JP2014507136A - NaV1.7ノックアウトマウスとそれらの使用 - Google Patents
NaV1.7ノックアウトマウスとそれらの使用 Download PDFInfo
- Publication number
- JP2014507136A JP2014507136A JP2013550571A JP2013550571A JP2014507136A JP 2014507136 A JP2014507136 A JP 2014507136A JP 2013550571 A JP2013550571 A JP 2013550571A JP 2013550571 A JP2013550571 A JP 2013550571A JP 2014507136 A JP2014507136 A JP 2014507136A
- Authority
- JP
- Japan
- Prior art keywords
- mouse
- pain
- knockout
- global
- mice
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 238000011813 knockout mouse model Methods 0.000 title claims abstract description 64
- 210000004027 cell Anatomy 0.000 claims abstract description 235
- 241000699666 Mus <mouse, genus> Species 0.000 claims abstract description 119
- 238000000034 method Methods 0.000 claims abstract description 115
- 238000012360 testing method Methods 0.000 claims abstract description 87
- 150000001875 compounds Chemical class 0.000 claims abstract description 43
- 238000003556 assay Methods 0.000 claims abstract description 32
- 210000004408 hybridoma Anatomy 0.000 claims abstract description 24
- 238000009395 breeding Methods 0.000 claims abstract description 16
- 230000001488 breeding effect Effects 0.000 claims abstract description 14
- 206010035226 Plasma cell myeloma Diseases 0.000 claims abstract description 11
- 201000000050 myeloid neoplasm Diseases 0.000 claims abstract description 11
- 230000004927 fusion Effects 0.000 claims abstract description 10
- 238000004113 cell culture Methods 0.000 claims abstract description 7
- 210000000056 organ Anatomy 0.000 claims abstract description 6
- 108090000623 proteins and genes Proteins 0.000 claims description 171
- 241000699670 Mus sp. Species 0.000 claims description 138
- 208000002193 Pain Diseases 0.000 claims description 108
- 230000036407 pain Effects 0.000 claims description 103
- 241000124008 Mammalia Species 0.000 claims description 95
- 230000004044 response Effects 0.000 claims description 63
- 102000004169 proteins and genes Human genes 0.000 claims description 59
- FVECELJHCSPHKY-UHFFFAOYSA-N Veratridine Natural products C1=C(OC)C(OC)=CC=C1C(=O)OC1C2(O)OC34CC5(O)C(CN6C(CCC(C)C6)C6(C)O)C6(O)C(O)CC5(O)C4CCC2C3(C)CC1 FVECELJHCSPHKY-UHFFFAOYSA-N 0.000 claims description 43
- FVECELJHCSPHKY-JLSHOZRYSA-N veratridine Chemical group C1=C(OC)C(OC)=CC=C1C(=O)O[C@@H]1[C@@]2(O)O[C@]34C[C@@]5(O)[C@H](CN6[C@@H](CC[C@H](C)C6)[C@@]6(C)O)[C@]6(O)[C@@H](O)C[C@@]5(O)[C@@H]4CC[C@H]2[C@]3(C)CC1 FVECELJHCSPHKY-JLSHOZRYSA-N 0.000 claims description 43
- 241000700159 Rattus Species 0.000 claims description 32
- 241000894007 species Species 0.000 claims description 24
- 239000005892 Deltamethrin Substances 0.000 claims description 20
- 229960002483 decamethrin Drugs 0.000 claims description 20
- OWZREIFADZCYQD-NSHGMRRFSA-N deltamethrin Chemical compound CC1(C)[C@@H](C=C(Br)Br)[C@H]1C(=O)O[C@H](C#N)C1=CC=CC(OC=2C=CC=CC=2)=C1 OWZREIFADZCYQD-NSHGMRRFSA-N 0.000 claims description 20
- 238000004519 manufacturing process Methods 0.000 claims description 20
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 claims description 19
- 239000013642 negative control Substances 0.000 claims description 19
- 239000012190 activator Substances 0.000 claims description 17
- 230000002829 reductive effect Effects 0.000 claims description 12
- 241000283973 Oryctolagus cuniculus Species 0.000 claims description 8
- 210000002569 neuron Anatomy 0.000 claims description 8
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 6
- 230000007423 decrease Effects 0.000 claims description 6
- 210000003323 beak Anatomy 0.000 claims description 3
- 235000016709 nutrition Nutrition 0.000 claims description 3
- 230000035764 nutrition Effects 0.000 claims description 3
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 2
- 230000001939 inductive effect Effects 0.000 claims description 2
- 230000009984 peri-natal effect Effects 0.000 claims description 2
- 206010013952 Dysphonia Diseases 0.000 claims 3
- 208000010473 Hoarseness Diseases 0.000 claims 3
- 241000131091 Lucanus cervus Species 0.000 claims 1
- 241000009328 Perro Species 0.000 claims 1
- 239000003112 inhibitor Substances 0.000 abstract description 17
- 238000012216 screening Methods 0.000 abstract description 7
- 241001465754 Metazoa Species 0.000 description 137
- 108700019146 Transgenes Proteins 0.000 description 99
- 108020004414 DNA Proteins 0.000 description 79
- 239000013598 vector Substances 0.000 description 60
- 108090000765 processed proteins & peptides Proteins 0.000 description 53
- 235000018102 proteins Nutrition 0.000 description 52
- 150000007523 nucleic acids Chemical class 0.000 description 46
- 125000003729 nucleotide group Chemical group 0.000 description 45
- 229920001184 polypeptide Polymers 0.000 description 43
- 102000004196 processed proteins & peptides Human genes 0.000 description 43
- 230000000694 effects Effects 0.000 description 41
- 108091028043 Nucleic acid sequence Proteins 0.000 description 40
- 230000014509 gene expression Effects 0.000 description 39
- 239000002773 nucleotide Substances 0.000 description 39
- 239000013615 primer Substances 0.000 description 38
- 230000006399 behavior Effects 0.000 description 36
- 102000039446 nucleic acids Human genes 0.000 description 36
- 108020004707 nucleic acids Proteins 0.000 description 36
- 102000018674 Sodium Channels Human genes 0.000 description 34
- 108010052164 Sodium Channels Proteins 0.000 description 34
- 238000003752 polymerase chain reaction Methods 0.000 description 34
- 101150080511 Scn9a gene Proteins 0.000 description 31
- 239000000203 mixture Substances 0.000 description 29
- WZUVPPKBWHMQCE-UHFFFAOYSA-N Haematoxylin Chemical compound C12=CC(O)=C(O)C=C2CC2(O)C1C1=CC=C(O)C(O)=C1OC2 WZUVPPKBWHMQCE-UHFFFAOYSA-N 0.000 description 28
- 210000001519 tissue Anatomy 0.000 description 27
- 102100031367 Sodium channel protein type 9 subunit alpha Human genes 0.000 description 26
- 230000027455 binding Effects 0.000 description 26
- 238000006243 chemical reaction Methods 0.000 description 26
- 210000001161 mammalian embryo Anatomy 0.000 description 26
- 239000003550 marker Substances 0.000 description 26
- 230000013011 mating Effects 0.000 description 23
- 230000009261 transgenic effect Effects 0.000 description 23
- 238000011282 treatment Methods 0.000 description 22
- 230000035772 mutation Effects 0.000 description 21
- 239000000047 product Substances 0.000 description 20
- 239000000243 solution Substances 0.000 description 20
- 230000000638 stimulation Effects 0.000 description 20
- 101000654386 Homo sapiens Sodium channel protein type 9 subunit alpha Proteins 0.000 description 19
- 239000000427 antigen Substances 0.000 description 19
- 108091007433 antigens Proteins 0.000 description 19
- 102000036639 antigens Human genes 0.000 description 19
- 239000003795 chemical substances by application Substances 0.000 description 19
- 239000002299 complementary DNA Substances 0.000 description 19
- 208000004296 neuralgia Diseases 0.000 description 19
- 239000003814 drug Substances 0.000 description 18
- 238000002347 injection Methods 0.000 description 18
- 239000007924 injection Substances 0.000 description 18
- 238000003780 insertion Methods 0.000 description 18
- 230000037431 insertion Effects 0.000 description 18
- 235000013336 milk Nutrition 0.000 description 18
- 239000008267 milk Substances 0.000 description 18
- 210000004080 milk Anatomy 0.000 description 18
- 208000021722 neuropathic pain Diseases 0.000 description 18
- 239000000523 sample Substances 0.000 description 18
- 230000006870 function Effects 0.000 description 17
- 230000036541 health Effects 0.000 description 17
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 16
- 239000012634 fragment Substances 0.000 description 16
- 238000000338 in vitro Methods 0.000 description 16
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 16
- 108060003951 Immunoglobulin Proteins 0.000 description 15
- 210000003169 central nervous system Anatomy 0.000 description 15
- 230000012010 growth Effects 0.000 description 15
- 210000001428 peripheral nervous system Anatomy 0.000 description 15
- 229940079593 drug Drugs 0.000 description 14
- YQGOJNYOYNNSMM-UHFFFAOYSA-N eosin Chemical compound [Na+].OC(=O)C1=CC=CC=C1C1=C2C=C(Br)C(=O)C(Br)=C2OC2=C(Br)C(O)=C(Br)C=C21 YQGOJNYOYNNSMM-UHFFFAOYSA-N 0.000 description 14
- 102000018358 immunoglobulin Human genes 0.000 description 14
- 238000010369 molecular cloning Methods 0.000 description 14
- 230000035897 transcription Effects 0.000 description 14
- 238000013518 transcription Methods 0.000 description 14
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 13
- 108091026890 Coding region Proteins 0.000 description 13
- 230000003321 amplification Effects 0.000 description 13
- 230000028161 membrane depolarization Effects 0.000 description 13
- 238000003199 nucleic acid amplification method Methods 0.000 description 13
- 238000002360 preparation method Methods 0.000 description 13
- 108091008146 restriction endonucleases Proteins 0.000 description 13
- 239000011734 sodium Substances 0.000 description 13
- VLPIATFUUWWMKC-SNVBAGLBSA-N (2r)-1-(2,6-dimethylphenoxy)propan-2-amine Chemical compound C[C@@H](N)COC1=C(C)C=CC=C1C VLPIATFUUWWMKC-SNVBAGLBSA-N 0.000 description 12
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 12
- 208000004454 Hyperalgesia Diseases 0.000 description 12
- 229930193140 Neomycin Natural products 0.000 description 12
- 108091034117 Oligonucleotide Proteins 0.000 description 12
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 12
- 201000010099 disease Diseases 0.000 description 12
- 239000012153 distilled water Substances 0.000 description 12
- 230000002068 genetic effect Effects 0.000 description 12
- 229960003404 mexiletine Drugs 0.000 description 12
- 229960004927 neomycin Drugs 0.000 description 12
- 230000002441 reversible effect Effects 0.000 description 12
- 210000003594 spinal ganglia Anatomy 0.000 description 12
- NTYJJOPFIAHURM-UHFFFAOYSA-N Histamine Chemical compound NCCC1=CN=CN1 NTYJJOPFIAHURM-UHFFFAOYSA-N 0.000 description 11
- 108010076504 Protein Sorting Signals Proteins 0.000 description 11
- 108010053752 Voltage-Gated Sodium Channels Proteins 0.000 description 11
- 102000016913 Voltage-Gated Sodium Channels Human genes 0.000 description 11
- 238000010367 cloning Methods 0.000 description 11
- 235000013305 food Nutrition 0.000 description 11
- 238000001727 in vivo Methods 0.000 description 11
- 102000040430 polynucleotide Human genes 0.000 description 11
- 108091033319 polynucleotide Proteins 0.000 description 11
- 239000002157 polynucleotide Substances 0.000 description 11
- 230000008569 process Effects 0.000 description 11
- 230000004913 activation Effects 0.000 description 10
- 230000003115 biocidal effect Effects 0.000 description 10
- 239000000090 biomarker Substances 0.000 description 10
- 239000005018 casein Substances 0.000 description 10
- BECPQYXYKAMYBN-UHFFFAOYSA-N casein, tech. Chemical compound NCCCCC(C(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(CC(C)C)N=C(O)C(CCC(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(C(C)O)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(COP(O)(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(N)CC1=CC=CC=C1 BECPQYXYKAMYBN-UHFFFAOYSA-N 0.000 description 10
- 235000021240 caseins Nutrition 0.000 description 10
- 238000011161 development Methods 0.000 description 10
- 230000018109 developmental process Effects 0.000 description 10
- 239000013604 expression vector Substances 0.000 description 10
- 230000001965 increasing effect Effects 0.000 description 10
- 230000009885 systemic effect Effects 0.000 description 10
- 230000000451 tissue damage Effects 0.000 description 10
- 231100000827 tissue damage Toxicity 0.000 description 10
- 210000001835 viscera Anatomy 0.000 description 10
- 108020004705 Codon Proteins 0.000 description 9
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 9
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 9
- 238000004458 analytical method Methods 0.000 description 9
- 210000002257 embryonic structure Anatomy 0.000 description 9
- 210000004209 hair Anatomy 0.000 description 9
- 230000001404 mediated effect Effects 0.000 description 9
- 238000009401 outcrossing Methods 0.000 description 9
- 238000002823 phage display Methods 0.000 description 9
- 239000003195 sodium channel blocking agent Substances 0.000 description 9
- 238000010561 standard procedure Methods 0.000 description 9
- 102000004190 Enzymes Human genes 0.000 description 8
- 108090000790 Enzymes Proteins 0.000 description 8
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 8
- 241000283984 Rodentia Species 0.000 description 8
- 239000003242 anti bacterial agent Substances 0.000 description 8
- 230000001419 dependent effect Effects 0.000 description 8
- 238000005516 engineering process Methods 0.000 description 8
- 239000003623 enhancer Substances 0.000 description 8
- 239000002609 medium Substances 0.000 description 8
- BQJCRHHNABKAKU-KBQPJGBKSA-N morphine Chemical compound O([C@H]1[C@H](C=C[C@H]23)O)C4=C5[C@@]12CCN(C)[C@@H]3CC5=CC=C4O BQJCRHHNABKAKU-KBQPJGBKSA-N 0.000 description 8
- 239000008188 pellet Substances 0.000 description 8
- 239000013612 plasmid Substances 0.000 description 8
- 238000000746 purification Methods 0.000 description 8
- 229910052708 sodium Inorganic materials 0.000 description 8
- 230000004083 survival effect Effects 0.000 description 8
- 238000001890 transfection Methods 0.000 description 8
- 241000219321 Caryophyllaceae Species 0.000 description 7
- 108091006146 Channels Proteins 0.000 description 7
- 102000053602 DNA Human genes 0.000 description 7
- 235000002845 Dianthus plumarius Nutrition 0.000 description 7
- 241000282412 Homo Species 0.000 description 7
- 101710097939 Sodium channel protein type 9 subunit alpha Proteins 0.000 description 7
- 238000002105 Southern blotting Methods 0.000 description 7
- 125000003275 alpha amino acid group Chemical group 0.000 description 7
- 210000003855 cell nucleus Anatomy 0.000 description 7
- 210000001638 cerebellum Anatomy 0.000 description 7
- 239000000306 component Substances 0.000 description 7
- 210000001671 embryonic stem cell Anatomy 0.000 description 7
- 210000004602 germ cell Anatomy 0.000 description 7
- 210000001320 hippocampus Anatomy 0.000 description 7
- 230000016784 immunoglobulin production Effects 0.000 description 7
- 210000003734 kidney Anatomy 0.000 description 7
- 230000004048 modification Effects 0.000 description 7
- 238000012986 modification Methods 0.000 description 7
- 210000000956 olfactory bulb Anatomy 0.000 description 7
- 230000007170 pathology Effects 0.000 description 7
- 239000002953 phosphate buffered saline Substances 0.000 description 7
- 238000011160 research Methods 0.000 description 7
- 239000003378 sodium channel stimulating agent Substances 0.000 description 7
- 210000000278 spinal cord Anatomy 0.000 description 7
- CFMYXEVWODSLAX-QOZOJKKESA-N tetrodotoxin Chemical compound O([C@@]([C@H]1O)(O)O[C@H]2[C@@]3(O)CO)[C@H]3[C@@H](O)[C@]11[C@H]2[C@@H](O)N=C(N)N1 CFMYXEVWODSLAX-QOZOJKKESA-N 0.000 description 7
- 229950010357 tetrodotoxin Drugs 0.000 description 7
- CFMYXEVWODSLAX-UHFFFAOYSA-N tetrodotoxin Natural products C12C(O)NC(=N)NC2(C2O)C(O)C3C(CO)(O)C1OC2(O)O3 CFMYXEVWODSLAX-UHFFFAOYSA-N 0.000 description 7
- 238000012546 transfer Methods 0.000 description 7
- 241000894006 Bacteria Species 0.000 description 6
- 206010015150 Erythema Diseases 0.000 description 6
- 241000588724 Escherichia coli Species 0.000 description 6
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 6
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 6
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 6
- 206010065390 Inflammatory pain Diseases 0.000 description 6
- 206010060862 Prostate cancer Diseases 0.000 description 6
- 208000000236 Prostatic Neoplasms Diseases 0.000 description 6
- 208000003251 Pruritus Diseases 0.000 description 6
- 206010053552 allodynia Diseases 0.000 description 6
- 229940024606 amino acid Drugs 0.000 description 6
- 235000001014 amino acid Nutrition 0.000 description 6
- 150000001413 amino acids Chemical class 0.000 description 6
- 238000012217 deletion Methods 0.000 description 6
- 230000037430 deletion Effects 0.000 description 6
- 238000004520 electroporation Methods 0.000 description 6
- 238000012224 gene deletion Methods 0.000 description 6
- 229960001340 histamine Drugs 0.000 description 6
- 230000003053 immunization Effects 0.000 description 6
- 238000002649 immunization Methods 0.000 description 6
- 230000002779 inactivation Effects 0.000 description 6
- 230000033001 locomotion Effects 0.000 description 6
- 108020004999 messenger RNA Proteins 0.000 description 6
- 230000003389 potentiating effect Effects 0.000 description 6
- 230000001105 regulatory effect Effects 0.000 description 6
- 230000002336 repolarization Effects 0.000 description 6
- 238000006748 scratching Methods 0.000 description 6
- 230000002393 scratching effect Effects 0.000 description 6
- 229940125794 sodium channel blocker Drugs 0.000 description 6
- 239000011780 sodium chloride Substances 0.000 description 6
- 239000000126 substance Substances 0.000 description 6
- 241000282472 Canis lupus familiaris Species 0.000 description 5
- 108091092195 Intron Proteins 0.000 description 5
- 206010028980 Neoplasm Diseases 0.000 description 5
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 5
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 5
- 230000001154 acute effect Effects 0.000 description 5
- 238000013459 approach Methods 0.000 description 5
- 230000005540 biological transmission Effects 0.000 description 5
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 5
- 230000000295 complement effect Effects 0.000 description 5
- 231100000321 erythema Toxicity 0.000 description 5
- 238000011156 evaluation Methods 0.000 description 5
- 210000003128 head Anatomy 0.000 description 5
- 229940088597 hormone Drugs 0.000 description 5
- 239000005556 hormone Substances 0.000 description 5
- 230000007803 itching Effects 0.000 description 5
- 210000004072 lung Anatomy 0.000 description 5
- 210000004962 mammalian cell Anatomy 0.000 description 5
- 239000012528 membrane Substances 0.000 description 5
- 238000000520 microinjection Methods 0.000 description 5
- 210000003928 nasal cavity Anatomy 0.000 description 5
- 210000000929 nociceptor Anatomy 0.000 description 5
- 108091008700 nociceptors Proteins 0.000 description 5
- 230000021368 organ growth Effects 0.000 description 5
- 230000008058 pain sensation Effects 0.000 description 5
- 210000000813 small intestine Anatomy 0.000 description 5
- 238000006467 substitution reaction Methods 0.000 description 5
- 230000001629 suppression Effects 0.000 description 5
- 238000003786 synthesis reaction Methods 0.000 description 5
- 230000001225 therapeutic effect Effects 0.000 description 5
- 230000005030 transcription termination Effects 0.000 description 5
- 238000013519 translation Methods 0.000 description 5
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 4
- 108700024394 Exon Proteins 0.000 description 4
- UGJMXCAKCUNAIE-UHFFFAOYSA-N Gabapentin Chemical compound OC(=O)CC1(CN)CCCCC1 UGJMXCAKCUNAIE-UHFFFAOYSA-N 0.000 description 4
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 4
- 241000238631 Hexapoda Species 0.000 description 4
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 4
- 108090000862 Ion Channels Proteins 0.000 description 4
- 102000004310 Ion Channels Human genes 0.000 description 4
- 108020004511 Recombinant DNA Proteins 0.000 description 4
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 4
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 4
- 208000005298 acute pain Diseases 0.000 description 4
- 239000002671 adjuvant Substances 0.000 description 4
- 229940088710 antibiotic agent Drugs 0.000 description 4
- 239000001961 anticonvulsive agent Substances 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 201000011510 cancer Diseases 0.000 description 4
- 230000008859 change Effects 0.000 description 4
- 238000001514 detection method Methods 0.000 description 4
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 4
- 208000035475 disorder Diseases 0.000 description 4
- 235000013601 eggs Nutrition 0.000 description 4
- 108020001507 fusion proteins Proteins 0.000 description 4
- 102000037865 fusion proteins Human genes 0.000 description 4
- 239000000499 gel Substances 0.000 description 4
- 239000011521 glass Substances 0.000 description 4
- 239000001963 growth medium Substances 0.000 description 4
- 230000028993 immune response Effects 0.000 description 4
- 230000002757 inflammatory effect Effects 0.000 description 4
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 4
- 238000007912 intraperitoneal administration Methods 0.000 description 4
- 238000001990 intravenous administration Methods 0.000 description 4
- 210000003292 kidney cell Anatomy 0.000 description 4
- 239000003446 ligand Substances 0.000 description 4
- 235000013384 milk substitute Nutrition 0.000 description 4
- 229960005181 morphine Drugs 0.000 description 4
- 230000001850 reproductive effect Effects 0.000 description 4
- 238000007920 subcutaneous administration Methods 0.000 description 4
- 239000006228 supernatant Substances 0.000 description 4
- 230000000699 topical effect Effects 0.000 description 4
- ZEUITGRIYCTCEM-KRWDZBQOSA-N (S)-duloxetine Chemical compound C1([C@@H](OC=2C3=CC=CC=C3C=CC=2)CCNC)=CC=CS1 ZEUITGRIYCTCEM-KRWDZBQOSA-N 0.000 description 3
- AYQMNFRCBKOMIA-UHFFFAOYSA-N 1-benzazepin-2-one Chemical class O=C1C=CC=C2C=CC=CC2=N1 AYQMNFRCBKOMIA-UHFFFAOYSA-N 0.000 description 3
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 3
- 206010000060 Abdominal distension Diseases 0.000 description 3
- 229920000936 Agarose Polymers 0.000 description 3
- UHOVQNZJYSORNB-UHFFFAOYSA-N Benzene Chemical compound C1=CC=CC=C1 UHOVQNZJYSORNB-UHFFFAOYSA-N 0.000 description 3
- 241000282693 Cercopithecidae Species 0.000 description 3
- 208000006561 Cluster Headache Diseases 0.000 description 3
- 241000699800 Cricetinae Species 0.000 description 3
- 108091029865 Exogenous DNA Proteins 0.000 description 3
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 3
- 239000007995 HEPES buffer Substances 0.000 description 3
- 208000035154 Hyperesthesia Diseases 0.000 description 3
- 102100029098 Hypoxanthine-guanine phosphoribosyltransferase Human genes 0.000 description 3
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 3
- 208000026350 Inborn Genetic disease Diseases 0.000 description 3
- PJAAESPGJOSQGZ-DZGBDDFRSA-N Isovelleral Chemical compound O=CC1=C[C@@H]2CC(C)(C)C[C@@H]2[C@@]2(C)C[C@]21C=O PJAAESPGJOSQGZ-DZGBDDFRSA-N 0.000 description 3
- NNJVILVZKWQKPM-UHFFFAOYSA-N Lidocaine Chemical compound CCN(CC)CC(=O)NC1=C(C)C=CC=C1C NNJVILVZKWQKPM-UHFFFAOYSA-N 0.000 description 3
- 238000012408 PCR amplification Methods 0.000 description 3
- KWYUFKZDYYNOTN-UHFFFAOYSA-M Potassium hydroxide Chemical compound [OH-].[K+] KWYUFKZDYYNOTN-UHFFFAOYSA-M 0.000 description 3
- 102000003566 TRPV1 Human genes 0.000 description 3
- 229940126422 TRPV1 antagonist Drugs 0.000 description 3
- 101150016206 Trpv1 gene Proteins 0.000 description 3
- 125000000539 amino acid group Chemical group 0.000 description 3
- 239000005557 antagonist Substances 0.000 description 3
- 230000000890 antigenic effect Effects 0.000 description 3
- 230000001580 bacterial effect Effects 0.000 description 3
- 108010005774 beta-Galactosidase Proteins 0.000 description 3
- 230000004071 biological effect Effects 0.000 description 3
- 210000002459 blastocyst Anatomy 0.000 description 3
- 210000004369 blood Anatomy 0.000 description 3
- 239000008280 blood Substances 0.000 description 3
- 238000009835 boiling Methods 0.000 description 3
- 230000001413 cellular effect Effects 0.000 description 3
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 3
- 239000013599 cloning vector Substances 0.000 description 3
- 208000018912 cluster headache syndrome Diseases 0.000 description 3
- 239000013065 commercial product Substances 0.000 description 3
- 230000002950 deficient Effects 0.000 description 3
- 239000005547 deoxyribonucleotide Substances 0.000 description 3
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 3
- 229910003460 diamond Inorganic materials 0.000 description 3
- 239000010432 diamond Substances 0.000 description 3
- 231100000673 dose–response relationship Toxicity 0.000 description 3
- 229960002866 duloxetine Drugs 0.000 description 3
- 239000012636 effector Substances 0.000 description 3
- 238000001962 electrophoresis Methods 0.000 description 3
- 201000011384 erythromelalgia Diseases 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- 210000003414 extremity Anatomy 0.000 description 3
- 230000004720 fertilization Effects 0.000 description 3
- 238000000684 flow cytometry Methods 0.000 description 3
- 208000016361 genetic disease Diseases 0.000 description 3
- 230000037308 hair color Effects 0.000 description 3
- 230000006801 homologous recombination Effects 0.000 description 3
- 238000002744 homologous recombination Methods 0.000 description 3
- 230000002163 immunogen Effects 0.000 description 3
- 230000000415 inactivating effect Effects 0.000 description 3
- 230000004941 influx Effects 0.000 description 3
- 230000005764 inhibitory process Effects 0.000 description 3
- 238000007918 intramuscular administration Methods 0.000 description 3
- 231100000518 lethal Toxicity 0.000 description 3
- 230000001665 lethal effect Effects 0.000 description 3
- 231100000225 lethality Toxicity 0.000 description 3
- 229960004194 lidocaine Drugs 0.000 description 3
- 210000004698 lymphocyte Anatomy 0.000 description 3
- 238000007726 management method Methods 0.000 description 3
- 230000002981 neuropathic effect Effects 0.000 description 3
- 235000015097 nutrients Nutrition 0.000 description 3
- 239000002751 oligonucleotide probe Substances 0.000 description 3
- 230000036961 partial effect Effects 0.000 description 3
- 230000002085 persistent effect Effects 0.000 description 3
- 230000008488 polyadenylation Effects 0.000 description 3
- NLKNQRATVPKPDG-UHFFFAOYSA-M potassium iodide Chemical compound [K+].[I-] NLKNQRATVPKPDG-UHFFFAOYSA-M 0.000 description 3
- 239000002987 primer (paints) Substances 0.000 description 3
- 238000012545 processing Methods 0.000 description 3
- 238000011084 recovery Methods 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- RPQXVSUAYFXFJA-HGRQIUPRSA-N saxitoxin Chemical compound NC(=O)OC[C@@H]1N=C(N)N2CCC(O)(O)[C@@]22N=C(N)N[C@@H]12 RPQXVSUAYFXFJA-HGRQIUPRSA-N 0.000 description 3
- RPQXVSUAYFXFJA-UHFFFAOYSA-N saxitoxin hydrate Natural products NC(=O)OCC1N=C(N)N2CCC(O)(O)C22NC(N)=NC12 RPQXVSUAYFXFJA-UHFFFAOYSA-N 0.000 description 3
- 230000003248 secreting effect Effects 0.000 description 3
- 230000028327 secretion Effects 0.000 description 3
- 230000035945 sensitivity Effects 0.000 description 3
- 210000002966 serum Anatomy 0.000 description 3
- 210000002784 stomach Anatomy 0.000 description 3
- 239000013589 supplement Substances 0.000 description 3
- 239000000725 suspension Substances 0.000 description 3
- 238000002560 therapeutic procedure Methods 0.000 description 3
- 230000001131 transforming effect Effects 0.000 description 3
- 238000011830 transgenic mouse model Methods 0.000 description 3
- 238000001262 western blot Methods 0.000 description 3
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 2
- HZAXFHJVJLSVMW-UHFFFAOYSA-N 2-Aminoethan-1-ol Chemical compound NCCO HZAXFHJVJLSVMW-UHFFFAOYSA-N 0.000 description 2
- HNXQXTQTPAJEJL-UHFFFAOYSA-N 2-aminopteridin-4-ol Chemical compound C1=CN=C2NC(N)=NC(=O)C2=N1 HNXQXTQTPAJEJL-UHFFFAOYSA-N 0.000 description 2
- VHPXWPAGHGHDCK-UHFFFAOYSA-N 5-chloro-4-[(3,4-dichlorophenoxy)methyl]-2-fluoro-n-methylsulfonylbenzamide Chemical compound C1=C(F)C(C(=O)NS(=O)(=O)C)=CC(Cl)=C1COC1=CC=C(Cl)C(Cl)=C1 VHPXWPAGHGHDCK-UHFFFAOYSA-N 0.000 description 2
- 206010001497 Agitation Diseases 0.000 description 2
- 208000031873 Animal Disease Models Diseases 0.000 description 2
- 206010003445 Ascites Diseases 0.000 description 2
- 241000228212 Aspergillus Species 0.000 description 2
- SPFYMRJSYKOXGV-UHFFFAOYSA-N Baytril Chemical compound C1CN(CC)CCN1C(C(=C1)F)=CC2=C1C(=O)C(C(O)=O)=CN2C1CC1 SPFYMRJSYKOXGV-UHFFFAOYSA-N 0.000 description 2
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 2
- 102000004506 Blood Proteins Human genes 0.000 description 2
- 108010017384 Blood Proteins Proteins 0.000 description 2
- 241000255789 Bombyx mori Species 0.000 description 2
- VTYYLEPIZMXCLO-UHFFFAOYSA-L Calcium carbonate Chemical compound [Ca+2].[O-]C([O-])=O VTYYLEPIZMXCLO-UHFFFAOYSA-L 0.000 description 2
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 2
- HEDRZPFGACZZDS-UHFFFAOYSA-N Chloroform Chemical compound ClC(Cl)Cl HEDRZPFGACZZDS-UHFFFAOYSA-N 0.000 description 2
- 108020004635 Complementary DNA Proteins 0.000 description 2
- 102000004594 DNA Polymerase I Human genes 0.000 description 2
- 108010017826 DNA Polymerase I Proteins 0.000 description 2
- 239000003155 DNA primer Substances 0.000 description 2
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 2
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 2
- 201000004624 Dermatitis Diseases 0.000 description 2
- 241000255925 Diptera Species 0.000 description 2
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 2
- 238000002965 ELISA Methods 0.000 description 2
- 241000196324 Embryophyta Species 0.000 description 2
- 102400001368 Epidermal growth factor Human genes 0.000 description 2
- 101800003838 Epidermal growth factor Proteins 0.000 description 2
- 241000206602 Eukaryota Species 0.000 description 2
- 108010008177 Fd immunoglobulins Proteins 0.000 description 2
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 2
- 206010019233 Headaches Diseases 0.000 description 2
- 101000654356 Homo sapiens Sodium channel protein type 10 subunit alpha Proteins 0.000 description 2
- 102000002265 Human Growth Hormone Human genes 0.000 description 2
- 108010000521 Human Growth Hormone Proteins 0.000 description 2
- 239000000854 Human Growth Hormone Substances 0.000 description 2
- XQFRJNBWHJMXHO-RRKCRQDMSA-N IDUR Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(I)=C1 XQFRJNBWHJMXHO-RRKCRQDMSA-N 0.000 description 2
- 206010061218 Inflammation Diseases 0.000 description 2
- 102000004877 Insulin Human genes 0.000 description 2
- 108090001061 Insulin Proteins 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- 241001529936 Murinae Species 0.000 description 2
- 241000699660 Mus musculus Species 0.000 description 2
- 229940124777 Nav1.7 inhibitor Drugs 0.000 description 2
- 208000028389 Nerve injury Diseases 0.000 description 2
- 108020005187 Oligonucleotide Probes Proteins 0.000 description 2
- 102000015636 Oligopeptides Human genes 0.000 description 2
- 108010038807 Oligopeptides Proteins 0.000 description 2
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Chemical compound OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 2
- 241000288906 Primates Species 0.000 description 2
- 239000012980 RPMI-1640 medium Substances 0.000 description 2
- VYGQUTWHTHXGQB-FFHKNEKCSA-N Retinol Palmitate Chemical compound CCCCCCCCCCCCCCCC(=O)OC\C=C(/C)\C=C\C=C(/C)\C=C\C1=C(C)CCCC1(C)C VYGQUTWHTHXGQB-FFHKNEKCSA-N 0.000 description 2
- 108091028664 Ribonucleotide Proteins 0.000 description 2
- 241000607142 Salmonella Species 0.000 description 2
- 206010039897 Sedation Diseases 0.000 description 2
- 241000607720 Serratia Species 0.000 description 2
- 102100031374 Sodium channel protein type 10 subunit alpha Human genes 0.000 description 2
- 108010062740 TRPV Cation Channels Proteins 0.000 description 2
- 102000011040 TRPV Cation Channels Human genes 0.000 description 2
- 108010025083 TRPV1 receptor Proteins 0.000 description 2
- 101710120037 Toxin CcdB Proteins 0.000 description 2
- 108091023040 Transcription factor Proteins 0.000 description 2
- 102000040945 Transcription factor Human genes 0.000 description 2
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 2
- 241000700605 Viruses Species 0.000 description 2
- 206010048010 Withdrawal syndrome Diseases 0.000 description 2
- 230000036982 action potential Effects 0.000 description 2
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 2
- 238000001042 affinity chromatography Methods 0.000 description 2
- 230000000202 analgesic effect Effects 0.000 description 2
- 229940035676 analgesics Drugs 0.000 description 2
- 238000011558 animal model by disease Methods 0.000 description 2
- 230000003042 antagnostic effect Effects 0.000 description 2
- 239000000730 antalgic agent Substances 0.000 description 2
- 210000000628 antibody-producing cell Anatomy 0.000 description 2
- 229940125681 anticonvulsant agent Drugs 0.000 description 2
- 229940105596 baytril Drugs 0.000 description 2
- 230000003542 behavioural effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 102000005936 beta-Galactosidase Human genes 0.000 description 2
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 2
- 238000010170 biological method Methods 0.000 description 2
- 230000033228 biological regulation Effects 0.000 description 2
- 230000037396 body weight Effects 0.000 description 2
- 230000004641 brain development Effects 0.000 description 2
- AIYUHDOJVYHVIT-UHFFFAOYSA-M caesium chloride Chemical compound [Cl-].[Cs+] AIYUHDOJVYHVIT-UHFFFAOYSA-M 0.000 description 2
- HUCVOHYBFXVBRW-UHFFFAOYSA-M caesium hydroxide Chemical compound [OH-].[Cs+] HUCVOHYBFXVBRW-UHFFFAOYSA-M 0.000 description 2
- 239000001506 calcium phosphate Substances 0.000 description 2
- 229910000389 calcium phosphate Inorganic materials 0.000 description 2
- 235000011010 calcium phosphates Nutrition 0.000 description 2
- YKPUWZUDDOIDPM-SOFGYWHQSA-N capsaicin Chemical compound COC1=CC(CNC(=O)CCCC\C=C\C(C)C)=CC=C1O YKPUWZUDDOIDPM-SOFGYWHQSA-N 0.000 description 2
- 230000010261 cell growth Effects 0.000 description 2
- 210000000170 cell membrane Anatomy 0.000 description 2
- 230000004663 cell proliferation Effects 0.000 description 2
- 238000005119 centrifugation Methods 0.000 description 2
- 239000003153 chemical reaction reagent Substances 0.000 description 2
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 2
- 230000002759 chromosomal effect Effects 0.000 description 2
- 230000021615 conjugation Effects 0.000 description 2
- 238000007796 conventional method Methods 0.000 description 2
- 230000007547 defect Effects 0.000 description 2
- 230000003111 delayed effect Effects 0.000 description 2
- 230000002999 depolarising effect Effects 0.000 description 2
- 238000013461 design Methods 0.000 description 2
- 238000000502 dialysis Methods 0.000 description 2
- 230000029087 digestion Effects 0.000 description 2
- 230000008034 disappearance Effects 0.000 description 2
- 229940116977 epidermal growth factor Drugs 0.000 description 2
- 206010015037 epilepsy Diseases 0.000 description 2
- 230000004634 feeding behavior Effects 0.000 description 2
- 238000000855 fermentation Methods 0.000 description 2
- 230000004151 fermentation Effects 0.000 description 2
- 239000012467 final product Substances 0.000 description 2
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 2
- 101150046339 fur gene Proteins 0.000 description 2
- 229960002870 gabapentin Drugs 0.000 description 2
- 102000054766 genetic haplotypes Human genes 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- 230000036449 good health Effects 0.000 description 2
- 239000003102 growth factor Substances 0.000 description 2
- 231100000869 headache Toxicity 0.000 description 2
- 238000009396 hybridization Methods 0.000 description 2
- 238000012872 hydroxylapatite chromatography Methods 0.000 description 2
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 2
- 230000005847 immunogenicity Effects 0.000 description 2
- 229940072221 immunoglobulins Drugs 0.000 description 2
- 208000019014 inability to feed Diseases 0.000 description 2
- 238000011534 incubation Methods 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 208000021267 infertility disease Diseases 0.000 description 2
- 230000004054 inflammatory process Effects 0.000 description 2
- 238000012966 insertion method Methods 0.000 description 2
- 229940125396 insulin Drugs 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 238000002955 isolation Methods 0.000 description 2
- 230000006651 lactation Effects 0.000 description 2
- 230000000670 limiting effect Effects 0.000 description 2
- 238000001638 lipofection Methods 0.000 description 2
- 230000004807 localization Effects 0.000 description 2
- 239000000463 material Substances 0.000 description 2
- 230000004060 metabolic process Effects 0.000 description 2
- MYWUZJCMWCOHBA-VIFPVBQESA-N methamphetamine Chemical compound CN[C@@H](C)CC1=CC=CC=C1 MYWUZJCMWCOHBA-VIFPVBQESA-N 0.000 description 2
- 244000005700 microbiome Species 0.000 description 2
- 210000005036 nerve Anatomy 0.000 description 2
- 230000008764 nerve damage Effects 0.000 description 2
- 210000000653 nervous system Anatomy 0.000 description 2
- 230000001537 neural effect Effects 0.000 description 2
- 208000015706 neuroendocrine disease Diseases 0.000 description 2
- 230000003040 nociceptive effect Effects 0.000 description 2
- 230000009437 off-target effect Effects 0.000 description 2
- 239000003921 oil Substances 0.000 description 2
- 235000019198 oils Nutrition 0.000 description 2
- 238000009400 out breeding Methods 0.000 description 2
- 230000002018 overexpression Effects 0.000 description 2
- 230000037324 pain perception Effects 0.000 description 2
- 238000004091 panning Methods 0.000 description 2
- 230000001314 paroxysmal effect Effects 0.000 description 2
- 210000005223 peripheral sensory neuron Anatomy 0.000 description 2
- 230000000144 pharmacologic effect Effects 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 230000004481 post-translational protein modification Effects 0.000 description 2
- 238000003127 radioimmunoassay Methods 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 238000012827 research and development Methods 0.000 description 2
- 230000031070 response to heat Effects 0.000 description 2
- 238000012552 review Methods 0.000 description 2
- 239000002336 ribonucleotide Substances 0.000 description 2
- 125000002652 ribonucleotide group Chemical group 0.000 description 2
- 230000036280 sedation Effects 0.000 description 2
- 230000035807 sensation Effects 0.000 description 2
- 230000001953 sensory effect Effects 0.000 description 2
- 238000012163 sequencing technique Methods 0.000 description 2
- 210000000717 sertoli cell Anatomy 0.000 description 2
- PUZPDOWCWNUUKD-UHFFFAOYSA-M sodium fluoride Chemical compound [F-].[Na+] PUZPDOWCWNUUKD-UHFFFAOYSA-M 0.000 description 2
- 210000000952 spleen Anatomy 0.000 description 2
- 210000004989 spleen cell Anatomy 0.000 description 2
- 238000010254 subcutaneous injection Methods 0.000 description 2
- 239000007929 subcutaneous injection Substances 0.000 description 2
- 238000001356 surgical procedure Methods 0.000 description 2
- 230000002889 sympathetic effect Effects 0.000 description 2
- 208000011580 syndromic disease Diseases 0.000 description 2
- XOAAWQZATWQOTB-UHFFFAOYSA-N taurine Chemical compound NCCS(O)(=O)=O XOAAWQZATWQOTB-UHFFFAOYSA-N 0.000 description 2
- 208000001608 teratocarcinoma Diseases 0.000 description 2
- 229940124597 therapeutic agent Drugs 0.000 description 2
- 229940104230 thymidine Drugs 0.000 description 2
- 230000002103 transcriptional effect Effects 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 230000026683 transduction Effects 0.000 description 2
- 230000009466 transformation Effects 0.000 description 2
- 238000002054 transplantation Methods 0.000 description 2
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 2
- 210000002700 urine Anatomy 0.000 description 2
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 description 2
- 210000004291 uterus Anatomy 0.000 description 2
- PHIQHXFUZVPYII-ZCFIWIBFSA-N (R)-carnitine Chemical compound C[N+](C)(C)C[C@H](O)CC([O-])=O PHIQHXFUZVPYII-ZCFIWIBFSA-N 0.000 description 1
- GVJHHUAWPYXKBD-UHFFFAOYSA-N (±)-α-Tocopherol Chemical compound OC1=C(C)C(C)=C2OC(CCCC(C)CCCC(C)CCCC(C)C)(C)CCC2=C1C GVJHHUAWPYXKBD-UHFFFAOYSA-N 0.000 description 1
- LEBVLXFERQHONN-UHFFFAOYSA-N 1-butyl-N-(2,6-dimethylphenyl)piperidine-2-carboxamide Chemical compound CCCCN1CCCCC1C(=O)NC1=C(C)C=CC=C1C LEBVLXFERQHONN-UHFFFAOYSA-N 0.000 description 1
- 101150114688 1.7 gene Proteins 0.000 description 1
- FALRKNHUBBKYCC-UHFFFAOYSA-N 2-(chloromethyl)pyridine-3-carbonitrile Chemical compound ClCC1=NC=CC=C1C#N FALRKNHUBBKYCC-UHFFFAOYSA-N 0.000 description 1
- BFSVOASYOCHEOV-UHFFFAOYSA-N 2-diethylaminoethanol Chemical compound CCN(CC)CCO BFSVOASYOCHEOV-UHFFFAOYSA-N 0.000 description 1
- RPERJPYDELTDMR-UHFFFAOYSA-K 2-hydroxyethyl(trimethyl)azanium;2-hydroxypropane-1,2,3-tricarboxylate Chemical compound C[N+](C)(C)CCO.C[N+](C)(C)CCO.C[N+](C)(C)CCO.[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O RPERJPYDELTDMR-UHFFFAOYSA-K 0.000 description 1
- NCGRTBNMEWDOSH-UHFFFAOYSA-N 2-hydroxypropane-1,2,3-tricarboxylic acid;tetrahydrate Chemical compound O.O.O.O.OC(=O)CC(O)(C(O)=O)CC(O)=O NCGRTBNMEWDOSH-UHFFFAOYSA-N 0.000 description 1
- AGFIRQJZCNVMCW-UAKXSSHOSA-N 5-bromouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(Br)=C1 AGFIRQJZCNVMCW-UAKXSSHOSA-N 0.000 description 1
- 241000238876 Acari Species 0.000 description 1
- 206010069754 Acquired gene mutation Diseases 0.000 description 1
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 1
- 102000006822 Agouti Signaling Protein Human genes 0.000 description 1
- 108010072151 Agouti Signaling Protein Proteins 0.000 description 1
- WSVLPVUVIUVCRA-KPKNDVKVSA-N Alpha-lactose monohydrate Chemical compound O.O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O WSVLPVUVIUVCRA-KPKNDVKVSA-N 0.000 description 1
- 206010002653 Anosmia Diseases 0.000 description 1
- 241001203868 Autographa californica Species 0.000 description 1
- 241000193830 Bacillus <bacterium> Species 0.000 description 1
- 244000063299 Bacillus subtilis Species 0.000 description 1
- 235000014469 Bacillus subtilis Nutrition 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 208000026310 Breast neoplasm Diseases 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- WSNMPAVSZJSIMT-UHFFFAOYSA-N COc1c(C)c2COC(=O)c2c(O)c1CC(O)C1(C)CCC(=O)O1 Chemical compound COc1c(C)c2COC(=O)c2c(O)c1CC(O)C1(C)CCC(=O)O1 WSNMPAVSZJSIMT-UHFFFAOYSA-N 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- UXVMQQNJUSDDNG-UHFFFAOYSA-L Calcium chloride Chemical compound [Cl-].[Cl-].[Ca+2] UXVMQQNJUSDDNG-UHFFFAOYSA-L 0.000 description 1
- 239000001736 Calcium glycerylphosphate Substances 0.000 description 1
- 241000222120 Candida <Saccharomycetales> Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 206010008025 Cerebellar ataxia Diseases 0.000 description 1
- 206010008342 Cervix carcinoma Diseases 0.000 description 1
- 241000282552 Chlorocebus aethiops Species 0.000 description 1
- 208000000094 Chronic Pain Diseases 0.000 description 1
- 241000699802 Cricetulus griseus Species 0.000 description 1
- 241000484025 Cuniculus Species 0.000 description 1
- QNAYBMKLOCPYGJ-UWTATZPHSA-N D-alanine Chemical compound C[C@@H](N)C(O)=O QNAYBMKLOCPYGJ-UWTATZPHSA-N 0.000 description 1
- QNAYBMKLOCPYGJ-UHFFFAOYSA-N D-alpha-Ala Natural products CC([NH3+])C([O-])=O QNAYBMKLOCPYGJ-UHFFFAOYSA-N 0.000 description 1
- HMFHBZSHGGEWLO-SOOFDHNKSA-N D-ribofuranose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H]1O HMFHBZSHGGEWLO-SOOFDHNKSA-N 0.000 description 1
- 235000001815 DL-alpha-tocopherol Nutrition 0.000 description 1
- 239000011627 DL-alpha-tocopherol Substances 0.000 description 1
- 108020003215 DNA Probes Proteins 0.000 description 1
- 239000003298 DNA probe Substances 0.000 description 1
- 230000007018 DNA scission Effects 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 230000006820 DNA synthesis Effects 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 102100024746 Dihydrofolate reductase Human genes 0.000 description 1
- 206010059866 Drug resistance Diseases 0.000 description 1
- 241000588914 Enterobacter Species 0.000 description 1
- 241000588921 Enterobacteriaceae Species 0.000 description 1
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 241000588698 Erwinia Species 0.000 description 1
- 241000620209 Escherichia coli DH5[alpha] Species 0.000 description 1
- 206010015548 Euthanasia Diseases 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 241000724791 Filamentous phage Species 0.000 description 1
- 241000192125 Firmicutes Species 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 206010064571 Gene mutation Diseases 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 1
- 244000068988 Glycine max Species 0.000 description 1
- 235000010469 Glycine max Nutrition 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 229930189130 Grayanotoxin Natural products 0.000 description 1
- JPEBAJKDWYGOHM-UHFFFAOYSA-N Grayanotoxin VIII Natural products C1C(O)C2(O)C(C)(C)C(O)CC2C(=C)C2CCC3C(=C)CC21C3O JPEBAJKDWYGOHM-UHFFFAOYSA-N 0.000 description 1
- IHEDDHMJFFWQJA-UHFFFAOYSA-N Grayanotoxin XI Natural products C1C(O)C2(O)C(C)(C)C(O)CC2C(=C)C2CC(O)C3C(C)(O)CC21C3O IHEDDHMJFFWQJA-UHFFFAOYSA-N 0.000 description 1
- BWMFRQKICHXLSH-ZZZNUFMVSA-N Grayanotoxin-III Natural products O[C@H]1[C@@H]2[C@@](O)(C)C[C@]31[C@H]([C@](O)(C)[C@H]1[C@](O)([C@H](O)C3)C(C)(C)[C@@H](O)C1)CC2 BWMFRQKICHXLSH-ZZZNUFMVSA-N 0.000 description 1
- 241001272567 Hominoidea Species 0.000 description 1
- 101100148835 Homo sapiens SCN9A gene Proteins 0.000 description 1
- 101100273566 Humulus lupulus CCL10 gene Proteins 0.000 description 1
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 1
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 1
- 206010020843 Hyperthermia Diseases 0.000 description 1
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 1
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 1
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 1
- 206010062016 Immunosuppression Diseases 0.000 description 1
- UGQMRVRMYYASKQ-KQYNXXCUSA-N Inosine Chemical class O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(O)=C2N=C1 UGQMRVRMYYASKQ-KQYNXXCUSA-N 0.000 description 1
- 241000588748 Klebsiella Species 0.000 description 1
- LEVWYRKDKASIDU-IMJSIDKUSA-N L-cystine Chemical compound [O-]C(=O)[C@@H]([NH3+])CSSC[C@H]([NH3+])C([O-])=O LEVWYRKDKASIDU-IMJSIDKUSA-N 0.000 description 1
- 235000019393 L-cystine Nutrition 0.000 description 1
- 239000004158 L-cystine Substances 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- 241000282553 Macaca Species 0.000 description 1
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 1
- 208000002720 Malnutrition Diseases 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- ABSPRNADVQNDOU-UHFFFAOYSA-N Menaquinone 1 Natural products C1=CC=C2C(=O)C(CC=C(C)C)=C(C)C(=O)C2=C1 ABSPRNADVQNDOU-UHFFFAOYSA-N 0.000 description 1
- 241000699729 Muridae Species 0.000 description 1
- 101100148836 Mus musculus Scn9a gene Proteins 0.000 description 1
- 241000282339 Mustela Species 0.000 description 1
- 241000237536 Mytilus edulis Species 0.000 description 1
- NQTADLQHYWFPDB-UHFFFAOYSA-N N-Hydroxysuccinimide Chemical compound ON1C(=O)CCC1=O NQTADLQHYWFPDB-UHFFFAOYSA-N 0.000 description 1
- CMWTZPSULFXXJA-UHFFFAOYSA-N Naproxen Natural products C1=C(C(C)C(O)=O)C=CC2=CC(OC)=CC=C21 CMWTZPSULFXXJA-UHFFFAOYSA-N 0.000 description 1
- 241001045988 Neogene Species 0.000 description 1
- 208000012902 Nervous system disease Diseases 0.000 description 1
- 208000025966 Neurological disease Diseases 0.000 description 1
- 238000000636 Northern blotting Methods 0.000 description 1
- 108020004711 Nucleic Acid Probes Proteins 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 235000019482 Palm oil Nutrition 0.000 description 1
- 206010033799 Paralysis Diseases 0.000 description 1
- 208000001300 Perinatal Death Diseases 0.000 description 1
- 241001325166 Phacelia congesta Species 0.000 description 1
- CXOFVDLJLONNDW-UHFFFAOYSA-N Phenytoin Chemical compound N1C(=O)NC(=O)C1(C=1C=CC=CC=1)C1=CC=CC=C1 CXOFVDLJLONNDW-UHFFFAOYSA-N 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- 241000235648 Pichia Species 0.000 description 1
- 239000004698 Polyethylene Substances 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 208000012287 Prolapse Diseases 0.000 description 1
- 108010029485 Protein Isoforms Proteins 0.000 description 1
- 102000001708 Protein Isoforms Human genes 0.000 description 1
- 241000588769 Proteus <enterobacteria> Species 0.000 description 1
- 241000589516 Pseudomonas Species 0.000 description 1
- 230000026279 RNA modification Effects 0.000 description 1
- 102000004879 Racemases and epimerases Human genes 0.000 description 1
- 108090001066 Racemases and epimerases Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- VYGQUTWHTHXGQB-UHFFFAOYSA-N Retinol hexadecanoate Natural products CCCCCCCCCCCCCCCC(=O)OCC=C(C)C=CC=C(C)C=CC1=C(C)CCCC1(C)C VYGQUTWHTHXGQB-UHFFFAOYSA-N 0.000 description 1
- PYMYPHUHKUWMLA-LMVFSUKVSA-N Ribose Natural products OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PYMYPHUHKUWMLA-LMVFSUKVSA-N 0.000 description 1
- 241000235070 Saccharomyces Species 0.000 description 1
- 241000235343 Saccharomycetales Species 0.000 description 1
- 241000831652 Salinivibrio sharmensis Species 0.000 description 1
- 241000235347 Schizosaccharomyces pombe Species 0.000 description 1
- 241000311088 Schwanniomyces Species 0.000 description 1
- 229920002684 Sepharose Polymers 0.000 description 1
- 241000607768 Shigella Species 0.000 description 1
- FKNQFGJONOIPTF-UHFFFAOYSA-N Sodium cation Chemical class [Na+] FKNQFGJONOIPTF-UHFFFAOYSA-N 0.000 description 1
- 229940089973 Sodium channel antagonist Drugs 0.000 description 1
- 208000027520 Somatoform disease Diseases 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 241000255588 Tephritidae Species 0.000 description 1
- 239000004098 Tetracycline Substances 0.000 description 1
- 108010034949 Thyroglobulin Proteins 0.000 description 1
- 102000009843 Thyroglobulin Human genes 0.000 description 1
- 108700009124 Transcription Initiation Site Proteins 0.000 description 1
- 102000004338 Transferrin Human genes 0.000 description 1
- 108090000901 Transferrin Proteins 0.000 description 1
- 206010044565 Tremor Diseases 0.000 description 1
- 241000223259 Trichoderma Species 0.000 description 1
- 101710162629 Trypsin inhibitor Proteins 0.000 description 1
- 229940122618 Trypsin inhibitor Drugs 0.000 description 1
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 description 1
- 108700005077 Viral Genes Proteins 0.000 description 1
- 108010046377 Whey Proteins Proteins 0.000 description 1
- 102000007544 Whey Proteins Human genes 0.000 description 1
- 208000027418 Wounds and injury Diseases 0.000 description 1
- 238000012452 Xenomouse strains Methods 0.000 description 1
- 241000235013 Yarrowia Species 0.000 description 1
- 230000002159 abnormal effect Effects 0.000 description 1
- 230000021736 acetylation Effects 0.000 description 1
- 238000006640 acetylation reaction Methods 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 238000000246 agarose gel electrophoresis Methods 0.000 description 1
- 230000004931 aggregating effect Effects 0.000 description 1
- HMFHBZSHGGEWLO-UHFFFAOYSA-N alpha-D-Furanose-Ribose Natural products OCC1OC(O)C(O)C1O HMFHBZSHGGEWLO-UHFFFAOYSA-N 0.000 description 1
- 229940037003 alum Drugs 0.000 description 1
- 238000012870 ammonium sulfate precipitation Methods 0.000 description 1
- 229960000723 ampicillin Drugs 0.000 description 1
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 238000005571 anion exchange chromatography Methods 0.000 description 1
- 230000008485 antagonism Effects 0.000 description 1
- 230000003110 anti-inflammatory effect Effects 0.000 description 1
- 230000003063 anti-neuropathic effect Effects 0.000 description 1
- 230000000692 anti-sense effect Effects 0.000 description 1
- 239000000074 antisense oligonucleotide Substances 0.000 description 1
- 238000012230 antisense oligonucleotides Methods 0.000 description 1
- 239000002249 anxiolytic agent Substances 0.000 description 1
- 230000000949 anxiolytic effect Effects 0.000 description 1
- 230000001363 autoimmune Effects 0.000 description 1
- 239000007640 basal medium Substances 0.000 description 1
- 230000006736 behavioral deficit Effects 0.000 description 1
- 238000009227 behaviour therapy Methods 0.000 description 1
- 238000005452 bending Methods 0.000 description 1
- WQZGKKKJIJFFOK-FPRJBGLDSA-N beta-D-galactose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-FPRJBGLDSA-N 0.000 description 1
- 230000001588 bifunctional effect Effects 0.000 description 1
- 238000010256 biochemical assay Methods 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 239000012620 biological material Substances 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 210000000601 blood cell Anatomy 0.000 description 1
- 239000012503 blood component Substances 0.000 description 1
- 230000036772 blood pressure Effects 0.000 description 1
- 230000036760 body temperature Effects 0.000 description 1
- 229940098773 bovine serum albumin Drugs 0.000 description 1
- 210000004556 brain Anatomy 0.000 description 1
- 239000000872 buffer Substances 0.000 description 1
- 229960003150 bupivacaine Drugs 0.000 description 1
- 239000011575 calcium Substances 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 229910000019 calcium carbonate Inorganic materials 0.000 description 1
- 239000001110 calcium chloride Substances 0.000 description 1
- 229910001628 calcium chloride Inorganic materials 0.000 description 1
- 229960002713 calcium chloride Drugs 0.000 description 1
- UHHRFSOMMCWGSO-UHFFFAOYSA-L calcium glycerophosphate Chemical compound [Ca+2].OCC(CO)OP([O-])([O-])=O UHHRFSOMMCWGSO-UHFFFAOYSA-L 0.000 description 1
- 229940095618 calcium glycerophosphate Drugs 0.000 description 1
- 235000019299 calcium glycerylphosphate Nutrition 0.000 description 1
- 229960002504 capsaicin Drugs 0.000 description 1
- 235000017663 capsaicin Nutrition 0.000 description 1
- FFGPTBGBLSHEPO-UHFFFAOYSA-N carbamazepine Chemical compound C1=CC2=CC=CC=C2N(C(=O)N)C2=CC=CC=C21 FFGPTBGBLSHEPO-UHFFFAOYSA-N 0.000 description 1
- 229960000623 carbamazepine Drugs 0.000 description 1
- 239000001569 carbon dioxide Substances 0.000 description 1
- 229910002092 carbon dioxide Inorganic materials 0.000 description 1
- 238000005277 cation exchange chromatography Methods 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 230000032823 cell division Effects 0.000 description 1
- 230000007910 cell fusion Effects 0.000 description 1
- 201000010881 cervical cancer Diseases 0.000 description 1
- 238000001311 chemical methods and process Methods 0.000 description 1
- 235000012000 cholesterol Nutrition 0.000 description 1
- 229960003257 choline citrate Drugs 0.000 description 1
- 239000013611 chromosomal DNA Substances 0.000 description 1
- 210000000349 chromosome Anatomy 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 239000003240 coconut oil Substances 0.000 description 1
- 235000019864 coconut oil Nutrition 0.000 description 1
- 239000003086 colorant Substances 0.000 description 1
- 238000004440 column chromatography Methods 0.000 description 1
- 230000001447 compensatory effect Effects 0.000 description 1
- 230000000536 complexating effect Effects 0.000 description 1
- 239000005289 controlled pore glass Substances 0.000 description 1
- JZCCFEFSEZPSOG-UHFFFAOYSA-L copper(II) sulfate pentahydrate Chemical compound O.O.O.O.O.[Cu+2].[O-]S([O-])(=O)=O JZCCFEFSEZPSOG-UHFFFAOYSA-L 0.000 description 1
- 235000005687 corn oil Nutrition 0.000 description 1
- 239000002285 corn oil Substances 0.000 description 1
- 239000003431 cross linking reagent Substances 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- 229960003067 cystine Drugs 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000034994 death Effects 0.000 description 1
- 231100000517 death Toxicity 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 230000007812 deficiency Effects 0.000 description 1
- 230000018044 dehydration Effects 0.000 description 1
- 238000006297 dehydration reaction Methods 0.000 description 1
- AAOVKJBEBIDNHE-UHFFFAOYSA-N diazepam Chemical compound N=1CC(=O)N(C)C2=CC=C(Cl)C=C2C=1C1=CC=CC=C1 AAOVKJBEBIDNHE-UHFFFAOYSA-N 0.000 description 1
- 229960003529 diazepam Drugs 0.000 description 1
- 235000005911 diet Nutrition 0.000 description 1
- 230000037213 diet Effects 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 108020001096 dihydrofolate reductase Proteins 0.000 description 1
- IGKWESNBSFVVHE-UHFFFAOYSA-N dihydrograyanotoxin ii Chemical compound C1CC2C(C)C3CC(O)C(C)(C)C3(O)C(O)CC22C(O)C1C(C)(O)C2 IGKWESNBSFVVHE-UHFFFAOYSA-N 0.000 description 1
- PGUYAANYCROBRT-UHFFFAOYSA-N dihydroxy-selanyl-selanylidene-lambda5-phosphane Chemical compound OP(O)([SeH])=[Se] PGUYAANYCROBRT-UHFFFAOYSA-N 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- NAGJZTKCGNOGPW-UHFFFAOYSA-K dioxido-sulfanylidene-sulfido-$l^{5}-phosphane Chemical compound [O-]P([O-])([S-])=S NAGJZTKCGNOGPW-UHFFFAOYSA-K 0.000 description 1
- ZZVUWRFHKOJYTH-UHFFFAOYSA-N diphenhydramine Chemical compound C=1C=CC=CC=1C(OCCN(C)C)C1=CC=CC=C1 ZZVUWRFHKOJYTH-UHFFFAOYSA-N 0.000 description 1
- 229960000520 diphenhydramine Drugs 0.000 description 1
- PYLIXCKOHOHGKQ-UHFFFAOYSA-L disodium;hydrogen phosphate;heptahydrate Chemical compound O.O.O.O.O.O.O.[Na+].[Na+].OP([O-])([O-])=O PYLIXCKOHOHGKQ-UHFFFAOYSA-L 0.000 description 1
- 230000007831 electrophysiology Effects 0.000 description 1
- 238000002001 electrophysiology Methods 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 150000002148 esters Chemical class 0.000 description 1
- 238000012869 ethanol precipitation Methods 0.000 description 1
- ZJXZSIYSNXKHEA-UHFFFAOYSA-N ethyl dihydrogen phosphate Chemical compound CCOP(O)(O)=O ZJXZSIYSNXKHEA-UHFFFAOYSA-N 0.000 description 1
- DEFVIWRASFVYLL-UHFFFAOYSA-N ethylene glycol bis(2-aminoethyl)tetraacetic acid Chemical compound OC(=O)CN(CC(O)=O)CCOCCOCCN(CC(O)=O)CC(O)=O DEFVIWRASFVYLL-UHFFFAOYSA-N 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 230000002964 excitative effect Effects 0.000 description 1
- 238000000605 extraction Methods 0.000 description 1
- 239000003885 eye ointment Substances 0.000 description 1
- 230000035558 fertility Effects 0.000 description 1
- 238000011049 filling Methods 0.000 description 1
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 1
- 230000004907 flux Effects 0.000 description 1
- 230000005714 functional activity Effects 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 230000002496 gastric effect Effects 0.000 description 1
- 238000001502 gel electrophoresis Methods 0.000 description 1
- 102000034356 gene-regulatory proteins Human genes 0.000 description 1
- 108091006104 gene-regulatory proteins Proteins 0.000 description 1
- 238000012214 genetic breeding Methods 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 238000003205 genotyping method Methods 0.000 description 1
- 229960002518 gentamicin Drugs 0.000 description 1
- 229930195712 glutamate Natural products 0.000 description 1
- 230000001279 glycosylating effect Effects 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 210000002149 gonad Anatomy 0.000 description 1
- BWMFRQKICHXLSH-FIRPSQKQSA-N grayanotoxin iii Chemical compound C1[C@@H](O)[C@]2(O)C(C)(C)[C@@H](O)C[C@H]2[C@](C)(O)[C@@H]2CC[C@H]3[C@](C)(O)C[C@]21[C@@H]3O BWMFRQKICHXLSH-FIRPSQKQSA-N 0.000 description 1
- 230000003648 hair appearance Effects 0.000 description 1
- 230000005802 health problem Effects 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 108060003552 hemocyanin Proteins 0.000 description 1
- 230000011132 hemopoiesis Effects 0.000 description 1
- 206010073071 hepatocellular carcinoma Diseases 0.000 description 1
- 230000001990 hyperexcitatory effect Effects 0.000 description 1
- 230000036031 hyperthermia Effects 0.000 description 1
- 230000002519 immonomodulatory effect Effects 0.000 description 1
- 230000008105 immune reaction Effects 0.000 description 1
- 238000010324 immunological assay Methods 0.000 description 1
- 238000001114 immunoprecipitation Methods 0.000 description 1
- 230000001506 immunosuppresive effect Effects 0.000 description 1
- 238000005462 in vivo assay Methods 0.000 description 1
- 238000010249 in-situ analysis Methods 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 239000003999 initiator Substances 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 208000014674 injury Diseases 0.000 description 1
- 150000002484 inorganic compounds Chemical class 0.000 description 1
- 229910010272 inorganic material Inorganic materials 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 229940126181 ion channel inhibitor Drugs 0.000 description 1
- SURQXAFEQWPFPV-UHFFFAOYSA-L iron(2+) sulfate heptahydrate Chemical compound O.O.O.O.O.O.O.[Fe+2].[O-]S([O-])(=O)=O SURQXAFEQWPFPV-UHFFFAOYSA-L 0.000 description 1
- 238000001155 isoelectric focusing Methods 0.000 description 1
- 230000009191 jumping Effects 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 229960001021 lactose monohydrate Drugs 0.000 description 1
- PYZRQGJRPPTADH-UHFFFAOYSA-N lamotrigine Chemical compound NC1=NC(N)=NN=C1C1=CC=CC(Cl)=C1Cl PYZRQGJRPPTADH-UHFFFAOYSA-N 0.000 description 1
- 229960001848 lamotrigine Drugs 0.000 description 1
- 231100000636 lethal dose Toxicity 0.000 description 1
- 108020001756 ligand binding domains Proteins 0.000 description 1
- 210000005229 liver cell Anatomy 0.000 description 1
- 208000004731 long QT syndrome Diseases 0.000 description 1
- 230000004777 loss-of-function mutation Effects 0.000 description 1
- 210000003141 lower extremity Anatomy 0.000 description 1
- 210000005265 lung cell Anatomy 0.000 description 1
- 210000001165 lymph node Anatomy 0.000 description 1
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 235000021073 macronutrients Nutrition 0.000 description 1
- 239000011777 magnesium Substances 0.000 description 1
- 229910052749 magnesium Inorganic materials 0.000 description 1
- 229940050906 magnesium chloride hexahydrate Drugs 0.000 description 1
- DHRRIBDTHFBPNG-UHFFFAOYSA-L magnesium dichloride hexahydrate Chemical compound O.O.O.O.O.O.[Mg+2].[Cl-].[Cl-] DHRRIBDTHFBPNG-UHFFFAOYSA-L 0.000 description 1
- 229940091250 magnesium supplement Drugs 0.000 description 1
- ISPYRSDWRDQNSW-UHFFFAOYSA-L manganese(II) sulfate monohydrate Chemical compound O.[Mn+2].[O-]S([O-])(=O)=O ISPYRSDWRDQNSW-UHFFFAOYSA-L 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 230000008883 metastatic behaviour Effects 0.000 description 1
- 208000010658 metastatic prostate carcinoma Diseases 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- 230000000813 microbial effect Effects 0.000 description 1
- 239000011785 micronutrient Substances 0.000 description 1
- 235000013369 micronutrients Nutrition 0.000 description 1
- 238000001823 molecular biology technique Methods 0.000 description 1
- 229940111688 monobasic potassium phosphate Drugs 0.000 description 1
- 235000019796 monopotassium phosphate Nutrition 0.000 description 1
- 230000000877 morphologic effect Effects 0.000 description 1
- 210000001611 motor endplate Anatomy 0.000 description 1
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 1
- 210000000663 muscle cell Anatomy 0.000 description 1
- 235000020638 mussel Nutrition 0.000 description 1
- 210000004165 myocardium Anatomy 0.000 description 1
- CMWTZPSULFXXJA-VIFPVBQESA-N naproxen Chemical compound C1=C([C@H](C)C(O)=O)C=CC2=CC(OC)=CC=C21 CMWTZPSULFXXJA-VIFPVBQESA-N 0.000 description 1
- 229960002009 naproxen Drugs 0.000 description 1
- 230000001338 necrotic effect Effects 0.000 description 1
- 101150091879 neo gene Proteins 0.000 description 1
- 230000008271 nervous system development Effects 0.000 description 1
- 210000004412 neuroendocrine cell Anatomy 0.000 description 1
- 230000000926 neurological effect Effects 0.000 description 1
- 230000008587 neuronal excitability Effects 0.000 description 1
- 239000002858 neurotransmitter agent Substances 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 239000002853 nucleic acid probe Substances 0.000 description 1
- 238000001668 nucleic acid synthesis Methods 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 230000000474 nursing effect Effects 0.000 description 1
- 235000018343 nutrient deficiency Nutrition 0.000 description 1
- 239000002674 ointment Substances 0.000 description 1
- 238000012346 open field test Methods 0.000 description 1
- 210000001672 ovary Anatomy 0.000 description 1
- 238000007500 overflow downdraw method Methods 0.000 description 1
- 210000003101 oviduct Anatomy 0.000 description 1
- 208000027753 pain disease Diseases 0.000 description 1
- 230000008052 pain pathway Effects 0.000 description 1
- 239000002540 palm oil Substances 0.000 description 1
- 208000019865 paroxysmal extreme pain disease Diseases 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 230000010412 perfusion Effects 0.000 description 1
- 230000000737 periodic effect Effects 0.000 description 1
- 210000002856 peripheral neuron Anatomy 0.000 description 1
- 210000001322 periplasm Anatomy 0.000 description 1
- 230000009038 pharmacological inhibition Effects 0.000 description 1
- 229960002036 phenytoin Drugs 0.000 description 1
- 239000003016 pheromone Substances 0.000 description 1
- 235000021317 phosphate Nutrition 0.000 description 1
- PTMHPRAIXMAOOB-UHFFFAOYSA-L phosphoramidate Chemical compound NP([O-])([O-])=O PTMHPRAIXMAOOB-UHFFFAOYSA-L 0.000 description 1
- 150000008300 phosphoramidites Chemical class 0.000 description 1
- 150000003013 phosphoric acid derivatives Chemical class 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- SHUZOJHMOBOZST-UHFFFAOYSA-N phylloquinone Natural products CC(C)CCCCC(C)CCC(C)CCCC(=CCC1=C(C)C(=O)c2ccccc2C1=O)C SHUZOJHMOBOZST-UHFFFAOYSA-N 0.000 description 1
- MBWXNTAXLNYFJB-NKFFZRIASA-N phylloquinone Chemical compound C1=CC=C2C(=O)C(C/C=C(C)/CCC[C@H](C)CCC[C@H](C)CCCC(C)C)=C(C)C(=O)C2=C1 MBWXNTAXLNYFJB-NKFFZRIASA-N 0.000 description 1
- 235000019175 phylloquinone Nutrition 0.000 description 1
- 239000011772 phylloquinone Substances 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 230000035479 physiological effects, processes and functions Effects 0.000 description 1
- 230000008288 physiological mechanism Effects 0.000 description 1
- 229960001898 phytomenadione Drugs 0.000 description 1
- SIOXPEMLGUPBBT-UHFFFAOYSA-N picolinic acid Chemical compound OC(=O)C1=CC=CC=N1 SIOXPEMLGUPBBT-UHFFFAOYSA-N 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- GNSKLFRGEWLPPA-UHFFFAOYSA-M potassium dihydrogen phosphate Chemical compound [K+].OP(O)([O-])=O GNSKLFRGEWLPPA-UHFFFAOYSA-M 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000009101 premedication Methods 0.000 description 1
- 238000003825 pressing Methods 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 230000009465 prokaryotic expression Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- XJMOSONTPMZWPB-UHFFFAOYSA-M propidium iodide Chemical compound [I-].[I-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CCC[N+](C)(CC)CC)=C1C1=CC=CC=C1 XJMOSONTPMZWPB-UHFFFAOYSA-M 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 238000001742 protein purification Methods 0.000 description 1
- 238000000734 protein sequencing Methods 0.000 description 1
- 230000012743 protein tagging Effects 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 238000001959 radiotherapy Methods 0.000 description 1
- 239000011541 reaction mixture Substances 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 230000000306 recurrent effect Effects 0.000 description 1
- 230000033458 reproduction Effects 0.000 description 1
- 239000011347 resin Substances 0.000 description 1
- 229920005989 resin Polymers 0.000 description 1
- 230000003938 response to stress Effects 0.000 description 1
- 230000000452 restraining effect Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 229940108325 retinyl palmitate Drugs 0.000 description 1
- 235000019172 retinyl palmitate Nutrition 0.000 description 1
- 239000011769 retinyl palmitate Substances 0.000 description 1
- 238000010839 reverse transcription Methods 0.000 description 1
- 238000004007 reversed phase HPLC Methods 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 210000001991 scapula Anatomy 0.000 description 1
- 238000013391 scatchard analysis Methods 0.000 description 1
- 238000010187 selection method Methods 0.000 description 1
- 239000006152 selective media Substances 0.000 description 1
- 230000008786 sensory perception of smell Effects 0.000 description 1
- 229960001153 serine Drugs 0.000 description 1
- 230000001568 sexual effect Effects 0.000 description 1
- 230000019491 signal transduction Effects 0.000 description 1
- 230000003584 silencer Effects 0.000 description 1
- 238000009097 single-agent therapy Methods 0.000 description 1
- 210000002027 skeletal muscle Anatomy 0.000 description 1
- 229940126121 sodium channel inhibitor Drugs 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- 239000011775 sodium fluoride Substances 0.000 description 1
- 235000013024 sodium fluoride Nutrition 0.000 description 1
- 229910001415 sodium ion Inorganic materials 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 230000037439 somatic mutation Effects 0.000 description 1
- 239000003549 soybean oil Substances 0.000 description 1
- 235000012424 soybean oil Nutrition 0.000 description 1
- 238000012421 spiking Methods 0.000 description 1
- 238000012453 sprague-dawley rat model Methods 0.000 description 1
- 238000003756 stirring Methods 0.000 description 1
- 239000000758 substrate Substances 0.000 description 1
- 229940014800 succinic anhydride Drugs 0.000 description 1
- 229940124530 sulfonamide Drugs 0.000 description 1
- 150000003456 sulfonamides Chemical class 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 230000002459 sustained effect Effects 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- 238000001308 synthesis method Methods 0.000 description 1
- 230000008685 targeting Effects 0.000 description 1
- 229960003080 taurine Drugs 0.000 description 1
- 229960002180 tetracycline Drugs 0.000 description 1
- 229930101283 tetracycline Natural products 0.000 description 1
- 235000019364 tetracycline Nutrition 0.000 description 1
- 150000003522 tetracyclines Chemical class 0.000 description 1
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 1
- 229960002175 thyroglobulin Drugs 0.000 description 1
- 229940042585 tocopherol acetate Drugs 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 231100000041 toxicology testing Toxicity 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- 239000012581 transferrin Substances 0.000 description 1
- QQOWHRYOXYEMTL-UHFFFAOYSA-N triazin-4-amine Chemical compound N=C1C=CN=NN1 QQOWHRYOXYEMTL-UHFFFAOYSA-N 0.000 description 1
- 210000000427 trigeminal ganglion Anatomy 0.000 description 1
- 206010044652 trigeminal neuralgia Diseases 0.000 description 1
- 239000002753 trypsin inhibitor Substances 0.000 description 1
- 229960004799 tryptophan Drugs 0.000 description 1
- 238000000108 ultra-filtration Methods 0.000 description 1
- 230000009452 underexpressoin Effects 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
- 230000003612 virological effect Effects 0.000 description 1
- 230000000007 visual effect Effects 0.000 description 1
- 239000011782 vitamin Substances 0.000 description 1
- 235000013343 vitamin Nutrition 0.000 description 1
- 229940088594 vitamin Drugs 0.000 description 1
- 229930003231 vitamin Natural products 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 230000003442 weekly effect Effects 0.000 description 1
- 235000021119 whey protein Nutrition 0.000 description 1
- RZLVQBNCHSJZPX-UHFFFAOYSA-L zinc sulfate heptahydrate Chemical compound O.O.O.O.O.O.O.[Zn+2].[O-]S([O-])(=O)=O RZLVQBNCHSJZPX-UHFFFAOYSA-L 0.000 description 1
Images




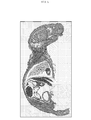








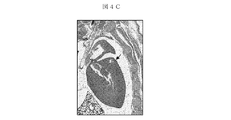








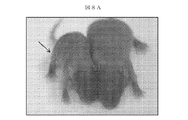














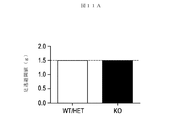






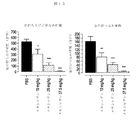






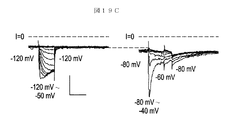


Classifications
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/8509—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells for producing genetically modified animals, e.g. transgenic
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
- A01K67/0275—Genetically modified vertebrates, e.g. transgenic
- A01K67/0276—Knock-out vertebrates
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K49/00—Preparations for testing in vivo
- A61K49/0004—Screening or testing of compounds for diagnosis of disorders, assessment of conditions, e.g. renal clearance, gastric emptying, testing for diabetes, allergy, rheuma, pancreas functions
- A61K49/0008—Screening agents using (non-human) animal models or transgenic animal models or chimeric hosts, e.g. Alzheimer disease animal model, transgenic model for heart failure
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0608—Germ cells
- C12N5/0609—Oocytes, oogonia
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0608—Germ cells
- C12N5/061—Sperm cells, spermatogonia
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/07—Animals genetically altered by homologous recombination
- A01K2217/075—Animals genetically altered by homologous recombination inducing loss of function, i.e. knock out
- A01K2217/077—Animals genetically altered by homologous recombination inducing loss of function, i.e. knock out heterozygous knock out animals displaying phenotype
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2227/00—Animals characterised by species
- A01K2227/10—Mammal
- A01K2227/105—Murine
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2267/00—Animals characterised by purpose
- A01K2267/03—Animal model, e.g. for test or diseases
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Zoology (AREA)
- Biomedical Technology (AREA)
- Organic Chemistry (AREA)
- Biotechnology (AREA)
- General Health & Medical Sciences (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Environmental Sciences (AREA)
- Veterinary Medicine (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Animal Behavior & Ethology (AREA)
- Microbiology (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Biodiversity & Conservation Biology (AREA)
- Animal Husbandry (AREA)
- Plant Pathology (AREA)
- Medicinal Chemistry (AREA)
- Endocrinology (AREA)
- Gastroenterology & Hepatology (AREA)
- Pathology (AREA)
- Rheumatology (AREA)
- Toxicology (AREA)
- Urology & Nephrology (AREA)
- Epidemiology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Public Health (AREA)
- Diabetes (AREA)
- Immunology (AREA)
- Physics & Mathematics (AREA)
- Cell Biology (AREA)
- Developmental Biology & Embryology (AREA)
- Reproductive Health (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
Abstract
生存可能なグローバルNaV1.7−/−ノックアウトマウス、およびグローバルNaV1.7−/−ノックアウトマウスの繁殖コロニーが開示される。また、NaV1.7−/−ノックアウトマウスにより産生された、機能性NaV1.7をコードしない単離マウス配偶子;NaV1.7−/−ノックアウトマウスから単離された単離NaV1.7−/−マウス細胞、またはその子孫細胞;および、NaV1.7−/−ノックアウトマウスから得た初代細胞培養物または二次細胞株および組織もしくは臓器外植片またはそれらの培養物、も開示される。さらに、最初、単離NaV1.7−/−マウス細胞および骨髄腫細胞の融合により形成されたハイブリドーマ、および、抗体を産生する方法も開示される。また、NaV1.7−/−ノックアウトマウスを使って検証された、有望なNaV1.7阻害剤の選別、および試験NaV1.7阻害剤化合物の用量範囲探索に有用なアッセイが開示される。
【選択図】図8B
【選択図】図8B
Description
本出願は、37C.F.R.セクション1.821(c)および1.821(e)で要求される、コンピュータ読み取り可能形式(CRF)および紙コピーの両方の役割を果たすASCII「txt」配列リストを含み、それは参照によってその全体が組み込まれる。2012年1月17日に作成されたその「txt」ファイルの名称は、A−1588−WO−PCT−SeqList011812.ST25.txtで、サイズは2kbである。
本出願全体を通し、種々の出版物が丸括弧または角括弧で囲って参照される。これらの出版物の開示は、本発明の属する最先端技術をさらに完全に記載するために、その全体が参照によって本出願に組み込まれる。
関連技術の考察
単離細胞株の使用(すなわち、インビトロ系)は、種々の遺伝子とタンパク質がもたらす生理的役割の理解には有用であるが、より完全な情報は、哺乳動物で(すなわち、インビボ系で)これらのタンパク質の役割の効果を直接研究することにより得ることができる。この目的のために、ある特定の遺伝子の発現レベルを変えた種々の哺乳動物が作られている。これらの哺乳動物の内の1つの種類は、いわゆる遺伝子導入哺乳動物である。これらの内の後者の哺乳動物は、新規遺伝子または複数遺伝子を有し、これらは異なる種由来であり、それらのインタクトゲノム中に挿入され、従って、「遺伝子導入」ということになる。別の種類は、ノックイン哺乳動物である。これらの動物は、自身の1つの遺伝子が削除され、その同じ遺伝子の変異体により置き換えられる。この手法は、選択遺伝子/タンパク質の発現過剰または発現低下を起こさせるために使用されることが多い。機能的リン酸化部位を有するキナーゼおよびタンパク質は、この手法での選択の標的である。最初の2つの技術の組み合わせを使用して、内在性宿主遺伝子の座位で外来性遺伝子、例えば、同等の遺伝子のマウス座位でヒト遺伝子、を発現させる「遺伝子導入−ノックイン」哺乳動物を生成できる。最後に挙げる手法は、グローバルヌル変異体、またはいわゆる、「ノックアウト」哺乳動物を生成することであり、そこでは内在性遺伝子の発現が、組換え技術か、または古典的遺伝子技術を使った遺伝子操作により抑制される。例えば、Nassarら(2006)は、電位依存性ナトリウムチャネルNaV1.3を発現できないグローバルヌル変異体マウス株を生成した(Nassar et al.、NaV1.3ヌル変異体マウスにおいて、神経損傷が強い異痛および異所性放電を誘導する(Nerve injury induces robust allodynia and ectopic discharges in NaV1.3 null mutant mice)、Mol.Pain 2:33(2006))。
単離細胞株の使用(すなわち、インビトロ系)は、種々の遺伝子とタンパク質がもたらす生理的役割の理解には有用であるが、より完全な情報は、哺乳動物で(すなわち、インビボ系で)これらのタンパク質の役割の効果を直接研究することにより得ることができる。この目的のために、ある特定の遺伝子の発現レベルを変えた種々の哺乳動物が作られている。これらの哺乳動物の内の1つの種類は、いわゆる遺伝子導入哺乳動物である。これらの内の後者の哺乳動物は、新規遺伝子または複数遺伝子を有し、これらは異なる種由来であり、それらのインタクトゲノム中に挿入され、従って、「遺伝子導入」ということになる。別の種類は、ノックイン哺乳動物である。これらの動物は、自身の1つの遺伝子が削除され、その同じ遺伝子の変異体により置き換えられる。この手法は、選択遺伝子/タンパク質の発現過剰または発現低下を起こさせるために使用されることが多い。機能的リン酸化部位を有するキナーゼおよびタンパク質は、この手法での選択の標的である。最初の2つの技術の組み合わせを使用して、内在性宿主遺伝子の座位で外来性遺伝子、例えば、同等の遺伝子のマウス座位でヒト遺伝子、を発現させる「遺伝子導入−ノックイン」哺乳動物を生成できる。最後に挙げる手法は、グローバルヌル変異体、またはいわゆる、「ノックアウト」哺乳動物を生成することであり、そこでは内在性遺伝子の発現が、組換え技術か、または古典的遺伝子技術を使った遺伝子操作により抑制される。例えば、Nassarら(2006)は、電位依存性ナトリウムチャネルNaV1.3を発現できないグローバルヌル変異体マウス株を生成した(Nassar et al.、NaV1.3ヌル変異体マウスにおいて、神経損傷が強い異痛および異所性放電を誘導する(Nerve injury induces robust allodynia and ectopic discharges in NaV1.3 null mutant mice)、Mol.Pain 2:33(2006))。
電位依存性ナトリウムチャネル(VGSC)は、興奮性細胞、例えば、中枢および末梢ニューロン、心臓ならびに骨格筋筋細胞、ならびに神経内分泌細胞における活動電位の開始と伝播に関与する糖タンパク質複合体である。哺乳類のナトリウムチャネルは、ヘテロ三量体であり、中心部の、気孔形成アルファ(α)サブユニットおよび補助的ベータ(β)サブユニットから構成される。アルファサブユニット遺伝子の変異は、発作性疾患、例えば、ヒトの癲癇、QT延長症候群、および高カリウム血性周期性四肢麻痺、ならびにマウスの運動終末板疾患および小脳失調に関連づけられている(Isom、ナトリウムチャネルβサブユニット:決して補助的役割などというようなものではない(Sodium channel beta subunits:anything but auxiliary)、Neuroscientist 7(1):42−54(2001))。β−サブユニットは、VGSC中のα−サブユニットの局在化、発現および機能的特性を調節する。
電位依存性ナトリウムチャネルは、9つの異なるサブタイプ(NaV1.1〜NaV1.9)から構成されるファミリーを含む。表1に示すように、これらのサブタイプは、組織特異的局在化および機能的差異を示す(Goldin、A.L.、ナトリウムチャネル研究の復活(Resurgence of sodium channel research)、Annu Rev Physiol 63:871−94(2001);Wilson et al.、電位依存性イオンチャネル阻害剤として有用な組成物(Compositions useful as inhibitors of voltage−gated ion channels)、米国特許公開第2005/0187217A1号、を参照)。その遺伝子ファミリーの3つのメンバー(NaV1.8、1.9、1.5)は、よく知られたナトリウムチャネル遮断薬テトロドトキシン(TTX)による遮断に対し抵抗性であり、これらは、この遺伝子ファミリー中でのサブタイプ特異性を示している。変異分析により、グルタメート387が、TTX結合に対し、極めて重要な残基であると特定された(Noda、M.、H.Suzuki、et al.、「ナトリウムチャネルIIに関し、ただ1つの点変異が、テトロドトキシンとサキシトキシンに不感受性を付与する(A single point mutation confers tetrodotoxin and saxitoxin insensitivity on the sodium channel II)」FEBS Lett 259(1):213−6(1989)、参照)。
VGSCファミリーとラットTTX IC50値。略号:CNS=中枢神経系、PNS=末梢神経系、DRG=脊髄後根神経節、TG=三叉神経節。(Wilson et al.、電位依存性イオンチャネル阻害剤として有用な組成物(Compositions useful as inhibitors of Voltage−gated ion channels)、米国特許公開第2005/0187217A1号;Goldin、ナトリウムチャネル研究の復活(Resurgence of Sodium Channel Research)、Annu Rev
Physiol 63:871−94(2001)、参照)。
Physiol 63:871−94(2001)、参照)。

一般的に、電位依存性ナトリウムチャネル(NaV)は、神経系の興奮性組織の活動電位の急速な上昇の開始に関与し、これは、正常および異常疼痛感覚を構成し、コードする電気信号を伝達する。NaVチャネルのアンタゴニストは、これらの疼痛信号を弱めることができ、限定されないが、急性、慢性、炎症性、神経障害性疼痛、等の種々の疼痛状態の治療に有用である。既知のNaVアンタゴニスト、例えば、TTX、リドカイン、ブピバカイン、フェニトイン、ラモトリギン、およびカルバマゼピンは、ヒトおよび動物モデルで、疼痛を弱めるのに有用であることが示されている(Mao、J.and L.L.Chen、神経障害性疼痛軽減のための全身性リドカイン(Systemic lidocaine for neuropathic pain relief)、Pain 87(1):7−17(2000);Jensen、T.S.、神経障害性疼痛における抗けいれん薬:理論的根拠と臨床的証拠(rationale and clinical evidence)、Eur J Pain 6(SupplA):61−68(2002);Rozen、T.D.、群発頭痛および三叉神経痛の管理における抗てんかん薬(Antiepileptic drugs in the management of cluster headache and trigeminal neuralgia)、Headache 41 Suppl 1:S25−32(2001);Backonja、M.M.、神経障害性疼痛の治療への抗けいれん薬の使用(Use of anticonvulsants for treatment of neuropathic pain)、Neurology 59(5Suppl2):S14−7(2002)、を参照)。
TTX感受性NaV1.7チャネルのαサブユニットは、SCN9A遺伝子によりコードされる。NaV1.7チャネルは、脊髄後根神経節の末梢感覚ニューロンで優先的に発現し、疼痛の知覚に関与する。ヒトでは、SCN9A遺伝子の変異は、過剰または低感受性で痛みを感ずる素因に関連づけられている。例えば、疼痛知覚でのNaV1.7チャネルの役割は、最近の臨床的遺伝子連鎖解析により確立され、これにより、SCN9A遺伝子の機能獲得型変異が、遺伝性疼痛症候群、例えば、原発性肢端紅痛症(PE)、遺伝性紅痛症(IEM)、および発作性激痛症(PEPD)の病因学的根拠として示された(例えば、Yang et al.、原発性肢端紅痛症の患者のナトリウムチャネルαサブユニットをコードするSCN9Aの変異(Mutations in SCN9A、encoding a sodium channel alpha subunit、in patients with primary erythermalgia)、J.Med.Genet.41:171−174(2004);Harty et al.、紅痛症におけるNaV1.7変異体A863P:侵害受容性脊髄後根神経節ニューロンの興奮性に対する、活性化および定常状態不活性化の変化の影響(NaV1.7 mutant A863P in erythromelalgia:effects of altered activation and steady−state inactivation on excitability of nociceptive dorsal root ganglion neurons)、J.Neurosci.26(48):12566−75(2006);Estacion et al.、連続体としてのNaV1.7機能獲得型変異のA1632Eは、紅痛症および発作性激痛症変異に関連する生理的変化を示し、両障害を発症する(NaV1.7 gain−of−function mutations as a continuum:A1632E displays physiological changes associated with erythromelalgia and paroxysmal extreme pain disorder mutations and produces symptoms of both disorders)、J.Neurosci.28(43):11079−88(2008)、を参照)。さらに、NaV1.7の過剰発現が、転移性の強い前立腺癌細胞株中で検出されている(Diss et al.、ヒト前立腺癌用の有望な新規マーカー:インビボ発現電位依存性ナトリウムチャネル(A potential novel marker for human prostate cancer:voltage−gated sodium channel expression in vivo)、Prostate Cancer and Prostatic Diseases 8:266−73(2005);Uysal−Onganer et al.、上皮増殖因子は、インビトロ転移性挙動のヒト前立腺癌PC−3M細胞を強化する:電位依存性ナトリウムチャネルの関与(Epidermal growth factor potentiates invitro metastatic behavior human prostate cancer PC−3M cells:involvement of voltage−gated sodium channel)、Molec.Cancer 6:76(2007)。
SCN9A遺伝子機能喪失型変異は、それ以外では健康な個人が、いかなる形の疼痛感覚も感じることが完全にできなくなる状態をもたらす(例えば、Ahmad et al.、SCN9Aの停止コドン変異は、疼痛感覚の欠落を引き起こす(A stop codon mutation in SCN9A causes lack of pain sensation)、Hum.Mol.Genet.16(17):2114−21(2007))。
条件付きノックアウトによるSCN9A遺伝子の細胞特異的欠失は、機械的、熱的または炎症性疼痛を知覚する能力を減らす(Nassar et al.、侵害受容器特異的遺伝子欠失は、急性および炎症性疼痛でのNaV1.7(PN1)の主要役割を明らかにする(Nociceptor−specific gene deletion reveals a major role for NaV1.7(PN1) in acute and inflammatory pain)、Proc.Natl.Acad.Sci、USA.101(34):12706−12711(2004))。
このような証拠に基づいて、脊髄後根神経節の末梢感覚ニューロン中でのNaV1.7チャネル活性または発現レベルの低減が、例えば、慢性痛、神経障害性疼痛、および神経痛のための有効な疼痛治療法として提案されている(例えば、Thakker et al.、疼痛の治療のためのSCN9A遺伝子発現および/または機能の抑制(Suppression of SCN9A gene expression and/or function for the treatment of pain)、国際公開第WO2009/033027A2号;Yeomans et al.、主要求心性神経中のNaV1.7ナトリウムチャネルのヘルペスベクター媒介ノックダウンによる炎症性痛覚過敏の減少(Decrease in inflammatory hyperalgesia by herpes vector−mediated knockdown of NaV1.7 sodium channels in primary afferents)、Hum.Gene Ther.16(2):271−7(2005);Fraser et al.、強力・選択的NaV1.7ナトリウムチャネル遮断薬(Potent and selective NaV1.7 sodium channel blockers)、国際公開第WO2007/109324A2号;Hoyt et al.、新規種類のベンゾアゼピノンNa(v)1.7遮断薬の発見:神経障害性疼痛用の有望な治療薬(Discovery of a novel class of benzazepinone Na(v)1.7 blockers:potential treatments for neuropathic pain)、Bioorg.Med.Chem.Lett.17(16):4630−34(2007);Hoyt et al.、ベンゾアゼピノンNaV 1.7遮断薬:神経障害性疼痛用の有望な治療薬(Benzazepinone NaV 1.7 blockers:Potential treatments for neuropathic pain)、Bioorg.Med.Chem.Lett.17(22):6172−77(2007))。
Nassarらは、マウス中の遺伝子切除を行い、疼痛経路におけるNaV1.7の機能を調べた;しかし、グローバルNaV1.7ヌル変異体は、摂食できなかったことが明らかな理由で、(ヒトとは異なり)出生後すぐに死ぬことが明らかになったと報告した(侵害受容器特異的遺伝子欠失は、急性および炎症性疼痛でのNaV1.7(PN1)の主要役割を明らかにする(Nociceptor−specific gene deletion reveals a major role for NaV1.7(PN1) in acute and inflammatory pain)、Proc Natl Acad Sci USA.101(34):12706−12711(2004))。実際、生き残った92匹の子の内で、72%は、ヘテロ接合体であり、残りはNaV1.7野性型であった。Nassarら(2004)は、「...全ての感覚および交感神経ニューロン中でのNaV1.7欠失は、周産期致死表現型を引き起こす」と述べた(Nassar et al.、同上、ページ12708)。 Nassarらは、認められた新生仔死亡を考慮して、Cre−loxP手法を使って、侵害受容器特異的ノックアウトを生成した。 これらの組織限定ノックアウトは、感覚および交感神経ニューロンの下位の集団では、もはやNav1.7の発現はないが、身体のその他の部位ではどこでもNav1.7の発現がある動物として記述された。それらのマウスは、NaV1.8CreデリーターマウスをfloxNaV1.7マウスと交配させ、組織限定NaV1.7−/−マウスおよび同腹子対照を生成させることにより作られた。これらの侵害受容器特異的動物は、その後、侵害受容および疼痛の機序を研究するために使用された。
Nassarらは、別の報告で、「NaV1.7のグローバル欠失は、P0で致死であるため、NaV1.8およびNaV1.7の両方のグローバルノックアウトを生成することはできない」と述べた(Nassar et al.、神経障害性疼痛は、NaV1.7およびNaV1.8の両方欠損マウスで通常通り発生する(Neuropathic pain develops normally in mice lacking both NaV1.7 and NaV1.8)、Mol.Pain 1−24(2005))。
上述の技術とは対照的に、本発明は、NaV1.7媒介生理機能の研究のための、および例えば、特に、疼痛および神経内分泌障害を標的とした医薬品の開発のための、特に、グローバルNaV1.7ヌル変異体マウスおよび繁殖性NaV1.7ノックアウトマウス株を提供する。
Nassar et al.、Nerve injury induces robust allodynia and ectopic discharges in NaV1.3 null mutant mice、Mol.Pain 2:33(2006)
Isom、Sodium channel beta subunits:anything but auxiliary、Neuroscientist 7(1):42−54(2001)
Goldin、A.L.、Resurgence of sodium channel research、Annu Rev Physiol 63:871−94(2001)
Wilson et al.、Compositions useful as inhibitors of voltage−gated ion channels
Noda、M.、H.Suzuki、et al.、A single point mutation confers tetrodotoxin and saxitoxin insensitivity on the sodium channel II、FEBS Lett 259(1):213−6(1989)
Mao、J.and L.L.Chen、Systemic lidocaine for neuropathic pain relief、Pain 87(1):7−17(2000)
Jensen、T.S.、rationale and clinical evidence、Eur J Pain 6(SupplA):61−68(2002)
Rozen、T.D.、Antiepileptic drugs in the management of cluster headache and trigeminal neuralgia、Headache 41 Suppl 1:S25−32(2001)
Backonja、M.M.、Use of anticonvulsants for treatment of neuropathic pain、Neurology 59(5Suppl2):S14−7(2002)
Yang et al.、Mutations in SCN9A、encoding a sodium channel alpha subunit、in patients with primary erythermalgia、J.Med.Genet.41:171−174(2004)
Harty et al.、NaV1.7 mutant A863P in erythromelalgia:effects of altered activation and steady−state inactivation on excitability of nociceptive dorsal root ganglion neurons、J.Neurosci.26(48):12566−75(2006)
Estacion et al.、NaV1.7 gain−of−function mutations as a continuum:A1632E displays physiological changes associated with erythromelalgia and paroxysmal extreme pain disorder mutations and produces symptoms of both disorders、J.Neurosci.28(43):11079−88(2008)
Diss et al.、A potential novel marker for human prostate cancer:voltage−gated sodium channel expression in vivo、Prostate Cancer and Prostatic Diseases 8:266−73(2005)
Uysal−Onganer et al.、Epidermal growth factor potentiates invitro metastatic behavior human prostate cancer PC−3M cells:involvement of voltage−gated sodium channel、Molec.Cancer 6:76(2007)
Ahmad et al.、A stop codon mutation in SCN9A causes lack of pain sensation、Hum.Mol.Genet.16(17):2114−21(2007)
Nassar et al.、Nociceptor−specific gene deletion reveals a major role for NaV1.7(PN1) in acute and inflammatory pain)、Proc.Natl.Acad.Sci、USA.101(34):12706−12711(2004)
Thakker et al.、Suppression of SCN9A gene expression and/or function for the treatment of pain
Yeomans et al.、Decrease in inflammatory hyperalgesia by herpes vector−mediated knockdown of NaV1.7 sodium channels in primary afferents、Hum.Gene Ther.16(2):271−7(2005)
Fraser et al.、Potent and selective NaV1.7 sodium channel blockers
Hoyt et al.、Discovery of a novel class of benzazepinone Na(v)1.7 blockers:potential treatments for neuropathic pain)、Bioorg.Med.Chem.Lett.17(16):4630−34(2007)
Hoyt et al.、Benzazepinone NaV 1.7 blockers:Potential treatments for neuropathic pain)、Bioorg.Med.Chem.Lett.17(22):6172−77(2007)
Nassar et al.、Neuropathic pain develops normally in mice lacking both NaV1.7 and NaV1.8)、Mol.Pain 1−24(2005)
本発明は、生存可能なグローバルNaV1.7−/−ノックアウトマウスに関し、このグローバルNaV1.7ノックアウト変異は、出生後0日目(P0)世代のマウスで早くも致死であるという当技術分野における教示とは驚くべき対比をなすものである(侵害受容器特異的遺伝子欠失は、急性および炎症性疼痛でのNaV1.7(PN1)の主要役割を明らかにする(Nociceptor−specific gene deletion reveals a major role for NaV1.7(PN1) in acute and inflammatory pain)、Proc.Natl.Acad.Sci、USA.101(34):12706−12711(2004)); (Nassar et al.、神経障害性疼痛は、NaV1.7およびNaV1.8の両方欠損マウスで通常通り発生する(Neuropathic pain develops normally in mice lacking both NaV1.7 and NaV1.8)、Mol.Pain 1−24(2005))。Nassar等により採用されたものと同じC57BL/6J背景下で、新生仔Nav1.7−/−マウスにより示された活力の欠如を注意深く観察し、C57BL/6Jに比べて高められた活力を有する株であり、さらに、その中の雌がC57BL/6Jに比べて強化された保育挙動を示す異なる株を計画的に選択することにより、我々は、これらのより活力のある株背景由来の、このような生存可能なグローバルNaV1.7−/−ノックアウトマウスを生成できた。
本発明の一実施形態では、マウスは、異系交配もしくは戻し交配されたグローバルNaV1.7−/−ノックアウトマウスであるか、または同様にNaV1.7−/−であるそれら由来の子孫マウスである。グローバルNaV1.7−/−ノックアウトマウスまたはそのNaV1.7−/−子孫は、また、同じ株または異なる株のNaV1.7+/+パートナーと交配させ、NaV1.7+/−の遺伝子型を有する他の子孫を生成させることができる。グローバルNaV1.7−/−ノックアウトマウスまたはそのNaV1.7−/−子孫は、また、同じ株または異なる株のNaV1.7+/−パートナーと交配させて、NaV1.7+/−またはNaV1.7−/−の遺伝子型を有する他の子孫を生成させることもできる。
本発明の別の実施形態では、グローバルNaV1.7−/−ノックアウトマウスは、成体である。
本発明の別の実施形態では、NaV1.7−/−マウス細胞(例えば、B−リンパ球、T細胞、または神経細胞)は、グローバルNaV1.7−/−ノックアウトマウスから単離でき、従って、子孫細胞、初代細胞培養または2次細胞株がグローバルNaV1.7−/−ノックアウトマウスから得られる。
本発明の他の実施形態では、組織または臓器植片、またはそれらの培養物もまた、グローバルNaV1.7−/−ノックアウトマウスから得られる。
本発明のグローバルNaV1.7−/−ノックアウトマウス成体は、繁殖性雄および雌をそれぞれ含むために、本発明の別の態様は、グローバルNaV1.7−/−ノックアウトマウスの繁殖コロニーに関し、成体グローバルNaV1.7−/−ノックアウトマウスの少なくとも1つの繁殖ペアを含む。
本発明の別の実施形態では、ハイブリドーマを、上述のNaV1.7−/−マウスB−リンパ球細胞、と骨髄腫細胞の融合により作ることができる。
本発明の別の態様では、NaV1.7に対する抗体の調製は、限定されないが、マウスまたはヒトNaV1.7を含む。本発明のNaV1.7−/−ノックアウトマウスにより産生された抗ヒトNaV1.7抗体のCDR配列に基づいて、これらのCDRを抗体中に組み込んだ、NaV1.7イオンチャネル活性に拮抗性のまたはそれを刺激するキメラまたはヒト化抗体を作り出すことができる。これは、治療に有効でありうる。
他の実施形態では、本発明のグローバルNaV1.7−/−ノックアウトマウスは、薬剤研究および開発、例えば、オンターゲット/オフターゲット効果の間の区別、または疼痛および鎮静効果の間の区別を行うためのインビボプロトコルに有用である。
例えば、本発明の一実施形態では、NaV1.7特異的生化学的刺激を伴うアッセイが、グローバルNaV1.7−/−ノックアウトマウスを使って検証された。このアッセイは、例えば、予期されるNaV1.7阻害剤の選別に対し有用であり、この選別は、研究または臨床的目的にとって有用でありうる。このアッセイは、下記を含む:
(a)哺乳動物(例えば、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒト)に、試験化合物(候補NaV1.7阻害剤)を投与し、続けて、
(b)投薬その哺乳動物に、陰性対照(試験化合物を受けていない)中で疼痛関連反応を誘導するのに有効な1回投与量のNaV1.7活性化因子(例えば、ベラトリジン、デルタメトリン、またはグラヤノトキシンIII)を投与し(このような投与は、全身性でも、局所的でもよい);および、次いで
(c)哺乳動物の疼痛関連反応が、陰性対照に比べて、減少するかどうかを判定する。記載のように、哺乳動物への試験化合物投与は、全身性(例えば、腹腔内、静脈内、筋肉内、または経口投与)であっても、または局所的(例えば、皮下、足底内、または局部投与)であってもよい。局所投与の場合は、Nav1.7活性化因子の局所投与と同じ部位またはその近くである必要がある。
(a)哺乳動物(例えば、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒト)に、試験化合物(候補NaV1.7阻害剤)を投与し、続けて、
(b)投薬その哺乳動物に、陰性対照(試験化合物を受けていない)中で疼痛関連反応を誘導するのに有効な1回投与量のNaV1.7活性化因子(例えば、ベラトリジン、デルタメトリン、またはグラヤノトキシンIII)を投与し(このような投与は、全身性でも、局所的でもよい);および、次いで
(c)哺乳動物の疼痛関連反応が、陰性対照に比べて、減少するかどうかを判定する。記載のように、哺乳動物への試験化合物投与は、全身性(例えば、腹腔内、静脈内、筋肉内、または経口投与)であっても、または局所的(例えば、皮下、足底内、または局部投与)であってもよい。局所投与の場合は、Nav1.7活性化因子の局所投与と同じ部位またはその近くである必要がある。
本発明の別の実施形態では、ナトリウムチャネル活性化因子は、NaV1.7特異的生化学的刺激を伴うアッセイで使用され、NaV1.7−遮断試験化合物の居所的または全身性の投与量を選択するのに有用である。臨床試験でのいずれかの試験化合物の適切な投与量の選択は、難しい仕事であり、理想的には、投与量を、ある種の、例えば、標的特異的であることがわかっているPETリガンドのバイオマーカーを置き換える量に較正することにより行われる。ナトリウムチャネル全般、および特に、NaV1.7には、今まで、このようなバイオマーカーがなかった。本明細書で、我々は、ベラトリジン、デルタメトリン、およびグラヤノトキシン、等のナトリウムチャネルの活性化因子は、適切な投与量をラットまたはマウスに注射すると、定量化可能な挙動による反応を生成することを開示する。これら3つの分子は、構造的に異なるが、それぞれがNaV1.7を活性化するという点で共通の生理的機序を共有する。実施例5で、我々は、これらのナトリウムチャネル活性化因子のいずれかを注射時に、ラットとマウスが、それぞれ、定量可能な用量依存性の足を引っ込めるおよび、足をなめる挙動を示すことを初めて明らかにする。これらの挙動は、モルヒネにより減少し、非選択的ナトリウムチャネルアンタゴニストのメキシレチンにより抑制されるため、これらの挙動が疼痛を反映し、ナトリウムチャネルにより媒介されていることを立証している。
最も重要なこととして、我々は、本明細書の実施例5で、NaV1.7活性化因子、例えば、ベラトリジンの1μgの、グローバルNav1.7−/−マウスの足へ注射が、足を引っ込める、または足をなめる挙動を起こさせず、他方、野性型マウスでは、この同じ1μg投与が、強い足を引っ込める挙動を起こさせることを示す。メキシレチン(NaV1.7ならびに他の全てのナトリウムチャネルを遮断する)の野性型マウスへの前投与が、1μg投与量のベラトリジンにより誘発される足を引っ込める挙動を抑制するので、NaV1.7の薬理学的遮断は、同じ効果を実現するはずである。従って、前臨床の試験で、ナトリウムチャネル活性化因子による刺激は、生きている動物に投与される所与の化合物がNaV1.7を遮断するか否かの有用な試験である。さらに、この試験は、臨床的疼痛症候群を治療するNaV1.7試験阻害剤の適正な投薬量を決定するために臨床的に使用でき得る。適正な臨床的投与量は、ナトリウムチャネル活性化因子の投与に対する疼痛反応を抑制する量であろう。
一実施形態では、本発明は、試験化合物(候補NaV1.7阻害剤)の用量範囲探索に有用な下記を含むアッセイを含む:
(a)第1の哺乳動物(例えば、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒト)に、試験化合物の第1の投与量を投与し、続けて、
(b)第1の哺乳動物に、陰性対照(試験化合物を受けていない)で疼痛関連反応を誘導するのに有効な(局所的または全身性)投与量のNaV1.7活性化因子(例えば、ベラトリジン、デルタメトリン、またはグラヤノトキシンIII)を投与し(このような投与は全身性でも局所的でもよい);次に
(c)疼痛関連反応が、陰性対照に比べて、第1の哺乳動物で減少するか否かを判定し;さらに
(d)疼痛関連反応が、陰性対照に比べて減少する試験化合物の最も少ない第2の投与量を特定する。哺乳動物への試験化合物投与は、全身性(例えば、腹腔内、静脈内、筋肉内、または経口投与)であっても、または局所的(例えば、皮下、足底内、局部投与)であってもよい。局所的な場合は、NaV1.7活性化因子の局所投与と同じ部位か、またはその近くである必要がある。回復期間が充分で、試験化合物およびNaV1.7活性化因子の効果が消滅し、いずれの残余試験化合物とNaV1.7活性化因子化合物の存在も哺乳動物で検出不可能になった後に、第1とは異なる第2の投与量レベルでの投与のために、第1の哺乳動物が後で再使用できる。あるいは、アッセイは、同じ種の第2の哺乳動物に試験化合物の第2の投与量を投与し、続いて、第2の哺乳動物に、陰性対照で疼痛関連反応を誘導するのに有効な投与量のNaV1.7活性化因子を投与し;さらに、疼痛関連反応が、陰性対照に比べて、第2の哺乳動物で減少するかどうかを判定することをさらに含むことができる。
(a)第1の哺乳動物(例えば、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒト)に、試験化合物の第1の投与量を投与し、続けて、
(b)第1の哺乳動物に、陰性対照(試験化合物を受けていない)で疼痛関連反応を誘導するのに有効な(局所的または全身性)投与量のNaV1.7活性化因子(例えば、ベラトリジン、デルタメトリン、またはグラヤノトキシンIII)を投与し(このような投与は全身性でも局所的でもよい);次に
(c)疼痛関連反応が、陰性対照に比べて、第1の哺乳動物で減少するか否かを判定し;さらに
(d)疼痛関連反応が、陰性対照に比べて減少する試験化合物の最も少ない第2の投与量を特定する。哺乳動物への試験化合物投与は、全身性(例えば、腹腔内、静脈内、筋肉内、または経口投与)であっても、または局所的(例えば、皮下、足底内、局部投与)であってもよい。局所的な場合は、NaV1.7活性化因子の局所投与と同じ部位か、またはその近くである必要がある。回復期間が充分で、試験化合物およびNaV1.7活性化因子の効果が消滅し、いずれの残余試験化合物とNaV1.7活性化因子化合物の存在も哺乳動物で検出不可能になった後に、第1とは異なる第2の投与量レベルでの投与のために、第1の哺乳動物が後で再使用できる。あるいは、アッセイは、同じ種の第2の哺乳動物に試験化合物の第2の投与量を投与し、続いて、第2の哺乳動物に、陰性対照で疼痛関連反応を誘導するのに有効な投与量のNaV1.7活性化因子を投与し;さらに、疼痛関連反応が、陰性対照に比べて、第2の哺乳動物で減少するかどうかを判定することをさらに含むことができる。
別の実施形態では、本発明は、試験化合物(候補NaV1.7阻害剤)の用量範囲探索に有用な下記を含むアッセイを含む:
(a)第1の哺乳動物(例えば、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒト)に、第1の投与量の試験化合物、および第2の哺乳動物(同じ種の)に、第1の投与量とは異なる第2の投与量の試験化合物を投与し、続けて、
(b)第1のおよび第2の哺乳動物に、陰性対照(試験化合物を受けていない)で疼痛関連反応を誘導するのに有効な量のNaV1.7活性化因子(例えば、ベラトリジン、デルタメトリン、またはグラヤノトキシンIII)を投与し(このような投与は、全身性でも局所的でもよい);その後、
(c)疼痛関連反応が、陰性対照に比べて、第1の哺乳動物と第2の哺乳動物で減少するか否かを判定し;さらに、
(d)疼痛関連反応が、陰性対照に比べて、低減する試験化合物の最も少ない第2の投与量を特定する。哺乳動物に投与される試験化合物投与は、全身性(例えば、腹腔内、静脈内、筋肉内、または経口投与)でも、または局所的(例えば、皮下、足底内、または局部投与)でもよい。局所的な場合は、NaV1.7活性化因子の局所投与と同じ部位か、またはその近くである必要がある。
(a)第1の哺乳動物(例えば、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒト)に、第1の投与量の試験化合物、および第2の哺乳動物(同じ種の)に、第1の投与量とは異なる第2の投与量の試験化合物を投与し、続けて、
(b)第1のおよび第2の哺乳動物に、陰性対照(試験化合物を受けていない)で疼痛関連反応を誘導するのに有効な量のNaV1.7活性化因子(例えば、ベラトリジン、デルタメトリン、またはグラヤノトキシンIII)を投与し(このような投与は、全身性でも局所的でもよい);その後、
(c)疼痛関連反応が、陰性対照に比べて、第1の哺乳動物と第2の哺乳動物で減少するか否かを判定し;さらに、
(d)疼痛関連反応が、陰性対照に比べて、低減する試験化合物の最も少ない第2の投与量を特定する。哺乳動物に投与される試験化合物投与は、全身性(例えば、腹腔内、静脈内、筋肉内、または経口投与)でも、または局所的(例えば、皮下、足底内、または局部投与)でもよい。局所的な場合は、NaV1.7活性化因子の局所投与と同じ部位か、またはその近くである必要がある。
このようにして、NaV1.7を抑制する投与量を決定し、推定上有害作用を生ずる可能性のあるより高い投与量と比較し、治療濃度域を決定できる。さらに、NaV1.7試験阻害剤の有効性の臨床試験では、このような投与量でのみ、治療の有効性をNaV1.7の結果と見なすことができる。グローバルNaV1.7−/−マウスにより得られる重要な知識は、ナトリウムチャネル活性化因子は、NaV1.7を介して、さらには、NaV1.7のみを介して、疼痛反応を生成することである。
以降の図および発明の詳細な説明を考慮すれば、多くの追加の本発明の態様および利点が明らかになるであろう。
実施形態の詳細な説明
本明細書で使われるセクション見出しは、構成上の目的のみのためであり、記載された主題を限定するものと解釈されるべきではない。
本明細書で使われるセクション見出しは、構成上の目的のみのためであり、記載された主題を限定するものと解釈されるべきではない。
定義
本明細書で別段の規定がない限り、本出願に関連して使われている科学的および技術的用語は、通常、当業者により理解されている意味を持つものとする。さらに、文脈から別義が要求されない限り、単数形の用語は、複数の対象物を含み、複数の用語は、反収の対象物を含むものとする。従って、文脈から別義が明確に指示されない限り、本明細書および添付請求項で使われる単数形「a」、「an」および「the」は、複数対象物を含む。例えば、「タンパク質(a protein)」への参照は、複数のタンパク質を含み;「細胞(a cell)」への参照は、複数の細胞集団を含む。
本明細書で別段の規定がない限り、本出願に関連して使われている科学的および技術的用語は、通常、当業者により理解されている意味を持つものとする。さらに、文脈から別義が要求されない限り、単数形の用語は、複数の対象物を含み、複数の用語は、反収の対象物を含むものとする。従って、文脈から別義が明確に指示されない限り、本明細書および添付請求項で使われる単数形「a」、「an」および「the」は、複数対象物を含む。例えば、「タンパク質(a protein)」への参照は、複数のタンパク質を含み;「細胞(a cell)」への参照は、複数の細胞集団を含む。
「ポリペプチド」および「タンパク質」は、本明細書では同義に使用され、ペプチド結合を介して共有結合で結合した2つ以上のアミノ酸分子鎖を含む。これらの用語は、特定の長さの生成物を意味しない。従って、「ペプチド」および「オリゴペプチド」は、ポリペプチドの定義の内に含まれる。この用語は、翻訳後修飾、例えば、グリコシル化、アセチル化、リン酸化、等、をされたポリペプチドを含む。さらに、タンパク質断片、類似体、変異または変種タンパク質、融合タンパク質、等は、ポリペプチドの意味の内に含まれる。この用語は、また、既知のタンパク質工学技術を使って、組換えにより発現されうる、1つまたは複数のアミノ酸類似体または非標準もしくは非天然アミノ酸が含まれる分子を含む。さらに、融合タンパク質は、周知の有機化学技術により本明細書記載のように誘導体化できる。
「組換え」という用語は、物質(例えば、核酸またはポリペプチド)がヒトの介入により、人工的にまたは合成により(すなわち、非天然的に)変えられていることを示す。変更は、その天然環境または状態内の、またはそこから取り出された物質に対し行うことができる。例えば、「組換え核酸」は、核酸を、例えば、クローニング、DNAシャフリングまたは他のよく知られた分子生物学的方法の間に、組み換えることにより作られるものである。このような分子生物学的方法の例は、Maniatis et al.、分子クローニング。実験マニュアル(Molecular Cloning.A Laboratory Manual).Cold Spring Harbor Laboratory、Cold Spring Harbor、N.Y(1982)、で見つけられる。「組換えDNA分子」は、このような分子生物学的技術により一緒に連結されたDNAのセグメントから構成される。本明細書で使われる「組換えタンパク質」または「組換えポリペプチド」という用語は、組換えDNA分子を使って発現されるタンパク質分子を意味する。「組換え宿主細胞」は、組換え核酸を含む、および/または発現する細胞である。
「ポリヌクレオチド」または「核酸」という用語は、2つ以上のヌクレオチド残基を含む単鎖および二重鎖ヌクレオチドポリマーの両方を含む。ヌクレオチド残基含有ポリヌクレオチドは、リボヌクレオチドもしくはデオキシリボヌクレオチドであってもよく、またはいずれかのヌクレオチドの型の修飾型であってもよい。前記修飾は、塩基修飾、例えば、ブロモウリジンおよびイノシン誘導体、リボース修飾、例えば、2’,3’−ジデオキシリボース、およびヌクレオチド間結合修飾、例えば、ホスホロチオアート、ホスホロジチオアート、ホスホロセレノアート、ホスホロジセレノアート、ホスホロアニロチオアート、ホスホラニラダートおよびホスホロアミダートを含む。
「オリゴヌクレオチド」という用語は、200以下のヌクレオチド残基を含むポリヌクレオチドを意味する。いくつかの実施形態では、オリゴヌクレオチドは、10〜60塩基長である。他の実施形態では、オリゴヌクレオチドは、12、13、14、15、16、17、18、19、または20〜40ヌクレオチド長である。オリゴヌクレオチドは、例えば、変異体遺伝子の作製での使用に対しては、単鎖でも、二重鎖でもよい。オリゴヌクレオチドは、センス、またはアンチセンスオリゴヌクレオチドのいずれでもよい。オリゴヌクレオチドは、標識を含んでもよく、これには、定量化または検出を容易にするための同位体標識(例えば、125I、14C、13C、35S、3H、2H、13N、15N、18O、17O、等);アッセイ検出用の蛍光標識、ハプテンまたは抗原性標識、が含まれる。オリゴヌクレオチドは、例えば、PCRプライマー、クローニングプライマーまたはハイブリダイゼーションプローブとして使用できる。
「ポリヌクレオチド配列」または「ヌクレオチド配列」または「核酸配列」は、本明細書で同義に使用され、ポリヌクレオチド中のヌクレオチド残基の一次配列であり、これには、オリゴヌクレオチド、DNA、およびRNA、核酸、または、文脈によっては、ヌクレオチド残基の一次配列を表す文字列を含む。いずれの特定ポリヌクレオチド配列からも、所与の核酸または相補的ポリヌクレオチド配列が決定可能である。ゲノムまたは合成由来のDNAまたはRNAが含まれ、単鎖でも二重鎖でも、また、センス鎖でもアンチセンス鎖でもよい。別段の指定がない限り、本明細書で考察されるいずれの単鎖ポリヌクレオチド配列の左側末端も、5’末端であり;二重鎖ポリヌクレオチド配列の左手方向は、5’方向と見なされる。5’から3’の新生RNA転写物の付加方向は、転写方向と呼ばれ;RNA転写物の5’から5’末端であるRNA転写物と同じ配列を有するDNA鎖上の配列領域は、「上流配列」と呼ばれ;RNA転写物の3’から3’末端であるRNA転写物と同じ配列を有するDNA鎖上の配列領域は、「下流配列」と呼ばれる。
本明細書で使用される「単離核酸分子」または「単離核酸配列」は、(1)特定され、通常自然の供給源において一緒に存在する少なくとも1つの混在核酸分子から分離されているか、または(2)クローン化され、増幅され、標識され、もしくは別の方法で背景核酸から区別され、それにより、目的の核酸の配列が特定できるようにされているか、いずれかの核酸分子である。単離核酸分子は、天然に見つかる形態または設定で存在するもの以外のものである。しかし、単離核酸分子は、通常、ポリペプチド(例えば、オリゴペプチドまたは抗体)を発現する細胞中に含まれる(例えば、核酸分子が天然の細胞とは異なる染色体の部位に存在する)核酸分子を含む。
本明細書で使用される「〜をコードする核酸分子」「〜をコードするDNA配列」および「〜をコードするDNA」という用語は、デオキシリボ核酸の鎖に沿ったデオキシリボヌクレオチドの順序または配列を指す。これらのデオキシリボヌクレオチドの順序は、mRNA鎖に沿ったリボヌクレオチドの順序を決定し、また、ポリペプチド(タンパク質)鎖に沿ったアミノ酸の順序も決定する。従って、DNA配列は、RNA配列およびアミノ酸配列をコードする。
「遺伝子」という用語は、広範に使用され、生物学的機能に関連するいずれの核酸をも意味する。遺伝子は、典型的には、コード配列、および/または、そのようなコード配列の発現に必要とされる調節配列を含む。「遺伝子」という用語は、特定のゲノムまたは組換え配列、ならびにその配列デコードされるcDNAまたはmRNAに適用される。「融合遺伝子」は、天然で一緒には見出されないか、または天然でそのコードされた融合タンパク質(すなわち、キメラタンパク質)中に存在する同じ配列中に一緒には見出されない、異なるタンパク質由来の部分を含むポリペプチドをコードするコーディング領域を含む。遺伝子は、また、例えば、他のタンパク質のための認識配列を形成する非発現核酸セグメントを含む。非発現核酸セグメントは、転写因子等の調節タンパク質が結合する転写調節領域であって、隣接または近くの配列の転写を生ずるものを含む。
「遺伝子の発現」または「核酸の発現」は、文脈に依存して、DNAのRNAへの転写(任意で、RNAの修飾、例えば、スプライシングを含む)、RNAのポリペプチドへの翻訳(場合により、その後のポリペプチドの翻訳後修飾を含む)、または転写と翻訳の両方を意味する。
本明細書で使用される「コーディング領域」または「コード配列」という用語は、構造遺伝子に関連して使われる場合、mRNA分子の翻訳の結果として新生ポリペプチドで見出されるアミノ酸をコードするヌクレオチド配列を意味する。真核生物では、コーディング領域は、5’側のイニシエータのメチオニンをコードするヌクレオチドトリプレット「ATG」および3’側の停止コドン(すなわち、TAA、TAG、TGA)を指定する3つのトリプレットの内の1つにより境界が形成されている。
「制御配列」または「制御シグナル」という用語は、特定の宿主細胞中で、それが連結されているコード配列の発現およびプロセッシングに影響を与えうるポリヌクレオチド配列を意味する。このような制御配列の性質は、宿主生物体に依存する可能性がある。特定の実施形態では、原核生物の制御配列は、プロモーター、リボソーム結合部、および転写終止配列を含むことができる。真核生物の制御配列は、1つまたは複数の転写因子用認識部位を含むプロモーター、転写エンハンサー配列または要素、ポリアデニル化部位、および転写終止配列を含むことができる。制御配列は、リーダー配列および/または融合パートナー配列を含むことができる。プロモーターおよびエンハンサーは、転写に関与する細胞タンパク質と特異的に相互作用する短い配列のDNAから構成される(Maniatis、et al.、Science 236:1237(1987))。プロモーターおよびエンハンサー配列は、酵母、昆虫および哺乳動物細胞ならびにウイルスの遺伝子、等の種々の真核生物の源から単離されている(類似の調節領域、すなわち、プロモーターは、原核生物でも見出される)。特定のプロモーターおよびエンハンサーの選択は、どの細胞型を使って目的のタンパク質を発現させる予定かに依存する。いくつかの真核生物のプロモーターおよびエンハンサーは、広い宿主範囲を有するが、他のものは、限られた細胞型サブセット中で機能する(概説は、Voss、et al.、Trends Biochem.Sci.、11:287(1986)、および、Maniatis、et al.、Science 236:1237(1987)、を参照)。
「ベクター」という用語は、タンパク質コード情報を宿主細胞中に移入するために使用されるいずれの分子または実体をも(例えば、核酸、プラスミド、バクテリオファージまたはウイルス)を意味する。
本明細書で使用される「発現ベクター」または「発現構築物」という用語は、特定の宿主細胞中で機能的に連結されたコード配列の発現に必要な所望のコード配列および適切な核酸制御配列を含む組換えDNA分子を意味する。発現ベクターは、限定されないが、転写、翻訳に影響するまたはこれを制御する配列、および、イントロンが存在する場合には、それに機能的に連結されたコーディング領域のRNAスプライシングに影響を与える配列を含むことができる。原核生物中の発現に必要な核酸配列は、プロモーター、任意選択でオペレーター配列、リボソーム結合部位および場合によっては他の配列を含む。真核細胞は、プロモーター、エンハンサー、ならびに終止およびポリアデニル化シグナルを利用することが知られている。分泌シグナルペプチド配列は、また、任意選択で、所望に応じ、その細胞から目的のポリペプチドのより簡単な単離を目的として、目的のコード配列に機能的に連結された発現ベクターによりコードされ、それにより、発現ポリペプチドが組換え宿主細胞により分泌されうる。このような技術は、当技術分野でよく知られている(例えば、Goodey、Andrew R.;et al.、ペプチドおよびDNA配列(Peptide and DNA sequences)、米国特許第5,302,697号;Weiner et al.、タンパク質分泌用組成物および方法(Compositions and methods for protein secretion)、米国特許第6,022,952号および米国特許第6,335,178号;Uemura et al.、タンパク質発現ベクターおよびその利用(Protein expression vector and utilization thereof)、米国特許第7,029,909号;Ruben et al.、27 ヒト分泌タンパク質(human secreted proteins)、米国特許公開第2003/0104400A1号)。
本明細書で使われる「操作可能な組み合わせで」、「操作可能な順に」および「機能的に連結された」という用語は、所与の遺伝子の転写、および/または所望のタンパク質分子の合成を指示できる核酸分子が産生されるような形式の核酸配列の結合を意味する。この用語は、また、機能的タンパク質が産生されるような形式でのアミノ酸配列の結合を意味する。例えば、タンパク質をコードする配列に「機能的に連結」されるベクター中の制御配列は、タンパク質をコードする配列の発現が、制御配列の転写活性に適合する条件下で実現されるように、それに対し連結される。
「宿主細胞」という用語は、核酸で形質転換されているか、または、形質転換されることが可能で、それにより、目的の遺伝子を発現する細胞を意味する。この用語は、目的の遺伝子が存在する限り、その子孫が形態学または遺伝的構成が元の親細胞に同じであっても同じでなくても、親細胞の子孫を含む。多数の入手可能な、よく知られた宿主細胞のいずれも、本発明の実施に使用できる。特定の宿主の選択は、技術分野によって認識されているいくつかの要因に依存する。これらには、例えば、選択発現ベクターとの適合性、DNA分子コードされるペプチドの毒性、形質転換速度、ペプチドの回収の容易さ、発現特性、バイオセーフティおよび費用、が含まれる。これらの要因の間のバランスは、全ての宿主が特定のDNA配列の発現に等しく有効なわけではないという理解と合致させなければならない。これらの一般ガイドライン内で、培養に有用な微生物の宿主細胞には、細菌(例えば、大腸菌種)、酵母(例えば、サッカロミセス種)および他の真菌細胞、昆虫細胞、植物細胞、哺乳類(ヒトを含む)細胞、例えば、CHO細胞およびHEK−293細胞、が含まれる。修飾は、DNAレベルでも同様に行うことができる。ペプチドをコードするDNA配列は、選択宿主細胞に適合性のより高いコドンに変更できる。大腸菌については、最適化コドンが、当技術分野で知られている。コドンを置換して、制限酵素部位を除去する、またはサイレント制限酵素部位を含めることができ、これは、選択宿主細胞中でのDNAプロセッシングを支援し得る。次いで、形質転換宿主は、培養され、精製される。宿主細胞は、従来の発酵条件下で培養し、所望の化合物を発現させることができる。このような発酵条件は、当技術分野でよく知られている。
「形質移入」という用語は、細胞による外来性または外因性DNAの取込を意味し、外因性のDNAが、細胞膜内に導入された場合に、細胞は「形質移入」されている。いくつかの形質移入技術が当技術分野でよく知られており、本明細書で開示されている。例えば、Graham et al.、1973、Virology 52:456;Sambrook et al.、2001、分子クローニング:実験マニュアル(Molecular Cloning:A Laboratory Manual)、同上;Davis et al.、1986、分子生物学における基礎的方法(Basic Methods in Molecular Biology)、Elsevier;Chu et al.、1981、Gene 13:197、を参照されたい。このような技術を使用して、1つまたは複数の外因性DNA成分を適切な宿主細胞中に導入することができる。
「形質転換」という用語は、細胞の遺伝的特性の変化を意味し、細胞は、改変されて新規DNAまたはRNAを含む場合、形質転換されている。例えば、形質移入、形質導入、または他の技術による新しい遺伝物質の導入により、その自然の状態から遺伝的に改変されている場合、細胞は形質転換されている。形質移入または形質導入後、形質転換DNAは、物理的にその細胞の染色体中に組み込むことにより、細胞のDNAと組み換えることができ、または、複製されることなく、エピソーム要素として一時的に維持でき、または、独立にプラスミドとして複製できる。形質転換DNAが細胞分裂により複製される場合、細胞は、「安定に形質転換」されていると見なされる。
同系交配マウス株(例えば、C57Bl/6J)を、異なる同系交配マウス株(例えば、BALB/c)に変化させるプロセスは、「戻し交配(backcrossing)」または「戻し交配(backcross)」と呼ばれる。これは、マウス株が意図した研究目的に適さない場合に有用であり;株は、育種計画により遺伝的に改変できる。遺伝子型を「株A」(例えば、C57Bl/6J)から「株B」(例えば、BALB/c)に完全に形質転換するために、マウスは、通常、少なくとも10回、戻し交配される必要があり;その後にのみ、それらは、「類遺伝子性」と呼ぶことができる。マウスを戻し交配するために、変異体マウスは、選ばれた同系交配株(例えば、BALB/c)と交配される。この交配の出生児は、「戻し交配#1」(または「N1」)と命名され、両方の株のハイブリッドである(例えば、約:50%C57Bl6/Jおよび50%BALB/c)。出生児は遺伝子型が特定でき、これらの目的の遺伝子に対するヘテロ接合体のみが、同系交配マウス株の野性型個体(例えば、野性型BALB/cマウス)と、再度交配される。通常、この手順は、類遺伝子性BALB/c系統が得られるまで約10回繰り返される。同系交配マウス株のいずれの組み合わせも、この手法を使用して選ばれた株の類遺伝子系統を作ることができる。例として、2010年12月までに、我々は、同系交配BALB/c背景の第5戻し交配世代(「N5」)を生成し、特徴付けした。ヘテロ接合の(「HET」)出生児は、約98.6%BALB/cの遺伝的背景である。2011年の5月には、我々は、最初のBALB/c類遺伝子性繁殖ペアを得た。我々は、この背景がNaV1.7KO新生児の生存率を改善するか否か(すなわち、ヒトケア/給餌の要求が減ったか否か)を評価するために現在交配している。今のところ、結果は、C57Bl6/J−BALB/cハイブリッドと同じであるように思われる。
同系交配マウス株(例えば、C57Bl/6J)を、非近交系マウス株(例えば、CD1)に変えるプロセスは、「異系交配(outcrossing)」または「異系交配(outcross)」と呼ばれる。これは、同系交配マウス株からの変異体(ノックアウト)動物が衰弱の徴候を示す場合に有用であり;目的の遺伝子に対するヘテロ接合出生児は、非近交系マウス株と交配して、遺伝的変異および活力を同系交配マウス株中に導入できる。マウスの異系交配を行うために、変異マウスは、選択された非近交系株(例えば、CD1)由来の野性型マウスと交配される。この交配の出生児は、ハイブリッドである(例えば、約:50%C57Bl6/Jおよび50%CD1)。非近交系マウスは、遺伝子プール中で変異が多すぎて類遺伝子性系統を作れないために、通常は、それ以上「異系交配」されない。
タンパク質の「ドメイン(domain)」または「領域(region)」(本明細書では同義に使用される)は、完全なタンパク質まで、およびこれを含む全体タンパク質のいずれかの部分であり、通常、完全なタンパク質より少ない部分を含む。ドメインは、(必要はないが)特定の生物学的、生化学的、または構造的機能または部位(例えば、リガンド結合ドメイン、またはサイトゾルの、膜貫通型の、もしくは細胞外のドメイン)を、残りのタンパク質鎖とは無関係に保持し、および/または、それらと関連付けられ得る。
「哺乳動物」は、哺乳類に分類されるいずれの動物をも意味し、ヒト、家畜(domestic and farm animal)、および動物園、スポーツ、またはペット動物を含み、例えば、イヌ、ウマ、ネコ、雌ウシ、ラット、マウス、非ヒト霊長類(例えば、サル、類人猿)、等である。
「げっ歯類(rodent)」および「複数げっ歯類(rodents)」という用語は、系統学的ネズミ目の全メンバーを指し、それら由来の全ての未来の世代のいずれかのおよび全ての子孫を含む。
「マウス」という用語は、ファミリーネズミ科のいずれかのおよび全てのメンバーを指し、ラットおよびマウスを含む。
生物学的物質、例えば、ポリペプチド、核酸、宿主細胞、等に関連して、本明細書全体を通して使用される「天然の」という用語は、自然で見出される物質を意味する。
動物、例えば、マウスまたは特に、グローバルNaV1.7−/−ノックアウトマウスに関し、「生存可能な」という用語は、動物が、成人期に達することができる(新生仔または幼動物の場合には)、または成人期に達していて、適切な栄養を摂ると自ら生きることができることを意味する。
「ノックアウト」という用語は、細胞中の内在性DNA配列によりコードされるタンパク質、例えば、サブユニットのナトリウムチャネル、電位依存性、タイプIX(「NaV1.7」としても知られる)の少なくとも一部の発現の部分的または完全抑制を意味する。「NaV1.7ノックアウト」、「NaV1.7KO」、「NaV1.7−/−」、「NaV1.7−/−ノックアウト」および「NaV1.7ヌル変異体」という用語は、本明細書では同義に使用され、機能性NaV1.7タンパク質の発現の完全抑制を示す細胞または哺乳動物を意味する。「hNaV1.7」という用語は、ヒトNaV1.7を意味する。
「ノックアウト構築物」という用語は、細胞中の内在性DNA配列によりコードされるタンパク質の発現を減少させるか、または抑制するように設計されている核酸配列を意味する。ノックアウト構築物として使用される核酸配列は、通常、(1)抑制される遺伝子のいくつかの部分(エキソン配列、イントロン配列、および/またはプロモーター配列)由来のDNA、および(2)細胞中のノックアウト構築物の存在を検出するために使用されるマーカー配列、からなる。ノックアウト構築物は、細胞中に挿入され、ネイティブDNA配列の転写を防ぐか、または阻止するような位置で、細胞のゲノムDNAに組み込まれる。このような挿入は、通常、相同組換えにより起こる(すなわち、ノックアウト構築物が細胞中に挿入され、組み換えられて、ノックアウト構築物が内在性DNAの対応する位置に組み込まれる時に、内在性DNA配列に相同なノックアウト構築物の領域が相互にハイブリダイズする)。ノックアウト構築物の核酸配列は、1)抑制される1つまたは複数の遺伝子のエキソンおよび/またはイントロンの完全または部分的配列、2)遺伝子の抑制されるプロモーターの完全または部分的配列、または3)これらの組み合わせ、を含むことができる。
典型的には、ノックアウト構築物は、胚性幹細胞(ES細胞)中に挿入され、通常、相同組換えプロセスにより、ES細胞ゲノムDNA中に統合される。このES細胞は、その後、発生過程の胚中に導入され、統合される。
「遺伝子の撹乱」および「遺伝子撹乱」という表現は、核酸配列のネイティブDNA配列の1つの領域(通常、1つまたは複数のエキソン)、および/または遺伝子のプロモーター領域中へ挿入し、遺伝子の野性型または天然配列に比べて、細胞中でのその遺伝子の発現を減らすか、または防ぐことを意味する。例として、撹乱されるDNA配列(プロモーターおよび/またはコーディング領域)に相補的なDNA配列中に挿入される抗生物質耐性遺伝子をコードするDNA配列を含む核酸構築物を調製できる。その後、この核酸構築物を細胞中に形質移入すると、構築物は、ゲノムDNAに統合される。従って、その時点でDNAが抗生物質抵抗性遺伝子により撹乱されているために、その細胞の多くの子孫は、少なくとも一部の細胞中で、もはやその遺伝子を発現しないか、または低いレベルで発現することになる。
「導入遺伝子」という用語は、哺乳動物または哺乳類胚の1つまたは複数の細胞中に挿入されうる、宿主とは異なる種に由来する単離ヌクレオチド配列を意味する。導入遺伝子は、任意選択で、直接、または間接的に、細胞機構と連携して、導入遺伝子の転写および/または発現を調節する機能を果たすことができる他の遺伝的要素(例えば、プロモーター、ポリA配列、等)に機能的に連結されていてもよい。あるいは、または追加として、導入遺伝子は、導入遺伝子の哺乳動物細胞または胚の核の染色体DNA中への統合を支援する(例えば、相同組換えで)ヌクレオチド配列に連結されてもよい。導入遺伝子は、哺乳動物の内在性遺伝物質中の特定のヌクレオチド配列に対し相同または異種であるか、またはハイブリッド配列(すなわち、哺乳動物の遺伝物質に対し、導入遺伝子の1つまたは複数の部分が相同で、1つまたは複数の部分が異種)であるヌクレオチド配列から構成されてもよい。導入遺伝子ヌクレオチド配列は、哺乳動物中で内因的に見出されるポリペプチドまたはポリペプチドの変異体をコードしてもよく、哺乳動物の天然に存在しないポリペプチド(すなわち、外因性のポリペプチド)をコードしてもよく、または、内在性および外因性ポリペプチドのハイブリッドをコードしてもよい。導入遺伝子がプロモーターに機能的に連結される場合は、そのプロモーターは、哺乳動物、および/または導入遺伝子に対し、相同であっても、異種であってもよい。あるいは、プロモーターは、内在性および外因性のプロモーター要素(エンハンサー、サイレンサー、サプレッサー、等)のハイブリッドであってもよい。
「疼痛関連反応」は、疼痛誘導刺激が加えられた場合、哺乳動物の特定の種で典型的に示される、例えば、マウスおよびラットでの足を持ち上げる、足をなめる、足を引っ込める、鳴く、またはこれらのいずれかの組み合わせの挙動のような、認識されるいずれかの挙動である。ヒト対象では、例えば、口頭または文書による疼痛の自己報告、または叫び声が「疼痛関連反応」といえる。
用語の「子孫」は、特定の哺乳動物、すなわち、そのゲノムDNAに挿入されたノックアウト構築物を含む哺乳動物由来で、それから派生するいずれかの、および全ての未来の世代を意味する。従って、いずれかの継続的世代の子孫が本明細書で含まれ、それにより、子孫のF1、F2、F3、世代、等は、どこまでもこの定義に含まれる。
追加の実施形態
1つ、2つ、またはそれ超の追加の目的遺伝子が、マウス由来の遺伝子(改変ヌクレオチド配列を有してもよい)または導入遺伝子の挿入により「ノックアウト」、または「ノックイン」されているグローバルNav1.7KOマウスが、本発明の範囲に含まれる。このような哺乳動物は、本明細書で各「ノックアウト」または遺伝子導入「ノックイン」構築物を生成するために記載された手続きを繰り返すことにより、または単一ノックアウト遺伝子を有する哺乳動物と相互に交配し、二重、または多重の、ノックアウトおよび/またはノックイン遺伝子型を有するものを選別することにより、生成できる。撹乱されるか、または組換えで発現されるDNA上の少なくとも一部の配列情報を入手でき、構築物および選別プローブの両方の調製に使用できるという条件で、ノックアウトまたはノックインされる遺伝子は、いずれの遺伝子でもよい。
1つ、2つ、またはそれ超の追加の目的遺伝子が、マウス由来の遺伝子(改変ヌクレオチド配列を有してもよい)または導入遺伝子の挿入により「ノックアウト」、または「ノックイン」されているグローバルNav1.7KOマウスが、本発明の範囲に含まれる。このような哺乳動物は、本明細書で各「ノックアウト」または遺伝子導入「ノックイン」構築物を生成するために記載された手続きを繰り返すことにより、または単一ノックアウト遺伝子を有する哺乳動物と相互に交配し、二重、または多重の、ノックアウトおよび/またはノックイン遺伝子型を有するものを選別することにより、生成できる。撹乱されるか、または組換えで発現されるDNA上の少なくとも一部の配列情報を入手でき、構築物および選別プローブの両方の調製に使用できるという条件で、ノックアウトまたはノックインされる遺伝子は、いずれの遺伝子でもよい。
ノックアウト遺伝子の選択
通常、ノックアウト構築物に使用されるDNAは、1つまたは複数のエキソンおよび/またはイントロン領域、および/またはプロモーター領域であるが、cDNAが充分に大きければ、cDNA配列であってもよい。一般的に、DNAは、少なくとも約1キロベース(kb)長、好ましくは、3〜4kbの長さであり、それにより、構築物がES細胞のゲノムDNAに導入される場合(下記で考察)、ハイブリダイゼーションに充分な相補的配列を提供する。典型的には、ノックアウトされる目的の遺伝子は、ノックアウトした場合、致死に繋がらない遺伝子であろう。
通常、ノックアウト構築物に使用されるDNAは、1つまたは複数のエキソンおよび/またはイントロン領域、および/またはプロモーター領域であるが、cDNAが充分に大きければ、cDNA配列であってもよい。一般的に、DNAは、少なくとも約1キロベース(kb)長、好ましくは、3〜4kbの長さであり、それにより、構築物がES細胞のゲノムDNAに導入される場合(下記で考察)、ハイブリダイゼーションに充分な相補的配列を提供する。典型的には、ノックアウトされる目的の遺伝子は、ノックアウトした場合、致死に繋がらない遺伝子であろう。
選択遺伝子をノックアウトするために使用されるDNA配列は、例えば、Sambrookら(分子クローニング:実験マニュアル(Molecular Cloning:A Laboratory Manual)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.[1989])により記載されたもの、等の当技術分野でよく知られた方法を使って得ることができる。このような方法には、例えば、少なくとも一部のゲノム配列を得るための、同じ遺伝子の少なくとも一部をコードするcDNAプローブを使ったゲノムライブラリーの選別が含まれる。あるいは、cDNA配列がノックアウト構築物で使用される場合は、そのcDNAは、オリゴヌクレオチドプローブまたは抗体を使ってcDNAライブラリーを選別することより(この場合、ライブラリーは、発現ベクター中に挿入される)得ることができる。プロモーター配列がノックアウト構築物で使用される場合は、合成DNAプローブは、プロモーター配列を含むゲノムライブラリーを選別するよう設計できる。
ノックアウト構築物で使用されるDNAを得る別の方法は、DNA合成装置を使って、DNA配列を合成により作製することである。
ノックアウト構築物をコードするDNA配列は、遺伝子操作およびES細胞への挿入に充分な量で生成されなければならない。増幅は、1)その配列を適切なベクターに配置し、急速にベクターを増幅できる細菌細胞または他の細胞を形質転換すること、2)PCR増幅により、または3)DNAシンセサイザーを使った合成により行うことができる。
ノックアウト構築物の調製
ノックアウト構築物の生成に使われるDNA配列は、典型的には、新しいマーカー遺伝子をコードするDNA配列が、このDNA配列内の適切な位置に挿入されうるような部位で切断されるように選択される特定の制限酵素で消化される。マーカー遺伝子挿入用の適切な位置は、自然遺伝子の発現を防ぐように機能する位置であり;この位置は、切断される配列中の制限酵素部位、およびエキソン配列、プロモーター配列または両方の内のどれが遮断されるのか(すなわち、プロモーター機能を抑制するまたはネイティブエキソンの合成を抑制するのに必要な正確な挿入位置)、等の種々の要因に依存する。典型的には、DNA切断用に選択される酵素は、より長いアームおよびより短いアームを生成し、より短いアームは、少なくとも約300ベース対(bp)である。いくつかの例では、マーカー遺伝子が、ノックアウト構築物に挿入される場合、元のゲノム配列と同程度のノックアウト構築物の長さを保持するために、1つまたは複数の抑制される遺伝子のエキソンの内の一部またはさらには全てを、実際に除去するのが望ましい。これらの場合では、ゲノムDNAは、適切な制限エンドヌクレアーゼで切断し、それにより、適切な大きさの断片が除去できる。
ノックアウト構築物の生成に使われるDNA配列は、典型的には、新しいマーカー遺伝子をコードするDNA配列が、このDNA配列内の適切な位置に挿入されうるような部位で切断されるように選択される特定の制限酵素で消化される。マーカー遺伝子挿入用の適切な位置は、自然遺伝子の発現を防ぐように機能する位置であり;この位置は、切断される配列中の制限酵素部位、およびエキソン配列、プロモーター配列または両方の内のどれが遮断されるのか(すなわち、プロモーター機能を抑制するまたはネイティブエキソンの合成を抑制するのに必要な正確な挿入位置)、等の種々の要因に依存する。典型的には、DNA切断用に選択される酵素は、より長いアームおよびより短いアームを生成し、より短いアームは、少なくとも約300ベース対(bp)である。いくつかの例では、マーカー遺伝子が、ノックアウト構築物に挿入される場合、元のゲノム配列と同程度のノックアウト構築物の長さを保持するために、1つまたは複数の抑制される遺伝子のエキソンの内の一部またはさらには全てを、実際に除去するのが望ましい。これらの場合では、ゲノムDNAは、適切な制限エンドヌクレアーゼで切断し、それにより、適切な大きさの断片が除去できる。
マーカー遺伝子は、検出可能な、および/またはアッセイ可能ないずれの核酸配列でもよいが、典型的には、それは、抗生物質耐性遺伝子またはゲノム中のその発現または存在が容易に検出できる他の遺伝子である。マーカー遺伝子は、通常、機能的に連結されたそれ自体のプロモーター、または挿入された細胞中で活性であるか、または容易に活性化できる任意のソース由来の別の強力プロモーターあるが、マーカー遺伝子は、抑制される遺伝子のプロモーターを使って転写でき得るので、それ自体のプロモーターが付随している必要はない。さらに、マーカー遺伝子は、通常、遺伝子の3’末端に結合したポリA配列を有し;この配列が遺伝子の転写を終止させる役目をする。好ましいマーカー遺伝子は、neo(ネオマイシン耐性遺伝子)およびベータgal(ベータガラクトシダーゼ)、等のいずれかの抗生物質耐性遺伝子である。
ゲノムDNA配列が適切な制限酵素により消化完了した後で、マーカー遺伝子配列は、当業者によく知られた、およびSambrookら(同上)に記載の方法を使ってゲノムDNA配列に連結される。連結されるDNA断片の末端は、適合性でなければならない;これは、適合性末端を生成する酵素で全ての断片を切断するか、または連結反応の前に平滑末端化することにより実現される。平滑末端化は、例えば、付着末端にはめ込まれるクレノー断片(DNAポリメラーゼI)の使用による、等の当技術分野でよく知られた方法を使って行える。
連結ノックアウト構築物は、胚性幹細胞中に直接挿入さても(下記で考察)、または、最初に、挿入に先立つ増幅のために適切なベクター中に置かれてもよい。好ましいベクターは、細菌細胞中で急速に増幅されるもの、例えば、pBluescript II SKベクター(Stratagene、San Diego、Calif.)またはpGEM7(Promega Corp.、Madison、Wis.)である。
胚性幹細胞の形質移入
本発明は、限定されないが、ウサギ、ラット、ハムスター、およびマウスを含む任意のげっ歯類の種由来のノックアウト哺乳動物の産生を意図している。好ましいげっ歯類には、ラットおよびマウスを含むネズミ科ファミリーのメンバーが含まれる。ノックアウト生成用のES細胞が導出できるマウス株には、C57BL/6、129SV、CD1、またはBALB/cが含まれる。一般的に、ノックアウト哺乳動物の産生に使われる胚性幹細胞(ES細胞)は、生成されるノックアウト哺乳動物と同じ種である。従って、例えば、マウス胚性幹細胞が、通常、ノックアウトマウスの生成に使用される。
本発明は、限定されないが、ウサギ、ラット、ハムスター、およびマウスを含む任意のげっ歯類の種由来のノックアウト哺乳動物の産生を意図している。好ましいげっ歯類には、ラットおよびマウスを含むネズミ科ファミリーのメンバーが含まれる。ノックアウト生成用のES細胞が導出できるマウス株には、C57BL/6、129SV、CD1、またはBALB/cが含まれる。一般的に、ノックアウト哺乳動物の産生に使われる胚性幹細胞(ES細胞)は、生成されるノックアウト哺乳動物と同じ種である。従って、例えば、マウス胚性幹細胞が、通常、ノックアウトマウスの生成に使用される。
胚性幹細胞(ES細胞)は、典型的には、ノックアウト構築物の生殖細胞系列伝達を生成するように、発生過程の胚の生殖細胞系列の一部中に組み込まれる能力とその一部になる能力で選択される。従って、この能力を有すると考えられるいずれのES細胞株も、本明細書での使用に適する。ES細胞の産生に通常使用される1つのマウス株は、129J株である。好ましいES細胞株は、マウス細胞株D3(アメリカ合衆国培養細胞株保存機関、カタログ番号:CRL 1934)である。細胞を培養し、下記に記載のもの等の、当業者よく知られた方法を使ってDNA挿入用に調製される:Robertson:奇形癌腫および胚性幹細胞:実践的手法(Teratocarcinomas and Embryonic Stem Cells:A Practical Approach)、E.J.Robertson、ed.IRL Press、Washington、D.C.[1987]、およびBradley et al.、Current Topics in Devel.Biol.、20:357−371[1986]、ならびにHogan et al.、マウス胚の操作:実験マニュアル(Manipulating the Mouse Embryo:A Laboratory Manual)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.[1986]。
ノックアウト構築物のES細胞中への挿入は、例えば、電気穿孔、微量注入、およびリン酸カルシウム処理(Robertson、ed.、同上中のLovell−Badgeを参照)等の当技術分野でよく知られた種々の方法を使って達成できる。好ましい挿入方法は、電気穿孔である。
細胞に挿入される各ノックアウト構築物DNAは、ノックアウト構築物がベクターに挿入されている場合は、最初に、直線化しなければならない。直線化は、ベクター配列内でのみ切断され、ノックアウト構築物配列内で切断されないように選択された適切な制限エンドヌクレアーゼでDNAを消化させることにより実現される。
DNA配列の挿入のために、ノックアウト構築物DNAは、選択された挿入方法に適切な条件下でES細胞に加えられる。2つ以上の構築物をES細胞中に導入する場合には、各構築物をコードするDNAを、同時にまたは1回で導入できる。
細胞を電気穿孔する場合は、ES細胞およびノックアウト構築物DNAを、電気穿孔装置を使い、メーカーの使用ガイドラインに従って電気パルスに曝す。電気穿孔後、細胞は、適切なインキュベーション条件下で回復させる。次に、細胞をノックアウト構築物の存在に関し選別する。
選別は、種々の方法を使って行える。マーカー遺伝子が抗生物質耐性遺伝子の場合は、それ以外は致死の濃度の抗生物質の存在下で細胞を培養する。生存細胞は、おそらく統合されたノックアウト構築物を有する。マーカー遺伝子が抗生物質耐性遺伝子以外である場合は、ES細胞ゲノムDNAのサザンブロットを使って、マーカー配列のみにハイブリダイズするように設計されたDNA配列で検出できる。最後に、マーカー遺伝子がその活性を検出できる酵素(例えば、ベータガラクトシダーゼ)をコードする遺伝子である場合は、酵素の基質を適切な条件下で細胞に添加し、酵素活性を解析できる。
ノックアウト構築物は、ES細胞ゲノム中のいくつかの部位に組み込むことができ、ランダム挿入イベントの発生に起因して、各細胞のゲノム中の異なる部位に組み込むことができ;所望の挿入部位は、ノックアウトされるDNA配列に相補的な位置にある。典型的には、ノックアウト構築物を取り込むES細胞の内の約1〜5パーセント未満が、望ましい部位にノックアウト構築物を実際に組み込まれる。ノックアウト構築物の適切な組込みを有するこれらの細胞を特定するために、例えば、Sambrook et al.、同上、に記載の方法、等の標準的な方法を使って、細胞からDNAを抽出できる。次に、DNAは、特定の制限酵素で消化されるゲノムDNAに、特異的パターンでハイブリダイズするように設計されたプローブまたは複数プローブを使って、サザンブロットで検出できる。あるいは、または追加として、ゲノムDNAは、特定のサイズと配列のDNA断片を増幅するように特別に設計されたプローブを使ってPCRにより増幅できる(すなわち、適切な位置のノックアウト構築物を含む細胞のみが、適切なサイズのDNA断片を生成する)。
胚の注入/移植
適切な部位にノックアウト構築物を含む適切なES細胞が特定された後で、細胞は、胚中に挿入される。挿入は、種々の方法で実現できるが、好ましい方法は、微量注入である。微量注入では、約10〜30細胞がマイクロピペット中に集められ、ES細胞を発生段階の胚中に組み込むのに適した発生段階にある胚中に注入される。
適切な部位にノックアウト構築物を含む適切なES細胞が特定された後で、細胞は、胚中に挿入される。挿入は、種々の方法で実現できるが、好ましい方法は、微量注入である。微量注入では、約10〜30細胞がマイクロピペット中に集められ、ES細胞を発生段階の胚中に組み込むのに適した発生段階にある胚中に注入される。
適切な胚の発生段階は、非常に種依存であるが、マウスでは、約3.5日である。胚は、妊娠雌の子宮を灌流することにより得られる。これを実現する適切な方法は、当業者には既知であり、Bradley(Robertson、ed.、同上)により説明されている。
適切な年齢/発生段階のいずれの胚でも使用に適するが、好ましい胚は、雄で、ES細胞遺伝子によりコードされる毛色とは異なる毛色をコードする遺伝子を有するものである。このようにして、出生児は、モザイク毛色を探す(ES細胞が発生段階の胚中に組み込まれたことを示す)ことにより、ノックアウト構築物の存在を容易に選別できる、従って、例えば、ES細胞株が白色毛皮の遺伝子を持つ場合、選択される胚は、黒または茶色の毛皮の遺伝子を持つものとなる。
ES細胞が胚中に導入された後、胚は、偽妊娠養母の子宮中に移植される。いずれの養母も使用可能であるが、典型的には、うまく交配し、繁殖する能力、および子どもの世話をする能力で選択される。このような養母は、典型的には、同じ種の精管切除された雄との交配により供給される。偽妊娠養母の段階は、移植の成功にとって重要であり、それは種依存である。マウスでは、この段階は、約2〜3日の偽妊娠である。
ノックアウト遺伝子の存在に関する選別
概して、養母の子として生まれた出生児は、最初に、毛色選択法(上述の)が採用された場合モザイク毛色で選別できる。さらに、または代替法として、出生児の尾部組織由来のDNAを、上述のように、サザンブロット、および/またはPCRを使ってノックアウト構築物の存在に関し選別できる。次に、モザイク模様に見える出生児が、それらが、生殖細胞系列中にノックアウト構築物を有し、ホモ接合のノックアウト動物を生成すると考えられる場合は、相互に交配される。出生児が生殖細胞系列伝達を有するか否かが不明確な場合は、それらは親株と、または他の株と交配でき、出生児をヘテロ接合性に関し選別できる。ヘテロ接合体は、上で説明のように、サザンブロット、および/またはDNAのPCR増幅により特定される。
概して、養母の子として生まれた出生児は、最初に、毛色選択法(上述の)が採用された場合モザイク毛色で選別できる。さらに、または代替法として、出生児の尾部組織由来のDNAを、上述のように、サザンブロット、および/またはPCRを使ってノックアウト構築物の存在に関し選別できる。次に、モザイク模様に見える出生児が、それらが、生殖細胞系列中にノックアウト構築物を有し、ホモ接合のノックアウト動物を生成すると考えられる場合は、相互に交配される。出生児が生殖細胞系列伝達を有するか否かが不明確な場合は、それらは親株と、または他の株と交配でき、出生児をヘテロ接合性に関し選別できる。ヘテロ接合体は、上で説明のように、サザンブロット、および/またはDNAのPCR増幅により特定される。
次いで、ヘテロ接合体が、相互に交配され、ホモ接合のノックアウト出生児を生成できる。ホモ接合体は、この交配の産物であるマウス、ならびにヘテロ接合体および野性型マウスであるとわかっているマウス由来の等量のゲノムDNAのサザンブロットにより特定できる。サザンブロットで選別するプローブは、上で説明のように設計できる。
他のノックアウト出生児を特定し、特徴付ける手段が利用できる。例えば、ノーザンブロット法を使って、ノックアウト遺伝子、マーカー遺伝子、または両方をコードする転写物の存在または非存在に関して、mRNAを調べることができる。さらに、ウェスタンブロットを使用して、ノックアウト遺伝子によりコードされたタンパク質に対する抗体を用い、またはマーカー遺伝子産物に対する抗体を用いて、ウェスタンブロットを探査することにより、これらの出生児の種々の組織中のノックアウトされた遺伝子の発現レベルを、この遺伝子が発現している場合に、評価できる。最後に、出生児由来の種々の細胞のインサイツ分析(例えば、細胞の固定および抗体で標識)および/またはFACS(蛍光標識細胞分取)分析を適切な抗体を使って行い、ノックアウト構築物遺伝子産物の存在または非存在を探索することができる。
新生仔のNaV1.7KOマウスは、明らかに摂食できないことを示すと報告された(Nassar et al.、2004)ために、本明細書の実施例1に記載のように、手による給餌を使って、栄養を新生仔のNaV1.7KOマウスに与えることができる。代わりの「手での給餌」の必要のない、静脈内給餌または胃挿入管およびシリンジポンプ等を使った給餌方法/人工飼育もまた、有用になりうる。しかし、必要な付随手術は、新生仔マウスに対しかなりのリスクを課す。あるいは、各マウスの子を単独でカップ中で飼育することを含む、いわゆる「pup in a Cup」を使用することができる(脳の発達の研究へのpup in a Cupモデルの使用(Use of pup in a Cup Model to Study Brain Development)、J.Nutr.、123:382−385(1993))。しかし、付随するシリンジポンプに関連する胃手術により引き起こされうる身体的傷害に加えて、この技術は、これらの社会性動物の行動障害を誘導し、従って、このようなマウスを使って得られるインビボデータの一部の信頼性に影響を与える可能性がある。上述のような合併症を避けて、ノックアウトの生存を増やす別の可能な方法は、ノックアウトを授乳雌親と一緒に置くことであり、そこでは、授乳がマウスの子の正常な同腹子により誘導される。すなわち、NaV1.7ノックアウトマウスの子は、NaV1.7ノックアウトマウスの子への給餌の必要性に応じ、正常な同腹子と交換される。雌親が外来性NaV1.7KOマウスの子を拒絶しないのを確実にするために、ノックアウトは、雌親の香りでマークされうる。時折、全ての遺伝子型の新生仔で腹部膨満が観察された。この腹部膨満は、腹部の膨満感の原因となる胃の空洞中の空気の存在を特徴とする。このような状況下では、永久取り付け型29ゲージ1/2インチ針を装着した極細インスリンシリンジを使って、空気を手作業で除去できる。
導入遺伝子技術
導入遺伝子の選択
典型的には、本発明で有用な導入遺伝子は、目的のポリペプチド、例えば、神経系、免疫反応、造血、炎症、細胞成長と増殖、細胞系列分化、および/またはストレス反応に関与するポリペプチドをコードするヌクレオチド配列であろう。本発明の範囲内に含まれるのは、1つ、2つ、またはそれ超の導入遺伝子の、本発明のNaV1.7ノックアウトマウスへの挿入である。
導入遺伝子の選択
典型的には、本発明で有用な導入遺伝子は、目的のポリペプチド、例えば、神経系、免疫反応、造血、炎症、細胞成長と増殖、細胞系列分化、および/またはストレス反応に関与するポリペプチドをコードするヌクレオチド配列であろう。本発明の範囲内に含まれるのは、1つ、2つ、またはそれ超の導入遺伝子の、本発明のNaV1.7ノックアウトマウスへの挿入である。
2つ以上の導入遺伝子が本発明で使用される場合は、導入遺伝子は、別々に調製、および挿入でき、または1つの挿入用構築物として一緒に生成できる。導入遺伝子は、各導入遺伝子の発現を進めるために選択された両方のプロモーターに対し、および/または哺乳動物に対し、相同であっても、異種であってもよい。さらに、導入遺伝子は、少なくといくらかの生物活性を有する、すなわち、同じ種の野性型、非遺伝子導入哺乳動物では容易に観察されないレベル(生化学的、細胞性および/または形態学的)で効果を示す、完全長cDNAであっても、ゲノムDNA配列であっても、またはそれらのいずれかの断片、サブユニットまたは変異体であってもよい。任意選択で、導入遺伝子は、ハイブリッドヌクレオチド配列、すなわち、相同および/または異種のcDNAおよび/またはゲノムDNA断片から構築された配列であってもよい。導入遺伝子は、また、任意選択で、天然cDNAおよび/またはゲノム配列、またはそれらの対立遺伝子変異体の内の1つまたは複数の変異体であってもよい。
それぞれの導入遺伝子は、当技術分野でよく知られた1つまたは複数の方法を使って、適切な量で単離し、得ることができる。導入遺伝子の単離に有用なこれらのおよび他の方法は、例えば、Sambrook et al.(分子クローニング:実験マニュアル(Molecular Cloning:A Laboratory Manual)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.[1989])およびBerger and Kimmel(酵素学における方法:分子クローニング技術ガイド(Methods in Enzymology:Guide to Molecular Cloning Techniques)、vol.152、Academic Press、Inc.、San Diego、Calif.[1987])、に記述されている。
それぞれの導入遺伝子のヌクレオチド配列が既知の場合は、導入遺伝子は、化学合成方法、例えば、Engels et al.(Angew.Chem.Int.Ed.Engl.、28:716−734[1989])に記載の方法を使って、全体または一部を合成できる。これらの方法には、特に、核酸合成ホスホトリエステル、ホスホラミダイトおよびH−ホスホナート法が含まれる。あるいは、導入遺伝子は、導入遺伝子DNAに選択的にハイブリダイズする1つまたは複数の核酸プローブ(クローン化される導入遺伝子に対し許容可能なレベルの相同性を有するオリゴヌクレオチド、cDNAまたは、ゲノムDNA断片、等)を使って、適切なcDNAまたはゲノムライブラリーを選別することにより得ることができる。別の適切な導入遺伝子を得る方法は、ポリメラーゼ連鎖反応(PCR)である。しかし、この方法をうまく使うには、導入遺伝子のヌクレオチド配列に関する充分な情報が、適切なヌクレオチド配列の増幅に有用な適切なオリゴヌクレオチドプライマーを設計するために利用できることが必要である。
選択の方法に、オリゴヌクレオチドプライマーまたはプローブの使用が必要な場合(例えば、PCR、cDNAまたはゲノムライブラリー選別)には、プローブまたはプライマーとして選択されるオリゴヌクレオチド配列は、ライブラリー選別またはPCRの間に起こる非特異的結合の量を最小限にするために、適切な長さであり、一義的である必要がある。プローブまたはプライマーの実際の配列は、通常、別の生物体からの同じまたは類似の遺伝子由来の保存または高相同配列もしくは領域に基づく。任意選択で、プローブまたはプライマーは、変性させてもよい。
導入遺伝子のアミノ酸配列のみがわかっている場合は、可能性のある、機能的核酸配列は、それぞれのアミノ酸残基に対する既知で好ましいコドンを使って導入遺伝子を推測できる。そこで、この配列を、化学的に合成できる。
本発明は、導入遺伝子変異体配列の使用を包含する。変異体導入遺伝子は、野性型配列と比較して、1つまたは複数のヌクレオチド置換、欠失、および/または挿入を含む導入遺伝子である。ヌクレオチド置換、欠失、および/または挿入により、野性型アミノ酸配列とはアミノ酸配列が異なる遺伝子産物(すなわち、タンパク質)を生成することができる。このような変異体の調製は、当技術分野でよく知られており、また、例えば、Wells et al.(Gene、34:315[1985])、およびSambrook et al、同上、に記載されている。
調節の要素の選択
導入遺伝子は、典型的には、プロモーターに機能的に連結されており、そこでプロモーターは、各導入遺伝子の発現を特定の様式で調節するように選択される。
導入遺伝子は、典型的には、プロモーターに機能的に連結されており、そこでプロモーターは、各導入遺伝子の発現を特定の様式で調節するように選択される。
2つ以上の導入遺伝子が使われる場合には、それぞれの導入遺伝子は、同じまたは異なるプロモーターにより調節できる。選択されたプロモーターは、相同(すなわち、導入遺伝子を形質移入される哺乳動物と同じ種由来)であっても、異種(すなわち、導入遺伝子を形質移入される哺乳動物の種以外の源由来)であってもよい。従って、各プロモーターの源は、いずれの単細胞、原核生物または真核生物の生物体からでもよく、またはいずれの脊椎動物または無脊椎動物生物体からのものでもよい。
他のベクター成分の選択
導入遺伝子およびプロモーターに加えて、本発明の導入遺伝子の調製に有用なベクターは、典型的には、(1)導入遺伝子が挿入される哺乳動物中での導入遺伝子の最適発現、および(2)細菌または哺乳類宿主細胞中でのベクターの増幅に有用な1つまたは複数の他の要素を含む。これらの要素のそれぞれは、ベクター中で、それぞれ他の要素に対して適切に配置され、それにより、それぞれの作用を最大化する。このような位置決めは、当業者にはよく知られている。下記の要素が、任意選択で、必要に応じ、ベクター中に含むことができる。
導入遺伝子およびプロモーターに加えて、本発明の導入遺伝子の調製に有用なベクターは、典型的には、(1)導入遺伝子が挿入される哺乳動物中での導入遺伝子の最適発現、および(2)細菌または哺乳類宿主細胞中でのベクターの増幅に有用な1つまたは複数の他の要素を含む。これらの要素のそれぞれは、ベクター中で、それぞれ他の要素に対して適切に配置され、それにより、それぞれの作用を最大化する。このような位置決めは、当業者にはよく知られている。下記の要素が、任意選択で、必要に応じ、ベクター中に含むことができる。
i.シグナル配列要素
導入遺伝子によりコードされるポリペプチドが分泌される本発明の実施形態にでは、シグナル配列と名付けられた小さなポリペプチドが存在する場合が多く、これは、導入遺伝子によりコードされたポリペプチドをそれが合成される細胞から外部へ誘導する。典型的には、シグナル配列は、導入遺伝子のコーディング領域中の5’末端の近く、または5’末端に配置される。多くのシグナル配列が特定されており、機能的で、従って、遺伝子導入組織と適合するそれらの内のいずれも、導入遺伝子と共に使用可能である。従って、シグナル配列をコードするヌクレオチド配列は、導入遺伝子に対し相同でも、異種でもよく、遺伝子導入哺乳動物に対し、相同でも、異種でもよい。さらに、シグナル配列をコードするヌクレオチド配列は、上述の方法を使って、化学的に合成できる。しかし、本明細書の目的のために好ましいシグナル配列は、導入遺伝子と共に天然で生成するもの(すなわち、導入遺伝子に相同である)である。
導入遺伝子によりコードされるポリペプチドが分泌される本発明の実施形態にでは、シグナル配列と名付けられた小さなポリペプチドが存在する場合が多く、これは、導入遺伝子によりコードされたポリペプチドをそれが合成される細胞から外部へ誘導する。典型的には、シグナル配列は、導入遺伝子のコーディング領域中の5’末端の近く、または5’末端に配置される。多くのシグナル配列が特定されており、機能的で、従って、遺伝子導入組織と適合するそれらの内のいずれも、導入遺伝子と共に使用可能である。従って、シグナル配列をコードするヌクレオチド配列は、導入遺伝子に対し相同でも、異種でもよく、遺伝子導入哺乳動物に対し、相同でも、異種でもよい。さらに、シグナル配列をコードするヌクレオチド配列は、上述の方法を使って、化学的に合成できる。しかし、本明細書の目的のために好ましいシグナル配列は、導入遺伝子と共に天然で生成するもの(すなわち、導入遺伝子に相同である)である。
ii.膜アンカードメイン要素
いくつかの例では、導入遺伝子を、特定の細胞膜の表面またはその細胞膜上に発現させることが好ましい場合がある。天然膜タンパク質は、ポリペプチドの一部として、タンパク質を膜に係留する役割をするアミノ酸の一続きの配列を含む。しかし、膜上の天然にはないタンパク質に対して、このような一続きの配列のアミノ酸を加えて、この特徴を付与できる。頻繁に、このアンカードメインは、ポリペプチド配列の内部部分であり、そのため、それをコードするヌクレオチド配列は、導入遺伝子ヌクレオチド配列の内部領域中へ操作される。しかし、他の場合では、アンカードメインをコードするヌクレオチド配列は、導入遺伝子ヌクレオチド配列の5’または3’末端に結合され得る。ここで、アンカードメインをコードするヌクレオチド配列は、最初に、導入遺伝子をコードするヌクレオチド配列とは別の成分として、ベクター中の適切な位置に配置できる。シグナル配列に関しては、アンカードメインは、いずれの源からであってもよく、従って、導入遺伝子および遺伝子導入哺乳動物の両方に対して、相同でも、異種でもよい。あるいは、アンカードメインは、前記の方法を使って化学的に合成できる。
いくつかの例では、導入遺伝子を、特定の細胞膜の表面またはその細胞膜上に発現させることが好ましい場合がある。天然膜タンパク質は、ポリペプチドの一部として、タンパク質を膜に係留する役割をするアミノ酸の一続きの配列を含む。しかし、膜上の天然にはないタンパク質に対して、このような一続きの配列のアミノ酸を加えて、この特徴を付与できる。頻繁に、このアンカードメインは、ポリペプチド配列の内部部分であり、そのため、それをコードするヌクレオチド配列は、導入遺伝子ヌクレオチド配列の内部領域中へ操作される。しかし、他の場合では、アンカードメインをコードするヌクレオチド配列は、導入遺伝子ヌクレオチド配列の5’または3’末端に結合され得る。ここで、アンカードメインをコードするヌクレオチド配列は、最初に、導入遺伝子をコードするヌクレオチド配列とは別の成分として、ベクター中の適切な位置に配置できる。シグナル配列に関しては、アンカードメインは、いずれの源からであってもよく、従って、導入遺伝子および遺伝子導入哺乳動物の両方に対して、相同でも、異種でもよい。あるいは、アンカードメインは、前記の方法を使って化学的に合成できる。
iii.複製起点要素
この成分は、典型的には、市販品購入される原核生物発現ベクターの一部であり、宿主細胞中でのベクターの増幅を支援する。選択したベクターが複製起点部位を含まない場合は、既知の配列に基づいて化学的に合成し、ベクターに連結できる。
この成分は、典型的には、市販品購入される原核生物発現ベクターの一部であり、宿主細胞中でのベクターの増幅を支援する。選択したベクターが複製起点部位を含まない場合は、既知の配列に基づいて化学的に合成し、ベクターに連結できる。
iv.転写終止要素
この要素は、ポリアデニル化またはポリA配列としても知られ、典型的には、ベクター中の導入遺伝子ヌクレオチド配列の3’に位置し、導入遺伝子の転写を終わらせる役割をする。この要素をコードするヌクレオチド配列は、ライブラリーから容易にクローン化されるか、またはベクターの一部として市販品の購入さえもできるが、上述のような、ヌクレオチド配列合成法を使って容易に合成することもできる。
この要素は、ポリアデニル化またはポリA配列としても知られ、典型的には、ベクター中の導入遺伝子ヌクレオチド配列の3’に位置し、導入遺伝子の転写を終わらせる役割をする。この要素をコードするヌクレオチド配列は、ライブラリーから容易にクローン化されるか、またはベクターの一部として市販品の購入さえもできるが、上述のような、ヌクレオチド配列合成法を使って容易に合成することもできる。
v.イントロン要素
多くの場合、導入遺伝子の転写は、クローニングベクター上の1つのイントロンまたは2つ以上のイントロンの存在(エキソンに連結された)により増加する。イントロンは、導入遺伝子がゲノムDNA配列の完全長または断片である場合は特に、導入遺伝子ヌクレオチド配列内の天然配列であり得る。イントロンがヌクレオチド配列内の天然配列ではない場合は(大抵のcDNAのように)、イントロンは、別のソースから入手できる。イントロンは、導入遺伝子、および/または遺伝子導入哺乳動物に対し相同であっても、異種であってもよい。イントロンが有効であるように転写される必要があるため、プロモーターおよび導入遺伝子に対するイントロンの位置は重要である。従って、導入遺伝子がcDNA配列である場合は、イントロンの好ましい位置は、転写開始部位に対し3’位置で、ポリA転写終止配列に対し5’位置である。好ましくは、cDNA導入遺伝子に対しては、イントロンは、導入遺伝子ヌクレオチド配列を妨害しないように、導入遺伝子ヌクレオチド配列の片側またはもう一方の側(すなわち、5’または3’)に配置される。挿入される宿主細胞に適合する限りにおいて、ウイルス、原核生物および真核生物(側物または動物)生命体を含む、いずれの源由来のいずれのイントロンも、本発明の実施に使用可能である。また、本明細書では、合成イントロンも含まれる。任意選択で、2つ以上のイントロンをベクター中で使用可能である。好ましいイントロンとエキソンのセットは、ヒト成長ホルモン(hGH)DNA配列である。
多くの場合、導入遺伝子の転写は、クローニングベクター上の1つのイントロンまたは2つ以上のイントロンの存在(エキソンに連結された)により増加する。イントロンは、導入遺伝子がゲノムDNA配列の完全長または断片である場合は特に、導入遺伝子ヌクレオチド配列内の天然配列であり得る。イントロンがヌクレオチド配列内の天然配列ではない場合は(大抵のcDNAのように)、イントロンは、別のソースから入手できる。イントロンは、導入遺伝子、および/または遺伝子導入哺乳動物に対し相同であっても、異種であってもよい。イントロンが有効であるように転写される必要があるため、プロモーターおよび導入遺伝子に対するイントロンの位置は重要である。従って、導入遺伝子がcDNA配列である場合は、イントロンの好ましい位置は、転写開始部位に対し3’位置で、ポリA転写終止配列に対し5’位置である。好ましくは、cDNA導入遺伝子に対しては、イントロンは、導入遺伝子ヌクレオチド配列を妨害しないように、導入遺伝子ヌクレオチド配列の片側またはもう一方の側(すなわち、5’または3’)に配置される。挿入される宿主細胞に適合する限りにおいて、ウイルス、原核生物および真核生物(側物または動物)生命体を含む、いずれの源由来のいずれのイントロンも、本発明の実施に使用可能である。また、本明細書では、合成イントロンも含まれる。任意選択で、2つ以上のイントロンをベクター中で使用可能である。好ましいイントロンとエキソンのセットは、ヒト成長ホルモン(hGH)DNA配列である。
vi.選択可能マーカー要素
選択可能マーカー遺伝子は、選択培養培地中で増殖される形質移入細胞の生存および増殖に必要なポリペプチドをコードする。典型的な選択マーカー遺伝子は、(a)抗生物質または他の毒素、例えば、原核生物宿主細胞に対してはアンピシリン、テトラサイクリン、またはカナマイシン、および哺乳動物細胞に対してはネオマイシン、ヒグロマイシン、またはメトトレキサート、に耐性を付与するか、(b)細胞の栄養要求欠陥を補足するか、または(c)複合培地から得られない重要な栄養素、例えば、桿菌培養用D−アラニンラセマーゼ、をコードする遺伝子を提供する、タンパク質をコードする。
選択可能マーカー遺伝子は、選択培養培地中で増殖される形質移入細胞の生存および増殖に必要なポリペプチドをコードする。典型的な選択マーカー遺伝子は、(a)抗生物質または他の毒素、例えば、原核生物宿主細胞に対してはアンピシリン、テトラサイクリン、またはカナマイシン、および哺乳動物細胞に対してはネオマイシン、ヒグロマイシン、またはメトトレキサート、に耐性を付与するか、(b)細胞の栄養要求欠陥を補足するか、または(c)複合培地から得られない重要な栄養素、例えば、桿菌培養用D−アラニンラセマーゼ、をコードする遺伝子を提供する、タンパク質をコードする。
本発明で有用な上述の全てのならびに他の要素は、当業者には、よく知られ、また、例えば、Sambrook et al.(分子クローニング:実験マニュアル(Molecular Cloning:A Laboratory Manual)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.[1989])およびBerger et al.、eds.(分子クローニング技術のガイド(Guide to Molecular Cloning Techniques)、Academic Press、Inc.、San Diego、Calif.[1987])、に記載されている。
クローニングベクターの構築
本発明の遺伝子導入哺乳動物の調製に有用な導入遺伝子カセットの増幅に最も役立つクローニングベクターは、原核細胞宿主に適合するものである。しかし、真核細胞宿主、およびこれらの細胞に適合するベクターは、本発明の範囲内に入る。
本発明の遺伝子導入哺乳動物の調製に有用な導入遺伝子カセットの増幅に最も役立つクローニングベクターは、原核細胞宿主に適合するものである。しかし、真核細胞宿主、およびこれらの細胞に適合するベクターは、本発明の範囲内に入る。
ある特定の場合では、クローニングベクター上に含められる種々の要素の一部が、市販のクローニングまたは増幅ベクター、例えば、pUC18、pUC19、pBR322、pGEMベクター(Promega Corp、Madison、Wis.)、pBluescript(登録商標)ベクター、例えば、pBIISK+/−(Stratagene Corp.、La Jolla、Calif.)、等、中に既に存在するものもある(これらの全ては、原核細胞宿主に対し安定である)。この場合、導入遺伝子をベクター中に挿入することが求められるだけである。
しかし、使用される1つまたは複数の要素が、クローニングまたは増幅ベクター上に予め存在しない場合は、個別に入手し、ベクターに連結することができる。それぞれの要素の入手および連結方法は、当業者にはよく知られており、上述の導入遺伝子(すなわち、DNA合成、ライブラリー選別、等)の入手方法に類似である。
導入遺伝子ヌクレオチド配列のクローニングまたは増幅用に、および/または哺乳類胚の形質移入用に使用されるベクターは、当技術分野でよく知られた方法を使って構築される。そのような方法には、Sambrook et al.、同上、に記載のように、例えば、制限エンドヌクレアーゼ消化の標準的な方法、連結反応、DNAおよび/またはRNAのアガロースおよびアクリルアミドゲル精製、DNAおよび/またはRNAのカラムクロマトグラフィー精製、DNAのフェノール/クロロホルム抽出、DNAシーケンシング、ポリメラーゼ連鎖反応増幅、等が含まれる。
本発明の実施に使用される最終的ベクターは、典型的には、市販のベクター等の出発クローニングまたは増幅ベクターから構築される。このベクターは、完成ベクターに含まれる要素の一部を含んでも、含まなくてもよい。出発ベクター中にいかなる所望の要素も存在しない場合は、各要素は、連結される要素の末端およびベクターの末端が連結反応に適合するように、適切な制限エンドヌクレアーゼでベクターを切断することにより、個別にベクターに連結できる。いくつかの場合では、満足のできる連結反応を行うために、連結される末端を一緒に「平滑末端化する」ことが必要となり得る。平滑末端化は、最初に、Klenow DNAポリメラーゼまたはT4DNAポリメラーゼを使って、4つ全てのヌクレオチドの存在下で、「付着末端」を埋めることにより実現される。この方法は、当技術分野でよく知られており、また、例えば、Sambrooketal.、同上、に記載されている。
あるいは、ベクターに挿入される2つ以上の要素を、最初に、一緒に連結し(相互に隣接して配置される場合は)、その後、ベクターに連結できる。
ベクターを作成する他の1つの方法は、種々の要素の全ての連結反応を、1つの反応混合物中で同時に行うことである。ここで、要素の不適切な連結反応または挿入のために、多くのナンセンスまたは非機能性ベクターが生成されるであろうが、機能性ベクターは、制限エンドヌクレアーゼ消化により特定、選択できる。
ベクターの作成後、増幅のために、原核生物宿主細胞中に形質移入できる。増幅に使われる細胞は、代表例としては、大腸菌DH5−アルファ(Gibco/BRL、Grand Island、N.Y.)およびDH5−アルファに類似の特性を有する他の大腸菌株である。
哺乳類宿主細胞が使用される場合は、例えば、チャイニーズハムスター卵巣(CHO細胞;Urlab et al.、Proc.Natl.Acad.Sci USA、77:4216[1980]))およびヒト胚性腎臓細胞株293(Graham et al.、J.Gen.Virol.、36:59[1977])等の細胞株、ならびに他の系統が適切である。
増幅のためのベクターの選択宿主細胞株への形質移入は、リン酸カルシウム、電気穿孔、微量注入、リポフェクションまたはDEAEデキストラン等の方法を使って実現される。選択された方法は、部分的に、形質移入される宿主細胞型に依存する。これらおよび他の適切な方法は、当業者にはよく知られており、また、Sambrook et al.、同上、に記載されている。
ベクターを十分に増幅されるのに充分長い時間(通常、大腸菌細胞の場合で一晩)培養の後、ベクター(この段階では、プラスミドと呼ばれることも多い)は、細胞から単離され、精製される。典型的には、細胞は、溶解され、プラスミドが他の細胞含有物から抽出される。プラスミド精製に適する方法には、特に、アルカリ溶解ミニプレップ法(Sambrook et al.、同上)が含まれる。
挿入用プラスミドの調製
典型的には、導入遺伝子を含むプラスミドは、直線化され、胚への挿入の前に、選択制限エンドヌクレアーゼを使って、その一部が除去される。いくつかの例では、ベクターの他の部分から直鎖断片として、導入遺伝子、プロモーター、および調節要素を単離し、それにより、導入遺伝子、プロモーター、イントロン(使われる場合は)、エンハンサー、ポリA配列、および任意選択で、シグナル配列または膜アンカードメインを含む直鎖ヌクレオチド配列のみを胚に注入することが好ましい場合がある。これは、これらの要素を含む核酸配列領域を取り出すようにプラスミドを切断し、アガロースゲル電気泳動法または他の適切な精製方法を使ってこの領域を精製することにより実現できる。
典型的には、導入遺伝子を含むプラスミドは、直線化され、胚への挿入の前に、選択制限エンドヌクレアーゼを使って、その一部が除去される。いくつかの例では、ベクターの他の部分から直鎖断片として、導入遺伝子、プロモーター、および調節要素を単離し、それにより、導入遺伝子、プロモーター、イントロン(使われる場合は)、エンハンサー、ポリA配列、および任意選択で、シグナル配列または膜アンカードメインを含む直鎖ヌクレオチド配列のみを胚に注入することが好ましい場合がある。これは、これらの要素を含む核酸配列領域を取り出すようにプラスミドを切断し、アガロースゲル電気泳動法または他の適切な精製方法を使ってこの領域を精製することにより実現できる。
遺伝子導入またはノックアウト哺乳動物の産生
遺伝子導入またはノックアウト(KO)哺乳動物は、当業者によく知られた方法を使って容易に調製できる。例えば、遺伝子導入げっ歯類を調製するために、Hogan et al.、eds.、(マウス胚の操作:実験マニュアル(Manipulating the Mouse Embryo:A Laboratory Manual)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.[1986])に記載のもの、等の方法を採用できる。
遺伝子導入またはノックアウト(KO)哺乳動物は、当業者によく知られた方法を使って容易に調製できる。例えば、遺伝子導入げっ歯類を調製するために、Hogan et al.、eds.、(マウス胚の操作:実験マニュアル(Manipulating the Mouse Embryo:A Laboratory Manual)、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.[1986])に記載のもの、等の方法を採用できる。
本発明を実施するのに使用されるいずれの哺乳類種の特定の系統も、一般的に良好な健康、良好な胚収量、良好な胚中での前核視感度、および良好な生殖適応度の観点から選択される。さらに、ハプロタイプが重要な要因である。例えば、遺伝子導入マウスが産生される場合、C57BL/6またはC57BL/6xDBA/2F1等の株、またはFVB系統がよく使われる(Charles River Labs、Boston、Mass.、The Jackson Laboratory、Bar Harbor、Me、またはTaconic Labs.、から市販品として入手可能)。好ましい株は、H−2b、H−2dまたはH−2qハプロタイプを有するもの、例えば、C57BL/6またはDBA/1である。本発明の実施に使われる系統は、それ自体遺伝子導入体であっても、および/またはノックアウト(すなわち、部分的に、または完全に抑制された1つまたは複数の遺伝子を有する哺乳動物)であってもよい。最初のノックアウト哺乳動物および遺伝子導入哺乳動物の両方の調製のために、同じ系統が使われるのが好ましいであろう。これは、その後の交配および戻し交配をより効率的にすることになろう。
ノックアウト(KO)マウス系統の生成(所望の遺伝子に対する)は、改変胚性幹(ES)細胞の、1つのマウス株「X」から異なるマウス株「Y」の胚盤胞中への移植から始まる。通常、異なる毛色のマウス株を選択して、両方の色からなる「キメラ」毛を有する動物を得るのが好都合である。例えば、改変ES細胞がアグーチ模様マウス系統(129/Sv)由来で、黒色マウス胚盤胞(C57/Bl6)に挿入される場合、アグーチ模様と黒毛を有するキメラが得られる。次の重要なステップは、改変ES細胞が生殖細胞系列伝達のための生殖腺を備えているかどうかを判定することである。これは、キメラマウスを、胚盤胞ソース(C57/Bl6)に対応する株の動物に交配することにより判定される。交配によりアグーチ模様(ES細胞色)の子を産生する場合は、C57Bl6/129Sv混合背景で、生殖系列伝達が実現されたことを示す。
異なるマウス株は違った挙動をするので、混合背景は、遺伝子調査で理想的なものではない。従って、1つの特定のマウス株から純粋なマウス系統を産生するのが好ましい。目的のマウスに交配する場合、戻し交配が同系交配マウス株との間で使われ、また、異系交配が非近交系マウス株との間で使われる。戻し交配は、選択同系交配マウス株に対し、凡そ99.99%の遺伝的同一性に達するまで、図1Aに図示するように行われ、これは約8〜10回の戻し交配を要し、研究目的のためには好ましい手法である。この新しい系統は、この時点で、類遺伝子性であると見なされる。異系交配(図1Bに図示)は、1回の交配の交配のみを含み、それぞれの親の系統の半分の遺伝コードを有するハイブリッドマウスが得られる。この系統は、類遺伝子性とは見なされず、また、系統のかなりの遺伝的変異性に起因して、類遺伝子性系統にすることができない。残念ながら、これは、近交系で特徴的なことであるように、戻し交配は、生成するマウスコロニーをより弱くする可能性があり、また、劣性遺伝性疾患になる傾向がある。異系交配は、1回(1x交配)のみ行われ、遺伝的多様性を高度同系交配マウス株中に挿入することが意図されているので、純粋な系統を生成できない。この方法は、育種系統に対し、繁殖性、活力および大きさを回復する場合が多い。
胚を得るために、および代用の宿主として役立たせるために使われる哺乳動物の年齢は、使われる種依存であるが、当業者なら簡単に判断できる。例えば、マウスが使われる場合は、より多くの胚を得ることができ、ホルモン注射よりよく反応することから、思春期前の雌が好ましい。
同様に、種馬として使用される雄哺乳動物は、通常、他の基準の中でも特に性的成熟年齢で選択される。
卵産生、交配、および/または胚の再移植用の雌を準備するのに、ホルモンまたは他の化学化合物の投与が必要な場合がある。使われるホルモン/補因子の型と量、ならびにホルモンの投与タイミングは、哺乳動物のそれぞれの種により変化する。このような配慮は、当業者には容易にわかることであろう。
典型的には、準備ができた雌(すなわち、受精できる卵子を産生している雌)は、種馬の雄と交配され、得られた受精胚は、次に、導入遺伝子導入用に取り出される。あるいは、卵子と精子を、適切な雌と雄から入手し、インビトロ受精用に使用して導入遺伝子の導入に適する胚を産生できる。
通常、受精胚は、前核が出現するまで適切な培地中でインキュベートされる。下記に記載のように、この頃に、導入遺伝子を含むヌクレオチド配列が雌または雄前核中に導入される。マウス等の一部の種では、雄前核が好ましい。
導入遺伝子ヌクレオチド配列の胚中への導入は、当技術分野で既知のいずれの手段によっても、例えば、微量注入、電気穿孔、またはリポフェクションにより実現できる。導入遺伝子ヌクレオチド配列の胚への導入後、胚は、様々な時間の間、インビトロでインキュベートされるか、または代用宿主に再移植されるか、またはこれらの両方がなされる。インビトロでの成熟までのインキュベーションは、本発明の範囲内に入る。1つのよく行われる方法は、胚をインビトロで、種に応じて約1〜7日間インキュベートし、その後、代用宿主中に再移植するものである。
再移植は、標準的な方法で行える。使用される雌「養母」株は、全般的耐久力および健康、ならびに、出生児の世話能力により選択される。マウスの場合、C57BL/6xDBA1またはCD1、またはBALB/c等の株が、通常、適切である。しかし、本発明のNav1.7−/−ノックアウト用には、C57BL/6背景は特に好適ではない。理由は、子が十分な活力に欠け、雌親が、通常、十分に熱心な母ではないからである。
通常、代用宿主は、麻酔され、胚が輸卵管中に挿入される。特定の宿主に移植される胚の数は、種により変化するが、通常は、種が天然で産出する出生児数と同程度である。
代用宿主の遺伝子導入出生児は、いずれの適切な方法を用いても、導入遺伝子の存在および/または発現により選別できる。選別は、少なくとも一部の導入遺伝子に相補的なプローブを使って、サザンブロットまたはノーザン法分析により行われる場合が多い。導入遺伝子によりコードされるタンパク質に対する抗体を使ったウェスタンブロット分析を、代わりの、または追加の、導入遺伝子産物の存在に関する選別方法として採用できる。典型的には、DNAが、尾部組織(尾先端から約1cmが取り出される)から調製され、導入遺伝子がサザン分析またはPCRにより分析される。あるいは、最高レベルで導入遺伝子を発現していると考えられる組織または細胞が導入遺伝子の存在および発現の観点から、サザン分析またはPCRを使って試験されるが、いずれの組織または細胞型も、この分析用に使用可能である。
代わりの、または追加の導入遺伝子の存在の評価方法には、限定されないが、適切な生化学的アッセイ、例えば、酵素、および/または免疫学的アッセイ、特定のマーカーまたは酵素活性用の組織学的株、フローサイトメトリー分析、等が含まれる。血液の分析もまた、血液中の導入遺伝子産物の検出、ならびに種々の型の血液細胞および他の血液成分のレベルに対する導入遺伝子の影響の評価に有用でありうる。
遺伝子導入哺乳動物の子孫は、遺伝子導入哺乳動物を適切なパートナーと交配することにより、または遺伝子導入哺乳動物から入手した卵子、および/または精子のインビトロ受精により、得ることができる。パートナーと交配を行う場合は、パートナーは、遺伝子導入および/またはノックアウトであっても、そうでなくてもよく、遺伝子導入パートナーの場合は、同じまたは異なる、または両方の導入遺伝子を含んでいてもよい。あるいは、パートナーは、親系統であってもよい。インビトロ受精が使われる場合は、受精胚は、代用宿主に移植して、またはインビトロでインキュベートしても、またはそれらの両方を行ってもよい。いずれかの方法を使って、上述の方法、または他の適切な方法により、導入遺伝子の存在について子孫を評価できる。
ノックアウト/遺伝子導入哺乳動物の調製
2つ以上のノックアウト構築物、および/または、2つ以上の導入遺伝子を含む哺乳動物が、いくつかの方法のいずれかを使って調製される。典型的には、調製方法は、本明細書記載の、所望のノックアウト構築物または導入遺伝子の1つをそれぞれ含む一連の哺乳動物、例えば、マウスを生成するものである。このような哺乳動物は、一連の交配、戻し交配および選択により育種され、最終的には、全体が所望のノックアウト構築物、および/または導入遺伝子を含む単一哺乳動物を生成する。この場合、哺乳動物は、ノックアウト構築物、および/または導入遺伝子の存在を除けば、野性型に類遺伝子性(遺伝的に同じ)である。
2つ以上のノックアウト構築物、および/または、2つ以上の導入遺伝子を含む哺乳動物が、いくつかの方法のいずれかを使って調製される。典型的には、調製方法は、本明細書記載の、所望のノックアウト構築物または導入遺伝子の1つをそれぞれ含む一連の哺乳動物、例えば、マウスを生成するものである。このような哺乳動物は、一連の交配、戻し交配および選択により育種され、最終的には、全体が所望のノックアウト構築物、および/または導入遺伝子を含む単一哺乳動物を生成する。この場合、哺乳動物は、ノックアウト構築物、および/または導入遺伝子の存在を除けば、野性型に類遺伝子性(遺伝的に同じ)である。
典型的には、交配および戻し交配は、育種プロセス中のそれぞれ特定のステップの目的に応じて、兄弟姉妹または親株を出生児と交配することで実現される。特定の例では、適切な染色体の部位にそれぞれのノックアウト構築物、および/または導入遺伝子を含む単一出生児を生成するために、多数の出生児を生成する必要がある場合もある。さらに、最終的に所望の遺伝子型を生成するために、数世代にわたり交配または戻し交配をする必要がある場合もある。
ノックアウト哺乳動物の用途
一般的に、ノックアウト哺乳動物は、抑制されている遺伝子または複数遺伝子に応じて様々な用途がある。例えば、遺伝子または複数遺伝子が、免疫抑制または炎症に関与すると考えられているコードタンパク質を抑制する場合、この哺乳動物は、免疫調節に有用な薬剤、すなわち、これらの活性を強化するか、または抑制する薬剤、を選別するために使用できる。
一般的に、ノックアウト哺乳動物は、抑制されている遺伝子または複数遺伝子に応じて様々な用途がある。例えば、遺伝子または複数遺伝子が、免疫抑制または炎症に関与すると考えられているコードタンパク質を抑制する場合、この哺乳動物は、免疫調節に有用な薬剤、すなわち、これらの活性を強化するか、または抑制する薬剤、を選別するために使用できる。
本発明のグローバルNaV1.7ノックアウトマウスは、疼痛、神経内分泌障害、または前立腺癌の治療に有望な薬剤を選別するために使用できる。有用な薬剤の選別は、一連の投与量でマウスに候補薬剤を投与し、種々の時点で評価される障害に対する薬剤の効果をアッセイすることを含む。さらに、本発明の哺乳動物は、神経系の発達の評価、および特定のNaV1.7遺伝子変異の効果の調査に役立つ可能性がある。本発明のNaV1.7ノックアウトマウスおよびその子孫の実施形態には、また、発現されうる追加の導入遺伝子、および/または含まれるノックアウト構築物に応じて、種々の用途が考えられる。有用な薬剤の選別には、最初に、哺乳動物で、疾患の誘導、または疾患モデルの誘導、次いで、一連の投与量による候補薬剤の哺乳動物への投与、および種々の時点での評価される疾患または障害に対する薬剤の効果のアッセイを含めることができる。代わりに、または追加で、薬剤は、疾患または疾患モデルの誘導を行う前に、または同時に投与でき得る。他の実施形態では、本発明のグローバルNaV1.7−/−ノックアウトマウスは、薬剤研究および開発、例えば、オンターゲット/オフターゲット効果を識別する、または疼痛および鎮静の効果を識別するためのインビボプロトコル用として、さらに有用である。
疾患または状態に使用される薬剤の選別に加えて、本発明の哺乳動物は、疾患または状態の予防または治療を目的とした治療法の設計に有用でありうる。例えば、哺乳動物は、疾患または状態の発症の前に、同時に、または後で、特定の食事、運動ルーチン、照射治療、および/または1つまたは複数の化合物または物質の組み合わせで治療でき得る。このような総合的療法または治療法は、疾患または状態と戦う上で、化合物単独による治療より、有効な場合がある。さらに、血圧、体温、体重、脈拍、挙動、毛の外観(ひだ状毛皮)、等の診断基準を評価でき得る。
本発明のグローバルNaV1.7−/−ノックアウトマウスは、また、1つまたは複数の細胞株を精製するのに使用できる。そのような細胞系統は、例えば、特定の組織または器官に対するノックアウトの影響の評価、および組織中でNaV1.7の活性レベルに影響する可能性のある化合物の選別、等の多くの用途がある。このような化合物は、治療薬として有用であり得る。
このような細胞系統の産生は、当業者に知られている種々の方法を使って実現できる。実際の培養条件は、培養される細胞の組織と型に依存する。必要以上の実験をせずに、細胞を使って、異なる濃度の多量および微量栄養素、増殖因子、血清、等を含む種々の培地が試験して、細胞の成長と増殖の最適条件を決定することができる。同様に、他の培養条件、例えば、インキュベーター中の細胞密度、培地温度、および二酸化炭素濃度も、容易に評価、最適化でき、また、このプロセスに影響する化合物を特定できる。
マウスまたはヒトNaV1.7等のNaV1.7に対する抗体の調製(NaV1.7KOマウスは、マウスNav1.7または他の種の密接に関連しているNav1.7配列、例えば、ヒトSCN9A遺伝子産物、に対し自己免疫寛容ではないため)を含む他の用途は、当業者には容易にわかるであろう。本発明のノックアウトマウスにより産生された抗ヒトNaV1.7抗体のCDR配列に基づいて、これらのCDRを抗体中に組み込んだ、NaV1.7イオンチャネル活性に対する拮抗または刺激用のキメラまたはヒト化抗体を開発できる。拮抗的または遮断的抗ヒトNaV1.7抗体の治療効果は、当業者には容易に明白となるであろう
抗体の産生
ポリクローナル抗体。ポリクローナル抗体は、典型的には、適切な抗原およびアジュバントの複数部位の皮下(sc)または腹腔内(ip)注射により動物中で産生される。あるいは、抗原は、動物のリンパ節に直接注入してもよい(Kilpatrick et al.、Hybridoma、16:381−389、1997、を参照)。改善された抗体反応は、二官能性または誘導体化薬剤、例えば、マレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介したコンジュゲーション)、N−ヒドロキシスクシンイミド(リジン残基を介したコンジュゲーション)、グルタルアルデヒド、コハク酸無水物または当技術分野で既知の他の薬剤を使って、適切な抗原を、免疫される種中で免疫原性のタンパク質、例えば、スカシ貝ヘモシアニン、血清アルブミン、ウシチログロブリン、または大豆トリプシン阻害剤、と複合体化させることにより得ることができる。
ポリクローナル抗体。ポリクローナル抗体は、典型的には、適切な抗原およびアジュバントの複数部位の皮下(sc)または腹腔内(ip)注射により動物中で産生される。あるいは、抗原は、動物のリンパ節に直接注入してもよい(Kilpatrick et al.、Hybridoma、16:381−389、1997、を参照)。改善された抗体反応は、二官能性または誘導体化薬剤、例えば、マレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介したコンジュゲーション)、N−ヒドロキシスクシンイミド(リジン残基を介したコンジュゲーション)、グルタルアルデヒド、コハク酸無水物または当技術分野で既知の他の薬剤を使って、適切な抗原を、免疫される種中で免疫原性のタンパク質、例えば、スカシ貝ヘモシアニン、血清アルブミン、ウシチログロブリン、または大豆トリプシン阻害剤、と複合体化させることにより得ることができる。
動物は、例えば、(マウスに対して)100μgのタンパク質または複合体を、3倍量のフロイント完全アジュバントと混合し、その溶液を皮内の複数部位に注射することにより、抗原、免疫原性コンジュゲート、または誘導体に対し免疫される。1ヶ月後、フロイント完全アジュバント中のペプチドまたはコンジュゲートの元の量の1/5〜1/10の複数部位の皮下注射により、動物は追加免疫される。追加免疫注射7〜14日後に、動物から採血し、血清の抗体力価がアッセイされる。動物は、力価がプラトーになるまで、追加免疫される。動物は、同じ抗原の複合体で追加免疫されるのが好ましいが、異なるタンパク質と、および/または異なる架橋結合試薬を介して複合体化されたものでもよい。複合体は、また、タンパク質融合体として組換え細胞培養でも作製可能である。また、ミョウバン等の凝集剤を適切に使って、免疫反応を適切に強化できる。
モノクローナル抗体。モノクローナル抗体は、当技術分野で知られているいずれの技術を用いても、例えば、免疫化計画の完結後に遺伝子導入動物から採取された脾臓細胞を不死化することによって、産生出来る。脾臓細胞は、当技術分野で知られているいずれの技術を用いても、例えば、それらを骨髄腫細胞と融合しハイブリドーマを産生することにより、不死化できる。例えば、モノクローナル抗体は、Kohler et al.、Nature、256:495(1975)、によって最初に報告されたハイブリドーマ法を使って作成できる。または、組換えDNA法(例えば、Cabilly et al.、免疫グロブリンの産生方法、およびその方法で使用するベクターと形質転換宿主細胞(Methods of producing immunoglobulins、vectors and transformed host cells for use therein)、米国特許第6、331、415号)により作製できる。この方法には、軽鎖および重鎖、任意選択で、抗体をグリコシル化できる哺乳動物細胞系統(例えば、CHO細胞)の全般的等モル産生を促進する「split DHFR」法が含まれる(例えば、抗体産生(Antibody production)、欧州特許第EP0481790A2号、および米国特許第5,545,403号)。
ハイブリドーマ法では、マウスまたは他の適切な宿主哺乳動物、例えば、ラット、ハムスターまたはマカクザルが、本明細書に記載のように免疫され、免疫化に使われるタンパク質に特異的に結合する抗体を産生するか、または産生が可能なリンパ球を誘発する。あるいは、リンパ球は、インビトロで免疫できる。次に、リンパ球は、適切な融合剤、例えば、ポリエチレングリコールを使って、骨髄腫細胞と融合され、ハイブリドーマ細胞を形成する(Goding、モノクローナル抗体:原理と実践(Monoclonal Antibodies:Principles and Practice)、pp.59−103(Academic Press、1986))。
ハイブリドーマ細胞は、調製されるとすぐに、好ましくは、未融合の親骨髄腫細胞の増殖または生存を抑制する1つまたは複数の物質を含む、適切な培地中に播種され、増殖される。例えば、親骨髄腫細胞が、酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠く場合は、ハイブリドーマ用の培地は、典型的に、HGPRT欠乏細胞の増殖を防ぐ物質である、ヒポキサンチン、アミノプテリン、およびチミジン(HAT培地)を含む。
好ましい骨髄腫細胞は、効率的に融合し、選択抗体産生細胞抗体の安定な高レベル産生を支援し、さらに培地に感受性の高いものである。ヒト骨髄腫およびマウスヒトヘテロミエローマ細胞系統は、また、ヒトモノクローナル抗体の産生用として記載されている(Kozbor、J.Immunol.、133:3001(1984);Brodeuretal.、モノクローナル抗体産生技術と適用(Monoclonal Antibody Production Techniques and Applications)、pp.51−63(Marcel Dekker、Inc.、NewYork、1987))。ハイブリドーマ産生融合体法で使われる骨髄腫細胞は、好ましくは非抗体産生性であり、高い融合効率、および酵素欠損を有し、この酵素欠陥は、特定の選択培地中での増殖を不能にし、所望の融合細胞(ハイブリドーマ)のみの増殖を支援する。マウス融合体に使われる適切な細胞系統の例には、Sp−20、P3−X63/Ag8、P3−X63−Ag8.653、NS1/1.Ag41、Sp210−Ag14、FO、NSO/U、MPC−11、MPC11−X45−GTG1.7およびS194/5XXOBul、が含まれ、ラット融合体に使われる細胞系統の例には、R210.RCY3、Y3−Ag1.2.3、IR983Fおよび4B210が含まれる。細胞融合体に有用な他の細胞系統は、U−266、GM1500−GRG2、LICR−LON−HMy2およびUC729−6である。
ハイブリドーマ細胞が成長している培地は、抗原に対するモノクローナル抗体の産生に関しアッセイされる。ハイブリドーマ細胞により産生されるモノクローナル抗体の結合特異性は、免疫沈降またはインビトロ結合アッセイ、例えば、ラジオイムノアッセイ(RIA)もしくは酵素免疫測定法(ELISA)により測定されるのが好ましい。モノクローナル抗体の結合親和性は、例えば、BIAcore(登録商標)またはスキャチャード解析により測定できる(Munsonetal.、Anal.Biochem.、107:220(1980);Fischer et al.、ペプチド免疫グロブリンコンジュゲート(peptide−immunoglobulin−conjugate)、国際公開第WO2007/045463A1号、実施例10、これらは、参照によりその全体が本明細書に組み込まれる)。
所望の特異性、親和性、および/または活性の抗体を産生するハイブリドーマ細胞が特定された後、クローンは、限界希釈法によりサブクローニングでき、標準的な方法で増殖できる(Goding、モノクローナル抗体:原理と実際(Monoclonal Antibodies:Principles and Practice)、pp.59−103(Academic Press、1986))。この目的のために適切な培地には、例えば、D−MEMまたはRPMI−1640培地が含まれる。さらに、ハイブリドーマ細胞は、動物中で腹水腫瘍としてインビボ増殖できる。
ハイブリドーマまたはmAbは、さらに選別されて、特定の特性、例えば、特異的抗原または標的との結合親和性を有するmAbを特定できる。サブクローンにより分泌されるモノクローナル抗体は、従来の免疫グロブリン精製法、例えば、タンパク質A−セファロース、ヒドロキシルアパタイトクロマトグラフィー、ゲル電気泳動法、透析、親和性クロマトグラフィー、またはいずれかの他の当技術分野で既知の他の適切な精製技術、を使って培地、腹水、または血清から、適切に分離される。
抗体および他のポリペプチドの組換え産生。免疫グロブリンまたは目的のポリペプチド由来の関連アミノ酸配列は、直接タンパク質配列決定により決定でき、適切なコード化ヌクレオチド配列がユニバーサルコドン表に従って設計できる。あるいは、ゲノムまたはモノクローナル抗体をコードするcDNAは、単離されて、このような抗体を産生している細胞から、従来の方法を使って配列決定される(例えば、モノクローナル抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合できるオリゴヌクレオチドプローブを使うことにより、)。関連DNA配列は、直接核酸配列決定により決定できる。
DNAのクローニングは標準的な方法を使って行われる(例えば、Sambrook et al.(1989)分子クローニング:研究室ガイド(Molecular Cloning:A Laboratory Guide)、Vols 1−3、Cold Spring Harbor Press、を参照。この文献は参照により本明細書に組み込まれる)。例えば、cDNAライブラリーを、ポリA+mRNAの、好ましくは、膜結合性mRNAの逆転写により生成でき、ライブラリーは、ヒト免疫グロブリンポリペプチド遺伝子配列に特異的なプローブを使って選別される。一実施形態では、しかしながら、ポリメラーゼ連鎖反応(PCR)を使って、目的の免疫グロブリン遺伝子セグメント(例えば、軽鎖または重鎖可変セグメント)をコードするcDNA(または完全長cDNAの一部)が増幅される。増幅された配列は、いずれの適切なベクター、例えば、発現ベクター、ミニ遺伝子ベクター、またはファージディスプレイベクター、にも容易に挿入できる。目的の免疫グロブリンポリペプチドの一部の配列を決定できる限り、使われるクローニングの特定の方法は、重大ではないことは、理解されよう。
抗体核酸の1つの源は、目的の抗原で免疫された動物由来のB細胞を得て、それを不死細胞に融合することにより産生されるハイブリドーマである。あるいは、核酸は、免疫された動物のB細胞(または全脾臓)から単離できる。また別の抗体をコードする核酸の源は、例えば、ファージディスプレイ技術により生成されるこのような核酸のライブラリーである。目的のペプチド、例えば、所望の結合特性を有する可変領域ペプチド、をコードするポリヌクレオチドは、標準的な方法、例えば、パニングにより特定できる。
免疫グロブリンポリペプチドの全可変領域をコードする配列が、決定できるが、一部の可変領域のみ、例えば、CDRコード部分のみを配列決定することで十分な場合もある。配列決定は、標準的な方法を使って行われる(例えば、Sambrook et al.(1989)分子クローニング:研究室ガイド(Molecular Cloning:A Laboratory Guide)、Vols 1−3、Cold Spring Harbor Press、およびSanger、F.et al.(1977)Proc.Natl.Acad.Sci.USA 74:5463−5467、を参照。これらの文献は、参照により本明細書に組み込まれる)。クローン化核酸の配列を、ヒト免疫グロブリン遺伝子およびcDNAの報告されている配列と比較することにより、当業者なら、配列決定された領域に応じて、容易に以下を決定できるであろう:(i)ハイブリドーマ免疫グロブリンポリペプチドの生殖系列セグメント使用(重鎖のアイソタイプを含む)、および(ii)N領域付加および体細胞変異のプロセスから得られた配列を含む、重鎖および軽鎖可変領域の配列。免疫グロブリン遺伝子配列情報の1つの源は、国立バイオテクノロジー情報センター、国立医学図書館、国立衛生研究所(メリーランド州Bethesda)である。
単離DNAは、制御配列に機能的に連結でき、または発現ベクター中に配置でき、次いで、発現ベクターを、それ以外の方法では免疫グロブリンタンパク質を産生しない宿主細胞中に形質移入し、組換え宿主細胞中のモノクローナル抗体の合成を指示できる。抗体の組換え産生は、当技術分野でよく知られている。
別の核酸配列と機能的関係に置かれる場合、核酸は、機能的に連結されている。例えば、プレ配列または分泌リーダー用のDNAは、ポリペプチドの分泌に関与するタンパク質前駆体として発現される場合、ポリペプチドのためのDNAに機能的に連結され、プロモーターまたはエンハンサーは、配列の転写に影響を与える場合、コード配列に機能的に連結され、またはリボソーム結合部位は、翻訳を促進するような位置にある場合、コード配列に機能的に連結されている。一般的に、機能的に連結されるは、連結されるDNA配列が近接しており、分泌リーダーの場合には、近接して、さらに読み相にあることを意味する。しかし、エンハンサーは、近接していなければならないことはない。連結は、好都合な制限酵素部位での連結反応により行われる。このような部位が存在しない場合は、合成オリゴヌクレオチドアダプターまたはリンカーが従来の方法に従い使用される。
多くのベクターが当技術分野で知られている。ベクター成分は、以下のうちの1つまたは複数を含んでもよい:シグナル配列(例えば、抗体の分泌を指示する;例えば、ATGGACATGAGGGTGCCCGCTCAGCTCCTGGGGCTCCTGCTGCTGTGGCTGAGAGGTGCGCGCTGT//配列番号4、これは、VK−1グナルペプチド配列MDMRVPAQLLGLLLLWLRGARC/配列番号5、をコードする)、複製起点、1つまたは複数の選択マーカー遺伝子(例えば、抗生物質または他の薬剤耐性を付与する、栄養要求欠陥を補充する、または重大な培地中にない重大な栄養素を供給することができる)、エンハンサー配列、プロモーター、および転写終止配列。これらは全て当技術分野でよく知られている。
細胞、細胞系統、および細胞培養物は、同義に使用されることが多く、本明細書の全てのこのような呼称は子孫を含む。形質転換体および形質転換細胞は、対象一次細胞および継代の数に関係なくそれ由来の培養物を含む。全ての子孫は、計画的または偶発性変異に起因して、DNA内容が正確に同じでない可能性があることも理解されよう。元の形質転換細胞での選別と同じ機能または生物活性を有する変異体子孫が含まれる。
代表的宿主細胞には、原核生物、酵母、または高等真核生物細胞が含まれる。原核生物宿主細胞には、真正細菌、例えば、グラム陰性またはグラム陽性菌、例えば、大腸菌属等の腸内細菌科、例えば、大腸菌、エンテロバクター、エルウィニア、クレブシエラ、プロテウス属、サルモネラ菌、例えば、ネズミチフス菌、セラチア、例えば、セラチア菌、および赤痢菌属、ならびに枯草菌およびB.リケニホルミス、等のバシラス属、シュードモナス属、およびストレプトマイセス属、が含まれる。真核生物の微生物、例えば、糸状菌類または酵母は、組換えポリペプチドまたは抗体用の、適切なクローニングまたは発現宿主である。出芽酵母、すなわち一般的パン酵母は、低級真核生物宿主微生物中では、最もよく使われる。しかし、いくつかの他の属、種、および株が、通常、利用可能で、本明細書で有用であり、これらは、下記のものがある:ピキア属、例えば、P.パストリス、分裂酵母;クリベロマイセス、ヤロウイア属;カンジダ;トリコデルマ・リージア;アカパンカビ;シュワニオミセスオキシデンタリス等のシュワニオミセス属;および糸状菌類、例えば、パンカビ、アオカビ類、トリポクラジウム、およびアスペルギルス宿主、例えば、偽巣性コウジ菌およびクロコウジカビ。
グリコシル化抗体の発現用の宿主細胞は、多細胞の生物体から誘導できる。無脊椎動物細胞の例には、植物および昆虫細胞が含まれる。多くのバキュロウイルス株および変異体ならびに、例えば、ヨトウガ(毛虫)、ネッタイシマカ(蚊)、ヒトスジシマカ(蚊)、キイロショウジョウバエ(果実蠅)、およびカイコ、等の宿主由来の対応する許容可能な昆虫宿主細胞が特定されている。このような細胞の形質移入用の種々のウイルス株、例えば、オートグラファ・カリフォルニカNPVのL−1変異体およびカイコNPVのBm−5株、等が公的に入手可能である。
脊椎動物宿主細胞は、また、適切な宿主であり、このような細胞からのポリペプチド(抗体を含む)の組換え産生が定型手段になっている。有用な哺乳類宿主細胞株の例は、チャイニーズハムスター卵巣細胞であり、これには、CHOK1細胞(ATCC CCL61)、DXB−11、DG−44、およびチャイニーズハムスター卵巣細胞/−DHFR(CHO、Urlaub et al.、Proc.Natl.Acad.Sci.USA 77:4216(1980));SV40で形質転換されたサル腎臓CV1系統(COS−7、ATCC CRL1651);ヒト胚性腎臓系統(浮遊培養による増殖用にサブクローニングされた293または293細胞、[Graham et al.、J.Gen Virol.36:59(1977)];ベビーハムスター腎臓細胞(BHK、ATCC CCL10);マウスセルトリ細胞(TM4、Mather、Biol.Reprod.23:243−251(1980));サル腎臓細胞(CV1 ATCC CCL70);アフリカミドリザル腎臓細胞(VERO−76、ATCC CRL−1587);ヒト子宮頚癌細胞(HELA、ATCC CCL2);イヌ腎臓細胞(MDCK、ATCC CCL34);BUFラット肝臓細胞(BRL3A、ATCC CRL1442);ヒト肺細胞(W138、ATCC CCL75);ヒト肝細胞腫細胞(HepG2、HB8065);マウス乳房腫瘍(MMT 060562、ATCC CCL51);TRI細胞(Mather et al.、Annals N.Y Acad.Sci.383:44−68(1982));MRC5細胞もしくはFS4細胞;または哺乳類骨髄腫細胞、が含まれる。
宿主細胞は、ポリペプチド(抗体を含む)産生のために形質転換されるか、または上記核酸またはベクターを形質移入され、プロモーターを誘導し、形質転換体を選択し、または所望の配列をコードする遺伝子を増幅するために、必要に応じて改善された従来の栄養素培地中で培養される。さらに、選択マーカーで分離された複数コピーの転写ユニットを有する新規ベクターおよび形質移入細胞系統が、抗体等のポリペプチドの発現に特に有用である。
本発明に有用なポリペプチドの産生に使用される宿主細胞は、種々の培地中で培養できる。市販の培地、例えば、ハムF10培地(Sigma)、基礎培地((MEM)、Sigma)、RPMI−1640(Sigma)、およびダルベッコ変法イーグル培地((DMEM)、Sigma)は、宿主細胞の培養に適する。さらに、下記に記載のいずれかの培地も、宿主細胞用培地として使用可能である:Ham et al.、Meth.Enz.58:44(1979)、Barnes et al.、Anal.Biochem.102:255(1980)、米国特許第4,767,704号;同4,657,866号;同4,927,762号;同4,560,655号;または同5,122,469号;国際公開第WO90103430号;同WO87/00195号;または米国再発行特許第30,985号。これらの培地のいずれも、必要に応じ、下記を補充できる:ホルモンおよび/または他の増殖因子(多例えば、インスリン、トランスフェリン、または上皮増殖因子)、塩(例えば、塩化ナトリウム、カルシウム、マグネシウム、およびリン酸塩)、緩衝剤(例えば、HEPES)、ヌクレオチド(例えば、アデノシンおよびチミジン)、抗生物質(例えば、ゲンタマイシン(登録商標)薬剤)、微量成分(μM範囲で最終濃度中に通常存在する無機化合物と定義される)、およびブドウ糖または等量のエネルギー源。他に必要ないずれかの補充剤も、当業者には既知の適切な濃度で含めることができる。培養条件、例えば、温度、pH、等は、発現用に選択された宿主細胞で以前使われたものであり、当業者には明らかであろう。
宿主細胞の培養の際には、組換えポリペプチドは、細胞内で、細胞周辺腔内で、または培地中へ直接分泌できる。最初のステップとして、抗体等のポリペプチドが細胞内で産生される場合は、宿主細胞または溶解断片、等の粒子状懐死組織片は、例えば、遠心分離または限外濾過により除去される。
抗体または抗体断片は、例えば、ヒドロキシルアパタイトクロマトグラフィー、陽イオンもしくは陰イオン交換クロマトグラフィー、または好ましくは、目的の抗原またはタンパク質Aもしくはタンパク質Gを親和性リガンドとして使用した親和性クロマトグラフィー、を使って、精製できる。タンパク質Aは、ヒトγ1、γ2、またはγ4重鎖に基づくポリペプチドを含むタンパク質の精製に使用できる(Lindmark et al.、J.Immunol.Meth.62:1−13(1983))。タンパク質Gは、全てのマウスアイソタイプおよびヒトγ3用に推奨されている(Guss et al.、EMBOJ.5:15671575(1986))。親和性リガンドが結合するマトリックスは、アガロースが最も多いが、他のマトリックスも利用可能である。機械的安定なマトリックス、例えば、気孔制御ガラスまたはポリ(スチレンジビニル)ベンゼンは、アガロースの場合より、高速の流量、および短い処理時間の実現を可能とする。タンパク質が、CH3ドメインを含む場合は、Bakerbond ABX(登録商標)樹脂(J.T.Baker、Phillipsburg、N.J.)が精製に有用である。回収される抗体にもよるが、エタノール沈殿、逆相HPLC、等電点電気泳動、SDS−PAGE、および硫酸アンモニウム沈殿法、等のタンパク質精製用の他の技術もまた可能である。
キメラ、ヒト化、Human Engineered(商標)、Xenomouse(登録商標)モノクローナル抗体。げっ歯類モノクローナル抗体の可変Igドメインがヒト定常Igドメインに融合されているキメラモノクローナル抗体は、当技術分野で既知の標準的方法を使って生成できる(Morrison、S.L.、et al.(1984)キメラヒト抗体分子;ヒト定常領域ドメインを有するマウス抗原結合ドメイン(Chimeric Human Antibody Molecules;Mouse Antigen Binding Domains with Human Constant Region Domains)、Proc.Natl.Acad.Sci.USA 81、6841−6855;および、Boulianne、G.L.、et al、Nature 312、643−646.(1984)、を参照)。例えば、(1)非ヒト相補性決定領域(CDR)をヒトフレームワークおよび定常領域上にグラフト化する(当技術分野で「CDRグラフティング」によるヒト化と呼ばれるプロセス)または(2)全非ヒト可変ドメインを移植するが、表面残基の置換によりひと様表面でそれらを「クローキング」する(当技術分野で「ベニアリング」と呼ばれるプロセス)または(3)それぞれの残基の抗原結合または抗体構造への関与の可能性およびその免疫原性に対する可能性に基づいて、選択非ヒトアミノ酸残基をよりヒト様に改変することによる、等の多くのヒト化または抗体配列のよりヒト様化改変技術に関して報告されている。例えば、Jones et al.、Nature 321:522 525(1986);Morrison et al.、Proc.Natl.Acad.Sci.、U.S.A.、81:6851 6855(1984);Morrison and Oi、Adv.Immunol.、44:65 92(1988);Verhoeyer et al.、Science 239:1534 1536(1988);Padlan、Molec.Immun.28:489 498(1991);Padlan、Molec.Immunol.31(3):169 217(1994);およびKettleborough、C.A.et al.、Protein Eng.4(7):773 83(1991);Co、M.S.、et al.(1994)、J.Immunol.152、2968−2976);Studnicka et al.Protein Engineering 7:805−814(1994);を参照されたい。これらのそれぞれは、参照により、その全体が、本明細書に組み込まれる。
例えば、(1)非ヒト相補性決定領域(CDR)をヒトフレームワークおよび定常領域上にグラフト化する(当技術分野で「CDRグラフティング」によるヒト化と呼ばれるプロセス)または(2)全非ヒト可変ドメインを移植するが、表面残基の置換によりひと様表面でそれらを「クローキング」する(当技術分野で「ベニアリング」と呼ばれるプロセス)または(3)それぞれの残基の抗原結合または抗体構造への関与の可能性およびその免疫原性に対する可能性に基づいて、選択非ヒトアミノ酸残基をよりヒト様に改変することによる、等の多くのヒト化または抗体配列のよりヒト様への改変に関する技術が、報告されている。例えば、Jones et al.、Nature 321:522 525(1986);Morrison et al.、Proc.Natl.Acad.Sci.、U.S.A.、81:6851 6855(1984);Morrison and Oi、Adv.Immunol.、44:65 92(1988);Verhoeyer et al.、Science 239:1534 1536(1988);Padlan、Molec.Immun.28:489 498(1991);Padlan、Molec.Immunol.31(3):169 217(1994);およびKettleborough、C.A.et al.、Protein Eng.4(7):773 83(1991);Co、M.S.、et al.(1994)、J.Immunol.152、2968−2976);Studnicka et al.Protein Engineering 7:805−814(1994);を参照されたい。これらのそれぞれは、参照により、その全体が、本明細書に組み込まれる。
抗体は、また、内在性免疫グロブリン産生のない遺伝子導入動物を使って産生でき、ヒト免疫グロブリン座位を含むように操作される(例えば、Mendez et al.、Nat.Genet.15:146−156(1997)、参照)。例えば、国際公開第WO98/24893号は、ヒトIg座位を有する遺伝子導入動物を開示しており、ここでは、動物は、内在性重鎖および軽鎖座位の不活化のために、機能的内在性免疫グロブリンを産生しない。国際公開第WO91/10741号は、また、免疫原に対し免疫反応を起こすことができる遺伝子導入非霊長類哺乳類宿主を開示し、ここでは、抗体は、霊長類の定常、および/または可変領域を有し、また、内在性免疫グロブリンをコードする座位が置換または不活化される。国際公開第WO96/30498号は、定常または可変領域の全てまたは一部を置換し、改変抗体分子を形成する、等の哺乳動物で免疫グロブリン座位を改変するためのCre/Lox系の使用を開示する。国際公開第WO94/02602号は、不活化内在性Ig座位および機能性ヒトIg座位を有する非ヒト哺乳類宿主を開示する。米国特許第5,939,598号は、マウスが内在性重鎖を欠き、1つまたは複数の異種定常領域を含む外因性の免疫グロブリン座位を発現する遺伝子導入マウスの作製方法を開示する。
上述の遺伝子導入動物を使って、選択された抗原性分子に対し免疫反応を生成でき、抗体産生細胞を動物から取り出して、ヒト由来モノクローナル抗体を分泌するハイブリドーマの産生に使用できる。免疫化プロトコル、アジュバント、等は、当技術分野で既知であり、例えば、国際公開第WO96/33735号に記載のように、遺伝子導入マウスの免疫化に使われる。モノクローナル抗体は、生物活性または対応するタンパク質の生理的な効果を抑制または中和する能力に関し試験できる。また、Jakobovits et al.、Proc.Natl.Acad.Sci.USA、90:2551(1993);Jakobovits et al.、Nature、362:255−258(1993);Bruggermann et al.、Year in Immuno.、7:33(1993);Mendez et al.、Nat.Genet.15:146−156(1997);および米国特許第5,591,669、同5,589,369号、同5,545,807号;ならびに米国特許公開第20020199213号、も参照されたい。米国特許公開第20030092125号は、動物の免疫反応を所望のエピトープに偏らせる方法を記載している。ヒト抗体は、また、インビトロ活性化B細胞によっても生成できる(米国特許第5,567,610号および同5,229,275号、参照)。
ファージディスプレイ技術による抗体産生
繊維状バクテリオファージ表面上のコード化抗体断片の組換えヒト抗体遺伝子レパートリー作成およびディスプレイ技術開発により、ヒト由来抗体生成の別の手段が提供されるようになった。ファージディスプレイについては、例えば、Dower et al.、国際公開第WO91/17271号、McCafferty et al.、国際公開第WO92/01047号、およびCaton and Koprowski、Proc.Natl.Acad.Sci.USA、87:6450−6454(1990)、に記載されている。これらは参照によりその全体が本明細書に組み込まれる。ファージ技術により産生された抗体は、通常、細菌中で抗原結合断片、例えば、FvまたはFab断片として産生され、従って、エフェクター機能に欠ける。エフェクター機能は、下記の2つの戦略の内の1つにより導入できる。断片を操作して、哺乳動物細胞中での発現のために完全抗体中に、またはエフェクター機能を誘発できる第2の結合部位を有する二重特異性抗体断片中に組み込むことができる。
繊維状バクテリオファージ表面上のコード化抗体断片の組換えヒト抗体遺伝子レパートリー作成およびディスプレイ技術開発により、ヒト由来抗体生成の別の手段が提供されるようになった。ファージディスプレイについては、例えば、Dower et al.、国際公開第WO91/17271号、McCafferty et al.、国際公開第WO92/01047号、およびCaton and Koprowski、Proc.Natl.Acad.Sci.USA、87:6450−6454(1990)、に記載されている。これらは参照によりその全体が本明細書に組み込まれる。ファージ技術により産生された抗体は、通常、細菌中で抗原結合断片、例えば、FvまたはFab断片として産生され、従って、エフェクター機能に欠ける。エフェクター機能は、下記の2つの戦略の内の1つにより導入できる。断片を操作して、哺乳動物細胞中での発現のために完全抗体中に、またはエフェクター機能を誘発できる第2の結合部位を有する二重特異性抗体断片中に組み込むことができる。
典型的には、抗体のFd断片(VH−CH1)および軽鎖(VL−CL)は、PCRにより別々にクローン化され、ランダムに組み合わされてコンビナトリアルファージディスプレイライブラリーを生成し、次に、これを、特定の抗原に対する結合に関し選択できる。抗体断片は、ファージ表面に発現され、FvまたはFab(従って、抗体断片をコードするDNAを含むファージ)の抗原結合による選択は、数ラウンドの抗原結合および再増幅により実現される。この方法は、パニングと呼ばれる。抗原に特異的な抗体断片が富化され、最終的に単離される。
ファージディスプレイ技術は、また、げっ歯類モノクローナル抗体のヒト化のための「誘導選択」と呼ばれる手法にも使うことができる(Jespers、L.S.、et al.、Bio/Technology 12、899−903(1994)、を参照)。このために、マウスモノクローナル抗体のFd断片を、ヒト軽鎖ライブラリーと組み合わせてディスプレイでき、得られたハイブリッドFabライブラリーを、次に、抗原で選択できる。マウスFd断片は、それにより、選択を誘導するテンプレートを提供する。その後、ヒト軽鎖は、ヒトFd断片ライブラリーと組み合わされる。生成されたライブラリーの選択により、完全ヒトFabが得られる。
ファージディスプレイライブラリーからヒト抗体を得るための種々の方法が報告されている(例えば、Hoogenboom et al.、J.Mol.Biol.、227:381(1991);Marks et al.、J.Mol.Biol、222:581−597(1991);米国特許第5,565,332号および同5,573,905号;Clackson、T.、and Wells、J.A.、TIBTECH 12、173−184(1994)、を参照)。特に、インビトロ選択および進化ofファージディスプレイライブラリー由来抗体の進展産物は強力なツールになっている(Burton、D.R.、and Barbas III、C.F.、Adv.Immunol.57、191−280(1994);および、Winter、G.、et al.、Annu.Rev.Immunol.12、433−455(1994);米国特許公開第20020004215号および国際公開第WO92/01047号;2003年10月9日公告の米国特許公開第20030190317号および米国特許第6,054,287号;米国特許第5,877,293号、を参照されたい)。Watkins、「ファージ発現抗体ライブラリーのキャプチャーリフト法による選別(Screening of Phage−Expressed Antibody Libraries by Capture Lift)」、分子生物学の方法、抗体ファージディスプレイ:方法とプロトコル(Methods in Molecular Biology、Antibody Phage Display:Methods and Protocols)178:187−193、および2003年3月6日公告の米国特許公開第20030044772号は、ファージ発現抗体ライブラリーまたは他の結合分子のキャプチャーリフト法による選別方法について記載している。この方法は、候補結合分子の固体支持上への固定化を含む方法である。
本発明は、下記の実施例への参照により、さらに完全に理解されるであろう。これらの実施例は、本発明の範囲を多少なりとも限定するものと解釈されるべきではない。
実施例1:戻し交配および異系交配
NaV1.7KO動物の報告されている明らかに摂食できないことによる新生仔の致死性(C57BL/6に対し少なくとも8世代戻し交配されたDeltagen、San Mateo、CA:B6.129P2−Scn9atm1Dgen/J由来ヘテロ接合のNav1.7+/−マウス)(Nassar et al.、2004)の理由で、我々は、これらの動物をCD1背景に異系交配し、その系統に活力を付与し、さらに、別の戻し交配をBALB/c系統で行い、類遺伝子性系統を生成した(2011年5月に受け取った最初の繁殖ペア)(図1A〜B参照)。
NaV1.7KO動物の報告されている明らかに摂食できないことによる新生仔の致死性(C57BL/6に対し少なくとも8世代戻し交配されたDeltagen、San Mateo、CA:B6.129P2−Scn9atm1Dgen/J由来ヘテロ接合のNav1.7+/−マウス)(Nassar et al.、2004)の理由で、我々は、これらの動物をCD1背景に異系交配し、その系統に活力を付与し、さらに、別の戻し交配をBALB/c系統で行い、類遺伝子性系統を生成した(2011年5月に受け取った最初の繁殖ペア)(図1A〜B参照)。
Scn9a−CD1KOまたはScn9a−BalbcKO動物に有意差は認められなかった。我々は、これらの動物を、NaV1.7KOと呼び、必要に応じ、この株を指定する。本明細書にに示される全データは、単一の異系交配および戻し交配で集められた。さらなる戻し交配(BALB/c系統のみに対し)は、現在実行中であり、今までもところ、得られた結果は、N4に対する戻し交配世代と同じである。生存のさらなる増加は認められなかった。戻し交配の最終結果は、N7(約99%BALB/c)で類遺伝子性系統に到達時に決定されなければならない。NaV1.7KOマウスは、それらの同腹子と大略同じサイズと重量で誕生した(例えば、図2B)が、差異は同腹子内ですぐに明らかになった(誕生後16時間以内;図2A)。NaV1.7KOマウスは、それらの同腹子ほど早く成長せず、少し弱かった。NaV1.7KO子は、資源獲得競争に苦労し、巣の底(食物供給源から離れている)までの到達で終わってしまい、ついには死亡する。明らかな摂食/ミルク哺乳の能力のために、我々は、NaV1.7KO子を野性型兄弟姉妹から離して養母と一緒に置くことが、より高い生存確率を与えるかどうかを考えた。残念ながら、それらの小さいサイズおよび相対的衰弱のために、雌親の適切な刺激が可能とはならなかった。我々は、すぐにミルク産生(授乳)の減少に気付き、新生仔のNaV1.7KO動物は死亡した。P4NaV1.7KO新生仔の徹底的解剖学的評価の後で、我々は、これらの動物は、機会があれば摂食することができたという結論に達した。図3A〜Gと図4A〜Eに示すように、全ての必要な臓器は、正しい位置にあった。
人工のマウスミルクを開発した(Auestad et al.、1989およびYajima et al.、2006、から変更したレシピ;本明細書の実施例2参照)。
最初に、我々は、誕生後24時間の体重に従って、同腹子中の全てのNaV1.7KO候補を特定した。大まかに、同腹子の最小1/4を、25%の予測メンデル比(Het x Het繁殖ペアに対し)に対応する有望なノックアウトとして特定した。より大きな「対照」動物を、下記式に従って巣から取り出した:
[候補KOの数]+2対照(KO数≦2の場合)
[候補KOの数]+3対照(KO数≧3の場合)
[候補KOの数]+2対照(KO数≦2の場合)
[候補KOの数]+3対照(KO数≧3の場合)
いくつかの対照動物の存在により、雌親の十分な授乳刺激を可能とし、その少ない数により、内部同腹子競合を減らした。対照動物は、標準的ポリメラーゼ連鎖反応(PCR)により測定して、両性の野性型およびヘテロ接合混合物であった。
簡単に説明すると、以下のプライマーを使って、コロニー中の全ての動物の遺伝子型の特定を行った:順方向Scn9a:5’−AGACTCTGCGTGCTGCTGGCAAAAAC−3’(配列番号1);順方向ネオマイシン:5’−GGGCCAGCTCATTCCTCCCACTCAT−3’(配列番号3)および逆方向Scn9a:5’−CGTGGAAAGACCTTTGTCCCACCTG−3’(配列番号2)。これらのプライマーは、267塩基対の内在性(E)バンド(プライマー順方向Scn9a+プライマー逆Scn9a);(図7Aのバンドを参照)、または389塩基対の標的(T)バンドを生じた(プライマー順方向ネオマイシン+プライマー逆方向Scn9a);図7Bのバンドを参照)。予想遺伝子型パターンは、以下の通りであった:
WT(+/+)動物 =内在性(E)バンドのみ
HET(+/−)動物 =内在性(E)+標的(T)バンド
KO(−/−)動物 =標的(T)バンドのみ。
HET(+/−)動物 =内在性(E)+標的(T)バンド
KO(−/−)動物 =標的(T)バンドのみ。
KO表現型は、内在性(E)バンドの「非存在」により決定されるので、我々は、最初に、一連のDNA濃度(25ng〜100ng)で試験し、産物を生成するのに十分なPCR反応用のインタクトDNAがあることを確認した。図7Aに示すように、わずか25ngのDNAで、野性型動物(AMA−50)中で視認できるPCR産物を生成するのに十分であった。同じDNA濃度の存在下で、KO動物(AMA−161)中では、PCR産物は存在しなかったが、このことは、この動物が内在性Scn9a遺伝子の無傷コピーを持っていることを示唆している。図7Bでは、標的プライマー対に対するPCR産物(バンド)の存在により、(i)NaV1.7KO DNA(AMA−161)は、良好な質の無傷であったこと、および(ii)動物AMA−161は、標的Neo遺伝子を持ち、従って、NaV1.7KOマウスと認められることが確認される。図7Cでは、PCR反応試料の対照は、DNAを含まず、従って、PCR産物を生成しなかった。
NaV1.7KOであると確認された動物は、2つの方法で再確認された。最初に、それらが独特の表現型(熱疼痛感覚の欠如、および嗅覚消失の存在;本明細書の実施例4参照)を示す、一連の挙動試験を受け、さらに遺伝子型が確認される。第2に、全動物は、安楽死の際に、同じPCR方法(上述の)により遺伝子型が再特定された。
NaV1.7KO候補に、1日3回を/7日間/週、これを14〜21日間続けて手で給餌した(図5Bのシリンジおよび図6の給餌を参照)。歯が生えると、追加のカロリー源として全ての新生仔マウスに対し柔らかい補助食物を利用できるようにした。合計28匹のNaV1.7KO動物を11ヶ月間の間に生成し、24匹はCD1背景を有し、4匹はBALB/c背景を有した。2つの背景の数の間の差異は、コロニーのサイズの反映であり、育種方法の困難さ、または結果ではない。我々は、動物は、どちらの背景のものでも非常に類似の方法で挙動を行うことを見出した(表2の「状態(status)」カラム参照)。2010年12月1日現在で18匹の動物が生存していた。8匹は、健康関連問題により失われ、試験(免疫化アッセイ)中の2匹の動物は、免疫化による健康上の合併症の後で、安楽死させる必要が生じた。それらの死は、特定の表現型に帰することはできなかった。雄雌両方の動物は、それぞれの株に対し予測比(約50%)で存在した。確認された遺伝子型の動物は、2010年12月1日現在、54日〜7.5ヶ月の年齢であった。表2(下記)は、それぞれのNaV1.7KOマウスのリストである。成人期に達すると、NaV1.7KO動物は、交配できた。NaV1.7KO動物の、雄(表2A)と雌(表2B)の両方が、同じ背景由来の逆の性のヘテロ接合の(HET)動物と交配された。雄雌両方共、繁殖可能であった。雄は、特に活気がなく、その仕事に非効率的であり、18の別々の交配イベントで1匹の同腹子の生成に繋がった(表2A)。いくつかの個体の場合に、泌尿性器の死後分析が行われ、精子数を測定し、全ての繁殖問題が調査された。繁殖性雄および不妊雄の両方が認められた。例えば、AMA−161は、分析され、不妊と判定された(セルトリ細胞および精原細胞が存在したが、成熟精子は存在しなかった。しかし、類似の表現型は、一部の対照動物中でも認められた)が、一方、AMA−380は、繁殖性であった。NaV1.7KO雄交配で観察された低い成功率は、NaV1.7KOマウスで観察された嗅覚消失(臭い感覚の欠如)表現型に関連付けられる可能性がある。NaV1.7KO雄は、ケージ中に置かれた雌のフェロモンを受け取ることができない可能性がある。両株の雌は、生殖の観点では成功であった(4交配イベントから3匹の同腹子)(表2B)。調査した全ての雌は、妊娠、同腹子の出産、および子への授乳/給餌ができた。ケージ中に置かれた雄は、対形成を妨害する可能性のある嗅覚消失の徴候のないNaV1.7HETであったので、雌にとって、臭い感覚は交配目的には必要ないであろう。
図8A〜Eは、NaV1.7KOマウスの外部表現型を示す。外部表現型は、図8A〜Dに示すように、サイズの顕著な差異を除いて、正常であった(図8A〜Dの矢印で示す、より小さいNaV1.7KOマウスを参照)。それらの目は、開かれ、歯は生え、さらに、毛はよく発達していた。それらは、同腹子とほぼ同時にケージ内で動くことができた。誕生後、16時間の早い時期に、NaV1.7KO動物は、同腹子よりサイズが小さいことが認められた。図8Eは、離乳後8週間にわたるサイズ比較を示す。離乳後野性型同腹子に比べて、NaV1.7KO動物は、約25%小さかった。このサイズ差は、成長の間に直面したカロリー制約と、従って、代償的供給源(完全な置換ではない)としての手による給餌に対する要求に起因する可能性がある。
Na V 1.7KOマウスで観察された臨床的イベント。NaV1.7KOコロニーを約1年間にわたりモニターした。これらの動物が要求する集中的新生仔ケアは別として、若干の再発性の離乳後の健康問題が認められた。これらが、遺伝的障害に関連するのか、または新生仔期間中の強い操作に起因するのかは、明らかでない。NaV1.7KOの約40%(集団の雌(8例)および雄(3例))で皮膚炎が観察された。この臨床的イベントは、下記治療により解決に成功した:週1回のBaytril(Western Medical Supply、Inc.、Arcadia、CA;cat.#2269)処置、続けて、三重抗生物質軟膏(Butler Schein Animal Health、Dublin、OH;cat.#031087)の毎日2回の局所的塗布を組み合わせた最適化治療。皮膚炎が目の近くに現れた場合には、局所の三重眼部軟膏(Webster Veterinary、Sterling、MA;cat.#07−805−4502)を代わりに使った。軽度の場合には、Baytril単独治療で問題を完全回復まで解決するのに十分であった。痒み行動アッセイ(本明細書の実施例4参照)から集めたデータに基づき、ジフェンヒドラミンは、Nav1.7KOマウスに対し投与されなかった。理由は、それらがヒスタミン誘導痒みに対し非感受性であるからである。NaV1.7KOマウスコロニーの約25%に拡張膀胱が認められた。これは、雌(2例)より雄(5例)で多かった。治療は、拡張膀胱領域に手で圧力をゆっくり加え、尿を放出させることから構成した。さらに、動物が脱水症の徴候を示す場合は、NaCl液の皮下注射を組み合わせた。陰茎脱が2、3匹の雄で報告され、器官を手でその腔中に戻し、発赤が観察される場合には、局所的三重抗生物質と組み合わせることにより解決した。臨床的徴候を示す全動物は、Teklad Diamond Soft Bedding(Harlan Laboratories、Indianapolis、IN;cat.#7089)に置き、標準的寝具とすり減らす作用を起こす接触を最小限にし、および/または妨害物が検出された場合には、尿産生をモニターした。
実施例2:人工マウスミルク(AMA)の調製
調製された人工マウスミルクの手順と組成は、Yajimaと共同研究者およびAuestadと共同研究者による記載に厳密に従って設計した(Yajima、M、et al.、「マウスの子人工飼育用のマウスミルクに同等の化学的に生成されたミルク代用品(A Chemically Derived Milk Substitute that is Compatible with Mouse Milk for Artificial Rearing of Mouse Pups)」Exp.Amin.55(4)、391−397、(2006);Auestad、N、et al.、ラットのミルクに同等のミルク代用品;その調製、組成および人工飼育ラットの成長および代謝に対する影響(Milk−substitutes comparable to rat’s milk;their preparation、composition and impact on development and metabolism in the artificially reared rat)、Br.J.Nutr.61:495−518(1989))。
調製された人工マウスミルクの手順と組成は、Yajimaと共同研究者およびAuestadと共同研究者による記載に厳密に従って設計した(Yajima、M、et al.、「マウスの子人工飼育用のマウスミルクに同等の化学的に生成されたミルク代用品(A Chemically Derived Milk Substitute that is Compatible with Mouse Milk for Artificial Rearing of Mouse Pups)」Exp.Amin.55(4)、391−397、(2006);Auestad、N、et al.、ラットのミルクに同等のミルク代用品;その調製、組成および人工飼育ラットの成長および代謝に対する影響(Milk−substitutes comparable to rat’s milk;their preparation、composition and impact on development and metabolism in the artificially reared rat)、Br.J.Nutr.61:495−518(1989))。
試薬と方法。入手元、使用量を含む使用試薬、溶剤と使用量、ならびに種々コメントが、表3(下記)に一覧表示されている。
方法。ステップ1:水酸化ナトリウムを秤量し、10Lビーカーに移した。蒸留水(1.3L)を加え、オーバーヘッドスターラーを適所に置いた。水酸化カリウムを秤量し、この溶液に加え、また、L−セリン、L−シスチンおよびL−トリプトファンを加えた。この溶液を水浴中、58℃まで加熱した。都合がよければ、この溶液または混合物を、ビンに移し、その後の使用の前に、5日間まで冷蔵庫中で貯蔵できる。次に、カゼインを温かい溶液にゆっくり加え、混合物を71℃まで90分間加熱した。この混合物を4Lのガラスビーカーに移し、50℃まで加熱した。オーバーヘッドスターラーを適所に置き、混合物を沸騰するまで加熱した(図5A)。ステップ2:グリセロ燐酸カルシウム、塩化マグネシウム六水和物および塩化カルシウムを秤量し、200mLの蒸留水に溶解し、20分間均一化した。この混合物を、カゼイン混合物に、攪拌を続けながらゆっくり添加した。ステップ3:炭酸カルシウムおよびクエン酸四水和物を秤量し、100mLの蒸留水に加え、1分間均一化し、カゼイン混合物にゆっくり加えた。ステップ4:リン酸ナトリウム二塩基性七水化物および一塩基性リン酸カリウムを秤量し、50mLの蒸留水に溶解し、カゼイン溶液にゆっくり添加した。ステップ5:ラクトース一水和物を秤量し、220mLの蒸留水中で均一化した後、カゼイン混合物に加えた。ステップ6:硫酸鉄七水化物およびクエン酸を秤量し、5mLの蒸留水に溶解し、カゼイン混合物に加えた。ステップ7:硫酸マンガン水和物、硫酸銅五水和物および硫酸亜鉛七水化物を秤量し、5mLの蒸留水に溶解後、カゼイン溶液に加えた。ステップ9:フッ化ナトリウムおよびヨウ化カリウムを秤量し、5mLの蒸留水に溶解し、カゼイン混合物に加えた。ステップ8および10:乳清タンパクを秤量し、600mLの蒸留水中で均一化した。この混合物に、L−カルニチン、アルファピコリン酸HCl、エタノールアミンおよびタウリンの10mL蒸留水中溶液を添加した。
ステップ11および12:パーム油、ココヤシ油、トウモロコシ油、MCT油、大豆油およびコレステロールの混合物をホットプレート上で60℃まで加熱した。二水素性クエン酸コリンとビタミンの混合物を秤量し、70mLの蒸留水中に懸濁した。水酸化ナトリウム(5N)を加えた。この混合物をカゼイン混合物にゆっくり加えた。オイル混合物をこのカゼイン混合物に加え、さらに蒸留水を加えて体積を4Lにした。混合物をビンに移し、3日以内に使用するまで冷蔵庫で貯蔵した。
貯蔵した人工ミルクを冷蔵庫から取り出し、水浴中の4Lビーカーに移し、沸騰するまで加熱した。水中で15分間煮沸することにより、ホモジナイザーチップを滅菌した。ステップ13:人工ミルク混合物を、均一化し、その間に、ビタミンK1、ビタミンAパルミチン酸塩およびDL−トコフェロール酢酸エステルを秤量し、添加した。均一化を続けながら人工ミルクを煮沸水浴中で保持し、その間に、無菌の15mLエッペンチューブ(図5B)に分注した。人工ミルクを使用するまで−80℃で貯蔵した。
実施例3:遺伝子型同定
下記のプライマーを使ってコロニー中の全ての動物の遺伝子型を特定した:順方向Scn9a5’AGACTCTGCGTGCTGCTGGCAAAAAC3’(配列番号1);逆方向Scn9a5’CGTGGAAAGACCTTTGTCCCACCTG3’(配列番号2)および順方向ネオマイシン5’GGGCCAGCTCATTCCTCCCACTCAT3’(配列番号3)。これらのプライマーは、267塩基対の内在性バンド(順方向Scn9a+逆方向Scn9a)または389塩基対の標的バンド(順方向ネオマイシン+逆方向Scn9a)生じた。
下記のプライマーを使ってコロニー中の全ての動物の遺伝子型を特定した:順方向Scn9a5’AGACTCTGCGTGCTGCTGGCAAAAAC3’(配列番号1);逆方向Scn9a5’CGTGGAAAGACCTTTGTCCCACCTG3’(配列番号2)および順方向ネオマイシン5’GGGCCAGCTCATTCCTCCCACTCAT3’(配列番号3)。これらのプライマーは、267塩基対の内在性バンド(順方向Scn9a+逆方向Scn9a)または389塩基対の標的バンド(順方向ネオマイシン+逆方向Scn9a)生じた。
PCRサイクル条件は、以下の通りであった:
(1)95℃で7分間、続けて、下記(2)〜(4)を35xサイクル、続けて、(5)〜(6):
(2)96℃で10秒間;
(3)60℃で30秒間;
(4)72℃で1.5分間、
(5)72℃で7分間;
(6)4℃に冷却。
(1)95℃で7分間、続けて、下記(2)〜(4)を35xサイクル、続けて、(5)〜(6):
(2)96℃で10秒間;
(3)60℃で30秒間;
(4)72℃で1.5分間、
(5)72℃で7分間;
(6)4℃に冷却。
同定遺伝子型パターンは以下の通りであった:
WTまたは+/+ =内在性(E)バンドのみ
HETまたは+/− =内在性(E)+標的(T)バンド
KOまたは−/− =標的にしている(T)バンドのみ
WTまたは+/+ =内在性(E)バンドのみ
HETまたは+/− =内在性(E)+標的(T)バンド
KOまたは−/− =標的にしている(T)バンドのみ
約25ng〜1μgのDNA濃度が、各試料で最初に試験された。内在性バンドは、AMA−161試料のいずれにも(または他のNav1.7KO試料のいずれにも)、最初確認した生存NaV1.7KO動物(CD1背景;図7A〜C)にも、存在しなかった。動物AMA−50は、確認済み野性型(WT)である。DNA不含の対照は、予想通りブランクであった。
実施例4:疼痛試験
熱試験。ポーサーマルスティミュレータは、研究者が不連続の熱刺激(放射熱)を特定の領域(例えば、足)に送出することを可能とする装置である。動物は、25℃に加熱されたガラス表面上の試験チャンバーに収容された。試験開始時、後肢の下のタイマーに連動した熱ビームのスイッチを入れる。刺激に反応した動物の足の動きが刺激を終わらせ、試験の終点としての役割を果たす。20秒の最大カットオフ時間を設定し、組織損傷を防いだ。典型的には、動物を、少なくとも5分の試験間隔をあけて、2回試験した。最初の2回の測定値が一致しない場合は、さらに1、2回の試験を行い、動物の真の熱閾値を明確にした。熱源から足を移動するまでの潜時を評価項目として記録した。
熱試験。ポーサーマルスティミュレータは、研究者が不連続の熱刺激(放射熱)を特定の領域(例えば、足)に送出することを可能とする装置である。動物は、25℃に加熱されたガラス表面上の試験チャンバーに収容された。試験開始時、後肢の下のタイマーに連動した熱ビームのスイッチを入れる。刺激に反応した動物の足の動きが刺激を終わらせ、試験の終点としての役割を果たす。20秒の最大カットオフ時間を設定し、組織損傷を防いだ。典型的には、動物を、少なくとも5分の試験間隔をあけて、2回試験した。最初の2回の測定値が一致しない場合は、さらに1、2回の試験を行い、動物の真の熱閾値を明確にした。熱源から足を移動するまでの潜時を評価項目として記録した。
図9Aは、NaV1.7KOマウス株Scn9a−CD1は、熱刺激(Hargreaves Apparatus)に対し、右足(n=3)では、明らかにごく僅かな反応があり、左足(n=5)では反応しなかったことを示す。図9Bは、NaV1.7KOマウス株Scn9a−BalbCは、両足ともに(n=1)反応しなかったことを示す。WTとHETの間では差は認められず(図9A〜B)、全て正常に反応し、熱刺激適用後10〜15秒以内に足を逃避させた。大部分のNaV1.7KO動物は、最大カットオフ時間(20秒)に到達するまで、熱刺激に耐えた。
ホットプレート装置は、所定の温度に設定された制御可能加熱表面を有する。マウスをその装置上に置き、熱に対する反応をモニターした。反応には、足を持ち上げる、足をなめる、足を引っ込める、および/またはジャンプする挙動が含まれる。最大時間制限を常に使用したが、刺激強度(すなわち、温度)に応じて変える。急性試験に対しては、装置に許容される最大時間は、温度を基準として次の通りとした:48℃=60秒;50℃=40秒;52.5℃=30秒;55℃=20秒。反復試験では、少なくとも10分の試験間隔をおき、試験から足を完全に回復させた。試験の開始時に、動物を試験チャンバー中に置き、タイマーをスタートさせた。反応、または最大カットオフ時間への到達のどちらかが先に発生した時点で、動物を装置から取り除いた。最初の反応までの潜時を評価項目として記録した。
図10A〜Hは、4種の異なる温度(48、50、52.5、および55℃)の試験で、両方のNaV1.7KOマウス株が熱疼痛に対し非感受性であったことを示す。Scn9a−CD1 NaV1.7−/−KOマウス(図10A〜D;n=14)およびScn9a−BalbCNaV1.7−/−KOマウス(図10E〜H;n=4)は、55℃の最高試験可能温度であっても、熱疼痛に対し非感受性であった。この最高温度では、設定カットオフ時間(20秒)でホットプレートから取り除き、組織損傷を避ける必要があった。
触覚アロディニアVon Frey試験。Von Freyフィラメントを使ってげっ歯類の機械的な感受性が評価される。マウスを、ワイヤメッシュフロアー上に置き、個別試験チャンバー中に収容し、落ち着くまで順化させる。次に、較正された種々の曲げ強度のフィラメントをマウスの足に押しつけ、無害触知(例えば、触覚)刺激に対する反応を測定した。一連のフィラメントに対する反応パターンおよび無反応により、動物の機械的な閾値を決定した。この閾値を、このアッセイの評価項目として使用した。
図11A〜Bは、NaV1.7KOマウス株Scn9a−CD1(図11A;n=16)およびScn9a−BalbC(図11B;n=4)が、Von Freyアロディニア刺激に対し、野性型/ヘテロ接合対照同腹子に類似の形式で反応したことを示す。全ての試験マウスは、1.5gのカットオフ閾値に達した。NaV1.7KO動物は、機械的な圧力を正常に認知すると思われ、この結果は、ヒトのNaV1.7欠失に関する報告された観察と一致する。
嗅覚消失試験。埋設食物試験(Buried Food Test)を開発して、マウスの嗅覚が損なわれていないかどうかを評価した。簡単に説明すると、1つのペレットをホームケージ中に置くことにより、動物が、ニャコラーダ香料入りの食物ペレットを鑑別できるように訓練した。各動物は、翌朝、ペレットが食べられ、食物として認識されたことを保証するためチェックされた。食物を試験日の前にケージ中に1日1個ずつ3日間の3回置いた。その後、試験日の前の18時間、動物を絶食させた。試験日に、マウスを、3cmのクリーン寝具を備えた標準的マウスケージに移し、5分間順化させた。5分後、マウスを、ランダムなコーナーの寝具の下1cmに埋設された食物ペレットを有する別のクリーンケージに移した。次に、マウスをケージに再導入した。食物を見つけ食べ始めるまでの潜時を記録し、試験の評価項目として使った。動物が15分後も食物を見つけられなかった場合は、試験は中止された。
図12A〜Bは、絶食NaV1.7KOマウス株Scn9a−CD1(図12A;n=14)およびScn9a−BalbC(図12B;n=4)が、アッセイの最初の200秒内にペレットを探し出した同年齢/同じ性別の対照(WT/HET)同腹子に比べて、15分の割り当て時間の間に隠された香料入りの食物ペレットの位置を探すことが困難であった(図12A)か、またはそれができなかった(図12B)ことを示す。
痒み試験。試験日に、刺激物注射の前に、全動物を観察チャンバーに30分間順化させた。150μgのヒスタミン二リン酸エステルの注射剤を、100μLの量で、動物の背中の肩甲骨の間の皮内に投与した。この領域は、試験の前日注射を配置しやすくするために剪毛した。マウスを手で優しく抑制しながら、皮内注射を行った。ヒスタミン注射後、動物を観察チャンバー中に置き、掻き動作を最大40分間数えた。痒み掻き動作の数を評価項目として記録した。
図13A〜Bは、野性型/ヘテロ接合の同腹子とは異なり、NaV1.7KOマウス株Scn9a−CD1(図13A;n=12)およびScn9a−BalbC(図13B;n=3)は、ヒスタミンに対し非感受性であり(ダイアモンド参照)、非常にわずかな掻き動作を示す(図13A)か、または掻き動作を全く示さなかった(図13B)ことを示す。NaV1.7KOマウスによる掻き動作の平均数は、野性型/ヘテロ接合の対照同腹子(逆三角形)の食塩水注射後の平均数の範囲内であった。特に、アッセイ最初の10分の間、野性型/ヘテロ接合の動物は、激しく掻いた(正位三角形)が、一方、NaV1.7KO(ダイアモンド)は、反応しなかった。
実施例5:NaV1.7阻害剤用バイオマーカーの特定のためのNaV1.7ノックアウトマウスの使用
Na V 1.7阻害剤用オンターゲット生化学的刺激。電位依存性ナトリウムチャネルは、神経細胞興奮性の一次決定因子であり、従って、疼痛を含む興奮性亢進の神経学的障害に対する新規治療の有望な標的である。ヒトの遺伝的障害からの臨床的証拠は、9つのナトリウムチャネルの内で、SCN9A遺伝子によりコードされるNaV1.7サブタイプは、疼痛の伝達に不可欠であることを示し、標的阻害剤の開発にとって魅力的なポイントとなっている。しかし、NaV1.7に対する療法の開発には、重要な課題がある。第1のハードルは、他のナトリウムチャネルサブタイプに比べて、NaV1.7に対する十分な選択性を有する分子を見つけることである;文献には、個別のNavサブタイプに対する選択性のある化合物の報告が含まれている(Jarvis、MF et al.、強力で、選択性のNav1.8ナトリウムチャネル遮断薬のA−803467が、ラットの神経障害性および炎症性疼痛を弱める(A−803467、a potent and selective Nav1.8 sodium channel blocker、attenuates neuropathic and inflammatory pain in the rat)、Proc Natl Acad Sci USA.104(20):8520−25(2007);Beaudoin、S et al.、スルホンアミド誘導体(Sulfonamide Derivatives)、国際公開第WO2010/079443A1号)。重要な追加のステップは、候補分子の送達モード、非特異的血漿タンパク質結合、半減期および他の薬物動態学的特性を含む薬剤様の特性をインビボで最適化することである。この後で、候補化合物は、動物疾患モデルの行動上の有効性に対する前臨床種で疼痛を含む試験ができる。その後の毒性学試験で、治療濃度域が規定され、投薬の方向付けがなされる。
Na V 1.7阻害剤用オンターゲット生化学的刺激。電位依存性ナトリウムチャネルは、神経細胞興奮性の一次決定因子であり、従って、疼痛を含む興奮性亢進の神経学的障害に対する新規治療の有望な標的である。ヒトの遺伝的障害からの臨床的証拠は、9つのナトリウムチャネルの内で、SCN9A遺伝子によりコードされるNaV1.7サブタイプは、疼痛の伝達に不可欠であることを示し、標的阻害剤の開発にとって魅力的なポイントとなっている。しかし、NaV1.7に対する療法の開発には、重要な課題がある。第1のハードルは、他のナトリウムチャネルサブタイプに比べて、NaV1.7に対する十分な選択性を有する分子を見つけることである;文献には、個別のNavサブタイプに対する選択性のある化合物の報告が含まれている(Jarvis、MF et al.、強力で、選択性のNav1.8ナトリウムチャネル遮断薬のA−803467が、ラットの神経障害性および炎症性疼痛を弱める(A−803467、a potent and selective Nav1.8 sodium channel blocker、attenuates neuropathic and inflammatory pain in the rat)、Proc Natl Acad Sci USA.104(20):8520−25(2007);Beaudoin、S et al.、スルホンアミド誘導体(Sulfonamide Derivatives)、国際公開第WO2010/079443A1号)。重要な追加のステップは、候補分子の送達モード、非特異的血漿タンパク質結合、半減期および他の薬物動態学的特性を含む薬剤様の特性をインビボで最適化することである。この後で、候補化合物は、動物疾患モデルの行動上の有効性に対する前臨床種で疼痛を含む試験ができる。その後の毒性学試験で、治療濃度域が規定され、投薬の方向付けがなされる。
これらの全てのステップで、候補分子が、実際に、標的に結合しているか否かを知ること(単なる有効性や暴露から把握するには困難なステップ)が重大である。例えば、非常に多くの血漿または脳暴露が標的結合に繋がらない場合もある;インビトロ透析または遠心分離実験により決定される平衡血漿タンパク質結合は、必ずしも候補分子の真の活性量を予測しない可能性がある;また、候補分子による個別または別々の多くのタンパク質の結合は、特に疼痛の動物疾患モデルの有効性を(真のまたは偽の)作り出すことができる。重大な課題は、オンターゲットバイオマーカーを介して、候補化合物がその意図した標的に到達し、機能的効果を行使したことをインビボ系で知ることである。これは、最も正確な治療濃度域を確立する上で重要である。候補分子の臨床的開発では、この課題は、さらに重要である。例えば、標的結合のないネガティブな臨床試験では、さらなる試験の必要性を示し、一方、試験された治療薬がそのバイオマーカーに結合しなかったネガティブな臨床試験では、通常、さらなる開発を終了する。
NaV1.7は、電位により活性化されるナトリウムイオンチャネルであり、バイオマーカーの基準として使われる可能性のあるどの化学神経伝達物質またはリガンドによっても活性化されない。わかっている限り、ニューロンの電気スパイクを産生するのは機能であるにすぎず、何ら信号伝達経路を開始または調節するものでない。これは、下流の生化学的影響の試験によるNaV1.7機能の抑制のモニタリングを非常に困難にする。従って、NaV1.7用の既知のバイオマーカー、特に、9つの全ての電位依存性ナトリウムチャネルの中でNaV1.7に対し特異的であることが知られているものは、存在しない。
我々は、電位依存性ナトリウムチャネル、および本明細書に記載の本発明のNaV1.7グローバルKOマウスの非特異的モジュレーターを使って、NaV1.7阻害剤に対しバイオマーカーとして機能するインビボアッセイを開発した。ナトリウムチャネルモジュレーターであるベラトリジンは、特徴的時間経過で成長したマウスとラットの、定量可能で強力な、用量依存性、および再現可能な、足をなめる、および足を引っ込める挙動を生み出した。これらの疼痛様挙動は、一部の薬物(例えば、モルヒネ、デュロキセチン)により幾分抑制されたが、全ての既存の疼痛用薬物(例えば、ガバペンチン)に抑制されるとは限らず、また、非特異的ナトリウムチャネル遮断薬(メキシレチン)により完全に抑制された。痛みによる足を引っ込める挙動が、2つのさらなる化学的に異なるナトリウムチャネルモジュレーターである、デルタメトリン(構造は下記II)およびグラヤノトキシンIII(「GRAY3」;構造は下記III)で観察され、さらに、このアッセイがナトリウムチャネルの機能を反映し、ベラトリジン固有の効果ではないという証拠が観察された。
ラットのベラトリジンに誘導された足を引っ込める挙動を測定するアッセイを開発した。重要なのは、NaV1.7のないマウス(グローバルノックアウトNaV1.7)の試験の場合は、ベラトリジンの効果がなく、ベラトリジンの効果は、完全にNaV1.7により媒介されるということである。従って、本発明のこの態様は、実験的NaV1.7遮断薬の、マウス、ラット、ウサギ、フェレット、イヌ、非ヒト霊長類、またはヒト等の哺乳類対象での、例えば、実験的VR1阻害剤の生化学的刺激マーカーとして機能するカプサイシン試験に類似の皮膚適用による試験に使われるオンターゲットおよびオンメカニズム生化学的刺激を示す(Chizh、BA et al.、TRPV1受容体媒介活性および炎症性のヒト痛覚過敏症に与えるTRPV1アンタゴニストSB−705498の効果(The effects of the TRPV1 antagonist SB−705498 on TRPV1 receptor−mediated activity and inflammatory hyperalgesia in humans)、Pain 132(1−2):132−141(2007);Gavva NR、Bannon AW、et al.、バニロイド受容体TRPV1アンタゴニストの反復投与がTRPV1遮断により誘発される高熱症を弱める(Repeated administration of vanilloid receptor TRPV1 antagonists attenuates hyper thermia elicited by TRPV1 blockade)、J Pharmacol Exp Ther 323:128−137(2007))。本発明のこの態様は、臨床試験用の適切な投与量の選択および臨床試験結果の解釈のための重要な利点、ならびに最良のナトリウムチャネル薬剤発見のためのツールを示している。
図14は、ベラトリジン(4α、9−エポキシ−3β−ベラトロイルオキシ−5β−セバン−4β、12、14、16β、17、20−ヘキサオール;化学構造は、下記Iに示す)によるNaV1.7のインビトロ調節を示す。図14に示す電流は、減算処理されていない。30μMベラトリジンの添加により、記載したように、ピーク電流が低下し、定常的、持続性内向き電流を生成した。この2つ目の効果は、正に帯電したナトリウムイオンのNaV1.7発現ニューロン中への連続内向き流束を生成し、後で疼痛として感知されるニューロンのスパイキングを生成することが予測できる。
HEK293細胞中で安定に発現したhNaV1.7を通る電流が、ホールセル構成のパッチクランプ法を使って、−100mVの保持電位から10mV間隔の一群の脱分極性電位パルスにより誘導された。
ホールセルパッチクランプ電気生理学。ホールセルパッチクランプ構成を使って室温(約22℃)で細胞を電位固定した。ピペット抵抗は、1.5〜2.0MΩであった。ホールセルキャパシタンスおよび直列抵抗は、補償しなかった。取込の間、電流を5kHzでフィルター(4極ベッセル型)にかけ、pClamp9.2を使って、20kHzでデジタイズした。細胞を培養皿から取り出し、化合物灌流用溶液交換マニホールドに連結されたマイクロピペットの前方に直接置いた。ナトリウム電流をモニターするために、−10mVまでの10msパルスを5秒毎に送出し、デルタメトリンおよびベラトリジンの場合には、外部化合物添加の前後で電流を記録した。あるいは、グラヤノトキシンIIIの場合には、細胞内の(ピペット)溶液中に含めたグラヤノトキシンIIIの電流を記録した。外部溶液は、次の構成とした:140mM NaCl、5.0mM KCl、2.0mM CaCl2、1.0mM MgCl2、10mM HEPES、および11mMブドウ糖、NaOHでpH7.4に調節。 内部溶液は次の構成とした:62.5mM CsCl、75mM CsF、2.5mM MgCl2、5mM EGTA、および10mM HEPES、CsOHでpH7.25に調節。結果を図19A〜Cに示す。これらの図は、デルタメトリンおよびグラヤノトキシンがhNaV1.7を活性化したことを示し、図14は、一群のステップ脱分極に反応して、ベラトリジンがhNaV1.7を活性化したことを示す。
ラット(Sprague−Dawley系、雄(別段の指定がなければ)、8〜10週齢)の足に注入したベラトリジンは、2つの別々の疼痛関連挙動を引き起こした。動物は、足を持ち上げ、足をなめ、また、動物は、足を引っ込めた(図15)。足底内にベラトリジン30μgを注射した。より高い投与量は、疼痛関連挙動に加えて激しい震えを含む不定の神経学的副作用を生じたが、さらに調査はしなかった。足を持ち上げるおよび足を引っ込める挙動は、ベラトリジン注射後20分間記録され、両方の挙動は、ベラトリジン注射の1時間前の経口投与(p.o.)によるナトリウムチャネル遮断薬メキシレチンにより用量依存的に抑制された。
既知の前臨床および臨床鎮痛剤を試験し、それらがベラトリジン誘導挙動を防げるかどうかを調べた。表4は、デュロキセチンおよびモルヒネの両方が、足を引っ込める挙動を回復させたが、足を持ち上げる挙動は回復させなかったことを示す。これは、ベラトリジン誘導反応は、疼痛を反映するが、モルヒネまたはデュロキセチンにより制御されない薬理学的に異なる構成要素を有する可能性があることを示唆している。さらなる鎮痛剤ガバペンチンおよび抗炎症性のナプロキセンは、足を持ち上げる、または足を引っ込める挙動を減らさず、ベラトリジン誘導された疼痛は、異なる経路で進むか、またはあまりに強すぎて、これらの比較的弱い薬剤では、軽減されないことを示唆している。「化合物52」は、インビボ使用に最適化されたアミノトリアジンNaV1.7遮断薬であり、Bregman et al.、ホルマリンモデルの持続性疼痛で経口有効性を有する強力な、状態依存性Nav1.7阻害剤の特定(Identification of a potent、state−dependent inhibitor of Nav1.7 with oral efficacy in the formalin model of persistent pain)、J.Med.Chem.54(13):4427−45、July 14、2011;epub June 2、2011)、に記載されている。この薬剤は、ホルマリンモデルの持続性疼痛に有効である。ジアゼパムは、前臨床で効果的な鎮痛剤ではなく、ベラトリジンモデルでは効果がなかった抗不安薬である。この結果は、観察された足を引っ込める疼痛を示す挙動と再度一致する。
表4.雄ラット(株)の疼痛試験に対する鎮痛剤化合物の効果。
化合物52は、Bregman et al.、ホルマリンモデルの持続性疼痛で経口有効性を有する強力な、状態依存性Nav1.7阻害剤の特定(Identification of a potent、state−dependent inhibitor of Nav1.7 with oral efficacy in the formalin model of persistent pain)、J.Med.Chem.54(13):4427−45、July 14、2011;epub June 2、2011)に記載されている。
化合物52は、Bregman et al.、ホルマリンモデルの持続性疼痛で経口有効性を有する強力な、状態依存性Nav1.7阻害剤の特定(Identification of a potent、state−dependent inhibitor of Nav1.7 with oral efficacy in the formalin model of persistent pain)、J.Med.Chem.54(13):4427−45、July 14、2011;epub June 2、2011)に記載されている。
雄マウス(CD1株、Harlan Laboratories、Inc.、Indianapolis、IN、から入手、35g〜45gの体重)に注射されたベラトリジンは、この場合も用量依存性的に、同等の足を持ち上げる挙動を起こした。図16は、1%エタノールのリン酸塩緩衝食塩水中溶液による表示投与量のベラトリジンのi.pl.注射30分後に記録した、合計足を持ち上げ挙動時間を示す。ベラトリジンの効果は、1μgベラトリジンの刺激の30分前の、30mg/kgの非特異的ナトリウムチャネル阻害剤メキシレチンのi.p.投与により遮断された。
2種の他のナトリウムチャネルモジュレーターを、雄マウス(CD1株、Harlanから入手)に別々に注射し、それぞれが、足を持ち上げる/足をなめる反応を生成した。図17A〜Bに、1%エタノールのリン酸塩緩衝食塩水溶液中のデルタメトリンの10μg懸濁液投与(図17A;n=6)またはグラヤノトキシンIIIの0.1μg投与(図17B;n=6)に反応した足を引っ込める挙動の合計時間を示す。食塩水溶液中のメキシレチンの30mg/kg i.p.事前投与は、デルタメトリンまたはグラヤノトキシンIIIに対する足を持ち上げる/足をなめる反応を抑制した。デルタメトリンで試験したマウスの体重は、30g〜45gで、グラヤノトキシンIIIで試験したマウスの体重は、30g〜45gであった。結果は、ベラトリジン関連痛み挙動は、ベラトリジンのみに特異的ではなく、ナトリウムチャネルに対する機能的影響を共有する3つの非関連分子が、同じ結果を生成したので、むしろ、1つまたは複数のナトリウムチャネルの活性化に特異的であることを支持している。
1つ、いくつか、または9つ全てのナトリウムチャネルアイソフォームの活性化は、痛み挙動を生成する可能性がある。しかし、最良のバイオマーカーは、NaV1.7(Scn9a)の特異的阻害剤を検出するものである。この目的のために、1μgのベラトリジンが、成体グローバルノックアウトNaV1.7マウス(n=5)および野性型ヘテロ接合体同腹子(n=6)の足に注射された。ノックアウトマウスは、誕生以降NaV1.7を欠いている;文献報告に反して、Nav1.7の除去は、必ずしも新生仔の致死に繋がらない。ベラトリジン注射は、野性型ヘテロ接合体マウスでは、強力な足を引っ込める反応を起こしたが、同量、同容量のベラトリジンは、NaV1.7ノックアウトでは反応を生じなかった(図18)。嗅覚および疼痛の欠如を除いて、この実験で使用されたような健康なNaV1.7ノックアウトマウスは、全体の動きに関する開放野外試験を含め、明確な欠陥を示さない。ノックアウトのカラム中に残されている少しの「足を上げる」時間は、マウスが、30分にわたる過程の中で行うであろう、足を持ち上げるまたは足をなめる挙動の正常な長さの時間に帰することができる。全ての挙動実験は、マウス遺伝子型または処理に関し、観察者盲検で行った。野生型ヘテロ接合体マウスのベラトリジン誘導の足を持ち上げる/足をなめる挙動は、また、メキシレチンの薬理学的投与により抑制された。
この実施例5で記載の結果は、ナトリウムチャネル活性化因子が、しっかりした、定量化可能な疼痛反応を生成し、この反応が、Nav1.7の活性化のみを介して媒介され、さらに、この反応が、ナトリウムチャネルの薬理学的抑制に対し感受性が高いことを示す。従って、本明細書に記載の本発明のベラトリジンバイオマーカーアッセイは、NaV1.7に特異的なオンターゲット生化学的刺激アッセイである。
実施例6:抗Nav1.7抗体を生成するためのNaV1.7ノックアウトマウスの使用
図20A〜Bは、本発明のNaV1.7ノックアウトマウスによる抗体生成を示す。NaV1.7ノックアウトマウスは、ヒトNaV1.7を発現している細胞で免疫された。抗体分泌ハイブリドーマ細胞を、マウス脾臓融合体から標準的手法で調製し、ハイブリドーマ細胞を培養し、さらに、上清をハイブリドーマ細胞のそれぞれのウエルから単離した。
図20A〜Bは、本発明のNaV1.7ノックアウトマウスによる抗体生成を示す。NaV1.7ノックアウトマウスは、ヒトNaV1.7を発現している細胞で免疫された。抗体分泌ハイブリドーマ細胞を、マウス脾臓融合体から標準的手法で調製し、ハイブリドーマ細胞を培養し、さらに、上清をハイブリドーマ細胞のそれぞれのウエルから単離した。
図20A〜Bは、代表的ハイブリドーマ上清の抗NaV1.7抗体の存在に関するフローサイトメトリーを使った試験を示す。HEK293細胞(親293細胞またはヒトNaV1.7を安定発現した細胞)を、試験上清および蛍光タグ抗マウス二次抗体と共にインキュベートした。各細胞を、健康な細胞集団でゲーティングしたフローサイトメトリー蛍光標識細胞分取(FACS)を使って、蛍光発光を測定した。抗体結合を測定するために、2x105ヒトNaV1.7発現細胞を1:20希釈IgG含有上清と共に4℃で1時間インキュベートした。非結合抗体を、PBS+2%FBSを使った2回の洗浄により除去した。細胞を、1μg/mLのフルオレセインイソチオシアナート(FITC)標識ヤギF(ab’)2抗マウスIgG二次抗体(Southern Biotech 1032−02)と共に、4℃で45分間、インキュベートした。PBS+2%FBSで2回洗浄後、細胞を0.5μg/mLのヨウ化プロピジウム溶液(Invitrogen P3566)中に再懸濁し、健康な細胞集団をゲーティングするフローサイトメトリー用BD−FACS Caliber(登録商標)選別装置に供した。
蛍光発光強度(下記プロットのx−軸;各点は単一細胞由来のデータを表す)は、293細胞発現hNav1.7の平均値(図20A)の方が、親HEK293細胞(図20B)からの平均値より大きかった。解釈は、ノックアウトマウスは抗体を産生し、これらの抗体は、hNaV1.7を標的としていた、ということである(Y−軸は、標識マーカーを反映していない)。
Claims (36)
- 生存可能なグローバルNaV1.7−/−ノックアウトマウス。
- グローバルNaV1.7−/−ノックアウトマウスが成体である、請求項1に記載のグローバルNaV1.7−/−ノックアウトマウス。
- 異系交配または戻し交配されたグローバルNaV1.7−/−ノックアウトマウス、もしくはそれら由来のNaV1.7−/−子孫マウスである、請求項1に記載のグローバルNaV1.7−/−ノックアウトマウス。
- 繁殖性である、請求項2に記載のグローバルNaV1.7−/−ノックアウトマウス。
- 雄である、請求項1に記載のグローバルNaV1.7−/−ノックアウトマウス。
- 雌である、請求項1に記載のグローバルNaV1.7−/−ノックアウトマウス。
- CD1マウス由来である、請求項1に記載のグローバルNaV1.7−/−ノックアウトマウス。
- BALB/cマウス由来である、請求項1に記載のグローバルNaV1.7−/−ノックアウトマウス。
- 請求項4に記載のNaV1.7−/−ノックアウトマウスにより産生された、機能性NaV1.7タンパク質をコードしない単離マウス配偶子。
- 雄配偶子である、請求項9に記載の単離マウス配偶子。
- 雌配偶子である、請求項9に記載の単離マウス配偶子。
- 請求項1のグローバルNaV1.7−/−ノックアウトマウスから単離された単離NaV1.7−/−マウス細胞、またはその子孫細胞。
- 請求項1のグローバルNaV1.7−/−ノックアウトマウス由来の初代細胞培養または二次細胞株。
- 請求項1のグローバルNaV1.7−/−ノックアウトマウス由来の組織または臓器外植片またはそれらの培養物。
- B−リンパ球、T細胞、または神経細胞である、請求項12に記載の単離マウス細胞、またはその子孫。
- 最初は、請求項15に記載のマウス細胞および骨髄腫細胞の融合により形成されたハイブリドーマ。
- 請求項4に記載のグローバルNaV1.7−/−ノックアウトマウスの少なくとも1つの繁殖ペアを含むグローバルNaV1.7−/−ノックアウトマウスの繁殖コロニー。
- グローバルNaV1.7−/−ノックアウトマウスの少なくとも1つの繁殖ペアがCD1背景を含む、請求項17に記載の繁殖コロニー。
- グローバルNaV1.7−/−ノックアウトマウスの少なくとも1つの繁殖ペアがBALB/c背景を含む、請求項17に記載の繁殖コロニー。
- 成体グローバルNaV1.7−/−ノックアウトマウスを生成する方法であって、
(a)生存可能な新生仔または周産期グローバルNaV1.7−/−ノックアウトマウスの子を得ることと、および
(b)その子が成人期に達するまで適切な栄養を与えることと、
を含む方法。 - 適切な栄養を与えることが、その子に手で餌を与えることを含む、請求項20に記載の方法。
- NaV1.7タンパク質で免疫された請求項1に記載のグローバルNaV1.7−/−ノックアウトマウスにより産生された、NaV1.7に対するマウス抗体をヒト化することを含む、抗体の産生方法。
- 請求項3に記載のグローバルNaV1.7−/−ノックアウトマウス由来の子孫であり、その遺伝子型がNaV1.7+/−である生存可能なマウス。
- (a)哺乳動物に試験化合物を投与することと、続けて、
(b)哺乳動物に、陰性対照に疼痛関連反応を誘導するのに有効な投与量のNaV1.7活性化因子を投与することと、および、次いで
(c)哺乳動物の疼痛関連反応が、陰性対照に比べて、減少するかどうかを判定すること、
を含むアッセイ。 - NaV1.7活性化因子が、ベラトリジン、デルタメトリン、またはグラヤノトキシンIIIである、請求項24に記載のアッセイ。
- 哺乳動物が、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒトである、請求項24に記載のアッセイ。
- 疼痛関連反応が、足を持ち上げること、足をなめること、足を引っ込めること、啼声、自己報告、またはこれらの反応のいずれかの組み合わせである、請求項24に記載のアッセイ。
- (a) 第1の哺乳動物に、第1の投与量の試験化合物を投与することと、続けて、
(b) 第1の哺乳動物に、陰性対照に疼痛関連反応を誘導するのに有効な投与量のNaV1.7活性化因子を投与することと、および、次いで、
(c) 陰性対照に比べて、疼痛関連反応が第1の哺乳動物で減少するかどうかを判定することと、ならびに
(d) 陰性対照に比べて、疼痛関連反応が減らされる、試験化合物の、最も低い第2の投与量を決定することと、
を含むアッセイ。 - NaV1.7活性化因子が、ベラトリジン、デルタメトリン、またはグラヤノトキシンIIIである、請求項28に記載のアッセイ。
- 哺乳動物が、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒトである、請求項28に記載のアッセイ。
- 疼痛関連反応が、足を持ち上げる、足をなめる、足を振り回す、啼声、自己報告、またはこれらの反応のいずれかの組み合わせである、請求項28に記載のアッセイ。
- (e)第2の同じ種の哺乳動物に、第2の投与量の試験化合物を投与することと、続けて、第2の哺乳動物に、陰性対照に疼痛関連反応を誘導するのに有効な投与量のNaV1.7活性化因子を投与することと、および、次いで、
(f)第2の哺乳動物の疼痛関連反応が、陰性対照に比べて、減少するかどうかを判定すること、
をさらに含む、請求項28に記載のアッセイ。 - (a)第1の哺乳動物に、第1の投与量の試験化合物および第2の同じ種の哺乳動物に、第1の投与量とは異なる第2の投与量の試験化合物を投与することと、続けて、
(b)第1のおよび第2の哺乳動物に、陰性対照で疼痛関連反応を誘導するのに有効な局所投与量のNaV1.7活性化因子を投与することと、および、次いで、
(c)疼痛関連反応が、陰性対照に比べて、第1の哺乳動物および第2の哺乳動物で減少するかどうかを判定することと、ならびに、
(d)陰性対照に比べて、疼痛関連反応が減らされる、試験化合物の、最も低い第2の投与量を特定することと、
を含むアッセイ。 - NaV1.7活性化因子が、ベラトリジン、デルタメトリン、またはグラヤノトキシンIIIである、請求項33に記載のアッセイ。
- 哺乳動物が、マウス、ラット、ウサギ、ケナガイタチ、イヌ、非ヒト霊長類、またはヒトである、請求項33に記載のアッセイ。
- 疼痛関連反応が、足を持ち上げること、足をなめること、足を引っ込めること、啼声、自己報告、またはこれらの反応のいずれかの組み合わせである、請求項33に記載のアッセイ。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201161433910P | 2011-01-18 | 2011-01-18 | |
| US61/433,910 | 2011-01-18 | ||
| US201161557877P | 2011-11-09 | 2011-11-09 | |
| US61/557,877 | 2011-11-09 | ||
| PCT/US2012/021757 WO2012099983A1 (en) | 2011-01-18 | 2012-01-18 | Nav1.7 knockout mice and uses thereof |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2014507136A true JP2014507136A (ja) | 2014-03-27 |
Family
ID=45561108
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013550571A Pending JP2014507136A (ja) | 2011-01-18 | 2012-01-18 | NaV1.7ノックアウトマウスとそれらの使用 |
Country Status (13)
| Country | Link |
|---|---|
| US (1) | US20120185956A1 (ja) |
| EP (1) | EP2665822A1 (ja) |
| JP (1) | JP2014507136A (ja) |
| KR (1) | KR20140009311A (ja) |
| CN (1) | CN103492575A (ja) |
| AU (1) | AU2012207366A1 (ja) |
| CA (1) | CA2823707A1 (ja) |
| CL (1) | CL2013002068A1 (ja) |
| EA (1) | EA201370161A1 (ja) |
| MX (1) | MX2013008282A (ja) |
| SG (1) | SG191993A1 (ja) |
| WO (1) | WO2012099983A1 (ja) |
| ZA (1) | ZA201305129B (ja) |
Families Citing this family (25)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US8871996B2 (en) | 2010-06-09 | 2014-10-28 | Regeneron Pharmaceuticals, Inc. | Mice expressing human voltage-gated sodium channels |
| US8486647B2 (en) | 2010-06-09 | 2013-07-16 | Regeneron Pharmaceuticals, Inc. | Neuropeptide release assay for sodium channels |
| EP3973981A1 (en) | 2012-05-18 | 2022-03-30 | Janssen Biotech, Inc. | Huwentoxin-iv variants and methods of use |
| US10414808B2 (en) | 2012-05-18 | 2019-09-17 | Janssen Biotech, Inc. | Huwentoxin-IV variants and methods of use |
| WO2014151472A1 (en) | 2013-03-15 | 2014-09-25 | Chromocell Corporation | Sodium channel modulators for the treatment of pain |
| FR3007763A1 (fr) * | 2013-06-28 | 2015-01-02 | Galderma Res & Dev | Anticorps diriges contre le canal sodique nav1.9 humain et leurs utilisations en diagnostic |
| FR3007656B1 (fr) * | 2013-06-28 | 2015-06-26 | Galderma Res & Dev | Modulateurs du canal sodique nav1.9 pour le traitement d'une maladie cutanee inflammatoire et methodes diagnostiques |
| FR3007762B1 (fr) * | 2013-06-28 | 2015-09-04 | Galderma Res & Dev | Anticorps diriges contre le canal sodique nav1.9 humain et leurs utilisations pour le diagnostic des maladies cutanees inflammatoires |
| CN105611923B (zh) | 2013-09-10 | 2019-08-23 | 卓莫赛尔公司 | 用于治疗疼痛和糖尿病的钠通道调节剂 |
| EP3418379B1 (en) | 2013-09-18 | 2020-12-09 | Kymab Limited | Methods, cells & organisms |
| BR112016007062A2 (pt) | 2013-10-03 | 2017-09-12 | Janssen Biotech Inc | variantes de protoxina-ii e métodos de uso |
| US9067998B1 (en) | 2014-07-15 | 2015-06-30 | Kymab Limited | Targeting PD-1 variants for treatment of cancer |
| US9914769B2 (en) | 2014-07-15 | 2018-03-13 | Kymab Limited | Precision medicine for cholesterol treatment |
| US8992927B1 (en) | 2014-07-15 | 2015-03-31 | Kymab Limited | Targeting human NAV1.7 variants for treatment of pain |
| US9045545B1 (en) | 2014-07-15 | 2015-06-02 | Kymab Limited | Precision medicine by targeting PD-L1 variants for treatment of cancer |
| US8986694B1 (en) | 2014-07-15 | 2015-03-24 | Kymab Limited | Targeting human nav1.7 variants for treatment of pain |
| GB201403775D0 (en) | 2014-03-04 | 2014-04-16 | Kymab Ltd | Antibodies, uses & methods |
| EP2918166A1 (en) | 2014-03-10 | 2015-09-16 | Westfälische Wilhelms-Universität Münster | TTP/MRP14 double knock out mouse model of psoriasis |
| US9139648B1 (en) | 2014-07-15 | 2015-09-22 | Kymab Limited | Precision medicine by targeting human NAV1.9 variants for treatment of pain |
| MA41642A (fr) | 2015-03-03 | 2018-01-09 | Janssen Biotech Inc | Variants de protoxine ii et méthodes d'utilisation |
| EP3277304B1 (en) | 2015-04-02 | 2021-08-04 | Janssen Biotech, Inc. | Protoxin-ii variants and methods of use |
| CN105684993B (zh) * | 2016-02-01 | 2018-06-01 | 江苏大学 | GABAergic神经元条件性敲除基因PGC-1α小鼠的制备 |
| TWI640536B (zh) | 2016-06-20 | 2018-11-11 | 克馬伯有限公司 | 抗體 |
| WO2018083248A1 (en) | 2016-11-03 | 2018-05-11 | Kymab Limited | Antibodies, combinations comprising antibodies, biomarkers, uses & methods |
| CN111560076B (zh) * | 2020-05-15 | 2021-05-07 | 广州百暨基因科技有限公司 | 一种嵌合抗原受体免疫细胞及其制备方法和应用 |
Family Cites Families (40)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4560655A (en) | 1982-12-16 | 1985-12-24 | Immunex Corporation | Serum-free cell culture medium and process for making same |
| US4657866A (en) | 1982-12-21 | 1987-04-14 | Sudhir Kumar | Serum-free, synthetic, completely chemically defined tissue culture media |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| US4767704A (en) | 1983-10-07 | 1988-08-30 | Columbia University In The City Of New York | Protein-free culture medium |
| GB8516415D0 (en) | 1985-06-28 | 1985-07-31 | Celltech Ltd | Culture of animal cells |
| US4927762A (en) | 1986-04-01 | 1990-05-22 | Cell Enterprises, Inc. | Cell culture medium with antioxidant |
| US5567610A (en) | 1986-09-04 | 1996-10-22 | Bioinvent International Ab | Method of producing human monoclonal antibodies and kit therefor |
| ATE135045T1 (de) | 1988-07-23 | 1996-03-15 | Delta Biotechnology Ltd | Sekretorische leader-sequenzen |
| WO1990003430A1 (en) | 1988-09-23 | 1990-04-05 | Cetus Corporation | Cell culture medium for enhanced cell growth, culture longevity and product expression |
| GB8823869D0 (en) | 1988-10-12 | 1988-11-16 | Medical Res Council | Production of antibodies |
| US5175384A (en) | 1988-12-05 | 1992-12-29 | Genpharm International | Transgenic mice depleted in mature t-cells and methods for making transgenic mice |
| SG48759A1 (en) | 1990-01-12 | 2002-07-23 | Abgenix Inc | Generation of xenogenic antibodies |
| US5229275A (en) | 1990-04-26 | 1993-07-20 | Akzo N.V. | In-vitro method for producing antigen-specific human monoclonal antibodies |
| US5427908A (en) | 1990-05-01 | 1995-06-27 | Affymax Technologies N.V. | Recombinant library screening methods |
| GB9014932D0 (en) | 1990-07-05 | 1990-08-22 | Celltech Ltd | Recombinant dna product and method |
| GB9015198D0 (en) | 1990-07-10 | 1990-08-29 | Brien Caroline J O | Binding substance |
| US5122469A (en) | 1990-10-03 | 1992-06-16 | Genentech, Inc. | Method for culturing Chinese hamster ovary cells to improve production of recombinant proteins |
| GB9022543D0 (en) | 1990-10-17 | 1990-11-28 | Wellcome Found | Antibody production |
| ES2136092T3 (es) | 1991-09-23 | 1999-11-16 | Medical Res Council | Procedimientos para la produccion de anticuerpos humanizados. |
| ATE244763T1 (de) | 1992-02-11 | 2003-07-15 | Cell Genesys Inc | Erzielen von homozygotem durch zielgerichtete genetische ereignisse |
| US5573905A (en) | 1992-03-30 | 1996-11-12 | The Scripps Research Institute | Encoded combinatorial chemical libraries |
| AU675661B2 (en) | 1992-07-24 | 1997-02-13 | Abgenix, Inc. | Generation of xenogeneic antibodies |
| US6054287A (en) | 1994-05-27 | 2000-04-25 | Methodist Hospital Of Indiana, Inc. | Cell-type-specific methods and devices for the low temperature preservation of the cells of an animal species |
| US6130364A (en) | 1995-03-29 | 2000-10-10 | Abgenix, Inc. | Production of antibodies using Cre-mediated site-specific recombination |
| EP0822830B1 (en) | 1995-04-27 | 2008-04-02 | Amgen Fremont Inc. | Human anti-IL-8 antibodies, derived from immunized xenomice |
| GB9712818D0 (en) | 1996-07-08 | 1997-08-20 | Cambridge Antibody Tech | Labelling and selection of specific binding molecules |
| PT1500329E (pt) | 1996-12-03 | 2012-06-18 | Amgen Fremont Inc | Anticorpos humanos que se ligam especificamente ao tnf alfa humano |
| US20020032315A1 (en) | 1997-08-06 | 2002-03-14 | Manuel Baca | Anti-vegf antibodies |
| US20030044772A1 (en) | 1997-08-04 | 2003-03-06 | Applied Molecular Evolution [Formerly Ixsys] | Methods for identifying ligand specific binding molecules |
| US6022952A (en) | 1998-04-01 | 2000-02-08 | University Of Alberta | Compositions and methods for protein secretion |
| US20020029391A1 (en) | 1998-04-15 | 2002-03-07 | Claude Geoffrey Davis | Epitope-driven human antibody production and gene expression profiling |
| ATE429504T1 (de) | 1998-11-20 | 2009-05-15 | Fuso Pharmaceutical Ind | Proteinexpressionsvektor und benutzung desselbigen |
| CA2368068A1 (en) | 1999-03-18 | 2000-09-21 | Steven M. Ruben | 27 human secreted proteins |
| CN101940189A (zh) | 2000-11-30 | 2011-01-12 | 米德列斯公司 | 用于生产人类抗体的转基因转染色体啮齿动物 |
| ATE548358T1 (de) | 2003-08-05 | 2012-03-15 | Vertex Pharma | Tetrahydrochinazolin-verbindungen als inhibitoren von spannungsabhängigen ionenkanälen |
| GB0512214D0 (en) * | 2005-06-15 | 2005-07-27 | Capsant Neurotechnologies Ltd | Method |
| TW200722436A (en) | 2005-10-21 | 2007-06-16 | Hoffmann La Roche | A peptide-immunoglobulin-conjugate |
| WO2007109324A2 (en) | 2006-03-21 | 2007-09-27 | Xenon Pharmaceuticals, Inc. | Potent and selective nav 1.7 sodium channel blockers |
| US8183221B2 (en) | 2007-09-05 | 2012-05-22 | Medtronic, Inc. | Suppression of SCN9A gene expression and/or function for the treatment of pain |
| MA32965B1 (fr) | 2009-01-12 | 2012-01-02 | Pfizer Ltd | Derives de sulfonamides |
-
2012
- 2012-01-18 JP JP2013550571A patent/JP2014507136A/ja active Pending
- 2012-01-18 WO PCT/US2012/021757 patent/WO2012099983A1/en active Application Filing
- 2012-01-18 SG SG2013054382A patent/SG191993A1/en unknown
- 2012-01-18 MX MX2013008282A patent/MX2013008282A/es unknown
- 2012-01-18 EA EA201370161A patent/EA201370161A1/ru unknown
- 2012-01-18 CA CA2823707A patent/CA2823707A1/en not_active Abandoned
- 2012-01-18 CN CN201280009502.XA patent/CN103492575A/zh active Pending
- 2012-01-18 EP EP12702103.8A patent/EP2665822A1/en not_active Withdrawn
- 2012-01-18 AU AU2012207366A patent/AU2012207366A1/en not_active Abandoned
- 2012-01-18 US US13/353,198 patent/US20120185956A1/en not_active Abandoned
- 2012-01-18 KR KR1020137021599A patent/KR20140009311A/ko not_active Application Discontinuation
-
2013
- 2013-07-08 ZA ZA2013/05129A patent/ZA201305129B/en unknown
- 2013-07-18 CL CL2013002068A patent/CL2013002068A1/es unknown
Also Published As
| Publication number | Publication date |
|---|---|
| SG191993A1 (en) | 2013-08-30 |
| EA201370161A1 (ru) | 2013-12-30 |
| AU2012207366A1 (en) | 2013-07-11 |
| US20120185956A1 (en) | 2012-07-19 |
| EP2665822A1 (en) | 2013-11-27 |
| CL2013002068A1 (es) | 2014-05-16 |
| CN103492575A (zh) | 2014-01-01 |
| MX2013008282A (es) | 2014-02-10 |
| KR20140009311A (ko) | 2014-01-22 |
| CA2823707A1 (en) | 2012-07-26 |
| ZA201305129B (en) | 2014-08-27 |
| WO2012099983A1 (en) | 2012-07-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2014507136A (ja) | NaV1.7ノックアウトマウスとそれらの使用 | |
| US20130115171A1 (en) | Nav1.7-related assays | |
| EP1573314B1 (en) | Transgenic mice expressing human cd20 | |
| US11154041B2 (en) | Genetically modified non-human animal with human or chimeric genes | |
| JP2021118745A (ja) | C9orf72遺伝子座における破壊を有する非ヒト動物 | |
| US11470827B2 (en) | Transgenic mice expressing human TREM proteins and methods of use thereof | |
| JP2022048157A (ja) | 上位及び下位運動ニューロン機能並びに知覚の減衰を示す非ヒト動物 | |
| JP4857450B2 (ja) | ヒト関節リウマチの病態を再現するトランスジェニック非ヒト哺乳動物 | |
| US9063123B2 (en) | Transgenic mice expressing human toll-like receptor 8 | |
| JPH10201396A (ja) | H2−m改変トランスジエニツク動物 | |
| JP2009511050A (ja) | バイオルミネセンス撮像のためのトランスジェニックrosa26−ルシフェラーゼマウス | |
| CA2471432C (en) | Nonhuman model animal lacking the ability to control lymphocyte migration | |
| US20050044581A1 (en) | Animals and cells containing a mutated alpha2/omega1 gene | |
| WO2006085673A1 (ja) | 無毛トランスジェニック動物 | |
| JP2004533802A (ja) | ヒトfadプレセニリン突然変異をもつ標的遺伝子組換え非ヒト哺乳動物および生殖による子孫 | |
| JP2015021853A (ja) | 精神症状モデル動物を用いた医薬品のスクリーニング | |
| JP2010527589A (ja) | エフリン受容体A6(EphA6)遺伝子の破壊と関連している記憶と学習の障害 | |
| Hitz | The role of MAP-Kinases in anxiety disorders and depression-studies with knockout and knockdown mouse models | |
| WO2002086131A1 (fr) | Unite de regulation de l'expression genetique et utilisation de cette unite | |
| Hitz | The role of MAP-Kinases in anxiety and depression: studies with knockout and knockdown mouse models | |
| JP2003009888A (ja) | 遺伝子発現制御ユニットおよびその利用 | |
| JP2014171413A (ja) | トランスジェニック非ヒト哺乳動物 |