KR20220038415A - 엔테로바이러스 d68에 대한 인간 모노클로날 항체 - Google Patents
엔테로바이러스 d68에 대한 인간 모노클로날 항체 Download PDFInfo
- Publication number
- KR20220038415A KR20220038415A KR1020227005641A KR20227005641A KR20220038415A KR 20220038415 A KR20220038415 A KR 20220038415A KR 1020227005641 A KR1020227005641 A KR 1020227005641A KR 20227005641 A KR20227005641 A KR 20227005641A KR 20220038415 A KR20220038415 A KR 20220038415A
- Authority
- KR
- South Korea
- Prior art keywords
- antibody
- fragment
- clone
- antibody fragment
- heavy chain
- Prior art date
Links
- 241000146324 Enterovirus D68 Species 0.000 title claims abstract description 250
- 238000000034 method Methods 0.000 claims abstract description 210
- 208000015181 infectious disease Diseases 0.000 claims description 293
- 102000008394 Immunoglobulin Fragments Human genes 0.000 claims description 173
- 108010021625 Immunoglobulin Fragments Proteins 0.000 claims description 173
- 230000027455 binding Effects 0.000 claims description 163
- 210000004027 cell Anatomy 0.000 claims description 162
- 238000009739 binding Methods 0.000 claims description 161
- 239000000427 antigen Substances 0.000 claims description 144
- 108091007433 antigens Proteins 0.000 claims description 143
- 102000036639 antigens Human genes 0.000 claims description 143
- 238000011282 treatment Methods 0.000 claims description 125
- 239000012634 fragment Substances 0.000 claims description 71
- 210000004369 blood Anatomy 0.000 claims description 65
- 239000008280 blood Substances 0.000 claims description 65
- 239000000203 mixture Substances 0.000 claims description 55
- 230000003612 virological effect Effects 0.000 claims description 55
- 229960005486 vaccine Drugs 0.000 claims description 48
- 210000004408 hybridoma Anatomy 0.000 claims description 45
- 238000003556 assay Methods 0.000 claims description 40
- 230000003993 interaction Effects 0.000 claims description 34
- 238000001514 detection method Methods 0.000 claims description 30
- 238000002965 ELISA Methods 0.000 claims description 29
- 230000013595 glycosylation Effects 0.000 claims description 28
- 238000006206 glycosylation reaction Methods 0.000 claims description 28
- 230000014509 gene expression Effects 0.000 claims description 27
- 230000008859 change Effects 0.000 claims description 26
- 230000035772 mutation Effects 0.000 claims description 26
- 238000009472 formulation Methods 0.000 claims description 25
- 238000004519 manufacturing process Methods 0.000 claims description 25
- 239000000126 substance Substances 0.000 claims description 24
- 230000001225 therapeutic effect Effects 0.000 claims description 24
- 230000000890 antigenic effect Effects 0.000 claims description 21
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 claims description 17
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 claims description 17
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 claims description 17
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 claims description 17
- 230000002255 enzymatic effect Effects 0.000 claims description 15
- 239000013598 vector Substances 0.000 claims description 14
- 238000001262 western blot Methods 0.000 claims description 13
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 11
- 239000007924 injection Substances 0.000 claims description 11
- 238000002347 injection Methods 0.000 claims description 11
- 210000002966 serum Anatomy 0.000 claims description 11
- 238000003127 radioimmunoassay Methods 0.000 claims description 10
- 239000013604 expression vector Substances 0.000 claims description 9
- 230000036541 health Effects 0.000 claims description 9
- 238000002560 therapeutic procedure Methods 0.000 claims description 9
- 230000000840 anti-viral effect Effects 0.000 claims description 8
- 102000020313 Cell-Penetrating Peptides Human genes 0.000 claims description 7
- 108010051109 Cell-Penetrating Peptides Proteins 0.000 claims description 7
- 230000000241 respiratory effect Effects 0.000 claims description 6
- 238000010186 staining Methods 0.000 claims description 6
- 210000002700 urine Anatomy 0.000 claims description 6
- 239000000443 aerosol Substances 0.000 claims description 5
- 208000032376 Lung infection Diseases 0.000 claims description 4
- 206010021888 Nervous system infections Diseases 0.000 claims description 4
- 206010036790 Productive cough Diseases 0.000 claims description 4
- 238000004520 electroporation Methods 0.000 claims description 4
- 210000003754 fetus Anatomy 0.000 claims description 4
- 210000002826 placenta Anatomy 0.000 claims description 4
- 210000003296 saliva Anatomy 0.000 claims description 4
- 210000000582 semen Anatomy 0.000 claims description 4
- 210000003802 sputum Anatomy 0.000 claims description 4
- 208000024794 sputum Diseases 0.000 claims description 4
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 claims description 4
- 241000710960 Sindbis virus Species 0.000 claims description 3
- 238000012575 bio-layer interferometry Methods 0.000 claims description 3
- 210000001124 body fluid Anatomy 0.000 claims description 3
- 230000007170 pathology Effects 0.000 claims description 3
- 230000035935 pregnancy Effects 0.000 claims description 3
- 238000007790 scraping Methods 0.000 claims description 3
- 210000004381 amniotic fluid Anatomy 0.000 claims description 2
- 238000011861 anti-inflammatory therapy Methods 0.000 claims description 2
- 210000003756 cervix mucus Anatomy 0.000 claims description 2
- 210000000416 exudates and transudate Anatomy 0.000 claims description 2
- 208000000509 infertility Diseases 0.000 claims description 2
- 230000036512 infertility Effects 0.000 claims description 2
- 231100000535 infertility Toxicity 0.000 claims description 2
- 210000003097 mucus Anatomy 0.000 claims description 2
- 210000005059 placental tissue Anatomy 0.000 claims description 2
- 210000001138 tear Anatomy 0.000 claims description 2
- 230000030414 genetic transfer Effects 0.000 claims 2
- 241000699670 Mus sp. Species 0.000 description 228
- 241000700605 Viruses Species 0.000 description 134
- 108090000623 proteins and genes Proteins 0.000 description 126
- 102000004169 proteins and genes Human genes 0.000 description 102
- 235000018102 proteins Nutrition 0.000 description 97
- 210000004072 lung Anatomy 0.000 description 88
- 235000001014 amino acid Nutrition 0.000 description 82
- 230000002829 reductive effect Effects 0.000 description 80
- 229940024606 amino acid Drugs 0.000 description 78
- 239000000523 sample Substances 0.000 description 76
- 108090000765 processed proteins & peptides Proteins 0.000 description 69
- 239000000902 placebo Substances 0.000 description 65
- 229940068196 placebo Drugs 0.000 description 65
- 150000001413 amino acids Chemical class 0.000 description 54
- 102000004196 processed proteins & peptides Human genes 0.000 description 51
- 229920001184 polypeptide Polymers 0.000 description 45
- 125000003275 alpha amino acid group Chemical group 0.000 description 40
- ZRALSGWEFCBTJO-UHFFFAOYSA-N Guanidine Chemical compound NC(N)=N ZRALSGWEFCBTJO-UHFFFAOYSA-N 0.000 description 38
- 125000005647 linker group Chemical group 0.000 description 35
- 230000006870 function Effects 0.000 description 33
- 210000001519 tissue Anatomy 0.000 description 32
- 230000000926 neurological effect Effects 0.000 description 30
- 230000000694 effects Effects 0.000 description 29
- 239000002245 particle Substances 0.000 description 28
- 238000012360 testing method Methods 0.000 description 27
- 241001465754 Metazoa Species 0.000 description 26
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 26
- 210000003719 b-lymphocyte Anatomy 0.000 description 26
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 24
- 230000004048 modification Effects 0.000 description 24
- 238000012986 modification Methods 0.000 description 24
- 108060003951 Immunoglobulin Proteins 0.000 description 21
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 21
- 102000018358 immunoglobulin Human genes 0.000 description 21
- 239000003446 ligand Substances 0.000 description 21
- 239000000243 solution Substances 0.000 description 21
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 20
- 241000709661 Enterovirus Species 0.000 description 20
- 102000004190 Enzymes Human genes 0.000 description 20
- 108090000790 Enzymes Proteins 0.000 description 20
- 238000013459 approach Methods 0.000 description 20
- 201000010099 disease Diseases 0.000 description 20
- 239000003814 drug Substances 0.000 description 20
- 229940088598 enzyme Drugs 0.000 description 20
- 229960004198 guanidine Drugs 0.000 description 20
- 102100021943 C-C motif chemokine 2 Human genes 0.000 description 19
- 101710155857 C-C motif chemokine 2 Proteins 0.000 description 19
- 102100032367 C-C motif chemokine 5 Human genes 0.000 description 19
- 108010055166 Chemokine CCL5 Proteins 0.000 description 19
- 210000001744 T-lymphocyte Anatomy 0.000 description 19
- CHJJGSNFBQVOTG-UHFFFAOYSA-N N-methyl-guanidine Natural products CNC(N)=N CHJJGSNFBQVOTG-UHFFFAOYSA-N 0.000 description 18
- 206010033799 Paralysis Diseases 0.000 description 18
- 230000015572 biosynthetic process Effects 0.000 description 18
- 239000003795 chemical substances by application Substances 0.000 description 18
- 230000034994 death Effects 0.000 description 18
- SWSQBOPZIKWTGO-UHFFFAOYSA-N dimethylaminoamidine Natural products CN(C)C(N)=N SWSQBOPZIKWTGO-UHFFFAOYSA-N 0.000 description 18
- 239000012636 effector Substances 0.000 description 18
- 230000004927 fusion Effects 0.000 description 18
- 230000003472 neutralizing effect Effects 0.000 description 18
- 108010002616 Interleukin-5 Proteins 0.000 description 17
- 108090001005 Interleukin-6 Proteins 0.000 description 17
- 102000004889 Interleukin-6 Human genes 0.000 description 17
- 238000011161 development Methods 0.000 description 17
- 230000018109 developmental process Effects 0.000 description 17
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 16
- 239000004472 Lysine Substances 0.000 description 16
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 15
- 239000000562 conjugate Substances 0.000 description 15
- 239000012528 membrane Substances 0.000 description 15
- 102000005962 receptors Human genes 0.000 description 15
- 108020003175 receptors Proteins 0.000 description 15
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 14
- 239000004475 Arginine Substances 0.000 description 14
- 108700012434 CCL3 Proteins 0.000 description 14
- 102000000013 Chemokine CCL3 Human genes 0.000 description 14
- 108020004414 DNA Proteins 0.000 description 14
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 14
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 14
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 14
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 14
- 238000000338 in vitro Methods 0.000 description 14
- 210000004379 membrane Anatomy 0.000 description 14
- 108090000174 Interleukin-10 Proteins 0.000 description 13
- 108010002350 Interleukin-2 Proteins 0.000 description 13
- 108010002386 Interleukin-3 Proteins 0.000 description 13
- 108090000978 Interleukin-4 Proteins 0.000 description 13
- 102100024952 Protein CBFA2T1 Human genes 0.000 description 13
- 208000010396 acute flaccid myelitis Diseases 0.000 description 13
- 238000007792 addition Methods 0.000 description 13
- 229940049595 antibody-drug conjugate Drugs 0.000 description 13
- 239000003431 cross linking reagent Substances 0.000 description 13
- 229940079593 drug Drugs 0.000 description 13
- 238000006386 neutralization reaction Methods 0.000 description 13
- 238000000746 purification Methods 0.000 description 13
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 12
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 12
- 102000013691 Interleukin-17 Human genes 0.000 description 12
- 108050003558 Interleukin-17 Proteins 0.000 description 12
- 206010035226 Plasma cell myeloma Diseases 0.000 description 12
- 238000001727 in vivo Methods 0.000 description 12
- 201000000050 myeloid neoplasm Diseases 0.000 description 12
- 244000052769 pathogen Species 0.000 description 12
- 238000005406 washing Methods 0.000 description 12
- -1 antibodies Proteins 0.000 description 11
- 210000004602 germ cell Anatomy 0.000 description 11
- 230000001976 improved effect Effects 0.000 description 11
- 230000001404 mediated effect Effects 0.000 description 11
- 239000002609 medium Substances 0.000 description 11
- 208000023504 respiratory system disease Diseases 0.000 description 11
- 208000024891 symptom Diseases 0.000 description 11
- 230000004580 weight loss Effects 0.000 description 11
- 108010019670 Chimeric Antigen Receptors Proteins 0.000 description 10
- 102000004127 Cytokines Human genes 0.000 description 10
- 108090000695 Cytokines Proteins 0.000 description 10
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 10
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 10
- 238000010171 animal model Methods 0.000 description 10
- 229960002685 biotin Drugs 0.000 description 10
- 235000020958 biotin Nutrition 0.000 description 10
- 239000011616 biotin Substances 0.000 description 10
- 239000002254 cytotoxic agent Substances 0.000 description 10
- 230000001419 dependent effect Effects 0.000 description 10
- 238000007912 intraperitoneal administration Methods 0.000 description 10
- 239000002773 nucleotide Substances 0.000 description 10
- 125000003729 nucleotide group Chemical group 0.000 description 10
- 230000003389 potentiating effect Effects 0.000 description 10
- 230000008569 process Effects 0.000 description 10
- 238000006467 substitution reaction Methods 0.000 description 10
- 230000004083 survival effect Effects 0.000 description 10
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 9
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 9
- 208000000474 Poliomyelitis Diseases 0.000 description 9
- 206010057190 Respiratory tract infections Diseases 0.000 description 9
- 239000002253 acid Substances 0.000 description 9
- 125000000539 amino acid group Chemical group 0.000 description 9
- 150000001720 carbohydrates Chemical class 0.000 description 9
- 231100000599 cytotoxic agent Toxicity 0.000 description 9
- 230000001472 cytotoxic effect Effects 0.000 description 9
- 239000000499 gel Substances 0.000 description 9
- 235000013922 glutamic acid Nutrition 0.000 description 9
- 239000004220 glutamic acid Substances 0.000 description 9
- 230000001965 increasing effect Effects 0.000 description 9
- 238000011534 incubation Methods 0.000 description 9
- 238000011081 inoculation Methods 0.000 description 9
- 239000000463 material Substances 0.000 description 9
- 150000007523 nucleic acids Chemical group 0.000 description 9
- 230000001717 pathogenic effect Effects 0.000 description 9
- 150000003839 salts Chemical class 0.000 description 9
- 210000000278 spinal cord Anatomy 0.000 description 9
- 108010073807 IgG Receptors Proteins 0.000 description 8
- 206010028980 Neoplasm Diseases 0.000 description 8
- 229940127089 cytotoxic agent Drugs 0.000 description 8
- 230000003247 decreasing effect Effects 0.000 description 8
- 230000036039 immunity Effects 0.000 description 8
- 229940127121 immunoconjugate Drugs 0.000 description 8
- 230000007246 mechanism Effects 0.000 description 8
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 7
- YZCKVEUIGOORGS-OUBTZVSYSA-N Deuterium Chemical group [2H] YZCKVEUIGOORGS-OUBTZVSYSA-N 0.000 description 7
- 241000991587 Enterovirus C Species 0.000 description 7
- 241000282412 Homo Species 0.000 description 7
- 102100037850 Interferon gamma Human genes 0.000 description 7
- 108010074328 Interferon-gamma Proteins 0.000 description 7
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 7
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 7
- 241000699666 Mus <mouse, genus> Species 0.000 description 7
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 7
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 7
- 239000012491 analyte Substances 0.000 description 7
- 238000004458 analytical method Methods 0.000 description 7
- 230000008901 benefit Effects 0.000 description 7
- 231100000433 cytotoxic Toxicity 0.000 description 7
- 229910052805 deuterium Inorganic materials 0.000 description 7
- 238000005516 engineering process Methods 0.000 description 7
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 7
- 230000004727 humoral immunity Effects 0.000 description 7
- 229910052739 hydrogen Inorganic materials 0.000 description 7
- 230000002209 hydrophobic effect Effects 0.000 description 7
- 238000003384 imaging method Methods 0.000 description 7
- 230000003053 immunization Effects 0.000 description 7
- 238000002649 immunization Methods 0.000 description 7
- 238000003018 immunoassay Methods 0.000 description 7
- 238000007918 intramuscular administration Methods 0.000 description 7
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 7
- 239000011159 matrix material Substances 0.000 description 7
- 238000002360 preparation method Methods 0.000 description 7
- 230000002265 prevention Effects 0.000 description 7
- 239000000047 product Substances 0.000 description 7
- 201000009410 rhabdomyosarcoma Diseases 0.000 description 7
- 235000004400 serine Nutrition 0.000 description 7
- 230000008685 targeting Effects 0.000 description 7
- 238000002255 vaccination Methods 0.000 description 7
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 7
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 6
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Natural products CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 6
- PNNNRSAQSRJVSB-SLPGGIOYSA-N Fucose Natural products C[C@H](O)[C@@H](O)[C@H](O)[C@H](O)C=O PNNNRSAQSRJVSB-SLPGGIOYSA-N 0.000 description 6
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 6
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 6
- SHZGCJCMOBCMKK-DHVFOXMCSA-N L-fucopyranose Chemical compound C[C@@H]1OC(O)[C@@H](O)[C@H](O)[C@@H]1O SHZGCJCMOBCMKK-DHVFOXMCSA-N 0.000 description 6
- MUBZPKHOEPUJKR-UHFFFAOYSA-N Oxalic acid Chemical compound OC(=O)C(O)=O MUBZPKHOEPUJKR-UHFFFAOYSA-N 0.000 description 6
- 108091008874 T cell receptors Proteins 0.000 description 6
- 239000004473 Threonine Substances 0.000 description 6
- 239000002671 adjuvant Substances 0.000 description 6
- 230000003321 amplification Effects 0.000 description 6
- 230000000903 blocking effect Effects 0.000 description 6
- 230000037396 body weight Effects 0.000 description 6
- 201000011510 cancer Diseases 0.000 description 6
- 235000014633 carbohydrates Nutrition 0.000 description 6
- 238000004113 cell culture Methods 0.000 description 6
- 238000006243 chemical reaction Methods 0.000 description 6
- 239000003153 chemical reaction reagent Substances 0.000 description 6
- 230000001186 cumulative effect Effects 0.000 description 6
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 6
- 238000002296 dynamic light scattering Methods 0.000 description 6
- 238000002474 experimental method Methods 0.000 description 6
- 239000012530 fluid Substances 0.000 description 6
- 125000000291 glutamic acid group Chemical group N[C@@H](CCC(O)=O)C(=O)* 0.000 description 6
- 150000004676 glycans Chemical class 0.000 description 6
- 230000028993 immune response Effects 0.000 description 6
- 210000000987 immune system Anatomy 0.000 description 6
- 230000002163 immunogen Effects 0.000 description 6
- 230000005847 immunogenicity Effects 0.000 description 6
- 238000001990 intravenous administration Methods 0.000 description 6
- 238000003199 nucleic acid amplification method Methods 0.000 description 6
- 108020004707 nucleic acids Proteins 0.000 description 6
- 102000039446 nucleic acids Human genes 0.000 description 6
- 239000002953 phosphate buffered saline Substances 0.000 description 6
- 238000000159 protein binding assay Methods 0.000 description 6
- 230000001105 regulatory effect Effects 0.000 description 6
- 239000011780 sodium chloride Substances 0.000 description 6
- 108090001008 Avidin Proteins 0.000 description 5
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 5
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 5
- 102000009109 Fc receptors Human genes 0.000 description 5
- 108010087819 Fc receptors Proteins 0.000 description 5
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 5
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 5
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 5
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 5
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 5
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 5
- 241001529936 Murinae Species 0.000 description 5
- 241000282339 Mustela Species 0.000 description 5
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 5
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 5
- 230000009471 action Effects 0.000 description 5
- 235000003704 aspartic acid Nutrition 0.000 description 5
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 5
- 229940098773 bovine serum albumin Drugs 0.000 description 5
- 239000000872 buffer Substances 0.000 description 5
- 239000002775 capsule Substances 0.000 description 5
- 238000003776 cleavage reaction Methods 0.000 description 5
- 230000000295 complement effect Effects 0.000 description 5
- 150000001875 compounds Chemical class 0.000 description 5
- 238000006471 dimerization reaction Methods 0.000 description 5
- 239000000284 extract Substances 0.000 description 5
- 239000001257 hydrogen Substances 0.000 description 5
- 230000033001 locomotion Effects 0.000 description 5
- 210000003141 lower extremity Anatomy 0.000 description 5
- 210000003205 muscle Anatomy 0.000 description 5
- 239000002105 nanoparticle Substances 0.000 description 5
- 229920002981 polyvinylidene fluoride Polymers 0.000 description 5
- 230000001681 protective effect Effects 0.000 description 5
- 230000005180 public health Effects 0.000 description 5
- 230000002285 radioactive effect Effects 0.000 description 5
- 238000011160 research Methods 0.000 description 5
- 238000009738 saturating Methods 0.000 description 5
- 230000007017 scission Effects 0.000 description 5
- 238000000926 separation method Methods 0.000 description 5
- 239000006228 supernatant Substances 0.000 description 5
- 229940124597 therapeutic agent Drugs 0.000 description 5
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 4
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 4
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 4
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 4
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 4
- 101150031278 MP gene Proteins 0.000 description 4
- 208000012902 Nervous system disease Diseases 0.000 description 4
- 208000025966 Neurological disease Diseases 0.000 description 4
- 239000000020 Nitrocellulose Substances 0.000 description 4
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 4
- 239000002202 Polyethylene glycol Substances 0.000 description 4
- 108010076504 Protein Sorting Signals Proteins 0.000 description 4
- 241000144282 Sigmodon Species 0.000 description 4
- 108010090804 Streptavidin Chemical class 0.000 description 4
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 4
- IEDXPSOJFSVCKU-HOKPPMCLSA-N [4-[[(2S)-5-(carbamoylamino)-2-[[(2S)-2-[6-(2,5-dioxopyrrolidin-1-yl)hexanoylamino]-3-methylbutanoyl]amino]pentanoyl]amino]phenyl]methyl N-[(2S)-1-[[(2S)-1-[[(3R,4S,5S)-1-[(2S)-2-[(1R,2R)-3-[[(1S,2R)-1-hydroxy-1-phenylpropan-2-yl]amino]-1-methoxy-2-methyl-3-oxopropyl]pyrrolidin-1-yl]-3-methoxy-5-methyl-1-oxoheptan-4-yl]-methylamino]-3-methyl-1-oxobutan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]-N-methylcarbamate Chemical compound CC[C@H](C)[C@@H]([C@@H](CC(=O)N1CCC[C@H]1[C@H](OC)[C@@H](C)C(=O)N[C@H](C)[C@@H](O)c1ccccc1)OC)N(C)C(=O)[C@@H](NC(=O)[C@H](C(C)C)N(C)C(=O)OCc1ccc(NC(=O)[C@H](CCCNC(N)=O)NC(=O)[C@@H](NC(=O)CCCCCN2C(=O)CCC2=O)C(C)C)cc1)C(C)C IEDXPSOJFSVCKU-HOKPPMCLSA-N 0.000 description 4
- 238000000540 analysis of variance Methods 0.000 description 4
- 239000000611 antibody drug conjugate Substances 0.000 description 4
- 230000005888 antibody-dependent cellular phagocytosis Effects 0.000 description 4
- 239000002246 antineoplastic agent Substances 0.000 description 4
- 239000003443 antiviral agent Substances 0.000 description 4
- 235000009582 asparagine Nutrition 0.000 description 4
- 229960001230 asparagine Drugs 0.000 description 4
- 208000006673 asthma Diseases 0.000 description 4
- 239000011324 bead Substances 0.000 description 4
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 4
- 210000000170 cell membrane Anatomy 0.000 description 4
- 238000005119 centrifugation Methods 0.000 description 4
- 238000012512 characterization method Methods 0.000 description 4
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 4
- 238000012258 culturing Methods 0.000 description 4
- 238000005520 cutting process Methods 0.000 description 4
- 238000000113 differential scanning calorimetry Methods 0.000 description 4
- 238000010790 dilution Methods 0.000 description 4
- 239000012895 dilution Substances 0.000 description 4
- 238000013401 experimental design Methods 0.000 description 4
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 4
- 230000012010 growth Effects 0.000 description 4
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 4
- 229940072221 immunoglobulins Drugs 0.000 description 4
- 238000003364 immunohistochemistry Methods 0.000 description 4
- 230000003834 intracellular effect Effects 0.000 description 4
- 150000002500 ions Chemical class 0.000 description 4
- 238000005259 measurement Methods 0.000 description 4
- 108020004999 messenger RNA Proteins 0.000 description 4
- 229960000485 methotrexate Drugs 0.000 description 4
- 238000012544 monitoring process Methods 0.000 description 4
- 229920001220 nitrocellulos Polymers 0.000 description 4
- 238000001543 one-way ANOVA Methods 0.000 description 4
- 229920001223 polyethylene glycol Polymers 0.000 description 4
- 102000040430 polynucleotide Human genes 0.000 description 4
- 108091033319 polynucleotide Proteins 0.000 description 4
- 239000002157 polynucleotide Substances 0.000 description 4
- 238000001556 precipitation Methods 0.000 description 4
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 4
- 238000003908 quality control method Methods 0.000 description 4
- 230000009257 reactivity Effects 0.000 description 4
- 238000006722 reduction reaction Methods 0.000 description 4
- 238000003757 reverse transcription PCR Methods 0.000 description 4
- 238000012216 screening Methods 0.000 description 4
- 239000006152 selective media Substances 0.000 description 4
- 230000035945 sensitivity Effects 0.000 description 4
- 238000012163 sequencing technique Methods 0.000 description 4
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 4
- 125000005629 sialic acid group Chemical group 0.000 description 4
- 241000894007 species Species 0.000 description 4
- 238000003786 synthesis reaction Methods 0.000 description 4
- 125000003396 thiol group Chemical group [H]S* 0.000 description 4
- 229940125575 vaccine candidate Drugs 0.000 description 4
- 230000029812 viral genome replication Effects 0.000 description 4
- AGNGYMCLFWQVGX-AGFFZDDWSA-N (e)-1-[(2s)-2-amino-2-carboxyethoxy]-2-diazonioethenolate Chemical compound OC(=O)[C@@H](N)CO\C([O-])=C\[N+]#N AGNGYMCLFWQVGX-AGFFZDDWSA-N 0.000 description 3
- TVZGACDUOSZQKY-LBPRGKRZSA-N 4-aminofolic acid Chemical compound C1=NC2=NC(N)=NC(N)=C2N=C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 TVZGACDUOSZQKY-LBPRGKRZSA-N 0.000 description 3
- LPMXVESGRSUGHW-UHFFFAOYSA-N Acolongiflorosid K Natural products OC1C(O)C(O)C(C)OC1OC1CC2(O)CCC3C4(O)CCC(C=5COC(=O)C=5)C4(C)CC(O)C3C2(CO)C(O)C1 LPMXVESGRSUGHW-UHFFFAOYSA-N 0.000 description 3
- 229920001817 Agar Polymers 0.000 description 3
- 102000019034 Chemokines Human genes 0.000 description 3
- 108010012236 Chemokines Proteins 0.000 description 3
- 206010011224 Cough Diseases 0.000 description 3
- 101710112752 Cytotoxin Proteins 0.000 description 3
- 241000588724 Escherichia coli Species 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- 101000840258 Homo sapiens Immunoglobulin J chain Proteins 0.000 description 3
- 101001133056 Homo sapiens Mucin-1 Proteins 0.000 description 3
- 102000009490 IgG Receptors Human genes 0.000 description 3
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 3
- 102100029571 Immunoglobulin J chain Human genes 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- 241000124008 Mammalia Species 0.000 description 3
- 102000018697 Membrane Proteins Human genes 0.000 description 3
- 108010052285 Membrane Proteins Proteins 0.000 description 3
- 102100034256 Mucin-1 Human genes 0.000 description 3
- QPCDCPDFJACHGM-UHFFFAOYSA-N N,N-bis{2-[bis(carboxymethyl)amino]ethyl}glycine Chemical group OC(=O)CN(CC(O)=O)CCN(CC(=O)O)CCN(CC(O)=O)CC(O)=O QPCDCPDFJACHGM-UHFFFAOYSA-N 0.000 description 3
- LPMXVESGRSUGHW-GHYGWZAOSA-N Ouabain Natural products O([C@@H]1[C@@H](O)[C@@H](O)[C@@H](O)[C@H](C)O1)[C@H]1C[C@@H](O)[C@@]2(CO)[C@@](O)(C1)CC[C@H]1[C@]3(O)[C@@](C)([C@H](C4=CC(=O)OC4)CC3)C[C@@H](O)[C@H]21 LPMXVESGRSUGHW-GHYGWZAOSA-N 0.000 description 3
- 206010057249 Phagocytosis Diseases 0.000 description 3
- 206010035664 Pneumonia Diseases 0.000 description 3
- 239000004365 Protease Substances 0.000 description 3
- 244000166550 Strophanthus gratus Species 0.000 description 3
- ZMANZCXQSJIPKH-UHFFFAOYSA-N Triethylamine Chemical compound CCN(CC)CC ZMANZCXQSJIPKH-UHFFFAOYSA-N 0.000 description 3
- 230000035508 accumulation Effects 0.000 description 3
- 238000009825 accumulation Methods 0.000 description 3
- 150000007513 acids Chemical class 0.000 description 3
- 230000004913 activation Effects 0.000 description 3
- 238000001042 affinity chromatography Methods 0.000 description 3
- 239000008272 agar Substances 0.000 description 3
- 235000004279 alanine Nutrition 0.000 description 3
- WQZGKKKJIJFFOK-PHYPRBDBSA-N alpha-D-galactose Chemical compound OC[C@H]1O[C@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-PHYPRBDBSA-N 0.000 description 3
- 229960003896 aminopterin Drugs 0.000 description 3
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 3
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 3
- 235000011130 ammonium sulphate Nutrition 0.000 description 3
- 102000025171 antigen binding proteins Human genes 0.000 description 3
- 108091000831 antigen binding proteins Proteins 0.000 description 3
- 229950011321 azaserine Drugs 0.000 description 3
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 3
- 239000012472 biological sample Substances 0.000 description 3
- 239000000969 carrier Substances 0.000 description 3
- 239000002738 chelating agent Substances 0.000 description 3
- 238000007385 chemical modification Methods 0.000 description 3
- 238000002983 circular dichroism Methods 0.000 description 3
- 239000011248 coating agent Substances 0.000 description 3
- 238000000576 coating method Methods 0.000 description 3
- 230000009260 cross reactivity Effects 0.000 description 3
- 239000002619 cytotoxin Substances 0.000 description 3
- 230000037430 deletion Effects 0.000 description 3
- 238000012217 deletion Methods 0.000 description 3
- 238000004925 denaturation Methods 0.000 description 3
- 230000036425 denaturation Effects 0.000 description 3
- 239000000032 diagnostic agent Substances 0.000 description 3
- 229940039227 diagnostic agent Drugs 0.000 description 3
- 230000029087 digestion Effects 0.000 description 3
- 239000000539 dimer Substances 0.000 description 3
- 238000009826 distribution Methods 0.000 description 3
- 231100000673 dose–response relationship Toxicity 0.000 description 3
- 210000003608 fece Anatomy 0.000 description 3
- 125000000524 functional group Chemical group 0.000 description 3
- 229930182830 galactose Natural products 0.000 description 3
- 230000002068 genetic effect Effects 0.000 description 3
- 229930195712 glutamate Natural products 0.000 description 3
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 3
- 239000003112 inhibitor Substances 0.000 description 3
- 230000005764 inhibitory process Effects 0.000 description 3
- 239000000543 intermediate Substances 0.000 description 3
- 238000001155 isoelectric focusing Methods 0.000 description 3
- 238000002955 isolation Methods 0.000 description 3
- 231100000518 lethal Toxicity 0.000 description 3
- 230000001665 lethal effect Effects 0.000 description 3
- 239000007788 liquid Substances 0.000 description 3
- 230000007935 neutral effect Effects 0.000 description 3
- LPMXVESGRSUGHW-HBYQJFLCSA-N ouabain Chemical compound O[C@@H]1[C@H](O)[C@@H](O)[C@H](C)O[C@H]1O[C@@H]1C[C@@]2(O)CC[C@H]3[C@@]4(O)CC[C@H](C=5COC(=O)C=5)[C@@]4(C)C[C@@H](O)[C@@H]3[C@@]2(CO)[C@H](O)C1 LPMXVESGRSUGHW-HBYQJFLCSA-N 0.000 description 3
- 229960003343 ouabain Drugs 0.000 description 3
- 238000007500 overflow downdraw method Methods 0.000 description 3
- 230000003647 oxidation Effects 0.000 description 3
- 238000007254 oxidation reaction Methods 0.000 description 3
- 230000005298 paramagnetic effect Effects 0.000 description 3
- 230000037361 pathway Effects 0.000 description 3
- 229960003330 pentetic acid Drugs 0.000 description 3
- 238000002823 phage display Methods 0.000 description 3
- 230000008782 phagocytosis Effects 0.000 description 3
- 210000002381 plasma Anatomy 0.000 description 3
- 239000004033 plastic Substances 0.000 description 3
- 229920003023 plastic Polymers 0.000 description 3
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 3
- 229920000136 polysorbate Polymers 0.000 description 3
- NLKNQRATVPKPDG-UHFFFAOYSA-M potassium iodide Chemical compound [K+].[I-] NLKNQRATVPKPDG-UHFFFAOYSA-M 0.000 description 3
- 239000000843 powder Substances 0.000 description 3
- 150000003141 primary amines Chemical group 0.000 description 3
- XOJVVFBFDXDTEG-UHFFFAOYSA-N pristane Chemical compound CC(C)CCCC(C)CCCC(C)CCCC(C)C XOJVVFBFDXDTEG-UHFFFAOYSA-N 0.000 description 3
- 230000002685 pulmonary effect Effects 0.000 description 3
- 238000011084 recovery Methods 0.000 description 3
- 230000009467 reduction Effects 0.000 description 3
- 230000004044 response Effects 0.000 description 3
- 230000009450 sialylation Effects 0.000 description 3
- 230000011664 signaling Effects 0.000 description 3
- 239000007787 solid Substances 0.000 description 3
- 210000001082 somatic cell Anatomy 0.000 description 3
- 125000006850 spacer group Chemical group 0.000 description 3
- 210000000952 spleen Anatomy 0.000 description 3
- 238000007920 subcutaneous administration Methods 0.000 description 3
- 208000011580 syndromic disease Diseases 0.000 description 3
- 238000012546 transfer Methods 0.000 description 3
- 230000009466 transformation Effects 0.000 description 3
- 230000032258 transport Effects 0.000 description 3
- GKSPIZSKQWTXQG-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-[1-(pyridin-2-yldisulfanyl)ethyl]benzoate Chemical group C=1C=C(C(=O)ON2C(CCC2=O)=O)C=CC=1C(C)SSC1=CC=CC=N1 GKSPIZSKQWTXQG-UHFFFAOYSA-N 0.000 description 2
- VBICKXHEKHSIBG-UHFFFAOYSA-N 1-monostearoylglycerol Chemical compound CCCCCCCCCCCCCCCCCC(=O)OCC(O)CO VBICKXHEKHSIBG-UHFFFAOYSA-N 0.000 description 2
- VGIRNWJSIRVFRT-UHFFFAOYSA-N 2',7'-difluorofluorescein Chemical compound OC(=O)C1=CC=CC=C1C1=C2C=C(F)C(=O)C=C2OC2=CC(O)=C(F)C=C21 VGIRNWJSIRVFRT-UHFFFAOYSA-N 0.000 description 2
- GANZODCWZFAEGN-UHFFFAOYSA-N 5-mercapto-2-nitro-benzoic acid Chemical compound OC(=O)C1=CC(S)=CC=C1[N+]([O-])=O GANZODCWZFAEGN-UHFFFAOYSA-N 0.000 description 2
- KDCGOANMDULRCW-UHFFFAOYSA-N 7H-purine Chemical compound N1=CNC2=NC=NC2=C1 KDCGOANMDULRCW-UHFFFAOYSA-N 0.000 description 2
- 206010069754 Acquired gene mutation Diseases 0.000 description 2
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 2
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 2
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 2
- 241000710929 Alphavirus Species 0.000 description 2
- QGZKDVFQNNGYKY-UHFFFAOYSA-O Ammonium Chemical compound [NH4+] QGZKDVFQNNGYKY-UHFFFAOYSA-O 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- 208000026310 Breast neoplasm Diseases 0.000 description 2
- 108010029697 CD40 Ligand Proteins 0.000 description 2
- 102100032937 CD40 ligand Human genes 0.000 description 2
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 2
- 108090000565 Capsid Proteins Proteins 0.000 description 2
- 102100023321 Ceruloplasmin Human genes 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- 208000035473 Communicable disease Diseases 0.000 description 2
- 241000709687 Coxsackievirus Species 0.000 description 2
- 239000004971 Cross linker Substances 0.000 description 2
- SRBFZHDQGSBBOR-IOVATXLUSA-N D-xylopyranose Chemical compound O[C@@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-IOVATXLUSA-N 0.000 description 2
- 241000702421 Dependoparvovirus Species 0.000 description 2
- FEWJPZIEWOKRBE-JCYAYHJZSA-N Dextrotartaric acid Chemical compound OC(=O)[C@H](O)[C@@H](O)C(O)=O FEWJPZIEWOKRBE-JCYAYHJZSA-N 0.000 description 2
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 241001466953 Echovirus Species 0.000 description 2
- 241000196324 Embryophyta Species 0.000 description 2
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 2
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 2
- 229910052688 Gadolinium Inorganic materials 0.000 description 2
- 239000004366 Glucose oxidase Substances 0.000 description 2
- 108010015776 Glucose oxidase Proteins 0.000 description 2
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 2
- 102000051366 Glycosyltransferases Human genes 0.000 description 2
- 108700023372 Glycosyltransferases Proteins 0.000 description 2
- 241000238631 Hexapoda Species 0.000 description 2
- 102100026122 High affinity immunoglobulin gamma Fc receptor I Human genes 0.000 description 2
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 2
- 101000611183 Homo sapiens Tumor necrosis factor Proteins 0.000 description 2
- 241000725303 Human immunodeficiency virus Species 0.000 description 2
- MHAJPDPJQMAIIY-UHFFFAOYSA-N Hydrogen peroxide Chemical compound OO MHAJPDPJQMAIIY-UHFFFAOYSA-N 0.000 description 2
- 102000018251 Hypoxanthine Phosphoribosyltransferase Human genes 0.000 description 2
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 2
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 2
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 description 2
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 description 2
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 description 2
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 description 2
- 206010061218 Inflammation Diseases 0.000 description 2
- 108010050904 Interferons Proteins 0.000 description 2
- 102000014150 Interferons Human genes 0.000 description 2
- 108010076561 Interleukin-23 Subunit p19 Proteins 0.000 description 2
- 102100036705 Interleukin-23 subunit alpha Human genes 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 2
- 108090001090 Lectins Proteins 0.000 description 2
- 102000004856 Lectins Human genes 0.000 description 2
- 206010028372 Muscular weakness Diseases 0.000 description 2
- 241000282341 Mustela putorius furo Species 0.000 description 2
- 208000003926 Myelitis Diseases 0.000 description 2
- OVRNDRQMDRJTHS-RTRLPJTCSA-N N-acetyl-D-glucosamine Chemical compound CC(=O)N[C@H]1C(O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-RTRLPJTCSA-N 0.000 description 2
- 230000004988 N-glycosylation Effects 0.000 description 2
- 238000005481 NMR spectroscopy Methods 0.000 description 2
- 230000004989 O-glycosylation Effects 0.000 description 2
- 108091005804 Peptidases Proteins 0.000 description 2
- 241000709664 Picornaviridae Species 0.000 description 2
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 2
- 241000700159 Rattus Species 0.000 description 2
- 102100037486 Reverse transcriptase/ribonuclease H Human genes 0.000 description 2
- 208000036071 Rhinorrhea Diseases 0.000 description 2
- 206010039101 Rhinorrhoea Diseases 0.000 description 2
- 206010061494 Rhinovirus infection Diseases 0.000 description 2
- 241000283984 Rodentia Species 0.000 description 2
- 108010071390 Serum Albumin Proteins 0.000 description 2
- 102000007562 Serum Albumin Human genes 0.000 description 2
- 206010041415 Spastic paralysis Diseases 0.000 description 2
- 229920002472 Starch Polymers 0.000 description 2
- NINIDFKCEFEMDL-UHFFFAOYSA-N Sulfur Chemical compound [S] NINIDFKCEFEMDL-UHFFFAOYSA-N 0.000 description 2
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 2
- FEWJPZIEWOKRBE-UHFFFAOYSA-N Tartaric acid Natural products [H+].[H+].[O-]C(=O)C(O)C(O)C([O-])=O FEWJPZIEWOKRBE-UHFFFAOYSA-N 0.000 description 2
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- 108060008683 Tumor Necrosis Factor Receptor Proteins 0.000 description 2
- 102100040247 Tumor necrosis factor Human genes 0.000 description 2
- 206010046306 Upper respiratory tract infection Diseases 0.000 description 2
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 2
- 108010046334 Urease Proteins 0.000 description 2
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 2
- 241000710959 Venezuelan equine encephalitis virus Species 0.000 description 2
- 108010067390 Viral Proteins Proteins 0.000 description 2
- 235000011054 acetic acid Nutrition 0.000 description 2
- 230000021736 acetylation Effects 0.000 description 2
- 238000006640 acetylation reaction Methods 0.000 description 2
- 230000003213 activating effect Effects 0.000 description 2
- 239000013543 active substance Substances 0.000 description 2
- 230000001154 acute effect Effects 0.000 description 2
- 239000007801 affinity label Substances 0.000 description 2
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 2
- 238000012867 alanine scanning Methods 0.000 description 2
- 230000004075 alteration Effects 0.000 description 2
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 2
- 238000012870 ammonium sulfate precipitation Methods 0.000 description 2
- 230000001093 anti-cancer Effects 0.000 description 2
- 230000002223 anti-pathogen Effects 0.000 description 2
- 230000000259 anti-tumor effect Effects 0.000 description 2
- 238000011091 antibody purification Methods 0.000 description 2
- 230000005875 antibody response Effects 0.000 description 2
- 229940009098 aspartate Drugs 0.000 description 2
- 125000004429 atom Chemical group 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000001588 bifunctional effect Effects 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 238000001574 biopsy Methods 0.000 description 2
- 239000011575 calcium Substances 0.000 description 2
- 229910052791 calcium Inorganic materials 0.000 description 2
- 238000005251 capillar electrophoresis Methods 0.000 description 2
- 229940055240 carimune Drugs 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 230000007541 cellular toxicity Effects 0.000 description 2
- 239000003638 chemical reducing agent Substances 0.000 description 2
- 230000003399 chemotactic effect Effects 0.000 description 2
- 239000003593 chromogenic compound Substances 0.000 description 2
- 229910017052 cobalt Inorganic materials 0.000 description 2
- 239000010941 cobalt Substances 0.000 description 2
- GUTLYIVDDKVIGB-UHFFFAOYSA-N cobalt atom Chemical compound [Co] GUTLYIVDDKVIGB-UHFFFAOYSA-N 0.000 description 2
- 239000002299 complementary DNA Substances 0.000 description 2
- 230000021615 conjugation Effects 0.000 description 2
- 238000010276 construction Methods 0.000 description 2
- 239000000356 contaminant Substances 0.000 description 2
- 239000007822 coupling agent Substances 0.000 description 2
- 238000004132 cross linking Methods 0.000 description 2
- 239000013078 crystal Substances 0.000 description 2
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 2
- 235000018417 cysteine Nutrition 0.000 description 2
- 230000009089 cytolysis Effects 0.000 description 2
- 230000003013 cytotoxicity Effects 0.000 description 2
- 231100000135 cytotoxicity Toxicity 0.000 description 2
- 230000006240 deamidation Effects 0.000 description 2
- 230000002950 deficient Effects 0.000 description 2
- 230000006735 deficit Effects 0.000 description 2
- 230000008021 deposition Effects 0.000 description 2
- 238000001212 derivatisation Methods 0.000 description 2
- 238000013461 design Methods 0.000 description 2
- 235000005911 diet Nutrition 0.000 description 2
- 230000037213 diet Effects 0.000 description 2
- 150000002019 disulfides Chemical class 0.000 description 2
- 150000004662 dithiols Chemical class 0.000 description 2
- 239000002552 dosage form Substances 0.000 description 2
- 239000006196 drop Substances 0.000 description 2
- 239000003937 drug carrier Substances 0.000 description 2
- 230000004064 dysfunction Effects 0.000 description 2
- 229960001484 edetic acid Drugs 0.000 description 2
- 238000001493 electron microscopy Methods 0.000 description 2
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 210000003414 extremity Anatomy 0.000 description 2
- 229960004887 ferric hydroxide Drugs 0.000 description 2
- 238000000684 flow cytometry Methods 0.000 description 2
- 108020001507 fusion proteins Proteins 0.000 description 2
- 102000037865 fusion proteins Human genes 0.000 description 2
- UIWYJDYFSGRHKR-UHFFFAOYSA-N gadolinium atom Chemical compound [Gd] UIWYJDYFSGRHKR-UHFFFAOYSA-N 0.000 description 2
- 108010074605 gamma-Globulins Proteins 0.000 description 2
- 238000001502 gel electrophoresis Methods 0.000 description 2
- 238000007429 general method Methods 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- 229940116332 glucose oxidase Drugs 0.000 description 2
- 235000019420 glucose oxidase Nutrition 0.000 description 2
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 2
- 229910052737 gold Inorganic materials 0.000 description 2
- 239000010931 gold Substances 0.000 description 2
- 210000004884 grey matter Anatomy 0.000 description 2
- 125000004435 hydrogen atom Chemical group [H]* 0.000 description 2
- 239000012216 imaging agent Substances 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 239000012642 immune effector Substances 0.000 description 2
- 230000002998 immunogenetic effect Effects 0.000 description 2
- 229940121354 immunomodulator Drugs 0.000 description 2
- 238000012744 immunostaining Methods 0.000 description 2
- 230000006872 improvement Effects 0.000 description 2
- APFVFJFRJDLVQX-AHCXROLUSA-N indium-111 Chemical compound [111In] APFVFJFRJDLVQX-AHCXROLUSA-N 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 230000004054 inflammatory process Effects 0.000 description 2
- 238000001802 infusion Methods 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- 230000002452 interceptive effect Effects 0.000 description 2
- 229940079322 interferon Drugs 0.000 description 2
- 210000000936 intestine Anatomy 0.000 description 2
- 229910052742 iron Inorganic materials 0.000 description 2
- IEECXTSVVFWGSE-UHFFFAOYSA-M iron(3+);oxygen(2-);hydroxide Chemical compound [OH-].[O-2].[Fe+3] IEECXTSVVFWGSE-UHFFFAOYSA-M 0.000 description 2
- 229960000310 isoleucine Drugs 0.000 description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 2
- QWTDNUCVQCZILF-UHFFFAOYSA-N isopentane Chemical compound CCC(C)C QWTDNUCVQCZILF-UHFFFAOYSA-N 0.000 description 2
- JJWLVOIRVHMVIS-UHFFFAOYSA-N isopropylamine Chemical compound CC(C)N JJWLVOIRVHMVIS-UHFFFAOYSA-N 0.000 description 2
- 108010045069 keyhole-limpet hemocyanin Proteins 0.000 description 2
- 238000002372 labelling Methods 0.000 description 2
- 239000008101 lactose Substances 0.000 description 2
- 239000002523 lectin Substances 0.000 description 2
- 150000002632 lipids Chemical class 0.000 description 2
- 210000001165 lymph node Anatomy 0.000 description 2
- 108010026228 mRNA guanylyltransferase Proteins 0.000 description 2
- HQKMJHAJHXVSDF-UHFFFAOYSA-L magnesium stearate Chemical compound [Mg+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O HQKMJHAJHXVSDF-UHFFFAOYSA-L 0.000 description 2
- 230000003211 malignant effect Effects 0.000 description 2
- 238000004949 mass spectrometry Methods 0.000 description 2
- 238000005399 mechanical ventilation Methods 0.000 description 2
- 229940126601 medicinal product Drugs 0.000 description 2
- 238000002844 melting Methods 0.000 description 2
- 230000008018 melting Effects 0.000 description 2
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 2
- 230000003278 mimic effect Effects 0.000 description 2
- 230000007659 motor function Effects 0.000 description 2
- 210000002161 motor neuron Anatomy 0.000 description 2
- 210000004877 mucosa Anatomy 0.000 description 2
- 239000013642 negative control Substances 0.000 description 2
- 108010068617 neonatal Fc receptor Proteins 0.000 description 2
- 210000000440 neutrophil Anatomy 0.000 description 2
- 239000003921 oil Substances 0.000 description 2
- 235000019198 oils Nutrition 0.000 description 2
- 229920001542 oligosaccharide Polymers 0.000 description 2
- 150000002482 oligosaccharides Chemical class 0.000 description 2
- 235000006408 oxalic acid Nutrition 0.000 description 2
- 239000007800 oxidant agent Substances 0.000 description 2
- 230000007030 peptide scission Effects 0.000 description 2
- 102000013415 peroxidase activity proteins Human genes 0.000 description 2
- 108040007629 peroxidase activity proteins Proteins 0.000 description 2
- 239000008194 pharmaceutical composition Substances 0.000 description 2
- 239000000546 pharmaceutical excipient Substances 0.000 description 2
- CTRLRINCMYICJO-UHFFFAOYSA-N phenyl azide Chemical compound [N-]=[N+]=NC1=CC=CC=C1 CTRLRINCMYICJO-UHFFFAOYSA-N 0.000 description 2
- 239000003016 pheromone Substances 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 229920000642 polymer Polymers 0.000 description 2
- 239000013641 positive control Substances 0.000 description 2
- 239000011591 potassium Substances 0.000 description 2
- 229910052700 potassium Inorganic materials 0.000 description 2
- 230000003449 preventive effect Effects 0.000 description 2
- MFDFERRIHVXMIY-UHFFFAOYSA-N procaine Chemical compound CCN(CC)CCOC(=O)C1=CC=C(N)C=C1 MFDFERRIHVXMIY-UHFFFAOYSA-N 0.000 description 2
- 229960004919 procaine Drugs 0.000 description 2
- 230000000770 proinflammatory effect Effects 0.000 description 2
- 238000011321 prophylaxis Methods 0.000 description 2
- 235000019419 proteases Nutrition 0.000 description 2
- 238000001742 protein purification Methods 0.000 description 2
- 230000006337 proteolytic cleavage Effects 0.000 description 2
- 238000003259 recombinant expression Methods 0.000 description 2
- 210000001533 respiratory mucosa Anatomy 0.000 description 2
- 208000020029 respiratory tract infectious disease Diseases 0.000 description 2
- PYWVYCXTNDRMGF-UHFFFAOYSA-N rhodamine B Chemical compound [Cl-].C=12C=CC(=[N+](CC)CC)C=C2OC2=CC(N(CC)CC)=CC=C2C=1C1=CC=CC=C1C(O)=O PYWVYCXTNDRMGF-UHFFFAOYSA-N 0.000 description 2
- 229920006395 saturated elastomer Polymers 0.000 description 2
- 230000028327 secretion Effects 0.000 description 2
- 238000002864 sequence alignment Methods 0.000 description 2
- 210000002027 skeletal muscle Anatomy 0.000 description 2
- 239000011734 sodium Substances 0.000 description 2
- 229910052708 sodium Inorganic materials 0.000 description 2
- FVAUCKIRQBBSSJ-UHFFFAOYSA-M sodium iodide Inorganic materials [Na+].[I-] FVAUCKIRQBBSSJ-UHFFFAOYSA-M 0.000 description 2
- 239000002904 solvent Substances 0.000 description 2
- 230000037439 somatic mutation Effects 0.000 description 2
- 239000008107 starch Substances 0.000 description 2
- 235000019698 starch Nutrition 0.000 description 2
- 238000007619 statistical method Methods 0.000 description 2
- 238000003860 storage Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 229910052717 sulfur Inorganic materials 0.000 description 2
- 239000011593 sulfur Substances 0.000 description 2
- 239000000725 suspension Substances 0.000 description 2
- 239000003826 tablet Substances 0.000 description 2
- 239000008399 tap water Substances 0.000 description 2
- 235000020679 tap water Nutrition 0.000 description 2
- 239000011975 tartaric acid Substances 0.000 description 2
- 235000002906 tartaric acid Nutrition 0.000 description 2
- ABZLKHKQJHEPAX-UHFFFAOYSA-N tetramethylrhodamine Chemical compound C=12C=CC(N(C)C)=CC2=[O+]C2=CC(N(C)C)=CC=C2C=1C1=CC=CC=C1C([O-])=O ABZLKHKQJHEPAX-UHFFFAOYSA-N 0.000 description 2
- 230000009258 tissue cross reactivity Effects 0.000 description 2
- 230000000699 topical effect Effects 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- GETQZCLCWQTVFV-UHFFFAOYSA-N trimethylamine Chemical compound CN(C)C GETQZCLCWQTVFV-UHFFFAOYSA-N 0.000 description 2
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 2
- 241000701161 unidentified adenovirus Species 0.000 description 2
- 241001529453 unidentified herpesvirus Species 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- 229910052720 vanadium Inorganic materials 0.000 description 2
- 239000003981 vehicle Substances 0.000 description 2
- 239000013603 viral vector Substances 0.000 description 2
- JWDFQMWEFLOOED-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 3-(pyridin-2-yldisulfanyl)propanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCSSC1=CC=CC=N1 JWDFQMWEFLOOED-UHFFFAOYSA-N 0.000 description 1
- QBYIENPQHBMVBV-HFEGYEGKSA-N (2R)-2-hydroxy-2-phenylacetic acid Chemical compound O[C@@H](C(O)=O)c1ccccc1.O[C@@H](C(O)=O)c1ccccc1 QBYIENPQHBMVBV-HFEGYEGKSA-N 0.000 description 1
- FFILOTSTFMXQJC-QCFYAKGBSA-N (2r,4r,5s,6s)-2-[3-[(2s,3s,4r,6s)-6-[(2s,3r,4r,5s,6r)-5-[(2s,3r,4r,5r,6r)-3-acetamido-4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-2-[(2r,3s,4r,5r,6r)-4,5-dihydroxy-2-(hydroxymethyl)-6-[(e)-3-hydroxy-2-(octadecanoylamino)octadec-4-enoxy]oxan-3-yl]oxy-3-hy Chemical compound O[C@@H]1[C@@H](O)[C@H](OCC(NC(=O)CCCCCCCCCCCCCCCCC)C(O)\C=C\CCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@@H]([C@@H](N)[C@H](O)C2)C(O)C(O)CO[C@]2(O[C@@H]([C@@H](N)[C@H](O)C2)C(O)C(O)CO)C(O)=O)C(O)=O)[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](CO)O1 FFILOTSTFMXQJC-QCFYAKGBSA-N 0.000 description 1
- KYBXNPIASYUWLN-WUCPZUCCSA-N (2s)-5-hydroxypyrrolidine-2-carboxylic acid Chemical compound OC1CC[C@@H](C(O)=O)N1 KYBXNPIASYUWLN-WUCPZUCCSA-N 0.000 description 1
- VYEWZWBILJHHCU-OMQUDAQFSA-N (e)-n-[(2s,3r,4r,5r,6r)-2-[(2r,3r,4s,5s,6s)-3-acetamido-5-amino-4-hydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-6-[2-[(2r,3s,4r,5r)-5-(2,4-dioxopyrimidin-1-yl)-3,4-dihydroxyoxolan-2-yl]-2-hydroxyethyl]-4,5-dihydroxyoxan-3-yl]-5-methylhex-2-enamide Chemical compound N1([C@@H]2O[C@@H]([C@H]([C@H]2O)O)C(O)C[C@@H]2[C@H](O)[C@H](O)[C@H]([C@@H](O2)O[C@@H]2[C@@H]([C@@H](O)[C@H](N)[C@@H](CO)O2)NC(C)=O)NC(=O)/C=C/CC(C)C)C=CC(=O)NC1=O VYEWZWBILJHHCU-OMQUDAQFSA-N 0.000 description 1
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 1
- XGIKILRODBEJIL-UHFFFAOYSA-N 1-(ethylamino)ethanol Chemical compound CCNC(C)O XGIKILRODBEJIL-UHFFFAOYSA-N 0.000 description 1
- UVBYMVOUBXYSFV-XUTVFYLZSA-N 1-methylpseudouridine Chemical compound O=C1NC(=O)N(C)C=C1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 UVBYMVOUBXYSFV-XUTVFYLZSA-N 0.000 description 1
- PNDPGZBMCMUPRI-HVTJNCQCSA-N 10043-66-0 Chemical compound [131I][131I] PNDPGZBMCMUPRI-HVTJNCQCSA-N 0.000 description 1
- WUAPFZMCVAUBPE-NJFSPNSNSA-N 188Re Chemical compound [188Re] WUAPFZMCVAUBPE-NJFSPNSNSA-N 0.000 description 1
- MIJDSYMOBYNHOT-UHFFFAOYSA-N 2-(ethylamino)ethanol Chemical compound CCNCCO MIJDSYMOBYNHOT-UHFFFAOYSA-N 0.000 description 1
- 125000000979 2-amino-2-oxoethyl group Chemical group [H]C([*])([H])C(=O)N([H])[H] 0.000 description 1
- ZOOGRGPOEVQQDX-UUOKFMHZSA-N 3',5'-cyclic GMP Chemical compound C([C@H]1O2)OP(O)(=O)O[C@H]1[C@@H](O)[C@@H]2N1C(N=C(NC2=O)N)=C2N=C1 ZOOGRGPOEVQQDX-UUOKFMHZSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- 229940117976 5-hydroxylysine Drugs 0.000 description 1
- IDLISIVVYLGCKO-UHFFFAOYSA-N 6-carboxy-4',5'-dichloro-2',7'-dimethoxyfluorescein Chemical compound O1C(=O)C2=CC=C(C(O)=O)C=C2C21C1=CC(OC)=C(O)C(Cl)=C1OC1=C2C=C(OC)C(O)=C1Cl IDLISIVVYLGCKO-UHFFFAOYSA-N 0.000 description 1
- BZTDTCNHAFUJOG-UHFFFAOYSA-N 6-carboxyfluorescein Chemical compound C12=CC=C(O)C=C2OC2=CC(O)=CC=C2C11OC(=O)C2=CC=C(C(=O)O)C=C21 BZTDTCNHAFUJOG-UHFFFAOYSA-N 0.000 description 1
- 208000004998 Abdominal Pain Diseases 0.000 description 1
- 101150044980 Akap1 gene Proteins 0.000 description 1
- 102000009027 Albumins Human genes 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 108091093088 Amplicon Proteins 0.000 description 1
- 108010032595 Antibody Binding Sites Proteins 0.000 description 1
- 208000002109 Argyria Diseases 0.000 description 1
- 240000003291 Armoracia rusticana Species 0.000 description 1
- 235000011330 Armoracia rusticana Nutrition 0.000 description 1
- 206010003445 Ascites Diseases 0.000 description 1
- 108091008875 B cell receptors Proteins 0.000 description 1
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 108700004676 Bence Jones Proteins 0.000 description 1
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 1
- BTBUEUYNUDRHOZ-UHFFFAOYSA-N Borate Chemical compound [O-]B([O-])[O-] BTBUEUYNUDRHOZ-UHFFFAOYSA-N 0.000 description 1
- 206010006187 Breast cancer Diseases 0.000 description 1
- 229940124293 CD30 monoclonal antibody Drugs 0.000 description 1
- 101100220616 Caenorhabditis elegans chk-2 gene Proteins 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 101710197658 Capsid protein VP1 Proteins 0.000 description 1
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 1
- 239000004215 Carbon black (E152) Substances 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010078791 Carrier Proteins Proteins 0.000 description 1
- 102000005600 Cathepsins Human genes 0.000 description 1
- 108010084457 Cathepsins Proteins 0.000 description 1
- 102000000844 Cell Surface Receptors Human genes 0.000 description 1
- 108010001857 Cell Surface Receptors Proteins 0.000 description 1
- 229940123150 Chelating agent Drugs 0.000 description 1
- ZAMOUSCENKQFHK-UHFFFAOYSA-N Chlorine atom Chemical compound [Cl] ZAMOUSCENKQFHK-UHFFFAOYSA-N 0.000 description 1
- VYZAMTAEIAYCRO-UHFFFAOYSA-N Chromium Chemical compound [Cr] VYZAMTAEIAYCRO-UHFFFAOYSA-N 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- 241000195493 Cryptophyta Species 0.000 description 1
- JPVYNHNXODAKFH-UHFFFAOYSA-N Cu2+ Chemical compound [Cu+2] JPVYNHNXODAKFH-UHFFFAOYSA-N 0.000 description 1
- 102000008130 Cyclic AMP-Dependent Protein Kinases Human genes 0.000 description 1
- 108010049894 Cyclic AMP-Dependent Protein Kinases Proteins 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 208000034423 Delivery Diseases 0.000 description 1
- 206010012310 Dengue fever Diseases 0.000 description 1
- 241000725619 Dengue virus Species 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 238000009007 Diagnostic Kit Methods 0.000 description 1
- 206010012735 Diarrhoea Diseases 0.000 description 1
- 101100072149 Drosophila melanogaster eIF2alpha gene Proteins 0.000 description 1
- 208000000059 Dyspnea Diseases 0.000 description 1
- 206010013975 Dyspnoeas Diseases 0.000 description 1
- 229910052692 Dysprosium Inorganic materials 0.000 description 1
- 231100000491 EC50 Toxicity 0.000 description 1
- 241001529459 Enterovirus A71 Species 0.000 description 1
- 229910052691 Erbium Inorganic materials 0.000 description 1
- 208000010201 Exanthema Diseases 0.000 description 1
- CWYNVVGOOAEACU-UHFFFAOYSA-N Fe2+ Chemical compound [Fe+2] CWYNVVGOOAEACU-UHFFFAOYSA-N 0.000 description 1
- 241000724791 Filamentous phage Species 0.000 description 1
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 1
- 208000007212 Foot-and-Mouth Disease Diseases 0.000 description 1
- 241000710198 Foot-and-mouth disease virus Species 0.000 description 1
- 108010001515 Galectin 4 Proteins 0.000 description 1
- 102100039556 Galectin-4 Human genes 0.000 description 1
- GYHNNYVSQQEPJS-OIOBTWANSA-N Gallium-67 Chemical compound [67Ga] GYHNNYVSQQEPJS-OIOBTWANSA-N 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- 102000006395 Globulins Human genes 0.000 description 1
- 108010044091 Globulins Proteins 0.000 description 1
- 108010024636 Glutathione Proteins 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 description 1
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 description 1
- 102000001398 Granzyme Human genes 0.000 description 1
- 108060005986 Granzyme Proteins 0.000 description 1
- 208000031886 HIV Infections Diseases 0.000 description 1
- 208000037357 HIV infectious disease Diseases 0.000 description 1
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 1
- 101000935040 Homo sapiens Integrin beta-2 Proteins 0.000 description 1
- 101000917826 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-a Proteins 0.000 description 1
- 101000917824 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-b Proteins 0.000 description 1
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 description 1
- 101000582320 Homo sapiens Neurogenic differentiation factor 6 Proteins 0.000 description 1
- 101000851434 Homo sapiens Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 1
- 101000851376 Homo sapiens Tumor necrosis factor receptor superfamily member 8 Proteins 0.000 description 1
- 241000414027 Human rhinovirus 87 Species 0.000 description 1
- 241000430519 Human rhinovirus sp. Species 0.000 description 1
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- 206010061598 Immunodeficiency Diseases 0.000 description 1
- 108010079585 Immunoglobulin Subunits Proteins 0.000 description 1
- 102000012745 Immunoglobulin Subunits Human genes 0.000 description 1
- 206010062016 Immunosuppression Diseases 0.000 description 1
- 206010022004 Influenza like illness Diseases 0.000 description 1
- 102100022297 Integrin alpha-X Human genes 0.000 description 1
- 102100025390 Integrin beta-2 Human genes 0.000 description 1
- 102000015696 Interleukins Human genes 0.000 description 1
- 108010063738 Interleukins Proteins 0.000 description 1
- 206010022678 Intestinal infections Diseases 0.000 description 1
- ZCYVEMRRCGMTRW-AHCXROLUSA-N Iodine-123 Chemical compound [123I] ZCYVEMRRCGMTRW-AHCXROLUSA-N 0.000 description 1
- 238000012313 Kruskal-Wallis test Methods 0.000 description 1
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- 102100038609 Lactoperoxidase Human genes 0.000 description 1
- 108010023244 Lactoperoxidase Proteins 0.000 description 1
- 241000713666 Lentivirus Species 0.000 description 1
- 239000000232 Lipid Bilayer Substances 0.000 description 1
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 1
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 108010046938 Macrophage Colony-Stimulating Factor Proteins 0.000 description 1
- 102000007651 Macrophage Colony-Stimulating Factor Human genes 0.000 description 1
- 102000009571 Macrophage Inflammatory Proteins Human genes 0.000 description 1
- 108010009474 Macrophage Inflammatory Proteins Proteins 0.000 description 1
- WAEMQWOKJMHJLA-UHFFFAOYSA-N Manganese(2+) Chemical compound [Mn+2] WAEMQWOKJMHJLA-UHFFFAOYSA-N 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 201000005505 Measles Diseases 0.000 description 1
- 201000009906 Meningitis Diseases 0.000 description 1
- 108010006519 Molecular Chaperones Proteins 0.000 description 1
- 102000005431 Molecular Chaperones Human genes 0.000 description 1
- 241000711408 Murine respirovirus Species 0.000 description 1
- 208000010428 Muscle Weakness Diseases 0.000 description 1
- 241000186359 Mycobacterium Species 0.000 description 1
- 208000009525 Myocarditis Diseases 0.000 description 1
- OVRNDRQMDRJTHS-CBQIKETKSA-N N-Acetyl-D-Galactosamine Chemical compound CC(=O)N[C@H]1[C@@H](O)O[C@H](CO)[C@H](O)[C@@H]1O OVRNDRQMDRJTHS-CBQIKETKSA-N 0.000 description 1
- NQTADLQHYWFPDB-UHFFFAOYSA-N N-Hydroxysuccinimide Chemical compound ON1C(=O)CCC1=O NQTADLQHYWFPDB-UHFFFAOYSA-N 0.000 description 1
- GXCLVBGFBYZDAG-UHFFFAOYSA-N N-[2-(1H-indol-3-yl)ethyl]-N-methylprop-2-en-1-amine Chemical compound CN(CCC1=CNC2=C1C=CC=C2)CC=C GXCLVBGFBYZDAG-UHFFFAOYSA-N 0.000 description 1
- OVRNDRQMDRJTHS-UHFFFAOYSA-N N-acelyl-D-glucosamine Natural products CC(=O)NC1C(O)OC(CO)C(O)C1O OVRNDRQMDRJTHS-UHFFFAOYSA-N 0.000 description 1
- MBLBDJOUHNCFQT-UHFFFAOYSA-N N-acetyl-D-galactosamine Natural products CC(=O)NC(C=O)C(O)C(O)C(O)CO MBLBDJOUHNCFQT-UHFFFAOYSA-N 0.000 description 1
- SQVRNKJHWKZAKO-PFQGKNLYSA-N N-acetyl-beta-neuraminic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)O[C@H]1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-PFQGKNLYSA-N 0.000 description 1
- MBLBDJOUHNCFQT-LXGUWJNJSA-N N-acetylglucosamine Natural products CC(=O)N[C@@H](C=O)[C@@H](O)[C@H](O)[C@H](O)CO MBLBDJOUHNCFQT-LXGUWJNJSA-N 0.000 description 1
- NXTVQNIVUKXOIL-UHFFFAOYSA-N N-chlorotoluene-p-sulfonamide Chemical compound CC1=CC=C(S(=O)(=O)NCl)C=C1 NXTVQNIVUKXOIL-UHFFFAOYSA-N 0.000 description 1
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 description 1
- 206010028740 Nasal dryness Diseases 0.000 description 1
- 208000006816 Neonatal Sepsis Diseases 0.000 description 1
- 206010029260 Neuroblastoma Diseases 0.000 description 1
- 102100030589 Neurogenic differentiation factor 6 Human genes 0.000 description 1
- VEQPNABPJHWNSG-UHFFFAOYSA-N Nickel(2+) Chemical compound [Ni+2] VEQPNABPJHWNSG-UHFFFAOYSA-N 0.000 description 1
- 244000061176 Nicotiana tabacum Species 0.000 description 1
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 1
- 241001263478 Norovirus Species 0.000 description 1
- AWZJFZMWSUBJAJ-UHFFFAOYSA-N OG-514 dye Chemical compound OC(=O)CSC1=C(F)C(F)=C(C(O)=O)C(C2=C3C=C(F)C(=O)C=C3OC3=CC(O)=C(F)C=C32)=C1F AWZJFZMWSUBJAJ-UHFFFAOYSA-N 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 206010068319 Oropharyngeal pain Diseases 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 240000007594 Oryza sativa Species 0.000 description 1
- 235000007164 Oryza sativa Nutrition 0.000 description 1
- 108010058846 Ovalbumin Proteins 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 208000033952 Paralysis flaccid Diseases 0.000 description 1
- 235000019483 Peanut oil Nutrition 0.000 description 1
- 102000057297 Pepsin A Human genes 0.000 description 1
- 108090000284 Pepsin A Proteins 0.000 description 1
- 102000004503 Perforin Human genes 0.000 description 1
- 108010056995 Perforin Proteins 0.000 description 1
- KHGNFPUMBJSZSM-UHFFFAOYSA-N Perforine Natural products COC1=C2CCC(O)C(CCC(C)(C)O)(OC)C2=NC2=C1C=CO2 KHGNFPUMBJSZSM-UHFFFAOYSA-N 0.000 description 1
- 102100022587 Peroxisomal multifunctional enzyme type 2 Human genes 0.000 description 1
- 101710125609 Peroxisomal multifunctional enzyme type 2 Proteins 0.000 description 1
- 201000007100 Pharyngitis Diseases 0.000 description 1
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 1
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 1
- OAICVXFJPJFONN-UHFFFAOYSA-N Phosphorus Chemical compound [P] OAICVXFJPJFONN-UHFFFAOYSA-N 0.000 description 1
- 241000276498 Pollachius virens Species 0.000 description 1
- 239000004793 Polystyrene Substances 0.000 description 1
- XBDQKXXYIPTUBI-UHFFFAOYSA-M Propionate Chemical compound CCC([O-])=O XBDQKXXYIPTUBI-UHFFFAOYSA-M 0.000 description 1
- 102000001253 Protein Kinase Human genes 0.000 description 1
- 206010037660 Pyrexia Diseases 0.000 description 1
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 1
- 206010037714 Quadriplegia Diseases 0.000 description 1
- IWYDHOAUDWTVEP-UHFFFAOYSA-N R-2-phenyl-2-hydroxyacetic acid Natural products OC(=O)C(O)C1=CC=CC=C1 IWYDHOAUDWTVEP-UHFFFAOYSA-N 0.000 description 1
- 238000002123 RNA extraction Methods 0.000 description 1
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 1
- 101710100968 Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 208000035415 Reinfection Diseases 0.000 description 1
- 208000004756 Respiratory Insufficiency Diseases 0.000 description 1
- 241000725643 Respiratory syncytial virus Species 0.000 description 1
- 206010062106 Respiratory tract infection viral Diseases 0.000 description 1
- 241000710122 Rhinovirus B14 Species 0.000 description 1
- 108010039491 Ricin Proteins 0.000 description 1
- 238000011579 SCID mouse model Methods 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 229910052772 Samarium Inorganic materials 0.000 description 1
- 108010084592 Saporins Proteins 0.000 description 1
- BUGBHKTXTAQXES-UHFFFAOYSA-N Selenium Chemical compound [Se] BUGBHKTXTAQXES-UHFFFAOYSA-N 0.000 description 1
- 229920005654 Sephadex Polymers 0.000 description 1
- 239000012507 Sephadex™ Substances 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- 241000144290 Sigmodon hispidus Species 0.000 description 1
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 1
- BQCADISMDOOEFD-UHFFFAOYSA-N Silver Chemical compound [Ag] BQCADISMDOOEFD-UHFFFAOYSA-N 0.000 description 1
- 108010003723 Single-Domain Antibodies Proteins 0.000 description 1
- 239000005708 Sodium hypochlorite Substances 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 108700005078 Synthetic Genes Proteins 0.000 description 1
- GKLVYJBZJHMRIY-OUBTZVSYSA-N Technetium-99 Chemical compound [99Tc] GKLVYJBZJHMRIY-OUBTZVSYSA-N 0.000 description 1
- 229910052771 Terbium Inorganic materials 0.000 description 1
- 101710120037 Toxin CcdB Proteins 0.000 description 1
- GYDJEQRTZSCIOI-UHFFFAOYSA-N Tranexamic acid Chemical compound NCC1CCC(C(O)=O)CC1 GYDJEQRTZSCIOI-UHFFFAOYSA-N 0.000 description 1
- 235000021307 Triticum Nutrition 0.000 description 1
- 244000098338 Triticum aestivum Species 0.000 description 1
- 102000004243 Tubulin Human genes 0.000 description 1
- 108090000704 Tubulin Proteins 0.000 description 1
- 102100036922 Tumor necrosis factor ligand superfamily member 13B Human genes 0.000 description 1
- 101710181056 Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 1
- 102100036857 Tumor necrosis factor receptor superfamily member 8 Human genes 0.000 description 1
- YJQCOFNZVFGCAF-UHFFFAOYSA-N Tunicamycin II Natural products O1C(CC(O)C2C(C(O)C(O2)N2C(NC(=O)C=C2)=O)O)C(O)C(O)C(NC(=O)C=CCCCCCCCCC(C)C)C1OC1OC(CO)C(O)C(O)C1NC(C)=O YJQCOFNZVFGCAF-UHFFFAOYSA-N 0.000 description 1
- 206010053614 Type III immune complex mediated reaction Diseases 0.000 description 1
- 101150117115 V gene Proteins 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 229940122803 Vinca alkaloid Drugs 0.000 description 1
- 230000010530 Virus Neutralization Effects 0.000 description 1
- 238000001793 Wilcoxon signed-rank test Methods 0.000 description 1
- 229910052769 Ytterbium Inorganic materials 0.000 description 1
- VWQVUPCCIRVNHF-OUBTZVSYSA-N Yttrium-90 Chemical compound [90Y] VWQVUPCCIRVNHF-OUBTZVSYSA-N 0.000 description 1
- PNDPGZBMCMUPRI-XXSWNUTMSA-N [125I][125I] Chemical compound [125I][125I] PNDPGZBMCMUPRI-XXSWNUTMSA-N 0.000 description 1
- 238000002835 absorbance Methods 0.000 description 1
- 238000010521 absorption reaction Methods 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 210000002534 adenoid Anatomy 0.000 description 1
- 230000001464 adherent effect Effects 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 238000003314 affinity selection Methods 0.000 description 1
- 230000002776 aggregation Effects 0.000 description 1
- 238000004220 aggregation Methods 0.000 description 1
- HAXFWIACAGNFHA-UHFFFAOYSA-N aldrithiol Chemical compound C=1C=CC=NC=1SSC1=CC=CC=N1 HAXFWIACAGNFHA-UHFFFAOYSA-N 0.000 description 1
- 230000000735 allogeneic effect Effects 0.000 description 1
- 229940037003 alum Drugs 0.000 description 1
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 1
- 230000009435 amidation Effects 0.000 description 1
- 238000007112 amidation reaction Methods 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 125000002344 aminooxy group Chemical group [H]N([H])O[*] 0.000 description 1
- 239000003708 ampul Substances 0.000 description 1
- 150000008064 anhydrides Chemical class 0.000 description 1
- 150000001450 anions Chemical class 0.000 description 1
- 230000036436 anti-hiv Effects 0.000 description 1
- 230000000852 anti-lentiviral effect Effects 0.000 description 1
- 229940044684 anti-microtubule agent Drugs 0.000 description 1
- 229940125644 antibody drug Drugs 0.000 description 1
- 210000000628 antibody-producing cell Anatomy 0.000 description 1
- 230000006907 apoptotic process Effects 0.000 description 1
- 239000012736 aqueous medium Substances 0.000 description 1
- 239000008346 aqueous phase Substances 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 1
- 125000000637 arginyl group Chemical group N[C@@H](CCCNC(N)=N)C(=O)* 0.000 description 1
- 125000000613 asparagine group Chemical group N[C@@H](CC(N)=O)C(=O)* 0.000 description 1
- 238000002820 assay format Methods 0.000 description 1
- RYXHOMYVWAEKHL-OUBTZVSYSA-N astatine-211 Chemical compound [211At] RYXHOMYVWAEKHL-OUBTZVSYSA-N 0.000 description 1
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 1
- 125000000852 azido group Chemical group *N=[N+]=[N-] 0.000 description 1
- 210000003323 beak Anatomy 0.000 description 1
- 230000006399 behavior Effects 0.000 description 1
- HFACYLZERDEVSX-UHFFFAOYSA-N benzidine Chemical compound C1=CC(N)=CC=C1C1=CC=C(N)C=C1 HFACYLZERDEVSX-UHFFFAOYSA-N 0.000 description 1
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 1
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 1
- VEZXCJBBBCKRPI-UHFFFAOYSA-N beta-propiolactone Chemical compound O=C1CCO1 VEZXCJBBBCKRPI-UHFFFAOYSA-N 0.000 description 1
- 238000013357 binding ELISA Methods 0.000 description 1
- 239000011230 binding agent Substances 0.000 description 1
- 230000000975 bioactive effect Effects 0.000 description 1
- 238000005842 biochemical reaction Methods 0.000 description 1
- 239000013060 biological fluid Substances 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 230000005540 biological transmission Effects 0.000 description 1
- 238000005415 bioluminescence Methods 0.000 description 1
- 230000029918 bioluminescence Effects 0.000 description 1
- 229960000074 biopharmaceutical Drugs 0.000 description 1
- 229910052797 bismuth Inorganic materials 0.000 description 1
- JCXGWMGPZLAOME-UHFFFAOYSA-N bismuth atom Chemical compound [Bi] JCXGWMGPZLAOME-UHFFFAOYSA-N 0.000 description 1
- 239000010839 body fluid Substances 0.000 description 1
- 210000001185 bone marrow Anatomy 0.000 description 1
- 210000004556 brain Anatomy 0.000 description 1
- 229960000455 brentuximab vedotin Drugs 0.000 description 1
- 239000006227 byproduct Substances 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 229940087373 calcium oxide Drugs 0.000 description 1
- 210000000234 capsid Anatomy 0.000 description 1
- 239000004202 carbamide Substances 0.000 description 1
- 150000001718 carbodiimides Chemical class 0.000 description 1
- 229910052799 carbon Inorganic materials 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 239000005018 casein Substances 0.000 description 1
- BECPQYXYKAMYBN-UHFFFAOYSA-N casein, tech. Chemical compound NCCCCC(C(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(CC(C)C)N=C(O)C(CCC(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(C(C)O)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(COP(O)(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(N)CC1=CC=CC=C1 BECPQYXYKAMYBN-UHFFFAOYSA-N 0.000 description 1
- 235000021240 caseins Nutrition 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 230000001364 causal effect Effects 0.000 description 1
- 238000000423 cell based assay Methods 0.000 description 1
- 230000011712 cell development Effects 0.000 description 1
- 230000032823 cell division Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 239000002771 cell marker Substances 0.000 description 1
- 239000013553 cell monolayer Substances 0.000 description 1
- 230000002032 cellular defenses Effects 0.000 description 1
- 229920002678 cellulose Polymers 0.000 description 1
- 239000001913 cellulose Substances 0.000 description 1
- 210000003169 central nervous system Anatomy 0.000 description 1
- 208000015114 central nervous system disease Diseases 0.000 description 1
- 239000013522 chelant Substances 0.000 description 1
- 238000012412 chemical coupling Methods 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 229940044683 chemotherapy drug Drugs 0.000 description 1
- 239000000460 chlorine Substances 0.000 description 1
- 229910052801 chlorine Inorganic materials 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 229910052804 chromium Inorganic materials 0.000 description 1
- 239000011651 chromium Substances 0.000 description 1
- BFGKITSFLPAWGI-UHFFFAOYSA-N chromium(3+) Chemical compound [Cr+3] BFGKITSFLPAWGI-UHFFFAOYSA-N 0.000 description 1
- 238000000978 circular dichroism spectroscopy Methods 0.000 description 1
- 238000001142 circular dichroism spectrum Methods 0.000 description 1
- 238000004140 cleaning Methods 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- 230000004186 co-expression Effects 0.000 description 1
- 239000000701 coagulant Substances 0.000 description 1
- XLJKHNWPARRRJB-UHFFFAOYSA-N cobalt(2+) Chemical compound [Co+2] XLJKHNWPARRRJB-UHFFFAOYSA-N 0.000 description 1
- AGVAZMGAQJOSFJ-WZHZPDAFSA-M cobalt(2+);[(2r,3s,4r,5s)-5-(5,6-dimethylbenzimidazol-1-yl)-4-hydroxy-2-(hydroxymethyl)oxolan-3-yl] [(2r)-1-[3-[(1r,2r,3r,4z,7s,9z,12s,13s,14z,17s,18s,19r)-2,13,18-tris(2-amino-2-oxoethyl)-7,12,17-tris(3-amino-3-oxopropyl)-3,5,8,8,13,15,18,19-octamethyl-2 Chemical compound [Co+2].N#[C-].[N-]([C@@H]1[C@H](CC(N)=O)[C@@]2(C)CCC(=O)NC[C@@H](C)OP(O)(=O)O[C@H]3[C@H]([C@H](O[C@@H]3CO)N3C4=CC(C)=C(C)C=C4N=C3)O)\C2=C(C)/C([C@H](C\2(C)C)CCC(N)=O)=N/C/2=C\C([C@H]([C@@]/2(CC(N)=O)C)CCC(N)=O)=N\C\2=C(C)/C2=N[C@]1(C)[C@@](C)(CC(N)=O)[C@@H]2CCC(N)=O AGVAZMGAQJOSFJ-WZHZPDAFSA-M 0.000 description 1
- 230000001447 compensatory effect Effects 0.000 description 1
- 230000009137 competitive binding Effects 0.000 description 1
- 230000024203 complement activation Effects 0.000 description 1
- 230000004154 complement system Effects 0.000 description 1
- 230000009850 completed effect Effects 0.000 description 1
- 239000008139 complexing agent Substances 0.000 description 1
- 239000012141 concentrate Substances 0.000 description 1
- 230000001268 conjugating effect Effects 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 238000001816 cooling Methods 0.000 description 1
- NKLPQNGYXWVELD-UHFFFAOYSA-M coomassie brilliant blue Chemical compound [Na+].C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=2C=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=C1 NKLPQNGYXWVELD-UHFFFAOYSA-M 0.000 description 1
- RYGMFSIKBFXOCR-AKLPVKDBSA-N copper-67 Chemical compound [67Cu] RYGMFSIKBFXOCR-AKLPVKDBSA-N 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- 230000000875 corresponding effect Effects 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000010168 coupling process Methods 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- 210000003792 cranial nerve Anatomy 0.000 description 1
- 238000012866 crystallographic experiment Methods 0.000 description 1
- UFULAYFCSOUIOV-UHFFFAOYSA-N cysteamine Chemical compound NCCS UFULAYFCSOUIOV-UHFFFAOYSA-N 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- 108010057085 cytokine receptors Proteins 0.000 description 1
- 102000003675 cytokine receptors Human genes 0.000 description 1
- 210000000805 cytoplasm Anatomy 0.000 description 1
- 210000005220 cytoplasmic tail Anatomy 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 238000002784 cytotoxicity assay Methods 0.000 description 1
- 231100000263 cytotoxicity test Toxicity 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000007547 defect Effects 0.000 description 1
- 230000018044 dehydration Effects 0.000 description 1
- 238000006297 dehydration reaction Methods 0.000 description 1
- YSMODUONRAFBET-UHFFFAOYSA-N delta-DL-hydroxylysine Natural products NCC(O)CCC(N)C(O)=O YSMODUONRAFBET-UHFFFAOYSA-N 0.000 description 1
- 208000025729 dengue disease Diseases 0.000 description 1
- 239000000645 desinfectant Substances 0.000 description 1
- 230000010460 detection of virus Effects 0.000 description 1
- 230000001627 detrimental effect Effects 0.000 description 1
- 239000008121 dextrose Substances 0.000 description 1
- 238000003745 diagnosis Methods 0.000 description 1
- 238000002050 diffraction method Methods 0.000 description 1
- 238000009792 diffusion process Methods 0.000 description 1
- 238000007865 diluting Methods 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- AFABGHUZZDYHJO-UHFFFAOYSA-N dimethyl butane Natural products CCCC(C)C AFABGHUZZDYHJO-UHFFFAOYSA-N 0.000 description 1
- 208000035475 disorder Diseases 0.000 description 1
- 238000010494 dissociation reaction Methods 0.000 description 1
- 230000005593 dissociations Effects 0.000 description 1
- 238000003110 dot immunobinding assay Methods 0.000 description 1
- 230000003828 downregulation Effects 0.000 description 1
- 241001493065 dsRNA viruses Species 0.000 description 1
- 230000002183 duodenal effect Effects 0.000 description 1
- KBQHZAAAGSGFKK-UHFFFAOYSA-N dysprosium atom Chemical compound [Dy] KBQHZAAAGSGFKK-UHFFFAOYSA-N 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 239000000839 emulsion Substances 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 230000009088 enzymatic function Effects 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- UYAHIZSMUZPPFV-UHFFFAOYSA-N erbium Chemical compound [Er] UYAHIZSMUZPPFV-UHFFFAOYSA-N 0.000 description 1
- YSMODUONRAFBET-UHNVWZDZSA-N erythro-5-hydroxy-L-lysine Chemical compound NC[C@H](O)CC[C@H](N)C(O)=O YSMODUONRAFBET-UHNVWZDZSA-N 0.000 description 1
- XJRPTMORGOIMMI-UHFFFAOYSA-N ethyl 2-amino-4-(trifluoromethyl)-1,3-thiazole-5-carboxylate Chemical compound CCOC(=O)C=1SC(N)=NC=1C(F)(F)F XJRPTMORGOIMMI-UHFFFAOYSA-N 0.000 description 1
- 201000005884 exanthem Diseases 0.000 description 1
- 238000002270 exclusion chromatography Methods 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 210000003722 extracellular fluid Anatomy 0.000 description 1
- 238000000605 extraction Methods 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 208000028331 flaccid paralysis Diseases 0.000 description 1
- 235000013312 flour Nutrition 0.000 description 1
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 1
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 1
- 238000001917 fluorescence detection Methods 0.000 description 1
- 238000005194 fractionation Methods 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 230000005021 gait Effects 0.000 description 1
- 210000001035 gastrointestinal tract Anatomy 0.000 description 1
- 238000001641 gel filtration chromatography Methods 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 238000010362 genome editing Methods 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-L glutamate group Chemical group N[C@@H](CCC(=O)[O-])C(=O)[O-] WHUUTDBJXJRKMK-VKHMYHEASA-L 0.000 description 1
- 229960003180 glutathione Drugs 0.000 description 1
- YQEMORVAKMFKLG-UHFFFAOYSA-N glycerine monostearate Natural products CCCCCCCCCCCCCCCCCC(=O)OC(CO)CO YQEMORVAKMFKLG-UHFFFAOYSA-N 0.000 description 1
- SVUQHVRAGMNPLW-UHFFFAOYSA-N glycerol monostearate Natural products CCCCCCCCCCCCCCCCC(=O)OCC(O)CO SVUQHVRAGMNPLW-UHFFFAOYSA-N 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- CBMIPXHVOVTTTL-UHFFFAOYSA-N gold(3+) Chemical compound [Au+3] CBMIPXHVOVTTTL-UHFFFAOYSA-N 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 229910052736 halogen Inorganic materials 0.000 description 1
- 150000002367 halogens Chemical class 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- 230000008821 health effect Effects 0.000 description 1
- 210000002216 heart Anatomy 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 229940022353 herceptin Drugs 0.000 description 1
- 231100000086 high toxicity Toxicity 0.000 description 1
- 238000001239 high-resolution electron microscopy Methods 0.000 description 1
- SCKNFLZJSOHWIV-UHFFFAOYSA-N holmium(3+) Chemical compound [Ho+3] SCKNFLZJSOHWIV-UHFFFAOYSA-N 0.000 description 1
- 229920001519 homopolymer Polymers 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 208000033519 human immunodeficiency virus infectious disease Diseases 0.000 description 1
- 244000052637 human pathogen Species 0.000 description 1
- 230000028996 humoral immune response Effects 0.000 description 1
- 230000008348 humoral response Effects 0.000 description 1
- 150000007857 hydrazones Chemical group 0.000 description 1
- 229930195733 hydrocarbon Natural products 0.000 description 1
- 150000002430 hydrocarbons Chemical class 0.000 description 1
- 235000011167 hydrochloric acid Nutrition 0.000 description 1
- 150000002431 hydrogen Chemical class 0.000 description 1
- WGCNASOHLSPBMP-UHFFFAOYSA-N hydroxyacetaldehyde Natural products OCC=O WGCNASOHLSPBMP-UHFFFAOYSA-N 0.000 description 1
- 125000002349 hydroxyamino group Chemical group [H]ON([H])[*] 0.000 description 1
- 229910052588 hydroxylapatite Inorganic materials 0.000 description 1
- 238000012872 hydroxylapatite chromatography Methods 0.000 description 1
- 239000005457 ice water Substances 0.000 description 1
- 150000003949 imides Chemical class 0.000 description 1
- 238000007654 immersion Methods 0.000 description 1
- 230000016178 immune complex formation Effects 0.000 description 1
- 230000008076 immune mechanism Effects 0.000 description 1
- 210000004201 immune sera Anatomy 0.000 description 1
- 229940042743 immune sera Drugs 0.000 description 1
- 238000003119 immunoblot Methods 0.000 description 1
- 238000013115 immunohistochemical detection Methods 0.000 description 1
- 230000003308 immunostimulating effect Effects 0.000 description 1
- 230000001506 immunosuppresive effect Effects 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 239000007943 implant Substances 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 229940029583 inactivated polio vaccine Drugs 0.000 description 1
- 230000001939 inductive effect Effects 0.000 description 1
- 230000002458 infectious effect Effects 0.000 description 1
- 230000001524 infective effect Effects 0.000 description 1
- 206010022000 influenza Diseases 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 230000030214 innervation Effects 0.000 description 1
- 150000007529 inorganic bases Chemical class 0.000 description 1
- 238000007689 inspection Methods 0.000 description 1
- 102000006495 integrins Human genes 0.000 description 1
- 108010044426 integrins Proteins 0.000 description 1
- 230000029225 intracellular protein transport Effects 0.000 description 1
- 230000004068 intracellular signaling Effects 0.000 description 1
- 230000010189 intracellular transport Effects 0.000 description 1
- 239000007927 intramuscular injection Substances 0.000 description 1
- 238000010255 intramuscular injection Methods 0.000 description 1
- 238000010253 intravenous injection Methods 0.000 description 1
- 229910052740 iodine Inorganic materials 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 238000006317 isomerization reaction Methods 0.000 description 1
- 150000002540 isothiocyanates Chemical class 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 210000003292 kidney cell Anatomy 0.000 description 1
- 230000002147 killing effect Effects 0.000 description 1
- 238000011005 laboratory method Methods 0.000 description 1
- 229940057428 lactoperoxidase Drugs 0.000 description 1
- CZMAIROVPAYCMU-UHFFFAOYSA-N lanthanum(3+) Chemical compound [La+3] CZMAIROVPAYCMU-UHFFFAOYSA-N 0.000 description 1
- 150000002605 large molecules Chemical class 0.000 description 1
- 238000012125 lateral flow test Methods 0.000 description 1
- 231100000225 lethality Toxicity 0.000 description 1
- 125000001909 leucine group Chemical group [H]N(*)C(C(*)=O)C([H])([H])C(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- 208000027905 limb weakness Diseases 0.000 description 1
- 231100000861 limb weakness Toxicity 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 238000011068 loading method Methods 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 239000008176 lyophilized powder Substances 0.000 description 1
- 230000002101 lytic effect Effects 0.000 description 1
- 229920002521 macromolecule Polymers 0.000 description 1
- ZLNQQNXFFQJAID-UHFFFAOYSA-L magnesium carbonate Chemical compound [Mg+2].[O-]C([O-])=O ZLNQQNXFFQJAID-UHFFFAOYSA-L 0.000 description 1
- 239000001095 magnesium carbonate Substances 0.000 description 1
- 229910000021 magnesium carbonate Inorganic materials 0.000 description 1
- 235000019359 magnesium stearate Nutrition 0.000 description 1
- 201000004792 malaria Diseases 0.000 description 1
- 238000007726 management method Methods 0.000 description 1
- 229960002510 mandelic acid Drugs 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 230000008774 maternal effect Effects 0.000 description 1
- 230000013011 mating Effects 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- WKPWGQKGSOKKOO-RSFHAFMBSA-N maytansine Chemical class CO[C@@H]([C@@]1(O)C[C@](OC(=O)N1)([C@H]([C@@H]1O[C@@]1(C)[C@@H](OC(=O)[C@H](C)N(C)C(C)=O)CC(=O)N1C)C)[H])\C=C\C=C(C)\CC2=CC(OC)=C(Cl)C1=C2 WKPWGQKGSOKKOO-RSFHAFMBSA-N 0.000 description 1
- 238000010297 mechanical methods and process Methods 0.000 description 1
- 238000002483 medication Methods 0.000 description 1
- 229960003151 mercaptamine Drugs 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- 229910021645 metal ion Inorganic materials 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- DYQNRMCKBFOWKH-UHFFFAOYSA-N methyl 4-hydroxybenzenecarboximidate Chemical compound COC(=N)C1=CC=C(O)C=C1 DYQNRMCKBFOWKH-UHFFFAOYSA-N 0.000 description 1
- 238000000386 microscopy Methods 0.000 description 1
- 230000005012 migration Effects 0.000 description 1
- 238000013508 migration Methods 0.000 description 1
- 235000013336 milk Nutrition 0.000 description 1
- 239000008267 milk Substances 0.000 description 1
- 210000004080 milk Anatomy 0.000 description 1
- 239000002480 mineral oil Substances 0.000 description 1
- 235000010446 mineral oil Nutrition 0.000 description 1
- 150000007522 mineralic acids Chemical class 0.000 description 1
- 210000003470 mitochondria Anatomy 0.000 description 1
- 238000002156 mixing Methods 0.000 description 1
- 102000035118 modified proteins Human genes 0.000 description 1
- 108091005573 modified proteins Proteins 0.000 description 1
- 239000003607 modifier Substances 0.000 description 1
- 239000002991 molded plastic Substances 0.000 description 1
- 238000001823 molecular biology technique Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 238000003032 molecular docking Methods 0.000 description 1
- 239000000178 monomer Substances 0.000 description 1
- 108010093470 monomethyl auristatin E Proteins 0.000 description 1
- 210000005087 mononuclear cell Anatomy 0.000 description 1
- 238000010172 mouse model Methods 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- 239000007923 nasal drop Substances 0.000 description 1
- 229940100662 nasal drops Drugs 0.000 description 1
- 210000001989 nasopharynx Anatomy 0.000 description 1
- 239000006199 nebulizer Substances 0.000 description 1
- QEFYFXOXNSNQGX-UHFFFAOYSA-N neodymium atom Chemical compound [Nd] QEFYFXOXNSNQGX-UHFFFAOYSA-N 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- 201000001119 neuropathy Diseases 0.000 description 1
- 230000007823 neuropathy Effects 0.000 description 1
- 244000309711 non-enveloped viruses Species 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 150000003833 nucleoside derivatives Chemical class 0.000 description 1
- 102000026415 nucleotide binding proteins Human genes 0.000 description 1
- 108091014756 nucleotide binding proteins Proteins 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 230000005868 ontogenesis Effects 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 150000007524 organic acids Chemical class 0.000 description 1
- 235000005985 organic acids Nutrition 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 230000003204 osmotic effect Effects 0.000 description 1
- 229940092253 ovalbumin Drugs 0.000 description 1
- 229910052760 oxygen Inorganic materials 0.000 description 1
- 239000001301 oxygen Substances 0.000 description 1
- 239000006179 pH buffering agent Substances 0.000 description 1
- VYNDHICBIRRPFP-UHFFFAOYSA-N pacific blue Chemical compound FC1=C(O)C(F)=C2OC(=O)C(C(=O)O)=CC2=C1 VYNDHICBIRRPFP-UHFFFAOYSA-N 0.000 description 1
- 210000002741 palatine tonsil Anatomy 0.000 description 1
- 238000004091 panning Methods 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 239000012188 paraffin wax Substances 0.000 description 1
- 230000001769 paralizing effect Effects 0.000 description 1
- 230000003071 parasitic effect Effects 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 230000008506 pathogenesis Effects 0.000 description 1
- 230000007918 pathogenicity Effects 0.000 description 1
- 230000001575 pathological effect Effects 0.000 description 1
- 239000000312 peanut oil Substances 0.000 description 1
- 230000006320 pegylation Effects 0.000 description 1
- 230000000149 penetrating effect Effects 0.000 description 1
- XYJRXVWERLGGKC-UHFFFAOYSA-D pentacalcium;hydroxide;triphosphate Chemical compound [OH-].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O XYJRXVWERLGGKC-UHFFFAOYSA-D 0.000 description 1
- 229940111202 pepsin Drugs 0.000 description 1
- 229930192851 perforin Natural products 0.000 description 1
- KHIWWQKSHDUIBK-UHFFFAOYSA-N periodic acid Chemical compound OI(=O)(=O)=O KHIWWQKSHDUIBK-UHFFFAOYSA-N 0.000 description 1
- 208000033808 peripheral neuropathy Diseases 0.000 description 1
- 239000003208 petroleum Substances 0.000 description 1
- 229940124531 pharmaceutical excipient Drugs 0.000 description 1
- 239000012071 phase Substances 0.000 description 1
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 235000011007 phosphoric acid Nutrition 0.000 description 1
- 229910052698 phosphorus Inorganic materials 0.000 description 1
- 239000011574 phosphorus Substances 0.000 description 1
- 238000006303 photolysis reaction Methods 0.000 description 1
- 230000015843 photosynthesis, light reaction Effects 0.000 description 1
- 230000035790 physiological processes and functions Effects 0.000 description 1
- 239000006187 pill Substances 0.000 description 1
- 210000003720 plasmablast Anatomy 0.000 description 1
- 239000013612 plasmid Substances 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- KQOXLKOJHVFTRN-UHFFFAOYSA-N pleconaril Chemical compound O1N=C(C)C=C1CCCOC1=C(C)C=C(C=2N=C(ON=2)C(F)(F)F)C=C1C KQOXLKOJHVFTRN-UHFFFAOYSA-N 0.000 description 1
- 229960000471 pleconaril Drugs 0.000 description 1
- 238000003752 polymerase chain reaction Methods 0.000 description 1
- 238000006116 polymerization reaction Methods 0.000 description 1
- 229920002223 polystyrene Polymers 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- DTBMTXYWRJNBGK-UHFFFAOYSA-L potassium;sodium;phthalate Chemical compound [Na+].[K+].[O-]C(=O)C1=CC=CC=C1C([O-])=O DTBMTXYWRJNBGK-UHFFFAOYSA-L 0.000 description 1
- 238000009597 pregnancy test Methods 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 230000001902 propagating effect Effects 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 229960000380 propiolactone Drugs 0.000 description 1
- QQONPFPTGQHPMA-UHFFFAOYSA-N propylene Natural products CC=C QQONPFPTGQHPMA-UHFFFAOYSA-N 0.000 description 1
- 125000004805 propylene group Chemical group [H]C([H])([H])C([H])([*:1])C([H])([H])[*:2] 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 108060006633 protein kinase Proteins 0.000 description 1
- 230000006920 protein precipitation Effects 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 239000002213 purine nucleotide Substances 0.000 description 1
- 230000006825 purine synthesis Effects 0.000 description 1
- 150000003212 purines Chemical class 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 238000004445 quantitative analysis Methods 0.000 description 1
- 238000012207 quantitative assay Methods 0.000 description 1
- 238000002708 random mutagenesis Methods 0.000 description 1
- 206010037844 rash Diseases 0.000 description 1
- 238000003753 real-time PCR Methods 0.000 description 1
- 230000008707 rearrangement Effects 0.000 description 1
- 230000007115 recruitment Effects 0.000 description 1
- 238000004064 recycling Methods 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 238000012827 research and development Methods 0.000 description 1
- 201000004193 respiratory failure Diseases 0.000 description 1
- 210000002345 respiratory system Anatomy 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 230000001177 retroviral effect Effects 0.000 description 1
- 238000004366 reverse phase liquid chromatography Methods 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- WUAPFZMCVAUBPE-IGMARMGPSA-N rhenium-186 Chemical compound [186Re] WUAPFZMCVAUBPE-IGMARMGPSA-N 0.000 description 1
- 210000003705 ribosome Anatomy 0.000 description 1
- 235000009566 rice Nutrition 0.000 description 1
- 238000010079 rubber tapping Methods 0.000 description 1
- CVHZOJJKTDOEJC-UHFFFAOYSA-N saccharin Chemical compound C1=CC=C2C(=O)NS(=O)(=O)C2=C1 CVHZOJJKTDOEJC-UHFFFAOYSA-N 0.000 description 1
- KZUNJOHGWZRPMI-UHFFFAOYSA-N samarium atom Chemical compound [Sm] KZUNJOHGWZRPMI-UHFFFAOYSA-N 0.000 description 1
- 238000005070 sampling Methods 0.000 description 1
- 238000003118 sandwich ELISA Methods 0.000 description 1
- 229910052711 selenium Inorganic materials 0.000 description 1
- 239000011669 selenium Substances 0.000 description 1
- 230000000405 serological effect Effects 0.000 description 1
- 206010040400 serum sickness Diseases 0.000 description 1
- 239000004017 serum-free culture medium Substances 0.000 description 1
- 239000008159 sesame oil Substances 0.000 description 1
- 235000011803 sesame oil Nutrition 0.000 description 1
- 230000009131 signaling function Effects 0.000 description 1
- 239000000741 silica gel Substances 0.000 description 1
- 229910002027 silica gel Inorganic materials 0.000 description 1
- 229910052709 silver Inorganic materials 0.000 description 1
- 239000004332 silver Substances 0.000 description 1
- 235000020183 skimmed milk Nutrition 0.000 description 1
- 206010041232 sneezing Diseases 0.000 description 1
- PTLRDCMBXHILCL-UHFFFAOYSA-M sodium arsenite Chemical compound [Na+].[O-][As]=O PTLRDCMBXHILCL-UHFFFAOYSA-M 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- SUKJFIGYRHOWBL-UHFFFAOYSA-N sodium hypochlorite Chemical compound [Na+].Cl[O-] SUKJFIGYRHOWBL-UHFFFAOYSA-N 0.000 description 1
- 235000009518 sodium iodide Nutrition 0.000 description 1
- RYYKJJJTJZKILX-UHFFFAOYSA-M sodium octadecanoate Chemical compound [Na+].CCCCCCCCCCCCCCCCCC([O-])=O RYYKJJJTJZKILX-UHFFFAOYSA-M 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 238000005063 solubilization Methods 0.000 description 1
- 230000007928 solubilization Effects 0.000 description 1
- 238000000527 sonication Methods 0.000 description 1
- 238000001179 sorption measurement Methods 0.000 description 1
- 239000003549 soybean oil Substances 0.000 description 1
- 235000012424 soybean oil Nutrition 0.000 description 1
- 238000001228 spectrum Methods 0.000 description 1
- 238000009987 spinning Methods 0.000 description 1
- 238000005507 spraying Methods 0.000 description 1
- 241000114864 ssRNA viruses Species 0.000 description 1
- 239000003381 stabilizer Substances 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 239000007858 starting material Substances 0.000 description 1
- 239000008227 sterile water for injection Substances 0.000 description 1
- 150000003431 steroids Chemical class 0.000 description 1
- 125000001424 substituent group Chemical group 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 230000009469 supplementation Effects 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 239000002344 surface layer Substances 0.000 description 1
- 239000004094 surface-active agent Substances 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 239000012730 sustained-release form Substances 0.000 description 1
- 238000002636 symptomatic treatment Methods 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 239000000454 talc Substances 0.000 description 1
- 229910052623 talc Inorganic materials 0.000 description 1
- 238000002626 targeted therapy Methods 0.000 description 1
- 229910052713 technetium Inorganic materials 0.000 description 1
- GKLVYJBZJHMRIY-UHFFFAOYSA-N technetium atom Chemical compound [Tc] GKLVYJBZJHMRIY-UHFFFAOYSA-N 0.000 description 1
- GZCRRIHWUXGPOV-UHFFFAOYSA-N terbium atom Chemical compound [Tb] GZCRRIHWUXGPOV-UHFFFAOYSA-N 0.000 description 1
- 238000010998 test method Methods 0.000 description 1
- MPLHNVLQVRSVEE-UHFFFAOYSA-N texas red Chemical compound [O-]S(=O)(=O)C1=CC(S(Cl)(=O)=O)=CC=C1C(C1=CC=2CCCN3CCCC(C=23)=C1O1)=C2C1=C(CCC1)C3=[N+]1CCCC3=C2 MPLHNVLQVRSVEE-UHFFFAOYSA-N 0.000 description 1
- 238000001757 thermogravimetry curve Methods 0.000 description 1
- 150000003568 thioethers Chemical class 0.000 description 1
- CNHYKKNIIGEXAY-UHFFFAOYSA-N thiolan-2-imine Chemical compound N=C1CCCS1 CNHYKKNIIGEXAY-UHFFFAOYSA-N 0.000 description 1
- 150000003573 thiols Chemical group 0.000 description 1
- 125000000341 threoninyl group Chemical group [H]OC([H])(C([H])([H])[H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 229940104230 thymidine Drugs 0.000 description 1
- 239000003104 tissue culture media Substances 0.000 description 1
- 238000003325 tomography Methods 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- 238000002627 tracheal intubation Methods 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 230000009752 translational inhibition Effects 0.000 description 1
- 230000005945 translocation Effects 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- 229960000575 trastuzumab Drugs 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 102000003298 tumor necrosis factor receptor Human genes 0.000 description 1
- MEYZYGMYMLNUHJ-UHFFFAOYSA-N tunicamycin Natural products CC(C)CCCCCCCCCC=CC(=O)NC1C(O)C(O)C(CC(O)C2OC(C(O)C2O)N3C=CC(=O)NC3=O)OC1OC4OC(CO)C(O)C(O)C4NC(=O)C MEYZYGMYMLNUHJ-UHFFFAOYSA-N 0.000 description 1
- 238000000108 ultra-filtration Methods 0.000 description 1
- 230000009677 vaginal delivery Effects 0.000 description 1
- LEONUFNNVUYDNQ-UHFFFAOYSA-N vanadium atom Chemical compound [V] LEONUFNNVUYDNQ-UHFFFAOYSA-N 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 238000009423 ventilation Methods 0.000 description 1
- 210000002845 virion Anatomy 0.000 description 1
- 238000001429 visible spectrum Methods 0.000 description 1
- 239000002699 waste material Substances 0.000 description 1
- 238000009736 wetting Methods 0.000 description 1
- 239000000080 wetting agent Substances 0.000 description 1
- NAWDYIZEMPQZHO-UHFFFAOYSA-N ytterbium Chemical compound [Yb] NAWDYIZEMPQZHO-UHFFFAOYSA-N 0.000 description 1
Images

































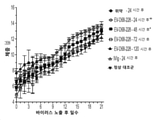
























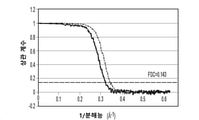




































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1009—Picornaviridae, e.g. hepatitis A virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
- A61K39/42—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum viral
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
- A61K45/06—Mixtures of active ingredients without chemical characterisation, e.g. antiphlogistics and cardiaca
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/569—Immunoassay; Biospecific binding assay; Materials therefor for microorganisms, e.g. protozoa, bacteria, viruses
- G01N33/56983—Viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/54—Medicinal preparations containing antigens or antibodies characterised by the route of administration
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/545—Medicinal preparations containing antigens or antibodies characterised by the dose, timing or administration schedule
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/55—Medicinal preparations containing antigens or antibodies characterised by the host/recipient, e.g. newborn with maternal antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/34—Identification of a linear epitope shorter than 20 amino acid residues or of a conformational epitope defined by amino acid residues
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/085—Picornaviridae, e.g. coxsackie virus, echovirus, enterovirus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2469/00—Immunoassays for the detection of microorganisms
- G01N2469/10—Detection of antigens from microorganism in sample from host
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Abstract
본 개시는 엔테로바이러스 D68(EV-D68)에 결합하는 항체 및 이의 사용 방법에 관한 것이다.
Description
우선권 주장
본 출원은, 2019년 7월 26일, 2019년 9월 12일 및 2020년 7월 2일에 각각 출원된 미국 가출원 일련번호 제62/878,955호, 제62/899,503호 및 제63/047,330호에 대한 우선권의 이익을 주장하고, 각 출원의 전체 내용은 참조에 의해 본 명세서에 도입된다.
배경
1. 개시의 분야
본 개시는 일반적으로 의학, 감염성 질환 및 면역학의 분야에 관한 것이다. 보다 구체적으로, 본 개시는 엔테로바이러스 D68에 결합하는 인간 항체 및 엔테로바이러스 D68 감염을 진단 및 치료하기 위해 이러한 항체를 사용하여 방법에 관한 것이다.
2. 배경
엔테로바이러스-D68(EV-D68)은 피코르나비리다에(Picornaviridae) 과의 엔테로바이러스 속의 포지티브-센스 단일-가닥 RNA 바이러스이다. 이 속 내의 기타 일반적 인간 병원체에는 폴리오바이러스, 에코바이러스, 콕사키바이러스, 라이노바이러스, 및 엔테로바이러스-A71 등의 기타 넘버링 엔테로바이러스가 포함된다[참조: Zell et al., 2017]. 이들 바이러스는, 구조적으로나 유전적으로는 유사하지만, 신생아 패혈증, 심근염, 소아마비, 뇌수막염, 호흡기 감염, 수족구병을 포함하는 다양한 소아 질환을 유발한다. 라이노바이러스와 마찬가지로, EV-D68은 주로 호흡기 병원체이고, 산에 불안정하고 33℃에서 가장 잘 복제된다[참조: Oberste et al., 2004]. 이러한 유사성은 EV-D68의 균주가 최초로 라이노바이러스 87로서 보고된 것과 같다[참조: Blomqvist et al., 2002].
1962년 캘리포니아의 폐렴에 걸린 어린이의 최초의 동정[참조: Schieble et al., 1967]부터 세기의 전환기에 이르기까지, EV-D68은 거의 검출되지 않았다[참조: Khetsurani et al., 2006]. 그 이래, EV-D68은 주로 소아에서 발생하는 호흡기 질환의 발생에서 세계적으로 검출됨에 따라 점점 중요성이 증가하는 병원체로서 인식되고 있다[참조: Xiang and Wang, 2016]. 이제까지 공지된 최대의 발생은 2014년 미국에서 발생하여 알래스카를 제외한 모든 주에 걸쳐 1,152건의 확인된 사례가 발생했다[참조: Oermann et al., 2015]. 이 수치는 2014년의 EV-D68의 실제 사례 수를 대폭 과소평가했을 가능성이 있고, 이는 경도의 상부 호흡기 감염이 이 바이러스의 검출에 필요한 특수한 검사를 수행하지 않을 가능성이 높기 때문이다. 천식 어린이는 특히 심각한 감염을 경험했지만[참조: Biggs et al., 2017], 한 기관의 연구에서는 천식 어린이에서 EV-D68이 비-EV-D68 엔테로바이러스/라이노바이러스 감염보다 심각하지 않은 것으로 나타났다[참조: Overdahl et al. , 2016]. 입원 환자의 절반 이상이 중환자실에 입원했고, 80%는 산소 보충을 받고, 23%는 비침습적 인공호흡을 필요로 하고, 8%는 삽관 및 기계적 인공호흡을 필요로 하고, 1%는 사망했다[참조: Midgley et al., 2015].
2014년의 발생과 동시에, 콜로라도주에서 급성 발병 이완성 마비와 뇌신경 기능 장애를 수반하는 소수의 소아 환자 모집단이 관찰되었다[참조: Messacar et al., 2015]. 이 증후군은 급성 이완성 척수염(AFM)으로 지정되었고, 척수의 화상 검사로 발견되는, 전형적으로 비대칭의 이완성 사지 쇠약 및 주로 회백질의 척수염을 수반하는 소아마비 유사 질환으로 정의되어 있다[참조: Div. of Viral Diseases, CDC, 2018]. 2014년의 발생 이후, 보다 많은 AFM 사례 및 클러스터가 EV-D68과 관련되었다. 현재까지, 임의의 환자 공급원으로부터 EV-D68 검사가 양성인 74건의 AFM 사례가 6개 대륙에서 확인되었다[참조: Messacar et al., 2018]. 질병 통제 예방 센터(Centers for Disease Control and Prevention; CDC)는 EV-D68을 AFM(Nat'l Center for Immun. Resp. Disease)의 입증된 원인으로서 공식적으로 인정하지 않지만, 다수의 전문가들은 인관관계가 있는 2개 사이의 관계를 검토하기에 충분한 설득력이 있는 증거의 우세를 발견한다[참조: Messacar et al. 2018]. 세계적 발생이 계속되기 때문에, 세계 보건 기구의 연구 개발 청사진에서는 EV-D68가 주요 공중 보건 위험으로서 수록되어 있다[참조: Ann. Rev. of Diseases, 2018].
EV-D68이 우선 병원체로서 매우 최근에 등장했기 때문에, 대부분의 초기 연구는 면역 반응을 특성화하는 것이 아니라, 바이러스의 역학을 정의하는 데 초점을 맞추었다. 따라서, EV-D68에 대한 체액성 면역의 연구는 초기 단계이다. 엔테로바이러스(Enterovirus) 속의 기타 바이러스로부터 보호하는 혈청 항체의 역할은 다양하다. 예를 들면, 비활성화된 폴리오바이러스 백신을 3회 접종하면, 혈청 중화 항체의 100% 유도에 근접하고, 마비성 소아마비의 예방에 80-96% 효과적이고; 그러나, 백신접종은 비강 또는 십이지장 IgA를 유도할 수 없기 때문에[참조: Sutter et al., 2018], 장 또는 비인두의 소아마비 바이러스 방출을 완전히 예방하지 못한다[참조: Vidor et al., 2018]. 라이노바이러스 감염의 연구에서는, 특정 혈청형 바이러스에 대한 체액성 면역이 수개월 이내에 동형 바이러스 재감염으로부터 확실히 보호하지 못하는 것을 나타낸다[참조: Howard et al., 2016]. 항체 반응과 관련된 보호의 정도의 이러한 차이는, 아마도 이러한 바이러스에 대한 병리의 상이한 부위: 폴리오바이러스의 최초 장내 감염 후의 이차 신경 확산 대 라이노바이러스의 국소 호흡기 감염 때문이다. EV-D68 감염은 호흡기 질환을 유발할 수 있고, 중추신경계 질환과 관련이 있기 때문에, 보호 및 질환 병인에서 항체의 역할은 복잡할 가능성이 있다. 이 역할을 보다 잘 이해하는 것은 EV-D68 감염에 대한 백신 및 치료법의 개발에 도움이 될 것이다.
따라서, 본 개시에 따르면, 대상체(subject)에서 엔테로바이러스 D68(EV-D68) 감염을 검출하는 방법이 제공되고, 이는 (a) 상기 대상체로부터의 샘플을 각각 표 3 및 4로부터의 클론-쌍(clone-paired) 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편과 접촉시키는 단계; 및 (b) 상기 항체 또는 항체 단편을 상기 샘플 중의 EV-D68 항원에 결합시킴으로써 상기 샘플에서 EV-D68을 검출하는 단계를 포함한다. 샘플은 혈액, 가래, 눈물, 타액, 점액 또는 혈청, 정액, 자궁경부 또는 질 분비물, 양수, 태반 조직, 소변, 삼출물, 누출물, 조직 스크래핑(tissue scraping), 호흡기 비말 또는 에어로졸, 대변 등의 체액일 수 있다. 검출은, 예를 들면, ELISA, RIA, 측방 유동 검정(lateral flow assay), 웨스턴 블롯(Western blot) 등을 포함할 수 있다. 이 방법은 단계 (a) 및 (b)를 2회 수행하고, 제1 검정과 비교하여 EV-D68 항원 수준의 변화를 결정하는 것을 추가로 포함할 수 있다.
항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성(identity)을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화(encoding)될 수 있다. 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(일본쇄(single chain) 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다.
또 다른 측면에서, 본 개시는 엔테로바이러스 D68(EV-D68)로 감염된 대상체를 치료하거나 EV-D68에 감염될 위험이 있는 대상체의 감염 가능성을 감소시키는 방법을 제공하고, 이는 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편을 상기 대상체에게 전달하는 것을 포함한다. 항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다. 항체는 임의의 아이소타입일 수 있고, 이로써 한정되지 않지만, IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)하고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된(engineered) 세포주의 발현을 변경(제거 또는 향상)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편을 포함한다. 항체는 키메라 항체(chimeric antibody) 또는 이중특이성 항체(bispecific antibody)일 수 있다.
항체 또는 항체 단편은 감염 전 또는 감염 후, 예를 들면, 감염 후 약 7일, 약 5일, 약 3일, 약 2일 또는 약 1일 이내에 투여될 수 있다. 치료는 폐 감염 및/또는 신경계 감염을 치료할 수 있거나, 치료는 폐 감염 및/또는 신경계 감염을 감소시킨다. 전달은 에어로졸 전달 등에 의한 항체 또는 항체 단편의 전신 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터를 사용한 유전 전달을 포함할 수 있다. 상기 방법은 항바이러스 요법 또는 항염증 요법 등의 제2 요법으로 상기 대상체를 치료하는 것을 추가로 포함할 수 있다.
또 다른 측면에서, 항체 또는 항체 단편이 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 특징으로 하는 모노클로날 항체(monoclonal antibody)가 제공된다. 항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다. 항체는 임의의 아이소타입일 수 있고, 이로써 한정되지 않지만, IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)하고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편을 포함한다. 항체는 키메라 항체 또는 이중특이성 항체일 수 있다. 항체 또는 항체 단편은 세포 투과 펩타이드(cell penetrating peptide)를 추가로 포함할 수 있고/있거나 인트라바디(intrabody)이다.
항체 또는 항체 단편을 암호화하는 하이브리도마 또는 조작된 세포가 제공되고, 여기서 항체 또는 항체 단편은 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 특징으로 한다. 하이브리도마 또는 조작된 세포는 표 1에 기재된 클론-쌍 가변 서열, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는 항체 또는 항체 단편을 암호화할 수 있다. 하이브리도마 또는 조작된 세포는, 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하거나, 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는 항체 또는 항체 단편을 함유할 수 있다. 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다. 항체는 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)하고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체는 키메라 항체 또는 이중특이성 항체일 수 있다. 항체 또는 항체 단편은 세포 투과 펩타이드를 추가로 포함하고/하거나 인트라바디이다.
또한, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 특징으로 하는 하나 이상의 항체 또는 항체 단편을 포함하는 백신 제형(vaccine formulation)이 제공된다. 항체 또는 항체 단편은 표 1에 제시된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다. 항체는 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)하고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체는 키메라 항체 또는 이중특이성 항체일 수 있다. 항체 또는 항체 단편은 세포 투과 펩타이드를 추가로 포함하고/하거나 인트라바디이다.
또 다른 실시양태는, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 특징으로 하는 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 포함하는 백신 제형을 포함한다. 항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다. 항체는 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)하고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체는 키메라 항체 또는 이중특이성 항체일 수 있다. 발현 벡터(들)는 신드비스(Sindbis) 바이러스 또는 VEE 벡터(들)이다. 백신 제형은 바늘 주사, 제트(jet) 주사 또는 전기천공에 의한 전달을 위해 제형화될 수 있다. 백신 제형은, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 특징으로 하는 별개의 항체 또는 항체 단편 등의 2차 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 추가로 포함할 수 있다. 항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다.
또 다른 실시양태는, 엔테로바이러스 D68로 감염되었거나 이의 감염 위험이 있는 임신 중의 대상체의 태반 및/또는 태아의 건강을 보호하는 방법을 포함하고, 상기 방법은 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편을 상기 대상체에게 전달하는 것을 포함한다. 항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다. 항체는 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)하고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체는 키메라 항체 또는 이중특이성 항체일 수 있다. 항체 또는 항체 단편은 세포 투과 펩타이드를 추가로 포함하고/하거나 인트라바디이다.
항체 또는 항체 단편은, 감염 전 또는 감염 후, 예를 들면, 감염 후 약 7일, 약 5일, 약 3일, 약 2일 또는 약 1일 이내에 투여될 수 있다. 대상체는 임신 중의 여성, 성적으로 활발한 여성, 또는 불임 치료를 받고 있는 여성일 수 있다. 전달은 항체 또는 항체 단편의 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터에 의한 유전적 전달을 포함할 수 있다. 항체 또는 항체 단편은 무처리 대조군과 비교하여 태반의 크기를 증가시킬 수 있다. 항체 또는 항체 단편은 무처리 대조군과 비교하여 태아의 바이러스 부하(viral load) 및/또는 병리를 감소시킬 수 있다.
또 다른 실시양태에서, 엔테로바이러스 D68 항원의 항원 완전성, 정확한 형태 및/또는 정확한 서열을 결정하는 방법이 제공되고, 이 방법은 (a) 상기 항원을 포함하는 샘플을, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 갖는 1차 항체 또는 항체 단편과 접촉시키는 단계; 및 (b) 상기 항원에 대한 상기 1차 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 완전성, 정확한 형태 및/또는 정확한 서열을 결정하는 단계를 포함한다. 샘플은 재조합적으로 생산된 항원을 포함할 수 있거나, 백신 제형 또는 백신 생산 배치(batch)를 포함할 수 있다. 검출은 ELISA, RIA, 웨스턴 블롯, 표면 플라즈몬 공명 또는 생물층 간섭계를 사용하는 바이오센서, 또는 유세포 분석 염색(flow cytometric staining)을 포함할 수 있다.
1차 항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. 1차 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 1차 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다. 이 방법은 단계 (a) 및 (b)를 2회 수행하여 시간 경과에 따른 항원의 항원 안정성을 결정하는 것을 추가로 포함할 수 있다.
상기 방법은 (c) 상기 항원을 포함하는 샘플을, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 갖는 2차 항체 또는 항체 단편과 접촉시키는 단계; 및 (d) 상기 항원에 대한 상기 2차 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 완전성을 결정하는 단계를 추가로 포함할 수 있다. 2차 항체 또는 항체 단편은 표 1에 기재된 클론-쌍 가변 서열에 의해, 표 1에 기재된 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. 2차 항체 또는 항체 단편은 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 2차 항체 단편은 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편일 수 있다. 상기 방법은 단계 (c) 및 (d)를 2회 수행하여 시간 경과에 따른 항원의 항원 안정성을 결정하는 것을 포함할 수 있다.
또한, 인간 모노클로날 항체 또는 항체 단편, 하이브리도마 또는 이를 생산하는 조작된 세포가 제공되고, 여기서 상기 항체는 적어도 2개의 바이러스 클레이드(viral clade)를 가로질러 EV-D68에 결합한다.
추가 실시양태는 인간 모노클로날 항체 또는 항체 단편, 하이브리도마 또는 이를 생성하는 조작된 세포를 포함하고, 여기서 상기 항체는 EV-D68 VP1, VP2 및 VP3에 결합한다. 항체는 EV-D68 VP1 DE 루프, 및/또는 EV-D68 VP2 EE 루프, 및/또는 EV-D68 VP3 N-말단 루프에 결합할 수 있다.
특허청구범위 및/또는 명세서에서 "포함하는"이라는 용어와 함께 사용되는 경우의 "a" 또는 "an"이라는 단어의 사용은 "하나"를 의미할 수 있지만, "하나 이상", "적어도 하나" 및 "하나 또는 하나 이상"의 의미와도 일치한다. "약"이라는 단어는 명시된 수치의 플러스 또는 마이너스 5%를 의미한다.
본 명세서에 기재된 임의의 방법 또는 조성물은 본 명세서에 기재된 임의의 기타 방법 또는 조성물과 관련하여 실시될 수 있음이 고려된다. 본 개시의 기타 목적, 특징 및 이점은 하기의 상세한 설명으로부터 명백해질 것이다. 그러나, 본 개시의 정신 및 범위 내의 다양한 변경 및 수정이 이러한 상세한 설명으로부터 당업자에게 명백해질 것이기 때문에, 상세한 설명 및 특정 예는, 본 개시의 특정 실시양태를 나타내면서, 예시로서만 제공되는 것으로 이해되어야 한다.
하기 도면은 본 명세서의 일부를 형성하고, 본 개시의 특정 측면을 추가로 입증하기 위해 포함된다. 본 개시는 본 명세서에 제시된 특정 실시양태의 상세한 설명과 조합하여 이들 도면 중 하나 이상을 참조함으로써 보다 잘 이해될 수 있다.
도 1. 항체 단리. 2014 EV-D68 발생으로부터 살아있는 바이러스 분리주를 ELISA 플레이트에 직접 코팅하는 간접 ELISA를 사용하여, 64개 인간 모노클로날 항체를 단리하고 스크리닝했다. 이어서, 임의의 히트는 골수종 세포와 융합하여 하이브리도마를 제조하고, 이는 이론적으로 영구적으로 계속 분열할 것이다. 항체를 정제한 후, 이들의 표현형을 정의하기 위해, 다수의 시험관내 특성화 단계가 수행된다.
도 2. 시험관내 항체 중화. 세포 배양 50% 감염 용량 또는 CCID50 검정을 사용하여 시험관내 중화를 수행한다. 일부 항체는 매우 강력하고, 일부는 여전히 매우 강력하고, 다른 항체는 약하고, mAb의 약 1/3이 일부 검출 가능한 중화 활성을 나타낸다.
도 3. 클레이드 사이의 교차 반응성 결합. 모든 mAb를 비교하기 위해, ELISA로부터 절반 최대 유효 농도 또는 EC50을 사용하여 히트 맵을 생성했다. 일부는 모든 클레이드에 매우 잘 결합되는 반면, 일부는 현저한 드롭아웃을 갖고, 통상은 멀리 떨어진 A2 클레이드를 수반한다.
도 4. 경쟁은 적어도 4개의 주요 표면 에피토프를 나타낸다. 피어슨 상관관계를 사용하여, 본 발명자는 클러스터에 대한 컬러 판독치로서 관련성 값을 생성했다. 이 관련성은 항체의 분류에 도움이 되지만, 실제 간섭 비율은 아니다. 본 발명자들은 이제 서로 관련될 수 있는 3 내지 4개의 표현형을 기술했다. 이를 통해, 임상 개발 후보를 특정할 수 있다.
도 5. 임상 후보. 중화에 추가하여, 추가 특성화를 통해, 추가 임상 후보가 밝혀졌다. mAb 159는, 믿을 수 없을 정도로 강력하고 바이러스의 모의 조제물에 결합하지 않지만, ELISA 또는 CCID50에서 교차 반응하지 않는다. mAb 105는 여전히 매우 강력하지만, 교차 반응이 매우 양호하다.
도 6A-C. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. EV-D68-228에 의한 치료는 감염 후 1일차(도 6A), 3일차(도 6B) 및 5일차(도 6C)에 폐 바이러스 역가를 유의하게 감소시켰다. 감염 후 1일, 3일 또는 5일차에 10, 3 또는 1mg/kg의 EV-D68-228로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. IVIg에 의한 치료는 감염 후 1일차에만 폐 바이러스 역가를 감소시킬 수 있었다. 구아니딘은 감염 후 1일차와 3일차에 폐 바이러스 역가를 유의하게 감소시켰지만, 5일차에는 감소시키지 않았다. 위약 치료 마우스와 비교하여 **P<0.01, ****P<0.0001.
도 7A-C. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. EV-D68-228 mAb에 의한 치료는 감염 후 1일차(도 7A), 3일차(도 7B) 및 5일차(도 7C)에 혈액 바이러스 역가를 감소시켰다. IVIg에 의한 치료는 감염 후 1, 3, 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. 구아니딘은 감염 후 1일차와 5일차에 혈액 바이러스 역가를 감소시켰지만, 3일차에는 감소시키지 않았다. 위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001.
도 8. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α 및 IL-1β의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 IL-1α 또는 IL-1β의 폐 농도를 유의하게 감소시키지 않았다. 위약 치료 마우스와 비교하여 ****P<0.0001. 그래프 내의 샘플의 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 9. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-6 및 MCP-1의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차에 IL-6 및 MCP-1의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 감염 후 3일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. 위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. 그래프 내의 샘플의 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 10. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 RANTES의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차 및 5일차에 RANTES의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 감염 후 어느 날자에도 RANTES의 폐 농도를 유의하게 감소시키지 않았다. 위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 11. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-2, IL-3 및 IL-4의 폐 농도. EV-D68 감염 후, IL-2, IL-3 또는 IL-4 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 12. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-5, IL-10 및 IL-12p70의 폐 농도. EV-D68 감염 후, IL-5, IL-10 또는 IL-12p70 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 13. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-17, IFNγ 및 TNFα의 폐 농도. EV-D68 감염 후, IL-17, IFNγ 또는 TNFα 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 14. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 MIP-1α 및 GM-CSF의 폐 농도. EV-D68 감염 후, MIP-1α 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 15. Expt. NIA-1850. EV-D68에 감염되고 EV-D68-228로 치료된 10일령 AG129 마우스의 생존. (n=6 마우스/그룹). 10, 3 또는 1mg/kg의 EV-D68-228에 의한 치료는 마우스를 사망으로부터 완전히 보호했다. 10mg/kg의 IVIg 용량은 6마리 중 4마리의 마우스를 사망으로부터 보호했다. 100mg/kg/일 용량의 구아니딘은 마우스를 사망으로부터 완전히 보호했다. 위약 치료된 마우스의 6마리 모두가 감염에 굴복했다. 위약 치료 마우스와 비교하여 *P<0.05, **P<0.01.
도 16. Expt. NIA-1850. EV-D68-228로 치료된 10일령의 EV-D68 감염 AG129 마우스의 평균 체중. 10, 3 또는 1mg/kg의 용량으로 처리하면 감염 관련 체중 감소로부터 마우스를 보호했다. 10mg/kg의 용량에서 IVIg를 사용한 치료는 또한 체중 감소로부터 마우스를 보호했다. 100mg/kg/일의 용량으로 구아니딘으로 처리된 마우스도 체중 감소로부터 보호되었다. 위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001.
도 17A-C. Expt. NIA-1850. EV-D68에 감염되고 EV-D68-228로 치료된 10일령의 AG129 마우스의 혈액 바이러스 역가. 10, 3 또는 1mg/kg의 EV-D68-228 용량으로 치료된 마우스에서는 감염 후 1일차(도 17A), 3일차(도 17B) 또는 5일차(도 17C)에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg의 IVIg에 의한 치료는 감염 후 1, 3 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. 혈액 바이러스 역가는 100mg/kg/일의 용량으로 구아니딘으로 치료함으로써 감염 후 1일, 3일 및 5일차에 유의하게 감소했다. 위약 처리 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
도 18. Expt. NIA-1850. EV-D68에 감염되고 EV-D68-228로 치료된 10일령의 AG129 마우스의 신경학적 스코어. 10, 3 또는 1mg/kg 용량으로 EV-D68-228에 의한 치료는 신경학적 스코어에 의해 측정한 임상 마비 징후를 예방했다. 10mg/kg의 IVIg에 의한 치료는 감염 후 3, 4 및 5일차에 신경학적 스코어를 감소시켰다. 100mg/kg/일의 구아니딘으로 치료된 마우스에서는 신경학적 스코어가 관찰되지 않았다. 위약 처리 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 19A-B. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. EV-D68-228에 의한 치료는 감염 후 3일차(도 19A) 및 5일차(도 19B)에 폐 바이러스 역가를 유의하게 감소시켰다. 감염 후 24시간 이내에, 10mg/kg의 EV-D68-228로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. 감염 후 48시간에 10 및 1mg/kg 용량으로 EV-D68-288에 의한 치료는 감염 후 3일차 및 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. IVIg에 의한 치료는 감염 후 24시간에 치료한 경우 폐 바이러스 역가를 감소시키지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 20A-B. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. EV-D68-228 mAb에 의한 치료는 감염 후 3일차(도 20A) 및 5일차(도 20B)에 혈액 바이러스 역가를 감소시켰다. 감염 후 48시간까지 EV-D68-228에 의한 치료는 감염 후 3일차와 5일차 모두에서 혈액 바이러스 역가를 검출 한계 미만으로 감소시켰다. IVIg에 의한 치료는 감염 후 3일차와 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, ****P<0.0001).
도 21. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α 및 IL-1β의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차 및 5일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IVIg에 의한 치료는 감염 후 3일차 및 5일차에 IL-1α 또는 IL-1β의 폐 농도를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 22. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-5 및 IL-6의 폐 농도. 감염 후 48시간까지 EV-D68-228에 의한 치료는 위약 치료된 마우스와 비교하여 감염 후 3일차에 IL-5 및 IL-6의 농도를 유의하게 감소시켰다. 감염 24시간 후 IVIg에 의한 치료는 감염 후 3일차에 IL-5 및 IL-6의 폐 농도를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 23. Expt. NIA-1849. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 MCP-1 및 RANTES의 폐 농도. 감염 후 48시간 이내에 EV-D68-228에 의한 치료는 위약 치료된 마우스와 비교하여 감염 후 3일차 및 5일차에 MCP-1 및 RANTES의 농도를 유의하게 감소시켰다. 감염 24시간 후 IVIg에 의한 치료는 감염 후 3일차에 MCP-1의 폐 농도를 유의하게 감소시켰고, 감염 후 3일차 및 5일차에 RANTES의 농도를 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 24. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-2, IL-3 및 IL-4의 폐 농도. EV-D68의 감염 후, IL-2, IL-3 또는 IL-4 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 25. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-10, IL-12p70 및 IL-17의 폐 농도. EV-D68의 감염 후, IL-5, IL-10 또는 IL-12p70 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 26. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IFNγ 및 TNFα의 폐 농도. EV-D68의 감염 후, IFNγ 또는 TNFα 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 27. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 MIP-1α 및 GM-CSF의 폐 농도. EV-D68 감염 후, MIP-1α 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 28. Expt. NIA-1870. EV-D68에 감염되고 감염 후 EV-D68-228로 치료된 10일령의 AG129 마우스의 생존. (n=6 마우스/그룹, n=7 마우스/그룹은 감염 후 120시간 치료됨). 10mg/kg의 EV-D68-228에 의한 치료는 감염 후 24시간에 마우스를 사망으로부터 완전히 보호했다. 감염 후 48시간에 EV-D68-228에 의한 치료는 6마리의 마우스 중 4마리를 사망으로부터 보호했다. EV-D68-228에 의한 감염 72시간 후에 치료한 6마리 마우스 중 1마리만이 감염에서 생존했다. 감염 120시간 후에 EV-D68-228로 치료된 그룹에서는 7마리의 마우스 중 어느 것도 생존하지 않았음에도 불구하고, 사망일의 지연으로 인해, 생존 곡선은 위약-치료된 마우스와는 상이했다. 10mg/kg의 IVIg 용량은, 감염 24시간 후에 치료된 경우, 6마리의 마우스 모두를 사망으로부터 보호했다. 위약 치료된 마우스 6마리 모두는 감염에 굴복했다(위약 치료된 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001).
도 29. Expt. NIA-1870. 감염 후 EV-D68-228로 치료된 EV-D68 감염 10일령의 AG129 마우스의 평균 체중. 감염 후 24시간 또는 48시간에 10mg/kg 용량의 EV-D68-228에 의한 치료는 마우스를 감염 관련 체중 감소로부터 보호했다. 감염 후 24시간에 10mg/kg 용량의 IVIg에 의한 치료는 마우스를 체중 감소로부터 보호하지 못했다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01).
도 30A-C. Expt. NIA-1870. EV-D68에 감염되고 감염 후 EV-D68-228로 치료된 10일령 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 30A), 3일차(도 30B) 또는 5일차(도 30C)에 제시된다. 10mg/kg 용량으로 EV-D68-228에 의한 치료는, 감염 후 24, 48 또는 72시간에 투여한 경우, 감염 후 3일차와 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. 감염 후 24시간에 투여된 10mg/kg 용량의 IVIg도 감염 후 3일차 및 5일차에 혈액 바이러스 역가를 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001).
도 31A-B. Expt. NIA-1870. EV-D68에 감염되고 감염 후 EV-D68-228로 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 2일차 및 3일차(도 31A) 및 감염 후 4일차 및 5일차(도 31B)에 제시된다. 감염 후 24시간에 10mg/kg 용량으로 EV-D68-228에 의한 치료는, 신경학적 스코어로 측정한 바와 같이, 3일, 4일 및 5일차의 임상적 마비 징후를 예방했다. 감염 24시간 후 10mg/kg의 IVIg에 의한 치료는 감염 후 3일, 4일 및 5일차에 신경학적 스코어를 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 32. EV-D68-면역 인간 대상체로부터의 mAb의 경쟁 결합 그룹. 관련성 스코어는 B1 클레이드 EV-D68 분리주를 사용하여 경쟁 결합 ELISA로부터 생성하고, mAb를 1~4로 지정된 4개의 경쟁 결합 그룹으로 클러스터링하는 데 사용했다. 적색 또는 청색으로 수록된 클론 번호는 강력하게 중화하는 mAb이고, 청색 클론 명칭은 이후 도면에서 상세히 연구된 2개의 mAb를 나타낸다.
도 33A-C. 인간 mAb의 중화 효능 및 결합 능력. (도 33A) MAb는 B1 클레이드 분리주를 사용하는 CCID50 중화 검정에서 IC50 값에 의해 경쟁-결합 그룹(비교 그룹, 그룹 5는 싱글톤의 잔류 컬렉션을 나타낸다) 내에서 순위가 매겨졌다. 본 발명자들은 또한 21개의 가장 강력하게 중화시키는 mAb에 대해 D 클레이드 및 페르몬(Fer.) 부리주의 중화를 시험했다. ">"는 최대 50μg/mL 농도에서 시험했을 때에 중화가 검출되지 않았음을 나타낸다. 공백 셀은 시험되지 않았음을 나타낸다. 적색 또는 청색으로 수록된 클론 번호는 강력하게 중화하는 mAb이고, 청색 클론 명칭은 이후 도면에서 상세히 연구된 2개의 mAb를 나타낸다. (도 33B) 생 바이러스 분리주 또는 모의 바이러스 조제물에 대한 결합 강도는 S자형 용량 반응 곡선에 적합하도록 정제된 mAb 희석액과 함께 간접 ELISA(도 33C)를 사용함으로써 생성된 EC50 값을 사용하여 표시된다. ">"는 EC50 값이 시험된 최대 농도인 100μg/mL를 초과하는 것을 나타내고, 이는 결합이 불충분하거나 없음을 시사한다.
도 34A-C. 2개 면역 복합체 사이의 구조적 특징 비교. (도 34A) EV-D68:Fab EV68-159(좌측) 또는 EV-D68:Fab EV68-228(우측)의 방사형 착색 크리오-EM 맵. 각 맵은 2개의 대칭 축을 따라 투영된다. 각 비대칭 단위의 5배, 3배 및 2배의 축은 각각 오각형 1개, 작은 삼각형 2개, 및 타원형 1개로 표지된 삼각형 윤곽을 사용하여 도시된다. (도 34B) 비대칭 단위에 대한 결합 위치 비교. 바이러스 단백질은 청색(VP1), 녹색(VP2) 및 적색(VP3)으로 착색된다. Fab 분자는 회색(EV68-159) 또는 자주색(EV68-228)으로 착색되고, 중쇄 또는 경쇄는 각각 암색 또는 명색 강도로 동일한 색상으로 표시된다. (도 34C) EV68-159 Fab(좌측) 또는 EV68-228 Fab(우측)의 풋프린트. 바이러스 표면의 방사형 착색된 2D 투영은 RIVEM을 사용하여 생성되었다. 4Å 거리 내에서 Fab 분자로부터의 임의의 원자와 대면하는 바이러스 표면 잔기는 밝은 청색(VP1), 밝은 녹색(VP2) 및 밝은 적색(VP3)으로 표시된다. 협곡 영역은 황색으로 도시된다. (도 34A) 및 (도 34C)의 스케일 바는 Å 단위로 측정된 반경 방향 거리를 나타낸다.
도 35. EV68-159 및 EV68-228의 결합 인터페이스의 확대도. 바이러스 캡시드는 표면으로 표시되고, Fab는 만화 표현으로 표시된다. VP1, VP2 및 VP3은 각각 백색, 짙은 회색 및 은색으로 착색되어 있다. 중쇄 또는 경쇄는 각각 주황색 또는 황색으로 착색된다. 상호작용을 수행하는 바이러스 잔류물은 중쇄와 경쇄를 기반으로 착색되고, 색상 강도는 VP 중 어느 것에 기초하여 변화한다. 결합 인터페이스에 관여하는 중쇄 및 경쇄 상보성 결정 영역(각각 HCDR 및 LCDR)은 화살표로 제시된다.
도 36A-D. 비리온-Fab 상호작용의 분자 상세. EV-D68:Fab EV68-159(도 36A-C) 및 EV-D68:Fab EV68-228(도 36D)의 결합 인터페이스에서의 대표적 상호작용. 수소 결합은 청록색으로 착색되고, 염 브릿지는 마젠타로 착색된다.
도 37A-D. MAb EV68-228은, 예방 또는 요법으로 사용되는 경우, EV-D68 유도된 호흡기 질환으로부터 마우스를 보호한다. 4주령의 AG129 계통 마우스(시점당 n = 4)에, 마우스-적응 B1 클레이드 EV-D68을 비강으로 접종하고, 항체를 복강내 투여하고; 표시된 조직에 대한 바이러스 역가를 CCID50 검정에 의해 측정했다. (도 37A-B) 지시된 항체 용량의 24시간 후에 마우스에게 바이러스를 접종하고, 이어서 지시된 시점에서 바이러스 역가를 측정했다. (도 37C-D) 마우스에게 바이러스를 접종하고, 4, 24 또는 48시간 후에 10mg/kg(지시된 경우 제외)의 항체를 접종하고, 이어서 바이러스 역가를 측정했다.
도 38A-F. MAb EV68-228은 EV-D68 감염 마우스에서 폐 염증을 감소시킨다. 4주령 AG129 마우스(시점당 n=4)(도 38A-C)은 지시된 항체 용량의 24시간 후에 비강내로 바이러스를 접종하거나, (도 38D-F)은 바이러스를 비강내로 접종하고, 이어서 4, 24 또는 48시간 후에 10mg/kg(지시된 경우 제외) 항체를 접종하고, 지시된 시점에서 사이토카인을 측정했다. ELISA를 사용하여 폐 균질액으로부터 사이토카인을 정량화했다. 둔네트(Dunnett)의 T3 다중 비교 검정을 사용한 일원 분산 분석을 사용하여, 각 시점에서 치료 그룹의 값을 위약 그룹과 비교했다(*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). IL-인터루킨; MCP-단핵 세포 주화성 단백질. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 39A-F. MAb EV68-228은, 예방 또는 요법으로 사용되는 경우, EV-D68 유도 신경계 질환으로부터 마우스를 보호한다. 10일령의 마우스에게 마우스-적응 B1 클레이드 EV-D68을 복강내 접종하고, 항체는 복강내 투여하고; 지시된 조직에 대한 바이러스 역가는 CCID50 검정에 의해 측정했다. (도 39A-C) 지시된 용량의 항체의 24시간 후에 마우스에게 바이러스를 접종하고, 이어서 (도 39A) 바이러스 역가를 측정하고(시점당 n = 3), (도 39B) 생존을 모니터링하고, (도 39C) 신경학적 스코어(시점당 n = 6)를 지시된 시점에서 기록했다. 스코어가 높으면, 운동 장애가 심각한 것을 나타낸다. (도 39D-F) 마우스에게 바이러스를 접종하고, 이어서 10mg/kg 용량의 항체를 접종하고, 이어서 (도 39D) 바이러스 역가를 측정하고(시점당 n = 3), (도 39E) 생존을 모니터링하고, (도 39F) 신경학적 스코어(시점당 n = 6, 120시간 후의 n = 9 제외)를 기록했다. 착색 수직 화살표는 치료 시간을 나타낸다.
도 40. 웨스턴 블롯 데이터. 모든 mAb는 웨스턴 블롯에 의해 B1 클레이드 바이러스를 염색하는 능력에 대해 시험되었다. 모든 긍정적 결과가 제시된다.
도 41. 인간 mAb의 상세한 특성. MAb는 B1 클레이드 분리주를 사용한 CCID50 중화 검정에서 IC50 값에 의해 경쟁 결합 그룹 내에서 순위가 매겨졌다. 본 발명자들은 또한 21개의 가장 강력하게 중화하는 mAb에 대해 D 클레이드 및 페르몬(Fer.) 분리주의 중화를 시험했다. ">"는 최대 50μg/mL 농도에서 시험할 때에 중화가 검출되지 않았음을 나타낸다. 공백 셀은 시험되지 않았음을 나타낸다. 적색 또는 청색 폰트로 수록된 클론 번호는 강력하게 중화하는 mAb이고, 청색 클론 명칭은 후속 도면에서 상세히 연구된 2개의 mAb를 나타낸다. 각 mAb에 대한 최종 IC50 값은 중복으로 수행된 3개의 개별 실험으로부터의 EC50 값의 로그 평균이다. 살아있는 바이러스 분리주에 대한 결합 강도는 S자형 용량 반응 곡선에 적합하도록 정제된 mAb 희석액과 함께 간접 ELISA를 사용하여 생성된 EC50 값을 기반으로 표시된다. ">"는 예측된 EC50 값이 시험된 최대 농도인 100μg/mL를 초과하는 것을 나타내고, 이는 결합이 불충분하거나 없음을 시사한다. 각 mAb에 대한 최종 EC50 값은 중복으로 수행된 3개의 개별 실험으로부터의 EC50 값의 로그 평균이다. IgG 서브타입은 표현형으로 결정되었다. 경쇄 유형은 카파(K) 또는 람다(L)이고, 유전적으로 결정되었다. WB는 웨스턴 블롯 반응성을 나타내고, 모든 mAb는 시험되었고, 공백 행은 반응성이 없음을 나타낸다.
도 42. 모든 mAb에 대한 간접 ELISA 데이터. 살아있는 바이러스 분리주에 대한 결합 강도는 S자형 용량 반응 곡선에 적합하도록 정제된 mAb 희석액과 함께 간접 ELISA를 사용하여 생성된 EC50 값으로 표시된다. 제시된 것은 3개의 실험 중 하나의 대표적 데이터이고, 에러 바는 중복 기술적 복제의 표준 편차를 나타낸다. 각 mAb에 대한 최종 EC50 값은 3개의 실험 모두로부터의 EC50 값의 로그 평균이다. 바이러스의 모의 조제물은 바이러스를 접종하지 않았지만 바이러스 감염 세포와 동일한 방식으로 제조된 RD 세포 단층으로부터의 것이다.
도 43. EV-D68로부터의 대표적 밀도: Fab EV68-228 전자 밀도 맵. 바이러스 단백질은 청색(VP1), 녹색(VP2), 적색(VP3) 및 마젠타색(VP4)으로 착색된다.
도 44. 면역 복합체 맵 해상도의 추정치. 맵 해상도는 금 표준 푸리에 쉘 상관관계(FSC) 컷오프 0.143을 기반으로 추정된다. EV68-159(적색 곡선) 또는 EV68-228(청색 곡선) 복합체의 최종적 해상도는 각각 2.9Å 또는 3.1Å이다.
도 45. Fab 결합 부위의 비교. EV68-159와 EV68-228은 각각 금색과 청색으로 착색되었고, 바이러스 표면은 청록색으로 착색되었다. 좌측 패널은 가변 도메인과 불변 도메인을 모두 표시하는 반면, 우측 패널은 가변 도메인만을 표시하고, 이는 2개 Fab의 풋프린트가 중첩하지 않음을 입증한다.
도 46. Fab 풋프린트의 확대도를 나타내는 로드맵. 바이러스 표면의 방사형 착색된 2D 투영은 RIVEM으로 생성되었다. 4Å 거리 내에서 Fab 분자로부터의 임의의 원자와 대면하는 바이러스 표면 잔기는 연한 청색(VP1), 연한 녹색(VP2) 및 연한 적색(VP3)으로 도시된다. 협곡 영역은 황색으로 도시된다.
도 47. EV68-228 Fab 중쇄의 벌키한 측쇄. 이 도면은 EV68-228 Fab를 안정화하기 위해 소수성 상호작용 네트워크를 형성하는 벌키한 측쇄의 예를 나타내고, 이는 EV68-159 Fab의 구조에서도 관찰된다.
도 48A-C. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. 폐 바이러스 역가는 감염 후 1일차(도 48A), 3일차(도 48B) 및 5일차(도 48C)에 제시된다. 감염 후 1일, 3일 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ****P<0.0001)
도 49A-C. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 49A), 3일차(도 49B) 및 5일차(도 49C)에 제시된다. 감염 후 1일, 3일 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 1일, 3일 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001)
도 50. 내선 NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 1일, 3일 및 5일차에 IL-1α의 농도를 유의하게 감소시켰다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료도 감염 후 3일차에 IL-1β의 농도를 감소시켰다. hIVIg에 의한 치료는 감염 후 3일차에 IL-1β의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-2 또는 IL-3의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 51. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-4, IL-5, IL-6 및 IL-10의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-5의 농도를, 감염 후 3일 및 5일차에 IL-6의 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차에 IL-5 및 IL-6의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-4 또는 IL-10의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 52. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFN-γ의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일 및 5일차에 MCP-1 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-12p70, IL-17 또는 IFN-γ의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 53. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 1일 및 3일차에 MIP-1α의 폐 농도를 유의하게 감소시켰고, 감염 후 3일 및 5일차에 RANTES의 농도를 감소시켰다. 10mg/kg 용량의 hIVIG는 감염 후 1일과 3일차에 MIP-1α의 농도를 유의하게 감소시켰고, 감염 후 3일차에 RANTES의 농도도 감소시켰다. EV-D68 감염 후 TNFα 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 54A-B. Expt. NIA-1930. 감염 24시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. 폐 바이러스 역가는 감염 후 3일차(도 54A) 및 5일차(도 54B)에 제시된다. 감염 후 3일차 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차에 폐 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001)
도 55A-B. Expt. NIA-1930. 감염 24시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 3일차(도 55A) 및 5일차(도 55B)에 제시된다. 감염 후 3일차 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001).
도 56. Expt. NIA-1930. 감염 24시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IL-3의 농도는 감염 후 3일차에 EV68-228-TP로 치료된 마우스 및 hIVIg로 치료된 마우스에서 유의하게 감소했다. 감염 후 IL-2 농도에서 유의한 변화는 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 57. Expt. NIA-1930. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-4, IL-5, IL-6 및 IL-10의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-5의 농도를, 감염 후 3일차 및 5일차에 IL-6의 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차에 IL-5의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-4 또는 IL-10의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 58. Expt. NIA-1930. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFN-γ의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차 및 5일차에 MCP-1 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-12p70, IL-17 또는 IFN-γ의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 59. Expt. NIA-1930. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 MIP-1α의 폐 농도를 유의하게 감소시켰고, 감염 후 3일차 및 5일차에 RANTES의 농도를 감소시켰다. EV-D68의 감염 후, TNFα 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 60A-B. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. 폐 바이러스 역가는 감염 후 3일차(도 60A) 및 5일차(도 60B)에 제시된다. EV68-228-CHO만이 감염 후 3일차 또는 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 61A-B. Expt. NIA-1930. 감염 48시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 3일차(도 61A) 및 5일차(도 61B)에 제시된다. 감염 후 3일차 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 62. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도. 10mg/kg의 EV68-228-CHO에 의한 치료만이 감염 후 3일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. 감염 후 IL-2 또는 IL-3의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 **P<0.01). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 63. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-4, IL-5, IL-6 및 IL-10의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-6 농도를 유의하게 감소시켰다. 감염 후 IL-4, IL-5 또는 IL-10의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료된 마우스와 비교하여 **P<0.01). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 64. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFN-γ의 폐 농도. 감염 후 IL-12p70, IL-17, MCP-1 또는 IFN-γ의 농도에서는 유의한 변화가 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 65. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도. EV-D68의 감염 후, TNFα, MIP-1α, GM-CSF 또는 RANTES 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 66. Expt. NIA-1931. EV-D68에 감염되고 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 생존. (n=10 마우스/그룹). 10mg/kg의 EV68-228-TP에 의한 치료는 사망으로부터 마우스를 완전히 보호했다. 10mg/kg의 EV68-228-CHO로 치료된 10마리의 마우스 모두 사망으로부터 보호되었다. 10mg/kg의 IVIg 용량은 10마리 중 8마리의 마우스를 사망으로부터 보호했다. 위약 치료된 마우스 10마리 중 9마리가 감염에 굴복했다(위약 치료 마우스와 비교하여 ***P<0.001, ****P<0.0001).
도 67. Expt. NIA-1931. 감염 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 10일령의 AG129 마우스의 초기 체중의 백분율. 위약 치료 마우스와 비교하여 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 마우스에서는 체중 감소의 유의한 차이가 관찰되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 체중 감소로부터 마우스를 보호하지 못했다.
도 68A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 1일차 및 3일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 68A) 및 3일차(도 68B)에 제시된다. 10mg/kg의 EV68-228-TP에 의한 감염 24시간 전의 치료는 감염 후 1일차 및 3일차에 바이러스 역가를 유의하게 감소시켰다. 10mg/kg의 EV68-228-CHO에 의한 감염 24시간 전의 치료는 감염 후 1일차 및 3일차에 바이러스 역가를 유의하게 감소시켰다. 감염 24시간 전에 10mg/kg의 IVIg에 의한 치료는 감염 후 3일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001).
도 69A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 5일차 및 7일차 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 5일차(도 69A) 및 7일차(도 69B)에 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 24시간 전의 치료는 감염 후 5일차에 바이러스 역가를 감소시켰다. 감염 후 7일차에는 바이러스가 검출되지 않았다.
도 70A-C. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 전에 EV68-228-TP 또는 감염 2 내지 10일 후에 EV68-228-CHO로 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 2-4일차(도 70A), 5-7일차(도 70B) 및 8-10일차(도 70C)에 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 24시간 전의 치료는 신경학적 스코어로 측정한 바와 같이 마비의 임상 징후를 완전히 예방했다. 감염 24시간 전에 10mg/kg 용량의 IVIg로 치료된 마우스에서는 신경학적 스코어가 관찰되지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 71. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 생존. (n=10 마우스/그룹). 10mg/kg의 EV68-228-TP에 의한 치료는 사망으로부터 마우스를 완전히 보호했다. 10mg/kg의 EV68-228-CHO로 치료된 10마리의 마우스 모두 사망으로부터 보호되었다. 10mg/kg의 IVIg는 10마리 중 10마리의 마우스를 사망으로부터 보호했다. 위약 치료된 마우스 10마리 중 8마리가 감염에 굴복했다(위약 치료된 마우스와 비교하여 ***P<0.001).
도 72. Expt. NIA-1931. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 10일령 AG129 마우스의 초기 체중의 백분율. 위약 치료 마우스와 비교하여 감염 24시간 후에 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 체중 감소의 유의한 차이가 관찰되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 체중 감소로부터 마우스를 보호하지 못했다.
도 73A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 후 1일 및 3일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 73A) 및 3일차(도 73B)에 제시된다. 감염 24시간 후에 10mg/kg의 EV68-228-TP에 의한 치료는 감염 후 3일차에 바이러스 역가를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg의 EV68-228-CHO에 의한 치료는 감염 후 3일차에 바이러스 역가를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg의 IVIg에 의한 치료는 감염 후 3일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001).
도 74A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 후 5일차 및 7일차에 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 5일차(도 74A) 및 7일차(도 74B)에 제시된다. 감염 후 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료한 후, 바이러스는 검출되지 않았다. 감염 후 7일차에서는 어떤 마우스로부터도 바이러스가 검출되지 않았다.
도 75A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 후 2-9일차에 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 2-5일차(도 75A) 및 6-9일차(도 75B)에 대해 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 24시간 후의 치료는 감염 후 2-9일차에 신경학적 스코어로 측정한 바와 같이 마비 징후를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg 용량의 hIVIg에 의한 치료는 감염 후 2일차 및 3일차에 신경학적 스코어만을 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 76A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 후 10-17일차에 EV68-228-TP 또는 EV68-228-CHO로 감염 후 24시간 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 10-13일차(도 76A) 및 14-17일차(도 76B)에 대해 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 후 24시간 치료는 감염 후 10-16일차에 신경학적 스코어로 측정한 바와 같이 마비 징후를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg 용량의 hIVIg 치료는 신경학적 스코어를 유의하게 감소시키지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 77. Expt. NIA-1931. EV-D68에 감염되고 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 생존. (n=10 마우스/그룹). 10mg/kg의 EV68-228-TP에 의한 치료는 10마리 중 9마리의 마우스를 사망으로부터 보호했다. 10mg/kg의 EV68-228-CHO로 치료된 마우스의 10마리 중 8마리가 사망으로부터 보호되었다. 10mg/kg의 IVIg 용량은 10마리 중 6마리의 마우스를 사망으로부터 보호했다. 위약 치료된 마우스의 10마리 중 9마리가 감염에 굴복했다(위약 치료된 마우스와 비교하여 **P<0.01, ***P<0.001).
도 78. Expt. NIA-1931. EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68 감염 10일령 AG129 마우스의 초기 체중 백분율. 위약 치료 마우스와 비교하여 감염 24시간 후에 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 처리된 마우스에서는 체중 감소의 유의한 차이가 관찰되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 체중 감소로부터 마우스를 보호하지 못했다.
도 79A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 1일차 및 3일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 79A) 및 3일차(도 79B)에 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 48시간 후의 치료는 혈액 바이러스 역가를 유의하게 감소시키지 않았다. 감염 48시간 후에 10mg/kg 용량의 IVIg에 의한 치료는 혈액 바이러스 역가를 유의하게 감소시키지 않았다.
도 80A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 5일차 및 7일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 5일차(도 80A) 및 7일차(도 80B)에 제시된다. 감염 후 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 감염 후 24시간 처리한 후, 바이러스는 검출되지 않았다. 감염 후 7일차에 어떤 마우스로부터도 바이러스는 검출되지 않았다.
도 81A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 후 3-10일차에 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 3-6일차(도 81A) 및 7-10일차(도 81B)에 대해 제시된다. 감염 48시간 후에 10mg/kg의 EV68-228-TP에 의한 치료는 감염 후 3-5일차에 신경학적 스코어를 유의하게 감소시켰다. 감염 48시간 후에 10mg/kg 용량의 EV68-228-CHO를 단일 투여하면, 감염 후 3-6일차에 신경학적 스코어로 측정한 바와 같이 마비 징후가 유의하게 감소했다. 감염 48시간 후에 10mg/kg 용량의 hIVIg에 의한 치료는 감염 후 신경학적 스코어를 유의하게 감소시키지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 1. 항체 단리. 2014 EV-D68 발생으로부터 살아있는 바이러스 분리주를 ELISA 플레이트에 직접 코팅하는 간접 ELISA를 사용하여, 64개 인간 모노클로날 항체를 단리하고 스크리닝했다. 이어서, 임의의 히트는 골수종 세포와 융합하여 하이브리도마를 제조하고, 이는 이론적으로 영구적으로 계속 분열할 것이다. 항체를 정제한 후, 이들의 표현형을 정의하기 위해, 다수의 시험관내 특성화 단계가 수행된다.
도 2. 시험관내 항체 중화. 세포 배양 50% 감염 용량 또는 CCID50 검정을 사용하여 시험관내 중화를 수행한다. 일부 항체는 매우 강력하고, 일부는 여전히 매우 강력하고, 다른 항체는 약하고, mAb의 약 1/3이 일부 검출 가능한 중화 활성을 나타낸다.
도 3. 클레이드 사이의 교차 반응성 결합. 모든 mAb를 비교하기 위해, ELISA로부터 절반 최대 유효 농도 또는 EC50을 사용하여 히트 맵을 생성했다. 일부는 모든 클레이드에 매우 잘 결합되는 반면, 일부는 현저한 드롭아웃을 갖고, 통상은 멀리 떨어진 A2 클레이드를 수반한다.
도 4. 경쟁은 적어도 4개의 주요 표면 에피토프를 나타낸다. 피어슨 상관관계를 사용하여, 본 발명자는 클러스터에 대한 컬러 판독치로서 관련성 값을 생성했다. 이 관련성은 항체의 분류에 도움이 되지만, 실제 간섭 비율은 아니다. 본 발명자들은 이제 서로 관련될 수 있는 3 내지 4개의 표현형을 기술했다. 이를 통해, 임상 개발 후보를 특정할 수 있다.
도 5. 임상 후보. 중화에 추가하여, 추가 특성화를 통해, 추가 임상 후보가 밝혀졌다. mAb 159는, 믿을 수 없을 정도로 강력하고 바이러스의 모의 조제물에 결합하지 않지만, ELISA 또는 CCID50에서 교차 반응하지 않는다. mAb 105는 여전히 매우 강력하지만, 교차 반응이 매우 양호하다.
도 6A-C. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. EV-D68-228에 의한 치료는 감염 후 1일차(도 6A), 3일차(도 6B) 및 5일차(도 6C)에 폐 바이러스 역가를 유의하게 감소시켰다. 감염 후 1일, 3일 또는 5일차에 10, 3 또는 1mg/kg의 EV-D68-228로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. IVIg에 의한 치료는 감염 후 1일차에만 폐 바이러스 역가를 감소시킬 수 있었다. 구아니딘은 감염 후 1일차와 3일차에 폐 바이러스 역가를 유의하게 감소시켰지만, 5일차에는 감소시키지 않았다. 위약 치료 마우스와 비교하여 **P<0.01, ****P<0.0001.
도 7A-C. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. EV-D68-228 mAb에 의한 치료는 감염 후 1일차(도 7A), 3일차(도 7B) 및 5일차(도 7C)에 혈액 바이러스 역가를 감소시켰다. IVIg에 의한 치료는 감염 후 1, 3, 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. 구아니딘은 감염 후 1일차와 5일차에 혈액 바이러스 역가를 감소시켰지만, 3일차에는 감소시키지 않았다. 위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001.
도 8. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α 및 IL-1β의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 IL-1α 또는 IL-1β의 폐 농도를 유의하게 감소시키지 않았다. 위약 치료 마우스와 비교하여 ****P<0.0001. 그래프 내의 샘플의 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 9. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-6 및 MCP-1의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차에 IL-6 및 MCP-1의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 감염 후 3일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. 위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. 그래프 내의 샘플의 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 10. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 RANTES의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차 및 5일차에 RANTES의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 감염 후 어느 날자에도 RANTES의 폐 농도를 유의하게 감소시키지 않았다. 위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 11. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-2, IL-3 및 IL-4의 폐 농도. EV-D68 감염 후, IL-2, IL-3 또는 IL-4 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 12. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-5, IL-10 및 IL-12p70의 폐 농도. EV-D68 감염 후, IL-5, IL-10 또는 IL-12p70 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 13. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-17, IFNγ 및 TNFα의 폐 농도. EV-D68 감염 후, IL-17, IFNγ 또는 TNFα 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 14. Expt. NIA-1849. EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 MIP-1α 및 GM-CSF의 폐 농도. EV-D68 감염 후, MIP-1α 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 15. Expt. NIA-1850. EV-D68에 감염되고 EV-D68-228로 치료된 10일령 AG129 마우스의 생존. (n=6 마우스/그룹). 10, 3 또는 1mg/kg의 EV-D68-228에 의한 치료는 마우스를 사망으로부터 완전히 보호했다. 10mg/kg의 IVIg 용량은 6마리 중 4마리의 마우스를 사망으로부터 보호했다. 100mg/kg/일 용량의 구아니딘은 마우스를 사망으로부터 완전히 보호했다. 위약 치료된 마우스의 6마리 모두가 감염에 굴복했다. 위약 치료 마우스와 비교하여 *P<0.05, **P<0.01.
도 16. Expt. NIA-1850. EV-D68-228로 치료된 10일령의 EV-D68 감염 AG129 마우스의 평균 체중. 10, 3 또는 1mg/kg의 용량으로 처리하면 감염 관련 체중 감소로부터 마우스를 보호했다. 10mg/kg의 용량에서 IVIg를 사용한 치료는 또한 체중 감소로부터 마우스를 보호했다. 100mg/kg/일의 용량으로 구아니딘으로 처리된 마우스도 체중 감소로부터 보호되었다. 위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001.
도 17A-C. Expt. NIA-1850. EV-D68에 감염되고 EV-D68-228로 치료된 10일령의 AG129 마우스의 혈액 바이러스 역가. 10, 3 또는 1mg/kg의 EV-D68-228 용량으로 치료된 마우스에서는 감염 후 1일차(도 17A), 3일차(도 17B) 또는 5일차(도 17C)에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg의 IVIg에 의한 치료는 감염 후 1, 3 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. 혈액 바이러스 역가는 100mg/kg/일의 용량으로 구아니딘으로 치료함으로써 감염 후 1일, 3일 및 5일차에 유의하게 감소했다. 위약 처리 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
도 18. Expt. NIA-1850. EV-D68에 감염되고 EV-D68-228로 치료된 10일령의 AG129 마우스의 신경학적 스코어. 10, 3 또는 1mg/kg 용량으로 EV-D68-228에 의한 치료는 신경학적 스코어에 의해 측정한 임상 마비 징후를 예방했다. 10mg/kg의 IVIg에 의한 치료는 감염 후 3, 4 및 5일차에 신경학적 스코어를 감소시켰다. 100mg/kg/일의 구아니딘으로 치료된 마우스에서는 신경학적 스코어가 관찰되지 않았다. 위약 처리 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 19A-B. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. EV-D68-228에 의한 치료는 감염 후 3일차(도 19A) 및 5일차(도 19B)에 폐 바이러스 역가를 유의하게 감소시켰다. 감염 후 24시간 이내에, 10mg/kg의 EV-D68-228로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. 감염 후 48시간에 10 및 1mg/kg 용량으로 EV-D68-288에 의한 치료는 감염 후 3일차 및 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. IVIg에 의한 치료는 감염 후 24시간에 치료한 경우 폐 바이러스 역가를 감소시키지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 20A-B. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. EV-D68-228 mAb에 의한 치료는 감염 후 3일차(도 20A) 및 5일차(도 20B)에 혈액 바이러스 역가를 감소시켰다. 감염 후 48시간까지 EV-D68-228에 의한 치료는 감염 후 3일차와 5일차 모두에서 혈액 바이러스 역가를 검출 한계 미만으로 감소시켰다. IVIg에 의한 치료는 감염 후 3일차와 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, ****P<0.0001).
도 21. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α 및 IL-1β의 폐 농도. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일차 및 5일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IVIg에 의한 치료는 감염 후 3일차 및 5일차에 IL-1α 또는 IL-1β의 폐 농도를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 22. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-5 및 IL-6의 폐 농도. 감염 후 48시간까지 EV-D68-228에 의한 치료는 위약 치료된 마우스와 비교하여 감염 후 3일차에 IL-5 및 IL-6의 농도를 유의하게 감소시켰다. 감염 24시간 후 IVIg에 의한 치료는 감염 후 3일차에 IL-5 및 IL-6의 폐 농도를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 23. Expt. NIA-1849. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 MCP-1 및 RANTES의 폐 농도. 감염 후 48시간 이내에 EV-D68-228에 의한 치료는 위약 치료된 마우스와 비교하여 감염 후 3일차 및 5일차에 MCP-1 및 RANTES의 농도를 유의하게 감소시켰다. 감염 24시간 후 IVIg에 의한 치료는 감염 후 3일차에 MCP-1의 폐 농도를 유의하게 감소시켰고, 감염 후 3일차 및 5일차에 RANTES의 농도를 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 24. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-2, IL-3 및 IL-4의 폐 농도. EV-D68의 감염 후, IL-2, IL-3 또는 IL-4 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 25. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-10, IL-12p70 및 IL-17의 폐 농도. EV-D68의 감염 후, IL-5, IL-10 또는 IL-12p70 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 26. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 IFNγ 및 TNFα의 폐 농도. EV-D68의 감염 후, IFNγ 또는 TNFα 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 27. Expt. NIA-1869. 감염 후 EV-D68-228로 치료된 EV-D68 감염 AG129 마우스로부터의 MIP-1α 및 GM-CSF의 폐 농도. EV-D68 감염 후, MIP-1α 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 28. Expt. NIA-1870. EV-D68에 감염되고 감염 후 EV-D68-228로 치료된 10일령의 AG129 마우스의 생존. (n=6 마우스/그룹, n=7 마우스/그룹은 감염 후 120시간 치료됨). 10mg/kg의 EV-D68-228에 의한 치료는 감염 후 24시간에 마우스를 사망으로부터 완전히 보호했다. 감염 후 48시간에 EV-D68-228에 의한 치료는 6마리의 마우스 중 4마리를 사망으로부터 보호했다. EV-D68-228에 의한 감염 72시간 후에 치료한 6마리 마우스 중 1마리만이 감염에서 생존했다. 감염 120시간 후에 EV-D68-228로 치료된 그룹에서는 7마리의 마우스 중 어느 것도 생존하지 않았음에도 불구하고, 사망일의 지연으로 인해, 생존 곡선은 위약-치료된 마우스와는 상이했다. 10mg/kg의 IVIg 용량은, 감염 24시간 후에 치료된 경우, 6마리의 마우스 모두를 사망으로부터 보호했다. 위약 치료된 마우스 6마리 모두는 감염에 굴복했다(위약 치료된 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001).
도 29. Expt. NIA-1870. 감염 후 EV-D68-228로 치료된 EV-D68 감염 10일령의 AG129 마우스의 평균 체중. 감염 후 24시간 또는 48시간에 10mg/kg 용량의 EV-D68-228에 의한 치료는 마우스를 감염 관련 체중 감소로부터 보호했다. 감염 후 24시간에 10mg/kg 용량의 IVIg에 의한 치료는 마우스를 체중 감소로부터 보호하지 못했다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01).
도 30A-C. Expt. NIA-1870. EV-D68에 감염되고 감염 후 EV-D68-228로 치료된 10일령 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 30A), 3일차(도 30B) 또는 5일차(도 30C)에 제시된다. 10mg/kg 용량으로 EV-D68-228에 의한 치료는, 감염 후 24, 48 또는 72시간에 투여한 경우, 감염 후 3일차와 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. 감염 후 24시간에 투여된 10mg/kg 용량의 IVIg도 감염 후 3일차 및 5일차에 혈액 바이러스 역가를 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001).
도 31A-B. Expt. NIA-1870. EV-D68에 감염되고 감염 후 EV-D68-228로 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 2일차 및 3일차(도 31A) 및 감염 후 4일차 및 5일차(도 31B)에 제시된다. 감염 후 24시간에 10mg/kg 용량으로 EV-D68-228에 의한 치료는, 신경학적 스코어로 측정한 바와 같이, 3일, 4일 및 5일차의 임상적 마비 징후를 예방했다. 감염 24시간 후 10mg/kg의 IVIg에 의한 치료는 감염 후 3일, 4일 및 5일차에 신경학적 스코어를 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 32. EV-D68-면역 인간 대상체로부터의 mAb의 경쟁 결합 그룹. 관련성 스코어는 B1 클레이드 EV-D68 분리주를 사용하여 경쟁 결합 ELISA로부터 생성하고, mAb를 1~4로 지정된 4개의 경쟁 결합 그룹으로 클러스터링하는 데 사용했다. 적색 또는 청색으로 수록된 클론 번호는 강력하게 중화하는 mAb이고, 청색 클론 명칭은 이후 도면에서 상세히 연구된 2개의 mAb를 나타낸다.
도 33A-C. 인간 mAb의 중화 효능 및 결합 능력. (도 33A) MAb는 B1 클레이드 분리주를 사용하는 CCID50 중화 검정에서 IC50 값에 의해 경쟁-결합 그룹(비교 그룹, 그룹 5는 싱글톤의 잔류 컬렉션을 나타낸다) 내에서 순위가 매겨졌다. 본 발명자들은 또한 21개의 가장 강력하게 중화시키는 mAb에 대해 D 클레이드 및 페르몬(Fer.) 부리주의 중화를 시험했다. ">"는 최대 50μg/mL 농도에서 시험했을 때에 중화가 검출되지 않았음을 나타낸다. 공백 셀은 시험되지 않았음을 나타낸다. 적색 또는 청색으로 수록된 클론 번호는 강력하게 중화하는 mAb이고, 청색 클론 명칭은 이후 도면에서 상세히 연구된 2개의 mAb를 나타낸다. (도 33B) 생 바이러스 분리주 또는 모의 바이러스 조제물에 대한 결합 강도는 S자형 용량 반응 곡선에 적합하도록 정제된 mAb 희석액과 함께 간접 ELISA(도 33C)를 사용함으로써 생성된 EC50 값을 사용하여 표시된다. ">"는 EC50 값이 시험된 최대 농도인 100μg/mL를 초과하는 것을 나타내고, 이는 결합이 불충분하거나 없음을 시사한다.
도 34A-C. 2개 면역 복합체 사이의 구조적 특징 비교. (도 34A) EV-D68:Fab EV68-159(좌측) 또는 EV-D68:Fab EV68-228(우측)의 방사형 착색 크리오-EM 맵. 각 맵은 2개의 대칭 축을 따라 투영된다. 각 비대칭 단위의 5배, 3배 및 2배의 축은 각각 오각형 1개, 작은 삼각형 2개, 및 타원형 1개로 표지된 삼각형 윤곽을 사용하여 도시된다. (도 34B) 비대칭 단위에 대한 결합 위치 비교. 바이러스 단백질은 청색(VP1), 녹색(VP2) 및 적색(VP3)으로 착색된다. Fab 분자는 회색(EV68-159) 또는 자주색(EV68-228)으로 착색되고, 중쇄 또는 경쇄는 각각 암색 또는 명색 강도로 동일한 색상으로 표시된다. (도 34C) EV68-159 Fab(좌측) 또는 EV68-228 Fab(우측)의 풋프린트. 바이러스 표면의 방사형 착색된 2D 투영은 RIVEM을 사용하여 생성되었다. 4Å 거리 내에서 Fab 분자로부터의 임의의 원자와 대면하는 바이러스 표면 잔기는 밝은 청색(VP1), 밝은 녹색(VP2) 및 밝은 적색(VP3)으로 표시된다. 협곡 영역은 황색으로 도시된다. (도 34A) 및 (도 34C)의 스케일 바는 Å 단위로 측정된 반경 방향 거리를 나타낸다.
도 35. EV68-159 및 EV68-228의 결합 인터페이스의 확대도. 바이러스 캡시드는 표면으로 표시되고, Fab는 만화 표현으로 표시된다. VP1, VP2 및 VP3은 각각 백색, 짙은 회색 및 은색으로 착색되어 있다. 중쇄 또는 경쇄는 각각 주황색 또는 황색으로 착색된다. 상호작용을 수행하는 바이러스 잔류물은 중쇄와 경쇄를 기반으로 착색되고, 색상 강도는 VP 중 어느 것에 기초하여 변화한다. 결합 인터페이스에 관여하는 중쇄 및 경쇄 상보성 결정 영역(각각 HCDR 및 LCDR)은 화살표로 제시된다.
도 36A-D. 비리온-Fab 상호작용의 분자 상세. EV-D68:Fab EV68-159(도 36A-C) 및 EV-D68:Fab EV68-228(도 36D)의 결합 인터페이스에서의 대표적 상호작용. 수소 결합은 청록색으로 착색되고, 염 브릿지는 마젠타로 착색된다.
도 37A-D. MAb EV68-228은, 예방 또는 요법으로 사용되는 경우, EV-D68 유도된 호흡기 질환으로부터 마우스를 보호한다. 4주령의 AG129 계통 마우스(시점당 n = 4)에, 마우스-적응 B1 클레이드 EV-D68을 비강으로 접종하고, 항체를 복강내 투여하고; 표시된 조직에 대한 바이러스 역가를 CCID50 검정에 의해 측정했다. (도 37A-B) 지시된 항체 용량의 24시간 후에 마우스에게 바이러스를 접종하고, 이어서 지시된 시점에서 바이러스 역가를 측정했다. (도 37C-D) 마우스에게 바이러스를 접종하고, 4, 24 또는 48시간 후에 10mg/kg(지시된 경우 제외)의 항체를 접종하고, 이어서 바이러스 역가를 측정했다.
도 38A-F. MAb EV68-228은 EV-D68 감염 마우스에서 폐 염증을 감소시킨다. 4주령 AG129 마우스(시점당 n=4)(도 38A-C)은 지시된 항체 용량의 24시간 후에 비강내로 바이러스를 접종하거나, (도 38D-F)은 바이러스를 비강내로 접종하고, 이어서 4, 24 또는 48시간 후에 10mg/kg(지시된 경우 제외) 항체를 접종하고, 지시된 시점에서 사이토카인을 측정했다. ELISA를 사용하여 폐 균질액으로부터 사이토카인을 정량화했다. 둔네트(Dunnett)의 T3 다중 비교 검정을 사용한 일원 분산 분석을 사용하여, 각 시점에서 치료 그룹의 값을 위약 그룹과 비교했다(*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). IL-인터루킨; MCP-단핵 세포 주화성 단백질. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 39A-F. MAb EV68-228은, 예방 또는 요법으로 사용되는 경우, EV-D68 유도 신경계 질환으로부터 마우스를 보호한다. 10일령의 마우스에게 마우스-적응 B1 클레이드 EV-D68을 복강내 접종하고, 항체는 복강내 투여하고; 지시된 조직에 대한 바이러스 역가는 CCID50 검정에 의해 측정했다. (도 39A-C) 지시된 용량의 항체의 24시간 후에 마우스에게 바이러스를 접종하고, 이어서 (도 39A) 바이러스 역가를 측정하고(시점당 n = 3), (도 39B) 생존을 모니터링하고, (도 39C) 신경학적 스코어(시점당 n = 6)를 지시된 시점에서 기록했다. 스코어가 높으면, 운동 장애가 심각한 것을 나타낸다. (도 39D-F) 마우스에게 바이러스를 접종하고, 이어서 10mg/kg 용량의 항체를 접종하고, 이어서 (도 39D) 바이러스 역가를 측정하고(시점당 n = 3), (도 39E) 생존을 모니터링하고, (도 39F) 신경학적 스코어(시점당 n = 6, 120시간 후의 n = 9 제외)를 기록했다. 착색 수직 화살표는 치료 시간을 나타낸다.
도 40. 웨스턴 블롯 데이터. 모든 mAb는 웨스턴 블롯에 의해 B1 클레이드 바이러스를 염색하는 능력에 대해 시험되었다. 모든 긍정적 결과가 제시된다.
도 41. 인간 mAb의 상세한 특성. MAb는 B1 클레이드 분리주를 사용한 CCID50 중화 검정에서 IC50 값에 의해 경쟁 결합 그룹 내에서 순위가 매겨졌다. 본 발명자들은 또한 21개의 가장 강력하게 중화하는 mAb에 대해 D 클레이드 및 페르몬(Fer.) 분리주의 중화를 시험했다. ">"는 최대 50μg/mL 농도에서 시험할 때에 중화가 검출되지 않았음을 나타낸다. 공백 셀은 시험되지 않았음을 나타낸다. 적색 또는 청색 폰트로 수록된 클론 번호는 강력하게 중화하는 mAb이고, 청색 클론 명칭은 후속 도면에서 상세히 연구된 2개의 mAb를 나타낸다. 각 mAb에 대한 최종 IC50 값은 중복으로 수행된 3개의 개별 실험으로부터의 EC50 값의 로그 평균이다. 살아있는 바이러스 분리주에 대한 결합 강도는 S자형 용량 반응 곡선에 적합하도록 정제된 mAb 희석액과 함께 간접 ELISA를 사용하여 생성된 EC50 값을 기반으로 표시된다. ">"는 예측된 EC50 값이 시험된 최대 농도인 100μg/mL를 초과하는 것을 나타내고, 이는 결합이 불충분하거나 없음을 시사한다. 각 mAb에 대한 최종 EC50 값은 중복으로 수행된 3개의 개별 실험으로부터의 EC50 값의 로그 평균이다. IgG 서브타입은 표현형으로 결정되었다. 경쇄 유형은 카파(K) 또는 람다(L)이고, 유전적으로 결정되었다. WB는 웨스턴 블롯 반응성을 나타내고, 모든 mAb는 시험되었고, 공백 행은 반응성이 없음을 나타낸다.
도 42. 모든 mAb에 대한 간접 ELISA 데이터. 살아있는 바이러스 분리주에 대한 결합 강도는 S자형 용량 반응 곡선에 적합하도록 정제된 mAb 희석액과 함께 간접 ELISA를 사용하여 생성된 EC50 값으로 표시된다. 제시된 것은 3개의 실험 중 하나의 대표적 데이터이고, 에러 바는 중복 기술적 복제의 표준 편차를 나타낸다. 각 mAb에 대한 최종 EC50 값은 3개의 실험 모두로부터의 EC50 값의 로그 평균이다. 바이러스의 모의 조제물은 바이러스를 접종하지 않았지만 바이러스 감염 세포와 동일한 방식으로 제조된 RD 세포 단층으로부터의 것이다.
도 43. EV-D68로부터의 대표적 밀도: Fab EV68-228 전자 밀도 맵. 바이러스 단백질은 청색(VP1), 녹색(VP2), 적색(VP3) 및 마젠타색(VP4)으로 착색된다.
도 44. 면역 복합체 맵 해상도의 추정치. 맵 해상도는 금 표준 푸리에 쉘 상관관계(FSC) 컷오프 0.143을 기반으로 추정된다. EV68-159(적색 곡선) 또는 EV68-228(청색 곡선) 복합체의 최종적 해상도는 각각 2.9Å 또는 3.1Å이다.
도 45. Fab 결합 부위의 비교. EV68-159와 EV68-228은 각각 금색과 청색으로 착색되었고, 바이러스 표면은 청록색으로 착색되었다. 좌측 패널은 가변 도메인과 불변 도메인을 모두 표시하는 반면, 우측 패널은 가변 도메인만을 표시하고, 이는 2개 Fab의 풋프린트가 중첩하지 않음을 입증한다.
도 46. Fab 풋프린트의 확대도를 나타내는 로드맵. 바이러스 표면의 방사형 착색된 2D 투영은 RIVEM으로 생성되었다. 4Å 거리 내에서 Fab 분자로부터의 임의의 원자와 대면하는 바이러스 표면 잔기는 연한 청색(VP1), 연한 녹색(VP2) 및 연한 적색(VP3)으로 도시된다. 협곡 영역은 황색으로 도시된다.
도 47. EV68-228 Fab 중쇄의 벌키한 측쇄. 이 도면은 EV68-228 Fab를 안정화하기 위해 소수성 상호작용 네트워크를 형성하는 벌키한 측쇄의 예를 나타내고, 이는 EV68-159 Fab의 구조에서도 관찰된다.
도 48A-C. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. 폐 바이러스 역가는 감염 후 1일차(도 48A), 3일차(도 48B) 및 5일차(도 48C)에 제시된다. 감염 후 1일, 3일 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ****P<0.0001)
도 49A-C. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 49A), 3일차(도 49B) 및 5일차(도 49C)에 제시된다. 감염 후 1일, 3일 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 1일, 3일 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001)
도 50. 내선 NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 1일, 3일 및 5일차에 IL-1α의 농도를 유의하게 감소시켰다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료도 감염 후 3일차에 IL-1β의 농도를 감소시켰다. hIVIg에 의한 치료는 감염 후 3일차에 IL-1β의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-2 또는 IL-3의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 51. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-4, IL-5, IL-6 및 IL-10의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-5의 농도를, 감염 후 3일 및 5일차에 IL-6의 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차에 IL-5 및 IL-6의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-4 또는 IL-10의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 52. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFN-γ의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일 및 5일차에 MCP-1 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-12p70, IL-17 또는 IFN-γ의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 53. Expt. NIA-1930. 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 1일 및 3일차에 MIP-1α의 폐 농도를 유의하게 감소시켰고, 감염 후 3일 및 5일차에 RANTES의 농도를 감소시켰다. 10mg/kg 용량의 hIVIG는 감염 후 1일과 3일차에 MIP-1α의 농도를 유의하게 감소시켰고, 감염 후 3일차에 RANTES의 농도도 감소시켰다. EV-D68 감염 후 TNFα 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 54A-B. Expt. NIA-1930. 감염 24시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. 폐 바이러스 역가는 감염 후 3일차(도 54A) 및 5일차(도 54B)에 제시된다. 감염 후 3일차 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 폐 바이러스 역가가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차에 폐 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 **P<0.01, ***P<0.001, ****P<0.0001)
도 55A-B. Expt. NIA-1930. 감염 24시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 3일차(도 55A) 및 5일차(도 55B)에 제시된다. 감염 후 3일차 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001).
도 56. Expt. NIA-1930. 감염 24시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IL-3의 농도는 감염 후 3일차에 EV68-228-TP로 치료된 마우스 및 hIVIg로 치료된 마우스에서 유의하게 감소했다. 감염 후 IL-2 농도에서 유의한 변화는 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 57. Expt. NIA-1930. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-4, IL-5, IL-6 및 IL-10의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-5의 농도를, 감염 후 3일차 및 5일차에 IL-6의 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차에 IL-5의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-4 또는 IL-10의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 58. Expt. NIA-1930. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFN-γ의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차 및 5일차에 MCP-1 농도를 유의하게 감소시켰다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. 감염 후 IL-12p70, IL-17 또는 IFN-γ의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 59. Expt. NIA-1930. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 MIP-1α의 폐 농도를 유의하게 감소시켰고, 감염 후 3일차 및 5일차에 RANTES의 농도를 감소시켰다. EV-D68의 감염 후, TNFα 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 60A-B. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 폐 바이러스 역가. 폐 바이러스 역가는 감염 후 3일차(도 60A) 및 5일차(도 60B)에 제시된다. EV68-228-CHO만이 감염 후 3일차 또는 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 61A-B. Expt. NIA-1930. 감염 48시간 후 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 3일차(도 61A) 및 5일차(도 61B)에 제시된다. 감염 후 3일차 또는 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았다. 10mg/kg의 hIVIg에 의한 치료는 감염 후 3일차 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 62. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도. 10mg/kg의 EV68-228-CHO에 의한 치료만이 감염 후 3일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. 감염 후 IL-2 또는 IL-3의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료 마우스와 비교하여 **P<0.01). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 63. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-4, IL-5, IL-6 및 IL-10의 폐 농도. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-6 농도를 유의하게 감소시켰다. 감염 후 IL-4, IL-5 또는 IL-10의 농도에서는 유의한 변화가 관찰되지 않았다. (위약 치료된 마우스와 비교하여 **P<0.01). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 64. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFN-γ의 폐 농도. 감염 후 IL-12p70, IL-17, MCP-1 또는 IFN-γ의 농도에서는 유의한 변화가 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 65. Expt. NIA-1930. 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도. EV-D68의 감염 후, TNFα, MIP-1α, GM-CSF 또는 RANTES 농도의 유의한 변화는 관찰되지 않았다. 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 66. Expt. NIA-1931. EV-D68에 감염되고 감염의 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 생존. (n=10 마우스/그룹). 10mg/kg의 EV68-228-TP에 의한 치료는 사망으로부터 마우스를 완전히 보호했다. 10mg/kg의 EV68-228-CHO로 치료된 10마리의 마우스 모두 사망으로부터 보호되었다. 10mg/kg의 IVIg 용량은 10마리 중 8마리의 마우스를 사망으로부터 보호했다. 위약 치료된 마우스 10마리 중 9마리가 감염에 굴복했다(위약 치료 마우스와 비교하여 ***P<0.001, ****P<0.0001).
도 67. Expt. NIA-1931. 감염 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 10일령의 AG129 마우스의 초기 체중의 백분율. 위약 치료 마우스와 비교하여 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 마우스에서는 체중 감소의 유의한 차이가 관찰되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 체중 감소로부터 마우스를 보호하지 못했다.
도 68A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 1일차 및 3일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 68A) 및 3일차(도 68B)에 제시된다. 10mg/kg의 EV68-228-TP에 의한 감염 24시간 전의 치료는 감염 후 1일차 및 3일차에 바이러스 역가를 유의하게 감소시켰다. 10mg/kg의 EV68-228-CHO에 의한 감염 24시간 전의 치료는 감염 후 1일차 및 3일차에 바이러스 역가를 유의하게 감소시켰다. 감염 24시간 전에 10mg/kg의 IVIg에 의한 치료는 감염 후 3일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001).
도 69A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 전에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 5일차 및 7일차 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 5일차(도 69A) 및 7일차(도 69B)에 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 24시간 전의 치료는 감염 후 5일차에 바이러스 역가를 감소시켰다. 감염 후 7일차에는 바이러스가 검출되지 않았다.
도 70A-C. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 전에 EV68-228-TP 또는 감염 2 내지 10일 후에 EV68-228-CHO로 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 2-4일차(도 70A), 5-7일차(도 70B) 및 8-10일차(도 70C)에 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 24시간 전의 치료는 신경학적 스코어로 측정한 바와 같이 마비의 임상 징후를 완전히 예방했다. 감염 24시간 전에 10mg/kg 용량의 IVIg로 치료된 마우스에서는 신경학적 스코어가 관찰되지 않았다. (위약 치료 마우스와 비교하여 ****P<0.0001).
도 71. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 생존. (n=10 마우스/그룹). 10mg/kg의 EV68-228-TP에 의한 치료는 사망으로부터 마우스를 완전히 보호했다. 10mg/kg의 EV68-228-CHO로 치료된 10마리의 마우스 모두 사망으로부터 보호되었다. 10mg/kg의 IVIg는 10마리 중 10마리의 마우스를 사망으로부터 보호했다. 위약 치료된 마우스 10마리 중 8마리가 감염에 굴복했다(위약 치료된 마우스와 비교하여 ***P<0.001).
도 72. Expt. NIA-1931. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 EV-D68 감염 10일령 AG129 마우스의 초기 체중의 백분율. 위약 치료 마우스와 비교하여 감염 24시간 후에 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 체중 감소의 유의한 차이가 관찰되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 체중 감소로부터 마우스를 보호하지 못했다.
도 73A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 후 1일 및 3일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 73A) 및 3일차(도 73B)에 제시된다. 감염 24시간 후에 10mg/kg의 EV68-228-TP에 의한 치료는 감염 후 3일차에 바이러스 역가를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg의 EV68-228-CHO에 의한 치료는 감염 후 3일차에 바이러스 역가를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg의 IVIg에 의한 치료는 감염 후 3일차에 혈액 바이러스 역가를 유의하게 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ****P<0.0001).
도 74A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 후 5일차 및 7일차에 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 5일차(도 74A) 및 7일차(도 74B)에 제시된다. 감염 후 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료한 후, 바이러스는 검출되지 않았다. 감염 후 7일차에서는 어떤 마우스로부터도 바이러스가 검출되지 않았다.
도 75A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 후 2-9일차에 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 2-5일차(도 75A) 및 6-9일차(도 75B)에 대해 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 24시간 후의 치료는 감염 후 2-9일차에 신경학적 스코어로 측정한 바와 같이 마비 징후를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg 용량의 hIVIg에 의한 치료는 감염 후 2일차 및 3일차에 신경학적 스코어만을 감소시켰다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 76A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 후 10-17일차에 EV68-228-TP 또는 EV68-228-CHO로 감염 후 24시간 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 10-13일차(도 76A) 및 14-17일차(도 76B)에 대해 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 후 24시간 치료는 감염 후 10-16일차에 신경학적 스코어로 측정한 바와 같이 마비 징후를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg 용량의 hIVIg 치료는 신경학적 스코어를 유의하게 감소시키지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, **P<0.01). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
도 77. Expt. NIA-1931. EV-D68에 감염되고 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 생존. (n=10 마우스/그룹). 10mg/kg의 EV68-228-TP에 의한 치료는 10마리 중 9마리의 마우스를 사망으로부터 보호했다. 10mg/kg의 EV68-228-CHO로 치료된 마우스의 10마리 중 8마리가 사망으로부터 보호되었다. 10mg/kg의 IVIg 용량은 10마리 중 6마리의 마우스를 사망으로부터 보호했다. 위약 치료된 마우스의 10마리 중 9마리가 감염에 굴복했다(위약 치료된 마우스와 비교하여 **P<0.01, ***P<0.001).
도 78. Expt. NIA-1931. EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68 감염 10일령 AG129 마우스의 초기 체중 백분율. 위약 치료 마우스와 비교하여 감염 24시간 후에 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 처리된 마우스에서는 체중 감소의 유의한 차이가 관찰되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 체중 감소로부터 마우스를 보호하지 못했다.
도 79A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 1일차 및 3일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 1일차(도 79A) 및 3일차(도 79B)에 제시된다. 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO에 의한 감염 48시간 후의 치료는 혈액 바이러스 역가를 유의하게 감소시키지 않았다. 감염 48시간 후에 10mg/kg 용량의 IVIg에 의한 치료는 혈액 바이러스 역가를 유의하게 감소시키지 않았다.
도 80A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 48시간 후에 EV68-228-TP 또는 EV68-228-CHO로 치료된 10일령 AG129 마우스의 감염 5일차 및 7일차의 혈액 바이러스 역가. 혈액 바이러스 역가는 감염 후 5일차(도 80A) 및 7일차(도 80B)에 제시된다. 감염 후 5일차에 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 감염 후 24시간 처리한 후, 바이러스는 검출되지 않았다. 감염 후 7일차에 어떤 마우스로부터도 바이러스는 검출되지 않았다.
도 81A-B. Expt. NIA-1931. EV-D68에 감염되고 감염 후 3-10일차에 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 10일령 AG129 마우스의 신경학적 스코어. 신경학적 스코어는 감염 후 3-6일차(도 81A) 및 7-10일차(도 81B)에 대해 제시된다. 감염 48시간 후에 10mg/kg의 EV68-228-TP에 의한 치료는 감염 후 3-5일차에 신경학적 스코어를 유의하게 감소시켰다. 감염 48시간 후에 10mg/kg 용량의 EV68-228-CHO를 단일 투여하면, 감염 후 3-6일차에 신경학적 스코어로 측정한 바와 같이 마비 징후가 유의하게 감소했다. 감염 48시간 후에 10mg/kg 용량의 hIVIg에 의한 치료는 감염 후 신경학적 스코어를 유의하게 감소시키지 않았다. (위약 치료 마우스와 비교하여 *P<0.05, ***P<0.001, ****P<0.0001). 그래프 내의 샘플 지정은 좌측에서 우측에 대응하고, 수직 범례는 상부에서 하부에 대응한다.
상기에서 논의한 바와 같이, 엔테로바이러스 D68(EV-D68)은 세계적으로 대부분 어린이에게 호흡기 질환의 발생을 유발할 수 있다. 소아마비-유사 신경근 약화 증후군인 급성 이완성 척수염의 클러스터는 종종 EV-D68 호흡기의 발생과 동시에 발생한다. 혈청역학 연구에 따르면, 2세 내지 5세 이상의 거의 모든 사람의 혈청에는 항-EV-D68 중화 항체가 포함되어 있고, 이는 EV-D68이 소아기의 유비쿼터스 병원체임을 시사한다. 그러나, 체액성 면역 반응이 지시되는 바이러스 에피토프에 대한 지식은 관련 바이러스에 대한 이전 연구로부터만 추론된다. 여기에서, 본 발명자들은 EV-D68에 대한 64개의 인간 모노클로날 항체를 생성했고, 그 중 일부는 강력하게 중화된다. 이들은 이 병원체에 대한 최초의 인간 mAb인 것으로 믿어지고, 본 발명자들은 EV-D68 감염의 검출, 치료 및 예방에서 이들의 사용을 제안한다. 본 개시의 이들 및 다른 측면은 하기에서 상세하게 설명된다.
I. 엔테로바이러스 D68
엔테로바이러스 68(EV68, EV-D68, HEV68)은 엔테로바이러스인 피코나비리다에(Picornaviridae) 과의 구성원이다. 1962년에 캘리포니아에서 처음 단리되고, 한때 희귀종으로 여겨졌지만, 21세기에는 전 세계적으로 급증했다. 급성 이완성 척수염이라는 소아마비-유사 장애를 유발하는 것으로 의심된다.
EV68은 100종류 초과의 엔테로바이러스 중의 하나이고, 폴리오바이러스, 콕사키바이러스 및 에코바이러스를 포함하는 ssRNA 바이러스 그룹이며, 외피가 없다. 다른 모든 엔테로바이러스와 달리, EV68은 산성 불안정성과 낮은 최적 성장 온도를 나타내고, 이들 둘 다는 인간 라이노바이러스의 특징이다. 이전에, 일부 연구자들은 인간 라이노바이러스 87이라고 불렀다. 천식과 면역억제를 갖는 성인의 질환도 보고되었지만, 5세 미만의 어린이와 천식을 갖는 어린이가 이 질환에 걸릴 위험이 가장 높은 것으로 보인다.
EV68이 발견된 이후로, EV68은 고립된 경우에 대부분 산발적으로 기술되었다. 2005년과 2011년 사이에 6개의 클러스터(10건 이상) 또는 발생이 필리핀, 일본, 네덜란드 및 미국의 조지아주, 펜실베니아주 및 애리조나주에서 보고되었다. EV68은 캘리포니아에서 2012/13년에 발생한 소아마비-유사 질환 클러스터의 5명의 어린이 중 2명에서 발견되었다. 2014년 8월, 이 바이러스는 미국에서 호흡기 질환의 클러스터를 유발했다. 10월 중순까지 46개 주와 컬럼비아 특별구에서 691명이 EV-D68에 의한 유발된 호흡기 질환에 걸렸고, 5명의 어린이가 사망했다. 2016년에는 유럽에서 29건의 사례가 보고되었다(프랑스와 스코틀랜드에서 5건, 스웨덴, 노르웨이 및 스페인에서 각각 3건).
2000년 이후, 원래의 바이러스 균주는 다양화하고, 유전적으로 상이한 발생 균주인 클레이드 B1을 진화시켰다. 이는 클레이드 B1이지만, AFM과 연관되어 있고 동물 모델에서 신경병증인 보다 오래된 균주는 아니다. 사례는 엔테로바이러스 시즌(대략 춘분과 추분 사이의 기간)에 발생하는 것으로 설명되었고, 이는 일반적으로 북반구에서 8월과 9월에 발생했다.
EV68은 거의 예외 없이 호흡기 질환을 유발하고, 이는 경증으로부터 중증까지 다양하지만, 전혀 증상이 없는 것으로부터 미묘한 독감 유사 증상, 쇠약성의 호흡기 질환 및 소아마비-유사 증후군의 증상에 드물게 관여하는 것으로 의심되는 관련 증상까지 다양한 증상을 유발할 수 있다. 모든 엔테로바이러스와 마찬가지로, 다양한 발진, 복통 및 연변을 유발할 수 있다. 초기 증상은 콧물, 인후통, 기침, 발열 등 감기의 증상과 유사하다. 질환이 진행됨에 따라, 폐렴과 같은 호흡곤란, 주의력 저하, 소변량 감소, 탈수 등의 보다 심각한 증상이 발생하고, 호흡부전을 유도할 수 있다.
경험한 증상의 중증도는 해당 인구통계학적 모집단에 따라 달라진다. 전문가들은 인구의 대다수가 실제로 엔테로바이러스에 노출됐지만, 건강한 성인에서는 증상이 나타나지 않는다고 추정한다. 대조적으로, EV-D68은 매우 약한 어린이뿐만 아니라, 매우 어린 어린이에게도 불균형적으로 쇠약해진다. 수백 명(472명)(대부분 청년)이 이 질환에 노출되었지만, 중증의 증상(예: 마비)으로 진단된 환자는 100명 미만이다.
이 바이러스는, 급성 이완 척수염의 희귀 근력 저하의 한 가지 원인으로 의심되고, 통상은 소아마비가 원인이고, 이는 바이러스의 검사에서 양성으로 된 2명의 캘리포니아 어린이는 발증으로부터 48시간 이내에 하나 이상의 사지가 마비되어 최고의 중증도에 도달했기 때문이다. 운동 기능의 회복은 6개월의 추적 관찰에서 불충분했다. 2014년 10월 시점에서, CDC는 엔테로바이러스 D68 활동의 증가와 일치하여, 콜로라도주 및 전국의 다른 보고서에서 마비 및/또는 두개 기능장애의 10건을 조사하고 있었다. 2014년 10월 현재, 실제의 사례 수는 100건 이상일 것으로 여겨졌다. 2018년 시점에서, EV-D68과 마비의 연관성은 강력하여, 브래드포드 힐(Bradford Hill)의 6개 기준을 완전히 충족하고, 2개를 부분적으로 충족한다. CDC는, 2018년 10월 17일에, "현재, 소아마비 바이러스가 이러한 AFM 사례의 원인이 아니라는 것을 알고 있다. CDC는 AFM 환자의 모든 대변 검체를 검사했지만, 소아마비 바이러스 양성의 검체는 없었다"고 말했다. 2014년에, 검출 속도를 높이기 위한 실시간 PCR 시험이 CDC에 의해 개발되었다.
특정한 치료법도 없고 백신도 없기 때문에, 질환은 이의 경과를 따라야 하고; 치료는 증상에 대해 수행된다(증상 치료). 대부분의 사람들은 완전히 회복되지만; 그러나, 일부는 입원해야 하고, 일부는 바이러스의 결과로서 사망했다. 5건의 EV68 마비 사례는 스테로이드, 정맥내 면역글로불린 및/또는 혈장 교환에 의한 치료에 실패했다. 운동 기능의 회복이 관찰되지 않았기 때문에, 치료는 명백한 이점이 없었다. 2015년 연구에서는 항바이러스제 플레코나릴이 EV-D68 치료에 유용할 수 있다고 시사했다.
이 바이러스는 대변뿐만 아니라 타액 및 가래를 통해 확산되기 때문에, 손을 씻는 것이 중요하다. 아픈 사람들은 재채기 또는 기침할 때에 코와 입을 가리는 것 등의 기본적 위생 대책에 의해 바이러스 확산을 감소시키려고 시도할 수 있다. 표면 및 장난감의 세정을 포함하는 기타 조치. EV-D68 감염으로 입원한 환자의 경우, CDC는 모든 엔테로바이러스에 권장되는 바와 같이 감염에 기초하는 예방 조치, 즉 표준 예방 조치, 접촉 예방 조치를 권장하고, 비말 예방 조치를 검토할 것을 권장한다. 2003년의 CDC에 따르면, 의료 시설의 표면은 몇몇 비외피 바이러스(예: 노로바이러스, 폴리오바이러스, 라이노바이러스)에 대해 EPA 라벨 표시된 병원 등급의 소독제로 세정해야 한다.
A. 바이러스 에피토프
혈청 항체 바이러스 중화 능력의 측정은 폴리클로날 항체 레퍼토리 전체의 활성을 반영한다. 그러나, 바이러스에 대한 체액성 반응을 완전히 이해하기 위해, 개별 항체가 결합하는 특정 바이러스 에피토프를 정의하고, 특정 에피토프에 결합하는 항체가 질환으로부터 보호하는지의 여부를 판단할 필요가 있다. 역사적으로, 엔테로바이러스 속의 바이러스에 대한 4개의 중화 면역에피토프(Nim)는 라이노바이러스-B14에 대해 생성된 뮤린 모노클로날 항체의 연구를 통해 동정되었다[참조: Rossmann et al., 1985]. EV-D68 특이적 모노클로날 항체 에피토프에 대한 연구는 현재까지 수행되지 않았다. 그러나, EV-D68의 결정 구조의 결정은 라이노바이러스와의 구조적 상동성을 관찰함으로써 EV-D68 비리온 상의 4개의 Nim의 위치의 가능성을 시사했다[참조: Liu et al., 2015a]. 미국[참조: Zhang et al., 2015] 또는 일본[참조: Imamura et al., 2014]로부터 EV-D68의 최근 인간 단리된 표면 단백질의 아미노산 서열을 페르몬(Fermon) 참조 바이러스 균주의 아미노산 서열과 비교한 연구는, Nim 및 근처의 인접 잔기에서 돌연변이가 특히 결정 구조가 무질서한 캡시드 단백질 VP1의 BC 및 DE 루프에서 발생했음을 시사한다[참조: Liu et al., 2015a]. 이러한 VP1 다형성은 기존의 체액성 면역을 회피함으로써 바이러스의 병원성의 증가에 기여하는 것이 가능하다. 뮤린 모노클로날 항체는 상업적으로 이용 가능하지만(출처에는 GeneTex, ThermoFisher 및 Sigma 포함), 면역원으로 사용된 특정 바이러스 표면 단백질의 목록을 제외하고, 이러한 항체의 에피토프에 대한 정보는 없다. 특히, 이러한 상용 항체 중 일부는 기타 엔테로바이러스 속 바이러스의 면역원으로부터 생성된 것이고, 이는 이형 항체가 EV-D68에 결합하는 능력을 시사한다. 이 가능성을 추가로 뒷받침하는 것으로서, 2개의 뮤린 mAb가 40개의 상이한 엔테로바이러스 종에 교차 반응했지만, 흥미롭게도 EV-D68이 반응하지 않은 유일한 시험 바이러스였다[참조: Miao et al., 2009]. 기타 유형의 엔테로바이러스에 의해 유도된 항체에 의한 EV-D68의 이형 분자 인식은 인생의 초기 단계에서 관찰된 보편적 EV-D68 혈청유병률에 기여할 수 있다. EV-D68 상의 인간 항체 에피토프의 미세한 특이성과 이러한 항체의 유형 특이성 또는 폭을 이해하는 것은 자연 감염에 의해 유도된 클론 인간 모노클로날 항체를 사용한 연구가 필요하다.
미국에서 2014년의 EV-D68이 발생하는 동안, 거의 모든 바이러스 분리주가 새로 출현한 B1 클레이드의 것이고, 밀접하게 관련된 B2 클레이드 또는 멀리 관련된 D 클레이드로부터의 바이러스의 검출은 보다 적었다[참조: Tan et al., 2016]. 이 연구의 1명을 제외한 모든 대상체는 B1 클레이드 분리주에 감염되었다(표 E). 2014년 이래, B3 클레이드 바이러스가 우세했고, B1 클레이드 바이러스는 더 이상 순환하지 않고[참조: Dyrdak et al., 2019], 2018년에, 미국 질병 예방 관리 센터에 의해 서열분석된 모든 EV-D68 분리주는 B3 클레이드로부터의 것이었다[참조: Kujawski et al., 2019]. 본 발명자들은 먼저 B1 클레이드 EV-D68 분리주를 사용하여 50% 세포 배양 감염 용량(CCID50) 검정에서 각 mAb의 시험관내 중화 능력을 측정했다(도 33A). 28개의 mAb는 50μg/mL 미만의 절반 최대 억제 농도(IC50)로 중화를 입증했고, mAb EV68-159는 0.32ng/mL의 IC50 값에서 가장 강력한 중화를 나타냈다(도 41). 본 발명자들은 D 클레이드 분리주에 대해 21개의 가장 강력하게 중화하는 mAb를 추가로 시험하고, 11개의 mAb가 그 바이러스를 중화시키고, 그 중 7개가 이종 바이러스의 IC50 값에 의해 적어도 10배의 효능 저하를 나타내는 것을 밝혀냈다. 페르몬(Fermon) 균주는 1962년으로부터의 분리주이고, 현대의 EV-D68 분리주와 매우 멀리 먼 관계에 있기 때문에, 클레이드 분류 체계에 적합하지 않다[참조: Tan et al., 2016]. 9개의 mAb는 페르몬 실험실 참조 균주를 중화했지만, 현대의 B1 클레이드 바이러스를 억제하는 것보다 덜 강력했다.
중화 검정은 교차 반응성을 과소평가할 수 있음을 인식하여, 본 발명자들은, 상기와 동일한 간접 ELISA 접근법을 사용하여, B1, B2 또는 D 클레이드으로부터의 대표적 EV-D68 분리주에 결합하기 위한 정제 mAb의 최대 유효 농도의 절반(EC50)을 생성했다(도 33B-C 및 도 42). ≤ 1 ㎍/mL의 B1 클레이드 분리주에 대한 결합에 대한 EC50 값을 갖는 mAb 중에서, 모두가 B2 클레이드 분리주에 결합하고, 약 절반은 D 클레이드 분리주에도 결합했다(도 33B 및 도 41). 일반적으로 약하게 결합하지만 시험된 모든 클레이드의 바이러스에 교차 반응하는 추가 부류의 mAb가 관찰되었다.
현재까지, 항체-EV-D68 상호작용의 구조적 연구는 마우스 mAb로 한정되었다[참조: Zheng et al., 2019]. 본 발명자들은 2개의 강력하게 중화하는 인간 mAb, 클레이드 특이적 mAb EV68-159 및 고도로 교차 반응성의 mAb EV68-228을 선택하여, 항원 결합 단편(Fab)과의 면역 복합체 및 크리오-전자 현미경(크리오-EM) 연구용의 B1 클레이드 EV-D68 분리주를 제조했다. 최종 밀도 맵은 2.9Å(EV68-159) 또는 3.1Å(EV68-228)의 해상도를 달성했다(도 34A, 도 43, 도 44 및 표 G). 구조는 2개의 상이한 결합 부위를 나타냈다: EV68-159는 3중 대칭 축 주위에 부착된 반면, EV68-228은 협곡 영역을 형성하는 함몰부 사이의 5중 축 주위에 결합했다(도 34A-C, 도 45). 따라서, 각 Fab에 대해, 총 60개의 카피가 바이러스 입자에 결합했다. 바이러스 표면과 상호작용하는 Fab 가변 도메인은 바이러스 캡시드 단백질과 유사한 강력한 밀도를 나타내고, 각 Fab의 원자 모델은 4개의 바이러스 캡시드 단백질과 함께 구축되었다. 대조적으로, 바이러스 표면에서 추가로 멀리 위치하는 Fab 불변 도메인은 보다 약한 밀도를 나타내고, 원자 모델의 구축으로부터 제외되었다. 폴리펩타이드 쇄의 백본과 아미노산 측쇄의 대부분은 밀도 맵에서 적절히 정렬되어 있고, 이는 바이러스 입자와 Fab 분자 사이의 결합 계면의 중요한 특징을 입증한다.
B. 신경 질환의 동물 모델
시험관내에서 항체에 의한 바이러스 중화를 측정하는 것은 통상 항체의 항바이러스 기능을 특성화하는 가장 신속하고 재현 가능한 방법이다. 그러나, 이 시험 방식은, 보체 활성화 또는 세포 표면 Fc 수용체와의 결합과 같이 생체내에서만 기능하는 항체의 Fc 부분에 의해 매개되는 이펙터 기능은 측정하지 않는다. 결과적으로, 시험관내 중화 능력의 크기는 바이러스 복제의 억제 수준 또는 생체내에서 관찰되는 질환에 대한 보호 수준과 완전하게 상관관계가 없을 수 있다[참조: (Lu et al., 2018)에서 검토]. 항체의 Fc 매개 이펙터 기능의 대부분은 완전한 보호 효과를 매개하기 위해 자연 면역계 또는 획득 면역계의 Fc 수용체 보유 세포를 필요로 한다. 전임상 연구에서 체액성 면역의 효능을 평가하기 위해서는 감염의 작은 동물 모델이 필요하다.
콜로라도[참조: Hixon et al., 2017a; 2017b], 유타[참조: Morrey et al., 2018] 및 중국[참조: Zhang et al., 2018]의 연구자는 2017년 초부터 EV-D68의 마우스 모델을 보고했다. 보고된 3개의 모델 모두에서, 10일령 이내의 마우스에게 바이러스를 접종한다. 이러한 모델 전체에서, EV-D68은 복강내[참조: Hixon et al., 2017a; 2017b; Morrey et al., 2018; Zhang et al., 2018], 근육내[참조: Hixon et al. 2017a; 2017b] 또는 뇌내 경로[참조: Hixon et al., 2017b] 투여될 때에 이완성 사지 마비 및 사망을 유발할 수 있는 것으로 나타났다. 특히, 비강내 접종은 시험된 73마리의 비근친교배 NIH 스위스 웹스터(Swiss Webster) 마우스 중 2마리에서만 마비를 유도하고, 척수에서 바이러스가 검출된 반면[참조: Hixon et al., 2017b], 인터페론 유형-I 및 -II 수용체를 결여하는 AG129 마우스의 비강내 접종은 척수가 아닌 근육에서 바이러스가 검출된 4마리 마우스 중 2마리에서 마비를 유발했다[참조: Morrey et al., 2018]. 그러나, 마우스에서 체액성 면역이 평가된 모든 연구는 바이러스 접종의 비생리학적 경로를 사용했다.
바이러스 항원은 골격근[참조: Morrey et al., 2018; Zhang et al., 2018] 및 척수[참조: Hixon et al., 2017b; Morrey et al., 2018]의 면역염색에 의해 조직에서 시각화되었다. 근육과 척수 둘 다의 관여는, EV-D68이 2개 메커니즘에 의해 마비를 유도할 수 있음을 시사한다: 1) 골격근에 대한 직접적 병리학적 효과 및 2) 중추 운동 뉴런의 상실. 척수의 운동 뉴런이 감염되었는데, 이는 소아마비의 병인과 유사하다. 면역염색 연구에서 뮤린 뉴런의 관여의 이러한 패턴은 AFM 환자의 척수 이미징에서 회백질 변화의 소견과도 상관관계가 있다[참조: Hixon et al., 2017b; Morrey et al., 2018]. 보다 민감한 실시간 정량적 RT-PCR 시험은 주로 근육과 척수뿐만 아니라, 뇌, 심장, 폐, 장, 간, 비장, 신장 및 혈액에서도 EV-D68 RNA가 검출되었다[참조: Zhang et al., 2018]. 그러나, 척수와 근육 외에는, 검출된 RNA의 양은 의심스러운 의미가 있었고, 이러한 조직의 진정한 지향성보다는 이러한 조직 내의 혈액에 존재하는 바이러스 게놈 카피를 양호하게 반영할 수 있다.
마비 및 치사의 이러한 모델에 대한 연구는 신경 질환으로부터 보호하는 항체의 능력에 대한 예비 연구를 촉진했다. EV-D68이 뇌내 경로를 통해 직접 접종된 경우에도, 나이브 마우스에게 수동적으로 전달된 면역 마우스로부터의 열-불활성화 혈청은 신경 질환을 예방했다[참조: Hixon et al., 2017b]. 또한, 인간 IVIG의 치료적 투여는 마우스 감염 후 6일까지 운동 장애를 감소시켰고[참조: Hixon et al., 2017a], 이는 IVIG에서 EV-D68 특이적 항체의 높은 역가의 소견과 일치한다[참조: Zhang et al., 2015]. 이 관찰은, 개별 환자가 EV-D68과 일시적으로 연관되는 AFM의 특징을 갖는 것으로 확인된 후, 백신 또는 인간 모노클로날 항체가 치료 효과를 매개할 수 있다는 희망을 제공한다. 또한, EV-D68의 공지된 발생 기간 동안 모체 체액성 면역이 약화된 영아 및 유아 등의 인구 모집단의 표적 백신접종 또는 면역예방은 AFM 사례를 예방할 수 있다.
C. 호흡기 질환의 동물 모델
EV-D68 감염에 관한 현재 마우스 연구의 주요 제한은, 마비 및 사망을 유도하기 위한 비생리학적 접종 경로의 사용과, 인간에서 EV-D68 감염의 주요 증상인 호흡기 질환의 부족이다. 코튼 랫트(Sigmodon hispidus)(Patel et al., 2016)와 페렛(Mustela putorius furo)(Zheng et al., 2017a) 모두에 대해, 각각의 단일 연구에 의해, EV-D68의 비강내 접종이 코와 폐에서 바이러스 복제를 초래하고 폐에서 전염증성 자연 면역 분자 전사체의 증가를 유도한다는 것이 입증되었다. 상기도 감염(기침, 콧물, 코 건조)과 일치하는 임상 증상도, 접종된 페렛의 약 25%에서 발증했다. 근육 또는 중추신경계 조직에서 EV-D68의 존재를 평가한 연구도, 명백한 사지 약화 또는 마비도 인식하지 않았다. 코튼 랫트의 백신접종은 일부 수준의 기존 체액성 면역이 호흡기 감염을 완전히 보호하지 못했고, 하기 추가로 기재되는 발견에서 실제로 유해할 수 있음을 나타낸다[참조: Patel et al., 2016].
인간 호흡기 바이러스 감염에 대한 모델로서 코튼 랫트와 페렛의 이론적 이점은 각각의 호흡기 상피가 특히 상피 표면의 글리칸의 시알화와 관련하여 인간 호흡 상피를 보다 밀접하게 모방할 수 있다는 것이고, 이는 인간에서는 강력하게 α2,6-결합되지만, 마우스에서는 결합하지 않는다[참조: Gagneux et al., 2003]. EV-D68은 시험관내 글리칸 어레이에서 α2,3-결합 시알산보다 α2,6-결합 시알산에 우선적으로 결합했다[참조: Imamura et al., 2014]. 형광 표지된 렉틴에 의한 기도의 염색은, 코튼 랫트가 하부 기도에 α2,6-결합 시알산만을 갖고, 상부 기도에 α2,6- 및 α2,3-결합 시알산의 혼합물을 갖는 것을 나타냈다[참조: Blanco et al., 2018]. 유사한 렉틴-기반 방법을 사용하여, 페렛은 상부 기도의 상피 글리칸에 α2,6-결합 시알산을 갖고[참조: Leigh et al., 1995], 하부 기도에 α2,6- 및 α2,3-결합 시알산의 혼합물을 갖는 것을 나타냈다[참조: Zheng et al., 2017a]. 그러나, 인간(Walther et al., 2013), 마우스(Bern et al., 2013) 및 페렛(Jia et al., 2014) 호흡기 조직에 대한 보다 최근의 정교한 글리코믹 분석은, 단순한 α2,6-결합 시알화의 밀도보다 일련의 글리칸 변형이 상이한 동물 종의 호흡기 바이러스의 친화성을 결정할 가능성이 있음을 나타냈다. 그럼에도 불구하고, 랫트와 페렛에서 인간 EV-D68 호흡기 질환의 모방에서 초기 연구의 성공은 인간 병인의 모델로서의 추가 개발에 고무적이다.
D. 백신
EV-D68에 대한 실험적 백신 후보는 아직 임상 개발에 들어가지 않았지만, 효모[참조: Zhang et al., 2018a] 또는 곤충 세포[참조: Dai et al., 2018]에서 생성된 바이러스 유사 입자(VLP) 백신 또는 베타-프로피오락톤 비활성화 EV-D68[참조: Zhang et al., 2018] 중의 어느 하나를 마우스에게 접종하는 초기 연구는 유망하다. VLP 백신은 이후의 치명적 복강내 챌린지로부터 마우스를 보호하는 항체 반응을 유도했다. 이러한 백신 후보 각각을 사용하여, 나이브 마우스에 대한 백신 면역 혈청의 수동적 전달은 치명적 복강내 챌린지로부터의 보호에 충분했다. VLP 기반 백신의 가능한 단점은 이러한 합성 작제물이 입자 표면의 모든 형태적으로 민감한 구조를 완전히 재현하는지의 여부에 대한 불확실성이다. 다른 바이러스에 대한 다수의 강력한 인간 바이러스 중화 항체는 엄격한 형태적 제약이 있는 복잡한 4차 항원 부위를 인식한다[참조: Crowe, JE, 2017]. VLP 상의 인간 항체 에피토프의 완전성은, 이러한 에피토프가 불명이기 않기 때문에, 현재 평가할 수 없다. 이러한 뮤린 백신 연구의 가능성을 추가로 완화하는 것은 살아있는 또는 UV 비활성화 EV-D68 중 어느 하나를 근육내 접종하고, 이어서, 나이브 동물의 감염과 비교하여 폐에서 염증의 향상이 관찰된 살아있는 동종 바이러스로 비강내 챌린지했다[참조: Patel et al., 2016]. 구체적으로, UV 비활성화 바이러스 백신접종은 폐 또는 코에서 바이러스 복제를 제한하지 않았고, 생 바이러스 백신접종으로부터의 면역에서 관찰된 균형 잡힌 Th1 및 Th2 표현형이 아닌 Th2 표현형을 향해 수득된 사이토카인 서명을 왜곡했다. 염증 및 질환의 향상은, 호흡기 합포체 바이러스[참조: Karron, R, 2018] 및 홍역[참조: Strebel et al., 2018]에 대한 비활성화 바이러스 백신 후보에서 주목되어 왔다. 이러한 혼합된 결과를 감안하면, EV-D68 백신 후보에 의해 유발된 보호 또는 향상의 면역 상관관계에 대한 추가의 신중한 연구가 필요하다.
II. 모노클로날 항체 및 이의 생산
"단리된 항체"는 이의 자연 환경의 성분으로부터 분리 및/또는 회수된 것이다. 이의 자연 환경의 오염 성분은 항체의 진단 또는 치료 용도를 방해하는 물질이고, 효소, 호르몬 및 기타 단백질 또는 비단백질 용질을 포함할 수 있다. 특정 실시양태에서, 항체는 (1) 로우리(Lowry) 방법에 의해 측정된 항체의 95중량% 초과, 및 가장 특히 99중량% 초과까지; (2) 스피닝 컵 서열분석기를 사용하여, N-말단 또는 내부 아미노산 서열의 적어도 15개 잔기를 수득하기에 충분한 정도까지; 또는 (3) 쿠마시에 블루 또는 은 염색을 사용하여 환원 또는 비환원 조건에서 SDS-PAGE에 의한 균질성까지 정제된다. 항체의 자연 환경의 적어도 하나의 성분이 존재하지 않을 것이기 때문에, 단리된 항체에는 재조합 세포 내의 제자리에서의 항체가 포함된다. 그러나, 일반적으로, 단리된 항체는 적어도 하나의 정제 단계에 의해 제조된다.
기본적 4개-쇄 항체 단위는 2개의 동일한 경쇄(L)와 2개의 동일한 중쇄(H)로 구성되는 헤테로사량체 당단백질이다. IgM 항체는 5개의 기본 헤테로사량체 단위와 J 쇄라고 하는 추가 폴리펩타이드로 구성되어 있기 때문에, 10개의 항원 결합 부위를 함유하는 반면, 분비된 IgA 항체는 중합하여, J 쇄와 함께 2-5개의 기본 4-쇄 단위를 포함하는 다가 집합체를 형성할 수 있다. IgG의 경우, 4-쇄 단위는 일반적으로 약 150,000 달톤이다. 각 L 쇄는 하나의 공유 디설파이드 결합에 의해 H 쇄에 결합되어 있는 반면, 2개의 H 쇄는 H 쇄 아이소타입에 따라 하나 이상의 디설파이드 결합에 의해 서로 연결되어 있다. 각 H 및 L 쇄는 또한 규칙적으로 간격을 둔 쇄 내 디설파이드 브릿지를 갖고 있다. 각 H 쇄는 N-말단에 가변 영역(VH)을 갖고, 이어서 알파 및 감마 쇄의 각각에 3개의 불변 도메인(CH)과 뮤 및 아이소타입에 4개의 CH 도메인을 갖는다. 각 L 쇄는 N-말단에 가변 영역(VL)을 갖고, 다른 쪽 끝에 불변 도메인(CL)을 갖는다. VL은 VH와 정렬되고, CL은 중쇄의 제1 불변 도메인(CH1)과 정렬된다. 특정 아미노산 잔기는 경쇄 및 중쇄 가변 영역 사이의 계면을 형성하는 것으로 여겨진다. VH와 VL의 쌍은 함께 단일 항원 결합 부위를 형성한다. 상이한 부류의 항체의 구조 및 특성에 대해서는, 예를 들면, 문헌[참조: Basic and Clinical Immunology, 8th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, Conn ., 1994, page 71, and Chapter 6]을 참조한다.
임의의 척추동물 종의 L 쇄는 불변 도메인(CL)의 아미노산 서열에 기초하여 카파와 람다라고 하는 2개의 명확하게 상이한 유형 중 어느 하나로 할당될 수 있다. 이들의 중쇄(CH)의 불변 도메인의 아미노산 서열에 따라, 면역글로불린은 상이한 부류 또는 아이소타입으로 할당될 수 있다. 면역글로불린에는 IgA, IgD, IgE, IgG 및 IgM의 5개 부류가 있고, 각각 알파, 델타, 엡실론, 감마 및 뮤로 지정된 중쇄를 갖는다. 감마 및 알파 부류는 CH 서열 및 기능의 비교적 사소한 차이에 기초하여 서브클래스로 분류되고, 인간은 IgG1, IgG2, IgG3, IgG4, IgA1 및 IgA2의 서브클래스를 발현한다.
"가변"이라는 용어는 V 도메인의 특정 세그먼트가 항체 사이에서 서열이 광범위하게 상이하다는 사실을 의미한다. V 도메인은 항원 결합을 매개하고, 특정 항원에 대한 특정 항체의 특이성을 정의한다. 그러나, 가변성은 가변 영역의 110개 아미노산 범위 전체에 걸쳐 균등하게 분포되지 않는다. 대신에, V 영역은 15-30개 아미노산의 프레임워크 영역(FR)이라고 하는 비교적 불변 스트레치로 구성되고, 각각 9-12개 아미노산 길이의 "초가변 영역"이라고 하는 극단의 가변성의 보다 짧은 영역으로 분리된다. 천연 중쇄 및 경쇄의 가변 영역은 각각 4개의 FR을 포함하고, 주로 베타 시트 구성을 채용하고, 3개의 초가변 영역에 의해 연결되고, 루프 연결을 형성하고, 일부 경우에는 베타 시트 구조의 일부를 형성한다. 각 쇄의 초가변 영역은 FR에 의해 근접하게 함께 유지되고, 다른 쇄의 초가변 영역과 함께, 항체의 항원 결합 부위의 형성에 기여한다[참조: Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)]. 불변 도메인은, 항체가 항원에 결합하는 데 직접적으로 관여하지 않지만, 항체 의존성 세포독성(ADCC), 항체 의존성 세포 식세포 작용(ADCP), 항체 의존성 호중구 식세포 작용(ADNP) 및 항체 의존적 보체 침착(ADCD)에서 항체의 참여 등의 다양한 이펙터 기능을 나타낸다.
본 명세서에 사용된 용어 "초가변 영역"은 항원 결합에 관여하는 항체의 아미노산 잔기를 지칭한다. 초가변 영역은 일반적으로 "상보성 결정 영역" 또는 "CDR"로부터의 아미노산 잔기[예를 들면, 카바트 넘버링 시스템에 따라 넘버링되는 경우, VL에서 약 잔기 24-34(L1), 50-56(L2) 및 89-97(L3), 및 VH에서 약 31-35(H1), 50-65(H2) 및 95-102(H3); Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)]; 및/또는 "초가변 루프"로부터의 잔기[예를 들면, 초티아 넘버링 시스템에 따라 넘버링되는 경우, VL에서 잔기 24-34(L1), 50-56(L2) 및 89-97(L3), 및 VH에서 26-32(H1), 52-56 (H2) 및 95-101(H3); Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987)]; 및/또는 "초가변 루프"/CDR로부터의 잔기[예를 들면, IMGT 넘버링 시스템에 따라 넘버링되는 경우, VL에서 잔기 27-38(L1), 56-65(L2) 및 105-120(L3), 및 VH에서 27-38(H1), 56-65(H2) 및 105-120(H3); Lefranc, MP et al. Nucl. Acids Res. 27:209-212(1999), Ruiz, M. et al. al. Nucl. Acids Res. 28:219-221(2000)]을 포함한다. 임의로, 항체는 하기 포인트 중의 하나 이상에서 대칭적 삽입물[예를 들면, AHo에 따라 넘버링되는 경우, VL에서 28, 36(L1), 63, 74-75(L2) 및 123(L3) 및 VsubH에서 28, 36(H1), 63, 74-75(H2) 및 123(H3); Honneger, A. and Plunkthun, A. J. Mol. Biol. 309:657-670(2001)]을 갖는다.
"생식세포계 핵산 잔기"는 불변 또는 가변 영역을 암호화하는 생식세포계 유전자에서 자연적으로 존재하는 핵산 잔기를 의미한다. "생식세포계 유전자"는 생식 세포(즉, 난자 또는 정자로 되는 운명의 세포)에서 발견되는 DNA이다. "생식세포계 돌연변이"는 단일 세포 단계에서 생식 세포 또는 접합자에서 발생한 특정 DNA의 유전적 변화를 지칭하고, 자손에게 전달되면, 이러한 돌연변이는 신체의 모든 세포에 도입된다. 생식세포계 돌연변이는 단일 체세포에서 획득되는 체세포 돌연변이와 대조적이다. 일부 경우에, 가변 영역을 암호화하는 생식세포계 DNA 서열의 뉴클레오티드는 돌연변이되고(즉, 체세포 돌연변이), 다른 뉴클레오티드로 치환된다.
본 명세서에 사용된 용어 "모노클로날 항체"는 실질적으로 균질한 항체 모집단으로부터 수득된 항체를 지칭하고, 즉 모집단을 포함하는 개별 항체는 소량으로 존재할 수 있는 가능한 자연 존재 돌연변이를 제외하고는 동일하다. 모노클로날 항체는 매우 특이적이고, 단일 항원 부위에 대해 지시된다. 추가로, 상이한 결정인자(에피토프)에 대해 지시된 상이한 항체를 포함하는 폴리클로날 항체 조제물과 대조적으로, 각 모노클로날 항체는 항원의 단일 결정인자에 대해 지시된다. 이들의 특이성 외에도, 모노클로날 항체는 이들이 다른 항체에 의해 오염되지 않은 상태로 합성될 수 있다는 점에서 유리하다. 수식어 "모노클로날"은 임의의 특정 방법에 의한 항체의 생산을 필요로 하는 것으로 해석되어서는 안 된다. 예를 들면, 본 개시에 유용한 모노클로날 항체는 문헌[참조: Kohler et al., Nature, 256:495 (1975)]에 의해 최초로 기재된 하이브리도마 방법론에 의해 제조될 수 있거나, 또는 항원 특이적 B 세포, 감염 또는 면역화에 응답하는 항원 특이적 형질아세포의 단일 세포 선별, 또는 벌크 선별된 항원 특이적 수집에서 단리 세포로부터 결합된 중쇄 및 경쇄의 포획 후에 박테리아, 진핵생물 동물 또는 식물 세포(참조: 예를 들면, 미국 특허 제4,816,567호)에서 재조합 DNA 방법을 사용하여 작제할 수 있다, "모노클로날 항체"는 또한, 예를 들면, 문헌[참조: Clackson et al., Nature, 352:624-628 (1991) and Marks et al., J. Mol. Biol., 222:581-597(1991)]에 기재된 기술을 사용하여 파지 항체 라이브러리로부터 단리할 수 있다.
A. 일반 방법
EV-D68에 결합하는 모노클로날 항체는 몇몇 용도가 있음을 이해할 것이다. 여기에는 EV-D68 감염의 검출 및 진단 및 이의 치료용 진단 키트의 생산이 포함된다. 이러한 맥락에서, 이러한 항체를 진단제 또는 치료제에 연결하거나, 이들을 포획제 또는 경쟁 검정의 경쟁물질로서 사용하거나, 추가 약제를 첨가하지 않고 개별적으로 사용할 수 있다. 항체는 하기에 추가로 논의되는 바와 같이 돌연변이시키거나 변형시킬 수 있다. 항체를 제조하고 특성화하는 방법은 당해 기술분야에 공지되어 있다[참조: 예를 들면, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988; 미국 특허 제4,196,265호].
모노클로날 항체(MAb)를 생성하는 방법은 일반적으로 폴리클로날 항체를 제조하는 방법과 동일한 방침에 따라 개시된다. 이 양 방법의 제1 단계는 적절한 숙주의 면역화, 또는 이전의 자연 감염 또는 허가된 백신 또는 실험적 백신에 의한 백신접종으로 인해 면역성을 갖는 대상체의 특정이다. 당해 기술분야에 공지되어 있는 바와 같이, 면역화를 위한 소정 조성물은 이의 면역원성이 상이할 수 있다. 따라서, 펩타이드 또는 폴리펩타이드 면역원을 담체에 커플링함으로써 달성될 수 있는 바와 같이, 숙주 면역계를 부스팅하는 것이 종종 필요하다. 예시적 및 바람직한 담체는 키홀 림펫 헤모시아닌(KLH) 및 소 혈청 알부민(BSA)이다. 오브알부민, 마우스 혈청 알부민 또는 래빗 혈청 알부민 등의 기타 알부민도 담체로서 사용할 수 있다. 폴리펩타이드를 담체 단백질에 접합시키는 수단은 당해 기술분야에 공지되어 있고, 글루타르알데히드, m-말레이미도벤코일-N-하이드록시석신이미드 에스테르, 카보디이미드 및 비스-비아조화 벤지딘을 포함한다. 또한, 당해 기술분야에 공지된 바와 같이, 특정 면역원 조성물의 면역원성은 애주번트로서 공지된 면역 반응의 비특이적 자극제를 사용함으로써 향상될 수 있다. 동물에서 예시적 및 바람직한 애주번트는 완전 프로인트 애주번트(사멸된 마이코박테리움을 함유하는 면역 반응의 비특이적 자극제), 불완전 프로인트 애주번트 및 수산화알루미늄 애주번트를 포함하고, 인간에서는 명반, CpG, MFP59 및 면역자극 분자의 조합("애주번트 시스템", 예컨대, AS01 또는 AS03)이 포함된다. EV-D68 특이적 B 세포를 유도하기 위한 추가의 실험 형태의 접종이 가능하다. 여기에는 나노입자 백신, 또는 물리적 전달 시스템(예: 지질 나노입자 또는 금 바이올리스틱 비드)에서 DNA 또는 RNA 유전자로서 전달되고 바늘, 유전자 총, 경피 전기천공 장치로 전달되는 유전자 암호화 항원이 포함된다. 항원 유전자는 또한, 아데노바이러스, 아데노-관련 바이러스, 폭스바이러스, 헤르페스바이러스 또는 알파바이러스 레플리콘 등의 복제 적격 또는 결함이 있는 바이러스 벡터, 또는 대안적으로 바이러스 유사 입자에 의해 암호화되는 바와 같이 운반될 수 있다.
자연 병원체에 대한 인간 항체의 경우, 적절한 접근법은 병원체에 노출된 대상체, 예를 들면, 질환에 걸린 것으로 진단된 대상체 또는 백신접종을 받아 병원체에 대해 보호 면역성을 생성하거나 실험적 백신의 안전성 또는 효능을 시험한 대상체를 특정하는 것이다. 순환하는 항-병원체 항체를 검출할 수 있고, 이어서 항체-양성 대상체로부터 B 세포를 암호화하거나 생산하는 항체를 수득할 수 있다.
폴리클로날 항체의 생산에 사용되는 면역원 조성물의 양은 면역원의 성질과 면역화에 사용되는 동물에 따라 상이하다. 면역원을 투여하기 위해 다양한 경로를 사용할 수 있다(피하, 근육내, 피내, 정맥내 및 복강내). 폴리클로날 항체의 생산은 면역화 후의 다양한 시점에서 면역화된 동물의 혈액을 샘플링함으로써 모니터링할 수 있다. 2차 부스팅 주사도 제공될 수 있다. 적절한 역가가 달성될 때까지, 부스팅 및 역가 측정의 프로세스를 반복한다. 원하는 수준의 면역원성이 수득되면, 면역화된 동물로부터 채혈하고, 혈청을 단리하여 저장하고/하거나, 동물을 사용하여 MAb를 생성할 수 있다.
면역화 후, 항체 생성 가능성이 있는 체세포, 특히 B 림프구(B 세포)가 MAb 생성 프로토콜에 사용하기 위해 선택된다. 이 세포는 생검된 비장, 림프절, 편도선 또는 아데노이드, 골수 흡인물 또는 생검, 폐 또는 위장관 등의 점막 기관으로부터의 조직 생검 또는 순환 혈액으로부터 수득할 수 있다. 이어서, 면역화된 동물 또는 면역화된 인간으로부터의 항체 생성 B 림프구를 면역화된 동물 또는 인간 또는 인간/마우스 키메라 세포와 일반적으로 동일한 종의 하나인 불사멸성 골수종 세포의 세포와 융합시킨다. 하이브리도마 생성 융합 절차에서의 사용에 적합한 골수종 세포주는 바람직하게는 비항체 생성이고, 높은 융합 효율을 갖고, 효소 결함에 의해, 따라서 원하는 융합 세포(하이브리도마)만의 성장을 서포트하는 특정 선택 배지에서 성장할 수 있게 된다. 당업자에게 공지된 바와 같이, 다수의 골수종 세포 중 어느 하나를 사용할 수 있다[참조: Goding, pp. 65-66, 1986; Campbell, pp. 75-83, 1984]. HMMA2.5 세포 또는 MFP-2 세포는 이러한 세포의 특히 유용한 예이다.
항체 생성 비장 또는 림프절 세포와 골수종 세포의 하이브리드를 생성하는 방법은 일반적으로 체세포와 골수종 세포를 2:1 비율로 혼합하는 것을 포함하지만, 비율은 세포막의 융합을 촉진하는 약제(들)(화학적 또는 전기적)의 존재하에 각각 약 20:1로부터 약 1:1까지 변화할 수 있다. 일부 경우에는, 초기 단계로서 엡스타인 바르 바이러스(EBV)로 인간 B 세포를 형질전환하면, B 세포의 크기가 증가하여, 비교적 큰 크기의 골수종 세포와의 융합이 향상된다. EBV에 의한 형질전환 효율은 형질전환 배지에서 CpG 및 Chk2 억제제 약물을 사용함으로써 향상된다. 대안적으로, 인간 B 세포는, IL-21 및 TNF 상과의 유형 II 구성원인 인간 B 세포 활성화 인자(BAFF) 등의 추가 가용성 인자를 함유하는 배지에서 CD40 리간드(CD154)를 발현하는 형질감염된 세포주와의 공-배양함으로써 활성화될 수 있다. 센다이 바이러스를 사용한 융합 방법은 문헌[참조: Kohler and Milstein(1975; 1976)]에 의해 기재되었고, 폴리에틸렌 글리콜(PEG)(예: 37%(v/v) PEG)을 사용한 융합 방법은 문헌[참조: Gefter et al. (1977)]에 기재되었다. 전기적으로 유도된 융합 방법의 사용도 적절하고[참조; Goding, pp. 71-74, 1986], 보다 양호한 효율성을 위한 프로세스가 있다[참조: Yu et al., 2008]. 융합 절차는 통상 약 1 × 10-6 내지 1 × 10-8의 낮은 빈도로 실행 가능한 하이브리드를 생산하지만, 최적화된 절차를 사용하면, 200분의 1에 근접하는 융합 효율을 달성할 수 있다[참조: Yu et al., 2008]. 그러나, 선택 배지에서 배양함으로써, 생존 가능한 융합된 하이브리드가 모체의 주입된 세포(특히 통상은 무기한으로 계속 분열하는 주입된 골수종 세포)로부터 분화되기 때문에, 융합 효율이 비교적 낮은 것은 문제를 일으키지 않는다. 선택 배지는 일반적으로 조직 배양 배지에서 뉴클레오티드의 드 노보 합성을 차단하는 약제를 포함하는 것이다. 예시적 및 바람직한 약제는 아미노프테린, 메토트렉세이트 및 아자세린이다. 아미노프테린 및 메토트렉세이트는 퓨린과 피리미딘 모두의 드 노보 합성을 차단하는 반면, 아자세린은 퓨린 합성만을 차단한다. 아미노프테린 또는 메토트렉세이트가 사용되는 경우, 배지에는 뉴클레오티드의 공급원으로서 하이포크산틴 및 티미딘이 보충된다(HAT 배지). 아자세린이 사용되는 경우, 배지에는 하이포크산틴이 보충된다. 골수종에 융합되지 않은 EBV 형질전환 세포주를 제거하기 위해, B 세포 공급원이 EBV 형질전환된 인간 B 세포주인 경우, 쿠아바인(Ouabain)이 추가된다.
바람직한 선택 매체는 HAT 또는 쿠아바인을 갖는 HAT이다. 뉴클레오티드 구조 경로를 조작할 수 있는 세포만이 HAT 배지에서 생존할 수 있다. 골수종 세포는 구조 경로의 중요한 효소(예: 하이포크산틴 포스포리보실 트랜스퍼라제(HPRT))에 결함이 있고, 생존할 수 없다. B 세포는 이 경로를 조작할 수 있지만, 배양 수명은 제한되어 있고, 통상은 약 2주 이내에 사망한다. 따라서, 선택적 배지에서 생존할 수 있는 유일한 세포는 골수종과 B 세포에서 형성된 하이브리드이다. 융합에 사용되는 B 세포의 공급원이 EBV-형질전환된 B 세포의 계통인 경우, 여기에서와 같이, EBV-형질전환된 B 세포는 약물 사멸에 감수성이 있기 때문에, 오우아바인은 하이브리드의 약물 선택에도 사용될 수 있지만, 사용되는 골수종 파트너는 오우아바인 내성으로 되도록 선택되었다.
배양은 특정 하이브리도마가 선택되는 하이브리도마 모집단을 제공한다. 일반적으로, 하이브리도마의 선택은 미세역가 플레이트에서 단일-클론 희석에 의해 세포를 배양하고, 이어서 개별 클론 상청액(약 2-3주 후)을 원하는 반응성에 대해 시험함으로써 수행된다. 분석은 방사면역분석, 효소 면역분석, 세포독성 분석, 플라크 분석 도트 면역결합 분석 등의 민감하고 간단하고 신속해야 한다. 이어서, 선택된 하이브리도마는 유세포 분석 분류에 의해 연속 희석하거나 단일 세포로 선별하고, 개별 항체 생산 세포주로 클로닝하고, 이어서 이 클론을 무기한으로 증식시켜 mAb를 제공할 수 있다. 세포주는 2개 기본 방식으로 MAb 생산에 이용될 수 있다. 하이브리도마 샘플은 동물(예: 마우스)에 (종종 복강으로) 주사할 수 있다. 임의로, 동물은 주사 전에 탄화수소, 특히 프리스탄(테트라메틸펜타데칸) 등의 오일로 프라이밍된다. 인간 하이브리도마를 이와 같이 사용하는 경우, 종양 거부반응을 방지하기 위해 SCID 마우스 등의 면역부전 마우스를 주사하는 것이 최적이다. 주사된 동물은 융합된 세포 하이브리드에 의해 생성된 특정 모노클로날 항체를 분비하는 종양을 발증시킨다. 이어서, 혈청 또는 복수액 등의 동물의 체액을 탭핑하여 고농도의 MAb를 제공할 수 있다. 개별 세포주는 또한 시험관내에서 배양될 수 있고, 여기서 MAb는 배양 배지로 자연적으로 분비되고, 이로부터 고농도로 용이하게 수득할 있다. 또는, 인간 하이브리도마 세포주는 세포 상청액에서 면역글로불린을 생성하기 위해 시험관내에서 사용할 수 있다. 세포주는 무혈청 배지에서의 성장에 적합시켜, 고순도의 인간 모노클로날 면역글로불린을 회수하는 능력을 최적화할 수 있다.
어느 하나의 수단에 의해 생성된 MAb는, 필요에 따라, 여과, 원심분리 및 FPLC 또는 친화성 크로마토그래피 등의 다양한 크로마토그래피 방법을 사용하여 추가로 정제할 수 있다. 본 개시의 모노클로날 항체의 단편은 펩신 또는 파파인 등의 효소에 의한 소화를 포함하는 방법에 의해 및/또는 화학적 환원에 의한 디설파이드 결합의 절단에 의해 정제된 모노클로날 항체로부터 수득될 수 있다. 또는, 본 개시에 포함되는 모노클로날 항체 단편은 자동화된 펩타이드 합성기를 사용하여 합성될 수 있다.
또한, 모노클로날 항체를 생성하기 위해 분자 클로닝 접근법을 사용할 수 있는 것으로 고려된다. 목적 항원으로 표지된 단일 B 세포는 상자성 비드 선택 또는 유세포 분석에 의한 선별을 사용하여 물리적으로 선별될 수 있고, 이어서 단일 세포로부터 RNA를 단리하고, RT-PCR에 의해 항체 유전자를 증폭시킬 수 있다. 또는, 항원 특이적 벌크 선별된 세포 모집단은 미세소포로 분리되고, 정합된 중쇄 및 경쇄 가변 유전자는 중쇄 및 경쇄 앰플리콘의 물리적 결합 또는 소포로부터의 중쇄 및 경쇄 유전자의 일반적 바코드를 사용하여 단일 세포에서 회수된다. 단일 세포를 형성하는 정합된 중쇄 및 경쇄 유전자는 RT-PCR 프라이머 및 세포당 하나의 바코드로 전사물을 마킹하기 위한 바코드를 함유하는 세포 투과성 나노입자로 세포를 처리함으로써 항원 특이적 B 세포의 모집단으로부터 수득할 수 있다. 항체 가변 유전자는 또한 하이브리도마 계통의 RNA 추출에 의해 단리할 수 있고, 항체 유전자는 RT-PCR에 의해 수득되고, 면역글로불린 발현 벡터에 클로닝될 수 있다. 또는, 조합 면역글로불린 파지미드 라이브러리는 세포주로부터 단리된 RNA로부터 제조되고, 적절한 항체를 발현하는 파지미드는 바이러스 항원을 사용하여 패닝함으로써 선택된다. 종래 하이브리도마 기술에 대한 이 접근법의 장점은 1회 라운드에서 약 104배 보다 많은 항체를 생산 및 스크리닝할 수 있고, H 및 L 쇄의 조합에 의해 새로운 특이성이 생성되어 적절한 항체를 발견할 가능성이 보다 높아진다는 것이다.
본 개시에 유용한 항체의 생산을 교시하는, 각각이 참조에 의해 본 명세서에 도입되는 기타 미국 특허에는 조합 접근법을 사용한 키메라 항체의 생산을 기재하는 미국 특허 제5,565,332호; 재조합 면역글로불린 조제물을 기재하는 미국 특허 제4,816,567호; 및 항체-치료제 접합체를 기재하는 미국 특허 제4,867,973호가 포함된다.
B. 본 개시의 항체
본 개시에 따른 항체는 우선 이들의 결합 특이성에 의해 정의될 수 있다. 당해 기술분야의 숙련가에게 공지된 기술을 사용하여 소정의 항체의 결합 특이성/친화성을 평가함으로써, 당해 기술분야의 숙련가는 이러한 항체가 본 청구범위의 범위 내에 속하는지의 여부를 결정할 수 있다. 예를 들면, 소정 항체가 결합하는 에피토프는 항원 분자 내에 위치한 3개 이상(예를 들면, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20)의 아미노산의 단일 연속 서열(예: 도메인 내의 선형 에피토프)로 이루어질 수 있다. 또는, 에피토프는 항원 분자 내에 위치한 복수의 비인접 아미노산(또는 아미노산 서열)으로 구성될 수 있다(예를 들면, 배좌 에피토프).
당업자에게 공지된 다양한 기술을 사용하여 항체가 폴리펩타이드 또는 단백질 내에서 "하나 이상의 아미노산과 상호작용"하는지의 여부를 결정할 수 있다. 예시적 기술은, 예를 들면, 문헌[참조: Antibodies, Harlow and Lane(Cold Spring Harbor Press, Cold Spring Harbor, N.Y.)]에 기재된 것 등의 일상적 교차-차단 검정을 포함한다. 교차 차단은 ELISA, 생물층 간섭계 또는 표면 플라즈몬 공명 등의 다양한 결합 검정에서 측정할 수 있다. 다른 방법에는 알라닌 스캐닝 돌연변이 분석, 펩타이드 블롯 분석[참조: Reineke (2004) Methods Mol. Biol. 248: 443-63], 펩타이드 절단 분석, 단일 입자 재구성을 사용한 고해상도 전자 현미경 기술, cryoEM 또는 단층 촬영, 결정학 연구 및 NMR 분석이 포함된다. 또한, 에피토프 절제, 에피토프 추출 및 항원의 화학적 변형 등의 방법을 사용할 수 있다[참조: Tomer (2000) Prot. Sci. 9: 487-496]. 항체가 상호작용하는 폴리펩타이드 내의 아미노산을 동정하기 위해 사용할 수 있는 또 다른 방법은 질량 분석에 의해 검출되는 수소/중수소 교환이다. 일반적으로, 수소/중수소 교환 방법은, 목적 단백질을 중수소로 표지한 후, 항체를 중수소로 표지된 단백질에 결합시키는 방식이다. 이어서, 단백질/항체 복합체는 물로 이동하고, 항체 복합체에 의해 보호되는 아미노산 내의 교환 가능한 양성자는 계면의 일부가 아닌 아미노산 내의 교환 가능한 양성자보다 느린 속도로 중수소로부터 수소로의 역교환을 받는다. 결과적으로, 단백질/항체 계면의 일부를 형성하는 아미노산은 중수소를 보유할 수 있기 때문에, 계면에 포함되지 않은 아미노산과 비교하여 비교적 높은 질량을 나타낼 수 있다. 항체의 해리 후, 표적 단백질은 프로테아제 절단 및 질량 분석 분석에 제공되고, 이에 의해 항체가 상호작용하는 특정 아미노산에 대응하는 중수소 표지된 잔기가 명백해진다[참조: 예를 들면, Ehring (1999) Analytical Biochemistry 267: 252-259; Engen and Smith(2001) Anal. Chem. 73: 256A-265A]. 항체가 EV-D68을 중화하는 경우, 고농도의 항체 존재하에 EV-D68을 시험관내 또는 동물 모델에서 EV-D68을 전파함으로써 항체 회피 돌연변이 변이체를 단리할 수 있다. 항체에 의해 표적화된 항원을 암호화하는 EV-D68 유전자의 서열 분석에 의해, 항체 회피를 제공하는 돌연변이(들)가 명백해지고, 에피토프 내의 잔기 또는 에피토프의 구조에 알로스테릭하게 영향을 미치는 것이 나타난다.
용어 "에피토프"는 B 및/또는 T 세포가 반응하는 항원 상의 부위를 지칭한다. B 세포 에피토프는 단백질의 3차 폴딩에 의해 병치된 인접 아미노산 또는 비인접 아미노산 모두로부터 형성될 수 있다. 인접한 아미노산으로부터 형성된 에피토프는 통상 변성 용매에 노출되어도 유지되는 반면, 3차 폴딩에 의해 형성된 에피토프는 통상 변성 용매로 처리하면 손실된다. 에피토프는 전형적으로 독특한 공간적 배좌로 적어도 3개, 보다 일반적으로 적어도 5개 또는 8-10개의 아미노산을 포함한다.
항원 구조-기반 항체 프로파일링(Antigen Structure-based Antibody Profiling; ASAP)이라고도 하는 변형-지원 프로파일링(Modification-Assisted Profiling; MAP)은 화학적으로 또는 효소적으로 변형된 항원 표면에 대한 각 항체의 결합 프로파일의 유사성에 따라 동일한 항원에 대해 지시된 다수의 모노클로날 항체(mAb)를 분류하는 방법이다[참조: US 2004/0101920, 참조에 의해 그 전체가 구체적으로 본 명세서에 도입됨). 각 카테고리는 다른 카테고리로 대표되는 에피토프와는 명백하게 상이하거나, 부분적으로 중복하는 고유한 에피토프를 반영할 수 있다. 이 기술에 의해, 유전적으로 동일한 항체를 신속하게 필터링할 수 있고, 유전적으로 상이한 항체에 초점을 맞추어 특성화를 수행할 수 있다. 하이브리도마 스크리닝에 적용하는 경우, MAP는 목적하는 특성을 갖는 mAb를 생성하는 희귀 하이브리도마 클론의 동정을 용이하게 할 수 있다. MAP를 사용하여, 본 개시의 항체를 상이한 에피토프에 결합하는 항체 그룹으로 분류할 수 있다.
본 개시는 동일한 에피토프, 또는 에피토프의 일부에 결합할 수 있는 항체를 포함한다. 마찬가지로, 본 개시는 또한, 표적 또는 이의 단편에 대한 결합에 대해 본 명세서에 기재된 임의의 특정 예시적 항체와 경쟁하는 항체를 포함한다. 당해 기술분야에 공지된 통상의 방법을 사용하여, 항체가 참조 항체와 동일한 에피토프에 결합하는지, 또는 결합에 대해 경쟁하는지의 여부를 용이하게 결정할 수 있다. 예를 들면, 시험 항체가 참조와 동일한 에피토프에 결합하는지를 결정하기 위해, 참조 항체는 포화 조건에서 표적에 결합시킨다. 이어서, 표적 분자에 결합하는 시험 항체의 능력이 평가된다. 시험 항체가 참조 항체와의 포화 결합 후에 표적 분자에 결합할 수 있는 경우, 시험 항체는 참조 항체와는 상이한 에피토프에 결합한다고 결론지을 수 있다. 반면에, 참조 항체와의 포화 결합 후에 시험 항체가 표적 분자에 결합할 수 없는 경우, 시험 항체는 참조 항체에 의해 결합된 에피토프와 동일한 에피토프에 결합할 수 있다.
항체가 참조 항-EV-D68 항체와의 결합에 대해 경쟁하는지의 여부를 결정하기 위해, 상기에서 설명된 결합 방법을 2개 방향으로 수행한다. 제1 방향에서, 참조 항체는 포화 조건하에서 EV-D68 항원에 결합하고, 이어서 EV-D68 항원에 대한 시험 항체의 결합을 평가한다. 제2 방향에서, 시험 항체를 포화 조건하에서 EV-D68 항원 분자에 결합시키고, 이어서 EV-D68 항원에 대한 참조 항체의 결합을 평가한다. 양 방향 모두에서, 제1(포화) 항체만이 EV-D68에 결합할 수 있는 경우, 시험 항체와 참조 항체는 EV-D68에 대한 결합에 대해 경쟁하는 것으로 결론지어진다. 당업자에 의해 이해되는 바와 같이, 참조 항체와의 결합을 위해 경쟁하는 항체는 반드시 참조 항체와 동일한 에피토프에 결합할 필요는 없지만, 중첩 또는 인접한 에피토프에 결합함으로써 참조 항체의 결합을 입체적으로 차단할 수 있다.
각각이 항원에 대한 다른 항체의 결합을 경쟁적으로 억제(차단)하는 경우, 2개의 항체는 동일하거나 중첩되는 에피토프에 결합한다. 즉, 1-, 5-, 10-, 20- 또는 100배 과량의 하나의 항체는, 경쟁 결합 검정에서 측정하는 경우, 다른 항체의 결합을 적어도 50%, 그러나 바람직하게는 75%, 90% 또는 심지어 99% 억제한다[참조: 예를 들면, Junghans et al., Cancer Res. 1990 50:1495-1502). 또는, 한 항체의 결합을 감소 또는 제거하는 항원의 본질적으로 모든 아미노산 돌연변이가 다른 항체의 결합을 감소 또는 제거하는 경우, 2개의 항체는 동일한 에피토프를 갖는다. 한 항체의 결합을 감소 또는 제거하는 일부 아미노산 돌연변이가 다른 항체의 결합을 감소 또는 제거하는 경우, 2개의 항체는 중첩 에피토프를 갖는다.
이어서, 추가의 통상의 실험(예를 들면, 펩타이드 돌연변이 및 결합 분석)을 실시하여, 관찰된 시험 항체의 결합의 결여가 실제로 참조 항체와 동일한 에피토프에 대한 결합에 의한 것인지, 또는 입체적 차단(또는 다른 현상)이 관찰된 결합의 결여의 원인지를 확인할 수 있다. 이러한 종류의 실험은 ELISA, RIA, 표면 플라즈몬 공명, 유세포 분석 또는 당해 기술분야에서 이용 가능한 임의의 기타 정량적 또는 정성적 항체 결합 검정을 사용하여 수행할 수 있다. EM 또는 결정학을 사용한 구조 연구는 결합을 위해 경쟁하는 2개의 항체가 동일한 에피토프를 인식하는지의 여부를 입증할 수도 있다.
또 다른 측면에서, 각각 표 3 및 4에 예시된 바와 같이, 중쇄 및 경쇄로부터 클론-쌍 CDR을 갖는 모노클로날 항체가 제공된다. 이러한 항체는 본 명세서에 기재된 방법을 사용하여 하기 실시예 섹션에서 논의된 클론에 의해 생성될 수 있다.
또 다른 측면에서, 항체는 추가의 "프레임워크" 영역을 포함하는 가변 서열에 의해 정의될 수 있다. 이들은 완전한 가변 영역을 암호화하거나 나타내는 표 1 및 2에 제공된다. 추가로, 항체 서열은, 임의로 하기에 보다 상세히 논의되는 방법을 사용하여 이들 서열과는 상이할 수 있다. 예를 들면, 핵산 서열은 (a) 가변 영역이 경쇄 및 중쇄의 불변 도메인으로부터 분리될 수 있고, (b) 핵산이 이에 의해 암호화된 잔기에 영향을 미치지 않고, 상기의 것과는 상이할 수 있고, (c) 핵산이 소정 백분율, 예를 들면, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 상동성으로 상기 핵산과는 상이할 수 있고, (d) 핵산이, 약 50℃ 내지 약 70℃의 온도에서 약 0.02M 내지 약 0.15M NaCl에 의해 제공되는 것과 같은 저염 및/또는 고온 조건하에서 예시되는 바와 같이, 높은 엄격성 조건하에서 하이브리드화하는 능력으로 인해 상기의 것과는 상이할 수 있고, (e) 아미노산이 소정 백분율, 예를 들면, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 상동성으로 상기 기재된 것과는 상이할 수 있고, 또는 (f) 아미노산이 보존적 치환(하기 설명됨)을 가능하게 함으로써 상기 기재된 것과는 상이할 수 있다는 점에서 상기 기재된 것들과는 상이할 수 있다. 상기의 각각은 표 1에 기재된 핵산 서열 및 표 2의 아미노산 서열에 적용된다.
폴리뉴클레오티드와 폴리펩타이드 서열을 비교하는 경우, 하기에 기재된 바와 같이, 최대 대응을 위해 정렬시킬 때에 2개의 서열의 뉴클레오티드 또는 아미노산 서열이 동일한 경우, 2개 서열은 "동일한" 것이라고 한다. 2개 서열 간의 비교는 통상 비교 윈도우에서 서열을 비교하여 서열 유사성의 국소 영역을 특정 및 비교함으로써 수행된다. 본 명세서에 사용된 "비교 윈도우"는 적어도 약 20개의 연속하는 위치, 통상 30 내지 약 75, 40 내지 약 50개의 세그먼트를 지칭하고, 여기서 서열은, 2개 서열이 최적으로 정렬된 후에 동일한 수의 연속하는 위치의 참조 서열과 비교할 수 있다.
비교를 위한 최적의 서열 정렬은 디펄트 파라미터를 사용하여 생물정보학 소프트웨어의 레이저진(Lasergene) 제품군(DNASTAR, Inc., Madison, WI)에서 메그얼라인(Megalign) 프로그램을 사용하여 수행할 수 있다. 이 프로그램은 하기 참고 문헌에 설명된 몇몇 정렬 방식을 구현한다: Dayhoff, M. O. (1978) A model of evolutionary change in proteins--Matrices for detecting distant relationships. In Dayhoff, M. O. (ed.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Washington D.C. Vol. 5, Suppl. 3, pp. 345-358; Hein J. (1990) Unified Approach to Alignment and Phylogeny pp. 626-645 Methods in Enzymology vol. 183, Academic Press, Inc., San Diego, Calif.; Higgins, D. G. and Sharp, P. M. (1989) CABIOS 5:151-153; Myers, E. W. and Muller W. (1988) CABIOS 4:11-17; Robinson, E. D. (1971) Comb. Theor 11:105; Santou, N. Nes, M. (1987) Mol. Biol. Evol. 4:406-425; Sneath, P. H. A. and Sokal, R. R. (1973) Numerical Taxonomy--the Principles and Practice of Numerical Taxonomy, Freeman Press, San Francisco, Calif.; Wilbur, W. J. and Lipman, D. J. (1983) Proc. Natl. Acad., Sci. USA 80:726-730.
또는, 비교를 위한 최적의 서열 정렬은 스미스 및 워터슨(Smith and Waterman (1981) Add. APL. Math 2:482)의 국소 동일성 알고리즘에 의해, 니들맨 및 운쉬(Needleman and Wunsch (1970) J. Mol. Biol. 48:443)의 동일성 정렬 알고리즘에 의해, 피어슨 및 립맨(Pearson and Lipman (1988) Proc. Natl. Acad. Sci. USA 85: 2444)의 유사성 검색 방법에 의해, 이들 알고리즘의 컴퓨터화 실장(GAP, BESTFIT, BLAST, FASTA, and TFASTA in the Wisconsin Genetics Software Package, Genetics Computer Group (GCG), 575 Science Dr., Madison, Wis.)에 의해 또는 검사에 의해 수행할 수 있다.
퍼센트 서열 동일성 및 서열 유사성을 결정하는 데 적합한 알고리즘의 한 특정 예는 문헌[참조: Altschul et al. (1977) Nucl. Acids Res. 25:3389-3402 and Altschul et al. (1990) J. Mol. Biol. 215:403-410]에 각각 기재되어 있는 BLAST 및 BLAST 2.0 알고리즘이다. BLAST 및 BLAST 2.0은, 예를 들면, 본 명세서에 기재된 파라미터와 함께, 본 개시의 폴리뉴클레오티드 및 폴리펩타이드에 대한 퍼센트 서열 동일성을 결정하기 위해 사용될 수 있다. BLAST 분석을 수행하기 위한 소프트웨어는 국립 생명공학 정보 센터를 통해 공개적으로 사용 가능하다. 항체 서열의 재배열된 특성과 각 유전자의 가변 길이는 단일 항체 서열에 대한 복수회 라운드의 BLAST 검색을 필요로 한다. 또한, 서로 상이한 유전자를 수동으로 조립하는 것은 곤란하고, 에러가 발생하기 쉽다. 서열 분석 도구 IgBLAST(월드 와이드 웹, ncbi.nlm.nih.gov/igblast/)는 생식세포계 V, D 및 J 유전자에 대한 정합, 재배열 접합부의 상세, Ig V 도메인 프레임워크 영역 상보성 결정 영역의 묘사를 식별한다. IgBLAST는 뉴클레오티드 또는 단백질 서열을 분석하고, 서열을 배치로 처리하고, 생식세포계 유전자 데이터베이스 및 기타 서열 데이터베이스를 동시에 검색하여, 가장 정합하는 생식세포계 V 유전자를 누락될 가능성을 최소화한다.
예시적 일례에서, 누적 스코어는 뉴클레오티드 서열에 대해, 파라미터 M(정합하는 잔기 쌍에 대한 보상 스코어; 항상 >0) 및 N(부정합하는 잔기에 대한 패널티 스코어; 항상 <0)을 사용하여 계산할 수 있다. 각 방향의 단어 히트의 확장은 다음과 같은 경우에 중지된다: 누적 정렬 스코어가 최대 달성 값으로부터 수량 X만큼 저하되거나; 하나 이상의 음수 스코어의 잔기 정렬의 누적에 의해, 누적 스코어가 0 이하가 되거나; 또는 어느 서열의 말단에 도달한 경우. BLAST 알고리즘 파라미터 W, T 및 X는 정렬의 감도 및 속도를 결정한다. BLASTN 프로그램(뉴클레오티드 서열용)은, 디펄트로서, 단어 길이(W) 11, 기대값(E) 10, 및 BLOSUM62 스코어링 매트릭스[참조: Henikoff 및 Henikoff(1989) Proc. Natl. Acad. Sci. USA 89:10915] 정렬, (B) 50, 기대치(E) 10, M=5, N=-4 및 양 가닥의 비교를 사용한다.
아미노산 서열의 경우, 스코어링 매트릭스를 사용하여 누적 스코어를 계산할 수 있다. 각 방향의 단어 히트 확장은 다음과 같은 경우에 중지된다: 누적 정렬 스코어가 최대 달성 값에서 수량 X만큼 저하되거나; 하나 이상의 음의 스코어 잔기 정렬의 누적으로 인해 누적 스코어가 0 이하가 되거나, 또는 어느 서열의 말단에 도달한 경우. BLAST 알고리즘 파라미터 W, T 및 X는 정렬의 감도와 속도를 결정한다.
하나의 접근법에서, "서열 동일성의 백분율"은 적어도 20개의 위치의 비교 윈도우에 걸쳐 2개의 최적으로 정렬된 서열을 비교함으로써 결정되고, 여기서 비교 윈도우에서 폴리뉴클레오티드 또는 폴리펩타이드 서열의 일부는 2개 서열의 최적 정렬을 위한 참조 서열(이는 부가 또는 결실을 포함하지 않음)과 비교하여 20% 이하, 통상 5 내지 15%, 또는 10 내지 12%의 부가 또는 결실(즉, 갭)을 포함할 수 있다. 백분율은, 양 서열에서 동일한 핵산 염기 또는 아미노산 잔기가 존재하는 위치의 수를 결정하여 정합된 위치의 수를 산출하고, 정합된 위치의 수를 참조 서열의 위치의 총 수(즉, 윈도위 크기)로 나누고, 결과에 100을 곱하여 서열 동일성의 백분율을 산출함으로써 계산된다.
항체를 정의하는 또 다른 방법은 하기에 기술된 항체 및 이의 항원 결합 단편의 임의의 "유도체"로 하는 것이다. 용어 "유도체"는 항원에 면역특이적으로 결합하지만, "모체"(또는 야생형) 분자와 비교하여 1, 2, 3, 4, 5 또는 그 이상의 아미노산 치환, 부가, 결실 또는 변형을 포함하는 항체 또는 이의 항원 결합 단편을 지칭한다. 이러한 아미노산 치환 또는 부가는 천연 존재(즉, DNA 암호화됨) 또는 비-천연 존재 아미노산 잔기를 도입할 수 있다. 용어 "유도체"는, 예를 들면, 향상된 또는 손상된 이펙터 또는 결합 특성을 나타내는 변이체 Fc 영역을 갖는, 예를 들면, 항체 등을 형성하기 위해 변경된 CH1, 힌지, CH2, CH3 또는 CH4 영역을 갖는 변이체를 포함한다. 용어 "유도체"는 비-아미노산 변형, 예를 들면, 글리코실화(예를 들면, 변화된 만노스, 2-N-아세틸글루코사민, 갈락토스, 푸코스, 글루코스, 시알산, 5-N-아세틸뉴라민산, 5-글리콜뉴라민산 등의 함량을 가짐), 아세틸화, 페길화, 인산화, 아미드화, 공지된 보호/차단 그룹에 의해 유도체화, 단백질 분해 절단, 세포 리간드 또는 다른 단백질 등에 연결될 수 있는 아미노산을 추가로 포함한다. 일부 실시양태에서, 변경된 탄수화물 변형은 하기 중 하나 이상을 조절한다: 항체의 가용화, 항체의 세포내 수송 및 분비의 촉진, 항체 조립의 촉진, 형태적 완전성 및 항체 매개 이펙터 기능. 특정 실시양태에서, 변경된 탄수화물 변형은 탄수화물 변형을 결여하는 항체와 비교하여 항체 매개 이펙터 기능을 향상시킨다. 변경된 항체 매개된 이펙터 기능을 유도하는 탄수화물 변형은 당해 기술분야에 공지되어 있다[참조: 예를 들면, Shields, R. L. et al. (2002) “Lack Of Fucose On Human IgG N-Linked Oligosaccharide Improves Binding To Human Fcgamma RIII And Antibody-Dependent Cellular Toxicity,” J. Biol. Chem. 277(30): 26733-26740; Davies J. et al. (2001) “Expression Of GnTIII In A Recombinant Anti-CD20 CHO Production Cell Line: Expression Of Antibodies With Altered Glycoforms Leads To An Increase In ADCC Through Higher Affinity For FC Gamma RIII,” Biotechnology & Bioengineering 74(4): 288-294]. 탄수화물 함량을 변경하는 방법은 당업자에게 공지되어 있다[참조: 예를 들면, Wallick, S. C. et al. (1988) “Glycosylation Of A VH Residue Of A Monoclonal Antibody Against Alpha (1----6) Dextran Increases Its Affinity For Antigen,” J. Exp. Med. 168(3): 1099-1109; Tao, M. H. et al. (1989) “Studies Of Aglycosylated Chimeric Mouse-Human IgG. Role Of Carbohydrate In The Structure And Effector Functions Mediated By The Human IgG Constant Region,” J. Immunol. 143(8): 2595-2601; Routledge, E. G. et al. (1995) “The Effect Of Aglycosylation On The Immunogenicity Of A Humanized Therapeutic CD3 Monoclonal Antibody,” Transplantation 60(8):847-53; Elliott, S. et al. (2003) “Enhancement Of Therapeutic Protein In Vivo Activities Through Glycoengineering,” Nature Biotechnol. 21:414-21; Shields, R. L. et al. (2002) “Lack Of Fucose On Human IgG N-Linked Oligosaccharide Improves Binding To Human Fcgamma RIII And Antibody-Dependent Cellular Toxicity,” J. Biol. Chem. 277(30): 26733-26740].
유도체 항체 또는 항체 단편은, 비드-기반 또는 세포-기반 검정 또는 동물 모델에서 생체내 연구에 의해 측정하는 경우, 항체 의존성 세포성 세포독성(ADCC), 항체 의존성 세포 식세포 작용(ADCP), 항체 의존성 호중구 식세포 작용(ADNP) 또는 항체 의존성 보체 침착(ADCD) 기능에서 바람직한 수준의 활성을 부여하기 위해 조작된 서열 또는 글리코실화 상태로 생성될 수 있다.
유도체 항체 또는 항체 단편은 특정의 화학적 절단, 아세틸화, 제형화, 투니카마이신의 대사 합성 등을 포함하지만 이들로 한정되지 않는 당업자에게 공지된 기술을 사용하여 화학적 변형에 의해 변형될 수 있다. 일 실시양태에서, 항체 유도체는 모 항체와 유사하거나 동일한 기능을 가질 것이다. 또 다른 실시양태에서, 항체 유도체는 모 항체와 비교하여 변화된 활성을 나타낼 것이다. 예를 들면, 유도체 항체(또는 이의 단편)는 모 항체보다도 이의 에피토프에 보다 강력하게 결합하거나, 또는 단백질 분해에 대해 보다 내성이 있을 수 있다.
C. 항체 서열의 조작
다양한 실시양태에서, 개선된 발현, 개선된 교차 반응성 또는 감소된 표적외 결합 등의 다양한 이유로 동정된 항체의 서열을 조작하도록 선택할 수 있다. 변형된 항체는 표준 분자 생물학적 기술을 통한 발현, 또는 폴리펩타이드의 화학적 합성을 포함하는, 당업자에게 공지된 임의의 기술에 의해 제조될 수 있다. 재조합 발현 방법은 이 문서의 다른 곳에서 취급한다. 하기는 항체 공학에 대한 관련 목표 기술에 대한 일반적 논의이다.
하이브리도마를 배양하고, 이어서 세포를 용해하고, 총 RNA를 추출할 수 있다. 랜덤 6량체를 RT와 함께 사용하여 RNA의 cDNA 카피를 생성하고, 이어서 모든 인간 가변 유전자 서열을 증폭할 것으로 예상되는 PCR 프라이머의 다중 혼합물을 사용하여 PCR을 수행할 수 있다. PCR 산물은 pGEM-T Easy 벡터에 클로닝하고, 표준 벡터 프라이머를 사용하여 자동화된 DNA 서열분석에 의해 서열분석할 수 있다. 결합 및 중화의 검정은 하이브리도마 상청액으로부터 수집된 항체를 사용하여 수행할 수 있고, 단백질 G 컬럼을 사용하여 FPLC로 정제할 수 있다.
재조합 전장 IgG 항체는 중쇄 및 경쇄 Fv DNA를 클로닝 벡터로부터 IgG 플라스미드 벡터로 서브클로닝하고, 293(예: Freestyle) 세포 또는 CHO 세포로 형질감염시킴으로써 생성할 수 있고, 항체는 293 또는 CHO 세포 상청액으로부터 수집 및 정제할 수 있다. 다른 적절한 숙주 세포 시스템에는 대장균 등의 세균, 곤충 세포(S2, Sf9, Sf29, 하이 파이브(High Five)), 식물 세포(예: 담배, 인간-유사 글리칸에 대한 조작의 존재 또는 부재), 조류 또는 마우스, 랫트, 염소 또는 소 등의 다양한 비인간 유전자도입 환경이 포함된다.
후속 항체 정제 및 숙주의 면역화 둘 다의 목적으로, 항체를 암호화하는 핵산의 발현이 또한 고려된다. 항체 코딩 서열은 천연 RNA 또는 변형된 RNA 등의 RNA일 수 있다. 변형된 RNA는 mRNA에 안정성 증가와 면역원성의 저하를 부여하고, 이에 의해 치료적으로 중요한 단백질의 발현을 촉진하는 특정 화학적 변형을 고려한다. 예를 들면, N1-메틸-슈도우리딘(N1mΨ)은 번역 능력의 점에서 기타 몇몇 뉴클레오사이드 변형 및 이들의 조합을 능가한다. 면역/eIF2α 인산화 의존성의 번역 억제를 오프로 하는 것에 추가하여, 도입된 N1mΨ 뉴클레오티드는 리보솜의 일시 정지 및 mRNA의 밀도를 증가시킴으로써 번역 프로세스의 역학을 극적으로 변화시킨다. 변형된 mRNA의 리보솜 로딩의 증가는 동일한 mRNA에서 리보솜 재순환 또는 새로운 리보솜 동원을 조장함으로써 이들을 개시에 대해 보다 관대하게 한다. 이러한 변형은 RNA 접종 후에 생체내에서 항체 발현을 향상시키기 위해 사용될 수 있다. RNA는, 천연 또는 변형된 것이든, 네이키드 RNA로서 또는 지질 나노입자 등의 전달 비히클에서 전달될 수 있다.
또는, 항체를 암호화하는 DNA를 동일한 목적으로 사용될 수 있다. DNA는 그것이 설계된 숙주 세포에서 활성인 프로모터를 포함하는 발현 카세트에 포함된다. 발현 카세트는 유리하게는 종래의 플라스미드 또는 미니벡터 등의 복제 가능한 벡터에 포함된다. 벡터는 폭스바이러스, 아데노바이러스, 헤르페스바이러스, 아데노-연관 바이러스 등의 바이러스 벡터를 포함하고, 렌티바이러스가 고려된다. VEE 바이러스 또는 신드비스(Sindbis) 바이러스에 기초한 알파바이러스 레플리콘 등의 항체 유전자를 암호화하는 레플리콘도 또한 고려된다. 이러한 벡터의 전달은 근육내, 피하 또는 피내 경로를 통한 바늘에 의해, 또는 생체내 발현이 요구되는 경우는 경피 전기천공에 의해 수행될 수 있다.
최종 cGMP 제조 프로세스와 동일한 숙주 세포 및 세포 배양 프로세스에서 생산된 항체의 신속한 가용성은 프로세스 개발 프로그램의 기간을 단축할 가능성이 있다. 론자(Lonza)는 CHO 세포에서 소량(최대 50g)의 항체를 신속하게 생산하기 위해 CDACF 배지에서 성장한 풀링된 형질감염체를 사용하는 일반적 방법을 개발했다. 진정한 일시적 시스템보다 약간 느리긴 하지만, 장점에는 보다 높은 제품 농도와 생산 세포주와 동일한 숙주 및 프로세스의 사용이 포함된다. 일회용 생물반응기에서 모델 항체를 발현하는 GS-CHO 풀의 성장 및 생산성의 예: 유가-배치 모드로 작동되는 일회용 백 생물반응기 배양물(5L 작업 용적)에서는 2g/L의 수확 항체 농도가 형질전환으로부터 9주 이내에 달성되었다.
항체 분자는, 예를 들면, mAb의 단백질분해 절단에 의해 생산되는 단편(예: F(ab'), F(ab')2), 또는, 예를 들면, 재조합 수단을 통해 생산 가능한 일본쇄 면역글로불린을 포함할 것이다. F(ab') 항체 유도체는 1가인 반면, F(ab')2 항체 유도체는 2가이다. 일 실시양태에서, 이러한 단편은 서로 조합되거나 다른 항체 단편 또는 수용체 리간드와 조합되어 "키메라" 결합 분자를 형성할 수 있다. 중요하게도, 이러한 키메라 분자는 동일한 분자의 상이한 에피토프에 결합할 수 있는 치환기를 함유할 수 있다.
관련 실시양태에서, 항체는 개시된 항체의 유도체, 예를 들면, 개시된 항체의 것과 동일한 CDR 서열을 포함하는 항체 (예를 들면, 키메라 또는 CDR-이식된 항체)이다. 또는, 항체 분자에 보존적 변화를 도입하는 것 등의 변형을 수행하는 것을 원할 수 있다. 이러한 변경을 수행할 때, 아미노산의 소수성 지수를 고려할 수 있다. 단백질에 상호작용적 생물학적 기능을 부여할 때의 수치료학적 아미노산 지수의 중요성은 당해 기술분야에서 일반적으로 이해된다[참조: Kyte and Doolittle, 1982]. 아미노산의 상대적 소수성 특성은 수득되는 단백질의 2차 구조에 기여하는 것으로 인정되고, 이에 의해 단백질과 다른 분자(예: 효소, 기질, 수용체, DNA, 항체, 항원, 등)와의 상호작용이 정의된다.
유사 아미노산의 치환은 친수성에 기초하여 효과적으로 수행될 수 있음이 또한 당해 기술분야에서 이해된다. 참조에 의해 본 명세서에서 도입된 미국 특허 제4,554,101호는 인접 아미노산의 친수성에 의해 지배되는 단백질의 최대 국소 평균 친수성이 단백질의 생물학적 특성과 상관관계가 있다고 서술하고 있다. 미국 특허 제4,554,101호에 상세히 기술된 바와 같이, 하기 친수성 값이 아미노산 잔기에 할당되었다: 염기성 아미노산: 아르기닌(+3.0), 라이신(+3.0) 및 히스티딘(-0.5); 산성 아미노산: 아스파르테이트(+3.0±1), 글루타메이트(+3.0±1), 아스파라긴(+0.2) 및 글루타민(+0.2); 친수성, 비이온성 아미노산: 세린(+0.3), 아스파라긴(+0.2), 글루타민(+0.2) 및 트레오닌(-0.4), 황 함유 아미노산: 시스테인(-1.0) 및 메티오닌(-1.3); 소수성, 비방향족 아미노산: 발린(-1.5), 류신(-1.8), 이소류신(-1.8), 프롤린(-0.5±1), 알라닌(-0.5) 및 글리신(0); 소수성, 방향족 아미노산: 트립토판(-3.4), 페닐알라닌(-2.5) 및 티로신(-2.3).
아미노산은 유사한 친수성을 갖는 또 다른 것으로 치환될 수 있고, 생물학적으로 또는 면역학적으로 변형된 단백질을 생성할 수 있는 것으로 이해된다. 이러한 변화에 있어서, 친수성 값이 ±2 이내인 아미노산의 치환이 바람직하고, ±1 이내인 것이 특히 바람직하고, ±0.5 이내인 것이 더욱 특히 바람직하다.
상기에서 개설한 바와 같이, 아미노산 치환은 일반적으로 아미노산 측쇄 치환기의 상대적 유사성, 예를 들면, 이들의 소수성, 친수성, 전하, 크기 등을 기반으로 한다. 전술한 다양한 특성을 고려하는 예시적 치환은 당업자에게 공지되어 있고, 하기를 포함한다: 아르기닌 및 라이신; 글루타메이트 및 아스파르테이트; 세린 및 트레오닌; 글루타민 및 아스파라긴; 및 발린, 류신 및 이소류신.
본 개시는 또한 아이소타입 변형을 고려한다. 상이한 이소타입을 갖도록 Fc 영역을 변형함으로써 상이한 기능을 달성할 수 있다. 예를 들면, IgG1으로 변경하면, 항체 의존성 세포독성이 증가할 수 있고, 클래스 A로 스위칭하면 조직 분포가 개선될 수 있으며, 클래스 M으로 스위칭하면 원자가가 개선될 수 있다.
대안적으로 또는 추가로, 아미노산 변형을, IL-23p19 결합 분자의 Fc 영역의 C1q 결합 및/또는 보체 의존성 세포독성(CDC) 기능을 변화시키는 하나 이상의 추가 아미노산 변형과 조합하는 것이 유용할 수 있다. 특정 목적의 결합 폴리펩타이드는, C1q에 결합하고 보체 의존성 세포독성을 나타내는 것일 수 있다. 기존의 C1q 결합 활성을 갖고 임의로 CDC를 매개하는 능력을 추가로 갖는 폴리펩타이드는 이들 활성 중 하나 또는 둘 모두가 향상되도록 변형될 수 있다. C1q를 변경하고/하거나 이의 보체 의존적 세포독성 기능을 변경하는 아미노산 변형은, 예를 들면, 참조에 의해 본 명세서에 도입되는 제WO/0042072호에 기재되어 있다.
예를 들면, C1q 결합 및/또는 FcγR 결합을 변형하고, 이에 의해 CDC 활성 및/또는 ADCC 활성을 변화시킴으로써 이펙터 기능이 변화된 항체의 Fc 영역을 설계할 수 있다. "이펙터 기능"은 생물학적 활성을 활성화 또는 감소시키는 역할을 한다(예: 대상체에서). 이펙터 기능의 예는 하기를 포함하지만 이들로 한정되지 않는다: C1q 결합; 보체 의존성 세포독성(CDC); Fc 수용체 결합; 항체 의존성 세포-매개 세포독성(ADCC); 식균 작용; 세포 표면 수용체(예: B 세포 수용체; BCR) 등의 하향조절. 이러한 이펙터 기능은 Fc 영역을 결합 도메인(예: 항체 가변 도메인)과 결합시킬 필요가 있고, 다양한 검정(예: Fc 결합 검정, ADCC 검정, CDC 검정 등)을 사용하여 평가할 수 있다.
예를 들면, 개선된 C1q 결합 및 개선된 FcγRIII 결합(예를 들면, 개선된 ADCC 활성 및 개선된 CDC 활성 둘 모두를 가짐)을 갖는 항체의 변이체 Fc 영역을 생성할 수 있다. 또는, 이펙터 기능을 저하시키거나 제거하는 것이 바람직한 경우, 변이체 Fc 영역은 CDC 활성의 저하 및/또는 ADCC 활성의 저하를 수반하여 조작될 수 있다. 다른 실시양태에서, 이들 활성 중 하나만을 증가시킬 수 있고, 임의로 다른 활성도 감소시킬 수 있다(예를 들면, 개선된 ADCC 활성을 갖지만, CDC 활성을 감소시킨 Fc 영역 변이체를 생성하기 위해 및 그 반대의 경우).
FcRn 결합. Fc 돌연변이는 또한 신생아 Fc 수용체(FcRn)와의 상호작용을 변화시키고 약동학적 특성을 개선하기 위해 도입 및 조작할 수 있다. FcRn에 대한 결합이 개선된 인간 Fc 변이체의 콜렉션이 기재되어 있다[참조: Shields et al., (2001)]. FcγRI, FcγRII, FcγRIII 및 FcRn에 대한 인간 IgG1의 결합 부위의 고해상도 맵핑 및 FcγR에 대한 결합이 개선된 IgG1 변이체의 설계[참조: J. Biol. Chem. 276:6591-6604]. 반감기를 증가시킬 수 있는 다수의 방법이 공지되어 있고[참조: Kuo and Aveson, (2011)], 아미노산 변형을 포함하는 것은, 알라닌 스캐닝 돌연변이유발, 랜덤 돌연변이유발, 및 신생아 Fc 수용체(FcRn)에 대한 결합 및/또는 생체내 거동을 평가하기 위한 스크리닝을 포함하는 기술을 통해 생성될 수 있다. 컴퓨터 전략 및 이어서 돌연변이유발을 사용하여, 돌연변이시킬 아미노산 돌연변이 중의 하나를 선택할 수 있다.
따라서, 본 개시는 FcRn에 대한 최적화된 결합을 갖는 항원 결합 단백질의 변이체를 제공한다. 특정 실시양태에서, 항원 결합 단백질의 상기 변이체는 상기 항원 결합 단백질의 Fc 영역에 적어도 하나의 아미노산 변형을 포함하고, 여기서 상기 변형은 상기 모 폴리펩타이드와 비교하여 Fc 영역의 226, 227, 228, 230, 231, 233, 234, 239, 241, 243, 246, 250, 252, 256, 259, 264, 265, 267, 269, 270, 276, 284, 285, 288, 289, 290, 291, 292, 294, 297, 298, 299, 301, 302, 303, 305, 307, 308, 309, 311, 315, 317, 320, 322, 325, 327, 330, 332, 334, 335, 338, 340, 342, 343, 345, 347, 350, 352, 354, 355, 356, 359, 360, 361, 362, 369, 370, 371, 375, 378, 380, 382, 384, 385, 386, 387, 389, 390, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401 403, 404, 408, 411, 412, 414, 415, 416, 418, 419, 420, 421, 422, 424, 426, 428, 433, 434, 438, 439, 440, 443, 444, 445, 446 및 447로 이루어진 그룹으로부터 선택되고, 여기서 Fc 영역의 아미노산 넘버링은 카바트(Kabat)의 EU 인덱스의 넘버링이다. 본 개시의 추가 측면에서, 변형은 M252Y/S254T/T256E이다.
또한, 다양한 간행물은, FcRn 결합 폴리펩타이드를 분자에 도입하거나, FcRn 결합 친화성이 보존되어 있지만 다른 Fc 수용체에 대한 친화성은 크게 감소되어 있는 항체와 분자를 융합시키거나, 항체의 FcRn 결합 도메인과 융합시킴으로써 반감기가 변형된 생리학적 활성 분자를 수득하는 방법을 설명한다[참조: 예를 들면, Kontermann (2009)].
유도체화된 항체는 포유동물, 특히 인간에서 모 항체의 반감기(예를 들면, 혈청 반감기)를 변경하기 위해 사용될 수 있다. 이러한 변경은 15일 초과, 바람직하게는 20일 초과, 25일 초과, 30일 초과, 35일 초과, 40일 초과, 45일 초과, 2개월 초과, 3개월 초과, 4개월 초과 또는 5개월 초과의 반감기를 초래할 수 있다. 포유동물, 바람직하게는 인간에서 본 개시의 항체 또는 이의 단편의 반감기의 증가는 포유동물에서 상기 항체 또는 항체 단편의 보다 높은 혈청 역가를 초래하고, 따라서 상기 항체 또는 항체 단편의 투여 빈도를 감소시키고/시키거나, 투여되는 상기 항체 또는 항체 단편의 농도를 감소시킨다. 생체내 반감기가 증가된 항체 또는 이의 단편은 당업자에게 공지된 기술에 의해 생성될 수 있다. 예를 들면, Fc 도메인과 FcRn 수용체 사이의 상호작용에 관여하는 것으로 동정된 아미노산 잔기를 변형(예를 들면, 치환, 결실 또는 부가)함으로써 생체내 반감기가 증가된 항체 또는 이의 단편을 생성할 수 있다.
문헌[참조: Beltramello et al. (2010)]은, CH2 도메인의 위치 1.3과 1.2의 류신 잔기(C-도메인에 대한 IMGT 고유 넘버링에 따름)가 알라닌 잔기로 치환되어 있는 것을 생성함으로써, 뎅기 바이러스 감염을 향상시키는 경향이 있기 때문에, 중화 mAb의 변형을 이전에 보고했다. "LALA" 돌연변이로서도 공지된 이 변형은 문헌[참조: Hessell et al. (2007)]에 기재되어 있는 바와 같이, FcγRI, FcγRII 및 FcγRIIIa에 대한 항체 결합을 무력화한다. 변이체 및 비변형 재조합 mAb를 4개 뎅기열 바이러스 혈청형에 의한 감염을 중화하고 향상하는 능력에 대해 비교했다. LALA 변이체는 변형되지 않은 mAb와 동일한 중화 활성을 유지했지만, 향상 활성을 완전히 결여하고 있었다. 따라서, 이러한 성질의 LALA 돌연변이는 현재 개시된 항체와 관련하여 고려된다.
변경된 글리코실화. 본 개시의 특정 실시양태는, 시알산, 갈락토스 또는 푸코스를 포함하지 않는 실질적으로 균질한 글리칸을 함유하는, 단리된 모노클로날 항체 또는 이의 항원 결합 단편이다. 모노클로날 항체는 중쇄 가변 영역 및 경쇄 가변 영역을 포함하고, 이들 둘 모두는 각각 중쇄 또는 경쇄 불변 영역에 부착될 수 있다. 전술한 실질적으로 균질한 글리칸은 중쇄 불변 영역에 공유 결합될 수 있다.
본 개시의 또 다른 실시양태는 신규 Fc 글리코실화 패턴을 갖는 mAb를 포함한다. 단리된 모노클로날 항체 또는 이의 항원 결합 단편은 GNGN 또는 G1/G2 글리코형에 의해 표시되는 실질적으로 균질한 조성물로 존재한다. Fc 글리코실화는 치료용 mAb의 항바이러스 및 항암 특성에서 중요한 역할을 한다. 본 개시는 시험관내에서 푸코스를 포함하지 않는 항-HIV mAb의 항-렌티바이러스 세포-매개 바이러스 억제의 증가를 나타내는 최근 연구와 일치한다. 코어 푸코스를 결여하는 균질한 글리칸을 사용한 본 개시의 이 실시양태는 특정 바이러스에 대한 보호가 2배 초과의 증가를 나타냈다. 코어 푸코스의 제거는 천연 킬러(NK) 세포에 의해 매개되는 mAb의 ADCC 활성을 극적으로 개선하지만, 다형핵 세포(PMN)의 ADCC 활성에는 역효과를 갖는 것으로 보인다.
GNGN 또는 G1/G2 글리코형으로 표시되는 실질적으로 균질한 조성물을 포함하는 단리된 모노클로날 항체 또는 이의 항원 결합 단편은, 실질적으로 균질한 GNGN 글리코형을 부재 및 G0, G1F, G2F, GNF, GNGNF 또는 GNGNFX 함유 글리코형의 존재하에 동일한 항체와 비교하여 Fc 감마 RI 및 Fc 감마 RIII에 대한 결합 친화성의 증가를 나타낸다. 본 개시의 일 실시양태에서, 항체는 1 × 10-8 M 이하의 Kd로 Fc 감마 RI로부터 해리되고, 1 × 10-7 M 이하의 Kd로 Fc 감마 RIII로부터 해리된다.
Fc 영역의 글리코실화는 통상 N-연결 또는 O-연결 중 어느 하나이다. N-연결은 아스파라긴 잔기의 측쇄에 대한 탄수화물 부분의 부착을 지칭한다. O-연결된 글리코실화는 당 N-아세틸갈락토사민, 갈락토오스 또는 자일로오스 중 하나가 하이드록시아미노산, 가장 일반적으로 세린 또는 트레오닌에 부착되는 것을 지칭하지만, 5-하이드록시프롤린 또는 5-하이드록시리신도 사용할 수 있다. 아스파라긴 측쇄 펩타이드 서열에 대한 탄수화물 모이어티의 효소적 부착에 대한 인식 서열은 아스파라긴-X-세린 및 아스파라긴-X-트레오닌이고, 여기서 X는 프롤린을 제외한 임의의 아미노산이다. 따라서, 폴리펩타이드에서 이들 펩타이드 서열 중 하나의 존재는 잠재적 글리코실화 부위를 생성한다.
글리코실화 패턴은, 예를 들면, 폴리펩타이드에서 발견되는 하나 이상의 글리코실화 부위(들)를 결실시키고/시키거나, 폴리펩타이드에 존재하지 않는 하나 이상의 글리코실화 부위(들)를 부가함으로써 변경될 수 있다. 항체의 Fc 영역에 대한 글리코실화 부위의 부가는 상기 기술된 트리펩타이드 서열(N-연결된 글리코실화 부위의 경우) 중 하나 이상을 함유하도록 아미노산 서열을 변경함으로써 편리하게 달성된다. 예시적 글리코실화 변이체는 중쇄의 잔기 Asn 297의 아미노산 치환을 갖는다. 변경은 또한, 원래의 폴리펩타이드의 서열(O-연결된 글리코실화 부위의 경우)에 하나 이상의 세린 또는 트레오닌 잔기를 부가하거나 치환함으로써 수행할 수 있다. 추가로, Asn 297을 Ala로 변경하면, 글리코실화 부위 중 하나를 제거할 수 있다.
특정 실시양태에서, 항체는 베타 (1,4)-N-아세틸글루코사미닐트랜스퍼라제 III(GnT III)를 발현하는 세포에서 발현되고, 그 결과, GnT III는 GlcNAc를 IL-23p19 항체에 부가한다. 이러한 방식으로 항체를 생산하는 방법은 WO/9954342, WO/03011878, 특허 공개 제20030003097A1호 및 문헌[참조: Umana et al., Nature Biotechnology, 17:176-180, February 1999]에 제공된다. 클러스터 정기적 산재된 짧은 팔린드롬 반복(Clustered Regularly Interspaced Short Palindromic Repeats; CRISPR) 등의 게놈 편집 기술을 사용하여, 글리코실화 등의 특정 번역 후 변형을 향상, 감소 또는 제거하도록 세포주를 변경할 수 있다. 예를 들면, CRISPR 기술을 사용하여, 재조합 모노클로날 항체의 발현에 사용되는 293 또는 CHO 세포에서 글리코실화 효소를 암호화하는 유전자를 제거할 수 있다.
모노클로날 항체 단백질 서열에 대한 책임의 제거. 인간 B 세포에서 수득된 항체 가변 유전자 서열을 조작하여, 이들의 제조 가능성과 안전성을 향상시키는 것이 가능하다. 잠재적 단백질 서열의 책임은 하기를 포함하는 부위와 관련된 서열 모티프를 검색함으로써 특정할 수 있다.
1) 쌍을 이루지 않은 Cys 잔기,
2) N-연결된 글리코실화,
3) Asn 탈아미드화,
4) Asp 이성질체화,
5) SYE 절단,
6) Met 산화,
7) Trp 산화,
8) N-말단 글루타메이트,
9) 인테그린 결합,
10) CD11c/CD18 결합, 또는
11) 단편화
이러한 모티프는 재조합 항체를 암호화하는 cDNA에 대한 합성 유전자를 변경하여 제거할 수 있다.
치료용 항체의 개발 분야에서 단백질 공학 노력은 특정 서열 또는 잔기가 용해도 차이와 관련되어 있음을 분명히 나타냈다[참조: Fernandez-Escamilla et al., Nature Biotech., 22(10), 1302-1306, 2004; Chennamsetty et al., PNAS, 106(29), 11937-11942, 2009, Voynov et al., Biocon. Chem., 21(2), 385-392, 2010]. 문헌에서 용해도-변경 돌연변이로부터의 증거는, 아스파르트산, 글루탐산 및 세린 등의 몇몇 친수성 잔기가 아스파라긴, 글루타민, 트레오닌, 리신 및 아르기닌 등의 기타 친수성 잔기보다 현저히 유리하게 단백질 용해도에 기여하는 것을 나타낸다.
안정성. 항체는 생물물리학적 특성을 향상시키기 위해 조작될 수 있다. 평균 겉보기 용융 온도를 사용하여, 고온을 사용하여 항체를 전개하고 상대적 안정성을 결정할 수 있다. 시차 주사 열량계(DSC)는 분자의 열용량(Cp)(1도당 분자의 가온에 필요한 열)을 온도의 함수로서 측정한다. DSC를 사용하여, 항체의 열 안정성을 연구할 수 있다. mAb에 대한 DSC 데이터는, 종종 mAb 구조 내에서 개별 도메인의 전개를 해결하고, 서모그램에서 최대 3개의 피크(Fab, CH2 및 CH3 도메인의 전개로부터)를 생성하기 때문에 특히 흥미롭다. 통상, Fab 도메인의 전개는 가장 강력한 피크를 생성한다. DSC 프로파일 및 Fc 부분의 상대적 안정성은 인간 IgG1, IgG2, IgG3 및 IgG4 서브클래스에 대한 특징적인 차이를 나타낸다[참조: Garber and Demarest, Biochem. Biophys. Res. Commun. 355, 751-757, 2007]. CD 분광기로 수행되는 원형 이색성(CD)을 사용하여, 평균 겉보기 용융 온도를 결정할 수도 있다. Far-UV CD 스펙트럼은 0.5nm 증분으로 200 내지 260nm 범위의 항체에 대해 측정된다. 최종 스펙트럼은 20회 누적의 평균으로 결정할 수 있다. 잔류 타원율 값은 배경을 차감한 후에 계산할 수 있다. 항체의 열 변성(0.1 mg/mL)은 235nm에서 25-95℃ 및 1℃/min의 가열 속도로 모니터링할 수 있다. 동적 광 산란(DLS)을 사용하여, 응집 경향을 평가할 수 있다. DLS는 단백질을 포함한 다양한 입자의 크기를 특성화하기 위해 사용된다. 시스템의 크기가 분산되지 않은 경우는 입자의 평균 유효 직경을 결정할 수 있다. 이 측정은 입자 코어의 크기, 표면 구조의 크기 및 입자 농도에 의존한다. DLS는 본질적으로 입자에 의한 산란 광 강도의 변동을 측정하기 위해 입자의 확산 계수를 결정할 수 있다. 상업용 DLA 기기의 DLS 소프트웨어는 다양한 직경의 입자 모집단을 표시한다. 안정성 연구는 DLS를 사용하여 편리하게 수행할 수 있다. 샘플의 DLS 측정은 입자의 유체역학적 반경이 증가하는지의 여부를 결정함으로써, 입자가 시간의 경과에 따라 응집되는지 또는 온도 변화에 따라 응집되는지의 여부를 나타낼 수 있다. 입자가 응집되면, 보다 큰 반경의 입자의 보다 큰 모집단을 볼 수 있다. 온도에 따른 안정성은 그 장소에서 온도를 제어하여 분석할 수 있다. 모세관 전기영동(CE) 기술에는 항체 안정성의 특징을 결정하기 위한 입증된 방법론이 포함된다. iCE 접근법을 사용하여, 탈아미드화, C-말단 라이신, 시알릴화, 산화, 글리코실화 및 단백질의 pI의 변화를 초래할 수 있는 단백질에 대한 기타 변화로 인한 항체 단백질 전하 변이체를 해결할 수 있다. 발현된 각각의 항체 단백질은, 단백질 심플 마우리스(Protein Simple Maurice) 기기를 사용하여, 모세관 컬럼(cIEF)에서 높은 처리량, 유리 용액 등전점 초점(IEF)에 의해 평가할 수 있다. 등전점(pIs)에 초점을 맞춘 분자를 실시간 모니터링하기 위해, 전체 컬럼 UV 흡수 검출을 30초마다 수행할 수 있다. 이 접근법은 종래의 겔 IEF의 고분해능과 컬럼 기반 분리에서 발견되는 정량 및 자동화의 장점을 조합하는 동시에 동원 단계의 필요성을 제거한다. 이 기술에 의해, 발현된 항체에 대한 동일성, 순도 및 불균일성 프로파일의 재현 가능한 정량 분석이 가능해진다. 이 결과는, 항체의 전하 불균일성 및 분자 크기를 특정하고, 흡광도 및 천연 형광 검출 모드로 0.7μg/mL까지의 검출 감도를 갖는다.
용해도. 항체 서열의 고유 용해도 스코어를 결정할 수 있다. 고유 용해도 스코어는 CamSol Intrinsic을 사용하여 계산할 수 있다[참조: Sormanni et al., J Mol Biol 427, 478-490, 2015]. scFv 등의 각 항체 단편의 HCDR3에서 잔기 95-102(Kabat 넘버링)에 대한 아미노산 서열은 용해도 스코어를 계산하기 위해 온라인 프로그램을 통해 평가될 수 있다. 또한, 실험실 기술을 사용하여 용해도를 결정할 수 있다. 용액이 포화되어 용해도 한계에 도달할 때까지 동결건조된 단백질을 용액에 첨가하거나, 적절한 분자량 컷오프를 갖는 미세농축기에서 한외여과에 의한 농축을 포함하는 다양한 기술이 존재한다. 가장 간단한 방법은 무정형 침전을 유도하는 것이고, 이는 황산암모늄을 사용하여 단백질 침전을 포함하는 방법을 사용하여 단백질의 용해도를 측정한다[참조: Trevino et al., J Mol Biol, 366: 449-460, 2007]. 황산암모늄 침전은 상대적 용해도 값에 대한 신속하고 정확한 정보를 제공한다. 황산암모늄 침전은 명확한 수성 및 고체상을 구비한 침전 용액을 생성하고, 비교적 소량의 단백질을 필요로 한다. 황산암모늄에 의한 무정형 침전의 유도를 사용하여 수행되는 용해도 측정은 다른 pH 값에서도 용이하게 수행할 수 있다. 단백질의 용해도는 pH에 크게 의존하고, pH는 용해도에 영향을 미치는 가장 중요한 외인성 요인으로 간주된다.
자가반응성. 일반적으로, 자가반응 클론은 개체발생 동안 음성 선택에 의해 제거되어야 한다고 생각되지만, 자가반응 특성을 갖는 다수의 인간 천연 항체가 성숙한 레퍼토리에 존속되고 자가반응성이 병원체에 대한 다수의 항체의 항바이러스 기능을 향상시킬 수 있다는 것이 명백해졌다. 초기 B 세포 발달 동안 항체의 HCDR3 루프는 종종 양전하가 풍부하고 자가반응성 패턴을 나타내는 것으로 주목되었다[참조: Wardemann et al., Science 301, 1374-1377, 2003]. 현미경(부착형 HeLa 또는 HEp-2 상피 세포를 사용) 및 유세포 측정 세포 표면 염색(현탁액 Jurkat T 세포 및 293S 인간 배아성 신장 세포를 사용)에서 인간 기원 세포에 대한 결합 수준을 평가함으로써, 소정 항체의 자가반응성을 시험할 수 있다. 자가반응성은 또한 조직 어레이 내의 조직에 대한 결합 평가를 사용하여 조사할 수 있다.
바람직한 잔기("인간 유사성"). 혈액 공여자의 인간 B 세포에 대한 B 세포 레퍼토리 심층 서열분석은 다수의 최근 연구에서 광범위하게 수행되고 있다. 인간 항체 레퍼토리의 상당 부분에 대한 서열 정보는 건강한 인간에게 공통하는 항체 서열 특징의 통계적 평가를 용이하게 한다. 인간 재조합 항체 가변 유전자 참조 데이터베이스에서 항체 서열의 특징에 대한 지식에 의해, 항체 서열의 "인간 유사성"(HL)의 위치 특이적 정도를 추정할 수 있다. HL은 치료용 항체 또는 백신으로서의 항체 등의 임상 용도에서 항체 개발에 유용한 것으로 나타났다. 목표는 항체의 인간 유사성을 증가시켜, 항체 약물의 효능을 현저히 감소시키거나, 심각한 건강 영향을 유발할 수 있는 잠재적 부작용 및 항-항체 면역 반응을 감소시키는 것이다. 총 약 4억개 서열의 3명의 건강한 인간 혈액 공여자의 조합된 항체 레퍼토리의 항체 특성을 평가하고, 항체의 초가변 영역에 초점을 맞춘 새로운 "상대적 인간 유사성"(rHL) 스코어를 작성할 수 있다. rHL 스코어를 통해, 인간(양성 스코어)과 비인간 서열(음성 스코어)을 용이하게 구별할 수 있다. 항체는 인간 레퍼토리에서 일반적이지 않은 잔기를 제거하도록 조작될 수 있다.
D. 일본쇄 항체
일본쇄 가변 단편(scFv)은 면역글로불린의 중쇄 및 경쇄 가변 영역의 융합이고, 짧은(통상 세린, 글리신) 링커로 함께 연결된다. 이 키메라 분자는 불변 영역의 제거와 링커 펩타이드의 도입에도 불구하고, 원래 면역글로불린의 특이성을 유지한다. 이 변형은 통상 특이성을 변경시키지 않는다. 이러한 분자는 항원 결합 도메인을 단일 펩타이드로서 발현시키는 것이 매우 편리한 파지 디스플레이를 용이하게 하기 위해 역사적으로 생성되었다. 또는, scFv는 하이브리도마 또는 B 세포로부터 유래하는 서브클로닝된 중쇄 및 경쇄로부터 직접 생성될 수 있다. 일본쇄 가변 단편은 완전한 항체 분자에서 발견되는 불변 Fc 영역을 결여하고 있으므로, 항체의 정제에 사용되는 공통 결합 부위(예: 단백질 A/G)가 없다. 단백질 L은 카파 경쇄의 가변 영역과 상호작용하기 때문에, 이러한 단편은 종종 단백질 L을 사용하여 정제/고정될 수 있다.
유연한 링커는 일반적으로 알라닌, 세린 및 글리신 등의 헬릭스 및 턴-촉진 아미노산 잔기로 구성된다. 그러나, 다른 잔류물도 기능할 수 있다. 문헌[참조: Tanget al. (1996)]은 단백질 링커 라이브러리로부터 일본쇄 항체(scFv)를 위해 조정된 링커를 신속하게 선택하는 수단으로서 파지 디스플레이를 사용했다. 중쇄 및 경쇄 가변 도메인에 대한 유전자가 가변 조성의 18-아미노산 폴리펩타이드를 암호화하는 세그먼트에 의해 연결된 랜덤 링커 라이브러리를 구축했다. scFv 레퍼토리(약 5 × 106개의 서로 상이한 구성원)를 필라멘트 파지에 표시하고, 합텐과의 친화성 선택을 수행했다. 선택된 변이체의 모집단은 결합 활성에서 유의한 증가를 나타냈지만, 상당한 서열 다양성을 유지했다. 이어서, 1054 개별 변이체를 스크리닝하면, 가용성 형태로 효율적으로 생성된 촉매 활성 scFv가 수득되었다. 서열 분석은 VH C 말단의 2개 잔기 후의 링커에 보존된 프롤린과, 선택된 테더의 유일한 공통 특징으로서 다른 위치에서 풍부한 아르기닌 및 프롤린을 나타냈다.
본 개시의 재조합 항체는 또한, 수용체의 이량체화 또는 다량체화를 가능하게 하는 서열 또는 모이어티를 포함할 수 있다. 이러한 서열은 J-쇄와 조합하여 다량체의 형성을 가능하게 하는 IgA로부터 유래하는 서열을 포함한다. 또 다른 다량체화 도메인은 Gal4 이량체화 도메인이다. 다른 실시양태에서, 쇄는 2개의 항체의 조합을 가능하게 하는 비오틴/아비딘 등의 약제로 변형시킬 수 있다.
별도의 실시양태에서, 일본쇄 항체는 비-펩타이드 링커 또는 화학 단위를 사용하여 수용체의 경쇄 및 중쇄를 연결함으로써 생성될 수 있다. 일반적으로, 경쇄 및 중쇄는 별개의 세포에서 생성되고, 정제되고, 후속적으로 적절한 방식으로 함께 연결될 것이다(즉, 중쇄의 N-말단이 적절한 화학 브릿지를 통해 경쇄의 C-말단에 부착됨).
가교 시약은 2개의 상이한 분자의 작용기를 연결하는 분자 브릿지를 형성하기 위해 사용된다(예: 안정화제 및 응고제). 그러나, 동일한 유사체의 이량체 또는 다량체 또는 상이한 유사체로 구성된 헤테로머 복합체가 생성될 수 있는 것으로 고려된다. 2개의 상이한 화합물을 단계적으로 연결하기 위해, 불필요한 단독중합체의 형성을 제거하는 헤테로-이작용성 가교결합제를 사용할 수 있다.
예시적 헤테로-이작용성 가교제는 2개의 반응성 그룹을 함유한다: 하나는 1급 아민 그룹(예: N-하이드록시 석신이미드)와 반응하고, 다른 하나는 티올 그룹(예: 피리딜 디설파이드, 말레이미드, 할로겐 등)와 반응한다. 1급 아민 반응성 그룹을 통해, 가교제는 하나의 단백질(예: 선택된 항체 또는 단편)의 라이신 잔기(들)와 반응할 수 있고, 티올 반응성 그룹을 통해, 이미 제1 단백질에 결합된 가교제는 다른 단백질(예: 선택제)의 시스테인 잔기(유리 설프하이드릴 그룹)와 반응한다.
혈액 내에서 적당한 안정성을 갖는 가교제를 사용하는 것이 바람직하다. 표적화제 및 치료제/예방제를 접합하기 위해 성공적으로 사용될 수 있는 다수 유형의 디설파이드 결합 함유 링커가 공지되어 있다. 입체 장애가 있는 디설파이드 결합을 포함하는 링커는 생체내에서 보다 큰 안정성을 제공하여, 작용 부위에 도달하기 전에 표적화 펩타이드의 방출을 방지하는 것을 증명할 수 있다. 따라서, 이러한 링커는 연결제의 하나의 그룹이다.
또 다른 가교제는 SMPT이고, 이는 인접한 벤젠 환 및 메틸 그룹에 의해 "입체 장애"되는 디설파이드 결합을 포함하는 이관능성 가교제이다. 디설파이드 결합의 입체 장애는 조직 및 혈액에 존재할 수 있는 글루타티온 등의 티올레이트 음이온에 의한 공격으로부터 결합을 보호하는 기능을 하고, 이에 의해 표적 부위로 결합물의 전달 전에 접합체의 디커플링을 방지하는 역할을 하는 것으로 여겨진다.
SMPT 가교 시약은, 기타 다수의 공지된 가교 시약과 마찬가지로, 시스테인의 SH 또는 1급 아민(예: 라이신의 엡실론 아미노 그룹) 등의 작용기를 가교하는 능력을 제공한다. 다른 가능한 유형의 가교제는 설포숙신이미딜-2-(p-아지도 살리실아미도) 에틸-1,3'-디티오프로피오네이트 등의 절단 가능한 디설파이드 결합을 함유하는 헤테로-이작용성 광반응성 페닐아지드를 포함한다. N-하이드록시-숙신이미딜 그룹은 1급 아미노 그룹과 반응하고, 페닐아지드(광분해시)는 임의의 아미노산 잔기와 비선택적으로 반응한다.
장애된 가교제에 추가하여, 장애되지 않은 링커도 또한, 본 명세서에 따라 사용될 수 있다. 보호된 디설파이드를 함유하거나 생성하는 것으로 간주되지 않는 기타 유용한 가교제에는 SATA, SPDP 및 2-이미노티올란이 포함된다[참조: Wawrzynczak & Thorpe, 1987]. 이러한 가교제의 사용은 당해 기술분야에서 잘 이해되어 있다. 또 다른 실시양태는 유연한 링커의 사용을 포함한다.
미국 특허 제4,680,338호는 아민-함유 중합체 및/또는 단백질과 리간드의 접합체를 생성하기 위해, 특히 킬레이트제, 약물, 효소, 검출 가능한 표지 등과 항체 접합체를 형성하는 데 유용한 이작용성 링커를 기재한다. 미국 특허 제5,141,648호 및 제5,563,250호는 다양한 온화한 조건하에서 절단가능한 불안정한 결합을 함유하는 절단 가능한 접합체를 개시하고 있다. 이 링커는 목적 약제가 링커에 직접 결합될 수 있고, 절단되어 활성제가 방출될 수 있다는 점에서 특히 유용한다. 특정 용도에는 항체 또는 약물 등의 단백질에 유리 아미노 또는 유리 설프하이드릴 그룹을 부가하는 것이 포함된다.
미국 특허 제5,856,456호는 폴리펩타이드 구성요소를 연결하여 융합 단백질, 예를 들면, 일본쇄 항체를 작제하기 위해 사용하기 위한 펩타이드 링커를 제공한다. 링커는 최대 약 50개 아미노산 길이이고, 하전된 아미노산(바람직하게는 아르기닌 또는 라이신)의 적어도 하나의 발생과 이에 계속하여 프롤린을 포함하고, 보다 큰 안정성과 감소된 응집을 특징으로 한다. 미국 특허 제5,880,270호는 다양한 면역진단 및 분리 기술에 유용한 아미노옥시-함유 링커를 개시하고 있다.
E. 다중특이성 항체
특정 실시양태에서, 본 개시의 항체는 이중특이성 또는 다중특이성이다. 이중특이성 항체는 적어도 2개의 상이한 에피토프에 대한 결합 특이성을 갖는 항체이다. 예시적 이중특이성 항체는 단일 항원의 2개의 상이한 에피토프에 결합할 수 있다. 기타 이러한 항체는 제1 항원 결합 부위를 제2 항원에 대한 결합 부위와 조합할 수 있다. 또는, 항-병원체 암(arm)은 T 세포 수용체 분자(예: CD3) 등의 백혈구 상의 유발 분자 또는 FcγRI(CD64), FcγRII(CD32) 및 Fc 감마 RIII(CD16) 등의 IgG의 Fc 수용체(FcγR)에 결합하는 암과 조합시킬 수 있고, 이에 의해 세포 방어 메커니즘을 감염된 세포에 집중시키고 국재화시킨다. 이중특이성 항체를 사용하여, 세포독성제를 감염된 세포에 국재화시킬 수 있다. 이 항체는 병원체 결합 암과 세포독성제(예: 사포린, 항-인터페론-α, 빈카 알칼로이드, 리신 A 쇄, 메토트렉세이트 또는 방사성 동위원소 합텐)에 결합하는 암을 갖고 있다. 이중특이성 항체는 전장 항체 또는 항체 단편(예: F(ab')2 이중특이성 항체)으로 제조할 수 있다. WO 제96/16673호는 이중특이성 항-ErbB2/항-Fc 감마 RIII 항체를 기재하고, 미국 특허 제5,837,234호는 이중특이성 항-ErbB2/항-Fc 감마 RI 항체를 개시한다. 이중특이성 항-ErbB2/Fc 알파 항체는 WO98/02463에 제시되어 있다. 미국 특허 제5,821,337호는 이중특이성 항-ErbB2/항-CD3 항체를 교시하고 있다.
이중특이성 항체의 제조 방법은 당해 기술분야에 공지되어 있다. 전장 이중특이성 항체의 종래의 생산은 2개의 면역글로불린 중쇄-경쇄 쌍의 공-발현을 기반으로 하고, 여기서 2개의 쇄는 상이한 특이성을 갖는다[참조: Millstein et al., Nature, 305:537-539 (1983)]. 면역글로불린 중쇄 및 경쇄는 랜덤으로 분류되어 있기 때문에, 이러한 하이브리도마(쿼드로마)는 10개의 상이한 항체 분자의 잠재적 혼합물을 생성하고, 이 중 하나만이 정확한 이중특이성 구조를 갖는다. 통상, 친화성 크로마토그래피 단계에 의해 수행되는 정확한 분자의 정제는 다소 번거롭고 생성물의 수율이 낮다. 유사한 절차는 WO 제93/08829호 및 문헌[참조: Traunecker et al., EMBO J., 10:3655-3659(1991)]에 개시되어 있다.
다른 접근법에 따르면, 목적하는 결합 특이성을 갖는 항체 가변 영역(항체-항원 결합 부위)을 면역글로불린 불변 도메인 서열에 융합시킨다. 바람직하게는, 융합은 힌지, CH2 및 CH3 영역의 적어도 일부를 포함하는 Ig 중쇄 불변 도메인과의 융합이다. 적어도 하나의 융합에 존재하는, 경쇄 결합에 필요한 부위를 포함하는 제1 중쇄 불변 영역(CH1)을 갖는 것이 바람직하다. 면역글로불린 중쇄 융합물 및 필요에 따라 면역글로불린 경쇄를 암호화하는 DNA는 별도의 발현 벡터에 삽입되고, 적합한 숙주 세포에 동시 형질감염된다. 이것은, 작제에 사용된 3개의 폴리펩타이드 쇄의 비균등한 비율이 목적하는 이중특이성 항체의 최적 수율을 제공하는 경우, 실시양태에서 3개의 폴리펩타이드 단편의 상호 비율을 조정할 때에 보다 큰 유연성을 제공한다. 그러나, 동일한 비율의 적어도 2개의 폴리펩타이드 쇄의 발현이 높은 수율을 초래하거나 비율이 목적하는 쇄 조합의 수율에 유의한 영향을 미치지 않는 경우, 2개 또는 3개의 모든 폴리펩타이드 쇄에 대한 코딩 서열을 단일 발현 벡터에 삽입하는 것이 가능하다.
이 접근법의 특정 실시양태에서, 이중특이성 항체는 한쪽 암에 제1 결합 특이성을 갖는 하이브리드 면역글로불린 중쇄 및 다른 쪽 암에 하이브리드 면역글로불린 중쇄-경쇄 쌍(제2 결합 특이성을 제공함)으로 구성된다. 이중특이성 분자의 절반에만 면역글로불린 경쇄가 존재함으로써 용이한 분리 방법이 제공되기 때문에, 이러한 비대칭 구조는 목적하지 않는 면역글로불린 쇄 조합으로부터 목적하는 이중특이성 화합물의 분리를 용이하게 하는 것으로 밝혀졌다. 이 접근법은 WO 제94/04690호에 개시되어 있다. 이중특이성 항체의 생성에 대한 추가 상세는, 예를 들면, 문헌[참조: Suresh et al., Methods in Enzymology, 121:210 (1986)]을 참조한다.
미국 특허 제5,731,168호에 기재된 또 다른 접근법에 따르면, 항체 분자의 쌍 사이의 계면은 재조합 세포 배양으로부터 회수되는 헤테로이량체의 백분율을 최대화하도록 조작할 수 있다. 선호되는 인터페이스는 CH3 도메인의 적어도 일부를 포함한다. 이 방법에서는, 1차 항체 분자의 계면으로부터 하나 이상의 작은 아미노산 측쇄가 보다 큰 측쇄(예: 티로신 또는 트립토판)로 치환된다. 큰 아미노산 측쇄를 보다 작은 측쇄(예: 알라닌 또는 트레오닌)로 치환함으로써, 큰 측쇄(들)와 동일하거나 유사한 크기의 보상성 "공동"은 2차 항체 분자의 계면에 생성된다. 이것은 호모이량체 등의 기타 불필요한 최종 생성물보다 헤테로이량체의 수율을 증가시키는 메커니즘을 제공한다.
이중특이성 항체에는 가교 또는 "헤테로접합체" 항체가 포함된다. 예를 들면, 헤테로접합체의 항체 중 하나는 아비딘에 결합되고, 다른 하나는 비오틴에 결합될 수 있다. 이러한 항체는, 예를 들면, 면역계 세포를 목적하지 않는 세포로 표적화하기 위해(미국 특허 제4,676,980호), 및 HIV 감염의 치료를 위해(WO 91/00360, WO 92/200373 및 EP 03089) 제안되었다. 헤테로접합체 항체는 임의의 편리한 가교 방법을 사용하여 제조할 수 있다. 적합한 가교제는 당해 기술분야에 공지되어 있고, 다수의 가교 기술과 함께 미국 특허 제4,676,980호에 개시되어 있다.
항체 단편으로부터 이중특이성 항체를 생성하는 기술도 문헌에 기재되어 있다. 예를 들면, 이중특이성 항체는 화학적 연결을 사용하여 제조할 수 있다. 문헌[참조: Brennan et al., Science, 229: 81 (1985)]은 무손상 항체를 단백질 분해적으로 절단하여 F(ab')2 단편을 생성하는 절차를 기재하고 있다. 이 단편은 디티올 착화제인 나트륨 아르세나이트의 존재하에서 환원되어, 인접 디티올을 안정화하고 분자간 디설파이드 형성을 방지한다. 이어서, 생성된 Fab' 단편을 티오니트로벤조에이트(TNB) 유도체로 전환시킨다. 이어서, Fab'-TNB 유도체 중 하나는 메르캅토에틸아민에 의한 환원에 의해 Fab'-티올로 재전환되고, 등몰량의 기타 Fab'-TNB 유도체와 혼합되어 이중특이성 항체를 형성한다. 생성된 이중특이성 항체는 효소의 선택적 고정화를 위한 약제로 사용될 수 있다.
이. 콜라이로부터 Fab'-SH 단편의 직접적 회수를 촉진하는 기술이 존재하고, 이는 화학적으로 결합하여 이중특이성 항체를 형성할 수 있다. 문헌[참조: Shalaby et al., J. Exp. Med., 175: 217-225(1992)]은 인간화 이중특이성 항체 F(ab')2 분자의 생산에 대해 기재되어 있다. 각각의 Fab' 단편은 이. 콜라이로부터 별도로 분비되었고, 시험관내에서 직접 화학적 커플링을 받아 이중특이성 항체를 형성했다. 이렇게 형성된 이중특이성 항체는 ErbB2 수용체를 과발현하는 세포 및 정상 인간 T 세포에 결합할 수 있을 뿐만 아니라, 인간 유방 종양 표적에 대한 인간 세포독성 림프구의 용해 활성을 유발할 수 있다.
재조합 세포 배양으로부터 직접 이중특이성 항체 단편을 제조하고 단리하기 위한 다양한 기술이 또한 기재되어 있다[참조: Merchant et al., Nat. Biotechnol. 16, 677-681 (1998) doi:10.1038/nbt0798-677pmid:9661204]. 예를 들면, 이중특이성 항체는 류신 지퍼를 사용하여 생산되었다[참조: Kostelny et al., J. Immunol., 148(5):1547-1553, 1992]. Fos 및 Jun 단백질로부터의 류신 지퍼 펩타이드는 유전자 융합에 의해 2개의 상이한 항체의 Fab' 부분에 연결되었다. 항체 호모이량체는 힌지 영역에서 환원되어 단량체를 형성하고, 이어서 재산화되어 항체 헤테로이량체를 형성했다. 이 방법은 또한 항체 호모이량체의 생산에도 이용될 수 있다. 문헌[참조: Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993)]에 의해 기재된 "디아바디" 기술은 이중특이성 항체 단편을 제조하기 위한 대체 메커니즘을 제공했다. 단편은, 동일한 쇄의 2개 도메인 사이의 페어링을 가능하게 하기에는 너무 짧은 링커에 의해 VL에 연결된 VH를 포함한다. 따라서, 하나의 단편의 VH 및 VL 도메인은 또 다른 단편의 상보적 VL 및 VH 도메인과 강제적으로 페어링되고, 이에 의해 2개의 항원 결합 부위가 형성된다. 일본쇄 Fv(sFv) 이량체를 사용하여 이중특이성 항체 단편을 제조하는 또 다른 전략도 보고되었다[참조: Gruber et al., J. Immunol., 152:5368(1994)].
특정 실시양태에서, 이중특이적 또는 다중특이적 항체는 DOCK-AND-LOCK™(DNL™) 복합체로서 형성될 수 있다(예를 들면, 미국 특허 제7,521,056호; 제7,527,787호; 제7,534,866호; 제7,550,143호 및 제7,666,400호 참조, 각각의 실시예의 섹션은 참조에 의해 본 명세서에 도입된다). 일반적으로, 이 기술은 cAMP 의존성 단백질 키나제(PKA)의 조절(R) 서브유닛의 이량체화 및 도킹 도메인(DDD) 서열과, 임의의 다양한 AKAP 단백질로부터 유래하는 앵커 도메인(AD) 서열 사이에서 발생하는 특이적으로 고친화성의 결합 상호작용을 이용한다[참조: Baillie et al., FEBS Letters. 2005; 579: 3264; Wong and Scott, Nat. Rev. Mol. Cell Biol. 2004; 5:959]. DDD 및 AD 펩타이드는 임의의 단백질, 펩타이드 또는 기타 분자에 부착될 수 있다. DDD 서열이 자발적으로 이량체화되어 AD 서열에 결합하기 때문에, 이 기술은 DDD 또는 AD 서열에 부착될 수 있는 선택된 분자 사이의 복합체의 형성이 가능해진다.
2가 이상의 원자가를 갖는 항체가 고려된다. 예를 들면, 삼중특이성 항체가 제조될 수 있다[참조: Tutt et al., J. Immunol. 147: 60, 1991; Xu et al., Science, 358(6359):85-90, 2017]. 다가 항체는 항체가 결합하는 항원을 발현하는 세포에 의해 2가 항체보다 신속하게 내재화(및/또는 이화)될 수 있다. 본 개시의 항체는 3개 이상의 항원 결합 부위를 갖는 다가 항체(예를 들면, 4가 항체)일 수 있고, 이는 항체의 폴리펩타이드 쇄를 암호화하는 핵산의 재조합 발현에 의해 용이하게 생성될 수 있다. 다가 항체는 이량체화 도메인 및 3개 이상의 항원 결합 부위를 포함할 수 있다. 바람직한 이량체화 도메인은 Fc 영역 또는 힌지 영역을 포함한다(또는 이로 구성된다). 이 시나리오에서는, 항체는 Fc 영역, 및 Fc 영역에 대한 아미노 말단의 3개 이상의 항원 결합 부위를 포함할 것이다. 본 명세서에서 바람직한 다가 항체는 3개 내지 약 8개, 그러나 바람직하게는 4개의 항원 결합 부위를 포함한다(또는 이들로 구성된다). 다가 항체는 적어도 하나의 폴리펩타이드 쇄(및 바람직하게는 2개의 폴리펩타이드 쇄)를 포함하고, 여기서 폴리펩타이드 쇄(들)는 2개 이상의 가변 영역을 포함한다. 예를 들면, 폴리펩타이드 쇄(들)는 VD1-(X1)n-VD2-(X2)n-Fc를 포함할 수 있고, 여기서 VD1은 제1 가변 영역이고, VD2는 제2 가변 영역이고, Fc는 Fc 영역의 하나의 폴리펩타이드 쇄이고, X1 및 X2는 아미노산 또는 폴리펩타이드를 나타내고, n은 0 또는 1이다. 예를 들면, 폴리펩타이드 쇄(들)는 하기를 포함할 수 있다: VH-CH1-유연한 링커-VH-CH1-Fc 영역 쇄; 또는 VH-CH1-VH-CH1-Fc 영역 쇄. 본 명세서에서 다가 항체는 바람직하게는 적어도 2개(및 바람직하게는 4개)의 경쇄 가변 영역 폴리펩타이드를 추가로 포함한다. 본 명세서의 다가 항체는, 예를 들면, 약 2 내지 약 8개의 경쇄 가변 영역 폴리펩타이드를 포함할 수 있다. 본 명세서에서 고려되는 경쇄 가변 영역 폴리펩타이드는 경쇄 가변 영역을 포함하고, 임의로 CL 도메인을 추가로 포함한다.
전하 변형은 다중특이성 항체의 상황에서 특히 유용하고, 여기서 Fab 분자의 아미노산 치환에 의해 경쇄와 부정합 중쇄(Bence-Jones 유형 부산물)와의 부정합이 감소하고, 이는 결합 아암 중 하나(또는 2개 이상의 항원 결합 Fab 분자를 포함하는 분자의 경우에는 하나 이상)에서 VH/VL 교환을 갖는 Fab 기반 이중/다중특이성 항원 결합 분자의 생성에서 발생할 수 있다(PCT 공개 번호 WO 제2015/150447호 참조, 특히 그 중의 예는 참조에 의해 그 전체가 본 명세서에 도입됨).
따라서, 특정 실시양태에서, 치료제에 포함된 항체는
(a) 제1 항원에 특이적으로 결합하는 제1 Fab 분자
(b) 제2 항원에 특이적으로 결합하는 제2 Fab 분자
를 포함하고,
여기서, Fab 경쇄 및 Fab 중쇄의 가변 도메인 VL 및 VH는 서로 치환되고,
제1 항원은 활성화 T 세포 항원이고 제2 항원은 표적 세포 항원이거나, 또는 제1 항원은 표적 세포 항원이고 제2 항원은 활성화 T 세포 항원이고;
여기서
i) a)하의 제1 Fab 분자의 불변 도메인 CL에서는, 위치 124의 아미노산이 양으로 하전된 아미노산(카바트에 따른 넘버링)으로 치환되고, a)하의 제1 Fab 분자의 불변 도메인 CH1에서는, 위치 147의 아미노산 또는 위치 213의 아미노산이 음으로 하전된 아미노산(카바트 EU 지수에 따른 넘버링)으로 치환되거나; 또는
ii) b)하의 제2 Fab 분자의 불변 도메인 CL에서는, 위치 124의 아미노산이 양으로 하전된 아미노산(카바트에 따른 넘버링)으로 치환되고, b)하의 제2 Fab 분자의 불변 도메인 CH1에서는, 위치 147의 아미노산 또는 위치 213의 아미노산이 음으로 하전된 아미노산(카바트 EU 지수에 따른 넘버링)으로 치환된다.
항체는 i) 및 ii)에 서술된 변형을 모두 포함하지 않을 수 있다. 제2 Fab 분자의 불변 도메인 CL 및 CH1은 서로 치환되지 않는다(즉, 교환되지 않은 상태로 유지됨).
항체의 또 다른 실시양태에서, a)하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 라이신(K), 아르기닌(R) 또는 히스티딘(H)(카바트에 따른 넘버링)(한 가지 바람직한 실시양태에서, 독립적으로 라이신(K) 또는 아르기닌(R)에 의해)에 의해 독립적으로 치환되고, a)하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산 또는 위치 213의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(카바트 EU 지수에 따른 넘버링)에 의해 독립적으로 치환된다.
추가 실시양태에서, a)하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K), 아르기닌(R) 또는 히스티딘(H)(카바트에 따른 넘버링)에 의해 독립적으로 치환되고, a)하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(카바트 EU 지수에 따른 넘버링)에 의해 독립적으로 치환된다.
특정 실시양태에서, a)하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 라이신(K), 아르기닌(R) 또는 히스티딘(H)(카바트에 따른 넘버링)(하나의 바람직한 실시양태에서는, 독립적으로 리신(K) 또는 아르기닌(R)에 의해)에 의해 독립적으로 치환되고, 위치 123의 아미노산은 리신(K), 아르기닌(R) 또는 히스티딘(H)(카바트에 따른 넘버링)(바람직한 일 실시양태에서는, 독립적으로 리신(K) 또는 아르기닌(R))에 의해 독립적으로 치환되고, a)하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(카바트 EU 지수에 따른 넘버링)에 의해 독립적으로 치환되고, 위치 213의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(카바트 EU 지수에 따른 넘버링)에 의해 독립적으로 치환된다.
보다 특정한 실시양태에서, a)하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K)(카바트에 따른 넘버링)에 의해 치환되고, 위치 123의 아미노산은 리신(K) 또는 아르기닌(R)(카바트에 따른 넘버링)에 의해 치환되고, a)하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E)(카바트 EU 지수에 따른 넘버링)에 의해 치환되고, 위치 213의 아미노산은 글루탐산(E)(카바트 EU 지수에 따른 넘버링)에 의해 치환된다.
추가로 특정한 실시양태에서, a)하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K)(카바트에 따른 넘버링)에 의해 치환되고, 위치 123의 아미노산은 아르기닌(R)(카바트에 따른 넘버링)에 의해 치환되고, a)하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E)(카바트 EU 지수에 따른 넘버링)에 의해 치환되고, 위치 213의 아미노산은 글루탐산(E)(카바트 EU 지수에 따른 넘버링)에 의해 치환된다.
F. 키메라 항원 수용체
인공 T 세포 수용체(키메라 T 세포 수용체, 키메라 면역수용체, 키메라 항원 수용체(CAR)라고도 공지됨)는 면역 이펙터 세포에 임의의 특이성을 이식하는 조작된 수용체이다. 통상, 이러한 수용체는 모노클로날 항체의 특이성을 T 세포에 이식하기 위해 사용되고, 레트로바이러스 벡터에 의해 코딩 서열의 전달이 촉진된다. 이러한 방식으로, 입양 세포 전달을 위해, 다수의 표적 특이적 T 세포가 생성될 수 있다. 이 접근법의 1상 임상 연구는 효능을 나타낸다.
이러한 분자의 가장 일반적 형태는 모노클로날 항체로부터 유래하는 일본쇄 가변 단편(scFv)의 융합물이고, CD3-제타 막관통 및 엔도도메인에 융합된다. 이러한 분자는 이의 표적의 scFv에 의한 인식에 대한 응답으로 제타 신호의 전달을 초래한다. 이러한 작제물의 예는 하이브리도마 14g2a(디시알로강글리오사이드 GD2를 인식함)로부터 유래하는 scFv의 융합물인 14g2a-제타이다. T 세포가 이 분자를 발현하면(통상은 온코레트로바이러스 벡터 형질도입에 의해 달성됨), GD2를 발현하는 표적 세포(예: 신경모세포종 세포)를 인식하고 사멸시킨다. 악성 B 세포를 표적화하기 위해, 연구자들은 B 계통 분자 CD19에 특이적인 키메라 면역수용체를 사용하여 T 세포의 특이성을 재지시했다.
면역글로불린 중쇄 및 경쇄의 가변 부분은 유연한 링커에 의해 융합되어 scFv를 형성한다. 이 scFv의 전에 신호 펩타이드가 있고, 신생 단백질을 소포체로 지시하고, 후속 표면 발현을 지시한다(이는 절단됨). 유연한 스페이서에 의해, scFv를 상이한 방향으로 배양시켜 항원 결합을 가능하게 한다. 막관통 도메인은 통상 세포 내로 돌출되어 목적하는 신호를 전달하는 신호전달 엔도도메인의 원래 분자로부터 유래하는 전형적 소수성 알파 헬릭스이다.
유형 I 단백질은 실제로 그 사이에 막관통 알파 헬릭스에 의해 연결된 2개의 단백질 도메인이다. 막관통 도메인이 통과하는 세포막 지질 이중층은 내측 부분(엔도도메인)을 외측 부분(엑토도메인)으로부터 단리하도록 작용한다. 한 단백질의 엑토도메인을 또 다른 단백질의 엔도도메인에 부착하면, 전자의 인식을 후자의 신호와 조합시킨 분자가 수득된다는 것은 놀라운 것은 아니다.
엑토도메인. 신호 펩타이드는 신생 단백질을 소포체로 유도한다. 이것은 수용체가 글리코실화되고 세포막에 고정되는 경우에 필수적이다. 모든 진핵생물 신호 펩타이드 서열은 통상 정상으로 작동한다. 일반적으로, 아미노 말단의 대부분 성분에 자연적으로 부착된 신호 펩타이드가 사용된다(예: 경쇄-링커-중쇄의 방향을 갖는 scFv에서는 경쇄의 천연 신호가 사용된다).
항원 인식 도메인은 통상 scFv이다. 그러나, 다수의 대안이 있다. 천연 T 세포 수용체(TCR) 알파 및 베타 일본쇄로부터의 항원 인식 도메인은, 단순한 엑토도메인(예: HIV 감염 세포를 인식하는 CD4 엑토도메인) 및 연결된 사이토카인 등의 보다 이국적 인식 성분(이는 사이토카인 수용체를 갖는 세포의 인식을 유도함)을 갖는 것으로 기재되어 있다. 사실, 소정 표적에 높은 친화성으로 결합하는 거의 모든 것을 항원 인식 영역으로 사용할 수 있다.
스페이서 영역은 항원 결합 도메인을 막관통 도메인에 연결한다. 항원 인식을 용이하게 하기 위해, 항원 결합 도메인이 상이한 방향으로 배향될 수 있도록 충분히 유연해야 한다. 가장 단순한 형태는 IgG1의 힌지 영역이다. 대안으로는 면역글로불린의 CH2CH3 영역과 CD3의 일부가 포함된다. 대부분의 scFv 기반 작제물에서는 IgG1 힌지로 충분하다. 그러나, 최상의 스페이서는 종종 경험적으로 결정되어야 한다.
막관통 도메인. 막관통 도메인은 막을 가로지르는 소수성 알파 헬릭스이다. 일반적으로, 엔도도메인의 가장 막 근위의 구성요소로부터의 막관통 도메인이 사용된다. 흥미롭게도, CD3-제타 막관통 도메인을 사용하면, 인공 TCR을 천연 TCR에 도입할 수 있고, 이는 천연 CD3-제타 막관통 하전 아스파라긴산 잔기의 존재에 의존하는 요인이다. 상이한 막관통 도메인은 상이한 수용체의 안정성을 초래한다. CD28 막관통 도메인은 밝게 표시된 안정한 수용체를 초래한다.
엔도도메인. 이것은 수용체의 "비즈니스-말단"이다. 항원 인식 후, 수용체가 클러스터링되고, 신호가 세포로 전달된다. 가장 일반적으로 사용되는 엔도도메인 성분은 3개의 ITAM을 포함하는 CD3-제타이다. 이것은, 항원이 결합된 후, 활성화 신호를 T 세포에 전달한다. CD3-제타는 완전히 유능한 활성화 신호를 제공하지 않을 수 있고, 추가 공-자극 신호가 필요하다.
"1세대" CAR은 통상 내인성 TCR로부터의 신호의 주요 전달물질인 CD3 ξ-쇄로부터의 세포내 도메인을 갖고 있다. "2세대" CAR은 다양한 공-자극 단백질 수용체(예: CD28, 41BB, ICOS)로부터의 세포내 신호전달 도메인을 CAR의 세포질 꼬리에 추가하여, T 세포에 추가 신호를 제공한다. 전임상 연구에 따르면, 2세대 CAR 디자인이 T 세포의 항종양 활성을 개선하는 것을 나타낸다. 보다 최근에는, "3세대" CAR은 CD3z-CD28-41BB 또는 CD3z-CD28-OX40 등의 복수의 신호전달 도메인을 조합하여 효능을 추가로 향상시킨다.
G. ADC
항체 약물 접합체 또는 ADC는 감염성 질환을 갖는 사람들의 치료를 위한 표적 요법으로서 설계된 새로운 부류의 매우 강력한 생물의약 약물이다. ADC는, 항체(mAb 전체 또는 일본쇄 가변 단편 또는 scFv)로 구성되는 복잡한 분자이고, 불안정한 결합을 갖는 안정한 화학적 링커를 통해 생물학적 활성 세포독성/항바이러스 페이로드 또는 약물에 연결된다. 항체 약물 접합체는 생체접합체 및 면역접합체의 예이다.
모노클로날 항체의 독자의 표적화 능력과 세포독성 약물의 암-사멸 능력을 조합함으로써, 항체-약물 접합체는 건강한 조직과 발병 조직을 민감하게 구별할 수 있다. 이는, 종래의 전신 접근법과 대조적으로, 항체-약물 접합체가 감염된 세포를 표적으로 하여 공격하기 때문에, 건강한 세포가 덜 심각하게 영향을 받는다는 것을 의미한다.
ADC 기반 항종양 요법의 개발에서, 항암제(예: 세포 독소 또는 세포독소)를, 특정 세포 마커를 특이적으로 표적화하는 항체(예: 이상적으로는, 감염된 세포 중 또는 상에서만 발견되는 단백질)에 커플링된다. 항체는 이러한 단백질을 체내에서 추적하여 암세포의 표면에 부착한다. 항체와 표적 단백질(항원) 사이의 생화학적 반응은 종양 세포에서 신호를 촉발하고, 이것이 세포독소와 함께 항체를 흡수하거나 내재화한다. ADC가 내재화된 후, 세포독성 약물이 방출되어 세포를 사멸하거나, 바이러스 복제를 손상시킨다. 이러한 표적화로 인해, 이상적으로는, 이 약물은 다른 약물보다 부작용이 적고 치료 윈도우가 더 넓어진다.
항체와 세포독성/항바이러스제 사이의 안정한 연결은 ADC의 중요한 측면이다. 링커는 디설파이드물, 하이드라존 또는 펩타이드(절단 가능) 또는 티오에테르(절단 불가능)를 포함한 화학적 모티프를 기반으로 하고, 세포독성제의 표적 세포로의 분포 및 전달을 제어한다. 절단 가능한 및 절단 불가능한 유형의 링커는 전임상 및 임상 시험에서 안전한 것으로 입증되었다. 브렌툭시맙 베토틴에는 강력하고 독성이 강한 항미세소관제인 모노메틸 오리스타틴 E 또는 MMAE(합성 항종양제)를 인간 특이적 CD30-양성 악성 세포에 전달하는 효소 감수성의 절단 가능한 링커가 포함되어 있다. 튜불린의 중합을 차단하여 세포 분열을 억제하는 MMAE의 독성이 높기 때문에, 단일제 화학요법 약물로서 사용할 수 없다. 그러나, 항-CD30 모노클로날 항체(cAC10, 종양 괴사 인자 또는 TNF 수용체의 세포막 단백질)에 연결된 MMAE의 조합은 세포외 유체에서 안정하고, 카텝신에 의해 절단 가능하고, 치료에 안전한 것으로 입증되었다. 또 다른 승인된 ADC인 트라스투주맙 엠타신은 마이탄신의 유도체인 미소관-형성 억제제 메르타신(DM-1)과, 안정한 비절단 링커에 의해 부착된 항체 트라투주맙(Herceptin®/Genentech/Roche)의 조합이다.
보다 우수하고 보다 안정한 링커의 가용성은 화학 결합의 기능을 변경했다. 링커 유형은 절단 가능 또는 절단 불가능하고, 세포독성(항암) 약물에 특정의 특성을 부여한다. 예를 들면, 절단 불가능한 링커는 약물을 세포 내에 유지한다. 그 결과, 전체 항체, 링커 및 세포독성제는 표적화된 암세포로 유입하고, 여기서 항체가 아미노산 수준으로 분해된다. 수득되는 복합체(아미노산, 링커 및 세포독성제)는 이제 활성 약물로 된다. 대조적으로, 절단 가능한 링커는 세포독성제를 방출하는 숙주 세포 내의 효소에 의해 촉매된다.
현재 개발 중의 또 다른 유형의 절단 가능한 링커는 세포독성/항바이러스 약물과 절단 부위 사이에 추가 분자를 추가한다. 이 링커 기술에 의해, 연구자는 절단 속도의 변화를 걱정하지 않고 보다 유연성의 ADC를 생성할 수 있다. 연구자들은 또한, 펩타이드 중의 아미노산을 서열분석하는 방법인 에드만(Edman) 분해를 기반으로 하는 펩타이드 절단의 신규 방법을 개발하고 있다. ADC의 개발에서 장래의 방향성에는 안정성과 치료 지수를 추가로 개선하기 위한 부위 특이적 접합(TDC)의 개발, 및 α 방출 면역접합체 및 항체 접합 나노입자의 개발도 포함된다.
H. BiTES
이중특이성 T-세포 인게이저(BiTE)는 항암제로서의 사용이 조사되는 인공 이중특이성 모노클로날 항체의 부류이다. 이들은 감염된 세포에 대하여 숙주의 면역계, 보다 구체적으로 T 세포의 세포독성 활성을 지시한다. BiTE는 마이크로메트 아게(Micromet AG)의 등록 상표이다.
BiTE는, 약 55킬로달톤의 단일 펩타이드 쇄 상의 서로 상이한 항체의 2개 일본쇄 가변 단편(scFv) 또는 4개의 상이한 유전자로부터의 아미노산 서열로 구성된 융합 단백질이다. scFv 중 하나는 CD3 수용체를 통해 T 세포에 결합하고, 다른 하나는 특정 분자를 통해 감염된 세포에 결합한다.
다른 이중특이성 항체와 마찬가지로 및 통상의 모노클로날 항체와는 달리, BiTE는 T 세포와 표적 세포 사이에 연결을 형성한다. 이것에 의해, T 세포는 MHC I 또는 공-자극 분자의 존재와 무관하게, 퍼포린 및 그랜자임 등의 단백질을 생성함으로써, 감염된 세포에 대해 세포독성/항바이러스 활성을 발휘한다. 이 단백질은 감염된 세포에 유입되어, 세포의 아폽토시스를 개시한다. 이 작용은 감염된 세포에 대한 T 세포 공격 동안 관찰되는 생리학적 프로세스를 모방한다.
I. 인트라바디
특정 실시양태에서, 항체는 세포 내에서의 작용에 적합한 재조합 항체이고, 이러한 항체는 "인트라바디"로서 공지되어 있다. 이러한 항체는 세포 내 단백질 수송의 변화, 효소 기능의 방해, 단백질-단백질 또는 단백질-DNA 상호작용의 차단 등, 다양한 메커니즘에 의해 표적 기능을 방해할 수 있다. 다수의 점에서, 이들의 구조는 상기 논의한 일본쇄 및 단일 도메인 항체의 구조를 모방 또는 유사하다. 실제로, 단일 전사물/일본쇄는 표적 세포에서의 세포내 발현을 가능하게 하고, 또한 세포막을 가로질러 단백질 수송을 보다 실현 가능하게 하는 중요한 기능이다. 그러나, 추가 기능이 필요하다.
인트라바디 치료의 실시에 영향을 미치는 2개의 주요 문제는 세포/조직의 표적화를 포함하는 전달 및 안정성이다. 전달과 관련하여, 조직-지시된 전달, 세포 유형 특이적 프로모터의 사용, 바이러스 기반의 전달 및 세포 투과성/막 전위 펩타이드의 사용 등, 다양한 접근법이 사용되었다. 안정성과 관련하여, 접근법은 일반적으로, 파지 디스플레이를 수반하고 컨센서스 서열의 서열 성숙 또는 발달을 포함할 수 있는 방법을 포함하여 무차별 힘에 의한 스크리닝, 또는 삽입 안정화 서열(예: Fc 영역, 샤페론 단백질 서열, 류신 지퍼) 및 디설파이드 치환/변형 등의 더욱 지시된 변형 중의 어느 하나이다.
인트라바디에 필요할 수 있는 추가 기능은 세포내 표적화를 위한 신호이다. 인트라바디(또는 기타 단백질)를 세포질, 핵, 미토콘드리아 및 ER 등의 세포내 영역에 표적화할 수 있는 벡터가 설계되었고, 상업적으로 입수 가능하다(Invitrogen Corp.; Persic et al., 1997).
세포에 유입될 수 있는 능력에 비추어, 인트라바디에는 기타 유형의 항체가 달성할 수 없는 추가 용도가 있다. 본 발명의 항체의 경우, 살아있는 세포에서 MUC1 세포질 도메인과 상호작용하는 능력은 신호전달 기능(다른 분자에 결합) 또는 올리고머 형성 등의 MUC1 CD와 관련된 기능을 방해할 수 있다. 특히, 이러한 항체는 MUC1 이량체 형성을 억제하기 위해 사용될 수 있는 것으로 고려된다.
J. 정제
특정 실시양태에서, 본 개시의 항체는 정제될 수 있다. 본 명세서에 사용된 용어 "정제된"은, 단백질이 이의 자연적으로 수득할 수 있는 상태에 대하여 임의의 정도까지 정제되는, 다른 성분으로부터 단리 가능한 조성물을 지칭하는 것을 의도한다. 따라서, 정제된 단백질은 자연적으로 발생할 수 있는 환경으로부터 해방된 단백질을 지칭한다. "실질적으로 정제된"이라는 용어가 사용되는 경우, 이 명칭은 단백질 또는 펩타이드가 조성물의 주요 성분을 형성하는, 예를 들면, 조성물에서 단백질의 약 50%, 약 60%, 약 70%, 약 80%, 약 90%, 약 95% 또는 그 이상을 구성하는 조성물을 지칭한다.
단백질 정제 기술은 당업자에게 공지되어 있다. 이러한 기술은 어느 수준에서는 세포 환경의 폴리펩타이드 및 비-폴리펩타이드 분획으로 조 분별하는 것을 포함한다. 폴리펩타이드를 다른 단백질로부터 분리한 후, 목적 폴리펩타이드는 부분적 또는 완전한 정제(또는 균질성을 위한 정제)를 달성하기 위해 크로마토그래피 및 전기영동 기술을 사용하여 추가로 정제할 수 있다. 순수한 펩타이드의 제조에 특히 적합한 분석 방법은 이온 교환 크로마토그래피, 배제 크로마토그래피; 폴리아크릴아미드 겔 전기영동; 등전점 초점이다. 단백질 정제를 위한 다른 방법에는 황산암모늄, PEG, 항체 등에 의한 침전 또는 열 변성과 그에 계속하여 원심분리; 겔 여과, 역상, 하이드록실아파타이트 및 친화성 크로마토그래피; 및 이러한 기술 및 기타 기술의 조합이 포함된다.
본 개시의 항체를 정제할 때에, 원핵생물 또는 진핵생물의 발현 시스템에서 폴리펩타이드를 발현시키고, 변성 조건을 사용하여 단백질을 추출하는 것이 바람직할 수 있다. 폴리펩타이드는 폴리펩타이드의 태깅된 부분에 결합하는 친화성 컬럼을 사용하여 기타 세포 성분으로부터 정제할 수 있다. 당해 기술분야에 일반적으로 공지된 바와 같이, 다양한 정제 단계를 수행하는 순서를 변경할 수 있거나, 또는 특정 단계를 생략하여도, 실질적으로 정제된 단백질 또는 펩타이드를 제조하기 위한 적합한 방법을 제공하는 것으로 믿어진다.
일반적으로 완전한 항체는 항체의 Fc 부분에 결합하는 약제(즉, 단백질 A)를 사용하여 분획화된다. 또는, 항원은 적절한 항체의 정제와 선택을 동시에 수행하기 위해 사용될 수 있다. 이러한 방법은 종종 컬럼, 필터 또는 비드 등의 지지체에 결합된 선택 약제를 이용한다. 항체는 지지체에 결합하고, 오염 물질을 제거하고(예: 세척), 조건(염, 열 등)을 적용함으로써 항체를 방출시킨다.
단백질 또는 펩타이드의 정제의 정도를 정량화하는 다양한 방법이 본 개시에 비추어 당업자에게 공지될 것이다. 여기에는, 예를 들면, 활성 분획의 비활성을 결정하는 것, 또는 SDS/PAGE 분석에 의해 분획 내의 폴리펩타이드의 양을 평가하는 것이 포함된다. 분획의 순도를 평가하는 또 다른 방법은 분획의 비활성을 계산하고, 이를 초기 추출물의 비활성과 비교하여 순도의 정도를 계산하는 것이다. 활성의 양을 나타내기 위해 사용되는 실제의 단위는 물론 정제를 추적하기 위해 선택된 특정 검정 기술 및 발현된 단백질 또는 펩타이드가 검출 가능한 활성을 나타내는지의 여부에 의존한다.
폴리펩타이드의 이동은, SDS/PAGE의 조건이 상이하면, 경우에 따라 현저히 변화할 수 있는 것으로 공지되어 있다[참조: Capaldi et al., 1977]. 따라서, 상이한 전기영동 조건하에서, 정제 또는 부분적으로 정제된 발현 생성물의 겉보기 분자량이 변화할 수 있음이 이해될 것이다.
III. 엔테로바이러스 D68 감염의 능동/수동 면역 및 치료/예방
A. 제형 및 투여
본 개시는 항-EV-D68 항체 및 이를 생성하기 위한 항원을 포함하는 약제학적 조성물을 제공한다. 이러한 조성물은 예방적 또는 치료적 유효량의 항체 또는 이의 단편, 또는 펩타이드 면역원, 및 약제학적으로 허용되는 담체를 포함한다. 특정 실시양태에서, 용어 "약제학적으로 허용되는"은 연방 또는 주 정부의 규제 당국에 의해 승인되거나, 동물, 보다 구체적으로는 인간에서의 사용에 대해 미국 약전 또는 기타 일반적으로 인정되는 약전에 수록되어 있는 것을 의미한다. 용어 "담체"는 치료제와 함께 투여되는 희석제, 부형제 또는 비히클을 지칭한다. 이러한 약제학적 담체는, 땅콩유, 대두유, 광유, 참기름 등의 석유, 동물성, 식물성 또는 합성 기원의 것을 포함하는, 물 및 오일 등의 멸균 액체일 수 있다. 약제학적 조성물이 정맥내 투여되는 경우, 물은 특정한 담체이다. 식염수, 덱스트로스 및 글리세롤 수용액도 특히 주사용 용액의 액체 담체로 사용할 수 있다. 기타 적합한 약제학적 부형제는 전분, 글루코스, 락토스, 수크로스, 젤라틴, 맥아, 쌀, 밀가루, 백악, 실리카겔, 스테아르산나트륨, 글리세롤 모노스테아레이트, 활석, 염화나트륨, 건조 탈지 분유, 글리세롤, 프로필렌, 글리콜, 물, 에탄올 등을 포함한다.
조성물은 또한, 필요에 따라, 소량의 습윤제 또는 유화제, 또는 pH 완충제를 함유할 수 있다. 이러한 조성물은 용액, 현탁액, 에멀젼, 정제, 환제, 캡슐, 분말, 지속 방출 제제 등의 형태를 취할 수 있다. 경구 제형은 약제학적 등급의 만니톨, 락토스, 전분, 스테아르산마그네슘, 사카린나트륨, 셀룰로오스, 탄산마그네슘 등의 표준 담체를 포함할 수 있다. 적합한 약제의 예는 문헌[참조: "Remington's Pharmaceutical Sciences"]에 기재되어 있다. 이러한 조성물은, 예방적 또는 치료적 유효량의 항체 또는 이의 단편을, 바람직하게는 정제된 형태로, 적절한 양의 담체와 함께, 환자에게 적절하게 투여하기 위한 형태를 제공한다. 제형은 경구, 정맥내, 동맥내, 협측, 비강내, 분무, 기관지 흡입, 직장내, 질, 국소 또는 기계적 인공호흡에 의해 전달될 수 있는 투여 방식에 적합해야 한다.
공개된 것과 같은 항체가 EV-D68 감염 위험이 있는 대상체에서 생체내에서 생성되는 경우의 활성 백신도 상정된다. 이러한 백신은 비경구 투여용으로 제형화될 수 있고, 예를 들면, 피내, 정맥내, 근육내, 피하, 에어로졸 또는 심지어 복강내 경로를 통한 주사용으로 제형화될 수 있다. 피내 및 근육내 경로에 의한 투여가 고려된다. 대안적으로, 백신은 점막에 국소 경로에 의해 점막에 직접 투여될 수 있고, 예를 들면, 점비제, 흡입, 분무기에 의해, 또는 직장내 또는 질 전달을 통해 투여될 수 있다. 약제학적으로 허용되는 염은 산성 염 및, 예를 들면, 염산 또는 인산 등의 무기산, 또는 아세트산, 옥살산, 타르타르산, 만델산 등의 유기산으로 형성된 염을 포함한다. 유리 카복실 그룹으로 형성된 염은 또한, 예를 들면, 나트륨, 칼륨, 암모늄, 칼슘 또는 수산화제2철 등의 무기 염기, 및 이소프로필아민, 트리메틸아민, 2-에틸아미노 에탄올, 히스티딘, 프로카인 등으로부터 유래할 수 있다.
인공적으로 획득한 수동 면역으로서 공지된 항체의 수동적 전달은 일반적으로 정맥내 주사 또는 근육내 주사의 사용을 수반한다. 항체의 형태는, 정맥내(IVIG) 또는 근육내(IG) 사용을 위한 풀링된 인간 면역글로불린으로서, 면역화되거나 질환으로부터 회복된 공여자로부터의 고역가 인간 IVIG 또는 IG으로서, 및 모노클로날 항체(MAb)로서, 인간 또는 동물의 혈장 또는 혈청일 수 있다. 이러한 면역은 일반적으로 짧은 기간 동안만 지속되고, 특히 비인간 기원의 감마 글로불린에 의한 과민 반응 및 혈청병의 잠재적 위험이 있다. 그러나, 수동 면역은 즉각적 보호를 제공한다. 항체는 주사에 적합한, 즉 멸균되고 주사 가능한 담체에서 제형화될 것이다.
일반적으로, 본 개시의 조성물의 성분은, 개별적으로 공급되거나, 예를 들면, 활성 물질의 양을 나타내는 앰플 또는 사셰 등의 밀봉된 용기에 건조 동결건조 분말 또는 물-비함유 농축물로서 단위 투여 형태로 혼합된다. 조성물이 주입에 의해 투여되는 경우, 이는 멸균 약제학적 등급의 물 또는 식염수를 포함하는 주입 병에 분배될 수 있다. 조성물이 주사에 의해 투여되는 경우, 주사용 멸균수 또는 식염수의 앰플을 제공하여 투여 전에 성분을 혼합할 수 있다.
본 개시의 조성물은 중성 또는 염 형태로 제형화될 수 있다. 약제학적으로 허용가능한 염은 염산, 인산, 아세트산, 옥살산, 타르타르산 등으로부터 유래된 것 등의 음이온으로 형성된 염, 및 나트륨, 칼륨, 암모늄, 칼슘, 수산화제이철, 이소프로필아민, 트리에틸아민, 2-에틸아미노에탄올, 히스티딘, 프로카인 등으로부터 유래된 것 등의 양이온으로 형성된 염을 포함한다.
2. ADCC
항체-의존성 세포-매개된 세포독성(ADCC)은 면역 이펙터 세포에 의해 항체 코팅된 표적 세포의 용해를 유도하는 면역 메커니즘이다. 표적 세포는 일반적으로 Fc 영역의 N-말단인 단백질 부분을 통해 Fc 영역을 포함하는 항체 또는 이의 단편이 특이적으로 결합하는 세포이다. "항체 의존성 세포-매개 세포독성(ADCC)의 증가/감소를 갖는 항체"는 당업자에게 공지된 임의의 적합한 방법에 의해 측정되는 ADCC의 증가/감소를 갖는 항체를 의미한다.
본원에 사용된 바와 같이, 용어 "증가/감소된 ADCC"는, 상기 정의된 ADCC의 메커니즘에 의해 표적 세포를 둘러싸고 있는 배지에서 소정 항체 농도에서 소정 시간에 용해되는 표적 세포 수의 증가/감소로서 정의되고/되거나, ADCC의 메커니즘에 의해 소정 시간에 소정 수의 표적 세포의 용해를 달성할 필요가 있는, 표적 세포를 둘러싸고 있는 배지에서 항체 농도의 감소/증가로서 정의된다. ADCC의 증가/감소는 동일한 표준적 생산, 정제, 제형 및 저장 방법(이는 당업자에게 공지됨)을 사용하여 동일한 유형의 숙주 세포에 의해 생성된 동일한 항체에 의해 매개되는 ADCC와 관련되지만, 이는 조작되지 않은 것이다. 예를 들면, 본 명세서에 기재된 방법에 의해, 변경된 글리코실화 패턴을 갖도록(예를 들면, 글리코실트랜스퍼라제, GnTIII 또는 기타 글리코실트랜스퍼라제를 발현하도록) 조작된 숙주 세포에 의해 생성된 항체에 의해 매개되는 ADCC의 증가는 동일한 유형의 조작되지 않은 숙주 세포에 의해 생성된 동일한 항체에 의해 매개되는 ADCC와 비교된다.
3. CDC
보체 의존성 세포독성(CDC)은 보체 시스템의 기능이다. 면역계의 항체 또는 세포의 관여 없이 병원체의 막을 손상시킴으로써 병원체를 사멸시키는 것은 면역계의 프로세스이다. 3개의 주요 프로세스가 있다. 3개 모두가 치명적 콜로이드 삼투압 팽창, 즉 CDC를 유발하는 병원체에 하나 이상의 막 공격 복합체(MAC)를 삽입한다. 이는 항체 또는 항체 단편이 항바이러스 효과를 발휘하는 메커니즘 중 하나이다.
IV. 항체 접합체
본 개시의 항체는 항체 접합체를 형성하기 위해 적어도 하나의 약제에 연결시킬 수 있다. 진단제 또는 치료제로서의 항체 분자의 효능을 증가시키기 위해, 적어도 하나의 목적하는 분자 또는 모이어티를 연결하거나, 공유 결합하거나, 또는 복합체를 형성하는 것이 통상적이다. 이러한 분자 또는 모이어티는 적어도 하나의 이펙터 또는 리포터 분자일 수 있지만, 이들로 한정되지 않는다. 이펙터 분자는 목적하는 활성, 예를 들면, 세포독성 활성을 갖는 분자를 포함한다. 항체에 부착된 이펙터 분자의 비제한적 예에는 독소, 항종양제, 치료용 효소, 방사성핵종, 항바이러스제, 킬레이트제, 사이토카인, 성장 인자 및 올리고- 또는 폴리뉴클레오티드가 포함된다. 대조적으로, 리포터 분자는 검정을 사용하여 검출될 수 있는 임의의 모이어티로서 정의된다. 항체에 접합된 리포터 분자의 비제한적 예에는 효소, 방사성 표지, 합텐, 형광 표지, 인광 분자, 화학발광 분자, 발색단, 광친화성 분자, 착색 입자 또는 리간드, 예를 들면, 비오틴이 포함된다.
항체 접합체는 일반적으로 진단제로서 사용하기에 바람직하다. 항체 진단은 일반적으로 2개 부류, 다양한 면역검정 등의 시험관내 진단에 사용되는 것과, 일반적으로 "항체 지시 이미징"으로 공지된 생체내 진단 프로토콜에 사용되는 것으로 분류된다. 항체에 대한 이들의 부착을 위한 방법과 마찬가지로, 다수의 적절한 이미징제가 당해 기술분야에 공지되어 있다(예를 들면, 미국 특허 제5,021,236호, 제4,938,948호 및 제4,472,509호 참조). 사용된 이미징 모이어티는 상자성 이온, 방사성 동위원소, 형광색소, NMR 검출 가능한 물질 및 X선 이미징제일 수 있다.
상자성 이온의 경우, 예로서, 크롬(III), 망간(II), 철(III), 철(II), 코발트(II), 니켈(II), 구리(II), 네오디뮴(III), 사마륨(III), 이테르븀(III), 가돌리늄(III), 바나듐(II), 테르븀(III), 디스프로슘(III), 홀뮴(III) 및/또는 에르븀(III) 등의 이온을 들 수 있고, 가돌리늄이 특히 바람직하다. X선 이미징 등의 기타 상황에서 유용한 이온에는 란탄(III), 금(III), 납(II), 특히 비스무트(III)가 포함되지만, 이들로 한정되지 않는다.
치료 및/또는 진단용의 방사성 동위원소의 경우, 아스타틴211, 14탄소, 51크롬, 36염소, 57코발트, 58코발트, 구리67, 152Eu, 갈륨67, 3수소, 요오드123, 요오드125, 요오드131, 인듐111, 59철, 32인, 레늄186, 레늄188, 75셀레늄, 35황, 테크니슘99m 및/또는 이트륨90을 들 수 있다. 125I는 특정 실시양태에서의 사용에 종종 바람직하고, 테크니시움99m 및/또는 인듐111은 또한 이들의 낮은 에너지 및 장거리 감지에 대한 적합성으로 인해 종종 바람직하다. 본 개시의 방사성 표지된 모노클로날 항체는 당해 기술분야에 공지된 방법에 따라 생성될 수 있다. 예를 들면, 모노클로날 항체는 요오드화나트륨 및/또는 요오드화칼륨 및 차아염소산나트륨 등의 화학적 산화제 또는 락토퍼옥시다제 등의 효소적 산화제와 접촉시킴으로써 요오드화할 수 있다. 본 개시에 따른 모노클로날 항체는, 예를 들면, 퍼테크네이트를 제1주석 용액으로 환원시키고, 환원된 테크네튬을 세파덱스 컬럼 상에 킬레이트화하고, 항체를 이 컬럼에 적용함으로써 테크네튬99m으로 표지될 수 있다. 또는, 예를 들면, 퍼테크네이트, SNCl2 등의 환원제, 나트륨-칼륨 프탈레이트 용액 등의 완충액 및 항체를 배양함으로써 직접 표지 기술을 사용할 수 있다. 금속 이온으로서 존재하는 방사성 동위원소를 항체에 결합시키기 위해 자주 사용되는 중간 작용기는 디에틸렌트리아민펜타아세트산(DTPA) 또는 에틸렌 디아민테트라아세트산(EDTA)이다.
접합체로서 사용하기 위해 고려되는 형광성 표지에는 알렉사(Alexa) 350, 알렉사(Alexa) 430, AMCA, BODIPY 630/650, BODIPY 650/665, BODIPY-FL, BODIPY-R6G, BODIPY-TMR, BODIPY-TRX, 카스카드 블루(Cascade Blue), Cy3, ,6-FAM, 플루오레세인 이소티오시아네이트, HEX, 6-JOE, 오레곤 그린(Oregon Green) 488, 오레곤 그린 500, 오레곤 그린 514, 퍼시픽 블루(Pacific Blue), REG, 로다민 그린(Rhodamine Green), 로다민 레드(Rhodamine Red), 레노그라핀(Renographin), ROX, TAMRA, TET, 테트라메틸로다민(Tetramethylrhodamine) 및/또는 텍사스 레드(Texas Red)가 포함된다.
본 개시에서 고려되는 추가 유형의 항체는 주로 시험관내에서의 사용을 위한 것이고, 여기서 항체는 2차 결합 리간드 및/또는 발색성 기질과의 접촉시 착색된 생성물을 생성하는 효소(효소 태그)에 연결된다. 적합한 효소의 예는 우레아제, 알칼리성 포스파타제, (서양고추냉이) 수소 퍼옥시다제 또는 글루코스 옥시다제를 포함한다. 바람직한 2차 결합 리간드는 비오틴 및 아비딘 및 스트렙트아비딘 화합물이다. 이러한 표지의 사용은 당업자에게 공지되어 있고, 예를 들면, 미국 특허 제3,817,837호, 제3,850,752호, 제3,939,350호, 제3,996,345호, 제4,277,437호, 제4,275,149호 및 제4,366,241호에 기재되어 있다.
항체에 대한 분자의 부위-특이적 부착의 또 다른 공지된 방법은 항체를 합텐 기반 친화성 표지와 반응시키는 것을 포함한다. 기본적으로, 합텐-기반 친화성 표지는 항원 결합 부위의 아미노산과 반응하고, 이에 의해 이 부위를 파괴하고, 특정 항원 반응을 차단한다. 그러나, 이는, 항체 접합체에 의한 항원 결합의 손실을 초래하기 때문에, 유리하지 않을 수 있다.
아지도 그룹을 포함하는 분자는 또한 낮은 강도의 자외선에 의해 생성되는 반응성 니트렌 중간체를 통해 단백질에 대한 공유 결합을 형성하기 위해 사용될 수 있다[참조: Potter and Haley, 1983]. 특히, 퓨린 뉴클레오티드의 2- 및 8-아지도 유사체는 조 세포 추출물 중의 뉴클레오티드 결합 단백질을 동정하기 위한 부위 특이적 광프로브로로서 사용되었다[참조: Owens & Haley, 1987; Atherton et al., 1985]. 2- 및 8-아지도 뉴클레오티드는 또한 정제된 단백질의 뉴클레오티드 결합 도메인을 맵핑하기 위해 사용되었고[참조: Khatoon et al., 1989; King et al., 1989; Dholakia et al., 1989], 항체 결합제로서 사용될 수 있다.
접합체 모이어티에 대한 항체의 부착 또는 접합을 위한 몇몇 방법이 당해 기술분야에 공지되어 있다. 일부 부착 방법은, 예를 들면, 디에틸렌트리아민펜타아세트산 무수물(DTPA) 등의 유기 킬레이트제; 에틸렌트리아민테트라아세트산; N-클로로-p-톨루엔설폰아미드; 및/또는 항체에 부착된 테트라클로로-3α-6α-디페닐글리코우릴-3을 사용하는 금속 킬레이트 착체의 사용을 수반한다(미국 특허 제4,472,509호 및 제4,938,948호). 모노클로날 항체는 또한 글루타르알데히드 또는 페리오데이트 등의 커플링제의 존재하에 효소와 반응시킬 수 있다. 플루오레세인 마커와의 접합체는 이러한 커플링제의 존재하에 또는 이소티오시아네이트와의 반응에 의해 제조된다. 미국 특허 제4,938,948호에서, 유방 종양의 이미징은 모노클로날 항체를 사용하여 달성되고, 검출 가능한 이미징 모이어티는 메틸-p-하이드록시벤즈이미데이트 또는 N-숙신이미딜-3-(4-하이드록시페닐)프로피오네이트 등의 링커를 사용하여 항체에 결합시킨다.
다른 실시양태에서, 항체 결합 부위를 변화시키지 않는 반응 조건을 사용하여, 면역글로불린의 Fc 영역에 설프하이드릴 그룹을 선택적으로 도입함으로써 면역글로불린의 유도체화가 고려된다. 이 방법에 따라 생성된 항체 접합체는 개선된 수명, 특이성 및 감도를 나타내는 것으로 개시되어 있다(미국 특허 제5,196,066호, 참조에 의해 본 명세서에 도입됨). 리포터 또는 이펙터 분자가 Fc 영역의 탄수화물 잔기에 접합되어 있는, 이펙터 또는 리포터 분자의 부위 특이적 부착도 문헌에 개시되어 있다[참조: O'Shannessy et al., 1987]. 이 접근법은 현재 임상 평가 중의 진단 및 치료적으로 유망한 항체를 생산하는 것으로 보고되었다.
V. 면역검출 방법
또 다른 실시양태에서, 본 개시는 EV-D68 및 이의 관련 항원을 결합, 정제, 제거, 정량화 및 달리 일반적으로 검출하기 위한 면역검출 방법에 관한 것이다. 이러한 방법은 종래의 의미에서 적용될 수 있지만, 백신 및 기타 바이러스 스톡의 품질 관리 및 모니터링에도 사용될 것이고, 여기서 본 개시에 따른 항체는 바이러스 중의 항원의 양 또는 완전성(즉, 장기 안정성)을 평가하기 위해 사용될 수 있다. 또는, 이 방법을 사용하여, 적절한/목적하는 반응성 프로파일에 대해 다양한 항체를 스크리닝할 수 있다.
다른 면역검출 방법에는 대상체에서 EV-D68의 존재를 결정하기 위한 특정 검정이 포함된다. 다양한 검정 형식이 고려되지만, 구체적으로는 타액, 혈액, 혈장, 가래, 정액, 소변, 호흡기 비말 또는 에어로졸 등의 대상체로부터 수득된 유체 중의 EV-D68을 검출하기 위해 사용되는 것들이다. 특히, 정액은 EV-D68을 검출하기 위한 실행 가능한 샘플로서 입증되었다[참조: Purpura et al., 2016; Mansuy et al., 2016; Barzon et al., 2016; Gornet et al., 2016; Duffy et al., 2009, CDC, 2016, Halfon et al., 2010, Elder et al. 2005]. 검정은 가정에서의 임신 검사와 유사한 회방향 유동 검정(하기 참조)을 포함하여 비의료(가정)에서의 사용을 위해 유리하게 포맷할 수 있다. 이러한 검정은 가족 구성원의 대상체에 의해 사용될 수 있도록 적절한 시약 및 지침을 포함하는 키트 형태로 팩키징될 수 있다.
일부 면역검출 방법에는 효소 결합 면역흡착 검정(ELISA), 방사선면역 검정(RIA), 면역방사성 검정, 형광면역 검정, 화학발광 검정, 생물발광 검정 및 웨스턴 블롯이 포함된다. 특히, 샘플 중의 특정 기생충 에피토프에 대해 지시된 EV-D68 항체의 검출 및 정량을 위한 경쟁 검정도 제공된다. 다양한 유용한 면역검출 방법의 단계는, 예를 들면, 문헌[참조: Doolittle 및 Ben-Zeev(1999), Gulbis and Galand(1993), De Jager et al. (1993), and Nakamura et al. (1987)]에 기재되어 있다. 일반적으로, 면역결합 방법은 EV-D68을 함유하는 것으로 의심되는 샘플을 수득하고, 경우에 따라, 면역복합체의 형성을 가능하게 하기에 효과적인 조건하에 본 개시에 따라 샘플을 제1 항체와 접촉시키는 것을 포함한다.
이러한 방법에는 샘플로부터 EV-D68 또는 관련 항원을 정제하는 방법이 포함된다. 항체는 바람직하게는 컬럼 매트릭스 형태 등의 고체 지지체에 연결되고, EV-D68 또는 항원 성분을 함유하는 것으로 의심되는 샘플은 고정된 항체에 적용된다. 불필요한 성분은 컬럼으로부터 세척되어 EV-D68 항원이 고정된 항체에 면역복합체로 잔류하고, 이어서 고정화된 항체는 컬럼으로부터 생물 또는 항원을 제거함으로써 수집된다.
면역 결합 방법에는 또한 샘플 중의 EV-D68 또는 관련 성분의 양을 검출 및 정량화하는 방법 및 결합 프로세스 동안 형성된 면역 복합체의 검출 및 정량화 방법이 포함된다. 여기에서는, EV-D68 또는 이의 항원을 포함하는 것으로 의심되는 샘플을 수득하고, 이 샘플을 EV-D68 또는 이의 성분에 결합하는 항체와 접촉시킨 후, 특정 조건하에서 형성된 면역 복합체의 양을 검출하고 정량화한다. 항원 검출과 관련하여, 분석된 생물학적 샘플은 조직 절편 또는 표본, 균질화된 조직 추출물, 혈액 및 혈청을 포함한 생물학적 유체, 또는 대변 또는 소변 등의 분비물 등의 EV-D68 또는 EV-D68 항원을 함유하는 것으로 의심되는 임의의 샘플일 수 있다.
선택된 생물학적 샘플을 효과적 조건하에서 면역 복합체(1차 면역 복합체)의 형성을 가능하게 하기에 충분한 시간 동안 항체와 접촉시키는 것은 일반적으로 항체 조성물을 샘플에 단순히 첨가하고, 항체가 EV-D68 또는 존재하는 항원과의 면역 복합체를 형성하기에 충분한 시간, 즉 EV-D68 또는 존재하는 항원에 결합하기에 충분한 시간 동안 혼합물을 인큐베이팅하는 것이다. 이 시간 후, 조직 절편, ELISA 플레이트, 도트 블롯 또는 웨스턴 블롯 등의 샘플 항체 조성물을 일반적으로 세척하여 비특이적으로 결합된 항체 종을 제거하여, 1차 면역 복합체 내에서 특이적으로 결합된 항체만이 검출되도록 한다.
일반적으로, 면역복합체 형성의 검출은 당해 기술분야에 공지되어 있고, 다수의 접근법을 적용함으로써 달성될 수 있다. 이러한 방법은 일반적으로 방사성, 형광성, 생물학적 및 효소적 태그 등의 표지 또는 마커의 검출을 기반으로 한다. 이러한 표지의 사용에 관한 특허에는 미국 특허 제3,817,837호, 제3,850,752호, 제3,939,350호, 제3,996,345호, 제4,277,437호, 제4,275,149호 및 제4,366,241호가 포함된다. 물론, 당해 기술분야에 공지된 바와 같이, 2차 항체 등의 2차 결합 리간드 및/또는 비오틴/아비딘 리간드 결합 배열을 사용함으로써 추가의 이점을 발견할 수 있다.
검출에 사용된 항체는 그 자체가 검출 가능한 표지에 연결될 수 있고, 이어서 이 표지를 단순히 검출하고, 이에 의해 조성물 중의 1차 면역 복합체의 양을 결정할 수 있다. 또는, 1차 면역 복합체 내에서 결합되는 1차 항체는 항체에 대한 결합 친화성을 갖는 2차 결합 리간드에 의해 검출할 수 있다. 이러한 경우, 2차 결합 리간드는 검출 가능한 표지에 연결될 수 있다. 2차 결합 리간드는 그 자체가 종종 항체이고, 따라서 "2차" 항체로 명명될 수 있다. 1차 면역 복합체는 효과적인 조건하에 2차 면역 복합체의 형성을 가능하게 하기에 충분한 시간 동안 표지된 2차 결합 리간드 또는 항체와 접촉시킨다. 이어서, 2차 면역 복합체를 일반적으로 세정하여, 비특이적으로 결합된 표지된 2차 항체 또는 리간드를 제거하고, 이어서 2차 면역 복합체의 잔류 표지를 검출한다.
추가 방법에는 2단계 접근법에 의한 1차 면역 복합체의 검출이 포함된다. 상기한 바와 같이, 항체에 대한 결합 친화성을 갖는 항체 등의 제2 결합 리간드를 사용하여 2차 면역 복합체를 형성한다. 세척 후, 2차 면역 복합체는 다시 효과적 조건하에서 면역 복합체(3차 면역 복합체)의 형성을 가능하게 하기에 충분한 기간 동안 2차 항체에 대해 결합 친화성을 갖는 3차 결합 리간드 또는 항체와 접촉시킨다. 3차 리간드 또는 항체는 검출 가능한 표지에 연결되어, 이렇게 형성된 3차 면역 복합체의 검출을 가능하게 한다. 이 시스템은, 이것이 바람직한 경우, 신호 증폭을 제공할 수 있다.
면역 검출의 한 가지 방법은 2개의 상이한 항체를 사용한다. 제1 비오틴화된 항체를 사용하여 표적 항원을 검출하고, 이어서 제2 항체를 사용하여 복합체화된 비오틴에 부착된 비오틴을 검출한다. 이 방법에서, 시험되는 샘플은 먼저 제1 단계의 항체를 포함하는 용액에서 인큐베이팅된다. 표적 항원이 존재하는 경우, 항체의 일부는 항원에 결합하여 비오틴화된 항체/항원 복합체를 형성한다. 이어서, 항체/항원 복합체는 스트렙트아비딘(또는 아비딘), 비오틴화된 DNA 및/또는 상보적 비오틴화된 DNA의 연속 용액에서 인큐베이션에 의해 증폭되고, 각 단계에서 항체/항원 복합체에 추가 비오틴 부위가 추가된다. 적절한 증폭 수준에 도달할 때까지 증폭 단계가 반복되고, 이 시점에서 샘플은 비오틴에 대한 제2 단계의 항체를 포함하는 용액에서 인큐베이팅된다. 이 제2 단계의 항체는, 예를 들면, 발색체 기질을 사용한 조직효소학에 의해 항체/항원 복합체의 존재를 검출하기 위해 사용할 수 있는 효소로 표지된다. 적절한 증폭에 의해, 육안으로 관찰 가능한 접합체를 생성할 수 있다.
면역검출의 또 다른 공지된 방법은 면역-PCR(폴리머라제 연쇄 반응) 방법론을 이용한다. PCR법은 비오틴화 DNA와의 인큐베이션까지는 캔토르(Cantor) 방법과 유사하지만, 스트렙트아비딘과 비오틴화 DNA의 인큐베이션을 복수회 사용하는 대신에, DNA/비오틴/스트렙트아비딘/항체 복합체를 낮은 pH 또는 높은 농도의 완충제로 세정하여 항체를 방출시킨다. 이어서, 수득된 세척 용액을 사용하여, 적절한 대조군을 갖는 적절한 프라이머를 사용하여 PCR 반응을 수행한다. 적어도 이론상으로는, PCR의 거대한 증폭 능력과 특이성을 이용하여 단일 항원 분자를 검출할 수 있다.
A. ELISA
면역검정은 가장 간단하고 직접적인 의미에서 결합 검정이다. 특정의 바람직한 면역검정은 당해 기술분야에 공지된 다양한 유형의 효소 결합 면역흡착 검정(ELISA) 및 방사선면역검정(RIA)이다. 조직 절편을 사용한 면역조직화학적 검출도 특히 유용하다. 그러나, 검출은 이러한 기술에 한정되지 않고, 웨스턴 블롯팅, 도트 블롯팅, FACS 분석 등도 또한 사용될 수 있음을 용이하게 이해할 것이다.
한 가지 예시적 ELISA에서, 본 개시의 항체는 폴리스티렌 미세적정 플레이트 내의 웰 등의 단백질 친화성을 나타내는 선택된 표면 상에 고정화시킨다. 이어서, EV-D68 또는 EV-D68 항원을 함유하는 것으로 의심되는 시험 조성물을 웰에 첨가한다. 비특이적으로 결합된 면역 복합체를 제거하기 위해 결합 및 세척한 후, 결합된 항원을 검출할 수 있다. 검출은, 검출 가능한 표지에 연결된 또 다른 항-EV-D68 항체를 추가함으로써 달성할 수 있다. 이러한 유형의 ELISA는 단순히 "샌드위치 ELISA"이다. 검출은 또한, 제2 항-EV-D68 항체를 첨가하고, 이어서 제2 항체에 대한 결합 친화성을 갖는 제3 항체를 첨가함으로써 달성할 수 있고, 제3 항체는 검출 가능한 표지에 연결된다.
또 다른 예시적 ELISA에서, EV-D68 또는 EV-D68 항원을 함유하는 것으로 의심되는 샘플은 웰 표면에 고정시키고, 이어서 본 개시의 항-EV-D68 항체와 접촉시킨다. 비특이적으로 결합된 면역 복합체를 제거하기 위해 결합 및 세척한 후, 결합된 항-EV-D68 항체가 검출된다. 초기 항-EV-D68 항체가 검출 가능한 표지에 연결된 경우, 면역 복합체를 직접 검출할 수 있다. 이 경우에도, 면역 복합체는 제1 항-EV-D68 항체에 대한 결합 친화성을 갖는 2차 항체를 사용하여 검출할 수 있고, 2차 항체는 검출 가능한 표지에 연결되어 있다.
사용된 포맷에 관계없이, ELISA는 코팅, 인큐베이션 및 결합, 비특이적으로 결합된 종을 제거하기 위한 세척, 결합된 면역 복합체의 검출 등 공통의 특정 기능을 갖고 있다. 이에 대해서 하기 설명한다.
플레이트를 항원 또는 항체로 코팅하는 경우, 일반적으로, 플레이트의 웰을 항원 또는 항체 용액과 함께 밤새 또는 지정된 시간 동안 인큐베이팅한다. 이어서, 플레이트의 웰을 세척하여 불완전하게 흡착된 물질을 제거한다. 이어서, 웰의 나머지 이용 가능한 표면은 시험 항혈청과 관련하여 항원적으로 중성인 비특이적 단백질로 "코팅"된다. 여기에는 소 혈청 알부민(BSA), 카제인 또는 분유 용액이 포함된다. 코팅은 고정화 표면의 비특이적 흡착 부위의 차단을 가능하게 하고, 따라서 표면에 대한 항혈청의 비특이적 결합에 의해 유발된 배경을 감소시킨다.
ELISA에서는, 직접적 절차가 아닌, 2차 또는 3차 검출 수단을 사용하는 것이 보다 일반적일 수 있다. 따라서, 단백질 또는 항체를 웰에 결합시키고, 비반응성 물질로 코팅하여 배경을 감소시키고, 세정하여 비결합된 물질을 제거한 후, 고정화 표면은 면역 복합체(항원/항체)의 형성을 가능하게 하기에 효과적인 조건하에서 시험되는 생물학적 샘플과 접촉시킨다. 이어서, 면역 복합체의 검출에는, 표지된 2차 결합 리간드 또는 항체, 및 표지된 3차 항체 또는 3차 결합 리간드와 조합하여 2차 결합 리간드 또는 항체가 필요하다.
"면역 복합체(항원/항체)의 형성을 가능하게 하기에 효과적인 조건하에서"는 조건이 바람직하게는 BSA, 소 감마 글로불린(BGG) 또는 인산 완충 식염수(PBS)/트윈 등의 용액으로 항원 및/또는 항체를 희석하는 것을 포함하는 것을 의미한다. 이러한 추가된 제제는 또한 비특이적 배경의 감소에도 역할을 하는 경향이 있다.
"적합한" 조건은 또한 인큐베이션이 효과적 결합을 가능하게 하기에 충분한 온도 또는 기간 동안 진행되는 것을 의미한다. 인큐베이션 단계는 일반적으로 약 1~2~4시간 정도, 바람직하게는 25℃ 내지 27℃ 정도의 온도에서 또는 약 4℃ 정도에서 밤새 수행할 수 있다.
ELISA의 모든 인큐베이션 단계에 계속하여, 접촉된 표면을 세척하여 복합체를 형성하지 않은 물질을 제거한다. 바람직한 세척 절차에는 PBS/트윈 등의 용액 또는 붕산염 완충액으로 세척하는 것을 포함한다. 시험 샘플과 원래 결합된 물질 사이에 특정의 면역 복합체의 형성 및 후속 세척에 계속하여, 미량의 면역 복합체의 발생이 결정될 수 있다.
검출 수단을 제공하기 위해, 2차 또는 3차 항체는 검출을 가능하게 하는 관련 표지를 가질 것이다. 바람직하게는, 이것은 적절한 발색성 기질과의 인큐베이션시에 발색을 생성하는 효소일 것이다. 따라서, 예를 들면, 추가 면역 복합체 형성의 발달을 조장하는 조건하에서 일정 기간 동안 우레아제, 글루코스 옥시다제, 알칼리성 포스파타제 또는 과산화수소-접합 항체와 1차 및 2차 면역 복합체를 접촉시키거나 인큐베이팅하는 것을 원할 것이다(예를 들면, PBS-트윈 등의 PBS 함유 용액에서 실온에서 2시간 동안 인큐베이션).
표지된 항체와 함께 인큐베이션한 후 및 결합되지 않은 물질을 제거하기 위해 세척한 후, 예를 들면, 요소, 브로모크레졸 퍼플 또는 2,2'-아지노-디-(3-에틸-벤즈티아졸린-6-설폰산(ABTS), 또는 효소 표지로서 퍼옥시다제의 경우에 H2O2에 의해 표지의 양을 정량화한다. 이어서, 예를 들면, 가시 스펙트럼 분광 광도계를 사용하여, 생성된 색상 정도를 측정함으로써 정량화를 달성한다.
또 다른 실시양태에서, 본 개시는 경쟁 포맷의 사용을 고려한다. 이것은 샘플에서 EV-D68 항체의 검출에 특히 유용하다. 경쟁 기반 검정에서, 미지의 양의 분석물 또는 항체는 공지된 양의 표지된 항체 또는 분석물을 치환하는 능력에 의해 결정된다. 따라서, 신호의 정량화 가능한 손실은 샘플에서 미지의 항체 또는 분석물의 양을 나타낸다.
여기에서, 본 발명자는, 샘플 중의 EV-D68 항체의 양을 결정하기 위해, 표지된 EV-D68 모노클로날 항체의 사용을 제안한다. 기본 포맷에는 미지 양의 EV-D68 모노클로날 항체(검출 가능한 표지에 연결됨)를 EV-D68 항원 또는 입자와 접촉시키는 것이 포함된다. EV-D68 항원 또는 생물은 바람직하게는 지지체에 부착된다. 표지된 모노클로날 항체를 지지체에 결합시킨 후, 샘플을 첨가하고, 샘플 중의 임의의 비표지 항체가 표지된 모노클로날 항체와 경쟁하고, 따라서 이를 치환할 수 있는 조건하에 인큐베이팅한다. 손실된 표지 또는 잔류하는 표지를 측정함으로써(및 결합된 표지의 원래 양으로부터 이를 차감), 비표지 항체의 양이 지지체에 결합되고, 따라서 샘플에 존재하는 항체의 양을 결정할 수 있다.
B. 웨스턴 블롯
웨스턴 블롯(또는 단백질 면역블롯)은 조직 균질물 또는 추출물의 소정 샘플에서 특정 단백질을 검출하기 위해 사용되는 분석 기술이다. 이는 겔 전기영동을 사용하여 폴리펩타이드의 길이(변성 조건) 또는 단백질의 3차원 구조(천연/비변성 조건)에 따라 천연 또는 변성된 단백질을 분리한다. 이어서, 단백질은 막(일반적으로 니트로셀룰로오스 또는 PVDF)으로 전송되고, 여기서 표적 단백질에 특이적인 항체를 사용하여 프로빙(검출)된다.
샘플은 전체 조직 또는 세포 배양으로부터 채취할 수 있다. 대부분의 경우, 고형 조직은 먼저 블렌더(샘플 용적이 큰 경우), 균질화기(용적이 작은 경우) 또는 초음파 처리를 사용하여 기계적으로 분해된다. 세포는 또한 상기 기계적 방법 중 하나에 의해 파괴되어 개방될 수 있다. 그러나, 세균, 바이러스 또는 환경 샘플이 단백질의 공급원으로 될 수 있으므로, 웨스턴 블롯팅은 세포 연구에만 한정되지 않음에 주의해야 한다. 세포의 용해를 촉진하고 단백질을 가용화하기 위해, 각종 계면활성제, 염 및 완충액을 사용할 수 있다. 프로테아제 및 포스파타제 억제제는 그 자체의 효소에 의한 샘플의 소화를 방지하기 위해 종종 첨가된다. 조직의 준비는 종종 단백질의 변성을 회피하기 위해 저온에서 수행된다.
샘플의 단백질은 겔 전기영동을 사용하여 분리된다. 단백질의 분리는 등전점(pI), 분자량, 전하 또는 이러한 요인의 조합에 의해 수행될 수 있다. 분리의 성질은 샘플의 처리와 겔의 성질에 따라 상이하다. 이것은 단백질을 결정하기 위한 매우 유용한 방법이다. 단일 샘플로부터의 단백질을 2차원으로 확산하는 2차원(2-D) 겔을 사용하는 것도 가능하다. 단백질은 1차원에서는 등전점(중성의 순 전하를 갖는 pH)에 따라, 2차원에서는 분자량에 따라 분리된다.
단백질을 항체 검출에 이용할 수 있도록 하기 위해, 이들은 겔 내로부터 니트로셀룰로오스 또는 폴리비닐리덴 디플루오라이드(PVDF)로 제조된 막으로 이동한다. 막을 겔 위에 놓고, 그 위에 여과지 스택을 놓는다. 전체 스택이 완충액에 배치되고, 모세관 작용에 의해 종이 위로 이동하고, 단백질이 함께 운반된다. 단백질을 전송하는 또 다른 방법은 전기블롯팅(electroblotting)이라고 하고, 전류를 사용하여 단백질을 겔로부터 PVDF 또는 니트로셀룰로오스 막으로 끌어당긴다. 단백질은 겔 내의 조직을 유지하면서 겔 내로부터 막으로 이동한다. 이 블롯팅 프로세스의 결과, 단백질은 검출을 위해 얇은 표면층에 노출된다(하기 참조). 두 종류의 막은 비특이적 단백질 결합 특성을 위해 선택된다(즉, 모든 단백질에 동등하게 결합함). 단백질 결합은 소수성 상호작용, 및 막과 단백질 사이의 하전 상호작용을 기반으로 한다. 니트로셀룰로오스 막은 PVDF보다 저렴하지만, 훨씬 더 깨지기 쉽고, 반복되는 프로빙에 충분히 견디지 못한다. 겔로부터 막으로의 단백질의 이동의 균일성과 전반적 효과는 쿠마시에 브릴리언트 블루 또는 폰소(Ponceau) S 염료로 막을 염색하여 확인할 수 있다. 전송되면, 단백질은 표지된 1차 항체 또는 비표지된 1차 항체를 사용하여 검출하고, 이어서 1차 항체의 Fc 영역에 결합하는 표지된 단백질 A 또는 2차 표지된 항체를 사용하여 간접적으로 검출한다.
C. 측면 유동 검정
측면 유동 면역크로마토그래피 검정으로도 공지된 측면 유동 검정은 특수한 고가의 장비 없이도 샘플(매트릭스) 중의 표적 분석물의 존재(또는 부재)를 검출하는 것을 목적으로 하는 단순한 장치이지만, 판독 장치에 의해 서포트되는 다수의 실험실 기반 적용이 존재한다. 통상, 이러한 시험은 재택 시험, 포인트 케어 시험 또는 실험실 사용을 위해 낮은 리소스의 의료 진단으로 사용된다. 널리 보급되고 공지된 적용은 가정용 임신 시험이다.
이 기술은 다공성 종이 또는 소결 폴리머 등의 일련의 모세관 베드를 기반으로 한다. 이들 요소의 각각은 자발적으로 체액(예: 소변)을 수송하는 능력을 갖고 있다. 제1 요소(샘플 패드)는 스폰지 역할을 하고, 과량의 샘플 유체를 보유한다. 침지되면, 유체는 제2 요소(접합체 패드)로 이동하고, 이 요소에는 제조업체가 소위 접합체, 즉 생리활성 입자의 건조된 포맷(하기 참조)을 염-당 매트릭스에 보존하고, 이는 입자의 표면에 고정화된 표적 분자(예: 항원) 및 이의 화학 파트너(예: 항체) 사이의 최적화된 화학적 반응을 보증하기 위한 모두를 함유한다. 샘플 유체는 염-당 매트릭스를 용해하지만, 입자도 용해하고, 하나의 조합된 수송 작용으로, 샘플과 접합체가 다공성 구조를 통해 유동하면서 혼합한다. 이러한 방식으로, 분석물은 제3 모세관 베드를 통해 추가로 이동하면서 입자에 결합한다. 이 물질에는, 제조업자에 의해 제3 분자를 고정화시킨 하나 이상의 영역(종종 스트립으로 지칭)이 있다. 샘플-접합체 혼합물이 이러한 스트립에 도달할 때까지, 분석물은 입자에 결합하고, 제3 '포착' 분자는 복합체에 결합한다. 잠시 후, 보다 많은 액체가 스트립을 통과하면, 입자가 축적되고, 스트립 영역의 색상이 변화된다. 통상, 적어도 2개의 스트립이 있다. 하나(대조군)는 임의의 입자를 포착하고, 이에 의해 반응 조건과 기술이 정상으로 기능하는 것을 나타내고, 다른 하나는 특정 포착 분자를 포함하고, 분석물 분자가 고정화된 입자만을 포착한다. 이러한 반응 구역을 통과한 후, 유체는 최종적 다공성 재료(심지)로 유입되고, 이는 단순히 폐기물 용기로서 기능한다. 측면 유동 시험은 경쟁 검정 또는 샌드위치 검정으로 기능할 수 있다. 측면 유동 검정은 미국 특허 제6,485,982호에 개시되어 있다.
D. 면역조직화학
본 개시의 항체는 또한, 면역조직화학(IHC)에 의한 연구를 위해 제조된, 신선-동결 및/또는 포르말린-고정, 파라핀-포매 조직 블록 둘 모두와 조합하여 사용될 수 있다. 이러한 미립자 표본으로부터 조직 블록을 제조하는 방법은 다양한 예후 인자에 대한 이전의 IHC 연구에서 성공적으로 사용되었고, 해당 분야의 숙련자에게 공지되어 있다[참조: Brown et al., 1990; Abbondanzo et al., 1990; Allred et al., 1990].
간단히 말하면, 동결 절편은, 작은 플라스틱 캡슐 내의 인산염 완충 식염수(PBS)에서 실온에서 50ng의 동결 "분쇄" 조직을 재수화하고; 원심분리에 의해 입자를 펠릿화하고; 점성 포매 매체(OCT)에 이들을 재현탁시키고; 캡슐을 반전시키고/시키거나 원심분리에 의해 다시 펠릿화하고; -70℃ 이소펜탄에서 급속 동결시키고; 플라스틱 캡슐을 절단하고/하거나 동결된 조직 실린더를 제거하고; 크라이오스탯 마이크로톰 척에 조직 실린더를 고정하고/하거나; 캡슐로부터 25-50개의 연속 절편을 절단함으로써 제조할 수 있다. 또는, 전체 냉동 조직 샘플을 연속 절편 절단에 사용할 수 있다.
영구 절편은, 플라스틱 마이크로퓨지 튜브에서 50mg 샘플을 재수화하고; 펠릿화하고; 10% 포르말린에 재현탁하여 4시간 고정시키고; 세척/펠릿화하고; 온화한 2.5% 아가에 재현탁시키고; 펠릿화하고; 빙수에서 냉각하여 아가를 고화시키고; 튜브로부터 조직/아가 블록을 제거하고; 파라핀에 블록을 침윤 및/또는 포매하고/하거나; 최대 50개의 연속 영구 절편을 절단하는 것을 수반하는 유사한 방법으로 제조할 수 있다. 또한, 전체 조직 샘플을 대용할 수 있다.
E. 면역검출 키트
또 다른 실시양태에서, 본 개시는 상기 면역검출 방법과 함께 사용하기 위한 면역검출 키트에 관한 것이다. 항체는 EV-D68 또는 EV-D68 항원을 검출하기 위해 사용될 수 있기 때문에, 항체는 키트에 포함될 수 있다. 따라서, 면역검출 키트는 적절한 용기 수단에서 EV-D68 또는 EV-D68 항원에 결합하는 1차 항체, 및 임의로 면역검출 시약을 포함할 것이다.
특정 실시양태에서, EV-D68 항체는 컬럼 매트릭스 및/또는 미세역가 플레이트의 웰 등의 고체 지지체에 사전에 결합시킬 수 있다. 키트의 면역검출 시약은 소정 항체와 연관되거나 연결된 검출 가능한 표지를 포함하여 다양한 형태 중 어느 하나를 취할 수 있다. 2차 결합 리간드와 연관되거나 부착된 검출 가능한 표지도 또한 고려된다. 예시적 2차 리간드는 1차 항체에 대해 결합 친화성을 갖는 2차 항체이다.
본 키트에 사용하기 위한 추가의 적합한 면역검출 시약은 1차 항체에 대해 결합 친화성을 갖는 2차 항체, 및 2차 항체에 대해 결합 친화성을 갖는 3차 항체를 포함하는 2-성분 시약을 포함하고, 제3 항체는 검출 가능한 표지에 연결된다. 상기 언급된 바와 같이, 다수의 예시적 표지가 당해 기술분야에 공지되어 있고, 이러한 모든 표지를 본 개시와 관련하여 사용할 수 있다.
키트는, 검출 검정을 위한 표준 곡선을 작성하기 위해 사용될 수 있는 바와 같이, 표지되거나 표지되지 않든지, EV-D68 또는 EV-D68 항원의 적절하게 분취된 조성물을 추가로 포함할 수 있다. 키트는, 완전히 접합된 형태로, 중간체의 형태로, 또는 키트 사용자에 의해 접합되는 별도의 모이어티로서 항체-표지 접합체를 포함할 수 있다. 키트의 구성요소는 수성 매질 또는 동결건조된 형태로 팩키징될 수 있다.
키트의 용기 수단은 일반적으로, 항체를 위치시킬 수 있거나, 또는 바람직하게는 적절하게 분취할 수 있는, 적어도 하나의 바이알, 시험관, 플라스크, 병, 주사기 또는 기타 용기 수단을 포함한다. 본 개시의 키트는 또한, 전형적으로, 상업적 판매를 위해 항체, 항원 및 기타 임의의 시약 용기를 긴밀하게 밀폐하여 수용하기 위한 수단을 포함할 것이다. 이러한 용기는 목적하는 바이알이 유지되는 주사 또는 취입 성형 플라스틱 용기를 포함할 수 있다.
F. 백신 및 항원 품질 관리 검정
본 개시는 또한, 샘플 중의 바이러스 항원의 항원성 완전성을 평가할 때에 사용하기 위한 본 명세서에 기재된 항체 및 항체 단편의 사용을 고려한다. 백신과 같은 생물학적 의약품은 통상 분자적으로 특성화될 수 없다는 점에서 화학 의약품과 상이하다. 항체는 상당히 복잡한 큰 분자이고, 제조마다 광범위하게 달라지는 능력을 갖는다. 이들은 또한, 인생 초기의 어린이를 포함하여 건강한 개인에게 투여되고, 따라서 그 자체가 해를 유발하지 않으면서, 생명을 위협하는 질환의 예방 또는 치료에 효과적인 것을 가능한 한 보장하기 위해, 이들의 품질에 중점을 두어야 한다.
백신의 생산 및 유통에서 세계화의 진전은 공중 위생상의 개념을 보다 적절하게 관리하기 위해 새로운 가능성을 열었지만, 다양한 공급원에서 조달된 백신의 동등성과 호환성에 대한 의문도 제기되었다. 따라서, 출발 물질, 생산 및 품질 관리 시험의 국제 표준화, 및 이러한 제품의 제조 및 사용 방식에 대한 규제 감독에 대한 높은 기대의 설정은 지속적 성공의 초석으로 되었다. 그러나, 이는 끊임없는 변화의 분야이고, 이 분야에서의 지속적 기술 발전은 가장 오래된 공중 위생의 위협, 및 새로운 위협(말라리아, 유행성 인플루엔자, HIV 등)에 대해 강력한 새로운 무기의 개발 약속을 제공하지만, 또한 제품이 달성 가능한 최고 수준의 품질을 지속적으로 충족하는 것을 보증하기 위해, 제조업체, 규제 당국 및 광범위한 의료계에 큰 압력을 가하고 있다.
따라서, 항원 또는 백신은 임의의 공급원으로부터 또는 제조 프로세스 중 임의의 시점에서 수득할 수 있다. 따라서, 품질 관리 프로세스는 바이러스 항원에 대한 본 명세서에 개시된 항체 또는 단편의 결합을 동정하는 면역검정을 위한 샘플을 조제하는 것으로부터 개시할 수 있다. 이러한 면역검정은 이 문서의 다른 부분에 개시되어 있고, 이들 중 어느 것을 사용하여, 항원의 구조적/항원성의 완전성을 평가할 수 있다. 허용 가능한 양의 항원적으로 정확한 및 무손상의 항원을 포함하는 샘플을 발견하기 위한 기준은 규제 당국에 의해 확립될 수 있다.
항원의 완전성이 평가되는 또 다른 중요한 실시양태는 저장 수명 및 저장 안정성을 결정하는 것이다. 백신을 포함한 대부분의 의약은 시간의 경과와 함께 악화될 수 있다. 따라서, 시간의 경과와 함께, 백신 등의 항원이 대상체에게 투여될 때 더 이상 항원성이 아니고/아니거나 면역 반응을 생성할 수 없도록, 분해 또는 불안정화하는 정도를 결정하는 것이 중요하다. 이 경우에도, 허용 가능한 양의 항원적으로 무손상 항원을 포함하는 샘플을 발견하기 위한 기준은 규제 당국에 의해 확립될 수 있다.
특정 실시양태에서, 바이러스 항원은 하나 이상의 보호 에피토프를 함유할 수 있다. 이러한 경우, 2, 3, 4, 5 또는 그 이상의 항체 등 하나 이상의 항체의 결합을 조사하는 검정을 사용하는 것이 유용할 수 있다. 이러한 항체는, 밀접하게 관련된 에피토프에 결합하기 때문에, 서로 인접하거나 심지어 중첩된다. 한편, 이들은 항원의 상이한 부분으로부터 별개의 에피토프를 나타낼 수 있다. 복수의 에피토프의 완전성을 조사함으로써, 항원의 전체적 완전성에 대한 보다 완전한 그림, 및 따라서 보호 면역 반응을 생성하는 능력을 결정할 수 있다.
본 개시에 기재된 바와 같은 항체 및 이의 단편은 또한 보호 EV-D68 항체의 존재를 검출함으로써 백신접종 절차의 효능을 모니터링하기 위한 키트에서 사용될 수 있다. 본 개시에 기재된 바와 같은 항체, 항체 단편, 또는 이의 변이체 및 유도체는 또한, 목적하는 면역원성을 갖는 백신 제조를 모니터링하기 위한 키트에서 사용될 수 있다.
VI. 실시예
하기 실시예는 바람직한 실시양태를 입증하기 위해 포함된다. 하기 실시예에 개시된 기술은 실시양태의 실시에서 양호하게 기능하기 위해 발명자에 의해 발견된 기술을 나타내고, 따라서 이의 실시를 위한 바람직한 방식을 구성하는 것으로 간주될 수 있다는 것이 당업자에 의해 이해되어야 한다. 그러나, 당업자는, 본 개시에 비추어, 개시되는 특정 실시양태에서 다수의 변경을 수행할 수 있고, 여전히 개시의 정신 및 범위를 벗어나지 않고서 유사하거나 유사한 결과를 수득할 수 있는 것을 인식해야 한다.
실시예 1 - 재료 및 방법
동물. 유타 주립 대학의 유타 과학기술연구소(USTAR) 빌딩에서 유지되고 있는 특정 병원체 비함유 콜로니로부터의 4주령 수컷 및 암컷 AG129 마우스. 마우스를 유타 주립 대학의 USTAR 빌딩에서 방사선 조사된 텍라드 설치류 다이어트(Harlan Teklad) 및 자동 급수된 수돗물로 사육 및 유지했다.
항체 및 화합물. 모노클로날 항체(mAb) EV-D68-228은 반더빌트 대학 의료 센터(Vanderbilt University Medical Center)의 제임스 크로우(James Crowe)로부터 제공되었다. EV-D68-228은 1.134mg/ml 농도의 용액으로 제공되었고, 치료를 위해 10, 3 및 1mg/kg의 용량으로 멸균 식염수에 희석되었다. rRSV-90은 5mg/ml 농도의 용액으로 제공되었고, 10mg/kg의 용량으로 음성 대조군 항체로 사용되었다. 정맥 면역글로불린(IVIg, Carimune, CSL Behring, King of Prussia, PA)은 지역 약국으로부터 구입하여 EV-D68-228 mAb에 대한 비교대조로서 사용되었다. 구아니딘 HCl(구아니딘)은 시그마-알드리히(Sigma-Aldrich)(St. Louis, MO)로부터 수득하고, 양성 대조군으로서 사용되었다.
바이러스. 엔테로바이러스 D68은 BEI 리소스, NIAID, NIH: 엔테로바이러스 D68, US/MO/14-18949, NR-49130로부터 수득했다. 이 바이러스는 4주령의 AG129 마우스의 폐에서 30회 연속 계대하고, 이어서 아메리칸 타입 컬쳐 컬렉션(American Type Culture Collection)(Manassas, VA)로부터 수득된 횡문근육종(RD) 세포에서 3회 플라크 정제되었다. 수득된 바이러스 스톡을 RD 세포에서 2회 증폭하여, 작업 스톡을 생성했다. 감염에 사용된 바이러스는 EV-D68 MP30 PP로 지정되었다.
실험 설계. 표 1에 제시된 바와 같이, 총 78마리의 마우스를 각각 12마리의 마우스로 이루어진 6개의 그룹으로 랜덤화하고, 6마리의 마우스 그룹을 정상 대조군으로 사용했다. 마우스는 감염 24시간 전에 EV-D68-228 mAb, IVIg 또는 위약 mAb의 복강내(IP) 투여에 의해 치료되었다. 마우스는, 90㎕ 용적의 MEM에서 EV-D68 MP30 PP의 1×104.5 CCID50을 비강내(IN) 주입하여 감염시켰다. 구아니딘에 의한 치료는 감염 4시간 후에 개시되었고, 5일 동안 1일 2회 계속되었다. 치료 전 및 그 후 매일 마우스의 체중을 측정했다. 폐 바이러스 역가, 혈액 바이러스 역가 및 폐 사이토카인 농도를 평가하기 위해, 감염 후 1일, 3일 및 5일차에 각 치료 그룹의 4마리 마우스를 안락사시켰다.
폐 사이토카인/케모카인 평가. 폐 균질액의 각 샘플은 제조업체의 지침(Quansys Biosciences Q-Plex™ Array, Logan, UT)에 따라 화학발광 ELISA 기반 검정을 사용하여 사이토카인 및 케모카인에 대해 시험되었다. 퀀시스 멀티플렉스(Quansys multiplex) ELISA는 16개의 상이한 포획 항체가 정의된 어레이의 96웰 플레이트의 각 웰에 적용된 정량적 시험이다. 각 샘플 상청액은 하기: IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-10, IL-12p70, IL-17, MCP-1, IFN-γ, TNFα, MIP-1α, GM-CSF 및 RANTES에 대해 시험되었다. 약어의 정의는 다음과 같다: IL - 인터루킨; MCP - 단핵 세포 주화성 단백질; IFN - 인터페론; TNF - 종양 괴사 인자, MIP - 마크로파지 염증성 단백질; GM-CSF - 과립구/마크로파지 콜로니 자극 인자; 및 RANTES - 활성화시에 조절되고, 정상 T 세포가 발현 및 분비된다.
통계 분석. 모든 수치 및 통계 분석은 프리즘(Prism) 8.0.2를 사용하여 완료되었다(GraphPad Software Inc.). 일원 분산 분석(ANOVA)을 사용하여, 감염 후 매일, 치료 그룹의 폐 및 혈액 바이러스 역가를 위약 치료 마우스의 폐 및 혈액 역가와 비교했다. 각 사이토카인/케모카인에 대해 이원 배치 분산 분석(ANOVA)을 사용하여 치료 마우스의 농도를 위약 치료 마우스와 비교했다.
실험 동물의 윤리 규칙. 이 연구는 2019년 3월 2일자의 유타 주립 대학(Utah State University)의 시설내 동물 관리 및 사용 위원회의 승인에 따라 수행되었다(2022년 3월 1일 만료). 이 작업은 유타 주립 대학의 AAALAC 인정의 실험 동물 연구 센터에서 수행되었다. 미국 정부(국립 위생 연구소(National Institutes of Health))의 승인이 실험 동물의 관리 및 사용에 관한 국립 위생 연구소 가이드(개정: 2011)에 따라 2018년 3월 9일에 갱신되었다(PHS 보증 번호 D16-00468[A3801-01]).
[표 I]
마우스에서 EV-D68 호흡기 감염의 치료를 위한 EV-D68-228의 효능을 텍스트화하기 위한 대한 실험 설계

실시예 2 - 결과
이 연구의 목적은 4주령 AG129 마우스에서 엔테로바이러스 D68(EV-D68) 호흡기 감염에 대한 EV-D68-228에 의한 치료의 효능을 결정하는 것이었다. 이 연구는 4주령 AG129 마우스에서 EV-D68 호흡기 감염의 치료에 대한 EV-D68-228 mAb의 효능을 결정했다.
도 6A-C는 EV-D68-228로 치료된 EV-D68-감염 AG129 마우스의 폐 바이러스 역가를 나타낸다. 10, 3 또는 1mg/kg 용량의 EV-D68-228로 치료한 마우스에서는 감염 후 1, 3 또는 5일차에 폐 바이러스 역가가 검출되지 않았다. 10mg/kg의 IVIg에 의한 치료는 감염 후 1일차에 폐 바이러스 역가를 유의하게 감소시켰지만, 감염 후 3일차 또는 5일차에는 감소시키지 않았다. 100mg/kg/일 용량의 구아니딘 치료는 감염 후 1일차 및 3일차에 폐 바이러스 역가를 유의하게 감소시켰지만, 감염 후 5일차에는 감소시키지 않았다.
도 7A-C는 EV-D68-228로 치료된 EV-D68-감염 AG129 마우스의 혈액 바이러스 역가를 나타낸다. 10, 3 또는 1mg/kg 용량의 EV-D68-228로 치료한 마우스에서는 감염 후 1일, 3일 또는 5일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 IVIg에 의한 치료는 또한 감염 후 1, 3, 5일차에 혈액 바이러스 역가를 감소시켰다. 구아니딘은 감염 후 1일 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰지만, 3일차에는 감소시키지 않았다.
도 8은 EV-D68-228로 치료된 EV-D68-감염 AG129 마우스로부터의 IL-1α 및 IL-1β의 폐 농도를 나타낸다. EV-D68-228에 의한 치료는 감염 후 3일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 IL-1α 또는 IL-1β의 폐 농도를 유의하게 감소시키지 않았다.
EV-D68-228로 치료된 EV-D68-감염 AG129 마우스로부터의 IL-6 및 MCP-1의 폐 농도는 도 9에 제시되어 있다. EV-D68-228에 의한 치료는 감염 후 3일차에 IL-6 및 MCP-1의 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 감염 후 3일차에 MCP-1의 폐 농도를 유의하게 감소시켰지만, IL-6은 감소시키지 않았다.
EV-D68-228로 치료된 EV-D68-감염 AG129 마우스로부터의 RANTES의 폐 농도는 도 10에 제시되어 있다. EV-D68-228에 의한 치료는 위약 치료 마우스와 비교하여 감염 후 3일 및 5일차에 RANTES 농도를 유의하게 감소시켰다. IVIg 또는 구아니딘에 의한 치료는 감염 후 1일, 3일 또는 5일차에 RANTES의 폐 농도를 유의하게 감소시키지 않았다.
도 11은 EV-D68-228로 치료된 EV-D68-감염 AG129 마우스로부터의 IL-2, IL-3 및 IL-4의 폐 농도를 나타낸다. EV-D68 감염 후, IL-2, IL-3 또는 IL-4 농도의 유의한 변화는 관찰되지 않았다.
도 12는 EV-D68-228로 치료된 EV-D68-감염 AG129 마우스로부터의 IL-5, IL-10 및 IL-12p70의 폐 농도를 나타낸다. EV-D68 감염 후, IL-5, IL-10 또는 IL-12p70 농도의 유의한 변화는 관찰되지 않았다.
EV-D68-228로 치료된 EV-D68-감염 AG129 마우스로부터의 IL-17, IFNγ 및 TNFα의 폐 농도는 도 13에 제시되어 있다. EV-D68 감염 후, IL-17, IFNγ 또는 TNFα 농도의 유의한 변화는 관찰되지 않았다.
EV-D68-228로 치료된 EV-D68-감염 AG129 마우스로부터의 MIP-1α 및 GM-CSF의 폐 농도는 도 14에 제시되어 있다. EV-D68 감염 후, MIP-1α 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다.
실시예 3 - 논의
이 연구는 4주령 AG129 마우스에서 EV-D68에 의해 유발된 호흡기 감염의 치료에 대한, EV-D68에 대한 mAb인 EV-D68-228의 효능을 결정했다.
10, 3 및 1mg/kg 용량의 EV-D68-228은 EV-D68 감염에 의해 유발되는 폐 바이러스 역가, 혈액 바이러스 역가 및 전염증성 사이토카인의 감소에 매우 효과적이었다. IL-1α, IL-1β, IL-6, MCP-1 및 RANTES의 폐 농도는 EV-D68-228에 의한 치료에 의해 유의하게 감소되었다.
EV-D68-228은 EV-D68 감염의 예방에 매우 효과적이었다.
실시예 4 - 재료 및 방법
동물. 유타 주립 대학의 유타 과학기술연구소(USTAR) 빌딩에서 유지되고 있는 특정 병원체 비함유 콜로니로부터의 10일령 AG129 마우스. 마우스를 유타 주립 대학의 USTAR 빌딩에서 방사선 조사된 텍라드 설치류 다이어트(Harlan Teklad) 및 자동 급수된 수돗물로 사육 및 유지했다.
항체 및 화합물. 모노클로날 항체(mAb) EV-D68-228은 반더빌트 대학 의료 센터(Vanderbilt University Medical Center)의 제임스 크로우(James Crowe)로부터 제공되었다. EV-D68-228은 1.134mg/ml 농도의 용액으로 제공되었고, 치료를 위해 10, 3 및 1mg/kg의 용량으로 멸균 식염수에 희석되었다. rRSV-90은 5mg/ml 농도의 용액으로 제공되었고, 10mg/kg의 용량으로 음성 대조군 항체로서 사용되었다. 정맥 면역글로불린(IVIg, Carimune, CSL Behring, King of Prussia, PA)은 지역 약국으로부터 구입하여 EV-D68-228 mAb에 대한 비교대조로서 사용되었다. 구아니딘 HCl(구아니딘)은 시그마-알드리히(St. Louis, MO)로부터 수득하고, 양성 대조군으로서 사용되었다.
바이러스. 엔테로바이러스 D68은 BEI 리소스, NIAID, NIH: 엔테로바이러스 D68, US/MO/14-18949, NR-49130로부터 수득했다. 바이러스는 4주령의 AG129 마우스의 폐에서 30회 연속 계대하고, 이어서 아메리칸 타입 컬쳐 컬렉션(Manassas, VA)으로부터 수득된 횡문근육종(RD) 세포에서 3회 플라크 정제되었다. 수득된 바이러스 스톡을 RD 세포에서 2회 증폭하여, 작업 스톡을 생성했다. 감염에 사용된 바이러스는 EV-D68 MP30 PP로 지정되었다.
실험 설계. 표 II에 제시된 바와 같이, 총 42마리의 마우스를 각각 6마리의 마우스로 이루어진 7개 그룹으로 랜덤화했다. 마우스는 감염 24시간 전에 EV-D68-228 mAb, IVIg 또는 위약 mAb의 복강내(IP) 투여에 의해 치료되었다. 마우스는, 100㎕ 용적의 MEM에서 EV-D68 MP30 PP의 1 × 106.5 CCID50을 복강내(IP) 주입하여 감염시켰다. 구아니딘에 의한 치료는 감염 4시간 후에 개시되었고, 5일 동안 1일 2회 계속되었다. 처리 전 및 그 후 매일 마우스의 체중을 측정했다. 21일차까지, 모든 마우스의 이환율, 사망률 및 신경학적 스코어를 매일 관찰했다. 신경학적 스코어(NS)는 다음과 같이 기록되었다: NS0 - 관찰 가능한 마비 없음, NS1 - 뒷다리가 비정상적으로 벌렸지만 정상 또는 약간 느린 보행, NS2 - 전진 운동에 사용하는 동안, 뒷다리가 부분적으로 붕괴하고 발이 끌림, NS3 - 뒷다리 및 뒷다리의 경직성 마비가 전진 운동에 사용되지 않음, NS4 - 뒷다리의 경직 마비 및 전진 운동 없음. NS4 스코어로 관찰된 모든 동물을 인도적으로 안락사시켰다.
통계 분석. 카플란-마이어(Kaplan-Meier) 생존 곡선은 프리즘 8.0.2로 생성되었다(GraphPad Software Inc.). 로그 순위(Mantel-Cox) 검정, 및 이어서 게한-브레슬로우-윌콕손(Gehan-Breslow-Wilcoxon) 검정을 사용하여 생존 곡선을 비교했다. 일원 분산 분석(ANOVA)을 사용하여, 감염 후 매일, 치료 그룹의 혈액 바이러스 역가를 위약 치료 마우스의 폐 및 혈액 역가와 비교했다. 일원 ANOVA를 사용하여 평균 체중을 비교했다. 크루스칼-왈리스(Kruskal-Wallis) 검정 및 이어서 둔(Dunn)의 다중 비교 검정을 사용하여 신경학적 스코어를 비교했다.
실험 동물의 윤리 규칙. 이 연구는 2019년 3월 2일자의 유타 주립 대학의 시설내 동물 관리 및 사용 위원회의 승인에 따라 수행되었다(2022년 3월 1일 만료). 이 작업은 유타 주립 대학의 AAALAC 인정의 실험 동물 연구 센터에서 수행되었다. 미국 정부(국립 위생 연구소)의 승인이 실험 동물의 관리 및 사용에 관한 국립 위생 연구소 가이드(개정: 2011)에 따라 2018년 3월 9일에 갱신되었다(PHS 보증 번호 D16-00468[A3801-01]).
[표 II]
마우스에서 EV-D68 신경 감염의 치료를 위한 EV-D68-228의 효능을 텍스트화하기 위한 실험 설계

실시예 5 - 결과
이 연구의 목적은 10일령의 AG129 마우스에서 엔테로바이러스 D68(EV-D68) 신경계 감염에 대한 EV-D68-228에 의한 치료의 효능을 결정하는 것이었다. 이 연구는 10일령의 AG129 마우스에서 EV-D68 신경 감염의 치료를 위한 EV-D68-228 mAb의 효능을 결정했다.
도 15는 EV-D68에 감염되고 EV-D68-228, IVIg 또는 구아니딘으로 치료된 10일령의 AG129 마우스에 대한 카플란-마이어 생존 곡선을 나타낸다. 10, 3 또는 1mg/kg 용량의 EV-D68-228은 EV-D68에 감염된 6마리 마우스의 각 그룹에서 사망으로부터 100% 보호를 제공했다. 10mg/kg 용량의 IVIg로 치료된 6마리의 마우스 중 2마리는 감염으로부터 생존하지 못했다. 100mg/kg/일 용량의 구아니딘 치료는 6마리의 마우스 모두를 사망으로부터 보호했다.
도 16은 EV-D68에 감염되고 EV-D68-228, IVIg 또는 구아니딘으로 치료된 10일령의 AG129 마우스에 대한 평균 체중을 나타낸다. 10, 3 또는 1mg/kg의 용량은 감염과 관련된 체중 감소로부터 마우스를 보호했다. 10mg/kg 용량의 IVIg는 또한 체중 감소로부터 마우스를 보호했다. 또한, 100mg/kg/일 용량의 구아니딘은 마우스를 체중 감소로부터 보호했다.
도 17A-C는 EV-D68-228로 치료된 EV-D68-감염 AG129 마우스의 혈액 바이러스 역가를 나타낸다. 10, 3 또는 1mg/kg 용량의 EV-D68-228로 치료한 마우스에서는 감염 후 1일, 3일 또는 5일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 IVIg에 의한 치료는 또한 감염 후 1, 3, 5일차에 혈액 바이러스 역가를 감소시켰다. 100mg/kg/일 용량의 구아니딘 치료는 감염 후 1일, 3일 및 5일차에 혈액 바이러스 역가를 유의하게 감소시켰다.
EV-D68에 감염되고 EV-D68-228, IVIg 또는 구아니딘으로 치료된 10일령의 AG129 마우스의 신경학적 스코어는 도 18에 제시되어 있다. 10, 3 또는 1mg/kg 용량의 EV-D68-228 Ab로 치료된 마우스에서는 신경학적 스코어가 관찰되지 않았다. 신경학적 스코어는 감염 후 4일, 5일 및 6일차에 10mg/kg 용량의 IVIg로 치료된 마우스에서 관찰되었다. 그러나, IVIg로 치료된 마우스의 신경학적 스코어는 위약 치료된 마우스와 비교하여 유의하게 감소했다. 100mg/kg/일 용량의 구아니딘으로 치료한 마우스에서는 신경학적 스코어가 관찰되지 않았다.
실시예 6 - 논의
이 연구에서는, 10일령의 AG129 마우스에서 EV-D68에 의해 유발된 신경 감염의 치료를 위한 EV-D68에 대한 mAb인 EV-D68-228의 효능을 결정했다.
10, 3, 및 1mg/kg 용량의 EV-D68-228은 EV-D68에 감염된 마우스의 사망률 및 체중 감소로부터 보호를 제공했다. 또한, EV-D68-228로 치료된 마우스에서는 감염 후 1일, 3일 또는 5일차에 혈액 바이러스 역가가 검출되지 않았다. 신경학적 스코어로 측정한 경우, 10, 3 또는 1mg/kg 용량의 EV-D68-228으로 치료된 마우스에서는 마비가 관찰되지 않았다.
EV-D68-228은, 10mg/kg 용량의 IVIg로 치료한 마우스에서 관찰된 생존율, 혈액 바이러스 역가 및 신경학적 스코어에 의해 나타난 바와 같이, 상업용 IVIg와 비교하여 EV-D68 감염으로부터 보호를 제공하는 것으로 나타났다.
EV-D68-228은, 양성 대조군인 구아니딘에 의한 치료와 비교한 경우, 사망률 및 마비로부터 동등한 보호를 제공했다. 그러나, 감염 후 3일차에 구아니딘 치료된 마우스에서는 혈액 바이러스 역가가 여전히 검출되었지만, 감염 후 1일, 3일 또는 5일차에 10, 3 또는 1mg/kg 용량의 EV-D68-228로 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았다.
실시예 7 - 마우스의 호흡기 감염에 대한 EV-D68-228의 효능
재료 및 방법
동물. 유타 주립 대학의 유타 과학기술연구소(USTAR) 빌딩에서 유지되고 있는 특정 병원체 비함유 콜로니로부터의 4주령의 수컷 및 암컷 AG129 마우스. 마우스를 유타 주립 대학의 USTAR 빌딩에서 방사선 조사된 텍라드 설치류 다이어트(Harlan Teklad) 및 자동 급수된 수돗물로 사육 및 유지했다.
항체 및 화합물. 모노클로날 항체(mAb) EV-D68-228은 반더빌트 대학 의료 센터의 제임스 크로우로부터 제공되었다. EV-D68-228은 1.134mg/ml 농도의 용액으로 제공되었고, 치료를 위해 10 또는 1mg/kg의 용량으로 멸균 식염수에 희석되었다. rRSV-90은 5mg/ml 농도의 용액으로 제공되었고, 10mg/kg의 용량으로 음성 대조군 항체로 사용되었다. 정맥 면역글로불린(IVIg, Carimune, CSL Behring, King of Prussia, PA)은 지역 약국으로부터 구입하여 EV-D68-228 mAb에 대한 비교대조로서 사용했다.
바이러스. 엔테로바이러스 D68은 BEI 리소스, NIAID, NIH: 엔테로바이러스 D68, US/MO/14-18949, NR-49130로부터 수득했다. 바이러스는 4주령의 AG129 마우스의 폐에서 30회 연속 계대하고, 이어서 아메리칸 타입 컬쳐 컬렉션(Manassas, VA)로부터 수득된 횡문근육종(RD) 세포에서 3회 플라크 정제되었다. 수득된 바이러스 스톡을 RD 세포에서 2회 증폭하여, 작업 스톡을 생성했다. 감염에 사용된 바이러스는 EV-D68 MP30 PP로 지정되었다.
실험 설계. 표 C에 제시된 바와 같이, 총 52마리의 마우스를 각각 8마리의 마우스로 이루어진 6개 그룹으로 랜덤화하고, 4마리의 마우스 그룹을 정상 대조군으로 사용했다. 마우스는 감염 후 4, 24 또는 48시간에 EV-D68-228 mAb, IVIg 또는 위약 mAb의 복강내(IP) 투여에 의해 치료되었다. 마우스는, 90㎕ 용적의 MEM에서 EV-D68 MP30 PP의 1×104.5 CCID50을 비강내(IN) 주입하여 감염시켰다. 치료 전 및 그 후 매일 마우스의 체중을 측정했다. 폐 바이러스 역가, 혈액 바이러스 역가 및 폐 사이토카인 농도의 평가를 위해, 감염 후 3일 및 5일차에 각 치료 그룹의 4마리 마우스를 안락사시켰다.
폐 사이토카인/케모카인 평가. 폐 균질액의 각 샘플은 제조업체의 지침(Quansys Biosciences Q-Plex™ Array, Logan, UT)에 따라 화학발광 ELISA 기반 검정을 사용하여 사이토카인 및 케모카인에 대해 시험되었다. 퀀시스 멀티플렉스(Quansys multiplex) ELISA는 16개의 상이한 포획 항체가 정의된 어레이의 96웰 플레이트의 각 웰에 적용된 정량적 시험이다. 각 샘플 상청액은 하기: IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-10, IL-12p70, IL-17, MCP-1, IFN-γ, TNFα, MIP-1α, GM-CSF 및 RANTES에 대해 시험되었다. 약어의 정의는 다음과 같다: IL - 인터루킨; MCP - 단핵 세포 주화성 단백질; IFN - 인터페론; TNF - 종양 괴사 인자, MIP - 마크로파지 염증성 단백질; GM-CSF - 과립구/마크로파지 콜로니 자극 인자; 및 RANTES - 활성화시에 조절되고, 정상 T 세포가 발현 및 분비된다.
통계 분석. 모든 수치 및 통계 분석은 프리즘(Prism) 8.2.0을 사용하여 완료되었다(GraphPad Software Inc.). 일원 분산 분석(ANOVA)을 사용하여, 감염 후 매일, 치료 그룹의 폐 및 혈액 바이러스 역가를 위약 치료 마우스의 폐 및 혈액 역가와 비교했다. 각 사이토카인/케모카인에 대해 이원 배치 분산 분석(ANOVA)을 사용하여 치료 마우스의 농도를 위약 치료 마우스와 비교했다.
실험 동물의 윤리 규칙. 이 연구는 유타 주립 대학의 시설내 동물 관리 및 사용 위원회의 승인에 따라 수행되었다. 이 작업은 유타 주립 대학의 AAALAC 인정의 실험 동물 연구 센터에서 수행되었다. 미국 국립 보건원(국립 위생 연구소(National Institutes of Health))의 승인은 실험 동물의 관리 및 사용에 관한 국립 위생 연구소 가이드(개정, 2011)에 따라 갱신되었다(PHS 보증 번호 D16-00468[A3801-01]).
[표 C]
Expt. NIA-1869. 실험 설계 - 마우스에서 EV-D68 호흡기 감염의 치료를 위한 EV-D68-228의 치료 효능.

결과 및 논의
이 연구에서는 4주령의 AG129 마우스에서 EV-D68 호흡기 감염의 치료를 위한 EV-D68-228 mAb의 치료 효능을 결정했다.
도 19는 EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스의 폐 바이러스 역가를 나타낸다. 감염 후 4시간 또는 24시간에서 치료를 개시한 경우, 10mg/kg 용량의 EV-D68-228로 치료한 마우스에서는 감염 후 3일 또는 5일에 폐 바이러스 역가가 검출되지 않았다. 감염 후 48시간에서 치료된 마우스에서는 10mg/kg 용량의 EV-D68-228은 감염 후 3일 및 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. 감염 후 48시간에서 1mg/kg의 용량도 감염 후 3일 및 5일차에 폐 바이러스 역가를 유의하게 감소시켰다. 감염 후 24시간에서 10mg/kg 용량의 IVIg에 의한 치료는 감염 후 3일 또는 5일차에 폐 바이러스 역가를 유의하게 감소시키지 않았다.
도 20은 EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스의 혈액 바이러스 역가를 나타낸다. 감염 후 4, 24 또는 48시간에 10mg/kg 용량의 EV-D68-228로 치료한 마우스에서는 감염 후 3일 또는 5일차에 혈액 바이러스 역가가 검출되지 않았다. 감염 후 48시간에 1mg/kg의 저용량의 EV-D68-228은 검출 한계 미만으로 혈액 바이러스 역가를 감소시켰다. 10mg/kg 용량의 IVIg에 의한 치료는 감염 후 3일 및 5일차에 혈액 바이러스 역가를 감소시켰다.
도 21은 EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-1α 및 IL-1β의 폐 농도를 나타낸다. EV-D68-228에 의한 치료적 치료는 감염 후 3일차 및 5일차에 IL-1α 및 IL-1β의 농도를 유의하게 감소시켰다. IVIg에 의한 치료는 IL-1α 또는 IL-1β의 폐 농도를 유의하게 감소시켰지만, 감염 후 3일차에만 감소시켰다.
EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-5 및 IL-6의 폐 농도는 도 22에 제시되어 있다. EV-D68-228에 의한 치료적 치료는 감염 후 3일차에 IL-5 및 IL-6의 농도를 유의하게 감소시켰다. 감염 24시간 후에 IVIg에 의한 치료는 감염 후 3일차에 IL-5 및 IL-6의 폐 농도를 유의하게 감소시켰다.
EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스로부터의 MCP-1 및 RANTES의 폐 농도가 도 23에 제시되어 있다. EV-D68-228에 의한 치료는, 위약 치료 마우스와 비교하여, 감염 후 3일차 및 5일차에 MCP-1 및 RANTES의 폐 농도를 유의하게 감소시켰다. IVIg에 의한 치료는 감염 후 3일차에 MCP-1의 폐 농도를 유의하게 감소시켰고, 감염 후 3일 및 5일차에 RANTES의 농도를 감소시켰다.
도 24는 EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-2, IL-3 및 IL-4의 폐 농도를 나타낸다. EV-D68 감염 후, IL-2, IL-3 또는 IL-4 농도의 유의한 변화는 관찰되지 않았다.
도 25는 EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-10, IL-12p70 및 IL-17의 폐 농도를 나타낸다. EV-D68 감염 후, IL-10, IL-12p70 또는 IL-17 농도의 유의한 변화는 관찰되지 않았다.
EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IFNγ 및 TNFα의 폐 농도는 도 26에 제시되어 있다. EV-D68 감염 후, IFNγ 또는 TNFα 농도의 유의한 변화는 관찰되지 않았다.
EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스로부터의 MIP-1α 및 GM-CSF의 폐 농도가 도 27에 제시되어 있다. EV-D68 감염 후, MIP-1α 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다.
따라서, 이 연구에서는 4주령의 AG129 마우스에서 EV-D68에 의해 유발된 호흡기 감염의 치료를 위한 EV-D68에 대한 mAb인 EV-D68-228의 치료 효능을 결정했다. 10 및 1mg/kg 용량의 EV-D68-228은, 감염 후 48시간 이내에 투여된 경우, 폐 바이러스 역가, 혈액 바이러스 역가 및 EV-D68 감염에 의해 유발된 전염증성 사이토카인의 감소에 매우 효과적이었다. IL-1α, IL-1β, IL-5, IL-6, MCP-1 및 RANTES의 폐 농도는 EV-D68-228에 의한 치료에 의해 유의하게 감소했다. EV-D68-228은 AG129 마우스의 EV-D68 호흡기 감염에 대한 치료적 치료로서 매우 효과적이었기 때문에, 폐 기능에 대한 EV-D68-228 치료의 영향을 결정하기 위해, 혈량측정법을 장래의 연구에서 사용할 수 있다.
실시예 8 - 마우스의 호흡기 감염에 대한 EV-D68-228의 효능
재료 및 방법
이 연구의 목적은 10일령의 AG129 마우스에서 엔테로바이러스 D68(EV-D68) 신경계 감염에 대한 EV-D68-228에 의한 치료의 치료 효능을 결정하는 것이었다.
동물. 유타 주립 대학의 유타 과학기술연구소(USTAR) 빌딩에서 유지되고 있는 특정 병원체 비함유 콜로니로부터의 10일령 AG129 마우스. 마우스를 유타 주립 대학의 USTAR 빌딩에서 방사선 조사된 텍라드 설치류 다이어트(Harlan Teklad) 및 자동 급수된 수돗물로 사육 및 유지했다.
항체 및 화합물. 모노클로날 항체(mAb) EV-D68-228은 반더빌트 대학 의료 센터의 제임스 크로우로부터 제공되었다. EV-D68-228은 1.134mg/ml 농도의 용액으로 제공되었고, 치료를 위해 멸균 식염수에서 10mg/kg의 용량으로 희석되었다. rRSV-90은 5mg/ml 농도의 용액으로 제공되었고, 10mg/kg의 용량으로 음성 대조군 항체로 사용되었다. 정맥 면역글로불린(IVIg, Carimune, CSL Behring, King of Prussia, PA)은 지역 약국으로부터 구입하여 EV-D68-228 mAb에 대한 비교대조로서 사용되었다.
바이러스. 엔테로바이러스 D68은 BEI 리소스, NIAID, NIH: 엔테로바이러스 D68, US/MO/14-18949, NR-49130로부터 수득했다. 이 바이러스는 4주령의 AG129 마우스의 폐에서 30회 연속 계대하고, 이어서 아메리칸 타입 컬쳐 컬렉션(Manassas, VA)으로부터 수득된 횡문근육종(RD) 세포에서 3회 플라크 정제되었다. 수득된 바이러스 스톡을 RD 세포에서 2회 증폭하여, 작업 스톡을 생성했다. 감염에 사용된 바이러스는 EV-D68 MP30 PP로 지정되었다.
실험 설계. 총 42마리의 마우스를 각각 6마리의 마우스로 이루어진 6개 그룹으로 랜덤화하고, 표 D에 제시된 바와 같이, 9마리 마우스의 한 그룹은 각각 치료 전에 사망을 설명하기 위한 것이다. 3마리의 마우스의 한 그룹을 정상 대조군으로 사용했다. 마우스는 100㎕ 용적의 MEM에서 EV-D68 MP30 PP의 1×106.5 CCID50의 복강내(IP) 투여에 의해 감염시켰다. 마우스는 감염 후 24, 48, 72, 또는 120시간에 EV-D68-228 mAb의 복강내(IP) 투여에 의해 치료되었다. IVIg 또는 위약 Ab에 의한 치료는 감염 후 24시간에 1회 수행되었다. 치료 전 및 그 후 매일 마우스의 체중을 측정했다. 21일차까지, 모든 마우스의 이환율, 사망률 및 신경학적 스코어를 매일 관찰했다. 신경학적 스코어(NS)는 다음과 같이 기록되었다: NS0 - 관찰 가능한 마비 없음, NS1 - 뒷다리가 비정상적으로 벌렸지만 정상 또는 약간 느린 보행, NS2 - 전진 운동에 사용하는 동안, 뒷다리가 부분적으로 붕괴하고 발이 끌림, NS3 - 뒷다리 및 뒷다리의 경직성 마비가 전진 운동에 사용되지 않음, NS4 - 뒷다리의 경직 마비 및 전진 운동 없음. NS4 스코어로 관찰된 모든 동물을 인도적으로 안락사시켰다.
통계 분석. 카플란-마이어 생존 곡선은 프리즘 8.2.0로 생성되었다(GraphPad Software Inc.). 로그 순위(Mantel-Cox) 검정, 및 이어서 게한-브레슬로우-윌콕손(Gehan-Breslow-Wilcoxon) 검정을 사용하여 생존 곡선을 비교했다. 일원 분산 분석(ANOVA)을 사용하여, 감염 후 매일, 치료 그룹의 혈액 바이러스 역가를 위약 치료 마우스의 폐 및 혈액 역가와 비교했다. 일원 ANOVA를 사용하여 평균 체중을 비교했다. 크루스칼-왈리스 검정 및 이어서 둔의 다중 비교 검정을 사용하여 신경학적 스코어를 비교했다.
실험 동물의 윤리 규칙. 이 연구는 유타 주립 대학의 시설내 동물 관리 및 사용 위원회의 승인에 따라 수행되었다. 이 작업은 유타 주립 대학의 AAALAC 인정의 실험 동물 연구 센터에서 수행되었다. 미국 국립 보건원(국립 위생 연구소)의 승인이 실험 동물의 관리 및 사용에 관한 국립 위생 연구소 가이드(개정, 2011)에 따라 갱신되었다(PHS 보증 번호 D16-00468[A3801-01]).
[표 D]
Expt. NIA-1870. 실험 설계 - 마우스에서 EV-D68 신경 감염 치료를 위한 EV-D68-228의 치료 효능.

결과 및 논의
이 연구에서는 10일령의 AG129 마우스에서 EV-D68 신경 감염의 치료적 치료를 위한 EV-D68-228 mAb의 효능을 결정했다.
도 28은 EV-D68로 감염되고 EV-D68-228로 감염 후에 치료된 10일령의 AG129 마우스에 대한 카플란-마이어 생존 곡선을 나타낸다. 10mg/kg 용량의 EV-D68-228을 단일 투여하면, 감염 후 24, 48 및 120시간에 치료된 마우스에서 유의한 생존 이점이 관찰되었다. 감염 후 120시간에 치료된 마우스의 생존 이점의 차이는 치료된 7마리의 마우스 모두가 여전히 감염에 굴복했기 때문에, 사망일의 증가에 의한 것이었다. 10mg/kg 용량의 IVIg는, 감염 후 24시간에 투여된 경우, 마우스를 완전히 사망으로부터 보호했다.
도 29는 EV-D68로 감염되고 EV-D68-228로 감염 후에 치료된 10일령의 AG129 마우스의 평균 체중을 나타낸다. 10mg/kg 단일 용량의 EV-D68-228은, 감염 후 24시간 또는 48시간에 투여된 경우, 감염 관련 체중 감소로부터 마우스를 보호했다. 감염 후 24시간에 투여된 IVIg의 단일 용량은 체중 감소로부터 마우스를 보호하지 못했다.
도 30A-C는 EV-D68-228로 감염 후에 치료된 EV-D68-감염 AG129 마우스의 혈액 바이러스 역가를 나타낸다. 감염 후 24시간, 48시간 또는 72시간에 10mg/kg의 EV-D68-228을 단일 투여하면, 감염 후 3일 및 5일차에 혈액 바이러스 역가가 유의하게 감소했다. 감염 후 24시간에 10mg/kg 용량의 IVIg에 의한 치료는 감염 후 3일차 및 5일차에 혈액 바이러스 역가를 감소시켰다. 감염 후 7일차에는 생존한 마우스의 어느 것에서도 바이러스 역가가 검출되지 않았다.
EV-D68로 감염되고 EV-D68-228로 감염 후에 치료된 10일령의 AG129 마우스의 신경학적 스코어는 도 31A-B에 제시되어 있다. 3, 4, 5일차의 신경학적 스코어는 10mg/kg 용량의 EV-D68-228을 감염 후 24시간에 치료한 마우스에서 유의하게 감소했다. 감염 후 24시간에 제공된 10mg/kg 용량의 IVIg는 감염 후 3, 4, 및 5일에서 신경학적 스코어를 감소시켰다. 위약 치료된 모든 마우스가 사망했기 때문에, 감염 후 5일 이후는 신경학적 스코어의 비교가 완료되지 않았다.
따라서, 이 연구에서는 10일령의 AG129 마우스에서 EV-D68에 의해 유발된 신경 감염의 치료를 위한, EV-D68에 대한 mAb인 EV-D68-228의 치료 효능을 결정했다. 감염 후 48시간 이내에 투여한 경우, 10mg/kg 단일 용량의 EV-D68-228은 사망 및 체중 감소로부터 마우스를 보호했다. 감염 후 72시간 이내에 치료된 마우스에서는 혈액 바이러스 역가의 유의한 감소가 관찰되었다. 신경학적 스코어에 의해 측정된 마비의 감소는 감염 후 24시간에 10mg/kg의 EV-D68로 치료된 마우스에서 관찰되었다. EV-D68-228은 EV-D68의 발생 동안 예방적 요법으로 사용될 수 있기 때문에, 감염 전의 다양한 시점에서 투여된 경우에 EV-D68-228의 효능을 결정하는 것이 유익할 것이다.
실시예 9 - 재료 및 방법
연구 설계. 본 발명자들은 EV-D68 감염에 반응하여 인간이 작제할 수 있는 항체를 동정하기 위해 이 연구를 설계했다. 따라서, 이들은 간접 ELISA 스크린에서 살아있는 바이러스 분리주를 사용하여 EV-D68 결합 항체를 분비하는 B 세포를 특정하고, 이들 B 세포를 골수종 세포와 전기 융합시켜 모노클로날 항체 분비 하이브리도마를 생성했다. 이어서, 본 발명자들은 CCID50, ELISA 및 크리오-EM 기반 기술을 사용하여 시험관내에서 이들 개별 mAb의 중화 및 결합 특성을 특성화했다. 이들은 인간에서 예방 및/또는 치료제로서 mAb EV68-228의 개발을 서포트하는 전임상 데이터를 생성하기 위해 생체내 실험을 추구했다. 이를 위해, 이들은 이론적 이점에 기초하여 AFM으로 인간을 치료하기 위해 광범위하게 사용되는 인간 IVIG와 비교하여 EV-D68 감염으로부터 마우스를 보호하는 mAb EV68-228의 효과를 연구했지만, 이 IVIG 치료는 지금까지 효과적인 것으로 입증되지 않았다. AG129 뮤린 감염 모델의 장점은 본 발명자들이 호흡기 및 신경학적 감염 모델 모두에서 항체 치료의 효과를 측정할 수 있다는 점이다.
세포주. RD 세포(인간, 여성 기원)는 아메리칸 타입 컬쳐 컬렉션(ATCC CCL-136)로부터 수득했다. RD 세포는, 10% 열 불활성화 소 태아 혈청(HI-FBS; HyClone), 1mM 나트륨 피루베이트 및 1% 페니실린-스트렙토마이신-암포테리신 B(ThermoFisher Scientific)이 보충된 둘베코 변형 이글 배지(DMEM)(ThermoFisher Scientific)에서 3℃의 5% CO2에서 배양했다. 구조 연구를 위해, RD 세포를 10% HI-FBS(Sigma-Aldrich) 및 비필수 아미노산(NEAA, Life Technologies)이 보충된 DMEM(Sigma-Aldrich)에서 배양했다. ExpiCHO(햄스터, 암컷 기원) 세포주는 써모피셔 사이언티픽(ThermoFisher Scientific)으로부터 구입하고, 제조업체의 프로토콜에 따라 배양했다. HMMA2.5 계통은 뮤린 골수종 세포주와 인간 골수종 세포주를 융합시킴으로써 생성된 비분비성 마우스-인간 헤테로골수종 세포주(성별 정보는 입수할 수 없음)이다[참조: Posner et al., 1987]. 이 세포주는 이전에 기재된 바와 같이 배양되었다[참조: Yu et al., 2008]. 모든 세포주는 마이코플라스마(Mycoplasma)에 대해 매월 시험되었고, 모든 경우에서 음성인 것으로 밝혀졌다.
바이러스. 이 연구에서 사용된 바이러스 분리주의 목록은 표 K를 참조한다. EV-D68 분리주는 하기 설명된 효소 결합 면역흡착 검정(ELISA)에서 사용하기 위해 RD 세포 단층 배양에서 2세대 증식시켰다. RD 세포 단층에 소정 바이러스 분리주를 접종하고, 70~90% 세포 사멸이 관찰될 때까지 모니터링했다. 이어서, 이 세포 배양 플라스크를 -80℃로 동결하고, 해동하고, 내용물을 스크래핑하고, 50mL 원뿔형 튜브에 수집했다. 이 조제물을 최대 출력 설정(Fisherbrand)의 도립 컵 초음파 분쇄기에서 20초 동안 3회 초음파 처리하고, 30초 동안 와동시키고, 이어서 추가로 20초 동안 2회 초음파 처리했다. 세포 파편을 펠렛화하고, 바이러스 함유 상청액을 PBS(w/v) 쿠션 중의 30% 수크로스 상에서 100,000 × g에서 4시간 동안 10℃에서 회전시켰다. 상청액을 버리고, 펠렛을 NTE 완충액(20mM 트리스, 120mM NaCl, 1mM EDTA pH 8.0) 중의 0.01%(w/v) 소 혈청 알부민(BSA)에 4℃에서 밤새 침지시켰다. 이어서, 재현탁된 펠릿을 10분 동안 10,000 × g에서 원심분리하여 추가로 정화시키고, 바이러스 분취량을 사용 준비가 될 때까지 -80℃에서 저장했다.
구조 연구에는, US/MO/14-18947 분리주를 사용했다. 바이러스는 RD 세포에서 계대되었고, 대규모 증식 전에 -80℃에서 보관되었다. RD 세포를 80% 컨플루언시로 성장시키고, 0.01의 감염 다중도에서 EV-D68로 감염시켰다. 감염 2일 후, 세포를 상청액과 함께 수집하고, 회전시켰다. 세포 펠릿을 수집하고, 복수의 동결/해동 사이클 후, 세포 파편을 제거하기 위해 회전시켰다. 모든 상청액을 합하고, 210,000 × g에서 2시간 동안 펠렛화했다. 펠렛을 인큐베이팅하고, 250mM HEPES(pH = 7.5), 250mM NaCl 완충액에 재현탁하고, 이어서 최종 농도까지 5mM MgCl2, 0.01mg/mL DNAse(Sigma-Aldrich), 0.8mg/mL 트립신, 15mM EDTA 및 1%(w/v) n-라우릴-사르코신을 보충했다. 이어서, 샘플을 210,000 × g에서 2시간 동안 펠렛화하고, 재현탁하고, 칼륨 타르트레이트 구배(10 내지 40%, w/v)에 로딩하여, 2시간 동안 160,000 × g에서 초원심분리의 최후 라운드를 수행했다. 튜브 중앙에 청색 밴드가 관찰되는 정제된 바이러스 샘플을 추출하고, 완충액을 20mM 트리스, 120mM NaCl, 1mM EDTA(pH=8.0) 완충액으로 교환하여 칼륨 타르트레이트를 제거했다.
CCID 50 검정에 의한 바이러스 부하 검출. 뮤린의 혈액 또는 폐 샘플 중의 바이러스 스톡 또는 바이러스의 적정은 RD 세포 배양 단층에서의 CCID50 검정에 의해 수행되었다. 간단히 말해서, 샘플의 10배 희석액을 증가시켜 96웰 플레이트의 3중 웰(50μL)에서 RD 세포 단층에 적용하고, 33℃에서 5% CO2에서 5일 동안 인큐베이팅하고, 이어서 1% 파라포름알데히드로 고정하여 크리스탈 바이올렛으로 염색했다. 세포변성 효과를 갖는 웰은 바이러스에 대해 양성으로 평가되고, 역가는 스피어만-케르버(Spearman-Kaerber) 방정식(Ramakrishnan, 2016)에 기반한 공식을 사용하여 결정했고; 검출 한계는 136 CCID50/mL였다.
바이러스 중화 검정. 바이러스 중화 검정은, 본질적으로 소아마비 바이러스에 대해 이전에 기재된 바와 같이[참조: Weldon et al., 2016], 표시된 바이러스를 사용하여 CCID50 포맷으로 수행되었다. 바이러스를 상승하는 농도의 mAb와 함께 33℃에서 1시간 동안 중복하여 인큐베이팅하고, 이어서 각 현탁액을 96웰 플레이트의 기술적 4중 웰(50μL)에서 RD 세포의 단층에 첨가했다. 33℃의 5% CO2에서 5일 동안 인큐베이팅한 후, 세포를 1% 파라포름알데히드로 고정하고, 크리스탈 바이올렛으로 염색했다. 세포변성 효과를 갖는 임의의 웰은 바이러스에 대해 양성으로 평가하고, 스피어만-케르버(Spearman-Kaerber) 방정식(Ramakrishnan, 2016)에 기반한 공식을 사용하여 최대 억제 농도의 절반(IC50)을 결정했고; 검출 한계는 57μg/mL 내지 4.8pg/mL였다.
마우스 모델. 10일령(신경학적 모델) 또는 4주령(호흡기 모델)의 수컷 및 암컷 AG129 마우스(인터페론 α/β 및 γ에 대한 수용체가 결핍됨)은 유타 주립 대학의 유타 과학기술연구소(Utah Science Technology) 빌딩에서 유지된 특이적 병원체 비함유 콜로니로부터 수득했다. 마우스는 유타 주립 대학의 USTAR 빌딩에서 방사선 조사된 텍라드 설치류 다이어트(Harlan Teklad) 및 자동 급수 수돗물로 사육 및 유지했다. 이 연구는 2019년 3월 2일자의 유타 주립 대학의 시설내 동물 관리 및 사용 위원회의 승인에 따라 수행되었다(2022년 3월 1일 만료). 이 작업은 유타 주립 대학의 AAALAC 인정의 실험 동물 연구 센터에서 수행되었다. 미국 정부(국립 위생 연구소) 승인이 실험 동물의 관리 및 사용에 관한 국립 위생 연구소 가이드(개정: 2011)에 따라 2018년 3월 9일에 갱신되었다(PHS 보증 번호 D16-00468[A3801-01]).
항체 및 대조군 치료는 PBS에 희석하고 EV-D68 접종 전후에 지시된 시점에서 복강내 주사로 투여했다. 구아니딘 HCl(Sigma-Aldrich)은 치료에 대한 양성 대조군으로 사용되었고[참조: Hurst et al., 2019], 감염 후 4시간에 개시하여 5일 동안 1일 2회 계속했다. 마우스 적응 EV-D68의 현탁액은 복강내 주사(신경학적 모델; 100μL 용적의 MEM에서 106.5 CCID50) 또는 비강내 주입(호흡 모델, 90μL 용적의 MEM에서 104.5 CCID50)에 의해 투여되었다. 처리 전 및 그 후 매일 마우스의 체중을 측정했다. 지시된 바와 같이, 폐 바이러스 역가, 혈액 바이러스 역가 또는 폐 사이토카인 농도의 측정을 위해, 감염 후의 지시된 시점에서 마우스를 인도적으로 안락사시켰다. 신경학적 모델의 경우, 21일까지 모든 마우스의 이환율, 사망률 및 신경학적 스코어에 대해 매일 관찰했다. 신경학적 스코어(NS)는 다음과 같이 기록되었다: NS0 - 관찰 가능한 마비 없음, NS1 - 뒷다리가 비정상적으로 벌렸지만 정상 또는 약간 느린 보행, NS2 - 전진 운동에 사용하는 동안, 뒷다리가 부분적으로 붕괴하고 발이 끌림, NS3 - 뒷다리 및 뒷다리의 경직성 마비가 전진 운동에 사용되지 않음, NS4 - 뒷다리의 경직 마비 및 전진 운동 없음. NS4 스코어로 관찰된 모든 동물은 인도적으로 안락사시켰다.
폐 사이토카인/케모카인 평가. 폐 균질액의 각 샘플은 제조업체의 지침(Quansys Biosciences Q-Plex™ Array, Logan, UT)에 따라 정량적 화학발광 ELISA 기반 검정을 사용하여 사이토카인 및 케모카인에 대해 시험되었다. 퀀시스 멀티플렉스(Quansys multiplex) ELISA는 16개의 별개의 포획 항체가 정의된 어레이의 96 웰 플레이트의 각 웰에 적용하는 정량적 시험이다.
인간 모노클로날 항체(mAb)의 생성. 대상체는 EV-D68 상부 호흡기 감염(Bochkov et al., 2014)을 실험실에서 기록한 천식의 소아 발증(COAST) 출생 코호트(Lemanske, 2002)로부터 특정되었다. 서면에 의한 동의가 수득된 후, 말초혈액을 수집하고, 말초혈액 단핵 세포(PBMC)가 SepMate 튜브(Stemcell Technologies)를 사용하여 정제될 수 있을 때까지 제조업자의 프로토콜에 따라 실온에서 보관하고, 이어서 소 태아 혈청(FBS) 중의 10%(v/v) 디메틸 설폭사이드(DMSO)에 동결 보존하고, 액체 질소의 기상으로 저장한다. 림프모구양 세포주(LCL)는 엡스타인-바르(Epstein-Barr) 바이러스, 세포 주기 체크포인트 키나제 2(chk2) 억제제(Sigma-Aldrich), CpG( Sigma-Aldrich) 및 배지 A(STEMCELL Technologies) 중의 사이클로스포린 A(Sigma-Aldrich)와 혼합함으로써 PBMC 내의 메모리 B 세포로부터 전술한 바와 같이 생성했다[참조: Smith and Crowe, 2015]. 1주일 후, LCL을 계수하고, 무관계 공여자로부터의 γ-조사된 인간 PBMC의 피더 층에서 확장했다. 추가로 1주일 후, 2014년 임상 분리주로부터 생성된 세포 배양으로 성장한 EV-D68 바이러스를 포함하는 살아있는 EV-D68 바이러스를 항원으로서 사용하는 간접 ELISA에 의해 CLC 상청액은 EV-D68 반응성 IgG의 존재에 대해 스크리닝했다. 이전에 기재된 바와 같이[참조: Yu et al., 2008], 바이러스 반응성 항체를 포함하는 웰로부터의 LCL을 전기 융합에 의해 HMMA2.5 골수종 세포에 융합시켰다. 융합 반응 후, 하이브리도마 계통을, HAT 배지 보충제(Sigma-Aldrich) 및 오아바인(Sigma-Aldrich)을 함유하는 선택 배지에서 384-웰 플레이트에서 배양한 후, 항체 생산을 위한 상청액을 스크리닝했다. 2주 후, 수득된 하이브리도마 세포주의 상청액을, 항원으로서 살아있는 바이러스를 사용하여 간접 ELISA로 스크리닝하고, EV-D68 반응성 항체를 갖는 웰의 세포주를 배양물에서 확장하고, 이어서 단일-세포 유세포 분석 선별에 의해 384-웰 세포 배양 플레이트로 클로닝했다. 이들 클로닝된 세포는 12웰 조직 배양 처리 플레이트(Corning)에서 약 50% 컨플루언트로 될 때까지 배지 E에서 확장하고, 이들의 상청액은 ELISA에 의해 바이러스 결합에 대해 스크리닝했다. ELISA에서 최고의 신호를 나타내는 웰을, mAb-생산 하이브리도마 세포주로서 선택하여, 추가로 사용했다.
MAb 아이소타입 및 유전자 서열 분석. 분비된 항체의 아이소타입 및 서브클래스는 서양고추냉이 퍼옥시다제(Southern Biotech)와 접합된 마우스 항-인간 IgG1, IgG2, IgG3 또는 IgG4 항체를 사용하여 결정되었다. 항체 중쇄 및 경쇄 가변 영역 유전자는 유세포 분석 선별에 의해 생물학적으로 클로닝된 하이브리도마 계통으로부터 수득된 RNA로부터 서열분석했다. RNeasy Mini 키트(Qiagen)를 사용하여 총 RNA를 추출했다. 수정된 5' RACE(cDNA 말단의 신속한 증폭) 접근법이 사용되었다[참조: Turchaninova et al., 2016]. 간단히 말해서, 5μL 총 RNA를 cDNA 합성 프라이머 믹스(각각 10μM)와 혼합하고, 70℃에서 2분 동안 인큐베이팅한 후, 인큐베이션 온도를 42℃로 저하시켜 합성 프라이머를 어닐링했다(1 내지 3분). 인큐베이션 후, 5× 제1-가닥 완충액(Clontech), DTT(20mM), 5' 주형 스위치 올리고(10μM), dNTP 용액(각각 10mM) 및 10× SMARTScribe 역전사 효소(Clontech)를 포함하는 혼합물을 프라이머 어닐링된 전체 RNA 반응에 첨가하고, 42℃에서 60분 동안 인큐베이팅했다. 제1-가닥 합성 반응은 암푸어 크기 선택 자기 비드 키트(Ampure Size Select Magnetic Bead Kit)를 1.8배의 비율로 사용하여 정제했다(Beckman Coulter). 이어서, 5μL의 제1-가닥 cDNA, 2× Q5 하이 피델리티 마스터믹스(High Fidelity Mastermix)(NEB), dNTP(각각 10mM), 전방향 범용 프라이머(10μM) 및 역방향 프라이머 믹스(중쇄 믹스에서 각각 0.2μM, 경쇄 믹스에서 각각 0.2μM)를 함유하는 단일 PCR 증폭 반응은 하기 조건으로 열 사이클링에 적용했다: 90초 동안 초기 변성, 이어서 98℃에서 10초 동안 30 사이클의 변성, 60℃에서 20초 동안 어닐링, 및 72℃에서 40초 동안 확장, 이어서 72℃에서 4분 동안 최종 확장 단계. 이 프로토콜에 사용된 모든 프라이머 서열은 이전에 기재되었다[참조: Turchaninova et al., 2016]. 제1 PCR 반응은 AMPure 크기 선택 자기 비드 키트를 0.6배의 비율로 사용하여 정제했다(Beckman Coulter). 이어서, 멀티플렉스 SMRT 서열분석 프로토콜(Pacific Biosciences)에 따라 앰플리콘 라이브러리를 제조하고, 세퀴얼 플랫폼 기기(Pacific Biosciences)에서 서열분석했다. 생 서열분석 데이터를 역다중화하고, 순환 컨센서스 서열(CCS)은 SMRT 분석 도구 쉬트(Pacific Biosciences)를 사용하여 결정했다. 유전자 절편, 상보성 결정 영역(CDR) 및 생식세포계 유전자로부터의 돌연변이의 동일성은 ImMunoGeneTics 데이터베이스[참조: Giudicelli and Lefranc, 2011]를 사용하여 정렬에 의해 결정되었다.
항체 생산 및 정제. 하이브리도마 유래의 mAb의 경우, 하이브리도마 세포를 하이브리도마 SFM(1X) 무혈청 배지(Gibco)에서 고갈될 때까지 성장시켰다. 재조합 mAb 생산을 위해, 중쇄 및 경쇄의 유전자를 암호화하는 cDNA를 합성하고, 전장 IgG1 단백질을 암호화하는 DNA 플라스미드 발현 벡터로 클로닝하고[참조: McLean et al., 2000], 이. 콜라이(E. coli) 세포로 형질전환시켰다. MAb 단백질은 제조사의 프로토콜에 따라 ExpiCHO 세포를 일시적으로 형질감염시킨 후에 생산되었다. 수득된 분비된 IgG는 단백질 G 컬럼(GE Healthcare Life Sciences)을 사용하는 AKTA 장치에서 고속 단백질 액체 크로마토그래피(FPLC)에 의해 여과된 배양 상청액으로부터 정제했다. 정제된 mAb를 PBS로 완충제 교환하고, 0.45μm의 멸균 기공 크기 필터 장치(Millipore)를 사용하여 여과하고, 농축하고, 사용할 때까지 -80℃에서 분취량으로 저장했다. 각 mAb의 분취량도 또한, mAb에 대해 20배 몰 과량의 비오틴과 함께 EZ-Link NHS-PEG4-비오틴 키트(ThermoFisher Scientific)를 사용하여 96웰 포맷으로 직접 비오틴화하고, 이어서 탈염 플레이트(Zeba, 7kDa 컷오프)를 사용하여 PBS로 다시 완충제 교환했다. 하이브리도마 유래 mAb는 시험관내 실험에서 사용되었고, 재조합 mAb는 생체내 실험에서 사용되었다. 풀링된 인간 면역글로불린은 정맥내 면역글로불린으로 구입했다(IVIG, Carimune, CSL Behring, King of Prussia, PA). RSV90은 본 발명자의 실험실에서 생산된 재조합 인간 IgG1 mAb이고, 마우스 실험에서 음성 대조군인 위약 mAb로 사용된다. 웨스턴 블롯에 사용된 폴리클로날 항-VP1, -VP2, -VP3 항체는 제네텍스(Genetex)에서 구입했다.
Fab 단편 생산. Fab 단편은 피어스(Pierce) Fab 제조 키트(ThermoFisher Scientific)를 통해 생성 및 정제했다. 고정된 파파인 바이알 스핀 컬럼 및 제바(Zeba) 스핀 탈염 컬럼은 사용 전에 소화 완충액(공급된 Fab 소화 완충액 10mL당 35mg 시스테인·HCl, pH 약 7.0)으로 평형화시켰다. NAb 단백질 A 플러스 스핀 컬럼은 사용 전에 PBS 완충액으로 평형화시켰다. 원래의 IgG 샘플을 제바 스핀 탈염 컬럼에 통과시키고, 제조된 IgG 샘플 0.5mL를 고정화 파파인 바이알에 적용하고, 5시간 Fab 소화 동안 37℃에서 인큐베이팅했다. 이어서, 최종 Fab 단편을 PBS로 완충액 교환하고, 4℃에서 저장했다.
EV-D68-특이적 ELISA. 배지 결합의 웰, 흑색 형광성 면역검정 미세역가 플레이트(Greiner Bio-One)를 100mM 중탄산염/탄산염 완충액, pH 9.6에 희석된 바이러스 스톡으로 코팅하고, 4℃에서 밤새 인큐베이팅했다. 플레이트는 0.05% 트윈-20(DPBS-T)을 함유하는 둘베코의 인산염 완충 식염수(DPBS) 중의 2% BSA로 1시간 동안 차단시켰다. mAb 스크리닝 검정을 위해, 하이브리도마 배양 상청액을 웰에 첨가하고, 주위 온도에서 2시간 동안 인큐베이팅했다. 결합된 항체는 HRP(Southern Biotech) 및 퀀타블루(QuantaBlu) 형광발생 퍼옥시다제 기질(ThermoFisher Scientific)과 접합된 Fc-특이적 염소 항-인간 IgG를 사용하여 검출되었다. 20분 후, 100mM 글리신(pH 10.5)을 첨가하여 반응을 켄칭하고, 시너지(Synergy) H1 마이크로플레이트 판독기(Biotek)를 사용하여 325nm에서 여기한 후 420nm에서 발광을 측정했다. 용량-반응 및 교차-반응성 검정을 위해, 정제된 mAb의 연속 희석액을 이중 기술적 복제로 웰에 적용하고, mAb 결합을 상기한 바와 같이 검출했고; 실험은 적어도 3회 수행되었다. 경쟁 ELISA의 경우, 미세역가 플레이트를 먼저 바이러스로 코팅하고, 이어서 정제된 mAb를 100μg/mL로 첨가하고, 33℃에서 3시간 동안 인큐베이팅했다. 이어서, 비오틴화된 mAb를 5㎍/mL의 최종 농도로 이 혼합물에 첨가하고, 주위 온도에서 1.5시간 동안 인큐베이팅했다. 세척 및 아비딘-퍼옥시다제(ThermoFisher Scientific)와의 30분간 인큐베이션한 후, mAb 결합이 상기한 바와 같이 검출되었다.
웨스턴 블롯. B1 클레이드 바이러스 조제물을 변성 및 환원 로딩 완충액과 혼합하고, 100℃에서 5분 동안 비등시킨 후, 노벡스(Novex) 샤프 사전-염색된 단백질 표준(ThermoFisher Scientific)과 함께 SDS-PAGE 겔에서 영동했다. 단백질을 막으로 전송하고, 차단 완충액(Li-Cor)으로 차단한 후, 스트립으로 절단하여, 개별 레인을 차단 완충액 중의 정제된 mAb로 염색할 수 있도록 했다. IRDye 800CW-접합된 염소 항-인간 2차 항체(Li-Cor)를 사용하여 mAb 결합을 검출했다. 분자량을 시각화하기 위해 스트립을 재조립하고, 오디세이(Odyssey) CLx 근적외선 이미저(Li-Cor)에서 이미지화했다.
크리오-EM 샘플의 준비 및 데이터 수집. EV-D68:Fab EV68-159 및 EV-D68:Fab EV68-228의 복합체 모두에 대해, 정제된 EV-D68 바이러스 및 Fab를 1:200의 몰비로 혼합했다. 실온에서 45 내지 60분 동안 인큐베이팅한 후, 3.5μL의 바이러스-Fab 혼합물 샘플을 글로우 방전된 400 메쉬의 레이시 탄소 필름 구리 그리드(Ted Pella Inc.)에 첨가했다. 그리드는 75 내지 80% 습도에서 3.5초 동안 블로팅한 후, 액체 에탄에서 플런지 동결(Cryoplunge 3 시스템, Gatan)했다. 크리오-EM 데이터 세트는 300kV 타이탄 크리오스 현미경(Titan Krios Microscope)(Thermo Fisher Scientific)에서 수집되었다. EV-D68:Fab EV68-228 데이터 세트의 경우, 64,000× 배율에서 K3 직접 검출 카메라(Gatan)를 구비한 레지논(Leginon)(Suloway et al., 2005) 프로그램을 사용하여 영상을 수집하고, 0.7 내지 2μm의 디포커스 범위와 함께 0.662Å의 초해상도 픽셀 크기를 수득했다. 2.6초의 노광 동안 44.2 전자/Å2의 총 전자 선량이 50프레임에 걸쳐 기록되었다. EV-D68:Fab EV68-159 데이터 세트는 K2 수미트(Summit) 직접 전자 검출기(Gatan)를 사용하여 81,000×의 공칭 배율에서 취득하여, 0.874Å의 초해상도 픽셀 크기, 0.7 내지 3.5μm의 디포커스 범위를 수득했다. 12초의 노광 동안 31.4 전자/Å2의 총 전자 선량이 60프레임으로 분할되었다. 전체적으로, EV-D68:Fab EV68-228 및 EV-D68:Fab EV68-159 데이터세트에 대해, 각각 462개의 무비 및 732개의 무비가 취득되었다.
이미지 처리. 양 데이터세트에 대해, 모션 보정은 데이터 수집 중에 아피온(Appion)(Lander et al., 2009)에서 실장된 MotionCor2(Zheng et al., 2017b)를 통해 생 무비 프레임에서 수행되었다. 콘트라스트 전달 함수(CTF)는 CTFFIND4를 사용하여 정렬된 프레임에서 추정했다[참조: Rohou and Grigorieff, 2015]. 입자 선택 주형은 아피온 매뉴얼 필커(Appion Manual Picker)[참조: Lander et al., 2009]를 사용하여 생성되었고, 자동 선택을 위한 주형은 XMIPP의 2차원(2D) 분류[참조: Sorzano et al., 2004]를 통해 수득되었다. 이어서, 이들 주형을 FindEM에서 자동 선택에 사용하고[참조: Roseman, 2004], RELION을 사용하여 입자를 추출했다. 이어서, 이들 입자는 RELION에서 복수회 라운드의 2D 및 3D 분류를 적용했다[참조: Scheres, 2012]. 그 결과, EV-D68:Fab EV68-228 및 EV-D68:Fab EV68-159 데이터세트에 대해 20,194개 및 30,554개의 입자가 생성되었고, 금 표준 정련 방법[참조: Guo and Jiang, 2014]에 따라 프로그램 JSPR을 사용하여 최종적 3D 20면체 재구성용으로 선택했다. 양 맵의 최종적 해상도는 0.143의 금 표준의 푸리에 쉘 상관관계 컷오프를 기반으로 추정되었다[참조: Scheres and Chen, 2012]. 맵 샤프닝은 RELION(Scheres, 2012)의 후처리에서 수행되었다. 데이터 수집 파라미터 및 관련 항목은 표 G에 요약되어 있다.
모델 구축, 정련 및 분석. EV-D68:Fab EV68-159 및 EV-D68:Fab EV68-228의 원자 구조에 동일한 방법이 사용되었다. EV-D68 페르몬(Fermon) 균주의 X선 결정학 모델(PDB 코드: 4WM8)이 모델 구축을 위한 개시 참조로서 선택되었고, 프로그램 키메라(Chimera)[참조: Pettersen et al., 2004]를 사용하여 밀도 맵에 수동으로 적합되었다. 초기 적합을 기본으로 사용하여, 모델을 쿠트(Coot)에서 재구축하고[참조: Emsley et al., 2010], PHENIX에서 실제 공간 정련[참조: Adams et al., 2010]을 사용하여 정련하여, 이상점과 불충분하게 적합한 로타머를 수정했다. 키메라[참조: Pettersen et al., 2004], 쿠트[참조: Emsley et al., 2010] 및 CCP4i2-PISA[참조: Potterton et al., 2018]를 사용하여 결합 계면 잔기를 결정했다. 최종 원자 모델은 MolProbity에서 검증되었다[참조: Chen et al., 2012]. 정련 통계는 표 G에 설명되어 있다.
중화 회피 돌연변이 바이러스의 선택. 클레이드 B1 EV-D68 분리주는 RD 세포에서 정제된 mAb의 양을 증가시키면서 선택하에 계대했다. mAb 및 바이러스를 33℃에서 1시간 동안 인큐베이팅한 후, 이 혼합물을 33℃에서 2시간 동안 세포 단층에 첨가했다. 이어서, 단층을 3회 세정하고, mAb 함유 배지를 다시 첨가했다. 이 배양물을 33℃에서 적어도 70%의 세포변성 효과가 관찰될 때까지 인큐베이팅하고(플레이트로부터 세포를 리프팅함), 이 시점에서 세포와 상청액을 함께 수집하고, -80℃로 동결했다. 이 샘플을 해동하고, 동일한 미세분리 튜브에서 도립 컵 초음파 처리기에서 최대 출력으로 30초 동안 초음파 처리하고, 다시 2×20초 초음파 처리했다. 세포 파편은 10,000×g에서 10분 동안 정화시켰다. 이어서, 바이러스 함유 상청액을 보다 높은 농도로 mAb를 함유하는 새로운 배지와 1:1로 혼합했다. 3회 계대에 걸쳐, mAb 농도는 5로부터 50, 500ng/mL로 증가했다. 바이러스 RNA는 TRI 시약 및 다이렉스-졸(Direct-zol) RNA MiniPrep 키트(Zymo Research)를 사용하여 회수했다. 3중으로, 본 발명자들은 cDNA 주형을 생성했고, 이로부터 바이러스 게놈의 P1 영역을 커버하는 3,080bp 앰플리콘을 PrimeScript 원 스텝(One Step) RT-PCR 키트 버젼 2(Takara) 및 프라이머 5'-CCTCCGGCCCCTGAAT(Fwd; 서열번호 641) 및 5'-CCATTGAATCCCTGGGCCTT(Rev; 서열번호 642)로 생성했다. 그들은 퍼시픽 바이오사이언시스(Pacific Biosciences)(PacBio) 차세대 서열분석 플랫폼을 사용하여 3개의 복제물 각각의 서열을 생성했다. 각 서열분석 실행의 2,000개 판독을 사용하여, 각 돌연변이가 관찰된 판독의 백분율을 정량화했다. 돌연변이는 음성 대조군 선택 mAb 선택으로부터의 모든 판독의 야생형 컨센서스 서열과 비교하여 결정되었다.
정량화 및 통계 분석. 기술 및 생물학적 복제는 방법 및 도면 범례에 제시되어 있다. 프리즘 v8(GraphPad)을 사용하여 통계 분석을 수행했다.
경쟁 결합 검정. ELISA 형광 값은, 관련 없는 사전 경쟁 mAb를 첨가하지 않고서 대조군 웰로부터 결정된 최대 결합의 백분율로 정규화되었다. 각각의 기타 비오틴화된 mAb에 대한 각각의 비오틴화된 mAb의 피어슨 상관관계는 판다스 파이톤(Pandas Python) 팩키지의 코르(corr) 방법을 사용하여 3개의 상이한 실험으로부터 중앙 억제 백분율을 사용하여 계산했다[참조: McKinney, 2010]. 이어서, 시본 파이톤(Seaborn Python) 팩키지의 클러스터맵(clustermap) 방법을 사용하여, 이러한 피어슨 상관관계에 대해 계층적 클러스터링을 수행했다. 클러스터링 정보는 뉴윅(newick) 형식으로 수출되고, 인터액티브 트리 오브 라이프(Interactive Tree of Life) v4[참조: Letunic and Bork, 2019]로 수입하고, 이는 최종 표시를 위해 엑셀(Microsoft)로 수입하기 전에 계층적으로 클러스터링된 히트맵을 표시하기 위해 사용되었다.
항체 ELISA 결합 실험. mAb 결합에 대한 EC50 값은, 하한치를 제로로 한정하고 상한치를 최고 포화 형광 값을 갖는 mAb의 최대 형광 값 미만으로 한정한 4-파라미터 S자형 용량-반응 비선형 회귀 분석을 사용하여 항체 농도의 로그 변환 후에 결정되었다.
바이러스 검정. MAb IC50 값은 스피어만-케르버(Spearman-Kaerber) 방정식에 기반한 공식을 사용하여 계산되었다[참조: Ramkrishnan, 2016]. 일원 분산 분석(ANOVA)과 둔네트(Dunnett)의 다중 비교 검정을 사용하여, 단일 풀링 분산을 사용하여, 뮤린의 혈장과 폐의 바이러스 역가를 비교했다. p < 0.05의 값은 유의한 것으로 간주되었다.
폐 사이토카인/케모카인 평가. 각 사이토카인/케모카인에 대해, 브라운-포르사이트(Brown-Forsythe) 일원 ANOVA 시험 및 둔네트(Dunnett)의 T3 다중 비교 시험을 사용하여, 치료된 마우스의 농도를 위약 치료된 마우스와 비교하고, 각 비교에 대해 개별 분산을 계산했다. 이 분석을 선택한 것은 본 발명자들이 각 측정에 대해 동일한 표준 편차를 상정하지 않았기 때문이다.
생체내 보호 연구. 생존 곡선은 카플란-마이어(Kaplan-Meier) 방법을 사용하여 생성되었고, 곡선은 로그 랭크 검정(Mantel-Cox)를 사용하여 비교되었다. 신경학적 스코어는 카이제곱 검정을 사용하여 비교되었다.
실시예 10 - 결과
사전 서면 동의를 획득한 후, 2014년 미국에서의 발병시에 EV-D68 호흡기 감염을 이전에 기록한 12명의 대상체가 혈액을 기증했고, 이로부터 본 발명자들은 말초 혈액 단핵 세포(PBMC)를 단리했다. 대상체는 감염 당시는 12~15세, 채혈 당시는 16~18세였다. 각 대상체는 EV-D68 관련 호흡기 질환의 병력이 있고, AFM의 증상은 없었다(표 E). 수집된 PBMC는 엡스타인-바 바이러스로 접종함으로써 시험관내에서 형질전환시켜, 항체를 분비하는 메모리 B 세포 유래 림프모구 세포주(LCL)를 생성했다, LCL 배양 상청액을 간접 ELISA에 사용하여 EV-D68 반응성 IgG의 존재를 스크리닝했다. 본 발명자들은, 2014년 임상 분리주로부터 생성된 EV-D68의 실험실 배양된 생 바이러스 조제물에 결합하지만 유사하게 제조된 비감염 세포 상청액에는 결합하지 않는 항체를 갖는 배양물을 선택했다. EV-D68 특이적 항체를 분비하는 LCL을 비분비성 골수종 세포주와 전기융합시킨 후, 수득된 하이브리도마 세포를 단일 세포로 선별하여, 완전한 인간 mAb를 분비하는 클론 하이브리도마를 생성했다(표 F).
본 발명자들은, 바이러스 표면 상의 얼마나 많은 주요 항원 부위가 자연 감염에 반응하여 제조된 인간 mAb에 의해 결합되는지를 결정하고자 했다. 유사한 에피토프를 인식하는 항체 그룹을 동정하기 위해, 간접 ELISA에서 mAb가 살아있는 바이러스에 대한 다른 mAb 각각의 결합과 경쟁할 수 있는지의 여부를 결정했다. 경쟁 결합 실험의 경우, 바이러스를 ELISA 플레이트에 직접 코팅하고, 이어서 고농도의 하나의 비표지 mAb와 함께 인큐베이팅했다. 이어서, 비오틴화에 의해 표지된 mAb를 보다 낮은 농도로 첨가하고, 1차 mAb의 존재하에 바이러스에 결합하는 2차 mAb의 능력을 결정했다. 이어서, 본 발명자들은, 억제 데이터와 피어슨 상관관계를 사용하여 항체 결합 패턴의 상호 관련성을 결정하고, 4개의 주요 경쟁-결합 그룹(도 32)을 동정했고, 이들을 그룹 1 내지 4로 명명했다. 이들은 각 mAb를 사용하여 EV-D68 조제물의 웨스턴 블롯을 염색하고, 경쟁 결합 그룹 2 및 3의 거의 모든 mAb가 VP1 단백질의 선형 에피토프에 결합한 반면, 기타 단일 mAb만이 바이러스 조제물 중의 임의의 단백질에 결합한다는 것을 발견했다(도 40).
2014년 미국에서 EV-D68의 발생 동안, 거의 모든 바이러스 분리주가 새롭게 출현한 B1 클레이드에 속했고, 밀접하게 관련된 B2 클레이드 또는 멀리 관련된 D 클레이드로부터의 바이러스의 검출은 보다 적었다[참조: Tan et al., 2016]. 이 연구의 대상자 중 1명을 제외한 모든 대상체는 B1 계통 분리주에 감염되었다(표 E). 2014년 이래, B3 클레이드 바이러스가 우세해졌고, B1 클레이드 바이러스는 더 이상 순환하지 않고[참조: Dyrdak et al., 2019]; 2018년에, 미국 질병 예방 관리 센터에 의해 서열분석된 모든 EV-D68 분리주는 B3 클레이드로부터의 것이었다[참조: Kujawski et al., 2019]. 본 발명자들은 먼저 B1 클레이드 EV-D68 분리주를 사용하여 50% 세포 배양 감염 용량(CCID50) 검정에서 각 mAb의 시험관내 중화 능력을 측정했다(도 33A). 28개의 mAb는 50μg/mL 미만의 절반 최대 억제 농도(IC50)로 중화를 입증했고, mAb EV68-159는 0.32ng/mL의 IC50 값에서 가장 강력한 중화를 나타냈다(도 41). 본 발명자들은 추가로, D 클레이드 분리주에 대해 21개의 가장 강력하게 중화하는 mAb를 시험하고, 11개의 mAb가 그 바이러스를 중화시키고, 이들 중의 7개가 이종 바이러스에 대한 IC50에 의해 적어도 10배의 효능 감소를 나타내는 것을 발견했다. 페르몬(Fermon) 균주는 1962년으로부터의 분리주이고, 현대 EV-D68 분리주와 너무 멀리 떨어져 있기 때문에, 클레이드 분류 체계에 적합하지 않다[참조: Tan et al., 2016]. 9개의 mAb는 페르몬 실험실 참조 균주를 중화했지만, 현대의 B1 클레이드 바이러스를 억제하는 것보다 덜 강력했다.
중화 검정은 교차 반응성을 과소평가할 수 있음을 인식하여, 본 발명자들은, 상기와 설명한 동일한 간접 ELISA 접근법을 사용하여, B1, B2 또는 D 클레이드로부터의 대표적 EV-D68 분리주에 결합하기 위해 정제된 mAb의 최대 유효 농도의 절반(EC50)을 생성했다(도 33B-C 및 도 42). B1 클레이드 분리주에 대한 ≤ 1㎍/mL의 결합에 대한 EC50 값을 갖는 mAb 중, 모두가 B2 클레이드 분리주에 결합된 반면, 약 절반은 D 클레이드 분리주에도 결합했다(도 33B 및 도 41). 일반적으로 약하게 결합하지만 시험된 모든 클레이드로부터의 바이러스에 교차 반응하는 추가 부류의 mAb가 관찰되었다.
현재까지, 항체-EV-D68 상호작용의 구조적 연구는 뮤린 mAb로 한정되었다[참조: Zheng et al., 2019]. 본 발명자들은, 2개의 강력하게 중화하는 인간 mAb, 클레이드 특이적 mAb EV68-159 및 고도 교차 반응성의 mAb EV68-228을 선택하여, 항원 결합 단편(Fabs)과의 면역 복합체 및 크리오-전자 현미경(cryo-EM) 연구용의 B1 클레이드 EV-D68 분리주를 제조했다. 최종 밀도 맵은 2.9Å(EV68-159) 또는 3.1Å(EV68-228)의 해상도를 달성했다(도 34A, 도 43, 도 44 및 표 G). 구조는 2개의 상이한 결합 부위를 나타냈다: EV68-159는 3중 대칭 축 주위에 부착된 반면, EV68-228은 협곡 영역을 형성하는 함몰부 사이의 5중 축 주위에 결합되었다(도 34A-C, 도 45). 따라서, 각 Fab에 대해, 총 60개 카피가 바이러스 입자에 결합된다. 바이러스 표면과 상호작용하는 Fab 가변 도메인은 바이러스 캡시드 단백질과 유사한 강력한 밀도를 나타내고, 각 Fab의 원자 모델은 4개의 바이러스 캡시드 단백질과 함께 구축되었다. 대조적으로, 바이러스 표면으로부터 추가로 멀리 위치한 Fab 불변 도메인은 보다 약한 밀도를 나타내고, 원자 모델의 구축으로부터 제외되었다. 폴리펩타이드 쇄의 백본과 대부분의 아미노산 측쇄는 밀도 맵에서 양호하게 정렬되어 있고, 이는 바이러스 입자와 Fab 분자 사이의 결합 계면의 중요한 특징을 입증한다.
양 모델 모두에 대해, Fab에 직면하고 Fab로부터 4Å의 거리 내에 있는 바이러스 표면 잔기가 풋프린트로서 식별되었다(도 34C, 도 46 및 표 H). 풋프린트는 양 Fab 분자가 1개의 프로토머 내에 있는 것을 나타낸다. EV-D68:Fab EV68-159 복합체에서는 각 Fab가 약 990Å2의 바이러스 표면적을 마스킹했다. 결합 계면(도 35)에서는, EV68-159 경쇄와 VP1의 C-말단의 3개 잔기: Glu271 및 Arg272(도 36A) 및 Asp285(도 36B) 사이에 본질적 상호작용이 발견되었다. 잔기 Glu271 및 Arg272는 CDR3 및 CDR1과 수소 결합을 형성했다. Arg272 및 Asp285는 각각 CDR3 및 CDR2 잔기와 염 브릿지를 형성했다. EV68-159의 중쇄는 마스킹된 표면적의 77%에 기여했다. 일련의 수소 결합이, 중쇄 상보성 결정 영역 2(CDR2) 및 CDR3과 B-β 가닥(βB) 전의 VP3 N-말단 루프 사이에서 발견되었다(도 36C).
EV-D68:Fab EV68-228 복합체에서는 각 Fab가 바이러스 캡시드 표면의 약 1,170Å2를 마스킹했다. EV68-159와 유사하게, EV68-228 Fab의 중쇄는 표면적의 약 84%를 마스킹함으로써 바이러스 캡시드와의 상호작용을 지배했다. 결합 계면(도 35)은 주로 중쇄 CDR과 VP1 βB 및 VP3 C-말단(도 36D) 사이에 형성된 수소 결합에 의해 안정화되었다. 경쇄 CDR1은 VP2 EF 루프와 상호작용했다. 또한, 중쇄 프레임워크 영역(FR) 3과 VP1 DE 루프 사이에 수소 결합이 형성되었다. 추가로, 경쇄 CDR3과 VP1의 C-말단 사이에 염 브릿지가 형성되었다. 전반적으로, EV68-228 Fab는 5중 축 주위의 바이러스 표면에 결합하고, 고전적 피코르나바이러스 중화 면역원성 부위(NIm) NIm-IB(VP1 DE 루프) 및 NIm-II(VP2 EF 루프)를 인식했다[참조: Rossmann et al., 1985].
벌키한 측쇄는 양 Fab의 계면에서 발견되었고, 소수성 상호작용 네트워크를 통해 구조를 안정화시키는 작용을 한다(도 47). 추가로, 중쇄 및 경쇄의 CDR1 및 CDR3 주변에서도 디설파이드 결합이 검출되었다. 또 다른 한 쌍의 시스테인인 Cys101 및 Cys106은 EV68-228 중쇄의 CDR3 내에서 발견되었고, 디설파이드 결합을 형성하기 위해 정확한 거리 및 방향에 있었다(도 36D). 구체적으로, 등고선 수준이 감소한 경우, 2개의 시스테인 측쇄의 밀도가 연결되었다. EV-D68:Fab EV68-228 복합체에 대해 상기에서 기재한 바와 같이, Cys101이 관여하는 협곡에 인접한 VP3 C-말단 잔기와 중쇄 CDR3 사이에 수소 결합이 관찰되었고, 이는 수소 결합과 디설파이드 결합을 모두 형성한다. 이러한 시스테인 잔기는 Fab 구조와 바이러스-Fab 결합 계면 모두를 안정화시키는 데 중요한 역할을 한다.
이어서, 본 발명자들은, 강력하게 중화되고 고도로 교차 반응성인 인간 mAb EV68-228이 EV-D68 감염의 작은 동물 모델에서 감염 및 질환을 예방하거나 치료할 수 있는지의 여부를 결정하고자 했다. 이들은 인터페론 α/β 및 γ에 대한 수용체가 결핍된 AG129 계통 마우스에서 호흡기 질환 또는 AFM-유사 신경계 질환을 유발하는 2개의 상이한 확립된 감염 모델을 사용하여 이 항바이러스 활성을 생체내에서 시험했다[참조: Evans et al., 2019; Hurst et al., 2019]. 먼저, 본 발명자들은 항체가 호흡기 감염 모델에서 바이러스혈증 및 폐 바이러스 복제를 감소시킬 수 있는지의 여부를 시험했다. 바이러스 접종 전일에 예방으로서 전신 투여된 MAb EV68-228은 시험된 각각의 농도에서 혈액(도 37A) 및 폐(도 37B)에 멸균 면역을 제공한 반면, 인간 IVIG는 혈액만을 멸균했다. 전염증성 사이토카인 분비의 유도는 EV68-228 치료된 마우스의 폐에서 억제되었다(도 38A-C). 바이러스 접종 후의 증가된 시간에 제공되는 치료제로 사용하는 경우, 다시 모든 치료제는 혈액 멸균에 매우 효과적이었지만(도 37C), EV68-228만이 폐에서 효능을 가졌다(도 37D). 본 발명자들은 유사하게 EV68-228-치료된 마우스의 폐에서 전염증성 사이토카인 수준의 감소를 관찰했다(도 38D-F).
이어서, 본 발명자들은 AFM 질환을 모방하는 감염의 신경학적 모델에서 항체의 수동적 전달의 효과를 평가했다. EV68-228 예방은 혈액의 멸균 면역(도 39A) 및 사망(도 39B) 또는 임의의 신경계 질환의 발병(도 39C)으로부터의 완전한 보호를 제공한 반면, IVIG 면역은 부분적으로만 보호되었다. 치료학적으로 제공되면, EV68-228 치료는 제공된 각각의 시점에서 투여 24시간 이내에 혈액을 멸균했다(도 39D). EV68-228은, 감염 후 48시간 이내에 제공되면, 생존율(도 39E) 및 신경 질환을 개선시켰고; 감염 후 72시간에 제공되었을 때, 생존한 마우스는 임상적으로 개선되었다(도 39F 및 표 I).
[표 E]
말초 혈액 단핵 세포를 제공한 대상체의 특성

1연령; 2F: 여성, M: 남성, 3Unk.: 불명, NHL: 히스패닉 또는 라틴계 아님; 4B: 흑인, W: 백인; 5C: 기침, H: 쉰 목소리, R: 비염, SP: 부비동 통증, W: 천명음, NR: 기록되지 않음; 6Mod.: 중등도, NR: 기록되지 않음.
[표 F]
인간 mAb의 서열 특성

월드 와이드 웹의 imgt.org에 있는 인터네셔널 ImMunoGeneTics(IMGT) 정보 시스템에 의해 동정된 서열 특성. IMGT가 복수의 유전자를 제외할 수 없는 경우, 제1 호출이 수록된다. ND: 미정.
[표 G]
크리오-EM 데이터 수집 파라미터 및 정제 통계

[표 H]
EV-D68 및 각각의 Fab의 구조적 접촉 아미노산 잔기

중쇄 및 경쇄는 각각 H 및 L로 표지된다. 특히, 고해상도의 전자 밀도 맵에서 명확하게 관찰되는 직접 상호작용만 수록되어 있다. 이것은, 최대 수소 결합 거리에서 전체 풋프린트를 표시하기 위해 4Å 컷오프를 사용하는 로드맵(도 34C 및 도 46)에서 강조 표시된 접촉 잔기와는 상이하다.
[표 I]
EV-68 접종 후에 항체로 치료된 개별 마우스의 신경학적 스코어


각 열은 도 39F의 그래프에 대응하는 지시된 치료 그룹 내의 개별 마우스로부터의 데이터이다. 일은 EV-D68 접종 후의 스코어가 결정된 일수를 나타낸다. 마우스는 4의 스코어에서 희생되거나 이미 사망했기 때문에, 4가 기록된 후의 일수에는 추가 스코어가 수록되지 않는다. 강조 표시된 세포는 임상적으로 개선되는 마우스의 예를 나타낸다.
[표 J]
중화 회피 아미노산 돌연변이

1 수록된 각 mAb는 RD 세포 배양에서 EV-D68의 3계대의 선택으로서 사용되었고, RSV-90은 생체내 실험을 위한 위약 치료로서 사용된 것과 동일한 음성 대조군 mAb이다. 각 치료에 대해, 구조 유전자는 3개의 기술적 복제에서 차세대 서열분석을 사용하여 서열분석되고, 복제당 2000개 서열이 분석된다. 수록된 수치는 이러한 복제의 평균이다. 2Fab 아미노산의 4Å 이내에 위치한 바이러스 아미노산인, 이 원고로부터의 크리오-EM 구조에 의해 결정된 접촉 잔기. 인접은 접촉 잔기에 직접 인접한 아미노산을 지칭한다. 3WT: 야생형. RSV-90 선택된 바이러스 서열의 컨센서스 서열을 나타낸다.
[표 K]
사용된 EV-68 분리주

ATCC: 아메리칸 타입 컬쳐 컬렉션; BEI: 생물방어 및 신흥 감염 연구 리소스 리포지토리, NIAID, NIH; USU: 유타 주립 대학; NA: 해당 없음.
실시예 11 - 논의
이러한 연구는 EV-D68 감염에 대한 인간 B 세포 반응의 다양한 특징을 나타낸다. 본 발명자들은 스크리닝 항원으로서 살아있는 바이러스 분리주를 사용하여 이러한 mAb를 단리하기 위한 접근법에서 편견이 없도록 시도했다. 본 발명자들은, 결합 및 중화 둘 모두에서 EV-D68의 클레이드 사이의 광범위한 교차 반응성을 관찰했기 때문에, 그들이 회수한 항체 표현형의 다양성은 이러한 전략의 결과일 수 있다. 흥미롭게도, 살아있는 바이러스 입자에 대한 강력한 결합은 반드시 높은 중화 효능을 예측하는 것은 아니다. 관찰된 경쟁 결합 그룹 중에서, 그룹 2 및 3만이 표현형의 균일성을 나타냈다. 양 그룹의 MAb는 교차 반응성이었지만, 그룹 2 mAb는 바이러스를 중화한 반면, 그룹 3 mAb는 중화하지 않았다. 웨스턴 블롯에서는, 그룹 2 및 3의 거의 모든 mAb가 VP1에 결합했고(도 40 및 41), 이는 이들이 선형 에피토프에 결합하는 것을 시사한다. 특히, 경쟁 결합 연구는 전장 IgG 분자를 사용했기 때문에, 관찰되는 경쟁은 인간 조직에서 발생하는 것과 같이 기능적이고, 반드시 바이러스 표면에 구조적으로 상이한 에피토프가 4개만 있다는 것을 나타내는 것은 아니다.
EV68-159 및 EV68-228의 웨스턴 블롯 반응성의 결여는 양 에피토프가 3개의 주요 바이러스 표면 단백질 모두에 걸쳐 있는 것을 나타내는 구조 연구의 발견과 상관관계가 있다. 펩타이드 마이크로어레이[참조: Mishra et al., 2019] 및 파지 라이브러리[참조: Schubert et al., 2019] 기술을 사용한 최근 진단 기술에 의해, 선형 에피토프를 인식하는 AF 환자의 뇌척수액에서 항체를 스캔했기 때문에, 이들 2개의 강력한 중화 mAb의 에피토프의 입체배좌-의존적 특성은 주목할 가치가 있다. 선형 에피토프를 인식하는 항체의 검출은 현재 귀중한 진단 도구에 사용될 수 있고, 그러나, 이러한 연구는 이러한 검사가 엔테로바이러스 감염에 대한 반응으로 이러한 환자가 수행하는 항체 반응의 품질에 대해 기껏해야 부분적으로만 정보를 제공한다는 것을 나타낸다. 구조는 또한 항체 매개 중화에 대한 분자적 기초를 시사한다. 프로토머 내에서 3개의 구조 단백질 모두와 접촉함으로써, 양 mAb 모두는 감염에 필요한 동적 구조 전이를 억제할 수 있는 것으로 보이고, 이는 잘 알려져 있지 않다.
EV68-228 중쇄의 CDR3에서 디설파이드 결합은 C형 간염[참조: Flyak et al., 2018] 및 인플루엔자 A 바이러스[참조: Bangaru et al., 2019] 둘 다를 포함하는 다수의 바이러스의 항바이러스 인간 mAb를 광범위하게 중화할 때에 본 발명자들이 관찰한 구조 모이어티이다. 시스테인 사이에 개재된 4개 내지 5개의 아미노산은 완전한 CDR3 루프의 가장 원위부에 작은 구조화 루프를 형성하여 바이러스 항원 결합에 최적인 사전 구성된 상태에서 CDR3을 안정화시킨다. 특히 EV68-228의 경우, Cys101은 수소 결합을 통해 VP3와도 직접 상호작용하기 때문에, 시스테인은 CDR3 루프 안정화 및 표적과의 상호작용에 모두 관여한다.
EV68-159 경쇄와 상호작용하는 3개의 VP1 잔기(Glu271, Arg272 및 Asp285) 및 EV68-159 중쇄와 상호작용하는 VP3의 N-말단 루프 상의 Glu59는 시알산 수용체 결합 부위[참조: Liu et al., 2015]에 인접하고, 이는 EV68-159 Fab가 시알산 수용체에 대한 바이러스의 결합을 차단할 수 있음을 시사한다. 특히, 이들 3개의 VP1 잔기는 AFM 환자의 뇌척수액에서 발견되는 항체에 의해 결합되는 22개 아미노산 VP1 C-말단 펩타이드에 위치한다[참조: Mishra et al., 2019; Schubert et al., 2019]. 추가로, EV68-159 Fab 중쇄와 VP3 N-말단 루프와의 상호작용에 의해, 4개의 VP의 N-말단이 캡시드 안정성에 기여하기 때문에, 바이러스의 탈코팅이 방지될 수 있다[참조: Filman et al., 1989]. EV68-228은 VP1 βB에 결합하여 바이러스의 탈코팅을 방지하여 진입에 필요한 VP1의 N-말단의 외재화를 억제할 수 있다. 또한, 항체 풋프린트에는 VP3의 C-말단에 있는 잔기가 포함되고, 이는 종래의 NIm의 일부가 아니다. 이러한 잔기는 협곡 수용체 결합 부위에 인접하여, mAb EV68-228이 수용체에 대한 바이러스의 결합을 차단할 수도 있음을 시사한다.
마지막으로, 소아마비 바이러스 2형 및 3형이 근절된 시기에, AFM이 증가하고 있고, 이 마비성 질환의 유행을 유발하는 EV-D68의 역할이 점점 더 명백해지고 있다. 인간 mAb EV68-228에 의한 예방이 생체내에서 얼마나 양호하게 기능하는지를 감안할 때, 이러한 데이터는 효과적인 EV-D68 백신이 AFM 질환을 예방할 수 있음을 시사한다. 실제로, 최근 연구에서는, 바이러스-유사 입자[참조: Zhang et al., 2018a; Dai et al., 2018] 및 불활성화된 EV-D68[참조: Patel et al., 2016] 백신 후보는 면역원성이고, 마우스의 감염을 보호하는 것이 제시되어 있다. 그러나, 불활성화된 EV-D68을 백신접종한 코튼 랫트에 대한 연구에서는, 후속 EV-D68 감염시에 호흡기 질환이 악화되었을 수 있음이 시사되었다[참조: Messacar et al., 2016]. 이 발견은 EV-D68 감염의 항체 의존성 향상(ADE) 가능성을 시사할 수 있지만, 마우스에서 본 발명자들은 시험한 항체 농도의 범위 내에서 폴리클로날 또는 모노클로날 항체에 의해 유발된 ADE를 관찰하지 못했다. 또한, 특히 이 항체가 인간 세포 재료에 대한 명백한 자가반응성 결합 없이 다양한 세트의 바이러스 분리주를 강력하게 중화시키기 때문에, EV-D68 감염의 초기에 요법으로서 mAb EV68-228을 사용하는 전망은 매력적이다(도 33C). IVIG는 이전의 생체내 연구에서 EV-D68에 의한 AFM-유사 질환으로부터 마우스를 보호했지만[참조: Hixon et al., 2017a], 지금까지의 IVIG는 AFM을 갖는 인간에게 이점을 부여하는 것으로 나타나지 않았다[참조: Messacar et al., 2016]. 그러나, IVIG는 EV-D68을 인식하는 작은 분획만을 갖는 폴리클로날 항체의 복잡한 혼합물이다. EV-D68 관련 AFM에 대한 MAb 예방 또는 요법은 사용할 수 있는 높은 특이성, 높은 효능 및 보다 낮은 항체 용량 때문에 IVIG보다 유망하다. 그러나, 복수의 에피토프에 대해 지시된 mAb 칵테일은 mAb 단독요법보다도 보호적일 수 있다. mAb 칵테일은 이론적으로 mAb 내성 바이러스의 출현에 대해 보다 높은 장벽을 제공할 것이지만, 본 발명자들은 생체내에서 내성을 관찰하지 않았다(도 37A-D 및 도 39A-F). 시험관내 EV68-228 내성 바이러스를 선택하기 위해 최적화된 조건하에서도, 본 발명자들은 중요성이 불명확한 돌연변이를 갖는 바이러스 게놈만을 동정할 수 있었다(표 J). 이러한 돌연변이를 갖는 재조합 바이러스를 제조하기 위한 역유전학 시스템이 없는 경우, 이들은 이러한 돌연변이가 중화로부터 탈출을 유발했는지의 여부를 구체적으로 확인할 수 없었다. 따라서, 잠재적 치료 사용 중에 내성이 출현할 가능성은 거의 없다. 이들 실험은 또한 EV-D68에 기인하는 중증 호흡기 질환 환자에서 치료 효능에 대한 희망을 제공하고, 이는 AFM과의 관련이 인정되기 이전에 2014 EV-D68의 발생을 공중 위생 당국의 주의를 환기시킨 임상 증후군이다[참조: Midgley et al., 2014]. 전반적으로, 여기에 제시된 연구는 인간의 자연적 EV-D68 감염이 호흡기와 신경계 모두에서 감염 및 질환을 예방 또는 치료할 수 있는 광범위하고 강력한 중화 항체를 암호화하는 B 세포를 유도한다는 것을 나타낸다.
실시예 12
이 연구의 목적은 4주령의 AG129 마우스에서 엔테로바이러스 D68(EV-D68) 호흡기 감염에 대한 담배 생산 EV68-228에 의한 치료의 효능을 결정하는 것이었다.
재료 및 방법
동물. 유타 주립 대학의 유타 과학기술연구소(USTAR) 빌딩에서 유지되고 있는 특정 병원체 비함유 콜로니로부터의 4주령 수컷 및 암컷 AG129 마우스. 마우스를 방사선 조사된 텍라드 설치류 다이어트(Harlan Teklad) 및 자동 급수된 수돗물에서 사육 및 유지했다.
항체 및 화합물. 담배 식물(EV68-228-TP) 및 차이니즈 햄스터 난소(CHO) 세포(EV68-228-CHO)에서 생산된 모노클로날 항체(mAb) EV68-228은 본 발명자들에 의해 제공되었다. 담배에서 생산된 항체와 CHO 항체는 모두 용액으로 제공되었고, 10mg/kg의 농도로 투여되었다. 담배 생산 항HIV(항-HIV-TP) 항체를 10mg/kg 용량으로 음성 대조군 항체로서 사용했다. 정맥 면역글로불린(HIVIg, Carimune, CSL Behring, King of Prussia, PA)은 지역 약국으로부터 구입하여 양성 대조군으로서 사용했다.
바이러스. 엔테로바이러스 D68은 BEI 리소스, NIAID, NIH: 엔테로바이러스 D68, US/MO/14-18949, NR-49130으로부터 수득했다. 이 바이러스는 4주령의 AG129 마우스의 폐에서 30회 연속 계대하고, 이어서 아메리칸 타입 컬쳐 컬렉션(Manassas, VA)로부터 수득된 횡문근육종(RD) 세포에서 3회 플라크 정제되었다. 수득된 바이러스 스톡을 RD 세포에서 2회 증폭하여, 작업 스톡을 생성했다. 감염에 사용된 바이러스는 EV-D68 MP30 PP로 지정되었다.
실험 설계. 표 1에 제시된 바와 같이, 총 118마리의 마우스를 각각 12마리로 이루어진 4개 그룹, 8마리 마우스로 이루어진 8개 그룹 및 정상 대조군에 사용되는 6마리 마우스로 이루어진 추가 그룹으로 랜덤화했다. 마우스는 감염 전 24시간 또는 감염 후 24시간 또는 48시간에 EV68-228 mAb, HIVIg 또는 위약 mAb의 복강내(IP) 투여에 의해 치료되었다. 90㎕ 용적의 MEM에서 1×104.5 CCID50의 EV-D68 MP30 PP를 비강내(IN) 주입함으로써 마우스를 감염시켰다. 처리 전 및 그 후 매일 마우스의 체중을 측정했다. 폐 바이러스 역가, 혈액 바이러스 역가 및 폐 사이토카인 농도의 평가를 위해, 감염 후 1일, 3일 및 5일차에 각 치료 그룹의 4마리의 마우스를 안락사시켰다. 감염 후 24시간 및 48시간 후에 치료된 마우스의 경우, 샘플은 감염 후 3일 및 5일차에만 수집되었다.
폐 사이토카인/케모카인 평가. 폐 균질액의 각 샘플은 제조업체의 지침(Quansys Biosciences Q-Plex™ Array, Logan, UT)에 따라 화학발광 ELISA 기반 검정을 사용하여 사이토카인 및 케모카인에 대해 시험되었다. 퀀시스 멀티플렉스 ELISA는 16개의 상이한 포획 항체가 정의된 어레이의 96웰 플레이트의 각 웰에 적용된 정량적 시험이다. 각 샘플 상청액은 하기: IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-10, IL-12p70, IL-17, MCP-1, IFN-γ, TNFα, MIP-1α, GM-CSF 및 RANTES에 대해 시험되었다. 약어의 정의는 다음과 같다: IL - 인터루킨; MCP - 단핵 세포 주화성 단백질; IFN - 인터페론; TNF - 종양 괴사 인자, MIP - 마크로파지 염증 단백질; GM-CSF - 과립구/마크로파지 콜로니 자극 인자; 및 RANTES - 활성화시에 조절되고, 정상 T 세포가 발현 및 분비된다.
통계 분석. 모든 수치 및 통계 분석은 프리즘 8.4.2를 사용하여 완료되었다(GraphPad Software Inc.). 일원 분산 분석(ANOVA)을 사용하여, 감염 후 매일, 치료 그룹의 폐 및 혈액 바이러스 역가를 위약 치료 마우스의 폐 및 혈액 역가와 비교했다. 각 사이토카인/케모카인에 대해 이원 배치 분산 분석(ANOVA)를 사용하여 치료 마우스의 농도를 위약 치료 마우스와 비교했다.
실험 동물의 윤리 규칙. 이 연구는 2019년 3월 2일자의 유타 주립 대학(Utah State University)의 시설내 동물 관리 및 사용 위원회의 승인에 따라 수행되었다(2022년 3월 1일 만료). 이 작업은 유타 주립 대학의 AAALAC 인정의 실험 동물 연구 센터에서 수행되었다. 미국 정부(국립 위생 연구소) 승인이 실험 동물의 관리 및 사용에 관한 국립 위생 연구소 가이드(개정: 2011)에 따라 2018년 3월 9일에 갱신되었다(PHS 보증 번호 D16-00468[A3801-01]).
[표 L]
Expt. NIA-1930/실험 설계: 마우스에서 EV-D68 호흡기 감염 치료에 대한 EV68-228-TP의 효능

결과 및 논의
이 연구는 4주령의 AG129 마우스에서 EV-D68 호흡기 감염 치료에 대한 담배 생산 EV68-228 mAb의 효능을 결정했다.
도 48A-C는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스의 폐 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 1일, 3일 또는 5일차에 폐 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 HIVIg에 의한 치료는 감염 후 5일차에 폐 바이러스 역가를 유의하게 감소시켰다.
도 49A-C는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스의 혈액 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 1일, 3일 또는 5일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 또한 감염 후 1, 3 및 5일차에 혈액 바이러스 역가를 감소시켰다.
도 50은 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도를 나타낸다. 감염 후 1일 및 3일차에, EV68-228-TP로 치료된 마우스와 EV68-228-CHO로 치료된 마우스에서 IL-1α의 폐 농도의 현저한 감소가 관찰되었다. 담배 생산 및 CHO 생산 치료는 감염 후 3일차에 IL-1β의 농도를 감소시켰다. hIVIg에 의한 치료는 IL-1α 또는 IL-1β의 폐 사이토카인 농도에 유의한 영향을 미치지 않았다. EV-D68 감염 후, IL-2 또는 IL-3의 폐 농도에 유의한 변화는 관찰되지 않았다.
EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스로부터의 Il-4, IL-5, IL-6 및 IL-10의 폐 농도는 도 51에 제시되어 있다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-5 농도를 유의하게 감소시켰고, 감염 후 3일차 및 5일차에 IL-6 농도를 감소시켰다. hIVIg에 의한 치료는 감염 후 3일차에 IL-5 및 IL-6의 폐 농도를 유의하게 감소시켰다. EV-D68의 감염 후, IL-4 또는 IL-10의 폐 농도에는 유의한 변화가 관찰되지 않았다.
도 52는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFNγ의 폐 농도를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. hIVIg에 의한 치료는 감염 후 3일 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. EV-D68의 감염 후, IL-12p70, IL-17 또는 IFNγ 농도의 유의한 변화는 관찰되지 않았다.
도 53은 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 1일 및 3일차에 MIP-1α의 폐 바이러스 농도를 감소시켰고, 감염 3일 및 5일차에 RANTES의 농도를 감소시켰다. 10mg/kg 용량의 hIVIg는 감염 후 1일 및 3일차에 MIP-1α의 폐 농도를 감소시켰고, 감염 후 3일차에 RANTES의 농도를 감소시켰다. EV-D68의 감염 후, TNFα 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다.
도 54A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 EV-D68-감염 AG129 마우스의 폐 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 3일 또는 5일차에 폐 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 HIVIg에 의한 치료는 감염 후 3일차에 폐 바이러스 역가를 유의하게 감소시켰다.
도 55A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 EV-D68-감염 AG129 마우스의 혈액 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 3일 또는 5일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 감염 후 3일 및 5일차에 혈액 바이러스 역가를 감소시켰다.
도 56은 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도를 나타낸다. 감염 후 3일차에, EV68-228-TP로 치료된 마우스 및 EV68-228-CHO로 치료된 마우스에서 IL-1α 및 IL-1β의 폐 농도의 유의한 감소가 관찰되었다. hIVIg에 의한 치료는 IL-1α 또는 IL-1β의 폐 사이토카인 농도에 유의한 영향을 미치지 않았다. IL-3의 농도는 EV68-228-TP로 치료된 마우스 및 감염 후 3일차에 hIVIg로 치료된 마우스에서 유의하게 감소했다. EV-D68의 감염 후, IL-2의 폐 농도에는 유의한 변화가 관찰되지 않았다.
EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 Il-4, IL-5, IL-6 및 IL-10의 폐 농도는 도 57에 제시되어 있다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-5 농도를 유의하게 감소시켰고, 감염 후 3일 및 5일차에 IL-6 농도를 감소시켰다. hIVIg에 의한 치료는 감염 후 3일차에 IL-5의 폐 농도를 유의하게 감소시켰다. EV-D68의 감염 후, 폐 조직에서 IL-4 또는 IL-10 농도의 유의한 변화는 관찰되지 않았다.
도 58은 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFNγ의 폐 농도를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. hIVIg에 의한 치료는 감염 후 3일 및 5일차에 MCP-1의 폐 농도를 유의하게 감소시켰다. EV-D68의 감염 후, IL-12p70, IL-17 또는 IFNγ 농도의 유의한 변화는 관찰되지 않았다.
도 59는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는 감염 후 3일차에 MIP-1α의 폐 바이러스 농도를 감소시켰고, 감염 후 3일 및 5일차에 RANTES의 농도를 감소시켰다. 10mg/kg 용량의 hIVIg는 감염 후 MIP-1α 또는 RANTES의 폐 농도를 유의하게 감소시키지 않았다. EV-D68의 감염 후, TNFα 또는 GM-CSF 농도의 유의한 변화는 관찰되지 않았다.
도 60A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스의 폐 바이러스 역가를 나타낸다. 감염 48시간 후에 치료를 수행한 경우, 10mg/kg 용량의 EV68-228-CHO에 의한 치료만이 폐 바이러스 역가를 감소시켰다. 감염 48시간 후에 치료를 투여한 경우, 10mg/kg 용량의 EV68-228-TP 또는 hIVIg에 의한 치료는 폐 바이러스 역가를 유의하게 감소시키지 않았다.
도 61A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스의 혈액 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 3일 또는 5일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 hIVIg에 의한 치료는 감염 후 3일 및 5일차에 혈액 바이러스 역가를 감소시켰다.
도 62는 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-1α, IL-1β, IL-2 및 IL-3의 폐 농도를 나타낸다. 감염 후 3일차에, EV68-228-CHO로 치료된 마우스에서는 IL-1α 및 IL-1β의 폐 농도의 현저한 감소가 관찰되었지만, EV68-228-TP로 치료된 마우스에서는 관찰되지 않았다. hIVIg에 의한 치료는 IL-1α 또는 IL-1β의 폐 사이토카인 농도에 유의한 영향을 미치지 않았다. EV-D68의 감염 후, IL-2 또는 IL-3의 폐 농도에 유의한 변화가 관찰되지 않았다.
EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 Il-4, IL-5, IL-6 및 IL-10의 폐 농도는 도 63에 제시되어 있다. 10mg/kg 용량의 EV68-228-CHO에 의한 치료는 감염 후 3일차에 IL-6 농도를 유의하게 감소시켰다. EV68-228-TP 또는 hIVIg에 의한 치료는 IL-6의 폐 농도를 유의하게 감소시키지 않았다. EV-D68의 감염 후, 폐 조직에서 IL-4, IL-5 또는 IL-10 농도의 유의한 변화는 관찰되지 않았다.
도 64는 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 IL-12p70, IL-17, MCP-1 및 IFNγ의 폐 농도를 나타낸다. 감염 48시간 후에 치료를 수행한 경우, EV-D68에 감염된 마우스에서 IL-12p70, IL-17, MCP-1 또는 IFNγ의 폐 농도에서 유의한 변화는 관찰되지 않았다.
도 65는 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스로부터의 TNFα, MIP-1α, GM-CSF 및 RANTES의 폐 농도를 나타낸다. 감염 48시간 후에 치료를 수행한 경우, EV-D68에 감염된 마우스에서 TNFα, MIP-1α, GM-CSF 또는 RANTES의 폐 농도에서 유의한 변화는 관찰되지 않았다.
결론
이 연구는 4주령의 AG129 마우스에서 EV-D68에 의해 유발된 호흡기 감염 치료에서 EV-D68에 대한 담배 생산 EV68-228 mAb의 효능을 결정했다. CHO에서 생산된 EV68-228 Ab를 비교대조로서 사용했다.
감염 전 24시간 또는 감염 후 24시간에 치료를 수행한 경우, 담배에서 생산된 Ab는 CHO에서 생산된 Ab와 비교하여 폐 바이러스 역가, 혈액 바이러스 역가 및 폐 사이토카인 농도에서 유사한 감소를 생성했다.
감염 후 48시간에 투여된 경우, Ab의 효능에서 약간의 차이가 관찰되었다. CHO 생산 Ab는 감염 후 3일 및 5일차에 폐 바이러스 역가를 유의하게 감소시킬 수 있었던 반면, 담배 생산 Ab는 감염 후 3일 또는 5일차에 폐 바이러스 역가를 감소시킬 수 없었다. 담배 생산 및 CHO 생산 Ab 모두는 감염 후 3일 및 5일차에 혈액 바이러스 역가를 감소시킬 수 있었다. 또한, CHO 생성 Ab는 IL-1α, IL-1β 및 IL-6의 폐 농도를 유의하게 감소시켰지만, 담배 생산 Ab는 사이토카인 농도에서 유사한 감소를 생성하지 않았다.
이러한 폐 바이러스 역가와 폐 사이토카인 농도의 차이는 감염 48시간 후에 치료가 수행된 경우에만 관찰되었고, 변동성은 감염 후의 Ab 치료의 한계로 인한 것일 수 있다. 그러나, CHO 생산 항체에 의한 EV-D68 호흡기 감염 후-치료를 평가한 이전의 연구(NIA-1869)에서, 1mg/kg의 용량은 IL-1α, IL-1β, IL-5, MCP-1 및 RANTES의 폐 농도를 감소시킬 수 있었다. 추가의 연구는, 담배 생산 항체와 CHO 생산 항체 사이에 유의한 차이가 있는지를 확인하기 위한 가치가 있을 것이다.
실시예 13
이 연구의 목적은 10일령의 AG129 마우스에서 엔테로바이러스 D68(EV-D68) 신경계 감염에 대한 담배 생산 EV68-228 항체에 의한 치료의 효능을 결정하는 것이었다. 차이니즈 햄스터 난소 세포에서 생산된 항체를 효능 비교대조로서 사용했다.
재료 및 방법
동물. 유타 주립 대학의 유타 과학기술연구소(USTAR) 빌딩에서 유지된 특정 병원체 비함유 콜로니로부터 10일령의 AG129 마우스. 마우스를 유타 주립 대학의 USTAR 빌딩에서 방사선 조사된 텍라드 설치류 다이어트(Harlan Teklad) 및 자동 급수된 수돗물로 사육 및 유지했다.
항체 및 화합물. 담배 식물(EV68-228-TP) 및 차이니즈 햄스터 난소(CHO) 세포(EV68-228-CHO)에서 생산된 모노클로날 항체(mAb) EV68-228은 반더빌드 대학 의료 센터의 발명자들에 의해 제공되었다. 담배 생산 항체와 CHO 항체는 모두 용액으로 제공되었고, 10mg/kg의 농도로 투여되었다. 담배 생산 항-HIV(항-HIV-TP) 항체는 10mg/kg의 용량에서 음성 대조군 항체로서 사용되었다. 정맥 면역글로불린(IVIg, Carimune, CSL Behring, King of Prussia, PA)은 지역 약국에서 구입하여 양성 대조군으로 사용했다.
바이러스. 엔테로바이러스 D68은 BEI 리소스, NIAID, NIH: 엔테로바이러스 D68, US/MO/14-18949, NR-49130에서 수득했다. 이 바이러스는 4주령의 AG129 마우스의 폐에서 30회 연속 계대하고, 이어서 아메리칸 타입 컬쳐 컬렉션(Manassas, VA)으로부터 수득된 횡문근육종(RD) 세포에서 3회 플라크 정제되었다. 수득된 바이러스 스톡을 RD 세포에서 2회 증폭하여, 작업 스톡을 생성했다. 감염에 사용된 바이러스는 EV-D68 MP30 PP로 지정되었다.
실험 설계. 표 1에 제시된 바와 같이, 총 125마리의 마우스를 각각 10마리씩 12개 그룹으로 랜덤화하고, 추가로 5마리의 마우스를 체중 증가에 대한 정상 대조군으로 사용했다. 마우스는 감염 24시간 전, 감염 24시간 또는 48시간 후에 EV68-228-TP 또는 EV68-228-CHO, IVIg 또는 위약을 복강내(IP) 투여하여 치료했다. 0.1mL 용적의 MEM에서 1×106.5 CCID50의 EV-D68 MP30 PP를 복강내(IP) 투여하여 마우스를 감염시켰다. 치료 전 및 그 후 매일 마우스의 체중을 측정했다. 혈액 바이러스 역가의 평가를 위해, 감염 후 1일, 3일, 5일 및 7일차에 5마리 마우스의 그룹으로부터 혈액을 수집했다. 21일까지, 모든 마우스의 이환율, 사망률 및 신경학적 스코어를 매일 관찰했다. 신경학적 스코어(NS)는 다음과 같이 기록되었다: NS0 - 관찰 가능한 마비 없음, NS1 - 뒷다리가 비정상적으로 벌렸지만 정상 또는 약간 느린 보행, NS2 - 전진 운동에 사용하는 동안, 뒷다리가 부분적으로 붕괴하고 발 끌림, NS3 - 뒷다리, 및 뒷다리의 경직 마비가 전진 운동에 사용되지 않음, NS4 - 뒷다리의 경직 마비 및 전진 운동 없음. NS4 스코어로 관찰된 모든 동물은 인도적으로 안락사시켰다.
통계 분석. 카플란-마이어 생존 곡선은 프리즘 8.4.2에서 생성되었다(GraphPad Software Inc.). 로그 랭크(Mantel-Cox) 검정, 및 이어서 게한-브레스로우-윌콕손(Gehan-Breslow-Wilcoxon) 검정을 사용하여 생존 곡선을 비교했다. 일원 분산 분석(ANOVA)을 사용하여, 감염 후 매일에 대해, 치료 그룹의 혈액 바이러스 역가를 위약 치료 마우스의 폐 및 혈액 역가와 비교했다. 일원 ANOVA를 사용하여 평균 체중을 비교했다. 크루스칼-왈리스(Kruskal-Wallis) 감정과 둔(Dunn)의 다중 비교 검정을 사용하여 신경학적 스코어를 비교했다.
실험 동물의 윤리 규칙. 이 연구는 2019년 3월 2일자의 유타 주립 대학(Utah State University)의 시설내 동물 관리 및 사용 위원회의 승인에 따라 수행되었다(2022년 3월 1일 만료). 이 작업은 유타 주립 대학의 AAALAC 인정의 실험 동물 연구 센터에서 수행되었다. 미국 정부(국립 위생 연구소) 승인이 실험 동물의 관리 및 사용에 관한 국립 위생 연구소 가이드(개정: 2011)에 따라 2018년 3월 9일에 갱신되었다(PHS 보증 번호 D16-00468[A3801-01]).
[표 M]
Expt. NIA-1931/실험 설계: 마우스에서 EV-D68 신경학적 감염의 치료에 대한 EV68-228-TP의 효능

결과 및 논의
이 연구는 10일령의 AG129 마우스에서 EV-D68 신경 감염 치료에 대한 담배 생산 EV68-228 mAb의 효능을 결정했다.
도 66은 EV-D68로 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 10일령의 AG129 마우스에 대한 카플란-마이어 생존 곡선을 나타낸다. 감염 24시간 전에 EV68-228-TP 또는 EV68-228-CHO 치료 모두는 사망으로부터 완전한 보호를 제공했다. 10mg/kg 단일 용량의 hIVIG는 10마리 중 8마리(80%)의 마우스를 사망으로부터 보호했다. 위약으로 치료된 10마리 중 9마리가 감염에 굴복했다.
도 67은 EV-D68로 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 10일령의 AG129 마우스에 대한 초기 체중의 백분율을 나타낸다. EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 마우스에서는, 위약 치료 그룹에서 생존한 마우스 1마리가 감염으로부터 회복된 후에 체중이 증가했기 때문에, 체중 감소로부터 유의한 보호가 관찰되지 않았다.
68A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스에 대한 감염 후 1일 및 3일차의 혈액 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 1일 또는 3일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 IVIg에 의한 치료는 또한 감염 후 3일차에 혈액 바이러스 역가를 감소시켰다.
도 69A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 EV-D68-감염 AG129 마우스에 대한 감염 후 5일 및 7일차의 혈액 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 5일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 IVIg에 의한 치료는 감염 후 5일차에 혈액 바이러스 역가를 유의하게 감소시키지 않았다. 감염 후 7일차에 어떤 치료 그룹의 혈액에서도 바이러스가 검출되지 않았다.
EV-D68로 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 10일령의 AG129 마우스에서 감염 후 2-10일차의 신경학적 스코어는 도 70A-B에 제시되어 있다. 감염 후 어느 일자에서든, 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 신경학적 스코어가 관찰되지 않았다. 또한, 감염 후 어느 시점에서든, 10mg/kg 용량의 IVIg로 치료한 마우스에서는 신경학적 스코어가 관찰되지 않았다. 유일한 생존 위약 치료 마우스는 마비의 징후를 나타내지 않았기 때문에, 감염 7일 후에 신경학적 스코어의 유의한 감소는 관찰되지 않았다.
EV-D68에 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령의 AG129 마우스의 카플란-마이어 생존 곡선이 도 71에 제시되어 있다. 감염 24시간 후에 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료하면, 마우스는 사망으로부터 완전히 보호되었다. 감염 24시간 후에 제공된 단일 용량의 hIVIg은 또한 10마리의 마우스 모두(100%)를 사망으로부터 보호했다. 위약으로 치료된 10마리 마우스 중 8마리가 감염에 굴복했다.
도 72는 EV-D68로 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령의 AG129 마우스에 대한 초기 체중의 백분율을 나타낸다. 위약 치료 그룹에서 생존한 2마리의 마우스가 체중 감염을 획득했기 때문에, EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 마우스에서는 체중 감소로부터 유의한 보호가 관찰되지 않았다.
EV-D68로 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령의 마우스로부터의 감염 후 1일 및 3일차의 혈액 바이러스 역가는 도 73A-B에 제시되어 있다. 치료가 감염 24시간 후에 수행되었기 때문에, 감염 후 1일차에는 혈액 바이러스 역가의 유의한 감소가 관찰되지 않았다. 감염 후 3일차에, EV68-228-TP 및 EV68-228-CHO에 의한 치료는 혈액 바이러스 역가를 유의하게 감소시켰다. 감염 24시간 후에 hIVIg에 의한 치료는 감염 후 3일차에 혈액 바이러스 역가를 유의하게 감소시켰다.
EV-D68에 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령의 마우스로부터의 감염 후 5일 및 7일차의 혈액 바이러스 역가는 도 74A-B에 제시되어 있다. EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 마우스의 혈액에서는 바이러스가 검출되지 않았지만, 위약 치료된 마우스는 2마리만이 혈액 수집을 위해 여전히 생존했고 어느 동물의 혈액에서도 바이러스가 검출되지 않았다. 위약 치료 동물에서는 바이러스 역가가 없기 때문에, 치료 그룹에 대해 통계적 유의성을 결정할 수 없었다. 감염 후 7일차에 어떤 그룹의 혈액에서도 바이러스가 검출되지 않았다.
도 75A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령의 AG129 마우스에서 감염 후 2-9일차의 신경학적 스코어를 나타낸다. 감염 24시간 후에 EV68-228-TP 또는 EV68-228-CHO에 의한 치료는, 감염 후 2-9일차에 신경학적 스코어의 감소에 의해 나타난 바와 같이, 마비 징후를 유의하게 감소시켰다. 감염 24시간 후에 10mg/kg 단일 용량의 hIVIg는 감염 후 2일 및 3일차에 신경학적 스코어를 감소시켰지만, 나머지 연구의 결과는 개선되지 않았다.
EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 후에 치료된 10일령의 AG129 마우스에서 감염 후 10-17일차의 신경학적 스코어는 도 76A-B에 제시되어 있다. EV68-228-TP 및 EV68-228-CHO는 감염 후 10-15일차에 신경학적 스코어를 유의하게 감소시켰다. hIVIg의 투여는 감염 후 10-17일차에 신경학적 스코어를 유의하게 감소시키지 않았다.
도 77은 EV-D68에 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 10일령의 AG129 마우스에 대한 카플란-마이어 생존 곡선을 나타낸다. 10mg/kg 용량의 EV68-228-TP에 의한 치료는 10마리 마우스 중 9마리(90%)의 마우스를 사망으로부터 보호했다. 10마리 마우스 중 8마리(80%)는 EV68-228-CHO로 감염 48시간 후에 치료된 감염에서 생존했다. 10mg/kg 단일 용량의 hIVIG는 10마리 마우스 중 6마리(60%)의 마우스를 사망으로부터 보호했다. 위약 치료된 10마리 마우스 중 1마리는 감염에서 생존했다.
도 78은 EV-D68에 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 10일령의 AG129 마우스에 대한 초기 체중의 백분율을 나타낸다. EV68-228-TP 또는 EV68-228-CHO로 감염 24시간 전에 치료된 마우스에서는, 위약 치료 그룹에서 생존한 1마리의 마우스가 감염 후에 체중을 획득했기 때문에, 체중 감소로부터 유의한 보호가 관찰되지 않았다.
도 79A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스에 대한 감염 후 1일 및 3일차의 혈액 바이러스 역가를 나타낸다. 감염 후 1일차에, 그 시점에서 치료가 수행되지 않았기 때문에, 치료 그룹 중 어느 것도 유의차는 없었다. 감염 후 3일차에, 10mg/kg의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서 혈액 바이러스 역가는 낮았지만, 통계적으로 유의차는 없었다. 이는 3일차의 혈액 수집의 24시간 전에 치료가 수행된 것 때문일 수 있다. 10mg/kg 용량의 IVIg에 의한 치료는 감염 후 1일차 또는 3일차에 혈액 바이러스 역가를 유의하게 감소시키지 않았다.
도 80A-B는 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후에 치료된 EV-D68-감염 AG129 마우스에 대한 감염 후 5일 및 7일차의 혈액 바이러스 역가를 나타낸다. 10mg/kg 용량의 EV68-228-TP 또는 EV68-228-CHO로 치료된 마우스에서는 감염 후 5일차에 혈액 바이러스 역가가 검출되지 않았다. 10mg/kg 용량의 IVIg에 의한 치료는 감염 후 5일차에 혈액 바이러스 역가를 유의하게 감소시키지 않았다. 감염 후 7일차에는 어떤 치료 그룹의 혈액에서도 바이러스가 검출되지 않았다.
EV-D68에 감염되고 EV68-228-TP 또는 EV68-228-CHO로 감염 48시간 후 치료된 10일령의 AG129 마우스에서 감염 후 3-10일차의 신경학적 스코어는 도 81A-B에 제시되어 있다. 10mg/kg의 EV68-228-TP에 의한 치료는 감염 후 3-5일차에 신경학적 스코어를 유의하게 감소시켰다. 10mg/kg의 EV68-228-CHO에 의한 치료는 감염 후 3-6일차에 신경학적 스코어를 유의하게 감소시켰다. 신경학적 스코어는 감염 후 3-5일차에 10mg/kg의 hIVIG로 치료된 마우스에서 유의하게 감소했다. 감염 후 7-10일차에는 어떤 치료 그룹에서도 신경학적 스코어의 유의한 감소가 관찰되지 않았다.
결론
이 연구는 EV-D68 신경 감염의 10일령 AG129 마우스에 대한 담배 생산 EV68-228 mAb의 효능을 결정했다.
담배 생산 mAb는 감염 24시간 전 및 감염 24시간 후 또는 48시간 후에 치료를 수행한 경우, CHO 생산 mAb와 비교하여 사망으로부터 유사한 보호를 제공했다.
또한, EV-D68에 감염된 10일령의 AG129 마우스에서, EV68-228-TP 및 EV68-228-CHO mAb 둘 다로 치료된 마우스에서 혈액 바이러스 역가의 유사한 감소가 관찰되었다.
신경학적 스코어의 감소에 의해 나타난 바와 같이 마비로부터 유사한 보호는 EV68-228-TP 및 EV68-228-CHO로 치료된 마우스에서 관찰되었다.
요약하면, 담배 생산 EV68-228은, CHO 생산 EV68-228과 비교하는 경우, 사망으로부터의 유사한 보호, 혈액 바이러스 역가의 감소 및 마비로부터 보호를 제공했다.
| 클론 | 서열번호 | 쇄 | 가변 서열 영역 |
| EV-D68-37 | 1 | 중쇄 | CAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCATGTGCAGCCTCTGGATTCATCTTCAGTCGCTACGCTCTGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGACTGGGTGGCAGTTATATCATATGATGCAAGAAATTCATATTACACAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAAAACACGCTGTTTCTGCAGATGAACAGTCTGAGAGCTGACGACACGGCTGTCTATTACTGTGCGAGACCGACTTTGCCCTACAGCAACAACTGGTACGCGCCTGAATACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 2 | 경쇄 | TCCTATGAGCTGACACAGCCACCCTCGGTGTCAGTGTCCCCAGGACAAACGGCCAGGATCACCTGCTCTGGAGATGCATTACCAAAAAAATATGCTTCTTGGTACCAGCAGAAGTCAGGCCAGGCCCCTGTGCTGGTCATCTATGAAGACACCAAACGACCCTCCGGGATCCCTGAGAGATTCTCTGGCTCCAGCTCAGGGACAATGGCCACCTTGACTATCAGTGGGGCCCAGGTGGAAGATGAAAGTGACTACTACTGTTCCTCGACAGACAGCAGTGGTAATCCTGTGCTATTCGGCGGAGGGACCAAGTTGACCGTCCTA | |
| EV-D68-40 | 3 | 중쇄 | GAGGTGCAGCTGGTAGAGTCTGGGGGAGACTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCTCCTTTAGCAGCTATGCCATGGCCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGCAGTGGGTCTCATCTATTAGTGGTAACGGTAATGGGAGATCCTATGCAGATTCTCTGAAGGGCCGGTTCACCACCTCCAAAGACCTTTCCAAGTATACCCTGTATCTGCAAATGAACAATCTGAGACCCGAGGACACGGCCATATATTACTGTGCGAAAGTTGTCCGTATAGCAGCTGTTTTGTATTACTTTGACTATTGGGGCCCGGGAACCCAGGTTACCGTCTCCTCA |
| 4 | 경쇄 | GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCACCTACTTAGCCTGGTACCAACAAAAGCCTGGCCAGGCTCCCAGGCTCCTCATCTATGAAGCATCCACCAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCATCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATCACTGTCAGCAGCGCAGCTCCTGGCCGATCACCTTCGGCCAAGGGACACGACTGGAGATTGAA | |
| EV-D68-41 | 5 | 중쇄 | GACGTGCAGCTGGTGGAGTCTGGGGGGGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCTGCCTCTGGATTCACTTTTAGCAACTATGCCATGACCTGGGTCCGCCAGGCTCTAGGGAAGGGGCTGGAGTGGGTCTCTTCTATTAGTGGTAGTGGTGGCCTCACATATTTCGCACACTCCGTGAAGGGCCGGCTCACCATCTCCAGAGACAACTCCAAGAATACCCTCTATCTGCAAATGAGCAGCCTGAGAGCCGAAGACACGGCCGTATATTACTGTGCGAGAGTGAAAAGTACAACTGGAACGACGGCGTTAGTTTTTGATATCTGGGGCCAAGGGACAATGGTCACCGTCTCTTCG |
| 6 | 경쇄 | CAGACTGTGGTGACCCAGGAGCCATCGTTCTCAGTGTCCCCTGGAGGGACAGTCACACTCACTTGTGGCTTGAGTTCTGGCTCAGTCTCTAGTAGTTACTACCCCAGTTGGTACCAGCAGACCCCAGGCCAGGCTCCACGCACGCTCATCTACAGTATAAACAGACGTTCTTCTGGGGTCCCTGATCGCTTCTCTGGCTTCATCCTTGGGAACAAAGCTGCCCTCACCATCAGGGGGGCCCAGGCAGATGATGAATCTGATTATTACTGTGGGCTGTATATGGGTAGTGGCATTTGGATCTTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-43 | 7 | 중쇄 | CAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCATTAACTATGGCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATCAAATGATGGAAGTTATAACTACGATGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACAAGGTTTATCTACAAATGAACAGCCTGAGACCTGAGGACACGGCTGTGTATTTCTGTGCGAAAGACAAACACGGTGACTTCGACTACTACGGTGTGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCCCA |
| 8 | 경쇄 | GACGTCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTATAGGAGACAGAGTCACCATCACTTGTCGGGCGGGTCAGGGAATTAGCAGCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAATTTGCAAAGTGGGGTCCCATCACGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGGAACTTACTATTGTCAACAGGCTGACAGTTTCCCTCGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA | |
| EV-D68-46 | 9 | 중쇄 | CAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTAGTTATGGCATACACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATCCTATGATGGAAGTGATAACACCTATGCACCTTTTGTGAACGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCTGACGACACGGCTGTGTATTACTGTGCGAGGCGTCGGCCTGGGAGCTTCCCAGGACTTTGCGACTACTGGGGCCAGGGAGCCCTGGTCACCGTCTCCTCA |
| 10 | 경쇄 | GACATCCAGATGACCCAGTCGCCTTCCACTATGTCTGCTTCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAAGTGGTTGGCCTGGTATCAGCAGAAGCCAGGGAAAGCCCCTAAACTCCTGATCTATAAGGCGTCTACCTTACAAACTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAACAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAACAGCATAATAGTTATTCGTACACTTTTGGCCAGGGGACCAAGGTGGAGATCAAG | |
| EV-D68-48 | 11 | 중쇄 | CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGTCCTCGGTGAAGGTCTCCTGCAAGGCTTCTGGAGGCAGCTTCAGCAGACTTACTATCATCTGGGTGCGGCAGGCCCCTGGACAAGGGCTTGAGTGGATGGGAGGGCACATCCCTATCTTTGGAACAACAAACTACGCACTGAAGTTCCAGGGCAGAGTCACGATTACCGCGGACAAAACCACGAGCACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCATATATTACTGTGCGAGAATGTATAGTGGCCATGACGGCGTTGATGTCTGGGGCCAAGGGACACTGGTCACCGTCTCTTCA |
| 12 | 경쇄 | GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGGAGCTACTTAGCCTGGTACCAACACAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCAAGGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCACCTGGCCTCCGGGAATGTTCGGCCAAGGGACCAGGGTGGAAATCAAA | |
| EV-D68-71 | 13 | 중쇄 | CAGCTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCATTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGTGGCTCCATCAGCAGTGGTTTTTACTACTGGGGCTGGATCCGCCAGCCCCCAGGGAAGGGGCTAGAGTGGATTGGGACTATCTATGATAGTGGGAGAACCTATGACAACCCGTCCCTCAAGAGTCGAGTCACCATATCCGCAGACACGTCCAAGAAGCAGTTCTCACTGACACTGAGGTCTGCGACCGCCGCGGACACGGCTGTGTATTTCTGTGCGAGACACCTTACCCACCTCTACGGTGACTACGTCACCCCTGATGCTTTAGATATCTGGGGCCAAGGGACAATGGTCACCGTCTCTTCA |
| 14 | 경쇄 | GAAATTGTGTTGACGCAGTCTCCAGGCACCCTGTCCTTGTCTCCAGGGGAAAGAGCCACCCTCTCTTGCAGGGCCAGTCAGAGCGTTAGTAGCAGCTTCTTAGCCTGGTACCAGCAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGGTGCGTCCAGCAGGGCCACCGGCATCCCAGACAGGTTCAGAGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAGCCTGAAGATTTTGCAGTATATTACTGTCAGCAGTATAGTAACTCACGTCTGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA | |
| EV-D68-72 | 15 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCACTACCTATAGCATGAACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGATTTCATACATTAGTAGTGGTAGTAGTAACATATACTACGCAGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAACTCACTGAATCTGCAAATGAGCAGCCTGAGAGACGAGGACACGGCTGTGTATTACTGTGCGAGAGCCCACGGACGTATAGTGAATTCTGGAGTGGTTATTAGTAGGTTCGACCCCTGGGGCCAGGGAATCCTGGTCACCGTCTCCTCA |
| 16 | 경쇄 | CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGACAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGGTGGTTATAACTATGTCTCCTGGTACCAACTGCACCCAGGCAAAGCCCCCAAACTCATGATTTTTGAGGTCACTTATCGGCCCTCAGGGGTTTCTAATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTTCTGCAGCTCCTATACAACCAGCAACACTCTCGTGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-74 | 17 | 중쇄 | CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGTAGGACTTCTGGATACACCTTCACGGCCTACTATATGCACTGGGTGCGACAGGCCCCCGGACAAGGGCCTGAGTGGATGGGAAGGATCAACCCGAGCAGTGGTGGCGCACAGTATGCACAGAAGTTTCAGGGCAGGGTCACCATGACCAGGGACACGTCCATCAGCACAACCTACATGACCCTGAGCGGGCTGACATCTGACGACACGGCCGTGTTTTACTGTGCGAGAATGGGTTGTCGTAGTGACCGGTGCTATTCGACCAACTACAACTTTGACCAGTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 18 | 경쇄 | TCCTATGTGCTGACTCAGCCACCCTCGGTGTCAGTGGCCCCAGGACAGACGGCCAGGATTCCCTGTGGGGGAAACAACATTGGAACTAAAAGTGTTCACTGGTACCAGCAGAAGCCAGGCCAGGCCCCTGTGCTGGTCGTCTCTAATGACAGCGACCGGCCCTCAGGGATCCCTGAGCGATTCTCTGGCTCCAAGTCTGGGAACACGGCCACCCTGACCATCAGCAGGGTCGAAGCCGGGGATGAGGCCGACTATTATTGTCAGGTGTGGGATAGTGGTATTGATGTCGTTTTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-75 | 19 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCCTGGTCAAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACTATCAGTCCCTATGGCATGAACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCATTCATTAGTAGTAGTAGTCGTTACACATATTACGCAGACTCAGTGAAGGGCCGTTTCACCATCTCCAGAGACAACGCCAAGAATTCACTGTCTCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCGAGAGAGAGGGGCCATAGCACCTCGTCCTCATACTTTGACTCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 20 | 경쇄 | TCCTATGTGCTGACTCAGCCACCCTCAGTGTCATTGGCCCCAGGAAAGACGGCCAGGATTACCTGTGGGGGAAACAACATTGGAACTAAAACTGTGAGCTGGTACCAGCAGAAGCCAGGCCAGGCCCCTGTGCTGGTCATGTATTATGATAGTGACCGGCCCTCAGGGATCCCTGAGCGATTCTCTGGCTCCAACTCTGGGAACACGGCCACCCTGACCATCAACAGGGTCGAGGCCGGGGATGAGGCCGACTATTACTGTCGGGTGTGGGATAGTGATACTGATCATCGAGTGTTCGGCGGGGGGACCAAGCTGACCGTCCTA | |
| EV-D68-76 | 21 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTAAAGCCTGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGATTCACTTTCAGTAACGCCTGGATTAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTTGGCCGTATTCAAACCAAAACTGATGGAGGGACAACAGACTACGCTGCACCCGTGAAAGGCAGATTCACCATCTCAAGAGATGATTCAAAAAACACGTTGTATCTGCAAATGAACAGCCTGAAAACCGAGGACACAGCCTTGTATTATTGTAGCACAGGACCGTATTACTATGATACTAGTGGTTACCCCCAACCCTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 22 | 경쇄 | TCCTATGAGCTGACTCAGCCACCCTCAGTGTCCGTGTCCCCAGGACAGACAGCCAGCATCACCTGCTCTGGAGATAAATTGGGAGATAAATATGCTTGCTGGTATCAGCAGAAGCCAGGCCAGTCCCCTGTGCTGGTCATCTATCAAGATACCAAGCGGCCCTCAGGGATCCCTGAGCGATTCTCTGGCTCCAACTCTGGGAACACAGCCACTCTGACCATCAGCGGGACCCAGGCTATGGATGAGGCTGACTATTACTGTCAGGCGTGGGACAGCAGCACTGTGGTATTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-80 | 23 | 중쇄 | GAGGTACAGATGGTGGAGTCTGGGGGAGGCCTGGTCAAGCCGGGGGGGTCCCTCAGACTCTCCTGTTCAGTCTCTGGATTCGACTTCAGTAGATACACCATGAACTGGTTCCGCCAGGCTCCAGGGGAGGGGCTGAAGTGGGTCTCGTCCATTAGTAGTACTAGTCTTTACACATTCTATGCGGACTCAGTGAAGGGCCGATTCTCCATCTCCAGAGACAACGCCCAGGGTTCCCTGTCTCTACAAATGAGCAGCCTGAGACCCGAAGATACGGCTGTCTATTATTGTGCGAGAGTCGTTGGTCCCGCCGAGTTAGATTACTGGGGCCAGGGAGTGCTGGTCACCGTCTCCTCA |
| 24 | 경쇄 | GTCACCCAGTTGACCCAGTCTCCATCCTCCCTGTCTGCTTCTGTCGGGGACAGAGTCACCATCACTTGCCGGGCAAGTCAGGACATTGGAGTTGACTTAGGTTGGTTTCAGCAGAGACCTGGGAAAGCCCCTAAACTCCTGATCTATGGTGCCTCCAGGTTGCAGAGTGGGGTCCCATCACGCTTCAGCGGGCGTGGATCTGGCACATTTTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTACAACTTACTTCTGTCTTCAAGATTATAATTACCCTTGGACGCTCGGCCAGGGGACCACGGTGGGAGTCAAA | |
| EV-D68-84 | 25 | 중쇄 | CAGGTGCACCTGGTGCAGTCTGGGTCTGCGGTGAGGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGCTTCTGGATACACCTTCACCGACTACTATATTCACTGGGTGCGACAGGCCCCTGGACAAGGCCTTGAGTGGATGGGATGGATCAACCCTAAAACTGGTGGCTCAAATTATACACAGAGGTTTCAGGCCAGGGTCACCATGACCTGGGACACGTCCATCAGTACAGCCTACATGGAGTTGAGCAGGCTGAGATCTGACGACACGGCCGTGTATTATTGTGCGAGGGCGGGCAGAAATGGCTACGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 26 | 경쇄 | TCCTATGAGCTGACACAGCCACCCTCGGTGTCAGTGTCCCCAGGACAGACGGCCAGGATCACCTGCTCTGCAGATGCATTGCCAAAGCAATATGCTTATTGGTACCAACAGAAGCCAGGCCAGGCCCCTGTGTTGATGATATATCAAGACACTGAGAGGCCCTCAGGGATCCCTGAGCGATTCTCTGGGTCCAGCTCAGGGACAACAGTCACGTTGACCATCAGTGGAGTCCAGGCAGAAGACGAGGCTGACTATTACTGTCAATCAGGAGACAGCAGTGGTACTTATCTAGTTTTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-85 | 27 | 중쇄 | GAGGTACAGCTCTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCAAATTTAGAAACTATGCCATGACCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAACTATTACTAGTGGTGGTAGTACAGAGTACGCAGACGCCGTGAAGGGCCGGTTCATCATCTCCAGAGACAATTCCAAGAACACGTTATATTTGCAAATGAACAGCCTGAGAGCCGACGACACGGCCGTATATTACTGTACAGTGCCGTGGGGTAACTACAATGACTACGTGTCTGACTACTGGGGCCAGGGAACCCTGGTCCCCGTCTCCTCA |
| 28 | 경쇄 | GAAGTTGTATTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGCAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGCGTGTTGGCAACTCCTTAGCCTGGTACCAACAAAAACCTGGCCAGGCTCCCAGCCTCCTCATCTATGATGCTTCCAAGAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCATCAGCCTAGAGTCTGAAGATTTTGCAGTTTATTACTGTCACCAACATAGCACCTGGCCTCGGGGGACCTTCGGCCAAGGGACACGACTGGAGATTAAA | |
| EV-D68-88 | 29 | 중쇄 | CAGGTGCACCTGGTTGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCATCTTCAGTAGATATCCTATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGTCTGGAGTGGGTGGCACTTATATCATATGATGGAAACAATAAATACTACGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTTTCTCCAAATGAACAGCCTGAGAGCTGAGGACACGGCTGTCTATTACTGTGCGAGACATTTCCTCCCATATAGCAGTAGTTGGTACCAGGGCTTTAACTACTGGGGCCAGGGAATCCTGGTCACCGTCTCCTCA |
| 30 | 경쇄 | AATTTTATGCTGACTCAGCCCCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCAGCATTGCCACCAACTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTTCCCCCACCCCTATAATCTTTGAAGATAGTCAAAGACCGTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAATTCTGCCTCCCTCACCATCTCTGGACTGAGGACTGACGACGAGGCTGACTACTACTGTCAGTCTTATGATAACAGCAATCGGGCTGTTGTATTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-89 | 31 | 중쇄 | CAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGAAGCGTCTGGATTCCTCTTCAGTCGCTATGGCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGACTGGGTGGCAGTTATATCGTATGATGGAAATAAGAAATATTATGCAGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCAAAGAACACGTTGTATCTGCAAGTGAACAGCCTGAGAGTCGAGGACACGGCTGTTTATTACTGTGCGAGAGGTGTCCCGTACGGTGACACCCTTACAGGGCTTGTCTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 32 | 경쇄 | AATTTTATGCTGACTCAGCCGCACTCTGTGTCGGAGTCTCCGGGGAAGACGGTAACCATCTCCTGCACCCGCAGCAGTGGCACCATTGCCAGCAACTATGTGCAGTGGTACCAGCAGCGCCCGGGCAGTGCCCCCACCACTGTAATCTATGAGGATAACCAAAGACCCTCTGGGGTCCCTGATCGGTTCTCTGGCTCCATCGACAGCTCCTCCAACTCTGCCTCCCTCACCATCTCTGGACTGAAGACTGAGGACGAGGCTGACTACTACTGTCAGTCTTATGATAACAGCGATCGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTT | |
| EV-D68-95 | 33 | 중쇄 | CAGGTCACCTTGAAGGAGTCTGGTCCTGTGCTGGTGAAACCCACAGAGACCCTCACGCTGACCTGCACCGTCTCTGGGTTCTCACTCAGGAACGCTAGAATGGGAGTGAGCTGGATCCGTCAGCCCCCAGGGAAGGCCCTGGAGTGGCTTGCACACATTTTTTCGAATGACGAAAAATCCTACAACACATCTCTGAAGAGCAGACTCTCCATCTCCAAGGACACCTCCAAAAGCCAGGTGGTCCTTACCATGACCAGCATGGACCCTTTGGACACAGCCACATATTTCTGTGCACGGCTACTGGTGGCTGGTACTTTCCTCCCCTCTCACTACTTTGACTACTGGGGCCAAGGAATCCTGGTCACCGTCTCCTCA |
| 34 | 경쇄 | TCCTATGTGCTGACTCAGCCACCCTCAGTGTCAGTGACCCCAGGAAAGACGGCCAGGATTACCTGTGGGGGAAACAACATTGGACTTAAAAGTGTCTTCTGGTACCAGGAGAGGCCAGACCAGGCCCCTGTGGTGGTCATCTATTATGATAGCGCCCGGCCCTCAGGGATCCCTGAGCGAATCTCTGGCTCCAAGTCTGGGAACACGGCCACCCTGACCATCACCAGGGTCGAAGCCGGGGATGAGGCCGACTATTTTTGTCAGGTGTGGGATAGTAGTCGTAATCATCCGGTCTTCGGCGGAGGGACCAAACTGACCGTCCTC | |
| EV-D68-97 | 35 | 중쇄 | GAGCTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTACCTATAGCATGAATTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTTTCATACATTAGTAGTAGTAGTAGTACCATACAGTACGCAGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAACTCACTGTATCTGCAAATGAATAGCCTGAGAGCCGAAGACACGGCCGTGTATTATTGTACGAGACAGGTCGGGGCGGATTTCAGTGGCCGCGGCTTTGACTACTGGGGCCAGGGAACCCTGCTCACCGTCTCCTCA |
| 36 | 경쇄 | TCCTATGAGCTGACTCAGCCACCCTCAGTGTCCGTGTCCCCAGGACAGACAGCCACCATCACCTGCTTTGGAGATAAATTGGGGGATAAATATGCTTGCTGGTATCAGCAGAAGCCAGGCCAGTCCCCTGTGTTGGTCATCTACCAAGATAGCAAGCGGCCCTCAGGGATCCCTGAGCGATTCTCTGGCTCCAAGTCTGGGAACACAGCCACTCTGACCATCAGCGGGACCCAGGCAATGGATGAGGCTGACTATTACT-GT-CAGGCGTGGGACAGCAGCACTGCAGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTC | |
| EV-D68-98 | 37 | 중쇄 | GAGGTACAACTAGTGGAGTCAGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCAAGTTTTCCGTCTATGCCTTGAGTTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGATTTCATATATTAGTAGTAGTGGTTCTACCATATATTATTCAGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAATGTCGGGAACTCACTGTTTGTGCAAATGAACAGCCTGAGAGCCGAGGACACGGGTATTTATTACTGTGCGACAGCCCGCCACATCACCAATGATGGTTTTGATATTTGGGGCCAAGGGACAATGGTCATCGTCTCTTCA |
| 38 | 경쇄 | GACATCCAGTTGACCCAGTCTCCATCCTTCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGGGCATTAGTAGGTTTTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATTCTGCGTCCACTTTACAAAGGGGGGTCCCATCAAGGTTCAGCGGCAGTGGATTTGGGACAGATTTCACTCTCACAATCAGTAGCCTTCAGCCTGAAGATTTTGCAACTTATTACTGTCAACAACTTAATAGTCACCCCCGAATGTTCACTTTCGGCCCTGGGACCACAGTGGATATCAAG | |
| EV-D68-105 | 39 | 중쇄 | CAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCGCTGTGTCTGGTTACTTAATCAGCAATGGTTACTACTGGGGCTGGATCCGGCAGCCCCCCGGGAAGGGGCTGGAGTGGATTGGGAGTATCTATCATACTAGAAGCACCTACTACAACCCGTCCCTCAAGAGTCGAGTCAGCATCTCAGTAGACACGTCCAAGAACCGGTTCTCCCTGAGGCTGAGGTCTGTGACCGCCGCAGACACGGCCTTTTATTACTGTGCGAGAGGCCCAGGCCACTGTTATGGTGATGACGACTGCTACGCGTACTACTTTGACCAGTGGGGCCAGGGAACCCCGGTCACCGTCTCCCCA |
| 40 | 경쇄 | GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTTCATCTGTAGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGGTATTAGCAACTGGTTAGCCTGGTATCAGCAGAACCCAGGGAAAGCTCCTAAACTCCTGATCTATGATGCCTCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAACAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAACAGGCTAACAGTTTCCCTTTCACTTTCGGCCCTGGGACCAAAGTGGATATCAAA | |
| EV-D68-110 | 41 | 중쇄 | CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGTCCTCGGTGAAGGTCTCCTGCAAGGCTTCTGGAGGCACCTTCAGCAGGTTTGCTATCAGCTGGGTGCGACAGGCCCCTGGACAAGGGCTTCAGTGGATGGGAGGGATCCTCCCTATCTTTGGTACAGCAAACTACGCACAGAAGTTCCAGGGCAGAGTCACGATTACCGCGGACACATCCACGAGCACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCGTGTATTACTGTGCGAGATCCCTCCCCTATTGTACTAATGATGTATGCTCAAACCAGAACACATTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 42 | 경쇄 | TCCTATGAGCTGACACAGCCACCCTCGGTGTCAGTGTCCCCAGGACAGGCGGCCAGGATCACCTGCTCTGGAGATGCATTGCCTAAGCAATATGCTTATTGGTACCAGCAGAAGCCAGGCCAGGCCCCTGTGTTGGTGATATATGAAGACAATAAGAGGCCCTCAGGGATCCCTGAGCGATTCTCTGGCTCCAGCTCAGGGACAACAGTCACGTTGACCATCAGTGGAGTCCAGGCAGAAGACGAGGCTGACTATTACTGTCAATCAGCAGACAGCAGTGGTACTTATGTGGTATTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-111 | 43 | 중쇄 | CAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCACAGACCCTGTCCCTCACCTGCACTGTCTCTGGTGGCTCCATCAGCAGTGGTGATTACTACTGGAGTTGGATCCGCCAGCCCCCAGGGAAGGGCCTGGAGTGGATTGGGTACATCTATTACAGTGGGAGCACCTACTACAACCCGTCCCTCAAGAGTCGAGTTACCATATCAGTAGACACGTCCAAGAACCAGTTCTCCCTGAAGCTGAGCTCTGTGACTGCCGCAGACACGGCCGTGTATTACTGTGCCAGCCGCTACGGTGACCCGATAGGGGACAACTGGTTCGACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 44 | 경쇄 | TCCTATGTGCTGACTCAGCCACCCTCAGTGTCAGTGACCCCAGGAAAGACGGCCAGGATTACCTGTGGGGGAAACAACATTGGACTTAAAAGTGTCTTCTGGTACCAGGAGAGGCCAGACCAGGCCCCTGTGGTGGTCATCTATTATGATAGCGCCCGGCCCTCAGGGATCCCTGAGCGAATCTCTGGCTCCAAGTCTGGGAACACGGCCACCCTGACCATCACCAGGGTCGAAGCCGGGGATGAGGCCGACTATTTTTGTCAGGTGTGGGATAGTAGTCGTAATCATCCGGTCTTCGGCGGAGGGACCAAACTGACCGTCCTC | |
| EV-D68-114 | 45 | 중쇄 | GAGGTGCAACTGTTGGAGTCGGGGGGAGGCTTGGTGCAGCCGGGGGGGTCCCTGAGACTCTCCTGCGCAGCCTCCGGATTCAGGTTTAGCTTCTATGGCATGACCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAAGTATTAGTGGTACTGGTGCTACCAGAAACTGCGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAACTCCAAGAACACGCTGTACCTGCAAATGGACAGCCTGAGAGTCGACGACACGGCCGTTTTTTATTGTGTGAGACGGTTCCCGATGACCACGGTGACAAGCTTTGACTCTTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 46 | 경쇄 | CAGTCTGCCCTGACTCAGCCTCGCTCAGTGTCCGGGTCTCCTGGACAGTCAGTCACCATCTCCTGCACTGGAACCAGCAGTGATGTTGGTGGTTATAACTTTGTCTCCTGGTACCAACAACACCCGGGCAAAGCCCCCAAACTCATGATTTTTGATGTCACTGGGCGGCCCTCAGGGGTCCCTGATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCATCGCTGGGCTCCAGGCTGAAGATGAGGCTGATTATTATTGTGGCGCATATGCGGGCTTTAACGCTCTTTTCGGCGGAGGGACCAAACTGACCGTCCTA | |
| EV-D68-116 | 47 | 중쇄 | GAGGTGCAGCTGGTGCAGTCGGGGGGAGGCTTGGTCCGGCCGGGGGGGTCCGTGAGACTCTCCTGTGTAGCCTCTGGATTCCCCTTCAATATGTTTTGGATGGGCTGGGTCCGCCAGACTCCAGGGAAGGGACTGGAGTGGGTGGCCAATATAAAACAGGATGGCAGTGAGAAATACTATGTCGACTCCGTGAAGGGCCGATTCGCCATCTCCAGAGACAATGCCAAGAACTCTCTCTTTCTTCAAATGGACAGCCTGAGTGTCGGGGACACGGCCATCTATTATTGTGTCAGAGAGGGGGTGCGAAGGGTCGTCGTACGTAGTACCGGTTACTTCGACTTTTGGGGCCAGGGACAGCTGGTCACCGTCTCCTCA |
| 48 | 경쇄 | TCCTATGAGCTGACACAGCCACCCTCGATGTCAGTGTCCCCAGGACAAACGGCCAGGATCACCTGCTCTGGAGATGCAGTGCCAATAAAATATGTTTATTGGTACCAACAGAGGTCAGGCCAGGCCCCTGTATTAGTCATCTATGAAGACGACAGACGACCCTCCGGGATCTCTGAGAGATTCTCTGGCTCCAGTTCAGGGACAACGGCCACCTTGACTATCACTGGGGCCCAGGTGGAGGATGAAGGTGACTACTATTGCTATTCAACAGACAGTAGTGGTTATCAGAGAGCGTTCGGCGGGGGGACCACGCTGACCGTCCTA | |
| EV-D68-150 | 49 | 중쇄 | CGGGTGCAGCTGCAGGAGTCGGCCCCAGGACTGGTGAGGCCTTCAGAGACCCTGTCCCTCACCTGCAGTGTCTCTGGTGGCCCCATCAGCAATGGTCCTTATTACTGGAGCTGGATCCGCCAGCACCCAGGGAAGGGCCTGGAGTGGATTGGATTCATCTTTTACAGTGGGAGCACCAACTACAACCCGTCCCTCCGGGGGCGCGTAACCATGGCAGTGGACACGTCTAAGAACCAGTTCTCCCTGAGGCTGAACTCTGTGACTGCCGCGGACACGGCCGTTTATTACTGTGCGAGACATGTGGTGACTGCGTCGGGGTGGTTCGACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 50 | 경쇄 | GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTGGCACCGACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGGTCCTCATCTATGATGCATTCAAGAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGAGTTCACTCTCACCATCAGCAGCCTCGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAGGTGGCCTCCCCCGTACACTTTTGGCCAGGGGACCAAGCTGGAGATCAAA | |
| EV-D68-151 | 51 | 중쇄 | CGGGTGCAGCTGCAGGAGTCGGCCCCAGGACTGGTGAGGCCTTCAGAGACCCTGTCCCTCACCTGCAGTGTCTCTGGTGGCCCCATCAGCAATGGTCCTTATTACTGGAGCTGGATCCGCCAGCACCCAGGGAAGGGCCTGGAGTGGATTGGATTCATCTTTTACAGTGGGAGCACCAACTACAACCCGTCCCTCCGGGGGCGCGTAACCATGGCAGTGGACACGTCTAAGAACCAGTTCTCCCTGAGGCTGAACTCTGTGACTGCCGCGGACACGGCCGTTTATTACTGTGCGAGACATGTGGTGACTGCGTCGGGGTGGTTCGACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 52 | 경쇄 | GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTGGCACCGACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGGTCCTCATCTATGATGCATTCAAGAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGAGTTCACTCTCACCATCAGCAGCCTCGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAGGTGGCCTCCCCCGTACACTTTTGGCCAGGGGACCAAGCTGGAGATCAAA | |
| EV-D68-152 | 53 | 중쇄 | CAGGTGCATCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGACTCTGGAGTCACCTTCAGTGACAATGCTTTGTACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTCGCAGTTATCTCATATGATGGAAGCAGTAGATACTACGCAGACTCCGTGAGGGGCCGGTTCACCATATCCAGAGACAATTCCAAGGACACGCTGTATCTGCAAATGAACAGACTGAGAGCTGAGGACACGGCTATTTATTACTGTGCGAGAGTCACAGCGGATTACTATGAGAGTAGTGGCAAGGTGTTTTGGGGCCAGGGAGCCCTGGTCGTCGTCTCCTCA |
| 54 | 경쇄 | GACATCCAGATGACCCAGTCTCCTTCCACCCTGTCTGCATCTGTGGGGGACAGAGTCTCCATCACTTGCCGGGCCAGTCAGAGTGTTAGGAGCTGGTTGGCCTGGTATCAGCACAAACCAGGGAAAGCCCCTAAACTCCTGATCTATAAGGCGTCTAGTTTAGAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAGCAGCCTGCAGGCTGATGATTTTGCAACTTATTACTGCCAACAGTATCAGACTTTTTCCTGGACGTTCGGCCAAGGGACCACGGTGGAAGTCAAA | |
| EV-D68-154 | 55 | 중쇄 | CAGGTGCAGCTGCAGGAGTCGGGCCCAGCACTGGTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGTGGCTCCATCAGTGATCACTACTGGAGCTGGATCCGGCAGCCCCCAGGGAAGGGACTGGAGTGGATTGGCTACATCTATACCAGTGGGACCACCAACTACAACCCCTCCCTCAAGAGTCGAGTCACCATATCAGTAGACACATCCAAGAAGCAGTTCTCCCTGAATCTGAGGTCTGTGACCGCCGCAGACACGGCCGTGTATTACTGTGCGAGAAGTCTAGAAACGGTGATCCGTTTCTACTACTACCACTACATGGACGTCTGGGGCAAAGGGACCACGGTGATCGTCTCATCA |
| 56 | 경쇄 | GATATTGTGATGACTCAGTCTCCACTCTCCCTGCCCGTCACCCCTGGAGAGCCGGCCTCCATCTCCTGCAGGTCTAGTCAGAGCCTCCTGCAGAGTGATGGGTACAGCTATTTGGATTGGTACCTGCAGAAGCCAGGGCAGTCTCCACAGCTCCTGATCTATTTGGGTTCTAATCGGGCCTCCGGGGTCCCTGACAGGTTCAGTGTCATTGGATCAGGCACATATTTTACACTGAAAATCAGCAGAGTGGAGGCTGAGGATGTTGGCGTTTATTTCTGCATGCAAGCTCTACAAACTCCGTGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA | |
| EV-D68-155 | 57 | 중쇄 | CAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCGCTGGCGGCTCCATCGGTGATTACCACTGGAACTGGATCCGGCAGCCCGCCGGGAAGGGGCTGGAGTGGATTGGGCGTATACATAGCAGTGGGAACACTGACTACAACCCCTCCCTCAAGAGTCGAGTCACCATGTCAGTAGACACGTCCAAGAACCAGTTCTCCCTGAAACTGAGGTCTGTGACCGCCGCGGACACGGCCGTGTATTACTGTGCGAGGCAAAATGTTTTTGATATCTGGGGCCAAGGGACAATGGTCACCGTCTCTTCA |
| 58 | 경쇄 | GACATCGTGATGACCCAGTCTCCAGACTCCCTGGCTCTGTCTCTGGGCGAGAGGGCCACCATCAACTGCAAGTCCAGCCAGAGTGTTTTATTCAGCTCCAACAATAAGAACTACTTAGCTTGGTACCAGCAGAAACCAGGACAGCCTCCTAAGTTGCTCATTTACTGGGCATCTACCCGGGAATCCGGGGTCCCTGACCGATTCAGTGGCAGCGGGTCTGAGACAGATTTCACTCTCACCATCAGCAGCCTGCAGGCTGAAGATGTGGCAGTTTATTTCTGTCAGCAATTTTATACTACTCCGCTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA | |
| EV-D68-156 | 59 | 중쇄 | CAGGTACAGATGCAGGAGTCGGGCCCAGGGCTGGTGAAGGCTTCGGAGACCCTGTCCCTCACCTGCAGTGTCTCTGGTATCTCCATCAATAACTACTATTGGAGTTGGTTCCGCCAGCCCCCCGGGAAGGGCCTGGAGTGGATCGGATATGTCTATTCTACTGGGAGTTCCAAGTACAATCCCTCCCTCGAGCGTCGAGCCACCATGTCAGTAGACACGTCCAACAACAACTTCTCCCTGAGGCTGACGTCTGTGACCACTGCGGACACGGCCGTCTACTACTGTGCGCGGGGGAGTATGCCGCATATCTGGGGCCAGGGCCTCCTGGTCACCGTCTCCTCA |
| 60 | 경쇄 | CAGTCTGTGCTGACTCAGCCACCCTCAGCGTCCGGGACCCCCGGACAGAGGGTCACCATCTCCTGTTCTGGTAGCACCTCCAACATCGAGACTAATTATGTATACTGGTACCAGCAGGTCCCAGGAACGGCCCCCAAGCCCCTCGTCTATAGGAATGATCAGCGCCCCTCGGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGCACCTCGGCCTCCCTGGTCATCAGTGGGCTCCGGACCGAGGATGAGGCTGCTTATTATTGTGCAGCTTGGGATGACAGTCTGAAAGCTCCGGTCTTCGGAGCTGGGACCAAGGTCGCCGTCCTC | |
| EV-D68-157 | 61 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGATAAAGCCGGGGGGGTCCCTTAGACTCTCCTGTGCAGCCTCTGGAATCACTTTCAGTAACGCCTGGATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAATGGGTTGGCCGTATTGAAAGCAAAATTGACGGTGGGACAATAGACTACGCTACACCCGTGAAAGGCAGATTCACCATCTCAAGAGATGATTCAAAAAACACGCTGTACCTGCAAATGAACAGCCTGAAAACCGAGGACACAGCCGTCTATTACTGTACCACAGACCAGGGCTACTATGATAGAAGTGGTTATTGGGTCGTCGGGAACCACTTTGACTACTGGGGCCAGGGAATCCTGGTCACCGTCTCCTCA |
| 62 | 경쇄 | CAGTCTGTGCTGACGCAGCCGCCCTCAGTGTCTGGGGCCCCAGGGCTGAGGGTCACCATCTCCTGTACTGGGAGCAGCACCAACATCGGGGCAGGTTATGATGTACACTGGTACCAGCACCTTCCAGGAACAGCCCCCAAACTCCTCATCTATGGTAACAGCAATCGGCCCTCAGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGCACCTCAGCCTCCCTGGCCATCACTGGGCTCCAGGCTGACGATGCGGCTGATTATTACTGCCAGTCCTATGACAGAAGCCTGAGTACTTATGTCTTCGGAACTGGGACCAAGGTCACCGTCCTA | |
| EV-D68-158 | 63 | 중쇄 | GAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGGTATTAGTGGTACTACAGGTAGCACATACTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGGTGCATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGATTCTCACTCCATGATAGTAGTTGATCATGCTTTTGATATCTGGGGCCAAGGGACAATGGTCACCGTCTCTTCA |
| 64 | 경쇄 | TCCTATGTGCTGACTCAGCCACCCTCGGTGTCAGTGGCCCCAGGACAGACGGCCAGGATTACCTGTGGGGGAAACAATATTGGAACTAAAAGTGTGCACTGGTACCAGCAGAGGCCAGGCCAGGCCCCTGTGCTGGTCGTCTATGATGATAGCGACCGGCCCTCAGGGATCCCTGAGCGATTCTCTGGCTCCAACTCTGGGAACACGGCCACCCTGACCATCAGCAGGGTCGAAGCCGGGGATGAGGCCGACTATTACTGTCAGGTGTGGGATAGTTATAATGTTCATTATGTCTTCGGAACAGGGACCAAGGTCACCGTCCTA | |
| EV-D68-159 | 65 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGGGGCCTGGTCAAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTACTTATATCATGACCTGGGTCCGCCAGGCCCCAGGGAGGGGGCTGGAGTGGGTCTCATCCATTAGTACCAGTAGTGTTTACACATTCTATGCAGATTCACTGAAGGGCCGGTTCACCATCTCCAGAGACAACGCCAAGAATTCAGTGTATCTGCAGATGAACAGCCTGAGAGCCGACGACACGGCTGTTTATTACTGTGCGAGGGAAGAAGGGTTTCGAGCTTATAACCTATACTGGGGCCAGGGAACCCTGGTCACTGTCTCCTCA |
| 66 | 경쇄 | CAGTCTGTGCTGACTCAGCCACCGTCAGCGTCTGGGACCCCCGGGCAGAGGGTCACCATCTCTTGTTCTGGAAGCAGCTCCAACATCGAATACAATTATGTTTACTGGTACCAGAAATTCCCAGGAACGGCCCCCAAACTCCTCATCTATAAAAATAATCAGCGGCCCTCAGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGCACCTCAGCCTCCCTGGCCATTAGTGGGCTCCGGTCCGAGGATGAGGGTGATTATTACTGTGCAGCATGGGATGACATCCTGAGTGGTGTGGTTTTCGGCGGGGGGACCAAGCTGACCGTCCTC | |
| EV-D68-160 | 67 | 중쇄 | CAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCAGAAACTCTCCTGTGCAGCCTCTGGATTCACGTTCAGTAGGTTTGGTATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATTTCGTTTGATGGAAGTAATAGATACTACGCAGACTCCGTGAAGGGGCGATTCACCATCACCAGAGACAATTCCAAGAACACATTGTATCTGCAAATGAACAACCTGAGACCTGAGGACACGGCTGTATATTACTGCGCGAGAGATTGGGATAGGCTGGTTCGTTCGGCGGTTGGCTACTGGGGCCAGGGAACCCTGGTCAGCGTCTCCTCA |
| 68 | 경쇄 | CAGTCTGCCCTGACTCAGCCTCGCTCAGTGTCCGGGTCTCCTGGACAGTCAGTCACCATCTCCTGCACTGGAACCAGTAATGATGTTGGTGGTTATAACTTTGTCTCCTGGTATCAGCAACACCCAGGCAAAGCCCCCAAACTAATGATTTTTGATGTCATTAGGCGGCCCTCAGGGGTCCCTGGTCGCTTCTCTGGCTCCAAGTCTGGCGACACGGCCTCCCTTATCATCTCTGGACTCCAGGCTGAGGATGAGGCTGATTATTACTGCTGCTCATATGCAGGCACCTACACCTGGGTATTCGGCGCAGGGACCACACTGACCGTCCTA | |
| EV-D68-161 | 69 | 중쇄 | CAGGTGCACCTGCAGGAGTCGGGCCCACGACTGGTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGTGGCTCCGTCAGCACTGCCACTTACTACTGGAGCTGGATCCGGCAGTCCCCAGGGAGGGGACTGGAGTGGATAGGATATATCTATTCCAGTGGTAACACCAACTACAACCCCTCCCTTAAGAGTCGAGTCACCATTTCTTTAGACACGCCCAACAACCAGCTCTCCCTGACGTTGACCTCTGTGACCGCTGCGGACACGGCCATTTATTATTGTGAGAGGCGCTTACGTATTCTGAGTATTGAGAGGAACTACTACGCTATGGACGTCTGGGGCCAAGGGACCCCGGTCACCGTCTCCTCA |
| 70 | 경쇄 | GAAGTTGTGTTGACGCAGACTCCAGGCACCCTGTCTTTGTCTCCGGGGGAAGGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGGGTTGTCAACAACTACTTAGCCTGGTACCAGCAGAGAGCTGGCCAGGCTCCCAGGCTCCTCATTTTTGGTGCATCCAACAGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGGAAGCTGGAGCCCGAAGATTTTGCAGTGTATTACTGTCAACAATATGGTAGCCCGTGGACGTTCGGCCACGGGACCAAGGTGGAAATGAAA | |
| EV-D68-162 | 71 | 중쇄 | CAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTAGTTATGCTATGCATTGGGTCCGCCAGGCTCCAGGCAGGGGGCTGGAGTGGGTGGCAGTTATATCATATGATGCAAGCAAGAAATACCACGCAGACTCCGTGAAGGGCCGATTCACAATCTCCAGAGACAGTTCCAAGAACACGCTGTTTCTGCAAATGAATAGCCTGAAACCTGAGGACACGGCTGTGTATTACTGTGCGAGAGATCATGTCCCCCCCAAGGATTGCAGTGATGGTAATTGCCACTCGGACTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA |
| 72 | 경쇄 | GACATCCAGATGACCCAGTCGCCTTCCACTATGTCTGCTTCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAAGTGGTTGGCCTGGTATCAGCAGAAGCCAGGGAAAGCCCCTAAACTCCTGATCTATAAGGCGTCTACCTTACAAACTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAACAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAACAGCATAATAGTTATTCGTACACTTTTGGCCAGGGGACCAAGGTGGAGATCAAG | |
| EV-D68-163 | 73 | 중쇄 | CAGGTCCAGCTTGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGATTTCTTGCAAGGCTTCTGGATACTCCTTCACTAACTTTGCTGTGCATTGGGTGCGCCAGGCCCCCGGACAAAGACTTGAGTGGATGGGATGGATCAACCCTGGCAATAGAAACACAAAGTATTCACACAACTTTCAGGGCAGAGTCACCATTACCAGGGACACATCCGCGAACACAGCCTACATGGAACTGAGCAGCCTGAGATCTCAAGACACGGCTGTGTATTACTGTGCGAGACTTCCGATAGCAGCAGCTGGCAGGGGCTGGTTCGACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCC |
| 74 | 경쇄 | GAAATTGTGTTGACGCAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTATCAGCACCTACTTAGCCTGGTACCAGCAGAGACCTGGCCAGGCTCCCAGGGTCCTCATCTATGATGTATCCACCAGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAGCCTGAAGATTTTGCAGTCTATTTCTGTCACCAGTATGGTAGTTCACCGGCGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA | |
| EV-D68-164 | 75 | 중쇄 | GAAGTGCAGCTGGTGGAGTCTGGGGGAGACTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAATACTTATGGCATGAACTGGGTCCGCCAGGCTCCAGGGAAGGGACTGGAGTGGGTTTCATACATTAGTAGTGCCACCACTACCTTCTACTACGCAGACTCTGTGAGGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAATTCACTATTTCTGCACATGAAGAGCCTGAGAGACGAAGACACGGCAGTTTATTACTGTGCGAGAGTCTATACTATGCTTCGCGGAGCGAGTATGGACGTCTGGGGCCACGGGACCACGGTCACCGTCTCCTCA |
| 76 | 경쇄 | GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTGGCACCTACCTGGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATTCAGCCAACAGGGCCACTGGCATCCCAGCCCGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCTGCGCATCACCTGGCCTCCTATATTCACTTTCGGCCCTGGGACCAAAGTGGATGTCAAA | |
| EV-D68-165 | 77 | 중쇄 | CAGGTGCACCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGTCTGGGGCCTCAGTGAAGGTCTCCTGCAAGACTTTTGGATACACCTTCACCGCCTACTATATGCACTGGGTGCGACAGGCCCCTGGACAGGGGCCTGAGTGGATGGGATGGATCAACCCTATCAGTGGTGGCACAAACTATGCACAGAAGTTTCAGGGCAGGGTCTCCATGACCAGGGACACGTCCATCAGCACAGCCTACATGGGCCTGAGCAGGCTGAGACCTGACGACACGGCCGTCTATTACTGTGCGAGAGTGAAGTGTAGTAGTGCCAACTGCTATGGGAACTTTGACTACTGGGGCCAGGGTACCCTGGTCACCGTCTCCTCA |
| 78 | 경쇄 | CAGTCTGCCCTGACTCAGCCTCCCTCCGCGTCCGGGTCTCCTGGAGAGTCAGTCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGGTGGTTATAACTATGTCTCCTGGTACCAACAACACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGGTCAGTAAGCGGCCCTCAGGGGTCCCTGATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCGTCTCTGGGCTCCAGGCTGAGGATGAGGCTGATTATTACTGCAGCTCATATGCAGGCAGCAACAATTTGGTATTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-166 | 79 | 중쇄 | GAGGTGCAGCTGGTGCAGTCTGGAGCAGAGGTGAAAAAGCCCGGTGAGTCTCTGAAGATCTCCTGTAAGGGTTCTGGATACAGGTTTACCAACTACCGGATCGGCTGGGTGCGCCAGATGCCCGGGAAAGGCCTGGAGTGGATGGGGATCATCTATCCCGGTGGCTCTGATACCAGATACAGTCCGTCCCTCCAAGGCCAGGTCACCATGTCAGTCGACAAGTCCATCAGCACCGCCTACCTGATGTGGAGCAGCCTGAAGGCCTCGGACACCGCCATGTATTACTGTGCTCGACAGACCACTCAAAATAGTGGCTACGATAGATGGTTTGACTCCTGGGGCCAGGGAACCCACGTCACCGTCTCCTCA |
| 80 | 경쇄 | CAGTCTGTACTGACTCAGCCACCCTCAGCGTCTGGGACCCCCGGGCAGAGGGTCACCATCTCTTGTTCTGGAAGCACCTCCAGCATCGGAAGTAATATTGTAAATTGGTACCAACACCTCCCAGGAACGGCCCCCAAACTCCTCATCTATATTAATAATCAGCGGCCCTCAGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGCACCTCAGCCTCCCTGGCCATCAGTGGGCTCCAGTCTGAGGATGAGGCTGACTATTACTGTGCAGCATGGGATGACAGCCTGAATGGTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTG | |
| EV-D68-181 | 81 | 중쇄 | GAGGTGCAGGTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGAATCACCTTTAGCAGGCATACTATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGGAGTGGTGGTAGCACATATCATGCAGACTCCGTGAAGGGCCGCTTCACCATCTCCAGAGACAGTTCCAAGAGCACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTTTATTACTGTGCGATCTCCGTGCCATTATTACGATTTTTGGAGTGGTTTCAACACCCTTTTGACTTCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 82 | 경쇄 | GAAATAGTGATGACGCAGTCTCCAGCCTCCCTGTCTGTGTCTCCAGGGGAAAGAGTCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTGGCAGCACCTTAGCCTGGTACCAGCACAAACCTGGCCAGGCTCCCAGGCTCCTCATCTCTGGTGCATCCACCAGGGCCACTGGTGTCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGAGTTCACTCTCACCATCAGCAGTCTGCAGTCTGAAGATTTTGCAGTTTACTACTGTCACCAGTATATTAACTGGCCTCCGTGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA | |
| EV-D68-183 | 83 | 중쇄 | GAGGTGCGGCTGGTGGAGTCTGGGGGAGGCCTGGTCAAACCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAATACATATTCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCGCCTCCATTAGTAGTACCGGAAGTTACATATACAATGCAGACTCACTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTGTTTCTGCAAATGAACAGCCTGAGAGTCGAAGACACGGCTGTGTATTACTGTGTGAGATTCACCATGACTACAGTGACTAACTTTGACTCATGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 84 | 경쇄 | CAGTCTGCCCTGACTCAGCCTCGCTCAGTGTCCGGGTCTCCTGGACAGTCAGTCACCATCTCCTGCACTGGAACCAGCAGTGATGTTGGTGCTTATAGCTATGTCTCCTGGTACCAACAACACCCAGGCAAAGCCCCCAAACTCATGATTTATGATGTCTACAGGCGGCCCTCAGGGGTCCCTGGTCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCCCTGACCGTCTCTGGGCTCCAGGCTGAGGATGAGGCTGATTACTACTGCTGCTCACATGCAGGCAGTCACACCTGGGTGTTCGGCGGAGGGACCAAGGTGACCGTCCTA | |
| EV-D68-185 | 85 | 중쇄 | CAGCTGCAGGTGGTGGCGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTAAAGCCTCTGGATTCACGTTCACCAATTATGGCATGCACTGGGTTCGCCAGGCGCCAGGCAAGGGGCTGGAGTGGGTGGCTTTTATATCATACGATGGAGGTAATAAATTTTATGCAGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAGGAACACGGTTTATCTGCAAATGAACAGCCTGAGAGTGGCGGACACGGCTATGTATTACTGTCCGAAGGTCATCCCCCACCCGTATTATGATAGTAGTGGTGATGATGCTTTTGATATCTGGGGCCAAGGGACAATGGTCGCCATTTCTTCA |
| 86 | 경쇄 | GAAATAGTGATGACGCAGTCTCCAGCCACCCTGTCTGTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCTGGGCCAGTCAGAGTATTAGCCGCAACTTAGCCTGGTATCAGCAAAAACCTGGCCAGGCTCCCCGACTCCTCATCTATGGTGCATCCACCAGGGCCACTGGTATCCCCGCCAAGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCGTCAGCAGCCTGCAGTCTGAAGACCTTGCAGTTTATTACTGTCAGCAGTATAGTAAGTTGCCTATCACCTTCGGCCAAGGGACACGACTGGAGATTAAA | |
| EV-D68-200 | 87 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCCCCTTCAGTAGTTATAGCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCATACATAAGTGGTAGTGGTGGTGACATATACTACGCAGACTCTGTGAAGGGCCGATTCACCATCTCCAGGGACAATGCCAGGAACTCACTGTCTCTGCAAATGAACAGCCTGAGAGCCGACGACACGGCTGTGTATTACTGTGCGAGAGGGCTGGTAGCAACAACTGGTACAAGGTACTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 88 | 경쇄 | GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGGAGTTACTTAGCCTGGTACCAACAGAGACCTGGCCAGGCTCCCAGGCTCCTCATCTACGATGCATCCAACAGGGCCACTGGCATCCCAGTCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCTACTGGCCTCCGTTCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA | |
| EV-D68-208 | 89 | 중쇄 | CAGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCGGGGTCCTCGGTGAAGGTCTCCTGCAAGGCTTCTGGAGGCACCTTCAGGAGGTTTGCTATCAGCTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGATGGGAGGGATCATCCCTATCCTAGGTAGAGGAAAGTACGCACAGAAGTTCCAGGGCAGAGTCAGGATTACCGCGGACGAATCCACGAGCACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAAGACACGGCCGTGTATTACTGTGCGAGATTTATTTCGACAGCCTCCTATGTTCCGGGGACCTTCGAGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCA |
| 90 | 경쇄 | GACATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCGCTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGAGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATGTTGCGGTTTATTACTGTCAGCAGCGTAGCGACTGGCCTCCGGGGACTTTTGGCCAGGGGACCAACGTGGAGATCAAA | |
| EV-D68-210 | 91 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCCTGGTCAAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGGAACTATAACATCAACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCATCCATTAGTAGTACTGGTAGTTACATACACTACGCAGATTTAGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCACTGTATCTGCAAATGAACAGCCTGAGAGTCGAGGACACGGCTGTATATTACTGTGCGCGAATGGTTAGGAATACGGTGACTGCCTTTGACTACTGGGGCCAGGGAACCCTGGTCTCCGTCTCCTCA |
| 92 | 경쇄 | CAGTCTGCCCTGACTCAGCCTGCCTCCGTGTCTGGGTCTCCTGGCCAGTCGATCACCATCTCCTGCACTGGAACCAGCAGTGATGTTGGTGGCTATAACTTTGTCTCCTGGTACCAACAACAGCCAGGCAGAGCCCCCAAACTCCTTATTTATGAAGTCATTAAGCGGCCCTCAGGGGTTTCTGATCGCTTCTCTGGCTCCAAGTCTGGCGACACGGCCTCCCTGACAATCTCTGGGCTCCAGGCTGAGGACGAGGCTGATTATTACTGCTGCTCATATGGGGGTAACAACTCTTGGATGTTCGGCGGAGGGACCATGCTGACCGTCCTA | |
| EV-D68-219 | 93 | 중쇄 | CAGGTCCAGGTGGTACAGTCTGGGGCTGAGATGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGATACAGGCTCATTGATTTACCCTTGCACTGGGTGCGACAGGCTCCTGGAAAAGGGCTTGAGTGGATGGGACTTTTTGATCCTGAAAAGGCTGAAGCCATCTACTCACAGAAATTCCAGGACAAGGTCACCATAAGCGAGGACACATCTATCGACACAGCCTACATGGAACTGAACAGCCTGCGCTCTGAAGACACGGCCGTCTACTACTGTGCAACTTGGGGAGTTGAGGTGGTGAATGGGAGAAGGGACTACTTTGACTCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 94 | 경쇄 | TCCTATGAGCTGACACAGCCACCCTCGGTGTCAGTGTCCCCAGGACAGACGGCCAGGATCACATGCTATGCAGATGTGTTGTCAAACCAATATACTTACTGGTATCAACAGAAGCCAGGCCAGGCCCCTGTGTTGGTGATATATAAAGACACTGAGAGGCCCTCAGGGATCCCTGAGCGATTTGCTGGCTCCAGCTCAGGGACAACAGTCACCTTAGTCATCAATGGAGTCCGGGCAGAGGACGAGGCTTACTATTACTGTCAATCAGCCGACAACACCAGAATTACGGTTTTCGGCGGAGGGACCAAGCTGTCCGTCCTA | |
| EV-D68-220 | 95 | 중쇄 | GGGGTGCAGCTGGTGGAGTCTGGGGGAGGATTGATACAGCCTGGAGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTAGTTTTGAAATGAACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTTTCATACATTAGTACTAGTGGTAGTACCATATACTACGCAGACTCTGTCAAGGGCCGATTCACCATCTCCAGAGACAACGCCAGGAACTCACTGTCTCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTTTATTACTGTGCGAGAGACGTGAGGGATTGTAGTGCTCTTACGTGCCCCCGAAGGGGAGATGCTTTTGATTTCTGGGGCCGTGGGACAAGGGTCACCGTCTCTTCA |
| 96 | 경쇄 | GATGTTGTGATGACTCAGTCTCCACTCTCCCTGCCCGTCACCCTTGGACAGTCGGCCTCCATCTCCTGCAGGTCTAGTCAAAGCCTCGTATACAGTGATGGAAACACCTACTTGAATTGGTTTCAGCAGAGGCCAGGCCAATCTCCAAGGCGCCTAATTTATAAGGTTTCTAACCGGGACTCTGGGGTCCCAGACAGATTCAGCGGCAGTGGGTCAGGCACTGATTTCACACTGAAAATCAGCAGGGTGGAGGCTGAGGATGTTGGGGTTTATTACTGCATGCAAGGTACACACTGGCCTCGCACTTTCGGCCCTGGGACCAAAGTGGATATCAAA | |
| EV-D68-221 | 97 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCCTGGTCAAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCAGTTTCAGTGTCTATCCCATGAACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCATCCATAAGTAGTAGTAGTCGTTACATATCCTACGCAGACTCACTGAGGGGTCGAATCACCATCTCCAGAGACAACGCCAAGAACTCACTGTATCTGCAAATGAACAGCCTGAGAGTCGAGGACACGGCTGTGTATTACTGTGTGAAAGTCGGGGGTAGCAAACACCAATACTACTTTGACTACTGGGGCCAGGGATCCCTGGTCACCGTCTCCTCA |
| 98 | 경쇄 | CAGTCTGTGCTGACTCAGCCACCCTCAACGTCTGGGATCCCCGGGCAGACGGTCACCATCTCTTGTTCTGGAAGCAGGTCCAACATCGGAAGTTATACTGTTAACTGGTACGAGCAACTCCCAGGAACGGCCCCCAAACTCGTCATCTTTAATAATAATCAGCGTCCCTCAGGGGTCCCTGACCGATTCTCTGGCTCCAAGTCTGGCACCTCAGCCTCCCTGGCCATCAGTGGGCTCCAGTCTGAGGGTGAAGCTGATTATTACTGTGCAGCATGGGATGACAGCCTGAATGGTGTGGTTTTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-224 | 99 | 중쇄 | CAGGTTCAGTTGGTGCAGTCTGGAGCTGAGGTGAAGAGGCCTGGGGCCTCAGTGAAGGTCTTCTGCAAGGCTTCTGGTTACACCTTTACGAATTATGACATCATCTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGGTGGGCTGGATCAGCACTTACAATGGTAACACAAACTATGAACAGAACCTCCAGGGCAGAGTCACCATGACCACAGACACATCCACGAGCACAGCCTACATGGAGCTGAGGAGCCTGAGATCTGACGACACGGCCGTTTACTATTGTGCGAGAGAGCGTTGTAGTACTAGTACCTGCTATAGTCGTTATGCTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 100 | 경쇄 | GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTGGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAATATCTATTTGAATTGGTATCAGCAAAAACCGGGGAAAGCCCCTAAGGTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCCGTGGCAGTGGGTCTGGGACAGATTTCATCCTCACCATCAGCAGTCTGCAACCTGAAGACTTTGCAACCTACTACTGTCAACAGAGTTACAGGTCCCCTCGGACGTTCGGCCAAGGGACCGAGGTGGAAATCAAA | |
| EV-D68-225 | 101 | 중쇄 | GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGCACAGCCTGGAGAGTCCATGAGACTCTCCTGTGTAGCCTCTGGATTCACCTTAAGTCGTTATGAAGTGAACTGGGTCCGCCAGGCTCCAGGGAAGGGGCTAGAGTGGCTTTCATACATTAGCAGTGGTGGTCCTTCCATATACTACGCAGACTCTGTGAAGGGCCGATTCACGATCTCCAGAAACAGCGCCGAGAACTCACTGGAACTACAAATGTCCACCCTGAGGACCGAGGACACGGCTGTTTATTATTGTATGAGAGAGGGTCTTACTTATTATGATAGTACTATTTGGGGCCAGGGAACCCTGGTCGCCGTCTCCTCA |
| 102 | 경쇄 | CAGAATGTGCTGACTCAATCGCCCTCTGCCTCTGCCTCCCTGGGAGCCTCGGTCAAACTCACCTGCACTCTGAACAGTGGGCACAGCAGATACGCCATCGCATGGCATCAACATCAGCCACAGAGGGGCCCTCGGTTCCTGATGAAGATTAATAGTGATGGCAGACACATCAGGGGGGACGGCATCTCTGATCGCTTCTCAGGCTCCGCCTCTGGGGCTGAGCGTCATCTCACCATCTCCAGCCTCCAGCCTGAGGATGAGGCTGACTATTATTGTCAGACCTGGGGCACTGGCTTTCGGGTGTTCGGCGGAGGGACCAAACTGACCGTCCTA | |
| EV-D68-227 | 103 | 중쇄 | GAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGAATATGCCATGCACTGGGTCCGGCAAGTTCCAGGGAAGGGCCTGGAGTGGGTCTCAGGTATTAGTTGGAATGGTGGTAGCAAAGGCTATGCGGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGATAACGCCAGGTATTCCCTGTCTCTGCAAATGAACAGTCTGAGAACTGAGGACACGGCCTTATATTACTGCGCAAAAGATGATTACGAGGGGGCTGGTTTTGATATCTGGGGCCAAGGGACAGTGGTCACCGTCTCTTCA |
| 104 | 경쇄 | GAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGGCTGGTACCAACAGAAATCTGGCCAGGCTCCCAGGCTCCTCATCTATGGTGCATCCAGCAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAATTTATTACTGTCAGCAGCGTAGCAACTGGCCTATCACCTTCGGCCAAGGGACACGACTGGAGATTAAA | |
| EV-D68-228 | 105 | 중쇄 | CAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGCTATTTAATTAGCAATGGTTACTACTGGGGCTGGATCCGGCAGTCCCCCGGGAAGGGGCTGGAGTGGATTGGGAGTATCTATTATACTAGGGACACCTACTACAACTGGTCCCTCAAGAGTCGAATCACCATATCAGTGGACACGTCCAAGAAACAGTTCTCCCTGAAGTTGTATTCTGTGACCGCCGCAGACACGGCCGTCTATTACTGTGTGAGACATGAGGGTTCTTGCAATGATGGAAGCTGTTACGGCTCGTTCGTTGACAACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 106 | 경쇄 | GACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGCATCTGTAGGAGACAGAGTGACCCTCACTTGTCGGGCGAGTCAGGATATTAGCAGCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACACATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTCGCAACTTACTTTTGTCAACAGGCTGACAGTTTCATCACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA | |
| EV-D68-231 | 107 | 중쇄 | GAGGTGCAACTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGATTTCACCTTCAGCAGCTACACCATGGCCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTTGAATGGGTCTCATCTATTAGTGGTGATGGTGTTAGCACAAAAAACGCAGACTCCGTGAAGGGCCGATTCTCCGTCTCCAGAGACAATTCCAAGAACACACTTTTTCTGCAACTGAACAGTCTGAGAGCCGAGGACACGGCCTTTTATTACTGTGCGAGGGGGGGGACCTTCCATAACTGGTACTTCGATCTCTGGGGCCGTGGCGTCTTGGTCACTGTCTCCTCA |
| 108 | 경쇄 | GAAATAGTGATGACGCAGTCTCCAGCCACCCTGTCTGTGTCTCCAGGGGAAACAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTATTGGCGACAACTTAGCCTGGTATCAGCAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTCTGGTGCATCCACAAGGGCCACTGATTTCCCAGCCAGGTTCCGTGGCAGTGGGTCTGGGACAGAGTTCACTCTCACCATCAGCAGCCTGCAGTCTGAAGACTTTGCAGTTTATTACTGTCAGCAGTATAAAAACTGGCCTCGGACGTTCGGCCGAGGGACCAAGGTGGAAGTCAGA | |
| EV-D68-234 | 109 | 중쇄 | GAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCGGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCACCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCCGGAGTGGGTCTCAGGTATTAGTGGTAGTGGTGGTAGCACAAACTACGCAGACTCTGCGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTATTGTGCGAAAGGGACCATTACTTACTCCTACTACTACATGGCCGTCTGGGGCAAAGGGACCACGGTCACCGTCTCCTCA |
| 110 | 경쇄 | GAAATTGTGTTGACGCAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGGCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGGAGCAGCTACTTAGCCTGGTACCAGCAGAGACCTGGCCAGGCTCCCAGGCTCCTCATCTATGGTGCATCCAGCAGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACGCTCACCATCGGCAGACTGGAGCCTGAAGATTTTGCAGTGTATTACTGTCAGCAGTATGGTACCTCAATAACTTTCGGCGGAGGGACCAAGGTGGAGATCAAA | |
| EV-D68-235 | 111 | 중쇄 | CAGGTCCAGGTGGTACAATCTGGGGCTGAGATGAGGAAGCCTGGGGCCTCAGTGAGGGTCTCCTGCAAGGTTTCCGGATACAGGCTCACTGATTTACCCTTGCACTGGGTGCGACAGGCTCCTGGAAAAGGGCTTGAATGGATTGGATTTGTTGATCTTGAAAAGCGCGAAATCATCTACGCACAGAAATTTCAGGGCAAAGTCACCATAACCGAGGACACATCTGCAGACACCGCCTACATGGAACTGAACAGCCTGCGATCTGAAGACACGGCCGTCTACTACTGTGCAACTTGGGGAATTGAGGTGGTGAATGGGAGGGACGAATTCTTTGACTCCTGGGGCCAGGGAACCCTGGTCTCCGTCTCCTCA |
| 112 | 경쇄 | TCCTATGAGCTGACACAGCCACCCTCGGTGTCAGTGTCCCCAGGACAGACGGCCAGGATCACATGCTATGCAGATGTGTTGTCAAAGCAATATACTTACTGGTATCAGCAGAAGCCAGGCCAGGCCCCTGTGTTGGTGATATATAAAGACACTGAGAGGCCCTCAGGGATCCCTGAGCGATTTGCTGGCTCCAGCTCAGGGACAACAGTCACCTTGATCATCAATGGAGTCCGGACAGAGGACGAGGCGTACTATTACTGTCAATCAGCCGACACCAGAATTACGGTTTTCGGCGGAGGGACCAAGCTGTCCGTCCTA | |
| EV-D68-236 | 113 | 중쇄 | CAGGTCCAGGTGGTCCAGTCTGGGGCTGAGATGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGATATAGTCTCAGTGATTTACCCTTGCACTGGGTGCGACAGGCTCCTGGAAAAGGGCTTGAGTGGATGGGACTTTTTGATCCTATAAACGGTGAAATCATCTACGCACAGACATTCCAGGGCAAAGTCACCATAAGCGAGGACACATCGATAGACACAGCCTACATGGAACTCAACAGCCTGCGATCTGAAGACACGGCCGTGTACTATTGTGCAACTTGGGGAGTTGCGGTGGTGAGTGGGAGAAGGGACTACTTTGACTCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 114 | 경쇄 | TCCTATGAGCTGACACAGCCACCCTCGGTGTCAGTGTCCCCAGGACAGACGGCCAGGATCACATGCTATGCAGCTGTATTGTCAAACCAATATACTTACTGGTATCAACAGAAGCCAGGCCAGGCCCCTGTTTTGGTGATATATAAAGACACTGAGAGGCCCTCAGGGATCCCTGAGCGATTTGCTGGCTCCAGCTCAGGGACAACAGTCACCTTGATCATCAATGGAGTCCGGACAGAGGACGAGGCTTACTATTACTGTCAAACAGCCGACACCAAATATACGGTTTTCGGCGGAGGGACCAAGCTGTCCGTCCTA | |
| EV-D68-241 | 115 | 중쇄 | CAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCACAGACCCTGTCCCTCACCTGCACTGTCTCTGGTGGCTCCATCACCAGTGGTGATTACTACTGGAATTGGATCCGCCAGCCCCCAGGGAAGGGCCTGGAGTGGATTGGGTACATCTATCACAGTGGGACCACCTACTACAACCCGTCCCTCAAGAGTCGAGTTACCATATCAGTAGACACGTCCAAGAACAGGTTCTCCCTGAAGTTGTCCTCTGTGACTGCCGCAGACACGGCCGTGTATTTTTGTGCCAGAGCCTACGCTTATGAATTTTGGAGCGGTTACCCTAACTGGTTCGACCCCTGGGGCCTGGGAACCCTGGTCACCGTCTCATCA |
| 116 | 경쇄 | GACATCCAGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTTGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGCGCATTAGTACCTATGTAAATTGGTATCAGGTGAAAGCAGGGACAGCCCCTAAGGTCCTGATCTATGCTGCGTCCAGTTTGCAAACTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATTGTCAGTCTACAACCTGAAGATTTTACAACCTACTTCTGTCAACAGAGTTACAGTCCCCCGTGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA | |
| EV-D68-247 | 117 | 중쇄 | CAGGTGCAGTTGGTGGAGTCTGGGGGAGGCTTGGTCCAGCCTGGTAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGGTTCATCTTCAATAGATATGCCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTCGAGTGGGTGGCTCTTATATCATATGATGGAATTAATAAATATTACGCAGACTCCGTGAAGGGCCGATTCTCCATCTCCAGAGACAATTCCAAGAGTACGCTGTATCTGCAAATGAACAGCCTCAGAGCTGAGGACACGGCTATCTTTTACTGTGCGAGAGGACTAGGATATTGTAGTGGTACCGGTGGTAGCTGTACACCCTTTGAATATTGGGGCCAGGGAATCCTGGTCACCGTCTCCTCA |
| 118 | 경쇄 | GACATCCAGATGACCCAGTCTCCACCCTCCCTGTCTGCATCTGTTGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGCATTAAGAAATATTTAAATTGGTATCAGCAGAAACCAGGGAATGCCCCTAAGCTCCTCATCTATGGCGCATCCAATTTGCAAACTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCGCTCTCTCCATCAGCAGTCTGGAAACTGAAGATTTTGCAACTTACTACTGTCAACAGAGTGACAGTGCCCCTCCCACTTTCGGCGGAGGGACCAAGGTGGAGTTCAAA | |
| EV-D68-254 | 119 | 중쇄 | CAGGTGCAGCTGCAACAGCGGGGCGCAGGGCTGTTGAAGCCCTCGGAGACCCTGTCCCTTACCTGCGAAATCTATGGTGCATCCCTCAATGATTACGACTGGACCTGGATCCGCCAGCCCCCAGGGAAGAGGCTGGAGTGGATTGGGGTCATCAATCGTCGTGACACTGTTGACTACAACCCGTCCCTCAAGAGTCGGGTCACCCTCTCACTTGACACGTCCAAGAACCAACTTTCCCTGAGTCTGAGTTCTGTGACCGCCGCGGACACGGCTGTTTATTACTGTGTGAGAGTCCCACGTCGGGGCTTTGAAGGGTCTTTCGGATTTTGTGATGATACTGCCTGCCGCTACGGGCATACCTGGTTCGACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 120 | 경쇄 | GACATTCAGATGACCCAGTCTCCTTCCTCCCTGTCTGCATCTGTTGGCGACAGAGTCACCATCACTTGCCGGGCAAGTCAGAGTATTAGAGATTATTTAAATTGGTATCAACAAAGACCAGGGAAAGCCCCTAAAGTCCTGATCTTTGCTGGTTCCCGTTTGGAAAGTGGGGTCCCATCGAGGTTTAGAGGCCGTGGATCTGGGACAGAATTCACTCTCACCATCAGCGATCTGCAACCTGAGGATTTTGCAACTTACTACTGTCAACAGAGTTACCTTACACCTCCGACATTCGGCCAAGGGACCACCGTCGATATCAAA | |
| EV-D68-260 | 121 | 중쇄 | CAGGTCACCTTGAAGGAGTCTGGTCCTGTGCTGGTGAAACCCACAGAGACCCTCACGCTGACCTGCACCGTCTCTGGATTCTCACTCCGCAATGGTAGAATGGGTGTGAGCTGGATCCGTCAGCCCCCAGGGAAGGCCCTGGAGTGGCTTGCACACATTTTTGCGAGTGACGAAAAATCTTACAGTACATCTCAGAGGACCAGGCTCTCCATCTCCAGGGACACCTCCAAAAGCCAAGTGGTCCTTAGCATGACCGACATGGACCCTGTGGACACAGCCACATATTACTGTGCGCGGATTTTGAAGTTTGGGACAATGAGGGCCGCATACTACTTTGACTACTGGGGCCAGGGAGCCCTGGTCCCCGTCTCCTCA |
| 122 | 경쇄 | TCCTATGTGCTGACTCAGCCACCCTCAGTGTCAGTGGCCCCAGGAAAGACGGCCAGGATTACCTGTGGGGGAATCAACATTGGAATTAGAACTGTACACTGGTACCAGCAGAAGCCAGGCCAGGCCCCTATGTTGGTCATCTATTATGATAGCGACCGGCCCTTAGGGATCCCTGAGCGATTCTCTGGCTCCAAGTCTGGGAACACGGCCACCCTGACCATCAGCAGGGTCGAAGCCGGGGATGAGGCCGACTATTACTGTCAGGTGTGGGATAGTAGTAGTGATCATGTTGTATTCGGCGGAGGGACCAAGCTGACCGCCCTA | |
| EV-D68-266 | 123 | 중쇄 | CAGGTGCACTTGGTGGAGTCTGGGGGAGGCTTGGTCAAGCCTGGAGGGTCCCTGAGACTCTCCTGTGCAGTTTCTGGATTCACCTTCAGTGACTACTACATGAGCTGGATCCGCCAGGCTCCGGGGAAGGGACTGGAGTGGCTTTCATACATTAGTAGTAGTGGTAGTACCATATACTACGCAGACTCTATGAAGGGCCGATTCACCATCTCCAGGGACAACGCCAGGAACTCACTCTATCTGCAAATGAACAGCCTGAGAGTCGAGGACACGGCCTTTTATTACTGTGCGGGGTCAAAGGTTGGCTATACTACTGGTCGAAGGAACTGGTACTTCGATCTCTGGGGCCGTGGCACCCTGGTCACTGTCTCCTCA |
| 124 | 경쇄 | CAGTCTGCCCTGACTCAGCCTCCCTCCGCGTCCGGGTCTCCTGGACAGTCAGTCACCATCTCCTGCACTGGAACCAGCAGTGACGTTGGTGCTTATAACTATGTCTCCTGGTACCAACAGCACCCAGGCAAAGCCCCCAAACTCATGATTTATGAGGTCACTAAGCGGCCCTCAGGGGTCCCTGATCGCTTCTCTGGCTCCAAGTCTGGCAACACGGCCTCTCTGACCGTCTCTGGGCTCCAGGCTGAGGATGAGGCTGATTATTACTGCAGCTCATATGCAGGCAACAACAATTTAGTCTTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-269 | 125 | 중쇄 | CAGGTGCAGTTGCTGCAGTCTGGGTCTGAGGTGAGGAAACCTGGGGCCTCAGTGAACATTCACTGTAAGGCATCTGGATTCACTTTCACCGACTTCTATTTACACTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGATGGGGATAATCAACCCTGAAACCGGTGAGACAACCTACTCACAGAAGTTTCAGGGCAGAGTCACCATGACCAGGGACACGTCCACGAGTGTAGTGAATCTGGAAGTGAGGAGCCTGAGATCTGAGGACACGGCCATATATTACTGTGCGAGAGATCTCGTTGTCGTAGTCCCCGTTGAAATGTCTCGGCGTGCCTTTGACATTTGGGGCCAAGGGATTATGGTCACAGTCTCCTCA |
| 126 | 경쇄 | TCCTATGTGCTGACTCAGCCACCCTCGGTGTCAGTGGCCCCAGGACAGACGGCCAGGATTCCCTGTGGGGGCAACAACATTGAACGTAAAAGTGTCCACTGGTACCAGCAGAGGCCAGGCCAGGCCCCTGTGTTGGTCGTCTATGATGATACTGTCCGGCCCTCAGGTATCCCTGAGCGATTCTCTGGCTCCAACTCCGGGAGCACGGCCACCCTGACCATCAGCAGGGTCGGAGCCGGGGATGAGGGCGACTATTATTGTCAGGTGTGGGACAGCACCACTGACCATGGGGTCTTCGGCGGAGGGACCAAGCTGACCGTCCTA | |
| EV-D68-271 | 127 | 중쇄 | CGGGTGCAGCTGGTGCAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGTCAGACTTCTGGATACATTTTCAGCGCCTACTACATCTATTGGGTGCGACAGGCCCCTGGACAAGGACTTGAGTGGATGGGACGGATGAACGCTAAGAGTGGAGGCGCAAACACTGCACAGCAGTTTCAGGGCAGACTCACCATGACCAGGGACATGTCCGTCAGCACAGCCTACATGGAACTGAGCAGGCTGCGATCGGACGACACGGCCGTCTATTATTGTGCGAGAGACTATAGGGATGACTACATGTGGGGGAGTTATCGGCCTTTAGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCA |
| 128 | 경쇄 | GAAGTTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCTCCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCGGCTACTTAGCCTGGTACCAACACAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCGCTGGCATCCCAGCCAGGTTCCGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTGGAGCCTGAAGATTTTGCAGTTTATTTCTGTCAGCAGCGTAGCAACGGGCTCACTTTCGGCGGAGGGACCAAGGTCGAGATCAAA |
| 클론 | 서열번호 | 쇄 | 가변 서열 |
| EV-D68-37 | 129 | 중쇄 | QVQLVESGGGVVQPGRSLRLSCAASGFIFSRYALHWVRQAPGKGLDWVAVISYDARNSYYTDSVKGRFTISRDNSKNTLFLQMNSLRADDTAVYYCARPTLPYSNNWYAPEYWGQGTLVTVSS |
| 130 | 경쇄 | SYELTQPPSVSVSPGQTARITCSGDALPKKYASWYQQKSGQAPVLVIYEDTKRPSGIPERFSGSSSGTMATLTISGAQVEDESDYYCSSTDSSGNPVLFGGGTKLTVL |
|
| EV-D68-40 | 131 | 중쇄 | EVQLVESGGDLVQPGGSLRLSCAASGFSFSSYAMAWVRQAPGKGLQWVSSISGNGNGRSYADSLKGRFTTSKDLSKYTLYLQMNNLRPEDTAIYYCAKVVRIAAVLYYFDYWGPGTQVTVSS |
| 132 | 경쇄 | EIVLTQSPATLSLSPGERATLSCRASQSVSTYLAWYQQKPGQAPRLLIYEASTRATGIPARFSGSGSGTDFTLIISSLEPEDFAVYHCQQRSSWPITFGQGTRLEIE |
|
| EV-D68-41 | 133 | 중쇄 | DVQLVESGGGLVQPGGSLRLSCAASGFTFSNYAMTWVRQALGKGLEWVSSISGSGGLTYFAHSVKGRLTISRDNSKNTLYLQMSSLRAEDTAVYYCARVKSTTGTTALVFDIWGQGTMVTVSS |
| 134 | 경쇄 | QTVVTQEPSFSVSPGGTVTLTCGLSSGSVSSSYYPSWYQQTPGQAPRTLIYSINRRSSGVPDRFSGFILGNKAALTIRGAQADDESDYYCGLYMGSGIWIFGGGTKLTVL |
|
| EV-D68-43 | 135 | 중쇄 | QVQLVESGGGVVQPGRSLRLSCAASGFTFINYGMHWVRQAPGKGLEWVAVISNDGSYNYDADSVKGRFTISRDNSKNKVYLQMNSLRPEDTAVYFCAKDKHGDFDYYGVDVWGQGTTVTVSP |
| 136 | 경쇄 | DVQMTQSPSSVSASIGDRVTITCRAGQGISSWLAWYQQKPGKAPKLLIYAASNLQSGVPSRFSGSGSGTDFTLTISSLQPEDFGTYYCQQADSFPRTFGQGTKVEIK |
|
| EV-D68-46 | 137 | 중쇄 | QVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGIHWVRQAPGKGLEWVAVISYDGSDNTYAPFVNGRFTISRDNSKNTLYLQMNSLRADDTAVYYCARRRPGSFPGLCDYWGQGALVTVSS |
| 138 | 경쇄 | DIQMTQSPSTMSASVGDRVTITCRASQSISKWLAWYQQKPGKAPKLLIYKASTLQTGVPSRFSGSGSGTEFTLTINSLQPDDFATYYCQQHNSYSYTFGQGTKVEIK |
|
| EV-D68-48 | 139 | 중쇄 | QVQLVQSGAEVKKPGSSVKVSCKASGGSFSRLTIIWVRQAPGQGLEWMGGHIPIFGTTNYALKFQGRVTITADKTTSTAYMELSSLRSEDTAIYYCARMYSGHDGVDVWGQGTLVTVSS |
| 140 | 경쇄 | EIVLTQSPATLSLSPGERATLSCRASQSVRSYLAWYQHKPGQAPRLLIYDASNRAKGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSTWPPGMFGQGTRVEIK |
|
| EV-D68-71 | 141 | 중쇄 | QLQLQESGPGLVKHSETLSLTCTVSGGSISSGFYYWGWIRQPPGKGLEWIGTIYDSGRTYDNPSLKSRVTISADTSKKQFSLTLRSATAADTAVYFCARHLTHLYGDYVTPDALDIWGQGTMVTVSS |
| 142 | 경쇄 | EIVLTQSPGTLSLSPGERATLSCRASQSVSSSFLAWYQQKPGQAPRLLIYGASSRATGIPDRFRGSGSGTDFTLTISRLEPEDFAVYYCQQYSNSRLTFGQGTKVEIK |
|
| EV-D68-72 | 143 | 중쇄 | EVQLVESGGGLVQPGGSLRLSCAASGFTFTTYSMNWVRQAPGKGLEWISYISSGSSNIYYADSVKGRFTISRDNAKNSLNLQMSSLRDEDTAVYYCARAHGRIVNSGVVISRFDPWGQGILVTVSS |
| 144 | 경쇄 | QSALTQPASVSGSPGQSITISCTGTSSDVGGYNYVSWYQLHPGKAPKLMIFEVTYRPSGVSNRFSGSKSGNTASLTISGLQAEDEADYFCSSYTTSNTLVVFGGGTKLTVL |
|
| EV-D68-74 | 145 | 중쇄 | QVQLVQSGAEVKKPGASVKVSCRTSGYTFTAYYMHWVRQAPGQGPEWMGRINPSSGGAQYAQKFQGRVTMTRDTSISTTYMTLSGLTSDDTAVFYCARMGCRSDRCYSTNYNFDQWGQGTLVTVSS |
| 146 | 경쇄 | SYVLTQPPSVSVAPGQTARIPCGGNNIGTKSVHWYQQKPGQAPVLVVSNDSDRPSGIPERFSGSKSGNTATLTISRVEAGDEADYYCQVWDSGIDVVFGGGTKLTVL |
|
| EV-D68-75 | 147 | 중쇄 | EVQLVESGGGLVKPGGSLRLSCAASGFTISPYGMNWVRQAPGKGLEWVSFISSSSRYTYYADSVKGRFTISRDNAKNSLSLQMNSLRAEDTAVYYCARERGHSTSSSYFDSWGQGTLVTVSS |
| 148 | 경쇄 | SYVLTQPPSVSLAPGKTARITCGGNNIGTKTVSWYQQKPGQAPVLVMYYDSDRPSGIPERFSGSNSGNTATLTINRVEAGDEADYYCRVWDSDTDHRVFGGGTKLTVL |
|
| EV-D68-76 | 149 | 중쇄 | EVQLVESGGGLVKPGGSLRLSCAASGFTFSNAWISWVRQAPGKGLEWVGRIQTKTDGGTTDYAAPVKGRFTISRDDSKNTLYLQMNSLKTEDTALYYCSTGPYYYDTSGYPQPFDYWGQGTLVTVSS |
| 150 | 경쇄 | SYELTQPPSVSVSPGQTASITCSGDKLGDKYACWYQQKPGQSPVLVIYQDTKRPSGIPERFSGSNSGNTATLTISGTQAMDEADYYCQAWDSSTVVFGGGTKLTVL |
|
| EV-D68-80 | 151 | 중쇄 | EVQMVESGGGLVKPGGSLRLSCSVSGFDFSRYTMNWFRQAPGEGLKWVSSISSTSLYTFYADSVKGRFSISRDNAQGSLSLQMSSLRPEDTAVYYCARVVGPAELDYWGQGVLVTVSS |
| 152 | 경쇄 | VTQLTQSPSSLSASVGDRVTITCRASQDIGVDLGWFQQRPGKAPKLLIYGASRLQSGVPSRFSGRGSGTFFTLTISSLQPEDFTTYFCLQDYNYPWTLGQGTTVGVK |
|
| EV-D68-84 | 153 | 중쇄 | QVHLVQSGSAVRKPGASVKVSCKASGYTFTDYYIHWVRQAPGQGLEWMGWINPKTGGSNYTQRFQARVTMTWDTSISTAYMELSRLRSDDTAVYYCARAGRNGYDYWGQGTLVTVSS |
| 154 | 경쇄 | SYELTQPPSVSVSPGQTARITCSADALPKQYAYWYQQKPGQAPVLMIYQDTERPSGIPERFSGSSSGTTVTLTISGVQAEDEADYYCQSGDSSGTYLVFGGGTKLTVL |
|
| EV-D68-85 | 155 | 중쇄 | EVQLLESGGGLVQPGGSLRLSCAASGFKFRNYAMTWVRQAPGKGLEWVSTITSGGSTEYADAVKGRFIISRDNSKNTLYLQMNSLRADDTAVYYCTVPWGNYNDYVSDYWGQGTLVPVSS |
| 156 | 경쇄 | EVVLTQSPATLSLSPGQRATLSCRASQRVGNSLAWYQQKPGQAPSLLIYDASKRATGIPARFSGSGSGTDFTLTIISLESEDFAVYYCHQHSTWPRGTFGQGTRLEIK |
|
| EV-D68-88 | 157 | 중쇄 | QVHLVESGGGVVQPGRSLRLSCAASGFIFSRYPMHWVRQAPGKGLEWVALISYDGNNKYYADSVKGRFTISRDNSKNTLFLQMNSLRAEDTAVYYCARHFLPYSSSWYQGFNYWGQGILVTVSS |
| 158 | 경쇄 | NFMLTQPHSVSESPGKTVTISCTRSSGSIATNYVQWYQQRPGSSPTPIIFEDSQRPSGVPDRFSGSIDSSSNSASLTISGLRTDDEADYYCQSYDNSNRAVVFGGGTKLTVL |
|
| EV-D68-89 | 159 | 중쇄 | QVQLVESGGGVVQPGRSLRLSCEASGFLFSRYGMHWVRQAPGKGLDWVAVISYDGNKKYYADSVKGRFTISRDNSKNTLYLQVNSLRVEDTAVYYCARGVPYGDTLTGLVYWGQGTLVTVSS |
| 160 | 경쇄 | NFMLTQPHSVSESPGKTVTISCTRSSGTIASNYVQWYQQRPGSAPTTVIYEDNQRPSGVPDRFSGSIDSSSNSASLTISGLKTEDEADYYCQSYDNSDRVFGGGTKLTVL |
|
| EV-D68-95 | 161 | 중쇄 | QVTLKESGPVLVKPTETLTLTCTVSGFSLRNARMGVSWIRQPPGKALEWLAHIFSNDEKSYNTSLKSRLSISKDTSKSQVVLTMTSMDPLDTATYFCARLLVAGTFLPSHYFDYWGQGILVTVSS |
| 162 | 경쇄 | SYVLTQPPSVSVTPGKTARITCGGNNIGLKSVFWYQERPDQAPVVVIYYDSARPSGIPERISGSKSGNTATLTITRVEAGDEADYFCQVWDSSRNHPVFGGGTKLTVL |
|
| EV-D68-97 | 163 | 중쇄 | ELQLVESGGGLVQPGGSLRLSCAASGFTFSTYSMNWVRQAPGKGLEWVSYISSSSSTIQYADSVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCTRQVGADFSGRGFDYWGQGTLLTVSS |
| 164 | 경쇄 | SYELTQPPSVSVSPGQTATITCFGDKLGDKYACWYQQKPGQSPVLVIYQDSKRPSGIPERFSGSKSGNTATLTISGTQAMDEADYYCQAWDSSTAVFGGGTKLTVL |
|
| EV-D68-98 | 165 | 중쇄 | EVQLVESGGGLVQPGGSLRLSCAASGFKFSVYALSWVRQAPGKGLEWISYISSSGSTIYYSDSVKGRFTISRDNVGNSLFVQMNSLRAEDTGIYYCATARHITNDGFDIWGQGTMVIVSS |
| 166 | 경쇄 | DIQLTQSPSFLSASVGDRVTITCRASQGISRFLAWYQQKPGKAPKLLIYSASTLQRGVPSRFSGSGFGTDFTLTISSLQPEDFATYYCQQLNSHPRMFTFGPGTTVDIK |
|
| EV-D68-105 | 167 | 중쇄 | QVQLQESGPGLVKPSETLSLTCAVSGYLISNGYYWGWIRQPPGKGLEWIGSIYHTRSTYYNPSLKSRVSISVDTSKNRFSLRLRSVTAADTAFYYCARGPGHCYGDDDCYAYYFDQWGQGTPVTVSP |
| 168 | 경쇄 | DIQMTQSPSSVSSSVGDRVTITCRASQGISNWLAWYQQNPGKAPKLLIYDASSLQSGVPSRFSGSGSGTDFTLTINSLQPEDFATYYCQQANSFPFTFGPGTKVDIK |
|
| EV-D68-110 | 169 | 중쇄 | QVQLVQSGAEVKKPGSSVKVSCKASGGTFSRFAISWVRQAPGQGLQWMGGILPIFGTANYAQKFQGRVTITADTSTSTAYMELSSLRSEDTAVYYCARSLPYCTNDVCSNQNTFDYWGQGTLVTVSS |
| 170 | 경쇄 | SYELTQPPSVSVSPGQAARITCSGDALPKQYAYWYQQKPGQAPVLVIYEDNKRPSGIPERFSGSSSGTTVTLTISGVQAEDEADYYCQSADSSGTYVVFGGGTKLTVL |
|
| EV-D68-111 | 171 | 중쇄 | QVQLQESGPGLVKPSQTLSLTCTVSGGSISSGDYYWSWIRQPPGKGLEWIGYIYYSGSTYYNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCASRYGDPIGDNWFDPWGQGTLVTVSS |
| 172 | 경쇄 | SYVLTQPPSVSVTPGKTARITCGGNNIGLKSVFWYQERPDQAPVVVIYYDSARPSGIPERISGSKSGNTATLTITRVEAGDEADYFCQVWDSSRNHPVFGGGTKLTVL |
|
| EV-D68-114 | 173 | 중쇄 | EVQLLESGGGLVQPGGSLRLSCAASGFRFSFYGMTWVRQAPGKGLEWVSSISGTGATRNCADSVKGRFTISRDNSKNTLYLQMDSLRVDDTAVFYCVRRFPMTTVTSFDSWGQGTLVTVSS |
| 174 | 경쇄 | QSALTQPRSVSGSPGQSVTISCTGTSSDVGGYNFVSWYQQHPGKAPKLMIFDVTGRPSGVPDRFSGSKSGNTASLTIAGLQAEDEADYYCGAYAGFNALFGGGTKLTVL |
|
| EV-D68-116 | 175 | 중쇄 | EVQLVQSGGGLVRPGGSVRLSCVASGFPFNMFWMGWVRQTPGKGLEWVANIKQDGSEKYYVDSVKGRFAISRDNAKNSLFLQMDSLSVGDTAIYYCVREGVRRVVVRSTGYFDFWGQGQLVTVSS |
| 176 | 경쇄 | SYELTQPPSMSVSPGQTARITCSGDAVPIKYVYWYQQRSGQAPVLVIYEDDRRPSGISERFSGSSSGTTATLTITGAQVEDEGDYYCYSTDSSGYQRAFGGGTTLTVL |
|
| EV-D68-150 | 177 | 중쇄 | RVQLQESAPGLVRPSETLSLTCSVSGGPISNGPYYWSWIRQHPGKGLEWIGFIFYSGSTNYNPSLRGRVTMAVDTSKNQFSLRLNSVTAADTAVYYCARHVVTASGWFDPWGQGTLVTVSS |
| 178 | 경쇄 | EIVLTQSPATLSLSPGERATLSCRASQSVGTDLAWYQQKPGQAPRVLIYDAFKRATGIPARFSGSGSGTEFTLTISSLEPEDFAVYYCQQRSRWPPPYTFGQGTKLEIK |
|
| EV-D68-151 | 179 | 중쇄 | RVQLQESAPGLVRPSETLSLTCSVSGGPISNGPYYWSWIRQHPGKGLEWIGFIFYSGSTNYNPSLRGRVTMAVDTSKNQFSLRLNSVTAADTAVYYCARHVVTASGWFDPWGQGTLVTVSS |
| 180 | 경쇄 | EIVLTQSPATLSLSPGERATLSCRASQSVGTDLAWYQQKPGQAPRVLIYDAFKRATGIPARFSGSGSGTEFTLTISSLEPEDFAVYYCQQRSRWPPPYTFGQGTKLEIK |
|
| EV-D68-152 | 181 | 중쇄 | QVHLVESGGGVVQPGRSLRLSCADSGVTFSDNALYWVRQAPGKGLEWVAVISYDGSSRYYADSVRGRFTISRDNSKDTLYLQMNRLRAEDTAIYYCARVTADYYESSGKVFWGQGALVVVSS |
| 182 | 경쇄 | DIQMTQSPSTLSASVGDRVSITCRASQSVRSWLAWYQHKPGKAPKLLIYKASSLESGVPSRFSGSGSGTEFTLTISSLQADDFATYYCQQYQTFSWTFGQGTTVEVK |
|
| EV-D68-154 | 183 | 중쇄 | QVQLQESGPALVKPSETLSLTCTVSGGSISDHYWSWIRQPPGKGLEWIGYIYTSGTTNYNPSLKSRVTISVDTSKKQFSLNLRSVTAADTAVYYCARSLETVIRFYYYHYMDVWGKGTTVIVSS |
| 184 | 경쇄 | DIVMTQSPLSLPVTPGEPASISCRSSQSLLQSDGYSYLDWYLQKPGQSPQLLIYLGSNRASGVPDRFSVIGSGTYFTLKISRVEAEDVGVYFCMQALQTPWTFGQGTKVEIK |
|
| EV-D68-155 | 185 | 중쇄 | QVQLQESGPGLVKPSETLSLTCTVAGGSIGDYHWNWIRQPAGKGLEWIGRIHSSGNTDYNPSLKSRVTMSVDTSKNQFSLKLRSVTAADTAVYYCARQNVFDIWGQGTMVTVSS |
| 186 | 경쇄 | DIVMTQSPDSLALSLGERATINCKSSQSVLFSSNNKNYLAWYQQKPGQPPKLLIYWASTRESGVPDRFSGSGSETDFTLTISSLQAEDVAVYFCQQFYTTPLTFGGGTKVEIK |
|
| EV-D68-156 | 187 | 중쇄 | QVQMQESGPGLVKASETLSLTCSVSGISINNYYWSWFRQPPGKGLEWIGYVYSTGSSKYNPSLERRATMSVDTSNNNFSLRLTSVTTADTAVYYCARGSMPHIWGQGLLVTVSS |
| 188 | 경쇄 | QSVLTQPPSASGTPGQRVTISCSGSTSNIETNYVYWYQQVPGTAPKPLVYRNDQRPSGVPDRFSGSKSGTSASLVISGLRTEDEAAYYCAAWDDSLKAPVFGAGTKVAVL |
|
| EV-D68-157 | 189 | 중쇄 | EVQLVESGGGLIKPGGSLRLSCAASGITFSNAWMSWVRQAPGKGLEWVGRIESKIDGGTIDYATPVKGRFTISRDDSKNTLYLQMNSLKTEDTAVYYCTTDQGYYDRSGYWVVGNHFDYWGQGILVTVSS |
| 190 | 경쇄 | QSVLTQPPSVSGAPGLRVTISCTGSSTNIGAGYDVHWYQHLPGTAPKLLIYGNSNRPSGVPDRFSGSKSGTSASLAITGLQADDAADYYCQSYDRSLSTYVFGTGTKVTVL |
|
| EV-D68-158 | 191 | 중쇄 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMNWVRQAPGKGLEWVSGISGTTGSTYYADSVKGRFTISRDNSKNTVHLQMNSLRAEDTAVYYCAKDSHSMIVVDHAFDIWGQGTMVTVSS |
| 192 | 경쇄 | SYVLTQPPSVSVAPGQTARITCGGNNIGTKSVHWYQQRPGQAPVLVVYDDSDRPSGIPERFSGSNSGNTATLTISRVEAGDEADYYCQVWDSYNVHYVFGTGTKVTVL |
|
| EV-D68-159 | 193 | 중쇄 | EVQLVESGGGLVKPGGSLRLSCAASGFTFSTYIMTWVRQAPGRGLEWVSSISTSSVYTFYADSLKGRFTISRDNAKNSVYLQMNSLRADDTAVYYCAREEGFRAYNLYWGQGTLVTVSS |
| 194 | 경쇄 | QSVLTQPPSASGTPGQRVTISCSGSSSNIEYNYVYWYQKFPGTAPKLLIYKNNQRPSGVPDRFSGSKSGTSASLAISGLRSEDEGDYYCAAWDDILSGVVFGGGTKLTVL |
|
| EV-D68-160 | 195 | 중쇄 | QVQLVESGGGVVQPGRSQKLSCAASGFTFSRFGMHWVRQAPGKGLEWVAVISFDGSNRYYADSVKGRFTITRDNSKNTLYLQMNNLRPEDTAVYYCARDWDRLVRSAVGYWGQGTLVSVSS |
| 196 | 경쇄 | QSALTQPRSVSGSPGQSVTISCTGTSNDVGGYNFVSWYQQHPGKAPKLMIFDVIRRPSGVPGRFSGSKSGDTASLIISGLQAEDEADYYCCSYAGTYTWVFGAGTTLTVL |
|
| EV-D68-161 | 197 | 중쇄 | QVHLQESGPRLVKPSETLSLTCTVSGGSVSTATYYWSWIRQSPGRGLEWIGYIYSSGNTNYNPSLKSRVTISLDTPNNQLSLTLTSVTAADTAIYYCERRLRILSIERNYYAMDVWGQGTPVTVSS |
| 198 | 경쇄 | EVVLTQTPGTLSLSPGEGATLSCRASQRVVNNYLAWYQQRAGQAPRLLIFGASNRATGIPDRFSGSGSGTDFTLTIRKLEPEDFAVYYCQQYGSPWTFGHGTKVEMK |
|
| EV-D68-162 | 199 | 중쇄 | QVQLVESGGGVVQPGRSLRLSCAASGFTFSSYAMHWVRQAPGRGLEWVAVISYDASKKYHADSVKGRFTISRDSSKNTLFLQMNSLKPEDTAVYYCARDHVPPKDCSDGNCHSDYGMDVWGQGTTVTVSS |
| 200 | 경쇄 | DIQMTQSPSTMSASVGDRVTITCRASQSISKWLAWYQQKPGKAPKLLIYKASTLQTGVPSRFSGSGSGTEFTLTINSLQPDDFATYYCQQHNSYSYTFGQGTKVEIK |
|
| EV-D68-163 | 201 | 중쇄 | QVQLVQSGAEVKKPGASVKISCKASGYSFTNFAVHWVRQAPGQRLEWMGWINPGNRNTKYSHNFQGRVTITRDTSANTAYMELSSLRSQDTAVYYCARLPIAAAGRGWFDPWGQGTLVTVSS |
| 202 | 경쇄 | EIVLTQSPGTLSLSPGERATLSCRASQSVISTYLAWYQQRPGQAPRVLIYDVSTRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYFCHQYGSSPATFGQGTKVEIK |
|
| EV-D68-164 | 203 | 중쇄 | EVQLVESGGDLVQPGGSLRLSCAASGFTFNTYGMNWVRQAPGKGLEWVSYISSATTTFYYADSVRGRFTISRDNAKNSLFLHMKSLRDEDTAVYYCARVYTMLRGASMDVWGHGTTVTVSS |
| 204 | 경쇄 | EIVLTQSPATLSLSPGERATLSCRASQSVGTYLAWYQQKPGQAPRLLIYDSANRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQLRITWPPIFTFGPGTKVDVK |
|
| EV-D68-165 | 205 | 중쇄 | QVHLVQSGAEVKKSGASVKVSCKTFGYTFTAYYMHWVRQAPGQGPEWMGWINPISGGTNYAQKFQGRVSMTRDTSISTAYMGLSRLRPDDTAVYYCARVKCSSANCYGNFDYWGQGTLVTVSS |
| 206 | 경쇄 | QSALTQPPSASGSPGESVTISCTGTSSDVGGYNYVSWYQQHPGKAPKLMIYEVSKRPSGVPDRFSGSKSGNTASLTVSGLQAEDEADYYCSSYAGSNNLVFGGGTKLTVL |
|
| EV-D68-166 | 207 | 중쇄 | EVQLVQSGAEVKKPGESLKISCKGSGYRFTNYRIGWVRQMPGKGLEWMGIIYPGGSDTRYSPSLQGQVTMSVDKSISTAYLMWSSLKASDTAMYYCARQTTQNSGYDRWFDSWGQGTHVTVSS |
| 208 | 경쇄 | QSVLTQPPSASGTPGQRVTISCSGSTSSIGSNIVNWYQHLPGTAPKLLIYINNQRPSGVPDRFSGSKSGTSASLAISGLQSEDEADYYCAAWDDSLNGWVFGGGTKLTVL |
|
| EV-D68-181 | 209 | 중쇄 | EVQVLESGGGLVQPGGSLRLSCAASGITFSRHTMSWVRQAPGKGLEWVSAISGSGGSTYHADSVKGRFTISRDSSKSTLYLQMNSLRAEDTAVYYCAISVPLLRFLEWFQHPFDFWGQGTLVTVSS |
| 210 | 경쇄 | EIVMTQSPASLSVSPGERVTLSCRASQSVGSTLAWYQHKPGQAPRLLISGASTRATGVPARFSGSGSGTEFTLTISSLQSEDFAVYYCHQYINWPPWTFGQGTKVEIK |
|
| EV-D68-183 | 211 | 중쇄 | EVRLVESGGGLVKPGGSLRLSCAASGFTFNTYSMSWVRQAPGKGLEWVASISSTGSYIYNADSLKGRFTISRDNAKNSLFLQMNSLRVEDTAVYYCVRFTMTTVTNFDSWGQGTLVTVSS |
| 212 | 경쇄 | QSALTQPRSVSGSPGQSVTISCTGTSSDVGAYSYVSWYQQHPGKAPKLMIYDVYRRPSGVPGRFSGSKSGNTASLTVSGLQAEDEADYYCCSHAGSHTWVFGGGTKVTVL |
|
| EV-D68-185 | 213 | 중쇄 | QLQVVASGGGVVQPGRSLRLSCKASGFTFTNYGMHWVRQAPGKGLEWVAFISYDGGNKFYADSVKGRFTISRDNSRNTVYLQMNSLRVADTAMYYCPKVIPHPYYDSSGDDAFDIWGQGTMVAISS |
| 214 | 경쇄 | EIVMTQSPATLSVSPGERATLSCWASQSISRNLAWYQQKPGQAPRLLIYGASTRATGIPAKFSGSGSGTDFTLTVSSLQSEDLAVYYCQQYSKLPITFGQGTRLEIK |
|
| EV-D68-200 | 215 | 중쇄 | EVQLVESGGGLVQPGGSLRLSCAASGFPFSSYSMSWVRQAPGKGLEWVSYISGSGGDIYYADSVKGRFTISRDNARNSLSLQMNSLRADDTAVYYCARGLVATTGTRYFDYWGQGTLVTVSS |
| 216 | 경쇄 | EIVLTQSPATLSLSPGERATLSCRASQSVRSYLAWYQQRPGQAPRLLIYDASNRATGIPVRFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSYWPPFTFGGGTKVEIK |
|
| EV-D68-208 | 217 | 중쇄 | QVQLVQSGAEVKKPGSSVKVSCKASGGTFRRFAISWVRQAPGQGLEWMGGIIPILGRGKYAQKFQGRVRITADESTSTAYMELSSLRSEDTAVYYCARFISTASYVPGTFEDVWGQGTTVTVSS |
| 218 | 경쇄 | DIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRAAGIPARFSGSGSETDFTLTISSLEPEDVAVYYCQQRSDWPPGTFGQGTNVEIK |
|
| EV-D68-210 | 219 | 중쇄 | EVQLVESGGGLVKPGGSLRLSCAASGFTFRNYNINWVRQAPGKGLEWVSSISSTGSYIHYADLVKGRFTISRDNAKNSLYLQMNSLRVEDTAVYYCARMVRNTVTAFDYWGQGTLVSVSS |
| 220 | 경쇄 | QSALTQPASVSGSPGQSITISCTGTSSDVGGYNFVSWYQQQPGRAPKLLIYEVIKRPSGVSDRFSGSKSGDTASLTISGLQAEDEADYYCCSYGGNNSWMFGGGTMLTVL |
|
| EV-D68-219 | 221 | 중쇄 | QVQVVQSGAEMKKPGASVKVSCKVSGYRLIDLPLHWVRQAPGKGLEWMGLFDPEKAEAIYSQKFQDKVTISEDTSIDTAYMELNSLRSEDTAVYYCATWGVEVVNGRRDYFDSWGQGTLVTVSS |
| 222 | 경쇄 | SYELTQPPSVSVSPGQTARITCYADVLSNQYTYWYQQKPGQAPVLVIYKDTERPSGIPERFAGSSSGTTVTLVINGVRAEDEAYYYCQSADNTRITVFGGGTKLSVL |
|
| EV-D68-220 | 223 | 중쇄 | GVQLVESGGGLIQPGGSLRLSCAASGFTFSSFEMNWVRQAPGKGLEWVSYISTSGSTIYYADSVKGRFTISRDNARNSLSLQMNSLRAEDTAVYYCARDVRDCSALTCPRRGDAFDFWGRGTRVTVSS |
| 224 | 경쇄 | DVVMTQSPLSLPVTLGQSASISCRSSQSLVYSDGNTYLNWFQQRPGQSPRRLIYKVSNRDSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCMQGTHWPRTFGPGTKVDIK |
|
| EV-D68-221 | 225 | 중쇄 | EVQLVESGGGLVKPGGSLRLSCAASGFSFSVYPMNWVRQAPGKGLEWVSSISSSSRYISYADSLRGRITISRDNAKNSLYLQMNSLRVEDTAVYYCVKVGGSKHQYYFDYWGQGSLVTVSS |
| 226 | 경쇄 | QSVLTQPPSTSGIPGQTVTISCSGSRSNIGSYTVNWYEQLPGTAPKLVIFNNNQRPSGVPDRFSGSKSGTSASLAISGLQSEGEADYYCAAWDDSLNGVVFGGGTKLTVL |
|
| EV-D68-224 | 227 | 중쇄 | QVQLVQSGAEVKRPGASVKVFCKASGYTFTNYDIIWVRQAPGQGLEWVGWISTYNGNTNYEQNLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARERCSTSTCYSRYADYWGQGTLVTVSS |
| 228 | 경쇄 | DIQMTQSPSSLSASVGDRVTITCRASQSINIYLNWYQQKPGKAPKVLIYAASSLQSGVPSRFRGSGSGTDFILTISSLQPEDFATYYCQQSYRSPRTFGQGTEVEIK |
|
| EV-D68-225 | 229 | 중쇄 | EVQLVESGGGLAQPGESMRLSCVASGFTLSRYEVNWVRQAPGKGLEWLSYISSGGPSIYYADSVKGRFTISRNSAENSLELQMSTLRTEDTAVYYCMREGLTYYDSTIWGQGTLVAVSS |
| 230 | 경쇄 | QNVLTQSPSASASLGASVKLTCTLNSGHSRYAIAWHQHQPQRGPRFLMKINSDGRHIRGDGISDRFSGSASGAERHLTISSLQPEDEADYYCQTWGTGFRVFGGGTKLTVL |
|
| EV-D68-227 | 231 | 중쇄 | EVQLVESGGGLVQPGRSLRLSCAASGFTFDEYAMHWVRQVPGKGLEWVSGISWNGGSKGYADSVKGRFTISRDNARYSLSLQMNSLRTEDTALYYCAKDDYEGAGFDIWGQGTVVTVSS |
| 232 | 경쇄 | EIVLTQSPATLSLSPGERATLSCRASQSVSSYLGWYQQKSGQAPRLLIYGASSRATGIPARFSGSGSGTDFTLTISSLEPEDFAIYYCQQRSNWPITFGQGTRLEIK |
|
| EV-D68-228 | 233 | 중쇄 | QVQLQESGPGLVKPSETLSLTCTVSGYLISNGYYWGWIRQSPGKGLEWIGSIYYTRDTYYNWSLKSRITISVDTSKKQFSLKLYSVTAADTAVYYCVRHEGSCNDGSCYGSFVDNWGQGTLVTVSS |
| 234 | 경쇄 | DIQMTQSPSSVSASVGDRVTLTCRASQDISSWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTHFTLTISSLQPEDFATYFCQQADSFITFGGGTKVEIK |
|
| EV-D68-231 | 235 | 중쇄 | EVQLLESGGGLVQPGGSLRLSCAASDFTFSSYTMAWVRQAPGKGLEWVSSISGDGVSTKNADSVKGRFSVSRDNSKNTLFLQLNSLRAEDTAFYYCARGGTFHNWYFDLWGRGVLVTVSS |
| 236 | 경쇄 | EIVMTQSPATLSVSPGETATLSCRASQSIGDNLAWYQQKPGQAPRLLISGASTRATDFPARFRGSGSGTEFTLTISSLQSEDFAVYYCQQYKNWPRTFGRGTKVEVR |
|
| EV-D68-234 | 237 | 중쇄 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSTYAMSWVRQAPGKGPEWVSGISGSGGSTNYADSAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGTITYSYYYMAVWGKGTTVTVSS |
| 238 | 경쇄 | EIVLTQSPGTLSLSPGERGTLSCRASQSVRSSYLAWYQQRPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTIGRLEPEDFAVYYCQQYGTSITFGGGTKVEIK |
|
| EV-D68-235 | 239 | 중쇄 | QVQVVQSGAEMRKPGASVRVSCKVSGYRLTDLPLHWVRQAPGKGLEWIGFVDLEKREIIYAQKFQGKVTITEDTSADTAYMELNSLRSEDTAVYYCATWGIEVVNGRDEFFDSWGQGTLVSVSS |
| 240 | 경쇄 | SYELTQPPSVSVSPGQTARITCYADVLSKQYTYWYQQKPGQAPVLVIYKDTERPSGIPERFAGSSSGTTVTLIINGVRTEDEAYYYCQSADTRITVFGGGTKLSVL |
|
| EV-D68-236 | 241 | 중쇄 | QVQVVQSGAEMKKPGASVKVSCKVSGYSLSDLPLHWVRQAPGKGLEWMGLFDPINGEIIYAQTFQGKVTISEDTSIDTAYMELNSLRSEDTAVYYCATWGVAVVSGRRDYFDSWGQGTLVTVSS |
| 242 | 경쇄 | SYELTQPPSVSVSPGQTARITCYAAVLSNQYTYWYQQKPGQAPVLVIYKDTERPSGIPERFAGSSSGTTVTLIINGVRTEDEAYYYCQTADTKYTVFGGGTKLSVL |
|
| EV-D68-241 | 243 | 중쇄 | QVQLQESGPGLVKPSQTLSLTCTVSGGSITSGDYYWNWIRQPPGKGLEWIGYIYHSGTTYYNPSLKSRVTISVDTSKNRFSLKLSSVTAADTAVYFCARAYAYEFWSGYPNWFDPWGLGTLVTVSS |
| 244 | 경쇄 | DIQMTQSPSSLSASVGDRVTITCRASQRISTYVNWYQVKAGTAPKVLIYAASSLQTGVPSRFSGSGSGTDFTLTIVSLQPEDFTTYFCQQSYSPPWTFGQGTKVEIK |
|
| EV-D68-247 | 245 | 중쇄 | QVQLVESGGGLVQPGRSLRLSCAASGFIFNRYAMHWVRQAPGKGLEWVALISYDGINKYYADSVKGRFSISRDNSKSTLYLQMNSLRAEDTAIFYCARGLGYCSGTGGSCTPFEYWGQGILVTVSS |
| 246 | 경쇄 | DIQMTQSPPSLSASVGDRVTITCRASQSIKKYLNWYQQKPGNAPKLLIYGASNLQTGVPSRFSGSGSGTDFALSISSLETEDFATYYCQQSDSAPPTFGGGTKVEFK |
|
| EV-D68-254 | 247 | 중쇄 | QVQLQQRGAGLLKPSETLSLTCEIYGASLNDYDWTWIRQPPGKRLEWIGVINRRDTVDYNPSLKSRVTLSLDTSKNQLSLSLSSVTAADTAVYYCVRVPRRGFEGSFGFCDDTACRYGHTWFDPWGQGTLVTVSS |
| 248 | 경쇄 | DIQMTQSPSSLSASVGDRVTITCRASQSIRDYLNWYQQRPGKAPKVLIFAGSRLESGVPSRFRGRGSGTEFTLTISDLQPEDFATYYCQQSYLTPPTFGQGTTVDIK |
|
| EV-D68-260 | 249 | 중쇄 | QVTLKESGPVLVKPTETLTLTCTVSGFSLRNGRMGVSWIRQPPGKALEWLAHIFASDEKSYSTSQRTRLSISRDTSKSQVVLSMTDMDPVDTATYYCARILKFGTMRAAYYFDYWGQGALVPVSS |
| 250 | 경쇄 | SYVLTQPPSVSVAPGKTARITCGGINIGIRTVHWYQQKPGQAPMLVIYYDSDRPLGIPERFSGSKSGNTATLTISRVEAGDEADYYCQVWDSSSDHVVFGGGTKLTAL |
|
| EV-D68-266 | 251 | 중쇄 | QVHLVESGGGLVKPGGSLRLSCAVSGFTFSDYYMSWIRQAPGKGLEWLSYISSSGSTIYYADSMKGRFTISRDNARNSLYLQMNSLRVEDTAFYYCAGSKVGYTTGRRNWYFDLWGRGTLVTVSS |
| 252 | 경쇄 | QSALTQPPSASGSPGQSVTISCTGTSSDVGAYNYVSWYQQHPGKAPKLMIYEVTKRPSGVPDRFSGSKSGNTASLTVSGLQAEDEADYYCSSYAGNNNLVFGGGTKLTVL |
|
| EV-D68-269 | 253 | 중쇄 | QVQLLQSGSEVRKPGASVNIHCKASGFTFTDFYLHWVRQAPGQGLEWMGIINPETGETTYSQKFQGRVTMTRDTSTSVVNLEVRSLRSEDTAIYYCARDLVVVVPVEMSRRAFDIWGQGIMVTVSS |
| 254 | 경쇄 | SYVLTQPPSVSVAPGQTARIPCGGNNIERKSVHWYQQRPGQAPVLVVYDDTVRPSGIPERFSGSNSGSTATLTISRVGAGDEGDYYCQVWDSTTDHGVFGGGTKLTVL |
|
| EV-D68-271 | 255 | 중쇄 | RVQLVQSGAEVKKPGASVKVSCQTSGYIFSAYYIYWVRQAPGQGLEWMGRMNAKSGGANTAQQFQGRLTMTRDMSVSTAYMELSRLRSDDTAVYYCARDYRDDYMWGSYRPLDYWGQGTLVTVSS |
| 256 | 경쇄 | EVVLTQSPATLSLSPGERASLSCRASQSVSGYLAWYQHKPGQAPRLLIYDASNRAAGIPARFRGSGSGTDFTLTISSLEPEDFAVYFCQQRSNGLTFGGGTKVEIK |
| 클론 |
CDRH1
(서열번호) |
CDRH2
(서열번호) |
CDRH3
(서열번호) |
| EV-D68-37 | GFIFSRYA | ISYDARNS | ARPTLPYSNNWYAPEY |
| (257) | (258) | (259) | |
| EV-D68-40 | GFSFSSYA | ISGNGNGR | AKVVRIAAVLYYFDY |
| (260) | (261) | (262) | |
| EV-D68-41 | GFTFSNYA | ISGSGGLT | ARVKSTTGTTALVFDI |
| (263) | (264) | (265) | |
| EV-D68-43 | GFTFINYG | ISNDGSYN | AKDKHGDFDYYGVDV |
| (266) | (267) | (268) | |
| EV-D68-46 | GFTFSSYG | ISYDGSDN | ARRRPGSFPGLCDY |
| (269) | (270) | (271) | |
| EV-D68-48 | GGSFSRLT | HIPIFGTT | ARMYSGHDGVDV |
| (272) | (273) | (274) | |
| EV-D68-71 | GGSISSGFYY | IYDSGRT | ARHLTHLYGDYVTPDALDI |
| (275) | (276) | (277) | |
| EV-D68-72 | GFTFTTYS | ISSGSSNI | ARAHGRIVNSGVVISRFDP |
| (278) | (279) | (280) | |
| EV-D68-74 | GYTFTAYY | INPSSGGA | ARMGCRSDRCYSTNYNFDQ |
| (281) | (282) | (283) | |
| EV-D68-75 | GFTISPYG | ISSSSRYT | ARERGHSTSSSYFDS |
| (284) | (285) | (286) | |
| EV-D68-76 | GFTFSNAW | IQTKTDGGTT | STGPYYYDTSGYPQPFDY |
| (287) | (288) | (289) | |
| EV-D68-80 | GFDFSRYT | ISSTSLYT | ARVVGPAELDY |
| (290) | (291) | (292) | |
| EV-D68-84 | GYTFTDYY | INPKTGGS | ARAGRNGYDY |
| (293) | (294) | (295) | |
| EV-D68-85 | GFKFRNYA | ITSGGST | TVPWGNYNDYVSDY |
| (296) | (297) | (298) | |
| EV-D68-88 | GFIFSRYP | ISYDGNNK | ARHFLPYSSSWYQGFNY |
| (299) | (300) | (301) | |
| EV-D68-89 | GFLFSRYG | ISYDGNKK | ARGVPYGDTLTGLVY |
| (302) | (303) | (304) | |
| EV-D68-95 | GFSLRNARMG | IFSNDEK | ARLLVAGTFLPSHYFDY |
| (305) | (306) | (307) | |
| EV-D68-97 | GFTFSTYS | ISSSSSTI | TRQVGADFSGRGFDY |
| (308) | (309) | (310) | |
| EV-D68-98 | GFKFSVYA | ISSSGSTI | ATARHITNDGFDI |
| (311) | (312) | (313) | |
| EV-D68-105 | GYLISNGYY | IYHTRST | ARGPGHCYGDDDCYAYYFDQ |
| (314) | (315) | (316) | |
| EV-D68-110 | GGTFSRFA | ILPIFGTA | ARSLPYCTNDVCSNQNTFDY |
| (317) | (318) | (319) | |
| EV-D68-111 | GGSISSGDYY | IYYSGST | ASRYGDPIGDNWFDP |
| (320) | (321) | (322) | |
| EV-D68-114 | GFRFSFYG | ISGTGATR | VRRFPMTTVTSFDS |
| (323) | (324) | (325) | |
| EV-D68-116 | GFPFNMFW | IKQDGSEK | VREGVRRVVVRSTGYFDF |
| (326) | (327) | (328) | |
| EV-D68-150 | GGPISNGPYY | IFYSGST | ARHVVTASGWFDP |
| (329) | (330) | (331) | |
| EV-D68-151 | GGPISNGPYY | IFYSGST | ARHVVTASGWFDP |
| (332) | (333) | (334) | |
| EV-D68-152 | GVTFSDNA | ISYDGSSR | ARVTADYYESSGKVF |
| (335) | (336) | (337) | |
| EV-D68-154 | GGSISDHY | IYTSGTT | ARSLETVIRFYYYHYMDV |
| (338) | (339) | (340) | |
| EV-D68-155 | GGSIGDYH | IHSSGNT | ARQNVFDI |
| (341) | (342) | (343) | |
| EV-D68-156 | GISINNYY | VYSTGSS | ARGSMPHI |
| (344) | (345) | (346) | |
| EV-D68-157 | GITFSNAW | IESKIDGGTI | TTDQGYYDRSGYWVVGNHFDY |
| (347) | (348) | (349) | |
| EV-D68-158 | GFTFSSYA | ISGTTGST | AKDSHSMIVVDHAFDI |
| (350) | (351) | (352) | |
| EV-D68-159 | GFTFSTYI | ISTSSVYT | AREEGFRAYNLY |
| (353) | (354) | (355) | |
| EV-D68-160 | GFTFSRFG | ISFDGSNR | ARDWDRLVRSAVGY |
| (356) | (357) | (358) | |
| EV-D68-161 | GGSVSTATYY | IYSSGNT | ERRLRILSIERNYYAMDV |
| (359) | (360) | (361) | |
| EV-D68-162 | GFTFSSYA | ISYDASKK | ARDHVPPKDCSDGNCHSDYGMDV |
| (362) | (363) | (364) | |
| EV-D68-163 | GYSFTNFA | INPGNRNT | ARLPIAAAGRGWFDP |
| (365) | (366) | (367) | |
| EV-D68-164 | GFTFNTYG | ISSATTTF | ARVYTMLRGASMDV |
| (368) | (369) | (370) | |
| EV-D68-165 | GYTFTAYY | INPISGGT | ARVKCSSANCYGNFDY |
| (371) | (372) | (373) | |
| EV-D68-166 | GYRFTNYR | IYPGGSDT | ARQTTQNSGYDRWFDS |
| (374) | (375) | (376) | |
| EV-D68-181 | GITFSRHT | ISGSGGST | AISVPLLRFLEWFQHPFDF |
| (377) | (378) | (379) | |
| EV-D68-183 | GFTFNTYS | ISSTGSYI | VRFTMTTVTNFDS |
| (380) | (381) | (382) | |
| EV-D68-185 | GFTFTNYG | ISYDGGNK | PKVIPHPYYDSSGDDAFDI |
| (383) | (384) | (385) | |
| EV-D68-200 | GFPFSSYS | ISGSGGDI | ARGLVATTGTRYFDY |
| (386) | (387) | (388) | |
| EV-D68-208 | GGTFRRFA | IIPILGRG | ARFISTASYVPGTFEDV |
| (389) | (390) | (391) | |
| EV-D68-210 | GFTFRNYN | ISSTGSYI | ARMVRNTVTAFDY |
| (392) | (393) | (394) | |
| EV-D68-219 | GYRLIDLP | FDPEKAEA | ATWGVEVVNGRRDYFDS |
| (395) | (396) | (397) | |
| EV-D68-220 | GFTFSSFE | ISTSGSTI | ARDVRDCSALTCPRRGDAFDF |
| (398) | (399) | (400) | |
| EV-D68-221 | GFSFSVYP | ISSSSRYI | VKVGGSKHQYYFDY |
| (401) | (402) | (403) | |
| EV-D68-224 | GYTFTNYD | ISTYNGNT | ARERCSTSTCYSRYADY |
| (404) | (405) | (406) | |
| EV-D68-225 | GFTLSRYE | ISSGGPSI | MREGLTYYDSTI |
| (407) | (408) | (409) | |
| EV-D68-227 | GFTFDEYA | ISWNGGSK | AKDDYEGAGFDI |
| (410) | (411) | (412) | |
| EV-D68-228 | GYLISNGYY | IYYTRDT | VRHEGSCNDGSCYGSFVDN |
| (413) | (414) | (415) | |
| EV-D68-231 | DFTFSSYT | ISGDGVST | ARGGTFHNWYFDL |
| (416) | (417) | (418) | |
| EV-D68-234 | GFTFSTYA | ISGSGGST | AKGTITYSYYYMAV |
| (419) | (420) | (421) | |
| EV-D68-235 | GYRLTDLP | VDLEKREI | ATWGIEVVNGRDEFFDS |
| (422) | (423) | (424) | |
| EV-D68-236 | GYSLSDLP | FDPINGEI | ATWGVAVVSGRRDYFDS |
| (425) | (426) | (427) | |
| EV-D68-241 | GGSITSGDYY | IYHSGTT | ARAYAYEFWSGYPNWFDP |
| (428) | (429) | (430) | |
| EV-D68-247 | GFIFNRYA | ISYDGINK | ARGLGYCSGTGGSCTPFEY |
| (431) | (432) | (433) | |
| EV-D68-254 | GASLNDYD | INRRDTV | VRVPRRGFEGSFGFCDDTACRYGHTWFDP |
| (434) | (435) | (436) | |
| EV-D68-260 | GFSLRNGRMG | IFASDEK | ARILKFGTMRAAYYFDY |
| (437) | (438) | (439) | |
| EV-D68-266 | GFTFSDYY | ISSSGSTI | AGSKVGYTTGRRNWYFDL |
| (440) | (441) | (442) | |
| EV-D68-269 | GFTFTDFY | INPETGET | ARDLVVVVPVEMSRRAFDI |
| (443) | (444) | (445) | |
| EV-D68-271 | GYIFSAYY | MNAKSGGA | ARDYRDDYMWGSYRPLDY |
| (446) | (447) | (448) |
| 클론 |
CDRL1
(서열번호) |
CDRL2
(서열번호) |
CDRL3
(서열번호) |
| EV-D68-37 | ALPKKY | EDT | SSTDSSGNPVL |
| (449) | (450) | (451) | |
| EV-D68-40 | QSVSTY | EAS | QQRSSWPIT |
| (452) | (453) | (454) | |
| EV-D68-41 | SGSVSSSYY | SIN | GLYMGSGIWI |
| (455) | (456) | (457) | |
| EV-D68-43 | QGISSW | AAS | QQADSFPRT |
| (458) | (459) | (460) | |
| EV-D68-46 | QSISKW | KAS | QQHNSYSYT |
| (461) | (462) | (463) | |
| EV-D68-48 | QSVRSY | DAS | QQRSTWPPGM |
| (464) | (465) | (466) | |
| EV-D68-71 | QSVSSSF | GAS | QQYSNSRLT |
| (467) | (468) | (469) | |
| EV-D68-72 | SSDVGGYNY | EVT | SSYTTSNTLVV |
| (470) | (471) | (472) | |
| EV-D68-74 | NIGTKS | NDS | QVWDSGIDVV |
| (473) | (474) | (475) | |
| EV-D68-75 | NIGTKT | YDS | RVWDSDTDHRV |
| (476) | (477) | (478) | |
| EV-D68-76 | KLGDKY | QDT | QAWDSSTVV |
| (479) | (480) | (481) | |
| EV-D68-80 | QDIGVD | GAS | LQDYNYPWT |
| (482) | (483) | (484) | |
| EV-D68-84 | ALPKQY | QDT | QSGDSSGTYLV |
| (485) | (486) | (487) | |
| EV-D68-85 | QRVGNS | DAS | HQHSTWPRGT |
| (488) | (489) | (490) | |
| EV-D68-88 | SGSIATNY | EDS | QSYDNSNRAVV |
| (491) | (492) | (493) | |
| EV-D68-89 | SGTIASNY | EDN | QSYDNSDRV |
| (494) | (495) | (496) | |
| EV-D68-95 | NIGLKS | YDS | QVWDSSRNHPV |
| (497) | (498) | (499) | |
| EV-D68-97 | KLGDKY | QDS | QAWDSSTAV |
| (500) | (501) | (502) | |
| EV-D68-98 | QGISRF | SAS | QQLNSHPRMFT |
| (503) | (504) | (505) | |
| EV-D68-105 | QGISNW | DAS | QQANSFPFT |
| (506) | (507) | (508) | |
| EV-D68-110 | ALPKQY | EDN | QSADSSGTYVV |
| (509) | (510) | (511) | |
| EV-D68-111 |
NIGLKS | YDS | QVWDSSRNHPV |
| (512) | (513) | (514) | |
| EV-D68-114 | SSDVGGYNF | DVT | GAYAGFNAL |
| (515) | (516) | (517) | |
| EV-D68-116 | AVPIKY | EDD | YSTDSSGYQRA |
| (518) | (519) | (520) | |
| EV-D68-150 | QSVGTD | DAF | QQRSRWPPPYT |
| (521) | (522) | (523) | |
| EV-D68-151 | QSVGTD | DAF | QQRSRWPPPYT |
| (524) | (525) | (526) | |
| EV-D68-152 | QSVRSW | KAS | QQYQTFSWT |
| (527) | (528) | (529) | |
| EV-D68-154 | QSLLQSDGYSY | LGS | MQALQTPWT |
| (530) | (531) | (532) | |
| EV-D68-155 | QSVLFSSNNKNY | WAS | QQFYTTPLT |
| (533) | (534) | (535) | |
| EV-D68-156 | TSNIETNY | RND | AAWDDSLKAPV |
| (536) | (537) | (538) | |
| EV-D68-157 | STNIGAGYD | GNS | QSYDRSLSTYV |
| (539) | (540) | (541) | |
| EV-D68-158 | NIGTKS | DDS | QVWDSYNVHYV |
| (542) | (543) | (544) | |
| EV-D68-159 | SSNIEYNY | KNN | AAWDDILSGVV |
| (545) | (546) | (547) | |
| EV-D68-160 | SNDVGGYNF | DVI | CSYAGTYTWV |
| (548) | (549) | (550) | |
| EV-D68-161 | QRVVNNY | GAS | QQYGSPWT |
| (551) | (552) | (553) | |
| EV-D68-162 | QSISKW | KAS | QQHNSYSYT |
| (554) | (555) | (556) | |
| EV-D68-163 | QSVISTY | DVS | HQYGSSPAT |
| (557) | (558) | (559) | |
| EV-D68-164 | QSVGTY | DSA | QLRITWPPIFT |
| (560) | (561) | (562) | |
| EV-D68-165 | SSDVGGYNY | EVS | SSYAGSNNLV |
| (563) | (564) | (565) | |
| EV-D68-166 | TSSIGSNI | INN | AAWDDSLNGWV |
| (566) | (567) | (568) | |
| EV-D68-181 | QSVGST | GAS | HQYINWPPWT |
| (569) | (570) | (571) | |
| EV-D68-183 | SSDVGAYSY | DVY | CSHAGSHTWV |
| (572) | (573) | (574) | |
| EV-D68-185 | QSISRN | GAS | QQYSKLPIT |
| (575) | (576) | (577) | |
| EV-D68-200 | QSVRSY | DAS | QQRSYWPPFT |
| (578) | (579) | (580) | |
| EV-D68-208 | QSVSSY | DAS | QQRSDWPPGT |
| (581) | (582) | (583) | |
| EV-D68-210 | SSDVGGYNF | EVI | CSYGGNNSWM |
| (584) | (585) | (586) | |
| EV-D68-219 | VLSNQY | KDT | QSADNTRITV |
| (587) | (588) | (589) | |
| EV-D68-220 | QSLVYSDGNTY | KVS | MQGTHWPRT |
| (590) | (591) | (592) | |
| EV-D68-221 | RSNIGSYT | NNN | AAWDDSLNGVV |
| (593) | (594) | (595) | |
| EV-D68-224 | QSINIY | AAS | QQSYRSPRT |
| (596) | (597) | (598) | |
| EV-D68-225 | SGHSRYA | INSDGRH | QTWGTGFRV |
| (599) | (600) | (601) | |
| EV-D68-227 | QSVSSY | GAS | QQRSNWPIT |
| (602) | (603) | (604) | |
| EV-D68-228 | QDISSW | AAS | QQADSFIT |
| (605) | (606) | (607) | |
| EV-D68-231 | QSIGDN | GAS | QQYKNWPRT |
| (608) | (609) | (610) | |
| EV-D68-234 | QSVRSSY | GAS | QQYGTSIT |
| (611) | (612) | (613) | |
| EV-D68-235 | VLSKQY | KDT | QSADTRITV |
| (614) | (615) | (616) | |
| EV-D68-236 | VLSNQY | KDT | QTADTKYTV |
| (617) | (618) | (619) | |
| EV-D68-241 | QRISTY | AAS | QQSYSPPWT |
| (620) | (621) | (622) | |
| EV-D68-247 | QSIKKY | GAS | QQSDSAPPT |
| (623) | (624) | (625) | |
| EV-D68-254 | QSIRDY | AGS | QQSYLTPPT |
| (626) | (627) | (628) | |
| EV-D68-260 | NIGIRT | YDS | QVWDSSSDHVV |
| (629) | (630) | (631) | |
| EV-D68-266 | SSDVGAYNY | EVT | SSYAGNNNLV |
| (632) | (633) | (634) | |
| EV-D68-269 | NIERKS | DDT | QVWDSTTDHGV |
| (635) | (636) | (637) | |
| EV-D68-271 | QSVSGY | DAS | QQRSNGLT |
| (638) | (639) | (640) |
*****************
본 명세서에 개시되고 청구된 모든 조성물 및 방법은 본 개시에 비추어 과도한 실험 없이 제조 및 실행될 수 있다. 본 개시의 조성물 및 방법은 바람직한 실시양태의 관점에서 설명되었지만, 당해 기술분야의 당업자에게는, 본 개시의 개념, 사상 및 범위를 벗어나지 않고도 본 명세서에 기재된 조성물 및 방법, 및 단계 또는 단계의 순서에 변경을 적용할 수 있음이 명백할 것이다. 보다 구체적으로, 화학적으로 및 생리학적으로 모두 관련된 특정 약제가 본 명세서에 기재된 약제를 대체하여도, 동일하거나 유사한 결과가 달성되는 것은 명백할 것이다. 당업자에게 자명한 이러한 모든 유사한 대체물 및 수정은 첨부된 청구범위에 의해 정의된 개시의 정신, 범위 및 개념의 범위 내에 있는 것으로 간주된다.
VII. 참조문헌
하기 참조문헌은 이들이 본 명세서에 기재된 것을 보충하는 예시적 절차 또는 기타 상세를 제공하는 범위에서 참조로 의애 본 명세서에 구체적으로 도입된다.
미국 특허 제3,817,837호
미국 특허 제3,850,752호
미국 특허 제3,939,350호
미국 특허 제3,996,345호
미국 특허 제4,196,265호
미국 특허 제4,275,149호
미국 특허 제4,277,437호
미국 특허 제4,366,241호
미국 특허 제4,472,509호
미국 특허 제4,554,101호
미국 특허 제4,680,338호
미국 특허 제4,816,567호
미국 특허 제4,867,973호
미국 특허 제4,938,948호
미국 특허 제5,021,236호
미국 특허 제5,141,648호
미국 특허 제5,196,066호
미국 특허 제5,563,250호
미국 특허 제5,565,332호
미국 특허 제5,856,456호
미국 특허 제5,880,270호
미국 특허 제6,485,982호




SEQUENCE LISTING
<110> VANDERBILT UNIVERSITY
<120> HUMAN MONOCLONAL ANTIBODIES TO ENTEROVIRUS D68
<130> VBLT.P0294WO
<140> Unknown
<141> 2020-07-24
<150> 62/878,955
<151> 2019-07-26
<150> 62/899,503
<151> 2019-09-12
<150> 63/047,330
<151> 2020-07-02
<160> 642
<170> PatentIn version 3.5
<210> 1
<211> 369
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 1
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcatgtgcag cctctggatt catcttcagt cgctacgctc tgcactgggt ccgccaggct 120
ccaggcaagg ggctggactg ggtggcagtt atatcatatg atgcaagaaa ttcatattac 180
acagactccg tgaagggccg attcaccatc tccagagaca attccaaaaa cacgctgttt 240
ctgcagatga acagtctgag agctgacgac acggctgtct attactgtgc gagaccgact 300
ttgccctaca gcaacaactg gtacgcgcct gaatactggg gccagggaac cctggtcacc 360
gtctcctca 369
<210> 2
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 2
tcctatgagc tgacacagcc accctcggtg tcagtgtccc caggacaaac ggccaggatc 60
acctgctctg gagatgcatt accaaaaaaa tatgcttctt ggtaccagca gaagtcaggc 120
caggcccctg tgctggtcat ctatgaagac accaaacgac cctccgggat ccctgagaga 180
ttctctggct ccagctcagg gacaatggcc accttgacta tcagtggggc ccaggtggaa 240
gatgaaagtg actactactg ttcctcgaca gacagcagtg gtaatcctgt gctattcggc 300
ggagggacca agttgaccgt ccta 324
<210> 3
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 3
gaggtgcagc tggtagagtc tgggggagac ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt ctcctttagc agctatgcca tggcctgggt ccgccaggct 120
ccagggaagg ggctgcagtg ggtctcatct attagtggta acggtaatgg gagatcctat 180
gcagattctc tgaagggccg gttcaccacc tccaaagacc tttccaagta taccctgtat 240
ctgcaaatga acaatctgag acccgaggac acggccatat attactgtgc gaaagttgtc 300
cgtatagcag ctgttttgta ttactttgac tattggggcc cgggaaccca ggttaccgtc 360
tcctca 366
<210> 4
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 4
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc acctacttag cctggtacca acaaaagcct 120
ggccaggctc ccaggctcct catctatgaa gcatccacca gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca tcatcagcag cctagagcct 240
gaagattttg cagtttatca ctgtcagcag cgcagctcct ggccgatcac cttcggccaa 300
gggacacgac tggagattga a 321
<210> 5
<211> 369
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 5
gacgtgcagc tggtggagtc tggggggggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgctg cctctggatt cacttttagc aactatgcca tgacctgggt ccgccaggct 120
ctagggaagg ggctggagtg ggtctcttct attagtggta gtggtggcct cacatatttc 180
gcacactccg tgaagggccg gctcaccatc tccagagaca actccaagaa taccctctat 240
ctgcaaatga gcagcctgag agccgaagac acggccgtat attactgtgc gagagtgaaa 300
agtacaactg gaacgacggc gttagttttt gatatctggg gccaagggac aatggtcacc 360
gtctcttcg 369
<210> 6
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 6
cagactgtgg tgacccagga gccatcgttc tcagtgtccc ctggagggac agtcacactc 60
acttgtggct tgagttctgg ctcagtctct agtagttact accccagttg gtaccagcag 120
accccaggcc aggctccacg cacgctcatc tacagtataa acagacgttc ttctggggtc 180
cctgatcgct tctctggctt catccttggg aacaaagctg ccctcaccat caggggggcc 240
caggcagatg atgaatctga ttattactgt gggctgtata tgggtagtgg catttggatc 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 7
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 7
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcatt aactatggca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcaaatg atggaagtta taactacgat 180
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa caaggtttat 240
ctacaaatga acagcctgag acctgaggac acggctgtgt atttctgtgc gaaagacaaa 300
cacggtgact tcgactacta cggtgtggac gtctggggcc aagggaccac ggtcaccgtc 360
tcccca 366
<210> 8
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 8
gacgtccaga tgacccagtc tccatcttcc gtgtctgcat ctataggaga cagagtcacc 60
atcacttgtc gggcgggtca gggaattagc agctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccaatt tgcaaagtgg ggtcccatca 180
cggttcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg gaacttacta ttgtcaacag gctgacagtt tccctcggac gttcggccaa 300
gggaccaagg tggaaatcaa a 321
<210> 9
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 9
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agttatggca tacactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcctatg atggaagtga taacacctat 180
gcaccttttg tgaacggccg attcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agctgacgac acggctgtgt attactgtgc gaggcgtcgg 300
cctgggagct tcccaggact ttgcgactac tggggccagg gagccctggt caccgtctcc 360
tca 363
<210> 10
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 10
gacatccaga tgacccagtc gccttccact atgtctgctt ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gagtattagt aagtggttgg cctggtatca gcagaagcca 120
gggaaagccc ctaaactcct gatctataag gcgtctacct tacaaactgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagaa ttcactctca ccatcaacag cctgcagcct 240
gatgattttg caacttatta ctgccaacag cataatagtt attcgtacac ttttggccag 300
gggaccaagg tggagatcaa g 321
<210> 11
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 11
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg cagcttcagc agacttacta tcatctgggt gcggcaggcc 120
cctggacaag ggcttgagtg gatgggaggg cacatcccta tctttggaac aacaaactac 180
gcactgaagt tccagggcag agtcacgatt accgcggaca aaaccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccatat attactgtgc gagaatgtat 300
agtggccatg acggcgttga tgtctggggc caagggacac tggtcaccgt ctcttca 357
<210> 12
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 12
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagg agctacttag cctggtacca acacaaacct 120
ggccaggctc ccaggctcct catctatgat gcatccaaca gggccaaggg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagagcct 240
gaagattttg cagtttatta ctgtcagcag cgtagcacct ggcctccggg aatgttcggc 300
caagggacca gggtggaaat caaa 324
<210> 13
<211> 381
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 13
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc attcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtggttttt actactgggg ctggatccgc 120
cagcccccag ggaaggggct agagtggatt gggactatct atgatagtgg gagaacctat 180
gacaacccgt ccctcaagag tcgagtcacc atatccgcag acacgtccaa gaagcagttc 240
tcactgacac tgaggtctgc gaccgccgcg gacacggctg tgtatttctg tgcgagacac 300
cttacccacc tctacggtga ctacgtcacc cctgatgctt tagatatctg gggccaaggg 360
acaatggtca ccgtctcttc a 381
<210> 14
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 14
gaaattgtgt tgacgcagtc tccaggcacc ctgtccttgt ctccagggga aagagccacc 60
ctctcttgca gggccagtca gagcgttagt agcagcttct tagcctggta ccagcagaaa 120
cctggccagg ctcccaggct cctcatctat ggtgcgtcca gcagggccac cggcatccca 180
gacaggttca gaggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtata ttactgtcag cagtatagta actcacgtct gacgttcggc 300
caagggacca aggtggaaat caaa 324
<210> 15
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 15
gaggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcact acctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg gatttcatac attagtagtg gtagtagtaa catatactac 180
gcagactctg tgaagggccg attcaccatc tccagagaca atgccaagaa ctcactgaat 240
ctgcaaatga gcagcctgag agacgaggac acggctgtgt attactgtgc gagagcccac 300
ggacgtatag tgaattctgg agtggttatt agtaggttcg acccctgggg ccagggaatc 360
ctggtcaccg tctcctca 378
<210> 16
<211> 333
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 16
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ggttataact atgtctcctg gtaccaactg 120
cacccaggca aagcccccaa actcatgatt tttgaggtca cttatcggcc ctcaggggtt 180
tctaatcgct tctctggctc caagtctggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttatttctgc agctcctata caaccagcaa cactctcgtg 300
gtgttcggcg gagggaccaa gctgaccgtc cta 333
<210> 17
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 17
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgtagga cttctggata caccttcacg gcctactata tgcactgggt gcgacaggcc 120
cccggacaag ggcctgagtg gatgggaagg atcaacccga gcagtggtgg cgcacagtat 180
gcacagaagt ttcagggcag ggtcaccatg accagggaca cgtccatcag cacaacctac 240
atgaccctga gcgggctgac atctgacgac acggccgtgt tttactgtgc gagaatgggt 300
tgtcgtagtg accggtgcta ttcgaccaac tacaactttg accagtgggg ccagggaacc 360
ctggtcaccg tctcctca 378
<210> 18
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 18
tcctatgtgc tgactcagcc accctcggtg tcagtggccc caggacagac ggccaggatt 60
ccctgtgggg gaaacaacat tggaactaaa agtgttcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcgt ctctaatgac agcgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaagtctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattattg tcaggtgtgg gatagtggta ttgatgtcgt tttcggcgga 300
gggaccaagc tgaccgtcct a 321
<210> 19
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 19
gaggtgcagc tggtggagtc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cactatcagt ccctatggca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcattc attagtagta gtagtcgtta cacatattac 180
gcagactcag tgaagggccg tttcaccatc tccagagaca acgccaagaa ttcactgtct 240
ctgcaaatga acagcctgag agccgaggac acggctgtgt attactgtgc gagagagagg 300
ggccatagca cctcgtcctc atactttgac tcctggggcc agggaaccct ggtcaccgtc 360
tcctca 366
<210> 20
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 20
tcctatgtgc tgactcagcc accctcagtg tcattggccc caggaaagac ggccaggatt 60
acctgtgggg gaaacaacat tggaactaaa actgtgagct ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat gtattatgat agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcaacagggt cgaggccggg 240
gatgaggccg actattactg tcgggtgtgg gatagtgata ctgatcatcg agtgttcggc 300
ggggggacca agctgaccgt ccta 324
<210> 21
<211> 381
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 21
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggatt cactttcagt aacgcctgga ttagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggttggccgt attcaaacca aaactgatgg agggacaaca 180
gactacgctg cacccgtgaa aggcagattc accatctcaa gagatgattc aaaaaacacg 240
ttgtatctgc aaatgaacag cctgaaaacc gaggacacag ccttgtatta ttgtagcaca 300
ggaccgtatt actatgatac tagtggttac ccccaaccct ttgactactg gggccaggga 360
accctggtca ccgtctcctc a 381
<210> 22
<211> 318
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 22
tcctatgagc tgactcagcc accctcagtg tccgtgtccc caggacagac agccagcatc 60
acctgctctg gagataaatt gggagataaa tatgcttgct ggtatcagca gaagccaggc 120
cagtcccctg tgctggtcat ctatcaagat accaagcggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacagcc actctgacca tcagcgggac ccaggctatg 240
gatgaggctg actattactg tcaggcgtgg gacagcagca ctgtggtatt cggcggaggg 300
accaagctga ccgtccta 318
<210> 23
<211> 354
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 23
gaggtacaga tggtggagtc tgggggaggc ctggtcaagc cgggggggtc cctcagactc 60
tcctgttcag tctctggatt cgacttcagt agatacacca tgaactggtt ccgccaggct 120
ccaggggagg ggctgaagtg ggtctcgtcc attagtagta ctagtcttta cacattctat 180
gcggactcag tgaagggccg attctccatc tccagagaca acgcccaggg ttccctgtct 240
ctacaaatga gcagcctgag acccgaagat acggctgtct attattgtgc gagagtcgtt 300
ggtcccgccg agttagatta ctggggccag ggagtgctgg tcaccgtctc ctca 354
<210> 24
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 24
gtcacccagt tgacccagtc tccatcctcc ctgtctgctt ctgtcgggga cagagtcacc 60
atcacttgcc gggcaagtca ggacattgga gttgacttag gttggtttca gcagagacct 120
gggaaagccc ctaaactcct gatctatggt gcctccaggt tgcagagtgg ggtcccatca 180
cgcttcagcg ggcgtggatc tggcacattt ttcactctca ccatcagcag cctgcagcct 240
gaagatttta caacttactt ctgtcttcaa gattataatt acccttggac gctcggccag 300
gggaccacgg tgggagtcaa a 321
<210> 25
<211> 351
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 25
caggtgcacc tggtgcagtc tgggtctgcg gtgaggaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggata caccttcacc gactactata ttcactgggt gcgacaggcc 120
cctggacaag gccttgagtg gatgggatgg atcaacccta aaactggtgg ctcaaattat 180
acacagaggt ttcaggccag ggtcaccatg acctgggaca cgtccatcag tacagcctac 240
atggagttga gcaggctgag atctgacgac acggccgtgt attattgtgc gagggcgggc 300
agaaatggct acgactactg gggccaggga accctggtca ccgtctcctc a 351
<210> 26
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 26
tcctatgagc tgacacagcc accctcggtg tcagtgtccc caggacagac ggccaggatc 60
acctgctctg cagatgcatt gccaaagcaa tatgcttatt ggtaccaaca gaagccaggc 120
caggcccctg tgttgatgat atatcaagac actgagaggc cctcagggat ccctgagcga 180
ttctctgggt ccagctcagg gacaacagtc acgttgacca tcagtggagt ccaggcagaa 240
gacgaggctg actattactg tcaatcagga gacagcagtg gtacttatct agttttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 27
<211> 360
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 27
gaggtacagc tcttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caaatttaga aactatgcca tgacctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaact attactagtg gtggtagtac agagtacgca 180
gacgccgtga agggccggtt catcatctcc agagacaatt ccaagaacac gttatatttg 240
caaatgaaca gcctgagagc cgacgacacg gccgtatatt actgtacagt gccgtggggt 300
aactacaatg actacgtgtc tgactactgg ggccagggaa ccctggtccc cgtctcctca 360
<210> 28
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 28
gaagttgtat tgacacagtc tccagccacc ctgtctttgt ctccagggca aagagccacc 60
ctctcctgca gggccagtca gcgtgttggc aactccttag cctggtacca acaaaaacct 120
ggccaggctc ccagcctcct catctatgat gcttccaaga gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcatcag cctagagtct 240
gaagattttg cagtttatta ctgtcaccaa catagcacct ggcctcgggg gaccttcggc 300
caagggacac gactggagat taaa 324
<210> 29
<211> 372
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 29
caggtgcacc tggttgagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt catcttcagt agatatccta tgcactgggt ccgccaggct 120
ccaggcaagg gtctggagtg ggtggcactt atatcatatg atggaaacaa taaatactac 180
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgttt 240
ctccaaatga acagcctgag agctgaggac acggctgtct attactgtgc gagacatttc 300
ctcccatata gcagtagttg gtaccagggc tttaactact ggggccaggg aatcctggtc 360
accgtctcct ca 372
<210> 30
<211> 336
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 30
aattttatgc tgactcagcc ccactctgtg tcggagtctc cggggaagac ggtaaccatc 60
tcctgcaccc gcagcagtgg cagcattgcc accaactatg tgcagtggta ccagcagcgc 120
ccgggcagtt cccccacccc tataatcttt gaagatagtc aaagaccgtc tggggtccct 180
gatcggttct ctggctccat cgacagctcc tccaattctg cctccctcac catctctgga 240
ctgaggactg acgacgaggc tgactactac tgtcagtctt atgataacag caatcgggct 300
gttgtattcg gcggagggac caagctgacc gtccta 336
<210> 31
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 31
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgaag cgtctggatt cctcttcagt cgctatggca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggactg ggtggcagtt atatcgtatg atggaaataa gaaatattat 180
gcagactctg tgaagggccg attcaccatc tccagagaca attcaaagaa cacgttgtat 240
ctgcaagtga acagcctgag agtcgaggac acggctgttt attactgtgc gagaggtgtc 300
ccgtacggtg acacccttac agggcttgtc tactggggcc agggaaccct ggtcaccgtc 360
tcctca 366
<210> 32
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 32
aattttatgc tgactcagcc gcactctgtg tcggagtctc cggggaagac ggtaaccatc 60
tcctgcaccc gcagcagtgg caccattgcc agcaactatg tgcagtggta ccagcagcgc 120
ccgggcagtg cccccaccac tgtaatctat gaggataacc aaagaccctc tggggtccct 180
gatcggttct ctggctccat cgacagctcc tccaactctg cctccctcac catctctgga 240
ctgaagactg aggacgaggc tgactactac tgtcagtctt atgataacag cgatcgggtg 300
ttcggcggag ggaccaagct gaccgtcctt 330
<210> 33
<211> 375
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 33
caggtcacct tgaaggagtc tggtcctgtg ctggtgaaac ccacagagac cctcacgctg 60
acctgcaccg tctctgggtt ctcactcagg aacgctagaa tgggagtgag ctggatccgt 120
cagcccccag ggaaggccct ggagtggctt gcacacattt tttcgaatga cgaaaaatcc 180
tacaacacat ctctgaagag cagactctcc atctccaagg acacctccaa aagccaggtg 240
gtccttacca tgaccagcat ggaccctttg gacacagcca catatttctg tgcacggcta 300
ctggtggctg gtactttcct cccctctcac tactttgact actggggcca aggaatcctg 360
gtcaccgtct cctca 375
<210> 34
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 34
tcctatgtgc tgactcagcc accctcagtg tcagtgaccc caggaaagac ggccaggatt 60
acctgtgggg gaaacaacat tggacttaaa agtgtcttct ggtaccagga gaggccagac 120
caggcccctg tggtggtcat ctattatgat agcgcccggc cctcagggat ccctgagcga 180
atctctggct ccaagtctgg gaacacggcc accctgacca tcaccagggt cgaagccggg 240
gatgaggccg actatttttg tcaggtgtgg gatagtagtc gtaatcatcc ggtcttcggc 300
ggagggacca aactgaccgt cctc 324
<210> 35
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 35
gagctgcagc tggtggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt acctatagca tgaattgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtttcatac attagtagta gtagtagtac catacagtac 180
gcagactctg tgaagggccg attcaccatc tccagagaca atgccaagaa ctcactgtat 240
ctgcaaatga atagcctgag agccgaagac acggccgtgt attattgtac gagacaggtc 300
ggggcggatt tcagtggccg cggctttgac tactggggcc agggaaccct gctcaccgtc 360
tcctca 366
<210> 36
<211> 318
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 36
tcctatgagc tgactcagcc accctcagtg tccgtgtccc caggacagac agccaccatc 60
acctgctttg gagataaatt gggggataaa tatgcttgct ggtatcagca gaagccaggc 120
cagtcccctg tgttggtcat ctaccaagat agcaagcggc cctcagggat ccctgagcga 180
ttctctggct ccaagtctgg gaacacagcc actctgacca tcagcgggac ccaggcaatg 240
gatgaggctg actattactg tcaggcgtgg gacagcagca ctgcagtgtt cggcggaggg 300
accaagctga ccgtcctc 318
<210> 37
<211> 360
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 37
gaggtacaac tagtggagtc agggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caagttttcc gtctatgcct tgagttgggt ccgccaggct 120
ccagggaagg ggctggagtg gatttcatat attagtagta gtggttctac catatattat 180
tcagactctg tgaagggccg attcaccatc tccagagaca atgtcgggaa ctcactgttt 240
gtgcaaatga acagcctgag agccgaggac acgggtattt attactgtgc gacagcccgc 300
cacatcacca atgatggttt tgatatttgg ggccaaggga caatggtcat cgtctcttca 360
<210> 38
<211> 327
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 38
gacatccagt tgacccagtc tccatccttc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gggcattagt aggtttttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctattct gcgtccactt tacaaagggg ggtcccatca 180
aggttcagcg gcagtggatt tgggacagat ttcactctca caatcagtag ccttcagcct 240
gaagattttg caacttatta ctgtcaacaa cttaatagtc acccccgaat gttcactttc 300
ggccctggga ccacagtgga tatcaag 327
<210> 39
<211> 381
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 39
caggtgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcgctg tgtctggtta cttaatcagc aatggttact actggggctg gatccggcag 120
ccccccggga aggggctgga gtggattggg agtatctatc atactagaag cacctactac 180
aacccgtccc tcaagagtcg agtcagcatc tcagtagaca cgtccaagaa ccggttctcc 240
ctgaggctga ggtctgtgac cgccgcagac acggcctttt attactgtgc gagaggccca 300
ggccactgtt atggtgatga cgactgctac gcgtactact ttgaccagtg gggccaggga 360
accccggtca ccgtctcccc a 381
<210> 40
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 40
gacatccaga tgacccagtc tccatcttcc gtgtcttcat ctgtaggaga cagagtcacc 60
atcacttgtc gggcgagtca gggtattagc aactggttag cctggtatca gcagaaccca 120
gggaaagctc ctaaactcct gatctatgat gcctccagtt tgcaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag cctgcagcct 240
gaagattttg caacttacta ttgtcaacag gctaacagtt tccctttcac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 41
<211> 381
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 41
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctgggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg caccttcagc aggtttgcta tcagctgggt gcgacaggcc 120
cctggacaag ggcttcagtg gatgggaggg atcctcccta tctttggtac agcaaactac 180
gcacagaagt tccagggcag agtcacgatt accgcggaca catccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtgc gagatccctc 300
ccctattgta ctaatgatgt atgctcaaac cagaacacat ttgactactg gggccaggga 360
accctggtca ccgtctcctc a 381
<210> 42
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 42
tcctatgagc tgacacagcc accctcggtg tcagtgtccc caggacaggc ggccaggatc 60
acctgctctg gagatgcatt gcctaagcaa tatgcttatt ggtaccagca gaagccaggc 120
caggcccctg tgttggtgat atatgaagac aataagaggc cctcagggat ccctgagcga 180
ttctctggct ccagctcagg gacaacagtc acgttgacca tcagtggagt ccaggcagaa 240
gacgaggctg actattactg tcaatcagca gacagcagtg gtacttatgt ggtattcggc 300
ggagggacca agctgaccgt ccta 324
<210> 43
<211> 369
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 43
caggtgcagc tgcaggagtc gggcccagga ctggtgaagc cttcacagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtggtgatt actactggag ttggatccgc 120
cagcccccag ggaagggcct ggagtggatt gggtacatct attacagtgg gagcacctac 180
tacaacccgt ccctcaagag tcgagttacc atatcagtag acacgtccaa gaaccagttc 240
tccctgaagc tgagctctgt gactgccgca gacacggccg tgtattactg tgccagccgc 300
tacggtgacc cgatagggga caactggttc gacccctggg gccagggaac cctggtcacc 360
gtctcctca 369
<210> 44
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 44
tcctatgtgc tgactcagcc accctcagtg tcagtgaccc caggaaagac ggccaggatt 60
acctgtgggg gaaacaacat tggacttaaa agtgtcttct ggtaccagga gaggccagac 120
caggcccctg tggtggtcat ctattatgat agcgcccggc cctcagggat ccctgagcga 180
atctctggct ccaagtctgg gaacacggcc accctgacca tcaccagggt cgaagccggg 240
gatgaggccg actatttttg tcaggtgtgg gatagtagtc gtaatcatcc ggtcttcggc 300
ggagggacca aactgaccgt cctc 324
<210> 45
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 45
gaggtgcaac tgttggagtc ggggggaggc ttggtgcagc cgggggggtc cctgagactc 60
tcctgcgcag cctccggatt caggtttagc ttctatggca tgacctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaagt attagtggta ctggtgctac cagaaactgc 180
gcagactccg tgaagggccg gttcaccatc tccagagaca actccaagaa cacgctgtac 240
ctgcaaatgg acagcctgag agtcgacgac acggccgttt tttattgtgt gagacggttc 300
ccgatgacca cggtgacaag ctttgactct tggggccagg gaaccctggt caccgtctcc 360
tca 363
<210> 46
<211> 327
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 46
cagtctgccc tgactcagcc tcgctcagtg tccgggtctc ctggacagtc agtcaccatc 60
tcctgcactg gaaccagcag tgatgttggt ggttataact ttgtctcctg gtaccaacaa 120
cacccgggca aagcccccaa actcatgatt tttgatgtca ctgggcggcc ctcaggggtc 180
cctgatcgct tctctggctc caagtctggc aacacggcct ccctgaccat cgctgggctc 240
caggctgaag atgaggctga ttattattgt ggcgcatatg cgggctttaa cgctcttttc 300
ggcggaggga ccaaactgac cgtccta 327
<210> 47
<211> 375
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 47
gaggtgcagc tggtgcagtc ggggggaggc ttggtccggc cgggggggtc cgtgagactc 60
tcctgtgtag cctctggatt ccccttcaat atgttttgga tgggctgggt ccgccagact 120
ccagggaagg gactggagtg ggtggccaat ataaaacagg atggcagtga gaaatactat 180
gtcgactccg tgaagggccg attcgccatc tccagagaca atgccaagaa ctctctcttt 240
cttcaaatgg acagcctgag tgtcggggac acggccatct attattgtgt cagagagggg 300
gtgcgaaggg tcgtcgtacg tagtaccggt tacttcgact tttggggcca gggacagctg 360
gtcaccgtct cctca 375
<210> 48
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 48
tcctatgagc tgacacagcc accctcgatg tcagtgtccc caggacaaac ggccaggatc 60
acctgctctg gagatgcagt gccaataaaa tatgtttatt ggtaccaaca gaggtcaggc 120
caggcccctg tattagtcat ctatgaagac gacagacgac cctccgggat ctctgagaga 180
ttctctggct ccagttcagg gacaacggcc accttgacta tcactggggc ccaggtggag 240
gatgaaggtg actactattg ctattcaaca gacagtagtg gttatcagag agcgttcggc 300
ggggggacca cgctgaccgt ccta 324
<210> 49
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 49
cgggtgcagc tgcaggagtc ggccccagga ctggtgaggc cttcagagac cctgtccctc 60
acctgcagtg tctctggtgg ccccatcagc aatggtcctt attactggag ctggatccgc 120
cagcacccag ggaagggcct ggagtggatt ggattcatct tttacagtgg gagcaccaac 180
tacaacccgt ccctccgggg gcgcgtaacc atggcagtgg acacgtctaa gaaccagttc 240
tccctgaggc tgaactctgt gactgccgcg gacacggccg tttattactg tgcgagacat 300
gtggtgactg cgtcggggtg gttcgacccc tggggccagg gaaccctggt caccgtctcc 360
tca 363
<210> 50
<211> 327
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 50
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttggc accgacttag cctggtacca acagaaacct 120
ggccaggctc ccagggtcct catctatgat gcattcaaga gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagag ttcactctca ccatcagcag cctcgagcct 240
gaagattttg cagtttatta ctgtcagcag cgtagcaggt ggcctccccc gtacactttt 300
ggccagggga ccaagctgga gatcaaa 327
<210> 51
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 51
cgggtgcagc tgcaggagtc ggccccagga ctggtgaggc cttcagagac cctgtccctc 60
acctgcagtg tctctggtgg ccccatcagc aatggtcctt attactggag ctggatccgc 120
cagcacccag ggaagggcct ggagtggatt ggattcatct tttacagtgg gagcaccaac 180
tacaacccgt ccctccgggg gcgcgtaacc atggcagtgg acacgtctaa gaaccagttc 240
tccctgaggc tgaactctgt gactgccgcg gacacggccg tttattactg tgcgagacat 300
gtggtgactg cgtcggggtg gttcgacccc tggggccagg gaaccctggt caccgtctcc 360
tca 363
<210> 52
<211> 327
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 52
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttggc accgacttag cctggtacca acagaaacct 120
ggccaggctc ccagggtcct catctatgat gcattcaaga gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagag ttcactctca ccatcagcag cctcgagcct 240
gaagattttg cagtttatta ctgtcagcag cgtagcaggt ggcctccccc gtacactttt 300
ggccagggga ccaagctgga gatcaaa 327
<210> 53
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 53
caggtgcatc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag actctggagt caccttcagt gacaatgctt tgtactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtcgcagtt atctcatatg atggaagcag tagatactac 180
gcagactccg tgaggggccg gttcaccata tccagagaca attccaagga cacgctgtat 240
ctgcaaatga acagactgag agctgaggac acggctattt attactgtgc gagagtcaca 300
gcggattact atgagagtag tggcaaggtg ttttggggcc agggagccct ggtcgtcgtc 360
tcctca 366
<210> 54
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 54
gacatccaga tgacccagtc tccttccacc ctgtctgcat ctgtggggga cagagtctcc 60
atcacttgcc gggccagtca gagtgttagg agctggttgg cctggtatca gcacaaacca 120
gggaaagccc ctaaactcct gatctataag gcgtctagtt tagaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagaa ttcactctca ccatcagcag cctgcaggct 240
gatgattttg caacttatta ctgccaacag tatcagactt tttcctggac gttcggccaa 300
gggaccacgg tggaagtcaa a 321
<210> 55
<211> 372
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 55
caggtgcagc tgcaggagtc gggcccagca ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagt gatcactact ggagctggat ccggcagccc 120
ccagggaagg gactggagtg gattggctac atctatacca gtgggaccac caactacaac 180
ccctccctca agagtcgagt caccatatca gtagacacat ccaagaagca gttctccctg 240
aatctgaggt ctgtgaccgc cgcagacacg gccgtgtatt actgtgcgag aagtctagaa 300
acggtgatcc gtttctacta ctaccactac atggacgtct ggggcaaagg gaccacggtg 360
atcgtctcat ca 372
<210> 56
<211> 336
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 56
gatattgtga tgactcagtc tccactctcc ctgcccgtca cccctggaga gccggcctcc 60
atctcctgca ggtctagtca gagcctcctg cagagtgatg ggtacagcta tttggattgg 120
tacctgcaga agccagggca gtctccacag ctcctgatct atttgggttc taatcgggcc 180
tccggggtcc ctgacaggtt cagtgtcatt ggatcaggca catattttac actgaaaatc 240
agcagagtgg aggctgagga tgttggcgtt tatttctgca tgcaagctct acaaactccg 300
tggacgttcg gccaagggac caaggtggaa atcaaa 336
<210> 57
<211> 342
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 57
caggtgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tcgctggcgg ctccatcggt gattaccact ggaactggat ccggcagccc 120
gccgggaagg ggctggagtg gattgggcgt atacatagca gtgggaacac tgactacaac 180
ccctccctca agagtcgagt caccatgtca gtagacacgt ccaagaacca gttctccctg 240
aaactgaggt ctgtgaccgc cgcggacacg gccgtgtatt actgtgcgag gcaaaatgtt 300
tttgatatct ggggccaagg gacaatggtc accgtctctt ca 342
<210> 58
<211> 339
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 58
gacatcgtga tgacccagtc tccagactcc ctggctctgt ctctgggcga gagggccacc 60
atcaactgca agtccagcca gagtgtttta ttcagctcca acaataagaa ctacttagct 120
tggtaccagc agaaaccagg acagcctcct aagttgctca tttactgggc atctacccgg 180
gaatccgggg tccctgaccg attcagtggc agcgggtctg agacagattt cactctcacc 240
atcagcagcc tgcaggctga agatgtggca gtttatttct gtcagcaatt ttatactact 300
ccgctcactt tcggcggagg gaccaaggtg gagatcaaa 339
<210> 59
<211> 342
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 59
caggtacaga tgcaggagtc gggcccaggg ctggtgaagg cttcggagac cctgtccctc 60
acctgcagtg tctctggtat ctccatcaat aactactatt ggagttggtt ccgccagccc 120
cccgggaagg gcctggagtg gatcggatat gtctattcta ctgggagttc caagtacaat 180
ccctccctcg agcgtcgagc caccatgtca gtagacacgt ccaacaacaa cttctccctg 240
aggctgacgt ctgtgaccac tgcggacacg gccgtctact actgtgcgcg ggggagtatg 300
ccgcatatct ggggccaggg cctcctggtc accgtctcct ca 342
<210> 60
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 60
cagtctgtgc tgactcagcc accctcagcg tccgggaccc ccggacagag ggtcaccatc 60
tcctgttctg gtagcacctc caacatcgag actaattatg tatactggta ccagcaggtc 120
ccaggaacgg cccccaagcc cctcgtctat aggaatgatc agcgcccctc gggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcggcctccc tggtcatcag tgggctccgg 240
accgaggatg aggctgctta ttattgtgca gcttgggatg acagtctgaa agctccggtc 300
ttcggagctg ggaccaaggt cgccgtcctc 330
<210> 61
<211> 390
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 61
gaggtgcagc tggtggagtc tgggggaggc ttgataaagc cgggggggtc ccttagactc 60
tcctgtgcag cctctggaat cactttcagt aacgcctgga tgagctgggt ccgccaggct 120
ccagggaagg ggctggaatg ggttggccgt attgaaagca aaattgacgg tgggacaata 180
gactacgcta cacccgtgaa aggcagattc accatctcaa gagatgattc aaaaaacacg 240
ctgtacctgc aaatgaacag cctgaaaacc gaggacacag ccgtctatta ctgtaccaca 300
gaccagggct actatgatag aagtggttat tgggtcgtcg ggaaccactt tgactactgg 360
ggccagggaa tcctggtcac cgtctcctca 390
<210> 62
<211> 333
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 62
cagtctgtgc tgacgcagcc gccctcagtg tctggggccc cagggctgag ggtcaccatc 60
tcctgtactg ggagcagcac caacatcggg gcaggttatg atgtacactg gtaccagcac 120
cttccaggaa cagcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactgggctc 240
caggctgacg atgcggctga ttattactgc cagtcctatg acagaagcct gagtacttat 300
gtcttcggaa ctgggaccaa ggtcaccgtc cta 333
<210> 63
<211> 369
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 63
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaggt attagtggta ctacaggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacggtgcat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gaaagattct 300
cactccatga tagtagttga tcatgctttt gatatctggg gccaagggac aatggtcacc 360
gtctcttca 369
<210> 64
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 64
tcctatgtgc tgactcagcc accctcggtg tcagtggccc caggacagac ggccaggatt 60
acctgtgggg gaaacaatat tggaactaaa agtgtgcact ggtaccagca gaggccaggc 120
caggcccctg tgctggtcgt ctatgatgat agcgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatagttata atgttcatta tgtcttcgga 300
acagggacca aggtcaccgt ccta 324
<210> 65
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 65
gaggtgcagc tggtggagtc tggggggggc ctggtcaagc cgggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt acttatatca tgacctgggt ccgccaggcc 120
ccagggaggg ggctggagtg ggtctcatcc attagtacca gtagtgttta cacattctat 180
gcagattcac tgaagggccg gttcaccatc tccagagaca acgccaagaa ttcagtgtat 240
ctgcagatga acagcctgag agccgacgac acggctgttt attactgtgc gagggaagaa 300
gggtttcgag cttataacct atactggggc cagggaaccc tggtcactgt ctcctca 357
<210> 66
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 66
cagtctgtgc tgactcagcc accgtcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcagctc caacatcgaa tacaattatg tttactggta ccagaaattc 120
ccaggaacgg cccccaaact cctcatctat aaaaataatc agcggccctc aggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccattag tgggctccgg 240
tccgaggatg agggtgatta ttactgtgca gcatgggatg acatcctgag tggtgtggtt 300
ttcggcgggg ggaccaagct gaccgtcctc 330
<210> 67
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 67
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc ccagaaactc 60
tcctgtgcag cctctggatt cacgttcagt aggtttggta tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atttcgtttg atggaagtaa tagatactac 180
gcagactccg tgaaggggcg attcaccatc accagagaca attccaagaa cacattgtat 240
ctgcaaatga acaacctgag acctgaggac acggctgtat attactgcgc gagagattgg 300
gataggctgg ttcgttcggc ggttggctac tggggccagg gaaccctggt cagcgtctcc 360
tca 363
<210> 68
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 68
cagtctgccc tgactcagcc tcgctcagtg tccgggtctc ctggacagtc agtcaccatc 60
tcctgcactg gaaccagtaa tgatgttggt ggttataact ttgtctcctg gtatcagcaa 120
cacccaggca aagcccccaa actaatgatt tttgatgtca ttaggcggcc ctcaggggtc 180
cctggtcgct tctctggctc caagtctggc gacacggcct cccttatcat ctctggactc 240
caggctgagg atgaggctga ttattactgc tgctcatatg caggcaccta cacctgggta 300
ttcggcgcag ggaccacact gaccgtccta 330
<210> 69
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 69
caggtgcacc tgcaggagtc gggcccacga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccgtcagc actgccactt actactggag ctggatccgg 120
cagtccccag ggaggggact ggagtggata ggatatatct attccagtgg taacaccaac 180
tacaacccct cccttaagag tcgagtcacc atttctttag acacgcccaa caaccagctc 240
tccctgacgt tgacctctgt gaccgctgcg gacacggcca tttattattg tgagaggcgc 300
ttacgtattc tgagtattga gaggaactac tacgctatgg acgtctgggg ccaagggacc 360
ccggtcaccg tctcctca 378
<210> 70
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 70
gaagttgtgt tgacgcagac tccaggcacc ctgtctttgt ctccggggga aggagccacc 60
ctctcctgca gggccagtca gagggttgtc aacaactact tagcctggta ccagcagaga 120
gctggccagg ctcccaggct cctcattttt ggtgcatcca acagggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag gaagctggag 240
cccgaagatt ttgcagtgta ttactgtcaa caatatggta gcccgtggac gttcggccac 300
gggaccaagg tggaaatgaa a 321
<210> 71
<211> 390
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 71
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agttatgcta tgcattgggt ccgccaggct 120
ccaggcaggg ggctggagtg ggtggcagtt atatcatatg atgcaagcaa gaaataccac 180
gcagactccg tgaagggccg attcacaatc tccagagaca gttccaagaa cacgctgttt 240
ctgcaaatga atagcctgaa acctgaggac acggctgtgt attactgtgc gagagatcat 300
gtccccccca aggattgcag tgatggtaat tgccactcgg actacggtat ggacgtctgg 360
ggccaaggga ccacggtcac cgtctcctca 390
<210> 72
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 72
gacatccaga tgacccagtc gccttccact atgtctgctt ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gagtattagt aagtggttgg cctggtatca gcagaagcca 120
gggaaagccc ctaaactcct gatctataag gcgtctacct tacaaactgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagaa ttcactctca ccatcaacag cctgcagcct 240
gatgattttg caacttatta ctgccaacag cataatagtt attcgtacac ttttggccag 300
gggaccaagg tggagatcaa g 321
<210> 73
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 73
caggtccagc ttgtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaagatt 60
tcttgcaagg cttctggata ctccttcact aactttgctg tgcattgggt gcgccaggcc 120
cccggacaaa gacttgagtg gatgggatgg atcaaccctg gcaatagaaa cacaaagtat 180
tcacacaact ttcagggcag agtcaccatt accagggaca catccgcgaa cacagcctac 240
atggaactga gcagcctgag atctcaagac acggctgtgt attactgtgc gagacttccg 300
atagcagcag ctggcagggg ctggttcgac ccctggggcc agggaaccct ggtcaccgtc 360
tcctcc 366
<210> 74
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 74
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttatc agcacctact tagcctggta ccagcagaga 120
cctggccagg ctcccagggt cctcatctat gatgtatcca ccagggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtcta tttctgtcac cagtatggta gttcaccggc gacgttcggc 300
caagggacca aggtggaaat caaa 324
<210> 75
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 75
gaagtgcagc tggtggagtc tgggggagac ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcaat acttatggca tgaactgggt ccgccaggct 120
ccagggaagg gactggagtg ggtttcatac attagtagtg ccaccactac cttctactac 180
gcagactctg tgaggggccg attcaccatc tccagagaca atgccaagaa ttcactattt 240
ctgcacatga agagcctgag agacgaagac acggcagttt attactgtgc gagagtctat 300
actatgcttc gcggagcgag tatggacgtc tggggccacg ggaccacggt caccgtctcc 360
tca 363
<210> 76
<211> 327
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 76
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttggc acctacctgg cctggtacca acagaaacct 120
ggccaggctc ccaggctcct catctatgat tcagccaaca gggccactgg catcccagcc 180
cggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagagcct 240
gaagattttg cagtttatta ctgtcagctg cgcatcacct ggcctcctat attcactttc 300
ggccctggga ccaaagtgga tgtcaaa 327
<210> 77
<211> 369
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 77
caggtgcacc tggtgcagtc tggggctgag gtgaagaagt ctggggcctc agtgaaggtc 60
tcctgcaaga cttttggata caccttcacc gcctactata tgcactgggt gcgacaggcc 120
cctggacagg ggcctgagtg gatgggatgg atcaacccta tcagtggtgg cacaaactat 180
gcacagaagt ttcagggcag ggtctccatg accagggaca cgtccatcag cacagcctac 240
atgggcctga gcaggctgag acctgacgac acggccgtct attactgtgc gagagtgaag 300
tgtagtagtg ccaactgcta tgggaacttt gactactggg gccagggtac cctggtcacc 360
gtctcctca 369
<210> 78
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 78
cagtctgccc tgactcagcc tccctccgcg tccgggtctc ctggagagtc agtcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ggttataact atgtctcctg gtaccaacaa 120
cacccaggca aagcccccaa actcatgatt tatgaggtca gtaagcggcc ctcaggggtc 180
cctgatcgct tctctggctc caagtctggc aacacggcct ccctgaccgt ctctgggctc 240
caggctgagg atgaggctga ttattactgc agctcatatg caggcagcaa caatttggta 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 79
<211> 369
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 79
gaggtgcagc tggtgcagtc tggagcagag gtgaaaaagc ccggtgagtc tctgaagatc 60
tcctgtaagg gttctggata caggtttacc aactaccgga tcggctgggt gcgccagatg 120
cccgggaaag gcctggagtg gatggggatc atctatcccg gtggctctga taccagatac 180
agtccgtccc tccaaggcca ggtcaccatg tcagtcgaca agtccatcag caccgcctac 240
ctgatgtgga gcagcctgaa ggcctcggac accgccatgt attactgtgc tcgacagacc 300
actcaaaata gtggctacga tagatggttt gactcctggg gccagggaac ccacgtcacc 360
gtctcctca 369
<210> 80
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 80
cagtctgtac tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcacctc cagcatcgga agtaatattg taaattggta ccaacacctc 120
ccaggaacgg cccccaaact cctcatctat attaataatc agcggccctc aggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tctgaggatg aggctgacta ttactgtgca gcatgggatg acagcctgaa tggttgggtg 300
ttcggcggag ggaccaagct gaccgtcctg 330
<210> 81
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 81
gaggtgcagg tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggaat cacctttagc aggcatacta tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggga gtggtggtag cacatatcat 180
gcagactccg tgaagggccg cttcaccatc tccagagaca gttccaagag cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgttt attactgtgc gatctccgtg 300
ccattattac gatttttgga gtggtttcaa cacccttttg acttctgggg ccagggaacc 360
ctggtcaccg tctcctca 378
<210> 82
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 82
gaaatagtga tgacgcagtc tccagcctcc ctgtctgtgt ctccagggga aagagtcacc 60
ctctcctgca gggccagtca gagtgttggc agcaccttag cctggtacca gcacaaacct 120
ggccaggctc ccaggctcct catctctggt gcatccacca gggccactgg tgtcccagcc 180
aggttcagtg gcagtgggtc tgggacagag ttcactctca ccatcagcag tctgcagtct 240
gaagattttg cagtttacta ctgtcaccag tatattaact ggcctccgtg gacgttcggc 300
caagggacca aggtggaaat caaa 324
<210> 83
<211> 360
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 83
gaggtgcggc tggtggagtc tgggggaggc ctggtcaaac ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcaat acatattcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtcgcctcc attagtagta ccggaagtta catatacaat 180
gcagactcac tgaagggccg attcaccatc tccagagaca acgccaagaa ctccctgttt 240
ctgcaaatga acagcctgag agtcgaagac acggctgtgt attactgtgt gagattcacc 300
atgactacag tgactaactt tgactcatgg ggccagggaa ccctggtcac cgtctcctca 360
<210> 84
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 84
cagtctgccc tgactcagcc tcgctcagtg tccgggtctc ctggacagtc agtcaccatc 60
tcctgcactg gaaccagcag tgatgttggt gcttatagct atgtctcctg gtaccaacaa 120
cacccaggca aagcccccaa actcatgatt tatgatgtct acaggcggcc ctcaggggtc 180
cctggtcgct tctctggctc caagtctggc aacacggcct ccctgaccgt ctctgggctc 240
caggctgagg atgaggctga ttactactgc tgctcacatg caggcagtca cacctgggtg 300
ttcggcggag ggaccaaggt gaccgtccta 330
<210> 85
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 85
cagctgcagg tggtggcgtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtaaag cctctggatt cacgttcacc aattatggca tgcactgggt tcgccaggcg 120
ccaggcaagg ggctggagtg ggtggctttt atatcatacg atggaggtaa taaattttat 180
gcagactctg tgaagggccg attcaccatc tccagagaca attccaggaa cacggtttat 240
ctgcaaatga acagcctgag agtggcggac acggctatgt attactgtcc gaaggtcatc 300
ccccacccgt attatgatag tagtggtgat gatgcttttg atatctgggg ccaagggaca 360
atggtcgcca tttcttca 378
<210> 86
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 86
gaaatagtga tgacgcagtc tccagccacc ctgtctgtgt ctccagggga aagagccacc 60
ctctcctgct gggccagtca gagtattagc cgcaacttag cctggtatca gcaaaaacct 120
ggccaggctc cccgactcct catctatggt gcatccacca gggccactgg tatccccgcc 180
aagttcagtg gcagtgggtc tgggacagac ttcactctca ccgtcagcag cctgcagtct 240
gaagaccttg cagtttatta ctgtcagcag tatagtaagt tgcctatcac cttcggccaa 300
gggacacgac tggagattaa a 321
<210> 87
<211> 366
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 87
gaggtgcagc tggtggagtc tgggggaggc ttggtacagc cgggggggtc cctgagactc 60
tcctgtgcag cctctggatt ccccttcagt agttatagca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatac ataagtggta gtggtggtga catatactac 180
gcagactctg tgaagggccg attcaccatc tccagggaca atgccaggaa ctcactgtct 240
ctgcaaatga acagcctgag agccgacgac acggctgtgt attactgtgc gagagggctg 300
gtagcaacaa ctggtacaag gtactttgac tactggggcc agggaaccct ggtcaccgtc 360
tcctca 366
<210> 88
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 88
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagg agttacttag cctggtacca acagagacct 120
ggccaggctc ccaggctcct catctacgat gcatccaaca gggccactgg catcccagtc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagagcct 240
gaagattttg cagtttatta ctgtcagcag cgtagctact ggcctccgtt cactttcggc 300
ggagggacca aggtggagat caaa 324
<210> 89
<211> 372
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 89
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc cggggtcctc ggtgaaggtc 60
tcctgcaagg cttctggagg caccttcagg aggtttgcta tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaggg atcatcccta tcctaggtag aggaaagtac 180
gcacagaagt tccagggcag agtcaggatt accgcggacg aatccacgag cacagcctac 240
atggagctga gcagcctgag atctgaagac acggccgtgt attactgtgc gagatttatt 300
tcgacagcct cctatgttcc ggggaccttc gaggacgtct ggggccaagg gaccacggtc 360
accgtctcct ca 372
<210> 90
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 90
gacattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc agctacttag cctggtacca acagaaacct 120
ggccaggctc ccaggctcct catctatgat gcatccaaca gggccgctgg catcccagcc 180
aggttcagtg gcagtgggtc tgagacagac ttcactctca ccatcagcag cctagagcct 240
gaagatgttg cggtttatta ctgtcagcag cgtagcgact ggcctccggg gacttttggc 300
caggggacca acgtggagat caaa 324
<210> 91
<211> 360
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 91
gaggtgcagc tggtggagtc tgggggaggc ctggtcaagc cgggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagg aactataaca tcaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc attagtagta ctggtagtta catacactac 180
gcagatttag tgaagggccg attcaccatc tccagagaca acgccaagaa ctcactgtat 240
ctgcaaatga acagcctgag agtcgaggac acggctgtat attactgtgc gcgaatggtt 300
aggaatacgg tgactgcctt tgactactgg ggccagggaa ccctggtctc cgtctcctca 360
<210> 92
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 92
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggccagtc gatcaccatc 60
tcctgcactg gaaccagcag tgatgttggt ggctataact ttgtctcctg gtaccaacaa 120
cagccaggca gagcccccaa actccttatt tatgaagtca ttaagcggcc ctcaggggtt 180
tctgatcgct tctctggctc caagtctggc gacacggcct ccctgacaat ctctgggctc 240
caggctgagg acgaggctga ttattactgc tgctcatatg ggggtaacaa ctcttggatg 300
ttcggcggag ggaccatgct gaccgtccta 330
<210> 93
<211> 372
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 93
caggtccagg tggtacagtc tggggctgag atgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg tttccggata caggctcatt gatttaccct tgcactgggt gcgacaggct 120
cctggaaaag ggcttgagtg gatgggactt tttgatcctg aaaaggctga agccatctac 180
tcacagaaat tccaggacaa ggtcaccata agcgaggaca catctatcga cacagcctac 240
atggaactga acagcctgcg ctctgaagac acggccgtct actactgtgc aacttgggga 300
gttgaggtgg tgaatgggag aagggactac tttgactcct ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 94
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 94
tcctatgagc tgacacagcc accctcggtg tcagtgtccc caggacagac ggccaggatc 60
acatgctatg cagatgtgtt gtcaaaccaa tatacttact ggtatcaaca gaagccaggc 120
caggcccctg tgttggtgat atataaagac actgagaggc cctcagggat ccctgagcga 180
tttgctggct ccagctcagg gacaacagtc accttagtca tcaatggagt ccgggcagag 240
gacgaggctt actattactg tcaatcagcc gacaacacca gaattacggt tttcggcgga 300
gggaccaagc tgtccgtcct a 321
<210> 95
<211> 384
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 95
ggggtgcagc tggtggagtc tgggggagga ttgatacagc ctggagggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agttttgaaa tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtttcatac attagtacta gtggtagtac catatactac 180
gcagactctg tcaagggccg attcaccatc tccagagaca acgccaggaa ctcactgtct 240
ctgcaaatga acagcctgag agccgaggac acggctgttt attactgtgc gagagacgtg 300
agggattgta gtgctcttac gtgcccccga aggggagatg cttttgattt ctggggccgt 360
gggacaaggg tcaccgtctc ttca 384
<210> 96
<211> 336
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 96
gatgttgtga tgactcagtc tccactctcc ctgcccgtca cccttggaca gtcggcctcc 60
atctcctgca ggtctagtca aagcctcgta tacagtgatg gaaacaccta cttgaattgg 120
tttcagcaga ggccaggcca atctccaagg cgcctaattt ataaggtttc taaccgggac 180
tctggggtcc cagacagatt cagcggcagt gggtcaggca ctgatttcac actgaaaatc 240
agcagggtgg aggctgagga tgttggggtt tattactgca tgcaaggtac acactggcct 300
cgcactttcg gccctgggac caaagtggat atcaaa 336
<210> 97
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 97
gaggtgcagc tggtggagtc tgggggaggc ctggtcaagc cgggggggtc cctgagactc 60
tcctgtgcag cctctggatt cagtttcagt gtctatccca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc ataagtagta gtagtcgtta catatcctac 180
gcagactcac tgaggggtcg aatcaccatc tccagagaca acgccaagaa ctcactgtat 240
ctgcaaatga acagcctgag agtcgaggac acggctgtgt attactgtgt gaaagtcggg 300
ggtagcaaac accaatacta ctttgactac tggggccagg gatccctggt caccgtctcc 360
tca 363
<210> 98
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 98
cagtctgtgc tgactcagcc accctcaacg tctgggatcc ccgggcagac ggtcaccatc 60
tcttgttctg gaagcaggtc caacatcgga agttatactg ttaactggta cgagcaactc 120
ccaggaacgg cccccaaact cgtcatcttt aataataatc agcgtccctc aggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tctgagggtg aagctgatta ttactgtgca gcatgggatg acagcctgaa tggtgtggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 99
<211> 372
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 99
caggttcagt tggtgcagtc tggagctgag gtgaagaggc ctggggcctc agtgaaggtc 60
ttctgcaagg cttctggtta cacctttacg aattatgaca tcatctgggt gcgacaggcc 120
cctggacaag ggcttgagtg ggtgggctgg atcagcactt acaatggtaa cacaaactat 180
gaacagaacc tccagggcag agtcaccatg accacagaca catccacgag cacagcctac 240
atggagctga ggagcctgag atctgacgac acggccgttt actattgtgc gagagagcgt 300
tgtagtacta gtacctgcta tagtcgttat gctgactact ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 100
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 100
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtgggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattaat atctatttga attggtatca gcaaaaaccg 120
gggaaagccc ctaaggtcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
aggttccgtg gcagtgggtc tgggacagat ttcatcctca ccatcagcag tctgcaacct 240
gaagactttg caacctacta ctgtcaacag agttacaggt cccctcggac gttcggccaa 300
gggaccgagg tggaaatcaa a 321
<210> 101
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 101
gaggtgcagc tggtggagtc tgggggaggc ttggcacagc ctggagagtc catgagactc 60
tcctgtgtag cctctggatt caccttaagt cgttatgaag tgaactgggt ccgccaggct 120
ccagggaagg ggctagagtg gctttcatac attagcagtg gtggtccttc catatactac 180
gcagactctg tgaagggccg attcacgatc tccagaaaca gcgccgagaa ctcactggaa 240
ctacaaatgt ccaccctgag gaccgaggac acggctgttt attattgtat gagagagggt 300
cttacttatt atgatagtac tatttggggc cagggaaccc tggtcgccgt ctcctca 357
<210> 102
<211> 333
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 102
cagaatgtgc tgactcaatc gccctctgcc tctgcctccc tgggagcctc ggtcaaactc 60
acctgcactc tgaacagtgg gcacagcaga tacgccatcg catggcatca acatcagcca 120
cagaggggcc ctcggttcct gatgaagatt aatagtgatg gcagacacat caggggggac 180
ggcatctctg atcgcttctc aggctccgcc tctggggctg agcgtcatct caccatctcc 240
agcctccagc ctgaggatga ggctgactat tattgtcaga cctggggcac tggctttcgg 300
gtgttcggcg gagggaccaa actgaccgtc cta 333
<210> 103
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 103
gaagtgcagc tggtggagtc tgggggaggc ttggtacagc ctggcaggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttgat gaatatgcca tgcactgggt ccggcaagtt 120
ccagggaagg gcctggagtg ggtctcaggt attagttgga atggtggtag caaaggctat 180
gcggactctg tgaagggccg attcaccatc tccagagata acgccaggta ttccctgtct 240
ctgcaaatga acagtctgag aactgaggac acggccttat attactgcgc aaaagatgat 300
tacgaggggg ctggttttga tatctggggc caagggacag tggtcaccgt ctcttca 357
<210> 104
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 104
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc agctacttag gctggtacca acagaaatct 120
ggccaggctc ccaggctcct catctatggt gcatccagca gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagagcct 240
gaagattttg caatttatta ctgtcagcag cgtagcaact ggcctatcac cttcggccaa 300
gggacacgac tggagattaa a 321
<210> 105
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 105
caggtgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggcta tttaattagc aatggttact actggggctg gatccggcag 120
tcccccggga aggggctgga gtggattggg agtatctatt atactaggga cacctactac 180
aactggtccc tcaagagtcg aatcaccata tcagtggaca cgtccaagaa acagttctcc 240
ctgaagttgt attctgtgac cgccgcagac acggccgtct attactgtgt gagacatgag 300
ggttcttgca atgatggaag ctgttacggc tcgttcgttg acaactgggg ccagggaacc 360
ctggtcaccg tctcctca 378
<210> 106
<211> 318
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 106
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtaggaga cagagtgacc 60
ctcacttgtc gggcgagtca ggatattagc agctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacacat ttcactctca ccatcagcag cctgcagcct 240
gaagatttcg caacttactt ttgtcaacag gctgacagtt tcatcacttt cggcggaggg 300
accaaggtgg agatcaaa 318
<210> 107
<211> 360
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 107
gaggtgcaac tgttggagtc tgggggaggc ttggtacagc cgggggggtc cctgagactc 60
tcctgtgcag cctctgattt caccttcagc agctacacca tggcctgggt ccgccaggct 120
ccagggaagg ggcttgaatg ggtctcatct attagtggtg atggtgttag cacaaaaaac 180
gcagactccg tgaagggccg attctccgtc tccagagaca attccaagaa cacacttttt 240
ctgcaactga acagtctgag agccgaggac acggcctttt attactgtgc gagggggggg 300
accttccata actggtactt cgatctctgg ggccgtggcg tcttggtcac tgtctcctca 360
<210> 108
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 108
gaaatagtga tgacgcagtc tccagccacc ctgtctgtgt ctccagggga aacagccacc 60
ctctcctgca gggccagtca gagtattggc gacaacttag cctggtatca gcagaaacct 120
ggccaggctc ccaggctcct catctctggt gcatccacaa gggccactga tttcccagcc 180
aggttccgtg gcagtgggtc tgggacagag ttcactctca ccatcagcag cctgcagtct 240
gaagactttg cagtttatta ctgtcagcag tataaaaact ggcctcggac gttcggccga 300
gggaccaagg tggaagtcag a 321
<210> 109
<211> 363
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 109
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc cgggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc acctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggccggagtg ggtctcaggt attagtggta gtggtggtag cacaaactac 180
gcagactctg cgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtat attattgtgc gaaagggacc 300
attacttact cctactacta catggccgtc tggggcaaag ggaccacggt caccgtctcc 360
tca 363
<210> 110
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 110
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga aagaggcacc 60
ctctcctgca gggccagtca gagtgttagg agcagctact tagcctggta ccagcagaga 120
cctggccagg ctcccaggct cctcatctat ggtgcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcacgc tcaccatcgg cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cagtatggta cctcaataac tttcggcgga 300
gggaccaagg tggagatcaa a 321
<210> 111
<211> 372
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 111
caggtccagg tggtacaatc tggggctgag atgaggaagc ctggggcctc agtgagggtc 60
tcctgcaagg tttccggata caggctcact gatttaccct tgcactgggt gcgacaggct 120
cctggaaaag ggcttgaatg gattggattt gttgatcttg aaaagcgcga aatcatctac 180
gcacagaaat ttcagggcaa agtcaccata accgaggaca catctgcaga caccgcctac 240
atggaactga acagcctgcg atctgaagac acggccgtct actactgtgc aacttgggga 300
attgaggtgg tgaatgggag ggacgaattc tttgactcct ggggccaggg aaccctggtc 360
tccgtctcct ca 372
<210> 112
<211> 318
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 112
tcctatgagc tgacacagcc accctcggtg tcagtgtccc caggacagac ggccaggatc 60
acatgctatg cagatgtgtt gtcaaagcaa tatacttact ggtatcagca gaagccaggc 120
caggcccctg tgttggtgat atataaagac actgagaggc cctcagggat ccctgagcga 180
tttgctggct ccagctcagg gacaacagtc accttgatca tcaatggagt ccggacagag 240
gacgaggcgt actattactg tcaatcagcc gacaccagaa ttacggtttt cggcggaggg 300
accaagctgt ccgtccta 318
<210> 113
<211> 372
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 113
caggtccagg tggtccagtc tggggctgag atgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg tttccggata tagtctcagt gatttaccct tgcactgggt gcgacaggct 120
cctggaaaag ggcttgagtg gatgggactt tttgatccta taaacggtga aatcatctac 180
gcacagacat tccagggcaa agtcaccata agcgaggaca catcgataga cacagcctac 240
atggaactca acagcctgcg atctgaagac acggccgtgt actattgtgc aacttgggga 300
gttgcggtgg tgagtgggag aagggactac tttgactcct ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 114
<211> 318
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 114
tcctatgagc tgacacagcc accctcggtg tcagtgtccc caggacagac ggccaggatc 60
acatgctatg cagctgtatt gtcaaaccaa tatacttact ggtatcaaca gaagccaggc 120
caggcccctg ttttggtgat atataaagac actgagaggc cctcagggat ccctgagcga 180
tttgctggct ccagctcagg gacaacagtc accttgatca tcaatggagt ccggacagag 240
gacgaggctt actattactg tcaaacagcc gacaccaaat atacggtttt cggcggaggg 300
accaagctgt ccgtccta 318
<210> 115
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 115
caggtgcagc tgcaggagtc gggcccagga ctggtgaagc cttcacagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcacc agtggtgatt actactggaa ttggatccgc 120
cagcccccag ggaagggcct ggagtggatt gggtacatct atcacagtgg gaccacctac 180
tacaacccgt ccctcaagag tcgagttacc atatcagtag acacgtccaa gaacaggttc 240
tccctgaagt tgtcctctgt gactgccgca gacacggccg tgtatttttg tgccagagcc 300
tacgcttatg aattttggag cggttaccct aactggttcg acccctgggg cctgggaacc 360
ctggtcaccg tctcatca 378
<210> 116
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 116
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgttggaga cagagtcacc 60
atcacttgcc gggcaagtca gcgcattagt acctatgtaa attggtatca ggtgaaagca 120
gggacagccc ctaaggtcct gatctatgct gcgtccagtt tgcaaactgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccattgtcag tctacaacct 240
gaagatttta caacctactt ctgtcaacag agttacagtc ccccgtggac gttcggccaa 300
gggaccaagg tggaaatcaa a 321
<210> 117
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 117
caggtgcagt tggtggagtc tgggggaggc ttggtccagc ctggtaggtc cctgagactc 60
tcctgtgcag cctctgggtt catcttcaat agatatgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctcgagtg ggtggctctt atatcatatg atggaattaa taaatattac 180
gcagactccg tgaagggccg attctccatc tccagagaca attccaagag tacgctgtat 240
ctgcaaatga acagcctcag agctgaggac acggctatct tttactgtgc gagaggacta 300
ggatattgta gtggtaccgg tggtagctgt acaccctttg aatattgggg ccagggaatc 360
ctggtcaccg tctcctca 378
<210> 118
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 118
gacatccaga tgacccagtc tccaccctcc ctgtctgcat ctgttggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattaag aaatatttaa attggtatca gcagaaacca 120
gggaatgccc ctaagctcct catctatggc gcatccaatt tgcaaactgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcgctctct ccatcagcag tctggaaact 240
gaagattttg caacttacta ctgtcaacag agtgacagtg cccctcccac tttcggcgga 300
gggaccaagg tggagttcaa a 321
<210> 119
<211> 405
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 119
caggtgcagc tgcaacagcg gggcgcaggg ctgttgaagc cctcggagac cctgtccctt 60
acctgcgaaa tctatggtgc atccctcaat gattacgact ggacctggat ccgccagccc 120
ccagggaaga ggctggagtg gattggggtc atcaatcgtc gtgacactgt tgactacaac 180
ccgtccctca agagtcgggt caccctctca cttgacacgt ccaagaacca actttccctg 240
agtctgagtt ctgtgaccgc cgcggacacg gctgtttatt actgtgtgag agtcccacgt 300
cggggctttg aagggtcttt cggattttgt gatgatactg cctgccgcta cgggcatacc 360
tggttcgacc cctggggcca gggaaccctg gtcaccgtct cctca 405
<210> 120
<211> 321
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 120
gacattcaga tgacccagtc tccttcctcc ctgtctgcat ctgttggcga cagagtcacc 60
atcacttgcc gggcaagtca gagtattaga gattatttaa attggtatca acaaagacca 120
gggaaagccc ctaaagtcct gatctttgct ggttcccgtt tggaaagtgg ggtcccatcg 180
aggtttagag gccgtggatc tgggacagaa ttcactctca ccatcagcga tctgcaacct 240
gaggattttg caacttacta ctgtcaacag agttacctta cacctccgac attcggccaa 300
gggaccaccg tcgatatcaa a 321
<210> 121
<211> 375
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 121
caggtcacct tgaaggagtc tggtcctgtg ctggtgaaac ccacagagac cctcacgctg 60
acctgcaccg tctctggatt ctcactccgc aatggtagaa tgggtgtgag ctggatccgt 120
cagcccccag ggaaggccct ggagtggctt gcacacattt ttgcgagtga cgaaaaatct 180
tacagtacat ctcagaggac caggctctcc atctccaggg acacctccaa aagccaagtg 240
gtccttagca tgaccgacat ggaccctgtg gacacagcca catattactg tgcgcggatt 300
ttgaagtttg ggacaatgag ggccgcatac tactttgact actggggcca gggagccctg 360
gtccccgtct cctca 375
<210> 122
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 122
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gaatcaacat tggaattaga actgtacact ggtaccagca gaagccaggc 120
caggccccta tgttggtcat ctattatgat agcgaccggc ccttagggat ccctgagcga 180
ttctctggct ccaagtctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatagtagta gtgatcatgt tgtattcggc 300
ggagggacca agctgaccgc ccta 324
<210> 123
<211> 375
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 123
caggtgcact tggtggagtc tgggggaggc ttggtcaagc ctggagggtc cctgagactc 60
tcctgtgcag tttctggatt caccttcagt gactactaca tgagctggat ccgccaggct 120
ccggggaagg gactggagtg gctttcatac attagtagta gtggtagtac catatactac 180
gcagactcta tgaagggccg attcaccatc tccagggaca acgccaggaa ctcactctat 240
ctgcaaatga acagcctgag agtcgaggac acggcctttt attactgtgc ggggtcaaag 300
gttggctata ctactggtcg aaggaactgg tacttcgatc tctggggccg tggcaccctg 360
gtcactgtct cctca 375
<210> 124
<211> 330
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 124
cagtctgccc tgactcagcc tccctccgcg tccgggtctc ctggacagtc agtcaccatc 60
tcctgcactg gaaccagcag tgacgttggt gcttataact atgtctcctg gtaccaacag 120
cacccaggca aagcccccaa actcatgatt tatgaggtca ctaagcggcc ctcaggggtc 180
cctgatcgct tctctggctc caagtctggc aacacggcct ctctgaccgt ctctgggctc 240
caggctgagg atgaggctga ttattactgc agctcatatg caggcaacaa caatttagtc 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 125
<211> 378
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 125
caggtgcagt tgctgcagtc tgggtctgag gtgaggaaac ctggggcctc agtgaacatt 60
cactgtaagg catctggatt cactttcacc gacttctatt tacactgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatggggata atcaaccctg aaaccggtga gacaacctac 180
tcacagaagt ttcagggcag agtcaccatg accagggaca cgtccacgag tgtagtgaat 240
ctggaagtga ggagcctgag atctgaggac acggccatat attactgtgc gagagatctc 300
gttgtcgtag tccccgttga aatgtctcgg cgtgcctttg acatttgggg ccaagggatt 360
atggtcacag tctcctca 378
<210> 126
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 126
tcctatgtgc tgactcagcc accctcggtg tcagtggccc caggacagac ggccaggatt 60
ccctgtgggg gcaacaacat tgaacgtaaa agtgtccact ggtaccagca gaggccaggc 120
caggcccctg tgttggtcgt ctatgatgat actgtccggc cctcaggtat ccctgagcga 180
ttctctggct ccaactccgg gagcacggcc accctgacca tcagcagggt cggagccggg 240
gatgagggcg actattattg tcaggtgtgg gacagcacca ctgaccatgg ggtcttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 127
<211> 375
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 127
cgggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgtcaga cttctggata cattttcagc gcctactaca tctattgggt gcgacaggcc 120
cctggacaag gacttgagtg gatgggacgg atgaacgcta agagtggagg cgcaaacact 180
gcacagcagt ttcagggcag actcaccatg accagggaca tgtccgtcag cacagcctac 240
atggaactga gcaggctgcg atcggacgac acggccgtct attattgtgc gagagactat 300
agggatgact acatgtgggg gagttatcgg cctttagact actggggcca gggaaccctg 360
gtcaccgtct cctca 375
<210> 128
<211> 318
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic oligonucleotide
<400> 128
gaagttgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagcctcc 60
ctctcctgca gggccagtca gagtgttagc ggctacttag cctggtacca acacaaacct 120
ggccaggctc ccaggctcct catctatgat gcatccaaca gggccgctgg catcccagcc 180
aggttccgtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctggagcct 240
gaagattttg cagtttattt ctgtcagcag cgtagcaacg ggctcacttt cggcggaggg 300
accaaggtcg agatcaaa 318
<210> 129
<211> 123
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 129
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Arg Tyr
20 25 30
Ala Leu His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Asp Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Ala Arg Asn Ser Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Thr Leu Pro Tyr Ser Asn Asn Trp Tyr Ala Pro Glu Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 130
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 130
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Ser Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Thr Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Met Ala Thr Leu Thr Ile Ser Gly Ala Gln Val Glu
65 70 75 80
Asp Glu Ser Asp Tyr Tyr Cys Ser Ser Thr Asp Ser Ser Gly Asn Pro
85 90 95
Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 131
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 131
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Ser Tyr
20 25 30
Ala Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ser Ser Ile Ser Gly Asn Gly Asn Gly Arg Ser Tyr Ala Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Thr Ser Lys Asp Leu Ser Lys Tyr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Asn Leu Arg Pro Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Lys Val Val Arg Ile Ala Ala Val Leu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Pro Gly Thr Gln Val Thr Val Ser Ser
115 120
<210> 132
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 132
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ile Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr His Cys Gln Gln Arg Ser Ser Trp Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Glu
100 105
<210> 133
<211> 123
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 133
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Thr Trp Val Arg Gln Ala Leu Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Gly Leu Thr Tyr Phe Ala His Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Lys Ser Thr Thr Gly Thr Thr Ala Leu Val Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 134
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 134
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Ser Ser
20 25 30
Tyr Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Ser Ile Asn Arg Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Phe Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Arg Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Gly Leu Tyr Met Gly Ser
85 90 95
Gly Ile Trp Ile Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 135
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 135
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ile Asn Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Asn Asp Gly Ser Tyr Asn Tyr Asp Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Lys Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Lys His Gly Asp Phe Asp Tyr Tyr Gly Val Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Pro
115 120
<210> 136
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 136
Asp Val Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Ile Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Gly Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Gly Thr Tyr Tyr Cys Gln Gln Ala Asp Ser Phe Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 137
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 137
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asp Asn Thr Tyr Ala Pro Phe Val
50 55 60
Asn Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Arg Pro Gly Ser Phe Pro Gly Leu Cys Asp Tyr Trp Gly
100 105 110
Gln Gly Ala Leu Val Thr Val Ser Ser
115 120
<210> 138
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 138
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Met Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Lys Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Asn Ser Tyr Ser Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 139
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 139
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Ser Phe Ser Arg Leu
20 25 30
Thr Ile Ile Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly His Ile Pro Ile Phe Gly Thr Thr Asn Tyr Ala Leu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Thr Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Met Tyr Ser Gly His Asp Gly Val Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 140
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 140
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Lys Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Thr Trp Pro Pro
85 90 95
Gly Met Phe Gly Gln Gly Thr Arg Val Glu Ile Lys
100 105
<210> 141
<211> 127
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 141
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys His Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Phe Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Thr Ile Tyr Asp Ser Gly Arg Thr Tyr Asp Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Ala Asp Thr Ser Lys Lys Gln Phe
65 70 75 80
Ser Leu Thr Leu Arg Ser Ala Thr Ala Ala Asp Thr Ala Val Tyr Phe
85 90 95
Cys Ala Arg His Leu Thr His Leu Tyr Gly Asp Tyr Val Thr Pro Asp
100 105 110
Ala Leu Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120 125
<210> 142
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 142
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Arg
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ser Asn Ser Arg
85 90 95
Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 143
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 143
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Thr Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ser Tyr Ile Ser Ser Gly Ser Ser Asn Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Asn
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala His Gly Arg Ile Val Asn Ser Gly Val Val Ile Ser Arg
100 105 110
Phe Asp Pro Trp Gly Gln Gly Ile Leu Val Thr Val Ser Ser
115 120 125
<210> 144
<211> 111
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 144
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Leu His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Phe Glu Val Thr Tyr Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Thr Thr Ser
85 90 95
Asn Thr Leu Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 145
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 145
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Arg Thr Ser Gly Tyr Thr Phe Thr Ala Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Arg Ile Asn Pro Ser Ser Gly Gly Ala Gln Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Thr Tyr
65 70 75 80
Met Thr Leu Ser Gly Leu Thr Ser Asp Asp Thr Ala Val Phe Tyr Cys
85 90 95
Ala Arg Met Gly Cys Arg Ser Asp Arg Cys Tyr Ser Thr Asn Tyr Asn
100 105 110
Phe Asp Gln Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 146
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 146
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Pro Cys Gly Gly Asn Asn Ile Gly Thr Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Ser
35 40 45
Asn Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Lys Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Gly Ile Asp Val
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 147
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 147
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Ile Ser Pro Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Phe Ile Ser Ser Ser Ser Arg Tyr Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Gly His Ser Thr Ser Ser Ser Tyr Phe Asp Ser Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 148
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 148
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Leu Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Thr Lys Thr Val
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Met Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Asn Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Arg Val Trp Asp Ser Asp Thr Asp His
85 90 95
Arg Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 149
<211> 127
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 149
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Gln Thr Lys Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Leu Tyr
85 90 95
Tyr Cys Ser Thr Gly Pro Tyr Tyr Tyr Asp Thr Ser Gly Tyr Pro Gln
100 105 110
Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 150
<211> 106
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 150
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Leu Gly Asp Lys Tyr Ala
20 25 30
Cys Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Thr Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Ser Thr Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 151
<211> 118
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 151
Glu Val Gln Met Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Val Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Thr Met Asn Trp Phe Arg Gln Ala Pro Gly Glu Gly Leu Lys Trp Val
35 40 45
Ser Ser Ile Ser Ser Thr Ser Leu Tyr Thr Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ser Ile Ser Arg Asp Asn Ala Gln Gly Ser Leu Ser
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Val Gly Pro Ala Glu Leu Asp Tyr Trp Gly Gln Gly Val
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 152
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 152
Val Thr Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Gly Val Asp
20 25 30
Leu Gly Trp Phe Gln Gln Arg Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Arg Gly Ser Gly Thr Phe Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Thr Thr Tyr Phe Cys Leu Gln Asp Tyr Asn Tyr Pro Trp
85 90 95
Thr Leu Gly Gln Gly Thr Thr Val Gly Val Lys
100 105
<210> 153
<211> 117
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 153
Gln Val His Leu Val Gln Ser Gly Ser Ala Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Lys Thr Gly Gly Ser Asn Tyr Thr Gln Arg Phe
50 55 60
Gln Ala Arg Val Thr Met Thr Trp Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Gly Arg Asn Gly Tyr Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 154
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 154
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Ala Asp Ala Leu Pro Lys Gln Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Met Ile Tyr
35 40 45
Gln Asp Thr Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Gly Asp Ser Ser Gly Thr Tyr
85 90 95
Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 155
<211> 120
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 155
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Lys Phe Arg Asn Tyr
20 25 30
Ala Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Thr Ser Gly Gly Ser Thr Glu Tyr Ala Asp Ala Val Lys
50 55 60
Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Asp Asp Thr Ala Val Tyr Tyr Cys Thr
85 90 95
Val Pro Trp Gly Asn Tyr Asn Asp Tyr Val Ser Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Pro Val Ser Ser
115 120
<210> 156
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 156
Glu Val Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Gln Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Gly Asn Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Ser Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Lys Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ile Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln His Ser Thr Trp Pro Arg
85 90 95
Gly Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 157
<211> 124
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 157
Gln Val His Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Arg Tyr
20 25 30
Pro Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Phe Leu Pro Tyr Ser Ser Ser Trp Tyr Gln Gly Phe Asn
100 105 110
Tyr Trp Gly Gln Gly Ile Leu Val Thr Val Ser Ser
115 120
<210> 158
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 158
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Ile Ala Thr Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ser Pro Thr Pro Ile
35 40 45
Ile Phe Glu Asp Ser Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Arg Thr Asp Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Asn
85 90 95
Ser Asn Arg Ala Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 159
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 159
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Glu Ala Ser Gly Phe Leu Phe Ser Arg Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Asp Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Asn Lys Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Val Asn Ser Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Val Pro Tyr Gly Asp Thr Leu Thr Gly Leu Val Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 160
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 160
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Thr Ile Ala Ser Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ala Pro Thr Thr Val
35 40 45
Ile Tyr Glu Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Asn
85 90 95
Ser Asp Arg Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 161
<211> 125
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 161
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Arg Asn Ala
20 25 30
Arg Met Gly Val Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Phe Ser Asn Asp Glu Lys Ser Tyr Asn Thr Ser
50 55 60
Leu Lys Ser Arg Leu Ser Ile Ser Lys Asp Thr Ser Lys Ser Gln Val
65 70 75 80
Val Leu Thr Met Thr Ser Met Asp Pro Leu Asp Thr Ala Thr Tyr Phe
85 90 95
Cys Ala Arg Leu Leu Val Ala Gly Thr Phe Leu Pro Ser His Tyr Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Ile Leu Val Thr Val Ser Ser
115 120 125
<210> 162
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 162
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Leu Lys Ser Val
20 25 30
Phe Trp Tyr Gln Glu Arg Pro Asp Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Tyr Asp Ser Ala Arg Pro Ser Gly Ile Pro Glu Arg Ile Ser Gly Ser
50 55 60
Lys Ser Gly Asn Thr Ala Thr Leu Thr Ile Thr Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Phe Cys Gln Val Trp Asp Ser Ser Arg Asn His
85 90 95
Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 163
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 163
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Ser Ser Thr Ile Gln Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Gln Val Gly Ala Asp Phe Ser Gly Arg Gly Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Leu Thr Val Ser Ser
115 120
<210> 164
<211> 106
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 164
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Thr Cys Phe Gly Asp Lys Leu Gly Asp Lys Tyr Ala
20 25 30
Cys Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Lys Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Ser Thr Ala Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 165
<211> 120
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 165
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Lys Phe Ser Val Tyr
20 25 30
Ala Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Gly Asn Ser Leu Phe
65 70 75 80
Val Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Ile Tyr Tyr Cys
85 90 95
Ala Thr Ala Arg His Ile Thr Asn Asp Gly Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Ile Val Ser Ser
115 120
<210> 166
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 166
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Arg Phe
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Leu Gln Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Phe Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Ser His Pro Arg
85 90 95
Met Phe Thr Phe Gly Pro Gly Thr Thr Val Asp Ile Lys
100 105
<210> 167
<211> 127
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 167
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Leu Ile Ser Asn Gly
20 25 30
Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Ser Ile Tyr His Thr Arg Ser Thr Tyr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Ser Ile Ser Val Asp Thr Ser Lys Asn Arg Phe Ser
65 70 75 80
Leu Arg Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Phe Tyr Tyr Cys
85 90 95
Ala Arg Gly Pro Gly His Cys Tyr Gly Asp Asp Asp Cys Tyr Ala Tyr
100 105 110
Tyr Phe Asp Gln Trp Gly Gln Gly Thr Pro Val Thr Val Ser Pro
115 120 125
<210> 168
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 168
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ser Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Asn Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 169
<211> 127
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 169
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Arg Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Gln Trp Met
35 40 45
Gly Gly Ile Leu Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Pro Tyr Cys Thr Asn Asp Val Cys Ser Asn Gln Asn
100 105 110
Thr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 170
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 170
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Ala Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Thr Tyr
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 171
<211> 123
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 171
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Asp Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Ser Arg Tyr Gly Asp Pro Ile Gly Asp Asn Trp Phe Asp Pro
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 172
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 172
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Leu Lys Ser Val
20 25 30
Phe Trp Tyr Gln Glu Arg Pro Asp Gln Ala Pro Val Val Val Ile Tyr
35 40 45
Tyr Asp Ser Ala Arg Pro Ser Gly Ile Pro Glu Arg Ile Ser Gly Ser
50 55 60
Lys Ser Gly Asn Thr Ala Thr Leu Thr Ile Thr Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Phe Cys Gln Val Trp Asp Ser Ser Arg Asn His
85 90 95
Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 173
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 173
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Arg Phe Ser Phe Tyr
20 25 30
Gly Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Thr Gly Ala Thr Arg Asn Cys Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Val Asp Asp Thr Ala Val Phe Tyr Cys
85 90 95
Val Arg Arg Phe Pro Met Thr Thr Val Thr Ser Phe Asp Ser Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 174
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 174
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Phe Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Phe Asp Val Thr Gly Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ala Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Tyr Ala Gly Phe
85 90 95
Asn Ala Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 175
<211> 125
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 175
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Arg Pro Gly Gly
1 5 10 15
Ser Val Arg Leu Ser Cys Val Ala Ser Gly Phe Pro Phe Asn Met Phe
20 25 30
Trp Met Gly Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Ala Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Ser Val Gly Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Val Arg Glu Gly Val Arg Arg Val Val Val Arg Ser Thr Gly Tyr Phe
100 105 110
Asp Phe Trp Gly Gln Gly Gln Leu Val Thr Val Ser Ser
115 120 125
<210> 176
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 176
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Met Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Val Pro Ile Lys Tyr Val
20 25 30
Tyr Trp Tyr Gln Gln Arg Ser Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Asp Arg Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Ala Thr Leu Thr Ile Thr Gly Ala Gln Val Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Tyr Ser Thr Asp Ser Ser Gly Tyr Gln
85 90 95
Arg Ala Phe Gly Gly Gly Thr Thr Leu Thr Val Leu
100 105
<210> 177
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 177
Arg Val Gln Leu Gln Glu Ser Ala Pro Gly Leu Val Arg Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Pro Ile Ser Asn Gly
20 25 30
Pro Tyr Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Phe Ile Phe Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Arg Gly Arg Val Thr Met Ala Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Arg Leu Asn Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg His Val Val Thr Ala Ser Gly Trp Phe Asp Pro Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 178
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 178
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Thr Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Val Leu Ile
35 40 45
Tyr Asp Ala Phe Lys Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Arg Trp Pro Pro
85 90 95
Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 179
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 179
Arg Val Gln Leu Gln Glu Ser Ala Pro Gly Leu Val Arg Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Pro Ile Ser Asn Gly
20 25 30
Pro Tyr Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Phe Ile Phe Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Arg Gly Arg Val Thr Met Ala Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Arg Leu Asn Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg His Val Val Thr Ala Ser Gly Trp Phe Asp Pro Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 180
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 180
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Thr Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Val Leu Ile
35 40 45
Tyr Asp Ala Phe Lys Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Arg Trp Pro Pro
85 90 95
Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 181
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 181
Gln Val His Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Asp Ser Gly Val Thr Phe Ser Asp Asn
20 25 30
Ala Leu Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Ser Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asp Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Arg Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Val Thr Ala Asp Tyr Tyr Glu Ser Ser Gly Lys Val Phe Trp
100 105 110
Gly Gln Gly Ala Leu Val Val Val Ser Ser
115 120
<210> 182
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 182
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Arg Ala Ser Gln Ser Val Arg Ser Trp
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gln Thr Phe Ser Trp
85 90 95
Thr Phe Gly Gln Gly Thr Thr Val Glu Val Lys
100 105
<210> 183
<211> 124
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 183
Gln Val Gln Leu Gln Glu Ser Gly Pro Ala Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Asp His
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Thr Ser Gly Thr Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Lys Gln Phe Ser Leu
65 70 75 80
Asn Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ser Leu Glu Thr Val Ile Arg Phe Tyr Tyr Tyr His Tyr Met Asp
100 105 110
Val Trp Gly Lys Gly Thr Thr Val Ile Val Ser Ser
115 120
<210> 184
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 184
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Gln Ser
20 25 30
Asp Gly Tyr Ser Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Val Ile Gly Ser Gly Thr Tyr Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Met Gln Ala
85 90 95
Leu Gln Thr Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 185
<211> 114
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 185
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ala Gly Gly Ser Ile Gly Asp Tyr
20 25 30
His Trp Asn Trp Ile Arg Gln Pro Ala Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile His Ser Ser Gly Asn Thr Asp Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Met Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gln Asn Val Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val
100 105 110
Ser Ser
<210> 186
<211> 113
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 186
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Leu Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Phe Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Glu Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Phe Cys Gln Gln
85 90 95
Phe Tyr Thr Thr Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 187
<211> 114
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 187
Gln Val Gln Met Gln Glu Ser Gly Pro Gly Leu Val Lys Ala Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Ile Ser Ile Asn Asn Tyr
20 25 30
Tyr Trp Ser Trp Phe Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Val Tyr Ser Thr Gly Ser Ser Lys Tyr Asn Pro Ser Leu Glu
50 55 60
Arg Arg Ala Thr Met Ser Val Asp Thr Ser Asn Asn Asn Phe Ser Leu
65 70 75 80
Arg Leu Thr Ser Val Thr Thr Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Ser Met Pro His Ile Trp Gly Gln Gly Leu Leu Val Thr Val
100 105 110
Ser Ser
<210> 188
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 188
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Glu Thr Asn
20 25 30
Tyr Val Tyr Trp Tyr Gln Gln Val Pro Gly Thr Ala Pro Lys Pro Leu
35 40 45
Val Tyr Arg Asn Asp Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Val Ile Ser Gly Leu Arg
65 70 75 80
Thr Glu Asp Glu Ala Ala Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Lys Ala Pro Val Phe Gly Ala Gly Thr Lys Val Ala Val Leu
100 105 110
<210> 189
<211> 130
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 189
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Glu Ser Lys Ile Asp Gly Gly Thr Ile Asp Tyr Ala Thr
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Asp Gln Gly Tyr Tyr Asp Arg Ser Gly Tyr Trp Val
100 105 110
Val Gly Asn His Phe Asp Tyr Trp Gly Gln Gly Ile Leu Val Thr Val
115 120 125
Ser Ser
130
<210> 190
<211> 111
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 190
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Leu
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Thr Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Asp Asp Ala Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Arg Ser
85 90 95
Leu Ser Thr Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 191
<211> 123
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 191
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Thr Thr Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val His
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Ser His Ser Met Ile Val Val Asp His Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 192
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 192
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Thr Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Tyr Asn Val His
85 90 95
Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105
<210> 193
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 193
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ile Met Thr Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Ser Ser Val Tyr Thr Phe Tyr Ala Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Glu Gly Phe Arg Ala Tyr Asn Leu Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 194
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 194
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Glu Tyr Asn
20 25 30
Tyr Val Tyr Trp Tyr Gln Lys Phe Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Lys Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Gly Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ile Leu
85 90 95
Ser Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 195
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 195
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Gln Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Asp Gly Ser Asn Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Thr Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Asn Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Trp Asp Arg Leu Val Arg Ser Ala Val Gly Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Ser Val Ser Ser
115 120
<210> 196
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 196
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Asn Asp Val Gly Gly Tyr
20 25 30
Asn Phe Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Phe Asp Val Ile Arg Arg Pro Ser Gly Val Pro Gly Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asp Thr Ala Ser Leu Ile Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Thr
85 90 95
Tyr Thr Trp Val Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105 110
<210> 197
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 197
Gln Val His Leu Gln Glu Ser Gly Pro Arg Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Thr Ala
20 25 30
Thr Tyr Tyr Trp Ser Trp Ile Arg Gln Ser Pro Gly Arg Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Ser Ser Gly Asn Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Leu Asp Thr Pro Asn Asn Gln Leu
65 70 75 80
Ser Leu Thr Leu Thr Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Glu Arg Arg Leu Arg Ile Leu Ser Ile Glu Arg Asn Tyr Tyr Ala
100 105 110
Met Asp Val Trp Gly Gln Gly Thr Pro Val Thr Val Ser Ser
115 120 125
<210> 198
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 198
Glu Val Val Leu Thr Gln Thr Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Gly Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Val Asn Asn
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Arg Ala Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Phe Gly Ala Ser Asn Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Lys Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Pro Trp
85 90 95
Thr Phe Gly His Gly Thr Lys Val Glu Met Lys
100 105
<210> 199
<211> 130
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 199
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Ala Ser Lys Lys Tyr His Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp His Val Pro Pro Lys Asp Cys Ser Asp Gly Asn Cys His
100 105 110
Ser Asp Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val
115 120 125
Ser Ser
130
<210> 200
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 200
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Met Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Lys Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Asn Ser Tyr Ser Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 201
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 201
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Asn Phe
20 25 30
Ala Val His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Gly Asn Arg Asn Thr Lys Tyr Ser His Asn Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Gln Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Pro Ile Ala Ala Ala Gly Arg Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 202
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 202
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ile Ser Thr
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln Ala Pro Arg Val Leu
35 40 45
Ile Tyr Asp Val Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Phe Cys His Gln Tyr Gly Ser Ser Pro
85 90 95
Ala Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 203
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 203
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ala Thr Thr Thr Phe Tyr Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Phe
65 70 75 80
Leu His Met Lys Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Tyr Thr Met Leu Arg Gly Ala Ser Met Asp Val Trp Gly
100 105 110
His Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 204
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 204
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ser Ala Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Leu Arg Ile Thr Trp Pro Pro
85 90 95
Ile Phe Thr Phe Gly Pro Gly Thr Lys Val Asp Val Lys
100 105
<210> 205
<211> 123
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 205
Gln Val His Leu Val Gln Ser Gly Ala Glu Val Lys Lys Ser Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Thr Phe Gly Tyr Thr Phe Thr Ala Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Pro Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Ile Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Ser Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Gly Leu Ser Arg Leu Arg Pro Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Lys Cys Ser Ser Ala Asn Cys Tyr Gly Asn Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 206
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 206
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Glu
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ala Gly Ser
85 90 95
Asn Asn Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 207
<211> 123
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 207
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Arg Phe Thr Asn Tyr
20 25 30
Arg Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Gly Ser Asp Thr Arg Tyr Ser Pro Ser Leu
50 55 60
Gln Gly Gln Val Thr Met Ser Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Met Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gln Thr Thr Gln Asn Ser Gly Tyr Asp Arg Trp Phe Asp Ser
100 105 110
Trp Gly Gln Gly Thr His Val Thr Val Ser Ser
115 120
<210> 208
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 208
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Ser Ile Gly Ser Asn
20 25 30
Ile Val Asn Trp Tyr Gln His Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ile Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 209
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 209
Glu Val Gln Val Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Thr Phe Ser Arg His
20 25 30
Thr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr His Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ile Ser Val Pro Leu Leu Arg Phe Leu Glu Trp Phe Gln His Pro
100 105 110
Phe Asp Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 210
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 210
Glu Ile Val Met Thr Gln Ser Pro Ala Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Ser Thr
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Ser Gly Ala Ser Thr Arg Ala Thr Gly Val Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Tyr Ile Asn Trp Pro Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 211
<211> 120
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 211
Glu Val Arg Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ser Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Thr Gly Ser Tyr Ile Tyr Asn Ala Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Phe Thr Met Thr Thr Val Thr Asn Phe Asp Ser Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 212
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 212
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Ala Tyr
20 25 30
Ser Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Tyr Arg Arg Pro Ser Gly Val Pro Gly Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser His Ala Gly Ser
85 90 95
His Thr Trp Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 213
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 213
Gln Leu Gln Val Val Ala Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Ser Tyr Asp Gly Gly Asn Lys Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Pro Lys Val Ile Pro His Pro Tyr Tyr Asp Ser Ser Gly Asp Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Ala Ile Ser Ser
115 120 125
<210> 214
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 214
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Trp Ala Ser Gln Ser Ile Ser Arg Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Lys Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Val Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln Tyr Ser Lys Leu Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 215
<211> 122
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 215
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Pro Phe Ser Ser Tyr
20 25 30
Ser Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Gly Ser Gly Gly Asp Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Ser Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Val Ala Thr Thr Gly Thr Arg Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 216
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 216
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Val Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Tyr Trp Pro Pro
85 90 95
Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 217
<211> 124
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 217
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Phe
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Leu Gly Arg Gly Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Arg Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Ile Ser Thr Ala Ser Tyr Val Pro Gly Thr Phe Glu Asp
100 105 110
Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 218
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 218
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Ala Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Glu Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asp Trp Pro Pro
85 90 95
Gly Thr Phe Gly Gln Gly Thr Asn Val Glu Ile Lys
100 105
<210> 219
<211> 120
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 219
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Asn Tyr
20 25 30
Asn Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Thr Gly Ser Tyr Ile His Tyr Ala Asp Leu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Val Arg Asn Thr Val Thr Ala Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Ser Val Ser Ser
115 120
<210> 220
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 220
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Phe Val Ser Trp Tyr Gln Gln Gln Pro Gly Arg Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Glu Val Ile Lys Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asp Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Gly Gly Asn
85 90 95
Asn Ser Trp Met Phe Gly Gly Gly Thr Met Leu Thr Val Leu
100 105 110
<210> 221
<211> 124
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 221
Gln Val Gln Val Val Gln Ser Gly Ala Glu Met Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Val Ser Gly Tyr Arg Leu Ile Asp Leu
20 25 30
Pro Leu His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Phe Asp Pro Glu Lys Ala Glu Ala Ile Tyr Ser Gln Lys Phe
50 55 60
Gln Asp Lys Val Thr Ile Ser Glu Asp Thr Ser Ile Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Trp Gly Val Glu Val Val Asn Gly Arg Arg Asp Tyr Phe Asp
100 105 110
Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 222
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 222
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Tyr Ala Asp Val Leu Ser Asn Gln Tyr Thr
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ala Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Val Ile Asn Gly Val Arg Ala Glu
65 70 75 80
Asp Glu Ala Tyr Tyr Tyr Cys Gln Ser Ala Asp Asn Thr Arg Ile Thr
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Ser Val Leu
100 105
<210> 223
<211> 128
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 223
Gly Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Thr Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Ser Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Val Arg Asp Cys Ser Ala Leu Thr Cys Pro Arg Arg Gly
100 105 110
Asp Ala Phe Asp Phe Trp Gly Arg Gly Thr Arg Val Thr Val Ser Ser
115 120 125
<210> 224
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 224
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Ser Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Tyr Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Lys Val Ser Asn Arg Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Thr His Trp Pro Arg Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105 110
<210> 225
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 225
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Val Tyr
20 25 30
Pro Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Arg Tyr Ile Ser Tyr Ala Asp Ser Leu
50 55 60
Arg Gly Arg Ile Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Lys Val Gly Gly Ser Lys His Gln Tyr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Ser Leu Val Thr Val Ser Ser
115 120
<210> 226
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 226
Gln Ser Val Leu Thr Gln Pro Pro Ser Thr Ser Gly Ile Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Ser Gly Ser Arg Ser Asn Ile Gly Ser Tyr
20 25 30
Thr Val Asn Trp Tyr Glu Gln Leu Pro Gly Thr Ala Pro Lys Leu Val
35 40 45
Ile Phe Asn Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Gly Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 227
<211> 124
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 227
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Phe Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Asp Ile Ile Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Val
35 40 45
Gly Trp Ile Ser Thr Tyr Asn Gly Asn Thr Asn Tyr Glu Gln Asn Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Arg Cys Ser Thr Ser Thr Cys Tyr Ser Arg Tyr Ala Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 228
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 228
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ile Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Ile Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Arg Ser Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Glu Val Glu Ile Lys
100 105
<210> 229
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 229
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ala Gln Pro Gly Glu
1 5 10 15
Ser Met Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Leu Ser Arg Tyr
20 25 30
Glu Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ser Tyr Ile Ser Ser Gly Gly Pro Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asn Ser Ala Glu Asn Ser Leu Glu
65 70 75 80
Leu Gln Met Ser Thr Leu Arg Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Met Arg Glu Gly Leu Thr Tyr Tyr Asp Ser Thr Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Ala Val Ser Ser
115
<210> 230
<211> 111
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 230
Gln Asn Val Leu Thr Gln Ser Pro Ser Ala Ser Ala Ser Leu Gly Ala
1 5 10 15
Ser Val Lys Leu Thr Cys Thr Leu Asn Ser Gly His Ser Arg Tyr Ala
20 25 30
Ile Ala Trp His Gln His Gln Pro Gln Arg Gly Pro Arg Phe Leu Met
35 40 45
Lys Ile Asn Ser Asp Gly Arg His Ile Arg Gly Asp Gly Ile Ser Asp
50 55 60
Arg Phe Ser Gly Ser Ala Ser Gly Ala Glu Arg His Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Trp Gly
85 90 95
Thr Gly Phe Arg Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 231
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 231
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Glu Tyr
20 25 30
Ala Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Asn Gly Gly Ser Lys Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Tyr Ser Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys Asp Asp Tyr Glu Gly Ala Gly Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Val Val Thr Val Ser Ser
115
<210> 232
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 232
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Ser Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Ile Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 233
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 233
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Leu Ile Ser Asn Gly
20 25 30
Tyr Tyr Trp Gly Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Ser Ile Tyr Tyr Thr Arg Asp Thr Tyr Tyr Asn Trp Ser Leu
50 55 60
Lys Ser Arg Ile Thr Ile Ser Val Asp Thr Ser Lys Lys Gln Phe Ser
65 70 75 80
Leu Lys Leu Tyr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg His Glu Gly Ser Cys Asn Asp Gly Ser Cys Tyr Gly Ser Phe
100 105 110
Val Asp Asn Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 234
<211> 106
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 234
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Leu Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Asp Ser Phe Ile Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 235
<211> 120
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 235
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Asp Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Asp Gly Val Ser Thr Lys Asn Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ser Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Leu Asn Ser Leu Arg Ala Glu Asp Thr Ala Phe Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Thr Phe His Asn Trp Tyr Phe Asp Leu Trp Gly Arg
100 105 110
Gly Val Leu Val Thr Val Ser Ser
115 120
<210> 236
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 236
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Thr Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Gly Asp Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Ser Gly Ala Ser Thr Arg Ala Thr Asp Phe Pro Ala Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Lys Asn Trp Pro Arg
85 90 95
Thr Phe Gly Arg Gly Thr Lys Val Glu Val Arg
100 105
<210> 237
<211> 121
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 237
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Ser Gly Gly Ser Thr Asn Tyr Ala Asp Ser Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Thr Ile Thr Tyr Ser Tyr Tyr Tyr Met Ala Val Trp Gly
100 105 110
Lys Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 238
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 238
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Gly Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Arg Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Arg Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Gly Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Thr Ser Ile
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 239
<211> 124
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 239
Gln Val Gln Val Val Gln Ser Gly Ala Glu Met Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Arg Val Ser Cys Lys Val Ser Gly Tyr Arg Leu Thr Asp Leu
20 25 30
Pro Leu His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Phe Val Asp Leu Glu Lys Arg Glu Ile Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Lys Val Thr Ile Thr Glu Asp Thr Ser Ala Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Trp Gly Ile Glu Val Val Asn Gly Arg Asp Glu Phe Phe Asp
100 105 110
Ser Trp Gly Gln Gly Thr Leu Val Ser Val Ser Ser
115 120
<210> 240
<211> 106
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 240
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Tyr Ala Asp Val Leu Ser Lys Gln Tyr Thr
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ala Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Ile Ile Asn Gly Val Arg Thr Glu
65 70 75 80
Asp Glu Ala Tyr Tyr Tyr Cys Gln Ser Ala Asp Thr Arg Ile Thr Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Ser Val Leu
100 105
<210> 241
<211> 124
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 241
Gln Val Gln Val Val Gln Ser Gly Ala Glu Met Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Val Ser Gly Tyr Ser Leu Ser Asp Leu
20 25 30
Pro Leu His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Phe Asp Pro Ile Asn Gly Glu Ile Ile Tyr Ala Gln Thr Phe
50 55 60
Gln Gly Lys Val Thr Ile Ser Glu Asp Thr Ser Ile Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Trp Gly Val Ala Val Val Ser Gly Arg Arg Asp Tyr Phe Asp
100 105 110
Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 242
<211> 106
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 242
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Tyr Ala Ala Val Leu Ser Asn Gln Tyr Thr
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ala Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Ile Ile Asn Gly Val Arg Thr Glu
65 70 75 80
Asp Glu Ala Tyr Tyr Tyr Cys Gln Thr Ala Asp Thr Lys Tyr Thr Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Ser Val Leu
100 105
<210> 243
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 243
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Thr Ser Gly
20 25 30
Asp Tyr Tyr Trp Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Thr Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Arg Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe
85 90 95
Cys Ala Arg Ala Tyr Ala Tyr Glu Phe Trp Ser Gly Tyr Pro Asn Trp
100 105 110
Phe Asp Pro Trp Gly Leu Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 244
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 244
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Arg Ile Ser Thr Tyr
20 25 30
Val Asn Trp Tyr Gln Val Lys Ala Gly Thr Ala Pro Lys Val Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Val Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Thr Thr Tyr Phe Cys Gln Gln Ser Tyr Ser Pro Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 245
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 245
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Asn Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Ser Tyr Asp Gly Ile Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ser Ile Ser Arg Asp Asn Ser Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Phe Tyr Cys
85 90 95
Ala Arg Gly Leu Gly Tyr Cys Ser Gly Thr Gly Gly Ser Cys Thr Pro
100 105 110
Phe Glu Tyr Trp Gly Gln Gly Ile Leu Val Thr Val Ser Ser
115 120 125
<210> 246
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 246
Asp Ile Gln Met Thr Gln Ser Pro Pro Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Lys Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Asn Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Ala Leu Ser Ile Ser Ser Leu Glu Thr
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asp Ser Ala Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Phe Lys
100 105
<210> 247
<211> 135
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 247
Gln Val Gln Leu Gln Gln Arg Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Glu Ile Tyr Gly Ala Ser Leu Asn Asp Tyr
20 25 30
Asp Trp Thr Trp Ile Arg Gln Pro Pro Gly Lys Arg Leu Glu Trp Ile
35 40 45
Gly Val Ile Asn Arg Arg Asp Thr Val Asp Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Leu Ser Leu Asp Thr Ser Lys Asn Gln Leu Ser Leu
65 70 75 80
Ser Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Val
85 90 95
Arg Val Pro Arg Arg Gly Phe Glu Gly Ser Phe Gly Phe Cys Asp Asp
100 105 110
Thr Ala Cys Arg Tyr Gly His Thr Trp Phe Asp Pro Trp Gly Gln Gly
115 120 125
Thr Leu Val Thr Val Ser Ser
130 135
<210> 248
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 248
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Arg Asp Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Arg Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Phe Ala Gly Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Arg Gly
50 55 60
Arg Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Leu Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Thr Val Asp Ile Lys
100 105
<210> 249
<211> 125
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 249
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Arg Asn Gly
20 25 30
Arg Met Gly Val Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Phe Ala Ser Asp Glu Lys Ser Tyr Ser Thr Ser
50 55 60
Gln Arg Thr Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val
65 70 75 80
Val Leu Ser Met Thr Asp Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Ile Leu Lys Phe Gly Thr Met Arg Ala Ala Tyr Tyr Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Ala Leu Val Pro Val Ser Ser
115 120 125
<210> 250
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 250
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Ile Asn Ile Gly Ile Arg Thr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Met Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Leu Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Lys Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp His
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Ala Leu
100 105
<210> 251
<211> 125
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 251
Gln Val His Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Met
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Phe Tyr Tyr Cys
85 90 95
Ala Gly Ser Lys Val Gly Tyr Thr Thr Gly Arg Arg Asn Trp Tyr Phe
100 105 110
Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 252
<211> 110
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 252
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Ala Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ala Gly Asn
85 90 95
Asn Asn Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 253
<211> 126
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 253
Gln Val Gln Leu Leu Gln Ser Gly Ser Glu Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Asn Ile His Cys Lys Ala Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Leu His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Glu Thr Gly Glu Thr Thr Tyr Ser Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Val Val Asn
65 70 75 80
Leu Glu Val Arg Ser Leu Arg Ser Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Val Val Val Val Pro Val Glu Met Ser Arg Arg Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Ile Met Val Thr Val Ser Ser
115 120 125
<210> 254
<211> 108
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 254
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Pro Cys Gly Gly Asn Asn Ile Glu Arg Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Thr Val Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Ser Thr Ala Thr Leu Thr Ile Ser Arg Val Gly Ala Gly
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Val Trp Asp Ser Thr Thr Asp His
85 90 95
Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 255
<211> 125
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 255
Arg Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Gln Thr Ser Gly Tyr Ile Phe Ser Ala Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Met Asn Ala Lys Ser Gly Gly Ala Asn Thr Ala Gln Gln Phe
50 55 60
Gln Gly Arg Leu Thr Met Thr Arg Asp Met Ser Val Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Arg Asp Asp Tyr Met Trp Gly Ser Tyr Arg Pro Leu
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 256
<211> 106
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic polypeptide
<400> 256
Glu Val Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Ser Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Gly Tyr
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Ala Gly Ile Pro Ala Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Arg Ser Asn Gly Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 257
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 257
Gly Phe Ile Phe Ser Arg Tyr Ala
1 5
<210> 258
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 258
Ile Ser Tyr Asp Ala Arg Asn Ser
1 5
<210> 259
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 259
Ala Arg Pro Thr Leu Pro Tyr Ser Asn Asn Trp Tyr Ala Pro Glu Tyr
1 5 10 15
<210> 260
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 260
Gly Phe Ser Phe Ser Ser Tyr Ala
1 5
<210> 261
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 261
Ile Ser Gly Asn Gly Asn Gly Arg
1 5
<210> 262
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 262
Ala Lys Val Val Arg Ile Ala Ala Val Leu Tyr Tyr Phe Asp Tyr
1 5 10 15
<210> 263
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 263
Gly Phe Thr Phe Ser Asn Tyr Ala
1 5
<210> 264
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 264
Ile Ser Gly Ser Gly Gly Leu Thr
1 5
<210> 265
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 265
Ala Arg Val Lys Ser Thr Thr Gly Thr Thr Ala Leu Val Phe Asp Ile
1 5 10 15
<210> 266
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 266
Gly Phe Thr Phe Ile Asn Tyr Gly
1 5
<210> 267
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 267
Ile Ser Asn Asp Gly Ser Tyr Asn
1 5
<210> 268
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 268
Ala Lys Asp Lys His Gly Asp Phe Asp Tyr Tyr Gly Val Asp Val
1 5 10 15
<210> 269
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 269
Gly Phe Thr Phe Ser Ser Tyr Gly
1 5
<210> 270
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 270
Ile Ser Tyr Asp Gly Ser Asp Asn
1 5
<210> 271
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 271
Ala Arg Arg Arg Pro Gly Ser Phe Pro Gly Leu Cys Asp Tyr
1 5 10
<210> 272
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 272
Gly Gly Ser Phe Ser Arg Leu Thr
1 5
<210> 273
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 273
His Ile Pro Ile Phe Gly Thr Thr
1 5
<210> 274
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 274
Ala Arg Met Tyr Ser Gly His Asp Gly Val Asp Val
1 5 10
<210> 275
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 275
Gly Gly Ser Ile Ser Ser Gly Phe Tyr Tyr
1 5 10
<210> 276
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 276
Ile Tyr Asp Ser Gly Arg Thr
1 5
<210> 277
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 277
Ala Arg His Leu Thr His Leu Tyr Gly Asp Tyr Val Thr Pro Asp Ala
1 5 10 15
Leu Asp Ile
<210> 278
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 278
Gly Phe Thr Phe Thr Thr Tyr Ser
1 5
<210> 279
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 279
Ile Ser Ser Gly Ser Ser Asn Ile
1 5
<210> 280
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 280
Ala Arg Ala His Gly Arg Ile Val Asn Ser Gly Val Val Ile Ser Arg
1 5 10 15
Phe Asp Pro
<210> 281
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 281
Gly Tyr Thr Phe Thr Ala Tyr Tyr
1 5
<210> 282
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 282
Ile Asn Pro Ser Ser Gly Gly Ala
1 5
<210> 283
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 283
Ala Arg Met Gly Cys Arg Ser Asp Arg Cys Tyr Ser Thr Asn Tyr Asn
1 5 10 15
Phe Asp Gln
<210> 284
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 284
Gly Phe Thr Ile Ser Pro Tyr Gly
1 5
<210> 285
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 285
Ile Ser Ser Ser Ser Arg Tyr Thr
1 5
<210> 286
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 286
Ala Arg Glu Arg Gly His Ser Thr Ser Ser Ser Tyr Phe Asp Ser
1 5 10 15
<210> 287
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 287
Gly Phe Thr Phe Ser Asn Ala Trp
1 5
<210> 288
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 288
Ile Gln Thr Lys Thr Asp Gly Gly Thr Thr
1 5 10
<210> 289
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 289
Ser Thr Gly Pro Tyr Tyr Tyr Asp Thr Ser Gly Tyr Pro Gln Pro Phe
1 5 10 15
Asp Tyr
<210> 290
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 290
Gly Phe Asp Phe Ser Arg Tyr Thr
1 5
<210> 291
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 291
Ile Ser Ser Thr Ser Leu Tyr Thr
1 5
<210> 292
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 292
Ala Arg Val Val Gly Pro Ala Glu Leu Asp Tyr
1 5 10
<210> 293
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 293
Gly Tyr Thr Phe Thr Asp Tyr Tyr
1 5
<210> 294
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 294
Ile Asn Pro Lys Thr Gly Gly Ser
1 5
<210> 295
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 295
Ala Arg Ala Gly Arg Asn Gly Tyr Asp Tyr
1 5 10
<210> 296
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 296
Gly Phe Lys Phe Arg Asn Tyr Ala
1 5
<210> 297
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 297
Ile Thr Ser Gly Gly Ser Thr
1 5
<210> 298
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 298
Thr Val Pro Trp Gly Asn Tyr Asn Asp Tyr Val Ser Asp Tyr
1 5 10
<210> 299
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 299
Gly Phe Ile Phe Ser Arg Tyr Pro
1 5
<210> 300
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 300
Ile Ser Tyr Asp Gly Asn Asn Lys
1 5
<210> 301
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 301
Ala Arg His Phe Leu Pro Tyr Ser Ser Ser Trp Tyr Gln Gly Phe Asn
1 5 10 15
Tyr
<210> 302
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 302
Gly Phe Leu Phe Ser Arg Tyr Gly
1 5
<210> 303
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 303
Ile Ser Tyr Asp Gly Asn Lys Lys
1 5
<210> 304
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 304
Ala Arg Gly Val Pro Tyr Gly Asp Thr Leu Thr Gly Leu Val Tyr
1 5 10 15
<210> 305
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 305
Gly Phe Ser Leu Arg Asn Ala Arg Met Gly
1 5 10
<210> 306
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 306
Ile Phe Ser Asn Asp Glu Lys
1 5
<210> 307
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 307
Ala Arg Leu Leu Val Ala Gly Thr Phe Leu Pro Ser His Tyr Phe Asp
1 5 10 15
Tyr
<210> 308
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 308
Gly Phe Thr Phe Ser Thr Tyr Ser
1 5
<210> 309
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 309
Ile Ser Ser Ser Ser Ser Thr Ile
1 5
<210> 310
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 310
Thr Arg Gln Val Gly Ala Asp Phe Ser Gly Arg Gly Phe Asp Tyr
1 5 10 15
<210> 311
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 311
Gly Phe Lys Phe Ser Val Tyr Ala
1 5
<210> 312
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 312
Ile Ser Ser Ser Gly Ser Thr Ile
1 5
<210> 313
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 313
Ala Thr Ala Arg His Ile Thr Asn Asp Gly Phe Asp Ile
1 5 10
<210> 314
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 314
Gly Tyr Leu Ile Ser Asn Gly Tyr Tyr
1 5
<210> 315
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 315
Ile Tyr His Thr Arg Ser Thr
1 5
<210> 316
<211> 20
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 316
Ala Arg Gly Pro Gly His Cys Tyr Gly Asp Asp Asp Cys Tyr Ala Tyr
1 5 10 15
Tyr Phe Asp Gln
20
<210> 317
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 317
Gly Gly Thr Phe Ser Arg Phe Ala
1 5
<210> 318
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 318
Ile Leu Pro Ile Phe Gly Thr Ala
1 5
<210> 319
<211> 20
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 319
Ala Arg Ser Leu Pro Tyr Cys Thr Asn Asp Val Cys Ser Asn Gln Asn
1 5 10 15
Thr Phe Asp Tyr
20
<210> 320
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 320
Gly Gly Ser Ile Ser Ser Gly Asp Tyr Tyr
1 5 10
<210> 321
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 321
Ile Tyr Tyr Ser Gly Ser Thr
1 5
<210> 322
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 322
Ala Ser Arg Tyr Gly Asp Pro Ile Gly Asp Asn Trp Phe Asp Pro
1 5 10 15
<210> 323
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 323
Gly Phe Arg Phe Ser Phe Tyr Gly
1 5
<210> 324
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 324
Ile Ser Gly Thr Gly Ala Thr Arg
1 5
<210> 325
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 325
Val Arg Arg Phe Pro Met Thr Thr Val Thr Ser Phe Asp Ser
1 5 10
<210> 326
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 326
Gly Phe Pro Phe Asn Met Phe Trp
1 5
<210> 327
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 327
Ile Lys Gln Asp Gly Ser Glu Lys
1 5
<210> 328
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 328
Val Arg Glu Gly Val Arg Arg Val Val Val Arg Ser Thr Gly Tyr Phe
1 5 10 15
Asp Phe
<210> 329
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 329
Gly Gly Pro Ile Ser Asn Gly Pro Tyr Tyr
1 5 10
<210> 330
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 330
Ile Phe Tyr Ser Gly Ser Thr
1 5
<210> 331
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 331
Ala Arg His Val Val Thr Ala Ser Gly Trp Phe Asp Pro
1 5 10
<210> 332
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 332
Gly Gly Pro Ile Ser Asn Gly Pro Tyr Tyr
1 5 10
<210> 333
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 333
Ile Phe Tyr Ser Gly Ser Thr
1 5
<210> 334
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 334
Ala Arg His Val Val Thr Ala Ser Gly Trp Phe Asp Pro
1 5 10
<210> 335
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 335
Gly Val Thr Phe Ser Asp Asn Ala
1 5
<210> 336
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 336
Ile Ser Tyr Asp Gly Ser Ser Arg
1 5
<210> 337
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 337
Ala Arg Val Thr Ala Asp Tyr Tyr Glu Ser Ser Gly Lys Val Phe
1 5 10 15
<210> 338
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 338
Gly Gly Ser Ile Ser Asp His Tyr
1 5
<210> 339
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 339
Ile Tyr Thr Ser Gly Thr Thr
1 5
<210> 340
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 340
Ala Arg Ser Leu Glu Thr Val Ile Arg Phe Tyr Tyr Tyr His Tyr Met
1 5 10 15
Asp Val
<210> 341
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 341
Gly Gly Ser Ile Gly Asp Tyr His
1 5
<210> 342
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 342
Ile His Ser Ser Gly Asn Thr
1 5
<210> 343
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 343
Ala Arg Gln Asn Val Phe Asp Ile
1 5
<210> 344
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 344
Gly Ile Ser Ile Asn Asn Tyr Tyr
1 5
<210> 345
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 345
Val Tyr Ser Thr Gly Ser Ser
1 5
<210> 346
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 346
Ala Arg Gly Ser Met Pro His Ile
1 5
<210> 347
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 347
Gly Ile Thr Phe Ser Asn Ala Trp
1 5
<210> 348
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 348
Ile Glu Ser Lys Ile Asp Gly Gly Thr Ile
1 5 10
<210> 349
<211> 21
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 349
Thr Thr Asp Gln Gly Tyr Tyr Asp Arg Ser Gly Tyr Trp Val Val Gly
1 5 10 15
Asn His Phe Asp Tyr
20
<210> 350
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 350
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 351
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 351
Ile Ser Gly Thr Thr Gly Ser Thr
1 5
<210> 352
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 352
Ala Lys Asp Ser His Ser Met Ile Val Val Asp His Ala Phe Asp Ile
1 5 10 15
<210> 353
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 353
Gly Phe Thr Phe Ser Thr Tyr Ile
1 5
<210> 354
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 354
Ile Ser Thr Ser Ser Val Tyr Thr
1 5
<210> 355
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 355
Ala Arg Glu Glu Gly Phe Arg Ala Tyr Asn Leu Tyr
1 5 10
<210> 356
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 356
Gly Phe Thr Phe Ser Arg Phe Gly
1 5
<210> 357
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 357
Ile Ser Phe Asp Gly Ser Asn Arg
1 5
<210> 358
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 358
Ala Arg Asp Trp Asp Arg Leu Val Arg Ser Ala Val Gly Tyr
1 5 10
<210> 359
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 359
Gly Gly Ser Val Ser Thr Ala Thr Tyr Tyr
1 5 10
<210> 360
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 360
Ile Tyr Ser Ser Gly Asn Thr
1 5
<210> 361
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 361
Glu Arg Arg Leu Arg Ile Leu Ser Ile Glu Arg Asn Tyr Tyr Ala Met
1 5 10 15
Asp Val
<210> 362
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 362
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 363
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 363
Ile Ser Tyr Asp Ala Ser Lys Lys
1 5
<210> 364
<211> 23
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 364
Ala Arg Asp His Val Pro Pro Lys Asp Cys Ser Asp Gly Asn Cys His
1 5 10 15
Ser Asp Tyr Gly Met Asp Val
20
<210> 365
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 365
Gly Tyr Ser Phe Thr Asn Phe Ala
1 5
<210> 366
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 366
Ile Asn Pro Gly Asn Arg Asn Thr
1 5
<210> 367
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 367
Ala Arg Leu Pro Ile Ala Ala Ala Gly Arg Gly Trp Phe Asp Pro
1 5 10 15
<210> 368
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 368
Gly Phe Thr Phe Asn Thr Tyr Gly
1 5
<210> 369
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 369
Ile Ser Ser Ala Thr Thr Thr Phe
1 5
<210> 370
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 370
Ala Arg Val Tyr Thr Met Leu Arg Gly Ala Ser Met Asp Val
1 5 10
<210> 371
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 371
Gly Tyr Thr Phe Thr Ala Tyr Tyr
1 5
<210> 372
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 372
Ile Asn Pro Ile Ser Gly Gly Thr
1 5
<210> 373
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 373
Ala Arg Val Lys Cys Ser Ser Ala Asn Cys Tyr Gly Asn Phe Asp Tyr
1 5 10 15
<210> 374
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 374
Gly Tyr Arg Phe Thr Asn Tyr Arg
1 5
<210> 375
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 375
Ile Tyr Pro Gly Gly Ser Asp Thr
1 5
<210> 376
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 376
Ala Arg Gln Thr Thr Gln Asn Ser Gly Tyr Asp Arg Trp Phe Asp Ser
1 5 10 15
<210> 377
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 377
Gly Ile Thr Phe Ser Arg His Thr
1 5
<210> 378
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 378
Ile Ser Gly Ser Gly Gly Ser Thr
1 5
<210> 379
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 379
Ala Ile Ser Val Pro Leu Leu Arg Phe Leu Glu Trp Phe Gln His Pro
1 5 10 15
Phe Asp Phe
<210> 380
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 380
Gly Phe Thr Phe Asn Thr Tyr Ser
1 5
<210> 381
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 381
Ile Ser Ser Thr Gly Ser Tyr Ile
1 5
<210> 382
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 382
Val Arg Phe Thr Met Thr Thr Val Thr Asn Phe Asp Ser
1 5 10
<210> 383
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 383
Gly Phe Thr Phe Thr Asn Tyr Gly
1 5
<210> 384
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 384
Ile Ser Tyr Asp Gly Gly Asn Lys
1 5
<210> 385
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 385
Pro Lys Val Ile Pro His Pro Tyr Tyr Asp Ser Ser Gly Asp Asp Ala
1 5 10 15
Phe Asp Ile
<210> 386
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 386
Gly Phe Pro Phe Ser Ser Tyr Ser
1 5
<210> 387
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 387
Ile Ser Gly Ser Gly Gly Asp Ile
1 5
<210> 388
<211> 15
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 388
Ala Arg Gly Leu Val Ala Thr Thr Gly Thr Arg Tyr Phe Asp Tyr
1 5 10 15
<210> 389
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 389
Gly Gly Thr Phe Arg Arg Phe Ala
1 5
<210> 390
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 390
Ile Ile Pro Ile Leu Gly Arg Gly
1 5
<210> 391
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 391
Ala Arg Phe Ile Ser Thr Ala Ser Tyr Val Pro Gly Thr Phe Glu Asp
1 5 10 15
Val
<210> 392
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 392
Gly Phe Thr Phe Arg Asn Tyr Asn
1 5
<210> 393
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 393
Ile Ser Ser Thr Gly Ser Tyr Ile
1 5
<210> 394
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 394
Ala Arg Met Val Arg Asn Thr Val Thr Ala Phe Asp Tyr
1 5 10
<210> 395
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 395
Gly Tyr Arg Leu Ile Asp Leu Pro
1 5
<210> 396
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 396
Phe Asp Pro Glu Lys Ala Glu Ala
1 5
<210> 397
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 397
Ala Thr Trp Gly Val Glu Val Val Asn Gly Arg Arg Asp Tyr Phe Asp
1 5 10 15
Ser
<210> 398
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 398
Gly Phe Thr Phe Ser Ser Phe Glu
1 5
<210> 399
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 399
Ile Ser Thr Ser Gly Ser Thr Ile
1 5
<210> 400
<211> 21
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 400
Ala Arg Asp Val Arg Asp Cys Ser Ala Leu Thr Cys Pro Arg Arg Gly
1 5 10 15
Asp Ala Phe Asp Phe
20
<210> 401
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 401
Gly Phe Ser Phe Ser Val Tyr Pro
1 5
<210> 402
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 402
Ile Ser Ser Ser Ser Arg Tyr Ile
1 5
<210> 403
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 403
Val Lys Val Gly Gly Ser Lys His Gln Tyr Tyr Phe Asp Tyr
1 5 10
<210> 404
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 404
Gly Tyr Thr Phe Thr Asn Tyr Asp
1 5
<210> 405
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 405
Ile Ser Thr Tyr Asn Gly Asn Thr
1 5
<210> 406
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 406
Ala Arg Glu Arg Cys Ser Thr Ser Thr Cys Tyr Ser Arg Tyr Ala Asp
1 5 10 15
Tyr
<210> 407
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 407
Gly Phe Thr Leu Ser Arg Tyr Glu
1 5
<210> 408
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 408
Ile Ser Ser Gly Gly Pro Ser Ile
1 5
<210> 409
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 409
Met Arg Glu Gly Leu Thr Tyr Tyr Asp Ser Thr Ile
1 5 10
<210> 410
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 410
Gly Phe Thr Phe Asp Glu Tyr Ala
1 5
<210> 411
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 411
Ile Ser Trp Asn Gly Gly Ser Lys
1 5
<210> 412
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 412
Ala Lys Asp Asp Tyr Glu Gly Ala Gly Phe Asp Ile
1 5 10
<210> 413
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 413
Gly Tyr Leu Ile Ser Asn Gly Tyr Tyr
1 5
<210> 414
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 414
Ile Tyr Tyr Thr Arg Asp Thr
1 5
<210> 415
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 415
Val Arg His Glu Gly Ser Cys Asn Asp Gly Ser Cys Tyr Gly Ser Phe
1 5 10 15
Val Asp Asn
<210> 416
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 416
Asp Phe Thr Phe Ser Ser Tyr Thr
1 5
<210> 417
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 417
Ile Ser Gly Asp Gly Val Ser Thr
1 5
<210> 418
<211> 13
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 418
Ala Arg Gly Gly Thr Phe His Asn Trp Tyr Phe Asp Leu
1 5 10
<210> 419
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 419
Gly Phe Thr Phe Ser Thr Tyr Ala
1 5
<210> 420
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 420
Ile Ser Gly Ser Gly Gly Ser Thr
1 5
<210> 421
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 421
Ala Lys Gly Thr Ile Thr Tyr Ser Tyr Tyr Tyr Met Ala Val
1 5 10
<210> 422
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 422
Gly Tyr Arg Leu Thr Asp Leu Pro
1 5
<210> 423
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 423
Val Asp Leu Glu Lys Arg Glu Ile
1 5
<210> 424
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 424
Ala Thr Trp Gly Ile Glu Val Val Asn Gly Arg Asp Glu Phe Phe Asp
1 5 10 15
Ser
<210> 425
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 425
Gly Tyr Ser Leu Ser Asp Leu Pro
1 5
<210> 426
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 426
Phe Asp Pro Ile Asn Gly Glu Ile
1 5
<210> 427
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 427
Ala Thr Trp Gly Val Ala Val Val Ser Gly Arg Arg Asp Tyr Phe Asp
1 5 10 15
Ser
<210> 428
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 428
Gly Gly Ser Ile Thr Ser Gly Asp Tyr Tyr
1 5 10
<210> 429
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 429
Ile Tyr His Ser Gly Thr Thr
1 5
<210> 430
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 430
Ala Arg Ala Tyr Ala Tyr Glu Phe Trp Ser Gly Tyr Pro Asn Trp Phe
1 5 10 15
Asp Pro
<210> 431
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 431
Gly Phe Ile Phe Asn Arg Tyr Ala
1 5
<210> 432
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 432
Ile Ser Tyr Asp Gly Ile Asn Lys
1 5
<210> 433
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 433
Ala Arg Gly Leu Gly Tyr Cys Ser Gly Thr Gly Gly Ser Cys Thr Pro
1 5 10 15
Phe Glu Tyr
<210> 434
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 434
Gly Ala Ser Leu Asn Asp Tyr Asp
1 5
<210> 435
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 435
Ile Asn Arg Arg Asp Thr Val
1 5
<210> 436
<211> 29
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 436
Val Arg Val Pro Arg Arg Gly Phe Glu Gly Ser Phe Gly Phe Cys Asp
1 5 10 15
Asp Thr Ala Cys Arg Tyr Gly His Thr Trp Phe Asp Pro
20 25
<210> 437
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 437
Gly Phe Ser Leu Arg Asn Gly Arg Met Gly
1 5 10
<210> 438
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 438
Ile Phe Ala Ser Asp Glu Lys
1 5
<210> 439
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 439
Ala Arg Ile Leu Lys Phe Gly Thr Met Arg Ala Ala Tyr Tyr Phe Asp
1 5 10 15
Tyr
<210> 440
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 440
Gly Phe Thr Phe Ser Asp Tyr Tyr
1 5
<210> 441
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 441
Ile Ser Ser Ser Gly Ser Thr Ile
1 5
<210> 442
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 442
Ala Gly Ser Lys Val Gly Tyr Thr Thr Gly Arg Arg Asn Trp Tyr Phe
1 5 10 15
Asp Leu
<210> 443
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 443
Gly Phe Thr Phe Thr Asp Phe Tyr
1 5
<210> 444
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 444
Ile Asn Pro Glu Thr Gly Glu Thr
1 5
<210> 445
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 445
Ala Arg Asp Leu Val Val Val Val Pro Val Glu Met Ser Arg Arg Ala
1 5 10 15
Phe Asp Ile
<210> 446
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 446
Gly Tyr Ile Phe Ser Ala Tyr Tyr
1 5
<210> 447
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 447
Met Asn Ala Lys Ser Gly Gly Ala
1 5
<210> 448
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 448
Ala Arg Asp Tyr Arg Asp Asp Tyr Met Trp Gly Ser Tyr Arg Pro Leu
1 5 10 15
Asp Tyr
<210> 449
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 449
Ala Leu Pro Lys Lys Tyr
1 5
<210> 450
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 450
Glu Asp Thr
1
<210> 451
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 451
Ser Ser Thr Asp Ser Ser Gly Asn Pro Val Leu
1 5 10
<210> 452
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 452
Gln Ser Val Ser Thr Tyr
1 5
<210> 453
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 453
Glu Ala Ser
1
<210> 454
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 454
Gln Gln Arg Ser Ser Trp Pro Ile Thr
1 5
<210> 455
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 455
Ser Gly Ser Val Ser Ser Ser Tyr Tyr
1 5
<210> 456
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 456
Ser Ile Asn
1
<210> 457
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 457
Gly Leu Tyr Met Gly Ser Gly Ile Trp Ile
1 5 10
<210> 458
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 458
Gln Gly Ile Ser Ser Trp
1 5
<210> 459
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 459
Ala Ala Ser
1
<210> 460
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 460
Gln Gln Ala Asp Ser Phe Pro Arg Thr
1 5
<210> 461
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 461
Gln Ser Ile Ser Lys Trp
1 5
<210> 462
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 462
Lys Ala Ser
1
<210> 463
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 463
Gln Gln His Asn Ser Tyr Ser Tyr Thr
1 5
<210> 464
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 464
Gln Ser Val Arg Ser Tyr
1 5
<210> 465
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 465
Asp Ala Ser
1
<210> 466
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 466
Gln Gln Arg Ser Thr Trp Pro Pro Gly Met
1 5 10
<210> 467
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 467
Gln Ser Val Ser Ser Ser Phe
1 5
<210> 468
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 468
Gly Ala Ser
1
<210> 469
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 469
Gln Gln Tyr Ser Asn Ser Arg Leu Thr
1 5
<210> 470
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 470
Ser Ser Asp Val Gly Gly Tyr Asn Tyr
1 5
<210> 471
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 471
Glu Val Thr
1
<210> 472
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 472
Ser Ser Tyr Thr Thr Ser Asn Thr Leu Val Val
1 5 10
<210> 473
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 473
Asn Ile Gly Thr Lys Ser
1 5
<210> 474
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 474
Asn Asp Ser
1
<210> 475
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 475
Gln Val Trp Asp Ser Gly Ile Asp Val Val
1 5 10
<210> 476
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 476
Asn Ile Gly Thr Lys Thr
1 5
<210> 477
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 477
Tyr Asp Ser
1
<210> 478
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 478
Arg Val Trp Asp Ser Asp Thr Asp His Arg Val
1 5 10
<210> 479
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 479
Lys Leu Gly Asp Lys Tyr
1 5
<210> 480
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 480
Gln Asp Thr
1
<210> 481
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 481
Gln Ala Trp Asp Ser Ser Thr Val Val
1 5
<210> 482
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 482
Gln Asp Ile Gly Val Asp
1 5
<210> 483
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 483
Gly Ala Ser
1
<210> 484
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 484
Leu Gln Asp Tyr Asn Tyr Pro Trp Thr
1 5
<210> 485
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 485
Ala Leu Pro Lys Gln Tyr
1 5
<210> 486
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 486
Gln Asp Thr
1
<210> 487
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 487
Gln Ser Gly Asp Ser Ser Gly Thr Tyr Leu Val
1 5 10
<210> 488
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 488
Gln Arg Val Gly Asn Ser
1 5
<210> 489
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 489
Asp Ala Ser
1
<210> 490
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 490
His Gln His Ser Thr Trp Pro Arg Gly Thr
1 5 10
<210> 491
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 491
Ser Gly Ser Ile Ala Thr Asn Tyr
1 5
<210> 492
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 492
Glu Asp Ser
1
<210> 493
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 493
Gln Ser Tyr Asp Asn Ser Asn Arg Ala Val Val
1 5 10
<210> 494
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 494
Ser Gly Thr Ile Ala Ser Asn Tyr
1 5
<210> 495
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 495
Glu Asp Asn
1
<210> 496
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 496
Gln Ser Tyr Asp Asn Ser Asp Arg Val
1 5
<210> 497
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 497
Asn Ile Gly Leu Lys Ser
1 5
<210> 498
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 498
Tyr Asp Ser
1
<210> 499
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 499
Gln Val Trp Asp Ser Ser Arg Asn His Pro Val
1 5 10
<210> 500
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 500
Lys Leu Gly Asp Lys Tyr
1 5
<210> 501
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 501
Gln Asp Ser
1
<210> 502
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 502
Gln Ala Trp Asp Ser Ser Thr Ala Val
1 5
<210> 503
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 503
Gln Gly Ile Ser Arg Phe
1 5
<210> 504
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 504
Ser Ala Ser
1
<210> 505
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 505
Gln Gln Leu Asn Ser His Pro Arg Met Phe Thr
1 5 10
<210> 506
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 506
Gln Gly Ile Ser Asn Trp
1 5
<210> 507
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 507
Asp Ala Ser
1
<210> 508
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 508
Gln Gln Ala Asn Ser Phe Pro Phe Thr
1 5
<210> 509
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 509
Ala Leu Pro Lys Gln Tyr
1 5
<210> 510
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 510
Glu Asp Asn
1
<210> 511
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 511
Gln Ser Ala Asp Ser Ser Gly Thr Tyr Val Val
1 5 10
<210> 512
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 512
Asn Ile Gly Leu Lys Ser
1 5
<210> 513
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 513
Tyr Asp Ser
1
<210> 514
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 514
Gln Val Trp Asp Ser Ser Arg Asn His Pro Val
1 5 10
<210> 515
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 515
Ser Ser Asp Val Gly Gly Tyr Asn Phe
1 5
<210> 516
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 516
Asp Val Thr
1
<210> 517
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 517
Gly Ala Tyr Ala Gly Phe Asn Ala Leu
1 5
<210> 518
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 518
Ala Val Pro Ile Lys Tyr
1 5
<210> 519
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 519
Glu Asp Asp
1
<210> 520
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 520
Tyr Ser Thr Asp Ser Ser Gly Tyr Gln Arg Ala
1 5 10
<210> 521
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 521
Gln Ser Val Gly Thr Asp
1 5
<210> 522
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 522
Asp Ala Phe
1
<210> 523
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 523
Gln Gln Arg Ser Arg Trp Pro Pro Pro Tyr Thr
1 5 10
<210> 524
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 524
Gln Ser Val Gly Thr Asp
1 5
<210> 525
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 525
Asp Ala Phe
1
<210> 526
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 526
Gln Gln Arg Ser Arg Trp Pro Pro Pro Tyr Thr
1 5 10
<210> 527
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 527
Gln Ser Val Arg Ser Trp
1 5
<210> 528
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 528
Lys Ala Ser
1
<210> 529
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 529
Gln Gln Tyr Gln Thr Phe Ser Trp Thr
1 5
<210> 530
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 530
Gln Ser Leu Leu Gln Ser Asp Gly Tyr Ser Tyr
1 5 10
<210> 531
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 531
Leu Gly Ser
1
<210> 532
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 532
Met Gln Ala Leu Gln Thr Pro Trp Thr
1 5
<210> 533
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 533
Gln Ser Val Leu Phe Ser Ser Asn Asn Lys Asn Tyr
1 5 10
<210> 534
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 534
Trp Ala Ser
1
<210> 535
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 535
Gln Gln Phe Tyr Thr Thr Pro Leu Thr
1 5
<210> 536
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 536
Thr Ser Asn Ile Glu Thr Asn Tyr
1 5
<210> 537
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 537
Arg Asn Asp
1
<210> 538
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 538
Ala Ala Trp Asp Asp Ser Leu Lys Ala Pro Val
1 5 10
<210> 539
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 539
Ser Thr Asn Ile Gly Ala Gly Tyr Asp
1 5
<210> 540
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 540
Gly Asn Ser
1
<210> 541
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 541
Gln Ser Tyr Asp Arg Ser Leu Ser Thr Tyr Val
1 5 10
<210> 542
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 542
Asn Ile Gly Thr Lys Ser
1 5
<210> 543
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 543
Asp Asp Ser
1
<210> 544
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 544
Gln Val Trp Asp Ser Tyr Asn Val His Tyr Val
1 5 10
<210> 545
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 545
Ser Ser Asn Ile Glu Tyr Asn Tyr
1 5
<210> 546
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 546
Lys Asn Asn
1
<210> 547
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 547
Ala Ala Trp Asp Asp Ile Leu Ser Gly Val Val
1 5 10
<210> 548
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 548
Ser Asn Asp Val Gly Gly Tyr Asn Phe
1 5
<210> 549
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 549
Asp Val Ile
1
<210> 550
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 550
Cys Ser Tyr Ala Gly Thr Tyr Thr Trp Val
1 5 10
<210> 551
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 551
Gln Arg Val Val Asn Asn Tyr
1 5
<210> 552
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 552
Gly Ala Ser
1
<210> 553
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 553
Gln Gln Tyr Gly Ser Pro Trp Thr
1 5
<210> 554
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 554
Gln Ser Ile Ser Lys Trp
1 5
<210> 555
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 555
Lys Ala Ser
1
<210> 556
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 556
Gln Gln His Asn Ser Tyr Ser Tyr Thr
1 5
<210> 557
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 557
Gln Ser Val Ile Ser Thr Tyr
1 5
<210> 558
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 558
Asp Val Ser
1
<210> 559
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 559
His Gln Tyr Gly Ser Ser Pro Ala Thr
1 5
<210> 560
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 560
Gln Ser Val Gly Thr Tyr
1 5
<210> 561
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 561
Asp Ser Ala
1
<210> 562
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 562
Gln Leu Arg Ile Thr Trp Pro Pro Ile Phe Thr
1 5 10
<210> 563
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 563
Ser Ser Asp Val Gly Gly Tyr Asn Tyr
1 5
<210> 564
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 564
Glu Val Ser
1
<210> 565
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 565
Ser Ser Tyr Ala Gly Ser Asn Asn Leu Val
1 5 10
<210> 566
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 566
Thr Ser Ser Ile Gly Ser Asn Ile
1 5
<210> 567
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 567
Ile Asn Asn
1
<210> 568
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 568
Ala Ala Trp Asp Asp Ser Leu Asn Gly Trp Val
1 5 10
<210> 569
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 569
Gln Ser Val Gly Ser Thr
1 5
<210> 570
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 570
Gly Ala Ser
1
<210> 571
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 571
His Gln Tyr Ile Asn Trp Pro Pro Trp Thr
1 5 10
<210> 572
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 572
Ser Ser Asp Val Gly Ala Tyr Ser Tyr
1 5
<210> 573
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 573
Asp Val Tyr
1
<210> 574
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 574
Cys Ser His Ala Gly Ser His Thr Trp Val
1 5 10
<210> 575
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 575
Gln Ser Ile Ser Arg Asn
1 5
<210> 576
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 576
Gly Ala Ser
1
<210> 577
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 577
Gln Gln Tyr Ser Lys Leu Pro Ile Thr
1 5
<210> 578
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 578
Gln Ser Val Arg Ser Tyr
1 5
<210> 579
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 579
Asp Ala Ser
1
<210> 580
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 580
Gln Gln Arg Ser Tyr Trp Pro Pro Phe Thr
1 5 10
<210> 581
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 581
Gln Ser Val Ser Ser Tyr
1 5
<210> 582
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 582
Asp Ala Ser
1
<210> 583
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 583
Gln Gln Arg Ser Asp Trp Pro Pro Gly Thr
1 5 10
<210> 584
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 584
Ser Ser Asp Val Gly Gly Tyr Asn Phe
1 5
<210> 585
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 585
Glu Val Ile
1
<210> 586
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 586
Cys Ser Tyr Gly Gly Asn Asn Ser Trp Met
1 5 10
<210> 587
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 587
Val Leu Ser Asn Gln Tyr
1 5
<210> 588
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 588
Lys Asp Thr
1
<210> 589
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 589
Gln Ser Ala Asp Asn Thr Arg Ile Thr Val
1 5 10
<210> 590
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 590
Gln Ser Leu Val Tyr Ser Asp Gly Asn Thr Tyr
1 5 10
<210> 591
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 591
Lys Val Ser
1
<210> 592
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 592
Met Gln Gly Thr His Trp Pro Arg Thr
1 5
<210> 593
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 593
Arg Ser Asn Ile Gly Ser Tyr Thr
1 5
<210> 594
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 594
Asn Asn Asn
1
<210> 595
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 595
Ala Ala Trp Asp Asp Ser Leu Asn Gly Val Val
1 5 10
<210> 596
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 596
Gln Ser Ile Asn Ile Tyr
1 5
<210> 597
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 597
Ala Ala Ser
1
<210> 598
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 598
Gln Gln Ser Tyr Arg Ser Pro Arg Thr
1 5
<210> 599
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 599
Ser Gly His Ser Arg Tyr Ala
1 5
<210> 600
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 600
Ile Asn Ser Asp Gly Arg His
1 5
<210> 601
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 601
Gln Thr Trp Gly Thr Gly Phe Arg Val
1 5
<210> 602
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 602
Gln Ser Val Ser Ser Tyr
1 5
<210> 603
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 603
Gly Ala Ser
1
<210> 604
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 604
Gln Gln Arg Ser Asn Trp Pro Ile Thr
1 5
<210> 605
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 605
Gln Asp Ile Ser Ser Trp
1 5
<210> 606
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 606
Ala Ala Ser
1
<210> 607
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 607
Gln Gln Ala Asp Ser Phe Ile Thr
1 5
<210> 608
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 608
Gln Ser Ile Gly Asp Asn
1 5
<210> 609
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 609
Gly Ala Ser
1
<210> 610
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 610
Gln Gln Tyr Lys Asn Trp Pro Arg Thr
1 5
<210> 611
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 611
Gln Ser Val Arg Ser Ser Tyr
1 5
<210> 612
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 612
Gly Ala Ser
1
<210> 613
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 613
Gln Gln Tyr Gly Thr Ser Ile Thr
1 5
<210> 614
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 614
Val Leu Ser Lys Gln Tyr
1 5
<210> 615
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 615
Lys Asp Thr
1
<210> 616
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 616
Gln Ser Ala Asp Thr Arg Ile Thr Val
1 5
<210> 617
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 617
Val Leu Ser Asn Gln Tyr
1 5
<210> 618
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 618
Lys Asp Thr
1
<210> 619
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 619
Gln Thr Ala Asp Thr Lys Tyr Thr Val
1 5
<210> 620
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 620
Gln Arg Ile Ser Thr Tyr
1 5
<210> 621
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 621
Ala Ala Ser
1
<210> 622
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 622
Gln Gln Ser Tyr Ser Pro Pro Trp Thr
1 5
<210> 623
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 623
Gln Ser Ile Lys Lys Tyr
1 5
<210> 624
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 624
Gly Ala Ser
1
<210> 625
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 625
Gln Gln Ser Asp Ser Ala Pro Pro Thr
1 5
<210> 626
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 626
Gln Ser Ile Arg Asp Tyr
1 5
<210> 627
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 627
Ala Gly Ser
1
<210> 628
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 628
Gln Gln Ser Tyr Leu Thr Pro Pro Thr
1 5
<210> 629
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 629
Asn Ile Gly Ile Arg Thr
1 5
<210> 630
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 630
Tyr Asp Ser
1
<210> 631
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 631
Gln Val Trp Asp Ser Ser Ser Asp His Val Val
1 5 10
<210> 632
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 632
Ser Ser Asp Val Gly Ala Tyr Asn Tyr
1 5
<210> 633
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 633
Glu Val Thr
1
<210> 634
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 634
Ser Ser Tyr Ala Gly Asn Asn Asn Leu Val
1 5 10
<210> 635
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 635
Asn Ile Glu Arg Lys Ser
1 5
<210> 636
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 636
Asp Asp Thr
1
<210> 637
<211> 11
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 637
Gln Val Trp Asp Ser Thr Thr Asp His Gly Val
1 5 10
<210> 638
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 638
Gln Ser Val Ser Gly Tyr
1 5
<210> 639
<211> 3
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 639
Asp Ala Ser
1
<210> 640
<211> 8
<212> PRT
<213> Artificial sequence
<220>
<223> Synthetic peptide
<400> 640
Gln Gln Arg Ser Asn Gly Leu Thr
1 5
<210> 641
<211> 16
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic primer
<400> 641
cctccggccc ctgaat 16
<210> 642
<211> 20
<212> DNA
<213> Artificial sequence
<220>
<223> Synthetic primer
<400> 642
ccattgaatc cctgggcctt 20
Claims (102)
- 엔테로바이러스 D68(EV-D68)에 감염된 대상체(subject)를 치료하거나 EV-D68에 감염될 위험이 있는 대상체의 감염 가능성을 감소시키는 방법으로서, 각각 표 3 및 4로부터의 클론-쌍(clone-paired)의 중쇄 및 경쇄 CDR 서열을 갖는 항체 단편을 상기 대상체에게 전달하는 것을 포함하는, 방법.
- 제1항에 있어서, 상기 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍의 경쇄 및 중쇄 가변 서열에 의해 암호화(encoding)되는, 방법.
- 제1항 또는 제2항에 있어서, 상기 항체 또는 항체 단편이 표 1에 기재된 바와 같이 95% 동일성(identity)을 갖는 클론-쌍의 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제1항 또는 제2항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성(identity)을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제1항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제1항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제1항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제1항 내지 제7항 중 어느 한 항에 있어서, 상기 항체 단편이 재조합 scFv(일본쇄(single chain) 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편이거나, 상기 항체가 키메라 항체(chimeric antibody) 또는 이중특이성 항체(bispecific antibody)인, 방법.
- 제1항 내지 제8항 중 어느 한 항에 있어서, 상기 항체가 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)시키고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된(engineered) 세포주의 발현을 변경(제거 또는 향상)시키도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편인, 방법.
- 제1항 내지 제9항 중 어느 한 항에 있어서, 제2 요법, 예컨대, 항바이러스 요법 또는 항염증 요법으로 상기 대상체를 치료하는 것을 추가로 포함하는, 방법.
- 제1항 내지 제10항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 감염 전 또는 감염 후에 투여되는, 방법.
- 제1항 내지 제11항 중 어느 한 항에 있어서, 상기 치료가 폐 감염 및/또는 신경계 감염을 치료하거나, 또는 치료가 폐 감염 및/또는 신경계 감염을 감소시키는, 방법.
- 제1항 내지 제12항 중 어느 한 항에 있어서, 전달이 항체 또는 항체 단편의 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터에 의한 유전적 전달을 포함하는, 방법.
- 대상체에서 엔테로바이러스 D68(EV-D68) 감염을 검출하는 방법으로서,
(a) 상기 대상체로부터의 샘플을 각각 표 3 및 4로부터의 클론-쌍의 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편과 접촉시키는 단계; 및
(b) 상기 항체 또는 항체 단편을 상기 샘플 중의 EV-D68 항원에 결합시킴으로써 상기 샘플 중의 EV-D68을 검출하는 단계를 포함하는, 방법. - 제14항에 있어서, 상기 샘플이 체액인, 방법.
- 제14항에 있어서, 상기 샘플이 혈액, 가래, 눈물, 타액, 점액 또는 혈청, 정액, 자궁경부 또는 질 분비물, 양수, 태반 조직, 소변, 삼출물, 누출물, 호흡기 비말 또는 에어로졸, 조직 스크래핑(tissue scraping) 또는 대변인, 방법.
- 제14항 내지 제16항 중 어느 한 항에 있어서, 상기 검출이 ELISA, RIA, 측방 유동 검정(lateral flow assay) 또는 웨스턴 블롯(western blot)을 포함하는, 방법.
- 제14항 내지 제17항 중 어느 한 항에 있어서, 단계 (a) 및 (b)를 2회 수행하고, 제1 검정과 비교하여 EV-D68 항원 수준의 변화를 결정하는 것을 추가로 포함하는, 방법.
- 제14항 내지 제18항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍의 가변 서열에 의해 암호화되는, 방법.
- 제14항 내지 제18항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍의 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제14항 내지 제18항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제14항 내지 제18항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제14항 내지 제18항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제14항 내지 제18항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제14항 내지 제24항 중 어느 한 항에 있어서, 상기 항체 단편이 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편인, 방법.
- 항체 또는 항체 단편이 각각 표 3 및 4로부터의 클론-쌍의 중쇄 및 경쇄 CDR 서열을 특징으로 하는, 모노클로날 항체(monoclonal antibody).
- 제26항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 모노클로날 항체.
- 제26항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 서열에 대해 적어도 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 모노클로날 항체.
- 제26항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 서열에 대해 적어도 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 모노클로날 항체.
- 제26항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 모노클로날 항체.
- 제26항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 모노클로날 항체.
- 제26항 내지 제31항 중 어느 한 항에 있어서, 상기 항체 단편이 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편인, 모노클로날 항체.
- 제26항 내지 제31항 중 어느 한 항에 있어서, 상기 항체가 키메라 항체이거나, 이중특이성 항체인, 모노클로날 항체.
- 제26항 내지 제33항 중 어느 한 항에 있어서, 상기 항체가 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)시키고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)시키도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편인, 모노클로날 항체.
- 제26항 내지 제34항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 세포 투과 펩타이드(cell penetrating peptide)를 추가로 포함하고/하거나 인트라바디(intrabody)인, 모노클로날 항체.
- 항체 또는 항체 단편을 암호화하는 하이브리도마 또는 조작된 세포로서, 상기 항체 또는 항체 단편이 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 특징으로 하는, 하이브리도마 또는 조작된 세포.
- 제36항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 하이브리도마 또는 조작된 세포.
- 제36항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 가변 서열에 대해 적어도 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 하이브리도마 또는 조작된 세포.
- 제36항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 가변 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 하이브리도마 또는 조작된 세포.
- 제36항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 하이브리도마 또는 조작된 세포.
- 제36에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 가변 서열에 대해 적어도 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 하이브리도마 또는 조작된 세포.
- 제36항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 하이브리도마 또는 조작된 세포.
- 제36항 내지 제42항 중 어느 한 항에 있어서, 상기 항체 단편이 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편인, 하이브리도마 또는 조작된 세포.
- 제36항 내지 제43항 중 어느 한 항에 있어서, 상기 항체가 키메라 항체 또는 이중특이성 항체인, 하이브리도마 또는 조작된 세포.
- 제36항 내지 제43항 중 어느 한 항에 있어서, 상기 항체가 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)시키고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)시키도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편인, 하이브리도마 또는 조작된 세포.
- 제36항 내지 제45항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 세포 투과 펩타이드를 추가로 포함하고/하거나 인트라바디인, 하이브리도마 또는 조작된 세포.
- 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 특징으로 하는 하나 이상의 항체 또는 항체 단편을 포함하는 백신 제형(vaccine formulation).
- 제47항에 있어서, 상기 항체 또는 항체 단편 중 적어도 하나가 표 1로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 백신 제형.
- 제47항에 있어서, 상기 항체 또는 항체 단편 중 적어도 하나가 표 1로부터의 클론-쌍 서열에 대해 적어도 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 백신 제형.
- 제47항에 있어서, 상기 항체 또는 항체 단편 중 적어도 하나가 표 1로부터의 클론-쌍 서열에 대해 적어도 95% 이상의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 백신 제형.
- 제47항에 있어서, 상기 항체 또는 항체 단편 중 적어도 하나가 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 백신 제형.
- 제47항에 있어서, 상기 항체 또는 항체 단편 중 적어도 하나가 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 백신 제형.
- 제47항 내지 제52항 중 어느 한 항에 있어서, 상기 항체 단편 중 적어도 하나가 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편인, 백신 제형.
- 제47항 내지 제52항 중 어느 한 항에 있어서, 상기 항체 중 적어도 하나가 키메라 항체이거나 이중특이성 항체인, 백신 제형.
- 제47항 내지 제54항 중 어느 한 항에 있어서, 상기 항체가 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)시키고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)시키도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편인, 백신 제형.
- 제47항 내지 제55항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편 중 적어도 하나가 세포 투과 펩타이드를 추가로 포함하고/하거나 인트라바디인, 백신 제형.
- 제26항 내지 제34항 중 어느 한 항에 따른 1차 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 포함하는 백신 제형.
- 제57항에 있어서, 상기 발현 벡터(들)가 신드비스(Sindbis) 바이러스 또는 VEE 벡터(들)인, 백신 제형.
- 제57항 내지 제58항 중 어느 한 항에 있어서, 바늘 주사, 제트(jet) 주사 또는 전기천공에 의한 전달용으로 제형화된, 백신 제형.
- 제57항에 있어서, 제26항 내지 제34항 중 어느 한 항의 별개의 항체 또는 항체 단편과 같은 2차 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 추가로 포함하는, 백신 제형.
- 엔테로바이러스 D68에 감염되었거나 이의 감염 위험이 있는 임신 중의 대상체의 태반 및/또는 태아의 건강을 보호하는 방법으로서, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편을 상기 대상체에게 전달하는 것을 포함하는, 방법.
- 제61항에 있어서, 상기 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제61항 또는 제62항에 있어서, 상기 항체 또는 항체 단편이 표 1에 기재된 바와 같이 95% 동일성을 갖는 클론-쌍 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제61항 또는 제62항에 있어서, 상기 항체 또는 항체 단편이 표 1로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제61항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제61항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제61항에 있어서, 상기 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제61항 내지 제67항 중 어느 한 항에 있어서, 상기 항체 단편이 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편인, 방법.
- 제61항 내지 제68항 중 어느 한 항에 있어서, 상기 항체가 IgG, 또는 FcR 상호작용을 변경(제거 또는 향상)시키고 반감기를 증가시키고/시키거나 치료 효능을 증가시키도록 돌연변이된 Fc 부분, 예컨대, LALA, N297, GASD/ALIE, YTE, DHS 또는 LS 돌연변이를 포함하거나, 또는 FcR 상호작용, 예컨대, 글리칸의 효소적 또는 화학적 부가 또는 제거, 또는 정의된 글리코실화 패턴으로 조작된 세포주의 발현을 변경(제거 또는 향상)시키도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편인, 방법.
- 제61항 내지 제67항 중 어느 한 항에 있어서, 상기 항체가 키메라 항체 또는 이중특이성 항체인, 방법.
- 제61항 내지 제70항 중 어느 한 항에 있어서, 상기 항체 또는 항체 단편이 감염 전 또는 감염 후에 투여되는, 방법.
- 제61항 내지 제71항 중 어느 한 항에 있어서, 상기 대상체가 임신 중의 여성, 성적으로 활발한 여성, 또는 불임 치료를 받고 있는 여성인, 방법.
- 제61항 내지 제72항 중 어느 한 항에 있어서, 전달이 항체 또는 항체 단편의 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터에 의한 유전적 전달을 포함하는, 방법.
- 제61항에 있어서, 상기 항체 또는 항체 단편이 미처리 대조군과 비교하여 태반의 크기를 증가시키는, 방법.
- 제61항에 있어서, 상기 항체 또는 항체 단편이 미처리 대조군과 비교하여 태아의 바이러스 부하(viral load) 및/또는 병리를 감소시키는, 방법.
- 엔테로바이러스 D68 항원의 항원 완전성, 정확한 형태 및/또는 정확한 서열을 결정하는 방법으로서,
(a) 상기 항원을 포함하는 샘플을, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 갖는 1차 항체 또는 항체 단편과 접촉시키는 단계; 및
(b) 상기 항원에 대한 상기 1차 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 완전성, 정확한 형태 및/또는 정확한 서열을 결정하는 단계
를 포함하는, 방법. - 제76항에 있어서, 상기 샘플이 재조합적으로 생산된 항원을 포함하는, 방법.
- 제76항에 있어서, 상기 샘플이 백신 제형 또는 백신 생산 배치(batch)를 포함하는, 방법.
- 제76항 내지 제78항 중 어느 한 항에 있어서, 검출이 ELISA, RIA, 웨스턴 블롯, 표면 플라즈몬 공명 또는 생물층 간섭계를 사용하는 바이오센서, 또는 유세포 분석 염색(flow cytometric staining)을 포함하는, 방법.
- 제76항 내지 제79항 중 어느 한 항에 있어서, 상기 1차 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 가변 서열에 의해 암호화되는, 방법.
- 제76항 내지 제79항 중 어느 한 항에 있어서, 상기 1차 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제76항 내지 제79항 중 어느 한 항에 있어서, 상기 1차 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제76항 내지 제79항 중 어느 한 항에 있어서, 상기 1차 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제76항 내지 제79항 중 어느 한 항에 있어서, 상기 1차 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제76항 내지 제79항 중 어느 한 항에 있어서, 상기 1차 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제76항 내지 제85항 중 어느 한 항에 있어서, 상기 1차 항체 단편이 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편인, 방법.
- 제76항 내지 제86항 중 어느 한 항에 있어서, 단계 (a) 및 (b)를 2회 수행하여, 시간 경과에 따른 항원의 항원 안정성을 결정하는 것을 추가로 포함하는, 방법.
- 제76항 내지 제87항 중 어느 한 항에 있어서,
(c) 상기 항원을 포함하는 샘플을, 각각 표 3 및 4로부터의 클론-쌍 중쇄 및 경쇄 CDR 서열을 갖는 2차 항체 또는 항체 단편과 접촉시키는 단계; 및
(d) 상기 항원에 대한 상기 2차 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 완전성을 결정하는 단계를 추가로 포함하는, 방법. - 제88항에 있어서, 상기 2차 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 가변 서열에 의해 암호화되는, 방법.
- 제89항에 있어서, 상기 2차 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 가변 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제89항에 있어서, 상기 2차 항체 또는 항체 단편이 표 1에 기재된 바와 같은 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화되는, 방법.
- 제89항에 있어서, 상기 2차 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 따른 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제89항에 있어서, 상기 2차 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 70%, 80% 또는 90% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제89항에 있어서, 상기 2차 항체 또는 항체 단편이 표 2로부터의 클론-쌍 서열에 대해 95% 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함하는, 방법.
- 제89항에 있어서, 상기 2차 항체 단편이 재조합 scFv(일본쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편 또는 Fv 단편인, 방법.
- 제89항에 있어서, 단계 (c) 및 (d)를 2회 수행하여 시간 경과에 따른 항원의 항원 안정성을 결정하는 것을 추가로 포함하는, 방법.
- 인간 모노클로날 항체 또는 항체 단편, 또는 하이브리도마 또는 이를 생산하는 조작된 세포로서, 상기 항체가 적어도 2개의 바이러스 클레이드(viral clade)를 가로질러 EV-D68에 결합하는, 인간 모노클로날 항체 또는 항체 단편, 또는 하이브리도마 또는 이를 생산하는 조작된 세포.
- 인간 모노클로날 항체 또는 항체 단편, 또는 하이브리도마 또는 이를 생산하는 조작된 세포로서, 상기 항체가 EV-D68 VP1, VP2 및 VP3에 결합하는, 인간 모노클로날 항체 또는 항체 단편, 또는 하이브리도마 또는 이를 생산하는 조작된 세포.
- 제98항에 있어서, 상기 항체가 EV-D68 VP1 DE 루프에 결합하는, 인간 모노클로날 항체 또는 항체 단편, 하이브리도마 또는 이를 생산하는 조작된 세포.
- 제98항에 있어서, 상기 항체가 EV-D68 VP2 EE 루프에 결합하는, 인간 모노클로날 항체 또는 항체 단편, 하이브리도마 또는 이를 생산하는 조작된 세포.
- 제98항에 있어서, 상기 항체가 EV-D68 VP3 N-말단 루프에 결합하는, 인간 모노클로날 항체 또는 항체 단편, 또는 하이브리도마 또는 이를 생산하는 조작된 세포.
- 제98항에 있어서, 상기 항체가 (a) EV-D68 VP3 N-말단 루프 및 EV-D68 VP2 EE 루프에 결합하거나, 또는 (b) EV-D68 VP2 EE 루프 및 EV-D68 VP1 DE 루프, 또는 (c) EV-D68 VP1 DE 루프 및 EV-D68 VP3 N-말단 루프, 또는 (d) EV-D68 VP3 N-말단 루프, EV-D68 VP1 DE 루프 및 EV-D68 VP2 EE 루프에 결합하는, 인간 모노클로날 항체 또는 항체 단편, 또는 하이브리도마 또는 이를 생산하는 조작된 세포.
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962878955P | 2019-07-26 | 2019-07-26 | |
| US62/878,955 | 2019-07-26 | ||
| US201962899503P | 2019-09-12 | 2019-09-12 | |
| US62/899,503 | 2019-09-12 | ||
| US202063047330P | 2020-07-02 | 2020-07-02 | |
| US63/047,330 | 2020-07-02 | ||
| PCT/US2020/043415 WO2021021605A1 (en) | 2019-07-26 | 2020-07-24 | Human monoclonal antibodies to enterovirus d68 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20220038415A true KR20220038415A (ko) | 2022-03-28 |
Family
ID=72039700
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020227005641A KR20220038415A (ko) | 2019-07-26 | 2020-07-24 | 엔테로바이러스 d68에 대한 인간 모노클로날 항체 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20220289828A1 (ko) |
| EP (1) | EP4004037A1 (ko) |
| JP (1) | JP2022541652A (ko) |
| KR (1) | KR20220038415A (ko) |
| CA (1) | CA3145278A1 (ko) |
| WO (1) | WO2021021605A1 (ko) |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB2605538A (en) | 2020-03-23 | 2022-10-05 | Hdt Bio Corp | Compositions and methods for delivery of RNA |
| WO2023039064A2 (en) * | 2021-09-08 | 2023-03-16 | The Scripps Research Institute | Broadly neutralizing antibodies against sars-like viruses |
| WO2023049753A2 (en) * | 2021-09-22 | 2023-03-30 | Hdt Bio Corp. | Compositions and methods for enhanced antigen binding proteins |
| LU103006B1 (de) * | 2022-09-15 | 2024-03-15 | Phaeosynt Gbr | Immunoassay zum nachweisen biologisch aktiver proteine |
Family Cites Families (44)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NL154598B (nl) | 1970-11-10 | 1977-09-15 | Organon Nv | Werkwijze voor het aantonen en bepalen van laagmoleculire verbindingen en van eiwitten die deze verbindingen specifiek kunnen binden, alsmede testverpakking. |
| US3817837A (en) | 1971-05-14 | 1974-06-18 | Syva Corp | Enzyme amplification assay |
| US3939350A (en) | 1974-04-29 | 1976-02-17 | Board Of Trustees Of The Leland Stanford Junior University | Fluorescent immunoassay employing total reflection for activation |
| US3996345A (en) | 1974-08-12 | 1976-12-07 | Syva Company | Fluorescence quenching with immunological pairs in immunoassays |
| US4196265A (en) | 1977-06-15 | 1980-04-01 | The Wistar Institute | Method of producing antibodies |
| FR2413974A1 (fr) | 1978-01-06 | 1979-08-03 | David Bernard | Sechoir pour feuilles imprimees par serigraphie |
| US4277437A (en) | 1978-04-05 | 1981-07-07 | Syva Company | Kit for carrying out chemically induced fluorescence immunoassay |
| US4275149A (en) | 1978-11-24 | 1981-06-23 | Syva Company | Macromolecular environment control in specific receptor assays |
| US4366241A (en) | 1980-08-07 | 1982-12-28 | Syva Company | Concentrating zone method in heterogeneous immunoassays |
| US4554101A (en) | 1981-01-09 | 1985-11-19 | New York Blood Center, Inc. | Identification and preparation of epitopes on antigens and allergens on the basis of hydrophilicity |
| US4957939A (en) | 1981-07-24 | 1990-09-18 | Schering Aktiengesellschaft | Sterile pharmaceutical compositions of gadolinium chelates useful enhancing NMR imaging |
| US4867973A (en) | 1984-08-31 | 1989-09-19 | Cytogen Corporation | Antibody-therapeutic agent conjugates |
| US4472509A (en) | 1982-06-07 | 1984-09-18 | Gansow Otto A | Metal chelate conjugated monoclonal antibodies |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| US4676980A (en) | 1985-09-23 | 1987-06-30 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Target specific cross-linked heteroantibodies |
| US4938948A (en) | 1985-10-07 | 1990-07-03 | Cetus Corporation | Method for imaging breast tumors using labeled monoclonal anti-human breast cancer antibodies |
| US4680338A (en) | 1985-10-17 | 1987-07-14 | Immunomedics, Inc. | Bifunctional linker |
| US5563250A (en) | 1987-12-02 | 1996-10-08 | Neorx Corporation | Cleavable conjugates for the delivery and release of agents in native form |
| US5141648A (en) | 1987-12-02 | 1992-08-25 | Neorx Corporation | Methods for isolating compounds using cleavable linker bound matrices |
| AU2684488A (en) | 1988-06-27 | 1990-01-04 | Carter-Wallace, Inc. | Test device and method for colored particle immunoassay |
| ES2096590T3 (es) | 1989-06-29 | 1997-03-16 | Medarex Inc | Reactivos biespecificos para la terapia del sida. |
| JPH049249A (ja) | 1990-04-27 | 1992-01-14 | Kusuda:Kk | 塗型剤吹き付け機 |
| EP0586505A1 (en) | 1991-05-14 | 1994-03-16 | Repligen Corporation | Heteroconjugate antibodies for treatment of hiv infection |
| LU91067I2 (fr) | 1991-06-14 | 2004-04-02 | Genentech Inc | Trastuzumab et ses variantes et dérivés immuno chimiques y compris les immotoxines |
| ES2136092T3 (es) | 1991-09-23 | 1999-11-16 | Medical Res Council | Procedimientos para la produccion de anticuerpos humanizados. |
| WO1993008829A1 (en) | 1991-11-04 | 1993-05-13 | The Regents Of The University Of California | Compositions that mediate killing of hiv-infected cells |
| DE69308573T2 (de) | 1992-08-17 | 1997-08-07 | Genentech Inc | Bispezifische immunoadhesine |
| AU5670194A (en) | 1992-11-20 | 1994-06-22 | Enzon, Inc. | Linker for linked fusion polypeptides |
| AU4289496A (en) | 1994-12-02 | 1996-06-19 | Chiron Corporation | Method of promoting an immune response with a bispecific antibody |
| US5731168A (en) | 1995-03-01 | 1998-03-24 | Genentech, Inc. | Method for making heteromultimeric polypeptides |
| WO1996040662A2 (en) | 1995-06-07 | 1996-12-19 | Cellpro, Incorporated | Aminooxy-containing linker compounds and their application in conjugates |
| US5837234A (en) | 1995-06-07 | 1998-11-17 | Cytotherapeutics, Inc. | Bioartificial organ containing cells encapsulated in a permselective polyether suflfone membrane |
| US5922845A (en) | 1996-07-11 | 1999-07-13 | Medarex, Inc. | Therapeutic multispecific compounds comprised of anti-Fcα receptor antibodies |
| DK1071700T3 (da) | 1998-04-20 | 2010-06-07 | Glycart Biotechnology Ag | Glykosylerings-modifikation af antistoffer til forbedring af antistofafhængig cellulær cytotoksicitet |
| EP1141024B1 (en) | 1999-01-15 | 2018-08-08 | Genentech, Inc. | POLYPEPTIDE COMPRISING A VARIANT HUMAN IgG1 Fc REGION |
| US7666400B2 (en) | 2005-04-06 | 2010-02-23 | Ibc Pharmaceuticals, Inc. | PEGylation by the dock and lock (DNL) technique |
| US7550143B2 (en) | 2005-04-06 | 2009-06-23 | Ibc Pharmaceuticals, Inc. | Methods for generating stably linked complexes composed of homodimers, homotetramers or dimers of dimers and uses |
| US7527787B2 (en) | 2005-10-19 | 2009-05-05 | Ibc Pharmaceuticals, Inc. | Multivalent immunoglobulin-based bioactive assemblies |
| US7534866B2 (en) | 2005-10-19 | 2009-05-19 | Ibc Pharmaceuticals, Inc. | Methods and compositions for generating bioactive assemblies of increased complexity and uses |
| CA2442801A1 (en) | 2001-04-02 | 2002-10-10 | Idec Pharmaceutical Corporation | Recombinant antibodies coexpressed with gntiii |
| KR20100018071A (ko) | 2001-08-03 | 2010-02-16 | 글리카트 바이오테크놀로지 아게 | 항체 의존적 세포 독성이 증가된 항체 글리코실화 변이체 |
| US20040101920A1 (en) | 2002-11-01 | 2004-05-27 | Czeslaw Radziejewski | Modification assisted profiling (MAP) methodology |
| CN101484182B (zh) | 2005-04-06 | 2014-06-11 | Ibc药品公司 | 由同二聚体、同四聚体或二聚体的二聚体组成的稳定连接复合体的生产方法及用途 |
| UA117289C2 (uk) | 2014-04-02 | 2018-07-10 | Ф. Хоффманн-Ля Рош Аг | Мультиспецифічне антитіло |
-
2020
- 2020-07-24 CA CA3145278A patent/CA3145278A1/en active Pending
- 2020-07-24 EP EP20754112.9A patent/EP4004037A1/en active Pending
- 2020-07-24 JP JP2022505261A patent/JP2022541652A/ja active Pending
- 2020-07-24 US US17/630,053 patent/US20220289828A1/en active Pending
- 2020-07-24 WO PCT/US2020/043415 patent/WO2021021605A1/en unknown
- 2020-07-24 KR KR1020227005641A patent/KR20220038415A/ko unknown
Also Published As
| Publication number | Publication date |
|---|---|
| EP4004037A1 (en) | 2022-06-01 |
| JP2022541652A (ja) | 2022-09-26 |
| CA3145278A1 (en) | 2021-02-04 |
| US20220289828A1 (en) | 2022-09-15 |
| WO2021021605A1 (en) | 2021-02-04 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR102535195B1 (ko) | 항-코로나바이러스 항체 및 사용 방법 | |
| CN108064240B (zh) | 针对流感血凝素的人抗体 | |
| KR102541905B1 (ko) | 치쿤군야 바이러스의 항체-매개 중화 | |
| US11345741B2 (en) | Human monoclonal antibodies to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) | |
| KR20220038415A (ko) | 엔테로바이러스 d68에 대한 인간 모노클로날 항체 | |
| KR20220140568A (ko) | 중증 급성 호흡기 증후군 코로나바이러스 2(SARS-CoV-2)에 대한 인간 단일클론 항체 | |
| US20210324050A1 (en) | Human monoclonal antibodies that neutralize pandemic gii.4 noroviruses | |
| KR20230142790A (ko) | 코로나바이러스 스파이크 단백질(spike protein)을 타겟팅하는 항체 | |
| US20230065377A1 (en) | Human antibodies to alphaviruses | |
| US20230122364A1 (en) | HUMAN MONOCLONAL ANTIBODIES TO SEVERE ACUTE RESPIRATORY SYNDROME CORONAVIRUS 2 (SARS-CoV-2) | |
| KR20220098003A (ko) | 한타바이러스에 대한 인간 모노클로날 항체 및 이의 사용 방법 | |
| KR20240006575A (ko) | SARS-CoV-2에 대한 사람 중화 모노클로날 항체 및 이의 용도 | |
| WO2021195385A1 (en) | HUMAN MONOCLONAL ANTIBODIES TO SEVERE ACUTE RESPIRATORY SYNDROME CORONAVIRUS 2 (SARS-GoV-2) | |
| RU2782256C2 (ru) | Антитела к коронавирусу и способы применения | |
| US20230181714A1 (en) | Human monoclonal antibodies to venezuelan equine encephalitis virus and uses therefor | |
| US20230085393A1 (en) | Human antibodies that neutralize zika virus and methods of use therefor | |
| WO2023187407A1 (en) | Human monoclonal antibodies binding to sars-cov-2 and methods of use thereof | |
| AU2020273365A1 (en) | Human antibodies to Ross River virus and methods of use therefor | |
| CN117836322A (zh) | 针对SARS-CoV-2的人中和单克隆抗体及其用途 | |
| KR20220113346A (ko) | 칸디다에 대한 항체 및 이의 용도 | |
| WO2024015760A2 (en) | Human monoclonal antibodies to omicron variant of severe acute respiratory syndrome coronavirus 2 (sars-cov- 2) | |
| CN117715930A (zh) | 抗体 |