CN1468312A - 嵌合GFP-水母发光蛋白作为单细胞水平的生物发光的Ca2+报道分子 - Google Patents
嵌合GFP-水母发光蛋白作为单细胞水平的生物发光的Ca2+报道分子 Download PDFInfo
- Publication number
- CN1468312A CN1468312A CNA018121047A CN01812104A CN1468312A CN 1468312 A CN1468312 A CN 1468312A CN A018121047 A CNA018121047 A CN A018121047A CN 01812104 A CN01812104 A CN 01812104A CN 1468312 A CN1468312 A CN 1468312A
- Authority
- CN
- China
- Prior art keywords
- seq
- ggc
- gly
- aequorin
- aag
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images











Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/06—Linear peptides containing only normal peptide links having 5 to 11 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K49/00—Preparations for testing in vivo
- A61K49/001—Preparation for luminescence or biological staining
- A61K49/0013—Luminescence
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/001—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof by chemical synthesis
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/43504—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates
- C07K14/43595—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from invertebrates from coelenteratae, e.g. medusae
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/08—Linear peptides containing only normal peptide links having 12 to 20 amino acids
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y10—TECHNICAL SUBJECTS COVERED BY FORMER USPC
- Y10S—TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y10S435/00—Chemistry: molecular biology and microbiology
- Y10S435/968—High energy substrates, e.g. fluorescent, chemiluminescent, radioactive
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y10—TECHNICAL SUBJECTS COVERED BY FORMER USPC
- Y10S—TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y10S930/00—Peptide or protein sequence
- Y10S930/01—Peptide or protein sequence
- Y10S930/31—Linker sequence
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Genetics & Genomics (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Gastroenterology & Hepatology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Public Health (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Toxicology (AREA)
- Zoology (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Epidemiology (AREA)
- Animal Behavior & Ethology (AREA)
- General Chemical & Material Sciences (AREA)
- Veterinary Medicine (AREA)
- Peptides Or Proteins (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Investigating Or Analysing Materials By The Use Of Chemical Reactions (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
Abstract
本发明涉及含共价连接光蛋白的荧光分子的经修饰的生物发光系统,其中两种蛋白质之间的所述连接具有稳定该经修饰的生物发光系统,且容许通过化学发光共振能量转移(CRET)进行能量转移的功能。
Description
相关申请的相互参照
本申请以2000年6月1日提交的美国临时申请第60/208,314号(代理人摘要编号03495.6051)、2000年6月6日提交的60/210,526(代理人摘要编号03495.6052)、和2000年12月14日提交的60/255,111(代理人摘要编号03495.6059)为基础,并要求它们的权益。本文依赖并收录这些申请的完整公开书作为参考。
发明背景
本发明涉及含共价连接光蛋白的荧光分子的经修饰的生物发光系统,所述光蛋白容许通过化学发光共振能量转移(CRET)进行能量转移。本发明还涉及经修饰的生物发光系统在体内和体外实验中的用途。
钙参与极其多种胞内过程的调控(1)。有几种技术最常用于胞内Ca2+监测。膜片钳和Ca2+选择性微电极能够在有限数目的细胞内进行Ca2+流累积测量。另一方面,可以通过荧光探针显现大量细胞的胞内Ca2+动力学(2)。遗传工具将为Ca2+监测提供新方法。
目前可以获得两组遗传Ca2+探针。第一类使用绿色荧光蛋白(GFP)的两种变体之间的荧光共振能量转移(FRET)原理。仅仅通过钙调蛋白结合序列或联合钙调蛋白使两种GFP共价相连,从而应答于Ca2+流发生(3)或不(4)发生分子内FRET。第二类包含生物发光蛋白质,诸如水母发光蛋白(5,6)。在存在氧分子时,阿朴水母发光蛋白(189个氨基酸)及其萤光素即腔肠吖嗪(coelenterazine)(Mr 423)形成有活性的蛋白质(7)。
Ca2+与水母发光蛋白(它具有三个Ca2+结合位点特征性的EF手结构域)的结合诱导构象变化,导致腔肠吖嗪经分子内反应而发生氧化。此外,由此生成的腔肠酰胺(coelenteramide)处于激发态,并在返回其基态时发出蓝光(最大:470nm)(8)。这种生物发光遗传标记展示超越Ca2+敏感性荧光染料的优势,即易于使用适当的调控元件和肽信号将其靶向特定的细胞和亚细胞区室(9)。生物发光过程不像荧光探针或蛋白质那样需要光激发,因而不诱导自发荧光、光漂白和生物降解问题。此外,水母发光蛋白无毒,不结合其它二价阳离子,而且甚至在进行高浓度显微注射时也不干扰[Ca2+]i缓冲系统。它与Ca2+的低亲和力(Kd=10μM)可能是其原因,并使得水母发光蛋白成为生物学Ca2+变化范围内的优良传感器。
尽管提供优良的信噪比,然而水母发光蛋白的信号因其低光量子产量(即每个结合Ca2+的蛋白质发射的光子数目)而非常难于检测。在用于分离水母发光蛋白的水母(Aequorea victoria)中(10),该蛋白质与GFP相关(11)。在Ca2+结合后,水母发光蛋白获得的能量由激活的氧合萤光素转移至GFP,而不发出蓝光。GFP受体荧光团通过无辐射能量转移由氧合腔肠吖嗪激发。然后,在激发态GFP返回其基态时发出绿光(最大:509nm)(12)。
这种分子间无辐射能量转移在生物发光中是常见的,而且早已显示在肾海鳃(Renilla,另一种腔肠动物)中提高生物发光过程的量子产量(13)。在体外测量得到的增益为3-5倍(14)。因为萤光素与绿色荧光蛋白结合到一起,所以有可能使用其低等摩尔浓度的成分在体外重建肾海鳃系统并获得光谱移动(14)。
在多管水母系统中,在溶液中不发生纯化的光蛋白与GFP之间的结合,甚至以高浓度存在时(15)。在体内,因高浓度的GFP而发生能量转移。在体外可以通过水母发光蛋白与GFP在DEAE纤维素膜上的共吸附来获得它(15)。Forster方程显示这种过程的效率依赖在FRET的情况中描述的几种条件。供体的放射谱与受体的激发谱必须具有最大交叠。能量转移还强烈依赖几何学,具体而言指两个偶极的相对取向和距离,并受到它们各自运动的调制(16)。
本发明的目的是开发联合水母发光蛋白的Ca2+敏感性与GFP的荧光特性的双重报道基因。可以通过经典表面荧光进行检测的融合蛋白可用于监测钙活动。本发明分子的构型提高了它们的整体周转,并容许有效的分子内化学发光共振能量转移(CRET)。结果,水母发光蛋白的量子产量显得提高了。本发明显示可以用加强型CCD相机在单个真核细胞中显现生理学钙信号。本文描述的其它构建物将融合蛋白靶向神经突膜。
发明概述
因而,本发明提供包含共价连接光蛋白的荧光分子的经修饰的生物发光系统,其中两种蛋白质之间的连接具有稳定经修饰的生物发光系统的功能,且容许通过化学发光共振能量转移(CRET)进行能量转移。在优选的实施方案中,生物发光系统包含共价连接水母发光蛋白的GFP蛋白,其中两种蛋白质之间的连接具有稳定该经修饰的生物发光系统的功能,且容许通过化学发光共振能量转移(CRET)进行能量转移。
在根据本发明的经修饰的生物发光系统的一个实施方案中,生物发光系统包含共价连接水母发光蛋白的GFP蛋白,其中两种蛋白质之间的连接由至少5个氨基酸、并可选至少5个氨基酸和至少一个拷贝的9个氨基酸构成。该连接具有稳定系统的功能,且容许通过化学发光共振能量转移(CRET)进行能量转移。
在优选的实施方案中,生物发光系统包含共价连接水母发光蛋白的GFP蛋白,其中两种蛋白质之间的连接优选由至少5个氨基酸和五个拷贝的9个氨基酸构成,并具有稳定系统的功能,且容许通过化学发光共振能量转移(CRET)进行能量转移。
两种蛋白质可以分开或一起起作用。另外,经修饰生物发光系统可以是对钙敏感的和/或对光敏感的。
本发明还提供了在体外筛选物理、化学、生化或生物学条件变化的方法,包括:
a)在不同样品中,在包含目的分析物的反应系统中提供根据本发明的生物发光系统;
b)量是否发光;并
c)根据发光检测变化。
此外,本发明提供在体内筛选物理、化学、生化或生物学条件变化的方法,包括:
a)对哺乳动物施用包含根据本发明的生物发光系统的可接受的组合物;
b)检测是否发光;并
c)可选的测量钙流的离子浓度。
另外,本发明提供包含纯化的多肽的组合物,其中组合物具有结合钙离子和容许可测量能量的功能特征,所述能量依赖在不存在任何光激发时在所述组合物中所结合的钙的数量和多肽的数量。
另外,本发明提供具有SEQ ID NO:1、SEQ ID NO:2、SEQ ID NO:3、SEQ ID NO:4、SEQ ID NO:5和SEQ ID NO:6的氨基酸序列的纯化的多肽。
在其它实施方案中,本发明提供具有SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11和SEQ ID NO:12的序列的多核苷酸。
本发明还提供保藏于C.N.C.M.且包含质粒号I-2507、质粒号I-2508、质粒号I-2509、质粒号I-2510、质粒号I-2511、质粒号I-2512或质粒号I-2513的培养物。
此外,本发明提供了肽接头,它在翻译后具有在最佳条件下使供体位点接近受体位点的功能,从而容许根据本发明的纯化多肽中通过化学发光的直接转移能量。核苷酸接头可以具有例如SEQ ID NO:13、SEQ ID NO:14、SEQ ID NO:15、SEQ ID NO:16或SEQ ID NO:17的核苷酸序列。肽接头可以包含至少5个氨基酸,且包含SEQ ID NO:18、SEQ ID NO:19、SEQ ID NO:20、SEQ ID NO:21或SEQ ID NO:22的氨基酸序列。
用于在体内或在体外测量能量转移的试剂盒包含至少一种根据本发明的多肽或根据本发明的多核苷酸,以及在存在或缺乏目的分子时显现或检测所述转移所必需的试剂。
在另一个实施方案中,本发明提供了具有如下通式的融合蛋白:
GFP-接头-AEQ,
其中GFP指绿色荧光蛋白;AEQ指水母发光蛋白;而接头指4-63个、优选14-50个氨基酸的多肽。接头可以包含如下氨基酸:
(Gly Gly Ser Gly Ser Gly Gly Gln Ser[SEQ ID NO:25])n,
其中n指1-5,优选为1或5。接头还可以包含氨基酸序列Ser GlyLeu Arg Ser[SEQ ID NO:26]。
用于存在Ca2+时在水母发光蛋白活化后通过化学发光共振能量转移(CRET)将能量由水母发光蛋白转移至绿色荧光蛋白的另一种融合蛋白具有如下通式:
GFP-接头-AEQ,
其中GFP指绿色荧光蛋白;AEQ指水母发光蛋白;而接头包含如下氨基酸:
(Gly Gly Ser Gly Ser Gly Gly Gln Ser[SEQ ID NO:25])n,
其中n指1-5;且其中融合蛋白具有Ca2+离子亲合力和至少24小时的半衰期。接头可以包含氨基酸序列Ser Gly Leu Arg Ser[SEQ IDNO:26]。另外,融合蛋白还可以包含将其靶向细胞或亚细胞区室的肽信号序列。
本发明还提供了编码上文所述融合蛋白的多核苷酸。
附图简述
下文将参照附图来描述本发明,其中:
图1是不同构建物的示意图。所有构建物都处于人类细胞肥大病毒启动子(PCMV)的控制之下。星号指示Val-163-Ala突变的位置。在pGA中,GFP与水母发光蛋白的编码序列由5个密码子分开。在pGiA中加入了1-5个接头(括弧中),其中i指接头的数目。设置接头的方向使之编码9个氨基酸的重复片段。将完整的突触结合蛋白1或其跨膜部分(tSyn)以符合读码框的方式融合G5A。
图2描绘了细胞提取物的Ca2+CRET活性。将水母发光蛋白和几种GFP-水母发光蛋白融合蛋白的发射光谱校准为最大强度的百分比。将CRET测量表述成绿色(500nm)对蓝色(450nm)光子的比率。
图3描绘了经pGm(A)、pGA(B)、pG2A(C)、和pG5A(D)转染的Neuro2A细胞中GFP-阿朴水母发光蛋白的GFP荧光。显示了表达pSG5A(E)或pStG5A(F)的细胞中GFP荧光的共焦重叠和突触结合蛋白的免疫染色。
图4描绘了在单细胞水平检测到的由Ca2+诱导的生物发光。将经pGA(A.1-4)或pSG5A(B)转染的Neuro2A细胞与5μM腔肠吖嗪一起在不含Ca2+的缓冲液中预保温。(A.3)GFP荧光使之有可能选择转染细胞。在加入CaCl2前记录的背景(A.2)对应于实验时间零点的相对光单位(RLU)水平(A.4,B)。将荧光强度和生物发光活性转化成标示的伪彩色。显示加入5mM CaCl2和5μM A23187后分别在开始采集后13秒钟和159秒钟时选定视野的代表性照片(A.1)。(A.4)每张图谱都指示单细胞的发光强度。
通过环绕个别细胞体定义了五个区域。对于pGA(数据未显示)或pSG5A(B)转染,在实验结束时(500秒钟)加入高浓度的CaCl2(100mM)以检查生物发光蛋白质是否仍有活性。(C)使用Fluo-3 AM在模拟转染的Neuro2A细胞上进行对照实验。
图5描绘了各种融合蛋白的蛋白质稳定性分析的结果。
图6描绘了水母发光蛋白和融合蛋白G5A的Ca2+亲合力测定的结果。
图7描绘了G5A、SG5A和水母发光蛋白的生物发光活性与Ca2+之间的校准曲线。
图8显示了在经腺病毒-G5A载体感染的培养物中游离神经元的荧光和由Ca2+诱导的生物发光活性。
图9显示了在经腺病毒-SG5A载体感染的培养物中游离神经元的荧光和由Ca2+诱导的生物发光活性。
图10显示了在爪蟾(Xenopus)胚的单细胞阶段注入GA质粒后发光活性的代表性图谱。
图11显示了具有GFP-水母发光蛋白的转基因爪蟾幼体。
发明详述
在腔肠动物中存在生物发光物种。大量研究显示生物发光是由对钙敏感的光蛋白产生的。这些蛋白质应答钙离子浓度的升高而发出闪光。在这些光蛋白中,水母发光蛋白是研究得最透彻的蛋白质之一(Blinks等人,1976)。
在水母Aequoria Victoria(Shimomura等人,1962)中分离得到的水母发光蛋白在结合三个钙离子后发出一闪蓝光,光谱最大波长470nm。与经典的萤光素酶-萤光素反应相反,发光不需要氧,而且光的总量与蛋白质的量成正比。然而,氧是通过阿朴水母发光蛋白(分子量21kDa)和腔肠吖嗪的作用而重建水母发光蛋白所必需的。光子发射是由三个钙离子结合水母发光蛋白后腔肠吖嗪中的过氧化反应引起的。已经为这种过程提出了两种假说:(i)水母发光蛋白与钙离子之间的结合通过蛋白质的构象变化使得氧能够与腔肠吖嗪发生反应从而诱导发光,和(ii)氧在腔肠吖嗪与阿朴水母发光蛋白的结合中发挥作用(Shimomura和Johnson,1978)。在体外和在体内可以通过存在β-巯基乙醇和氧时去除氧合萤光素,加上萤光素(腔肠吖嗪)来再造水母发光蛋白(Shimomura和Johnson,1978)。为了重建水母发光蛋白而加入β-巯基乙醇或还原剂是必需的,这可能是由于在带负电荷的微环境中所包含的第145位半胱氨酸存在至少一个巯基(Charbonneau等人,1985)。
根据结合蛋白质的腔肠吖嗪的类型,已经表征了超过三十种具有不同钙离子亲合力的半合成水母发光蛋白(Shimomura,1991;收入本文作为参考)。水母发光蛋白与钙离子之间的解离常数估计在0.1mM(Allen等人,1997)与1mM(Prasher等人,1985)之间。尽管发光与钙离子浓度之间的关系可能不是线性的,然而已经测定了发光与钙离子浓度之间的对数关系(Johnson和Shimomura,1978)。事实上,当Ca2+浓度由10-7M升至10-6时,信噪比升高了200倍;而由10-6M升至10-5时,是1000倍(Cobbold和Rink,1987)。此外,信号发射的动力学快得足以检测Ca2+离子浓度的瞬间升高。已经显示,在钙饱和条件下,时间常数为6毫秒的光强度升高(Blinks等人,1978)。因而,水母发光蛋白是非常适于在生理学条件下测量快速Ca2+离子增加的光蛋白。
Prasher等人(1985)和Inouye等人(1985)克隆了阿朴水母发光蛋白基因,导致表达载体的生成,从而有可能通过与细胞核、细胞质、线粒体、内质网或质膜信号肽融合而将它靶向特定的细胞区室(Kendall等人,1992;Di Giorgio等人,1996)。另外,蛋白质的体内表达使其低水平的检测成为可能,而不干扰钙的胞内生理学。
本质上,光蛋白活性常常与第二种蛋白质有关。最常见的就是“绿色荧光蛋白”即GFP。在这种情况中发光事实上是绿色的。Johnson等人(1962)在二十世纪六十年代提出了水母发光蛋白与GFP之间通过辐射机制进行能量转移的假说。GFP可能吸收水母发光蛋白在存在Ca2+时发出的蓝光并再次发出最大波长509nm的光谱。其它研究显示通过无辐射机制发生这种能量转移,而这种机制是通过在GFP与水母发光蛋白之间形成异型四聚体而成为可能的。Morise等人(1974)成功的在体外显现了这种能量转移,而且两种分子在DEAE-纤维素膜上的共吸附促进该过程。因而,通过这种机制似乎有可能提高系统的量子效率(Ward和Cormier,1976)。
最近克隆的GFP(同样是在水母Aequoria Victoria中分离得到的)(Prasher等人,1992),在不同的生物学系统中用作细胞表达和谱系的标记(Cubitt等人,1995)。使用经典荧光显微镜检术在活生物体和固定组织中检测这种蛋白质相对容易。另外,荧光发射不需要添加辅助因子或辅酶,且依赖自身催化的翻译后加工。由九个氨基酸构成的荧光团的特征是在第65位丝氨酸与第67位甘氨酸之间形成环,从而产生中间物咪唑烷5,随后是第66位酪氨酸的氧化将其转变成脱氢酪氨酸(Heim等人,1994)。该基团发现于由11个β层构成的圆筒内侧,它构成了与发色团直接作用的环境(Yang等人,1996)。
实时监测钙流有助于理解中枢神经系统的发育、可塑性和机能。在水母中,化学发光、钙结合、水母发光蛋白与绿色荧光蛋白(GFP)有关,而绿色生物发光信号是在Ca2+刺激后发出的。由于激发后发射的光子较弱,因此在细胞和亚细胞水平难以检测单独的水母发光蛋白。
因此开始开发对钙敏感、具有更高量子产量的新标记。本发明利用两种分子之间的化学发光共振能量转移(CRET)。通过融合GFP与水母发光蛋白而构建了对钙敏感的生物发光报道基因,使得发光多得多。
已经在哺乳动物中评估了这些融合蛋白的化学发光和荧光活性。使用冷却加强型CCD(偶联电荷装置)相机在单细胞水平显像胞质(cystosolic)Ca2+的升高。这种双功能报道基因应当能够在转基因动物中调查神经元网络和特定亚细胞区室中的钙活动。
GFP-水母发光蛋白融合蛋白作为由Ca2+激活的报道基因
根据本发明,用水母发光蛋白与GFP构建了融合蛋白,以提高由Ca2+诱导的生物发光的量子产量。仅仅通过GFP与水母发光蛋白的共表达不能提高这种活性(数据未显示)。因为阿朴水母发光蛋白的C端脯氨酸显示参与由Ca2+激活的生物发光过程(20),所以将耐热GFP(Gm)以符合读码框的方式融合在其N端(图1)。
在GFP与阿朴水母发光蛋白之间使用不同大小的接头而生成了不同构建物。最短的间隔物是由5个氨基酸构成的,而最长的含50个氨基酸(图1)。所有融合蛋白都显示比单独的水母发光蛋白更好的由Ca2+触发的生物发光活性。发光活性的增益范围是19-65倍(表1),这可能是由于蛋白质稳定性更好。表1:由Ca2+诱导的化学发光活性
*当进行的测量超过两次时,给出SEM。否则给出两次测量。
| 名称 平均值±SEM*(RLUx106/10U βgal) |
| PA 0.15(0.10;0.21)PGa 10.01±4.4pG1A 2.96(3.39;2.53)pG2A 8.39(9.54;7.23)pG4A 7.78(12.02、3.53)pG5A 8.15±1.72 |
下文详细描述了表1中鉴定的质粒。下列序列标识符用于描述每种质粒插入片段的氨基酸和核苷酸序列。表2:序列标识符
*阿朴水母发光蛋白的核苷酸序列包含于美国专利号5,422,266。
| 质粒插入片段 | 氨基酸序列 | 核苷酸序列 |
| AGAG1AG2AG4AG5ASeG5A | *SEQ ID NO:1SEQ ID NO:2SEQ ID NO:3SEQ ID NO:4SEQ ID NO:5SEQ ID NO:6 | *SEQ ID NO:7SEQ ID NO:8SEQ ID NO:9SEQ ID NO:10SEQ ID NO:11SEQ ID NO:12 |
这些构建物中所用接头的鉴定如下:GFP-水母发光蛋白接头的DNA序列pGA(菌株I-2507) TCC GGC CTC AGA TCT[SEQ ID NO:13]pG1A(菌株I-2508)TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
CTC AGA TCT[SEQ ID NO:14]pG2A(菌株I-2509)TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
GGG AGC GGA TCC GGC GGC CAG TCC GGC CTC AGA
TCT[SEQ ID NO:15]pG4A(菌株I-2510)TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
GGG AGC GGA TCC GGC GGC CAG TCC GGC GGG AGC
GGA TCC GGC GGC CAG TCC GGC GGG AGC GGA TCC
GGC GGC CAG TCC GGC CTC AGA TCT[SEQ ID NO:
16]pG5A(菌株I-2511)TCC GGC GGG AGC GGA
TCC GGC GGC CAG
TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
GGG AGC GGA
TCC GGC GGC CAG TCC GGC GGG AGC GGA TCC GGC
GGC CAG
TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
CTC AGA TCT[SEQ ID NO:17]pSeG5A菌株(I-2512)和pStG5A(菌株I-2513)与pG5A的接头序列相同接头的肽序列pGA Ser Gly Leu Arg Ser[SEQ ID NO:18]pG1A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Leu Arg Ser
[SEQ ID NO:19]pG2A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly
Ser Gly Gly Gln Ser Gly Leu Arg Ser[SEQ ID NO:20]pG4A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly
Ser Gly Gly Gln Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser
Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Leu Arg Ser[SEQ
ID NO:21]pG5A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly
Ser Gly Gly Gln Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser
Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly Ser
Gly Gly Gln Ser Gly Leu Arg Ser[SEQ ID NO:22]pSeG5A和pStG5A与pGSA相同
包含上述多核苷酸的质粒已经保藏于巴斯德研究院(InstitutPasteur,28,rue du Docteur Roux,75724,巴黎Cedex 15,法国)的国家微生物培养物收藏中心(Collection Nationale de Culturesde Microorganismes即C.N.C.M.),如下:
| 插入片段 | 质粒 | 编号 | 保藏日期 |
| AGAG1AG2AG4AG5ASeG5AStG5A | pAeq+PGapG1ApG2ApG4ApG5ApSeG5ApStG5A | I-2506I-2507I-2508I-2509I-2510I-2511I-2512I-2513 | 2000年6月22日2000年6月22日2000年6月22日2000年6月22日2000年6月22日2000年6月22日2000年6月22日2000年6月22日 |
重组阿朴水母发光蛋白在胞质中不稳定,半衰期大约20分钟(21)。相反,GFP是非常稳定的蛋白质,而且在嵌合蛋白中可能稳定阿朴水母发光蛋白。通过用嘌呤霉素(50μg/ml)处理6小时而以COS7细胞中的瞬时表达评估了不同胞质蛋白质的周转时间。此后,大部分融合蛋白展示活性降低30%;比较而言,仅有阿朴水母发光蛋白时,损失80%(图5)。观察到在体外本发明的融合蛋白比单独的水母发光蛋白更灵敏。在Ca2+浓度低至38nM时,G5A仍给出高于背景的显著信号;而水母发光蛋白需要多28倍的钙(1μM)才能产生类似信号(图6)。能量转移还可能改进GFP-水母发光蛋白的量子产量,从而能够进行更有效的钙离子检测。为了鉴别有助于更高发光的因素,有必要在纯化的杂合蛋白上研究GFP荧光激发态的弛豫机制。
更一般的说,本发明的一个实施方案由GFP和水母发光蛋白的基因着手而提供了嵌合蛋白。改进的量子产量将依赖蛋白质通过化学发光共振能量转移(CRET)机制进行的功能偶联。因而,在重建水母发光蛋白且其结合钙离子后,激活后的水母发光蛋白将其能量发送至GFP,GFP继而发出绿光而返回其基态。两种蛋白质之间功能偶联的优化集中于三点:
1.改进37℃时GFP构象变化的诱导,导致GFP在哺乳动物细胞中进行更多发射;
2.改变水母发光蛋白使用的密码子以适应哺乳动物细胞,从而增强其表达;和
3.在两种蛋白质之间加入连接肽。
关于第三点,首先完成了以五个氨基酸分开两种蛋白质的最初分子构建物。然后加入1-5个拷贝的九个氨基酸的序列。将这些构建物置于真核细胞表达载体中,处于CMV(细胞肥大病毒)启动子的控制之下,从而能够确认它们的功能。可以如下鉴定这些融合蛋白:(i)用波长470nm的光激发生物学制剂,并通过荧光显微镜检术(FITC滤镜)检测GFP信号;(ii)在结合Ca2+离子后通过蓝光发射检测水母发光蛋白活性。
下列术语在用于本文时具有如下含义:发光
由原子或分子发出UV、可见光或IR范围内的电磁辐射。这种发射源自从电子激发态向能量较弱的状态、通常是基态的跃迁。荧光
荧光由单线态(singlet)产生,很短,受电子激发。在激发源离开的同时,这种发光就消失了。化学发光
由化学反应引起的发光。生物发光
由活的生物体产生的可见的化学发光。本发明模拟在水母中天然存在的系统,而不固定于支持物。生物发光系统
根据本发明的生物发光系统是三组分嵌合分子,其中部是肽接头和辅酶(即腔肠吖嗪)。与接头共价连接的第一种分子和第二种分子可以是任何分子,只要第一种分子上具有供体位点而第二种分子上具有受体位点(受体-接头-配体、抗体-接头-抗原)。可以将嵌合蛋白融合破伤风毒素片段以获得它在轴突上的逆向和跨突触运输(Coen,L.、Osta,R.、Maury,M.和Brulet,P.,1997,Construction ofhybrid proteins that migrate retrogradely andtranssynaptically into the central nervous system即构建能够逆向和跨突触迁移至中枢神经系统的杂合蛋白,Proc.Natl.Acad.Sci.USA,94:9400-9405),或融合膜受体。无辐射
当水母发光蛋白受到钙离子的结合时,不发射光子至GFP(因此本发明中不存在蓝光发射,两种蛋白质之间直接进行能量转移)。FRET系统
通过荧光共振进行的能量转移(即GFP的两种变体之间)。
参考文献:
Fluorescent indicators for Ca2+ based on green fluorescentproteins and calmodulin.即基于绿色荧光蛋白和钙调蛋白的荧光Ca2+指示剂。
Miyawaki,A.、Llopis,J.、Heim,R.、McCaffery,J.M.、Adams,J.A.、Ikura,M.和Tsien,R.Y.,1997,Nature,388:882-887。
Detection in living cells of Ca2+-dependent changes in thefluorescence emission of an indicator composed of two greenfluorescent protein variants linked by a calmodulin-bindingsequence.Anew class of fluorescent indicators.即使用由两种绿色荧光蛋白变体经钙调蛋白结合序列相连而构成的指示剂在活细胞中检测依赖Ca2+的荧光发射变化。一类新的荧光指示剂。
Romoser,V.A.、Hinkle,P.M.和Persechini,A.,1997,J.Biol.Chem.,272:13270-13274。CRET
通过化学发光共振进行的能量转移(即包含GFP-水母发光蛋白(水母Aequorea)但不含接头或GFP-obeline的融合蛋白)。
参考文献:
Chemiluminescence energy transfer.即化学发光能量转移。
Campbell,A.K.,1988,《Chemiluminescence,Principles andApplication in Biology and Medecin》(化学发光的原理及在生物学和医学中的应用),Ellis Horwood有限公司编,英国奇切斯特市,第475-534页。BRET
通过生物发光共振进行的能量转移(即GFP与萤光素酶(水母Renilla)之间的相互作用)。
参考文献:
A bioluminescence resonance energy transfer(BRET)system:application to interacting circadiant clock protein.即生物发光共振能量转移(BRET)系统:用于与昼夜节律钟蛋白相互作用的应用。
Xu,Y.、Piston,D.W.和Johnson,C.H.,1999,Proc.Natl.Acad.Sci.USA,96:151-156。应用1:在真核生物体中研究细胞群体的钙信号
可以通过同源重组或在特异性启动子控制下的基因转移而实现将对钙敏感的生物发光蛋白靶向细胞群体或特定组织。在小鼠胚细胞中通过同源重组用这种新标记取代诸如Hoxc-8和Otxl基因,使之有可能获得突变型小鼠的新系。这种方法将能够检测一组神经细胞中的电活动,而且还将使之有可能完成通过取代LacZ基因而获得的突变体的表型研究(Le Mouéllic等人,1990,1992;Acampora等人,1996)。对于Hoxc-8基因座,标记的表达应当位于由C7节段开始的脊髓腹角(ventral horns)(Le Mouéllic等人,1990)。已经发现了支配这些肌肉的运动神经元在躯体定位组构中的异常(Tiret等人,1998),而且可能因此着手研究关于发育过程中钙流在建立这些神经连接中的作用。在Otxl模型中,考虑到已经显示了定位于大脑皮层第5和6层及间脑、中脑和小脑区域的表达,转基因应当在前额的特定区域中表达(Frantz等人,1994)。通过用LacZ基因取代该基因而获得的突变型小鼠显示大脑皮层厚度减小及海马、中脑和小脑异常(Acampora等人,1996)。在这些小鼠中观察到的丧失平衡和旋转运动可能归咎于感觉器官异常,特别是眼和内耳。这些小鼠还发生全身化癫痫发作。缺陷连接和/或异常电活动的建立可能牵涉这些病理学过程的发生(McNamara,1992)。一方面,这种新标记的使用使之有可能通过功能和动力学方法来确证这些假设;另一方面,使之有可能阐释癫痫症在成人中及在发育过程中的产生。应用2:关于胞内钙的作用的研究
钙参与大量的细胞机制,诸如细胞迁移、膜兴奋、线粒体代谢、分泌、有丝分裂和突触可塑性(Berridge等人,1998)。在细胞和亚细胞水平编码钙信息是复杂的,涉及空间、时间和数量因素。有可能通过融合肽信号例如突触结合蛋白而将本发明的标记靶向不同亚细胞区室。
范例A:靶向细胞核区室将使之有可能研究钙在转录激活机制和涉及程序化细胞死亡(凋亡)的机制中的作用。
范例B:将包含产生不同发射光谱的GFP的两种融合蛋白靶向两种细胞区室例如细胞质和内质网,将使之有可能研究细胞激活过程中的钙流调控。
范例C:将融合蛋白靶向突触将使之有可能研究神经细胞中神经递质释放过程中与电活动有关的钙活动。第一种可能性由突触蛋白质诸如突触结合蛋白或SNAP25、GFP与水母发光蛋白之间的三组分融合实现。胞吐过程中存在的蛋白质-蛋白质相互作用使之有可能考虑第二种可能性:GFP与水母发光蛋白之间的功能偶联,其一融合囊泡蛋白质,另一融合胞质蛋白质。只有在钙离子浓度升高而不同蛋白质相互作用的过程中获得信号。应用3:在细胞群体水平研究钙信号
具有胞间转运特性的蛋白质诸如破伤风毒素片段C(TTC)或疱疹病毒VP22蛋白质与GFP及水母发光蛋白的三组分融合将使之有可能观察到相连细胞群体例如神经网络中的钙活动。关于对钙离子敏感的生物发光标记表达载体的构建的描述第一步:pEGFP-C1dKS(KpnI-SmaI删除)
用KpnI和SmaI酶对pEGFP-C1质粒(Clontech,见附图)进行双酶切。用绿豆核酸酶切平KpnI的粘端后,将两个末端连接起来。
5'GTC GAC GGT ACC GCG GGC CCG GGA TCC 3'
3'CAG CTG
CCA TGG CGC CC
G GGC CCT AGG 5'
KpnI SmaI
GTC GAC GGT AC G GGA TCC
CAG CTG C C CCT AGG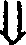
GTC GAC G G GGA TCC
CAG CTG C C CCT AGG
GTC GAC GGG GAT CC
CAG CTG CC
C CTA GG
SalI BamHI第二步:pEGFP-Clmut(GFP诱变)
在由pEGFP-CldKS制备的单链分子中使用四种诱变寡核苷酸。每种寡核苷酸包含一个或几个错配(下文以小写字母表示),从而引起期望的突变。在选择的pEGFP-Clmut质粒(能够被SacII酶切开,但不能被AgeI酶切开)中,通过测序确认了所有突变。-破坏AgeI位点,导入SacII位点,并删除“野生型”GFP中通常不存在的缬氨酸密码子(Prasher,D.C.、Eckenrode,R.K.、Ward,W.W.、Prendergast,F.G.和Cormier,M.J.,1992,Primary structure of theAequorea Victoria green fluorescent protein即Aequoreavictoria绿色荧光蛋白的一级结构,Gene,111,229-233。)
SacII Met Ser Lys Gly AspoGM1: 5-′GCGCTACCGcggGCCACC ATG AGC AAG GGC GAG 3'pEGFP-CldKS: 5'GCGCT
ACCGGTCGCCACC ATG GTG AGC AAG GGC GAG 3'
AgeI Val-用丙氨酸密码子替代第163位缬氨酸密码子,以增加在37℃采取正确构象的GFP的数量(Siemering,K.R.、Golbik,R.、Sever,R.和Haseloff,J.,1996,Mutations that suppress the thermosensitityof green fluorescent protein即遏制绿色荧光蛋白的热敏感性的突变,Current Biol.,6:1653-1663。)
Ile Lys Ala Asn Phe LysoGM2: 5'GC ATC AAG Gcc AAC TTC AAG 3'pEGFP-CldKS 5'GC ATC AAG GTG AAG TTC AAG 3'
Val-用“野生型”GFP中通常存在的组氨酸密码子替代第231位亮氨酸密码子(Prasher,D.C.、Eckenrode,V.K.、Ward,W.W.、Prendergast,F.G.和Cormier,M.J.,1992,Primary structure of the AequoreaVictoria green fluorescent protein即Aequorea victoria绿色荧光蛋白的一级结构,Gene,111:229-233。)
Ile Thr His Asn MetoGM3: 5'GG ACT ATC CaC GGC ATG GA 3'pEGFP-CldKS: 5'GG ACT ATC CTC GGC ATC GA 3'
Leu第三步:pEGFPmut-Aeq(GFP-水母发光蛋白融合蛋白)
在包含水母发光蛋白(Aeq)编码序列的载体上进行四个PCR(聚合酶链式反应),从而能够分别使用引物oAE5A与oAE3A、oAE5B与oAE3B、oAE5C与oAE3C、oAE5D与oAE3D扩增A、B、C和D片段。将交叠区用于在连续PCR(Ho,S.N.、Hunt,H.D.、Horton,R.M.、Pullen,J.K.、和Pease,L.R.,1989,Site-directed mutagenesis by overlapextension using the polymerase chain reaction即使用聚合酶链式反应通过交叠延伸进行的定点诱变,Gene,77:51-59)过程中装配不同部分。使用引物oAE5A与oAE3B由A与B片段的混合物扩增A+B片段。相似的,使用引物oAESA与oAE3D由C与D片段的混合物扩增C+D片段。最后,使用引物oAE5A与oAE3D获得完整编码序列即A+B+C+D。
每种寡核苷酸包含一个或几个错配,下文以小写字母表示。相反,“野生型”序列以大写字母表示。引物oAE5A抑制原始的翻译起始密码子(ATG)并导入了BglII位点。引物oAE3D在紧挨翻译终止密码子(TAA)的后面导入了XhoI位点。用BglII和XhoI酶消化最后的PCR产物,并将其克隆到pEGFP-Clmut质粒的BglTI-SalI位点中,使得缬氨酸密码子(GTC)即水母发光蛋白的第一个密码子与GFP处于相同的读码框中(见附图)。其它引物导入了“沉默”突变,不改变蛋白质序列,但更改水母Aequoria Victoria的六个密码子,以改进它们在哺乳动物中的表达(Wada,K-N.、Aota,S.-I.、Tsuchiya,R.、Ishibashi,F.、Gojobori,T.和Ikemura,T.,1990,Codon usage tabulated from the GenBankgenetic sequence data即根据GenBank遗传序列数据获得的密码子使用率列表,Nucleic Acids Res.,18(增刊):2367-2411)。通过测序确证了整个序列的完整性。oAE5A CCATG5'AGCTTCAgatct GTC AAA CTT ACA TCA GAC TTC GAC AAC CCA AGA TGG ATT GGA CGA3'TCGAAGTctaca CAG TTT GAA TGT AGT CTG AAG CTG TTG GGT TCT ACC TAA CCT GCT
BGlIICAC AAG CAT ATG TTC AAT TTC CTT GAT GTC AAC CAC AAT GGA AAA ATC TCT CTT GAC GAGTG TTC GTA TAC AAG TTA AAG GAA CTA CAG TTG GTG TTA CCT TTT TAG AGA GAA CTG CTATG GTC TAC AAG GCA TCT GAT ATT GTC ATC AAT AAC CTT GGA GCA ACA CCT GAG CAA GCTAC CAG ATG TTC CGT AGA CTA TAA CAG TAG TTA TTG GAA CCT CGT TGT GGA CTC GTT CG
oAE5B AAAA CGA CAC AAA GAT GCT GTg GAA GCC TTC TTC GGA GGA GCT GGA ATG AAA TAT GGT GTGTTT GCT GTG
TTT CTA CGA CAc CTT CGG AAG AAG CCT CCT CGA CCT TAC TTT ATA CCA CAC
T oAE3AGAA ACT GAT TGG CCT GCA TAT ATT GAA GGA TGG AAA AAA TTG GCT ACT GAT GAA TTG GAGCTT TGA CTA ACC GGA CGT ATA TAA CTT CCT ACC TTT TTT AAC CGA TGA CTA CTT AAC CTC
oAE5C G T AAAA TAC GCC AAA AAC GAA CCA ACC CTC ATC CGc ATc TGG GGT GAT GCT TTG TTT GAT ATCTTT ATG CGG TTT TTG CTT GGT TGg GAG TAG GCg TAg ACC CCA CTA CGA AAC AAA CTA TAG
C A T oAE3BGTT GAC AAA GAT CAA AAT GGA GCT ATT ACA CTG GAT GAA TGG AAA GCA TAC ACC AAA GCTCAA CTG TTT CTA GTT TTA CCT CGA TAA TGT GAC CTA CTT ACC TTT CGT ATG TGG TTT CGAGCT GGT ATC ATC CAA TCA TCA GAA GAT TGC GAG GAA ACA TTC AGA GTG TGC GAT ATT GATCGA CCA TAG TAG GTT AGT AGT CTT CTA ACG CTC CTT TGT AAG TCT CAC ACG CTA TAA CTA
oAE5D A T AGAA AGT GGA CAA CTC GAT GTT GAT GAG ATG ACA AGA CAg CAT cTg GGA TTT TGG TAC ACCCTT TCA CCT GTT GAG CTA CAA CTA CTC TAC TGT TCT GTc GTA gAc CCT AAA ACC ATG TGG
T A T oAE3D
XhoIATG GAT CCT GCT TGC GAA AAG CTC TAC GGT GGA GCT GTC CCC TAA TCTcGAGGATCTTT 3'TAC CTA GGA CGA ACG CTT TTC GAG ATG CCA CCT CGA CAG GGG ATT AGAgCTCCTAGAAA 5'
T oAE3D第四步:pGCA(插入间隔序列)
在pEGFPmur-Aeq质粒中,在GFP与水母发光蛋白的编码序列之间存在五个氨基酸的序列。观察结果引导我们通过在BspEL位点插入间隔序列而延长该区域。编码九个氨基酸的序列的两种互补寡核苷酸赋予组合物以很多柔性,因为其中富含甘氨酸和丝氨酸。插入后,只在一侧保留BspEI位点,但是仍可以连续添加新的间隔序列。通过BspEI酶控制每个阶段的取向。需要两个拷贝的该序列以恢复GFP的正常荧光,但是水母发光蛋白与GFP之间的能量转移在五个拷贝时是最佳的。通过测序确认了pGCA质粒(5×9个氨基酸+五个最初氨基酸=50个氨基酸)的整个间隔序列。Lys Ser Gly Leu Arg Ser Val5'AAG TCC GGA CTC AGA TCT GTC 3'3'TTC
AGG CCT GAG
TCT AGA CAG 5'GFP BspEI BGlII Aeq 5'AAG T GC GGA CTC AGA TCT GTC 3'3'TTC AGG CC T GAG TCT AGA CAG 5'
5'AAG T GC GGA CTC AGA TCT GTC 3'3'TTC AGG CC T GAG TCT AGA CAG 5'
+
Gly Gly Ser Gly Ser Gly Gly Gln Ser5'CC GGC GGG AGC GGA TCC GGC GGC CAG T 3'3'
G CCC TCG
CCT AGG CCG CCG GTC
AGG CC 5'
BamHI BspEI
通过在GFP与水母发光蛋白之间插入间隔物来优化能量转移
激发态氧合萤光素与GFP发色团之间的无辐射能量转移强烈依赖它们的整体几何学和它们各自的运动。因此,设计了主要由丝氨酸和甘氨酸残基构成的接头,以插入具有可变长度的柔性元件。
在瞬时转染Neuro2A细胞后48小时制备的细胞提取物上测量了Ca2+触发后发射的绿色与蓝色光子的比率。在透过适当滤镜后对透过分光镜发射的光子进行计数。GFP与水母发光蛋白的共价连接(GA)显著改变了最大发光的波长(图2),从而证明了分子内能量转移。通过加入1-5个接头,绿光对蓝光(500/450nm)的比率进一步由3升至大约7(图2,CRET)。初步测量指示在使用SG5A时该比率可以达到差不多11,可能是由于锚定在膜上的融合蛋白的累积(见材料和方法)。
还使用单色器分析了不同构建物的发射光谱。水母发光蛋白显示广阔的光谱,最大波长474±6.9nm,带宽(对应于具有最大发射值50%的低与高波长之间的距离)108.3±20.1nm(图2)。在范围为506.7±1.2nm至514.1+3.4nm的GFP-水母发光蛋白构建物的峰值发射中存在明显趋向绿色的峰移。增加接头的长度进一步影响光谱的锐度,正如较窄的带宽所示,pGA和pG5A分别是88.4+9.4nm和56.0±3.3nm。没有证据表明任何pG1A-pG5A构建物具有双峰光谱,指示在pGA的情况中可能是不完全的最佳转移。
当GFP与水母发光蛋白之间的间隔物长度超过14个氨基酸时,供体与受体偶极可能具有更多自由,以处于有利于最佳分子内能量转移的构型。本发明系统产生的效率与在体内测量到的分子间CRET相当(22,23),并为关于无辐射能量转移机制的生物物理研究提供方便模型。GFP-阿朴水母发光蛋白的细胞定位和靶向
检验了GFP-阿朴水母发光蛋白构建物的细胞定位。图3图示了瞬时转染Neuro2A细胞后48小时的GFP活性。正如预计的,突变型GFP的单独表达(Gm)在胞质以及细胞核中显示均匀的荧光,因为GFP是能够扩散到细胞核中的小型蛋白质。与原始EGFP(数据未显示)相比,突变V163A显著改进荧光信号并降低光漂白,这可能是因为正确折叠的蛋白质的浓度较高。还观察到所有GFP-阿朴水母发光蛋白构建物在Neuro2A细胞(图3A-D)以及COS-7细胞中均匀分布。在包含具有最短接头:GA、G1A和G2A的融合蛋白的胞质中常常出现亮点。这些斑点在使用G4A时不常发生,而在使用Gm和G5A时从未观察到。在瞬时转染过程中高浓度表达的蛋白质能够诱导GFP的聚集(24),这也将受到水母发光蛋白的存在和它们分开的距离的影响。
还用完整或部分突触结合蛋白1分子将GFP-阿朴水母发光蛋白靶向神经递质囊泡。突触结合蛋白1是突触囊泡的跨膜蛋白,并参与神经递质的胞吐(25)。为了显现突触前区室中的钙微域,信号应当比神经元细胞质中的均匀分布时更精确。在三组分融合蛋白SG5A(图1)中,将突触结合蛋白1的完整编码序列以符合读码框的方式置于G5A的上游。在这种情况中,GFP荧光可以与突触结合蛋白的免疫染色叠加,但是在细胞表面也看得见(图3E)。在神经元(26)和Neuro2A细胞中,突触结合蛋白1位于神经元突起,但是在质膜中检测不到,这可能是因为胞吐的动态机制之后跟随着快速胞吞。当将GFP-阿朴水母发光蛋白只与包含跨膜结构域但缺乏胞质结构域的突触结合蛋白N端部分融合时(tSG5A,图1),强烈荧光限制于胞质内(图3F)。点状标记说明这种蛋白质限制于高尔基体外侧系统。本发明三组分融合分子的正确靶向不发生于tSG5A,而且在SG5A情况中显得变慢。在融合完整突触结合蛋白时,生物发光标记被阻滞在质膜中,但是标记了Neuro2A细胞中存在的所有神经突突起。单细胞中的Ca2+检测
对Neuro2A细胞进行pA、pGA、pG2A、pG5A的瞬时转染或用pA与pGm的共转染(图1)。在不含Ca++的缓冲液中用天然腔肠吖嗪重建水母发光蛋白后,使用经典加强型CCD相机在加入CaCl2溶液(5mM)后测量光子发射(图4A.1和4A.4)。背景可忽略(图4A.2)的条件下,整合时间1秒足以记录表达任何融合蛋白的单细胞中的信号(图4A.1)。只用水母发光蛋白或联合共表达的游离GFP不能显现任何信号(数据未出示)。正如我们在体外所观察到的,未结合GFP的存在不能改进水母发光蛋白的化学发光。由于发光水平较低,因此除了靶向线粒体时从未在单细胞中检测到原位表达的水母发光蛋白。使用冷却的加强型CCD相机,在将水母发光蛋白融合细胞色素c氧化物后,Rutter等人(1996)(27)成功的检测到线粒体内Ca2+信号。只有在使用更加灵敏但对单细胞分析缺乏空间分辨率的光电倍增器(PMT)时,编码细胞质水母发光蛋白的转基因才能报道单层细胞中的钙活动。本发明GFP-水母发光蛋白融合蛋白的稳定性和改进的发光使之有可能在单细胞水平检测生理学Ca2+信号。
测量前的钙缺乏或使用的转染条件可能诱导细胞去极化,使得依赖电压的Ca2+通道的打开有可能对针对加入CaCl2的快速生物发光应答负有责任(图4A)。然后,由于Ca2+通道的脱敏和通过依赖Ca2+的K+通道的膜去极化(28),发光回到背景水平。Fluo-3在Neuro2A细胞的模拟转染中显示相似的图谱(图4C)。随后加入的Ca2+离子载体(A23187)诱导强度相当但动力学不同的二次光子发射。在用pSG5A转染的Neuro2A细胞中能够检测到较低光强度(图4B)。当荧光钙探针锚定于膜的内表面时,应答动力学比非靶向的探针要快得多(29)。生物发光报道分子SG5A的使用可能需要具有更高空间和时间分辨率的系统。在任何情况中,观察到的应答都不是由于水母发光蛋白的完全消耗,因为在将浓Ca2+溶液(100mM)应用于细胞时仍能观察到更多的生物发光(例如图4B)。对于每种构建物,测量至少重复4次。观察到了个体细胞应答的可变性,这可能是由于细胞群体的异质性。需要进行进一步研究以校准相对光单位(RLU)与Ca2+浓度。膜片钳技术将能够鉴定参与这些应答的钙通道的类型和细胞转染对膜电位的影响。
本发明的转基因应当能够显现完整动物的神经网络的电活动。在体外,直到最近还使用两种方法。第一种方法基于在神经细胞中将胞吐偶联于synaptolucin的发光(30)。当靶向突触囊泡内部的萤光素酶在胞外空间与ATP发生反应时发生发光。作者使用这种系统获得了与神经递质释放相关的信号,但是需要很长采集时间(超过30秒钟)的低光水平。在第二种方法中,通过FRET将荧光Ca2+敏感性标记用于测量胞内Ca2+(3,4,31)。对于单细胞检测,这种技术需要足够浓度的探针以区别信号同由生物学化合物的自身荧光产生的背景以及两个GFP之间不依赖钙的能量转移的可能性。整合时间也相对较长,在4与20秒之间。
本发明因而提供新的双功能杂合物,其中可以通过GFP荧光来追踪表达模式,而水母发光蛋白部分是Ca2+活动的报道分子。此外,遵循CRET原理的两种成分的功能偶联导致更多的发光量和更高的Ca2+敏感性。已经使用冷却的加强型CCD相机以1秒整合时间在单细胞中评估了这些遗传标记的生物发光活性。最近开发的低水平光检测系统应当能够以更短的整合时间、和更高的空间分辨率检测CRET信号。可以在转基因动物中在体内研究胞内和胞间Ca2+信号,其中GFP-水母发光蛋白靶向特定的细胞群体和/或特定的亚细胞区室。具体而言,从而能够实时且同时显现完整神经回路的细胞中的钙振荡。
下面的实施例将更详细的描述本发明。实施例1:GFP-水母发光蛋白融合蛋白的构建
所有构建物都在pEGFP-C1载体(Clontech)中生成。为了在哺乳动物细胞中获得最大表达,对EGFP基因进行了密码子优化。它还在发色团中包含2处突变,F64L和S65T,它们改变了激发光谱且增强了荧光强度(17)。通过单链诱变,用丙氨酸替代EGFP的第163位缬氨酸,从而改进蛋白质的正确折叠并增强处于37℃时的荧光(18,19)。将水母发光蛋白编码序列(由M.-T.Nicolas慷慨惠赠)以符合读码框的方式融合在EGFP基因的3’端,即插入pEGFP-C1的BgIII/SalI位点。为了在哺乳动物细胞中获得更好的表达,通过使用交叠延伸PCR(聚合酶链式反应)的定点诱变手段,更改了七个密码子。然后,将互补寡核苷酸5’-CCGGCGGGAGCGGATCCGGCGGCCAGT-3’[SEQ ID NO:23]和5’-CCGGACTGGCCGCCGGATCCGCTCCCG-3’[SEQ ID NO:24]插入GFP与水母发光蛋白之间15bp序列中的BspEI位点。只在-端保留BspEI位点,从而能够连续加入1-5个接头序列(pG1A-pG5A)。
使用突触蛋白质突触结合蛋白1(其cDNA质粒由M.Fukuda慷慨惠赠)在pG5A中制作两个额外的融合构建物。将编码完整开放读码框或前134个N端氨基酸(包含蛋白质的跨膜结构域)的序列以符合读码框的方式融合在GFP-水母发光蛋白基因的5’端。实施例2:细胞培养和转染
在添加10%(v/v)热处理胎牛血清、2mm谷氨酰胺(LifeTechnologeis-Gibco,英国)和100单位链霉素-青霉素(LifeTechnologies-Gibco,英国)的Dulbecco氏Eagle培养基(LifeTechnologies-Gibco,英国)中培养成神经细胞瘤细胞(Neuro2A,小鼠)。将培养物于37℃在含8%CO2的潮湿空气中进行保温,并使用磷酸钙技术或FuGENE 6TM转染试剂(Roche)进行瞬时转染。实施例3:体外对Ca2+敏感的化学发光和CRET活性
转染后48小时在250μl含10mM β-巯基乙醇、4mM EDTA、5μM腔肠吖嗪的PBS中于4℃在2-4小时内收获细胞。在含溶于PBS的1mM EDTA中漂洗细胞,并在400μl低渗缓冲液(20mM Tris-HCl pH7.5/5mMEDTA/5mM β-巯基乙醇及蛋白酶抑制剂混合物,依照制造商,Roche)中于4℃在30分钟-1小时内收获细胞.使细胞通过30号针头从而破坏细胞膜,并以13000rmp、于4℃微量离心1小时后获得细胞提取物。收获所有构建物的上清液;但是,对于SGSA,进一步重悬膜沉淀。在光度计(Lumat LB95501 E&EG Berthold)中测量钙敏感性化学发光活性。取一部分(10μl)置于光度计的样品管(装有90μl 10mM Tris-HClpH7.5)中,并在注入100μl 50mM CaCl2/10mM Tris-HCl pH7.5溶液后测量光强度,表述成相对光单位(R.L.U.)。
对于CRET测量,取一部分转染细胞的提取物置于贮液槽中,并使之接触连接光子计数相机(Photek三微通道板加强型CCD相机:Photek216)的光纤束。在捕捉信号前,让光透过单色器,从而能够对发射光子进行光谱分析。在注入CaCl2前20秒钟开始进行采集,并持续至注入CaCl2溶液(50mM)后40秒钟。为了测定绿色/蓝色光子比率,进行相同流程,但是在这种情况中,系统测量透过分光镜后又穿过蓝色(450nm)和绿色(500nm)滤镜的发光。实施例4:GFP荧光和免疫定位
转染后48小时在含4%低聚甲醛的PBS pH7.4中固定Neuro2A细胞,在PBS中漂洗,并封固。在使用氩-氪激光并以多线模式进行操作的共聚焦激光扫描显微镜(Zeiss,海德尔堡,德国)或配备表面发光系统(Zeiss,海德尔堡,德国)的Axiophot显微镜下显现GFP荧光。为了荧光定位被靶向的GFP-水母发光蛋白的,将固定细胞用溶于PBS pH7.4的50mM NH4Cl于室温预处理5分钟,并在含2%BSA/0.02%屈立通/山羊血清溶液的PBS中渗透1小时。然后应用针对突触结合蛋白的抗体(StressGen SYA-130)达2-4小时。然后在PBS中漂洗细胞,并在含2%BSA/0.02%屈立通及以1/100稀释的二抗(TRITC缀合抗体)的PBS中孵育。然后在PBS中清洗细胞并封固。实施例5:单细胞生物发光检测
转染后48小时,在124mM NaCl/5mM KCl/15mM Hepes pH7.4/5mMNaHCO3/1mM NaH2PO4/0.5mM MgSO4/1.5mM CaCl2/5.5mM葡萄糖中漂洗细胞,稍后在不含CaCl2但包含5μM腔肠吖嗪的相同缓冲液中于37℃保温2-4小时以重建水母发光蛋白,然后漂洗。用配备BH2-RFCA表面荧光单元的修改型Olympus竖式显微镜(BHS)显现钙信号,并通过平面40倍Olympus长工作距离水浸透镜(N.A.O.7)记录。GFP荧光容许选择在转染细胞上的记录区域。关闭激发光源,而相机的增益增加了。使用冷却Photonic Science extended ISIS视频相机整合每秒钟的图象。图4中的每张图谱都展示我们使用Axon Imaging Workbench 2214软件围绕单个细胞体定义的区域内的发光量。将荧光强度和CRET活性转化成标示的伪彩色。使用Fluo-3 AM在模拟转染的NeuroA细胞上进行对照以检查实验条件。实施例6:蛋白质稳定性
通过用嘌呤霉素(50μg/ml)处理6小时而以COS7细胞中的瞬时表达评估不同胞质蛋白质的周转时间。以存在5μM腔肠吖嗪重建水母发光蛋白后所获得的细胞提取物进行Ca2+诱导的化学发光活性。在光度计(Lumat LB95501 E&EG Berthold)中测量钙敏感性化学发光活性。取一部分(10μl)置于光度计的样品管(装有90μl 10mM Tris-HClpH7.5)中,并在注入100μl 50mM CaCl2/10mM Tris-HCl pH7.5溶液后测量光强度,表述成相对光单位(RLU)。将相对化学发光活性表述成相对于时间零点时活性(100%)的百分比。结果显示于图5。在图5中可以看出,此后,大部分融合蛋白展示活性降低30%,比较而言,仅有阿朴水母发光蛋白时损失80%。实施例7:水母发光蛋白和G5A的Ca2+亲合力的测定
以存在5μM腔肠吖嗪重建水母发光蛋白后所获得的细胞提取物进行由Ca2+诱导的化学发光活性。在光度计(Lumat LB95501 E&EGBerthold)中测量钙敏感性化学发光活性。取一部分(10μl)置于光度计的样品管(装有90μl 10mM Tris-HCl pH7.5)中,并在注入100μl不同的Ca/EGTA溶液后测量光强度,表述成相对光单位(RLU)。结果显示于图6。在图6中可以看出,在Ca2+浓度低至38nM时,G5A仍给出高于背景的显著信号;而水母发光蛋白需要多28倍(1μM)的钙才能产生类似信号。嵌合GFP-水母发光蛋白作为单细胞水平的生物发光Ca2+报道分子
关于嵌合GFP-水母发光蛋白钙敏感性生物发光报道分子的发明,已经开发新的应用,而且已经获得了一些关于GFP-水母发光蛋白融合蛋白对Ca2+离子敏感性的初步数据。实施例8:G5A和SG5A的Ca2+敏感性:生物发光信号与Ca2+浓度之间的校准曲线
以存在5μM腔肠吖嗪重建水母发光蛋白后所获得的细胞提取物进行两种构建物G5A和SG5A的Ca2+敏感性的测量。在光度计(LumatLB95501 E&EG Berthold)中测量钙敏感性化学发光活性。取一部分(10μl)置于光度计的样品管(装有90μl 10mM Tris-HCl pH7.5)中,并在注入100ml不同的Ca/EGTA溶液(分子探针钙校准缓冲液试剂盒)后测量光强度,表述成相对光单位(RLU)。图7显示了G5A、SG5A和水母发光蛋白的Ca2+反应曲线。曲线展示了比率L/Lmax与Ca2+之间的关系。L指任何指定[Ca2+]处的RLS率,而Lmax指饱和[Ca2+]处的RLU率。这些结果显示各种形式的GFP-水母发光蛋白的Ca2+亲合力要比水母发光蛋白高得多。实施例9:GFP-水母发光蛋白报道分子的新应用
开发了包含GFP-水母发光蛋白的腺病毒载体。使用这些新构建物,可以以较高效率转染培养物中来自大鼠脊髓的游离神经元。图8和图9描绘了在游离神经元细胞的单细胞水平检测到的由Ca2+诱导的生物发光信号。将由含G5A(图8)或SG5A(图9)的腺病毒载体感染的神经元细胞与5μM腔肠吖嗪一起在不含Ca2+的缓冲液中预保温。将荧光强度和生物发光活性转化成伪彩色。图8a-c和9a分别显示了加入5mM和2.5mM CaCl2后12和9秒钟选定视野的代表性照片。图8d-e和9b是在加入离子霉素和高浓度的CaCl2(100mM)后获得的。实施例10:GFP-水母发光蛋白报道分子在爪蟾中的体内表达和钙活动的测量
研究了爪蟾早期和晚期胚胎发生过程中的钙信号。图10显示了发光活性的代表性图谱,图解了在爪蟾胚单细胞阶段注入GA质粒后神经诱导过程中胞内钙的变化。图11显示了包含GFP-水母发光蛋白的转基因爪蟾幼体。这些技术也可用于转基因斑马鱼和小鼠。这些结果显示这些钙报道分子可用于极其多种生物体或组织,以显现钙活动和测量钙浓度。
总之,可用于生物发光系统中通过CRET系统的能量转移的新接头具有如下特性:形式:
描述了接头的不同氨基酸序列和肽序列。它的长度包括最小值4-9个氨基酸,可以通过一组7-12个氨基酸(在一个优选的实施方案中是9个氨基酸)进行延伸。所述氨基酸组可延伸至63个氨基酸,即9×6次。例如使用由5个氨基酸及随后9氨基酸的1-5次重复组成的肽接头进行了实验。功能:
它的第一种功能是使进行直接能量传送的两个分子的供体位点接近受体位点。该接头为通过CRET的能量转移提供最佳环境。
第二种功能是通过与GFP的融合来提高水母发光蛋白的半衰期从而稳定所述系统。将水母发光蛋白连接半衰期超过24小时的GFP。应用:
在生物发光系统中,蛋白质-蛋白质相互作用的趋势。
具有接头的生物发光系统的应用:癫痫形成、SNC病(在发育过程中和在成人中显现神经元细胞活动)、脊髓中同源异型基因HOX-C8参与的神经肌肉联系。
包含根据本发明的接头的嵌合蛋白在凋亡中的应用,显现胞内钙池的变化。
活器官如脾中钙波(胞内和胞间钙波)作用的显现和准确确定。结果:
嵌合蛋白因分子半衰期的增加而更稳定。钙敏感性的增加是重要的。
本发明的接头具有令人惊讶的特性。包含水母发光蛋白和接头的嵌合分子的钙离子敏感性与单独的水母发光蛋白不同。本发明提供了更好的敏感性。
该接头使之有可能将水母发光蛋白与GFP连在一起。下面的参考文献证明了两种分子在一起但无接头时不发生相互作用:Morise,H.、Shimomura,O.、Johonson,F.H.和Winant,J.,1974,IntermolecularEnergy Transfer in the bioluminescent system of Aequoria(多管水母生物发光系统中的分子间能量转移),Biochemistry,13:2656-2662。
这是第一次在活的单细胞系统(或活的动物)中获得水母发光蛋白信号的显现。
总之,实时监测钙流将有助于理解中枢神经系统的发育、可塑性和机能。在水母中,化学发光钙结合水母发光蛋白与绿色荧光蛋白(GFP)相关,并在Ca2+刺激后发出绿色生物发光信号。我们决定使用两种分子之间的这种化学发光共振能量转移(CRET)。已经通过融合GFP与水母发光蛋白而构建了钙敏感性生物发光报道基因,使得发光多得多。已经在哺乳动物细胞中评估了这些融合蛋白的化学发光和荧光活性。使用冷却加强型CCD相机在单细胞水平显像了胞质Ca2+的升高。这种双功能报道基因应当能够在转基因动物中调查神经元网络和特定亚细胞区室中的钙活动。
下面是本文提到的序列和相应的序列标识符:肽序列:M S K G E E L F T G V V P I L V E L D G D V N G H K F S V S G E G E G D A TY G K L T L K F I C T T G K L P V P W·P T L V T T L T Y G V Q C F S R Y P DH M K Q H D F F K S A M P E G Y V Q E R T I F F K D D G N Y K T R A E V K FE G D T L V N R I E L K G I D F K E D G N I L G H K L E Y N Y N S H N V Y IM A D K Q K N G I K A N F K I R H N I E D G S V Q L A D H Y Q Q N T P I G DG P V L L P D N H Y L S T Q S A L S K D P N E K R D H M V L L E F V T A A GI T H G M D E L Y K S G L R S V K L T S D F D N P R W I G R H K H M F N F LD V N H N G K I S L D E M V Y K A S D I V I N N L G A T P E Q A K R H K D AV E k F F G G A G M K Y G V E T D W P A Y I E G W K K L A T D E L E K Y A KN E P T L I R I W G D A L F D I V D K D Q N G A I T L D E W K A Y T K A A GI I Q S S E D C E E T F R V C D I D E S G Q L D V D E M T R Q H L G F W Y TM D P A C E K L Y G G A V P [SEQ ID NO:1]G1AM S K G E E L F T G V V P I L V E L D G D V N G H K F S V S G E G E G D A TY G K L T L K F I C T T G K L P V P W P T L V T T L T Y G V Q C F S R Y P DH M K Q H D F F K S A M P E G Y V Q E R T I F F K D D G N Y K T R A E V K FE G D T L V N R I E L K G I D F K E D G N I L G H K L E Y N Y N S H N V Y IM A D K Q K N G I K A N F K I R H N I E D G S V Q L A D H Y Q Q N T P I G DG P V L L P D N H Y L S T Q S A L S K D P N E K R D H M V L L E F V T A A GI T H G M D E L Y K S G G S G S G G Q S G L R S V K L T S D F D N P R W I GR H K H M F N F L D V N H N G K I S L D E M V Y K A S D I V I N N L G A T PE Q A K R H K D A V E A F F G G A G M K Y G V E T D W P A Y I E G W K K L AT D E L E K Y A K N E P T L I R I W G D A L F D I V D K D Q N G A I T L D EW K A Y T K A A G I I Q S S E D C E E T F R V C D I D E S G Q L D V D E M TR Q H L G F W Y T M D P A C E K L Y G G A V P [SEQ ID NO:2]G2AM S K G E E L F T G V V P I L V E L D G D V N G H K F S V S G E G E G D A TY G K L T L K F I C T T G K L P V P W P T L V T T L T Y G V Q C F S R Y P DH M K Q H D F F K S A M P E G Y V Q E R T I F F K D D G N Y K T R A E V K FE G D T L V N R I E L K G I D F K E D G N I L G H K L E Y N Y N S H N V Y IM A D K Q K N G I K A N F K I R H N I E D G S V Q L A D H Y Q Q N T P I G DG P V L L P D N H Y L S T Q S A L S K D P N E K R D H M V L L E F V T A A GI T H G M D E L Y K S G G S G S G G Q S G G S G S G G Q S G L R S V K L T SD F D N P R W I G R H K H M F N F L D V N H N G K I S L D E M V Y K A S D IV I N N L G A T P E Q A K R H K D A V E A F F G G A G M K Y G V E T D W P AY I E G W K K L A T D E L E K Y A K N E P T L I R I W G D A L F D I V D K DQ N G A I T L D E W K A Y T K A A G I I Q S S E D C E E T F R V C D I D E SG Q L D V D E M T R Q H L G F W Y T M D P A C E K L Y G G A V P[SEQ ID NO:3]G4AM S K G E E L F T G V V P I L V E L D G D V N G H K F S V S G E G E G D A TY G K L T L K F I C T T G K L P V P W P T L V T T L T Y G V Q C F S R Y P DH M K Q H D F F K S A M P E G Y V Q E R T I F F K D D G N Y K T R A E V K FE G D T L V N R I E L K G I D F K E D G N I L G H K L E Y N Y N S H N V Y IM A D K Q K N G I K A N F K I R H N I E D G S V Q L A D H Y Q Q N T P I G DG P V L L P D N H Y L S T Q S A L S K D P N E K R D H M V L L E F V T A A GI T H G M D E L Y K S G G S G S G G Q S G G S G S G G Q S G G S G S G G Q SG G S G S G G Q S G L R S V K L T S D F D N P R W I G R H K H M F N F L D VN H N G K I S L D E M V Y K A S D I V I N N L G A T P E Q A K R H K D A V EA F F G G A G M K Y G V E T D W P A Y I E G W K K L A T D E L E K Y A K N EP T L I R I W G D A L F D I V D K D Q N G A I T L D E W K A Y T K A A G I IQ S S E D C E E T F R V C D I D E S G Q L D V D E M T R Q H L G F W Y T M DP A C E K L Y G G A V P [SEQ ID NO:4]G5AM S K G E E L F T G V V P I L V E L D G D V N G H K F S V S G E G E G D A TY G K L T L K F I C T T G K L P V P W P T L V T T L T Y G V Q C F S R Y P DH M K Q H D F F K S A M P E G Y V Q E R T I F F K D D G N Y K T R A E V K FE G D T L V N R I E L K G I D F K E D G N I L G H K L E Y N Y N S H N V Y IM A D K Q K N G I K A N F K I R H N I E D G S V Q L A D H Y Q Q N T P I G DG P V L L P D N H Y L S T Q S A L S K D P N E K R D H M V L L E F V T A A GI T H G M D E L Y K S G G S G S G G Q S G G S G S G G Q S G G S G S G G Q SG G S G S G G Q S G G S G S G G Q S G L R S V K L T S D F D N P R W I G R HK H M F N F L D V N H N G K I S L D E M V Y K A S D I V I N N L G A T P E QA K R H K D A V E A F F G G A G M K Y G V E T D W P A Y I E G W K K L A T DE L E K Y A K N E P T L I R I W G D A L F D I V D K D Q N G A I T L D E W KA Y T K A A G I I Q S S E D C E E T F R V C D I D E S G Q L D V D E M T R QH L G F W Y T M D P A C E K L Y G G A V P [SEQ ID NO:5]SeG5AM V S A S R P E A L A A P V T T V A T L V P H N A T E P A S P G E G K E D AF S K L K Q K F M N E L H K I P L P P W A L I A I A I V A V L L V V T C C FC V C K K C L F K K K N K K K G K E K G G K N A I N M K D V K D L G K T M KD Q A L K D D D A E T G L T D G E E K E E P K E E E K L G K L Q Y S L D Y DF Q N N Q L L V G I I Q A A E L P A L D M G G T S D P Y V K V F L L P D K KK K F E T K V H R K T L N P V F N E Q F T F K V P Y S E L G G K T L V M A VY D F D R F S K H D I I G E F K V P M N T V D F G H V T E E W R D L Q S A EK E E Q E K L G D I C F S L R Y V P T A G K L T V V I L E A K N L K K M D VG G L S D P Y V K I H L M Q N G K R L K K K K T T I K K N T L N P Y Y N E SF S F E V P F E Q I Q K V Q V V V T V L D Y D K I G K N D A I G K V F V G YN S T G A E L R H W S D M L A N P R R P I A Q W H T L Q V E E E V D A M L AV K R S G N S G R A T M S K G E E L F T G V V P I L V E L D G D V N G H K FS V S G E G E G D A T Y G K L T L K F I C T T G K L p V P W P T L V T T L TY G V Q C F S R Y P D H M K Q H D F F K S A M P E G Y V Q E R T I F F K D DG N Y K T R A E V K F E G D T L V N R I E L K G I D F K E D G N I L G H K LE Y N Y N S H N V Y I M A D K Q K N G I K A N F K I R H N I E D G S V Q L AD H Y Q Q N T P I G D G P V L L P D N H Y L S T Q S A L S K D P N E K R D HM V L L E F V T A A G I T H G M D E L Y K S G G S G S G G Q S G G S G S G GQ S G G S G S G G Q S G G S G S G G Q S G G S G S G G Q S G L R S V K L T SD F D N P R W I G R H K H M F N F L D V N H N G K I S L D E M V Y K A S D IV I N N L G A T P E Q A K R H K D A V E A F F G G A G M K Y G V E T D W P AY I E G W K K L A T D E L E K Y A K N E P T L I R I W G D A L F D I V D K DQ N G A I T L D E W K A Y T K A A G I I Q S S E D C E E T F R V C D I D E SG Q L D V D E M T R Q H L G F W Y T M D P A C E K L Y G G A V P[SEQ ID NO:6)GAAtg agc aag ggc gag gag ctg ttc acc ggg gtg gtg ccc atc ctg gtc gag ctg gacggc gac gta aac ggc cac aag ttc agc gtg tcc ggc gag ggc gag ggc gat gcc acctac ggc aag ctg acc ctg aag ttc atc tgc acc acc ggc aag ctg ccc gtg ccc tggccc acc ctc gtg acc acc ctg acc tac ggc gtg cag tgc ttc agc cgc tac ccc gaccac atg aag cag cac gac ttc ttc aag tcc gcc atg ccc gaa ggc tac gtc cag gagcgc acc atc ttc ttc aag gac gac ggc aac tac aag acc cgc gcc gag gtg aag ttcgag ggc gac acc ctg gtg aac cgc atc gag ctg aag ggc atc gac ttc aag gag gacggc aac atc ctg ggg cac aag ctg gag tac aac tac aac agc cac aac gtc tat atcatg gcc gac aag cag aag aac ggc atc aag gCC aac ttc aag atc cqc cac aac atcgag gac ggc agc gtg cag ctc qcc gac cac tac cag cag aac acc ccc atc ggc gacggc ccc gtg ctg ctg ccc gac aac cac tac ctg agc acc cag tcc gcc ctg agc aaagac ccc aac gag aag cgc gat cac atg gtc ctg ctg gag ttc gtg acc gcc gcc gggatc act cAc ggc atg gac gag ctg tac aag tcc gga ctc aGA TCT gtc aaa ctt acatca gac ttc gac aac cca aga tgg att gga cga cac aag cat atg ttc aat ttc cttgat gtc aac cac aat gga aaa atc tct ctt gac gag atg gtc tac aag gca tct gatatt gtc atc aat aac ctt gga gca aca cct gag caa gcc aaa cga cac aaa gat gctgtG gaa gcc ttc ttc gga gga gct gga atg aaa tat ggt gtg gaa act gat tgg cctgca tat att gaa gga tgg aaa aaa ttg gct act gat gaa ttg gag aaa tac gcc aaaaac gaa cca acC ctc atc cgC ata tgg ggt gat gct ttg ttt gat atc gtt gac aaagat caa aat gga gct att aca ctg gat gaa tgg aaa gca tac acc aaa gct gct ggtatc atc caa tca tca gaa gat tgc gag gaa aca ttc aga gtg tgc gat att gat gaaagt gga caa ctc gat gtt gat gag atg aca aga caG cat CtG gga ttt tgg tac accatg gat cct gct tgc gaa aag ctc tac ggt gga gct gtc ccc[SEQ ID NO:7]G1AAtg agc aag ggc gag gag ctg ttc acc ggg gtg gtg ccc atc ctg gtc gag ctg gacggc gac gta aac ggc cac aag ttc agc gtg tcc ggc gag ggc gag ggc gat gcc acctac ggc aag ctg acc ctg aag ttc atc tgc acc acc ggc aag ctg ccc gtg ccc tggccc acc ctc gtg acc acc ctg acc tac ggc gtg cag tgc ttc agc cgc tac ccc gaccac atg aag cag cac gac ttc ttc aag tcc gcc atg ccc gaa ggc tac gtc cag gagcgc acc atc ttc ttc aag gac gac ggc aac tac aag acc cgc gcc gag gtg aag ttcgag ggc gac acc ctg gtg aac cgc atc gag ctg aag ggc atc gac ttc aag gag gacggc aac atc ctg ggg cac aag ctg gag tac aac tac aac agc cac aac gtc tat atcatg gcc gac aag cag aag aac ggc atc aag gCC aac ttc aag atc cgc cac aac atcgag gac ggc agc gtg cag ctc gcc gac cac tac cag cag aac acc ccc atc ggc gacggc ccc gtg ctg ctg ccc gac aac cac tac ctg agc acc cag tcc gcc ctg agc aaagac ccc aac gag aag cgc gat cac atg gtc ctg ctg gag ttc gtg acc gcc gcc gggatc act cAc ggc atg gac gag ctg tac aag tcc ggc ggg agc gga tcc ggc ggc cagtcc ggc ctc aGA TCT gtc aaa ctt aca tca gac ttc gac aac cca aga tgg att ggacga cac aag cat atg ttc aat ttc ctt gat gtc aac cac aat gga aaa atc tct cttgac gag atg gtc tac aag gca tct gat att gtc atc aat aac ctt gga gca aca cctgag caa gcc aaa cga cac aaa gat gct gtG gaa gcc ttc ttc gga gga gct gga atgaaa tat ggt gtg gaa act gat tgg cct gca tat att gaa gga tgg aaa aaa ttg gctact gat gaa ttg gag aaa tac gcc aaa aac gaa cca acC ctc atc cgC ata tgg ggtgat gct ttg ttt gat atc gtt gac aaa gat caa aat gga gct att aca ctg gat gaatgg aaa gca tac acc aaa gct gct ggt atc atc caa tca tca gaa gat tgc gag gaaaca ttc aga gtg tgc gat att gat gaa agt gga caa ctc gat gtt gat gag atg acaaga caG cat CtG gga ttt tgg tac acc atg gat cct gct tgc gaa aag ctc tac ggtgga gct gtc ccc [SEQID NO:8]G2AAtg agc aag ggc gag gag ctg ttc acc ggg gtg gtg ccc atc ctg gtc gag ctg gacggc gac gta aac ggc cac aag ttc agc gtg tcc ggc gag ggc gag ggc gat gcc acctac ggc aag ctg acc ctg aag ttc atc tgc acc acc ggc aag ctg ccc gtg ccc tggccc acc ctc gtg acc acc ctg acc tac ggc gtg cag tgc ttc agc cgc tac ccc gaccac atg aag cag cac gac ttc ttc aag tcc gcc atg ccc gaa ggc tac gtc cag gagcgc acc atc ttc ttc aag gac gac ggc aac tac aag acc cgc gcc gag gtg aag ttcgag ggc gac acc ctg gtg aac cgc atc gag ctg aag ggc atc gac ttc aag gag gacggc aac atc ctg ggg cac aag ctg gag tac aac tac aac agc cac aac gtc tat atcatg gcc gac aag cag aag aac ggc atc aag gCC aac ttc aag atc cgc cac aac atcgag gac ggc agc gtg cag ctc gcc gac cac tac cag cag aac acc ccc atc ggc gacggc ccc gtg ctg ctg ccc gac aac cac tac ctg agc acc cag tcc gcc ctg agc aaagac ccc aac gag aag cgc gat cac atg gtc ctg ctg gag ttc gtg acc gcc gcc gggatc act cAc ggc atg gac gag ctg tac aag tcc ggc ggg agc gga tcc ggc ggc cagtcc ggc ggg agc gga tcc ggc ggc cag tcc ggc ctc aGA TCT gtc aaa ctt aca tcagac ttc gac aac cca aga tgg att gga cga cac aag cat atg ttc aat ttc ctt gatgtc aac cac aat gga aaa atc tct ctt gac gag atg gtc tac aag gca tct gat attgtc atc aat aac ctt gga gca aca cct gag caa gcc aaa cga cac aaa gat gct gtGgaa gcc ttc ttc gga gga gct gga atg aaa tat ggt gtg gaa act gat tgg cct gcatat att gaa gga tgg aaa aaa ttg gct act gat gaa ttg gag aaa tac gcc aaa aacgaa cca acC ctc atc cgC ata tgg ggt gat gct ttg ttt gat atc gtt gac aaa gatcaa aat gga gct att aca ctg gat gaa tgg aaa gca tac acc aaa gct gct ggt atcatc caa tca tca gaa gat tgc gag gaa aca ttc aga gtg tgc gat att gat gaa agtgga caa ctc gat gtt gat gag atg aca aga caG cat CtG gga ttt tgg tac acc atggat cct gct tgc gaa aag ctc tac ggt gga gct gtc ccc [SEQ ID NO:9]G4AAtg agc aag ggc gag gag ctg ttc acc ggg gtg gtg ccc atc ctg gtc gag ctg gacggc gac gta aac ggc cac aag ttc agc gtg tcc ggc gag ggc gag ggc gat gcc acctac ggc aag ctg acc ctg aag ttc atc tgc acc acc ggc aag ctg ccc gtg ccc tggccc acc ctc gtg acc acc ctg acc tac ggc gtg cag tgc ttc agc cgc tac ccc gaccac atg aag cag cac gac ttc ttc aag tcc gcc atg ccc gaa ggc tac gtc cag gagcgc acc atc ttc ttc aag gac gac ggc aac tac aag acc cgc gcc gag gtg aag ttcgag ggc gac acc ctg gtg aac cgc atc gag ctg aag ggc atc gac ttc aag gag gacggc aac atc ctg ggg cac aag ctg gag tac aac tac aac agc cac aac gtc tat atcatg gcc gac aag cag aag aac ggc atc aag gCC aac ttc aag atc cgc cac aac atcgag gac ggc agc gtg cag ctc gcc gac cac tac cag cag aac acc ccc atc ggc gacggc ccc gtg ctg ctg ccc gac aac cac tac ctg agc acc cag tcc gcc ctg agc aaagac ccc aac gag aag cgc gat cac atg gtc ctg ctg gag ttc gtg acc gcc gcc gggatc act cAc ggc atg gac gag ctg tac aag tcc ggc ggg agc gga tcc ggc ggc cagtcc ggc ggg agc gga tcc ggc ggc cag tcc ggc ggg agc gga tcc ggc ggc cag tccggc ggg agc gga tcc ggc ggc cag tcc ggc ctc aGA TCT gtc aaa ctt aca tca gacttc gac aac cca aga tgg att gga cga cac aag cat atg ttc aat ttc ctt gat gtcaac cac aat gga aaa atc tct ctt gac gag atg gtc tac aag gca tct gat att gtcatc aat aac ctt gga gca aca cct gag caa gcc aaa cga cac aaa gat gct gtG gaagcc ttc ttc gga gga gct gga atg aaa tat ggt gtg gaa act gat tgg cct gca tatatt gaa gga tgg aaa aaa ttg gct act gat gaa ttg gag aaa tac gcc aaa aac gaacca acC ctc atc cgC ata tgg ggt gat gct ttg ttt gat atc gtt gac aaa gat caaaat qga gct att aca ctg gat gaa tgg aaa gca tac acc aaa gct gct ggt atc atccaa tca tca gaa gat tgc gag gaa aca ttc aga gtg tgc gat att gat gaa agt ggacaa ctc gat gtt gat gag atg aca aga caG cat CtG gga ttt tgg tac acc atg gatcct gct tgc gaa aag ctc tac ggt gga gct gtc ccc [SEQ ID NO:10]G5AAtg agc aag ggc gag gag ctg ttc acc ggg gtg gtg ccc atc ctg gtc gag ctg gacggc gac gta aac ggc cac aag ttc agc gtg tcc ggc gag ggc gag ggc gat gcc acctac ggc aag ctg acc ctg aag ttc atc tgc acc acc ggc aag ctg ccc gtg ccc tggccc acc ctc gtg acc acc ctg acc tac ggc gtg cag tgc ttc agc cgc tac ccc gaccac atg aag cag cac gac ttc ttc aag tcc gcc atg ccc gaa ggc tac gtc cag gagcgc acc atc ttc ttc aag gac gac ggc aac tac aag acc cgc gcc gag gtg aag ttcgag ggc gac acc ctg gtg aac cgc atc gag ctg aag ggc atc gac ttc aag gag gacggc aac atc ctg ggg cac aag ctg gag tac aac tac aac agc cac aac gtc tat atcatg gcc gac aag cag aag aac ggc atc aag gCC aac ttc aag atc cgc cac aac atcgag gac ggc agc gtg cag ctc gcc gac cac tac cag cag aac acc ccc atc ggc gacggc ccc gtg ctg ctg ccc gac aac cac tac ctg agc acc cag tcc gcc ctg agc aaagac ccc aac gag aag cgc gat cac atg gtc ctg ctg gag ttc gtg acc gcc gcc gggatc act cAc ggc atg gac gag ctg tac aag tcc ggc ggg agc gga tcc ggc ggc cagtcc ggc ggg agc gga tcc ggc ggc cag tcc ggc ggg agc gga tcc ggc ggc cag tccggc ggg agc gga tcc ggc ggc cag tcc ggc ggg agc gga tcc ggc ggc cag tcc ggcctc aGA TCT gtc aaa ctt aca tca gac ttc gac aac cca aga tgg att gga cga cacaag cat atg ttc aat ttc ctt gat gtc aac cac aat gga aaa atc tct ctt gac gagatg gtc tac aag gca tct gat att gtc atc aat aac ctt gga gca aca cct gag caagcc aaa cga cac aaa gat gct gtG gaa gcc ttc ttc gga gga gct gga atg aaa tatggt gtg gaa act gat tgg cct gca tat att gaa gga tgg aaa aaa ttg gct act gatgaa ttg gag aaa tac gcc aaa aac gaa cca acC ctc atc cgC ata tgg ggt gat gctttg ttt gat atc gtt gac aaa gat caa aat gga gct att aca ctg gat gaa tgg aaagca tac acc aaa gct gct ggt atc atc caa tca tca gaa gat tgc gag gaa aca ttcaga gtg tgc gat att gat gaa aqt gga caa ctc gat gtt gat gag atg aca aga caGcat CtG gga ttt tgg tac acc atg gat cct gct tgc gaa aag ctc tac ggt gga gctgtc ccc [SEQ ID NO: 11]SeG5AAtg gtg agt gcc agt cgt cct gag gcc ctg gct gcc cct gtc acc act gtt gcg accctt gtc cca cac aac gcc act gag cca gcc agt cct ggg gaa ggg aag gaa gat gccttt tcc aag ctg aag cag aag ttt atg aat gaa ctg cat aaa atc cca ttg cca ccgtgg gcc tta att gcc ata gcc ata gtt gcg gtc ctt cta gtc gtg acc tgc tgc ttctgt gtc tgt aag aaa tgt ttg ttc aaa aag aaa aac aag aag aag gga aag gaa aaggga ggg aag aac gcc att aac atg aaa gac gtg aaa gac tta ggg aag acc atg aaggat cag gcc ctt aag gat gac gat gct gaa act gga ctg act gat gga gaa gaa aaggag gag ccc aag gaa gag gag aaa ctg gga aag ctt caa tat tca ctg gac tat gacttc cag aat aac cag ctg ctg gtg gga atc atc cag gct gct gaa ctg ccc gcc ctggac atg gga ggc aca tct gat cca tac gtc aaa gtc ttc ctg ctg ccc gac aaa aagaag aag ttt gag aca aaa gtc cac cgg aaa acc ctc aat cca gtc ttc aat gaa cagttt act ttc aag gtg cca tac tcg gaa tta ggt ggc aag aca ctg gtg atg gct gtgtat gat ttt gac cgc ttc tcc aag cac gac atc att gga gag ttc aaa gtt cct atgaac acc gtg gat ttt ggc cac gtc acc gag gag tgg cgc gat ctc cag agt gct gagaaa gaa gag caa gag aaa ctg ggt gac atc tgc ttc tcc ctc cgc tac gtc cct actgcc ggc aag ctg act gtt gtc att ctg gaa gcc aag aac ctg aag aag atg gat gtgqgt ggc tta tct gat ccc tat gta aag att cac ctg atg cag aac ggc aag aga ctgaag aag aaa aag aca acg att aag aag aac aca ctt aac ccc tac tac aat gag tccttc agc ttt gaa gtt ccg ttc gag caa atc cag aaa gtg caa gtg gtg gta act gttttg gac tat gac aag att ggc aag aac gac gcc atc ggc aaa gtc ttt gtg ggc tacaac agc acc ggc gca gag ctg cga cac tgg tca gac atg ctg gcc aac ccc cgg cgcccc atc gcc cag tgg cac act ctg cag gta gag gag gag gtt gat gcc atg ctg gctgtc aag aGA tCC GGG AAT TCC GGG CGG gcc acc atg agc aag ggc gag gag ctg ttcacc ggg gtg gtg ccc atc ctg gtc gag ctg gac ggc gac gta aac ggc cac aag ttcagc gtg tcc ggc gag ggc gag ggc gat gcc acc tac ggc aag ctg acc ctg aag ttcatc tgc acc acc ggc aag ctg ccc gtg ccc tgg ccc acc ctc gtg acc acc ctg acctac ggc gtg cag tgc ttc agc cgc tac ccc gac cac atg aag cag cac gac ttc ttcaag tcc gcc atg ccc gaa ggc tac gtc cag gag cgc acc atc ttc ttc aag gac gacggc aac tac aag acc cgc gcc gag gtg aag ttc gag ggc gac acc ctg gtg aac cgcatc gag ctg aag ggc atc gac ttc aag gag gac ggc aac atc ctg ggg cac aag ctggag tac aac tac aac agc cac aac gtc tat atc atg gcc gac aag cag aag aac ggcatc aag gCC aac ttc aag atc cgc cac aac atc gag gac ggc agc gtg cag ctc gccgac cac tac cag cag aac acc ccc atc ggc gac ggc ccc gtg ctq ctg ccc gac aaccac tac ctg agc acc cag tcc gcc ctg agc aaa gac ccc aac gag aag cgc gat cacatg gtc ctg ctg gag ttc gtg acc gcc gcc ggg atc act cAc ggc atg gac gag ctgtac aag tcc ggc ggg agc gga tcc ggc ggc cag tcc ggc ggg agc gga tcc ggc ggccag tcc ggc ggg agc gga tcc ggc ggc cag tcc ggc ggg agc gga tcc ggc ggc cagtcc ggc ggg agc gga tcc ggc ggc cag tcc ggc ctc aGA TCT gtc aaa ctt aca tcagac ttc gac aac cca aga tgg att gga cga cac aag cat atg ttc aat ttc ctt gatgtc aac cac aat gga aaa atc tct ctt gac gag atg gtc tac aag gca tct gat attgtc atc aat aac ctt gga gca aca cct gag caa gcc aaa cga cac aaa gat gct gtGgaa gcc ttc ttc gga gga gct gga atg aaa tat ggt gtg gaa act gat tgg cct gcatat att gaa gga tgg aaa aaa ttg gct act gat gaa ttg gag aaa tac gcc aaa aacgaa cca acC ctc atc cgC ata tgg ggt gat gct ttg ttt gat atc gtt gac aaa gatcaa aat gga gct att aca ctg gat gaa tgg aaa gca tac acc aaa gct gct ggt atcatc caa tca tca gaa gat tgc gag gaa aca ttc aga gtq tgc gat att gat gaa agtgga caa ctc gat gtt gat gag atg aca aga caG cat CtG gga ttt tgg tac acc atggat cct gct tgc gaa aag ctc tac ggt gga gct gtc ccc [SEQ ID NO:12]GFP-水母发光蛋白接头的DNA序列:pGA(菌株I2507) TCC GGC CTC AGA TCT[SEQ ID NO:13]pG1A(菌株I2508) TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
CTC AGA TCT[SEQ ID NO:14]pG2A(菌株I2509) TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
GGG AGC GGA TCC GGC GGC CAG TCC GGC CTC AGA
TCT[SEQ ID NO:15]pG4A(菌株I2510) TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
GGG AGC GGA TCC GGC GGC CAG TCC GGC GGG AGC
GGA TCC GGC GGC CAG TCC GGC GGG AGC GGA TCC
GGC GGC CAG TCC GGC CTC AGA TCT[SEQ ID NO:
16]pG5A(菌株I-2511)TCC GGC GGG AGC GGA TCC GGC GGC CAG TCC GGC
GGG AGC GGA TCC GGC GGC CAG TCC GGC GGG AGC
GGA TCC GGC GGC CAG TCC GGC GGG AGC GGA TCC
GGC GGC CAG TCC GGC GGG AGC GGA TCC GGC GGC
CAG TCC GGC CTC AGA TCT[SEQ ID NO:17]pSeG5A(菌株I2512)和pStG5A(菌株I2513)与pG5A相同接头的肽序列:pGA Ser Gly Leu Arg Ser[SEQ ID NO:18]pG1A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Leu Arg Ser
[SEQ ID NO:19]pG2A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly
Ser Gly Gly Gln Ser Gly Leu Arg Ser[SEQ ID NO:20]pG4A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly
Ser Gly Gly Gln Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser
Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Leu Arg Ser[SEQ
ID NO:21]pG5A Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly
Ser Gly Gly Gln Ser Gly Gly Ser Gly Ser Gly Gly Gln Ser
Gly Gly Ser Gly Ser Gly Gly Gln Ser Gly Gly Ser Gly Ser
Gly Gly Gln Ser Gly Leu Arg Ser[SEQ ID NO:22]PseG5A和pStGSA与pG5A相同参考文献
本文引用了下列发表物。本文依赖并收入这些发表物的完整公开书作为参考。
1.Berridge,M.J.(1998)Neuron 21,13-26.
2.Cobbold,P.H.,& Rink,T.J.(1987)Biochem.J.248,313-323.
3.Miyawaki,A.,Griesbeck,O.,Heim,R.,& Tsien,R.Y.(1999)Proc.Natl.Acad.Sci.USA 96,2135-2140.
4.Romoser,V.A.,Hinkle,P.M.,& Persechini,A.(1997)J.Biol.Chem.272,13270-13274.
5.Inui,S.,Noguchi,M.,Sack,Y.,Takagi,Y.,Miyata,T.,Awing,S.,Miyata,T,&Tsuji,F.I.(1985)Proc.Natl.Acad.Sci.USA 82,3154-3158.
6.Prasher,D.,McCann,R.O.,& Cormier,M.J.(1985)Biochem.Biophys.Res.Comm.126,1259-1268.
7.Tsuji,F.I.,Inouye,S.,Goto,T.,& Sakaki,Y.(1986)Proc.Natl.Acad.Sci.USA83,8107-8111.
8.Shimomura,O.,& Johnson,F.H.(1978)Proc.Natl.Acad.Sci.USA 75,2611-2615.
9.Sala-Newby,G.B.,Badminton,M N.,Evans,W.H.,Georges,C.H.,Jones,H.E.,Kendal,J.M.,Ribeiro,A.R.,& Campbell,A.K.(2000)Methods Enzymol.305,479-498.
10.Shimomura,O.,Johnson,F.H.,& Saiga,Y.(1962)J.Cell Comp.Physiol.59,223-239.
11.Johnson,F.H.,Shimomura,O.,Saiga,Y.,Gershman,L.C.,Reynolds,G.T.,&Waters,J.R.(1962)J.Cell Comp.Physiol 60,85-103.
12.Cubitt,A.B.,Heim,R.,Adams,S.R.,Boyd,A.E.,Gross,L.A.,& Tsien,R.Y.(1995)Trends Biochem.Sci.,20,448-455.
13.Ward,W.W.,& Cormier,M.J.(1976)J.Phys.Chem.80,2289-2291.
14.Ward,W.W.1 & Cormier,M.J.(1978)Photochem.Photobiol.27,389-396.
15.Morise,H.,Shimomura,O.,Johnson,F.H.,& WinNT,J.(1974)Biochemistry13,2656-2662.
16.Campbell,A.K.,1988,《Chemiluminescence,Principles andApplication in Biology and Medecin》(化学发光的原理及在生物学和医学中的应用),Ellis Horwood有限公司(奇切斯特市)编,
17.Cormack,B.P.,Valdivia,R.H.,& Falkow,S.(1996)Gene 173,33-38.
18.Crameri,A.,Whitehom,E.A.,Tate,E.,Stemmer,W.P.C.(1996)NatureBiotech.14,315-319.
19.Siemering,K.R.,Golbik,R.,Sever,R.,& Haseloff,J.(1996)Curr.Biol.6,1653-1663.
20.Watkins,N.J.,& Campbell,A.K.(1993)Biochem.J.,293,181-185.
21.Badminton,M.N.,Sala-Newby,G.B.,Kendall,J.M.,& Campbell,A.K.(1995)Biochem.Biophys.Res.Comm.217,950-957.
22.Morin,J.G.,& Hastings,J.W.(1970)J.Cell.Physiol.77,313-318.
23.Campbell,A.K.,& Hallett,M.B.(1978)Proc.Physiol.Soc.,287,4-5.
24.Yang,F.,Moss,L.G.,& Phillips,Jr.,G.N.(1996)Nature Biotech.14,1246-1251.
25.Brose,N.,Petrenko,A.G.,Sladhof,T,C.,& Jahm,R.(1992)Science 256,1021-1025.
26.Coco,S.,Verderio,C.,De Camilli,P.,& Matteoli,M(1998)J.Neurochem.71,1987-1992.
27.Rutter,G.A.,Burnett,P.,Rizzuto,R.,Brini,M.,Murgia,M.,Pozzan,T.,TavaréJ.M.,& Denton,R.M.(1996)Proc.Natl.Acad.Sci.USA 93,5489-5494.
28.Sah,P.(1996)Trends Neurosci.19,150-154.
29.Etter,E.F.,Minta,A.,Poenie,M.,& Fay,F.S.(1996)Proc.Natl.Acad.Sci.USA 93,5368-5373.
30.Miesenbck,G.,& Rothman,J.E.(1997)Prod.Natl.Acad.Sci.USA 94,3402-3407.
31.Miyawaki,A.,Llopis,J.,Heim,R.,McCaffery,J.M.,Adams,J.A.,Ikura,M.,&Tsien,R.Y,(1997)Nature 388,882-887.
本文还完整收入美国专利号5,683,888作为参考。
Claims (52)
1.包含纯化的多肽的组合物,其中所述组合物具有结合钙离子和容许可测量的能量的功能性特征,所述能量在不存在任何光激发时依赖所述组合物中所结合的钙的数量和多肽的数量。
2.具有SEQ ID NO:1的氨基酸序列的纯化的多肽。
3.具有SEQ ID NO:2的氨基酸序列的纯化的多肽。
4.具有SEQ ID NO:3的氨基酸序列的纯化的多肽。
5.具有SEQ ID NO:4的氨基酸序列的纯化的多肽。
6.具有SEQ ID NO:5的氨基酸序列的纯化的多肽。
7.具有SEQ ID NO:6的氨基酸序列的纯化的多肽。
8.具有SEQ ID NO:7的序列的纯化的多核苷酸。
9.具有SEQ ID NO:8的序列的纯化的多核苷酸。
10.具有SEQ ID NO:9的序列的纯化的多核苷酸。
11.具有SEQ ID NO:10的序列的纯化的多核苷酸。
12.具有SEQ ID NO:11的序列的纯化的多核苷酸。
13.具有SEQ ID NO:12的序列的纯化的多核苷酸。
14.具有SEQ ID NO:13的多核苷酸序列的多核苷酸接头。
15.具有SEQ ID NO:14的多核苷酸序列的多核苷酸接头。
16.具有SEQ ID NO:15的多核苷酸序列的多核苷酸接头。
17.具有SEQ ID NO:16的多核苷酸序列的多核苷酸接头。
18.具有SEQ ID NO:17的多核苷酸序列的多核苷酸接头。
19.根据权利要求14-18任一项的多核苷酸接头,它在翻译后具有在最佳条件下使供体位点接近受体位点的功能,从而容许根据权利要求1的纯化的多肽中通过化学发光共振能量转移(CRET)进行直接的能量转移。
20.具有至少5个氨基酸且包含SEQ ID NO:18的氨基酸序列的肽接头。
21.具有至少5个氨基酸且包含SEQ ID NO:19的氨基酸序列的肽接头。
22.具有至少5个氨基酸且包含SEQ ID NO:20的氨基酸序列的肽接头。
23.具有至少5个氨基酸且包含SEQ ID NO:21的氨基酸序列的肽接头。
24.具有至少5个氨基酸且包含SEQ ID NO:22的氨基酸序列的肽接头。
25.肽接头,它具有在最佳条件下使供体位点接近受体位点的功能,从而容许根据权利要求2-7的纯化的多肽中通过化学发光进行直接的能量转移。
26.根据权利要求20-25任一项的肽接头,它具有在最佳条件下使供体位点接近受体位点的功能,从而容许在根据权利要求1的纯化的多肽存在时进行直接的能量转移。
27.根据权利要求20-26任一项的肽接头,它具有在体内和/或在体外稳定经修饰的生物发光系统的能力。
28.包含两个生物发光蛋白质和根据权利要求20-27任一项的肽接头的经修饰的生物发光系统。
29.根据权利要求28的经修饰的生物发光系统,其中所述两个生物发光蛋白质至少包含水母发光蛋白。
30.根据权利要求28或29的经修饰的生物发光系统,它包含如下成分:水母发光蛋白和GFP蛋白。
31.用于在体内或在体外测量能量转移的试剂盒,它至少包含一种根据权利要求2-7的多肽或根据权利要求8-13的多核苷酸,以及在存在或缺乏目的分子时显现或检测所述转移所必需的试剂。
32.具有如下通式的融合蛋白:
GFP-接头-AEQ,
其中GFP指绿色荧光蛋白;AEQ指水母发光蛋白;而接头指4-63个氨基酸的多肽。
33.权利要求32的融合蛋白,其中接头包含14-50个氨基酸。
34.权利要求32和33的融合蛋白,其中接头包含如下氨基酸:
(Gly Gly Ser Gly Ser Gly Gly Gln Ser[SEQ ID NO:251])n,
其中n是1-5。
35.权利要求34的融合蛋白,其中n是1。
36.权利要求34的融合蛋白,其中n是5。
37.用于在Ca2+存在时活化水母发光蛋白后通过化学发光共振能量转移(CRET)将能量由水母发光蛋白转移至绿色荧光蛋白的融合蛋白,其中融合蛋白具有如下通式:
GFP-接头-AEQ,
其中GFP指绿色荧光蛋白;AEQ指水母发光蛋白;而接头包含如下氨基酸:
(Gly Gly Ser Gly Ser Gly Gly Gln Ser[SEQ ID NO:25])n,
其中n是1-5;且其中融合蛋白具有Ca2+离子亲合力和至少24小时的半衰期。
38.权利要求32-37的融合蛋白,其中接头包含氨基酸序列SerGly Leu Arg Ser[SEQ ID NO:26]。
39.权利要求32-38的融合蛋白,它还包含将融合蛋白靶向细胞或亚细胞区室的肽信号序列。
40.编码权利要求32-39任一项的融合蛋白的多核苷酸。
41.根据权利要求1的组合物,其中所述纯化的多肽是根据权利要求2-7任一项的纯化的多肽、或根据权利要求28-30任一项的经修饰的生物发光系统、或根据权利要求32-39任一项的融合蛋白。
42.保藏于C.N.C.M.且包含质粒I-2507的培养物。
43.保藏于C.N.C.M.且包含质粒I-2508的培养物。
44.保藏于C.N.C.M.且包含质粒I-2509的培养物。
45.保藏于C.N.C.M.且包含质粒I-2510的培养物。
46.保藏于C.N.C.M.且包含质粒I-2511的培养物。
47.保藏于C.N.C.M.且包含质粒I-2512的培养物。
48.保藏于C.N.C.M.且包含质粒I-2513的培养物。
49.在体内筛选物理、化学、生化或生物学状态变化的方法,包括:
a)对哺乳动物施用根据权利要求1或41的组合物;
b)检测是否发光;并
c)可选地测量钙流的离子浓度。
50.在体外筛选物理、化学、生化或生物学条件变化的方法,包
(a)在存在或缺乏待测目的分子时向包含目的分析物的反应系统中加入根据权利要求1或41的组合物;并
(b)显现步骤(a)中产生的能量的发射。
51.筛选导致体内物理、化学、生化或生物学条件变化的产物的方法,包括:
(a)在存在或缺乏待测目的分子时对脊椎动物施用包含根据权利要求1或41的组合物的药学可接受介质;
(b)在存在所述组合物时检测产生的能量;并
(c)可选地测量检测步骤(b)中的能量所必需的所述目的分子的有效浓度。
52.在体外筛选能够在根据权利要求1或41的组合物中调节能量的分子的方法,包括:
(a)向包含待测分子的反应系统中提供在生物学样品中的根据权利要求1或41的组合物;
(b)通过与包含根据权利要求1-41的所述组合物但不含待测分子的对照样品比较来检测能量的调节;并
(c)可选测定能够抑制或提高所述组合物的能量转移的所述分子的最低有效浓度。
Applications Claiming Priority (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US20831400P | 2000-06-01 | 2000-06-01 | |
| US60/208,314 | 2000-06-01 | ||
| US21052600P | 2000-06-09 | 2000-06-09 | |
| US60/210,526 | 2000-06-09 | ||
| US25511100P | 2000-12-14 | 2000-12-14 | |
| US60/255,111 | 2000-12-14 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN1468312A true CN1468312A (zh) | 2004-01-14 |
| CN100467604C CN100467604C (zh) | 2009-03-11 |
Family
ID=27395180
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN01812104.7A Expired - Fee Related CN100467604C (zh) | 2000-06-01 | 2001-06-01 | 嵌合GFP-水母发光蛋白作为单细胞水平的生物发光的Ca2+报道分子 |
Country Status (14)
| Country | Link |
|---|---|
| US (3) | US6936475B2 (zh) |
| EP (1) | EP1292613B9 (zh) |
| JP (1) | JP5389307B2 (zh) |
| CN (1) | CN100467604C (zh) |
| AT (1) | ATE407142T1 (zh) |
| AU (1) | AU784887B2 (zh) |
| CA (1) | CA2411304C (zh) |
| CY (1) | CY1110438T1 (zh) |
| DE (1) | DE60135652D1 (zh) |
| DK (1) | DK1292613T3 (zh) |
| ES (1) | ES2312454T3 (zh) |
| HK (1) | HK1060370A1 (zh) |
| PT (1) | PT1292613E (zh) |
| WO (1) | WO2001092300A2 (zh) |
Families Citing this family (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| PT1292613E (pt) * | 2000-06-01 | 2008-11-10 | Pasteur Institut | Gpf-aequorina quiméria como repórteres de ca++ bioluminiscentes, ao nível da célula singular |
| CA2421858A1 (en) * | 2000-09-07 | 2002-03-14 | Bjorn A. Olde | Reporter system for cell surface receptor-ligand binding |
| DE03779067T1 (de) * | 2002-11-12 | 2006-06-22 | Zakrytoe Aktsionernoe Obschestvo "Evrogen" | Fluoreszenzproteine und chromoproteine aus nicht-aequorea-hydrozoa-spezies sowie verfahren zur verwendung davon |
| CN1304571C (zh) * | 2003-10-16 | 2007-03-14 | 清华大学 | 一种培育转基因斑马鱼的方法及其专用质粒 |
| EP1714155A1 (en) * | 2004-02-12 | 2006-10-25 | Institut Pasteur | Non-invasive real-time in vivo bioluminescence imaging of local ca2+ dynamics in living organisms |
| GB0625678D0 (en) | 2006-12-21 | 2007-01-31 | Lux Biotechnology Ltd | Composition and method for detection of demineralisation |
| ES2427854B1 (es) * | 2012-03-29 | 2014-09-15 | Universidad De Valladolid | Sensores de calcio y métodos para la detección de calcio libre intracelular |
| ES2452485B1 (es) * | 2012-07-13 | 2015-03-09 | Univ Valladolid | Mutantes de apoacuorina y metodos para su uso |
| ES2445018B1 (es) * | 2012-07-27 | 2014-12-12 | Universidad De Castilla La Mancha | VARIANTES DE LA PROTEÍNA DE FUSIÓN SENSIBLE AL CALCIO tdTOMATO-AEQUORINA |
| FR2996225B1 (fr) | 2012-10-03 | 2014-10-03 | Centre Nat Rech Scient | Analogues de la coelenterazine emettant une lumiere decalee vers le rouge visant l'imagerie in vivo |
| GB201700317D0 (en) | 2017-01-09 | 2017-02-22 | Calcivis Ltd | Detection device |
| WO2019241318A1 (en) * | 2018-06-13 | 2019-12-19 | Cornell University | Bioluminescent markers of neural activity |
| USD1042180S1 (en) | 2020-05-28 | 2024-09-17 | Calcivis Limited | Dental detection device |
Family Cites Families (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB8916806D0 (en) * | 1989-07-22 | 1989-09-06 | Univ Wales Medicine | Modified proteins |
| US5498521A (en) * | 1990-01-24 | 1996-03-12 | President And Fellows Of Harvard College | Diagnosis of hereditary retinal degenerative diseases |
| US5614184A (en) * | 1992-07-28 | 1997-03-25 | New England Deaconess Hospital | Recombinant human erythropoietin mutants and therapeutic methods employing them |
| US5714666A (en) * | 1993-02-09 | 1998-02-03 | Children's Hospital Of Philadelphia | Measurement of intracellular calcium using bioluminescent apoaequorin expressed in mammalian cells |
| US6818439B1 (en) * | 1994-12-30 | 2004-11-16 | Chiron Corporation | Methods for administration of recombinant gene delivery vehicles for treatment of hemophilia and other disorders |
| US5744326A (en) * | 1996-03-11 | 1998-04-28 | The Immune Response Corporation | Use of viral CIS-acting post-transcriptional regulatory sequences to increase expression of intronless genes containing near-consensus splice sites |
| AU765703B2 (en) * | 1998-03-27 | 2003-09-25 | Bruce J. Bryan | Luciferases, fluorescent proteins, nucleic acids encoding the luciferases and fluorescent proteins and the use thereof in diagnostics, high throughput screening and novelty items |
| DK1071809T3 (da) * | 1998-04-17 | 2003-12-01 | Rigel Pharmaceuticals Inc | Multiparameter FACS test til påvisning af ændringer i cellulære parametre |
| US6800492B2 (en) | 2000-06-01 | 2004-10-05 | Institute Pasteur | Chimeric GFP-aequorin as bioluminescent Ca++ reporters at the single cell level |
| PT1292613E (pt) | 2000-06-01 | 2008-11-10 | Pasteur Institut | Gpf-aequorina quiméria como repórteres de ca++ bioluminiscentes, ao nível da célula singular |
-
2001
- 2001-06-01 PT PT01949426T patent/PT1292613E/pt unknown
- 2001-06-01 AT AT01949426T patent/ATE407142T1/de active
- 2001-06-01 DE DE60135652T patent/DE60135652D1/de not_active Expired - Lifetime
- 2001-06-01 ES ES01949426T patent/ES2312454T3/es not_active Expired - Lifetime
- 2001-06-01 EP EP01949426A patent/EP1292613B9/en not_active Expired - Lifetime
- 2001-06-01 AU AU70583/01A patent/AU784887B2/en not_active Ceased
- 2001-06-01 CN CN01812104.7A patent/CN100467604C/zh not_active Expired - Fee Related
- 2001-06-01 DK DK01949426T patent/DK1292613T3/da active
- 2001-06-01 CA CA2411304A patent/CA2411304C/en not_active Expired - Fee Related
- 2001-06-01 WO PCT/EP2001/007057 patent/WO2001092300A2/en active IP Right Grant
- 2001-06-01 JP JP2002500911A patent/JP5389307B2/ja not_active Expired - Fee Related
-
2002
- 2002-12-02 US US10/307,389 patent/US6936475B2/en not_active Expired - Fee Related
-
2004
- 2004-05-14 HK HK04103420.0A patent/HK1060370A1/xx not_active IP Right Cessation
-
2005
- 2005-06-10 US US11/149,177 patent/US7763443B2/en not_active Expired - Fee Related
-
2008
- 2008-12-02 CY CY20081101381T patent/CY1110438T1/el unknown
-
2010
- 2010-06-04 US US12/794,147 patent/US20100330667A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| US20030175807A1 (en) | 2003-09-18 |
| US20100330667A1 (en) | 2010-12-30 |
| AU7058301A (en) | 2001-12-11 |
| PT1292613E (pt) | 2008-11-10 |
| JP5389307B2 (ja) | 2014-01-15 |
| WO2001092300A2 (en) | 2001-12-06 |
| DE60135652D1 (de) | 2008-10-16 |
| US6936475B2 (en) | 2005-08-30 |
| US20080213879A1 (en) | 2008-09-04 |
| CN100467604C (zh) | 2009-03-11 |
| ATE407142T1 (de) | 2008-09-15 |
| US7763443B2 (en) | 2010-07-27 |
| ES2312454T3 (es) | 2009-03-01 |
| CA2411304C (en) | 2014-07-29 |
| CA2411304A1 (en) | 2001-12-06 |
| AU784887B2 (en) | 2006-07-20 |
| CY1110438T1 (el) | 2015-04-29 |
| EP1292613A2 (en) | 2003-03-19 |
| WO2001092300A3 (en) | 2002-11-07 |
| EP1292613B1 (en) | 2008-09-03 |
| JP2004532001A (ja) | 2004-10-21 |
| EP1292613B9 (en) | 2009-04-01 |
| DK1292613T3 (da) | 2008-11-24 |
| HK1060370A1 (en) | 2004-08-06 |
| WO2001092300A9 (en) | 2009-01-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US7858844B2 (en) | Non aggregating fluorescent proteins and methods for using the same | |
| AU730040B2 (en) | Renilla luciferase and green fluorescent protein fusion genes | |
| CN1317299C (zh) | 生物发光提高的发光蛋白质 | |
| JP5963391B2 (ja) | Anthozoa綱の非生物発光性種由来の蛍光タンパク質、そのようなタンパク質をコードする遺伝子、およびそれらの使用 | |
| JP3810791B2 (ja) | 緑色蛍光タンパク質の使用 | |
| US7763443B2 (en) | Polynucleotides encoding chimeric GFP-aequorin as bioluminescent Ca++ reporters | |
| JP2017051208A (ja) | 花虫類に由来する発色団/蛍光体、およびそれらの使用法 | |
| JP4330338B2 (ja) | 大きく赤側にシフトした蛍光タンパク質 | |
| US9102750B2 (en) | Branchiostoma derived fluorescent proteins | |
| US6800492B2 (en) | Chimeric GFP-aequorin as bioluminescent Ca++ reporters at the single cell level | |
| CA2434737C (en) | Non aggregating fluorescent proteins and methods for using the same | |
| JP2010017186A (ja) | 連結された色素/蛍光性ドメインをコードする核酸ならびにその使用法 | |
| CN1438239A (zh) | 一种新型荧光蛋白,编码其的多核苷酸序列及其用途 | |
| Tsien et al. | Red-shifted fluorescent proteins mPlum and mRaspberry and polynucleotides encoding the same |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| REG | Reference to a national code |
Ref country code: HK Ref legal event code: DE Ref document number: 1060370 Country of ref document: HK |
|
| C14 | Grant of patent or utility model | ||
| GR01 | Patent grant | ||
| REG | Reference to a national code |
Ref country code: HK Ref legal event code: GR Ref document number: 1060370 Country of ref document: HK |
|
| CF01 | Termination of patent right due to non-payment of annual fee | ||
| CF01 | Termination of patent right due to non-payment of annual fee |
Granted publication date: 20090311 Termination date: 20160601 |