CN108064308B - 细菌中产生双链蛋白的方法 - Google Patents
细菌中产生双链蛋白的方法 Download PDFInfo
- Publication number
- CN108064308B CN108064308B CN201580059973.5A CN201580059973A CN108064308B CN 108064308 B CN108064308 B CN 108064308B CN 201580059973 A CN201580059973 A CN 201580059973A CN 108064308 B CN108064308 B CN 108064308B
- Authority
- CN
- China
- Prior art keywords
- coli
- protein
- antibody
- production
- growth
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images




























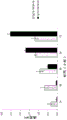
























Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/24—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against cytokines, lymphokines or interferons
- C07K16/244—Interleukins [IL]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/46—Hybrid immunoglobulins
- C07K16/468—Immunoglobulins having two or more different antigen binding sites, e.g. multifunctional antibodies
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/10—Immunoglobulins specific features characterized by their source of isolation or production
- C07K2317/14—Specific host cells or culture conditions, e.g. components, pH or temperature
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/31—Immunoglobulins specific features characterized by aspects of specificity or valency multispecific
Abstract
本文提供了产生含有两条链的多肽(如包括轻链和重链的抗体)的方法。具体地说,提供了通过利用优化的表达载体和培养过程在细菌中产生异源分泌性蛋白的方法。
Description
相关申请的交叉引用
本申请要求2015年8月20日提交的美国临时申请号62/207,882和2014年11月5提交的美国临时申请号62/075792的权益,所述文献通过引用的方式完整并入本文。
提交的ASCII文本文件的序列表
以下在ASCII文本文件上提交的内容通过引用方式完整并入本文:序列表的计算机可读形式(文件名:146392024040SEQLIST.TXT,记录日期:2015年11月5日,大小:49KB)。
发明领域
本公开涉及产生重组多肽(如抗体)的方法。更具体地,本公开涉及通过利用优化的表达载体和培养过程在细菌中产生异源分泌性蛋白的方法。
背景
自1978年在大肠杆菌中生产人胰岛素以来,在原核宿主细胞中生产重组蛋白已经成为许多重要治疗药的来源。随着分子生物学工具和知识已经取得进展,重组治疗药的复杂性也已经增加。这些重组蛋白的生产要求产物显示多种特性如正确翻译、折叠、装配、形成二硫键和转运至周质。已知许多重组蛋白、尤其具有的那些二硫键(例如,双链蛋白,包括而不限于抗体和抗体片段)的表达导致原核宿主细胞中形成包涵体(Spadiut等人,Trendsin Biotechnology,32:54,2014)。因此,需要在原核宿主细胞中以产业规模重组生产正确折叠和装配的双链蛋白的表达系统和过程。
单克隆抗体代表增长最快的重组治疗药类型之一,其中多种单克隆抗体已经获准治疗多种疾病或就此接受审查(Nelson等人,Nature Review Drug Discovery,9:767,2010)。传统的单克隆抗体结合单一靶抗原。对于许多疾病,可能有利的是利用结合多于一种靶抗原的抗体,即,多特异性抗体。这类抗体可以用于针对多种治疗靶的组合方法中(参见,例如,Bostrom等人,Science 323:1610,2009;和Wu等人,Nature Biotechnology,25:1290,2007)。例如,可以产生这样的双特异性抗体,它们同时结合癌细胞表面上表达的表位和T细胞上表达的表位以诱导T细胞介导的肿瘤细胞杀伤作用(Shalaby等人,ClinicalImmunology,74:185,1995)。
临床使用双特异性抗体要求以有产业意义的量生产双链蛋白的能力。尽管已经描述改善原核宿主细胞中重组蛋白生产的载体组件(参见,例如,Schlapschy等人,ProteinEngineering,Design and Selection,19:385,2006;和Simmon等人,Journal ofImmunological Methods 263:133,2002),但本文所述的结果显示修饰表达载体并未解决生产双链蛋白期间遇到的全部生产问题。仍需要以制备规模高效产生重组双链蛋白(如抗体片段和半抗体)的最佳方法。
本文援引的全部参考文献,包括专利申请、专利出版物和UniProtKB/Swiss-Prot登录号,均通过引用方式完整并入本文,如同特别和单独指明每份独立的参考文献通过引用的方式并入。
概述
在一个方面,本文中提供在原核宿主细胞中产生包含两条链的多肽的方法,所述方法包括:(a)培养宿主细胞以表达多肽的两条链,因而当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞包含多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;和(3)编码选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合的至少一种伴侣蛋白的第三翻译单位;其中在培养基中在包含以下的条件下培养宿主细胞:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收生物活性多肽。还提供在原核宿主细胞中产生包含两条链的多肽的方法,所述方法包括:(a)在培养基中在包含以下的条件下培养宿主细胞以表达多肽的两条链:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,因而当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞包含多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;和(3)编码选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合的至少一种伴侣蛋白的第三翻译单位;其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收生物活性多肽。还提供在原核宿主细胞中产生包含两条链的多肽的方法,所述方法包括:(a)培养宿主细胞以表达多肽的两条链,因而当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞包含多核苷酸,所述多核苷酸包含:(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合;其中在培养基中在包含以下的条件下培养宿主细胞:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收生物活性多肽。还提供在原核宿主细胞中产生包含两条链的多肽的方法,所述方法包括:(a)在培养基中在包含以下的条件下培养宿主细胞以表达多肽的两条链:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,因而当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞包含多核苷酸,所述多核苷酸包含:(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合;其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收生物活性多肽。在一些实施方案中,该多肽包含三、四或五条链。在一些实施方案中,pH培养基在生产阶段期间维持在6.7和7.3之间的pH范围内。在一些实施方案中,多核苷酸还包含启动子的三个副本,其中第一副本与第一翻译单位有效组合,第二副本与第二翻译单位有效组合并且第三副本与第三翻译单位有效组合,以驱动第一链、第二链和伴侣蛋白的转录。
在一些实施方案中,编码三种伴侣蛋白中两者的两个翻译单位是单一转录单位(双顺反子单位)的组成部分。在一些实施方案中,多核苷酸还包含与每个翻译单位有效组合的启动子。在一些实施方案中,启动子是诱导型启动子。在一些实施方案中,诱导型启动子是在IPTG诱导作用不存在的情况下驱动第一链、第二链和伴侣蛋白转录的IPTG诱导型启动子。在一些实施方案中,诱导型启动子是当培养基中的磷酸盐已经耗尽时驱动第一链、第二链和伴侣蛋白转录的Pho启动子。在一些实施方案中,多核苷酸还包含选择标记并且培养基包含由引起宿主细胞保留所述多核苷酸的单一抗生素组成的选择剂。在一些实施方案中,第一翻译单位包含与第一链编码区有效组合的第一翻译起始区域(TIR),并且第二翻译单位包含与第二链编码区有效组合的第二翻译起始区域(TIR),其中第一和第二TIR的相对翻译强度是约1.0至约3.0。在一些实施方案中,至少一种伴侣蛋白或第一伴侣蛋白包括肽酰基-脯氨酰异构酶。在一些实施方案中,肽酰基-脯氨酰异构酶是FkpA蛋白。在一些实施方案中,FkpA是大肠杆菌(E.coli)FkpA。在一些实施方案中,至少一种伴侣蛋白还包括或第二伴侣蛋白和第三伴侣蛋白之一或两者包括蛋白质二硫键氧化还原酶。在一些实施方案中,蛋白质二硫键氧化还原酶是DsbA蛋白和DsbC蛋白之一或两者。在一些实施方案中,至少一种蛋白质二硫键氧化还原酶是大肠杆菌DsbA和大肠杆菌DsbC之一或两者。在一些实施方案中,原核宿主细胞是革兰氏阴性细菌。在一些实施方案中,革兰氏阴性细菌是大肠杆菌。在一些实施方案中,大肠杆菌是内源蛋白酶活性缺陷的菌株。在一些实施方案中,大肠杆菌是具有degpS210A突变的菌株。在一些实施方案中,大肠杆菌是具有基因型W3110 ΔfhuA ΔphoA ilvG2096(Valr)Δprc spr43H1 ΔdegP ΔmanA lacIQ ΔompT ΔmenE degpS210A的菌株。在一些实施方案中,多肽对宿主细胞为异源。在一些实施方案中,多肽是异二聚体(例如,双特异性抗体)的单体。在一些实施方案中,多肽的两条链彼此通过至少一个二硫键连接。在一些实施方案中,多肽的两条链彼此通过多肽接头连接。在一些实施方案中,多肽是其中第一链和第二链包含免疫球蛋白重链和免疫球蛋白轻链的单价抗体。在一些实施方案中,免疫球蛋白重链是IgG1或IgG4同种型。在一些实施方案中,单价抗体能够特异性结合抗原。在一些实施方案中,抗原是细胞因子。在一些实施方案中,细胞因子选自趋化因子、干扰素、白介素、淋巴因子和肿瘤坏死因子。在一些实施方案中,生长因子是血管内皮生长因子。在一些实施方案中,抗原选自IL-4、IL13、IL-14、IL-17、VEGFA和VEGFC。在一些实施方案中,多肽是分泌性蛋白。在一些实施方案中,分泌性蛋白从宿主细胞的周质回收。在一些实施方案中,抗体从宿主细胞的周质回收。在一些实施方案中,生长温度在生长阶段期间处于约30℃至约34℃的范围内,并且生产温度在生产阶段期间处于约25℃至约29℃的范围内。在一些实施方案中,生长搅拌速率在生长阶段期间处于约600至800转/分钟的范围内,并且生产搅拌速率在生产阶段期间处于约300至约500转/分钟的范围内。在一些实施方案中,生长搅拌速率足以在生长阶段期间在宿主细胞中实现在生产阶段期间宿主细胞中最大氧摄取速率之上0.5至2.5mmol/L/分钟的氧摄取速率。在一些实施方案中,宿主细胞的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且宿主细胞的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内。在一些实施方案中,生长搅拌速率在生产搅拌速率之上约10%至约40%。
在另一个方面,本文中提供在原核宿主细胞中产生包含重链和轻链的半抗体(half antibody)的方法,所述方法包括:(a)在培养基中在包含以下的条件下培养宿主细胞以表达重链和轻链:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,因而当表达时,重链和轻链装配以在宿主细胞中形成半抗体;其中宿主细胞包含多核苷酸,所述多核苷酸包含:(1)编码半抗体的重链的第一翻译单位;(2)编码半抗体的轻链的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合;其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收半抗体。在一些实施方案中,半抗体包含至少一个成臼(hole)突变或至少一个成杵(knob)突变。还提供在原核宿主细胞中产生包含重链和轻链的抗IL13半抗体的方法,所述方法包括:(a)在培养基中在包含以下的条件下培养宿主细胞以表达重链和轻链:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,其中(i)重链包含重链可变结构域,所述重链可变结构域包含SEQ ID NO:9的HVR-H1、SEQID NO:10的HVR-H2和SEQ ID NO:11的HVR-H3;并且(ii)轻链包含轻链可变结构域,所述轻链可变结构域包含SEQ ID NO:12的HVR-L1、SEQ ID NO:13的HVR-L2和SEQ ID NO:14的HVR-L3,因而当表达时,重链和轻链装配以在宿主细胞中形成抗IL13半抗体;其中宿主细胞包含多核苷酸,所述多核苷酸包含:(1)编码半抗体的重链的第一翻译单位;(2)编码轻链半抗体的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合;其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收抗IL13半抗体。在一些实施方案中,抗IL13半抗体包含至少一个成杵突变。还提供在原核宿主细胞中产生包含重链和轻链的抗IL17半抗体的方法,所述方法包括:(a)在培养基中在包含以下的条件下培养宿主细胞以表达重链和轻链:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,其中(i)重链包含重链可变结构域,所述重链可变结构域包含SEQ ID NO:20的HVR-H1、SEQ ID NO:21的HVR-H2和SEQ ID NO:22的HVR-H3;并且(ii)轻链包含轻链可变结构域,所述轻链可变结构域包含SEQ ID NO:23的HVR-L1、SEQ ID NO:24的HVR-L2和SEQ ID NO:25的HVR-L3,因而当表达时,重链和轻链装配以在宿主细胞中形成抗IL17半抗体;其中宿主细胞包含多核苷酸,所述多核苷酸包含:(1)编码半抗体的重链的第一翻译单位;(2)编码半抗体的轻链的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合;其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收抗IL17半抗体。在一些实施方案中,抗IL17半抗体包含至少一个成臼突变。在一些实施方案中,多核苷酸还包含启动子的三个副本,其中第一副本与第一翻译单位有效组合,第二副本与第二翻译单位有效组合并且第三副本与第三翻译单位有效组合,以驱动重链、轻链和伴侣蛋白的转录。在一些实施方案中,启动子是诱导型启动子。在一些实施方案中,诱导型启动子是在IPTG诱导作用不存在的情况下驱动重链、轻链和伴侣蛋白转录的IPTG诱导型启动子。在一些实施方案中,诱导型启动子是当培养基中的磷酸盐已经耗尽时驱动重链、轻链和伴侣蛋白转录的Pho启动子。在一些实施方案中,多核苷酸还包含选择标记并且培养基包含由引起宿主细胞保留单价抗体的单一抗生素组成的选择剂。在一些实施方案中,第一翻译单位包含与重链可变结构域的编码区有效组合的第一翻译起始区域(TIR),并且第二翻译单位包含与轻链可变结构域的编码区有效组合的第二翻译起始区域(TIR),其中第一和第二TIR的相对翻译强度是约1.0至约3.0。在一些实施方案中,至少一种伴侣蛋白或第一伴侣蛋白包括肽酰基-脯氨酰异构酶。在一些实施方案中,肽酰基-脯氨酰异构酶是FkpA蛋白。在一些实施方案中,FkpA是大肠杆菌FkpA。在一些实施方案中,至少一种伴侣蛋白还包括或第二伴侣蛋白和第三伴侣蛋白之一或两者包括蛋白质二硫键氧化还原酶。在一些实施方案中,蛋白质二硫键氧化还原酶是DsbA蛋白和DsbC蛋白之一或两者。在一些实施方案中,至少一种蛋白质二硫键氧化还原酶是大肠杆菌DsbA和大肠杆菌DsbC之一或两者。在一些实施方案中,原核宿主细胞是革兰氏阴性细菌。在一些实施方案中,革兰氏阴性细菌是大肠杆菌。在一些实施方案中,大肠杆菌是内源蛋白酶活性缺陷的菌株。在一些实施方案中,大肠杆菌是具有degpS210A突变的菌株。在一些实施方案中,大肠杆菌是具有基因型W3110 ΔfhuA ΔphoA ilvG2096(Valr)Δprc spr43H1 ΔdegP ΔmanA lacIQ ΔompT ΔmenE degpS210A的菌株。在一些实施方案中,生长温度在生长阶段期间处于约30℃至约34℃的范围内,并且生产温度在生产阶段期间处于约25℃至约29℃的范围内。在一些实施方案中,生长搅拌速率足以在生长阶段期间在宿主细胞中实现在生产阶段期间宿主细胞中最大氧摄取速率之上0.5至2.5mmol/L/分钟的氧摄取速率。在一些实施方案中,宿主细胞的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且宿主细胞的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内。在一些实施方案中,生长搅拌速率在生产搅拌速率之上约10%至约40%。还提供产生双特异性抗体的方法,所述双特异性抗体包含能够结合第一抗原的第一半抗体和能够结合第二抗原的第二半抗体,所述方法包括:在还原条件下将第一半抗体与第二半抗体组合以产生双特异性抗体,其中第一半抗体包含至少一个成杵突变并且第二半抗体包含至少一个成臼突变,并且其中第一半抗体和第二半抗体均通过前述实施方案中任一项的方法产生。在一些实施方案中,第一半部分抗原和第二半部分抗原是不同的抗原。在一些实施方案中,第一半抗体能够结合IL-13。在一些实施方案中,第二半抗体能够结合IL-17。在一些实施方案中,该方法还包括添加还原剂以实现还原条件的步骤。在一些实施方案中,还原剂是谷胱甘肽。在又一个方面,本文中提供产生双特异性抗体的方法,所述双特异性抗体包含能够结合IL13的第一半抗体和能够结合IL17的第二半抗体,所述方法包括:(a)培养第一原核宿主细胞以表达第一半抗体的第一重链和第一轻链,其中(i)第一重链包含第一重链可变结构域,所述第一重链可变结构域包含SEQ ID NO:9的HVR-H1、SEQ IDNO:10的HVR-H2和SEQ ID NO:11的HVR-H3;和(ii)第一轻链包含第一轻链可变结构域,所述第一轻链可变结构域包含SEQ ID NO:12的HVR-L1、SEQ ID NO:13的HVR-L2和SEQ ID NO:14的HVR-L3,因而当表达时,第一重链和第一轻链装配以在宿主细胞中形成第一半抗体;并且(a’)培养第二原核宿主细胞以表达第二半抗体的第二重链和第二轻链,其中(i)第二重链包含第二重链可变结构域,所述第二重链可变结构域包含SEQ ID NO:20的HVR-H1、SEQ IDNO:21的HVR-H2和SEQ ID NO:22的HVR-H3;和(ii)第二轻链包含第二轻链可变结构域,所述第二轻链可变结构域包含SEQ ID NO:23的HVR-L1、SEQ ID NO:24的HVR-L2和SEQ ID NO:25的HVR-L3,因而当表达时,第二重链和第二轻链装配以在宿主细胞中形成第二半抗体;其中第一宿主细胞包含第一多核苷酸,所述第一多核苷酸包含:(1)编码第一重链的第一翻译单位;(2)编码第一轻链的第二翻译单位;并且第二宿主细胞包含第二多核苷酸,所述第二多核苷酸包含:(1’)编码第二重链的第三翻译单位;(2’)编码第二轻链的第四翻译单位;其中第一多核苷酸和第二多核苷酸均还包含:(3)编码第一伴侣蛋白的第五翻译单位;(4)编码第二伴侣蛋白的第六翻译单位;和(5)编码第三伴侣蛋白的第七翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合;其中将第一宿主细胞和第二宿主细胞分别地在培养基中在包含以下的条件下培养:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;(b)从第一宿主细胞回收第一半抗体;(b’)从第二宿主细胞回收第二半抗体;和)将第一半抗体与第二半抗体在还原条件下组合以产生能够与IL-13和IL-17同时结合的双特异性抗体。在一些实施方案中,第一半抗体包含至少一个成杵突变,并且第二半抗体包含至少一个成臼突变。在一些实施方案中,该方法还包括添加还原剂以实现还原条件的步骤。在一些实施方案中,还原剂是谷胱甘肽。还提供包含前述实施方案任一项的双特异性抗体的组合物。在一些实施方案中,抗IL13半抗体的重链可变结构域包含SEQ ID NO:7的氨基酸序列和抗IL13抗体的轻链可变结构域包含SEQ ID NO:8的氨基酸序列。在一些实施方案中,抗IL13半抗体的重链包含SEQ ID NO:15或SEQ ID NO:16的氨基酸序列。在一些实施方案中,抗IL13半抗体的轻链包含SEQ ID NO:17的氨基酸序列。在一些实施方案中,抗IL17半抗体的重链可变结构域包含SEQ ID NO:18的氨基酸序列和抗IL17半抗体的轻链可变结构域包含SEQ ID NO:19的氨基酸序列。在一些实施方案中,抗IL17半抗体的重链包含SEQ ID NO:26或SEQ IDNO:27的氨基酸序列。在一些实施方案中,抗IL17半抗体的轻链包含SEQ ID NO:28的氨基酸序列。
应该理解,本文所述的各个实施方案的一个、一些或全部特征可以组合以形成本公开的其它实施方案。本公开的这些方面和其他方面将对本领域技术人员而言变得显而易见。本公开的这些和其他实施方案由后续详述进一步描述。
附图简述
图1A-C显示从xIL13半抗体(hAb)生产载体产生总轻链(LC)亚单位和重链(HC)亚单位。图1A提供从TIR1,1(黑色柱)和TIR2,2(条纹柱)生产载体产生的LC和HC总亚单位的图形,如通过RP-HPLC所测量。图1B提供从TIR1,1生产载体产生的LC和HC(黑色柱)或可溶性LC和HC(黑色柱)总亚单位的图形。图1C提供从TIR2,2生产载体产生的总LC和HC(黑色柱)或可溶性LC和HC(黑色柱)亚单位的图形。
图2显示使用TIR1,1(黑色柱)或TIR2,2(条纹柱)hAb生产载体时xIL13 hAb的滴度,如通过双柱RP-HPLC所测量。
图3A-B显示细菌宿主细胞中蛋白质的折叠和装配。图3A是描述细菌蛋白产生的示意图,其显示使用伴侣蛋白时,周质中蛋白质的折叠和装配。图3B是伴侣蛋白(包括肽酰基-脯氨酰异构酶(“Ppiases”)、氧化还原酶和其他伴侣蛋白)的列表。
图4A显示用来筛选FkpA变体的相容性系统。图4B显示编码抗体LC、HC和FkpA的单一xIL13质粒(pxIL13.2.2.FkpAc13)的产生。
图5A-B显示当滴定FkpA表达时xVEGF IgG1 hAb的产生。图5A描述了显示当表达不同水平的FkpA时hAb和可溶性单体重链积累的蛋白质印迹结果,而考马斯染色的凝胶显示每种条件下可溶性总蛋白产生。图5B图形显示当表达不同水平的FkpA时产生的hAb的滴度。
图6A-B显示当表达不同水平的FkpA时xIL13 IgG4 hAb的产生。图6A提供了显示使用不同载体系统时所产生xIL13 hAb的滴度的图形,并且伴随显示在每种条件下hAb和可溶性单体重链积累的蛋白质印迹结果。图6B提供了显示使用不同载体系统时所产生FkpA的量的图形,并且伴随显示在每种条件下FkpA表达的蛋白质印迹结果。在两个小图中,“内源FkpA水平”指不含有编码FkpA的质粒的细菌宿主细胞;“相容性FkpA水平”指xIL13和FkpA从独立(相容)的质粒表达;并且“单一FkpA水平”指同时表达xIL13和FkpA的单一载体。
图7A-B显示当FkpA诱导型表达时xIL4 IgG4 hAb的产生。图7A描述了显示hAb和可溶性单体重链积累的蛋白质印迹结果。图7B提供了显示使用FkpA诱导型表达时所产生xIL4hAb的滴度的图形,并且伴随显示FkpA表达的蛋白质印迹结果。在两个小图中,样品1使用TIR1,1载体产生xIL4 hAb并且不过量表达FkpA的;样品2使用TIR2,2载体产生xIL4 hAb并且不过量表达FkpA;样品3使用TIR1,1产生xIL4 hAb并使用IPTG诱导FkpA表达;并且样品4使用TIR2,2产生xIL4 hAb并使用IPTG诱导FkpA表达。
图8A-C显示当FkpA表达时xVEGFC IgG1 hAb的产生。图8A提供了显示使用不同载体系统时所产生xVEGFC hAb的滴度的图形。图8B描述了显示在两种条件下产生的可溶性总蛋白的凝胶,以FkpA条带作为标记。图8C描述了显示xVEGFC hAb和可溶性单体重链积累的蛋白质印迹结果。在全部小图中,样品1使用TIR2,2载体产生xVEGFC hAb并且不含有表达FkpA的质粒;样品2使用TIR2,2载体产生xIL4 hAb并使用IPTG诱导FkpA表达。
图9显示了使用表达xIL13 hAb(pxIL13.2.2.FkpAc13)的第一质粒和表达DsbA和DsbC(pJJ247)的第二质粒的相容性质粒系统。
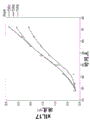
图10提供了显示随时间推移在具有或没有IPTG诱导作用的情况下,使用xIL13.2.2.FkpAc13生产质粒和表达DsbA和DsbC的相容性质粒时xIL13 hAb产生的图形。
图11A显示xIL14hAb相容性质粒系统。图11B显示了编码xIL14hAb LC和HC、FkpA、DsbA和DsbC的单一质粒的产生。
图12A-B显示在FkpA、DsbA和DsbC表达不存在的情况下(1和2,如标记);在IPTG诱导的FkpA表达存在下(3和4,如标记);和在IPTG不存在时在含FkpA、DsbA和DsbC的质粒存在下(5和6,如标记)用TIR1,1或TIR2,2载体产生xIL4 hAb。图12A描述了显示在多种条件下xIL4 hAb和可溶性单体重链积累的蛋白质印迹结果。图12B提供了显示在多种条件下所产生xIL4 hAb的滴度的图形,并且伴随显示FkpA表达的蛋白质印迹结果。
图13显示在FkpA、DsbA和DsbC表达不存在的情况下(柱1);在IPTG诱导的FkpA表达存在下(柱2);和在IPTG不存在时在含FkpA、DsbA和DsbC的质粒存在下(柱3)用TIR1,1载体产生xVEGFC hAb。
图14显示在FkpA、DsbA和DsbC表达不存在的情况下(1和3,如标记);和在IPTG不存在时在含FkpA、DsbA和DsbC的质粒存在下(2和4,如标记)用TIR1,1或TIR2,2载体产生xVEGFA IgG1 hAb。
图15显示当FkpA、DsbA、和DsbC从相同载体(“单一”)表达时和当FkpA,DsbA和DsbC从第二相容性载体(“相容性”)表达时,连同阴性对照在没有抗体表达载体的情况下和在没有DsbA、DsbC、和FkpA过量表达的情况下(“对照”),用TIR2,2载体产生xIL4 hAb。蛋白质印迹结果显示DsbA、DsbC和FkpA表达。
图16A显示利用前述pxIL13.2.2.FkpAc13生产质粒和相容性氧化还原酶质粒(pJJ247)的xIL13相容性质粒系统。图16B显示了并入来自pxIL13.2.2.FkpAc13和pJJ247的可读框(ORF)的单一质粒(MD157)的产生。
图17显示当FkpA、DsbA、和DsbC从相同载体(“单一”)表达时和当DsbA和DsbC从相容性载体表达并且FkpA从xIL13.2.2.FkpAc13载体(“相容性”)表达时,随时间推移用TIR2,2载体产生xIL3 hAb。这些载体使用phoA启动子驱动FkpA表达。
图18显示随时间推移在固定的细胞搅拌速率下培育的培养物中的平均氧摄取速率(OUR),所述细胞携带两种载体:表达xIL13 hAb和FkpA的TIR2,2载体和在IPTG诱导型启动子下表达DsbA和DsbC的载体。显示在IPTG存在或不存在下培育的培养物的OUR。提供用于每种条件的样品的数目(“N”)。
图19显示在固定的细胞搅拌速率下培育的培养物中的平均重量摩尔渗透压浓度,所述细胞携带两种载体:表达xIL13 hAb和FkpA的TIR2,2载体和在IPTG诱导型启动子下表达DsbA和DsbC的载体。显示在IPTG存在或不存在下培育的培养物的重量摩尔渗透压浓度。提供用于每种条件的样品的数目(“N”)。
图20显示随时间推移从下述细胞产生的xIL13 hAb的平均滴度,所述细胞携带两种载体:表达xIL13 hAb和FkpA的TIR2,2载体和在IPTG诱导型启动子下表达DsbA和DsbC的载体。显示在IPTG存在或不存在下培育的培养物所产生抗体的滴度。提供用于每种条件的样品的数目(“N”)。
图21显示随时间推移在650转/分钟搅拌26小时下培育、随后变换至足以实现标记OUR设定点的较低搅拌速率的培养物中的平均OUR。细胞携带两种载体:表达xIL13 hAb和FkpA的TIR2,2载体和在IPTG不存在下表达DsbA和DsbC的载体。提供用于每种条件的样品的数目(“N”)。
图22显示随时间推移在650转/分钟搅拌26小时下培育、随后变换至足以实现标记OUR设定点的较低搅拌速率的培养物中的平均重量摩尔渗透压浓度。细胞携带两种载体:表达xIL13 hAb和FkpA的TIR2,2载体和在IPTG不存在下表达的载体。提供用于每种条件的样品的数目(“N”)。
图23显示在两个时间点(54小时和72小时)在650转/分钟搅拌26小时下培育、随后变换至足以实现标记OUR设定点的较低搅拌速率的培养物中xIL13 hAb产生的平均滴度。细胞携带两种载体:表达xIL13 hAb和FkpA的TIR2,2载体和在IPTG不存在下表达DsbA和DsbC的载体。提供用于每种条件的样品的数目(“N”)。
图24显示在IPTG不存在的情况下从TIR2,2载体产生xIL13 hAb的培养物随时间推移的平均细胞密度(OD550nm),所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。在生长阶段和生产阶段(Tg/Tp),将培养物在28℃或30℃的恒定温度培育。搅拌变换进行26小时进入发酵。提供用于每种条件的样品的数目(“N”)。
图25显示在IPTG不存在的情况下从TIR2,2载体产生xIL13 hAb的细胞培养物随时间推移的平均OUR,所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。在生长阶段和生产阶段(Tg/Tp),将培养物在28℃或30℃的恒定温度培育。提供用于每种条件的样品的数目(“N”)。
图26显示在IPTG不存在的情况下从TIR2,2载体产生xIL13 hAb的细胞培养物中随时间推移的平均磷酸盐浓度,所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。在生长阶段和生产阶段(Tg/Tp),将培养物在28℃或30℃的恒定温度培育。提供用于每种条件的样品的数目(“N”)。
图27显示在IPTG不存在的情况下细胞培养物中从TIR2,2载体产生的xIL13 hAb的平均滴度,所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。在生长阶段和生产阶段(Tg/Tp),将培养物在28℃或30℃的恒定温度培育。
图28显示在IPTG不存在的情况下从TIR2,2载体产生xIL13 hAb的培养物随时间推移的平均细胞密度(OD550nm),所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。培养物在28℃或30℃恒定温度培育(分别是Tg/Tp 28℃或Tg/Tp 30℃),或在生长阶段期间在30℃培育,随后变换至生产阶段的28℃(Tg 30 Tp 28)。提供用于每种条件的样品的数目(“N”)。
图29显示在IPTG不存在的情况下从TIR2,2载体产生xIL13 hAb的细胞培养物中随时间推移的平均磷酸盐浓度,所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。培养物在28℃或30℃恒定温度培育(分别是Tg/Tp 28℃或Tg/Tp 30℃),或在生长阶段期间在30℃培育,随后变换至生产阶段的28℃(Tg 30 Tp 28)。提供用于每种条件的样品的数目(“N”)。
图30显示在IPTG不存在的情况下从TIR2,2载体产生xIL13 hAb的细胞培养物随时间推移的OUR,所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。培养物在28℃或30℃恒定温度培育(分别是Tg/Tp 28℃或Tg/Tp 30℃),或在生长阶段期间在30℃培育,随后变换至生产阶段的28℃(Tg 30 Tp 28)。提供用于每种条件的样品的数目(“N”)。
图31显示在IPTG不存在的情况下从TIR2,2载体产生的xIL13 hAb随时间推移的平均滴度,所述载体还编码受phoA启动子驱动的FkpA和受tacII启动子驱动的DsbA和DsbC。培养物在如标记的28℃或30℃恒定温度培育(分别是Tg/Tp 28℃或Tg/Tp 30℃),或在生长阶段期间在30℃培育,随后变换至生产阶段的28℃(Tg 30 Tp 28)。
图32显示在依据附表中样式确定的不同工艺条件下采用单一质粒(MD157)时xIL13 hAb滴度的部分析因设计实验(DoE)分析的结果。
图33显示在IPTG不存在的情况下从TIR2,2载体产生的xIL4 hAb的滴度,所述载体还编码受tacII启动子驱动的FkpA、DsbA和DsbC。培养物在30℃恒定温度培育(Tg/Tp 30℃),或在生长阶段期间在34℃培育,随后变换至生产阶段的25℃(Tg 34 Tp 25)。
图34显示在依据附表中样式确定的不同工艺条件下采用单一质粒(MD341)时xIL17 hAb滴度的部分析因设计实验(DoE)分析的结果。
图35显示第一优化伴侣蛋白共表达并且随后优化工艺步骤(例如,搅拌速率、Tg和Tp)对xIL13 hAb滴度的影响。
图36A显示来自66F8和67A6宿主菌株中进行的发酵的可溶性xIL13 hAb滴度。图36B显示在66F8和67A6宿主菌株中72小时处的xIL13轻链和重链总浓度。两种条件的N=2。
图37A显示在恒定发酵温度,来自66F8和67A6宿主菌株中进行的发酵的可溶性xIL14 hAb滴度。图37B显示在使用温度变换的发酵条件下,来自66F8和67A6宿主菌株中进行的发酵的可溶性xIL14 hAb总滴度。两种条件的N=2。
图38A和图38B分别显示在使用温度变换的发酵条件下,来自66F8和67A6宿主菌株中进行的发酵的xIL4轻链滴度和xIL4重链滴度。两种条件的N=2。
图39提供xIL33 hAb分泌质粒图。LC和HC可读框独立地置于与TIR2有效组合。
图40显示在恒定温度30℃不存在伴侣蛋白DsbA、DsbC、FkpA共表达的情况下进行的发酵(基础情况)中和在相同工艺条件下存在伴侣蛋白DsbA、DsbC和FkpA共表达时进行的发酵(w/伴侣蛋白)中xIL33hAb的积累。
图41显示用采用含有伴侣蛋白FkpA、DsbA、DsbC的xIL33 hAb单一质粒进行的实验设计(DoE)的aIL33hAb滴度差异。DoE因素包括pH、生长温度(Tg)、生产温度(Tp)和生产阶段目标氧摄取速率(OUR)。
图42提供TIR1(SEQ ID NO:42)、TIR2(SEQ ID NO:43)和TIR3(SEQ ID NO:44)FkpA信号序列变体的核苷酸序列。单核苷酸置换在特定密码子的第三位置进行并且代表不改变FkpA信号肽序列(SEQ ID NO:45)的氨基酸序列的同义密码子变化。
图43显示FkpA TIR变体相对于TIR1 FkpA变体(质粒19)的定量性强度。
图44显示用TIR1、TIR2和TIR3 FkpA TIR变体进行的发酵中xIL13hAb的积累。每种条件下产生的TIR1、TIR2和TIR3变体滴度分别是1.5g/L、2.5g/L和4.0g/L。
图45显示用TIR1、TIR2和TIR3 FkpA TIR变体进行的发酵中xIL33hAb的积累。
图46显示用TIR1、TIR2和TIR3 FkpA变体进行的发酵中xIL17 hAb的积累曲线。
图47A显示用FkpA TIR变体进行的发酵中xIL13 hAb的积累曲线。图47B显示在发酵结束时来自xIL13 hAb过程的可溶性部分中存在的FkpA的水平。
图48A显示改变的氧转移速率(OTR)条件和对照发酵条件的氧摄取速率(OUR)。改变的OTR发酵和对照发酵实现相似的最高OUR约5mmol/L/分钟及相似的搅拌变换后目标OUR2.75mmol/L/分钟。图48B显示改变的OTR发酵条件和对照发酵条件的生长曲线。改变的OTR发酵和对照发酵具有相似的生长曲线并且都实现250的最高OD550。xIL13 hAb对照(Ctrl)最佳条件=1巴背压(BP),20标准升/分钟(SLPM),和650至475转/分钟的搅拌速率变换。xIL13hAb改变的OTR条件=0.3巴背压,13SLPM和880至650转/分钟的搅拌速率变换。
图49显示改变的OTR条件和对照条件的xIL13 hAb积累曲线。改变的条件和对照条件在发酵期间具有相似的积累曲线并且分别实现72小时处4.1g/L和4.2g/L的最大平均滴度。
详细描述
本文中提供的例示显示,共表达与编码一种多链蛋白的每条链的翻译单位(例如,半抗体的轻链和重链)组合的一种或多种特定伴侣蛋白增加了原核宿主细胞系统中已装配的多链蛋白的产生。例子还显示后续工艺改善,如发酵的某些阶段的特定温度和搅拌速率,导致超出表达载体改善之外明显增强的产量和稳健性。总体上,本文所述的方法实现示例性双链多肽(例如,半抗体)的产量增长至少10倍。
在一个方面,本文中提供在原核宿主细胞中通过培养宿主细胞以表达多肽的两条链而产生含有两条链的多肽的方法,其中当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞含有多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;和(3)编码选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合的至少一种伴侣蛋白的第三翻译单位;其中在培养基中在包含以下的条件下培养宿主细胞:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收生物活性多肽。在一个方面,该多肽由两条链组成,而在另一个方面,该多肽包含三、四、五或更多条链。
在另一个方面,本文中提供在原核宿主细胞中通过培养宿主细胞以表达多肽的两条链而产生含有两条链的多肽的方法,其中当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞含有多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合;其中在培养基中在包含以下的条件下培养宿主细胞:包含生产温度和生产搅拌速率的生产阶段,和包含生产温度和生产搅拌速率的生产阶段,其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟;和(b)从宿主细胞回收生物活性多肽。在一个方面,该多肽由两条链组成,而在另一个方面,该多肽包含三、四、五或更多条链。
I.定义
在详细描述本公开之前,本发明不限于具体的组合物或生物系统,它们当然可以变动。还应当理解本文中所用的术语其目的仅在于描述具体实施例,并且不意图是限制性的。
除非上下文另外清楚地说明,否则如本说明书和所附权利要求书中所用,单数形式“一个(a)”、“一种(an)”和“该(the)”包括复数指称。例如,对“一个分子”的指称任选地包括两个或更多个此类分子的组合等。
术语“约”如本文所用,指本技术领域的技术人员轻易已知的相应值的寻常误差范围。本文中对“约”某个值或参数的谈及包括(并且描述)涉及该值或参数本身的实施方案。在最大值时,如本文用于指示某值的术语“约”涵盖该值的90%至110%(例如,约1.0至约3.0的第一和第二TIR的相对翻译强度指处于0.9和3.3之间范围内的相对翻译强度)。
可以理解本文中所述的本公开的方面和实施方案包括“包括”、“由……组成”和“基本上由……组成”的方面和实施方案。
如本文所用的术语“包含两条链的多肽”(术语“双链蛋白”和“双链多肽”也可以在本文中互换使用)意指含有多于一条不同多肽链的任何多肽。在一些实施方案中,双链蛋白可以包括通过一个或多个分子间键(包括而不限于二硫键)连接在一起的两条或更多条多肽的大分子复合体。在一些实施方案中,双链蛋白可以包括其中氨基酸序列属于多肽接头连接的两条不同多肽链(例如,抗体重链和抗体轻链)的单一多肽。在这种情况下,双链蛋白可以实体上代表单链,但是单链的两个或更多个部分可以在功能上如同它们作为两个独立蛋白质链那样发挥作用。例如,单链抗体可以包含有功能的重链和有功能的轻链,然而在通过多肽接头连接时,所述重链和轻链如同它们作为仅通过分子间键(例如,一个或多个二硫键)缔合的独立多肽那样折叠并装配。
如本文所用,术语“载体”意指能够运输已经与之连接的另一个核酸的核酸分子。一种类型的载体是“质粒“,其指可以向其中连接额外DNA区段的环状双链DNA环。另一个类型载体是噬菌体载体。另一类型的载体是病毒载体,其中额外的DNA区段可以连入病毒基因组中。某些载体能够在其导入的宿主细胞中自主复制(例如,具有细菌复制起点的细菌载体和附加体型哺乳动物载体)。其他载体(例如非附加体型哺乳动物载体)可以在导入宿主细胞时整合到该宿主细胞的基因组中,并因而随宿主基因组一起复制。另外,某些载体能够指导与它们有效连接的基因表达。这类载体在本文中称作“重组表达载体”(或简单地,“重组载体”)。通常,重组DNA技术中使用的表达载体经常处于质粒形式。在本说明书中,“质粒”和“载体”可以互换地使用,因为质粒是最常用形式的载体。
如本文所用,术语“顺反子”意指广义上等同于翻译单位的遗传元件,所述翻译单位包含编码多肽链的核苷酸序列和毗邻的控制区。一个“顺反子”可以包括,例如,一个或多个可读框、翻译起始区(TIR;如下文定义)、信号序列和终止区。
“多顺反子”表达载体指含有并表达在一个单一启动子调节控制下的多个顺反子的单一载体。多顺反子载体的常见例子是含有和表达处于一个启动子控制下的两条不同多肽的“双顺反子”载体。当双顺反子或多顺反子载体表达时,多个基因首先作为单一转录单位转录,并且随后分别翻译。
“转录单位”指作为单一RNA转录物转录的多核苷酸。“翻译单位”指编码并且当翻译时产生多肽的多核苷酸。如上文描述,多顺反子多核苷酸可以含有带多个翻译单位的单一转录单位。
根据本公开的“单独顺反子”表达载体指包含至少两个单独启动子-顺反子对的单一载体,其中每个顺反子受其自身启动子控制。当单独顺反子表达载体表达时,不同基因的转录和翻译过程是分离和单独的。
如本文所用的“伴侣蛋白”指辅助其他大分子(包括而不限于双链蛋白)折叠或装配的任何蛋白质。通常,伴侣蛋白可以通过许多不同机制发挥作用以促进蛋白质折叠或装配。例如,伴侣蛋白可以促进蛋白质折叠和/或装配、催化链内二硫键形成、促进蛋白质(例如,聚集的或错误折叠的蛋白质或多蛋白复合体)解折叠和/或解装配、防止聚集、辅助蛋白质降解等。
如本文所用,“翻译起始区”或TIR或翻译起始区或翻译起始序列指提供目的基因翻译起始效率的核酸区域。通常,特定顺反子内部的TIR涵盖核糖体结合位点(RBS)和RBS5'和3'的序列。定义RBS最少含有Shine-Dalgarno区域和起始密码子(AUG)。因此,TIR还包含待翻译的核酸序列的至少一部分。优选地,本公开的TIR包括编码信号肽的分泌信号序列,所述分泌信号序列位于顺反子内部编码轻链或重链的序列之前。TIR变体在TIR区域内部含有改变TIR性能(如下文定义的其翻译强度)的序列变体(尤其是置换)。优选地,本公开的TIR变体在分泌信号序列的前2个至约14个、优选地约4个至12个、更优选地约6个密码子范围内含有序列置换,所述分泌信号序列位于顺反子内部编码轻链或重链的序列之前。
如本文所用的术语“翻译强度”指对照系统中分泌型多肽的量度,其中一种或多种TIR变体用来指导多肽的分泌并且结果在相同培养和分析条件下与野生型TIR或某些其他对照比较。不限于任一项理论,如本文所用的“翻译强度”可以包括例如并且不限于,mRNA稳定性量度、核糖体与核糖体结合位点结合的效率等。
“分泌信号序列”或“信号序列”指编码短信号肽的核酸序列,所述短信号肽可以用来指导新合成的目的蛋白穿过细胞膜,通常是原核生物的内膜或其内膜和外膜。因而,目的蛋白如免疫球蛋白轻链或重链多肽分泌入原核宿主细胞的周质或分泌入培养基。由分泌信号序列编码的信号肽可以相对于宿主细胞为内源,或它们可以是外源,包含对待表达多肽为天然的信号肽。分泌信号序列一般存在于待表达多肽的氨基端,并且一般在多肽的生物合成及其从细胞质分泌之间经酶促移除。因此,信号肽通常不存在于成熟的蛋白产物中。
“有效连接”指两个或更多个组件的并列(juxtaposition),其中如此描述的组件处于允许它们以其预期方式发挥功能的关系中。例如,如果启动子顺式发挥作用以控制或调节所连接序列的转录,则启动子与编码序列有效连接。通常但不必然地,“有效连接”的DNA序列是连续的,并且在需要连接两个蛋白质编码区或在分泌性前导序列的情况下,是连续并且符合可读框的。然而,虽然有效连接的启动子是通常位于编码序列的上游,但是它不必然地与编码序列连续。有效连接的增强子可以位于编码序列上游、其内部或下游并且远离启动子。通过本领域已知的重组方法,例如,使用PCR方法学、通过复性或通过在便利的限制性位点连接,完成连接。如果便利的限制性位点不存在,则根据常规惯例使用合成性寡核苷酸衔接子或接头。
如本文所用,“调节元件”指顺式存在的、编码异源多肽的多核苷酸转录和翻译成多肽必需的核苷酸序列。转录调节元件正常情况下包含待表达基因序列5'的启动子、转录起始位点和终止位点及多聚腺苷化信号序列。术语“转录起始位点”指在构建体中与并入初级转录物(即,mRNA前体)的第一核酸相对应的核酸;转录起始位点可以与启动子序列重叠。
“启动子”指控制与其有效连接的基因或序列转录的多核苷酸序列。启动子包括RNA聚合酶结合和转录启动的信号。所用的启动子将在其中构思表达所选择序列的宿主细胞的细胞类型中有功能。许多启动子,包括来自多种不同来源的组成型、诱导型和阻遏型启动子,是本领域熟知的(并且在数据库如GenBank中鉴定)并且是原样或在克隆的多核苷酸内部可获得的(例如,来自保藏机构如ATCC以及其他商业来源或个人来源)。采用诱导型启动子,启动子的活性响应于信号(例如,存在IPTG或磷酸盐耗尽)而增加或减少。
如本文所用,术语“宿主细胞”(或“重组宿主细胞”)意指在指通过引入外源多核苷酸(例如重组质粒或载体)已经按遗传方式改变或能够按遗传方式改变的细胞。应当理解这类术语不仅意指具体的主题细胞,还意指这种细胞的子代。因为某些修饰可以因突变或环境影响而出现于后续世代中,所以这类子代实际上可以与亲代细胞不完全相同,但仍包含于如本文中所用的术语“宿主细胞”的范围内。
术语“药物制剂”指一种制备物,所述制备物处于这类形式从而允许有效成分的生物学活性有效,并且不含有对于将会施用该制剂的受试者不可接受地有毒的额外组分。这类制剂是无菌的。“可药用的”赋形剂(载体、添加物)是可以合理地施用至主题哺乳动物以提供有效剂量的所用有效成分的那些赋形剂。
出于治疗目的,“受试者”或“个体”指划归为哺乳动物的任何动物,包括人、家养动物和家畜,和动物园、体育用或宠物动物,如犬、马、猫、奶牛等。优选地,哺乳动物是人。
术语“抗体”在本文中以最广意义使用并且特别涵盖单克隆抗体(包括全长单克隆抗体)、多克隆抗体、多特异性抗体(例如双特异性抗体)和抗体片段,只要它们显示出所需的生物学活性即可。
“分离”的抗体是已经鉴定并且与其自然环境的组分分离和/或从其回收的一种抗体。其自然环境的杂质组分是将干扰该抗体的研究用途、诊断用途或治疗用途的物质并且可以包括酶、激素和其他蛋白质性或非蛋白质性溶质。在某些实施方案中,将该抗体(1)纯化至以重量计大于95%的抗体,如通过例如Lowry法测定,并且在一些实施方案中,纯化至以重量计大于99%;(2)纯化至足以通过利用例如转杯测序仪(spinning cup sequenator)获得至少15个N端残基或内部氨基酸序列的程度或纯化(3)至通过还原或非还原条件下使用考马斯蓝或银染SDS-PAGE所确定的均一性。分离的抗体包括在重组细胞内部在原位的抗体,因为该抗体的自然环境的至少一种组分将不存在。然而,一般将通过至少一个纯化步骤制备分离的抗体。
“天然抗体”通常是由两条相同轻链(L)和两条相同重链(H)组成的约150,000道尔顿的异四聚体糖蛋白。每条轻链通过一个共价二硫键与重链连接,而二硫键的数目在不同免疫球蛋白同种型的重链之间不同。每条重链和轻链还具有规则间隔的链内二硫键。每条重链在一端具有可变结构域(VH),随后是多个恒定结构域。每条轻链在一端具有具有可变结构域(VL)并在另一端具有恒定结构域;轻链的恒定结构域与重链的第一恒定结构域对齐,并且轻链可变结构域与重链的可变结构域对齐。认为特殊氨基酸残基形成轻链可变结构域和重链可变结构域之间的界面。
术语“恒定结构域”指免疫球蛋白分子的部分,所述部分相对于含有抗原结合位点的免疫球蛋白其余部分-可变结构域而具有更保守的氨基酸序列。恒定结构域含有重链的CH1、CH2和CH3结构域(总称为,CH)和轻链的CHL(或CL)结构域。
抗体的“可变区”或“可变结构域”指抗体重链或轻链的氨基端结构域。重链的可变结构域可以称作“VH”。可变结构域轻链可以称作“VL”。这些结构域通常是抗体的最可变部分并且含有抗原结合位点。
术语“可变的”指这样的事实:可变结构域的某些部分在序列方面在抗体之间明显不同并且用于每个特殊抗体对其特殊抗原的结合作用和特异性中。然而,变异性并不在抗体的可变结构域范围内均匀分布。它在轻链可变结构域和重链可变结构域中均集中于三个区段(称作高变区(HVR))内。可变结构域的较高度保守部分称作构架区(FR)。天然重链和轻链的可变结构域各自包含由3个HVR连接的大体上采取β折叠构型的4个FR区,所述HVR形成环,所述环连接该β折叠结构和在一些情况下形成该β折叠结构的部分。每条链中的HVR由FR区密切接近地固定在一起,并且与来自其他链的HVR形成抗体的抗原-结合部位(参见Kabat等人,Sequences of Proteins of Immunological Interest,第五版,NationalInstitute of Health,Bethesda,Md.(1991))。恒定结构不直接参与抗体与抗原结合,但是展示多种效应子功能,如使抗体参与抗体依赖的细胞毒性(ADCC)。
来自任何哺乳动物物种的抗体(免疫球蛋白)的“轻链”可以基于其恒定域的氨基酸序列划分成两个明显不同类型(称作κ(“κ”)和λ(“λ”))之一。
如本文所用的术语IgG“同种型”或“亚类”指免疫球蛋白的由其恒定区的化学特征和抗原性特征限定的任何亚类。
取决于其重链恒定域的氨基酸序列,抗体(免疫球蛋白)可以归属至不同“类别”。存在五个主要类别的免疫球蛋白:IgA、IgD、IgE、IgG和IgM,并且这些类别中的几种可以进一步划分成亚类(同种型),例如,IgG1、IgG2、IgG3、IgG4、IgA1和IgA2。与不同类别免疫球蛋白相对应的重链恒定结构域分别称作α、γ、ε、γ和μ。不同类别的免疫球蛋白的亚单位结构和三维构型是熟知的并且总体上例如在Abbas等人,Cellular and Mol.Immunology,第四版(W.B.Saunders,Co.,2000)中描述。抗体可以是通过抗体与一种或多种其他蛋白质或肽共价或非共价缔合所形成的较大融合分子的组成部分。
术语“全长抗体”、“完好抗体”和“完整抗体”在本文中可互换地用来指处于其基本上完好形式的抗体,而不是如下文定义的抗体片段。该术语特别指具有重链的抗体,所述重链含有Fc区。
出于本文目的,“裸抗体”指不与细胞毒部分或放射标记物缀合的抗体。
“抗体片段”包含完整抗体的一部分,优选地包括其抗原结合区。在一些实施方案中,本文所述的抗体片段是抗原结合片段。抗体片段的例子包括Fab、Fab'、F(ab')2和Fv片段;双体抗体;线性抗体;单链抗体分子;和从抗体片段形成的多特异性抗体。
木瓜蛋白酶消化抗体产生了两个相同的抗原结合片段,称作“Fab片段,各自具有单个抗原结合位点,和一个残余“Fc片段,该片段的名称反映其容易结晶的能力。胃蛋白酶处理产生了具有两个抗原结合位点并且仍能够交联抗原的F(ab')2片段。
“Fv”是含有完整抗原结合位点的最小抗体片段。在一个实施方案中,双链Fv种类由处于紧密非共价缔合的一个重链可变结构域和一个轻链可变结构域的二聚体组成。在单链Fv(scFv)种类中,一个重链可变结构域和一个轻链可变结构域可以通过柔性肽接头如此共价连接,从而轻链和重链可以按照与双链Fv种类中类似的“二聚体”结构结合。正是在这种格局中每个可变结构域的3个HVR相互作用以限定VH-VL二聚体表面上的抗原结合位点。总体上,这6个HVR向抗体赋予抗原结合特异性。然而,甚至单个可变结构域(或仅包含3个对抗原特异的HVR的半个Fv)具有识别并结合抗原的能力,虽然以比完整结合部位更低的亲和力进行。
Fab片段含有重链可变结构域和轻链可变结构域并且还含有轻链的恒定结构域和重链的第一恒定结构域(CH1)。Fab'片段因在重链CH1结构域的羧基端处附加额外几个残基(包括来自抗体铰链区的一个或多个半胱氨酸)而与Fab片段不同。Fab'-SH在本文是Fab'的名称,其中恒定结构域的半胱氨酸残基携带游离巯基。F(ab')2抗体片段最初作为成对的Fab片段产生,所述成对的Fab片段在它们之间具有铰合半胱氨酸。抗体片段的其他化学偶联也是已知的。
“单链Fv”或“scFv”抗体片段包括抗体的VH和VL结构域,其中这些结构域存在于单一多肽链中。通常,scFv多肽还包含在VH结构域和VL结构域之间的多肽接头,所述多肽接头能够使scFv形成用于抗原结合的所需结构。关于scFv的综述,例如参见Pluckthün,ThePharmacology of Monoclonal Antibodies,第113卷,Rosenburg和Moore编著,,(Springer-Verlag,New York,1994),第269-315。
术语“双体抗体”指具有两个抗原结合位点的抗体片段,所述片段包括在相同的多肽链(VH-VL)中与轻链可变结构域(VL)连接的重链可变结构域(VH)。通过使用太短以至于不能引起相同链上两个结构域之间配对的接头,这些结构域被迫与另一条链的互补性结构域配对并且产生两个抗原结合位点。双体抗体可以是双价或双特异性的。双体抗体更充分例如在以下文献中描述:EP 404,097;WO 1993/01161;Hudson等人,Nat.Med.9:129-134(2003);和Hollinger等人,Proc.Natl.Acad.Sci.USA 90:6444-6448(1993)。三体抗体和四体抗体还在Hudson等人,Nat.Med.9:129-134(2003)中描述。
如本文所用的术语“单克隆抗体”指从基本上同质的抗体群体中获得的抗体,例如,构成这个群体的各个抗体是基本上相同的,例外是可能少量存在的可能突变,例如天然存在的突变。因此,修饰语“单克隆”表明该抗体的特征为不是单独抗体的混合物。在某些实施方案中,这种单克隆抗体一般包括了包含结合某靶的多肽序列的抗体,其中通过以下方法获得靶结合的多肽序列,所述方法包括从多个多肽序列中选择单个靶结合的多肽序列。例如,该选择过程可以是从多个克隆(杂交瘤克隆、噬菌体克隆或重组DNA克隆的汇集物)选择独特克隆。应当理解,可以进一步改变所选择的靶结合序列,例如以便改善对该靶的亲和力、以便人源化靶结合序列、以便改善其在细胞培养中的产生、以便降低其体内免疫原性、以便产生多特异性抗体等,并且包含改变的靶结合序列的抗体也是本公开的单克隆抗体。与一般包括针对不同决定簇(表位)的不同抗体的多克隆抗体制备物相反,单克隆抗体制备物的每种单克隆抗体针对抗原上的单一决定簇。除了它们的特异性外,单克隆抗体制备物也是有利的,在于它们一般不混杂有其他免疫球蛋白。
修饰语“单克隆的”表明该抗体的特征为从基本上同质的抗体群体获得,并且不得解释为要求通过任何特定方法产生该抗体。例如,根据本发明待使用的单克隆抗体可以通过多种技术产生,所述技术例如包括在原核宿主细胞中表达、杂交瘤方法(例如,Kohler和Milstein,Nature,256:495-97(1975);Hongo等人,Hybridoma,14(3):253-260(1995);Harlow等人,Antibodies:A Laboratory Manual,(Cold Spring Harbor LaboratoryPress,第2版,1988);Hammerling等人,引自:Monoclonal Antibodies and T-CellHybridomas 563-681(Elsevier,N.Y.,1981)、重组DNA方法(见,例如,美国专利号4,816,567)、噬菌体展示技术(参见,例如,Clackson等人,Nature,352:624-628(1991);Marks等人,J.Mol.Biol.222:581-597(1992);Sidhu等人,J.Mol.Biol.338(2):299-310(2004);Lee等人,J.Mol.Biol.340(5):1073-1093(2004);Fellouse,Proc.Natl.Acad.Sci.USA 101(34):12467-12472(2004);和Lee等人,J.Immunol.Methods 284(1-2):119-132(2004)、和在具有编码人免疫球蛋白序列的部分或全部人免疫球蛋白基因座或基因的动物中产生人抗体或人样抗体的技术(见,例如,WO 1998/24893;WO 1996/34096;WO 1996/33735;WO1991/10741;Jakobovits等人,Proc.Natl.Acad.Sci.USA 90:2551(1993);Jakobovits等人,Nature 362:255-258(1993);Bruggemann等人,Year in Immunol.7:33(1993);美国专利号5,545,807;5,545,806;5,569,825;5,625,126;5,633,425;和5,661,016;Marks等人,Bio/Technology 10:779-783(1992);Lonberg等人,Nature 368:856-859(1994);Morrison,Nature 368:812-813(1994);Fishwild等人,Nature Biotechnol.14:845-851(1996);Neuberger,Nature Biotechnol.14:826(1996)以及Lonberg和Huszar,Intern.Rev.Immunol.13:65-93(1995)。
本文中的“单克隆抗体”特别包括这些“嵌合抗体”,其中重链和/或轻链的部分与源自特定物种或属于特定抗体类别或亚类的抗体以及这种抗体的片段中的相应序列相同或同源,同时所述链的其余部分与源自另一个物种或属于另一个抗体类别或亚类的抗体以及这种抗体的片段中的相应序列相同或同源,只要它们显示所需的生物学活性(参见,例如,美国专利号4,816,567;和Morrison等人,Proc.Natl.Acad.Sci.USA 81:6851-6855(1984))。嵌合抗体包括 抗体,其中抗体的抗原结合区衍生自通过例如用目的抗原免疫接种猕猴所产生的抗体。
抗体,其中抗体的抗原结合区衍生自通过例如用目的抗原免疫接种猕猴所产生的抗体。
非人(例如,小鼠)抗体的“人源化”形式是含有从非人免疫球蛋白衍生的最少序列的嵌合抗体。在一个实施例中,人源化抗体是这样的人免疫球蛋白(受者抗体),其中来自受者的HVR中的残基由来自非人类物种(供者抗体)(如小鼠、大鼠、兔或非人类灵长类)的具有所需特异性、亲和力和能力的HVR中的残基替换。在一些情况下,人免疫球蛋白的FR残基由相应的非人残基替换。另外,人源化抗体可以包含在受体抗体中或在供体抗体中不存在的残基。可以作出这些修饰以进一步完善抗体性能。通常,人源化抗体将包含至少1个、并且一般2个可变结构域的基本上全部,其中全部或基本上全部的高变环与非人免疫球蛋白的那些高变环对应并且全部或基本上全部的FR区是人免疫球蛋白序列的那些FR。人源化抗体任选地还将包含免疫球蛋白恒定区(Fc)的至少部分,通常是人免疫球蛋白恒定区的部分。对于进一步细节,例如参见Jones等人,Nature 321:522-525(1986);Reichmann等人,Nature332:323-329(1988);和Presta,Curr.Op.Struct.Biol.2:593-596(1992)。还参见,例如,Vaswani和Hamilton,Ann.Allergy,Asthma&Immunol.1:105-115(1998);Harris,Biochem.Soc.Transactions 23:1035-1038(1995);Hurle和Gross,Curr.Op.Biotech.5:428-433(1994);和美国专利号6,982,321和7,087,409。
“人抗体”是一种抗体,所述抗体拥有与人产生的抗体的氨基酸序列相对应的氨基酸序列和/或已经使用如本文中公开的产生人抗体的任何技术产生。人抗体的这种定义特别排除包含非人抗原结合残基的人源化抗体。可以使用本领域已知的多种技术(包括噬菌体展示文库)产生人抗体。Hoogenboom和Winter,J.Mol.Biol.,227:381(1991);Marks等人,J.Mol.Biol.,222:581(1991)。在Cole等人,Monoclonal Antibodies and CancerTherapy,Alan R.Liss,第77页(1985);Boerner等人,J.Immunol.,147(1):86-95(1991)中描述的方法也可用于制备人单克隆抗体。还参见van Dijk和van de Winkel,Curr.Opin.Pharmacol.,5:368-74(2001)。可以通过施用抗原至转基因动物制备人抗体,其中所述转基因动物已经被修饰成响应于抗原攻击而产生此类抗体,但是其内源基因座已经失能,例如,免疫接种的xenomice(关于XENOMOUSETM技术,例如参见美国专利号6,075,181和6,150,584)。关于借助人B-细胞杂交瘤技术产生的人抗体,还例如参见Li等人,Proc.Natl.Acad.Sci.USA,103:3557-3562(2006)。
“物种依赖性抗体”是这样的抗体,该抗体对来自第一哺乳动物物种的抗原的亲和力比它对来自第二哺乳动物物种的抗原的同源物的亲合力更强。正常情况下,物种依赖性抗体“特异性结合”于人抗原(例如,具有不多于约1x10-7M,优选地不多于约1x10-8M和优选地不多于约1x10-9M的结合亲和力(Kd)值),但是对来自第二非人哺乳动物物种的这种抗原的同源物的结合亲和力比它对人抗原的亲和力弱至少约50倍或至少约500倍或至少约1000倍。物种依赖性抗体可以是如上文定义的多种类型抗体中的任一种,但优选地是人源化或人抗体。
术语“超变区”、“HVR”或“HV”指抗体可变结构域中在序列方上高度变化和/或形成结构上确定的环的区域。通常,抗体包含六个HVR;VH中三个(H1、H2、H3)和VL中三个(L1、L2、L3)。在天然抗体中,H3和L3显示这六个HVR的大部分多样性,并且尤其认为H3在赋予抗体精细特异性方面发挥独特作用。见,例如,Xu等人,Immunity 13:37-45(2000);Johnson和Wu,在Methods in Molecular Biology 248:1-25中(Lo,编著,Human Press,Totowa,N.J.,2003)。实际上,仅由重链组成的天然存在的camelid抗体在轻链不存在的情况下是有功能和稳定的。见,例如,Hamers-Casterman等人,Nature 363:446-448(1993);Sheriff等人,Nature Struct.Biol.3:733-736(1996)。
本文中使用和包括了众多的HVR描述。Kabat互补性决定区(CDR)基于序列变异性并且是最常使用的(Kabat等人,Sequences of Proteins of Immunological Interest,第5版.Public Health Service,National Institutes of Health,Bethesda,Md.(1991))。相反,Chothia指结构环的位置(Chothia和Lesk J.Mol.Biol.196:901-917(1987))。AbMHVR代表了Kabat HVR和Chothia结构环之间的折中方案,并且由Oxford Molecular's AbM建模软件使用。“接触(contact)”HVR基于可获得的复合物晶体结构的分析结果。下文指出来自这些HVR的每一者中的残基。
表1a.抗体高变区

HVR可以包含如下“延长的HVR”:VL中的24-36或24-34(L1)、46-56或50-56(L2)和89-97或89-96(L3)和VH中的26-35(H1)、50-65或49-65(H2)和93-102、94-102或95-102(H3)。对于这些定义中的每一者,可变结构域残基根据上文Kabat等人编号。
“构架”残基或“FR”残基是除如本文中定义的HVR残基之外的那些可变结构域残基。
术语“如Kabat中编号的可变结构域残基”或“如Kabat中编号的氨基酸位置”和其变型,指用于上文Kabat等人中编纂抗体的重链可变结构域或轻链可变结构域的编号体系。使用这种编号体系,实际线性氨基酸序列可以含有与缩短可变结构域的FR或HVR或插入其中相对应的较少或额外的氨基酸。例如,重链可变结构域可以在H2的残基52后包含单个氨基酸插入物(根据Kabat,残基52a)及在重链FR残基82后包含插入的残基(例如,根据Kabat,残基82a、82b和82c等)。可以通过在抗体的同源性序列的区域与“标准”Kabat编号的序列比对,确定给定抗体的残基的Kabat编号。
当提到可变结构域中的残基(大约轻链第1-107位残基和重链第1-113位残基)时,通常使用Kabat编号体系(例如,Kabat等人,Sequences of Immunological Interest.第5版Public Health Service,National Institutes of Health,Bethesda,Md.(1991))。当提到免疫球蛋白重链恒定区中的残基时,通常使用“EU编号体系”或“EU index”(例如,上文Kabat等人中报道的EU index)。“如Kabat中那样的EU index”指人IgG1EU抗体的残基编号法。
表述“线性抗体”指在Zapata等人(1995 Protein Eng,8(10):1057-1062)中描述的抗体。简而言之,这些抗体包含一对串联Fd区段(VH-CH1-VH-CH1),它们与互补性轻链多肽一起形成一对抗原结合区。线性抗体可以是双特异或单特异的。
II.分子优化
文中提供在原核宿主细胞中通过培养宿主细胞以表达多肽的两条链而产生含有两条链的多肽的方法,其中当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞含有多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;和(3)编码选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合的至少一种伴侣蛋白的第三翻译单位。本文中还提供在原核宿主细胞中通过培养宿主细胞以表达多肽的两条链而产生含有两条链的多肽的方法,其中当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽;其中宿主细胞含有多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合。
在一些实施方案中,将宿主细胞培养以表达多肽的两条链,其中当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽。如本文所用,两条链折叠和装配可以指促进最终采取正确的三维双链蛋白构象、双链蛋白装配或两者的任何或全部步骤。折叠和装配可以指每条链折叠和装配成其正确的构象和折叠,或它可以指通过两条蛋白质链的分子间键产生的复合体的折叠和装配。类似地,每条链可以折叠并装配以形成生物活性多肽,或通过两条蛋白质链的分子间键产生的复合体可以折叠并装配以形成整体上的生物活性多肽。
生物活性多肽可以指能够实施赋予该多肽的功能的任何多肽。生物活性多肽的功能可以包括,而不限于,正确的折叠或装配、与另一个大分子结合或其他相互作用和酶活性。以说明的方式,生物活性抗体可以指能够实施赋予该抗体的至少一种功能的抗体,包括而不限于与表位结合或拥有抗体Fc区的特性,如下文进一步详述。
伴侣蛋白
在一些实施方案中,本公开的多核苷酸含有编码至少一种伴侣蛋白的翻译单位。如上文描述,伴侣蛋白可以指辅助其他大分子(包括而不限于双链蛋白)折叠或装配的任何蛋白质。伴侣蛋白的例子可以包括而不限于肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶和热休克蛋白(如Hsp60、Hsp70、Hsp90和Hsp100蛋白)。伴侣蛋白还可以辅助蛋白质跨膜转运,例如,多肽链跨质膜或内质网膜转位。
在一些实施方案中,伴侣蛋白可以是肽酰基-脯氨酰异构酶。肽酰基-脯氨酰异构酶(术语“脯氨酰异构酶”、“旋转异构酶(rotamase)”和“PPiase”可以在本文互换使用)可以指催化脯氨酸或脯氨酰-亚氨基肽键的顺式和反式异构体互变的任何酶。这种反应的EC编号是EC 5.2.1.8。已知或预测催化由这个EC编号描述的反应的任何蛋白质可以是本公开的肽酰基-脯氨酰异构酶。肽酰基-脯氨酰异构酶活性也可以依据GO术语ID GO:0003755描述。已知或预测拥有由这个GO术语ID描述的分子功能的任何蛋白质可以是本公开的肽酰基-脯氨酰异构酶。
本领域已知肽酰基-脯氨酰异构酶活性促进蛋白质折叠和装配。在一些实施方案中,肽酰基-脯氨酰异构酶可以通过将蛋白质反式脯氨酰键转化成的顺式脯氨酰键,辅助蛋白质折叠和装配,其中所述蛋白质的正确折叠的结构包含顺式脯氨酰键。还已知一些肽酰基-脯氨酰异构酶增强缺少顺式脯氨酰键的蛋白质折叠和装配(Bothmann H andPluckthun A 2000J.Biol.Chem.275:17100)。在一些实施方案中,肽酰基-脯氨酰异构酶可以辅助缺少顺式脯氨酰键的蛋白质发生蛋白质折叠和装配。因此,尽管肽酰基-脯氨酰异构酶活性可以充当鉴定可用于本文所述方法的伴侣蛋白的功能特征,但是使用肽酰基-脯氨酰异构酶并不必然限于其催化活性本身。
在一些实施方案中,肽酰基-脯氨酰异构酶是FkpA蛋白。在一些实施方案中,FkpA蛋白是大肠杆菌FkpA。大肠杆菌FkpA可以指由属于大肠杆菌物种的细菌的任何菌株或分离株中的fkpA基因编码的任何多肽。在一些实施方案中,大肠杆菌FkpA指由依据EcoGene登录号EG12900描述的fkpA基因编码的蛋白质。在一些实施方案中,大肠杆菌FkpA指具有依据NCBI参考序列登录号NP_417806描述的序列的蛋白质。
其他FkpA蛋白是本领域已知的。FkpA蛋白的例子可以包括,而不限于鲍氏志贺氏菌(S.boydii)肽酰基-脯氨酰异构酶(NCBI参考序列号WP_000838252)、杨格枸橼酸杆菌(C.youngae)肽酰基-脯氨酰异构酶(NCBI参考序列号WP_006687366)、产酸克雷伯氏菌(K.oxytoca)肽酰基-脯氨酰异构酶(NCBI参考序列号WP_004125943)、肠道沙门氏菌(S.enterica)肽酰基-脯氨酰异构酶(NCBI参考序列号WP_000838233)、肺炎克雷伯氏菌(K.pneumoniae)肽酰基-脯氨酰异构酶(NCBI参考序列号WP_019704642)、酿酒酵母(S.cerevisiae)FPR3p(NCBI参考序列号NP_013637)、小家鼠(M.musculus)Fkpb1a(NCBI参考序列号NP_032045)、小家鼠Fkpb2(NCBI参考序列号NP_032046)、智人(H.sapiens)FKBP2(NCBI参考序列号NP_001128680)和黑腹果蝇(D.melanogaster)CG14715(NCBI参考序列号NP_650101)。在一些实施方案中,本公开的FkpA蛋白与大肠杆菌FkpA具有至少约80%、至少约81%、至少约82%、至少约83%、至少约84%、至少约85%、至少约86%、至少约87%、至少约88%、至少约89%、至少约90%、至少约91%、至少约92%、至少约93%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性。
在一些实施方案中,伴侣蛋白可以是蛋白质二硫键氧化还原酶。蛋白质二硫化物氧化还原酶(术语“蛋白质二硫化物异构酶”和“巯基-二硫键异构酶”可以在本文互换使用)可以指催化蛋白质中二硫键重排的任何酶。例如,蛋白质二硫键氧化还原酶可以催化半胱氨酸的氧化以形成蛋白质中的二硫键。蛋白质二硫键氧化还原酶还可以催化蛋白质中错配的二硫键异构化。这个反应的EC编号是EC 5.3.4.1。已知或预测催化由这个EC编号描述的反应的任何蛋白质可以是本公开的蛋白质二硫键氧化还原酶。蛋白质二硫键氧化还原酶活性也可以依据GO术语ID GO:0015035描述。已知或预测拥有由这个GO术语ID描述的分子功能的任何蛋白质可以是本公开的蛋白质二硫键氧化还原酶。
本领域已知蛋白质二硫键氧化还原酶活性促进蛋白质折叠和装配。例如,蛋白质二硫键氧化还原酶活性促进蛋白质折叠和装配期间正确的分子内和分子间二硫键形成。具体地说,蛋白质二硫键氧化还原酶活性对原核细胞周质中表达的带二硫键的蛋白质是重要的。
在一些实施方案中,蛋白质二硫键氧化还原酶是DsbA蛋白。在一些实施方案中,DsbA蛋白是大肠杆菌DsbA。大肠杆菌DsbA可以指由属于大肠杆菌物种的细菌的任何菌株或分离株中的dsbA基因编码的任何多肽。在一些实施方案中,大肠杆菌DsbA指由依据EcoGene登录号EG11297描述的dsbA基因编码的蛋白质。在一些实施方案中,大肠杆菌DsbA指具有依据NCBI参考序列登录号NP_418297描述的序列的蛋白质。
其他DsbA蛋白是本领域已知的。DsbA蛋白的例子可以包括,而不限于福氏志贺氏菌(S.flexneri)巯基-二硫键异构酶(NCBI参考序列号WP_000725335)、痢疾志贺氏菌(S.dysenteriae)巯基-二硫键异构酶(NCBI参考序列号WP_000725348)、杨格枸橼酸杆菌巯基-二硫键异构酶(NCBI参考序列号WP_006686108)和肠道沙门氏菌巯基-二硫键异构酶(NCBI参考序列号WP_023240584)。在一些实施方案中,本公开的DsbA蛋白与大肠杆菌DsbA具有至少约80%、至少约81%、至少约82%、至少约83%、至少约84%、至少约85%、至少约86%、至少约87%、至少约88%、至少约89%、至少约90%、至少约91%、至少约92%、至少约93%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性。
在一些实施方案中,蛋白质二硫键氧化还原酶是DsbC蛋白。在一些实施方案中,DsbC蛋白是大肠杆菌DsbC。大肠杆菌DsbC可以指由属于大肠杆菌物种的细菌的任何菌株或分离株中的dsbC基因编码的任何多肽。在一些实施方案中,大肠杆菌DsbC指由依据EcoGene登录号EG11070描述的dsbC基因编码的蛋白质。在一些实施方案中,大肠杆菌DsbC指具有依据NCBI参考序列登录号NP_417369描述的序列的蛋白质。
其他DsbC蛋白是本领域已知的。DsbC蛋白的例子可以包括,而不限于,宋氏志贺氏菌(S.sonnei)蛋白质-二硫键异构酶(NCBI参考序列号WP_000715206)、痢疾志贺氏菌蛋白质-二硫键异构酶(NCBI参考序列号WP_000715209)、弗格森埃希菌(E.fergusonii)蛋白质-二硫键异构酶(NCBI参考序列号WP_000715225)、邦戈尔沙门氏菌(S.bongori)巯基:二硫键相互作用蛋白DsbC(NCBI参考序列号WP_020845161)和肠道沙门氏菌蛋白质二硫化物异构酶DsbC(NCBI参考序列号WP_023183515)。在一些实施方案中,本公开的DsbC蛋白与大肠杆菌DsbC具有至少约80%、至少约81%、至少约82%、至少约83%、至少约84%、至少约85%、至少约86%、至少约87%、至少约88%、至少约89%、至少约90%、至少约91%、至少约92%、至少约93%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%同一性。
为确定两个氨基酸序列或两个核酸序列的同一性百分数,将所述序列出于最佳比较目的比对(例如,可以为最佳比对而在第一和第二氨基酸序列或核酸序列之一或二者中引入空位或可以为比较目的而抛弃非同源序列)。在一个实施例中,为比较目的所比对的参考序列的长度是至少50%、一般至少75%、和甚至更一般地至少80%、85%、90%、95%或100%的参考序列长度。随后比较在对应氨基酸位置或核苷酸位置处的氨基酸残基或核苷酸。当第一序列中的位置由第二序列中对应位置处的相同氨基酸残基或核苷酸占据时,则所述分子在这个位置处是相同的(如本文所用,氨基酸或核酸“同一性”等同于氨基酸或核酸“同源性”)。
考虑到为最佳比对这两个序列而需要引入的空位的数目和每个空位的长度,两个序列之间的同一性百分数随所述序列共有的相同位置变化而变化。对于序列比较,一般地一个序列充当与检验序列比较的参考序列。当使用序列比较算法时,将检验序列和参考序列输入计算机,如果需要,指定子序列坐标,并指定序列算法程序参数。可以使用默认程序参数或可以指定备选参数。基于程序参数,序列比较算法随后相对于参考序列计算检验序列的序列同一性百分数。当比较两个序列的同一性时,不需要序列是连续的,但是任何空位将为其带来将减少总体同一性百分数的罚分。对于blastn,默认参数是空位开口罚分=5并且空位延伸罚分=2。对于blastp,默认参数是空位开口罚分=11并且空位延伸罚分=1。
如本文所用,“比较窗口”包括涉及具有任一连续位置数的节段的参考,所述的连续位置数选自20至600、通常约50至约200、更通常约100至约150,其中一个序列可以与具有相同连续位置数的参考序列在最佳比对这两个序列后进行比较。用于比较的序列比对方法是本领域熟知的。可以使用已知算法(例如通过Smith和Waterman,Adv Appl Math,2:482,1981的局部同源性算法、通过Needleman和Wunsch,J Mol Biol,48:443,1970的同源性比对算法、通过Pearson和Lipman,Proc Natl Acad Sci USA,85:2444,1988的相似性检索方法、通过这些算法的计算机化执行-威斯康星遗传学软件包(Wisconsin Genetics SoftwarePackage)(遗传学计算机组(Genetics Computer Group),Madison,WI)中的FASTDB(Intelligenetics)、BLAST(美国国家生物医学信息中心)、GAP、BESTFIT、FASTA和TFASTA或通过手工比对和目视检查,实施用于比较的最佳序列比对。
适于确定序列同一性百分数和序列相似性的优选算法例子是FASTA算法(Pearson和Lipman,Proc Natl Acad Sci USA,85:2444,1988;和Pearson,Methods Enzymol,266:227-258,1996)。优化在DNA序列的FASTA比对中用来计算同一性百分数的优选参数,BL50矩阵15:-5,k-tuple=2;连接罚分=40,优化=28;空位罚分=-12,空位长度罚分=-2;和宽度=16。
适用于确定序列同一性和序列相似性百分数的另一个优选的算法例子是BLAST算法和BLAST 2.0算法(分别是Altschul等人,Nuc Acids Res,25:3389-3402,1977;和Altschul等人,J Mol Biol,215:403-410,1990)。采用本文所述的参数,使用BLAST和BLAST2.0以确定本公开的核酸和蛋白质的序列同一性百分数。进行BLAST分析的软件是通过美国国家生物技术信息中心网站可公开获得的。这种算法涉及首先通过确定查询序列中长度为W的短字,鉴定高评分序列对(HSP),其中与数据库序列中具有相同长度的字比对时,所述短字匹配或满足某些正值阈评分T。将T称作相邻字评分阈值。这些初始相邻字命中充当种子,所述种子用于启动检索以找到含有这些种子的更长HSP。所述字命中在两个方向沿每个序列尽可能远地延伸,只要可以提高累积比对评分。对于核苷酸序列,使用参数M(一对匹配残基的报酬评分;总是>0)和N(错配残基的惩罚评分;总是<0)计算累积评分。对于氨基酸序列,使用评分矩阵计算累积评分。字命中在每个方向上的延伸在以下情况时停止:累积比对评分从其最大实现值跌落达量X;累积评分因积累一个或更多负评分残基比对比对结果而达到或低于零;或抵达两个序列中任一序列的末端。BLAST算法参数W、T和X决定比对的灵敏度和速度。BLASTN程序(对于核苷酸序列)使用字长度(W)11、期望(E)10、M=5、N=-4和两条链比较作为默认。对于氨基酸序列,BLASTP程序使用字长度3和期望(E)10,以及BLOSUM62评分矩阵(Henikoff和Henikoff,Proc.Natl.Acad.Sci.USA 89:10915,1989)、比对结果(B)50、期望(E)10、M=5、N=-4和两条链的比较作为默认。
BLAST算法也进行两个序列之间相似性的统计分析(参见,例如,Karlin和Altschul,Proc.Natl.Acad.Sci.USA 90:5873-5787,1993)。由BLAST算法提供的相似性的一种度量是最小总和概率(P(N)),其提供两个核苷酸序列或氨基酸序列之间的匹配会因偶然发生的概率指示。例如,如果最小总和概率在测试核酸与参考核酸的比较中小于约0.2、更优选小于约0.01并且最优选小于约0.001,则将认为核酸相似于参考核酸。
另一个有用算法的例子是PILEUP。使用累进配对比对法以显示关系和序列同一性百分数,PILEUP从一组相关的序列中产生多重序列比对结果。它还绘制进化树或树状聚类图(dendogram),其中所述进化树或树状聚类图显示用来产生该比对结果的聚类关系。PILEUP使用累进比对法(Feng和Doolittle,J.Mol.Evol.,35:351-360[1987])的简化形式,其使用与一种公开方法类似的方法(Higgins和Sharp,CABIOS 5:151-153,1989)。该程序可以比对直至300个序列,各自最大长度为5,000个核苷酸或氨基酸。多重比对程序始于两个最相似序列的两两比对,产生两个所比对序列的聚类。这个聚类随后与下一个最相关的序列或所比对序列的聚类比对。通过简单延伸两个单独序列的两两比对结果,比对序列的两个聚类。通过一系列累进性两两比对,实现最终比对结果。通过对序列比较的区域指定特定序列及其氨基酸或核苷酸坐标并且通过指定程序参数,运行该程序。使用PILEUP时,将参考序列与其他检验序列比较以使用以下参数确定序列同一性百分数关系:默认空位权重(3.00)、默认空位长度权重(0.10)和加权的末端空位。PILEUP可以从GCG序列分析软件包获得,例如,版本7.0(Devereaux等人,Nuc Acids Res,12:387-395,1984)。
适于多重DNA和氨基酸序列比对的另一个优选算法例子结果是CLUSTALW程序(Thompson等人,Nucl Acids.Res,22:4673-4680,1994)。ClustalW执行成组序列之间的多重配对比较并且基于同源性,将它们装配成多重比对结果。空位开口罚分和空位延伸罚分分别是10和0.05。对于氨基酸比对,BLOSUM算法可以用作蛋白质权重矩阵(Henikoff和Henikoff,Proc Natl Acad Sci USA,89:10915-10919,1992)。
表达盒和载体
在一些实施方案中,宿主细胞含有多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;和(3)编码选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合的至少一种伴侣蛋白的第三翻译单位。在一些实施方案中,宿主细胞含有多核苷酸,所述多核苷酸包含(1)编码多肽第一链的第一翻译单位;(2)编码多肽第二链的第二翻译单位;(3)编码第一伴侣蛋白的第三翻译单位;(4)编码第二伴侣蛋白的第四翻译单位;和(5)编码第三伴侣蛋白的第五翻译单位,其中第一、第二和第三伴侣蛋白选自肽酰基-脯氨酰异构酶、蛋白质二硫键氧化还原酶及其组合。本公开发现,可以使用单一质粒系统(即,含有编码双链蛋白中每条链的翻译单位和一个或多个编码一种或多种伴侣蛋白的翻译单位的单一多核苷酸)或相容性质粒系统(即,含有编码双链蛋白中每条链的翻译单位的第一多核苷酸和含有一个或多个编码一种或多种伴侣蛋白的翻译单位的第二多核苷酸)实现正确折叠和装配的双链蛋白增加的产量。
在一些实施方案中,多核苷酸还含有启动子的三个副本,其中第一副本与第一翻译单位有效组合,第二副本与第二翻译单位有效组合并且第三副本与第三翻译单位有效组合,以驱动第一链、第二链和伴侣蛋白的转录。在一些实施方案中,编码三种伴侣蛋白中两者的两个翻译单位是单一转录单位的组成部分。在一些实施方案中,多核苷酸还含有与每个翻译单位有效组合的启动子。
在一些实施方案中,启动子是诱导型启动子。如上文描述,诱导型启动子的活性响应于信号而增加或减少。例如,诱导型启动子可以响应于信号(如IPTG)存在而促进转录。诱导型启动子可以响应于信号(如磷酸盐)缺乏而促进转录。在这些场景的任一者之下,转录的量可以或可以不与信号的量或其缺乏成正比。适于原核宿主细胞的诱导型启动子的众多例子是本领域已知的。这些可以包括,而不限于lac、tac、trc、trp、pho、recA、tetA、nar、phage PL、cspA、T7和PBAD启动子(关于更详细的描述,参见Terpe K.2006Appl.Microbiol.Biotechnol.72:211)。在一些实施方案中,使用诱导型启动子的三个副本以协调的方式驱动单独翻译单位(例如,双链蛋白的两条链和伴侣蛋白)表达。
在一些实施方案中,诱导型启动子是IPTG诱导型启动子。IPTG诱导型启动子可以指以响应于异丙基β-D-1-硫代半乳糖吡喃糖苷(IPTG)或能够促进从lac操纵子转录的任何其他乳糖衍生物(例如,异乳糖)的方式促进转录的任何多核苷酸序列。IPTG诱导型启动子的许多例子是本领域已知的,包括而不限于tac(例如,tacI、tacII等)启动子、lac启动子及其衍生物(例如,lacUV5、taclac等)。
在一些实施方案中,诱导型启动子是驱动第一链、第二链和伴侣蛋白转录的IPTG诱导型启动子。本公开令人惊讶地发现,一种调节伴侣蛋白表达的IPTG诱导型启动子可以在不受IPTG诱导的情况下,与该IPTG诱导型启动子由IPTG诱导时伴侣蛋白的表达相比,以促进更高产物滴度的水平促进伴侣蛋白表达。
在一些实施方案中,诱导型启动子是在培养基中磷酸盐已经耗尽时驱动第一链、第二链和伴侣蛋白转录的pho启动子。pho启动子可以指以响应于胞外磷酸盐(例如,无机磷酸盐)的方式促进转录的任何多核苷酸序列。例如,大肠杆菌中的磷酸盐(Pho)调节子包括感知胞外磷酸盐并且响应于磷酸盐水平通过Pho启动子调节多种下游基因表达的蛋白质组分(关于更详细的描述,参见Hsieh YJ和Wanner BL,2010Curr.Opin.Microbiol.13(2):198)。当细菌生长在培养基中时,已知这种Pho调节子的表达在培养基中可获得磷酸盐(例如,无机磷酸盐,Pi)时被阻遏并且在磷酸盐已经耗尽时被诱导。在本文所述方法中使用的pho启动子的一个非限制性例子是大肠杆菌phoA启动子。这种启动子是广泛已知的并且在本领域用来以依赖细胞培养基中磷酸盐浓度的方式调节原核宿主细胞中重组蛋白表达(关于更详细的描述,参见Lubke C等人,1995 Enzyme Microb.Technol.17(10):923)。
在一些实施方案中,多核苷酸还含有选择标记并且培养基包含了具有单一抗生素的选择剂,所述单一抗生素引起宿主细胞保留所述多核苷酸。有利地,本文所述的方法允许产生双链蛋白的链以及一种或多种伴侣蛋白的共表达,从而编码这些组分中每者的翻译单位均包含于单一多核苷酸(例如,如本文所述的表达质粒或单一质粒系统)中。这种系统优点是,由于这些组分全部由相同质粒编码,仅需要一个选择标记以便在原核宿主细胞中维持这些多核苷酸。
选择标记可以指编码蛋白质的任何多核苷酸,所述蛋白质促进宿主细胞经历选择(即,用来相对于缺少选择标记的细胞的丰度,偏好地增加携带选择标记的细胞的丰度的任何条件)时该细胞的存活。选择标记的例子是促进宿主细胞在抗生素存在下存活的基因。众多选择标记和含单一抗生素的相应选择剂是本领域已知的。例如并且不限于,许多选择标记和相应的抗生素在Jang CW和Magnuson T 2013 PLoS ONE 8(2):e57075中描述和援引。在一些实施方案中,选择标记可以指补足宿主细胞基因组中存在的基因缺失的基因(例如,从质粒表达的基因)。在这些例子中,当细胞经历选择时(即,在需要从宿主基因组缺失的基因的活性的条件下生长),质粒供应的基因副本补足宿主基因组的缺乏,因而选出携带外源补足基因的细胞。这类基因可以包括缺陷型标记或为产生细胞培养基中缺少的特定养分所需要的基因,本文进一步描述其例子。下文进一步描述选择标记和抗生素的几个例子。
在一些实施方案中,第一翻译单位包含与第一链编码区有效组合的第一翻译起始区域(TIR),并且第二翻译单位包含与第二链编码区有效组合的第二翻译起始区域(TIR)。已知翻译起始区(TIRs)对原核宿主细胞中重组蛋白的翻译重要(参见,例如,Simmons LC和Yansura DG 1996 Nat.Biotechnol.14:629和Vimberg V等人,2007 BMC Mol.Biol.8:100)。TIR可以决定翻译单位的翻译效率。TIR一般包含多个翻译单位特征,如起始密码子、Shine-Dalgarno(SD)序列和翻译增强子。TIR还可以包含编码信号肽的分泌信号序列。TIR的序列和及其各特征之间的间距可以调节翻译起始效率。
在一些实施方案中,第一和第二TIR的相对翻译强度是约1.0至约3.0。本文中描述了包含编码双链蛋白中每条链的翻译单位的载体,并且每个翻译单位可以包含TIR。如本文所用,“翻译强度”可以指通过翻译单位的翻译产生的多肽。这种产生可以取决于众多特征,包括而不限于mRNA翻译、mRNA稳定性、核糖体与mRNA结合的效率、和折叠、装配和/或因而编码的多肽转位。相对翻译强度可以指当实验性TIR和对照TIR由培养在相同条件下的相似原核宿主细胞(例如,相同病毒属和物种)表达时,与具有野生型或对照TIR的翻译单位编码的多肽的产量相比,由具有特定或实验性TIR的翻译单位编码的多肽的产量。可以在美国专利号8,361,744中找到TIR的进一步描述。
重组多肽
本公开的某些方面涉及产生带两条链的多肽的方法。有利地,本文所述的方法可以用于促进许多不同类型的蛋白质(尤其具有二硫键的那些,如上文所述的双链蛋白)表达、折叠和装配。下文描述特定双链蛋白,但是本文所述的方法不限于这些具体实施方案。如本文所用,双链蛋白可以包括含有多于一条不同多肽链的蛋白质。尽管本文所述的许多实施方案包括具有两条多肽链的双链蛋白,但是构思并且可以通过本文所述的方法产生具有多于两条多肽链(例如,三条或更多条多肽)的双链蛋白。如上文描述,还构思并且可以通过本文所述的方法产生由单条多肽链组成的双链蛋白,其中所述单条多肽链像其是两条不同的多肽链一样缔合(例如,单链抗体、单链可变片段等)。
在一些实施方案中,本公开双链多肽的两条链彼此通过至少一个二硫键连接。二硫键可以指连接两个巯基的任何共价键。多肽中的二硫键一般在半胱氨酸残基的巯基之间形成。本领域已知多肽二硫键对多种多肽(如本公开的双链蛋白)的折叠和装配重要。多肽二硫键可以包括单条多肽链中半胱氨酸残基之间的二硫键(即,分子内或链内二硫键)。多肽二硫键还可以包括分离的多肽链上存在的半胱氨酸残基之间的二硫键(即,分子间或链间二硫键)。因此,在一些实施方案中,双链多肽的两条链彼此通过至少一个二硫键连接。
本领域已知二硫键对抗体和抗体片段的折叠和装配重要。已知不同抗体同种型和某个同种型内部的不同亚类拥有不同的二硫键样式。例如,IgG抗体可以含有12个链内二硫键、一个在每条轻链和其相应重链之间的链间二硫键和重链之间2个和11个之间的链间二硫键,这取决于具体的IgG亚类(关于更详细的描述,参见Liu H和May K 2012 MAbs.4(1):17)。还已知IgM(参见,例如,Wiersma EJ和Shulman MJ 1995J.Immunol.154(10):5265)、IgE(参见,例如,Helm BA等人,1991 Eur.J.Immunol.21(6):1543)、IgA(参见,例如,Chintalacharuvu KR等人,2002J.Immunol.169(9):5072)和IgD(参见,例如,Shin SU等人,1992 Hum.Antibodies Hybridomas 3(2):65)在折叠和装配期间形成二硫键。
在一些实施方案中,本公开的双链多肽对宿主细胞为异源。如本文所用,当提到宿主细胞使用时,异源多肽可以指在宿主细胞中不内源表达(即,当宿主细胞与自然界分离时)的任何多肽。异源多肽还可以指可以由宿主细胞内源表达,但在不同于宿主细胞与自然界分离时的调节作用下表达的多肽。不同调节作用的例子可以包括而不限于不同的表达量、响应于不同刺激而表达,或任何其他改变的表达情景,如因使用异源启动子如诱导型启动子所致。
在一些实施方案中,本公开的双链多肽是异二聚体的单体。如本文所用,异二聚体可以指含有两种不同多肽的任何多肽复合体或处于有效链接的多肽复合体。异二聚体的非限制性例子是由两个不同抗体单体组成(即,处于有效链接的轻链-重链对)的双特异性抗体或双价抗体。在这个例子中,识别第一抗原的第一重链-轻链对的折叠和装配产生第一抗体单体。识别第二抗原的第二重链-轻链对的折叠和装配产生第二抗体单体。这些单体可以通过本领域已知的任何手段(下文相对于双特异性抗体更详细地描述)装配以形成异二聚体。关于异二聚抗体样式的示意性例子的更多详情,参见Ridgway JBB等人,1996 ProteinEng.9(7):617。
在一些实施方案中,本公开的双链多肽是其中第一链和第二链代表免疫球蛋白重链和免疫球蛋白轻链的单价抗体。如本文所用,单价抗体可以指从抗体重链和抗体轻链产生的任何多肽复合体,所述的抗体重链和抗体轻链有效连接在一起以形成其中该重链-轻链对不与第二重链-轻链对有效连接的重链-轻链对。术语“半抗体(hAb)”可以在本文互换使用。
在一些实施方案中,本公开的单价抗体能够特异性结合抗原。如本文所用,术语“结合”、“特异性结合”或是“特异的”指可度量和可重复的相互作用如靶和抗体之间结合作用,所述相互作用在分子(包括生物分子)的不均一群体存在时决定靶的存在。例如,与靶(其可以是表位)结合或特异性结合的抗体是这样的抗体,与它结合其他靶相比,所述抗体按照更大亲和力、亲合力、更轻易地和/或以更长持续时间结合这种靶。在一个实施方案中,抗体与不相关靶结合的程度小于该抗体与靶结合的程度约10%,如通过放射免疫测定(RIA)所测量。在某些实施方案中,与靶特异性结合的抗体具有≤1μM、≤100nM、≤10nM、≤1nM或≤0.1nM的解离常数(Kd)。在某些实施方案中,抗体与某蛋白质上来自不同物种的该蛋白质之间保守的表位特异性结合。在另一个实施方案中,特异性结合可以包括,但不要求排他性结合。
在一些实施方案中,本公开的双链多肽是分泌性蛋白。如本文所用,分泌性蛋白可以指是由宿主细胞分泌入宿主细胞周质或胞外环境的任何蛋白质。分泌性蛋白可以是宿主细胞内源分泌的蛋白质,或分泌性蛋白可以是并非由宿主细胞内源分泌、但以促进其分泌的方式修饰的蛋白质。例如,一般在多肽N末端找得到的信号序列的存在可以指导多肽至分泌途径供分泌。众多信号序列是本领域已知的并且可以用于促进分泌性蛋白分泌或允许宿主细胞并不天然分泌的蛋白质分泌;参见,例如,Picken等人,Infect.Immun.42:269-275(1983);Simmons和Yansura,Nature Biotechnology 14:629-634(1996);和Humphreys DP等人,2000 Protein Expr.Purif.20(2):252。信号序列的一个非限制性例子是热稳定性肠毒素II(STII)信号序列。
在一些实施方案中,本公开的分泌性蛋白从宿主细胞的周质回收。本领域已知周质指革兰氏阴性细菌细胞的内膜或胞质膜和外膜之间的间隙。在不希望受理论约束的情况下,认为周质是支持二硫键形成的氧化环境。因此,可能有利的是,使得具有二硫键作为其正确折叠及装配的结构的组成部分的多肽(例如,本公开的双链蛋白)定位至周质(关于更详细的描述,参见Schlapschy M等人,2006 Protein Eng.Des.Sel.19(8):385)。
用于回收周质蛋白的众多方法是本领域已知的。大规模纯化周质蛋白的一个非限制性例子在欧洲专利号EP1356052 B1中描述(参见,例如,实施例4)。可以通过从球形质体制备物提取周质级分,回收周质蛋白(参见,例如,Schlapschy M等人,2006 ProteinEng.Des.Sel.19(8):385)。一旦已经产生周质提取物,可以通过本领域已知的任何标准蛋白质纯化技术(如亲和纯化、色谱等)纯化周质蛋白。
抗体
本文所述的双链蛋白可以通过本领域已知的任何合适技术制备。一个示例类型的双链蛋白是抗体。如下文描述,使用本领域可用于产生抗体的技术制备抗体,所述技术的示例性方法在以下部分中更详细地描述。本领域技术人员会认识到,下文描述的许多方法可以适用于除抗体之外的双链蛋白。
抗体针对目的抗原(例如,并且不限于,PD-L1(如人PD-L1)、HER2、或CD3(如人CD3)、IL13、IL4、VEGFC、VEGFA和VEGF)。优选地,抗原是生物上重要的多肽并且向患有疾病的哺乳动物的施用抗体可以在哺乳动物中产生治疗益处。
IL13抗体和IL17抗体
在一些实施方案中,本公开的抗体针对白介素-13(在本文中称作IL-13或IL13)。例如,该抗体可以是针对IL13的单价抗体或“半抗体”、包含两个针对IL13的单价重链-轻链对的完整抗体(例如,两个相同单价重链-轻链对;两个单价重链-轻链对,各自包含识别相同IL13表位的不同HVR或CDR;或两个单价重链-轻链对,各自包含识别IL13的非重叠表位或部分重叠表位的不同HVR或CDR),或包含针对IL13的重链-轻链对和针对不同抗原的重链-轻链对的双特异性抗体。
IL13多肽的例子是本领域已知的。在一些实施方案中,IL13多肽是人IL13多肽。在一些实施方案中,IL13多肽是IL13的前体形式。IL13多肽的前体形式的非限制性例子是人IL13前体,如Swiss-Prot登录号P35225.2所代表。在一些实施方案中,IL13多肽包含序列:

在其他实施方案中,IL13是IL13的成熟形式(例如,缺少信号序列)。在一些实施方案中,IL13多肽包含序列:

在一些实施方案中,本公开的抗体是针对白介素-17(在本文中称作IL-17或IL17)。例如,该抗体可以是针对IL17的单价抗体或“半抗体”、包含两个针对IL17的单价重链-轻链对的完整抗体(例如,两个相同单价重链-轻链对;两个单价重链-轻链对,各自包含识别相同IL17表位的不同HVR或CDR;或两个单价重链-轻链对,各自包含识别IL17的非重叠表位或部分重叠表位的不同HVR或CDR),或包含针对IL17的重链-轻链对和针对不同抗原的重链-轻链对的双特异性抗体。
在IL17家族中存在六个成员:IL17A、IL17B、IL17C、IL7D、IL17E和IL17F。IL17有时在本领域中用来指IL17A,IL17家族的原型。在全部IL17家族成员之间,IL17A和IL17F具有最高的序列同源性。IL17家族成员形成同型二聚体,例如,IL17AA和IL17FF。此外,IL17A和IL17F形成异二聚体(例如,IL17AF)。参见,例如Gaffen,S.L.,2009,Nature Review 9:556-567;Hymowitz等人,2001,EMBO J.20:5332-41;和WO 2005/010044,所述文献通过引用方式完整并入本文。在某些实施方案中,本公开的抗体针对IL17A。在某些实施方案中,本公开的抗体针对IL17F。在某些实施方案中,抗IL17抗体或结合IL17的抗体指结合IL17AA、FF和AF的抗体(或IL17A F交叉反应性抗体)。在某些实施方案中,本公开的抗体与IL17A和IL17F均交叉反应(例如,抗IL17AA、AF和FF)。关于IL17AA抗体、IL17FF抗体和IL17AF抗体的进一步描述,参见,例如,美国专利号8,715,669。
IL17多肽(例如,IL17A或IL17F)的例子是本领域已知的。在一些实施方案中,IL17多肽是人IL17多肽。在一些实施方案中,IL17多肽是IL17A的前体形式。IL17A多肽的前体形式的非限制性例子是人IL17A前体,如Swiss-Prot登录号Q16552.1所代表。在一些实施方案中,IL17A多肽包含序列:
MTPGKTSLVSLLLLLSLEAIVKAGITIPRNPGCPNSEDKNFPRTVNVNLNIHNRNTNTNPKRSSDYYNRSTSPWNLHRNEDPERYPSVIWEAKCRHLGCINADGNVDYHMNSVPIQQEILVLRREPPHCPNSFRLEKILVSVGCTCVTPIVHHVA(SEQ ID NO:3)
在其他实施方案中,IL17A是IL17A的成熟形式(例如,缺少信号序列)。在一些实施方案中,IL17A多肽包含序列:
GITIPRNPGCPNSEDKNFPRTVMVNLNIHNRNTNTNPKRSSDYYNRSTSPWNLHRNEDPERYPSVIWEAKCRHLGCINADGNVDYHMNSVPIQQEILVLRREPPHCPNSFRLEKILVSVGCTCVTPIVHHVA(SEQ ID NO:4).
在一些实施方案中,IL17F多肽是人IL17F多肽。在一些实施方案中,IL17F多肽是IL17F的前体形式。IL17F多肽的前体形式的非限制性例子是人IL17F前体,如SWiss-Prot登录号Q96PD4.3所代表。在一些实施方案中,IL17F多肽包含序列:
MTVKTLHGPAMVKYLLLSILGLAFLSEAAARKIPKVGHTFFQKPESCPPVPGGSMKLDIGIINENQRVSMSRNIESRSTSPWNYTVTWDPNRYPSEVVQAQCRNLGCINAQGKEDISMNSVPIQQETLVVRRKHQGCSVSFQLEKVLVTVGCTCVTPVIHHVQ(SEQ ID NO:5)
在其他实施方案中,IL17F是IL17F的成熟形式(例如,缺少信号序列)。在一些实施方案中,IL17F多肽包含序列:
RKIPKVGHTFFQKPESCPPVPGGSMKLDIGIINENQRVSMSRNIESRSTSPWNYTVTWDPNRYPSEVVQAQCRNLGCINAQGKEDISMNSVPIQQETLVVRRKHQGCSVSFQLEKVLVTVGCTCVTPVIHHVQ(SEQ ID NO:6).
在一些实施方案中,本文中提供了包含重链可变结构域和轻链可变结构域的抗IL13抗体,其中:
(a)重链可变结构域包含分别与AYSVN(SEQ ID NO:9)、MIWGDGKIVYNSALKS(SEQ IDNO:10)和DGYYPYAMDN(SEQ ID NO:11)具有至少85%序列同一性的HVR-H1、HVR-H2和HVR-H3序列,和/或
(b)轻链可变结构域包含分别与RASKSVDSYGNSFMH(SEQ ID NO:12),LASNLES(SEQID NO:13)和QQNNEDPRT(SEQ ID NO:14)具有至少85%序列同一性的HVR-L1、HVR-L2和HVR-L3序列。
在一个具体方面,与参考序列相比,序列同一性是至少86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。
在一些实施方案中,抗IL13抗体包含SEQ ID NO:7的重链可变结构域序列和/或SEQ ID NO:8的轻链可变结构域序列。在又一个实施方案中,提供包含重链和/或轻链序列的分离的抗IL13抗体,其中:
(a)重链可变结构域序列与下述参考重链序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%序列同一性:
EVTLRESGPALVKPTQTLTLTCTVSGFSLSAYSVNWIRQPPGKALEWLAMIWGDGKIVYNSALKSRLTISKDTSKNQVVLTMTNMDPVDTATYYCAGDGYYPYAMDNWGQGSLVTVSS(SEQ ID NO:7),和/或
(b)轻链可变结构域序列与下述参考重链序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%序列同一性:
DIVLTQSPDSLSVSLGERATINCRASKSVDSYGNSFMHWYQQKPGQPPKLLIYLASNLESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQNNEDPRTFGGGTKVEIKR(SEQ ID NO:8).
在一些实施方案中,本文中提供了包含重链可变结构域和轻链可变结构域的抗IL17A F交叉反应性抗体,其中:
(a)重链可变结构域包含分别与DYAMH(SEQ ID NO:20)、GINWSSGGIGYADSVKG(SEQID NO:21)和DIGGFGEFYWNFGL(SEQ ID NO:22)具有至少85%序列同一性的HVR-H1、HVR-H2和HVR-H3序列,和/或
(b)轻链可变结构域包含分别与RASQSVRSYLA(SEQ ID NO:23),DASNRAT(SEQ IDNO:24)和QQRSNWPPAT(SEQ ID NO:25)具有至少85%序列同一性的HVR-L1、HVR-L2和HVR-L3序列。
在一个具体方面,与参考序列相比,序列同一性是至少86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。
在一些实施方案中,抗IL17A F交叉反应性抗体包含SEQ ID NO:18的重链可变结构域序列和/或SEQ ID NO:19的轻链可变结构域序列。在又一个实施方案中,提供包含重链和/或轻链序列的分离的抗IL17A F交叉反应性抗体,其中:
(a)重链序列与下述参考重链序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%序列同一性:
EVQLVESGGGLVQPGRSLRLSCAASGFTFDDYAMHWVRQAPGKGLEWVSGINWSSGGIGYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCARDIGGFGEFYWNFGLWGRGTLVTVSS(SEQ ID NO:18),和/或
(b)轻链序列与下述参考重链序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%序列同一性:
EIVLTQSPATLSLSPGERATLSCRASQSVRSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPPATFGGGTKVEIK(SEQ ID NO:19).
在本发明的一个方面,提供多特异性抗体,其中抗体包含第一单价或半抗体和第二单价或半抗体,其中第一半抗体包含结合IL-17的第一VH/VL单元并且第二半抗体包含结合IL-13的第二VH/VL单元。在一些实施方案中,第一半抗体不结合IL-13,并且第二半抗体不结合IL-17。在某些具体实施方案中,本文中提供的多特异性抗体与IL-17AA、IL-17AF和IL-17FF结合,抑制IL-17AA诱导的、IL-17AF诱导和IL-17FF诱导的活性,并且抑制IL-13诱导的活性。在某些这类实施例中,与单一IL-17A或IL-17F诱导的活性对比,抗IL-13/IL-17AA、AF和FF双特异性抗体有利地阻断由全部IL-17A和IL-17F细胞因子诱导的活性。在一些实施方案中,本文中提供的多特异性抗体与IL-17AA、IL-17AF和IL-17FF结合。在一些实施方案中,IL-17AA诱导的活性是IL-17AA诱导的体内或体外基因表达和/或细胞的增殖。在一些实施方案中,IL-17AF诱导的活性是IL-17AF诱导的体内或体外基因表达和/或细胞的增殖。在一些此类实施方案中,IL-17FF诱导的活性是IL-17FF诱导的体内或体外基因表达和/或细胞的增殖。在一些实施方案中,IL-13诱导的活性是IL-13诱导的体内或体外基因表达和/或细胞的增殖。在一些实施方案中,本文中提供的多特异性抗体不抑制IL-13与IL-13Rα1的结合。
在一些实施方案中,抗IL13/IL17AA AF FF双特异性抗体(或抗IL13/IL17双特异性抗体)包含第一半抗体和第二半抗体,其中第一半抗体包含了结合IL-17AA、AF和FF的第一VH/VL单元并且第二半抗体包含了结合IL-13的第二VH/VL单元,其中第一VH/VL单元包含了包含SEQ ID NO:20的氨基酸序列的HVR-H1、包含SEQ ID NO:21的氨基酸序列的HVR-H2、包含SEQ ID NO:22的氨基酸序列的HVR-H3、包含SEQ ID NO:23的氨基酸序列的HVR-L1、包含SEQ ID NO:24的氨基酸序列的HVR-L2和包含SEQ ID NO:25的氨基酸序列的HVR-L3,并且其中第二VH/VL单元包含了包含SEQ ID NO:9的氨基酸序列的HVR-H1、包含SEQ ID NO:10的氨基酸序列的HVR-H2、包含SEQ ID NO:11的氨基酸序列的HVR-H3、包含SEQ ID NO:12的氨基酸序列的HVR-L1、包含SEQ ID NO:13的氨基酸序列的HVR-L2和包含SEQ ID NO:14的氨基酸序列的HVR-L3。在一些实施方案中,第一半抗体不结合IL-13,并且第二半抗体不结合IL-17。参见美国专利号8,715,669;美国专利号8,771,697;美国专利号8,088,618;和PCT/US2015/017168。
在一些实施方案中,第一VH/VL单元包含与SEQ ID NO:18的氨基酸序列具有至少95%、至少96%、至少97%、至少98%或至少99%序列同一性的VH序列和与SEQ ID NO:19的氨基酸序列具有至少95%、至少96%、至少97%、至少98%或至少99%序列同一性的VL序列,并且其中第二VH/VL单元包含与SEQ ID NO:7的氨基酸序列具有至少95%、至少96%、至少97%、至少98%、或至少99%序列同一性的VH序列和与SEQ ID NO:8的氨基酸序列具有至少95%、至少96%、至少97%、至少98%、或至少99%序列同一性VL序列。在一些实施方案中,提供多特异性抗体,其中第一VH/VL单元包含具有SEQ ID NO:18的氨基酸序列的VH序列和具有SEQ ID NO:19的氨基酸序列的VL序列,并且其中第二VH/VL单元包含具有SEQ IDNO:7的氨基酸序列的VH序列和具有SEQ ID NO:8的氨基酸序列的VL序列。
在某些实施方案中,结合IL17AA AF和FF的第一半抗体包含了包含SEQ ID NO:26的氨基酸序列的重链和包含SEQ ID NO:28的氨基酸序列的轻链;并且结合IL13的第二半抗体包含了包含SEQ ID NO:15的氨基酸序列的重链和包含SEQ ID NO:17的氨基酸序列的轻链。在某些实施方案中,结合IL17AA AF和FF的第一半抗体包含了包含SEQ ID NO:27的氨基酸序列的重链和包含SEQ ID NO:28的氨基酸序列的轻链;并且结合IL13的第二半抗体包含了包含SEQ ID NO:16的氨基酸序列的重链和包含SEQ ID NO:17的氨基酸序列的轻链。
在一些实施方案中,本公开抗体的CH3和/或CH2结构域来自IgG(例如,IgG1亚型、IgG2亚型、IgG2A亚型、IgG2B亚型、IgG3亚型或IgG4亚型)。在一些实施方案中,本公开抗体的CH3和/或CH2结构域可以包含一个或多个成杵或成臼突变,如下表2中描述的那些。
在某些实施方案中,本公开抗体的CH3和/或CH2结构域来自IgG4亚型。在一些实施方案中,本公开抗体的IgG4CH3和/或CH2结构域可以包含一个或多个额外的突变,包括而不限于S228P突变(EU编号)。
本文中另外提供编码本公开的抗体重链可变结构域/轻链可变结构域或重链/轻链的多核苷酸序列。在某些实施方案中,本公开的抗IL13抗体(例如,结合IL13的半抗体)包含由包含SEQ ID NO:29的多核苷酸序列编码的重链可变结构域和/或由包含SEQ ID NO:30的多核苷酸序列编码的轻链可变结构域。在某些实施方案中,本公开的抗IL13抗体(例如,结合IL13的半抗体)包含由包含SEQ ID NO:31或SEQ ID NO:32的多核苷酸序列编码的重链和/或由包含SEQ ID NO:33的多核苷酸序列编码的轻链可变结构域。在某些实施方案中,本公开的抗IL17A F交叉反应性抗体(例如,结合IL17AA、AF和FF的半抗体)包含由包含SEQ IDNO:34的多核苷酸序列编码的重链可变结构域和/或由包含SEQ ID NO:35的多核苷酸序列编码的轻链可变结构域。在某些实施方案中,本公开的抗IL17A F交叉反应性抗体(例如,结合IL17AA、AF和FF的半抗体)包含由包含SEQ ID NO:36或SEQ ID NO:37的多核苷酸序列编码的重链和/或由包含SEQ ID NO:38的多核苷酸序列编码的轻链可变结构域。
在一些实施方案中,本公开的抗体可以含有经密码子优化以便在特定宿主细胞(例如原核宿主细胞,如大肠杆菌)中表达的多肽序列。在某些实施方案中,本公开的抗IL17AF抗体(例如,结合IL17AA、AF和FF的半抗体)包含由包含SEQ ID NO:39或SEQ ID NO:40的多核苷酸序列编码的重链和/或由包含SEQ ID NO:41的多核苷酸序列编码的轻链可变结构域。
抗体特性
在某些实施方案中,本文提供的抗体具有≤1μM、≤150nM,≤100nM、≤50nM、≤10nM、≤1nM、≤0.1nM、≤0.01nM或≤0.001nM(例如10-8M或更小,例如从10-8M至10-13M,例如,从10-9M至10-13M)的解离常数(Kd)。
在一个实施方案中,通过放射标记抗原结合测定法(RIA)测量Kd,所述测定法用目的抗体的Fab形式和其抗原如以下测定法所述那样进行。通过以下方式测量Fab对抗原的溶液结合亲和力:在未标记抗原的滴定系列存在下用最低浓度的(125I)的标记抗原平衡Fab,随后用抗Fab抗体包被的平板捕获结合的抗原(参见,例如,Chen等人,J.Mol.Biol.293:865-881(1999))。为了确立该测定法的条件,将 多孔板(ThermoScientific)用50mM碳酸钠(pH 9.6)中的5μg/mL抗Fab捕获抗体(Cappel Labs)包被过夜并随后用PBS中的2%(w/v)牛血清白蛋白在室温(大约23℃)封闭2至5小时。在不吸附性平板(Nunc#269620)中,将100pM或26pM[125I]-抗原与目的Fab的连续稀释物混合。随后将目的Fab孵育过夜;然而,孵育可以持续较长时间(例如,约65小时)以确保达到平衡。此后,将混合物转移至捕获平板以便在室温孵育(例如,1小时)。随后移除溶液并将平板用PBS中的0.1%聚山梨醇酯/>
多孔板(ThermoScientific)用50mM碳酸钠(pH 9.6)中的5μg/mL抗Fab捕获抗体(Cappel Labs)包被过夜并随后用PBS中的2%(w/v)牛血清白蛋白在室温(大约23℃)封闭2至5小时。在不吸附性平板(Nunc#269620)中,将100pM或26pM[125I]-抗原与目的Fab的连续稀释物混合。随后将目的Fab孵育过夜;然而,孵育可以持续较长时间(例如,约65小时)以确保达到平衡。此后,将混合物转移至捕获平板以便在室温孵育(例如,1小时)。随后移除溶液并将平板用PBS中的0.1%聚山梨醇酯/> 洗涤8次。当平板已经干燥时,添加150μl/孔闪烁体(MICROSCINT-20TM;Packard),并且将平板在TOPCOUNTTMγ计数器(Packard)上计数10分钟。选择产生小于或等于20%最大结合作用的每种Fab的浓度用于竞争性结合测定法中。
洗涤8次。当平板已经干燥时,添加150μl/孔闪烁体(MICROSCINT-20TM;Packard),并且将平板在TOPCOUNTTMγ计数器(Packard)上计数10分钟。选择产生小于或等于20%最大结合作用的每种Fab的浓度用于竞争性结合测定法中。
根据另一个实施方案,使用表面等离子体共振测定法,使用 或/>
或/> (BIAcore,Inc.,Piscataway,NJ)在25℃采用固定抗原CM5芯片以约10个响应单位(RU),测量Kd。简而言之,根据供应商说明书,以盐酸N-乙基-N'-(3-二甲氨基丙基)-碳二亚胺(EDC)和N-羟基琥珀酰亚胺(NHS)活化羧甲基化葡聚糖生物传感器芯片(CM5,BIACORE,Inc.)。抗原用10mM乙酸钠,pH 4.8稀释至5μg/ml(约0.2μM),随后以5μl/分钟的流速上样以实现偶联蛋白的大约10个响应单位(RU)。注射抗原后,注入1M乙醇胺以封闭未反应的基团。对于动力学测量,在25℃以大约25μl/分钟的流速注入含0.05%聚山梨醇酯20(TWEEN-20TM)表面活性剂的PBS(PBST)中Fab的2倍连续稀释物(0.78nM至500nM)。使用简单一对一Langmuir结合模型(/>
(BIAcore,Inc.,Piscataway,NJ)在25℃采用固定抗原CM5芯片以约10个响应单位(RU),测量Kd。简而言之,根据供应商说明书,以盐酸N-乙基-N'-(3-二甲氨基丙基)-碳二亚胺(EDC)和N-羟基琥珀酰亚胺(NHS)活化羧甲基化葡聚糖生物传感器芯片(CM5,BIACORE,Inc.)。抗原用10mM乙酸钠,pH 4.8稀释至5μg/ml(约0.2μM),随后以5μl/分钟的流速上样以实现偶联蛋白的大约10个响应单位(RU)。注射抗原后,注入1M乙醇胺以封闭未反应的基团。对于动力学测量,在25℃以大约25μl/分钟的流速注入含0.05%聚山梨醇酯20(TWEEN-20TM)表面活性剂的PBS(PBST)中Fab的2倍连续稀释物(0.78nM至500nM)。使用简单一对一Langmuir结合模型(/> 评价软件3.2版),通过同时拟合缔合和解离传感图(sensorgram),计算缔合速率(kon)和解离速率(koff)。将平衡解离常数(Kd)计算为koff/kon比。参见,例如,Chen等人,J.Mol.Biol.293:865-881(1999)。如果通过以上表面等离子体共振测定法显示缔合速率超过106M-1s-1,则可以使用荧光猝灭技术测定缔合速率,其中在如光谱仪(如配备止流法(stop-flow)的分光光度计(Aviv Instruments)或具有搅拌型比色皿的8000-系列SLM-AMINCOTM分光光度计(ThermoSpectronic))中所测量的增加浓度的抗原存在下,所述荧光猝灭技术在25℃测量在PBS,pH 7.2中的20nM抗抗原抗体(Fab形式)的荧光发射强度(激发=295nm;发射=340nm,16nm带通)增加或减少。
评价软件3.2版),通过同时拟合缔合和解离传感图(sensorgram),计算缔合速率(kon)和解离速率(koff)。将平衡解离常数(Kd)计算为koff/kon比。参见,例如,Chen等人,J.Mol.Biol.293:865-881(1999)。如果通过以上表面等离子体共振测定法显示缔合速率超过106M-1s-1,则可以使用荧光猝灭技术测定缔合速率,其中在如光谱仪(如配备止流法(stop-flow)的分光光度计(Aviv Instruments)或具有搅拌型比色皿的8000-系列SLM-AMINCOTM分光光度计(ThermoSpectronic))中所测量的增加浓度的抗原存在下,所述荧光猝灭技术在25℃测量在PBS,pH 7.2中的20nM抗抗原抗体(Fab形式)的荧光发射强度(激发=295nm;发射=340nm,16nm带通)增加或减少。
(i)抗原制备
可溶性抗原或其片段,任选地与其他分子缀合,可以用作产生抗体的免疫原。对于跨膜分子如受体,这些分子的片段(例如受体的胞外结构域)可以用作免疫原。备选地,表达所述跨膜分子的细胞可以用作免疫原。此类细胞可以源自天然来源(例如癌细胞系)或可以是已经通过重组技术转化以表达所述跨膜分子的细胞。其他抗原及其用于制备抗体的形式对本领域技术人员是显而易见的。
(ii)某些基于抗体的方法
优选地在动物中通过多次皮下(sc)或腹膜内(ip)注射相关抗原和佐剂产生多克隆抗体。在一些实施方案中,可以有用的是,使用双官能剂或衍化剂,例如,马来酰亚胺苯甲酰硫代琥珀酰亚胺酯(通过半胱氨酸残基缀合)、N-羟琥珀酰亚胺(通过赖氨酸残基)、戊二醛、琥珀酐、SOCl2或其中R和R1是不同烷基的R1N=C=NR,使相关抗原缀合至在待免疫接种的物种中具有免疫原性的蛋白质,例如,匙孔槭血蓝蛋白、血清白蛋白、牛甲状腺球蛋白或大豆胰蛋白酶抑制蛋白。
通过将例如100μg或5μg蛋白质或缀合物(分别用于兔或小鼠)与3体积弗氏完全佐剂组合并且在多个部位皮内注射该溶液,将动物针对抗原、免疫原性缀合物或衍生物免疫。1个月后,这些动物用弗氏完全佐剂中1/5至1/10初始量的肽或缀合物,通过在多个部位皮下注射进行强化免疫。7日至14日后,将动物采血并且测定血清的抗体滴度。对动物强化免疫直至滴度进入平台期。优选地,动物用相同抗原的缀合物强化免疫,但是所述抗原与不同的蛋白质缀合和/或借助不同的交联试剂缀合。缀合物也可以在重组细胞培养物中作为蛋白质融合物产生。另外,适当地使用聚集剂如明矾来增强免疫反应。
可以使用Kohler等人,Nature,256:495(1975)首次描述及在Hongo等人,Hybridoma,14(3):253-260(1995);Harlow等人,Antibodies:A Laboratory Manual,(ColdSpring Harbor Laboratory Press,2nd ed.1988);Hammerling等人,引自MonoclonalAntiodies and T-Cell Hybridomas 563-681(Elsevier,N.Y.,1981),和Ni,XiandaiMianyixue,26(4):265-268(2006)进一步就人-人杂交瘤描述的杂交瘤方法产生单克隆抗体。额外的方法包括例如美国专利号7,189,826中关于从杂交瘤细胞系产生人天然单克隆IgM抗体所描述的那些方法。人杂交瘤技术(三体瘤技术)在Vollmers和Brandlein,Histology and Histopathology,20(3):927-937(2005)以及Vollmers和Brandlein,Methods and Findings in Experimental and Clinical Pharmacology,27(3):185-91(2005)中描述。一旦所需的单克隆抗体已经从杂交瘤分离,编码它们的多核苷酸可以亚克隆入原核表达载体,并且可以通过本文所述的任何方法在原核宿主细胞中表达,产生抗体。
(iii)文库衍生的抗体
可以通过对组合文库筛选具有所需活性或多种活性的抗体,分离本公开的抗体。例如,本领域已知用于产生噬菌体展示文库并对这类文库筛选拥有所需结合特征的抗体的多种方法,如实施例3中描述的方法。额外的方法例如综述于Hoogenboom等人,引自Methodsin Molecular Biology 178:1-37(O'Brien等人编著,Human Press,Totowa,NJ,2001)中并且例如还在McCafferty等人,Nature 348:552-554;Clackson等人,Nature 352:624-628(1991);Marks等人,J.Mol.Biol.222:581-597(1992);Marks和Bradbury,引自Methods inMolecular Biology 248:161-175(Lo编著,Human Press,Totowa,NJ,2003);Sidhu等人,J.Mol.Biol.338(2):299-310(2004);Lee等人,J.Mol.Biol.340(5):1073-1093(2004);Fellouse,Proc.Natl.Acad.Sci.USA 101(34):12467-12472(2004);和Lee等人,J.Immunol.Methods 284(1-2):119-132(2004)。
在某些噬菌体展示法中,VH基因和VL基因库分别由聚合酶链反应(PCR)克隆并且在噬菌体文库中随机重组,其中随后可以对所述噬菌体文库筛选结合抗原的噬菌体,如Winter等人,Ann.Rev.Immunol.,12:433-455(1994)中所述。噬菌体一般将抗体片段展示为单链Fv(scFv)片段或展示为Fab片段。来自免疫来源的文库在不要求构建杂交瘤的情况下提供针对免疫原的高亲和力抗体。备选地,可以(例如,从人)克隆天然库以在不进行任何免疫的情况下,提供针对广泛类型非自身抗原的抗体和还针对自身抗原的抗体的单一来源,如Griffiths等人,EMBO J,12:725-734(1993)所述。最后,也可以通过从干细胞克隆未重排的V-基因区段并使用含有随机序列以编码高度可变的CDR3区并实现体外重排的PCR引物,合成地产生天然文库,如Hoogenboom和Winter,J.Mol.Biol.,227:381-388(1992)所述。描述人抗体噬菌体文库的专利公布例如包括:美国专利号5,750,373和美国专利公开号2005/0079574、2005/0119455、2005/0266000、2007/0117126、2007/0160598、2007/0237764、2007/0292936和2009/0002360。
将从人抗体文库分离的抗体或抗体片段视为本文中的人抗体或人抗体片段。
(iv)嵌合抗体、人源化抗体和人抗体
在某些实施方案中,本文提供的抗体是嵌合抗体。某些嵌合抗体例如在美国专利号4,816,567;和Morrison等人,Proc.Natl.Acad.Sci.USA,81:6851-6855(1984))中描述。在一个例子中,嵌合抗体包含非人类可变区(例如,源自小鼠、大鼠、仓鼠、兔或非人灵长类如猴的可变区)和人类恒定区。在又一个例子中,嵌合抗体是“类转换”抗体,其中类或亚类已经相对亲本抗体发生改变。嵌合抗体包括其抗原结合其片段。
在某些实施方案中,嵌合抗体是人源化抗体。一般,将非人抗体人源化以降低对人的免疫原性,同时保留亲本非人抗体的特异性和亲和力。通常,人源化抗体包含其中HVR(例如,CDR)(或其部分)衍生自非人类抗体并且FR(或其部分)衍生自人抗体序列的一个或多个可变结构域。人源化抗体任选地也将包含人恒定区的至少一部分。在一些实施方案中,将人源化抗体中的一些FR残基置换为来自非人类抗体(例如,从中衍生HVR残基的抗体)的相应残基,例如,以恢复或改善抗体特异性或亲和力。
人源化抗体和制造它们的方法在例如Almagro和Fransson,Front.Biosci.13:1619-1633(2008)中,并且例如还在Riechmann等人,Nature 332:323-329(1988);Queen等人,Proc.Nat’l Acad.Sci.USA 86:10029-10033(1989);美国专利号5,821,337,7,527,791,6,982,321,和7,087,409;Kashmiri等人,Methods 36:25-34(2005)(描述SDR(a-CDR)移植);Padlan,Mol.Immunol.28:489-498(1991)(描述“表面重塑”);Dall'Acqua等人,Methods 36:43-60(2005)(描述“FR改组”);和Osbourn等人,Methods 36:61-68(2005)和Klimka等人,Br.J.Cancer,83:252-260(2000)(描述针对FR改组的“导向选择”方案)中综述。
可以用于人源化的人构架区包括但不限于:使用“最佳配合”法选择的构架区(参见,例如,Sims等人,J.Immunol.151:2296(1993));从具有特定亚组的轻链可变区或重链可变区的人抗体的共有序列衍生的构架区(参见,例如,Carter等人,Proc.Natl.Acad.Sci.USA,89:4285(1992);和Presta等人,J.Immunol.,151:2623(1993));人成熟(体细胞突变)构架区或人种系构架区(参见,例如,Almagro和Fransson,Front.Biosci.13:1619-1633(2008));和从筛选FR文库衍生的构架区(参见,例如,Baca等人,J.Biol.Chem.272:10678-10684(1997)和Rosok等人,J.Biol.Chem.271:22611-22618(1996))。
在某些实施方案中,本文提供的抗体是人抗体。可以使用本领域已知的多种技术产生人抗体。人抗体总体上在van Dijk和van de Winkel,Curr.Opin.Pharmacol.5:368-74(2001)和Lonberg,Curr.Opin.Immunol.20:450-459(2008)中描述。可以例如并且不限于通过本文所述任何方法在原核宿主细胞中从原核表达载体表达,产生人抗体。
也可以通过分离从人衍生的噬菌体展示文库中选择的Fv克隆可变结构域序列产生人抗体。这类可变结构域序列随后可以与所需的人恒定结构域组合。下文描述用于从抗体文库选出人抗体的技术。
(v)抗体片段
抗体片段可以通过传统手段如酶消化法或通过重组技术产生。在某些情况下,使用抗体片段而非完整抗体存在优势。所述片段的更小尺寸允许迅速清除,并且可以导致进抵实体肿瘤的改善。关于某些抗体片段的综述,参见Hudson等人(2003)Nat.Med.9:129-134。
已经开发了用于产生抗体片段的多种技术。传统上,借助蛋白酶解消化完整抗体衍生这些片段(参见,例如,Morimoto等人,Journal of Biochemical and BiophysicalMethods 24:107-117(1992);和Brennan等人,Science,229:81(1985))。然而,现在可以通过重组宿主细胞直接产生这些片段。Fab、Fv和ScFv抗体片段均可以在大肠杆菌(E.coil)中表达并从中分泌出来,因而允许轻易产生大量的这些片段。抗体片段可以从上文讨论的抗体噬菌体文库分离。备选地,Fab'-SH片段可以从大肠杆菌直接回收并且化学地偶联以形成F(ab')2片段(Carter等人,Bio/Technology 10:163-167(1992))。根据另一种方法,F(ab')2片段可以从重组宿主细胞培养物直接分离。具有增加的体内半寿期的Fab和F(ab')2片段包含救援受体(salvage receptor)结合表位残基在美国专利号5,869,046中描述。用于产生抗体片段的其他技术对于技术人员将是显而易见的。在某些实施方案中,抗体是单链Fv片段(scFv)。参见WO 93/16185;美国专利号5,571,894和5,587,458。Fv和scFv是具有恒定区缺乏的完好结合部位的仅有种类;因而,它们可以适合用于体内使用期间减少的非特异性结合作用。可以构建scFv融合蛋白以导致效应子蛋白在scFv氨基端或羧基端处融合。见上文的Antibody Engineering,Borrebaeck编著。抗体片段也可以是“线性抗体”,例如,如美国专利号5,641,870中所述。此类线型抗体可以是单特异性或双特异性的。
(vi)多特异性抗体
多特异性抗体对至少两个不同表位具有结合特异性,其中表位通常来自不同抗原。尽管这类分子正常情况下将仅结合两个不同的表位(即双特异性抗体,BsAb),但在本文中使用时,这个表述涵盖具有额外特异性的抗体如三特异性抗体。可以将双特异性抗体制备为全长抗体或抗体片段(例如,F(ab’)2双特异性抗体)。
用于产生双特异性抗体的方法是本领域已知的。全长双特异性抗体的传统生产基于共表达两个免疫球蛋白重链-轻链对,其中所述两种链具有不同特异性(Millstein等人,Nature,305:537-539(1983))。因为免疫球蛋白重链和轻链的随机分配,这些杂交瘤(四合瘤(quadroma)产生10种不同抗体分子的可能混合物,其中仅一种分子具有正确的双特异性结构。正确分子的纯化相当繁琐,通常借助亲和色谱步骤进行,并且产物产率低。在WO 93/0882中和Traunecker等人,EMBO J.,10:3655-3659(1991)中公开了相似的方法。
本领域已知的产生双特异性抗体的一个方法是“杵入臼”或“突出部分入空穴”方法(参见,例如,美国专利号5,731,168)。在这种方法中,两个免疫球蛋白多肽(例如,重链多肽)各自包含界面。一条免疫球蛋白多肽的界面与另一条免疫球蛋白多肽上的相应界面相互作用,因而允许两条免疫球蛋白多肽结合。这些界面可以如此工程化,从而位于一条免疫球蛋白多肽的界面中的“杵”或“突出部分”(这些术语可以在本文互换使用)对应于位于另一条免疫球蛋白多肽的界面中的“臼”或“空穴”(这些术语可以在本文互换使用)。在一些实施方案中,臼具有与杵相同或相似的大小并且适当地安置,从而当两个界面相互作用时,一个界面的杵可布置在另一个界面的相应臼中。在不希望受理论约束的情况下,认为这种情况令异源多聚体稳定并且有利于异源多聚体超过其他种类(如同型多聚体)形成。在一些实施方案中,这种方法可以用来促进两条不同的免疫球蛋白多肽的异源多聚化,产生针对不同表位具有结合特异性的包含两条免疫球蛋白多肽的双特异性抗体。
在一些实施方案中,可以通过将小氨基酸侧链替换为较大侧链,构建杵。在一些实施方案中,可以通过将大氨基酸侧链替换为较小侧链,构建臼。杵或臼可以存在于初始界面中,或它们可以合成地引入。例如,可以通过改变编码界面的核酸序列以将至少一个“原始氨基酸残基替换为至少一个“输入”氨基酸残基,合成地引入杵或臼。用于改变核酸序列的方法可以包括本领域熟知的标准分子生物学技术。下表中显示各种氨基酸残基的侧链体积。在一些实施方案中,原始残基具有小的侧链体积(例如,丙氨酸、天冬酰胺、天冬氨酸、甘氨酸、丝氨酸、苏氨酸或缬氨酸),并且用于形成杵的输入残基是天然存在的氨基酸并且可以包括精氨酸、苯丙氨酸、酪氨酸和色氨酸。在一些实施方案中,原始残基具有大的侧链体积(例如,精氨酸、苯丙氨酸、酪氨酸和色氨酸),并且用于形成臼的输入残基是天然存在的氨基酸并且可以包括丙氨酸、丝氨酸、苏氨酸和缬氨酸。
表1b.氨基酸残基的特性


a扣减水分子量的氨基酸分子量。值来自Handbook of Chemistry and Physics,第43版,Cleveland,Chemical Rubber Publishing Co.,1961。
b值来自A.A.Zamyatnin,Prog.Biophys.Mol.Biol.24:107-123,1972。
c值来自C.Chothia,J.Mol.Biol.105:1-14,1975。该参考文献的图6-20中定义可及表面区域。
在一些实施方案中,基于异源多聚体的三维结构,确定形成杵或臼的原始残基。本领域已知获得三维结构的技术可以包括X射线结晶学和NMR。在一些实施方案中,该界面是免疫球蛋白恒定结构域的CH3结构域。在这些实施方案中,人IgG1的CH3/CH3界面涉及每结构域中位于四条反平行β-链上的16个残基。在不希望受理论约束的情况下,突变的残基优选地位于两个中央反平行β-链上,以最大限度降低杵可能被周围溶剂而非伴侣CH3结构域中的互补臼容纳的风险。在一些实施方案中,在两条免疫球蛋白多肽中形成相应杵和孔的突变对应于下表中提供的一个或多个对子。
表2.相应成杵和成臼突变的示例性集合


突变由原始残基表示,随后是使用Kabat编号体系时的位置,并且随后是输入残基(全部残基均以单字母氨基酸代码给出)。多个突变由冒号分隔。
在一些实施方案中,免疫球蛋白多肽包含一个CH3杵构域,所述CH3结构域包含上表2中列出的一个或多个氨基酸置换。在一些实施方案中,双特异性抗体包含第一免疫球蛋白多肽和第二免疫球蛋白多肽,所述第一免疫球蛋白多肽包含了包含表2左栏中列出的一个或多个氨基酸置换的CH3结构,所述第二免疫球蛋白多肽包含了包含表2右栏中列出的一个或多个氨基酸置换的CH3结构。作为成杵和成臼对子的一个非限制性例子,在一些实施方案中,双特异性抗体包含第一免疫球蛋白多肽和第二免疫球蛋白多肽,所述第一免疫球蛋白多肽包含了包含T366W突变的CH3结构,所述第二免疫球蛋白多肽包含了包含T366S、L368A和Y407V突变的CH3结构。
在如上文讨论那样突变DNA后,可以使用本领域已知的标准重组技术和细胞系统,表达和纯化编码具有一个或多个相应成杵或成臼突变的修饰的免疫球蛋白多肽的多核苷酸。参见,例如,美国专利号5,731,168;5,807,706;5,821,333;7,642,228;7,695,936;8,216,805;美国公开号2013/0089553;和Spies等人,Nature Biotechnology 31:753-758,2013。可以使用原核宿主细胞,如大肠杆菌,产生修饰的免疫球蛋白多肽。相应的携带杵和携带臼的免疫球蛋白多肽可以在宿主细胞中以共培养方式表达并且作为异源多聚体一起纯化,或它们可以在单一培养物中表达、分别纯化并且在体外装配。在一些实施方案中,使用本领域已知的标准细菌培养技术,共培养细菌宿主细胞的两个菌株(一个菌株表达带杵的免疫球蛋白多肽,并且另一个菌株表达带臼的免疫球蛋白多肽)。在一些实施方案中,两个菌株可以按特定比率混合,例如,从而在培养物中实现相等的表达水平。在一些实施方案中,两个菌株可以按50:50、60:40或70:30的比率混合。在表达多肽后,可以将细胞一起裂解,并且可以提取蛋白质。允许测量同型多聚体种类对异源多聚体种类的丰度的本领域已知的标准技术可以包括大小排阻色谱。在一些实施方案中,将每种修饰的免疫球蛋白多肽使用标准重组技术分别表达、回收并且体外装配在一起。如下文更详细地描述,装配可以例如通过以下方式实现:纯化每种修饰的免疫球蛋白多肽、将其按相等质量混合并且一起孵育,还原二硫键(例如,通过二硫苏糖醇处理)、浓缩和使多肽再氧化。形成的双特异性抗体可以使用标准技术纯化,包括阳离子交换色谱,并且使用标准技术(包括大小排阻色谱)测量。关于这些方法的更详细描述,参见Speis等人,Nat Biotechnol 31:753-8,2013。
每个半抗体可以具有工程化入重链的杵(突出部分)或臼(空穴),如美国专利号7,642,228中所述。简而言之,CH3杵突变体可以首先产生。可以随后通过随机化靠近在伴侣CH3结构域上的杵的残基366、368和407,产生CH3臼突变体的文库。在某些实施方案中,杵突变包含T366W,并且臼突变包含IgG1或IgG4主链中的T366S、L368和Y407V。其他免疫球蛋白同种型中的等价突变可以由本领域技术人员产生。另外,技术人员将轻易地理解,优选用于双特异性抗体的两个半抗体应当是相同的同种型。
通过在细菌宿主细胞(例如,大肠杆菌)中表达重链构建体和轻链构建体,在分立的培养物中产生含有杵突变或臼突变的半抗体。每个半抗体可以通过蛋白A亲和色谱分别纯化。可以通过HiTrap MabSelect SuReTM柱,纯化来自杵半抗体和臼半抗体的澄清的细胞提取物。可以装配蛋白A纯化的具有不同特异性的半抗体,以在体外在氧化还原反应中在还原剂存在下形成双特异性抗体。
任何合适的方法可以用来准备所需的还原条件。例如,可以通过向反应(如本发明的装配混合物)添加还原剂/还原物质,准备所需的还原条件。合适的还原剂包括而不限于二硫苏糖醇(DTT)、三(2-羧乙基)膦)(TCEP)、硫代乙醇酸、抗坏血酸(维生素C)、巯基乙酸、谷胱甘肽(GSH)、β-巯基乙胺、半胱氨酸/胱氨酸、GSH/氧化型谷胱甘肽(GSSG)、半胱胺/胱胺、glycylcysteine和β-巯基乙醇、优选地GSH。在某些具体实施方案中,还原剂是弱还原剂,包括而不限于GSH、β-巯基乙胺、半胱氨酸/胱氨酸、GSH/GSSG、半胱胺/胱胺、甘氨酰半胱氨酸和β-巯基乙醇、优选地GSH。在某些优选的实施方案中,还原剂是GSH。在某些实施方案中,还原剂不是DTT。按合适浓度并且在合适实验条件下选择合适的还原剂以在反应中实现所需还原条件,处于本领域普通技术人员的能力范围内。例如,在20℃的具有10g/L双特异性抗体蛋白质浓度的溶液中10mM L-还原型谷胱甘肽将产生约-400mV的起始氧化还原电势。例如,添加至装配混合物的谷胱甘肽产生有利于杵入臼双特异性装配的微弱还原性条件。相似类型的其他还原剂如BMEA(β-巯基乙胺)可以具有相似的作用。参见WO 2013/055958,所述文献通过引用方式完整并入本文。可以使用本领域已知的任何合适方法,估计和测量反应的还原条件。例如,可以使用刃天青指示物(还原条件下从蓝色褪色至无色)测量还原条件。为了更精确测量,可以使用氧化还原电势计(如BROADLEY 制作的ORP电极)。
制作的ORP电极)。
在某些具体实施方案中,还原条件是弱还原条件。如本文所用的术语“弱还原剂”或“弱还原条件”指在25℃具有负氧化电势的还原剂或通过这种还原剂准备的还原条件。当pH在7和9之间及温度在15℃和39℃之间时,还原剂的氧化电势优选地在-50至-600mV、-100至-600mV、-200至-600mV、-100至-500mV、-150至-300mV之间,更优选地在约-300至-500mV之间,最优选地是约-400mV。本领域技术人员将能够选择合适的还原剂以准备所需的还原条件。熟练研究者会认识到,强还原剂,即,对于相同浓度、pH和温度比上文提到的还原剂具有更负氧化电势的还原剂,可以在更低的浓度使用。在一个优选实施方案中,蛋白质将能够在还原剂存在下在上述条件下孵育时形成二硫键。弱还原剂的例子包括而不限于谷胱甘肽、β-巯基乙胺、胱氨酸/半胱氨酸、GSH/GSSG、半胱胺/胱胺、甘氨酰半胱氨酸和β-巯基乙醇。在某些实施方案中,与200X GSH:抗体摩尔比类似的氧化电势可以作为使用其他还原剂时可以预期高效装配的弱还原条件的参考点使用。
谷胱甘肽浓度可以根据术语“体积摩尔浓度”或依据相对于装配混合物中存在的半抗体的量而言的摩尔比或摩尔过量表述。对装配混合物中的蛋白质浓度使用还原剂对照的目标摩尔比;这防止因可变动的蛋白质浓度所致的还原过度或还原不足。在某些其他实施方案中,将还原剂相对于半抗体总量以2-600X、2-200X、2-300X、2-400X、2-500X、2-20X、2-8X、20-50X、50-600X、50-200X、或100-300X摩尔过量,优选地50-400X、更优选地100-300X并且最优选地200X摩尔过量添加至装配混合物。在某些实施方案中,装配混合物具有7和9之间的pH,优选地pH 8.5。
在某些实施方案中,第一半抗体的培养物和第二半抗体的培养物可以合并并且随后在合并的培养物中裂解。合并的释放的第一半抗体和第二半抗体可以在还原条件下形成双特异性抗体。参见WO 2011/133886,所述文献通过引用方式完整并入本文。
根据一种不同方法,具有所需杵合特异性的抗体可变结构域(抗体-抗原结合位点)与免疫球蛋白恒定结构域序列融合。这种融合物优选地具有免疫球蛋白重链恒定结域,包括铰链区、CH2区和CH3区的至少部分。常见的是使得含有轻链结合作用所必需的位点的第一重链恒定区(CH1)在所述融合物至少一个中存在。将编码免疫球蛋白重链融合物和(如果所需)免疫球蛋白轻链的DNA插入单独的表达载体中并且共转染至合适的宿主生物中。在构建种所用的比率不等的3多肽链提供最佳产率时的实施方案中,这在调节3种多肽片段的相互比例方面提供灵活性。然而,当至少两条多肽链以等比率的表达导致高产率时或当该比率没有特定意义时,可能将2条或全部3条多肽链的编码序列在一个表达载体中插入。
在这种方案的一个实施方案中,双特异性抗体由一条臂中具有第一结合特异性的杂合免疫球蛋白重链和另一条臂中的杂合免疫球蛋白重链-轻链对(提供第二结合特异性)组成。发现这种非对称结构促进所需的双特异性化合物与不想要的免疫球蛋白链组合分离,因为免疫球蛋白轻链在所述双特异性分子的仅一半中的存在提供了便利的分离方式。这种方法在WO 94/04690中公开。关于产生双特异性抗体的进一步细节,见,例如Suresh等人,Methods in Enzymology,121:210(1986)。
根据WO 96/27011中描述的另一个方法,可以将一对抗体分子之间的界面工程化,以使得从重组细胞培养物回收的异二聚体的百分数最大化。该界面包含抗体恒定结构域的CH3结构域的至少一部分。在这种方法中,来自第一抗体分子界面的一个或多个小氨基酸侧链由较大侧链(例如,酪氨酸或色氨酸)替换。通过将大氨基酸侧链替换为较小侧链(例如,丙氨酸或苏氨酸),在第二抗体分子的界面上产生与大体积侧链具有相同或相似尺寸的补偿“空穴”。这提供了相对于其他不想要的产物(如同型二聚体)而言增加异二聚体产率的机制。
双特异性抗体包括交联抗体或“异质缀合”抗体。例如,异质缀合下的抗体之一可以偶联于抗生物素蛋白,另一种抗体偶联于生物素。已经例如提出这类抗体将免疫系统细胞导引至不想要的细胞(美国专利号4,676,980)并用于治疗HIV感染(WO 91/00360、WO 92/200373和EP 3089)。可以使用任何便利的交联方法产生异质缀合抗体。合适的交联剂是本领域熟知的并且连同许多交联技术一起在美国专利号4,676,980中公开。
也已经在文献中描述从抗体片段产生双特异性抗体的技术。例如,可以利用化学键制备双特异性抗体。Brennan等人,Science,229:81(1985)描述了其中完整抗体经蛋白酶解方式切割以产生F(ab’)2片段的方法。这些片段在稳定邻位二硫醇并防止分子间二硫化物形成的二硫醇络合剂亚砷酸钠存在下还原。产生的Fab'片段随后转化成硫代硝基苯甲酸酯(TNB)衍生物。随后将Fab'-TNB衍生物之一通过用巯基乙胺还原再转化成Fab'-巯基并且与等摩尔量的其他Fab'-TNB衍生物混合以形成双特异性抗体。产生的双特异性抗体可以用作用于选择性固定酶的物质。
最近的进展已经促进从大肠杆菌直接回收Fab’-SH片段,其中可以化学地偶联所述片段以形成双特异性抗体。Shalaby等人,J.Exp.Med.,175:217-225(1992)描述了全人双特异性抗体F(ab’)2分子的产生。每种Fab'片段分别从大肠杆菌分泌并经历体外定向化学偶联以形成双特异性抗体。
也已经描述了从重组细胞培养物直接产生并分离双特异性抗体片段的多种技术。例如,已经使用亮氨酸拉链产生双特异性抗体。Kostelny等人,J.Immunol.,148(5):1547-1553(1992)。来自Fos和Jun蛋白的亮氨酸拉链肽通过基因融合与两个不同抗体的Fab'部分连接。抗体同型二聚体在铰链区经还原以形成单体和随后再氧化以形成抗体异二聚体。也可以将这种方法用于抗体同型二聚体的产生。Hollinger等人,Proc.Natl.Acad.Sci.USA,90:6444-6448(1993)描述的“双体抗体”技术已经为产生双特异性抗体片段提供替代性机制。这些片段包含通过接头与轻链可变结构域(VL)连接的重链可变结构域(VH),其中所述接头太短,以至于不允许相同链上的这两个结构域之间配对。因而,迫使一个片段的VH结构域和VL结构域与另一个片段的互补性VL结构域和VH结构域配对,因而形成两个抗原结合位点。还已经报道了通过利用单链Fv(sFv)二聚体产生双特异性抗体片段的另一种策略。参见Gruber等人,J.Immunol.152:5368(1994)。
另一项产生双特异性抗体片段的技术是“双特异性T细胞接合者”或 法(参见,例如,WO 2004/106381、WO 2005/061547、WO 2007/042261和WO 2008/119567)。这种方法利用布置在单一多肽上的两个抗体可变结构域。例如,单条多肽链包含两个单链Fv(scFv片段,各自具有可变重链(VH)和可变轻链(VL)结构域,所述结构域由长度足以在这两个结构域之间引起分子内缔合的多肽接头分隔。这条单一多肽还在两个scFv片段之间包含多肽间隔序列。每个scFv识别不同的表位,并且这些表位可以对不同的细胞类型特异,从而当每个scFv与其同族表位接合时,令两个不同细胞类型的细胞紧邻或系留。这种方法的一个具体实施方案包括识别由免疫细胞表达的细胞表面抗原(例如,T细胞上的CD3多肽)的scFv,所述scFv与识别由靶细胞(如恶性细胞或肿瘤细胞)表达的细胞表面抗原的另一个scFv连接。
法(参见,例如,WO 2004/106381、WO 2005/061547、WO 2007/042261和WO 2008/119567)。这种方法利用布置在单一多肽上的两个抗体可变结构域。例如,单条多肽链包含两个单链Fv(scFv片段,各自具有可变重链(VH)和可变轻链(VL)结构域,所述结构域由长度足以在这两个结构域之间引起分子内缔合的多肽接头分隔。这条单一多肽还在两个scFv片段之间包含多肽间隔序列。每个scFv识别不同的表位,并且这些表位可以对不同的细胞类型特异,从而当每个scFv与其同族表位接合时,令两个不同细胞类型的细胞紧邻或系留。这种方法的一个具体实施方案包括识别由免疫细胞表达的细胞表面抗原(例如,T细胞上的CD3多肽)的scFv,所述scFv与识别由靶细胞(如恶性细胞或肿瘤细胞)表达的细胞表面抗原的另一个scFv连接。
由于它是单一多肽,所以可以使用本领域已知的任何原核细胞表达系统表达双特异性T细胞接合者。但是,可能必需特定纯化技术(参见,例如,EP1691833)以分离单体双特异性T细胞接合者与其他多聚体种类,所述多聚体种类可能具有除单体的意图活性之外的生物学活性。在一个示例性纯化方案中,含有分泌型多肽的溶液首先经历金属亲和色谱,并且用咪唑浓度梯度洗脱多肽。使用阴离子交换色谱进一步纯化这种洗脱物,并且使用氯化钠浓度梯度洗脱多肽。最后,这种洗脱物经历大小排阻色谱以分离单体与多聚体种类。
构思了大于二价的抗体。例如,可以制备三特异性抗体。Tuft等人,J.Immunol.147:60(1991)。
在一些实施方案中,双链蛋白是多特异性抗体或双特异性抗体的组成部分。多特异性抗体或双特异性抗体可以含有本公开的两种或更多种单价抗体。
在一些实施方案中,双特异性抗体的第一抗原结合结构域包含一个或多个重链恒定结构域,其中一个或多个重链恒定结构域选自第一CH1(CH11)结构域、第一CH2(CH21)结构域、第一CH3(CH31)结构域;并且双特异性抗体的第二抗原结合结构域包含一个或多个重链恒定结构域,其中一个或多个重链恒定结构域选自第二CH1(CH12)结构域、第二CH2(CH22)结构域和第二CH3(CH32)结构域。在一些实施方案中,第一抗原结合结构域的一个或多个重链恒定结构域至少之一与第二抗原结合结构域的另一个重链恒定结构域配对。在一些实施方案中,CH31结构域和CH32结构域各自包含突出部分或空穴,并且其中CH31结构域中的突出部分或空穴分别可布置在CH32结构域的空穴或突出部分中。在一些实施方案中,CH31结构域和CH32结构域在界面处所述突出部分和空穴之间相遇。本文表2中显示CH31结构域和CH32结构域中的示例性氨基酸置换集合。在一些实施方案中,CH21结构域和CH22结构域各自包含突出部分或空穴,并且其中CH21结构域中的突出部分或空穴分别可布置在CH22结构域的空穴或突出部分中。在一些实施方案中,CH21结构域和CH22结构域在界面处所述突出部分和空穴之间相遇。在一些实施方案中,IgG的CH31结构域和/或CH32结构域在根据如美国专利号8,216,805的图5中所示的氨基酸编号法选自347、349、350、351、366、368、370、392、394、395、398、399、405、407和409的残基处含有一个或多个氨基酸置换。在一些实施方案中,突出部分包含一种或多种引入的残基,所述残基选自精氨酸(R)残基、苯丙氨酸(F)残基、酪氨酸(Y)残基和色氨酸(W)残基。在一些实施方案中,空穴包含一个或多个引入的残基,所述残基选自丙氨酸(A)残基、丝氨酸(S)残基、苏氨酸(T)残基和缬氨酸(V)残基。在一些实施方案中,CH3结构域和/或CH2结构域来自IgG(例如,IgG1亚型、IgG2亚型、IgG2A亚型、IgG2B亚型、IgG3亚型或IgG4亚型)。在一些实施方案中,双特异性抗体的一个CH3结构域包含氨基酸置换T366Y,并且另一个CH3结构域包含氨基酸置换Y407T。在一些实施方案中,一个CH3结构域包含氨基酸置换T366W,并且另一个CH3结构域包含氨基酸置换Y407A。在一些实施方案中,一个CH3结构域包含氨基酸置换F405A,并且另一个CH3结构域包含氨基酸置换T394W。在一些实施方案中,一个CH3结构域包含氨基酸置换T366Y和F405A,并且另一个CH3结构域包含氨基酸置换T394W和Y407T。在一些实施方案中,一个CH3结构域包含氨基酸置换T366W和F405W,并且另一个CH3结构域包含氨基酸置换T394S和Y407A。在一些实施方案中,一个CH3结构域包含氨基酸置换F405W和Y407A,并且另一个CH3结构域包含氨基酸置换T366W和T394S。在一些实施方案中,一个CH3结构域包含氨基酸置换F405W,并且另一个CH3结构域包含氨基酸置换T394S。突变由原始残基表示,随后是使用Kabat编号体系时的位置,并且随后是输入残基。还参见在美国专利号8,216,805的图5中的编号法。
(vii)单结构域抗体
在一些实施方案中,本公开的抗体是单结构域抗体。单结构域抗体是一种单条多肽链,其包含抗体的全部或一部分重链可变结构域或全部或一部分轻链可变结构域。在某些实施方案中,单结构域抗体是人单结构域抗体(Domantis,Inc.,Waltham,Mass.;参见,例如,美国专利号6,248,516B1)。在一个实施例中,单结构域抗体由抗体的全部或一部分的重链可变结构域组成。
(viii)抗体变体
在一些实施方案中,构思了本文所述的抗体的氨基酸序列修饰。例如,可能想要改善该抗体的结合亲和力和/或其他生物学特性。可以通过向编码抗体的核苷酸序列引入适宜的改变或通过肽合成制备该抗体的氨基酸序列变体。此类修饰包括例如,从抗体的氨基酸序列内部缺失残基和/或将残基插入所述氨基酸序列中和/或置换所述氨基酸序列中的残基。可以产生缺失、插入和置换的任意组合以获得最终构建体,只要所述最终构建体拥有想要的特征。可以在产生序列之时在主题抗体氨基酸序列中引入氨基酸改变。
(ix)置换变体、插入变体和缺失变体
在某些实施方案中,提供具有一个或多个氨基酸置换的抗体变体。用于置换诱变的目的位点包括HVR和FR。表1中在“保守性置换”标题下显示保守性置换。表1中在“示例性置换”标题下并且参考如下文进一步描述的氨基酸侧链类别显示更明显的变化。可以将氨基酸置换引入目的抗体中并且对产物筛选所需的活性,例如,保留/改善的抗原结合作用、降低的免疫原性或改善的ADCC或CDC。
表3.示例性置换。


氨基酸可以根据常见的侧链特性分组:
a.疏水性:正亮氨酸、Met、Ala、Val、Leu;Ile;
b.中性亲水:Cys、Ser、Thr、Asn;Gln;
c.酸性:Asp、Glu;
d.碱性:His、Lys、Arg;
e.影响链方向的残基:Gly、Pro;
f.芳族:Trp、Tyr、Phe。
非保守性置换将使这些类别之一的成员交换为另一个类别的成员。
一个置换变体类型涉及置换亲本抗体(例如,人源化或人抗体)的一个或多个高变区残基。通常,经选择用于进一步研究的所得变体将相对于亲本抗体在某些生物学特性方面(例如,增加的亲和力、降低的免疫原性)具有修饰(例如,改善)和/或将基本上保留亲本抗体的某些生物学特性。示例性置换变体是亲和力成熟抗体,所述抗体可以例如使用基于噬菌体展示的亲和力成熟技术(如本文所述的那些技术)便利地产生。简而言之,将一个或多个HVR残基突变并且将变体抗体在噬菌体上展示并筛选特定生物学活性(例如结合亲和力)。
可以在HVR中做出改变(例如,置换),例如以改善抗体亲和力。这类改变可以在HVR“热点”(即,在体细胞成熟过程期间以高频率经历突变的密码子所编码的残基(参见,例如,Chowdhury,Methods Mol.Biol.207:179-196(2008))和/或SDR(a-CDR)中做出,同时对所产生的变体VH或VL测试结合亲和力。已经例如在Hoogenboom等人,引自Methods inMolecular Biology 178:1-37(O'Brien等人编著,Human Press,Totowa,NJ,(2001))中描述了通过构建次级文库并从中重新选择实现亲和力成熟。在亲和力成熟的一些实施方案中,通过多种方法(例如,易错PCR、链改组或寡核苷酸定向诱变)的任一种,将多样性引入所选择用于成熟的可变基因中。随后产生次级文库。随后筛选该文库以鉴定具有所需亲和力的任何抗体变体。另一种引入多样性的方法涉及HVR指导的方法,其中将几个HVR残基(例如,一次4-6个残基)随机分组。可以特别地鉴定参与抗原结合的HVR残基,例如,使用丙氨酸扫描法诱变或建模。特别地经常靶向CDR-H3和CDR-L3。
在某些实施方案中,置换、插入或缺失可以在一个或多个HVR内部出现,只要这类改变不实质降低抗体结合抗原的能力。例如,可以在HVR中做出不实质降低结合亲和力的保守性改变(例如,如本文中提供的保守性置换)。这类改变可以在HVR“热点”或SDR的外部。在上文提供的变体VH和VL序列的某些实施方案中,每个HVR或者不予改变或含有不多于一个、两个或三个氨基酸置换。
一种用于鉴定可以被靶向以便诱变的抗体残基或区域的有用方法称作“丙氨酸扫描法诱变法”,如Cunningham和Wells(1989)Science,244:1081-1085所述。在这种方法中,鉴定一个残基或靶残基组(例如,带电荷残基如arg、asp、his、lys和glu)并且用中性或带负电荷的氨基酸(例如,丙氨酸或聚丙氨酸)替换以确定该抗体与抗原的相互作用是否受影响。可以在对于初始置换显示功能敏感的氨基酸位置处引入其他置换。备选地或另外,测定抗原-抗体复合物的晶体结构以鉴定抗体和抗原之间的接触点。可以将这类接触残基和邻近残基作为置换候选物打靶或消除。可以筛选变体以确定它们是否含有所需的特性。
氨基酸序列插入包括长度从1个残基至含有成百个或更多个残基的多肽间变动的氨基端和/或羧基端融合,以及单个或多个氨基酸残基的序列内插入。末端插入的例子包括具有N末端甲硫氨酰基残基的抗体。抗体分子的其他插入性变体包括抗体的N末端或C末端与酶(例如,针对ADEPT的酶)或增加该抗体血清半寿期的多肽融合。
(x)Fc区变体
在某些实施方案中,可以将一个或多个氨基酸修饰引入本文提供的抗体的Fc区中,因而产生Fc区变体。Fc区变体可以包含人Fc区序列(例如,人IgG1、IgG2、IgG3或IgG4 Fc区),所述的人Fc区序列在一个或多个氨基酸位置处包含氨基酸修饰(例如,置换)。
在某些实施方案中,本公开构思了拥有一些但非全部效应子功能的抗体变体,这使所述抗体变体成为下述应用的有利候选物,其中抗体的体内半寿期重要,而某些效应子功能(如补体和ADCC)是不必要或有害的。可以实施体外和/或体内细胞毒性测定法以证实CDC和/或ADCC活性降低/耗尽。例如,可以实施Fc受体(FcR)结合测定法以确保抗体缺少FcγR结合作用(因此可能缺少ADCC活性),但是保留FcRn结合能力。介导ADCC的原代细胞-NK细胞仅表达Fc(RIII,而单核细胞表达Fc(RI、Fc(RII和Fc(RIII。Ravetch和Kinet,Annu.Rev.Immunol.9:457-492(1991)的第464页上的表3中总结了造血细胞上的FcR表达。评估目的分子的ADCC活性的体外测定法的非限制性例子在美国专利号5,500,362(参见,例如,Hellstrom,I.等人,Proc.Nat’l Acad.Sci.USA 83:7059-7063(1986))和Hellstrom,I等人,Proc.Nat’l Acad.Sci.USA 82:1499-1502(1985);5,821,337(参见Bruggemann,M.等人,J.Exp.Med.166:1351-1361(1987))中描述。备选地,可以使用非放射性分析方法(见,例如,用于流式细胞术的ACTITM非放射性细胞毒性测定法(CellTechnology,Inc.MountainView,CA;和CytoTox 非放射性细胞毒性测定法(Promega,Madison,WI)。用于此类测定法的效应细胞包括外周血单核细胞(PBMC)和天然杀伤(NK)细胞。备选或额外地,可以在体内,例如,在动物模型中,如在Clynes等人,Proc.Nat’l Acad.Sci.USA 95:652-656(1998)公开中的那种动物模型中评估目的分子的ADCC活性。也可以实施C1q结合测定法以证实抗体不能结合C1q并因此缺少CDC活性。参见,例如,WO 2006/029879和WO 2005/100402中的C1q和C3c结合ELISA。为了评估补体激活,可以进行CDC测定法(见,例如,Gazzano-Santoro等人,J.Immunol.Methods 202:163(1996);Cragg,M.S.等人,Blood 101:1045-1052(2003);和Cragg,M.S.和M.J.Glennie,Blood 103:2738-2743(2004))。也可以使用本领域已知的方法确定FcRn结合作用和体内清除率/半寿期(见,例如,Petkova,S.B.等人,Int’l.Immunol.18(12):1759-1769(2006))。
非放射性细胞毒性测定法(Promega,Madison,WI)。用于此类测定法的效应细胞包括外周血单核细胞(PBMC)和天然杀伤(NK)细胞。备选或额外地,可以在体内,例如,在动物模型中,如在Clynes等人,Proc.Nat’l Acad.Sci.USA 95:652-656(1998)公开中的那种动物模型中评估目的分子的ADCC活性。也可以实施C1q结合测定法以证实抗体不能结合C1q并因此缺少CDC活性。参见,例如,WO 2006/029879和WO 2005/100402中的C1q和C3c结合ELISA。为了评估补体激活,可以进行CDC测定法(见,例如,Gazzano-Santoro等人,J.Immunol.Methods 202:163(1996);Cragg,M.S.等人,Blood 101:1045-1052(2003);和Cragg,M.S.和M.J.Glennie,Blood 103:2738-2743(2004))。也可以使用本领域已知的方法确定FcRn结合作用和体内清除率/半寿期(见,例如,Petkova,S.B.等人,Int’l.Immunol.18(12):1759-1769(2006))。
效应子功能减少的抗体包括置换了一个或多个Fc区残基238、265、269、270、297、327和329(美国专利号6,737,056)的那些。这类Fc突变体包括在氨基酸位置265、269、270、297和327的两个或更多个位置处具有置换的Fc突变体,包括残基265和297置换成丙氨酸的所谓“DANA”Fc突变体(美国专利号7,332,581)。
描述了具有FcR结合作用改善或削弱的某些抗体变体。(参见,例如,美国专利号6,737,056;WO 2004/056312和Shields等人,J.Biol.Chem.9(2):6591-6604(2001))。
在某些实施方案中,抗体变体包含具有改善ADCC的一个或多个氨基酸置换(例如,在Fc区的位置298、333和/或334处的置换)(残基的EU编号)的Fc区。在一个示例性实施方案中,抗体在其Fc区中包含以下氨基酸置换:S298A、E333A和K334A。
在一些实施方案中,在Fc区内做出改变,所述改变导致变更(即改善或削弱)的C1q结合作用和/或补体依赖细胞毒性(CDC),例如,如美国专利号6,194,551、WO 99/51642和Idusogie等人,J.Immunol.164:4178-4184(2000)中所述。
在US 2005/0014934 A1(Hinton等人)中描述了半寿期增加和新生Fc受体(FcRn)结合作用改善的抗体,所述新生Fc受体负责转移母源IgG至胎儿(Guyer等人,J.Immunol.117:587(1976)和Kim等人,J.Immunol.24:249(1994)。这些抗体包含其中具有一个或多个置换的Fc区,其中所述置换改善Fc区与FcRn的结合。这类Fc变体包括在一个或多个Fc区残基:238、256、265、272、286、303、305、307、311、312、317、340、356、360、362、376、378、380、382、413、424或434处具有置换(例如,Fc区残基434置换)的那些(美国专利号7,371,826)。还参见涉及Fc区变体的其他例子的Duncan和Winter,Nature 322:738-40(1988);美国专利号5,648,260;美国专利号5,624,821;和WO 94/29351。
(xi)抗体衍生物
可以进一步修饰本公开的抗体,以含有本领域已知并轻易可获得的额外的非蛋白质部分。在某些实施方案中,适于抗体衍生化的这些部分是水溶性聚合物。水溶性聚合物的非限制性例子包括但不限于聚乙二醇(PEG)、乙二醇/丙二醇的共聚物、羧甲基纤维素、葡聚糖、聚乙烯醇、聚乙烯吡咯烷酮、聚1,3-二氧戊环、聚1,3,6-三噁烷、亚乙基/马来酐共聚物、聚氨基酸(均聚物或无规共聚物)、和葡聚糖或聚(n-乙烯基吡咯烷酮)聚乙二醇、丙二醇均聚物、聚环氧丙烷/环氧乙烷共聚物、聚氧乙基化多元醇(例如、丙三醇)、聚乙烯醇和它们的混合物。聚乙二醇丙醛可以具有制造方面的优点,原因在于其在水中的稳定性。这种聚合物可以具有任何分子量,并可以是分枝或不分枝的。与抗体连接的聚合物的数目可以变动,并且如果连接多于一个聚合物,它们可以是相同或不同的分子。通常,用于衍生化的聚合物的数目和/或类型可以基于以下考虑事项确定,包括但不限于待改善的抗体特定特性或功能、抗体衍生物是否将用于限定情况下的疗法中等等。
(xii)载体、宿主细胞和重组方法
也可以使用重组方法产生抗体。为了重组产生抗抗原抗体,将编码该抗体的核酸分离并插入复制型载体以进一步克隆(DNA扩增)或表达。可以使用常规方法(例如,通过使用能够与编码抗体重链和轻链的基因特异性结合的寡核苷酸探针),容易地分离出编码抗体的DNA。许多载体是可用的。载体组件通常包括但不限于以下一者或多者:信号序列、复制起点、一个或多个标记基因、增强子元件、启动子和转录终止序列。
(a)信号序列组件
本公开的抗体可以不仅直接重组地产生,还作为具有异源多肽的融合多肽重组地产生,所述异源多肽优选地是在成熟蛋白或多肽的N末端具有特定切割位点的信号序列或其他多肽。选择的异源信号序列优选地是宿主细胞识别并加工(例如,由信号肽酶切割)的一个异源信号序列。对于不识别并加工天然抗体信号序列的原核宿主细胞,将信号序列由选择自碱性磷酸酶前导序列、青霉素酶前导序列、lpp前导序列或热稳定肠毒素II前导序列的原核信号序列置换。
(b)复制起点
表达载体和克隆载体均含有能够使载体在一种或多种选择的宿主细胞中复制的核酸序列。通常,在克隆载体中,这个序列是令载体能够独立于宿主染色体DNA而复制的一个序列并且包括复制起点或自主复制序列。对于多种原核宿主细胞,这类序列是熟知的。例如,来自质粒pBR322的复制起点适用于大部分革兰氏阴性细菌。
(c)选择基因组件
表达和克隆载体可以含有选择基因,也称作选择标记。常见的选择基因编码这样的蛋白质,这些蛋白质(a)赋予针对抗生素或其他毒素的抗性,例如,氨苄青霉素、新霉素、甲氨蝶呤或四环素,(b)互补性营养缺陷型,或(c)供应从复合培养基不可获得的关键养分,例如,编码杆菌用D-丙氨酸消旋酶的基因。
选择方案的一个例子利用使宿主细胞生长停滞的药物。用异源基因成功转化的那些细胞产生赋予耐药性的蛋白质并且因此实现选择方案。这种显性选择的例子使用药物新霉素、麦考酚酸和潮霉素。
另一个选择方案使用具有移除下述基因的染色体缺失的原核宿主细胞,所述基因的基因产物对特定培养基中生长是必需的。在这些例子中,在特定培养基中培育时,成功用补足宿主细胞的染色体缺失的异源基因转化的那些细胞将存活。可用于这个方案中的基因的例子可以包括营养缺陷型标记基因或在特定培养基中培育宿主细胞时为产生必需养分所要求的其他基因。
(d)启动子组件
表达载体和克隆载体通常含有由宿主生物识别并且与编码抗体的核酸有效连接的启动子。适合随原核宿主一起使用启动子包括phoA启动子、β-内酰胺酶和乳糖启动子系统、碱性磷酸酶启动子、色氨酸(trp)启动子系统和杂合启动子例如tac启动子。但是,其他已知的细菌启动子是合适的。用于细菌系统中的启动子还将含有与编码抗体的DNA有效连接的Shine-Dalgarno(S.D.)序列。
(e)翻译起始区域组件
如上文描述,已知翻译起始区(TIRs)对原核宿主细胞中重组蛋白的翻译重要(参见,例如,Simmons LC和Yansura DG 1996 Nat.Biotechnol.14:629和Vimberg V等人,2007BMC Mol.Biol.8:100)。TIR可以决定翻译单位的翻译效率。TIR一般包含多个翻译单位特征,如起始密码子、Shine-Dalgarno(SD)序列和翻译增强子。TIR还可以包含编码信号肽的分泌信号序列。TIR的序列和及其各特征之间的间距可以调节翻译起始效率。关于蛋白质生产中TIR用途的进一步描述,参见,例如,美国专利号5,840,523(Simmon等人),所述专利描述了用于优化表达和分泌的翻译起始区域(TIR)和信号序列。
(f)转录终止组件
原核宿主细胞中所用的表达载体还可以含有为终止mRNA转录和稳定mRNA必需的序列。在原核细胞中,终止子可以包括Rho依赖性或Rho非依赖性终止子。可用于原核宿主细胞中的终止子的一个例子包括而不限于λt0终止子(Scholtissek和Grosse,Nucleic AcidsRes.15:3185,1987)。
III.工艺优化
本文中提供在原核宿主细胞中通过培养宿主细胞以表达多肽的两条链而产生含有两条链的多肽的方法,其中当表达时,两条链折叠并装配以在宿主细胞中形成生物活性多肽,其中在培养基中在包含以下的条件下培养宿主细胞:包含生长温度和生长搅拌速率的生长阶段,和包含生产温度和生产搅拌速率的生产阶段,其中生长温度在生产温度之上2℃至10℃,并且生长搅拌速率在生产搅拌速率之上50至250转/分钟。本公开令人惊讶地发现,某些生产工艺优化引起双链蛋白产率的大幅度增加,如通过本文所述的数据展示。
(g)宿主细胞的选择和转化
本公开的某些方面涉及原核宿主细胞。克隆或表达在载体中的DNA的合适原核生物在此包括真细菌,如革兰氏阴性或革兰氏阳性生物,例如,肠杆菌科(Enterobacteriaceae)如埃希氏菌属(Escherichia)(例如,大肠杆菌)、肠杆菌属(Enterobacter)、欧文氏菌属(Erwinia)、克雷伯氏菌属(Klebsiella)、变形菌属(Proteus)、沙门氏菌属(Salmonella)(例如,鼠伤寒沙门氏菌(Salmonellatyphimurium))、沙雷菌属(Serratia)(例如,粘质沙雷菌(Serratia marcescans))和志贺氏菌(Shigella)、以及芽孢杆菌纲(Bacilli)如枯草芽孢杆菌(B.subtilis)和地衣芽孢杆菌(B.licheniformis)(例如,在1989年4月12日公开的DD 266,710的地衣芽孢杆菌41P)、假单胞菌属(Pseudomonas)(如铜绿假单胞菌(P.aeruginosa))和链霉菌属(Streptomyces)。一个优选的大肠杆菌克隆宿主是大肠杆菌294(ATCC31,446),不过其他菌株如大肠杆菌B、大肠杆菌X1776(ATCC31,537)和大肠杆菌W3110(ATCC27,325)是合适的。这些例子是示意性的而非限制性的
在一些实施方案中,原核宿主细胞是革兰氏阴性细菌。革兰氏阴性细菌指通过革兰氏染色检测到的含有外膜的任何细菌,所述外膜包围肽聚糖层。许多革兰氏阴性细菌宿主细胞是本领域已知的。例如,已知革兰氏阴性细菌包括而不限于变形菌门(Proteobacteria),如α-变形菌纲(Alphaproteobacteria),β-变形菌纲(Betaproteobacteria),γ-变形菌纲(Gammaproteobacteria)、ζ-变形菌纲(Zetaproteobacteria)、ε-变形菌纲(Epsilonproteobacteria),δ-变形菌纲(Deltaproteobacteria),和酸杆菌门(Acidobacteria);蓝细菌(Cyanobacteria);和螺旋体。熟知的革兰氏阴性细菌可以包括来自以下属的物种,如埃希氏属、沙门氏菌属、志贺氏菌属、假单胞菌属、螺杆菌属(Heliobacter)、军团菌属(Legionella)、奈瑟氏菌属(Neisseria)和克雷伯氏菌属。
在一些实施方案中,本公开的革兰氏阴性细菌是大肠杆菌。如本文所用,大肠杆菌可以指属于物种大肠杆菌的细菌的任何菌株或分离株。大肠杆菌可以包括天然存在的菌株或已经过基因修饰(如通过突变或用如本文所述的质粒转化)的菌株。
在一些实施方案中,本公开的大肠杆菌是内源蛋白酶活性缺陷的菌株。在不希望受理论约束的情况下,认为内源蛋白酶活性缺陷的菌株可以引起增强的重组蛋白生产,如本公开的周质蛋白,因为某些内源蛋白酶对重组表达的底物具有活性(关于这样一个例子,参见Baneyx F和Georgiu G 1990 J.Bacteriol.172(1):491)。内源蛋白酶活性缺陷的菌株可以包括其中编码内源蛋白酶的基因突变、缺失或否则失活的菌株。这类基因的例子可以包括而不限于degP、prc和ompT。用于多种类型原核宿主细胞中引入突变(例如,用于工程化内源蛋白酶活性缺陷的菌株)的方法是本领域熟知的;参见,例如,Snyder L等人,2013Molecular Genetics of Bacteria第4版ASM Press)。
双链蛋白如全长抗体、抗体融合蛋白和抗体片段可以在细菌中产生,尤其当不需要糖基化和Fc效应子功能时,如当治疗性抗体与本身在肿瘤细胞毁灭中显示有效性的细胞毒药物(例如,毒素)缀合时。全长抗体具有在循环中更长的半寿期。大肠杆菌中产生是更迅速和更为成本有效的。关于在细菌中表达抗体片段和多肽,参见,例如,美国专利号5,648,237(Carter等人)、美国专利号5,789,199(Joly等人)、美国专利号5,840,523(Simmon等人),所述专利描述了用于优化表达和分泌的翻译起始区域(TIR)和信号序列。还参见Charlton,Methods in Molecular Biology,第248卷(B.K.C.Lo编著,Humana Press,Totowa,N.J.,2003),第245-254页,其描述在大肠杆菌中表达抗体片段。在表达后,抗体可以从大肠杆菌细胞糊状物在可溶性级分中分离并且可以例如通过蛋白A柱或蛋白G柱(取决于同种型)纯化抗体。可以类似于用于纯化例如在CHO细胞中表达的抗体的方法,实施最终纯化。
将宿主细胞用上述用于双链蛋白生产的表达载体或克隆载体转化并且在常规营养培养基中培养,所述常规营养培养基经调整以诱导启动子,选择转化体或扩增编码所需序列的基因。
(h)培养宿主细胞
本公开的某些方面涉及在培养基中在包括生长阶段和生产阶段的条件下培养宿主细胞。这些阶段各自还可以指特定相期间培育宿主细胞的条件。例如,如本文所用,生长阶段可以包含生长温度和生长搅拌速率,并且生产阶段可以包含生产温度和生产搅拌速率。
生长阶段可以指期间宿主细胞培养物正在指数生长的任何时间。如本文所用的生长温度可以指在宿主细胞的生长阶段期间含有本公开的宿主细胞的培养基的温度。宿主细胞培养物的生长阶段可以通过本领域共知的方法确定,例如,通过随时间推移测量培养物的光密度(例如,在约550nm、约600nm波长,或前者之间的波长)并且确定在什么时间指数生长阶段停止。如果宿主细胞含有带pho启动子的载体,则生长阶段可以指期间宿主细胞培养物正在指数生长并且培养基中磷酸盐浓度足以防止诱导pho启动子介导的基因转录的任何时间。
生产阶段可以指期间宿主细胞培养物正在产生产物的任何时间。如本文所用的生产温度可以指在宿主细胞的生产阶段期间含有本公开的宿主细胞的培养基的温度。如果宿主细胞含有带驱动某产物表达的pho启动子的载体,则生产阶段可以指期间培养基中磷酸盐浓度足够低以至诱导pho启动子介导的产物基因转录的任何时间。
在一些实施方案中,将本公开的宿主细胞在生产温度之上2℃至10℃的生长温度培养。在一些实施方案中,将本公开的宿主细胞在生产温度之上2℃、3℃、4℃、5℃、6℃、7℃、8℃、9℃或10℃的生长温度培养。在一些实施方案中,生长温度在生产温度之上约小于以下任何量(以℃计):10、9.5、9、8.5、8、7.5、7、6.5、6、5.5、5、4.5、4、3.5、3或2.5。在一些实施方案中,生长温度在生产温度之上约大于以下任何量(以℃计):2、2.5、3、3.5、4、4.5、5、5.5、6、6.5、7、7.5、8、8.5、9或9.5。即,生长温度在生产温度之上达下述量范围中的任一者(以℃计),所述量范围具有上限10、9.5、9、8.5、8、7.5、7、6.5、6、5.5、5、4.5、4、3.5、3或2.5和独立选择的下限2、2.5、3、3.5、4、4.5、5、5.5、6、6.5、7、7.5、8、8.5、9或9.5,其中下限小于上限。
在一些实施方案中,生长温度在生长阶段期间处于约30℃至约34℃的范围内。在一些实施方案中,生长温度在生长阶段期间是约30℃、约30.5℃、约31℃、约31.5℃、约32℃、约32.5℃、约33℃、约33.5℃或约34℃。在一些实施方案中,生长温度在生长阶段期间小于约以下温度的任一者(以℃计):34、33.5、33、32.5、32、31.5、31或30.5。在一些实施方案中,生长温度在生长阶段期间大于约以下温度的任一者(以℃计):30、30.5、31、31.5、32、32.5、33或33.5。即,生长温度在生长阶段期间可以是下述温度范围的任一者(以℃计),所述的温度范围具有上限34、33.5、33、32.5、32、31.5、31或30.5和独立选择的下限30、30.5、31、31.5、32、32.5、33或33.5,其中下限小于上限。
在一些实施方案中,生产温度在生产阶段期间处于约25℃至约29℃的范围内。在一些实施方案中,生产温度在生产阶段期间是约25℃、约25.5℃、约26℃、约26.5℃、约27℃、约27.5℃、约28℃、约28.5℃或约29℃。在一些实施方案中,生产温度在生产阶段期间小于约以下温度的任一者(以℃计):29、28.5、28、27.5、27、26.5、26或25.5。在一些实施方案中,生产温度在生产阶段期间大于约以下温度的任一者(以℃计):25、25.5、26、26.5、27、27.5、28或28.5。即,生产温度在生产阶段期间可以是下述温度范围的任一者(以℃计),所述的温度范围具有上限29、28.5、28、27.5、27、26.5、26或25.5和单独选择的下限25、25.5、26、26.5、27、27.5、28或28.5,其中下限小于上限。
搅拌速率指(例如,通过振摇)搅拌细胞培养物的速率。如本文所用的生长搅拌速率可以指在宿主细胞的生长阶段期间搅拌含有本公开的宿主细胞的培养基的速率。如本文所用的生产搅拌速率可以指在宿主细胞的生产阶段期间搅拌含有本公开的宿主细胞的培养基的速率。可以搅拌细胞培养物以维持细胞培养物的通气。
在一些实施方案中,将本公开的宿主细胞在生产搅拌速率之上50至250转/分钟的生长搅拌速率培养。在一些实施方案中,将本公开的宿主细胞在生产搅拌速率之上50转/分钟、75转/分钟、100转/分钟、125转/分钟、150转/分钟、175转/分钟、200转/分钟、225转/分钟或250转/分钟的生长搅拌速率培养。在一些实施方案中,生长搅拌速率在生产搅拌速率之上约小于以下速率的任一者(以转/分钟计):250、225、200、175、150、125、100或75。在一些实施方案中,生长搅拌速率在生产搅拌速率之下约大于以下速率的任一者(以转/分钟计):50、75、100、125、150、175、200或225。即,生长搅拌速率在生产搅拌速率之上达下述速率范围的任一者(以转/分钟计),所述的速率范围具有上限250、225、200、175、150、125、100或75和独立选择的下限50、75、100、125、150、175、200或225,其中下限小于上限。
在一些实施方案中,生长搅拌速率在生长阶段期间处于约600至800转/分钟的范围内。在一些实施方案中,生长搅拌速率在生长阶段期间是约600转/分钟、约625转/分钟、约650转/分钟、约675转/分钟、约700转/分钟、约725转/分钟、约750转/分钟、约775转/分钟或约800转/分钟。在一些实施方案中,生长搅拌速率小于约以下速率的任一者(以转/分钟计):800、775、750、725、700、675、650或625。在一些实施方案中,生长搅拌速率在生长阶段期间大于约以下速率的任一者(以转/分钟计):600、625、650、675、700、725、750或775。即,生长搅拌速率在生长阶段期间是下述速率范围的任一者(以转/分钟计),所述的速率范围具有上限800、775、750、725、700、675、650或625和独立选择的下限600、625、650、675、700、725、750或775,其中下限小于上限。
在一些实施方案中,生产搅拌速率在生产阶段期间处于约300至500转/分钟的范围内。在一些实施方案中,生产搅拌速率在生产阶段期间是约300转/分钟、约325转/分钟、约350转/分钟、约375转/分钟、约400转/分钟、约425转/分钟、约450转/分钟、约475转/分钟或约500转/分钟。在一些实施方案中,生产搅拌速率在生产阶段期间是小于约以下速率的任一者(以转/分钟计):500、475、450、425、400、375、350或325。在一些实施方案中,生产搅拌速率在生产阶段期间大于约以下速率的任一者(以转/分钟计):300、325、350、375、400、425、450或475。即,生产搅拌速率在生产阶段期间是下述速率范围的任一者(以转/分钟计),所述的速率范围具有上限500、475、450、425、400、375、350或325和独立选择的下限300、325、350、375、400、425、450或475,其中下限小于上限。
在不希望受理论约束的情况下,认为当氧浓度有限时,氧转移速率(OTR)等于细胞的氧摄取速率(OUR)或代谢率。可以通过调节搅拌速率促进对OTR的操纵,因此操纵OUR。OUR和其与OTR关系的更多详细描述可以在Ochoa-Garci等人,Biotechnol.Adv.27:153,2009中找到。用于测量细胞培养物的OUR的技术是本领域已知的并且包括,而不限于,使用质谱仪监测来自细胞培养物的尾气的组成并计算细胞培养物的氧摄取速率和二氧化碳释放速率。
在一些实施方案中,将本公开的宿主细胞在这样的生长搅拌速率培养,所述生长搅拌速率足以在生长阶段期间在宿主细胞中实现在生产阶段期间宿主细胞中最大氧摄取速率之上0.5至2.5mmol/L/分钟的氧摄取速率。本公开发现,在生产阶段期间减少细胞培养物的搅拌速率至如与生长阶段相比,足以在宿主细胞中实现氧摄取速率小于约2.5mmol/L/分钟的速率,大大增强产物(如本公开的双链多肽)的产生。
在一些实施方案中,将本公开的宿主细胞在这样的生长搅拌速率培养,所述生长搅拌速率足以在生长阶段期间在宿主细胞中实现在生产阶段期间宿主细胞中最大氧摄取速率之上0.5mmol/L/分钟、1.0mmol/L/分钟、1.5mmol/L/分钟、2.0mmol/L/分钟或2.5mmol/L/分钟的氧摄取速率。在一些实施方案中,将本公开的宿主细胞在这样的生长搅拌速率培养,所述生长搅拌速率足以在生长阶段期间在宿主细胞中实现在生产阶段期间宿主细胞最大氧摄取速率之上小于约以下氧摄取速率的任一者(以mmol/L/分钟计)的氧摄取速率:2.5、2.0、1.5或1.0。在一些实施方案中,将本公开的宿主细胞在这样的生长搅拌速率培养,所述生长搅拌速率足以在生长阶段期间在宿主细胞中实现在生产阶段期间宿主细胞最大氧摄取速率之上大于约以下氧摄取速率的任一者(以mmol/L/分钟计)的氧摄取速率:0.5、1.0、1.5、2.0或2.5。即,将本公开的宿主细胞在这样的生长搅拌速率培养,所述生长搅拌速率足以在生长阶段期间在宿主细胞中实现在生产阶段期间宿主细胞最大氧摄取速率之上下述氧摄取速率范围中任一者(以mmol/L/分钟计)的氧摄取速率,所述的氧摄取速率范围具有上限2.5、2.0、1.5、或1.0和单独选择的下限0.5、1.0、1.5、2.0、或2.5,其中下限小于上限。
在一些实施方案中,宿主细胞的最大氧摄取速率在生长阶段期间处于3.5mmol/L/分钟至4.5mmol/L/分钟的范围内。在一些实施方案中,宿主细胞的最大氧摄取速率在生长阶段期间是3.5mmol/L/分钟、3.75mmol/L/分钟、4.0mmol/L/分钟、4.25mmol/L/分钟或4.5mmol/L/分钟。在一些实施方案中,宿主细胞的氧摄取速率在生产阶段期间处于1.0mmol/L/分钟至3.0mmol/L/分钟的范围内。在一些实施方案中,宿主细胞的氧摄取速率在生产阶段期间是1.0mmol/L/分钟、1.25mmol/L/分钟、1.5mmol/L/分钟、1.75mmol/L/分钟、2.0mmol/L/分钟、2.25mmol/L/分钟、2.5mmol/L/分钟、2.75mmol/L/分钟或3.0mmol/L/分钟。
在一些实施方案中,将本公开的宿主细胞在比生产搅拌速率高约10%至约40%(rpm/rpm)的生长搅拌速率培养。在一些实施方案中,将宿主细胞在这样的生长搅拌速率培养,所述生长搅拌速率具有至少10%、15%、20%、25%、30%或35%(rpm/rpm)生产搅拌速率的下限和不超过生产搅拌速率40%、35%、30%、25%、20%或15%(rpm/rpm)的独立选择的上限。在一个优选实施方案中,将本公开的宿主细胞在10L发酵罐中在1巴背压和20L/分钟的通气速率培养。
可以在多种培养基中培养本公开的宿主细胞。如本文所用的“培养基”指支持本公开的细菌生长的任何组合物或培养液。合适的培养基可以为液态或固态并含有支持细胞生长和活力的任何养分、盐、缓冲剂、元素和其他化合物。培养基的常见养分可以包括氮源、碳源、氨基酸源、糖源、微量元素源、维生素源和矿物质源。这些养分可以作为单独组分(如在限定成分培养基中)或作为复杂提取物组分(例如,酵母提取物)添加。培养基可以富含养分以支持快速生长或最起码支持较慢的生长。培养基还可以含有用来抑制污染性生物生长或将其杀死的任何物质(例如,抗生素)。培养基还可以含有用来控制诱导型启动子或酶的活性的任何化合物(作为一个例子,可以包含IPTG以诱导lac操纵子或功能上相似的启动子控制的任何多核苷酸表达)。合适培养基的许多例子是本领域熟知的并且包括而不限于M9培养基、溶源性培养液(LB)、Terrific培养液(TB)、NZY培养液、SOB培养基和YT培养液。
这些培养基的任一者可以根据需要补充以盐(如氯化钠、钙、镁和磷酸盐)、缓冲剂(如HEPES)、核苷(如腺苷和胸苷)、抗生素、抗真菌剂、微量元素(定义为通常以微摩尔范围的终浓度存在的无机化合物)、葡萄糖和/或适宜的能量源。存在于原核细胞培养基中的常见成分包括酵母提取物、盐(例如,NaCl)、胰蛋白胨、缓冲液(例如,磷酸盐缓冲液)、甘油等。也可以按本领域技术人员已知的适宜浓度包括任何其他的必需补充物。培养条件如温度、pH等是先前随经选择的原核宿主细胞表达一起所用的那些培养条件,并且将是普通技术人员是显而易见的。
(i)纯化生物活性多肽
本公开的某些方面涉及从宿主细胞回收生物活性多肽。通常,回收(术语“纯化”或“纯化”可以在本文互换使用)本公开的生物活性多肽涉及从宿主细胞(或如果多肽分泌入培养基,从细胞培养基)分离多肽并且从其他缔合的大分子(例如,细胞残片和其他多肽)纯化多肽。从多种宿主细胞区间纯化多种蛋白质的众多技术是本领域已知的(参见,例如,Evans,Jr.,TC和Xu MQ(编著)Heterologous Gene Expression in E.coli(2011)Methodsin Molecular Biology第705卷,Humana Presss)。下文描述示例性技术,但是这些技术仅出于补充技术人员理解的示意性目的纳入并且无论如何不意在起到限制作用。
当使用重组技术时,双链蛋白如分泌性蛋白可以在胞内、在周质间隙产生或直接分泌入培养基。如果分泌性蛋白以胞内方式产生,则作为第一步骤,移除颗粒状残片(宿主细胞或裂解的片段),例如,通过离心或超滤移除。
在一些实施方案中,分泌性蛋白从宿主细胞的周质回收。Carter等人,Bio/Technology 10:163-167(1992)描述了用于分离分泌到大肠杆菌周质间隙的分泌性蛋白的方法。简而言之,将细胞糊状物在乙酸钠(pH 3.5)、EDTA和苯甲基磺酰氟(PMSF)存在下经约30分钟融化。通过离心移除细胞残片。在分泌性蛋白分泌入培养基的情况下,首先通常使用市售的蛋白质浓缩滤器,例如,Amicon或Millipore Pellicon超滤装置,浓缩来自这类表达系统的上清液。可以在前述任意步骤中包含蛋白酶抑制剂如PMSF以抑制蛋白酶解,并且可以包含抗生素以防止外来污染物增长。
可以例如使用羟基磷灰石色谱、疏水相互作用色谱、凝胶电泳、透析和亲和色谱,纯化从细胞制备的分泌蛋白质组合物,亲和色谱属于通常优选的纯化步骤之一。就抗体而言,蛋白A作为亲和力配体的适用性取决于抗体中存在的任何免疫球蛋白Fc结构域的种类和同种型。蛋白A可以用来纯化基于人γ1、γ2或γ4重链的抗体(Lindmark等人,J.Immunol.Meth.,62:1-13(1983))。推荐蛋白G用于全部小鼠同种型和用于人γ3(Guss等人,EMBO J.5:15671575(1986))。亲和力配体与之连接的基质最经常是琼脂糖,但是其他基质是可用的。力学稳定的基质如可控孔度玻璃或聚(苯乙烯-二乙烯)苯允许比用琼脂糖所实现的流速和处理时间更快的流速和更短的处理时间。在抗体包含CH3结构域的情况下,Bakerbond ABXTM树脂(J.T.Baker,Phillipsburg,N.J.)可用于纯化。根据待回收的抗体,用于蛋白纯化的其他技术如在离子交换柱上分级、乙醇沉淀、反相HPLC、硅胶上色谱、肝素SEPHAROSETM上色谱、阴离子或阳离子交换树脂(如聚天冬氨酸柱)上色谱、聚焦色谱,SDS-PAGE和硫酸铵沉淀法也可用的。本领域技术人员会认识到,可用于抗体回收的这些技术中许多可以容易地适用于回收其他双链蛋白,如分泌性蛋白。
实施例
公开内容通过参考以下实施例将更充分理解。然而,它们不应当解释为限制本公开的范围。可以理解本文所述的实施例和实施方案的目的仅在于说明,并且在其影响下的多种修改或改变将会启发本领域技术人员并且将纳入本申请的精神与权限范围内和所附权利要求书的范围内。
缩写:Ab(抗体)、hAb(半抗体)、HC(重链)、IPTG(异丙基β-D-1-硫代半乳糖吡喃糖苷)、LC(轻链)、OD(光密度)、ORF(可读框)、OTR(氧转移速率)、OUR(氧摄取速率)、Tg(生长温度)、Tp(生产温度)、TIR(翻译起始区域)、xIL4(抗白介素-4)、xIL13(抗白介素-13)、xIL17(抗白介素-17)、和xIL33(抗白介素-33)。
实施例1:伴侣蛋白水平和氧化还原酶水平对半抗体生产滴度的效应
使用重组技术生产异源分泌型蛋白对许多治疗性分子重要。多特异性抗体(例如,双特异性抗体)是具有多种重要治疗性用途(如癌症和其他应用中免疫治疗)的异源分泌型蛋白的一个非限制性实施例。产生治疗用多特异性抗体需要以产业规模生产这些抗体的结构单元(如半抗体(hAb))的能力。为满足这种需求,本文描述了导致产量相对于标准方法显著增加的优化表达载体和工艺步骤。重要地,发现,与伴侣蛋白FkpA、DsbA和DsbC组合而共表达hAb重链(HC)和轻链(LC)的单一质粒或相容性质粒系统显著增强已装配的hAb的产量。这个系统经测试并且发现改善多种hAb的生产,显示其产生多种分泌型蛋白的广泛实用性。工艺步骤(包含,例如,搅拌速率、pH、FkpA启动子和在不同培养阶段的培养温度)的后续优化导致甚至产物产量的进一步显著增加。
材料与方法
半抗体(hAb)载体构建
载体以EP1356052中所描述相似的方式构建。具体地说,产生用于表达hAb的多种表达载体。对于每种载体,将表达盒在EcoRI位点克隆入大肠杆菌质粒pBR322的构架(Sutcliffe,Cold Spring Harbor Symp.Quant.Biol.43:77-90,1978)。每个表达盒含有至少以下组件:(1)用于控制转录的phoA启动子(Kikuchi等人,Nucleic Acids Res.9:5671-5678,1981);(2)用于翻译起始的来自大肠杆菌trp的Shine-Dalgarno序列、热稳定性肠毒素II(STII)信号序列或二者组合(Chang等人,Gene 55:189-196,1987);和(3)结束转录的λt0终止子(Scholtissek和Grosse,Nucleic Acids Res.15:3185,1987)。额外地,STII信号序列或该信号序列的沉默密码子变体位于轻链或重链的编码序列之前。这个序列指导多肽分泌入周质(Picken等人,Infect.Immun.42:269-275,1983;和Simmon和Yansura,NatureBiotechnology 14:629-634,1996)。
带有单独顺反子的载体设计成提供免疫球蛋白轻链基因和重链基因的单独表达。在这类载体中,每条链的顺反子单元处于其自身Pho启动子控制下并且后接λt0终止子。另外,每个顺反子并入TIR(翻译起始区域)以调节轻链和重链的表达(Simmon等人,J.Immunol.Meth.,263:133-147,2002)。在一个示例性实施方案中,该表达盒从5’至3’含有第一PhoA启动子,随后是针对轻链的顺反子(TIR-L+轻链)和第一λt0终止子,及第二PhoA启动子,随后是针对重链的顺反子(TIR-H+重链)和第二λt0终止子。备选地,该表达盒从5’至3’含有第一PhoA启动子,随后是针对重链的顺反子(TIR-H+重链)和第一λt0终止子,及第二PhoA启动子,随后是针对轻链的顺反子(TIR-L+轻链)和第二λt0终止子。TIR-L和TIR-H含于STII信号序列或其变体内部。
对于xIL13和xIL4 IgG4 hAb表达载体,评价1,1和2,2的TIR组合。对于xIL17和xIL33 IgG4 hAb表达载体,评价TIR 2,2。第一数字代表轻链的TIR强度并且第二数字代表重链的TIR强度。除xIL13、xIL4、xIL17和xIL33 IgG4 hAb之外,还构建并测试了其他IgG1同种型hAb载体。
伴侣蛋白表达质粒构建
为了确定与上文所述载体组合以产生最高xIL13、xIL4、xIL17和xIL33 hAb滴度的伴侣蛋白表达,提供用于共表达的质粒。测试了多种已知的伴侣蛋白,包括FkpA蛋白,一种具有伴侣蛋白活性的肽酰脯氨酰顺-反异构酶。为此,构建了含有FkpA的ORF的相容性质粒(pACYC,Novagen,Madison,WI),如EP1356052中所述(参见,例如,实施例9-10,尤其第[0207]段)。
基于这项工作针对本发明过程扩充,类似地生成一组FkpA相容性载体以调节FkpA共表达水平。如先前所描述通过优化信号肽实现对FkpA水平的调节(Simmon和Yansura,上文,1996)。简言之,FkpA ORF的5'末端含有FkpA信号肽(天然或变体)或STII信号肽。全部FkpA变体基因构建体均处于tacII启动子控制下。
这些质粒随后用上文描述的hAb表达质粒共转化入菌株66F8。宿主菌株66F8的基因型是W3110 ΔfhuA ΔphoA ilvG2096(Valr)Δprc spr43H1 ΔdegP ΔmanA lacIQ ΔompT ΔmenE。
对于xIL13 hAb,构建编码半抗体的LC和HC及FkpA的TIR1,1和TIR2,2质粒并且用来转化66F8。在这些质粒构建体中,FkpA的表达通过受LC的ORF上游的phoA启动子控制。pBR322质粒一般维持在大约30个副本/细胞(Bolivar等人,Gene,2:95-113,1977)并且pACYC质粒一般维持在大约15个副本/细胞(Chang和Cohen,J.Bacteriol.,134:1141-1156,1978)。在不希望受理论约束的情况下,认为将FkpA移动到Ab表达质粒上时,副本数的增加可以导致产生的FkpA的量增加。
氧化还原酶质粒构建
类似于描述用于共表达FkpA的相容性质粒系统,相容性质粒用来筛选与并入前述FkpA TIR变体之一的hAb表达质粒组合的多种已知的氧化还原酶。这项工作用标示为pxIL13.2.2.FkpAc13的TIR2,2质粒进行。此外,相容性质粒用来筛选与xIL4和其他IgG1hAb组合的FkpA和氧化还原酶。
原始氧化还原酶相容性质粒构建如EP1356052中所述(参见,例如,实施例9)。氧化还原酶的筛选包括从相容性质粒JJ247连同hAb表达质粒xIL13.2.2.FkpAc13表达。在xIL13hAb实施例中,氧化还原酶表达用1mM IPTG诱导或保持不诱导以调节氧化还原酶水平。
还构建了编码hAb的LC和HC和伴侣蛋白FkpA、DsbA和DsbC的单一质粒并用来转化66F8。在xIL13 hAb实施例中,构建两个TIR2,2单一质粒,差异在于用来驱动FkpA表达的启动子。单一质粒MD157含有用于表达FkpA的phoA启动子并且质粒KA01含有tacII启动子。利用不同的启动子允许进一步调节FkpA表达。在xIL17 hAb实施例中,构建TIR2,2单一质粒(MD341),所述质粒利用phoA启动子表达FkpA。在全部单一质粒条件下,DsbA和DsbC的表达以多顺反子方式处于tacII启动子控制下。在xIL4 hAb实施例中,构建TIR2,2单一质粒,所述质粒并入下文描述的相容性伴侣蛋白质粒AH8145的ORF并标示为CB1。
除上文描述的单一质粒系统之外,还评价了具有多种IgG1同种型和IgG4同种型hAb的三伴侣蛋白相容性质粒系统。在相容性质粒系统中,将先前描述的FkpA TIR变体之一克隆入多顺反子DsbA和DsbC相容性质粒(JJ247)并标示为AH8145。
发酵过程
大规模生产是基本上如EP1356052中所述(参见,例如,实施例4和第[0159]-[160]段)。对于每个10升发酵,0.5mL冷冻的种子培养物(含有10-15%DMSO)解冻并用来接种含有500ml大豆LB培养基的2L摇瓶,所述培养基补充有0.5ml四环素溶液(5mg/ml)和/或2mL卡那霉素溶液(5mg/mL)和2.5ml 1M磷酸二氢钠溶液。将这个种子培养物在振摇下在30℃培育大约16小时并随后用来接种10升发酵罐。
该发酵罐最初含有大约7.0升培养基,所述培养基含有1.1g葡萄糖、100ml 1M硫酸镁、10ml微量元素溶液(终体积1升中100ml盐酸、27g氯化铁六水合物、8g硫酸锌七水合物、7g氯化钴六水合物、7g钼酸钠二水合物、8g硫酸铜五水合物、2g硼酸、5g硫酸锰一水合物),或者20ml四环素溶液(乙醇中5mg/ml)或250mL氨苄青霉素溶液(2mg/mL)、1袋HCD盐(37.5g硫酸铵、19.5g磷酸氢二钾、9.75g磷酸二氢钠二水合物、7.5g柠檬酸钠二水合物、11.3g磷酸二氢钾)、200g BL4大豆(大豆蛋白水解物)和100克酵母提取物。发酵最初在30℃以20标准升/分钟(slpm)通气量进行并且被控制在pH 7.0±0.2(不过在一些情况下发生超出这个范围的偶尔波动)。发酵罐的背压维持在表压1巴并且搅拌速率最初设定至650转/分钟。如下文实施例2中详细讨论,也可以变动搅拌速率以操纵发酵罐中的氧转移速率并且因此控制细胞呼吸速率。此外如下文实施例3中详细讨论,可以调节在生长阶段和生产阶段期间的温度以最大化产物产量。
发酵罐用来自摇瓶的含有细胞的培养基接种后,使用基于计算机的算法向发酵罐输入浓葡萄糖溶液时,将培养物在发酵罐中培育至高细胞密度。根据需要,还将氢氧化铵(58%溶液)和硫酸(24%溶液)输入发酵罐以控制pH。在一些情况下还添加L-61消泡剂以控制起泡。
当培养物达到大约40OD550的细胞密度时,将额外的100ml 1M硫酸镁添加至发酵罐。额外地,将浓盐进料(终体积1250ml中12.5g硫酸铵、32.5g磷酸氢二钾、16.25g磷酸二氢钠二水合物、2.5g二水合柠檬酸钠、18.75g磷酸二氢钾、10ml 2.7%氯化铁和10ml微量元素)添加至发酵罐并且当培养物达到大约20OD550时以2.5ml/分钟的速率启动并且持续直至向发酵罐添加大约1250ml。发酵一般持续70-80小时。
供电泳、免疫印迹和HPLC分析的样品制备
未还原的可溶性样品制备类似于EP1356052中所描述的情况(参见,例如,实施例4,尤其第[0162]段)。具体地说,未还原的可溶性样品如下制备:在发酵过程期间取得的1mL完整培养液样品的冷冻样品在室温解冻。将100μL解冻的完整培养液添加至500μL提取缓冲液。(提取缓冲液:10mM Tris,pH 6.8,5mM EDTA,新鲜添加的0.2mg/mL鸡蛋溶菌酶,和新鲜配制的碘乙酸至终浓度5-10mM)。将完整培养液样品加提取缓冲液在冰上孵育5-10分钟,随后超声处理2x 10个脉冲,随后在4℃和14,000转/分钟离心15-20分钟。将上清液作为可溶性部分移除。为了通过SDS-PAGE和免疫印迹结果分析,将可溶性级分1:10稀释入无还原剂的2X Novex Tricine样品缓冲液。将10μL这种制备物加载于10孔Novex 10%Bis-TrisNuPage凝胶上和在150V以MES缓冲液电泳。随后将凝胶用于免疫印迹或用考马斯蓝染色。
提交可溶性级分的样品以通过LC-κ/RP测定法分析。这种测定法是二维HPLC测定法,其中第一柱是捕获含有κ-轻链的IgG组分的亲和柱并且第二柱是反相柱。一体化HPLC工作站以双柱模式设置。溶剂贮库是:溶剂1A,亲和力上样缓冲液;溶剂1B,亲和力洗脱缓冲液,水中0.2%TFA;溶剂2A,反相水性缓冲液,水中0.1%TFA;溶剂2B,反相有机洗脱缓冲液、80%乙腈中0.1%TFA。第一柱是购自Life Technologies(Carlsbad,CA)的 CaptureSelectTM LCκ亲和柱(2.1x 30mm)。涉及亲和柱的全部程序均在环境温度进行。
CaptureSelectTM LCκ亲和柱(2.1x 30mm)。涉及亲和柱的全部程序均在环境温度进行。
第二柱是购自Life Technologies(Carlsbad,CA)的 R2 20μm反相柱(2.1x 30mm)。反相柱温度维持在-80℃。
R2 20μm反相柱(2.1x 30mm)。反相柱温度维持在-80℃。
亲和柱在上样缓冲液中平衡,并且样品以2ml/分钟流量加载。指引通流至废物瓶。在加载样品后,将亲和柱用上样缓冲液(3ml)洗涤以减少非特异性结合的组分。随后,通过阀门转换,使亲和柱与反相柱连通并用洗脱缓冲液(5ml)以2ml/分钟的流量洗脱,以转移亲和捕获的组分至反相柱。在这个转移步骤期间,令一体化UV检测器位于亲和柱后反相柱之前,因此监测亲和柱的洗脱物,该洗脱物变成到达反相柱的负载。在洗脱和从反相柱断开后,将亲和柱通过水(2ml)洗涤并且随后用上样缓冲液(4ml)再平衡。
加载的反相柱用0.1%TFA水溶液(1.1ml)洗涤。流量设定至2ml/分钟并且快速梯度(0.25分钟)运行至35%溶剂2B(0.1%TFA/80%乙腈),随后经3分钟内至50%溶剂2B的浅梯度。通过0.5分钟内至100%溶剂2B的梯度完成洗脱并保持1.5分钟。随后使反相柱在0.05分钟内返回初始条件并保持2分钟以再平衡。在280nm和214nm监测柱洗脱物。基于来自反相柱的分离,通过将积分的峰面积与浓度已知的那些标准品比较,进行定量。
还定量发酵过程期间产生的LC和HC的可溶物量和总量。为了进行总RP-HPLC定量,将发酵液样品用6M HCl胍、360mM TRIS、2mM EDTA pH 8.6连同200mM中的100mM DL-二硫苏糖醇(Sigm目录号#43816)稀释10倍。将样品涡旋混合,在60℃孵育20分钟并在4℃以13,000转/分钟离心15分钟。使用0.22um滤器过滤可溶性部分,之后上样在HPLC上。使用的HPLC系统是Agilent Technologies 1290 Infinity系统。将样品上样到Zorbax 300SB-C3 RapidResolution(4.6x 150mm 3.5微米)分析柱(目录号#863973-909)上。流动相A由SQH2O中的0.1%三氟醋酸(Thermo目录号#28901)组成并且流动相B由HPLC级乙腈(Honeywell目录号#AH015-4)中的0.08%三氟乙酸组成。
为了进行总可溶物RP-HPLC定量,将发酵液样品用Pro-Scientific DPS-20均化(10,000转/分钟)及超声处理(88%振幅)10秒4次。随后将样品在4℃和14,000转/分钟离心15-20分钟。取出上清液作为可溶性部分并在RP-HPLC上如上文所述那样变性和加工。
双特异性抗IL13/IL17抗体装配
亲和纯化的抗IL13和抗IL17半抗体按1:1质量比合并,随后滴定至pH 8.5。相对于从装配过程期望的双特异性抗体理论最大量,将100x过量摩尔比的还原型L-谷胱甘肽(GSH)添加至混合物,并且将温度从室温调整至35℃。两个半抗体在体外在氧化还原反应中装配以形成抗IL13/抗IL17双特异性抗体。氧化还原反应在35℃继续12-24小时。通过调节pH至6.5,猝灭装配反应,并且将汇集物用水稀释,之后双特异性抗体经历单独或组合的通过离子交换剂纯化或疏水性柱色谱纯化。
结果
FkpA表达对hAb滴度的影响
如图1A中所示,轻链和重链亚单位从xIL13 hAb TIR1,1载体的产生分别导致2.9g/L和1.2g/L的轻链生产总量和重链生产总量。对于TIR2,2,产生的轻链总量和重链总量分别是6.4g/L和4.1g/L。
但是,总亚单位生产的量不导致明显的可溶性亚单位积累。使用xIL13 TIR1,1hAb质粒时,产生的轻链可溶性总量和重链可溶性总量分别是0.2g/L和小于0.1g/L(图1B)。对于TIR2,2 hAb,产生的轻链可溶性总量和重链可溶性总量分别是0.3g/L和0.3g/L(图1C)。对于TIR1,1发酵,产生的已装配xIL13 hAb的滴度仅是0.1g/L,并且TIR2,2发酵滴度小于0.2g/L(图2)。这些结果表示,存在明显的hAb产物无效折叠和/或装配。因此,执行进一步工作以评价使用共表达伴侣蛋白的TIR1,1质粒和TIR2,2质粒对滴度的影响。
已知几个类性的伴侣蛋白促进蛋白质折叠和装配(图3A-B)。特定目的伴侣蛋白包括已知其作为肽酰基-脯氨酰顺-反异构酶和伴侣蛋白发挥作用的FkpA和已知其作为氧化还原酶发挥作用的DsbA和DsbC。实施实验以检验FkpA、DsbA和DsbC的表达是否影响hAb产生。
具体地说,检查了使用单独质粒(相容性质粒系统)或编码hAb HC ORF和LC ORF的相同质粒(单一质粒系统)表达FkpA的影响(图4A-B)。单一表达质粒的使用消除了使用多种抗生素或其他选择性压力手段以确保质粒保留的需求。在用于xIL13 hAb的单一质粒系统中,驱动FkpA表达的启动子从IPTG诱导型tacII启动子变成phoA启动子。因此,培养物中磷酸盐耗尽导致HC、LC和FkpA表达。
对于IgG1同种型hAb(xVEGF),除了装配的hAb增加之外,FkpA伴侣蛋白共表达水平增加与可溶性单体重链积累的量增加相关(图5A)。在这个实验中,用1mM IPTG诱导作用筛选一组如先前所描述的FkpA表达变体。在无FkpA共表达的情况下,xVEGF hAb的滴度是0.2g/L,在FkpA共表达(FkpA(wt))水平最高的条件下,hAb滴度是大约0.4g/L(图5B)。
为了产生xIL13 hAb,测试了使用TIR1,1或TIR2,2抗体表达载体的相容性质粒系统(从一个质粒表达xIL13HC和LC并且从另一个质粒表达FkpA,如图4A中所示)。对于采用1mM IPTG诱导以驱动FkpA共表达的相容性质粒系统,TIR1,1质粒的滴度是0.2g/L并且TIR2,2质粒的滴度是0.4g/L(图6A,泳道3和4)。与具有内源FkpA水平的TIR1,1条件和TIR2,2条件相比,这导致滴度增加大约两倍(图6A,泳道1和2)。图6B显示如与内源表达相比,相容性系统中诱导的FkpA表达量,如通过超高效液相色谱(UPLC)以百万吸附单位(mAu)计和蛋白质印迹结果所测量。
还测试了图4B中所示的用于表达FkpA和产生hAb的单一质粒系统。对于xIL13hAb,单一质粒TIR1,1和TIR2,2的滴度分别是0.4g/L和0.7g/L(图6A)。这代表滴度超过相容性质粒系统增加大约两倍。单一质粒系统中的FkpA表达水平也比相容性质粒系统中诱导的FkpA表达水平高大约两倍(图6B)。在两种TIR条件下,FkpA表达水平增加均与可溶性单体重链积累的量增加相关。
从相容性质粒系统诱导FkpA共表达的情况下,用TIR1,1条件和TIR2,2条件测试第三种hAb(xIL4 IgG4)。在这个实验中,FkpA(wt)共表达增加了两种TIR条件下可溶性单体重链的量(图7A),但是不导致产生的hAb的滴度增加(图7B)。
从相容性质粒系统诱导FkpA共表达的情况下,测试第四种hAb(xVEGFC IgG1)。在这个实验中,FkpA(wt)共表达增加了可溶性单体重链的量并且将滴度从0.5g/L增加至0.8g/L(图8A)。如通过考马斯染色所确定的FkpA表达增加(图8B)与可溶性单体HC链积累增加相关(图8C)。总之,这些结果表明,FkpA表达增强可溶性单体重链的积累,但是对已装配的hAb滴度的影响不定。这表明需要进一步优化。
DsbA和DsbC表达对hAb滴度的影响
通过上文描述的pxIL13.2.2.FkpAc13(单一质粒)与表达氧化还原酶DsbA和DsbC的相容性质粒组合进一步优化xIL13 hAb产生(图9)。表达DsbA和DsbC(JJ247)的质粒连同pxIL13.2.2.FkpAc13增加hAb滴度。图10显示在IPTG诱导DsbA和DsbC存在和不存在的情况下,随时间推移产生的xIL13 hAb的滴度。在进入发酵的52小时实现具有JJ247的pxIL13.2.2.FkpAc13的最高滴度。在这个时间点,在无IPTG的条件(非诱导条件)下,xIL13滴度是1.2±0.2g/L,并且在含IPTG的条件(诱导条件)下,xIL13滴度是1.0±0.2g/L(图10)。对于测试的两种条件,滴度从52小时至72小时明显降低并且这归因于发酵过程期间氧摄取速率(OUR)降低和重量摩尔渗透压浓度升高,如实施例2中所述那样。
还用xIL4TIR1,1和TIR2,2Ab表达质粒评价了用于表达DsbA、DsbC和FkpA的相容性系统(AH8145)(图11A)。全部三种伴侣蛋白的非诱导型共表达分别产生0.8g/L和1.2g/L的TIR1,1滴度和TIR2,2滴度(图12,泳道5和6)。这代表如与前述的FkpA相容性TIR2,2条件相比,TIR2,2条件的滴度增加大约六倍(图12,泳道3和4)。
用另一种hAb进一步评价非诱导的相容性AH8145质粒系统。使用基于TIR2,2的载体产生xVEGFC IgG1 hAb,如与仅FkpA共表达相比,未诱导的相容性AH8145条件将滴度从0.8g/L增加至1.0g/L(图13)。用AH8145相容性质粒,测试第五hAb(xVEGFA IgG1)连同TIR1,1和TIR2,2条件。在无伴侣蛋白共表达的情况下,TIR1,1条件和TIR2,2条件的滴度相似,约0.9g/L(图14,泳道1和3)。在未诱导的AH8145条件下采用相容性质粒时,TIR1,1质粒和TIR2,2质粒的滴度分别是1.2g/L和1.7g/L(图14,泳道2和4)。
生成了并入AH8145的伴侣蛋白ORF的xIL4TIR2,2单一质粒(CB1)并且图11B中显示该质粒。在不添加IPTG的情况下,CB1质粒产生比随相容性质粒系统所观察到的滴度略高的滴度(图15)。应当指出,这种比较中报告的hAb滴度从利用如实施例2中所述的优化搅拌策略的发酵中产生。通过蛋白质印迹结果,采用CB1时全部三种伴侣蛋白的水平均略高(图15)。在不希望受理论约束的情况下,单一质粒系统可以允许更高的质粒拷贝数,这可以导致更高的产量。
将xIL13单一质粒系统MD157与非诱导的相容性pxIL13.2.2.FkpAc13和JJ247相容性质粒系统比较(图16)。与相容性系统中的1.9±0.04g/L相比,MD157单一质粒条件产生2.1±0.3g/L的滴度(图17)。应当指出,这种比较中报告的滴度从利用如实施例2中所述的优化搅拌策略的发酵中产生。
综上,这些结果显示,FkpA与DsbA和DsbC一起共表达增加了已装配hAb的产生,使用多种hAb构建体证实这些影响。
实施例2:氧摄取对半抗体产生的影响
上述结果显示,通过FkpA、DsbA、和DsbC与hAb HC和LC一起共表达实现hAb生产改善。但是,即便具有这种增强的产量,发现滴度在52小时生产后多个时间点进入停滞或甚至削弱。因此,进行额外的实验以进一步优化hAb生产。
如前述,发酵最初在30℃以20标准升/分钟(slpm)通气量进行并且被控制在pH7.0±0.2。发酵罐的背压维持在表压1巴并且搅拌速率最初设定至650转/分钟。额外的表背压产生200%的初始溶解氧浓度(dO2)。在dO2信号下降低于起始水平的2%后,将浓葡萄糖溶液输入生产培养物,一般约两小时,并且在发酵期间持续输入,从而葡萄糖是非限制性养分。
在12小时,培养物中的细胞密度足以维持dO2浓度在零百分数处或其附近。在不希望受理论约束的情况下,认为当氧浓度有限时,氧转移速率(OTR)等于细胞的氧摄取速率(OUR)或代谢率。可以通过调节搅拌速率促进对OTR的操纵,并且这种操纵直接影响培养物的OUR。质谱仪用于监测来自发酵的尾气的组成并使得计算发酵中的氧摄取速率和二氧化碳释放速率成为可能。
如图18中所示,在上文描述的具有JJ247的pxIL13.2.2.FkpAc13质粒系统中,诱导的和未诱导的TIR2,2培养物均在第12小时具有4.25mmol/L/分钟的最高OUR,在该时刻后O2变得有限。将这些培养物在以650转/分钟的恒定速率搅拌下培育。图19显示这些培养物随时间推移的重量摩尔渗透压浓度。综上,图18-19显示,在50小时后,细胞培养物的OUR急剧下降并且重量摩尔渗透压浓度升高。
图20显示这些培养物中随时间推移产生的xIL13 hAb的滴度。观察到滴度从52小时处的1.2g/L±0.2g/L降至72小时处的0.7±0.04g/L。类似地,在诱导条件下,观察到从52小时处的1.0±0.2g/L降至72小时处的0.5±0.05g/L。令人好奇地,这些培养物中生产的这种降低在OUR下降和重量摩尔渗透压浓度升高的相同时间出现。
为了缓和滴度降低并消除重量摩尔渗透压浓度升高,采用非诱导的TIR2,2条件评价在26小时进行的搅拌变换。搅拌速率从650转/分钟变换为实现目标OUR设定点的水平。测试了四个不同的OUR目标设定点:大约1.5、1.75、2.0和2.5mmol/L/分钟。OUR设定点在大约1.5和2.0mmol/L/分钟之间的搅拌变换消除了OUR降低(图21)和重量摩尔渗透压浓度升高(图22)。
重要地,在全部搅拌变换条件中,滴度高于54小时处未变换的条件下的滴度(图23)。在大约2.5mmol/L/分钟条件下,OUR在约60小时下降(图21)并且重量摩尔渗透压浓度在72小时升高至最高1200mOsm(图22)。在72小时,大约2.5mmol/L/分钟条件的滴度(0.7g/L)降至与未变换的条件相似的水平(0.6±0.1g/L)。大约2.0mmol/L/分钟条件下的平均滴度是四个所测试条件中最高的(2.4±0.6g/L);然而,滴度存在某种变异性并且似乎存在OUR特征的轻微下降。大约1.5mmol/L/分钟条件具有重现的平均滴度2.1±0.3g/L,但是在发酵后期,似乎再次存在OUR特征的某种变异。大约1.75mmol/L/分钟条件既有可重现的滴度(1.8±0.2g/L),又具有始终如一的OUR趋势,因此选择它作为优选的设定点。
这些结果显示,变换培养物的搅拌速率缓和了观察到的OUR下降和重量摩尔渗透压浓度升高。重要地,这些搅拌变换还允许增强hAb生产滴度,特别地在稍后的生产时间点。
实施例3:温度对半抗体产生的影响
尽管先前实施例显示hAb生产明显增长,但进行额外的试验以仍然进一步优化生产过程。因此,对于xIL13 hAb生产,使用在实施例1中描述的高效单一质粒系统和在实施例2中描述的大约1.75mmol/L/分钟的OUR设定点作为起点。该过程仍然进一步优化以产生显著更大的生产产量,如下文描述。
本公开的优选实施方案的发酵过程可以划分成两个不同部分:生长阶段和生产阶段。在生长阶段,大部分养分处于过量,并且培养物密度快速地增加。在生产阶段中,磷酸盐变得有限,生长停止,并且目的产物的表达开始。对这些阶段分别测试温度对hAb生产的影响。
与温度实验组合的情况下,评价第二宿主。在这些实验中,将MD157质粒转化入67A6生产性宿主。67A6菌株的基因型是W3110 ΔfhuA ΔphoA ilvG+ Δprc spr43H1 ΔdegP ΔmanA lacIQ ΔompT ΔmenE742 degPS210A。
67A6菌株含有蛋白酶缺陷型等位基因degP基因(也称作htr和ptd,其编码DegP)的敲入。DegP(也称作蛋白酶Do)是在高温存活所要求的周质丝氨酸蛋白酶。另外,DegP充当分子伴侣。丙氨酸置换丝氨酸消除了DegP的蛋白酶解活性,但不影响其伴侣蛋白功能(Spies等人,Cell,97:339-347,1999)。
对xIL13TIR2,2单一质粒系统的生长阶段(Tg)和生产阶段(Tp)均进行温度优化。图24显示以30℃或28℃的恒定Tg/Tp培育的培养物的生长。如与30℃的恒定Tg/Tp相比,28℃的恒定Tg/Tp导致生长速率迟滞。图25显示在这些温度培育的培养物的OUR。类似于生长速率,如与30℃相比,当培养物在28℃的恒定Tg/Tp培育时OUR延迟。
如图26中所示,使用30℃的恒定Tg/Tp时,磷酸盐在22±2小时耗尽。当Tg/Tp向下变动至28℃时,培养物的生长速率如上文所述受阻,并且磷酸盐耗尽变动至26±2小时。
图27显示这些培养物的xIL13 hAb生产。对于每种条件,产物表达在磷酸盐耗尽出现时的时间开始。如与恒定30℃Tg/Tp条件的1.3±0.2g/L相比,恒定28℃Tg/Tp条件实现2.5±0.2g/L的最终滴度。
在增加生产阶段时间量的尝试中,测试温度变换策略。在这个实验中,生长阶段温度设定在30℃并且在20小时,温度变动至28℃。对于Tg30/Tp 28℃变换和恒定Tg/Tp30℃条件,在0和20小时之间的生长特征(图28)、磷酸盐特征(图29)和OUR特征(图30)相似。
如图31中所示,比较Tg30/Tp 28℃条件下产生的3.1g/L最终滴度与恒定Tg/Tp28℃条件下产生的2.5g/L最终滴度,在产物诱导时使用变换至28℃的温度引起滴度进一步增加0.6g/L。这些结果显示在较高温度培育培养物,随后在生产时降低培养温度可以导致产物形成明显增加。
进行一项部分析因设计实验(DoE)以确定利用67A6宿主菌株时xIL13 hAb的最佳运行条件。该DoE关注三个运行参数和FkpA水平。
表3-1.xIL13 hAb参数


如表3-1中所示,运行参数包括Tg和Tp以及pH。样式指特定参数的运行范围;(-)指低值参数,(+)指高值参数,并且0001和0002指中心点参数。Tg为30℃至34℃,Tp为25℃至28℃,和pH在6.7和7.0之间。产生的FkpA的量受所用启动子调节。高水平的FkpA从MD157上的pho启动子产生并且低水平FkpA从KA01上未诱导的tac启动子产生。通过进行10个实验进行部分析因分析。
如图32中所示,这些因素显著影响所产生的xIL13 hAb的滴度。通过表3-1中所述用于DoE实验的条件产生的hAb的滴度为大约1.5g/L至大约4.5g/L。
采用从xIL13DoE确定的最佳发酵条件(Tg34℃,Tp28℃,pH 7.0),测试xIL4单一质粒(CB1)。但是在CB1条件下,tacII启动子在无IPTG诱导作用时用来驱动FkpA表达。将xIL14hAb滴度与其中Tg/Tp保持在30℃恒定及pH维持在7.0的发酵条件比较。在不添加IPTG的情况下采用所述的新工艺条件,xIL4 hAb滴度从1.3g/L增加至3.1g/L(图33)。
进行一项部分析因设计实验(DoE)以确定xIL17 hAb的最佳运行条件。通过将xIL13 hAb单一质粒(MD157)LC和HC的ORF更换为xIL17LC和HC的ORF,构建MD341单一质粒。用于表达抗体片段和伴侣蛋白的启动子与xIL13(MD157)质粒相同。DoE关注三个运行参数。
表3-2.xIL17 hAb参数
| 样式 | 生长温度(Tg) | 生产温度(Tp) | pH | 滴度(g/l) |
| -+- | 30 | 30 | 6.7 | 0.38 |
| 0001 | 32 | 27.5 | 7.0 | 2.0 |
| --- | 30 | 25 | 6.7 | 2.5 |
| --+ | 30 | 25 | 7.3 | 2.6 |
| +-+ | 34 | 25 | 7.3 | 2.6 |
| 0002 | 32 | 27.5 | 7.0 | 1.9 |
| ++- | 34 | 30 | 6.7 | 0.34 |
| +-- | 34 | 25 | 6.7 | 2.0 |
| -++ | 30 | 30 | 7.3 | 0.2 |
| +++ | 34 | 30 | 7.3 | 0.12 |
如表3-2中所示,运行参数包括Tg和Tp以及pH。Tg为30℃至34℃,Tp为25℃至30℃,和pH在6.7和7.3之间。图34显示在Tp为25℃的条件下实现了最显著的滴度积累。
图35显示第一优化伴侣蛋白共表达并且随后优化工艺步骤(例如,搅拌速率、Tg和Tp)对xIL13 hAb滴度产量的影响。在一个示例性实施方案中,分子优化(例如,伴侣蛋白表达和载体特征)引起滴度增加大约16倍。通过工艺开发(例如,搅拌速率、Tg和Tp),进一步增强这种生产水平约3.5倍。综上,这些结果显示,可以通过优化多个变量(包括伴侣蛋白表达、载体系、搅拌速率、pH和生长/生产温度)实现生产和稳健性的显著增长。最终确定,其中Tg在34℃、Tp在25℃,pH 6.7的条件和由phoA启动子产生的FkpA的水平导致xIL13 hAb最高滴度。
实施例4:宿主菌株对半抗体产生的影响
尽管先前实施例显示hAb生产明显增长,但进行额外的试验以表征宿主菌株66F8和67A6之间生产hAb的潜在差别。因此,对于hAb生产,在两个大肠杆菌宿主菌株:66F8和67A6中评价实施例1中描述的高效单一质粒系统、在实施例2中描述的OUR变换和实施例3中描述的温度变换。菌株66F8的基因型是W3110 ΔfhuA ΔphoA ilvG+ Δprc spr43H1 ΔdegP ΔmanA lacIQ ΔompT ΔmenE742。67A6菌株的基因型是W3110 ΔfhuA ΔphoA ilvG+ Δprc spr43H1 ΔdegP ΔmanA lacIQ ΔompT ΔmenE742 degPS210A。
xIL13 hAb在66F8和67A6宿主菌株中均表达。发酵在使用如上文所述的温度变换和搅拌速率变换的条件下进行。如与66F8菌株相比,67A6菌株的使用导致可溶性xIL13 hAb滴度增加(图36A)及LC和HC的总亚单位积累增加(图36B)。
xIL4 hAb也在66F8和67A6宿主菌株中均表达。当xIL4 hAb发酵在30℃的恒定温度进行时,约1.5g/L的相似滴度从两种菌株获得(图37A)。但是,在其中Tg从34℃降至Tp 25℃的发酵条件下,67A6菌株产生3.0g/L平均数并且66F8菌株产生2.0g/L平均数(图37B)。此外,在使用温度变换的条件下67A6发酵中LC和HC的总亚单位积累大于66F8发酵中(图38A和图38B)。
因此,xIL13 hAb和xIL4 hAb是由67A6宿主菌株和相对于生长温度生产温度降低提供的滴度益处的两个例子。不受理论约束,重组蛋白积累似乎在缺少DegP的宿主菌株中进入停滞,而在具有突变DegPS210A的宿主菌株中,重组蛋白似乎积累直至发酵结束。
实施例5:使用优化的表达载体和优化的培养条件生产xIL33 hAb
通过将xIL13 hAb单一质粒(MD157)LC和HC的ORF更换为xIL33 LC和HC的ORF,构建xIL33 hAb(MD501质粒)。用于表达抗体片段和伴侣蛋白的启动子与xIL13(MD157)质粒相同(图39)。用仅含有xIL33 LC和HC的ORF的xIL33 hAb表达载体(MD481)进行单一发酵。MD481质粒不含有分子伴侣DsbA、DsbC或FkpA的ORF。在30℃恒定温度、pH 7.0和搅拌速率650转/分钟(图40的基础情况)进行发酵。接下来,用含有xIL33 LC和HC的ORF以及分子伴侣DsbA、DsbC或FkpA的ORF的xIL33 hAb表达载体(MD501)进行单一发酵。与MD481发酵相同的运行条件用于MD501发酵。如与基础情况相比,单一质粒(MD501)的使用导致xIL-33hAb滴度增加大约10倍。
进行一项部分析因设计实验(DoE)以确定xIL33 hAbMD501单一质粒的最佳培养条件。DoE关注采用10个实验的部分析因中的四个参数,所述部分析因包含67A6宿主中的两个中心点重复(表5-1)。在OD550 150进行搅拌速率变换和温度变换。
表5-1.xIL33 hAb参数

如表5-1中所示,Tg为30℃至34℃,Tp为25℃至30℃,并且pH在6.7和7.3之间,以及OUR设定点为1.9至2.8mmol O2/L/分钟。图41显示中心点条件向滴度提供最明显的获益,实现的最高滴度为4.0±0.05g/L,这解释了如与基础情况运行条件相比hAb滴度的进一步增加(图40)。最佳培养条件包括pH 7.0,Tg 32℃,Tp 27.5℃(2小时爬坡)和目标OUR大约2.3mmol/L分钟(2小时搅拌速率爬坡)。
实施例6:FkpA优化
在优化FkpA表达水平的尝试中,两种额外的FkpA TIR变体在xIL13、xIL17和xIL33hAb的单一质粒(MD157、MD341和MD501质粒)中测试。如所述,将FkpA TIR变体与内源FkpA信号序列相比时表征(图42)。MD157、MD341和MD501质粒并入FkpAc13的ORF,所述ORF与这三种质粒的TIR强度相关(图43)。在MD157、MD341和MD501单一质粒中,将FkpAc13ORF替换为FkpATIR1或TIR2ORF并且在先前对每种hAb确定的优化发酵条件下测试。
FkpA表达水平增加与xIL13 hAb积累增加相关(图44)。FkpA TIR1条件和TIR2条件分别导致0.5g/L和2.5g/L的FkpA最终量及1.5g/L和2.5g/L的xIL13 hAb滴度。TIR3条件产生4g/L FkpA和3.8g/L hAb。FkpA表达水平还影响生产阶段OUR特征。总体上,FkpA的滴定增加所产生hAb的量约3倍。
在xIL33 hAb发酵中,积累曲线的最低滴度与FkpA TIR1条件相关,试验结束滴度为2.4g/L(图45)。与TIR1条件相比时,xIL33 TIR2条件和TIR3条件导致xIL33 hAb滴度增加大约2倍(图45)。数据表明,FkpA水平增加有益于hAb产生。
在xIL17 hAb发酵中,积累曲线的最低滴度与FkpA TIR1条件相关,试验结束滴度为2.0g/L(图46)。
使用xIL13 hAb的前述最佳条件,测试了两个额外的FkpA TIR变体(TIR2.3和TIR6)。TIR2.3滴度曲线趋向高于先前测试的TIR2条件,但是仍然低于对照TIR3条件(图47A)。TIR6条件产生类似于TIR2.3条件的滴度积累曲线,但再次低于TIR3对照。如通过蛋白质印迹分析所测定的FkpA积累显示了各TIR变体之间FkpA的滴定(图47B)。数据显示,存在与hAb产生相关的最佳FkpA表达量,在xIL13 hAb(MD157单一质粒)情况下,这是TIR3。
实施例7:氧转移速率(OTR)条件的影响
还用第二OTR策略测试了实施例3中确定的xIL13 hAb最佳条件。在实验(N=3)中,容器背压(BP)和鼓泡速率分别从1.0巴减少至0.3巴及从20SLPM减少至13SLPM。在恢复因背压和鼓泡速率减少所致OTR损失的尝试中,生长阶段的搅拌速率和生产阶段的搅拌速率分别从650转/分钟增加至880转/分钟和从475转/分钟增加至650转/分钟。容器背压、鼓泡速率和搅拌速率的不同组合可以用来实现与先前实施例中所提及的那些OTR条件相似的OTR条件。
对于整个发酵过程,用恒定BP进行发酵。在改变的OTR条件下,背压维持在0.3巴(N=3),并且在对照条件中,背压维持在1.0巴(N=5)。对于整个发酵过程,用恒定通气量进行发酵。在改变的OTR条件下,通气量维持在13SLPM(N=3),并且在对照条件中,通气量维持在20SLPM(N=5)。在改变的OTR条件和对照条件下,发酵在150OD550实施搅拌变换。在改变的OTR条件下,初始搅拌速率设定至880转/分钟并且变动至650转/分钟,并且在对照条件下,初始搅拌速率是设定至650转/分钟并且变动至475转/分钟。
搅拌速率增加补偿了原因是鼓泡和背压减少的OTR降低。改变的OTR条件产生与对照条件相似的最高OUR和生产阶段OUR(图48A)和生长曲线(图48B)。改变的OTR条件产生与对照条件相似的积累曲线和最大滴度(4.1±0.4g/L)(图49)。
序列
除非另外指出,否则从N末端至C末端展示全部多肽序列。
除非另外指出,否则从5’至3’展示全部多核苷酸序列。
人IL13前体多肽
MALLLTTVIALTCLGGFASPGPVPPSTALRELIEELVNITQNQKAPLCNGSMVWSINLTAGMYCAALESLINVSGCSAIEKTQRMLSGFCPHKVSAGQFSSLHVRDTKIEVAQFVKDLLLHLKKLFREGRFN(SEQ ID NO:1)
人IL13成熟多肽
SPGPVPPSTALRELIEELVNITQNQKAPLCNGSMVWSINLTAGMYCAALESLINVSGCSAIEKTQRMLSGFCPHKVSAGQFSSLHVRDTKIEVAQFVKDLLLHLKKLFREGRFN(SEQ ID NO:2)
人IL17A前体多肽
MTPGKTSLVSLLLLLSLEAIVKAGITIPRNPGCPNSEDKNFPRTVMVNLNIHNRNTNTNPKRSSDYYNRSTSPWNLHRNEDPERYPSVIWEAKCRHLGCINADGNVDYHMNSVPIQQEILVLRREPPHCPNSFRLEKILVSVGCTCVTPIVHHVA(SEQ ID NO:3)
人IL17A成熟多肽
GITIPRNPGCPNSEDKNFPRTVMVNLNIHNRNTNTNPKRSSDYYNRSTSPWNLHRNEDPERYPSVIWEAKCRHLGCINADGNVDYHMNSVPIQQEILVLRREPPHCPNSFRLEKILVSVGCTCVTPIVHHVA(SEQ ID NO:4)
人IL17F前体多肽
MTVKTLHGPAMVKYLLLSILGLAFLSEAAARKIPKVGHTFFQKPESCPPVPGGSMKLDIGIINENQRVSMSRNIESRSTSPWNYTVTWDPNRYPSEVVQAQCRNLGCINAQGKEDISMNSVPIQQETLVVRRKHQGCSVSFQLEKVLVTVGCTCVTPVIHHVQ(SEQ ID NO:5)
人IL17F成熟多肽
RKIPKVGHTFFQKPESCPPVPGGSMKLDIGIINENQRVSMSRNIESRSTSPWNYTVTWDPNRYPSEVVQAQCRNLGCINAQGKEDISMNSVPIQQETLVVRRKHQGCSVSFQLEKVLVTVGCTCVTPVIHHVQ(SEQ ID NO:6)
抗IL13重链可变结构域
EVTLRESGPALVKPTQTLTLTCTVSGFSLSAYSVNWIRQPPGKALEWLAMIWGDGKIVYNSALKSRLTISKDTSKNQVVLTMTNMDPVDTATYYCAGDGYYPYAMDNWGQGSLVTVSS(SEQ ID NO:7)
抗IL13轻链可变结构域
DIVLTQSPDSLSVSLGERATINCRASKSVDSYGNSFMHWYQQKPGQPPKLLIYLASNLESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQNNEDPRTFGGGTKVEIKR(SEQ ID NO:8)
抗IL13 HVR-H1
AYSVN(SEQ ID NO:9)
抗IL13 HVR-H2
MIWGDGKIVYNSALKS(SEQ ID NO:10)
抗IL13 HVR-H3
DGYYPYAMDN(SEQ ID NO:11)
抗IL13 HVR-L1
RASKSVDSYGNSFMH(SEQ ID NO:12)
抗IL13 HVR-L2
LASNLES(SEQ ID NO:13)
抗IL13 HVR-L3
QQNNEDPRT(SEQ ID NO:14)
具有形成杵的T366W突变的抗IL13 IgG4重链
EVTLRESGPALVKPTQTLTLTCTVSGFSLSAYSVNWIRQPPGKALEWLAMIWGDGKIVYNSALKSRLTISKDTSKNQVVLTMTNMDPVDTATYYCAGDGYYPYAMDNWGQGSLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLG(SEQID NO:15)
具有形成杵的T366W突变的抗IL13 IgG4重链

抗IL13轻链
DIVLTQSPDSLSVSLGERATINCRASKSVDSYGNSFMHWYQQKPGQPPKLLIYLASNLESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYCQQNNEDPRTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC(SEQ ID NO:17)
抗IL17A F交叉反应性Ab重链可变结构域
EVQLVESGGGLVQPGRSLRLSCAASGFTFDDYAMHWVRQAPGKGLEWVSGINWSSGGIGYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCARDIGGFGEFYWNFGLWGRGTLVTVSS(SEQ ID NO:18)
抗IL17A F交叉反应性Ab轻链可变结构域
EIVLTQSPATLSLSPGERATLSCRASQSVRSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPPATFGGGTKVEIK(SEQ ID NO:19)
抗IL17A F交叉反应性Ab HVR-H1
DYAMH(SEQ ID NO:20)
抗IL17A F交叉反应性Ab HVR-H2
GINWSSGGIGYADSVKG(SEQ ID NO:21)
抗IL17A F交叉反应性Ab HVR-H3
DIGGFGEFYWNFGL(SEQ ID NO:22)
抗IL17A F交叉反应性Ab HVR-L1
RASQSVRSYLA(SEQ ID NO:23)
抗IL17A F交叉反应性Ab HVR-L2
DASNRAT(SEQ ID NO:24)
抗IL17A F交叉反应性Ab HVR-L3
QQRSNWPPAT(SEQ ID NO:25)
具有形成臼的T366S/L368A/Y407V突变的抗IL17A F交叉反应性Ab IgG4重链
EVQLVESGGGLVQPGRSLRLSCAASGFTFDDYAMHWVRQAPGKGLEWVSGINWSSGGIGYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCARDIGGFGEFYWNFGLWGRGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLG(SEQ ID NO:26)
具有形成臼的T366S/L368A/Y407V突变的抗IL17A F交叉反应性Ab IgG4重链
EVQLVESGGGLVQPGRSLRLSCAASGFTFDDYAMHWVRQAPGKGLEWVSGINWSSGGIGYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCARDIGGFGEFYWNFGLWGRGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK(SEQ ID NO:27)
抗IL17A F交叉反应性Ab轻链
EIVLTQSPATLSLSPGERATLSCRASQSVRSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPPATFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC(SEQID NO:28)
抗IL13重链可变结构域的多核苷酸序列
GAAGTTACCCTGCGCGAGAGCGGCCCAGCCCTGGTGAAGCCAACCCAGACCCTGACCCTGACCTGCACCGTCAGCGGCTTCAGCCTGAGCGCCTACAGCGTGAACTGGATCCGCCAGCCACCAGGCAAGGCCCTGGAGTGGCTGGCCATGATCTGGGGCGACGGCAAGATCGTGTACAACAGCGCCCTGAAGAGCCGCCTGACCATCAGCAAGGACACCAGCAAGAACCAGGTGGTGCTGACCATGACCAACATGGACCCAGTGGACACCGCCACCTACTACTGCGCCGGCGACGGCTACTACCCATACGCCATGGACAACTGGGGCCAGGGCAGCCTGGTGACCGTGAGCAGC(SEQ ID NO:29)
抗IL13轻链可变结构域的多核苷酸序列
GATATCGTGCTGACCCAGAGCCCAGACAGCCTGTCTGTGAGCCTGGGCGAGCGCGCCACCATCAACTGCCGCGCCAGCAAAAGCGTGGACAGCTACGGCAACAGCTTCATGCACTGGTATCAGCAGAAGCCAGGCCAGCCACCCAAGCTGCTGATCTACCTGGCCAGCAACCTGGAGAGCGGCGTGCCAGACCGCTTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGACCATCAGCTCTCTGCAGGCCGAGGATGTGGCCGTGTACTACTGCCAGCAGAACAACGAGGACCCACGCACCTTCGGTGGCGGTACCAAGGTGGAGATCAAA(SEQ ID NO:30)
具有形成杵的T366W突变的抗IL13 IgG4重链多核苷酸序列
GAAGTTACCCTGCGCGAGAGCGGCCCAGCCCTGGTGAAGCCAACCCAGACCCTGACCCTGACCTGCACCGTCAGCGGCTTCAGCCTGAGCGCCTACAGCGTGAACTGGATCCGCCAGCCACCAGGCAAGGCCCTGGAGTGGCTGGCCATGATCTGGGGCGACGGCAAGATCGTGTACAACAGCGCCCTGAAGAGCCGCCTGACCATCAGCAAGGACACCAGCAAGAACCAGGTGGTGCTGACCATGACCAACATGGACCCAGTGGACACCGCCACCTACTACTGCGCCGGCGACGGCTACTACCCATACGCCATGGACAACTGGGGCCAGGGCAGCCTGGTGACCGTGAGCAGCGCTTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTGCTCCCGCAGTACTTCTGAGTCCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCAAGACCTACACGTGCAACGTGGATCACAAGCCCAGCAACACCAAGGTGGACAAACGCGTTGAGTCCAAATATGGTCCCCCATGCCCACCATGCCCAGCACCTGAGTTCCTGGGGGGACCATCAGTCTTCCTGTTCCCCCCAAAACCCAAGGACACTCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCAGGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGATGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTTCAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCGTCCTCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAGCCACAGGTGTACACCCTGCCCCCATCCCAGGAGGAGATGACCAAGAACCAGGTCAGCCTGTGGTGCCTGGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAGGCTAACCGTGGACAAGAGCAGGTGGCAGGAGGGGAATGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCTGGGT(SEQ ID NO:31)
具有形成杵的T366W突变的抗IL13 IgG4重链多核苷酸序列
GAAGTTACCCTGCGCGAGAGCGGCCCAGCCCTGGTGAAGCCAACCCAGACCCTGACCCTGACCTGCACCGTCAGCGGCTTCAGCCTGAGCGCCTACAGCGTGAACTGGATCCGCCAGCCACCAGGCAAGGCCCTGGAGTGGCTGGCCATGATCTGGGGCGACGGCAAGATCGTGTACAACAGCGCCCTGAAGAGCCGCCTGACCATCAGCAAGGACACCAGCAAGAACCAGGTGGTGCTGACCATGACCAACATGGACCCAGTGGACACCGCCACCTACTACTGCGCCGGCGACGGCTACTACCCATACGCCATGGACAACTGGGGCCAGGGCAGCCTGGTGACCGTGAGCAGCGCTTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTGCTCCCGCAGTACTTCTGAGTCCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCAAGACCTACACGTGCAACGTGGATCACAAGCCCAGCAACACCAAGGTGGACAAACGCGTTGAGTCCAAATATGGTCCCCCATGCCCACCATGCCCAGCACCTGAGTTCCTGGGGGGACCATCAGTCTTCCTGTTCCCCCCAAAACCCAAGGACACTCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCAGGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGATGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTTCAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCGTCCTCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAGCCACAGGTGTACACCCTGCCCCCATCCCAGGAGGAGATGACCAAGAACCAGGTCAGCCTGTGGTGCCTGGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAGGCTAACCGTGGACAAGAGCAGGTGGCAGGAGGGGAATGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCTGGGTAAATAA(SEQ ID NO:32)
抗IL13轻链多核苷酸
GATATCGTGCTGACCCAGAGCCCAGACAGCCTGTCTGTGAGCCTGGGCGAGCGCGCCACCATCAACTGCCGCGCCAGCAAAAGCGTGGACAGCTACGGCAACAGCTTCATGCACTGGTATCAGCAGAAGCCAGGCCAGCCACCCAAGCTGCTGATCTACCTGGCCAGCAACCTGGAGAGCGGCGTGCCAGACCGCTTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGACCATCAGCTCTCTGCAGGCCGAGGATGTGGCCGTGTACTACTGCCAGCAGAACAACGAGGACCCACGCACCTTCGGTGGCGGTACCAAGGTGGAGATCAAACGAACTGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCTTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCCGTGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA(SEQ ID NO:33)
抗IL17A F交叉反应性Ab重链可变结构域多核苷酸
GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGATTATGCCATGCACTGGGTCCGGCAAGCTCCAGGGAAGGGCCTGGAGTGGGTCTCAGGTATTAATTGGAGCAGTGGTGGCATAGGCTATGCGGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTGTATCTGCAAATGAACAGTCTGAGAGCTGAGGACACGGCCTTGTATTACTGTGCAAGAGATATCGGGGGGTTCGGGGAGTTTTACTGGAACTTCGGTCTCTGGGGCCGTGGCACCCTGGTCACTGTCTCCTCA(SEQID NO:34)
抗IL17A F交叉反应性Ab轻链可变结构域多核苷酸
GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGAAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAACTGGCCTCCGGCCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA(SEQ ID NO:35)
具有形成臼的T366S/L368A/Y407V突变的抗IL17A F交叉反应性Ab IgG4重链多核苷酸
GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGATTATGCCATGCACTGGGTCCGGCAAGCTCCAGGGAAGGGCCTGGAGTGGGTCTCAGGTATTAATTGGAGCAGTGGTGGCATAGGCTATGCGGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTGTATCTGCAAATGAACAGTCTGAGAGCTGAGGACACGGCCTTGTATTACTGTGCAAGAGATATCGGGGGGTTCGGGGAGTTTTACTGGAACTTCGGTCTCTGGGGCCGTGGCACCCTGGTCACTGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTGCTCCCGCAGTACTTCTGAGTCCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCAAGACCTACACGTGCAACGTGGATCACAAGCCCAGCAACACCAAGGTGGACAAACGCGTTGAGTCCAAATATGGTCCCCCATGCCCACCATGCCCAGCACCTGAGTTCCTGGGGGGACCATCAGTCTTCCTGTTCCCCCCAAAACCCAAGGACACTCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCAGGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGATGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTTCAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCGTCCTCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAGCCACAGGTGTACACCCTGCCCCCATCCCAGGAGGAGATGACCAAGAACCAGGTCAGCCTGAGCTGCGCTGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCGTTAGCAGGCTAACCGTGGACAAGAGCAGGTGGCAGGAGGGGAATGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCTGGGT(SEQ ID NO:36)
具有形成臼的T366S/L368A/Y407V突变的抗IL17A F交叉反应性Ab IgG4重链多核苷酸
GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGATTATGCCATGCACTGGGTCCGGCAAGCTCCAGGGAAGGGCCTGGAGTGGGTCTCAGGTATTAATTGGAGCAGTGGTGGCATAGGCTATGCGGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTGTATCTGCAAATGAACAGTCTGAGAGCTGAGGACACGGCCTTGTATTACTGTGCAAGAGATATCGGGGGGTTCGGGGAGTTTTACTGGAACTTCGGTCTCTGGGGCCGTGGCACCCTGGTCACTGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTGCTCCCGCAGTACTTCTGAGTCCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCAAGACCTACACGTGCAACGTGGATCACAAGCCCAGCAACACCAAGGTGGACAAACGCGTTGAGTCCAAATATGGTCCCCCATGCCCACCATGCCCAGCACCTGAGTTCCTGGGGGGACCATCAGTCTTCCTGTTCCCCCCAAAACCCAAGGACACTCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCAGGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGATGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTTCAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCGTCCTCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAGCCACAGGTGTACACCCTGCCCCCATCCCAGGAGGAGATGACCAAGAACCAGGTCAGCCTGAGCTGCGCTGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCGTTAGCAGGCTAACCGTGGACAAGAGCAGGTGGCAGGAGGGGAATGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCTGGGTAAATAA(SEQ IDNO:37)
抗IL17A F交叉反应性Ab轻链多核苷酸
GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGAAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAACTGGCCTCCGGCCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAACGAACTGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCTTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCCGTGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA(SEQ ID NO:38)
具有形成臼的T366S/L368A/Y407V突变的密码子优化的抗IL17A F交叉反应性AbIgG4重链多核苷酸
GAAGTTCAGCTGGTTGAAAGCGGTGGTGGTCTGGTTCAGCCTGGTCGTAGCCTGCGTCTGAGCTGTGCAGCAAGCGGTTTTACCTTTGATGATTATGCCATGCATTGGGTTCGTCAGGCACCGGGTAAAGGTCTGGAATGGGTTAGCGGTATTAATTGGAGCAGCGGTGGTATTGGTTATGCAGATAGCGTTAAAGGTCGTTTTACCATTAGCCGTGATAATGCCAAAAATAGCCTGTACCTGCAGATGAATAGTCTGCGTGCAGAAGATACCGCACTGTATTATTGTGCACGTGATATTGGTGGTTTTGGCGAATTCTATTGGAATTTTGGTCTGTGGGGTCGTGGCACCCTGGTTACCGTTAGCAGCGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTGCTCCCGCAGTACTTCTGAGTCCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCAAGACCTACACGTGCAACGTGGATCACAAGCCCAGCAACACCAAGGTGGACAAACGCGTTGAGTCCAAATATGGTCCCCCATGCCCACCATGCCCAGCACCTGAGTTCCTGGGGGGACCATCAGTCTTCCTGTTCCCCCCAAAACCCAAGGACACTCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCAGGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGATGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTTCAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCGTCCTCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAGCCACAGGTGTACACCCTGCCCCCATCCCAGGAGGAGATGACCAAGAACCAGGTCAGCCTGAGCTGCGCTGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCGTTAGCAGGCTAACCGTGGACAAGAGCAGGTGGCAGGAGGGGAATGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCTGGGT(SEQ ID NO:39)
具有形成臼的T366S/L368A/Y407V突变的密码子优化的抗IL17A F交叉反应性AbIgG4重链多核苷酸
GAAGTTCAGCTGGTTGAAAGCGGTGGTGGTCTGGTTCAGCCTGGTCGTAGCCTGCGTCTGAGCTGTGCAGCAAGCGGTTTTACCTTTGATGATTATGCCATGCATTGGGTTCGTCAGGCACCGGGTAAAGGTCTGGAATGGGTTAGCGGTATTAATTGGAGCAGCGGTGGTATTGGTTATGCAGATAGCGTTAAAGGTCGTTTTACCATTAGCCGTGATAATGCCAAAAATAGCCTGTACCTGCAGATGAATAGTCTGCGTGCAGAAGATACCGCACTGTATTATTGTGCACGTGATATTGGTGGTTTTGGCGAATTCTATTGGAATTTTGGTCTGTGGGGTCGTGGCACCCTGGTTACCGTTAGCAGCGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTGCTCCCGCAGTACTTCTGAGTCCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACTGTGCCCTCTAGCAGCTTGGGCACCAAGACCTACACGTGCAACGTGGATCACAAGCCCAGCAACACCAAGGTGGACAAACGCGTTGAGTCCAAATATGGTCCCCCATGCCCACCATGCCCAGCACCTGAGTTCCTGGGGGGACCATCAGTCTTCCTGTTCCCCCCAAAACCCAAGGACACTCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCAGGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGATGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTTCAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCGTCCTCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAGCCACAGGTGTACACCCTGCCCCCATCCCAGGAGGAGATGACCAAGAACCAGGTCAGCCTGAGCTGCGCTGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCGTTAGCAGGCTAACCGTGGACAAGAGCAGGTGGCAGGAGGGGAATGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCTGGGTAAATAA(SEQ IDNO:40)
密码子优化的抗IL17A F交叉反应性Ab轻链多核苷酸
GAAATTGTTCTGACCCAGAGTCCGGCAACCCTGAGCCTGAGTCCGGGTGAACGTGCCACCCTGAGCTGTCGTGCAAGCCAGAGCGTTCGTAGCTATCTGGCATGGTATCAGCAGAAACCGGGTCAGGCACCGCGTCTGCTGATTTATGATGCAAGCAATCGTGCAACCGGTATTCCGGCACGTTTTAGCGGTAGCGGTAGTGGCACCGATTTTACCCTGACCATTAGCAGCCTGGAACCGGAAGATTTTGCAGTGTATTATTGTCAGCAGCGTAGCAATTGGCCACCGGCAACCTTTGGTGGTGGCACCAAAGTTGAAATTAAACGAACTGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCTTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCCGTGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAA(SEQ ID NO:41)
FkpA TIR1

FkpA TIR2

FkpA TIR3(c13)

FkpA信号肽
MKSLFKVTLLATTMAVALHAPITFA
(SEQ ID NO:45)



























Claims (38)
1.一种在大肠杆菌中产生包含两条链的多肽的方法,所述方法包括:
(a)在培养基中在包含以下的条件下培养大肠杆菌以表达多肽的两条链:
包含生长温度和生长搅拌速率的生长阶段,
从生长温度至较低的生产温度的变换,从生长搅拌速率至较低的生产搅拌速率的变换,和
包含生产温度和生产搅拌速率的生产阶段,
因而当表达时,两条链折叠并装配以在大肠杆菌中形成生物活性多肽;
其中大肠杆菌是具有degp S210A突变的菌株,且包含多核苷酸,所述多核苷酸包含
(1)编码多肽第一链的第一翻译单位;
(2)编码多肽第二链的第二翻译单位;和
(3)编码伴侣蛋白的第三翻译单位,其中所述伴侣蛋白是FkpA蛋白,DsbA蛋白和DsbC蛋白的组合;
其中大肠杆菌的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且大肠杆菌的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内;并且
(b)从大肠杆菌回收生物活性多肽。
2.一种在大肠杆菌中产生包含两条链的多肽的方法,所述方法包括:
(a)在培养基中在包含以下的条件下培养大肠杆菌以表达多肽的两条链:
包含生长温度和生长搅拌速率的生长阶段,
从生长温度至较低的生产温度的变换,从生长搅拌速率至较低的生产搅拌速率的变换,和
包含生产温度和生产搅拌速率的生产阶段,
因而当表达时,两条链折叠并装配以在大肠杆菌中形成生物活性多肽;
其中大肠杆菌是具有degp S210A突变的菌株,且包含多核苷酸,所述多核苷酸包含
(1)编码多肽第一链的第一翻译单位;
(2)编码多肽第二链的第二翻译单位;
(3)编码第一伴侣蛋白的第三翻译单位;
(4)编码第二伴侣蛋白的第四翻译单位;和
(5)编码第三伴侣蛋白的第五翻译单位,
其中第一、第二和第三伴侣蛋白是FkpA蛋白,DsbA蛋白和DsbC;
其中大肠杆菌的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且大肠杆菌的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内;并且
(b)从大肠杆菌回收生物活性多肽。
3.根据权利要求1或权利要求2所述的方法,其中多核苷酸还包含启动子的三个副本,其中第一副本与第一翻译单位有效组合,第二副本与第二翻译单位有效组合并且第三副本与第三翻译单位有效组合,以驱动第一链、第二链和伴侣蛋白的转录。
4.根据权利要求3所述的方法,其中启动子是诱导型启动子。
5.根据权利要求4所述的方法,其中诱导型启动子是在IPTG诱导作用不存在的情况下驱动第一链、第二链和伴侣蛋白转录的IPTG诱导型启动子。
6.根据权利要求4所述的方法,其中诱导型启动子是当培养基中的磷酸盐已经耗尽时驱动第一链、第二链和伴侣蛋白转录的PhoA启动子。
7.根据权利要求1或2所述的方法,其中多核苷酸还包含选择标记并且培养基包含由引起大肠杆菌保留所述多核苷酸的单一抗生素组成的选择剂。
8.根据权利要求1或2所述的方法,其中第一翻译单位包含与第一链编码区有效组合的第一翻译起始区域(TIR),并且第二翻译单位包含与第二链编码区有效组合的第二翻译起始区域(TIR),其中第一和第二TIR的相对翻译强度是1.0至3.0。
9.根据权利要求1或2所述的方法,其中FkpA是大肠杆菌(E.coli)FkpA。
10.根据权利要求1或2所述的方法,其中DsbA蛋白和DsbC蛋白是大肠杆菌DsbA蛋白和大肠杆菌DsbC蛋白。
11.根据权利要求1所述的方法,其中大肠杆菌是具有基因型W3110ΔfhuAΔphoAilvG2096(Valr)Δprc spr43H1ΔdegPΔmanA lacIQΔompTΔmenE degp S210A的菌株。
12.根据权利要求1或2所述的方法,其中多肽是异二聚体的单体。
13.根据权利要求1所述的方法,其中多肽的两条链彼此通过至少一个二硫键连接。
14.根据权利要求1所述的方法,其中多肽是其中第一链和第二链包含免疫球蛋白重链和免疫球蛋白轻链的单价抗体。
15.根据权利要求14所述的方法,其中单价抗体能够特异性结合抗原。
16.根据权利要求1所述的方法,其中多肽是分泌性蛋白。
17.根据权利要求16所述的方法,其中分泌性蛋白从大肠杆菌的周质回收。
18.根据权利要求1所述的方法,其中生长搅拌速率足以在生长阶段期间在大肠杆菌中实现在生产阶段期间大肠杆菌中最大氧摄取速率之上0.5至2.5mmol/L/分钟的氧摄取速率。
19.在大肠杆菌中产生包含重链和轻链的半抗体的方法,所述方法包括:
(a)在培养基中在包含以下的条件下培养大肠杆菌以表达重链和轻链:
包含生长温度和生长搅拌速率的生长阶段,
从生长温度至较低的生产温度的变换,从生长搅拌速率至较低的生产搅拌速率的变换,和
包含生产温度和生产搅拌速率的生产阶段,
因而当表达时,重链和轻链装配以在大肠杆菌中形成半抗体;
其中大肠杆菌是具有degp S210A突变的菌株,且包含多核苷酸,所述多核苷酸包含
(1)编码半抗体的重链的第一翻译单位;
(2)编码半抗体的轻链的第二翻译单位;
(3)编码第一伴侣蛋白的第三翻译单位;
(4)编码第二伴侣蛋白的第四翻译单位;和
(5)编码第三伴侣蛋白的第五翻译单位,
其中第一、第二和第三伴侣蛋白是FkpA蛋白,DsbA蛋白和DsbC蛋白;
其中大肠杆菌的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且大肠杆菌的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内;并且
(b)从大肠杆菌回收半抗体。
20.根据权利要求19所述的方法,其中半抗体包含至少一个成臼突变或至少一个成杵突变。
21.在大肠杆菌中产生包含重链和轻链的抗IL13半抗体的方法,所述方法包括:
(a)在培养基中在包含以下的条件下培养大肠杆菌以表达重链和轻链:
包含生长温度和生长搅拌速率的生长阶段,
从生长温度至较低的生产温度的变换,从生长搅拌速率至较低的生产搅拌速率的变换,和
包含生产温度和生产搅拌速率的生产阶段,
其中(i)重链包含重链可变结构域,所述重链可变结构域包含SEQ ID NO:9的HVR-H1、SEQ ID NO:10的HVR-H2和SEQ ID NO:11的HVR-H3;和
(ii)轻链包含轻链可变结构域,所述轻链可变结构域包含SEQ IDNO:12的HVR-L1、SEQID NO:13的HVR-L2和SEQ ID NO:14的HVR-L3,因而当表达时,重链和轻链装配以在大肠杆菌中形成抗IL13半抗体;
其中大肠杆菌是具有degp S210A突变的菌株,且包含多核苷酸,所述多核苷酸包含
(1)编码半抗体的重链的第一翻译单位;
(2)编码半抗体的轻链的第二翻译单位;
(3)编码第一伴侣蛋白的第三翻译单位;
(4)编码第二伴侣蛋白的第四翻译单位;和
(5)编码第三伴侣蛋白的第五翻译单位,
其中第一、第二和第三伴侣蛋白是FkpA蛋白,DsbA蛋白和DsbC蛋白;
其中大肠杆菌的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且大肠杆菌的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内;并且
(b)从大肠杆菌回收抗IL13半抗体。
22.根据权利要求21所述的方法,其中抗IL13半抗体包含至少一个成杵突变。
23.在大肠杆菌中产生包含重链和轻链的抗IL17半抗体的方法,所述方法包括:
(a)在培养基中在包含以下的条件下培养大肠杆菌以表达重链和轻链:
包含生长温度和生长搅拌速率的生长阶段,
从生长温度至较低的生产温度的变换,从生长搅拌速率至较低的生产搅拌速率的变换,和
包含生产温度和生产搅拌速率的生产阶段,
其中(i)重链包含重链可变结构域,所述重链可变结构域包含SEQ ID NO:20的HVR-H1、SEQ ID NO:21的HVR-H2和SEQ ID NO:22的HVR-H3;并且(ii)轻链包含轻链可变结构域,所述轻链可变结构域包含SEQ ID NO:23的HVR-L1、SEQ ID NO:24的HVR-L2和SEQ ID NO:25的HVR-L3,因而当表达时,重链和轻链装配以在大肠杆菌中形成抗IL17半抗体;
其中大肠杆菌是具有degp S210A突变的菌株,且包含多核苷酸,所述多核苷酸包含
(1)编码半抗体的重链的第一翻译单位;
(2)编码半抗体的轻链的第二翻译单位;
(3)编码第一伴侣蛋白的第三翻译单位;
(4)编码第二伴侣蛋白的第四翻译单位;和
(5)编码第三伴侣蛋白的第五翻译单位,
其中第一、第二和第三伴侣蛋白是FkpA蛋白,DsbA蛋白和DsbC蛋白;
其中大肠杆菌的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且大肠杆菌的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内;并且
(b)从大肠杆菌回收抗IL17半抗体。
24.根据权利要求23所述的方法,其中抗IL17半抗体包含至少一个成臼突变。
25.根据权利要求19-24中任一项所述的方法,其中多核苷酸还包含启动子的三个副本,其中第一副本与第一翻译单位有效组合,第二副本与第二翻译单位有效组合并且第三副本与第三翻译单位有效组合,以驱动重链、轻链和伴侣蛋白的转录。
26.根据权利要求25所述的方法,其中启动子是诱导型启动子。
27.根据权利要求26所述的方法,其中诱导型启动子是在IPTG诱导作用不存在的情况下驱动重链、轻链和伴侣蛋白转录的IPTG诱导型启动子。
28.根据权利要求26所述的方法,其中诱导型启动子是当培养基中的磷酸盐已经耗尽时驱动重链、轻链和伴侣蛋白转录的phoA启动子。
29.根据权利要求19-24中任一项所述的方法,其中多核苷酸还包含选择标记并且培养基包含由引起大肠杆菌保留单价抗体的单一抗生素组成的选择剂。
30.根据权利要求19-24中任一项所述的方法,其中第一翻译单位包含与重链可变结构域的编码区有效组合的第一翻译起始区域(TIR),并且第二翻译单位包含与轻链可变结构域的编码区有效组合的第二翻译起始区域(TIR),其中第一和第二TIR的相对翻译强度是1.0至3.0。
31.根据权利要求19-24中任一项所述的方法,其中FkpA是大肠杆菌FkpA。
32.根据权利要求19-24中任一项所述的方法,其中DsbA和DsbC是大肠杆菌DsbA和大肠杆菌DsbC。
33.根据权利要求19-24中任一项所述的方法,其中生长搅拌速率足以在生长阶段期间在大肠杆菌中实现在生产阶段期间大肠杆菌中最大氧摄取速率之上0.5至2.5mmol/L/分钟的氧摄取速率。
34.产生双特异性抗体的方法,所述双特异性抗体包含能够结合IL13的第一半抗体和能够结合IL17的第二半抗体,所述方法包括:
(a)培养第一大肠杆菌以表达第一半抗体的第一重链和第一轻链,其中(i)第一重链包含第一重链可变结构域,所述第一重链可变结构域包含SEQ ID NO:9的HVR-H1、SEQ ID NO:10的HVR-H2和SEQ ID NO:11的HVR-H3;和(ii)第一轻链包含第一轻链可变结构域,所述第一轻链可变结构域包含SEQ ID NO:12的HVR-L1、SEQ ID NO:13的HVR-L2和SEQ ID NO:14的HVR-L3,因而当表达时,第一重链和第一轻链装配以在大肠杆菌中形成第一半抗体;并且
(a’)培养第二大肠杆菌以表达第二半抗体的第二重链和第二轻链,其中(i)第二重链包含第二重链可变结构域,所述第二重链可变结构域包含SEQ ID NO:20的HVR-H1、SEQ IDNO:21的HVR-H2和SEQ ID NO:22的HVR-H3;和(ii)第二轻链包含第二轻链可变结构域,所述第二轻链可变结构域包含SEQ ID NO:23的HVR-L1、SEQ ID NO:24的HVR-L2和SEQ ID NO:25的HVR-L3,因而当表达时,第二重链和第二轻链装配以在大肠杆菌中形成第二半抗体;
其中第一大肠杆菌包含第一多核苷酸,所述第一多核苷酸包含:
(1)编码第一重链的第一翻译单位;
(2)编码第一轻链的第二翻译单位;和
第二大肠杆菌包含第二多核苷酸,所述第二多核苷酸包含:
(1’)编码第二重链的第三翻译单位;
(2’)编码第二轻链的第四翻译单位;
其中第一多核苷酸和第二多核苷酸均还包含:
(3)编码第一伴侣蛋白的第五翻译单位;
(4)编码第二伴侣蛋白的第六翻译单位;和
(5)编码第三伴侣蛋白的第七翻译单位,
其中第一、第二和第三伴侣蛋白是FkpA蛋白,DsbA蛋白和DsbC蛋白;
其中将第一大肠杆菌和第二大肠杆菌分别地在培养基中在包含以下的条件下培养:
包含生长温度和生长搅拌速率的生长阶段,
从生长温度至较低的生产温度的变换,从生长搅拌速率至较低的生产搅拌速率的变换,和
包含生产温度和生产搅拌速率的生产阶段,
其中大肠杆菌的最大氧摄取速率在生长阶段期间处于3.5至4.5mmol/L/分钟的范围内,并且大肠杆菌的氧摄取速率在生产阶段期间处于1.0至3.0mmol/L/分钟的范围内;
(b)从第一大肠杆菌回收第一半抗体;
(b’)从第二大肠杆菌回收第二半抗体;并且
(c)在还原条件下将第一半抗体与第二半抗体组合以产生能够与IL-13和IL-17同时结合的双特异性抗体。
35.根据权利要求34所述的方法,其中第一半抗体包含至少一个成杵突变,并且第二半抗体包含至少一个成臼突变。
36.根据权利要求34或35所述的方法,还包括添加还原剂以实现还原条件的步骤。
37.根据权利要求36所述的方法,其中还原剂是谷胱甘肽。
38.根据权利要求1、2、19、21、23和34中任一项所述的方法,其中生长搅拌速率比生产搅拌速率高10%至40%。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201462075792P | 2014-11-05 | 2014-11-05 | |
| US62/075,792 | 2014-11-05 | ||
| US201562207882P | 2015-08-20 | 2015-08-20 | |
| US62/207,882 | 2015-08-20 | ||
| PCT/US2015/059339 WO2016073791A1 (en) | 2014-11-05 | 2015-11-05 | Methods of producing two chain proteins in bacteria |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN108064308A CN108064308A (zh) | 2018-05-22 |
| CN108064308B true CN108064308B (zh) | 2023-06-09 |
Family
ID=54754738
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201580059973.5A Active CN108064308B (zh) | 2014-11-05 | 2015-11-05 | 细菌中产生双链蛋白的方法 |
Country Status (13)
| Country | Link |
|---|---|
| US (2) | US10112994B2 (zh) |
| EP (1) | EP3215526A1 (zh) |
| JP (3) | JP6875276B2 (zh) |
| KR (1) | KR20170075793A (zh) |
| CN (1) | CN108064308B (zh) |
| AU (1) | AU2015342961B2 (zh) |
| BR (1) | BR112017009152A2 (zh) |
| CA (1) | CA2966558C (zh) |
| HK (1) | HK1255201A1 (zh) |
| IL (1) | IL252027B (zh) |
| MX (1) | MX2017005930A (zh) |
| RU (1) | RU2705274C2 (zh) |
| WO (1) | WO2016073791A1 (zh) |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN108064308B (zh) | 2014-11-05 | 2023-06-09 | 豪夫迈·罗氏有限公司 | 细菌中产生双链蛋白的方法 |
| KR102544705B1 (ko) | 2014-11-05 | 2023-06-15 | 제넨테크, 인크. | 박테리아 내 2쇄 단백질을 생산하는 방법 |
| TW202031899A (zh) | 2018-11-05 | 2020-09-01 | 美商建南德克公司 | 在原核宿主細胞中產生二鏈蛋白質之方法 |
| SG11202106713UA (en) * | 2019-01-23 | 2021-07-29 | Genentech Inc | Methods of producing multimeric proteins in eukaryotic host cells |
Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2002061090A2 (en) * | 2000-12-14 | 2002-08-08 | Genentech, Inc. | Prokaryotically produced antibodies and uses thereof |
Family Cites Families (86)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DE266710C (zh) | ||||
| FR2413974A1 (fr) | 1978-01-06 | 1979-08-03 | David Bernard | Sechoir pour feuilles imprimees par serigraphie |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| DD266710A3 (de) | 1983-06-06 | 1989-04-12 | Ve Forschungszentrum Biotechnologie | Verfahren zur biotechnischen Herstellung van alkalischer Phosphatase |
| US4676980A (en) | 1985-09-23 | 1987-06-30 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Target specific cross-linked heteroantibodies |
| US6548640B1 (en) | 1986-03-27 | 2003-04-15 | Btg International Limited | Altered antibodies |
| IL85035A0 (en) | 1987-01-08 | 1988-06-30 | Int Genetic Eng | Polynucleotide molecule,a chimeric antibody with specificity for human b cell surface antigen,a process for the preparation and methods utilizing the same |
| JP3101690B2 (ja) | 1987-03-18 | 2000-10-23 | エス・ビィ・2・インコーポレイテッド | 変性抗体の、または変性抗体に関する改良 |
| CA1338518C (en) | 1987-09-23 | 1996-08-13 | Joyce M. Zarling | Antibody heteroconjugates for the killing of hiv-infected cells |
| GB8823869D0 (en) | 1988-10-12 | 1988-11-16 | Medical Res Council | Production of antibodies |
| EP0368684B2 (en) | 1988-11-11 | 2004-09-29 | Medical Research Council | Cloning immunoglobulin variable domain sequences. |
| DE3920358A1 (de) | 1989-06-22 | 1991-01-17 | Behringwerke Ag | Bispezifische und oligospezifische, mono- und oligovalente antikoerperkonstrukte, ihre herstellung und verwendung |
| ES2096590T3 (es) | 1989-06-29 | 1997-03-16 | Medarex Inc | Reactivos biespecificos para la terapia del sida. |
| US6075181A (en) | 1990-01-12 | 2000-06-13 | Abgenix, Inc. | Human antibodies derived from immunized xenomice |
| ATE139258T1 (de) | 1990-01-12 | 1996-06-15 | Cell Genesys Inc | Erzeugung xenogener antikörper |
| US6150584A (en) | 1990-01-12 | 2000-11-21 | Abgenix, Inc. | Human antibodies derived from immunized xenomice |
| US5545806A (en) | 1990-08-29 | 1996-08-13 | Genpharm International, Inc. | Ransgenic non-human animals for producing heterologous antibodies |
| US5633425A (en) | 1990-08-29 | 1997-05-27 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| EP0546073B1 (en) | 1990-08-29 | 1997-09-10 | GenPharm International, Inc. | production and use of transgenic non-human animals capable of producing heterologous antibodies |
| US5625126A (en) | 1990-08-29 | 1997-04-29 | Genpharm International, Inc. | Transgenic non-human animals for producing heterologous antibodies |
| US5661016A (en) | 1990-08-29 | 1997-08-26 | Genpharm International Inc. | Transgenic non-human animals capable of producing heterologous antibodies of various isotypes |
| DK0564531T3 (da) | 1990-12-03 | 1998-09-28 | Genentech Inc | Berigelsesfremgangsmåde for variantproteiner med ændrede bindingsegenskaber |
| US5571894A (en) | 1991-02-05 | 1996-11-05 | Ciba-Geigy Corporation | Recombinant antibodies specific for a growth factor receptor |
| EP0586505A1 (en) | 1991-05-14 | 1994-03-16 | Repligen Corporation | Heteroconjugate antibodies for treatment of hiv infection |
| LU91067I2 (fr) | 1991-06-14 | 2004-04-02 | Genentech Inc | Trastuzumab et ses variantes et dérivés immuno chimiques y compris les immotoxines |
| GB9114948D0 (en) | 1991-07-11 | 1991-08-28 | Pfizer Ltd | Process for preparing sertraline intermediates |
| CA2116774C (en) | 1991-09-19 | 2003-11-11 | Paul J. Carter | Expression in e. coli antibody fragments having at least a cysteine present as a free thiol. use for the production of bifunctional f(ab') 2 antibodies |
| US5587458A (en) | 1991-10-07 | 1996-12-24 | Aronex Pharmaceuticals, Inc. | Anti-erbB-2 antibodies, combinations thereof, and therapeutic and diagnostic uses thereof |
| WO1993008829A1 (en) | 1991-11-04 | 1993-05-13 | The Regents Of The University Of California | Compositions that mediate killing of hiv-infected cells |
| CA2372813A1 (en) | 1992-02-06 | 1993-08-19 | L.L. Houston | Biosynthetic binding protein for cancer marker |
| DE69308573T2 (de) | 1992-08-17 | 1997-08-07 | Genentech Inc | Bispezifische immunoadhesine |
| WO1994029351A2 (en) | 1993-06-16 | 1994-12-22 | Celltech Limited | Antibodies |
| US5789199A (en) | 1994-11-03 | 1998-08-04 | Genentech, Inc. | Process for bacterial production of polypeptides |
| US5840523A (en) | 1995-03-01 | 1998-11-24 | Genetech, Inc. | Methods and compositions for secretion of heterologous polypeptides |
| US5731168A (en) | 1995-03-01 | 1998-03-24 | Genentech, Inc. | Method for making heteromultimeric polypeptides |
| US5641870A (en) | 1995-04-20 | 1997-06-24 | Genentech, Inc. | Low pH hydrophobic interaction chromatography for antibody purification |
| US5869046A (en) | 1995-04-14 | 1999-02-09 | Genentech, Inc. | Altered polypeptides with increased half-life |
| JP4312259B2 (ja) | 1995-04-27 | 2009-08-12 | アムジェン フレモント インク. | 免疫したゼノマウス(XenoMouse)に由来するヒト抗体 |
| AU2466895A (en) | 1995-04-28 | 1996-11-18 | Abgenix, Inc. | Human antibodies derived from immunized xenomice |
| KR100643058B1 (ko) | 1996-12-03 | 2006-11-13 | 아브게닉스, 인크. | 복수의 Vн 및 Vк 영역을 함유하는 인간 면역글로불린 유전자좌를 갖는 트랜스제닉 포유동물 및 이로부터 생성된 항체 |
| US6610833B1 (en) | 1997-11-24 | 2003-08-26 | The Institute For Human Genetics And Biochemistry | Monoclonal human natural antibodies |
| IL136544A0 (en) | 1997-12-05 | 2001-06-14 | Scripps Research Inst | Humanization of murine antibody |
| US6194551B1 (en) | 1998-04-02 | 2001-02-27 | Genentech, Inc. | Polypeptide variants |
| CA2323757C (en) | 1998-04-02 | 2011-08-02 | Genentech, Inc. | Antibody variants and fragments thereof |
| US6737056B1 (en) | 1999-01-15 | 2004-05-18 | Genentech, Inc. | Polypeptide variants with altered effector function |
| EP1141024B1 (en) | 1999-01-15 | 2018-08-08 | Genentech, Inc. | POLYPEPTIDE COMPRISING A VARIANT HUMAN IgG1 Fc REGION |
| AU3305100A (en) | 1999-03-17 | 2000-10-04 | Medical Research Council | Method for refolding molecules of polypeptides containing ig domains |
| GB9911569D0 (en) * | 1999-05-18 | 1999-07-21 | Oxford Biomedica Ltd | Antibodies |
| EP1240319A1 (en) | 1999-12-15 | 2002-09-18 | Genentech, Inc. | Shotgun scanning, a combinatorial method for mapping functional protein epitopes |
| JP4878724B2 (ja) * | 2000-11-03 | 2012-02-15 | ジェネンテック, インコーポレイテッド | 組み換えタンパク質を発現する発酵における代謝速度シフト |
| CA2454731C (en) | 2001-08-27 | 2010-11-02 | Genentech, Inc. | A system for antibody expression and assembly |
| CA2457652C (en) | 2001-08-31 | 2012-08-07 | Avidex Limited | Soluble t cell receptor |
| NZ556507A (en) | 2002-06-03 | 2010-03-26 | Genentech Inc | Synthetic antibody phage libraries |
| PT1944322E (pt) * | 2002-07-19 | 2015-07-01 | Abbvie Biotechnology Ltd | Tratamento de distúrbios relacionados com tnf-alfa |
| US7361740B2 (en) | 2002-10-15 | 2008-04-22 | Pdl Biopharma, Inc. | Alteration of FcRn binding affinities or serum half-lives of antibodies by mutagenesis |
| ES2347241T3 (es) | 2002-12-16 | 2010-10-27 | Genentech, Inc. | Variantes de inmunoglobulina y sus utilizaciones. |
| AU2004205631A1 (en) | 2003-01-16 | 2004-08-05 | Genentech, Inc. | Synthetic antibody phage libraries |
| EP1629012B1 (en) | 2003-05-31 | 2018-11-28 | Amgen Research (Munich) GmbH | Pharmaceutical compositions comprising bispecific anti-cd3, anti-cd19 antibody constructs for the treatment of b-cell related disorders |
| EP2277908A3 (en) | 2003-07-08 | 2011-12-14 | Genentech, Inc. | IL-17A/F heterologous polypeptides, antibodies and therapeutic uses thereof |
| EP2186527A1 (en) | 2003-11-28 | 2010-05-19 | Micromet AG | Compositions comprising polypeptides |
| US7235641B2 (en) | 2003-12-22 | 2007-06-26 | Micromet Ag | Bispecific antibodies |
| DK1711528T3 (da) * | 2003-12-23 | 2012-08-20 | Genentech Inc | Behandling af cancer med hidtil ukendte anti-il 13 monoklonale antistoffer |
| JP4501451B2 (ja) | 2004-02-18 | 2010-07-14 | 株式会社豊田自動織機 | 塗料組成物、塗料組成物を用いた透明性保護膜の製造方法および透明性保護膜を有する有機ガラス |
| CN1961003B (zh) | 2004-03-31 | 2013-03-27 | 健泰科生物技术公司 | 人源化抗TGF-β抗体 |
| US7785903B2 (en) | 2004-04-09 | 2010-08-31 | Genentech, Inc. | Variable domain library and uses |
| SG172616A1 (en) | 2004-04-13 | 2011-07-28 | Hoffmann La Roche | Anti-p-selectin antibodies |
| CA2577082A1 (en) | 2004-09-02 | 2006-03-16 | Genentech, Inc. | Heteromultimeric molecules |
| TWI380996B (zh) | 2004-09-17 | 2013-01-01 | Hoffmann La Roche | 抗ox40l抗體 |
| EP1940881B1 (en) | 2005-10-11 | 2016-11-30 | Amgen Research (Munich) GmbH | Compositions comprising cross-species-specific antibodies and uses thereof |
| EP2465870A1 (en) | 2005-11-07 | 2012-06-20 | Genentech, Inc. | Binding polypeptides with diversified and consensus VH/VL hypervariable sequences |
| EP1973951A2 (en) | 2005-12-02 | 2008-10-01 | Genentech, Inc. | Binding polypeptides with restricted diversity sequences |
| EP2016101A2 (en) | 2006-05-09 | 2009-01-21 | Genentech, Inc. | Binding polypeptides with optimized scaffolds |
| CN101687915B8 (zh) | 2007-04-03 | 2018-08-03 | 安进研发(慕尼黑)股份有限公司 | 跨物种特异性CD3-ε结合结构域 |
| CN100592373C (zh) | 2007-05-25 | 2010-02-24 | 群康科技(深圳)有限公司 | 液晶显示面板驱动装置及其驱动方法 |
| AU2009245440C1 (en) * | 2008-05-05 | 2013-03-14 | Novimmune Sa | Anti-IL-17A/IL-17F cross-reactive antibodies and methods of use thereof |
| SG175427A1 (en) | 2009-05-01 | 2011-12-29 | Abbott Lab | Dual variable domain immunoglobulins and uses thereof |
| SI2496601T1 (sl) | 2009-11-05 | 2017-09-29 | F. Hoffmann-La Roche Ag | Tehnike in sestavek za sekrecijo heterolognih polipeptidov |
| CA2796633C (en) | 2010-04-23 | 2020-10-27 | Genentech, Inc. | Production of heteromultimeric proteins |
| US9732143B2 (en) * | 2011-02-03 | 2017-08-15 | Xoma Technology Ltd. | Methods and materials for enhancing functional protein expression in bacteria |
| US9158890B2 (en) | 2011-07-27 | 2015-10-13 | At&T Mobility Ii Llc | Mobile applications and methods for conveying performance information of a cardiac pacemaker |
| WO2013055958A1 (en) * | 2011-10-11 | 2013-04-18 | Genentech, Inc. | Improved assembly of bispecific antibodies |
| CN105307676A (zh) * | 2013-04-05 | 2016-02-03 | 豪夫迈·罗氏有限公司 | 抗il-4抗体和双特异性抗体及其用途 |
| ES2922358T3 (es) | 2013-04-19 | 2022-09-13 | Sutro Biopharma Inc | Expresión de proteínas biológicamente activas en un sistema bacteriano de síntesis sin células utilizando extractos celulares con niveles elevados de chaperonas exógenas |
| MX2016010729A (es) * | 2014-02-21 | 2016-10-26 | Genentech Inc | Anticuerpos biespecificos anti-il-13 / il-17 y sus usos. |
| KR102544705B1 (ko) | 2014-11-05 | 2023-06-15 | 제넨테크, 인크. | 박테리아 내 2쇄 단백질을 생산하는 방법 |
| CN108064308B (zh) | 2014-11-05 | 2023-06-09 | 豪夫迈·罗氏有限公司 | 细菌中产生双链蛋白的方法 |
-
2015
- 2015-11-05 CN CN201580059973.5A patent/CN108064308B/zh active Active
- 2015-11-05 MX MX2017005930A patent/MX2017005930A/es unknown
- 2015-11-05 WO PCT/US2015/059339 patent/WO2016073791A1/en active Application Filing
- 2015-11-05 JP JP2017523894A patent/JP6875276B2/ja active Active
- 2015-11-05 AU AU2015342961A patent/AU2015342961B2/en active Active
- 2015-11-05 CA CA2966558A patent/CA2966558C/en active Active
- 2015-11-05 RU RU2017119403A patent/RU2705274C2/ru active
- 2015-11-05 US US14/934,100 patent/US10112994B2/en active Active
- 2015-11-05 BR BR112017009152A patent/BR112017009152A2/pt active Search and Examination
- 2015-11-05 KR KR1020177014850A patent/KR20170075793A/ko not_active Application Discontinuation
- 2015-11-05 EP EP15802233.5A patent/EP3215526A1/en active Pending
-
2017
- 2017-04-30 IL IL252027A patent/IL252027B/en unknown
-
2018
- 2018-09-21 US US16/138,180 patent/US11299539B2/en active Active
- 2018-11-08 HK HK18114324.8A patent/HK1255201A1/zh unknown
-
2021
- 2021-04-20 JP JP2021070858A patent/JP2021119777A/ja not_active Withdrawn
-
2023
- 2023-03-14 JP JP2023039366A patent/JP2023088935A/ja active Pending
Patent Citations (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2002061090A2 (en) * | 2000-12-14 | 2002-08-08 | Genentech, Inc. | Prokaryotically produced antibodies and uses thereof |
Also Published As
| Publication number | Publication date |
|---|---|
| JP6875276B2 (ja) | 2021-05-19 |
| RU2017119403A (ru) | 2018-12-05 |
| AU2015342961B2 (en) | 2021-08-12 |
| MX2017005930A (es) | 2017-06-30 |
| CA2966558C (en) | 2024-03-12 |
| AU2015342961A1 (en) | 2017-06-15 |
| IL252027A0 (en) | 2017-06-29 |
| RU2017119403A3 (zh) | 2019-03-27 |
| WO2016073791A1 (en) | 2016-05-12 |
| CN108064308A (zh) | 2018-05-22 |
| RU2705274C2 (ru) | 2019-11-06 |
| JP2023088935A (ja) | 2023-06-27 |
| US20190112368A1 (en) | 2019-04-18 |
| US11299539B2 (en) | 2022-04-12 |
| HK1255201A1 (zh) | 2019-08-09 |
| JP2021119777A (ja) | 2021-08-19 |
| CA2966558A1 (en) | 2016-05-12 |
| KR20170075793A (ko) | 2017-07-03 |
| US20160159898A1 (en) | 2016-06-09 |
| JP2018500008A (ja) | 2018-01-11 |
| US10112994B2 (en) | 2018-10-30 |
| EP3215526A1 (en) | 2017-09-13 |
| IL252027B (en) | 2021-10-31 |
| BR112017009152A2 (pt) | 2018-03-06 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6997273B2 (ja) | 細菌における2鎖タンパク質の生成方法 | |
| JP2021119777A (ja) | 細菌における2鎖タンパク質の生成方法 | |
| US20210261973A1 (en) | Methods of producing two chain proteins in prokaryotic host cells |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| REG | Reference to a national code |
Ref country code: HK Ref legal event code: DE Ref document number: 1255201 Country of ref document: HK |
|
| GR01 | Patent grant | ||
| GR01 | Patent grant |