KR20220115995A - 인간 염색체 9 개방 판독 프레임 72 (C9orf72) iRNA 제제 조성물 및 이의 사용 방법 - Google Patents
인간 염색체 9 개방 판독 프레임 72 (C9orf72) iRNA 제제 조성물 및 이의 사용 방법 Download PDFInfo
- Publication number
- KR20220115995A KR20220115995A KR1020227023988A KR20227023988A KR20220115995A KR 20220115995 A KR20220115995 A KR 20220115995A KR 1020227023988 A KR1020227023988 A KR 1020227023988A KR 20227023988 A KR20227023988 A KR 20227023988A KR 20220115995 A KR20220115995 A KR 20220115995A
- Authority
- KR
- South Korea
- Prior art keywords
- nucleotides
- c9orf72
- dsrna agent
- strand
- antisense strand
- Prior art date
Links
Images




































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/70—Carbohydrates; Sugars; Derivatives thereof
- A61K31/7088—Compounds having three or more nucleosides or nucleotides
- A61K31/713—Double-stranded nucleic acids or oligonucleotides
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/28—Drugs for disorders of the nervous system for treating neurodegenerative disorders of the central nervous system, e.g. nootropic agents, cognition enhancers, drugs for treating Alzheimer's disease or other forms of dementia
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/14—Type of nucleic acid interfering N.A.
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/31—Chemical structure of the backbone
- C12N2310/315—Phosphorothioates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/31—Chemical structure of the backbone
- C12N2310/318—Chemical structure of the backbone where the PO2 is completely replaced, e.g. MMI or formacetal
- C12N2310/3183—Diol linkers, e.g. glycols or propanediols
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/32—Chemical structure of the sugar
- C12N2310/321—2'-O-R Modification
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/32—Chemical structure of the sugar
- C12N2310/322—2'-R Modification
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/35—Nature of the modification
- C12N2310/351—Conjugate
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/30—Chemical structure
- C12N2310/35—Nature of the modification
- C12N2310/351—Conjugate
- C12N2310/3515—Lipophilic moiety, e.g. cholesterol
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Neurology (AREA)
- Neurosurgery (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Public Health (AREA)
- Medicinal Chemistry (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Hospice & Palliative Care (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Psychiatry (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Epidemiology (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Saccharide Compounds (AREA)
- Investigating Or Analysing Biological Materials (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicinal Preparation (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
본 개시내용은 인간 염색체 9 개방 판독 프레임 72(C9orf72) 유전자를 표적으로 하는 이중 가닥 리보핵산(dsRNAi) 작용제 및 조성물, 뿐만 아니라 이러한 dsRNAi 제제 및 조성물을 사용하여 C9orf72 유전자의 발현을 억제하는 방법 및 C9orf72-관련 질환 또는 장애, 예를 들어 C9orf72 근위축성 측삭 경화증/전두측두엽 치매 또는 C9orf72 확장으로 인한 헌팅턴-유사 증후군을 갖는 대상체를 치료하는 방법에 관한 것이다.
Description
관련 출원
본 출원은 2019년 12월 13일에 출원된 미국 가출원 제62/947,605호 및 2019년 12월 13일에 출원된 미국 가출원 제62/947,768호에 대한 우선권의 이익을 주장한다. 전술한 각 출원의 전체 내용은 본원에 참고로 포함된다.
서열 목록
본 출원은 ASCII 형식으로 전자적으로 제출된 서열 목록을 함유하며 이의 전문이 본원에 참고로 포함된다. 2020년 XXX에 생성된 상기 ASCII 사본은 이름이 121301_10220_SL.txt이고 크기가 XXX바이트이다.
인간 염색체 9 개방 판독 프레임 72(C9orf72)는 c9orf72 유전자에 의해 암호화된 단백질이다. C9orf72는 대뇌 피질, 뉴런의 세포질 뿐만 아니라 시냅스 전 말단과 같은 뇌의 많은 영역에서 발견된다.
전사 대체 시작 및 종료 부위의 차등 사용은 C9orf72 DNA로부터 3개의 RNA 전사체를 생성한다. 이들은 변이체 2(NM_018325.4) 및 3(NM_001256054.2)으로부터 유래된 약 54kDa의 긴 이소형(이소형 A) 및 변이체 1 (NM_145005.6)로부터 유래된 약 24kDa의 짧은 이소형(이소형 B)으로 구성된 두 개의 단백질 이소형을 암호화한다(예를 들어, 참조; 문헌 [Barker, et al. (2017) Frontiers Cell Neurosci 11:1-15]의 도 1).
C9orf72 유전자의 두 가지 대안적으로 사용되는 첫 번째 엑손은 엑손 1a 및 1b이다(예를 들어, 참조; 상기 [Barker, et al.]의 도 1). 엑손 1a와 1b 사이의 C9orf72 유전자의 첫 번째 인트론에서 큰 GGGGCC(G4C2) 헥사뉴클레오티드 반복부 확장(약 2-22개 카피에서 700-1600개 카피로)은 C9orf72 단백질의 기능을 방해하고 뚜렷한 임상적 특징을 갖지만 공통적인 병리학적 특징 및 유전적 원인을 갖는 여러 신경퇴행성 질환을 초래하는 병원성인 것으로 나타났다(Ling, et al. (2013) Neuron 79:416-438). 특히, C9orf72 유전자에서 헥사뉴클레오티드 반복부 확장의 존재는 뇌와 척수에서 운동 뉴런의 치명적인 퇴행성 질환인 가족성 및 산발성 근위축성 측삭 경화증(ALS)의 가장 흔한 유전적 원인이다. 실제로, C9orf72 돌연변이 헥사뉴클레오티드 반복부 확장은 가족성 ALS의 약 40% 및 산발성 ALS 대상체의 8-10%에 존재한다. C9orf72 유전자의 헥사뉴클레오티드 반복부 확장은 행동 및 언어 결함을 특징으로 하고 뇌의 전두엽 및 전두측두엽의 신경 위축으로 병리학적으로 나타나는 알츠하이머병 다음으로 가장 흔한 형태의 초로성 치매인 전두측두엽 치매(FTD)의 가장 흔한 가족성 원인이다. 근긴장 이상, 무도증, 간대성 근경련, 떨림 및 강직, 인지 및 기억 장애, 초기 정신 장애 및 행동 문제를 포함하는 운동 장애를 특징으로 하는, C9orf72 확장으로 인한 헌팅턴-유사 증후군이 또한 C9orf72 유전자의 헥사뉴클레오티드 반복부 확장과 관련이 있다.
C9orf72 단백질의 기능이 여전히 조사되고 있지만, C9orf72는 세포골격, 자가포식 및 식작용 전달을 조절하는데 관여하는 Rab 단백질과 상호작용하고 이를 활성화시키는 것으로 나타났다. 또한, 수많은 세포 경로가 C9orf72 헥사뉴클레오티드 반복부 확장과 관련된 신경퇴행성 질환에서 잘못 조절되는 것으로 입증되었다. 예를 들면, 변경된 RNA 프로세싱은 C9orf72 질환에 대한 연구의 최전선에서 일관되게 나타났다. 이것은 반복부 서열의 양방향 전사, 핵 병소 격리 특이적 RNA-결합 단백질(RBP)로의 반복부 RNA의 축적 및 반복부-관련 비-AUG(RAN)-개시 번역에 의한 디펩티드 반복부 단백질(DPR)로의 RNA 반복부의 번역을 포함한다. 추가로, RNA 중합효소 II로부터의 C9orf72 RNA의 방출의 중단, 세포질에서의 번역 및 퇴화는 C9orf72 헥사뉴클레오티드 반복부 확장에 의해 중단되는 것으로 나타났다. 게다가, 전사, 스플라이싱 및 국소화 측면에서 C9orf72 RNA 자체의 프로세싱에서 여러 변경이 확인되었다(예를 들어, 참조; 상기 [Barker, et al.]).
메커니즘과 상관없이, 여러 그룹에서 센스 및 안티센스 C9orf72-함유 병소의 존재 뿐만 아니라 C9orf72-관련 질환을 갖는 대상체의 신경계에서 여러 세포 유형에서 반복부-관련 비-AUG-의존성(RAN) 번역을 통해 센스 또는 안티센스 반복부-함유 C9orf72 RNA의 모든 판독 프레임으로부터 생성된 비정상적인 디펩티드-반복부(DPR) 단백질(폴리(GA), 폴리(GR), 폴리(GP), 폴리(PA) 및 폴리(PR))의 존재를 확인하였다(Lagier-Tourenne, et al. (2013) Proc Natl Acad Sci USA doi/10.1073/pnas.1318835110; Jiang, et al. (2016) Neuron 90:535-550). 게다가, C9orf72 대립형질 중 하나가 비활성화된 마우스에서는 질환이 유발되지 않았지만 C9orf72 대립형질 둘 다가 비활성화된 마우스에서는 비장비대, 림프절 비대 및 경미한 사회적 상호작용 결손이 관찰되었지만 운동 기능 장애는 관찰되지 않았다. 또한, 최대 450개의 GGGGCC 반복부가 있는 인간 C9orf72 RNA를 발현하는 마우스에서 헥사뉴클레오티드 확장이 해마 뉴런 손실, 불안 증가, 및 인지 기능 손상을 동반한 AUG-비의존적 번역에 의해 합성된 센스 및 안티센스 RNA-함유 방소 및 디펩티드-반복부 단백질의 연령-, 반복부-길이- 및 발현-수준-의존적 축적을 유발하는 것으로 나타났다(Jiang, et al. (2016) Neuron 90:535-550).
C9orf72-관련 질환, 예를 들어, C9orf72 근위축성 측삭 경화증, C9orf72 전두측두엽 치매 또는 헌팅턴병, 예를 들어 C9orf72 확장으로 인한 헌팅턴-유사 증후군을 갖는 대상체에 대한 치료법은 현재 없으며, 치료는 단지 질환이 진행됨에 따라 증상을 완화하고 환자의 삶의 질을 개선하는 것을 목표로 한다.
따라서, 예를 들어, C9orf72-관련 장애를 갖는 대상체의 치료를 위해 C9orf72 유전자의 발현을 선택적이고 효율적으로 억제할 수 있는 제제가 당업계에 필요하다.
본 개시내용은 확장된 GGGGCC (G4C2) 반복부를 갖는 C9orf72 유전자와 같은 C9orf72 유전자의 RNA 전사체의 RNA-유도 침묵 복합체(RISC)-매개된 절단에 영향을 미치는 RNAi 조성물을 제공한다. C9orf72 유전자는 세포, 예를 들어 인간과 같은 대상체 내의 세포 내에 있을 수 있다. 이들 iRNA의 사용은 포유동물에서 상응하는 유전자(C9orf72 유전자)의 mRNA의 표적화된 분해를 가능하게 한다.
본 발명의 iRNA는 C9orf72 유전자, 예를 들어 유전자의 인트론에서 확장된 GGGGCC 헥사뉴클레오티드 반복부를 갖고 뉴클레오티드 변형의 조합을 갖는 C9orf72 유전자를 표적으로 하도록 설계되었다. 제제는 성숙 C9orf72 mRNA(인트론이 스플라이싱된 mRNA) 또는 헥사뉴클레오티드 반복부를 함유하는 C9orf72 RNA(C9orf72 인트론 1A를 함유하는 RNA)를 표적으로 할 수 있다. 본 발명의 iRNA는 헥사뉴클레오티드 반복부를 함유하는 C9orf72 RNA의 수준을 감소시키는 것보다 덜 C9orf72 성숙 mRNA의 수준을 감소시킬 수 있다. 예를 들면, 본 발명의 iRNA는 C9orf72 성숙 mRNA의 수준을 약 50% 이하로 감소시키고, 센스- 및 안티센스-함유 병소 및 비정상적인 디펩티드-반복부(DPR) 단백질(폴리(GA), 폴리(GR), 폴리(GP), 폴리(PA) 및 폴리(PR))의 수준을 낮추고/낮추키거나 헥사뉴클레오티드-반복부를 함유하는 C9orf72 RNA의 수준을 약 50% 이상 감소시키고, 센스- 및 안티센스-함유 병소 및 비정상적인 디펩티드 반복부(DPR) 단백질(폴리(GA), 폴리(GR), 폴리(GP), 폴리(PA) 및 폴리(PR))의 수준을 낮추고/낮출 수 있다. 이론에 의해 제한하려는 것은 아니지만, 전술한 특성 및 특이 표적 부위의 조합 또는 하위-조합, 또는 이들 iRNA의 특정 변형은 본 발명의 iRNA에 개선된 효능, 안정성, 역가, 내구성 및 안전성을 부여하는 것으로 여겨진다.
하나의 측면에서, 본 발명은 세포에서 C9orf72 표적 RNA를 녹다운하기 위한 이중 가닥 리보핵산(dsRNA) 제제를 제공한다.
하나의 양태에서, dsRNA 제제는 헥사뉴클레오티드 반복부를 함유하는 C9orf72 표적 RNA의 영역을 표적으로 한다.
하나의 측면에서, 본 발명은 헥사뉴클레오티드-반복부를 함유하는 C9orf72 RNA와 같은 C9orf72의 발현을 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제를 제공하며, 여기서 dsRNA 제제는 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고, 여기서 안티센스 가닥은 서열번호 125의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고, 여기서 센스 가닥, 안티센스 가닥, 또는 센스 가닥과 안티센스 가닥 둘 다는 하나 이상의 친유성 모이어티에 접합된다.
또 다른 측면에서, 본 발명은 헥사뉴클레오티드-반복부를 함유하는 C9orf72 RNA와 같은 C9orf72의 발현을 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제를 제공하며, 여기서 dsRNA 제제는 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고, 여기서 센스 가닥은 서열번호 133의 뉴클레오티드 200-290과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고, 안티센스 가닥은 서열번호 134의 상응하는 뉴클레오티드 서열의 적어도 15개의 연속 뉴클레오티드를 포함하고, 여기서 센스 가닥, 안티센스 가닥, 또는 센스 가닥과 안티센스 가닥 둘 다는 하나 이상의 친유성 모이어티에 접합된다.
하나의 측면에서, 본 발명은 헥사뉴클레오티드-반복부를 함유하는 C9orf72 RNA와 같은 C9orf72의 발현을 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제를 제공하며, 여기서 dsRNA 제제는 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고, 여기서 센스 가닥은 표 10A, 10B, 12-15, 19, 20, 또는 21 중 어느 하나의 안티센스 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함한다.
하나의 양태에서, 센스 및 안티센스 가닥의 뉴클레오티드 서열은 표 12 또는 13 중 어느 하나의 센스 및 안티센스 가닥 뉴클레오티드 서열 중 어느 하나를 포함한다. 하나의 양태에서, 안티센스 가닥의 뉴클레오티드 서열은 표 12 또는 13 중 어느 하나의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나를 포함한다.
하나의 양태에서, 센스 가닥은 서열번호 121의 뉴클레오티드 230-270, 233-262, 800-840, 800-830, 802-828, 1240-1290, 1240-1280, 1247-1288, 1590-1645, 1590-1620, 및 1594-1642의 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고, 안티센스 가닥은 서열번호 125의 상응하는 뉴클레오티드 서열로부터의 적어도 15개의 연속 뉴클레오티드를 포함한다.
하나의 양태에서, 센스 가닥은 서열번호 121의 뉴클레오티드 1594-1616, 802-824, 239-261, 1308-1330, 233-255, 1595-1617, 240-262, 1532-1554, 237-259, 3268-3290, 806-828, 1620-1642, 526-548, 1169-1191, 1266-1288, 1247-1269, 586-608, 1257-1279, 및 400-422의 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고, 안티센스 가닥은 서열번호 125의 상응하는 뉴클레오티드 서열로부터의 적어도 15개의 연속 뉴클레오티드를 포함한다.
하나의 양태에서, 안티센스 가닥은 AD-348904.1, AD-348136.1, AD-347612.1, AD-348639.1, AD-347606.1, AD-348905.1, AD-347613.1, AD-348842.1, AD-347610.1, AD-350329.1, AD-348140.1, AD-348930.1, AD-347863.1, AD-348500.1, AD-348597.1, AD-348578.1, AD-347923.1, AD-348588.1, 및 AD-347773.1로 이루어진 그룹으로부터 선택된 이중체(duplex)의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나와는 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함한다.
하나의 양태에서, 센스 및 안티센스 가닥의 뉴클레오티드 서열은 표 14, 15, 19, 20 또는 21 중 어느 하나의 센스 및 안티센스 가닥 뉴클레오티드 서열 중 어느 하나를 포함한다.
하나의 측면에서, 본 발명은 서열번호 1의 다중 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하는 C9orf72의 발현을 선택적으로 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제를 제공하며, 여기서 dsRNA 제제는 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고, 여기서 안티센스 가닥은 표 14, 15 및 21 중 어느 하나의 안티센스 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함한다.
하나의 양태에서, 안티센스 가닥은 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함한다.
하나의 양태에서, 안티센스 가닥은 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15, 16, 17, 18, 19, 또는 20개의 연속 뉴클레오티드를 포함한다. 일부 양태에서, 센스 및 안티센스 가닥은 각각 독립적으로 19, 20, 21, 23, 23 24, 또는 25개의 뉴클레오티드 길이이다.
하나의 양태에서, 안티센스 가닥은 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 적어도 하나와 2개 이하의 뉴클레오티드가 다른 적어도 15, 16, 17, 18, 19, 또는 20개의 연속 뉴클레오티드를 포함한다. 일부 양태에서, 센스 및 안티센스 가닥은 각각 독립적으로 19, 20, 21, 23, 23 24, 또는 25개의 뉴클레오티드 길이이다.
하나의 양태에서, 안티센스 가닥은 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나와 1개 이하의 뉴클레오티드가 다른 적어도 15, 16, 17, 18, 19, 또는 20개의 연속 뉴클레오티드를 포함한다. 일부 양태에서, 센스 및 안티센스 가닥은 각각 독립적으로 19, 20, 21, 23, 23 24, 또는 25개의 뉴클레오티드 길이이다.
하나의 양태에서, 안티센스 가닥은 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나로부터의 적어도 15, 16, 17, 18, 19, 또는 20개의 연속 뉴클레오티드를 포함한다. 일부 양태에서, 센스 및 안티센스 가닥은 각각 독립적으로 19, 20, 21, 23, 23 24, 또는 25개의 뉴클레오티드 길이이다.
하나의 양태에서, C9orf72 표적 RNA는 서열번호 1의 다중 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하고, dsRNA 제제는 센스 가닥 및 안티센스 가닥을 포함하며, 안티센스 가닥은 C9orf72 표적 RNA에 대한 상보성 영역을 포함하고, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 뉴클레오티드 서열의 역 상보체와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함한다.
하나의 양태에서, C9orf72 표적 RNA는 서열번호 1의 다중 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하고, dsRNA 제제는 센스 가닥 및 안티센스 가닥을 포함하고, 안티센스 가닥은 C9orf72 표적 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 서열에 존재하는 15-25개의 연속 뉴클레오티드의 서열과 적어도 80% 상보성을 갖는 15-25개의 연속 뉴클레오티드의 서열을 포함한다.
하나의 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A와 엑손 1B 사이의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드의 역 상보체를 포함한다.
하나의 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A와 헥사뉴클레오티드 반복부 사이의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드의 역 상보체를 포함한다.
하나의 양태에서, 센스 가닥은 서열번호 115의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드를 포함하고, 안티센스 가닥은 서열번호 116의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드를 포함한다.
하나의 양태에서, 센스 가닥은 서열번호 117의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드를 포함하고, 안티센스 가닥은 서열번호 118의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드를 포함한다.
하나의 양태에서, 안티센스 가닥은 서열번호 3, 5, 7, 9, 11, 및 13의 안티센스 가닥 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드를 포함하고/하거나 센스 가닥은 서열번호 2, 4, 6, 8, 10, 및 12의 센스 가닥 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 인접 뉴클레오티드를 포함한다.
하나의 양태에서, C9orf72 표적 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 뉴클레오티드 서열은 성숙 C9orf72 메신저 RNA에 존재하지 않는다.
하나의 양태에서, 센스 가닥, 안티센스 가닥, 또는 센스 가닥과 안티센스 가닥 둘 다는 하나 이상의 친유성 모이어티에 접합된다.
하나의 양태에서, 친유성 모이어티는 dsRNA 제제의 이중 가닥 영역에서 하나 이상의 내부 위치에 접합된다.
하나의 양태에서, 친유성 모이어티는 링커 또는 담체를 통해 접합된다.
하나의 양태에서, logKow로 측정되는, 친유성 모이어티의 친유성은 0을 초과한다.
하나의 양태에서, 이중 가닥 RNAi 제제의 혈장 단백질 결합 분석에서 결합되지 않은 분획으로 측정되는, 이중 가닥 RNAi 제제의 소수성은 0.2를 초과한다.
하나의 양태에서, 혈장 단백질 결합 분석은 인간 혈청 알부민 단백질을 사용하는 전기영동 이동성 변화 분석이다.
일부 양태에서, dsRNA 제제는 적어도 하나의 변형된 뉴클레오티드를 포함한다.
하나의 양태에서, 5개 이하의 센스 가닥 뉴클레오티드 및 5개 이하의 안티센스 가닥 뉴클레오티드는 변형되지 않은 뉴클레오티드이다.
하나의 양태에서, 센스 가닥의 모든 뉴클레오티드 및 안티센스 가닥의 모든 뉴클레오티드는 변형된 뉴클레오티드이다.
하나의 양태에서, 변형된 뉴클레오티드 중 적어도 하나는 데옥시-뉴클레오티드, 3'-말단 데옥시-티민(dT) 뉴클레오티드, 2'-O-메틸 변형된 뉴클레오티드, 2'-플루오로 변형된 뉴클레오티드, 2'-데옥시-변형된 뉴클레오티드, 고정 뉴클레오티드(locked nucleotide), 비고정 뉴클레오티드(unlocked nucleotide), 형태적으로 제한된 뉴클레오티드, 제한된 에틸 뉴클레오티드, 무염기 뉴클레오티드, 2'-아미노-변형된 뉴클레오티드, 2'-O-알릴-변형된 뉴클레오티드, 2'-C-알킬-변형된 뉴클레오티드, 2'-하이드록실-변형된 뉴클레오티드, 2'-메톡시에틸 변형된 뉴클레오티드, 2'-O-알킬-변형된 뉴클레오티드, 모르폴리노 뉴클레오티드, 포스포르아미데이트, 뉴클레오티드를 포함하는 비천연 염기, 테트라하이드로피란 변형된 뉴클레오티드, 1,5-안하이드로헥시톨 변형된 뉴클레오티드, 사이클로헥세닐 변형된 뉴클레오티드, 5'-포스포로티오에이트 그룹을 포함하는 뉴클레오티드, 5'-메틸포스포네이트 그룹을 포함하는 뉴클레오티드, 5' 포스페이트 또는 5' 포스페이트 모방체를 포함하는 뉴클레오티드, 비닐 포스포네이트를 포함하는 뉴클레오티드, 아데노신-글리콜 핵산(GNA)을 포함하는 뉴클레오티드, 티미딘-글리콜 핵산(GNA) S-이성체를 포함하는 뉴클레오티드, 2-하이드록시메틸-테트라하이드로푸란-5-포스페이트를 포함하는 뉴클레오티드, 2'-데옥시티미딘-3'포스페이트를 포함하는 뉴클레오티드, 2'-데옥시구아노신-3'-포스페이트를 포함하는 뉴클레오티드, 및 콜레스테릴 유도체 및 도데칸산 비스데실아미드 그룹에 연결된 말단 뉴클레오티드; 및 이들의 조합의 그룹으로부터 선택된다.
하나의 양태에서, 변형된 뉴클레오티드는 2'-데옥시-2'-플루오로 변형된 뉴클레오티드, 2'-데옥시-변형된 뉴클레오티드, 3'-말단 데옥시-티민 뉴클레오티드(dT), 고정 뉴클레오티드, 무염기 뉴클레오티드, 2'-아미노-변형된 뉴클레오티드, 2'-알킬-변형된 뉴클레오티드, 모르폴리노 뉴클레오티드, 포스포르아미데이트, 및 뉴클레오티드를 포함하는 비천연 염기로 이루어진 그룹으로부터 선택된다.
하나의 양태에서, 변형된 뉴클레오티드는 3'-말단 데옥시-티민 뉴클레오티드(dT)의 짧은 서열을 포함한다.
하나의 양태에서, 뉴클레오티드에 대한 변형은 2'-O-메틸, GNA 및 2' 플루오로 변형이다.
하나의 양태에서, 센스 가닥의 뉴클레오티드의 실질적으로 전부는 2'-O-메틸 변형 및 2'-플루오로 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다. 일부 양태에서, 센스 가닥의 뉴클레오티드의 전부는 2'-O-메틸 변형 및 2'-플루오로 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다.
하나의 양태에서, 안티센스 가닥의 뉴클레오티드의 실질적으로 전부는 2'-O-메틸 변형 및 2'-플루오로 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다. 일부 양태에서, 안티센스 가닥의 뉴클레오티드의 전부는 2'-O-메틸 변형 및 2'-플루오로 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다.
하나의 양태에서, 센스 가닥의 뉴클레오티드의 실질적으로 전부는 2'-O-메틸 변형, 2'-플루오로 변형, 및 글리콜 핵산(GNA) 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다. 일부 양태에서, 센스 가닥의 뉴클레오티드의 전부는 2'-O-메틸 변형, 2'-플루오로 변형, 및 글리콜 핵산 (GNA) 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다.
하나의 양태에서, 안티센스 가닥의 뉴클레오티드의 실질적으로 전부는 2'-O-메틸 변형, 2'-플루오로 변형, 및 글리콜 핵산(GNA) 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다. 일부 양태에서, 안티센스 가닥의 뉴클레오티드의 전부는 2'-O-메틸 변형, 2'-플루오로 변형, 및 글리콜 핵산(GNA) 변형으로 이루어진 그룹으로부터 선택된 변형을 포함한다.
일부 양태에서, dsRNA 제제는 적어도 하나의 포스포로티오에이트 뉴클레오티드간 결합을 추가로 포함한다.
하나의 양태에서, dsRNA 제제는 6-8개의 포스포로티오에이트 뉴클레오티드간 결합을 포함한다.
하나의 양태에서, 센스 가닥은 적어도 하나의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 포함하고 안티센스 가닥은 적어도 하나의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 포함한다.
하나의 양태에서, 센스 가닥은 적어도 2개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 포함한다.
하나의 양태에서, 안티센스 가닥은 적어도 2개, 적어도 3개, 또는 적어도 4개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 포함한다.
하나의 양태에서, 적어도 하나의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합은 한 가닥의 5'-말단에 있거나, 한 가닥의 3'-말단에 있거나, 한 가닥의 5'-말단과 3'-말단 둘 다에 있다.
하나의 양태에서, 적어도 하나의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합은 센스 가닥의 5'-말단에 있다. 일부 양태에서, 센스 가닥은 5'-말단에 2개의 포스포로티오에이트 뉴클레오티드간 결합을 포함한다.
하나의 양태에서, 적어도 하나의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합은 안티센스 가닥의 5' 말단과 3' 말단 둘 다에 있다. 일부 양태에서, 안티센스 가닥은 5'-말단에 2개의 포스포로티오에이트 뉴클레오티드간 결합 및 3'-말단에 2개의 포스포로티오에이트 뉴클레오티드간 결합을 포함한다.
하나의 양태에서, 센스 가닥의 뉴클레오티드의 전부는 2'-O-메틸 변형 및 2'-플루오로 변형으로 이루어진 그룹으로부터 선택된 변형을 포함하고, 안티센스 가닥의 뉴클레오티드의 전부는 2'-O-메틸 변형 및 2'-플루오로 변형으로 이루어진 그룹으로부터 선택된 변형을 포함하고, 센스 가닥은 5'-말단에 2개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고, 안티센스 가닥은 5'-말단에 2개의 포스포로티오에이트 뉴클레오티드간 결합 및 3'-말단에 2개의 포스포로티오에이트 뉴클레오티드간 결합을 포함한다.
하나의 양태에서, (I) 센스 가닥은 (a) 다중 뉴클레오티드에 2'-O-메틸 변형; (b) 다중 뉴클레오티드에 2'-플루오로 변형; 및 (c) 다중 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (II) 안티센스 가닥은 (a) 다중 뉴클레오티드에 2'-O-메틸 변형; (c) 다중 뉴클레오티드에 2'-플루오로 변형; 및 (d) 다중 뉴클레오티드 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함한다. 임의로, dsRNA 제제는 안티센스 가닥의 3'-말단에 돌출부(overhang) 및 안티센스 가닥의 5' 말단에 평활 말단(blunt end)을 포함한다.
하나의 양태에서, (I) 센스 가닥은 (a) 21개의 뉴클레오티드의 길이; (b) 5' 말단으로부터 계수하여 뉴클레오티드 위치 1-6, 8, 및 12-21에 2'-O-메틸 변형; (c) 5' 말단으로부터 계수하여 뉴클레오티드 위치 7 및 9-11에 2'-플루오로 변형; 및 (d) 5' 말단으로부터 계수하여 뉴클레오티드 위치 1 및 2 사이에 및 뉴클레오티드 위치 2 및 3 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (II) 안티센스 가닥은 (a) 23개의 뉴클레오티드의 길이; (b) 5' 말단으로부터 계수하여 위치 1, 3-5, 7, 10-13, 15 및 17-23에 2'-O-메틸 변형; (c) 5' 말단으로부터 계수하여 위치 2, 6, 8, 9, 14, 및 16에 2'-플루오로 변형; 및 (d) (5' 말단으로부터 계수하여) 뉴클레오티드 위치 1 및 2 사이에, 뉴클레오티드 위치 2 및 3 사이에, 뉴클레오티드 위치 21 및 22 사이에, 및 뉴클레오티드 위치 22 및 23 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함하며, 여기서 dsRNA 제제는 안티센스 가닥의 3'-말단에 2-뉴클레오티드 돌출부 및 안티센스 가닥의 5' 말단에 평활 말단을 포함한다.
하나의 양태에서, 안티센스 가닥은 서열번호 3, 5, 7, 9, 11, 및 13의 안티센스 가닥 서열 중 어느 하나를 포함하고/하거나 센스 가닥은 서열번호 2, 4, 6, 8, 10, 및 12의 센스 가닥 서열 중 어느 하나를 포함한다.
하나의 양태에서, 센스 가닥은 30개 이하 뉴클레오티드 길이이다. 또 다른 양태에서, 안티센스 가닥은 30개 이하 뉴클레오티드 길이이다. 하나의 양태에서, 센스 가닥 및 안티센스 가닥은 각각 독립적으로 30개 이하 뉴클레오티드 길이이다.
하나의 양태에서, 적어도 하나의 가닥은 적어도 1개의 뉴클레오티드의 3'-돌출부를 포함한다. 또 다른 양태에서, 적어도 하나의 가닥은 적어도 2개의 뉴클레오티드의 3'-돌출부를 포함한다. 하나의 양태에서, 안티센스 가닥은 3'-돌출부를 포함한다.
이중 가닥 영역은 15-30개의 뉴클레오티드 쌍 길이; 17-23개의 뉴클레오티드 쌍 길이; 17-25개의 뉴클레오티드 쌍 길이; 23-27개의 뉴클레오티드 쌍 길이; 19-21개의 뉴클레오티드 쌍 길이; 21-23개의 뉴클레오티드 쌍 길이, 또는 21개의 뉴클레오티드 쌍 길이일 수 있다.
센스 가닥 및 안티센스 가닥은 각각 독립적으로 19-30개의 뉴클레오티드; 19-25개의 뉴클레오티드; 19-23개의 뉴클레오티드; 또는 21-23개의 뉴클레오티드 길이, 또는 21개의 뉴클레오티드 길이일 수 있다.
하나의 양태에서, 상보성 영역은 적어도 17개의 뉴클레오티드 길이이다. 다른 양태에서, 상보성 영역은 19-30개의 뉴클레오티드 길이; 19-25개의 뉴클레오티드 길이; 또는 21-23개의 뉴클레오티드 길이이다.
하나의 양태에서, 상보성 영역은 적어도 17개의 뉴클레오티드 길이이다. 다른 양태에서, 상보성 영역은 19-30개의 뉴클레오티드 길이; 19-25개의 뉴클레오티드 길이; 또는 21-23개의 뉴클레오티드 길이이다.
하나의 양태에서, 상보성 영역은 C9orf72 유전자의 엑손 1A의 시작과 엑손 2의 시작 사이의 서열에 대해 적어도 85% 상보적이다. 일부 양태에서, 안티센스 가닥은 C9orf72 표적 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 서열에 존재하는 15-25개의 연속 뉴클레오티드의 서열에 대해 적어도 85% 상보성을 갖는 15-25개의 연속 뉴클레오티드의 서열을 포함한다. 다른 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 서열에 대해 적어도 90% 상보적이다. 하나의 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 서열에 대해 적어도 95% 상보적이다. 일부 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 서열에 대해 100% 상보적이다. 일부 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 말단과 헥사뉴클레오티드 반복부 영역의 시작 사이의 서열에 대해 100% 상보적이다.
하나의 양태에서, 상보성 영역은 C9orf72 유전자의 엑손 1A의 말단과 인트론 1A의 헥사뉴클레오티드 반복부의 시작 사이의 서열에 대해 적어도 85% 상보적이다. 일부 양태에서, 안티센스 가닥은 C9orf72 표적 RNA의 엑손 1A의 말단과 인트론 1A의 헥사뉴클레오티드 반복부의 시작 사이의 서열에 존재하는 15-25개의 연속 뉴클레오티드의 서열에 대해 적어도 85% 상보성을 갖는 15-25개의 연속 뉴클레오티드의 서열을 포함한다. 다른 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 말단과 인트론 1A의 헥사뉴클레오티드 반복부의 시작 사이의 서열에 대해 적어도 90% 상보적이다. 하나의 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 말단과 인트론 1A의 헥사뉴클레오티드 반복부의 시작 사이의 서열에 대해 적어도 95% 상보적이다. 일부 양태에서, 상보성 영역은 C9orf72 표적 RNA의 엑손 1A의 말단과 인트론 1A의 헥사뉴클레오티드 반복부의 시작 사이의 서열에 대해 100% 상보적이다.
하나의 양태에서, 하나 이상의 친유성 모이어티는 예를 들어 링커 또는 담체를 통해 하나 이상의 가닥 상의 하나 이상의 내부 위치에 접합된다.
하나의 양태에서, 내부 위치는 적어도 하나의 가닥의 각 끝에서 말단 두 개 위치를 제외한 모든 위치를 포함한다.
또 다른 양태에서, 내부 위치는 적어도 하나의 가닥의 각 끝에서 말단 세 개 위치를 제외한 모든 위치를 포함한다.
하나의 양태에서, 내부 위치는 센스 가닥의 절단 부위를 제외한다.
하나의 양태에서, 내부 위치는 센스 가닥의 5'-말단으로부터 계수하여 위치 9-12를 제외한 모든 위치를 포함한다.
또 다른 양태에서, 내부 위치는 센스 가닥의 3'-말단으로부터 계수하여 위치 11-13을 제외한 모든 위치를 포함한다.
하나의 양태에서, 내부 위치는 안티센스 가닥의 절단 부위 영역을 제외한다.
하나의 양태에서, 내부 위치는 안티센스 가닥의 5'-말단으로부터 계수하여 위치 12-14를 제외한 모든 위치를 포함한다.
하나의 양태에서, 내부 위치는 3'-말단으로부터 계수하여 센스 가닥 상의 위치 11-13 및 5'-말단으로부터 계수하여 안티센스 가닥 상의 위치 12-14를 제외한 모든 위치를 포함한다.
하나의 양태에서, 하나 이상의 친유성 모이어티는 각 가닥의 5' 말단으로부터 계수하여 센스 가닥 상의 위치 4-8 및 13-18, 및 안티센스 가닥 상의 위치 6-10 및 15-18로 이루어진 그룹으로부터 선택된 내부 위치 중 하나 이상에 접합된다.
또 다른 양태에서, 하나 이상의 친유성 모이어티는 각 가닥의 5' 말단으로부터 계수하여 센스 가닥 상의 위치 5, 6, 7, 15, 및 17, 및 안티센스 가닥 상의 위치 15 및 17로 이루어진 그룹으로부터 선택된 내부 위치 중 하나 이상에 접합된다.
하나의 양태에서, 이중 가닥 영역의 내부 위치는 센스 가닥의 절단 부위 영역을 제외한다.
하나의 양태에서, 센스 가닥은 21개의 뉴클레오티드 길이이고, 안티센스 가닥은 23개의 뉴클레오티드 길이이며, 친유성 모이어티는 센스 가닥의 위치 21, 위치 20, 위치 15, 위치 1, 위치 7, 위치 6, 또는 위치 2 또는 안티센스 가닥의 위치 16에 접합된다.
하나의 양태에서, 친유성 모이어티는 센스 가닥의 위치 21, 위치 20, 위치 15, 위치 1, 또는 위치 7에 접합된다.
또 다른 양태에서, 친유성 모이어티는 센스 가닥의 위치 21, 위치 20, 또는 위치 15에 접합된다.
또 다른 양태에서, 친유성 모이어티는 센스 가닥의 위치 20 또는 위치 15에 접합된다.
하나의 양태에서, 친유성 모이어티는 안티센스 가닥의 위치 16에 접합된다.
하나의 양태에서, 친유성 모이어티는 지방족, 지환족 또는 다지환족 화합물이다.
하나의 양태에서, 친유성 모이어티는 지질, 콜레스테롤, 레티노산, 콜산, 아다만탄 아세트산, 1-피렌 부티르산, 디하이드로테스토스테론, 1,3-비스-O(헥사데실)글리세롤, 게라닐옥시헥시아놀, 헥사데실글리세롤, 보르네올, 멘톨, 1,3-프로판디올, 헵타데실 그룹, 팔미트산, 미리스트산, O3-(올레오일)리토콜산, O3-(올레오일)콜렌산, 디메톡시트리틸, 또는 페녹사진으로 이루어진 그룹으로부터 선택된다.
하나의 양태에서, 친유성 모이어티는 포화 또는 불포화 C4-C30 탄화수소 쇄, 및 하이드록실, 아민, 카복실산, 설포네이트, 포스페이트, 티올, 아지드 및 알킨으로 이루어진 그룹으로부터 선택된 임의의 작용 그룹을 함유한다.
하나의 양태에서, 친유성 모이어티는 포화 또는 불포화 C6-C18 탄화수소 쇄를 함유한다.
하나의 양태에서, 친유성 모이어티는 포화 또는 불포화 C16 탄화수소 쇄를 함유한다.
하나의 양태에서, 포화 또는 불포화 C16 탄화수소 쇄는 가닥의 5'-말단으로부터 계수하여 위치 6에 접합된다.
하나의 양태에서, 친유성 모이어티는 내부 위치(들) 또는 이중 가닥 영역에서 하나 이상의 뉴클레오티드(들)를 대체하는 담체를 통해 접합된다.
하나의 양태에서, 담체는 피롤리디닐, 피라졸리닐, 피라졸리디닐, 이미다졸리닐, 이미다졸리디닐, 피페리디닐, 피페라지닐, [1,3]디옥솔라닐, 옥사졸리디닐, 이속사졸리디닐, 모르폴리닐, 티아졸리디닐, 이소티아졸리디닐, 퀴녹살리닐, 피리다지노닐, 테트라하이드로푸라닐 및 데칼리닐로 이루어진 그룹으로부터 선택된 사이클릭 그룹이거나; 세리놀 백본 또는 디에탄올아민 백본을 기반으로 하는 비사이클릭 모이어티이다.
하나의 양태에서, 친유성 모이어티는 에테르, 티오에테르, 요소, 카보네이트, 아민, 아미드, 말레이미드-티오에테르, 디설파이드, 포스포디에스테르, 설폰아미드 결합, 클릭 반응의 생성물, 또는 카바메이트를 함유하는 링커를 통해 이중 가닥 iRNA 제제에 접합된다.
하나의 양태에서, 친유성 모이어티는 핵염기, 당 모이어티, 또는 뉴클레오시드간 결합에 접합된다.
하나의 양태에서, 친유성 모이어티 또는 표적화 리간드는 DNA, RNA, 디설파이드, 아미드; 갈락토사민, 글루코사민, 글루코스, 갈락토스, 만노스의 작용화된 단당류 또는 올리고당류, 및 이들의 조합으로 이루어진 그룹으로부터 선택된 생체-절단 가능한 링커를 통해 접합된다.
하나의 양태에서, 센스 가닥의 3' 말단은 아민을 갖는 사이클릭 그룹인 말단 캡을 통해 보호되고, 상기 사이클릭 그룹은 피롤리디닐, 피라졸리닐, 피라졸리디닐, 이미다졸리닐, 이미다졸리디닐, 피페리디닐, 피페라지닐, [1,3]디옥솔라닐, 옥사졸리디닐, 이속사졸리디닐, 모르폴리닐, 티아졸리디닐, 이소티아졸리디닐, 퀴녹살리닐, 피리다지노닐, 테트라하이드로푸라닐 및 데칼리닐로 이루어진 그룹으로부터 선택된다.
하나의 양태에서, dsRNA 제제는 간 조직을 표적으로 하는 표적화 리간드를 추가로 포함한다.
하나의 양태에서, 표적화 리간드는 GalNAc 접합체이다.
하나의 양태에서, dsRNA 제제는 Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및 Rp 배위 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함한다.
또 다른 양태에서, dsRNA 제제는 Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및 Rp 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함한다.
또 다른 양태에서, dsRNA 제제는 Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째, 두 번째 및 세 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및 Rp 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함한다.
또 다른 양태에서, dsRNA 제제는 Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 세 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및 Rp 배위 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함한다.
또 다른 양태에서, dsRNA 제제는 Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및 Rp 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함한다.
하나의 양태에서, dsRNA 제제는 안티센스 가닥의 5'-말단에 포스페이트 또는 포스페이트 모방체를 추가로 포함한다.
하나의 양태에서, 포스페이트 모방체는 5'-비닐 포스포네이트(VP)이다.
하나의 양태에서, 이중체의 안티센스 가닥의 5'-말단의 1 위치의 염기 쌍은 AU 염기 쌍이다.
하나의 양태에서, 센스 가닥은 총 21개의 뉴클레오티드를 갖고 안티센스 가닥은 총 23개의 뉴클레오티드를 갖는다.
하나의 양태에서, dsRNA 제제는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 24-48시간 이내에 적어도 10%, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 또는 적어도 60%까지 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 억제한다.
하나의 양태에서, dsRNA 제제는 성숙 C9orf72 메신저 RNA의 발현에 비해 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 선택적으로 억제한다.
하나의 양태에서, dsRNA 제제는 성숙 C9orf72 메신저 RNA를 발현하는 세포에 투여한 후 24-48시간 이내에 50% 미만, 40% 미만, 30% 미만, 20% 미만, 또는 10% 미만까지 성숙 C9orf72 메신저 RNA의 발현을 억제한다.
하나의 양태에서, dsRNA 제제는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 24-48시간 이내에 디펩티드 반복부 단백질 합성을 감소시킨다. 일부 양태에서, dsRNA 제제는 세포에 투여한 후 24-48시간 이내에 적어도 10%, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 또는 적어도 80%까지 디펩티드 반복부 단백질 합성을 감소시킨다.
본 발명은 또한 본 발명의 dsRNA 제제 등을 포함하는 C9orf72를 암호화하는 유전자의 발현을 억제하기 위한 세포 및 약제학적 조성물을 제공한다.
하나의 양태에서, dsRNA 제제는 염수 또는 물과 같은 완충되지 않은 용액에 존재한다.
또 다른 양태에서, dsRNA 제제는 아세테이트, 시트레이트, 프롤라민, 카보네이트, 또는 포스페이트 또는 이들의 임의의 조합을 포함하는 완충 용액과 같은 완충 용액; 또는 인산염 완충 염수(PBS)에 존재한다.
하나의 측면에서, 본 발명은 세포, 예를 들어 운동 뉴런과 같은 뉴런에서 헥사뉴클레오티드 반복부의 다중 연속 카피를 포함하는 C9orf72 유전자와 같은 헥사뉴클레오티드-반복부를 함유하는 C9orf72 RNA와 같은 C9orf72의 발현을 억제하는 방법을 제공하며, 상기 방법은 세포를 본 발명의 dsRNA 제제 또는 본 발명의 약제학적 조성물과 접촉시켜 세포에서 C9orf72 유전자의 발현을 억제함을 포함한다.
또 다른 측면에서, 본 발명은 세포에서 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집을 감소시키는 방법을 제공한다. 상기 방법은 본 발명의 dsRNA 제제를 세포에 도입함으로써 세포에서 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집을 감소시킴을 포함한다.
또 다른 측면에서, 본 발명은 세포에서 핵 및/또는 세포질 센스 및/또는 안티센스 C9orf72 RNA 병소 (RNA foci)를 감소시키는 방법을 제공한다. 상기 방법은 본 발명의 dsRNA 제제를 세포에 도입함으로써 세포에서 핵 및/또는 세포질 센스 및/또는 안티센스 C9orf72 RNA 병소를 감소시킴을 포함한다.
하나의 양태에서, 세포는 대상체 내에 있다.
하나의 양태에서, 대상체는 인간이다.
하나의 양태에서, 대상체는 C9orf72-헥사뉴클레오티드-반복부-확장-관련 질환, 상태 또는 장애와 같은 C9orf72-관련 장애를 갖거나 발병할 위험이 있다.
하나의 양태에서, C9orf72-관련 장애는 C9orf72 근위축성 측삭 경화증/전두측두엽 치매 또는 C9orf72 확장으로 인한 헌팅턴-유사 증후군으로 이루어진 그룹으로부터 선택된다.
하나의 양태에서, 세포를 dsRNA 제제와 접촉시키면 C9orf72의 발현이 50% 이하로 억제된다.
하나의 양태에서, dsRNA 제제는, 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 24-48시간 이내에, 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 mRNA의 발현을 적어도 10%, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 또는 적어도 60%까지 억제한다.
일부 양태에서, dsRNA 제제는 성숙 C9orf72 메신저 RNA의 발현에 비해 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 선택적으로 억제한다. 다른 양태에서, dsRNA 제제는, 성숙 C9orf72 메신저 RNA를 발현하는 세포에 투여한 후 24-48시간 이내에, 성숙 C9orf72 메신저 RNA의 발현을 50% 미만, 40% 미만, 30% 미만, 20% 미만, 또는 10% 미만까지 억제한다.
일부 양태에서, dsRNA 제제는 세포에서 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집을 감소시킨다.
일부 양태에서, dsRNA 제제는 세포에서 핵 및/또는 세포질 센스 및/또는 안티센스 C9orf72 RNA 병소를 감소시킨다.
하나의 양태에서, C9orf72의 발현을 억제시키면 대상체의 혈청에서 C9orf72 단백질 수준이 50% 이하까지 감소된다.
일부 양태에서, dsRNA 제제는, 세포에 투여한 후 24-48시간 이내에, 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집을 적어도 10%, 적어도 20%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60%, 적어도 70%, 또는 적어도 80%까지 감소시킨다.
하나의 측면에서, 본 발명은 대상체에게 치료학적 유효량의 본 발명의 dsRNA 제제, 또는 본 발명의 약제학적 조성물을 투여하여 C9orf72 발현의 감소로부터 혜택을 받는 장애를 갖는 대상체를 치료함을 포함하여, C9orf72-헥사뉴클레오티드-반복부-확장-관련 질환, 상태 또는 장애와 같은 표적 C9orf72 RNA를 녹다운함으로써 혜택을 받는 장애를 갖는 대상체를 치료하는 방법을 제공한다.
또 다른 측면에서, 본 발명은 대상체에게 예방학적 유효량의 본 발명의 dsRNA 제제, 또는 본 발명의 약제학적 조성물을 투여하여 C9orf72 발현의 감소로부터 혜택을 받는 장애를 갖는 대상체에서 적어도 하나의 증상을 예방함을 포함하여, C9orf72-헥사뉴클레오티드-반복부-확장-관련 질환, 상태 또는 장애와 같은 C9orf72 발현의 감소로부터 혜택을 받는 장애를 갖는 대상체에서 적어도 하나의 증상을 예방하는 방법을 제공한다.
하나의 양태에서, 장애는 C9orf72-관련 장애이다.
하나의 양태에서, C9orf72-관련 장애는 C9orf72 근위축성 측삭 경화증/전두측두엽 치매 또는 C9orf72 확장으로 인한 헌팅턴-유사 증후군으로 이루어진 그룹으로부터 선택된다.
하나의 양태에서, 대상체는 인간이다.
하나의 양태에서, 대상체에 대한 제제의 투여는 C9orf72 단백질 축적의 감소를 야기한다.
일부 양태에서, 상기 방법은 대상체에서 디펩티드 반복부 단백질 합성을 감소시키거나 디펩티드 반복부 단백질 응집을 감소시킨다. 일부 양태에서, 상기 방법은 대상체에서 서열번호 1의 다중 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 감소시킨다.
하나의 양태에서, dsRNA 제제는 약 0.01 mg/kg 내지 약 50mg/kg의 용량으로 대상체에게 투여된다.
하나의 양태에서, dsRNA 제제는 피하로 대상체에게 투여된다.
또 다른 양태에서, dsRNA 제제는 척수강내로 대상체에게 투여된다.
하나의 양태에서, 본 발명의 방법은 대상체로부터의 샘플(들)에서 C9orf72의 수준을 측정함을 추가로 포함한다.
하나의 양태에서, 대상체 샘플(들)에서의 C9orf72의 수준은 혈액, 혈청 또는 뇌척수액 샘플(들)의 C9orf72 단백질 수준이다.
하나의 양태에서, 본 발명의 방법은 추가 치료제를 대상체에게 투여함을 추가로 포함한다.
하나의 측면에서, 본 발명은 본 발명의 dsRNA 제제, 또는 본 발명의 약제학적 조성물을 포함하는 키트를 제공한다.
또 다른 측면에서, 본 발명은 본 발명의 dsRNA 제제, 또는 본 발명의 약제학적 조성물을 포함하는 바이알을 제공한다.
또 다른 측면에서, 본 발명은 본 발명의 dsRNA 제제, 또는 본 발명의 약제학적 조성물을 포함하는 시린지를 제공한다.
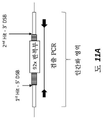
도 1(축척되지 않음)은 야생형 마우스 C9orf72 유전자좌, 야생형 인간 C9orf72 유전자좌, 및 서열번호 1(GGGGCC)(MAID8029a)에 제시된 헥사뉴클레오티드 서열의 92개 반복부를 포함하는 인간화 마우스 C9orf72 유전자좌의 개략도이다. 마우스 서열은 점선 및 점선이 있는 상자로 표시되고 인간 서열은 검은색 실선으로 표시된다. 엑손은 상자로 표시된다. 헥사뉴클레오티드 반복부 서열의 위치가 표시된다.
도 2(축척되지 않음)는 표적 게놈 유전자좌에서 반복부 서열(예를 들어 C9orf72 유전자좌에서 서열번호 1에 제시된 헥사뉴클레오티드 서열)을 확장하기 위해 재조합 기구를 사용하기 위해 반복부 서열 부근에 이중-가닥 절단을 도입하기 위해 뉴클레아제 제제를 사용하는 개념의 개략도이다.
도 3(축척되지 않음)은 C9orf72 인간화, 92x-반복부-함유 ES 세포(MAID8029a)에서 헥사뉴클레오티드 반복부 확장 서열의 말단 부근에 위치한 8개의 가이드 RNA 표적 서열의 위치가 있는 개략도이다.
도 4는 인간화, 92x-반복부-함유 대립유전자(MAID8029a)와 동일한 서열을 포함하는 플라스미드(8028 Stvec)를 사용하여 무세포 시스템에서 평가된 8개의 가이드 RNA의 절단 효율을 도시한다.
도 5(축척되지 않음)는 92x 반복부 확장 서열의 5' 말단 부근(상자 2), 3' 말단 부근(상자 3) 또는 5' 및 3' 말단 둘 다의 부근(상자 1)에 이중-가닥 절단을 도입하여 92x 반복부를 확장하기 위한 계획의 개략도이다. 검은색 상자는 내인성 마우스 서열을 나타내고; 흰색 상자는 인간화 영역을 나타낸다.
도 6A(축척되지 않음)는 내인성 C9orf72 ES 세포 클론에서 서열번호 1에 제시된 헥사뉴클레오티드 서열의 인스턴스의 수를 평가하기 위해 사용되는 종래의 2-프라이머 PCR의 개략도이다.
도 6B(축척되지 않음)는 내인성 C9orf72 ES 세포 클론에서 서열번호 1에 제시된 헥사뉴클레오티드 서열의 인스턴스의 수를 평가하기 위해 3개의 프라이머를 사용하는 프라임 PCR의 개략도이다.
도 7A는 92x 반복부 확장 서열의 5' 말단 부근의 절단 후 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래의 PCR의 결과를 보여준다. 확장된 반복부는 별표로 표시된다.
도 7B는 92x 반복부 확장 서열의 3' 말단 부근의 절단 후 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래의 PCR의 결과를 보여준다. 확장된 반복부는 별표로 표시된다.
도 8은 도 7A로부터의 클론 중 하나 및 도 7B로부터의 클론 중 하나에서 반복부의 수를 확인하기 위한 프라임 PCR의 결과를 보여준다. 모 8029a(92x 반복부) 클론을 대조군으로 사용하였다. 도 8은 모세관 전기영동으로부터의 결과를 보여준다. 신호 강도는 Y축에, PCR 산물 크기는 X축에 있다. 판독값은 피크의 수이다.
도 9는 도 7A로부터의 클론 중 하나 및 도 7B로부터의 클론 중 하나에서 반복부의 수를 확인하기 위한 프라임 PCR의 결과를 보여준다.
도 10은 92x 반복부 확장 서열의 5' 말단 부근의 절단, 92x 반복부 확장 서열의 3' 말단 부근의 절단 또는 92x 반복부 확장 서열의 5' 및 3' 말단 둘 다 부근의 절단 후 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래 PCR의 결과를 보여준다. 확장된 반복부는 화살표로 표시된다.
도 11A(축척되지 않음)는 첫 번째 확장된 클론을 생성하기 위해 92x 반복부 확장 서열의 5' 말단 부근에 이중-가닥 절단을 먼저 도입한 다음 두 번째 확장된 클론을 생성하기 위해 첫 번째 확장된 클론에서 반복부 확장 서열의 3' 말단 부근에 이중-가닥 절단을 도입함으로써 마우스 ES 세포에서 92x 반복부를 확장하기 위한 계획의 개략도를 보여준다. 인간화 영역은 라벨 및 흰색 상자로 표시된다. 5' DSB 및 3' DSB의 위치가 PCR에 의한 검출용 프라이머의 위치와 함께 표시된다.
도 11B는 250x 반복부 확장 서열의 3' 말단 부근의 250x 반복부 확장 서열 클론의 절단 후 두 번째 확장에 따른 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래의 PCR의 결과를 보여준다. 확장된 반복부 및 모 반복부는 표지된 화살표로 표시된다.
도 12A 내지 12D는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 250, 또는 300개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손 1A-엑손 2 스플라이싱된 전사체인(도 12A), 엑손 1B-엑손 2 스플라이싱된 전사체인(도 12B), 엑손 1A 부근에 인트론 서열을 함유하는(도 12C), 엑손 1B 부근에 인트론 서열을 보유하는(도 12D) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 12E 내지 12H는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 500, 또는 500개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손 1A-엑손 2 스플라이싱된 전사체인(도 12E), 엑손 1B-엑손 2 스플라이싱된 전사체인(도 12F), 엑손 1A 부근에 인트론 서열을 함유하는(도 12G), 엑손 1B 부근에 인트론 서열을 보유하는(도 12H) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 13(상단)은 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500, 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)의 용해물의 웨스턴 슬롯 블롯 이미지이다. 0㎍, 1.25㎍, 2.5㎍, 5㎍, 10㎍, 또는 20㎍ 총 단백질을 함유하는 용해물을 항-폴리-GlyPro 항체 또는 항-폴리-GlyAla 항체로 블롯팅하였다. 도 13(하단)은 도면의 상단 부분의 웨스턴 슬롯 블롯의 정량화를 보여준다.
도 14A 내지 14B는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500, 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서(도 14A) 또는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 마우스로부터의 뇌간 및 척수 샘플에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3 또는 300개 반복부를 포함하는 변형된 (인간화) C9orf72 유전자좌에 대해 이종접합성인 마우스로부터의 뇌간 및 척수 샘플에서(도 14B) 액손 1A 부근에 인트론 서열을 함유하는 C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 15A 내지 15D는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 축하형(hypaxial-like) 운동 뉴런 또는 사지형(limb-like) 운동 뉴런인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손 1A-엑손 2 스플라이싱된 전사체인(도 15A), 엑손 1B-엑손 2 스플라이싱된 전사체인(도 15B), 엑손 1A 부근에 인트론 서열을 함유하는(도 15C), 엑손 1B 부근에 인트론 서열을 보유하는(도 15D) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 15E 내지 15F는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 축하형 운동 뉴런 또는 사지형 운동 뉴런인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 스플라이싱되지 않은 전구체 전사체(도 15E) 또는 스플라이싱된 C9orf72 mRNA(도 15F)인 C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 16A 내지 16B는 처리 후 24시간 및 48시간에서의, 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 C9orf72의 인트론 1을 표적화하는 siRNA, 스플라이싱된 C9orf72 mRNA를 표적화하는 siRNA(엑손 11(siRNA 1), 엑손 5(siRNAs 2 및 6), 또는 엑손 2(siRNAs 3 및 5), 또는 엑손 9(siRNA4)), 또는 대조군 GAPDH siRNA로 처리된 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손2-3 스플라이싱된 전사체이거나(도 16A) 엑손 1A 부근에 인트론 서열을 함유하는(도 16B) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. 별표는 도 17A 및 17B의 웨스턴 블롯에 해당하는 샘플을 나타낸다.
도 17A는 처리 후 48시간에서의, 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 C9orf72의 인트론 1을 표적화하는 siRNA, 스플라이싱된 C9orf72 mRNA를 표적화하는 siRNA(엑손 11(siRNA 1), 엑손 5(siRNAs 2), 또는 엑손 2(siRNAs 3)), 또는 대조군 GAPDH siRNA로 처리된 배아 줄기 세포 유래된 운동 뉴런(ESMN)의 용해물의 웨스턴 슬롯 블롯 이미지이다. 1.25㎍ 또는 2.5㎍ 총 단백질을 함유하는 용해물을 항-폴리-GlyAla 항체로 블롯팅하였다. 도 17B는 도 17A의 웨스턴 슬롯 블롯의 정량화를 보여준다.
도 18A는 C9orf72의 인트론 1A를 표적화하는 siRNA의 개략도(축척되지 않음)이다.
도 18B는 C9orf72 pre-mRNA의 상이한 섹션에 걸쳐 있는 TAQMAN® 정성적 PCR 분석 A, B, C, D, E, F, G, H, I 및 J의 개략도(축척되지 않음)이다. 프라이머 및 프로브의 서열은 표 8에 나타내어져 있다.
도 19A 내지 19C는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 엑손 1A를 표적화하는 siRNA(siRNA E1A #1, E1A #2 및 E1A #3), 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA I1A #2, I1A #4, I1A #6 및 I1A #10), 또는 헥사뉴클레오티드 반복부의 하류 및 엑손 1B의 하류에 있는 인트론 1B(siRNAs I1B #1 및 I1B #6)로 형질감염시킨지 24시간 후 C9orf72 인트론 1A 서열을 함유하는(도 19A; 분석 F), C9orf72 엑손 2-3 스플라이싱된 전이체인(도 19B), 또는 C9orf72 엑손 1a-2 스플라이싱된 전이체인(도 19C; 분석 B) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA는 대조군으로 사용되었고 mock은 음성 대조군으로 사용되었다.
도 20A 내지 20C는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 엑손 1A를 표적화하는 siRNA(siRNA E1A #1, E1A #2 및 E1A #3), 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA I1A #2, I1A #4, I1A #6 및 I1A #10), 또는 헥사뉴클레오티드 반복부의 하류 및 엑손 1B의 하류에 있는 인트론 1B(siRNAs I1B #1 및 I1B #6)로 형질감염시킨지 48시간 후 C9orf72 인트론 1A 서열을 함유하는(도 20A; 분석 F), C9orf72 엑손 2-3 스플라이싱된 전이체인(도 20B), 또는 C9orf72 엑손 1a-2 스플라이싱된 전이체인(도 20C; 분석 B) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA는 대조군으로 사용되었고 mock은 음성 대조군으로 사용되었다.
도 21A 내지 21C는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 엑손 1A를 표적화하는 siRNA(siRNA E1A #1, E1A #2 및 E1A #3), 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA I1A #2, I1A #4, I1A #6 및 I1A #10), 또는 헥사뉴클레오티드 반복부의 하류 및 엑손 1B의 하류에 있는 인트론 1B(siRNAs I1B #1 및 I1B #6)로 형질감염시킨지 72시간 후 C9orf72 인트론 1A 서열을 함유하는(도 21A; 분석 F), C9orf72 엑손 2-3 스플라이싱된 전이체인(도 21B), 또는 C9orf72 엑손 1a-2 스플라이싱된 전이체인(도 21C; 분석 B) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA는 대조군으로 사용되었고 mock은 음성 대조군으로 사용되었다.
도 22A 내지 22E는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 성숙 C9orf72 mRNA를 표적화하는 siRNA(siRNA #1) 또는 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA #12)로 전기천공한지 24시간 후 C9orf72 인트론 1A 서열을 함유하는(도 22A; 분석 F), 엑손 1A에 결합된 C9orf72 인트론 1A 서열을 함유하는(도 22B; 분석 E), C9orf72 엑손 1a-2 스플라이싱된 전사체인(도 22C; 분석 B), C9orf72 엑손 1b-2 스플라이싱된 전사체인(도 22D; 분석 C), 또는 C9orf72 엑손 5-6 스플라이싱된 전사체인(도 22E; 분석 D) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA 및 mock은 음성 대조군으로 사용되었다.
도 23A 내지 23E는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 성숙 C9orf72 mRNA를 표적화하는 siRNA(siRNA #1) 또는 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA #12)로 전기천공한지 48시간 후 C9orf72 인트론 1A 서열을 함유하는(도 23A; 분석 F), 엑손 1A에 결합된 C9orf72 인트론 1A 서열을 함유하는(도 23B; 분석 E), C9orf72 엑손 1a-2 스플라이싱된 전사체인(도 23C; 분석 B), C9orf72 엑손 1b-2 스플라이싱된 전사체인(도 23D; 분석 C), 또는 C9orf72 엑손 5-6 스플라이싱된 전사체인(도 23E; 분석 D) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA 및 mock은 음성 대조군으로 사용되었다.
도 2(축척되지 않음)는 표적 게놈 유전자좌에서 반복부 서열(예를 들어 C9orf72 유전자좌에서 서열번호 1에 제시된 헥사뉴클레오티드 서열)을 확장하기 위해 재조합 기구를 사용하기 위해 반복부 서열 부근에 이중-가닥 절단을 도입하기 위해 뉴클레아제 제제를 사용하는 개념의 개략도이다.
도 3(축척되지 않음)은 C9orf72 인간화, 92x-반복부-함유 ES 세포(MAID8029a)에서 헥사뉴클레오티드 반복부 확장 서열의 말단 부근에 위치한 8개의 가이드 RNA 표적 서열의 위치가 있는 개략도이다.
도 4는 인간화, 92x-반복부-함유 대립유전자(MAID8029a)와 동일한 서열을 포함하는 플라스미드(8028 Stvec)를 사용하여 무세포 시스템에서 평가된 8개의 가이드 RNA의 절단 효율을 도시한다.
도 5(축척되지 않음)는 92x 반복부 확장 서열의 5' 말단 부근(상자 2), 3' 말단 부근(상자 3) 또는 5' 및 3' 말단 둘 다의 부근(상자 1)에 이중-가닥 절단을 도입하여 92x 반복부를 확장하기 위한 계획의 개략도이다. 검은색 상자는 내인성 마우스 서열을 나타내고; 흰색 상자는 인간화 영역을 나타낸다.
도 6A(축척되지 않음)는 내인성 C9orf72 ES 세포 클론에서 서열번호 1에 제시된 헥사뉴클레오티드 서열의 인스턴스의 수를 평가하기 위해 사용되는 종래의 2-프라이머 PCR의 개략도이다.
도 6B(축척되지 않음)는 내인성 C9orf72 ES 세포 클론에서 서열번호 1에 제시된 헥사뉴클레오티드 서열의 인스턴스의 수를 평가하기 위해 3개의 프라이머를 사용하는 프라임 PCR의 개략도이다.
도 7A는 92x 반복부 확장 서열의 5' 말단 부근의 절단 후 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래의 PCR의 결과를 보여준다. 확장된 반복부는 별표로 표시된다.
도 7B는 92x 반복부 확장 서열의 3' 말단 부근의 절단 후 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래의 PCR의 결과를 보여준다. 확장된 반복부는 별표로 표시된다.
도 8은 도 7A로부터의 클론 중 하나 및 도 7B로부터의 클론 중 하나에서 반복부의 수를 확인하기 위한 프라임 PCR의 결과를 보여준다. 모 8029a(92x 반복부) 클론을 대조군으로 사용하였다. 도 8은 모세관 전기영동으로부터의 결과를 보여준다. 신호 강도는 Y축에, PCR 산물 크기는 X축에 있다. 판독값은 피크의 수이다.
도 9는 도 7A로부터의 클론 중 하나 및 도 7B로부터의 클론 중 하나에서 반복부의 수를 확인하기 위한 프라임 PCR의 결과를 보여준다.
도 10은 92x 반복부 확장 서열의 5' 말단 부근의 절단, 92x 반복부 확장 서열의 3' 말단 부근의 절단 또는 92x 반복부 확장 서열의 5' 및 3' 말단 둘 다 부근의 절단 후 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래 PCR의 결과를 보여준다. 확장된 반복부는 화살표로 표시된다.
도 11A(축척되지 않음)는 첫 번째 확장된 클론을 생성하기 위해 92x 반복부 확장 서열의 5' 말단 부근에 이중-가닥 절단을 먼저 도입한 다음 두 번째 확장된 클론을 생성하기 위해 첫 번째 확장된 클론에서 반복부 확장 서열의 3' 말단 부근에 이중-가닥 절단을 도입함으로써 마우스 ES 세포에서 92x 반복부를 확장하기 위한 계획의 개략도를 보여준다. 인간화 영역은 라벨 및 흰색 상자로 표시된다. 5' DSB 및 3' DSB의 위치가 PCR에 의한 검출용 프라이머의 위치와 함께 표시된다.
도 11B는 250x 반복부 확장 서열의 3' 말단 부근의 250x 반복부 확장 서열 클론의 절단 후 두 번째 확장에 따른 반복부 영역의 크기를 평가하기 위한 C9orf72 유전자좌의 종래의 PCR의 결과를 보여준다. 확장된 반복부 및 모 반복부는 표지된 화살표로 표시된다.
도 12A 내지 12D는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 250, 또는 300개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손 1A-엑손 2 스플라이싱된 전사체인(도 12A), 엑손 1B-엑손 2 스플라이싱된 전사체인(도 12B), 엑손 1A 부근에 인트론 서열을 함유하는(도 12C), 엑손 1B 부근에 인트론 서열을 보유하는(도 12D) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 12E 내지 12H는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 500, 또는 500개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손 1A-엑손 2 스플라이싱된 전사체인(도 12E), 엑손 1B-엑손 2 스플라이싱된 전사체인(도 12F), 엑손 1A 부근에 인트론 서열을 함유하는(도 12G), 엑손 1B 부근에 인트론 서열을 보유하는(도 12H) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 13(상단)은 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500, 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)의 용해물의 웨스턴 슬롯 블롯 이미지이다. 0㎍, 1.25㎍, 2.5㎍, 5㎍, 10㎍, 또는 20㎍ 총 단백질을 함유하는 용해물을 항-폴리-GlyPro 항체 또는 항-폴리-GlyAla 항체로 블롯팅하였다. 도 13(하단)은 도면의 상단 부분의 웨스턴 슬롯 블롯의 정량화를 보여준다.
도 14A 내지 14B는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500, 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서(도 14A) 또는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 마우스로부터의 뇌간 및 척수 샘플에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3 또는 300개 반복부를 포함하는 변형된 (인간화) C9orf72 유전자좌에 대해 이종접합성인 마우스로부터의 뇌간 및 척수 샘플에서(도 14B) 액손 1A 부근에 인트론 서열을 함유하는 C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 15A 내지 15D는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 축하형(hypaxial-like) 운동 뉴런 또는 사지형(limb-like) 운동 뉴런인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손 1A-엑손 2 스플라이싱된 전사체인(도 15A), 엑손 1B-엑손 2 스플라이싱된 전사체인(도 15B), 엑손 1A 부근에 인트론 서열을 함유하는(도 15C), 엑손 1B 부근에 인트론 서열을 보유하는(도 15D) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 15E 내지 15F는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3개 반복부를 포함하는 ESMN에 비해 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 3, 92, 300, 500 또는 600개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 축하형 운동 뉴런 또는 사지형 운동 뉴런인 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 스플라이싱되지 않은 전구체 전사체(도 15E) 또는 스플라이싱된 C9orf72 mRNA(도 15F)인 C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다.
도 16A 내지 16B는 처리 후 24시간 및 48시간에서의, 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 C9orf72의 인트론 1을 표적화하는 siRNA, 스플라이싱된 C9orf72 mRNA를 표적화하는 siRNA(엑손 11(siRNA 1), 엑손 5(siRNAs 2 및 6), 또는 엑손 2(siRNAs 3 및 5), 또는 엑손 9(siRNA4)), 또는 대조군 GAPDH siRNA로 처리된 배아 줄기 세포 유래된 운동 뉴런(ESMN)에서, 엑손2-3 스플라이싱된 전사체이거나(도 16A) 엑손 1A 부근에 인트론 서열을 함유하는(도 16B) C9orf72 유전자좌로부터의 전사체의 발현 수준(y-축)(각 도면 상단의 C9orf72 유전자좌의 묘사에 나타난 TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. 별표는 도 17A 및 17B의 웨스턴 블롯에 해당하는 샘플을 나타낸다.
도 17A는 처리 후 48시간에서의, 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌에 대해 이종접합성이고 C9orf72의 인트론 1을 표적화하는 siRNA, 스플라이싱된 C9orf72 mRNA를 표적화하는 siRNA(엑손 11(siRNA 1), 엑손 5(siRNAs 2), 또는 엑손 2(siRNAs 3)), 또는 대조군 GAPDH siRNA로 처리된 배아 줄기 세포 유래된 운동 뉴런(ESMN)의 용해물의 웨스턴 슬롯 블롯 이미지이다. 1.25㎍ 또는 2.5㎍ 총 단백질을 함유하는 용해물을 항-폴리-GlyAla 항체로 블롯팅하였다. 도 17B는 도 17A의 웨스턴 슬롯 블롯의 정량화를 보여준다.
도 18A는 C9orf72의 인트론 1A를 표적화하는 siRNA의 개략도(축척되지 않음)이다.
도 18B는 C9orf72 pre-mRNA의 상이한 섹션에 걸쳐 있는 TAQMAN® 정성적 PCR 분석 A, B, C, D, E, F, G, H, I 및 J의 개략도(축척되지 않음)이다. 프라이머 및 프로브의 서열은 표 8에 나타내어져 있다.
도 19A 내지 19C는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 엑손 1A를 표적화하는 siRNA(siRNA E1A #1, E1A #2 및 E1A #3), 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA I1A #2, I1A #4, I1A #6 및 I1A #10), 또는 헥사뉴클레오티드 반복부의 하류 및 엑손 1B의 하류에 있는 인트론 1B(siRNAs I1B #1 및 I1B #6)로 형질감염시킨지 24시간 후 C9orf72 인트론 1A 서열을 함유하는(도 19A; 분석 F), C9orf72 엑손 2-3 스플라이싱된 전이체인(도 19B), 또는 C9orf72 엑손 1a-2 스플라이싱된 전이체인(도 19C; 분석 B) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA는 대조군으로 사용되었고 mock은 음성 대조군으로 사용되었다.
도 20A 내지 20C는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 엑손 1A를 표적화하는 siRNA(siRNA E1A #1, E1A #2 및 E1A #3), 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA I1A #2, I1A #4, I1A #6 및 I1A #10), 또는 헥사뉴클레오티드 반복부의 하류 및 엑손 1B의 하류에 있는 인트론 1B(siRNAs I1B #1 및 I1B #6)로 형질감염시킨지 48시간 후 C9orf72 인트론 1A 서열을 함유하는(도 20A; 분석 F), C9orf72 엑손 2-3 스플라이싱된 전이체인(도 20B), 또는 C9orf72 엑손 1a-2 스플라이싱된 전이체인(도 20C; 분석 B) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA는 대조군으로 사용되었고 mock은 음성 대조군으로 사용되었다.
도 21A 내지 21C는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 엑손 1A를 표적화하는 siRNA(siRNA E1A #1, E1A #2 및 E1A #3), 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA I1A #2, I1A #4, I1A #6 및 I1A #10), 또는 헥사뉴클레오티드 반복부의 하류 및 엑손 1B의 하류에 있는 인트론 1B(siRNAs I1B #1 및 I1B #6)로 형질감염시킨지 72시간 후 C9orf72 인트론 1A 서열을 함유하는(도 21A; 분석 F), C9orf72 엑손 2-3 스플라이싱된 전이체인(도 21B), 또는 C9orf72 엑손 1a-2 스플라이싱된 전이체인(도 21C; 분석 B) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA는 대조군으로 사용되었고 mock은 음성 대조군으로 사용되었다.
도 22A 내지 22E는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 성숙 C9orf72 mRNA를 표적화하는 siRNA(siRNA #1) 또는 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA #12)로 전기천공한지 24시간 후 C9orf72 인트론 1A 서열을 함유하는(도 22A; 분석 F), 엑손 1A에 결합된 C9orf72 인트론 1A 서열을 함유하는(도 22B; 분석 E), C9orf72 엑손 1a-2 스플라이싱된 전사체인(도 22C; 분석 B), C9orf72 엑손 1b-2 스플라이싱된 전사체인(도 22D; 분석 C), 또는 C9orf72 엑손 5-6 스플라이싱된 전사체인(도 22E; 분석 D) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA 및 mock은 음성 대조군으로 사용되었다.
도 23A 내지 23E는 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 300개 반복부를 포함하는 변형된 C9orf72 유전자좌를 포함하는 마우스 배아 줄기 세포에서 성숙 C9orf72 mRNA를 표적화하는 siRNA(siRNA #1) 또는 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A(siRNA #12)로 전기천공한지 48시간 후 C9orf72 인트론 1A 서열을 함유하는(도 23A; 분석 F), 엑손 1A에 결합된 C9orf72 인트론 1A 서열을 함유하는(도 23B; 분석 E), C9orf72 엑손 1a-2 스플라이싱된 전사체인(도 23C; 분석 B), C9orf72 엑손 1b-2 스플라이싱된 전사체인(도 23D; 분석 C), 또는 C9orf72 엑손 5-6 스플라이싱된 전사체인(도 23E; 분석 D) C9orf72 유전자좌로부터의 전사체의 상대적 발현 수준(y-축)(TAQMAN® 정량적 역전사-결합 PCR(RT-qPCR) 분석에 의해 결정됨)을 보여주는 막대 그래프이다. Gapdh siRNA 및 mock은 음성 대조군으로 사용되었다.
본 개시내용은 확장된 GGGGCC (G4C2) 반복부를 갖는 C9orf72 유전자와 같은 C9orf72 유전자의 RNA 전사체의 RNA-유도 침묵 복합체(RISC)-매개된 절단에 영향을 미치는 RNAi 조성물을 제공한다. C9orf72 유전자는 세포, 예를 들어 인간과 같은 대상체 내의 세포 내에 있을 수 있다. 이들 iRNA의 사용은 포유동물에서 상응하는 유전자(C9orf72 유전자)의 mRNA의 표적화된 분해를 가능하게 한다.
본 발명의 iRNA는 C9orf72 표적 RNA, 예를 들어 유전자의 인트론에서 확장된 GGGGCC 헥사뉴클레오티드 반복부를 갖고 뉴클레오티드 변형의 조합을 갖는 C9orf72 표적 RNA를 표적으로 하도록 설계되었다. 제제는 성숙 C9orf72 mRNA(인트론이 스플라이싱된 mRNA) 또는 C9orf7 mRNA 전구체(인트론을 함유하는 mRNA)를 표적으로 할 수 있다. 본 발명의 iRNA는 C9orf7유전자(예를 들어, 성숙 mRNA)의 발현을 약 50% 이하까지 억제하고 센스- 및 안티센스-함유 병소 및 비정상적인 디펩티드-반복부(DPR) 단백질(폴리(GA), 폴리(GR), 폴리(GP), 폴리(PA) 및 폴리(PR))의 수준을 감소시킬 수 있고/있거나 C9orf7유전자(예를 들어, 전구체 mRNA)의 발현을 약 50% 초과까지 억제하고 센스- 및 안티센스-함유 병소 및 비정상적인 디펩티드-반복부(DPR) 단백질(폴리(GA), 폴리(GR), 폴리(GP), 폴리(PA) 및 폴리(PR))의 수준을 감소시킬 수 있다. 이론에 의해 제한하려는 것은 아니지만, 전술한 특성 및 특이 표적 부위의 조합 또는 하위-조합, 또는 이들 iRNA의 특정 변형은 본 발명의 iRNA에 개선된 효능, 안정성, 역가, 내구성 및 안전성을 부여하는 것으로 여겨진다.
따라서, 본 개시내용은 또한 C9orf72 유전자의 발현을 억제하기 위해 또는 C9orf72 유전자의 발현을 억제하거나 감소시키는 것으로부터 혜택을 받는 장애, 예를 들어, C9orf72-관련 질환, 예를 들면, C9orf72 근위축성 측삭 경화증/전두측두엽 치매, 또는 헌팅턴 질환, 예를 들어, C9orf72 확장으로 인한 헌팅턴-유사 증후군과 같은 C9orf72 유전자의 인트론에서 확장된 GGGGCC 헥사뉴클레오티드 반복부와 관련된 질환을 갖는 대상체를 치료하기 위해 본 개시내용의 RNAi 조성물을 사용하는 방법을 제공한다.
본 개시내용의 RNAi 제제는 약 30개 이하 뉴클레오티드 길이, 예를 들어, 15-30, 15-29, 15-28, 15-27, 15-26, 15-25, 15-24, 15-23, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 18-30, 18-29, 18-28, 18-27, 18-26, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 19-30, 19-29, 19-28, 19-27, 19-26, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-30, 20-29, 20-28, 20-27, 20-26, 20-25, 20-24,20-23, 20-22, 20-21, 21-30, 21-29, 21-28, 21-27, 21-26, 21-25, 21-24, 21-23, 또는 21-22개의 뉴클레오티드 길이인 영역을 갖는 RNA 가닥(안티센스 가닥)을 포함하며, 이 영역은 C9orf72 유전자의 표적 RNA 전사체의 적어도 일부, 예를 들어, C9orf72 인트론에 실질적으로 상보적이다. 특정 양태에서, 본 개시내용의 RNAi 제제는 약 21-23개의 뉴클레오티드 길이인 영역을 갖는 RNA 가닥(안티센스 가닥)을 포함하며, 이 영역은 C9orf72 유전자의 표적 RNA 전사체의 적어도 일부, 예를 들어, C9orf72 인트론에 실질적으로 상보적이다.
특정 양태에서, 본 개시내용의 RNAi 제제는 C9orf72 유전자의 mRNA 전사체의 적어도 일부에 실질적으로 상보적인 적어도 19개의 연속 뉴클레오티드의 영역을 갖는 더 긴 길이, 예를 들면 66개 이하 뉴클레오티드, 예를 들어, 36-66, 26-36, 25-36, 31-60, 22-43, 27-53개의 뉴클레오티드 길이를 포함할 수 있는 RNA 가닥(안티센스 가닥)을 포함한다. 더 긴 길이 안티센스 가닥을 갖는 이러한 RNAi 제제는 바람직하게는 센스 및 안티센스 가닥이 18-30개의 연속 뉴클레오티드의 이중체를 형성하는 20-60개의 뉴클레오티드 길이의 두 번째 RNA 가닥(센스 가닥)을 포함한다.
이러한 RNAi 제제의 사용은 포유동물에서 C9orf72 유전자의 표적 RNA의 표적화된 분해를 가능하게 한다. 따라서, 이들 RNAi 제제를 포함하는 방법 및 조성물은 표적 C9orf72 RNA의 녹다운, 정상 C9orf72 단백질의 감소 및/또는 병원성 헥사뉴클레오티드 반복부 확장으로부터 생성된 병원성 디펩티드 반복부 단백질의 감소에 의해 혜택을 받는 대상체, 예를 들어 C9orf72 근위축성 측삭 경화증/전두측두엽 치매 또는 현팅턴 질환, 예를 들어, C9orf72 확장으로 인한 헌팅턴-유사 증후군과 같은 C9orf72-관련 질환을 갖는 대상체를 치료하는데 유용하다.
하기 상세한 설명은 C9orf72 유전자의 발현을 억제하기 위해 RNAi 제제를 함유하는 조성물을 제조하고 사용하는 방법 뿐만 아니라 유전자 발현의 억제 또는 감소로부터 혜택을 받는 질환 및 장애를 갖는 대상체를 치료하기 위한 조성물 및 방법을 개시한다.
I. 정의
본 개시내용이 보다 쉽게 이해될 수 있도록 하기 위해, 먼저 특정한 용어를 정의한다. 또한, 매개변수의 값 또는 값의 범위가 인용될 때마다, 인용된 값의 중간 값 및 범위도 본 개시내용의 일부로 의도된다는 것을 주지해야 한다.
관사 "a" 및 "an"은 본원에서 관사의 문법적 대상의 하나 또는 하나 초과(즉, 적어도 하나)를 지칭하는데 사용된다. 예를 들면, "요소(an element)"는 하나의 요소 또는 하나 초과의 요소, 예를 들어, 복수의 요소를 의미한다.
용어 "포함하는"은 본원에서 어구 "포함하지만 이에 제한되지 않는"을 의미하고 이와 상호교환적으로 사용된다. 용어 "또는"은 문맥이 명백하게 달리 나타내지 않는 한 용어 "및/또는"을 의미하고 이와 상호교환적으로 사용된다.
용어 "약"은 당업계에서 전형적인 허용 범위 내를 의미하기 위해 본원에서 사용된다. 예를 들면, "약"은 평균에서 약 2 표준 편차로 이해될 수 있다. 특정 양태에서, 약은 ±10%를 의미한다. 특정 양태에서, 약은 ±5%를 의미한다. 일련의 숫자 또는 범위 앞에 약이 있는 경우 "약"이 일련의 또는 범위의 각 숫자를 수식할 수 있는 것으로 이해된다.
숫자 또는 일련의 숫자 앞에 있는 용어 "적어도"는 용어 "적어도"에 인접한 숫자 및 문맥상 분명한 바와 같이 논리적으로 포함될 수 있는 모든 후속 숫자 또는 정수를 포함하는 것으로 이해된다. 예를 들면, 핵산 분자에서 뉴클레오티드의 수는 정수여야 한다. 예를 들면, "21개의 뉴클레오티드 핵산 분자의 적어도 18개의 뉴클레오티드"는 18개, 19개, 20개 또는 21개의 뉴클레오티드가 표시된 특성을 가짐을 의미한다. 일련의 숫자 또는 범위 앞에 적어도가 존재하는 경우, "적어도"는 일련의 또는 범위의 각 숫자를 수식할 수 있는 것으로 이해된다.
본원에 사용된 바와 같이, "이하(no more than)" 또는 "미만(less than)"은 어구에 인접한 값 및 0까지의, 문맥으로부터 논리적인 바와 같이, 논리적 하위 값 또는 정수로서 이해된다. 예를 들면, "2개 이하의 뉴클레오티드"의 돌출부를 갖는 이중체는 2개, 1개 또는 0개의 뉴클레오티드 돌출부를 갖는다. 일련의 숫자 또는 범위 앞에 "이하"가 존재하는 경우 "이하"가 일련 또는 범위의 각 숫자를 수식할 수 있는 것으로 이해된다.
본원에 사용된 바와 같이, 검출 방법은 존재하는 분석물의 양이 방법의 검출 수준 미만인지 결정하는 것을 포함할 수 있다.
센스 또는 안티센스 가닥에 대해 표시된 표적 부위와 뉴클레오티드 서열 사이가 상충하는 경우, 표시된 서열이 우선한다.
화학 구조와 화학명 사이가 상충하는 경우, 화학 구조가 우선한다.
하나 이상의 인용된 요소를 "포함하는(comprising)" 또는 "포함하는(including)" 조성물 또는 방법은 구체적으로 언급되지 않은 다른 요소를 포함할 수 있다. 예를 들면, 단백질을 "포함한다(comprises)" 또는 "포함한다(includes)" 조성물은 단백질을 단독으로 또는 다른 성분과 조합하여 함유할 수 있다. "본질적으로 구성되는"이라는 과도적 어구는 청구 범위가 청구 범위에 인용된 특정 요소 및 청구된 발명의 기본적이고 신규한 특성(들)에 실질적으로 영향을 미치지 않는 요소를 포함하는 것으로 해석되어야 함을 의미한다. 따라서, "본질적으로 구성되는"이라는 용어는 본 발명의 청구범위에서 사용될 때 "포함하는"과 동등한 것으로 해석되도록 의도되지 않는다.
"임의의" 또는 "임의로"는 후속적으로 기술된 사건 또는 상황이 발생하거나 발생하지 않을 수 있으며 설명은 사건 또는 상황이 발생하는 경우 및 사건 또는 상황이 발생하지 않는 경우를 포함함을 의미한다.
"C9orf72-SMCR8 복합체 서브유닛", 구아닌 뉴클레오티드 교환 C9orf72", "염색체 9 개방 판독 프레임 72", "단백질 C9orf72", "DENNL72", "FTDALS1", "ALSFTD" 및 "FTDALS"로도 알려진 "C9orf72" 유전자는 엔도솜 트래피킹의 조절에 관여하는 잘 알려진 단백질인 C9orf72를 암호화하는 유전자를 지칭한다. C9orf72 단백질은 자가포식 및 식작용 전달에 관여하는 Rab 단백질과 상호작용하는 것으로 나타났다. 이 유전자로부터의 전사체의 교호 5' 엑손 사이의 인트론 서열에서 약 2 내지 약 22개 카피에서 약 700 내지 약 1600개 카피로의 GGGGCC 반복부의 확장은 C9orf72 근위축성 측삭 경화증/전두측두엽 치매 및 현팅턴 질환, 예를 들어, C9orf72 확장으로 인한 헌팅턴-유사 증후군과 연관된다. 대체 스플라이싱은 상이한 이소형을 암호화하는 다중 전사체 변이체를 야기한다.
C9orf72의 예시적인 뉴클레오티드 및 아미노산 서열은, 예를 들면, GenBank 수탁번호 NM_001256054.2 (호모 사피엔스 C9orf72, 서열번호 121, 역 상보체 서열번호 125; GenBank 수탁번호: XM_005581570.2 (필리핀 원숭이 C9orf72, 서열번호 122, 역 상보체 서열번호 126); GenBank 수탁번호 NM_001081343.2 (생쥐 C9orf72, 서열번호 123, 역 상보제 서열번호 127); 및 GenBank 수탁번호: NM_001007702.1 (시궁쥐 C9orf72, 서열번호 124, 역 상보체 서열번호 128)에서 찾을 수 있다.
인간 C9orf72의 추가 뉴클레오티드 및 아미노산 서열은, 예를 들면, GenBank 수탁번호 NM_145005.6, 전사체 변이체 1 (서열번호 129, 역 상보체 서열번호 130); 및 NM_018325.5, 전사체 변이체 2 (서열번호 131, 역 상보체 서열번호 132)에서 찾을 수 있다.
C9orf72 유전자를 지닌 인간 염색체 9의 게놈 영역의 뉴클레오티드 서열은, 예를 들면, GenBank에서 이용 가능한 Genome Reference Consortium Human Build 38(인간 게놈 빌드 38 또는 GRCh38이라고도 함)에서 찾을 수 있다. C9orf72 유전자를 지닌 인간 염색체 9의 게놈 영역의 뉴클레오티드 서열은 또한, 예를 들면, 인간 염색체 9의 뉴클레오티드 27546545 내지 27573866에 상응하는 GenBank 수탁번호 NC_000009.12 (서열번호 133, 약 상보체 서열번호 134)에서 찾을 수 있다. 인간 c9orf72 유전자의 뉴클레오티드 서열은, 예를 들면, GenBank 수탁번호 NG_031977.1에서 찾을 수 있다.
C9orf72 서열의 추가 예는 공개적으로 이용 가능한 데이터베이스, 예를 들면, GenBank, OMIM 및 UniProt에서 찾을 수 있다.
C9orf72에 대한 추가 정보는, 예를 들면, www.ncbi.nlm.nih.gov/gene/203228에서 찾을 수 있다. 본원에 사용된 용어 C9orf72는 또한 임상 변이 데이터베이스, 예를 들면, www.ncbi.nlm.nih.gov/clinvar/?term=NM_001256054.2에 제공된 변이체를 포함하여 C9orf72 유전자의 변이를 지칭한다.
상기 GenBank 수탁번호 및 Gene 데이터베이스 번호 각각의 전체 내용은 본 출원을 제출한 날짜를 기준으로 본원에 참고로 포함된다.
본원에서 사용된 "표적 서열"은 1차 전사 산물의 RNA 프로세싱의 산물인 mRNA를 포함하는 C9orf72 유전자의 전사 동안 형성된 mRNA 분자의 뉴클레오티드 서열의 연속 부분을 지칭한다. 하나의 양태에서, 서열의 표적 부분은 적어도 C9orf72 유전자의 전사 동안 형성된 mRNA 분자의 뉴클레오티드 서열의 그 부분에서 또는 근처에서 RNAi-지시된 절단을 위한 기질로 작용하기에 충분할 정도로 길 것이다.
C9orf72 mRNA(표적 C9orf72 RNA)는 C9orf72 유전자로부터 전사된 RNA이다. C9orf72 mRNA는 C9orf72 성숙 mRNA, C9orf72 전구체 mRNA, 또는 이의 임의의 부분(예를 들어, 스플라이싱된 인트론성 영역 또는 대안적으로 스플라이싱된 RNA)을 포함한다. C9orf72 성숙 mRNA는 인트론이 제거(스플라이싱)되고 C9orf72 단백질이 번역된 C9orf72 mRNA이다. C9orf72 전구체 mRNA는 적어도 1개의 인트론, 특히 첫 번째 인트론(인트론 1)이 제거되지 않은 C9orf72 mRNA이다.
C9orf72 단백질은 C9orf72 mRNA로부터 발현된 임의의 단백질을 포함한다. C9orf72 단백질은 C9orf72 성숙 mRNA로부터 발현된 단백질 뿐만 아니라 헥사뉴클레오티드 반복부를 함유하는 C9orf72 RNA로부터의 반복부-관련 비-AUG (AUG) 번역으로부터 야기되는 디펩티드 반복부 단백질(예를 들어, 폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌))을 포함한다.
C9orf72 표적 RNA는 서열번호 1의 다중 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함할 수 있다. C9orf72 표적 RNA는, 예를 들면, 병원성 헥사뉴클레오티드 반복부 확장을 갖는 (예를 들면, 적어도 약 30, 적어도 약 35, 적어도 약 40, 적어도 약 50, 적어도 약 60, 적어도 약 70, 적어도 약 80, 적어도 약 100, 적어도 약 200, 적어도 약 300, 적어도 약 400, 또는 적어도 약 500개 카피의 헥사뉴클레오티드 반복부를 갖는) 것일 수 있다.
표적 서열은 약 15-30개의 뉴클레오티드 길이일 수 있다. 예를 들면, 표적 서열은 약 15-30개의 뉴클레오티드, 15-29, 15-28, 15-27, 15-26, 15-25, 15-24, 15-23, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 18-30, 18-29, 18-28, 18-27, 18-26, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 19-30, 19-29, 19-28, 19-27, 19-26, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-30, 20-29, 20-28, 20-27, 20-26, 20-25, 20-24,20-23, 20-22, 20-21, 21-30, 21-29, 21-28, 21-27, 21-26, 21-25, 21-24, 21-23, 또는 21-22개의 뉴클레오티드 길이일 수 있다. 특정 양태에서, 표적 서열은 19-23개의 뉴클레오티드 길이, 임의로 21-23개의 뉴클레오티드 길이이다. 상기 인용된 범위 및 길이의 중간 범위 및 길이 또한 본 개시내용의 일부로 고려된다.
본원에 사용된 용어 "서열을 포함하는 가닥"은 표준 뉴클레오티드 명명법을 사용하여 언급된 서열에 의해 기술되는 뉴클레오티드의 쇄를 포함하는 올리고뉴클레오티드를 지칭한다.
"G," "C," "A," "T" 및 "U"는 각각 일반적으로 변형되거나 변형되지 않은 뉴클레오티드의 맥락에서 각각 염기로서 구아닌, 시토신, 아데닌, 티미딘 및 우라실을 함유하는 뉴클레오티드를 대표한다. 그러나, 용어 "리보뉴클레오티드" 또는 "뉴클레오티드"는 또한 하기에 추가로 상세히 설명된 바와 같이 변형된 뉴클레오티드 또는 대리 치환 모이어티를 지칭할 수 있음이 이해될 것이다(예를 들어, 참조; 표 11). 숙련가는 구아닌, 시토신, 아데닌, 티미딘 및 우라실이 이러한 대체 모이어티를 지닌 뉴클레오티드를 포함하는 올리고뉴클레오티드의 염기 쌍 형성 특성을 실질적으로 변경하지 않으면서 다른 모이어티에 의해 대체될 수 있음을 잘 알고 있다. 예를 들면, 제한 없이, 염기로서 이노신을 포함하는 뉴클레오티드는 아데닌, 시토신 또는 우라실을 포함하는 뉴클레오티드와 염기 쌍을 이룰 수 있다. 따라서, 우라실, 구아닌 또는 아데닌을 함유하는 뉴클레오티드는 본 개시내용에 특징화된 dsRNA의 뉴클레오티드 서열에서, 예를 들면, 이노신을 함유하는 뉴클레오티드에 의해 대체될 수 있다. 또 다른 예에서, 올리고뉴클레오티드의 어느 곳에서나 아데닌 및 시토신은 각각 구아닌 및 우라실로 대체되어 표적 mRNA와 G-U 동요 염기 쌍을 형성할 수 있다. 이러한 대체 모이어티를 함유하는 서열은 본 개시내용에 특성화된 조성물 및 방법에 적합하다.
본원에서 상호교환적으로 사용되는 용어 "iRNA", "RNAi 제제", "iRNA 제제", "RNA 간섭 제제"는 용어가 본원에 정의된 바와 같은 RNA를 함유하고 RNA-유도 침묵 복합체(RISC) 경로를 통해 RNA 전사체의 표적화된 절단을 매개하는 제제를 지칭한다. RNA 간섭(RNAi)은 mRNA의 서열-특이적 분해를 지시하는 과정이다. RNAi는, 예를 들어, 포유동물 대상체와 같은 대상체 내의 세포에서 C9orf72, C9orf72-관련 전사체 또는 C9orf72-관련 펩티드의 발현을 조절, 예를 들어, 억제한다.
하나의 양태에서, 본 개시내용의 RNAi 제제는 표적 RNA 서열, 예를 들어 C9orf72 표적 mRNA 서열과 상호작용하여 표적 RNA의 절단을 지시하는 단일 가닥 RNAi를 포함한다. 이론에 결부시키고자 함이 없이, 세포 내로 도입된 긴 이중 가닥 RNA는 Dicer로 알려진 유형 III 엔도뉴클레아제에 의해 센스 가닥 및 안티센스 가닥을 포함하는 이중 가닥 짧은 간섭 RNA(siRNA)로 분해되는 것으로 믿어진다(Sharp et al. (2001) Genes Dev. 15:485). 리보뉴클레아제-III-유사 효소인 Dicer는 이러한 dsRNA를 특징적인 2개의 염기 3' 돌출부가 있는 19-23개 염기 쌍 짧은 간섭 RNA로 되게 프로세싱한다(Bernstein, et al., (2001) Nature 409:363). 그후 이러한 siRNA는 RNA-유도 침묵 복합체(RISC)에 통합되어 하나 이상의 헬리카제가 siRNA 이중체를 풀어 상보적 안티센스 가닥이 표적 인식을 가이드할 수 있게 한다(Nykanen, et al., (2001) Cell 107:309). 적절한 표적 mRNA에 결합시, RISC 내의 하나 이상의 엔도뉴클레아제가 표적을 절단하여 침묵을 유도한다(Elbashir, et al., (2001) Genes Dev. 15:188). 따라서, 하나의 측면에서 본 개시내용은 표적 유전자, 즉, C9orf72 유전자의 침묵을 수행하기 위해 RISC 복합체의 형성을 촉진하고 세포 내에서 생성되는 단일 가닥 RNA(ssRNA)(siRNA 이중체의 안티센스 가닥)에 관한 것이다. 따라서, 용어 "siRNA"는 또한 상기 기술된 바와 같은 RNAi를 지칭하기 위해 본원에서 사용된다.
또 다른 양태에서, RNAi 제제는 표적 mRNA를 억제하기 위해 세포 또는 유기체에 도입되는 단일 가닥 RNA일 수 있다. 단일 가닥 RNAi 제제는 RISC 엔도뉴클레아제인 아르고너트(Argonaute) 2에 결합하여 표적 mRNA를 절단한다. 단일 가닥 siRNA는 일반적으로 15-30개의 뉴클레오티드이며 화학적으로 변형된다. 단일 가닥 RNA의 설계 및 시험은 미국 특허 제8,101,348호 및 문헌[Lima et al., (2012) Cell 150:883-894]에 기술되어 있으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다. 본원에 기술된 안티센스 뉴클레오티드 서열 중 어느 것은 본원에 기술된 바와 같은 단일 가닥 siRNA로서 또는 문헌[Lima et al., (2012) Cell 150:883-894]에 기술된 방법에 의해 화학적으로 변형된 대로 사용될 수 있다.
또 다른 양태에서, 본 개시내용의 조성물 및 방법에 사용하기 위한 "RNAi 제제"는 이중 가닥 RNA이고 본원에서 "이중 가닥 RNAi 제제", "이중 가닥 RNA(dsRNA) 분자", "dsRNA 제제" 또는 "dsRNA"라고 한다. 용어 "dsRNA"는 표적 RNA, 즉 C9orf72 유전자에 대해 "센스" 및 "안티센스" 배향을 갖는 것으로 지칭되는 2개의 역평행 및 실질적으로 상보적인 핵산 가닥을 포함하는 이중체 구조를 갖는 리보핵산 분자의 복합체를 지칭한다. 본 개시내용의 일부 양태에서, 이중 가닥 RNA(dsRNA)는 본원에서 RNA 간섭 또는 RNAi로 지칭되는 전사후 유전자-침묵 기전을 통해 표적 RNA, 즉 mRNA의 분해를 유발한다.
본원에 기술된 dsRNA 제제는 안티센스 올리고뉴클레오티드(ASO) 또는 갭머(gapmer) 안티센스 올리고뉴클레오티드(ASO)와 상이할 수 있다(즉, 포함하지 않음).
일반적으로, dsRNA 분자는 리보뉴클레오티드를 포함할 수 있지만, 본원에 상세히 기술된 바와 같이, 각각 또는 양쪽 가닥은 또한 하나 이상의 비-리보뉴클레오티드, 예를 들어, 데옥시리보뉴클레오티드, 변형된 뉴클레오티드를 포함할 수 있다. 또한, 본 명세서에 사용된 바와 같이, "RNAi 제제"는 화학적 변형을 갖는 리보뉴클레오티드를 포함할 수 있으며; RNAi 제제는 다중 뉴클레오티드에 실질적인 변형을 포함할 수 있다. 본원에 사용된 용어 "변형된 뉴클레오티드"는 독립적으로 변형된 당 모이어티, 변형된 뉴클레오티드간 결합 또는 변형된 핵염기를 갖는 뉴클레오티드를 지칭한다. 따라서, 변형된 뉴클레오티드라는 용어는 뉴클레오시드간 결합, 당 모이어티 또는 핵염기에 대한, 예를 들어, 작용 그룹 또는 원자의 치환, 부가 또는 제거를 포함한다. 본 개시내용의 제제에 사용하기에 적합한 변형은 본원에 개시되거나 당업계에 공지된 모든 유형의 변형을 포함한다. siRNA 유형 분자에서 사용되는 바와 같은 임의의 이러한 변형은 본 명세서 및 청구범위의 목적을 위해 "RNAi 제제"에 의해 포함된다.
본 개시내용의 특정 양태에서, RNAi 제제 내에 존재하는 경우 데옥시-뉴클레오티드의 포함은 변형된 뉴클레오티드를 구성하는 것으로 간주될 수 있다.
이중체 영역은 RISC 경로를 통해 원하는 표적 RNA의 특이적 분해를 허용하는 임의의 길이일 수 있으며, 길이는 약 15-36개 염기 쌍, 예를 들면, 약 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 또는 36개 염기 쌍 길이, 예를 들어 약 15-30, 15-29, 15-28, 15-27, 15-26, 15-25, 15-24, 15-23, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 18-30, 18-29, 18-28, 18-27, 18-26, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 19-30, 19-29, 19-28, 19-27, 19-26, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-30, 20-29, 20-28, 20-27, 20-26, 20-25, 20-24,20-23, 20-22, 20-21, 21-30, 21-29, 21-28, 21-27, 21-26, 21-25, 21-24, 21-23, 또는 21-22개 염기 쌍 길이에 이를 수 있다. 특정 양태에서, 이중체 영역은 19-21개 염기 쌍 길이, 예를 들어, 21개 염기 쌍 길이이다. 상기 인용된 범위 및 길이의 중간 범위 및 길이 또한 본 개시내용의 일부로 고려된다.
이중체 구조를 형성하는 두 가닥은 하나의 더 큰 RNA 분자의 상이한 부분일 수 있거나 별도의 RNA 분자일 수 있다. 두 가닥이 하나의 더 큰 분자의 일부이고 따라서 한 가닥의 3'-말단과 각각의 다른 가닥의 5'-말단 사이에 뉴클레오티드의 중단되지 않은 사슬로 연결되어 이중체 구조를 형성하는 경우, 결합 RNA 쇄는 "헤어핀 루프"라고 한다. 헤어핀 루프는 적어도 하나의 쌍을 이루지 않은 뉴클레오티드를 포함할 수 있다. 일부 양태에서, 헤어핀 루프는 적어도 4개, 적어도 5개, 적어도 6개, 적어도 7개, 적어도 8개, 적어도 9개, 적어도 10개, 적어도 20개, 적어도 23개 이상의 쌍을 이루지 않은 뉴클레오티드 또는 dsRNA의 표적 부위에 지시되지 않는 뉴클레오티드를 포함할 수 있다. 일부 양태에서, 헤어핀 루프는 10개 이하의 뉴클레오티드일 수 있다. 일부 양태에서, 헤어핀 루프는 8개 이하의 쌍을 이루지 않은 뉴클레오티드일 수 있다. 일부 양태에서, 헤어핀 루프는 4-10개의 쌍을 이루지 않은 뉴클레오티드일 수 있다. 일부 양태에서, 헤어핀 루프는 4-8개의 뉴클레오티드일 수 있다.
dsRNA의 2개의 실질적으로 상보적인 가닥이 별도의 RNA 분자로 구성되는 경우, 이들 분자는 공유적으로 연결될 필요는 없지만 공유적으로 연결될 수 있다. 특정 양태에서 (본원의 다른 곳에 정의된 특정 다른 구조도 "링커"로 지칭될 수 있는 것으로 주지되지만) 두 가닥이 이중체 구조를 형성하는 한 가닥의 3'-말단과 다른 가닥의 5'-말단 사이에 중단되지 않은 뉴클레오티드 사슬 이외의 수단에 의해 공유적으로 연결되는 경우, 연결 구조를 "링커"라고 한다. RNA 가닥은 동일하거나 다른 수의 뉴클레오티드를 가질 수 있다. 염기 쌍의 최대 수는 dsRNA의 가장 짧은 가닥에 있는 뉴클레오티드 수에서 이중체에 존재하는 돌출부를 뺀 것이다. 이중체 구조에 더하여, RNAi는 하나 이상의 뉴클레오티드 돌출부를 포함할 수 있다. RNAi 제제의 하나의 양태에서, 적어도 하나의 가닥은 적어도 1개의 뉴클레오티드의 3' 돌출부를 포함한다. 또 다른 양태에서, 적어도 하나의 가닥은 적어도 2개의 뉴클레오티드, 예를 들어, 2, 3, 4, 5, 6, 7, 9, 10, 11, 12, 13, 14, 또는 15개의 뉴클레오티드의 3' 돌출부를 포함한다. 다른 양태에서, RNAi 제제의 적어도 하나의 가닥은 적어도 1개의 뉴클레오티드의 5' 돌출부를 포함한다. 특정 양태에서, 적어도 하나의 가닥은 적어도 2개의 뉴클레오티드, 예를 들어, 2, 3, 4, 5, 6, 7, 9, 10, 11, 12, 13, 14, 또는 15개의 뉴클레오티드의 5' 돌출부를 포함한다. 여전히 또 다른 양태에서, RNAi 제제의 한 가닥의 3' 및 5' 말단 모두는 적어도 1개의 뉴클레오티드의 돌출부를 포함한다.
하나의 양태에서, 본 개시내용의 RNAi 제제는 표적 RNA 서열, 예를 들어, C9orf72 표적 mRNA 서열과 상호작용하여 표적 RNA의 절단을 지시하는, 각각의 가닥이 독립적으로 19-23개의 뉴클레오티드를 포함하는 dsRNA이다.
일부 양태에서, 본 발명의 iRNA는 표적 RNA 서열, 예를 들어, C9orf72 표적 mRNA 서열과 상호작용하여 표적 RNA의 절단을 지시하는 24-30개의 뉴클레오티드의 dsRNA이다.
본원에 사용된 용어 "뉴클레오티드 돌출부"는 RNAi 제제, 예를 들어, dsRNA의 이중체 구조로부터 돌출된 적어도 하나의 쌍을 이루지 않은 뉴클레오티드를 지칭한다. 예를 들면, dsRNA의 한 가닥의 3'-말단이 다른 가닥의 5'-말단을 넘어 확장되거나 그 반대의 경우, 뉴클레오티드 돌출부가 있다. dsRNA는 적어도 하나의 뉴클레오티드의 돌출부를 포함할 수 있고; 대안적으로, 돌출부는 적어도 2개의 뉴클레오티드, 적어도 3개의 뉴클레오티드, 적어도 4개의 뉴클레오티드, 적어도 5개의 뉴클레오티드 또는 그 이상을 포함할 수 있다. 뉴클레오티드 돌출부는 데옥시뉴클레오티드/뉴클레오시드를 포함하는 뉴클레오티드/뉴클레오시드 유사체를 포함하거나 이로 이루어질 수 있다. 돌출부(들)는 센스 가닥, 안티센스 가닥 또는 이들의 임의의 조합에 있을 수 있다. 또한, 돌출부의 뉴클레오티드(들)는 dsRNA의 안티센스 또는 센스 가닥의 5'-말단, 3'-말단 또는 양쪽 말단에 존재할 수 있다.
하나의 양태에서, dsRNA의 안티센스 가닥은 3'-말단 또는 5'-말단에 돌출부, 1-10개의 뉴클레오티드, 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 또는 10개의 뉴클레오티드를 갖는다. 하나의 양태에서, dsRNA의 센스 가닥은 3'-말단 또는 5'-말단에 돌출부, 1-10개의 뉴클레오티드, 예를 들어, a 1, 2, 3, 4, 5, 6, 7, 8, 9, 또는 10개의 뉴클레오티드를 갖는다. 또 다른 양태에서, 돌출부에 있는 하나 이상의 뉴클레오티드는 뉴클레오시드 티오포스페이트로 대체된다.
특정 양태에서, dsRNA의 안티센스 가닥은 3'-말단 또는 5'-말단에 돌출부, 1-10개의 뉴클레오티드, 예를 들어, 0-3, 1-3, 2-4, 2-5, 4-10, 5-10, 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 또는 10개의 뉴클레오티드를 갖는다. 하나의 양태에서, dsRNA의 센스 가닥은 3'-말단 또는 5'-말단에 돌출부, 1-10개의 뉴클레오티드, 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 또는 10개의 뉴클레오티드를 갖는다. 또 다른 양태에서, 돌출부에 있는 하나 이상의 뉴클레오티드는 뉴클레오시드 티오포스페이트로 대체된다.
특정 양태에서, 센스 가닥 또는 안티센스 가닥 상의 돌출부는 10개의 뉴클레오티드보다 긴 연장된 길이, 예를 들어, 1-30개의 뉴클레오티드, 2-30개의 뉴클레오티드, 10-30개의 뉴클레오티드, 또는 10-15개의 뉴클레오티드 길이를 포함할 수 있다. 특정 양태에서, 연장된 돌출부는 이중체의 센스 가닥에 있다. 특정 양태에서, 연장된 돌출부는 이중체의 센스 가닥의 3' 말단에 존재한다. 특정 양태에서, 연장된 돌출부는 이중체의 센스 가닥의 5' 말단에 존재한다. 특정 양태에서, 연장된 돌출부는 이중체의 안티센스 가닥에 있다. 특정 양태에서, 연장된 돌출부는 이중체의 안티센스 가닥의 3' 말단에 존재한다. 특정 양태에서, 연장된 돌출부는 이중체의 안티센스 가닥의 5' 말단에 존재한다. 특정 양태에서, 돌출부에 있는 하나 이상의 뉴클레오티드는 뉴클레오시드 티오포스페이트로 대체된다. 특정 양태에서, 돌출부는 생리학적 조건하에서 안정한 헤어핀 구조를 형성할 수 있도록 자가-상보적인 부분을 포함한다.
특정 양태에서, 가닥 중 하나가 열역학적으로 안정화되는 테트라루프 구조를 포함하는 구조를 포함하여, 적어도 하나의 가닥의 적어도 하나의 말단이 이중체 표적화 영역 너머로 연장된다(예를 들어, 참조; 미국 특허 제8,513,207호 및 제8,927,705호 뿐만 아니라 제W02010033225호, 이들 각각의 전체 내용은 본원에 참고로 포함됨). 이러한 구조는 단일 가닥 연장부(분자의 한쪽 또는 양쪽에 있음) 및 이중 가닥 연장부를 포함할 수 있다.
특정 양태에서, 센스 가닥의 3' 말단 및 안티센스 가닥의 5' 말단은 리보뉴클레오티드, 데옥시리보뉴클레오티드 또는 둘 다를 포함하는 폴리뉴클레오티드 서열에 의해 연결되고, 임의로 여기서 폴리뉴클레오티드 서열은 테트라루프 서열을 포함한다. 특정 양태에서, 센스 가닥은 25-35개의 뉴클레오티드 길이이다.
테트라루프는 리보뉴클레오티드, 데옥시리보뉴클레오티드, 변형된 뉴클레오티드 및 이들의 조합을 함유할 수 있다. 전형적으로, 테트라루프는 4 내지 5개의 뉴클레오티드를 갖는다. 일부 양태에서, 루프는 GAAA로서 제시된 서열을 포함한다. 일부 양태에서, 루프(GAAA)의 뉴클레오티드 중 적어도 하나는 뉴클레오티드 변형을 포함한다. 일부 양태에서, 변형된 뉴클레오티드는 2'-변형을 포함한다. 일부 양태에서, 2'-변형은 2'-아미노에틸, 2'-플루오로, 2'-O-메틸, 2'-O-메톡시에틸, 2'-아미노디에톡시메탄올, 2'-adem, 및 2'-데옥시-2'-플루오로-d-아라비노핵산으로 이루어진 그룹으로부터 선택된 변형이다. 일부 양태에서, 루푸의 뉴클레오티드 모두는 변형된다. 일부 양태에서, GAAA 서열의 G는 2'-OH를 포함한다. 일부 양태에서, GAAA 서열의 뉴클레오티드 각각은 2'-O-메틸 변형을 포함한다. 일부 양태에서, GAAA 서열의 A 각각은 2'-OH를 포함하고 GAAA 서열의 G는 2'-O-메틸 변형을 포함한다. 바람직한 양태에서, 일부 양태에서, GAAA 서열의 A 각각은 2'-O-메톡시에틸(MOE) 변형을 포함하고 GAAA 서열의 G는 2'-O-메틸 변형을 포함하거나; GAAA 서열의 A 각각은 2'-adem 변형을 포함하고 GAAA 서열의 G는 2'-O-메틸 변형을 포함한다. 예를 들어, PCT 공개 제WO 2020/206350호를 참조하며, 이의 전체 내용은 본원에 참고로 포함된다.
예시적인 2'adem 변형된 뉴클레오티드가 아래에 나타내어져 있다:

dsRNA와 관련하여 본원에 사용된 용어 "평활" 또는 "평활 말단"은 dsRNA의 주어진 말단 끝에 쌍을 이루지 않은 뉴클레오티드 또는 뉴클레오티드 유사체가 존재하지 않음, 즉 뉴클레오티드 돌출부가 없음을 의미한다. dsRNA의 한쪽 또는 양쪽 말단은 평활할 수 있다. dsRNA의 양쪽 말단이 평활한 경우, dsRNA는 평활 말단이라고 한다. 명확히 하자면, "평활 말단" dsRNA는 양쪽 말단이 평활한 dsRNA, 즉, 분자의 어느 말단에 뉴클레오티드 돌출부가 없다. 가장 자주 이러한 분자는 이의 전체 길이에 걸쳐 이중 가닥이 될 것이다.
용어 "안티센스 가닥" 또는 "가이드 가닥"은 표적 서열, 예를 들어, C9orf72 mRNA에 실질적으로 상보적인 영역을 포함하는 RNAi 제제, 예를 들어, dsRNA의 가닥을 지칭한다.
본원에 사용된 용어 "상보성 영역"은 본원에 정의된 바와 같은 표적 서열, 예를 들어, C9orf72 뉴클레오티드 서열에 실질적으로 상보적인 안티센스 가닥 상의 영역을 지칭한다. 상보성 영역이 표적 서열에 완전히 상보적이지 않은 경우, 미스매치는 분자의 내부 또는 말단 영역에 있을 수 있다. 일반적으로, 가장 허용되는 미스매치는 말단 영역에, 예를 들어, RNAi 제제의 5'- 또는 3'-말단의 5, 4, 3, 또는 2개의 뉴클레오티드 내에 있다. 일부 양태에서, 본 발명의 이중 가닥 RNA 제제는 안티센스 가닥에 뉴클레오티드 미스매치를 포함한다. 일부 양태에서, 본 발명의 이중 가닥 RNA 제제의 안티센스 가닥은 표적 mRNA와 4개 이하의 미스매치를 포함하고, 예를 들어, 안티센스 가닥은 표적 mRNA와 4, 3, 2, 1, 또는 0개 미스매치를 포함한다. 일부 양태에서, 본 발명의 안티센스 가닥 이중 가닥 RNA 제제는 센스 가닥과 4개 이하의 미스매치를 포함하고, 예를 들어, 안티센스 가닥은 센스 가닥과 4, 3, 2, 1, 또는 0개 미스매치를 포함한다. 일부 양태에서, 본 발명의 이중 가닥 RNA 제제는 센스 가닥에 뉴클레오티드 미스매치를 포함한다. 일부 양태에서, 본 발명의 이중 가닥 RNA 제제의 센스 가닥은 안티센스 가닥과 4개 이하의 미스매치를 포함하고, 예를 들어, 센스 가닥은 안티센스 가닥과 4, 3, 2, 1, 또는 0개 미스매치를 포함한다. 일부 양태에서, 뉴클레오티드 미스매치는, 예를 들면, iRNA의 3'-말단으로부터 5, 4, 3개의 뉴클레오티드 내에 있다. 또 다른 양태에서, 뉴클레오티드 미스매치는, 예를 들면, iRNA 제제의 3'-말단 뉴클레오티드에 있다. 일부 양태에서, 미스매치(들)는 시드 영역에 있지 않다.
따라서, 본원에 기술된 바와 같은 RNAi 제제는 표적 서열에 대한 하나 이상의 미스매치를 함유할 수 있다. 하나의 양태에서, 본원에 기술된 바와 같은 RNAi 제제는 3개 이하의 미스매치(즉, 3, 2, 1, 또는 0개의 미스매치)를 함유한다. 하나의 양태에서, 본원에 기술된 바와 같은 RNAi 제제는 2개 이하의 미스매치를 함유한다. 하나의 양태에서, 본원에 기술된 바와 같은 RNAi 제제는 1개 이하의 미스매치를 함유한다. 하나의 양태에서, 본원에 기술된 바와 같은 RNAi 제제는 0개의 미스매치를 함유한다. 특정 양태에서, RNAi 제제의 안티센스 가닥이 표적 서열에 대한 미스매트를 함유하는 경우, 미스매치는 임의로 상보성 영역의 5'- 또는 3'-말단으로부터 마지막 5개의 뉴클레오티드 내로 제한될 수 있다. 예를 들면, 이러한 양태에서, 23개의 뉴클레오티드 RNAi 제제의 경우, C9orf72 유전자의 영역에 상보적인 가닥은 일반적으로 중심 13개의 뉴클레오티드 내에 임의의 미스매치를 함유하지 않는다. 본원에 기술된 방법 또는 당업계에 공지된 방법을 사용하여 표적 서열에 대한 미스매치를 함유하는 RNAi 제제가 C9orf72 유전자의 발현을 억제하는데 효과적인지 여부를 결정할 수 있다. 특히 C9orf72 유전자의 특정 상보성 영역이 집단 내에서 다형성 서열 변이를 갖는 것으로 알려져 있다면, C9orf72 유전자의 발현을 억제하는데 있어서의 미스매치를 갖는 RNAi 제제의 효능을 고려하는 것이 중요하다.
본원에 사용된 바와 같이, "실질적으로 모든 뉴클레오티드가 변형된다"는 전적으로 변형된 것은 아니지만 대부분 변형되고 5, 4, 3, 2 또는 1개 이하의 비변형 뉴클레오티드를 포함할 수 있다.
본원에 사용된 용어 "센스 가닥" 또는 "운송 가닥(passenger strand)"은 그 용어가 본원에 정의된 바와 같이 안티센스 가닥의 영역에 실질적으로 상보적인 영역을 포함하는 RNAi 제제의 가닥을 지칭한다.
본원에 사용된 용어 "절단 영역"은 절단 부위에 바로 인접하여 위치하는 영역을 지칭한다. 절단 부위는 절단이 일어나는 표적 상의 부위이다. 일부 양태에서, 절단 영역은 절단 부위의 말단 및 절단 부위에 바로 인접한 3개의 염기를 포함한다. 일부 양태에서, 절단 영역은 절단 부위의 말단 및 절단 부위에 바로 인접한 2개의 염기를 포함한다. 일부 양태에서, 절단 부위는 안티센스 가닥의 뉴클레오티드 10 및 11에 의해 결합된 부위에서 특이적으로 발생하고, 절단 영역은 뉴클레오티드 11, 12 및 13을 포함한다.
본원에 사용된 바와 같이, 달리 나타내지 않는 한, 제2 뉴클레오티드 서열과 관련하여 제1 뉴클레오티드 서열을 기술하기 위해 사용될 때 용어 "상보적"은 숙련가에 의해 이해되는 바와 같이, 제2 뉴클레오티드 서열을 포함하는 올리고뉴클레오티드 또는 폴리뉴클레오티드와 특정 조건 하에서 혼성화하고 이중체 구조를 형성하는 제1 뉴클레오티드 서열을 포함하는 올리고뉴클레오티드 또는 폴리뉴클레오티드의 능력을 지칭한다. 이러한 조건은, 예를 들면, 엄격한 조건일 수 있으며, 여기서 엄격한 조건은 다음을 포함할 수 있다: 400mM NaCl, 40mM PIPES pH 6.4, 1mM EDTA, 50℃ 또는 70℃에서 12-16시간에 이은 세척(예를 들어, 참조; "Molecular Cloning: A Laboratory Manual, Sambrook, et al. (1989) Cold Spring Harbor Laboratory Press). 유기체 내부에서 직면할 수 있는 생리학적으로 관련된 조건과 같은 다른 조건이 적용될 수 있다. 숙련가는 혼성화된 뉴클레오티드의 궁극적인 적용에 따라 2개의 서열의 상보성 시험에 가장 적합한 조건 세트를 결정할 수 있을 것이다.
RNAi 제제 내의, 예를 들어, 본원에 기술된 바와 같은 dsRNA 내의 상보적 서열은 하나 또는 둘 모두의 뉴클레오티드 서열의 전체 길이에 걸쳐 제1 뉴클레오티드 서열을 포함하는 올리고뉴클레오티드 또는 폴리뉴클레오티드를 제2 뉴클레오티드 서열을 포함하는 올리고뉴클레오티드 또는 폴리뉴클레오티드로 염기쌍을 형성하는 것을 포함한다. 이러한 서열은 본원에서 서로에 대해 "완전히 상보적인" 것으로 지칭될 수 있다. 그러나, 본원에서 첫 번째 서열이 두 번째 서열과 관련하여 "실질적으로 상보적"인 것으로 언급되는 경우, 두 서열은 완전히 상보적일 수 있거나, 이들은 이들의 궁극적인 용도, 예를 들어 RISC 경로를 통한 유전자 발현의 억제와 가장 관련이 있는 조건하에서 혼성화할 수 있는 능력을 유지하면서 최대 30개 염기쌍 이중체에 대한 혼성화시 하나 이상, 그러나 일반적으로 5, 4, 3 또는 2개 이하의 미스매치 염기쌍을 형성할 수 있다. 그러나, 2개의 올리고뉴클레오티드가 혼성화시 하나 이상의 단일 가닥 돌출부를 형성하도록 설계된 경우, 이러한 돌출부는 상보성 결정과 관련하여 미스매치로서 간주되지 않는다. 예를 들면, 하나의 올리고뉴클레오티드 21개의 뉴클레오티드 길이 및 또 다른 올리고뉴클레오티드 23개의 뉴클레오티드 길이를 포함하는 dsRNA(여기서 더 긴 올리고뉴클레오티드는 더 짧은 올리고뉴클레오티드에 완전히 상보적인 21개의 뉴클레오티드의 서열을 포함한다)는 본원에 기술된 목적을 위해 "완전히 상보적"인 것으로 지칭될 수 있다.
본원에 사용된 "상보적" 서열은 또한, 혼성화하는 이들의 능력에 대한 상기 요건이 이행되는 한, 비-왓슨-크릭 염기쌍 또는 비-천연 및 변형 뉴클레오티드로부터 형성된 염기쌍을 포함하거나 이들로부터 완전히 형성될 수 있다. 이러한 비-왓슨-크릭 염기쌍은 G:U Wobble 또는 Hoogstein 염기쌍을 포함하지만, 이에 제한되지 않는다.
본원에서 용어 "상보적인", "완전히 상보적인" 및 "실질적으로 상보적인"은 이들의 사용 맥락으로부터 이해되는 바와 같이 dsRNA의 센스 가닥과 안티센스 가닥 사이 또는 RNAi 제제의 안티센스 가닥과 표적 서열 사이의 염기 매칭과 관련하여 사용될 수 있다.
본원에서 사용되는 바와 같이, 메신저 RNA(mRNA)의 "적어도 일부에 실질적으로 상보적인" 폴리뉴클레오티드는 관심 mRNA(예를 들어, C9orf72를 암호화하는 mRNA)의 인접 부분에 실질적으로 상보적인 폴리뉴클레오티드를 지칭한다. 예를 들면, 폴리뉴클레오티드는 서열이 C9orf72를 암호화하는 mRNA의 중단되지 않은 부분에 실질적으로 상보적인 경우 C9orf72 mRNA의 적어도 일부에 상보적이다.
따라서, 일부 양태에서, 본원에 개시된 안티센스 폴리뉴클레오티드는 표적 C9orf72 서열에 완전히 상보적이다. 다른 양태에서, 본원에 개시된 안티센스 폴리뉴클레오티드는 표적 C9orf72 서열에 실질적으로 상보적이며, 서열번호 121-124 및 133 중 어느 하나의 뉴클레오티드 서열의 등가 서열에 대해 이의 전체 길이에 걸쳐서 적어도 80% 상보적인, 예를 들어 약 85%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 또는 약 99% 상보적인 연속 뉴클레오티드 서열을 포함한다.
일부 양태에서, 본원에 개시된 안티센스 폴리뉴클레오티드는 표적 C9orf72 서열의 단편에 실질적으로 상보적이며, 서열번호 121의 뉴클레오티드 1594-1616, 802-824, 239-261, 1308-1330, 233-255, 1595-1617, 240-262, 1532-1554, 237-259, 3268-3290, 806-828, 1620-1642, 526-548, 1169-1191, 1266-1288, 1247-1269, 586-608, 1257-1279, 및 400-422의 그룹으로부터 선택된 서열번호 121의 단편에 대해 이의 전체 길이에 걸쳐서 적어도 80% 상보적인, 예를 들어 약 85%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 또는 약 99% 상보적인 연속 뉴클레오티드 서열을 포함한다.
일부 양태에서, 본원에 개시된 안티센스 폴리뉴클레오티드는 표적 C9orf72 서열의 단편에 실질적으로 상보적이며, 서열번호 133의 단편, 예를 들어 뉴클레오티드 200-290, 예를 들어, 뉴클레오티드 200-290, 200-280, 200-270, 200-260, 200-250, 200-240, 200-230, 200-225, 210-290, 210-280, 210-270, 210-260, 210-250, 210-240, 210-235, 220-290, 220-280, 220-270, 220-260, 220-250, 220-245, 225-250, 225-245, 230-290, 230-280, 230-270, 230-260, 230-255, 235-260, 240-265, 240-290, 240-280, 240-270, 240-265, 250-290, 250-280, 250-275, 260-290, 260-285)에 대해 이의 전체 길이에 걸쳐서 적어도 80% 상보적인, 예를 들어 약 85%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 또는 약 99% 상보적인 연속 뉴클레오티드 서열을 포함한다. 상기 인용된 범위의 중간 범위도 본 개시내용의 일부로 고려된다.
다른 양태에서, 본원에 개시된 안티센스 폴리뉴클레오티드는 표적 C9orf72 서열에 실질적으로 상보적이며, 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나의 센스 가닥 뉴클레오티드 서열 중 어느 하나 또는 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나의 센스 가닥 뉴클레오티드 서열 중 어느 하나의 단편에 대해 이의 전체 길이에 걸쳐서 적어도 80% 상보적인, 예를 들어 약 85%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 약 99% 또는 100% 상보적인 연속 뉴클레오티드 서열을 포함한다.
하나의 양태에서, 본 개시내용의 RNAi 제제는 차례로 표적 C9orf72 서열과 동일한 안티센스 폴리뉴클레오티드에 실질적으로 상보적인 센스 가닥을 포함하며, 여기서 센스 가닥 뉴클레오티드는 서열번호 125-128 및 134의 뉴클레오티드 서열의 등가 영역에 대해 이의 전체 길이에 걸쳐서 적어도 80% 상보적인, 예를 들어 약 85%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 약 99% 또는 100% 상보적인 연속 뉴클레오티드 서열을 포함한다.
일부 양태에서, 본 발명의 iRNA는 차례로 표적 C9orf72 서열에 상보적인 안티센스 폴리뉴클레오티드에 실질적으로 상보적인 센스 가닥을 포함하며, 여기서 센스 가닥 뉴클레오티드는 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나, 또는 표 10A, 10B, 12-25, 19 또는 20 중 어느 하나의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나의 단편에 대해 이의 전체 길이에 걸쳐서 적어도 80% 상보적인, 예를 들어 약 85%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 약 99% 또는 100% 상보적인 연속 뉴클레오티드 서열을 포함한다.
특정 양태에서, 센스 및 안티센스 가닥은 이중체 AD-348904.1, AD-348136.1, AD-347612.1, AD-348639.1, AD-347606.1, AD-348905.1, AD-347613.1, AD-348842.1, AD-347610.1, AD-350329.1, AD-348140.1, AD-348930.1, AD-347863.1, AD-348500.1, AD-348597.1, AD-348578.1, AD-347923.1, AD-348588.1, AD-347773.1, 및 AD-348136.1 중 어느 하나로부터 선택된다.
특정 양태에서, 센스 및 안티센스 가닥은 이중체 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1 중 어느 하나로부터 선택된다.
하나의 양태에서, C9orf72 유전자의 발현의 적어도 부분적인 억제는, C9orf72 유전자가 전사되는 첫 번째 세포 또는 세포 그룹에서 단리되거나 검출될 수 있고 첫 번째 세포 또는 세포 그룹과 실질적으로 동일하지만 그렇게 처리되지 않은 두 번째 세포 또는 세포 그룹(대조군 세포)과 비교하여 C9orf72 유전자의 발현이 억제되도록 처리된, C9orf72 mRNA, 예를 들어, 센스 mRNA, 안티센스 mRNA, 총 C9orf72 mRNA, 센스 C9orf72 반복부-함유 mRNA, 및/또는 안티센스 C9orf72 반복부-함유 mRNA의 양의 감소에 의해 평가된다. 억제 정도는 다음과 같이 표현될 수 있다:

본원에 사용된 어구 "세포를 RNAi 제제, 예를 들어 dsRNA와 접촉시키는 것"은 임의의 가능한 수단에 의해 세포를 접촉시키는 것을 포함한다. 세포를 RNAi 제제와 접촉시키는 것은 시험관내에서 세포를 RNAi 제제와 접촉시키거나 생체내에서 세포를 RNAi 제제와 접촉시키는 것을 포함한다. 접촉은 직접 또는 간접적으로 수행될 수 있다. 따라서, 예를 들면, RNAi 제제는 방법을 수행하는 개인에 의해 세포와 물리적 접촉에 놓일 수 있거나, 대안적으로 RNAi 제제는 세포와 후속적으로 접촉하도록 허용하거나 야기하는 상황에 놓일 수 있다.
시험관내에서 세포를 접촉시키는 것은, 예를 들면, 세포를 RNAi 제제와 함께 배양함으로써 수행될 수 있다. 생체내에서 세포를 접촉시키는 것은, 예를 들면, RNAi 제제를 세포가 위치한 조직 내로 또는 그 근처에 주입함으로써, 또는 RNAi 제제를 다른 영역, 예를 들어 중추신경계(CNS)에, 임의로 척수강내, 유리체내 또는 기타 주사를 통해, 또는 제제가 후속적으로 접촉하고자 하는 세포가 위치하는 조직에 도달하도록 혈류 또는 피하 공간에 주사함으로써 수행될 수 있다. 예를 들면, RNAi 제제는 리간드, 예를 들어, 하기에 기술되고, 예를 들어, 본원에 참고로 포함된 제PCT/US2019/031170호에 추가로 상세하게 기술되어 있는 친유성 모이어티 또는 모이어티들을 함유하거나 이에 결합될 수 있으며, 이것은 관심 부위, 예를 들어, CNS에서 RNAi 제제를 지시하거나 달리 안정화시킨다. 접촉시키는 시험관내 및 생체내 방법의 조합이 또한 가능하다. 예를 들면, 세포는 또한 시험관내에서 RNAi 제제와 접촉되고 후속적으로 대상체로 이식될 수 있다.
하나의 양태에서, 세포를 RNAi 제제와 접촉시키는 것은 세포 내로의 섭취 또는 흡수를 촉진하거나 수행함으로써 "RNAi 제제를 세포 내로 도입" 또는 "전달하는 것"을 포함한다. RNAi 제제의 흡수 또는 섭취는 비보조 확산 또는 활성 세포 과정을 통해, 또는 보조 제제 또는 장치에 의해 발생할 수 있다. RNAi 제제를 세포 내로 도입하는 것은 시험관내 또는 생체내일 수 있다. 예를 들면, 생체내 도입을 위해, RNAi 제제는 조직 부위에 주사되거나 전신적으로 투여될 수 있다. 세포로의 시험관내 도입은 전기천공법 및 리포펙션과 같은 당업계에 공지된 방법을 포함한다. 추가 접근법은 본원에서 이하에 기술되거나 당업계에 공지되어 있다.
용어 "친유성" 또는 "친유성 모이어티"는 광범위하게는 지질에 대한 친화성을 갖는 임의의 화합물 또는 화학적 모이어티를 지칭한다. 친유성 모이어티의 친유성을 특성화하는 한 가지 방법은 옥탄올-물 분배 계수, logKow이며, 여기서 Kow는 평형 상태에서 2상 시스템의 수성 상의 농도에 대한 옥탄올 상의 화학물질 농도의 비율이다. 옥탄올-물 분배 계수는 물질의 실험실 측정된 특성이다. 그러나, 이것은 또한 제1-원칙 또는 경험적 방법을 사용하여 계산된 화학 물질의 구조적 구성 요소에 기인한 계수를 사용하여 예측할 수 있다(예를 들면, 참조; Tetko et al., J. Chem . Inf . Comput . Sci . 41:1407-21 (2001), 이것은 전문이 본원에 참고로 포함됨). 이것은 물보다 비수성 또는 유성 환경을 선호하는 물질의 경향에 대한 열역학적 척도를 제공한다(즉, 친수성/친유성 균형). 원칙적으로, 화학 물질은 logKow가 0을 초과할 때 특성에 있어서 친유성이다. 전형적으로, 친유성 모이어티는 1 초과, 1.5 초과, 2 초과, 3 초과, 4 초과, 5 초과, 또는 10 초과의 logKow를 보유한다. 예를 들어, 6-아미노 헥산올의 logKow는, 예를 들어 대략 0.7인 것으로 예측된다. 동일한 방법을 사용하여, 콜레스테릴 N-(헥산-6-올) 카바메이트의 logKow는 10.7인 것으로 예측된다.
분자의 친유성은 이것이 운반하는 작용 그룹에 따라 변할 수 있다. 예를 들어, 친유성 모이어티의 말단에 하이드록실 그룹 또는 아민 그룹을 첨가하면 친유성 모이어티의 분배 계수(예를 들어, logKow) 값을 증가 또는 감소할 수 있다.
대안적으로, 하나 이상의 친유성 모이어티에 접합된 이중 가닥 RNAi 제제의 소수성은 이의 단백질 결합 특성에 의해 측정될 수 있다. 예를 들어, 특정 양태에서, 이중 가닥 RNAi 제제의 혈장 단백질 결합 분석에서 비결합 분획은 이중 가닥 RNAi 제제의 상대적 소수성과 양의 상관관계가 있는 것으로 결정될 수 있으며, 이는 이중 가닥 RNAi 제제의 침묵 활성과 양의 상관관계가 있을 수 있다.
하나의 양태에서, 결정된 혈장 단백질 결합 분석은 인간 혈청 알부민 단백질을 사용한 전기영동 이동성 변화 분석(EMSA)이다. 이러한 결합 분석의 예시적인 프로토콜은, 예를 들어, 제PCT/US2019/031170호에 상세히 예시되어 있다. 결합 분석에서 비결합 siRNA의 분획에 의해 측정된 이중 가닥 RNAi 제제의 소수성은 향상된 생체내 siRNA의 전달을 위해 0.15를 초과, 0.2를 초과, 0.25를 초과, 0.3을 초과, 0.35를 초과, 0.4를 초과, 0.45를 초과 또는 0.5를 초과한다.
따라서, 이중 가닥 RNAi 제제의 내부 위치(들)에 친유성 모이어티를 접합하는 것은 siRNA의 향상된 생체내 전달을 위한 최적의 소수성을 제공한다.
용어 "지질 나노입자" 또는 "LNP"는 약제학적 활성 분자, 예를 들어 핵산 분자, 예를 들어, RNAi 제제가 전사되는 RNAi 제제 또는 플라스미드를 캡슐화하는 지질층을 포함하는 소포이다. LNP는, 예를 들면, 미국 특허 제6,858,225호, 제6,815,432호, 제8,158,601호, 및 제8,058,069호에 기술되어 있으며, 이의 전체 내용은 본원에 참고로 포함된다.
본원에 사용된 바와 같이, "대상체"는 영장류(예를 들어, 인간, 비인간 영장류, 예를 들어, 원숭이 및 침팬지), 또는 비영장류(예를 들어, 래트 또는 마우스)를 포함하는 포유동물과 같은 동물이다. 바람직한 양태에서, 대상체는 인간, 예를 들어, C9orf72 발현의 감소로부터 혜택을 받는 질환, 장애 또는 상태에 대해 치료 또는 평가되는 인간; 표적 C9orf72 RNA 수준의 감소로부터 혜택을 받는 질환, 장애 또는 상태에 대한 위험이 있는 인간; C9orf72 발현의 감소로부터 혜택을 받는 질환, 장애 또는 상태를 가진 인간; 또는 본원에 기술된 바와 같은 C9orf72 발현의 감소로부터 혜택을 받는 질환, 장애 또는 상태에 대해 치료받는 인간이다. 일부 양태에서, 대상체는 여성 인간이다. 다른 양태에서, 대상체는 남성 인간이다. 하나의 양태에서, 대상체는 성인 대상체이다. 하나의 양태에서, 대상체는 소아 대상체이다. 또 다른 양태에서, 대상체는 청소년 대상체, 즉 20세 미만의 대상체이다.
본원에 사용된 용어 "치료하는" 또는 "치료"는 C9orf72 유전자 발현 또는 C9orf72 단백질 생산과 관련된 하나 이상의 징후 또는 증상, 예를 들어, C9orf72-관련 질환, 예를 들어 C9orf72-관련 질환의 완화 또는 개선을 포함하지만 이에 제한되지 않는 유익한 또는 원하는 결과를 지칭한다. "치료"는 또한 치료의 부재하에서 예상되는 생존과 비교하여 생존을 연장하는 것을 의미할 수 있다.
대상체에서 C9orf72 수준 또는 질환 마커 또는 증상과 관련하여 "낮은(lower)"이라는 용어는 이러한 수준의 통계적으로 유의한 감소를 지칭한다. 감소는, 예를 들면, 적어도 10%, 15%, 20%, 25%, 30%, %, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 또는 그 이상일 수 있다. 특정 양태에서, 감소는 적어도 20%이다. 특정 양태에서, 감소는 질환 마커, 예를 들어, 센스- 또는 안티센스-함유 병소의 수준 및/또는 비정상적인 디펩티드 반복부 단백질의 수준의 적어도 50%, 예를 들어, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 또는 그 이상의 감소이다. 일부 양태에서, 감소는 질환 마커, 예를 들어, C9orf72 단백질 및/또는 유전자 발현 수준의 50% 이하, 예를 들어, 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 또는 5% 이하이다. 대상체의 C9orf72 수준의 맥락에서 "낮은"은 바람직하게는 이러한 장애가 없는 개인에 대해 정상 범위 내로 허용되는 수준까지 내려간다. 특정 양태에서, "낮은"은 질환을 앓고 있는 대상체에 대한 마커 또는 증상의 수준과 개인에 대해 정상 범위 내에서 허용되는 수준 사이의 차이의 감소, 예를 들어, 비만한 개인과 정상 범위 내에서 허용되는 체중을 가진 개인 사이의 체중 감소 수준이다.
본원에 사용된 "예방" 또는 "예방하는"은, C9orf72 유전자의 발현 또는 C9orf72 단백질 생성의 감소로부터 혜택을 받는 질환, 장애 또는 이의 상태와 관련하여 사용될 때, 대상체가 이러한 질환, 장애 또는 상태와 관련된 증상, 예를 들어, C9orf72-관련 질환의 증상을 발병할 가능성의 감소를 지칭한다. 질환, 장애 또는 상태의 발병 실패 또는 이러한 질환, 장애 또는 상태와 관련된 증상의 발달 감소(예를 들어, 해당 질환 또는 장애에 대해 임상적으로 허용되는 규모에서 적어도 약 10%까지) 또는 지연된(예를 들어, 수 일, 수 주, 수 개월 또는 수 년) 증상의 발현이 효과적인 예방으로 간주된다.
본원에 사용된 용어 "C9orf72-관련 질환" 또는 "C9orf72-관련 장애"는 C9orf72의 발현 및/또는 활성의 감소로부터 혜택을 받는 임의의 질환 또는 장애를 포함한다. 예시적인 C9orf72-관련 질환은 대상체가 C9orf72 유전자의 엑손 1a와 1b 사이의 인트론에 헥사뉴클레오티드 반복부(GGGCC) 확장을 지닌 질환, 예를 들어, 근위축성 측삭 경화증(ALS) 또는 전두측두엽 치매(FTD) 및 C9orf72 확장으로 인한 헌팅턴-유사 증후군을 포함한다.
정상 G4C2 반복부는 ~25 단위 이하이고, 고침투성 질병 대립유전자는 전형적으로 4,000 단위 이상에 이르는 ~60 반복부 단위 이상이고; 드물게 47 내지 60개 반복부가 가족의 질병으로 분리된다. 반복부-프라이밍된 PCR 분석은 전형적으로 더 작은 확장(<80)을 검출하는데 사용되지만 더 큰 반복부의 크기를 정확하게 결정하려면 길이 추정치를 제공하는 다른 기술(예를 들어, 서던 블롯 혼성화)이 필요하다.
C9orf72 유전자의 인트론에 GGGGCC (또는 G4C2) 헥사뉴클레오티드 확장을 갖는 대상체는 동일한 계열에서도 근위축성 측삭 경화증(ALS) 또는 전두측두엽 치매(FTD)로 나타날 수 있으며, 따라서, 이러한 확장과 관련된 신경변성을 본원에서는 "C9orf72 근위축성 측삭 경화증/전두측두엽 치매" 또는 C9orf72 ALS/FTD"라고 한다. 이것은 상염색체 우성 질환이며, ALS 가족의 약 3분의 1을 차지하고 ALS 클리닉에서 산발적인 사례의 5-10%를 차지하는 가장 흔한 형태의 가족성 ALS이다. 이것은 또한 가족성 FTD의 약 1/4을 설명하는 FTD의 일반적인 원인이다. 증상 발현 연령은 30세에서 70세 사이이며 평균 발병 연령은 50대 후반이다. C9orf72-매개 ALS는 가장 자주 전형적인 ALS와 유사하고, 구근 또는 사지 발병일 수 있고, 빠르게 진행될 수 있고(항상 그런 것은 아니지만) 나중에 인지 증상과 연관될 수 있다. 따라서, C9orf72-매개 ALS는 임의의 ALS 환자에서와 마찬가지로 평가되고 치료된다. C9orf72-매개 FTD의 패턴은 가장 일반적으로 탈억제, 무관심 및 실행 기능 장애를 포함한 모든 범위의 행동 및 인지 증상을 갖는 행동 변이형 FTD이다. 덜 일반적으로, C9orf72-매개 FTD는 어의 변이형 원발성 진행성 실어증(semantic variant primary progressive aphasia)(PPA) 또는 비달변성 변이형 PPA를 나타내며, 매우 드물게, 피질기저 증후군, 진행성 핵상 마비 또는 HD 유사-증후군과 유사할 수 있다. 때때로 파킨슨병 특징이 C9orf72-매개 ALS 또는 FTD에서 보여진다.
대상체는 행동의 점진적인 변화, 실행 기능 장애 및/또는 언어 장애를 특징으로 하는 전두측두엽 변성(frontotemporal lobar degeneration)(FTLD)을 나타낼 수 있다. 세 가지 FTLD 임상 증후군 중에서, 행동 변이형 FTD(bvFTD)가 가장 흔하지만 독점적인 것은 아니다. 이것은 뇌 MRI에서의 우세한 전두엽 위축과 함께 진행성 행동 장애와 실행 기능의 감소를 특징으로 한다. 전체 ALS 표현형에 대한 기준을 충족하거나 충족하지 않을 수 있는 상부 또는 하부 운동 뉴런 기능장애(또는 둘 다)를 포함하는 운동 뉴런 질환이 또한 존재할 수 있다. C9orf72-관련 bvFTD를 가진 많은 개인에게 나타나는 어느 정도의 파킨슨증은 일반적으로 떨림이 없는 무운동-강직 유형이며 레보도파에 반응이 없다.
현팅턴병-유사 증후군(HD-유사 증후군, 또는 HDL 증후군)은 전형적으로 무도병, 인지 쇠퇴 또는 치매 및 행동적 또는 정신과적 문제가 복합적으로 나타난다는 점에서 현팅턴병(HD)과 매우 유사한 유전성 신경퇴행성 질환의 계열이다.
C9orf72 확장으로 인한 헌팅턴병-유사 증후군을 갖는 대상체는 근긴장이상, 무도증, 간대성 근경련, 떨림 및 강직을 포함한 운동 장애를 갖는 것으로 특징지어진다. 관련된 특징으로는 또한 인지 및 기억 장애, 초기 정신 장애 및 행동 문제가 있다. 발병시 평균 연령은 약 43세(8-60세 범위)이다. 초기의 정신과 및 행동 문제(우울증, 무관심, 강박 행동 및 정신병 포함)가 일반적이다. 인지 증상은 실행 기능 장애로 나타난다. 운동 장애가 두드러진다: 파킨슨병 특징(parkinsonian feature) 및 삼지골신근돌기염 특징(pyramidal feature)이 또한 나타날 수 있다. 본원에 사용된 "치료학적 유효량"은 C9orf72-관련 질환을 갖는 대상체에게 투여될 때 (예를 들어, 기존 질환 또는 질환의 하나 이상의 증상을 감소, 개선 또는 유지함으로써) 질병의 치료를 수행하기에 충분한 RNAi 제제의 양을 포함하는 것으로 의도된다. "치료학적 유효량"은 RNAi 제제, 제제가 투여되는 방법, 질환 및 이의 중증도 및 병력, 연령, 체중, 가족력, 유전적 구성, 선행 또는 수반하는 치료의 유형(있는 경우) 및 치료될 대상의 기타 개별적인 특성에 따라 달라질 수 있다.
본원에 사용된 "예방학적 유효량"은 C9orf72-관련 장애를 갖는 대상체에게 투여될 때 질환 또는 질환의 하나 이상의 증상을 예방하거나 개선하기에 충분한 RNAi 제제의 양을 포함하도록 의도된다. 질환을 개선하는 것은 질환의 경과를 늦추거나 나중에 발병하는 질환의 중증도를 감소시키는 것을 포함한다. "예방학적 유효량"은 RNAi 제제, 제제가 투여되는 방식, 질환의 위험 정도, 병력, 연령, 체중, 가족력, 유전적 구성, 선행 또는 수반되는 치료의 유형(있는 경우), 및 치료할 환자의 기타 개별적인 특성에 따라 달라질 수 있다.
"치료학적 유효량" 또는 "예방학적 유효량"은 또한 임의의 치료에 적용할 수 있는 합리적인 이익/위험 비율로 일부 원하는 국소 또는 전신 효과를 생성하는 RNAi 제제의 양을 포함한다. 본 개시내용의 방법에 사용된 RNAi 제제는 이러한 치료에 적용할 수 있는 합리적인 이익/위험 비율을 생성하기에 충분한 양으로 투여될 수 있다.
어구 "약제학적으로 허용되는"은 건전한 의학적 판단의 범위 내에서 과도한 독성, 자극, 알레르기 반응, 또는 기타 문제 또는 합병증 없이 인간 대상체 및 동물 대상체의 조직과 접촉하여 사용하기에 적합하고 합당한 이익/위험 비율에 상응하는 화합물, 물질, 조성물 또는 투여 형태를 지칭하기 위해 본원에서 사용된다.
본원에 사용된 어구 "약제학적으로 허용되는 담체"는 대상 화합물을 신체의 한 기관 또는 일부에서 신체의 다른 기관 또는 일부로 운반하거나 수송하는데 관여하는 약제학적으로 허용되는 물질, 조성물 또는 비히클, 예를 들어 액체 또는 고체 충전제, 희석제, 부형제, 제조 보조제(예를 들어, 윤활제, 활석 마그네슘, 칼슘 또는 아연 스테아레이트, 또는 스테아르산), 또는 용매 캡슐화 물질을 의미한다. 각 담체는 제형의 다른 성분과 사용성이고 치료되는 대상체에 해를 끼치지 않는다는 의미에서 "허용"되어야 한다. 약제학적으로 허용되는 담체로서 역할을 할 수 있는 물질의 몇 가지 예는 다음을 포함한다: (1) 락토스, 글루코스 및 수크로스와 같은 당; (2) 옥수수 전분 및 감자 전분과 같은 전분; (3) 셀룰로스, 및 나트륨 카복시메틸셀룰로스, 에틸 셀룰로스 및 셀룰로스 아세테이트와 같은 이의 유도체; (4) 분말상 트라가칸트; (5) 맥아; (6) 젤라틴; (7) 마그네슘 상태, 나트륨 라우릴 설페이트 및 활석과 같은 윤활제; (8) 코코아 버터 및 좌약 왁스와 같은 부형제; (9) 땅콩유, 면실유, 홍화유, 호마유, 올리브유, 옥수수유 및 대두유와 같은 오일; (10) 프로필렌 글리콜과 같은 글리콜; (11) 글리세린, 소르비톨, 만니톨 및 폴리에틸렌 글리콜과 같은 폴리올; (12) 에틸 올레이트 및 에틸 라우레이트와 같은 에스테르; (13) 한천; (14) 수산화마그네슘 및 수산화알루미늄과 같은 완충제; (15) 알긴산; (16) 피로겐-비함유 물; (17) 등장성 염수; (18) 링거액; (19) 에틸 알코올; (20) pH 완충 용액; (21) 폴리에스테르, 폴리카보네이트 또는 폴리무수물; (22) 폴리펩티드 및 아미노산과 같은 증량제, (23) 혈청 알부민, HDL 및 LDL과 같은 혈청 성분; 및 (22) 약제학적 제형에 사용되는 기타 무독성 상용성 물질.
본원에 사용된 용어 "샘플"은 대상체로부터 단리된 유사한 체액, 세포 또는 조직의 집합체 뿐만 아니라 대상체 내에 존재하는 체액, 세포 또는 조직을 포함한다. 생물학적 유체의 예는 혈액, 혈청 및 장액, 혈장, 뇌척수액, 안액, 림프, 소변, 타액 등을 포함한다. 조직 샘플은 조직, 장기 또는 국소 영역의 샘플을 포함할 수 있다. 예를 들면, 샘플은 특정 기관, 기관의 일부 또는 이들 기관 내의 체액이나 세포로부터 유래될 수 있다. 특정 양태에서, 샘플은 뇌(예를 들어, 전체 뇌 또는 뇌의 특정 분절, 예를 들어, 선조체, 또는 뇌의 특정 유형의 세포, 예를 들어, 뉴런 및 신경교세포(성상세포, 희돌기아교세포, 소교세포))로부터 유래될 수 있다. 일부 양태에서, "대상체로부터 유래된 샘플"은 대상체로부터 채취한 혈액 또는 이로부터 유래된 혈장 또는 혈청을 지칭한다. 추가 양태에서, "대상체로부터 유래된 샘플"은 대상체로부터 유래된 뇌 조직(또는 그의 하위 구성요소) 또는 망막 조직(또는 그의 하위 구성요소)을 지칭한다.
II. 본 개시내용의 RNAi 제제
본원의 다른 곳에서 기술된 바와 같이, C9orf72의 돌연변이는 가족성 전두측두엽 치매(FTD) 및 근위축성 측삭 경화증(ALS)과 관련된다. 돌연변이는 C9orf72 유전자의 엑손 1A 및 엑손 1B 사이의 인트론 내에 위치한 G4C2 (서열번호 1) 헥사뉴클레오티드 반복부의 확장의 결과이다. 헥사뉴클레오티드 반복부는 비-AUG-개시된 메커니즘을 통해 번역될 수 있다. 반복부 확장-함유 RNA(표적 RNA)의 축적 또는 반복부 서열의 번역은 FTD 및/또는 ALS 또는 FTD 및/또는 ALS와 관련된 질환 증상을 유발하거나 기여할 수 있다.
따라서, 본 발명은 헥사뉴클레오티드 반복부 확장과 관련된, C9orf72-관련 발현 산물, RNA 및/또는 번역된 폴리펩티드의 발현을 선택적으로 및 효율적으로 감소시키는 dsRNA RNAi 제제를 제공한다. 일부 양태에서, dsRNA 제제는 헥사뉴클레오티드-반복체-함유 RNA(표적 RNA)를 표적화(예를 들어, 선택적으로 표적화)하고 헥사뉴클레오티드-반복체-함유 RNA로부터 발현된 표적 RNA 및 폴리펩티드를 녹다운시킨다. dsRNA 제제는 RNA 병소의 반복부-길이-의존적 형성, 특정 RNA-결합 단백질의 격리, 및 뉴런에서 반복부-관련 비-AUG (AUG) 번역으로부터 야기된 디펩티드 반복부 단백질(예를 들어, 폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌))의 축적 및 응집을 포함하지만 이에 제한되지 않는 FTD 및/또는 ALS와 관련된 징후 또는 증상의 치료적 치료 및/또는 예방을 위한 방법에 사용될 수 있다. dsRNA 제제는 운동 뉴런 질환의 징후 및 증상 및 치매의 징후 및 증상을 포함하지만 이에 제한되지 않는 FTD 및/또는 ALS와 관련된 징후 또는 증상의 치료적 치료 및/또는 예방을 위한 방법에 사용될 수 있다. 운동 뉴런 질환의 징후 및 증상은, 예를 들면, 넘어짐, 물건 떨어뜨림, 팔 및/또는 다리의 비정상적인 피로, 불분명한 발음, 근육 경축 및 경련, 제어할 수 없는 웃음 또는 울음 시간, 및 호흡 곤란을 포함할 수 있다. 치매의 징후 및 증상은 행동 변화, 성격 변화, 발음 및 언어 문제, 및 운동-관련 문제를 포함할 수 있다. 이러한 방법은 본원에 기술된 하나 이상의 dsRNA 제제를 대상체(예를 들면, 인간 또는 동물 대상체)에게 투여하는 것을 포함한다.
본원에 기술된 dsRNA 제제는 반복부-함유 C9orf72 RNA(예를 들어, RNA 병소로 분석됨)의 축적을 중단 또는 감소시킬 수 있고, 이에 의해 RAN 번역에 의한 디펩티드 반복부 단백질의 합성을 방지할 수 있다.
일부 양태에서, 본 발명의 dsRNA 제제는 성숙한 C9orf72 mRNA(즉, 인트론이 스플라이싱된 mRNA)를 표적으로 한다. 다른 양태에서, 본 발명의 dsRNA 제제는 인트론 1A(즉, 인트론이 스플라이싱되지 않은 mRNA, 전구체 mRNA에서 스플라이싱된 RNA 영역, 또는 대안적으로 스플라이싱된 RNA)와 같은 인트론을 함유하는 C9orf72 RNA를 표적으로 한다.
일부 양태에서, 본 발명의 dsRNA 제제는 반복부의 상류에서 비암호화 엑손 1A에서 개시되는 C9orf72 전사체를 표적으로 한다. C9orf72 반복부 확장은 본질적으로 항상 이형접합 돌연변이로서 발생하는 우성 돌연변이이다. 따라서, 일부 양태에서, 본원에 기술된 dsRNA 제제는 엑손 1A에서 개시되며 주로 병원성 대립유전자에만 영향을 미치는 전사체를 표적으로 하며; 정상 대립유전자는 반복부 하류의 엑손 1B에서 거의 독점적으로 전사를 개시한다. 따라서 정상적인(비병원성) 전사체는 보존되므로 dsRNA 제제는 모든 C9orf72 단백질 생산을 없애지 않는다.
하나의 양태에서, RNAi 제제는 세포, 예를 들어, 대상체, 포유동물, 예를 들어 C9orf72-관련 질환을 갖는 인간 내의 세포에서 C9orf72 유전자의 발현을 억제하기 위한 이중 가닥 리보핵산(dsRNA) 분자를 포함한다. dsRNA는 C9orf72 유전자의 발현에서 형성된 mRNA의 적어도 일부에 상보적인 상보성 영역을 갖는 안티센스 가닥을 포함한다. 상보성 영역은 약 15-30개의 뉴클레오티드 이하의 길이이다. C9orf72 유전자를 발현하는 세포와 접촉시, RNAi 제제는 C9orf72 유전자(예를 들면, 인간 유전자, 영장류 유전자, 비영장류 유전자)의 발현을, 예를 들면, PCR 또는 분지형 DNA(bDNA)-기반 방법, 또는 예를 들어 웨스턴 블롯팅 또는 유세포 분석 기술을 사용하는 면역형광 분석과 같은 단백질-기반 방법에 의해 분석하여 적어도 50%까지 억제한다. 바람직한 양태에서, 녹다운 수준은 하기 실시예 1에 제공된 이중-루시페라제 검정 방법을 사용하여 인간 신경모세포종 BE(2)C 세포에서 분석된다.
일부 양태에서, 본원에 기술된 dsRNA 제제는 반복부-함유 C9orf72 RNA(예를 들어, RNA 병소로서 분석됨)의 축적을 중단 또는 감소시키고 이에 의해 RAN 번역에 의한 디펩티드 반복부 단백질의 합성을 방지하도록 설계된다. 이를 달성하기 위해, 특정 양태에서, dsRNA 제제는 C9orf72 mRNA 단백질 암호화 서열을 표적으로 한다(예를 들어, 표 12 및 13 참조). 다른 양태에서, dsRNA 제제는 반복부의 상류에 있는 비암호화 엑손 1A에서 개시되는 C9orf72 전사체를 표적으로 한다. 반복부의 하류에서 개시되는 전사체는 병원성 RNA를 함유하지 않을 것이다. C9orf72 반복부 확장은 본질적으로 항상 이형접합 돌연변이로서 발생하는 우성 돌연변이이다. 따라서, 본원에 기술된 dsRNA 제제가 엑손 1A에서 개시되는 전사체를 표적으로 할 때, 이들은 주로 병원성 대립유전자에만 영향을 미치며; 정상 대립유전자는 반복부의 하류에 있는 엑손 1B에서 거의 독점적으로 전사를 개시하므로 정상(비병원성) 전사체는 보존된다.
dsRNA는 dsRNA가 사용될 조건하에서 상보적이고 혼성화되어 이중체 구조를 형성하는 2개의 RNA 가닥을 포함한다. dsRNA의 한 가닥(안티센스 가닥)은 표적 서열에 실질적으로 상보적이고 일반적으로 완전히 상보적인 상보성 영역을 포함한다. 표적 서열은 C9orf72 유전자의 발현 동안 형성된 mRNA의 서열로부터 유래될 수 있다. 다른 가닥(센스 가닥)은 두 가닥이 적절한 조건하에 결합될 때 혼성화되고 이중체 구조를 형성하도록 안티센스 가닥에 상보적인 영역을 포함한다. 본원의 다른 곳에서 기술되고 당업계에 공지된 바와 같이, dsRNA의 상보적 서열은 또한 별개의 올리고뉴클레오티드 상에 존재하는 것과는 대조적으로 단일 핵산 분자의 자가-상보적 영역으로서 포함될 수 있다.
일반적으로, 이중체 구조는 15 내지 30개 염기 쌍 길이, 예를 들어, 15-29, 15-28, 15-27, 15-26, 15-25, 15-24, 15-23, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 18-30, 18-29, 18-28, 18-27, 18-26, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 19-30, 19-29, 19-28, 19-27, 19-26, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-30, 20-29, 20-28, 20-27, 20-26, 20-25, 20-24,20-23, 20-22, 20-21, 21-30, 21-29, 21-28, 21-27, 21-26, 21-25, 21-24, 21-23, 또는 21-22개 염기 쌍 길이이다. 특정의 바람직한 양태에서, 이중체 구조는 18 내지 25개 염기 쌍 길이, 예를 들어, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-25, 20-24,20-23, 20-22, 20-21, 21-25, 21-24, 21-23, 21-22, 22-25, 22-24, 22-23, 23-25, 23-24 또는 24-25개 염기 쌍 길이, 예를 들면, 19-21개 염기쌍 길이이다. 상기 인용된 범위 및 길이의 중간 범위 및 길이 또한 본 개시내용의 일부로 고려된다.
유사하게, 표적 서열에 대한 상보성 영역은 15 내지 30개의 뉴클레오티드 길이, 예를 들어, 15-29, 15-28, 15-27, 15-26, 15-25, 15-24, 15-23, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 18-30, 18-29, 18-28, 18-27, 18-26, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 19-30, 19-29, 19-28, 19-27, 19-26, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-30, 20-29, 20-28, 20-27, 20-26, 20-25, 20-24,20-23, 20-22, 20-21, 21-30, 21-29, 21-28, 21-27, 21-26, 21-25, 21-24, 21-23, 또는 21-22개의 뉴클레오티드 길이, 예를 들면 19-23개의 뉴클레오티드 길이 또는 21-23개의 뉴클레오티드 길이이다. 상기 인용된 범위 및 길이의 중간 범위 및 길이 또한 본 개시내용의 일부로 고려된다.
일부 양태에서, 이중체 구조는 19 내지 30개 염기 쌍 길이이다. 유사하게, 표적 서열에 대한 상보성 영역은 19 내지 30개의 뉴클레오티드 길이이다.
일부 양태에서, dsRNA는 15 내지 23개의 뉴클레오티드 길이, 19 내지 23개의 뉴클레오티드 길이, 또는 25 내지 30개의 뉴클레오티드 길이이다. 일반적으로, dsRNA는 Dicer 효소의 기질로서 역할을 하기에 충분히 길다. 예를 들면, 약 21 내지 23개의 뉴클레오티드보다 긴 dsRNA가 Dicer에 대한 기질로 작용할 수 있다는 것은 당업계에 잘 알려져 있다. 통상의 숙련가가 또한 인식하는 바와 같이, 절단을 위해 표적화된 RNA의 영역은 가장 흔히 더 큰 RNA 분자, 종종 mRNA 분자의 일부일 것이다. 관련되는 경우, mRNA 표적의 "일부"는 이것이 RNAi-유도 절단(즉, RISC 경로를 통한 절단)을 위한 기질이 되도록 하기에 충분한 길이의 mRNA 표적의 연속 서열이다.
당업계의 숙련가는 또한 이중체 영역이 dsRNA의 1차 기능적 부분, 예를 들어약 15 내지 36개 염기 쌍, 예를 들어, 15-36, 15-35, 15-34, 15-33, 15-32, 15-31, 15-30, 15-29, 15-28, 15-27, 15-26, 15-25, 15-24, 15-23, 15-22, 15-21, 15-20, 15-19, 15-18, 15-17, 18-30, 18-29, 18-28, 18-27, 18-26, 18-25, 18-24, 18-23, 18-22, 18-21, 18-20, 19-30, 19-29, 19-28, 19-27, 19-26, 19-25, 19-24, 19-23, 19-22, 19-21, 19-20, 20-30, 20-29, 20-28, 20-27, 20-26, 20-25, 20-24,20-23, 20-22, 20-21, 21-30, 21-29, 21-28, 21-27, 21-26, 21-25, 21-24, 21-23, 또는 21-22개 염기 쌍, 예를 들면, 19-21개 염기 쌍의 이중체 영역이라는 것을 인식할 것이다. 따라서, 하나의 양태에서, 절단을 위한 원하는 RNA를 표적으로 하는, 예를 들어, 15-30개 염기 쌍의 기능적 이중체로 처리될 정도로, 30개 이상 염기 쌍의 이중체 영역을 갖는 RNA 분자 또는 RNA 분자의 복합체는 dsRNA이다. 따라서, 통상의 숙련가는 하나의 양태에서, miRNA가 dsRNA임을 인식할 것이다. 또 다른 양태에서, dsRNA는 자연 발생 miRNA가 아니다. 또 다른 양태에서, C9orf72 발현을 표적화하는데 유용한 RNAi 제제는 더 큰 dsRNA의 절단에 의해 표적 세포에서 생성되지 않는다.
본원에 기술된 dsRNA는 하나 이상의 단일-가닥 뉴클레오티드 돌출부, 예를 들어, 1, 2, 3, 또는 4개의 뉴클레오티드를 추가로 포함할 수 있다. 뉴클레오티드 돌출부는 데옥시뉴클레오티드/뉴클레오시드를 포함하는 뉴클레오티드/뉴클레오시드 유사체를 포함하거나 이들로 이루어질 수 있다. 돌출부는 센스 가닥, 안티센스 가닥 또는 이들의 임의의 조합에 있을 수 있다. 또한, 돌출부의 뉴클레오티드(들)는 dsRNA의 안티센스 또는 센스 가닥의 5'-말단, 3'-말단 또는 양쪽 말단에 존재할 수 있다.
dsRNA는 당업계에 공지된 표준 방법에 의해 합성될 수 있다. 본 발명의 이중 가닥 RNAi 화합물은 2단계 절차를 사용하여 제조될 수 있다. 먼저, 이중 가닥 RNA 분자의 개별 가닥을 별도로 제조한다. 그후, 구성 요소 가닥을 어닐링한다. siRNA 화합물의 개별 가닥은 용액상 또는 고체상 유기 합성 또는 둘 다를 사용하여 제조될 수 있다. 유기 합성은 비천연 또는 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 가닥을 쉽게 제조할 수 있다는 이점을 제공한다. 유사하게, 본 발명의 단일-가닥 올리고뉴클레오티드는 용액상 또는 고체상 유기 합성 또는 둘 다를 사용하여 제조할 수 있다.
특정 양태에서, 본 발명의 dsRNA 제제는 서열번호 1의 다중 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA, 예를 들면, (예를 들면, 헥사뉴클레오티드 반복부의 적어도 약 30, 적어도 약 35, 적어도 약 40, 적어도 약 50, 적어도 약 60, 적어도 약 70, 적어도 약 80, 적어도 약 100, 적어도 약 200, 적어도 약 300, 적어도 약 400, 또는 적어도 약 500개 카피를 갖는) 병원성 헥사뉴클레오티드 반복부 확장을 갖는 C9orf72 표적 RNA를 표적으로 한다.
본원에 기술된 dsRNA 제제는 헥사뉴클레오티드-반복체 함유 C9orf72 RNA(C9orf72 표적 RNA)의 임의의 영역을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 시작과 엑손 2의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1A는 포함하지만 엑손 2는 포함하지 않음)을 표적으로 할 수 있다(즉, 상응하는 C9orf72 유전자에서 엑손 1A의 시작부터 엑손 2의 시작까지의 영역에 해당). 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 시작과 엑손 1B의 끝 사이의 뉴클레오티드 서열(즉, 엑손 1A 및 엑손 1B를 포함함)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 시작과 엑손 1B의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1A는 포함하지만 엑손 1B는 포함하지 않음)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 시작과 헥사뉴클레오티드 반복부 서열의 끝 사이의 뉴클레오티드 서열(즉, 엑손 1A 및 헥사뉴클레오티드 반복부 서열를 포함함)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 시작과 헥사뉴클레오티드 반복부 서열의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1A는 포함하지만 헥사뉴클레오티드 반복부 서열은 포함하지 않음)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 시작과 엑손 1A의 끝 사이의 뉴클레오티드 서열을 표적으로 할 수 있다.
특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 끝과 엑손 2의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1A를 포함하지 않고 엑손 2를 포함하지 않음)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 끝과 엑손 1B의 끝 사이의 뉴클레오티드 서열(즉, 엑손 1A를 포함하지 않지만 엑손 1B를 포함함)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 끝과 엑손 1B의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1A를 포함하지 않고 엑손 1B를 포함하지 않음)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1A의 끝과 헥사뉴클레오티드 반복부 서열의 끝 사이의 뉴클레오티드 서열(즉, 엑손 1A를 포함하지 않지만 헥사뉴클레오티드 반복부 서열을 포함함)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 엑손 1A의 끝과 헥사뉴클레오티드 반복부 서열의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1A를 포함하지 않고 헥사뉴클레오티드 반복부 서열을 포함하지 않음)을 표적으로 할 수 있다.
특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 헥사뉴클레오티드 반복부 서열의 시작과 엑손 2의 시작 사이의 뉴클레오티드 서열(즉, 헥사뉴클레오티드 반복부 서열를 포함하지만 엑손 2를 포함하지 않음)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 헥사뉴클레오티드 반복부 서열의 시작과 엑손 1B의 끝 사이의 뉴클레오티드 서열(즉, 헥사뉴클레오티드 반복부 서열 및 엑손 1B를 포함함)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 헥사뉴클레오티드 반복부 서열의 시작과 엑손 1B의 시작 사이의 뉴클레오티드 서열(즉, 헥사뉴클레오티드 반복부 서열을 포함하지만 엑손 1B는 포함하지 않음)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 헥사뉴클레오티드 반복부 서열의 시작과 헥사뉴클레오티드 반복부 서열의 끝 사이의 뉴클레오티드 서열을 표적으로 할 수 있다.
특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 헥사뉴클레오티드 반복부 서열의 끝과 엑손 2의 시작 사이의 뉴클레오티드 서열(즉, 헥사뉴클레오티드 반복부 서열을 포함하지 않고 엑손 2를 포함하지 않음)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 헥사뉴클레오티드 반복부 서열의 끝과 엑손 1B의 끝 사이의 뉴클레오티드 서열(즉, 헥사뉴클레오티드 반복부 서열을 포함하지 않지만 엑손 1B를 포함함)을 표적으로 할 수 있다. 특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 헥사뉴클레오티드 반복부 서열의 끝과 엑손 1B의 시작 사이의 뉴클레오티드 서열(즉, 헥사뉴클레오티드 반복부 서열을 포함하지 않고 엑손 1B를 포함하지 않음)을 표적으로 할 수 있다.
특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1B의 시작의 끝과 엑손 2의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1B는 포함하고 엑손 2는 포함하지 않음)을 표적으로 할 수 있다.
특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1B의 시작의 끝과 엑손 1B의 끝 사이의 뉴클레오티드 서열을 표적으로 할 수 있다.
특정 양태에서, 본원에 기술된 dsRNA 제제는 C9orf72 RNA의 엑손 1B의 끝의 끝과 엑손 2의 시작 사이의 뉴클레오티드 서열(즉, 엑손 1B를 포함하지 않고 엑손 2를 포함하지 않음)을 표적으로 할 수 있다.
일부 dsRNA 제제는 C9orf72 RNA의 인트론 영역을 표적으로 한다. 예를 들면, 본원에 기술된 dsRNA 제제는 C9orf72 유전자의 엑손 1A와 엑손 1B 사이의 인트론(즉, C9orf72 표적 RNA의 엑손 1A의 끝과 엑손 1B의 시작 사이의 인트론)을 표적으로 할 수 있다. dsRNA 제제에 의해 표적화된 영역은 C9orf72 표적 RNA의 엑손 1A와 헥사뉴클레오티드 반복부 서열 사이의 인트론 서열에서 헥사뉴클레오티드 반복부의 (5') 상류 또는 (3') 하류에 있을 수 있다. 특정 예에서, dsRNA 제제는 C9orf72 표적 RNA에서 헥사뉴클레오티드 반복부의 (5') 상류의 영역을 표적으로 한다. 예를 들면, dsRNA 제제는 엑손 1A의 끝과 헥사뉴클레오티드 반복부 서열의 시작 사이의 인트론 서열을 표적으로 할 수 있다.
하나의 양태에서, dsRNA 제제는 인간 게놈 어셈블리 GRCh38/hg38 배위 chr9:27,567,165-27,573,866 사이의 인간 C9orf72 유전자의 영역에 상응하는 C9orf72 표적 RNA의 영역을 표적으로 한다(엑손 1A의 시작부터 엑손 2의 시작(즉, 엑손 2 이전의 인트론의 끝)까지의 영역에 상응함; 이 영역으로부터 전사된 세 개의 GGGGCC 반복부를 갖는 예시적인 RNA 서열이 서열번호 111에 제시되어 있으며, 역 상보체는 서열번호 112에 제시되어 있다). 또 다른 특정 예에서, dsRNA 제제는 인간 게놈 어셈블리 GRCh38/hg38 배위 chr9:27,573,494-27,573,708 사이의 인간 C9orf72 유전자의 영역에 상응하는 C9orf72 표적 RNA의 영역을 표적으로 한다(엑손 1A와 엑손 1B 사이(즉, 엑손 1A의 끝과 엑손 1B의 시작 사이)의 인트론 서열의 영역에 상응함; 이 영역으로부터 전사된 세 개의 GGGGCC 반복부를 갖는 예시적인 RNA 서열이 서열번호 113에 제시되어 있으며, 역 상보체는 서열번호 114에 제시되어 있다). 또 다른 특정 예에서, dsRNA 제제는 인간 게놈 어셈블리 GRCh38/hg38 배위 chr9:27,573,547-27,573,708 사이의 인간 C9orf72 유전자의 영역에 상응하는 C9orf72 표적 RNA의 영역을 표적으로 한다(엑손 1A와 헥사뉴클레오티드 반복부 서열의 시작 사이(즉, 엑손 1A의 끝과 헥사뉴클레오티드 반복부 서열의 시작 사이)의 인트론 서열의 영역에 상응함; 이 영역으로부터 전사된 예시적인 RNA 서열이 서열번호 115에 제시되어 있으며, 역 상보체는 서열번호 116에 제시되어 있다). 또 다른 특정 예에서, dsRNA 제제는 인간 게놈 어셈블리 GRCh38/hg38 배위 chr9:27,573,605-27,573,640 사이의 인간 C9orf72 유전자의 영역에 상응하는 C9orf72 표적 RNA의 영역을 표적으로 한다(엑손 1A와 헥사뉴클레오티드 반복부 서열의 시작 사이의 인트론 서열의 영역으로부터의 단편에 상응함; 이 영역으로부터 저사된 예시적인 RNA 서열이 서열번호 117에 제시되어 있으며, 역 상보체는 서열번호 118에 제시되어 있다).
특정 양태에서, 본 발명의 dsRNA 제제는 서열번호 133의 dsRNA 제제 표적 뉴클레오티드 200-290을 표적으로 한다.
특정 양태에서, dsRNA 제제는 서열번호 121의 뉴클레오티드 230-270, 233-262, 800-840, 800-830, 802-828, 1240-1290, 1240-1280, 1247-1288, 1590-1645, 1590-1620, 및 1594-1642의 뉴클레오티드 서열 중 어느 하나로부터의 뉴클레오티드를 표적으로 한다.
특정 양태에서, dsRNA 제제는 서열번호 121의 뉴클레오티드 1594-1616, 802-824, 239-261, 1308-1330, 233-255, 1595-1617, 240-262, 1532-1554, 237-259, 3268-3290, 806-828, 1620-1642, 526-548, 1169-1191, 1266-1288, 1247-1269, 586-608, 1257-1279, 및 400-422의 뉴클레오티드 서열 중 어느 하나로부터의 뉴클레오티드를 표적으로 한다.
하나의 측면에서, 본 개시내용의 dsRNA는 적어도 두 개의 뉴클레오티드 서열, 센스 서열 및 안티센스 서열을 포함한다. C9orf72에 대한 센스 가닥 서열은 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나에 제공된 서열의 그룹으로부터 선택될 수 있고 센스 가닥의 안티센스 가닥의 상응하는 뉴클레오티드 서열은 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나의 서열의 그룹으로부터 선택될 수 있다. 이러한 측면에서, 두 개의 서열 중 하나는 두 개의 서열 중 다른 하나에 상보적이고, 서열 중 하나는 C9orf72 유전자의 발현에서 생성된 mRNA의 서열에 실질적으로 상보적이다. 이와 같이, 이러한 측면에서, dsRNA는 두 개의 올리고뉴클레오티드를 포함할 것이며, 여기서 하나의 올리고뉴클레오티드는 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나에서 센스 가닥(운송 가닥)으로 기술되고 두 번째 올리고뉴클레오티드는 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나에서 센스 가닥의 안티센스 가닥(가이드 가닥)으로 기술된다.
하나의 양태에서, dsRNA의 실질적으로 상보적인 서열은 별도의 올리고뉴클레오티드에 포함된다. 또 다른 양태에서, dsRNA의 실질적으로 상보적인 서열은 단일 올리고뉴클레오티드에 포함된다.
표 10A, 10B, 12-15, 19 또는 20의 서열이 변형되거나 접합된 서열로서 기술되어 있지만, 본 개시내용의 RNAi 제제의 RNA, 예를 들어, 본 개시내용의 dsRNA는 비-변형, 비-접합되거나, 또는 본원에 기술된 것과 상이하게 변형 또는 접합된, 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나에 제시된 서열 중 어느 하나를 포함할 수 있는 것으로 이해될 것이다. 예를 들면, 표 13 및 15에 나타낸 본 발명의 제제의 센스 가닥이 GalNAc 리간드에 접합되어 있지만, 이들 제제는 본원에 기술된 바와 같이 CNS, 예를 들어, C16 리간드로의 전달을 지시하는 모이어티에 접합될 수 있다. 친유성 리간드는 본 출원에서 제공된 임의의 위치에 포함될 수 있다.
통상의 숙련가는 약 20 내지 23개 염기 쌍, 예를 들어, 21개 염기 쌍의 이중체 구조를 갖는 dsRNA가 RNA 간섭을 유도하는데 특히 효과적인 것으로 환영받고 있음을 잘 알고 있다(Elbashir et al., (2001) EMBO J., 20:6877-6888). 그러나, 다른 사람들은 더 짧거나 더 긴 RNA 이중체 구조도 효과적일 수 있음을 발견하였다(Chu and Rana (2007) RNA 14:1714-1719; Kim et al. (2005) Nat Biotech 23:222-226). 위에 기술된 양태에서, 본원에 제공된 올리고뉴클레오티드 서열의 특성 덕분에, 본원에 기술된 dsRNA는 최소 21개의 뉴클레오티드 길이의 적어도 하나의 가닥을 포함할 수 있다. 더 짧은 이중체에서 한쪽 또는 양쪽 말단에 단지 몇 개의 뉴클레오티드를 뺀 것이 위에 기술된 dsRNA와 비교하여 유사하게 효과적일 수 있음을 합리적으로 예상할 수 있다. 따라서, 본원에 제공된 서열 중 하나로부터 유래된 적어도 15, 16, 17, 18, 19, 20개 이상의 연속 뉴클레오티드의 서열을 갖고, 예를 들어, Be(2)c 세포 및 10nM 농도의 RNA 제제를 갖는 시험관내 검정 및 본원의 실시예에 제공된 바와 같은 PCR 검정을 사용하여 전체 서열을 포함하는 dsRNA로부터 10, 15, 20, 25 또는 30% 이하 억제만큼 C9orf72 유전자의 발현을 억제하는 능력이 상이한 dsRNA는 본 개시내용의 범위 내에 있는 것으로 고려된다.
또한, 본원에 기술된 RNA는 RISC-매개 절단에 민감한 C9orf72 전사체의 부위(들)를 확인한다. 이와 같이, 본 개시내용은 이 부위(들) 내에서 표적화하는 RNAi 제제를 추가로 특징으로 한다. 본원에 사용된 바와 같이, RNAi 제제는 RNAi 제제가 그 특정 부위 내의 어느 곳에서나 전사체의 절단을 촉진하는 경우 RNA 전사체의 특정 부위 내를 표적화한다고 한다. 이러한 RNAi 제제는 일반적으로 C9orf72 유전자에서 선택된 서열에 인접한 영역으로부터 취해진 추가의 뉴클레오티드 서열에 커플링된 본원에 제공된 서열 중 하나로부터의 적어도 15개의 연속 뉴클레오티드, 바람직하게는 적어도 19개의 뉴클레오티드를 포함할 것이다.
본원에 개시된 dsRNA 제제는 헥사뉴클레오티드 반복부을 포함하는 C9orf72 표적 RNA의 발현을 억제한다. 발현 억제는 임의의 수준의 억제(예를 들어, 발현의 부분적 억제)를 포함한다. 예를 들면, dsRNA 제제는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 적어도 약 10%, 적어도 약 20%, 적어도 약 30%, 적어도 약 40%, 적어도 약 50%, 적어도 약 60%, 적어도 약 70%, 적어도 약 80%, 또는 적어도 약 90%까지 (또는 C9orf72 표적 RNA가 검출되지 않는 지점까지) 억제할 수 있다. 예를 들면, 이러한 수준의 억제는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 24-48시간 이내에 이루어질 수 있다. 감소는, 예를 들면, dsRNA 제제로 처리하기 전의 세포에 비해 또는 dsRNA 제제로 처리되지 않은 대조군 세포에 비해서 일 수 있다.
본원에 개시된 dsRNA 제제는 또한, 예를 들면, 성숙 C9orf72 메신저 RNA의 발현에 비해 인트론성 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 선택적으로 억제할 수 있다. 이러한 맥락에서 성숙 C9orf72 메신저 RNA는 스플라이싱되고 프로세싱된 C9orf72 RNA 전사체이다. 성숙 C9orf72 메신저 RNA는 엑손으로만 구성되며 모든 인트론이 제거된다. dsRNA 제제는 C9orf72 표적 RNA의 발현의 상대적 감소가 C9orf72 표적 RNA를 발현하는 세포에 dsRNA 제제를 투여한 후 성숙 C9orf72 메신저 RNA의 발현의 상대적 감소보다 큰 경우 성숙 C9orf72 메신저 RNA의 발현에 비해 인트로성 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 선택적으로 억제할 수 있다. 예를 들면, dsRNA 제제는 성숙 C9orf72 메신저 RNA의 발현을 약 50% 미만, 약 40% 미만, 약 30% 미만, 약 20% 미만, 약 10% 미만, 또는 약 5% 미만까지 억제할 수 있다(또는, 예를 들면, 발현에 통계적으로 유의하거나 기능적으로 유의한 영향을 미치지 않는다). 예를 들면, 이러한 수준의 억제는 성숙 C9orf72 메신저 RNA를 발현하는 세포에 투여한 후 24-48시간 이내에 이루어질 수 있다.
본원에 개시된 dsRNA 제제는 또한, 예를 들면, 세포에서 (예를 들어, 세포에 투여 후 24-48시간 이내에) 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 수준을 감소시킬 수 있다. 예를 들면, dsRNA 제제는 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 수준을 적어도 약 10%, 적어도 약 20%, 적어도 약 30%, 적어도 약 40%, 적어도 약 50%, 적어도 약 60%, 적어도 약 70%, 적어도 약 80%, 또는 적어도 약 90%까지 감소시킬 수 있다. 감소는, 예를 들면, dsRNA 제제로 처리하기 전의 세포에 비해 또는 dsRNA 제제로 처리되지 않은 대조군 세포에 비해서 일 수 있다.
III. 본 개시내용의 변형된 RNAi 제제
하나의 양태에서, 본 개시내용의 RNAi 제제의 RNA, 예를 들어, dsRNA는 변형되지 않으며, 예를 들어, 당업계에 공지되고 본원에 기술된 화학적 변형 또는 접합을 포함하지 않는다. 바람직한 양태에서, 본 개시내용의 RNAi 제제의 RNA, 예를 들어, dsRNA는 안정성 또는 기타 유익한 특성을 향상시키기 위해 화학적으로 변형된다. 본 개시내용의 특정 양태에서, 본 개시내용의 RNAi 제제의 뉴클레오티드의 실질적으로 전부는 변형된다. 본 개시내용의 다른 양태에서, 본 개시내용의 RNAi 제제의 뉴클레오티드의 전부는 변형된다. "뉴클레오티드의 실질적으로 전부가 변형된" 본 개시내용의 RNAi 제제는 전부는 아니지만 대부분 변형되며 5, 4, 3, 2개 이하의 변형되지 않은 뉴클레오티드를 포함할 수 있다. 본 개시내용의 여전히 또 다른 양태에서, 본 개시내용의 RNAi 제제는 5, 4, 3, 2 또는 1개 이하의 변형된 뉴클레오티드를 포함할 수 있다.
본 개시내용에서 특징으로 하는 핵산은 본원에 참고로 포함된 문헌["Current protocols in nucleic acid chemistry," Beaucage, S.L. et al. (Edrs.), John Wiley & Sons, Inc., New York, NY, USA]에 기술된 것과 같은 당업계에 잘 확립된 방법에 의해 합성되거나 변형될 수 있다. 변형은, 예를 들면, 말단 변형, 예를 들어, 5'-말단 변형(인산화, 접합, 역 결합) 또는 3'-말단 변형(접합, DNA 뉴클레오티드, 역 결합 등); 염기 변형, 예를 들어, 안정화 염기, 불안정화 염기 또는 파트너의 확장된 레퍼토리와 염기쌍을 이루는 염기로 대체, 염기(무염기 뉴클레오티드) 또는 접합된 염기의 제거; 당 변형(예를 들어, 2'-위치 또는 4'-위치에서) 또는 당의 대체; 또는 포스포디에스테르 결합의 변형 또는 대체를 포함하는 백본 변형을 포함한다. 본원에 기술된 양태에서 유용한 RNAi 재제의 특정 예는 변형된 백본을 함유하거나 천연 뉴클레오시드간 결합을 함유하지 않는 RNA를 포함하지만, 이에 제한되지 않는다. 변형된 백본을 갖는 RNA는 특히 백본에 인 원자를 갖지 않는 것을 포함한다. 본 명세서의 목적을 위해, 그리고 때때로 당업계에서 참조되는 바와 같이, 뉴클레오시드간 백본에 인 원자를 갖지 않는 변형된 RNA는 또한 올리고뉴클레오시드인 것으로 간주될 수 있다. 일부 양태에서, 변형된 RNAi 제제는 이의 뉴클레오시드간 백본에 인 원자를 가질 것이다.
변형된 RNA 백본은, 예를 들면, 포스포로티오에이트, 키랄 포스포로티오에이트, 포스포로디티오에이트, 포스포트리에스테르, 아미노알킬포스포트리에스테르, 3'-알킬렌 포스포네이트 및 키랄 포스포네이트를 포함한 메틸 및 기타 알킬 포스포네이트, 포스피네이트, 3'-아미노 포스포르아미데이트 및 아미노알킬포스포르아미데이트를 포함한 포스포르아미데이트, 티오노포스포르아미데이트, 티오노알킬포스포네이트, 티오노알킬포스포트리에스테르, 및 정상적인 3'-5' 결합을 갖는 보라노포스페이트, 이의 2'-5'-결합된 유사체, 및 뉴클레오시드 단위의 인접한 쌍이 3'-5'에서 5'-3'으로 또는 2'-5'에서 5'-2'로 연결된 역 극성을 갖는 것을 포함한다. 다양한 염, 혼합된 염 및 유리 산 형태가 또한 포함된다. 본 발명의 일부 양태에서, 본 발명의 dsRNA 제제는 유리 산 형태이다. 본 발명의 다른 양태에서, 본 발명의 dsRNA 제제는 염 형태이다. 하나의 양태에서, 본 발명의 dsRNA 제제는 나트륨 염 형태이다. 특정 양태에서, 본 발명의 dsRNA 제제가 나트륨 염 형태인 경우, 나트륨 이온은 제제에 존재하는 실질적으로 모든 포스포디에스테르 및/또는 포스포로티오테이트 그룹에 대한 짝이온으로서 제제에 존재한다. 실질적으로 모든 포스포디에스테르 및/또는 포스포로티오에이트 결합이 나트륨 짝이온을 갖는 제제는 나트륨 짝이온 없이 5, 4, 3, 2, 또는 1개 이하의 포스포디에스테르 및/또는 포스포로티오에이트 결합을 포함한다. 일부 양태에서, 본 발명의 dsRNA 제제가 나트륨 염 형태인 경우, 나트륨 이온은 제제에 존재하는 모든 포스포디에스테르 및/또는 포스포로티오테이트 그룹에 대한 짝이온으로서 제제에 존재한다.
상기 인-함유 결합의 제조를 교시하는 대표적인 미국 특허는 미국 특허 제3,687,808호; 제4,469,863호; 제4,476,301호; 제5,023,243호; 제5,177,195호; 제5,188,897호; 제5,264,423호; 제5,276,019호; 제5,278,302호; 제5,286,717호; 제5,321,131호; 제5,399,676호; 제5,405,939호; 제5,453,496호; 제5,455,233호; 제5,466,677호; 제5,476,925호; 제5,519,126호; 제5,536,821호; 제5,541,316호; 제5,550,111호; 제5,563,253호; 제5,571,799호; 제5,587,361호; 제5,625,050호; 제6,028,188호; 제6,124,445호; 제6,160,109호; 제6,169,170호; 제6,172,209호; 제6,239,265호; 제6,277,603호; 제6,326,199호; 제6,346,614호; 제6,444,423호; 제6,531,590호; 제6,534,639호; 제6,608,035호; 제6,683,167호; 제6,858,715호; 제6,867,294호; 제6,878,805호; 제7,015,315호; 제7,041,816호; 제7,273,933호; 제7,321,029호; 및 미국 특허 제RE39464호를 포함하지만, 이에 제한되지 않으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
그 안에 인 원자를 포함하지 않는 변형된 RNA 백본은 단쇄 알킬 또는 사이클로알킬 뉴클레오시드간 결합, 혼합된 헤테로원자 및 알킬 또는 사이클로알킬 뉴클레오시드간 결합, 또는 하나 이상의 단쇄 헤테로원자 또는 헤테로사이클릭 뉴클레오시드간 결합에 의해 형성된 백본을 갖는다. 이들은 모르폴리노 결합(뉴클레오시드의 당 부분에서 부분적으로 형성됨); 실록산 백본; 설파이드, 설폭사이드 및 설폰 백본; 포름아세틸 및 티오포름아세틸 백본; 메틸렌 포름아세틸 및 티오포름아세틸 백본; 알켄 함유 백본; 설파메이트 백본; 메틸렌이미노 및 메틸렌하이드라지노 백본; 설포네이트 및 설폰아미드 백본; 아미드 백본을 갖는 것; 및 혼합된 N, O, S 및 CH2 구성 부분을 갖는 기타를 포함한다.
상기 올리고뉴클레오시드의 제조를 교시하는 대표적인 미국 특허는 미국 특허 제5,034,506호; 제5,166,315호; 제5,185,444호; 제5,214,134호; 제5,216,141호; 제5,235,033호; 제5,64,562호; 제5,264,564호; 제5,405,938호; 제5,434,257호; 제5,466,677호; 제5,470,967호; 제5,489,677호; 제5,541,307호; 제5,561,225호; 제5,596,086호; 제5,602,240호; 제5,608,046호; 제5,610,289호; 제5,618,704호; 제5,623,070호; 제5,663,312호; 제5,633,360호; 제5,677,437호; 및 제5,677,439호를 포함하지만, 이에 제한되지 않으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
다른 양태에서, 뉴클렐오티드의 당 및 뉴클레오시드간 결합, 즉 백본 둘 다가 신규 그룹으로 대체된 적절한 RNA 모방체가 RNAi 제제에 사용하기 위해 고려된다. 염기 단위는 적절한 핵산 표적 화합물과의 혼성화를 위해 유지된다. 탁월한 혼성화 특성을 갖는 것으로 나타난 RNA 모방체인 이러한 올리고머 화합물 중 하나를 펩티드 핵산(PNA)이라고 한다. PNA 화합물에서, RNA의 당 백본은 아미드를 포함하는 백본, 특히 아미노에틸글리신 백본으로 대체된다. 핵염기는 유지되고 백본의 아미드 부분의 아자 질소 원자에 직접 또는 간접적으로 결합된다. PNA 화합물의 제조를 교시하는 대표적인 미국 특허는 미국 특허 제5,539,082호; 제5,714,331호; 및 제5,719,262호를 포함하지만 이에 제한되지 않으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다. 본 개시내용의 RNAi 제제에 사용하기에 적합한 추가의 PNA 화합물은, 예를 들면, 문헌[Nielsen et al., Science, 1991, 254, 1497-1500]에 기술되어 있다.
본 개시내용에서 특징으로 하는 일부 양태는 포스포로티오에이트 백본을 갖는 RNA 및 헤테로원자 백본, 특히 상기 참고된 미국 특허 제5,489,677호의 --CH2--NH--CH2-, --CH2--N(CH3)--O--CH2--[메틸렌(메틸이미노) 또는 MMI 백본으로 알려짐], --CH2--O--N(CH3)--CH2--, --CH2--N(CH3)--N(CH3)--CH2-- 및 --N(CH3)--CH2--CH2--[여기서 천연 포스포디에스테르 백본은 --O--P--O--CH2--로 나타내어진다], 및 상기 참고된 미국 특허 제5,602,240호의 아미드 백본을 갖는 올리고뉴클레오시드를 포함한다.
변형된 RNA는 또한 하나 이상의 치환된 당 모이어티를 함유할 수 있다. 본원에 특징지어진 RNAi 제제, 예를 들어, dsRNA는 2'-위치에 다음 중 하나를 포함할 수 있다: OH; F; O-, S-, 또는 N-알킬; O-, S-, 또는 N-알케닐; O-, S- 또는 N-알키닐; 또는 O-알킬-O-알킬, 여기서 알킬, 알케닐 및 알키닐은 치환되거나 치환되지 않은 C1 내지 C10 알킬 또는 C2 내지 C10 알케닐 및 알키닐일 수 있다. 예시적인 적합한 변형은 O[(CH2)nO] mCH3, O(CH2).nOCH3, O(CH2)nNH2, O(CH2) nCH3, O(CH2)nONH2, 및 O(CH2)nON[(CH2)nCH3)]2를 포함하고, 여기서 n 및 m은 1 내지 약 10이다. 다른 양태에서, dsRNA는 2' 위치에 다음 중 하나를 포함한다: C1 내지 C10 저급 알킬, 치환된 저급 알킬, 알크아릴, 아르알킬, O-알크아릴 또는 O-아르알킬, SH, SCH3, OCN, Cl, Br, CN, CF3, OCF3, SOCH3, SO2CH3, ONO2, NO2, N3, NH2, 헤테로사이클로알킬, 헤테로사이클로알크아릴, 아미노알킬아미노, 폴리알킬아미노, 치환된 실릴, RNA 절단 그룹, 리포터 그룹, 인터칼레이터, RNAi 제제의 약동학 특성을 개선하기 위한 그룹, 또는 RNAi 재제의 약력학적 특성을 개선하기 위한 그룹, 및 유사한 특성을 갖는 다른 치환체. 일부 양태에서, 변형은 2'-메톡시에톡시(2'-O--CH2CH2OCH3, 2'-O-(2-메톡시에틸) 또는 2'-MOE로도 알려짐)(Martin et al., Helv . Chim . Acta, 1995, 78:486-504) 즉, 알콕시-알콕시 그룹을 포함한다. 또 다른 예시적인 변형은 아래 본원 실시예에 기술된 바와 같이 2'-DMAOE로도 알려진 2'-디메틸아미노옥시에톡시, 즉 O(CH2)2ON(CH3)2 그룹, 및 2'-디메틸아미노에톡시에톡시(2'-O-디메틸아미노에톡시에틸 또는 2'-DMAEOE로도 당업계에 알려짐), 즉, 2'-O--CH2--O--CH2--N(CH2)2이다. 추가의 예시적인 변형은 다음을 포함한다: 5'-Me-2'-F 뉴클레오티드, 5'-Me-2'-OMe 뉴클레오티드, 5'-Me-2'-데옥시뉴클레오티드, (이러한 세 가지 계열에서 R 및 S 이성체 둘 다); 2'-알콕시알킬; 및 2'-NMA(N-메틸아세트아미드).
또 다른 변형은 2'-메톡시(2'-OCH3), 2'-아미노프로폭시(2'-OCH2CH2CH2NH2), 2'-O-헥사데실, 및 2'-플루오로(2'-F)를 포함한다. 유사한 변형이 또한 RNAi 제제의 RNA 상의 다른 위치에서, 특히 3' 말단 뉴클레오티드 상의 당의 3' 위치 또는 2'-5' 결합된 dsRNA 및 5' 말단 뉴클레오티드의 5' 위치에서 이루어질 수 있다. RNAi 제제는 또한 펜토푸라노실 당 대신에 사이클로부틸 모이어티와 같은 당 모방체를 가질 수 있다. 이러한 변형된 당 구조의 제조를 교시하는 대표적인 미국 특허는 미국 특허 제4,981,957호; 제5,118,800호; 제5,319,080호; 제5,359,044호; 제5,393,878호; 제5,446,137호; 제5,466,786호; 제5,514,785호; 제5,519,134호; 제5,567,811호; 제5,576,427호; 제5,591,722호; 제5,597,909호; 제5,610,300호; 제5,627,053호; 제5,639,873호; 제5,646,265호; 제5,658,873호; 제5,670,633호; 및 제5,700,920호를 포함하지만, 이에 제한되지 않고, 그 중 일부는 본 출원과 공동으로 소유된다. 상기 각각의 전체 내용은 본원에 참고로 포함된다.
본 개시내용의 RNAi 제제는 또한 핵염기(종종 단순히 "염기"로 당업계에서 지칭됨) 변형 또는 치환을 포함할 수 있다. 본원에 사용된 바와 같이, "비변형" 또는 "천연" 핵염기는 퓨린 염기 아데닌(A) 및 구아닌(G), 및 피리미딘 염기 티민(T), 시토신(C) 및 우라실(U)을 포함한다. 변형된 핵염기는 기타 합성 및 천연 핵염기, 예를 들어 5-메틸시토신(5-me-C), 5-하이드록시메틸 시토신, 크산틴, 하이포크산틴, 2-아미노아데닌, 아데닌 및 구아닌의 6-메틸 및 기타 알킬 유도체, 아데닌 및 구아닌의 2-프로필 및 기타 알킬 유도체, 2-티오우라실, 2-티오티민 및 2-티오시토신, 5-할로우라실 및 시토신, 5-프로피닐 우라실 및 시토신, 6-아조 우라실, 시토신 및 티민, 5-우라실(슈도우라실), 4-티오우라실, 8-할로, 8-아미노, 8-티올, 8-티오알킬, 8-하이드록실 및 기타 8-치환된 아데닌 및 구아닌, 5-할로, 특히 5-브로모, 5-트리플루오로메틸 및 기타 5-치환된 우라실 및 시토신, 7-메틸구아닌 및 7-메틸아데닌, 8-아자구아닌 및 8-아자아데닌, 7-데아자구아닌 및 7-다아자아데닌 및 3-데아자구아닌 및 3-데아자아데닌을 포함한다. 추가의 핵염기는 미국 특허 제3,687,808호에 개시된 것, 문헌[Modified Nucleosides in Biochemistry, Biotechnology and Medicine, Herdewijn, P. ed. Wiley-VCH, 2008]에 개시된 것; 문헌[The Concise Encyclopedia Of Polymer Science And Engineering, pages 858-859, Kroschwitz, J. L, ed. John Wiley & Sons, 1990]에 개시된 것, 문헌[Englisch et al., (1991) Angewandte Chemie , International Edition, 30:613]에 개시된 것, 및 문헌[Sanghvi, Y S., Chapter 15, dsRNA Research and Applications, pages 289-302, Crooke, S. T. and Lebleu, B., Ed., CRC Press, 1993]에 개시된 것을 포함한다. 이들 핵염기 중 일부는 본 개시내용에 특징지어진 올리고머 화합물의 결합 친화도를 증가시키는데 특히 유용하다. 이들은 2-아미노프로필아데닌, 5-프로피닐우라실 및 5-프로피닐시토신을 포함한 5-치환된 피리미딘, 6-아자피리미딘 및 N-2, N-6 및 0-6 치환된 퓨린을 포함한다. 5-메틸시토신 치환은 핵산 이중체 안정성을 0.6-1.2℃까지 증가시키는 것으로 나타났으며(Sanghvi, Y. S., Crooke, S. T. and Lebleu, B., Eds., dsRNA Research and Applications, CRC Press, Boca Raton, 1993, pp. 276-278), 더욱 특히 염기 치환이 2'-O-메톡시에틸 당 변형과 조합될 때 예시적인 염 치환이다.
상기 언급된 변형된 핵염기 중 일부 및 기타 변형된 핵염기의 제조를 교시하는 대표적인 미국 특허는 상기 언급된 미국 특허 제3,687,808호, 제4,845,205호; 제5,130,30호; 제5,134,066호; 제5,175,273호; 제5,367,066호; 제5,432,272호; 제5,457,187호; 제5,459,255호; 제5,484,908호; 제5,502,177호; 제5,525,711호; 제5,552,540호; 제5,587,469호; 제5,594,121호, 제5,596,091호; 제5,614,617호; 제5,681,941호; 제5,750,692호; 제6,015,886호; 제6,147,200호; 제6,166,197호; 제6,222,025호; 제6,235,887호; 제6,380,368호; 제6,528,640호; 제6,639,062호; 제6,617,438호; 제7,045,610호; 제7,427,672호; 및 제7,495,088호를 포함하지만 이에 제한되지 않으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
본 개시내용의 RNAi 제제는 또한 하나 이상의 고정 핵산(LNA)을 포함하도록 변형될 수 있다. 잠금 핵산은 리보스 모이어티가 2' 및 4' 탄소를 연결하는 추가 브릿지를 포함하는 변형된 리보스 모이어티를 갖는 뉴클레오티드이다. 이 구조는 3'-엔도 구조적 형태에서 리보스를 효과적으로 "고정"한다. siRNA에 고정 핵산을 추가하면 혈청내 siRNA 안정성이 증가하고 오프-타겟 효과(off-target effect)가 감소하는 것으로 나타났다(Elmen, J. et al., (2005) Nucleic Acids Research 33(1):439-447; Mook, OR. et al., (2007) Mol Canc Ther 6(3):833-843; Grunweller, A. et al., (2003) Nucleic Acids Research 31(12):3185-3193).
본 개시내용의 RNAi 제제는 또한 하나 이상의 바이사이클릭 당 모이어티를 포함하도록 변형될 수 있다. "바이사이클릭 당"은 두 원자의 브릿징에 의해 변형된 푸라노실 환이다. "바이사이클릭 뉴클레오사이드"("BNA")는 당 고리의 2개의 탄소 원자를 연결하여 바이사이클릭 환 시스템을 형성하는 브릿지를 포함하는 당 모이어티를 갖는 뉴클레오시드이다. 특정 양태에서, 브릿지는 당 고리의 4'-탄소와 2'-탄소를 연결한다. 따라서, 일부 양태에서, 본 개시내용의 제제는 하나 이상의 고정 핵산(LNA)을 포함할 수 있다. 고정 핵산은 리보스 모이어티가 2' 및 4' 탄소를 연결하는 추가 브릿지를 포함하는 변형된 리보스 모이어티를 갖는 뉴클레오티드이다. 즉, LNA는 4'-CH2-O-2' 브릿지를 포함하는 바이사이클릭 당 모이어티를 포함하는 뉴클레오티드이다. 이것은 3'-엔도 구조적 형태에서 리보스를 효과적으로 "고정"한다. siRNA에 고정 핵산을 추가하면 혈청내 siRNA 안정성이 증가하고 오프-타겟 효과가 감소하는 것으로 나타났다(Elmen, J. et al., (2005) Nucleic Acids Research 33(1):439-447; Mook, OR. et al., (2007) Mol Canc Ther 6(3):833-843; Grunweller, A. et al., (2003) Nucleic Acids Research 31(12):3185-3193). 본 개시내용의 폴리뉴클레오티드에 사용하기 위한 바이사이클릭 뉴클레오시드의 예는 4' 및 2' 리보실 환 원자 사이에 브릿지를 포함하는 뉴클레오사이드를 제한 없이 포함한다. 특정 양태에서, 본 개시내용의 안티센스 폴리뉴클레오티드 제제는 4'에서 2'로의 브릿지를 포함하는 하나 이상의 바이사이클릭 뉴클레오시드를 포함한다. 이러한 4'에서 2'로의 브릿징된 바이사이클릭 뉴클레오시드의 예는 4'-(CH2)―O-2' (LNA); 4'-(CH2)―S-2'; 4'-(CH2)2―O-2' (ENA); 4'-CH(CH3)―O-2' ("제한된 에틸" 또는 "cEt"라고도 함) 및 4'-CH(CH2OCH3)―O-2' (및 이의 유사체; 예를 들어, 참조; 미국 특허 제7,399,845호 참조); 4'―C(CH3)(CH3)-O-2' (및 이의 유사체; 예를 들어, 미국 특허 제8,278,283호 참조); 4'―CH2-N(OCH3)-2' (및 이의 유사체; 예를 들어, 미국 특허 제8,278,425호 참조); 4'―CH2-O―N(CH3)-2' (예를 들어, 미국 특허 공개 제2004/0171570호); 4'―CH2-N(R)―O-2' (여기서 R은 H, C1-C12 알킬, 또는 보호 그룹이다) (예를 들어, 미국 특허 제7,427,672호 참조); 4'―CH2-C(H)(CH3)-2' (예를 들어, 참조; Chattopadhyaya et al., J. Org . Chem., 2009, 74, 118-134); 및 4'―CH2-C(=CH2)-2' (및 이의 유사체; 예를 들어, 미국 특허 제8,278,426호 참조)를 포함하지만, 이에 제한되지 않는다. 상기 각각의 전체 내용은 본원에 참고로 포함된다.
고정 핵산 뉴클레오티드의 제조를 교시하는 추가의 대표적인 미국 특허 및 미국 특허 공개는 다음을 포함하지만 이에 제한되지 않는다: 미국 특허 제6,268,490호; 제6,525,191호; 제6,670,461호; 제6,770,748호; 제6,794,499호; 제6,998,484호; 제7,053,207호; 제7,034,133호; 제7,084,125호; 제7,399,845호; 제7,427,672호; 제7,569,686호; 제7,741,457호; 제8,022,193호; 제8,030,467호; 제8,278,425호; 제8,278,426호; 제8,278,283호; 제US 2008/0039618호; 및 제US 2009/0012281호, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
예를 들면 α-L-리보푸라노스 및 β-D-리보푸라노스를 포함하는 하나 이상의 입체화학적 당 배열을 갖는 전술한 바이사이클릭 뉴클레오시드 중 어느 것이 제조될 수 있다(제WO 99/14226호 참조).
본 개시내용의 RNAi 제제는 또한 하나 이상의 제한된 에틸 뉴클레오티드를 포함하도록 변형될 수 있다. 본원에 사용된 "제한된 에틸 뉴클레오티드(constrained ethyl nucleotide)" 또는 "cEt"는 4'-CH(CH3)-O-2' 브릿지를 포함하는 바이사이클릭 당 모이어티를 포함하는 고정 핵산이다. 하나의 양태에서, 제한된 에틸 뉴클레오티드는 본원에서 "S-cEt"로 지칭되는 S 형태이다.
본 개시내용의 RNAi 제제는 또한 하나 이상의 "형태적으로 제한된 뉴클레오티드"(conformationally restricted nucleotide; "CRN")를 포함할 수 있다. CRN은 리보스의 C2' 및 C4' 탄소 또는 리보스의 C3 및 -C5' 탄소를 연결하는 링커를 갖는 뉴클레오티드 유사체이다. CRN은 리보스 환을 안정적인 형태로 고정하고 mRNA에 대한 혼성화 친화도를 증가시킨다. 링커는 안정성과 친화성을 위한 최적의 위치에 산소를 배치하여 리보스 환 주름(ring puckering)을 덜 야기하기에 충분한 길이이다.
위에 언급된 특정 CRN의 제조를 교시하는 대표적인 공보는 제US 2013/0190383호; 및 제WO 2013/036868호를 포함하지만, 이에 제한되지 않으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
일부 양태에서, 본 개시내용의 RNAi 제제는 UNA(비고정 핵산) 뉴클레오티드인 하나 이상의 단량체를 포함한다. UNA는 당의 결합 중 어느 것이 제거되어 비고정된 "당" 잔기를 형성하는 비고정된 비사이클릭 핵산이다. 하나의 예에서, UNA는 또한 C1'-C4' 사이의 결합(즉, C1' 및 C4' 탄소 사이의 공유 탄소-산소-탄소 결합)이 제거된 단량체를 포함한다. 또 다른 예에서, 당의 C2'-C3' 결합(즉, C2'과 C3' 탄소 사이의 공유 탄소-탄소 결합)이 제거되었다(참조; Nuc . Acids Symp . Series, 52, 133-134 (2008) and Fluiter et al., Mol . Biosyst ., 2009, 10, 1039, 본원에 참고로 포함됨).
UNA의 제조를 교시하는 대표적인 미국 공보는 제US8,314,227호; 및 미국 특허 공개 제2013/0096289호; 제2013/0011922호; 및 제2011/0313020호를 포함하지만, 이에 제한되지 않으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
RNA 분자의 말단에 대한 잠재적인 안정화 변형은 N-(아세틸아미노카프로일)-4-하이드록시프롤리놀(Hyp-C6-NHAc), N-(카프로일-4-하이드록시프롤리놀(Hyp-C6), N-(아세틸-4-하이드록시프롤리놀(Hyp-NHAc), 티미딘-2'-0-데옥시티미딘(에테르), N-(아미노카프로일)-4-하이드록시프롤리놀(Hyp-C6-아미노), 2-도코사노일-우리딘-3"-포스페이트, 역 염기 dT(idT) 및 기타를 포함할 수 있다. 이러한 변형의 개시내용은 제WO 2011/005861호에서 찾을 수 있다.
본 개시내용의 RNAi 제제의 또 다른 변형은 RNAi 제제의 안티센스 가닥 상에 5' 포스페이트 또는 5' 포스페이트 모방체, 예를 들어, 5'-말단 포스페이트 또는 포스페이트 모방체를 포함한다. 적합한 포스페이트 모방체는, 예를 들면, 제US 2012/0157511호에 개시되어 있으며, 이의 전체 내용은 본원에 참고로 포함된다.
A. 본 개시내용의 모티프를 포함하는 변형된 RNAi 제제
개시내용의 특정 측면에서, 본 개시내용의 이중 가닥 RNAi 제제는, 예를 들면, 제WO 2013/075035호에 개시된 바와 같은 화학적 변형을 갖는 제제를 포함하며, 이의 전체 내용은 본원에 참고로 포함된다. 본원에 및 제WO 2013/075035호에 나타낸 바와 같이, 특히 절단 부위 또는 그 부근에서 RNAi 제제의 센스 가닥 또는 안티센스 가닥에 3개의 연속 뉴클레오티드에 대한 3개의 동일한 변형의 하나 이상의 모티프를 도입함으로써 보다 우수한 결과가 수득될 수 있다. 일부 양태에서, RNAi 제제의 센스 가닥 및 안티센스 가닥은 달리 완전히 변형될 수 있다. 이러한 모티프의 도입은, 존재하는 경우, 센스 또는 안티센스 가닥의 변형 패턴을 방해한다. RNAi 제제는, 예를 들어, 센스 가닥 상에서, 친유성 리간드, 예를 들어 C16 리간드와 임의로 접합될 수 있다. RNAi 제제는 예를 들어 안티센스 가닥의 하나 이상의 잔기 상에서 (S)-글리콜 핵산(GNA) 변형으로 임의로 변형될 수 있다. 생성된 RNAi 제제는 보다 우수한 유전자 침묵 활성을 나타낸다.
따라서, 본 개시내용은 생체내에서 표적 유전자(즉, C9orf72 유전자)의 발현을 억제할 수 있는 이중 가닥 RNAi 제제를 제공한다. RNAi 제제는 센스 가닥 및 안티센스 가닥을 포함한다. RNAi 제제의 각 가닥은 15-30개의 뉴클레오티드 길이일 수 있다. 예를 들면, 각 가닥은 16-30개의 뉴클레오티드 길이, 17-30개의 뉴클레오티드 길이, 25-30개의 뉴클레오티드 길이, 27-30개의 뉴클레오티드 길이, 17-23개의 뉴클레오티드 길이, 17-21개의 뉴클레오티드 길이, 17-19개의 뉴클레오티드 길이, 19-25개의 뉴클레오티드 길이, 19-23개의 뉴클레오티드 길이, 19-21개의 뉴클레오티드 길이, 21-25개의 뉴클레오티드 길이, 또는 21-23개의 뉴클레오티드 길이일 수 있다. 특정 양태에서, 각 가닥은 19-23개의 뉴클레오티드 길이이다.
센스 가닥 및 안티센스 가닥은 전형적으로 본원에서 "RNAi 제제"라고도 하는 이중체 이중 가닥 RNA("dsRNA")를 형성한다. RNAi 제제의 이중체 영역은 15-30개의 뉴클레오티드 쌍 길이일 수 있다. 예를 들면, 이중체 영역은 16-30개의 뉴클레오티드 쌍 길이, 17-30개의 뉴클레오티드 쌍 길이, 27-30개의 뉴클레오티드 쌍 길이, 17-23개의 뉴클레오티드 쌍 길이, 17-21개의 뉴클레오티드 쌍 길이, 17-19개의 뉴클레오티드 쌍 길이, 19-25개의 뉴클레오티드 쌍 길이, 19-23개의 뉴클레오티드 쌍 길이, 19- 21개의 뉴클레오티드 쌍 길이, 21-25개의 뉴클레오티드 쌍 길이, 또는 21-23개의 뉴클레오티드 쌍 길이일 수 있다. 또 다른 예에서, 이중체 영역은 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 및 27개의 뉴클레오티드 길이로부터 선택된다. 바람직한 양태에서, 이중체 영역은 19-21개의 뉴클레오티드 쌍 길이이다.
하나의 양태에서, RNAi 제제는 3'-말단, 5'-말단, 또는 한 가닥 또는 두 가닥 모두의 양 말단에 하나 이상의 돌출 영역 또는 캡핑 그룹을 함유할 수 있다. 돌출부는 1-6개의 뉴클레오티드 길이, 예를 들어 2-6개의 뉴클레오티드 길이, 1-5개의 뉴클레오티드 길이, 2-5개의 뉴클레오티드 길이, 1-4개의 뉴클레오티드 길이, 2-4개의 뉴클레오티드 길이, 1-3개의 뉴클레오티드 길이, 2-3개의 뉴클레오티드 길이, 또는 1-2개의 뉴클레오티드 길이. 바람직한 양태에서, 뉴클레오티드 돌출부 영역은 2개의 뉴클레오티드 길이일 수 있다. 돌출부는 한 가닥이 다른 가닥보다 긴 결과이거나 동일한 길이의 두 가닥이 엇갈려서 생긴 결과일 수 있다. 돌출부는 표적 mRNA와 미스매치를 형성할 수 있거나 표적화된 유전자 서열에 상보적일 수 있거나 다른 서열일 수 있다. 첫 번째 및 두 번째 가닥은 또한, 예를 들어, 헤어핀을 형성하기 위한 추가 염기에 의해, 또는 다른 비염기 링커에 의해 연결될 수 있다.
하나의 양태에서, RNAi 제제의 돌출부 영역에 있는 뉴클레오티드는 각각 독립적으로 2'-당 변형된, 예를 들어 2-F, 2'-O-메틸, 티미딘(T), 및 이들의 임의의 조합을 포함하지만 이에 제한되지 않는 변형되거나 비변형된 뉴클레오티드일 수 있다.
예를 들면, TT는 어느 하나의 가닥 상의 어느 하나의 말단에 대한 돌출부 서열일 수 있다. 돌출부는 표적 mRNA와 미스매치를 형성할 수 있거나 표적화된 유전자 서열에 상보적일 수 있거나 다른 서열일 수 있다.
RNAi 제제의 센스 가닥, 안티센스 가닥 또는 양 가닥에서 5'- 또는 3'-돌출부는 인산화될 수 있다. 일부 양태에서, 돌출부 영역(들)은 2개의 뉴클레오티드 사이에 포스포로티오에이트를 갖는 2개의 뉴클레오티드를 함유하며, 여기서 2개의 뉴클레오티드는 동일하거나 상이할 수 있다. 하나의 양태에서, 돌출부는 센스 가닥, 안티센스 가닥, 또는 양 가닥의 3'-말단에 존재한다. 하나의 양태에서, 이러한 3'-돌출부는 안티센스 가닥에 존재한다. 하나의 양태에서, 이러한 3'-돌출부는 센스 가닥에 존재한다.
RNAi 제제는 이의 전체 안정성에 영향을 주지 않으면서 RNAi의 간섭 활성을 강화할 수 있는 단일 돌출부만 함유할 수 있다. 예를 들면, 단일 가닥 돌출부는 센스 가닥의 3'-말단 끝에 또는, 대안적으로, 안티센스 가닥의 3'말단 끝에 위치할 수 있다. RNAi는 또한 안티센스 가닥의 5'-말단(또는 센스 가닥의 3'-말단)에 위치한 평활 말단을 가질 수 있으며, 그 반대도 가능하다. 일반적으로, RNAi의 안티센스 가닥은 3'-말단에 뉴클레오티드 돌출부가 있고 5'-말단이 평활하다. 이론에 결부시키고자 하는 것은 아니지만, 안티센스 가닥의 5'-말단 및 안티센스 가닥의 3'-말단 돌출부에 있는 비대칭 평활 말단은 가이드 가닥을 RISC 공정에 부하하는 것을 유리하게 한다.
하나의 양태에서, RNAi 제제는 19개의 뉴클레오티드 길이의 이중 말단 블런트머(bluntmer)이며, 여기서 센스 가닥은 5' 말단으로부터 위치 7, 8, 9에서 3개의 연속 뉴클레오티드 상에 3개의 2'-F 변형의 적어도 하나의 모티프를 함유한다. 안티센스 가닥은 5' 말단으로부터 위치 11, 12, 13에서 3개의 연속 뉴클레오티드 상에 3개의 2'-O-메틸 변형의 적어도 하나의 모티프를 함유한다.
또 다른 양태에서, RNAi 제제는 20개의 뉴클레오티드 길이의 이중 말단 블런트머이며, 여기서 센스 가닥은 5' 말단으로부터 위치 8, 9, 10에서 3개의 연속 뉴클레오티드 상에 3개의 2'-F 변형의 적어도 하나의 모티프를 함유한다. 안티센스 가닥은 5' 말단으로부터 위치 11, 12, 13에서 3개의 연속 뉴클레오티드 상에 3개의 2'-O-메틸 변형의 적어도 하나의 모티프를 함유한다.
또 다른 양태에서, RNAi 제제는 21개의 뉴클레오티드 길이의 이중 말단 블런트머이며, 여기서 센스 가닥은 5' 말단으로부터 위치 9, 10, 11에서 3개의 연속 뉴클레오티드 상에 3개의 2'-F 변형의 적어도 하나의 모티프를 함유한다. 안티센스 가닥은 5' 말단으로부터 위치 11, 12, 13에서 3개의 연속 뉴클레오티드 상에 3개의 2'-O-메틸 변형의 적어도 하나의 모티프를 함유한다.
하나의 양태에서, RNAi 제제는 21개의 뉴클레오티드 센스 가닥 및 23개의 뉴클레오티드 안티센스 가닥을 포함하며, 여기서 센스 가닥은 5' 말단으로부터 위치 9, 10, 11에서 3개의 연속 뉴클레오티드 상에 3개의 2'-F 변형의 적어도 하나의 모티프를 함유하고; 안티센스 가닥은 5' 말단으로부터 위치 11, 12, 13에서 3개의 연속 뉴클레오티드 상에 3개의 2'-O-메틸 변형의 적어도 하나의 모티프를 함유하며, 여기서 RNAi 제제의 한쪽 말단은 평활한 반면 다른 쪽 말단은 2개의 뉴클레오티드 돌출부를 포함한다. 바람직하게는, 2개의 뉴클레오티드 돌출부는 안티센스 가닥의 3'-말단에 있다. 2개의 뉴클레오티드 돌출부가 안티센스 가닥의 3' 말단에 있는 경우, 말단 3개의 뉴클레오티드 사이에 2개의 포스포로티오에이트 뉴클레오티드간 결합이 있을 수 있으며, 여기서 3개의 뉴클레오티드 중 2개는 돌출부 뉴클레오티드이고, 세 번째 뉴클레오티드는 돌출부 뉴클레오티드 옆에 있는 쌍을 이루는 뉴클레오티드이다. 하나의 양태에서, RNAi 제제는 추가로 센스 가닥의 5'-말단과 안티센스 가닥의 5'-말단 둘 다에서 말단 3개의 뉴클레오티드 사이에 2개의 포스포로티오에이트 뉴클레오티드간 결합을 갖는다. 하나의 양태에서, 모티프의 일부인 뉴클레오티드를 포함하는, RNAi 제제의 센스 가닥 및 안티센스 가닥의 모든 뉴클레오티드는 변형된 뉴클레오티드이다. 하나의 양태에서 각 잔기는 독립적으로, 예를 들어, 교대 모티프에서 2'-O-메틸 또는 3'-플루오로로 변형된다. 임의로, RNAi 제제는 리간드(예를 들어, 친유성 리간드, 임의로 C16 리간드)를 추가로 포함한다.
하나의 양태에서, RNAi 제제는 센스 및 안티센스 가닥을 포함하고, 여기서 센스 가닥은 25-30개의 뉴클레오티드 잔기 길이이고, 여기서 5' 말단 뉴클레오티드(위치 1)에서 시작하여 첫 번째 가닥의 위치 1 내지 23은 적어도 8개의 리보뉴클레오티드를 포함하고; 안티센스 가닥은 36-66개의 뉴클레오티드 잔기 길이이고, 3' 말단 뉴클레오티드에서 시작하여, 센스 가닥의 위치 1-23과 쌍을 이루는 위치에 적어도 8개의 리보뉴클레오티드를 포함하여 이중체를 형성하고; 여기서 안티센스 가닥의 적어도 3' 말단 뉴클레오티드는 센스 가닥과 쌍을 이루지 않고, 최대 6개의 연속적인 3' 말단 뉴클레오티드는 센스 가닥과 쌍을 이루지 않아, 1-6개의 뉴클레오티드의 3' 단일 가닥 돌출부를 형성하며; 여기서 안티센스 가닥의 5' 말단은 센스 가닥과 쌍을 이루지 않은 10-30개의 연속 뉴클레오티드를 포함하여 10-30개의 뉴클레오티드 단일 가닥 5' 돌출부를 형성하고; 여기서 적어도 센스 가닥 5' 말단 및 3' 말단 뉴클레오티드는 센스 및 안티센스 가닥이 최대 상보성을 위해 정렬될 때 안티센스 가닥의 뉴클레오티드와 염기쌍을 이루어 센스 가닥과 안티센스 가닥 사이에 실질적으로 이중체화된 영역을 형성하고; 안티센스 가닥은 이중 가닥 핵산이 포유동물 세포에 도입될 때 표적 유전자 발현을 감소시키기 위해 안티센스 가닥 길이의 적어도 19개의 리보뉴클레오티드를 따라 표적 RNA에 대해 충분히 상보적이고; 여기서 센스 가닥은 3개의 연속적인 뉴클레오티드에 3개의 2'-F 변형의 적어도 하나의 모티프를 함유하고, 여기서 모티프 중 적어도 하나는 절단 부위 또는 그 부근에서 발생한다. 안티센스 가닥은 절단 부위에 또는 그 부근에서 3개의 연속 뉴클레오티드 상에 3개의 2'-O-메틸 변형의 적어도 하나의 모티프를 함유한다.
하나의 양태에서, RNAi 제제는 센스 및 안티센스 가닥을 포함하고, 여기서 RNAi 제제는 적어도 25개 및 최대 29개의 뉴클레오티드인 길이를 갖는 제1 가닥 및 5' 말단으로부터 위치 11, 12, 13에서 3개의 연속 뉴클레오티드 상에 3개의 2'-O-메틸 변형의 적어도 하나의 모티프를 갖는 최대 30개의 뉴클레오티드인 길이를 갖는 제2 가닥을 포함하고; 여기서 첫 번째 가닥의 3' 말단과 두 번째 가닥의 5' 말단은 평활 말단을 형성하고 두 번째 가닥은 첫 번째 가닥보다 3' 말단에서 1-4개의 뉴클레오티드 더 길고, 여기서 이중체 영역은 적어도 25개의 뉴클레오티드 길이이고, 두 번째 가닥은 RNAi 제제가 포유동물 세포에 도입될 때 표적 유전자 발현을 감소시키기 위해 두 번째 가닥 길이의 적어도 19개의 뉴클레오티드를 따라 표적 mRNA에 충분히 상보적이고, 여기서 RNAi 제제의 다이서 절단은 우선적으로 두 번째 가닥의 3' 말단을 포함하는 siRNA를 초래하여 포유동물에서 표적 유전자의 발현을 감소시킨다. 임의로, RNAi 제제는 리간드를 추가로 포함한다.
하나의 양태에서, RNAi 제제의 센스 가닥은 3개의 연속 뉴클레오티드 상에 세 개의 동일한 변형의 적어도 하나의 모티프를 함유하며, 여기서 모티프 중 하나는 센스 가닥의 절단 부위에서 발생한다.
하나의 양태에서, RNAi 제제의 안티센스 가닥 또한 3개의 연속 뉴클레오티드 상에 세 개의 동일한 변형의 적어도 하나의 모티프를 함유할 수 있며, 여기서 모티프 중 하나는 안티센스 가닥의 절단 부위에 또는 그 부근에서 발생한다.
17-23개의 뉴클레오티드 길이의 이중체 영역을 갖는 RNAi 제제의 경우, 안티센스 가닥의 절단 부위는 전형적으로 5'-말단으로부터 대략 10, 11 및 12 위치이다. 따라서, 3개의 동일한 변형의 모티프는 안티센스 가닥의 9, 10, 11 위치; 10, 11, 12 위치; 11, 12, 13 위치; 12, 13, 14 위치; 또는 13, 14, 15 위치에서 발생할 수 있으며, 개수는 안티센스 가닥의 5'-말단으로부터의 1st 뉴클레오티드부터 시작하거나, 개수는 안티센스 가닥의 5'-말단으로부터의 이중체 영역 내의 1st 쌍을 이룬 뉴클레오티드로부터 시작한다. 안티센스 가닥의 절단 부위는 또한 5'-말단으로부터 RNAi의 이중체 영역의 길이에 따라 변할 수 있다.
RNAi 제제의 센스 가닥은 가닥의 절단 부위에서 3개의 연속 뉴클레오티드에 3개의 동일한 변형의 적어도 하나의 모티프를 함유할 수 있고; 안티센스 가닥은 가닥의 절단 부위 또는 그 부근에서 3개의 연속 뉴클레오티드에 3개의 동일한 변형의 적어도 하나의 모티프를 가질 수 있다. 센스 가닥 및 안티센스 가닥이 dsRNA 이중체를 형성할 때, 센스 가닥 및 안티센스 가닥은 센스 가닥 상의 3개의 뉴클레오티드 중 하나의 모티프 및 안티센스 가닥 상의 3개의 뉴클레오티드 중 하나의 모티프가 적어도 하나의 뉴클레오티드 중첩을 갖도록 정렬될 수 있으며, 즉 센스 가닥에서 모티프의 세 개의 뉴클레오티드 중 적어도 하나는 안티센스 가닥에서 모티프의 세 개의 뉴클레오티드 중 적어도 하나와 염기 쌍을 형성한다. 대안적으로, 적어도 2개의 뉴클레오티드가 중첩할 수 있거나, 3개의 뉴클레오티드 모두가 중첩할 수 있다.
하나의 양태에서, RNAi 제제의 센스 가닥은 3개의 연속 뉴클레오티드에 3개의 동일한 변형의 하나 이상의 모티프를 함유할 수 있다. 첫 번째 모티프는 가닥의 절단 부위 또는 그 부근에서 발생할 수 있고 다른 모티프는 윙 변형(wing modification)일 수 있다. 본원에서 용어 "윙 변형"은 동일한 가닥의 절단 부위 또는 그 부근에서 모티프로부터 분리된 가닥의 다른 부분에서 발생하는 모티프를 지칭한다. 윙 변형은 첫 번째 모티프에 인접하거나 적어도 하나 이상의 뉴클레오티드에 의해 분리된다. 모티프가 서로 바로 인접해 있는 경우 모티프의 화학적 성질은 서로 구별되며, 모티프가 하나 이상의 뉴클레오티드에 의해 분리되는 경우 화학적 성질은 동일하거나 상이할 수 있다. 두 개 이상의 윙 변형이 존재할 수 있다. 예를 들어, 2개의 윙 변형이 존재하는 경우, 각 윙 변형은 절단 부위 또는 그 부근에 있는 첫 번째 모티프에 대한 한쪽 말단에서 또는 리드 모티프의 어느 한 쪽에서 발생할 수 있다.
센스 가닥과 같이, RNAi 제제의 안티센스 가닥은 가닥의 절단 부위 또는 그 부근에서 발생하는 모티프 중 적어도 하나와 함께 3개의 연속 뉴클레오티드에 3개의 동일한 변형의 하나 이상의 모티프를 함유할 수 있다. 이러한 안티센스 가닥은 또한 센스 가닥에 존재할 수 있는 윙 변형과 유사한 정렬에서 하나 이상의 윙 변형을 포함할 수 있다.
하나의 양태에서, RNAi 제제의 센스 가닥 또는 안티센스 가닥에 대한 윙 변형은 전형적으로 가닥의 3'-말단, 5'-말단 또는 양 말단에 처음 1개 또는 2개의 말단 뉴클레오티드를 포함하지 않는다.
또 다른 양태에서, RNAi 제제의 센스 가닥 또는 안티센스 가닥에 대한 윙 변형은 전형적으로 가닥의 3'-말단, 5'-말단 또는 양 말단에서 이중체 영역 내에 처음 1개 또는 2개의 쌍을 이루는 뉴클레오티드를 포함하지 않는다.
RNAi 제제의 센스 가닥 및 안티센스 가닥이 각각 적어도 하나의 윙 변형을 함유하는 경우, 윙 변형은 이중체 영역의 동일한 말단에 있을 수 있으며 1개, 2개 또는 3개의 뉴클레오티드의 중첩을 가질 수 있다.
RNAi 제제의 센스 가닥 및 안티센스 가닥이 각각 적어도 두 개의 윙 변형을 함유하는 경우, 센스 가닥 및 안티센스 가닥은 각각 하나의 가닥으로부터의 2개의 변형이 1개, 2개 또는 3개의 뉴클레오티드의 중첩을 갖는 이중체 영역의 한쪽 말단에 있고; 각각 하나의 가닥으로부터의 2개의 변형이 1개, 2개 또는 3개의 뉴클레오티드의 중첩을 갖는 이중체 영역의 다른 말단에 있고; 하나의 가닥으로부터의 2개의 변형이 이중체 영역에 1개, 2개 또는 3개의 뉴클레오티드의 중첩을 갖는 리드 모티프의 각 측면에 있도록 정렬될 수 있다.
하나의 양태에서, RNAi 제제는 표적과, 이중체 내에, 또는 이들의 조합에 미스매치(들)를 포함한다. 미스매치는 돌출부 영역 또는 이중체 영역에서 발생할 수 있다. 염기 쌍은 해리 또는 용융을 촉진하는 이들의 경향을 기준으로 하여 순위가 매겨질 수 있다(예를 들어, 특정 쌍의 결합 또는 해리의 자유 에너지에 대해, 가장 간단한 접근법은 개별 쌍을 기반으로 쌍을 조사하는 것이지만, 다음 이웃하거나 유사한 분석도 사용될 수 있다). 해리 촉진 측면에서: A:U가 G:C보다 바람직하고; G:U가 G:C보다 바람직하며; I:C가 G:C보다 바람직하다(I=이노신). 미스매치, 예를 들어, 비-정준 또는 정준 이외의 쌍(본원의 다른 곳에 기술된 바와 같음)이 정준 (A:T, A:U, G:C) 쌍보다 바람직하고; 보편적인 염기를 포함하는 쌍이 정준 쌍보다 바람직하다.
하나의 양태에서, RNAi 제제는 이중체의 5'-말단에서 안티센스 가닥의 해리를 촉진하기 위해 A:U, G:U, I:C의 그룹으로부터 독립적으로 선택된 안티센스 가닥의 5'-말단으로부터의 이중체 영역 내에 처음 1, 2, 3, 4, 또는 5개 염기 쌍 중 적어도 하나, 및 미스매칭된 쌍, 예를 들어, 보편적인 염기를 포함하는 비-정준 또는 정준 이외의 쌍을 포함한다.
하나의 양태에서, 안티센스 가닥의 5'-말단으로부터 이중체 영역 내의 1 위치에 있는 뉴클레오티드는 A, dA, dU, U, 및 dT로 이루어진 그룹으로부터 선택된다. 대안적으로, 안티센스 가닥의 5'-말단으로부터 이중체 영역 내의 처음 1, 2 또는 3개 염기 쌍 중 적어도 하나는 AU 염기 쌍이다. 예를 들면, 안티센스 가닥의 5'-말단으로부터 이중체 영역 내의 첫 번째 염기 쌍은 AU 염기 쌍이다.
또 다른 양태에서, 센스 가닥의 3'-말단에 있는 뉴클레오티드는 데옥시-티민(dT)이다. 또 다른 양태에서, 안티센스 가닥의 3'-말단에 있는 뉴클레오티드는 데옥시-티민(dT)이다. 하나의 양태에서, 센스 또는 안티센스 가닥의 3'-말단에 데옥시-티민 뉴클레오티드의 짧은 서열, 예를 들면, 2개의 dT 뉴클레오티드가 있다.
하나의 양태에서, 센스 가닥 서열은 화학식 I로 나타내어질 수 있다:
[화학식 I]
5' np-Na-(X X X )i-Nb-Y Y Y -Nb-(Z Z Z )j-Na-nq 3'
여기서:
i 및 j는 각각 독립적으로 0 또는 1이고;
p 및 q는 각각 독립적으로 0-6이고;
각각의 Na는 독립적으로 0-25개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타내고, 각 서열은 적어도 두 개의 상이하게 변형된 뉴클레오티드를 포함하며;
각각의 Nb는 독립적으로 0-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타내고;
각각의 np 및 nq는 독립적으로 돌출부 뉴클레오티드를 나타내며;
Nb 및 Y는 동일한 변형을 갖지 않고;
XXX, YYY 및 ZZZ는 각각 독립적으로 3개의 연속 뉴클레오티드에 대한 3개의 동일한 변형의 하나의 모티프를 나타낸다. 바람직하게는 YYY는 모두 2'-F 변형된 뉴클레오티드이다.
하나의 양태에서, Na 및 Nb는 교대 패턴의 변형을 포함한다.
하나의 양태에서, YYY 모티프는 센스 가닥의 절단 부위 또는 그 부군에서 발생한다. 예를 들면, RNAi 제제가 17-23개의 뉴클레오티드 길이의 이중체 영역을 갖는 경우, YYY 모티프는 센스 가닥의 절단 부위 또는 그 부근에서 발생할 수 있으며(예를 들어: 위치 6, 7, 8, 7, 8, 9, 8, 9, 10, 9, 10, 11, 10, 11, 12 또는 11, 12, 13), 개수는 5'-말단으로부터 1st 뉴클레오티드로부터 시작하거나; 또는 임의로, 개수는 5'-말단으로부터 이중체 영역 내의 1st 쌍을 이루는 뉴클레오티드에서 시작한다.
하나의 양태에서, i는 1이고 j는 0이거나, i는 0이고 j는 1이거나, i 및 j 둘 다 1이다. 따라서 센스 가닥은 다음 화학식으로 나타낼 수 있다:
[화학식 Ib]
5' np-Na-YYY-Nb-ZZZ-Na-nq 3';
[화학식 Ic]
5' np-Na-XXX-Nb-YYY-Na-nq 3'; 또는
[화학식 Id]
5' np-Na-XXX-Nb-YYY-Nb-ZZZ-Na-nq 3'.
센스 가닥이 화학식 Ib로 나타내어질 때, Nb는 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다.
각각의 Na는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낼 수 있다.
센스 가닥이 화학식 Ic로 나타내어질 때, Nb는 0-10, 0-7, 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낼 수 있다.
센스 가닥이 화학식 Id로 나타내어질 때, 각각의 Nb는 독립적으로 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 바람직하게는, Nb는 0, 1, 2, 3, 4, 5 또는 6이다. 각각의 Na는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낼 수 있다.
각각의 X, Y 및 Z는 서로 동일하거나 상이할 수 있다.
다른 양태에서, i는 0이고 j는 0이며, 센스 가닥은 하기 화학식으로 나타낼 수 있다:
[화학식 Ia]
5' np-Na-YYY- Na-nq 3'.
센스 가닥이 화학식 Ia로 나타내어질 때, 각각의 Na는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낼 수 있다.
하나의 양태에서, RNAi의 안티센스 가닥 서열은 화학식 II로 나타낼 수 있다:
[화학식 II]
5' nq '-Na'-(Z'Z'Z')k-Nb'-Y'Y'Y'-Nb'-(X'X'X')l-N'a-np' 3'
여기서:
k 및 l은 각각 독립적으로 0 또는 1이고;
p' 및 q'는 각각 독립적으로 0-6이고;
각각의 Na'는 독립적으로 0-25개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타내고, 각각의 서열은 적어도 두 개의 상이하게 변형된 뉴클레오티드를 포함하며;
각각의 Nb'는 독립적으로 0-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타내고;
각각의 np' 및 nq'는 독립적으로 돌출부 뉴클레오티드를 나타내고;
여기서 Nb' 및 Y'는 동일한 변형을 갖지 않고;
X'X'X', Y'Y'Y' 및 Z'Z'Z'는 각각 독립적으로 3개의 연속 뉴클레오티드에 대한 3개의 동일한 변형의 하나의 모티프를 나타낸다.
하나의 양태에서, Na' 또는 Nb'는 교대 패턴의 변형을 포함한다.
Y'Y'Y' 모티프는 안티센스 가닥의 절단 부위 또는 그 부근에서 발생한다. 예를 들면, RNAi 제제가 17-23개의 뉴클레오티드 길이의 이중체 영역을 갖는 경우, Y'Y'Y' 모티프는 안티센스 가닥의 위치 9, 10, 11; 10, 11, 12; 11, 12, 13; 12, 13, 14; 또는 13, 14, 15에서 발생할 수 있으며, 개수는 5'-말단으로부터 1st 뉴클레오티드에서 시작하거나; 임의로, 개수는 5'-말단으로부터 이중체 영역 내의 1st 쌍을 이루는 뉴클레오티드에서 시작한다. 바람직하게는, Y'Y'Y' 모티프는 위치 11, 12, 13에서 발생한다.
하나의 양태에서, Y'Y'Y' 모티프는 모두 2'-OMe 변형된 뉴클레오티드이다.
하나의 양태에서, k는 1이고 l은 0이거나, k는 0이고 l은 1이거나, k 및 l 둘 다 1이다.
따라서 안티센스 가닥은 하기 화학식으로 나타낼 수 있다:
[화학식 IIb]
5' nq '-Na'-Z'Z'Z'-Nb'-Y'Y'Y'-Nb'-np ' 3';
[화학식 IIc]
5' nq '-Na'-Y'Y'Y'-Nb'-X'X'X'-np ' 3'; 또는
[화학식 IId]
5' nq '-Na'-Z'Z'Z'-Nb'-Y'Y'Y'-Nb'-X'X'X'-N'a-np ' 3'.
안티센스 가닥이 화학식 IIb로 나타내어질 때, Nb'는 0-10, 0-7, 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na'는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다.
안티센스 가닥이 화학식 IIc로 나타내어질 때, Nb'는 0-10, 0-7, 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na'는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다.
안티센스 가닥이 화학식 IId로 나타내어질 때, 각각의 Nb'는 독립적으로 0-10, 0-7, 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na'는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 바람직하게는, Nb는 0, 1, 2, 3, 4, 5 또는 6이다.
다른 양태에서, k는 0이고 l은 0이며 안티센스 가닥은 다음 화학식으로 나타낼 수 있다:
화학식 Ia
5' np '-Na'-Y'Y'Y'- Na'-nq ' 3'.
안티센스 가닥이 화학식 IIa로 나타내어질 때, 각각의 Na'는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다.
각각의 X', Y' 및 Z'는 서로 동일하거나 상이할 수 있다.
센스 가닥 및 안티센스 가닥의 각 뉴클레오티드는 독립적으로 LNA, HNA, CeNA, 2'-메톡시에틸, 2'-O-메틸, 2'-O-알릴, 2'-C-알릴, 2'-하이드록실, 또는 2'-플루오로로 변형될 수 있다. 예를 들면, 센스 가닥 및 안티센스 가닥의 각 뉴클레오티드는 독립적으로 2'-O-메틸 또는 2'-플루오로로 변형된다. 각각의 X, Y, Z, X', Y' 및 Z'는 특히 2'-O-메틸 변형 또는 2'-플루오로 변형을 나타낼 수 있다.
하나의 양태에서, RNAi 제제의 센스 가닥은 이중체 영역이 21 nt인 경우 가닥의 9, 10 및 11 위치에서 발생하는 YYY 모티프를 함유할 수 있으며, 개수는 5'-말단으로부터의 1st 뉴클레오티드부터 시작하거나, 임의로, 개수는 5'-말단으로부터의 이중체 영역 내의 1st 쌍을 이룬 뉴클레오티드로부터 시작하고; Y는 2'-F 변형을 나타낸다. 센스 가닥은 이중체 영역의 반대쪽 말단에 윙 변형으로서 XXX 모티프 또는 ZZZ 모티프를 추가로 함유할 수 있으며; XXX 및 ZZZ는 각각 독립적으로 2'OMe 변형 또는 2'-F 변형을 나타낸다.
하나의 양태에서 안티센스 가닥은 가닥의 위치 11, 12, 13에서 발생하는 Y'Y'Y' 모티프를 함유할 수 있으며, 개수는 5'-말단으로부터의 1st 뉴클레오티드부터 시작하거나, 임의로, 개수는 5'-말단으로부터의 이중체 영역 내의 1st 쌍을 이룬 뉴클레오티드로부터 시작하고; Y'는 2'-O-메틸 변형을 나타낸다. 안티센스 가닥은 이중체 영역의 반대쪽 말단에 윙 변형으로서 X'X'X' 모티프 또는 Z'Z'Z' 모티프를 추가로 함유할 수 있으며; X'X'X' 및 Z'Z'Z'는 각각 독립적으로 2'-OMe 변형 또는 2'-F 변형을 나타낸다.
상기 화학식 Ia, Ib, Ic, 및 Id 중 어느 하나로 나타내어지는 센스 가닥은 각각 화학식 IIa, IIb, IIc, 및 IId 중 어느 하나로 나타내어지는 안티센스 가닥과 이중체를 형성한다.
따라서, 본 개시내용의 방법에서 사용하기 위한 RNAi 제제는 센스 가닥 및 안티센스 가닥을 포함할 수 있고, 각 가닥은 14 내지 30개의 뉴클레오티드를 가지며, RNAi 이중체는 화학식 III으로 나타내어진다:
[화학식 III]
센스: 5' np -Na-(X X X)i -Nb- Y Y Y -Nb -(Z Z Z)j-Na-nq 3'
안티센스: 3' np'-Na'-(X' X' X')k -Nb'- Y' Y' Y' -Nb' -(Z' Z' Z')l-Na'-nq' 5'
여기서:
i, j, k, 및 l은 각각 독립적으로 0 또는 1이고;
p, p', q, 및 q'는 각각 독립적으로 0-6이고;
각각의 Na 및 Na '는 독립적으로 0-25개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타내고, 각 서열은 적어도 두 개의 상이하게 변형된 뉴클레오티드를 포함하며;
각각의 Nb 및 Nb '는 독립적으로 0-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타내고;
여기서
각각의 np', np, nq', 및 nq는, 이들 각각이 존재하거나 존재하지 않을 수 있지만, 독립적으로 돌출부 뉴클레오티드를 나타내고;
XXX, YYY, ZZZ, X'X'X', Y'Y'Y', Z'Z'Z'는 각각 독립적으로 3개의 연속 뉴클레오티드에 대한 3개의 동일한 변형의 하나의 모티프를 나타낸다.
하나의 양태에서, i는 0이고 j는 0이거나; i는 1이고 j는 0이거나; i는 0이고 j는 1이거나; i 및 j 둘 다는 0이거나; i 및 j 둘 다 1이다. 또 다른 양태에서, k는 0이고 l은 0이거나; k는 1이고 l은 0이거나; k는 0이고 l은 1이거나; k 및 l 둘 다 0이거나; k 및 l 둘 다 1이다.
RNAi 이중체를 형성하는 센스 가닥 및 안티센스 가닥의 예시적인 조합은 아래 화학식을 포함한다:
[화학식 IIIa]
5' np - Na -Y Y Y -Na-nq 3'
3' np '-Na '-Y' Y' Y' -Na 'nq ' 5'
[화학식 IIIb]
5' np -Na -Y Y Y -Nb -Z Z Z -Na-nq 3'
3' np '-Na '-Y' Y' Y' -Nb '-Z' Z' Z' -Na 'nq ' 5'
[화학식 IIIc]
5' np-Na- X X X -Nb -Y Y Y - Na-nq 3'
3' np '-Na '-X' X' X' -Nb '-Y' Y' Y'-Na '-nq ' 5'
[화학식 IIId]
5' np -Na -X X X -Nb-Y Y Y -Nb- Z Z Z -Na-nq 3'
3' np '-Na '-X' X' X' -Nb '-Y' Y' Y' -Nb '-Z' Z' Z'-Na-nq ' 5'
RNAi 제제가 화학식 IIIa로 나타내어질 때, 각각의 Na는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다.
RNAi 제제가 화학식 IIIb로 나타내어질 때, 각각의 Nb는 독립적으로 1-10, 1-7, 1-5 또는 1-4개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다.
RNAi 제제가 화학식 IIIc로 나타내어질 때, 각각의 Nb, Nb'는 독립적으로 0-10, 0-7, 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다.
RNAi 제제가 화학식 IIId로 나타내어질 때, 각각의 Nb, Nb'는 독립적으로 0-10, 0-7, 0-10, 0-7, 0-5, 0-4, 0-2 또는 0개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na, Na '는 독립적으로 2-20, 2-15, 또는 2-10개 변형된 뉴클레오티드를 포함하는 올리고뉴클레오티드 서열을 나타낸다. 각각의 Na, Na', Nb 및 Nb '는 독립적으로 교대 패턴의 변형을 포함한다.
하나의 양태에서, RNAi 제제가 화학식 IIId로 나타내어질 때, Na 변형은 2'-O-메틸 또는 2'-플루오로 변형이다. 또 다른 양태에서, RNAi 제제가 화학식 IIId로 나타내어질 때, Na 변형은 2'-O-메틸 또는 2'-플루오로 변형이고 np' >0 및 적어도 하나의 np'는 포스포로티오에이트 결합을 통해 이웃하는 뉴클레오티드에 연결된다. 또 다른 양태에서, RNAi 제제가 화학식 IIId로 나타내어질 때, Na 변형은 2'-O-메틸 또는 2'-플루오로 변형이고 np' >0 및 적어도 하나의 np'는 포스포로티오에이트 결합을 통해 이웃하는 뉴클레오티드에 연결되고, 센스 가닥은 2가 또는 3가 분지형 링커(아래에 기술됨)를 통해 부착된 하나 이상의 C16 (또는 관련) 모이어티에 접합된다. 또 다른 양태에서, RNAi 제제가 화학식 IIId로 나타내어질 때, Na 변형은 2'-O-메틸 또는 2'-플루오로 변형이고 np' >0 및 적어도 하나의 np'는 포스포로티오에이트 결합을 통해 이웃하는 뉴클레오티드에 연결되고, 센스 가닥은 적어도 하나의 포스포로티오에이트 결합을 포함하고, 센스 가닥은 2가 또는 3가 분지형 링커를 통해 임의로 부착된 하나 이상의 친유성, 예를 들어, C16 (또는 관련) 모이어티에 접합된다.
하나의 양태에서, RNAi 제제가 화학식 IIIa로 나타내어질 때, Na 변형은 2'-O-메틸 또는 2'-플루오로 변형이고, np' >0 및 적어도 하나의 np'는 포스포로티오에이트 결합을 통해 이웃하는 뉴클레오티드에 연결되고, 센스 가닥은 적어도 하나의 포스포로티오에이트 결합을 포함하고, 센스 가닥은 2가 또는 3가 분지형 링커를 통해 부착된 하나 이상의 친유성, 예를 들어, C16 (또는 관련) 모이어티에 접합된다.
하나의 양태에서, RNAi 제제는 화학식 III, IIIa, IIIb, IIIc, 및 IIId로 나타내어지는 적어도 두 개의 이중체를 함유하는 다량체이며, 여기서 이중체는 링커에 의해 연결된다. 링커는 절단 가능하거나 절단 불가능할 수 있다. 임의로, 다량체는 리간드를 추가로 포함한다. 각각의 이중체는 동일한 유전자 또는 2개의 상이한 유전자를 표적으로 할 수 있거나; 각각의 이중체는 2개의 상이한 표적 부위에서 동일한 유전자를 표적으로 할 수 있다.
하나의 양태에서, RNAi 제제는 화학식 III, IIIa, IIIb, IIIc, 및 IIId로 나타내어지는 3, 4, 5, 6개 이상의 이중체를 함유하는 다량체이며, 여기서 이중체는 링커에 의해 연결된다. 링커는 절단 가능하거나 절단 불가능할 수 있다. 임의로, 다량체는 리간드를 추가로 포함한다. 각각의 이중체는 동일한 유전자 또는 2개의 상이한 유전자를 표적으로 할 수 있거나; 각각의 이중체는 2개의 상이한 표적 부위에서 동일한 유전자를 표적으로 할 수 있다.
하나의 양태에서, 화학식 III, IIIa, IIIb, IIIc, 및 IIId로 나타내어지는 두 개의 RNAi 제제는 5' 말단, 및 3' 말단 중 하나 또는 둘 다에서 서로 연결되며 리간드에 임의로 접합된다. 각각의 제제는 동일한 유전자 또는 2개의 상이한 유전자를 표적으로 할 수 있거나; 각각의 제제는 2개의 상이한 표적 부위에서 동일한 유전자를 표적으로 할 수 있다.
다양한 공보가 본 개시내용의 방법에 사용될 수 있는 다량체 RNAi 제제를 기술한다. 이러한 공보는 제WO2007/091269호, 제WO2010/141511호, 제WO2007/117686호, 제WO2009/014887호, 및 제WO2011/031520호; 및 제US 7858769호를 포함하고, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
특정 양태에서, 본 개시내용의 조성물 및 방법은 본원에 기술된 바와 같은 RNAi 제제의 비닐 포스포네이트(VP) 변형을 포함한다. 예시적인 양태에서, 본 개시내용의 비닐 포스포네이트는 하기 구조를 갖는다:

본 개시내용의 비닐 포스포네이트는 본 개시내용의 dsRNA의 안티센스 또는 센스 가닥에 부착될 수 있다. 특정의 바람직한 양태에서, 본 개시내용의 비닐 포스포네이트는 임의로 dsRNA의 안티센스 가닥의 5' 말단에서 dsRNA의 안티센스 가닥에 부착된다.
비닐 포스페이트 변형이 또한 본 개시내용의 조성물 및 방법에 대해 고려된다. 예시적인 비닐 포스페이트 구조는 다음과 같다:

i. 열적으로 불안정한 변형
특정 양태에서, dsRNA 분자는 오프-타겟 유전자 침묵을 감소시키거나 억제하기 위해 안티센스 가닥의 시드 영역에(즉, 안티센스 가닥의 5'-말단의 위치 2-9에서) 열적으로 불안정한 변형을 통합함으로써 RNA 간섭에 대해 최적화될 수 있다. 안티센스 가닥의, 5' 말단부터 개수하여, 처음 9개의 뉴클레오티드 위치 내에 이중체의 적어도 하나의 열적으로 불안정한 변형을 포함하는 안티센스 가닥을 가진 dsRNA가 오프-타겟 유전자 침묵 활성을 감소시킨 것으로 밝혀졌다. 따라서, 일부 양태에서, 안티센스 가닥은 안티센스 가닥의 5' 영역의 처음 9개의 뉴클레오티드 위치 내에서 이중체의 적어도 하나(예를 들어, 1, 2, 3, 4, 5개 또는 그 이상)의 열적으로 불안정한 변형을 포함한다. 일부 양태에서, 이중체의 하나 이상의 열적으로 불안정한 변형(들)은 안티센스 가닥의 5'-말단으로부터 위치 2-9, 또는 바람직하게는 위치 4-8에 위치한다. 일부 추가 양태에서, 이중체의 열적으로 불안정한 변형(들)은 안티센스 가닥의 5'-말단으로부터 위치 6, 7 또는 8에 위치한다. 여전히 일부 추가 양태에서, 이중체의 열적으로 불안정한 변형은 안티센스 가닥의 5'-말단으로부터 위치 7에 위치한다. 용어 "열적으로 불안정한 변형(들)"은 더 낮은 전체 용융 온도(Tm)(바람직하게는 이러한 변형(들)이 없는 dsRNA의 Tm보다 1, 2, 3 또는 4도 더 낮은 Tm)를 갖는 dsRNA로 야기되는 변형(들)을 포함한다. 일부 양태에서, 이중체의 열적으로 불안정한 변형은 안티센스 가닥의 5'-말단으로부터 위치 2, 3, 4, 5 또는 9에 위치한다.
열적으로 불안정한 변형은 무염기 변형; 반대 가닥의 반대 뉴클레오티드와의 미스매치; 및 당 변형, 예를 들어 2'-데옥시 변형 또는 비사이클릭 뉴클레오티드, 예를 들어, 비고정 핵산(UNA) 또는 글리콜 핵산(GNA)을 포함할 수 있지만, 이에 제한되지 않는다.
예시된 무염기 변형은 다음을 포함하지만, 이에 제한되지 않는다:

여기서 R = H, Me, Et 또는 OMe; R' = H, Me, Et 또는 OMe; R" = H, Me, Et 또는 OMe

여기서 B는 변형되거나 변형되지 않은 핵염기이다.
예시된 당 변형은 다음을 포함하지만, 이에 제한되지 않는다:


여기서 B는 변형되거나 변형되지 않은 핵염기이다.
일부 양태에서 이중체의 열적으로 불안정한 변형은 다음으로 이루어진 그룹으로부터 선택된다:

여기서 B는 변형되거나 변형되지 않은 핵염기이고 각 구조의 별표는 R, S 또는 라세미체를 나타낸다.
용어 "비사이클릭 뉴클레오티드"는 비사이클릭 리보스 당을 갖는 임의의 뉴클레오티드를 지칭하며, 예를 들면, 여기서 리보스 탄소 간의 임의이 결합(예를 들어, C1'-C2', C2'-C3', C3'-C4', C4'-O4' 또는 C1'-O4')이 없거나 리보스 탄소 또는 산소(예를 들어, C1', C2', C3', C4' 또는 O4') 중 적어도 하나가 독립적으로 또는 조합하여 뉴클레오티드로부터 부재하다. 일부 양태에서, 비사이클릭 뉴클레오티드는  ,
,  ,
,  ,
,  또는
또는  이고, 여기서 B는 변형되거나 변형되지 않은 핵염기이고, R1 및 R2는 독립적으로 H, 할로겐, OR3, 또는 알킬이고; R3은 H, 알킬, 사이클로알킬, 아릴, 아르알킬, 헤테로아릴 또는 당이다). 용어 "UNA"는 비고정된 비사이클릭 핵산을 지칭하며, 여기서 당의 결합 중 어느 것이 제거되어 비고정된 "당" 잔기를 형성한다. 하나의 예에서, UNA는 또한 C1'-C4' 사이의 결합(즉, C1'과 C4' 탄소 사이의 공유 탄소-산소-탄소 결합)이 제거된 단량체를 포함한다. 또 다른 예에서, 당의 C2'-C3' 결합(즉, C2' 및 C3' 탄소 사이의 공유 탄소-탄소 결합)이 제거된다(참조; Mikhailov et. al., Tetrahedron Letters, 26 (17): 2059 (1985); and Fluiter et al., Mol. Biosyst., 10: 1039 (2009), 전문이 본원에 참고로 포함됨). 비사이클릭 유도체는 Watson-Crick 쌍에 영향을 주지 않고 더 큰 백본 유연성을 제공한다. 비사이클릭 뉴클레오티드는 2'-5' 또는 3'-5' 결합을 통해 연결될 수 있다.
이고, 여기서 B는 변형되거나 변형되지 않은 핵염기이고, R1 및 R2는 독립적으로 H, 할로겐, OR3, 또는 알킬이고; R3은 H, 알킬, 사이클로알킬, 아릴, 아르알킬, 헤테로아릴 또는 당이다). 용어 "UNA"는 비고정된 비사이클릭 핵산을 지칭하며, 여기서 당의 결합 중 어느 것이 제거되어 비고정된 "당" 잔기를 형성한다. 하나의 예에서, UNA는 또한 C1'-C4' 사이의 결합(즉, C1'과 C4' 탄소 사이의 공유 탄소-산소-탄소 결합)이 제거된 단량체를 포함한다. 또 다른 예에서, 당의 C2'-C3' 결합(즉, C2' 및 C3' 탄소 사이의 공유 탄소-탄소 결합)이 제거된다(참조; Mikhailov et. al., Tetrahedron Letters, 26 (17): 2059 (1985); and Fluiter et al., Mol. Biosyst., 10: 1039 (2009), 전문이 본원에 참고로 포함됨). 비사이클릭 유도체는 Watson-Crick 쌍에 영향을 주지 않고 더 큰 백본 유연성을 제공한다. 비사이클릭 뉴클레오티드는 2'-5' 또는 3'-5' 결합을 통해 연결될 수 있다.
용어 'GNA'는 DNA 또는 RNA와 유사하지만 포스포디에스테르 결합에 의해 연결된 반복 글리세롤 단위로 구성되어 있다는 점에서 이의 "백분"의 구성이 상이한 글리콜 핵산을 지칭한다:

이중체의 열적으로 불안정한 변형은 열적으로 불안정한 뉴클레오티드와 dsRNA 이중체 내의 반대 가닥에 있는 반대 뉴클레오티드 사이의 미스매치(즉, 비상보적 염기쌍)일 수 있다. 예시적인 미스매치 염기 쌍은 G:G, G:A, G:U, G:T, A:A, A:C, C:C, C:U, C:T, U:U, T:T, U:T, 또는 이들의 조합을 포함한다. 당업계에 공지된 다른 미스매치 염기 쌍도 본 발명에 적합하다. 자연 발생 뉴클레오티드 또는 변형된 뉴클레오티드인 뉴클레오티드 간에 미스매치가 발생할 수 있으며, 즉 미스매치 염기 쌍은 뉴클레오티드의 리보스 당의 변형과 무관하게 각 뉴클레오티드로부터의 핵염기 간에 발생할 수 있다. 특정 양태에서, dsRNA 분자는 2'-데옥시 핵염기인 미스매치 불일치 쌍에 적어도 하나의 핵염기를 함유하며; 예를 들어, 2'-데옥시 핵염기는 센스 가닥에 있다.
일부 양태에서, 안티센스 가닥의 시드 영역에서 이중체의 열적으로 불안정한 변형은 다음과 같은 표적 mRNA의 상보적 염기에 대한 손상된 W-C H-결합을 갖는 뉴클레오티드를 포함한다:

무염기 뉴클레오티드, 비사이클릭 뉴클레오티드 변형(UNA 및 GNA 포함) 및 미스매치 변형의 더 많은 예가 제WO 2011/133876호에 상세히 기술되어 있으며, 이는 전문이 본원에 참고로 포함된다.
열적으로 불안정한 변형은 또한 반대 염기와 수소 결합을 형성하는 능력이 감소되거나 폐지된 보편적 염기, 및 포스페이트 변형을 포함할 수 있다.
일부 양태에서, 이중체의 열적으로 불안정한 변형은 반대 가닥의 염기와 수소 결합을 형성하는 능력이 손상되거나 완전히 폐지된 핵염기 변형과 같지만 이에 제한되지 않는 비-정준 염기를 가진 뉴클레오티드를 포함한다. 이들 핵염기 변형은 제WO 2010/0011895호에 기술된 바와 같이 dsRNA 이중체의 중심 영역의 불안정화에 대해 평가되었으며, 이것은 전문이 본원에 참고로 포함된다. 예시적인 핵염기 변형은 다음과 같다:

일부 양태에서, 안티센스 가닥의 시드 영역에서 이중체의 열적으로 불안정한 변형은 다음과 같은 표적 mRNA의 염기에 상보적인 하나 이상의 α-뉴클레오티드를 포함한다:

여기서 R은 H, OH, OCH3, F, NH2, NHMe, NMe2 또는 O-알킬이다.
천연 포스포디에스테르 결합과 비교하여 dsRNA 이중체의 열 안정성을 감소시키는 것으로 알려진 예시적인 포스페이트 변형은 다음과 같다:

R 그룹에 대한 알킬은 C1-C6알킬일 수 있다. R 그룹에 대한 구체적인 알킬은 메틸, 에틸, 프로필, 이소프로필, 부틸, 펜틸 및 헥실을 포함하지만, 이에 제한되지 않는다.
숙련가가 인식하는 바와 같이, 핵염기의 기능적 역할이 본 개시내용의 RNAi 제제의 특이성을 정의하고 있다는 측면에서, 핵염기 변형이, 예를 들어, 오프-타겟 효과에 비해 온-타겟 효과를 향상시킬 목적으로, 예를 들어 본 발명의 RNAi 제제에 불안정 변형을 도입하기 위해, 본원에 기술된 바와 같은 다양한 방식으로 수행될 수 있지만, 이용 가능한 및, 일반적으로, 본 개시내용의 RNAi 제제에 존재하는 변형의 범위는 비-핵염기 변형, 예를 들어 폴리리보뉴클레오티드의 당 그룹 또는 포스페이트 백본에 대한 변형에 대해 훨씬 더 큰 경향이 있다. 이러한 변형은 본 개시내용의 다른 섹션에 더욱 상세히 기술되어 있으며 상기 또는 본원의 다른 곳에 기술된 바와 같이 천연 핵염기 또는 변형된 핵염기를 보유하는 본 개시내용의 RNAi 제제에 대해 명시적으로 고려된다.
열적으로 불안정한 변형을 포함하는 안티센스 가닥에 더하여, dsRNA는 또한 하나 이상의 안정화 변형을 포함할 수 있다. 예를 들면, dsRNA는 적어도 2개(예를 들어, 2개, 3개, 4개, 5개, 6개, 7개, 8개, 9개, 10개 또는 그 이상)의 안정화 변형을 포함할 수 있다. 제한 없이, 안정화 변형은 모두 하나의 가닥에 존재할 수 있다. 일부 양태에서, 센스 및 안티센스 가닥 둘 다는 적어도 두 개의 안정화 변형을 포함한다. 안정화 변형은 센스 가닥 또는 안티센스 가닥의 임의의 뉴클레오티드에서 발생할 수 있다. 예를 들어, 안정화 변형은 센스 가닥 또는 안티센스 가닥의 모든 뉴클레오티드에서 발생할 수 있거나; 각 안정화 변형은 센스 가닥 또는 안티센스 가닥에서 교대 패턴으로 발생할 수 있거나; 센스 가닥 또는 안티센스 가닥은 교대 패턴으로 안정화 변형을 둘 다 포함한다. 센스 가닥에 대한 안정화 변경의 교대 패턴은 안티센스 가닥과 동일하거나 상이할 수 있고, 센스 가닥에 대한 안정화 변형의 교대 패턴은 안티센스 가닥에 대한 안정화 변형의 교대 패턴에 비해 이동을 가질 수 있다.
일부 양태에서, 안티센스 가닥은 적어도 2개(예를 들어, 2개, 3개, 4개, 5개, 6개, 7개, 8개, 9개, 10개 또는 그 이상)의 안정화 변형을 포함한다. 제한 없이, 안티센스 가닥의 안정화 변형은 임의의 위치에 존재할 수 있다. 일부 양태에서, 안티센스는 5'-말단으로부터 위치 2, 6, 8, 9, 14, 및 16에 안정화 변형을 포함한다. 일부 다른 양태에서, 안티센스는 5'-말단으로부터 위치 2, 6, 14, 및 16에 안정화 변형을 포함한다. 여전히 일부 다른 양태에서, 안티센스는 5'-말단으로부터 위치 2, 14, 및 16에 안정화 변형을 포함한다.
일부 양태에서, 안티센스 가닥은 불안정화 변형에 인접한 적어도 하나의 안정화 변형을 포함한다. 예를 들면, 안정화 변형은 불안정화 변형의 5'-말단 또는 3'-말단에, 즉 불안정화 변형의 위치로부터 위치 -1 또는 +1에 있는 뉴클레오티드일 수 있다. 일부 양태에서, 안티센스 가닥은 불안정화 변형의 5'-말단 및 3'-말단 각각에, 즉 불안정화 변형의 위치로부터 위치 -1 및 +1에 안정화 변형을 포함한다.
일부 양태에서, 안티센스 가닥은 불안정화 변형의 3'-말단에, 즉 불안정화 변형의 위치로부터 위치 +1 및 +2에 적어도 2개의 안정화 변형을 포함한다.
일부 양태에서, 센스 가닥은 적어도 2개(예를 들어, 2개, 3개, 4개, 5개, 6개, 7개, 8개, 9개, 10개 또는 그 이상)의 안정화 변형을 포함한다. 제한 없이, 센스 가닥의 안정화 변형은 임의의 위치에 존재할 수 있다. 일부 양태에서, 센스 가닥은 5'-말단으로부터 위치 7, 10 및 11에 안정화 변형을 포함한다. 일부 다른 양태에서, 센스 가닥은 5'-말단으로부터 위치 7, 9, 10, 및 11에 안정화 변형을 포함한다. 일부 양태에서, 센스 가닥은 안티센스 가닥의 5'-말단으로부터 개수하여 안티센스 가닥의 위치 11, 12, 및 15에 상보적인 또는 반대인 위치에 안정화 변형을 포함한다. 일부 다른 양태에서, 센스 가닥은 안티센스 가닥의 5'-말단으로부터 개수하여 안티센스 가닥의 위치 11, 12, 13 및 15에 상보적인 또는 반대인 위치에 안정화 변형을 포함한다. 일부 양태에서, 센스 가닥은 2, 3 또는 4개의 안정화 변형의 블록을 포함한다.
일부 양태에서, 센스 가닥은 안티센스 가닥에서 이중체의 열적으로 불안정한 변형에 상보적인 또는 반대인 위치에 안정화 변형을 포함하지 않는다.
예시적인 열적으로 안정한 변형은 2'-플루오로 변형을 포함하지만 이에 제한되지 않는다. 기타 열적으로 안정한 변형은 LNA를 포함하지만 이에 제한되지 않는다.
일부 양태에서, 본 개시내용의 dsRNA는 적어도 4개(예를 들어, 4개, 5개, 6개, 7개, 8개, 9개, 10개 또는 그 이상)의 2'-플루오로 뉴클레오티드를 포함한다. 제한 없이, 2'-플루오로 뉴클레오티드는 모두 하나의 가닥에 존재할 수 있다. 일부 양태에서, 센스 및 안티센스 가닥 둘 다는 적어도 두 개의 2'-플루오로 뉴클레오티드를 포함한다. 2'-플루오로 변형은 센스 가닥 또는 안티센스 가닥의 임의의 뉴클레오티드에서 발생할 수 있다. 예를 들어, 2'-플루오로 변형은 센스 가닥 또는 안티센스 가닥의 모든 뉴클레오티드에서 발생할 수 있거나; 각각의 2'-플루오로 변형은 센스 가닥 또는 안티센스 가닥의 교대 패턴에서 발생할 수 있거나; 센스 가닥 또는 안티센스 가닥은 교대 패턴에 2'-플루오로 변형 둘 다를 포함한다. 센스 가닥의 2'-플루오로 변형의 교대 패턴은 안티센스 가닥과 동일하거나 상이할 수 있고, 센스 가닥의 2'-플루오로 변형의 교대 패턴은 안티센스 가닥의 2'-플루오로 변형의 교대 패턴에 비해 이동을 가질 수 있다.
일부 양태에서, 안티센스 가닥은 적어도 2개(예를 들어, 2개, 3개, 4개, 5개, 6개, 7개, 8개, 9개, 10개 또는 그 이상)의 2'-플루오로 뉴클레오티드를 포함한다. 제한 없이, 안티센스 가닥의 2'-플루오로 변형은 임의의 위치에 존재할 수 있다. 일부 양태에서, 안티센스는 5'-말단으로부터 위치 2, 6, 8, 9, 14, 및 16에 2'-플루오로 뉴클레오티드를 포함한다. 일부 다른 양태에서, 안티센스는 5'-말단으로부터 위치 2, 6, 14, 및 16에 2'-플루오로 뉴클레오티드를 포함한다. 여전히 일부 다른 양태에서, 안티센스는 5'-말단으로부터 위치 2, 14, 및 16에 2'-플루오로 뉴클레오티드를 포함한다.
일부 양태에서, 안티센스 가닥은 불안정화 변형에 인접한 적어도 하나의 2'-플루오로 뉴클레오티드를 변형을 포함한다. 예를 들면, 2'-플루오로 뉴클레오티드는 불안정화 변형의 5'-말단 또는 3'-말단에, 즉 불안정화 변형의 위치로부터 위치 -1 또는 +1에 있는 뉴클레오티드일 수 있다. 일부 양태에서, 안티센스 가닥은 불안정화 변형의 5'-말단 및 3'-말단 각각에, 즉 불안정화 변형의 위치로부터 위치 -1 및 +1에 2'-플루오로 뉴클레오티드를 포함한다.
일부 양태에서, 안티센스 가닥은 불안정화 변형의 3'-말단에, 즉 불안정화 변형의 위치로부터 위치 +1 및 +2에 적어도 2개의 2'-플루오로 뉴클레오티드를 포함한다.
일부 양태에서, 센스 가닥은 적어도 2개(예를 들어, 2개, 3개, 4개, 5개, 6개, 7개, 8개, 9개, 10개 또는 그 이상)의 2'-플루오로 뉴클레오티드를 포함한다. 제한 없이, 센스 가닥의 2'-플루오로 변형은 임의의 위치에 존재할 수 있다. 일부 양태에서, 안티센스는 5'-말단으로부터 위치 7, 10, 및 11에 2'-플루오로 뉴클레오티드를 포함한다. 일부 다른 양태에서, 센스 가닥은 5'-말단으로부터 위치 7, 9, 10, 및 11에 2'-플루오로 뉴클레오티드를 포함한다. 일부 양태에서, 센스 가닥은 안티센스 가닥의 5'-말단으로부터 개수하여 안티센스 가닥의 위치 11, 12, 및 15에 상보적인 또는 반대인 위치에 2'-플루오로 뉴클레오티드를 포함한다. 일부 다른 양태에서, 센스 가닥은 안티센스 가닥의 5'-말단으로부터 개수하여 안티센스 가닥의 위치 11, 12, 13, 및 15에 상보적인 또는 반대인 위치에 2'-플루오로 뉴클레오티드를 포함한다. 일부 양태에서, 센스 가닥은 2개, 3개 또는 4개의 2'-플루오로 뉴클레오티드의 블록을 포함한다.
일부 양태에서, 센스 가닥은 안티센스 가닥에서 이중체의 열적으로 불안정한 변형에 상보적인 또는 반대인 위치에 2'-플루오로 뉴클레오티드를 포함하지 않는다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 21개의 뉴클레오티드 (nt) 센스 가닥 및 23의 뉴클레오티드 (nt) 안티세스를 포함하고, 여기서 안티센스 가닥은 적어도 하나의 열적으로 불안정한 뉴클레오티드를 함유하고, 여기서 적어도 하나의 열적으로 불안정한 뉴클레오티드는 안티센스 가닥의 시드 영역에서 (즉, 안티센스 가닥의 5'-말단의 위치 2-9에) 발생하며, 여기서 dsRNA의 한쪽 말단은 평활한 반면 다른 쪽 말단은 2 nt 돌출부를 포함하고, 여기서 dsRNA는 임의로 다음 특징들 중 적어도 하나(예를 들어, 1개, 2개, 3개, 4개, 5개, 6개 또는 7개 모두)를 추가로 갖는다: (i) 안티센스는 2, 3, 4, 5 또는 6개 2'-플루오로 변형을 포함하고; (ii) 안티센스는 1, 2, 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (iii) 센스 가닥은 리간드와 접합되고; (iv) 센스 가닥은 2, 3, 4 또는 5개 2'-플루오로 변형을 포함하고; (v) 센스 가닥은 1, 2, 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (vi) dsRNA는 적어도 4개의 2'-플루오로 변형을 포함하고; (vii) dsRNA는 안티센스 가닥의 5'-말단에 평활 말단을 포함한다. 바람직하게는, 2 nt 돌출부는 안티센스의 3'-말단에 있다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 및 안티센스 가닥을 포함하고, 여기서: 센스 가닥은 25-30개의 뉴클레오티드 잔기 길이이고, 여기서 5' 말단 뉴클레오티드(위치 1)로부터 시작하여, 상기 센스 가닥의 위치 1 내지 23은 적어도 8개의 리보뉴클레오티드를 포함하고; 안티센스 가닥은 36-66개의 뉴클레오티드 잔기 길이이고, 3' 말단 뉴클레오티드로부터 시작하여, 센스 가닥의 위치 1-23과 쌍을 이루는 위치에서 적어도 8개의 리보뉴클레오티드가 이중체를 형성하고; 여기서 안티센스 가닥의 적어도 3' 말단 뉴클레오티드는 센스 가닥과 쌍을 이루지 않고, 최대 6개의 연속 3' 말단 뉴클레오티드가 센스 가닥과 쌍을 이루지 않아 1-6개의 뉴클레오티드의 3' 단일 가닥 돌출부를 형성하고; 여기서 안티센스 가닥의 5' 말단은 센스 가닥과 쌍을 이루지 않은 10-30개의 연속 뉴클레오티드를 포함하여 10-30개의 뉴클레오티드 단일 가닥 5' 돌출부를 형성하고; 여기서 적어도 센스 가닥 5' 말단 및 3' 말단 뉴클레오티드는 센스 및 안티센스 가닥이 최대 상보성을 위해 정렬될 때 안티센스 가닥의 뉴클레오티드와 염기 쌍을 이루어 센스 가닥과 안티센스 가닥 사이에 실질적으로 이중체화된 영역을 형성하고; 안티센스 가닥은 상기 이중 가닥 핵산이 포유동물 세포에 도입될 때 표적 유전자 발현을 감소시키기 위해 안티센스 가닥 길이의 적어도 19개 리보뉴클레오티드를 따라 표적 RNA에 충분히 상보적이고; 여기서 안티센스 가닥은 적어도 하나의 열적으로 불안정한 뉴클레오티드를 함유하고, 여기서 적어도 하나의 열적으로 불안정한 뉴클레오티드는 안티센스 가닥의 시드 영역에 (즉 안티센스 가닥의 5'-말단의 위치 2-9에) 있다. 예를 들면, 열적으로 불안정한 뉴클레오티드는 센스 가닥의 5'-말단의 위치 14-17에 상보적인 또는 반대인 위치 사이에서 발생하며, 여기서 dsRNA는 임의로 다음 특징들 중 적어도 하나(예를 들어, 1개, 2개, 3개, 4개, 5개, 6개 또는 7개 모두)를 추가로 갖는다: (i) 안티센스는 2, 3, 4, 5, 또는 6개 2'-플루오로 변형을 포함하고; (ii) 안티센스는 1, 2, 3, 4, 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (iii) 센스 가닥은 리간드와 접합되며; (iv) 센스 가닥은 2, 3, 4, 또는 5개 2'-플루오로 변형을 포함하고; (v) 센스 가닥은 1, 2, 3, 4, 또는 5 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (vi) dsRNA는 적어도 4개의 2'-플루오로 변형을 포함하고; (vii) dsRNA는 12-30개의 뉴클레오티드 쌍 길이의 이중체 영역을 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 및 안티센스 가닥을 포함하고, 여기서 상기 dsRNA 분자는 적어도 25개 및 최대 29개의 뉴클레오티드인 길이를 갖는 센스 가닥 및 최대 30개의 뉴클레오티드인 길이를 갖는 안티센스 가닥을 포함하고 센스 가닥은 5' 말단으로부터 위치 11에 효소 분해에 민감한 변형된 뉴클레오티드를 포함하고, 여기서 상기 센스 가닥의 3' 말단 및 상기 안티센스 가닥의 5' 말단은 평활 말단을 형성하고 상기 안티센스 가닥은 센스 가닥보다 이의 3' 말단에서 1-4개의 뉴클레오티드가 더 길며, 여기서 이중체 영역은 적어도 25개의 뉴클레오티드 길이이고, 상기 안티센스 가닥은 상기 dsRNA 분자가 포유동물 세포 내로 도입될 때 표적 유전자 발현을 감소시키기 위해 상기 안티센스 가닥 길이의 적어도 19 nt를 따라 표적 mRNA에 충분히 상보적이고, 여기서 상기 dsRNA의 다이서 절단은 상기 안티센스 가닥의 상기 3' 말단을 포함하는 siRNA를 우선적으로 생성하여 포유동물에서 표적 유전자의 발현을 감소시키며, 여기서 안티센스 가닥은 적어도 하나의 열적으로 불안정한 뉴클레오티드를 함유하고, 적어도 하나의 열적으로 불안정한 뉴클레오티드는 안티센스 가닥의 시드 영역에 (즉, 안티센스 가닥의 5'-말단의 위치 2-9에) 있고, 여기서 dsRNA는 임의로 다음 특징들 중 적어도 하나(예를 들어, 1개, 2개, 3개, 4개, 5개, 6개 또는 7개 모두)를 추가로 갖는다: (i) 안티센스는 2, 3, 4, 5, 또는 6개 2'-플루오로 변형을 포함하고; (ii) 안티센스는 1, 2, 3, 4, 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (iii) 센스 가닥은 리간드와 접합되고; (iv) 센스 가닥은 2, 3, 4, 또는 5개 2'-플루오로 변형을 포함하고; (v) 센스 가닥은 1, 2, 3, 4, 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (vi) dsRNA는 적어도 4개의 2'-플루오로 변형을 포함하고; (vii) dsRNA는 12-29개의 뉴클레오티드 쌍 길이의 이중체 영역을 갖는다.
일부 양태에서, dsRNA 분자의 센스 가닥 및 안티센스 가닥의 모든 뉴클레오티드는 변형될 수 있다. 각각의 뉴클레오티드는 비-결합 포스페이트 산소 중 하나 또는 둘 다 또는 하나 이상의 결합 포스페이트 산소의 하나 이상의 변경; 리보스 당, 예를 들어 리보스 당의 2' 하이드록실의 구성분의 변경; "데포스포" 링커로의 포스페이트 모이어티의 대량 대체; 자연 발생 염기의 변형 또는 대체; 및 리보스-포스페이트 백본의 대체 또는 변형을 포함할 수 있는 동일하거나 상이한 변형으로 변형될 수 있다.
핵산은 서브유닛의 중합체이기 때문에, 많은 변형이 핵산 내에서 반복되는 위치, 예를 들어, 염기의 변형, 또는 포스페이트 모이어티, 또는 포스페이트 모이어티의 비결합 O에서 발생한다. 일부 경우에, 변형은 핵산의 모든 대상 위치에서 발생하지만 많은 경우에 발생하지 않는다. 예를 들자면, 변형은 3' 또는 5' 말단 위치에서만 발생할 수 있고, 단지 말단 영역에서, 예를 들어, 말단 뉴클레오티드 상의 위치에 또는 가닥의 마지막 2, 3, 4, 5, 또는 10개의 뉴클레오티드에서 발생할 수 있다. 변형은 이중 가닥 영역, 단일 가닥 영역 또는 둘 다에서 발생할 수 있다. 변형은 RNA의 이중 가닥 영역에서만 발생하거나 RNA의 단일 가닥 영역에서만 발생할 수 있다. 예를 들어, 비-결합 O 위치에서의 포스포로티오에이트 변형은 한쪽 또는 양쪽 말단에서만 발생할 수 있고, 단지 말단 영역에서, 예를 들어, 말단 뉴클레오티드 상의 위치에 또는 가닥의 마지막 2, 3, 4, 5, 또는 10개의 뉴클레오티드에서 발생할 수 있거나, 이중 가닥 및 단일 가닥 영역, 특히 말단에서 발생할 수 있다. 5' 말단 또는 말단들은 인산화될 수 있다.
안정성을 향상시키거나, 돌출부에 특정 염기를 포함시키거나, 변형된 뉴클레오티드 또는 뉴클레오티드 대용물을 단일 가닥 돌출부에, 예를 들어, 5' 또는 3' 돌출부에, 또는 둘 다에 포함시키는 것이 가능할 수 있다. 예를 들어, 돌출부에 퓨린 뉴클레오티드를 포함시키는 것이 바람직할 수 있다. 일부 양태에서 3' 또는 5' 돌출부의 염기 전부 또는 일부는, 예를 들어, 본원에 기술된 변형으로 변형될 수 있다. 변형은, 예를 들어, 당업계에 공지된 변형으로의 리보스 당의 2' 위치에서의 변형의 사용, 예를 들어, 핵염기의 리보당 대신에 변형된 데옥시리보뉴클레오티드, 2'-데옥시-2'-플루오로(2'-F) 또는 2'-O-메틸의 사용, 및 포스페이트 그룹에서의 변형, 예를 들어, 포스포로티오에이트 변형을 포함할 수 있다. 돌출부는 표적 서열과 상동성일 필요는 없다.
일부 양태에서, 센스 가닥 및 안티센스 가닥의 각 잔기는 독립적으로 LNA, HNA, CeNA, 2'-메톡시에틸, 2'-O-메틸, 2'-O-알킬, 2'-C-알릴, 2'-데옥시, 또는 2'-플루오로로 변형된다. 가닥은 하나 이상의 변형을 함유할 수 있다. 일부 양태에서, 센스 가닥 및 안티센스 가닥의 각 가닥은 독립적으로 2'-O-메틸 또는 2'-플루오로로 변형된다. 이러한 변형은 안티센스 가닥에 존재하는 이중체의 적어도 하나의 열적으로 불안정한 변형에 더하여 존재하는 것으로 이해해야 한다.
적어도 2개의 상이한 변형이 전형적으로 센스 가닥 및 안티센스 가닥에 존재한다. 이러한 2개의 변형은 2'-데옥시, 2'- O-메틸 또는 2'-플루오로 변형, 비사이클릭 뉴클레오티드 등일 수 있다. 일부 양태에서, 센스 가닥 및 안티센스 가닥 각각은 2'-O-메틸 또는 2'-데옥시로부터 선택된 두 개의 상이하게 변형된 뉴클레오티드를 포함한다. 일부 양태에서, 센스 가닥 및 안티센스 가닥의 각 잔기는 독립적으로 2'-O-메틸 뉴클레오티드, 2'-데옥시 뉴클레오티드, 2'-데옥시-2'-플루오로 뉴클레오티드, 2'-O-N-메틸아세트아미도 (2'-O-NMA) 뉴클레오티드, 2'-O-디메틸아미노에톡시에틸 (2'-O-DMAEOE) 뉴클레오티드, 2'-O-아미노프로필 (2'-O-AP) 뉴클레오티드, 또는 2'-아라-F 뉴클레오티드로 변형된다. 다시, 이러한 변형은 안티센스 가닥에 존재하는 이중체의 적어도 하나의 열적으로 불안정한 변형에 더하여 존재하는 것으로 이해해야 한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 특히 B1, B2, B3, B1', B2', B3', B4' 영역에 교대 패턴의 변형을 포함한다. 본원에 사용된 용어 "교대 모티프" 또는 "대체 패턴"은 하나 이상의 변형을 갖는 모티프를 지칭하며, 각각의 변형은 하나의 가닥의 교대 뉴클레오티드 상에서 발생한다. 교대 뉴클레오티드는 다른 뉴클레오티드마다 1개 또는 3개의 뉴클레오티드마다 1개, 또는 유사한 패턴을 나타낼 수 있다. 예를 들면, A, B 및 C 각각이 뉴클레오티드에 대한 한 가지 유형의 변형을 나타낸다면, 교대 모티프는 "ABABABABABAB...", "AABBAABBAABB...", "AABAABAABAAB...", "AAABAAABAAAB...", "AAABBBAAABBB..." 또는 "ABCABCABCABC..." 등일 수 있다. 교대 모티프에 함유된 변형의 유형은 동일하거나 상이할 수 있다. 예를 들면, A, B, C, D 각각이 뉴클레오티드에 대한 한 가지 유형의 변형을 나타낸다면, 교대 패턴, 즉 모든 다른 뉴클레오티드에 대한 변형은 동일할 수 있지만, 각각의 센스 가닥 또는 안티센스 가닥은 "ABABAB...", "ACACAC..." "BDBDBD..." 또는 "CDCDCD..." 등과 같은 교대 모티프 내에서 변형의 여러 가능성으로부터 선택될 수 있다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 안티센스 가닥의 교대 모티프에 대한 변형 패턴에 비해 센스 가닥의 교대 모티프에 대한 변형 패턴이 이동된 것을 포함한다. 이동은 센스 가닥의 뉴클레오티드의 변형된 그룹이 안티센스 가닥의 뉴클레오티드의 상이하게 변형된 그룹에 상응하고 그 반대로도 마찬가지이도록 이루어질 수 있다. 예를 들면, dsRNA 이중체에서 센스 가닥이 안티센스 가닥과 쌍을 이룰 때, 이중체 영역 내에서 센스 가닥의 교대 모티프는 가닥의 5'-3'로부터 "ABABAB"로 시작할 수 있고 안티센스 가닥의 교대 모티프는 가닥의 3'-5'로부터 "BABABA"로 시작할 수 있다. 또 다른 예로서, 이중체 영역 내에서 센스 가닥의 교대 모티프는 가닥의 5'-3'로부터 "AABBAABB"로 시작할 수 있고 안티센스 가닥의 교대 모티프는 가닥의 3'-5'로부터 "BBAABBAA"로 시작할 수 있어서, 센스 가닥과 안티센스 가닥 사이에 변형 패턴의 완전한 또는 부분적인 이동이 있다.
본 개시내용의 dsRNA 분자는 적어도 하나의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 추가로 포함할 수 있다. 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형은 센스 가닥 또는 안티센스 가닥의 임의의 뉴클레오티드에서 또는 가닥의 임의의 위치 둘 다에서 발생할 수 있다. 예를 들어, 뉴클레오티드간 결합 변형은 센스 가닥 또는 안티센스 가닥의 모든 뉴클레오티드에서 발생할 수 있거나; 각 뉴클레오티드간 결합 변형은 센스 가닥 또는 안티센스 가닥의 교대 패턴에서 발생할 수 있거나; 센스 가닥 또는 안티센스 가닥은 교대 패턴에 뉴클레오티드간 결합 변형 둘 다를 포함한다. 센스 가닥에 대한 뉴클레오티드간 결합 변형의 교대 패턴은 안티센스 가닥과 동일하거나 상이할 수 있으며, 센스 가닥에 대한 뉴클레오티드간 결합 변형의 교대 패턴은 안티센스 가닥에 대한 뉴클레오티드간 결합 변형의 교대 패턴에 비해 이동을 가질 수 있다.
일부 양태에서, dsRNA 분자는 돌출부 영역에 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형을 포함한다. 예를 들면, 돌출부 영역은 두 개의 뉴클레오티드 간에 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 갖는 두 개의 뉴클레오티드를 포함한다. 뉴클레오티드간 결합 변형은 또한 이중체 영역 내에서 돌출부 뉴클레오티드를 말단 쌍을 이루는 뉴클레오티드와 연결하도록 만들 수 있다. 예를 들면, 적어도 2, 3, 4, 또는 모든 돌출부 뉴클레오티드는 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 통해 연결될 수 있으며, 임의로, 돌출부 뉴클레오티드 다음에는 돌출부 뉴클레오티드를 쌍을 이루는 뉴클레오티드와 결합하는 추가의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합이 있을 수 있다. 예를 들어, 말단 3개의 뉴클레이티드 간에 적어도 2개의 포스포로티오에이트 뉴클레오티드간 결합이 있을 수 있으며, 여기서 3개의 뉴클레오티드 중 2개는 돌출부 뉴클레오티드이고, 세 번째는 돌출부 뉴클레오티드 다음의 쌍을 이루는 뉴클레오티드이다. 바람직하게는, 이러한 말단 3개의 뉴클레오티드는 안티센스 가닥의 3' 말단에 있을 수 있다.
일부 양태에서, dsRNA 분자의 센스 가닥은 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 또는 16개 포스페이트 뉴클레오티드간 결합에 의해 분리된 2개 내지 10개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 1-10개 블록을 포함하며, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 안티센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 또는 18개 포스페이트 뉴클레오티드간 결합에 의해 분리된 2개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하며, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 또는 16개 포스페이트 뉴클레오티드간 결합에 의해 분리된 3개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하고, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 또는 14개 포스페이트 뉴클레오티드간 결합에 의해 분리된 4개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하고, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 또는 12개 포스페이트 뉴클레오티드간 결합에 의해 분리된 5개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하고, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 4, 5, 6, 7, 8, 9, 또는 10개 포스페이트 뉴클레오티드간 결합에 의해 분리된 6개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하고, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 4, 5, 6, 7, 또는 8개 포스페이트 뉴클레오티드간 결합에 의해 분리된 7개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하고, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 4, 5, 또는 6개 포스페이트 뉴클레오티드간 결합에 의해 분리된 8개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하고, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, dsRNA 분자의 안티센스 가닥은 1, 2, 3, 또는 4개 포스페이트 뉴클레오티드간 결합에 의해 분리된 9개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합의 두 개의 블록을 포함하고, 여기서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 중 하나는 올리고뉴클레오티드 서열의 임의의 위치에 배치되고 상기 안티센스 가닥은 포스포로티오에이트, 메틸포스포네이트 및 포스페이트 뉴클레오티드간 결합의 임의의 조합을 포함하는 센스 가닥 또는 포스포로티오에이트 또는 메틸포스포네이트 또는 포스페이트 결합을 포함하는 안티센스 가닥과 쌍을 이룬다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 또는 안티센스 가닥의 말단 위치(들)의 1-10 내에 하나 이상의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형을 추가로 포함한다. 예를 들면, 적어도 2, 3, 4, 5, 6, 7, 8, 9, 또는 10개의 뉴클레오티드가 센스 또는 안티센스 가닥의 한쪽 말단 또는 양 말단에서 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합을 통해 연결될 수 있다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 또는 안티센스 가닥의 각각의 이중체의 내부 영역의 1-10 내에 하나 이상의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형을 추가로 포함한다. 예를 들면, 적어도 2, 3, 4, 5, 6, 7, 8, 9, 또는 10개의 뉴클레오티드가 센스 가닥의 5'-말단으로부터 개수하여 이중체 영역의 위치 8-16에서 포스포로티오에이트 메틸포스포네이트 뉴클레오티드간 결합을 통해 연결될 수 있고; dsRNA 분자는 임의로 말단 위치(들)의 1-10 내에 하나 이상의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형을 추가로 포함할 수 있다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 1 내지 5개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형(들) 및 위치 18-23 내에 1 내지 5개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형(들), 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 1 내지 5개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1 내지 5개를 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 또는 메틸포스포네이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1-5 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 18-23 내에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 20 및 21에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21에 1개를 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 20 및 21에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21 및 22에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21 및 22에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 22 및 23에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 센스 가닥의 (5'-말단으로부터 개수하여) 위치 1에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 21에 1개의 포스포로티오에이트 뉴클레오티드간 결합 변형, 및 안티센스 가닥의 (5'-말단으로부터 개수하여) 위치 1 및 2에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형 및 위치 23 및 23에 2개의 포스포로티오에이트 뉴클레오티드간 결합 변형을 추가로 포함한다.
일부 양태에서, 본 개시내용의 화합물은 백본 키랄 중심의 패턴을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 5개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 6개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 7개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 8개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 9개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 10개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 11개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 12개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 13개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 14개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 15개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 16개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 17개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 18개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 19개의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 8개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 7개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 6개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 5개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 4개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 3개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 2개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Rp 배위의 1개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 8개 이하의 뉴클레오티드간 결합을 포함한다(비제한적인 예로서, 포스포디에스테르). 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 7개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 6개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 5개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 4개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 3개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 2개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 키랄이 아닌 1개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 10개의 뉴클레오티드간 결합, 및 키랄이 아닌 8개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 11개의 뉴클레오티드간 결합, 및 키랄이 아닌 7개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 12개의 뉴클레오티드간 결합, 및 키랄이 아닌 6개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 13개의 뉴클레오티드간 결합, 및 키랄이 아닌 6개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 14개의 뉴클레오티드간 결합, 및 키랄이 아닌 5개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, 백본 키랄 중심의 일반적인 패턴은 Sp 배위의 적어도 15개의 뉴클레오티드간 결합, 및 키랄이 아닌 4개 이하의 뉴클레오티드간 결합을 포함한다. 일부 양태에서, Sp 배위의 뉴클레오티드간 결합은 임의로 인접하거나 인접하지 않는다. 일부 양태에서, Rp 배위의 뉴클레오티드간 결합은 임의로 인접하거나 인접하지 않는다. 일부 양태에서, 키랄이 아닌 뉴클레오티드간 결합은 임의로 인접하거나 인접하지 않는다.
일부 양태에서, 본 개시내용의 화합물은 입체화학 블록인 블록을 포함한다. 일부 양태에서, 블록은 블록의 각 뉴클레오티드간 결합이 Rp라는 점에서 Rp 블록이다. 일부 양태에서, 5'-블록은 Rp 블록이다. 일부 양태에서, 3'-블록은 Rp 블록이다. 일부 양태에서, 블록은 블록의 각 뉴클레오티드간 결합이 Sp라는 점에서 Sp 블록이다. 일부 양태에서, 5'-블록은 Sp 블록이다. 일부 양태에서, 3'-블록은 Sp 블록이다. 일부 양태에서, 제공된 올리고뉴클레오티드는 Rp 및 Sp 블록 모두를 포함한다. 일부 양태에서, 제공된 올리고뉴클레오티드는 하나 이상의 Rp를 포함하지만 Sp 블록은 포함하지 않는다. 일부 양태에서, 제공된 올리고뉴클레오티드는 하나 이상의 Sp를 포함하지만 Rp 블록은 포함하지 않는다. 일부 양태에서, 제공된 올리고뉴클레오티드는 각각의 뉴클레오티드간 결합이 천연 포스페이트 결합인 하나 이상의 PO 블록을 포함한다.
일부 양태에서, 본 개시내용의 화합물은 5'-블록이 Sp 블록인 것을 포함하며, 여기서 각 당 모이어티는 2'-F 변형을 포함한다. 일부 양태에서, 5'-블록은 Sp 블록이며, 여기서 각 뉴클레오티드간 결합은 변형된 뉴클레오티드간 결합이고 각 당 모이어티는 2'-F 변형을 포함한다. 일부 양태에서, 5'-블록은 Sp 블록이며, 여기서 각 뉴클레오티드간 결합은 포스포로티오에이트 결합이고 각 당 모이어티는 2'-F 변형을 포함한다. 일부 양태에서, 5'-블록은 4개 이상의 뉴클레오시드 단위를 포함한다. 일부 양태에서, 5'-블록은 5개 이상의 뉴클레오시드 단위를 포함한다. 일부 양태에서, 5'-블록은 6개 이상의 뉴클레오시드 단위를 포함한다. 일부 양태에서, 5'-블록은 7개 이상의 뉴클레오시드 단위를 포함한다. 일부 양태에서, 3'-블록은 Sp 블록이고, 여기서 각 당 모이어티는 2'-F 변형을 포함한다. 일부 양태에서, 3'-블록은 Sp 블록이고, 여기서 각 뉴클레오티드간 결합은 변형된 뉴클레오티드간 결합이고 각 당 모이어티는 2'-F 변형을 포함한다. 일부 양태에서, 3'-블록은 Sp 블록이며, 여기서 각 뉴클레오티드간 결합은 포스포로티오에이트 결합이고 각 당 모이어티는 2'-F 변형을 포함한다. 일부 양태에서, 3'-블록은 4개 이상의 뉴클레오시드 단위를 포함한다. 일부 양태에서, 3'-블록은 5개 이상의 뉴클레오시드 단위를 포함한다. 일부 양태에서, 3'-블록은 6개 이상의 뉴클레오시드 단위를 포함한다. 일부 양태에서, 3'-블록은 7개 이상의 뉴클레오시드 단위를 포함한다.
일부 양태에서, 본 개시내용의 화합물은 영역의 한 유형의 뉴클레오시드 또는 올리고뉴클레오티드 다음에 특정 유형의 뉴클레오티드간 결합, 예를 들어, 천연 포스페이트 결합, 변형된 뉴클레오티드간 결합, Rp 키랄 뉴클레오티드간 결합, Sp 키랄 뉴클레오티드간 결합 등이 뒤따르는 것을 포함한다. 일부 양태에서, A 다음에는 Sp가 뒤따른다. 일부 양태에서, A 다음에는 Rp가 뒤따른다. 일부 양태에서, A 다음에는 천연 포스페이트 결합(PO)이 뒤따른다. 일부 양태에서, U 다음에는 Sp가 뒤따른다. 일부 양태에서, U 다음에는 Rp가 뒤따른다. 일부 양태에서, U 다음에는 천연 포스페이트 결합(PO)이 뒤따른다. 일부 양태에서, C 다음에는 Sp가 뒤따른다. 일부 양태에서, C 다음에는 Rp가 뒤따른다. 일부 양태에서, C 다음에는 천연 포스페이트 결합(PO)이 뒤따른다. 일부 양태에서, G 다음에는 Sp가 뒤따른다. 일부 양태에서, G 다음에는 Rp가 뒤따른다. 일부 양태에서, G 다음에는 천연 포스페이트 결합(PO)이 뒤따른다. 일부 양태에서, C 및 U 다음에는 Sp가 뒤따른다. 일부 양태에서, C 및 U 다음에는 Rp가 뒤따른다. 일부 양태에서, C 및 U 다음에는 천연 포스페이트 결합(PO)이 뒤따른다. 일부 양태에서, A 및 G 다음에는 Sp가 뒤따른다. 일부 양태에서, A 및 G 다음에는 Rp가 뒤따른다.
일부 양태에서, 안티센스 가닥은 뉴클레오티드 위치 21 및 22 사이에 및 뉴클레오티드 위치 22 및 23 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함하며, 여기서 안티센스 가닥은 안티센스 가닥의 시드 영역에 (즉, 안티센스 가닥의 5'-말단의 위치 2-9에) 위치한 이중체의 적어도 하나의 열적으로 불안정한 변형을 함유하고, 여기서 dsRNA는 임의로 다음 특징들 중 적어도 하나(예를 들어, 1개, 2개, 3개, 4개, 5개, 6개, 7개 또는 8개 모두)를 추가로 갖는다: (i) 안티센스는 2, 3, 4, 5 또는 6개 2'-플루오로 변형을 포함하고; (ii) 안티센스는 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (iii) 센스 가닥은 리간드와 접합되고; (iv) 센스 가닥은 2, 3, 4 또는 5개 2'-플루오로 변형을 포함하고; (v) 센스 가닥은 1, 2, 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (vi) dsRNA는 적어도 4개의 2'-플루오로 변형을 포함하고; (vii) dsRNA는 12-40개의 뉴클레오티드 쌍 길이의 이중체 영역을 포함하고; (viii) dsRNA는 안티센스 가닥의 5'-말단에 평활 말단을 갖는다.
일부 양태에서, 안티센스 가닥은 뉴클레오티드 위치 1 및 2 사이에, 뉴클레오티드 위치 2 및 3 사이에, 뉴클레오티드 위치 21 및 22 사이에, 및 뉴클레오티드 위치 22 및 23 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함하며, 여기서 안티센스 가닥은 안티센스 가닥의 시드 영역에 (즉, 안티센스 가닥의 5'-말단의 위치 2-9에) 위치한 이중체의 적어도 하나의 열적으로 불안정한 변형을 함유하고, 여기서 dsRNA는 임의로 다음 특징들 중 적어도 하나(예를 들어, 1개, 2개, 3개, 4개, 5개, 6개, 7개 또는 8개 모두)를 추가로 갖는다: (i) 안티센스는 2, 3, 4, 5 또는 6개 2'-플루오로 변형을 포함하고; (ii) 센스 가닥은 리간드와 접합되고; (iii) 센스 가닥은 2, 3, 4 또는 5개 2'-플루오로 변형을 포함하고; (iv) 센스 가닥은 1, 2, 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (v) dsRNA는 적어도 4개의 2'-플루오로 변형을 포함하고; (vi) dsRNA는 12-40개의 뉴클레오티드 쌍 길이의 이중체 영역을 포함하고; (vii) dsRNA는 12-40개의 뉴클레오티드 쌍 길이의 이중체 영역을 포함하고; (viii) dsRNA는 안티센스 가닥의 5'-말단에 평활 말단을 갖는다.
일부 양태에서, 센스 가닥은 뉴클레오티드 위치 1 및 2 사이에, 및 뉴클레오티드 위치 2 및 3 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함하고, 여기서 안티센스 가닥은 안티센스 가닥의 시드 영역에 (즉, 안티센스 가닥의 5'-말단의 위치 2-9에) 위치한 이중체의 적어도 하나의 열적으로 불안정한 변형을 함유하고, 여기서 dsRNA는 임의로 다음 특징들 중 적어도 하나(예를 들어, 1개, 2개, 3개, 4개, 5개, 6개, 7개 또는 8개 모두)를 추가로 갖는다: (i) 안티센스는 2, 3, 4, 5 또는 6개 2'-플루오로 변형을 포함하고; (ii) 안티센스는 1, 2, 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (iii) 센스 가닥은 리간드와 접합되고; (iv) 센스 가닥은 2, 3, 4 또는 5개 2'-플루오로 변형을 포함하고; (v) 센스 가닥은 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (vi) dsRNA는 적어도 4개의 2'-플루오로 변형을 포함하고; (vii) dsRNA는 12-40개의 뉴클레오티드 쌍 길이의 이중체 영역을 포함하고; (viii) dsRNA는 안티센스 가닥의 5'-말단에 평활 말단을 갖는다.
일부 양태에서, 센스 가닥은 뉴클레오티드 위치 1 및 2 사이에 및 뉴클레오티드 위치 2 및 3 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함하고, 안티센스 가닥은 뉴클레오티드 위치 1 및 2 사이에, 뉴클레오티드 위치 2 및 3 사이에, 뉴클레오티드 위치 21 및 22 사이에, 및 뉴클레오티드 위치 22 및 23 사이에 포스포로티오에이트 뉴클레오티드간 결합을 포함하며, 여기서 안티센스 가닥은 안티센스 가닥의 시드 영역에 (즉, 안티센스 가닥의 5'-말단의 위치 2-9에) 위치한 이중체의 적어도 하나의 열적으로 불안정한 변형을 함유하고, 여기서 dsRNA는 임의로 다음 특징들 중 적어도 하나(예를 들어, 1개, 2개, 3개, 4개, 5개, 6개 또는 7개 모두)를 추가로 갖는다: (i) 안티센스는 2, 3, 4, 5 또는 6개 2'-플루오로 변형을 포함하고; (ii) 센스 가닥은 리간드와 접합되고; (iii) 센스 가닥은 2, 3, 4 또는 5개 2'-플루오로 변형을 포함하고; (iv) 센스 가닥은 3, 4 또는 5개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하고; (v) dsRNA는 적어도 4개의 2'-플루오로 변형을 포함하고; (vi) dsRNA는 12-40개의 뉴클레오티드 쌍 길이의 이중체 영역을 포함하고; (vii) dsRNA는 안티센스 가닥의 5'-말단에 평활 말단을 갖는다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 표적과, 이중체 내에, 또는 이들의 조합에 미스매치(들)를 포함한다. 미스매치는 돌출부 영역 또는 이중체 영역에서 발생할 수 있다. 염기 쌍은 해리 또는 용융을 촉진하는 이들의 경향을 기준으로 하여 순위가 매겨질 수 있다(예를 들어, 특정 쌍의 결합 또는 해리의 자유 에너지에 대해, 가장 간단한 접근법은 개별 쌍을 기반으로 쌍을 조사하는 것이지만, 다음 이웃하거나 유사한 분석도 사용될 수 있다). 해리 촉진 측면에서: A:U가 G:C보다 바람직하고; G:U가 G:C보다 바람직하며; I:C가 G:C보다 바람직하다(I=이노신). 미스매치, 예를 들어, 비-정준 또는 정준 이외의 쌍(본원의 다른 곳에 기술된 바와 같음)이 정준 (A:T, A:U, G:C) 쌍보다 바람직하고; 보편적인 염기를 포함하는 쌍이 정준 쌍보다 바람직하다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 이중체의 5'-말단에서 안티센스 가닥의 해리를 촉진하기 위해 A:U, G:U, I:C의 그룹으로부터 독립적으로 선택될 수 있는 안티센스 가닥의 5'-말단으로부터의 이중체 영역 내에 처음 1, 2, 3, 4, 또는 5개 염기 쌍 중 적어도 하나, 및 미스매칭된 쌍, 예를 들어, 보편적인 염기를 포함하는 비-정준 또는 정준 이외의 쌍을 포함한다.
일부 양태에서, 안티센스 가닥의 5'-말단으로부터 이중체 영역 내의 1 위치에 있는 뉴클레오티드는 A, dA, dU, U, 및 dT로 이루어진 그룹으로부터 선택된다. 대안적으로, 안티센스 가닥의 5'-말단으로부터 이중체 영역 내의 처음 1, 2 또는 3개 염기 쌍 중 적어도 하나는 AU 염기 쌍이다. 예를 들면, 안티센스 가닥의 5'-말단으로부터 이중체 영역 내의 처음 염기 쌍은 AU 염기 쌍이다.
단일 가닥 또는 이중 가닥 올리고뉴클레오티드의 임의의 위치에서 디뉴클레오티드의 포스포디에스테르(PO), 포스포로티오에이트(PS), 또는 포스포로디티오에이트(PS2) 결합의 3' 말단에 4'-변형된 또는 5'-변형된 뉴클레오티드를 도입하면 뉴클레오티드간 결합에 입체 효과(steric effect)를 발휘하여 뉴클레아제에 대해 이를 보호하거나 안정화할 수 있는 것으로 밝혀졌다.
일부 양태에서, 5'-변형된 뉴클레오시드가 단일 가닥 또는 이중 가닥 siRNA의 임의의 위치에서 디뉴클레오티드의 3'-말단에 도입된다. 예를 들어, 5'-알킬화된 뉴클레오시드가 단일 가닥 또는 이중 가닥 siRNA의 임의의 위치에서 디뉴클레오티드의 3'-말단에 도입될 수 있다. 리보스 당의 5' 위치의 알킬 그룹은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다. 예시적인 5'-알킬화된 뉴클레오시드는 5'-메틸 뉴클레오시드이다. 5'-메틸은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다.
일부 양태에서, 4'-변형된 뉴클레오시드가 단일 가닥 또는 이중 가닥 siRNA의 임의의 위치에서 디뉴클레오티드의 3'-말단에 도입된다. 예를 들어, 4'-알킬화된 뉴클레오시드가 단일 가닥 또는 이중 가닥 siRNA의 임의의 위치에서 디뉴클레오티드의 3'-말단에 도입될 수 있다. 리보스 당의 4' 위치의 알킬 그룹은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다. 예시적인 4'-알킬화된 뉴클레오시드는 4'-메틸 뉴클레오시드이다. 4'-메틸은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다. 대안적으로, 4'-O-알킬화된 뉴클레오시드가 단일 가닥 또는 이중 가닥 siRNA의 임의의 위치에서 디뉴클레오티드의 3'-말단에 도입될 수 있다. 리보스 당의 4'-O-알킬은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다. 예시적인 4'-O-알킬화된 뉴클레오시드는 4'-O-메틸 뉴클레오시드이다. 4'-O-메틸은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다.
일부 양태에서, 5'-알킬화된 뉴클레오시드가 dsRNA의 센스 가닥 또는 안티센스 가닥의 임의의 위치에 도입되며, 이러한 변형은 dsRNA의 효능을 유지하거나 개선시킨다. 5'-알킬은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다. 예시적인 5'-알킬화된 뉴클레오시드는 5'-메틸 뉴클레오시드이다. 5'-메틸은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다.
일부 양태에서, 4'-알킬화된 뉴클레오시드가 dsRNA의 센스 가닥 또는 안티센스 가닥의 임의의 위치에 도입되며, 이러한 변형은 dsRNA의 효능을 유지하거나 개선시킨다. 4'-알킬은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다. 예시적인 4'-알킬화된 뉴클레오시드는 4'-메틸 뉴클레오시드이다. 4'-메틸은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다.
일부 양태에서, 4'-O-알킬화된 뉴클레오시드가 dsRNA의 센스 가닥 또는 안티센스 가닥의 임의의 위치에 도입되며, 이러한 변형은 dsRNA의 효능을 유지하거나 개선시킨다. 5'-알킬은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다. 예시적인 4'-O-알킬화된 뉴클레오시드는 4'-O-메틸 뉴클레오시드이다. 4'-O-메틸은 라세미 또는 키랄성으로 순수한 R 또는 S 이성체일 수 있다.
일부 양태에서, 본 개시내용의 dsRNA 분자는 (2'-H, 2'-OH 및 2'-OMe를 갖는 및 P=O 또는 P=S를 갖는) 2'-5' 결합을 포함할 수 있다. 예를 들면, 2'-5' 결합 변형은 뉴클레아제 내성을 촉진하거나 안티센스 가닥에 대한 센스 가닥의 결합을 억제하는데 사용될 수 있거나, RISC에 의한 센스 가닥 활성화를 피하기 위해 센스 가닥의 5' 말단에서 사용될 수 있다.
또 다른 양태에서, 본 개시내용의 dsRNA 분자는 L 당(예를 들어, L 리보스, 2'-H, 2'-OH 및 2'-OMe를 갖는 L-아라비노스)을 포함할 수 있다. 예를 들면, 이러한 L 당 변형은 뉴클레아제 내성을 촉진하거나 안티센스 가닥에 대한 센스 가닥의 결합을 억제하는데 사용될 수 있거나, RISC에 의한 센스 가닥 활성화를 피하기 위해 센스 가닥의 5' 말단에서 사용될 수 있다.
다양한 공보가 본 개시내용의 dsRNA와 함께 사용될 수 있는 다량체 siRNA를 기술하고 있다. 이러한 공보는 전문이 본원에 참고로 포함된 제WO2007/091269호, 제US 7858769호, 제WO2010/141511호, 제WO2007/117686호, 제WO2009/014887호, 및 제WO2011/031520호를 포함한다.
아래에 보다 상세히 기술된 바와 같이, RNAi 제제에 대한 하나 이상의 탄수화물 모이어티의 접합을 함유하는 RNAi 제제는 RNAi 제제의 하나 이상의 특성을 최적화할 수 있다. 다수의 경우에, 탄수화물 모이어티는 RNAi 제제의 변형된 서브유닛에 부착될 것이다. 예를 들면, dsRNA 제제의 하나 이상의 리보뉴클레오티드 서브유닛의 리보스 당은 또 다른 모이어티, 예를 들어, 탄수화물 리간드가 부착된 비-탄수화물(바람직하게는 사이클릭) 담체로 대체될 수 있다. 서브유닛의 리보스 당이 그렇게 대체된 리보뉴클레오티드 서브유닛을 본원에서 리보스 대체 변형 서브유닛(RRMS)이라고 한다. 사이클릭 담체는 카보사이클릭 환 시스템, 즉 모든 환 원자가 탄소 원자일 수 있거나, 또는 헤테로사이클릭 환 시스템, 즉 하나 이상의 환 원자가 헤테로원자, 예를 들어, 질소, 산소, 황일 수 있다. 사이클릭 담체는 모노사이클릭 환 시스템일 수 있거나, 둘 이상의 환, 예를 들어 융합된 환을 함유할 수 있다. 사이클릭 담체는 완전 포화된 환 시스템일 수 있거나, 하나 이상의 이중 결합을 함유할 수 있다.
리간드는 담체를 통해 폴리뉴클레오티드에 부착될 수 있다. 담체는 (i) 적어도 하나의 "백본 부착점", 바람직하게는 두 개의 "백본 부착점" 및 (ii) 적어도 하나의 "테더링 부착점"을 포함한다. 본원에 사용된 "백본 부착점"은 리보핵산의 백본, 예를 들어, 포스페이트, 또는 변형된 포스페이트, 예를 들어, 황 함유, 백본에 담체를 통합하는데 사용될 수 있고 이에 적합한 작용 그룹, 예를 들어, 하이드록실 그룹, 또는 일반적으로 결합을 지칭한다. "테더링 부착점"(TAP)은 일부 양태에서 사이클릭 담체의 구성 환 원자, 예를 들어, 선택된 모이어티를 연결하는 탄소 원자 또는 헤테로원자(백본 부착점을 제공하는 원자와 구별됨)를 지칭한다. 모이어티는, 예를 들어, 탄수화물, 예를 들어 단당류, 이당류, 삼당류, 사당류, 올리고당류 및 다당류일 수 있다. 임의로, 선택된 모이어티는 개재 테더(intervening tether)에 의해 사이클릭 담체에 연결된다. 따라서, 사이클릭 담체는 종종 작용 그룹, 예를 들어 아미노 그룹을 포함하거나, 일반적으로 구성 환에 대한 다른 화학적 실체, 예를 들어 리간드의 통합 또는 테더링에 적합한 결합을 제공할 것이다.
RNAi 제제는 담체를 통해 리간드에 접합될 수 있으며, 여기서 담체는 사이클릭 그룹 또는 비사이클릭 그룹일 수 있고; 바람직하게는, 사이클릭 그룹은 피롤리디닐, 피라졸리닐, 피라졸리디닐, 이미다졸리닐, 이미다졸리디닐, 피페리디닐, 피페라지닐, [1,3]디옥솔란, 옥사졸리디닐, 이속사졸리디닐, 모르폴리닐, 티아졸리디닐, 이소티아졸리디닐, 퀴녹살리디닐, 피리다지노닐, 테트라하이드로푸릴 및 데칼린으로부터 선택되고; 바람직하게는, 비사이클릭 그룹은 세리놀 백본 또는 디에탄올아민 백본으로부터 선택된다.
특정의 구체적인 태에서, 본 개시내용의 방법에 사용하기 위한 RNAi 제제는 표 10A, 10B, 12-15, 19 또는 20 중 어느 하나에 열거된 제제의 그룹으로부터 선택된 제제이다. 이들 제제는 리간드를 추가로 포함할 수 있다.
IV. 리간드에
접합된
iRNA
본 발명의 iRNA의 RNA의 또 다른 변형은, 예를 들어, 세포 내로의 iRNA의 활성, 세포 분포 또는 세포 흡수를 향상시키는 하나 이상의 리간드, 모이어티 또는 접합체를 iRNA에 화학적으로 연결하는 것을 포함한다. 이러한 모이어티는 지질 모이어티, 예를 들어 콜레스테롤 모이어티(Letsinger et al., Proc . Natl . Acid. Sci. USA, 1989, 86: 6553-6556), 콜산(Manoharan et al., Biorg . Med . Chem . Let., 1994, 4:1053-1060), 티오에테르, 예를 들어, 베릴-S-트리틸티올(Manoharan et al., Ann. N.Y . Acad . Sci ., 1992, 660:306-309; Manoharan et al., Biorg . Med. Chem . Let., 1993, 3:2765-2770), 티오콜레스테롤(Oberhauser et al., Nucl . Acids Res., 1992, 20:533-538), 지방족 쇄, 예를 들어, 도데칸디올 또는 운데실 잔기(Saison-Behmoaras et al., EMBO J, 1991, 10:1111-1118; Kabanov et al., FEBS Lett ., 1990, 259:327-330; Svinarchuk et al., Biochimie, 1993, 75:49-54), 인지질, 예를 들어, 디-헥사데실-rac-글리세롤 또는 트리에틸-암모늄 1,2-디-O-헥사데실-rac-글리세로-3-포스포네이트(Manoharan et al., Tetrahedron Lett ., 1995, 36:3651-3654; Shea et al., Nucl . Acids Res., 1990, 18:3777-3783), 폴리아민 또는 폴리에틸렌 글리콜 쇄(Manoharan et al., Nucleosides & Nucleotides, 1995, 14:969-973), 또는 아다만탄 아세트산(Manoharan et al., Tetrahedron Lett ., 1995, 36:3651-3654), 팔미틸 모이어티(Mishra et al., Biochim . Biophys . Acta, 1995, 1264:229-237), 또는 옥타데실아민 또는 헥실아미노-카보닐옥시콜레스테롤 모이어티(Crooke et al., J. Pharmacol . Exp . Ther ., 1996, 277:923-937)를 포함하지만, 이에 제한되지 않는다.
특정 양태에서, 리간드는 이것이 통합되는 iRNA 제제의 분포, 표적화 또는 수명을 변경한다. 일부 양태에서, 리간드는, 예를 들어, 이러한 리간드가 부재하는 종에 비해, 선택된 표적, 예를 들어, 분자, 세포 또는 세포 유형, 구획, 예를 들어, 세포 또는 기관 구획, 조직, 기관 또는 신체의 부위에 대해 향상된 친화성을 제공한다. 전형적인 리간드는 이중체화된 핵산에서 이중체 쌍형성에 참여할 것이다.
리간드는 단백질(예를 들어, 인간 혈청 알부민(HSA), 저밀도 지단백질(LDL) 또는 글로불린); 탄수화물(예를 들어, 덱스트란, 풀루란, 키틴, 키토산, 이눌린, 사이클로덱스트린 또는 히알루론산); 또는 지질과 같은 자연 발생 물질을 포함할 수 있다. 리간드는 또한 합성 중합체, 예를 들어 합성 폴리아미노산과 같은 재조합 또는 합성 분자일 수 있다. 폴리아미노산의 예는 폴리리신(PLL), 폴리 L-아스파르트산, 폴리 L-글루탐산, 스티렌-말레산 무수물 공중합체, 폴리(L-락티드-코-글리콜리드) 공중합체, 디비닐 에테르-말레산 무수물 공중합체, N-(2-하이드록시프로필)메타크릴아미드 공중합체(HMPA), 폴리에틸렌 글리콜(PEG), 폴리비닐 알코올(PVA), 폴리우레탄, 폴리(2-에틸아크릴산), N-이소프로필아크릴아미드 중합체 또는 폴리포스파진이다. 폴리아민의 예는 폴리에틸렌이민, 폴리리신(PLL), 스페르민, 스페르미딘, 폴리아민, 슈도펩티드-폴리아민, 펩티드모방체 폴리아민, 덴드리머 폴리아민, 아르기닌, 아미딘, 프로타민, 양이온성 지질, 양이온성 포르피린, 폴리아민의 4급 염, 또는 α 나선형 펩티드를 포함한다.
리간드는 또한 표적화 그룹, 예를 들어, 세포 또는 조직 표적화제, 예를 들어, 렉틴, 당단백질, 지질 또는 단백질, 예를 들어, 신장 세포와 같은 특정 세포 유형에 결합하는 항체를 포함할 수 있다. 표적화 그룹은 티로트로핀, 멜라노트로핀, 렉틴, 당단백질, 계면활성제 단백질 A, 뮤신 탄수화물, 다가 락토스, 다가 갈락토스, N-아세틸-갈락토사민, N-아세틸-글루코사민 다가 만노스, 다가 푸코스, 글리코실화된 폴리아미노산, 다가 갈락토스, 트랜스페린, 비스포스포네이트, 폴리글루타메이트, 폴리아스파르테이트, 지질, 콜레스테롤, 스테로이드, 담즙산, 엽산, 비타민 B12, 비오틴, 또는 RGD 펩티드 또는 RGD 펩티드 모방체일 수 있다. 특정 양태에서, 리간드는 다가 갈락토스, 예를 들어, N-아세틸-갈락토사민이다.
리간드의 또 다른 예는 염료, 인터칼레이션제(예를 들어, 아크리딘), 가교결합제(예를 들어, 소랄렌, 미토마이신 C), 포르피린(TPPC4, 텍사피린, 사피린), 폴리사이클릭 방향족 탄화수소(예를 들어, 페나진, 디하이드로페나진), 인공 엔도뉴클레아제(예를 들어, EDTA), 친유성 분자, 예를 들어, 콜레스테롤, 콜산, 아다만탄 아세트산, 1-피렌 부티르산, 디하이드로테스토스테론, 1,3-비스-O(헥사데실)글리세롤, 게라닐옥시헥실 그룹, 헥사데실글리세롤, 보르네올, 멘톨, 1,3-프로판디올, 헵타데실 그룹, 팔미트산, 미리스트산, O3-(올레오일)리토콜산, O3-(올레오일)콜렌산, 디메톡시트리틸, 또는 페녹사진) 및 펩티드 접합체(예를 들어, 안테나페디아 펩티드, Tat 펩티드), 알킬화제, 포스페이트, 아미노, 머캅토, PEG(예를 들어, PEG-40K), MPEG, [MPEG]2, 폴리아미노, 알킬, 치환된 알킬, 방사성 표지 마커, 효소, 합텐(예를 들어, 비오틴) , 수송/흡수 촉진제(예를 들어, 아스피린, 비타민 E, 엽산), 합성 리보뉴클레아제(예를 들어, 이미다졸, 비스이미다졸, 히스타민, 이미다졸 클러스터, 아크리딘-이미다졸 접합체, 테트라아자마크로사이클의 Eu3+ 복합체), 디니트로페닐, HRP, 또는 AP를 포함한다.
리간드는 단백질, 예를 들어, 당단백질 또는 펩티드, 예를 들어, 공동-리간드에 대해 특이적 친화성을 갖는 분자, 또는 항체, 예를 들어 암세포, 내피 세포, 또는 뼈 세포와 같은 특정 세포 유형에 결합하는 항체일 수 있다. 리간드는 또한 호르몬 및 호르몬 수용체를 포함할 수 있다. 이들은 또한 지질, 렉틴, 탄수화물, 비타민, 보조인자, 다가 락토스, 다가 갈락토스, N-아세틸-갈락토사민, N-아세틸-글루코사민 다가 만노스, 또는 다가 푸코스와 같은 비-펩티드 종을 포함할 수 있다. 리간드는, 예를 들면, 지질다당류, p38 MAP 키나제의 활성화제, 또는 NF-κB의 활성화제일 수 있다.
리간드는, 예를 들면, 세포의 세포골격을 파괴함으로써, 예를 들어, 세포의 미세소관, 미세섬유 또는 중간 섬유를 파괴함으로써 세포 내로의 iRNA 제제의 흡수를 증가시킬 수 있는 물질, 예를 들어, 약물일 수 있다. 약물은, 예를 들면, 탁손, 빈크리스틴, 빈블라스틴, 사이토칼라신, 노코다졸, 자플라키놀리드, 라트룬쿨린 A, 팔로이딘, 스윈홀리드 A, 인다노신 또는 미오세르빈일 수 있다.
일부 양태에서, 본원에 기술된 바와 같은 iRNA에 부착된 리간드는 약동학적 조절제(PK 조절제)로서 작용한다. PK 조절제는 친유체, 담즙산, 스테로이드, 인지질 유사체, 펩티드, 단백질 결합제, PEG, 비타민 등을 포함한다. 예시적인 PK 조절제는 콜레스테롤, 지방산, 콜산, 리토콜산, 디알킬글리세리드, 디아실글리세리드, 인지질, 스핑고지질, 나프록센, 이부프로펜, 비타민 E, 비오틴 등을 포함하지만, 이에 제한되지 않는다. 다수의 포스포로티오에이트 결합을 포함하는 올리고뉴클레오티드는 또한 혈청 단백질에 결합하는 것으로 알려져 있으므로 백본에 다수의 포스포로티오에이트 결합을 포함하는 짧은 올리고뉴클레오티드, 예를 들어, 약 5개 염기, 10개 염기, 15개 염기 또는 20개 염기의 올리고뉴클레오티드가 또한 리간드로서(예를 들어, PK 조절 리간드로서) 본 발명에 적용할 수 있다. 또한, 혈청 성분(예를 들어, 혈청 단백질)에 결합하는 앱타머가 또한 본원에 기술된 양태에서 PK 조절 리간드로서 사용하기에 적합하다.
본 발명의 리간드-접합된 iRNA는 올리고뉴클레오티드에 대한 결합 분자의 부착으로부터 유래된 것과 같은 펜던트 반응성 작용기를 보유하는 올리고뉴클레오티드의 사용에 의해 합성될 수 있다(아래 기술됨). 이러한 반응성 올리고뉴클레오티드는 상업적으로 이용 가능한 리간드, 다양한 보호 그룹 중 임의의 것을 함유하는 합성된 리간드, 또는 이에 부착된 결합 모이어티를 갖는 리간드와 직접 반응할 수 있다.
본 발명의 접합체에 사용되는 올리고뉴클레오티드는 공지된 고체상 합성 기술을 통해 편리하고 통상적으로 제조될 수 있다. 이러한 합성을 위한 장비는, 예를 들면, Applied Biosystems®(Foster City, CA)를 포함한 여러 공급업체에서 판매된다. 당업계에 공지된 이러한 합성을 위한 임의의 다른 수단이 추가로 또는 대안적으로 사용될 수 있다. 포스포로티오에이트 및 알킬화 유도체와 같은 다른 올리고뉴클레오티드를 제조하기 위해 유사한 기술을 사용하는 것이 또한 알려져 있다.
본 발명의 리간드-접합된 올리고뉴클레오티드 및 리간드-분자 보유 서열-특이 결합된 뉴클레오시드에서, 올리고뉴클레오티드 및 올리고뉴클레오시드는 표준 뉴클레오티드 또는 뉴클레오시드 전구체, 또는 이미 결합 모이어티를 보유하는 뉴클레오티드 또는 뉴클레오시드 접합체 전구체, 이미 리간드 분자를 보유하는 리간드-뉴클레오티드 또는 뉴클레오시드-접합체 전구체, 또는 비-뉴클레오시드 리간드-보유 구성 블록을 사용하여 적합한 DNA 합성기 상에서 조립될 수 있다.
이미 결합 모이어티를 보유하는 뉴클레오티드-접합체 전구체를 사용하는 경우, 서열-특이 결합된 뉴클레오시드의 합성이 전형적으로 완료되고, 그후 리간드 분자가 결합 모이어티와 반응하여 리간드-접합된 올리고뉴클레오티드를 형성한다. 일부 양태에서, 본 발명의 올리고뉴클레오티드 또는 결합된 뉴클레오시드는 상업적으로 이용 가능하고 올리고뉴클레오티드 합성에 일상적으로 사용되는 표준 포스포르아미다이트 및 비표준 포스포아르미다이트 외에 리간드-뉴클레오시드 접합체로부터 유도된 포스포르아미다이트를 사용하여 자동화 합성기에 의해 합성된다.
A. 지질 접합체
특정 양태에서, 리간드 또는 접합체는 지질 또는 지질-기반 분자이다. 이러한 지질 또는 지질-기반 분자는 전형적으로 인간 혈청 알부민(HSA)과 같은 혈청 단백질에 결합할 수 있다. HSA 결합 리간드는 표적 조직, 예를 들어, 신체의 비-신장 표적 조직에 접합체가 분포되는 것을 허용한다. 예를 들면, 표적 조직은 간의 실질 세포를 포함한 간일 수 있다. HSA에 결합할 수 있는 다른 분자가 또한 리간드로서 사용될 수 있다. 예를 들면, 나프록센 또는 아스피린이 사용될 수 있다. 지질 또는 지질-기반 리간드는 (a) 접합체의 분해에 대한 저항성을 증가시키거나, (b) 표적 세포 또는 세포막으로의 표적화 또는 수송을 증가시키거나, (c) 혈청 단백질, 예를 들면, HSA에 대한 결합을 조정하는데 사용될 수 있다.
지질-기반 리간드는 표적 조직에 대한 접합체의 결합을 조절, 예를 들어, 제어(예를 들어, 억제)하는데 사용될 수 있다. 예를 들면, HSA에 더 강하게 결합하는 지질 또는 지질-기반 리간드는 신장을 표적으로 할 가능성이 적어서 신체에서 제거될 가능성이 덜하다. HSA에 덜 강하게 결합하는 지질 또는 지질-기반 리간드를 사용하여 접합체를 신장에 표적화할 수 있다.
특정 양태에서, 지질-기반 리간드는 HSA에 결합한다. 예를 들면, 리간드는 비-신장 조직에 대한 접합체의 분포가 향상되도록 충분한 친화도로 HSA에 결합할 수 있다. 그러나, 친화도는 전형적으로 HSA-리간드 결합을 되돌릴 수 없을 정도로 강하지 않다.
특정 양태에서, 지질-기반 리간드는 신장으로의 접합체의 분포가 향상되도록 HSA에 약하게 결합하거나 전혀 결합하지 않는다. 신장 세포를 표적으로 하는 다른 모이어티가 또한 지질-기반 리간드 대신에 또는 이에 더하여 사용될 수 있다.
또 다른 측면에서, 리간드는 모이어티, 예를 들어, 표적 세포, 예를 들어, 증식 세포에 의해 흡수되는 비타민이다. 이들은 악성 또는 비악성 유형의, 예를 들어, 암세포의 원치 않는 세포 증식을 특징으로 하는 장애를 치료하는데 특히 유용하다. 예시적인 비타민은 비타민 A, E 및 K를 포함한다. 다른 예시적인 비타민은 비타민 B, 예를 들어, 엽산, B12, 리보플라빈, 비오틴, 피리독살 또는 암세포에 의해 흡수되는 기타 비타민 또는 영양소를 포함한다. HSA 및 저밀도 지단백(LDL)도 포함된다.
B. 세포
투과제
또 다른 측면에서, 리간드는 나선형 세포 투과제와 같은 세포 투과제이다. 특정 양태에서, 제제는 양친매성이다. 예시적인 제제는 tat 또는 안테노페디아(antennopedia)와 같은 펩티드이다. 제제가 펩티드인 경우, 이것은 펩티드 모방체, 인버토머(invertomer), 비펩티드 또는 유사-펩티드 결합, 및 D-아미노산의 사용을 포함하여 변형될 수 있다. 나선형 제제는 전형적으로 α-나선형 제제이고 친유성 및 소유성 상을 가질 수 있다.
리간드는 펩티드 또는 펩티드 모방체일 수 있다. 펩티드 모방체(본원에서 올리고펩티드 모방체로도 지칭됨)는 천연 펩티드와 유사한 정의된 3차원 구조로 접힐 수 있는 분자이다. iRNA 제제에 대한 펩티드 및 펩티드 모방체의 부착은 세포 인식 및 흡수를 향상시키는 것과 같이 iRNA의 약동학적 분포에 영향을 미칠 수 있다. 펩티드 또는 펩티드 모방체 모이어티는 약 5-50개 아미노산 길이, 예를 들어, 약 5, 10, 15, 20, 25, 30, 35, 40, 45, 또는 50개 아미노산 길이일 수 있다.
펩티드 또는 펩티드 모방체는, 예를 들면, 세포 투과 펩티드, 양이온성 펩티드, 양친매성 펩티드 또는 소수성 펩티드(예를 들어, 주로 Tyr, Trp, 또는 Phe로 구성됨)일 수 있다. 펩티드 모이어티는 덴드리머 펩티드, 제한된 펩티드 또는 가교결합된 펩티드일 수 있다. 또 다른 대안에서, 펩티드 모이어티는 소수성 막 전좌 서열(MTS)을 포함할 수 있다. 예시적인 소수성 MTS-함유 펩티드는 아미노산 서열 AAVALLPAVLLALLAP (서열번호 ____)을 갖는 RFGF이다. 소수성 MTS를 함유하는 RFGF 유사체(예를 들어, 아미노산 서열 AALLPVLLAAP (서열번호 ____))가 또한 표적화 모이어티일 수 있다. 펩티드 모이어티는 세포막을 가로질러 펩티드, 올리고뉴클레오티드 및 단백질을 포함하는 큰 극성 분자를 운반할 수 있는 "전달" 펩티드일 수 있다. 예를 들어, HIV Tat 단백질(GRKKRRQRRRPPQ (서열번호 ____)) 및 드로소필라 안테나페디아 ( Drosophila Antennapedia ) 단백질(RQIKIWFQNRRMKWKK (서열번호 ____))로부터의 서열이 전달 펩티드로서 기능할 수 있는 것으로 밝혀졌다. 펩티드 또는 펩티드 모방체는 파지-디스플레이 라이브러리 또는 OBOC(one-bead-one-compound) 조합 라이브러리로부터 식별된 펩티드와 같은 DNA의 랜덤 서열에 의해 암호화될 수 있다(Lam et al., Nature, 354:82-84, 1991). 전형적으로, 통합된 단량체 단위를 통해 dsRNA 제제에 테더링된 펩티드 또는 펩티드 모방체는 아르기닌-글리신-아스파르트산(RGD)-펩티드 또는 RGD 모방체와 같은 세포 표적화 펩티드이다. 펩티드 모이어티는 약 5개 아미노산 내지 약 40개 아미노산의 길이 범위일 수 있다. 펩티드 모이어티는 안정성을 증가시키거나 구조적 특성을 지시하기 위해서와 같이 구조적 변형을 가질 수 있다. 아래에 기술된 임의의 구조적 변형이 사용될 수 있다.
본 발명의 조성물 및 방법에 사용하기 위한 RGD 펩티드는 선형 또는 고리형일 수 있고, 특정 조직(들)에 대한 표적화를 촉진하기 위해 변형, 예를 들어 글리코실화 또는 메틸화될 수 있다. RGD-함유 펩티드 및 펩티드 모방체는 D-아미노산 및 합성 RGD 모방체를 포함할 수 있다. RGD 외에도, 인테그린 리간드를 표적으로 하는 다른 모이어티를 사용할 수 있다. 이 리간드의 바람직한 접합체는 PECAM-1 또는 VEGF를 표적으로 한다.
RGD 펩티드 모이어티는 특정 세포 유형, 예를 들어 내피 종양 세포 또는 유방암 종양 세포와 같은 종양 세포를 표적화하는데 사용될 수 있다(Zitzmann et al., Cancer Res., 62:5139-43, 2002). RGD 펩티드는 폐, 신장, 비장 또는 간을 포함한 다양한 다른 조직의 종양에 대한 dsRNA 제제의 표적화를 촉진할 수 있다(Aoki et al., Cancer Gene Therapy 8:783-787, 2001). 전형적으로, RGD 펩티드는 신장에 대한 iRNA 제제의 표적화를 촉진할 것이다. RGD 펩티드는 선형 또는 고리형일 수 있고, 특정 조직에 대한 표적화를 용이하게 하기 위해 변형, 예를 들어, 글리코실화 또는 메틸화될 수 있다. 예를 들면, 글리코실화된 RGD 펩티드는 αVß3을 발현하는 종양 세포에 iRNA 작용제를 전달할 수 있다(Haubner et al., Jour. Nucl. Med ., 42:326-336, 2001).
"세포 투과 펩티드"는 세포, 예를 들어 박테리아 또는 진균 세포와 같은 미생물 세포, 또는 인간 세포와 같은 포유동물 세포를 투과할 수 있다. 미생물 세포-투과 펩티드는, 예를 들면, α-나선형 선형 펩티드(예를 들어, LL-37 또는 세로핀 P1), 이황화 결합-함유 펩티드(예를 들어, α-디펜신, β-디펜신 또는 박테네신), 또는 1개 또는 2개의 주요 아미노산만을 함유하는 펩티드(예를 들어, PR-39 또는 인돌리시딘)일 수 있다. 세포 투과 펩티드는 또한 핵 국소화 신호(NLS)를 포함할 수 있다. 예를 들면, 세포 투과 펩티드는 HIV-1 gp41의 융합 펩티드 도메인 및 SV40 대형 T 항원의 NLS로부터 유래된 MPG와 같은 이분형 양친매성 펩티드일 수 있다(Simeoni et al., Nucl. Acids Res. 31:2717-2724, 2003).
C. 탄수화물 접합체
본 발명의 조성물 및 방법의 일부 양태에서, iRNA는 탄수화물을 추가로 포함한다. 탄수화물 접합된 iRNA는 핵산의 생체내 전달에 유용할 뿐만 아니라 본원에 기술된 바와 같이 생체내 치료 용도에 적합한 조성물이다. 본원에 사용된 "탄수화물"은 각 탄소 원자에 결합된 산소, 질소 또는 황 원자와 함께 적어도 6개의 탄소 원자(선형, 분지형 또는 고리형일 수 있음)를 갖는 하나 이상의 단당류 단위로 구성된 탄수화물 자체인 화합물; 또는 각 탄소 원자에 결합된 산소, 질소 또는 황 원자와 함께 각각 적어도 6개의 탄소 원자(선형, 분지형 또는 고리형일 수 있음)를 갖는 하나 이상의 단당류 단위로 구성된 탄수화물 모이어티를 그 일부로 갖는 화합물을 지칭한다. 대표적인 탄수화물은 당(약 4, 5, 6, 7, 8, 또는 9개의 단당류 단위를 함유하는 단당류, 이당류, 삼당류 및 올리고당류), 및 전분, 글리코겐, 셀룰로스 및 다당류 검과 같은 다당류를 포함한다. 특정 단당류는 C5 이상(예를 들어, C5, C6, C7, 또는 C8) 당을 포함하고; 이당류 및 삼당류는 2개 또는 3개의 단당류 단위(예를 들어, C5, C6, C7 또는 C8)를 갖는 당을 포함한다.
특정 양태에서, 탄수화물 접합체는 단당류를 포함한다.
특정 양태에서, 단당류는 N-아세틸갈락토사민(GalNAc)이다. 하나 이상의 N-아세틸갈락토사민(GalNAc) 유도체를 포함하는 GalNAc 접합체는, 예를 들면, 미국 제8,106,022호에 기술되어 있으며, 이의 전체 내용은 본원에 참고로 포함된다. 일부 양태에서, GalNAc 접합체는 iRNA를 특정 세포로 표적화하는 리간드로서 역할을 한다. 일부 양태에서, GalNAc 접합체는, 예를 들어, 간 세포(예를 들어, 간세포)의 아시알로당단백질 수용체에 대한 리간드로서 역할을 함으로써 iRNA를 간 세포에 표적화한다.
일부 양태에서, 탄수화물 접합체는 하나 이상의 GalNAc 유도체를 포함한다. GalNAc 유도체는 링커, 예를 들어, 2가 또는 3가 분지형 링커를 통해 부착될 수 있다. 일부 양태에서 GalNAc 접합체는 센스 가닥의 3' 말단에 접합된다. 일부 양태에서, GalNAc 접합체는 링커, 예를 들어, 본원에 기술된 바와 같은 링커를 통해 iRNA 제제에 (예를 들어, 센스 가닥의 3' 말단에) 접합된다. 일부 양태에서 GalNAc 접합체는 센스 가닥의 5' 말단에 접합된다. 일부 양태에서, GalNAc 접합체는 링커, 예를 들어, 본원에 기술된 바와 같은 링커를 통해 iRNA 제제에 (예를 들어, 센스 가닥의 5' 말단에) 접합된다.
본 발명의 특정 양태에서, GalNAc 또는 GalNAc 유도체는 1가 링커를 통해 본 발명의 iRNA 제제에 부착된다. 일부 양태에서, GalNAc 또는 GalNAc 유도체는 2가 링커를 통해 본 발명의 iRNA 제제에 부착된다. 본 발명의 또 다른 양태에서, GalNAc 또는 GalNAc 유도체는 3가 링커를 통해 본 발명의 iRNA 제제에 부착된다. 본 발명의 다른 양태에서, GalNAc 또는 GalNAc 유도체는 4가 링커를 통해 본 발명의 iRNA 제제에 부착된다.
특정 양태에서, 본 발명의 이중 가닥 RNAi 제제는 iRNA 제제에 부착된 하나의 GalNAc 또는 GalNAc 유도체를 포함한다. 특정 양태에서, 본 발명의 이중 가닥 RNAi 제제는 복수의 1가 링커를 통해 이중 가닥 RNAi 제제의 다수의 뉴클레오티드에 각각 독립적으로 부착된 복수의(예를 들면, 2, 3, 4, 5, 또는 6개) GalNAc 또는 GalNAc 유도체를 포함한다.
일부 양태에서, 예를 들면, 본 발명의 iRNA 제제의 2개의 가닥이 한 가닥의 3'-말단과 각각의 다른 가닥의 5'-말단 사이에 중단되지 않은 뉴클레오티드 쇄에 의해 연결되어 복수의 쌍을 이루지 않은 뉴클레오티드를 포함하는 헤어핀 루프를 형성하는 하나의 더 큰 분자의 일부인 경우, 헤어핀 루프 내의 각각의 쌍을 이루지 않은 뉴클레오티드는 독립적으로 1가 링커를 통해 부착된 GalNAc 또는 GalNAc 유도체를 포함할 수 있다. 헤어핀 루프는 또한 이중체의 한 가닥에서 연장된 돌출부에 의해 형성될 수 있다.
일부 양태에서, 예를 들면, 본 발명의 iRNA 제제의 2개의 가닥이 한 가닥의 3'-말단과 각각의 다른 가닥의 5'-말단 사이에 중단되지 않은 뉴클레오티드 쇄에 의해 연결되어 복수의 쌍을 이루지 않은 뉴클레오티드를 포함하는 헤어핀 루프를 형성하는 하나의 더 큰 분자의 일부인 경우, 헤어핀 루프 내의 각각의 쌍을 이루지 않은 뉴클레오티드는 독립적으로 1가 링커를 통해 부착된 GalNAc 또는 GalNAc 유도체를 포함할 수 있다. 헤어핀 루프는 또한 이중체의 한 가닥에서 연장된 돌출부에 의해 형성될 수 있다.
일부 양태에서, GalNAc 접합체는 화학식 II이다.
[화학식 II]

일부 양태에서, RNAi 제제는 하기 개략도에 나타낸 바와 같이 링커를 통해 탄수화물 접합체에 부착되며, 여기서 X는 O 또는 S이다.

일부 양태에서, RNAi 제제는 표 11에 정의되고 아래에 나타낸 바와 같이 L96에 접합된다:

특정 양태에서, 본 발명의 조성물 및 방법에 사용하기 위한 탄수화물 접합체는 다음으로 이루어진 그룹으로부터 선택된다:
화학식 II

[화학식 III]

[화학식 IV]

[화학식 V]

[화학식 VI]

[화학식 VII]

[화학식 VIII]

[화학식 IX]

[화학식 X]

[화학식 XI]

[화학식 XII]

[화학식 XIII]

[화학식 XIV]

[화학식 XV]

[화학식 XVI]

[화학식 XVII]

[화학식 XVIII]

[화학식 XIX]

[화학식 XX]

[화학식 XXI]

[화학식 XXII]

[화학식 XXIII]

[화학식 XXIV]

[화학식 XXV]

[화학식 XXVI]

[화학식 XXVII]

[화학식 XXVIII, XXIX]

[화학식 XXX, XXXI]

[화학식 XXXII, XXXIII]

[화학식 XXXIV]

특정 양태에서, 본 발명의 조성물 및 방법에 사용하기 위한 탄수화물 접합체는 단당류이다. 특정 양태에서, 단당류는 화학식 II와 같은 N-아세틸갈락토사민이다:
화학식 II

본원에 기술된 양태에서 사용하기 위한 또 다른 대표적인 탄수화물 접합체는 다음을 포함하지만, 이에 제한되지는 않는다:
[화학식 XXXVI]

X 또는 Y 중 하나가 올리고뉴클레오티드일 때, 다른 하나는 수소이다.
일부 양태에서, 적합한 리간드는 제WO 2019/055633호에 개시된 리간드이며, 이의 전체 내용은 본원에 참고로 포함된다. 하나의 양태에서 리간드는 아래 구조를 포함한다:

특정 양태에서, 본 개시내용의 RNAi 제제는 GalNAc 리간드가 현재 본 개시내용의 바람직한 척수강내/CNS 전달 경로(들)에 대해 제한된 가치를 갖는 것으로 예상되더라도, GalNAc 리간드를 포함할 수 있다.
본 발명의 특정 양태에서, GalNAc 또는 GalNAc 유도체는 1가 링커를 통해 본 발명의 iRNA 제제에 부착된다. 일부 양태에서, GalNAc 또는 GalNAc 유도체는 2가 링커를 통해 본 발명의 iRNA 제제에 부착된다. 본 발명의 또 다른 양태에서, GalNAc 또는 GalNAc 유도체는 3가 링커를 통해 본 발명의 iRNA 제제에 부착된다.
하나의 양태에서, 본 발명의 이중 가닥 RNAi 제제는 iRNA 제제에 부착된 하나 이상의 GalNAc 또는 GalNAc 유도체를 포함한다. GalNAc는 센스 가닥 또는 안티센스 가닥에서 링커를 통해 임의의 뉴클레오티드에 부착될 수 있다. GalNac는 센스 가닥의 5'-말단, 센스 가닥의 3' 말단, 안티센스 가닥의 5'-말단, 또는 안티센스 가닥의 3'-말단에 부착될 수 있다. 하나의 양태에서, GalNAc는, 예를 들어, 3가 링커를 통해 센스 가닥의 3' 말단에 부착된다.
다른 양태에서, 본 발명의 이중 가닥 RNAi 제제는 각각 독립적으로 복수의 링커, 예를 들어, 1가 링커를 통해 이중 가닥 RNAi 제제의 복수의 뉴클레오티드에 부착된 복수의(예를 들면, 2, 3, 4, 5, 또는 6개) GalNAc 또는 GalNAc 유도체를 포함한다.
일부 양태에서, 예를 들면, 본 발명의 iRNA 제제의 2개의 가닥이 한 가닥의 3'-말단과 각각의 다른 가닥의 5'-말단 사이에 중단되지 않은 뉴클레오티드 쇄에 의해 연결되어 복수의 쌍을 이루지 않은 뉴클레오티드를 포함하는 헤어핀 루프를 형성하는 하나의 더 큰 분자의 일부인 경우, 헤어핀 루프 내의 각각의 쌍을 이루지 않은 뉴클레오티드는 독립적으로 1가 링커를 통해 부착된 GalNAc 또는 GalNAc 유도체를 포함할 수 있다.
일부 양태에서, 탄수화물 접합체는 PK 조절제 또는 세포 투과 펩티드와 같지만 이에 제한되지 않는 상기 기술된 바와 같은 하나 이상의 추가 리간드를 추가로 포함한다.
본 발명에 사용하기에 적합한 추가의 탄수화물 접합체 및 링커는 제WO 2014/179620호 및 제WO 2014/179627호에 기술된 것들을 포함하며, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
D. 링커
일부 양태에서, 본원에 기술된 접합체 또는 리간드는 절단 가능하거나 절단 불가능할 수 있는 다양한 링커를 사용하여 iRNA 올리고뉴클레오티드에 부착될 수 있다.
용어 "링커" 또는 "연결 그룹"은 화합물의 두 부분을 연결하고, 예를 들어, 화합물의 두 부분을 공유적으로 부착시키는 유기 모이어티를 의미한다. 링커는 전형적으로 직접 결합 또는 산소 또는 황과 같은 원자, NR8, C(O), C(O)NH, SO, SO2, SO2NH와 같은 단위 또는 치환되거나 비치환된 알킬, 치환되거나 비치환된 알케닐, 치환되거나 비치환된 알키닐, 아릴알킬, 아릴알케닐, 아릴알키닐, 헤테로아릴알킬, 헤테로아릴알케닐, 헤테로아릴알키닐, 헤테로사이클릴알킬, 헤테로사이클릴알케닐, 헤테로사이클릴알키닐, 아릴, 헤테로아릴, 헤테로사이클릴, 사이클로알킬, 사이클로알케닐, 알킬아릴알킬, 알킬아릴알케닐, 알킬아릴알키닐, 알케닐아릴알킬, 알케닐아릴알케닐, 알케닐아릴알키닐, 알키닐아릴알킬, 알키닐아릴알케닐, 알키닐아릴알키닐, 알킬헤테로아릴알킬, 알킬헤테로아릴알케닐, 알킬헤테로아릴알키닐, 알케닐헤테로아릴알킬, 알케닐헤테로아릴알케닐, 알케닐헤테로아릴알키닐, 알키닐헤테로아릴알킬, 알키닐헤테로아릴알케닐, 알키닐헤테로아릴알키닐, 알킬헤테로사이클릴알킬, 알킬헤테로사이클릴알케닐, 알킬헤테로사이클릴알키닐, 알케닐헤테로사이클릴알킬, 알케닐헤테로사이클릴알케닐, 알케닐헤테로사이클릴알키닐, 알키닐헤테로사이클릴알킬, 알키닐헤테로사이클릴알케닐, 알키닐헤테로사이클릴알키닐, 알킬아릴, 알케닐아릴, 알키닐아릴, 알킬헤테로아릴, 알케닐헤테로아릴, 알키닐헤테로아릴과 같지만 이에 제한되지 않는 원소의 쇄를 포함하고, 여기서 하나 이상의 메틸렌은 O, S, S(O), SO2, N(R8), C(O), 치환되거나 비치환된 아릴, 치환되거나 비치환된 헤테로아릴, 치환되거나 비치환된 헤테로사이클릭에 의해 차단되거나 말단화될 수 있고; 여기서 R8은 수소, 아실, 지방족 또는 치환된 지방족이다. 특정 양태에서, 링커는 약 1-24개 원자, 2-24, 3-24, 4-24, 5-24, 6-24, 6-18, 7-18, 8-18개 원자, 7-17, 8-17, 6-16, 7-16, 또는 8-16개 원자이다.
절단 가능한 연결 그룹은 세포 외부에서 충분히 안정하지만 표적 세포에 진입시 절단되어 링커가 함께 유지하고 있는 두 부분을 해제하는 그룹이다. 바람직한 양태에서, 절단 가능한 연결 그룹은 대상체의 혈액에서 또는 제2 참조 조건(예를 들어, 혈액 또는 혈청에서 발견되는 조건을 모방하거나 나타내도록 선택될 수 있음) 하에서보다 표적 세포에서 또는 제1 참조 조건(예를 들어, 세포내 조건을 모방하거나 나타내도록 선택될 수 있음) 하에서 적어도 약 10배, 20배, 30배, 40배, 50배, 60배, 70배, 80배, 90배 또는 그 이상, 또는 적어도 약 100배 더 빨리 절단된다.
절단 가능한 연결 그룹은 절단제, 예를 들어, pH, 산화환원 전위 또는 분해성 분자의 존재에 민감하다. 일반적으로, 절단제는 혈청 또는 혈액에서보다 세포 내에서 더 널리 퍼져 있거나 더 높은 수준 또는 활성으로 발견된다. 이러한 분해제의 예는 다음을 포함한다: 예를 들어, 산화 또는 환원 효소 또는 환원에 의해 산화 환원 절단 가능한 연결 그룹을 분해할 수 있는, 세포에 존재하는, 머캅탄과 같은 환원제를 포함하여 특정 기질에 대해 선택되거나 기질 특이성이 없는 산화 환원제; 에스테라제; 산성 환경을 생성할 수 있는 엔도솜 또는 제제, 예를 들어, 5 이하의 pH를 야기하는 것; 일반 산, 펩티다제(기질 특이적일 수 있음) 및 포스파타아제로 작용함으로써 산 절단 가능한 연결 그룹을 가수분해하거나 분해할 수 있는 효소.
이황화 결합과 같은 절단 가능한 연결 그룹은 pH에 민감할 수 있다. 인간 혈청의 pH는 7.4인 반면, 평균 세포내 pH는 약 7.1-7.3 범위로 약간 더 낮다. 엔도솜은 5.5-6.0 범위의 더 산성인 pH를 가지며, 리소좀은 약 5.0에서 훨씬 더 산성인 pH를 갖는다. 일부 링커는 바람직한 pH에서 절단되어 세포 내부의 리간드로부터 또는 세포의 원하는 구획으로 양이온성 지질을 방출하는 절단 가능한 연결 그룹을 가질 것이다.
링커는 특정 효소에 의해 절단할 수 있는 절단 가능한 연결 그룹을 포함할 수 있다. 링커에 통합된 절단 가능한 연결 그룹의 유형은 표적화될 세포에 따라 달라질 수 있다. 예를 들면, 간-표적화 리간드는 에스테르 그룹을 포함하는 링커를 통해 양이온성 지질에 연결될 수 있다. 간 세포에는 에스테라제가 풍부하며, 따라서 링커는 에스테라제가 풍부하지 않은 세포 유형에서보다 간 세포에서 더 효율적으로 절단된다. 에스테라제가 풍부한 다른 세포 유형은 폐, 신장 피질 및 고환의 세포를 포함한다.
펩티드 결합을 함유하는 링커는 간 세포 및 활막 세포와 같은 펩티다제가 풍부한 세포 유형을 표적화할 때 사용될 수 있다.
일반적으로, 후보 절단 가능한 연결 그룹의 적합성은 후보 연결 그룹을 절단하는 분해제(또는 조건)의 능력을 시험함으로써 평가될 수 있다. 또한 혈액에서 또는 다른 비표적 조직과 접촉할 때 절단에 저항하는 능력에 대해 후보 절단 가능한 연결 그룹을 시험하는 것이 바람직할 것이다. 따라서, 첫 번째 조건과 두 번째 조건 사이의 절단에 대한 상대적인 민감성을 결정할 수 있으며, 여기서 첫 번째 조건은 표적 세포에서 절단을 나타내도록 선택되고 두 번째 조건은 다른 조직 또는 생물학적 유체, 예를 들어 혈액 또는 혈청에서 절단을 나타내도록 선택된다. 평가는 무세포 시스템, 세포, 세포 배양물, 장기 또는 조직 배양물, 또는 전체 동물에서 수행될 수 있다. 무세포 또는 배양 조건에서 초기 평가를 수행하고 전체 동물에서 추가 평가에 의해 확인하는 것이 유용할 수 있다. 바람직한 양태에서, 유용한 후보 화합물은 혈액 또는 혈청(또는 세포외 조건을 모방하도록 선택된 시험관내 조건하에서)과 비교하여 세포에서(또는 세포내 조건을 모방하도록 선택된 시험관내 조건하에서) 적어도 약 2, 4, 10, 20, 30, 40, 50, 60, 70, 80, 90, 또는 약 100배 더 빠르게 절단된다.
i. 산화환원 절단 가능한 연결 그룹
특정 양태에서, 절단 가능한 연결 그룹은 환원 또는 산화시 절단되는 산화환원 절단 가능한 연결 그룹이다. 환원적으로 절단 가능한 연결 그룹의 예는 이황화물 연결 그룹(-S-S-)이다. 후보 절단 가능한 연결 그룹이 적합한 "환원적으로 절단 가능한 연결 그룹"인지 또는 예를 들어 특정 iRNA 모이어티 및 특정 표적화제와 함께 사용하기에 적합한지 여부를 결정하기 위해, 본원에 기술된 방법을 살펴볼 수 있다. 예를 들면, 후보물질은 세포, 예를 들어, 표적 세포에서 관찰될 절단 속도를 모방하는 당업계에 공지된 시약을 사용하여 디티오트레이톨(DTT) 또는 기타 환원제와의 배양에 의해 평가될 수 있다. 후보물질은 또한 혈액 또는 혈청 조건을 모방하도록 선택된 조건하에서 평가될 수 있다. 하나의 양태에서, 후보 화합물은 혈액에서 최대 약 10%까지 절단된다. 다른 양태에서, 유용한 후보 화합물은 혈액(또는 세포외 조건을 모방하도록 선택된 시험관내 조건하에서)과 비교하여 세포에서(또는 세포내 조건을 모방하도록 선택된 시험관내 조건하에서) 적어도 약 2, 4, 10, 20, 30, 40, 50, 60, 70, 80, 90, 또는 약 100배 더 빠르게 분해된다. 후보 화합물의 절단 속도는 세포외 배지를 모방하도록 선택된 조건과 비교하여 세포내 배지를 모방하도록 선택된 조건하에서 표준 효소 동역학 분석을 사용하여 결정될 수 있다.
ii. 포스페이트-기반 절단 가능한 연결 그룹
특정 양태에서, 절단 가능한 링커는 포스페이트-기반 절단 가능한 연결 그룹을 포함한다. 포스페이트-기반 절단 가능한 연결 그룹은 포스페이트 그룹을 분해하거나 가수분해하는 제제에 의해 절단된다. 세포에서 포스페이트 그룹을 절단하는 제제의 예는 세포에서 포스파타제와 같은 효소이다. 포스페이트-기반 연결 그룹의 예는 -O-P(O)(ORk)-O-, -O-P(S)(ORk)-O-, -O-P(S)(SRk)-O-, -S-P(O)(ORk)-O-, -O-P(O)(ORk)-S-, -S-P(O)(ORk)-S-, -O-P(S)(ORk)-S-, -S-P(S)(ORk)-O-, -O-P(O)(Rk)-O-, -O-P(S)(Rk)-O-, -S-P(O)(Rk)-O-, -S-P(S)(Rk)-O-, -S-P(O)(Rk)-S-, -O-P(S)(Rk)-S-이다. 바람직한 양태는 -O-P(O)(OH)-O-, -O-P(S)(OH)-O-, -O-P(S)(SH)-O-, -S-P(O)(OH)-O-, -O-P(O)(OH)-S-, -S-P(O)(OH)-S-, -O-P(S)(OH)-S-, -S-P(S)(OH)-O-, -O-P(O)(H)-O-, -O-P(S)(H)-O-, -S-P(O)(H)-O, -S-P(S)(H)-O-, -S-P(O)(H)-S-, -O-P(S)(H)-S-이다. 바람직한 양태는 -O-P(O)(OH)-O-이다. 이러한 후보물질은 위에 기술된 것과 유사한 방법을 사용하여 평가될 수 있다.
iii. 산 절단 가능한 연결 그룹
특정 양태에서, 절단 가능한 링커는 산 절단 가능한 연결 그룹을 포함한다. 산 절단 가능한 연결 그룹은 산성 조건하에서 절단되는 연결 그룹이다. 바람직한 양태에서 산 절단 가능한 연결 그룹은 pH가 약 6.5 이하(예를 들어, 약 6.0, 5.75, 5.5, 5.25, 5.0 이하)인 산성 환경에서 또는 일반 산으로서 작용할 수 있는 효소와 같은 제제에 의해 절단된다. 세포에서, 엔도솜 및 리소좀과 같은 특정의 낮은 pH 소기관은 산 절단 가능한 연결 그룹에 대한 절단 환경을 제공할 수 있다. 산 절단가능한 연결 그룹의 예는 하이드라존, 에스테르, 및 아미노산의 에스테르를 포함하지만 이에 제한되지 않는다. 산 절단 가능한 그룹은 일반 화학식 -C=NN-, C(O)O 또는 -OC(O)를 가질 수 있다. 바람직한 양태는 에스테르의 산소에 부착된 탄소(알콕시 그룹)가 아릴 그룹, 치환된 알킬 그룹, 또는 3급 알킬 그룹, 예를 들어 디메틸 펜틸 또는 t-부틸일 때이다. 이러한 후보물질은 위에 기술된 것과 유사한 방법을 사용하여 평가될 수 있다.
iv. 에스테르-기반 절단 가능한 연결 그룹
특정 양태에서, 절단 가능한 링커는 에스테르-기반 절단 가능한 연결 그룹을 포함한다. 에스테르-기반 절단 가능한 연결 그룹은 세포에서 에스테라제 및 아미다제와 같은 효소에 의해 절단된다. 에스테르-기반 절단 가능한 연결 그룹의 예는 알킬렌, 알케닐렌 및 알키닐렌 그룹의 에스테르를 포함하지만, 이에 제한되지 않는다. 에스테르 절단 가능한 연결 그룹은 일반 화학식 -C(O)O-, 또는 -OC(O)-를 갖는다. 이러한 후보물질은 위에 기술된 것과 유사한 방법을 사용하여 평가될 수 있다.
v. 펩티드-기반 절단 가능한 연결 그룹
또 다른 양태에서, 절단 가능한 링커는 펩티드-기반 절단 가능한 연결 그룹을 포함한다. 펩티드-기반 절단 가능한 연결 그룹은 세포에서 펩티다제 및 프로테아제와 같은 효소에 의해 절단된다. 펩티드-기반 절단 가능한 연결 그룹은 올리고펩티드(예를 들어, 디펩티드, 트리펩티드 등) 및 폴리펩티드를 생성하기 위해 아미노산 사이에 형성되는 펩티드 결합이다. 펩티드-기반 절단 가능한 그룹은 아미드 그룹(-C(O)NH-)을 포함하지 않는다. 아미드 그룹은 임의의 알킬렌, 알케닐렌 또는 알키넬렌 사이에 형성될 수 있다. 펩티드 결합은 펩티드와 단백질을 생성하기 위해 아미노산 사이에 형성되는 특수한 유형의 아미드 결합이다. 펩티드 기반 절단 그룹은 일반적으로 펩티드와 단백질을 생성하는 아미노산 사이에 형성된 펩티드 결합(즉, 아미드 결합)으로 제한되며 전체 아미드 작용 그룹을 포함하지 않는다. 펩티드-기반 절단 가능한 연결 그룹은 일반 화학식 -NHCHRAC(O)NHCHRBC(O)- (서열번호 __)를 가지며, 여기서 RA 및 RB는 두 개의 인접한 아미노산의 R 그룹이다. 이러한 후보물질은 위에 기술된 것과 유사한 방법을 사용하여 평가될 수 있다.
일부 양태에서, 본 발명의 iRNA는 링커를 통해 탄수화물에 접합된다. 본 발명의 조성물 및 방법의 링커를 갖는 iRNA 탄수화물 접합체의 비제한적인 예는 다음을 포함하지만, 이에 제한되지 않는다:
[화학식 XXXVII]

[화학식 XXXVIII]

[화학식 XXXIX]

[화학식 XL]

[화학식 XLI]

[화학식 XLII]

[화학식 XLIII]

[화학식 XLIV]

X 또는 Y 중 하나가 올리고뉴클레오티드일 때, 다른 하나는 수소이다.
본 발명의 조성물 및 방법의 특정 양태에서, 리간드는 2가 또는 3가 분지형 링커를 통해 부착된 하나 이상의 "GalNAc"(N-아세틸갈락토사민) 유도체이다.
특정 양태에서, 본 발명의 dsRNA는 화학식 XLV 내지 XLVI 중 어느 하나로 나타낸 구조의 그룹으로부터 선택된 2가 또는 3가 분지형 링커에 접합된다:
[화학식 XLV]

[화학식 XLVI]

[화학식 XLVII]

[화학식 XLVIII]

여기서:
q2A, q2B, q3A, q3B, q4A, q4B, q5A, q5B 및 q5C는 각각의 발생에 대해 독립적으로 0-20을 나타내고 여기서 반복 단위는 동일하거나 상이할 수 있으며;
P2A, P2B, P3A, P3B, P4A, P4B, P5A, P5B, P5C, T2A, T2B, T3A, T3B, T4A, T4B, T4A, T5B, T5C는 각각의 발생에 대해 각각 독립적으로 부재하거나, CO, NH, O, S, OC(O), NHC(O), CH2, CH2NH 또는 CH2O이고;
Q2A, Q2B, Q3A, Q3B, Q4A, Q4B, Q5A, Q5B, Q5C는 각각의 발생에 대해 독립적으로 부재하거나, 알킬렌, 치환된 알킬렌이고, 여기서 하나 이상의 메틸렌은 O, S, S(O), SO2, N(RN), C(R')=C(R''), C≡C 또는 C(O) 중 하나 이상에 의해 차단되거나 말단화될 수 있고;
R2A, R2B, R3A, R3B, R4A, R4B, R5A, R5B, R5C는 각각의 발생에 대해 각각 독립적으로 부재하거나, NH, O, S, CH2, C(O)O, C(O)NH, NHCH(Ra)C(O), -C(O)-CH(Ra)-NH-, CO, CH=N-O,  ,
,  ,
,  ,
,  ,
,  또는 헤테로사이클릴이고;
또는 헤테로사이클릴이고;
L2A, L2B, L3A, L3B, L4A, L4B, L5A, L5B 및 L5C는 리간드를 나타내고; 즉 각각의 발생에 대해 각각 독립적으로 단당류(예를 들어 GalNAc), 이당류, 삼당류, 사당류, 올리고당류 또는 다당류를 나타내고; Ra는 H 또는 아미노산 측쇄이다. 화학식 XLIX와 같은 3가 접합 GalNAc 유도체는 표적 유전자의 발현을 억제하기 위한 RNAi 제제와 함께 사용하는데 특히 유용하다:
[화학식 XLIX]

여기서 L5A, L5B 및 L5C는 단당류, 예를 들어 GalNAc 유도체를 나타낸다.
GalNAc 유도체를 접합하는 적합한 2가 및 3가 분지형 링커 그룹의 예는 화학식 II, VII, XI, X, 및 XIII으로서 위에 인용된 구조를 포함하지만, 이에 제한되지 않는다.
RNA 접합체의 제조를 교시하는 대표적인 미국 특허는 미국 특허 제4,828,979호; 제4,948,882호; 제5,218,105호; 제5,525,465호; 제5,541,313호; 제5,545,730호; 제5,552,538호; 제5,578,717호; 제5,580,731호; 제5,591,584호; 제5,109,124호; 제5,118,802호; 제5,138,045호; 제5,414,077호; 제5,486,603호; 제5,512,439호; 제5,578,718호; 제5,608,046호; 제4,587,044호; 제4,605,735호; 제4,667,025호; 제4,762,779호; 제4,789,737호; 제4,824,941호; 제4,835,263호; 제4,876,335호; 제4,904,582호; 제4,958,013호; 제5,082,830호; 제5,112,963호; 제5,214,136호; 제5,082,830호; 제5,112,963호; 제5,214,136호; 제5,245,022호; 제5,254,469호; 제5,258,506호; 제5,262,536호; 제5,272,250호; 제5,292,873호; 제5,317,098호; 제5,371,241호, 제5,391,723호; 제5,416,203호, 제5,451,463호; 제5,510,475호; 제5,512,667호; 제5,514,785호; 제5,565,552호; 제5,567,810호; 제5,574,142호; 제5,585,481호; 제5,587,371호; 제5,595,726호; 제5,597,696호; 제5,599,923호; 제5,599,928; 제5,688,941; 제6,294,664; 제6,320,017; 제6,576,752; 제6,783,931호; 제6,900,297호; 제7,037,646호; 및 제8,106,022호를 포함하지만, 이에 제한되지 않으며, 이들 각각의 전체 내용은 본원에 참고로 포함된다.
주어진 화합물의 모든 위치가 균일하게 변형될 필요는 없으며, 실제로 전술한 변형 중 하나 이상이 단일 화합물에 또는 심지어 iRNA 내의 단일 뉴클레오시드에 통합될 수 있다. 본 발명은 또한 키메라 화합물인 iRNA 화합물을 포함한다.
본 발명의 맥락에서 "키메라" iRNA 화합물 또는 "키메라"는 각각 적어도 하나의 단량체 단위, 즉, dsRNA 화합물의 경우 뉴클레오티드로 구성된 2개 이상의 화학적으로 구별되는 영역을 함유하는 iRNA 화합물, 바람직하게는 dsRNA 제제이다. 이들 iRNA는 전형적으로 뉴클레아제 분해에 대한 증가된 내성, 증가된 세포 흡수, 또는 표적 핵산에 대한 증가된 결합 친화성을 iRNA에 부여하도록 RNA가 변형된 적어도 하나의 영역을 함유한다. iRNA의 추가 영역은 RNA:DNA 또는 RNA:RNA 하이브리드를 절단할 수 있는 효소에 대하 기질로서 역할을 할 수 있다. 예를 들자면, RNase H는 RNA:DNA 이중체의 RNA 가닥을 절단하는 세포 엔도뉴클레아제이다. 따라서 RNase H의 활성화는 RNA 표적의 절단을 야기하여 iRNA의 유전자 발현 억제 효율을 크게 향상시킨다. 결과적으로, 동일한 표적 영역에 혼성화하는 포스포로티오에이트 데옥시 dsRNA와 비교하여 키메라 dsRNA가 사용될 때 더 짧은 iRNA로 유사한 결과를 종종 수득할 수 있다. RNA 표적의 절단은 겔 전기영동 및, 필요한 경우, 당업계에 공지된 관련 핵산 혼성화 기술에 의해 일상적으로 검출될 수 있다.
특정 예에서, iRNA의 RNA는 비-리간드 그룹에 의해 변형될 수 있다. iRNA의 활성, 세포 분포 또는 세포 흡수를 향상시키기 위해 다수의 비-리간드 분자가 iRNA에 접합되었으며, 이러한 접합을 수행하는 절차는 과학 문헌에서 이용 가능하다. 이러한 비-리간드 모이어티는 지질 모이어티, 예를 들어 콜레스테롤(Kubo, T. et al., Biochem . Biophys . Res. Comm., 2007, 365(1):54-61; Letsinger et al., Proc. Natl . Acad . Sci . USA, 1989, 86:6553), 콜산(Manoharan et al., Bioorg . Med. Chem . Lett., 1994, 4:1053), 티오에테르, 예를 들어, 헥실-S-트리틸티올(Manoharan et al., Ann. N.Y . Acad . Sci., 1992, 660:306; Manoharan et al., Bioorg. Med . Chem . Let., 1993, 3:2765), 티오콜레스테롤(Oberhauser et al., Nucl. Acids Res., 1992, 20:533), 지방족 쇄, 예를 들어, 도데칸디올 또는 운데실 잔기(Saison-Behmoaras et al., EMBO J., 1991, 10:111; Kabanov et al., FEBS Lett., 1990, 259:327; Svinarchuk et al., Biochimie, 1993, 75:49), 인지질, 예를 들어, 디-헥사데실-rac-글리세롤 또는 트리에틸암모늄 1,2-디-O-헥사데실-rac-글리세로-3-H-포스포네이트(Manoharan et al., Tetrahedron Lett., 1995, 36:3651; Shea et al., Nucl . Acids Res., 1990, 18:3777), 폴리아민 또는 폴리에틸렌 글리콜 쇄(Manoharan et al., Nucleosides & Nucleotides, 1995, 14:969), 또는 아다만탄 아세트산(Manoharan et al., Tetrahedron Lett., 1995, 36:3651), 팔미틸 모이어티(Mishra et al., Biochim . Biophys . Acta, 1995, 1264:229), 또는 옥타데실아민 또는 헥실아미노-카보닐-옥시콜레스테롤 모이어티(Crooke et al., J. Pharmacol. Exp . Ther., 1996, 277:923)를 포함하였다. 이러한 RNA 접합체의 제조를 교시하는 대표적인 미국 특허가 위에 열거되어 있다. 전형적인 접합 프로토콜은 서열의 하나 이상의 위치에서 아미노링커를 보유하는 RNA의 합성을 포함한다. 그후 아미노 그룹을 적절한 커플링 또는 활성화 시약을 사용하여 접합되는 분자와 반응시킨다. 접합 반응은 RNA가 여전히 고체 지지체에 결합되어 있거나 RNA가 절단된 후 용액상에서 수행될 수 있다. HPLC에 의한 RNA 접합체의 정제는 전형적으로 순수 접합체를 제공한다.
V. 본 개시내용의 RNAi 제제의 전달
세포, 예를 들어, 인간 대상체(예를 들어, 이를 필요로 하는 대상체, 예를 들어 C9orf72-관련 장애, 예를 들어, C9orf72-관련 질환을 갖는 대상체)와 같은 대상체 내의 세포로의 본 개시내용의 RNAi 제제의 전달은 다양한 방식으로 달성될 수 있다. 예를 들면, 전달은 시험관내 또는 생체내에서 세포를 본 개시내용의 RNAi 제제와 접촉시킴으로써 수행될 수 있다. 생체내 전달은 또한 RNAi 제제, 예를 들어, dsRNA를 포함하는 조성물을 대상체에게 투여함으로써 직접 수행될 수 있다. 대안적으로, 생체내 전달은 RNAi 제제의 발현을 암호화하고 지시하는 하나 이상의 벡터를 투여함으로써 간접적으로 수행될 수 있다. 이러한 대안은 아래에서 추가로 논의된다.
일반적으로, 핵산 분자를 전달하는 임의의 방법(시험관내 또는 생체내)이 본 개시내용의 RNAi 제제와 함께 사용하기 위해 채택될 수 있다(예를 들어, 참조; Akhtar S. and Julian RL., (1992) Trends Cell. Biol . 2(5):139-144 및 제WO94/02595호, 이들은 전문이 본원에 참고로 포함됨). 생체내 전달의 경우, RNAi 제제를 전달하기 위해 고려해야 할 요소는, 예를 들면, 전달되는 제제의 생물학적 안정성, 비특이적 효과의 방지, 및 표적 조직에서 전달된 제제의 축적을 포함한다. RNAi 제제의 비특이적 효과는 국소 투여에 의해, 예를 들면, 조직에 직접 주사 또는 이식하거나 제제를 국부적으로 투여함으로써 최소화될 수 있다. 치료 부위에의 국소 투여는 제제의 국소 농도를 최대화하고, 달리 제제에 의해 손상될 수 있거나 제제를 분해할 수 있는 전신 조직에의 제제의 노출을 제한하고, 투여될 RNAi 제제의 총 용량을 더욱 낮춘다. 몇몇 연구에서는 RNAi 제제가 국소적으로 투여될 때 유전자 산물의 성공적인 녹다운이 나타났다. 예를 들면, 시노몰구스 원숭이에서 유리체내 주사(Tolentino, MJ. et al., (2004) Retina 24:132-138) 및 마우스에서 망막하 주사(Reich, SJ. et al. (2003) Mol . Vis . 9:210-216)에 의한 VEGF dsRNA의 안내 전달은 둘 다 연령-관련 황반 변성의 실험 모델에서 신생혈관 형성을 방지하는 것으로 나타났다. 또한, 마우스에서 dsRNA의 직접 종양내 주사는 종양 체적을 감소시키고(Pille, J. et al. (2005) Mol . Ther . 11:267-274) 종양-보유 마우스의 생존을 연장할 수 있다(Kim, WJ. et al., (2006) Mol . Ther . 14:343-350; Li, S. et al., (2007) Mol . Ther . 15:515-523). RNA 간섭은 직접 주사에 의한 CNS로의(Dorn, G. et al., (2004) Nucleic Acids 32:e49; Tan, PH. et al. (2005) Gene Ther. 12:59-66; Makimura, H. et a.l (2002) BMC Neurosci . 3:18; Shishkina, GT., et al. (2004) Neuroscience 129:521-528; Thakker, ER., et al. (2004) Proc. Natl . Acad . Sci . U.S.A . 101:17270-17275; Akaneya,Y., et al. (2005) J. Neurophysiol. 93:594-602) 및 비내 투여에 의한 폐로의(Howard, KA. et al., (2006) Mol . Ther . 14:476-484; Zhang, X. et al., (2004) J. Biol . Chem . 279:10677-10684; Bitko, V. et al., (2005) Nat. Med . 11:50-55) 국소 전달에서도 성공을 나타내었다. 질환의 치료를 위해 RNAi 제제를 전신적으로 투여하기 위해, RNA는 변형되거나 대안적으로 약물 전달 시스템을 사용하여 전달될 수 있고; 두 방법 모두 생체내에서 엔도- 및 엑소-뉴클레아제에 의한 dsRNA의 급속한 분해를 방지하는 작용을 한다. RNA 또는 약제학적 담체의 변형은 또한 표적 조직에 대한 RNAi 제제의 표적화를 허용하고 바람직하지 않은 오프-타겟 효과를 피할 수 있다(예를 들어, 이론에 결부시키고자 함이 없이, 본원에 기술된 바와 같은 GNA의 사용은 dsRNA의 시드 영역을 불안정화하는 것으로 확인되었으며, 그 결과 이러한 시드 영역 불안정화에 의해 오프-타겟 효과가 상당히 약해지기 때문에 오프-타겟 효과에 비해 온-타겟 효과에 대한 이러한 dsRNA의 선호도를 향상시켰다). RNAi 제제는 콜레스테롤과 같은 친유성 그룹에 대한 화학적 접합에 의해 변형되어 세포 흡수를 향상시키고 분해를 방지할 수 있다. 예를 들면, 친유성 콜레스테롤 모이어티에 접합된 ApoB에 대해 지시된 RNAi 제제를 마우스에 전신 주사하여 간과 공장 모두에서 apoB mRNA의 녹다운을 초래하였다(Soutschek, J. et al., (2004) Nature 432:173-178). RNAi 제제를 앱타머에 접합하면 전립선암의 마우스 모델에서 종양 성장을 억제하고 종양 퇴행을 매개하는 것으로 나타났다(McNamara, JO. et al., (2006) Nat. Biotechnol . 24:1005-1015). 대안적인 양태에서, RNAi 제제는 나노입자, 덴드리머, 중합체, 리포솜 또는 양이온 전달 시스템과 같은 약물 전달 시스템을 사용하여 전달될 수 있다. 양으로 하전된 양이온 전달 시스템은 분자 RNAi 제제(음으로 하전됨)의 결합을 촉진하고 또한 음으로 하전된 세포막에서 상호작용을 향상시켜 세포에 의한 RNAi 제제의 효율적인 흡수를 가능하게 한다. 양이온성 지질, 덴드리머 또는 중합체는 RNAi 제제에 결합되거나 RNAi 제제를 둘러싸는 소포 또는 미셀을 형성하도록 유도될 수 있다(예를 들어, 참조; Kim SH. et al., (2008) Journal of Controlled Release 129(2):107-116). 소포 또는 미셀의 형성은 전신으로 투여될 때 RNAi 제제의 분해를 추가로 방지한다. 양이온성-RNAi 제제 복합체를 제조하고 투여하는 방법은 당업계의 숙련가의 능력 범위 내에 있다(예를 들어, 참조; Sorensen, DR., et al. (2003) J. Mol . Biol 327:761-766; Verma, UN. et al., (2003) Clin . Cancer Res. 9:1291-1300; Arnold, AS et al. (2007) J. Hypertens. 25:197-205, 이는 전문이 본원에 참고로 포함됨). RNAi 제제의 전신 전달에 유용한 약물 전달 시스템의 일부 비제한적 예는 DOTAP(Sorensen, DR., et al (2003), supra; Verma, UN. et al., (2003), supra), Oligofectamine, "solid nucleic acid lipid particles" (Zimmermann, TS. et al., (2006) Nature 441:111-114), 카르디오리핀(Chien, PY. et al., (2005) Cancer Gene Ther . 12:321-328; Pal, A. et al., (2005) Int J. Oncol . 26:1087-1091), 폴리에틸렌이민(Bonnet ME. et al., (2008) Pharm . Res. Aug 16 Epub ahead of print; Aigner, A. (2006) J. Biomed. Biotechnol . 71659), Arg-Gly-Asp (RGD) 펩티드(Liu, S. (2006) Mol . Pharm. 3:472-487), 및 폴리아미도아민(Tomalia, DA. et al., (2007) Biochem . Soc. Trans. 35:61-67; Yoo, H. et al., (1999) Pharm . Res. 16:1799-1804)을 포함한다. 일부 양태에서, RNAi 제제는 전신 투여를 위해 사이클로덱스트린과 복합체를 형성한다. RNAi 제제 및 사이클로덱스트린의 투여 방법 및 약제학적 조성물은 미국 특허 제7,427,605호에서 찾을 수 있으며, 이는 전문이 본원에 참고로 포함된다.
본 개시내용의 특정 측면은 세포를 본 개시내용의 이중 가닥 RNAi 제제와 접촉시킴을 포함하여, 세포에서 C9orf72 표적 유전자의 발현을 감소시키는 방법에 관한 것이다. 하나의 양태에서, 세포는 간외 세포, 임의로 CNS 세포이다.
본 개시내용의 또 다른 측면은 대상체에게 본 개시내용의 이중 가닥 RNAi 제제를 투여함을 포함하여, 대상체에서 C9orf72 표적 유전자의 발현을 감소시키는 방법에 관한 것이다.
본 개시내용의 또 다른 측면은 대상체에게 본 개시내용의 이중 가닥 C9orf72-표적화 RNAi 제제의 치료학적 유효량을 투여함으로써 대상체를 치료함을 포함하여, CNS 장애를 갖는 대상체를 치료하는 방법에 관한 것이다. 본 개시내용의 방법에 의해 치료될 수 있는 예시적인 CNS 장애는 C9orf72-관련 질환을 포함한다.
하나의 양태에서, 이중 가닥 RNAi 제제는 척수강내 투여된다. 이중 가닥 RNAi 제제의 척수강내 투여에 의해, 상기 방법은 뇌(예를 들어, 선조체) 또는 척추 조직, 예를 들어, 피질, 소뇌, 경추, 요추 및 흉추에서 C9orf72 표적 유전자의 발현을 감소시킬 수 있다.
설명을 쉽게 하기 위해, 이 섹션에서 제형, 조성물 및 방법은 변형된 siRNA 화합물과 관련하여 주로 논의된다. 그러나, 이러한 제형, 조성물 및 방법은 다른 siRNA 화합물, 예를 들어, 변형되지 않은 siRNA 화합물로 실시될 수 있고, 이러한 실시는 본 개시내용 내에 있는 것으로 이해될 수 있다. RNAi 제제를 포함하는 조성물은 다양한 경로에 의해 대상체에게 전달될 수 있다. 예시적인 경로는 척수강내, 정맥내, 국소, 직장, 항문, 질, 비강, 폐 및 안구를 포함한다.
본 개시내용의 RNAi 제제는 투여에 적합한 약제학적 조성물에 혼입될 수 있다. 이러한 조성물은 전형적으로 하나 이상의 종의 RNAi 제제 및 약제학적으로 허용되는 담체를 포함한다. 본원에 사용된 "약제학적으로 허용되는 담체"라는 용어는 약제학적 투여와 상용성인 임의의 및 모든 용매, 분산 매질, 코팅제, 항균제 및 항진균제, 등장제 및 흡수 지연제 등을 포함하도록 의도된다. 약제학적 활성 물질에 대한 이러한 매질 및 제제의 사용은 당업계에 잘 알려져 있다. 임의의 통상적인 매질 또는 제제가 활성 화합물과 비상용성인 한을 제외하고, 조성물에서 이의 사용이 고려된다. 보충 활성 화합물이 또한 조성물에 포함될 수 있다.
본 개시내용의 약제학적 조성물은 국소 또는 전신 치료가 바람직한지 여부 및 치료될 부위에 따라 다양한 방식으로 투여될 수 있다. 투여는 국소(안과, 질, 직장, 비내, 경피 포함), 경구 또는 비경구일 수 있다. 비경구 투여는 정맥내 점적, 피하, 복강내 또는 근육내 주사, 또는 척수강내 또는 뇌실내 투여를 포함한다.
투여 경로 및 부위는 표적화를 향상시키기 위해 선택될 수 있다. 예를 들면, 근육 세포를 표적화하기 위해서는, 관심 근육에 근육내 주사하는 것이 논리적인 선택이 될 것이다. 폐 세포는 에어로졸 형태의 RNAi 제제를 투여함으로써 표적화될 수 있다. 혈관 내피 세포는 풍선 카테터를 RNAi 제제로 코팅하고 RNA를 기계적으로 도입함으로써 표적화될 수 있다.
국소 투여를 위한 제형은 경피 패치, 연고, 로션, 크림, 젤, 점적제, 좌약, 스프레이, 액체 및 분말을 포함할 수 있다. 통상적인 약제학적 담체, 수성, 분말 또는 유성 기재, 증점제 등이 필요하거나 바람직할 수 있다. 코팅된 콘돔, 장갑 등도 유용할 수 있다.
경구 투여용 조성물은 분말 또는 과립, 물 중의 현탁액 또는 용액, 시럽, 엘릭시르 또는 비수성 매질, 정제, 캡슐, 로젠지 또는 트로키를 포함한다. 정제의 경우, 사용될 수 있는 담체는 락토스, 나트륨 시트레이트 및 인산의 염을 포함한다. 전분과 같은 다양한 붕해제 및 마그네슘 스테아레이트, 나트륨 라우릴 설페이트 및 활석과 같은 윤활제가 정제에서 통상적으로 사용된다. 캡슐 형태의 경구 투여를 위해, 유용한 희석제는 락토스 및 고분자량 폴리에틸렌 글리콜이다. 경구 사용을 위해 수성 현탁액이 필요한 경우, 핵산 조성물은 유화제 및 현탁제와 조합될 수 있다. 원하는 경우, 특정 감미제 또는 방향제가 첨가될 수 있다.
척수강내 또는 뇌실내 투여용 조성물은 완충제, 희석제 및 기타 적합한 첨가제를 또한 함유할 수 있는 멸균 수용액을 포함할 수 있다.
비경구 투여용 제형은 완충제, 희석제 및 기타 적절한 첨가제를 또한 함유할 수 있는 멸균 수용액을 포함할 수 있다. 심실내 주사는, 예를 들면, 저장소에 부착된 심실내 카테터에 의해 촉진될 수 있다. 정맥내 사용의 경우, 용질의 총 농도를 조절하여 제제를 등장성으로 만들 수 있다.
하나의 양태에서, siRNA 화합물, 예를 들어 이중 가닥 siRNA 화합물, 또는 ssiRNA 화합물, 조성물의 투여는 비경구, 예를 들어, 정맥내(예를 들어, 볼루스 또는 확산성 주입으로서), 피내, 복강내, 근육내, 척수강내, 뇌실내, 두개내, 피하, 경점막, 협측, 설하, 내시경, 직장, 경구, 질, 국소, 폐, 비내, 요도 또는 안구이다. 투여는 대상체에 의해 또는 다른 사람, 예를 들어 의료 제공자에 의해 제공될 수 있다. 약물은 측정된 용량으로 또는 계량된 용량을 전달하는 디스펜서로 제공될 수 있다. 선택된 전달 방식은 아래에서 보다 상세히 논의된다.
A. 척수강내 투여.
하나의 양태에서, 이중 가닥 RNAi 제제는 척수강내 주사(즉, 뇌 및 척수 조직을 감싸는 척수액으로의 주사)에 의해 전달된다. 척수액에의 RNAi 제제의 척수강내 주사는 볼루스 주사로서 또는 피부 아래에 이식될 수 있는 미니펌프를 통해 수행되어, 척수액으로의 siRNA의 규칙적이고 일정한 전달을 제공할 수 있다. 척수액이 생성되는 맥락막 신경총으로부터의 척수액의 순환은 척수 및 후근 신경절 주위를 따라 아래로, 이어서 소뇌를 지나 피질을 거쳐 지주막 과립으로 흐르며, 여기서 유체가 CNS를 빠져나갈 수 있고, 주입된 화합물의 크기, 안정성 및 용해도에 따라, 척수강내로 전달된 분자는 CNS 전체에 걸쳐 표적과 부딪힐 수 있다.
일부 양태에서, 척수강내 투여는 펌프를 통해 이루어진다. 펌프는 외과적으로 이식된 삼투 펌프일 수 있다. 하나의 양태에서, 삼투 펌프는 척추관의 지주막하 공간에 이식되어 척수강내 투여를 용이하게 한다.
일부 양태에서, 척수강내 투여는 일정 용적의 약제를 함유하는 저장소, 및 저장소에 함유된 약제의 일부를 전달하도록 구성된 펌프를 포함하는 약제용 척수강내 전달 시스템을 통해 이루어진다. 이러한 척수강내 전달 시스템에 대한 보다 상세한 사항은 제WO 2015/116658호에서 찾을 수 있으며, 이는 전문이 참고로 포함된다.
척수강내 주사된 RNAi 제제의 양은 표적 유전자마다 다를 수 있으며 적용되어야 하는 적절한 양은 각 표적 유전자에 대해 개별적으로 결정되어야 할 수 있다. 전형적으로, 이 양은 10㎍ 내지 2mg, 바람직하게는 50㎍ 내지 1500㎍, 보다 바람직하게는 100㎍ 내지 1000㎍ 범위이다.
B. 본 개시내용의 벡터 암호화된 RNAi 제제
C9orf72 유전자를 표적으로 하는 RNAi 제제는 DNA 또는 RNA 벡터에 삽입된 전사 단위로부터 발현될 수 있다(예를 들어, 참조; Couture, A, et al., TIG . (1996), 12:5-10; 제WO 00/22113호, 제WO 00/22114호, 및 제US 6,054,299호). 발현은 사용된 특정 작제물 및 표적 조직 또는 세포 유형에 따라 바람직하게는 (수개월 이상) 지속된다. 이러한 이식유전자는 통합 또는 비통합 벡터일 수 있는 선형 작제물, 원형 플라스미드 또는 바이러스 벡터로서 도입될 수 있다. 이식유전자는 또한 염색체외 플라스미드로 유전될 수 있도록 작제될 수 있다(Gassmann, et al., (1995) Proc. Natl. Acad. Sci. USA 92:1292).
RNAi 제제의 개별 가닥 또는 가닥들은 발현 벡터 상의 프로모터로부터 전사될 수 있다. 예를 들면, dsRNA를 생성하기 위해 2개의 별도의 가닥이 발현되어야 하는 경우, 2개의 별도의 발현 벡터가 표적 세포 내로 (예를 들어, 형질감염 또는 감염에 의해) 공동-도입될 수 있다. 대안적으로, dsRNA의 각 개별 가닥은 동일한 발현 플라스미드에 위치한 프로모터에 의해 전사될 수 있다. 하나의 양태에서, dsRNA는 dsRNA가 스템 및 루프 구조를 갖도록 링커 폴리뉴클레오티드 서열에 의해 연결된 역반복 폴리뉴클레오티드로서 발현된다.
RNAi 제제 발현 벡터는 일반적으로 DNA 플라스미드 또는 바이러스 벡터이다. 진핵 세포와 상용성인 발현 벡터, 바람직하게는 척추동물 세포와 상용성인 것들이 본원에 기술된 바와 같은 RNAi 제제의 발현을 위한 재조합 작제물을 생성하는데 사용될 수 있다. RNAi 제제 발현 벡터의 전달은 예를 들어 정맥내 또는 근육내 투여에 의해, 환자로부터 외식된 표적 세포에 투여한 다음 환자에게 재도입함으로써, 또는 원하는 표적 세포로의 도입을 허용하는 임의의 다른 수단에 의해 전신적일 수 있다.
본원에 기술된 방법 및 조성물에 사용될 수 있는 바이러스 벡터 시스템은 다음을 포함하지만, 이에 제한되지 않는다: (a) 아데노바이러스 벡터; (b) 렌티바이러스 벡터, 몰로니 뮤린 백혈병 바이러스 등을 포함하지만 이에 제한되지 않는 레트로바이러스 벡터; (c) 아데노-관련 바이러스 벡터; (d) 단순 포진 바이러스 벡터; (e) SV 40 벡터; (f) 폴리오마 바이러스 벡터; (g) 유두종 바이러스 벡터; (h) 피코르나바이러스 벡터; (i) 수두 바이러스 벡터, 예를 들어 오르소폭스(orthopox), 예를 들어, 백시니아 바이러스 벡터 또는 아비폭스(avipox), 예를 들어, 카나리아 수두 또는 가금류 수두; 및 (j) 헬퍼-의존성 또는 소화관이 없는 아데노바이러스. 복제-결함 바이러스도 유리할 수 있다. 상이한 벡터가 세포의 게놈에 혼입되거나 혼입되지 않을 것이다. 작제물은 원하는 경우 형질감염을 위한 바이러스 서열을 포함할 수 있다. 대안적으로, 작제물은 에피솜 복제가 가능한 벡터, 예를 들어 EPV 및 EBV 벡터에 혼입될 수 있다. RNAi 제제의 재조합 발현을 위한 작제물은 일반적으로 표적 세포에서 RNAi 제제의 발현을 보장하기 위해 조절 요소, 예를 들어, 프로모터, 인핸서 등을 필요로 할 것이다. 벡터 및 작제물에 대해 고려해야 할 또 다른 측면은 당업계에 공지되어 있다.
VI. 본 발명의 약제학적 조성물
본 개시내용은 또한 본 개시내용의 RNAi 제제를 포함하는 약제학적 조성물 및 제형을 포함한다. 하나의 양태에서, 본원에 기술된 바와 같은 RNAi 제제 및 약제학적으로 허용되는 담체를 함유하는 약제학적 조성물이 본원에 제공된다. RNAi 제제를 함유하는 약제학적 조성물은 C9orf72의 발현 또는 활성과 관련된 질환 또는 장애, 예를 들어, C9orf72-관련 질환을 치료하는데 유용하다.
일부 양태에서, 본 발명의 약제학적 조성물은 무균이다. 또 다른 양태에서, 본 발명의 약제학적 조성물은 피로겐이 없다.
이러한 약제학적 조성물은 전달 방식에 기반하여 제형화된다. 한 가지 예는, 예를 들어, 정맥내(IV), 근육내(IM) 또는 피하(subQ) 전달에 의한 비경구 전달을 통한 전신 투여용으로 제형화된 조성물이다. 또 다른 예는 척수강내 또는 유리체내 주사 경로에 의해, 임의로 뇌(예를 들어, 선조체)로의 주입에 의해, 예를 들어 연속 펌프 주입에 의해 CNS로의 직접 전달용으로 제형화된 조성물이다.
본 개시내용의 약제학적 조성물은 C9orf72 유전자의 발현을 억제하기에 충분한 투여량으로 투여될 수 있다. 일반적으로, 본 개시내용의 RNAi 제제의 적합한 용량은 1일당 수용자의 체중 킬로그램당 약 0.001 내지 약 200.0 밀리그램의 범위, 일반적으로 1일당 체중 킬로그램당 약 1 내지 50mg의 범위일 것이다.
반복-투여 섭생은 RNAi 제제의 치료량을 정기적으로, 예를 들어 매월 내지 6개월마다 1회 투여하는 것을 포함할 수 있다. 특정 양태에서, RNAi 제제는 약 분기당 1회(즉, 약 3개월마다 1회) 내지 약 1년에 2회 투여된다.
초기 치료 섭생(예를 들어, 부하 용량) 후에, 치료는 덜 빈번하게 투여될 수 있다.
다른 양태에서, 약제학적 조성물의 단일 용량은 후속 용량이 1, 2, 3 또는 4개월 이상보다 많지 않은 간격으로 투여될 수 있도록 장기간 지속될 수 있다. 본 개시내용의 일부 양태에서, 본 개시내용의 약제학적 조성물의 단일 용량은 1개월에 1회 투여된다. 본 개시내용의 다른 양태에서, 본 개시내용의 약제학적 조성물의 단일 용량은 분기당 1회 내지 1년에 2회 투여된다.
숙련된 기술자는 질환 또는 장애의 중증도, 이전 치료, 대상체의 일반적인 건강 또는 연령, 및 존재하는 기타 질환을 포함하지만 이에 제한되지 않는 특정 요인이 대상체를 효과적으로 치료하는데 필요한 투여량 및 시기에 영향을 미칠 수 있음을 이해할 것이다. 더욱이, 치료학적 유효량의 조성물로 대상체를 치료하는 것은 단일 치료 또는 일련의 치료를 포함할 수 있다.
마우스 유전학의 발전으로 C9orf72의 발현 감소로 이익을 얻을 수 있는 ALS 및 FTD와 같은 다양한 인간 질환 연구를 위한 많은 마우스 모델이 생성되었다. 이러한 모델은 RNAi 제제의 생체내 시험 뿐만 아니라 치료학적 유효 용량을 결정하는데에도 사용될 수 있다. 적합한 설치류 모델이 당업계에 공지되어 있으며, 예를 들면, 문헌[Cepeda, et al. (ASN Neuro (2010) 2(2):e00033) and Pouladi, et al. (Nat Reviews (2013) 14:708)]에 기술된 것들을 포함한다.
본 개시내용의 약제학적 조성물은 국소 또는 전신 치료가 바람직한지 여부 및 치료될 부위에 따라 다양한 방식으로 투여될 수 있다. 투여는 국소(예를 들어, 경피 패치에 의해), 폐, 예를 들어, 네뷸라이저를 포함하여 분말 또는 에어로졸의 흡입 또는 통기에 의해; 기관내, 비내, 표피 및 경피, 경구 또는 비경구일 수 있다. 비경구 투여는 정맥내, 동맥내, 피하, 복강내 또는 근육내 주사 또는 주입; 예를 들어, 이식된 장치를 통해 피하; 또는 예를 들어 실질내, 척수강내 또는 뇌실내 투여에 의해 두개내를 포함한다.
RNAi 제제는 CNS(예를 들어, 뇌의 뉴런, 신경교 또는 혈관 조직)와 같은 특정 조직을 표적화하는 방식으로 전달될 수 있다.
국소 투여를 위한 약제학적 조성물 및 제형은 경피 패치, 연고, 로션, 크림, 겔, 점적제, 좌약, 스프레이, 액체 및 분말을 포함할 수 있다. 통상적인 약제학적 담체, 수성, 분말 또는 유성 기재, 증점제 등이 필요하거나 바람직할 수 있다. 코팅된 콘돔, 장갑 등도 유용할 수 있다. 적합한 국소 제형은 본 개시내용에서 특성화된 RNAi 제제가 지질, 리포솜, 지방산, 지방산 에스테르, 스테로이드, 킬레이트화제 및 계면활성제와 같은 국소 전달제와 혼합되어 있는 것을 포함한다. 적합한 지질 및 리포솜은 중성(예를 들어, 디올레오일포스파티딜 DOPE 에탄올아민, 디미리스토일포스파티딜 콜린 DMPC, 디스테아로일포스파티딜 콜린) 음성(예를 들면, 디미리스토일포스파티딜 글리세롤 DMPG) 및 양이온성(예를 들어, 디올레오일테트라메틸아미노 프로필 DOTAP 및 디올레오일포스파티딜 에탄올아민 DOTMA)을 포함한다. 본 개시내용에서 특성화된 RNAi 제제는 리포솜 내에 캡슐화될 수 있거나 이에 대한 복합체, 특히 양이온성 리포솜에 대한 복합체를 형성할 수 있다. 대안적으로, RNAi 제제는 지질, 특히 양이온성 지질과 복합체화될 수 있다. 적합한 지방산 및 에스테르는 아라키돈산, 올레산, 에이코산산, 라우르산, 카프릴산, 카프르산, 미리스트산, 팔미트산, 스테아르산, 리놀레산, 리놀렌산, 디카프레이트, 트리카프레이트, 모노올레인, 디라우린, 글리세릴 1-모노카프레이트, 1-도데실아자사이클로헵탄-2-온, 아실카르니틴, 아실콜린 또는 C1-20 알킬 에스테르(예를 들어, 이소프로필미리스테이트 IPM), 모노글리세리드, 디글리세리드 또는 이들의 약제학적으로 허용되는 염을 포함하지만, 이에 제한되지 않는다. 국소 제형은 본원에 참고로 포함되는 미국 제6,747,014호에 상세히 기술되어 있다.
A. 막 분자 어셈블리를 포함하는 RNAi 제제 제형
본 개시내용의 조성물 및 방법에 사용하기 위한 RNAi 제제는 막 분자 어셈블리, 예를 들어, 리포솜 또는 미셀에서의 전달을 위해 제형화될 수 있다. 본원에 사용된 용어 "리포솜"은 적어도 하나의 이중층, 예를 들어, 하나의 이중층 또는 복수의 이중층으로 배열된 양친매성 지질로 구성된 소포를 지칭한다. 리포솜은 친유성 물질로부터 형성된 막 및 수성 내부를 갖는 단층 및 다중층 소포를 포함한다. 수성 부분은 RNAi 제제 조성물을 함유한다. 친유성 물질은 수성 내부를 수성 외부로부터 분리하며, 이것은 전형적으로 RNAi 제제 조성물을 포함하지 않지만 일부 예에서는 포함할 수 있다. 리포솜은 활성 성분을 작용 부위로 이송 및 전달하는데 유용하다. 리포솜 막은 구조적으로 생물학적 막과 유사하기 때문에, 리포솜을 조직에 적용하는 경우, 리포솜 이중층이 세포막의 이중층과 융합된다. 리포솜과 세포의 병합이 진행됨에 따라, RNAi 제제를 포함하는 내부 수성 내용물은 세포로 전달되고, 여기서 RNAi 제제는 표적 RNA에 특이적으로 결합할 수 있고 RNAi를 매개할 수 있다. 일부 경우에 리포솜은 또한, 예를 들어, 특정 세포 유형으로 RNAi 제제를 지시하기 위해 특이적으로 표적화된다.
RNAi 제제를 함유하는 리포솜은 다양한 방법으로 제조될 수 있다. 하나의 예에서, 리포솜의 지질 성분은 미셀이 지질 성분으로 형성되도록 세제에 용해된다. 예를 들면, 지질 성분은 양친매성 양이온성 지질 또는 지질 접합체일 수 있다. 세제는 높은 임계 미셀 농도를 가질 수 있으며 비이온성일 수 있다. 예시적인 세제는 콜레이트, CHAPS, 옥틸글루코시드, 데옥시콜레이트 및 라우로일 사르코신을 포함한다. 그후 RNAi 제제 조제물을 지질 성분을 포함하는 미셀에 첨가한다. 지질 상의 양이온성 그룹은 RNAi 제제와 상호작용하고 RNAi 제제 주위에 축합되어 리포솜을 형성한다. 축합 후, 세제를, 예를 들어, 투석에 의해 제거하여 RNAi 제제의 리포솜 제제를 생성한다.
필요한 경우 축합을 돕는 담체 화합물을 축합 반응 동안, 예를 들어, 제어 첨가에 의해 첨가할 수 있다. 예를 들면, 담체 화합물은 핵산 이외의 중합체(예를 들어, 스페르민 또는 스페르미딘)일 수 있다. pH를 또한 축합에 유리하도록 조정할 수 있다.
폴리뉴클레오티드/양이온성 지질 복합체를 전달 비히클의 구조적 성분으로 포함하는 안정한 폴리뉴클레오티드 전달 비히클을 제조하는 방법은, 예를 들어, 제WO 96/37194호에 추가로 기술되어 있으며, 이의 전체 내용은 본원에 참고로 포함된다. 리포솜 형성은 또한 문헌[Felgner, P. L. et al., (1987) Proc . Natl . Acad . Sci. USA 8:7413-7417; United States Patent No. 4,897,355; United States Patent No. 5,171,678; Bangham et al., (1965) M. Mol . Biol . 23:238; Olson et al., (1979) Biochim . Biophys . Acta 557:9; Szoka et al., (1978) Proc . Natl . Acad. Sci . 75: 4194; Mayhew et al., (1984) Biochim . Biophys . Acta 775:169; Kim et al., (1983) Biochim . Biophys . Acta 728:339; and Fukunaga et al., (1984) Endocrinol . 115:757]에 기술된 예시적인 방법의 하나 이상의 측면을 포함할 수 있다. 전달 비히클로서 사용하기에 적절한 크기의 지질 응집체를 제조하기 위해 일반적으로 사용되는 기술은 초음파 처리 및 동결-해동 및 압출을 포함한다(예를 들어, 참조; Mayer et al., (1986) Biochim . Biophys . Acta 858:161). 미세유동화는 일관되게 작고(50 내지 200nm) 비교적 균일한 응집체가 요구되는 경우 사용될 수 있다(Mayhew et al., (1984) Biochim . Biophys . Acta 775:169). 이러한 방법은 RNAi 제제 조제물을 리포솜으로 포장하는데 용이하게 채택된다.
리포솜은 크게 두 부류로 나뉜다. 양이온성 리포솜은 음으로 하전된 핵산 분자와 상호작용하여 안정한 복합체를 형성하는 양으로 하전된 리포솜이다. 양으로 하전된 핵산/리포솜 복합체는 음으로 하전된 세포 표면에 결합하여 엔도솜에 내재화된다. 엔도솜 내의 산성 pH로 인해, 리포솜이 파열되어 이들의 내용물이 세포질로 방출된다(Wang et al. (1987) Biochem . Biophys . Res. Commun ., 147:980-985).
pH에 민감하거나 음으로 하전된 리포솜은 핵산과 복합체화되기 보다는 핵산을 포획한다. 핵산과 지질 모두 유사하게 하전되기 때문에, 복합체 형성보다는 반발이 일어난다. 그럼에도 불구하고, 일부 핵산은 이러한 리포솜의 수성 내부 내에 포획된다. pH 민감성 리포솜은 티미딘 키나제 유전자를 암호화하는 핵산을 배양물의 세포 단층에 전달하는데 사용되었다. 외인성 유전자의 발현이 표적 세포에서 검출되었다(Zhou et al. (1992) Journal of Controlled Release, 19:269-274).
리포솜 조성물의 한 가지 주요 유형은 천연 유래 포스파티딜콜린 이외의 인지질을 포함한다. 예를 들면, 중성 리포솜 조성물은 디미리스토일 포스파티딜콜린(DMPC) 또는 디팔미토일 포스파티딜콜린(DPPC)으로부터 형성될 수 있다. 음이온성 리포솜 조성물은 일반적으로 디미리스토일 포스파티딜글리세롤로부터 형성되는 반면, 음이온성 융해성 리포솜은 주로 디올레오일 포스파티딜에탄올아민(DOPE)으로부터 형성된다. 또 다른 유형의 리포솜 조성물은, 예를 들면, 대두 PC 및 계란 PC와 같은 포스파티딜콜린(PC)으로부터 형성된다. 또 다른 유형은 인지질 또는 포스파티딜콜린 또는 콜레스테롤의 혼합물로부터 형성된다.
시험관내 및 생체내에서 세포 내로 리포솜을 도입하는 또 다른 방법의 예는 미국 특허 제5,283,185호; 미국 특허 제5,171,678호; 제WO 94/00569호; 제WO 93/24640호; 제WO 91/16024호; 문헌[Felgner, (1994) J. Biol. Chem. 269:2550; Nabel, (1993) Proc. Natl. Acad. Sci. 90:11307; Nabel, (1992) Human Gene Ther. 3:649; Gershon, (1993) Biochem. 32:7143; and Strauss, (1992) EMBO J. 11:417]을 포함한다.
비이온성 리포솜 시스템, 특히 비이온성 계면활성제 및 콜레스테롤을 포함하는 시스템을 또한 피부로의 약물의 전달에서의 유용성을 결정하기 위해 조사하였다. NovasomeTM I(글리세릴 디라우레이트/콜레스테롤/폴리옥시에틸렌-10-스테아릴 에테르) 및 NovasomeTM II(글리세릴 디스테아레이트/콜레스테롤/폴리옥시에틸렌-10-스테아릴 에테르)를 포함하는 비이온성 리포솜 제형이 마우스 피부의 진피에 사이클로스포린-A를 전달하는데 사용되었다. 결과는 이러한 비이온성 리포솜 시스템이 피부의 다른 층으로 사이클로스포린 A의 침착을 촉진하는데 효과적임을 나타내었다(Hu et al., (1994) S.T.P.Pharma. Sci., 4(6):466).
리포솜은 또한 "입체적으로 안정화된" 리포솜을 포함하며, 이 용어는 본원에 사용되는 바와 같이, 리포솜에 혼입될 때, 이러한 특수화된 지질이 결여된 리포솜에 비해 향상된 순환 수명을 야기하는 하나 이상의 특수화된 지질을 포함하는 리포솜을 지칭한다. 입체적으로 안정화된 리포솜의 예는 리포솜의 소포-형성 지질 부분의 일부가 (A) 모노시알로강글리오시드 GM1과 같은 하나 이상의 당지질을 포함하거나 (B) 폴리에틸렌 글리콜(PEG) 모이어티와 같은 하나 이상의 친수성 중합체로 유도체화된 것이다. 임의의 특정 이론에 결부시키고자 하는 것은 아니지만, 당업계에서는 적어도 강글리오시드, 스핑고미엘린 또는 PEG-유도체화된 지질을 함유하는 입체적으로 안정화된 리포솜의 경우, 이러한 입체적으로 안정화된 리포솜의 향상된 순환 반감기는 세망내피계(RES)의 세포로의 감소된 흡수로부터 유래하는 것으로 생각된다(Allen et al., (1987) FEBS Letters, 223:42; Wu et al., (1993) Cancer Research, 53:3765).
하나 이상의 당지질을 포함하는 다양한 리포솜이 당업계에 공지되어 있다. Papahadjopoulos 등(Ann. N.Y. Acad. Sci., (1987), 507:64)은 모노시알로강글리오시드 GM1, 갈락토세레브로시드 설페이트 및 포스파티딜이노시톨이 리포솜의 혈액 반감기를 개선하는 능력을 보고하였다. 이러한 발견은 Gabizon 등(Proc . Natl . Acad. Sci . U.S.A ., (1988), 85,:6949)에 의해 설명되었다. Allen 등의 미국 특허 제4,837,028호와 제WO 88/04924호는 (1) 스핑고미엘린 및 (2) 강글리오시드 GM1 또는 갈락토세레브로시드 설페이트 에스테르를 포함하는 리포솜을 개시하고 있다. 미국 특허 제5,543,152호(Webb 등)는 스핑고미엘린을 포함하는 리포솜을 개시하고 있다. 1,2-sn-디미리스토일포스파티딜콜린을 포함하는 리포솜은 제WO 97/13499호(Lim 등)에 개시되어 있다.
하나의 양태에서, 양이온성 리포솜이 사용된다. 양이온성 리포솜은 세포막에 융합할 수 있다는 장점을 갖는다. 비-양이온성 리포솜은, 원형질막과 효율적으로 융합할 수는 없지만, 생체내에서 대식세포에 의해 흡수되고 RNAi 제제를 대식세포에 전달하는데 사용될 수 있다.
리포솜의 추가 장점은 다음을 포함한다: 천연 인지질로부터 수득된 리포솜은 생체적합성 및 생분해성이고; 리포솜은 광범위한 물 및 지질 가용성 약물을 통합할 수 있으며; 리포솜은 이들 내부 구획의 캡슐화된 RNAi 제제를 대사 및 분해로부터 보호할 수 있다(Rosoff, in "Pharmaceutical Dosage Forms," Lieberman, Rieger and Banker (Eds.), 1988, volume 1, p. 245). 리포솜 제형의 제조에서 중요한 고려 사항은 지질 표면 전하, 소포 크기 및 리포솜의 수성 용적이다.
양으로 하전된 합성 양이온성 지질, N-[1-(2,3-디올레일옥시)프로필]-N,N,N-트리메틸암모늄 클로라이드(DOTMA)는 핵산과 자발적으로 상호작용하여 조직 배양 세포의 세포막의 음으로 하전된 지질과 융합하여 RNAi 제제를 전달할 수 있는 지질-핵산 복합체를 형성하는 작은 리포솜을 형성하는데 사용될 수 있다(예를 들어, DOTMA 및 DNA에서의 이의 용도의 설명에 대해서는 문헌[Felgner, P. L. et al., (1987) Proc . Natl . Acad . Sci . USA 8:7413-7417], 및 미국 특허 제4,897,355호 참조).
DOTMA 유사체, 1,2-비스(올레오일옥시)-3-(트리메틸암모니아)프로판(DOTAP)은 인지질과 함께 사용되어 DNA-복합체 소포를 형성할 수 있다. Lipofectin™(Bethesda Research Laboratories, Gaithersburg, Md.)는 음으로 하전된 폴리뉴클레오티드와 자발적으로 상호작용하여 복합체를 형성하는 양으로 하전된 DOTMA 리포솜을 포함하는 살아있는 조직 배양 세포로의 고도의 음이온성 핵산의 전달을 위한 효과적인 제제이다. 충분히 양으로 하전된 리포솜이 사용되는 경우, 생성된 복합체의 순전하도 양전하이다. 이러한 방식으로 제조된 양으로 하전된 복합체는 음으로 하전된 세포 표면에 자발적으로 부착되고, 원형질막과 융합되어 기능적 핵산을, 예를 들어, 조직 배양 세포로 효율적으로 전달한다. 상업적으로 이용 가능한 또 다른 양이온성 지질, 1,2-비스(올레오일옥시)-3,3-(트리메틸암모니아)프로판("DOTAP")(Boehringer Mannheim, Indianapolis, Indiana)은 올레오일 모이어티가 에테르 결합보다는 에스테르에 의해 결합된다는 점에서 DOTMA와 다르다.
보고된 다른 양이온성 지질 화합물은, 예를 들면, 2가지 유형의 지질 중 하나에 접합된 카복시스페르민을 포함한 다양한 모이어티에 접합된 것들을 포함하고 5-카복시스페르밀글리신 디옥타올레오일아미드("DOGS")(Transfectam™, Promega, Madison, Wisconsin) 및 디팔미토일포스파티딜에탄올아민 5-카복시스페르밀-아미드("DPPES")를 포함한다(예를 들어, 미국 특허 제5,171,678호 참조).
또 다른 양이온성 지질 접합체는 DOPE와 조합하여 리포솜으로 제형화된 콜레스테롤을 사용한 지질의 유도체화("DC-Chol")를 포함한다(참조; Gao, X. and Huang, L., (1991) Biochim . Biophys . Res. Commun . 179:280). DOPE에 폴리리신을 접합시켜 만든 리포폴리리신은 혈청의 존재하에서 형질감염에 효과적인 것으로 보고되었다(Zhou, X. et al., (1991) Biochim . Biophys . Acta 1065:8). 특정 세포주의 경우, 접합된 양이온성 지질을 함유하는 이러한 리포솜은 DOTMA-함유 조성물보다 더 낮은 독성을 나타내고 더 효율적인 형질감염을 제공한다고 한다. 기타 상업적으로 이용 가능한 양이온성 지질 제품은 DMRIE 및 DMRIE-HP(Vical, La Jolla, California) 및 리포펙타민(DOSPA)(Life Technology, Inc., Gaithersburg, Maryland)을 포함한다. 올리고뉴클레오티드의 전달에 적합한 다른 양이온성 지질은 제WO 98/39359호 및 제WO 96/37194호에 기술되어 있다.
리포솜 제형은 특히 국소 투여에 적합하며, 리포솜은 다른 제형에 비해 몇 가지 장점을 나타낸다. 이러한 장점은 투여된 약물의 높은 전신 흡수, 원하는 표적에서 투여된 약물의 증가된 축적 및 RNAi 제제를 피부에 투여하는 능력과 관련된 부작용 감소를 포함한다. 일부 구현에서, 리포솜은 RNAi 제제를 표피 세포에 전달하고 또한 RNAi 제제의 진피 조직, 예를 들어 피부로의 침투를 향상시키기 위해 사용된다. 예를 들면, 리포솜은 국소적으로 적용될 수 있다. 리포솜으로 제형화된 약물의 피부에의 국소 전달이 기록되었다(예를 들어, 참조; Weiner et al., (1992) Journal of Drug Targeting, vol. 2,405-410 and du Plessis et al., (1992) Antiviral Research, 18:259-265; Mannino, R. J. and Fould-Fogerite, S., (1998) Biotechniques 6:682-690; Itani, T. et al., (1987) Gene 56:267-276; Nicolau, C. et al. (1987) Meth . Enzymol . 149:157-176; Straubinger, R. M. and Papahadjopoulos, D. (1983) Meth . Enzymol . 101:512-527; Wang, C. Y. and Huang, L., (1987) Proc. Natl. Acad. Sci. USA 84:7851-7855).
비이온성 리포솜 시스템, 특히 비이온성 계면활성제 및 콜레스테롤을 포함하는 시스템을 또한 피부로의 약물의 전달에 있어서의 이들의 유용성을 결정하기 위해 조사하였다. Novasome I(글리세릴 디라우레이트/콜레스테롤/폴리옥시에틸렌-10-스테아릴 에테르) 및 Novasome II(글리세릴 디스테아레이트/콜레스테롤/폴리옥시에틸렌-10-스테아릴 에테르)를 포함하는 비이온성 리포솜 제형을 사용하여 약물을 마우스 피부의 진피에 전달하였다. RNAi 제제를 갖는 이러한 제형이 피부 장애를 치료하는데 유용하다.
RNAi 제제를 포함하는 리포솜은 고도로 변형 가능하게 만들어질 있다. 이러한 변형성은 리포솜이 리포솜의 평균 반경보다 작은 기공을 통해 침투할 수 있게 한다. 예를 들면, 트랜스퍼솜(transfersome)이 변형 가능한 리포솜의 한 유형이다. 트랜스퍼솜은 표면 가장자리 활성화제, 통상적으로 계면활성제를 표준 리포솜 조성에 추가하여 만들 수 있다. RNAi 제제를 포함하는 트랜스퍼솜은, 예를 들면, 피부의 각질형성세포에 RNAi 제제를 전달하기 위해 감염에 의해 피하로 전달될 수 있다. 온전한 포유동물 피부를 통과하기 위해서는, 지질 소포가 적절한 경피 구배의 영향하에 각각 직경이 50nm 미만인 일련의 미세 기공을 통과해야 한다. 또한, 지질 특성으로 인해, 이러한 트랜스퍼솜은 자가-최적화(예를 들어, 피부에서 모공의 모양에 적응), 자가-복구할 수 있으며, 자주 단편화 및 종종 자가-부하 없이 이들의 표적에 도달할 수 있다.
본 개시내용에 적용할 수 있는 다른 제형은 2008년 1월 2일 출원된 미국 가출원 일련번호 제61/018,616호; 2008년 1월 2일 출원된 제61/018,611호; 2008년 3월 26일 출원된 제61/039,748호; 2008년 4월 22일 출원된 제61/047,087호 및 2008년 5월 8일 출원된 제61/051,528호에 기술되어 있다. 2007년 10월 3일 출원된 PCT 출원 제PCT/US2007/080331호 또한 본 개시내용에 적용할 수 있는 제형을 기술하고 있다.
리포솜의 또 다른 유형인 트랜스퍼솜은 약물 전달 비히클의 매력적인 후보인 고도로 변형 가능한 지질 응집체이다. 트랜스퍼솜은 매우 변형 가능하여 소적보다 작은 기공을 통해 쉽게 침투할 수 있는 지질 소적으로 설명할 수 있다. 트랜스퍼솜은 이들이 사용되는 환경에 적응할 수 있으며, 예를 들어, 이들은 자가-최적화(피부의 모공 모양에 적응), 자가-복구하며, 자주 단편화 및 종종 자가-부하 없이 이들의 표적에 도달한다. 트랜스퍼솜을 만들기 위해, 표면 가장자리 활성화제, 통상적으로 계면활성제를 표준 리포솜 조성에 추가할 수 있다. 트랜스퍼솜은 혈청 알부민을 피부에 전달하는데 사용되었다. 혈청 알부민의 트랜스퍼솜-매개된 전달은 혈청 알부민을 함유한 용액의 피하 주사만큼 효과적인 것으로 나타났다.
계면활성제는 본원에 기술된 것과 같은 제형, 특히 에멀젼(마이크로에멀젼 포함) 및 리포솜에 광범위하게 적용된다. 천연 및 합성 계면활성제의 여러 다양한 유형의 특성을 분류하고 순위를 매기는 가장 일반적인 방법은 친수성/친유성 평형(hydrophile/lipophile balance; HLB)을 사용하는 것이다. 친수성 그룹("헤드"라고도 함)의 특성은 제형에 사용되는 상이한 계면활성제를 분류하는데 가장 유용한 수단을 제공한다(Rieger, in Pharmaceutical Dosage Forms, Marcel Dekker, Inc., New York, N.Y., 1988, p. 285).
계면활성제 분자가 이온화되지 않으면, 비이온성 계면활성제로 분류된다. 비이온성 계면활성제는 제약 및 화장품에 광범위하게 적용되며 광범위한 pH 값에 걸쳐 사용 가능하다. 일반적으로, 이들의 HLB 값은 이들의 구조에 따라 2 내지 약 18 범위이다. 비이온성 계면활성제는 비이온성 에스테르, 예를 들어, 에틸렌 글리콜 에스테르, 프로필렌 글리콜 에스테르, 글리세릴 에스테르, 폴리글리세릴 에스테르, 소르비탄 에스테르, 수크로스 에스테르, 및 에톡실화 에스테르를 포함한다. 지방 알코올 에톡실레이트, 프로폭실화 알코올 및 에톡실화/프로폭실화 블록 중합체와 같은 비이온성 알칸올아미드 및 에테르도 이 부류에 포함된다. 폴리옥시에틸렌 계면활성제가 비이온성 계면활성제 부류 중 가장 인기있는 구성원이다.
계면활성제 분자가 물에 용해되거나 분산될 때 음전하를 띠면, 계면활성제는 음이온성으로 분류된다. 음이온성 계면활성제는 카복실레이트, 예를 들어 비누, 아실 락틸레이트, 아미노산의 아실 아미드, 황산의 에스테르, 예를 들어 알킬 설페이트 및 에톡실화 알킬 설페이트, 설포네이트, 예를 들어 벤젠 설포네이트, 아실 이세티오네이트, 아실 타우레이트 및 설포석시네이트, 및 포스페이트를 포함한다. 음이온성 계면활성제 부류의 가장 중요한 구성원은 알킬 설페이트 및 비누이다.
계면활성제 분자가 물에 용해되거나 분산될 때 양전하를 띠면, 계면활성제는 양이온성으로 분류된다. 양이온성 계면활성제는 4급 암모늄 염 및 에톡실화 아민을 포함한다. 4급 암모늄 염이 이 부류 중 가장 많이 사용되는 구성원이다.
계면활성제 분자가 양전하 또는 음전하를 띠는 능력을 갖는다면, 계면활성제는 양쪽성으로 분류된다. 양쪽성 계면활성제는 아크릴산 유도체, 치환된 알킬아미드, N-알킬베타인 및 포스파티드를 포함한다.
의약품, 제형 및 에멀젼에서의 계면활성제의 사용이 검토되었다(Rieger, in Pharmaceutical Dosage Forms, Marcel Dekker, Inc., New York, N.Y., 1988, p. 285).
본 개시내용의 방법에 사용하기 위한 RNAi 제제는 또한 미셀 제형으로서 제공될 수 있다. "미셀"은 본원에서, 분자의 모든 소수성 부분이 안쪽으로 향하여 친수성 부분이 주변 수성 상과 접촉하도록 양친매성 분자가 구형 구조로 배열되는 분자 조립체의 특정 유형으로서 정의된다. 환경이 소수성인 경우 반대 배열이 존재한다.
경피 막을 통한 전달에 적합한 혼합 미셀 제형은 siRNA 조성물의 수용액, 알칼리 금속 C8 내지 C22 알킬 설페이트, 및 미셀 형성 화합물을 혼합함으로써 제조될 수 있다. 예시적인 미셀 형성 화합물은 레시틴, 히알루론산, 히알루론산의 약제학적으로 허용되는 염, 글리콜산, 락트산, 카모마일 추출물, 오이 추출물, 올레산, 리놀레산, 리놀렌산, 모노올레인, 모노올레에이트, 모노라우레이트, 보리지 오일, 달맞이꽃 오일, 멘톨, 트리하이드록시 옥소 콜라닐 글리신 및 이의 약제학적으로 허용되는 염, 글리세린, 폴리글리세린, 리신, 폴리리신, 트리올레인, 폴리옥시에틸렌 에테르 및 이들의 유사체, 폴리도카놀 알킬 에테르 및 이들의 유사체, 케노데옥시콜레이트, 데옥시콜레이트, 및 이들의 혼합물을 포함한다. 미셀 형성 화합물은 알칼리 금속 알킬 설페이트의 첨가와 동시에 또는 후에 첨가될 수 있다. 혼합 미셀은 실질적으로 모든 종류의 성분 혼합으로 형성되지만 더 작은 크기의 미셀을 제공하기 위해서는 격렬한 혼합으로 형성될 것이다.
하나의 방법에서, siRNA 조성물 및 적어도 알칼리 금속 알킬 설페이트를 함유하는 제1 미셀 조성물을 제조한다. 그후 제1 미셀 조성물을 적어도 3개의 미셀 형성 화합물과 혼합하여 혼합 미셀 조성물을 형성한다. 또 다른 방법에서, 미셀 조성물은 siRNA 조성물, 알칼리 금속 알킬 설페이트, 및 미셀 형성 화합물 중 적어도 하나를 혼합한 다음 나머지 미셀 형성 화합물을 첨가하여 격렬하게 혼합함으로써 제조된다.
페놀 또는 m-크레졸을 혼합 미셀 조성물에 첨가하여 제형을 안정화하고 박테리아 성장을 방지할 수 있다. 대안적으로, 페놀 또는 m-크레졸이 미셀 형성 성분과 함께 첨가될 수 있다. 혼합 미셀 조성물의 형성 후에 글리세린과 같은 등장제를 첨가할 수도 있다.
미셀 제형을 스프레이로서 전달하기 위해, 제형을 에어로졸 디스펜서에 넣고 디스펜서에 추진제를 충전할 수 있다. 압력하에 있는 추진제는 디스펜서에서 액체 형태이다. 성분의 비율은 수성 상 및 추진제 상이 하나가 되도록. 즉, 하나의 상이 존재하도록 조정된다. 두 개의 상이 있다면, 예를 들어 계량 밸브를 통해, 내용물의 일부를 분배하기 전에 디스펜서를 진탕시킬 필요가 있다. 약제의 분배된 용량은 미세 스프레이에서 계량 밸브로부터 추진된다.
추진제는 수소-함유 클로로플루오로카본, 수소-함유 플루오로카본, 디메틸 에테르 및 디에틸 에테르를 포함할 수 있다. 특정 양태에서, HFA 134a(1,1,1,2 테트라플루오로에탄)이 사용될 수 있다.
필수 성분의 특정 농도는 비교적 간단한 실험으로 결정할 수 있다. 구강을 통한 흡수를 위해서는, 주사 또는 위장관을 통한 투여에 대한 투여량을, 예를 들어, 적어도 2배 또는 3배로 증가시키는 것이 종종 바람직하다.
B. 지질 입자
본 개시내용의 RNAi 제제, 예를 들어, dsRNA는 지질 제형, 예를 들어, LNP, 또는 다른 핵산-지질 입자에 완전히 캡슐화될 수 있다.
본원에 사용된 용어 "LNP"는 안정한 핵산-지질 입자를 지칭한다. LNP는 전형적으로 양이온성 지질, 비-양이온성 지질, 및 입자의 응집을 방지하는 지질(예를 들어, PEG-지질 접합체)을 함유한다. LNP는 정맥 주사(i.v.) 후 연장된 순환 수명을 나타내고 말단 부위(예를 들어, 투여 부위와 물리적으로 분리된 부위)에 축적되기 때문에 전신 적용에 매우 유용하다. LNP는 제WO 00/03683호에 제시된 바와 같이 캡슐화된 축합제-핵산 복합체를 포함하는 "pSPLP"를 포함한다. 본 개시내용의 입자는 전형적으로 약 50nm 내지 약 150nm, 보다 전형적으로 약 60nm 내지 약 130nm, 보다 전형적으로 약 70nm 내지 약 110nm, 가장 전형적으로 약 70nm 내지 약 90nm의 평균 직경을 가지며, 실질적으로 무독성이다. 또한, 본 개시내용의 핵산-지질 입자에 존재할 때 핵산은 뉴클레아제에 의한 분해에 대해 수용액에서 내성이 있다. 핵산-지질 입자 및 이의 제조 방법은, 예를 들어, 미국 특허 제5,976,567호; 제5,981,501호; 제6,534,484호; 제6,586,410호; 제6,815,432호; 미국 특허 공개 제2010/0324120호 및 제WO 96/40964호에 개시되어 있다.
하나의 양태에서, 지질 대 약물 비(질량/질량 비)(예를 들어, 지질 대 dsRNA 비)는 약 1:1 내지 약 50:1, 약 1:1 내지 약 25:1, 약 3:1 내지 약 15:1, 약 내지 약 10:1, 약 5:1 내지 약 9:1, 또는 약 6:1 내지 약 9:1 범위일 것이다. 상기 인용된 범위의 중간 범위도 본 개시내용의 일부로 고려된다.
RNAi 제제의 전달을 위한 특정 LNP 제형은, 예를 들어, 본원에 참고로 포함된 제WO 2008/042973호에 기술된 "LNP01" 제형을 포함하여 당업계에 기술되어 있다.
추가의 예시적인 지질-dsRNA 제형은 하기 표 2에서 확인된다.


DSPC: 디스테아로일포스파티딜콜린
DPPC: 디팔미토일포스파티딜콜린
PEG-DMG: PEG-디디미리스토일 글리세롤 (C14-PEG, 또는 PEG-C14) (2000의 평균 분자량을 갖는 PEG)
PEG-DSG: PEG-디스티릴 글리세롤(C18-PEG, 또는 PEG-C18) (2000의 평균 분자량을 갖는 PEG)
PEG-cDMA: PEG-카바모일-1,2-디미리스틸옥시프로필아민 (2000의 평균 분자량을 갖는 PEG)
SNALP(1,2-디리놀레닐옥시-N,N-디메틸아미노프로판(DLinDMA)) 포함 제형은 본원에 참고로 포함된 제WO 2009/127060호에 기술되어 있다.
XTC 포함 제형은 전체 내용이 본원에 참고로 포함된 제WO 2010/088537호에 기술되어 있다.
MC3 포함 제형은, 예를 들어, 전체 내용이 본원에 참고로 포함된 미국 특허 공개 제2010/0324120호에 기술되어 있다.
ALNY-100 포함 제형은 전체 내용이 본원에 참고로 포함된 제WO 2010/054406호에 기술되어 있다.
C12-200 포함 제형은 전체 내용이 본원에 참고로 포함된 제WO 2010/129709호에 기술되어 있다.
경구 투여를 위한 조성물 및 제형은 분말 또는 과립, 미세미립자, 나노미립자, 물 또는 비수성 매질 중 현탁액 또는 용액, 캡슐, 겔 캡슐, 샤쉐, 정제 또는 미니정제를 포함한다. 증점제, 방향제, 희석제, 유화제, 분산 보조제 또는 결합제가 바람직할 수 있다. 일부 양태에서, 경구 제형은 본 개시내용에 특성화된 dsRNA가 하나 이상의 침투 증강제 계면활성제 및 킬레이트제와 함께 투여되는 제형이다. 적합한 계면활성제는 지방산 또는 이의 에스테르 또는 염, 담즙산 또는 이의 염을 포함한다. 적합한 담즙산/염은 케노데옥시콜산(CDCA) 및 우르소데옥시케노데옥시콜산(UDCA), 콜산, 데하이드로콜산, 데옥시콜산, 글루콜산, 글리콜산, 글리코데옥시콜산, 타우로콜산, 타우로데옥시콜산, 나트륨 타우로-24,25-디하이드로-퓨시데이트 및 나트륨 글리코디하이드로퓨시데이트를 포함한다. 적합한 지방산은 아라키돈산, 운데칸산, 올레산, 라우르산, 카프릴산, 카프르산, 미리스트산, 팔미트산, 스테아르산, 리놀레산, 리놀렌산, 디카프레이트, 트리카프레이트, 모노올레인, 디라우린, 글리세릴 1-모노카프레이트, 1-도데실아자사이클로헵탄-2-온, 아실카르니틴, 아실콜린, 또는 모노글리세리드, 디글리세리드 또는 이의 약제학적으로 허용되는 염(예를 들어, 나트륨)을 포함한다. 일부 양태에서, 침투 증강제의 조합, 예를 들어 담즙산/염과 조합된 지방산/염이 사용된다. 한 가지 예시적인 조합은 라우르산, 카프르산 및 UDCA의 나트륨 염이다. 추가 침투 증강제는 폴리옥시에틸렌-9-라우릴 에테르, 폴리옥시에틸렌-20-세틸 에테르를 포함한다. 본 개시내용에서 특성화된 dsRNA는 분무 건조된 입자를 포함하는 과립 형태로 경구로 전달될 수 있거나, 마이크로 또는 나노입자를 형성하도록 복합체화될 수 있다. dsRNA 착화제는 폴리아미노산; 폴리이민; 폴리아크릴레이트; 폴리알킬아크릴레이트, 폴리옥세테인, 폴리알킬시아노아크릴레이트; 양이온화된 젤라틴, 알부민, 전분, 아크릴레이트, 폴리에틸렌글리콜(PEG) 및 전분; 폴리알킬시아노아크릴레이트; DEAE-유도체화된 폴리이민, 풀루란, 셀룰로스 및 전분을 포함한다. 적합한 착화제는 키토산, N-트리메틸키토산, 폴리-L-리신, 폴리히스티딘, 폴리오르니틴, 폴리스페르민, 프로타민, 폴리비닐피리딘, 폴리티오디에틸아미노메틸에틸렌 P(TDAE), 폴리아미노스티렌(예를 들어, p-아미노), 폴리(메틸시아노아크릴레이트), 폴리(에틸시아노아크릴레이트), 폴리(부틸시아노아크릴레이트), 폴리(이소부틸시아노아크릴레이트), 폴리(이소헥실시아노아크릴레이트), DEAE-메타크릴레이트, DEAE-헥실아크릴레이트, DEAE-아크릴아미드, DEAE-알부민 및 DEAE-덱스트란, 폴리메틸아크릴레이트, 폴리헥실아크릴레이트, 폴리(D,L-락트산), 폴리(DL-락트산-코-글리콜산(PLGA), 알기네이트, 및 폴리에틸렌글리콜(PEG)을 포함한다. dsRNA에 대한 경구 제형 및 이의 제조는 미국 특허 제6,887,906호, 미국 제2003/0027780호 및 미국 특허 제6,747,014호에 상세히 기술되어 있으며, 이들 각각은 본원에 참고로 포함된다.
비경구, 실질내(뇌로), 척수강내, 뇌실내 또는 간내 투여를 위한 조성물 및 제형은 완충제, 희석제, 및 침투 증강제, 담체 화합물 및 기타 약제학적으로 허용되는 담체 또는 부형제와 같은 기타 적합한 첨가제를 또한 함유할 수 있는 멸균 수용액을 포함할 수 있다.
본 개시내용의 약제학적 조성물은 용액, 에멀젼, 및 리포솜-함유 제형을 를 포함하지만, 이에 제한되지 않는다. 이러한 조성물은 미리 형성된 액체, 자가-유화 고체 및 자가-유화 반고체를 포함하지만 이에 제한되지 않는 다양한 성분으로부터 생성될 수 있다. APP-관련 질환 또는 장애를 치료할 때 뇌를 표적으로 하는 제형이 특히 바람직하다.
단위 투여 형태로 통상적으로 제시될 수 있는 본 개시내용의 약제학적 제형은 제약 산업에 널리 공지된 통상적인 기술에 따라 제조될 수 있다. 이러한 기술은 활성 성분을 약제학적 담체(들) 또는 부형제(들)와 회합시키는 단계를 포함한다. 일반적으로, 제형은 활성 성분을 액체 담체 또는 미분된 고체 담체 또는 둘 다와 균일하고 친밀하게 회합시킨 다음, 필요한 경우 생성물을 성형함으로써 제조된다.
본 개시내용의 조성물은 정제, 캡슐, 젤 캡슐, 액체 시럽, 연질 젤, 좌약 및 관장제와 같지만 이에 제한되지 않는 임의의 많은 가능한 투여 형태로 제형화될 수 있다. 본 개시내용의 조성물은 또한 수성, 비수성 또는 혼합 매질에서 현탁액으로서 제형화될 수 있다. 수성 현탁액은, 예를 들면, 나트륨 카복시메틸셀룰로스, 소르비톨 또는 덱스트란을 포함하는 현탁액의 점도를 증가시키는 물질을 추가로 함유할 수 있다. 현탁액은 안정제를 또한 함유할 수 있다.
C. 추가 제형
i. 에멀젼
본 개시내용의 조성물은 에멀젼으로서 제조 및 제형화될 수 있다. 에멀젼은 전형적으로 직경이 0.1mm를 초과하는 액적 형태로 한 액체가 다른 액체에 분산되어 있는 이종 시스템이다(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199; Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., Volume 1, p. 245; Block in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 2, p. 335; Higuchi et al., in Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa., 1985, p. 301). 에멀젼은 종종 서로 밀접하게 혼합되고 분산된 2개의 비혼화성 액체 상을 포함하는 2상 시스템이다. 일반적으로, 에멀젼은 유중수(w/o) 또는 수중유(o/w) 종류일 수 있다. 수성 상이 미세한 액적으로서 벌크 오일 상에 미분되고 분산되는 경우, 생성된 조성물을 유중수(w/o) 에멀젼이라고 한다. 대안적으로, 오일 상이 미세한 액적으로서 벌크 수성 상에 미분되고 분산되는 경우, 생성된 조성물을 수중유(o/w) 에멀젼이라고 한다. 에멀젼은 분산된 상 외에 추가 성분을 함유할 수 있으며, 수성 상, 오일 상 또는 별도의 상으로 자체적으로 용액으로서 존재할 수 있는 활성 약물을 함유할 수 있다. 유화제, 안정제, 염료 및 항산화제와 같은 약제학적 부형제가 또한 필요에 따라 에멀젼에 존재할 수 있다. 약제학적 에멀젼은 또한, 예를 들면, 유중수중유(o/w/o) 및 수중유중수(w/o/w) 에멀젼의 경우와 같이 2개 이상의 상으로 구성된 다중 에멀젼일 수 있다. 이러한 복합체 제형은 종종 단순 이원 에멀젼이 제공하지 않는 특정 이점을 제공한다. o/w 에멀젼의 개별 오일 소적이 작은 물방울을 둘러싸고 있는 다중 에멀젼이 w/o/w 에멀젼을 구성한다. 마찬가지로, 유성 연속 상으로 안정화된 물의 소구체에 둘러싸인 오일 소적의 시스템은 o/w/o 에멀젼을 제공한다.
에멀젼은 열역학적 안정성이 거의 또는 전혀 없는 것을 특징으로 한다. 종종, 에멀젼의 분산된 또는 불연속 상은 외부 또는 연속 상으로 잘 분산되고 유화제의 수단 또는 제형의 점도를 통해 이러한 형태로 유지된다. 에멀젼의 상 중 하나는 에멀젼-스타일 연고 기재 및 크림의 경우와 같이 반고체 또는 고체일 수 있다. 에멀젼을 안정화시키는 다른 수단은 에멀젼의 어느 상에 혼입될 수 있는 유화제의 사용을 수반한다. 유화제는 크게 4개 범주로 분류될 수 있다: 합성 계면활성제, 자연 발생 유화제, 흡수 기재, 미분된 고체(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199).
계면활성제로도 알려진 합성 계면활성제는 에멀젼의 제형화에 폭넓게 적용할 수 있으며 문헌에서 검토되었다(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY; Rieger, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 285; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), Marcel Dekker, Inc., New York, N.Y., 1988, volume 1, p. 199). 계면활성제는 전형적으로 양친매성이며 친수성 부분과 소수성 부분을 포함한다. 계면활성제의 친수성 대 소수성 특성의 비율을 친수성/친유성 평형(HLB)이라고 하며 이것은 제형의 제조에서 계면활성제를 분류하고 선택하는데 유용한 도구이다. 계면활성제는 친수성 그룹의 성질에 기반하여 상이한 부류로 분류될 수 있다: 비이온성, 음이온성, 양이온성 및 양쪽성(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY Rieger, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 285).
에멀젼 제형에 사용되는 자연 발생 유화제는 라놀린, 밀랍, 포스파티드, 레시틴 및 아카시아를 포함한다. 무수 라놀린 및 친수성 바셀린과 같은 흡수 기재는 물을 빨아들여 w/o 에멀젼을 형성하면서도 이들의 반고체 조도를 유지할 수 있도록 친수성을 가지고 있다. 미분된 고체는 또한 특히 계면활성제 및 점성 제제와 조합하여 우수한 유화제로서 사용되었다. 이들은 중금속 수산화물과 같은 극성 무기 고체, 벤토나이트, 아타풀자이트, 헥토라이트, 카올린, 몬모릴로나이트, 콜로이드성 알루미늄 실리케이트 및 콜로이드성 마그네슘 알루미늄 실리케이트와 같은 비팽윤성 점토, 안료 및 탄소 또는 글리세릴 트리스테아레이트와 같은 비극성 고체를 포함한다.
매우 다양한 비유화 물질이 또한 에멀젼 제형에 포함되며 에멀젼의 특성에 기여한다. 이들은 지방, 오일, 왁스, 지방산, 지방 알코올, 지방 에스테르, 보습제, 친수성 콜로이드, 방부제 및 항산화제를 포함한다(Block, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 335; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199).
친수성 콜로이드 또는 하이드로콜로이드는 자연 발생 검 및 합성 중합체, 예를 들어 다당류(예를 들면, 아카시아, 한천, 알긴산, 카라기난, 구아 검, 카라야 검 및 트라가칸트), 셀룰로스 유도체(예를 들면, 카복시메틸셀룰로스 및 카복시프로필셀룰로스), 및 합성 중합체(예를 들면, 카보머, 셀룰로스 에테르 및 카복시비닐 중합체)를 포함한다. 이들은 물에 분산되거나 팽윤되어 분산상 액적 주위에 강한 계면 필름을 형성함으로써 및 외부 상의 점도를 증가시킴으로써 에멀젼을 안정화시키는 콜로이드성 용액을 형성한다.
에멀젼은 종종 미생물의 성장을 쉽게 지원할 수 있는 탄수화물, 단백질, 스테롤 및 포스파티드와 같은 다수의 성분을 포함하기 때문에, 이러한 제형은 종종 방부제를 포함한다. 에멀젼 제형에 포함되어 일반적으로 사용되는 방부제는 메틸 파라벤, 프로필 파라벤, 4급 암모늄 염, 벤잘코늄 클로라이드, p-하이드록시벤조산 의 에스테르, 및 붕산을 포함한다. 항산화제는 또한 제형의 악화를 방지하기 위해 일반적으로 에멀젼 제형에 첨가된다. 사용되는 항산화제는 토코페롤, 알킬 갈레이트, 부틸화 하이드록시아니솔, 부틸화 하이드록시톨루엔과 같은 자유 라디칼 제거제, 또는 아스코르브산 및 나트륨 메타비설파이트와 같은 환원제, 및 시트르산, 타르타르산, 및 레시틴과 같은 항산화제 상승제일 수 있다.
피부과, 경구 및 비경구 경로를 통한 에멀젼 제형의 적용 및 이들의 제조 방법은 문헌에서 검토되었다(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199). 경구 전달용 에멀젼 제형은 제형화의 용이성 뿐만 아니라 흡수 및 생체이용효율 측면으로부터의 효능 때문에 매우 널리 사용되어 왔다(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY; Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 245; Idson, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 199). 광유-기반 완하제, 지용성 비타민 및 고지방 영양 제제는 일반적으로 o/w 에멀젼으로서 경구 투여되는 물질이다.
ii. 마이크로에멀젼
본 개시내용의 하나의 양태에서, RNAi 제제 및 핵산의 조성물은 마이크로에멀젼으로 제형화된다. 마이크로에멀젼은 광학적으로 등방성이며 열역학적으로 안정적인 단일 액체 용액인 물, 오일 및 양친매성 물질의 시스템으로서 정의될 수 있다(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY; Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 245). 전형적으로, 마이크로에멀젼은 먼저 수성 계면활성제 용액에 오일을 분산시킨 다음 충분한 양의 제4 성분, 일반적으로 중간 쇄 길이의 알코올을 첨가하여 투명한 시스템을 형성함으로써 제조되는 시스템이다. 따라서, 마이크로에멀젼은 또한 표면 활성 분자의 계면 필름에 의해 안정화된 두 개의 비혼화성 액체의 열역학적으로 안정하고 등방성으로 투명한 분산액으로서 기술되었다(Leung and Shah, in: Controlled Release of Drugs: Polymers and Aggregate Systems, Rosoff, M., Ed., 1989, VCH Publishers, New York, pages 185-215). 마이크로에멀젼은 일반적으로 오일, 물, 계면활성제, 보조계면활성제 및 전해질을 포함하는 3 내지 5가지 성분의 조합을 통해 제조된다. 마이크로에멀젼이 유중수(w/o) 또는 수중유(o/w) 유형인지 여부는 사용된 오일 및 계면활성제의 특성, 및 계면활성제 분자의 극성 머리와 탄화수소 꼬리의 구조 및 기하학적 패킹에 따라 좌우된다(Schott, in Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa., 1985, p. 271).
상태도를 활용하는 현상학적 접근법이 광범위하게 연구되었으며 마이크로에멀젼을 제형화하는 방법에 대한 포괄적인 지식을 당업계의 숙련가에게 제공하였다(예를 들어, 참조; Ansel's Pharmaceutical Dosage Forms and Drug Delivery Systems, Allen, LV., Popovich NG., and Ansel HC., 2004, Lippincott Williams & Wilkins (8th ed.), New York, NY; Rosoff, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 245; Block, in Pharmaceutical Dosage Forms, Lieberman, Rieger and Banker (Eds.), 1988, Marcel Dekker, Inc., New York, N.Y., volume 1, p. 335). 기존의 에멀젼과 비교하여, 마이크로에멀젼은 자발적으로 형성되는 열역학적으로 안정적인 액적의 제형에서 수불용성 약물을 가용화하는 이점을 제공한다.
마이크로에멀젼의 제조에 사용되는 계면활성제는 이온성 계면활성제, 비이온성 계면활성제, Brij 96, 폴리옥시에틸렌 올레일 에테르, 폴리글리세롤 지방산 에스테르, 테트라글리세롤 모노라우레이트(ML310), 테트라글리세롤 모노올레에이트(MO310), 헥사글리세롤 모노올레에이트(PO310), 헥사글리세롤 펜타올레에이트(PO500), 데카글리세롤 모노카프레이트(MCA750), 데카글리세롤 모노올레에이트(MO750), 데카글리세롤 세퀴올레에이트(SO750), 데카글리세롤 데카올레레이트(DAO750)를 단독으로 또는 보조계면활성제와 조합하여 포함하지만, 이에 제한되지 않는다. 일반적으로 에탄올, 1-프로판올, 및 1-부탄올과 같은 단쇄 알코올인 보조계면활성제는 계면활성제 필름에 침투하여 계면활성제 분자 사이에 생성된 공극 공간으로 인해 무질서한 필름을 생성함으로써 계면 유동성을 증가시키는 역할을 한다. 그러나, 마이크로에멀젼은 보조계면활성제를 사용하지 않고 제조될 수 있으며 무알코올 자가-유화 마이크로에멀젼 시스템은 당업계에 공지되어 있다. 수성 상은 전형적으로 물, 약물의 수용액, 글리세롤, PEG300, PEG400, 폴리글리세롤, 프로필렌 글리콜, 및 에틸렌 글리콜의 유도체일 수 있지만, 이에 제한되지 않는다. 오일 상은 Captex 300, Captex 355, Capmul MCM, 지방산 에스테르, 중쇄(C8-C12) 모노, 디, 및 트리-글리세리드, 폴리옥시에틸화 글리세릴 지방산 에스테르, 지방 알코올, 폴리글리콜화 글리세리드, 포화 폴리글리콜화 C8-C10 글리세리드, 식물성 오일 및 실리콘 오일과 같은 물질을 포함할 수 있지만, 이에 제한되지 않는다.
마이크로에멀젼은 약물 가용화 및 약물의 향상된 흡수의 관점에서 특히 관심이 있다. 지질 기반 마이크로에멀젼(o/w 및 w/o 둘 다)은 펩티드를 포함한 약물의 경구 생체이용효율을 향상시키기 위해 제안되었다(예를 들어, 참조; 미국 특허 제6,191,105호; 제7,063,860호; 제7,070,802호; 제7,157,099호; Constantinides et al., Pharmaceutical Research, 1994, 11, 1385-1390; Ritschel, Meth. Find. Exp. Clin. Pharmacol., 1993, 13, 205). 마이크로에멀젼은 개선된 약물 가용화, 효소 가수분해로부터의 약물의 보호, 막 유동성과 투과성에 있어서의 계면활성제-유도된 변경으로 인한 약물 흡수의 가능한 향상, 제조 용이성, 고체 투여 형태를 능가하는 경구 투여의 용이성, 개선된 임상 효능, 및 감소된 독성의 이점을 제공한다(예를 들어, 참조; 미국 특허 제6,191,105호; 제7,063,860호; 제7,070,802호; 제7,157,099호; Constantinides et al., Pharmaceutical Research, 1994, 11, 1385; Ho et al., J. Pharm. Sci., 1996, 85, 138-143). 종종 마이크로에멀젼은 이들의 성분이 주위 온도에서 합쳐질 때 자발적으로 형성될 수 있다. 이것은 열 불안정성 약물, 펩티드 또는 RNAi 제제를 제형화할 때 특히 유리할 수 있다. 마이크로에멀젼은 또한 화장품 및 제약 용도 모두에서 활성 성분의 경피 전달에 효과적이었다. 본 개시내용의 마이크로에멀젼 조성물 및 제형은 위장관으로부터의 RNAi 제제 및 핵산의 증가된 전신 흡수를 촉진할 뿐만 아니라 RNAi 제제 및 핵산의 국소 세포 흡수를 개선할 것으로 예상된다.
본 개시내용의 마이크로에멀젼은 또한 제제의 특성을 개선하고 본 개시내용의 RNAi 제제 및 핵산의 흡수를 향상시키기 위해 소르비탄 모노스테아레이트(Grill 3), 라브라솔 및 침투 증강제와 같은 추가 성분 및 첨가제를 함유할 수 있다. 본 발명의 마이크로에멀젼에 사용되는 침투 증강제는 5가지 광범위한 범주--계면활성제, 지방산, 담즙산 염, 킬레이트화제, 및 비-킬레이트화 비-계면활성제--중 하나로 분류될 수 있다(Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, p. 92). 이러한 부류 각각은 상기 논의되어 있다.
iii. 미세입자
본 개시내용의 RNAi 제제는 입자, 예를 들어, 마이크로입자에 혼입될 수 있다. 미세입자는 분무 건조에 의해 생성될 수 있지만 동결건조, 증발, 유동상 건조, 진공 건조 또는 이러한 기술의 조합을 포함한 다른 방법에 의해 생성될 수도 있다.
iv. 침투 증강제
하나의 양태에서, 본 개시내용은 핵산, 특히 RNAi 제제를 동물의 피부에 효율적으로 전달하기 위해 다양한 침투 증강제를 사용한다. 대부분의 약물은 이온화된 형태와 비이온화된 형태로 용액에 존재한다. 그러나, 일반적으로 지용성 또는 친유성 약물만이 세포막을 쉽게 통과한다. 비친유성 약물이라도 통과하고자 하는 막을 침투 증강제로 처리하면 세포막을 통과할 수 있다는 것이 발견되었다. 세포막을 가로지른 비친유성 약물의 확산을 돕는 것 외에도, 침투 증강제는 또한 친유성 약물의 투과성을 향상시킨다.
침투 증강제는 5가지 광범위한 범주, 즉, 계면활성제, 지방산, 담즙염, 킬레이트화제 및 비-킬레이트화 비-계면활성제 중 하나에 속하는 것으로 분류될 수 있다(예를 들어, 참조; Malmsten, M. Surfactants and polymers in drug delivery, Informa Health Care, New York, NY, 2002; Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, p.92). 위에서 언급한 각 부류의 침투 증강제는 아래에서 보다 상세하게 기술된다.
계면활성제(또는 "표면 활성제")는 수용액에 용해될 때 용액의 표면 장력 또는 수용액과 다른 액체 사이의 계면 장력을 감소시켜 그 결과 점막을 통한 RNAi 제제의 흡수가 향상된다. 담즙산 염 및 지방산 이외에, 이러한 침투 증강제는, 예를 들면, 나트륨 라우릴 설페이트, 폴리옥시에틸렌-9-라우릴 에테르 및 폴리옥시에틸렌-20-세틸 에테르를 포함한다(예를 들어, 참조; Malmsten, M. Surfactants and polymers in drug delivery, Informa Health Care, New York, NY, 2002; Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, p.92); and perfluorochemical emulsions, such as FC-43. Takahashi et al., J. Pharm. Pharmacol., 1988, 40, 252).
침투 증강제로서 작용하는 다양한 지방산 및 이들의 유도체는, 예를 들면, 올레산, 라우르산, 카프르산(n-데칸산), 미리스트산, 팔미트산, 스테아르산, 리놀레산, 리놀렌산, 디카프레이트, 트리카프레이트, 모노올레인(1-모노올레오일-rac-글리세롤), 디라우린, 카프릴산, 아라키돈산, 글리세롤 1-모노카프레이트, 1-도데실아자사이클로헵탄-2-온, 아실카르니틴, 아실콜린, 이의 C1-20 알킬 에스테르(예를 들어, 메틸, 이소프로필 및 t-부틸), 및 이의 모노- 및 디-글리세리드(즉, 올레에이트, 라우레이트, 카프레이트, 미리스테이트, 팔미테이트, 스테아레이트, 리놀레이트 등)을 포함한다(예를 들어, 참조; Touitou, E., et al. Enhancement in Drug Delivery, CRC Press, Danvers, MA, 2006; Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, p.92; Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33; El Hariri et al., J. Pharm. Pharmacol., 1992, 44, 651-654).
담즙의 생리학적 역할은 지질 및 지용성 비타민의 분산 및 흡수의 촉진을 포함한다(예를 들어, 참조; Malmsten, M. Surfactants and polymers in drug delivery, Informa Health Care, New York, NY, 2002; Brunton, Chapter 38 in: Goodman & Gilman's The Pharmacological Basis of Therapeutics, 9th Ed., Hardman et al. Eds., McGraw-Hill, New York, 1996, pp. 934-935). 다양한 천연 담즙산 염 및 이들의 합성 유도체가 침투 촉진제로서 작용한다. 따라서 용어 "담즙산 염"은 담즙의 자연 발생 성분 중 어느 것 뿐만 아니라 이들의 합성 유도체 중 어느 것을 포함한다. 적합한 담즙산 염은, 예를 들면, 콜산(또는 이의 약제학적으로 허용되는 나트륨 염, 나트륨 콜레이트), 데하이드로콜산(나트륨 데하이드로콜레이트), 데옥시콜산(나트륨 데옥시콜레이트), 글루콜산(나트륨 글루콜레이트), 글리콜산(나트륨 글리코콜레이트), 글리코데옥시콜산(나트륨 글리코데옥시콜레이트), 타우로콜산(나트륨 타우로콜레이트), 타우로데옥시콜산(나트륨 타우로데옥시콜레이트), 케노데옥시콜산(나트륨 케노데옥시콜레이트), 우르소데옥시콜산(UDCA), 나트륨 타우로-24,25-디하이드로-퓨시데이트(STDHF), 나트륨 글리코디하이드로퓨시데이트 및 폴리옥시에틸렌-9-라우릴 에테르(POE)를 포함한다(예를 들어, 참조; Malmsten, M. Surfactants and polymers in drug delivery, Informa Health Care, New York, NY, 2002; Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, page 92; Swinyard, Chapter 39 In: Remington's Pharmaceutical Sciences, 18th Ed., Gennaro, ed., Mack Publishing Co., Easton, Pa., 1990, pages 782-783; Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33; Yamamoto et al., J. Pharm. Exp. Ther., 1992, 263, 25; Yamashita et al., J. Pharm. Sci., 1990, 79, 579-583).
본 개시내용과 관련하여 사용되는 킬레이트화제는 금속 이온과 복합체를 형성하여 용액으로부터 금속 이온을 제거하여 그 결과 점막을 통한 RNAi 제제의 흡수가 향상되는 화합물로서 정의될 수 있다. 본 개시내용에서 침투 증강제로서의 이들의 사용과 관련하여, 대부분의 특징화된 DNA 뉴클레아제는 촉매작용을 위해 2가 금속 이온을 필요로 하고 따라서 킬레이트제에 의해 억제되기 때문에, 킬레이트화제는 DNase 억제제로도 작용한다는 추가 이점을 갖는다(Jarrett, J. Chromatogr., 1993, 618, 315-339). 적합한 킬레이트화제는 이나트륨 에틸렌디아민테트라아세테이트(EDTA), 시트르산, 살리실레이트(예를 들어, 나트륨 살리실레이트, 5-메톡시살리실레이트 및 호모바닐레이트), 콜라겐의 N-아실 유도체, 라우레트-9- 및 베타-디케톤(엔아민)의 N-아미노 아실 유도체를 포함하지만 이에 제한되지 않는다(예를 들어, 참조; Katdare, A. et al., Excipient development for pharmaceutical, biotechnology, and drug delivery, CRC Press, Danvers, MA, 2006; Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, page 92; Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33; Buur et al., J. Control Rel., 1990, 14, 43-51).
본원에 사용된 바와 같이, 비-킬레이트화 비-계면활성제 침투 증진 화합물은 킬레이트화제로서 또는 계면활성제로서 미미한 활성을 입증하지만 그럼에도 불구하고 소화 점막을 통한 RNAi 제제의 흡수를 향상시키는 화합물로서 정의될 수 있다(예를 들어, 참조; Muranishi, Critical Reviews in Therapeutic Drug Carrier Systems, 1990, 7, 1-33). 이러한 종류의 침투 증강제는, 예를 들면, 불포화 사이클릭 요소, 1-알킬- 및 1-알케닐아자사이클로-알카논 유도체를 포함한다(Lee et al., Critical Reviews in Therapeutic Drug Carrier Systems, 1991, page 92); and non-steroidal anti-inflammatory agents such as diclofenac sodium, indomethacin and phenylbutazone (Yamashita et al., J. Pharm. Pharmacol., 1987, 39, 621-626).
세포 수준에서 RNAi 제제의 흡수를 향상시키는 제제가 또한 본 개시내용의 약제학적 및 기타 조성물에 첨가될 수 있다. 리포펙틴(Junichi 등, 미국 특허 제5,705,188호)과 같은 양이온성 지질, 양이온성 글리세롤 유도체, 및 폴리리신(제WO 97/30731호)과 같은 폴리양이온성 분자도 dsRNA의 세포 흡수를 향상시키는 것으로 알려져 있다.
에틸렌 글리콜 및 프로필렌 글리콜과 같은 글리콜, 2-피롤과 같은 피롤, 아존, 및 리모넨 및 멘톤과 같은 테르펜을 포함한 기타 제제가 투여된 핵산의 침투를 향상시키기 위해 사용될 수 있다.
vi. 부형제
담체 화합물과 대조적으로, "약제학적 담체" 또는 "부형제"는 하나 이상의 핵산을 동물에게 전달하기 위한 약제학적으로 허용되는 용매, 현탁제 또는 임의의 기타 약리학적으로 불활성인 비히클이다. 부형제는 액체 또는 고체일 수 있고 주어진 약제학적 조성물의 핵산 및 기타 성분과 조합될 때 원하는 부피, 조도 등을 제공하도록 계획된 투여 방식을 염두에 두고 선택된다. 전형적인 약제학적 담체는 결합제(예를 들어, 예비젤라틴화 옥수수 전분, 폴리비닐피롤리돈 또는 하이드록시프로필 메틸셀룰로스 등); 충전제(예를 들어, 락토스 및 기타 당류, 미세결정질 셀룰로스, 펙틴, 젤라틴, 황산칼슘, 에틸 셀룰로스, 폴리아크릴레이트 또는 인산수소칼슘 등); 윤활제(예를 들어, 마그네슘 스테아레이트, 활석, 실리카, 콜로이드성 이산화규소, 스테아르산, 금속성 스테아레이트, 수소화 식물성 오일, 옥수수 전분, 폴리에틸렌 글리콜, 나트륨 벤조에이트, 나트륨 아세테이트 등); 붕해제(예를 들어, 전분, 나트륨 전분 글리콜레이트 등); 및 습윤제(예를 들어, 나트륨 라우릴 설페이트 등)를 포함하지만, 이에 제한되지 않는다.
핵산과 유해하게 반응하지 않는 비경구 투여에 적합한 약제학적으로 허용되는 유기 또는 무기 부형제가 또한 본 개시내용의 조성물을 제형화하는데 사용될 수 있다. 적합한 약제학적으로 허용가능한 담체는 물, 염 용액, 알코올, 폴리에틸렌 글리콜, 젤라틴, 락토스, 아밀로스, 마그네슘 스테아레이트, 활석, 규산, 점성 파라핀, 하이드록시메틸셀룰로스, 폴리비닐피롤리돈 등을 포함하지만, 이에 제한되지 않는다.
핵산의 국소 투여를 위한 제형은 멸균 및 비멸균 수용액, 알코올과 같은 일반적인 용매 중의 비수성 용액, 또는 액체 또는 고체 오일 기재 중의 핵산의 용액을 포함할 수 있다. 용액은 또한 완충제, 희석제 및 기타 적합한 첨가제를 포함할 수 있다. 핵산과 유해하게 반응하지 않는 비경구 투여에 적합한 약제학적으로 허용되는 유기 또는 무기 부형제가 사용될 수 있다.
적합한 약제학적으로 허용되는 부형제는 물, 염 용액, 알코올, 폴리에틸렌 글리콜, 젤라틴, 락토스, 아밀로스, 마그네슘 스테아레이트, 활석, 규산, 점성 파라핀, 하이드록시메틸셀룰로스, 폴리비닐피롤리돈 등을 포함하지만, 이에 제한되지 않는다.
vii. 기타 성분
본 개시내용의 조성물은 약제학적 조성물에서 통상적으로 발견되는 기타 보조 성분을 당업계에 확립된 사용 수준으로 추가로 함유할 수 있다. 띠라서, 예를 들면, 조성물은, 예를 들면, 항소양제, 수렴제, 국소 마취제 또는 항염증제와 같은 추가의 상용성 약제학적 활성 물질을 함유할 수 있거나, 염료, 방향제, 방부제, 항산화제, 불투명화제, 증점제 및 안정제와 같은 본 개시내용의 조성물의 다양한 투여 형태를 물리적으로 제형화하는데 유용한 추가 물질을 함유할 수 있다. 그러나, 이러한 물질은, 첨가될 때, 본 개시내용의 조성물의 성분의 생물학적 활성을 과도하게 방해해서는 안 된다. 제형은 멸균될 수 있고, 원하는 경우, 제형의 핵산(들)과 유해하게 상호작용하지 않는 보조제, 예를 들어, 윤활제, 방부제, 안정제, 습윤제, 유화제, 삼투압에 영향을 미치는 염, 완충제, 착색제, 방향제 또는 방향 물질 등과 혼합될 수 있다.
수성 현탁액은, 예를 들면, 나트륨 카복시셀룰로스, 소르비톨 또는 덱스트란을 포함하는 현탁액의 점도를 증가시키는 물질을 함유할 수 있다. 현탁액은 또한 안정제를 함유할 수 있다.
일부 양태에서, 본 개시내용에 특성화된 약제학적 조성물은 (a) 하나 이상의 RNAi 제제 및 (b) 비-RNAi 메카니즘에 의해 기능하고 C9orf72-관련 장애를 치료하는데 유용한 하나 이상의 제제를 포함한다. 이러한 제제의 예는 모노아민 억제제, 레세르핀, 항경련제, 항정신병제 및 항우울제를 포함하지만, 이에 제한되지 않는다.
이러한 화합물의 독성 및 치료 효능은, 예를 들어, LD50(집단의 50%에 치명적인 용량) 및 ED50(집단의 50%에서 치료적으로 효과적인 용량)을 결정하기 위해 세포 배양물 또는 실험 동물에서 표준 약제학적 절차에 의해 결정될 수 있다. 독성 효과와 치료 효과 사이의 용량 비가 치료 지수이며 LD50/ED50의 비율로 표현될 수 있다. 높은 치료 지수를 나타내는 화합물이 바람직하다.
세포 배양물 분석 및 동물 연구로부터 수득된 데이터는 인간에 사용하기 위한 투여량 범위를 수립하는데 사용될 수 있다. 본 개시내용에서 본원에 특성화된 조성물의 투여량은 일반적으로 독성이 거의 또는 전혀 없는 ED50을 포함하는 순환 농도 범위 내에 있다. 투여량은 사용된 투여 형태 및 사용된 투여 경로에 따라 이 범위 내에서 변할 수 있다. 본 개시내용에 특성화된 방법에 사용되는 임의의 화합물에 대해, 치료학적 유효량은 초기에 세포 배양물 검정으로부터 추정될 수 있다. 용량은 화합물의 순환 혈장 농도 범위, 또는 적절한 경우 세포 배양물에서 결정된 바와 같은 IC50(즉, 증상의 최대-절반 억제를 달성하는 시험 화합물의 농도)을 포함하는 표적 서열의 폴리펩티드 산물의 순환 혈장 농도 범위(예를 들어 감소된 농도의 폴리펩티드 달성)를 달성하기 위해 동물 모델에서 수립될 수 있다. 이러한 정보는 인간에서 유용한 용량을 보다 정확하게 결정하는데 사용될 수 있다. 혈장 중 수준은, 예를 들면, 고성능 액체 크로마토그래피에 의해 측정될 수 있다.
이들의 투여에 더하여, 상기 논의된 바와 같이, 본 개시내용에 특성화된 RNAi 제제는 뉴클레오티드 반복부 발현에 의해 매개되는 병리학적 과정의 치료에 효과적인 기타 공지된 제제와 조합하여 투여될 수 있다. 어떠한 경우에도, 투여 의사는 당업계에 공지되거나 본원에 기술된 효능의 표준 측정을 사용하여 관찰된 결과에 기초하여 RNAi 제제 투여의 양 및 시기를 조정할 수 있다.
VII. 키트
특정 측면에서, 본 개시내용은 siRNA 화합물, 예를 들어, 이중 가닥 siRNA 화합물, 또는 ssiRNA 화합물(예를 들어, 전구체, 예를 들어, ssiRNA 화합물로 프로세싱될 수 있는 더 큰 siRNA 화합물, 또는 siRNA 화합물을 암호화하는 DNA, 예를 들어, 이중 가닥 siRNA 화합물, 또는 ssiRNA 화합물, 또는 이들의 전구체)의 약제학적 제형을 함유하는 적합한 용기를 포함하는 키트를 제공한다.
이러한 키트는 하나 이상의 dsRNA 제제 및 사용 지침서, 예를 들어, 예방적 또는 치료학적 유효량의 dsRNA 제제를 투여하기 위한 지침서를 포함한다. dsRNA 제제는 바이알 또는 사전-충전된 주사기에 있을 수 있다. 키트는 임의로 dsRNA 제제를 투여하기 위한 수단(예를 들어, 사전-충전된 주사기와 같은 주사 장치), 또는 C3의 억제를 측정하기 위한 수단(예를 들어, C9orf72 mRNA, C9orf72 단백질 및/또는 C9orf72 활성의 억제를 측정하기 위한 수단)을 추가로 포함할 수 있다. C9orf72의 억제를 측정하기 위한 이러한 수단은, 예를 들어, CSF 및/또는 혈장 샘플과 같은 대상체로부터 샘플을 수득하기 위한 수단을 포함할 수 있다. 본 발명의 키트는 임의로 치료학적 유효량 또는 예방학적 유효량을 결정하기 위한 수단을 추가로 포함할 수 있다.
특정 양태에서 약제학적 제형의 개별 성분은 하나의 용기에 제공될 수 있다. 대안적으로, 약제학적 제형의 성분을 2개 이상의 용기, 즉 siRNA 화합물 제제를 위한 하나의 용기 및 담체 화합물을 위한 적어도 또 다른 용기에 별도로 제공하는 것이 바람직할 수 있다. 키트는 단일 상자에 하나 이상의 용기와 같은 다수의 상이한 구성으로 포장될 수 있다. 상이한 성분들은, 예를 들어, 키트와 함께 제공된 지침서에 따라 조합될 수 있다. 성분들은, 예를 들어, 약제학적 조성물을 제조 및 투여하기 위해 본원에 기술된 방법에 따라 조합될 수 있다. 키트는 또한 전달 장치를 포함할 수 있다.
VII. C9orf72 발현을 억제하는 방법
본 개시내용은 또한 세포에서 C9orf72 유전자의 발현을 억제하는 방법을 제공한다. 상기 방법은 세포에서 C9orf72의 발현을 억제하기에 효과적인 양의 RNAi 제제, 예를 들어 이중 가닥 RNAi 제제와 세포를 접촉시켜 세포에서 C9orf72의 발현을 억제함을 포함한다. 본 개시내용의 특정 양태에서, C9orf72는 CNS(예를 들면, 뇌) 세포에서 우선적으로 억제된다.
세포를 RNAi 제제, 예를 들어 이중 가닥 RNAi 제제와 접촉시키는 것은 시험관내 또는 생체내에서 수행될 수 있다. 세포를 생체내에서 RNAi 제제와 접촉시키는 것은 대상체, 예를 들어 인간 대상체 내의 세포 또는 세포 그룹을 RNAi 제제와 접촉시키는 것을 포함한다. 세포를 접촉시키는 시험관내 및 생체내 방법의 조합도 가능하다.
세포를 접촉시키는 것은 위에서 논의한 바와 같이 직접적이거나 간접적일 수 있다. 또한, 세포를 접촉시키는 것은 본원에 기술되거나 당업계에 공지된 임의의 리간드를 포함하는 표적화 리간드를 통해 달성될 수 있다. 일부 양태에서, 표적화 리간드는 탄수화물 모이어티, 예를 들어, GalNAc 리간드, 또는 RNAi 제제를 관심 부위로 지시하는 임의의 다른 리간드이다.
본원에 사용된 용어 "억제하는"은 "감소", "침묵", "하향조절", "억제" 및 기타 유사한 용어와 상호교환적으로 사용되며, 임의의 억제 수준을 포함한다. 특정 양태에서, 예를 들어, 본 개시내용의 RNAi 제제에 대한 억제 수준은 세포 배양 조건에서 평가될 수 있으며, 예를 들어, 여기서 세포 배양물 중 세포는 세포 부근에서 10nM 이하, 1nM 이하 등의 농도에서 리포펙타민™-매개된 형질감염을 통해 형질감염된다. 주어진 RNAi 제제의 녹다운은 세포 배양물에서 전처리된 수준 대 세포 배양물에서 후처리된 수준의 비교를 통해 결정될 수 있으며, 임의로 스크램블된 또는 다른 형태의 대조군 RNAi 제제와 병행하여 처리된 세포와 비교하기도 한다. 이에 따라 세포 배양에서, 예를 들어, 약 50%의 녹다운이 "억제" 또는 "감소", "하향 조절" 또는 "억제" 등이 발생했음을 나타내는 것으로 식별될 수 있다. 표적화된 mRNA 또는 암호화된 단백질 수준(및 따라서 본 개시내용의 RNAi 제제에 의해 야기되는 "억제" 정도 등)의 평가는 또한 당업계에 기술된 바와 같이 적절하게 제어된 조건하에서 본 개시내용의 RNAi 제제에 대한 생체내 시스템에서 평가될 수 있음이 명백히 고려된다.
본원에 사용된 문구 "C9orf72 유전자의 발현을 억제하는" 또는 "C9orf72의 발현을 억제하는"은 임의의 C9orf72 유전자(예를 들어, 마우스 C9orf72 유전자, 래트 C9orf72 유전자, 원숭이 C9orf72 유전자, 또는 인간 C9orf72 유전자) 뿐만 아니라, C9orf72 단백질을 암호화하는 C9orf72 유전자의 변이체 또는 돌연변이체, 예를 들어, 유전자의 인트론에 확장된 헥사뉴클레오티드 반복부를 갖는 C9orf72 유전자의 발현의 억제를 포함한다. 따라서, C9orf72 유전자는 야생형 C9orf72 유전자, 돌연변이 C9orf72 유전자, 또는 유전자 조작된 세포, 세포 그룹 또는 유기체의 맥락에서 유전자이식 C9orf72 유전자일 수 있다.
"C9orf72 유전자의 발현을 억제"하는 것은 C9orf72 유전자의 임의의 수준의 억제, 예를 들어, C9orf72 유전자의 발현의 적어도 부분적 억제, 예를 들어 적어도 20% 억제를 포함한다. 특정 양태에서, 억제는 적어도 30%, 적어도 40%, 또는 바람직하게는 적어도 50%이다. 다른 양태에서, 억제는 50% 이하, 예를 들어, 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 또는 5% 이하이다.
C9orf72 유전자의 발현은 C9orf72 유전자 발현과 관련된 임의의 변수의 수준, 예를 들어, C9orf72 mRNA 수준(예를 들어, 센스 mRNA, 안티센스 mRNA, 전체 C9orf72 mRNA, 센스 C9orf72 반복부-함유 mRNA, 및/또는 안티센스 C9orf72 반복부-함유 mRNA) 또는 C9orf72 단백질 수준(예를 들어, 전체 C9orf72 단백질, 야생형 C9orf72 단백질, 또는 확장된 반복부-함유 단백질), 또는, 예를 들면, 센스- 또는 안티센스-함유 병소의 수준 및/또는 비정상적인 디펩티드 반복부 단백질의 수준에 기반하여 평가될 수 있다.
억제는 대조군 수준과 비교하여 이러한 변수 중 하나 이상의 절대적 또는 상대적 수준의 감소에 의해 평가될 수 있다. 대조군 수준은 당업계에서 사용되는 임의의 유형의 대조군 수준, 예를 들어, 투여-전 기준선 수준, 또는 대조군으로 처리되지 않거나 처리된(예를 들어, 완충제 단독 대조군 및 불활성 제제 대조군) 유사한 대상체, 세포 또는 샘플로부터 결정된 수준일 수 있다.
예를 들면, 본 개시내용의 방법의 일부 양태에서, C9orf72 유전자의 발현(센스- 또는 안티센스-함유 병소 및/또는 비정상적인 디펩티드 반복부 단백질 수준에 의해 평가됨)이 적어도 20%, 30%, 40%, 50%, 60%, 70%, 80%, 85%, 90% 또는 95%, 또는 분석의 검출 수준 이하까지 억제된다. 본 개시내용의 방법의 다른 양태에서, C9orf72 유전자의 발현(예를 들어, mRNA 또는 단백질 발현 수준에 의해 평가됨)은 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 또는 5% 이하까지 억제된다. 특정 양태에서, 상기 방법은, 예를 들어, C9orf72의 발현을 감소시키는 제제로 대상체를 치료한 후 임상적으로 관련된 결과에 의해 입증된 바와 같이, C9orf72의 발현의 임상적으로 관련된 억제를 포함한다.
C9orf72 유전자의 발현의 억제는, 첫 번째 세포 또는 세포 그룹과 실질적으로 동일하지만 그렇게 처리되지 않았거나 처리되지 않은 두 번째 세포 또는 세포 그룹(RNAi 제제로 처리되지 않았거나 관심 유전자를 표적으로 하는 RNAi 제제로 처리되지 않은 대조군 세포)과 비교하여, C9orf72 유전자가 전사되고 C9orf72 유전자의 발현이 억제되도록 (예를 들어, 세포 또는 세포들을 본 개시내용의 RNAi 제제와 접촉시킴으로써 또는 세포가 존재하거나 존재했던 대상체에게 본 개시내용의 RNAi 제제를 투여함으로써) 처리된 첫 번째 세포 또는 세포 그룹(이러한 세포는, 예를 들면, 대상체로부터 유래된 샘플에 존재할 수 있음)에 의해 발현되는 mRNA의 양의 감소로 나타날 수 있다. 억제 정도는 다음과 같이 표현될 수 있다:

다른 양태에서, C9orf72 유전자의 발현 억제는 C9orf72 유전자 발현, 예를 들어, C9orf72 단백질 발현, 센스- 또는 안티센스-함유 병소 및/또는 비정상적인 디펩티드 반복질 단백질의 수준과 기능적으로 연결된 매개변수의 감소 측면에서 평가될 수 있다. C9orf72 유전자 침묵은 발현 작제물로부터 내인성 또는 이종성인 C9orf72를 발현하는 임의의 세포에서, 및 당업계에 공지된 임의의 검정에 의해 결정될 수 있다.
C9orf72 단백질의 발현의 억제는 세포 또는 세포 그룹에 의해 발현된 C9orf72 단백질(또는, 예를 들어, 본원에 기술된 바와 같은 기능적 매개변수)의 수준(예를 들어, 대상체로부터 유래된 샘플에서 발현되는 단백질 수준)의 감소로 나타날 수 있다. 상기 설명된 바와 같이, mRNA 억제의 평가를 위해, 처리된 세포 또는 세포 그룹에서의 단백질 발현 수준의 억제는 대조군 세포 또는 세포 그룹에서의 단백질 수준의 백분율로서 유사하게 표현될 수 있다.
C9orf72 유전자의 발현의 억제를 평가하기 위해 사용될 수 있는 대조군 세포 또는 세포 그룹은 본 개시내용의 RNAi 제제와 아직 접촉되지 않은 세포 또는 세포 그룹을 포함한다. 예를 들면, 대조군 세포 또는 세포 그룹은 RNAi 제제로 대상체를 치료하기 전에 개별 대상체(예를 들어, 인간 또는 동물 대상체)로부터 유래할 수 있다.
세포 또는 세포 그룹에 의해 발현되는 C9orf72 mRNA의 수준은 mRNA 발현을 평가하기 위해 당업계에 공지된 임의의 방법을 사용하여 결정될 수 있다. 하나의 양태에서, 샘플에서 C9orf72의 발현의 수준은 전사된 폴리뉴클레오티드 또는 이의 일부, 예를 들어 C9orf72 유전자의 mRNA를 검출함으로써 결정된다. RNA는, 예를 들면, 산 페놀/구아니딘 이소티오시아네이트 추출(RNAzol B; Biogenesis), RNeasy™ RNA 제조 키트(Qiagen®) 또는 PAXgene(PreAnalytix, Switzerland)을 사용함을 포함하여 RNA 추출 기술을 사용하여 세포로부터 추출될 수 있다. 리보핵산 혼성화를 사용하는 전형적인 검정 형식은 핵 실행 검정, RT-PCR, RNase 보호 검정, 노던 블롯팅, 제자리 혼성화, 및 마이크로어레이 분석을 포함한다. 가닥 특이적 C9orf72 mRNA는, 예를 들면, 문헌[Jiang, et al. supra, Lagier-Tourenne, et al., supra and Jiang, et al., supra]에 기술된 정량적 RT-PCR 및.또는 액적 디지털 PCR 방법을 사용하여 검출될 수 있다. 순환 C9orf72 mRNA는 제WO2012/177906호에 기술된 방법을 사용하여 검출될 수 있으며, 이의 전체 내용은 본원에 참고로 포함된다.
일부 양태에서, C9orf72의 발현의 수준은 핵산 프로브를 사용하여 결정된다. 본원에 사용된 용어 "프로브"는 특정 C9orf72 핵산 또는 단백질, 또는 이의 단편에 선택적으로 결합할 수 있는 임의의 분자를 지칭한다. 프로브는 당업계의 숙련가에 의해 합성되거나 적절한 생물학적 제제로부터 유래될 수 있다. 프로브는 표지화되도록 특별히 설계될 수 있다. 프로브로서 사용될 수 있는 분자의 예는 RNA, DNA, 단백질, 항체 및 유기 분자를 포함하지만, 이에 제한되지 않는다.
단리된 mRNA는 서던 또는 노던 분석, 중합효소 연쇄 반응(PCR) 분석 및 프로브 어레이를 포함하지만 이에 제한되지 않는 혼성화 또는 증폭 분석에 사용될 수 있다. mRNA 수준의 측정을 위한 한 가지 방법은 단리된 mRNA를 C9orf72 mRNA에 혼성화할 수 있는 핵산 분자(프로브)와 접촉시키는 것을 포함한다. 하나의 양태에서, mRNA를 고체 표면에 고정시키고, 예를 들면, 단리된 mRNA를 아가로스 겔에 유동시키고 mRNA를 겔에서 니트로셀룰로스와 같은 막으로 전달함으로써 프로브와 접촉시킨다. 대안적인 양태에서, 프로브(들)를 고체 표면에 고정시키고 mRNA를, 예를 들면, Affymetrix® 유전자 칩 어레이에서 프로브(들)와 접촉시킨다. 숙련가는 C9orf72 mRNA의 수준을 결정하는데 사용하기 위해 공지된 mRNA 검출 방법을 쉽게 조정할 수 있다.
샘플에서 C9orf72의 발현의 수준을 결정하기 위한 대안적인 방법은, 예를 들어, RT-PCR(실험적 양태는 문헌[Mullis, 1987, US Patent No. 4,683,202), ligase chain reaction (Barany (1991) Proc . Natl . Acad . Sci . USA 88:189-193]에 제시됨), 자가 지속 서열 복제(Guatelli et al. (1990) Proc . Natl . Acad . Sci . USA 87:1874-1878), 전사 증폭 시스템(Kwoh et al. (1989) Proc . Natl . Acad . Sci . USA 86:1173-1177), Q-베타 복제효소(Lizardi et al. (1988) Bio/Technology 6:1197), 롤링 서클 복제(Lizardi et al., US Patent No. 5,854,033) 또는 임의의 다른 핵산 증폭 방법에 이은 당업계의 숙련가들에게 잘 알려진 기술을 사용한 증폭된 분자의 검출에 의한, 예를 들면, 샘플 내의 mRNA의 핵산 증폭 또는 역전사효소(cDNA를 제조하기 위한) 과정을 포함한다. 이러한 검출 체계는 핵산 분자가 매우 적은 수로 존재하는 경우 핵산 분자의 검출에 특히 유용하다. 본 개시내용의 특정 측면에서, C9orf72의 발현 수준은 정량적 형광성 RT-PCR(즉, TaqMan™ 시스템)에 의해, Dual-Glo® 루시페라제 검정에 의해, 또는 C9orf72 발현 또는 mRNA 수준의 측정을 위해 당업계에서 인정되는 기타 방법에 의해 결정된다.
C9orf72 mRNA의 발현 수준은 막 블롯(예를 들어 노던, 서던, 도트 등과 같은 혼성화 분석에 사용됨), 또는 마이크로웰, 샘플 튜브, 겔, 비드 또는 섬유(또는 결합된 핵산을 포함하는 임의의 고체 지지체)을 사용하여 모니터링될 수 있다. 본원에 참고로 포함된 미국 특허 제5,770,722호, 제5,874,219호, 제5,744,305호, 제5,677,195호 및 제5,445,934호를 참조한다. C9orf72 발현 수준의 결정은 또한 용액 중의 핵산 프로브를 사용하는 것을 포함할 수 있다.
일부 양태에서, mRNA 발현의 수준은 분기형 DNA(bDNA) 분석 또는 실시간 PCR(qPCR)을 사용하여 평가된다. 이러한 PCR 방법의 사용은 본원에 제시된 실시예에 기술되고 예시된다. 이러한 방법은 또한 C9orf72 핵산의 검출에 사용될 수 있다.
C9orf72 단백질 발현의 수준은 단백질 수준의 측정을 위해 당업계에 공지된 임의의 방법을 사용하여 결정될 수 있다. 이러한 방법은, 예를 들면, 전기영동, 모세관 전기영동, 고성능 액체 크로마토그래피(HPLC), 박층 크로마토그래피(TLC), 과확산 크로마토그래피, 유체 또는 겔 침강소 반응, 흡수 분광법, 비색 분석, 분광 광도 분석, 유세포 분석, 면역확산(단일 또는 이중), 면역전기영동, 웨스턴 블롯팅, 방사선면역분석(RIA), 효소-결합 면역흡착 분석(ELISA), 면역형광 분석, 전기화학발광 분석 등을 포함한다. 이러한 분석은 또한 C9orf72 단백질의 존재 또는 복제를 나타내는 단백질의 검출에 사용될 수 있다.
센스- 또는 안티센스-함유 병소의 수준 및 비정상적인 디펩티드 반복부 단백질의 수준은, 예를 들면, 형광 제자리 혼성화(FISH), 면역조직화학 및 면역분석을 포함하는 당업계의 통상의 숙련가에게 잘 알려진 방법을 사용하여 평가될 수 있다(예를 들어, 참조; Jiang, et al. supra). 일부 양태에서, C9orf72-관련 질환의 치료에서 본 개시내용의 방법의 효능은 C9orf72 mRNA 수준의 감소에 의해 (예를 들어, C9orf72 수준에 대한 CSF 샘플 및/또는 혈장 샘플의 평가, 뇌 생검 등에 의해) 평가된다.
본 개시내용의 방법의 일부 양태에서, RNAi 제제는 RNAi 제제가 대상체 내의 특정 부위에 전달되도록 대상체에 투여된다. C9orf72의 발현의 억제는 대상체 내의 특정 부위로부터 유래된 샘플, 예를 들어, CNS 세포에서 C9orf72 mRNA(예를 들어, 센스 mRNA, 안티센스 mRNA, 전체 C9orf72 mRNA, 센스 C9orf72 반복부-함유 mRNA, 및/또는 안티센스 C9orf72 반복부-함유 mRNA), C9orf72 단백질(예를 들어, 전체 C9orf72 단백질, 야생형 C9orf72 단백질, 또는 확장된 반복부-함유 단백질), 센스-함유 병소, 안티센스-함유 병소, 비정상적인 디펩티드 반복부 단백질 수준의 변화 또는 수준 측정을 사용하여 평가될 수 있다. 특정 양태에서, 방법은, 예를 들면, 꼬리 위축의 안정화 또는 억제(예를 들어, 체적 MRI(vMRI)에 의해 평가됨), 대상체로부터의 CSF 샘플에서 신경필라멘트 경쇄(Nfl) 수준의 안정화 또는 감소, 돌연변이 C9orf72 mRNA 또는 절단 돌연변이 C9orf72 단백질, 예를 들어, 전장 돌연변이 C9orf72 mRNA 또는 단백질 및 절단 돌연변이 C9orf72 mRNA 또는 단백질 중 하나 또는 둘 다의 감소, 및 통합된 C9orf72-관련 질환 평가 척도(UHDRS) 점수의 안정화 또는 개선과 같은 C9orf72의 발현을 감소시키는 제제로 대상체를 치료한 후 임상적으로 관련된 결과에 의해 입증된 바와 같이 C9orf72 발현의 임상적으로 관련된 억제를 포함한다.
본원에 사용된 바와 같이, 분석물의 수준을 검출하거나 결정하는이라는 용어는 물질, 예를 들어, 단백질, RNA가 존재하는지 여부를 결정하는 단계를 수행함을 의미하는 것으로 이해된다. 본원에 사용된 바와 같이, 검출하거나 결정하는 방법은 사용된 방법에 대한 검출 수준 미만인 분석물 수준의 검출 또는 결정을 포함한다.
IX. C9orf72-관련 질환을 치료 또는 예방하는 방법
본원에 기술된 바와 같이, C9orf72 반복부-함유 RNA를 감소시키는 것은 RNA 간섭의 자연적 과정을 통해 달성될 수 있으며, 여기서 dsRNA는 RNA-유도된 침묵 복합체에 의한 표적 RNA의 절단에 이은 세포 뉴클레아제에 의한 RNA 절단 단편의 분해를 지시한다. 그러나, RNA 간섭은 핵에 보유된 RNA에 작용할 것으로 예상되지 않는 우세한 세포질 과정이다. 인트론-함유 RNA는 성숙 mRNA로 빠르게 스플라이싱되는 mRNA 전구체로서 또는 핵에서 빠르게 분해되는 스플라이싱된 인트론으로서 일반적으로 수명이 짧다. 따라서, 인트론-함유 RNA는 RNA 간섭에 의한 표적화에 사용할 수 없을 것이라고 예상하는 것이 합리적이다.
그러나, 본원에서 입증된 바와 같이, GGGGCC 반복부 확장에 인접한 인트론 서열은 표적으로 하는 siRNA는 C9orf72 성숙 mRNA에 거의 또는 전혀 영향을 미치지 않으면서 인트론-함유 C9orf72 RNA의 축적을 감소시켰다. 인트론-표적화 siRNA는 또한 디펩티드 반복부 단백질의 생산을 감소시켰다. 놀랍게도, 본 발명자들은 이 영역을 표적으로 하는 개시된 siRNA가 인트론-함유 C9orf72 RNA 수준을 감소시키는데 효과적임을 보여주었다. 결과는 디펩티드 반복부 단백질 합성을 담당하는 인트론-함유 C9orf72 RNA의 상당 부분이 세포질에 존재함을 보여준다. 대조적으로, C9orf72 mRNA 단백질 코딩 서열을 표적으로 하는 siRNA는 mRNA의 강한 녹다운을 생성하지만 인트론-함유 전사체에는 영향을 미치지 않았고 디펩티드 반복부 단백질 합성을 눈에 띄게 감소시키지 않았다. 인트론-표적화 siRNA 및 mRNA-표적화 siRNA 사이의 결과 차이는 표적화된 서열의 두 부류가 공유적으로 연결되지 않은 별도의 RNA에 존재함을 시사한다.
본원에 개시된 방법은 C9orf72 반복부 확장 질환의 주요 병원성 성분인 디펩티드 반복부 단백질의 합성의 치료적 감소를 제공하면서 C9orf72 mRNA는 보존함으로써, 핵에서 1차 C9orf72 전사체를 표적으로 하는 안티센스 올리고뉴클레오티드의 사용과 같은 치료 전략으로 발생할 수 있는 C9orf72 단백질의 감소의 가능한 역효과를 피할 수 있다.
본원에 개시된 방법 중 일부는 세포에서 서열번호 1의 다수의 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 억제하기 위한 것이다. C9orf72 표적 RNA는, 예를 들면, 병원성 헥사뉴클레오티드 반복부 확장(예를 들면, 헥사뉴클레오티드 반복부의 적어도 약 30, 적어도 약 35, 적어도 약 40, 적어도 약 50, 적어도 약 60, 적어도 약 70, 적어도 약 80, 적어도 약 100, 적어도 약 200, 적어도 약 300, 적어도 약 400, 또는 적어도 약 500개 카피를 가짐)을 가진 것일 수 있다. 이러한 방법은 본원에 개시된 임의의 dsRNA 제제를 세포 내로 도입함으로써 세포에서 C9orf72 표적 RNA의 발현을 억제함을 포함할 수 있다.
따라서, 본 개시내용은 또한 세포에서 C9orf72 발현을 감소 또는 억제하기 위해 본 개시내용의 RNAi 제제 또는 본 개시내용의 RNAi 제제를 함유하는 조성물을 사용하는 방법을 제공한다. 상기 방법은 세포를 본 개시내용의 dsRNA와 접촉시키는 단계 및 C9orf72 유전자의 mRNA 전사체의 분해를 수득하기에 충분한 시간 동안 세포를 유지함으로써 세포에서 C9orf72 유전자의 발현을 억제하는 단계를 포함한다.
또한, 본 개시내용은 또한 세포에서 센스- 및 안티센스-함유 병소의 수준을 감소시키고/시키거나 형성을 억제하기 위해 본 개시내용의 RNAi 제제 또는 본 개시내용의 RNAi 제제를 함유하는 조성물을 사용하는 방법을 제공한다. 상기 방법은 세포를 본 개시내용의 dsRNA와 접촉시켜, 세포에서 C9orf72 센스- 및 안티센스-함유 병소의 수준을 감소시키는 것을 포함한다.
본 개시내용은 또한 세포에서 비정상적인 디펩티드 반복부 단백질의 수준을 감소시키고/시키거나 형성을 억제하기 위해 본 개시내용의 RNAi 제제 또는 본 개시내용의 RNAi 제제를 함유하는 조성물을 사용하는 방법을 제공한다. 상기 방법은 세포를 본 개시내용의 dsRNA와 접촉시켜 세포에서 비정상적인 디펩티드 반복부 단백질의 수준을 감소시키는 것을 포함한다.
이러한 방법은 세포에서 C9orf72 표적 RNA의 발현을 평가하는 단계 및/또는 세포에서 성숙 C9orf72 mRNA의 발현을 평가하는 단계를 추가로 포함할 수 있다. 평가는, 예를 들면, C9orf72 표적 RNA를 검출하기 위해 역전사 정량적 중합효소 연쇄 반응에 의해 수행될 수 있다. 그러나 임의의 다른 적절한 방법이 사용될 수 있다.
본 개시내용의 방법에서 세포는 시험관내 또는 생체내에서 접촉될 수 있으며, 즉 세포는 대상체 내에 있을 수 있다.
본 개시내용의 방법을 사용한 치료에 적합한 세포는 C9orf72 유전자를 발현하는 임의의 세포 또는 확장된 GGGGCC 반복부를 갖는 C9orf72 유전자를 발현하는 세포일 수 있다. 본 개시내용의 방법에 사용하기에 적합한 세포는 포유동물 세포, 예를 들어, 영장류 세포(예를 들어, 인간 세포 또는 비인간 영장류 세포, 예를 들어, 원숭이 세포 또는 침팬지 세포), 비영장류 세포(예를 들어, 래트 세포 또는 마우스 세포)일 수 있다. 하나의 양태에서, 세포는 인간 세포, 예를 들어, 인간 CNS 세포이다. 일부 양태에서, 세포는 비인간 동물 단세포 단계 배아, 비인간 동물 배아 줄기 세포, 배아 줄기 세포 유래 운동 뉴런, 뇌 세포, 피질 세포, 뉴런 세포, 근육 세포, 심장 세포 또는 생식 세포이다.
일부 양태에서, 세포는 병원성 헥사뉴클레오티드 반복부 확장을 포함하는 C9orf72 유전자좌를 포함할 수 있다. 병원성 헥사뉴클레오티드 반복부 확장은 다음 병리학적 판독 중 하나 또는 둘 다와 관련된 C9orf72 유전자에서 첫 번째로 추정되는 2개의 비암호화 엑손(엑손 1A 및 1B)을 분리하는 개재 서열에서 GGGGCC(서열번호 1)의 다수의 반복으로 구성된 확장이다: (1) 센스 및 안티센스 반복부-함유 RNA는 뉴런 및 기타 세포에서 별개의 병소로서 시각화될 수 있고; (2) 센스 및 안티센스 반복부-함유 RNA로부터의 반복부-관련 비-AUG-의존적 번역에 의해 합성된 디펩티드 반복부 단백질―폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌)―이 세포에서 검출될 수 있다. 반복부의 수는 C9orf72 ALS 또는 C9orf72 FTD를 갖지 않는 사람의 유전자좌에서 일반적으로 볼 수 있는 것보다 더 많은 반복부의 수일 수 있다. 대안적으로, 병원성 헥사뉴클레오티드 반복부 확장은 C9orf72 ALS 또는 C9orf72 FTD를 갖는 대상체로부터의 C9orf72 유전자좌에서의 확장(즉, 반복부의 수)일 수 있다. 병원성 헥사뉴클레오티드 반복부 확장은 GGGGCC(서열번호 1)의 복수의 반복을 갖는다. 예를 들면, 병원성 헥사뉴클레오티드 반복부 확장은, 예를 들면, 헥사뉴클레오티드 반복부의 적어도 약 30, 적어도 약 35, 적어도 약 40, 적어도 약 50, 적어도 약 60, 적어도 약 70, 적어도 약 80, 적어도 약 100, 적어도 약 200, 적어도 약 300, 적어도 약 400, 또는 적어도 약 500개 카피를 가질 수 있다.
세포는, 예를 들면, C9orf72 ALS 또는 C9orf72 FTD를 포함하는 C9orf72-헥사뉴클레오티드-반복부-확장 관련 질환을 갖거나 발병 위험이 있는 대상체로부터의 세포(예를 들어, 뉴런 또는 운동 뉴런)일 수 있다.
본원에 개시된 방법의 세포는 C9orf72 유전자좌를 포함하는 임의의 유형의 세포일 수 있다. C9orf72 유전자좌는 헥사뉴클레오티드 반복부 확장 서열 또는 본원의 다른 곳에 기술된 바와 같은 병원성 헥사뉴클레오티드 반복부 확장 서열을 포함할 수 있다. 헥사뉴클레오티드 반복부 확장 서열은 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 100개 이상의 반복부를 포함할 수 있다.
C9orf72 헥사뉴클레오티드 반복부 확장 서열은 일반적으로 서열번호 1로서 제시된 헥사뉴클레오티드 서열 GGGGCC의 적어도 두 개의 인스턴스(즉, 두 개의 반복부)를 포함하는 뉴클레오티드 서열이다. 일부 헥사뉴클레오티드 반복부 확장 서열에서, 반복부는 연속적이다(개재 서열 없이 서로 인접함). 반복부 확장 서열은, 예를 들면, 내인성 C9orf72 유전자좌의 첫 번째 비암호화 내인성 엑손과 엑손 2 사이에 위치할 수 있다.
헥사뉴클레오티드 반복부 확장 서열은 임의의 수의 반복부를 가질 수 있다. 예를 들면, 반복 확장 서열은 약 95개 이상의 반복부, 96개 이상의 반복부, 약 97개 이상의 반복부, 약 98개 이상의 반복부, 약 99개 이상의 반복부, 약 100개 이상의 반복부, 약 101개 이상의 반복부, 약 102개 이상의 반복부, 약 103개 이상의 반복부, 약 104개 이상의 반복부, 약 105개 이상의 반복부, 약 150개 이상의 반복부, 약 200개 이상의 반복부, 약 250개 이상의 반복부, 약 295개 이상의 반복부, 약 296개 이상의 반복부, 약 297개 이상의 반복부, 약 298개 이상의 반복부, 약 299개 이상의 반복부, 약 300개 이상의 반복부, 약 301개 이상의 반복부, 약 302개 이상의 반복부, 약 303개 이상의 반복부, 약 304개 이상의 반복부, 약 305개 이상의 반복부, 약 350개 이상의 반복부, 약 400개 이상의 반복부, 약 450개 이상의 반복부, 약 500개 이상의 반복부, 약 550개 이상의 반복부, 약 595개 이상의 반복부, 약 596개 이상의 반복부, 약 597개 이상의 반복부, 약 598개 이상의 반복부, 약 599개 이상의 반복부, 약 600개 이상의 반복부, 약 601개 이상의 반복부, 약 602개 이상의 반복부, 약 603개 이상의 반복부, 약 604개 이상의 반복부, 또는 약 605개 이상의 반복부를 포함할 수 있다. 대안적으로, 반복부 확장 서열은 적어도 약 95개 반복부, 적어도 약 96개 반복부, 적어도 약 97개 반복부, 적어도 약 98개 반복부, 적어도 약 99개 반복부, 적어도 약 100개 반복부, 적어도 약 101개 반복부, 적어도 약 102개 반복부, 적어도 약 103개 반복부, 적어도 약 104개 반복부, 적어도 약 105개 반복부, 적어도 약 150개 반복부, 적어도 약 200개 반복부, 적어도 약 250개 반복부, 적어도 약 295개 반복부, 적어도 약 296개 반복부, 적어도 약 297개 반복부, 적어도 약 298개 반복부, 적어도 약 299개 반복부, 적어도 약 300개 반복부, 적어도 약 301개 반복부, 적어도 약 302개 반복부, 적어도 약 303개 반복부, 적어도 약 304개 반복부, 적어도 약 305개 반복부, 적어도 약 350개 반복부, 적어도 약 400개 반복부, 적어도 약 450개 반복부, 적어도 약 500개 반복부, 적어도 약 550개 반복부, 적어도 약 595개 반복부, 적어도 약 596개 반복부, 적어도 약 597개 반복부, 적어도 약 598개 반복부, 적어도 약 599개 반복부, 적어도 약 600개 반복부, 적어도 약 601개 반복부, 적어도 약 602개 반복부, 적어도 약 603개 반복부, 적어도 약 604개 반복부, 또는 적어도 약 605개 반복부를 포함할 수 있다. 특정 예에서, 헥사뉴클레오티드 반복부 확장 서열은 약 100개 이상의 반복부, 약 300개 이상의 반복부, 약 600개 이상의 반복부, 적어도 약 100개 반복부, 적어도 약 300개 반복부, 또는 적어도 약 600개 반복부를 포함한다.
비-인간 세포인 세포에서, 반복부 확장 서열은 이종 반복부 확장 서열일 수 있다. 대안적으로, 반복부 확장 서열은 내인성 반복부 확장 서열일 수 있다. 핵산과 관련하여 사용될 때 이종이라는 용어는 핵산이 동일한 분자에서 자연적으로 함께 발생하지 않는 적어도 2개의 분절을 포함함을 나타낸다. 예를 들면, 핵산의 분절과 관련하여 사용될 때 이종이라는 용어는 핵산이 자연에서 서로 동일한 관계(예를 들어, 함께 연결된)에서 발견되지 않는 2개 이상의 하위서열을 포함함을 나타낸다. 하나의 예로서, 핵산의 이종 영역은 자연에서 다른 분자와 관련하여 발견되지 않는 또 다른 핵산 분자 내에 있거나 이에 부착된 핵산의 분절이다. 예를 들면, 핵산의 이종성 영역은 자연에서 C9orf72 서열과 관련하여 발견되지 않는 반복부 확장 서열이 측면에 있는 C9orf72 서열을 포함할 수 있다. 대안적으로, 핵산의 이종 영역은 내인성 비인간 핵산 서열이 측면에 있는 인간 핵산 서열을 포함할 수 있다.
일부 예에서, 변형된 내인성 비-인간 C9orf72 유전자좌는 또한 추가의 이종상동성 C9orf72 서열을 포함할 수 있다. 한 예로서, 비-인간 C9orf72 유전자좌는 인간 C9orf72 뉴클레오티드 서열을 포함할 수 있다. 예를 들면, 비-인간 동물 C9orf72 유전자좌는 내인성 C9orf72 유전자좌의 5' 비번역 및/또는 비-코딩 내인성 비-인간 서열의 헥사뉴클레오티드 반복부 확장 서열 및 측면 이종상동성 인간 C9orf72 서열(즉, 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 반복부가 측면에 있는 이종(예를 들어, 인간) 서열)로의 대체를 포함할 수 있다. 이종 반복부 확장 서열 및/또는 측면 서열은 자연 발생 게놈 서열(예를 들어, 자연 발생 인간 게놈 서열)일 수 있다. 구체적인 예로서, 내인성 엑손 1(예를 들어, 엑손 1A 및/또는 1B)과 내인성 비인간 C9orf72 유전자좌의 ATG 시작 코돈 또는 이의 일부 사이에 걸쳐 있는(그리고 임의로 이의 적어도 일부를 포함하는) 비번역된 및/또는 비코딩 서열은 이종 헥사뉴클레오티드 반복부 확장 서열로 대체될 수 있고, 임의로 이종상동 인간 C9orf72 서열과 함께 측면에 있을 수 있다. 예를 들면, 내인성 C9orf72 유전자좌에 삽입된 서열은 5' 내지 3': 제1 이종 헥사뉴클레오티드 측면 서열, 서열번호 1에 제시된 헥사뉴클레오티드의 복수 반복부, 및 제2 이종 헥사뉴클레오티드 측면 서열을 포함할 수 있다.
하나의 양태에서, 인간 C9orf72 유전자의 엑손 1A 및/또는 엑손 1B의 전부 또는 일부에 걸쳐 있는 (및 임의로 포함하는) 이종 인간 C9orf72 서열이 내인성 C9orf72 유전자좌에 삽입된다. 예를 들면, 제1 이종 헥사뉴클레오티드 측면 서열은 서열번호 46으로 제시된 서열 또는 이의 일부를 포함할 수 있고/있거나 제2 이종 헥사뉴클레오티드 측면 서열은 서열번호 47로 제시된 서열 또는 이의 일부를 포함할 수 있다. 따라서, 일부 예에서, 이종 헥사뉴클레오티드 반복부 확장 서열이 삽입되는 내인성 C9orf72 유전자좌는 서열번호 46 및/또는 서열번호 47에 제시된 서열을 포함하는 인간 서열을 포함할 수 있다.
또 다른 양태 예에서, 내인성 C9orf72 유전자좌는 마우스 C9orf72 유전자좌의 비-코딩 서열의 일부를 마우스 C9orf72 프로모터 및/또는 인간 조절 요소(예를 들어, 인간 C9orf72 유전자의 엑손 1A 및/또는 1B에서 발견될 수 있는 것)를 갖는 작동 가능한 결합에 배치된 이종 인간 헥사뉴클레오티드 반복부 확장 서열로 대체하는 것을 포함한다. 제US 2018/0094267호 및 제WO 2018/064600호를 참조하며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다. 내인성 비인간 C9orf72 유전자좌에 삽입할 목적으로, 이종 헥사뉴클레오티드 반복부 확장 서열은 서열번호 1로서 제시된 복수의(즉, 적어도 두 개의 인스턴스(반복부))의 헥사뉴클레오티드 서열을 포함할 수 있고 인간 염색체 9 개방 판독 프레임 72(C9orf72)의 비 코딩 엑손 1A 및 1B, 또는 이의 일부에 걸쳐 있는(및 임의로 포함하는) 게놈 핵산 서열과 동일하거나 실질적으로 동일할 수 있다.
세포는 시험관내, 생체외 또는 생체내일 수 있다. 예를 들면, 세포는 동물 내에서 생체내일 수 있다. 세포 또는 동물은 수컷 또는 암컷일 수 있다. 세포 또는 동물은 내인성 C9orf72 유전자좌에 삽입된 헥사뉴클레오티드 반복부 확장 서열에 대해 이형접합성 또는 동형접합성일 수 있다. 이배체 유기체는 각 유전자 좌에 두 개의 대립유전자를 갖는다. 대립유전자의 각 쌍은 특정 유전자 좌의 유전자형을 나타낸다. 유전자형은 특정 유전자좌에 2개의 동일한 대립유전자가 있으면 동형접합으로, 두 개의 대립유전자가 상이하면 이형접합으로 기술된다. 비-인간 동물은 이들의 생식계열 게놈의 내인성 C9orf72 유전자좌에 삽입된 헥사뉴클레오티드 반복부 확장 서열을 포함할 수 있다.
C9orf72 발현(예를 들어, 센스 mRNA, 안티센스 mRNA, 전체 C9orf72 mRNA, 센스 C9orf72 반복부-함유 mRNA, 안티센스 C9orf72 반복부-함유 mRNA 수준, 전체 C9orf72 단백질, 및/또는 C9orf72 반복부-함유 단백질에 의해 평가됨)은 세포에서 약 20, 25, 30, 35, 40, 45, 또는 50%까지 억제된다. 바람직한 양태에서, C9orf72 발현은 50% 이하, 예를 들어, 50%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 또는 5% 이하까지 억제된다.
C9orf72 표적 RNA의 발현 감소는 어느 정도일 수 있다. 센스- 또는 안티센스-함유 병소 및/또는 비정상적인 디펩티드 반복부 단백질 수준에 의해 평가되는 억제는 세포에서 적어도 20%, 30%, 40%, 바람직하게는 적어도 50%, 60%, 70%, 80%, 85%, 90% 또는 95%, 또는 분석의 검출 수준 이하로 억제된다.
일부 양태에서, dsRNA 제제는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA와 같은 C9orf72 표적 RNA의 발현을 적어도 약 10%, 적어도 약 20%, 적어도 약 30%, 적어도 약 40%, 적어도 약 50%, 적어도 약 60%, 적어도 약 70%, 적어도 약 80%, 또는 적어도 약 90%(또는 C9orf72 표적 RNA가 검출되지 않는 지점까지) 억제할 수 있다. 예를 들면, 이러한 억제 수준은 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 약 1일 이내, 약 2일 이내, 약 3일 이내, 약 4일 이내, 약 5일 이내, 약 6일 이내, 약 일주일 이내, 또는 약 24 내지 약 48시간 이내일 수 있다. 감소는, 예를 들면, dsRNA 제제로 처리하기 전의 세포를 기준으로 하거나 dsRNA 제제로 처리되지 않은 대조군 세포를 기준으로 할 수 있다.
방법의 일부에서, 본 발명의 dsRNA 제제는 성숙 C9orf72 메신저 RNA의 발현에 비해 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA와 같은 C9orf72 표적 RNA의 발현을 선택적으로 억제한다. 이러한 맥락에서 성숙 C9orf72 메신저 RNA는 스플라이싱되고 처리된 C9orf72 RNA 전사체이다. 성숙 C9orf72 메신저 RNA는 엑손으로만 구성되며 모든 인트론이 제거된다. dsRNA 제제는 C9orf72 표적 RNA의 발현의 상대적 감소가 dsRNA 제제를 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 성숙 C9orf72 메신저 RNA의 발현의 상대적 감소보다 크다면 성숙 C9orf72 메신저 RNA의 발현에 비해 인트론 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현을 선택적으로 억제한다. 예를 들면, 특정 양태에서, 본 발명의 dsRNA 제제는 성숙 C9orf72 메신저 RNA의 발현을 약 50% 미만, 약 40% 미만, 약 30% 미만, 약 20% 미만, 약 10% 미만, 또는 약 5% 미만까지 억제한다(또는, 예를 들면, 발현에 대해 통계적으로 유의하거나 기능적으로 유의한 영향을 미치지 않는다). 예를 들면, 이러한 억제 수준은 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 약 1일 이내, 약 2일 이내, 약 3일 이내, 약 4일 이내, 약 5일 이내, 약 6일 이내, 약 일주일 이내, 또는 약 24 내지 약 48시간 이내일 수 있다. 감소는, 예를 들면, dsRNA 제제로 처리하기 전의 세포를 기준으로 하거나 dsRNA 제제로 처리되지 않은 대조군 세포를 기준으로 할 수 있다.
본원에 개시된 방법 중 일부는 세포에서 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집체를 감소시키기 위한 것이다. 이러한 방법은 본원에 개시된 임의의 dsTNA 제제를 세포 내로 도입함으로써 세포에서 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집체를 감소시키는 것을 포함할 수 있다.
이러한 방법은 세포에서 디펩티드 반복부 단백질 응집체(예를 들어, 폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌))의 존재를 평가함을 추가로 포함할 수 있다. 특정 예에서, 디펩티드 반복부 단백질은 폴리(글리신-알라닌) 및/또는 폴리(글리신-프롤린)일 수 있다. 평가는, 예를 들면, 디펩티드 반복부 단백질 응집체를 검출하기 위한 면역조직화학 또는 웨스턴 블롯 분석에 의해 수행될 수 있다. 그러나, 임의의 다른 적절한 방법이 사용될 수 있다.
디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집체의 감소는 어느 정도일 수 있다. 예를 들면, dsRNA 제제는 디펩티드 반복부 단백질 합성 또는 디펩티드 반복부 단백질 응집체를 적어도 약 10%, 적어도 약 20%, 적어도 약 30%, 적어도 약 40%, 적어도 약 50%, 적어도 약 60%, 적어도 약 70%, 적어도 약 80%, 또는 적어도 약 90% (또는 디펩티드 반복부 단백질 응집체가 검출되지 않는 지점까지) 감소시킬 수 있다. 예를 들면, 이러한 억제 수준은 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 약 1일 이내, 약 2일 이내, 약 3일 이내, 약 4일 이내, 약 5일 이내, 약 6일 이내, 약 일주일 이내, 또는 약 24 내지 약 48시간 이내일 수 있다. 감소는, 예를 들면, dsRNA 제제로 처리하기 전의 세포를 기준으로 하거나 dsRNA 제제로 처리되지 않은 대조군 세포를 기준으로 할 수 있다.
이러한 방법은 세포에서 핵 및/또는 세포질 센스 및/또는 안티센스 C9orf72 RNA 병소의 존재를 평가함을 추가로 포함할 수 있다.
핵 및/또는 세포질 센스 및/또는 안티센스 C9orf72 RNA 병소의 존재의 감소는 어느 정도일 수 있다. 예를 들면, dsRNA 제제는 핵 및/또는 세포질 센스 및/또는 안티센스 C9orf72 RNA 병소의 존재를 적어도 약 10%, 적어도 약 20%, 적어도 약 30%, 적어도 약 40%, 적어도 약 50%, 적어도 약 60%, 적어도 약 70%, 적어도 약 80%, 또는 적어도 약 90% (또는 핵 및/또는 세포질 센스 및/또는 안티센스 C9orf72 RNA 병소가 검출되지 않는 지점까지) 감소시킬 수 있다. 예를 들면, 이러한 억제 수준은 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 발현하는 세포에 투여한 후 약 1일 이내, 약 2일 이내, 약 3일 이내, 약 4일 이내, 약 5일 이내, 약 6일 이내, 약 일주일 이내, 또는 약 24 내지 약 48시간 이내일 수 있다. 감소는, 예를 들면, dsRNA 제제로 처리하기 전의 세포를 기준으로 하거나 dsRNA 제제로 처리되지 않은 대조군 세포를 기준으로 할 수 있다.
본 개시내용의 생체내 방법은 RNAi 제제를 함유하는 조성물을 대상체에게 투여함을 포함할 수 있으며, 여기서 RNAi 제제는 치료될 포유동물의 C9orf72 유전자의 RNA 전사체의 적어도 일부에 상보적인 뉴클레오티드 서열을 포함한다. 치료될 유기체가 인간과 같은 포유동물인 경우, 조성물은 두개내(예를 들어, 뇌실내, 실질내 및 척수강내), 정맥내, 근육내, 유리체내, 피하, 경피, 기도(에어로졸), 비강, 직장 및 국소(협측 및 설하 포함) 투여를 포함한 경구, 복강내, 또는 비경구 경로를 포함하지만 이에 제한되지 않는 당업계에 알려진 임의의 수단에 의해 투여될 수 있다. 특정 양태에서, 조성물은 정맥내 주입 또는 주사에 의해 투여된다. 특정 양태에서, 조성물은 피하 주사에 의해 투여된다. 특정 양태에서, 조성물은 척수강내 주사에 의해 투여된다.
일부 양태에서, 투여는 데포트 주사를 통해 이루어진다. 데포트 주사는 장기간에 걸쳐 일관된 방식으로 RNAi 제제를 방출할 수 있다. 따라서, 데포트 주사는 원하는 효과, 예를 들어, C9orf72의 원하는 억제, 또는 치료적 또는 예방적 효과를 수득하기 위해 필요한 투여 빈도를 감소시킬 수 있다. 데포트 주사는 또한 보다 일관된 혈청 농도를 제공할 수 있다. 데포트 주사는 피하 주사 또는 근육내 주사를 포함할 수 있다. 바람직한 양태에서, 데포트 주사는 피하 주사이다.
일부 양태에서, 투여는 펌프를 통해 이루어진다. 펌프는 외부 펌프 또는 외과적으로 이식된 펌프일 수 있다. 특정 양태에서, 펌프는 피하 이식된 삼투압 펌프이다. 다른 양태에서, 펌프는 주입 펌프이다. 주입 펌프는 두개내, 정맥내, 피하, 동맥 또는 경막외 주입에 사용될 수 있다. 바람직한 양태에서, 주입 펌프는 피하 주입 펌프이다. 다른 양태에서, 펌프는 RNAi 제제를 CNS에 전달하는 외과적으로 이식된 펌프이다.
투여 방식은 국소 치료 또는 전신 치료가 필요한지 여부 및 치료할 부위에 기반하여 선택될 수 있다. 투여 경로 및 부위는 표적화를 향상시키기 위해 선택될 수 있다.
하나의 측면에서, 본 개시내용은 또한 포유동물에서 C9orf72 유전자의 발현을 억제하는 방법을 제공한다. 상기 방법은 포유동물의 세포에서 C9orf72 유전자를 표적으로 하는 dsRNA를 포함하는 조성물을 포유동물에게 투여하여 세포에서 C9orf72 유전자의 발현을 억제하는 것을 포함한다. 유전자 발현의 감소는 당업계에 공지된 임의의 방법에 의해 및 본원에 기술된 방법, 예를 들어, qRT-PCR에 의해 평가될 수 있다. 단백질 생산의 감소는 당업계에 공지된 임의의 방법에 의해 및 본원에 기술된 방법, 예를 들어, ELISA에 의해 평가될 수 있다. 하나의 양태에서, CNS 생검 샘플 또는 뇌척수액(CSF) 샘플은 C9orf72 유전자 또는 단백질 발현(또는 따라서 프록시)의 감소를 모니터링하기 위한 조직 재료로서 역할을 한다.
본 개시내용은 이를 필요로 하는 대상체의 치료 방법을 추가로 제공한다. 본 개시내용의 치료 방법은 본 개시내용의 RNAi 제제를 대상체, 예를 들어, C9orf72 발현의 억제로부터 혜택을 받는 대상체, 예를 들어 C9orf72 유전자의 인트론에 GGGGCC 확장된 뉴클레오티드 반복부를 갖는 대상체에게 C9orf72 유전자를 표적으로 하는 RNAi 제제 또는 aC9orf72 유전자를 표적으로 하는 RNAi 제제를 포함하는 약제학적 조성물의 치료학적 유효량으로 투여함을 포함한다.
또한, 본 개시내용은 대상체에서 C9orf72-관련 질환 또는 장애(예를 들어, C9orf72-관련 장애)의 진행을 예방, 치료 또는 억제하는 방법을 제공한다. 상기 방법은 치료학적 유효량의 임의의 RNAi 제제, 예를 들어, dsRNA 제제, 또는 본원에 제공된 약제학적 조성물을 대상체에게 투여함으로써, 대상체에서 C9orf72-관련 질환 또는 장애의 진행을 예방, 치료 또는 억제하는 것을 포함한다.
일부 양태에서, 방법은 C9orf72-헥사뉴클레오티드-반복부-확장-관련 질환, 상태 또는 장애를 앓고 있는 대상체를 치료하기 위한 것이다. 이러한 방법은 또한 서열번호 1의 다수의 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA의 발현 감소로부터 혜택을 받는 질환, 장애 또는 상태를 갖는 대상체(예를 들어, C9orf72-헥사뉴클레오티드-반복부-확장-관련 질환, 상태 또는 장애를 갖거나 발병 위험이 있는 대상체)에서 적어도 하나의 증상을 예방하거나 개선하기 위한 것일 수 있다. C9orf72 표적 RNA는, 예를 들면, (예를 들면, 헥사뉴클레오티드 반복부의 적어도 약 30, 적어도 약 35, 적어도 약 40, 적어도 약 50, 적어도 약 60, 적어도 약 70, 적어도 약 80, 적어도 약 100, 적어도 약 200, 적어도 약 300, 적어도 약 400, 또는 적어도 약 500개 카피를 갖는) 병원성 헥사뉴클레오티드 반복부 확장을 갖는 것일 수 있다. C9orf72-헥사뉴클레오티드-반복부-확장-관련 질환, 상태 또는 장애는 C9orf72 유전자의 5' 비-코딩 부분에 있는 헥사뉴클레오티드 반복부 (GGGGCC; 서열번호 1)의 확장에 의해 유발되거나 이와 관련된 것이다. 예는 근위축성 측삭 경화증(ALS) 및 전두측두엽 치매(FTD)를 포함한다. FTD 및/또는 ALS와 관련된 징후 또는 증상은 RNA 병소의 반복부-길이-의존적 형성, 특정 RNA-결합 단백질의 격리, 및 뉴런에서의 반복부-관련 비-AUG (AUG) 번역으로부터 야기되는 디펩티드 반복부 단백질(예를 들어, 폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌))의 축적 및 응집을 포함하지만, 이에 제한되지 않는다. 본 발명의 dsRNA 제제는 운동 뉴런 질환의 징후 및 증상 및 치매의 징후 및 증상을 포함하지만 이에 제한되지 않는 FTD 및/또는 ALS와 관련된 징후 또는 증상의 치료적 치료 및/또는 예방을 위한 방법에 사용될 수 있다. 운동 뉴런 질환의 징후 및 증상은, 예를 들면, 넘어짐, 물건 떨어뜨림, 팔 및/또는 다리의 비정상적인 피로, 불분명한 발음, 근육 경축 및 경련, 제어할 수 없는 웃음 또는 울음 시간, 및 호흡 곤란을 포함할 수 있다. 치매의 징후 및 증상은, 예를 들면, 행동 변화, 성격 변화, 발음 및 언어 문제, 및 운동-관련 문제를 포함할 수 있다.
본 개시내용의 RNAi 제제는 "유리 RNAi 제제"로서 투여될 수 있다. 유리 RNAi 제제는 약제학적 조성물의 부재하에 투여된다. 네이키드 RNAi 제제는 적절한 완충 용액에 있을 수 있다. 완충 용액은 아세테이트, 시트레이트, 프롤라민, 카보네이트 또는 포스페이트, 또는 이들의 임의의 조합을 포함할 수 있다. 하나의 양태에서, 완충 용액은 인산염 완충 염수(PBS)이다. RNAi 제제를 함유하는 완충 용액의 pH 및 삼투압은 대상체에게 투여하기에 적합하도록 조정될 수 있다.
대안적으로, 본 개시내용의 RNAi 제제는 dsRNA 리포솜 제형과 같은 약제학적 조성물로서 투여될 수 있다.
C9orf72 유전자 발현의 감소 또는 억제로부터 헤택을 받는 대상체는 C9orf72-관련 질환, 예를 들어, C9orf72-관련 질환을 앓고 있는 대상체이다. 예시적인 C9orf72-관련 질환은 ALS, FTD, C9ALS/FTD 및 C9orf72 확장으로 인한 헌팅턴-유사 증후군, 예를 들어, C9orf72 유전자의 인트론에 확장된 GGGGCC 헥사뉴클레오티드 반복부를 갖는 대상체를 포함하지만, 이에 제한되지 않는다.
본 개시내용은, 예를 들어, C9orf72 발현의 감소 또는 억제로부터 혜택을 받는 대상체, 예를 들어, C9orf72-관련 장애를 갖는 대상체를 치료하기 위해 다른 약제 또는 다른 치료적 방법, 예를 들어, 공지된 약제 또는 공지된 치료적 방법, 예를 들면, 이러한 질환을 치료하기 위해 현재 사용되는 것들과 조합하여 RNAi 제제 또는 이의 약제학적 조성물을 사용하기 위한 방법을 추가로 제공한다. 예를 들면, 특정 양태에서, C9orf72를 표적으로 하는 RNAi 제제는 본원의 다른 곳에 기술된 바와 같은 또는 당업계에 달리 공지된 바와 같은 C9orf72-관련 장애의 치료에 유용한 제제와 조합하여 투여된다. 예를 들면, C9orf72 발현의 감소로부터 혜택을 받는 대상체, 예를 들어, C9orf72-관련 장애를 갖는 대상체를 치료하는데 적합한 추가 제제는 C9orf72-관련 질환의 증상을 치료하기 위해 현재 사용되는 제제를 포함할 수 있다. RNAi 제제 및 추가 치료제는 동시에 또는 동일한 조합으로, 예를 들어, 척수강내로 투여될 수 있거나, 추가 치료제는 별도의 조성물의 일부로서 또는 별도의 시간에 또는 당업계에 공지되거나 본원에 기술된 다른 방법에 의해 투여될 수 있다.
예시적인 추가 치료제는, 예를 들면, 모노아민 억제제, 예를 들어, 테트라베나진(Xenazine), 듀테트라베나진(Austedo), 및 레세르핀, 항경련제, 예를 들어, 발프로산(Depakote, Depakene, Depacon), 및 클로나제팜(Klonopin), 항정신병제, 예를 들어, 리스페리돈(Risperdal) 및 할로페리돌(Haldol), 및 항우울제, 예를 들어, 파록세틴(Paxil)을 포함한다.
하나의 양태에서, 상기 방법은 표적 C9orf72 유전자의 발현이 적어도 1개월 동안 감소되도록 본원에 특성화된 조성물을 투여함을 포함한다. 바람직한 양태에서, 발현은 적어도 2개월, 3개월, 6개월 동안 감소된다.
바람직하게는, 본원에 특성화된 방법 및 조성물에 유용한 RNAi 제제는 표적 C9orf72 유전자의 (일차 또는 프로세싱된) RNA를 특이적으로 표적화한다. RNAi 제제를 사용하여 이들 유전자의 발현을 억제하기 위한 조성물 및 방법은 본원에 기술된 바와 같이 제조 및 수행될 수 있다.
본 개시내용의 방법에 따른 dsRNA의 투여는 C9orf72-관련 장애를 갖는 환자에서 이러한 질환 또는 장애의 중증도, 징후, 증상 또는 마커의 감소를 초래할 수 있다. 이러한 맥락에서 "감소"는 이러한 수준의 통계적으로 유의하거나 임상적으로 유의한 감소를 의미한다. 감소는, 예를 들면, 적어도 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 또는 약 100%일 수 있다.
질환의 치료 또는 예방의 효능은, 예를 들면, 질환 진행, 질환 완화, 증상 중증도, 통증 감소, 삶의 질, 치료 효과를 유지하는데 필요한 약물의 투여량, 질환 마커의 수준 또는 예방을 위해 치료되거나 표적이 되는 주어진 질환에 적절한 다른 측정 가능한 매개변수를 측정함으로써 평가될 수 있다. 이러한 매개변수 중 어느 하나 또는 매개변수의 임의의 조합을 측정함으로써 치료 또는 예방의 효능을 모니터링하는 것은 당업계의 숙련가의 능력 범위 내에 있다. 예를 들면, C9orf72-관련 장애의 치료 효능은, 예를 들어 대상체의 주기적인 모니터링에 의해 평가될 수 있다. 이후 판독값과 초기 판독값의 비교는 치료가 효과적인지 여부의 표시를 의사에게 제공한다. 이러한 매개변수 중 어느 하나 또는 매개변수의 임의의 조합을 측정함으로써 치료 또는 예방의 효능을 모니터링하는 것은 당업계의 숙련가의 능력 범위 내에 있다. C9orf72를 표적으로 하는 RNAi 제제 또는 이의 약제학적 조성물의 투여와 관련하여, C9orf72-관련 장애"에 대해 "효과적"이라는 것은 임상적으로 적절한 방식으로의 투여가 증상의 개선, 치료, 질환의 감소, 수명 연장, 삶의 질 개선 또는 C9orf72-관련 장애 및 관련 원인 치료에 정통한 의사가 일반적으로 긍정적으로 인정하는 기타 효과와 같은, 환자의 적어도 통계적으로 유의한 부분에 대해 유익한 효과를 초래한다는 것을 나타낸다.
치료 또는 예방 효과는 질환 상태의 하나 이상의 매개변수에서 통계적으로 유의한 개선이 있을 때, 또는 달리 예상할 수 있는 증상을 악화시키거나 발병하지 않음으로써 명백하다. 예를 들자면, 질환의 측정 가능한 매개변수에서 적어도 10%, 바람직하게는 적어도 20%, 30%, 40%, 50% 또는 그 이상의 유리한 변화가 효과적인 치료를 나타낼 수 있다. 주어진 RNAi 제제 약물 또는 그 약물의 제형에 대한 효능은 또한 당업계에 공지된 주어진 질환에 대한 실험 동물 모델을 사용하여 판단할 수 있다. 실험 동물 모델을 사용하는 경우, 마커 또는 증상의 통계적으로 유의한 감소가 관찰될 때 치료의 효능이 입증된다.
대안적으로, 효능은 임상적으로 허용되는 질환 중증도 등급 척도에 기초하여 진단 분야의 숙련자에 의해 결정된 바와 같이 질환의 중증도의 감소에 의해 측정될 수 있다. 예를 들어, 적절한 척도를 사용하여 측정된 질환의 중증도의 감소를 초래하는 임의의 긍정적인 변화는 본원에 기술된 바와 같은 RNAi 제제 또는 RNAi 제제 제형을 사용한 적절한 치료를 나타낸다.
대상체는 약 0.01mg/kg 내지 약 200mg/kg과 같은 치료량의 dsRNA를 투여받을 수 있다.
RNAi 제제는 일정 기간에 걸쳐 정기적으로, 척수강내, 유리체내 주사 또는 정맥내 주입을 통해 투여될 수 있다. 특정 양태에서, 초기 치료 섭생 후에, 치료를 덜 빈번하게 투여할 수 있다. RNAi 제제의 투여는, 예를 들어, 세포, 조직, 혈액, CSF 샘플 또는 환자의 다른 구획에서 C9orf72 수준을 감소시킬 수 있다. 하나의 양태에서, RNAi 제제의 투여는, 예를 들어, 세포, 조직, 혈액, CSF 샘플 또는 환자의 다른 구획에서 C9orf72 수준을 50% 이하까지 감소시킬 수 있다.
전체 용량의 RNAi 제제를 투여하기 전에, 환자는 5% 주입 반응과 같은 더 적은 용량을 투여받고 알레르기 반응과 같은 부작용에 대해 모니터링될 수 있다. 또 다른 예에서, 환자는 증가된 사이토카인(예를 들어, TNF-알파 또는 INF-알파) 수준과 같은 원치 않는 면역자극 효과에 대해 모니터링될 수 있다.
대안적으로, RNAi 제제는 피하, 즉 피하 주사에 의해 투여될 수 있다. 1회 이상의 주사를 사용하여 원하는, 예를 들어 월 용량의 RNAi 제제를 대상체에게 전달할 수 있다. 주사는 일정 기간에 걸쳐 반복될 수 있다. 투여는 정기적으로 반복될 수 있다. 특정 양태에서, 초기 치료 섭생 후, 치료는 덜 빈번하게 투여될 수 있다. 반복-용량 섭생은 매월 또는 분기 1회로 연장하여, 1년에 2회, 1년에 1회와 같이 정기적으로 치료량의 RNAi 제제의 투여를 포함할 수 있다. 특정 양태에서, RNAi 제제는 약 월 1회 또는 약 분기에 1회(즉, 약 3개월에 1회) 투여된다.
달리 정의되지 않는 한, 본원에 사용된 모든 기술 용어 및 과학 용어는 본 발명이 속하는 기술 분야의 통상의 숙련가에 의해 일반적으로 이해하는 것과 동일한 의미를 갖는다. 본원에 기술된 것과 유사하거나 동등한 방법 및 재료가 본 발명에 특성화된 RNAi 제제 및 방법의 실시 또는 시험에 사용될 수 있지만, 적합한 방법 및 재료는 하기에 기술되어 있다. 본원에 언급된 모든 간행물, 특허 출원, 특허 및 기타 참고 문헌은 전문이 본원에 참고로 포함된다. 충돌하는 경우, 정의를 포함한 본 명세서가 우선할 것이다. 또한, 재료, 방법 및 예는 예시일 뿐이며 제한하려는 의도가 아니다.
서열 목록이 함께 제출되며 제출된 바와 같은 명세서의 일부를 형성한다.
첨부된 서열 목록에 나열된 뉴클레오티드 및 아미노산 서열은 뉴클레오티드 염기의 경우 표준 문자 약어를 사용하고 아미노산의 경우 3자리 코드를 사용하여 표시된다. 뉴클레오티드 서열은 서열의 5' 말단에서 시작하여 3' 말단까지 앞으로(즉, 각 줄의 왼쪽에서 오른쪽으로) 진행하는 표준 관례를 따른다. 각 뉴클레오티드 서열의 한 가닥이 표시되지만, 표시된 가닥을 참조하여 상보적 가닥이 포함되는 것으로 이해된다. 아미노산 서열을 암호화하는 뉴클레오티드 서열이 제공되는 경우, 동일한 아미노산 서열을 암호화하는 이의 코돈 축퇴 변이체도 제공되는 것으로 이해된다. 아미노산 서열은 서열의 아미노 말단에서 시작하여 카복시 말단까지 앞으로(즉, 각 줄의 왼쪽에서 오른쪽으로) 진행하는 표준 관례를 따른다. 하기 표 3은 본원에 개시된 특정 서열에 대한 간략한 설명을 제공한다.




실시예
실시예
1.
C9orf72
유전자좌에서의
헥사뉴클레오티드
반복부
확장
I. 개요
근위축성 측삭 경화증(ALS) 및 전두측두엽 치매(FTD)는 ALS의 경우 운동 뉴런 질환을, FTD의 경우 치매를 유발하는 치명적인 신경퇴행성 질환이다. 둘 다는 항상 치명적이다. ALS 및 FTD는 자발적 또는 가족성(즉, 유전적) 질환으로 나타날 수 있다. ALS와 FTD의 가장 흔한 유전적 원인은 기능이 완전히 이해되지 않은 단백질을 암호화하는 C9orf72 유전자의 5' 비암호화 부분에 있는 헥사뉴클레오티드 반복부 (GGGGCC; 서열번호 1)의 확장이다. 영향을 받지 않은 사람들은 보통 이들의 C9orf72 유전자에 수 개 내지 수십 개의 헥사뉴클레오티드 반복부를 갖는 반면, ALS 및 FTD가 발병한 사람들은 부모 중 한 명에게서만 헥사뉴클레오티드 반복부의 수백 내지 수천 개의 카피의 반복 확장을 물려받는다. 유전적 관찰은 C9orf72 ALS 및 FTD가 우성 유전 질환이며 병리학적 기능의 획득으로 인한 것임을 시사한다.
어떻게 C9orf72 헥사뉴클레오티드 반복부 확장이 운동 뉴런 질환 및 치매를 일으키는지는 알려져 있지 않지만, C9orf72 ALS 및 FTLD 환자에서의 두 가지 보편적인 사후 병리학적 소견이 반복부 확장과 관련이 있다: (1) 센스 및 안티센스 반복부-함유 RNA는 형광 제자리 혼성화에 의해 뉴런 및 기타 세포에서 별개의 병소로 시각화될 수 있고; (2) 센스 및 안티센스 반복부-함유 RNA로부터 반복부-관련 비-AUG-의존성 번역에 의해 합성된 디펩티드 반복부 단백질―폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌)―이 면역조직화학에 의해 세포에서 검출될 수 있다. 질병 가설 중 하나에서는 병소로서 시각화된 반복체-함유 RNA가 RNA 결합 단백질을 격리함으로써 세포 RNA 대사를 방해한다고 제안한다. 두 번째 질병 가설은 디펩티드 반복부 단백질이 RNA 대사, 단백질 항상성 및 핵세포질 수송에 광범위한 독성 영향을 미친다고 상정한다. 두 병원성 메커니즘 모두 질병에 기여할 수 있다. C9orf72 반복부-함유 RNA 전사체가 그 자체로 또는 디펩티드 반복부 단백질의 번역을 위한 주형으로서 ALS 및 FTLD에서 발병을 촉진하는 경우, 일반적인 치료 전략은 GGGGCC 반복부-함유 RNA를 파괴하거나 디펩티드 반복부 단백질로 번역되는 이들의 능력을 폐지하는 것일 것이다.
C9orf72 유전자는 2개의 전사 개시 부위로부터 전사체를 생성한다. 상류 부위는 대체 비-코딩 엑손 1A로 전사를 개시하는 반면 하류 부위는 대체 엑손 1B로 전사를 개시한다. 엑손 1A 및 1B는 모두 단백질-코딩 서열의 시작을 포함하는 엑손 2에 스플라이싱될 수 있다. 병원성 헥사뉴클레오티드 반복부 확장은 엑손 1A와 1B 사이에 위치한다. 따라서, 엑손 1A로부터 개시된 전사는 반복부-함유 RNA를 생성할 수 있는 반면, 엑손 1B로부터 개시된 전사는 그렇게 할 수 없다.
마우스에서 C9orf72 반복부 확장 질환을 모델링하기 위해, 아래에 기술된 바와 같이, 마우스 배아 줄기(ES) 세포에서 대립유전자 시리즈를 작제하였으며, 여기서 엑손 1A의 일부, 1A와 1B 사이의 인트론 서열, 모든 엑손 1B 및 하류 인트론의 일부를 포함하는 인간 C9orf72 유전자의 단편을 마우스 C9orf72 유전자의 한 대립유전자의 상동 위치에 정확하게 배치하였다. 예를 들어, 제US 2018/0094267호 및 제WO 2018/064600호를 참조하며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다. 정상적인 3개의 반복부 내지 병리학적인 600개의 반복부까지의 범위인 헥사뉴클레오티드 반복부 확장을 인간 유전자에서 발견된 위치에 배치하였다.
ALS와 관련된 세포 유형에 대한 확장의 효과를 연구하기 위해 상이한 반복부 확장을 갖는 마우스 ES 세포 클론을 배양물에서 운동 뉴런으로 분화시켰다. 유전자 변형된 인간화 C9orf72 대립유전자로부터 생성된 전사체를 조사한 결과, 3개 반복부 정상 대조군에서 우세한 엑손 1B 스플라이싱된 전사체로부터 더 긴 반복부 확장을 갖는 대립유전자에서 엑손 1A 스플라이싱된 전사체의 출현 증가로의 전환이 있는 것이 발견되었다. 또한, 스플라이싱되지 않은 인트론-함유 전사체의 축적이 관찰되었으며, 이의 풍부도는 헥사뉴클레오티드 반복부 확장의 길이와 직접적인 상관관계가 있으며, 이는 더 긴 반복부 확장, 더 많은 반복부-함유 전사체가 C9orf72 유전자로부터 생산되는 이기적 피드-포워드 루프(feed-forward loop)를 시사한다. 정상적인 C9orf72 mRNA 및 단백질의 합성을 보존하면서 디펩티드 반복부 단백질 합성을 위한 주형으로서 파괴 또는 비활성화를 위한 반복부-함유 인트론 전사체를 표적화하는 것은 C9orf72 반복부 확장 질환에 대한 안전하고 효과적인 치료 전략이 될 것으로 예상된다.
C9orf72 반복부-함유 RNA를 감소시키는 한 가지 가능한 접근법은 siRNA가 RNA-유도된 침묵 복합체에 의한 표적 RNA의 절단을 지시한 다음 세포 뉴클레아제에 의한 RNA 절단 단편의 분해를 지시하는 RNA 간섭의 자연적 과정을 통한 것이다. 그러나 RNA 간섭은 핵에 보유된 RNA에 작용할 것으로 예상되지 않는 주로 세포질 과정이다. 인트론-함유 RNA는 성숙한 mRNA로 빠르게 스플라이싱되는 mRNA 전구체로서 또는 핵에서 빠르게 분해되는 스플라이싱된 인트론으로서 일반적으로 수명이 짧다. 따라서 인트론-함유 RNA는 RNA 간섭에 의한 표적화에 사용할 수 없을 것이라고 예상하는 것이 합리적이다.
그러나, GGGGCC 반복부 확장에 인접한 인트론 서열을 표적으로 하는 siRNA는 C9orf72 성숙 mRNA에 거의 또는 전혀 영향을 미치지 않으면서 인트론-함유 C9orf72 RNA의 감소된 축적을 촉진하는 것으로 입증되었다. 인트론-표적화 siRNA는 또한 디펩티드 반복부 단백질의 생산을 감소시켰다. 이러한 예상치 못한 실험 결과는 C9orf72 C9orf72 헥사뉴클레오티드 반복부 확장을 갖는 세포에 축적되는 인트론-함유 RNA가 RNA 간섭에 민감함을 나타낸다. 결과는 디펩티드 반복부 단백질 합성을 담당하는 인트론-함유 C9orf72 RNA의 상당 부분이 세포질에 존재함을 보여준다. 대조적으로, C9orf72 mRNA 단백질 코딩 서열을 표적으로 하는 siRNA는 mRNA의 강한 녹다운을 생성하지만 인트론-함유 전사체에는 영향을 미치지 않았고 디펩티드 반복부 단백질 합성을 눈에 띄게 감소시키지 않았다. 인트론-표적화 siRNA와 mRNA-표적화 siRNA 사이의 결과 차이는 표적화된 서열의 두 부류가 공유적으로 연결되지 않은 별도의 RNA에 존재함을 시사한다.
본원에 개시된 방법 및 조성물은 C9orf72 mRNA를 보존함으로써 핵의 1차 C9orf72 전사체를 표적으로 하는 안티센스 올리고뉴클레오티드의 사용과 같은 치료 전략에서 발생할 수 있는 C9orf72 단백질 감소의 가능한 부작용을 피면서 C9orf72 반복부 확장 질환의 주요 병원성 성분인 디펩티드 반복부 단백질의 합성의 치료적 감소를 제공한다.
II. C9orf72
,
헥사뉴클레오티드
반복부
확장 서열, 및 관련 질환
루게릭병이라고도 하는 근위축성 측삭 경화증(ALS)은 상부 및/또는 하부 운동 뉴런의 상실을 특징으로 하는 가장 흔한 성인-발병 마비 장애이다. ALS는 미국 전역에서 최대 20,000명에서 발생하며 매년 약 5,000명의 새로운 사례가 발생한다. 전두측두엽 치매(FTD; 픽병, 전두측두엽 변성 또는 FTLD라고도 함)는 뇌의 전두엽 또는 측두엽의 진행성 세포 퇴화에 의해 유발되는 장애 그룹이다. FTD는 모든 치매 사례의 10%-15%를 차지하는 것으로 보고된다. 인간 C9orf72 유전자의 두 개의 비-코딩 엑손인 엑손 1A와 1B 사이의 (및 임의로 이에 걸친) 헥사뉴클레오티드 반복부 확장 서열은 ALS 및 FTD 둘 다와 관련이 있다. GGGGCC (서열번호 1) 헥사뉴클레오티드 반복부 확장이 가족성 및 다수의 비-가족성 ALS 사례의 약 50%를 차지하는 것으로 추정된다. C9orf72 반복부 확장은 본질적으로 항상 이형 돌연변이로 발생하는 우성 돌연변이이기 때문이다. 이것은 가족성 FTD 사례의 약 25% 및 산발성 FTD 사례의 약 8%에서 존재한다.
예를 들면, RNA 병소의 반복부-길이-의존적 형성, 특정 RNA-결합 단백질의 격리, 및 뉴런에서의 반복부-관련 비-AUG (AUG) 번역으로부터 야기된 디펩티드 반복부 단백질(예를 들어, 폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌))의 축적 및 응집과 같은, C9orf72에서 헥사뉴클레오티드 반복부 확장 서열과 관련된 다수의 병리학적 측면이 보고되었다.
C9orf72가 엔도솜 트래피킹을 조절하는 것으로 보고되었지만, C9orf72의 많은 세포 기능은 아직 알려지지 않다. 실제로, C9orf72는 알려지지 않은 기능을 가진 특성화되지 않은 단백질을 암호화하는 유전자이다.
마우스 C9orf72 전사체 변이체가 보고되었다. 예를 들어, 문헌[Koppers et al. (2015) Ann. Neurol . 78:426-438 and Atkinson et al. (2015) Acta Neuropathologica Communications 3:59]을 참조하며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다. 3개의 보고된 마우스 C9orf72 전사체 변이체에 대한 게놈 정보는 또한 Ensembl 웹사이트에서 ENSMUST00000108127(V1), ENSMUST00000108126(V2) 및 ENSMUST00000084724(V3)의 명칭하에 이용 가능하다. 예시적인 비-인간(예를 들어, 설치류) C9orf72 mRNA 및 아미노산 서열은 서열번호 40-43, 123, 및 124에 제시되어 있다. 마우스 C9orf72의 mRNA 및 아미노산 서열은 각각 GenBank 수탁번호 NM_001081343 및 NP_001074812에서 찾을 수 있으며, 이것은 모든 목적을 위해 전문이 본원에 참고로 포함된다. NM_001081343.1 및 NP_001074812.1의 서열은 각각 서열번호 40 (및 서열번호 123) 및 41에 제시되어 있다. 래트 C9orf72의 mRNA 및 아미노산 서열은 각각 GenBank 수탁번호 NM_001007702 및 NP_001007703에서 찾을 수 있으며, 이것은 모든 목적을 위해 전문이 본원에 참고로 포함된다. NM_001007702.1 및 NP_001007703.1의 서열은 각각 서열번호 42 (및 서열번호 124) 및 43에 제시되어 있다.
인간 C9orf72 전사체 변이체가 또한 알려져 있다. 하나의 인간 C9orf72 전사체 변이체는 중심 및 3' 코딩 영역에 다중 엑손이 없으며 이의 3' 말단 엑손은 변이체 3(아래 참조)에서 사용되는 스플라이스 부위를 넘어 확장되어, 변이체 3에 비해 새로운 3' 비번역 영역(UTR)이 생성된다. 이 변이체는 상당히 더 짧은 폴리펩티드를 암호화하고 이의 C-말단 아미노산은 2개의 다른 변이체에 의해 암호화되는 것과 비교하여 구별된다. 이 변이체의 mRNA 및 아미노산 서열은 각각 GenBank 수탁번호 NM_145005.6 및 NP_659442.2에서 찾을 수 있으며, 이것은 모든 목적을 위해 전문이 본원에 참고로 포함된다. NM_145005.6 및 NP_659442.2의 서열은 각각 서열번호 94 및 서열번호 110에 제시되어 있다. 두 번째 인간 C9orf72 전사체 변이체(2)는 변이체 3과 비교하여 5' 비번역 영역(UTR)이 다르다. 이 변이체의 mRNA 및 아미노산 서열은 각각 GenBank 수탁번호 NM_018325.4 및 NP_060795.1에서 찾을 수 있으며, 이것은 모든 목적을 위해 전문이 본원에 참고로 포함된다. NM_018325.4 및 NP_060795.1의 서열은 각각 서열번호 26 및 서열번호 27에 제시되어 있다. 세 번째 인간 C9orf72 전사체 변이체(3)는 3개의 보고된 변이체 중 가장 긴 서열을 포함하고 더 긴 이소형을 암호화한다. 이 변이체의 mRNA 및 아미노산 서열은 각각 GenBank 수탁번호 NM_001256054.2 및 NP_001242983.1에서 찾을 수 있으며, 이것은 모든 목적을 위해 전문이 본원에 참고로 포함된다. NM_001256054.2 및 NP_001242983.1의 서열은 각각 서열번호 28 (및 서열번호 121) 및 서열번호 29에 제시되어 있다. 변이체 2 및 3은 동일한 단백질을 암호화한다.
III.
C9orf72
유전자좌에
헥사뉴클레오티드
반복부
확장을 포함하는
ES
세포 클론의 생성
위의 논의된 바와 같이, 근위축성 측삭 경화증(ALS)은 운동 뉴런 사멸을 유발하여 마비를 일으키는 진행성 신경퇴행성 장애이다. 미국에서는 매년 5,000명이 ALS로 진단된다. 질환의 10%는 가족에서 전염되며, 이 범주 중 가장 흔한 원인은 C9orf72 유전자 좌에서의 GGGGCC (서열번호 1) 헥사뉴클레오티드 반복부 확장이다. 건강한 개인은 전형적으로 반복부의 30개 미만의 카피를 가지며, 영향을 받은 환자는 종종 1000개 이상의 카피를 갖는다. 이러한 긴 반복 스트레치가 어떻게 질환을 유발하는지에 대한 정확한 메커니즘은 유용한 동물 모델의 부족 때문에 아직 규명되지 않았다. 헥사뉴클레오티드 반복부 서열에서 GC 함량이 높으면 미생물에서 반복부를 유지하는 것 뿐만 아니라 반복부를 갖는 DNA 단편을 합성하는 것도 어려워진다. 따라서, 유전자이식 동물을 생성하기 위한 표적 벡터와 같은 물질을 제조하는 것은 매우 어렵다. 이것이 현재까지 ALS의 C9orf72 반복부 확장 유형에 대한 유용한 동물 모델이 거의 없는 주요 이유 중 하나이며 현장에서 더 나은 동물 모델에 대한 강한 요구가 있다.
표적 벡터 작제를 위한 이러한 어려운 중간 단계를 우회하기 위해, 데 노보(de novo) 표적 벡터 생산 및 ES 세포 표적화보다는 게놈의 올바른 위치에 이미 삽입된 비교적 짧은 헥사뉴클레오티드 반복부를 확장하는 접근법이 취해졌다. 반복부 확장을 위한 출발 물질로서, 마우스 C9orf72 유전자 좌의 일부가 GGGGCC(서열번호 1) 헥사뉴클레오티드의 92x 반복부를 함유하는 인간 대응물로 대체된 이형 ES 세포 클론을 사용하였다. 도 1 참조. 이 대립유전자는 이전에 개발되었다. 제US 2018/0094267호 및 제WO 2018/064600호를 참조하며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다.
포유류 세포 염색체에서 DNA 이중-가닥 파손(DSB)이 발생하는 경우, 세포는 DSB를 인식하고 상동 재조합(HR) 또는 비상동 말단 연결(NHEJ) 또는 단일 가닥 어닐링(SSA)과 같은 비상동 재조합 경로를 통해 손상을 복구한다. HR에서, 손상된 염색분체는 손상되지 않은 염색분체를 이의 복구를 위한 주형으로서 사용한다. 이 경로에서, 일단 DSB가 발생하면, 3' 가닥 절제를 포함한 많은 과정을 통해 DSB의 가장자리가 변형된다. 노출된 5' 단일 가닥은 Rad51의 다중 분자에 의해 코팅되고, 그후 Rad51 필라멘트가 게놈에서 상동성 서열을 찾기 시작한다. Rad51 필라멘트는 노출된 5' 단일 가닥이 상동성을 갖는 이중 가닥 DNA에 자신을 삽입할 수 있다. 침범된 5' 가닥은 Rad51 필라멘트가 들어가는 바로 그 서열을 사용하여 수복을 개시한다. Rad51 필라멘트가 상동성 서열을 찾는 정확도가 정확한 DNA 서열을 보존하는 열쇠이다. 반복부를 확장하는 접근법은 노출된 5' 가닥이 반복적인 서열만 함유하도록 헥사뉴클레오티드 반복부 근처에 DSB를 도입함으로써, Rad51 필라멘트가 잘못된 위치로 들어갈 수 있고 이것이 Rad51 필라멘트에 의한 상동성 검색을 헛갈리게 하여 반복부의 확장 또는 수축으로 이어질 수 있다는 가설에 근거하였다. 도 2 참조.
구체적으로, 본원에 사용된 접근법은 CRISPR/Cas9 뉴클레아제를 사용하여 헥사뉴클레오티드 반복부의 말단 부근에 DSB를 도입함으로써 인간화 마우스 배아 줄기세포의 C9f72 인트론 1에 있는 GGGGCC (서열번호 1) 헥사뉴클레오티드 반복부를 확장하는 것이었다. 몇몇 가이드 RNA(gRNA)는 절제 후 5' 단일 가닥에서 반복적인 서열을 노출시키기 위해 가능한 한 가깝게 헥사뉴클레오티드 반복부 부근에 설계되었다. 도 3 및 표 4 참조.

목표는 반복부에 가능한 한 가깝게 그리고 높은 절단 효율로 절단하는 gRNA를 수득하는 것이었다. DNA 절단 효율은 인간화된 92x-반복부-함유 대립유전자와 동일한 서열을 함유하는 플라스미드(8028 Stvec)를 사용하여 시험관내에서 시험하였다. 가장 높은 절단 효율을 갖는 gRNA를 반복부의 5' 측과 3' 측 둘 다로부터 선택하였다. 이들은 5' gRNA-1 및 3' gRNA-2였다. 도 4 참조.
ES 세포에서 DNA를 변형하기 위해, 전기천공법에 의해 SpCas9(스트렙토코커스 피오게네스 (Streptococcus pyogenes ) Cas9; Thermo Fisher) 및 gRNA를 RNA 및 단백질 복합체(RNP)로서 C9orf72 인간화, 92x-반복부-함유 ES 세포(MAID8029a)에 도입하였다. ES 세포 콜로니를 선택하고, 콜로니를 96-웰 배양 플레이트에서 성장시키고, 분석을 위해 게놈 DNA를 정제하였다. 세 가지 실험을 수행하였다: (1) 반복부의 5' 측에 단일 DSB를 도입하고; (2) 반복부의 3' 측에 단일 DSB를 도입하며; 및 (3) 반복부의 5' 및 3' 측 둘 다에 DSB를 도입한다. 도 5 참조. 기존의 2-프라이머 PCR(도 6A) 및 AmplideX PCR/CE C9orf72 키트를 사용하여 3개의 프라이머를 사용하는 프라임 PCR(도 6B)을 사용하여 분석을 수행하였다. 프라임 PCR의 경우, ES 세포로부터의 게놈 DNA를 주형으로 사용하였다. 3개의 프라이머가 사용되었다: 반복부의 5' 외부에 위치한 프라이머, 반복부의 3' 외부에 위치한 프라이머, 및 도 6B에 나타낸 바와 같이 반복부 내부의 반복 서열에 어닐링하는 세 번째 프라이머. 이러한 PCR 반응은 도 6B에 나타낸 바와 같이 여러 상이한 크기의 PCR 산물을 생성한다. 세 번째 프라이머는 반복부의 주어진 단위에서 중합효소 반응을 프라이밍할 수 있기 때문에, 수득된 PCR 산물의 수는 기본적으로 클론이 갖는 반복부의 수이다. 이러한 PCR 산물에 대해 형광 신호를 검출하는 모세관 전기영동을 실행하였다(프라이머 중 하나는 형광 표지됨).
AmplideX PCR/CE C9orf72 키트(Asuragen)는 내인성 C9orf72 ES 세포 클론에서 서열번호 1로서 제시된 헥사뉴클레오티드 서열의 인스턴스 수를 확인하기 위해 제조업체의 지침에 따라 사용되었다. 3x 반복 클론, 92x 반복 클론 및 30x 반복 클론으로부터의 정제된 mESC 게놈 전체 DNA를 대조군으로 사용하였다. 표 5의 프라이머 및 AmplideX PCR/CE C9orf72 키트로부터의 반복-특이 프라이머를 사용한 PCR은 ABI 9700 열 순환기(Thermo Fisher)에서 수행하였다. 앰플리콘은 POP-7 중합체(Thermo Fisher) 및 NuSieve 아가로스 겔(Lonza)을 사용하여 ABI 3500xL GeneScan 상에서 모세관 전기영동으로 크기를 조정하였다. 2-log DNA 사다리(New England BioLabs) 분자량 마커를 비교를 위해 아가로스 겔에 부하하고, 밴드를 SYBR Gold Nucleic Acid Stain(Thermo Fisher)으로 시각화하였다.

반복부의 어느 한 쪽에 DSB를 도입하면 기존의 2-프라이머 PCR을 사용하여 입증된 바와 같이 반복부 영역에서 크기 변경이 야기되었다. 도 7A 및 7B 참조. 각 조건에 대한 88개 콜로니 중 16개에 대한 결과가 도 7A 및 7B에 나타내어져 있다. 반복부 확장을 확인하기 위해 프라임 PCR을 사용한 추가 분석을 위해 가장 큰 확장을 갖는 5' 단일 DSB 클론 및 3' 단일 DSB 클론을 선택하였다. 프라임 PCR은 클론 9253D-B2(5' 단일 DSB)는 145개 반복을 갖고 클론 9253A-B1(3' 단일 DSB)은 에 130개의 반복과 92개의 반복을 가짐을 확인시켜 주었다. 도 8 및 도 9 참조. 도 8은 모세관 전기영동으로부터의 결과를 보여준다. 신호 강도는 Y축에, PCR 산물 크기는 X축에 있다. 판독값은 피크의 수이다. 상단 패널은 92x 상위 클론에서 가져온 것이며 92개의 피크가 있다. 50개의 피크는 중간 패널에서 계수되었고 30x 피크는 하단 패널에서 계수되었다. 하단 패널에서 두 개의 높은 피크가 관찰되었다. 이것은 본 발명자들이 고른 ES 세포 콜로니가 균일한 클론이 아니기 때문일 수 있다.
동일한 실험을 유사한 결과로 다시 반복하였다. 도 10 참조. 각 조건(단일 5' DSB, 단일 3' DSB 또는 5' 및 3' DSB 둘 다)에 대해, 88개 클론을 시험하였다. 반복부에서 DSB는 반복 불안정을 야기하였다. 반복부의 어느 한 쪽 또는 양쪽에 있는 DSB는 반복 확장을 야기하였다(92x에서 최대 ~300x). 상이한 클론에서의 확장 또는 수축에 대한 완전한 분석이 표 6에 제공되어 있다. 완전 수축은 TAQMAN 분석 또는 Asuragen AmplideX PCR/CE C9orf72 키트에 의해 호출되었다.

가장 큰 반복부는 5' 및 3' DSB를 함께 도입함으로써 수득되었다. 단일 DSB를 도입함으로써 수득된 가장 큰 반복부는 약 250X였으며, 여기서 3' gRNA 부위는 여전히 온전하였으며 반복 확장 가능하였다. 그 반복부를 더욱 확장하기 위해, 250x 반복 클론(MAID9253D-C5)을 사용하고 상기한 바와 같이 Cas9/gRNA RNP의 도입을 통해 3' 말단에 DSB를 도입하였다. 250x 대립유전자를 사용하여, 반복부를 대략 ~600x의 크기로 추가로 확장시켰다. 도 11A 및 11B 및 표 7 참조.

실시예
2.
C9orf72
유전자 좌에
헥사뉴클레오티드
반복부
확장을 포함하는 운동 뉴런 또는 뇌 조직의 분석
마우스에서 헥사뉴클레오티드 반복부의 크기의 안정성을 상기한 바와 같이 AmplideX PCR/CE C9orf72 키트(Asuragen)를 사용하여 확인하였다.
야생형 C9orf72 유전자좌(대조군) 또는 실시예 1로부터의 유전자 변형된 C9orf72 유전자좌를 포함하는 마우스 배아-줄기-세포-유래된 운동 뉴런(ESMN)의 RNA 전사체를 각각 제US 2018/0094267호 및 제WO 2018/064600호에 기술된 바와 같이 시험하였으며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다. RNA 병소 및 디펩티드 반복부 단백질 수준은 야생형 C9orf72 유전자좌(대조군) 또는 실시예 1로부터의 유전자 변형된 C9orf72 유전자좌를 포함하는 모 배아 줄기 세포로부터 유래된 ESMN에서 평가하였다. 재료 및 방법은 아래에 기술되어 있다.
추가로, 야생형 C9orf72 유전자좌(대조군) 또는 실시예 1로부터의 유전자 변형된 C9orf72 유전자좌를 포함하는 마우스 뇌 조직 및 모 배아 줄기 세포의 RNA 전사체를 각각 제US 2018/0094267호 및 제WO 2018/064600호에 기술된 바와 같이 시험하였으며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다. RNA 병소 및 디펩티드 반복부 단백질 수준은 야생형 C9orf72 유전자좌(대조군) 또는 실시예 1로부터의 유전자 변형된 C9orf72 유전자좌를 포함하는 모 배아 줄기 세포로부터 유래된 ESMN에서 평가하였다. 재료 및 방법은 아래에 기술되어 있다.
배아-줄기-세포-
유래된
운동 뉴런
실시예 1로부터의 배아 줄기 세포(ESC)를 배아 줄기 세포 배지(ESM; DMEM + 15% 소 태아 혈청 + 페니실린/스트렙토마이신 + 글루타민 + 비필수 아미노산 + 뉴클레오시드 + β-머캅토에탄올 + 나트륨 피루베이트 + LIF)에서 2일 동안 배양하였으며, 그 동안 배지를 매일 교체하였다. 트립신 처리 1시간 전에 ES 배지를 ADFNK 배지(고급 DMEM/F12 + 신경기저 배지 + 10% 녹아웃 혈청 + 페니실린/스트렙토마이신 + 글루타민 + β-머캅토에탄올) 7mL로 교체하였다. ADFNK 배지를 흡인하고, ESC를 0.05% 트립신-EDTA로 트립신 처리하였다. 펠렛화된 세포를 12mL의 ADFNK에 재현탁시키고 현탁액에서 2일 동안 성장시켰다. 세포를 레티노산(RA), 평활화 효능제 및 푸르모르파민이 보충된 ADFNK에서 추가 4일 동안 배양하여 사지-유사 운동 뉴런(ESMN)을 수득하였다. 분리된 운동 뉴런을 배아-줄기-세포-유래 운동 뉴런 배지(ESMN; 신경기저 배지 + 2% 말 혈청 + B27 + 글루타민 + 페니실린/스트렙토마이신 + β-머캅토에탄올 + 10ng/mL GDNF, BDNF, CNTF)에 플레이팅하고 성숙시켰다.
정량적 중합효소 연쇄 반응
각 샘플로부터 총 RNA를 추출하고 다양한 영역의 측면에 있는 프라이머 및 변형된 C9orf72 유전자좌의 영역을 검출하는 프로브를 사용하여 역전사시켰다. 검출 가능한 영역은 마우스 및 인간 서열의 접합부에 걸쳐 있는 영역, 인간 서열만 또는 마우스 서열만을 포함한다. GAPDH, DROSHA 또는 β2-마이크로글로불린의 qPCR은 쉽게 이용 가능한 키트의 프로브 및 프라이머를 사용하여 수행하였다.
구체적으로, RNA를 야생형(WT) C9orf72 유전자좌(대조군) 또는 유전자 변형된 C9orf72 유전자좌를 포함하는 배아-줄기-세포-유래 운동 뉴런(ESMN)으로부터 단리하였다. 또 다른 실험에서, RNA를 또한 야생형(WT) C9orf72 유전자좌(대조군) 또는 유전자 변형된 C9orf72 유전자좌를 포함하는 마우스로부터 단리된 모 배아 줄기(ES) 세포 또는 전체 뇌로부터 단리한다.
총 RNA를 제조사의 프로토콜(Zymo Research)에 따라 Direct-zol RNA Miniprep 플러스 키트를 사용하여 단리하였다. 총 RNA를 제조사의 프로토콜(Invitrogen)에 따라 Turbo DNA-비함유 키트를 사용하여 DNase로 처리하고 20ng/μL로 희석하였다. 역전사(RT) 및 PCR은 Quantitect 프로브 RT-PCR 키트(Qiagen)를 사용한 1단계 반응으로 수행하였다. qRT-PCR 반응은 10μL의 최종 용적을 만들도록 2μL RNA 및 RT-PCR 마스터 믹스, ROX 염료, RT-믹스 및 20X 유전자 특이적 프라이머-프로브 믹스를 함유하는 8μL 혼합물을 함유하였다.
달리 명시되지 않는 한, 최종 프라이머 및 프로브 농도는 각각 0.5μM 및 0.25μM였다. qRT-PCR은 ViiA™ 7 실시간 PCR 검출 시스템(ThermoFisher)에서 수행하였다. PCR 반응은 광학 384웰 플레이트에서 45회 사이클 동안 45℃ 10분에서 RT-단계 후 95℃ 10분 및 2단계 사이클링 95℃ 5초, 60℃ 30초로 반복하여 사중으로 수행하였다. 각 분석(A, B, C, D, E, F, G, H, I, 및 J)에서 사용된 프라이머 및 프로브의 서열 및 서열번호는 표 8에 제공되어 있다.


웨스턴
블롯
분석
분화된 배양체(EB)를 수집하고 SDS 샘플 완충액(2% SDS, 10% 글리세롤, 5% β-머캅토에탄올, 60mM TrisHCl, pH 6.8, 브로모페놀 블루)에서 균질화하였다. 단백질 추출물은 RC DC 단백질 분석(BioRad)을 사용하여 정량하였다. 추출물(10㎍)을 4-20% SDS-PAGE 겔(ThermoFisher)에서 실행하고 iBLOT 전송 장치(ThermoFisher)를 사용하여 니트로셀룰로스 막으로 옮겼다. 면역블롯은 C9orf72 및 GAPDH(Millipore)에 대한 1차 항체로 프로빙하였다. 결합된 항체는 호스래디쉬 퍼옥시다제(Abcam)에 접합된 이차 항체와 함께 배양한 다음 SuperSignal West Pico 화학발광 기질(Thermo Scientific)을 사용하여 화학발광하여 검출하였다. 신호는 Full Speed Blue 민감성 의료용 X선 필름(Ewen Parker XRay Corporation)을 사용하여 자가방사선 촬영으로 검출하였다. 상대적 단백질 수준은 ImageJ를 사용하여 계산하였다. 데이터는 표시되지 않는다.
센스 또는
안티센스
RNA 전사 산물의 검출을 위한 형광 제자리
혼성화
(FISH)
형광 제자리 혼성화(FISH)를 사용하여 상기한 바와 같이 생성된 배아-줄기-세포-유래 운동 뉴런(ESMN)에서 서열번호 1로서 제시된 헥사뉴클레오티드 반복부 서열로부터 전사된 RNA의 위치를 결정하였다. 간단히 말해서, ESMN을 4-웰 챔버 슬라이드(Lab-Tek II 챔버 슬라이드 시스템, ThermoFisher Scientific)에서 성장시키고 PBS에서 4% PFA(Electron Microscopy Sciences)로 고정하였다. 그후 세포를 디에틸 피로카보네이트(DEPC) PBS/0.2% Triton X-100(Fisher Scientific, 카탈로그 #BP151)으로 투과시키고 DEPC-PBS로 세척하고, 아래 기술된 바와 같이 RNA 전사 산물의 검출을 위해 차단하고 LNA 올리고뉴클레오티드로 염색하였다. 염색 후, 슬라이드를 적절한 형광 염료와 함께 배양하고, Fluoromount G(Southern Biotech)를 장착하고 공초점 현미경을 사용하여 시각화하였다.
슬라이드를 50% 포름아미드(IBI Scientific, 카탈로그 #IB72020), DEPC 2X SSC[300mM 염화나트륨, 30mM 시트르산나트륨(pH 7.0)], 10%(w/v) 덱스트란 설페이트(Sigma-Aldrich, 카탈로그 #D8960) 및 DEPC 50mM 인산나트륨(pH 7.0)로 구성된 완충액으로 LNA 프로브의 경우 66℃에서 30분 동안 예비-혼성화하였다. 그후, 혼성화 완충액을 배출하고, 혼성화 완충액 중 40nM LNA 프로브 믹스 400μL를 각 슬라이드에 첨가하고 암실에서 (LNA 프로브의 경우) 66℃에서 3시간 동안 배양하였다. LNA 프로브와 함께 배양된 슬라이드를 실온에서 DEPC 2×SSC/0.1% Tween 20(Fisher Scientific, 카탈로그 번호 BP337)에서 한 번, DEPC 0.1×SSC에서 65℃에서 세 번 세정하였다. 후속적으로 슬라이드를 1㎍/mL DAPI(Molecular Probes Inc.)와 함께 배양하였다.
또 다른 실험에서, 슬라이드를 50% 포름아미드(IBI Scientific, 카탈로그 #IB72020), DEPC 2X SSC[300mM 염화나트륨, 30mM 시트르산나트륨(pH 7.0)], 10%(w/v) 덱스트란 설페이트(Sigma-Aldrich, 카탈로그 #D8960) 및 DEPC 50mM 인산나트륨(pH 7.0)로 구성된 완충액으로 66℃(LNA 프로브의 경우) 또는 55℃(DNA 프로브의 경우)에서 30분 동안 예비-혼성화하였다. 그후, 혼성화 완충액을 배출하고, 혼성화 완충액 중 200ng/mL DNA 프로브 믹스 400μL를 각 슬라이드에 첨가하고 암실에서 55℃에서 3시간 동안 배양하였다. DNA 프로브와 함께 배양된 슬라이드를 2×SSC 중 40% 포름아미드로 3회 세척하고 PBS에서 한 번 간단하게 세척하였다. 후속적으로 슬라이드를 1㎍/mL DAPI(Molecular Probes Inc.)와 함께 배양하였다.
본 실시예에서 사용된 LNA 및 DNA 올리고뉴클레오티드 프로브의 서열 및 서열번호 뿐만 아니라 프로브의 혼성화 조건은 하기 표 9에 제공되어 있다. 고정 핵산(LNA)은 리보스 모이어티가 2' 산소와 4' 탄소를 연결하는 추가 브릿지로 변형된 핵산 유사체이다.

디펩티드
반복부
단백질 산물의 검출(
면역형광
및
웨스턴
슬롯
블롯
)
면역형광을 사용하여 상기한 바와 같이 생성된 배아-줄기-세포-유래 운동 뉴런(ESMN)에서 디펩티드 반복부 단백질 생산을 평가하였다. 간단히 말해서, ESMN을 4-웰 챔버 슬라이드(Lab-Tek II 챔버 슬라이드 시스템, ThermoFisher Scientific)에서 성장시키고 PBS에서 4% PFA(Electron Microscopy Sciences)로 고정하였다. 그후 세포를 디에틸 피로카보네이트(DEPC) PBS/0.2% Triton X-100(Fisher Scientific, 카탈로그 #BP151)으로 투과시키고 DEPC-PBS로 세척하고, 아래 기술된 바와 같이 RNA 전사 산물의 검출을 위해 차단하고 항-폴리GA 항체로 염색하였다. 염색 후, 슬라이드를 적절한 형광 염료와 함께 배양하고, Fluoromount G(Southern Biotech)를 장착하고 공초점 현미경을 사용하여 시각화하였다.
투과화 후, 슬라이드를 0.2% Triton X100을 갖는 Tris 완충 염수(pH 7.4)(TBS-T)에 희석된 5% 정상 당나귀 혈청으로 차단하였다. 슬라이드를 5% 정상 당나귀 혈청을 갖는 TBS-T에 희석된 폴리-GA(Millipore)에 대한 1차 항체와 함께 4℃에서 밤새 배양하였다. TBS-T로 3회 세척한 후, 슬라이드를 Alexa 488 또는 555(TBS-T에서 1:1000)에 결합된 종-특이 이차 항체 및 DAPI(1㎍/mL)(Molecular Probes Inc.)와 함께 실온에서 1시간 동안 배양하였다. TBS-T로 3회 세척한 후, 슬라이드에 Fluoromount G(Southern Biotech)를 장착하고 공초점 현미경을 사용하여 시각화하였다.
슬롯 블롯 분석을 위해, 분화된 배양체(EB)를 수집하고 SDS 샘플 완충액(2% SDS, 10% 글리세롤, 5% β-머캅토에탄올, 60mM TrisHCl, pH 6.8, 브로모페놀 블루)에서 균질화하였다. 단백질 추출물을 RC DC 단백질 분석(BioRad)을 사용하여 정량하였다. 0㎍, 1.25㎍, 2.5㎍, 5㎍, 10㎍ 또는 20㎍을 함유하는 용해물을 진공하에 Bio-Slot 48-웰 미세여과 시스템(Bio-Rad)을 사용하여 니트로셀룰로오스 막에 고정하였다. 막을 TBS-T로 세척하고, 폴리(GP)(1:5,000, Novus biologicals) 및 폴리 GA(1:5000, Millipore)에 대한 항체로 블롯팅하였다. 막을 HRP 접합된 이차 항체와 함께 배양한 후, 밴드는 ECL과 웨스턴 블로팅 검출 시스템(Pierce)에 의해 시각화하였다.
결과
G4C2 헥사뉴클레오티드 서열의 3개 반복부, 92개 반복부, 250개 반복부, 300개 반복부, 500개 반복부, 또는 600개 반복부를 갖는 대립유전자 시리즈의 인간화 C9orf72 대립유전자를 포함하는 배아-줄기-세포-유래 운동 뉴런(ESMN)을 시험하였다. 도 12C, 도 12D, 도 12G, 및 도 12H에 나타낸 바와 같이, C9orf72 유전자좌에 헥사뉴클레오티드 반복부 확장 서열을 포함하는 ESMN은 인트론 1 서열을 보유하는 C9orf72 mRNA 전사체의 발현 증가를 보여주었다. 또한, 더 큰 G4C2 반복부 확장은 엑손 1A의 사용을 증가시키고 엑손 1B의 사용을 감소시켰다. 각각 도 12A와 12B, 및 도 12E와 12F를 참조한다. G4C2 반복부 확장을 함유하는 ESMN은 또한 핵 및 세포질 센스 및 안티센스 C9orf72 RNA 병소를 함유하였다(데이터는 표시되지 않음). 센스 G4C2 병소가 더 크며 핵소체에 분포되어 있다(데이터는 표시되지 않음). 300개 이상의 반복부 확장을 갖는 ESMN에서의 핵소체를 갖는 RNA 병소의 국소화는 ALS-환자-유래 세포에서 병리학적 발견을 재현한다. 또한, G4C2 헥사뉴클레오티드 서열의 반복부의 증가된 수는 헥사뉴클레오티드 반복부 서열의 전사체로부터 (RAN 번역, 비AUG 기전을 통해) 번역된 증가된 디펩티드 반복부 단백질(polyGA 및 polyGP)의 존재와 직접적인 상관관계가 있다. 예를 들어, 도 13을 참조한다. 요약하면, 대립유전자 시리즈 ES 세포로부터 유래된 운동 뉴런은 ALS 질환의 분자적 특징(센스 및 안티센스 반복부 RNA 병소, 핵소체에 국소화된 반복 RNA 병소, 디펩티드 반복부 단백질의 5가지 형태 중 적어도 2개, 및 인트론-함유 전사체의 축적 증가)을 재현하여, 신경퇴행성 질환에 대한 질병 모델로서 본원에 개시된 비인간 동물의 사용을 뒷받침한다.
표 8의 분석 C, B, F, H, E 및 D에 대한 정량적 PCR 반응은 2가지 유형의 ESMN에서 반복되었다: 축하형 운동 뉴런(MN) 및 사지형 운동 뉴런(MN). 축하형 MN는 모델에서 축하 MN을 자극한다. 축하 MN에 의해 자극된 근육의 예는 늑간근, 횡격막 및 복벽 근육을 포함한다. 사지형 MN은 운동 측주로부터 발생하고 앞다리와 뒷다리에 있는 것과 같은 말단 사지 근육(전경골, 비복근 및 둔근)을 자극하는 것이다. 주로 축하 MN은 레티노산(RA)과 소닉 헤지호그 효능제(SAG)를 추가하는 프로토콜에 의해 생성된다. 사지형 MN은 본 발명자들이 사지형 MN를 생성할 수 있는 RA 및 SAG의 상단에 1μM 푸르모르파민을 추가함으로써 생성될 수 있다.
G4C2 헥사뉴클레오티드 서열의 3개 반복부, 92개 반복부, 300개 반복부, 500개 반복부, 또는 600개 반복부를 갖는 대립유전자 시리즈의 인간화 C9orf72 대립유전자를 포함하는 운동 뉴런을 시험하였다. 도 15A 및 도 15B에 나타낸 바와 같이, 더 큰 G4C2 반복부 확장은 엑손 1A의 사용을 증가시키고 엑손 1B의 사용을 감소시켰다. 도 15C 및 도 15D에 나타낸 바와 같이, C9orf72 유전자좌에 헥사뉴클레오티드 반복부 확장 서열을 포함하는 ESMN은 인트론 1 서열을 보유하는 C9orf72 mRNA 전사체의 발현 증가를 보여주었다. 도 15E에 나타낸 바와 같이, 더 큰 G4C2 반복부 확장은 비스플라이싱된 전구체의 발현을 증가시켰다. 도 15F에 나타낸 바와 같이, 스플라이싱된 C9orf72 mRNA의 발현은 반복부 크기에 따라 크게 변하지 않았다.
실시예
3.
C9orf72
유전자 좌에
헥사뉴클레오티드
반복부
확장을 포함하는 마우스의 생성
F0 마우스는 실시예 1에 기술된 300x 반복 ES 세포 클론을 사용하여 VELOCIMOUSE® 방법을 사용하여 생성하였다. 예를 들어, 제US 7,576,259; 제US 7,659,442; 제US 7,294,754; 제US 2008/0078000; 및 문헌[Poueymirou et al. (2007) Nat. Biotechnol . 25(1):91-99]을 참조하며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다. VELOCIMOUSE® 방법에서, 표적 마우스 배아 줄기(ES) 세포를 레이저-보조 주사를 통해 상실기 이전 배아, 즉 8-세포 단계 배아에 주입되어 완전히 ES-세포-유래인 F0 세대 마우스를 효율적으로 생산한다.
C9orf72 300x 반복 마우스(즉, G4C2 헥사뉴클레오티드 서열의 300개 반복부를 갖는 인간화 C9orf72 대립유전자) 및 대조군 C9orf72 3x 반복 마우스(즉, G4C2 헥사뉴클레오티드 서열의 3개 반복부를 갖는 인간화 C9orf72)로부터의 뇌간 및 척수 샘플의 RNA 전사체를 실시예 3에서와 같이 시험하였다. RNA 병소 및 디펩티드 반복부 단백질 수준은 실시예 3에서와 같이 C9orf72 300x 반복 마우스로부터의 뇌간 및 척수 샘플에서 평가하였다. 도 14B에 나타낸 바와 같이, C9orf72 300x 반복 마우스로부터의 뇌간 및 척수 샘플은 인트론 1 서열을 보유하는 C9orf72 mRNA 전사체의 발현 증가를 보여주었다. 또한, 척수 샘플(즉, L4/L5 요추 척수 운동 뉴런)을 시험하였으며 핵 및 세포질 센스 및 안티센스 C9orf72 RNA 병소를 함유하는 것으로 나타났다(데이터는 표시되지 않음). 유사하게, 이들 샘플은 헥사뉴클레오티드 반복부 서열의 전사체로부터 (RAN 번역, 비AUG 기전을 통해) 번역된 디펩티드 반복부 단백질(polyGA)의 존재가 증가하였다(데이터는 표시되지 않음).
마찬가지로, G4C2 헥사뉴클레오티드 서열의 3개 반복부, 92개 반복부, 300개 반복부, 500개 반복부, 또는 600개 반복부를 갖는 대립유전자 시리즈의 인간화 C9orf72 대립유전자를 포함하는 배아-줄기-세포-유래 운동 뉴런(ESMN)을 시험하였다. 도 14A에 나타낸 바와 같이, C9orf72 유전자 좌에 헥사뉴클레오티드 반복부 확장 서열을 포함하는 ESMN은 인트론 1 서열을 보유하는 C9orf72 mRNA 전사체의 발현 증가를 보여주었다. G4C2 반복부 확장을 함유하는 ESMN은 또한 핵 및 세포질 센스 및 안티센스 C9orf72 RNA 병소 및 헥사뉴클레오티드 반복부 서열의 전사체로부터 (RAN 번역, 비AUG 기전을 통해) 번역된 디펩티드 반복부 단백질(polyGA)의 증가된 존재를 함유하였다(데이터는 표시되지 않음).
또한, 공지된 방법을 사용하여 GGGGCC(서열번호 1) 헥사뉴클레오티드의 대략 500개 반복부 또는 대략 600개 반복부를 함유하는 인간 대응물로 대체된 마우스 C9orf72 유전자 좌를 갖는 F0 마우스를 생성하였다.
실시예
4.
C9orf72
-
반복부
-인접
인트론
서열을 표적으로 하는
siRNA는
인트론
-함유 RNA 및
디펩티드
반복부
단백질을
감소시킨다
근위축성 측삭 경화증(ALS) 및 전두측두엽 변성(FTLD)는 ALS의 경우 운동 뉴런 질환을, FTD의 경우 치매를 유발하는 치명적인 신경퇴행성 질환이다. 둘 다는 항상 치명적이다. ALS 및 FTLD는 자발적 또는 가족성(즉, 유전적) 질환으로 나타날 수 있다. ALS와 FTLD의 가장 흔한 유전적 원인은 기능이 완전히 이해되지 않은 단백질을 암호화하는 C9orf72 유전자의 5' 비암호화 부분에 있는 헥사뉴클레오티드 반복부 (GGGGCC; 서열번호 1)의 확장이다. 영향을 받지 않은 사람들은 보통 이들의 C9orf72 유전자에 수 개 내지 수십 개의 헥사뉴클레오티드 반복부를 갖는 반면, ALS 및 FTLD가 발병한 사람들은 부모 중 한 명에게서만 헥사뉴클레오티드 반복부의 수백 내지 수천 개의 카피의 반복 확장을 물려받는다. C9orf72 단백질의 기능 상실을 유발할 것으로 예상되는 돌연변이는 ALS 및 FTLD와 관련이 없다. 이러한 유전적 관찰은 C9orf72 ALS 및 FTD가 우성 유전 질환이며 병리학적 기능의 획득으로 인한 것임을 시사한다.
어떻게 C9orf72 헥사뉴클레오티드 반복부 확장이 운동 뉴런 질환 및 치매를 일으키는지는 알려져 있지 않지만, C9orf72 ALS 및 FTLD 환자에서의 두 가지 보편적인 사후 병리학적 소견이 반복 확장과 관련이 있다: (1) 센스 및 안티센스 반복부-함유 RNA는 형광 제자리 혼성화에 의해 뉴런 및 기타 세포에서 별개의 병소로 시각화될 수 있고; (2) 센스 및 안티센스 반복부-함유 RNA로부터 반복부-관련 비-AUG-의존성 번역에 의해 합성된 디펩티드 반복부 단백질―폴리(글리신-알라닌), 폴리(글리신-프롤린), 폴리(글리신-아르기닌), 폴리(알라닌-프롤린), 및 폴리(프롤린-아르기닌)―이 면역조직화학에 의해 세포에서 검출될 수 있다. 질병 가설 중 하나에서는 병소로서 시각화된 반복체-함유 RNA가 RNA 결합 단백질을 격리함으로써 세포 RNA 대사를 방해한다고 제안한다. 두 번째 질병 가설은 디펩티드 반복부 단백질이 RNA 대사, 단백질 항상성 및 핵세포질 수송에 광범위한 독성 영향을 미친다고 상정한다. 두 병원성 메커니즘 모두 질병에 기여할 수 있다. C9orf72 반복부-함유 RNA 전사체가 그 자체로 또는 디펩티드 반복부 단백질의 번역을 위한 주형으로서 ALS 및 FTLD에서 발병을 촉진하는 경우, 일반적인 치료 전략은 GGGGCC-반복부-함유 RNA를 파괴하거나 디펩티드 반복부 단백질로 번역되는 이들의 능력을 폐지하는 것일 것이다.
C9orf72 유전자는 2개의 전사 개시 부위로부터 전사체를 생성한다. 상류 부위는 대체 비-코딩 엑손 1A로 전사를 개시하는 반면 하류 부위는 대체 엑손 1B로 전사를 개시한다. 엑손 1A 및 1B는 모두 단백질-코딩 서열의 시작을 포함하는 엑손 2에 스플라이싱될 수 있다. 병원성 헥사뉴클레오티드 반복부 확장은 엑손 1A와 1B 사이에 위치하기 때문에, 엑손 1A로부터 개시된 전사는 반복부-함유 RNA를 생성할 수 있는 반면, 엑손 1B로부터의 개시는 그렇게 할 수 없다.
마우스에서 C9orf72 반복부 확장 질환을 모델링하기 위해, 마우스 배아 줄기(ES) 세포에서 대립유전자 시리즈를 생성하였으며, 여기서 엑손 1A의 일부, 1A와 1B 사이의 인트론 서열, 모든 엑손 1B 및 하류 인트론의 일부를 포함하는 인간 C9orf72 유전자의 단편을 마우스 C9orf72 유전자의 한 대립유전자의 상동 위치에 정확하게 배치하였다. 예를 들어, 제US 2018/0094267호 및 제WO 2018/064600호를 참조하며, 이들 각각은 모든 목적을 위해 전문이 본원에 참고로 포함된다. 정상적인 3개의 반복부 내지 병리학적인 600개의 반복부까지의 범위인 헥사뉴클레오티드 반복 확장부를 인간 유전자에서 발견된 위치에 배치하였다. 실시예 1 참조.
ALS와 관련된 세포 유형에 대한 확장의 효과를 연구하기 위해 상이한 반복부 확장을 갖는 마우스 ES 세포 클론을 배양물에서 운동 뉴런으로 분화시켰다. 실시예 3 참조. 유전자 변형된 인간화 C9orf72 대립유전자로부터 생성된 전사체를 조사한 결과, 3개 반복부 정상 대조군에서 우세한 엑손 1B 스플라이싱된 전사체로부터 더 긴 반복부 확장을 갖는 대립유전자에서 엑손 1A 스플라이싱된 전사체의 출현 증가로의 전환이 있는 것이 발견되었다. 도 12A, 도 12B, 도 12E, 및 도 12F 참조. 또한, 스플라이싱되지 않은 인트론-함유 전사체의 축적이 관찰되었으며, 이의 풍부도는 헥사뉴클레오티드 반복부의 길이와 직접적인 상관관계가 있으며, 이는 더 긴 반복부 확장, 더 많은 반복부-함유 전사체가 C9orf72 유전자로부터 생산되는 이기적 피드-포워드 루프(feed-forward loop)를 시사한다. 도 12C, 도 12D, 도 12G, 및 도 12H 참조. 정상적인 C9orf72 mRNA 및 단백질의 합성을 보존하면서 디펩티드 반복부 단백질 합성을 위한 주형으로서 파괴 또는 비활성화를 위한 반복부-함유 인트론 전사체를 표적화하는 것은 C9orf72 반복부 확장 질환에 대한 안전하고 효과적인 치료 전략이 될 것으로 예상된다.
C9orf72 반복부-함유 RNA를 감소시키는 한 가지 가능한 접근법은 siRNA가 RNA-유도된 침묵 복합체에 의한 표적 RNA의 절단을 지시한 다음 세포 뉴클레아제에 의한 RNA 절단 단편의 분해를 지시하는 RNA 간섭의 자연적 과정을 통한 것이다. 그러나 RNA 간섭은 핵에 보유된 RNA에 작용할 것으로 예상되지 않는 주로 세포질 과정이다. 인트론-함유 RNA는 성숙한 mRNA로 빠르게 스플라이싱되는 mRNA 전구체로서 또는 핵에서 빠르게 분해되는 스플라이싱된 인트론으로서 일반적으로 수명이 짧다. 따라서 인트론-함유 RNA는 RNA 간섭에 의한 표적화에 사용할 수 없을 것이라고 예상하는 것이 합리적이다.
그러나, GGGGCC 헥사뉴클레오티드의 300개 반복부를 갖는 인간화된 C9orf72 유전자를 보유하는 마우스 배아 줄기 세포를 C9orf72 RNA 서열을 표적으로 하는 siRNA로 처리한 실험에서, GGGGCC 반복부 확장에 인접한 인트론 서열을 표적으로 하는 siRNA가 C9orf72 성숙 mRNA에 영향을 미치지 않으면서(도 16A 참조) 인트론-함유 C9orf72 RNA의 축적을 적어도 60%까지 감소시키는 것(도 16B 참조)을 촉진하였다.
엑손 1A 부근의 인트론 서열을 함유하는 전사체를 측정하는 인트론-함유 RT-qPCR 분석에 사용된 프라이머 및 프로브는 표 8로부터의 분석 F이다. 엑손2-3 RT-qPCR 부석에 사용된 프라이머 및 프로브는 ThermoFisher(분석 Mm01216829_m1)로부터 구입하였다. 대조군 GAPDH siRNA는 ThermoFisher(제품 번호 4390849)로부터 구입하였다.
헥사뉴클레오티드 반복부의 C9orf72 성숙 mRNA 또는 C9orf72 인트론 5'를 표적으로 하는 siRNA는 실시예 5 및 6에 기술된 바와 같이 Neuro2a 또는 BE(2)C 세포에서 단일 용량 스크린에서의 활성에 대해 설계, 합성 및 분석되었다.
본 실시예에서 GGGGCC 헥사뉴클레오티드의 300개 반복부를 갖는 인간화 C9orf72 유전자를 보유하는 마우스 배아 줄기 세포를 처리하는데 사용되는 siRNA의 센스 가닥 및 안티센스 가닥의 비변형 뉴클레오티드 서열이 표 10A에 나타내어져 있다. 본 실시예에서 GGGGCC 헥사뉴클레오티드의 300개 반복부를 갖는 인간화 C9orf72 유전자를 보유하는 마우스 배아 줄기 세포를 처리하는데 사용되는 siRNA의 센스 가닥 및 안티센스 가닥의 비변형 뉴클레오티드 서열은 표 10A에 나타내어져 있다. 본 실시예에서 GGGGCC 헥사뉴클레오티드의 300개 반복부를 갖는 인간화 C9orf72 유전자를 보유하는 마우스 배아 줄기 세포를 처리하는데 사용되는 siRNA의 센스 가닥 및 안티센스 가닥의 변형 뉴클레오티드 서열은 표 10B에 나타내어져 있다.
인트론-표적화 siRNA는 또한 디펩티드 반복부 단백질의 생산을 75%만큼 감소시켰다. 도 17A-17B 참조. 이러한 예상치 못한 실험 결과는 C9orf72 헥사뉴클레오티드 반복부 확장을 갖는 세포에 축적되는 인트론-함유 RNA가 RNA 간섭에 민감함을 나타낸다. 결과는 디펩티드 반복부 단백질 합성을 담당하는 인트론-함유 C9orf72 RNA의 상당 부분이 세포질에 존재함을 입증한다. 대조적으로, C9orf72 mRNA 단백질 코딩 서열을 표적으로 하는 siRNA는 mRNA의 강한 녹다운을 생성하지만 인트론-함유 전사체에는 영향을 미치지 않았고 디펩티드 반복부 단백질 합성을 눈에 띄게 감소시키지 않았다. 도 16A-16B 참조. 인트론-표적화 siRNA와 mRNA-표적화 siRNA 사이의 결과 차이는 표적화된 서열의 두 부류가 공유적으로 연결되지 않은 별도의 RNA에 존재함을 시사한다.


서열번호 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 및 24의 센스 가닥은 다음의 특징/변형을 모두 포함하였다: (a) 21개의 뉴클레오티드 길이, (b) (5' 말단에서 계수하여) 위치 1-6, 8, 및 12-21에 2'-O-메틸 변형; (c) (5' 말단에서 계수하여) 위치 7 및 9-11에 2'-F 변형; (d) (5' 말단에서 계수하여) 뉴클레오티드 위치 1 및 2 및 뉴클레오티드 위치 2 및 3 사이의 포스포로티오에이트 뉴클레오티드간 결합, 및 (e) 3'-말단에 부착된 아시알로당단백질 수용체(ASGPR) 리간드, 여기서 ASGPR 리간드는 3가 분지형 링커를 통해 부착된 3개의 GalNAc 유도체를 포함한다.
서열번호 3, 5, 7, 9, 11, 및 13의 안티센스 가닥은 다음의 특징/변형을 모두 포함하였다: (a) 23개의 뉴클레오티드 길이, (b) (5' 말단에서 계수하여) 위치 1, 3-5, 7, 10-13, 15, 및 17-23에 2'-O-메틸 변형; (c) (5' 말단에서 계수하여) 위치 2, 6, 8, 9, 14, 및 16에 2'-F 변형; 및 (d) (5' 말단에서 계수하여) (d) 뉴클레오티드 위치 1 및 2, 뉴클레오티드 위치 2 및 3, 뉴클레오티드 위치 21 및 22, 및 뉴클레오티드 위치 22 및 23 사이의 포스포로티오에이트 뉴클레오티드간 결합.
서열번호 15, 17, 19, 21, 23, 및 25의 안티센스 가닥은 다음의 특징/변형을 모두 포함하였다: (a) 23개의 뉴클레오티드 길이, (b) 위치 1, 3-6, 8-13, 15, 및 17-23에 2'-O-메틸 변형; (c) 위치 2, 14, 및 16에 2'-F 변형; (d) 위치 7에 글리콜 핵산(GNA); 및 (e) (5' 말단에서 계수하여) 뉴클레오티드 위치 1 및 2, 뉴클레오티드 위치 2 및 3, 뉴클레오티드 위치 21 및 22, 및 뉴클레오티드 위치 22 및 23 사이의 포스포로티오에이트 뉴클레오티드간 결합.
표 10A 및 10B의 이중 가닥 RNAi 제제 각각은 안티센스 가닥의 3'-말단에 2-뉴클레오티드 돌출부 및 안티센스 가닥의 5' 말단에 평활 말단을 포함하였다.
실시예
5.
RNAi
제제 설계, 합성, 선택, 및
시험관내
평가
당해 실시예는 C9orf72 RNAi 제제의 설계, 합성, 선택, 및 시험관내 평가를 위한 방법을 기술한다.
시약의 공급원
시약의 공급원이 본원에 구체적으로 제공되지 않은 경우, 이러한 시약은 분자 생물학에 적용하기 위한 품질/순도 표준으로 분자 생물학용 시약 공급업체로부터 수득할 수 있다.
생물정보학
인간 C9orf72 전사체(C9orf72; 인간 NCBI refseqID NM_001256054; NCBI GeneID: 203228)를 표적으로 하는 siRNA는 맞춤형 R 및 Python 스크립트를 사용하여 설계되었다. 인간 NM_001256054 mRNA는 3356개 염기의 길이를 갖는다.
비변형된 C9orf72 센스 및 안티센스 가닥 뉴클레오티드 서열의 상세 목록은 표 12에 나타내어져 있다. 변형된 C9orf72 센스 및 안티센스 가닥 뉴클레오티드 서열의 상세 목록은 표 13에 나타내어져 있다.
인간 C9orf72 유전자(GenBank 수탁번호 NC_000009.12)에서 엑손 1a와 1b 사이의 인트론을 표적으로 하는 siRNA는 맞춤형 R 및 Python 스크립트를 사용하여 설계되었다. 비변형된 C9orf72 센스 및 안티센스 가닥 뉴클레오티드 서열의 상세 목록은 표 14 및 19에 나타내어져 있다. 변형된 C9orf72 센스 및 안티센스 가닥 뉴클레오티드 서열의 사세 목록은 표 15 및 20에 나타내어져 있다.
본 출원 전반에 걸쳐, 소수가 없는 이중체 명칭은 단지 이중체의 배치 번호를 참조하는 소수가 있는 이중체 명칭과 동등한 것으로 이해해야 한다. 예를 들면, AD-347430은 AD-347430.1과 동등하다.
시험관내
Cos
-7(이중-
루시퍼라제
psiCHECK2
벡터), BE(2)-C, 및
Neuro
-2a 스크리닝
세포 배양 및 형질감염
:
Cos-7(ATCC)은 웰당 Opti-MEM에 희석된 1ng/ul의 C9orf72 인트론 1 psiCHECK2 벡터(Blue Heron Biotechnology) 5μl, 4.9μl의 Opti-MEM + 0.1μl의 리포펙타민 2000(Invitrogen, Carlsbad CA. cat #11668-019)을 384-웰 플레이트 내로, 각 siRNA 이중체의 4개 레플리케이트로 하여, 웰당 5μl의 siRNA 이중체에 첨가함으로써 형질감염시키고, 실온에서 15분 동안 배양하였다. 그후 ~5x103개 세포를 함유하는 35μl의 둘베코 개질된 이글 배지(ThermoFisher)를 siRNA 혼합물에 첨가하였다. 세포를 48시간 동안 배양한 다음 Firefly(형질감염 대조군) 및 Renilla(표적 서열에 융합) 루시페라제 측정을 수행하였다. 10nM, 1nM 및 0.1nM에서 3가지 용량 실험을 수행하였다.
BE(2)-C(ATCC)는 웰당 4.9μl의 Opti-MEM + 0.1μl의 RNAiMAX(Invitrogen, Carlsbad CA. cat # 13778-150)을 384-웰 플레이트 내로, 각 siRNA 이중체의 4개 레플리케이트로 하여, 웰당 5μl의 siRNA 이중체에 첨가함으로써 형질감염시키고, 실온에서 15분 동안 배양하였다. 그후 ~5x103개 세포를 함유하는 최소 필수 배지와 F12 배지의 1:1 혼합물(ThermoFisher) 40μl를 siRNA 혼합물에 첨가하였다. RNA 정제 전에 세포를 48시간 동안 배양하였다. 10nM 및 0.1nM에서 2가지 용량 실험을 수행하였다.
Neuro-2a(ATCC)는 웰당 4.9μl의 Opti-MEM + 0.1μl의 RNAiMAX(Invitrogen, Carlsbad CA. cat # 13778-150)을 384-웰 플레이트 내로, 각 siRNA 이중체의 4개 레플리케이트로 하여, 웰당 5μl의 siRNA 이중체에 첨가함으로써 형질감염시키고, 실온에서 15분 동안 배양하였다. 그후 ~5x103개 세포를 함유하는 최소 필수 배지(ThermoFisher) 40μl를 siRNA 혼합물에 첨가하였다. RNA 정제 전에 세포를 48시간 동안 배양하였다. 10nM 및 0.1nM에서 2가지 용량 실험을 수행하였다.
DYNABEADS
mRNA
단리
키트를
사용한 총 RNA 단리:
RNA는 DYNABEAD(Invitrogen, cat#61012)를 사용하는 BioTek-EL406 플랫폼에서 자동화된 프로토콜을 사용하여 단리하였다. 간단히 말해서, 용해/결합 완충액 70ul 및 3μl의 자기 비드를 함유하는 용해 완충액 10ul를 세포가 있는 플레이트에 첨가하였다. 플레이트를 실온에서 10분 동안 전자기 진탕기에서 배양한 다음 자기 비드를 포획하고 상청액을 제거하였다. 그후 비드-결합된 RNA를 150μl 세척 완충액 A로 2회 세척하고 세척 완충액 B로 1회 세척하였다. 그후, 비드를 150μl 용출 완충액으로 세척하고, 재포획하고, 상청액을 제거하였다.
ABI
고용량 cDNA
역전사
키트
(Applied
Biosystems
, Foster City, CA, Cat #4368813)를 사용한 cDNA 합성:
반응당 1μl 10X 완충액, 0.4ul 25X dNTP, 1μl 10x 랜덤 프라이머, 0.5μl 역전사효소, 0.5μl RNase 억제제 및 6.6μl의 H2O를 함유하는 마스터 믹스 10μl를 상기 단리된 RNA에 첨가하였다. 플레이트를 밀봉하고, 혼합하고, 전자기 진탕기에서 실온에서 10분 동안 배양한 다음 37℃에서 2시간 동안 배양하였다.
실시간
PCR
:
2μl의 cDNA 및 5μl Lightcycler 480 프로브 마스터 믹스(Roche Cat # 04887301001)를 384 웰 플레이트(Roche cat # 04887301001)에서 웰당 0.5μl의 인간 GAPDH TaqMan 프로브(4326317E) 및 0.5μl C9orf72 인간 프로브(Hs00376619_m1, Thermo) 또는 0.5μl 마우스 GAPDH TaqMan 프로브(4352339E) 및 0.5μl C9orf72 마우스 프로브(Mm01216837_m1, Thermo)에 첨가하였다. 실시간 PCR은 LightCycler480 실시간 PCR 시스템(Roche)에서 수행하였다. 각 이중체를 적어도 2회 시험하였으며 데이터를 비표적화 대조군 siRNA로 형질감염된 세포에 대해 정규화하였다. 상대적 배수 변화를 계산하기 위해, △△Ct 방법을 사용하여 실시간 데이터를 분석하고 비-표적화 대조군 siRNA로 형질감염된 세포로 수행한 분석에 대해 정규화하였다.
Neuro2a 세포에서 표 12 및 13에 나열된 dsRNA 제제의 스크리닝의 결과는 표 16에 나타내어져 있다. BE(2)c 세포에서 표 12 및 13에 나열된 dsRNA 제제의 스크리닝의 결과는 표 17에 나타내어져 있다. Cos7 세포에서 표 14 및 15에 나열된 dsRNA 제제의 스크리닝의 결과는 표 18에 나타내어져 있다.





























실시예
6. 유전자이식 마우스의
생체내
평가
당해 실시예는 최대, 예를 들어, 450개의 GGGGCC 반복부를 갖는 인간 C9orf72 RNA를 발현하는 유전자이식 마우스에서 C9orf72 RNAi 제제의 생체내 평가를 위한 방법을 기술한다(예를 들어, 참조; Jiang, et al. (supra).
실시예 5에서 설계되고 분석된 선택된 dsRNA 제제의 능력은 최대 450개의 GGGGCC 반복부를 갖는 인간 C9orf72 RNA를 발현하는 마우스에서 센스- 또는 안티센스-함유 병소의 수준을 감소시키는 능력에 대해 평가된다.
간단히 말해서, 대조군 한배 새끼, 최대 450개의 GGGGCC 반복부를 갖는 인간 C9orf72 RNA에 대해 이형접합성인 마우스, 및 최대 450개의 GGGGCC 반복부를 갖는 인간 C9orf72 RNA에 대해 동형접합성인 마우스에 이중체 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1 또는 AD-463860.1, 또는 위약을 포함하는 관심 dsRNA 제제의 단일 용량을 척수강내 또는 피하로 투여한다. 투여한지 2주 후, 동물을 희생시키고, 대뇌 피질, 척수, 간, 비장 및 경부 림프절을 포함한 혈액 및 조직 샘플을 수집한다.
상기 논의된 바와 같이, 전사 대체 시작 및 종료 부위 생성의 차등 사용에 의해 생성된 3개의 C9orf72 전사체가 있다. 따라서, 유해한 반복부-함유 mRNA의 수준에 대한 dsRNA 제제의 투여 효과를 결정하기 위해, 반복부-함유 C9orf72 mRNA, 총 C9orf72 mRNA, 및 엑손 1b-함유 mRNA 수준을 qRT-PCR에 의해 피질 및 척수에서 결정한다(예를 들어, 참조; above and Jiang, supra).
결과는 단일 용량의 dsRNA 제제의 투여가 반복부-함유 C9orf72 mRNA의 생성을 억제하지만 총 C9orf72 mRNA 수준에 최소한의 영향만 미치고 엑손 1b-함유 mRNA 수준에는 영향을 미치지 않는다는 것을 입증한다.
C9orf72 센스 가닥- 및 안티센스 가닥-함유 병소 둘 다의 수 및/또는 형성을 감소시키는 dsRNA 제제의 효과를 결정하기 위해, 상기 Jiang에 기술된 FISH 방법을 상기로부터의 관심 이중체가 투여된 동물로부터 수득된 샘플에 사용한다. 사용되는 프로브는 센스 및 안티센스 RNA 헥사뉴클레오티드 반복부에 대한 것을 포함한다(Exiqon, Inc.). 모든 혼성화 단계는 RNase-비함유 조건하에서 수행된다. 15 마이크로미터 뇌 및 척수 OCT 동결 절편을 투과화하고 절편을 차단한다. 그후 절편을 변성 프로브와 혼성화한다. 혼성화 후, 슬라이드를 세척한다. 리포푸신의 자가형광을 켄칭시키고 세포핵을 DAPI로 염색한다. 마우스 전두엽 피질, 해마 치상회, 후비장 피질 및 소뇌 분자층에서 센스 및 안티센스 RNA 병소의 정량은 맹검 조사자에 의해 수행된다. 3 내지 6장의 무작위 사진을 100X 배율하에 공초점 현미경으로 촬영하고 200-400개의 세포를 계수한다.
결과는 단일 용량의 dsRNA 제제의 투여가 전두엽 피질, 해마 치상회, 후비장 피질 및 소뇌 분자층에서 센스 가닥- 및 안티센스 가닥-함유 병소의 수준을 감소시킨다는 것을 입증한다.
비정상적인 디펩티드 반복부 단백질 수준 및 폴리(GP) 및 폴리(GA) 부담 및 크기의 수준에 대한 제제의 투여 효과는 또한 상기 관심 이중체가 투여된 동물에서, 예를 들면, 상기 Jiang에 기술된 바와 같이 평가된다.
면역조직화학이 마우스 반뇌 및 척수에서 비정상적인 디펩티드 반복부 단백질 수준을 식별하고 평가하는데 사용된다. 간단히 말해서, 마우스 반뇌의 8 내지 10 마이크론 두께 시상 조각 또는 척수의 관상 조각을 포르말린-고정된, 파라핀-매봉된 블록으로부터 절단하고 유리 슬라이드에 장착한다. 건조 후, 슬라이드는 파라핀을 제거하고 크실렌에 재수화하고 세척하기 전에 알코올로 세척한다. 그후 슬라이드가 스팀처리하고 차단한다. 폴리(GP), 폴리(GA), 폴리(GR), 폴리(PA), 폴리(PR), GFAP, IBA-1, CD3, F4/80, 및 CD45R/B220에 대해 상업적으로 이용 가능한 항체로 밤새 염색한 후, HRP-접합된 이차 항체를 적용하고 기질과 함께 퍼옥시다제 활성을 발현시킨다. 절편을 Harris의 개질된 헤마톡실린으로 대조염색하고 커버슬립한다.
폴리(GP) 및 폴리(GA) 봉입체 부담 및 크기를 정량하기 위해, 폴리(GP) 또는 폴리(GA)에 대해 면역염색된 마우스 반뇌 절편을 40X 배율로 스캔하여 고해상도 디지털 이미지를 수득한다. 적절한 소프트웨어를 사용하여, 해마의 봉입체의 수 또는 팽대후부 피질의 묘사된 영역을 계수한다. 이 영역에서 봉입체의 크기를 측정하기 위해, 63X 배율하에 현미경으로 이미지를 촬영한다. 주어진 필드의 각 봉입체가 한 번만 분석되지만, 병소에 있는 봉입체에 대해서만 분석이 수행되도록 보장하기 위해 필드의 다중 이미지를 촬영할 수 있다. 이미지를 개방하고 확대하며, 윤곽 도구를 사용하여 각 봉입체를 추적하여 이의 면적(μm2)을 결정한다. 각 마우스에 대해, 각 시험 영역 내에서 μm2 단위의 봉입체의 평균 크기를 계산한다.
데이터는 단일 용량의 dsRNA 제제의 투여가 비정상적인 디펩티드 반복부 단백질 수준, 특히 폴리(GP) 및 폴리(GA) 봉입체 부담 및 크기의 수준을 감소시키는지 여부를 결정하는데 사용된다.













실시예
7. 엑손 1A,
인트론
1A, 및
인트론
1B를 표적으로 하는
C9orf72
siRNA
실시예 4에서 관찰된 현상을 추가로 조사하기 위해, GGGGCC 헥사뉴클레오티드의 300개 반복부를 갖는 인간화 C9orf72 유전자를 보유하는 마우스 배아 줄기 세포를 엑손 1A, 인트론 1A(헥사뉴클레오티드 반복부 확장의 상류), 및 인트론 1B(헥사뉴클레오티드 반복부 확장의 하류 및 엑손 1B의 하류)의 C9orf72 RNA 서열을 표적으로 하는 siRNA로 처리하였다. 3개의 siRNA는 엑손 1A를 표적으로 하고(siRNA E1A #1, E1A #2, 및 E1A #3), 4개의 siRNA는 헥사뉴클레오티드 반복부의 상류에 있는 인트론 1A를 표적으로 하고(siRNA I1A #2, I1A #4, I1A #6, 및 I1A #10), 2개의 siRNA는 헥사뉴클레오티드 반복부의 하류 및 엑손 1B의 하류에 있는 인트론 1B를 표적으로 하였다(siRNA I1B #1 및 I1B #6). 표 21은 이들 제제의 센스 가닥 및 안티센스 가닥 뉴클레오티드 서열을 제공한다. 인트론 1A를 표적으로 하는 4개의 siRNA에 의해 표적화된 C9orf72 영역은 실시예 4에서 사용된 siRNA와 함께 도 18A에 나타내어져 있다. Gapdh siRNA는 양성 대조군으로 사용되었으며, Gapdh 발현에 대한 TAQMAN® 정성적 PCR 분석은 Gapdh 발현의 녹다운을 확인시켜 주었다(데이터는 표시되지 않음). Mock은 음성 대조군으로 사용되었다. 25nM siRNA로 형질감염시킨 후 24시간, 48시간 및 72시간에 유전자 발현을 평가하였다. 그후 TAQMAN® 정성적 PCR 분석을 변형된 C9orf72 유전자좌의 이러한 영역을 검출하는 프로브 및 다양한 영역의 측면에 있는 프라이머를 사용하여 수행하였다. 프라이머 및 프로브는 도 18B 및 표 8에 나타내어져 있다. 인트론 1A(GGGGCC 반복부 확장의 상류)에서 인트론 서열을 표적으로 하는 siRNA는 C9orf72 성숙 mRNA에는 거의 영향을 미치지 않으면서(Thermo Fisher Scientific로부터의 엑손 2-3 분석 ID Mm01216829 m1; 24시간, 48시간, 및 72시간 시점에 대해 도 19B, 20B, 및 21B 참조) 인트론-1A-함유 C9orf72 RNA의 감소된 축적을 촉진하는 것으로 입증되었다(분석 F; 각각 24시간, 48시간, 및 72시간 시점에 대해 도 19A, 20A 및 21A 참조). 대조적으로, 엑손 1A 또는 인트론 1B(GGGGCC 반복부 확장의 하류 및 엑손 1B의 하류)를 표적으로 하는 siRNA는 인트론-1A-함유 C9orf72 RNA의 축적을 감소시키지 않았다(각각 24시간, 48시간, 및 72시간 시점에 대해 도 19A, 20A, 및 21A 참조). Gapdh siRNA 대조군은 C9orf72 RNA의 특정 녹다운이 단순히 세포에서 일반적인 RNA 간섭 활성의 결과가 아님을 보여준다.
C9orf72 성숙 RNA를 표적으로 하는 siRNA의 효과를 GGGGCC 헥사뉴클레오티드의 300개 반복부를 갖는 인간화 C9orf72 유전자를 지닌 마우스 배아 줄기 세포에서 C9orf72 인트론 1A를 표적으로 하는 siRNA의 효과와 비교하였다. siRNA를 세포에 전기천공하고, 전기천공 후 24시간 및 48시간에 유전자 발현을 평가하였다. C9orf72 성숙 RNA를 표적으로 하는 하나의 siRNA를 사용하고(siRNA #1), C9orf72 인트론 1A를 표적으로 하는 하나의 siRNA를 사용하였다(siRNA #12). 그후 TAQMAN® 정성적 PCR 분석을 변형된 C9orf72 유전자좌의 이러한 영역을 검출하는 프로브 및 다양한 영역의 측면에 있는 프라이머를 사용하여 수행하였다. 프라이머 및 프로브는 도 18B 및 표 8에 나타내어져 있다. siRNA 데이터는 엄격하였으며 C9orf72 성숙 RNA에 거의 영향을 미치지 않으면서(분석 D)(도 22E 및 23E의 siRNA #12 참조) 인트론 1A를 표적화할 때 인트론-1A-함유 C9orf72 RNA(분석 F)의 선택적 녹다운의 이전 결과를 재현하였다(도 22A 및 23A의 siRNA #12 참조). C9orf72 성숙 RNA를 표적으로 하는 siRNA는 반대 효과를 가졌다.
또한, 성숙 mRNA의 본체를 표적으로 하는 siRNA는 C9orf72 엑손 1A-엑손 2 스플라이싱된 mRNA를 녹다운시켰으며(분석 B), 이는 이들이 공유 결합되어 있음을 나타낸다. 대조적으로, C9orf7 2 인트론 1A(헥사뉴클레오티드 반복의 상류)를 표적으로 하는 siRNA는 엑손 1A-엑손 2 스플라이싱된 mRNA의 발현에 유의한 영향을 미치지 않았다. 도 22C 및 23C 참조 .
인트론-1A-표적화 siRNA는 또한 분석 C 결과에 의해 입증된 바와 같이 엑손 1B에서 개시되는 우세 정상 C9orf72 mRNA를 녹다운시키지 않았다. 도 22D 및 23D 참조. 분석 C(엑손 1B-엑손 2 스플라이싱)는 헥사뉴클레오티드 반복부에 인접한 인트론-1A-함유 RNA(아마도 병원성 반복부-함유 RNA)를 녹다운시키는 siRNA가 정상 mRNA를 보존하며, 따라서, C9orf72 mRNA의 가장 풍부한 형태로부터 번역된 C9orf72 단백질을 눈에 띄게 감소시키지 않는다는 것을 입증하였다.
분석 E는 인트론-1A-표적화 siRNA가 헥사뉴클레오티드 반복부의 상부에 있는 인트론 서열을 보유하고 엑손 1A에 연결된 RNA도 녹다운한다는 것을 보여준다. 도 22B 및 23B 참조. 이러한 유형의 전사체는 전사가 엑손 1A에서 시작되고 반복 서열을 통해 엑손 1B로 확장된 다음, 엑손 1B에서 엑손 2 및 mRNA의 본체로 스플라이싱되는 것일 수 있다. 이러한 RNA는 헥사뉴클레오티드 반복부에 의해 암호화된 디펩티드 반복부 단백질 합성을 위한 주형으로 작용할 수 있다. 본 발명자들의 결과는 인트론-1A-표적화 siRNA가 이러한 전사체를 파괴하고 본 발명자들이 이 siRNA로 처리할 때 관찰한 적어도 폴리(Gly-Ala)에 대해 DPR 단백질 합성의 감소에 기여할 것임을 보여준다. 요약하면, 헥사뉴클레오티드 반복부의 상류에 있는 인트론을 표적으로 하는 siRNA는 성숙 C9orf72 mRNA를 보존한다. 이들은 인접한 인트론에 연결된 엑손 1A를 갖는 RNA를 포함하여 인트론-함유 C9orf72 전사체에 대해 매우 특이적이다.

































<110> ALNYLAM PHARMACEUTICALS, INC.
REGENERON PHARMACEUTICALS, INC.
<120> HUMAN CHROMOSOME 9 OPEN READING FRAME 72 (C9ORF72) IRNA AGENT
COMPOSITIONS AND METHODS OF USE THEREOF
<130> 121301-10220
<140> PCT/US2020/064159
<141> 2020-12-10
<150> 62/947,605
<151> 2019-12-13
<150> 62/947,768
<151> 2019-12-13
<160> 1074
<170> PatentIn version 3.5
<210> 1
<211> 6
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1
ggggcc 6
<210> 2
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 2
aaagaccuga uaaagauuaa u 21
<210> 3
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 3
auuaaucuuu aucaggucuu uuc 23
<210> 4
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 4
aaaagaccug auaaagauua a 21
<210> 5
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 5
uuaaucuuua ucaggucuuu ucu 23
<210> 6
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 6
cugauaaaga uuaaccagaa u 21
<210> 7
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 7
auucugguua aucuuuauca ggu 23
<210> 8
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 8
aaagauuaac cagaagaaaa u 21
<210> 9
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 9
auuuucuucu gguuaaucuu uau 23
<210> 10
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 10
auaaagauua accagaagaa a 21
<210> 11
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 11
uuucuucugg uuaaucuuua uca 23
<210> 12
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 12
agaaaagacc ugauaaagau u 21
<210> 13
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 13
aaucuuuauc aggucuuuuc uug 23
<210> 14
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 14
gaagaccuuu cuacacuagu u 21
<210> 15
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 15
aacuagtgua gaaaggucuu cca 23
<210> 16
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 16
uagcugauac aguacucaau u 21
<210> 17
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 17
aauugaguac uguaucagcu aua 23
<210> 18
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 18
caagacagag auugcuuuaa u 21
<210> 19
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 19
auuaaagcaa ucucugucuu ggc 23
<210> 20
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 20
gucuuacaca gagacacucu a 21
<210> 21
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 21
uagagugucu cuguguaaga cau 23
<210> 22
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 22
uguugccaag acagagauug u 21
<210> 23
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 23
acaauctcug ucuuggcaac agc 23
<210> 24
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 24
ugauacagua cucaaugaug a 21
<210> 25
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 25
ucaucatuga guacuguauc agc 23
<210> 26
<211> 3261
<212> DNA
<213> Homo sapiens
<400> 26
gggcggggct gcggttgcgg tgcctgcgcc cgcggcggcg gaggcgcagg cggtggcgag 60
tggatatctc cggagcattt ggataatgtg acagttggaa tgcagtgatg tcgactcttt 120
gcccaccgcc atctccagct gttgccaaga cagagattgc tttaagtggc aaatcacctt 180
tattagcagc tacttttgct tactgggaca atattcttgg tcctagagta aggcacattt 240
gggctccaaa gacagaacag gtacttctca gtgatggaga aataactttt cttgccaacc 300
acactctaaa tggagaaatc cttcgaaatg cagagagtgg tgctatagat gtaaagtttt 360
ttgtcttgtc tgaaaaggga gtgattattg tttcattaat ctttgatgga aactggaatg 420
gggatcgcag cacatatgga ctatcaatta tacttccaca gacagaactt agtttctacc 480
tcccacttca tagagtgtgt gttgatagat taacacatat aatccggaaa ggaagaatat 540
ggatgcataa ggaaagacaa gaaaatgtcc agaagattat cttagaaggc acagagagaa 600
tggaagatca gggtcagagt attattccaa tgcttactgg agaagtgatt cctgtaatgg 660
aactgctttc atctatgaaa tcacacagtg ttcctgaaga aatagatata gctgatacag 720
tactcaatga tgatgatatt ggtgacagct gtcatgaagg ctttcttctc aatgccatca 780
gctcacactt gcaaacctgt ggctgttccg ttgtagtagg tagcagtgca gagaaagtaa 840
ataagatagt cagaacatta tgcctttttc tgactccagc agagagaaaa tgctccaggt 900
tatgtgaagc agaatcatca tttaaatatg agtcagggct ctttgtacaa ggcctgctaa 960
aggattcaac tggaagcttt gtgctgcctt tccggcaagt catgtatgct ccatatccca 1020
ccacacacat agatgtggat gtcaatactg tgaagcagat gccaccctgt catgaacata 1080
tttataatca gcgtagatac atgagatccg agctgacagc cttctggaga gccacttcag 1140
aagaagacat ggctcaggat acgatcatct acactgacga aagctttact cctgatttga 1200
atatttttca agatgtctta cacagagaca ctctagtgaa agccttcctg gatcaggtct 1260
ttcagctgaa acctggctta tctctcagaa gtactttcct tgcacagttt ctacttgtcc 1320
ttcacagaaa agccttgaca ctaataaaat atatagaaga cgatacgcag aagggaaaaa 1380
agccctttaa atctcttcgg aacctgaaga tagaccttga tttaacagca gagggcgatc 1440
ttaacataat aatggctctg gctgagaaaa ttaaaccagg cctacactct tttatctttg 1500
gaagaccttt ctacactagt gtgcaagaac gagatgttct aatgactttt taaatgtgta 1560
acttaataag cctattccat cacaatcatg atcgctggta aagtagctca gtggtgtggg 1620
gaaacgttcc cctggatcat actccagaat tctgctctca gcaattgcag ttaagtaagt 1680
tacactacag ttctcacaag agcctgtgag gggatgtcag gtgcatcatt acattgggtg 1740
tctcttttcc tagatttatg cttttgggat acagacctat gtttacaata taataaatat 1800
tattgctatc ttttaaagat ataataatag gatgtaaact tgaccacaac tactgttttt 1860
ttgaaataca tgattcatgg tttacatgtg tcaaggtgaa atctgagttg gcttttacag 1920
atagttgact ttctatcttt tggcattctt tggtgtgtag aattactgta atacttctgc 1980
aatcaactga aaactagagc ctttaaatga tttcaattcc acagaaagaa agtgagcttg 2040
aacataggat gagctttaga aagaaaattg atcaagcaga tgtttaattg gaattgatta 2100
ttagatccta ctttgtggat ttagtccctg ggattcagtc tgtagaaatg tctaatagtt 2160
ctctatagtc cttgttcctg gtgaaccaca gttagggtgt tttgtttatt ttattgttct 2220
tgctattgtt gatattctat gtagttgagc tctgtaaaag gaaattgtat tttatgtttt 2280
agtaattgtt gccaactttt taaattaatt ttcattattt ttgagccaaa ttgaaatgtg 2340
cacctcctgt gccttttttc tccttagaaa atctaattac ttggaacaag ttcagatttc 2400
actggtcagt cattttcatc ttgttttctt cttgctaagt cttaccatgt acctgctttg 2460
gcaatcattg caactctgag attataaaat gccttagaga atatactaac taataagatc 2520
tttttttcag aaacagaaaa tagttccttg agtacttcct tcttgcattt ctgcctatgt 2580
ttttgaagtt gttgctgttt gcctgcaata ggctataagg aatagcagga gaaattttac 2640
tgaagtgctg ttttcctagg tgctactttg gcagagctaa gttatctttt gttttcttaa 2700
tgcgtttgga ccattttgct ggctataaaa taactgatta atataattct aacacaatgt 2760
tgacattgta gttacacaaa cacaaataaa tattttattt aaaattctgg aagtaatata 2820
aaagggaaaa tatatttata agaaagggat aaaggtaata gagcccttct gccccccacc 2880
caccaaattt acacaacaaa atgacatgtt cgaatgtgaa aggtcataat agctttccca 2940
tcatgaatca gaaagatgtg gacagcttga tgttttagac aaccactgaa ctagatgact 3000
gttgtactgt agctcagtca tttaaaaaat atataaatac taccttgtag tgtcccatac 3060
tgtgtttttt acatggtaga ttcttattta agtgctaact ggttattttc tttggctggt 3120
ttattgtact gttatacaga atgtaagttg tacagtgaaa taagttatta aagcatgtgt 3180
aaacattgtt atatatcttt tctcctaaat ggagaatttt gaataaaata tatttgaaat 3240
tttaaaaaaa aaaaaaaaaa a 3261
<210> 27
<211> 481
<212> PRT
<213> Homo sapiens
<400> 27
Met Ser Thr Leu Cys Pro Pro Pro Ser Pro Ala Val Ala Lys Thr Glu
1 5 10 15
Ile Ala Leu Ser Gly Lys Ser Pro Leu Leu Ala Ala Thr Phe Ala Tyr
20 25 30
Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala Pro Lys
35 40 45
Thr Glu Gln Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu Ala Asn
50 55 60
His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly Ala Ile
65 70 75 80
Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile Val Ser
85 90 95
Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr Gly Leu
100 105 110
Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro Leu His
115 120 125
Arg Val Cys Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly Arg Ile
130 135 140
Trp Met His Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Ile Leu Glu
145 150 155 160
Gly Thr Glu Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro Met Leu
165 170 175
Thr Gly Glu Val Ile Pro Val Met Glu Leu Leu Ser Ser Met Lys Ser
180 185 190
His Ser Val Pro Glu Glu Ile Asp Ile Ala Asp Thr Val Leu Asn Asp
195 200 205
Asp Asp Ile Gly Asp Ser Cys His Glu Gly Phe Leu Leu Asn Ala Ile
210 215 220
Ser Ser His Leu Gln Thr Cys Gly Cys Ser Val Val Val Gly Ser Ser
225 230 235 240
Ala Glu Lys Val Asn Lys Ile Val Arg Thr Leu Cys Leu Phe Leu Thr
245 250 255
Pro Ala Glu Arg Lys Cys Ser Arg Leu Cys Glu Ala Glu Ser Ser Phe
260 265 270
Lys Tyr Glu Ser Gly Leu Phe Val Gln Gly Leu Leu Lys Asp Ser Thr
275 280 285
Gly Ser Phe Val Leu Pro Phe Arg Gln Val Met Tyr Ala Pro Tyr Pro
290 295 300
Thr Thr His Ile Asp Val Asp Val Asn Thr Val Lys Gln Met Pro Pro
305 310 315 320
Cys His Glu His Ile Tyr Asn Gln Arg Arg Tyr Met Arg Ser Glu Leu
325 330 335
Thr Ala Phe Trp Arg Ala Thr Ser Glu Glu Asp Met Ala Gln Asp Thr
340 345 350
Ile Ile Tyr Thr Asp Glu Ser Phe Thr Pro Asp Leu Asn Ile Phe Gln
355 360 365
Asp Val Leu His Arg Asp Thr Leu Val Lys Ala Phe Leu Asp Gln Val
370 375 380
Phe Gln Leu Lys Pro Gly Leu Ser Leu Arg Ser Thr Phe Leu Ala Gln
385 390 395 400
Phe Leu Leu Val Leu His Arg Lys Ala Leu Thr Leu Ile Lys Tyr Ile
405 410 415
Glu Asp Asp Thr Gln Lys Gly Lys Lys Pro Phe Lys Ser Leu Arg Asn
420 425 430
Leu Lys Ile Asp Leu Asp Leu Thr Ala Glu Gly Asp Leu Asn Ile Ile
435 440 445
Met Ala Leu Ala Glu Lys Ile Lys Pro Gly Leu His Ser Phe Ile Phe
450 455 460
Gly Arg Pro Phe Tyr Thr Ser Val Gln Glu Arg Asp Val Leu Met Thr
465 470 475 480
Phe
<210> 28
<211> 3356
<212> DNA
<213> Homo sapiens
<400> 28
acgtaaccta cggtgtcccg ctaggaaaga gaggtgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg gtgtgtcagc cgtccctgct gcccggttgc ttctcttttg 120
ggggcggggt ctagcaagag caggtgtggg tttaggagat atctccggag catttggata 180
atgtgacagt tggaatgcag tgatgtcgac tctttgccca ccgccatctc cagctgttgc 240
caagacagag attgctttaa gtggcaaatc acctttatta gcagctactt ttgcttactg 300
ggacaatatt cttggtccta gagtaaggca catttgggct ccaaagacag aacaggtact 360
tctcagtgat ggagaaataa cttttcttgc caaccacact ctaaatggag aaatccttcg 420
aaatgcagag agtggtgcta tagatgtaaa gttttttgtc ttgtctgaaa agggagtgat 480
tattgtttca ttaatctttg atggaaactg gaatggggat cgcagcacat atggactatc 540
aattatactt ccacagacag aacttagttt ctacctccca cttcatagag tgtgtgttga 600
tagattaaca catataatcc ggaaaggaag aatatggatg cataaggaaa gacaagaaaa 660
tgtccagaag attatcttag aaggcacaga gagaatggaa gatcagggtc agagtattat 720
tccaatgctt actggagaag tgattcctgt aatggaactg ctttcatcta tgaaatcaca 780
cagtgttcct gaagaaatag atatagctga tacagtactc aatgatgatg atattggtga 840
cagctgtcat gaaggctttc ttctcaatgc catcagctca cacttgcaaa cctgtggctg 900
ttccgttgta gtaggtagca gtgcagagaa agtaaataag atagtcagaa cattatgcct 960
ttttctgact ccagcagaga gaaaatgctc caggttatgt gaagcagaat catcatttaa 1020
atatgagtca gggctctttg tacaaggcct gctaaaggat tcaactggaa gctttgtgct 1080
gcctttccgg caagtcatgt atgctccata tcccaccaca cacatagatg tggatgtcaa 1140
tactgtgaag cagatgccac cctgtcatga acatatttat aatcagcgta gatacatgag 1200
atccgagctg acagccttct ggagagccac ttcagaagaa gacatggctc aggatacgat 1260
catctacact gacgaaagct ttactcctga tttgaatatt tttcaagatg tcttacacag 1320
agacactcta gtgaaagcct tcctggatca ggtctttcag ctgaaacctg gcttatctct 1380
cagaagtact ttccttgcac agtttctact tgtccttcac agaaaagcct tgacactaat 1440
aaaatatata gaagacgata cgcagaaggg aaaaaagccc tttaaatctc ttcggaacct 1500
gaagatagac cttgatttaa cagcagaggg cgatcttaac ataataatgg ctctggctga 1560
gaaaattaaa ccaggcctac actcttttat ctttggaaga cctttctaca ctagtgtgca 1620
agaacgagat gttctaatga ctttttaaat gtgtaactta ataagcctat tccatcacaa 1680
tcatgatcgc tggtaaagta gctcagtggt gtggggaaac gttcccctgg atcatactcc 1740
agaattctgc tctcagcaat tgcagttaag taagttacac tacagttctc acaagagcct 1800
gtgaggggat gtcaggtgca tcattacatt gggtgtctct tttcctagat ttatgctttt 1860
gggatacaga cctatgttta caatataata aatattattg ctatctttta aagatataat 1920
aataggatgt aaacttgacc acaactactg tttttttgaa atacatgatt catggtttac 1980
atgtgtcaag gtgaaatctg agttggcttt tacagatagt tgactttcta tcttttggca 2040
ttctttggtg tgtagaatta ctgtaatact tctgcaatca actgaaaact agagccttta 2100
aatgatttca attccacaga aagaaagtga gcttgaacat aggatgagct ttagaaagaa 2160
aattgatcaa gcagatgttt aattggaatt gattattaga tcctactttg tggatttagt 2220
ccctgggatt cagtctgtag aaatgtctaa tagttctcta tagtccttgt tcctggtgaa 2280
ccacagttag ggtgttttgt ttattttatt gttcttgcta ttgttgatat tctatgtagt 2340
tgagctctgt aaaaggaaat tgtattttat gttttagtaa ttgttgccaa ctttttaaat 2400
taattttcat tatttttgag ccaaattgaa atgtgcacct cctgtgcctt ttttctcctt 2460
agaaaatcta attacttgga acaagttcag atttcactgg tcagtcattt tcatcttgtt 2520
ttcttcttgc taagtcttac catgtacctg ctttggcaat cattgcaact ctgagattat 2580
aaaatgcctt agagaatata ctaactaata agatcttttt ttcagaaaca gaaaatagtt 2640
ccttgagtac ttccttcttg catttctgcc tatgtttttg aagttgttgc tgtttgcctg 2700
caataggcta taaggaatag caggagaaat tttactgaag tgctgttttc ctaggtgcta 2760
ctttggcaga gctaagttat cttttgtttt cttaatgcgt ttggaccatt ttgctggcta 2820
taaaataact gattaatata attctaacac aatgttgaca ttgtagttac acaaacacaa 2880
ataaatattt tatttaaaat tctggaagta atataaaagg gaaaatatat ttataagaaa 2940
gggataaagg taatagagcc cttctgcccc ccacccacca aatttacaca acaaaatgac 3000
atgttcgaat gtgaaaggtc ataatagctt tcccatcatg aatcagaaag atgtggacag 3060
cttgatgttt tagacaacca ctgaactaga tgactgttgt actgtagctc agtcatttaa 3120
aaaatatata aatactacct tgtagtgtcc catactgtgt tttttacatg gtagattctt 3180
atttaagtgc taactggtta ttttctttgg ctggtttatt gtactgttat acagaatgta 3240
agttgtacag tgaaataagt tattaaagca tgtgtaaaca ttgttatata tcttttctcc 3300
taaatggaga attttgaata aaatatattt gaaattttaa aaaaaaaaaa aaaaaa 3356
<210> 29
<211> 481
<212> PRT
<213> Homo sapiens
<400> 29
Met Ser Thr Leu Cys Pro Pro Pro Ser Pro Ala Val Ala Lys Thr Glu
1 5 10 15
Ile Ala Leu Ser Gly Lys Ser Pro Leu Leu Ala Ala Thr Phe Ala Tyr
20 25 30
Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala Pro Lys
35 40 45
Thr Glu Gln Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu Ala Asn
50 55 60
His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly Ala Ile
65 70 75 80
Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile Val Ser
85 90 95
Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr Gly Leu
100 105 110
Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro Leu His
115 120 125
Arg Val Cys Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly Arg Ile
130 135 140
Trp Met His Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Ile Leu Glu
145 150 155 160
Gly Thr Glu Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro Met Leu
165 170 175
Thr Gly Glu Val Ile Pro Val Met Glu Leu Leu Ser Ser Met Lys Ser
180 185 190
His Ser Val Pro Glu Glu Ile Asp Ile Ala Asp Thr Val Leu Asn Asp
195 200 205
Asp Asp Ile Gly Asp Ser Cys His Glu Gly Phe Leu Leu Asn Ala Ile
210 215 220
Ser Ser His Leu Gln Thr Cys Gly Cys Ser Val Val Val Gly Ser Ser
225 230 235 240
Ala Glu Lys Val Asn Lys Ile Val Arg Thr Leu Cys Leu Phe Leu Thr
245 250 255
Pro Ala Glu Arg Lys Cys Ser Arg Leu Cys Glu Ala Glu Ser Ser Phe
260 265 270
Lys Tyr Glu Ser Gly Leu Phe Val Gln Gly Leu Leu Lys Asp Ser Thr
275 280 285
Gly Ser Phe Val Leu Pro Phe Arg Gln Val Met Tyr Ala Pro Tyr Pro
290 295 300
Thr Thr His Ile Asp Val Asp Val Asn Thr Val Lys Gln Met Pro Pro
305 310 315 320
Cys His Glu His Ile Tyr Asn Gln Arg Arg Tyr Met Arg Ser Glu Leu
325 330 335
Thr Ala Phe Trp Arg Ala Thr Ser Glu Glu Asp Met Ala Gln Asp Thr
340 345 350
Ile Ile Tyr Thr Asp Glu Ser Phe Thr Pro Asp Leu Asn Ile Phe Gln
355 360 365
Asp Val Leu His Arg Asp Thr Leu Val Lys Ala Phe Leu Asp Gln Val
370 375 380
Phe Gln Leu Lys Pro Gly Leu Ser Leu Arg Ser Thr Phe Leu Ala Gln
385 390 395 400
Phe Leu Leu Val Leu His Arg Lys Ala Leu Thr Leu Ile Lys Tyr Ile
405 410 415
Glu Asp Asp Thr Gln Lys Gly Lys Lys Pro Phe Lys Ser Leu Arg Asn
420 425 430
Leu Lys Ile Asp Leu Asp Leu Thr Ala Glu Gly Asp Leu Asn Ile Ile
435 440 445
Met Ala Leu Ala Glu Lys Ile Lys Pro Gly Leu His Ser Phe Ile Phe
450 455 460
Gly Arg Pro Phe Tyr Thr Ser Val Gln Glu Arg Asp Val Leu Met Thr
465 470 475 480
Phe
<210> 30
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 30
gctatgcgat cgccgtctcg 20
<210> 31
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 31
ccccggcccc ggccccgaga 20
<210> 32
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 32
cgccgtctcg gggccggggc 20
<210> 33
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 33
ccgtctcggg gccggggccg 20
<210> 34
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 34
cggccggccc tcgagggtct 20
<210> 35
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 35
ggccggggcc gagaccctcg 20
<210> 36
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 36
gagggtctcg gccccggccc 20
<210> 37
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 37
ctcggccccg gccccggccc 20
<210> 38
<211> 16
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 38
ccccggcccc ggcccc 16
<210> 39
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 39
ggggccgggg ccggggggcc cc 22
<210> 40
<211> 3198
<212> DNA
<213> Mus musculus
<400> 40
gtgtccgggg cggggcggtc ccggggcggg gcccggagcg ggctgcggtt gcggtccctg 60
cgccggcggt gaaggcgcag cagcggcgag tggctattgc aagcgttcgg ataatgtgag 120
acctggaatg cagtgagacc tgggatgcag ggatgtcgac tatctgcccc ccaccatctc 180
ctgctgttgc caagacagag attgctttaa gtggtgaatc acccttgttg gcggctacct 240
ttgcttactg ggataatatt cttggtccta gagtaaggca tatttgggct ccaaagacag 300
accaagtgct tctcagtgat ggagaaataa cttttcttgc caaccacact ctaaatggag 360
aaattcttcg aaatgcagag agtggggcta tagatgtaaa attttttgtc ttatctgaaa 420
aaggggtaat tattgtttca ttaatcttcg acggaaactg gaatggagat cggagcactt 480
atggactatc aattatactg ccgcagacag agctgagctt ctacctccca cttcacagag 540
tgtgtgttga caggctaaca cacattattc gaaaaggaag aatatggatg cataaggaaa 600
gacaagaaaa tgtccagaaa attgtcttgg aaggcacaga gaggatggaa gatcagggtc 660
agagtatcat tcccatgctt actggggaag tcattcctgt aatggagctg cttgcatcta 720
tgaaatccca cagtgttcct gaagacattg atatagctga tacagtgctc aatgatgatg 780
acattggtga cagctgtcac gaaggctttc ttctcaatgc catcagctca cacctgcaga 840
cctgtggctg ttccgttgta gttggcagca gtgcagagaa agtaaataag atagtaagaa 900
cgctgtgcct ttttctgaca ccagcagaga ggaaatgctc caggctgtgt gaagcagaat 960
cgtcctttaa gtacgaatcg ggactctttg tgcaaggctt gctaaaggat gcaacaggca 1020
gttttgtcct acccttccgg caagttatgt atgccccgta ccccaccacg cacattgatg 1080
tggatgtcaa cactgtcaag cagatgccac cgtgtcatga acatatttat aatcaacgca 1140
gatacatgag gtcagagctg acagccttct ggagggcaac ttcagaagag gacatggcgc 1200
aggacaccat catctacaca gatgagagct tcactcctga tttgaatatt ttccaagatg 1260
tcttacacag agacactcta gtgaaagcct tcctggatca ggtcttccat ttgaagcctg 1320
gcctgtctct caggagtact ttccttgcac agttcctcct cattcttcac agaaaagcct 1380
tgacactaat caagtacatc gaggatgata cgcagaaggg gaaaaagccc tttaagtctc 1440
ttcggaacct gaagatagat cttgatttaa cagcagaggg cgatcttaac ataataatgg 1500
ctctagctga gaaaattaag ccaggcctac actctttcat ctttgggaga cctttctaca 1560
ctagtgtaca agaacgtgat gttctaatga ccttttgacc gtgtggtttg ctgtgtctgt 1620
ctcttcacag tcacacctgc tgttacagtg tctcagcagt gtgtgggcac atccttcctc 1680
ccgagtcctg ctgcaggaca gggtacacta cacttgtcag tagaagtctg tacctgatgt 1740
caggtgcatc gttacagtga atgactcttc ctagaataga tgtactcttt tagggcctta 1800
tgtttacaat tatcctaagt actattgctg tcttttaaag atatgaatga tggaatatac 1860
acttgaccat aactgctgat tggttttttg ttttgttttg tttgttttct tggaaactta 1920
tgattcctgg tttacatgta ccacactgaa accctcgtta gctttacaga taaagtgtga 1980
gttgacttcc tgcccctctg tgttctgtgg tatgtccgat tacttctgcc acagctaaac 2040
attagagcat ttaaagtttg cagttcctca gaaaggaact tagtctgact acagattagt 2100
tcttgagaga agacactgat agggcagagc tgtaggtgaa atcagttgtt agcccttcct 2160
ttatagacgt agtccttcag attcggtctg tacagaaatg ccgaggggtc atgcatgggc 2220
cctgagtatc gtgacctgtg acaagttttt tgttggttta ttgtagttct gtcaaagaaa 2280
gtggcatttg tttttataat tgttgccaac ttttaaggtt aattttcatt atttttgagc 2340
cgaattaaaa tgcgcacctc ctgtgccttt cccaatcttg gaaaatataa tttcttggca 2400
gagggtcaga tttcagggcc cagtcacttt catctgacca ccctttgcac ggctgccgtg 2460
tgcctggctt agattagaag tccttgttaa gtatgtcaga gtacattcgc tgataagatc 2520
tttgaagagc agggaagcgt cttgcctctt tcctttggtt tctgcctgta ctctggtgtt 2580
tcccgtgtca cctgcatcat aggaacagca gagaaatctg acccagtgct atttttctag 2640
gtgctactat ggcaaactca agtggtctgt ttctgttcct gtaacgttcg actatctcgc 2700
tagctgtgaa gtactgatta gtggagttct gtgcaacagc agtgtaggag tatacacaaa 2760
cacaaatatg tgtttctatt taaaactgtg gacttagcat aaaaagggag aatatattta 2820
ttttttacaa aagggataaa aatgggcccc gttcctcacc caccagattt agcgagaaaa 2880
agctttctat tctgaaaggt cacggtggct ttggcattac aaatcagaac aacacacact 2940
gaccatgatg gcttgtgaac taactgcaag gcactccgtc atggtaagcg agtaggtccc 3000
acctcctagt gtgccgctca ttgctttaca cagtagaatc ttatttgagt gctaattgtt 3060
gtctttgctg ctttactgtg ttgttataga aaatgtaagc tgtacagtga ataagttatt 3120
gaagcatgtg taaacactgt tatatatctt ttctcctaga tggggaattt tgaataaaat 3180
acctttgaaa ttctgtgt 3198
<210> 41
<211> 481
<212> PRT
<213> Mus musculus
<400> 41
Met Ser Thr Ile Cys Pro Pro Pro Ser Pro Ala Val Ala Lys Thr Glu
1 5 10 15
Ile Ala Leu Ser Gly Glu Ser Pro Leu Leu Ala Ala Thr Phe Ala Tyr
20 25 30
Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala Pro Lys
35 40 45
Thr Asp Gln Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu Ala Asn
50 55 60
His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly Ala Ile
65 70 75 80
Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile Val Ser
85 90 95
Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr Gly Leu
100 105 110
Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro Leu His
115 120 125
Arg Val Cys Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly Arg Ile
130 135 140
Trp Met His Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Val Leu Glu
145 150 155 160
Gly Thr Glu Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro Met Leu
165 170 175
Thr Gly Glu Val Ile Pro Val Met Glu Leu Leu Ala Ser Met Lys Ser
180 185 190
His Ser Val Pro Glu Asp Ile Asp Ile Ala Asp Thr Val Leu Asn Asp
195 200 205
Asp Asp Ile Gly Asp Ser Cys His Glu Gly Phe Leu Leu Asn Ala Ile
210 215 220
Ser Ser His Leu Gln Thr Cys Gly Cys Ser Val Val Val Gly Ser Ser
225 230 235 240
Ala Glu Lys Val Asn Lys Ile Val Arg Thr Leu Cys Leu Phe Leu Thr
245 250 255
Pro Ala Glu Arg Lys Cys Ser Arg Leu Cys Glu Ala Glu Ser Ser Phe
260 265 270
Lys Tyr Glu Ser Gly Leu Phe Val Gln Gly Leu Leu Lys Asp Ala Thr
275 280 285
Gly Ser Phe Val Leu Pro Phe Arg Gln Val Met Tyr Ala Pro Tyr Pro
290 295 300
Thr Thr His Ile Asp Val Asp Val Asn Thr Val Lys Gln Met Pro Pro
305 310 315 320
Cys His Glu His Ile Tyr Asn Gln Arg Arg Tyr Met Arg Ser Glu Leu
325 330 335
Thr Ala Phe Trp Arg Ala Thr Ser Glu Glu Asp Met Ala Gln Asp Thr
340 345 350
Ile Ile Tyr Thr Asp Glu Ser Phe Thr Pro Asp Leu Asn Ile Phe Gln
355 360 365
Asp Val Leu His Arg Asp Thr Leu Val Lys Ala Phe Leu Asp Gln Val
370 375 380
Phe His Leu Lys Pro Gly Leu Ser Leu Arg Ser Thr Phe Leu Ala Gln
385 390 395 400
Phe Leu Leu Ile Leu His Arg Lys Ala Leu Thr Leu Ile Lys Tyr Ile
405 410 415
Glu Asp Asp Thr Gln Lys Gly Lys Lys Pro Phe Lys Ser Leu Arg Asn
420 425 430
Leu Lys Ile Asp Leu Asp Leu Thr Ala Glu Gly Asp Leu Asn Ile Ile
435 440 445
Met Ala Leu Ala Glu Lys Ile Lys Pro Gly Leu His Ser Phe Ile Phe
450 455 460
Gly Arg Pro Phe Tyr Thr Ser Val Gln Glu Arg Asp Val Leu Met Thr
465 470 475 480
Phe
<210> 42
<211> 3435
<212> DNA
<213> Rattus norvegicus
<400> 42
cgtttgtagt gtcagccatc ccaattgcct gttccttctc tgtgggagtg gtgtctagac 60
agtccaggca gggtatgcta ggcaggtgcg ttttggttgc ctcagatcgc aacttgactc 120
cataacggtg accaaagaca aaagaaggaa accagattaa aaagaaccgg acacagaccc 180
ctgcagaatc tggagcggcc gtggttgggg gcggggctac gacggggcgg actcgggggc 240
gtgggagggc ggggccgggg cggggcccgg agccggctgc ggttgcggtc cctgcgccgg 300
cggtgaaggc gcagcggcgg cgagtggcta ttgcaagcgt ttggataatg tgagacctgg 360
gatgcaggga tgtcgactat ctgcccccca ccatctcctg ctgttgccaa gacagagatt 420
gctttaagtg gtgaatcacc cttgttggcg gctacctttg cttactggga taatattctt 480
ggtcctagag taaggcacat ttgggctcca aagacagacc aagtactcct cagtgatgga 540
gaaatcactt ttcttgccaa ccacactctg aatggagaaa ttcttcggaa tgcggagagt 600
ggggcaatag atgtaaagtt ttttgtctta tctgaaaagg gcgtcattat tgtttcatta 660
atcttcgacg ggaactggaa cggagatcgg agcacttacg gactatcaat tatactgccg 720
cagacggagc tgagtttcta cctcccactg cacagagtgt gtgttgacag gctaacgcac 780
atcattcgaa aaggaaggat atggatgcac aaggaaagac aagaaaatgt ccagaaaatt 840
gtcttggaag gcaccgagag gatggaagat cagggtcaga gtatcatccc tatgcttact 900
ggggaggtca tccctgtgat ggagctgctt gcgtctatga gatcacacag tgttcctgaa 960
gacctcgata tagctgatac agtactcaat gatgatgaca ttggtgacag ctgtcatgaa 1020
ggctttcttc tcaatgccat cagctcacat ctgcagacct gcggctgttc tgtggtggta 1080
ggcagcagtg cagagaaagt aaataagata gtaagaacac tgtgcctttt tctgacacca 1140
gcagagagga agtgctccag gctgtgtgaa gccgaatcgt cctttaaata cgaatctgga 1200
ctctttgtac aaggcttgct aaaggatgcg actggcagtt ttgtactacc tttccggcaa 1260
gttatgtatg ccccttatcc caccacacac atcgatgtgg atgtcaacac tgtcaagcag 1320
atgccaccgt gtcatgaaca tatttataat caacgcagat acatgaggtc agagctgaca 1380
gccttctgga gggcaacttc agaagaggac atggctcagg acaccatcat ctacacagat 1440
gagagcttca ctcctgattt gaatattttc caagatgtct tacacagaga cactctagtg 1500
aaagcctttc tggatcaggt cttccatttg aagcctggcc tgtctctcag gagtactttc 1560
cttgcacagt tcctcctcat tcttcacaga aaagccttga cactaatcaa gtacatagag 1620
gatgacacgc agaaggggaa aaagcccttt aagtctcttc ggaacctgaa gatagatctt 1680
gatttaacag cagagggcga ccttaacata ataatggctc tagctgagaa aattaagcca 1740
ggcctacact ctttcatctt cgggagacct ttctacacta gtgtccaaga acgtgatgtt 1800
ctaatgactt tttaaacatg tggtttgctc cgtgtgtctc atgacagtca cacttgctgt 1860
tacagtgtct cagcgctttg gacacatcct tcctccaggg tcctgccgca ggacacgtta 1920
cactacactt gtcagtagag gtctgtacca gatgtcaggt acatcgttgt agtgaatgtc 1980
tcttttccta gactagatgt accctcgtag ggacttatgt ttacaaccct cctaagtact 2040
agtgctgtct tgtaaggata cgaatgaagg gatgtaaact tcaccacaac tgctggttgg 2100
ttttgttgtt tttgtttttt gaaacttata attcatggtt tacatgcatc acactgaaac 2160
cctagttagc tttttacagg taagctgtga gttgactgcc tgtccctgtg ttctctggcc 2220
tgtacgatct gtggcgtgta ggatcacttt tgcaacaact aaaaactaaa gcactttgtt 2280
tgcagttcta cagaaagcaa cttagtctgt ctgcagattc gtttttgaaa gaagacatga 2340
gaaagcggag ttttaggtga agtcagttgt tggatcttcc tttatagact tagtccttta 2400
gatgtggtct gtatagacat gcccaaccat catgcatggg cactgaatat cgtgaactgt 2460
ggtatgcttt ttgttggttt attgtacttc tgtcaaagaa agtggcattg gtttttataa 2520
ttgttgccaa gttttaaggt taattttcat tatttttgag ccaaattaaa atgtgcacct 2580
cctgtgcctt tcccaatctt ggaaaatata atttcttggc agaaggtcag atttcagggc 2640
ccagtcactt tcgtctgact tccctttgca cagtccgcca tgggcctggc ttagaagttc 2700
ttgtaaacta tgccagagag tacattcgct gataaaatct tctttgcaga gcaggagagc 2760
ttcttgcctc tttcctttca tttctgcctg gactttggtg ttctccacgt tccctgcatc 2820
ctaaggacag caggagaact ctgaccccag tgctatttct ctaggtgcta ttgtggcaaa 2880
ctcaagcggt ccgtctctgt ccctgtaacg ttcgtacctt gctggctgtg aagtactgac 2940
tggtaaagct ccgtgctaca gcagtgtagg gtatacacaa acacaagtaa gtgttttatt 3000
taaaactgtg gacttagcat aaaaagggag actatattta ttttttacaa aagggataaa 3060
aatggaaccc tttcctcacc caccagattt agtcagaaaa aaacattcta ttctgaaagg 3120
tcacagtggt tttgacatga cacatcagaa caacgcacac tgtccatgat ggcttatgaa 3180
ctccaagtca ctccatcatg gtaaatgggt agatccctcc ttctagtgtg ccacaccatt 3240
gcttcccaca gtagaatctt atttaagtgc taagtgttgt ctctgctggt ttactctgtt 3300
gttttagaga atgtaagttg tatagtgaat aagttattga agcatgtgta aacactgtta 3360
tacatctttt ctcctagatg gggaatttgg aataaaatac ctttaaaatt caaaaaaaaa 3420
aaaaaaaaaa aaaaa 3435
<210> 43
<211> 481
<212> PRT
<213> Rattus norvegicus
<400> 43
Met Ser Thr Ile Cys Pro Pro Pro Ser Pro Ala Val Ala Lys Thr Glu
1 5 10 15
Ile Ala Leu Ser Gly Glu Ser Pro Leu Leu Ala Ala Thr Phe Ala Tyr
20 25 30
Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala Pro Lys
35 40 45
Thr Asp Gln Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu Ala Asn
50 55 60
His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly Ala Ile
65 70 75 80
Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile Val Ser
85 90 95
Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr Gly Leu
100 105 110
Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro Leu His
115 120 125
Arg Val Cys Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly Arg Ile
130 135 140
Trp Met His Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Val Leu Glu
145 150 155 160
Gly Thr Glu Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro Met Leu
165 170 175
Thr Gly Glu Val Ile Pro Val Met Glu Leu Leu Ala Ser Met Arg Ser
180 185 190
His Ser Val Pro Glu Asp Leu Asp Ile Ala Asp Thr Val Leu Asn Asp
195 200 205
Asp Asp Ile Gly Asp Ser Cys His Glu Gly Phe Leu Leu Asn Ala Ile
210 215 220
Ser Ser His Leu Gln Thr Cys Gly Cys Ser Val Val Val Gly Ser Ser
225 230 235 240
Ala Glu Lys Val Asn Lys Ile Val Arg Thr Leu Cys Leu Phe Leu Thr
245 250 255
Pro Ala Glu Arg Lys Cys Ser Arg Leu Cys Glu Ala Glu Ser Ser Phe
260 265 270
Lys Tyr Glu Ser Gly Leu Phe Val Gln Gly Leu Leu Lys Asp Ala Thr
275 280 285
Gly Ser Phe Val Leu Pro Phe Arg Gln Val Met Tyr Ala Pro Tyr Pro
290 295 300
Thr Thr His Ile Asp Val Asp Val Asn Thr Val Lys Gln Met Pro Pro
305 310 315 320
Cys His Glu His Ile Tyr Asn Gln Arg Arg Tyr Met Arg Ser Glu Leu
325 330 335
Thr Ala Phe Trp Arg Ala Thr Ser Glu Glu Asp Met Ala Gln Asp Thr
340 345 350
Ile Ile Tyr Thr Asp Glu Ser Phe Thr Pro Asp Leu Asn Ile Phe Gln
355 360 365
Asp Val Leu His Arg Asp Thr Leu Val Lys Ala Phe Leu Asp Gln Val
370 375 380
Phe His Leu Lys Pro Gly Leu Ser Leu Arg Ser Thr Phe Leu Ala Gln
385 390 395 400
Phe Leu Leu Ile Leu His Arg Lys Ala Leu Thr Leu Ile Lys Tyr Ile
405 410 415
Glu Asp Asp Thr Gln Lys Gly Lys Lys Pro Phe Lys Ser Leu Arg Asn
420 425 430
Leu Lys Ile Asp Leu Asp Leu Thr Ala Glu Gly Asp Leu Asn Ile Ile
435 440 445
Met Ala Leu Ala Glu Lys Ile Lys Pro Gly Leu His Ser Phe Ile Phe
450 455 460
Gly Arg Pro Phe Tyr Thr Ser Val Gln Glu Arg Asp Val Leu Met Thr
465 470 475 480
Phe
<210> 44
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 44
ccccggcccc ggccccgg 18
<210> 45
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 45
ggggccgggg ccggggc 17
<210> 46
<211> 198
<212> DNA
<213> Homo sapiens
<400> 46
gggtctagca agagcaggtg tgggtttagg aggtgtgtgt ttttgttttt cccaccctct 60
ctccccacta cttgctctca cagtactcgc tgagggtgaa caagaaaaga cctgataaag 120
attaaccaga agaaaacaag gagggaaaca accgcagcct gtagcaagct ctggaactca 180
ggagtcgcgc gctatgcg 198
<210> 47
<211> 118
<212> DNA
<213> Homo sapiens
<400> 47
gcgatcgcgg ggcgtggtcg gggcgggccc gggggcgggc ccggggcggg gctgcggttg 60
cggtgcctgc gcccgcggcg gcggaggcgc aggcggtggc gagtgggtga gtgaggag 118
<210> 48
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 48
ggccccggcc ccggcccc 18
<210> 49
<211> 12
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 49
agtcagtcag tc 12
<210> 50
<211> 49
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 50
ggggccgggg ccggggccgg ggccggggcc ggggccagtc agtcagtca 49
<210> 51
<211> 9
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 51
tgactgact 9
<210> 52
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 52
ccccggcccc ggccccgaga 20
<210> 53
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 53
cggccggccc tcgagggtct 20
<210> 54
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 54
gagggtctcg gccccggccc 20
<210> 55
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 55
ctcggccccg gccccggccc 20
<210> 56
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 56
gcuaugcgau cgccgucucg guuuuagagc uaugcuguuu ug 42
<210> 57
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 57
ccccggcccc ggccccgaga guuuuagagc uaugcuguuu ug 42
<210> 58
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 58
gcgcgctatg cgatcgccgt ctcggggccg gggccggggc cgg 43
<210> 59
<211> 62
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 59
ccggggccgg ggccggggcc gagaccctcg agggccggcc gctagcgcga tcgcggggcg 60
tg 62
<210> 60
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 60
ggggccgggg ccggggccgg ggccggggcc ggggcc 36
<210> 61
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 61
catcccaatt gccctttcc 19
<210> 62
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 62
cccacacctg ctcttgctag a 21
<210> 63
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 63
tctaggtgga aagtggg 17
<210> 64
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 64
gagcaggtgt gggtttagga 20
<210> 65
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 65
ccaggtctca ctgcattcca 20
<210> 66
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 66
attgcaagcg ttcggataat gtgaga 26
<210> 67
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 67
gctgtcacga aggctttctt c 21
<210> 68
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 68
gcactgctgc caactacaac 20
<210> 69
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 69
tcaatgccat cagctcacac ctgc 24
<210> 70
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 70
aagaggcgcg ggtagaa 17
<210> 71
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 71
cagcttcggt cagagaaatg ag 22
<210> 72
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 72
ctctcctcag agctcgacgc attt 24
<210> 73
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 73
ctgcacaatt tcagcccaag 20
<210> 74
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 74
caggtcatgt cccacagaat 20
<210> 75
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 75
catatgaggg cagcaatgca agtc 24
<210> 76
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 76
cgccgucucg gggccggggc guuuuagagc uaugcuguuu ug 42
<210> 77
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 77
ccgucucggg gccggggccg guuuuagagc uaugcuguuu ug 42
<210> 78
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 78
cggccggccc ucgagggucu guuuuagagc uaugcuguuu ug 42
<210> 79
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 79
ggccggggcc gagacccucg guuuuagagc uaugcuguuu ug 42
<210> 80
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 80
gagggucucg gccccggccc guuuuagagc uaugcuguuu ug 42
<210> 81
<211> 42
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 81
cucggccccg gccccggccc guuuuagagc uaugcuguuu ug 42
<210> 82
<211> 89
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 82
guuggaacca uucaaaacag cauagcaagu uaaaauaagg cuaguccguu aucaacuuga 60
aaaaguggca ccgagucggu gcuuuuuuu 89
<210> 83
<211> 5
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 83
ggggu 5
<210> 84
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 84
gcuaugcgau cgccgucucg 20
<210> 85
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 85
ccccggcccc ggccccgaga 20
<210> 86
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 86
cgccgucucg gggccggggc 20
<210> 87
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 87
ccgucucggg gccggggccg 20
<210> 88
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 88
cggccggccc ucgagggucu 20
<210> 89
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 89
ggccggggcc gagacccucg 20
<210> 90
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 90
gagggucucg gccccggccc 20
<210> 91
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 91
cucggccccg gccccggccc 20
<210> 92
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 92
tgcgcctccg ccgccgcggg cgcaggcacc gcaaccgca 39
<210> 93
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 93
cgcagcctgt agcaagctct ggaactcagg agtcg 35
<210> 94
<211> 1957
<212> DNA
<213> Homo sapiens
<400> 94
acgtaaccta cggtgtcccg ctaggaaaga gaggtgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg atatctccgg agcatttgga taatgtgaca gttggaatgc 120
agtgatgtcg actctttgcc caccgccatc tccagctgtt gccaagacag agattgcttt 180
aagtggcaaa tcacctttat tagcagctac ttttgcttac tgggacaata ttcttggtcc 240
tagagtaagg cacatttggg ctccaaagac agaacaggta cttctcagtg atggagaaat 300
aacttttctt gccaaccaca ctctaaatgg agaaatcctt cgaaatgcag agagtggtgc 360
tatagatgta aagttttttg tcttgtctga aaagggagtg attattgttt cattaatctt 420
tgatggaaac tggaatgggg atcgcagcac atatggacta tcaattatac ttccacagac 480
agaacttagt ttctacctcc cacttcatag agtgtgtgtt gatagattaa cacatataat 540
ccggaaagga agaatatgga tgcataagga aagacaagaa aatgtccaga agattatctt 600
agaaggcaca gagagaatgg aagatcaggg tcagagtatt attccaatgc ttactggaga 660
agtgattcct gtaatggaac tgctttcatc tatgaaatca cacagtgttc ctgaagaaat 720
agatatagct gatacagtac tcaatgatga tgatattggt gacagctgtc atgaaggctt 780
tcttctcaag taagaatttt tcttttcata aaagctggat gaagcagata ccatcttatg 840
ctcacctatg acaagatttg gaagaaagaa aataacagac tgtctactta gattgttcta 900
gggacattac gtatttgaac tgttgcttaa atttgtgtta tttttcactc attatatttc 960
tatatatatt tggtgttatt ccatttgcta tttaaagaaa ccgagtttcc atcccagaca 1020
agaaatcatg gccccttgct tgattctggt ttcttgtttt acttctcatt aaagctaaca 1080
gaatcctttc atattaagtt gtactgtaga tgaacttaag ttatttaggc gtagaacaaa 1140
attattcata tttatactga tctttttcca tccagcagtg gagtttagta cttaagagtt 1200
tgtgccctta aaccagactc cctggattaa tgctgtgtac ccgtgggcaa ggtgcctgaa 1260
ttctctatac acctatttcc tcatctgtaa aatggcaata atagtaatag tacctaatgt 1320
gtagggttgt tataagcatt gagtaagata aataatataa agcacttaga acagtgcctg 1380
gaacataaaa acacttaata atagctcata gctaacattt cctatttaca tttcttctag 1440
aaatagccag tatttgttga gtgcctacat gttagttcct ttactagttg ctttacatgt 1500
attatcttat attctgtttt aaagtttctt cacagttaca gattttcatg aaattttact 1560
tttaataaaa gagaagtaaa agtataaagt attcactttt atgttcacag tcttttcctt 1620
taggctcatg atggagtatc agaggcatga gtgtgtttaa cctaagagcc ttaatggctt 1680
gaatcagaag cactttagtc ctgtatctgt tcagtgtcag cctttcatac atcattttaa 1740
atcccatttg actttaagta agtcacttaa tctctctaca tgtcaatttc ttcagctata 1800
aaatgatggt atttcaataa ataaatacat taattaaatg atattatact gactaattgg 1860
gctgttttaa ggctcaataa gaaaatttct gtgaaaggtc tctagaaaat gtaggttcct 1920
atacaaataa aagataacat tgtgcttata aaaaaaa 1957
<210> 95
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 95
gatagtcgac atccctgcat c 21
<210> 96
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 96
ggtggcgagt ggctattg 18
<210> 97
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 97
aagcgttcgg ataatgtgag acctgg 26
<210> 98
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 98
tctcacagta ctcgctgagg gtga 24
<210> 99
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 99
aagagcaggt gtgggtttag 20
<210> 100
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 100
cggttgtttc cctccttgt 19
<210> 101
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 101
cccactactt gctctcacag 20
<210> 102
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 102
tacaggctgc ggttgttt 18
<210> 103
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 103
actcgctgag ggtgaacaag aaa 23
<210> 104
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 104
cgagtgggtg agtgagga 18
<210> 105
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 105
ttctacccgc gcctctt 17
<210> 106
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 106
atcctggcgg gtggctgttt 20
<210> 107
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 107
cggataatgt gagacctgga at 22
<210> 108
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 108
aaaggtagcc gccaacaa 18
<210> 109
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 109
accatctcct gctgttgcca aga 23
<210> 110
<211> 222
<212> PRT
<213> Homo sapiens
<400> 110
Met Ser Thr Leu Cys Pro Pro Pro Ser Pro Ala Val Ala Lys Thr Glu
1 5 10 15
Ile Ala Leu Ser Gly Lys Ser Pro Leu Leu Ala Ala Thr Phe Ala Tyr
20 25 30
Trp Asp Asn Ile Leu Gly Pro Arg Val Arg His Ile Trp Ala Pro Lys
35 40 45
Thr Glu Gln Val Leu Leu Ser Asp Gly Glu Ile Thr Phe Leu Ala Asn
50 55 60
His Thr Leu Asn Gly Glu Ile Leu Arg Asn Ala Glu Ser Gly Ala Ile
65 70 75 80
Asp Val Lys Phe Phe Val Leu Ser Glu Lys Gly Val Ile Ile Val Ser
85 90 95
Leu Ile Phe Asp Gly Asn Trp Asn Gly Asp Arg Ser Thr Tyr Gly Leu
100 105 110
Ser Ile Ile Leu Pro Gln Thr Glu Leu Ser Phe Tyr Leu Pro Leu His
115 120 125
Arg Val Cys Val Asp Arg Leu Thr His Ile Ile Arg Lys Gly Arg Ile
130 135 140
Trp Met His Lys Glu Arg Gln Glu Asn Val Gln Lys Ile Ile Leu Glu
145 150 155 160
Gly Thr Glu Arg Met Glu Asp Gln Gly Gln Ser Ile Ile Pro Met Leu
165 170 175
Thr Gly Glu Val Ile Pro Val Met Glu Leu Leu Ser Ser Met Lys Ser
180 185 190
His Ser Val Pro Glu Glu Ile Asp Ile Ala Asp Thr Val Leu Asn Asp
195 200 205
Asp Asp Ile Gly Asp Ser Cys His Glu Gly Phe Leu Leu Lys
210 215 220
<210> 111
<211> 6702
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<220>
<221> misc_feature
<222> (1)..(158)
<223> /note="Exon 1A"
<220>
<221> misc_feature
<222> (321)..(338)
<223> /note="3x Hexanucleotide Repeat"
<220>
<221> misc_feature
<222> (374)..(436)
<223> /note="Exon 1B"
<400> 111
acguaaccua cggugucccg cuaggaaaga gaggugcguc aaacagcgac aaguuccgcc 60
cacguaaaag augacgcuug gugugucagc cgucccugcu gcccgguugc uucucuuuug 120
ggggcggggu cuagcaagag cagguguggg uuuaggaggu guguguuuuu guuuuuccca 180
cccucucucc ccacuacuug cucucacagu acucgcugag ggugaacaag aaaagaccug 240
auaaagauua accagaagaa aacaaggagg gaaacaaccg cagccuguag caagcucugg 300
aacucaggag ucgcgcgcua ggggccgggg ccggggccgg ggcguggucg gggcgggccc 360
gggggcgggc ccggggcggg gcugcgguug cggugccugc gcccgcggcg gcggaggcgc 420
aggcgguggc gaguggguga gugaggaggc ggcauccugg cggguggcug uuugggguuc 480
ggcugccggg aagaggcgcg gguagaagcg ggggcucucc ucagagcucg acgcauuuuu 540
acuuucccuc ucauuucucu gaccgaagcu gggugucggg cuuucgccuc uagcgacugg 600
uggaauugcc ugcauccggg ccccgggcuu cccggcggcg gcggcggcgg cggcggcgca 660
gggacaaggg auggggaucu ggccucuucc uugcuuuccc gcccucagua cccgagcugu 720
cuccuucccg gggacccgcu gggagcgcug ccgcugcggg cucgagaaaa gggagccucg 780
gguacugaga ggccucgccu gggggaaggc cggagggugg gcggcgcgcg gcuucugcgg 840
accaagucgg gguucgcuag gaacccgaga cggucccugc cggcgaggag aucaugcggg 900
augagauggg gguguggaga cgccugcaca auuucagccc aagcuucuag agagugguga 960
ugacuugcau augagggcag caaugcaagu cggugugcuc cccauucugu gggacaugac 1020
cugguugcuu cacagcuccg agaugacaca gacuugcuua aaggaaguga cuauugugac 1080
uugggcauca cuugacugau gguaaucagu ugucuaaaga agugcacaga uuacaugucc 1140
gugugcucau ugggucuauc uggccgcguu gaacaccacc aggcuuugua uucagaaaca 1200
ggagggaggu ccugcacuuu cccaggaggg guggcccuuu cagaugcaau cgagauuguu 1260
aggcucuggg agaguaguug ccugguugug gcaguuggua aauuucuauu caaacaguug 1320
ccaugcacca guuguucaca acaaggguac guaaucuguc uggcauuacu ucuacuuuug 1380
uacaaaggau caaaaaaaaa aaagauacug uuaagauaug auuuuucuca gacuuuggga 1440
aacuuuuaac auaaucugug aauaucacag aaacaagacu aucauauagg ggauauuaau 1500
aaccuggagu cagaauacuu gaaauacggu gucauuugac acgggcauug uugucaccac 1560
cucugccaag gccugccacu uuaggaaaac ccugaaucag uuggaaacug cuacaugcug 1620
auaguacauc ugaaacaaga acgagaguaa uuaccacauu ccagauuguu cacuaagcca 1680
gcauuuaccu gcuccaggaa aaaauuacaa gcaccuuaug aaguugauaa aauauuuugu 1740
uuggcuaugu uggcacucca caauuugcuu ucagagaaac aaaguaaacc aaggaggacu 1800
ucuguuuuuc aagucugccc ucggguucua uucuacguua auuagauagu ucccaggagg 1860
acuagguuag ccuaccuauu gucugagaaa cuuggaacug ugagaaaugg ccagauagug 1920
auaugaacuu caccuuccag ucuucccuga uguugaagau ugagaaagug uugugaacuu 1980
ucugguacug uaaacaguuc acuguccuug aagugguccu gggcagcucc uguuguggaa 2040
aguggacggu uuaggauccu gcuucucuuu gggcugggag aaaauaaaca gcaugguuac 2100
aaguauugag agccagguug gagaaggugg cuuacaccug uaaugccaga gcuuugggag 2160
gcggaggcaa gaggaucacu ugaagccagg aguucaagcu caaccugggc aacguagacc 2220
cugucucuac aaaaaauuaa aaacuuagcc gggcguggug augugcaccu guaguccuag 2280
cuacuuggga ggcugaggca ggagggucau uugagcccaa gaguuugaag uuaccgagag 2340
cuaugauccu gccagugcau uccagccugg augacaaaac gagacccugu cucuaaaaaa 2400
caagaaguga gggcuuuaug auuguagaau uuucacuaca auagcagugg accaaccacc 2460
uuucuaaaua ccaaucaggg aagagauggu ugauuuuuua acagacguuu aaagaaaaag 2520
caaaaccuca aacuuagcac ucuacuaaca guuuuagcag auguuaauua auguaaucau 2580
gucugcaugu augggauuau uuccagaaag uguauuggga aaccucucau gaacccugug 2640
agcaagccac cgucucacuc aauuugaauc uuggcuuccc ucaaaagacu ggcuaauguu 2700
ugguaacucu cuggaguaga cagcacuaca uguacguaag auagguacau aaacaacuau 2760
ugguuuugag cugauuuuuu ucagcugcau uugcauguau ggauuuuucu caccaaagac 2820
gaugacuuca aguauuagua aaauaauugu acagcucucc ugauuauacu ucucugugac 2880
auuucauuuc ccaggcuauu ucuuuuggua ggauuuaaaa cuaagcaauu caguaugauc 2940
uuuguccuuc auuuucuuuc uuauucuuuu uguuuguuug uuuguuuguu uuuuucuuga 3000
ggcagagucu cucucugucg cccaggcugg agugcagugg cgccaucuca gcucauugca 3060
accucugcca ccuccggguu caagagauuc uccugccuca gccucccgag uagcugggau 3120
uacagguguc caccaccaca cccggcuaau uuuuuguauu uuuaguagag gugggguuuc 3180
accauguugg ccaggcuggu cuugagcucc ugaccucagg ugauccaccu gccucggccu 3240
accaaagagc ugggauaaca ggugugaccc accaugcccg gcccauuuuu uuuuucuuau 3300
ucuguuagga gugagagugu aacuagcagu auaauaguuc aauuuucaca acgugguaaa 3360
aguuucccua uaauucaauc agauuuugcu ccaggguuca guucuguuuu aggaaauacu 3420
uuuauuuuca guuuaaugau gaaauauuag aguuguaaua uugccuuuau gauuauccac 3480
cuuuuuaacc uaaaagaaug aaagaaaaau auguuugcaa uauaauuuua ugguuguaug 3540
uuaacuuaau ucauuauguu ggccuccagu uugcuguugu uaguuaugac agcaguagug 3600
ucauuaccau uucaauucag auuacauucc uauauuugau cauuguaaac ugacugcuua 3660
cauuguauua aaaacagugg auauuuuaaa gaagcuguac ggcuuauauc uagugcuguc 3720
ucuuaagacu auuaaauuga uacaacauau uuaaaaguaa auauuaccua aaugaauuuu 3780
ugaaauuaca aauacacgug uuaaaacugu cguuguguuc aaccauuucu guacauacuu 3840
agaguuaacu guuuugccag gcucuguaug ccuacucaua auaugauaaa agcacucauc 3900
uaaugcucug uaaauagaag ucagugcuuu ccaucagacu gaacucucuu gacaagaugu 3960
ggaugaaauu cuuuaaguaa aauuguuuac uuugucauac auuuacagau caaauguuag 4020
cucccaaagc aaucauaugg caaagauagg uauaucauag uuugccuauu agcugcuuug 4080
uauugcuauu auuauaaaua gacuucacag uuuuagacuu gcuuagguga aauugcaauu 4140
cuuuuuacuu ucagucuuag auaacaaguc uucaauuaua guacaaucac acauugcuua 4200
ggaaugcauc auuaggcgau uuugucauua ugcaaacauc auagagugua cuuacacaaa 4260
ccuagauagu auagccuuua uguaccuagg ccguauggua uagucuguug cuccuaggcc 4320
acaaaccugu acaacuguua cuguacugaa uacuauagac aguuguaaca cagugguaaa 4380
uauuuaucua aauauaugca aacagagaaa agguacagua aaaguauggu auaaaagaua 4440
augguauacc uguguaggcc acuuaccacg aauggagcuu gcaggacuag aaguugcucu 4500
gggugaguca gugagugagu ggugaauuaa ugugaaggcc uagaacacug uacaccacug 4560
uagacuauaa acacaguacg cugaagcuac accaaauuua ucuuaacagu uuuucuucaa 4620
uaaaaaauua uaacuuuuua acuuuguaaa cuuuuuaauu uuuuaacuuu uaaaauacuu 4680
agcuugaaac acaaauacau uguauagcua uacaaaaaua uuuuuucuuu guauccuuau 4740
ucuagaagcu uuuuucuauu uucuauuuua aauuuuuuuu uuuacuuguu agucguuuuu 4800
guuaaaaacu aaaacacaca cacuuucacc uaggcauaga caggauuagg aucaucagua 4860
ucacucccuu ccaccucacu gccuuccacc uccacaucuu gucccacugg aagguuuuua 4920
ggggcaauaa cacacaugua gcugucaccu augauaacag ugcuuucugu ugaauaccuc 4980
cugaaggacu ugccugaggc uguuuuacau uuaacuuaaa aaaaaaaaaa guagaaggag 5040
ugcacucuaa aauaacaaua aaaggcauag uauagugaau acauaaacca gcaauguagu 5100
aguuuauuau caaguguugu acacuguaau aauuguaugu gcuauacuuu aaauaacuug 5160
caaaauagua cuaagaccuu augaugguua cagugucacu aaggcaauag cauauuuuca 5220
gguccauugu aaucuaaugg gacuaccauc auauaugcag ucuaccauug acugaaacgu 5280
uacauggcac auaacuguau uugcaagaau gauuuguuuu acauuaauau cacauaggau 5340
guaccuuuuu agagugguau guuuaugugg auuaagaugu acaaguugag caaggggacc 5400
aagagcccug gguucugucu uggaugugag cguuuauguu cuucuccuca ugucuguuuu 5460
cucauuaaau ucaaaggcuu gaacgggccc uauuuagccc uucuguuuuc uacguguucu 5520
aaauaacuaa agcuuuuaaa uucuagccau uuaguguaga acucucuuug cagugaugaa 5580
augcuguauu gguuucuugg cuagcauauu aaauauuuuu aucuuugucu ugauacuuca 5640
augucguuuu aaacaucagg aucgggcuuc aguauucuca uaaccagaga guucacugag 5700
gauacaggac uguuugccca uuuuuuguua uggcuccaga cuugugguau uuccaugucu 5760
uuuuuuuuuu uuuuuuuuuu gaccuuuuag cggcuuuaaa guauuucugu uguuaggugu 5820
uguauuacuu uucuaagauu acuuaacaaa gcaccacaaa cugaguggcu uuaaacaaca 5880
gcaauuuauu cucucacaau ucuagaagcu agaaguccga aaucaaagug uugacagggg 5940
caugaucuuc aagagagaag acucuuuccu ugccucuucc uggcuucugg ugguuaccag 6000
caauccugag uguuccuuuc uugccuugua guuucaacaa uccaguaucu gccuuuuguc 6060
uucacauggc ugucuaccau uugucucugu gucuccaaau cucucuccuu auaaacacag 6120
caguuauugg auuaggcccc acucuaaucc aguaugaccc cauuuuaaca ugauuacacu 6180
uauuucuaga uaaggucaca uucacguaca ccaaggguua ggaauugaac auaucuuuuu 6240
gggggacaca auucaaccca caagugucag ucucuagcug agccuuuccc uuccuguuuu 6300
ucuccuuuuu aguugcuaug gguuaggggc caaaucucca gucauacuag aauugcacau 6360
ggacuggaua uuugggaaua cugcgggucu auucuaugag cuuuaguaug uaacauuuaa 6420
uaucagugua aagaagcccu uuuuuaaguu auuucuuuga auuucuaaau guaugcccug 6480
aauauaagua acaaguuacc augucuugua aaaugaucau aucaacaaac auuuaaugug 6540
caccuacugu gcuaguugaa ugucuuuauc cugauaggag auaacaggau uccacaucuu 6600
ugacuuaaga ggacaaacca aauaugucua aaucauuugg gguuuugaug gauaucuuua 6660
aauugcugaa ccuaaucauu gguuucauau gucauuguuu ag 6702
<210> 112
<211> 6702
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<220>
<221> misc_feature
<222> (6267)..(6329)
<223> /note="Exon 1B"
<220>
<221> misc_feature
<222> (6365)..(6382)
<223> /note="3x Hexanucleotide Repeat"
<220>
<221> misc_feature
<222> (6545)..(6702)
<223> /note="Exon 1A"
<400> 112
cuaaacaaug acauaugaaa ccaaugauua gguucagcaa uuuaaagaua uccaucaaaa 60
ccccaaauga uuuagacaua uuugguuugu ccucuuaagu caaagaugug gaauccuguu 120
aucuccuauc aggauaaaga cauucaacua gcacaguagg ugcacauuaa auguuuguug 180
auaugaucau uuuacaagac augguaacuu guuacuuaua uucagggcau acauuuagaa 240
auucaaagaa auaacuuaaa aaagggcuuc uuuacacuga uauuaaaugu uacauacuaa 300
agcucauaga auagacccgc aguauuccca aauauccagu ccaugugcaa uucuaguaug 360
acuggagauu uggccccuaa cccauagcaa cuaaaaagga gaaaaacagg aagggaaagg 420
cucagcuaga gacugacacu uguggguuga auuguguccc ccaaaaagau auguucaauu 480
ccuaacccuu gguguacgug aaugugaccu uaucuagaaa uaaguguaau cauguuaaaa 540
uggggucaua cuggauuaga guggggccua auccaauaac ugcuguguuu auaaggagag 600
agauuuggag acacagagac aaaugguaga cagccaugug aagacaaaag gcagauacug 660
gauuguugaa acuacaaggc aagaaaggaa cacucaggau ugcugguaac caccagaagc 720
caggaagagg caaggaaaga gucuucucuc uugaagauca ugccccuguc aacacuuuga 780
uuucggacuu cuagcuucua gaauugugag agaauaaauu gcuguuguuu aaagccacuc 840
aguuuguggu gcuuuguuaa guaaucuuag aaaaguaaua caacaccuaa caacagaaau 900
acuuuaaagc cgcuaaaagg ucaaaaaaaa aaaaaaaaaa aaagacaugg aaauaccaca 960
agucuggagc cauaacaaaa aaugggcaaa caguccugua uccucaguga acucucuggu 1020
uaugagaaua cugaagcccg auccugaugu uuaaaacgac auugaaguau caagacaaag 1080
auaaaaauau uuaauaugcu agccaagaaa ccaauacagc auuucaucac ugcaaagaga 1140
guucuacacu aaauggcuag aauuuaaaag cuuuaguuau uuagaacacg uagaaaacag 1200
aagggcuaaa uagggcccgu ucaagccuuu gaauuuaaug agaaaacaga caugaggaga 1260
agaacauaaa cgcucacauc caagacagaa cccagggcuc uugguccccu ugcucaacuu 1320
guacaucuua auccacauaa acauaccacu cuaaaaaggu acauccuaug ugauauuaau 1380
guaaaacaaa ucauucuugc aaauacaguu augugccaug uaacguuuca gucaauggua 1440
gacugcauau augaugguag ucccauuaga uuacaaugga ccugaaaaua ugcuauugcc 1500
uuagugacac uguaaccauc auaaggucuu aguacuauuu ugcaaguuau uuaaaguaua 1560
gcacauacaa uuauuacagu guacaacacu ugauaauaaa cuacuacauu gcugguuuau 1620
guauucacua uacuaugccu uuuauuguua uuuuagagug cacuccuucu acuuuuuuuu 1680
uuuuuaaguu aaauguaaaa cagccucagg caaguccuuc aggagguauu caacagaaag 1740
cacuguuauc auaggugaca gcuacaugug uguuauugcc ccuaaaaacc uuccaguggg 1800
acaagaugug gagguggaag gcagugaggu ggaagggagu gauacugaug auccuaaucc 1860
ugucuaugcc uaggugaaag uguguguguu uuaguuuuua acaaaaacga cuaacaagua 1920
aaaaaaaaaa uuuaaaauag aaaauagaaa aaagcuucua gaauaaggau acaaagaaaa 1980
aauauuuuug uauagcuaua caauguauuu guguuucaag cuaaguauuu uaaaaguuaa 2040
aaaauuaaaa aguuuacaaa guuaaaaagu uauaauuuuu uauugaagaa aaacuguuaa 2100
gauaaauuug guguagcuuc agcguacugu guuuauaguc uacaguggug uacaguguuc 2160
uaggccuuca cauuaauuca ccacucacuc acugacucac ccagagcaac uucuaguccu 2220
gcaagcucca uucgugguaa guggccuaca cagguauacc auuaucuuuu auaccauacu 2280
uuuacuguac cuuuucucug uuugcauaua uuuagauaaa uauuuaccac uguguuacaa 2340
cugucuauag uauucaguac aguaacaguu guacagguuu guggccuagg agcaacagac 2400
uauaccauac ggccuaggua cauaaaggcu auacuaucua gguuugugua aguacacucu 2460
augauguuug cauaaugaca aaaucgccua augaugcauu ccuaagcaau gugugauugu 2520
acuauaauug aagacuuguu aucuaagacu gaaaguaaaa agaauugcaa uuucaccuaa 2580
gcaagucuaa aacugugaag ucuauuuaua auaauagcaa uacaaagcag cuaauaggca 2640
aacuaugaua uaccuaucuu ugccauauga uugcuuuggg agcuaacauu ugaucuguaa 2700
auguaugaca aaguaaacaa uuuuacuuaa agaauuucau ccacaucuug ucaagagagu 2760
ucagucugau ggaaagcacu gacuucuauu uacagagcau uagaugagug cuuuuaucau 2820
auuaugagua ggcauacaga gccuggcaaa acaguuaacu cuaaguaugu acagaaaugg 2880
uugaacacaa cgacaguuuu aacacgugua uuuguaauuu caaaaauuca uuuagguaau 2940
auuuacuuuu aaauauguug uaucaauuua auagucuuaa gagacagcac uagauauaag 3000
ccguacagcu ucuuuaaaau auccacuguu uuuaauacaa uguaagcagu caguuuacaa 3060
ugaucaaaua uaggaaugua aucugaauug aaaugguaau gacacuacug cugucauaac 3120
uaacaacagc aaacuggagg ccaacauaau gaauuaaguu aacauacaac cauaaaauua 3180
uauugcaaac auauuuuucu uucauucuuu uagguuaaaa agguggauaa ucauaaaggc 3240
aauauuacaa cucuaauauu ucaucauuaa acugaaaaua aaaguauuuc cuaaaacaga 3300
acugaacccu ggagcaaaau cugauugaau uauagggaaa cuuuuaccac guugugaaaa 3360
uugaacuauu auacugcuag uuacacucuc acuccuaaca gaauaagaaa aaaaaaaugg 3420
gccgggcaug gugggucaca ccuguuaucc cagcucuuug guaggccgag gcagguggau 3480
caccugaggu caggagcuca agaccagccu ggccaacaug gugaaacccc accucuacua 3540
aaaauacaaa aaauuagccg gguguggugg uggacaccug uaaucccagc uacucgggag 3600
gcugaggcag gagaaucucu ugaacccgga gguggcagag guugcaauga gcugagaugg 3660
cgccacugca cuccagccug ggcgacagag agagacucug ccucaagaaa aaaacaaaca 3720
aacaaacaaa caaaaagaau aagaaagaaa augaaggaca aagaucauac ugaauugcuu 3780
aguuuuaaau ccuaccaaaa gaaauagccu gggaaaugaa augucacaga gaaguauaau 3840
caggagagcu guacaauuau uuuacuaaua cuugaaguca ucgucuuugg ugagaaaaau 3900
ccauacaugc aaaugcagcu gaaaaaaauc agcucaaaac caauaguugu uuauguaccu 3960
aucuuacgua cauguagugc ugucuacucc agagaguuac caaacauuag ccagucuuuu 4020
gagggaagcc aagauucaaa uugagugaga cgguggcuug cucacagggu ucaugagagg 4080
uuucccaaua cacuuucugg aaauaauccc auacaugcag acaugauuac auuaauuaac 4140
aucugcuaaa acuguuagua gagugcuaag uuugagguuu ugcuuuuucu uuaaacgucu 4200
guuaaaaaau caaccaucuc uucccugauu gguauuuaga aaggugguug guccacugcu 4260
auuguaguga aaauucuaca aucauaaagc ccucacuucu uguuuuuuag agacaggguc 4320
ucguuuuguc auccaggcug gaaugcacug gcaggaucau agcucucggu aacuucaaac 4380
ucuugggcuc aaaugacccu ccugccucag ccucccaagu agcuaggacu acaggugcac 4440
aucaccacgc ccggcuaagu uuuuaauuuu uuguagagac agggucuacg uugcccaggu 4500
ugagcuugaa cuccuggcuu caagugaucc ucuugccucc gccucccaaa gcucuggcau 4560
uacaggugua agccaccuuc uccaaccugg cucucaauac uuguaaccau gcuguuuauu 4620
uucucccagc ccaaagagaa gcaggauccu aaaccgucca cuuuccacaa caggagcugc 4680
ccaggaccac uucaaggaca gugaacuguu uacaguacca gaaaguucac aacacuuucu 4740
caaucuucaa caucagggaa gacuggaagg ugaaguucau aucacuaucu ggccauuucu 4800
cacaguucca aguuucucag acaauaggua ggcuaaccua guccuccugg gaacuaucua 4860
auuaacguag aauagaaccc gagggcagac uugaaaaaca gaaguccucc uugguuuacu 4920
uuguuucucu gaaagcaaau uguggagugc caacauagcc aaacaaaaua uuuuaucaac 4980
uucauaaggu gcuuguaauu uuuuccugga gcagguaaau gcuggcuuag ugaacaaucu 5040
ggaauguggu aauuacucuc guucuuguuu cagauguacu aucagcaugu agcaguuucc 5100
aacugauuca ggguuuuccu aaaguggcag gccuuggcag agguggugac aacaaugccc 5160
gugucaaaug acaccguauu ucaaguauuc ugacuccagg uuauuaauau ccccuauaug 5220
auagucuugu uucugugaua uucacagauu auguuaaaag uuucccaaag ucugagaaaa 5280
aucauaucuu aacaguaucu uuuuuuuuuu ugauccuuug uacaaaagua gaaguaaugc 5340
cagacagauu acguacccuu guugugaaca acuggugcau ggcaacuguu ugaauagaaa 5400
uuuaccaacu gccacaacca ggcaacuacu cucccagagc cuaacaaucu cgauugcauc 5460
ugaaagggcc accccuccug ggaaagugca ggaccucccu ccuguuucug aauacaaagc 5520
cugguggugu ucaacgcggc cagauagacc caaugagcac acggacaugu aaucugugca 5580
cuucuuuaga caacugauua ccaucaguca agugaugccc aagucacaau agucacuucc 5640
uuuaagcaag ucugugucau cucggagcug ugaagcaacc aggucauguc ccacagaaug 5700
gggagcacac cgacuugcau ugcugcccuc auaugcaagu caucaccacu cucuagaagc 5760
uugggcugaa auugugcagg cgucuccaca cccccaucuc aucccgcaug aucuccucgc 5820
cggcagggac cgucucgggu uccuagcgaa ccccgacuug guccgcagaa gccgcgcgcc 5880
gcccacccuc cggccuuccc ccaggcgagg ccucucagua cccgaggcuc ccuuuucucg 5940
agcccgcagc ggcagcgcuc ccagcggguc cccgggaagg agacagcucg gguacugagg 6000
gcgggaaagc aaggaagagg ccagaucccc aucccuuguc ccugcgccgc cgccgccgcc 6060
gccgccgccg ggaagcccgg ggcccggaug caggcaauuc caccagucgc uagaggcgaa 6120
agcccgacac ccagcuucgg ucagagaaau gagagggaaa guaaaaaugc gucgagcucu 6180
gaggagagcc cccgcuucua cccgcgccuc uucccggcag ccgaacccca aacagccacc 6240
cgccaggaug ccgccuccuc acucacccac ucgccaccgc cugcgccucc gccgccgcgg 6300
gcgcaggcac cgcaaccgca gccccgcccc gggcccgccc ccgggcccgc cccgaccacg 6360
ccccggcccc ggccccggcc ccuagcgcgc gacuccugag uuccagagcu ugcuacaggc 6420
ugcgguuguu ucccuccuug uuuucuucug guuaaucuuu aucaggucuu uucuuguuca 6480
cccucagcga guacugugag agcaaguagu ggggagagag ggugggaaaa acaaaaacac 6540
acaccuccua aacccacacc ugcucuugcu agaccccgcc cccaaaagag aagcaaccgg 6600
gcagcaggga cggcugacac accaagcguc aucuuuuacg ugggcggaac uugucgcugu 6660
uugacgcacc ucucuuuccu agcgggacac cguagguuac gu 6702
<210> 113
<211> 215
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<220>
<221> misc_feature
<222> (163)..(180)
<223> /note="3x Hexanucleotide Repeat"
<400> 113
guguguguuu uuguuuuucc cacccucucu ccccacuacu ugcucucaca guacucgcug 60
agggugaaca agaaaagacc ugauaaagau uaaccagaag aaaacaagga gggaaacaac 120
cgcagccugu agcaagcucu ggaacucagg agucgcgcgc uaggggccgg ggccggggcc 180
ggggcguggu cggggcgggc ccgggggcgg gcccg 215
<210> 114
<211> 215
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<220>
<221> misc_feature
<222> (36)..(53)
<223> /note="3x Hexanucleotide Repeat"
<400> 114
cgggcccgcc cccgggcccg ccccgaccac gccccggccc cggccccggc cccuagcgcg 60
cgacuccuga guuccagagc uugcuacagg cugcgguugu uucccuccuu guuuucuucu 120
gguuaaucuu uaucaggucu uuucuuguuc acccucagcg aguacuguga gagcaaguag 180
uggggagaga gggugggaaa aacaaaaaca cacac 215
<210> 115
<211> 162
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 115
guguguguuu uuguuuuucc cacccucucu ccccacuacu ugcucucaca guacucgcug 60
agggugaaca agaaaagacc ugauaaagau uaaccagaag aaaacaagga gggaaacaac 120
cgcagccugu agcaagcucu ggaacucagg agucgcgcgc ua 162
<210> 116
<211> 162
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 116
uagcgcgcga cuccugaguu ccagagcuug cuacaggcug cgguuguuuc ccuccuuguu 60
uucuucuggu uaaucuuuau caggucuuuu cuuguucacc cucagcgagu acugugagag 120
caaguagugg ggagagaggg ugggaaaaac aaaaacacac ac 162
<210> 117
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 117
caagaaaaga ccugauaaag auuaaccaga agaaaa 36
<210> 118
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 118
uuuucuucug guuaaucuuu aucaggucuu uucuug 36
<210> 119
<400> 119
000
<210> 120
<400> 120
000
<210> 121
<211> 3356
<212> DNA
<213> Homo sapiens
<400> 121
acgtaaccta cggtgtcccg ctaggaaaga gaggtgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg gtgtgtcagc cgtccctgct gcccggttgc ttctcttttg 120
ggggcggggt ctagcaagag caggtgtggg tttaggagat atctccggag catttggata 180
atgtgacagt tggaatgcag tgatgtcgac tctttgccca ccgccatctc cagctgttgc 240
caagacagag attgctttaa gtggcaaatc acctttatta gcagctactt ttgcttactg 300
ggacaatatt cttggtccta gagtaaggca catttgggct ccaaagacag aacaggtact 360
tctcagtgat ggagaaataa cttttcttgc caaccacact ctaaatggag aaatccttcg 420
aaatgcagag agtggtgcta tagatgtaaa gttttttgtc ttgtctgaaa agggagtgat 480
tattgtttca ttaatctttg atggaaactg gaatggggat cgcagcacat atggactatc 540
aattatactt ccacagacag aacttagttt ctacctccca cttcatagag tgtgtgttga 600
tagattaaca catataatcc ggaaaggaag aatatggatg cataaggaaa gacaagaaaa 660
tgtccagaag attatcttag aaggcacaga gagaatggaa gatcagggtc agagtattat 720
tccaatgctt actggagaag tgattcctgt aatggaactg ctttcatcta tgaaatcaca 780
cagtgttcct gaagaaatag atatagctga tacagtactc aatgatgatg atattggtga 840
cagctgtcat gaaggctttc ttctcaatgc catcagctca cacttgcaaa cctgtggctg 900
ttccgttgta gtaggtagca gtgcagagaa agtaaataag atagtcagaa cattatgcct 960
ttttctgact ccagcagaga gaaaatgctc caggttatgt gaagcagaat catcatttaa 1020
atatgagtca gggctctttg tacaaggcct gctaaaggat tcaactggaa gctttgtgct 1080
gcctttccgg caagtcatgt atgctccata tcccaccaca cacatagatg tggatgtcaa 1140
tactgtgaag cagatgccac cctgtcatga acatatttat aatcagcgta gatacatgag 1200
atccgagctg acagccttct ggagagccac ttcagaagaa gacatggctc aggatacgat 1260
catctacact gacgaaagct ttactcctga tttgaatatt tttcaagatg tcttacacag 1320
agacactcta gtgaaagcct tcctggatca ggtctttcag ctgaaacctg gcttatctct 1380
cagaagtact ttccttgcac agtttctact tgtccttcac agaaaagcct tgacactaat 1440
aaaatatata gaagacgata cgcagaaggg aaaaaagccc tttaaatctc ttcggaacct 1500
gaagatagac cttgatttaa cagcagaggg cgatcttaac ataataatgg ctctggctga 1560
gaaaattaaa ccaggcctac actcttttat ctttggaaga cctttctaca ctagtgtgca 1620
agaacgagat gttctaatga ctttttaaat gtgtaactta ataagcctat tccatcacaa 1680
tcatgatcgc tggtaaagta gctcagtggt gtggggaaac gttcccctgg atcatactcc 1740
agaattctgc tctcagcaat tgcagttaag taagttacac tacagttctc acaagagcct 1800
gtgaggggat gtcaggtgca tcattacatt gggtgtctct tttcctagat ttatgctttt 1860
gggatacaga cctatgttta caatataata aatattattg ctatctttta aagatataat 1920
aataggatgt aaacttgacc acaactactg tttttttgaa atacatgatt catggtttac 1980
atgtgtcaag gtgaaatctg agttggcttt tacagatagt tgactttcta tcttttggca 2040
ttctttggtg tgtagaatta ctgtaatact tctgcaatca actgaaaact agagccttta 2100
aatgatttca attccacaga aagaaagtga gcttgaacat aggatgagct ttagaaagaa 2160
aattgatcaa gcagatgttt aattggaatt gattattaga tcctactttg tggatttagt 2220
ccctgggatt cagtctgtag aaatgtctaa tagttctcta tagtccttgt tcctggtgaa 2280
ccacagttag ggtgttttgt ttattttatt gttcttgcta ttgttgatat tctatgtagt 2340
tgagctctgt aaaaggaaat tgtattttat gttttagtaa ttgttgccaa ctttttaaat 2400
taattttcat tatttttgag ccaaattgaa atgtgcacct cctgtgcctt ttttctcctt 2460
agaaaatcta attacttgga acaagttcag atttcactgg tcagtcattt tcatcttgtt 2520
ttcttcttgc taagtcttac catgtacctg ctttggcaat cattgcaact ctgagattat 2580
aaaatgcctt agagaatata ctaactaata agatcttttt ttcagaaaca gaaaatagtt 2640
ccttgagtac ttccttcttg catttctgcc tatgtttttg aagttgttgc tgtttgcctg 2700
caataggcta taaggaatag caggagaaat tttactgaag tgctgttttc ctaggtgcta 2760
ctttggcaga gctaagttat cttttgtttt cttaatgcgt ttggaccatt ttgctggcta 2820
taaaataact gattaatata attctaacac aatgttgaca ttgtagttac acaaacacaa 2880
ataaatattt tatttaaaat tctggaagta atataaaagg gaaaatatat ttataagaaa 2940
gggataaagg taatagagcc cttctgcccc ccacccacca aatttacaca acaaaatgac 3000
atgttcgaat gtgaaaggtc ataatagctt tcccatcatg aatcagaaag atgtggacag 3060
cttgatgttt tagacaacca ctgaactaga tgactgttgt actgtagctc agtcatttaa 3120
aaaatatata aatactacct tgtagtgtcc catactgtgt tttttacatg gtagattctt 3180
atttaagtgc taactggtta ttttctttgg ctggtttatt gtactgttat acagaatgta 3240
agttgtacag tgaaataagt tattaaagca tgtgtaaaca ttgttatata tcttttctcc 3300
taaatggaga attttgaata aaatatattt gaaattttaa aaaaaaaaaa aaaaaa 3356
<210> 122
<211> 3339
<212> DNA
<213> Macaca fascicularis
<400> 122
acgtaaccta cggtgtcccg ctaggaaaga gaggcgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg gtgcgtcagc cgtccctgct gcccggttcc ttctctctgg 120
gggcggggcc tggctagagc aggtgtgggt ttaggagata tctcaggagc atttggataa 180
tgtgacagtt ggaatgcagt gatgtcgact ctttgcccac cgccatctcc agctgttgcc 240
aagacagaga ttgctttaag tggtgaatca cctttattag cagctacttt tgcttactgg 300
gacaatattc ttggtcctag agtaaggcac atttgggctc caaagacaga acaggtactt 360
ctcagtgacg gagaaataac ttttcttgcc aaccacactc taaatggaga aatccttcga 420
aatgcagaga gtggtgctat agatgtaaag ttttttgtct tgtctgaaaa gggagtgatt 480
attgtttcat taatctttga tggaaactgg aatggggatc gcagcacata cggactatca 540
attatacttc cacagacaga acttagtttc tacctcccac ttcatagagt gtgtgttgat 600
agattaacac atataatccg gaaaggaaga atatggatgc ataaggaaag acaagaaaat 660
gtccagaaga ttatcttaga aggcacagag agaatggaag atcagggtca gagtattatt 720
ccaatgctta ctggagaagt gattcctgta atggaactgc tttcatctat gaaatcacac 780
agtgttcctg aagaaataga tatagctgat acagtactca atgatgatga tattggtgac 840
agttgtcatg aaggctttct tctcaatgcc atcagctcac acttgcaaac ctgtggctgt 900
tccgttgtag taggtagcag tgcagagaaa gtaaataaga tagtcagaac attatgcctt 960
tttctgactg cagcagagag aaaatgctcc aggttatgtg aagcagaatc atcatttaaa 1020
tatgagtcag ggctctttgt acagggcctg ctaaaggatt caactggaag ctttgtgctg 1080
cctttccggc aagtcatgta tgctccatat cccaccacac acatagatgt ggatgtcaat 1140
actgtgaagc agatgccacc ctgtcatgaa catatttata atcagcgtag atacatgaga 1200
tccgagctga cagccttctg gagagccact tcagaagaag acatggctca ggatacgatc 1260
atctacactg acgaaagctt tactcctgat ttgaatattt ttcaagatgt cttacacaga 1320
gacactctag tgaaagcctt cctggatcag gtctttcagc tgaaacctgg cttatctctc 1380
aggagtactt tccttgcaca gtttttactt gtccttcaca gaaaagcctt gacactaata 1440
aaatatatag aagatgatac gcagaaggga aaaaagccct ttaaatctct tcggaacctg 1500
aagatagacc ttgatttaac agcagagggc gatcttaaca taataatggc tctggctgag 1560
aaaattaaac caggcctaca ctcttttatc tttggaagac ctttctacac tagtgtacaa 1620
gaacgagatg ttctaatgac tttttaaatg tgtaacttaa taagcctatt ccatcacaat 1680
cgtgatcgct gctaaagtag ctcggtggtg tggggaaaca ttcccctgga tcatactcca 1740
gagctctgct cggcagttgc agttaagtta gttacactac agttctcaca agagtctgtg 1800
aggggatgtc aggtgcatca ttacattgga tgtctctttt cctagattta tgcttttggg 1860
atacagacct atgtttacaa tataataggt attattgctg tcttttaaat atataataat 1920
aggatataaa cttgaccaca actgctgttt ttttgaaata tatgattcat ggtttacatg 1980
tattaaggtg aaatccgagt tcgcttttac agatattagt tgactttcta tcttttggca 2040
ttctttggtg tgtggaatta ctgtaatact tctgcaatca actgaaaatt agagccttta 2100
aatgatttca gttccacaga aagaaagtga gcttcaacat aggataagct ttagaaagag 2160
aattgatcaa gcagatgttt aattggaatt gattattaga tcctgctttg tggatttagc 2220
cctcgggatt cagtctgtag aaatgtctga tagttctcta tagtccctgc tcatggtgaa 2280
ccacagttag gatgttttgt ttgttttatt gttgttgcta ttgttgatgt tctatatagt 2340
tgagctctgt aaaaggaaat tgtattttat gttttagtag ttgttgccaa ctttttaaat 2400
taattttcat tatttttgag ccaaattgaa atgtgcacct cctgtgcctt ttttttcctt 2460
ggaaaatcga attacttgga agaagttcag atttcactgg tcagtcgttt tcatcttgtt 2520
ttcttcttgc agagtcttac catgtacctg ctttggcaat cattgtaact ctgagattat 2580
aaaatgcatt agagaatata ttaactaata agatcttttt tttcaggaac agaaaatagt 2640
tccttgagta cttccttctt acatttctgc ccatgttttt gaagttgttg ccatttgcct 2700
gcaataggct ataaggaata gcaggagaaa ttttactgaa gtgctatttt tctaggtgct 2760
actttggcag agctaagtgg tctgtttctt ttgtttcctt aatgcgtttg gaccattttg 2820
ctggctgtaa aataactgat taatataatt ctaacacaat attgacattg tagtgtacac 2880
aaacacaaat attttattta aaactggaag taacataaaa gggaaaatat atttataaga 2940
aaggaataaa ggtaatagag ctcttctgtc ccccagccac caaatttaca caacaaaatg 3000
atatgttcta atgtgaaagg tcataatagc tttcccatca ttaatcagaa agatgtggca 3060
gcttgatttt tcagacaacc cctgaactag atgactgttg tactgtagct cagtcattta 3120
aaaaatatat aaatactatc tcgtagtgtc ccatactatg ttttttacat gatagattct 3180
tatttaagtg ctacctggtt attttctttg gctggtttat tgtactgtta tatagaatgt 3240
aagttgtaca gtgaaataag ttattaaagc atgtgtaaac attgttatat atcttttctc 3300
ctagatggag aattttgaat aaaatatatt tgaaatttt 3339
<210> 123
<211> 3151
<212> DNA
<213> Mus musculus
<400> 123
gcggttgcgg tccctgcgcc ggcggtgaag gcgcagcagc ggcgagtggc tattgcaagc 60
gttcggataa tgtgagacct ggaatgcagt gagacctggg atgcagggat gtcgactatc 120
tgccccccac catctcctgc tgttgccaag acagagattg ctttaagtgg tgaatcaccc 180
ttgttggcgg ctacctttgc ttactgggat aatattcttg gtcctagagt aaggcatatt 240
tgggctccaa agacagacca agtgcttctc agtgatggag aaataacttt tcttgccaac 300
cacactctaa atggagaaat tcttcgaaat gcagagagtg gggctataga tgtaaaattt 360
tttgtcttat ctgaaaaagg ggtaattatt gtttcattaa tcttcgacgg aaactggaat 420
ggagatcgga gcacttatgg actatcaatt atactgccgc agacagagct gagcttctac 480
ctcccacttc acagagtgtg tgttgacagg ctaacacaca ttattcgaaa aggaagaata 540
tggatgcata aggaaagaca agaaaatgtc cagaaaattg tcttggaagg cacagagagg 600
atggaagatc agggtcagag tatcattccc atgcttactg gggaagtcat tcctgtaatg 660
gagctgcttg catctatgaa atcccacagt gttcctgaag acattgatat agctgataca 720
gtgctcaatg atgatgacat tggtgacagc tgtcacgaag gctttcttct caatgccatc 780
agctcacacc tgcagacctg tggctgttcc gttgtagttg gcagcagtgc agagaaagta 840
aataagatag taagaacgct gtgccttttt ctgacaccag cagagaggaa atgctccagg 900
ctgtgtgaag cagaatcgtc ctttaagtac gaatcgggac tctttgtgca aggcttgcta 960
aaggatgcaa caggcagttt tgtcctaccc ttccggcaag ttatgtatgc cccgtacccc 1020
accacgcaca ttgatgtgga tgtcaacact gtcaagcaga tgccaccgtg tcatgaacat 1080
atttataatc aacgcagata catgaggtca gagctgacag ccttctggag ggcaacttca 1140
gaagaggaca tggcgcagga caccatcatc tacacagatg agagcttcac tcctgatttg 1200
aatattttcc aagatgtctt acacagagac actctagtga aagccttcct ggatcaggtc 1260
ttccatttga agcctggcct gtctctcagg agtactttcc ttgcacagtt cctcctcatt 1320
cttcacagaa aagccttgac actaatcaag tacatcgagg atgatacgca gaaggggaaa 1380
aagcccttta agtctcttcg gaacctgaag atagatcttg atttaacagc agagggcgat 1440
cttaacataa taatggctct agctgagaaa attaagccag gcctacactc tttcatcttt 1500
gggagacctt tctacactag tgtacaagaa cgtgatgttc taatgacctt ttgaccgtgt 1560
ggtttgctgt gtctgtctct tcacagtcac acctgctgtt acagtgtctc agcagtgtgt 1620
gggcacatcc ttcctcccga gtcctgctgc aggacagggt acactacact tgtcagtaga 1680
agtctgtacc tgatgtcagg tgcatcgtta cagtgaatga ctcttcctag aatagatgta 1740
ctcttttagg gccttatgtt tacaattatc ctaagtacta ttgctgtctt ttaaagatat 1800
gaatgatgga atatacactt gaccataact gctgattggt tttttgtttt gttttgtttg 1860
ttttcttgga aacttatgat tcctggttta catgtaccac actgaaaccc tcgttagctt 1920
tacagataaa gtgtgagttg acttcctgcc cctctgtgtt ctgtggtatg tccgattact 1980
tctgccacag ctaaacatta gagcatttaa agtttgcagt tcctcagaaa ggaacttagt 2040
ctgactacag attagttctt gagagaagac actgataggg cagagctgta ggtgaaatca 2100
gttgttagcc cttcctttat agacgtagtc cttcagattc ggtctgtaca gaaatgccga 2160
ggggtcatgc atgggccctg agtatcgtga cctgtgacaa gttttttgtt ggtttattgt 2220
agttctgtca aagaaagtgg catttgtttt tataattgtt gccaactttt aaggttaatt 2280
ttcattattt ttgagccgaa ttaaaatgcg cacctcctgt gcctttccca atcttggaaa 2340
atataatttc ttggcagagg gtcagatttc agggcccagt cactttcatc tgaccaccct 2400
ttgcacggct gccgtgtgcc tggcttagat tagaagtcct tgttaagtat gtcagagtac 2460
attcgctgat aagatctttg aagagcaggg aagcgtcttg cctctttcct ttggtttctg 2520
cctgtactct ggtgtttccc gtgtcacctg catcatagga acagcagaga aatctgaccc 2580
agtgctattt ttctaggtgc tactatggca aactcaagtg gtctgtttct gttcctgtaa 2640
cgttcgacta tctcgctagc tgtgaagtac tgattagtgg agttctgtgc aacagcagtg 2700
taggagtata cacaaacaca aatatgtgtt tctatttaaa actgtggact tagcataaaa 2760
agggagaata tatttatttt ttacaaaagg gataaaaatg ggccccgttc ctcacccacc 2820
agatttagcg agaaaaagct ttctattctg aaaggtcacg gtggctttgg cattacaaat 2880
cagaacaaca cacactgacc atgatggctt gtgaactaac tgcaaggcac tccgtcatgg 2940
taagcgagta ggtcccacct cctagtgtgc cgctcattgc tttacacagt agaatcttat 3000
ttgagtgcta attgttgtct ttgctgcttt actgtgttgt tatagaaaat gtaagctgta 3060
cagtgaataa gttattgaag catgtgtaaa cactgttata tatcttttct cctagatggg 3120
gaattttgaa taaaatacct ttgaaattct g 3151
<210> 124
<211> 3435
<212> DNA
<213> Rattus norvegicus
<400> 124
cgtttgtagt gtcagccatc ccaattgcct gttccttctc tgtgggagtg gtgtctagac 60
agtccaggca gggtatgcta ggcaggtgcg ttttggttgc ctcagatcgc aacttgactc 120
cataacggtg accaaagaca aaagaaggaa accagattaa aaagaaccgg acacagaccc 180
ctgcagaatc tggagcggcc gtggttgggg gcggggctac gacggggcgg actcgggggc 240
gtgggagggc ggggccgggg cggggcccgg agccggctgc ggttgcggtc cctgcgccgg 300
cggtgaaggc gcagcggcgg cgagtggcta ttgcaagcgt ttggataatg tgagacctgg 360
gatgcaggga tgtcgactat ctgcccccca ccatctcctg ctgttgccaa gacagagatt 420
gctttaagtg gtgaatcacc cttgttggcg gctacctttg cttactggga taatattctt 480
ggtcctagag taaggcacat ttgggctcca aagacagacc aagtactcct cagtgatgga 540
gaaatcactt ttcttgccaa ccacactctg aatggagaaa ttcttcggaa tgcggagagt 600
ggggcaatag atgtaaagtt ttttgtctta tctgaaaagg gcgtcattat tgtttcatta 660
atcttcgacg ggaactggaa cggagatcgg agcacttacg gactatcaat tatactgccg 720
cagacggagc tgagtttcta cctcccactg cacagagtgt gtgttgacag gctaacgcac 780
atcattcgaa aaggaaggat atggatgcac aaggaaagac aagaaaatgt ccagaaaatt 840
gtcttggaag gcaccgagag gatggaagat cagggtcaga gtatcatccc tatgcttact 900
ggggaggtca tccctgtgat ggagctgctt gcgtctatga gatcacacag tgttcctgaa 960
gacctcgata tagctgatac agtactcaat gatgatgaca ttggtgacag ctgtcatgaa 1020
ggctttcttc tcaatgccat cagctcacat ctgcagacct gcggctgttc tgtggtggta 1080
ggcagcagtg cagagaaagt aaataagata gtaagaacac tgtgcctttt tctgacacca 1140
gcagagagga agtgctccag gctgtgtgaa gccgaatcgt cctttaaata cgaatctgga 1200
ctctttgtac aaggcttgct aaaggatgcg actggcagtt ttgtactacc tttccggcaa 1260
gttatgtatg ccccttatcc caccacacac atcgatgtgg atgtcaacac tgtcaagcag 1320
atgccaccgt gtcatgaaca tatttataat caacgcagat acatgaggtc agagctgaca 1380
gccttctgga gggcaacttc agaagaggac atggctcagg acaccatcat ctacacagat 1440
gagagcttca ctcctgattt gaatattttc caagatgtct tacacagaga cactctagtg 1500
aaagcctttc tggatcaggt cttccatttg aagcctggcc tgtctctcag gagtactttc 1560
cttgcacagt tcctcctcat tcttcacaga aaagccttga cactaatcaa gtacatagag 1620
gatgacacgc agaaggggaa aaagcccttt aagtctcttc ggaacctgaa gatagatctt 1680
gatttaacag cagagggcga ccttaacata ataatggctc tagctgagaa aattaagcca 1740
ggcctacact ctttcatctt cgggagacct ttctacacta gtgtccaaga acgtgatgtt 1800
ctaatgactt tttaaacatg tggtttgctc cgtgtgtctc atgacagtca cacttgctgt 1860
tacagtgtct cagcgctttg gacacatcct tcctccaggg tcctgccgca ggacacgtta 1920
cactacactt gtcagtagag gtctgtacca gatgtcaggt acatcgttgt agtgaatgtc 1980
tcttttccta gactagatgt accctcgtag ggacttatgt ttacaaccct cctaagtact 2040
agtgctgtct tgtaaggata cgaatgaagg gatgtaaact tcaccacaac tgctggttgg 2100
ttttgttgtt tttgtttttt gaaacttata attcatggtt tacatgcatc acactgaaac 2160
cctagttagc tttttacagg taagctgtga gttgactgcc tgtccctgtg ttctctggcc 2220
tgtacgatct gtggcgtgta ggatcacttt tgcaacaact aaaaactaaa gcactttgtt 2280
tgcagttcta cagaaagcaa cttagtctgt ctgcagattc gtttttgaaa gaagacatga 2340
gaaagcggag ttttaggtga agtcagttgt tggatcttcc tttatagact tagtccttta 2400
gatgtggtct gtatagacat gcccaaccat catgcatggg cactgaatat cgtgaactgt 2460
ggtatgcttt ttgttggttt attgtacttc tgtcaaagaa agtggcattg gtttttataa 2520
ttgttgccaa gttttaaggt taattttcat tatttttgag ccaaattaaa atgtgcacct 2580
cctgtgcctt tcccaatctt ggaaaatata atttcttggc agaaggtcag atttcagggc 2640
ccagtcactt tcgtctgact tccctttgca cagtccgcca tgggcctggc ttagaagttc 2700
ttgtaaacta tgccagagag tacattcgct gataaaatct tctttgcaga gcaggagagc 2760
ttcttgcctc tttcctttca tttctgcctg gactttggtg ttctccacgt tccctgcatc 2820
ctaaggacag caggagaact ctgaccccag tgctatttct ctaggtgcta ttgtggcaaa 2880
ctcaagcggt ccgtctctgt ccctgtaacg ttcgtacctt gctggctgtg aagtactgac 2940
tggtaaagct ccgtgctaca gcagtgtagg gtatacacaa acacaagtaa gtgttttatt 3000
taaaactgtg gacttagcat aaaaagggag actatattta ttttttacaa aagggataaa 3060
aatggaaccc tttcctcacc caccagattt agtcagaaaa aaacattcta ttctgaaagg 3120
tcacagtggt tttgacatga cacatcagaa caacgcacac tgtccatgat ggcttatgaa 3180
ctccaagtca ctccatcatg gtaaatgggt agatccctcc ttctagtgtg ccacaccatt 3240
gcttcccaca gtagaatctt atttaagtgc taagtgttgt ctctgctggt ttactctgtt 3300
gttttagaga atgtaagttg tatagtgaat aagttattga agcatgtgta aacactgtta 3360
tacatctttt ctcctagatg gggaatttgg aataaaatac ctttaaaatt caaaaaaaaa 3420
aaaaaaaaaa aaaaa 3435
<210> 125
<211> 3356
<212> DNA
<213> Homo sapiens
<400> 125
tttttttttt ttttttttaa aatttcaaat atattttatt caaaattctc catttaggag 60
aaaagatata taacaatgtt tacacatgct ttaataactt atttcactgt acaacttaca 120
ttctgtataa cagtacaata aaccagccaa agaaaataac cagttagcac ttaaataaga 180
atctaccatg taaaaaacac agtatgggac actacaaggt agtatttata tattttttaa 240
atgactgagc tacagtacaa cagtcatcta gttcagtggt tgtctaaaac atcaagctgt 300
ccacatcttt ctgattcatg atgggaaagc tattatgacc tttcacattc gaacatgtca 360
ttttgttgtg taaatttggt gggtgggggg cagaagggct ctattacctt tatccctttc 420
ttataaatat attttccctt ttatattact tccagaattt taaataaaat atttatttgt 480
gtttgtgtaa ctacaatgtc aacattgtgt tagaattata ttaatcagtt attttatagc 540
cagcaaaatg gtccaaacgc attaagaaaa caaaagataa cttagctctg ccaaagtagc 600
acctaggaaa acagcacttc agtaaaattt ctcctgctat tccttatagc ctattgcagg 660
caaacagcaa caacttcaaa aacataggca gaaatgcaag aaggaagtac tcaaggaact 720
attttctgtt tctgaaaaaa agatcttatt agttagtata ttctctaagg cattttataa 780
tctcagagtt gcaatgattg ccaaagcagg tacatggtaa gacttagcaa gaagaaaaca 840
agatgaaaat gactgaccag tgaaatctga acttgttcca agtaattaga ttttctaagg 900
agaaaaaagg cacaggaggt gcacatttca atttggctca aaaataatga aaattaattt 960
aaaaagttgg caacaattac taaaacataa aatacaattt ccttttacag agctcaacta 1020
catagaatat caacaatagc aagaacaata aaataaacaa aacaccctaa ctgtggttca 1080
ccaggaacaa ggactataga gaactattag acatttctac agactgaatc ccagggacta 1140
aatccacaaa gtaggatcta ataatcaatt ccaattaaac atctgcttga tcaattttct 1200
ttctaaagct catcctatgt tcaagctcac tttctttctg tggaattgaa atcatttaaa 1260
ggctctagtt ttcagttgat tgcagaagta ttacagtaat tctacacacc aaagaatgcc 1320
aaaagataga aagtcaacta tctgtaaaag ccaactcaga tttcaccttg acacatgtaa 1380
accatgaatc atgtatttca aaaaaacagt agttgtggtc aagtttacat cctattatta 1440
tatctttaaa agatagcaat aatatttatt atattgtaaa cataggtctg tatcccaaaa 1500
gcataaatct aggaaaagag acacccaatg taatgatgca cctgacatcc cctcacaggc 1560
tcttgtgaga actgtagtgt aacttactta actgcaattg ctgagagcag aattctggag 1620
tatgatccag gggaacgttt ccccacacca ctgagctact ttaccagcga tcatgattgt 1680
gatggaatag gcttattaag ttacacattt aaaaagtcat tagaacatct cgttcttgca 1740
cactagtgta gaaaggtctt ccaaagataa aagagtgtag gcctggttta attttctcag 1800
ccagagccat tattatgtta agatcgccct ctgctgttaa atcaaggtct atcttcaggt 1860
tccgaagaga tttaaagggc ttttttccct tctgcgtatc gtcttctata tattttatta 1920
gtgtcaaggc ttttctgtga aggacaagta gaaactgtgc aaggaaagta cttctgagag 1980
ataagccagg tttcagctga aagacctgat ccaggaaggc tttcactaga gtgtctctgt 2040
gtaagacatc ttgaaaaata ttcaaatcag gagtaaagct ttcgtcagtg tagatgatcg 2100
tatcctgagc catgtcttct tctgaagtgg ctctccagaa ggctgtcagc tcggatctca 2160
tgtatctacg ctgattataa atatgttcat gacagggtgg catctgcttc acagtattga 2220
catccacatc tatgtgtgtg gtgggatatg gagcatacat gacttgccgg aaaggcagca 2280
caaagcttcc agttgaatcc tttagcaggc cttgtacaaa gagccctgac tcatatttaa 2340
atgatgattc tgcttcacat aacctggagc attttctctc tgctggagtc agaaaaaggc 2400
ataatgttct gactatctta tttactttct ctgcactgct acctactaca acggaacagc 2460
cacaggtttg caagtgtgag ctgatggcat tgagaagaaa gccttcatga cagctgtcac 2520
caatatcatc atcattgagt actgtatcag ctatatctat ttcttcagga acactgtgtg 2580
atttcataga tgaaagcagt tccattacag gaatcacttc tccagtaagc attggaataa 2640
tactctgacc ctgatcttcc attctctctg tgccttctaa gataatcttc tggacatttt 2700
cttgtctttc cttatgcatc catattcttc ctttccggat tatatgtgtt aatctatcaa 2760
cacacactct atgaagtggg aggtagaaac taagttctgt ctgtggaagt ataattgata 2820
gtccatatgt gctgcgatcc ccattccagt ttccatcaaa gattaatgaa acaataatca 2880
ctcccttttc agacaagaca aaaaacttta catctatagc accactctct gcatttcgaa 2940
ggatttctcc atttagagtg tggttggcaa gaaaagttat ttctccatca ctgagaagta 3000
cctgttctgt ctttggagcc caaatgtgcc ttactctagg accaagaata ttgtcccagt 3060
aagcaaaagt agctgctaat aaaggtgatt tgccacttaa agcaatctct gtcttggcaa 3120
cagctggaga tggcggtggg caaagagtcg acatcactgc attccaactg tcacattatc 3180
caaatgctcc ggagatatct cctaaaccca cacctgctct tgctagaccc cgcccccaaa 3240
agagaagcaa ccgggcagca gggacggctg acacaccaag cgtcatcttt tacgtgggcg 3300
gaacttgtcg ctgtttgacg cacctctctt tcctagcggg acaccgtagg ttacgt 3356
<210> 126
<211> 3339
<212> DNA
<213> Macaca fascicularis
<400> 126
aaaatttcaa atatatttta ttcaaaattc tccatctagg agaaaagata tataacaatg 60
tttacacatg ctttaataac ttatttcact gtacaactta cattctatat aacagtacaa 120
taaaccagcc aaagaaaata accaggtagc acttaaataa gaatctatca tgtaaaaaac 180
atagtatggg acactacgag atagtattta tatatttttt aaatgactga gctacagtac 240
aacagtcatc tagttcaggg gttgtctgaa aaatcaagct gccacatctt tctgattaat 300
gatgggaaag ctattatgac ctttcacatt agaacatatc attttgttgt gtaaatttgg 360
tggctggggg acagaagagc tctattacct ttattccttt cttataaata tattttccct 420
tttatgttac ttccagtttt aaataaaata tttgtgtttg tgtacactac aatgtcaata 480
ttgtgttaga attatattaa tcagttattt tacagccagc aaaatggtcc aaacgcatta 540
aggaaacaaa agaaacagac cacttagctc tgccaaagta gcacctagaa aaatagcact 600
tcagtaaaat ttctcctgct attccttata gcctattgca ggcaaatggc aacaacttca 660
aaaacatggg cagaaatgta agaaggaagt actcaaggaa ctattttctg ttcctgaaaa 720
aaaagatctt attagttaat atattctcta atgcatttta taatctcaga gttacaatga 780
ttgccaaagc aggtacatgg taagactctg caagaagaaa acaagatgaa aacgactgac 840
cagtgaaatc tgaacttctt ccaagtaatt cgattttcca aggaaaaaaa aggcacagga 900
ggtgcacatt tcaatttggc tcaaaaataa tgaaaattaa tttaaaaagt tggcaacaac 960
tactaaaaca taaaatacaa tttcctttta cagagctcaa ctatatagaa catcaacaat 1020
agcaacaaca ataaaacaaa caaaacatcc taactgtggt tcaccatgag cagggactat 1080
agagaactat cagacatttc tacagactga atcccgaggg ctaaatccac aaagcaggat 1140
ctaataatca attccaatta aacatctgct tgatcaattc tctttctaaa gcttatccta 1200
tgttgaagct cactttcttt ctgtggaact gaaatcattt aaaggctcta attttcagtt 1260
gattgcagaa gtattacagt aattccacac accaaagaat gccaaaagat agaaagtcaa 1320
ctaatatctg taaaagcgaa ctcggatttc accttaatac atgtaaacca tgaatcatat 1380
atttcaaaaa aacagcagtt gtggtcaagt ttatatccta ttattatata tttaaaagac 1440
agcaataata cctattatat tgtaaacata ggtctgtatc ccaaaagcat aaatctagga 1500
aaagagacat ccaatgtaat gatgcacctg acatcccctc acagactctt gtgagaactg 1560
tagtgtaact aacttaactg caactgccga gcagagctct ggagtatgat ccaggggaat 1620
gtttccccac accaccgagc tactttagca gcgatcacga ttgtgatgga ataggcttat 1680
taagttacac atttaaaaag tcattagaac atctcgttct tgtacactag tgtagaaagg 1740
tcttccaaag ataaaagagt gtaggcctgg tttaattttc tcagccagag ccattattat 1800
gttaagatcg ccctctgctg ttaaatcaag gtctatcttc aggttccgaa gagatttaaa 1860
gggctttttt cccttctgcg tatcatcttc tatatatttt attagtgtca aggcttttct 1920
gtgaaggaca agtaaaaact gtgcaaggaa agtactcctg agagataagc caggtttcag 1980
ctgaaagacc tgatccagga aggctttcac tagagtgtct ctgtgtaaga catcttgaaa 2040
aatattcaaa tcaggagtaa agctttcgtc agtgtagatg atcgtatcct gagccatgtc 2100
ttcttctgaa gtggctctcc agaaggctgt cagctcggat ctcatgtatc tacgctgatt 2160
ataaatatgt tcatgacagg gtggcatctg cttcacagta ttgacatcca catctatgtg 2220
tgtggtggga tatggagcat acatgacttg ccggaaaggc agcacaaagc ttccagttga 2280
atcctttagc aggccctgta caaagagccc tgactcatat ttaaatgatg attctgcttc 2340
acataacctg gagcattttc tctctgctgc agtcagaaaa aggcataatg ttctgactat 2400
cttatttact ttctctgcac tgctacctac tacaacggaa cagccacagg tttgcaagtg 2460
tgagctgatg gcattgagaa gaaagccttc atgacaactg tcaccaatat catcatcatt 2520
gagtactgta tcagctatat ctatttcttc aggaacactg tgtgatttca tagatgaaag 2580
cagttccatt acaggaatca cttctccagt aagcattgga ataatactct gaccctgatc 2640
ttccattctc tctgtgcctt ctaagataat cttctggaca ttttcttgtc tttccttatg 2700
catccatatt cttcctttcc ggattatatg tgttaatcta tcaacacaca ctctatgaag 2760
tgggaggtag aaactaagtt ctgtctgtgg aagtataatt gatagtccgt atgtgctgcg 2820
atccccattc cagtttccat caaagattaa tgaaacaata atcactccct tttcagacaa 2880
gacaaaaaac tttacatcta tagcaccact ctctgcattt cgaaggattt ctccatttag 2940
agtgtggttg gcaagaaaag ttatttctcc gtcactgaga agtacctgtt ctgtctttgg 3000
agcccaaatg tgccttactc taggaccaag aatattgtcc cagtaagcaa aagtagctgc 3060
taataaaggt gattcaccac ttaaagcaat ctctgtcttg gcaacagctg gagatggcgg 3120
tgggcaaaga gtcgacatca ctgcattcca actgtcacat tatccaaatg ctcctgagat 3180
atctcctaaa cccacacctg ctctagccag gccccgcccc cagagagaag gaaccgggca 3240
gcagggacgg ctgacgcacc aagcgtcatc ttttacgtgg gcggaacttg tcgctgtttg 3300
acgcgcctct ctttcctagc gggacaccgt aggttacgt 3339
<210> 127
<211> 3151
<212> DNA
<213> Mus musculus
<400> 127
cagaatttca aaggtatttt attcaaaatt ccccatctag gagaaaagat atataacagt 60
gtttacacat gcttcaataa cttattcact gtacagctta cattttctat aacaacacag 120
taaagcagca aagacaacaa ttagcactca aataagattc tactgtgtaa agcaatgagc 180
ggcacactag gaggtgggac ctactcgctt accatgacgg agtgccttgc agttagttca 240
caagccatca tggtcagtgt gtgttgttct gatttgtaat gccaaagcca ccgtgacctt 300
tcagaataga aagctttttc tcgctaaatc tggtgggtga ggaacggggc ccatttttat 360
cccttttgta aaaaataaat atattctccc tttttatgct aagtccacag ttttaaatag 420
aaacacatat ttgtgtttgt gtatactcct acactgctgt tgcacagaac tccactaatc 480
agtacttcac agctagcgag atagtcgaac gttacaggaa cagaaacaga ccacttgagt 540
ttgccatagt agcacctaga aaaatagcac tgggtcagat ttctctgctg ttcctatgat 600
gcaggtgaca cgggaaacac cagagtacag gcagaaacca aaggaaagag gcaagacgct 660
tccctgctct tcaaagatct tatcagcgaa tgtactctga catacttaac aaggacttct 720
aatctaagcc aggcacacgg cagccgtgca aagggtggtc agatgaaagt gactgggccc 780
tgaaatctga ccctctgcca agaaattata ttttccaaga ttgggaaagg cacaggaggt 840
gcgcatttta attcggctca aaaataatga aaattaacct taaaagttgg caacaattat 900
aaaaacaaat gccactttct ttgacagaac tacaataaac caacaaaaaa cttgtcacag 960
gtcacgatac tcagggccca tgcatgaccc ctcggcattt ctgtacagac cgaatctgaa 1020
ggactacgtc tataaaggaa gggctaacaa ctgatttcac ctacagctct gccctatcag 1080
tgtcttctct caagaactaa tctgtagtca gactaagttc ctttctgagg aactgcaaac 1140
tttaaatgct ctaatgttta gctgtggcag aagtaatcgg acataccaca gaacacagag 1200
gggcaggaag tcaactcaca ctttatctgt aaagctaacg agggtttcag tgtggtacat 1260
gtaaaccagg aatcataagt ttccaagaaa acaaacaaaa caaaacaaaa aaccaatcag 1320
cagttatggt caagtgtata ttccatcatt catatcttta aaagacagca atagtactta 1380
ggataattgt aaacataagg ccctaaaaga gtacatctat tctaggaaga gtcattcact 1440
gtaacgatgc acctgacatc aggtacagac ttctactgac aagtgtagtg taccctgtcc 1500
tgcagcagga ctcgggagga aggatgtgcc cacacactgc tgagacactg taacagcagg 1560
tgtgactgtg aagagacaga cacagcaaac cacacggtca aaaggtcatt agaacatcac 1620
gttcttgtac actagtgtag aaaggtctcc caaagatgaa agagtgtagg cctggcttaa 1680
ttttctcagc tagagccatt attatgttaa gatcgccctc tgctgttaaa tcaagatcta 1740
tcttcaggtt ccgaagagac ttaaagggct ttttcccctt ctgcgtatca tcctcgatgt 1800
acttgattag tgtcaaggct tttctgtgaa gaatgaggag gaactgtgca aggaaagtac 1860
tcctgagaga caggccaggc ttcaaatgga agacctgatc caggaaggct ttcactagag 1920
tgtctctgtg taagacatct tggaaaatat tcaaatcagg agtgaagctc tcatctgtgt 1980
agatgatggt gtcctgcgcc atgtcctctt ctgaagttgc cctccagaag gctgtcagct 2040
ctgacctcat gtatctgcgt tgattataaa tatgttcatg acacggtggc atctgcttga 2100
cagtgttgac atccacatca atgtgcgtgg tggggtacgg ggcatacata acttgccgga 2160
agggtaggac aaaactgcct gttgcatcct ttagcaagcc ttgcacaaag agtcccgatt 2220
cgtacttaaa ggacgattct gcttcacaca gcctggagca tttcctctct gctggtgtca 2280
gaaaaaggca cagcgttctt actatcttat ttactttctc tgcactgctg ccaactacaa 2340
cggaacagcc acaggtctgc aggtgtgagc tgatggcatt gagaagaaag ccttcgtgac 2400
agctgtcacc aatgtcatca tcattgagca ctgtatcagc tatatcaatg tcttcaggaa 2460
cactgtggga tttcatagat gcaagcagct ccattacagg aatgacttcc ccagtaagca 2520
tgggaatgat actctgaccc tgatcttcca tcctctctgt gccttccaag acaattttct 2580
ggacattttc ttgtctttcc ttatgcatcc atattcttcc ttttcgaata atgtgtgtta 2640
gcctgtcaac acacactctg tgaagtggga ggtagaagct cagctctgtc tgcggcagta 2700
taattgatag tccataagtg ctccgatctc cattccagtt tccgtcgaag attaatgaaa 2760
caataattac ccctttttca gataagacaa aaaattttac atctatagcc ccactctctg 2820
catttcgaag aatttctcca tttagagtgt ggttggcaag aaaagttatt tctccatcac 2880
tgagaagcac ttggtctgtc tttggagccc aaatatgcct tactctagga ccaagaatat 2940
tatcccagta agcaaaggta gccgccaaca agggtgattc accacttaaa gcaatctctg 3000
tcttggcaac agcaggagat ggtggggggc agatagtcga catccctgca tcccaggtct 3060
cactgcattc caggtctcac attatccgaa cgcttgcaat agccactcgc cgctgctgcg 3120
ccttcaccgc cggcgcaggg accgcaaccg c 3151
<210> 128
<211> 3435
<212> DNA
<213> Rattus norvegicus
<400> 128
tttttttttt tttttttttt ttttgaattt taaaggtatt ttattccaaa ttccccatct 60
aggagaaaag atgtataaca gtgtttacac atgcttcaat aacttattca ctatacaact 120
tacattctct aaaacaacag agtaaaccag cagagacaac acttagcact taaataagat 180
tctactgtgg gaagcaatgg tgtggcacac tagaaggagg gatctaccca tttaccatga 240
tggagtgact tggagttcat aagccatcat ggacagtgtg cgttgttctg atgtgtcatg 300
tcaaaaccac tgtgaccttt cagaatagaa tgtttttttc tgactaaatc tggtgggtga 360
ggaaagggtt ccatttttat cccttttgta aaaaataaat atagtctccc tttttatgct 420
aagtccacag ttttaaataa aacacttact tgtgtttgtg tataccctac actgctgtag 480
cacggagctt taccagtcag tacttcacag ccagcaaggt acgaacgtta cagggacaga 540
gacggaccgc ttgagtttgc cacaatagca cctagagaaa tagcactggg gtcagagttc 600
tcctgctgtc cttaggatgc agggaacgtg gagaacacca aagtccaggc agaaatgaaa 660
ggaaagaggc aagaagctct cctgctctgc aaagaagatt ttatcagcga atgtactctc 720
tggcatagtt tacaagaact tctaagccag gcccatggcg gactgtgcaa agggaagtca 780
gacgaaagtg actgggccct gaaatctgac cttctgccaa gaaattatat tttccaagat 840
tgggaaaggc acaggaggtg cacattttaa tttggctcaa aaataatgaa aattaacctt 900
aaaacttggc aacaattata aaaaccaatg ccactttctt tgacagaagt acaataaacc 960
aacaaaaagc ataccacagt tcacgatatt cagtgcccat gcatgatggt tgggcatgtc 1020
tatacagacc acatctaaag gactaagtct ataaaggaag atccaacaac tgacttcacc 1080
taaaactccg ctttctcatg tcttctttca aaaacgaatc tgcagacaga ctaagttgct 1140
ttctgtagaa ctgcaaacaa agtgctttag tttttagttg ttgcaaaagt gatcctacac 1200
gccacagatc gtacaggcca gagaacacag ggacaggcag tcaactcaca gcttacctgt 1260
aaaaagctaa ctagggtttc agtgtgatgc atgtaaacca tgaattataa gtttcaaaaa 1320
acaaaaacaa caaaaccaac cagcagttgt ggtgaagttt acatcccttc attcgtatcc 1380
ttacaagaca gcactagtac ttaggagggt tgtaaacata agtccctacg agggtacatc 1440
tagtctagga aaagagacat tcactacaac gatgtacctg acatctggta cagacctcta 1500
ctgacaagtg tagtgtaacg tgtcctgcgg caggaccctg gaggaaggat gtgtccaaag 1560
cgctgagaca ctgtaacagc aagtgtgact gtcatgagac acacggagca aaccacatgt 1620
ttaaaaagtc attagaacat cacgttcttg gacactagtg tagaaaggtc tcccgaagat 1680
gaaagagtgt aggcctggct taattttctc agctagagcc attattatgt taaggtcgcc 1740
ctctgctgtt aaatcaagat ctatcttcag gttccgaaga gacttaaagg gctttttccc 1800
cttctgcgtg tcatcctcta tgtacttgat tagtgtcaag gcttttctgt gaagaatgag 1860
gaggaactgt gcaaggaaag tactcctgag agacaggcca ggcttcaaat ggaagacctg 1920
atccagaaag gctttcacta gagtgtctct gtgtaagaca tcttggaaaa tattcaaatc 1980
aggagtgaag ctctcatctg tgtagatgat ggtgtcctga gccatgtcct cttctgaagt 2040
tgccctccag aaggctgtca gctctgacct catgtatctg cgttgattat aaatatgttc 2100
atgacacggt ggcatctgct tgacagtgtt gacatccaca tcgatgtgtg tggtgggata 2160
aggggcatac ataacttgcc ggaaaggtag tacaaaactg ccagtcgcat cctttagcaa 2220
gccttgtaca aagagtccag attcgtattt aaaggacgat tcggcttcac acagcctgga 2280
gcacttcctc tctgctggtg tcagaaaaag gcacagtgtt cttactatct tatttacttt 2340
ctctgcactg ctgcctacca ccacagaaca gccgcaggtc tgcagatgtg agctgatggc 2400
attgagaaga aagccttcat gacagctgtc accaatgtca tcatcattga gtactgtatc 2460
agctatatcg aggtcttcag gaacactgtg tgatctcata gacgcaagca gctccatcac 2520
agggatgacc tccccagtaa gcatagggat gatactctga ccctgatctt ccatcctctc 2580
ggtgccttcc aagacaattt tctggacatt ttcttgtctt tccttgtgca tccatatcct 2640
tccttttcga atgatgtgcg ttagcctgtc aacacacact ctgtgcagtg ggaggtagaa 2700
actcagctcc gtctgcggca gtataattga tagtccgtaa gtgctccgat ctccgttcca 2760
gttcccgtcg aagattaatg aaacaataat gacgcccttt tcagataaga caaaaaactt 2820
tacatctatt gccccactct ccgcattccg aagaatttct ccattcagag tgtggttggc 2880
aagaaaagtg atttctccat cactgaggag tacttggtct gtctttggag cccaaatgtg 2940
ccttactcta ggaccaagaa tattatccca gtaagcaaag gtagccgcca acaagggtga 3000
ttcaccactt aaagcaatct ctgtcttggc aacagcagga gatggtgggg ggcagatagt 3060
cgacatccct gcatcccagg tctcacatta tccaaacgct tgcaatagcc actcgccgcc 3120
gctgcgcctt caccgccggc gcagggaccg caaccgcagc cggctccggg ccccgccccg 3180
gccccgccct cccacgcccc cgagtccgcc ccgtcgtagc cccgccccca accacggccg 3240
ctccagattc tgcaggggtc tgtgtccggt tctttttaat ctggtttcct tcttttgtct 3300
ttggtcaccg ttatggagtc aagttgcgat ctgaggcaac caaaacgcac ctgcctagca 3360
taccctgcct ggactgtcta gacaccactc ccacagagaa ggaacaggca attgggatgg 3420
ctgacactac aaacg 3435
<210> 129
<211> 1957
<212> DNA
<213> Homo sapiens
<400> 129
acgtaaccta cggtgtcccg ctaggaaaga gaggtgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg atatctccgg agcatttgga taatgtgaca gttggaatgc 120
agtgatgtcg actctttgcc caccgccatc tccagctgtt gccaagacag agattgcttt 180
aagtggcaaa tcacctttat tagcagctac ttttgcttac tgggacaata ttcttggtcc 240
tagagtaagg cacatttggg ctccaaagac agaacaggta cttctcagtg atggagaaat 300
aacttttctt gccaaccaca ctctaaatgg agaaatcctt cgaaatgcag agagtggtgc 360
tatagatgta aagttttttg tcttgtctga aaagggagtg attattgttt cattaatctt 420
tgatggaaac tggaatgggg atcgcagcac atatggacta tcaattatac ttccacagac 480
agaacttagt ttctacctcc cacttcatag agtgtgtgtt gatagattaa cacatataat 540
ccggaaagga agaatatgga tgcataagga aagacaagaa aatgtccaga agattatctt 600
agaaggcaca gagagaatgg aagatcaggg tcagagtatt attccaatgc ttactggaga 660
agtgattcct gtaatggaac tgctttcatc tatgaaatca cacagtgttc ctgaagaaat 720
agatatagct gatacagtac tcaatgatga tgatattggt gacagctgtc atgaaggctt 780
tcttctcaag taagaatttt tcttttcata aaagctggat gaagcagata ccatcttatg 840
ctcacctatg acaagatttg gaagaaagaa aataacagac tgtctactta gattgttcta 900
gggacattac gtatttgaac tgttgcttaa atttgtgtta tttttcactc attatatttc 960
tatatatatt tggtgttatt ccatttgcta tttaaagaaa ccgagtttcc atcccagaca 1020
agaaatcatg gccccttgct tgattctggt ttcttgtttt acttctcatt aaagctaaca 1080
gaatcctttc atattaagtt gtactgtaga tgaacttaag ttatttaggc gtagaacaaa 1140
attattcata tttatactga tctttttcca tccagcagtg gagtttagta cttaagagtt 1200
tgtgccctta aaccagactc cctggattaa tgctgtgtac ccgtgggcaa ggtgcctgaa 1260
ttctctatac acctatttcc tcatctgtaa aatggcaata atagtaatag tacctaatgt 1320
gtagggttgt tataagcatt gagtaagata aataatataa agcacttaga acagtgcctg 1380
gaacataaaa acacttaata atagctcata gctaacattt cctatttaca tttcttctag 1440
aaatagccag tatttgttga gtgcctacat gttagttcct ttactagttg ctttacatgt 1500
attatcttat attctgtttt aaagtttctt cacagttaca gattttcatg aaattttact 1560
tttaataaaa gagaagtaaa agtataaagt attcactttt atgttcacag tcttttcctt 1620
taggctcatg atggagtatc agaggcatga gtgtgtttaa cctaagagcc ttaatggctt 1680
gaatcagaag cactttagtc ctgtatctgt tcagtgtcag cctttcatac atcattttaa 1740
atcccatttg actttaagta agtcacttaa tctctctaca tgtcaatttc ttcagctata 1800
aaatgatggt atttcaataa ataaatacat taattaaatg atattatact gactaattgg 1860
gctgttttaa ggctcaataa gaaaatttct gtgaaaggtc tctagaaaat gtaggttcct 1920
atacaaataa aagataacat tgtgcttata aaaaaaa 1957
<210> 130
<211> 1957
<212> DNA
<213> Homo sapiens
<400> 130
ttttttttat aagcacaatg ttatctttta tttgtatagg aacctacatt ttctagagac 60
ctttcacaga aattttctta ttgagcctta aaacagccca attagtcagt ataatatcat 120
ttaattaatg tatttattta ttgaaatacc atcattttat agctgaagaa attgacatgt 180
agagagatta agtgacttac ttaaagtcaa atgggattta aaatgatgta tgaaaggctg 240
acactgaaca gatacaggac taaagtgctt ctgattcaag ccattaaggc tcttaggtta 300
aacacactca tgcctctgat actccatcat gagcctaaag gaaaagactg tgaacataaa 360
agtgaatact ttatactttt acttctcttt tattaaaagt aaaatttcat gaaaatctgt 420
aactgtgaag aaactttaaa acagaatata agataataca tgtaaagcaa ctagtaaagg 480
aactaacatg taggcactca acaaatactg gctatttcta gaagaaatgt aaataggaaa 540
tgttagctat gagctattat taagtgtttt tatgttccag gcactgttct aagtgcttta 600
tattatttat cttactcaat gcttataaca accctacaca ttaggtacta ttactattat 660
tgccatttta cagatgagga aataggtgta tagagaattc aggcaccttg cccacgggta 720
cacagcatta atccagggag tctggtttaa gggcacaaac tcttaagtac taaactccac 780
tgctggatgg aaaaagatca gtataaatat gaataatttt gttctacgcc taaataactt 840
aagttcatct acagtacaac ttaatatgaa aggattctgt tagctttaat gagaagtaaa 900
acaagaaacc agaatcaagc aaggggccat gatttcttgt ctgggatgga aactcggttt 960
ctttaaatag caaatggaat aacaccaaat atatatagaa atataatgag tgaaaaataa 1020
cacaaattta agcaacagtt caaatacgta atgtccctag aacaatctaa gtagacagtc 1080
tgttattttc tttcttccaa atcttgtcat aggtgagcat aagatggtat ctgcttcatc 1140
cagcttttat gaaaagaaaa attcttactt gagaagaaag ccttcatgac agctgtcacc 1200
aatatcatca tcattgagta ctgtatcagc tatatctatt tcttcaggaa cactgtgtga 1260
tttcatagat gaaagcagtt ccattacagg aatcacttct ccagtaagca ttggaataat 1320
actctgaccc tgatcttcca ttctctctgt gccttctaag ataatcttct ggacattttc 1380
ttgtctttcc ttatgcatcc atattcttcc tttccggatt atatgtgtta atctatcaac 1440
acacactcta tgaagtggga ggtagaaact aagttctgtc tgtggaagta taattgatag 1500
tccatatgtg ctgcgatccc cattccagtt tccatcaaag attaatgaaa caataatcac 1560
tcccttttca gacaagacaa aaaactttac atctatagca ccactctctg catttcgaag 1620
gatttctcca tttagagtgt ggttggcaag aaaagttatt tctccatcac tgagaagtac 1680
ctgttctgtc tttggagccc aaatgtgcct tactctagga ccaagaatat tgtcccagta 1740
agcaaaagta gctgctaata aaggtgattt gccacttaaa gcaatctctg tcttggcaac 1800
agctggagat ggcggtgggc aaagagtcga catcactgca ttccaactgt cacattatcc 1860
aaatgctccg gagatatcaa gcgtcatctt ttacgtgggc ggaacttgtc gctgtttgac 1920
gcacctctct ttcctagcgg gacaccgtag gttacgt 1957
<210> 131
<211> 3231
<212> DNA
<213> Homo sapiens
<400> 131
ggttgcggtg cctgcgcccg cggcggcgga ggcgcaggcg gtggcgagtg gatatctccg 60
gagcatttgg ataatgtgac agttggaatg cagtgatgtc gactctttgc ccaccgccat 120
ctccagctgt tgccaagaca gagattgctt taagtggcaa atcaccttta ttagcagcta 180
cttttgctta ctgggacaat attcttggtc ctagagtaag gcacatttgg gctccaaaga 240
cagaacaggt acttctcagt gatggagaaa taacttttct tgccaaccac actctaaatg 300
gagaaatcct tcgaaatgca gagagtggtg ctatagatgt aaagtttttt gtcttgtctg 360
aaaagggagt gattattgtt tcattaatct ttgatggaaa ctggaatggg gatcgcagca 420
catatggact atcaattata cttccacaga cagaacttag tttctacctc ccacttcata 480
gagtgtgtgt tgatagatta acacatataa tccggaaagg aagaatatgg atgcataagg 540
aaagacaaga aaatgtccag aagattatct tagaaggcac agagagaatg gaagatcagg 600
gtcagagtat tattccaatg cttactggag aagtgattcc tgtaatggaa ctgctttcat 660
ctatgaaatc acacagtgtt cctgaagaaa tagatatagc tgatacagta ctcaatgatg 720
atgatattgg tgacagctgt catgaaggct ttcttctcaa tgccatcagc tcacacttgc 780
aaacctgtgg ctgttccgtt gtagtaggta gcagtgcaga gaaagtaaat aagatagtca 840
gaacattatg cctttttctg actccagcag agagaaaatg ctccaggtta tgtgaagcag 900
aatcatcatt taaatatgag tcagggctct ttgtacaagg cctgctaaag gattcaactg 960
gaagctttgt gctgcctttc cggcaagtca tgtatgctcc atatcccacc acacacatag 1020
atgtggatgt caatactgtg aagcagatgc caccctgtca tgaacatatt tataatcagc 1080
gtagatacat gagatccgag ctgacagcct tctggagagc cacttcagaa gaagacatgg 1140
ctcaggatac gatcatctac actgacgaaa gctttactcc tgatttgaat atttttcaag 1200
atgtcttaca cagagacact ctagtgaaag ccttcctgga tcaggtcttt cagctgaaac 1260
ctggcttatc tctcagaagt actttccttg cacagtttct acttgtcctt cacagaaaag 1320
ccttgacact aataaaatat atagaagacg atacgcagaa gggaaaaaag ccctttaaat 1380
ctcttcggaa cctgaagata gaccttgatt taacagcaga gggcgatctt aacataataa 1440
tggctctggc tgagaaaatt aaaccaggcc tacactcttt tatctttgga agacctttct 1500
acactagtgt gcaagaacga gatgttctaa tgacttttta aatgtgtaac ttaataagcc 1560
tattccatca caatcatgat cgctggtaaa gtagctcagt ggtgtgggga aacgttcccc 1620
tggatcatac tccagaattc tgctctcagc aattgcagtt aagtaagtta cactacagtt 1680
ctcacaagag cctgtgaggg gatgtcaggt gcatcattac attgggtgtc tcttttccta 1740
gatttatgct tttgggatac agacctatgt ttacaatata ataaatatta ttgctatctt 1800
ttaaagatat aataatagga tgtaaacttg accacaacta ctgttttttt gaaatacatg 1860
attcatggtt tacatgtgtc aaggtgaaat ctgagttggc ttttacagat agttgacttt 1920
ctatcttttg gcattctttg gtgtgtagaa ttactgtaat acttctgcaa tcaactgaaa 1980
actagagcct ttaaatgatt tcaattccac agaaagaaag tgagcttgaa cataggatga 2040
gctttagaaa gaaaattgat caagcagatg tttaattgga attgattatt agatcctact 2100
ttgtggattt agtccctggg attcagtctg tagaaatgtc taatagttct ctatagtcct 2160
tgttcctggt gaaccacagt tagggtgttt tgtttatttt attgttcttg ctattgttga 2220
tattctatgt agttgagctc tgtaaaagga aattgtattt tatgttttag taattgttgc 2280
caacttttta aattaatttt cattattttt gagccaaatt gaaatgtgca cctcctgtgc 2340
cttttttctc cttagaaaat ctaattactt ggaacaagtt cagatttcac tggtcagtca 2400
ttttcatctt gttttcttct tgctaagtct taccatgtac ctgctttggc aatcattgca 2460
actctgagat tataaaatgc cttagagaat atactaacta ataagatctt tttttcagaa 2520
acagaaaata gttccttgag tacttccttc ttgcatttct gcctatgttt ttgaagttgt 2580
tgctgtttgc ctgcaatagg ctataaggaa tagcaggaga aattttactg aagtgctgtt 2640
ttcctaggtg ctactttggc agagctaagt tatcttttgt tttcttaatg cgtttggacc 2700
attttgctgg ctataaaata actgattaat ataattctaa cacaatgttg acattgtagt 2760
tacacaaaca caaataaata ttttatttaa aattctggaa gtaatataaa agggaaaata 2820
tatttataag aaagggataa aggtaataga gcccttctgc cccccaccca ccaaatttac 2880
acaacaaaat gacatgttcg aatgtgaaag gtcataatag ctttcccatc atgaatcaga 2940
aagatgtgga cagcttgatg ttttagacaa ccactgaact agatgactgt tgtactgtag 3000
ctcagtcatt taaaaaatat ataaatacta ccttgtagtg tcccatactg tgttttttac 3060
atggtagatt cttatttaag tgctaactgg ttattttctt tggctggttt attgtactgt 3120
tatacagaat gtaagttgta cagtgaaata agttattaaa gcatgtgtaa acattgttat 3180
atatcttttc tcctaaatgg agaattttga ataaaatata tttgaaattt t 3231
<210> 132
<211> 3231
<212> DNA
<213> Homo sapiens
<400> 132
aaaatttcaa atatatttta ttcaaaattc tccatttagg agaaaagata tataacaatg 60
tttacacatg ctttaataac ttatttcact gtacaactta cattctgtat aacagtacaa 120
taaaccagcc aaagaaaata accagttagc acttaaataa gaatctacca tgtaaaaaac 180
acagtatggg acactacaag gtagtattta tatatttttt aaatgactga gctacagtac 240
aacagtcatc tagttcagtg gttgtctaaa acatcaagct gtccacatct ttctgattca 300
tgatgggaaa gctattatga cctttcacat tcgaacatgt cattttgttg tgtaaatttg 360
gtgggtgggg ggcagaaggg ctctattacc tttatccctt tcttataaat atattttccc 420
ttttatatta cttccagaat tttaaataaa atatttattt gtgtttgtgt aactacaatg 480
tcaacattgt gttagaatta tattaatcag ttattttata gccagcaaaa tggtccaaac 540
gcattaagaa aacaaaagat aacttagctc tgccaaagta gcacctagga aaacagcact 600
tcagtaaaat ttctcctgct attccttata gcctattgca ggcaaacagc aacaacttca 660
aaaacatagg cagaaatgca agaaggaagt actcaaggaa ctattttctg tttctgaaaa 720
aaagatctta ttagttagta tattctctaa ggcattttat aatctcagag ttgcaatgat 780
tgccaaagca ggtacatggt aagacttagc aagaagaaaa caagatgaaa atgactgacc 840
agtgaaatct gaacttgttc caagtaatta gattttctaa ggagaaaaaa ggcacaggag 900
gtgcacattt caatttggct caaaaataat gaaaattaat ttaaaaagtt ggcaacaatt 960
actaaaacat aaaatacaat ttccttttac agagctcaac tacatagaat atcaacaata 1020
gcaagaacaa taaaataaac aaaacaccct aactgtggtt caccaggaac aaggactata 1080
gagaactatt agacatttct acagactgaa tcccagggac taaatccaca aagtaggatc 1140
taataatcaa ttccaattaa acatctgctt gatcaatttt ctttctaaag ctcatcctat 1200
gttcaagctc actttctttc tgtggaattg aaatcattta aaggctctag ttttcagttg 1260
attgcagaag tattacagta attctacaca ccaaagaatg ccaaaagata gaaagtcaac 1320
tatctgtaaa agccaactca gatttcacct tgacacatgt aaaccatgaa tcatgtattt 1380
caaaaaaaca gtagttgtgg tcaagtttac atcctattat tatatcttta aaagatagca 1440
ataatattta ttatattgta aacataggtc tgtatcccaa aagcataaat ctaggaaaag 1500
agacacccaa tgtaatgatg cacctgacat cccctcacag gctcttgtga gaactgtagt 1560
gtaacttact taactgcaat tgctgagagc agaattctgg agtatgatcc aggggaacgt 1620
ttccccacac cactgagcta ctttaccagc gatcatgatt gtgatggaat aggcttatta 1680
agttacacat ttaaaaagtc attagaacat ctcgttcttg cacactagtg tagaaaggtc 1740
ttccaaagat aaaagagtgt aggcctggtt taattttctc agccagagcc attattatgt 1800
taagatcgcc ctctgctgtt aaatcaaggt ctatcttcag gttccgaaga gatttaaagg 1860
gcttttttcc cttctgcgta tcgtcttcta tatattttat tagtgtcaag gcttttctgt 1920
gaaggacaag tagaaactgt gcaaggaaag tacttctgag agataagcca ggtttcagct 1980
gaaagacctg atccaggaag gctttcacta gagtgtctct gtgtaagaca tcttgaaaaa 2040
tattcaaatc aggagtaaag ctttcgtcag tgtagatgat cgtatcctga gccatgtctt 2100
cttctgaagt ggctctccag aaggctgtca gctcggatct catgtatcta cgctgattat 2160
aaatatgttc atgacagggt ggcatctgct tcacagtatt gacatccaca tctatgtgtg 2220
tggtgggata tggagcatac atgacttgcc ggaaaggcag cacaaagctt ccagttgaat 2280
cctttagcag gccttgtaca aagagccctg actcatattt aaatgatgat tctgcttcac 2340
ataacctgga gcattttctc tctgctggag tcagaaaaag gcataatgtt ctgactatct 2400
tatttacttt ctctgcactg ctacctacta caacggaaca gccacaggtt tgcaagtgtg 2460
agctgatggc attgagaaga aagccttcat gacagctgtc accaatatca tcatcattga 2520
gtactgtatc agctatatct atttcttcag gaacactgtg tgatttcata gatgaaagca 2580
gttccattac aggaatcact tctccagtaa gcattggaat aatactctga ccctgatctt 2640
ccattctctc tgtgccttct aagataatct tctggacatt ttcttgtctt tccttatgca 2700
tccatattct tcctttccgg attatatgtg ttaatctatc aacacacact ctatgaagtg 2760
ggaggtagaa actaagttct gtctgtggaa gtataattga tagtccatat gtgctgcgat 2820
ccccattcca gtttccatca aagattaatg aaacaataat cactcccttt tcagacaaga 2880
caaaaaactt tacatctata gcaccactct ctgcatttcg aaggatttct ccatttagag 2940
tgtggttggc aagaaaagtt atttctccat cactgagaag tacctgttct gtctttggag 3000
cccaaatgtg ccttactcta ggaccaagaa tattgtccca gtaagcaaaa gtagctgcta 3060
ataaaggtga tttgccactt aaagcaatct ctgtcttggc aacagctgga gatggcggtg 3120
ggcaaagagt cgacatcact gcattccaac tgtcacatta tccaaatgct ccggagatat 3180
ccactcgcca ccgcctgcgc ctccgccgcc gcgggcgcag gcaccgcaac c 3231
<210> 133
<211> 27321
<212> DNA
<213> Homo sapiens
<400> 133
acgtaaccta cggtgtcccg ctaggaaaga gaggtgcgtc aaacagcgac aagttccgcc 60
cacgtaaaag atgacgcttg gtgtgtcagc cgtccctgct gcccggttgc ttctcttttg 120
ggggcggggt ctagcaagag caggtgtggg tttaggaggt gtgtgttttt gtttttccca 180
ccctctctcc ccactacttg ctctcacagt actcgctgag ggtgaacaag aaaagacctg 240
ataaagatta accagaagaa aacaaggagg gaaacaaccg cagcctgtag caagctctgg 300
aactcaggag tcgcgcgcta ggggccgggg ccggggccgg ggcgtggtcg gggcgggccc 360
gggggcgggc ccggggcggg gctgcggttg cggtgcctgc gcccgcggcg gcggaggcgc 420
aggcggtggc gagtgggtga gtgaggaggc ggcatcctgg cgggtggctg tttggggttc 480
ggctgccggg aagaggcgcg ggtagaagcg ggggctctcc tcagagctcg acgcattttt 540
actttccctc tcatttctct gaccgaagct gggtgtcggg ctttcgcctc tagcgactgg 600
tggaattgcc tgcatccggg ccccgggctt cccggcggcg gcggcggcgg cggcggcgca 660
gggacaaggg atggggatct ggcctcttcc ttgctttccc gccctcagta cccgagctgt 720
ctccttcccg gggacccgct gggagcgctg ccgctgcggg ctcgagaaaa gggagcctcg 780
ggtactgaga ggcctcgcct gggggaaggc cggagggtgg gcggcgcgcg gcttctgcgg 840
accaagtcgg ggttcgctag gaacccgaga cggtccctgc cggcgaggag atcatgcggg 900
atgagatggg ggtgtggaga cgcctgcaca atttcagccc aagcttctag agagtggtga 960
tgacttgcat atgagggcag caatgcaagt cggtgtgctc cccattctgt gggacatgac 1020
ctggttgctt cacagctccg agatgacaca gacttgctta aaggaagtga ctattgtgac 1080
ttgggcatca cttgactgat ggtaatcagt tgtctaaaga agtgcacaga ttacatgtcc 1140
gtgtgctcat tgggtctatc tggccgcgtt gaacaccacc aggctttgta ttcagaaaca 1200
ggagggaggt cctgcacttt cccaggaggg gtggcccttt cagatgcaat cgagattgtt 1260
aggctctggg agagtagttg cctggttgtg gcagttggta aatttctatt caaacagttg 1320
ccatgcacca gttgttcaca acaagggtac gtaatctgtc tggcattact tctacttttg 1380
tacaaaggat caaaaaaaaa aaagatactg ttaagatatg atttttctca gactttggga 1440
aacttttaac ataatctgtg aatatcacag aaacaagact atcatatagg ggatattaat 1500
aacctggagt cagaatactt gaaatacggt gtcatttgac acgggcattg ttgtcaccac 1560
ctctgccaag gcctgccact ttaggaaaac cctgaatcag ttggaaactg ctacatgctg 1620
atagtacatc tgaaacaaga acgagagtaa ttaccacatt ccagattgtt cactaagcca 1680
gcatttacct gctccaggaa aaaattacaa gcaccttatg aagttgataa aatattttgt 1740
ttggctatgt tggcactcca caatttgctt tcagagaaac aaagtaaacc aaggaggact 1800
tctgtttttc aagtctgccc tcgggttcta ttctacgtta attagatagt tcccaggagg 1860
actaggttag cctacctatt gtctgagaaa cttggaactg tgagaaatgg ccagatagtg 1920
atatgaactt caccttccag tcttccctga tgttgaagat tgagaaagtg ttgtgaactt 1980
tctggtactg taaacagttc actgtccttg aagtggtcct gggcagctcc tgttgtggaa 2040
agtggacggt ttaggatcct gcttctcttt gggctgggag aaaataaaca gcatggttac 2100
aagtattgag agccaggttg gagaaggtgg cttacacctg taatgccaga gctttgggag 2160
gcggaggcaa gaggatcact tgaagccagg agttcaagct caacctgggc aacgtagacc 2220
ctgtctctac aaaaaattaa aaacttagcc gggcgtggtg atgtgcacct gtagtcctag 2280
ctacttggga ggctgaggca ggagggtcat ttgagcccaa gagtttgaag ttaccgagag 2340
ctatgatcct gccagtgcat tccagcctgg atgacaaaac gagaccctgt ctctaaaaaa 2400
caagaagtga gggctttatg attgtagaat tttcactaca atagcagtgg accaaccacc 2460
tttctaaata ccaatcaggg aagagatggt tgatttttta acagacgttt aaagaaaaag 2520
caaaacctca aacttagcac tctactaaca gttttagcag atgttaatta atgtaatcat 2580
gtctgcatgt atgggattat ttccagaaag tgtattggga aacctctcat gaaccctgtg 2640
agcaagccac cgtctcactc aatttgaatc ttggcttccc tcaaaagact ggctaatgtt 2700
tggtaactct ctggagtaga cagcactaca tgtacgtaag ataggtacat aaacaactat 2760
tggttttgag ctgatttttt tcagctgcat ttgcatgtat ggatttttct caccaaagac 2820
gatgacttca agtattagta aaataattgt acagctctcc tgattatact tctctgtgac 2880
atttcatttc ccaggctatt tcttttggta ggatttaaaa ctaagcaatt cagtatgatc 2940
tttgtccttc attttctttc ttattctttt tgtttgtttg tttgtttgtt tttttcttga 3000
ggcagagtct ctctctgtcg cccaggctgg agtgcagtgg cgccatctca gctcattgca 3060
acctctgcca cctccgggtt caagagattc tcctgcctca gcctcccgag tagctgggat 3120
tacaggtgtc caccaccaca cccggctaat tttttgtatt tttagtagag gtggggtttc 3180
accatgttgg ccaggctggt cttgagctcc tgacctcagg tgatccacct gcctcggcct 3240
accaaagagc tgggataaca ggtgtgaccc accatgcccg gcccattttt tttttcttat 3300
tctgttagga gtgagagtgt aactagcagt ataatagttc aattttcaca acgtggtaaa 3360
agtttcccta taattcaatc agattttgct ccagggttca gttctgtttt aggaaatact 3420
tttattttca gtttaatgat gaaatattag agttgtaata ttgcctttat gattatccac 3480
ctttttaacc taaaagaatg aaagaaaaat atgtttgcaa tataatttta tggttgtatg 3540
ttaacttaat tcattatgtt ggcctccagt ttgctgttgt tagttatgac agcagtagtg 3600
tcattaccat ttcaattcag attacattcc tatatttgat cattgtaaac tgactgctta 3660
cattgtatta aaaacagtgg atattttaaa gaagctgtac ggcttatatc tagtgctgtc 3720
tcttaagact attaaattga tacaacatat ttaaaagtaa atattaccta aatgaatttt 3780
tgaaattaca aatacacgtg ttaaaactgt cgttgtgttc aaccatttct gtacatactt 3840
agagttaact gttttgccag gctctgtatg cctactcata atatgataaa agcactcatc 3900
taatgctctg taaatagaag tcagtgcttt ccatcagact gaactctctt gacaagatgt 3960
ggatgaaatt ctttaagtaa aattgtttac tttgtcatac atttacagat caaatgttag 4020
ctcccaaagc aatcatatgg caaagatagg tatatcatag tttgcctatt agctgctttg 4080
tattgctatt attataaata gacttcacag ttttagactt gcttaggtga aattgcaatt 4140
ctttttactt tcagtcttag ataacaagtc ttcaattata gtacaatcac acattgctta 4200
ggaatgcatc attaggcgat tttgtcatta tgcaaacatc atagagtgta cttacacaaa 4260
cctagatagt atagccttta tgtacctagg ccgtatggta tagtctgttg ctcctaggcc 4320
acaaacctgt acaactgtta ctgtactgaa tactatagac agttgtaaca cagtggtaaa 4380
tatttatcta aatatatgca aacagagaaa aggtacagta aaagtatggt ataaaagata 4440
atggtatacc tgtgtaggcc acttaccacg aatggagctt gcaggactag aagttgctct 4500
gggtgagtca gtgagtgagt ggtgaattaa tgtgaaggcc tagaacactg tacaccactg 4560
tagactataa acacagtacg ctgaagctac accaaattta tcttaacagt ttttcttcaa 4620
taaaaaatta taacttttta actttgtaaa ctttttaatt ttttaacttt taaaatactt 4680
agcttgaaac acaaatacat tgtatagcta tacaaaaata ttttttcttt gtatccttat 4740
tctagaagct tttttctatt ttctatttta aatttttttt tttacttgtt agtcgttttt 4800
gttaaaaact aaaacacaca cactttcacc taggcataga caggattagg atcatcagta 4860
tcactccctt ccacctcact gccttccacc tccacatctt gtcccactgg aaggttttta 4920
ggggcaataa cacacatgta gctgtcacct atgataacag tgctttctgt tgaatacctc 4980
ctgaaggact tgcctgaggc tgttttacat ttaacttaaa aaaaaaaaaa gtagaaggag 5040
tgcactctaa aataacaata aaaggcatag tatagtgaat acataaacca gcaatgtagt 5100
agtttattat caagtgttgt acactgtaat aattgtatgt gctatacttt aaataacttg 5160
caaaatagta ctaagacctt atgatggtta cagtgtcact aaggcaatag catattttca 5220
ggtccattgt aatctaatgg gactaccatc atatatgcag tctaccattg actgaaacgt 5280
tacatggcac ataactgtat ttgcaagaat gatttgtttt acattaatat cacataggat 5340
gtaccttttt agagtggtat gtttatgtgg attaagatgt acaagttgag caaggggacc 5400
aagagccctg ggttctgtct tggatgtgag cgtttatgtt cttctcctca tgtctgtttt 5460
ctcattaaat tcaaaggctt gaacgggccc tatttagccc ttctgttttc tacgtgttct 5520
aaataactaa agcttttaaa ttctagccat ttagtgtaga actctctttg cagtgatgaa 5580
atgctgtatt ggtttcttgg ctagcatatt aaatattttt atctttgtct tgatacttca 5640
atgtcgtttt aaacatcagg atcgggcttc agtattctca taaccagaga gttcactgag 5700
gatacaggac tgtttgccca ttttttgtta tggctccaga cttgtggtat ttccatgtct 5760
tttttttttt tttttttttt gaccttttag cggctttaaa gtatttctgt tgttaggtgt 5820
tgtattactt ttctaagatt acttaacaaa gcaccacaaa ctgagtggct ttaaacaaca 5880
gcaatttatt ctctcacaat tctagaagct agaagtccga aatcaaagtg ttgacagggg 5940
catgatcttc aagagagaag actctttcct tgcctcttcc tggcttctgg tggttaccag 6000
caatcctgag tgttcctttc ttgccttgta gtttcaacaa tccagtatct gccttttgtc 6060
ttcacatggc tgtctaccat ttgtctctgt gtctccaaat ctctctcctt ataaacacag 6120
cagttattgg attaggcccc actctaatcc agtatgaccc cattttaaca tgattacact 6180
tatttctaga taaggtcaca ttcacgtaca ccaagggtta ggaattgaac atatcttttt 6240
gggggacaca attcaaccca caagtgtcag tctctagctg agcctttccc ttcctgtttt 6300
tctccttttt agttgctatg ggttaggggc caaatctcca gtcatactag aattgcacat 6360
ggactggata tttgggaata ctgcgggtct attctatgag ctttagtatg taacatttaa 6420
tatcagtgta aagaagccct tttttaagtt atttctttga atttctaaat gtatgccctg 6480
aatataagta acaagttacc atgtcttgta aaatgatcat atcaacaaac atttaatgtg 6540
cacctactgt gctagttgaa tgtctttatc ctgataggag ataacaggat tccacatctt 6600
tgacttaaga ggacaaacca aatatgtcta aatcatttgg ggttttgatg gatatcttta 6660
aattgctgaa cctaatcatt ggtttcatat gtcattgttt agatatctcc ggagcatttg 6720
gataatgtga cagttggaat gcagtgatgt cgactctttg cccaccgcca tctccagctg 6780
ttgccaagac agagattgct ttaagtggca aatcaccttt attagcagct acttttgctt 6840
actgggacaa tattcttggt cctagagtaa ggcacatttg ggctccaaag acagaacagg 6900
tacttctcag tgatggagaa ataacttttc ttgccaacca cactctaaat ggagaaatcc 6960
ttcgaaatgc agagagtggt gctatagatg taaagttttt tgtcttgtct gaaaagggag 7020
tgattattgt ttcattaatc tttgatggaa actggaatgg ggatcgcagc acatatggac 7080
tatcaattat acttccacag acagaactta gtttctacct cccacttcat agagtgtgtg 7140
ttgatagatt aacacatata atccggaaag gaagaatatg gatgcataag gtaagtgatt 7200
tttcagctta ttaatcatgt taacctatct gttgaaagct tattttctgg tacatataaa 7260
tcttattttt ttaattatat gcagtgaaca tcaaacaata aatgttattt attttgcatt 7320
taccctatta gatacaaata catctggtct gatacctgtc atcttcatat taactgtgga 7380
aggtacgaaa tggtagctcc acattataga tgaaaagcta aagcttagac aaataaagaa 7440
acttttagac cctggattct tcttgggagc ctttgactct aatacctttt gtttcccttt 7500
cattgcacaa ttctgtcttt tgcttactac tatgtgtaag tataacagtt caaagtaata 7560
gtttcataag ctgttggtca tgtagccttt ggtctcttta acctctttgc caagttccca 7620
ggttcataaa atgaggaggt tgaatggaat ggttcccaag agaattcctt ttaatcttac 7680
agaaattatt gttttcctaa atcctgtagt tgaatatata atgctattta catttcagta 7740
tagttttgat gtatctaaag aacacattga attctccttc ctgtgttcca gtttgatact 7800
aacctgaaag tccattaagc attaccagtt ttaaaaggct tttgcccaat agtaaggaaa 7860
aataatatct tttaaaagaa taatttttta ctatgtttgc aggcttactt ccttttttct 7920
cacattatga aactcttaaa atcaggagaa tcttttaaac aacatcataa tgtttaattt 7980
gaaaagtgca agtcattctt ttcctttttg aaactatgca gatgttacat tgactgtttt 8040
ctgtgaagtt atcttttttt cactgcagaa taaaggttgt tttgatttta ttttgtattg 8100
tttatgagaa catgcatttg ttgggttaat ttcctacccc tgcccccatt ttttccctaa 8160
agtagaaagt atttttcttg tgaactaaat tactacacaa gaacatgtct attgaaaaat 8220
aagcaagtat caaaatgttg tgggttgttt ttttaaataa attttctctt gctcaggaaa 8280
gacaagaaaa tgtccagaag attatcttag aaggcacaga gagaatggaa gatcaggtat 8340
atgcaaattg catactgtca aatgtttttc tcacagcatg tatctgtata aggttgatgg 8400
ctacatttgt caaggccttg gagacatacg aataagcctt taatggagct tttatggagg 8460
tgtacagaat aaactggagg aagatttcca tatcttaaac ccaaagagtt aaatcagtaa 8520
acaaaggaaa atagtaattg catctacaaa ttaatatttg ctcccttttt ttttctgttt 8580
gcccagaata aattttggat aacttgttca tagtaaaaat aaaaaaaatt gtctctgata 8640
tgttctttaa ggtactactt ctcgaacctt tccctagaag tagctgtaac agaaggagag 8700
catatgtacc cctgaggtat ctgtctgggg tgtaggccca ggtccacaca atatttcttc 8760
taagtcttat gttgtatcgt taagactcat gcaatttaca ttttattcca taactatttt 8820
agtattaaaa tttgtcagtg atatttctta ccctctcctc taggaaaatg tgccatgttt 8880
atcccttggc tttgaatgcc cctcaggaac agacactaag agtttgagaa gcatggttac 8940
aagggtgtgg cttcccctgc ggaaactaag tacagactat ttcactgtaa agcagagaag 9000
ttcttttgaa ggagaatctc cagtgaagaa agagttcttc acttttactt ccatttcctc 9060
ttgtgggtga ccctcaatgc tccttgtaaa actccaatat tttaaacatg gctgttttgc 9120
ctttctttgc ttctttttag catgaatgag acagatgata ctttaaaaaa gtaattaaaa 9180
aaaaaaactt gtgaaaatac atggccataa tacagaaccc aatacaatga tctcctttac 9240
caaattgtta tgtttgtact tttgtagata gctttccaat tcagagacag ttattctgtg 9300
taaaggtctg acttaacaag aaaagatttc cctttaccca aagaatccca gtccttattt 9360
gctggtcaat aagcagggtc cccaggaatg gggtaacttt cagcaccctc taacccacta 9420
gttattagta gactaattaa gtaaacttat cgcaagttga ggaaacttag aaccaactaa 9480
aattctgctt ttactgggat tttgtttttt caaaccagaa acctttactt aagttgacta 9540
ctattaatga attttggtct ctcttttaag tgctcttctt aaaaatgtta tcttactgct 9600
gagaagttca agtttgggaa gtacaaggag gaatagaaac ttaagagatt ttcttttaga 9660
gcctcttctg tatttagccc tgtaggattt tttttttttt tttttttttt ggtgttgttg 9720
agcttcagtg aggctattca ttcacttata ctgataatgt ctgagatact gtgaatgaaa 9780
tactatgtat gcttaaacct aagaggaaat attttcccaa aattattctt cccgaaaagg 9840
aggagttgcc ttttgattga gttcttgcaa atctcacaac gactttattt tgaacaatac 9900
tgtttgggga tgatgcatta gtttgaaaca acttcagttg tagctgtcat ctgataaaat 9960
tgcttcacag ggaaggaaat ttaacacgga tctagtcatt attcttgtta gattgaatgt 10020
gtgaattgta attgtaaaca ggcatgataa ttattacttt aaaaactaaa aacagtgaat 10080
agttagttgt ggaggttact aaaggatggt ttttttttaa ataaaacttt cagcattatg 10140
caaatgggca tatggcttag gataaaactt ccagaagtag catcacattt aaattctcaa 10200
gcaacttaat aatatggggc tctgaaaaac tggttaaggt tactccaaaa atggccctgg 10260
gtctgacaaa gattctaact taaagatgct tatgaagact ttgagtaaaa tcatttcata 10320
aaataagtga ggaaaaacaa ctagtattaa attcatctta aataatgtat gatttaaaaa 10380
atatgtttag ctaaaaatgc atagtcattt gacaatttca tttatatctc aaaaaattta 10440
cttaaccaag ttggtcacaa aactgatgag actggtggtg gtagtgaata aatgagggac 10500
catccatatt tgagacactt tacatttgtg atgtgttata ctgaattttc agtttgattc 10560
tatagactac aaatttcaaa attacaattt caagatgtaa taagtagtaa tatcttgaaa 10620
tagctctaaa gggaattttt ctgttttatt gattcttaaa atatatgtgc tgattttgat 10680
ttgcatttgg gtagattata cttttatgag tatggaggtt aggtattgat tcaagttttc 10740
cttacctatt tggtaaggat ttcaaagtct ttttgtgctt ggttttcctc atttttaaat 10800
atgaaatata ttgatgacct ttaacaaatt ttttttatct caaattttaa aggagatctt 10860
ttctaaaaga ggcatgatga cttaatcatt gcatgtaaca gtaaacgata aaccaatgat 10920
tccatactct ctaaagaata aaagtgagct ttagggccgg gcatggtcag aaatttgaca 10980
ccaacctggc caacatggcg aaaccccgtc tctactaaaa atacaaaaat cagccgggca 11040
tggtggcggc acctatagtc ccagctactt gggaggatga gacaggagag tcacttgaac 11100
ctgggaggag aggttgcagt gagctgagat cacgccattg cactccagcc tgagcaatga 11160
aagcaaaact ccatctcaaa aaaaaaaaaa gaaaagaaag aataaaagtg agctttggat 11220
tgcatataaa tcctttagac atgtagtaga cttgtttgat actgtgtttg aacaaattac 11280
gaagtatttt catcaaagaa tgttattgtt tgatgttatt tttatttttt attgcccagc 11340
ttctctcata ttacgtgatt ttcttcactt catgtcactt tattgtgcag ggtcagagta 11400
ttattccaat gcttactgga gaagtgattc ctgtaatgga actgctttca tctatgaaat 11460
cacacagtgt tcctgaagaa atagatgtaa gtttaaatga gagcaattat acactttatg 11520
agttttttgg ggttatagta ttattatgta tattattaat attctaattt taatagtaag 11580
gactttgtca tacatactat tcacatacag tattagccac tttagcaaat aagcacacac 11640
aaaatcctgg attttatggc aaaacagagg catttttgat cagtgatgac aaaattaaat 11700
tcattttgtt tatttcatta cttttataat tcctaaaagt gggaggatcc cagctcttat 11760
aggagcaatt aatatttaat gtagtgtctt ttgaaacaaa actgtgtgcc aaagtagtaa 11820
ccattaatgg aagtttactt gtagtcacaa atttagtttc cttaatcatt tgttgaggac 11880
gttttgaatc acacactatg agtgttaaga gataccttta ggaaactatt cttgttgttt 11940
tctgattttg tcatttaggt tagtctcctg attctgacag ctcagaagag gaagttgttc 12000
ttgtaaaaat tgtttaacct gcttgaccag ctttcacatt tgttcttctg aagtttatgg 12060
tagtgcacag agattgtttt ttggggagtc ttgattctcg gaaatgaagg cagtgtgtta 12120
tattgaatcc agacttccga aaacttgtat attaaaagtg ttatttcaac actatgttac 12180
agccagacta atttttttat tttttgatgc attttagata gctgatacag tactcaatga 12240
tgatgatatt ggtgacagct gtcatgaagg ctttcttctc aagtaagaat ttttcttttc 12300
ataaaagctg gatgaagcag ataccatctt atgctcacct atgacaagat ttggaagaaa 12360
gaaaataaca gactgtctac ttagattgtt ctagggacat tacgtatttg aactgttgct 12420
taaatttgtg ttatttttca ctcattatat ttctatatat atttggtgtt attccatttg 12480
ctatttaaag aaaccgagtt tccatcccag acaagaaatc atggcccctt gcttgattct 12540
ggtttcttgt tttacttctc attaaagcta acagaatcct ttcatattaa gttgtactgt 12600
agatgaactt aagttattta ggcgtagaac aaaattattc atatttatac tgatcttttt 12660
ccatccagca gtggagttta gtacttaaga gtttgtgccc ttaaaccaga ctccctggat 12720
taatgctgtg tacccgtggg caaggtgcct gaattctcta tacacctatt tcctcatctg 12780
taaaatggca ataatagtaa tagtacctaa tgtgtagggt tgttataagc attgagtaag 12840
ataaataata taaagcactt agaacagtgc ctggaacata aaaacactta ataatagctc 12900
atagctaaca tttcctattt acatttcttc tagaaatagc cagtatttgt tgagtgccta 12960
catgttagtt cctttactag ttgctttaca tgtattatct tatattctgt tttaaagttt 13020
cttcacagtt acagattttc atgaaatttt acttttaata aaagagaagt aaaagtataa 13080
agtattcact tttatgttca cagtcttttc ctttaggctc atgatggagt atcagaggca 13140
tgagtgtgtt taacctaaga gccttaatgg cttgaatcag aagcacttta gtcctgtatc 13200
tgttcagtgt cagcctttca tacatcattt taaatcccat ttgactttaa gtaagtcact 13260
taatctctct acatgtcaat ttcttcagct ataaaatgat ggtatttcaa taaataaata 13320
cattaattaa atgatattat actgactaat tgggctgttt taaggctcaa taagaaaatt 13380
tctgtgaaag gtctctagaa aatgtaggtt cctatacaaa taaaagataa cattgtgctt 13440
atagcttcgg tgtttatcat ataaagctat tctgagttat ttgaagagct cacctacttt 13500
tttttgtttt tagtttgtta aattgtttta taggcaatgt ttttaatctg ttttctttaa 13560
cttacagtgc catcagctca cacttgcaaa cctgtggctg ttccgttgta gtaggtagca 13620
gtgcagagaa agtaaataag gtagtttatt ttataatcta gcaaatgatt tgactcttta 13680
agactgatga tatatcatgg attgtcattt aaatggtagg ttgcaattaa aatgatctag 13740
tagtataagg aggcaatgta atctcatcaa attgctaaga caccttgtgg caacagtgag 13800
tttgaaataa actgagtaag aatcatttat cagtttattt tgatagctcg gaaataccag 13860
tgtcagtagt gtataaatgg ttttgagaat atattaaaat cagatatata aaaaaaatta 13920
ctcttctatt tcccaatgtt atctttaaca aatctgaaga tagtcatgta cttttggtag 13980
tagttccaaa gaaatgttat ttgtttattc atcttgattt cattgtcttc gctttccttc 14040
taaatctgtc ccttctaggg agctattggg attaagtggt cattgattat tatactttat 14100
tcagtaatgt ttctgaccct ttccttcagt gctacttgag ttaattaagg attaatgaac 14160
agttacattt ccaagcatta gctaataaac taaaggattt tgcacttttc ttcactgacc 14220
attagttaga aagagttcag agataagtat gtgtatcttt caatttcagc aaacctaatt 14280
ttttaaaaaa agttttacat aggaaatatg ttggaaatga tactttacaa agatattcat 14340
aatttttttt tgtaatcagc tactttgtat atttacatga gccttaattt atatttctca 14400
tataaccatt tatgagagct tagtatacct gtgtcattat attgcatcta cgaactagtg 14460
accttattcc ttctgttacc tcaaacaggt ggctttccat ctgtgatctc caaagcctta 14520
ggttgcacag agtgactgcc gagctgcttt atgaagggag aaaggctcca tagttggagt 14580
gttttttttt ttttttttaa acatttttcc catcctccat cctcttgagg gagaatagct 14640
taccttttat cttgttttaa tttgagaaag aagttgccac cactctaggt tgaaaaccac 14700
tcctttaaca taataactgt ggatatggtt tgaatttcaa gatagttaca tgccttttta 14760
tttttcctaa tagagctgta ggtcaaatat tattagaatc agatttctaa atcccaccca 14820
atgacctgct tattttaaat caaattcaat aattaattct cttctttttg gaggatctgg 14880
acattctttg atatttctta caacgaattt catgtgtaga cccactaaac agaagctata 14940
aaagttgcat ggtcaaataa gtctgagaaa gtctgcagat gatataattc acctgaagag 15000
tcacagtatg tagccaaatg ttaaaggttt tgagatgcca tacagtaaat ttaccaagca 15060
ttttctaaat ttatttgacc acagaatccc tattttaagc aacaactgtt acatcccatg 15120
gattccaggt gactaaagaa tacttatttc ttaggatatg ttttattgat aataacaatt 15180
aaaatttcag atatctttca taagcaaatc agtggtcttt ttacttcatg ttttaatgct 15240
aaaatatttt cttttataga tagtcagaac attatgcctt tttctgactc cagcagagag 15300
aaaatgctcc aggttatgtg aagcagaatc atcatttaaa tatgagtcag ggctctttgt 15360
acaaggcctg ctaaaggtat agtttctagt tatcacaagt gaaaccactt ttctaaaatc 15420
atttttgaga ctctttatag acaaatctta aatattagca tttaatgtat ctcatattga 15480
catgcccaga gactgacttc ctttacacag ttctgcacat agactatatg tcttatggat 15540
ttatagttag tatcatcagt gaaacaccat agaataccct ttgtgttcca ggtgggtccc 15600
tgttcctaca tgtctagcct caggactttt ttttttttaa cacatgctta aatcaggttg 15660
cacatcaaaa ataagatcat ttctttttaa ctaaatagat ttgaatttta ttgaaaaaaa 15720
attttaaaca tctttaagaa gcttatagga tttaagcaat tcctatgtat gtgtactaaa 15780
atatatatat ttctatatat aatatatatt agaaaaaaat tgtatttttc ttttatttga 15840
gtctactgtc aaggagcaaa acagagaaat gtaaattagc aattatttat aatacttaaa 15900
gggaagaaag ttgttcacct tgttgaatct attattgtta tttcaattat agtcccaaga 15960
cgtgaagaaa tagctttcct aatggttatg tgattgtctc atagtgacta ctttcttgag 16020
gatgtagcca cggcaaaatg aaataaaaaa atttaaaaat tgttgcaaat acaagttata 16080
ttaggctttt gtgcattttc aataatgtgc tgctatgaac tcagaatgat agtatttaaa 16140
tatagaaact agttaaagga aacgtagttt ctatttgagt tatacatatc tgtaaattag 16200
aacttctcct gttaaaggca taataaagtg cttaatactt ttgtttcctc agcaccctct 16260
catttaatta tataatttta gttctgaaag ggacctatac cagatgccta gaggaaattt 16320
caaaactatg atctaatgaa aaaatattta atagttctcc atgcaaatac aaatcatata 16380
gttttccaga aaataccttt gacattatac aaagatgatt atcacagcat tataatagta 16440
aaaaaatgga aatagcctct ttcttctgtt ctgttcatag cacagtgcct catacgcagt 16500
aggttattat tacatggtaa ctggctaccc caactgatta ggaaagaagt aaatttgttt 16560
tataaaaata catactcatt gaggtgcata gaataattaa gaaattaaaa gacacttgta 16620
attttgaatc cagtgaatac ccactgttaa tatttggtat atctctttct agtctttttt 16680
tcccttttgc atgtattttc tttaagactc ccacccccac tggatcatct ctgcatgttc 16740
taatctgctt ttttcacagc agattctaag cctctttgaa tatcaacaca aacttcaaca 16800
acttcatcta tagatgccaa ataataaatt catttttatt tacttaacca cttcctttgg 16860
atgcttaggt cattctgatg ttttgctatt gaaaccaatg ctatactgaa cacttctgtc 16920
actaaaactt tgcacacact catgaatagc ttcttaggat aaatttttag agatggattt 16980
gctaaatcag agaccatttt ttaaaattaa aaaacaatta ttcatatcgt ttggcatgta 17040
agacagtaaa ttttcctttt attttgacag gattcaactg gaagctttgt gctgcctttc 17100
cggcaagtca tgtatgctcc atatcccacc acacacatag atgtggatgt caatactgtg 17160
aagcagatgc caccctgtca tgaacatatt tataatcagc gtagatacat gagatccgag 17220
ctgacagcct tctggagagc cacttcagaa gaagacatgg ctcaggatac gatcatctac 17280
actgacgaaa gctttactcc tgatttgtac gtaatgctct gcctgctggt actgtagtca 17340
agcaatatga aattgtgtct tttacgaata aaaacaaaac agaagttgca tttaaaaaga 17400
aagaaatatt accagcagaa ttatgcttga agaaacattt aatcaagcat ttttttctta 17460
aatgttcttc tttttccata caattgtgtt taccctaaaa taggtaagat taacccttaa 17520
agtaaatatt taactatttg tttaataaat atatattgag ctcctaggca ctgttctagg 17580
taccgggctt aatagtggcc aaccagacag ccccagcccc agcccctaca ttgtgtatag 17640
tctattatgt aacagttatt gaatggactt attaacaaaa ccaaagaagt aattctaagt 17700
cttttttttc ttgacatatg aatataaaat acagcaaaac tgttaaaata tattaatgga 17760
acattttttt actttgcatt ttatattgtt attcacttct tatttttttt taaaaaaaaa 17820
agcctgaaca gtaaattcaa aaggaaaagt aatgataatt aattgttgag catggaccca 17880
acttgaaaaa aaaaatgatg atgataaatc tataatccta aaaccctaag taaacactta 17940
aaagatgttc tgaaatcagg aaaagaatta tagtatactt ttgtgtttct cttttatcag 18000
ttgaaaaaag gcacagtagc tcatgcctgt aagaacagag ctttgggagt gcaaggcagg 18060
cggatcactt gaggccagga gttccagacc agcctgggca acatagtgaa accccatctc 18120
tacaaaaaat aaaaaagaat tattggaatg tgtttctgtg tgcctgtaat cctagctatt 18180
ccgaaagctg aggcaggagg atcttttgag cccaggagtt tgaggttaca gggagttatg 18240
atgtgccagt gtactccagc ctggggaaca ccgagactct gtcttattta aaaaaaaaaa 18300
aaaaaaaatg cttgcaataa tgcctggcac atagaaggta acagtaagtg ttaactgtaa 18360
taacccaggt ctaagtgtgt aaggcaatag aaaaattggg gcaaataagc ctgacctatg 18420
tatctacaga atcagtttga gcttaggtaa cagacctgtg gagcaccagt aattacacag 18480
taagtgttaa ccaaaagcat agaataggaa tatcttgttc aagggacccc cagccttata 18540
catctcaagg tgcagaaaga tgacttaata taggacccat tttttcctag ttctccagag 18600
tttttattgg ttcttgagaa agtagtaggg gaatgtttta gaaaatgaat tggtccaact 18660
gaaattacat gtcagtaagt ttttatatat tggtaaattt tagtagacat gtagaagttt 18720
tctaattaat ctgtgccttg aaacattttc ttttttccta aagtgcttag tattttttcc 18780
gttttttgat tggttacttg ggagcttttt tgaggaaatt tagtgaactg cagaatgggt 18840
ttgcaaccat ttggtatttt tgttttgttt tttagaggat gtatgtgtat tttaacattt 18900
cttaatcatt tttagccagc tatgtttgtt ttgctgattt gacaaactac agttagacag 18960
ctattctcat tttgctgatc atgacaaaat aatatcctga atttttaaat tttgcatcca 19020
gctctaaatt ttctaaacat aaaattgtcc aaaaaatagt attttcagcc actagattgt 19080
gtgttaagtc tattgtcaca gagtcatttt acttttaagt atatgttttt acatgttaat 19140
tatgtttgtt atttttaatt ttaacttttt aaaataattc cagtcactgc caatacatga 19200
aaaattggtc actggaattt tttttttgac ttttatttta ggttcatgtg tacatgtgca 19260
ggtgtgttat acaggtaaat tgcgtgtcat gagggtttgg tgtacaggtg atttcattac 19320
ccaggtaata agcatagtac ccaataggta gttttttgat cctcaccctt ctcccaccct 19380
caagtaggcc ctggtgttgc tgtttccttc tttgtgtcca tgtatactca gtgtttagct 19440
cccacttaga agtgagaaca tgcggtagtt ggttttctgt tcctggatta gttcacttag 19500
gataatgacc tctagctcca tctggttttt atggctgcat agtattccat ggtgtatatg 19560
tatcacattt tctttatcca gtctaccatt gataggcatt taggttgatt ccctgtcttt 19620
gttatcatga atagtgctgt gatgaacata cacatgcatg tgtctttatg gtagaaaaat 19680
ttgtattcct ttaggtacat atagaataat ggggttgcta gggtgaatgg tagttctatt 19740
ttcagttatt tgagaaatct tcaaactgct tttcataata gctaaactaa tttacagtcc 19800
cgccagcagt gtataagtgt tcccttttct ccacaacctt gccaacatct gtgatttttt 19860
gactttttaa taatagccat tcctagagaa ttgatttgca attctctatt agtgatatta 19920
agcatttttt catatgcttt ttagctgtct gtatatattc ttctgaaaaa ttttcatgtc 19980
ctttgcccag tttgtagtgg ggtgggttgt tttttgcttg ttaattagtt ttaagttcct 20040
tccagattct gcatatccct ttgttggata catggtttgc agatattttt ctcccattgt 20100
gtaggttgtc ttttactctg ttgatagttt cttttgccat gcaggagctc gttaggtccc 20160
atttgtgttt gtttttgttg cagttgcttt tggcgtcttc atcataaaat ctgtgccagg 20220
gcctatgtcc agaatggtat ttcctaggtt gtcttccagg gtttttacaa ttttagattt 20280
tacgtttatg tctttaatcc atcttgagtt gatttttgta tatggcacaa ggaaggggtc 20340
cagtttcact ccaattccta tggctagcaa ttatcccagc accatttatt gaatacggag 20400
tcctttcccc attgcttgtt ttttgtcaac tttgttgaag atcagatggt tgtaagtgtg 20460
tggctttatt tcttggctct ctattctcca ttggtctatg tgtctgtttt tataacagta 20520
ccctgctgtt caggttccta tagcctttta gtataaaatc ggctaatgtg atgcctccag 20580
ctttgttctt tttgcttagg attgctttgg ctatttgggc tcctttttgg gtccatatta 20640
attttaaaac agttttttct ggttttgtga aggatatcat tggtagttta taggaatagc 20700
attgaatctg tagattgctt tgggcagtat ggccatttta acaatattaa ttcttcctat 20760
ctatgaatat ggaatgtttt tccatgtgtt tgtgtcatct ctttatacct gatgtataaa 20820
gaaaagctgg tattattcct actcaatctg ttccaaaaaa ttgaggagga ggaactcttc 20880
cctaatgagg ccagcatcat tctgatacca aaacctggca gagacacaac agaaaaaaga 20940
aaacttcagg ccaatatcct tgatgaatat agatgcaaaa atcctcaaca aaatactagc 21000
aaaccaaatc cagcagcaca tcaaaaagct gatctacttt gatcaagtag gctttatccc 21060
tgggatgcaa ggttggttca acatacacaa atcaataagt gtgattcatc acataaacag 21120
agctaaaaac aaaaaccaca agattatctc aataggtaga gaaaaggttg tcaataaaat 21180
ttaacatcct ccatgttaaa aaccttcagt aggtcaggtg tagtgactca cacctgtaat 21240
cccagcactt tgggaggcca aggcgggcat atctcttaag cccaggagtt caagacgagc 21300
ctaggcagca tggtgaaacc ccatctctac aaaaaaaaaa aaaaaaaaaa attagcttgg 21360
tatggtgaca tgcacctata gtcccagcta ttcaggaggt tgaggtggga ggattgtttg 21420
agcccgggag gcagaggttg gcagcgagct gagatcatgc caccgcactc cagcctgggc 21480
aacggagtga gaccctgtct caaaaaagaa aaatcacaaa caatcctaaa caaactaggc 21540
attgaaggaa catgcctcaa aaaaataaga accatctatg acagacccat agccaatatc 21600
ttaccaaatg ggcaaaagct ggaagtattc tccttgagaa ccgtaacaag acaaggatgt 21660
ccactctcac cactcctttt cagcatagtt ctggaagtcc tagccagagc aatcaggaaa 21720
gagaaagaaa gaaagacatt cagataggaa gagaagaagt caaactattt ctgtttgcag 21780
gcagtataat tctgtaccta gaaaatctca tagtctctgc ccagaaactc ctaaatctgt 21840
taaaaatttc agcaaagttt tggcattctc tatactccaa caccttccaa agtgagagca 21900
aaatcaagaa cacagtccca ttcacaatag ccgcaaaacg aataaaatac ctaggaatcc 21960
agctaaccag ggaggtgaaa gatctctatg agaattacaa aacactgctg aaagaaatca 22020
gagatgacac aaacaaatgg aaatgttctt ttttaacacc ttgctttatc taattcactt 22080
atgatgaaga tactcattca gtggaacagg tataataagt ccactcgatt aaatataagc 22140
cttattctct ttccagagcc caagaagggg cactatcagt gcccagtcaa taatgacgaa 22200
atgctaatat ttttcccctt tacggtttct ttcttctgta gtgtggtaca ctcgtttctt 22260
aagataagga aacttgaact accttcctgt ttgcttctac acatacccat tctctttttt 22320
tgccactctg gtcaggtata ggatgatccc taccactttc agttaaaaac tcctcctctt 22380
actaaatgtt ctcttaccct ctggcctgag tagaacctag ggaaaatgga agagaaaaag 22440
atgaaaggga ggtggggcct gggaagggaa taagtagtcc tgtttgtttg tgtgtttgct 22500
ttagcacctg ctatatccta ggtgctgtgt taggcacaca ttattttaag tggccattat 22560
attactacta ctcactctgg tcgttgccaa ggtaggtagt actttcttgg atagttggtt 22620
catgttactt acagatggtg ggcttgttga ggcaaaccca gtggataatc atcggagtgt 22680
gttctctaat ctcactcaaa tttttcttca cattttttgg tttgttttgg tttttgatgg 22740
tagtggctta tttttgttgc tggtttgttt tttgtttttt tttgagatgg caagaattgg 22800
tagttttatt tattaattgc ctaagggtct ctactttttt taaaagatga gagtagtaaa 22860
atagattgat agatacatac atacccttac tggggactgc ttatattctt tagagaaaaa 22920
attacatatt agcctgacaa acaccagtaa aatgtaaata tatccttgag taaataaatg 22980
aatgtatatt ttgtgtctcc aaatatatat atctatattc ttacaaatgt gtttatatgt 23040
aatatcaatt tataagaact taaaatgttg gctcaagtga gggattgtgg aaggtagcat 23100
tatatggcca tttcaacatt tgaacttttt tcttttcttc attttcttct tttcttcagg 23160
aatatttttc aagatgtctt acacagagac actctagtga aagccttcct ggatcaggta 23220
aatgttgaac ttgagattgt cagagtgaat gatatgacat gttttctttt ttaatatatc 23280
ctacaatgcc tgttctatat atttatattc ccctggatca tgccccagag ttctgctcag 23340
caattgcagt taagttagtt acactacagt tctcagaaga gtctgtgagg gcatgtcaag 23400
tgcatcatta cattggttgc ctcttgtcct agatttatgc ttcgggaatt cagacctttg 23460
tttacaatat aataaatatt attgctatct tttaaagata taataataag atataaagtt 23520
gaccacaact actgtttttt gaaacataga attcctggtt tacatgtatc aaagtgaaat 23580
ctgacttagc ttttacagat ataatatata catatatata tcctgcaatg cttgtactat 23640
atatgtagta caagtatata tatatgtttg tgtgtgtata tatatatagt acgagcatat 23700
atacatatta ccagcattgt aggatatata tatgtttata tattaaaaaa aagttataaa 23760
cttaaaaccc tattatgtta tgtagagtat atgttatata tgatatgtaa aatatataac 23820
atatactcta tgatagagtg taatatattt tttatatata ttttaacatt tataaaatga 23880
tagaattaag aattgagtcc taatctgttt tattaggtgc tttttgtagt gtctggtctt 23940
tctaaagtgt ctaaatgatt tttccttttg acttattaat ggggaagagc ctgtatatta 24000
acaattaaga gtgcagcatt ccatacgtca aacaacaaac attttaattc aagcattaac 24060
ctataacaag taagtttttt tttttttttt gagaaaggga ggttgtttat ttgcctgaaa 24120
tgactcaaaa atatttttga aacatagtgt acttatttaa ataacatctt tattgtttca 24180
ttcttttaaa aaatatctac ttaattacac agttgaagga aatcgtagat tatatggaac 24240
ttatttctta atatattaca gtttgttata ataacattct ggggatcagg ccaggaaact 24300
gtgtcataga taaagctttg aaataatgag atccttatgt ttactagaaa ttttggattg 24360
agatctatga ggtctgtgac atattgcgaa gttcaaggaa aattcgtagg cctggaattt 24420
catgcttctc aagctgacat aaaatccctc ccactctcca cctcatcata tgcacacatt 24480
ctactcctac ccacccactc caccccctgc aaaagtacag gtatatgaat gtctcaaaac 24540
cataggctca tcttctagga gcttcaatgt tatttgaaga tttgggcaga aaaaattaag 24600
taatacgaaa taacttatgt atgagtttta aaagtgaagt aaacatggat gtattctgaa 24660
gtagaatgca aaatttgaat gcatttttaa agataaatta gaaaacttct aaaaactgtc 24720
agattgtctg ggcctggtgg cttatgcctg taatcccagc actttgggag tccgaggtgg 24780
gtggatcaca aggtcaggag atcgagacca tcctgccaac atggtgaaac cccgtctcta 24840
ctaagtatac aaaaattagc tgggcgtggc agcgtgtgcc tgtaatccca gctacctggg 24900
aggctgaggc aggagaatcg cttgaaccca ggaggtgtag gttgcagtga gtcaagatcg 24960
cgccactgca ctttagcctg gtgacagagc tagactccgt ctcaaaaaaa aaaaaaaata 25020
tcagattgtt cctacaccta gtgcttctat accacactcc tgttaggggg catcagtgga 25080
aatggttaag gagatgttta gtgtgtattg tctgccaagc actgtcaaca ctgtcataga 25140
aacttctgta cgagtagaat gtgagcaaat tatgtgttga aatggttcct ctccctgcag 25200
gtctttcagc tgaaacctgg cttatctctc agaagtactt tccttgcaca gtttctactt 25260
gtccttcaca gaaaagcctt gacactaata aaatatatag aagacgatac gtgagtaaaa 25320
ctcctacacg gaagaaaaac ctttgtacat tgtttttttg ttttgtttcc tttgtacatt 25380
ttctatatca taatttttgc gcttcttttt tttttttttt tttttttttt tccattattt 25440
ttaggcagaa gggaaaaaag ccctttaaat ctcttcggaa cctgaagata gaccttgatt 25500
taacagcaga gggcgatctt aacataataa tggctctggc tgagaaaatt aaaccaggcc 25560
tacactcttt tatctttgga agacctttct acactagtgt gcaagaacga gatgttctaa 25620
tgacttttta aatgtgtaac ttaataagcc tattccatca caatcatgat cgctggtaaa 25680
gtagctcagt ggtgtgggga aacgttcccc tggatcatac tccagaattc tgctctcagc 25740
aattgcagtt aagtaagtta cactacagtt ctcacaagag cctgtgaggg gatgtcaggt 25800
gcatcattac attgggtgtc tcttttccta gatttatgct tttgggatac agacctatgt 25860
ttacaatata ataaatatta ttgctatctt ttaaagatat aataatagga tgtaaacttg 25920
accacaacta ctgttttttt gaaatacatg attcatggtt tacatgtgtc aaggtgaaat 25980
ctgagttggc ttttacagat agttgacttt ctatcttttg gcattctttg gtgtgtagaa 26040
ttactgtaat acttctgcaa tcaactgaaa actagagcct ttaaatgatt tcaattccac 26100
agaaagaaag tgagcttgaa cataggatga gctttagaaa gaaaattgat caagcagatg 26160
tttaattgga attgattatt agatcctact ttgtggattt agtccctggg attcagtctg 26220
tagaaatgtc taatagttct ctatagtcct tgttcctggt gaaccacagt tagggtgttt 26280
tgtttatttt attgttcttg ctattgttga tattctatgt agttgagctc tgtaaaagga 26340
aattgtattt tatgttttag taattgttgc caacttttta aattaatttt cattattttt 26400
gagccaaatt gaaatgtgca cctcctgtgc cttttttctc cttagaaaat ctaattactt 26460
ggaacaagtt cagatttcac tggtcagtca ttttcatctt gttttcttct tgctaagtct 26520
taccatgtac ctgctttggc aatcattgca actctgagat tataaaatgc cttagagaat 26580
atactaacta ataagatctt tttttcagaa acagaaaata gttccttgag tacttccttc 26640
ttgcatttct gcctatgttt ttgaagttgt tgctgtttgc ctgcaatagg ctataaggaa 26700
tagcaggaga aattttactg aagtgctgtt ttcctaggtg ctactttggc agagctaagt 26760
tatcttttgt tttcttaatg cgtttggacc attttgctgg ctataaaata actgattaat 26820
ataattctaa cacaatgttg acattgtagt tacacaaaca caaataaata ttttatttaa 26880
aattctggaa gtaatataaa agggaaaata tatttataag aaagggataa aggtaataga 26940
gcccttctgc cccccaccca ccaaatttac acaacaaaat gacatgttcg aatgtgaaag 27000
gtcataatag ctttcccatc atgaatcaga aagatgtgga cagcttgatg ttttagacaa 27060
ccactgaact agatgactgt tgtactgtag ctcagtcatt taaaaaatat ataaatacta 27120
ccttgtagtg tcccatactg tgttttttac atggtagatt cttatttaag tgctaactgg 27180
ttattttctt tggctggttt attgtactgt tatacagaat gtaagttgta cagtgaaata 27240
agttattaaa gcatgtgtaa acattgttat atatcttttc tcctaaatgg agaattttga 27300
ataaaatata tttgaaattt t 27321
<210> 134
<211> 27321
<212> DNA
<213> Homo sapiens
<400> 134
aaaatttcaa atatatttta ttcaaaattc tccatttagg agaaaagata tataacaatg 60
tttacacatg ctttaataac ttatttcact gtacaactta cattctgtat aacagtacaa 120
taaaccagcc aaagaaaata accagttagc acttaaataa gaatctacca tgtaaaaaac 180
acagtatggg acactacaag gtagtattta tatatttttt aaatgactga gctacagtac 240
aacagtcatc tagttcagtg gttgtctaaa acatcaagct gtccacatct ttctgattca 300
tgatgggaaa gctattatga cctttcacat tcgaacatgt cattttgttg tgtaaatttg 360
gtgggtgggg ggcagaaggg ctctattacc tttatccctt tcttataaat atattttccc 420
ttttatatta cttccagaat tttaaataaa atatttattt gtgtttgtgt aactacaatg 480
tcaacattgt gttagaatta tattaatcag ttattttata gccagcaaaa tggtccaaac 540
gcattaagaa aacaaaagat aacttagctc tgccaaagta gcacctagga aaacagcact 600
tcagtaaaat ttctcctgct attccttata gcctattgca ggcaaacagc aacaacttca 660
aaaacatagg cagaaatgca agaaggaagt actcaaggaa ctattttctg tttctgaaaa 720
aaagatctta ttagttagta tattctctaa ggcattttat aatctcagag ttgcaatgat 780
tgccaaagca ggtacatggt aagacttagc aagaagaaaa caagatgaaa atgactgacc 840
agtgaaatct gaacttgttc caagtaatta gattttctaa ggagaaaaaa ggcacaggag 900
gtgcacattt caatttggct caaaaataat gaaaattaat ttaaaaagtt ggcaacaatt 960
actaaaacat aaaatacaat ttccttttac agagctcaac tacatagaat atcaacaata 1020
gcaagaacaa taaaataaac aaaacaccct aactgtggtt caccaggaac aaggactata 1080
gagaactatt agacatttct acagactgaa tcccagggac taaatccaca aagtaggatc 1140
taataatcaa ttccaattaa acatctgctt gatcaatttt ctttctaaag ctcatcctat 1200
gttcaagctc actttctttc tgtggaattg aaatcattta aaggctctag ttttcagttg 1260
attgcagaag tattacagta attctacaca ccaaagaatg ccaaaagata gaaagtcaac 1320
tatctgtaaa agccaactca gatttcacct tgacacatgt aaaccatgaa tcatgtattt 1380
caaaaaaaca gtagttgtgg tcaagtttac atcctattat tatatcttta aaagatagca 1440
ataatattta ttatattgta aacataggtc tgtatcccaa aagcataaat ctaggaaaag 1500
agacacccaa tgtaatgatg cacctgacat cccctcacag gctcttgtga gaactgtagt 1560
gtaacttact taactgcaat tgctgagagc agaattctgg agtatgatcc aggggaacgt 1620
ttccccacac cactgagcta ctttaccagc gatcatgatt gtgatggaat aggcttatta 1680
agttacacat ttaaaaagtc attagaacat ctcgttcttg cacactagtg tagaaaggtc 1740
ttccaaagat aaaagagtgt aggcctggtt taattttctc agccagagcc attattatgt 1800
taagatcgcc ctctgctgtt aaatcaaggt ctatcttcag gttccgaaga gatttaaagg 1860
gcttttttcc cttctgccta aaaataatgg aaaaaaaaaa aaaaaaaaaa aaaaaagaag 1920
cgcaaaaatt atgatataga aaatgtacaa aggaaacaaa acaaaaaaac aatgtacaaa 1980
ggtttttctt ccgtgtagga gttttactca cgtatcgtct tctatatatt ttattagtgt 2040
caaggctttt ctgtgaagga caagtagaaa ctgtgcaagg aaagtacttc tgagagataa 2100
gccaggtttc agctgaaaga cctgcaggga gaggaaccat ttcaacacat aatttgctca 2160
cattctactc gtacagaagt ttctatgaca gtgttgacag tgcttggcag acaatacaca 2220
ctaaacatct ccttaaccat ttccactgat gccccctaac aggagtgtgg tatagaagca 2280
ctaggtgtag gaacaatctg atattttttt ttttttttga gacggagtct agctctgtca 2340
ccaggctaaa gtgcagtggc gcgatcttga ctcactgcaa cctacacctc ctgggttcaa 2400
gcgattctcc tgcctcagcc tcccaggtag ctgggattac aggcacacgc tgccacgccc 2460
agctaatttt tgtatactta gtagagacgg ggtttcacca tgttggcagg atggtctcga 2520
tctcctgacc ttgtgatcca cccacctcgg actcccaaag tgctgggatt acaggcataa 2580
gccaccaggc ccagacaatc tgacagtttt tagaagtttt ctaatttatc tttaaaaatg 2640
cattcaaatt ttgcattcta cttcagaata catccatgtt tacttcactt ttaaaactca 2700
tacataagtt atttcgtatt acttaatttt ttctgcccaa atcttcaaat aacattgaag 2760
ctcctagaag atgagcctat ggttttgaga cattcatata cctgtacttt tgcagggggt 2820
ggagtgggtg ggtaggagta gaatgtgtgc atatgatgag gtggagagtg ggagggattt 2880
tatgtcagct tgagaagcat gaaattccag gcctacgaat tttccttgaa cttcgcaata 2940
tgtcacagac ctcatagatc tcaatccaaa atttctagta aacataagga tctcattatt 3000
tcaaagcttt atctatgaca cagtttcctg gcctgatccc cagaatgtta ttataacaaa 3060
ctgtaatata ttaagaaata agttccatat aatctacgat ttccttcaac tgtgtaatta 3120
agtagatatt ttttaaaaga atgaaacaat aaagatgtta tttaaataag tacactatgt 3180
ttcaaaaata tttttgagtc atttcaggca aataaacaac ctccctttct caaaaaaaaa 3240
aaaaaaactt acttgttata ggttaatgct tgaattaaaa tgtttgttgt ttgacgtatg 3300
gaatgctgca ctcttaattg ttaatataca ggctcttccc cattaataag tcaaaaggaa 3360
aaatcattta gacactttag aaagaccaga cactacaaaa agcacctaat aaaacagatt 3420
aggactcaat tcttaattct atcattttat aaatgttaaa atatatataa aaaatatatt 3480
acactctatc atagagtata tgttatatat tttacatatc atatataaca tatactctac 3540
ataacataat agggttttaa gtttataact tttttttaat atataaacat atatatatcc 3600
tacaatgctg gtaatatgta tatatgctcg tactatatat atatacacac acaaacatat 3660
atatatactt gtactacata tatagtacaa gcattgcagg atatatatat gtatatatta 3720
tatctgtaaa agctaagtca gatttcactt tgatacatgt aaaccaggaa ttctatgttt 3780
caaaaaacag tagttgtggt caactttata tcttattatt atatctttaa aagatagcaa 3840
taatatttat tatattgtaa acaaaggtct gaattcccga agcataaatc taggacaaga 3900
ggcaaccaat gtaatgatgc acttgacatg ccctcacaga ctcttctgag aactgtagtg 3960
taactaactt aactgcaatt gctgagcaga actctggggc atgatccagg ggaatataaa 4020
tatatagaac aggcattgta ggatatatta aaaaagaaaa catgtcatat cattcactct 4080
gacaatctca agttcaacat ttacctgatc caggaaggct ttcactagag tgtctctgtg 4140
taagacatct tgaaaaatat tcctgaagaa aagaagaaaa tgaagaaaag aaaaaagttc 4200
aaatgttgaa atggccatat aatgctacct tccacaatcc ctcacttgag ccaacatttt 4260
aagttcttat aaattgatat tacatataaa cacatttgta agaatataga tatatatatt 4320
tggagacaca aaatatacat tcatttattt actcaaggat atatttacat tttactggtg 4380
tttgtcaggc taatatgtaa ttttttctct aaagaatata agcagtcccc agtaagggta 4440
tgtatgtatc tatcaatcta ttttactact ctcatctttt aaaaaaagta gagaccctta 4500
ggcaattaat aaataaaact accaattctt gccatctcaa aaaaaaacaa aaaacaaacc 4560
agcaacaaaa ataagccact accatcaaaa accaaaacaa accaaaaaat gtgaagaaaa 4620
atttgagtga gattagagaa cacactccga tgattatcca ctgggtttgc ctcaacaagc 4680
ccaccatctg taagtaacat gaaccaacta tccaagaaag tactacctac cttggcaacg 4740
accagagtga gtagtagtaa tataatggcc acttaaaata atgtgtgcct aacacagcac 4800
ctaggatata gcaggtgcta aagcaaacac acaaacaaac aggactactt attcccttcc 4860
caggccccac ctccctttca tctttttctc ttccattttc cctaggttct actcaggcca 4920
gagggtaaga gaacatttag taagaggagg agtttttaac tgaaagtggt agggatcatc 4980
ctatacctga ccagagtggc aaaaaaagag aatgggtatg tgtagaagca aacaggaagg 5040
tagttcaagt ttccttatct taagaaacga gtgtaccaca ctacagaaga aagaaaccgt 5100
aaaggggaaa aatattagca tttcgtcatt attgactggg cactgatagt gccccttctt 5160
gggctctgga aagagaataa ggcttatatt taatcgagtg gacttattat acctgttcca 5220
ctgaatgagt atcttcatca taagtgaatt agataaagca aggtgttaaa aaagaacatt 5280
tccatttgtt tgtgtcatct ctgatttctt tcagcagtgt tttgtaattc tcatagagat 5340
ctttcacctc cctggttagc tggattccta ggtattttat tcgttttgcg gctattgtga 5400
atgggactgt gttcttgatt ttgctctcac tttggaaggt gttggagtat agagaatgcc 5460
aaaactttgc tgaaattttt aacagattta ggagtttctg ggcagagact atgagatttt 5520
ctaggtacag aattatactg cctgcaaaca gaaatagttt gacttcttct cttcctatct 5580
gaatgtcttt ctttctttct ctttcctgat tgctctggct aggacttcca gaactatgct 5640
gaaaaggagt ggtgagagtg gacatccttg tcttgttacg gttctcaagg agaatacttc 5700
cagcttttgc ccatttggta agatattggc tatgggtctg tcatagatgg ttcttatttt 5760
tttgaggcat gttccttcaa tgcctagttt gtttaggatt gtttgtgatt tttctttttt 5820
gagacagggt ctcactccgt tgcccaggct ggagtgcggt ggcatgatct cagctcgctg 5880
ccaacctctg cctcccgggc tcaaacaatc ctcccacctc aacctcctga atagctggga 5940
ctataggtgc atgtcaccat accaagctaa tttttttttt tttttttttt tgtagagatg 6000
gggtttcacc atgctgccta ggctcgtctt gaactcctgg gcttaagaga tatgcccgcc 6060
ttggcctccc aaagtgctgg gattacaggt gtgagtcact acacctgacc tactgaaggt 6120
ttttaacatg gaggatgtta aattttattg acaacctttt ctctacctat tgagataatc 6180
ttgtggtttt tgtttttagc tctgtttatg tgatgaatca cacttattga tttgtgtatg 6240
ttgaaccaac cttgcatccc agggataaag cctacttgat caaagtagat cagctttttg 6300
atgtgctgct ggatttggtt tgctagtatt ttgttgagga tttttgcatc tatattcatc 6360
aaggatattg gcctgaagtt ttcttttttc tgttgtgtct ctgccaggtt ttggtatcag 6420
aatgatgctg gcctcattag ggaagagttc ctcctcctca attttttgga acagattgag 6480
taggaataat accagctttt ctttatacat caggtataaa gagatgacac aaacacatgg 6540
aaaaacattc catattcata gataggaaga attaatattg ttaaaatggc catactgccc 6600
aaagcaatct acagattcaa tgctattcct ataaactacc aatgatatcc ttcacaaaac 6660
cagaaaaaac tgttttaaaa ttaatatgga cccaaaaagg agcccaaata gccaaagcaa 6720
tcctaagcaa aaagaacaaa gctggaggca tcacattagc cgattttata ctaaaaggct 6780
ataggaacct gaacagcagg gtactgttat aaaaacagac acatagacca atggagaata 6840
gagagccaag aaataaagcc acacacttac aaccatctga tcttcaacaa agttgacaaa 6900
aaacaagcaa tggggaaagg actccgtatt caataaatgg tgctgggata attgctagcc 6960
ataggaattg gagtgaaact ggaccccttc cttgtgccat atacaaaaat caactcaaga 7020
tggattaaag acataaacgt aaaatctaaa attgtaaaaa ccctggaaga caacctagga 7080
aataccattc tggacatagg ccctggcaca gattttatga tgaagacgcc aaaagcaact 7140
gcaacaaaaa caaacacaaa tgggacctaa cgagctcctg catggcaaaa gaaactatca 7200
acagagtaaa agacaaccta cacaatggga gaaaaatatc tgcaaaccat gtatccaaca 7260
aagggatatg cagaatctgg aaggaactta aaactaatta acaagcaaaa aacaacccac 7320
cccactacaa actgggcaaa ggacatgaaa atttttcaga agaatatata cagacagcta 7380
aaaagcatat gaaaaaatgc ttaatatcac taatagagaa ttgcaaatca attctctagg 7440
aatggctatt attaaaaagt caaaaaatca cagatgttgg caaggttgtg gagaaaaggg 7500
aacacttata cactgctggc gggactgtaa attagtttag ctattatgaa aagcagtttg 7560
aagatttctc aaataactga aaatagaact accattcacc ctagcaaccc cattattcta 7620
tatgtaccta aaggaataca aatttttcta ccataaagac acatgcatgt gtatgttcat 7680
cacagcacta ttcatgataa caaagacagg gaatcaacct aaatgcctat caatggtaga 7740
ctggataaag aaaatgtgat acatatacac catggaatac tatgcagcca taaaaaccag 7800
atggagctag aggtcattat cctaagtgaa ctaatccagg aacagaaaac caactaccgc 7860
atgttctcac ttctaagtgg gagctaaaca ctgagtatac atggacacaa agaaggaaac 7920
agcaacacca gggcctactt gagggtggga gaagggtgag gatcaaaaaa ctacctattg 7980
ggtactatgc ttattacctg ggtaatgaaa tcacctgtac accaaaccct catgacacgc 8040
aatttacctg tataacacac ctgcacatgt acacatgaac ctaaaataaa agtcaaaaaa 8100
aaaattccag tgaccaattt ttcatgtatt ggcagtgact ggaattattt taaaaagtta 8160
aaattaaaaa taacaaacat aattaacatg taaaaacata tacttaaaag taaaatgact 8220
ctgtgacaat agacttaaca cacaatctag tggctgaaaa tactattttt tggacaattt 8280
tatgtttaga aaatttagag ctggatgcaa aatttaaaaa ttcaggatat tattttgtca 8340
tgatcagcaa aatgagaata gctgtctaac tgtagtttgt caaatcagca aaacaaacat 8400
agctggctaa aaatgattaa gaaatgttaa aatacacata catcctctaa aaaacaaaac 8460
aaaaatacca aatggttgca aacccattct gcagttcact aaatttcctc aaaaaagctc 8520
ccaagtaacc aatcaaaaaa cggaaaaaat actaagcact ttaggaaaaa agaaaatgtt 8580
tcaaggcaca gattaattag aaaacttcta catgtctact aaaatttacc aatatataaa 8640
aacttactga catgtaattt cagttggacc aattcatttt ctaaaacatt cccctactac 8700
tttctcaaga accaataaaa actctggaga actaggaaaa aatgggtcct atattaagtc 8760
atctttctgc accttgagat gtataaggct gggggtccct tgaacaagat attcctattc 8820
tatgcttttg gttaacactt actgtgtaat tactggtgct ccacaggtct gttacctaag 8880
ctcaaactga ttctgtagat acataggtca ggcttatttg ccccaatttt tctattgcct 8940
tacacactta gacctgggtt attacagtta acacttactg ttaccttcta tgtgccaggc 9000
attattgcaa gcattttttt tttttttttt ttaaataaga cagagtctcg gtgttcccca 9060
ggctggagta cactggcaca tcataactcc ctgtaacctc aaactcctgg gctcaaaaga 9120
tcctcctgcc tcagctttcg gaatagctag gattacaggc acacagaaac acattccaat 9180
aattcttttt tattttttgt agagatgggg tttcactatg ttgcccaggc tggtctggaa 9240
ctcctggcct caagtgatcc gcctgccttg cactcccaaa gctctgttct tacaggcatg 9300
agctactgtg ccttttttca actgataaaa gagaaacaca aaagtatact ataattcttt 9360
tcctgatttc agaacatctt ttaagtgttt acttagggtt ttaggattat agatttatca 9420
tcatcatttt ttttttcaag ttgggtccat gctcaacaat taattatcat tacttttcct 9480
tttgaattta ctgttcaggc tttttttttt aaaaaaaaat aagaagtgaa taacaatata 9540
aaatgcaaag taaaaaaatg ttccattaat atattttaac agttttgctg tattttatat 9600
tcatatgtca agaaaaaaaa gacttagaat tacttctttg gttttgttaa taagtccatt 9660
caataactgt tacataatag actatacaca atgtaggggc tggggctggg gctgtctggt 9720
tggccactat taagcccggt acctagaaca gtgcctagga gctcaatata tatttattaa 9780
acaaatagtt aaatatttac tttaagggtt aatcttacct attttagggt aaacacaatt 9840
gtatggaaaa agaagaacat ttaagaaaaa aatgcttgat taaatgtttc ttcaagcata 9900
attctgctgg taatatttct ttctttttaa atgcaacttc tgttttgttt ttattcgtaa 9960
aagacacaat ttcatattgc ttgactacag taccagcagg cagagcatta cgtacaaatc 10020
aggagtaaag ctttcgtcag tgtagatgat cgtatcctga gccatgtctt cttctgaagt 10080
ggctctccag aaggctgtca gctcggatct catgtatcta cgctgattat aaatatgttc 10140
atgacagggt ggcatctgct tcacagtatt gacatccaca tctatgtgtg tggtgggata 10200
tggagcatac atgacttgcc ggaaaggcag cacaaagctt ccagttgaat cctgtcaaaa 10260
taaaaggaaa atttactgtc ttacatgcca aacgatatga ataattgttt tttaatttta 10320
aaaaatggtc tctgatttag caaatccatc tctaaaaatt tatcctaaga agctattcat 10380
gagtgtgtgc aaagttttag tgacagaagt gttcagtata gcattggttt caatagcaaa 10440
acatcagaat gacctaagca tccaaaggaa gtggttaagt aaataaaaat gaatttatta 10500
tttggcatct atagatgaag ttgttgaagt ttgtgttgat attcaaagag gcttagaatc 10560
tgctgtgaaa aaagcagatt agaacatgca gagatgatcc agtgggggtg ggagtcttaa 10620
agaaaataca tgcaaaaggg aaaaaaagac tagaaagaga tataccaaat attaacagtg 10680
ggtattcact ggattcaaaa ttacaagtgt cttttaattt cttaattatt ctatgcacct 10740
caatgagtat gtatttttat aaaacaaatt tacttctttc ctaatcagtt ggggtagcca 10800
gttaccatgt aataataacc tactgcgtat gaggcactgt gctatgaaca gaacagaaga 10860
aagaggctat ttccattttt ttactattat aatgctgtga taatcatctt tgtataatgt 10920
caaaggtatt ttctggaaaa ctatatgatt tgtatttgca tggagaacta ttaaatattt 10980
tttcattaga tcatagtttt gaaatttcct ctaggcatct ggtataggtc cctttcagaa 11040
ctaaaattat ataattaaat gagagggtgc tgaggaaaca aaagtattaa gcactttatt 11100
atgcctttaa caggagaagt tctaatttac agatatgtat aactcaaata gaaactacgt 11160
ttcctttaac tagtttctat atttaaatac tatcattctg agttcatagc agcacattat 11220
tgaaaatgca caaaagccta atataacttg tatttgcaac aatttttaaa tttttttatt 11280
tcattttgcc gtggctacat cctcaagaaa gtagtcacta tgagacaatc acataaccat 11340
taggaaagct atttcttcac gtcttgggac tataattgaa ataacaataa tagattcaac 11400
aaggtgaaca actttcttcc ctttaagtat tataaataat tgctaattta catttctctg 11460
ttttgctcct tgacagtaga ctcaaataaa agaaaaatac aatttttttc taatatatat 11520
tatatataga aatatatata ttttagtaca catacatagg aattgcttaa atcctataag 11580
cttcttaaag atgtttaaaa ttttttttca ataaaattca aatctattta gttaaaaaga 11640
aatgatctta tttttgatgt gcaacctgat ttaagcatgt gttaaaaaaa aaaaagtcct 11700
gaggctagac atgtaggaac agggacccac ctggaacaca aagggtattc tatggtgttt 11760
cactgatgat actaactata aatccataag acatatagtc tatgtgcaga actgtgtaaa 11820
ggaagtcagt ctctgggcat gtcaatatga gatacattaa atgctaatat ttaagatttg 11880
tctataaaga gtctcaaaaa tgattttaga aaagtggttt cacttgtgat aactagaaac 11940
tataccttta gcaggccttg tacaaagagc cctgactcat atttaaatga tgattctgct 12000
tcacataacc tggagcattt tctctctgct ggagtcagaa aaaggcataa tgttctgact 12060
atctataaaa gaaaatattt tagcattaaa acatgaagta aaaagaccac tgatttgctt 12120
atgaaagata tctgaaattt taattgttat tatcaataaa acatatccta agaaataagt 12180
attctttagt cacctggaat ccatgggatg taacagttgt tgcttaaaat agggattctg 12240
tggtcaaata aatttagaaa atgcttggta aatttactgt atggcatctc aaaaccttta 12300
acatttggct acatactgtg actcttcagg tgaattatat catctgcaga ctttctcaga 12360
cttatttgac catgcaactt ttatagcttc tgtttagtgg gtctacacat gaaattcgtt 12420
gtaagaaata tcaaagaatg tccagatcct ccaaaaagaa gagaattaat tattgaattt 12480
gatttaaaat aagcaggtca ttgggtggga tttagaaatc tgattctaat aatatttgac 12540
ctacagctct attaggaaaa ataaaaaggc atgtaactat cttgaaattc aaaccatatc 12600
cacagttatt atgttaaagg agtggttttc aacctagagt ggtggcaact tctttctcaa 12660
attaaaacaa gataaaaggt aagctattct ccctcaagag gatggaggat gggaaaaatg 12720
tttaaaaaaa aaaaaaaaaa cactccaact atggagcctt tctcccttca taaagcagct 12780
cggcagtcac tctgtgcaac ctaaggcttt ggagatcaca gatggaaagc cacctgtttg 12840
aggtaacaga aggaataagg tcactagttc gtagatgcaa tataatgaca caggtatact 12900
aagctctcat aaatggttat atgagaaata taaattaagg ctcatgtaaa tatacaaagt 12960
agctgattac aaaaaaaaat tatgaatatc tttgtaaagt atcatttcca acatatttcc 13020
tatgtaaaac tttttttaaa aaattaggtt tgctgaaatt gaaagataca catacttatc 13080
tctgaactct ttctaactaa tggtcagtga agaaaagtgc aaaatccttt agtttattag 13140
ctaatgcttg gaaatgtaac tgttcattaa tccttaatta actcaagtag cactgaagga 13200
aagggtcaga aacattactg aataaagtat aataatcaat gaccacttaa tcccaatagc 13260
tccctagaag ggacagattt agaaggaaag cgaagacaat gaaatcaaga tgaataaaca 13320
aataacattt ctttggaact actaccaaaa gtacatgact atcttcagat ttgttaaaga 13380
taacattggg aaatagaaga gtaatttttt ttatatatct gattttaata tattctcaaa 13440
accatttata cactactgac actggtattt ccgagctatc aaaataaact gataaatgat 13500
tcttactcag tttatttcaa actcactgtt gccacaaggt gtcttagcaa tttgatgaga 13560
ttacattgcc tccttatact actagatcat tttaattgca acctaccatt taaatgacaa 13620
tccatgatat atcatcagtc ttaaagagtc aaatcatttg ctagattata aaataaacta 13680
ccttatttac tttctctgca ctgctaccta ctacaacgga acagccacag gtttgcaagt 13740
gtgagctgat ggcactgtaa gttaaagaaa acagattaaa aacattgcct ataaaacaat 13800
ttaacaaact aaaaacaaaa aaaagtaggt gagctcttca aataactcag aatagcttta 13860
tatgataaac accgaagcta taagcacaat gttatctttt atttgtatag gaacctacat 13920
tttctagaga cctttcacag aaattttctt attgagcctt aaaacagccc aattagtcag 13980
tataatatca tttaattaat gtatttattt attgaaatac catcatttta tagctgaaga 14040
aattgacatg tagagagatt aagtgactta cttaaagtca aatgggattt aaaatgatgt 14100
atgaaaggct gacactgaac agatacagga ctaaagtgct tctgattcaa gccattaagg 14160
ctcttaggtt aaacacactc atgcctctga tactccatca tgagcctaaa ggaaaagact 14220
gtgaacataa aagtgaatac tttatacttt tacttctctt ttattaaaag taaaatttca 14280
tgaaaatctg taactgtgaa gaaactttaa aacagaatat aagataatac atgtaaagca 14340
actagtaaag gaactaacat gtaggcactc aacaaatact ggctatttct agaagaaatg 14400
taaataggaa atgttagcta tgagctatta ttaagtgttt ttatgttcca ggcactgttc 14460
taagtgcttt atattattta tcttactcaa tgcttataac aaccctacac attaggtact 14520
attactatta ttgccatttt acagatgagg aaataggtgt atagagaatt caggcacctt 14580
gcccacgggt acacagcatt aatccaggga gtctggttta agggcacaaa ctcttaagta 14640
ctaaactcca ctgctggatg gaaaaagatc agtataaata tgaataattt tgttctacgc 14700
ctaaataact taagttcatc tacagtacaa cttaatatga aaggattctg ttagctttaa 14760
tgagaagtaa aacaagaaac cagaatcaag caaggggcca tgatttcttg tctgggatgg 14820
aaactcggtt tctttaaata gcaaatggaa taacaccaaa tatatataga aatataatga 14880
gtgaaaaata acacaaattt aagcaacagt tcaaatacgt aatgtcccta gaacaatcta 14940
agtagacagt ctgttatttt ctttcttcca aatcttgtca taggtgagca taagatggta 15000
tctgcttcat ccagctttta tgaaaagaaa aattcttact tgagaagaaa gccttcatga 15060
cagctgtcac caatatcatc atcattgagt actgtatcag ctatctaaaa tgcatcaaaa 15120
aataaaaaaa ttagtctggc tgtaacatag tgttgaaata acacttttaa tatacaagtt 15180
ttcggaagtc tggattcaat ataacacact gccttcattt ccgagaatca agactcccca 15240
aaaaacaatc tctgtgcact accataaact tcagaagaac aaatgtgaaa gctggtcaag 15300
caggttaaac aatttttaca agaacaactt cctcttctga gctgtcagaa tcaggagact 15360
aacctaaatg acaaaatcag aaaacaacaa gaatagtttc ctaaaggtat ctcttaacac 15420
tcatagtgtg tgattcaaaa cgtcctcaac aaatgattaa ggaaactaaa tttgtgacta 15480
caagtaaact tccattaatg gttactactt tggcacacag ttttgtttca aaagacacta 15540
cattaaatat taattgctcc tataagagct gggatcctcc cacttttagg aattataaaa 15600
gtaatgaaat aaacaaaatg aatttaattt tgtcatcact gatcaaaaat gcctctgttt 15660
tgccataaaa tccaggattt tgtgtgtgct tatttgctaa agtggctaat actgtatgtg 15720
aatagtatgt atgacaaagt ccttactatt aaaattagaa tattaataat atacataata 15780
atactataac cccaaaaaac tcataaagtg tataattgct ctcatttaaa cttacatcta 15840
tttcttcagg aacactgtgt gatttcatag atgaaagcag ttccattaca ggaatcactt 15900
ctccagtaag cattggaata atactctgac cctgcacaat aaagtgacat gaagtgaaga 15960
aaatcacgta atatgagaga agctgggcaa taaaaaataa aaataacatc aaacaataac 16020
attctttgat gaaaatactt cgtaatttgt tcaaacacag tatcaaacaa gtctactaca 16080
tgtctaaagg atttatatgc aatccaaagc tcacttttat tctttctttt cttttttttt 16140
ttttgagatg gagttttgct ttcattgctc aggctggagt gcaatggcgt gatctcagct 16200
cactgcaacc tctcctccca ggttcaagtg actctcctgt ctcatcctcc caagtagctg 16260
ggactatagg tgccgccacc atgcccggct gatttttgta tttttagtag agacggggtt 16320
tcgccatgtt ggccaggttg gtgtcaaatt tctgaccatg cccggcccta aagctcactt 16380
ttattcttta gagagtatgg aatcattggt ttatcgttta ctgttacatg caatgattaa 16440
gtcatcatgc ctcttttaga aaagatctcc tttaaaattt gagataaaaa aaatttgtta 16500
aaggtcatca atatatttca tatttaaaaa tgaggaaaac caagcacaaa aagactttga 16560
aatccttacc aaataggtaa ggaaaacttg aatcaatacc taacctccat actcataaaa 16620
gtataatcta cccaaatgca aatcaaaatc agcacatata ttttaagaat caataaaaca 16680
gaaaaattcc ctttagagct atttcaagat attactactt attacatctt gaaattgtaa 16740
ttttgaaatt tgtagtctat agaatcaaac tgaaaattca gtataacaca tcacaaatgt 16800
aaagtgtctc aaatatggat ggtccctcat ttattcacta ccaccaccag tctcatcagt 16860
tttgtgacca acttggttaa gtaaattttt tgagatataa atgaaattgt caaatgacta 16920
tgcattttta gctaaacata ttttttaaat catacattat ttaagatgaa tttaatacta 16980
gttgtttttc ctcacttatt ttatgaaatg attttactca aagtcttcat aagcatcttt 17040
aagttagaat ctttgtcaga cccagggcca tttttggagt aaccttaacc agtttttcag 17100
agccccatat tattaagttg cttgagaatt taaatgtgat gctacttctg gaagttttat 17160
cctaagccat atgcccattt gcataatgct gaaagtttta tttaaaaaaa aaccatcctt 17220
tagtaacctc cacaactaac tattcactgt ttttagtttt taaagtaata attatcatgc 17280
ctgtttacaa ttacaattca cacattcaat ctaacaagaa taatgactag atccgtgtta 17340
aatttccttc cctgtgaagc aattttatca gatgacagct acaactgaag ttgtttcaaa 17400
ctaatgcatc atccccaaac agtattgttc aaaataaagt cgttgtgaga tttgcaagaa 17460
ctcaatcaaa aggcaactcc tccttttcgg gaagaataat tttgggaaaa tatttcctct 17520
taggtttaag catacatagt atttcattca cagtatctca gacattatca gtataagtga 17580
atgaatagcc tcactgaagc tcaacaacac caaaaaaaaa aaaaaaaaaa aaaatcctac 17640
agggctaaat acagaagagg ctctaaaaga aaatctctta agtttctatt cctccttgta 17700
cttcccaaac ttgaacttct cagcagtaag ataacatttt taagaagagc acttaaaaga 17760
gagaccaaaa ttcattaata gtagtcaact taagtaaagg tttctggttt gaaaaaacaa 17820
aatcccagta aaagcagaat tttagttggt tctaagtttc ctcaacttgc gataagttta 17880
cttaattagt ctactaataa ctagtgggtt agagggtgct gaaagttacc ccattcctgg 17940
ggaccctgct tattgaccag caaataagga ctgggattct ttgggtaaag ggaaatcttt 18000
tcttgttaag tcagaccttt acacagaata actgtctctg aattggaaag ctatctacaa 18060
aagtacaaac ataacaattt ggtaaaggag atcattgtat tgggttctgt attatggcca 18120
tgtattttca caagtttttt tttttaatta cttttttaaa gtatcatctg tctcattcat 18180
gctaaaaaga agcaaagaaa ggcaaaacag ccatgtttaa aatattggag ttttacaagg 18240
agcattgagg gtcacccaca agaggaaatg gaagtaaaag tgaagaactc tttcttcact 18300
ggagattctc cttcaaaaga acttctctgc tttacagtga aatagtctgt acttagtttc 18360
cgcaggggaa gccacaccct tgtaaccatg cttctcaaac tcttagtgtc tgttcctgag 18420
gggcattcaa agccaaggga taaacatggc acattttcct agaggagagg gtaagaaata 18480
tcactgacaa attttaatac taaaatagtt atggaataaa atgtaaattg catgagtctt 18540
aacgatacaa cataagactt agaagaaata ttgtgtggac ctgggcctac accccagaca 18600
gatacctcag gggtacatat gctctccttc tgttacagct acttctaggg aaaggttcga 18660
gaagtagtac cttaaagaac atatcagaga caattttttt tatttttact atgaacaagt 18720
tatccaaaat ttattctggg caaacagaaa aaaaaaggga gcaaatatta atttgtagat 18780
gcaattacta ttttcctttg tttactgatt taactctttg ggtttaagat atggaaatct 18840
tcctccagtt tattctgtac acctccataa aagctccatt aaaggcttat tcgtatgtct 18900
ccaaggcctt gacaaatgta gccatcaacc ttatacagat acatgctgtg agaaaaacat 18960
ttgacagtat gcaatttgca tatacctgat cttccattct ctctgtgcct tctaagataa 19020
tcttctggac attttcttgt ctttcctgag caagagaaaa tttatttaaa aaaacaaccc 19080
acaacatttt gatacttgct tatttttcaa tagacatgtt cttgtgtagt aatttagttc 19140
acaagaaaaa tactttctac tttagggaaa aaatgggggc aggggtagga aattaaccca 19200
acaaatgcat gttctcataa acaatacaaa ataaaatcaa aacaaccttt attctgcagt 19260
gaaaaaaaga taacttcaca gaaaacagtc aatgtaacat ctgcatagtt tcaaaaagga 19320
aaagaatgac ttgcactttt caaattaaac attatgatgt tgtttaaaag attctcctga 19380
ttttaagagt ttcataatgt gagaaaaaag gaagtaagcc tgcaaacata gtaaaaaatt 19440
attcttttaa aagatattat ttttccttac tattgggcaa aagcctttta aaactggtaa 19500
tgcttaatgg actttcaggt tagtatcaaa ctggaacaca ggaaggagaa ttcaatgtgt 19560
tctttagata catcaaaact atactgaaat gtaaatagca ttatatattc aactacagga 19620
tttaggaaaa caataatttc tgtaagatta aaaggaattc tcttgggaac cattccattc 19680
aacctcctca ttttatgaac ctgggaactt ggcaaagagg ttaaagagac caaaggctac 19740
atgaccaaca gcttatgaaa ctattacttt gaactgttat acttacacat agtagtaagc 19800
aaaagacaga attgtgcaat gaaagggaaa caaaaggtat tagagtcaaa ggctcccaag 19860
aagaatccag ggtctaaaag tttctttatt tgtctaagct ttagcttttc atctataatg 19920
tggagctacc atttcgtacc ttccacagtt aatatgaaga tgacaggtat cagaccagat 19980
gtatttgtat ctaatagggt aaatgcaaaa taaataacat ttattgtttg atgttcactg 20040
catataatta aaaaaataag atttatatgt accagaaaat aagctttcaa cagataggtt 20100
aacatgatta ataagctgaa aaatcactta ccttatgcat ccatattctt cctttccgga 20160
ttatatgtgt taatctatca acacacactc tatgaagtgg gaggtagaaa ctaagttctg 20220
tctgtggaag tataattgat agtccatatg tgctgcgatc cccattccag tttccatcaa 20280
agattaatga aacaataatc actccctttt cagacaagac aaaaaacttt acatctatag 20340
caccactctc tgcatttcga aggatttctc catttagagt gtggttggca agaaaagtta 20400
tttctccatc actgagaagt acctgttctg tctttggagc ccaaatgtgc cttactctag 20460
gaccaagaat attgtcccag taagcaaaag tagctgctaa taaaggtgat ttgccactta 20520
aagcaatctc tgtcttggca acagctggag atggcggtgg gcaaagagtc gacatcactg 20580
cattccaact gtcacattat ccaaatgctc cggagatatc taaacaatga catatgaaac 20640
caatgattag gttcagcaat ttaaagatat ccatcaaaac cccaaatgat ttagacatat 20700
ttggtttgtc ctcttaagtc aaagatgtgg aatcctgtta tctcctatca ggataaagac 20760
attcaactag cacagtaggt gcacattaaa tgtttgttga tatgatcatt ttacaagaca 20820
tggtaacttg ttacttatat tcagggcata catttagaaa ttcaaagaaa taacttaaaa 20880
aagggcttct ttacactgat attaaatgtt acatactaaa gctcatagaa tagacccgca 20940
gtattcccaa atatccagtc catgtgcaat tctagtatga ctggagattt ggcccctaac 21000
ccatagcaac taaaaaggag aaaaacagga agggaaaggc tcagctagag actgacactt 21060
gtgggttgaa ttgtgtcccc caaaaagata tgttcaattc ctaacccttg gtgtacgtga 21120
atgtgacctt atctagaaat aagtgtaatc atgttaaaat ggggtcatac tggattagag 21180
tggggcctaa tccaataact gctgtgttta taaggagaga gatttggaga cacagagaca 21240
aatggtagac agccatgtga agacaaaagg cagatactgg attgttgaaa ctacaaggca 21300
agaaaggaac actcaggatt gctggtaacc accagaagcc aggaagaggc aaggaaagag 21360
tcttctctct tgaagatcat gcccctgtca acactttgat ttcggacttc tagcttctag 21420
aattgtgaga gaataaattg ctgttgttta aagccactca gtttgtggtg ctttgttaag 21480
taatcttaga aaagtaatac aacacctaac aacagaaata ctttaaagcc gctaaaaggt 21540
caaaaaaaaa aaaaaaaaaa aagacatgga aataccacaa gtctggagcc ataacaaaaa 21600
atgggcaaac agtcctgtat cctcagtgaa ctctctggtt atgagaatac tgaagcccga 21660
tcctgatgtt taaaacgaca ttgaagtatc aagacaaaga taaaaatatt taatatgcta 21720
gccaagaaac caatacagca tttcatcact gcaaagagag ttctacacta aatggctaga 21780
atttaaaagc tttagttatt tagaacacgt agaaaacaga agggctaaat agggcccgtt 21840
caagcctttg aatttaatga gaaaacagac atgaggagaa gaacataaac gctcacatcc 21900
aagacagaac ccagggctct tggtcccctt gctcaacttg tacatcttaa tccacataaa 21960
cataccactc taaaaaggta catcctatgt gatattaatg taaaacaaat cattcttgca 22020
aatacagtta tgtgccatgt aacgtttcag tcaatggtag actgcatata tgatggtagt 22080
cccattagat tacaatggac ctgaaaatat gctattgcct tagtgacact gtaaccatca 22140
taaggtctta gtactatttt gcaagttatt taaagtatag cacatacaat tattacagtg 22200
tacaacactt gataataaac tactacattg ctggtttatg tattcactat actatgcctt 22260
ttattgttat tttagagtgc actccttcta cttttttttt ttttaagtta aatgtaaaac 22320
agcctcaggc aagtccttca ggaggtattc aacagaaagc actgttatca taggtgacag 22380
ctacatgtgt gttattgccc ctaaaaacct tccagtggga caagatgtgg aggtggaagg 22440
cagtgaggtg gaagggagtg atactgatga tcctaatcct gtctatgcct aggtgaaagt 22500
gtgtgtgttt tagtttttaa caaaaacgac taacaagtaa aaaaaaaaat ttaaaataga 22560
aaatagaaaa aagcttctag aataaggata caaagaaaaa atatttttgt atagctatac 22620
aatgtatttg tgtttcaagc taagtatttt aaaagttaaa aaattaaaaa gtttacaaag 22680
ttaaaaagtt ataatttttt attgaagaaa aactgttaag ataaatttgg tgtagcttca 22740
gcgtactgtg tttatagtct acagtggtgt acagtgttct aggccttcac attaattcac 22800
cactcactca ctgactcacc cagagcaact tctagtcctg caagctccat tcgtggtaag 22860
tggcctacac aggtatacca ttatctttta taccatactt ttactgtacc ttttctctgt 22920
ttgcatatat ttagataaat atttaccact gtgttacaac tgtctatagt attcagtaca 22980
gtaacagttg tacaggtttg tggcctagga gcaacagact ataccatacg gcctaggtac 23040
ataaaggcta tactatctag gtttgtgtaa gtacactcta tgatgtttgc ataatgacaa 23100
aatcgcctaa tgatgcattc ctaagcaatg tgtgattgta ctataattga agacttgtta 23160
tctaagactg aaagtaaaaa gaattgcaat ttcacctaag caagtctaaa actgtgaagt 23220
ctatttataa taatagcaat acaaagcagc taataggcaa actatgatat acctatcttt 23280
gccatatgat tgctttggga gctaacattt gatctgtaaa tgtatgacaa agtaaacaat 23340
tttacttaaa gaatttcatc cacatcttgt caagagagtt cagtctgatg gaaagcactg 23400
acttctattt acagagcatt agatgagtgc ttttatcata ttatgagtag gcatacagag 23460
cctggcaaaa cagttaactc taagtatgta cagaaatggt tgaacacaac gacagtttta 23520
acacgtgtat ttgtaatttc aaaaattcat ttaggtaata tttactttta aatatgttgt 23580
atcaatttaa tagtcttaag agacagcact agatataagc cgtacagctt ctttaaaata 23640
tccactgttt ttaatacaat gtaagcagtc agtttacaat gatcaaatat aggaatgtaa 23700
tctgaattga aatggtaatg acactactgc tgtcataact aacaacagca aactggaggc 23760
caacataatg aattaagtta acatacaacc ataaaattat attgcaaaca tatttttctt 23820
tcattctttt aggttaaaaa ggtggataat cataaaggca atattacaac tctaatattt 23880
catcattaaa ctgaaaataa aagtatttcc taaaacagaa ctgaaccctg gagcaaaatc 23940
tgattgaatt atagggaaac ttttaccacg ttgtgaaaat tgaactatta tactgctagt 24000
tacactctca ctcctaacag aataagaaaa aaaaaatggg ccgggcatgg tgggtcacac 24060
ctgttatccc agctctttgg taggccgagg caggtggatc acctgaggtc aggagctcaa 24120
gaccagcctg gccaacatgg tgaaacccca cctctactaa aaatacaaaa aattagccgg 24180
gtgtggtggt ggacacctgt aatcccagct actcgggagg ctgaggcagg agaatctctt 24240
gaacccggag gtggcagagg ttgcaatgag ctgagatggc gccactgcac tccagcctgg 24300
gcgacagaga gagactctgc ctcaagaaaa aaacaaacaa acaaacaaac aaaaagaata 24360
agaaagaaaa tgaaggacaa agatcatact gaattgctta gttttaaatc ctaccaaaag 24420
aaatagcctg ggaaatgaaa tgtcacagag aagtataatc aggagagctg tacaattatt 24480
ttactaatac ttgaagtcat cgtctttggt gagaaaaatc catacatgca aatgcagctg 24540
aaaaaaatca gctcaaaacc aatagttgtt tatgtaccta tcttacgtac atgtagtgct 24600
gtctactcca gagagttacc aaacattagc cagtcttttg agggaagcca agattcaaat 24660
tgagtgagac ggtggcttgc tcacagggtt catgagaggt ttcccaatac actttctgga 24720
aataatccca tacatgcaga catgattaca ttaattaaca tctgctaaaa ctgttagtag 24780
agtgctaagt ttgaggtttt gctttttctt taaacgtctg ttaaaaaatc aaccatctct 24840
tccctgattg gtatttagaa aggtggttgg tccactgcta ttgtagtgaa aattctacaa 24900
tcataaagcc ctcacttctt gttttttaga gacagggtct cgttttgtca tccaggctgg 24960
aatgcactgg caggatcata gctctcggta acttcaaact cttgggctca aatgaccctc 25020
ctgcctcagc ctcccaagta gctaggacta caggtgcaca tcaccacgcc cggctaagtt 25080
tttaattttt tgtagagaca gggtctacgt tgcccaggtt gagcttgaac tcctggcttc 25140
aagtgatcct cttgcctccg cctcccaaag ctctggcatt acaggtgtaa gccaccttct 25200
ccaacctggc tctcaatact tgtaaccatg ctgtttattt tctcccagcc caaagagaag 25260
caggatccta aaccgtccac tttccacaac aggagctgcc caggaccact tcaaggacag 25320
tgaactgttt acagtaccag aaagttcaca acactttctc aatcttcaac atcagggaag 25380
actggaaggt gaagttcata tcactatctg gccatttctc acagttccaa gtttctcaga 25440
caataggtag gctaacctag tcctcctggg aactatctaa ttaacgtaga atagaacccg 25500
agggcagact tgaaaaacag aagtcctcct tggtttactt tgtttctctg aaagcaaatt 25560
gtggagtgcc aacatagcca aacaaaatat tttatcaact tcataaggtg cttgtaattt 25620
tttcctggag caggtaaatg ctggcttagt gaacaatctg gaatgtggta attactctcg 25680
ttcttgtttc agatgtacta tcagcatgta gcagtttcca actgattcag ggttttccta 25740
aagtggcagg ccttggcaga ggtggtgaca acaatgcccg tgtcaaatga caccgtattt 25800
caagtattct gactccaggt tattaatatc ccctatatga tagtcttgtt tctgtgatat 25860
tcacagatta tgttaaaagt ttcccaaagt ctgagaaaaa tcatatctta acagtatctt 25920
tttttttttt gatcctttgt acaaaagtag aagtaatgcc agacagatta cgtacccttg 25980
ttgtgaacaa ctggtgcatg gcaactgttt gaatagaaat ttaccaactg ccacaaccag 26040
gcaactactc tcccagagcc taacaatctc gattgcatct gaaagggcca cccctcctgg 26100
gaaagtgcag gacctccctc ctgtttctga atacaaagcc tggtggtgtt caacgcggcc 26160
agatagaccc aatgagcaca cggacatgta atctgtgcac ttctttagac aactgattac 26220
catcagtcaa gtgatgccca agtcacaata gtcacttcct ttaagcaagt ctgtgtcatc 26280
tcggagctgt gaagcaacca ggtcatgtcc cacagaatgg ggagcacacc gacttgcatt 26340
gctgccctca tatgcaagtc atcaccactc tctagaagct tgggctgaaa ttgtgcaggc 26400
gtctccacac ccccatctca tcccgcatga tctcctcgcc ggcagggacc gtctcgggtt 26460
cctagcgaac cccgacttgg tccgcagaag ccgcgcgccg cccaccctcc ggccttcccc 26520
caggcgaggc ctctcagtac ccgaggctcc cttttctcga gcccgcagcg gcagcgctcc 26580
cagcgggtcc ccgggaagga gacagctcgg gtactgaggg cgggaaagca aggaagaggc 26640
cagatcccca tcccttgtcc ctgcgccgcc gccgccgccg ccgccgccgg gaagcccggg 26700
gcccggatgc aggcaattcc accagtcgct agaggcgaaa gcccgacacc cagcttcggt 26760
cagagaaatg agagggaaag taaaaatgcg tcgagctctg aggagagccc ccgcttctac 26820
ccgcgcctct tcccggcagc cgaaccccaa acagccaccc gccaggatgc cgcctcctca 26880
ctcacccact cgccaccgcc tgcgcctccg ccgccgcggg cgcaggcacc gcaaccgcag 26940
ccccgccccg ggcccgcccc cgggcccgcc ccgaccacgc cccggccccg gccccggccc 27000
ctagcgcgcg actcctgagt tccagagctt gctacaggct gcggttgttt ccctccttgt 27060
tttcttctgg ttaatcttta tcaggtcttt tcttgttcac cctcagcgag tactgtgaga 27120
gcaagtagtg gggagagagg gtgggaaaaa caaaaacaca cacctcctaa acccacacct 27180
gctcttgcta gaccccgccc ccaaaagaga agcaaccggg cagcagggac ggctgacaca 27240
ccaagcgtca tcttttacgt gggcggaact tgtcgctgtt tgacgcacct ctctttccta 27300
gcgggacacc gtaggttacg t 27321
<210> 135
<211> 2700
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 135
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 60
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 120
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 180
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 240
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 300
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 360
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 420
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 480
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 540
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 600
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 660
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 720
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 780
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 840
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 900
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 960
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1020
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1080
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1140
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1200
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1260
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1320
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1380
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1440
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1500
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1560
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1620
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1680
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1740
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1800
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1860
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1920
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1980
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2040
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2100
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2160
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2220
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2280
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2340
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2400
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2460
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2520
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2580
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2640
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2700
<210> 136
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
RFGF peptide"
<400> 136
Ala Ala Val Ala Leu Leu Pro Ala Val Leu Leu Ala Leu Leu Ala Pro
1 5 10 15
<210> 137
<211> 11
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
RFGF analogue peptide"
<400> 137
Ala Ala Leu Leu Pro Val Leu Leu Ala Ala Pro
1 5 10
<210> 138
<211> 13
<212> PRT
<213> Human immunodeficiency virus
<400> 138
Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Pro Pro Gln
1 5 10
<210> 139
<211> 16
<212> PRT
<213> Drosophila sp.
<400> 139
Arg Gln Ile Lys Ile Trp Phe Gln Asn Arg Arg Met Lys Trp Lys Lys
1 5 10 15
<210> 140
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 140
ggggccgggg ccggggcc 18
<210> 141
<211> 552
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 141
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 60
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 120
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 180
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 240
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 300
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 360
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 420
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 480
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 540
ggggccgggg cc 552
<210> 142
<211> 1800
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 142
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 60
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 120
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 180
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 240
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 300
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 360
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 420
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 480
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 540
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 600
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 660
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 720
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 780
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 840
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 900
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 960
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1020
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1080
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1140
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1200
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1260
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1320
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1380
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1440
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1500
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1560
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1620
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1680
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1740
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1800
<210> 143
<211> 3000
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 143
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 60
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 120
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 180
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 240
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 300
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 360
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 420
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 480
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 540
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 600
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 660
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 720
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 780
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 840
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 900
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 960
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1020
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1080
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1140
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1200
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1260
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1320
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1380
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1440
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1500
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1560
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1620
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1680
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1740
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1800
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1860
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1920
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1980
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2040
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2100
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2160
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2220
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2280
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2340
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2400
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2460
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2520
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2580
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2640
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2700
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2760
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2820
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2880
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2940
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3000
<210> 144
<211> 3600
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 144
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 60
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 120
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 180
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 240
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 300
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 360
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 420
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 480
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 540
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 600
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 660
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 720
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 780
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 840
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 900
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 960
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1020
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1080
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1140
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1200
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1260
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1320
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1380
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1440
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1500
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1560
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1620
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1680
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1740
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1800
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1860
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1920
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1980
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2040
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2100
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2160
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2220
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2280
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2340
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2400
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2460
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2520
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2580
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2640
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2700
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2760
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2820
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2880
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 2940
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3000
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3060
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3120
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3180
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3240
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3300
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3360
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3420
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3480
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3540
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 3600
<210> 145
<211> 1500
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 145
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 60
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 120
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 180
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 240
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 300
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 360
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 420
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 480
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 540
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 600
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 660
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 720
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 780
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 840
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 900
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 960
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1020
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1080
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1140
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1200
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1260
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1320
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1380
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1440
ggggccgggg ccggggccgg ggccggggcc ggggccgggg ccggggccgg ggccggggcc 1500
<210> 146
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 146
uguugccaag acagagauug u 21
<210> 147
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 147
acaauctcug ucuuggcaac agc 23
<210> 148
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 148
caagacagag auugcuuuaa u 21
<210> 149
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 149
auuaaagcaa ucucugucuu ggc 23
<210> 150
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 150
uagcugauac aguacucaau u 21
<210> 151
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 151
aauugaguac uguaucagcu aua 23
<210> 152
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 152
ugauacagua cucaaugaug a 21
<210> 153
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 153
ucaucatuga guacuguauc agc 23
<210> 154
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 154
gucuuacaca gagacacucu a 21
<210> 155
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 155
uagagugucu cuguguaaga cau 23
<210> 156
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 156
gaagaccuuu cuacacuagu u 21
<210> 157
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 157
aacuagtgua gaaaggucuu cca 23
<210> 158
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 158
agaaaagacc ugauaaagau u 21
<210> 159
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 159
aaucuuuauc aggucuuuuc uug 23
<210> 160
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 160
aaaagaccug auaaagauua a 21
<210> 161
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 161
uuaaucuuua ucaggucuuu ucu 23
<210> 162
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 162
aaagaccuga uaaagauuaa u 21
<210> 163
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 163
auuaaucuuu aucaggucuu uuc 23
<210> 164
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 164
cugauaaaga uuaaccagaa u 21
<210> 165
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 165
auucugguua aucuuuauca ggu 23
<210> 166
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 166
auaaagauua accagaagaa a 21
<210> 167
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 167
uuucuucugg uuaaucuuua uca 23
<210> 168
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 168
aaagauuaac cagaagaaaa u 21
<210> 169
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 169
auuuucuucu gguuaaucuu uau 23
<210> 170
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 170
ugcgucaaac agcgacaagu u 21
<210> 171
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 171
aacuugtcgc uguuugacgc acc 23
<210> 172
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 172
cgucaaacag cgacaaguuc u 21
<210> 173
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 173
agaacutguc gcuguuugac gca 23
<210> 174
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 174
cccacguaaa agaugacgcu u 21
<210> 175
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 175
aagcgucauc uuuuacgugg gcg 23
<210> 176
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 176
ccacguaaaa gaugacgcuu u 21
<210> 177
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 177
aaagcgtcau cuuuuacgug ggc 23
<210> 178
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 178
agugaugucg acucuuugcc u 21
<210> 179
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 179
aggcaaagag ucgacaucac ugc 23
<210> 180
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 180
cagcuguugc caagacagag a 21
<210> 181
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 181
ucucugtcuu ggcaacagcu gga 23
<210> 182
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 182
agcuguugcc aagacagaga u 21
<210> 183
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 183
aucucugucu uggcaacagc ugg 23
<210> 184
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 184
gccaagacag agauugcuuu a 21
<210> 185
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 185
uaaagcaauc ucugucuugg caa 23
<210> 186
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 186
aagacagaga uugcuuuaag u 21
<210> 187
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 187
acuuaaagca aucucugucu ugg 23
<210> 188
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 188
agacagagau ugcuuuaagu u 21
<210> 189
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 189
aacuuaaagc aaucucuguc uug 23
<210> 190
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 190
gacagagauu gcuuuaagug u 21
<210> 191
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 191
acacuuaaag caaucucugu cuu 23
<210> 192
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 192
uagcagcuac uuuugcuuac u 21
<210> 193
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 193
aguaagcaaa aguagcugcu aau 23
<210> 194
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 194
acaauauucu ugguccuaga u 21
<210> 195
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 195
aucuaggacc aagaauauug ucc 23
<210> 196
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 196
aauauucuug guccuagagu a 21
<210> 197
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 197
uacucuagga ccaagaauau ugu 23
<210> 198
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 198
auauucuugg uccuagagua a 21
<210> 199
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 199
uuacuctagg accaagaaua uug 23
<210> 200
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 200
agacagaaca gguacuucuc a 21
<210> 201
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 201
ugagaaguac cuguucuguc uuu 23
<210> 202
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 202
uaaauggaga aauccuucga a 21
<210> 203
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 203
uucgaaggau uucuccauuu aga 23
<210> 204
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 204
ggagaaaucc uucgaaaugc a 21
<210> 205
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 205
ugcauutcga aggauuucuc cau 23
<210> 206
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 206
cauuaaucuu ugauggaaac u 21
<210> 207
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 207
aguuuccauc aaagauuaau gaa 23
<210> 208
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 208
auuaaucuuu gauggaaacu u 21
<210> 209
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 209
aaguuuccau caaagauuaa uga 23
<210> 210
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 210
cauauggacu aucaauuaua u 21
<210> 211
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 211
auauaatuga uaguccauau gug 23
<210> 212
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 212
uauggacuau caauuauacu u 21
<210> 213
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 213
aaguauaauu gauaguccau aug 23
<210> 214
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 214
uggacuauca auuauacuuc u 21
<210> 215
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 215
agaaguauaa uugauagucc aua 23
<210> 216
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 216
ucaauuauac uuccacagac a 21
<210> 217
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 217
ugucugtgga aguauaauug aua 23
<210> 218
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 218
acagaacuua guuucuaccu u 21
<210> 219
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 219
aagguagaaa cuaaguucug ucu 23
<210> 220
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 220
cagaacuuag uuucuaccuc u 21
<210> 221
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 221
agagguagaa acuaaguucu guc 23
<210> 222
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 222
gagugugugu ugauagauua a 21
<210> 223
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 223
uuaauctauc aacacacacu cua 23
<210> 224
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 224
aguguguguu gauagauuaa u 21
<210> 225
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 225
auuaaucuau caacacacac ucu 23
<210> 226
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 226
uguguguuga uagauuaaca u 21
<210> 227
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 227
auguuaaucu aucaacacac acu 23
<210> 228
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 228
aacacauaua auccggaaag u 21
<210> 229
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 229
acuuuccgga uuauaugugu uaa 23
<210> 230
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 230
uguccagaag auuaucuuag a 21
<210> 231
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 231
ucuaagauaa ucuucuggac auu 23
<210> 232
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 232
gaacugcuuu caucuaugaa a 21
<210> 233
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 233
uuucauagau gaaagcaguu cca 23
<210> 234
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 234
cugucaugaa ggcuuucuuc u 21
<210> 235
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 235
agaagaaagc cuucaugaca gcu 23
<210> 236
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 236
uguuccguug uaguagguag u 21
<210> 237
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 237
acuacctacu acaacggaac agc 23
<210> 238
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 238
aucauuuaaa uaugagucag u 21
<210> 239
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 239
acugactcau auuuaaauga uga 23
<210> 240
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 240
ccugcuaaag gauucaacug u 21
<210> 241
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 241
acaguugaau ccuuuagcag gcc 23
<210> 242
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 242
cacccuguca ugaacauauu u 21
<210> 243
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 243
aaauauguuc augacagggu ggc 23
<210> 244
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 244
ugaacauauu uauaaucagc u 21
<210> 245
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 245
agcugatuau aaauauguuc aug 23
<210> 246
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 246
acauauuuau aaucagcgua u 21
<210> 247
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 247
auacgctgau uauaaauaug uuc 23
<210> 248
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 248
auauuuauaa ucagcguaga u 21
<210> 249
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 249
aucuacgcug auuauaaaua ugu 23
<210> 250
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 250
ucaggauacg aucaucuaca u 21
<210> 251
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 251
auguagauga ucguauccug agc 23
<210> 252
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 252
aucaucuaca cugacgaaag u 21
<210> 253
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 253
acuuucguca guguagauga ucg 23
<210> 254
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 254
caucuacacu gacgaaagcu u 21
<210> 255
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 255
aagcuutcgu caguguagau gau 23
<210> 256
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 256
aucuacacug acgaaagcuu u 21
<210> 257
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 257
aaagcutucg ucaguguaga uga 23
<210> 258
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 258
cacugacgaa agcuuuacuc u 21
<210> 259
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 259
agaguaaagc uuucgucagu gua 23
<210> 260
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 260
acugacgaaa gcuuuacucc u 21
<210> 261
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 261
aggaguaaag cuuucgucag ugu 23
<210> 262
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 262
cugacgaaag cuuuacuccu u 21
<210> 263
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 263
aaggagtaaa gcuuucguca gug 23
<210> 264
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 264
uuucaagaug ucuuacacag a 21
<210> 265
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 265
ucugugtaag acaucuugaa aaa 23
<210> 266
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 266
cuggaucagg ucuuucagcu u 21
<210> 267
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 267
aagcugaaag accugaucca gga 23
<210> 268
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 268
cucagaagua cuuuccuugc a 21
<210> 269
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 269
ugcaaggaaa guacuucuga gag 23
<210> 270
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 270
ucuacuuguc cuucacagaa a 21
<210> 271
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 271
uuucugtgaa ggacaaguag aaa 23
<210> 272
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 272
gaaaagccuu gacacuaaua a 21
<210> 273
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 273
uuauuagugu caaggcuuuu cug 23
<210> 274
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 274
ccuuuaaauc ucuucggaac u 21
<210> 275
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 275
aguuccgaag agauuuaaag ggc 23
<210> 276
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 276
cuuuaaaucu cuucggaacc u 21
<210> 277
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 277
agguuccgaa gagauuuaaa ggg 23
<210> 278
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 278
uuuaaaucuc uucggaaccu u 21
<210> 279
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 279
aagguuccga agagauuuaa agg 23
<210> 280
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 280
gggcgaucuu aacauaauaa u 21
<210> 281
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 281
auuauuaugu uaagaucgcc cuc 23
<210> 282
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 282
ucuuaacaua auaauggcuc u 21
<210> 283
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 283
agagccauua uuauguuaag auc 23
<210> 284
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 284
uuaucuuugg aagaccuuuc u 21
<210> 285
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 285
agaaaggucu uccaaagaua aaa 23
<210> 286
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 286
aagaccuuuc uacacuagug u 21
<210> 287
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 287
acacuagugu agaaaggucu ucc 23
<210> 288
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 288
agaccuuucu acacuagugu u 21
<210> 289
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 289
aacacuagug uagaaagguc uuc 23
<210> 290
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 290
gcaagaacga gauguucuaa u 21
<210> 291
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 291
auuagaacau cucguucuug cac 23
<210> 292
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 292
gaacgagaug uucuaaugac u 21
<210> 293
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 293
agucautaga acaucucguu cuu 23
<210> 294
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 294
aacgagaugu ucuaaugacu u 21
<210> 295
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 295
aagucatuag aacaucucgu ucu 23
<210> 296
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 296
acgagauguu cuaaugacuu u 21
<210> 297
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 297
aaagucauua gaacaucucg uuc 23
<210> 298
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 298
auguguaacu uaauaagccu a 21
<210> 299
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 299
uaggcutauu aaguuacaca uuu 23
<210> 300
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 300
guguaacuua auaagccuau u 21
<210> 301
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 301
aauaggcuua uuaaguuaca cau 23
<210> 302
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 302
guaacuuaau aagccuauuc u 21
<210> 303
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 303
agaauaggcu uauuaaguua cac 23
<210> 304
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 304
uaacuuaaua agccuauucc a 21
<210> 305
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 305
uggaauaggc uuauuaaguu aca 23
<210> 306
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 306
aacuuaauaa gccuauucca u 21
<210> 307
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 307
auggaatagg cuuauuaagu uac 23
<210> 308
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 308
guuaaguaag uuacacuaca u 21
<210> 309
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 309
auguagtgua acuuacuuaa cug 23
<210> 310
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 310
uaaguuacac uacaguucuc a 21
<210> 311
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 311
ugagaacugu aguguaacuu acu 23
<210> 312
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 312
guuacacuac aguucucaca a 21
<210> 313
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 313
uugugagaac uguaguguaa cuu 23
<210> 314
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 314
cagaccuaug uuuacaauau a 21
<210> 315
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 315
uauauuguaa acauaggucu gua 23
<210> 316
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 316
agaccuaugu uuacaauaua a 21
<210> 317
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 317
uuauautgua aacauagguc ugu 23
<210> 318
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 318
gaucaagcag auguuuaauu u 21
<210> 319
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 319
aaauuaaaca ucugcuugau caa 23
<210> 320
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 320
ugggauucag ucuguagaaa u 21
<210> 321
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 321
auuucuacag acugaauccc agg 23
<210> 322
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 322
ugucuaauag uucucuauag u 21
<210> 323
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 323
acuauagaga acuauuagac auu 23
<210> 324
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 324
cuaauaguuc ucuauagucc u 21
<210> 325
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 325
aggacuauag agaacuauua gac 23
<210> 326
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 326
agccaaauug aaaugugcac u 21
<210> 327
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 327
agugcacauu ucaauuuggc uca 23
<210> 328
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 328
uucuugcuaa gucuuaccau u 21
<210> 329
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 329
aaugguaaga cuuagcaaga aga 23
<210> 330
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 330
ugcaauaggc uauaaggaau a 21
<210> 331
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 331
uauucctuau agccuauugc agg 23
<210> 332
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 332
gcaauaggcu auaaggaaua u 21
<210> 333
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 333
auauuccuua uagccuauug cag 23
<210> 334
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 334
gaaaggucau aauagcuuuc u 21
<210> 335
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 335
agaaagcuau uaugaccuuu cac 23
<210> 336
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 336
aaaggucaua auagcuuucc u 21
<210> 337
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 337
aggaaagcua uuaugaccuu uca 23
<210> 338
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 338
aacuagauga cuguuguacu u 21
<210> 339
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 339
aaguacaaca gucaucuagu uca 23
<210> 340
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 340
cuguagcuca gucauuuaaa a 21
<210> 341
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 341
uuuuaaauga cugagcuaca gua 23
<210> 342
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 342
cugguuuauu guacuguuau a 21
<210> 343
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 343
uauaacagua caauaaacca gcc 23
<210> 344
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 344
auguguaaac auuguuauau a 21
<210> 345
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 345
uauauaacaa uguuuacaca ugc 23
<210> 346
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 346
ugcgucaaac agcgacaagu u 21
<210> 347
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 347
aacuugtcgc uguuugacgc acc 23
<210> 348
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 348
ggugcgucaa acagcgacaa guu 23
<210> 349
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 349
cgucaaacag cgacaaguuc u 21
<210> 350
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 350
agaacutguc gcuguuugac gca 23
<210> 351
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 351
ugcgucaaac agcgacaagu ucc 23
<210> 352
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 352
cccacguaaa agaugacgcu u 21
<210> 353
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 353
aagcgucauc uuuuacgugg gcg 23
<210> 354
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 354
cgcccacgua aaagaugacg cuu 23
<210> 355
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 355
ccacguaaaa gaugacgcuu u 21
<210> 356
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 356
aaagcgtcau cuuuuacgug ggc 23
<210> 357
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 357
gcccacguaa aagaugacgc uug 23
<210> 358
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 358
agugaugucg acucuuugcc u 21
<210> 359
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 359
aggcaaagag ucgacaucac ugc 23
<210> 360
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 360
gcagugaugu cgacucuuug ccc 23
<210> 361
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 361
cagcuguugc caagacagag a 21
<210> 362
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 362
ucucugtcuu ggcaacagcu gga 23
<210> 363
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 363
uccagcuguu gccaagacag aga 23
<210> 364
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 364
agcuguugcc aagacagaga u 21
<210> 365
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 365
aucucugucu uggcaacagc ugg 23
<210> 366
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 366
ccagcuguug ccaagacaga gau 23
<210> 367
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 367
gcuguugcca agacagagau ugc 23
<210> 368
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 368
gccaagacag agauugcuuu a 21
<210> 369
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 369
uaaagcaauc ucugucuugg caa 23
<210> 370
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 370
uugccaagac agagauugcu uua 23
<210> 371
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 371
gccaagacag agauugcuuu aag 23
<210> 372
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 372
aagacagaga uugcuuuaag u 21
<210> 373
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 373
acuuaaagca aucucugucu ugg 23
<210> 374
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 374
ccaagacaga gauugcuuua agu 23
<210> 375
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 375
agacagagau ugcuuuaagu u 21
<210> 376
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 376
aacuuaaagc aaucucuguc uug 23
<210> 377
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 377
caagacagag auugcuuuaa gug 23
<210> 378
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 378
gacagagauu gcuuuaagug u 21
<210> 379
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 379
acacuuaaag caaucucugu cuu 23
<210> 380
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 380
aagacagaga uugcuuuaag ugg 23
<210> 381
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 381
uagcagcuac uuuugcuuac u 21
<210> 382
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 382
aguaagcaaa aguagcugcu aau 23
<210> 383
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 383
auuagcagcu acuuuugcuu acu 23
<210> 384
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 384
acaauauucu ugguccuaga u 21
<210> 385
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 385
aucuaggacc aagaauauug ucc 23
<210> 386
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 386
ggacaauauu cuugguccua gag 23
<210> 387
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 387
aauauucuug guccuagagu a 21
<210> 388
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 388
uacucuagga ccaagaauau ugu 23
<210> 389
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 389
acaauauucu ugguccuaga gua 23
<210> 390
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 390
auauucuugg uccuagagua a 21
<210> 391
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 391
uuacuctagg accaagaaua uug 23
<210> 392
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 392
caauauucuu gguccuagag uaa 23
<210> 393
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 393
agacagaaca gguacuucuc a 21
<210> 394
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 394
ugagaaguac cuguucuguc uuu 23
<210> 395
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 395
aaagacagaa cagguacuuc uca 23
<210> 396
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 396
uaaauggaga aauccuucga a 21
<210> 397
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 397
uucgaaggau uucuccauuu aga 23
<210> 398
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 398
ucuaaaugga gaaauccuuc gaa 23
<210> 399
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 399
ggagaaaucc uucgaaaugc a 21
<210> 400
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 400
ugcauutcga aggauuucuc cau 23
<210> 401
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 401
auggagaaau ccuucgaaau gca 23
<210> 402
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 402
cauuaaucuu ugauggaaac u 21
<210> 403
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 403
aguuuccauc aaagauuaau gaa 23
<210> 404
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 404
uucauuaauc uuugauggaa acu 23
<210> 405
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 405
auuaaucuuu gauggaaacu u 21
<210> 406
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 406
aaguuuccau caaagauuaa uga 23
<210> 407
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 407
ucauuaaucu uugauggaaa cug 23
<210> 408
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 408
cauauggacu aucaauuaua u 21
<210> 409
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 409
auauaatuga uaguccauau gug 23
<210> 410
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 410
cacauaugga cuaucaauua uac 23
<210> 411
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 411
uauggacuau caauuauacu u 21
<210> 412
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 412
aaguauaauu gauaguccau aug 23
<210> 413
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 413
cauauggacu aucaauuaua cuu 23
<210> 414
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 414
uggacuauca auuauacuuc u 21
<210> 415
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 415
agaaguauaa uugauagucc aua 23
<210> 416
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 416
uauggacuau caauuauacu ucc 23
<210> 417
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 417
ucaauuauac uuccacagac a 21
<210> 418
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 418
ugucugtgga aguauaauug aua 23
<210> 419
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 419
uaucaauuau acuuccacag aca 23
<210> 420
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 420
acagaacuua guuucuaccu u 21
<210> 421
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 421
aagguagaaa cuaaguucug ucu 23
<210> 422
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 422
agacagaacu uaguuucuac cuc 23
<210> 423
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 423
cagaacuuag uuucuaccuc u 21
<210> 424
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 424
agagguagaa acuaaguucu guc 23
<210> 425
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 425
gacagaacuu aguuucuacc ucc 23
<210> 426
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 426
gagugugugu ugauagauua a 21
<210> 427
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 427
uuaauctauc aacacacacu cua 23
<210> 428
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 428
uagagugugu guugauagau uaa 23
<210> 429
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 429
aguguguguu gauagauuaa u 21
<210> 430
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 430
auuaaucuau caacacacac ucu 23
<210> 431
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 431
agagugugug uugauagauu aac 23
<210> 432
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 432
uguguguuga uagauuaaca u 21
<210> 433
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 433
auguuaaucu aucaacacac acu 23
<210> 434
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 434
aguguguguu gauagauuaa cac 23
<210> 435
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 435
aacacauaua auccggaaag u 21
<210> 436
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 436
acuuuccgga uuauaugugu uaa 23
<210> 437
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 437
uuaacacaua uaauccggaa agg 23
<210> 438
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 438
uguccagaag auuaucuuag a 21
<210> 439
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 439
ucuaagauaa ucuucuggac auu 23
<210> 440
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 440
aauguccaga agauuaucuu aga 23
<210> 441
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 441
gaacugcuuu caucuaugaa a 21
<210> 442
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 442
uuucauagau gaaagcaguu cca 23
<210> 443
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 443
uggaacugcu uucaucuaug aaa 23
<210> 444
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 444
uauagcugau acaguacuca aug 23
<210> 445
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 445
gcugauacag uacucaauga uga 23
<210> 446
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 446
cugucaugaa ggcuuucuuc u 21
<210> 447
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 447
agaagaaagc cuucaugaca gcu 23
<210> 448
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 448
agcugucaug aaggcuuucu ucu 23
<210> 449
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 449
uguuccguug uaguagguag u 21
<210> 450
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 450
acuacctacu acaacggaac agc 23
<210> 451
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 451
gcuguuccgu uguaguaggu agc 23
<210> 452
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 452
aucauuuaaa uaugagucag u 21
<210> 453
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 453
acugactcau auuuaaauga uga 23
<210> 454
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 454
ucaucauuua aauaugaguc agg 23
<210> 455
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 455
ccugcuaaag gauucaacug u 21
<210> 456
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 456
acaguugaau ccuuuagcag gcc 23
<210> 457
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 457
ggccugcuaa aggauucaac ugg 23
<210> 458
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 458
cacccuguca ugaacauauu u 21
<210> 459
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 459
aaauauguuc augacagggu ggc 23
<210> 460
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 460
gccacccugu caugaacaua uuu 23
<210> 461
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 461
ugaacauauu uauaaucagc u 21
<210> 462
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 462
agcugatuau aaauauguuc aug 23
<210> 463
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 463
caugaacaua uuuauaauca gcg 23
<210> 464
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 464
acauauuuau aaucagcgua u 21
<210> 465
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 465
auacgctgau uauaaauaug uuc 23
<210> 466
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 466
gaacauauuu auaaucagcg uag 23
<210> 467
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 467
auauuuauaa ucagcguaga u 21
<210> 468
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 468
aucuacgcug auuauaaaua ugu 23
<210> 469
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 469
acauauuuau aaucagcgua gau 23
<210> 470
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 470
ucaggauacg aucaucuaca u 21
<210> 471
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 471
auguagauga ucguauccug agc 23
<210> 472
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 472
gcucaggaua cgaucaucua cac 23
<210> 473
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 473
aucaucuaca cugacgaaag u 21
<210> 474
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 474
acuuucguca guguagauga ucg 23
<210> 475
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 475
cgaucaucua cacugacgaa agc 23
<210> 476
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 476
caucuacacu gacgaaagcu u 21
<210> 477
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 477
aagcuutcgu caguguagau gau 23
<210> 478
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 478
aucaucuaca cugacgaaag cuu 23
<210> 479
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 479
aucuacacug acgaaagcuu u 21
<210> 480
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 480
aaagcutucg ucaguguaga uga 23
<210> 481
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 481
ucaucuacac ugacgaaagc uuu 23
<210> 482
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 482
cacugacgaa agcuuuacuc u 21
<210> 483
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 483
agaguaaagc uuucgucagu gua 23
<210> 484
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 484
uacacugacg aaagcuuuac ucc 23
<210> 485
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 485
acugacgaaa gcuuuacucc u 21
<210> 486
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 486
aggaguaaag cuuucgucag ugu 23
<210> 487
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 487
acacugacga aagcuuuacu ccu 23
<210> 488
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 488
cugacgaaag cuuuacuccu u 21
<210> 489
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 489
aaggagtaaa gcuuucguca gug 23
<210> 490
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 490
cacugacgaa agcuuuacuc cug 23
<210> 491
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 491
uuucaagaug ucuuacacag a 21
<210> 492
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 492
ucugugtaag acaucuugaa aaa 23
<210> 493
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 493
uuuuucaaga ugucuuacac aga 23
<210> 494
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 494
augucuuaca cagagacacu cua 23
<210> 495
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 495
cuggaucagg ucuuucagcu u 21
<210> 496
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 496
aagcugaaag accugaucca gga 23
<210> 497
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 497
uccuggauca ggucuuucag cug 23
<210> 498
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 498
cucagaagua cuuuccuugc a 21
<210> 499
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 499
ugcaaggaaa guacuucuga gag 23
<210> 500
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 500
cucucagaag uacuuuccuu gca 23
<210> 501
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 501
ucuacuuguc cuucacagaa a 21
<210> 502
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 502
uuucugtgaa ggacaaguag aaa 23
<210> 503
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 503
uuucuacuug uccuucacag aaa 23
<210> 504
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 504
gaaaagccuu gacacuaaua a 21
<210> 505
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 505
uuauuagugu caaggcuuuu cug 23
<210> 506
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 506
cagaaaagcc uugacacuaa uaa 23
<210> 507
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 507
ccuuuaaauc ucuucggaac u 21
<210> 508
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 508
aguuccgaag agauuuaaag ggc 23
<210> 509
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 509
gcccuuuaaa ucucuucgga acc 23
<210> 510
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 510
cuuuaaaucu cuucggaacc u 21
<210> 511
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 511
agguuccgaa gagauuuaaa ggg 23
<210> 512
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 512
cccuuuaaau cucuucggaa ccu 23
<210> 513
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 513
uuuaaaucuc uucggaaccu u 21
<210> 514
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 514
aagguuccga agagauuuaa agg 23
<210> 515
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 515
ccuuuaaauc ucuucggaac cug 23
<210> 516
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 516
gggcgaucuu aacauaauaa u 21
<210> 517
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 517
auuauuaugu uaagaucgcc cuc 23
<210> 518
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 518
gagggcgauc uuaacauaau aau 23
<210> 519
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 519
ucuuaacaua auaauggcuc u 21
<210> 520
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 520
agagccauua uuauguuaag auc 23
<210> 521
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 521
gaucuuaaca uaauaauggc ucu 23
<210> 522
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 522
uuaucuuugg aagaccuuuc u 21
<210> 523
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 523
agaaaggucu uccaaagaua aaa 23
<210> 524
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 524
uuuuaucuuu ggaagaccuu ucu 23
<210> 525
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 525
uggaagaccu uucuacacua gug 23
<210> 526
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 526
aagaccuuuc uacacuagug u 21
<210> 527
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 527
acacuagugu agaaaggucu ucc 23
<210> 528
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 528
ggaagaccuu ucuacacuag ugu 23
<210> 529
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 529
agaccuuucu acacuagugu u 21
<210> 530
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 530
aacacuagug uagaaagguc uuc 23
<210> 531
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 531
gaagaccuuu cuacacuagu gug 23
<210> 532
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 532
gcaagaacga gauguucuaa u 21
<210> 533
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 533
auuagaacau cucguucuug cac 23
<210> 534
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 534
gugcaagaac gagauguucu aau 23
<210> 535
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 535
gaacgagaug uucuaaugac u 21
<210> 536
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 536
agucautaga acaucucguu cuu 23
<210> 537
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 537
aagaacgaga uguucuaaug acu 23
<210> 538
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 538
aacgagaugu ucuaaugacu u 21
<210> 539
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 539
aagucatuag aacaucucgu ucu 23
<210> 540
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 540
agaacgagau guucuaauga cuu 23
<210> 541
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 541
acgagauguu cuaaugacuu u 21
<210> 542
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 542
aaagucauua gaacaucucg uuc 23
<210> 543
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 543
gaacgagaug uucuaaugac uuu 23
<210> 544
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 544
auguguaacu uaauaagccu a 21
<210> 545
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 545
uaggcutauu aaguuacaca uuu 23
<210> 546
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 546
aaauguguaa cuuaauaagc cua 23
<210> 547
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 547
guguaacuua auaagccuau u 21
<210> 548
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 548
aauaggcuua uuaaguuaca cau 23
<210> 549
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 549
auguguaacu uaauaagccu auu 23
<210> 550
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 550
guaacuuaau aagccuauuc u 21
<210> 551
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 551
agaauaggcu uauuaaguua cac 23
<210> 552
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 552
guguaacuua auaagccuau ucc 23
<210> 553
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 553
uaacuuaaua agccuauucc a 21
<210> 554
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 554
uggaauaggc uuauuaaguu aca 23
<210> 555
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 555
uguaacuuaa uaagccuauu cca 23
<210> 556
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 556
aacuuaauaa gccuauucca u 21
<210> 557
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 557
auggaatagg cuuauuaagu uac 23
<210> 558
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 558
guaacuuaau aagccuauuc cau 23
<210> 559
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 559
guuaaguaag uuacacuaca u 21
<210> 560
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 560
auguagtgua acuuacuuaa cug 23
<210> 561
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 561
caguuaagua aguuacacua cag 23
<210> 562
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 562
uaaguuacac uacaguucuc a 21
<210> 563
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 563
ugagaacugu aguguaacuu acu 23
<210> 564
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 564
aguaaguuac acuacaguuc uca 23
<210> 565
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 565
guuacacuac aguucucaca a 21
<210> 566
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 566
uugugagaac uguaguguaa cuu 23
<210> 567
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 567
aaguuacacu acaguucuca caa 23
<210> 568
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 568
cagaccuaug uuuacaauau a 21
<210> 569
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 569
uauauuguaa acauaggucu gua 23
<210> 570
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 570
uacagaccua uguuuacaau aua 23
<210> 571
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 571
agaccuaugu uuacaauaua a 21
<210> 572
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 572
uuauautgua aacauagguc ugu 23
<210> 573
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 573
acagaccuau guuuacaaua uaa 23
<210> 574
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 574
gaucaagcag auguuuaauu u 21
<210> 575
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 575
aaauuaaaca ucugcuugau caa 23
<210> 576
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 576
uugaucaagc agauguuuaa uug 23
<210> 577
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 577
ugggauucag ucuguagaaa u 21
<210> 578
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 578
auuucuacag acugaauccc agg 23
<210> 579
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 579
ccugggauuc agucuguaga aau 23
<210> 580
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 580
ugucuaauag uucucuauag u 21
<210> 581
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 581
acuauagaga acuauuagac auu 23
<210> 582
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 582
aaugucuaau aguucucuau agu 23
<210> 583
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 583
cuaauaguuc ucuauagucc u 21
<210> 584
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 584
aggacuauag agaacuauua gac 23
<210> 585
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 585
gucuaauagu ucucuauagu ccu 23
<210> 586
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 586
agccaaauug aaaugugcac u 21
<210> 587
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 587
agugcacauu ucaauuuggc uca 23
<210> 588
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 588
ugagccaaau ugaaaugugc acc 23
<210> 589
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 589
uucuugcuaa gucuuaccau u 21
<210> 590
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 590
aaugguaaga cuuagcaaga aga 23
<210> 591
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 591
ucuucuugcu aagucuuacc aug 23
<210> 592
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 592
ugcaauaggc uauaaggaau a 21
<210> 593
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 593
uauucctuau agccuauugc agg 23
<210> 594
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 594
ccugcaauag gcuauaagga aua 23
<210> 595
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 595
gcaauaggcu auaaggaaua u 21
<210> 596
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 596
auauuccuua uagccuauug cag 23
<210> 597
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 597
cugcaauagg cuauaaggaa uag 23
<210> 598
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 598
gaaaggucau aauagcuuuc u 21
<210> 599
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 599
agaaagcuau uaugaccuuu cac 23
<210> 600
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 600
gugaaagguc auaauagcuu ucc 23
<210> 601
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 601
aaaggucaua auagcuuucc u 21
<210> 602
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 602
aggaaagcua uuaugaccuu uca 23
<210> 603
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 603
ugaaagguca uaauagcuuu ccc 23
<210> 604
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 604
aacuagauga cuguuguacu u 21
<210> 605
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 605
aaguacaaca gucaucuagu uca 23
<210> 606
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 606
ugaacuagau gacuguugua cug 23
<210> 607
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 607
cuguagcuca gucauuuaaa a 21
<210> 608
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 608
uuuuaaauga cugagcuaca gua 23
<210> 609
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 609
uacuguagcu cagucauuua aaa 23
<210> 610
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 610
cugguuuauu guacuguuau a 21
<210> 611
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 611
uauaacagua caauaaacca gcc 23
<210> 612
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 612
ggcugguuua uuguacuguu aua 23
<210> 613
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 613
auguguaaac auuguuauau a 21
<210> 614
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 614
uauauaacaa uguuuacaca ugc 23
<210> 615
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 615
gcauguguaa acauuguuau aua 23
<210> 616
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 616
gcugagggug aacaagaaaa u 21
<210> 617
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 617
auuuucuugu ucacccucag cga 23
<210> 618
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 618
gauaaagauu aaccagaaga a 21
<210> 619
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 619
uucuucuggu uaaucuuuau cag 23
<210> 620
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 620
aaccagaaga aaacaaggag u 21
<210> 621
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 621
acuccuuguu uucuucuggu uaa 23
<210> 622
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 622
auuaaccaga agaaaacaag u 21
<210> 623
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 623
acuuguuuuc uucugguuaa ucu 23
<210> 624
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 624
uaaccagaag aaaacaagga u 21
<210> 625
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 625
auccuuguuu ucuucugguu aau 23
<210> 626
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 626
aacaagaaaa gaccugauaa a 21
<210> 627
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 627
uuuaucaggu cuuuucuugu uca 23
<210> 628
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 628
gaaaagaccu gauaaagauu a 21
<210> 629
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 629
uaaucuuuau caggucuuuu cuu 23
<210> 630
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 630
acaagaaaag accugauaaa u 21
<210> 631
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 631
auuuaucagg ucuuuucuug uuc 23
<210> 632
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 632
aagaccugau aaagauuaac u 21
<210> 633
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 633
aguuaaucuu uaucaggucu uuu 23
<210> 634
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 634
gaccugauaa agauuaacca u 21
<210> 635
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 635
augguuaauc uuuaucaggu cuu 23
<210> 636
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 636
ugauaaagau uaaccagaag a 21
<210> 637
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 637
ucuucugguu aaucuuuauc agg 23
<210> 638
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 638
accugauaaa gauuaaccag a 21
<210> 639
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 639
ucugguuaau cuuuaucagg ucu 23
<210> 640
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 640
aagaaaagac cugauaaaga u 21
<210> 641
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 641
aucuuuauca ggucuuuucu ugu 23
<210> 642
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 642
cugaggguga acaagaaaag a 21
<210> 643
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 643
ucuuuucuug uucacccuca gcg 23
<210> 644
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 644
ugaacaagaa aagaccugau a 21
<210> 645
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 645
uaucaggucu uuucuuguuc acc 23
<210> 646
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 646
gagggugaac aagaaaagac u 21
<210> 647
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 647
agucuuuucu uguucacccu cag 23
<210> 648
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 648
gugaacaaga aaagaccuga u 21
<210> 649
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 649
aucaggucuu uucuuguuca ccc 23
<210> 650
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 650
aagauuaacc agaagaaaac a 21
<210> 651
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 651
uguuuucuuc ugguuaaucu uua 23
<210> 652
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 652
agggugaaca agaaaagacc u 21
<210> 653
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 653
aggucuuuuc uuguucaccc uca 23
<210> 654
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 654
ccugauaaag auuaaccaga a 21
<210> 655
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 655
uucugguuaa ucuuuaucag guc 23
<210> 656
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 656
ugcucucaca guacucgcug a 21
<210> 657
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 657
ucagcgagua cugugagagc aag 23
<210> 658
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 658
ggugaacaag aaaagaccug a 21
<210> 659
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 659
ucaggucuuu ucuuguucac ccu 23
<210> 660
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 660
cucgcugagg gugaacaaga a 21
<210> 661
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 661
uucuuguuca cccucagcga gua 23
<210> 662
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 662
cgcugagggu gaacaagaaa a 21
<210> 663
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 663
uuuucuuguu cacccucagc gag 23
<210> 664
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 664
caguacucgc ugagggugaa u 21
<210> 665
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 665
auucacccuc agcgaguacu gug 23
<210> 666
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 666
ucgcugaggg ugaacaagaa a 21
<210> 667
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 667
uuucuuguuc acccucagcg agu 23
<210> 668
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 668
cuccccacua cuugcucuca u 21
<210> 669
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 669
augagagcaa guagugggga gag 23
<210> 670
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 670
acaaccgcag ccuguagcaa u 21
<210> 671
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 671
auugcuacag gcugcgguug uuu 23
<210> 672
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 672
uugcucucac aguacucgcu u 21
<210> 673
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 673
aagcgaguac ugugagagca agu 23
<210> 674
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 674
uacucgcuga gggugaacaa u 21
<210> 675
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 675
auuguucacc cucagcgagu acu 23
<210> 676
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 676
guagcaagcu cuggaacuca u 21
<210> 677
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 677
augaguucca gagcuugcua cag 23
<210> 678
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 678
agcucuggaa cucaggaguc u 21
<210> 679
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 679
agacuccuga guuccagagc uug 23
<210> 680
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 680
ccccacuacu ugcucucaca u 21
<210> 681
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 681
augugagagc aaguaguggg gag 23
<210> 682
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 682
cucucacagu acucgcugag u 21
<210> 683
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 683
acucagcgag uacugugaga gca 23
<210> 684
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 684
uguguuuuug uuuuucccac u 21
<210> 685
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 685
agugggaaaa acaaaaacac aca 23
<210> 686
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 686
gaacaagaaa agaccugaua a 21
<210> 687
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 687
uuaucagguc uuuucuuguu cac 23
<210> 688
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 688
accagaagaa aacaaggagg u 21
<210> 689
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 689
accuccuugu uuucuucugg uua 23
<210> 690
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 690
uagcaagcuc uggaacucag u 21
<210> 691
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 691
acugaguucc agagcuugcu aca 23
<210> 692
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 692
agaccugaua aagauuaacc a 21
<210> 693
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 693
ugguuaaucu uuaucagguc uuu 23
<210> 694
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 694
guacucgcug agggugaaca a 21
<210> 695
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 695
uuguucaccc ucagcgagua cug 23
<210> 696
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 696
acuugcucuc acaguacucg u 21
<210> 697
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 697
acgaguacug ugagagcaag uag 23
<210> 698
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 698
gccuguagca agcucuggaa u 21
<210> 699
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 699
auuccagagc uugcuacagg cug 23
<210> 700
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 700
caagcucugg aacucaggag u 21
<210> 701
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 701
acuccugagu uccagagcuu gcu 23
<210> 702
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 702
guguguuuuu guuuuuccca u 21
<210> 703
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 703
augggaaaaa caaaaacaca cac 23
<210> 704
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 704
cacuacuugc ucucacagua u 21
<210> 705
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 705
auacugugag agcaaguagu ggg 23
<210> 706
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 706
aagcucugga acucaggagu u 21
<210> 707
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 707
aacuccugag uuccagagcu ugc 23
<210> 708
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 708
cuacuugcuc ucacaguacu u 21
<210> 709
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 709
aaguacugug agagcaagua gug 23
<210> 710
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 710
uacuugcucu cacaguacuc u 21
<210> 711
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 711
agaguacugu gagagcaagu agu 23
<210> 712
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 712
caaccgcagc cuguagcaag u 21
<210> 713
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 713
acuugcuaca ggcugcgguu guu 23
<210> 714
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 714
gggugaacaa gaaaagaccu u 21
<210> 715
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 715
aaggucuuuu cuuguucacc cuc 23
<210> 716
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 716
cuugcucuca caguacucgc u 21
<210> 717
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 717
agcgaguacu gugagagcaa gua 23
<210> 718
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 718
uuaaccagaa gaaaacaagg a 21
<210> 719
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 719
uccuuguuuu cuucugguua auc 23
<210> 720
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 720
ucuccccacu acuugcucuc a 21
<210> 721
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 721
ugagagcaag uaguggggag aga 23
<210> 722
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 722
gcucucacag uacucgcuga u 21
<210> 723
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 723
aucagcgagu acugugagag caa 23
<210> 724
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 724
gcucuggaac ucaggagucg u 21
<210> 725
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 725
acgacuccug aguuccagag cuu 23
<210> 726
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 726
acaguacucg cugaggguga a 21
<210> 727
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 727
uucacccuca gcgaguacug uga 23
<210> 728
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 728
aaccgcagcc uguagcaagc u 21
<210> 729
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 729
agcuugcuac aggcugcggu ugu 23
<210> 730
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 730
uccccacuac uugcucucac a 21
<210> 731
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 731
ugugagagca aguagugggg aga 23
<210> 732
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 732
acuacuugcu cucacaguac u 21
<210> 733
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 733
aguacuguga gagcaaguag ugg 23
<210> 734
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 734
ggaaacaacc gcagccugua u 21
<210> 735
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 735
auacaggcug cgguuguuuc ccu 23
<210> 736
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 736
ccuguagcaa gcucuggaac u 21
<210> 737
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 737
aguuccagag cuugcuacag gcu 23
<210> 738
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 738
ugagggugaa caagaaaaga u 21
<210> 739
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 739
aucuuuucuu guucacccuc agc 23
<210> 740
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 740
uguagcaagc ucuggaacuc a 21
<210> 741
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 741
ugaguuccag agcuugcuac agg 23
<210> 742
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 742
aguacucgcu gagggugaac a 21
<210> 743
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 743
uguucacccu cagcgaguac ugu 23
<210> 744
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 744
cccacuacuu gcucucacag u 21
<210> 745
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 745
acugugagag caaguagugg gga 23
<210> 746
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 746
agccuguagc aagcucugga a 21
<210> 747
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 747
uuccagagcu ugcuacaggc ugc 23
<210> 748
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 748
cucacaguac ucgcugaggg u 21
<210> 749
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 749
acccucagcg aguacuguga gag 23
<210> 750
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 750
ccacuacuug cucucacagu a 21
<210> 751
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 751
uacugugaga gcaaguagug ggg 23
<210> 752
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 752
gcaagcucug gaacucagga u 21
<210> 753
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 753
auccugaguu ccagagcuug cua 23
<210> 754
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 754
cuguagcaag cucuggaacu u 21
<210> 755
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 755
aaguuccaga gcuugcuaca ggc 23
<210> 756
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 756
ccgcagccug uagcaagcuc u 21
<210> 757
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 757
agagcuugcu acaggcugcg guu 23
<210> 758
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 758
acucgcugag ggugaacaag a 21
<210> 759
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 759
ucuuguucac ccucagcgag uac 23
<210> 760
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 760
ucaggagucg cgcgcuaggg u 21
<210> 761
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 761
acccuagcgc gcgacuccug agu 23
<210> 762
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 762
agcaagcucu ggaacucagg a 21
<210> 763
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 763
uccugaguuc cagagcuugc uac 23
<210> 764
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 764
guuuuuguuu uucccacccu u 21
<210> 765
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 765
aaggguggga aaaacaaaaa cac 23
<210> 766
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 766
guguuuuugu uuuucccacc u 21
<210> 767
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 767
aggugggaaa aacaaaaaca cac 23
<210> 768
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 768
aacaaccgca gccuguagca a 21
<210> 769
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 769
uugcuacagg cugcgguugu uuc 23
<210> 770
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 770
caggagucgc gcgcuagggg u 21
<210> 771
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 771
accccuagcg cgcgacuccu gag 23
<210> 772
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 772
ggagucgcgc gcuaggggcc u 21
<210> 773
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 773
aggccccuag cgcgcgacuc cug 23
<210> 774
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 774
cacaguacuc gcugagggug a 21
<210> 775
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 775
ucacccucag cgaguacugu gag 23
<210> 776
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 776
gggaaacaac cgcagccugu a 21
<210> 777
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 777
uacaggcugc gguuguuucc cuc 23
<210> 778
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 778
aggagucgcg cgcuaggggc u 21
<210> 779
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 779
agccccuagc gcgcgacucc uga 23
<210> 780
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 780
aaacaaccgc agccuguagc a 21
<210> 781
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 781
ugcuacaggc ugcgguuguu ucc 23
<210> 782
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 782
gagucgcgcg cuaggggccg u 21
<210> 783
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 783
acggccccua gcgcgcgacu ccu 23
<210> 784
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 784
agucgcgcgc uaggggccgg u 21
<210> 785
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 785
accggccccu agcgcgcgac ucc 23
<210> 786
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 786
gcugagggug aacaagaaaa u 21
<210> 787
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 787
auuuucuugu ucacccucag cga 23
<210> 788
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 788
gauaaagauu aaccagaaga a 21
<210> 789
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 789
uucuucuggu uaaucuuuau cag 23
<210> 790
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 790
aaccagaaga aaacaaggag u 21
<210> 791
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 791
acuccuuguu uucuucuggu uaa 23
<210> 792
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 792
auuaaccaga agaaaacaag u 21
<210> 793
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 793
acuuguuuuc uucugguuaa ucu 23
<210> 794
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 794
uaaccagaag aaaacaagga u 21
<210> 795
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 795
auccuuguuu ucuucugguu aau 23
<210> 796
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 796
aacaagaaaa gaccugauaa a 21
<210> 797
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 797
uuuaucaggu cuuuucuugu uca 23
<210> 798
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 798
gaaaagaccu gauaaagauu a 21
<210> 799
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 799
uaaucuuuau caggucuuuu cuu 23
<210> 800
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 800
acaagaaaag accugauaaa u 21
<210> 801
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 801
auuuaucagg ucuuuucuug uuc 23
<210> 802
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 802
aagaccugau aaagauuaac u 21
<210> 803
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 803
aguuaaucuu uaucaggucu uuu 23
<210> 804
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 804
gaccugauaa agauuaacca u 21
<210> 805
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 805
augguuaauc uuuaucaggu cuu 23
<210> 806
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 806
ugauaaagau uaaccagaag a 21
<210> 807
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 807
ucuucugguu aaucuuuauc agg 23
<210> 808
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 808
accugauaaa gauuaaccag a 21
<210> 809
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 809
ucugguuaau cuuuaucagg ucu 23
<210> 810
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 810
aagaaaagac cugauaaaga u 21
<210> 811
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 811
aucuuuauca ggucuuuucu ugu 23
<210> 812
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 812
cugaggguga acaagaaaag a 21
<210> 813
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 813
ucuuuucuug uucacccuca gcg 23
<210> 814
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 814
ugaacaagaa aagaccugau a 21
<210> 815
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 815
uaucaggucu uuucuuguuc acc 23
<210> 816
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 816
gagggugaac aagaaaagac u 21
<210> 817
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 817
agucuuuucu uguucacccu cag 23
<210> 818
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 818
gugaacaaga aaagaccuga u 21
<210> 819
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 819
aucaggucuu uucuuguuca ccc 23
<210> 820
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 820
aagauuaacc agaagaaaac a 21
<210> 821
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 821
uguuuucuuc ugguuaaucu uua 23
<210> 822
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 822
agggugaaca agaaaagacc u 21
<210> 823
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 823
aggucuuuuc uuguucaccc uca 23
<210> 824
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 824
ccugauaaag auuaaccaga a 21
<210> 825
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 825
uucugguuaa ucuuuaucag guc 23
<210> 826
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 826
ugcucucaca guacucgcug a 21
<210> 827
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 827
ucagcgagua cugugagagc aag 23
<210> 828
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 828
ggugaacaag aaaagaccug a 21
<210> 829
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 829
ucaggucuuu ucuuguucac ccu 23
<210> 830
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 830
cucgcugagg gugaacaaga a 21
<210> 831
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 831
uucuuguuca cccucagcga gua 23
<210> 832
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 832
cgcugagggu gaacaagaaa a 21
<210> 833
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 833
uuuucuuguu cacccucagc gag 23
<210> 834
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 834
caguacucgc ugagggugaa u 21
<210> 835
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 835
auucacccuc agcgaguacu gug 23
<210> 836
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 836
ucgcugaggg ugaacaagaa a 21
<210> 837
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 837
uuucuuguuc acccucagcg agu 23
<210> 838
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 838
cuccccacua cuugcucuca u 21
<210> 839
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 839
augagagcaa guagugggga gag 23
<210> 840
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 840
acaaccgcag ccuguagcaa u 21
<210> 841
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 841
auugcuacag gcugcgguug uuu 23
<210> 842
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 842
uugcucucac aguacucgcu u 21
<210> 843
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 843
aagcgaguac ugugagagca agu 23
<210> 844
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 844
uacucgcuga gggugaacaa u 21
<210> 845
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 845
auuguucacc cucagcgagu acu 23
<210> 846
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 846
guagcaagcu cuggaacuca u 21
<210> 847
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 847
augaguucca gagcuugcua cag 23
<210> 848
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 848
agcucuggaa cucaggaguc u 21
<210> 849
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 849
agacuccuga guuccagagc uug 23
<210> 850
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 850
ccccacuacu ugcucucaca u 21
<210> 851
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 851
augugagagc aaguaguggg gag 23
<210> 852
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 852
cucucacagu acucgcugag u 21
<210> 853
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 853
acucagcgag uacugugaga gca 23
<210> 854
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 854
uguguuuuug uuuuucccac u 21
<210> 855
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 855
agugggaaaa acaaaaacac aca 23
<210> 856
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 856
gaacaagaaa agaccugaua a 21
<210> 857
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 857
uuaucagguc uuuucuuguu cac 23
<210> 858
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 858
accagaagaa aacaaggagg u 21
<210> 859
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 859
accuccuugu uuucuucugg uua 23
<210> 860
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 860
uagcaagcuc uggaacucag u 21
<210> 861
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 861
acugaguucc agagcuugcu aca 23
<210> 862
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 862
agaccugaua aagauuaacc a 21
<210> 863
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 863
ugguuaaucu uuaucagguc uuu 23
<210> 864
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 864
guacucgcug agggugaaca a 21
<210> 865
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 865
uuguucaccc ucagcgagua cug 23
<210> 866
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 866
acuugcucuc acaguacucg u 21
<210> 867
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 867
acgaguacug ugagagcaag uag 23
<210> 868
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 868
gccuguagca agcucuggaa u 21
<210> 869
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 869
auuccagagc uugcuacagg cug 23
<210> 870
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 870
caagcucugg aacucaggag u 21
<210> 871
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 871
acuccugagu uccagagcuu gcu 23
<210> 872
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 872
guguguuuuu guuuuuccca u 21
<210> 873
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 873
augggaaaaa caaaaacaca cac 23
<210> 874
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 874
cacuacuugc ucucacagua u 21
<210> 875
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 875
auacugugag agcaaguagu ggg 23
<210> 876
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 876
aagcucugga acucaggagu u 21
<210> 877
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 877
aacuccugag uuccagagcu ugc 23
<210> 878
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 878
cuacuugcuc ucacaguacu u 21
<210> 879
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 879
aaguacugug agagcaagua gug 23
<210> 880
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 880
uacuugcucu cacaguacuc u 21
<210> 881
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 881
agaguacugu gagagcaagu agu 23
<210> 882
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 882
caaccgcagc cuguagcaag u 21
<210> 883
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 883
acuugcuaca ggcugcgguu guu 23
<210> 884
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 884
gggugaacaa gaaaagaccu u 21
<210> 885
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 885
aaggucuuuu cuuguucacc cuc 23
<210> 886
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 886
cuugcucuca caguacucgc u 21
<210> 887
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 887
agcgaguacu gugagagcaa gua 23
<210> 888
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 888
uuaaccagaa gaaaacaagg a 21
<210> 889
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 889
uccuuguuuu cuucugguua auc 23
<210> 890
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 890
ucuccccacu acuugcucuc a 21
<210> 891
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 891
ugagagcaag uaguggggag aga 23
<210> 892
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 892
gcucucacag uacucgcuga u 21
<210> 893
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 893
aucagcgagu acugugagag caa 23
<210> 894
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 894
gcucuggaac ucaggagucg u 21
<210> 895
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 895
acgacuccug aguuccagag cuu 23
<210> 896
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 896
acaguacucg cugaggguga a 21
<210> 897
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 897
uucacccuca gcgaguacug uga 23
<210> 898
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 898
aaccgcagcc uguagcaagc u 21
<210> 899
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 899
agcuugcuac aggcugcggu ugu 23
<210> 900
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 900
uccccacuac uugcucucac a 21
<210> 901
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 901
ugugagagca aguagugggg aga 23
<210> 902
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 902
acuacuugcu cucacaguac u 21
<210> 903
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 903
aguacuguga gagcaaguag ugg 23
<210> 904
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 904
ggaaacaacc gcagccugua u 21
<210> 905
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 905
auacaggcug cgguuguuuc ccu 23
<210> 906
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 906
ccuguagcaa gcucuggaac u 21
<210> 907
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 907
aguuccagag cuugcuacag gcu 23
<210> 908
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 908
ugagggugaa caagaaaaga u 21
<210> 909
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 909
aucuuuucuu guucacccuc agc 23
<210> 910
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 910
uguagcaagc ucuggaacuc a 21
<210> 911
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 911
ugaguuccag agcuugcuac agg 23
<210> 912
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 912
aguacucgcu gagggugaac a 21
<210> 913
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 913
uguucacccu cagcgaguac ugu 23
<210> 914
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 914
cccacuacuu gcucucacag u 21
<210> 915
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 915
acugugagag caaguagugg gga 23
<210> 916
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 916
agccuguagc aagcucugga a 21
<210> 917
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 917
uuccagagcu ugcuacaggc ugc 23
<210> 918
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 918
cucacaguac ucgcugaggg u 21
<210> 919
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 919
acccucagcg aguacuguga gag 23
<210> 920
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 920
ccacuacuug cucucacagu a 21
<210> 921
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 921
uacugugaga gcaaguagug ggg 23
<210> 922
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 922
gcaagcucug gaacucagga u 21
<210> 923
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 923
auccugaguu ccagagcuug cua 23
<210> 924
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 924
cuguagcaag cucuggaacu u 21
<210> 925
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 925
aaguuccaga gcuugcuaca ggc 23
<210> 926
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 926
ccgcagccug uagcaagcuc u 21
<210> 927
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 927
agagcuugcu acaggcugcg guu 23
<210> 928
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 928
acucgcugag ggugaacaag a 21
<210> 929
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 929
ucuuguucac ccucagcgag uac 23
<210> 930
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 930
ucaggagucg cgcgcuaggg u 21
<210> 931
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 931
acccuagcgc gcgacuccug agu 23
<210> 932
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 932
agcaagcucu ggaacucagg a 21
<210> 933
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 933
uccugaguuc cagagcuugc uac 23
<210> 934
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 934
guuuuuguuu uucccacccu u 21
<210> 935
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 935
aaggguggga aaaacaaaaa cac 23
<210> 936
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 936
guguuuuugu uuuucccacc u 21
<210> 937
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 937
aggugggaaa aacaaaaaca cac 23
<210> 938
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 938
aacaaccgca gccuguagca a 21
<210> 939
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 939
uugcuacagg cugcgguugu uuc 23
<210> 940
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 940
caggagucgc gcgcuagggg u 21
<210> 941
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 941
accccuagcg cgcgacuccu gag 23
<210> 942
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 942
ggagucgcgc gcuaggggcc u 21
<210> 943
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 943
aggccccuag cgcgcgacuc cug 23
<210> 944
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 944
cacaguacuc gcugagggug a 21
<210> 945
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 945
ucacccucag cgaguacugu gag 23
<210> 946
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 946
gggaaacaac cgcagccugu a 21
<210> 947
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 947
uacaggcugc gguuguuucc cuc 23
<210> 948
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 948
aggagucgcg cgcuaggggc u 21
<210> 949
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 949
agccccuagc gcgcgacucc uga 23
<210> 950
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 950
aaacaaccgc agccuguagc a 21
<210> 951
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 951
ugcuacaggc ugcgguuguu ucc 23
<210> 952
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 952
gagucgcgcg cuaggggccg u 21
<210> 953
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 953
acggccccua gcgcgcgacu ccu 23
<210> 954
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 954
agucgcgcgc uaggggccgg u 21
<210> 955
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 955
accggccccu agcgcgcgac ucc 23
<210> 956
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 956
ccucucuccc cacuacuugc u 21
<210> 957
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 957
agcaaguagu ggggagagag ggu 23
<210> 958
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 958
ucacaguacu cgcugagggu u 21
<210> 959
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 959
aacccucagc gaguacugug aga 23
<210> 960
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 960
guguguguuu uuguuuuucc cac 23
<210> 961
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 961
uguguguuuu uguuuuuccc acc 23
<210> 962
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 962
guguguuuuu guuuuuccca ccc 23
<210> 963
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 963
guguuuuugu uuuucccacc cuc 23
<210> 964
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 964
acccucucuc cccacuacuu gcu 23
<210> 965
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 965
agcaaguagu ggggagagag ggu 23
<210> 966
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 966
ccucucuccc cacuacuugc u 21
<210> 967
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 967
ucucucccca cuacuugcuc uca 23
<210> 968
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 968
cucuccccac uacuugcucu cac 23
<210> 969
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 969
ucuccccacu acuugcucuc aca 23
<210> 970
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 970
cuccccacua cuugcucuca cag 23
<210> 971
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 971
uccccacuac uugcucucac agu 23
<210> 972
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 972
ccccacuacu ugcucucaca gua 23
<210> 973
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 973
cccacuacuu gcucucacag uac 23
<210> 974
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 974
ccacuacuug cucucacagu acu 23
<210> 975
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 975
cacuacuugc ucucacagua cuc 23
<210> 976
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 976
acuacuugcu cucacaguac ucg 23
<210> 977
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 977
cuacuugcuc ucacaguacu cgc 23
<210> 978
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 978
uacuugcucu cacaguacuc gcu 23
<210> 979
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 979
acuugcucuc acaguacucg cug 23
<210> 980
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 980
cuugcucuca caguacucgc uga 23
<210> 981
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 981
uugcucucac aguacucgcu gag 23
<210> 982
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 982
ugcucucaca guacucgcug agg 23
<210> 983
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 983
cucucacagu acucgcugag ggu 23
<210> 984
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 984
ucucacagua cucgcugagg gug 23
<210> 985
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 985
aacccucagc gaguacugug aga 23
<210> 986
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 986
ucacaguacu cgcugagggu u 21
<210> 987
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 987
cucacaguac ucgcugaggg uga 23
<210> 988
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 988
ucacaguacu cgcugagggu gaa 23
<210> 989
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 989
cacaguacuc gcugagggug aac 23
<210> 990
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 990
acaguacucg cugaggguga aca 23
<210> 991
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 991
caguacucgc ugagggugaa caa 23
<210> 992
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 992
aguacucgcu gagggugaac aag 23
<210> 993
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 993
guacucgcug agggugaaca aga 23
<210> 994
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 994
uacucgcuga gggugaacaa gaa 23
<210> 995
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 995
acucgcugag ggugaacaag aaa 23
<210> 996
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 996
cucgcugagg gugaacaaga aaa 23
<210> 997
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 997
ucgcugaggg ugaacaagaa aag 23
<210> 998
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 998
cgcugagggu gaacaagaaa aga 23
<210> 999
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 999
gcugagggug aacaagaaaa gac 23
<210> 1000
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1000
cugaggguga acaagaaaag acc 23
<210> 1001
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1001
ugagggugaa caagaaaaga ccu 23
<210> 1002
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1002
gagggugaac aagaaaagac cug 23
<210> 1003
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1003
agggugaaca agaaaagacc uga 23
<210> 1004
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1004
gggugaacaa gaaaagaccu gau 23
<210> 1005
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1005
ggugaacaag aaaagaccug aua 23
<210> 1006
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1006
gugaacaaga aaagaccuga uaa 23
<210> 1007
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1007
ugaacaagaa aagaccugau aaa 23
<210> 1008
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1008
gaacaagaaa agaccugaua aag 23
<210> 1009
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1009
acaagaaaag accugauaaa gau 23
<210> 1010
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1010
caagaaaaga ccugauaaag auu 23
<210> 1011
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1011
aagaaaagac cugauaaaga uua 23
<210> 1012
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1012
agaaaagacc ugauaaagau uaa 23
<210> 1013
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1013
gaaaagaccu gauaaagauu aac 23
<210> 1014
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1014
aaaagaccug auaaagauua acc 23
<210> 1015
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1015
aaagaccuga uaaagauuaa cca 23
<210> 1016
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1016
aagaccugau aaagauuaac cag 23
<210> 1017
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1017
agaccugaua aagauuaacc aga 23
<210> 1018
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1018
gaccugauaa agauuaacca gaa 23
<210> 1019
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1019
accugauaaa gauuaaccag aag 23
<210> 1020
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1020
ccugauaaag auuaaccaga aga 23
<210> 1021
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1021
cugauaaaga uuaaccagaa gaa 23
<210> 1022
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1022
ugauaaagau uaaccagaag aaa 23
<210> 1023
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1023
auaaagauua accagaagaa aac 23
<210> 1024
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1024
uaaagauuaa ccagaagaaa aca 23
<210> 1025
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1025
agauuaacca gaagaaaaca agg 23
<210> 1026
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1026
gauuaaccag aagaaaacaa gga 23
<210> 1027
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1027
auuaaccaga agaaaacaag gag 23
<210> 1028
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1028
uuaaccagaa gaaaacaagg agg 23
<210> 1029
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1029
uaaccagaag aaaacaagga ggg 23
<210> 1030
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1030
gagggaaaca accgcagccu gua 23
<210> 1031
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1031
agggaaacaa ccgcagccug uag 23
<210> 1032
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1032
ggaaacaacc gcagccugua gca 23
<210> 1033
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1033
gaaacaaccg cagccuguag caa 23
<210> 1034
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1034
aaacaaccgc agccuguagc aag 23
<210> 1035
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1035
aacaaccgca gccuguagca agc 23
<210> 1036
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1036
acaaccgcag ccuguagcaa gcu 23
<210> 1037
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1037
aaccgcagcc uguagcaagc ucu 23
<210> 1038
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1038
gcagccugua gcaagcucug gaa 23
<210> 1039
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1039
cagccuguag caagcucugg aac 23
<210> 1040
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1040
agccuguagc aagcucugga acu 23
<210> 1041
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1041
gccuguagca agcucuggaa cuc 23
<210> 1042
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1042
ccuguagcaa gcucuggaac uca 23
<210> 1043
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1043
cuguagcaag cucuggaacu cag 23
<210> 1044
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1044
uguagcaagc ucuggaacuc agg 23
<210> 1045
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1045
guagcaagcu cuggaacuca gga 23
<210> 1046
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1046
uagcaagcuc uggaacucag gag 23
<210> 1047
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1047
agcaagcucu ggaacucagg agu 23
<210> 1048
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1048
gcaagcucug gaacucagga guc 23
<210> 1049
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1049
caagcucugg aacucaggag ucg 23
<210> 1050
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1050
aagcucugga acucaggagu cgc 23
<210> 1051
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1051
acucaggagu cgcgcgcuag ggg 23
<210> 1052
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1052
cucaggaguc gcgcgcuagg ggc 23
<210> 1053
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1053
ucaggagucg cgcgcuaggg gcc 23
<210> 1054
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1054
caggagucgc gcgcuagggg ccg 23
<210> 1055
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1055
aggagucgcg cgcuaggggc cgg 23
<210> 1056
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1056
ggagucgcgc gcuaggggcc ggg 23
<210> 1057
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1057
ggaaaguggg gucuagcaau u 21
<210> 1058
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1058
uugcuagacc ccacuuuccu u 21
<210> 1059
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1059
caagagcagg uguggguuuu u 21
<210> 1060
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1060
aaacccacac cugcucuugu u 21
<210> 1061
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1061
ccuucuaggu ggaaaguggu u 21
<210> 1062
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1062
ccacuuucca ccuagaaggu u 21
<210> 1063
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1063
caagaaaaga ccugauaaau u 21
<210> 1064
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1064
uuuaucaggu cuuuucuugu u 21
<210> 1065
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1065
gaaaagaccu gauaaagauu u 21
<210> 1066
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1066
aucuuuauca ggucuuuucu u 21
<210> 1067
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1067
cgcugagggu gaacaagaau u 21
<210> 1068
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1068
uucuuguuca cccucagcgu u 21
<210> 1069
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1069
gaagaaaaca aggagggaau u 21
<210> 1070
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1070
uucccuccuu guuuucuucu u 21
<210> 1071
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1071
ccucagagcu cgacgcauuu u 21
<210> 1072
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1072
aaugcgucga gcucugaggu u 21
<210> 1073
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1073
ucuagcgacu gguggaauuu u 21
<210> 1074
<211> 21
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1074
aauuccacca gucgcuagau u 21
Claims (118)
- C9orf72의 발현을 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제로서,
상기 dsRNA 제제가 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고,
상기 센스 가닥이 표 10A, 10B, 12 내지 15, 19, 20, 또는 21 중 어느 하나의 안티센스 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제. - C9orf72의 발현을 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제로서,
상기 dsRNA 제제가 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고,
상기 센스 가닥이 서열번호 133의 뉴클레오티드 200 내지 290과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고, 상기 안티센스 가닥이 서열번호 134의 상응하는 뉴클레오티드 서열의 적어도 15개의 연속 뉴클레오티드를 포함하며,
상기 센스 가닥, 안티센스 가닥, 또는 센스 가닥과 안티센스 가닥이 모두 하나 이상의 친유성 모이어티에 접합되는, dsRNA 제제. - C9orf72의 발현을 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제로서,
상기 dsRNA 제제가 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고,
상기 안티센스 가닥이 서열번호 125의 뉴클레오티드 서열과 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고,
상기 센스 가닥, 안티센스 가닥, 또는 센스 가닥과 안티센스 가닥이 모두 하나 이상의 친유성 모이어티에 접합되는, dsRNA 제제. - 제1항 또는 제2항에 있어서, 상기 센스 및 안티센스 가닥의 뉴클레오티드 서열이 표 14, 15, 19 또는 20 중 어느 하나의 센스 및 안티센스 가닥 뉴클레오티드 서열 중 어느 하나를 포함하는, dsRNA 제제.
- 제1항, 제2항 및 제4항 중 어느 한 항에 있어서, 상기 안티센스 가닥이 AD-348904.1, AD-348136.1, AD-347612.1, AD-348639.1, AD-347606.1, AD-348905.1, AD-347613.1, AD-348842.1, AD-347610.1, AD-350329.1, AD-348140.1, AD-348930.1, AD-347863.1, AD-348500.1, AD-348597.1, AD-348578.1, AD-347923.1, AD-348588.1, 및 AD-347773.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제.
- 서열번호 1의 다중 연속 카피를 포함하는 헥사뉴클레오티드 반복부를 포함하는 C9orf72 표적 RNA를 선택적으로 억제하기 위한 이중 가닥 리보핵산(dsRNA) 제제로서,
상기 dsRNA 제제가 이중 가닥 영역을 형성하는 센스 가닥 및 안티센스 가닥을 포함하고,
상기 안티센스 가닥이 표 14, 15, 및 21 중 어느 하나의 안티센스 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제. - 제1항, 제2항, 제4항, 제5항 및 제6항 중 어느 한 항에 있어서, 상기 안티센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제.
- 제7항에 있어서, 상기 안티센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나의 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제.
- 제7항에 있어서, 상기 안티센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 뉴클레오티드 서열을 포함하는, dsRNA 제제.
- 제7항에 있어서, 상기 안티센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나의 뉴클레오티드 서열을 포함하는, dsRNA 제제.
- 제7항에 있어서, 상기 안티센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 안티센스 가닥 뉴클레오티드 서열 중 어느 하나의 뉴클레오티드 서열로 이루어지는, dsRNA 제제.
- 제7항 내지 제11항 중 어느 한 항에 있어서, 상기 센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 센스 가닥 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제.
- 제7항 내지 제12항 중 어느 한 항에 있어서, 상기 센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 센스 가닥 뉴클레오티드 서열 중 어느 하나로부터 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제.
- 제7항 내지 제13항 중 어느 한 항에 있어서, 상기 센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 센스 가닥 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 뉴클레오티드 서열을 포함하는, dsRNA 제제.
- 제7항 내지 제14항 중 어느 한 항에 있어서, 상기 센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 센스 가닥 뉴클레오티드 서열 중 어느 하나의 뉴클레오티드 서열을 포함하는, dsRNA 제제.
- 제7항 내지 제15항 중 어느 한 항에 있어서, 상기 센스 가닥이 AD-463863.1, AD-463862.1, AD-463869.1, AD-463873.1, AD-463872.1, 및 AD-463860.1로 이루어진 그룹으로부터 선택된 이중체의 센스 가닥 뉴클레오티드 서열 중 어느 하나의 뉴클레오티드 서열로 이루어지는, dsRNA 제제.
- 제1항 또는 제3항에 있어서, 상기 센스 및 안티센스 가닥의 뉴클레오티드 서열이 표 12 또는 표 13 중 어느 하나의 센스 및 안티센스 가닥 뉴클레오티드 서열 중 어느 하나를 포함하는, dsRNA 제제.
- 제1항, 제3항 및 제17항 중 어느 한 항에 있어서, 상기 센스 가닥이 서열번호 121의 뉴클레오티드 230-270, 233-262, 800-840, 800-830, 802-828, 1240-1290, 1240-1280, 1247-1288, 1590-1645, 1590-1620, 및 1594-1642의 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고, 상기 안티센스 가닥이 서열번호 125의 상응하는 뉴클레오티드 서열의 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제.
- 제1항, 제3항, 제17항 및 제18항 중 어느 한 항에 있어서, 상기 센스 가닥이 서열번호 121의 뉴클레오티드 1594-1616, 802-824, 239-261, 1308-1330, 233-255, 1595-1617, 240-262, 1532-1554, 237-259, 3268-3290, 806-828, 1620-1642, 526-548, 1169-1191, 1266-1288, 1247-1269, 586-608, 1257-1279, 및 400-422의 뉴클레오티드 서열 중 어느 하나와 3개 이하의 뉴클레오티드가 다른 적어도 15개의 연속 뉴클레오티드를 포함하고, 안티센스 가닥이 서열번호 125의 상응하는 뉴클레오티드 서열의 적어도 15개의 연속 뉴클레오티드를 포함하는, dsRNA 제제.
- 제1항 및 제4항 내지 제19항 중 어느 한 항에 있어서, 상기 센스 가닥, 안티센스 가닥, 또는 센스 가닥과 안티센스 가닥이 모두 하나 이상의 친유성 모이어티에 접합되는, dsRNA 제제.
- 제20항에 있어서, 상기 친유성 모이어티가 dsRNA 제제의 이중 가닥 영역에서 하나 이상의 내부 위치에 접합되는, dsRNA 제제.
- 제20항 또는 제21항에 있어서, 상기 친유성 모이어티가 링커 또는 담체를 통해 접합되는, dsRNA 제제.
- 제20항 내지 제22항 중 어느 한 항에 있어서, logKow로 측정되는, 친유성 모이어티의 친유성이 0을 초과하는, dsRNA 제제.
- 제1항 내지 제23항 중 어느 한 항에 있어서, 상기 이중 가닥 RNAi 제제의 혈장 단백질 결합 분석에서 결합되지 않은 분획으로 측정되는, 이중 가닥 RNAi 제제의 소수성이 0.2를 초과하는, dsRNA 제제.
- 제24항에 있어서, 상기 혈장 단백질 결합 분석이 인간 혈청 알부민 단백질을 사용하는 전기영동 이동성 변화 분석인, dsRNA 제제.
- 제1항 내지 제25항 중 어느 한 항에 있어서, 적어도 하나의 변형된 뉴클레오티드를 포함하는, dsRNA 제제.
- 제26항에 있어서, 상기 5개 이하의 센스 가닥 뉴클레오티드 및 5개 이하의 안티센스 가닥 뉴클레오티드가 변형되지 않은 뉴클레오티드인, dsRNA 제제.
- 제26항에 있어서, 상기 센스 가닥의 모든 뉴클레오티드 및 안티센스 가닥의 모든 뉴클레오티드가 변형된 뉴클레오티드인, dsRNA 제제.
- 제26항 내지 제28항 중 어느 한 항에 있어서, 상기 변형된 뉴클레오티드 중 적어도 하나가 데옥시-뉴클레오티드, 3'-말단 데옥시-티민(dT) 뉴클레오티드, 2'-O-메틸 변형된 뉴클레오티드, 2'-플루오로 변형된 뉴클레오티드, 2'-데옥시-변형된 뉴클레오티드, 고정 뉴클레오티드(locked nucleotide), 비고정 뉴클레오티드(unlocked nucleotide), 형태적으로 제한된 뉴클레오티드, 제한된 에틸 뉴클레오티드, 무염기 뉴클레오티드, 2'-아미노-변형된 뉴클레오티드, 2'-O-알릴-변형된 뉴클레오티드, 2'-C-알킬-변형된 뉴클레오티드, 2'-하이드록실-변형된 뉴클레오티드, 2'-메톡시에틸 변형된 뉴클레오티드, 2'-O-알킬-변형된 뉴클레오티드, 모르폴리노 뉴클레오티드, 포스포르아미데이트, 뉴클레오티드를 포함하는 비천연 염기, 테트라하이드로피란 변형된 뉴클레오티드, 1,5-안하이드로헥시톨 변형된 뉴클레오티드, 사이클로헥세닐 변형된 뉴클레오티드, 5'-포스포로티오에이트 그룹을 포함하는 뉴클레오티드, 5'-메틸포스포네이트 그룹을 포함하는 뉴클레오티드, 5' 포스페이트 또는 5' 포스페이트 모방체를 포함하는 뉴클레오티드, 비닐 포스포네이트를 포함하는 뉴클레오티드, 아데노신-글리콜 핵산(GNA)을 포함하는 뉴클레오티드, 티미딘-글리콜 핵산(GNA) S-이성체를 포함하는 뉴클레오티드, 2-하이드록시메틸-테트라하이드로푸란-5-포스페이트를 포함하는 뉴클레오티드, 2'-데옥시티미딘-3'포스페이트를 포함하는 뉴클레오티드, 2'-데옥시구아노신-3'-포스페이트를 포함하는 뉴클레오티드, 및 콜레스테릴 유도체 및 도데칸산 비스데실아미드 그룹에 연결된 말단 뉴클레오티드; 및 이들의 조합의 그룹으로부터 선택되는, dsRNA 제제.
- 제29항에 있어서, 상기 변형된 뉴클레오티드가 2'-데옥시-2'-플루오로 변형된 뉴클레오티드, 2'-데옥시-변형된 뉴클레오티드, 3'-말단 데옥시-티민 뉴클레오티드(dT), 고정 뉴클레오티드, 무염기 뉴클레오티드, 2'-아미노-변형된 뉴클레오티드, 2'-알킬-변형된 뉴클레오티드, 모르폴리노 뉴클레오티드, 포스포르아미데이트, 및 뉴클레오티드를 포함하는 비천연 염기로 이루어진 그룹으로부터 선택되는, dsRNA 제제.
- 제29항에 있어서, 상기 변형된 뉴클레오티드가 3'-말단 데옥시-티민 뉴클레오티드(dT)의 짧은 서열을 포함하는, dsRNA 제제.
- 제29항에 있어서, 상기 뉴클레오티드에 대한 변형이 2'-O-메틸, GNA 및 2' 플루오로 변형인, dsRNA 제제.
- 제1항 내지 제32항 중 어느 한 항에 있어서, 적어도 1개의 포스포로티오에이트 뉴클레오티드간 결합을 추가로 포함하는, dsRNA 제제.
- 제33항에 있어서, 6 내지 8개의 포스포로티오에이트 뉴클레오티드간 결합을 포함하는, dsRNA 제제.
- 제1항 내지 제34항 중 어느 한 항에 있어서, 각 가닥이 30개 이하의 뉴클레오티드 길이인, dsRNA 제제.
- 제1항 내지 제35항 중 어느 한 항에 있어서, 적어도 하나의 가닥이 적어도 1개의 뉴클레오티드의 3' 돌출부를 포함하는, dsRNA 제제.
- 제1항 내지 제36항 중 어느 한 항에 있어서, 적어도 하나의 가닥이 적어도 2개의 뉴클레오티드의 3' 돌출부를 포함하는, dsRNA 제제.
- 제1항 내지 제37항 중 어느 한 항에 있어서, 상기 이중 가닥 영역이 15 내지 30개의 뉴클레오티드 쌍 길이인, dsRNA 제제.
- 제38항에 있어서, 상기 이중 가닥 영역이 17 내지 23개의 뉴클레오티드 쌍 길이인, dsRNA 제제.
- 제38항에 있어서, 상기 이중 가닥 영역이 17 내지 25개의 뉴클레오티드 쌍 길이인, dsRNA 제제.
- 제38항에 있어서, 상기 이중 가닥 영역이 23 내지 27개의 뉴클레오티드 쌍 길이인, dsRNA 제제.
- 제38항에 있어서, 상기 이중 가닥 영역이 19 내지 21개의 뉴클레오티드 쌍 길이인, dsRNA 제제.
- 제38항에 있어서, 상기 이중 가닥 영역이 21 내지 23개의 뉴클레오티드 쌍 길이인, dsRNA 제제.
- 제1항 내지 제43항 중 어느 한 항에 있어서, 각 가닥이 19 내지 30개의 뉴클레오티드 길이인, dsRNA 제제.
- 제1항 내지 제43항 중 어느 한 항에 있어서, 각 가닥이 19 내지 23개의 뉴클레오티드 길이인, dsRNA 제제.
- 제1항 내지 제43항 중 어느 한 항에 있어서, 각 가닥이 21 내지 23개의 뉴클레오티드 길이인, dsRNA 제제.
- 제21항 내지 제46항 중 어느 한 항에 있어서, 하나 이상의 친유성 모이어티가 적어도 하나의 가닥 상의 하나 이상의 내부 위치에 접합되는, dsRNA 제제.
- 제47항에 있어서, 하나 이상의 친유성 모이어티가 링커 또는 담체를 통해 적어도 하나의 가닥 상의 하나 이상의 내부 위치에 접합되는, dsRNA 제제.
- 제48항에 있어서, 상기 내부 위치가 적어도 하나의 가닥의 각 끝 말단의 두 위치를 제외한 모든 위치를 포함하는, dsRNA 제제.
- 제48항에 있어서, 상기 내부 위치가 적어도 하나의 가닥의 각 끝 말단의 세 위치를 제외한 모든 위치를 포함하는, dsRNA 제제.
- 제48항 내지 제50항 중 어느 한 항에 있어서, 상기 내부 위치가 센스 가닥의 절단 부위 영역을 제외하는, dsRNA 제제.
- 제51항에 있어서, 상기 내부 위치가 센스 가닥의 5'-말단으로부터 계수하여 위치 9 내지 12를 제외한 모든 위치를 포함하는, dsRNA 제제.
- 제51항에 있어서, 상기 내부 위치가 센스 가닥의 3'-말단으로부터 계수하여 위치 11 내지 13을 제외한 모든 위치를 포함하는, dsRNA 제제.
- 제48항 내지 제50항 중 어느 한 항에 있어서, 상기 내부 위치가 안티센스 가닥의 절단 부위 영역을 제외하는, dsRNA 제제.
- 제54항에 있어서, 상기 내부 위치가 안티센스 가닥의 5'-말단으로부터 계수하여 위치 12 내지 14를 제외한 모든 위치를 포함하는, dsRNA 제제.
- 제48항 내지 제50항 중 어느 한 항에 있어서, 상기 내부 위치가 3'-말단으로부터 계수하여 센스 가닥 상의 위치 11 내지 13, 및 5'-말단으로부터 계수하여 안티센스 가닥 상의 위치 12 내지 14를 제외한 모든 위치를 포함하는, dsRNA 제제.
- 제21항 내지 제56항 중 어느 한 항에 있어서, 상기 하나 이상의 친유성 모이어티가 각 가닥의 5' 말단으로부터 계수하여 센스 가닥 상의 위치 4 내지 8 및 13 내지 18, 및 안티센스 가닥 상의 위치 6 내지 10 및 15 내지 18로 이루어진 그룹으로부터 선택된 내부 위치 중 하나 이상에 접합되는, dsRNA 제제.
- 제57항에 있어서, 상기 하나 이상의 친유성 모이어티가 각 가닥의 5' 말단으로부터 계수하여 센스 가닥 상의 위치 5, 6, 7, 15, 및 17, 및 안티센스 가닥 상의 위치 15 및 17로 이루어진 그룹으로부터 선택된 내부 위치 중 하나 이상에 접합되는, dsRNA 제제.
- 제21항에 있어서, 상기 이중 가닥 영역의 내부 위치가 센스 가닥의 절단 부위 영역을 제외하는, dsRNA 제제.
- 제20항 내지 제59항 중 어느 한 항에 있어서, 상기 센스 가닥이 21개의 뉴클레오티드 길이이고, 상기 안티센스 가닥이 23개의 뉴클레오티드 길이이며, 상기 친유성 모이어티가 센스 가닥의 위치 21, 위치 20, 위치 15, 위치 1, 위치 7, 위치 6, 또는 위치 2 또는 안티센스 가닥의 위치 16에 접합되는, dsRNA 제제.
- 제60항에 있어서, 상기 친유성 모이어티가 센스 가닥의 위치 21, 위치 20, 위치 15, 위치 1, 또는 위치 7에 접합되는, dsRNA 제제.
- 제60항에 있어서, 상기 친유성 모이어티가 센스 가닥의 위치 21, 위치 20, 또는 위치 15에 접합되는, dsRNA 제제.
- 제60항에 있어서, 상기 친유성 모이어티가 센스 가닥의 위치 20 또는 위치 15에 접합되는, dsRNA 제제.
- 제60항에 있어서, 상기 친유성 모이어티가 안티센스 가닥의 위치 16에 접합되는, dsRNA 제제.
- 제20항 내지 제64항 중 어느 한 항에 있어서, 상기 친유성 모이어티가 지방족, 지환족 또는 다지환족 화합물인, dsRNA 제제.
- 제65항에 있어서, 상기 친유성 모이어티가 지질, 콜레스테롤, 레티노산, 콜산, 아다만탄 아세트산, 1-피렌 부티르산, 디하이드로테스토스테론, 1,3-비스-O(헥사데실)글리세롤, 게라닐옥시헥시아놀, 헥사데실글리세롤, 보르네올, 멘톨, 1,3-프로판디올, 헵타데실 그룹, 팔미트산, 미리스트산, O3-(올레오일)리토콜산, O3-(올레오일)콜렌산, 디메톡시트리틸, 또는 페녹사진으로 이루어진 그룹으로부터 선택되는, dsRNA 제제.
- 제65항에 있어서, 상기 친유성 모이어티가 포화 또는 불포화 C4-C30 탄화수소 쇄, 및 하이드록실, 아민, 카복실산, 설포네이트, 포스페이트, 티올, 아지드 및 알킨으로 이루어진 그룹으로부터 선택된 임의의 작용 그룹을 함유하는, dsRNA 제제.
- 제67항에 있어서, 상기 친유성 모이어티가 포화 또는 불포화 C6-C18 탄화수소 쇄를 함유하는, dsRNA 제제.
- 제67항에 있어서, 상기 친유성 모이어티가 포화 또는 불포화 C16 탄화수소 쇄를 함유하는, dsRNA 제제.
- 제69항에 있어서, 상기 포화 또는 불포화 C16 탄화수소 쇄가 가닥의 5'-말단으로부터 계수하여 위치 6에 접합되는, dsRNA 제제.
- 제20항 내지 제70항 중 어느 한 항에 있어서, 상기 친유성 모이어티가 내부 위치(들) 또는 이중 가닥 영역에서 하나 이상의 뉴클레오티드(들)를 대체하는 담체를 통해 접합되는, dsRNA 제제.
- 제71항에 있어서, 상기 담체가 피롤리디닐, 피라졸리닐, 피라졸리디닐, 이미다졸리닐, 이미다졸리디닐, 피페리디닐, 피페라지닐, [1,3]디옥솔라닐, 옥사졸리디닐, 이속사졸리디닐, 모르폴리닐, 티아졸리디닐, 이소티아졸리디닐, 퀴녹살리닐, 피리다지노닐, 테트라하이드로푸라닐 및 데칼리닐로 이루어진 그룹으로부터 선택된 사이클릭 그룹이거나; 세리놀 백본 또는 디에탄올아민 백본을 기반으로 하는 비사이클릭 모이어티인, dsRNA 제제.
- 제20항 내지 제70항 중 어느 한 항에 있어서, 상기 친유성 모이어티가 에테르, 티오에테르, 요소, 카보네이트, 아민, 아미드, 말레이미드-티오에테르, 디설파이드, 포스포디에스테르, 설폰아미드 결합, 클릭 반응의 생성물, 또는 카바메이트를 함유하는 링커를 통해 이중 가닥 iRNA 제제에 접합되는, dsRNA 제제.
- 제20항 내지 제73항 중 어느 한 항에 있어서, 상기 친유성 모이어티가 핵염기, 당 모이어티, 또는 뉴클레오시드간 결합에 접합되는, dsRNA 제제.
- 제20항 내지 제74항 중 어느 한 항에 있어서, 상기 친유성 모이어티 또는 표적화 리간드가 DNA, RNA, 디설파이드, 아미드; 갈락토사민, 글루코사민, 글루코스, 갈락토스, 만노스의 작용화된 단당류 또는 올리고당류, 및 이들의 조합으로 이루어진 그룹으로부터 선택된 생체-절단 가능한 링커를 통해 접합되는, dsRNA 제제.
- 제20항 내지 제75항 중 어느 한 항에 있어서, 상기 센스 가닥의 3' 말단이 아민을 갖는 사이클릭 그룹인 말단 캡을 통해 보호되고, 상기 사이클릭 그룹이 피롤리디닐, 피라졸리닐, 피라졸리디닐, 이미다졸리닐, 이미다졸리디닐, 피페리디닐, 피페라지닐, [1,3]디옥솔라닐, 옥사졸리디닐, 이속사졸리디닐, 모르폴리닐, 티아졸리디닐, 이소티아졸리디닐, 퀴녹살리닐, 피리다지노닐, 테트라하이드로푸라닐 및 데칼리닐로 이루어진 그룹으로부터 선택되는, dsRNA 제제.
- 제1항 내지 제19항 중 어느 한 항에 있어서, 간 조직을 표적으로 하는 표적화 리간드를 추가로 포함하는, dsRNA 제제.
- 제77항에 있어서, 상기 표적화 리간드가 GalNAc 접합체인, dsRNA 제제.
- 제1항 내지 제78항 중 어느 한 항에 있어서,
Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형,
Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및
Rp 배위 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함하는, dsRNA 제제. - 제1항 내지 제78항 중 어느 한 항에 있어서,
Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형,
Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및
Rp 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함하는, dsRNA 제제. - 제1항 내지 제78항 중 어느 한 항에 있어서,
Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째, 두 번째 및 세 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형,
Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및
Rp 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함하는, dsRNA 제제. - 제1항 내지 제78항 중 어느 한 항에 있어서,
Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형,
Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 세 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형,
Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및
Rp 배위 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함하는, dsRNA 제제. - 제1항 내지 제78항 중 어느 한 항에 있어서,
Sp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 3' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형,
Rp 배위의 결합 인 원자를 갖는, 안티센스 가닥의 5' 말단에 첫 번째 및 두 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형, 및
Rp 또는 Sp 배위의 결합 인 원자를 갖는, 센스 가닥의 5' 말단에 첫 번째 뉴클레오티드간 결합에서 발생하는 말단 키랄 변형을 추가로 포함하는, dsRNA 제제. - 제1항 내지 제83항 중 어느 한 항에 있어서, 상기 안티센스 가닥의 5'-말단에 포스페이트 또는 포스페이트 모방체를 추가로 포함하는, dsRNA 제제.
- 제84항에 있어서, 상기 포스페이트 모방체가 5'-비닐 포스포네이트(VP)인, dsRNA 제제.
- 제1항 내지 제83항 중 어느 한 항에 있어서, 상기 이중체의 안티센스 가닥의 5'-말단의 1 위치의 염기 쌍이 AU 염기 쌍인, dsRNA 제제.
- 제1항 내지 제83항 중 어느 한 항에 있어서, 상기 센스 가닥이 총 21개의 뉴클레오티드를 갖고 안티센스 가닥이 총 23개의 뉴클레오티드를 갖는, dsRNA 제제.
- 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제를 함유하는 세포.
- 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제를 포함하는, C9orf72의 발현을 억제하기 위한 약제학적 조성물.
- 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제 및 지질 제제를 포함하는 약제학적 조성물.
- 제89항 또는 제90항에 있어서, 상기 dsRNA 제제가 완충되지 않은 용액에 존재하는, 약제학적 조성물.
- 제88항에 있어서, 상기 완충되지 않은 용액이 염수 또는 물인, 약제학적 조성물.
- 제89항 또는 제90항에 있어서, 상기 dsRNA 제제가 완충 용액에 존재하는, 약제학적 조성물.
- 제93항에 있어서, 상기 완충 용액이 아세테이트, 시트레이트, 프롤라민, 카보네이트, 또는 포스페이트 또는 이들의 임의의 조합을 포함하는, 약제학적 조성물.
- 제93항에 있어서, 상기 완충 용액이 인산염 완충 염수(PBS)인, 약제학적 조성물.
- 세포에서 C9orf72의 발현을 억제하는 방법으로서, 세포를 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제, 또는 제89항 내지 제95항 중의 어느 한 항의 약제학적 조성물과 접촉시켜, 세포에서 C9orf72의 발현을 억제하는 단계를 포함하는, 방법.
- 제96항에 있어서, 상기 세포가 대상체 내에 있는, 방법.
- 제97항에 있어서, 상기 대상체가 인간인, 방법.
- 제98항에 있어서, 상기 대상체가 C9orf72-관련 장애를 갖는, 방법.
- 제99항에 있어서, 상기 C9orf72-관련 장애가 C9orf72 근위축성 측삭 경화증/전두측두엽 치매 또는 C9orf72 확장으로 인한 헌팅턴-유사 증후군으로 이루어진 그룹으로부터 선택되는, 방법.
- 제96항 내지 제100항 중 어느 한 항에 있어서, 세포를 dsRNA 제제와 접촉시키는 것이 C9orf72의 발현을 50% 이하로 억제하는, 방법.
- 제96항 내지 제101항 중 어느 한 항에 있어서, 상기 C9orf72의 발현을 억제시키는 것이 대상체의 혈청에서 C9orf72 단백질 수준을 50% 이하로 감소시키는, 방법.
- C9orf72 발현의 감소로부터 혜택을 받는 장애를 갖는 대상체를 치료하는 방법으로서, 대상체에게 치료학적 유효량의 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제, 또는 제89항 내지 제95항 중의 어느 한 항의 약제학적 조성물을 투여하여 C9orf72 발현의 감소로부터 혜택을 받는 장애를 갖는 대상체를 치료함을 포함하는, 방법.
- C9orf72 발현의 감소로부터 혜택을 받는 장애를 갖는 대상체에서 적어도 하나의 증상을 예방하는 방법으로서, 대상체에게 예방학적 유효량의 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제, 또는 제89항 내지 제95항 중의 어느 한 항의 약제학적 조성물을 투여하여 C9orf72 발현의 감소로부터 혜택을 받는 장애를 갖는 대상체에서 적어도 하나의 증상을 예방함을 포함하는, 방법.
- 제103항 또는 제104항에 있어서, 상기 장애가 C9orf72-관련 장애인, 방법.
- 제105항에 있어서, 상기 C9orf723-관련 장애가 C9orf72 근위축성 측삭 경화증/전두측두엽 치매 및 C9orf72 확장으로 인한 헌팅턴-유사 증후군으로 이루어진 그룹으로부터 선택되는, 방법.
- 제106항에 있어서, 상기 대상체가 인간인, 방법.
- 제106항 또는 제107항에 있어서, 상기 대상체로의 상기 제제의 투여가 RNA 병소 (RNA foci)의 반복부-길이-의존적 형성, 특정 RNA-결합 단백질의 격리, 또는 폴리(글리신-알라닌) 펩티드, 폴리(글리신-프롤린) 펩티드, 폴리(글리신-아르기닌) 펩티드, 폴리(알라닌-프롤린) 펩티드, 또는 폴리(프롤린-아르기닌) 펩티드의 축적 또는 응집의 감소를 야기하는, 방법.
- 제108항에 있어서, 상기 RNA 병소 (RNA foci)의 반복부-길이-의존적 형성, 특정 RNA-결합 단백질의 격리, 또는 폴리(글리신-알라닌) 펩티드, 폴리(글리신-프롤린) 펩티드, 폴리(글리신-아르기닌) 펩티드, 폴리(알라닌-프롤린) 펩티드, 또는 폴리(프롤린-아르기닌) 펩티드의 축적 또는 응집이 50% 이상 감소하고 C9orf72 성숙 RNA의 발현이 50% 미만으로 억제되는, 방법.
- 제103항 내지 제109항 중 어느 한 항에 있어서, 상기 dsRNA 제제가 약 0.01 mg/kg 내지 약 50mg/kg의 용량으로 대상체에게 투여되는, 방법.
- 제103항 내지 제110항 중 어느 한 항에 있어서, 상기 dsRNA 제제가 피하로 대상체에게 투여되는, 방법.
- 제93항 내지 제110항 중 어느 한 항에 있어서, 상기 dsRNA 제제가 척수강내로 대상체에게 투여되는, 방법.
- 제93항 내지 제112항 중 어느 한 항에 있어서, 대상체의 샘플(들)에서 C9orf72의 수준을 측정하는 단계를 추가로 포함하는, 방법.
- 제113항에 있어서, 상기 대상체 샘플(들)에서의 C9orf72의 수준이 혈액, 혈청 또는 뇌척수액 샘플(들)의 C9orf72 단백질 수준인, 방법.
- 제103항 내지 제114항 중 어느 한 항에 있어서, 추가 치료제를 대상체에게 투여하는 단계를 추가로 포함하는, 방법.
- 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제 또는 제89항 내지 제95항 중의 어느 한 항의 약제학적 조성물을 포함하는 키트.
- 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제 또는 제89항 내지 제95항 중의 어느 한 항의 약제학적 조성물을 포함하는 바이알.
- 제1항 내지 제87항 중의 어느 한 항의 dsRNA 제제 또는 제89항 내지 제95항 중의 어느 한 항의 약제학적 조성물을 포함하는 시린지.
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962947768P | 2019-12-13 | 2019-12-13 | |
| US201962947605P | 2019-12-13 | 2019-12-13 | |
| US62/947,605 | 2019-12-13 | ||
| US62/947,768 | 2019-12-13 | ||
| PCT/US2020/064159 WO2021119226A1 (en) | 2019-12-13 | 2020-12-10 | Human chromosome 9 open reading frame 72 (c9orf72) irna agent compositions and methods of use thereof |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20220115995A true KR20220115995A (ko) | 2022-08-19 |
Family
ID=74004172
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020227023988A KR20220115995A (ko) | 2019-12-13 | 2020-12-10 | 인간 염색체 9 개방 판독 프레임 72 (C9orf72) iRNA 제제 조성물 및 이의 사용 방법 |
Country Status (14)
| Country | Link |
|---|---|
| US (1) | US20230114649A1 (ko) |
| EP (1) | EP4073251A1 (ko) |
| JP (1) | JP2023506181A (ko) |
| KR (1) | KR20220115995A (ko) |
| CN (1) | CN115151641A (ko) |
| AU (1) | AU2020402885A1 (ko) |
| BR (1) | BR112022011417A2 (ko) |
| CA (1) | CA3164599A1 (ko) |
| CL (1) | CL2022001503A1 (ko) |
| CO (1) | CO2022009787A2 (ko) |
| IL (1) | IL293824A (ko) |
| MX (1) | MX2022006433A (ko) |
| TW (1) | TW202140509A (ko) |
| WO (1) | WO2021119226A1 (ko) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2020252020A1 (en) * | 2019-03-29 | 2021-11-11 | University Of Massachusetts | Oligonucleotide-based modulation of C90RF72 |
| US20210340535A1 (en) * | 2020-03-27 | 2021-11-04 | University Of Massachusetts | DUAL-ACTING siRNA BASED MODULATION OF C9orf72 |
| BR112023025224A2 (pt) | 2021-06-04 | 2024-02-27 | Alnylam Pharmaceuticals Inc | Quadro de leitura aberto 72 do cromossomo humano 9 (c9orf72) composições de agente de irna e métodos de uso dos mesmos |
| WO2024036343A2 (en) * | 2022-08-12 | 2024-02-15 | University Of Miami | Synergistic nucleic acid based therapeutics and methods of use for treating genetic disorders |
| WO2024107765A2 (en) | 2022-11-14 | 2024-05-23 | Regeneron Pharmaceuticals, Inc. | Compositions and methods for fibroblast growth factor receptor 3-mediated delivery to astrocytes |
Family Cites Families (251)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3687808A (en) | 1969-08-14 | 1972-08-29 | Univ Leland Stanford Junior | Synthetic polynucleotides |
| US4469863A (en) | 1980-11-12 | 1984-09-04 | Ts O Paul O P | Nonionic nucleic acid alkyl and aryl phosphonates and processes for manufacture and use thereof |
| US5023243A (en) | 1981-10-23 | 1991-06-11 | Molecular Biosystems, Inc. | Oligonucleotide therapeutic agent and method of making same |
| US4476301A (en) | 1982-04-29 | 1984-10-09 | Centre National De La Recherche Scientifique | Oligonucleotides, a process for preparing the same and their application as mediators of the action of interferon |
| JPS5927900A (ja) | 1982-08-09 | 1984-02-14 | Wakunaga Seiyaku Kk | 固定化オリゴヌクレオチド |
| FR2540122B1 (fr) | 1983-01-27 | 1985-11-29 | Centre Nat Rech Scient | Nouveaux composes comportant une sequence d'oligonucleotide liee a un agent d'intercalation, leur procede de synthese et leur application |
| US4605735A (en) | 1983-02-14 | 1986-08-12 | Wakunaga Seiyaku Kabushiki Kaisha | Oligonucleotide derivatives |
| US4948882A (en) | 1983-02-22 | 1990-08-14 | Syngene, Inc. | Single-stranded labelled oligonucleotides, reactive monomers and methods of synthesis |
| US4824941A (en) | 1983-03-10 | 1989-04-25 | Julian Gordon | Specific antibody to the native form of 2'5'-oligonucleotides, the method of preparation and the use as reagents in immunoassays or for binding 2'5'-oligonucleotides in biological systems |
| US4587044A (en) | 1983-09-01 | 1986-05-06 | The Johns Hopkins University | Linkage of proteins to nucleic acids |
| US5118800A (en) | 1983-12-20 | 1992-06-02 | California Institute Of Technology | Oligonucleotides possessing a primary amino group in the terminal nucleotide |
| US5118802A (en) | 1983-12-20 | 1992-06-02 | California Institute Of Technology | DNA-reporter conjugates linked via the 2' or 5'-primary amino group of the 5'-terminal nucleoside |
| US5550111A (en) | 1984-07-11 | 1996-08-27 | Temple University-Of The Commonwealth System Of Higher Education | Dual action 2',5'-oligoadenylate antiviral derivatives and uses thereof |
| FR2567892B1 (fr) | 1984-07-19 | 1989-02-17 | Centre Nat Rech Scient | Nouveaux oligonucleotides, leur procede de preparation et leurs applications comme mediateurs dans le developpement des effets des interferons |
| US5258506A (en) | 1984-10-16 | 1993-11-02 | Chiron Corporation | Photolabile reagents for incorporation into oligonucleotide chains |
| US5430136A (en) | 1984-10-16 | 1995-07-04 | Chiron Corporation | Oligonucleotides having selectably cleavable and/or abasic sites |
| US5367066A (en) | 1984-10-16 | 1994-11-22 | Chiron Corporation | Oligonucleotides with selectably cleavable and/or abasic sites |
| US4828979A (en) | 1984-11-08 | 1989-05-09 | Life Technologies, Inc. | Nucleotide analogs for nucleic acid labeling and detection |
| US4897355A (en) | 1985-01-07 | 1990-01-30 | Syntex (U.S.A.) Inc. | N[ω,(ω-1)-dialkyloxy]- and N-[ω,(ω-1)-dialkenyloxy]-alk-1-yl-N,N,N-tetrasubstituted ammonium lipids and uses therefor |
| FR2575751B1 (fr) | 1985-01-08 | 1987-04-03 | Pasteur Institut | Nouveaux nucleosides de derives de l'adenosine, leur preparation et leurs applications biologiques |
| US5034506A (en) | 1985-03-15 | 1991-07-23 | Anti-Gene Development Group | Uncharged morpholino-based polymers having achiral intersubunit linkages |
| US5405938A (en) | 1989-12-20 | 1995-04-11 | Anti-Gene Development Group | Sequence-specific binding polymers for duplex nucleic acids |
| US5166315A (en) | 1989-12-20 | 1992-11-24 | Anti-Gene Development Group | Sequence-specific binding polymers for duplex nucleic acids |
| US5185444A (en) | 1985-03-15 | 1993-02-09 | Anti-Gene Deveopment Group | Uncharged morpolino-based polymers having phosphorous containing chiral intersubunit linkages |
| US5235033A (en) | 1985-03-15 | 1993-08-10 | Anti-Gene Development Group | Alpha-morpholino ribonucleoside derivatives and polymers thereof |
| US4683202A (en) | 1985-03-28 | 1987-07-28 | Cetus Corporation | Process for amplifying nucleic acid sequences |
| US4762779A (en) | 1985-06-13 | 1988-08-09 | Amgen Inc. | Compositions and methods for functionalizing nucleic acids |
| US5317098A (en) | 1986-03-17 | 1994-05-31 | Hiroaki Shizuya | Non-radioisotope tagging of fragments |
| JPS638396A (ja) | 1986-06-30 | 1988-01-14 | Wakunaga Pharmaceut Co Ltd | ポリ標識化オリゴヌクレオチド誘導体 |
| US4920016A (en) | 1986-12-24 | 1990-04-24 | Linear Technology, Inc. | Liposomes with enhanced circulation time |
| US4837028A (en) | 1986-12-24 | 1989-06-06 | Liposome Technology, Inc. | Liposomes with enhanced circulation time |
| US5264423A (en) | 1987-03-25 | 1993-11-23 | The United States Of America As Represented By The Department Of Health And Human Services | Inhibitors for replication of retroviruses and for the expression of oncogene products |
| US5276019A (en) | 1987-03-25 | 1994-01-04 | The United States Of America As Represented By The Department Of Health And Human Services | Inhibitors for replication of retroviruses and for the expression of oncogene products |
| US4904582A (en) | 1987-06-11 | 1990-02-27 | Synthetic Genetics | Novel amphiphilic nucleic acid conjugates |
| WO1988010264A1 (en) | 1987-06-24 | 1988-12-29 | Howard Florey Institute Of Experimental Physiology | Nucleoside derivatives |
| US5585481A (en) | 1987-09-21 | 1996-12-17 | Gen-Probe Incorporated | Linking reagents for nucleotide probes |
| US5188897A (en) | 1987-10-22 | 1993-02-23 | Temple University Of The Commonwealth System Of Higher Education | Encapsulated 2',5'-phosphorothioate oligoadenylates |
| US4924624A (en) | 1987-10-22 | 1990-05-15 | Temple University-Of The Commonwealth System Of Higher Education | 2,',5'-phosphorothioate oligoadenylates and plant antiviral uses thereof |
| US5525465A (en) | 1987-10-28 | 1996-06-11 | Howard Florey Institute Of Experimental Physiology And Medicine | Oligonucleotide-polyamide conjugates and methods of production and applications of the same |
| DE3738460A1 (de) | 1987-11-12 | 1989-05-24 | Max Planck Gesellschaft | Modifizierte oligonukleotide |
| US5082830A (en) | 1988-02-26 | 1992-01-21 | Enzo Biochem, Inc. | End labeled nucleotide probe |
| WO1989009221A1 (en) | 1988-03-25 | 1989-10-05 | University Of Virginia Alumni Patents Foundation | Oligonucleotide n-alkylphosphoramidates |
| US5278302A (en) | 1988-05-26 | 1994-01-11 | University Patents, Inc. | Polynucleotide phosphorodithioates |
| US5109124A (en) | 1988-06-01 | 1992-04-28 | Biogen, Inc. | Nucleic acid probe linked to a label having a terminal cysteine |
| US5216141A (en) | 1988-06-06 | 1993-06-01 | Benner Steven A | Oligonucleotide analogs containing sulfur linkages |
| US5175273A (en) | 1988-07-01 | 1992-12-29 | Genentech, Inc. | Nucleic acid intercalating agents |
| US5262536A (en) | 1988-09-15 | 1993-11-16 | E. I. Du Pont De Nemours And Company | Reagents for the preparation of 5'-tagged oligonucleotides |
| US5512439A (en) | 1988-11-21 | 1996-04-30 | Dynal As | Oligonucleotide-linked magnetic particles and uses thereof |
| US5599923A (en) | 1989-03-06 | 1997-02-04 | Board Of Regents, University Of Tx | Texaphyrin metal complexes having improved functionalization |
| US5457183A (en) | 1989-03-06 | 1995-10-10 | Board Of Regents, The University Of Texas System | Hydroxylated texaphyrins |
| FR2645866B1 (fr) | 1989-04-17 | 1991-07-05 | Centre Nat Rech Scient | Nouvelles lipopolyamines, leur preparation et leur emploi |
| US5391723A (en) | 1989-05-31 | 1995-02-21 | Neorx Corporation | Oligonucleotide conjugates |
| US4958013A (en) | 1989-06-06 | 1990-09-18 | Northwestern University | Cholesteryl modified oligonucleotides |
| US5143854A (en) | 1989-06-07 | 1992-09-01 | Affymax Technologies N.V. | Large scale photolithographic solid phase synthesis of polypeptides and receptor binding screening thereof |
| US5744101A (en) | 1989-06-07 | 1998-04-28 | Affymax Technologies N.V. | Photolabile nucleoside protecting groups |
| US5451463A (en) | 1989-08-28 | 1995-09-19 | Clontech Laboratories, Inc. | Non-nucleoside 1,3-diol reagents for labeling synthetic oligonucleotides |
| US5134066A (en) | 1989-08-29 | 1992-07-28 | Monsanto Company | Improved probes using nucleosides containing 3-dezauracil analogs |
| US5254469A (en) | 1989-09-12 | 1993-10-19 | Eastman Kodak Company | Oligonucleotide-enzyme conjugate that can be used as a probe in hybridization assays and polymerase chain reaction procedures |
| US5591722A (en) | 1989-09-15 | 1997-01-07 | Southern Research Institute | 2'-deoxy-4'-thioribonucleosides and their antiviral activity |
| US5399676A (en) | 1989-10-23 | 1995-03-21 | Gilead Sciences | Oligonucleotides with inverted polarity |
| ATE269870T1 (de) | 1989-10-24 | 2004-07-15 | Isis Pharmaceuticals Inc | 2'-modifizierte oligonukleotide |
| US5264564A (en) | 1989-10-24 | 1993-11-23 | Gilead Sciences | Oligonucleotide analogs with novel linkages |
| US5292873A (en) | 1989-11-29 | 1994-03-08 | The Research Foundation Of State University Of New York | Nucleic acids labeled with naphthoquinone probe |
| US5177198A (en) | 1989-11-30 | 1993-01-05 | University Of N.C. At Chapel Hill | Process for preparing oligoribonucleoside and oligodeoxyribonucleoside boranophosphates |
| CA2029273A1 (en) | 1989-12-04 | 1991-06-05 | Christine L. Brakel | Modified nucleotide compounds |
| US5486603A (en) | 1990-01-08 | 1996-01-23 | Gilead Sciences, Inc. | Oligonucleotide having enhanced binding affinity |
| US5670633A (en) | 1990-01-11 | 1997-09-23 | Isis Pharmaceuticals, Inc. | Sugar modified oligonucleotides that detect and modulate gene expression |
| US5587361A (en) | 1991-10-15 | 1996-12-24 | Isis Pharmaceuticals, Inc. | Oligonucleotides having phosphorothioate linkages of high chiral purity |
| WO1993013121A1 (en) | 1991-12-24 | 1993-07-08 | Isis Pharmaceuticals, Inc. | Gapped 2' modified oligonucleotides |
| US5459255A (en) | 1990-01-11 | 1995-10-17 | Isis Pharmaceuticals, Inc. | N-2 substituted purines |
| US5681941A (en) | 1990-01-11 | 1997-10-28 | Isis Pharmaceuticals, Inc. | Substituted purines and oligonucleotide cross-linking |
| US5852188A (en) | 1990-01-11 | 1998-12-22 | Isis Pharmaceuticals, Inc. | Oligonucleotides having chiral phosphorus linkages |
| US7037646B1 (en) | 1990-01-11 | 2006-05-02 | Isis Pharmaceuticals, Inc. | Amine-derivatized nucleosides and oligonucleosides |
| US6783931B1 (en) | 1990-01-11 | 2004-08-31 | Isis Pharmaceuticals, Inc. | Amine-derivatized nucleosides and oligonucleosides |
| US5587470A (en) | 1990-01-11 | 1996-12-24 | Isis Pharmaceuticals, Inc. | 3-deazapurines |
| US5646265A (en) | 1990-01-11 | 1997-07-08 | Isis Pharmceuticals, Inc. | Process for the preparation of 2'-O-alkyl purine phosphoramidites |
| US5578718A (en) | 1990-01-11 | 1996-11-26 | Isis Pharmaceuticals, Inc. | Thiol-derivatized nucleosides |
| AU7579991A (en) | 1990-02-20 | 1991-09-18 | Gilead Sciences, Inc. | Pseudonucleosides and pseudonucleotides and their polymers |
| US5214136A (en) | 1990-02-20 | 1993-05-25 | Gilead Sciences, Inc. | Anthraquinone-derivatives oligonucleotides |
| US5321131A (en) | 1990-03-08 | 1994-06-14 | Hybridon, Inc. | Site-specific functionalization of oligodeoxynucleotides for non-radioactive labelling |
| US5470967A (en) | 1990-04-10 | 1995-11-28 | The Dupont Merck Pharmaceutical Company | Oligonucleotide analogs with sulfamate linkages |
| US5264618A (en) | 1990-04-19 | 1993-11-23 | Vical, Inc. | Cationic lipids for intracellular delivery of biologically active molecules |
| GB9009980D0 (en) | 1990-05-03 | 1990-06-27 | Amersham Int Plc | Phosphoramidite derivatives,their preparation and the use thereof in the incorporation of reporter groups on synthetic oligonucleotides |
| EP0745689A3 (en) | 1990-05-11 | 1996-12-11 | Microprobe Corporation | A dipstick for a nucleic acid hybridization assay |
| US5608046A (en) | 1990-07-27 | 1997-03-04 | Isis Pharmaceuticals, Inc. | Conjugated 4'-desmethyl nucleoside analog compounds |
| US5623070A (en) | 1990-07-27 | 1997-04-22 | Isis Pharmaceuticals, Inc. | Heteroatomic oligonucleoside linkages |
| US5618704A (en) | 1990-07-27 | 1997-04-08 | Isis Pharmacueticals, Inc. | Backbone-modified oligonucleotide analogs and preparation thereof through radical coupling |
| US5489677A (en) | 1990-07-27 | 1996-02-06 | Isis Pharmaceuticals, Inc. | Oligonucleoside linkages containing adjacent oxygen and nitrogen atoms |
| US5541307A (en) | 1990-07-27 | 1996-07-30 | Isis Pharmaceuticals, Inc. | Backbone modified oligonucleotide analogs and solid phase synthesis thereof |
| US5677437A (en) | 1990-07-27 | 1997-10-14 | Isis Pharmaceuticals, Inc. | Heteroatomic oligonucleoside linkages |
| DE69126530T2 (de) | 1990-07-27 | 1998-02-05 | Isis Pharmaceutical, Inc., Carlsbad, Calif. | Nuklease resistente, pyrimidin modifizierte oligonukleotide, die die gen-expression detektieren und modulieren |
| US5602240A (en) | 1990-07-27 | 1997-02-11 | Ciba Geigy Ag. | Backbone modified oligonucleotide analogs |
| US5138045A (en) | 1990-07-27 | 1992-08-11 | Isis Pharmaceuticals | Polyamine conjugated oligonucleotides |
| US5610289A (en) | 1990-07-27 | 1997-03-11 | Isis Pharmaceuticals, Inc. | Backbone modified oligonucleotide analogues |
| US5218105A (en) | 1990-07-27 | 1993-06-08 | Isis Pharmaceuticals | Polyamine conjugated oligonucleotides |
| US5688941A (en) | 1990-07-27 | 1997-11-18 | Isis Pharmaceuticals, Inc. | Methods of making conjugated 4' desmethyl nucleoside analog compounds |
| US5245022A (en) | 1990-08-03 | 1993-09-14 | Sterling Drug, Inc. | Exonuclease resistant terminally substituted oligonucleotides |
| BR9106729A (pt) | 1990-08-03 | 1993-07-20 | Sterling Winthrop Inc | Composto,processos para inibir a degradacao por nuclease de compostos e para estabilizar sequencias de nicleotideos ou oligonucleosideos,composicao utilizavel para inibir expressao de genes e processo para inibir expressao de genes em um mamifero necessitando de tal tratamento |
| US5512667A (en) | 1990-08-28 | 1996-04-30 | Reed; Michael W. | Trifunctional intermediates for preparing 3'-tailed oligonucleotides |
| US5214134A (en) | 1990-09-12 | 1993-05-25 | Sterling Winthrop Inc. | Process of linking nucleosides with a siloxane bridge |
| US5561225A (en) | 1990-09-19 | 1996-10-01 | Southern Research Institute | Polynucleotide analogs containing sulfonate and sulfonamide internucleoside linkages |
| CA2092002A1 (en) | 1990-09-20 | 1992-03-21 | Mark Matteucci | Modified internucleoside linkages |
| US5432272A (en) | 1990-10-09 | 1995-07-11 | Benner; Steven A. | Method for incorporating into a DNA or RNA oligonucleotide using nucleotides bearing heterocyclic bases |
| KR930702373A (ko) | 1990-11-08 | 1993-09-08 | 안토니 제이. 페이네 | 합성 올리고누클레오티드에 대한 다중 리포터(Reporter)그룹의 첨합 |
| GB9100304D0 (en) | 1991-01-08 | 1991-02-20 | Ici Plc | Compound |
| US7015315B1 (en) | 1991-12-24 | 2006-03-21 | Isis Pharmaceuticals, Inc. | Gapped oligonucleotides |
| US5714331A (en) | 1991-05-24 | 1998-02-03 | Buchardt, Deceased; Ole | Peptide nucleic acids having enhanced binding affinity, sequence specificity and solubility |
| US5719262A (en) | 1993-11-22 | 1998-02-17 | Buchardt, Deceased; Ole | Peptide nucleic acids having amino acid side chains |
| US5539082A (en) | 1993-04-26 | 1996-07-23 | Nielsen; Peter E. | Peptide nucleic acids |
| US5371241A (en) | 1991-07-19 | 1994-12-06 | Pharmacia P-L Biochemicals Inc. | Fluorescein labelled phosphoramidites |
| US5571799A (en) | 1991-08-12 | 1996-11-05 | Basco, Ltd. | (2'-5') oligoadenylate analogues useful as inhibitors of host-v5.-graft response |
| US5283185A (en) | 1991-08-28 | 1994-02-01 | University Of Tennessee Research Corporation | Method for delivering nucleic acids into cells |
| DE59208572D1 (de) | 1991-10-17 | 1997-07-10 | Ciba Geigy Ag | Bicyclische Nukleoside, Oligonukleotide, Verfahren zu deren Herstellung und Zwischenprodukte |
| US5594121A (en) | 1991-11-07 | 1997-01-14 | Gilead Sciences, Inc. | Enhanced triple-helix and double-helix formation with oligomers containing modified purines |
| ATE293011T1 (de) | 1991-11-22 | 2005-04-15 | Affymetrix Inc A Delaware Corp | Kombinatorische strategien für die polymersynthese |
| US5484908A (en) | 1991-11-26 | 1996-01-16 | Gilead Sciences, Inc. | Oligonucleotides containing 5-propynyl pyrimidines |
| US6235887B1 (en) | 1991-11-26 | 2001-05-22 | Isis Pharmaceuticals, Inc. | Enhanced triple-helix and double-helix formation directed by oligonucleotides containing modified pyrimidines |
| US5359044A (en) | 1991-12-13 | 1994-10-25 | Isis Pharmaceuticals | Cyclobutyl oligonucleotide surrogates |
| US6277603B1 (en) | 1991-12-24 | 2001-08-21 | Isis Pharmaceuticals, Inc. | PNA-DNA-PNA chimeric macromolecules |
| US5595726A (en) | 1992-01-21 | 1997-01-21 | Pharmacyclics, Inc. | Chromophore probe for detection of nucleic acid |
| US5565552A (en) | 1992-01-21 | 1996-10-15 | Pharmacyclics, Inc. | Method of expanded porphyrin-oligonucleotide conjugate synthesis |
| FR2687679B1 (fr) | 1992-02-05 | 1994-10-28 | Centre Nat Rech Scient | Oligothionucleotides. |
| DE4203923A1 (de) | 1992-02-11 | 1993-08-12 | Henkel Kgaa | Verfahren zur herstellung von polycarboxylaten auf polysaccharid-basis |
| US5633360A (en) | 1992-04-14 | 1997-05-27 | Gilead Sciences, Inc. | Oligonucleotide analogs capable of passive cell membrane permeation |
| US5434257A (en) | 1992-06-01 | 1995-07-18 | Gilead Sciences, Inc. | Binding compentent oligomers containing unsaturated 3',5' and 2',5' linkages |
| EP0646178A1 (en) | 1992-06-04 | 1995-04-05 | The Regents Of The University Of California | expression cassette with regularoty regions functional in the mammmlian host |
| AU4541093A (en) | 1992-06-18 | 1994-01-24 | Genpharm International, Inc. | Methods for producing transgenic non-human animals harboring a yeast artificial chromosome |
| EP0577558A2 (de) | 1992-07-01 | 1994-01-05 | Ciba-Geigy Ag | Carbocyclische Nukleoside mit bicyclischen Ringen, Oligonukleotide daraus, Verfahren zu deren Herstellung, deren Verwendung und Zwischenproduckte |
| US5272250A (en) | 1992-07-10 | 1993-12-21 | Spielvogel Bernard F | Boronated phosphoramidate compounds |
| EP1251170A3 (en) | 1992-07-17 | 2002-10-30 | Ribozyme Pharmaceuticals, Inc. | Method and reagent for treatment of NF-kappaB dependent animal diseases |
| US6346614B1 (en) | 1992-07-23 | 2002-02-12 | Hybridon, Inc. | Hybrid oligonucleotide phosphorothioates |
| US5574142A (en) | 1992-12-15 | 1996-11-12 | Microprobe Corporation | Peptide linkers for improved oligonucleotide delivery |
| US5476925A (en) | 1993-02-01 | 1995-12-19 | Northwestern University | Oligodeoxyribonucleotides including 3'-aminonucleoside-phosphoramidate linkages and terminal 3'-amino groups |
| DE69424406T2 (de) | 1993-02-19 | 2000-10-26 | Nippon Shinyaku Co., Ltd. | Arzneistoffzusammensetzung, die ein nukleinsäurecopolymer enthält |
| GB9304618D0 (en) | 1993-03-06 | 1993-04-21 | Ciba Geigy Ag | Chemical compounds |
| WO1994022864A1 (en) | 1993-03-30 | 1994-10-13 | Sterling Winthrop Inc. | Acyclic nucleoside analogs and oligonucleotide sequences containing them |
| AU6412794A (en) | 1993-03-31 | 1994-10-24 | Sterling Winthrop Inc. | Oligonucleotides with amide linkages replacing phosphodiester linkages |
| DE4311944A1 (de) | 1993-04-10 | 1994-10-13 | Degussa | Umhüllte Natriumpercarbonatpartikel, Verfahren zu deren Herstellung und sie enthaltende Wasch-, Reinigungs- und Bleichmittelzusammensetzungen |
| US6191105B1 (en) | 1993-05-10 | 2001-02-20 | Protein Delivery, Inc. | Hydrophilic and lipophilic balanced microemulsion formulations of free-form and/or conjugation-stabilized therapeutic agents such as insulin |
| US5955591A (en) | 1993-05-12 | 1999-09-21 | Imbach; Jean-Louis | Phosphotriester oligonucleotides, amidites and method of preparation |
| US6015886A (en) | 1993-05-24 | 2000-01-18 | Chemgenes Corporation | Oligonucleotide phosphate esters |
| US6294664B1 (en) | 1993-07-29 | 2001-09-25 | Isis Pharmaceuticals, Inc. | Synthesis of oligonucleotides |
| US5502177A (en) | 1993-09-17 | 1996-03-26 | Gilead Sciences, Inc. | Pyrimidine derivatives for labeled binding partners |
| WO1995014030A1 (en) | 1993-11-16 | 1995-05-26 | Genta Incorporated | Synthetic oligomers having chirally pure phosphonate internucleosidyl linkages mixed with non-phosphonate internucleosidyl linkages |
| US5457187A (en) | 1993-12-08 | 1995-10-10 | Board Of Regents University Of Nebraska | Oligonucleotides containing 5-fluorouracil |
| US5446137B1 (en) | 1993-12-09 | 1998-10-06 | Behringwerke Ag | Oligonucleotides containing 4'-substituted nucleotides |
| US5519134A (en) | 1994-01-11 | 1996-05-21 | Isis Pharmaceuticals, Inc. | Pyrrolidine-containing monomers and oligomers |
| US5599922A (en) | 1994-03-18 | 1997-02-04 | Lynx Therapeutics, Inc. | Oligonucleotide N3'-P5' phosphoramidates: hybridization and nuclease resistance properties |
| US5596091A (en) | 1994-03-18 | 1997-01-21 | The Regents Of The University Of California | Antisense oligonucleotides comprising 5-aminoalkyl pyrimidine nucleotides |
| US5627053A (en) | 1994-03-29 | 1997-05-06 | Ribozyme Pharmaceuticals, Inc. | 2'deoxy-2'-alkylnucleotide containing nucleic acid |
| US5625050A (en) | 1994-03-31 | 1997-04-29 | Amgen Inc. | Modified oligonucleotides and intermediates useful in nucleic acid therapeutics |
| US6054299A (en) | 1994-04-29 | 2000-04-25 | Conrad; Charles A. | Stem-loop cloning vector and method |
| US5525711A (en) | 1994-05-18 | 1996-06-11 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Pteridine nucleotide analogs as fluorescent DNA probes |
| US5543152A (en) | 1994-06-20 | 1996-08-06 | Inex Pharmaceuticals Corporation | Sphingosomes for enhanced drug delivery |
| US5597696A (en) | 1994-07-18 | 1997-01-28 | Becton Dickinson And Company | Covalent cyanine dye oligonucleotide conjugates |
| US5597909A (en) | 1994-08-25 | 1997-01-28 | Chiron Corporation | Polynucleotide reagents containing modified deoxyribose moieties, and associated methods of synthesis and use |
| US5580731A (en) | 1994-08-25 | 1996-12-03 | Chiron Corporation | N-4 modified pyrimidine deoxynucleotides and oligonucleotide probes synthesized therewith |
| US5556752A (en) | 1994-10-24 | 1996-09-17 | Affymetrix, Inc. | Surface-bound, unimolecular, double-stranded DNA |
| US6608035B1 (en) | 1994-10-25 | 2003-08-19 | Hybridon, Inc. | Method of down-regulating gene expression |
| US6166197A (en) | 1995-03-06 | 2000-12-26 | Isis Pharmaceuticals, Inc. | Oligomeric compounds having pyrimidine nucleotide (S) with 2'and 5 substitutions |
| AU5359496A (en) | 1995-03-06 | 1996-09-23 | Isis Pharmaceuticals, Inc. | Improved process for the synthesis of 2'-o-substituted pyrimidines and oligomeric compounds therefrom |
| JPH11508231A (ja) | 1995-05-26 | 1999-07-21 | ソマティックス セラピー コーポレイション | 安定な脂質/核酸複合体を含む送達ビヒクル |
| US5981501A (en) | 1995-06-07 | 1999-11-09 | Inex Pharmaceuticals Corp. | Methods for encapsulating plasmids in lipid bilayers |
| US7422902B1 (en) | 1995-06-07 | 2008-09-09 | The University Of British Columbia | Lipid-nucleic acid particles prepared via a hydrophobic lipid-nucleic acid complex intermediate and use for gene transfer |
| CA2222328C (en) | 1995-06-07 | 2012-01-10 | Inex Pharmaceuticals Corporation | Lipid-nucleic acid particles prepared via a hydrophobic lipid-nucleic acid complex intermediate and use for gene transfer |
| US5545531A (en) | 1995-06-07 | 1996-08-13 | Affymax Technologies N.V. | Methods for making a device for concurrently processing multiple biological chip assays |
| US5858397A (en) | 1995-10-11 | 1999-01-12 | University Of British Columbia | Liposomal formulations of mitoxantrone |
| US6160109A (en) | 1995-10-20 | 2000-12-12 | Isis Pharmaceuticals, Inc. | Preparation of phosphorothioate and boranophosphate oligomers |
| US5854033A (en) | 1995-11-21 | 1998-12-29 | Yale University | Rolling circle replication reporter systems |
| US5858401A (en) | 1996-01-22 | 1999-01-12 | Sidmak Laboratories, Inc. | Pharmaceutical composition for cyclosporines |
| US5994316A (en) | 1996-02-21 | 1999-11-30 | The Immune Response Corporation | Method of preparing polynucleotide-carrier complexes for delivery to cells |
| US6444423B1 (en) | 1996-06-07 | 2002-09-03 | Molecular Dynamics, Inc. | Nucleosides comprising polydentate ligands |
| US6576752B1 (en) | 1997-02-14 | 2003-06-10 | Isis Pharmaceuticals, Inc. | Aminooxy functionalized oligomers |
| US6172209B1 (en) | 1997-02-14 | 2001-01-09 | Isis Pharmaceuticals Inc. | Aminooxy-modified oligonucleotides and methods for making same |
| US6639062B2 (en) | 1997-02-14 | 2003-10-28 | Isis Pharmaceuticals, Inc. | Aminooxy-modified nucleosidic compounds and oligomeric compounds prepared therefrom |
| US6034135A (en) | 1997-03-06 | 2000-03-07 | Promega Biosciences, Inc. | Dimeric cationic lipids |
| JP3756313B2 (ja) | 1997-03-07 | 2006-03-15 | 武 今西 | 新規ビシクロヌクレオシド及びオリゴヌクレオチド類縁体 |
| US6770748B2 (en) | 1997-03-07 | 2004-08-03 | Takeshi Imanishi | Bicyclonucleoside and oligonucleotide analogue |
| EP1027033B1 (en) | 1997-05-14 | 2009-07-22 | The University Of British Columbia | High efficiency encapsulation of nucleic acids in lipid vesicles |
| JP2002510319A (ja) | 1997-07-01 | 2002-04-02 | アイシス・ファーマシューティカルス・インコーポレーテッド | オリゴヌクレオチドの消化管を介したデリバリーのための組成物及び方法 |
| EP2341058A3 (en) | 1997-09-12 | 2011-11-23 | Exiqon A/S | Oligonucleotide Analogues |
| US6794499B2 (en) | 1997-09-12 | 2004-09-21 | Exiqon A/S | Oligonucleotide analogues |
| US6528640B1 (en) | 1997-11-05 | 2003-03-04 | Ribozyme Pharmaceuticals, Incorporated | Synthetic ribonucleic acids with RNAse activity |
| US6617438B1 (en) | 1997-11-05 | 2003-09-09 | Sirna Therapeutics, Inc. | Oligoribonucleotides with enzymatic activity |
| US6320017B1 (en) | 1997-12-23 | 2001-11-20 | Inex Pharmaceuticals Corp. | Polyamide oligomers |
| US7273933B1 (en) | 1998-02-26 | 2007-09-25 | Isis Pharmaceuticals, Inc. | Methods for synthesis of oligonucleotides |
| US7045610B2 (en) | 1998-04-03 | 2006-05-16 | Epoch Biosciences, Inc. | Modified oligonucleotides for mismatch discrimination |
| US6531590B1 (en) | 1998-04-24 | 2003-03-11 | Isis Pharmaceuticals, Inc. | Processes for the synthesis of oligonucleotide compounds |
| US6867294B1 (en) | 1998-07-14 | 2005-03-15 | Isis Pharmaceuticals, Inc. | Gapped oligomers having site specific chiral phosphorothioate internucleoside linkages |
| DE69906977T2 (de) | 1998-07-20 | 2004-05-19 | Protiva Biotherapeutics Inc., Burnaby | In liposomen verkapselte nukleinsäurekomplexe |
| WO2000022113A1 (en) | 1998-10-09 | 2000-04-20 | Ingene, Inc. | ENZYMATIC SYNTHESIS OF ssDNA |
| BR9914773A (pt) | 1998-10-09 | 2002-02-05 | Ingene Inc | Conjunto de elementos genéricos, método para a produção de dna de cordão único, transcrição de mrna, construção de ácido nucléico, transcrição de ssdna, vetor, sistema vetor, célula hospedeira, conjunto para a produção de uma sequência de ácido nucléico de cordão único, método para a produção in vivo ou in vitro de uma sequência de ácido nucléico de cordão único, transcrição de cdna de cordão único, ácido nucléico inibidor, molécula heteroduplex, e composição farmacêutica |
| US6465628B1 (en) | 1999-02-04 | 2002-10-15 | Isis Pharmaceuticals, Inc. | Process for the synthesis of oligomeric compounds |
| WO2000050050A1 (en) | 1999-02-23 | 2000-08-31 | Isis Pharmaceuticals, Inc. | Multiparticulate formulation |
| US7084125B2 (en) | 1999-03-18 | 2006-08-01 | Exiqon A/S | Xylo-LNA analogues |
| US7053207B2 (en) | 1999-05-04 | 2006-05-30 | Exiqon A/S | L-ribo-LNA analogues |
| US6525191B1 (en) | 1999-05-11 | 2003-02-25 | Kanda S. Ramasamy | Conformationally constrained L-nucleosides |
| US6593466B1 (en) | 1999-07-07 | 2003-07-15 | Isis Pharmaceuticals, Inc. | Guanidinium functionalized nucleotides and precursors thereof |
| US6147200A (en) | 1999-08-19 | 2000-11-14 | Isis Pharmaceuticals, Inc. | 2'-O-acetamido modified monomers and oligomers |
| WO2001053307A1 (en) | 2000-01-21 | 2001-07-26 | Geron Corporation | 2'-arabino-fluorooligonucleotide n3'→p5'phosphoramidates: their synthesis and use |
| IT1318539B1 (it) | 2000-05-26 | 2003-08-27 | Italfarmaco Spa | Composizioni farmaceutiche a rilascio prolungato per lasomministrazione parenterale di sostanze idrofile biologicamente |
| JP4413493B2 (ja) | 2000-10-04 | 2010-02-10 | サンタリス ファーマ アー/エス | プリンlna類似体の改善された合成方法 |
| WO2005078097A2 (en) | 2004-02-10 | 2005-08-25 | Sirna Therapeutics, Inc. | RNA INTERFERENCE MEDIATED INHIBITION OF GENE EXPRESSION USING MULTIFUNCTIONAL SHORT INTERFERING NUCLEIC ACID (Multifunctional siNA) |
| WO2003015698A2 (en) | 2001-08-13 | 2003-02-27 | University Of Pittsburgh | Application of lipid vehicles and use for drug delivery |
| EP1520022B1 (en) | 2002-07-10 | 2015-07-22 | Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. | Rna-interference by single-stranded rna molecules |
| US6878805B2 (en) | 2002-08-16 | 2005-04-12 | Isis Pharmaceuticals, Inc. | Peptide-conjugated oligomeric compounds |
| AU2003291753B2 (en) | 2002-11-05 | 2010-07-08 | Isis Pharmaceuticals, Inc. | Polycyclic sugar surrogate-containing oligomeric compounds and compositions for use in gene modulation |
| EP1560840B1 (en) | 2002-11-05 | 2015-05-06 | Isis Pharmaceuticals, Inc. | Compositions comprising alternating 2'-modified nucleosides for use in gene modulation |
| US7427672B2 (en) | 2003-08-28 | 2008-09-23 | Takeshi Imanishi | Artificial nucleic acids of n-o bond crosslinkage type |
| PL1802193T3 (pl) | 2004-10-19 | 2014-09-30 | Regeneron Pharma | Sposób wytwarzania myszy homozygotycznej pod względem modyfikacji genetycznej |
| CA2603730A1 (en) | 2005-03-31 | 2006-10-05 | Calando Pharmaceuticals, Inc. | Inhibitors of ribonucleotide reductase subunit 2 and uses thereof |
| EP2314594B1 (en) | 2006-01-27 | 2014-07-23 | Isis Pharmaceuticals, Inc. | 6-modified bicyclic nucleic acid analogs |
| US7569686B1 (en) | 2006-01-27 | 2009-08-04 | Isis Pharmaceuticals, Inc. | Compounds and methods for synthesis of bicyclic nucleic acid analogs |
| WO2007091269A2 (en) | 2006-02-08 | 2007-08-16 | Quark Pharmaceuticals, Inc. | NOVEL TANDEM siRNAS |
| KR101221589B1 (ko) | 2006-04-07 | 2013-01-15 | 이데라 파마슈티칼즈, 인코포레이티드 | Tlr7 및 tlr8에 대한 안정화된 면역 조절성 rna〔simra〕 화합물 |
| WO2007134181A2 (en) | 2006-05-11 | 2007-11-22 | Isis Pharmaceuticals, Inc. | 5'-modified bicyclic nucleic acid analogs |
| KR101129509B1 (ko) | 2006-10-03 | 2012-04-13 | 알닐람 파마슈티칼스 인코포레이티드 | 지질 함유 조성물 |
| US20100105134A1 (en) | 2007-03-02 | 2010-04-29 | Mdrna, Inc. | Nucleic acid compounds for inhibiting gene expression and uses thereof |
| US8314227B2 (en) | 2007-05-22 | 2012-11-20 | Marina Biotech, Inc. | Hydroxymethyl substituted RNA oligonucleotides and RNA complexes |
| DK2170917T3 (da) | 2007-05-30 | 2012-10-08 | Isis Pharmaceuticals Inc | N-Substituerede bicycliske nukleinsyreanaloge med aminomethylenbro |
| ES2386492T3 (es) | 2007-06-08 | 2012-08-21 | Isis Pharmaceuticals, Inc. | Análogos de ácidos nucleicos bicíclicos carbocíclicos |
| CA2692579C (en) | 2007-07-05 | 2016-05-03 | Isis Pharmaceuticals, Inc. | 6-disubstituted bicyclic nucleic acid analogs |
| EP2357231A2 (en) | 2007-07-09 | 2011-08-17 | Idera Pharmaceuticals, Inc. | Stabilized immune modulatory RNA (SIMRA) compounds |
| CA3043911A1 (en) | 2007-12-04 | 2009-07-02 | Arbutus Biopharma Corporation | Targeting lipids |
| HUE034483T2 (en) | 2008-04-15 | 2018-02-28 | Protiva Biotherapeutics Inc | New lipid preparations for introducing a nucleic acid |
| CA2732229C (en) | 2008-07-25 | 2023-10-17 | Alnylam Pharmaceuticals, Inc. | Enhancement of sirna silencing activity using universal bases or mismatches in the sense strand |
| JP2012504389A (ja) | 2008-09-22 | 2012-02-23 | ダイセルナ ファーマシューティカルズ, インコーポレイテッド | 修飾を有するdsRNAによる遺伝子発現の特異的な阻害のための組成物および方法 |
| SG10201901089TA (en) | 2008-11-10 | 2019-03-28 | Arbutus Biopharma Corp | Novel lipids and compositions for the delivery of therapeutics |
| CN102239259A (zh) | 2008-12-03 | 2011-11-09 | 玛瑞纳生物技术有限公司 | UsiRNA复合物 |
| US20100249214A1 (en) | 2009-02-11 | 2010-09-30 | Dicerna Pharmaceuticals | Multiplex dicer substrate rna interference molecules having joining sequences |
| KR101728655B1 (ko) | 2008-12-18 | 2017-04-19 | 다이서나 파마수이티컬, 인크. | 유전자 발현의 특이적 억제를 위한 연장된 다이서 기질 제제 및 방법 |
| EP3243504A1 (en) | 2009-01-29 | 2017-11-15 | Arbutus Biopharma Corporation | Improved lipid formulation |
| SG10201402054UA (en) | 2009-05-05 | 2014-09-26 | Muthiah Manoharan | Lipid compositions |
| CN102575252B (zh) | 2009-06-01 | 2016-04-20 | 光环生物干扰疗法公司 | 用于多价rna干扰的多核苷酸、组合物及其使用方法 |
| HUE056773T2 (hu) | 2009-06-10 | 2022-03-28 | Arbutus Biopharma Corp | Továbbfejlesztett lipid készítmény |
| WO2011005860A2 (en) | 2009-07-07 | 2011-01-13 | Alnylam Pharmaceuticals, Inc. | 5' phosphate mimics |
| US9512164B2 (en) | 2009-07-07 | 2016-12-06 | Alnylam Pharmaceuticals, Inc. | Oligonucleotide end caps |
| DK2470656T3 (da) | 2009-08-27 | 2015-06-22 | Idera Pharmaceuticals Inc | Sammensætning til hæmning af genekspression og anvendelser heraf |
| US10913767B2 (en) | 2010-04-22 | 2021-02-09 | Alnylam Pharmaceuticals, Inc. | Oligonucleotides comprising acyclic and abasic nucleosides and analogs |
| WO2011139710A1 (en) | 2010-04-26 | 2011-11-10 | Marina Biotech, Inc. | Nucleic acid compounds with conformationally restricted monomers and uses thereof |
| US20140275211A1 (en) | 2011-06-21 | 2014-09-18 | Alnylam Pharmaceuticals, Inc. | Assays and methods for determining activity of a therapeutic agent in a subject |
| SG11201401314PA (en) | 2011-09-07 | 2014-09-26 | Marina Biotech Inc | Synthesis and uses of nucleic acid compounds with conformationally restricted monomers |
| WO2013075035A1 (en) | 2011-11-18 | 2013-05-23 | Alnylam Pharmaceuticals | Rnai agents, compositions and methods of use thereof for treating transthyretin (ttr) associated diseases |
| EP2906696B2 (en) * | 2012-10-15 | 2022-12-14 | Ionis Pharmaceuticals, Inc. | Methods for modulating c9orf72 expression |
| CN111593051A (zh) | 2013-05-01 | 2020-08-28 | Ionis制药公司 | 组合物和方法 |
| US10549035B2 (en) | 2014-01-31 | 2020-02-04 | University Hospitals Health System, Inc. | Systems and methods for intrathecal delivery of a pharmaceutical agent |
| WO2018064600A1 (en) | 2016-09-30 | 2018-04-05 | Regeneron Pharmaceuticals, Inc. | Non-human animals having a hexanucleotide repeat expansion in a c9orf72 locus |
| WO2019051025A2 (en) * | 2017-09-06 | 2019-03-14 | The Trustees Of Columbia University In The City Of New York | TREATMENT OF AGGRESSIVE CANCERS BY TARGETING C9ORF72 |
| CN111343994B (zh) | 2017-09-14 | 2023-11-21 | 箭头药业股份有限公司 | 用于抑制血管生成素-样3 (ANGPTL3)的表达的RNAi剂和组合物以及使用方法 |
| KR20210018267A (ko) * | 2018-05-07 | 2021-02-17 | 알닐람 파마슈티칼스 인코포레이티드 | 간외 전달 |
| EP3947683A1 (en) | 2019-04-04 | 2022-02-09 | Dicerna Pharmaceuticals, Inc. | Compositions and methods for inhibiting gene expression in the central nervous system |
-
2020
- 2020-12-10 WO PCT/US2020/064159 patent/WO2021119226A1/en unknown
- 2020-12-10 AU AU2020402885A patent/AU2020402885A1/en active Pending
- 2020-12-10 TW TW109143671A patent/TW202140509A/zh unknown
- 2020-12-10 MX MX2022006433A patent/MX2022006433A/es unknown
- 2020-12-10 IL IL293824A patent/IL293824A/en unknown
- 2020-12-10 JP JP2022535508A patent/JP2023506181A/ja active Pending
- 2020-12-10 EP EP20829269.8A patent/EP4073251A1/en active Pending
- 2020-12-10 CA CA3164599A patent/CA3164599A1/en active Pending
- 2020-12-10 CN CN202080096562.4A patent/CN115151641A/zh active Pending
- 2020-12-10 KR KR1020227023988A patent/KR20220115995A/ko unknown
- 2020-12-10 BR BR112022011417A patent/BR112022011417A2/pt unknown
-
2022
- 2022-06-08 CL CL2022001503A patent/CL2022001503A1/es unknown
- 2022-06-09 US US17/836,020 patent/US20230114649A1/en active Pending
- 2022-07-13 CO CONC2022/0009787A patent/CO2022009787A2/es unknown
Also Published As
| Publication number | Publication date |
|---|---|
| CL2022001503A1 (es) | 2023-03-10 |
| MX2022006433A (es) | 2022-06-23 |
| WO2021119226A1 (en) | 2021-06-17 |
| US20230114649A1 (en) | 2023-04-13 |
| CO2022009787A2 (es) | 2022-07-19 |
| TW202140509A (zh) | 2021-11-01 |
| AU2020402885A1 (en) | 2022-06-16 |
| CA3164599A1 (en) | 2021-06-17 |
| BR112022011417A2 (pt) | 2022-08-30 |
| JP2023506181A (ja) | 2023-02-15 |
| EP4073251A1 (en) | 2022-10-19 |
| CN115151641A (zh) | 2022-10-04 |
| IL293824A (en) | 2022-08-01 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20240294911A1 (en) | INSULIN-LIKE GROWTH FACTOR BINDING PROTEIN, ACID LABILE SUBUNIT (IGFALS) AND INSULIN-LIKE GROWTH FACTOR 1 (IGF-1) iRNA COMPOSITIONS AND METHODS OF USE THEREOF | |
| KR20200133373A (ko) | 17β-하이드록시스테로이드 데하이드로게나제 유형 13 (HSD17B13) iRNA 조성물 및 그것의 사용 방법 | |
| KR20220115995A (ko) | 인간 염색체 9 개방 판독 프레임 72 (C9orf72) iRNA 제제 조성물 및 이의 사용 방법 | |
| KR20210111780A (ko) | 아밀로이드 전구체 단백질(APP) RNAi 제제 조성물 및 그것의 사용 방법 | |
| KR20230005194A (ko) | 미세소관 관련된 단백질 TAU (MAPT) iRNA 제제 조성물 및 이의 사용 방법 | |
| US20240011032A1 (en) | Snca irna compositions and methods of use thereof for treating or preventing snca-associated neurodegenerative diseases | |
| US20220380762A1 (en) | HUNTINGTIN (HTT) iRNA AGENT COMPOSITIONS AND METHODS OF USE THEREOF | |
| US20240294906A1 (en) | Atxn2 irna compositions and methods of use thereof for treating or preventing atxn2-associated neurodegenerative diseases | |
| KR20230018377A (ko) | 아포지질단백질 e (apoe) irna 제제 조성물 및 이의 사용 방법 | |
| JP2024534961A (ja) | 拡大したエンドソームによって特徴付けられる疾患を処置または予防するためのAPP iRNA組成物およびその使用方法 | |
| KR20230162024A (ko) | 헌팅틴(HTT) iRNA 제제 조성물 및 이의 사용 방법 | |
| KR20240017911A (ko) | 인간 염색체 9 개방 해독 프레임 72(C9orf72) iRNA 제제 조성물 및 이의 사용 방법 | |
| KR20220133246A (ko) | 류신 풍부 반복 키나아제 2 (LRRK2) iRNA 작용제 조성물 및 이의 사용 방법 | |
| US20230056569A1 (en) | Ataxin3 (atxn3) rnai agent compositions and methods of use thereof | |
| US20240301426A1 (en) | Leucine-rich repeat kinase 2 (lrrk2) irna agent compositions and methods of use thereof | |
| US20230032974A1 (en) | SMALL RIBOSOMAL PROTEIN SUBUNIT 25 (RPS25) iRNA AGENT COMPOSITIONS AND METHODS OF USE THEREOF | |
| WO2023154900A2 (en) | Microtubule associated protein tau (mapt) irna agent compositions and methods of use thereof | |
| EA046043B1 (ru) | КОМПОЗИЦИИ иРНК 17B-ГИДРОКСИСТЕРОИДДЕГИДРОГЕНАЗЫ ТИПА 13 (HSD17B13) И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | |
| CN117561335A (zh) | 富含亮氨酸的重复激酶2(LRRK2)iRNA药剂组合物和其使用方法 |