KR20170133507A - 인간화된 인플루엔자 모노클로날 항체 및 그의 사용 방법 - Google Patents
인간화된 인플루엔자 모노클로날 항체 및 그의 사용 방법 Download PDFInfo
- Publication number
- KR20170133507A KR20170133507A KR1020177032200A KR20177032200A KR20170133507A KR 20170133507 A KR20170133507 A KR 20170133507A KR 1020177032200 A KR1020177032200 A KR 1020177032200A KR 20177032200 A KR20177032200 A KR 20177032200A KR 20170133507 A KR20170133507 A KR 20170133507A
- Authority
- KR
- South Korea
- Prior art keywords
- antibody
- seq
- acid sequence
- virus
- amino acid
- Prior art date
Links
- 238000000034 method Methods 0.000 title claims abstract description 94
- 206010022000 influenza Diseases 0.000 title claims abstract description 38
- 230000027455 binding Effects 0.000 claims abstract description 146
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 101
- 241000712461 unidentified influenza virus Species 0.000 claims abstract description 70
- 102000004169 proteins and genes Human genes 0.000 claims abstract description 68
- 241000282414 Homo sapiens Species 0.000 claims description 103
- 241000700605 Viruses Species 0.000 claims description 87
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 51
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 42
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 36
- 239000000203 mixture Substances 0.000 claims description 36
- 206010057190 Respiratory tract infections Diseases 0.000 claims description 33
- 239000012634 fragment Substances 0.000 claims description 33
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 33
- 239000013598 vector Substances 0.000 claims description 32
- 229920001184 polypeptide Polymers 0.000 claims description 23
- 238000006386 neutralization reaction Methods 0.000 claims description 22
- 241000712431 Influenza A virus Species 0.000 claims description 19
- 201000010099 disease Diseases 0.000 claims description 18
- 208000035475 disorder Diseases 0.000 claims description 18
- 150000007523 nucleic acids Chemical group 0.000 claims description 17
- 150000001413 amino acids Chemical class 0.000 claims description 15
- 101710154606 Hemagglutinin Proteins 0.000 claims description 12
- 101710093908 Outer capsid protein VP4 Proteins 0.000 claims description 12
- 101710135467 Outer capsid protein sigma-1 Proteins 0.000 claims description 12
- 101710176177 Protein A56 Proteins 0.000 claims description 12
- 239000003814 drug Substances 0.000 claims description 12
- 239000000185 hemagglutinin Substances 0.000 claims description 10
- 108020004707 nucleic acids Proteins 0.000 claims description 9
- 102000039446 nucleic acids Human genes 0.000 claims description 9
- 230000002265 prevention Effects 0.000 claims description 9
- 230000001850 reproductive effect Effects 0.000 claims description 8
- 101001037139 Homo sapiens Immunoglobulin heavy variable 3-30 Proteins 0.000 claims description 7
- 102100040219 Immunoglobulin heavy variable 3-30 Human genes 0.000 claims description 7
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 6
- 150000003384 small molecules Chemical class 0.000 claims description 6
- 229940124597 therapeutic agent Drugs 0.000 claims description 6
- 239000003053 toxin Substances 0.000 claims description 5
- 231100000765 toxin Toxicity 0.000 claims description 5
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 4
- 102000004127 Cytokines Human genes 0.000 claims description 4
- 108090000695 Cytokines Proteins 0.000 claims description 4
- 101001054838 Homo sapiens Immunoglobulin lambda variable 1-44 Proteins 0.000 claims description 4
- 102100026921 Immunoglobulin lambda variable 1-44 Human genes 0.000 claims description 4
- 239000003443 antiviral agent Substances 0.000 claims description 4
- 108020004459 Small interfering RNA Proteins 0.000 claims description 2
- 238000005520 cutting process Methods 0.000 claims description 2
- 239000000427 antigen Substances 0.000 abstract description 80
- 108091007433 antigens Proteins 0.000 abstract description 69
- 102000036639 antigens Human genes 0.000 abstract description 69
- 230000003472 neutralizing effect Effects 0.000 abstract description 34
- 229960005486 vaccine Drugs 0.000 abstract description 13
- 238000004519 manufacturing process Methods 0.000 abstract description 11
- 230000009385 viral infection Effects 0.000 abstract description 8
- 229940124873 Influenza virus vaccine Drugs 0.000 abstract 2
- 230000004043 responsiveness Effects 0.000 abstract 1
- 210000004027 cell Anatomy 0.000 description 110
- 235000018102 proteins Nutrition 0.000 description 66
- 150000001875 compounds Chemical class 0.000 description 55
- 238000012360 testing method Methods 0.000 description 29
- 238000003556 assay Methods 0.000 description 27
- 108020004414 DNA Proteins 0.000 description 23
- 241000699670 Mus sp. Species 0.000 description 22
- 239000003795 chemical substances by application Substances 0.000 description 22
- 238000002474 experimental method Methods 0.000 description 22
- 238000011282 treatment Methods 0.000 description 22
- 108060003951 Immunoglobulin Proteins 0.000 description 20
- 238000004458 analytical method Methods 0.000 description 20
- 102000018358 immunoglobulin Human genes 0.000 description 20
- 238000000338 in vitro Methods 0.000 description 20
- 102100035824 Unconventional myosin-Ig Human genes 0.000 description 19
- 210000001806 memory b lymphocyte Anatomy 0.000 description 19
- 239000000126 substance Substances 0.000 description 18
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 17
- 208000015181 infectious disease Diseases 0.000 description 17
- 230000003993 interaction Effects 0.000 description 17
- 239000000523 sample Substances 0.000 description 17
- 241001465754 Metazoa Species 0.000 description 16
- 108010006232 Neuraminidase Proteins 0.000 description 16
- 235000001014 amino acid Nutrition 0.000 description 16
- 102000005348 Neuraminidase Human genes 0.000 description 15
- 108020001507 fusion proteins Proteins 0.000 description 15
- 102000037865 fusion proteins Human genes 0.000 description 15
- 230000005847 immunogenicity Effects 0.000 description 15
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 14
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 14
- 102000004190 Enzymes Human genes 0.000 description 14
- 108090000790 Enzymes Proteins 0.000 description 14
- 108090000631 Trypsin Proteins 0.000 description 14
- 102000004142 Trypsin Human genes 0.000 description 14
- 229940024606 amino acid Drugs 0.000 description 14
- 210000003719 b-lymphocyte Anatomy 0.000 description 14
- 150000001720 carbohydrates Chemical class 0.000 description 14
- 229940088598 enzyme Drugs 0.000 description 14
- 210000004408 hybridoma Anatomy 0.000 description 14
- 239000012588 trypsin Substances 0.000 description 14
- 239000012472 biological sample Substances 0.000 description 13
- 230000035772 mutation Effects 0.000 description 13
- 239000005089 Luciferase Substances 0.000 description 12
- 238000003776 cleavage reaction Methods 0.000 description 12
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 12
- 239000002609 medium Substances 0.000 description 12
- 239000002953 phosphate buffered saline Substances 0.000 description 12
- 230000008707 rearrangement Effects 0.000 description 12
- 230000007017 scission Effects 0.000 description 12
- 239000006228 supernatant Substances 0.000 description 12
- 206010069754 Acquired gene mutation Diseases 0.000 description 11
- 108060001084 Luciferase Proteins 0.000 description 11
- 241000699666 Mus <mouse, genus> Species 0.000 description 11
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 11
- 230000004927 fusion Effects 0.000 description 11
- -1 glutaraldehyde) Chemical class 0.000 description 11
- 238000001727 in vivo Methods 0.000 description 11
- 230000037439 somatic mutation Effects 0.000 description 11
- 230000008859 change Effects 0.000 description 10
- 230000001419 dependent effect Effects 0.000 description 10
- 238000001514 detection method Methods 0.000 description 10
- 238000010494 dissociation reaction Methods 0.000 description 10
- 230000005593 dissociations Effects 0.000 description 10
- 230000000694 effects Effects 0.000 description 10
- 230000003053 immunization Effects 0.000 description 10
- 230000001965 increasing effect Effects 0.000 description 10
- 208000037798 influenza B Diseases 0.000 description 10
- 239000002502 liposome Substances 0.000 description 10
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 10
- 210000002966 serum Anatomy 0.000 description 10
- 239000013638 trimer Substances 0.000 description 10
- 241000282412 Homo Species 0.000 description 9
- 238000005859 coupling reaction Methods 0.000 description 9
- 210000004602 germ cell Anatomy 0.000 description 9
- 238000002649 immunization Methods 0.000 description 9
- 238000011534 incubation Methods 0.000 description 9
- 238000011081 inoculation Methods 0.000 description 9
- 238000003032 molecular docking Methods 0.000 description 9
- 230000004224 protection Effects 0.000 description 9
- 230000002285 radioactive effect Effects 0.000 description 9
- 238000012216 screening Methods 0.000 description 9
- 239000000243 solution Substances 0.000 description 9
- 230000001225 therapeutic effect Effects 0.000 description 9
- 210000001519 tissue Anatomy 0.000 description 9
- 230000003612 virological effect Effects 0.000 description 9
- 108010090804 Streptavidin Proteins 0.000 description 8
- 230000000890 antigenic effect Effects 0.000 description 8
- 239000012131 assay buffer Substances 0.000 description 8
- 230000008878 coupling Effects 0.000 description 8
- 238000010168 coupling process Methods 0.000 description 8
- 239000012636 effector Substances 0.000 description 8
- 230000002209 hydrophobic effect Effects 0.000 description 8
- 108700010900 influenza virus proteins Proteins 0.000 description 8
- 238000002372 labelling Methods 0.000 description 8
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 7
- 241001112090 Pseudovirus Species 0.000 description 7
- 241000700584 Simplexvirus Species 0.000 description 7
- 239000011324 bead Substances 0.000 description 7
- 230000015572 biosynthetic process Effects 0.000 description 7
- 229960002685 biotin Drugs 0.000 description 7
- 235000020958 biotin Nutrition 0.000 description 7
- 239000011616 biotin Substances 0.000 description 7
- 210000000170 cell membrane Anatomy 0.000 description 7
- 239000013604 expression vector Substances 0.000 description 7
- 238000009472 formulation Methods 0.000 description 7
- 230000001404 mediated effect Effects 0.000 description 7
- 230000001717 pathogenic effect Effects 0.000 description 7
- 239000013612 plasmid Substances 0.000 description 7
- 230000009467 reduction Effects 0.000 description 7
- 238000007423 screening assay Methods 0.000 description 7
- 239000007790 solid phase Substances 0.000 description 7
- 239000002904 solvent Substances 0.000 description 7
- LMDZBCPBFSXMTL-UHFFFAOYSA-N 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide Chemical compound CCN=C=NCCCN(C)C LMDZBCPBFSXMTL-UHFFFAOYSA-N 0.000 description 6
- CSCPPACGZOOCGX-UHFFFAOYSA-N Acetone Chemical compound CC(C)=O CSCPPACGZOOCGX-UHFFFAOYSA-N 0.000 description 6
- 208000002979 Influenza in Birds Diseases 0.000 description 6
- DNIAPMSPPWPWGF-UHFFFAOYSA-N Propylene glycol Chemical compound CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 6
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 6
- 230000004913 activation Effects 0.000 description 6
- 238000001042 affinity chromatography Methods 0.000 description 6
- 239000012491 analyte Substances 0.000 description 6
- 206010064097 avian influenza Diseases 0.000 description 6
- 230000008901 benefit Effects 0.000 description 6
- 238000006243 chemical reaction Methods 0.000 description 6
- 230000000295 complement effect Effects 0.000 description 6
- 210000004443 dendritic cell Anatomy 0.000 description 6
- 230000001900 immune effect Effects 0.000 description 6
- 238000003018 immunoassay Methods 0.000 description 6
- 230000002163 immunogen Effects 0.000 description 6
- 230000001976 improved effect Effects 0.000 description 6
- 230000002458 infectious effect Effects 0.000 description 6
- 230000002401 inhibitory effect Effects 0.000 description 6
- 239000000463 material Substances 0.000 description 6
- 230000034217 membrane fusion Effects 0.000 description 6
- 239000008194 pharmaceutical composition Substances 0.000 description 6
- 239000002243 precursor Substances 0.000 description 6
- 238000002360 preparation method Methods 0.000 description 6
- 102000005962 receptors Human genes 0.000 description 6
- 108020003175 receptors Proteins 0.000 description 6
- 102200007903 rs796065047 Human genes 0.000 description 6
- RPENMORRBUTCPR-UHFFFAOYSA-M sodium;1-hydroxy-2,5-dioxopyrrolidine-3-sulfonate Chemical compound [Na+].ON1C(=O)CC(S([O-])(=O)=O)C1=O RPENMORRBUTCPR-UHFFFAOYSA-M 0.000 description 6
- 230000004083 survival effect Effects 0.000 description 6
- 230000001960 triggered effect Effects 0.000 description 6
- 241000272525 Anas platyrhynchos Species 0.000 description 5
- 241000271566 Aves Species 0.000 description 5
- 108010041986 DNA Vaccines Proteins 0.000 description 5
- 229940021995 DNA vaccine Drugs 0.000 description 5
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 5
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 5
- 206010035226 Plasma cell myeloma Diseases 0.000 description 5
- 229920002684 Sepharose Polymers 0.000 description 5
- WLJPJRGQRNCIQS-ZLUOBGJFSA-N Ser-Ser-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O WLJPJRGQRNCIQS-ZLUOBGJFSA-N 0.000 description 5
- 239000002671 adjuvant Substances 0.000 description 5
- 210000000612 antigen-presenting cell Anatomy 0.000 description 5
- 230000000903 blocking effect Effects 0.000 description 5
- 210000004369 blood Anatomy 0.000 description 5
- 239000008280 blood Substances 0.000 description 5
- 239000000969 carrier Substances 0.000 description 5
- 239000007822 coupling agent Substances 0.000 description 5
- 239000013078 crystal Substances 0.000 description 5
- 229940079593 drug Drugs 0.000 description 5
- 238000005516 engineering process Methods 0.000 description 5
- XKUKSGPZAADMRA-UHFFFAOYSA-N glycyl-glycyl-glycine Natural products NCC(=O)NCC(=O)NCC(O)=O XKUKSGPZAADMRA-UHFFFAOYSA-N 0.000 description 5
- 230000028993 immune response Effects 0.000 description 5
- 208000037797 influenza A Diseases 0.000 description 5
- 230000005764 inhibitory process Effects 0.000 description 5
- 231100000636 lethal dose Toxicity 0.000 description 5
- 239000003550 marker Substances 0.000 description 5
- 238000005259 measurement Methods 0.000 description 5
- 230000007246 mechanism Effects 0.000 description 5
- 239000012528 membrane Substances 0.000 description 5
- 201000000050 myeloid neoplasm Diseases 0.000 description 5
- 239000000047 product Substances 0.000 description 5
- 230000000069 prophylactic effect Effects 0.000 description 5
- 238000002818 protein evolution Methods 0.000 description 5
- 230000004044 response Effects 0.000 description 5
- 238000002864 sequence alignment Methods 0.000 description 5
- 238000013207 serial dilution Methods 0.000 description 5
- 238000010561 standard procedure Methods 0.000 description 5
- 108010061238 threonyl-glycine Proteins 0.000 description 5
- 238000002255 vaccination Methods 0.000 description 5
- 108010073969 valyllysine Proteins 0.000 description 5
- 239000013603 viral vector Substances 0.000 description 5
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 4
- 241000196324 Embryophyta Species 0.000 description 4
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 4
- POJJAZJHBGXEGM-YUMQZZPRSA-N Gly-Ser-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)CN POJJAZJHBGXEGM-YUMQZZPRSA-N 0.000 description 4
- 101150039660 HA gene Proteins 0.000 description 4
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 4
- 241000880493 Leptailurus serval Species 0.000 description 4
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 4
- 206010028980 Neoplasm Diseases 0.000 description 4
- 239000012980 RPMI-1640 medium Substances 0.000 description 4
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 4
- ANHVRCNNGJMJNG-BZSNNMDCSA-N Tyr-Tyr-Cys Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)N[C@@H](CS)C(=O)O)N)O ANHVRCNNGJMJNG-BZSNNMDCSA-N 0.000 description 4
- 208000036142 Viral infection Diseases 0.000 description 4
- 238000010521 absorption reaction Methods 0.000 description 4
- 108010086434 alanyl-seryl-glycine Proteins 0.000 description 4
- 108010047857 aspartylglycine Proteins 0.000 description 4
- 230000037396 body weight Effects 0.000 description 4
- 239000000872 buffer Substances 0.000 description 4
- 201000011510 cancer Diseases 0.000 description 4
- 239000003153 chemical reaction reagent Substances 0.000 description 4
- 229940127089 cytotoxic agent Drugs 0.000 description 4
- 230000003013 cytotoxicity Effects 0.000 description 4
- 231100000135 cytotoxicity Toxicity 0.000 description 4
- 238000010586 diagram Methods 0.000 description 4
- 239000006185 dispersion Substances 0.000 description 4
- 239000003937 drug carrier Substances 0.000 description 4
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 4
- 239000001963 growth medium Substances 0.000 description 4
- 210000000987 immune system Anatomy 0.000 description 4
- 238000001114 immunoprecipitation Methods 0.000 description 4
- 239000004615 ingredient Substances 0.000 description 4
- 239000003112 inhibitor Substances 0.000 description 4
- 239000007924 injection Substances 0.000 description 4
- 238000002347 injection Methods 0.000 description 4
- 238000001990 intravenous administration Methods 0.000 description 4
- 238000002955 isolation Methods 0.000 description 4
- 108010054155 lysyllysine Proteins 0.000 description 4
- 210000004962 mammalian cell Anatomy 0.000 description 4
- 239000011159 matrix material Substances 0.000 description 4
- 239000003094 microcapsule Substances 0.000 description 4
- 239000002773 nucleotide Substances 0.000 description 4
- 239000002245 particle Substances 0.000 description 4
- 244000052769 pathogen Species 0.000 description 4
- 108010051242 phenylalanylserine Proteins 0.000 description 4
- 238000000746 purification Methods 0.000 description 4
- 230000009257 reactivity Effects 0.000 description 4
- 238000003757 reverse transcription PCR Methods 0.000 description 4
- 239000007787 solid Substances 0.000 description 4
- 208000024891 symptom Diseases 0.000 description 4
- 108700012359 toxins Proteins 0.000 description 4
- 238000001890 transfection Methods 0.000 description 4
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 4
- 230000004580 weight loss Effects 0.000 description 4
- GKSPIZSKQWTXQG-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-[1-(pyridin-2-yldisulfanyl)ethyl]benzoate Chemical compound C=1C=C(C(=O)ON2C(CCC2=O)=O)C=CC=1C(C)SSC1=CC=CC=N1 GKSPIZSKQWTXQG-UHFFFAOYSA-N 0.000 description 3
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 description 3
- YYSWCHMLFJLLBJ-ZLUOBGJFSA-N Ala-Ala-Ser Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O YYSWCHMLFJLLBJ-ZLUOBGJFSA-N 0.000 description 3
- KQFRUSHJPKXBMB-BHDSKKPTSA-N Ala-Ala-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](C)NC(=O)[C@@H](N)C)C(O)=O)=CNC2=C1 KQFRUSHJPKXBMB-BHDSKKPTSA-N 0.000 description 3
- VNYMOTCMNHJGTG-JBDRJPRFSA-N Ala-Ile-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O VNYMOTCMNHJGTG-JBDRJPRFSA-N 0.000 description 3
- RTZCUEHYUQZIDE-WHFBIAKZSA-N Ala-Ser-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O RTZCUEHYUQZIDE-WHFBIAKZSA-N 0.000 description 3
- 235000006576 Althaea officinalis Nutrition 0.000 description 3
- ATABBWFGOHKROJ-GUBZILKMSA-N Arg-Pro-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O ATABBWFGOHKROJ-GUBZILKMSA-N 0.000 description 3
- QLSRIZIDQXDQHK-RCWTZXSCSA-N Arg-Val-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QLSRIZIDQXDQHK-RCWTZXSCSA-N 0.000 description 3
- WONGRTVAMHFGBE-WDSKDSINSA-N Asn-Gly-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CC(=O)N)N WONGRTVAMHFGBE-WDSKDSINSA-N 0.000 description 3
- MKJBPDLENBUHQU-CIUDSAMLSA-N Asn-Ser-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O MKJBPDLENBUHQU-CIUDSAMLSA-N 0.000 description 3
- MRQQMVZUHXUPEV-IHRRRGAJSA-N Asp-Arg-Phe Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O MRQQMVZUHXUPEV-IHRRRGAJSA-N 0.000 description 3
- WVDDGKGOMKODPV-UHFFFAOYSA-N Benzyl alcohol Chemical compound OCC1=CC=CC=C1 WVDDGKGOMKODPV-UHFFFAOYSA-N 0.000 description 3
- 101800001415 Bri23 peptide Proteins 0.000 description 3
- 101800000655 C-terminal peptide Proteins 0.000 description 3
- 102400000107 C-terminal peptide Human genes 0.000 description 3
- 108091026890 Coding region Proteins 0.000 description 3
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 3
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 3
- ZBKUIQNCRIYVGH-SDDRHHMPSA-N Gln-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)N)N ZBKUIQNCRIYVGH-SDDRHHMPSA-N 0.000 description 3
- WZZSKAJIHTUUSG-ACZMJKKPSA-N Glu-Ala-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(O)=O WZZSKAJIHTUUSG-ACZMJKKPSA-N 0.000 description 3
- JWNZHMSRZXXGTM-XKBZYTNZSA-N Glu-Ser-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JWNZHMSRZXXGTM-XKBZYTNZSA-N 0.000 description 3
- TWTPDFFBLQEBOE-IUCAKERBSA-N Gly-Leu-Gln Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O TWTPDFFBLQEBOE-IUCAKERBSA-N 0.000 description 3
- FFJQHWKSGAWSTJ-BFHQHQDPSA-N Gly-Thr-Ala Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O FFJQHWKSGAWSTJ-BFHQHQDPSA-N 0.000 description 3
- YGHSQRJSHKYUJY-SCZZXKLOSA-N Gly-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)CN YGHSQRJSHKYUJY-SCZZXKLOSA-N 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 3
- 102000018251 Hypoxanthine Phosphoribosyltransferase Human genes 0.000 description 3
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 3
- CDGLBYSAZFIIJO-RCOVLWMOSA-N Ile-Gly-Gly Chemical compound CC[C@H](C)[C@H]([NH3+])C(=O)NCC(=O)NCC([O-])=O CDGLBYSAZFIIJO-RCOVLWMOSA-N 0.000 description 3
- FBGXMKUWQFPHFB-JBDRJPRFSA-N Ile-Ser-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O)N FBGXMKUWQFPHFB-JBDRJPRFSA-N 0.000 description 3
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 3
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 3
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 description 3
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 description 3
- LZDNBBYBDGBADK-UHFFFAOYSA-N L-valyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)C(C)C)C(O)=O)=CNC2=C1 LZDNBBYBDGBADK-UHFFFAOYSA-N 0.000 description 3
- YVKSMSDXKMSIRX-GUBZILKMSA-N Leu-Glu-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O YVKSMSDXKMSIRX-GUBZILKMSA-N 0.000 description 3
- LCNASHSOFMRYFO-WDCWCFNPSA-N Leu-Thr-Gln Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CCC(N)=O LCNASHSOFMRYFO-WDCWCFNPSA-N 0.000 description 3
- 241001529936 Murinae Species 0.000 description 3
- 101100068676 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) gln-1 gene Proteins 0.000 description 3
- YMIZSYUAZJSOFL-SRVKXCTJSA-N Phe-Ser-Asn Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O YMIZSYUAZJSOFL-SRVKXCTJSA-N 0.000 description 3
- 239000002202 Polyethylene glycol Substances 0.000 description 3
- RMODQFBNDDENCP-IHRRRGAJSA-N Pro-Lys-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O RMODQFBNDDENCP-IHRRRGAJSA-N 0.000 description 3
- FDMKYQQYJKYCLV-GUBZILKMSA-N Pro-Pro-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 FDMKYQQYJKYCLV-GUBZILKMSA-N 0.000 description 3
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 3
- YQHZVYJAGWMHES-ZLUOBGJFSA-N Ser-Ala-Ser Chemical compound OC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O YQHZVYJAGWMHES-ZLUOBGJFSA-N 0.000 description 3
- HJEBZBMOTCQYDN-ACZMJKKPSA-N Ser-Glu-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O HJEBZBMOTCQYDN-ACZMJKKPSA-N 0.000 description 3
- UIGMAMGZOJVTDN-WHFBIAKZSA-N Ser-Gly-Ser Chemical compound OC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O UIGMAMGZOJVTDN-WHFBIAKZSA-N 0.000 description 3
- SFTZWNJFZYOLBD-ZDLURKLDSA-N Ser-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CO SFTZWNJFZYOLBD-ZDLURKLDSA-N 0.000 description 3
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 3
- 241000282898 Sus scrofa Species 0.000 description 3
- MXDOAJQRJBMGMO-FJXKBIBVSA-N Thr-Pro-Gly Chemical compound C[C@@H](O)[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O MXDOAJQRJBMGMO-FJXKBIBVSA-N 0.000 description 3
- 108010087924 alanylproline Proteins 0.000 description 3
- 125000000539 amino acid group Chemical group 0.000 description 3
- 239000003242 anti bacterial agent Substances 0.000 description 3
- 239000002246 antineoplastic agent Substances 0.000 description 3
- 238000013459 approach Methods 0.000 description 3
- 108010068265 aspartyltyrosine Proteins 0.000 description 3
- DQXBYHZEEUGOBF-UHFFFAOYSA-N but-3-enoic acid;ethene Chemical compound C=C.OC(=O)CC=C DQXBYHZEEUGOBF-UHFFFAOYSA-N 0.000 description 3
- 150000001718 carbodiimides Chemical class 0.000 description 3
- 235000014633 carbohydrates Nutrition 0.000 description 3
- 238000005119 centrifugation Methods 0.000 description 3
- 238000010367 cloning Methods 0.000 description 3
- 230000009918 complex formation Effects 0.000 description 3
- 239000002254 cytotoxic agent Substances 0.000 description 3
- 231100000599 cytotoxic agent Toxicity 0.000 description 3
- 238000002784 cytotoxicity assay Methods 0.000 description 3
- 231100000263 cytotoxicity test Toxicity 0.000 description 3
- 230000029087 digestion Effects 0.000 description 3
- 125000005442 diisocyanate group Chemical group 0.000 description 3
- 239000002612 dispersion medium Substances 0.000 description 3
- 238000009510 drug design Methods 0.000 description 3
- 239000005038 ethylene vinyl acetate Substances 0.000 description 3
- 230000002349 favourable effect Effects 0.000 description 3
- 238000000684 flow cytometry Methods 0.000 description 3
- 239000012530 fluid Substances 0.000 description 3
- 230000006870 function Effects 0.000 description 3
- 239000000499 gel Substances 0.000 description 3
- XBGGUPMXALFZOT-UHFFFAOYSA-N glycyl-L-tyrosine hemihydrate Natural products NCC(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 XBGGUPMXALFZOT-UHFFFAOYSA-N 0.000 description 3
- 108010010147 glycylglutamine Proteins 0.000 description 3
- 108010050848 glycylleucine Proteins 0.000 description 3
- 108010087823 glycyltyrosine Proteins 0.000 description 3
- 238000003384 imaging method Methods 0.000 description 3
- 229940124452 immunizing agent Drugs 0.000 description 3
- 230000001939 inductive effect Effects 0.000 description 3
- 230000002452 interceptive effect Effects 0.000 description 3
- 239000010410 layer Substances 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 150000002632 lipids Chemical class 0.000 description 3
- 210000004698 lymphocyte Anatomy 0.000 description 3
- 108010017391 lysylvaline Proteins 0.000 description 3
- 125000003729 nucleotide group Chemical group 0.000 description 3
- PGZUMBJQJWIWGJ-ONAKXNSWSA-N oseltamivir phosphate Chemical compound OP(O)(O)=O.CCOC(=O)C1=C[C@@H](OC(CC)CC)[C@H](NC(C)=O)[C@@H](N)C1 PGZUMBJQJWIWGJ-ONAKXNSWSA-N 0.000 description 3
- 108010012581 phenylalanylglutamate Proteins 0.000 description 3
- 229920001200 poly(ethylene-vinyl acetate) Polymers 0.000 description 3
- 229920001223 polyethylene glycol Polymers 0.000 description 3
- 239000000843 powder Substances 0.000 description 3
- 230000001681 protective effect Effects 0.000 description 3
- 208000009305 pseudorabies Diseases 0.000 description 3
- 238000011160 research Methods 0.000 description 3
- 230000002441 reversible effect Effects 0.000 description 3
- 230000001932 seasonal effect Effects 0.000 description 3
- 238000012163 sequencing technique Methods 0.000 description 3
- 125000003607 serino group Chemical group [H]N([H])[C@]([H])(C(=O)[*])C(O[H])([H])[H] 0.000 description 3
- 239000011780 sodium chloride Substances 0.000 description 3
- 238000003860 storage Methods 0.000 description 3
- 239000003826 tablet Substances 0.000 description 3
- 229940061367 tamiflu Drugs 0.000 description 3
- 230000009261 transgenic effect Effects 0.000 description 3
- 238000003146 transient transfection Methods 0.000 description 3
- 241000701161 unidentified adenovirus Species 0.000 description 3
- 229940125575 vaccine candidate Drugs 0.000 description 3
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 2
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 2
- VPFUWHKTPYPNGT-UHFFFAOYSA-N 3-(3,4-dihydroxyphenyl)-1-(5-hydroxy-2,2-dimethylchromen-6-yl)propan-1-one Chemical compound OC1=C2C=CC(C)(C)OC2=CC=C1C(=O)CCC1=CC=C(O)C(O)=C1 VPFUWHKTPYPNGT-UHFFFAOYSA-N 0.000 description 2
- LSLIRHLIUDVNBN-CIUDSAMLSA-N Ala-Asp-Lys Chemical compound C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCCCN LSLIRHLIUDVNBN-CIUDSAMLSA-N 0.000 description 2
- HYIDEIQUCBKIPL-CQDKDKBSSA-N Ala-Phe-His Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N HYIDEIQUCBKIPL-CQDKDKBSSA-N 0.000 description 2
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 2
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 2
- PPMTUXJSQDNUDE-CIUDSAMLSA-N Asn-Glu-Arg Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N PPMTUXJSQDNUDE-CIUDSAMLSA-N 0.000 description 2
- ALHMNHZJBYBYHS-DCAQKATOSA-N Asn-Lys-Arg Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O ALHMNHZJBYBYHS-DCAQKATOSA-N 0.000 description 2
- YUUIAUXBNOHFRJ-IHRRRGAJSA-N Asn-Phe-Met Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCSC)C(O)=O YUUIAUXBNOHFRJ-IHRRRGAJSA-N 0.000 description 2
- GHWWTICYPDKPTE-NGZCFLSTSA-N Asn-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(=O)N)N GHWWTICYPDKPTE-NGZCFLSTSA-N 0.000 description 2
- QXHVOUSPVAWEMX-ZLUOBGJFSA-N Asp-Asp-Ser Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O QXHVOUSPVAWEMX-ZLUOBGJFSA-N 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- 241000972773 Aulopiformes Species 0.000 description 2
- 108090001008 Avidin Proteins 0.000 description 2
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 2
- 241000894006 Bacteria Species 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- 102100027207 CD27 antigen Human genes 0.000 description 2
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 2
- TVYMKYUSZSVOAG-ZLUOBGJFSA-N Cys-Ala-Ala Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O TVYMKYUSZSVOAG-ZLUOBGJFSA-N 0.000 description 2
- 108020003215 DNA Probes Proteins 0.000 description 2
- 239000003298 DNA probe Substances 0.000 description 2
- 101710088194 Dehydrogenase Proteins 0.000 description 2
- 238000002965 ELISA Methods 0.000 description 2
- 241000233866 Fungi Species 0.000 description 2
- 241000287828 Gallus gallus Species 0.000 description 2
- 108010010803 Gelatin Proteins 0.000 description 2
- IHSGESFHTMFHRB-GUBZILKMSA-N Gln-Lys-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCC(N)=O IHSGESFHTMFHRB-GUBZILKMSA-N 0.000 description 2
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 2
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 2
- 108010024636 Glutathione Proteins 0.000 description 2
- YMUFWNJHVPQNQD-ZKWXMUAHSA-N Gly-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)CN YMUFWNJHVPQNQD-ZKWXMUAHSA-N 0.000 description 2
- JVWPPCWUDRJGAE-YUMQZZPRSA-N Gly-Asn-Leu Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O JVWPPCWUDRJGAE-YUMQZZPRSA-N 0.000 description 2
- QPDUVFSVVAOUHE-XVKPBYJWSA-N Gly-Gln-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CCC(N)=O)NC(=O)CN)C(O)=O QPDUVFSVVAOUHE-XVKPBYJWSA-N 0.000 description 2
- HQRHFUYMGCHHJS-LURJTMIESA-N Gly-Gly-Arg Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N HQRHFUYMGCHHJS-LURJTMIESA-N 0.000 description 2
- UQJNXZSSGQIPIQ-FBCQKBJTSA-N Gly-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)CN UQJNXZSSGQIPIQ-FBCQKBJTSA-N 0.000 description 2
- NSTUFLGQJCOCDL-UWVGGRQHSA-N Gly-Leu-Arg Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N NSTUFLGQJCOCDL-UWVGGRQHSA-N 0.000 description 2
- WNZOCXUOGVYYBJ-CDMKHQONSA-N Gly-Phe-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)CN)O WNZOCXUOGVYYBJ-CDMKHQONSA-N 0.000 description 2
- WNGHUXFWEWTKAO-YUMQZZPRSA-N Gly-Ser-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)CN WNGHUXFWEWTKAO-YUMQZZPRSA-N 0.000 description 2
- BZKDJRSZWLPJNI-SRVKXCTJSA-N His-His-Ser Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CO)C(O)=O BZKDJRSZWLPJNI-SRVKXCTJSA-N 0.000 description 2
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 2
- 101000914511 Homo sapiens CD27 antigen Proteins 0.000 description 2
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 2
- 241000725303 Human immunodeficiency virus Species 0.000 description 2
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 2
- PARSHQDZROHERM-NHCYSSNCSA-N Ile-Lys-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)NCC(=O)O)N PARSHQDZROHERM-NHCYSSNCSA-N 0.000 description 2
- AYLAAGNJNVZDPY-CYDGBPFRSA-N Ile-Met-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCSC)C(=O)O)N AYLAAGNJNVZDPY-CYDGBPFRSA-N 0.000 description 2
- NGKPIPCGMLWHBX-WZLNRYEVSA-N Ile-Tyr-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N NGKPIPCGMLWHBX-WZLNRYEVSA-N 0.000 description 2
- 101900222562 Influenza A virus Nucleoprotein Proteins 0.000 description 2
- 108010002350 Interleukin-2 Proteins 0.000 description 2
- IBMVEYRWAWIOTN-UHFFFAOYSA-N L-Leucyl-L-Arginyl-L-Proline Natural products CC(C)CC(N)C(=O)NC(CCCN=C(N)N)C(=O)N1CCCC1C(O)=O IBMVEYRWAWIOTN-UHFFFAOYSA-N 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 2
- XBBKIIGCUMBKCO-JXUBOQSCSA-N Leu-Ala-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XBBKIIGCUMBKCO-JXUBOQSCSA-N 0.000 description 2
- RVVBWTWPNFDYBE-SRVKXCTJSA-N Leu-Glu-Arg Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O RVVBWTWPNFDYBE-SRVKXCTJSA-N 0.000 description 2
- RXGLHDWAZQECBI-SRVKXCTJSA-N Leu-Leu-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O RXGLHDWAZQECBI-SRVKXCTJSA-N 0.000 description 2
- BJWKOATWNQJPSK-SRVKXCTJSA-N Leu-Met-Glu Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N BJWKOATWNQJPSK-SRVKXCTJSA-N 0.000 description 2
- NJNRBRKHOWSGMN-SRVKXCTJSA-N Lys-Leu-Asn Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O NJNRBRKHOWSGMN-SRVKXCTJSA-N 0.000 description 2
- OIQSIMFSVLLWBX-VOAKCMCISA-N Lys-Leu-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OIQSIMFSVLLWBX-VOAKCMCISA-N 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 241000282341 Mustela putorius furo Species 0.000 description 2
- 102100038895 Myc proto-oncogene protein Human genes 0.000 description 2
- 101710135898 Myc proto-oncogene protein Proteins 0.000 description 2
- WUGMRIBZSVSJNP-UHFFFAOYSA-N N-L-alanyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)C)C(O)=O)=CNC2=C1 WUGMRIBZSVSJNP-UHFFFAOYSA-N 0.000 description 2
- XMBSYZWANAQXEV-UHFFFAOYSA-N N-alpha-L-glutamyl-L-phenylalanine Natural products OC(=O)CCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XMBSYZWANAQXEV-UHFFFAOYSA-N 0.000 description 2
- 230000004988 N-glycosylation Effects 0.000 description 2
- 108010066427 N-valyltryptophan Proteins 0.000 description 2
- 108091061960 Naked DNA Proteins 0.000 description 2
- 102000011931 Nucleoproteins Human genes 0.000 description 2
- 108010061100 Nucleoproteins Proteins 0.000 description 2
- CSYVXYQDIVCQNU-QWRGUYRKSA-N Phe-Asp-Gly Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O CSYVXYQDIVCQNU-QWRGUYRKSA-N 0.000 description 2
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Chemical compound OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 2
- 206010035664 Pneumonia Diseases 0.000 description 2
- WCNVGGZRTNHOOS-ULQDDVLXSA-N Pro-Lys-Tyr Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)O WCNVGGZRTNHOOS-ULQDDVLXSA-N 0.000 description 2
- 239000004365 Protease Substances 0.000 description 2
- REFJWTPEDVJJIY-UHFFFAOYSA-N Quercetin Chemical compound C=1C(O)=CC(O)=C(C(C=2O)=O)C=1OC=2C1=CC=C(O)C(O)=C1 REFJWTPEDVJJIY-UHFFFAOYSA-N 0.000 description 2
- 108020004511 Recombinant DNA Proteins 0.000 description 2
- QYSFWUIXDFJUDW-DCAQKATOSA-N Ser-Leu-Arg Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O QYSFWUIXDFJUDW-DCAQKATOSA-N 0.000 description 2
- KCNSGAMPBPYUAI-CIUDSAMLSA-N Ser-Leu-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O KCNSGAMPBPYUAI-CIUDSAMLSA-N 0.000 description 2
- LGIMRDKGABDMBN-DCAQKATOSA-N Ser-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CO)N LGIMRDKGABDMBN-DCAQKATOSA-N 0.000 description 2
- XXNLGZRRSKPSGF-HTUGSXCWSA-N Thr-Gln-Phe Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N)O XXNLGZRRSKPSGF-HTUGSXCWSA-N 0.000 description 2
- NJGMALCNYAMYCB-JRQIVUDYSA-N Thr-Tyr-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(N)=O)C(O)=O NJGMALCNYAMYCB-JRQIVUDYSA-N 0.000 description 2
- CURFABYITJVKEW-QTKMDUPCSA-N Thr-Val-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N)O CURFABYITJVKEW-QTKMDUPCSA-N 0.000 description 2
- 108090000190 Thrombin Proteins 0.000 description 2
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 2
- 102220470359 Thymosin beta-10_H14A_mutation Human genes 0.000 description 2
- 101710150448 Transcriptional regulator Myc Proteins 0.000 description 2
- ZHDQRPWESGUDST-JBACZVJFSA-N Trp-Phe-Gln Chemical compound C([C@H](NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)N)C(=O)N[C@@H](CCC(N)=O)C(O)=O)C1=CC=CC=C1 ZHDQRPWESGUDST-JBACZVJFSA-N 0.000 description 2
- TZXFLDNBYYGLKA-BZSNNMDCSA-N Tyr-Asp-Tyr Chemical compound C([C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=C(O)C=C1 TZXFLDNBYYGLKA-BZSNNMDCSA-N 0.000 description 2
- WVGKPKDWYQXWLU-BZSNNMDCSA-N Tyr-His-Lys Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)N[C@@H](CCCCN)C(=O)O)N)O WVGKPKDWYQXWLU-BZSNNMDCSA-N 0.000 description 2
- KZKMBGXCNLPYKD-YEPSODPASA-N Val-Gly-Thr Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(O)=O KZKMBGXCNLPYKD-YEPSODPASA-N 0.000 description 2
- CKTMJBPRVQWPHU-JSGCOSHPSA-N Val-Phe-Gly Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)NCC(=O)O)N CKTMJBPRVQWPHU-JSGCOSHPSA-N 0.000 description 2
- 238000002835 absorbance Methods 0.000 description 2
- 239000004480 active ingredient Substances 0.000 description 2
- 230000009824 affinity maturation Effects 0.000 description 2
- 239000000556 agonist Substances 0.000 description 2
- 108010024078 alanyl-glycyl-serine Proteins 0.000 description 2
- 108010047495 alanylglycine Proteins 0.000 description 2
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 2
- 238000010171 animal model Methods 0.000 description 2
- 230000000844 anti-bacterial effect Effects 0.000 description 2
- 210000000628 antibody-producing cell Anatomy 0.000 description 2
- 229940121375 antifungal agent Drugs 0.000 description 2
- 239000003429 antifungal agent Substances 0.000 description 2
- 230000030741 antigen processing and presentation Effects 0.000 description 2
- 229940041181 antineoplastic drug Drugs 0.000 description 2
- 108010069926 arginyl-glycyl-serine Proteins 0.000 description 2
- 235000010323 ascorbic acid Nutrition 0.000 description 2
- 229960005070 ascorbic acid Drugs 0.000 description 2
- 239000011668 ascorbic acid Substances 0.000 description 2
- 108010077245 asparaginyl-proline Proteins 0.000 description 2
- CKLJMWTZIZZHCS-REOHCLBHSA-N aspartic acid group Chemical class N[C@@H](CC(=O)O)C(=O)O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- 239000011230 binding agent Substances 0.000 description 2
- 239000013060 biological fluid Substances 0.000 description 2
- 230000005540 biological transmission Effects 0.000 description 2
- 108091006004 biotinylated proteins Proteins 0.000 description 2
- 210000001124 body fluid Anatomy 0.000 description 2
- 239000010839 body fluid Substances 0.000 description 2
- 239000002775 capsule Substances 0.000 description 2
- 238000012512 characterization method Methods 0.000 description 2
- 239000002738 chelating agent Substances 0.000 description 2
- OSASVXMJTNOKOY-UHFFFAOYSA-N chlorobutanol Chemical compound CC(C)(O)C(Cl)(Cl)Cl OSASVXMJTNOKOY-UHFFFAOYSA-N 0.000 description 2
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 2
- 238000004587 chromatography analysis Methods 0.000 description 2
- 210000000349 chromosome Anatomy 0.000 description 2
- 238000000576 coating method Methods 0.000 description 2
- 230000021615 conjugation Effects 0.000 description 2
- 229920001577 copolymer Polymers 0.000 description 2
- 230000009260 cross reactivity Effects 0.000 description 2
- 239000012228 culture supernatant Substances 0.000 description 2
- 210000000805 cytoplasm Anatomy 0.000 description 2
- 230000034994 death Effects 0.000 description 2
- 239000003405 delayed action preparation Substances 0.000 description 2
- 239000003599 detergent Substances 0.000 description 2
- 238000003745 diagnosis Methods 0.000 description 2
- 239000003085 diluting agent Substances 0.000 description 2
- 125000002228 disulfide group Chemical group 0.000 description 2
- 238000012377 drug delivery Methods 0.000 description 2
- 210000001671 embryonic stem cell Anatomy 0.000 description 2
- 210000001163 endosome Anatomy 0.000 description 2
- 230000002255 enzymatic effect Effects 0.000 description 2
- 239000013613 expression plasmid Substances 0.000 description 2
- 238000001914 filtration Methods 0.000 description 2
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 2
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 2
- 239000008273 gelatin Substances 0.000 description 2
- 229920000159 gelatin Polymers 0.000 description 2
- 235000019322 gelatine Nutrition 0.000 description 2
- 235000011852 gelatine desserts Nutrition 0.000 description 2
- 108010078144 glutaminyl-glycine Proteins 0.000 description 2
- 108010055341 glutamyl-glutamic acid Proteins 0.000 description 2
- 229960003180 glutathione Drugs 0.000 description 2
- 235000011187 glycerol Nutrition 0.000 description 2
- 108010090037 glycyl-alanyl-isoleucine Proteins 0.000 description 2
- 108010062266 glycyl-glycyl-argininal Proteins 0.000 description 2
- 108010067216 glycyl-glycyl-glycine Proteins 0.000 description 2
- 108010066198 glycyl-leucyl-phenylalanine Proteins 0.000 description 2
- 108010048994 glycyl-tyrosyl-alanine Proteins 0.000 description 2
- 108010015792 glycyllysine Proteins 0.000 description 2
- 230000012010 growth Effects 0.000 description 2
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical compound O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 2
- NAQMVNRVTILPCV-UHFFFAOYSA-N hexane-1,6-diamine Chemical compound NCCCCCCN NAQMVNRVTILPCV-UHFFFAOYSA-N 0.000 description 2
- 230000028996 humoral immune response Effects 0.000 description 2
- 210000004754 hybrid cell Anatomy 0.000 description 2
- 239000000017 hydrogel Substances 0.000 description 2
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 2
- 230000036039 immunity Effects 0.000 description 2
- 238000010166 immunofluorescence Methods 0.000 description 2
- 230000016784 immunoglobulin production Effects 0.000 description 2
- 229940072221 immunoglobulins Drugs 0.000 description 2
- 230000002637 immunotoxin Effects 0.000 description 2
- 239000002596 immunotoxin Substances 0.000 description 2
- 231100000608 immunotoxin Toxicity 0.000 description 2
- 229940051026 immunotoxin Drugs 0.000 description 2
- 230000006872 improvement Effects 0.000 description 2
- 229960003971 influenza vaccine Drugs 0.000 description 2
- 238000007912 intraperitoneal administration Methods 0.000 description 2
- 108010027338 isoleucylcysteine Proteins 0.000 description 2
- 210000003734 kidney Anatomy 0.000 description 2
- 239000008101 lactose Substances 0.000 description 2
- 108010083708 leucyl-aspartyl-valine Proteins 0.000 description 2
- 108010057821 leucylproline Proteins 0.000 description 2
- 210000000265 leukocyte Anatomy 0.000 description 2
- RGLRXNKKBLIBQS-XNHQSDQCSA-N leuprolide acetate Chemical compound CC(O)=O.CCNC(=O)[C@@H]1CCCN1C(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](CC(C)C)NC(=O)[C@@H](NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H]1NC(=O)CC1)CC1=CC=C(O)C=C1 RGLRXNKKBLIBQS-XNHQSDQCSA-N 0.000 description 2
- 239000003446 ligand Substances 0.000 description 2
- 210000002809 long lived plasma cell Anatomy 0.000 description 2
- 238000003670 luciferase enzyme activity assay Methods 0.000 description 2
- HQKMJHAJHXVSDF-UHFFFAOYSA-L magnesium stearate Chemical compound [Mg+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O HQKMJHAJHXVSDF-UHFFFAOYSA-L 0.000 description 2
- 238000013507 mapping Methods 0.000 description 2
- 108020004999 messenger RNA Proteins 0.000 description 2
- OSWPMRLSEDHDFF-UHFFFAOYSA-N methyl salicylate Chemical compound COC(=O)C1=CC=CC=C1O OSWPMRLSEDHDFF-UHFFFAOYSA-N 0.000 description 2
- LXCFILQKKLGQFO-UHFFFAOYSA-N methylparaben Chemical compound COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 2
- 230000005012 migration Effects 0.000 description 2
- 238000013508 migration Methods 0.000 description 2
- 239000000178 monomer Substances 0.000 description 2
- 210000002569 neuron Anatomy 0.000 description 2
- 229910052757 nitrogen Inorganic materials 0.000 description 2
- 239000000346 nonvolatile oil Substances 0.000 description 2
- 238000011275 oncology therapy Methods 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 238000002823 phage display Methods 0.000 description 2
- 239000000546 pharmaceutical excipient Substances 0.000 description 2
- 239000012071 phase Substances 0.000 description 2
- 108010070409 phenylalanyl-glycyl-glycine Proteins 0.000 description 2
- 238000013081 phylogenetic analysis Methods 0.000 description 2
- 210000004180 plasmocyte Anatomy 0.000 description 2
- 229920003023 plastic Polymers 0.000 description 2
- 239000004033 plastic Substances 0.000 description 2
- 231100000614 poison Toxicity 0.000 description 2
- 239000002574 poison Substances 0.000 description 2
- 229920000747 poly(lactic acid) Polymers 0.000 description 2
- 230000003389 potentiating effect Effects 0.000 description 2
- 244000144977 poultry Species 0.000 description 2
- 230000003449 preventive effect Effects 0.000 description 2
- 108700042769 prolyl-leucyl-glycine Proteins 0.000 description 2
- 230000002829 reductive effect Effects 0.000 description 2
- PYWVYCXTNDRMGF-UHFFFAOYSA-N rhodamine B Chemical compound [Cl-].C=12C=CC(=[N+](CC)CC)C=C2OC2=CC(N(CC)CC)=CC=C2C=1C1=CC=CC=C1C(O)=O PYWVYCXTNDRMGF-UHFFFAOYSA-N 0.000 description 2
- 235000019515 salmon Nutrition 0.000 description 2
- 108010048397 seryl-lysyl-leucine Proteins 0.000 description 2
- 239000002356 single layer Substances 0.000 description 2
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 2
- 239000012064 sodium phosphate buffer Substances 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 238000010186 staining Methods 0.000 description 2
- 238000007920 subcutaneous administration Methods 0.000 description 2
- 238000006467 substitution reaction Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 235000000346 sugar Nutrition 0.000 description 2
- 239000000725 suspension Substances 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 230000008685 targeting Effects 0.000 description 2
- AURFVNDXGLQSNN-UHFFFAOYSA-K trisodium 2-hydroxypropane-1,2,3-tricarboxylic acid phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O.OC(=O)CC(O)(C(O)=O)CC(O)=O AURFVNDXGLQSNN-UHFFFAOYSA-K 0.000 description 2
- GPRLSGONYQIRFK-MNYXATJNSA-N triton Chemical compound [3H+] GPRLSGONYQIRFK-MNYXATJNSA-N 0.000 description 2
- 108010077037 tyrosyl-tyrosyl-phenylalanine Proteins 0.000 description 2
- 241001529453 unidentified herpesvirus Species 0.000 description 2
- 229940124856 vaccine component Drugs 0.000 description 2
- 210000002845 virion Anatomy 0.000 description 2
- 238000001262 western blot Methods 0.000 description 2
- NLEBIOOXCVAHBD-YHBSTRCHSA-N (2r,3r,4s,5s,6r)-2-[(2r,3s,4r,5r,6s)-6-dodecoxy-4,5-dihydroxy-2-(hydroxymethyl)oxan-3-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol Chemical compound O[C@@H]1[C@@H](O)[C@@H](OCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 NLEBIOOXCVAHBD-YHBSTRCHSA-N 0.000 description 1
- CWFMWBHMIMNZLN-NAKRPEOUSA-N (2s)-1-[(2s)-2-[[(2s,3s)-2-amino-3-methylpentanoyl]amino]propanoyl]pyrrolidine-2-carboxylic acid Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O CWFMWBHMIMNZLN-NAKRPEOUSA-N 0.000 description 1
- XMQUEQJCYRFIQS-YFKPBYRVSA-N (2s)-2-amino-5-ethoxy-5-oxopentanoic acid Chemical compound CCOC(=O)CC[C@H](N)C(O)=O XMQUEQJCYRFIQS-YFKPBYRVSA-N 0.000 description 1
- 125000004066 1-hydroxyethyl group Chemical group [H]OC([H])([*])C([H])([H])[H] 0.000 description 1
- IIZPXYDJLKNOIY-JXPKJXOSSA-N 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphocholine Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCC\C=C/C\C=C/C\C=C/C\C=C/CCCCC IIZPXYDJLKNOIY-JXPKJXOSSA-N 0.000 description 1
- ASNTZYQMIUCEBV-UHFFFAOYSA-N 2,5-dioxo-1-[6-[3-(pyridin-2-yldisulfanyl)propanoylamino]hexanoyloxy]pyrrolidine-3-sulfonic acid Chemical compound O=C1C(S(=O)(=O)O)CC(=O)N1OC(=O)CCCCCNC(=O)CCSSC1=CC=CC=N1 ASNTZYQMIUCEBV-UHFFFAOYSA-N 0.000 description 1
- GVJXGCIPWAVXJP-UHFFFAOYSA-N 2,5-dioxo-1-oxoniopyrrolidine-3-sulfonate Chemical compound ON1C(=O)CC(S(O)(=O)=O)C1=O GVJXGCIPWAVXJP-UHFFFAOYSA-N 0.000 description 1
- FATXTKJILXPNJL-UHFFFAOYSA-N 2-[[2-[2-[(2-amino-3-methylpentanoyl)amino]propanoylamino]acetyl]amino]-3-phenylpropanoic acid Chemical compound CCC(C)C(N)C(=O)NC(C)C(=O)NCC(=O)NC(C(O)=O)CC1=CC=CC=C1 FATXTKJILXPNJL-UHFFFAOYSA-N 0.000 description 1
- FZDFGHZZPBUTGP-UHFFFAOYSA-N 2-[[2-[bis(carboxymethyl)amino]-3-(4-isothiocyanatophenyl)propyl]-[2-[bis(carboxymethyl)amino]propyl]amino]acetic acid Chemical compound OC(=O)CN(CC(O)=O)C(C)CN(CC(O)=O)CC(N(CC(O)=O)CC(O)=O)CC1=CC=C(N=C=S)C=C1 FZDFGHZZPBUTGP-UHFFFAOYSA-N 0.000 description 1
- 125000000979 2-amino-2-oxoethyl group Chemical group [H]C([*])([H])C(=O)N([H])[H] 0.000 description 1
- BFSVOASYOCHEOV-UHFFFAOYSA-N 2-diethylaminoethanol Chemical compound CCN(CC)CCO BFSVOASYOCHEOV-UHFFFAOYSA-N 0.000 description 1
- XBBVURRQGJPTHH-UHFFFAOYSA-N 2-hydroxyacetic acid;2-hydroxypropanoic acid Chemical compound OCC(O)=O.CC(O)C(O)=O XBBVURRQGJPTHH-UHFFFAOYSA-N 0.000 description 1
- FPQQSJJWHUJYPU-UHFFFAOYSA-N 3-(dimethylamino)propyliminomethylidene-ethylazanium;chloride Chemical compound Cl.CCN=C=NCCCN(C)C FPQQSJJWHUJYPU-UHFFFAOYSA-N 0.000 description 1
- UMCMPZBLKLEWAF-BCTGSCMUSA-N 3-[(3-cholamidopropyl)dimethylammonio]propane-1-sulfonate Chemical compound C([C@H]1C[C@H]2O)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(=O)NCCC[N+](C)(C)CCCS([O-])(=O)=O)C)[C@@]2(C)[C@@H](O)C1 UMCMPZBLKLEWAF-BCTGSCMUSA-N 0.000 description 1
- GUQQBLRVXOUDTN-XOHPMCGNSA-N 3-[dimethyl-[3-[[(4r)-4-[(3r,5s,7r,8r,9s,10s,12s,13r,14s,17r)-3,7,12-trihydroxy-10,13-dimethyl-2,3,4,5,6,7,8,9,11,12,14,15,16,17-tetradecahydro-1h-cyclopenta[a]phenanthren-17-yl]pentanoyl]amino]propyl]azaniumyl]-2-hydroxypropane-1-sulfonate Chemical compound C([C@H]1C[C@H]2O)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(=O)NCCC[N+](C)(C)CC(O)CS([O-])(=O)=O)C)[C@@]2(C)[C@@H](O)C1 GUQQBLRVXOUDTN-XOHPMCGNSA-N 0.000 description 1
- TVZGACDUOSZQKY-LBPRGKRZSA-N 4-aminofolic acid Chemical compound C1=NC2=NC(N)=NC(N)=C2N=C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 TVZGACDUOSZQKY-LBPRGKRZSA-N 0.000 description 1
- CJIJXIFQYOPWTF-UHFFFAOYSA-N 7-hydroxycoumarin Natural products O1C(=O)C=CC2=CC(O)=CC=C21 CJIJXIFQYOPWTF-UHFFFAOYSA-N 0.000 description 1
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 1
- 108010022752 Acetylcholinesterase Proteins 0.000 description 1
- 102000012440 Acetylcholinesterase Human genes 0.000 description 1
- FXKNPWNXPQZLES-ZLUOBGJFSA-N Ala-Asn-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O FXKNPWNXPQZLES-ZLUOBGJFSA-N 0.000 description 1
- DECCMEWNXSNSDO-ZLUOBGJFSA-N Ala-Cys-Ala Chemical compound C[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@@H](C)C(O)=O DECCMEWNXSNSDO-ZLUOBGJFSA-N 0.000 description 1
- AWAXZRDKUHOPBO-GUBZILKMSA-N Ala-Gln-Lys Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCCN)C(O)=O AWAXZRDKUHOPBO-GUBZILKMSA-N 0.000 description 1
- KXEVYGKATAMXJJ-ACZMJKKPSA-N Ala-Glu-Asp Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O KXEVYGKATAMXJJ-ACZMJKKPSA-N 0.000 description 1
- HXNNRBHASOSVPG-GUBZILKMSA-N Ala-Glu-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O HXNNRBHASOSVPG-GUBZILKMSA-N 0.000 description 1
- LMFXXZPPZDCPTA-ZKWXMUAHSA-N Ala-Gly-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@H](C)N LMFXXZPPZDCPTA-ZKWXMUAHSA-N 0.000 description 1
- SIGTYDNEPYEXGK-ZANVPECISA-N Ala-Gly-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)CNC(=O)[C@@H](N)C)C(O)=O)=CNC2=C1 SIGTYDNEPYEXGK-ZANVPECISA-N 0.000 description 1
- IFKQPMZRDQZSHI-GHCJXIJMSA-N Ala-Ile-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O IFKQPMZRDQZSHI-GHCJXIJMSA-N 0.000 description 1
- CFPQUJZTLUQUTJ-HTFCKZLJSA-N Ala-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@H](C)N CFPQUJZTLUQUTJ-HTFCKZLJSA-N 0.000 description 1
- SDZRIBWEVVRDQI-CIUDSAMLSA-N Ala-Lys-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(O)=O SDZRIBWEVVRDQI-CIUDSAMLSA-N 0.000 description 1
- PMQXMXAASGFUDX-SRVKXCTJSA-N Ala-Lys-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](C)N)CCCCN PMQXMXAASGFUDX-SRVKXCTJSA-N 0.000 description 1
- IORKCNUBHNIMKY-CIUDSAMLSA-N Ala-Pro-Glu Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O IORKCNUBHNIMKY-CIUDSAMLSA-N 0.000 description 1
- VRTOMXFZHGWHIJ-KZVJFYERSA-N Ala-Thr-Arg Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O VRTOMXFZHGWHIJ-KZVJFYERSA-N 0.000 description 1
- VHAQSYHSDKERBS-XPUUQOCRSA-N Ala-Val-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O VHAQSYHSDKERBS-XPUUQOCRSA-N 0.000 description 1
- REWSWYIDQIELBE-FXQIFTODSA-N Ala-Val-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O REWSWYIDQIELBE-FXQIFTODSA-N 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 102000009027 Albumins Human genes 0.000 description 1
- HULHGJZIZXCPLD-FXQIFTODSA-N Arg-Ala-Cys Chemical compound C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N HULHGJZIZXCPLD-FXQIFTODSA-N 0.000 description 1
- RVDVDRUZWZIBJQ-CIUDSAMLSA-N Arg-Asn-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O RVDVDRUZWZIBJQ-CIUDSAMLSA-N 0.000 description 1
- PQWTZSNVWSOFFK-FXQIFTODSA-N Arg-Asp-Asn Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)CN=C(N)N PQWTZSNVWSOFFK-FXQIFTODSA-N 0.000 description 1
- OQCWXQJLCDPRHV-UWVGGRQHSA-N Arg-Gly-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O OQCWXQJLCDPRHV-UWVGGRQHSA-N 0.000 description 1
- WVNFNPGXYADPPO-BQBZGAKWSA-N Arg-Gly-Ser Chemical compound NC(N)=NCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O WVNFNPGXYADPPO-BQBZGAKWSA-N 0.000 description 1
- HIMXTOIXVXWHTB-DCAQKATOSA-N Arg-Met-Gln Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N HIMXTOIXVXWHTB-DCAQKATOSA-N 0.000 description 1
- SLQQPJBDBVPVQV-JYJNAYRXSA-N Arg-Phe-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](C(C)C)C(O)=O SLQQPJBDBVPVQV-JYJNAYRXSA-N 0.000 description 1
- FVBZXNSRIDVYJS-AVGNSLFASA-N Arg-Pro-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCN=C(N)N FVBZXNSRIDVYJS-AVGNSLFASA-N 0.000 description 1
- AWMAZIIEFPFHCP-RCWTZXSCSA-N Arg-Pro-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O AWMAZIIEFPFHCP-RCWTZXSCSA-N 0.000 description 1
- RYQSYXFGFOTJDJ-RHYQMDGZSA-N Arg-Thr-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O RYQSYXFGFOTJDJ-RHYQMDGZSA-N 0.000 description 1
- XHFXZQHTLJVZBN-FXQIFTODSA-N Asn-Arg-Asn Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N)CN=C(N)N XHFXZQHTLJVZBN-FXQIFTODSA-N 0.000 description 1
- RCENDENBBJFJHZ-ACZMJKKPSA-N Asn-Asn-Gln Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O RCENDENBBJFJHZ-ACZMJKKPSA-N 0.000 description 1
- KXFCBAHYSLJCCY-ZLUOBGJFSA-N Asn-Asn-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O KXFCBAHYSLJCCY-ZLUOBGJFSA-N 0.000 description 1
- VKCOHFFSTKCXEQ-OLHMAJIHSA-N Asn-Asn-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VKCOHFFSTKCXEQ-OLHMAJIHSA-N 0.000 description 1
- CUQUEHYSSFETRD-ACZMJKKPSA-N Asn-Asp-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CC(=O)N)N CUQUEHYSSFETRD-ACZMJKKPSA-N 0.000 description 1
- BHQQRVARKXWXPP-ACZMJKKPSA-N Asn-Asp-Glu Chemical compound C(CC(=O)O)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CC(=O)N)N BHQQRVARKXWXPP-ACZMJKKPSA-N 0.000 description 1
- KLKHFFMNGWULBN-VKHMYHEASA-N Asn-Gly Chemical compound NC(=O)C[C@H](N)C(=O)NCC(O)=O KLKHFFMNGWULBN-VKHMYHEASA-N 0.000 description 1
- PLVAAIPKSGUXDV-WHFBIAKZSA-N Asn-Gly-Cys Chemical compound C([C@@H](C(=O)NCC(=O)N[C@@H](CS)C(=O)O)N)C(=O)N PLVAAIPKSGUXDV-WHFBIAKZSA-N 0.000 description 1
- PBSQFBAJKPLRJY-BYULHYEWSA-N Asn-Gly-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CC(=O)N)N PBSQFBAJKPLRJY-BYULHYEWSA-N 0.000 description 1
- RAQMSGVCGSJKCL-FOHZUACHSA-N Asn-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(N)=O RAQMSGVCGSJKCL-FOHZUACHSA-N 0.000 description 1
- BZWRLDPIWKOVKB-ZPFDUUQYSA-N Asn-Leu-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O BZWRLDPIWKOVKB-ZPFDUUQYSA-N 0.000 description 1
- PBFXCUOEGVJTMV-QXEWZRGKSA-N Asn-Met-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C(C)C)C(O)=O PBFXCUOEGVJTMV-QXEWZRGKSA-N 0.000 description 1
- QXOPPIDJKPEKCW-GUBZILKMSA-N Asn-Pro-Arg Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CC(=O)N)N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O QXOPPIDJKPEKCW-GUBZILKMSA-N 0.000 description 1
- GMUOCGCDOYYWPD-FXQIFTODSA-N Asn-Pro-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O GMUOCGCDOYYWPD-FXQIFTODSA-N 0.000 description 1
- UGXYFDQFLVCDFC-CIUDSAMLSA-N Asn-Ser-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O UGXYFDQFLVCDFC-CIUDSAMLSA-N 0.000 description 1
- HNXWVVHIGTZTBO-LKXGYXEUSA-N Asn-Ser-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O HNXWVVHIGTZTBO-LKXGYXEUSA-N 0.000 description 1
- NCXTYSVDWLAQGZ-ZKWXMUAHSA-N Asn-Ser-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O NCXTYSVDWLAQGZ-ZKWXMUAHSA-N 0.000 description 1
- VBKIFHUVGLOJKT-FKZODXBYSA-N Asn-Thr Chemical compound C[C@@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)N)O VBKIFHUVGLOJKT-FKZODXBYSA-N 0.000 description 1
- JBDLMLZNDRLDIX-HJGDQZAQSA-N Asn-Thr-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O JBDLMLZNDRLDIX-HJGDQZAQSA-N 0.000 description 1
- AMGQTNHANMRPOE-LKXGYXEUSA-N Asn-Thr-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O AMGQTNHANMRPOE-LKXGYXEUSA-N 0.000 description 1
- UXHYOWXTJLBEPG-GSSVUCPTSA-N Asn-Thr-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O UXHYOWXTJLBEPG-GSSVUCPTSA-N 0.000 description 1
- XZFONYMRYTVLPL-NHCYSSNCSA-N Asn-Val-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CC(=O)N)N XZFONYMRYTVLPL-NHCYSSNCSA-N 0.000 description 1
- LMIWYCWRJVMAIQ-NHCYSSNCSA-N Asn-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)N)N LMIWYCWRJVMAIQ-NHCYSSNCSA-N 0.000 description 1
- XOQYDFCQPWAMSA-KKHAAJSZSA-N Asn-Val-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XOQYDFCQPWAMSA-KKHAAJSZSA-N 0.000 description 1
- IXIWEFWRKIUMQX-DCAQKATOSA-N Asp-Arg-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H](N)CC(O)=O IXIWEFWRKIUMQX-DCAQKATOSA-N 0.000 description 1
- HTOZUYZQPICRAP-BPUTZDHNSA-N Asp-Arg-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(=O)O)N HTOZUYZQPICRAP-BPUTZDHNSA-N 0.000 description 1
- KNMRXHIAVXHCLW-ZLUOBGJFSA-N Asp-Asn-Ser Chemical compound C([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CO)C(=O)O)N)C(=O)O KNMRXHIAVXHCLW-ZLUOBGJFSA-N 0.000 description 1
- RDRMWJBLOSRRAW-BYULHYEWSA-N Asp-Asn-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O RDRMWJBLOSRRAW-BYULHYEWSA-N 0.000 description 1
- WCFCYFDBMNFSPA-ACZMJKKPSA-N Asp-Asp-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCC(O)=O WCFCYFDBMNFSPA-ACZMJKKPSA-N 0.000 description 1
- VHQOCWWKXIOAQI-WDSKDSINSA-N Asp-Gln-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O VHQOCWWKXIOAQI-WDSKDSINSA-N 0.000 description 1
- PMEHKVHZQKJACS-PEFMBERDSA-N Asp-Gln-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O PMEHKVHZQKJACS-PEFMBERDSA-N 0.000 description 1
- QCVXMEHGFUMKCO-YUMQZZPRSA-N Asp-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(O)=O QCVXMEHGFUMKCO-YUMQZZPRSA-N 0.000 description 1
- KHGPWGKPYHPOIK-QWRGUYRKSA-N Asp-Gly-Phe Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O KHGPWGKPYHPOIK-QWRGUYRKSA-N 0.000 description 1
- SNDBKTFJWVEVPO-WHFBIAKZSA-N Asp-Gly-Ser Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](CO)C(O)=O SNDBKTFJWVEVPO-WHFBIAKZSA-N 0.000 description 1
- NRIFEOUAFLTMFJ-AAEUAGOBSA-N Asp-Gly-Trp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O NRIFEOUAFLTMFJ-AAEUAGOBSA-N 0.000 description 1
- KRQFMDNIUOVRIF-KKUMJFAQSA-N Asp-Phe-His Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CC(=O)O)N KRQFMDNIUOVRIF-KKUMJFAQSA-N 0.000 description 1
- ZBYLEBZCVKLPCY-FXQIFTODSA-N Asp-Ser-Arg Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O ZBYLEBZCVKLPCY-FXQIFTODSA-N 0.000 description 1
- XXAMCEGRCZQGEM-ZLUOBGJFSA-N Asp-Ser-Asn Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O XXAMCEGRCZQGEM-ZLUOBGJFSA-N 0.000 description 1
- JSHWXQIZOCVWIA-ZKWXMUAHSA-N Asp-Ser-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O JSHWXQIZOCVWIA-ZKWXMUAHSA-N 0.000 description 1
- MNQMTYSEKZHIDF-GCJQMDKQSA-N Asp-Thr-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O MNQMTYSEKZHIDF-GCJQMDKQSA-N 0.000 description 1
- UTLCRGFJFSZWAW-OLHMAJIHSA-N Asp-Thr-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)O)N)O UTLCRGFJFSZWAW-OLHMAJIHSA-N 0.000 description 1
- KNDCWFXCFKSEBM-AVGNSLFASA-N Asp-Tyr-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O KNDCWFXCFKSEBM-AVGNSLFASA-N 0.000 description 1
- VHUKCUHLFMRHOD-MELADBBJSA-N Asp-Tyr-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=C(C=C2)O)NC(=O)[C@H](CC(=O)O)N)C(=O)O VHUKCUHLFMRHOD-MELADBBJSA-N 0.000 description 1
- GYNUXDMCDILYIQ-QRTARXTBSA-N Asp-Val-Trp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CC(=O)O)N GYNUXDMCDILYIQ-QRTARXTBSA-N 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 241000416162 Astragalus gummifer Species 0.000 description 1
- 241000700663 Avipoxvirus Species 0.000 description 1
- 230000003844 B-cell-activation Effects 0.000 description 1
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 1
- 102100026189 Beta-galactosidase Human genes 0.000 description 1
- 102100021277 Beta-secretase 2 Human genes 0.000 description 1
- 208000031648 Body Weight Changes Diseases 0.000 description 1
- YFSNBQBDMHKKQP-UHFFFAOYSA-N C(C)(=O)O.C(C)(=O)O.C(C)(=O)O.C(C)(=O)O.C(C)(=O)O.N(=C=S)C1(CNCC(NCCN)C)CC=CC=C1 Chemical compound C(C)(=O)O.C(C)(=O)O.C(C)(=O)O.C(C)(=O)O.C(C)(=O)O.N(=C=S)C1(CNCC(NCCN)C)CC=CC=C1 YFSNBQBDMHKKQP-UHFFFAOYSA-N 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 102100032937 CD40 ligand Human genes 0.000 description 1
- 101100505161 Caenorhabditis elegans mel-32 gene Proteins 0.000 description 1
- 241000282465 Canis Species 0.000 description 1
- 101710158575 Cap-specific mRNA (nucleoside-2'-O-)-methyltransferase Proteins 0.000 description 1
- OKTJSMMVPCPJKN-NJFSPNSNSA-N Carbon-14 Chemical compound [14C] OKTJSMMVPCPJKN-NJFSPNSNSA-N 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 102100035882 Catalase Human genes 0.000 description 1
- 108010053835 Catalase Proteins 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 102000001327 Chemokine CCL5 Human genes 0.000 description 1
- 108010055166 Chemokine CCL5 Proteins 0.000 description 1
- 102000019034 Chemokines Human genes 0.000 description 1
- 108010012236 Chemokines Proteins 0.000 description 1
- VEXZGXHMUGYJMC-UHFFFAOYSA-M Chloride anion Chemical compound [Cl-] VEXZGXHMUGYJMC-UHFFFAOYSA-M 0.000 description 1
- KRKNYBCHXYNGOX-UHFFFAOYSA-K Citrate Chemical compound [O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O KRKNYBCHXYNGOX-UHFFFAOYSA-K 0.000 description 1
- 102000008186 Collagen Human genes 0.000 description 1
- 108010035532 Collagen Proteins 0.000 description 1
- 208000035473 Communicable disease Diseases 0.000 description 1
- 108010060123 Conjugate Vaccines Proteins 0.000 description 1
- 229920002261 Corn starch Polymers 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- 241000699802 Cricetulus griseus Species 0.000 description 1
- 108700032819 Croton tiglium crotin II Proteins 0.000 description 1
- 241000195493 Cryptophyta Species 0.000 description 1
- DCJNIJAWIRPPBB-CIUDSAMLSA-N Cys-Ala-Lys Chemical compound C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CS)N DCJNIJAWIRPPBB-CIUDSAMLSA-N 0.000 description 1
- LHLSSZYQFUNWRZ-NAKRPEOUSA-N Cys-Arg-Ile Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O LHLSSZYQFUNWRZ-NAKRPEOUSA-N 0.000 description 1
- KIHRUISMQZVCNO-ZLUOBGJFSA-N Cys-Asp-Asp Chemical compound SC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O KIHRUISMQZVCNO-ZLUOBGJFSA-N 0.000 description 1
- UCMIKRLLIOVDRJ-XKBZYTNZSA-N Cys-Gln-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CS)N)O UCMIKRLLIOVDRJ-XKBZYTNZSA-N 0.000 description 1
- BDWIZLQVVWQMTB-XKBZYTNZSA-N Cys-Glu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CS)N)O BDWIZLQVVWQMTB-XKBZYTNZSA-N 0.000 description 1
- VBIIZCXWOZDIHS-ACZMJKKPSA-N Cys-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H](N)CS VBIIZCXWOZDIHS-ACZMJKKPSA-N 0.000 description 1
- ZMWOJVAXTOUHAP-ZKWXMUAHSA-N Cys-Ile-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CS)N ZMWOJVAXTOUHAP-ZKWXMUAHSA-N 0.000 description 1
- IDFVDSBJNMPBSX-SRVKXCTJSA-N Cys-Lys-Leu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O IDFVDSBJNMPBSX-SRVKXCTJSA-N 0.000 description 1
- QVLKXRMFNGHDRO-FXQIFTODSA-N Cys-Met-Asn Chemical compound SC[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(N)=O)C(O)=O QVLKXRMFNGHDRO-FXQIFTODSA-N 0.000 description 1
- OZSBRCONEMXYOJ-AVGNSLFASA-N Cys-Phe-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CS)N OZSBRCONEMXYOJ-AVGNSLFASA-N 0.000 description 1
- HMWBPUDETPKSSS-DCAQKATOSA-N Cys-Pro-Lys Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CS)N)C(=O)N[C@@H](CCCCN)C(=O)O HMWBPUDETPKSSS-DCAQKATOSA-N 0.000 description 1
- KVCJEMHFLGVINV-ZLUOBGJFSA-N Cys-Ser-Asn Chemical compound SC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC(N)=O KVCJEMHFLGVINV-ZLUOBGJFSA-N 0.000 description 1
- SRZZZTMJARUVPI-JBDRJPRFSA-N Cys-Ser-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CS)N SRZZZTMJARUVPI-JBDRJPRFSA-N 0.000 description 1
- CLEFUAZULXANBU-MELADBBJSA-N Cys-Tyr-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=C(C=C2)O)NC(=O)[C@H](CS)N)C(=O)O CLEFUAZULXANBU-MELADBBJSA-N 0.000 description 1
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 1
- IGXWBGJHJZYPQS-SSDOTTSWSA-N D-Luciferin Chemical compound OC(=O)[C@H]1CSC(C=2SC3=CC=C(O)C=C3N=2)=N1 IGXWBGJHJZYPQS-SSDOTTSWSA-N 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 1
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Natural products CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 1
- 238000011238 DNA vaccination Methods 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- CYCGRDQQIOGCKX-UHFFFAOYSA-N Dehydro-luciferin Natural products OC(=O)C1=CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 CYCGRDQQIOGCKX-UHFFFAOYSA-N 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 1
- 102000016607 Diphtheria Toxin Human genes 0.000 description 1
- 108010053187 Diphtheria Toxin Proteins 0.000 description 1
- 206010059866 Drug resistance Diseases 0.000 description 1
- 241000792859 Enema Species 0.000 description 1
- 108090000371 Esterases Proteins 0.000 description 1
- 101710082714 Exotoxin A Proteins 0.000 description 1
- 108010087819 Fc receptors Proteins 0.000 description 1
- 102000009109 Fc receptors Human genes 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 101710189104 Fibritin Proteins 0.000 description 1
- 108090000331 Firefly luciferases Proteins 0.000 description 1
- BJGNCJDXODQBOB-UHFFFAOYSA-N Fivefly Luciferin Natural products OC(=O)C1CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 BJGNCJDXODQBOB-UHFFFAOYSA-N 0.000 description 1
- PNNNRSAQSRJVSB-SLPGGIOYSA-N Fucose Natural products C[C@H](O)[C@@H](O)[C@H](O)[C@H](O)C=O PNNNRSAQSRJVSB-SLPGGIOYSA-N 0.000 description 1
- OYTPNWYZORARHL-XHNCKOQMSA-N Gln-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)N)N OYTPNWYZORARHL-XHNCKOQMSA-N 0.000 description 1
- DTMLKCYOQKZXKZ-HJGDQZAQSA-N Gln-Arg-Thr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O DTMLKCYOQKZXKZ-HJGDQZAQSA-N 0.000 description 1
- PCKOTDPDHIBGRW-CIUDSAMLSA-N Gln-Cys-Arg Chemical compound C(C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CCC(=O)N)N)CN=C(N)N PCKOTDPDHIBGRW-CIUDSAMLSA-N 0.000 description 1
- GNMQDOGFWYWPNM-LAEOZQHASA-N Gln-Gly-Ile Chemical compound CC[C@H](C)[C@H](NC(=O)CNC(=O)[C@@H](N)CCC(N)=O)C(O)=O GNMQDOGFWYWPNM-LAEOZQHASA-N 0.000 description 1
- DAAUVRPSZRDMBV-KBIXCLLPSA-N Gln-Ile-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCC(=O)N)N DAAUVRPSZRDMBV-KBIXCLLPSA-N 0.000 description 1
- HXOLDXKNWKLDMM-YVNDNENWSA-N Gln-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CCC(=O)N)N HXOLDXKNWKLDMM-YVNDNENWSA-N 0.000 description 1
- FTIJVMLAGRAYMJ-MNXVOIDGSA-N Gln-Ile-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCC(N)=O FTIJVMLAGRAYMJ-MNXVOIDGSA-N 0.000 description 1
- HWEINOMSWQSJDC-SRVKXCTJSA-N Gln-Leu-Arg Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O HWEINOMSWQSJDC-SRVKXCTJSA-N 0.000 description 1
- MSHXWFKYXJTLEZ-CIUDSAMLSA-N Gln-Met-Asn Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCC(=O)N)N MSHXWFKYXJTLEZ-CIUDSAMLSA-N 0.000 description 1
- FALJZCPMTGJOHX-SRVKXCTJSA-N Gln-Met-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(O)=O FALJZCPMTGJOHX-SRVKXCTJSA-N 0.000 description 1
- FQCILXROGNOZON-YUMQZZPRSA-N Gln-Pro-Gly Chemical compound NC(=O)CC[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O FQCILXROGNOZON-YUMQZZPRSA-N 0.000 description 1
- OTQSTOXRUBVWAP-NRPADANISA-N Gln-Ser-Val Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O OTQSTOXRUBVWAP-NRPADANISA-N 0.000 description 1
- ZFBBMCKQSNJZSN-AUTRQRHGSA-N Gln-Val-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O ZFBBMCKQSNJZSN-AUTRQRHGSA-N 0.000 description 1
- NCWOMXABNYEPLY-NRPADANISA-N Glu-Ala-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(O)=O NCWOMXABNYEPLY-NRPADANISA-N 0.000 description 1
- WOMUDRVDJMHTCV-DCAQKATOSA-N Glu-Arg-Arg Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O WOMUDRVDJMHTCV-DCAQKATOSA-N 0.000 description 1
- CVPXINNKRTZBMO-CIUDSAMLSA-N Glu-Arg-Asn Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)CN=C(N)N CVPXINNKRTZBMO-CIUDSAMLSA-N 0.000 description 1
- OBIHEDRRSMRKLU-ACZMJKKPSA-N Glu-Cys-Asp Chemical compound C(CC(=O)O)[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)O)C(=O)O)N OBIHEDRRSMRKLU-ACZMJKKPSA-N 0.000 description 1
- ZXLZWUQBRYGDNS-CIUDSAMLSA-N Glu-Cys-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CCC(=O)O)N ZXLZWUQBRYGDNS-CIUDSAMLSA-N 0.000 description 1
- BUZMZDDKFCSKOT-CIUDSAMLSA-N Glu-Glu-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O BUZMZDDKFCSKOT-CIUDSAMLSA-N 0.000 description 1
- IQACOVZVOMVILH-FXQIFTODSA-N Glu-Glu-Ser Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O IQACOVZVOMVILH-FXQIFTODSA-N 0.000 description 1
- OAGVHWYIBZMWLA-YFKPBYRVSA-N Glu-Gly-Gly Chemical compound OC(=O)CC[C@H](N)C(=O)NCC(=O)NCC(O)=O OAGVHWYIBZMWLA-YFKPBYRVSA-N 0.000 description 1
- HPJLZFTUUJKWAJ-JHEQGTHGSA-N Glu-Gly-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(O)=O HPJLZFTUUJKWAJ-JHEQGTHGSA-N 0.000 description 1
- ATVYZJGOZLVXDK-IUCAKERBSA-N Glu-Leu-Gly Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O ATVYZJGOZLVXDK-IUCAKERBSA-N 0.000 description 1
- MWMJCGBSIORNCD-AVGNSLFASA-N Glu-Leu-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O MWMJCGBSIORNCD-AVGNSLFASA-N 0.000 description 1
- IVGJYOOGJLFKQE-AVGNSLFASA-N Glu-Leu-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N IVGJYOOGJLFKQE-AVGNSLFASA-N 0.000 description 1
- BCYGDJXHAGZNPQ-DCAQKATOSA-N Glu-Lys-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O BCYGDJXHAGZNPQ-DCAQKATOSA-N 0.000 description 1
- ILWHFUZZCFYSKT-AVGNSLFASA-N Glu-Lys-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O ILWHFUZZCFYSKT-AVGNSLFASA-N 0.000 description 1
- ZGEJRLJEAMPEDV-SRVKXCTJSA-N Glu-Lys-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(=O)O)N ZGEJRLJEAMPEDV-SRVKXCTJSA-N 0.000 description 1
- QDMVXRNLOPTPIE-WDCWCFNPSA-N Glu-Lys-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QDMVXRNLOPTPIE-WDCWCFNPSA-N 0.000 description 1
- GMAGZGCAYLQBKF-NHCYSSNCSA-N Glu-Met-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C(C)C)C(O)=O GMAGZGCAYLQBKF-NHCYSSNCSA-N 0.000 description 1
- FGSGPLRPQCZBSQ-AVGNSLFASA-N Glu-Phe-Ser Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O FGSGPLRPQCZBSQ-AVGNSLFASA-N 0.000 description 1
- QCMVGXDELYMZET-GLLZPBPUSA-N Glu-Thr-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O QCMVGXDELYMZET-GLLZPBPUSA-N 0.000 description 1
- DTLLNDVORUEOTM-WDCWCFNPSA-N Glu-Thr-Lys Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(O)=O DTLLNDVORUEOTM-WDCWCFNPSA-N 0.000 description 1
- HAGKYCXGTRUUFI-RYUDHWBXSA-N Glu-Tyr-Gly Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CCC(=O)O)N)O HAGKYCXGTRUUFI-RYUDHWBXSA-N 0.000 description 1
- 239000004366 Glucose oxidase Substances 0.000 description 1
- 108010015776 Glucose oxidase Proteins 0.000 description 1
- OVSKVOOUFAKODB-UWVGGRQHSA-N Gly-Arg-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCN=C(N)N OVSKVOOUFAKODB-UWVGGRQHSA-N 0.000 description 1
- GGEJHJIXRBTJPD-BYPYZUCNSA-N Gly-Asn-Gly Chemical compound NCC(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O GGEJHJIXRBTJPD-BYPYZUCNSA-N 0.000 description 1
- XRTDOIOIBMAXCT-NKWVEPMBSA-N Gly-Asn-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)N)NC(=O)CN)C(=O)O XRTDOIOIBMAXCT-NKWVEPMBSA-N 0.000 description 1
- SUDUYJOBLHQAMI-WHFBIAKZSA-N Gly-Asp-Cys Chemical compound NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CS)C(O)=O SUDUYJOBLHQAMI-WHFBIAKZSA-N 0.000 description 1
- QGZSAHIZRQHCEQ-QWRGUYRKSA-N Gly-Asp-Tyr Chemical compound NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 QGZSAHIZRQHCEQ-QWRGUYRKSA-N 0.000 description 1
- PNMUAGGSDZXTHX-BYPYZUCNSA-N Gly-Gln Chemical compound NCC(=O)N[C@H](C(O)=O)CCC(N)=O PNMUAGGSDZXTHX-BYPYZUCNSA-N 0.000 description 1
- BIRKKBCSAIHDDF-WDSKDSINSA-N Gly-Glu-Cys Chemical compound NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CS)C(O)=O BIRKKBCSAIHDDF-WDSKDSINSA-N 0.000 description 1
- HFXJIZNEXNIZIJ-BQBZGAKWSA-N Gly-Glu-Gln Chemical compound NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O HFXJIZNEXNIZIJ-BQBZGAKWSA-N 0.000 description 1
- YWAQATDNEKZFFK-BYPYZUCNSA-N Gly-Gly-Ser Chemical compound NCC(=O)NCC(=O)N[C@@H](CO)C(O)=O YWAQATDNEKZFFK-BYPYZUCNSA-N 0.000 description 1
- BHPQOIPBLYJNAW-NGZCFLSTSA-N Gly-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)CN BHPQOIPBLYJNAW-NGZCFLSTSA-N 0.000 description 1
- HAXARWKYFIIHKD-ZKWXMUAHSA-N Gly-Ile-Ser Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O HAXARWKYFIIHKD-ZKWXMUAHSA-N 0.000 description 1
- YIFUFYZELCMPJP-YUMQZZPRSA-N Gly-Leu-Cys Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CS)C(O)=O YIFUFYZELCMPJP-YUMQZZPRSA-N 0.000 description 1
- TVUWMSBGMVAHSJ-KBPBESRZSA-N Gly-Leu-Phe Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 TVUWMSBGMVAHSJ-KBPBESRZSA-N 0.000 description 1
- GMTXWRIDLGTVFC-IUCAKERBSA-N Gly-Lys-Glu Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O GMTXWRIDLGTVFC-IUCAKERBSA-N 0.000 description 1
- PDUHNKAFQXQNLH-ZETCQYMHSA-N Gly-Lys-Gly Chemical compound NCCCC[C@H](NC(=O)CN)C(=O)NCC(O)=O PDUHNKAFQXQNLH-ZETCQYMHSA-N 0.000 description 1
- MHXKHKWHPNETGG-QWRGUYRKSA-N Gly-Lys-Leu Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O MHXKHKWHPNETGG-QWRGUYRKSA-N 0.000 description 1
- RVGMVLVBDRQVKB-UWVGGRQHSA-N Gly-Met-His Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)CN RVGMVLVBDRQVKB-UWVGGRQHSA-N 0.000 description 1
- FJWSJWACLMTDMI-WPRPVWTQSA-N Gly-Met-Val Chemical compound [H]NCC(=O)N[C@@H](CCSC)C(=O)N[C@@H](C(C)C)C(O)=O FJWSJWACLMTDMI-WPRPVWTQSA-N 0.000 description 1
- IBYOLNARKHMLBG-WHOFXGATSA-N Gly-Phe-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CC=CC=C1 IBYOLNARKHMLBG-WHOFXGATSA-N 0.000 description 1
- JPVGHHQGKPQYIL-KBPBESRZSA-N Gly-Phe-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CC=CC=C1 JPVGHHQGKPQYIL-KBPBESRZSA-N 0.000 description 1
- JJGBXTYGTKWGAT-YUMQZZPRSA-N Gly-Pro-Glu Chemical compound NCC(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O JJGBXTYGTKWGAT-YUMQZZPRSA-N 0.000 description 1
- GAAHQHNCMIAYEX-UWVGGRQHSA-N Gly-Pro-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)CN GAAHQHNCMIAYEX-UWVGGRQHSA-N 0.000 description 1
- YOBGUCWZPXJHTN-BQBZGAKWSA-N Gly-Ser-Arg Chemical compound NCC(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCN=C(N)N YOBGUCWZPXJHTN-BQBZGAKWSA-N 0.000 description 1
- JSLVAHYTAJJEQH-QWRGUYRKSA-N Gly-Ser-Phe Chemical compound NCC(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 JSLVAHYTAJJEQH-QWRGUYRKSA-N 0.000 description 1
- FKYQEVBRZSFAMJ-QWRGUYRKSA-N Gly-Ser-Tyr Chemical compound NCC(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 FKYQEVBRZSFAMJ-QWRGUYRKSA-N 0.000 description 1
- FFALDIDGPLUDKV-ZDLURKLDSA-N Gly-Thr-Ser Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O FFALDIDGPLUDKV-ZDLURKLDSA-N 0.000 description 1
- FOKISINOENBSDM-WLTAIBSBSA-N Gly-Thr-Tyr Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O FOKISINOENBSDM-WLTAIBSBSA-N 0.000 description 1
- PYFHPYDQHCEVIT-KBPBESRZSA-N Gly-Trp-Gln Chemical compound [H]NCC(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CCC(N)=O)C(O)=O PYFHPYDQHCEVIT-KBPBESRZSA-N 0.000 description 1
- SFOXOSKVTLDEDM-HOTGVXAUSA-N Gly-Trp-Leu Chemical compound C1=CC=C2C(C[C@@H](C(=O)N[C@@H](CC(C)C)C(O)=O)NC(=O)CN)=CNC2=C1 SFOXOSKVTLDEDM-HOTGVXAUSA-N 0.000 description 1
- IROABALAWGJQGM-OALUTQOASA-N Gly-Trp-Tyr Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)O)NC(=O)CN IROABALAWGJQGM-OALUTQOASA-N 0.000 description 1
- UIQGJYUEQDOODF-KWQFWETISA-N Gly-Tyr-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)CN)CC1=CC=C(O)C=C1 UIQGJYUEQDOODF-KWQFWETISA-N 0.000 description 1
- IZVICCORZOSGPT-JSGCOSHPSA-N Gly-Val-Tyr Chemical compound [H]NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O IZVICCORZOSGPT-JSGCOSHPSA-N 0.000 description 1
- KSOBNUBCYHGUKH-UWVGGRQHSA-N Gly-Val-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)CN KSOBNUBCYHGUKH-UWVGGRQHSA-N 0.000 description 1
- JZNWSCPGTDBMEW-UHFFFAOYSA-N Glycerophosphorylethanolamin Natural products NCCOP(O)(=O)OCC(O)CO JZNWSCPGTDBMEW-UHFFFAOYSA-N 0.000 description 1
- 108060003393 Granulin Proteins 0.000 description 1
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 1
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 1
- JBCLFWXMTIKCCB-UHFFFAOYSA-N H-Gly-Phe-OH Natural products NCC(=O)NC(C(O)=O)CC1=CC=CC=C1 JBCLFWXMTIKCCB-UHFFFAOYSA-N 0.000 description 1
- 208000031886 HIV Infections Diseases 0.000 description 1
- 241000711549 Hepacivirus C Species 0.000 description 1
- 241000700721 Hepatitis B virus Species 0.000 description 1
- MJNWEIMBXKKCSF-XVYDVKMFSA-N His-Ala-Asn Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC1=CN=CN1)N MJNWEIMBXKKCSF-XVYDVKMFSA-N 0.000 description 1
- NOQPTNXSGNPJNS-YUMQZZPRSA-N His-Asn-Gly Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O NOQPTNXSGNPJNS-YUMQZZPRSA-N 0.000 description 1
- JBSLJUPMTYLLFH-MELADBBJSA-N His-His-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CN=CN2)NC(=O)[C@H](CC3=CN=CN3)N)C(=O)O JBSLJUPMTYLLFH-MELADBBJSA-N 0.000 description 1
- YAALVYQFVJNXIV-KKUMJFAQSA-N His-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC1=CN=CN1 YAALVYQFVJNXIV-KKUMJFAQSA-N 0.000 description 1
- YAEKRYQASVCDLK-JYJNAYRXSA-N His-Phe-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CC2=CN=CN2)N YAEKRYQASVCDLK-JYJNAYRXSA-N 0.000 description 1
- NBWATNYAUVSAEQ-ZEILLAHLSA-N His-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CC1=CN=CN1)N)O NBWATNYAUVSAEQ-ZEILLAHLSA-N 0.000 description 1
- 102000018713 Histocompatibility Antigens Class II Human genes 0.000 description 1
- 101100005713 Homo sapiens CD4 gene Proteins 0.000 description 1
- 101000815628 Homo sapiens Regulatory-associated protein of mTOR Proteins 0.000 description 1
- 101000652747 Homo sapiens Target of rapamycin complex 2 subunit MAPKAP1 Proteins 0.000 description 1
- 101000648491 Homo sapiens Transportin-1 Proteins 0.000 description 1
- 108090000144 Human Proteins Proteins 0.000 description 1
- 102000003839 Human Proteins Human genes 0.000 description 1
- 102000008100 Human Serum Albumin Human genes 0.000 description 1
- 108091006905 Human Serum Albumin Proteins 0.000 description 1
- 241000701024 Human betaherpesvirus 5 Species 0.000 description 1
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 1
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 1
- CYHYBSGMHMHKOA-CIQUZCHMSA-N Ile-Ala-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N CYHYBSGMHMHKOA-CIQUZCHMSA-N 0.000 description 1
- ZZHGKECPZXPXJF-PCBIJLKTSA-N Ile-Asn-Phe Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 ZZHGKECPZXPXJF-PCBIJLKTSA-N 0.000 description 1
- MTFVYKQRLXYAQN-LAEOZQHASA-N Ile-Glu-Gly Chemical compound [H]N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O MTFVYKQRLXYAQN-LAEOZQHASA-N 0.000 description 1
- SPQWWEZBHXHUJN-KBIXCLLPSA-N Ile-Glu-Ser Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O SPQWWEZBHXHUJN-KBIXCLLPSA-N 0.000 description 1
- LEHPJMKVGFPSSP-ZQINRCPSSA-N Ile-Glu-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)[C@@H](C)CC)C(O)=O)=CNC2=C1 LEHPJMKVGFPSSP-ZQINRCPSSA-N 0.000 description 1
- LPFBXFILACZHIB-LAEOZQHASA-N Ile-Gly-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)N[C@@H](CCC(=O)O)C(=O)O)N LPFBXFILACZHIB-LAEOZQHASA-N 0.000 description 1
- LBRCLQMZAHRTLV-ZKWXMUAHSA-N Ile-Gly-Ser Chemical compound CC[C@H](C)[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O LBRCLQMZAHRTLV-ZKWXMUAHSA-N 0.000 description 1
- UAQSZXGJGLHMNV-XEGUGMAKSA-N Ile-Gly-Tyr Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N UAQSZXGJGLHMNV-XEGUGMAKSA-N 0.000 description 1
- GLLAUPMJCGKPFY-BLMTYFJBSA-N Ile-Ile-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)[C@@H](C)CC)C(O)=O)=CNC2=C1 GLLAUPMJCGKPFY-BLMTYFJBSA-N 0.000 description 1
- HPCFRQWLTRDGHT-AJNGGQMLSA-N Ile-Leu-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O HPCFRQWLTRDGHT-AJNGGQMLSA-N 0.000 description 1
- PMMMQRVUMVURGJ-XUXIUFHCSA-N Ile-Leu-Pro Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(O)=O PMMMQRVUMVURGJ-XUXIUFHCSA-N 0.000 description 1
- ZUPJCJINYQISSN-XUXIUFHCSA-N Ile-Met-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCCN)C(=O)O)N ZUPJCJINYQISSN-XUXIUFHCSA-N 0.000 description 1
- SAVXZJYTTQQQDD-QEWYBTABSA-N Ile-Phe-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N SAVXZJYTTQQQDD-QEWYBTABSA-N 0.000 description 1
- ZDNNDIJTUHQCAM-MXAVVETBSA-N Ile-Ser-Phe Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N ZDNNDIJTUHQCAM-MXAVVETBSA-N 0.000 description 1
- SAEWJTCJQVZQNZ-IUKAMOBKSA-N Ile-Thr-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N SAEWJTCJQVZQNZ-IUKAMOBKSA-N 0.000 description 1
- WKSHBPRUIRGWRZ-KCTSRDHCSA-N Ile-Trp-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)NCC(=O)O)N WKSHBPRUIRGWRZ-KCTSRDHCSA-N 0.000 description 1
- DTPGSUQHUMELQB-GVARAGBVSA-N Ile-Tyr-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](C)C(O)=O)CC1=CC=C(O)C=C1 DTPGSUQHUMELQB-GVARAGBVSA-N 0.000 description 1
- GVEODXUBBFDBPW-MGHWNKPDSA-N Ile-Tyr-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=C(O)C=C1 GVEODXUBBFDBPW-MGHWNKPDSA-N 0.000 description 1
- ZGKVPOSSTGHJAF-HJPIBITLSA-N Ile-Tyr-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CO)C(=O)O)N ZGKVPOSSTGHJAF-HJPIBITLSA-N 0.000 description 1
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 1
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 1
- 102000018071 Immunoglobulin Fc Fragments Human genes 0.000 description 1
- 108010091135 Immunoglobulin Fc Fragments Proteins 0.000 description 1
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 1
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 description 1
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 description 1
- 206010062016 Immunosuppression Diseases 0.000 description 1
- 238000012695 Interfacial polymerization Methods 0.000 description 1
- PWWVAXIEGOYWEE-UHFFFAOYSA-N Isophenergan Chemical compound C1=CC=C2N(CC(C)N(C)C)C3=CC=CC=C3SC2=C1 PWWVAXIEGOYWEE-UHFFFAOYSA-N 0.000 description 1
- PMGDADKJMCOXHX-UHFFFAOYSA-N L-Arginyl-L-glutamin-acetat Natural products NC(=N)NCCCC(N)C(=O)NC(CCC(N)=O)C(O)=O PMGDADKJMCOXHX-UHFFFAOYSA-N 0.000 description 1
- SHZGCJCMOBCMKK-DHVFOXMCSA-N L-fucopyranose Chemical compound C[C@@H]1OC(O)[C@@H](O)[C@H](O)[C@@H]1O SHZGCJCMOBCMKK-DHVFOXMCSA-N 0.000 description 1
- LHSGPCFBGJHPCY-UHFFFAOYSA-N L-leucine-L-tyrosine Natural products CC(C)CC(N)C(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 LHSGPCFBGJHPCY-UHFFFAOYSA-N 0.000 description 1
- QPRQGENIBFLVEB-BJDJZHNGSA-N Leu-Ala-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O QPRQGENIBFLVEB-BJDJZHNGSA-N 0.000 description 1
- VCSBGUACOYUIGD-CIUDSAMLSA-N Leu-Asn-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O VCSBGUACOYUIGD-CIUDSAMLSA-N 0.000 description 1
- POJPZSMTTMLSTG-SRVKXCTJSA-N Leu-Asn-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N POJPZSMTTMLSTG-SRVKXCTJSA-N 0.000 description 1
- XVSJMWYYLHPDKY-DCAQKATOSA-N Leu-Asp-Met Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCSC)C(O)=O XVSJMWYYLHPDKY-DCAQKATOSA-N 0.000 description 1
- QCSFMCFHVGTLFF-NHCYSSNCSA-N Leu-Asp-Val Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O QCSFMCFHVGTLFF-NHCYSSNCSA-N 0.000 description 1
- AXZGZMGRBDQTEY-SRVKXCTJSA-N Leu-Gln-Met Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCSC)C(O)=O AXZGZMGRBDQTEY-SRVKXCTJSA-N 0.000 description 1
- QVFGXCVIXXBFHO-AVGNSLFASA-N Leu-Glu-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O QVFGXCVIXXBFHO-AVGNSLFASA-N 0.000 description 1
- HQUXQAMSWFIRET-AVGNSLFASA-N Leu-Glu-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CCCCN HQUXQAMSWFIRET-AVGNSLFASA-N 0.000 description 1
- FEHQLKKBVJHSEC-SZMVWBNQSA-N Leu-Glu-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC(C)C)C(O)=O)=CNC2=C1 FEHQLKKBVJHSEC-SZMVWBNQSA-N 0.000 description 1
- FMEICTQWUKNAGC-YUMQZZPRSA-N Leu-Gly-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O FMEICTQWUKNAGC-YUMQZZPRSA-N 0.000 description 1
- LAPSXOAUPNOINL-YUMQZZPRSA-N Leu-Gly-Asp Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC(O)=O LAPSXOAUPNOINL-YUMQZZPRSA-N 0.000 description 1
- PDQDCFBVYXEFSD-SRVKXCTJSA-N Leu-Leu-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O PDQDCFBVYXEFSD-SRVKXCTJSA-N 0.000 description 1
- QNBVTHNJGCOVFA-AVGNSLFASA-N Leu-Leu-Glu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CCC(O)=O QNBVTHNJGCOVFA-AVGNSLFASA-N 0.000 description 1
- YOKVEHGYYQEQOP-QWRGUYRKSA-N Leu-Leu-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O YOKVEHGYYQEQOP-QWRGUYRKSA-N 0.000 description 1
- REPBGZHJKYWFMJ-KKUMJFAQSA-N Leu-Lys-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N REPBGZHJKYWFMJ-KKUMJFAQSA-N 0.000 description 1
- LZHJZLHSRGWBBE-IHRRRGAJSA-N Leu-Lys-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O LZHJZLHSRGWBBE-IHRRRGAJSA-N 0.000 description 1
- YRRCOJOXAJNSAX-IHRRRGAJSA-N Leu-Pro-Lys Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)O)N YRRCOJOXAJNSAX-IHRRRGAJSA-N 0.000 description 1
- CHJKEDSZNSONPS-DCAQKATOSA-N Leu-Pro-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O CHJKEDSZNSONPS-DCAQKATOSA-N 0.000 description 1
- KIZIOFNVSOSKJI-CIUDSAMLSA-N Leu-Ser-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O)N KIZIOFNVSOSKJI-CIUDSAMLSA-N 0.000 description 1
- SVBJIZVVYJYGLA-DCAQKATOSA-N Leu-Ser-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O SVBJIZVVYJYGLA-DCAQKATOSA-N 0.000 description 1
- LJBVRCDPWOJOEK-PPCPHDFISA-N Leu-Thr-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O LJBVRCDPWOJOEK-PPCPHDFISA-N 0.000 description 1
- DAYQSYGBCUKVKT-VOAKCMCISA-N Leu-Thr-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(O)=O DAYQSYGBCUKVKT-VOAKCMCISA-N 0.000 description 1
- ISSAURVGLGAPDK-KKUMJFAQSA-N Leu-Tyr-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O ISSAURVGLGAPDK-KKUMJFAQSA-N 0.000 description 1
- VHTIZYYHIUHMCA-JYJNAYRXSA-N Leu-Tyr-Gln Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O VHTIZYYHIUHMCA-JYJNAYRXSA-N 0.000 description 1
- YIRIDPUGZKHMHT-ACRUOGEOSA-N Leu-Tyr-Tyr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O YIRIDPUGZKHMHT-ACRUOGEOSA-N 0.000 description 1
- QESXLSQLQHHTIX-RHYQMDGZSA-N Leu-Val-Thr Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QESXLSQLQHHTIX-RHYQMDGZSA-N 0.000 description 1
- 108010000817 Leuprolide Proteins 0.000 description 1
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 1
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- DDWFXDSYGUXRAY-UHFFFAOYSA-N Luciferin Natural products CCc1c(C)c(CC2NC(=O)C(=C2C=C)C)[nH]c1Cc3[nH]c4C(=C5/NC(CC(=O)O)C(C)C5CC(=O)O)CC(=O)c4c3C DDWFXDSYGUXRAY-UHFFFAOYSA-N 0.000 description 1
- 102000008072 Lymphokines Human genes 0.000 description 1
- 108010074338 Lymphokines Proteins 0.000 description 1
- GAOJCVKPIGHTGO-UWVGGRQHSA-N Lys-Arg-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O GAOJCVKPIGHTGO-UWVGGRQHSA-N 0.000 description 1
- NQCJGQHHYZNUDK-DCAQKATOSA-N Lys-Arg-Ser Chemical compound NCCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CO)C(O)=O)CCCN=C(N)N NQCJGQHHYZNUDK-DCAQKATOSA-N 0.000 description 1
- FACUGMGEFUEBTI-SRVKXCTJSA-N Lys-Asn-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CCCCN FACUGMGEFUEBTI-SRVKXCTJSA-N 0.000 description 1
- FLCMXEFCTLXBTL-DCAQKATOSA-N Lys-Asp-Arg Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N FLCMXEFCTLXBTL-DCAQKATOSA-N 0.000 description 1
- WGCKDDHUFPQSMZ-ZPFDUUQYSA-N Lys-Asp-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CCCCN WGCKDDHUFPQSMZ-ZPFDUUQYSA-N 0.000 description 1
- QIJVAFLRMVBHMU-KKUMJFAQSA-N Lys-Asp-Phe Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O QIJVAFLRMVBHMU-KKUMJFAQSA-N 0.000 description 1
- MLLKLNYPZRDIQG-GUBZILKMSA-N Lys-Cys-Gln Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N MLLKLNYPZRDIQG-GUBZILKMSA-N 0.000 description 1
- ZXEUFAVXODIPHC-GUBZILKMSA-N Lys-Glu-Asn Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ZXEUFAVXODIPHC-GUBZILKMSA-N 0.000 description 1
- GCMWRRQAKQXDED-IUCAKERBSA-N Lys-Glu-Gly Chemical compound [NH3+]CCCC[C@H]([NH3+])C(=O)N[C@@H](CCC([O-])=O)C(=O)NCC([O-])=O GCMWRRQAKQXDED-IUCAKERBSA-N 0.000 description 1
- DCRWPTBMWMGADO-AVGNSLFASA-N Lys-Glu-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O DCRWPTBMWMGADO-AVGNSLFASA-N 0.000 description 1
- DUTMKEAPLLUGNO-JYJNAYRXSA-N Lys-Glu-Phe Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O DUTMKEAPLLUGNO-JYJNAYRXSA-N 0.000 description 1
- GQZMPWBZQALKJO-UWVGGRQHSA-N Lys-Gly-Arg Chemical compound [H]N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(O)=O GQZMPWBZQALKJO-UWVGGRQHSA-N 0.000 description 1
- FHIAJWBDZVHLAH-YUMQZZPRSA-N Lys-Gly-Ser Chemical compound NCCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O FHIAJWBDZVHLAH-YUMQZZPRSA-N 0.000 description 1
- ZMMDPRTXLAEMOD-BZSNNMDCSA-N Lys-His-Phe Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O ZMMDPRTXLAEMOD-BZSNNMDCSA-N 0.000 description 1
- PRSBSVAVOQOAMI-BJDJZHNGSA-N Lys-Ile-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCCCN PRSBSVAVOQOAMI-BJDJZHNGSA-N 0.000 description 1
- PINHPJWGVBKQII-SRVKXCTJSA-N Lys-Leu-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCCN)N PINHPJWGVBKQII-SRVKXCTJSA-N 0.000 description 1
- WRODMZBHNNPRLN-SRVKXCTJSA-N Lys-Leu-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O WRODMZBHNNPRLN-SRVKXCTJSA-N 0.000 description 1
- LJADEBULDNKJNK-IHRRRGAJSA-N Lys-Leu-Val Chemical compound CC(C)C[C@H](NC(=O)[C@@H](N)CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O LJADEBULDNKJNK-IHRRRGAJSA-N 0.000 description 1
- GOVDTWNJCBRRBJ-DCAQKATOSA-N Lys-Met-Asn Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCCCN)N GOVDTWNJCBRRBJ-DCAQKATOSA-N 0.000 description 1
- ZCWWVXAXWUAEPZ-SRVKXCTJSA-N Lys-Met-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCC(O)=O)C(O)=O ZCWWVXAXWUAEPZ-SRVKXCTJSA-N 0.000 description 1
- JMNRXRPBHFGXQX-GUBZILKMSA-N Lys-Ser-Glu Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O JMNRXRPBHFGXQX-GUBZILKMSA-N 0.000 description 1
- JHNOXVASMSXSNB-WEDXCCLWSA-N Lys-Thr-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O JHNOXVASMSXSNB-WEDXCCLWSA-N 0.000 description 1
- XYLSGAWRCZECIQ-JYJNAYRXSA-N Lys-Tyr-Glu Chemical compound NCCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CCC(O)=O)C(O)=O)CC1=CC=C(O)C=C1 XYLSGAWRCZECIQ-JYJNAYRXSA-N 0.000 description 1
- FPQMQEOVSKMVMA-ACRUOGEOSA-N Lys-Tyr-Tyr Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)O)NC(=O)[C@H](CCCCN)N)O FPQMQEOVSKMVMA-ACRUOGEOSA-N 0.000 description 1
- OHXUUQDOBQKSNB-AVGNSLFASA-N Lys-Val-Arg Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O OHXUUQDOBQKSNB-AVGNSLFASA-N 0.000 description 1
- MDDUIRLQCYVRDO-NHCYSSNCSA-N Lys-Val-Asn Chemical compound NC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCCN MDDUIRLQCYVRDO-NHCYSSNCSA-N 0.000 description 1
- NYTDJEZBAAFLLG-IHRRRGAJSA-N Lys-Val-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(O)=O NYTDJEZBAAFLLG-IHRRRGAJSA-N 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 108091054438 MHC class II family Proteins 0.000 description 1
- 239000004907 Macro-emulsion Substances 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 241000712079 Measles morbillivirus Species 0.000 description 1
- 102000018897 Membrane Fusion Proteins Human genes 0.000 description 1
- 108010027796 Membrane Fusion Proteins Proteins 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 244000246386 Mentha pulegium Species 0.000 description 1
- 235000016257 Mentha pulegium Nutrition 0.000 description 1
- 235000004357 Mentha x piperita Nutrition 0.000 description 1
- QGQGAIBGTUJRBR-NAKRPEOUSA-N Met-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCSC QGQGAIBGTUJRBR-NAKRPEOUSA-N 0.000 description 1
- MTBVQFFQMXHCPC-CIUDSAMLSA-N Met-Glu-Asp Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O MTBVQFFQMXHCPC-CIUDSAMLSA-N 0.000 description 1
- RNAGAJXCSPDPRK-KKUMJFAQSA-N Met-Glu-Phe Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 RNAGAJXCSPDPRK-KKUMJFAQSA-N 0.000 description 1
- OSZTUONKUMCWEP-XUXIUFHCSA-N Met-Leu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC OSZTUONKUMCWEP-XUXIUFHCSA-N 0.000 description 1
- LCPUWQLULVXROY-RHYQMDGZSA-N Met-Lys-Thr Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LCPUWQLULVXROY-RHYQMDGZSA-N 0.000 description 1
- HNQXYIVNRUXQLU-BPUTZDHNSA-N Met-Trp-Asp Chemical compound CSCC[C@H](N)C(=O)N[C@@H](Cc1c[nH]c2ccccc12)C(=O)N[C@@H](CC(O)=O)C(O)=O HNQXYIVNRUXQLU-BPUTZDHNSA-N 0.000 description 1
- OVTOTTGZBWXLFU-QXEWZRGKSA-N Met-Val-Asp Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(O)=O OVTOTTGZBWXLFU-QXEWZRGKSA-N 0.000 description 1
- OTKQHDPECKUDSB-SZMVWBNQSA-N Met-Val-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCSC)C(O)=O)=CNC2=C1 OTKQHDPECKUDSB-SZMVWBNQSA-N 0.000 description 1
- 229920000168 Microcrystalline cellulose Polymers 0.000 description 1
- 244000302512 Momordica charantia Species 0.000 description 1
- 235000009811 Momordica charantia Nutrition 0.000 description 1
- 208000016285 Movement disease Diseases 0.000 description 1
- 229920000715 Mucilage Polymers 0.000 description 1
- 241000711386 Mumps virus Species 0.000 description 1
- 241000699660 Mus musculus Species 0.000 description 1
- YBAFDPFAUTYYRW-UHFFFAOYSA-N N-L-alpha-glutamyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=O YBAFDPFAUTYYRW-UHFFFAOYSA-N 0.000 description 1
- XZFYRXDAULDNFX-UHFFFAOYSA-N N-L-cysteinyl-L-phenylalanine Natural products SCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XZFYRXDAULDNFX-UHFFFAOYSA-N 0.000 description 1
- AJHCSUXXECOXOY-UHFFFAOYSA-N N-glycyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)CN)C(O)=O)=CNC2=C1 AJHCSUXXECOXOY-UHFFFAOYSA-N 0.000 description 1
- 206010060860 Neurological symptom Diseases 0.000 description 1
- 108090001074 Nucleocapsid Proteins Proteins 0.000 description 1
- 230000004989 O-glycosylation Effects 0.000 description 1
- 108020005187 Oligonucleotide Probes Proteins 0.000 description 1
- 108010038807 Oligopeptides Proteins 0.000 description 1
- 102000015636 Oligopeptides Human genes 0.000 description 1
- 241000712464 Orthomyxoviridae Species 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 206010033799 Paralysis Diseases 0.000 description 1
- 208000002606 Paramyxoviridae Infections Diseases 0.000 description 1
- 102000057297 Pepsin A Human genes 0.000 description 1
- 108090000284 Pepsin A Proteins 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 108010067902 Peptide Library Proteins 0.000 description 1
- OJUMUUXGSXUZJZ-SRVKXCTJSA-N Phe-Asp-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O OJUMUUXGSXUZJZ-SRVKXCTJSA-N 0.000 description 1
- CUMXHKAOHNWRFQ-BZSNNMDCSA-N Phe-Asp-Tyr Chemical compound C([C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=CC=C1 CUMXHKAOHNWRFQ-BZSNNMDCSA-N 0.000 description 1
- OYQBFWWQSVIHBN-FHWLQOOXSA-N Phe-Glu-Phe Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O OYQBFWWQSVIHBN-FHWLQOOXSA-N 0.000 description 1
- JJHVFCUWLSKADD-ONGXEEELSA-N Phe-Gly-Ala Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)NCC(=O)N[C@@H](C)C(O)=O JJHVFCUWLSKADD-ONGXEEELSA-N 0.000 description 1
- RVRRHFPCEOVRKQ-KKUMJFAQSA-N Phe-His-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)N[C@@H](CC(=O)N)C(=O)O)N RVRRHFPCEOVRKQ-KKUMJFAQSA-N 0.000 description 1
- FXYXBEZMRACDDR-KKUMJFAQSA-N Phe-His-Asp Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(O)=O)C(O)=O FXYXBEZMRACDDR-KKUMJFAQSA-N 0.000 description 1
- ZUQACJLOHYRVPJ-DKIMLUQUSA-N Phe-Lys-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CC1=CC=CC=C1 ZUQACJLOHYRVPJ-DKIMLUQUSA-N 0.000 description 1
- OXKJSGGTHFMGDT-UFYCRDLUSA-N Phe-Phe-Arg Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C1=CC=CC=C1 OXKJSGGTHFMGDT-UFYCRDLUSA-N 0.000 description 1
- GMWNQSGWWGKTSF-LFSVMHDDSA-N Phe-Thr-Ala Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O GMWNQSGWWGKTSF-LFSVMHDDSA-N 0.000 description 1
- ZVJGAXNBBKPYOE-HKUYNNGSSA-N Phe-Trp-Gly Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)NCC(O)=O)C1=CC=CC=C1 ZVJGAXNBBKPYOE-HKUYNNGSSA-N 0.000 description 1
- WSAPMHXTQAOAQQ-BVSLBCMMSA-N Phe-Trp-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CC3=CC=CC=C3)N WSAPMHXTQAOAQQ-BVSLBCMMSA-N 0.000 description 1
- XBCOOBCTVMMQSC-BVSLBCMMSA-N Phe-Val-Trp Chemical compound C([C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)C1=CC=CC=C1 XBCOOBCTVMMQSC-BVSLBCMMSA-N 0.000 description 1
- 108010053210 Phycocyanin Proteins 0.000 description 1
- 101100413173 Phytolacca americana PAP2 gene Proteins 0.000 description 1
- 241000276498 Pollachius virens Species 0.000 description 1
- 229920002732 Polyanhydride Polymers 0.000 description 1
- 229920000954 Polyglycolide Polymers 0.000 description 1
- 108010039918 Polylysine Proteins 0.000 description 1
- 229920001710 Polyorthoester Polymers 0.000 description 1
- 229920002685 Polyoxyl 35CastorOil Polymers 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 241000288906 Primates Species 0.000 description 1
- SSSFPISOZOLQNP-GUBZILKMSA-N Pro-Arg-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O SSSFPISOZOLQNP-GUBZILKMSA-N 0.000 description 1
- QCARZLHECSFOGG-CIUDSAMLSA-N Pro-Glu-Cys Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CS)C(=O)O QCARZLHECSFOGG-CIUDSAMLSA-N 0.000 description 1
- WVOXLKUUVCCCSU-ZPFDUUQYSA-N Pro-Glu-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WVOXLKUUVCCCSU-ZPFDUUQYSA-N 0.000 description 1
- ZTVCLZLGHZXLOT-ULQDDVLXSA-N Pro-Glu-Trp Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC2=CNC3=CC=CC=C32)C(=O)O ZTVCLZLGHZXLOT-ULQDDVLXSA-N 0.000 description 1
- AQGUSRZKDZYGGV-GMOBBJLQSA-N Pro-Ile-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O AQGUSRZKDZYGGV-GMOBBJLQSA-N 0.000 description 1
- HFNPOYOKIPGAEI-SRVKXCTJSA-N Pro-Leu-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 HFNPOYOKIPGAEI-SRVKXCTJSA-N 0.000 description 1
- FXGIMYRVJJEIIM-UWVGGRQHSA-N Pro-Leu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 FXGIMYRVJJEIIM-UWVGGRQHSA-N 0.000 description 1
- VTFXTWDFPTWNJY-RHYQMDGZSA-N Pro-Leu-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VTFXTWDFPTWNJY-RHYQMDGZSA-N 0.000 description 1
- GNADVDLLGVSXLS-ULQDDVLXSA-N Pro-Phe-His Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CNC=N1)C(O)=O GNADVDLLGVSXLS-ULQDDVLXSA-N 0.000 description 1
- SNGZLPOXVRTNMB-LPEHRKFASA-N Pro-Ser-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CO)C(=O)N2CCC[C@@H]2C(=O)O SNGZLPOXVRTNMB-LPEHRKFASA-N 0.000 description 1
- QDDJNKWPTJHROJ-UFYCRDLUSA-N Pro-Tyr-Tyr Chemical compound C([C@@H](C(=O)O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H]1NCCC1)C1=CC=C(O)C=C1 QDDJNKWPTJHROJ-UFYCRDLUSA-N 0.000 description 1
- XDKKMRPRRCOELJ-GUBZILKMSA-N Pro-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H]1CCCN1 XDKKMRPRRCOELJ-GUBZILKMSA-N 0.000 description 1
- 241001415846 Procellariidae Species 0.000 description 1
- 102000007327 Protamines Human genes 0.000 description 1
- 108010007568 Protamines Proteins 0.000 description 1
- 241000589517 Pseudomonas aeruginosa Species 0.000 description 1
- ZVOLCUVKHLEPEV-UHFFFAOYSA-N Quercetagetin Natural products C1=C(O)C(O)=CC=C1C1=C(O)C(=O)C2=C(O)C(O)=C(O)C=C2O1 ZVOLCUVKHLEPEV-UHFFFAOYSA-N 0.000 description 1
- 241000711798 Rabies lyssavirus Species 0.000 description 1
- 208000035415 Reinfection Diseases 0.000 description 1
- 241000725643 Respiratory syncytial virus Species 0.000 description 1
- 108091027981 Response element Proteins 0.000 description 1
- HWTZYBCRDDUBJY-UHFFFAOYSA-N Rhynchosin Natural products C1=C(O)C(O)=CC=C1C1=C(O)C(=O)C2=CC(O)=C(O)C=C2O1 HWTZYBCRDDUBJY-UHFFFAOYSA-N 0.000 description 1
- 102000004389 Ribonucleoproteins Human genes 0.000 description 1
- 108010081734 Ribonucleoproteins Proteins 0.000 description 1
- 108010039491 Ricin Proteins 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 1
- 241000710799 Rubella virus Species 0.000 description 1
- QEDMOZUJTGEIBF-FXQIFTODSA-N Ser-Arg-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O QEDMOZUJTGEIBF-FXQIFTODSA-N 0.000 description 1
- VAUMZJHYZQXZBQ-WHFBIAKZSA-N Ser-Asn-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O VAUMZJHYZQXZBQ-WHFBIAKZSA-N 0.000 description 1
- VGNYHOBZJKWRGI-CIUDSAMLSA-N Ser-Asn-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CO VGNYHOBZJKWRGI-CIUDSAMLSA-N 0.000 description 1
- TYYBJUYSTWJHGO-ZKWXMUAHSA-N Ser-Asn-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O TYYBJUYSTWJHGO-ZKWXMUAHSA-N 0.000 description 1
- HEQPKICPPDOSIN-SRVKXCTJSA-N Ser-Asp-Tyr Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 HEQPKICPPDOSIN-SRVKXCTJSA-N 0.000 description 1
- MIJWOJAXARLEHA-WDSKDSINSA-N Ser-Gly-Glu Chemical compound OC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(O)=O MIJWOJAXARLEHA-WDSKDSINSA-N 0.000 description 1
- YMTLKLXDFCSCNX-BYPYZUCNSA-N Ser-Gly-Gly Chemical compound OC[C@H](N)C(=O)NCC(=O)NCC(O)=O YMTLKLXDFCSCNX-BYPYZUCNSA-N 0.000 description 1
- IOVHBRCQOGWAQH-ZKWXMUAHSA-N Ser-Gly-Ile Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H]([C@@H](C)CC)C(O)=O IOVHBRCQOGWAQH-ZKWXMUAHSA-N 0.000 description 1
- OQPNSDWGAMFJNU-QWRGUYRKSA-N Ser-Gly-Tyr Chemical compound OC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 OQPNSDWGAMFJNU-QWRGUYRKSA-N 0.000 description 1
- SFTZTYBXIXLRGQ-JBDRJPRFSA-N Ser-Ile-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O SFTZTYBXIXLRGQ-JBDRJPRFSA-N 0.000 description 1
- FUMGHWDRRFCKEP-CIUDSAMLSA-N Ser-Leu-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O FUMGHWDRRFCKEP-CIUDSAMLSA-N 0.000 description 1
- GJFYFGOEWLDQGW-GUBZILKMSA-N Ser-Leu-Gln Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CO)N GJFYFGOEWLDQGW-GUBZILKMSA-N 0.000 description 1
- YUJLIIRMIAGMCQ-CIUDSAMLSA-N Ser-Leu-Ser Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O YUJLIIRMIAGMCQ-CIUDSAMLSA-N 0.000 description 1
- HDBOEVPDIDDEPC-CIUDSAMLSA-N Ser-Lys-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(O)=O HDBOEVPDIDDEPC-CIUDSAMLSA-N 0.000 description 1
- XUDRHBPSPAPDJP-SRVKXCTJSA-N Ser-Lys-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CO XUDRHBPSPAPDJP-SRVKXCTJSA-N 0.000 description 1
- VXYQOFXBIXKPCX-BQBZGAKWSA-N Ser-Met-Gly Chemical compound CSCC[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CO)N VXYQOFXBIXKPCX-BQBZGAKWSA-N 0.000 description 1
- GDUZTEQRAOXYJS-SRVKXCTJSA-N Ser-Phe-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CO)N GDUZTEQRAOXYJS-SRVKXCTJSA-N 0.000 description 1
- UGTZYIPOBYXWRW-SRVKXCTJSA-N Ser-Phe-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O UGTZYIPOBYXWRW-SRVKXCTJSA-N 0.000 description 1
- RRVFEDGUXSYWOW-BZSNNMDCSA-N Ser-Phe-Phe Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O RRVFEDGUXSYWOW-BZSNNMDCSA-N 0.000 description 1
- XZKQVQKUZMAADP-IMJSIDKUSA-N Ser-Ser Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(O)=O XZKQVQKUZMAADP-IMJSIDKUSA-N 0.000 description 1
- SRSPTFBENMJHMR-WHFBIAKZSA-N Ser-Ser-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O SRSPTFBENMJHMR-WHFBIAKZSA-N 0.000 description 1
- AABIBDJHSKIMJK-FXQIFTODSA-N Ser-Ser-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCSC)C(O)=O AABIBDJHSKIMJK-FXQIFTODSA-N 0.000 description 1
- VGQVAVQWKJLIRM-FXQIFTODSA-N Ser-Ser-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O VGQVAVQWKJLIRM-FXQIFTODSA-N 0.000 description 1
- PURRNJBBXDDWLX-ZDLURKLDSA-N Ser-Thr-Gly Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CO)N)O PURRNJBBXDDWLX-ZDLURKLDSA-N 0.000 description 1
- FLMYSKVSDVHLEW-SVSWQMSJSA-N Ser-Thr-Ile Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O FLMYSKVSDVHLEW-SVSWQMSJSA-N 0.000 description 1
- NADLKBTYNKUJEP-KATARQTJSA-N Ser-Thr-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O NADLKBTYNKUJEP-KATARQTJSA-N 0.000 description 1
- NERYDXBVARJIQS-JYBASQMISA-N Ser-Trp-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CO)N)O NERYDXBVARJIQS-JYBASQMISA-N 0.000 description 1
- PQEQXWRVHQAAKS-SRVKXCTJSA-N Ser-Tyr-Asn Chemical compound NC(=O)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CO)N)CC1=CC=C(O)C=C1 PQEQXWRVHQAAKS-SRVKXCTJSA-N 0.000 description 1
- UBTNVMGPMYDYIU-HJPIBITLSA-N Ser-Tyr-Ile Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O UBTNVMGPMYDYIU-HJPIBITLSA-N 0.000 description 1
- MFQMZDPAZRZAPV-NAKRPEOUSA-N Ser-Val-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CO)N MFQMZDPAZRZAPV-NAKRPEOUSA-N 0.000 description 1
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 1
- DWAQJAXMDSEUJJ-UHFFFAOYSA-M Sodium bisulfite Chemical compound [Na+].OS([O-])=O DWAQJAXMDSEUJJ-UHFFFAOYSA-M 0.000 description 1
- 238000002105 Southern blotting Methods 0.000 description 1
- 241000269319 Squalius cephalus Species 0.000 description 1
- 229920002472 Starch Polymers 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 230000024932 T cell mediated immunity Effects 0.000 description 1
- 108091008874 T cell receptors Proteins 0.000 description 1
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 1
- 102100036011 T-cell surface glycoprotein CD4 Human genes 0.000 description 1
- 210000001744 T-lymphocyte Anatomy 0.000 description 1
- 102100030904 Target of rapamycin complex 2 subunit MAPKAP1 Human genes 0.000 description 1
- ZMZDMBWJUHKJPS-UHFFFAOYSA-M Thiocyanate anion Chemical compound [S-]C#N ZMZDMBWJUHKJPS-UHFFFAOYSA-M 0.000 description 1
- XSLXHSYIVPGEER-KZVJFYERSA-N Thr-Ala-Val Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(O)=O XSLXHSYIVPGEER-KZVJFYERSA-N 0.000 description 1
- JTEICXDKGWKRRV-HJGDQZAQSA-N Thr-Asn-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N)O JTEICXDKGWKRRV-HJGDQZAQSA-N 0.000 description 1
- OJRNZRROAIAHDL-LKXGYXEUSA-N Thr-Asn-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O OJRNZRROAIAHDL-LKXGYXEUSA-N 0.000 description 1
- GUZGCDIZVGODML-NKIYYHGXSA-N Thr-Gln-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N)O GUZGCDIZVGODML-NKIYYHGXSA-N 0.000 description 1
- GKWNLDNXMMLRMC-GLLZPBPUSA-N Thr-Glu-Gln Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N)O GKWNLDNXMMLRMC-GLLZPBPUSA-N 0.000 description 1
- SHOMROOOQBDGRL-JHEQGTHGSA-N Thr-Glu-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O SHOMROOOQBDGRL-JHEQGTHGSA-N 0.000 description 1
- HJOSVGCWOTYJFG-WDCWCFNPSA-N Thr-Glu-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N)O HJOSVGCWOTYJFG-WDCWCFNPSA-N 0.000 description 1
- SIMKLINEDYOTKL-MBLNEYKQSA-N Thr-His-Ala Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](C)C(=O)O)N)O SIMKLINEDYOTKL-MBLNEYKQSA-N 0.000 description 1
- VUSAEKOXGNEYNE-PBCZWWQYSA-N Thr-His-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)O VUSAEKOXGNEYNE-PBCZWWQYSA-N 0.000 description 1
- PAXANSWUSVPFNK-IUKAMOBKSA-N Thr-Ile-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N PAXANSWUSVPFNK-IUKAMOBKSA-N 0.000 description 1
- ADPHPKGWVDHWML-PPCPHDFISA-N Thr-Ile-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N ADPHPKGWVDHWML-PPCPHDFISA-N 0.000 description 1
- FQPDRTDDEZXCEC-SVSWQMSJSA-N Thr-Ile-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O FQPDRTDDEZXCEC-SVSWQMSJSA-N 0.000 description 1
- IMDMLDSVUSMAEJ-HJGDQZAQSA-N Thr-Leu-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O IMDMLDSVUSMAEJ-HJGDQZAQSA-N 0.000 description 1
- RRRRCRYTLZVCEN-HJGDQZAQSA-N Thr-Leu-Asp Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O RRRRCRYTLZVCEN-HJGDQZAQSA-N 0.000 description 1
- VTVVYQOXJCZVEB-WDCWCFNPSA-N Thr-Leu-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O VTVVYQOXJCZVEB-WDCWCFNPSA-N 0.000 description 1
- IJVNLNRVDUTWDD-MEYUZBJRSA-N Thr-Leu-Tyr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O IJVNLNRVDUTWDD-MEYUZBJRSA-N 0.000 description 1
- XKWABWFMQXMUMT-HJGDQZAQSA-N Thr-Pro-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O XKWABWFMQXMUMT-HJGDQZAQSA-N 0.000 description 1
- DEGCBBCMYWNJNA-RHYQMDGZSA-N Thr-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)[C@@H](C)O DEGCBBCMYWNJNA-RHYQMDGZSA-N 0.000 description 1
- BBPCSGKKPJUYRB-UVOCVTCTSA-N Thr-Thr-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O BBPCSGKKPJUYRB-UVOCVTCTSA-N 0.000 description 1
- COYHRQWNJDJCNA-NUJDXYNKSA-N Thr-Thr-Thr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O COYHRQWNJDJCNA-NUJDXYNKSA-N 0.000 description 1
- OGOYMQWIWHGTGH-KZVJFYERSA-N Thr-Val-Ala Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O OGOYMQWIWHGTGH-KZVJFYERSA-N 0.000 description 1
- BKIOKSLLAAZYTC-KKHAAJSZSA-N Thr-Val-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O BKIOKSLLAAZYTC-KKHAAJSZSA-N 0.000 description 1
- 101710120037 Toxin CcdB Proteins 0.000 description 1
- 229920001615 Tragacanth Polymers 0.000 description 1
- 102000004357 Transferases Human genes 0.000 description 1
- 108090000992 Transferases Proteins 0.000 description 1
- 235000021307 Triticum Nutrition 0.000 description 1
- 244000098338 Triticum aestivum Species 0.000 description 1
- IQGJAHMZWBTRIF-UBHSHLNASA-N Trp-Asp-Asn Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N IQGJAHMZWBTRIF-UBHSHLNASA-N 0.000 description 1
- VEYXZZGMIBKXCN-UBHSHLNASA-N Trp-Asp-Asp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N VEYXZZGMIBKXCN-UBHSHLNASA-N 0.000 description 1
- NKUIXQOJUAEIET-AQZXSJQPSA-N Trp-Asp-Thr Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@H](O)C)C(O)=O)=CNC2=C1 NKUIXQOJUAEIET-AQZXSJQPSA-N 0.000 description 1
- WPSYJHFHZYJXMW-JSGCOSHPSA-N Trp-Gln-Gly Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O WPSYJHFHZYJXMW-JSGCOSHPSA-N 0.000 description 1
- UKWSFUSPGPBJGU-VFAJRCTISA-N Trp-Leu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N)O UKWSFUSPGPBJGU-VFAJRCTISA-N 0.000 description 1
- KYWBVMKEYAEDIX-BPUTZDHNSA-N Trp-Met-Cys Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CS)C(O)=O)=CNC2=C1 KYWBVMKEYAEDIX-BPUTZDHNSA-N 0.000 description 1
- LVTKHGUGBGNBPL-UHFFFAOYSA-N Trp-P-1 Chemical compound N1C2=CC=CC=C2C2=C1C(C)=C(N)N=C2C LVTKHGUGBGNBPL-UHFFFAOYSA-N 0.000 description 1
- ABRICLFKFRFDKS-IHPCNDPISA-N Trp-Ser-Tyr Chemical compound C([C@H](NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)N)C(O)=O)C1=CC=C(O)C=C1 ABRICLFKFRFDKS-IHPCNDPISA-N 0.000 description 1
- JTMZSIRTZKLBOA-NWLDYVSISA-N Trp-Thr-Gln Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(O)=O JTMZSIRTZKLBOA-NWLDYVSISA-N 0.000 description 1
- HTGJDTPQYFMKNC-VFAJRCTISA-N Trp-Thr-Leu Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)[C@@H](C)O)=CNC2=C1 HTGJDTPQYFMKNC-VFAJRCTISA-N 0.000 description 1
- PKZIWSHDJYIPRH-JBACZVJFSA-N Trp-Tyr-Gln Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O PKZIWSHDJYIPRH-JBACZVJFSA-N 0.000 description 1
- MBLJBGZWLHTJBH-SZMVWBNQSA-N Trp-Val-Arg Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O)=CNC2=C1 MBLJBGZWLHTJBH-SZMVWBNQSA-N 0.000 description 1
- NOXKHHXSHQFSGJ-FQPOAREZSA-N Tyr-Ala-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 NOXKHHXSHQFSGJ-FQPOAREZSA-N 0.000 description 1
- JWHOIHCOHMZSAR-QWRGUYRKSA-N Tyr-Asp-Gly Chemical compound OC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 JWHOIHCOHMZSAR-QWRGUYRKSA-N 0.000 description 1
- JFDGVHXRCKEBAU-KKUMJFAQSA-N Tyr-Asp-Lys Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N)O JFDGVHXRCKEBAU-KKUMJFAQSA-N 0.000 description 1
- CRHFOYCJGVJPLE-AVGNSLFASA-N Tyr-Gln-Asn Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)O CRHFOYCJGVJPLE-AVGNSLFASA-N 0.000 description 1
- WVRUKYLYMFGKAN-IHRRRGAJSA-N Tyr-Glu-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 WVRUKYLYMFGKAN-IHRRRGAJSA-N 0.000 description 1
- NOOMDULIORCDNF-IRXDYDNUSA-N Tyr-Gly-Phe Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O NOOMDULIORCDNF-IRXDYDNUSA-N 0.000 description 1
- KEANSLVUGJADPN-LKTVYLICSA-N Tyr-His-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CC2=CC=C(C=C2)O)N KEANSLVUGJADPN-LKTVYLICSA-N 0.000 description 1
- SZEIFUXUTBBQFQ-STQMWFEESA-N Tyr-Pro-Gly Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O SZEIFUXUTBBQFQ-STQMWFEESA-N 0.000 description 1
- MWUYSCVVPVITMW-IGNZVWTISA-N Tyr-Tyr-Ala Chemical compound C([C@@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@@H](N)CC=1C=CC(O)=CC=1)C1=CC=C(O)C=C1 MWUYSCVVPVITMW-IGNZVWTISA-N 0.000 description 1
- TYGHOWWWMTWVKM-HJOGWXRNSA-N Tyr-Tyr-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=C(O)C=C1 TYGHOWWWMTWVKM-HJOGWXRNSA-N 0.000 description 1
- RVGVIWNHABGIFH-IHRRRGAJSA-N Tyr-Val-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O RVGVIWNHABGIFH-IHRRRGAJSA-N 0.000 description 1
- ASQFIHTXXMFENG-XPUUQOCRSA-N Val-Ala-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O ASQFIHTXXMFENG-XPUUQOCRSA-N 0.000 description 1
- RUCNAYOMFXRIKJ-DCAQKATOSA-N Val-Ala-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN RUCNAYOMFXRIKJ-DCAQKATOSA-N 0.000 description 1
- COYSIHFOCOMGCF-WPRPVWTQSA-N Val-Arg-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@H](C(=O)NCC(O)=O)CCCN=C(N)N COYSIHFOCOMGCF-WPRPVWTQSA-N 0.000 description 1
- COYSIHFOCOMGCF-UHFFFAOYSA-N Val-Arg-Gly Natural products CC(C)C(N)C(=O)NC(C(=O)NCC(O)=O)CCCN=C(N)N COYSIHFOCOMGCF-UHFFFAOYSA-N 0.000 description 1
- UDNYEPLJTRDMEJ-RCOVLWMOSA-N Val-Asn-Gly Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)NCC(=O)O)N UDNYEPLJTRDMEJ-RCOVLWMOSA-N 0.000 description 1
- JLFKWDAZBRYCGX-ZKWXMUAHSA-N Val-Asn-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CO)C(=O)O)N JLFKWDAZBRYCGX-ZKWXMUAHSA-N 0.000 description 1
- DHINLYMWMXQGMQ-IHRRRGAJSA-N Val-His-His Chemical compound C([C@H](NC(=O)[C@@H](N)C(C)C)C(=O)N[C@@H](CC=1NC=NC=1)C(O)=O)C1=CN=CN1 DHINLYMWMXQGMQ-IHRRRGAJSA-N 0.000 description 1
- YTUABZMPYKCWCQ-XQQFMLRXSA-N Val-His-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N2CCC[C@@H]2C(=O)O)N YTUABZMPYKCWCQ-XQQFMLRXSA-N 0.000 description 1
- VXDSPJJQUQDCKH-UKJIMTQDSA-N Val-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](C(C)C)N VXDSPJJQUQDCKH-UKJIMTQDSA-N 0.000 description 1
- OVBMCNDKCWAXMZ-NAKRPEOUSA-N Val-Ile-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](C(C)C)N OVBMCNDKCWAXMZ-NAKRPEOUSA-N 0.000 description 1
- XXWBHOWRARMUOC-NHCYSSNCSA-N Val-Lys-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(=O)N)C(=O)O)N XXWBHOWRARMUOC-NHCYSSNCSA-N 0.000 description 1
- QRVPEKJBBRYISE-XUXIUFHCSA-N Val-Lys-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C(C)C)N QRVPEKJBBRYISE-XUXIUFHCSA-N 0.000 description 1
- ZRSZTKTVPNSUNA-IHRRRGAJSA-N Val-Lys-Leu Chemical compound CC(C)C[C@H](NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)C(C)C)C(O)=O ZRSZTKTVPNSUNA-IHRRRGAJSA-N 0.000 description 1
- JAKHAONCJJZVHT-DCAQKATOSA-N Val-Lys-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)O)N JAKHAONCJJZVHT-DCAQKATOSA-N 0.000 description 1
- RYQUMYBMOJYYDK-NHCYSSNCSA-N Val-Pro-Glu Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(=O)O)C(=O)O)N RYQUMYBMOJYYDK-NHCYSSNCSA-N 0.000 description 1
- UGFMVXRXULGLNO-XPUUQOCRSA-N Val-Ser-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O UGFMVXRXULGLNO-XPUUQOCRSA-N 0.000 description 1
- NZYNRRGJJVSSTJ-GUBZILKMSA-N Val-Ser-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O NZYNRRGJJVSSTJ-GUBZILKMSA-N 0.000 description 1
- TVGWMCTYUFBXAP-QTKMDUPCSA-N Val-Thr-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](C(C)C)N)O TVGWMCTYUFBXAP-QTKMDUPCSA-N 0.000 description 1
- HTONZBWRYUKUKC-RCWTZXSCSA-N Val-Thr-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O HTONZBWRYUKUKC-RCWTZXSCSA-N 0.000 description 1
- DOBHJKVVACOQTN-DZKIICNBSA-N Val-Tyr-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)CC1=CC=C(O)C=C1 DOBHJKVVACOQTN-DZKIICNBSA-N 0.000 description 1
- 240000001866 Vernicia fordii Species 0.000 description 1
- 229940118555 Viral entry inhibitor Drugs 0.000 description 1
- 238000012452 Xenomouse strains Methods 0.000 description 1
- 239000002250 absorbent Substances 0.000 description 1
- 230000002745 absorbent Effects 0.000 description 1
- 239000003070 absorption delaying agent Substances 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- OFLXLNCGODUUOT-UHFFFAOYSA-N acetohydrazide Chemical class C\C(O)=N\N OFLXLNCGODUUOT-UHFFFAOYSA-N 0.000 description 1
- 229940022698 acetylcholinesterase Drugs 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 230000006978 adaptation Effects 0.000 description 1
- 239000000853 adhesive Substances 0.000 description 1
- 239000000443 aerosol Substances 0.000 description 1
- 108010008685 alanyl-glutamyl-aspartic acid Proteins 0.000 description 1
- 108010069020 alanyl-prolyl-glycine Proteins 0.000 description 1
- 108010011559 alanylphenylalanine Proteins 0.000 description 1
- 150000001299 aldehydes Chemical class 0.000 description 1
- 239000000783 alginic acid Substances 0.000 description 1
- 235000010443 alginic acid Nutrition 0.000 description 1
- 229920000615 alginic acid Polymers 0.000 description 1
- 229960001126 alginic acid Drugs 0.000 description 1
- 150000004781 alginic acids Chemical class 0.000 description 1
- 125000001931 aliphatic group Chemical group 0.000 description 1
- 125000005599 alkyl carboxylate group Chemical group 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 229960003896 aminopterin Drugs 0.000 description 1
- 230000000840 anti-viral effect Effects 0.000 description 1
- 230000005875 antibody response Effects 0.000 description 1
- 239000003963 antioxidant agent Substances 0.000 description 1
- 235000006708 antioxidants Nutrition 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- 108010008355 arginyl-glutamine Proteins 0.000 description 1
- 108010072041 arginyl-glycyl-aspartic acid Proteins 0.000 description 1
- 108010068380 arginylarginine Proteins 0.000 description 1
- 210000001106 artificial yeast chromosome Anatomy 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 108010069205 aspartyl-phenylalanine Proteins 0.000 description 1
- 108010038633 aspartylglutamate Proteins 0.000 description 1
- 108010092854 aspartyllysine Proteins 0.000 description 1
- 230000002238 attenuated effect Effects 0.000 description 1
- DMLAVOWQYNRWNQ-UHFFFAOYSA-N azobenzene Chemical compound C1=CC=CC=C1N=NC1=CC=CC=C1 DMLAVOWQYNRWNQ-UHFFFAOYSA-N 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 210000003323 beak Anatomy 0.000 description 1
- 235000019445 benzyl alcohol Nutrition 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 108010005774 beta-Galactosidase Proteins 0.000 description 1
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 1
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 239000003833 bile salt Substances 0.000 description 1
- 229940093761 bile salts Drugs 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 238000004166 bioassay Methods 0.000 description 1
- 229920000249 biocompatible polymer Polymers 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 238000001574 biopsy Methods 0.000 description 1
- 238000007413 biotinylation Methods 0.000 description 1
- 230000006287 biotinylation Effects 0.000 description 1
- 238000009534 blood test Methods 0.000 description 1
- 230000004579 body weight change Effects 0.000 description 1
- 210000004556 brain Anatomy 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 239000001569 carbon dioxide Substances 0.000 description 1
- 229910002092 carbon dioxide Inorganic materials 0.000 description 1
- 238000000423 cell based assay Methods 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 230000030833 cell death Effects 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 230000001413 cellular effect Effects 0.000 description 1
- 230000036755 cellular response Effects 0.000 description 1
- 239000003638 chemical reducing agent Substances 0.000 description 1
- 229960004926 chlorobutanol Drugs 0.000 description 1
- 235000012000 cholesterol Nutrition 0.000 description 1
- 230000004087 circulation Effects 0.000 description 1
- 238000005354 coacervation Methods 0.000 description 1
- 229940110456 cocoa butter Drugs 0.000 description 1
- 235000019868 cocoa butter Nutrition 0.000 description 1
- 229920001436 collagen Polymers 0.000 description 1
- 229940075614 colloidal silicon dioxide Drugs 0.000 description 1
- 238000012875 competitive assay Methods 0.000 description 1
- 238000010668 complexation reaction Methods 0.000 description 1
- 239000002131 composite material Substances 0.000 description 1
- 238000013329 compounding Methods 0.000 description 1
- 238000009833 condensation Methods 0.000 description 1
- 230000005494 condensation Effects 0.000 description 1
- 235000009508 confectionery Nutrition 0.000 description 1
- 229940031670 conjugate vaccine Drugs 0.000 description 1
- 230000001268 conjugating effect Effects 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 238000013270 controlled release Methods 0.000 description 1
- 239000008120 corn starch Substances 0.000 description 1
- 239000006071 cream Substances 0.000 description 1
- 238000004132 cross linking Methods 0.000 description 1
- 239000003431 cross linking reagent Substances 0.000 description 1
- 238000012136 culture method Methods 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- 108010060199 cysteinylproline Proteins 0.000 description 1
- 108010069495 cysteinyltyrosine Proteins 0.000 description 1
- 238000007405 data analysis Methods 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- 230000001934 delay Effects 0.000 description 1
- 230000003111 delayed effect Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- 239000008121 dextrose Substances 0.000 description 1
- 238000002405 diagnostic procedure Methods 0.000 description 1
- 238000000502 dialysis Methods 0.000 description 1
- 229930191339 dianthin Natural products 0.000 description 1
- UGMCXQCYOVCMTB-UHFFFAOYSA-K dihydroxy(stearato)aluminium Chemical compound CCCCCCCCCCCCCCCCCC(=O)O[Al](O)O UGMCXQCYOVCMTB-UHFFFAOYSA-K 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 125000000118 dimethyl group Chemical group [H]C([H])([H])* 0.000 description 1
- IYNJILVYWCLNBH-UHFFFAOYSA-N dimethylazanium;2-hydroxypropane-1-sulfonate Chemical compound C[NH2+]C.CC(O)CS([O-])(=O)=O IYNJILVYWCLNBH-UHFFFAOYSA-N 0.000 description 1
- 206010013023 diphtheria Diseases 0.000 description 1
- 150000002016 disaccharides Chemical class 0.000 description 1
- 239000007884 disintegrant Substances 0.000 description 1
- 238000004090 dissolution Methods 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- ZWIBGKZDAWNIFC-UHFFFAOYSA-N disuccinimidyl suberate Chemical compound O=C1CCC(=O)N1OC(=O)CCCCCCC(=O)ON1C(=O)CCC1=O ZWIBGKZDAWNIFC-UHFFFAOYSA-N 0.000 description 1
- 231100000673 dose–response relationship Toxicity 0.000 description 1
- 238000009509 drug development Methods 0.000 description 1
- 241001493065 dsRNA viruses Species 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 239000007920 enema Substances 0.000 description 1
- 229940079360 enema for constipation Drugs 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- 150000002148 esters Chemical class 0.000 description 1
- RTZKZFJDLAIYFH-UHFFFAOYSA-N ether Substances CCOCC RTZKZFJDLAIYFH-UHFFFAOYSA-N 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 238000001704 evaporation Methods 0.000 description 1
- 239000000284 extract Substances 0.000 description 1
- 239000012997 ficoll-paque Substances 0.000 description 1
- 239000000796 flavoring agent Substances 0.000 description 1
- ZFKJVJIDPQDDFY-UHFFFAOYSA-N fluorescamine Chemical compound C12=CC=CC=C2C(=O)OC1(C1=O)OC=C1C1=CC=CC=C1 ZFKJVJIDPQDDFY-UHFFFAOYSA-N 0.000 description 1
- 239000007850 fluorescent dye Substances 0.000 description 1
- 150000002222 fluorine compounds Chemical class 0.000 description 1
- 235000013355 food flavoring agent Nutrition 0.000 description 1
- 235000003599 food sweetener Nutrition 0.000 description 1
- 238000004108 freeze drying Methods 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- IECPWNUMDGFDKC-MZJAQBGESA-M fusidate Chemical class O[C@@H]([C@@H]12)C[C@H]3\C(=C(/CCC=C(C)C)C([O-])=O)[C@@H](OC(C)=O)C[C@]3(C)[C@@]2(C)CC[C@@H]2[C@]1(C)CC[C@@H](O)[C@H]2C IECPWNUMDGFDKC-MZJAQBGESA-M 0.000 description 1
- 230000000799 fusogenic effect Effects 0.000 description 1
- 108010006664 gamma-glutamyl-glycyl-glycine Proteins 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 238000001502 gel electrophoresis Methods 0.000 description 1
- 239000007903 gelatin capsule Substances 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 239000011521 glass Substances 0.000 description 1
- 229940116332 glucose oxidase Drugs 0.000 description 1
- 235000019420 glucose oxidase Nutrition 0.000 description 1
- 229930182478 glucoside Natural products 0.000 description 1
- 229960002989 glutamic acid Drugs 0.000 description 1
- 125000005456 glyceride group Chemical group 0.000 description 1
- 150000002337 glycosamines Chemical group 0.000 description 1
- VPZXBVLAVMBEQI-UHFFFAOYSA-N glycyl-DL-alpha-alanine Natural products OC(=O)C(C)NC(=O)CN VPZXBVLAVMBEQI-UHFFFAOYSA-N 0.000 description 1
- 108010089804 glycyl-threonine Proteins 0.000 description 1
- 108010081551 glycylphenylalanine Proteins 0.000 description 1
- 108010037850 glycylvaline Proteins 0.000 description 1
- 239000003966 growth inhibitor Substances 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 208000002672 hepatitis B Diseases 0.000 description 1
- 239000000833 heterodimer Substances 0.000 description 1
- SYECJBOWSGTPLU-UHFFFAOYSA-N hexane-1,1-diamine Chemical compound CCCCCC(N)N SYECJBOWSGTPLU-UHFFFAOYSA-N 0.000 description 1
- FUZZWVXGSFPDMH-UHFFFAOYSA-N hexanoic acid Chemical compound CCCCCC(O)=O FUZZWVXGSFPDMH-UHFFFAOYSA-N 0.000 description 1
- 108010040030 histidinoalanine Proteins 0.000 description 1
- 125000000487 histidyl group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C([H])=N1 0.000 description 1
- 108010025306 histidylleucine Proteins 0.000 description 1
- 108010085325 histidylproline Proteins 0.000 description 1
- 235000001050 hortel pimenta Nutrition 0.000 description 1
- 230000007236 host immunity Effects 0.000 description 1
- 230000004727 humoral immunity Effects 0.000 description 1
- 230000008348 humoral response Effects 0.000 description 1
- 238000009396 hybridization Methods 0.000 description 1
- ZMZDMBWJUHKJPS-UHFFFAOYSA-N hydrogen thiocyanate Natural products SC#N ZMZDMBWJUHKJPS-UHFFFAOYSA-N 0.000 description 1
- 230000007062 hydrolysis Effects 0.000 description 1
- 238000006460 hydrolysis reaction Methods 0.000 description 1
- OUUQCZGPVNCOIJ-UHFFFAOYSA-N hydroperoxyl Chemical compound O[O] OUUQCZGPVNCOIJ-UHFFFAOYSA-N 0.000 description 1
- 229920001600 hydrophobic polymer Polymers 0.000 description 1
- 238000012872 hydroxylapatite chromatography Methods 0.000 description 1
- 229920003063 hydroxymethyl cellulose Polymers 0.000 description 1
- 229940031574 hydroxymethyl cellulose Drugs 0.000 description 1
- 230000003100 immobilizing effect Effects 0.000 description 1
- 210000004201 immune sera Anatomy 0.000 description 1
- 229940042743 immune sera Drugs 0.000 description 1
- 239000002955 immunomodulating agent Substances 0.000 description 1
- 230000002584 immunomodulator Effects 0.000 description 1
- 229940121354 immunomodulator Drugs 0.000 description 1
- 230000002480 immunoprotective effect Effects 0.000 description 1
- 230000001506 immunosuppresive effect Effects 0.000 description 1
- 230000001024 immunotherapeutic effect Effects 0.000 description 1
- 238000009169 immunotherapy Methods 0.000 description 1
- 239000007943 implant Substances 0.000 description 1
- 238000000126 in silico method Methods 0.000 description 1
- 238000007901 in situ hybridization Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 239000003701 inert diluent Substances 0.000 description 1
- 230000004941 influx Effects 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 239000007972 injectable composition Substances 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 230000002687 intercalation Effects 0.000 description 1
- 238000009830 intercalation Methods 0.000 description 1
- 229940079322 interferon Drugs 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 239000007928 intraperitoneal injection Substances 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- 108010031424 isoleucyl-prolyl-proline Proteins 0.000 description 1
- 108010044374 isoleucyl-tyrosine Proteins 0.000 description 1
- 239000007951 isotonicity adjuster Substances 0.000 description 1
- MWDZOUNAPSSOEL-UHFFFAOYSA-N kaempferol Natural products OC1=C(C(=O)c2cc(O)cc(O)c2O1)c3ccc(O)cc3 MWDZOUNAPSSOEL-UHFFFAOYSA-N 0.000 description 1
- 238000012933 kinetic analysis Methods 0.000 description 1
- 150000002605 large molecules Chemical class 0.000 description 1
- 230000002045 lasting effect Effects 0.000 description 1
- 239000000787 lecithin Substances 0.000 description 1
- 235000010445 lecithin Nutrition 0.000 description 1
- 229940067606 lecithin Drugs 0.000 description 1
- 231100000518 lethal Toxicity 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 108010012058 leucyltyrosine Proteins 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- 229960004338 leuprorelin Drugs 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 239000007788 liquid Substances 0.000 description 1
- 230000005923 long-lasting effect Effects 0.000 description 1
- 239000007937 lozenge Substances 0.000 description 1
- 239000000314 lubricant Substances 0.000 description 1
- 238000004020 luminiscence type Methods 0.000 description 1
- HWYHZTIRURJOHG-UHFFFAOYSA-N luminol Chemical compound O=C1NNC(=O)C2=C1C(N)=CC=C2 HWYHZTIRURJOHG-UHFFFAOYSA-N 0.000 description 1
- 210000002751 lymph Anatomy 0.000 description 1
- 210000001165 lymph node Anatomy 0.000 description 1
- 108010003700 lysyl aspartic acid Proteins 0.000 description 1
- 108010064235 lysylglycine Proteins 0.000 description 1
- 229920002521 macromolecule Polymers 0.000 description 1
- 235000019359 magnesium stearate Nutrition 0.000 description 1
- 238000012423 maintenance Methods 0.000 description 1
- VZCYOOQTPOCHFL-UPHRSURJSA-N maleic acid Chemical compound OC(=O)\C=C/C(O)=O VZCYOOQTPOCHFL-UPHRSURJSA-N 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 230000010534 mechanism of action Effects 0.000 description 1
- 108010034507 methionyltryptophan Proteins 0.000 description 1
- DFTAZNAEBRBBKP-UHFFFAOYSA-N methyl 4-sulfanylbutanimidate Chemical compound COC(=N)CCCS DFTAZNAEBRBBKP-UHFFFAOYSA-N 0.000 description 1
- STZCRXQWRGQSJD-GEEYTBSJSA-M methyl orange Chemical compound [Na+].C1=CC(N(C)C)=CC=C1\N=N\C1=CC=C(S([O-])(=O)=O)C=C1 STZCRXQWRGQSJD-GEEYTBSJSA-M 0.000 description 1
- 229940012189 methyl orange Drugs 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 229960001047 methyl salicylate Drugs 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- 239000004530 micro-emulsion Substances 0.000 description 1
- 230000000813 microbial effect Effects 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 230000004089 microcirculation Effects 0.000 description 1
- 235000019813 microcrystalline cellulose Nutrition 0.000 description 1
- 239000008108 microcrystalline cellulose Substances 0.000 description 1
- 229940016286 microcrystalline cellulose Drugs 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 239000004005 microsphere Substances 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 150000002772 monosaccharides Chemical class 0.000 description 1
- 238000010172 mouse model Methods 0.000 description 1
- 239000002324 mouth wash Substances 0.000 description 1
- 229940051866 mouthwash Drugs 0.000 description 1
- ZTLGJPIZUOVDMT-UHFFFAOYSA-N n,n-dichlorotriazin-4-amine Chemical compound ClN(Cl)C1=CC=NN=N1 ZTLGJPIZUOVDMT-UHFFFAOYSA-N 0.000 description 1
- SBWGZAXBCCNRTM-CTHBEMJXSA-N n-methyl-n-[(2s,3r,4r,5r)-2,3,4,5,6-pentahydroxyhexyl]octanamide Chemical compound CCCCCCCC(=O)N(C)C[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO SBWGZAXBCCNRTM-CTHBEMJXSA-N 0.000 description 1
- 239000002088 nanocapsule Substances 0.000 description 1
- 239000002105 nanoparticle Substances 0.000 description 1
- 239000007922 nasal spray Substances 0.000 description 1
- 229940097496 nasal spray Drugs 0.000 description 1
- 239000006218 nasal suppository Substances 0.000 description 1
- 230000004719 natural immunity Effects 0.000 description 1
- 239000006199 nebulizer Substances 0.000 description 1
- 239000002687 nonaqueous vehicle Substances 0.000 description 1
- HEGSGKPQLMEBJL-RKQHYHRCSA-N octyl beta-D-glucopyranoside Chemical compound CCCCCCCCO[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O HEGSGKPQLMEBJL-RKQHYHRCSA-N 0.000 description 1
- 239000002674 ointment Substances 0.000 description 1
- KVWDHTXUZHCGIO-UHFFFAOYSA-N olanzapine Chemical compound C1CN(C)CCN1C1=NC2=CC=CC=C2NC2=C1C=C(C)S2 KVWDHTXUZHCGIO-UHFFFAOYSA-N 0.000 description 1
- 229960005017 olanzapine Drugs 0.000 description 1
- 239000002751 oligonucleotide probe Substances 0.000 description 1
- 229920001542 oligosaccharide Polymers 0.000 description 1
- 150000002482 oligosaccharides Chemical class 0.000 description 1
- 150000002894 organic compounds Chemical class 0.000 description 1
- VSZGPKBBMSAYNT-RRFJBIMHSA-N oseltamivir Chemical compound CCOC(=O)C1=C[C@@H](OC(CC)CC)[C@H](NC(C)=O)[C@@H](N)C1 VSZGPKBBMSAYNT-RRFJBIMHSA-N 0.000 description 1
- 229960003752 oseltamivir Drugs 0.000 description 1
- 210000001672 ovary Anatomy 0.000 description 1
- QUANRIQJNFHVEU-UHFFFAOYSA-N oxirane;propane-1,2,3-triol Chemical compound C1CO1.OCC(O)CO QUANRIQJNFHVEU-UHFFFAOYSA-N 0.000 description 1
- 238000004806 packaging method and process Methods 0.000 description 1
- 208000021090 palsy Diseases 0.000 description 1
- 238000004091 panning Methods 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 229940111202 pepsin Drugs 0.000 description 1
- 229940023041 peptide vaccine Drugs 0.000 description 1
- 210000005259 peripheral blood Anatomy 0.000 description 1
- 239000011886 peripheral blood Substances 0.000 description 1
- 210000005105 peripheral blood lymphocyte Anatomy 0.000 description 1
- 229960003742 phenol Drugs 0.000 description 1
- 108010083476 phenylalanyltryptophan Proteins 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- WTJKGGKOPKCXLL-RRHRGVEJSA-N phosphatidylcholine Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCCCCCCC=CCCCCCCCC WTJKGGKOPKCXLL-RRHRGVEJSA-N 0.000 description 1
- 150000008104 phosphatidylethanolamines Chemical class 0.000 description 1
- 230000004962 physiological condition Effects 0.000 description 1
- 239000002504 physiological saline solution Substances 0.000 description 1
- 239000006187 pill Substances 0.000 description 1
- 210000002381 plasma Anatomy 0.000 description 1
- 229920003229 poly(methyl methacrylate) Polymers 0.000 description 1
- 229920000728 polyester Polymers 0.000 description 1
- 239000008389 polyethoxylated castor oil Substances 0.000 description 1
- 239000004633 polyglycolic acid Substances 0.000 description 1
- 229920002338 polyhydroxyethylmethacrylate Polymers 0.000 description 1
- 239000004626 polylactic acid Substances 0.000 description 1
- 229920000656 polylysine Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 239000004926 polymethyl methacrylate Substances 0.000 description 1
- 229920005862 polyol Polymers 0.000 description 1
- 150000003077 polyols Chemical class 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 150000004804 polysaccharides Chemical class 0.000 description 1
- 229920002451 polyvinyl alcohol Polymers 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 230000008092 positive effect Effects 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 230000037452 priming Effects 0.000 description 1
- 238000003672 processing method Methods 0.000 description 1
- 230000002035 prolonged effect Effects 0.000 description 1
- 108010029020 prolylglycine Proteins 0.000 description 1
- 108010053725 prolylvaline Proteins 0.000 description 1
- 239000003380 propellant Substances 0.000 description 1
- 229940048914 protamine Drugs 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 235000004252 protein component Nutrition 0.000 description 1
- 210000001938 protoplast Anatomy 0.000 description 1
- 230000005180 public health Effects 0.000 description 1
- 235000011962 puddings Nutrition 0.000 description 1
- 229960001285 quercetin Drugs 0.000 description 1
- 235000005875 quercetin Nutrition 0.000 description 1
- 230000005855 radiation Effects 0.000 description 1
- 239000012857 radioactive material Substances 0.000 description 1
- 239000000941 radioactive substance Substances 0.000 description 1
- 239000002287 radioligand Substances 0.000 description 1
- 239000000376 reactant Substances 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000000306 recurrent effect Effects 0.000 description 1
- 239000012925 reference material Substances 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 230000000241 respiratory effect Effects 0.000 description 1
- 230000008593 response to virus Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 230000001177 retroviral effect Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 102200015462 rs121912296 Human genes 0.000 description 1
- CVHZOJJKTDOEJC-UHFFFAOYSA-N saccharin Chemical compound C1=CC=C2C(=O)NS(=O)(=O)C2=C1 CVHZOJJKTDOEJC-UHFFFAOYSA-N 0.000 description 1
- 229940081974 saccharin Drugs 0.000 description 1
- 235000019204 saccharin Nutrition 0.000 description 1
- 239000000901 saccharin and its Na,K and Ca salt Substances 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 108010026333 seryl-proline Proteins 0.000 description 1
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 1
- 238000004088 simulation Methods 0.000 description 1
- 235000010267 sodium hydrogen sulphite Nutrition 0.000 description 1
- 239000006104 solid solution Substances 0.000 description 1
- 210000001082 somatic cell Anatomy 0.000 description 1
- 239000000600 sorbitol Substances 0.000 description 1
- 230000009870 specific binding Effects 0.000 description 1
- 210000004989 spleen cell Anatomy 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 239000007921 spray Substances 0.000 description 1
- 235000019698 starch Nutrition 0.000 description 1
- 230000003068 static effect Effects 0.000 description 1
- 239000007929 subcutaneous injection Substances 0.000 description 1
- 238000010254 subcutaneous injection Methods 0.000 description 1
- 229940031626 subunit vaccine Drugs 0.000 description 1
- KZNICNPSHKQLFF-UHFFFAOYSA-N succinimide Chemical class O=C1CCC(=O)N1 KZNICNPSHKQLFF-UHFFFAOYSA-N 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 150000005846 sugar alcohols Polymers 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 239000000829 suppository Substances 0.000 description 1
- 239000002511 suppository base Substances 0.000 description 1
- 238000005211 surface analysis Methods 0.000 description 1
- 239000004094 surface-active agent Substances 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 239000012730 sustained-release form Substances 0.000 description 1
- 239000003765 sweetening agent Substances 0.000 description 1
- 230000002194 synthesizing effect Effects 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 238000011285 therapeutic regimen Methods 0.000 description 1
- 229940021747 therapeutic vaccine Drugs 0.000 description 1
- RTKIYNMVFMVABJ-UHFFFAOYSA-L thimerosal Chemical compound [Na+].CC[Hg]SC1=CC=CC=C1C([O-])=O RTKIYNMVFMVABJ-UHFFFAOYSA-L 0.000 description 1
- 229940033663 thimerosal Drugs 0.000 description 1
- 150000007970 thio esters Chemical class 0.000 description 1
- 150000003568 thioethers Chemical class 0.000 description 1
- CNHYKKNIIGEXAY-UHFFFAOYSA-N thiolan-2-imine Chemical compound N=C1CCCS1 CNHYKKNIIGEXAY-UHFFFAOYSA-N 0.000 description 1
- 108010031491 threonyl-lysyl-glutamic acid Proteins 0.000 description 1
- 229960004072 thrombin Drugs 0.000 description 1
- 229940104230 thymidine Drugs 0.000 description 1
- 238000004448 titration Methods 0.000 description 1
- 230000000699 topical effect Effects 0.000 description 1
- 239000000196 tragacanth Substances 0.000 description 1
- 235000010487 tragacanth Nutrition 0.000 description 1
- 229940116362 tragacanth Drugs 0.000 description 1
- VZCYOOQTPOCHFL-UHFFFAOYSA-N trans-butenedioic acid Natural products OC(=O)C=CC(O)=O VZCYOOQTPOCHFL-UHFFFAOYSA-N 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 230000002463 transducing effect Effects 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 238000011830 transgenic mouse model Methods 0.000 description 1
- 108010080629 tryptophan-leucine Proteins 0.000 description 1
- 108010038745 tryptophylglycine Proteins 0.000 description 1
- 108010044292 tryptophyltyrosine Proteins 0.000 description 1
- ORHBXUUXSCNDEV-UHFFFAOYSA-N umbelliferone Chemical compound C1=CC(=O)OC2=CC(O)=CC=C21 ORHBXUUXSCNDEV-UHFFFAOYSA-N 0.000 description 1
- HFTAFOQKODTIJY-UHFFFAOYSA-N umbelliferone Natural products Cc1cc2C=CC(=O)Oc2cc1OCC=CC(C)(C)O HFTAFOQKODTIJY-UHFFFAOYSA-N 0.000 description 1
- 238000001291 vacuum drying Methods 0.000 description 1
- 108010009962 valyltyrosine Proteins 0.000 description 1
- 239000003981 vehicle Substances 0.000 description 1
- 229960004854 viral vaccine Drugs 0.000 description 1
- 230000024058 virion binding Effects 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 239000008215 water for injection Substances 0.000 description 1
- ARAIBEBZBOPLMB-UFGQHTETSA-N zanamivir Chemical compound CC(=O)N[C@@H]1[C@@H](N=C(N)N)C=C(C(O)=O)O[C@H]1[C@H](O)[C@H](O)CO ARAIBEBZBOPLMB-UFGQHTETSA-N 0.000 description 1
- 229960001028 zanamivir Drugs 0.000 description 1
Images







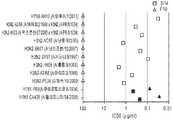











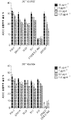
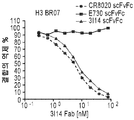













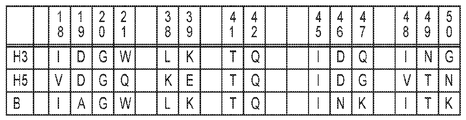


















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1018—Orthomyxoviridae, e.g. influenza virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/145—Orthomyxoviridae, e.g. influenza virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/16—Antivirals for RNA viruses for influenza or rhinoviruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/33—Crossreactivity, e.g. for species or epitope, or lack of said crossreactivity
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/34—Identification of a linear epitope shorter than 20 amino acid residues or of a conformational epitope defined by amino acid residues
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
- C07K2317/732—Antibody-dependent cellular cytotoxicity [ADCC]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
Abstract
본 발명은 인플루엔자 바이러스의 HA 단백질의 스템 도메인에 결합하기 위해 중요한 구조적 결정자, 및 상기 결정자에 기초하여 고친화도 인플루엔자 중화 항체의 제조를 위한 그의 사용 방법을 제공한다. 본 발명은 또한 인플루엔자 바이러스 백신의 효능을 결정하기 위한 수단을 제공한다. 또한, 본 발명은 대상체에서 인플루엔자 바이러스 백신의 효능을 결정하기 위해, 또는 백신 또는 인플루엔자 바이러스 감염에 대한 사전 면역 노출 또는 항원 반응성을 예측하기 위해 유용한 분자 시그너쳐를 제공한다.
Description
관련 출원
본원은 그 내용 전체가 본원에 참고로 포함된, 2016년 4월 8일 출원된 미국 가출원 제62/144,729호의 우선권 및 이익을 주장한다.
발명의 분야
본 발명은 일반적으로 인플루엔자 중화 항체 및 그의 사용 방법에 관한 것이다.
정부 이익
본 발명은 미국 정부의 지원에 의해 이루어졌다. 미국 정부는 본 발명에 대해 특정 권리를 갖는다.
인플루엔자 대유행은 인체 건강에 대한 가장 심각한 급성 감염 위협 중 하나이다. 백신 접종은 계절성 및 대유행성 인플루엔자 및 그의 합병증을 예방하는 주요 수단으로 남아 있다. 인플루엔자 바이러스의 여러 아형에 대한 광범위한 면역을 유도하는 "보편적인" 인플루엔자 백신은 의학 연구에서 오랫동안 추구된 목표였다. HA의 스템(stem) 상의 고도로 보존된 소수성 포켓에 결합하는, 광범위하게 중화하는 인간 "이종아형(heterosubtypic)" 항체(sBnAb)의 최근 발견은 상기 백신 개발 노력을 재점화하였다. 그러나, 계절성 인플루엔자 또는 H5N1 백신의 혈청에서 또는 상업용 정맥내 이뮤노글로불린(IVIG) 제제에서 sBnAb의 매우 낮은 농도만이 검출된다.
인플루엔자 바이러스에 대한 면역요법을 위한 모노클로날 항체(mAb) 및 약물을 생산하기 위한 지속적인 노력이 있다. 구체적으로, 다양한 인플루엔자 균주 모두를 중화하는 치료 화합물의 개발에 대한 노력이 진행되고 있다. 현재, 이 목표를 달성할 수 있는 소수의 mAb만이 보고되어 있다. 이들 mAb는 파지 항체 라이브러리를 패닝(panning)하고 백신 접종된 자원자로부터 B 세포를 스크리닝함으로써 단리하였다. 그러나, 광범위한 인플루엔자 중화 항체의 특성에 대한 이해의 증가는 인플루엔자 중화 항체의 광범위한 패널의 발견 및 생산을 위한 보다 합리적인 설계 방법에 특정 구조적 결정 요인을 도입하는데 유용할 수 있다.
따라서, 인플루엔자 바이러스를 광범위하게 중화할 수 있는 추가의 모노클로날 항체 및 합리적인 설계 방법을 통해 상기 항체의 친화도 또는 효능을 증가시키는 방법에 대한 필요성이 존재한다.
발명의 개요
본 발명은 다음 특징을 갖는, 재조합 생산된, 단리된 모노클로날 항체를 특징으로 한다: IGHV3-30 생식계열 유전자에 의해 코딩되는 중쇄 가변 영역; 헤마글루티닌(HA) 단백질의 스템 영역의 에피토프에 대한 결합; 및 그룹 1 및 그룹 2 인플루엔자 A 바이러스의 중화.
한 측면에서, 모노클로날 항체는 다음 특징 중 하나 이상을 갖는다: IGLV1-44 생식계열 유전자에 의해 코딩되는 경쇄 가변 영역; 서열 번호 9의 아미노산 서열을 포함하는 중쇄 CDR3; 절단되지 않은 HA0에 대한 결합; HA0 절단 방지; 및/또는 서열 번호 18에 따라 넘버링될 때 HA2 폴리펩티드의 아미노산 18, 19, 20, 21, 36, 38, 39, 41, 42, 45, 45, 49 및 53에 의해 규정되는 구조적 에피토프(conformational epitope)에 대한 결합.
본 발명은 서열 번호 7의 아미노산 서열을 포함하는 CDR1; 서열 번호 8의 아미노산 서열을 포함하는 CDR2 및 서열 번호 9의 아미노산 서열을 포함하는 CDR3을 포함하는 중쇄; 및 서열 번호 10 또는 서열 번호 14의 아미노산 서열을 포함하는 CDR1; 서열 번호 11의 아미노산 서열을 포함하는 CDR2 및 서열 번호 12 또는 서열 번호 13의 아미노산 서열을 포함하는 CDR3을 포함하는 경쇄를 포함하는, 인플루엔자 바이러스를 중화하는 단리된 모노클로날 항체를 특징으로 한다.
본 발명은 또한 서열 번호 2의 VH 아미노산 서열 및 서열 번호 4 또는 서열 번호 6의 VL 아미노산 서열; 서열 번호 2의 VH 아미노산 서열 및 서열 번호 24의 VL 아미노산 서열; 또는 서열 번호 22의 VH 아미노산 서열 및 서열 번호 4 또는 6의 VL 아미노산 서열을 포함하는 단리된 모노클로날 항체를 특징으로 한다.
항체는 HA2 폴리펩티드의 아미노산 18, 19, 20, 21, 36, 38, 39, 41, 42, 45, 45, 49 및 53에 의해 규정된 구조적 에피토프에 결합한다.
한 측면에서, 항체는 단일쇄 Fv 항체, Fab 단편, Fab' 단편, 또는 F(ab')2 단편이다. 또 다른 측면에서, 항체는 치료제에 연결된다. 예를 들어, 치료제는 독소, 방사성 표지, siRNA, 소분자 또는 사이토카인이다.
본 발명은 또한 본원에 개시된 임의의 항체 및 담체를 포함하는 조성물을 특징으로 한다.
본 발명은 서열 번호 1, 3 및 5로부터 선택된 핵산 서열을 포함하는 핵산 서열을 제공한다. 또 다른 실시양태에서, 서열 번호 2, 4 및 6으로부터 선택된 아미노산 서열을 포함하는 폴리펩티드를 코딩하는 핵산 서열이 제공된다. 한 측면에서, 서열 번호 2, 4 및 6으로부터 선택된 아미노산 서열을 포함하는 폴리펩티드가 제공된다.
본 발명은 서열 번호 1, 2, 3, 4, 5 및 6으로부터 선택된 핵산 서열을 포함하는 벡터를 제공한다. 한 측면에서, 본 발명은 핵산 서열 1, 2, 3, 4 또는 6을 포함하는 벡터를 포함하는 세포를 제공한다.
본 발명은 또한 본원에 개시된 임의의 항체를 생산하는 세포를 제공한다.
본 발명은 추가로 인플루엔자 바이러스에 의해 유발된 질환 또는 장애를 앓을 위험이 있는 인간에게 본원에서 설명되는 임의의 모노클로날 항체의 치료 유효량을 투여함으로써 상기 질환 또는 장애를 치료하는 방법을 제공한다. 선택적으로, 상기 방법은 항바이러스제를 투여하는 단계를 추가로 포함한다.
본 발명의 다른 특징 및 이점은 다음의 상세한 설명 및 청구범위로부터 명백할 것이고, 그에 의해 포함될 것이다.
도 1a는 인간 기억 B 세포 레퍼토리로부터 인플루엔자 바이러스에 대해 광범위하게 중화하는 Ab의 단리를 나타내는 개략도이다. 대표적인 형광 표지 세포 분류(FACS) 데이터도 제공된다. FACS 데이터는 총 PBMC로부터 단리된 H3-반응성 기억 B 세포의 빈도를 나타낸다. 도 1b는 아미노산 서열에 기초한 인플루엔자 A 바이러스의 18 HA 아형의 계통수(phylogenetic tree)를 도시한 것이다. 그룹 1 아형 및 그룹 2 아형이 개략도에 표시된다. 아미노산 거리 규모 막대는 0.1의 거리를 나타낸다.
도 2는 넓은 범위의 그룹 1 및 그룹 2 HA에 대한 3I14 결합을 보여주는 일련의 FACS 그래프이다. 293T 세포를 상이한 HA 발현 플라스미드로 일시적으로 형질감염시키고, 정제된 scFvFc 항체 및 APC 표지된 마우스 항-인간 Fc 항체로 염색하였다. 3I14 (적색 선), F10 (그룹 1 특이적, 녹색 선), CR8020 (그룹 2 특이적, 청색 선), FI6v3 (그룹 1 및 2 특이적, 보라색 선), CR9114 (그룹 1 & 2 특이적, 오렌지색 선) 및 무관한 mAb Fm-6 (항-SARS 바이러스, 회색의 채워진 히스토그램)의 결합을 유동 세포 계측법으로 분석하였다.
도 3은 그룹 1(적색) 또는 그룹 2(청색) 아형을 나타내는 재조합 HA에 대한 3I14 IgG1(A) 결합(Kd 값) 또는 scFvFc Ab(B) 결합(Kd 값)을 보여주는 일련의 그래프이다.
도 4는 그룹 1(적색) 또는 그룹 2(청색) 아형의 감염성 바이러스의 3I14 IgG1 중화(A)(IC50 값) 또는 3I14 scFvFc Ab 중화(IC50 값)을 보여주는 일련의 그래프이다. 3I14는 정사각형으로 표시하고; 항-그룹 1 mAb F10은 삼각형으로 나타내었다. IC50 값에 사용된 그래프는 2-3회의 독립적인 실험의 평균 중화 역가에 의해 결정하였다.
도 5는 그룹 1 또는 그룹 2 아형의 슈도바이러스(pseudovirus)의 3I14 IgG1 중화(A)(IC50 값) 또는 3I14 scFvFc Ab 중화(B)(IC50 값)을 보여주는 일련의 그래프이다. 이들 데이터는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다. 도 5b에서, 항-그룹 1 mAb F10 scFvFc를 참고용으로 사용하였다.
도 6은 인플루엔자 바이러스에 노출되고 5, 20 또는 25 mg 중 하나의 농도에서 정제된 IgG를 복강내 투여한 마우스의 생존율(A)을 보여주는 일련의 그래프이다. 도 6b는 인플루엔자 바이러스에 노출되고 지시된 특정 항체 농도를 투여한 마우스에서 체중 조성의 변화를 보여주는 일련의 그래프이다. 이러한 분석을 위해, 5마리의 마우스의 그룹을, H7N7-NL219, H7N9-AH13, H3N2-BR07 또는 H5N1-VN04 인플루엔자 바이러스 (~10 LD50)의 i.n. 접종에 의한 치사 시험접종(challenge) 24시간 전에 20/25 또는 5 mg/kg 투여량의 정제된 IgG의 복강내 투여로 처리하였다. (a) bnAb 3I14(적색) 및 그룹 1 대조군 mAb F10(흑색)으로 처리한 마우스의 생존율(%) 및(b) 체중 변화(%).
도 7은 항체가 사용되지 않거나, 3I14 또는 Fm-6(IgG1 대조군)이 사용된 트립신 절단 억제 분석으로부터의 결과를 보여주는 겔이다. 이 분석을 위해, 0.4 ㎍의 재조합 H3-히스티딘(H3-BR07)을 100 ㎍/mL 트립신-울트라(New England Biolabs, 미국)를 함유하는 pH 8.0의 Tris-HCl 완충제 내에서 2.5 ㎍ 3I14 또는 Fm-6 IgG1의 존재 하에 또는 항체의 부재 하에 37℃에서 인큐베이팅하였다. 트립신 소화는 1% BSA의 첨가에 의해 여러 시점에서 억제되었다. 샘플을 12% 환원 SDS-PAGE에서 이동시키고, HisProbe-HRP Ab를 사용하여 블로팅하였다.
도 8은 3I14 IgG1이 표면 발현된 H3-A268 및 H3-BR07에서 낮은 pH에 의해 촉발된 구조적 재배열을 방지함을 보여주는 일련의 개략도 및 그래프이다. 표면 발현된 H3의 구조적 재배열은 3I14(솔리드 막대) 및 헤드 결합 대조군 mAb E730(오픈 막대)의 FACS 염색에 의해 검출되었다. 결합은 미처리된 HA(HA0)에 결합하는 백분율로 표현된다. 이 항체 억제 분석을 위해, H3를 mAb 처리 없이, 3I14로, 또는 대조군 Ab인 Fm-6 IgG1로 전처리한 후, 절단된 HA를 pH 4.9에 노출시켰다. 데이터는 3회의 독립적인 실험의 평균 + SD를 나타낸다.
도 9a 및 9b는 3I14가 항체 의존성 세포독성(ADCC)을 매개한다는 것을 입증한다. 도 9a는 ADCC 기반 분석의 결과를 보여주는 그래프이다. 3I14 및 다른 항-스템 bnAb인 FI6v3, CR9114, 39.29, F10 및 CR8020은 H3- 및 H5-발현 293T 세포에서 ADCC를 유도하였다. 이들 분석을 위해, 분석하기 전에 플레이트에 1x104/웰 H3 또는 H5-발현 293T 세포를 부착시킨 다음, 배지를 낮은 IgG 혈청 분석 완충제(0.5%의 낮은 IgG FBS를 갖는 RPMI 1640)으로 대체하였다. 상이한 bnAb를 5, 1, 0.2 및 0.04 ㎍/㎖의 농도로 각각의 웰에 첨가하였다. 1시간 후에, 낮은 IgG 혈청 분석 완충제에서 플레이트를 분석하기 위해 Jurkat 이펙터 세포를 6.0x104 세포/웰로 첨가하고, 6시간 동안 인큐베이팅하였다. 상청액을 수거하고, 루시페라제 활성을 Bio-GloTM 루시페라제 분석 키트(Promega)를 사용하여 측정하였다. 도 9b는 H3- 및 H5-발현 293T 세포에서 3I14 및 다른 항-스템 bnAb인 FI6v3, CR9114, 39.29, F10 및 CR8020 유도 ADCC를 보여주는 일련의 그래프이다. 2x104/웰 H3 또는 H5-발현 293T 세포를 분석 전에 플레이트에 부착시킨 다음, 배지를 낮은 IgG 혈청 분석 완충제(0.5%의 낮은 IgG FBS를 갖는 RPMI 1640)으로 교체하였다. 상이한 bnAb를 10, 5, 2.5 및 1.25 ㎍/ml의 농도로 각각의 웰에 첨가하였다. 1시간 후, 낮은 IgG 혈청 분석 완충제에서 플레이트를 분석하기 위해 PBMC를 1.2x105 세포/웰로 첨가하고, 6시간 동안 인큐베이팅하였다. 상청액을 수거하고, 루시페라제 활성을 LDH 세포독성 분석 키트(Pierce)를 사용하여 측정하였다. 데이터는 평균 ± S.E.M.을 나타낸다. 실험은 3회 수행하였고, 비슷한 결과를 얻었다. 데이터는 그룹당 3회 반복한 하나의 독립적인 실험을 나타낸다.
도 10은 3I14가 H3 또는 H5에 대한 결합을 위해 다른 항-스템 bnAb인 FI6v3, CR9114, 39.29, F10 및 CR8020과 교차 경쟁함을 보여주는 일련의 그래프이다. 이들 분석을 위해, 5 ㎍/ml H3-BR07 또는 H5-VN04 단백질을 ELISA 플레이트 상에 고정하고, 80 nM 내지 0.3 nM의 농도 범위의 3I14 Fab의 2배 연속 희석액과 함께 인큐베이팅하였고; 이들을 5 nM의 다른 scFvFc Ab와 추가로 혼합하였다. scFvFc Ab의 결합은 HRP 접합 항-인간 CH2 항체를 사용하여 검출되었다.
도 11은 3I14 및 다른 생식계열 중쇄(a, c) 또는 경쇄 영역(b, d)의 아미노산 서열 정렬을 도시한 것이다. 대응하는 V, D 및 J 서열은 IMGT 데이터베이스를 사용하여 결정하였고 비교를 위해 나타내었다.
도 12a 및 도 12b는 3개의 VH3-30 BnAb인 39.29(4KVN으로부터), FI6(3ZTJ로부터) 및 Mab3.1(4PY8)을 사용한 3I14 모델의 중첩을 보여주는 일련의 개략도이다. 3I14 항체의 중쇄 CDR뿐만 아니라 중쇄 및 경쇄의 상대적인 위치를 보여주는 단백질 모델 개략도가 제시된다. 중쇄는 녹색으로 표시되고, 경쇄는 회색으로 표시된다. 도 12의 중쇄 CDR은 다음과 같이 색상으로 구분된다: 3I14 HCDR은 청색, 다른 BnAb의 HCDR1은 적색, HCDR2는 자홍색, HCDR3은 청록색이다.
bnAb는 리본 방식으로 표시된다. 도 12b에서, 중쇄는 청색이고, 경쇄는 청록색이다. HCDR3은 3I14에서 적색으로, FI6v3에 자홍색으로, 39.29에서 황색으로, MAb 3.1에서 녹색으로 강조 표시된다.
도 13a-d는 3I14의 모델링 및 H3/H5과의 도킹을 보여주는 일련의 개략도이다. H3 삼량체 모델의 줄기 상의 3I14 에피토프 구조가 도 13a에 도시되어 있다. 도 13b는 HA와 함께 IGVH3-30 유래 bnAb의 복합체 구조를 도시한 것이다. HA상의 에피토프 잔기는 표면 제시로 도시되고, bnAb의 CDR 루프는 리본으로 표시된다. HA 단량체의 HA1은 밀에, HA2는 연어살색에 존재하고, 에피토프 잔기는 적색으로 표지된다. 핵심 잔기 L38 및 K39는 황색으로 착색되어 있다. BnAb의 중쇄 CDR은 청색이고, 경쇄 CDR은 청록색이다. 3I14는 바이오루미네이트(BioLuminate)의 항체 모델링 모듈을 사용하여 상동성을 모델링하였고, 모델은 로제타독(RosettaDock)과 도킹하기 전에 H3/FI6v3 복합체 구조에 중첩되었다. 다른 3개의 IGHV3-30 bnAb인 FI6v3, 39.29 및 MAb 3.1은 프로테인 데이터 뱅크(Protein Data Bank)에서 다운로드하였다. 도 13c는 3I14 내의 D94와 H3/H5의 상호작용을 도시한 것이다. H3은 청록색으로 표시되고 여기서 K39는 스틱으로 제시되고; H5는 녹색으로 표시되고 여기서 E39는 스틱으로 제시되고; 3I14는 H3/3I14 모델에서 오렌지색으로, H5/3I14 모델에서는 황색으로 표시되고 여기서 D94는 스틱으로 제시된다. 도 13d는 H3/3I14 복합 모델에서 3I14 경쇄의 G31과 H3의 상호작용을 도시한 것이다. H3의 HA2 도메인의 나선 A는 청록색 리본으로 표시되고; 3I14의 경쇄는 오렌지색으로 리본으로 표시되고; G31의 주쇄 원자는 스틱으로 표시되고 H3 HA2의 Q42 및 D46의 측쇄 원자는 막대기로 표시되고; G31과 H3 사이의 거리는 녹색 점선으로 표시되고 흑색으로 표지된다(PyMOL Molecular Graphics System, Version 0.99 rc6 Schroedinger, LLC).
도 14는 H3/3I14 및 H5/3I14 모델의 서열 정렬(a) 및 구조 중첩(b)을 도시한 것이다.
또한, 15a-15f는 재조합 H5-VN04(a) 및 H3-PE09(b)에 결합하는 3I14 WT 및 VLD94N IgG1 변이체의 Kd 결합 값을 보여주는 그래프 및 표이다. 녹색 또는 청색 곡선은 생물층 간섭 측정 실험으로부터 얻은 실험 기록이고, 적색 곡선은 Kd 값을 계산하기 위해 사용되는 데이터에 대한 전반적인 최적 적합도(best global fit)이다. 결합 곡선 친화도 측정채(Kd 값)는 표 4에 보고된다. 3I14 WT는 1.15 nm의 Kd 값으로 정제된 H5-VN04에 결합한 반면, 3I14 VLD94N 돌연변이체는 0.19 nM에서 10배 더 높은 친화도로 H5-VN04에 결합하였다. 도 15c-f는 재조합 Ha에 결합하는 3I14 scFvFc를 나타내는 일련의 그래프이다.
도 16a-d는 슈도타이핑된(pseudotyped) 바이러스 H5N1-VN04 및 감염성 바이러스 H3N2-BR07과 함께 인큐베이팅한 후, 3I14 WT 및 VLD94N 돌연변이체 IgG1의 중화 값을 나타내는 일련의 그래프이다. 도 16c 및 16d에서, 3I14(흑색) 및 VLD94N 변이체(적색)는 슈도타이핑된 바이러스 H5N1-VN04(c) 및 H3N2-BR07 바이러스(d)를 중화하였다. 이 데이터는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다.
도 17은 3I14 scFvFc Ab가 인플루엔자 바이러스 감염 및 HA-슈도타이핑된 루시페라제 리포터 바이러스를 중화함을 나타내는 일련의 그래프이다. MAb 3I14(흑색) 및 항-그룹 1 mAb F10(적색)은 감염성 바이러스 및 슈도타이핑된 바이러스의 상이한 균주를 중화하였다. 데이터는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다.
도 18은 조작된 3I14 효모-CDR 라이브러리의 H5에 대한 결합과 비교하여 3I14-WT 효모의 결합을 나타내는 일련의 유동 세포 계측 그래프이다. 3I14 효모-CDR 라이브러리는 H5에 대한 결합을 증가시키는 변이체에 대한 효모 디스플레이를 사용하여 조작되었다.
도 19는 재조합 H3-PE09의 전장 또는 HA1에 결합하는 3I14 scFvFc Ab를 나타내는 그래프이다.
도 20은 3I14 IgG1 변이체의 재조합 H1, H3 및 H5에 대한 결합을 나타내는 일련의 그래프이다. 청색 곡선은 생물층 간섭 측정 실험으로부터 얻은 실험 기록이고, 적색 곡선은 표 7에 제시된 Kd를 계산하기 위해 사용된 데이터에 대한 전반적인 최적 적합도이다.
도 2는 넓은 범위의 그룹 1 및 그룹 2 HA에 대한 3I14 결합을 보여주는 일련의 FACS 그래프이다. 293T 세포를 상이한 HA 발현 플라스미드로 일시적으로 형질감염시키고, 정제된 scFvFc 항체 및 APC 표지된 마우스 항-인간 Fc 항체로 염색하였다. 3I14 (적색 선), F10 (그룹 1 특이적, 녹색 선), CR8020 (그룹 2 특이적, 청색 선), FI6v3 (그룹 1 및 2 특이적, 보라색 선), CR9114 (그룹 1 & 2 특이적, 오렌지색 선) 및 무관한 mAb Fm-6 (항-SARS 바이러스, 회색의 채워진 히스토그램)의 결합을 유동 세포 계측법으로 분석하였다.
도 3은 그룹 1(적색) 또는 그룹 2(청색) 아형을 나타내는 재조합 HA에 대한 3I14 IgG1(A) 결합(Kd 값) 또는 scFvFc Ab(B) 결합(Kd 값)을 보여주는 일련의 그래프이다.
도 4는 그룹 1(적색) 또는 그룹 2(청색) 아형의 감염성 바이러스의 3I14 IgG1 중화(A)(IC50 값) 또는 3I14 scFvFc Ab 중화(IC50 값)을 보여주는 일련의 그래프이다. 3I14는 정사각형으로 표시하고; 항-그룹 1 mAb F10은 삼각형으로 나타내었다. IC50 값에 사용된 그래프는 2-3회의 독립적인 실험의 평균 중화 역가에 의해 결정하였다.
도 5는 그룹 1 또는 그룹 2 아형의 슈도바이러스(pseudovirus)의 3I14 IgG1 중화(A)(IC50 값) 또는 3I14 scFvFc Ab 중화(B)(IC50 값)을 보여주는 일련의 그래프이다. 이들 데이터는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다. 도 5b에서, 항-그룹 1 mAb F10 scFvFc를 참고용으로 사용하였다.
도 6은 인플루엔자 바이러스에 노출되고 5, 20 또는 25 mg 중 하나의 농도에서 정제된 IgG를 복강내 투여한 마우스의 생존율(A)을 보여주는 일련의 그래프이다. 도 6b는 인플루엔자 바이러스에 노출되고 지시된 특정 항체 농도를 투여한 마우스에서 체중 조성의 변화를 보여주는 일련의 그래프이다. 이러한 분석을 위해, 5마리의 마우스의 그룹을, H7N7-NL219, H7N9-AH13, H3N2-BR07 또는 H5N1-VN04 인플루엔자 바이러스 (~10 LD50)의 i.n. 접종에 의한 치사 시험접종(challenge) 24시간 전에 20/25 또는 5 mg/kg 투여량의 정제된 IgG의 복강내 투여로 처리하였다. (a) bnAb 3I14(적색) 및 그룹 1 대조군 mAb F10(흑색)으로 처리한 마우스의 생존율(%) 및(b) 체중 변화(%).
도 7은 항체가 사용되지 않거나, 3I14 또는 Fm-6(IgG1 대조군)이 사용된 트립신 절단 억제 분석으로부터의 결과를 보여주는 겔이다. 이 분석을 위해, 0.4 ㎍의 재조합 H3-히스티딘(H3-BR07)을 100 ㎍/mL 트립신-울트라(New England Biolabs, 미국)를 함유하는 pH 8.0의 Tris-HCl 완충제 내에서 2.5 ㎍ 3I14 또는 Fm-6 IgG1의 존재 하에 또는 항체의 부재 하에 37℃에서 인큐베이팅하였다. 트립신 소화는 1% BSA의 첨가에 의해 여러 시점에서 억제되었다. 샘플을 12% 환원 SDS-PAGE에서 이동시키고, HisProbe-HRP Ab를 사용하여 블로팅하였다.
도 8은 3I14 IgG1이 표면 발현된 H3-A268 및 H3-BR07에서 낮은 pH에 의해 촉발된 구조적 재배열을 방지함을 보여주는 일련의 개략도 및 그래프이다. 표면 발현된 H3의 구조적 재배열은 3I14(솔리드 막대) 및 헤드 결합 대조군 mAb E730(오픈 막대)의 FACS 염색에 의해 검출되었다. 결합은 미처리된 HA(HA0)에 결합하는 백분율로 표현된다. 이 항체 억제 분석을 위해, H3를 mAb 처리 없이, 3I14로, 또는 대조군 Ab인 Fm-6 IgG1로 전처리한 후, 절단된 HA를 pH 4.9에 노출시켰다. 데이터는 3회의 독립적인 실험의 평균 + SD를 나타낸다.
도 9a 및 9b는 3I14가 항체 의존성 세포독성(ADCC)을 매개한다는 것을 입증한다. 도 9a는 ADCC 기반 분석의 결과를 보여주는 그래프이다. 3I14 및 다른 항-스템 bnAb인 FI6v3, CR9114, 39.29, F10 및 CR8020은 H3- 및 H5-발현 293T 세포에서 ADCC를 유도하였다. 이들 분석을 위해, 분석하기 전에 플레이트에 1x104/웰 H3 또는 H5-발현 293T 세포를 부착시킨 다음, 배지를 낮은 IgG 혈청 분석 완충제(0.5%의 낮은 IgG FBS를 갖는 RPMI 1640)으로 대체하였다. 상이한 bnAb를 5, 1, 0.2 및 0.04 ㎍/㎖의 농도로 각각의 웰에 첨가하였다. 1시간 후에, 낮은 IgG 혈청 분석 완충제에서 플레이트를 분석하기 위해 Jurkat 이펙터 세포를 6.0x104 세포/웰로 첨가하고, 6시간 동안 인큐베이팅하였다. 상청액을 수거하고, 루시페라제 활성을 Bio-GloTM 루시페라제 분석 키트(Promega)를 사용하여 측정하였다. 도 9b는 H3- 및 H5-발현 293T 세포에서 3I14 및 다른 항-스템 bnAb인 FI6v3, CR9114, 39.29, F10 및 CR8020 유도 ADCC를 보여주는 일련의 그래프이다. 2x104/웰 H3 또는 H5-발현 293T 세포를 분석 전에 플레이트에 부착시킨 다음, 배지를 낮은 IgG 혈청 분석 완충제(0.5%의 낮은 IgG FBS를 갖는 RPMI 1640)으로 교체하였다. 상이한 bnAb를 10, 5, 2.5 및 1.25 ㎍/ml의 농도로 각각의 웰에 첨가하였다. 1시간 후, 낮은 IgG 혈청 분석 완충제에서 플레이트를 분석하기 위해 PBMC를 1.2x105 세포/웰로 첨가하고, 6시간 동안 인큐베이팅하였다. 상청액을 수거하고, 루시페라제 활성을 LDH 세포독성 분석 키트(Pierce)를 사용하여 측정하였다. 데이터는 평균 ± S.E.M.을 나타낸다. 실험은 3회 수행하였고, 비슷한 결과를 얻었다. 데이터는 그룹당 3회 반복한 하나의 독립적인 실험을 나타낸다.
도 10은 3I14가 H3 또는 H5에 대한 결합을 위해 다른 항-스템 bnAb인 FI6v3, CR9114, 39.29, F10 및 CR8020과 교차 경쟁함을 보여주는 일련의 그래프이다. 이들 분석을 위해, 5 ㎍/ml H3-BR07 또는 H5-VN04 단백질을 ELISA 플레이트 상에 고정하고, 80 nM 내지 0.3 nM의 농도 범위의 3I14 Fab의 2배 연속 희석액과 함께 인큐베이팅하였고; 이들을 5 nM의 다른 scFvFc Ab와 추가로 혼합하였다. scFvFc Ab의 결합은 HRP 접합 항-인간 CH2 항체를 사용하여 검출되었다.
도 11은 3I14 및 다른 생식계열 중쇄(a, c) 또는 경쇄 영역(b, d)의 아미노산 서열 정렬을 도시한 것이다. 대응하는 V, D 및 J 서열은 IMGT 데이터베이스를 사용하여 결정하였고 비교를 위해 나타내었다.
도 12a 및 도 12b는 3개의 VH3-30 BnAb인 39.29(4KVN으로부터), FI6(3ZTJ로부터) 및 Mab3.1(4PY8)을 사용한 3I14 모델의 중첩을 보여주는 일련의 개략도이다. 3I14 항체의 중쇄 CDR뿐만 아니라 중쇄 및 경쇄의 상대적인 위치를 보여주는 단백질 모델 개략도가 제시된다. 중쇄는 녹색으로 표시되고, 경쇄는 회색으로 표시된다. 도 12의 중쇄 CDR은 다음과 같이 색상으로 구분된다: 3I14 HCDR은 청색, 다른 BnAb의 HCDR1은 적색, HCDR2는 자홍색, HCDR3은 청록색이다.
bnAb는 리본 방식으로 표시된다. 도 12b에서, 중쇄는 청색이고, 경쇄는 청록색이다. HCDR3은 3I14에서 적색으로, FI6v3에 자홍색으로, 39.29에서 황색으로, MAb 3.1에서 녹색으로 강조 표시된다.
도 13a-d는 3I14의 모델링 및 H3/H5과의 도킹을 보여주는 일련의 개략도이다. H3 삼량체 모델의 줄기 상의 3I14 에피토프 구조가 도 13a에 도시되어 있다. 도 13b는 HA와 함께 IGVH3-30 유래 bnAb의 복합체 구조를 도시한 것이다. HA상의 에피토프 잔기는 표면 제시로 도시되고, bnAb의 CDR 루프는 리본으로 표시된다. HA 단량체의 HA1은 밀에, HA2는 연어살색에 존재하고, 에피토프 잔기는 적색으로 표지된다. 핵심 잔기 L38 및 K39는 황색으로 착색되어 있다. BnAb의 중쇄 CDR은 청색이고, 경쇄 CDR은 청록색이다. 3I14는 바이오루미네이트(BioLuminate)의 항체 모델링 모듈을 사용하여 상동성을 모델링하였고, 모델은 로제타독(RosettaDock)과 도킹하기 전에 H3/FI6v3 복합체 구조에 중첩되었다. 다른 3개의 IGHV3-30 bnAb인 FI6v3, 39.29 및 MAb 3.1은 프로테인 데이터 뱅크(Protein Data Bank)에서 다운로드하였다. 도 13c는 3I14 내의 D94와 H3/H5의 상호작용을 도시한 것이다. H3은 청록색으로 표시되고 여기서 K39는 스틱으로 제시되고; H5는 녹색으로 표시되고 여기서 E39는 스틱으로 제시되고; 3I14는 H3/3I14 모델에서 오렌지색으로, H5/3I14 모델에서는 황색으로 표시되고 여기서 D94는 스틱으로 제시된다. 도 13d는 H3/3I14 복합 모델에서 3I14 경쇄의 G31과 H3의 상호작용을 도시한 것이다. H3의 HA2 도메인의 나선 A는 청록색 리본으로 표시되고; 3I14의 경쇄는 오렌지색으로 리본으로 표시되고; G31의 주쇄 원자는 스틱으로 표시되고 H3 HA2의 Q42 및 D46의 측쇄 원자는 막대기로 표시되고; G31과 H3 사이의 거리는 녹색 점선으로 표시되고 흑색으로 표지된다(PyMOL Molecular Graphics System, Version 0.99 rc6 Schroedinger, LLC).
도 14는 H3/3I14 및 H5/3I14 모델의 서열 정렬(a) 및 구조 중첩(b)을 도시한 것이다.
또한, 15a-15f는 재조합 H5-VN04(a) 및 H3-PE09(b)에 결합하는 3I14 WT 및 VLD94N IgG1 변이체의 Kd 결합 값을 보여주는 그래프 및 표이다. 녹색 또는 청색 곡선은 생물층 간섭 측정 실험으로부터 얻은 실험 기록이고, 적색 곡선은 Kd 값을 계산하기 위해 사용되는 데이터에 대한 전반적인 최적 적합도(best global fit)이다. 결합 곡선 친화도 측정채(Kd 값)는 표 4에 보고된다. 3I14 WT는 1.15 nm의 Kd 값으로 정제된 H5-VN04에 결합한 반면, 3I14 VLD94N 돌연변이체는 0.19 nM에서 10배 더 높은 친화도로 H5-VN04에 결합하였다. 도 15c-f는 재조합 Ha에 결합하는 3I14 scFvFc를 나타내는 일련의 그래프이다.
도 16a-d는 슈도타이핑된(pseudotyped) 바이러스 H5N1-VN04 및 감염성 바이러스 H3N2-BR07과 함께 인큐베이팅한 후, 3I14 WT 및 VLD94N 돌연변이체 IgG1의 중화 값을 나타내는 일련의 그래프이다. 도 16c 및 16d에서, 3I14(흑색) 및 VLD94N 변이체(적색)는 슈도타이핑된 바이러스 H5N1-VN04(c) 및 H3N2-BR07 바이러스(d)를 중화하였다. 이 데이터는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다.
도 17은 3I14 scFvFc Ab가 인플루엔자 바이러스 감염 및 HA-슈도타이핑된 루시페라제 리포터 바이러스를 중화함을 나타내는 일련의 그래프이다. MAb 3I14(흑색) 및 항-그룹 1 mAb F10(적색)은 감염성 바이러스 및 슈도타이핑된 바이러스의 상이한 균주를 중화하였다. 데이터는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다.
도 18은 조작된 3I14 효모-CDR 라이브러리의 H5에 대한 결합과 비교하여 3I14-WT 효모의 결합을 나타내는 일련의 유동 세포 계측 그래프이다. 3I14 효모-CDR 라이브러리는 H5에 대한 결합을 증가시키는 변이체에 대한 효모 디스플레이를 사용하여 조작되었다.
도 19는 재조합 H3-PE09의 전장 또는 HA1에 결합하는 3I14 scFvFc Ab를 나타내는 그래프이다.
도 20은 3I14 IgG1 변이체의 재조합 H1, H3 및 H5에 대한 결합을 나타내는 일련의 그래프이다. 청색 곡선은 생물층 간섭 측정 실험으로부터 얻은 실험 기록이고, 적색 곡선은 표 7에 제시된 Kd를 계산하기 위해 사용된 데이터에 대한 전반적인 최적 적합도이다.
인플루엔자 A는 음성 단일 가닥 RNA 바이러스이고, 10개의 단백질을 코딩하는 8 개 분절의 게놈을 갖는다. 이것은 뉴클레오캡시드 및 매트릭스 단백질의 항원성에 의해 정의된 인플루엔자 바이러스 A, B 및 C의 속을 포함하는 오르토믹소비리대(Orthomyxoviridae)에 속한다. 일반적으로, 인플루엔자 A 바이러스는 인간의 보다 심각한 질환과 연관된다. 인플루엔자 A 바이러스는 2개의 표면 단백질, 즉 세포 진입을 위해 숙주 세포에 비리온을 부착시키는 헤마글루티닌(HA: hemagglutinin) 및 자손체 바이러스 또는 세포 표면에 부착된 숙주 시알산을 절단함으로써 자손체 바이러스의 전파를 촉진하는 뉴라미니다제(NA: neuraminidase)에 의해 추가로 하위 분류된다.
HA 및 NA의 다양한 조합에 의해 인플루엔자 A 바이러스의 모든 아형을 구성하는 16개의 HA 아형 및 9개의 NA 아형이 존재한다. 16개의 HA 및 9개의 NA 바이러스 아형의 모든 조합은 물새에서 발견된다. 수 백종의 조류 인플루엔자 A 바이러스 균주 중, 인간의 감염을 일으킨 것으로 알려진 바이러스는 오직 H5N1, H7N3, H7N7 및 H9N2 뿐이다. 일반적으로, 이들 바이러스에 의한 인간의 감염은 경증 증상 및 극히 적은 심각한 병을 일으켰고. H7N7에 의한 폐렴의 유일한 치명적인 사례가 있었다. 그러나, 인간에게 자연적인 면역이 없는 고병원성 H5N1 바이러스는 예외이다. RNA 폴리머라제의 부정확(infidelity) 및 숙주 면역의 선택 압력은 돌연변이의 축적 및 이들 단백질의 표면 항원성의 변화로 이어질 수 있다. 이 항원 변화는 항원 드리프트(antigenic drift)라고한다. 또한, 그의 분절화된 게놈의 결과로서, 인플루엔자 A 바이러스의 2개의 상이한 아형이 동일한 세포를 감염시키면 유전자 분절이 뒤섞일 수 있다. 예를 들어, 인간 H3N2 바이러스 및 조류 H5N1 바이러스가 인간 또는 포유동물 종의 다른 구성원을 동시에 감염시키면, 이러한 사건은 새로운 H5N2를 생성할 수 있다. 이 새로운 바이러스는 대부분의 유전자 분절이 인간의 바이러스로부터 유래하기 때문에 인간으로부터 인간으로 효율적으로 전염될 수 있다. 그러한 유전자 재편성(reassortment)은 항원 변위(antigenic shift)라고 불리는 중요한 항원 변화를 일으키는데, 이것은 전체 집단의 대부분이 재편성 바이러스에 대한 어떠한 중화 항체도 갖지 않을 것임을 의미한다. 이러한 상황은 인플루엔자 H5N1 폐렴의 높은 사망률과 함께, 공중 보건 분야의 가장 두려운 시나리오 중의 하나이다.
인플루엔자 바이러스 헤마글루티닌(HA)은 인플루엔자 바이러스의 가장 가변적 인 항원이고, 바이러스의 세포 내로의 유입을 담당한다. 이는 단일 디술피드 결합에 의해 연결된 2개의 폴리펩티드 HA1 및 HA2로 번역 후 절단되는 삼량체 전구체 폴리펩티드 HA0으로서 합성된다. HA의 HA1 사슬은 바이러스의 세포 표면에 대한 부착을 담당한다. HA2는 엔도솜에서 바이러스와 세포막의 융합을 매개하여, 리보핵단백질 복합체가 세포질로 방출되도록 한다. HA1과 달리, HA2 분자는 HA의 비교적 보존된 부분을 나타낸다. 제2 면역원성 인플루엔자 단백질은 뉴라미니다제(NA)이다. 이 사량체 당단백질은 생산자 세포에서 표면 시알산으로부터 비리온을 방출하는 역할을 하며, 기도의 표적 세포에 대한 접근을 촉진하는 역할을 할 수도 있다. NA에 대한 중화 항체가 동물 및 인간을 보호하지만, 그 작용 메커니즘에 대한 데이터가 부족하다. N 뉴라미니다제의 결정 구조에 관한 최근 보고서는 항체를 비롯하여 새로운 항인플루엔자 약물을 개발하기 위해 이용될 수 있는 활성 부위에 인접한 공동의 존재를 입증하였다. 이 발견은 H5N1 바이러스에 대한 약물인 오셀타미비르(타미플루) 및 자나미비르(렐렌자)에 대한 약물 내성의 출현에 비추어 특히 중요하다.
20여 년 전에, H3 아형의 HA 분자는 항원 드리프트 변이체의 HA의 서열을 결정함으로써 특성화되었고, 항원 에피토프는 분자의 3차원 구조 상에 매핑되었다. 그 이후, 조류 병원성 바이러스의 H1, H2, H5에 있는 항원 부위가 H3의 3차원 구조에 매핑되었다. 1997년 홍콩에서 인간에게 H5N1 감염이 발생하고 1999년에 인간 환자로부터 H9N2 바이러스가 단리된 후, 두 단백질의 X선 구조가 밝혀졌다. 그러나, 구조를 해결하기 위해 사용된 1997년 돼지 단리물(A/오리/싱가폴/3/97), 및 보다 최근에 단리된 고병원성 균주의 항원 드리프트가 중요하다. 실제로, 돼지 단리물(A/오리/싱가폴/3/97)과 HPAI H5N1 균주(A/베트남1203/04) 사이에는 28건의 사소한 변화 및 2건의 잠재적으로 큰 변화가 존재한다.
2004-2005년 발생한 H5 HA 유전자의 계통발생 분석은 클레이드(clade) 1 및 2로 지칭되는 HA 유전자의 2개의 상이한 계통을 보여주었다. HPAI H5N1 균주(A/베트남1203/04)는 클레드 1의 구성원이다. 이들 클레이드 각각의 바이러스는 아시아의 중첩되지 않는 지리적 영역에 분포한다. 인도차이나의 H5N1 바이러스는 클레이드 1 내에 밀집되어 있는 반면, 여러 주변 국가로부터 단리된 H5N1은 클레이드 1 단리물과 구별되며, 보다 분기하는 클레이드 2에 속한다. 클레이드 1 바이러스는 베트남, 태국, 캄보디아의 인간 및 조류로부터 단리되었지만, 라오스 및 말레이시아에서는 조류로부터만 단리되었다. 클레이드 2 바이러스는 중국, 인도네시아, 일본 및 한국의 조류로부터만 단리된 바이러스에서 발견되었다. 가장 최근의 역학 연구는 바이러스 도입, 유행성 및 진화와 관련된 질문을 해결하기 위해 공공 데이터베이스에서 이용 가능한 서열 데이터와 함께, 인도네시아 및 베트남 전역의 가금류로부터 단리된 82개의 H5N1 바이러스, 및 베트남 남부의 11개의 인간 단리물을 분석하였다36. 계통발생 분석은 인도네시아의 모든 바이러스가 H5N1 유전자형 Z 바이러스의 차별적인 방계(sublineage)를 형성함을 보여주었고, 이것은 상기 발생이 지난 2년 동안 전국에 걸친 확산을 통한 단일 도입으로부터 비롯된 것일 가능성이 있음을 시사한다. 인도네시아에서 계속된 바이러스 활동은 조류 이동에 의한 반복적인 도입보다는 전국적인 가금류 이동을 통한 전염에 기인하였다. 인도네시아 및 베트남 내에서, H5N1 바이러스는 시간이 지남에 따라 각국 내의 지리적으로 다른 그룹으로 진화하였다.
최근에, A/베트남1203/4의 HA 구조가 해결되었다. 클레이드 1 및 2 바이러스로부터의 HPAI 2004 및 2005 단리물의 HA 유전자와 그의 아미노산 서열의 비교는 수용체 결합 도메인 주위에 주로 클러스터링된 항원 변이의 13개 위치를 확인하고, 나머지는 잔존 에스테라제 도메인 내에 존재하였다. 항원 변이의 영역은 H1 및 H3 혈청형에서 확인되었다. H1의 경우, 이들 부위는 Sa, Sb, Ca 및 Cb로 지정되고, H3의 경우, 부위는 A, B, C 및 D로 지정된다. H5 HA의 탈출 돌연변이체는 3개의 에피토프로 클러스터링될 수 있다; 부위 1: H3의 항원 부위 A 및 H2의 Ca2에 중첩되는 노출 루프(HA1 140-145); 부위 2: H3 혈청형에서 항원 부위 B에 상응하는 HA1 잔기 156 및 157; 부위 3: H1 HA 및 H9 혈청형에서 Sa 부위로 제한되는 HA1 129-133. 스미스(Smith)에 의한 최근의 연구에서, 아미노산 수준에서의 양성 선택의 검출은 HA 단백질 내의 8개의 잔기가 양성 선택 하에 있음을 보여주었다. 이들 잔기는 항원 부위 A 및 E 내의 5개(위치 83, 86, 138, 140 및 141)를 포함하고; 2개는 수용체 결합에 관여하고(위치 129 및 175); 위치 156은 수용체 결합 부위 근처에 잠재적인 N-연결된 글리코실화를 위한 부위이다. 그 결과는 HA 내의 3개의 잔기(Val 86, Ser 129 및 Thr 156)가 닭 또는 오리 단리물에서보다 인간 단리물에서 더 자주 관찰되었고, 인간에 대한 H5N1 유전자형 Z의 조기 적응을 나타낼 가능성이 있음을 보여주었다. 상기 연구로부터 또 다른 중요한 발견은 인도네시아와 베트남 방계 사이의 계통발생적 차이가 또한 상기 두 그룹의 바이러스 사이의 항원 교차 반응성의 유의한 차이에 반영된다는 것이다. 구체적으로, 인도네시아의 바이러스는 A/베트남1203/04에 대한 흰족제비 항혈청에 반응하지 않았고, 베트남의 대표적인 바이러스는 인도네시아 바이러스 IDN/5/06 및 Dk/IDN/MS/04에 대한 흰족제비 항혈청과 반응하지 않았다. 이러한 발견은 균주가 계통발생학적으로뿐만 아니라 항원 상으로 구분된다는 면역 인간 혈청 및 인간 1997 및 2003 H5N1 바이러스에 대한 초기 연구들과 일치한다. 따라서, 자연적 변이 및 탈출 돌연변이체는 어떤 균주(들)가 수동 및 능동 면역화를 위해 사용되어야 하는지에 대한 결정에 바이러스의 지속적인 진화가 영향을 주어야 함을 시사한다.
scFV
및
모노클로날
항체의 확인 및 특성 규명
고친화도, 교차-아형, 광범위 중화 인간 항-HA mAb가 확인되었다. 항원 특이적 기억 B 세포는 사량체화된 H3(A/브리즈번/10/2007) 헤마글루티닌(HA) 삼량체를 사용하여 인간 PBMC로부터 단리되었다. H3 반응성 단일 기억 B 세포를 플레이트 내에 분류하고, 시험관 내에서 자극하였다. 40% 이상의 분류된 B 세포는 14일 후에 배양 상청액에서 평균 200 ng/ml IgG를 생산하였다. 팽창된 B 세포로부터의 상청액은 MSD 또는 고도로 민감한 중화 분석에 의해 그의 이종아형 결합 특이성 및 중화 활성에 대해 측정되었다. 선택적 클론으로부터의 항체 유전자를 단일 세포 RT-PCR에 의해 회수하였다.
7명의 개체로부터 2688개의 기억 B 클론의 스크리닝을 통해, 11%의 클론 가능한 기억 B 세포가 H3 헤마글루티닌과 반응하였다. 그 중, H3/H7, H3/H7/H1 및 H3/H7/H1/인플루엔자 B 이종아형 결합 집단은 각각 16%, 6.9% 및 0.35%이었다. 광범위하게 중화하는 새로운 Ab 3I14가 확인되었다. 3I14는 특성화되었고, 그룹 1 및 그룹 2 인플루엔자 A 바이러스 둘 모두에 대해 교차 반응성 결합 및 중화 활성을 갖는 것으로 나타났다. 이것은 그룹 1 또는 그룹 2 인플루엔자 A 바이러스를 중화하는 다른 알려진 항인플루엔자 항체, 예컨대 F10, CR6261, MAb 3.1 및 CR8020와는 대조적이다. 단지, 항인플루엔자 항체 FI6v3, CR9114, 39.39, MAb 1.12 및 CT149만이 그룹 1 및 그룹 2의 인간 인플루엔자 A 바이러스를 중화할 수 있다. 인간 형질 세포, 형질모세포 및 CD138+ HA 특이적 항체 분비 세포의 배양액으로부터 단리된 FI6v3, CR9114, 39.39, MAb 1.12 및 CT149와 대조적으로, 본 발명의 항체, 예를 들어 3I14는 기억 B 세포로부터 단리되었다. 바이러스 재감염 및 백신 접종에 대한 반응으로, 오래 생존하는 형질 세포는 특히 원래의 바이러스를 기억하는 중화 항체를 생산하는 반면, 기억 B 세포는 생식 센터에 다시 들어가 변이체 바이러스에 특이적인 고친화도 중화 항체를 생산함으로써 기여한다. 또한, 기억 B 세포의 체세포 돌연변이는 항체 분기(divergence) 및 선택의 반복된 사이클을 통해 노인에 축적될 수 있다. 따라서, 기억 B 세포는 오래 생존하는 형질 세포보다 항원 특이성의 보다 넓은 레퍼토리를 갖는다. 오래 지속되고 광범위하게 효과가 있는 백신이 안정적인 기억 B 세포 집단을 개발하고 강력한 bnAb 반응을 이끌어내는 것이 필수적인 것으로 생각된다. 따라서, 본 발명의 항체는 다른 공지된 항인플루엔자 항체보다 더 큰 치료 유용성을 가질 것이다.
본 발명의 항체는 그룹 2(H3, H4, H7, H14 및 H15) 및 그룹 1(H1, H2, H5, H6, H8, H9, H11, H12 및 H16) 인플루엔자 A 둘 모두의 혈청형에 걸친 표면 발현된 HA에 결합한다. 구체적으로, 본 발명의 항체의 결합 친화도(Kd)는 약 1 pM 내지 1 μM, 약 1 pM 내지 1 nM 또는 약 1 nM 내지 1 nM이다. 예를 들어, 항체는 그룹 1(H1, H5 및 H9) 및 그룹 2(H3, H4, H7 및 H17)에 대해 약 1 pM 내지 1 μM의 결합 친화도를 갖는다. 바람직하게는, 그룹 1(H1, H5 및 H9) 및 그룹 2(H3, H4, H7 및 H17)에 대한 결합 친화도(Kd)는 약 0.01 nM 내지 10 nM이다. 일부 실시양태에서, 항체는 그룹 2 인플루엔자 A 바이러스 HA(H3, H4, H7 및 H14)에 대해 약 1 pM 내지 1 μM의 결합 친화도를 갖는다. 바람직하게는, 그룹 2 HA(H3, H4, H7 및 H14)에 대한 결합 친화도(Kd)는 <1 nM이다.
구체적으로, 3I14는 그룹 2(H3, H4, H7 및 H14) 및 그룹 1(H1, H5 및 H9)에 속하는 상이한 아형의 정제된 HA 단백질에 0.01 nM 내지 10 nM의 해리 상수(Kd)로 및 시험된 모든 그룹 2 HA(H3, H4, H7 및 H14)에 높은 친화도(평균 Kd < 0.1 nM)로 결합하였다. 또한, 3I14는 높은 친화도로 그룹 1 H1 아형(H1-CA09, H1-SI06 및 H1-PR8)에 결합한 반면, 다른 그룹 1 아형(H5-VN04, H5-IN05 및 H9-HK99)에 대한 그의 친화도는 더 낮았다(각각 평균 Kd = 1.02, 1.05 및 5.23 nM). H5 인플루엔자 바이러스 아형에 대한 상기 더 낮은 결합 친화도는 FI6v3 및 39.29와 같이 이전에 설명된 다른 광범위하게 중화하는 항체와는 다르다.
본 발명의 항체는 인플루엔자 A 바이러스를 중화한다. "중화하다" 또는 "중화"는 바이러스 입자에 대한 항체의 결합에 의한 바이러스 감염성의 감소를 유도하고, 따라서 바이러스에 의해 코딩되는 전사 또는 합성에 선행하는 바이러스의 복제 사이클의 단계를 차단함을 의미한다. 항체는 다양한 메커니즘에 의해 바이러스를 중화할 수 있고, 예를 들어, 항체는 수용체에 대한 비리온 결합을 방해하여 바이러스를 중화하거나, 세포 내로의 흡수를 차단하거나, 엔도솜에서 게놈의 탈각(coating)을 방지하거나, 바이러스 입자를 응집 또는 용해시킬 수 있다.
본 발명의 항체는 그룹 2 및 그룹 1 인플루엔자 A 바이러스 둘 모두의 혈청형을 중화한다. 본 발명의 항체는 약 0.001 내지 5 ㎍/mL, 약 0.001 내지 4 ㎍/mL, 또는 약 0.001 내지 3 ㎍/mL의 50% 최대 억제 농도(IC50)를 갖는다. 바람직하게는, 항체는 약 0.03 내지 2 ㎍/mL, 약 0.03 내지 1.0 ㎍/mL의 IC50을 갖는다. 훨씬 더 바람직하게는, 항체는 약 0.001 내지 0.5 ㎍/mL, 약 0.001 내지 0.05 ㎍/mL, 또는 약 0.001 내지 0.03 ㎍/mL의 IC50을 갖는다. 훨씬 더 바람직하게는, 항체는 약 0.01 내지 0.5 ㎍/mL, 약 0.1 내지 0.5 ㎍/mL, 또는 약 0.2 내지 0.5 ㎍/mL의 IC50을 갖는다. 바람직하게는, 항체는 약 0.05 내지 0.008 ㎍/mL, 또는 약 0.04 내지 0.008 ㎍/mL의 IC50을 갖는다. 가장 바람직하게는, 항체는 약 0.03 내지 1.08 ㎍/mL, 약 0.007 내지 0.027 ㎍/mL, 약 0.225 내지 0.413 ㎍/mL, 또는 약 0.040 내지 0.008 ㎍/mL의 IC50을 갖는다.
구체적으로, 본 발명의 항체는 그룹 2 바이러스(예를 들어, H3, H7, A/위스콘신/67/05(HA, NA) x A/푸에르토리코/8/34 및 A/아이치/2/68(HA, NA) x A/푸에르토리코/8/34 및 H7N9-AH13)를 중화한다. 본 발명의 항체는 또한 슈도바이러스 H7N1-FPN 및 H7N1-NL219 균주를 중화한다. 또한, 본 발명의 항체는 그룹 1 H1 균주(H1-CA09 및 H1-PR8) 및 슈도바이러스 H5-VN04 및 H5-HK97을 중화한다.
본 발명의 항체는 생체 내에서 그룹 1 및 그룹 2 인플루엔자 A 바이러스 둘 모두에 대한 예방 효능을 갖는다. 본 발명의 항체는 바이러스 감염에 대한 50%, 60%, 70%, 80%, 90%, 95% 또는 100%의 예방적 보호를 제공한다. 구체적으로, 본 발명의 항체는 H7N7-NL219 또는 H7N9-AH13 시험접종으로부터 완전한 보호, H3N2-BR07 시험접종에 대해 80%의 보호 및 H5N1-VN04 시험접종에 대해 60%의 보호를 제공한다.
본 발명의 항체는 미성숙 HA0의 절단을 방지한다. HA0 단백질이 HA1 및 HA2를 형성하기 위해 절단되지 않으면, 바이러스-숙주 막 융합은 발생할 수 없다. 따라서, HA가 절단되지 않은 인플루엔자 바이러스는 전염성이 없다. 따라서, 본 발명의 항체는 인플루엔자 감염을 차단하는데 유용하며, 따라서 예를 들어 타미플루와 같은 다른 항바이러스제와 조합하여 사용될 수 있다.
중요하게는, 본 발명의 항체는 절단되지 않은 HA 전구체(HA0) 단백질 및 2개의 성숙한 형태의 HA1 단백질 및 HA2 단백질에 결합한다.
추가로, 본 발명의 항체는 낮은 pH에 의해 촉발된 HA 구조적 재배열을 방지한다.
항체는 Fc 의존성 바이러스 청소(clearance)를 매개한다. 일부 실시양태에서, 항체는 항체 의존성 세포독성(ADCC)을 증가시킨다. 대안으로, 항체는 생체내 보호를 위한 Fc 의존성 면역 매개 메커니즘에 참여한다.
본 발명의 항체의 가변 중쇄는 IGHV3-30 생식계열 유전자에 의해 코딩된다. 항체의 가변 경쇄는 IGLV1-44 생식계열 유전자에 의해 코딩된다. IGHV3-30 항체는 HA 결합에 기여하는 소수성 코어를 형성하기 위해 HCDR3을 사용한다. 항체는 예컨대 긴 상보성 결정 영역 3(HCDR3)을 생성하기 위해 재배열된 중쇄를 갖는다. 긴 HCDR3의 길이는 약 12 내지 30개의 아미노산(예를 들어, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30개)일 수 있다. 바람직한 실시양태에서, 긴 HCDR3은 약 23개 아미노산 길이이다. 일부 실시양태에서, 긴 HCDR3은 VH 및 IGHJ4*02 접합부 둘 모두에서 큰 N-부가물이 측면에 있는 IGHD3-22*01 DH 분절을 사용한다.
항체는 가변 중쇄 및/또는 가변 경쇄에 체세포 돌연변이를 갖는다. 가변 중쇄의 체세포 돌연변이의 수는 약 2 내지 30(예를 들어, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30)일 수 있다. 일부 실시양태에서, 가변 중쇄의 체세포 돌연변이의 수는 약 15이다. 가변 경쇄의 체세포 돌연변이의 수는 약 2 내지 15(예를 들어, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15)일 수 있다. 일부 실시양태에서, 체세포 돌연변이의 수는 약 7이다.
에피토프 매핑 및 경쟁 분석은 HA 스템에 위치하는 고도로 보존된 에피토프를 보여주었다. 예를 들어, 본 발명의 항체는 서열 번호 18에 따라 넘버링될 때 HA2의 아미노산 잔기 18, 19, 20, 21, 36, 38, 39, 41, 42, 45, 46, 49 및 53에 의해 규정되는 구조적 에피토프에 결합한다. 대안으로, 본 발명의 항체는 서열 번호 18에 따라 넘버링될 때 HA2의 아미노산 잔기 18, 19, 20, 21, 38, 39, 41, 42, 45, 46, 47, 48, 49 및 50에 의해 규정되는 구조적 에피토프에 결합한다. 선택적으로, 항체는 HA1에 결합한다.
보존된 에피토프 잔기 서열은 펩티드 잔기에 의해 규정된다.
중등도의 아형 HA 균주에 대한 그의 효능을 개선하기 위해 3I14를 최적화하기 위해 구조-기반 항체 공학이 사용되어 왔다. 이 3I14의 고친화도 변이체는 본원에서 3I14VLD94N으로 지칭되고, 아미노산 위치 9에서 3I14 VH 내의 아스파르트산(D)의 아스파라긴(N)으로의 아미노산 치환에 의해 생성되었다.
VLD94L 치환은 H5에 대한 항체의 결합을 허용하거나 증가시킨다. H5에 대한 결합 친화도의 증가는 야생형 3I14에 비해 약 5 내지 15배이다. H5-VN04에 대한 3I14VLD94N의 Kd는 약 0.2 nM 미만이다.
추가의 구조 기반 공학은 H5에 대한 결합 친화도를 증가시킬 수 있다. 구체적으로, H5에 대한 결합 친화도의 증가는 LCDR1의 잔기 31에서의 글리신(G)을 또 다른 아미노산으로 치환함으로써 달성된다. 예를 들어, 잔기 31의 글리신(G)은 세린(S)으로 치환될 수 있다.
본 발명에 따른 중화 인플루엔자 항체의 핵산 및 아미노산 서열을 아래에 제시한다:




<표 8> IGHV3-03*18 및 IGLV1-44*01의 핵산 서열

3I14 및 3A14VLD94N 중화 인플루엔자 항체의 중쇄 및 경쇄 상보성 결정 영역의 아미노산 서열을 하기 표 2에 제시한다.
<표 2>

1X는 글리신 이외의 다른 임의의 아미노산일 수 있다. 바람직하게는, X는 세린이다.
<표 9> 헤마글루티닌 핵산 및 아미노산 서열



본원에서 사용되는 바와 같이, 용어 "항체"는 이뮤노글로불린 분자 및 이뮤노글로불린(Ig) 분자의 면역학적 활성 부분, 즉 항원에 특이적으로 결합(항원과 면역 반응)하는 항원 결합 부위를 함유하는 분자를 의미한다. "특이적으로 결합하다" 또는 "와 면역반응하다"는 항체가 원하는 항원의 하나 이상의 항원 결정자와 반응하고 다른 폴리펩티드와는 반응하지 않음을 의미한다. 항체는 폴리클로날, 모노클로날, 키메라, dAb(도메인 항체), 단일쇄, Fab, Fab' 및 F(ab')2 단편, scFv 및 Fab 발현 라이브러리를 포함하고, 이로 제한되지 않는다.
단일쇄 Fv("scFv") 폴리펩티드 분자는 펩티드 코딩 링커에 의해 연결된 VH- 및 VL-코딩 유전자를 포함하는 유전자 융합체로부터 발현될 수 있는 공유 연결된 VH:VL 이종이량체이다(문헌 [Huston et al. (1988) Proc Nat Acad Sci USA 85(16):5879-5883] 참조). 항체 V 영역으로부터 자연적으로 응집되지만 화학적으로 분리된 경쇄 및 중쇄 폴리펩티드를, 항원 결합 부위의 구조와 실질적으로 유사한 3차원 구조로 폴딩될 scFv 분자로 전환하는 화학 구조를 식별하기 위한 많은 방법이 기술되어 있다. 예를 들어, 미국 특허 제5,091,513호; 제5,132,405호; 및 제4,946,778호를 참조한다.
매우 큰 미자극(naive) 인간 scFv 라이브러리가 존재하고, 과다한 표적 분자에 대하여 재배열된 항체 유전자의 큰 공급원을 제공하기 위해 생성될 수 있다. 질환에 특이적인 항체를 단리하기 위해 감염성 질환이 있는 개체로부터 더 작은 라이브러리를 구축할 수 있다([Barbas et al., Proc. Natl. Acad. Sci. USA 89:9339-43 (1992)]; [Zebedee et al., Proc. Natl. Acad. Sci. USA 89:3175-79 (1992)] 참조).
일반적으로, 인간으로부터 수득된 항체 분자는 분자 내에 존재하는 중쇄의 특성에 의해 서로 상이한 클래스 IgG, IgM, IgA, IgE 및 IgD 중 임의의 것에 관련된다. 특정 클래스는 IgG1, IgG2, IgG3, IgG4 등과 같은 하위클래스를 또한 갖는다. 또한, 인간에서, 경쇄는 카파 사슬 또는 람다 사슬일 수 있다. 바람직하게는, 항체는 IgG1 또는 IgG4이다.
항체는 키메라 항체이다. 키메라 항체는 성숙 항체 중쇄와 생식계열 경쇄(mHgL)를 페어링하거나 또는 생식계열 중쇄와 성숙 경쇄(gHmL)를 페어링함으로써 생산된다. 키메라 항체는 야생형(WT) 항체와 비교하여 증가된 결합 친화도(Kd)을 갖는다. 예를 들어, 특정 바이러스(예를 들어, H1-CA09)에 대한 mHgL 및 gHmL 키메라 변이체의 결합 친화도는 약 0.001 nM 미만일 수 있다. 대안으로, mHgL 및 gHmL 키메라 변이체의 결합 친화도(Kd)는 WT 항체(예를 들어, 바이러스 H5-VN04 및 H3-PE09에 대한)에서 발견된 것보다 더 작다. 선택적으로, mHgL 및 gHmL 키메라 변이체의 결합 친화도는 WT 항체의 결합 친화도와 거의 동일하다.
"항원 결합 부위" 또는 "결합 부분"이라는 용어는 항원 결합에 참여하는 이뮤노글로불린 분자의 부분을 의미한다. 항원 결합 부위는 중쇄("H") 및 경쇄("L")의 N 말단 가변("V") 영역의 아미노산 잔기에 의해 형성된다. "초가변 영역"이라 불리는 중쇄 및 경쇄의 V 영역 내의 3개의 고도로 분기하는 스트레치(stretch)는 "프레임워크 영역" 또는 "FR"로 알려진 보다 보존된 인접 스트레치 사이에 삽입된다. 따라서, 용어 "FR"은 이뮤노글로불린에서 초가변 영역 사이에서 및 이에 인접하여 자연적으로 발견되는 아미노산 서열을 의미한다. 항체 분자에서, 경쇄의 초가변 영역 및 중쇄의 3개의 초가변 영역은 3차원 공간에서 서로에 대해 배치되어 항원 결합 표면을 형성한다. 항원 결합 표면은 결합된 항원의 3차원 표면에 상보성이고, 중쇄 및 경쇄 각각의 3개의 초가변 영역은 "상보성 결정 영역" 또는 "CDR"로 지칭된다.
본원에서 사용되는 바와 같이, 용어 "에피토프"는 이뮤노글로불린, scFv 또는 T 세포 수용체에 특이적으로 결합할 수 있는 임의의 단백질 결정자를 포함한다. 에피토프 결정자는 보통 아미노산 또는 당 측쇄와 같은 분자의 화학적 활성 표면 군으로 이루어지며, 일반적으로 특이적 전하 특성뿐만 아니라 특이적인 3차원 구조적 특성을 갖는다. 예를 들어, 항체는 폴리펩티드의 N-말단 또는 C-말단 펩티드에 대해 생성될 수 있다.
본원에서 사용되는 바와 같이, 용어 "면역학적 결합" 및 "면역학적 결합 특성"은 이뮤노글로불린 분자와 이뮤노글로불린이 특이적인 항원 사이에서 발생하는 유형의 비공유 상호작용을 나타낸다. 면역학적 결합 상호작용의 강도 또는 친화도는 상호작용의 해리 상수(Kd)로 표현될 수 있고, 여기서 보다 작은 Kd는 보다 큰 친화도를 나타낸다. 선택된 폴리펩티드의 면역학적 결합 특성은 관련 기술 분야에 널리 알려된 방법을 사용하여 정량될 수 있다. 그러한 방법 중 하나는 항원 결합 부위/항원 복합체 형성 및 해리의 속도를 측정하는 것을 수반하며, 이들 속도는 복합체 파트너의 농도, 상호작용의 친화도 및 양 방향의 속도에 동등하게 영향을 미치는 기하학적 파라미터에 의존한다. 따라서, "온 레이트(on rate) 상수"(Kon) 및 "오프 레이트(off rate) 상수"(Koff)는 농도 및 회합 및 해리의 실제 속도를 계산함으로써 결정될 수 있다(문헌[Nature 361:186-87(1993)] 참조). Koff/Kon의 비율은 친화도와 관련이 없는 모든 파라미터의 취소를 가능하게 하며, 해리 상수 Kd와 같다(일반적으로 문헌 [Davies et al. (1990) Annual Rev Biochem 59:439-473] 참조). 본 발명의 항체는 방사성 리간드 결합 분석 또는 관련 기술 분야의 통상의 기술자에게 공지된 유사한 분석과 같은 분석에 의해 측정될 때, 평형 결합 상수(Kd)가 1 μM, 바람직하게는 100 nM, 보다 바람직하게는 10 nM, 가장 바람직하게는 100 pM 내지 약 1 pM 일 때 인플루엔자 에피토프에 특이적으로 결합한다고 언급된다.
본 발명의 인플루엔자 단백질(예를 들어, HA 또는 뉴라미니다제), 또는 그의 유도체, 단편, 유사체, 상동체 또는 오르톨로그(ortholog)는 이들 단백질 성분에 면역특이적으로 결합하는 항체의 생성에서 면역원으로서 이용될 수 있다.
관련 기술 분야의 통상의 기술자는 본 발명의 인간 모노클로날 항체가 인플루엔자 바이러스의 HA 단백질에 결합하는 것을 인간 모노클로날 항체가 막는지의 여부를 확인함으로써, 인간 모노클로날 항체가 본 발명의 인간 모노클로날 항체와 동일한 특이성을 갖는지를 과도한 실험을 수행하지 않으면서 결정할 수 있다는 것을 인식할 것이다. 본 발명의 인간 모노클로날 항체에 의한 결합의 감소에 의해 제시되는 바와 같이, 시험되는 인간 모노클로날 항체가 본 발명의 인간 모노클로날 항체와 경쟁하는 경우, 2개의 모노클로날 항체는 동일하거나 또는 밀접하게 관련된 에피토프에 결합하는 것으로 보인다.
인간 모노클로날 항체가 본 발명의 인간 모노클로날 항체의 특이성을 갖는지의 여부를 결정하는 또 다른 방법은 본 발명의 인간 모노클로날 항체를 통상적으로 반응성 인 인플루엔자 HA 단백질과 미리 인큐베이팅한 후, 시험되는 인간 모노클로날 항체를 첨가하여 시험되는 인간 모노클로날 항체가 HA 단백질에 결합하는 그의 능력이 억제되는지의 여부를 결정하는 것이다. 시험되는 인간 모노클로날 항체가 억제된다면, 이 항체는 거의 틀림없이 본 발명의 모노클로날 항체와 동일하거나 기능적으로 동등한 에피토프 특이성을 갖는다. 본 발명의 인간 모노클로날 항체의 스크리닝은 또한 인플루엔자 바이러스를 사용하고 시험 모노클로날 항체가 인플루엔자 바이러스를 중화할 수 있는지의 여부를 결정함으로써 수행될 수 있다.
관련 기술 분야에 공지된 다양한 절차가 본 발명의 단백질, 또는 그의 유도체, 단편, 유사체, 상동체 또는 오르톨로그에 대해 작용하는 폴리클로날 또는 모노클로날 항체의 제조에 사용될 수 있다(예를 들어, 본원에 참고로 포함된 문헌 [Antibodies: A Laboratory Manual, Harlow E, and Lane D, 1988, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY] 참조).
항체는 주로 면역 혈청의 IgG 분획을 제공하는 단백질 A 또는 단백질 G를 사용하는 친화도 크로마토그래피와 같은 잘 알려진 기술에 의해 정제될 수 있다. 후속적으로 또는 대안으로, 추구되는 이뮤노글로불린의 표적인 특정 항원 또는 그의 에피토프를 면역친화도 크로마토그래피에 의해 면역 특이적 항체를 정제하기 위해 컬럼에 고정할 수 있다. 이뮤노글로불린의 정제는 예를 들어 문헌 [D. Wilkinson, The Scientist, published by The Scientist, Inc., Philadelphia PA, Vol. 14, No. 8 (April 17, 2000), pp. 25-28]에서 논의된다.
본원에서 사용되는 용어 "모노클로날 항체" 또는 "MAb" 또는 "모노클로날 항체 조성물"은 특유한 경쇄 유전자 산물 및 특유한 중쇄 유전자 산물로 이루어진 항체 분자의 단지 하나의 분자 종만을 함유하는 항체 분자의 집단을 지칭한다. 특히, 모노클로날 항체의 상보성 결정 영역(CDR)은 집단의 모든 분자에서 동일하다. MAb는 특유한 결합 친화도를 특징으로 하는 항원의 특정 에피토프와 면역반응할 수 있는 항원 결합 부위를 함유한다.
모노클로날 항체는 문헌 [Kohler and Milstein, Nature, 256:495 (1975)]에 기재된 것과 같은 하이브리도마 방법을 사용하여 제조할 수 있다. 하이브리도마 방법에서, 마우스, 햄스터 또는 다른 적절한 숙주 동물은 전형적으로 면역화제에 특이적으로 결합할 항체를 생산하거나 생산할 수 있는 림프구를 유도하기 위해 면역화제로 면역화된다. 다르게는, 림프구는 시험관 내에서 면역화할 수 있다.
면역화제는 전형적으로 단백질 항원, 그의 단편 또는 그의 융합 단백질을 포함할 것이다. 일반적으로, 인간 기원 세포를 원할 경우 말초 혈액 림프구를 사용하거나, 비인간 포유동물 공급원을 원할 경우에는 비장 세포 또는 림프절 세포를 사용한다. 이어서, 림프구를 폴리에틸렌 글리콜과 같은 적절한 융합제를 사용하여 불멸화된 세포주와 융합시켜 하이브리도마 세포를 형성시킨다(Goding, Monoclonal Antibodies: Principles and Practice, Academic Press,(1986) pp. 59-103). 불멸화된 세포주는 일반적으로 형질전환된 포유동물 세포, 특히 설치류, 소 및 인간 기원의 골수종 세포이다. 보통, 래트 또는 마우스 골수종 세포주가 사용된다. 하이브리도마 세포는 바람직하게는 융합되지 않은 불멸화된 세포의 성장 또는 생존을 억제하는 하나 이상의 물질을 함유하는 적합한 배양 배지에서 배양될 수 있다. 예를 들어, 모 세포에 효소 히포크산틴 구아닌 포스포리보실 트랜스퍼라제(HGPRT 또는 HPRT)가 결여되면, 하이브리도마에 대한 배양 배지에는 전형적으로 HGPRT-결핍된 세포의 성장을 억제하는 히포크산틴, 아미노프테린 및 티미딘("HAT 배지")을 포함할 것이다.
바람직한 불멸화된 세포주는 효율적으로 융합되고 선택된 항체 생산 세포에 의한 항체의 안정한 고수준 발현을 지지하며 HAT 배지와 같은 배지에 민감한 것이다. 보다 바람직한 불멸화된 세포주는 뮤린 골수종 세포주이며, 예를 들어 미국 캘리포니아주 샌 디에고 소재의 솔크 인스티튜트 셀 디스프리뷰션 센터(Salk Institute Cell Distribution Center) 및 미국 버지니아주 매나사스 소재의 아메리칸 타입 컬쳐 컬렉션(American Type Culture Collection)으로부터 얻을 수 있다. 인간 골수종 및 마우스-인간 이종골수종 세포주가 또한 인간 모노클로날 항체의 생산을 위해 설명되었다(문헌 [Kozbor, J. Immunol., 133:3001 (1984)]; [Brodeur et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63)] 참조).
하이브리도마 세포가 배양된 배양 배지를 항원에 대해 작용하는 모노클로날 항체의 존재에 대해 분석할 수 있다. 바람직하게는, 하이브리도마 세포에 의해 생성된 모노클로날 항체의 결합 특이성은 면역침강 또는 시험관내 결합 분석, 예컨대 방사성 면면분석(RIA) 또는 효소 연결 면역흡착 분석(ELISA)에 의해 결정된다. 이러한 기술 및 분석은 관련 기술 분야에 알려져 있다. 모노클로날 항체의 결합 친화도는 예를 들어 문헌[Munson and Pollard, Anal. Biochem, 107:220(1980)]의 스캐챠드(Scatchard) 분석에 의해 결정될 수 있다. 또한, 모노클로날 항체의 치료적 적용에서, 표적 항원에 대한 고도의 특이성 및 높은 결합 친화도를 갖는 항체를 확인하는 것이 중요하다.
목적하는 하이브리도마 세포가 확인된 후, 클론은 제한 희석 절차에 의해 서브클로닝되고 표준 방법에 의해 성장될 수 있다(문헌 [Goding, Monoclonal Antibodies: Principles and Practice, Academic Press,(1986) pp. 59-103] 참조). 이를 위해 적절한 배지는 예를 들어, 둘베코 변형 이글 배지(Dulbecco's Modified Eagle's Medium) 및 RPMI-1640 배지를 포함한다. 다르게는, 하이브리도마 세포는 포유동물에서 복수로서 생체 내에서 성장할 수 있다.
서브클론에 의해 분비된 모노클로날 항체는 통상적인 이뮤노글로불린 정제 절차, 예컨대 단백질 A-세파로스, 히드록실아파타이트 크로마토그래피, 겔 전기영동, 투석 또는 친화도 크로마토그래피에 의해 배양 배지 또는 복수액으로부터 단리되거나 정제될 수 있다.
모노클로날 항체는 또한 미국 특허 제4,816,567호에 기재된 것과 같은 재조합 DNA 방법에 의해 제조될 수 있다. 본 발명의 모노클로날 항체를 코딩하는 DNA는 통상적 인 절차를 사용하여(예를 들어, 뮤린 항체의 중쇄 및 경쇄를 코딩하는 유전자에 특이적으로 결합할 수 있는 올리고뉴클레오티드 프로브를 사용함으로써) 용이하게 단리 및 서열결정될 수 있다. 본 발명의 하이브리도마 세포는 이러한 DNA의 바람직한 공급원으로 작용한다. 일단 단리된 DNA를 발현 벡터에 도입한 다음, 그렇지 않으면 이뮤노글로불린 단백질을 생산하지 않는 숙주 세포, 예컨대 원숭이 COS 세포, 차이니즈 햄스터 난소(CHO) 세포 또는 골수종 세포 내로 형질감염시켜 재조합 숙주 세포에서 모노클로날 항체를 수득한다. DNA는 또한 예를 들어 상동성 뮤린 서열 대신에 인간 중쇄 및 경쇄 불변 도메인에 대한 코딩 서열로 치환하거나(미국 특허 제4,816,567호; 문헌 [Morrison, Nature 368, 812-13(1994)] 참조) 또는 비-이뮤노글로불린 폴리펩티드의 코딩 서열의 전부 또는 일부를 이뮤노글로불린 코딩 서열에 공유 연결함으로써 변형될 수 있다. 상기 비-이뮤노글로불린 폴리펩티드는 본 발명의 항체의 불변 도메인을 치환할 수 있거나, 또는 키메라 2가 항체를 제조하기 위해 본 발명의 항체의 한 항원 조합 부위의 가변 도메인을 치환할 수 있다.
완전 인간 항체는 CDR을 비롯한 경쇄 및 중쇄 둘 모두의 전체 서열이 인간 유전자로부터 유래하는 항체 분자이다. 이러한 항체는 본원에서 "인간 항체" 또는 "완전 인간 항체"로 지칭된다. 인간 모노클로날 항체는 트리오(trioma) 기술; 인간 B 세포 하이브리도마 기술(문헌 [Kozbor, et al., 1983 Immunol Today 4: 72] 참조); 및 인간 모노클로날 항체를 생산하는 EBV 하이브리도마 기술(문헌 [Cole, et al., 1985 In: MONOCLONAL ANTIBODIES AND CANCER THERAPY, Alan R. Liss, Inc., pp 77-96] 참조)을 사용하여 제조할 수 있다. 인간 모노클로날 항체는 인간 하이브리도마(문헌 [Cote, et al., 1983, Proc Natl Acad Sci USA 80: 2026-2030] 참조)를 사용하거나 또는 시험관 내에서 엡스타인 바 바이러스로 인간 B 세포를 형질전환함으로써 생산될 수 있다 (문헌 [Cole, et al., 1985 In: Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96] 참조).
또한, 인간 항체는 또한 파지 디스플레이 라이브러리를 포함하는 추가의 기술을 사용하여 생산될 수 있다(문헌 [Hoogenboom and Winter, J. Mol. Biol, 227:381(1991)]; [Marks et al., J. Mol. Biol, 222:581 (1991)] 참조). 유사하게, 인간 항체는 트랜스제닉(transgenic) 동물, 예를 들어, 내인성 이뮤노글로불린 유전자가 부분적으로 또는 완전히 불활성화된 마우스 내로 인간 이뮤노글로불린 유전자좌를 도입함으로써 제조될 수 있다. 시험접종시에, 인간 항체 생산이 관찰되고, 이것은 유전자 재배열, 조립 및 항체 레퍼토리를 포함하여 모든 측면에서 인간과 매우 유사하다. 이 방법은 예를 들어 미국 특허 번호 제5,545,807호; 제5,545,806호; 제5,569,825호; 제5,625,126호; 제5,633,425호; 제5,661,016호 및 문헌 [Marks et al., Bio/Technology 10, 779-783 (1992)]; [Lonberg et al., Nature 368 856-859 (1994)]; [Morrison, Nature 368, 812-13 (1994)]; [Fishwild et al., Nature Biotechnology 14, 845-51 (1996)]; [Neuberger, Nature Biotechnology 14, 826 (1996)]; 및 [Lonberg and Huszar, Intern. Rev. Immunol. 13 65-93 (1995)]에 기재되어 있다.
인간 항체는 항원 시험접종에 대한 반응으로 동물의 내인성 항체가 아닌 완전 인간 항체를 생산하도록 변형된 트랜스제닉 비인간 동물을 사용하여 추가로 생산될 수 있다(PCT 공개 WO94/02602 참조). 비인간 숙주에서 중쇄 및 경쇄 이뮤노글로불린을 코딩하는 내인성 유전자는 무능화되고, 인간 중쇄 및 경쇄 이뮤노글로불린을 코딩하는 활성 유전자좌가 숙주의 게놈에 삽입된다. 인간 유전자는 예를 들어 필요한 인간 DNA 분절을 포함하는 효모 인공 염색체를 사용하여 통합된다. 모든 원하는 변형을 제공하는 동물은 이어서 변형의 전체 보완에 미치지 않는 변형을 함유하는 중간체 트랜스제닉 동물을 교배시킴으로써 자손체로서 얻어진다. 상기 비인간 동물의 바람직한 실시양태는 마우스이고, PCT 공개 WO 96/33735 및 WO 96/34096에 개시된 바와 같이 제노마우스(Xenomouse)™로 불린다. 이 동물은 완전 인간 이뮤노글로불린을 분비하는 B 세포를 생성한다. 항체는 예를 들어 폴리클로날 항체의 제제로서 관심 면역원에 의한 면역화 후 동물로부터 직접 또는 모노클로날 항체를 생성하는 하이브리도마와 같은 동물 유래의 불멸화된 B 세포로부터 수득될 수 있다. 추가로, 인간 가변 영역을 갖는 이뮤노글로불린을 코딩하는 유전자는 직접 항체를 얻기 위해 회수 및 발현되거나, 또는 예를 들어 단일쇄 Fv(scFv) 분자와 같은 항체의 유사체를 수득하기 위해 추가로 변형될 수 있다.
내인성 이뮤노글로불린 중쇄의 발현이 결여된 마우스로서 예시된 비인간 숙주를 생산하는 방법의 일례가 미국 특허 제5,939,598호에 개시되어 있다. 이것은 유전자좌의 재배치를 방지하고 재배열된 이뮤노글로불린 중쇄 유전자좌의 전사체의 형성을 방지하기 위해 배아 줄기세포에서 적어도 하나의 내인성 중쇄 유전자좌로부터 J 분절 유전자를 결실시키고, 여기서 결실은 선택가능 마커를 코딩하는 유전자를 함유하는 표적 벡터에 의해 실행되는 것인 단계; 그의 체세포 및 생식 세포가 선택가능 마커를 코딩하는 유전자를 함유하는 트랜스제닉 마우스를 배아 줄기세포로부터 생산하는 단계를 포함하는 방법에 의해 수득될 수 있다.
관심 항체, 예컨대 인간 항체를 생산하는 한 방법은 미국 특허 제5,916,771호에 개시되어 있다. 이 방법은 중쇄를 코딩하는 뉴클레오티드 서열을 함유하는 발현 벡터를 배양 중의 하나의 포유동물 숙주 세포에 도입하는 단계, 경쇄를 코딩하는 뉴클레오티드 서열을 함유하는 발현 벡터를 또 다른 포유동물 숙주 세포에 도입하는 단계 및 두 세포를 융합시켜 하이브리드 세포를 형성하는 단계를 포함한다. 하이브리드 세포는 중쇄 및 경쇄를 함유하는 항체를 발현한다.
이 절차의 추가의 개선으로, 면역원에서 임상적으로 관련있는 에피토프를 확인하는 방법, 및 높은 친화도로 관련 에피토프에 면역 특이적으로 결합하는 항체를 선택하는 상관관계가 있는 방법이 PCT 공개 WO 99/53049에 개시되어 있다.
항체는 상기 설명한 단일쇄 항체를 코딩하는 DNA 분절을 포함하는 벡터에 의해 발현될 수 있다.
이들은 벡터, 리포솜, 네이키드(naked) DNA, 보조제-보조 DNA, 유전자 총, 카테테르 등을 포함할 수 있다. 벡터는 표적화 모이어티(moiety)(예를 들어, 세포 표면에 대한 리간드) 및 핵산 결합 모이어티(예를 들어, 폴리라이신)을 갖는 WO 93/64701에 기재된 바와 같은 화학적 접합체, 바이러스 벡터(예를 들어, DNA 또는 RNA 바이러스 벡터), 표적 모이어티(예를 들어, 표적 세포에 특이적인 항체) 및 핵산 결합 모이어티(예를 들어, 프로타민)을 포함하는 융합 단백질인 PCT/US 95/02140(WO 95/22618)에 기재된 융합 단백질, 플라스미드, 파지 등을 포함할 수 있다. 벡터는 염색체, 비-염색체 또는 합성 벡터일 수 있다.
바람직한 벡터는 바이러스 벡터, 융합 단백질 및 화학적 접합체를 포함한다. 레트로바이러스 벡터는 몰로니 뮤린 백혈병 바이러스를 포함한다. DNA 바이러스 벡터가 바람직하다. 이러한 벡터는 오르토폭스(orthopox) 또는 아비폭스(avipox) 벡터와 같은 폭스 벡터, 단순 포진 바이러스(HSV: herpes simplex I virus) 벡터와 같은 헤르페스바이러스 벡터(문헌 [Geller, A. I. et al., J. Neurochem, 64:487 (1995)]; [Lim, F., et al., in DNA Cloning: Mammalian Systems, D. Glover, Ed. (Oxford Univ. Press, Oxford England) (1995)]; [Geller, A. I. et al., Proc Natl. Acad. Sci.: U.S.A. 90:7603 (1993)]; [Geller, A. I., et al., Proc Natl. Acad. Sci USA 87:1149 (1990)] 참조), 아데노 바이러스 벡터(문헌 [LeGal LaSalle et al., Science, 259:988 (1993)]; [Davidson, et al., Nat. Genet 3:219 (1993)]; [Yang, et al., J. Virol. 69:2004 (1995)] 참조) 및 아데노 연관 바이러스 벡터(문헌 [Kaplitt, M. G., et al., Nat. Genet. 8:148 (1994)] 참조)를 포함한다.
폭스 바이러스 벡터는 유전자를 세포질에 도입한다. 아비폭스 바이러스 벡터는 단지 핵산의 단기적인 발현만을 유도한다. 아데노바이러스 벡터, 아데노 연관 바이러스 벡터 및 단순 포진 바이러스(HSV) 벡터가 핵산을 신경 세포에 도입하기에 바람직하다. 아데노 바이러스 벡터는 아데노 연관 바이러스(약 4개월; HSV 벡터보다 더 짧음)보다 더 짧은 기간의 발현(약 2개월)을 유도한다. 선택된 특정 벡터는 표적 세포 및 치료되는 병태에 따라 결정될 것이다. 도입은 표준 기술, 예를 들어 감염, 형질감염, 형질도입 또는 형질전환에 의한 것일 수 있다. 유전자 전달 방식의 예는 예를 들어 네이키드 DNA, CaPO4 침전, DEAE 덱스트란, 전기천공, 원형질체 융합, 리포펙션(lipofection), 세포 미세주입 및 바이러스 벡터를 포함한다.
벡터는 본질적으로 임의의 목적하는 표적 세포를 표적화하는데 사용될 수 있다. 예를 들어, 정위적(stereotaxic) 주사는 벡터(예를 들어, 아데노바이러스, HSV)를 원하는 위치로 유도하기 위해 사용될 수 있다. 추가로, 입자는 신크로메드 인퓨젼 시스템(SynchroMed Infusion System)과 같은 미니펌프 주입 시스템을 사용한 뇌실내(icv) 주입에 의해 전달될 수 있다. 대류라는 대량 유동을 기반으로 한 방법은 뇌의 확장된 영역에 큰 분자를 전달하는 데 효과적인 것으로 입증되었고, 표적 세포에 벡터를 전달하는 데 유용할 수 있다([Bobo et al., Proc. Natl. Acad. Sci. USA 91:2076-2080 (1994)]; [Morrison et al., Am. J. Physiol. 266:292-305 (1994)]). 사용될 수 있는 다른 방법은 카테테르, 정맥내, 비경구, 복강내 및 피하 주사, 및 경구 또는 다른 공지된 투여 경로를 포함한다.
이들 벡터는 다양한 방법으로 사용될 수 있는 다량의 항체를 발현하기 위해 사용될 수 있다. 예를 들어, 샘플에서 인플루엔자 바이러스의 존재를 감지한다. 항체는 또한 인플루엔자 바이러스 세포막 융합체에 결합하여 파괴시키려는 시도에 사용될 수 있다.
본 발명의 항원 단백질에 특이적인 단일쇄 항체의 제조를 위해 기술을 적용할 수 있다(예를 들어, 미국 특허 제4,946,778호 참조). 또한, 단백질 또는 그의 유도체, 단편, 유사체 또는 상동체에 대해 원하는 특이성을 갖는 모노클로날 Fab 단편의 신속하고 효과적인 확인을 가능하게 하기 위해, Fab 발현 라이브러리의 구축을 위해 방법을 적용할 수 있다(예를 들어, 문헌 [Huse, et al., 1989 Science 246: 1275-1281] 참조). 다음을 포함하고 이로 제한되지 않는, 단백질 항원에 대한 이디오타입(idiotype)을 함유하는 항체 단편은 관련 기술 분야에 공지된 기술에 의해 생산될 수 있다: (i) 항체 분자의 펩신 소화에 의해 생성된 F(ab')2 단편; (ii) F(ab')2 단편의 디술피드 다리를 환원시킴으로써 생성된 Fab 단편; (iii) 파파인 및 환원제로 항체 분자를 처리함으로써 생성된 Fab 단편 및 (iv) Fv 단편.
이종접합체 항체가 또한 본 발명의 범위 내에 포함된다. 이종접합체 항체는 2개의 공유 연결된 항체로 이루어진다. 이러한 항체는 예를 들어, 면역계 세포를 원하지 않는 세포로 표적화하기 위해(미국 특허 제4,676,980호 참조) 및 HIV 감염의 치료를 위해(WO 91/00360; WO 92/200373; EP 03089 참조) 제안되었다. 항체는 가교결합제를 수반하는 것을 포함하는 합성 단백질 화학에 공지된 방법을 사용하여 시험관 내에서 제조될 수 있는 것이 고려된다. 예를 들어, 면역독소는 디술피드 교환 반응을 이용하거나 티오에테르 결합을 형성함으로써 제조될 수 있다. 이를 위해 적합한 시약의 예는 이미노티올레이트 및 메틸-4-머캅토부티르이미데이트, 및 예를 들어 미국 특허 제4,676,980호에 개시된 것을 포함한다.
예를 들어, 인플루엔자의 치료에서 항체의 유효성을 향상시키기 위해, 이펙터 기능에 관하여 본 발명의 항체를 변형시키는 것이 바람직할 수 있다. 예를 들어, 시스테인 잔기(들)가 Fc 영역 내로 도입될 수 있고, 이에 의해 상기 영역에서 사슬간 디술피드 결합의 형성이 가능해진다. 이렇게 생성된 동종이량체 항체는 개선된 내재화 능력 및/또는 증가된 보체 매개 세포 사멸 및 항체 의존성 세포독성(ADCC)을 가질 수 있다(문헌 [Caron et al., J. Exp Med., 176: 1191-1195 (1992)] 및 [Shopes, J. Immunol., 148: 2918-2922 (1992)] 참조). 대안으로, 이중 Fc 영역을 갖고 이에 의해 향상된 보체 용해 및 ADCC 능력을 가질 수 있는 항체를 조작할 수 있다(문헌 [Stevenson et al., Anti-Cancer Drug Design, 3: 219-230 (1989)] 참조).
본 발명은 또한 세포독성제, 예컨대 독소(예를 들어, 박테리아, 진균, 식물 또는 동물 기원의 효소 활성을 갖는 독소, 또는 그의 단편) 또는 방사성 동위원소(즉, 방사성 접합체)에 접합된 항체를 포함하는 면역접합체에 관한 것이다.
사용할 수 있는 효소 활성을 갖는 독소 및 그의 단편은 디프테리아 A 사슬, 디프테리아 독소의 비결합 활성 단편, 외독소 A 사슬(슈도모나스 아에루기노사(Pseudomonas aeruginosa) 유래), 리신 A 사슬, 아브린 A 사슬, 모데신 A 사슬, 알파-사르신, 알류리테스 포르디이(Aleurites fordii) 단백질, 디안틴 단백질, 피톨라카 아메리카나(Phytolaca americana) 단백질(PAPI, PAPII 및 PAP-S), 모모르디카 카란티아(momordica charantia) 억제제, 쿠르친, 크로틴, 사파오나리아 오피시날리스(sapaonaria officinalis) 억제제, 겔로닌, 미토겔린, 레스트릭토신, 페노마이신, 에노마이신 및 트리코테센을 포함한다. 다양한 방사성 핵종이 방사성 접합된 항체의 생산을 위해 이용가능하다. 그 예는 212Bi, 131I, 131In, 90Y 및 186Re를 포함한다.
항체 및 세포독성제의 접합체는 다양한 이기능성 단백질-커플링제, 예컨대 N-숙신이미딜-3-(2-피리딜디티올)프로피오네이트(SPDP), 이미노티올란(IT), 이미도에스테르의 이기능성 유도체(예컨대, 디메틸 아디피미데이트 HCL), 활성 에스테르(예컨대, 디숙신이미딜 수베레이트), 알데히드(예컨대, 글루타르알데히드), 비스-아지도 화합물(예컨대, 비스(p-아지도벤조일)헥산디아민), 비스-디아조늄 유도체(예컨대, 비스-(p-디아조늄벤조일)-에틸렌디아민), 디이소시아네이트(예컨대, 톨리엔 2,6-디이소시아네이트) 및 비스-활성 불소 화합물(예컨대, 1,5-디플루오로-2,4-디니트로벤젠)을 사용하여 제조된다. 예를 들어, 리신 면역독소는 문헌 [Vitetta et al., Science 238: 1098(1987)]에 기재된 바와 같이 제조될 수 있다. 탄소 14 표지된 1-이소티오시아나토벤질-3-메틸디에틸렌 트리아민펜타아세트산(MX-DTPA)은 방사성 뉴클레오티드를 항체에 접합시키기 위한 예시적인 킬레이팅제이다(WO94/11026 참조).
관련 기술 분야의 통상의 기술자는 매우 다양한 가능한 모이어티가 생성되는 항체 또는 본 발명의 다른 분자에 커플링될 수 있음을 인식할 것이다(예를 들어, 그 전체 내용이 본원에 참고로 포함된 문헌 ["Conjugate Vaccines", Contributions to Microbiology and Immunology, J. M. Cruse and R. E. Lewis, Jr (eds), Carger Press, New York, (1989)] 참조).
커플링은 항체 및 다른 모이어티가 그 각각의 활성을 유지하는 한, 두 분자를 결합시킬 수 있는 임의의 화학 반응에 의해 달성될 수 있다. 이 연결은 예를 들어 공유 결합, 친화도 결합, 삽입(intercalation), 배위 결합 및 복합체화와 같은 많은 화학적 메커니즘을 포함할 수 있다. 그러나, 바람직한 결합은 공유 결합이다. 공유 결합은 기존 측쇄의 직접 축합 또는 외부 가교 분자의 도입에 의해 달성될 수 있다. 많은 2가 또는 다가 연결제가 본 발명의 항체와 같은 단백질 분자를 다른 분자에 결합시키는데 유용하다. 예를 들어, 대표적인 커플링제는 티오에스테르, 카르보디이미드, 숙신이미드 에스테르, 디이소시아네이트, 글루타르알데히드, 디아조벤젠 및 헥사메틸렌 디아민과 같은 유기 화합물을 포함할 수 있다. 이 목록은 관련 기술 분야에 공지된 다양한 클래스의 커플링제를 포괄하고자 하는 것이 아니라, 보다 일반적인 커플링제의 예를 제시하는 것이다(문헌 [Killen and Lindstrom, Jour. Immun. 133:1335-2549 (1984)]; [Jansen et al., Immunological Reviews 62:185-216 (1982)]; 및 [Vitetta et al., Science 238:1098 (1987)] 참조).
바람직한 링커는 문헌에 기재되어 있다(예를 들어, MBS(M-말레이미도벤조일-N-히드록시숙신이미드 에스테르)의 사용을 기술하는 문헌 [Ramakrishnan, S. et al., Cancer Res. 44:201-208 (1984)] 참조). 또한, 올리고펩티드 링커에 의해 항체에 커플 링된 할로겐화된 아세틸 히드라지드 유도체의 사용을 기술한 미국 특허 제5,030,719호를 참조한다. 특히 바람직한 링커는 다음을 포함한다: (i) EDC(1-에틸-3-(3-디메틸아미노-프로필) 카르보디이미드 히드로클로라이드, (ii) SMPT(4-숙신이미딜옥시카르보닐-알파-메틸-알파-(2-피리딜-디티오)-톨루엔(Pierce Chem. Co., Cat. (21558G)); (iii) SPDP(숙신이미딜-6[3-(2-피리딜디티오)프로피온아미도]헥사노에이트(Pierce Chem. Co., Cat. #21558G); (iv) 술포-LC-SPDP(술포숙신이미딜 6 [3-(2-피리딜디티오)-프로피온아미드]헥사노에이트(Pierce Chem. Co. Cat. #2165-G) 및 (v) EDC에 접합된 술포-NHS(N-히드록시술포-숙신이미드: Pierce Chem. Co., Cat. # 24510).
상기 설명된 링커는 상이한 속성을 갖는 성분을 함유하고, 따라서 상이한 물리화학적 특성을 갖는 접합체를 유도한다. 예를 들어, 알킬 카르복실레이트의 술포-NHS 에스테르는 방향족 카르복실레이트의 술포-NHS 에스테르보다 더 안정하다. NHS-에스테르 함유 링커는 술포-NHS 에스테르보다 덜 가용성이다. 또한, 링커 SMPT는 입체 장애 디술피드 결합을 함유하고, 증가된 안정성을 갖는 접합체를 형성할 수 있다. 디술피드 연결은 일반적으로 시험관 내에서 절단되기 때문에 다른 결합보다 덜 안정하여 이용가능한 접합체가 더 적게 된다. 특히 Sulfo-NHS는 카르보디이드 커플링의 안정성을 향상시킬 수 있다. 술포-NHS와 함께 사용되는 경우 카르보디이미드 커플링(예를 들어, EDC)는 카르보디이미드 커플링 반응 단독보다 가수분해에 대한 내성이 더 강한 에스테르를 형성한다.
본원에 개시된 항체는 또한 면역리포솜으로서 제제화될 수 있다. 항체를 함유하는 리포솜은 관련 기술 분야에 알려진 방법, 예를 들어 문헌 [Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688 (1985)]; [Hwang et al., Proc. Natl. Acad. Sci. USA, 77: 4030 (1980)]; 및 미국 특허 제4,485,045호 및 제4,544,545호에 기재된 방법에 의해 제조된다. 순환 시간이 향상된 리포솜은 미국 특허 제5,013,556호에 개시되어 있다.
특히 유용한 리포솜은 포스파티딜콜린, 콜레스테롤 및 PEG-유도체화 포스파티딜에탄올아민(PEG-PE)을 포함하는 지질 조성물로 역상 증발 방법에 의해 생성될 수 있다. 리포솜은 소정의 공극 크기의 필터를 통해 압출되어 원하는 직경의 리포솜을 생성한다. 본 발명의 항체의 Fab' 단편은 디술피드 교환 반응을 통해 문헌 [Martin et al., J. Biol. Chem., 257: 286-288 (1982)]에 기재된 바와 같이리포솜에 접합될 수 있다.
인플루엔자 바이러스에 대한 항체의 용도
원하는 특이성을 갖는 항체의 스크리닝 방법은 효소 연결 면역흡착 분석(ELISA) 및 관련 기술 분야에 알려진 다른 면역학적 매개 기술을 포함하고 이로 제한되지 않는다.
HA(또는 그의 단편)와 같은 인플루엔자 바이러스 단백질에 대해 작용하는 항체는 인플루엔자 바이러스 단백질의 국재화 및/또는 정량에 관한 관련 기술 분야에 알려진 방법에 (예를 들어, 적절한 생리학적 샘플 내의 인플루엔자 바이러스 단백질의 수준 측정에 사용하기 위해, 진단 방법에 사용하기 위해, 단백질의 영상화에 사용하기 위해 등) 사용될 수 있다. 제시된 실시양태에서, 항체 유래 항원 결합 도메인을 함유하는, 인플루엔자 바이러스 단백질, 또는 그의 유도체, 단편, 유사체 또는 상동체에 특이적인 항체는 약리학적 활성 화합물(이하 "치료제"라 함)로서 이용된다.
본 발명의 인플루엔자 바이러스 단백질에 특이적인 항체는 표준 기술, 예컨대 면역친화도, 크로마토그래피 또는 면역침전에 의해 인플루엔자 바이러스 폴리펩티드를 단리하기 위해 사용될 수 있다. 인플루엔자 바이러스 단백질(또는 그의 단편)에 대해 작용하는 항체는 예를 들어, 주어진 치료 요법의 효능을 결정하기 위한 임상 시험 절차의 일부로서 조직 내 단백질 수준을 모니터링하기 위해 진단학적으로 사용될 수 있다. 항체를 검출 가능한 물질에 커플링시킴으로써(즉, 물리적으로 연결함으로써) 검출을 용이하게 할 수 있다. 검출 가능한 물질의 예는 다양한 효소, 보결 분자단, 형광 물질, 발광 물질, 생물 발광 물질 및 방사성 물질을 포함한다. 적합한 효소의 예는 양고추냉이 퍼옥시다제, 알칼리성 포스파타제, β-갈락토시다제 또는 아세틸콜린에스테라제를 포함하고; 적합한 보결 분자단 복합체의 예는 스트렙타비딘/비오틴 및 아비딘/비오틴을 포함하고; 적합한 형광 물질의 예는 움벨리페론, 플루오레세인, 플루오레세인 이소티오시아네이트, 로다민, 디클로로트리아지닐아민 플루오레세인, 단실 클로라이드 또는 피코에리트린을 포함하고; 발광 물질의 예는 루미놀을 포함하고; 생물 발광 물질의 예는 루시페라제, 루시페린 및 에쿼린을 포함하고, 적합한 방사성 물질의 예는 125I, 131I, 35S 또는 3H를 포함한다.
폴리클로날, 모노클로날, 인간화 및 완전 인간 항체를 비롯한 본 발명의 항체는 치료제로서 사용될 수 있다. 그러한 작용제는 일반적으로 대상체에서 인플루엔자 바이러스 관련 질환 또는 병상(예를 들어, 조류 독감)을 치료 또는 예방하기 위해 사용될 것이다. 항체 제제, 바람직하게는 그의 표적 항원에 대해 높은 특이성 및 높은 친화도을 갖는 제제가 대상에게 투여되고, 일반적으로 그의 표적에 대한 결합으로 인해 효과를 나타낼 것이다. 항체의 투여는 바이러스의 세포 내로의 내재화를 폐기, 억제 또는 방해할 수 있다. 이 경우, 항체는 표적에 결합하고, 자연 발생 리간드의 결합 부위를 가려서 바이러스의 세포막에 대한 융합을 차단함으로써 바이러스의 내재화를 억제한다.
본 발명의 항체의 치료 유효량은 일반적으로 치료 목적을 달성하는데 필요한 양에 관련된다. 상기한 바와 같이, 이것은 특정 경우에 표적의 기능을 방해하는 항체와 그의 표적 항원 사이의 결합 상호작용일 수 있다. 투여될 필요가 있는 양은 또한 그의 특이적 항원에 대한 항체의 결합 친화도에 의존할 것이고, 또한 항체가 투여되는 다른 대상체로부터 투여된 항체가 고갈되는 속도에 의해 좌우될 것이다. 본 발명의 항체 또는 항체 단편의 치료상 효과적인 투여를 위한 통상적인 범위는 비제한적인 예로서 약 0.1 mg/kg(체중) 내지 약 50 mg/kg(체중)일 수 있다. 일반적인 투여 빈도는 예를 들어 1일 2회 내지 1주 1회일 수 있다.
본 발명의 인플루엔자 바이러스 단백질 또는 그의 단편에 특이적으로 결합하는 항체뿐만 아니라 본원에 개시된 스크리닝 분석에 의해 확인된 다른 분자는 인플루엔자 바이러스 관련 장애를 치료하기 위해 약학 조성물의 형태로 투여될 수 있다. 그러한 조성물의 제조에 관한 원리 및 고려 사항뿐만 아니라 성분의 선택에 대한 지침은 예를 들어 문헌 [Remington: The Science And Practice Of Pharmacy 19th ed. (Alfonso R. Gennaro, et al., editors) Mack Pub. Co., Easton, Pa., 1995]; [Drug Absorption Enhancement: Concepts, Possibilities, Limitations, And Trends, Harwood Academic Publishers, Langhorne, Pa., 1994]; 및 [Peptide And Protein Drug Delivery (Advances In Parenteral Sciences, Vol. 4), 1991, M. Dekker, New York]에 제시되어 있다.
항체 단편이 사용되는 경우, 표적 단백질의 결합 도메인에 특이적으로 결합하는 가장 작은 억제 단편이 바람직하다. 예를 들어, 항체의 가변 영역 서열에 기초하여, 펩티드 분자는 표적 단백질 서열을 결합하는 능력을 보유하도록 설계될 수 있다. 이러한 펩티드는 화학적으로 합성되고/되거나 재조합 DNA 기술에 의해 생산될 수 있다(예를 들어, 문헌 [Marasco et al., Proc. Natl. Acad. Sci. USA, 90: 7889-7893 (1993)] 참조). 제제는 또한 치료되는 특정 징후에 필요한 하나 초과의 활성 화합물, 바람직하게는 서로 유해한 영향을 주지 않는 보완 활성을 갖는 활성 화합물을 함유할 수 있다. 대안으로, 또는 부가적으로, 조성물은 예를 들어 세포독성제, 사이토카인, 화학치료제 또는 성장 억제제와 같은 그의 기능을 향상시키는 작용제를 포함할 수 있다. 이러한 분자는 목적하는 목적에 효과적인 양으로 적절하게 조합하여 존재한다.
활성 성분은 또한 예를 들어 코아세르베이션(coacervation) 기술에 의해 또는 계면 중합에 의해 제조된 마이크로캡슐, 예를 들어 각각 히드록시메틸셀룰로스 또는 젤라틴-마이크로캡슐 및 폴리(메틸메타크릴레이트) 마이크로캡슐에, 콜로이드성 약물 전달 시스템(예를 들어, 리포솜, 알부민 미세구, 미세에멀젼, 나노 입자 및 나노캡슐)에 또는 거대에멀젼에 포획될 수 있다.
생체내 투여에 사용되는 제제는 멸균되어야 한다. 이것은 멸균 여과막을 통한 여과에 의해 용이하게 달성된다.
서방성 제제를 제조할 수 있다. 서방성 제제의 적합한 예는 항체를 함유하는 고체 소수성 중합체의 반투과성 매트릭스를 포함하고, 이 매트릭스는 필름 또는 미세캡슐과 같은 성형된 물품의 형태이다. 서방성 매트릭스의 예는 다음을 포함한다: 폴리에스테르, 히드로겔(예를 들어, 폴리(2-히드록시에틸-메타크릴레이트) 또는 폴리(비닐알콜)), 폴리락티드(미국 특허 제3,773,919호), L-글루탐산과 γ 에틸-L-글루타메이트, 비분해성 에틸렌-비닐 아세테이트, 분해성 락트산-글리콜산 공중합체, 예컨대 루프론 데포(LUPRON DEPOT)™(락트산-글리콜산 공중합체 및 류프롤리드 아세테이트로 이루어진 주사가능한 미소구) 및 폴리-D-(-)-3-히드록시 부티르산. 에틸렌-비닐 아세테이트 및 락트산-글리콜산과 같은 중합체는 100일 초과 동안 분자를 방출할 수 있지만, 특정 히드로겔은 보다 짧은 시간 동안 단백질을 방출한다.
본 발명에 따른 항체는 샘플에서 인플루엔자 바이러스(또는 그의 단백질 또는 단백질 단편)의 존재를 검출하기 위한 작용제로서 사용될 수 있다. 바람직하게는, 항체는 검출 가능한 표지를 함유한다. 항체는 폴리클로날 또는 보다 바람직하게는 모노클로날 항체일 수 있다. 무손상 항체 또는 그의 단편(예를 들어, Fab, scFv 또는 F(ab)2)을 사용할 수 있다. 프로브 또는 항체와 관련하여 용어 "표지된"은 프로브 또는 항체에 대한 검출 가능한 물질의 커플링(즉, 물리적 연결)에 의한 프로브 또는 항체의 직접 표지뿐만 아니라 직접 표지된 또 다른 시약과의 반응성에 의한 프로브 또는 항체의 간접 표지를 포함하는 것이 의도된다. 간접 표지의 예는 형광 표지된 2차 항체를 사용하는 1차 항체의 검출 및 형광 표지된 스트렙타비딘으로 검출될 수 있도록 비오틴을 사용한 DNA 프로브의 말단 표지를 포함한다. "생물학적 샘플"이라는 용어는 대상체로부터 단리된 조직, 세포 및 생물학적 유체뿐만 아니라 대상체 내에 존재하는 조직, 세포 및 체액을 포함하도록 의도된다. 따라서 "생물학적 샘플"이라는 용어의 사용에는 혈액, 및 혈청, 혈장 또는 림프를 포함하는 혈액의 분획 또는 성분이 포함된다. 즉, 본 발명의 검출 방법은 시험관 내뿐만 아니라 생체 내에서 생물학적 샘플 내의 분석물 mRNA, 단백질 또는 게놈 DNA를 검출하기 위해 사용될 수 있다. 예를 들어, 분석물 mRNA를 검출하기 위한 시험관내 기술은 노던(Northern) 혼성화 및 인 시츄(in situ) 혼성화를 포함한다. 분석물 단백질의 검출을 위한 시험관내 기술은 효소 연결 면역흡착 분석(ELISA), 웨스턴(Western) 블롯, 면역침전 및 면역형광을 포함한다. 분석물 게놈 DNA를 검출하기 위한 시험관내 기술은 서던(Southern) 혼성화를 포함한다. 면역분석을 수행하는 절차는 예를 들어 문헌 ["ELISA: Theory and Practice: Methods in Molecular Biology", Vol. 42, J. R. Crowther (Ed.) Human Press, Totowa, NJ, 1995]; ["Immunoassay", E. Diamandis and T. Christopoulus, Academic Press, Inc., San Diego, CA, 1996]; 및 ["Practice and Theory of Enzyme Immunoassays", P. Tijssen, Elsevier Science Publishers, Amsterdam, 1985]에 기재되어 있다. 또한, 분석물 단백질의 검출을 위한 생체내 기술은 표지된 항-분석물 단백질 항체를 대상체에 도입하는 것을 포함한다. 예를 들어, 항체는 대상체 내의 그의 존재 및 위치가 표준 영상화 기술에 의해 검출될 수 있는 방사성 마커로 표지될 수 있다.
약학 조성물
본 발명의 항체 또는 작용제(본원에서 "활성 화합물"로도 지칭함) 및 그의 유도체, 단편, 유사체 및 상동체는 투여에 적합한 약학 조성물에 혼입될 수 있다. 이러한 조성물은 전형적으로 항체 또는 작용제 및 약학적으로 허용되는 담체를 포함한다. 본원에서 사용되는 바와 같이, 용어 "약학적으로 허용되는 담체"는 약학적 투여와 상용성인 임의의 및 모든 용매, 분산매, 코팅, 항박테리아 및 항진균제, 등장성 흡수 지연제 등을 포함하는 것이 의도된다. 적합한 담체는 본원에 참고로 포함된, 이 분야의 표준 참고문헌 [Remington's Pharmaceutical Sciences]의 최신판에 기재되어 있다. 이러한 담체 또는 희석제의 바람직한 예는 물, 염수, 링거 용액, 덱스트로스 용액 및 5% 인간 혈청 알부민을 포함하고, 이로 제한되지 않는다. 리포솜 및 비수성 비히클, 예컨대 고정유가 또한 사용될 수 있다. 약학적 활성 물질에 대한 상기 매질 및 작용제의 사용은 관련 기술 분야에 잘 알려져 있다. 임의의 통상적인 매질 또는 작용제가 활성 화합물과 비상용성인 경우를 제외하고, 조성물에서 그의 사용이 고려된다. 보충 활성 화합물이 또한 조성물에 혼입될 수 있다.
본 발명의 약학 조성물은 그의 의도된 투여 경로에 적합할 수 있도록 제제화된다. 투여 경로의 예는 비경구, 예를 들어 정맥내, 피내, 피하, 경구(예를 들어, 흡입), 경피(즉, 국소), 경점막 및 직장 투여를 포함한다. 비경구, 피내 또는 피하 적용에 사용되는 용액 또는 현탁액은 다음 성분을 포함할 수 있다: 멸균 희석제, 예컨대 주사용수, 염수 용액, 고정유, 폴리에틸렌 글리콜, 글리세린, 프로필렌 글리콜 또는 다른 합성 용매; 항박테리아제, 예컨대 벤질 알콜 또는 메틸 파라벤; 항산화제, 예컨대 아스코르브산 또는 아황산수소나트륨; 킬레이팅제, 예컨대 에틸렌디아민테트라아세트산(EDTA); 완충제, 예컨대 아세테이트, 시트레이트 또는 포스페이트, 및 긴장성 조절제, 예컨대 염화나트륨 또는 덱스트로스. pH는 산 또는 염기, 예컨대 염산 또는 수산화나트륨로 조절할 수 있다. 비경구 제제는 유리 또는 플라스틱으로 제조된 앰퓰, 일회용 주사기 또는 다중 용량 바이알에 넣을 수 있다.
주사가능한 용도에 적합한 약학 조성물은 멸균 주사가능 용액 또는 분산액의 즉석 제조를 위한 멸균 수용액(수용성인 경우) 또는 분산액 및 멸균 분말을 포함한다. 정맥내 투여를 위해, 적합한 담체는 생리학적 염수, 정균수, 크레모포르(Cremophor) EL(BASF, 미국 뉴저지주 파시파니) 또는 인산 완충 염수(PBS)를 포함한다. 모든 경우에, 조성물은 멸균되어야 하고, 용이한 주사를 실시할 수 있을 정도로 유동적이어야 한다. 이는 제조 및 저장 조건 하에서 안정해야 하고, 미생물, 예컨대 박테리아 및 진균의 오염 작용에 대해 보존되어야 한다. 담체는 예를 들어 물, 에탄올, 폴리올(예를 들어, 글리세롤, 프로필렌 글리콜 및 액체 폴리에틸렌 글리콜 등) 및 이들의 적합한 혼합물을 함유하는 용매 또는 분산매일 수 있다. 적절한 유동성은 예를 들어 레시틴과 같은 코팅의 사용, 분산액의 경우 요구되는 입자 크기의 유지 및 계면활성제의 사용에 의해 유지될 수 있다. 미생물 작용의 예방은 다양한 항박테리아 및 항진균제, 예를 들어 파라벤, 클로로부탄올, 페놀, 아스코르브산, 티메로살 등에 의해 달성될 수 있다. 많은 경우에, 등장성 물질, 예를 들어 당, 폴리알콜, 예컨대 만니톨, 소르비톨, 염화나트륨을 조성물에 포함시키는 것이 바람직할 것이다. 주사가능한 조성물의 장기 흡수는 조성물에 흡수를 지연시키는 작용제, 예를 들어 모노스테아르산알루미늄 및 젤라틴을 포함시킴으로써 달성될 수 있다.
멸균 주사가능 용액은 적절한 용매 내의 필요한 양의 활성 화합물을 필요에 따라 상기 열거된 성분 중 하나 또는 조합물과 혼입하고, 이어서 여과 멸균함으로써 제조할 수 있다. 일반적으로, 분산액은 활성 화합물을 염기성 분산매 및 상기 열거된 것들 중 필요한 다른 성분을 함유하는 멸균 비히클에 혼입함으로써 제조된다. 멸균 주사가능 용액의 제조를 위한 멸균 분말의 경우, 제조 방법은 이전에 제조된 멸균 여과된 그의 용액으로부터 활성 성분 및 임의의 추가의 원하는 성분의 분말을 수득하는 진공 건조 및 동결건조이다.
경구 조성물은 일반적으로 불활성 희석제 또는 식용 담체를 포함한다. 이들은 젤라틴 캡슐 내에 넣거나 압축하여 정제로 만들 수 있다. 경구 치료 투여를 위해, 활성 화합물은 부형제와 함께 혼입될 수 있고, 정제, 트로키제 또는 캡슐의 형태로 사용될 수 있다. 경구 조성물은 또한 구강 세정제로서 사용하기 위해 유체 담체를 사용하여 제조될 수 있고, 여기서 유체 담체 내의 화합물은 경구 적용되고 입에서 씻어내고 뱉어내거나 삼켜진다. 약학적으로 상용성인 결합제 및/또는 보조제 물질은 조성물의 일부로서 포함될 수 있다. 정제, 환제, 캡슐, 트로키제 등은 하기 성분 또는 유사한 성질의 화합물 중 임의의 것을 함유할 수 있다: 결합제, 예컨대 미정질 셀룰로스, 트라가칸트 고무 또는 젤라틴; 부형제, 예컨대 전분 또는 락토스, 붕해제, 예컨대 알긴산, 프리모겔(Primogel) 또는 옥수수 전분; 윤활제, 예컨대 스테아르산마그네슘 또는 스테로테트(Sterotes); 활주제, 예컨대 콜로이드성 이산화규소; 감미제, 예컨대 수크로스 또는 사카린; 또는 향미제, 예컨대 페퍼민트, 메틸 살리실레이트 또는 오렌지 향료.
흡입에 의한 투여를 위해, 화합물은 적절한 추진제, 예를 들어, 기체, 예컨대 이산화탄소를 함유한 가압된 용기 또는 분배기, 또는 네뷸라이저(nebulizer)로부터 에어로졸 스프레이 형태로 전달된다.
전신 투여는 또한 경점막 또는 경피 수단에 의해서 시행될 수 있다. 경점막 또는 경피 투여를 위해, 침투되는 장벽에 적절한 침투제가 제제에 사용된다. 이러한 침투제는 일반적으로 관련 기술 분야에 알려져 있고, 예를 들어 경점막 투여를 위해 세제, 담즙산염, 및 푸시드산 유도체를 포함한다. 경점막 투여는 비강 스프레이 또는 좌제의 사용을 통해 달성될 수 있다. 경피 투여를 위해, 활성 화합물은 관련 기술 분야에 일반적으로 알려진 연고, 고약, 겔 또는 크림으로 제제화된다.
화합물은 또한 직장 전달용 좌제(예를 들어, 코코아 버터 및 다른 글리세라이드와 같은 통상의 좌약 기제를 사용한) 또는 정체 관장제 형태로 제조될 수 있다.
한 실시양태에서, 활성 화합물은 임플란트 및 미세캡슐화된 전달 시스템을 포함하는 제어 방출 제제와 같이 신체로부터의 신속한 제거로부터 화합물을 보호하는 담체로 제조된다. 에틸렌 비닐 아세테이트, 폴리안히드라이드, 폴리글리콜산, 콜라겐, 폴리오르토에스테르 및 폴리락트산과 같은 생분해성 생체적합성 중합체가 사용될 수 있다. 이러한 제제의 제조 방법은 통상의 기술자에게 명백할 것이다. 상기 물질은 또한 알자 코포레이션(Alza Corporation) 및 노바 파마슈티칼스, 인크.(Nova Pharmaceuticals, Inc.)로부터 상업적으로 입수할 수 있다. 현탁액(바이러스 항원에 대한 모노클로날 항체를 사용하여 감염된 세포로 표적화되는 리포솜 포함)을 또한 약학적으로 허용되는 담체로 사용할 수 있다. 이들은 미국 특허 4,522,811에 기재된 바와 같이 관련 기술 분야의 통상의 기술자에게 알려진 방법에 따라 제조될 수 있다.
투여의 용이성 및 투여량의 균일성을 위해 투여 단위 형태로 경구 또는 비경구 조성물을 제제화하는 것이 특히 바람직하다. 본원에서 사용된 투여 단위 형태는 치료될 대상체에 대한 단일 투여량으로 적합한 물리적으로 별개의 단위를 의미하고; 각각의 단위는 필요한 약학 담체와 함께 원하는 치료 효과를 생성하도록 계산된 미리 결정된 양의 활성 화합물을 함유한다. 본 발명의 투여 단위 형태에 대한 명세는 활성 화합물의 고유한 특성 및 달성되어야 할 특정 치료 효과, 및 개체의 치료를 위한 활성 화합물을 배합하는 기술에 내재된 한계에 의해 좌우되고 직접적으로 의존한다.
약학 조성물은 투여 지침과 함께 용기, 팩 또는 분배기에 포함될 수 있다.
스크리닝 방법
본 발명은 인플루엔자 바이러스의 세포막에 대한 융합을 조절하거나 달리 방해하는 조절자, 즉 후보 또는 시험 화합물 또는 작용제(예를 들어, 펩티드, 펩티드 모방체, 소분자 또는 다른 약물)를 확인하기 위한 방법(본원에서 "스크리닝 분석"으로도 불림)을 제공한다. 또한, 인플루엔자 감염의 치료에 유용한 화합물을 확인하는 방법도 제공된다. 본 발명은 또한 본원에서 설명되는 스크리닝 분석을 사용하여 확인된 화합물을 포함한다.
예를 들어, 본 발명은 인플루엔자 바이러스와 세포막 사이의 상호작용을 조절하는 후보 또는 시험 화합물을 스크리닝하기 위한 분석을 제공한다. 본 발명의 시험 화합물은 생물학적 라이브러리; 공간적으로 다룰 수 있는 평행 고체상 또는 용액상 라이브러리; 디컨볼루션(deconvolution)을 필요로 하는 합성 라이브러리 방법; "하나의 비드 하나의 화합물(one-bead one-compound)" 라이브러리 방법; 및 친화도 크로마토그래피 선택을 이용한 합성 라이브러리 방법을 비롯하여 관련 기술 분야에 알려진 조합 라이브러리 방법에서의 많은 접근 방법 중 임의의 것을 사용하여 수득할 수 있다. 생물학적 라이브러리 방법은 펩티드 라이브러리로 제한되지만, 다른 4개의 방법은 펩티드, 비펩티드 올리고머 또는 화합물의 소분자 라이브러리에 적용할 수 있다(예를 들어, 문헌 [Lam, 1997. Anticancer Drug Design 12: 145] 참조).
본원에서 사용되는 "소분자"는 약 5 kD 미만, 가장 바람직하게는 약 4 kD 미만의 분자량을 갖는 조성물을 의미한다. 소분자는 예를 들어 핵산, 펩티드, 폴리펩티드, 펩티드 모방체, 탄수화물, 지질 또는 다른 유기 또는 무기 분자일 수 있다. 진균, 박테리아 또는 조류 추출물과 같은 화학적 및/또는 생물학적 혼합물의 라이브러리는 관련 기술 분야에 공지되어 있으며, 본 발명의 임의의 분석법으로 스크리닝될 수 있다.
분자 라이브러리의 합성 방법의 예는 관련 기술 분야에서, 예를 들어 다음 문헌에서 볼 수 있다: [DeWitt, et al., 1993. Proc. Natl. Acad. Sci. U.S.A. 90: 6909]; [Erb, et al., 1994. Proc. Natl. Acad. Sci. U.S.A. 91: 11422]; [Zuckermann, et al., 1994. J. Med. Chem. 37: 2678]; [Cho, et al., 1993. Science 261: 1303]; [Carrell, et al., 1994. Angew. Chem. Int. Ed. Engl. 33: 2059]; [Carell, et al., 1994. Angew. Chem. Int. Ed. Engl. 33: 2061]; 및 [Gallop, et al., 1994. J. Med. Chem. 37: 1233].
화합물의 라이브러리는 용액 내에(예를 들어, 문헌 [Houghten, 1992. Biotechniques 13: 412-421] 참조), 또는 비드 상에 (문헌 [Lam, 1991. Nature 354: 82-84] 참조), 칩 상에 (문헌 [Fodor, 1993. Nature 364: 555-556] 참조), 박테리아(미국 특허 제5,223,409호 참조), 포자(미국 특허 제5,233,409호 참조), 플라스미드(문헌 [Cull, et al., 1992. Proc. Natl. Acad. Sci. USA 89: 1865-1869] 참조) 또는 파지(문헌 [Scott and Smith, 1990. Science 249: 386-390]; [Devlin, 1990. Science 249: 404-406]; [Cwirla, et al., 1990. Proc. Natl. Acad. Sci. U.S.A. 87: 6378-6382]; [Felici, 1991. J. Mol. Biol. 222: 301-310]; 및 미국 특허 제5,233,409호 참조) 상에 제시될 수 있다.
한 실시양태에서, 후보 화합물을 항체-항원 복합체에 도입하고, 후보 화합물이 항체-항원 복합체를 파괴하는지 결정하는데, 이 복합체의 파괴는 후보 화합물이 인플루엔자 바이러스와 세포막 사이의 상호작용을 조절함을 나타낸다.
또 다른 실시양태에서, 적어도 하나의 중화 모노클로날 항체에 노출된 적어도 하나의 HA 단백질이 제공된다. 항체-항원 복합체의 형성이 검출되고, 하나 이상의 후보 화합물이 복합체에 도입된다. 하나 이상의 후보 화합물의 도입 후에 항체-항원 복합체가 파괴되면, 후보 화합물은 인플루엔자 바이러스 관련 질환 또는 장애, 예를 들어 조류 독감을 치료하는데 유용하다. 예를 들어, 적어도 하나의 인플루엔자 바이러스 단백질은 인플루엔자 바이러스 분자로서 제공될 수 있다.
항체-항원 복합체를 방해하거나 파괴하는 시험 화합물의 능력을 결정하는 것은 예를 들어 시험 화합물의 항원 또는 그의 생물학적 활성 부분에 대한 결합이 복합체 내의 표지된 화합물을 검출함으로써 결정될 수 있도록 시험 화합물을 방사성 동위 원소 또는 효소 표지와 커플링시킴으로써 달성될 수 있다. 예를 들어, 시험 화합물은 125I, 35S, 14C 또는 3H를 직접 또는 간접적으로 표지하고, 방사성 동위원소를 방사선 방출의 직접 계산 또는 섬광(scintillation) 계수에 의해 검출한다. 대안으로, 시험 화합물은 예를 들어 양고추냉이 퍼옥시다제, 알칼리성 포스파타제 또는 루시페라제로 효소적으로 표지될 수 있고, 효소 표지는 적합한 기질의 생성물로의 전환을 측정함으로써 검출될 수 있다.
한 실시양태에서, 분석은 항체-항원 복합체를 시험 화합물과 접촉시키는 단계 및 항원과 상호작용하거나 기존의 항체-항원 복합체를 파괴하는 시험 화합물의 능력을 결정하는 단계를 포함한다. 이 실시양태에서, 시험 화합물이 항원과 상호작용하고/하거나 항체-항원 복합체를 파괴하는 능력을 결정하는 것은 시험 화합물이 항체에 비해 항원 또는 그의 생물학적 활성 부분에 우선적으로 결합하는 능력을 결정하는 것을 포함한다.
또 다른 실시양태에서, 분석은 항체-항원 복합체를 시험 화합물과 접촉시키는 단계 및 항체-항원 복합체를 조절하는 시험 화합물의 능력을 측정하는 단계를 포함한다. 시험 화합물이 항체-항원 복합체를 조절하는 능력을 결정하는 것은 예를 들어 시험 화합물의 존재 하에 항원이 항체에 결합하거나 항체와 상호작용하는 능력을 결정함으로써 달성될 수 있다.
관련 기술 분야의 통상의 기술자는 본원에 개시된 임의의 스크리닝 방법에서 항체가 인플루엔자 바이러스 중화 항체일 수 있음을 인식할 것이다. 또한, 항원은 HA 단백질 또는 그의 일부일 수 있다. 본원에 기재된 임의의 분석에서, 후보 화합물이 본 발명의 모노클로날 항체와 HA 단백질 사이의 결합을 방해하는 능력은 후보 화합물이 인플루엔자 바이러스와 세포막의 융합을 방해하거나 조절할 수 있음을 나타낸다. 또한, 세포에 대한 HA 단백질의 결합에 의해 인플루엔자 바이러스가 세포 내로 유입되기 때문에, 상기 후보 화합물은 또한 인플루엔자 바이러스 관련 질환 또는 장애, 예를 들어 조류 독감의 치료에 유용할 것이다.
본원에 개시된 스크리닝 방법은 세포-기반 분석 또는 무세포 분석으로서 수행될 수 있다. 본 발명의 무세포 분석에는 가용성 형태 또는 막 결합 형태의 HA 단백질 및 그의 단편 둘 모두의 사용이 가능하다. 막 결합 형태의 HA 단백질을 포함하는 무세포 분석의 경우, 막 결합 형태의 단백질이 용액 중에 유지되도록 가용화제를 이용하는 것이 바람직할 수 있다. 이러한 가용화제의 예는 비이온성 세제, 예컨대 n-옥틸글루코시드, n-도데실글루코시드, N-도데실말토시드, 옥타노일-N-메틸글루카미드, 데카노일-N-메틸글루카미드, 트리톤(Triton)® X-100, 트리톤® X-114, 테시트(Thesit)®, 이소트리데시폴리(에틸렌 글리콜 에테르)n, N-도데실-N,N-디메틸-3-암모니오-1-프로판 술포네이트, 3-(3-콜라미도프로필)디메틸암모니올-1-프로판 술포네이트(CHAPS), 또는 3-(3-콜라미도프로필)디메틸암모니올-2-히드록시-1-프로판 술포네이트(CHAPSO)를 포함한다.
하나 초과의 실시양태에서, 후보 화합물의 도입 후에 하나 또는 둘 모두의 복합체화되지 않은 형태로부터 복합체화된 형태의 분리를 용이하게 하고 분석의 자동화를 수용하기 위해 항체 또는 항원을 고정하는 것이 바람직할 수 있다. 후보 화합물의 존재 및 부재 하에서의 항체-항원 복합체의 관찰은 반응물을 포함하기에 적합한 임의의 용기에서 수행될 수 있다. 상기 용기의 예는 미량역가 플레이트, 시험관 및 미세 원심분리관을 포함한다. 한 실시양태에서, 단백질 중 하나 또는 둘 모두가 매트릭스에 결합되는 것을 허용하는 도메인을 부가하는 융합 단백질이 제공될 수 있다. 예를 들어, GST-항체 융합 단백질 또는 GST-항원 융합 단백질을 글루타티온 세파로스 비드(Sigma Chemical, 미국 미주리주 세이트 루이스 소재) 또는 글루타티온 유도체화된 미량역가 플레이트에 흡착시킨 후, 시험 화합물과 조합하고, 혼합물을 복합체 형성에 도움이 되는 조건(예를 들어, 염 및 pH의 생리학적 조건)에서 인큐베이팅한다. 인큐베이팅 후, 비드 또는 미량역가 플레이트 웰을 세척하여 결합되지 않은 임의의 성분, 비드의 경우에 고정된 매트릭스를 제거하고, 복합체는 직접 또는 간접적으로 결정된다. 다르게는, 복합체는 매트릭스로부터 해리될 수 있고, 항체-항원 복합체 형성 수준은 표준 기술을 사용하여 결정될 수 있다.
매트릭스 상에 단백질을 고정하는 다른 기술도 또한 본 발명의 스크리닝 분석에 사용될 수 있다. 예를 들어, 항체 또는 항원은(예를 들어, 비오틴과 스트렙타비딘의 접합을 이용하여 고정될 수 있다. 비오티닐화된 항체 또는 항원 분자는 관련 기술 분야에 공지된 기술(비오티닐화 키트, Pierce Chemicals, 미국 일리노이주 록포드 소재)을 사용하여 비오틴-NHS(N-히드록시-숙신이미드)로부터 제조되고, 스트렙타비딘 코팅된 96웰 플레이트(Pierce Chemical)의 웰에 고정된다. 다르게는, 관심 항체 또는 항원과 반응하지만 관심 항체-항원 복합체의 형성은 방해하지 않는 다른 항체가 플레이트의 웰에 유도체화될 수 있고, 결합되지 않은 항체 또는 항원은 항체 접합에 의해 웰에 포획될 수 있다. GST 고정된 복합체에 대해 상기 설명된 방법에 추가하여, 상기 복합체를 검출하는 방법은 항체 또는 항원과 반응하는 상기 다른 항체를 사용한 복합체의 면역검출을 포함한다.
본 발명은 추가로 상기 언급된 스크리닝 분석 중 임의의 것으로 확인된 신규한 작용제 및 본원에서 설명되는 치료를 위한 그의 용도에 관련된다.
진단 분석
본 발명의 항체는 적절한 분석, 예를 들어, 통상적인 유형의 면역분석법에 의해 검출될 수 있다. 예를 들어, 인플루엔자 단백질(HA1, HA2 또는 뉴라미니다제) 또는 그의 단편이 고상에 부착되는 분석을 수행할 수 있다. 샘플 중의 항체가 고상에 고정된 폴리펩티드에 결합할 수 있기에 충분한 시간 동안 인큐베이팅을 유지한다. 이 제1 인큐베이팅 후, 고상을 샘플로부터 분리한다. 고상을 세척하여, 샘플에 또한 존재할 수 있 비결합 물질 및 방해 물질, 예컨대 비특이적 단백질을 제거한다. 이어서, 고정된 폴리펩티드에 결합된 관심 항체를 함유하는 고상을 제2의 표지된 항체 또는 비오틴 또는 아비딘과 같은 커플링제에 결합된 항체와 함께 인큐베이팅한다. 상기 제2 항체는 또 다른 항인플루엔자 항체 또는 또 다른 항체일 수 있다. 항체에 대한 표지는 관련 기술 분야에 공지되어 있고, 방사성 핵종, 효소(예를 들어, 말레에이트 데히드로게나제, 양고추냉이 퍼옥시다제, 글루코스 옥시다제, 카탈라제), 형광 물질(플루오레세인 이소티오시아네이트, 로다민, 피코시아닌, 플루오레스카민), 비오틴 등을 포함한다. 표지된 항체를 고체와 함께 인큐베이팅되고, 고상에 결합된 표지가 측정된다. 이러한 면역분석 및 다른 면역분석은 관련 기술 분야의 통상의 기술자에 의해 용이하게 수행될 수 있다.
생물학적 샘플에서 인플루엔자 바이러스의 존재 또는 부재를 검출하기 위한 예시적인 방법은 시험 대상체로부터 생물학적 샘플을 수득하고, 인플루엔자 바이러스가 생물학적 샘플에서 검출되도록 생물학적 샘플을 본 발명에 따른 표지된 모노클로날 또는 scFv 항체와 접촉시키는 것을 수반한다.
프로브 또는 항체와 관련하여 본원에서 사용되는 용어 "표지된"은 프로브 또는 항체에 대한 검출 가능한 물질의 커플링(즉, 물리적 연결)에 의한 프로브 또는 항체의 직접 표지뿐만 아니라, 직접 표지된 또 다른 시약과의 반응성에 의한 프로브 또는 항체의 간접적인 표지를 포함하는 것이 의도된다. 간접 표지의 예는 형광 표지된 2차 항체를 사용하는 1차 항체의 검출 및 형광 표지된 스트렙타비딘으로 검출될 수 있도록 비오틴을 사용한 DNA 프로브의 말단 표지를 포함한다. "생물학적 샘플"이라는 용어는 대상체로부터 단리된 조직, 세포 및 생물학적 유체뿐만 아니라, 대상체 내에 존재하는 조직, 세포 및 체액을 포함하는 것이 의도된다. 즉, 본 발명의 검출 방법은 시험관 내에서뿐만 아니라 생체 내에서 생물학적 샘플 내의 인플루엔자 바이러스를 검출하기 위해 사용될 수 있다. 예를 들어, 인플루엔자 바이러스를 검출하기 위한 시험관내 기술은 효소 연결 면역흡착 분석(ELISA), 웨스턴 블롯, 면역침전 및 면역형광을 포함한다. 또한, 인플루엔자 바이러스의 검출을 위한 생체내 기술은 표지된 항인플루엔자 바이러스 항체를 대상체 내에 도입하는 것을 포함한다. 예를 들어, 항체는 대상체 내의 그의 존재 및 위치가 표준 영상화 기술에 의해 검출될 수 있는 방사성 마커로 표지될 수 있다.
한 실시양태에서, 생물학적 샘플은 시험 대상체로부터의 단백질 분자를 함유한다. 하나의 바람직한 생물학적 샘플은 대상체로부터 통상적인 수단에 의해 단리된 말초 혈액 백혈구 샘플이다.
본 발명은 또한 생물학적 샘플에서 인플루엔자 바이러스의 존재를 검출하기 위한 키트를 포함한다. 예를 들어, 키트는 생물학적 샘플에서 인플루엔자 바이러스를 검출할 수 있는 표지된 화합물 또는 작용제(예를 들어, 항인플루엔자 scFv 또는 모노클로날 항체); 샘플에서 인플루엔자 바이러스의 양을 결정하는 수단; 및 샘플 중의 인플루엔자 바이러스의 양을 표준물질과 비교하는 수단을 포함한다. 화합물 또는 작용제는 적절한 용기에 포장될 수 있다. 키트는 샘플에서 인플루엔자 바이러스를 검출하기 위해 키트를 사용하기 위한 지침을 추가로 포함할 수 있다.
수동 면역
수동 면역은 바이러스 질환의 예방 및 치료를 위한 효과적이고 안전한 전략으로 입증되었다 (각각 본원에 참고로 포함된 문헌 [Keller et al., Clin. Microbiol. Rev. 13:602-14 (2000)]; [Casadevall, Nat. Biotechnol. 20:114 (2002)]; [Shibata et al., Nat. Med. 5:204-10 (1999)]; 및 [Igarashi et al., Nat. Med. 5:211-16 (1999)] 참조). 대안적이고 더 많은 시간이 소용되는 백신 및 신약 개발이 진행 중이지만, 중화 인간 모노클로날 항체를 사용하는 수동 면역은 조류 독감과 같은 인플루엔자의 긴급 예방 및 치료를 위한 즉각적인 치료 전략을 제공할 수 있다.
서브유닛 백신은 종래의 면역원에 비해 유의한 이점을 잠재적으로 제공한다. 이들은 전통적인 사멸된 또는 약독화된 전체 병원체 백신의 생산, 유통 및 전달에 내재된 안전 위험을 회피한다. 또한, 이들은 확인된 보호 에피토프만을 포함하도록 합리적으로 설계될 수 있기 때문에, 억제성 T 에피토프(문헌 [Steward et al., J. Virol. 69:7668 (1995)] 참조) 또는 쓸데없는 비보호 반응을 유도함으로써 면역계를 파괴하는 면역우성 B 에피토프(예를 들어, "디코이(decoy)" 에피토프)(문헌 [Garrity et al., J. Immunol. 159:279 (1997)] 참조)를 회피할 수 있다.
또한, 관련 기술 분야의 통상의 기술자는 많은 상이한 바이러스, 시험접종 경로 및 동물 모델에 대해 시험관 내에서의 항체 중화 활성과 생체 내에서의 보호 사이에 양호한 상관관계가 존재함을 인식할 것이다(문헌 [Burton, Natl. Rev. Immunol. 2:706-13 (2002)]; [Parren et al., Adv. Immunol. 77:195-262 (2001)] 참조). 본원에서 제시되는 데이터는 D7, D8, F10, G17, H40, A66, D80, E88, E90 및 H98 인간 모노클로날 항체가 인플루엔자의 긴급 예방 및 치료를 위한 강력한 바이러스 진입 억제제로서의 그의 임상 유용성을 결정하기 위해 생체내 동물 실험에서 추가로 개발되어 시험될 수 있음을 보여준다.
백신 접종에서의 항원-Ig
키메라
처음 항체가 항원 결정자를 면역계에 효율적으로 제시하기 위한 스캐폴드로서 사용된 지 10년이 넘는 시간이 흘렀다(문헌 [Zanetti, Nature 355:476-77 (1992)]; [Zaghouani et al., Proc. Natl. Acad. Sci. USA 92:631-35 (1995)] 참조). 펩티드가 IgG 분자(예를 들어, 본원에 기재된 11A 또는 256 IgG 모노클로날 항체)의 필수 부분으로 포함되는 경우, 펩티드 에피토프의 항원성 및 면역원성은 자유 펩티드에 비해 크게 향상된다. 이러한 향상은 항원-IgG 키메라의 더 긴 반감기, 더 우수한 제시 및 그들의 천연 구조를 모방한 속박된(constrained) 구조에 기인한 것일 수 있다.
또한, 항원-Ig 키메라를 사용하는 추가의 이점은 항원-Ig 키메라의 가변 또는 Fc 영역이 전문 항원 제시 세포(APC)를 표적화하기 위해 사용될 수 있다는 것이다. 현재까지, 중쇄 가변 유전자(VH)의 상보성 결정 영역(CDR)이 B 또는 T 세포에 의해 인식되는 다양한 항원 펩티드로 대체된 재조합 Ig가 생성되었다. 이러한 항원-Ig 키메라는 체액성 및 세포성 면역 반응 둘 모두를 유도하기 위해 사용되어 왔다(문헌 [Bona et al., Immunol. Today 19:126-33 (1998)] 참조).
CDR3 루프에 이식된 특정 에피토프를 갖는 키메라는 HIV-1 gp120 V3-루프 또는 인간 CD4 수용체의 제1 세포외 도메인(D1)에 대한 체액성 반응을 유도하기 위해 사용되어왔다(문헌 [Lanza et al., Proc. Natl. Acad. Sci. USA 90:11683-87 (1993)]; [Zaghouani et al., Proc. Natl. Acad. Sci. USA 92:631-35 (1995)] 참조). 면역 혈청은 HIV-1MN에 의한 CD4 SupT1 세포의 감염을 예방하거나(항-gp120 V3C) 합포체(syncytia) 형성을 억제할 수 있다(항-CD4-D1). CDR2 및 CDR3은 펩티드 에피토프로 동시에 대체될 수 있고, 삽입되는 펩티드의 길이는 최대 19개의 아미노산일 수 있다.
대안으로, 한 그룹은 펩티드 항원이 Ig 불변(C) 영역의 루프 내에 제시되고 키메라의 가변 영역이 B-세포의 표면상의 IgD 또는 B 세포, 수지상 세포(DC: dendritic cell) 및 대식세포를 포함하는 전문 APC 상의 MHC 클래스 II 분자를 표적화하기 위해 사용될 수 있는 "트로이바디(troybody)" 전략을 개발하였다(문헌 [Lunde et al., Biochem. Soc. Trans. 30:500-6 (2002)] 참조).
항원-Ig 키메라는 또한 항원을 IgG 분자의 Fc 부분과 직접 융합시킴으로써 제조될 수 있다. 문헌 [You et al., Cancer Res. 61:3704-1121(2001)]에서는 이 방법을 사용하여 B형 간염 바이러스 코어 항원에 대한 매우 높은 수준의 항체를 비롯하여 특정 면역 반응의 모든 아암(arm)을 얻을 수 있었다.
DNA 백신 접종
DNA 백신은 안정하고, 항원에게 자연적으로 처리될 기회를 제공할 수 있고, 보다 오래 지속되는 반응을 유도할 수 있다. 매우 매력적인 면역 전략이지만, DNA 백신은 종종 면역 반응을 유도할 수 있는 가능성이 매우 제한적이다. 수지상 세포(DC)와 같은 직업적인 APC에 의한 주입된 DNA의 불충분한 흡수가 이러한 제한의 주요 원인일 수 있다. 항원-Ig 키메라 백신과 조합하여, APC 항원 제시의 향상에 기초한 유망한 새로운 DNA 백신 전략이 보고되었고(문헌 [Casares, et al., Viral Immunol. 10:129-36 (1997)]; [Gerloni et al., Nat. Biotech. 15:876-81 (1997)]; [Gerloni et al., DNA Cell Biol. 16:611-25 (1997)]; [You et al., Cancer Res. 61:3704-11 (2001)] 참조), 이것은 DC 표면 상의 Fc 수용체(FcγR)의 존재를 이용한다.
항원(Ag)-Ig 키메라를 코딩하는 DNA 백신을 생성하는 것이 가능하다. 면역화시에, Ag-Ig 융합 단백질은 DNA 분자를 흡수하는 세포에 의해 발현되고 분비될 것이다. 분비된 Ag-Ig 융합 단백질은 B 세포 반응을 유도하는 동안, DC 표면 상의 FcγR과 Fc 단편의 상호작용에 의해 포획되고 내재화될 수 있으며, 이것은 효율적인 항원 제시를 촉진하고 항원 특이적인 면역 반응을 크게 향상시킬 것이다. 동일한 원리를 적용하여, 기능성 항-MHC II 특이적 scFv 영역 유전자를 보유하는 항원-Ig 키메라를 코딩하는 DNA는 또한 면역원이 3가지 유형의 모든 APC를 표적화하도록 할 수 있다. 면역 반응은 필요한 경우, 시험관 내에서 생성된 동일한 단백질 항원을 사용하여 추가로 부스팅될 수 있다(즉, "프라이밍 및 부스팅"). 이 전략을 사용하면, 인플루엔자 바이러스의 감염에 대한 특이적인 세포성 및 체액성 면역 반응은 DNA 백신의 근내(i.m.) 주사를 통해 달성되었다(문헌 [Casares et al., Viral. Immunol. 10:129-36 (1997)] 참조).
백신 조성물
하나 이상의 모노클로날 항체 또는 ScFv 및 이들의 조합물의 혼합물을 일반적으로 포함하는 치료 또는 예방 조성물이 본원에서 제공된다. 예방 백신은 인플루엔자 바이러스 감염을 예방하기 위해 사용할 수 있고, 치료 백신은 인플루엔자 바이러스 감염 후에 개체를 치료하기 위해 사용할 수 있다. 예방적 사용은 예방 접종 대상체에서 인플루엔자 바이러스에 대한 증가된 항체 역가를 제공하는 것을 포함한다. 이러한 방식으로, 인플루엔자를 보유할 위험이 높은 대상체에게는 인플루엔자 바이러스에 대한 수동 면역이 제공될 수 있다.
이들 백신 조성물은 보조 면역조절제, 예를 들어 IL-2, 변형된 IL-2(Cys125 → Ser125), GM-CSF, IL-12, γ-인터페론, IP-10, MIP1β, 및 RANTES를 포함하고 이로 제한되지 않는 사이토카인, 림포카인 및 케모카인과 함께 투여될 수 있다.
면역화 방법
본 발명의 백신은 다른 항바이러스 백신보다 더 우수한 면역보호 및 면역치료 특성을 갖는다.
본 발명은 대상체의 면역화, 예를 들어, 면역 반응의 유도 방법을 제공한다. 대상체는 병원성 외피 바이러스의 막 융합 단백질을 함유하는 조성물을 대상체에게 투여함으로써 면역화된다. 상기 융합 단백질은 생물학적으로 적합한 매트릭스에 코팅되거나 포매된다.
융합 단백질은 글리코실화되고, 예를 들어, 탄수화물 모이어티를 함유한다. 탄수화물 모이어티는 단당류, 이당류(들), 올리고당(들), 다당류(들), 또는 이들의 유도체(예를 들어, 술포- 또는 포스포-치환된)의 형태일 수 있다. 탄수화물은 선형 또는 분 지형이다. 탄수화물 모이어티는 폴리펩티드에 N-연결 또는 O-연결한다. N-연결된 글리코실화는 아스파라긴 측쇄의 아미드 질소에 대한 것이고, O-연결된 글리코실화는 세린 및 트레오닌 측쇄의 히드록시 산소에 대한 것이다.
탄수화물 모이어티는 백신 접종되는 대상체에 내인성이다. 대안으로, 탄수화물 모이어티는 백신 접종되는 대상체에 외인성이다. 탄수화물 모이어티는 전형적으로 백신 접종되는 대상체의 폴리펩티드 상에 발현되지 않는 탄수화물 모이어티이다. 예를 들어, 탄수화물 모이어티는 식물 특이적 탄수화물이다. 식물 특이적 탄수화물 모이어티는 예를 들어 코어 결합된 α1,3 푸코스 또는 코어 결합된 β1,2 크실로스를 갖는 N-연결된 글리칸을 포함한다. 대안으로, 탄수화물 모이어티는 백신 접종되는 대상체의 폴리펩티드 또는 지질 상에 발현되는 탄수화물 모이어티이다. 예를 들어, 많은 숙주 세포가 인간 유사 당 부착물을 갖는 인간 단백질을 생산하도록 유전적으로 조작되었다.
예를 들어, 융합 단백질은 삼량체 헤마글루티닌 단백질이다. 선택적으로, 헤마글루티닌 단백질은 식물 세포와 같은 비포유동물 세포에서 생산된다.
대상체는 바이러스 감염이 발생할 위험이 있거나 바이러스 감염을 앓을 위험이 있다. 외피 바이러스는 엡스타인-바 바이러스, 단순 포진 바이러스 1형 및 2형, 인간 사이토메갈로바이러스, 인간 헤르페스바이러스 8형, 수두 대상포진 바이러스, B형 간염 바이러스, C형 간염 바이러스, 인간 면역결핍 바이러스, 인플루엔자 바이러스, 홍역 바이러스, 유행성 이하선염 바이러스, 파라인플루엔자 바이러스, 호흡기 세포융합 바이러스, 광견병 바이러스 및 풍진 바이러스를 포함한다.
본원에서 설명되는 방법은 바이러스 감염의 하나 이상의 증상의 중증도 감소 또는 완화를 유도한다. 감염은 표준 방법을 사용하여 의사에 의해 진단되고/되거나 모니터링된다. 면역화를 필요로 하는 대상체는 관련 기술 분야에 공지된 방법에 의해 확인된다. 예를 들어, 대상체는 CDC의 면역화에 대한 일반적인 권고안(General Recommendation on Immunization)(51(RR02) pp1-36)에 개관된 바와 같이 면역화된다. 암은 신체 검사, 생검, 혈액 검사 또는 엑스레이로 진단된다.
대상체는 예를 들어 임의의 포유동물, 예를 들어 인간, 영장류, 마우스, 래트, 개, 고양이, 소, 말, 돼지, 어류 또는 조류이다. 치료는 감염 진단 전에 시행된다. 다르게는, 치료는 진단 후에 시행된다.
치료의 효능은 특정 장애 또는 감염을 진단 또는 치료하기 위한 임의의 공지된 방법과 관련하여 결정된다. 장애의 하나 이상의 징상의 완화는 화합물이 임상적 이익을 부여함을 나타낸다.
백신 잠재력에 대한 항원 단백질 단편(
APF
)의 평가
체액성 면역을 목표로 하는 백신 후보는 성공하기 위해서 다음과 같은 적어도 3가지 기준을 충족시켜야 한다: 강력한 항체 반응을 촉발하여야 하고("면역원성"); 백신 후보가 촉발하는 항체의 상당 분획은 병원체와 교차 반응을 가져야 하고("면역원성 적합성"); 백신 후보가 촉발하는 항체는 보호성이어야 한다. 면역원성은 종종 보조제 또는 담체를 사용하여 향상될 수 있지만, 면역원성 적합성 및 보호 유도 능력(중화에 의해 입증되는)은 백신 성분으로서의 항원의 성공을 최종적으로 결정할 항원의 고유한 특성이다.
면역원성 평가
"면역원성 적합성"은 병원체와 교차 반응하는 항원에 의해 유도된 항체의 분획으로서 정의된다(문헌 [Matthews et al., J. Immunol.169:837 (2002)] 참조). 이것은 병원체와 교차 반응하지 않는 항체를 포함하는, 항원에 의해 유도된 모든 항체의 역가에 의해 측정되는 면역원성과는 구별된다. 부적절한 면역원성 적합성은 아마도 현재까지 펩티드 백신의 실망스러운 실적에 기여했을 것이다. 항체에 높은 친화도로 결합하여 높은 항체 역가를 촉발하는 펩티드는 종종 적절한 면역원성 적합성이 결여되고, 따라서 잠재적인 백신 성분으로서 실패하게 된다. 따라서, 인플루엔자 백신 후보를 선택하는 기준의 하나로서 면역원성 적합성을 포함시키는 것이 중요한다.
낮은 면역원성 적합성에 대한 공통적인 설명은 대부분의 짧은 펩티드의 구조적 유연성이다. 특히, 유연한 펩티드는 환자의 항체와 잘 결합할 수 있고, 미자극 대상체에서 실질적인 항체 역가를 유도할 수 있다. 그러나, 펩티드가 구조의 큰 레퍼토리를 갖는다면, 펩티드가 미자극 대상체에서 유도하는 항체의 우세는 무손상 병원체 상의 상응하는 천연 에피토프와 교차 반응하지 못할 수 있다.
짧은 펩티드와 마찬가지로, 일부 APF는 매우 유연할 수 있고, 따라서 백신 성분으로서 실패할 수 있다. 가장 면역학적으로 적합한 APF는 전체 단백질의 구조(context) 외부에서 본질적으로 속박되는 자가 폴딩 단백질 하위도메인으로 이루어질 수 있다.
면역원성 적합성은 1차적으로 APF 자체의 특성이고 반응하는 면역계의 특성이 아니기 때문에, 궁극적으로 APF가 인간에서 수행되어야 하더라도 면역원성 적합성은 동물 모델(예를 들어, 마우스)에서 평가할 수 있다.
APF에 의해 달성된 면역원성 적합성은 문헌 [Matthews et al., J. Immunol. 169:837 (2002)]에 기재된 것과 유사한 절차를 통해 정제된 스파이크 또는 막 단백질로 항-APF 혈청을 면역흡수시킴으로써 평가된다. IgG는 면역화된 마우스로부터 수집된 혈청으로부터 정제된다. 정제된 비오티닐화된 단백질(마우스가 면역화된 특정 APF에 따라 적절한)을 마우스 IgG와 혼합하고, 인큐베이팅한다. 이어서, 스트렙타비딘으로 코팅된 세파로스 비드를 임의의 결합된 IgG와 함께 모든 비오티닐화된 단백질을 포획하기에 충분한 양으로 첨가한다. 스트렙타비딘으로 코팅된 비드를 미세 원심분리기에서 13,000 rpm으로 원심분리함으로써 제거하여, 각각 단백질에 대해 작용하는 항체들이 고갈된 IgG를 얻는다. 모의 면역흡수는 비오티닐화된 BSA가 모의 흡수제로서 인플루엔자 단백질을 대체하는 것을 제외하고는 동일한 방식으로 병행하여 수행된다.
APF의 면역원성 적합성을 측정하기 위해, 흡수된 항체 및 모의 흡수된 항체의 역가를 면역화 APF에 대한 ELISA에서 동시에 측정한다. 파지 디스플레이 NPL로부터 선택된 APF 친화도에 대해, 이들 ELISA에 대한 항원은 정제된 APF-GST 융합 단백질일 것이다. 포유동물 세포 디스플레이 NPL로부터의 잠재적으로 글리코실화된 APF에 대한 이들 ELISA에 대한 항원은 포유동물 세포에 의해 분비된 APF-Fc 융합 단백질이고 단백질 A로 정제될 것이다. 모의 흡수된 항체에 비해 흡수된 항체의 항-APF 역가의 감소 비율은 APF의 면역원성 적합성에 대한 척도를 제공할 것이다.
처리 방법
본 발명은 인플루엔자 바이러스 관련 질환 또는 장애의 위험이 있는(또는 감염되기 쉬운) 대상체를 처리하는 예방 및 치료 방법을 제공한다. 그러한 질환 또는 장애는 예를 들어 조류 독감을 포함하고, 이로 제한되지 않는다.
예방 방법
한 측면에서, 본 발명은 본 발명의 모노클로날 항체 또는 scFv 항체 또는 본 발명의 방법에 따라 확인된 작용제를 대상체에 투여함으로써 대상체에서 인플루엔자 바이러스 관련 질환 또는 장애를 예방하는 방법을 제공한다. 예를 들어, scFv 및/또는 모노클로날 항체는 치료 유효량으로 투여될 수 있다. 선택적으로, 두 가지 이상의 항인플루엔자 항체가 공동 투여된다.
인플루엔자 바이러스 관련 질환 또는 장애의 위험이 있는 대상체는 감염된 인간과 접촉하거나 일부 다른 방식으로 인플루엔자 바이러스에 노출된 환자를 포함한다. 예방제의 투여는 질환 또는 장애가 예방되거나, 대안으로 그의 진행이 지연되도록 인플루엔자 바이러스 관련 질환 또는 장애의 특징적인 증상이 나타나기 전에 시행될 수 있다.
적절한 작용제는 본원에서 설명되는 스크리닝 분석에 기초하여 결정될 수 있다. 대안으로, 또는 부가적으로, 투여되는 작용제는 본 발명의 방법에 따라 확인된 인플루엔자 바이러스를 중화하는 scFv 또는 모노클로날 항체이다.
치료 방법
본 발명의 또 다른 측면은 환자에서 인플루엔자 바이러스 관련 질환 또는 장애를 치료하는 방법에 관한 것이다. 한 실시양태에서, 상기 방법은 작용제(예를 들어, 본원에서 설명되는 스크리닝 분석에 의해 확인된 작용제 및/또는 본 발명의 방법에 따라 확인된 scFv 항체 또는 모노클로날 항체), 또는 인플루엔자를 중화하는 작용제의 조합물을 질환 또는 장애를 앓고 있는 환자에게 투여하는 것을 수반한다. 예를 들어, 본 발명의 항체는 다른 항바이러스제, 예를 들어 타미플루와 조합하여 사용될 수 있다.
본 발명은 청구범위에 기재된 본 발명의 범위를 제한하지 않는 하기 실시예에서 추가로 설명될 것이다.
실시예
실시예
1: 단일 기억 B 세포의 배양액으로부터 인플루엔자 바이러스에 대한 BnAb의 단리
광범위한 중화 항체(BnAb) 단리 과정의 개략도가 도 1에 도시되어 있다. 인간 기억 B 세포 레퍼토리로부터 인플루엔자 바이러스에 대한 bnAb를 단리하기 위해, 본 발명자들은 시험관 내에서 인간 기억 B 세포 활성화 및 분화를 허용하는 신속하고 신뢰할 수 있는 배양 방법을 확립하였다. 항원 특이적 인간 기억 B 세포(CD19+CD27+)는 사량체화된 H3(A/브리즈번/10/07) 삼량체를 사용하여 7명의 건강한 공여자의 말초 혈액 단핵 세포(PBMC)로부터 단리되었고; 기억 B 세포의 단지 0.19%-1.08%만이 H3와 반응성이었다(표 3). 이들 B 세포를 웰당 하나의 세포의 밀도로 384웰 플레이트 내에 분류하고, 방사선 조사된 CD40L 형질감염된 세포의 존재 하에 배양하였다. 14일 후, 7명의 공여자로부터 유래된 1051개(2688 배앵약 내의 39.1%)의 배양 상청액은 IgG 또는 IgM을 분비하는 것으로 밝혀졌고, H3(A/브리즈번/10/07), H7(A/캐나다/RV444/04), H1(A/캘리포니아/04/09) 및 인플루엔자 B(B/말레이지아/2506/04)의 HA와의 반응성에 대해 순차적으로 시험되었다. 상기 스크린을 통해, 237개(22.55%)의 팽창된 기억 B 세포가 H3에 결합된 Ig를 분비하는 것으로 밝혀졌다(표 3). 이 시험관내 팽창 단계는 분류 및 RT-PCR만으로 얻은 0.61%의 회수에 비해 37배 증가한 H3 반응성 B 세포의 회수를 보였다(데이터는 나타내지 않았다). 그룹 2 균주 H3/H7 내의 교차 반응성 클론의 평균 백분율은 18.14%이었다. 놀랍게도, H3 결합 클론의 13.08% 및 8.44%는 각각 그룹 1 H1 균주 및 H7/H1 균주에 이종아형 결합을 나타내었다. 단지 3.38%의 H3 반응성(H3+) 클론만이 또한 인플루엔자 B에 결합하는 것으로 밝혀졌다. 다음으로, 이종아형 결합을 보인 기억 B 세포 클론의 상청액을 H3N2(A/브리즈번/10/07)에 대한 미세중화를 시험하였다. H3/H7/H1 교차반응성 및 중화를 나타낸 공여자 3으로부터 유래된 하나의 bnAb인 3I14에 대한 특성이 추가로 분석되었다.
<표 3> 7명의 건강한 공여자에서 팽창된 기억 B 세포(mB)

3I14는
고도로
돌연변이된
IGHV3
-30 코딩 항체이다.
친화도 성숙에 대한 체세포 돌연변이의 기여를 평가하기 위해, 본 발명자들은 3I14 VH 및 VL 생식계열 버전(3I14-GL), 및 생식계열(g) 경쇄와 짝을 이루는 성숙한(m) 3I14 중쇄에 의해 형성된 키메라 항체(3I14-mHgL) 및 그 역으로 형성된 항체(3I14-gHmL)를 생성하였다(도 11). 3I14 변이체 항체를 인간 IgG1로 발현시키고, H1, H5 및 H3에 대한 이들의 결합 친화도를 평가하였다(표 4 및 도 20). 놀랍게도, 3I14-GL 변이체는 각각 H3에 대한 결합 친화도는 >15배 감소하고 H1에 대한 결합 친화도는 4.7배 증가하면서, 여전히 H3 및 H1에 대해 nM 및 nM 미만 범위로 결합하였다(표 4). H3 및 H1에 대한 3I14-GL 결합 친화도의 이러한 변화는 Koff의 각각 13.9배 또는 7.5배의 증가 및 감소에 주로 기인하였다. 흥미로운 것은, 3I14-GL은 이러한 분석 조건 하에서 H5에 결합하지 않았다는 것이다.

2개의 키메라 형태를 야생형(WT) 3I14와 비교하면, 3I14의 VH 및 VL 모두에 존재하는 체세포 돌연변이가 H3 결합에 동등하게 기여하는 것으로 보인다(Kd: 0.658 nM 대 0.733 nM). 또한, 중쇄 및 경쇄 키메라 둘 모두 Koff < 1.0E-7s-1로 H1에 대한 본질적 비가역적인 결합을 유도하였다. 그러나, H5의 경우, VL 돌연변이는 각각 Koff의 5.2배 및 2.2배 감소 때문에 VH 돌연변이보다 친화도 향상에 더 많이 기여한다(7.5배 대 1.9배). 이 연구에서, 본 발명자들은 3I14-GL이 H1에 대한 더 높은 친화도 결합 및 H3에 대한 중등도의 친화도를 나타내었으면, Koff의 변화가 3I14-WT와 비교시의 동역학 차이의 큰 원인이 되는 것으로 결론내렸다. H5 결합의 경우, 3I14의 체세포 돌연변이는 VL 돌연변이와의 결합에 절대적으로 필요하며, VH 변화보다 결합에 더 큰 기여를 제공한다. H1, H3 및 H5에 대한 결합 친화도의 모든 변화는 주로 해리 속도(Koff) 상수의 변화의 결과이다.
중쇄(VH) 및 경쇄(VL)의 가변 영역의 서열을 RT-PCR을 사용하여 팽창된 단일 세포 배양물로부터 회수하였다. 3I14는 IGHV3-30*18 및 IGLV1-44*01 생식계열 유전자에 의해 코딩된다. 재배열된 중쇄는 긴 상보성 결정 영역 3(HCDR3)(23개 아미노산)을 보유하고, VH 및 IGHJ4*02 접합부 둘 모두에 큰 N-부가가 측면에 있는 IGHD3-22*01 DH 분절을 사용한다(도 11c 및 11d). 3I14 mAb는 프레임워크 및 CDR 둘 모두에서 관찰되는 프라이머 인접 영역을 제외한 15개의 가변 중쇄 및 7개의 가변 경쇄 체세포 돌연변이를 갖는다.
실시예
2: 3I14는
그룹 1 및 그룹 2 인플루엔자 바이러스 둘
모두에 대한 결합 및
중화를 제시한다.
3I14는 유동 세포 계측법에 의해 그룹 2(H3, H4, H7, H14 및 H15) 및 그룹 1(H1, H2, H5, H6, H8, H9, H11, H12 및 H16) 인플루엔자 A 바이러스의 혈청형에 걸쳐 세포 표면 발현된 HA에 결합하였다(도 2). 3I14는 또한 그룹 2(H3, H4, H7 및 H14) 및 그룹 1(H1, H5 및 H9)에 속하는 상이한 아형의 정제된 HA 단백질에 0.01 nM 내지 10 nM의 해리 상수(Kd)로 결합하였다(도 3 및 도 15c). 3I14는 높은 친화도(평균 Kd < 0.1 nM)로 시험된 모든 그룹 2 HA(H3, H4, H7 및 H14)에 결합하였다. 또한, 3I14는 그룹 1 H1 아형(H1-CA09, H1-SI06 및 H1-PR8)에는 높은 친화도로 결합한 반면, 다른 그룹 1 아형(H5-VN04, H5-IN05 및 H9-HK99)에 대한 친화도는 낮았다(평균 Kd = 각각 1.02, 1.05 및 5.23 nM).
3I14는 2개의 재조합 바이러스 균주(A/위스콘신/67/05(HA, NA) x A/푸에르토리코/8/34 및 A/아이치/2/68(HA, NA) x A/푸에르토리코/8/34) 및 새로운 H7N9-AH13 균주를 포함하는 다수의 그룹 2(H3 및 H7) 바이러스를 강력하게 중화하였고, 50% 최대 억제 농도(IC50) 값은 0.032 내지 1.074 ㎍/ml이었다(도 4 및 도 17). 이것은 또한 슈도바이러스 H7N1-FPN 및 H7N1-NL219 균주를 0.007 내지 0.027 ㎍/ml 범위의 IC50 값으로 중화하였다(도 5b 및 도 17). 또한, 3I14는 그룹 1 H1 균주(H1-CA09 및 H1-PR8)을 0.225 및 0.413 ㎍/ml의 IC50 값으로 중화하고(도 5b 및 도 10), 슈도바이러스 H5-VN04 및 H5-HK97을 각각 0.040 및 0.008 ㎍/ml의 IC50 값으로 중화하였다(도 5b 및 도 17).
실시예
3: 3I14
IgG1
결합(
Kd
값)
재조합 HA 삼량체에 결합하는 bnAb의 동역학 분석은 옥테트(Octet)® RED96 기기(ForteBio, Inc.)를 사용하여 25℃에서 생물층 간섭 측정기에서 수행되었다. 5 nM의 bnAb IgG1은 피어스 단백질(Pierce Protein) 미함유 차단 완충제(Tween-20 함유 PBS) 내에서 항인간 IgG Fc 바이오센서에 180초 동안 포획되었다. 재조합 전장 HA는 6.25 내지 100 nM 범위의 농도에서 로딩되었다. 모든 실험은 HA와 항-인간 IgG Fc 사이의 잠재적인 비특이적 상호작용에 대해 시험하는 추가적인 항-인간 IgG Fc 항체 바이오센서를 포함하였다. kon의 측정을 위해, 센서를 최대 20 농도의 HA에 노출시킴으로써 3I14 IgG1의 회합을 600초 동안 측정하였다. koff의 측정을 위해, 3I14 IgG1의 해리를 900 초 동안 측정하였다. 친화도 상수(Kd)는 포르테바이오(ForteBio) 데이터 분석 7을 사용하여 계산하였다.
3I14는 0.01 nM 내지 10 nM 범위의 Kd 값으로 그룹 1(H1 및 H5) 및 그룹 2(H3, H4, H7 및 H14)에 속하는 상이한 아형의 정제된 HA 단백질에 결합하였다(도 3). 3I14는 높은 친화도(평균 Kd < 0.1 nM)로 대부분의 그룹 2 HA(H3, H4, H7 및 H14)에 결합하였다. 이와 대조적으로, 3I14는 높은 친화도로 H1 아형(H1-CA409, H1-SI06 및 H1-PR8)에 결합한 반면, 다른 그룹 1 아형(H5-VN04 및 H5-IN05)에 대한 그룹 친화도는 상당히 더 낮았다(평균 Kd > 1 n M).
H1 아형 A/캘리포니아/04/09(H1-CA409), A/솔로몬 아일랜드/3/06(H1-SI06) 및 A/푸에르토리코/8/34(H1-PR8); H3 아형 A/퍼쓰/16/09(H3-PE09), A/우루과이/716/07(H3-UY07), A/위스콘신/67/05(H3-WI05), A/브리즈번/10/07(H3-BR07), A/뉴욕/55/04(H3-NY04) 및 A/빅토리아/341/11(H3-VIC11); H5 A/베트남/1203/04(H5-VN04), A/홍콩/213/03(H5-HK03) 및 A/인도네시아/05/05(H5-ID05); H7 A/네덜란드/219/03(H7-NL219), A/캐나다/RV444/04(H7-CA444) 및 A/안후이/1/13(H7-AH13)의 재조합 전장 HA 단백질(rHA)은 NIH BEIR 보관소(NIAID, NIH)로부터 얻었다. 재조합 전장 H3 A/위스콘신/12/2010(H3-WI10)은 인플루엔자 리에이전트 리소시즈(IRR: Influenza Reagent Resources, 미국 매나사스 소재)을 통해 입수하였다. 아형 H3 A/아이치/2/68(H3-A268), H4 A/청둥 오리/네덜란드/2/05(H4-NL05) 및 H14 A/청둥오리/아스트라칸/263/82(H14-AS82)의 재조합 전장 HA는 리딩턴(R.C. Liddington) 박사(Burnham Institute for Medical Research, 미국 캘리포니아주 소재)로부터 친절하게 기부받았다.
실시예
4.
3I14
IgG1
중화(IC50 값)
IC50 그래프는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다. 3I14를 정사각형으로 나타내고, 항-그룹 1 mAb F10 IgG1(삼각형으로 표시)을 대조군으로 사용하였다(도 4 및 도 5).
MDCK 세포(1.5 x 104 세포/웰)를 96웰 조직 배양 플레이트에 접종하고, PBS로 2회 세척한 후, 2 ㎍/mL 트립신 및 0.5% BSA로 보충된 DMEM에서 인큐베이팅하였다. 바이러스의 100 TCID50(중간 조직 배양 감염량)을 96웰 플레이트에서 Ab 또는 항체 함유 상청액의 2배 연속 희석액과 동일한 부피로 혼합하고, 37℃에서 1시간 동안 인큐베이팅하였다. 인큐베이팅 후, Ab-바이러스 혼합물을 융합성(confluent) MDCK 단일층에 이중으로 옮기고, 37℃에서 21시간 동안 인큐베이팅하였다. 세포를 PBS로 세척하고, 아세톤에 고정하고, 인플루엔자 A 바이러스 핵단백질(NP)(클론 A3, BEI)에 대한 mAb를 사용한 간접 ELISA에 의해 바이러스 항원을 검출하였다. 50% 최대 억제 농도(IC50)는 배경 차감 후 바이러스 대조군을 포함하는 웰에 비해 효과가 50% 감소한 Ab 농도이다.
3I14는 시험관 내에서 그룹 1(H1) 및 그룹 2(H3 및 H7) 바이러스를 중화하였다. 3I14는 0.032 내지 1.336 ㎍/ml의 IC50 값으로 상이한 H1, H3 및 H7 바이러스를 강력하게 중화하였다(도 4 및 5).
실시예
5. 그룹 1 및 그룹 2 아형을 나타내는
슈도바이러스의
3I14
IgG1
중화(IC50 값)
IC50 그래프는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다. 3I14는 정사각형으로 나타내었고, 항-그룹 1 mAb F10 IgG1(삼각형으로 나타냄)을 대조군으로 사용하였다.
3I14는 0.032 내지 1.336 ㎍/ml 범위의 IC50 값으로, 둘 모두 그룹 2 슈도바이러스인 H7N1-FPN 및 H7N1-NL219를 강력하게 중화하였다. 또한, 각각 2.137 및 4.601 ㎍/ml의 IC50 값으로 그룹 1 슈도바이러스 H5-VN04 및 H5-HK97을 중화하였다(도 5).
실시예
6. 마우스에서 그룹 2 및 1 인플루엔자 바이러스에 대한
3I14의
예방 효능
본 발명자들은 BALB/c 마우스 모델에서 H5N1, H3N2, H7N7 및 H7N9 감염에 대한 보호 효능을 평가하기 위해 3I14를 전장 인간 IgG1로 전환하였다(도 6). 항-그룹 1 Ab인 F1012가 균주 특이적 대조군으로서 사용되었다. H7N7-NL219, H7N9-AH13, H3N2-BR07-ma 및 H5N1-VN04 바이러스의 치사 용량으로 시험접종하기 1일 전에 3I14 및 F10 IgG1을 다양한 용량으로 처리하였다. 5 mg/kg의 3I14 IgG1을 사용한 예방은 H7N7-NL219 또는 H7N9-AH13 시험접종 후에 14일 내지 18일에 최소 체중 감소를 보이면서 마우스를 완전히 보호하였다(도 6a). 25 mg/kg 3I14의 용량에서, IgG1은 H3N2-BR07에 대해 80%의 보호를, H5N1-VN04에 대해 60%의 보호를 나타냈다. 모든 생존 마우스는 관찰 기간이 끝날 때까지 체중 감소의 역전을 보였다(도 6b). H3N2 BR07, H5N1 VN04, H7N9 AU13 또는 H7N7 NL219 인플루엔자 바이러스의 i.n. 접종에 의해 시험접종되어 치사 전에 24시간 동안 5마리의 마우스 그룹을 25 또는 5 mg/kg의 정제된 IgG로 복강 내로 처리하였다. bnAb 3I14(적색), 그룹 1 대조군 mAb F10(흑색) 및 그룹 2 대조군 mAb A533(청색)으로 처리한 마우스의 생존율(%)(A) 및 체중 변화(%)(B).
실험 1일 전에, 5마리의 암컷 8-10주령 BALB/c 마우스 그룹에게 각각 0.5 mL 부피로 복강내(i.p.) 경로에 의해 3I14, F10 및 A533-IgG1을 5 mg/kg의 저농도 및 20 또는 25 mg/kg의 고농도로 주입하였다. 6개의 마우스 그룹을 마우스 적합화된 그룹 A/베트남/1203/04(H5N1), A/브리즈번/10/07(H3N2), A/네덜란드/219/03(H7N7) 또는 A/안후이/1/13(H7N9)의 10 LD50을 비내 감염시켰다. 마우스는 바이러스 시험접종 당일에 체중을 측정한 다음, 생존을 모니터링하고, 14일 또는 18일 동안 매일 체중을 측정하였다. 동물 연구는 승인된 기관 동물 관리 및 사용 위원회 규약(Institutional Animal Care and Use Committee protocols)에 따라 수행되었다.
치사량의 H5N1-VN04, H3N2-BR07, H7N7-NL219 및 H7N9-AU13 바이러스로 시험접종하기 1일 전에 마우스를 3I14, F10(그룹 1 대조군 Ab) 및 A533(그룹 2 대조군 Ab) IgG1의 다양한 용량으로 처리하였다. 5 mg/kg 이상의 3I14 IgG1을 사용한 예방은 관찰 기간 동안 최소 체중 감소와 함께 H7N7-NL219 또는 H7N9-AU13 투여 후에 사망으로부터 마우스를 완전히 보호하였다. 이러한 결과는 3I14 IgG1이 치사량의 H3N2-BR07로 시험접종된 마우스를 효과적으로 보호하고(60-80%), 치사량의 H5N1-VN04로 시험접종된 마우스를 부분적으로 보호함(20-60%)을 나타낸다(도 6a). 5 mg/kg의 3I14 IgG1의 용량은 H3N2 및 H5N1에 의한 사망을 예방하기 위해 단지 부분적으로만 보호하지만, 모든 생존 마우스는 5 또는 25 mg/kg의 용량에서 관찰 기간 종료시에 체중 감소의 역전을 보였다 (도 6b).
실시예
7: 3I14는
트립신 매개 HA 성숙 및 pH 의존성
구고적
변화를 차단한다.
스템에 대해 작용하는 bnAb는 HA의 pH 의존성 구조적 변화 및 막 융합을 방해하는 것으로 알려져 있다12 , 14, 16. 전구체 HA0의 절단은 산성 엔도솜 환경에서의 막 융합의 후속 활성화를 위해 HA를 프라이밍한다. 미성숙 HA0은 일반적으로 호흡 상피 세포 상의 표면 프로테아제에 의해 HA1 및 HA2로 처리되고28 , 29, 이것은 트립신에 의한 HA0 처리에 의해 실험적으로 모방된다30. 3I14가 HA0 절단 부위 및 HA2 N-말단 융합 펩티드를 포함하는 HA의 스템 도메인을 표적으로 하기 때문에, 본 발명자들은 3I14가 또한 HA0의 트립신 절단 활성화를 차단할 수 있는지 또는 HA 매개 바이러스-숙주 막 융합을 방해하는지의 여부를 시험하였다. 도 7은 대조군 항-HSS IgG1(Fm-6)은 그렇지 않지만 3I14 IgG1은 미성숙 HA0의 절단을 방지함을 보여준다. 본 발명자들은 또한 표면 발현된 H3-A2/68 및 H3-BR07을 이용하여 3I14가 낮은 pH에 의해 촉발되는 구조적 재배열을 방지함을 분석하였다. 도 8(상부)은 3I14가 트립신 활성화 단독 후에(좌측 중간) 또는 낮은 pH 촉발이 이어질 때(우측 중간) 미절단된 HA 전구체(HA0)(좌측)와 2개의 성숙 형태(HA) 둘 모두에 결합함을 보여주다. 대조적으로, 3I14는 DTT 환원에 의해 매개되는 해리된 HA2에는 결합하지 않았다(우측). 3I14가 낮은 pH 촉발 전에 성숙한 HA에 미리 결합될 때, 항체는 DTT 처리 후에 결합을 유지하였고, 이것은 3I14가 pH 의존성 HA 재배열을 억제함을 나타낸다(도 8, 하부). 또한, 3I14의 사전 결합은, E730 Ab(항-HA1)의 결합이 DTT 처리 후에 보존되었기 때문에 HA1-HA2 해리를 방지하였다(도 8, 하부). 이러한 데이터로부터, 본 발명자들은 HA 스템 에피토프에 대한 3I14 결합이 HA0 절단 및 pH 의존성 구조적 변화의 억제를 유도한다고 결론을 내렸다.
실시예
8: 3I14
IgG1은
표면 발현된 H3-A268 및 H3-BR07에 대한, 낮은 pH에 의해 촉발된 구조적 재배열을 억제한다.
표면 발현된 H3의 구조적 재배열은 3I14(솔리드 막대) 및 헤드 결합 대조군 mAb E730(오픈 막대)의 FACS 염색에 의해 검출되었다(도 8). 다양한 구조는 상응하는 그래프 위에 표시되고, 다음과 같았다: 절단되지 않은 전구체(HA0); 트립신 활성화, 절단(HA); 낮은 pH 촉발, 절단(pH 4.9); 및 DTT 환원, 삼량체 HA2(tHA2). 결합은 미처리된 HA(HA0)에 결합하는 백분율로 표현된다. 항체 억제 분석을 위해, H3를 mAb 없이, 3I14로, 또는 대조군 Ab인 Fm-6으로 전처리한 후, 절단된 HA를 pH 4.9에 노출시켰다. 데이터는 3회의 독립적인 실험의 평균 + SD를 나타낸다.
도 8에 도시된 바와 같이, 3I14는 트립신 활성화 및 낮은 pH 촉발 후에 미절단된 HA 전구체(HA0) 및 성숙 형태(HA) 둘 모두에 결합하였지만, DTT 환원에 의해 매개되는 해리된 HA2에는 결합하지 않았다. 3I14가 낮은 pH 촉발 전에 성숙 HA에 미리 결합되는 동안, 항체는 DTT 처리 후에도 결합된 상태로 유지되고, 이것은 3I14가 pH 의존성 HA 재배열 및 후속 막 융합을 억제함을 나타낸다. 또한, 3I14의 사전 결합은, DTT 처리 후에 E730 Ab(항-HA1)의 결합이 보존되었기 때문에 HA1이 HA2로부터 해리되는 것을 방지하였다.
MDCK 세포를 전장 재조합 인플루엔자 A pcDNA3.1-H3-A268 및 H3-BR07 플라스미드로 형질감염시켰다. 형질감염 30시간 후, 세포를 0.2% EDTA/PBS를 사용하여 플라스틱 지지체로부터 분리하였다. 상이한 HA 구조 형태 및 구조에 대한 mAb 결합을 측정하기 위해, 세포 샘플을 분할하고, 각각의 처리 단계 후에 3I14 또는 E730 IgG1(항-H3 헤드)로 염색하였다. 분리된 세포는 이후 실온에서 5분 동안 트립신(TrypLETM Select Enzyme, Gibco)으로 처리하고, 1% BSA/PBS로 세척하고, 시트르산-인산나트륨 완충제(pH 4.9)에서 15분 동안 인큐베이팅하고, 세척한 후, PBS 내의 50 mM 디티오트레이톨(DTT)과 함께 실온에서 20분 동안 인큐베이팅하였다. 다르게는, 낮은 pH 단계 전에 5 ㎍ 3I14 또는 Fm-6 IgG1을 첨가하였다. 후속 처리의 샘플을 APC 접합된 항-인간 Fc(BioLegend, Inc.)로 염색하였다. 염색된 세포는 FACS Diva 소프트웨어(Becton Dickinson)로 BD FACSAriaTM II를 사용하여 분석하였다.
실시예
9: 3I14은
FC
의존성 바이러스 청소를 매개한다.
항-스템 bnAb는 mAb 매개 항바이러스 청소의 주요 메커니즘으로 여겨지는, 인플루엔자 바이러스 감염 세포의 FcγR 의존성 세포독성을 효율적으로 매개하는 것으로 보고되었다31. 3I14 및 다른 항-스템 bnAb에 의한 항체 의존성 세포독성(ADCC)의 특성을 조사하기 위해, 본 발명자들은 표적으로서 HA 발현 293T 세포와 함께 인간 FcγIIIa 및 활성화된 T 세포의 핵 인자(NFAT) 유도 루시페라제32를 안정적으로 발현하는 조작된 Jurkat 이펙터 세포를 사용하여 시험관 내에서 대리 리포터(surrogate reporter) 기반 ADCC 분석을 수행하였다. H3 발현 293T 표적 세포와 함께 인큐베이팅한 후, 3I14는 FI6v3, CR9114, 39.29 및 그룹 2 mAb CR8020을 포함하는 다른 항-스템 bnAb와 대등한 수준으로 용량 의존적 방식으로 Jurkat 리포터 세포에서 유의한 루시페라제 반응을 유도하였다(도 9). 이 분석의 특이성은 항-그룹 1 mAb인 F10으로부터의 반응의 부재에 의해 증명되었다. 3I14는 또한 H5 발현 293T 세포에 대한 루시페라제 반응을 특이적으로 유도하였지만, FI6v3, CR9114, 39.29 및 F10보다는 낮은 수준으로 유도하였다. 본 발명자들은 H5 발현 293T 표적 세포에 대한 CR8020의 낮은 반응성을 관찰하였다(도 9). 이들 데이터는 3I14가 또한 생체내 보호를 위한 Fc 의존성 면역 매개 메커니즘을 관여할 가능성이 있음을 지지한다.
실시예
10.
3I14는
H3 또는 H5와 결합하기 위해 다른 항-줄기
bnAb인
FI6, CR9114, 39.29, F10 및 CR8020과 교차
경쟁한다.
ELISA 플레이트 상에 고정된 5 ㎍/ml H3-BR07 또는 H5-VN04 단백질을 80 nM 내지 0.3 nM의 3I14 Fab의 2배 연속 희석액과 함께 인큐베이팅하고, 5 nM에서 다른 scFvFc Ab와 혼합하였다. 1시간 동안 동시 인큐베이팅한 후, HRP 접합된 항-인간 CH2 항체를 사용하여 scFvFc Ab의 결합을 검출하였다. 3I14 Fab는 H3-BR07에 결합하기 위해 CR8020, CR9114, FI6 및 39.29를 비롯한 다른 항-줄기 Ab와 교차 경쟁하지만, 항-HA1 항체인 E730과는 경쟁하지 않았다(도 10a-d). 3I14는 또한 H5-VN04에 대한 39.29 및 F10의 결합을 억제하지만, 항-헤드 항체 2A의 결합은 억제하지 않았다(도 10e-f). 이러한 결과는 3I14가 다른 항-줄기 bnAb의 알려진 에피토프와 중복되거나 매우 근접한 HA 스템 영역 내의 에피토프를 표적화함을 제시한다.
실시예
11. 시험관 내에서의 구조 기반 친화도 성숙
H3 및 H5에 대한 동일하지 않은 결합 강도에 대한 분자 기초를 특성화하고 H5N1 균주에 대한 개선된 친화도를 갖는 3I14를 조작하기 위해, 본 발명자들은 먼저 3I14 구조의 인 실리코(in silico) 시뮬레이션을 위한 항체 구조 예측 프로그램인 바이오루미네이트(BioLuminate)33를 사용하였다. 다른 3개의 IGHV3-30 bnAb인 FI6v3, 39.29 및 MAb 3.1과 3I14 모델의 중첩은 도 12b에 도시된다. 이러한 항체 간의 주요 차이점은 HA와 접촉하는 루프 구조를 형성하는 FI6v3의 LCDR1이 더 긴 것을 제외하고는 HCDR3의 구조임이 분명하다.
다음으로, 3I14 모델을 로제타독 서버34를 사용하여 H3 삼량체 구조에 도킹시켰다. 3I14는 H3 및 H5에 결합하기 위해 FI6v3 및 39.29와 경쟁하고, MAb 3.1은 FI6v3 및 39.29와 동일한 보존된 에피토프를 점유하기 때문에18, 본 발명자들은 3I14가 H3/H5와 상호작용하기 위해 FI6v3, 39.29 및 MAb 3.1과 유사한 결합 방식을 채택할 것이라는 가설을 세웠다. 상기 3개의 Ab-HA 공결정(co-crystal) 구조에 대해서, HCDR3은 융합 펩티드 및 나선 A와 소수성 코어를 형성하는데 중요한 역할을 수행한다15 , 17, 18. HA와 유의한 상호작용을 하기보다는, HCDR1 및 HCDR2는 결합을 촉진하기 위해 HCDR3 루프를 안정화하는 것으로 보인다. 친수성 경쇄 CDR 잔기는 또한 HA와 상호작용하고 소수성 코어를 둘러싸지만, 경쇄의 배향은 보존되지 않고 결합에 관련된 잔기도 아니다. 이러한 관찰은 HCDR3을 에피토프와 상호작용하는 최적의 위치로 배향시킴으로써 경쇄가 주로 결합에 기여한다는 것을 제시한다.
이러한 해결된 공결정 구조에 기초하여, 본 발명자들은 3I14/H3(도 13b) 및 3I14/H5 복합체(제시되지 않음)의 가장 유사한 결합 모델을 1000개의 디코이로부터 선택하였다. 두 복합체의 경면에 대한 철저한 분석은 3I14가 H5보다 H3/H1에 더 강하게 결합하는 이유를 이해하기 위해 수행되었다(표 5). 활발한 계산35은 전기적 반발로 인해 E39가 H5/3I14 모델에서 D94로부터 멀리 회전하면서 염 다리를 형성할 수 있고 H5 결합에 유리하지 않을 수 있는, H3 모델에서 3I14 경쇄의 D94와 K39 사이의 매우 유리한 결합 기여를 보여준다(도 13c). H5 이외에, E39 아미노산 변화는 또한 그룹 1 H2, H6, H11 및 H13 균주에서도 발견된다(표 6). 또 다른 놀라운 변이는 H3 위치 L38에 있고, 여기서 상기 잔기는 일부 그룹 1 균주에 대해 K/R38로 변경된다(표 6). 그러나, 결합 기여는 두 모델에서 L/K38이 유리한 내지 매우 유리한 결합(총 유리한 자유 에너지의 ~70%)으로 HCDR3 잔기 Y104, F105 및 F109에 접촉함을 보여주고, 따라서 본 발명자들은 이들 잔기가 두 HA에 대한 결합에 긍정적인 영향을 갖는다고 간주하였다


실시예
12: H3
삼량체
모델의 줄기 상의
3I14
에피토프의
구조.
도 13은 H3의 줄기 상의 3I14 에피토프의 개략도이다. 도면에서, 3I14의 중쇄는 청색으로 표시되고, 경쇄는 자홍색으로 표시된다. H3 삼량체의 줄기는 연어살색, 녹색 및 청록색이다. 잔기 넘버링은 H3 또는 Ab 서열에 기초하여 철저히 이루어진다.
3I14
/H3 도킹
3I14 모델은 본 발명자들의 리눅스 머신에 설치된 독립형 소프트웨어로서 로제타독을 사용하여 H3 삼량체 구조에 도킹되었다. 로제타독은 국소 고해상도 도킹을 처리하고 여분의 회전이성질체(rotamer) 및 루프 재배열을 허용하는 그의 기능 때문에 선택된다. 도킹 전에 3I14 모델은 H3/39.29 복합체 구조 내에서 39.29에 중첩되었다. 여분의 측쇄 회전이성질체가 첨가되었고, 고해상도 단독 프로토콜이 수행되었다. 1000개의 디코이가 생성되었고, 최상의 점수를 가진 클러스터링된 모델을 PyMol로 철저히 분석하였다. 3I14가 H3에 결합하기 위해 FI6과 경쟁한다는 것을 감안하여, 본 발명자들은 3I14가 H3와 상호작용하기 위해 39.29, FI6 및 Mab3.1과 동일한 방식을 채택할 것이라는 가설을 세웠다. 따라서, 최종 모델을 선택할 때 다음의 기준이 적용되었다: HCDR3 및 H3의 HA2의 융합 펩티드 및 나선 A 상의 소수성 잔기는 계면에서 소수성 코어를 형성하기 위해 긴밀하게 접촉하여야 하고; HCDR2 및 HCDR1 잔기는 다른 복합체와 마찬가지로 HCDR3와 유사한 상호작용을 하고; 경쇄 CDR은 H3과 주로 친수성 상호작용을 한다. 1000개의 디코이로부터 상위 10개 모델 중에서, 6개 모델이 이 기준에 적합하며, 서로 매우 유사하다. 따라서, 6점 이내의 최고 점수를 갖는 모델이 추가의 분석을 위해 선택되었다.
실시예
13 H3/
3I14
및 H5/
3I14
모델의 서열 정렬 및 구조 중첩.
도 14a는 H3, H5 및 인플루엔자 B의 스템 에피토프의 서열 정렬을 도시한 것이다. 도 14b는 잔기 38 및 39에서 H3/3I14 및 H5/3I14 모델의 구조 중첩을 도시한 것이다. H3은 청록색으로 표시되고, H5는 황색으로 표시되고; H3/3I14 모델의 3I14는 청색(중쇄) 및 황색(경쇄)으로 표시되며, H5/3I14 모델의 3I14는 오렌지색으로 표시된다. H3 잔기 Leu38 및 Lys39는 표지된다. 중쇄의 잔기 F100F 및 경쇄의 D93은 각각 38 및 39와 상호작용하며, 이들 또한 표지된다.
3I14
/H5 도킹
H5/3I14 및 인플루엔자 B/3I14 복합체를 3I14/H3 복합체와 동일한 방식으로 모델링하였다. 3I14 모델과 H5 삼량체 또는 H3 삼량체 모두 H3/39.29 복합체 구조와 중첩되고, 2개의 구조 파일은 도킹의 초기 모델로서 하나의 3I14/H5 복합체 또는 3I14/인플루엔자 B 복합체로 병합되었다. 흥미롭게도, H3/3I14 복합 모델을 위해 선택된 모델과 유사한 모델은 H5/3I14 및 인플루엔자 B/3I14 복합체 둘 모두를 위한 최상의 모델 중 하나이다. 따라서, 이러한 유사한 모델을 추가 분석을 위한 최종 모델로 선택하였다.
보다 양호한
H5 결합을 위한
3I14
조작
H3/3I14 및 H5/3I14 계면의 철저한 분석은 3I14가 H5보다 H3에 더 강력하게 결합하는 이유를 이해하기 위해 수행되었다. 모든 에피토프 잔기의 서열 정렬은 가장 현저한 변이가 H3 내의 L38-K39가 H5에서 K38-39E로 되는 것임을 보여주었다(도 14a). L38은 H3/3I14 모델에서 소수성 코어의 일부이며, 3I14의 HCDR3으로부터의 F100F와 상호작용한다(도 14b). 놀랍게도, H5/3I14 모델의 K38은 하전된 아민기를 용매로 향하게 하고 지방족 사슬을 F100F에 대면하게 함으로써 동일한 접촉을 이룰 수 있다. 모델에 따르면,이 돌연변이는 결합 친화도에 영향을 줄 수 있는 것으로 보이지 않는다. 이와 반대로, K39는 H3/3I14 모델에서 3I14의 경쇄로부터의 D94와 접촉하고, E39는 전기적 반발로 인해 H5/3I14 모델에서 D94로부터 멀리 회전한다(도 14b). 명백하게, K39E 돌연변이는 H5 결합에 유리하지 않고, 아마도 이것이 3I14가 H3에 비해 H5에 더 약하게 결합하는 이유이다. 이 가설을 시험하기 위해, 상이한 아형으로부터의 HA의 위치 38-39의 잔기가 3I14에 결합하는 그의 능력과 비교하여 조사되었다. L38 및 K39를 갖는 HA가 3I14에 강하게 결합하고 K38 및 E39를 갖는 HA는 보다 약한 결합을 보이는 강한 상관 관계가 밝혀질 수 있다. 종합적으로, 본 발명자들은 3I14의 경쇄 내의 D94K 돌연변이가 H5에 대한 3I14의 결합 선호도를 역전시키고 H3에 대한 결합 친화도를 낮출 것이라는 가설을 제시한다. 또한, 본 발명자들은 D94N 3I14 변이체가 H3 및 H5 둘 모두에 똑같이 잘 결합할 것이라는 가설을 제시한다. 본 발명자들은 상호작용을 위한 주요 원동력이 HCDR3으로부터의 소수성 상호작용이기 때문에, D94N 돌연변이가 H3 결합의 약화를 유발할 것으로 기대하지 않는다. 이 위치에서의 상호작용이 반발적이지 않으면, 그것은 친화도에 유의한 영향을 분명히 주지 않는다.
실시예
14: 재조합 H5-VN04(A) 및 H3-PE09(B)에 대한
3I14
WT 및
VLD94N
돌연변이 IgG1의 결합(Kd 값).
3I14
VLD94N
변이체는
H5에 대한 결합 및
중화 활성을 개선한다.
E39 및 D94의 제안된 반발 효과를 제거하기 위해, 본 발명자들은 부위에서 음전하 상실을 유도하는 단일 Asp→Asn(D→N) 돌연변이가 H3 및 H5 둘 모두에 동일하게 잘 결합할 것이라는 가설을 세웠다. 이러한 구조 기반 변형을 조사하기 위해, 본 발명자들은 먼저 WT 3I14 및 VLD94N 변이체 IgG1 둘 모두의 결합 친화도를 평가하였다. 표 3에 나타낸 바와 같이, VLD94N 변이체는 H5에 대한 결합 친화도를 거의 10배 증가시켰지만, H3에 대한 결합에서 어떠한 유의한 변화도 일으키지 않았다. 흥미롭게도, H5에 대한 더 높은 친화도는 또한 해리 속도의 감소 때문인 반면, 회합률은 동일하였다(표 7 및 도 15).

본 발명자들은 또한 H5 슈도타이핑된 또는 H3 감염성 바이러스에 대한 3I14 VLD94N 변이체의 활성을 평가하기 위해 중화 분석을 수행하였다(도 16c 및 16d). 3I14에 비해, VLD94N 변이체는 10배 더 높은 효능으로 H5-VN04의 슈도바이러스를 중화하였다(IC5O: 8.65 ng/ml 대 81.58 ng/ml)(도 16c). 한편, H3-BR07에 대한 중화 활성은 VLD94N 변이체에서 그대로 유지되었다(IC5O: 336.6 ng/ml 대 305.4 ng/ml)(도 16d). 이러한 결과는 최적화된 3I14 VLD94N 변이체가 H3에 대한 그의 효능을 유지하면서 H5에 대한 결합 및 중화 능력을 증가시킨다는 것을 입증한다.
추가의 실험은 3I14가 1.96 nM의 Kd로 H3-PE09에 결합한 반면, H3-PE09에 대한 3I14 VLD94N 돌연변이체의 친화도는 유사함(평균 Kd = 2.34 nM)을 입증하였다. D94N 돌연변이는 H3 결합의 약화를 유도하지는 않지만, H5에 대한 결합 친화도의 증가를 유도한다(도 15a 및 15b).
실시예
15: 3I14
WT 및
VLD94N
돌연변이체는
IgG1이
슈도타이핑
바이러스
H5N1
-VN04 및 감염성 바이러스
H3N2
-BR07을 중화한다.
도 16은 H5N1-VN04 및 H3N2-BR07 감염 바이러스의 중화를 나타내는 일련의 그래프이다. 3I14 WT(흑색) 및 VLD94N 돌연변이체(적색)는 슈도타이핑된 바이러스 H5N1-VN04(A) 및 H3N2 BR07 바이러스(B)를 중화하였다. 항-그룹 1 mAb F10(청색)을 대조군으로 사용하였다. 이들 데이터는 2-3회의 독립적인 실험의 평균 중화 역가를 나타낸다.
3I14 VLD94N 돌연변이체 IgG1은 3I14 WT보다 더 높은 IC50 값으로 슈도바이러스 H5N1-VN04를 중화하고, 또한 유사한 IC50 값으로 H3N2 바이러스도 중화하였다.
실시예
16: H5에 대한 결합이
증가된
3I14
변이체의
단리를 위한 조작된 효모 디스플레이.
7개의 효모 디스플레이 라이브러리는 HCDR1, HCDR2, HCDR3, LCDR1, LCDR2, LCDR3 및 LCDR4의 잔기를 무작위로 선발함으로써 생성되었다. 이들 효모 디스플레이 라이브러리는, 선택되고 효소 디스플레이 벡터 pCTCON2 내로 클로닝되는 단일쇄 3I14 변이체의 풀을 생성하기 위해 사용되었다. 단리된 구축물은 제시용 마커로서 작용하기 위해 항체의 C-말단에 c-Myc 태그를 부착할 것이다. 항체 발현 및 표면 디스플레이는 24-48시간 동안 20도에서 SGCAA 배지에서 라이브러리를 성장시킴으로써 유도될 것이다. 3I14 변이체의 성공적인 제시는 항-c-Myc FITC 표지에 의해 검출될 것이다. H5 HA는 형광 표지로 표지되고, 1시간 동안 염색에 첨가된다. 결합되지 않은 시약은 씻어내고, 표지된 라이브러리는 H5 HA 양성 클론에 대해 분류될 것이다.
도 18에 도시된 바와 같이, FACS 분석에 의해 입증된 바와 같이, 3I14 WT 효모 및 CDR 라이브러리 둘 모두는 C-myc 및 H5에 대해 양성이었다. 이중 양성 H5 및 c-myc 양성 집단은 3I14-WT-효모 라이브러리와 비교하여 3I14 효모 CDR 라이브러리에서 0.039%로부터 0.090%로 증가되었다.
향후 연구에서, 양성 클론을 배양하고, 양성 집단을 풍부하게 하기 위해 3회 다시 분류할 것이다. 양성 클론은 FACS 분석에 의해 확인되고, 서열결정으로 확인될 것이다. FACS 분류와 조합한 효모 디스플레이는 항체공학을 위해 성공적인 것으로 입증되었고, 인플루엔자 B HA에 결합할 수 있는 3I14 변이체 클론을 단리하기 위해 사용될 것이다. 인플루엔자 B HA에 대한 초기 결합제가 확인되면, 후속 라운드의 스크리닝이 다색 분류를 사용하여 수행될 것이고, 즉 H3, H5 및 인플루엔자 B HA가 상이한 형광 표지로 표지되고 삼중 양성 3I14 변이체가 분류될 것이다.
실시예
17:
에피토프
매핑
및 결합 경쟁
3I14 인식을 위한 HA의 에피토프를 조사하기 위해, 본 발명자들은 옥테트® RED96 기기에서 HA 또는 HA1 서브유닛의 전장에 대한 그의 결합 활성을 평가하였다. 3I14는 삼량체 전장 H3 균주인 A/퍼쓰/16/09(PE-09)에 결합하지만, 그의 HA1 서브유닛에는 결합하지 않았다(도 19). 본 발명자들은 3I14와 다른 스템에 대해 작용하는 bnAb인 FI6v3, CR9114, 39.29, F10 및 CR8020 사이의 결합 경쟁 분석을 추가로 수행하였다(도 2). 3I14 Fab는 H3-BR07에 대한 다른 항-스템 Ab인 CR9114, FI6v3 및 39.29의 결합을 강하게 억제하지만, 헤드에 대해 작용하는 항-H3 mAb E730(미공개된 항체 서열)은 억제하지 않는다(도 10). 또한, 3I14는 보다 막에 근접한 에피토프 14에 대해 작용하는 CR8020과 경쟁한다. 3I14는 H5-VN04에 대한 39.29 및 F10의 결합을 부분적으로 억제하지만, 항-H5 헤드 항체 2A12의 결합은 억제하지 않는다(도 2e, f). 이 결과는 3I14가 다른 bnAb의 알려진 스템 에피토프와 겹치거나 매우 가깝다는 것을 입증한다. 또한, 3I14는 H3의 강력한 억제제이며 H5의 중등도의 억제제이다. 이러한 결과는 H3 및 H5에 대한 3I14 결합의 친화도 측정치와 일치한다.
실시예
18: 물질 및 방법
세포
최근의 계절성 인플루엔자 백신 접종을 보고하지 않은 7명의 건강한 성인의 신선한 PBMC는 IRB가 승인한 인간 프로토콜에 따라 2012년 12월 DFCI 크래프트 패밀리 블러드 도너 센터(Kraft Family Blood Donor Center)에서 백혈구 성분 채집술 동안 수집된 버려진 "칼라(collar)"를 사용하여 얻었다. 아메리칸 타입 컬쳐 컬렉션(미국 버지니아주 매나사스 소재)으로부터 마딘-다비 개 신장(MDCK: Madin-Darby canine kidney) 세포, 293T 및 293F 세포를 얻었다.
재조합
헤마글루티닌의
제조
H3(A/브리즈번/10/2007)의 세포외 도메인, 잔기 17 내지 531은 아비태그(Avitag)(아미노산 서열: GGGLNDIFEAQKIEWHE), 트롬빈 절단 부위, 삼량체화 T4 피브리틴 폴돈(fibritin foldon) 및 6개의 히스티딘 잔기를 함유하는 C 말단 펩티드를 포함하는 융합 단백질로서 발현되었다. 융합 단백질 H3-ATTH는 293F 세포에서 발현되고, Ni-NTA 친화도 크로마토그래피에 의해 상청액으로부터 정제되었다. 정제된 재조합 HA 단백질은 트롬빈 효소(Novagen, 독일 다름슈타트 소재)에 의해 절단되고, 제조사의 지시에 따라 BirA 효소(Avidity, 미국 콜로라도주 오로라 소재)로 비오티닐화하였다.
A/뉴욕/18/09(H1-NY09), A/텍사스/05/09(H1-TX09), A/일본/305/57(H2-JP57), A/아이치/2/68(H3-A2/68), A/브리즈번/10/07(H3-BR07), A/네덜란드/2/2005(H4-NL05), A/베트남/1203/04(H5-VN04), A/홍콩/1073/99(H5-HK99), A/닭/뉴욕/14677-13/98(H6-NY98), A/네덜란드/219/03(H7-NL219), A/칠면조/온타리오/6118/68(H8-ON68), A/홍콩/1073/99(H9-HK99), A/오리/멤피스/546/74(H11-MEM74), A/오리/알버타/60/76(H12-AB76), A/청둥오리/아스트라칸/263/1982(H14-AS82), A/쉬어워터/웨스트 오스트레일리아/2576/79(H15-WA79) 및 A/붉은부리 갈매기/스웨덴/2/99(H16-SE06)의 전장 HA 유전자를 pcDNA3.1 벡터에 클로닝하고, 세포 표면 발현된 HA를 생산하기 위해 293T/17 세포에 형질감염시켰다.
H1 아형 A/캘리포니아/04/09(H1-CA09), A/솔로몬 아일랜드/3/06(H1-SI06) 및 A/푸에르토리코/8/34(H1-PR8); H3 A/퍼쓰/16/09(H3-PE09), A/우루과이/716/07(H3-UY07) 및 A/빅토리아/341/11(H3-VIC11); H5 A/베트남/1203/04(H5-VN04) 및 A/인도네시아/05/05(H5-ID05); H7 A/네덜란드/219/03(H7-NL219), A/캐나다/RV444/04(H7-CA444) 및 A/안후이/1/13(H7-AH13); H9 A/홍콩/1073/99(H9-HK99)의 재조합 전장 HA 단백질은 NIH BEIR 보관소(NIH, 미국 버지니아주 매나사스 소재)에서 입수하였다. 아형 H4 A/청둥오리/네덜란드/2/05(H4-NL05) 및 H14 A/청둥오리/아스트라칸/263/82(H14-AS82)의 재조합 전장 HA는 리딩턴 박사(Burnham Institute for Medical Research, 미국 캘리포니아주 소재)로부터 친절하게 기부받았다.
인플루엔자 바이러스 및 HA
슈도타이핑된
바이러스의 제조
야생형 인플루엔자 바이러스 A/캘리포니아/4/09(H1N1-CA09), A/푸에르토리코/8/34(H1N1-PR8), A/퍼쓰/16/09(H3N2-PE09), A/아이치/2/68(H3N2-A2/68), A/홍콩/8/68(H3N2-HK68), A/시드니/5/97(H3N2-SY97), A/브리즈번/10/07(H3N2-BR07), A/위스콘신/67/05(HA, NA) x A/푸에르토리코/8/34(H3N2), A/아이치/2/68(HA, NA) x A/푸에르토리코/8/34(H3N2) 및 A/난창/993/95(H3N2-NC95)는 NIH BEIR 보관소(NIH, 미국 버지니아주 매나사스 소재)에서 얻은 후, 마딘-다비 개 신장(MDCK) 세포에서 표준 바이러스 배양 기술에 의해 성장시켰다. 동물 시험접종 연구에 사용된 A/브리즈번/10/2007-ma(H3N2)는 PR8 재조합(reassortant) 바이러스 x-17146에서 유래된 마우스 적응 바이러스이다.
A/베트남/1203/04(H5-VN04), A/홍콩/156/97(H5-HK97), A/네덜란드/219/07(H7-NL219), A/FPV/로스톡/1934(H7-FPV)의 전장 HA 유전자 및 H5-VN04의 뉴라미니다제 유전자 N1(Genbank 수탁 번호 AAW80723)을 별개로 pcDNA3.1 플라스미드에 클로닝하였다. Env-슈도타이핑된-루시페라제 리포터 바이러스는 이전에 설명된 바와 같이 293T/17 세포에서 생산되었다12. 간단히 설명하면, pcDNA3.1-H5-VN04, H5-HK97, H7-NL219 또는 H7-FPV 플라스미드를 N1 발현 플라스미드 pcDNA3.1-N1-VN04, HIV 패키징 벡터 pCMVR8.2 및 리포터 벡터 pHIV-Luc와 함께 293T/17 세포에 별개로 동시 형질감염시켰다 . 바이러스 상청액을 형질감염 48시간 후에 수거하였다. 바이러스 적정은 POLARstar 오메가(Omega) 미세플레이트 판독기(BMG LABTECH, 독일 오르텐베르크 소재)를 사용하여 루시페라제 활성을 측정함으로써 평가하였다.
H3 결합 기억 세포의
FACS
분류
피콜-파크(Ficoll-Paque) 구배(GE HealthCare)를 사용하여 수집된 혈액으로부터 신선한 PBMC를 단리하였다. CD19+/CD27+ B 세포는 비오티닐화 H3-ATTH 및 알로피코시아닌(APC) 표지된 스트렙타비딘으로 염색되었다. 단일 H3 반응성 기억 B 세포를 384웰 플레이트 내에 분류하였다. 14일간의 팽창 후, 상청액을 재조합 H1(H1-CA09), H3(H3-BR07) 및 H7(H7-CA444) HA 단백질에 대한 반응성에 대해 시험하고, 메조 스케일 디스커버리 멀티플렉스(MSD: Meso Scale Discovery multiplex, 미국 메릴랜드주 록빌 소재)에 의해 분석하였다. 이어서, 반응 상청액에 대해 H3N2-BR07에 대한 시험관내 중화 활성을 측정하였다. 모든 H3N2 중화 항체는 이전에 설명된 바와 같이47 프라이머를 사용한 단일 세포 RT-PCR에 의해 구조되었다.
3I14
scFv
및
IgG
항체의 발현 및 정제
본 발명자들은 3I14 Ab를 pcDNA3.1-힌지 scFvFc 미니바디(minibody) 발현 벡터 내로 초기에 이동시켜, scFv를 인간 IgG1의 힌지, CH2 및 CH3 도메인과의 융합 생성물로서 생성하는 신속한 단일 단계 클로닝 절차를 사용하였다12. 정제된 3I14 scFvFc를 사용하여 상이한 아형의 다중 HA 및 바이러스에 대한 결합 및 중화 활성을 평가하였다(도 1a-b 및 도 2). 전체 인간 IgG1에 대해, scFv의 유전자 단편을 인간 IgG1 발현 벡터 TCAE648에 별개로 서브클로닝하였다. scFvFc 또는 IgG1은 일시적인 형질감염에 의해 293F 세포에서 발현되고, 단백질 A 세파로스 친화도 크로마토그래피에 의해 정제되었다.
동역학 및
K
d
결정
본 발명자들은 3I14 Ab를 pcDNA3.1-힌지 scFvFc 미니바디 발현 벡터 내로 초기에 이동시켜, scFv를 인간 IgG1의 힌지, CH2 및 CH3 도메인과의 융합 생성물로서 생성하는 신속한 단일 단계 클로닝 절차를 사용하였다12. 정제된 3I14 scFvFc를 사용하여 상이한 아형의 다중 HA 및 바이러스에 대한 결합 및 중화 활성을 평가하였다(도 1a-b 및 도 2). 전체 인간 IgG1에 대해, scFv의 유전자 단편을 인간 IgG1 발현 벡터 TCAE648에 별개로 서브클로닝하였다. scFvFc 또는 IgG1은 일시적인 형질감염에 의해 293F 세포에서 발현되고, 단백질 A 세파로스 친화도 크로마토그래피에 의해 정제되었다.
미세중화 분석
실험 전에, MDCK 세포(1.5 x 104 세포/웰)를 96웰 조직 배양 플레이트에 접종하고, PBS로 2회 세척한 후, 2 ㎍/㎖의 트립신 및 0.5% BSA로 보충된 DMEM 배지에서 인큐베이팅하였다. 바이러스의 100 TCID50(중간 조직 배양 감염량)을 96웰 플레이트에서 Ab 또는 항체 함유 상청액의 2배 연속 희석액과 동일한 부피로 혼합하고, 37℃에서 1시간 동안 인큐베이팅하였다. 인큐베이팅 후, Ab-바이러스 혼합물을 융합성 MDCK 단일층에 이중으로 옮기고, 37℃에서 21시간 동안 인큐베이팅하였다. 세포를 PBS로 세척하고, 80% 아세톤에 고정하고, 인플루엔자 A 바이러스 핵단백질(NP)(클론 A3, BEI)에 대한 mAb를 사용한 간접 ELISA에 의해 바이러스 항원을 검출하였다.
마우스에서의 예방 연구
바이러스 시험접종 24시간 전에 5마리의 암컷 8-10주령 BALB/c 마우스의 그룹에게 3I14 및 F10 IgG1을 저용량 (5 mg/kg) 또는 고용량(20 또는 25 mg/kg)으로 각각 0.5 mL 부피로 복강내(i.p.) 경로로 투여하였다. 마우스의 모든 그룹(n=6)은 10 LD50 (중간 치사 용량)의 A/베트남/1203/04(H5N1), A/브리즈번/10/07-ma(H3N2), A/네덜란드/219/03(H7N7) 또는 A/안후이/1/13(H7N9)로 비내 감염시켰다. 마우스는 바이러스 시험접종 당일에 체중을 측정한 후, 14일 또는 18일 동안 매일 임상 징후를 모니터링하고 체중을 기록하였다. 초기 체중에 비해 체중 25% 이상 감소하거나 임상 징후(무반응 또는 심각한 신경학적 증상, 예컨대 뒷다리 마비, 운동 장애) 지수에 대한 점수 4를 생존 종점으로 사용하였다. 동물 연구는 승인된 기관 동물 관리 및 사용 위원회 규약에 따라 수행되었다.
항체 결합 경쟁
ELISA 플레이트에 고정된 5 ㎍/ml H3-BR07 또는 H5-VN04 단백질을 80 nM 내지 0.3 nM의 3I14 Fab의 2배 연속 희석액과 함께 인큐베이팅하고, 5 nM에서 다른 scFvFc Ab와 혼합하였다. 1시간 동안 동시 인큐베이팅한 후, HRP 접합된 항-인간 CH2 항체(Life Technologies, 미국 뉴욕주 그랜드 아일랜드 소재)를 사용하여 scFvFc Ab의 결합을 검출하고, POLARstar 오메가 마이크로 플레이트 리더(BMG LABTECH, 독일 오르텐베르크 소재)에서 수퍼 아쿠아블루(Super AquaBlue) ELISA 기질(ebioscience, 미국 캘리포니아주 샌디에고 소재)을 사용하여 측정하였다.
트립신 절단 억제 분석
0.4 ㎍의 재조합 H3-히스티딘(H3-ATTH) 단백질을 37℃에서 100 ㎍/ml 트립신-울트라(New England Biolabs, 미국 매사추세츠주 입스위치)를 함유하는 pH 8.0의 트리스-HCl 완충제 내에서 2.5 ㎍의 3I14 또는 항-SARS Fm-6 IgG1의 존재 하에, 또는 항체의 부재 하에 인큐베이팅하였다. 트립신 소화는 1% BSA의 첨가에 의해 여러 시점에서 억제되었다. 샘플을 환원 조건 하에서 12% 환원 SDS-PAGE 겔 상에서 이동시키고, 히스프로브(HisProbe)-HRP 및 수퍼시그날 웨스트 히스프로브(SuperSignal West HisProbe) 키트(Pierce Biotechnology, 미국 일리노이주 록포드 소재)를 사용하여 블로팅하였다.
구조적 변화
FACS
분석
293T/17 세포를 전장 재조합 인플루엔자 A pcDNA3.1-H3-A2/68 및 H3-BR07 플라스미드로 형질감염시켰다. 형질감염 후 ~30시간에, 세포를 0.2% 에틸렌디아민테트라아세트산(EDTA)을 사용하여 배양 용기로부터 분리하였다. 상이한 HA 구조적 입체형태에 대한 mAb 결합을 측정하기 위해, 세포 샘플을 상이한 처리에 노출시키고, 분취하고, 3I14 또는 E730 scFvFc Ab로 염색하였다. 분리된 세포를 트립신(Gibco, 미국 뉴욕주 그랜드 아일랜드 소재)으로 연속적으로 5분간 실온에서 처리하고, 1% BSA/PBS로 세척하고, 시트르산-인산나트륨 완충제(pH 4.9)에서 15분 동안 인큐베이팅하고, 세척한 후, 실온에서 PBS 내의 50 mM 디티오트레이톨(DTT)과 함께 20분 동안 인큐베이팅하였다. 대안으로, 5 ㎍의 3I14 또는 항-SARS Ab Fm-6 IgG1을 낮은 pH 단계 전에 첨가하였다. 연속 처리 샘플을 APC 접합된 항-인간 Fc(BioLegend, 미국 캘리포니아주 샌디에고 소재)로 염색하였다. 염색된 세포는 FACS 디바(Diva) 소프트웨어(Becton Dickinson, 미국 뉴욕주 프랭클린 레이크스 소재)로 BD FACSAria™ II를 사용하여 분석하였다.
항체 의존성 세포독성 분석
ADCC 리포터 생물학적 분석은 이펙터 세포(Promega)로서 FcγRIIIa 수용체, V158(고친화도) 변이체, 및 반딧불이 루시페라제의 발현을 유도하는 NFAT 반응 요소를 안정하게 발현하는 조작된 Jurkat 세포를 사용한다. ADCC에서의 항체 생물학적 활성은 NFAT 경로 활성화의 결과로서 생성된 루시페라제를 통해 정량되고; 이펙터 세포에서 루시페라제 활성은 발광 판독값으로 정량된다. 표적 세포로서, 1 x 104/웰 H3- 또는 H5-발현 293T 세포를 분석하기 전에 평평 바닥 96웰 플레이트에 부착한 후, 배지를 낮은 IgG 혈청 분석 완충제(0.5%의 낮은 IgG FBS를 함유하는 RPMI 1640)로 교체하였다. scFvFc 항체를 각각의 웰에 1, 0.2 및 0.04 ㎍/ml의 최종 농도로 첨가하였다. 1시간 후에, Jurkat 이펙터 세포를 낮은 IgG 혈청 분석 완충제 내에서 분석 플레이트에 6.0x104/웰로 첨가하고, 6시간 동안 인큐베이팅하였다. 300 x g에서 원심분리하여 상청액을 회수하고, POLARstar 오메가 마이크로 플레이트 리더(BMG LABTECH, 독일 오르텐베르크 소재)에 의해 490 nm에서 바이오-글로™ 루시페라제(Bio-Glo Luciferase) 분석 키트(Promega, 미국 위스콘신주 매디슨 소재)를 사용하여 측정하였다.
도 9b의 추가의 ADCC 방법이 바로 설명된다. ADCC 분석은 건강한 인간 공여자로부터의 신선한 PBMC를 사용하여 HA 발현 293T 세포에서 수행되었다. ADCC 활성은 락토스 데히드로게나제(LDH) 방출 분석(Pierce Biotechnology, 미국 일리노이주 록포드 소재)에 의해 결정되었다. 피콜-파크 구배(GE HealthCare)를 사용하여 수집된 혈액으로부터 이펙터 세포로서 신선한 PBMC를 단리하였다. 표적 세포로서, 분석 전에 2x104/웰의 H3- 또는 H5-발현 293T 세포를 단단한 둥근 바닥 96웰 플레이트에 부착한 후, 배지를 낮은 IgG 혈청 분석 완충제(0.5% 낮은 IgG FBS를 함유하는 RPMI 1640)로 교체하였다. scFvFc 항체를 각각의 웰에 10, 5, 2.5 및 1.25 ㎍/ml의 최종 농도로 첨가하였다. 1시간 후에, PBMC를 낮은 IgG 혈청 분석 완충제 내에서 분석 플레이트에 1.2x105/웰로 첨가하고, 6시간 동안 인큐베이팅하였다. 300 x g에서 원심분리하여 상청액을 회수하고, 벤치마크 플러스 리더(Benchmark Plus Reader)(Bio-Rad, 미국 캘리포니아주 허큘레스 소재)에 의해 490 nm 및 680 nm에서 LDH 세포독성 분석 키트(Pierce Biotechnology, 미국 일리노이주 록포드 소재)를 사용하여 측정하였다. LDH 활성은 490 nm 흡광도 판독값으로부터 680 nm 흡광도 값(배경)을 차감함으로써 결정되었다. 세포독성 %는 다음과 같이 계산되었다: 세포독성 % = 100 x (E - SE - ST)/(M - ST) (여기서, E는 항체와 함께 배양한 E/T 배양물로부터 방출된 LDH이고; SE는 이펙터로부터 자연 방출된 LDH이고; ST는 표적으로부터 자연 방출된 LDH이고; M은 용해된 표적으로부터 방출된 최대 LDH임). 데이터는 3회의 독립적인 실험으로부터 대표적인 실험을 나타내고, 모든 시험은 3중으로 수행되었다. 데이터는 3회의 독립적인 실험으로부터의 대표적인 실험을 나타내고, 모든 시험은 삼중으로 수행되었다.
서열 분석
전장 인플루엔자 A HA 서열은 NCBI(National Center for Biotechnology Information) 데이터베이스의 인플루엔자 바이러스 리소스(Influenza Virus Resource)에서 다운로드되었다. 계통수(PHYML)는 Geneious 소프트웨어를 사용한 그들의 아미노산 서열 비교를 기초로 한다. 새로운 bnAb인 3I14는 IMGT 데이터베이스(http://imgt.cines.fr)를 사용하여 생식계열 유전자 사용, 체세포 돌연변이, N-뉴클레오티드 삽입 및 동족체 가변 중쇄(VH) 및 경쇄(VL) 유전자 쌍에 대해 분석되었다. 단일 또는 다중 생식계열 돌연변이가 생식계열로 복귀한 항체 변이체는 합성(Genewiz, 미국 뉴저지주 사우쓰 플레인필드 소재)에 의해 생성되고 서열결정에 의해 확인되었다. F10, FI6v3, CR9114, CR8020 및 39.29의 VH 및 VK 서열은 프로테인 데이터 뱅크(PDB 접근 코드)를 통해 얻었고, 상응하는 유전자를 합성하고 일시적 형질감염에 의해 발현시켰다.
인
실리코
구조
모델링
3I14는 바이오루미네이트의 항체 모델링 모듈을 사용하여 상동성을 모델링하였다. 모델은 로제타독과 도킹하기 전에 H3/FI6 복합체 구조에 중첩되었다. 측쇄 및 루프 재배열을 허용하면서 고해상도 도킹만 수행하였다. 1000개의 디코이가 각각의 도킹에 대해 생성되었고, RMSD 값을 기초로 하여 클러스터링되었다. 최종 모델은 결과 세션에서 설명된 클러스터 크기 및 기준에 따라 선택되었다.
다른 실시양태
본 발명은 그의 상세한 설명과 관련하여 설명되었지만, 상기 설명은 첨부된 청구범위에 의해 한정되는 본 발명의 범위를 예시하기 위한 것이고 그 범위를 제한하는 것이 아니다. 다른 측면, 이점 및 변형은 하기 청구범위의 범주 내에 포함된다.
SEQUENCE LISTING
<110> Dana-Farber Cancer Institute, Inc.
<120> HUMANIZED INFLUENZA MONOCLONAL ANTIBODIES AND METHODS OF USE
THEREOF
<130> DFCI-109/01WO 322270-2476
<150> US 62/144,729
<151> 2016-04-08
<160> 25
<170> PatentIn version 3.5
<210> 1
<211> 390
<212> DNA
<213> Homo sapiens
<400> 1
caggtgcagc tgttggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt aactatggca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcaatt atatcatttg atggaagtaa aaaatattat 180
gcaaactccg tgaagggccg atccaccatc tccagagaca attccaagaa cacgctgtct 240
ctgcaaatga acagcctggg acctgaggac acggctctat attactgtgc gaaactgccc 300
tccccgtatt actttgatag tcggttcgtg tgggtcgccg ccagcgcatt tcacttctgg 360
ggccagggaa tcctggtcac cgtctcttca 390
<210> 2
<211> 130
<212> PRT
<213> Homo sapiens
<400> 2
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Ser Phe Asp Gly Ser Lys Lys Tyr Tyr Ala Asn Ser Val
50 55 60
Lys Gly Arg Ser Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Gly Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys Leu Pro Ser Pro Tyr Tyr Phe Asp Ser Arg Phe Val Trp Val
100 105 110
Ala Ala Ser Ala Phe His Phe Trp Gly Gln Gly Ile Leu Val Thr Val
115 120 125
Ser Ser
130
<210> 3
<211> 330
<212> DNA
<213> Homo sapiens
<400> 3
aattttatgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgctctg gaagcagctc caacatcgga ggtaatactg tacactggtt ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat actaatagtc tgcggccctc aggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tctgaggatg aggctgatta ttactgtgca gcatgggatg acagcctaaa tggtcaggtg 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 4
<211> 110
<212> PRT
<213> Homo sapiens
<400> 4
Asn Phe Met Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Gly Asn
20 25 30
Thr Val His Trp Phe Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Thr Asn Ser Leu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 5
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Modified VL chain of 3I14VLD94N nucleic acid
<400> 5
aattttatgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgctctg gaagcagctc caacatcgga ggtaatactg tacactggtt ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat actaatagtc tgcggccctc aggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tctgaggatg aggctgatta ttactgtgca gcatgggata acagcctaaa tggtcaggtg 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 6
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Modified VL chain of 3I14VLD94N amino acid
<400> 6
Asn Phe Met Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Gly Asn
20 25 30
Thr Val His Trp Phe Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Thr Asn Ser Leu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asn Ser Leu
85 90 95
Asn Gly Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 7
<211> 8
<212> PRT
<213> Homo sapiens
<400> 7
Gly Phe Thr Phe Ser Asn Tyr Gly
1 5
<210> 8
<211> 8
<212> PRT
<213> Homo sapiens
<400> 8
Ile Ser Phe Asp Gly Ser Lys Lys
1 5
<210> 9
<211> 25
<212> PRT
<213> Homo sapiens
<400> 9
Cys Ala Lys Leu Pro Ser Pro Tyr Tyr Phe Asp Ser Arg Phe Val Trp
1 5 10 15
Val Ala Ala Ser Ala Phe His Phe Trp
20 25
<210> 10
<211> 8
<212> PRT
<213> Homo sapiens
<400> 10
Ser Ser Asn Ile Gly Gly Asn Thr
1 5
<210> 11
<211> 3
<212> PRT
<213> Homo sapiens
<400> 11
Thr Asn Ser
1
<210> 12
<211> 13
<212> PRT
<213> Homo sapiens
<400> 12
Cys Ala Ala Trp Asp Asp Ser Leu Asn Gly Gln Val Phe
1 5 10
<210> 13
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Modified 3I14VLD94N Light CDR3
<400> 13
Cys Ala Ala Trp Asp Asn Ser Leu Asn Gly Gln Val Phe
1 5 10
<210> 14
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Modified 3I14VLD94N Light CDR1
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa = Any amino acid other the glycine; preferably X is serine
<400> 14
Ser Ser Asn Ile Gly Xaa Asn Thr
1 5
<210> 15
<211> 324
<212> PRT
<213> Influenza A Virus
<400> 15
Asp Gln Ile Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Glu Lys Val
1 5 10 15
Asp Thr Asn Leu Glu Arg Asn Val Thr Val Thr His Ala Lys Asp Ile
20 25 30
Leu Glu Lys Thr His Asn Gly Lys Leu Cys Lys Leu Asn Gly Ile Pro
35 40 45
Pro Leu Glu Leu Gly Asp Cys Ser Ile Ala Gly Trp Leu Leu Gly Asn
50 55 60
Pro Glu Cys Asp Arg Leu Leu Ser Val Pro Glu Trp Ser Tyr Ile Met
65 70 75 80
Glu Lys Glu Asn Pro Arg Asp Gly Leu Cys Tyr Pro Gly Ser Phe Asn
85 90 95
Asp Tyr Glu Glu Leu Lys His Leu Leu Ser Ser Val Lys His Phe Glu
100 105 110
Lys Val Lys Ile Leu Pro Lys Asp Arg Trp Thr Gln His Thr Thr Thr
115 120 125
Gly Gly Ser Arg Ala Cys Ala Val Ser Gly Asn Pro Ser Phe Phe Arg
130 135 140
Asn Met Val Trp Leu Thr Lys Glu Gly Ser Asp Tyr Pro Val Ala Lys
145 150 155 160
Gly Ser Tyr Asn Asn Thr Ser Gly Glu Gln Met Leu Ile Ile Trp Gly
165 170 175
Val His His Pro Ile Asp Glu Thr Glu Gln Arg Thr Leu Tyr Gln Asn
180 185 190
Val Gly Thr Tyr Val Ser Val Gly Thr Ser Thr Leu Asn Lys Arg Ser
195 200 205
Thr Pro Glu Ile Ala Thr Arg Pro Lys Val Asn Gly Gln Gly Gly Arg
210 215 220
Met Glu Phe Ser Trp Thr Leu Leu Asp Met Trp Asp Thr Ile Asn Phe
225 230 235 240
Glu Ser Thr Gly Asn Leu Ile Ala Pro Glu Tyr Gly Phe Lys Ile Ser
245 250 255
Lys Arg Gly Ser Ser Gly Ile Met Lys Thr Glu Gly Thr Leu Glu Asn
260 265 270
Cys Glu Thr Lys Cys Gln Thr Pro Leu Gly Ala Ile Asn Thr Thr Leu
275 280 285
Pro Phe His Asn Val His Pro Leu Thr Ile Gly Glu Cys Pro Lys Tyr
290 295 300
Val Lys Ser Glu Lys Leu Val Leu Ala Thr Gly Leu Arg Asn Val Pro
305 310 315 320
Gln Ile Glu Ser
<210> 16
<211> 974
<212> DNA
<213> Influenza A Virus
<400> 16
gaccagatat gcattggata ccatgccaat aattccacag agatggtcga cacaattcta 60
gagcggaacg tcactgtgac tcatgccaag gacattcttg agaagaccca taacggaaag 120
ttatgcaaac taaacggaat ccctccactt gaactagggg actgtagcat tgccggatgg 180
ctccttggaa atccagaatg tgataggctt ctaagtgtgc cagaatggtc ctatataatg 240
gagaaagaaa acccgagaga cggtttgtgt tatccaggca gcttcaatga ttatgaagaa 300
ttgaaacatc tcctcagcag cgtgaaacat ttcgagaaag taaagattct gcccaaagat 360
agatggacac agcatacaac aactggaggt tcacgggcct gcgcggtgtc tggtaatcca 420
tcattcttca ggaacatggt ctggctgaca aagaaaggat cagattatcc ggttgccaaa 480
ggatcgtaca acaatacaag cggagaacaa atgctaataa tttggggggt gcaccatccc 540
aatgatgaga cagaacaaag aacattgtac cagaatgtgg gaacctatgt ttccgtaggc 600
acatcaacat tgaacaaaag gtcaacccca gaaatagcaa caaggcttaa agtgaatgga 660
caaggaggta gaatggaatt ctcttggacc ctcttggata tgtgggacac cataaatttt 720
gagagtactg gtaatctaat tgcaccagag tatggattca aaatatcgaa aagaggtagt 780
tcagggatca tgaaaacaga aggaacactt gagaactgtg agaccaaatg ccaaactcct 840
ttgggagcaa taaatacaac attgcctttt cacaatgtcc acccactgac aataggtgag 900
tgccccaaat atgtaaaatc ggagaagttg gtcttagcaa caggactaag gaatgttccc 960
cagattgaat caag 974
<210> 17
<211> 222
<212> PRT
<213> Influenza A Virus
<400> 17
Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile Glu Gly Gly Trp Gln Gly
1 5 10 15
Met Val Asp Gly Trp Tyr Gly Tyr His His Ser Asn Asp Gln Gly Ser
20 25 30
Gly Tyr Ala Ala Asp Lys Glu Ser Thr Gln Lys Ala Phe Asp Gly Ile
35 40 45
Thr Asn Lys Val Asn Ser Val Ile Glu Lys Met Asn Thr Gln Phe Glu
50 55 60
Ala Val Gly Lys Glu Phe Gly Asn Leu Glu Arg Arg Leu Glu Asn Leu
65 70 75 80
Asn Lys Arg Met Glu Asp Gly Phe Leu Asp Val Trp Thr Tyr Asn Ala
85 90 95
Glu Leu Leu Val Leu Met Glu Asn Glu Arg Thr Leu Asp Phe His Asp
100 105 110
Ser Asn Val Lys Asn Leu Tyr Asp Lys Val Arg Met Gln Leu Arg Asp
115 120 125
Asn Val Lys Glu Leu Gly Asn Gly Cys Phe Glu Phe Tyr His Lys Cys
130 135 140
Asp Asp Glu Cys Met Asn Ser Val Lys Asn Gly Thr Tyr Asp Tyr Pro
145 150 155 160
Lys Tyr Glu Glu Glu Ser Lys Leu Asn Arg Asn Glu Ile Lys Gly Val
165 170 175
Lys Leu Ser Ser Met Gly Val Tyr Gln Ile Leu Ala Ile Tyr Ala Thr
180 185 190
Val Ala Gly Ser Leu Ser Leu Ala Ile Met Met Ala Gly Ile Ser Phe
195 200 205
Trp Met Cys Ser Asn Gly Ser Leu Gln Cys Arg Ile Cys Ile
210 215 220
<210> 18
<211> 668
<212> DNA
<213> Influenza A Virus
<400> 18
gattgtttgg ggcaatagct ggttttatag aaggaggatg gcaaggaatg gttgatggtt 60
ggtatggata ccatcacagc aatgaccagg gatcagggta tgcagcagac aaagaatcca 120
ctcaaaaggc atttgatgga atcaccaaca aggtaaattc tgtgattgaa aagatgaaca 180
cccaatttga agctgttggg aaagaattca gtaacttaga gagaagactg gagaacttga 240
acaaaaagat ggaagacggg tttctagatg tgtggacata caatgctgag cttctagttc 300
tgatggaaaa tgagaggaca cttgactttc atgattctaa tgtcaagaat ctgtatgata 360
aagtcagaat gcagttgaga gacaacgtca aagaactagg aaatggatgt tttgaatttt 420
atcacaaatg tgatgatgaa tgcatgaata gtgtgaaaac cgggacgtat gattatccca 480
agtatgaaga agagtctaaa ctaaatagaa atgaaatcaa aggggtaaaa ttgagcagca 540
tgggggttta tcaaatcctt gccatttatg ctacagtagc aggttctctg tcactggcaa 600
tcatgatggc tgggatctct ttctggatgt gctccaacgg gtctctgcag tgcaggatct 660
gcatatga 668
<210> 19
<211> 562
<212> PRT
<213> Influenza A Virus
<400> 19
Met Ala Ile Ile Tyr Leu Ile Leu Leu Phe Thr Ala Val Arg Gly Asp
1 5 10 15
Gln Ile Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Glu Met Val Asp
20 25 30
Thr Ile Leu Glu Arg Asn Val Thr Val Thr His Ala Lys Asp Ile Leu
35 40 45
Glu Lys Thr His Asn Gly Lys Leu Cys Lys Leu Asn Gly Ile Pro Pro
50 55 60
Leu Glu Leu Gly Asp Cys Ser Ile Ala Gly Trp Leu Leu Gly Asn Pro
65 70 75 80
Glu Cys Asp Arg Leu Leu Ser Val Pro Glu Trp Ser Tyr Ile Met Glu
85 90 95
Lys Glu Asn Pro Arg Asp Gly Leu Cys Tyr Pro Gly Ser Phe Asn Asp
100 105 110
Tyr Glu Glu Leu Lys His Leu Leu Ser Ser Val Lys His Phe Glu Lys
115 120 125
Val Lys Ile Leu Pro Lys Asp Arg Trp Thr Gln His Thr Thr Thr Gly
130 135 140
Gly Ser Arg Ala Cys Ala Val Ser Gly Asn Pro Ser Phe Phe Arg Asn
145 150 155 160
Met Val Trp Leu Thr Lys Lys Gly Ser Asp Tyr Pro Val Ala Lys Gly
165 170 175
Ser Tyr Asn Asn Thr Ser Gly Glu Gln Met Leu Ile Ile Trp Gly Val
180 185 190
His His Pro Asn Asp Glu Thr Glu Gln Arg Thr Leu Tyr Gln Asn Val
195 200 205
Gly Thr Tyr Val Ser Val Gly Thr Ser Thr Leu Asn Lys Arg Ser Thr
210 215 220
Pro Glu Ile Ala Thr Arg Leu Lys Val Asn Gly Gln Gly Gly Arg Met
225 230 235 240
Glu Phe Ser Trp Thr Leu Leu Asp Met Trp Asp Thr Ile Asn Phe Glu
245 250 255
Ser Thr Gly Asn Leu Ile Ala Pro Glu Tyr Gly Phe Lys Ile Ser Lys
260 265 270
Arg Gly Ser Ser Gly Ile Met Lys Thr Glu Gly Thr Leu Glu Asn Cys
275 280 285
Glu Thr Lys Cys Gln Thr Pro Leu Gly Ala Ile Asn Thr Thr Leu Pro
290 295 300
Phe His Asn Val His Pro Leu Thr Ile Gly Glu Cys Pro Lys Tyr Val
305 310 315 320
Lys Ser Glu Lys Leu Val Leu Ala Thr Gly Leu Arg Asn Val Pro Gln
325 330 335
Ile Glu Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile Glu Gly
340 345 350
Gly Trp Gln Gly Met Val Asp Gly Trp Tyr Gly Tyr His His Ser Asn
355 360 365
Asp Gln Gly Ser Gly Tyr Ala Ala Asp Lys Glu Ser Thr Gln Lys Ala
370 375 380
Phe Asp Gly Ile Thr Asn Lys Val Asn Ser Val Ile Glu Lys Met Asn
385 390 395 400
Thr Gln Phe Glu Ala Val Gly Lys Glu Phe Ser Asn Leu Glu Arg Arg
405 410 415
Leu Glu Asn Leu Asn Lys Lys Met Glu Asp Gly Phe Leu Asp Val Trp
420 425 430
Thr Tyr Asn Ala Glu Leu Leu Val Leu Met Glu Asn Glu Arg Thr Leu
435 440 445
Asp Phe His Asp Ser Asn Val Lys Asn Leu Tyr Asp Lys Val Arg Met
450 455 460
Gln Leu Arg Asp Asn Val Lys Glu Leu Gly Asn Gly Cys Phe Glu Phe
465 470 475 480
Tyr His Lys Cys Asp Asp Glu Cys Met Asn Ser Val Lys Thr Gly Thr
485 490 495
Tyr Asp Tyr Pro Lys Tyr Glu Glu Glu Ser Lys Leu Asn Arg Asn Glu
500 505 510
Ile Lys Gly Val Lys Leu Ser Ser Met Gly Val Tyr Gln Ile Leu Ala
515 520 525
Ile Tyr Ala Thr Val Ala Gly Ser Leu Ser Leu Ala Ile Met Met Ala
530 535 540
Gly Ile Ser Phe Trp Met Cys Ser Asn Gly Ser Leu Gln Cys Arg Ile
545 550 555 560
Cys Ile
<210> 20
<211> 1689
<212> DNA
<213> Influenza A Virus
<400> 20
atggccatca tttatctcat tctcctgttc acagcagtga gaggggacca gatatgcatt 60
ggataccatg ccaataattc cacagagatg gtcgacacaa ttctagagcg gaacgtcact 120
gtgactcatg ccaaggacat tcttgagaag acccataacg gaaagttatg caaactaaac 180
ggaatccctc cacttgaact aggggactgt agcattgccg gatggctcct tggaaatcca 240
gaatgtgata ggcttctaag tgtgccagaa tggtcctata taatggagaa agaaaacccg 300
agagacggtt tgtgttatcc aggcagcttc aatgattatg aagaattgaa acatctcctc 360
agcagcgtga aacatttcga gaaagtaaag attctgccca aagatagatg gacacagcat 420
acaacaactg gaggttcacg ggcctgcgcg gtgtctggta atccatcatt cttcaggaac 480
atggtctggc tgacaaagaa aggatcagat tatccggttg ccaaaggatc gtacaacaat 540
acaagcggag aacaaatgct aataatttgg ggggtgcacc atcccaatga tgagacagaa 600
caaagaacat tgtaccagaa tgtgggaacc tatgtttccg taggcacatc aacattgaac 660
aaaaggtcaa ccccagaaat agcaacaagg cttaaagtga atggacaagg aggtagaatg 720
gaattctctt ggaccctctt ggatatgtgg gacaccataa attttgagag tactggtaat 780
ctaattgcac cagagtatgg attcaaaata tcgaaaagag gtagttcagg gatcatgaaa 840
acagaaggaa cacttgagaa ctgtgagacc aaatgccaaa ctcctttggg agcaataaat 900
acaacattgc cttttcacaa tgtccaccca ctgacaatag gtgagtgccc caaatatgta 960
aaatcggaga agttggtctt agcaacagga ctaaggaatg ttccccagat tgaatcaaga 1020
ggattgtttg gggcaatagc tggttttata gaaggaggat ggcaaggaat ggttgatggt 1080
tggtatggat accatcacag caatgaccag ggatcagggt atgcagcaga caaagaatcc 1140
actcaaaagg catttgatgg aatcaccaac aaggtaaatt ctgtgattga aaagatgaac 1200
acccaatttg aagctgttgg gaaagaattc agtaacttag agagaagact ggagaacttg 1260
aacaaaaaga tggaagacgg gtttctagat gtgtggacat acaatgctga gcttctagtt 1320
ctgatggaaa atgagaggac acttgacttt catgattcta atgtcaagaa tctgtatgat 1380
aaagtcagaa tgcagttgag agacaacgtc aaagaactag gaaatggatg ttttgaattt 1440
tatcacaaat gtgatgatga atgcatgaat agtgtgaaaa ccgggacgta tgattatccc 1500
aagtatgaag aagagtctaa actaaataga aatgaaatca aaggggtaaa attgagcagc 1560
atgggggttt atcaaatcct tgccatttat gctacagtag caggttctct gtcactggca 1620
atcatgatgg ctgggatctc tttctggatg tgctccaacg ggtctctgca gtgcaggatc 1680
tgcatatga 1689
<210> 21
<211> 755
<212> DNA
<213> Homo sapiens
<400> 21
catggagttt gggctgagct gggttttcct cgttgctctt ttaagaggtg attcatggag 60
aaatagagag actgagtgtg agtgaacatg agtgagaaaa actggatttg tgtggcattt 120
tctgataacg gtgtccttct gtttgcaggt gtccagtgtc aggtgcagct ggtggagtct 180
gggggaggcg tggtccagcc tgggaggtcc ctgagactct cctgtgcagc ctctggattc 240
accttcagta gctatggcat gcactgggtc cgccaggctc caggcaaggg gctggagtgg 300
gtggcagtta tatcatatga tggaagtaat aaatactatg cagactccgt gaagggccga 360
ttcaccatct ccagagacaa ttccaagaac acgctgtatc tgcaaatgaa cagcctgaga 420
gctgaggaca cggctgtgta ttactgtgcg aaagacacag tgaggggaag tcattgtgcg 480
cccagacaca aacctccctg caggaacgct ggcgggaaat cagcggcagg gggcgctcag 540
gagccactga tcagagtcag ccctagaggc aggtgcagat ggaggctgtt tcctgtcagg 600
atgtgggact ttgtcttctt ctgacagttc cccaaggaac ctcttaaatt tagaaaactg 660
tgcctaacaa tgtcttctct atgcatatga ggaccttttc tccctagcac aaaatgcaga 720
ttgacgctga cacggatgaa aattcctcaa ccatg 755
<210> 22
<211> 66
<212> PRT
<213> Homo sapiens
<400> 22
Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly Arg Leu Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
20 25 30
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
35 40 45
Lys Asp Phe Gly Pro Lys Arg Pro Thr Gly Asp Tyr Phe Asp Tyr Trp
50 55 60
Gly Gln
65
<210> 23
<211> 296
<212> DNA
<213> Homo sapiens
<400> 23
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcagctc caacatcgga agtaatactg taaactggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat agtaataatc agcggccctc aggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tctgaggatg aggctgatta ttactgtgca gcatgggatg acagcctgaa tggtcc 296
<210> 24
<211> 98
<212> PRT
<213> Homo sapiens
<400> 24
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly
<210> 25
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> C-terminal peptide tag
<400> 25
Gly Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His
1 5 10 15
Glu
Claims (20)
- 재조합 생산된, 단리된 모노클로날 항체로서, 상기 항체는
a. IGHV3-30 생식계열 유전자에 의해 코딩되는 중쇄 가변 영역;
b. 헤마글루티닌(HA) 단백질의 스템 영역의 에피토프에 대한 결합; 및
c. 그룹 1 및 그룹 2 인플루엔자 A 바이러스의 중화
의 특징을 갖는 것인 재조합 생산된, 단리된 모노클로날 항체. - 제1항에 있어서, 항체가
d. IGLV1-44 생식계열 유전자에 의해 코딩되는 경쇄 가변 영역;
e. 서열 번호 9의 아미노산 서열을 포함하는 중쇄 CDR3;
f. 절단되지 않은 HA0에 대한 결합;
g. HA0 절단 방지; 및/또는
h. 서열 번호 18에 따라 넘버링될 때 HA2 폴리펩티드의 아미노산 18, 19, 20, 21, 36, 38, 39, 41, 42, 45, 45, 49 및 53에 의해 규정되는 구조적 에피토프에 대한 결합
의 특징 중 하나 이상을 갖는 것인 모노클로날 항체. - a. 서열 번호 7의 아미노산 서열을 포함하는 CDR1; 서열 번호 8의 아미노산 서열을 포함하는 CDR2 및 서열 번호 9의 아미노산 서열을 포함하는 CDR3을 포함하는 중쇄; 및
b. 서열 번호 10 또는 서열 번호 14의 아미노산 서열을 포함하는 CDR1; 서열 번호 11의 아미노산 서열을 포함하는 CDR2 및 서열 번호 12 또는 서열 번호 13의 아미노산 서열을 포함하는 CDR3을 포함하는 경쇄
를 포함하는, 인플루엔자 바이러스를 중화하는 단리된 모노클로날 항체. - 단리된 모노클로날 항체로서, 상기 항체는
a. 서열 번호 2의 VH 아미노산 서열 및 서열 번호 4 또는 서열 번호 6의 VL 아미노산 서열;
b. 서열 번호 2의 VH 아미노산 서열 및 서열 번호 24의 VL 아미노산 서열; 또는
c. 서열 번호 22의 VH 아미노산 서열 및 서열 번호 4 또는 6의 VL 아미노산 서열
을 포함하는 것인 단리된 모노클로날 항체. - 제3항 또는 제4항에 있어서, 상기 항체가 인플루엔자 바이러스의 HA의 스템 영역에 결합하는 것인 항체.
- 제3항 내지 제5항 중 어느 한 항에 있어서, 상기 인플루엔자 바이러스가 인플루엔자 A 바이러스인 항체.
- 제3항 내지 제6항 중 어느 한 항에 있어서, 상기 항체가 인플루엔자 A 바이러스 그룹 I 및 그룹 II를 중화하는 것인 항체.
- 제3항 내지 제7항 중 어느 한 항에 있어서, 상기 항체가 HA2 폴리펩티드의 아미노산 18, 19, 20, 21, 36, 38, 39, 41, 42, 45, 45, 49 및 53에 의해 규정되는 구조적 에피토프에 결합하는 것인 항체.
- 제1항 내지 제8항 중 어느 한 항에 있어서, 상기 항체가 단일쇄 Fv 항체, Fab 단편, Fab' 단편, 또는 F(ab')2 단편인 항체.
- 제1항 내지 제9항 중 어느 한 항에 있어서, 치료제에 연결된 항체.
- 제7항에 있어서, 상기 치료제가 독소, 방사성 표지, siRNA, 소분자 또는 사이토카인인 항체.
- 제1항 내지 제11항 중 어느 한 항에 따른 항체를 생산하는 세포.
- 제1항 내지 제11항 중 어느 한 항에 따른 항체 및 담체를 포함하는 조성물.
- 서열 번호 1, 3 및 5로부터 선택된 핵산 서열을 포함하는 핵산 서열.
- 서열 번호 2, 4 및 6으로부터 선택된 아미노산 서열을 포함하는 폴리펩티드를 코딩하는 핵산 서열.
- 서열 번호 2, 4 및 6으로부터 선택된 아미노산 서열을 포함하는 폴리펩티드.
- 제14항 또는 제15항의 핵산을 포함하는 벡터.
- 제17항의 벡터를 포함하는 세포.
- 인플루엔자 바이러스에 의해 유발되는 질환 또는 장애를 예방 또는 치료하는 방법으로서, 상기 방법은 상기 질환 또는 장애를 앓을 위험이 있는 인간에게 치료 유효량의 제13항의 조성물을 투여하는 단계를 포함하는 것인 방법.
- 제19항에 있어서, 항바이러스제를 투여하는 단계를 추가로 포함하는 방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201562144729P | 2015-04-08 | 2015-04-08 | |
| US62/144,729 | 2015-04-08 | ||
| PCT/US2016/026800 WO2016164835A1 (en) | 2015-04-08 | 2016-04-08 | Humanized influenza monoclonal antibodies and methods of use thereof |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20170133507A true KR20170133507A (ko) | 2017-12-05 |
Family
ID=55754481
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020177032200A KR20170133507A (ko) | 2015-04-08 | 2016-04-08 | 인간화된 인플루엔자 모노클로날 항체 및 그의 사용 방법 |
Country Status (9)
| Country | Link |
|---|---|
| US (2) | US11135282B2 (ko) |
| EP (1) | EP3280730B1 (ko) |
| JP (3) | JP6960856B2 (ko) |
| KR (1) | KR20170133507A (ko) |
| CN (1) | CN107750253B (ko) |
| AU (2) | AU2016246065B2 (ko) |
| CA (1) | CA2981969C (ko) |
| IL (2) | IL254951B (ko) |
| WO (1) | WO2016164835A1 (ko) |
Families Citing this family (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP4043489A1 (en) * | 2014-03-19 | 2022-08-17 | Dana-Farber Cancer Institute, Inc. | Immunogenetic restriction on elicitation of antibodies |
| JP7269167B2 (ja) | 2016-10-14 | 2023-05-08 | デイナ ファーバー キャンサー インスティチュート,インコーポレイテッド | モジュラー四価二重特異性抗体プラットフォーム |
| SG11202006379UA (en) * | 2018-01-26 | 2020-07-29 | Regeneron Pharma | Human antibodies to influenza hemagglutinin |
| BR112021002432A2 (pt) | 2018-08-13 | 2021-05-04 | Regeneron Pharmaceuticals, Inc. | seleção de proteína terapêutica em condições in vivo simuladas |
| CN109448781B (zh) * | 2018-11-06 | 2021-09-14 | 云南大学 | 一种流感病毒抗原变化的预测方法 |
| CN111423507B (zh) * | 2019-01-10 | 2022-04-15 | 中国科学院分子细胞科学卓越创新中心 | 广谱性中和流感病毒的全人抗体 |
| EP3946613A1 (en) | 2019-03-25 | 2022-02-09 | Visterra, Inc. | Compositions and methods for treating and preventing influenza |
| KR20220113749A (ko) * | 2019-12-11 | 2022-08-16 | 비스테라, 인크. | 인플루엔자를 치료하고 예방하는 조성물 및 방법 |
| WO2023081471A1 (en) | 2021-11-05 | 2023-05-11 | Dana-Farber Cancer Institute, Inc. | Human broadly crossreactive influenza monoclonal antibodies and methods of use thereof |
| CN114366809A (zh) * | 2022-01-12 | 2022-04-19 | 广东粤港澳大湾区国家纳米科技创新研究院 | 一种铝纳米晶递送系统及其结合疫苗抗原分子自组装颗粒佐剂疫苗 |
Family Cites Families (40)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3773919A (en) | 1969-10-23 | 1973-11-20 | Du Pont | Polylactide-drug mixtures |
| FR2413974A1 (fr) | 1978-01-06 | 1979-08-03 | David Bernard | Sechoir pour feuilles imprimees par serigraphie |
| US4485045A (en) | 1981-07-06 | 1984-11-27 | Research Corporation | Synthetic phosphatidyl cholines useful in forming liposomes |
| US4522811A (en) | 1982-07-08 | 1985-06-11 | Syntex (U.S.A.) Inc. | Serial injection of muramyldipeptides and liposomes enhances the anti-infective activity of muramyldipeptides |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| US4544545A (en) | 1983-06-20 | 1985-10-01 | Trustees University Of Massachusetts | Liposomes containing modified cholesterol for organ targeting |
| US4676980A (en) | 1985-09-23 | 1987-06-30 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Target specific cross-linked heteroantibodies |
| WO1988001513A1 (en) | 1986-08-28 | 1988-03-10 | Teijin Limited | Cytocidal antibody complex and process for its preparation |
| US4946778A (en) | 1987-09-21 | 1990-08-07 | Genex Corporation | Single polypeptide chain binding molecules |
| US5132405A (en) | 1987-05-21 | 1992-07-21 | Creative Biomolecules, Inc. | Biosynthetic antibody binding sites |
| US5091513A (en) | 1987-05-21 | 1992-02-25 | Creative Biomolecules, Inc. | Biosynthetic antibody binding sites |
| US5223409A (en) | 1988-09-02 | 1993-06-29 | Protein Engineering Corp. | Directed evolution of novel binding proteins |
| GB8823869D0 (en) | 1988-10-12 | 1988-11-16 | Medical Res Council | Production of antibodies |
| EP0739904A1 (en) | 1989-06-29 | 1996-10-30 | Medarex, Inc. | Bispecific reagents for aids therapy |
| US5013556A (en) | 1989-10-20 | 1991-05-07 | Liposome Technology, Inc. | Liposomes with enhanced circulation time |
| DE69133566T2 (de) | 1990-01-12 | 2007-12-06 | Amgen Fremont Inc. | Bildung von xenogenen Antikörpern |
| EP0547065B1 (en) | 1990-06-29 | 2001-08-29 | Large Scale Biology Corporation | Melanin production by transformed microorganisms |
| DK0814159T3 (da) | 1990-08-29 | 2005-10-24 | Genpharm Int | Transgene, ikke-humane dyr, der er i stand til at danne heterologe antistoffer |
| US5625126A (en) | 1990-08-29 | 1997-04-29 | Genpharm International, Inc. | Transgenic non-human animals for producing heterologous antibodies |
| US5633425A (en) | 1990-08-29 | 1997-05-27 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| US5661016A (en) | 1990-08-29 | 1997-08-26 | Genpharm International Inc. | Transgenic non-human animals capable of producing heterologous antibodies of various isotypes |
| US5545806A (en) | 1990-08-29 | 1996-08-13 | Genpharm International, Inc. | Ransgenic non-human animals for producing heterologous antibodies |
| WO1992020373A1 (en) | 1991-05-14 | 1992-11-26 | Repligen Corporation | Heteroconjugate antibodies for treatment of hiv infection |
| US5233409A (en) | 1992-02-25 | 1993-08-03 | Schwab Karl W | Color analysis of organic constituents in sedimentary rocks for thermal maturity |
| ES2301158T3 (es) | 1992-07-24 | 2008-06-16 | Amgen Fremont Inc. | Produccion de anticuerpos xenogenicos. |
| PT752248E (pt) | 1992-11-13 | 2001-01-31 | Idec Pharma Corp | Aplicacao terapeutica de anticorpos quimericos e marcados radioactivamente contra antigenios de diferenciacao restrita de linfocitos b humanos para o tratamento do linfoma de celulas b |
| AU1925195A (en) | 1994-02-22 | 1995-09-04 | Dana-Farber Cancer Institute | Nucleic acid delivery system, method of synthesis and uses thereof |
| EP0822830B1 (en) | 1995-04-27 | 2008-04-02 | Amgen Fremont Inc. | Human anti-IL-8 antibodies, derived from immunized xenomice |
| AU2466895A (en) | 1995-04-28 | 1996-11-18 | Abgenix, Inc. | Human antibodies derived from immunized xenomice |
| US5916771A (en) | 1996-10-11 | 1999-06-29 | Abgenix, Inc. | Production of a multimeric protein by cell fusion method |
| US20020029391A1 (en) | 1998-04-15 | 2002-03-07 | Claude Geoffrey Davis | Epitope-driven human antibody production and gene expression profiling |
| JP5813629B2 (ja) * | 2009-05-11 | 2015-11-17 | クルセル ホランド ベー ヴェー | インフルエンザウイルスh3n2を中和可能なヒト抗体及び/または抗原結合フラグメントおよびその使用 |
| US20130022608A1 (en) | 2010-03-26 | 2013-01-24 | Pomona Ricerca S.R.L. | FULL-LENGTH IgG IMMUNOGLOBULINS CAPABLE OF RECOGNIZING A HETEROSUBTYPE NEUTRALIZING EPITOPE ON THE HEMAGGLUTININ STEM REGION AND USES THEREOF |
| US9534042B2 (en) * | 2010-09-03 | 2017-01-03 | Fujita Health University | Influenza virus-neutralizing antibody and screening method therefor |
| KR101941724B1 (ko) * | 2011-07-14 | 2019-01-23 | 얀센 백신스 앤드 프리벤션 비.브이. | 계통발생군 1과 계통발생군 2의 인플루엔자 a형 바이러스 및 인플루엔자 b형 바이러스를 중화시킬 수 있는 인간 결합분자 |
| SI3418300T1 (sl) * | 2011-07-18 | 2021-03-31 | Institute For Research In Biomedicine | Nevtralizirajoča protitelesa proti virusu influence A in njihove uporabe |
| AU2012316930B2 (en) * | 2011-09-30 | 2016-06-09 | Celltrion Inc. | Binding molecule having influenza A virus-neutralizing activity produced from human B cell |
| US9969794B2 (en) * | 2012-05-10 | 2018-05-15 | Visterra, Inc. | HA binding agents |
| WO2014078268A2 (en) | 2012-11-13 | 2014-05-22 | Genentech, Inc. | Anti-hemagglutinin antibodies and methods of use |
| US9111624B2 (en) | 2013-03-22 | 2015-08-18 | Katsuyuki Fujita | Semiconductor memory device |
-
2016
- 2016-04-08 CA CA2981969A patent/CA2981969C/en active Active
- 2016-04-08 CN CN201680033270.XA patent/CN107750253B/zh active Active
- 2016-04-08 EP EP16716806.1A patent/EP3280730B1/en active Active
- 2016-04-08 WO PCT/US2016/026800 patent/WO2016164835A1/en active Application Filing
- 2016-04-08 US US15/564,897 patent/US11135282B2/en active Active
- 2016-04-08 AU AU2016246065A patent/AU2016246065B2/en active Active
- 2016-04-08 JP JP2017553107A patent/JP6960856B2/ja active Active
- 2016-04-08 KR KR1020177032200A patent/KR20170133507A/ko active IP Right Grant
-
2017
- 2017-10-09 IL IL254951A patent/IL254951B/en unknown
-
2021
- 2021-05-18 JP JP2021083700A patent/JP2021121204A/ja active Pending
- 2021-08-18 US US17/405,748 patent/US20220054624A1/en active Pending
- 2021-09-03 AU AU2021225240A patent/AU2021225240A1/en not_active Abandoned
- 2021-09-09 IL IL286208A patent/IL286208A/en unknown
-
2023
- 2023-07-18 JP JP2023116535A patent/JP2023130499A/ja not_active Withdrawn
Also Published As
| Publication number | Publication date |
|---|---|
| EP3280730C0 (en) | 2024-01-03 |
| AU2021225240A1 (en) | 2021-09-30 |
| AU2016246065A1 (en) | 2017-10-26 |
| EP3280730B1 (en) | 2024-01-03 |
| JP2023130499A (ja) | 2023-09-20 |
| IL254951B (en) | 2021-09-30 |
| JP2021121204A (ja) | 2021-08-26 |
| EP3280730A1 (en) | 2018-02-14 |
| JP6960856B2 (ja) | 2021-11-05 |
| IL254951A0 (en) | 2017-12-31 |
| US11135282B2 (en) | 2021-10-05 |
| AU2016246065B2 (en) | 2021-06-03 |
| US20220054624A1 (en) | 2022-02-24 |
| CA2981969C (en) | 2023-12-05 |
| JP2018518940A (ja) | 2018-07-19 |
| CN107750253B (zh) | 2022-10-04 |
| IL286208A (en) | 2021-10-31 |
| US20180099040A1 (en) | 2018-04-12 |
| WO2016164835A1 (en) | 2016-10-13 |
| CN107750253A (zh) | 2018-03-02 |
| CA2981969A1 (en) | 2016-10-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20220054624A1 (en) | Humanized influenza monoclonal antibodies and methods of use thereof | |
| JP2021176841A (ja) | インフルエンザウイルスに対する抗体およびその使用方法 | |
| KR20160145813A (ko) | 중동 호흡기 증후군 코로나바이러스 중화 항체 및 이의 사용 방법 | |
| JP2011506344A5 (ko) | ||
| AU2019204310B2 (en) | Antibodies against influenza virus and methods of use thereof | |
| WO2023081471A1 (en) | Human broadly crossreactive influenza monoclonal antibodies and methods of use thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| E902 | Notification of reason for refusal | ||
| E701 | Decision to grant or registration of patent right |