KR20130132936A - 구강 관리 제품에 사용하기 위한 효소에 의한 과산 생성 - Google Patents
구강 관리 제품에 사용하기 위한 효소에 의한 과산 생성 Download PDFInfo
- Publication number
- KR20130132936A KR20130132936A KR20137019091A KR20137019091A KR20130132936A KR 20130132936 A KR20130132936 A KR 20130132936A KR 20137019091 A KR20137019091 A KR 20137019091A KR 20137019091 A KR20137019091 A KR 20137019091A KR 20130132936 A KR20130132936 A KR 20130132936A
- Authority
- KR
- South Korea
- Prior art keywords
- enzyme
- seq
- oral
- group
- peptide
- Prior art date
Links
- 150000004965 peroxy acids Chemical class 0.000 title claims abstract description 100
- 230000002255 enzymatic effect Effects 0.000 title description 13
- 102000004190 Enzymes Human genes 0.000 claims abstract description 386
- 108090000790 Enzymes Proteins 0.000 claims abstract description 386
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 244
- 239000000203 mixture Substances 0.000 claims abstract description 158
- 238000000034 method Methods 0.000 claims abstract description 120
- 230000000694 effects Effects 0.000 claims abstract description 117
- 239000000758 substrate Substances 0.000 claims abstract description 104
- 108020001507 fusion proteins Proteins 0.000 claims abstract description 62
- 102000037865 fusion proteins Human genes 0.000 claims abstract description 62
- 108090000371 Esterases Proteins 0.000 claims abstract description 61
- 239000003795 chemical substances by application Substances 0.000 claims abstract description 47
- 230000008901 benefit Effects 0.000 claims abstract description 45
- -1 CE-7 carbohydrate Chemical class 0.000 claims abstract description 40
- 230000002087 whitening effect Effects 0.000 claims abstract description 36
- 238000004061 bleaching Methods 0.000 claims abstract description 25
- 230000009467 reduction Effects 0.000 claims abstract description 16
- 238000004659 sterilization and disinfection Methods 0.000 claims abstract description 13
- 238000011033 desalting Methods 0.000 claims abstract description 8
- 238000004332 deodorization Methods 0.000 claims abstract description 6
- 238000006722 reduction reaction Methods 0.000 claims abstract description 4
- 230000027455 binding Effects 0.000 claims description 149
- KFSLWBXXFJQRDL-UHFFFAOYSA-N Peracetic acid Chemical compound CC(=O)OO KFSLWBXXFJQRDL-UHFFFAOYSA-N 0.000 claims description 143
- 210000003298 dental enamel Anatomy 0.000 claims description 120
- URAYPUMNDPQOKB-UHFFFAOYSA-N triacetin Chemical compound CC(=O)OCC(OC(C)=O)COC(C)=O URAYPUMNDPQOKB-UHFFFAOYSA-N 0.000 claims description 101
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 93
- 238000006243 chemical reaction Methods 0.000 claims description 93
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 91
- 108090000623 proteins and genes Proteins 0.000 claims description 72
- 239000003054 catalyst Substances 0.000 claims description 68
- 125000002887 hydroxy group Chemical group [H]O* 0.000 claims description 67
- 239000000047 product Substances 0.000 claims description 57
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 56
- 239000000843 powder Substances 0.000 claims description 55
- 150000001413 amino acids Chemical class 0.000 claims description 54
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 claims description 54
- RTZKZFJDLAIYFH-UHFFFAOYSA-N Diethyl ether Chemical compound CCOCC RTZKZFJDLAIYFH-UHFFFAOYSA-N 0.000 claims description 50
- 235000013773 glyceryl triacetate Nutrition 0.000 claims description 50
- 229960002622 triacetin Drugs 0.000 claims description 49
- 150000001720 carbohydrates Chemical class 0.000 claims description 47
- 239000001087 glyceryl triacetate Substances 0.000 claims description 47
- 125000000229 (C1-C4)alkoxy group Chemical group 0.000 claims description 39
- 125000000217 alkyl group Chemical group 0.000 claims description 39
- 230000003301 hydrolyzing effect Effects 0.000 claims description 37
- 125000001072 heteroaryl group Chemical group 0.000 claims description 34
- 150000002148 esters Chemical class 0.000 claims description 32
- 238000004519 manufacturing process Methods 0.000 claims description 30
- 125000004185 ester group Chemical group 0.000 claims description 28
- 102000004169 proteins and genes Human genes 0.000 claims description 28
- 210000000214 mouth Anatomy 0.000 claims description 26
- 238000011282 treatment Methods 0.000 claims description 24
- 125000003118 aryl group Chemical group 0.000 claims description 23
- 108010009043 arylesterase Proteins 0.000 claims description 23
- 102000028848 arylesterase Human genes 0.000 claims description 23
- 229920001184 polypeptide Polymers 0.000 claims description 22
- 239000007788 liquid Substances 0.000 claims description 21
- 230000008569 process Effects 0.000 claims description 21
- 125000006850 spacer group Chemical group 0.000 claims description 20
- 125000004432 carbon atom Chemical group C* 0.000 claims description 19
- 229910052799 carbon Inorganic materials 0.000 claims description 18
- 229910052739 hydrogen Inorganic materials 0.000 claims description 16
- 229920001282 polysaccharide Polymers 0.000 claims description 16
- 239000005017 polysaccharide Substances 0.000 claims description 16
- 150000002016 disaccharides Chemical class 0.000 claims description 15
- 238000002156 mixing Methods 0.000 claims description 14
- 150000002772 monosaccharides Chemical class 0.000 claims description 14
- 102000035195 Peptidases Human genes 0.000 claims description 13
- 108091005804 Peptidases Proteins 0.000 claims description 13
- 239000004365 Protease Substances 0.000 claims description 13
- 125000003342 alkenyl group Chemical group 0.000 claims description 13
- 125000002877 alkyl aryl group Chemical group 0.000 claims description 13
- 125000005213 alkyl heteroaryl group Chemical group 0.000 claims description 13
- 125000000304 alkynyl group Chemical group 0.000 claims description 13
- 108060003951 Immunoglobulin Proteins 0.000 claims description 12
- 241000187480 Mycobacterium smegmatis Species 0.000 claims description 12
- 125000002843 carboxylic acid group Chemical group 0.000 claims description 12
- 102000018358 immunoglobulin Human genes 0.000 claims description 12
- 108010021625 Immunoglobulin Fragments Proteins 0.000 claims description 11
- 102000008394 Immunoglobulin Fragments Human genes 0.000 claims description 11
- 239000000499 gel Substances 0.000 claims description 11
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 claims description 10
- 239000012634 fragment Substances 0.000 claims description 10
- 229910052760 oxygen Inorganic materials 0.000 claims description 10
- 239000001301 oxygen Substances 0.000 claims description 10
- 208000002925 dental caries Diseases 0.000 claims description 9
- XJJPVDONOLMSOS-UHFFFAOYSA-N 3-[(2-chlorophenyl)-(4-hydroxy-2-oxochromen-3-yl)methyl]-4-hydroxychromen-2-one Chemical compound O=C1OC=2C=CC=CC=2C(O)=C1C(C=1C(OC2=CC=CC=C2C=1O)=O)C1=CC=CC=C1Cl XJJPVDONOLMSOS-UHFFFAOYSA-N 0.000 claims description 8
- 102000004882 Lipase Human genes 0.000 claims description 8
- 239000004367 Lipase Substances 0.000 claims description 8
- 108090001060 Lipase Proteins 0.000 claims description 8
- 235000019421 lipase Nutrition 0.000 claims description 8
- 108700016155 Acyl transferases Proteins 0.000 claims description 7
- 108700014220 acyltransferase activity proteins Proteins 0.000 claims description 6
- 230000000249 desinfective effect Effects 0.000 claims description 6
- 239000006072 paste Substances 0.000 claims description 6
- 239000000606 toothpaste Substances 0.000 claims description 6
- 206010016228 Fasciitis Diseases 0.000 claims description 5
- 208000007027 Oral Candidiasis Diseases 0.000 claims description 5
- 208000007565 gingivitis Diseases 0.000 claims description 5
- 239000002324 mouth wash Substances 0.000 claims description 5
- 230000001877 deodorizing effect Effects 0.000 claims description 4
- 229940034610 toothpaste Drugs 0.000 claims description 4
- 102000057234 Acyl transferases Human genes 0.000 claims description 3
- 238000010612 desalination reaction Methods 0.000 claims description 3
- 229940051866 mouthwash Drugs 0.000 claims description 3
- 230000002265 prevention Effects 0.000 claims description 3
- 239000003826 tablet Substances 0.000 claims description 3
- 241000282832 Camelidae Species 0.000 claims description 2
- 241000628997 Flos Species 0.000 claims description 2
- 239000006071 cream Substances 0.000 claims description 2
- 229940023486 oral product Drugs 0.000 claims description 2
- 239000013588 oral product Substances 0.000 claims description 2
- 150000004676 glycans Chemical class 0.000 claims 5
- 239000008376 breath freshener Substances 0.000 claims 1
- 239000002674 ointment Substances 0.000 claims 1
- 150000001733 carboxylic acid esters Chemical class 0.000 abstract description 52
- 229940088598 enzyme Drugs 0.000 description 359
- 239000000243 solution Substances 0.000 description 127
- MHAJPDPJQMAIIY-UHFFFAOYSA-N Hydrogen peroxide Chemical compound OO MHAJPDPJQMAIIY-UHFFFAOYSA-N 0.000 description 92
- 108010080434 cephalosporin-C deacetylase Proteins 0.000 description 81
- 238000009472 formulation Methods 0.000 description 67
- 150000007523 nucleic acids Chemical group 0.000 description 59
- 108010093941 acetylxylan esterase Proteins 0.000 description 54
- 210000004027 cell Anatomy 0.000 description 51
- 235000001014 amino acid Nutrition 0.000 description 45
- 229960002163 hydrogen peroxide Drugs 0.000 description 45
- 229940024606 amino acid Drugs 0.000 description 44
- XYJRXVWERLGGKC-UHFFFAOYSA-D pentacalcium;hydroxide;triphosphate Chemical compound [OH-].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O XYJRXVWERLGGKC-UHFFFAOYSA-D 0.000 description 42
- 229910052588 hydroxylapatite Inorganic materials 0.000 description 41
- 239000000872 buffer Substances 0.000 description 39
- 108091028043 Nucleic acid sequence Proteins 0.000 description 34
- SCKXCAADGDQQCS-UHFFFAOYSA-N Performic acid Chemical compound OOC=O SCKXCAADGDQQCS-UHFFFAOYSA-N 0.000 description 33
- 239000008363 phosphate buffer Substances 0.000 description 33
- 235000014633 carbohydrates Nutrition 0.000 description 31
- 230000004927 fusion Effects 0.000 description 30
- 102000039446 nucleic acids Human genes 0.000 description 27
- 108020004707 nucleic acids Proteins 0.000 description 27
- 235000018102 proteins Nutrition 0.000 description 27
- 239000000546 pharmaceutical excipient Substances 0.000 description 23
- 239000000126 substance Substances 0.000 description 23
- 230000014509 gene expression Effects 0.000 description 22
- 239000000463 material Substances 0.000 description 22
- 244000063299 Bacillus subtilis Species 0.000 description 20
- 235000014469 Bacillus subtilis Nutrition 0.000 description 20
- 239000000523 sample Substances 0.000 description 19
- 230000008685 targeting Effects 0.000 description 19
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 18
- 102000004157 Hydrolases Human genes 0.000 description 18
- 108090000604 Hydrolases Proteins 0.000 description 18
- VZCYOOQTPOCHFL-UHFFFAOYSA-N trans-butenedioic acid Natural products OC(=O)C=CC(O)=O VZCYOOQTPOCHFL-UHFFFAOYSA-N 0.000 description 18
- 241000283690 Bos taurus Species 0.000 description 17
- 108091026890 Coding region Proteins 0.000 description 17
- 108020004414 DNA Proteins 0.000 description 17
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 17
- 239000002245 particle Substances 0.000 description 17
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 16
- 239000002253 acid Substances 0.000 description 16
- 238000001514 detection method Methods 0.000 description 16
- 229920001542 oligosaccharide Polymers 0.000 description 16
- 150000002482 oligosaccharides Chemical class 0.000 description 16
- 241000589540 Pseudomonas fluorescens Species 0.000 description 14
- 238000007792 addition Methods 0.000 description 14
- 230000003647 oxidation Effects 0.000 description 14
- 238000007254 oxidation reaction Methods 0.000 description 14
- 238000003860 storage Methods 0.000 description 14
- VZCYOOQTPOCHFL-UPHRSURJSA-N maleic acid Chemical compound OC(=O)\C=C/C(O)=O VZCYOOQTPOCHFL-UPHRSURJSA-N 0.000 description 13
- 230000000813 microbial effect Effects 0.000 description 13
- 239000002773 nucleotide Substances 0.000 description 13
- 125000003729 nucleotide group Chemical group 0.000 description 13
- 239000006228 supernatant Substances 0.000 description 13
- 150000007513 acids Chemical class 0.000 description 12
- 108010036413 histidylglycine Proteins 0.000 description 12
- 238000005259 measurement Methods 0.000 description 12
- 230000001105 regulatory effect Effects 0.000 description 12
- 238000006467 substitution reaction Methods 0.000 description 12
- 108020004705 Codon Proteins 0.000 description 11
- DBMJMQXJHONAFJ-UHFFFAOYSA-M Sodium laurylsulphate Chemical compound [Na+].CCCCCCCCCCCCOS([O-])(=O)=O DBMJMQXJHONAFJ-UHFFFAOYSA-M 0.000 description 11
- UYXTWWCETRIEDR-UHFFFAOYSA-N Tributyrin Chemical compound CCCC(=O)OCC(OC(=O)CCC)COC(=O)CCC UYXTWWCETRIEDR-UHFFFAOYSA-N 0.000 description 11
- 150000001721 carbon Chemical group 0.000 description 11
- 238000004140 cleaning Methods 0.000 description 11
- UQLDLKMNUJERMK-UHFFFAOYSA-L di(octadecanoyloxy)lead Chemical compound [Pb+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O UQLDLKMNUJERMK-UHFFFAOYSA-L 0.000 description 11
- 238000002474 experimental method Methods 0.000 description 11
- 238000006460 hydrolysis reaction Methods 0.000 description 11
- 235000019333 sodium laurylsulphate Nutrition 0.000 description 11
- 108010053835 Catalase Proteins 0.000 description 10
- 241000588724 Escherichia coli Species 0.000 description 10
- 229910019142 PO4 Inorganic materials 0.000 description 10
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 10
- 239000003153 chemical reaction reagent Substances 0.000 description 10
- UYAAVKFHBMJOJZ-UHFFFAOYSA-N diimidazo[1,3-b:1',3'-e]pyrazine-5,10-dione Chemical compound O=C1C2=CN=CN2C(=O)C2=CN=CN12 UYAAVKFHBMJOJZ-UHFFFAOYSA-N 0.000 description 10
- 235000021317 phosphate Nutrition 0.000 description 10
- 230000019612 pigmentation Effects 0.000 description 10
- 150000004804 polysaccharides Chemical class 0.000 description 10
- 229940116423 propylene glycol diacetate Drugs 0.000 description 10
- 239000007787 solid Substances 0.000 description 10
- KMZHZAAOEWVPSE-UHFFFAOYSA-N 2,3-dihydroxypropyl acetate Chemical compound CC(=O)OCC(O)CO KMZHZAAOEWVPSE-UHFFFAOYSA-N 0.000 description 9
- 125000000539 amino acid group Chemical group 0.000 description 9
- 238000005119 centrifugation Methods 0.000 description 9
- 150000001875 compounds Chemical class 0.000 description 9
- 238000009396 hybridization Methods 0.000 description 9
- 244000005700 microbiome Species 0.000 description 9
- 239000008188 pellet Substances 0.000 description 9
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 9
- 239000010452 phosphate Substances 0.000 description 9
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 8
- JTXMVXSTHSMVQF-UHFFFAOYSA-N 2-acetyloxyethyl acetate Chemical compound CC(=O)OCCOC(C)=O JTXMVXSTHSMVQF-UHFFFAOYSA-N 0.000 description 8
- 102000016938 Catalase Human genes 0.000 description 8
- 239000004348 Glyceryl diacetate Substances 0.000 description 8
- 238000004458 analytical method Methods 0.000 description 8
- 239000007864 aqueous solution Substances 0.000 description 8
- 230000015572 biosynthetic process Effects 0.000 description 8
- 238000010790 dilution Methods 0.000 description 8
- 239000012895 dilution Substances 0.000 description 8
- 235000019443 glyceryl diacetate Nutrition 0.000 description 8
- 238000004128 high performance liquid chromatography Methods 0.000 description 8
- 230000007062 hydrolysis Effects 0.000 description 8
- DNIAPMSPPWPWGF-UHFFFAOYSA-N monopropylene glycol Natural products CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 8
- 239000003921 oil Substances 0.000 description 8
- 235000019198 oils Nutrition 0.000 description 8
- 150000002978 peroxides Chemical class 0.000 description 8
- QZAYGJVTTNCVMB-UHFFFAOYSA-N serotonin Chemical compound C1=C(O)C=C2C(CCN)=CNC2=C1 QZAYGJVTTNCVMB-UHFFFAOYSA-N 0.000 description 8
- 239000011734 sodium Substances 0.000 description 8
- 108010073969 valyllysine Proteins 0.000 description 8
- 239000013598 vector Substances 0.000 description 8
- 241000894006 Bacteria Species 0.000 description 7
- BVKZGUZCCUSVTD-UHFFFAOYSA-M Bicarbonate Chemical compound OC([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-M 0.000 description 7
- KRKNYBCHXYNGOX-UHFFFAOYSA-K Citrate Chemical compound [O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O KRKNYBCHXYNGOX-UHFFFAOYSA-K 0.000 description 7
- 241000282326 Felis catus Species 0.000 description 7
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 7
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 7
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 7
- ZIJKGAXBCRWEOL-SAXBRCJISA-N Sucrose octaacetate Chemical compound CC(=O)O[C@H]1[C@H](OC(C)=O)[C@@H](COC(=O)C)O[C@@]1(COC(C)=O)O[C@@H]1[C@H](OC(C)=O)[C@@H](OC(C)=O)[C@H](OC(C)=O)[C@@H](COC(C)=O)O1 ZIJKGAXBCRWEOL-SAXBRCJISA-N 0.000 description 7
- 239000001344 [(2S,3S,4R,5R)-4-acetyloxy-2,5-bis(acetyloxymethyl)-2-[(2R,3R,4S,5R,6R)-3,4,5-triacetyloxy-6-(acetyloxymethyl)oxan-2-yl]oxyoxolan-3-yl] acetate Substances 0.000 description 7
- 230000003197 catalytic effect Effects 0.000 description 7
- 235000011180 diphosphates Nutrition 0.000 description 7
- 102220500059 eIF5-mimic protein 2_S54V_mutation Human genes 0.000 description 7
- 239000000284 extract Substances 0.000 description 7
- 230000012010 growth Effects 0.000 description 7
- 125000001183 hydrocarbyl group Chemical group 0.000 description 7
- 230000000670 limiting effect Effects 0.000 description 7
- 108010064235 lysylglycine Proteins 0.000 description 7
- 238000002360 preparation method Methods 0.000 description 7
- 229910052708 sodium Inorganic materials 0.000 description 7
- 229940083542 sodium Drugs 0.000 description 7
- 229940013883 sucrose octaacetate Drugs 0.000 description 7
- 239000004094 surface-active agent Substances 0.000 description 7
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 6
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 6
- 102000004316 Oxidoreductases Human genes 0.000 description 6
- 108090000854 Oxidoreductases Proteins 0.000 description 6
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Chemical compound CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 6
- 239000004480 active ingredient Substances 0.000 description 6
- 230000001580 bacterial effect Effects 0.000 description 6
- 230000003115 biocidal effect Effects 0.000 description 6
- 239000006172 buffering agent Substances 0.000 description 6
- XPPKVPWEQAFLFU-UHFFFAOYSA-J diphosphate(4-) Chemical compound [O-]P([O-])(=O)OP([O-])([O-])=O XPPKVPWEQAFLFU-UHFFFAOYSA-J 0.000 description 6
- 239000003814 drug Substances 0.000 description 6
- VZCYOOQTPOCHFL-OWOJBTEDSA-L fumarate(2-) Chemical compound [O-]C(=O)\C=C\C([O-])=O VZCYOOQTPOCHFL-OWOJBTEDSA-L 0.000 description 6
- 235000011187 glycerol Nutrition 0.000 description 6
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Natural products C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 6
- YACKEPLHDIMKIO-UHFFFAOYSA-N methylphosphonic acid Chemical compound CP(O)(O)=O YACKEPLHDIMKIO-UHFFFAOYSA-N 0.000 description 6
- 239000013612 plasmid Substances 0.000 description 6
- 108091033319 polynucleotide Proteins 0.000 description 6
- 102000040430 polynucleotide Human genes 0.000 description 6
- 239000002157 polynucleotide Substances 0.000 description 6
- 238000011002 quantification Methods 0.000 description 6
- 239000011541 reaction mixture Substances 0.000 description 6
- 230000001954 sterilising effect Effects 0.000 description 6
- KDYFGRWQOYBRFD-UHFFFAOYSA-L succinate(2-) Chemical compound [O-]C(=O)CCC([O-])=O KDYFGRWQOYBRFD-UHFFFAOYSA-L 0.000 description 6
- YZWRNSARCRTXDS-UHFFFAOYSA-N tripropionin Chemical compound CCC(=O)OCC(OC(=O)CC)COC(=O)CC YZWRNSARCRTXDS-UHFFFAOYSA-N 0.000 description 6
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 5
- 241000193830 Bacillus <bacterium> Species 0.000 description 5
- 241000194103 Bacillus pumilus Species 0.000 description 5
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 5
- 239000005913 Maltodextrin Substances 0.000 description 5
- 229920002774 Maltodextrin Polymers 0.000 description 5
- 102000003992 Peroxidases Human genes 0.000 description 5
- MLHOXUWWKVQEJB-UHFFFAOYSA-N Propyleneglycol diacetate Chemical compound CC(=O)OC(C)COC(C)=O MLHOXUWWKVQEJB-UHFFFAOYSA-N 0.000 description 5
- 241000589516 Pseudomonas Species 0.000 description 5
- 244000057717 Streptococcus lactis Species 0.000 description 5
- 235000014897 Streptococcus lactis Nutrition 0.000 description 5
- GWEVSGVZZGPLCZ-UHFFFAOYSA-N Titan oxide Chemical compound O=[Ti]=O GWEVSGVZZGPLCZ-UHFFFAOYSA-N 0.000 description 5
- 238000002835 absorbance Methods 0.000 description 5
- 238000004422 calculation algorithm Methods 0.000 description 5
- 230000008859 change Effects 0.000 description 5
- 238000004040 coloring Methods 0.000 description 5
- 239000000356 contaminant Substances 0.000 description 5
- 239000006184 cosolvent Substances 0.000 description 5
- 230000007423 decrease Effects 0.000 description 5
- 210000004268 dentin Anatomy 0.000 description 5
- 108010025153 lysyl-alanyl-alanine Proteins 0.000 description 5
- 229940035034 maltodextrin Drugs 0.000 description 5
- 239000002609 medium Substances 0.000 description 5
- RIEABXYBQSLTFR-UHFFFAOYSA-N monobutyrin Chemical compound CCCC(=O)OCC(O)CO RIEABXYBQSLTFR-UHFFFAOYSA-N 0.000 description 5
- NLKNQRATVPKPDG-UHFFFAOYSA-M potassium iodide Substances [K+].[I-] NLKNQRATVPKPDG-UHFFFAOYSA-M 0.000 description 5
- 239000008057 potassium phosphate buffer Substances 0.000 description 5
- 159000000001 potassium salts Chemical class 0.000 description 5
- 239000002243 precursor Substances 0.000 description 5
- 238000011084 recovery Methods 0.000 description 5
- 210000003296 saliva Anatomy 0.000 description 5
- 239000011780 sodium chloride Substances 0.000 description 5
- 239000002904 solvent Substances 0.000 description 5
- 239000007921 spray Substances 0.000 description 5
- 238000012546 transfer Methods 0.000 description 5
- 238000013519 translation Methods 0.000 description 5
- AQLJVWUFPCUVLO-UHFFFAOYSA-N urea hydrogen peroxide Chemical class OO.NC(N)=O AQLJVWUFPCUVLO-UHFFFAOYSA-N 0.000 description 5
- 125000005023 xylyl group Chemical group 0.000 description 5
- PUPZLCDOIYMWBV-UHFFFAOYSA-N (+/-)-1,3-Butanediol Chemical compound CC(O)CCO PUPZLCDOIYMWBV-UHFFFAOYSA-N 0.000 description 4
- KBPLFHHGFOOTCA-UHFFFAOYSA-N 1-Octanol Chemical compound CCCCCCCCO KBPLFHHGFOOTCA-UHFFFAOYSA-N 0.000 description 4
- XJFPXLWGZWAWRQ-UHFFFAOYSA-N 2-[[2-[[2-[[2-[[2-[(2-azaniumylacetyl)amino]acetyl]amino]acetyl]amino]acetyl]amino]acetyl]amino]acetate Chemical compound NCC(=O)NCC(=O)NCC(=O)NCC(=O)NCC(=O)NCC(O)=O XJFPXLWGZWAWRQ-UHFFFAOYSA-N 0.000 description 4
- LNNSWWRRYJLGNI-NAKRPEOUSA-N Ala-Ile-Val Chemical compound C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O LNNSWWRRYJLGNI-NAKRPEOUSA-N 0.000 description 4
- DPNZTBKGAUAZQU-DLOVCJGASA-N Ala-Leu-His Chemical compound C[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N DPNZTBKGAUAZQU-DLOVCJGASA-N 0.000 description 4
- MDNAVFBZPROEHO-DCAQKATOSA-N Ala-Lys-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O MDNAVFBZPROEHO-DCAQKATOSA-N 0.000 description 4
- MDNAVFBZPROEHO-UHFFFAOYSA-N Ala-Lys-Val Natural products CC(C)C(C(O)=O)NC(=O)C(NC(=O)C(C)N)CCCCN MDNAVFBZPROEHO-UHFFFAOYSA-N 0.000 description 4
- SYIFFFHSXBNPMC-UWJYBYFXSA-N Ala-Ser-Tyr Chemical compound C[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N SYIFFFHSXBNPMC-UWJYBYFXSA-N 0.000 description 4
- IYKVSFNGSWTTNZ-GUBZILKMSA-N Ala-Val-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N IYKVSFNGSWTTNZ-GUBZILKMSA-N 0.000 description 4
- NLYYHIKRBRMAJV-AEJSXWLSSA-N Ala-Val-Pro Chemical compound C[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N NLYYHIKRBRMAJV-AEJSXWLSSA-N 0.000 description 4
- NKNILFJYKKHBKE-WPRPVWTQSA-N Arg-Gly-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O NKNILFJYKKHBKE-WPRPVWTQSA-N 0.000 description 4
- HJDNZFIYILEIKR-OSUNSFLBSA-N Arg-Ile-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O HJDNZFIYILEIKR-OSUNSFLBSA-N 0.000 description 4
- JEOCWTUOMKEEMF-RHYQMDGZSA-N Arg-Leu-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JEOCWTUOMKEEMF-RHYQMDGZSA-N 0.000 description 4
- RAUPFUCUDBQYHE-AVGNSLFASA-N Asn-Phe-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O RAUPFUCUDBQYHE-AVGNSLFASA-N 0.000 description 4
- WSGVTKZFVJSJOG-RCOVLWMOSA-N Asp-Gly-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O WSGVTKZFVJSJOG-RCOVLWMOSA-N 0.000 description 4
- RPUYTJJZXQBWDT-SRVKXCTJSA-N Asp-Phe-Ser Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC(=O)O)N RPUYTJJZXQBWDT-SRVKXCTJSA-N 0.000 description 4
- GCACQYDBDHRVGE-LKXGYXEUSA-N Asp-Thr-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H]([C@H](O)C)NC(=O)[C@@H](N)CC(O)=O GCACQYDBDHRVGE-LKXGYXEUSA-N 0.000 description 4
- XWKBWZXGNXTDKY-ZKWXMUAHSA-N Asp-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC(O)=O XWKBWZXGNXTDKY-ZKWXMUAHSA-N 0.000 description 4
- 241000194110 Bacillus sp. (in: Bacteria) Species 0.000 description 4
- LGIKBBLQVSWUGK-DCAQKATOSA-N Gln-Leu-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O LGIKBBLQVSWUGK-DCAQKATOSA-N 0.000 description 4
- QDMVXRNLOPTPIE-WDCWCFNPSA-N Glu-Lys-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QDMVXRNLOPTPIE-WDCWCFNPSA-N 0.000 description 4
- YUXIEONARHPUTK-JBACZVJFSA-N Glu-Phe-Trp Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CNC3=CC=CC=C32)C(=O)O)NC(=O)[C@H](CCC(=O)O)N YUXIEONARHPUTK-JBACZVJFSA-N 0.000 description 4
- 239000004366 Glucose oxidase Substances 0.000 description 4
- 108010015776 Glucose oxidase Proteins 0.000 description 4
- UXJHNZODTMHWRD-WHFBIAKZSA-N Gly-Asn-Ala Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(O)=O UXJHNZODTMHWRD-WHFBIAKZSA-N 0.000 description 4
- XLFHCWHXKSFVIB-BQBZGAKWSA-N Gly-Gln-Gln Chemical compound NCC(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O XLFHCWHXKSFVIB-BQBZGAKWSA-N 0.000 description 4
- STVHDEHTKFXBJQ-LAEOZQHASA-N Gly-Glu-Ile Chemical compound [H]NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O STVHDEHTKFXBJQ-LAEOZQHASA-N 0.000 description 4
- FQKKPCWTZZEDIC-XPUUQOCRSA-N Gly-His-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)CN)CC1=CN=CN1 FQKKPCWTZZEDIC-XPUUQOCRSA-N 0.000 description 4
- UESJMAMHDLEHGM-NHCYSSNCSA-N Gly-Ile-Leu Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O UESJMAMHDLEHGM-NHCYSSNCSA-N 0.000 description 4
- IROABALAWGJQGM-OALUTQOASA-N Gly-Trp-Tyr Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)O)NC(=O)CN IROABALAWGJQGM-OALUTQOASA-N 0.000 description 4
- UIQGJYUEQDOODF-KWQFWETISA-N Gly-Tyr-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)CN)CC1=CC=C(O)C=C1 UIQGJYUEQDOODF-KWQFWETISA-N 0.000 description 4
- YJDALMUYJIENAG-QWRGUYRKSA-N Gly-Tyr-Asn Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)CN)O YJDALMUYJIENAG-QWRGUYRKSA-N 0.000 description 4
- NLZVTPYXYXMCIP-XUXIUFHCSA-N Ile-Pro-Lys Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(O)=O NLZVTPYXYXMCIP-XUXIUFHCSA-N 0.000 description 4
- PXKACEXYLPBMAD-JBDRJPRFSA-N Ile-Ser-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)O)N PXKACEXYLPBMAD-JBDRJPRFSA-N 0.000 description 4
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 4
- RCFDOSNHHZGBOY-UHFFFAOYSA-N L-isoleucyl-L-alanine Natural products CCC(C)C(N)C(=O)NC(C)C(O)=O RCFDOSNHHZGBOY-UHFFFAOYSA-N 0.000 description 4
- KFKWRHQBZQICHA-STQMWFEESA-N L-leucyl-L-phenylalanine Natural products CC(C)C[C@H](N)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 KFKWRHQBZQICHA-STQMWFEESA-N 0.000 description 4
- KVMULWOHPPMHHE-DCAQKATOSA-N Leu-Glu-Gln Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O KVMULWOHPPMHHE-DCAQKATOSA-N 0.000 description 4
- HVJVUYQWFYMGJS-GVXVVHGQSA-N Leu-Glu-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O HVJVUYQWFYMGJS-GVXVVHGQSA-N 0.000 description 4
- JRJLGNFWYFSJHB-HOCLYGCPSA-N Leu-Gly-Trp Chemical compound [H]N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O JRJLGNFWYFSJHB-HOCLYGCPSA-N 0.000 description 4
- IZPVWNSAVUQBGP-CIUDSAMLSA-N Leu-Ser-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O IZPVWNSAVUQBGP-CIUDSAMLSA-N 0.000 description 4
- LJBVRCDPWOJOEK-PPCPHDFISA-N Leu-Thr-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O LJBVRCDPWOJOEK-PPCPHDFISA-N 0.000 description 4
- FBNPMTNBFFAMMH-AVGNSLFASA-N Leu-Val-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N FBNPMTNBFFAMMH-AVGNSLFASA-N 0.000 description 4
- FBNPMTNBFFAMMH-UHFFFAOYSA-N Leu-Val-Arg Natural products CC(C)CC(N)C(=O)NC(C(C)C)C(=O)NC(C(O)=O)CCCN=C(N)N FBNPMTNBFFAMMH-UHFFFAOYSA-N 0.000 description 4
- KWUKZRFFKPLUPE-HJGDQZAQSA-N Lys-Asp-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O KWUKZRFFKPLUPE-HJGDQZAQSA-N 0.000 description 4
- PSVAVKGDUAKZKU-BZSNNMDCSA-N Lys-Tyr-His Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CCCCN)N)O PSVAVKGDUAKZKU-BZSNNMDCSA-N 0.000 description 4
- HMZPYMSEAALNAE-ULQDDVLXSA-N Lys-Val-Tyr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O HMZPYMSEAALNAE-ULQDDVLXSA-N 0.000 description 4
- 239000004472 Lysine Substances 0.000 description 4
- VOOINLQYUZOREH-SRVKXCTJSA-N Met-Gln-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCSC)N VOOINLQYUZOREH-SRVKXCTJSA-N 0.000 description 4
- IHRFZLQEQVHXFA-RHYQMDGZSA-N Met-Thr-Lys Chemical compound CSCC[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CCCCN IHRFZLQEQVHXFA-RHYQMDGZSA-N 0.000 description 4
- SITLTJHOQZFJGG-UHFFFAOYSA-N N-L-alpha-glutamyl-L-valine Natural products CC(C)C(C(O)=O)NC(=O)C(N)CCC(O)=O SITLTJHOQZFJGG-UHFFFAOYSA-N 0.000 description 4
- DDYIRGBOZVKRFR-AVGNSLFASA-N Phe-Asp-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N DDYIRGBOZVKRFR-AVGNSLFASA-N 0.000 description 4
- RIYZXJVARWJLKS-KKUMJFAQSA-N Phe-Asp-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1 RIYZXJVARWJLKS-KKUMJFAQSA-N 0.000 description 4
- VJLLEKDQJSMHRU-STQMWFEESA-N Phe-Gly-Met Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)NCC(=O)N[C@@H](CCSC)C(O)=O VJLLEKDQJSMHRU-STQMWFEESA-N 0.000 description 4
- 241000235648 Pichia Species 0.000 description 4
- KPDRZQUWJKTMBP-DCAQKATOSA-N Pro-Asp-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@@H]1CCCN1 KPDRZQUWJKTMBP-DCAQKATOSA-N 0.000 description 4
- YTWNSIDWAFSEEI-RWMBFGLXSA-N Pro-His-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC2=CN=CN2)C(=O)N3CCC[C@@H]3C(=O)O YTWNSIDWAFSEEI-RWMBFGLXSA-N 0.000 description 4
- CLJLVCYFABNTHP-DCAQKATOSA-N Pro-Leu-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O CLJLVCYFABNTHP-DCAQKATOSA-N 0.000 description 4
- YHUBAXGAAYULJY-ULQDDVLXSA-N Pro-Tyr-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(O)=O YHUBAXGAAYULJY-ULQDDVLXSA-N 0.000 description 4
- GYDFRTRSSXOZCR-ACZMJKKPSA-N Ser-Ser-Glu Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O GYDFRTRSSXOZCR-ACZMJKKPSA-N 0.000 description 4
- MQBTXMPQNCGSSZ-OSUNSFLBSA-N Thr-Arg-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)[C@@H](C)O)CCCN=C(N)N MQBTXMPQNCGSSZ-OSUNSFLBSA-N 0.000 description 4
- CJEHCEOXPLASCK-MEYUZBJRSA-N Thr-Tyr-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)[C@H](O)C)CC1=CC=C(O)C=C1 CJEHCEOXPLASCK-MEYUZBJRSA-N 0.000 description 4
- BSCBBPKDVOZICB-KKUMJFAQSA-N Tyr-Leu-Asp Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O BSCBBPKDVOZICB-KKUMJFAQSA-N 0.000 description 4
- PGEFRHBWGOJPJT-KKUMJFAQSA-N Tyr-Lys-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(O)=O PGEFRHBWGOJPJT-KKUMJFAQSA-N 0.000 description 4
- CDBXVDXSLPLFMD-BPNCWPANSA-N Tyr-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CC1=CC=C(O)C=C1 CDBXVDXSLPLFMD-BPNCWPANSA-N 0.000 description 4
- VPEFOFYNHBWFNQ-UFYCRDLUSA-N Tyr-Pro-Tyr Chemical compound C([C@H](N)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=C(O)C=C1 VPEFOFYNHBWFNQ-UFYCRDLUSA-N 0.000 description 4
- RMRFSFXLFWWAJZ-HJOGWXRNSA-N Tyr-Tyr-Tyr Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=C(O)C=C1 RMRFSFXLFWWAJZ-HJOGWXRNSA-N 0.000 description 4
- FZSPNKUFROZBSG-ZKWXMUAHSA-N Val-Ala-Asp Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC(O)=O FZSPNKUFROZBSG-ZKWXMUAHSA-N 0.000 description 4
- NMPXRFYMZDIBRF-ZOBUZTSGSA-N Val-Asn-Trp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N NMPXRFYMZDIBRF-ZOBUZTSGSA-N 0.000 description 4
- VLOYGOZDPGYWFO-LAEOZQHASA-N Val-Asp-Glu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O VLOYGOZDPGYWFO-LAEOZQHASA-N 0.000 description 4
- YQYFYUSYEDNLSD-YEPSODPASA-N Val-Thr-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O YQYFYUSYEDNLSD-YEPSODPASA-N 0.000 description 4
- LLPWGHLVUPBSLP-IJLUTSLNSA-N [(2r,3r,4r)-3,4-diacetyloxy-3,4-dihydro-2h-pyran-2-yl]methyl acetate Chemical compound CC(=O)OC[C@H]1OC=C[C@@H](OC(C)=O)[C@H]1OC(C)=O LLPWGHLVUPBSLP-IJLUTSLNSA-N 0.000 description 4
- LLPWGHLVUPBSLP-UTUOFQBUSA-N [(2r,3s,4r)-3,4-diacetyloxy-3,4-dihydro-2h-pyran-2-yl]methyl acetate Chemical compound CC(=O)OC[C@H]1OC=C[C@@H](OC(C)=O)[C@@H]1OC(C)=O LLPWGHLVUPBSLP-UTUOFQBUSA-N 0.000 description 4
- 102000045404 acyltransferase activity proteins Human genes 0.000 description 4
- 239000004599 antimicrobial Substances 0.000 description 4
- 230000000903 blocking effect Effects 0.000 description 4
- WERYXYBDKMZEQL-UHFFFAOYSA-N butane-1,4-diol Chemical compound OCCCCO WERYXYBDKMZEQL-UHFFFAOYSA-N 0.000 description 4
- 238000005251 capillar electrophoresis Methods 0.000 description 4
- 238000004042 decolorization Methods 0.000 description 4
- 238000012217 deletion Methods 0.000 description 4
- 230000037430 deletion Effects 0.000 description 4
- 238000013461 design Methods 0.000 description 4
- 239000006185 dispersion Substances 0.000 description 4
- 238000010494 dissociation reaction Methods 0.000 description 4
- 230000005593 dissociations Effects 0.000 description 4
- 238000004090 dissolution Methods 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 238000000855 fermentation Methods 0.000 description 4
- 230000004151 fermentation Effects 0.000 description 4
- 108010063718 gamma-glutamylaspartic acid Proteins 0.000 description 4
- 229940116332 glucose oxidase Drugs 0.000 description 4
- 235000019420 glucose oxidase Nutrition 0.000 description 4
- XKUKSGPZAADMRA-UHFFFAOYSA-N glycyl-glycyl-glycine Natural products NCC(=O)NCC(=O)NCC(O)=O XKUKSGPZAADMRA-UHFFFAOYSA-N 0.000 description 4
- 108010026364 glycyl-glycyl-leucine Proteins 0.000 description 4
- 238000011065 in-situ storage Methods 0.000 description 4
- 238000011534 incubation Methods 0.000 description 4
- 108010053037 kyotorphin Proteins 0.000 description 4
- 108010044056 leucyl-phenylalanine Proteins 0.000 description 4
- 108010057821 leucylproline Proteins 0.000 description 4
- 230000014759 maintenance of location Effects 0.000 description 4
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 4
- 238000002823 phage display Methods 0.000 description 4
- 229920000642 polymer Polymers 0.000 description 4
- 229920000136 polysorbate Polymers 0.000 description 4
- 235000013772 propylene glycol Nutrition 0.000 description 4
- 238000000746 purification Methods 0.000 description 4
- 102200082948 rs33916412 Human genes 0.000 description 4
- 150000003839 salts Chemical class 0.000 description 4
- 238000002864 sequence alignment Methods 0.000 description 4
- 229940076279 serotonin Drugs 0.000 description 4
- 239000012064 sodium phosphate buffer Substances 0.000 description 4
- 239000003381 stabilizer Substances 0.000 description 4
- 239000011550 stock solution Substances 0.000 description 4
- 229940124597 therapeutic agent Drugs 0.000 description 4
- 108010061238 threonyl-glycine Proteins 0.000 description 4
- 238000013518 transcription Methods 0.000 description 4
- 230000035897 transcription Effects 0.000 description 4
- 230000009466 transformation Effects 0.000 description 4
- 238000005406 washing Methods 0.000 description 4
- 229920001221 xylan Polymers 0.000 description 4
- KBWFWZJNPVZRRG-UHFFFAOYSA-N 1,3-dibutyrin Chemical compound CCCC(=O)OCC(O)COC(=O)CCC KBWFWZJNPVZRRG-UHFFFAOYSA-N 0.000 description 3
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 3
- CSCPPACGZOOCGX-UHFFFAOYSA-N Acetone Chemical compound CC(C)=O CSCPPACGZOOCGX-UHFFFAOYSA-N 0.000 description 3
- 229920000936 Agarose Polymers 0.000 description 3
- PBAMJJXWDQXOJA-FXQIFTODSA-N Ala-Asp-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N PBAMJJXWDQXOJA-FXQIFTODSA-N 0.000 description 3
- GKAZXNDATBWNBI-DCAQKATOSA-N Ala-Met-Lys Chemical compound C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCCN)C(=O)O)N GKAZXNDATBWNBI-DCAQKATOSA-N 0.000 description 3
- BTRULDJUUVGRNE-DCAQKATOSA-N Ala-Pro-Lys Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(O)=O BTRULDJUUVGRNE-DCAQKATOSA-N 0.000 description 3
- AOAKQKVICDWCLB-UWJYBYFXSA-N Ala-Tyr-Asn Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N AOAKQKVICDWCLB-UWJYBYFXSA-N 0.000 description 3
- BAVDUESNGSMLPI-CIUDSAMLSA-N Arg-Asn-Gly-Ser Chemical compound NC(=N)NCCC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(=O)N[C@@H](CO)C(O)=O BAVDUESNGSMLPI-CIUDSAMLSA-N 0.000 description 3
- UVTGNSWSRSCPLP-UHFFFAOYSA-N Arg-Tyr Natural products NC(CCNC(=N)N)C(=O)NC(Cc1ccc(O)cc1)C(=O)O UVTGNSWSRSCPLP-UHFFFAOYSA-N 0.000 description 3
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical group [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 3
- 229940123748 Catalase inhibitor Drugs 0.000 description 3
- 108700010070 Codon Usage Proteins 0.000 description 3
- 208000002064 Dental Plaque Diseases 0.000 description 3
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 3
- 241000588722 Escherichia Species 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- RZSLYUUFFVHFRQ-FXQIFTODSA-N Gln-Ala-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O RZSLYUUFFVHFRQ-FXQIFTODSA-N 0.000 description 3
- IWUFOVSLWADEJC-AVGNSLFASA-N Gln-His-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(C)C)C(O)=O IWUFOVSLWADEJC-AVGNSLFASA-N 0.000 description 3
- YPFFHGRJCUBXPX-NHCYSSNCSA-N Gln-Pro-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCC(N)=O)C(O)=O YPFFHGRJCUBXPX-NHCYSSNCSA-N 0.000 description 3
- DUGYCMAIAKAQPB-GLLZPBPUSA-N Gln-Thr-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O DUGYCMAIAKAQPB-GLLZPBPUSA-N 0.000 description 3
- MUSGDMDGNGXULI-DCAQKATOSA-N Glu-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O MUSGDMDGNGXULI-DCAQKATOSA-N 0.000 description 3
- IVGJYOOGJLFKQE-AVGNSLFASA-N Glu-Leu-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N IVGJYOOGJLFKQE-AVGNSLFASA-N 0.000 description 3
- YQPFCZVKMUVZIN-AUTRQRHGSA-N Glu-Val-Gln Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O YQPFCZVKMUVZIN-AUTRQRHGSA-N 0.000 description 3
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 3
- UXDDRFCJKNROTO-UHFFFAOYSA-N Glycerol 1,2-diacetate Chemical compound CC(=O)OCC(CO)OC(C)=O UXDDRFCJKNROTO-UHFFFAOYSA-N 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- ZYDYEPDFFVCUBI-SRVKXCTJSA-N His-Glu-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CC1=CN=CN1)N ZYDYEPDFFVCUBI-SRVKXCTJSA-N 0.000 description 3
- WGHJXSONOOTTCZ-JYJNAYRXSA-N His-Glu-Tyr Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O WGHJXSONOOTTCZ-JYJNAYRXSA-N 0.000 description 3
- OQDLKDUVMTUPPG-AVGNSLFASA-N His-Leu-Glu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O OQDLKDUVMTUPPG-AVGNSLFASA-N 0.000 description 3
- HDODQNPMSHDXJT-GHCJXIJMSA-N Ile-Asn-Ser Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O HDODQNPMSHDXJT-GHCJXIJMSA-N 0.000 description 3
- QSPLUJGYOPZINY-ZPFDUUQYSA-N Ile-Asp-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N QSPLUJGYOPZINY-ZPFDUUQYSA-N 0.000 description 3
- NYEYYMLUABXDMC-NHCYSSNCSA-N Ile-Gly-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)O)N NYEYYMLUABXDMC-NHCYSSNCSA-N 0.000 description 3
- RVNOXPZHMUWCLW-GMOBBJLQSA-N Ile-Met-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(=O)N)C(=O)O)N RVNOXPZHMUWCLW-GMOBBJLQSA-N 0.000 description 3
- IITVUURPOYGCTD-NAKRPEOUSA-N Ile-Pro-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O IITVUURPOYGCTD-NAKRPEOUSA-N 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- 125000000998 L-alanino group Chemical group [H]N([*])[C@](C([H])([H])[H])([H])C(=O)O[H] 0.000 description 3
- FEWJPZIEWOKRBE-JCYAYHJZSA-L L-tartrate(2-) Chemical class [O-]C(=O)[C@H](O)[C@@H](O)C([O-])=O FEWJPZIEWOKRBE-JCYAYHJZSA-L 0.000 description 3
- CQGSYZCULZMEDE-UHFFFAOYSA-N Leu-Gln-Pro Natural products CC(C)CC(N)C(=O)NC(CCC(N)=O)C(=O)N1CCCC1C(O)=O CQGSYZCULZMEDE-UHFFFAOYSA-N 0.000 description 3
- HDHQQEDVWQGBEE-DCAQKATOSA-N Leu-Met-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CO)C(O)=O HDHQQEDVWQGBEE-DCAQKATOSA-N 0.000 description 3
- MYZMQWHPDAYKIE-SRVKXCTJSA-N Lys-Leu-Ala Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O MYZMQWHPDAYKIE-SRVKXCTJSA-N 0.000 description 3
- WRODMZBHNNPRLN-SRVKXCTJSA-N Lys-Leu-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O WRODMZBHNNPRLN-SRVKXCTJSA-N 0.000 description 3
- 241000186359 Mycobacterium Species 0.000 description 3
- 108091034117 Oligonucleotide Proteins 0.000 description 3
- OXKJSGGTHFMGDT-UFYCRDLUSA-N Phe-Phe-Arg Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C1=CC=CC=C1 OXKJSGGTHFMGDT-UFYCRDLUSA-N 0.000 description 3
- RBRNEFJTEHPDSL-ACRUOGEOSA-N Phe-Phe-Lys Chemical compound C([C@@H](C(=O)N[C@@H](CCCCN)C(O)=O)NC(=O)[C@@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 RBRNEFJTEHPDSL-ACRUOGEOSA-N 0.000 description 3
- UEHYFUCOGHWASA-HJGDQZAQSA-N Pro-Glu-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H]1CCCN1 UEHYFUCOGHWASA-HJGDQZAQSA-N 0.000 description 3
- FDMKYQQYJKYCLV-GUBZILKMSA-N Pro-Pro-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 FDMKYQQYJKYCLV-GUBZILKMSA-N 0.000 description 3
- 241000316848 Rhodococcus <scale insect> Species 0.000 description 3
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 3
- 108090000787 Subtilisin Proteins 0.000 description 3
- 241000999856 Thermotoga maritima MSB8 Species 0.000 description 3
- YOOAQCZYZHGUAZ-KATARQTJSA-N Thr-Leu-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O YOOAQCZYZHGUAZ-KATARQTJSA-N 0.000 description 3
- SPIFGZFZMVLPHN-UNQGMJICSA-N Thr-Val-Phe Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O SPIFGZFZMVLPHN-UNQGMJICSA-N 0.000 description 3
- LMKKMCGTDANZTR-BZSNNMDCSA-N Tyr-Phe-Asp Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CC(O)=O)C(O)=O)C1=CC=C(O)C=C1 LMKKMCGTDANZTR-BZSNNMDCSA-N 0.000 description 3
- OKDNSNWJEXAMSU-IRXDYDNUSA-N Tyr-Phe-Gly Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)NCC(O)=O)C1=CC=C(O)C=C1 OKDNSNWJEXAMSU-IRXDYDNUSA-N 0.000 description 3
- VPGCVZRRBYOGCD-AVGNSLFASA-N Val-Lys-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O VPGCVZRRBYOGCD-AVGNSLFASA-N 0.000 description 3
- JXCOEPXCBVCTRD-JYJNAYRXSA-N Val-Tyr-Arg Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N JXCOEPXCBVCTRD-JYJNAYRXSA-N 0.000 description 3
- IHNHAHWGVLXCCI-FDYHWXHSSA-N [(2r,3r,4r,5s)-3,4,5-triacetyloxyoxolan-2-yl]methyl acetate Chemical compound CC(=O)OC[C@H]1O[C@@H](OC(C)=O)[C@H](OC(C)=O)[C@@H]1OC(C)=O IHNHAHWGVLXCCI-FDYHWXHSSA-N 0.000 description 3
- FEQXFAYSNRWXDW-RKQHYHRCSA-N [(2r,3r,4s,5r,6s)-4,5,6-triacetyloxy-2-(hydroxymethyl)oxan-3-yl] acetate Chemical compound CC(=O)O[C@@H]1O[C@H](CO)[C@@H](OC(C)=O)[C@H](OC(C)=O)[C@H]1OC(C)=O FEQXFAYSNRWXDW-RKQHYHRCSA-N 0.000 description 3
- LPTITAGPBXDDGR-OWYFMNJBSA-N [(2r,3r,4s,5s,6r)-3,4,5,6-tetraacetyloxyoxan-2-yl]methyl acetate Chemical compound CC(=O)OC[C@H]1O[C@H](OC(C)=O)[C@@H](OC(C)=O)[C@@H](OC(C)=O)[C@@H]1OC(C)=O LPTITAGPBXDDGR-OWYFMNJBSA-N 0.000 description 3
- 239000013543 active substance Substances 0.000 description 3
- 230000002411 adverse Effects 0.000 description 3
- 235000004279 alanine Nutrition 0.000 description 3
- 125000001931 aliphatic group Chemical group 0.000 description 3
- 230000002882 anti-plaque Effects 0.000 description 3
- 238000003556 assay Methods 0.000 description 3
- 238000002819 bacterial display Methods 0.000 description 3
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 3
- 239000003139 biocide Substances 0.000 description 3
- 239000007844 bleaching agent Substances 0.000 description 3
- 230000003139 buffering effect Effects 0.000 description 3
- 210000004899 c-terminal region Anatomy 0.000 description 3
- 150000001732 carboxylic acid derivatives Chemical class 0.000 description 3
- 239000000969 carrier Substances 0.000 description 3
- 239000000919 ceramic Substances 0.000 description 3
- 239000013043 chemical agent Substances 0.000 description 3
- 239000003086 colorant Substances 0.000 description 3
- 238000013329 compounding Methods 0.000 description 3
- 239000002826 coolant Substances 0.000 description 3
- 239000002537 cosmetic Substances 0.000 description 3
- 239000000645 desinfectant Substances 0.000 description 3
- 235000014113 dietary fatty acids Nutrition 0.000 description 3
- 239000012149 elution buffer Substances 0.000 description 3
- 235000019441 ethanol Nutrition 0.000 description 3
- 239000013604 expression vector Substances 0.000 description 3
- 239000000194 fatty acid Substances 0.000 description 3
- 229930195729 fatty acid Natural products 0.000 description 3
- 229960001031 glucose Drugs 0.000 description 3
- 239000008103 glucose Substances 0.000 description 3
- 108010020688 glycylhistidine Proteins 0.000 description 3
- 239000004615 ingredient Substances 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- 230000002147 killing effect Effects 0.000 description 3
- 108010076756 leucyl-alanyl-phenylalanine Proteins 0.000 description 3
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 3
- 108010003700 lysyl aspartic acid Proteins 0.000 description 3
- 238000002824 mRNA display Methods 0.000 description 3
- 238000012423 maintenance Methods 0.000 description 3
- 239000011159 matrix material Substances 0.000 description 3
- 238000005374 membrane filtration Methods 0.000 description 3
- OSWPMRLSEDHDFF-UHFFFAOYSA-N methyl salicylate Chemical compound COC(=O)C1=CC=CC=C1O OSWPMRLSEDHDFF-UHFFFAOYSA-N 0.000 description 3
- 238000002703 mutagenesis Methods 0.000 description 3
- 231100000350 mutagenesis Toxicity 0.000 description 3
- QBQNDWBZRZGJTF-JOAYCEEESA-N n-[(2s,3r,4r,5r,6r)-2,4,5-triacetyl-2,4,5-trihydroxy-6-(1-hydroxy-2-oxopropyl)oxan-3-yl]acetamide Chemical compound CC(=O)N[C@H]1[C@@](O)(C(C)=O)O[C@H](C(O)C(C)=O)[C@](O)(C(C)=O)[C@@]1(O)C(C)=O QBQNDWBZRZGJTF-JOAYCEEESA-N 0.000 description 3
- 235000015097 nutrients Nutrition 0.000 description 3
- 108010024607 phenylalanylalanine Proteins 0.000 description 3
- 239000000049 pigment Substances 0.000 description 3
- 239000004033 plastic Substances 0.000 description 3
- 229920003023 plastic Polymers 0.000 description 3
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 3
- XAEFZNCEHLXOMS-UHFFFAOYSA-M potassium benzoate Chemical compound [K+].[O-]C(=O)C1=CC=CC=C1 XAEFZNCEHLXOMS-UHFFFAOYSA-M 0.000 description 3
- 238000012545 processing Methods 0.000 description 3
- 108010053725 prolylvaline Proteins 0.000 description 3
- 238000000159 protein binding assay Methods 0.000 description 3
- 238000002818 protein evolution Methods 0.000 description 3
- 239000000376 reactant Substances 0.000 description 3
- 239000011347 resin Substances 0.000 description 3
- 229920005989 resin Polymers 0.000 description 3
- 102200010049 rs869025189 Human genes 0.000 description 3
- 238000012163 sequencing technique Methods 0.000 description 3
- 108010026333 seryl-proline Proteins 0.000 description 3
- 210000004872 soft tissue Anatomy 0.000 description 3
- 241000894007 species Species 0.000 description 3
- 238000001694 spray drying Methods 0.000 description 3
- 235000000346 sugar Nutrition 0.000 description 3
- 229940095064 tartrate Drugs 0.000 description 3
- 238000012360 testing method Methods 0.000 description 3
- 210000001519 tissue Anatomy 0.000 description 3
- 235000010215 titanium dioxide Nutrition 0.000 description 3
- 230000000699 topical effect Effects 0.000 description 3
- 239000003981 vehicle Substances 0.000 description 3
- 150000004823 xylans Chemical class 0.000 description 3
- HDTRYLNUVZCQOY-UHFFFAOYSA-N α-D-glucopyranosyl-α-D-glucopyranoside Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(O)C(O)C(CO)O1 HDTRYLNUVZCQOY-UHFFFAOYSA-N 0.000 description 2
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- DNIAPMSPPWPWGF-VKHMYHEASA-N (+)-propylene glycol Chemical compound C[C@H](O)CO DNIAPMSPPWPWGF-VKHMYHEASA-N 0.000 description 2
- DNIAPMSPPWPWGF-GSVOUGTGSA-N (R)-(-)-Propylene glycol Chemical compound C[C@@H](O)CO DNIAPMSPPWPWGF-GSVOUGTGSA-N 0.000 description 2
- 229940015975 1,2-hexanediol Drugs 0.000 description 2
- YPFDHNVEDLHUCE-UHFFFAOYSA-N 1,3-propanediol Substances OCCCO YPFDHNVEDLHUCE-UHFFFAOYSA-N 0.000 description 2
- CKCRUEKHEVDQDC-DSVABHIPSA-N 1-hydroxy-1-[(2r,3r,4s,5r,6r)-3,4,5-triacetyl-3,4,5,6-tetrahydroxyoxan-2-yl]propan-2-one Chemical compound CC(=O)C(O)[C@H]1O[C@@H](O)[C@@](O)(C(C)=O)[C@](O)(C(C)=O)[C@@]1(O)C(C)=O CKCRUEKHEVDQDC-DSVABHIPSA-N 0.000 description 2
- ARXJGSRGQADJSQ-UHFFFAOYSA-N 1-methoxypropan-2-ol Chemical compound COCC(C)O ARXJGSRGQADJSQ-UHFFFAOYSA-N 0.000 description 2
- CPOBTYJRKAJERX-UHFFFAOYSA-N 3-ethyl-n-[(3-ethyl-1,3-benzothiazol-2-ylidene)amino]-1,3-benzothiazol-2-imine Chemical compound S1C2=CC=CC=C2N(CC)C1=NN=C1SC2=CC=CC=C2N1CC CPOBTYJRKAJERX-UHFFFAOYSA-N 0.000 description 2
- DBTMGCOVALSLOR-UHFFFAOYSA-N 32-alpha-galactosyl-3-alpha-galactosyl-galactose Natural products OC1C(O)C(O)C(CO)OC1OC1C(O)C(OC2C(C(CO)OC(O)C2O)O)OC(CO)C1O DBTMGCOVALSLOR-UHFFFAOYSA-N 0.000 description 2
- HGINCPLSRVDWNT-UHFFFAOYSA-N Acrolein Chemical compound C=CC=O HGINCPLSRVDWNT-UHFFFAOYSA-N 0.000 description 2
- 241000589158 Agrobacterium Species 0.000 description 2
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 2
- QNIACYURSSCLRP-GUBZILKMSA-N Asp-Lys-Gln Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(O)=O QNIACYURSSCLRP-GUBZILKMSA-N 0.000 description 2
- VSMYBNPOHYAXSD-GUBZILKMSA-N Asp-Lys-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O VSMYBNPOHYAXSD-GUBZILKMSA-N 0.000 description 2
- 241000228212 Aspergillus Species 0.000 description 2
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 2
- 241000186146 Brevibacterium Species 0.000 description 2
- 241000222120 Candida <Saccharomycetales> Species 0.000 description 2
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical compound O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 2
- GHXZTYHSJHQHIJ-UHFFFAOYSA-N Chlorhexidine Chemical compound C=1C=C(Cl)C=CC=1NC(N)=NC(N)=NCCCCCCN=C(N)N=C(N)NC1=CC=C(Cl)C=C1 GHXZTYHSJHQHIJ-UHFFFAOYSA-N 0.000 description 2
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 2
- SRBFZHDQGSBBOR-IOVATXLUSA-N D-xylopyranose Chemical compound O[C@@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-IOVATXLUSA-N 0.000 description 2
- KRHYYFGTRYWZRS-UHFFFAOYSA-M Fluoride anion Chemical compound [F-] KRHYYFGTRYWZRS-UHFFFAOYSA-M 0.000 description 2
- 241000233866 Fungi Species 0.000 description 2
- AALUCPRYHRPMAG-UHFFFAOYSA-N Glycerol 1-propanoate Chemical compound CCC(=O)OCC(O)CO AALUCPRYHRPMAG-UHFFFAOYSA-N 0.000 description 2
- 108010093096 Immobilized Enzymes Proteins 0.000 description 2
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 2
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 2
- 241000235649 Kluyveromyces Species 0.000 description 2
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 2
- 229930195725 Mannitol Natural products 0.000 description 2
- ALQSHHUCVQOPAS-UHFFFAOYSA-N Pentane-1,5-diol Chemical compound OCCCCCO ALQSHHUCVQOPAS-UHFFFAOYSA-N 0.000 description 2
- 108700020962 Peroxidase Proteins 0.000 description 2
- 241001542817 Phaffia Species 0.000 description 2
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 2
- 229920001214 Polysorbate 60 Polymers 0.000 description 2
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 2
- MUPFEKGTMRGPLJ-RMMQSMQOSA-N Raffinose Natural products O(C[C@H]1[C@@H](O)[C@H](O)[C@@H](O)[C@@H](O[C@@]2(CO)[C@H](O)[C@@H](O)[C@@H](CO)O2)O1)[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 MUPFEKGTMRGPLJ-RMMQSMQOSA-N 0.000 description 2
- 241000193448 Ruminiclostridium thermocellum Species 0.000 description 2
- 241000235070 Saccharomyces Species 0.000 description 2
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 2
- 241000607142 Salmonella Species 0.000 description 2
- 238000012300 Sequence Analysis Methods 0.000 description 2
- IUXGJEIKJBYKOO-SRVKXCTJSA-N Ser-Leu-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CO)N IUXGJEIKJBYKOO-SRVKXCTJSA-N 0.000 description 2
- QPPYAWVLAVXISR-DCAQKATOSA-N Ser-Pro-His Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CO)N)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O QPPYAWVLAVXISR-DCAQKATOSA-N 0.000 description 2
- 241000736131 Sphingomonas Species 0.000 description 2
- 241000194019 Streptococcus mutans Species 0.000 description 2
- 241000187747 Streptomyces Species 0.000 description 2
- 108010056079 Subtilisins Proteins 0.000 description 2
- 102000005158 Subtilisins Human genes 0.000 description 2
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 2
- 229930006000 Sucrose Natural products 0.000 description 2
- 244000269722 Thea sinensis Species 0.000 description 2
- 241001137870 Thermoanaerobacterium Species 0.000 description 2
- 241000204666 Thermotoga maritima Species 0.000 description 2
- HJOSVGCWOTYJFG-WDCWCFNPSA-N Thr-Glu-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N)O HJOSVGCWOTYJFG-WDCWCFNPSA-N 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 239000004473 Threonine Substances 0.000 description 2
- 206010044032 Tooth discolouration Diseases 0.000 description 2
- HDTRYLNUVZCQOY-WSWWMNSNSA-N Trehalose Natural products O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-WSWWMNSNSA-N 0.000 description 2
- 241000223259 Trichoderma Species 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- MUPFEKGTMRGPLJ-UHFFFAOYSA-N UNPD196149 Natural products OC1C(O)C(CO)OC1(CO)OC1C(O)C(O)C(O)C(COC2C(C(O)C(O)C(CO)O2)O)O1 MUPFEKGTMRGPLJ-UHFFFAOYSA-N 0.000 description 2
- 239000001083 [(2R,3R,4S,5R)-1,2,4,5-tetraacetyloxy-6-oxohexan-3-yl] acetate Substances 0.000 description 2
- IHNHAHWGVLXCCI-PFGBXZAXSA-N [(2r,3r,4r)-3,4,5-triacetyloxyoxolan-2-yl]methyl acetate Chemical compound CC(=O)OC[C@H]1OC(OC(C)=O)[C@H](OC(C)=O)[C@@H]1OC(C)=O IHNHAHWGVLXCCI-PFGBXZAXSA-N 0.000 description 2
- SHBHJRVMGYVXKK-DYPLGBCKSA-N [(2r,3r,4r,5s)-3,4,6-triacetyloxy-5-hydroxyoxan-2-yl]methyl acetate Chemical compound CC(=O)OC[C@H]1OC(OC(C)=O)[C@@H](O)[C@@H](OC(C)=O)[C@@H]1OC(C)=O SHBHJRVMGYVXKK-DYPLGBCKSA-N 0.000 description 2
- UAOKXEHOENRFMP-ZJIFWQFVSA-N [(2r,3r,4s,5r)-2,3,4,5-tetraacetyloxy-6-oxohexyl] acetate Chemical compound CC(=O)OC[C@@H](OC(C)=O)[C@@H](OC(C)=O)[C@H](OC(C)=O)[C@@H](OC(C)=O)C=O UAOKXEHOENRFMP-ZJIFWQFVSA-N 0.000 description 2
- MJOQJPYNENPSSS-PFGBXZAXSA-N [(3r,4r,5r)-4,5,6-triacetyloxyoxan-3-yl] acetate Chemical compound CC(=O)O[C@@H]1COC(OC(C)=O)[C@H](OC(C)=O)[C@@H]1OC(C)=O MJOQJPYNENPSSS-PFGBXZAXSA-N 0.000 description 2
- 125000002777 acetyl group Chemical group [H]C([H])([H])C(*)=O 0.000 description 2
- 150000001298 alcohols Chemical class 0.000 description 2
- HDTRYLNUVZCQOY-LIZSDCNHSA-N alpha,alpha-trehalose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 HDTRYLNUVZCQOY-LIZSDCNHSA-N 0.000 description 2
- 230000000692 anti-sense effect Effects 0.000 description 2
- 239000012062 aqueous buffer Substances 0.000 description 2
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 2
- 238000000889 atomisation Methods 0.000 description 2
- 239000002585 base Substances 0.000 description 2
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 2
- AFYNADDZULBEJA-UHFFFAOYSA-N bicinchoninic acid Chemical compound C1=CC=CC2=NC(C=3C=C(C4=CC=CC=C4N=3)C(=O)O)=CC(C(O)=O)=C21 AFYNADDZULBEJA-UHFFFAOYSA-N 0.000 description 2
- 230000033228 biological regulation Effects 0.000 description 2
- 229940098773 bovine serum albumin Drugs 0.000 description 2
- 239000007853 buffer solution Substances 0.000 description 2
- BMRWNKZVCUKKSR-UHFFFAOYSA-N butane-1,2-diol Chemical compound CCC(O)CO BMRWNKZVCUKKSR-UHFFFAOYSA-N 0.000 description 2
- OWBTYPJTUOEWEK-UHFFFAOYSA-N butane-2,3-diol Chemical compound CC(O)C(C)O OWBTYPJTUOEWEK-UHFFFAOYSA-N 0.000 description 2
- 229940078916 carbamide peroxide Drugs 0.000 description 2
- 239000011203 carbon fibre reinforced carbon Substances 0.000 description 2
- 230000015556 catabolic process Effects 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 229920002678 cellulose Polymers 0.000 description 2
- 235000010980 cellulose Nutrition 0.000 description 2
- HOKIDJSKDBPKTQ-GLXFQSAKSA-N cephalosporin C Chemical compound S1CC(COC(=O)C)=C(C(O)=O)N2C(=O)[C@@H](NC(=O)CCC[C@@H](N)C(O)=O)[C@@H]12 HOKIDJSKDBPKTQ-GLXFQSAKSA-N 0.000 description 2
- 229960003260 chlorhexidine Drugs 0.000 description 2
- 239000010941 cobalt Substances 0.000 description 2
- 229910017052 cobalt Inorganic materials 0.000 description 2
- GUTLYIVDDKVIGB-UHFFFAOYSA-N cobalt atom Chemical compound [Co] GUTLYIVDDKVIGB-UHFFFAOYSA-N 0.000 description 2
- 239000002299 complementary DNA Substances 0.000 description 2
- 238000012136 culture method Methods 0.000 description 2
- 125000004122 cyclic group Chemical group 0.000 description 2
- JHIVVAPYMSGYDF-UHFFFAOYSA-N cyclohexanone Chemical compound O=C1CCCCC1 JHIVVAPYMSGYDF-UHFFFAOYSA-N 0.000 description 2
- 230000006378 damage Effects 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- 239000008367 deionised water Substances 0.000 description 2
- 229910021641 deionized water Inorganic materials 0.000 description 2
- 230000008021 deposition Effects 0.000 description 2
- 150000005690 diesters Chemical class 0.000 description 2
- OSVXSBDYLRYLIG-UHFFFAOYSA-N dioxidochlorine(.) Chemical compound O=Cl=O OSVXSBDYLRYLIG-UHFFFAOYSA-N 0.000 description 2
- 239000011363 dried mixture Substances 0.000 description 2
- 239000000839 emulsion Substances 0.000 description 2
- 239000006167 equilibration buffer Substances 0.000 description 2
- LZCLXQDLBQLTDK-UHFFFAOYSA-N ethyl 2-hydroxypropanoate Chemical compound CCOC(=O)C(C)O LZCLXQDLBQLTDK-UHFFFAOYSA-N 0.000 description 2
- OMSUIQOIVADKIM-UHFFFAOYSA-N ethyl 3-hydroxybutyrate Chemical compound CCOC(=O)CC(C)O OMSUIQOIVADKIM-UHFFFAOYSA-N 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 230000001747 exhibiting effect Effects 0.000 description 2
- 239000000796 flavoring agent Substances 0.000 description 2
- 229940091249 fluoride supplement Drugs 0.000 description 2
- 238000011010 flushing procedure Methods 0.000 description 2
- 235000013305 food Nutrition 0.000 description 2
- 235000013355 food flavoring agent Nutrition 0.000 description 2
- 238000004817 gas chromatography Methods 0.000 description 2
- 108010078144 glutaminyl-glycine Proteins 0.000 description 2
- 125000003976 glyceryl group Chemical group [H]C([*])([H])C(O[H])([H])C(O[H])([H])[H] 0.000 description 2
- 239000001963 growth medium Substances 0.000 description 2
- 150000004820 halides Chemical class 0.000 description 2
- 238000010438 heat treatment Methods 0.000 description 2
- FHKSXSQHXQEMOK-UHFFFAOYSA-N hexane-1,2-diol Chemical compound CCCCC(O)CO FHKSXSQHXQEMOK-UHFFFAOYSA-N 0.000 description 2
- XXMIOPMDWAUFGU-UHFFFAOYSA-N hexane-1,6-diol Chemical compound OCCCCCCO XXMIOPMDWAUFGU-UHFFFAOYSA-N 0.000 description 2
- OHMBHFSEKCCCBW-UHFFFAOYSA-N hexane-2,5-diol Chemical compound CC(O)CCC(C)O OHMBHFSEKCCCBW-UHFFFAOYSA-N 0.000 description 2
- 230000036571 hydration Effects 0.000 description 2
- 238000006703 hydration reaction Methods 0.000 description 2
- 125000004435 hydrogen atom Chemical group [H]* 0.000 description 2
- WQYVRQLZKVEZGA-UHFFFAOYSA-N hypochlorite Chemical compound Cl[O-] WQYVRQLZKVEZGA-UHFFFAOYSA-N 0.000 description 2
- ZCZCOXLLICTZAH-UHFFFAOYSA-N hypothiocyanous acid Chemical compound OSC#N ZCZCOXLLICTZAH-UHFFFAOYSA-N 0.000 description 2
- 239000003622 immobilized catalyst Substances 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 2
- 230000003834 intracellular effect Effects 0.000 description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 2
- 229960000310 isoleucine Drugs 0.000 description 2
- 239000008101 lactose Substances 0.000 description 2
- 230000007774 longterm Effects 0.000 description 2
- 239000000594 mannitol Substances 0.000 description 2
- 235000010355 mannitol Nutrition 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- 108020004999 messenger RNA Proteins 0.000 description 2
- 229910052751 metal Inorganic materials 0.000 description 2
- 239000002184 metal Substances 0.000 description 2
- VNWKTOKETHGBQD-UHFFFAOYSA-N methane Chemical compound C VNWKTOKETHGBQD-UHFFFAOYSA-N 0.000 description 2
- 229930182817 methionine Natural products 0.000 description 2
- 230000003278 mimic effect Effects 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 238000007899 nucleic acid hybridization Methods 0.000 description 2
- 150000002894 organic compounds Chemical class 0.000 description 2
- 239000003960 organic solvent Substances 0.000 description 2
- 238000004806 packaging method and process Methods 0.000 description 2
- WCVRQHFDJLLWFE-UHFFFAOYSA-N pentane-1,2-diol Chemical compound CCCC(O)CO WCVRQHFDJLLWFE-UHFFFAOYSA-N 0.000 description 2
- GLOBUAZSRIOKLN-UHFFFAOYSA-N pentane-1,4-diol Chemical compound CC(O)CCCO GLOBUAZSRIOKLN-UHFFFAOYSA-N 0.000 description 2
- 108040007629 peroxidase activity proteins Proteins 0.000 description 2
- XNGIFLGASWRNHJ-UHFFFAOYSA-N phthalic acid Chemical compound OC(=O)C1=CC=CC=C1C(O)=O XNGIFLGASWRNHJ-UHFFFAOYSA-N 0.000 description 2
- 229920005862 polyol Polymers 0.000 description 2
- 150000003077 polyols Chemical class 0.000 description 2
- 229920000166 polytrimethylene carbonate Polymers 0.000 description 2
- 239000003755 preservative agent Substances 0.000 description 2
- MUPFEKGTMRGPLJ-ZQSKZDJDSA-N raffinose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO[C@@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)O)O1 MUPFEKGTMRGPLJ-ZQSKZDJDSA-N 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 210000003705 ribosome Anatomy 0.000 description 2
- 230000003248 secreting effect Effects 0.000 description 2
- 239000007974 sodium acetate buffer Substances 0.000 description 2
- PUZPDOWCWNUUKD-UHFFFAOYSA-M sodium fluoride Chemical compound [F-].[Na+] PUZPDOWCWNUUKD-UHFFFAOYSA-M 0.000 description 2
- 238000005507 spraying Methods 0.000 description 2
- 239000005720 sucrose Substances 0.000 description 2
- 150000008163 sugars Chemical class 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- MGSRCZKZVOBKFT-UHFFFAOYSA-N thymol Chemical compound CC(C)C1=CC=C(C)C=C1O MGSRCZKZVOBKFT-UHFFFAOYSA-N 0.000 description 2
- 239000004408 titanium dioxide Substances 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- PZSJOBKRSVRODF-UHFFFAOYSA-N vanillin acetate Chemical compound COC1=CC(C=O)=CC=C1OC(C)=O PZSJOBKRSVRODF-UHFFFAOYSA-N 0.000 description 2
- 238000011179 visual inspection Methods 0.000 description 2
- UAOKXEHOENRFMP-UHFFFAOYSA-N (2,3,4,5-tetraacetyloxy-6-oxohexyl) acetate Chemical compound CC(=O)OCC(OC(C)=O)C(OC(C)=O)C(OC(C)=O)C(OC(C)=O)C=O UAOKXEHOENRFMP-UHFFFAOYSA-N 0.000 description 1
- OMDQUFIYNPYJFM-XKDAHURESA-N (2r,3r,4s,5r,6s)-2-(hydroxymethyl)-6-[[(2r,3s,4r,5s,6r)-4,5,6-trihydroxy-3-[(2s,3s,4s,5s,6r)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyoxan-2-yl]methoxy]oxane-3,4,5-triol Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1OC[C@@H]1[C@@H](O[C@H]2[C@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)[C@H](O)[C@H](O)[C@H](O)O1 OMDQUFIYNPYJFM-XKDAHURESA-N 0.000 description 1
- PPLUPPGMFOKIQT-UHFFFAOYSA-N (3-hydroxy-2-propanoyloxypropyl) propanoate Chemical compound CCC(=O)OCC(CO)OC(=O)CC PPLUPPGMFOKIQT-UHFFFAOYSA-N 0.000 description 1
- BHQCQFFYRZLCQQ-UHFFFAOYSA-N (3alpha,5alpha,7alpha,12alpha)-3,7,12-trihydroxy-cholan-24-oic acid Natural products OC1CC2CC(O)CCC2(C)C2C1C1CCC(C(CCC(O)=O)C)C1(C)C(O)C2 BHQCQFFYRZLCQQ-UHFFFAOYSA-N 0.000 description 1
- FQVLRGLGWNWPSS-BXBUPLCLSA-N (4r,7s,10s,13s,16r)-16-acetamido-13-(1h-imidazol-5-ylmethyl)-10-methyl-6,9,12,15-tetraoxo-7-propan-2-yl-1,2-dithia-5,8,11,14-tetrazacycloheptadecane-4-carboxamide Chemical compound N1C(=O)[C@@H](NC(C)=O)CSSC[C@@H](C(N)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C)NC(=O)[C@@H]1CC1=CN=CN1 FQVLRGLGWNWPSS-BXBUPLCLSA-N 0.000 description 1
- QAVITTVTXPZTSE-UHFFFAOYSA-N (5-formylfuran-2-yl)methyl acetate Chemical compound CC(=O)OCC1=CC=C(C=O)O1 QAVITTVTXPZTSE-UHFFFAOYSA-N 0.000 description 1
- AWHAUPZHZYUHOM-UHFFFAOYSA-N 1,2-dibutyrin Chemical compound CCCC(=O)OCC(CO)OC(=O)CCC AWHAUPZHZYUHOM-UHFFFAOYSA-N 0.000 description 1
- DSVGICPKBRQDDX-UHFFFAOYSA-N 1,3-diacetoxypropane Chemical compound CC(=O)OCCCOC(C)=O DSVGICPKBRQDDX-UHFFFAOYSA-N 0.000 description 1
- LDVVTQMJQSCDMK-UHFFFAOYSA-N 1,3-dihydroxypropan-2-yl formate Chemical compound OCC(CO)OC=O LDVVTQMJQSCDMK-UHFFFAOYSA-N 0.000 description 1
- XUKSWKGOQKREON-UHFFFAOYSA-N 1,4-diacetoxybutane Chemical compound CC(=O)OCCCCOC(C)=O XUKSWKGOQKREON-UHFFFAOYSA-N 0.000 description 1
- WEEGYLXZBRQIMU-UHFFFAOYSA-N 1,8-cineole Natural products C1CC2CCC1(C)OC2(C)C WEEGYLXZBRQIMU-UHFFFAOYSA-N 0.000 description 1
- RWNUSVWFHDHRCJ-UHFFFAOYSA-N 1-butoxypropan-2-ol Chemical compound CCCCOCC(C)O RWNUSVWFHDHRCJ-UHFFFAOYSA-N 0.000 description 1
- SOFBKAHFJMHMPJ-UHFFFAOYSA-N 1-hydroxyethyl hypochlorite Chemical compound CC(O)OCl SOFBKAHFJMHMPJ-UHFFFAOYSA-N 0.000 description 1
- IIZPXYDJLKNOIY-JXPKJXOSSA-N 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphocholine Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCC\C=C/C\C=C/C\C=C/C\C=C/CCCCC IIZPXYDJLKNOIY-JXPKJXOSSA-N 0.000 description 1
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 1
- OAYXUHPQHDHDDZ-UHFFFAOYSA-N 2-(2-butoxyethoxy)ethanol Chemical compound CCCCOCCOCCO OAYXUHPQHDHDDZ-UHFFFAOYSA-N 0.000 description 1
- SBASXUCJHJRPEV-UHFFFAOYSA-N 2-(2-methoxyethoxy)ethanol Chemical compound COCCOCCO SBASXUCJHJRPEV-UHFFFAOYSA-N 0.000 description 1
- CUDYYMUUJHLCGZ-UHFFFAOYSA-N 2-(2-methoxypropoxy)propan-1-ol Chemical compound COC(C)COC(C)CO CUDYYMUUJHLCGZ-UHFFFAOYSA-N 0.000 description 1
- WAEVWDZKMBQDEJ-UHFFFAOYSA-N 2-[2-(2-methoxypropoxy)propoxy]propan-1-ol Chemical compound COC(C)COC(C)COC(C)CO WAEVWDZKMBQDEJ-UHFFFAOYSA-N 0.000 description 1
- MWWXARALRVYLAE-UHFFFAOYSA-N 2-acetyloxybut-3-enyl acetate Chemical compound CC(=O)OCC(C=C)OC(C)=O MWWXARALRVYLAE-UHFFFAOYSA-N 0.000 description 1
- LJCNDNBULVLKSG-UHFFFAOYSA-N 2-aminoacetic acid;butane Chemical compound CCCC.CCCC.NCC(O)=O LJCNDNBULVLKSG-UHFFFAOYSA-N 0.000 description 1
- IVLXQGJVBGMLRR-UHFFFAOYSA-N 2-aminoacetic acid;hydron;chloride Chemical compound Cl.NCC(O)=O IVLXQGJVBGMLRR-UHFFFAOYSA-N 0.000 description 1
- ICPWFHKNYYRBSZ-UHFFFAOYSA-M 2-methoxypropanoate Chemical compound COC(C)C([O-])=O ICPWFHKNYYRBSZ-UHFFFAOYSA-M 0.000 description 1
- XGRSAFKZAGGXJV-UHFFFAOYSA-N 3-azaniumyl-3-cyclohexylpropanoate Chemical compound OC(=O)CC(N)C1CCCCC1 XGRSAFKZAGGXJV-UHFFFAOYSA-N 0.000 description 1
- GDBUZIKSJGRBJP-UHFFFAOYSA-N 4-acetoxy benzoic acid Chemical compound CC(=O)OC1=CC=C(C(O)=O)C=C1 GDBUZIKSJGRBJP-UHFFFAOYSA-N 0.000 description 1
- OSDLLIBGSJNGJE-UHFFFAOYSA-N 4-chloro-3,5-dimethylphenol Chemical compound CC1=CC(O)=CC(C)=C1Cl OSDLLIBGSJNGJE-UHFFFAOYSA-N 0.000 description 1
- COYZTUXPHRHKGV-UHFFFAOYSA-N 4-methoxy-5-[3-(2-methoxy-4-nitro-5-sulfophenyl)-5-(phenylcarbamoyl)-1h-tetrazol-2-yl]-2-nitrobenzenesulfonic acid Chemical compound COC1=CC([N+]([O-])=O)=C(S(O)(=O)=O)C=C1N1N(C=2C(=CC(=C(C=2)S(O)(=O)=O)[N+]([O-])=O)OC)N=C(C(=O)NC=2C=CC=CC=2)N1 COYZTUXPHRHKGV-UHFFFAOYSA-N 0.000 description 1
- 101710163881 5,6-dihydroxyindole-2-carboxylic acid oxidase Proteins 0.000 description 1
- PIJBVCVBCQOWMM-UHFFFAOYSA-N 5-acetyloxypentyl acetate Chemical compound CC(=O)OCCCCCOC(C)=O PIJBVCVBCQOWMM-UHFFFAOYSA-N 0.000 description 1
- ODEHMIGXGLNAKK-OESPXIITSA-N 6-kestotriose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)OC[C@@H]1[C@@H](O)[C@H](O)[C@](CO)(O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)O1 ODEHMIGXGLNAKK-OESPXIITSA-N 0.000 description 1
- LPEKGGXMPWTOCB-UHFFFAOYSA-N 8beta-(2,3-epoxy-2-methylbutyryloxy)-14-acetoxytithifolin Natural products COC(=O)C(C)O LPEKGGXMPWTOCB-UHFFFAOYSA-N 0.000 description 1
- 240000000073 Achillea millefolium Species 0.000 description 1
- 235000007754 Achillea millefolium Nutrition 0.000 description 1
- 241000589291 Acinetobacter Species 0.000 description 1
- 241000588986 Alcaligenes Species 0.000 description 1
- 102100034035 Alcohol dehydrogenase 1A Human genes 0.000 description 1
- 108010025188 Alcohol oxidase Proteins 0.000 description 1
- 102100036826 Aldehyde oxidase Human genes 0.000 description 1
- 229920001450 Alpha-Cyclodextrin Polymers 0.000 description 1
- 229920000945 Amylopectin Polymers 0.000 description 1
- 241000192542 Anabaena Species 0.000 description 1
- 108700042778 Antimicrobial Peptides Proteins 0.000 description 1
- 102000044503 Antimicrobial Peptides Human genes 0.000 description 1
- 108020005544 Antisense RNA Proteins 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- PDECQIHABNQRHN-GUBZILKMSA-N Asp-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC(O)=O PDECQIHABNQRHN-GUBZILKMSA-N 0.000 description 1
- BSYNRYMUTXBXSQ-UHFFFAOYSA-N Aspirin Chemical compound CC(=O)OC1=CC=CC=C1C(O)=O BSYNRYMUTXBXSQ-UHFFFAOYSA-N 0.000 description 1
- 241001328122 Bacillus clausii Species 0.000 description 1
- 241000936493 Bacillus clausii KSM-K16 Species 0.000 description 1
- 241000423334 Bacillus halodurans C-125 Species 0.000 description 1
- 239000002028 Biomass Substances 0.000 description 1
- 208000035985 Body Odor Diseases 0.000 description 1
- 239000004343 Calcium peroxide Substances 0.000 description 1
- 101710128063 Carbohydrate oxidase Proteins 0.000 description 1
- BVKZGUZCCUSVTD-UHFFFAOYSA-L Carbonate Chemical compound [O-]C([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-L 0.000 description 1
- 229920002134 Carboxymethyl cellulose Polymers 0.000 description 1
- 108050004290 Cecropin Proteins 0.000 description 1
- 229930186147 Cephalosporin Natural products 0.000 description 1
- 229920001661 Chitosan Polymers 0.000 description 1
- ZAMOUSCENKQFHK-UHFFFAOYSA-N Chlorine atom Chemical compound [Cl] ZAMOUSCENKQFHK-UHFFFAOYSA-N 0.000 description 1
- 239000004155 Chlorine dioxide Substances 0.000 description 1
- 241000191366 Chlorobium Species 0.000 description 1
- 239000004380 Cholic acid Substances 0.000 description 1
- 241000190831 Chromatium Species 0.000 description 1
- 241000605056 Cytophaga Species 0.000 description 1
- GUBGYTABKSRVRQ-CUHNMECISA-N D-Cellobiose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-CUHNMECISA-N 0.000 description 1
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 1
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 1
- RXVWSYJTUUKTEA-UHFFFAOYSA-N D-maltotriose Natural products OC1C(O)C(OC(C(O)CO)C(O)C(O)C=O)OC(CO)C1OC1C(O)C(O)C(O)C(CO)O1 RXVWSYJTUUKTEA-UHFFFAOYSA-N 0.000 description 1
- 241000192093 Deinococcus Species 0.000 description 1
- 238000002965 ELISA Methods 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 241000588698 Erwinia Species 0.000 description 1
- 241000190844 Erythrobacter Species 0.000 description 1
- DBVJJBKOTRCVKF-UHFFFAOYSA-N Etidronic acid Chemical compound OP(=O)(O)C(O)(C)P(O)(O)=O DBVJJBKOTRCVKF-UHFFFAOYSA-N 0.000 description 1
- WEEGYLXZBRQIMU-WAAGHKOSSA-N Eucalyptol Chemical compound C1C[C@H]2CC[C@]1(C)OC2(C)C WEEGYLXZBRQIMU-WAAGHKOSSA-N 0.000 description 1
- 229920002444 Exopolysaccharide Polymers 0.000 description 1
- 241000589565 Flavobacterium Species 0.000 description 1
- 229920000855 Fucoidan Polymers 0.000 description 1
- 206010017533 Fungal infection Diseases 0.000 description 1
- 101150094690 GAL1 gene Proteins 0.000 description 1
- 101150038242 GAL10 gene Proteins 0.000 description 1
- 229920000926 Galactomannan Polymers 0.000 description 1
- 102100028501 Galanin peptides Human genes 0.000 description 1
- 102100024637 Galectin-10 Human genes 0.000 description 1
- 240000001238 Gaultheria procumbens Species 0.000 description 1
- 235000007297 Gaultheria procumbens Nutrition 0.000 description 1
- 241000193385 Geobacillus stearothermophilus Species 0.000 description 1
- 101000892220 Geobacillus thermodenitrificans (strain NG80-2) Long-chain-alcohol dehydrogenase 1 Proteins 0.000 description 1
- IOFDDSNZJDIGPB-GVXVVHGQSA-N Gln-Leu-Val Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O IOFDDSNZJDIGPB-GVXVVHGQSA-N 0.000 description 1
- KASDBWKLWJKTLJ-GUBZILKMSA-N Glu-Glu-Met Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCSC)C(O)=O KASDBWKLWJKTLJ-GUBZILKMSA-N 0.000 description 1
- 229920001503 Glucan Polymers 0.000 description 1
- 102100031181 Glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 description 1
- 239000004347 Glyceryl monoacetate Substances 0.000 description 1
- 102000005744 Glycoside Hydrolases Human genes 0.000 description 1
- 108010031186 Glycoside Hydrolases Proteins 0.000 description 1
- 101150009006 HIS3 gene Proteins 0.000 description 1
- 101100246753 Halobacterium salinarum (strain ATCC 700922 / JCM 11081 / NRC-1) pyrF gene Proteins 0.000 description 1
- 241000282412 Homo Species 0.000 description 1
- 101000780443 Homo sapiens Alcohol dehydrogenase 1A Proteins 0.000 description 1
- 101000928314 Homo sapiens Aldehyde oxidase Proteins 0.000 description 1
- 101100121078 Homo sapiens GAL gene Proteins 0.000 description 1
- 101001046426 Homo sapiens cGMP-dependent protein kinase 1 Proteins 0.000 description 1
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- 108090001061 Insulin Proteins 0.000 description 1
- 102000004877 Insulin Human genes 0.000 description 1
- 102000004195 Isomerases Human genes 0.000 description 1
- 108090000769 Isomerases Proteins 0.000 description 1
- 241000588748 Klebsiella Species 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- JVTAAEKCZFNVCJ-UHFFFAOYSA-M Lactate Chemical compound CC(O)C([O-])=O JVTAAEKCZFNVCJ-UHFFFAOYSA-M 0.000 description 1
- 241001147746 Lactobacillus delbrueckii subsp. lactis Species 0.000 description 1
- PVMPDMIKUVNOBD-CIUDSAMLSA-N Leu-Asp-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O PVMPDMIKUVNOBD-CIUDSAMLSA-N 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 102000004317 Lyases Human genes 0.000 description 1
- 108090000856 Lyases Proteins 0.000 description 1
- FZIJIFCXUCZHOL-CIUDSAMLSA-N Lys-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN FZIJIFCXUCZHOL-CIUDSAMLSA-N 0.000 description 1
- DIBZLYZXTSVGLN-CIUDSAMLSA-N Lys-Ser-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O DIBZLYZXTSVGLN-CIUDSAMLSA-N 0.000 description 1
- 108060003100 Magainin Proteins 0.000 description 1
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 1
- 229920000057 Mannan Polymers 0.000 description 1
- 241000589195 Mesorhizobium loti Species 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- 241000202974 Methanobacterium Species 0.000 description 1
- LDLDJEAVRNAEBW-UHFFFAOYSA-N Methyl 3-hydroxybutyrate Chemical compound COC(=O)CC(C)O LDLDJEAVRNAEBW-UHFFFAOYSA-N 0.000 description 1
- 241000589350 Methylobacter Species 0.000 description 1
- 241000589345 Methylococcus Species 0.000 description 1
- 241000589966 Methylocystis Species 0.000 description 1
- 241001533203 Methylomicrobium Species 0.000 description 1
- 241000589344 Methylomonas Species 0.000 description 1
- 241000589354 Methylosinus Species 0.000 description 1
- 239000004909 Moisturizer Substances 0.000 description 1
- 208000031888 Mycoses Diseases 0.000 description 1
- 241000863420 Myxococcus Species 0.000 description 1
- 208000025157 Oral disease Diseases 0.000 description 1
- 240000007594 Oryza sativa Species 0.000 description 1
- 235000007164 Oryza sativa Nutrition 0.000 description 1
- CBENFWSGALASAD-UHFFFAOYSA-N Ozone Chemical compound [O-][O+]=O CBENFWSGALASAD-UHFFFAOYSA-N 0.000 description 1
- 101150012394 PHO5 gene Proteins 0.000 description 1
- 241000520272 Pantoea Species 0.000 description 1
- 241000233805 Phoenix Species 0.000 description 1
- 108091007643 Phosphate carriers Proteins 0.000 description 1
- OAICVXFJPJFONN-UHFFFAOYSA-N Phosphorus Chemical compound [P] OAICVXFJPJFONN-UHFFFAOYSA-N 0.000 description 1
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 1
- 239000004698 Polyethylene Substances 0.000 description 1
- 229920002593 Polyethylene Glycol 800 Polymers 0.000 description 1
- 239000004743 Polypropylene Substances 0.000 description 1
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 1
- 102000029797 Prion Human genes 0.000 description 1
- 108091000054 Prion Proteins 0.000 description 1
- 108010076504 Protein Sorting Signals Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 241000191025 Rhodobacter Species 0.000 description 1
- 101100394989 Rhodopseudomonas palustris (strain ATCC BAA-98 / CGA009) hisI gene Proteins 0.000 description 1
- 101100434411 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) ADH1 gene Proteins 0.000 description 1
- 101001000154 Schistosoma mansoni Phosphoglycerate kinase Proteins 0.000 description 1
- 206010040904 Skin odour abnormal Diseases 0.000 description 1
- 244000061458 Solanum melongena Species 0.000 description 1
- 235000002597 Solanum melongena Nutrition 0.000 description 1
- UQZIYBXSHAGNOE-USOSMYMVSA-N Stachyose Natural products O(C[C@H]1[C@@H](O)[C@H](O)[C@H](O)[C@@H](O[C@@]2(CO)[C@H](O)[C@@H](O)[C@@H](CO)O2)O1)[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@H](CO[C@@H]2[C@@H](O)[C@@H](O)[C@@H](O)[C@H](CO)O2)O1 UQZIYBXSHAGNOE-USOSMYMVSA-N 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- NINIDFKCEFEMDL-UHFFFAOYSA-N Sulfur Chemical compound [S] NINIDFKCEFEMDL-UHFFFAOYSA-N 0.000 description 1
- 241000192707 Synechococcus Species 0.000 description 1
- 241000192584 Synechocystis Species 0.000 description 1
- 108700005078 Synthetic Genes Proteins 0.000 description 1
- 241000123734 Thermoanaerobacterium sp. Species 0.000 description 1
- 241000204652 Thermotoga Species 0.000 description 1
- 241000204664 Thermotoga neapolitana Species 0.000 description 1
- 241000334089 Thermotoga petrophila Species 0.000 description 1
- 241000605118 Thiobacillus Species 0.000 description 1
- MGJLBZFUXUGMML-VOAKCMCISA-N Thr-Lys-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N)O MGJLBZFUXUGMML-VOAKCMCISA-N 0.000 description 1
- NYQIZWROIMIQSL-VEVYYDQMSA-N Thr-Pro-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O NYQIZWROIMIQSL-VEVYYDQMSA-N 0.000 description 1
- 239000005844 Thymol Substances 0.000 description 1
- ATJFFYVFTNAWJD-UHFFFAOYSA-N Tin Chemical compound [Sn] ATJFFYVFTNAWJD-UHFFFAOYSA-N 0.000 description 1
- 208000004509 Tooth Discoloration Diseases 0.000 description 1
- 108700019146 Transgenes Proteins 0.000 description 1
- XEFQLINVKFYRCS-UHFFFAOYSA-N Triclosan Chemical compound OC1=CC(Cl)=CC=C1OC1=CC=C(Cl)C=C1Cl XEFQLINVKFYRCS-UHFFFAOYSA-N 0.000 description 1
- 229920004890 Triton X-100 Polymers 0.000 description 1
- 101150050575 URA3 gene Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- 241000235013 Yarrowia Species 0.000 description 1
- 241000588901 Zymomonas Species 0.000 description 1
- IEOLRPPTIGNUNP-RKQHYHRCSA-N [(2r,3r,4s,5r,6r)-3,4,5-triacetyloxy-6-hydroxyoxan-2-yl]methyl acetate Chemical compound CC(=O)OC[C@H]1O[C@@H](O)[C@H](OC(C)=O)[C@@H](OC(C)=O)[C@@H]1OC(C)=O IEOLRPPTIGNUNP-RKQHYHRCSA-N 0.000 description 1
- NJVBTKVPPOFGAT-XMTFNYHQSA-N [(2s,3r,4r,5r)-2,3,4,5,6-pentaacetyloxyhexyl] acetate Chemical compound CC(=O)OC[C@@H](OC(C)=O)[C@@H](OC(C)=O)[C@H](OC(C)=O)[C@@H](OC(C)=O)COC(C)=O NJVBTKVPPOFGAT-XMTFNYHQSA-N 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 239000003082 abrasive agent Substances 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- 150000001242 acetic acid derivatives Chemical class 0.000 description 1
- 230000021736 acetylation Effects 0.000 description 1
- 238000006640 acetylation reaction Methods 0.000 description 1
- 125000003668 acetyloxy group Chemical group [H]C([H])([H])C(=O)O[*] 0.000 description 1
- 229960001138 acetylsalicylic acid Drugs 0.000 description 1
- 238000010306 acid treatment Methods 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 239000012190 activator Substances 0.000 description 1
- 125000002252 acyl group Chemical group 0.000 description 1
- 101150102866 adc1 gene Proteins 0.000 description 1
- 239000000853 adhesive Substances 0.000 description 1
- 230000001070 adhesive effect Effects 0.000 description 1
- 230000032683 aging Effects 0.000 description 1
- 150000001299 aldehydes Chemical class 0.000 description 1
- 239000003513 alkali Substances 0.000 description 1
- DBTMGCOVALSLOR-VXXRBQRTSA-N alpha-D-Glcp-(1->3)-alpha-D-Glcp-(1->3)-D-Glcp Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](O)[C@@H](O[C@H]2[C@@H]([C@@H](CO)OC(O)[C@@H]2O)O)O[C@H](CO)[C@H]1O DBTMGCOVALSLOR-VXXRBQRTSA-N 0.000 description 1
- WQZGKKKJIJFFOK-UHFFFAOYSA-N alpha-D-glucopyranose Natural products OCC1OC(O)C(O)C(O)C1O WQZGKKKJIJFFOK-UHFFFAOYSA-N 0.000 description 1
- HFHDHCJBZVLPGP-RWMJIURBSA-N alpha-cyclodextrin Chemical compound OC[C@H]([C@H]([C@@H]([C@H]1O)O)O[C@H]2O[C@@H]([C@@H](O[C@H]3O[C@H](CO)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](CO)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](CO)[C@H]([C@@H]([C@H]3O)O)O3)[C@H](O)[C@H]2O)CO)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O)[C@@H]3O[C@@H]1CO HFHDHCJBZVLPGP-RWMJIURBSA-N 0.000 description 1
- 229940043377 alpha-cyclodextrin Drugs 0.000 description 1
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 1
- 150000001408 amides Chemical class 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 230000006229 amino acid addition Effects 0.000 description 1
- 238000012197 amplification kit Methods 0.000 description 1
- 230000000181 anti-adherent effect Effects 0.000 description 1
- 230000000844 anti-bacterial effect Effects 0.000 description 1
- 239000002260 anti-inflammatory agent Substances 0.000 description 1
- 229940121363 anti-inflammatory agent Drugs 0.000 description 1
- 239000003911 antiadherent Substances 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 239000003963 antioxidant agent Substances 0.000 description 1
- 229940027983 antiseptic and disinfectant quaternary ammonium compound Drugs 0.000 description 1
- 238000013459 approach Methods 0.000 description 1
- PYMYPHUHKUWMLA-WDCZJNDASA-N arabinose Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)C=O PYMYPHUHKUWMLA-WDCZJNDASA-N 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 150000007860 aryl ester derivatives Chemical class 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 125000004429 atom Chemical group 0.000 description 1
- 239000003899 bactericide agent Substances 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QUYVBRFLSA-N beta-maltose Chemical compound OC[C@H]1O[C@H](O[C@H]2[C@H](O)[C@@H](O)[C@H](O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@@H]1O GUBGYTABKSRVRQ-QUYVBRFLSA-N 0.000 description 1
- 235000013361 beverage Nutrition 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 238000005415 bioluminescence Methods 0.000 description 1
- 230000029918 bioluminescence Effects 0.000 description 1
- 230000001851 biosynthetic effect Effects 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- OWMVSZAMULFTJU-UHFFFAOYSA-N bis-tris Chemical compound OCCN(CCO)C(CO)(CO)CO OWMVSZAMULFTJU-UHFFFAOYSA-N 0.000 description 1
- 229920001400 block copolymer Polymers 0.000 description 1
- 239000004067 bulking agent Substances 0.000 description 1
- BJUFAVXYJRJBLQ-UHFFFAOYSA-N butanoic acid;propane-1,2,3-triol Chemical compound CCCC(O)=O.OCC(O)CO BJUFAVXYJRJBLQ-UHFFFAOYSA-N 0.000 description 1
- 125000000484 butyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])C([H])([H])[H] 0.000 description 1
- 239000006227 byproduct Substances 0.000 description 1
- 102220359928 c.229A>G Human genes 0.000 description 1
- 102100022422 cGMP-dependent protein kinase 1 Human genes 0.000 description 1
- LHJQIRIGXXHNLA-UHFFFAOYSA-N calcium peroxide Chemical compound [Ca+2].[O-][O-] LHJQIRIGXXHNLA-UHFFFAOYSA-N 0.000 description 1
- 235000019402 calcium peroxide Nutrition 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 229940041514 candida albicans extract Drugs 0.000 description 1
- 229940077731 carbohydrate nutrients Drugs 0.000 description 1
- 239000001569 carbon dioxide Substances 0.000 description 1
- 229910002092 carbon dioxide Inorganic materials 0.000 description 1
- 239000011545 carbonate/bicarbonate buffer Substances 0.000 description 1
- 239000001768 carboxy methyl cellulose Substances 0.000 description 1
- 235000010948 carboxy methyl cellulose Nutrition 0.000 description 1
- 239000008112 carboxymethyl-cellulose Substances 0.000 description 1
- 239000012876 carrier material Substances 0.000 description 1
- 239000004359 castor oil Substances 0.000 description 1
- 235000019438 castor oil Nutrition 0.000 description 1
- 125000002091 cationic group Chemical group 0.000 description 1
- 238000007444 cell Immobilization Methods 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 239000013592 cell lysate Substances 0.000 description 1
- 239000001913 cellulose Substances 0.000 description 1
- 229920003086 cellulose ether Polymers 0.000 description 1
- 229940124587 cephalosporin Drugs 0.000 description 1
- 150000001780 cephalosporins Chemical class 0.000 description 1
- 229960001927 cetylpyridinium chloride Drugs 0.000 description 1
- NFCRBQADEGXVDL-UHFFFAOYSA-M cetylpyridinium chloride monohydrate Chemical compound O.[Cl-].CCCCCCCCCCCCCCCC[N+]1=CC=CC=C1 NFCRBQADEGXVDL-UHFFFAOYSA-M 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 239000002738 chelating agent Substances 0.000 description 1
- 230000009920 chelation Effects 0.000 description 1
- 239000003638 chemical reducing agent Substances 0.000 description 1
- 229940112822 chewing gum Drugs 0.000 description 1
- 235000015218 chewing gum Nutrition 0.000 description 1
- 239000000460 chlorine Substances 0.000 description 1
- 229910052801 chlorine Inorganic materials 0.000 description 1
- 235000019398 chlorine dioxide Nutrition 0.000 description 1
- 229960005443 chloroxylenol Drugs 0.000 description 1
- 235000019416 cholic acid Nutrition 0.000 description 1
- BHQCQFFYRZLCQQ-OELDTZBJSA-N cholic acid Chemical compound C([C@H]1C[C@H]2O)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(O)=O)C)[C@@]2(C)[C@@H](O)C1 BHQCQFFYRZLCQQ-OELDTZBJSA-N 0.000 description 1
- 229960002471 cholic acid Drugs 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 210000000349 chromosome Anatomy 0.000 description 1
- 229960005233 cineole Drugs 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 150000001860 citric acid derivatives Chemical class 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 238000004737 colorimetric analysis Methods 0.000 description 1
- 230000000295 complement effect Effects 0.000 description 1
- 239000003184 complementary RNA Substances 0.000 description 1
- 230000000536 complexating effect Effects 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 239000012141 concentrate Substances 0.000 description 1
- 239000003636 conditioned culture medium Substances 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 150000001879 copper Chemical class 0.000 description 1
- 239000003431 cross linking reagent Substances 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 125000001995 cyclobutyl group Chemical group [H]C1([H])C([H])([H])C([H])(*)C1([H])[H] 0.000 description 1
- HPXRVTGHNJAIIH-UHFFFAOYSA-N cyclohexanol Chemical compound OC1CCCCC1 HPXRVTGHNJAIIH-UHFFFAOYSA-N 0.000 description 1
- 125000000113 cyclohexyl group Chemical group [H]C1([H])C([H])([H])C([H])([H])C([H])(*)C([H])([H])C1([H])[H] 0.000 description 1
- 125000001511 cyclopentyl group Chemical group [H]C1([H])C([H])([H])C([H])([H])C([H])(*)C1([H])[H] 0.000 description 1
- 125000001559 cyclopropyl group Chemical group [H]C1([H])C([H])([H])C1([H])* 0.000 description 1
- 231100000599 cytotoxic agent Toxicity 0.000 description 1
- 229940127089 cytotoxic agent Drugs 0.000 description 1
- 239000002254 cytotoxic agent Substances 0.000 description 1
- 230000006196 deacetylation Effects 0.000 description 1
- 238000003381 deacetylation reaction Methods 0.000 description 1
- 230000000593 degrading effect Effects 0.000 description 1
- 239000000551 dentifrice Substances 0.000 description 1
- 239000003975 dentin desensitizing agent Substances 0.000 description 1
- KXGVEGMKQFWNSR-UHFFFAOYSA-N deoxycholic acid Natural products C1CC2CC(O)CCC2(C)C2C1C1CCC(C(CCC(O)=O)C)C1(C)C(O)C2 KXGVEGMKQFWNSR-UHFFFAOYSA-N 0.000 description 1
- 230000035614 depigmentation Effects 0.000 description 1
- 239000003599 detergent Substances 0.000 description 1
- 229940099371 diacetylated monoglycerides Drugs 0.000 description 1
- GUJOJGAPFQRJSV-UHFFFAOYSA-N dialuminum;dioxosilane;oxygen(2-);hydrate Chemical compound O.[O-2].[O-2].[O-2].[Al+3].[Al+3].O=[Si]=O.O=[Si]=O.O=[Si]=O.O=[Si]=O GUJOJGAPFQRJSV-UHFFFAOYSA-N 0.000 description 1
- 229910003460 diamond Inorganic materials 0.000 description 1
- 239000010432 diamond Substances 0.000 description 1
- 150000002009 diols Chemical class 0.000 description 1
- 238000007598 dipping method Methods 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 1
- VTIIJXUACCWYHX-UHFFFAOYSA-L disodium;carboxylatooxy carbonate Chemical compound [Na+].[Na+].[O-]C(=O)OOC([O-])=O VTIIJXUACCWYHX-UHFFFAOYSA-L 0.000 description 1
- 239000002270 dispersing agent Substances 0.000 description 1
- 239000012153 distilled water Substances 0.000 description 1
- 230000003828 downregulation Effects 0.000 description 1
- ODQWQRRAPPTVAG-GZTJUZNOSA-N doxepin Chemical compound C1OC2=CC=CC=C2C(=C/CCN(C)C)/C2=CC=CC=C21 ODQWQRRAPPTVAG-GZTJUZNOSA-N 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 238000001035 drying Methods 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 239000000975 dye Substances 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 230000007515 enzymatic degradation Effects 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- 238000001952 enzyme assay Methods 0.000 description 1
- 150000002168 ethanoic acid esters Chemical class 0.000 description 1
- ZANNOFHADGWOLI-UHFFFAOYSA-N ethyl 2-hydroxyacetate Chemical compound CCOC(=O)CO ZANNOFHADGWOLI-UHFFFAOYSA-N 0.000 description 1
- JLEKJZUYWFJPMB-UHFFFAOYSA-N ethyl 2-methoxyacetate Chemical compound CCOC(=O)COC JLEKJZUYWFJPMB-UHFFFAOYSA-N 0.000 description 1
- 125000001495 ethyl group Chemical group [H]C([H])([H])C([H])([H])* 0.000 description 1
- 229940116333 ethyl lactate Drugs 0.000 description 1
- DEFVIWRASFVYLL-UHFFFAOYSA-N ethylene glycol bis(2-aminoethyl)tetraacetic acid Chemical compound OC(=O)CN(CC(O)=O)CCOCCOCCN(CC(O)=O)CC(O)=O DEFVIWRASFVYLL-UHFFFAOYSA-N 0.000 description 1
- 238000001704 evaporation Methods 0.000 description 1
- 230000008020 evaporation Effects 0.000 description 1
- 230000005284 excitation Effects 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 239000012527 feed solution Substances 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- 230000037406 food intake Effects 0.000 description 1
- 235000003599 food sweetener Nutrition 0.000 description 1
- 238000007710 freezing Methods 0.000 description 1
- 230000008014 freezing Effects 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 238000012239 gene modification Methods 0.000 description 1
- 238000007429 general method Methods 0.000 description 1
- 230000005017 genetic modification Effects 0.000 description 1
- 235000013617 genetically modified food Nutrition 0.000 description 1
- 239000011521 glass Substances 0.000 description 1
- 230000009477 glass transition Effects 0.000 description 1
- 108010046301 glucose peroxidase Proteins 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 1
- 125000005456 glyceride group Chemical group 0.000 description 1
- 108010090622 glycerol oxidase Proteins 0.000 description 1
- ZEMPKEQAKRGZGQ-XOQCFJPHSA-N glycerol triricinoleate Natural products CCCCCC[C@@H](O)CC=CCCCCCCCC(=O)OC[C@@H](COC(=O)CCCCCCCC=CC[C@@H](O)CCCCCC)OC(=O)CCCCCCCC=CC[C@H](O)CCCCCC ZEMPKEQAKRGZGQ-XOQCFJPHSA-N 0.000 description 1
- 235000019442 glyceryl monoacetate Nutrition 0.000 description 1
- 239000008187 granular material Substances 0.000 description 1
- 230000005484 gravity Effects 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 231100000206 health hazard Toxicity 0.000 description 1
- 125000004051 hexyl group Chemical group [H]C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])* 0.000 description 1
- 239000003906 humectant Substances 0.000 description 1
- DLINORNFHVEIFE-UHFFFAOYSA-N hydrogen peroxide;zinc Chemical compound [Zn].OO DLINORNFHVEIFE-UHFFFAOYSA-N 0.000 description 1
- 230000002209 hydrophobic effect Effects 0.000 description 1
- 125000001165 hydrophobic group Chemical group 0.000 description 1
- 229920003063 hydroxymethyl cellulose Polymers 0.000 description 1
- 229940031574 hydroxymethyl cellulose Drugs 0.000 description 1
- 239000001866 hydroxypropyl methyl cellulose Substances 0.000 description 1
- 235000010979 hydroxypropyl methyl cellulose Nutrition 0.000 description 1
- 229920003088 hydroxypropyl methyl cellulose Polymers 0.000 description 1
- UFVKGYZPFZQRLF-UHFFFAOYSA-N hydroxypropyl methyl cellulose Chemical compound OC1C(O)C(OC)OC(CO)C1OC1C(O)C(O)C(OC2C(C(O)C(OC3C(C(O)C(O)C(CO)O3)O)C(CO)O2)O)C(CO)O1 UFVKGYZPFZQRLF-UHFFFAOYSA-N 0.000 description 1
- 238000005286 illumination Methods 0.000 description 1
- 210000001822 immobilized cell Anatomy 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 239000005414 inactive ingredient Substances 0.000 description 1
- 210000003000 inclusion body Anatomy 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 230000002401 inhibitory effect Effects 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 229910001410 inorganic ion Inorganic materials 0.000 description 1
- 229910052500 inorganic mineral Inorganic materials 0.000 description 1
- 229940125396 insulin Drugs 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 239000013067 intermediate product Substances 0.000 description 1
- 239000002563 ionic surfactant Substances 0.000 description 1
- 229910052742 iron Inorganic materials 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- 230000007794 irritation Effects 0.000 description 1
- 125000000959 isobutyl group Chemical group [H]C([H])([H])C([H])(C([H])([H])[H])C([H])([H])* 0.000 description 1
- 125000001449 isopropyl group Chemical group [H]C([H])([H])C([H])(*)C([H])([H])[H] 0.000 description 1
- 150000002576 ketones Chemical class 0.000 description 1
- 108010004902 lactose oxidase Proteins 0.000 description 1
- JCQLYHFGKNRPGE-FCVZTGTOSA-N lactulose Chemical compound OC[C@H]1O[C@](O)(CO)[C@@H](O)[C@@H]1O[C@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 JCQLYHFGKNRPGE-FCVZTGTOSA-N 0.000 description 1
- 229960000511 lactulose Drugs 0.000 description 1
- PFCRQPBOOFTZGQ-UHFFFAOYSA-N lactulose keto form Natural products OCC(=O)C(O)C(C(O)CO)OC1OC(CO)C(O)C(O)C1O PFCRQPBOOFTZGQ-UHFFFAOYSA-N 0.000 description 1
- 238000011031 large-scale manufacturing process Methods 0.000 description 1
- 239000007942 layered tablet Substances 0.000 description 1
- 239000000787 lecithin Substances 0.000 description 1
- 235000010445 lecithin Nutrition 0.000 description 1
- 229940067606 lecithin Drugs 0.000 description 1
- 231100000225 lethality Toxicity 0.000 description 1
- AIHDCSAXVMAMJH-GFBKWZILSA-N levan Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)OC[C@@H]1[C@@H](O)[C@H](O)[C@](CO)(CO[C@@H]2[C@H]([C@H](O)[C@@](O)(CO)O2)O)O1 AIHDCSAXVMAMJH-GFBKWZILSA-N 0.000 description 1
- 239000012669 liquid formulation Substances 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 239000006210 lotion Substances 0.000 description 1
- 239000007937 lozenge Substances 0.000 description 1
- 239000006166 lysate Substances 0.000 description 1
- 150000002688 maleic acid derivatives Chemical class 0.000 description 1
- 150000004701 malic acid derivatives Chemical class 0.000 description 1
- FGPATWVHNYVVEE-SKPZHCOCSA-N maltotriulose Chemical compound OC[C@H]1OC(O)(CO)[C@@H](O)[C@@H]1O[C@@H]1[C@H](O)[C@@H](O)[C@H](O[C@@H]2[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)[C@@H](CO)O1 FGPATWVHNYVVEE-SKPZHCOCSA-N 0.000 description 1
- FYGDTMLNYKFZSV-UHFFFAOYSA-N mannotriose Natural products OC1C(O)C(O)C(CO)OC1OC1C(CO)OC(OC2C(OC(O)C(O)C2O)CO)C(O)C1O FYGDTMLNYKFZSV-UHFFFAOYSA-N 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 238000004949 mass spectrometry Methods 0.000 description 1
- 238000000691 measurement method Methods 0.000 description 1
- 239000001525 mentha piperita l. herb oil Substances 0.000 description 1
- 239000001683 mentha spicata herb oil Substances 0.000 description 1
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 1
- 229940057867 methyl lactate Drugs 0.000 description 1
- 229960001047 methyl salicylate Drugs 0.000 description 1
- 230000003641 microbiacidal effect Effects 0.000 description 1
- 244000000010 microbial pathogen Species 0.000 description 1
- 229940124561 microbicide Drugs 0.000 description 1
- 244000005706 microflora Species 0.000 description 1
- 239000011785 micronutrient Substances 0.000 description 1
- 235000013369 micronutrients Nutrition 0.000 description 1
- 235000010755 mineral Nutrition 0.000 description 1
- 239000011707 mineral Substances 0.000 description 1
- 230000001333 moisturizer Effects 0.000 description 1
- 239000012778 molding material Substances 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 208000030194 mouth disease Diseases 0.000 description 1
- 230000035772 mutation Effects 0.000 description 1
- 239000002105 nanoparticle Substances 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 238000006386 neutralization reaction Methods 0.000 description 1
- 239000002736 nonionic surfactant Substances 0.000 description 1
- 235000019645 odor Nutrition 0.000 description 1
- 230000003287 optical effect Effects 0.000 description 1
- 238000005457 optimization Methods 0.000 description 1
- 239000008375 oral care agent Substances 0.000 description 1
- 150000007524 organic acids Chemical class 0.000 description 1
- 235000005985 organic acids Nutrition 0.000 description 1
- 150000002898 organic sulfur compounds Chemical class 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 239000007800 oxidant agent Substances 0.000 description 1
- 230000001590 oxidative effect Effects 0.000 description 1
- 150000002927 oxygen compounds Chemical class 0.000 description 1
- 239000006174 pH buffer Substances 0.000 description 1
- 238000010422 painting Methods 0.000 description 1
- 238000004091 panning Methods 0.000 description 1
- 238000005192 partition Methods 0.000 description 1
- 230000001717 pathogenic effect Effects 0.000 description 1
- 239000001814 pectin Substances 0.000 description 1
- 235000010987 pectin Nutrition 0.000 description 1
- 229920001277 pectin Polymers 0.000 description 1
- 230000006320 pegylation Effects 0.000 description 1
- 125000001147 pentyl group Chemical group C(CCCC)* 0.000 description 1
- 235000019477 peppermint oil Nutrition 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 125000000864 peroxy group Chemical group O(O*)* 0.000 description 1
- 239000012071 phase Substances 0.000 description 1
- 239000007981 phosphate-citrate buffer Substances 0.000 description 1
- 150000003013 phosphoric acid derivatives Chemical class 0.000 description 1
- 239000011574 phosphorus Substances 0.000 description 1
- 229910052698 phosphorus Inorganic materials 0.000 description 1
- 230000029553 photosynthesis Effects 0.000 description 1
- 238000010672 photosynthesis Methods 0.000 description 1
- 230000000704 physical effect Effects 0.000 description 1
- 229920001983 poloxamer Polymers 0.000 description 1
- 229920000573 polyethylene Polymers 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229940085675 polyethylene glycol 800 Drugs 0.000 description 1
- 239000000244 polyoxyethylene sorbitan monooleate Substances 0.000 description 1
- 235000010482 polyoxyethylene sorbitan monooleate Nutrition 0.000 description 1
- 235000013824 polyphenols Nutrition 0.000 description 1
- 229920001155 polypropylene Polymers 0.000 description 1
- 229920000053 polysorbate 80 Polymers 0.000 description 1
- 229940068968 polysorbate 80 Drugs 0.000 description 1
- 239000011591 potassium Substances 0.000 description 1
- 229960003975 potassium Drugs 0.000 description 1
- 229910052700 potassium Inorganic materials 0.000 description 1
- 239000001508 potassium citrate Substances 0.000 description 1
- 229960002635 potassium citrate Drugs 0.000 description 1
- QEEAPRPFLLJWCF-UHFFFAOYSA-K potassium citrate (anhydrous) Chemical compound [K+].[K+].[K+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O QEEAPRPFLLJWCF-UHFFFAOYSA-K 0.000 description 1
- 235000011082 potassium citrates Nutrition 0.000 description 1
- 239000002244 precipitate Substances 0.000 description 1
- 238000004321 preservation Methods 0.000 description 1
- 230000002335 preservative effect Effects 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 230000035755 proliferation Effects 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- WHMQHKUIPUYTKP-UHFFFAOYSA-N propane-1,2,3-triol;propanoic acid Chemical compound CCC(O)=O.OCC(O)CO WHMQHKUIPUYTKP-UHFFFAOYSA-N 0.000 description 1
- 238000011321 prophylaxis Methods 0.000 description 1
- 125000001436 propyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])[H] 0.000 description 1
- LLHKCFNBLRBOGN-UHFFFAOYSA-N propylene glycol methyl ether acetate Chemical compound COCC(C)OC(C)=O LLHKCFNBLRBOGN-UHFFFAOYSA-N 0.000 description 1
- 150000003856 quaternary ammonium compounds Chemical class 0.000 description 1
- 150000003242 quaternary ammonium salts Chemical class 0.000 description 1
- 238000002708 random mutagenesis Methods 0.000 description 1
- 230000035484 reaction time Effects 0.000 description 1
- 238000003259 recombinant expression Methods 0.000 description 1
- 235000020095 red wine Nutrition 0.000 description 1
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 230000000241 respiratory effect Effects 0.000 description 1
- 230000029058 respiratory gaseous exchange Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 238000002702 ribosome display Methods 0.000 description 1
- 235000009566 rice Nutrition 0.000 description 1
- 102220148016 rs200944017 Human genes 0.000 description 1
- 229940016590 sarkosyl Drugs 0.000 description 1
- 108700004121 sarkosyl Proteins 0.000 description 1
- 229930195734 saturated hydrocarbon Natural products 0.000 description 1
- 210000004761 scalp Anatomy 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 230000028327 secretion Effects 0.000 description 1
- 239000002453 shampoo Substances 0.000 description 1
- 229910052710 silicon Inorganic materials 0.000 description 1
- 239000010703 silicon Substances 0.000 description 1
- 239000000377 silicon dioxide Substances 0.000 description 1
- 239000002002 slurry Substances 0.000 description 1
- APSBXTVYXVQYAB-UHFFFAOYSA-M sodium docusate Chemical compound [Na+].CCCCC(CC)COC(=O)CC(S([O-])(=O)=O)C(=O)OCC(CC)CCCC APSBXTVYXVQYAB-UHFFFAOYSA-M 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- 239000011775 sodium fluoride Substances 0.000 description 1
- 235000013024 sodium fluoride Nutrition 0.000 description 1
- KSAVQLQVUXSOCR-UHFFFAOYSA-M sodium lauroyl sarcosinate Chemical compound [Na+].CCCCCCCCCCCC(=O)N(C)CC([O-])=O KSAVQLQVUXSOCR-UHFFFAOYSA-M 0.000 description 1
- 229960004711 sodium monofluorophosphate Drugs 0.000 description 1
- 229960001922 sodium perborate Drugs 0.000 description 1
- 229940045872 sodium percarbonate Drugs 0.000 description 1
- PFUVRDFDKPNGAV-UHFFFAOYSA-N sodium peroxide Chemical compound [Na+].[Na+].[O-][O-] PFUVRDFDKPNGAV-UHFFFAOYSA-N 0.000 description 1
- 159000000000 sodium salts Chemical class 0.000 description 1
- MWNQXXOSWHCCOZ-UHFFFAOYSA-L sodium;oxido carbonate Chemical compound [Na+].[O-]OC([O-])=O MWNQXXOSWHCCOZ-UHFFFAOYSA-L 0.000 description 1
- YKLJGMBLPUQQOI-UHFFFAOYSA-M sodium;oxidooxy(oxo)borane Chemical compound [Na+].[O-]OB=O YKLJGMBLPUQQOI-UHFFFAOYSA-M 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 239000000600 sorbitol Substances 0.000 description 1
- 235000019721 spearmint oil Nutrition 0.000 description 1
- UNFWWIHTNXNPBV-WXKVUWSESA-N spectinomycin Chemical compound O([C@@H]1[C@@H](NC)[C@@H](O)[C@H]([C@@H]([C@H]1O1)O)NC)[C@]2(O)[C@H]1O[C@H](C)CC2=O UNFWWIHTNXNPBV-WXKVUWSESA-N 0.000 description 1
- 229960000268 spectinomycin Drugs 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- UQZIYBXSHAGNOE-XNSRJBNMSA-N stachyose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO[C@@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO[C@@H]3[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O3)O)O2)O)O1 UQZIYBXSHAGNOE-XNSRJBNMSA-N 0.000 description 1
- 229910001220 stainless steel Inorganic materials 0.000 description 1
- 239000010935 stainless steel Substances 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 238000003756 stirring Methods 0.000 description 1
- 239000012089 stop solution Substances 0.000 description 1
- 150000003900 succinic acid esters Chemical class 0.000 description 1
- 229910052717 sulfur Inorganic materials 0.000 description 1
- 239000011593 sulfur Substances 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 239000003765 sweetening agent Substances 0.000 description 1
- 229920002994 synthetic fiber Polymers 0.000 description 1
- 235000018553 tannin Nutrition 0.000 description 1
- 229920001864 tannin Polymers 0.000 description 1
- 239000001648 tannin Substances 0.000 description 1
- 239000013077 target material Substances 0.000 description 1
- 150000003899 tartaric acid esters Chemical class 0.000 description 1
- 238000005496 tempering Methods 0.000 description 1
- 125000000999 tert-butyl group Chemical group [H]C([H])([H])C(*)(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- 239000012085 test solution Substances 0.000 description 1
- 239000002562 thickening agent Substances 0.000 description 1
- 150000003567 thiocyanates Chemical class 0.000 description 1
- 229960000790 thymol Drugs 0.000 description 1
- 238000004448 titration Methods 0.000 description 1
- 235000019505 tobacco product Nutrition 0.000 description 1
- 230000036367 tooth discoloration Effects 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 230000032258 transport Effects 0.000 description 1
- 150000003626 triacylglycerols Chemical class 0.000 description 1
- 229960003500 triclosan Drugs 0.000 description 1
- WEAPVABOECTMGR-UHFFFAOYSA-N triethyl 2-acetyloxypropane-1,2,3-tricarboxylate Chemical compound CCOC(=O)CC(C(=O)OCC)(OC(C)=O)CC(=O)OCC WEAPVABOECTMGR-UHFFFAOYSA-N 0.000 description 1
- 239000012137 tryptone Substances 0.000 description 1
- 210000000689 upper leg Anatomy 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- MWOOGOJBHIARFG-UHFFFAOYSA-N vanillin Chemical compound COC1=CC(C=O)=CC=C1O MWOOGOJBHIARFG-UHFFFAOYSA-N 0.000 description 1
- 235000012141 vanillin Nutrition 0.000 description 1
- 229940117960 vanillin Drugs 0.000 description 1
- FGQOOHJZONJGDT-UHFFFAOYSA-N vanillin Natural products COC1=CC(O)=CC(C=O)=C1 FGQOOHJZONJGDT-UHFFFAOYSA-N 0.000 description 1
- 239000007762 w/o emulsion Substances 0.000 description 1
- 239000011534 wash buffer Substances 0.000 description 1
- 239000002699 waste material Substances 0.000 description 1
- 239000000080 wetting agent Substances 0.000 description 1
- 239000008096 xylene Substances 0.000 description 1
- 150000003738 xylenes Chemical class 0.000 description 1
- 239000012138 yeast extract Substances 0.000 description 1
- 229940105296 zinc peroxide Drugs 0.000 description 1
- 229910052845 zircon Inorganic materials 0.000 description 1
- GFQYVLUOOAAOGM-UHFFFAOYSA-N zirconium(iv) silicate Chemical compound [Zr+4].[O-][Si]([O-])([O-])[O-] GFQYVLUOOAAOGM-UHFFFAOYSA-N 0.000 description 1
- FYGDTMLNYKFZSV-BYLHFPJWSA-N β-1,4-galactotrioside Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@H](CO)O[C@@H](O[C@@H]2[C@@H](O[C@@H](O)[C@H](O)[C@H]2O)CO)[C@H](O)[C@H]1O FYGDTMLNYKFZSV-BYLHFPJWSA-N 0.000 description 1
Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/30—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds
- A61K8/33—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds containing oxygen
- A61K8/37—Esters of carboxylic acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61Q—SPECIFIC USE OF COSMETICS OR SIMILAR TOILETRY PREPARATIONS
- A61Q11/00—Preparations for care of the teeth, of the oral cavity or of dentures; Dentifrices, e.g. toothpastes; Mouth rinses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/19—Cosmetics or similar toiletry preparations characterised by the composition containing inorganic ingredients
- A61K8/22—Peroxides; Oxygen; Ozone
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/30—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds
- A61K8/33—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds containing oxygen
- A61K8/35—Ketones, e.g. benzophenone
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/30—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds
- A61K8/33—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds containing oxygen
- A61K8/37—Esters of carboxylic acids
- A61K8/375—Esters of carboxylic acids the alcohol moiety containing more than one hydroxy group
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/30—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds
- A61K8/33—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds containing oxygen
- A61K8/38—Percompounds, e.g. peracids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/30—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds
- A61K8/60—Sugars; Derivatives thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/30—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds
- A61K8/64—Proteins; Peptides; Derivatives or degradation products thereof
- A61K8/65—Collagen; Gelatin; Keratin; Derivatives or degradation products thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K8/00—Cosmetics or similar toiletry preparations
- A61K8/18—Cosmetics or similar toiletry preparations characterised by the composition
- A61K8/30—Cosmetics or similar toiletry preparations characterised by the composition containing organic compounds
- A61K8/64—Proteins; Peptides; Derivatives or degradation products thereof
- A61K8/66—Enzymes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- General Health & Medical Sciences (AREA)
- Animal Behavior & Ethology (AREA)
- Epidemiology (AREA)
- Birds (AREA)
- Emergency Medicine (AREA)
- Chemical & Material Sciences (AREA)
- Oral & Maxillofacial Surgery (AREA)
- Inorganic Chemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Oncology (AREA)
- Medicinal Chemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Organic Chemistry (AREA)
- Communicable Diseases (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Cosmetics (AREA)
- Enzymes And Modification Thereof (AREA)
- Peptides Or Proteins (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
과산계 유익제로 구강 표면을 처리하기 위한 조성물 및 방법이 본 명세서에 개시된다. 과산 유익제는 구강 표면 탈색, 미백, 소독, 탈염, 탈취, 생물막의 감소 또는 제거 및 그들의 조합을 위해 사용될 수 있다. 과산은 과산소원의 존재 하에 과가수분해 활성을 갖는 CE-7 탄수화물 에스테라제(과가수분해효소)를 사용하여 카르복실산 에스테르 기질로부터 효소에 의해 생성된다. 구강 표면에 대하여 친화성을 갖는 펩티드 성분에 직접적으로 또는 임의의 링커를 통하여 결합된 과가수분해효소를 포함하는 융합 단백질을 사용하여, 과가수분해 활성을 구강 표면에 표적화시킬 수 있다.
Description
관련-출원과의 상호 참조
본 출원은 본 명세서에 그 전문이 참고로 포함된 미국 가특허 출원 제61/424,903호(출원일: 2010년 12월 20일)에 대한 우선권을 청구한다.
본 발명은 구강 관리 유익제(benefit agent)로서 적어도 하나의 과산을 포함하는 개인 관리 제품의 분야에 관한 것이다. 과산은 적어도 하나의 적절한 카르복실산 에스테르 기질 및 과산소원의 존재 하에 효소에 의해 생성된다. 구체적으로, 과가수분해(perhydrolytic) 활성을 갖는 효소 촉매를 사용하여, 구강 관리 제품에 사용하기 위한 과산 유익제를 생성한다. 과가수분해 효소는 효소에 의해 생성된 과산이 원하는 표면 상에서 또는 그 근처에서 생성되도록 구강 표면에 대하여 친화성을 갖는 적어도 하나의 펩티드 성분을 함유하도록 엔지니어링된 융합 단백질("표적화된 과가수분해효소")의 형태일 수 있다.
퍼옥시카르복실산("과산")은 효과적인 항미생물제이다. 원치 않는 미생물 성장에 대하여 경질 표면, 식품, 살아있는 식물 조직 및 의료 장치를 세정하고/거나 소독하고/거나 살균하는 방법이 기재되어 있다(예를 들어, 미국 특허 제6,545,047호; 미국 특허 제6,183,807호; 미국 특허 제6,518,307호; 미국 특허 제5,683,724호; 및 미국 특허 제6,635,286호). 또한, 과산은 세탁용 세제 응용을 위한 표백(bleaching) 조성물을 제조하는데 유용한 것으로 보고되어 있다(예를 들어, 미국 특허 제3,974,082호; 미국 특허 제5,296,161호; 및 미국 특허 제5,364,554호).
또한, 과산을 포함하는 구강 관리 조성물이 개시되어 있다. 미국 특허 제5,302,375호(비시오, 디.(Viscio, D.))에는 비히클 중에 용해된 과아세트산을 포함하는 치아를 미백시키기 위한 구강 조성물이 개시되며, 여기서, 과아세트산은 물, 아세틸살리실산 및 수용성 알칼리 금속 과탄산염을 배합함으로써 동소에서 비히클 내에서 생성된다. 미국 특허 제5,279,816호(처치(Church) 등)에는 착색되거나 변색된 치아를 미백시키기 위한 과아세트산을 포함하는 조성물의 용도가 개시된다. 미국 특허 제6,221,341호 및 제7,189,385호(몬트고메리, 알.(Montgomery, R.))에는 치아를 미백시키기 위한 방법에서 사용하기에 적절한 퍼옥시산 치아 미백 조성물이 개시되어 있다. 더욱 구체적으로, 과아세트산 조성물은 과산화수소 전구체, 글리세린의 아세트산 에스테르 및 물을 배합하여, 화학적 과가수분해를 통하여, 과아세트산을 생성함에 의해 생성된다. 효소에 의한 과가수분해는 기재되어 있지 않다.
미국 특허 출원 공개 제2009-0311198호(콘카르(Concar) 등)에는 치아를 탈색시키기 위한, 과가수분해 활성을 갖는 마이코박테리움 스메그마티스(M. smegmatis) 효소를 포함하는 구강 조성물이 개시되어 있다. 과산 유익제를 생성하기 위한 CE-7 과가수분해효소의 이용은 개시되지 않았다. 또한, 콘카르 등은 구강 관리 조성물에서 표적화된 과가수분해 효소의 이용에 대해서는 언급하지 않았다.
바디 케어(body care) 제품에 과가수분해 활성을 갖는 특정 변이체 서브틸리신 칼스베르그(subtilisin Carlsberg) 프로테아제를 포함하는 것이 빌란트(Wieland) 등의 미국 특허 제7,510,859호에 개시되어 있다. 특정 변이체 프로테아제를 뛰어넘는 과가수분해 효소가 기재되어 있지 않거나, 개인 관리 유익제로서 과산의 효소에 의한 생성을 설명하는 어떠한 실험예도 존재하지 않는다.
미국 특허 출원 공개 제2008-0176783 A1호; 제2008-0176299 A1호; 제2009-0005590 A1호; 및 제2010-0041752 A1호(디코시모(DiCosimo) 등)에는 카르복실산 에스테르 기질을 (적절한 과산소원, 예컨대 과산화수소의 존재 하에) 소독제 및/또는 탈색제로 사용하기에 충분한 농도로 퍼옥시카르복실산으로 전환시키기 위한 상당한 과가수분해 활성을 특징으로 하는, 구조적으로 탄수화물 에스테라제의 CE-7 패밀리의 구성원으로 분류된 효소(즉, 세팔로스포린 C 데아세틸라제[CAH] 및 아세틸 자일란 에스테라제[AXE])가 개시되어 있다. 탄수화물 에스테라제의 CE-7 패밀리의 일부 구성원은 일단 반응 성분이 혼합되면, 알코올, 다이올 및 글리세롤의 아세틸 에스테르로부터 1분 내에 4000 내지 5000 ppm, 그리고 5분 내지 30분에 최대 9000 ppm의 과아세트산을 생성하기에 충분한 과가수분해 활성을 갖는 것으로 입증되었다(디코시모 등, 미국 특허 출원 공개 제2009-0005590 A1호). 미국 특허 출원 공개 제2010-0087529 A1호에는 과가수분해 활성이 향상된 변이체 CE-7 효소가 기재되어 있다. 비록 CE-7 과가수분해효소(perhydrolase)가 특출한 과가수분해 활성을 갖지만, 개인 관리 제품에서의 그들의 이용은 개시된 적이 없다. 이와 같이, 해결해야 할 문제는 과산 유익제의 생성을 위해 적어도 하나의 CE-7 과가수분해효소의 이용을 포함하는 개인 관리 조성물 및 방법을 제공하는 것이다.
과산은 원하는 혜택에 대하여 표적화되지 않은 물질을 포함하는 다양한 물질에 대하여 반응성일 수 있는 강력한 산화제이다. 이와 같이, 특정 개인 관리 응용은 원하는 표적 체표면 상에 또는 그 근처에 과산 생성을 국소화시킴으로써 과산 유익제를 원하는 체표면에 표적화/집중시키는 능력으로부터 유리할 수 있다. 효소에 의한 과산 생성은 과가수분해효소를 체표면에 표적화시킴으로써 유리할 수 있다. 과가수분해효소를 운반 물질에 표적화시켜, 효소 농도 및 사용자의 노출을 제한함으로써 추가의 이익이 달성될 수 있다.
구강 물질과 결합되는 효소를 사용하는 구강 관리 표면을 처리하는 구강 관리 조성물 및/또는 방법이 보고되어 있다. 미국 특허 제4,138,476호(심슨(Simonson) 등)에는 복합화 시약(complexing reagent)을 통하여 치아의 표면에 대하여 친화성을 갖는 포스페이트 캐리어 기(carrier group)에 공유 결합되는 글루칸-분해 효소의 사용을 포함하는 플라크의 처리 방법이 개시되어 있다. 글루칸 침착물의 효소에 의한 분해는 플라크 물질의 용해 및 분산을 증진시키는 것을 나타낸다.
미국 특허 출원 공개 제2005-0158253호, 미국 특허 제6,830,745호(버드니(Budny) 등)에는 생물막(biofilm) 구조를 효소에 의해 분해하기 위한 고정(anchor) 효소 복합체 및 박테리아 상에서 직접적으로 작용할 수 있는 제2 고정 효소 성분을 포함하는 2-성분 조성물이 개시되어 있다. 생물막 분해 효소는 엑소폴리사카라이드(exopolysaccharide) 백본(backbone) 구조를 직접적으로 분해하는 것이다.
미국 특허 제5,871,714호(버드니, 제이.)에는 고정 분자에 결합된 플라크 매트릭스를 분해하는 효소를 포함하는 박테리아 성장/콜로니화의 조절(예를 들어, 치아 플라크 감소)을 위한 조성물이 개시된다. 표적화된 과가수분해효소의 이용이 기재되어 있지 않다.
미국 특허 제5,490,988호 및 EP 0479,600 B1호(베그스(Beggs) 등)에는 표적 부위에 대한 결합을 위한 수단으로서의 항체 단편의 용도가 개시되며, 여기서, 치료제는 치료제를 표적 부위에 부착시키기 위한 항체 단편에 첨부되는 추가의 펩티드를 통하여 연결된다. 치아 플라크에서 박테리아의 항원 성분에 대하여 친화성을 갖는 변형된 항체 단편을 포함하여 치료제를 전달하는 구강 관리 제품이 개시된다. 치료제는 효소 또는 효소의 조합, 예를 들어, 산화된 할라이드를 형성하기 위하여 과산화효소와 병용한 산화효소에 의해 생성되는 세포독성제일 수 있다. 과산 유익제를 생성하기 위한 표적화된 과가수분해효소의 이용이 기재되어 있지 않다.
EP 0450,800 B1호(베그스 등)에는 함께 작용하여 구강 미생물총에서 발생하는 종을 공격하는 2가지 상이한 효소의 이용이 개시된다. 제1 효소는 제2 효소에 대한 기질로 사용되어 입안의 표적에 대하여 활성인 제제를 생성하는 중간 생성물을 생성한다. 각 효소는 입안의 표적 표면에 대하여 친화성을 갖는 항체 또는 항체 단편에 부착되며, 이에 의해, 사용 중에, 효소는 서로 인접한 표적 부위에 결합된다. 할라이드 또는 티오시아네이트의 존재 하에서 과산화효소에 의해 전환되어, 각각 하이포할라이트 또는 하이포티오시아네이트를 생성할 수 있는 과산화수소를 생성하기 위한 글루코스 산화효소의 조합이 예시된다. 과산 유익제를 생성하기 위한 표적화된 과가수분해효소의 이용은 기재되어 있지 않다.
EP 0451,972 B1호(베그스 등)에는 2가지 효소를 포함하는 제품이 기재되며, 상기 제품은 표적에 대한제를 생성하기 위한 제1 효소 및 제1 효소에 대한 기질인 중간체를 생성하기 위한 제2 효소를 포함하며; 상기 제품은 추가로 둘 모두의 효소에 부착되거나 부착될 수 있어 효소를 함께 결합시키며, 이에 의해 표적 세포에 결합하는 복합체를 형성하는 연결 수단(다시 말하면, 항체 또는 항체 단편)을 포함한다. 하이포할라이트 또는 하이포티오시아네이트 활성제의 형성을 촉매작용시키는 과산화효소에 결합된 산화효소(과산화수소를 생성할 수 있는)가 예시된다.
EP 0453,097 B1호(베그스 등)에는 자가 조립가능하여, 활성제와 표적 부위 간에 가교를 형성할 수 있는 복수의 항체 또는 항체 단편을 사용하는 활성제의 표적 부위로의 전달이 기재된다. 활성제는 글루코스 산화효소 또는 글루코스 산화효소 및 과산화효소의 조합이다. 과산 유익제를 생성하기 위한 표적화된 과가수분해효소의 이용은 기재되어 있지 않다.
펩티드 친화성 물질로서의 항체, 항체 단편(Fab), 단쇄 융합 가변 영역 항체(scFc), 카멜리대(Camelidae) 항체 및 라지 스캐폴드 디스플레이 단백질(large scaffold display protein)의 이용은 그들의 크기와 비용 때문에 일부 개인 관리 응용에 적절하지 않을 수 있다. 이와 같이, 소정의 저비용 미용 응용에서, 표적화된 유익제의 전달을 위해 더 짧거나 비용이 보다 낮은 펩티드 친화성 물질을 사용하는 것이 필요하다.
미용 유익제를 체표면에 표적화시키기 위한 체표면에 대하여 강력한 친화성을 갖는 더 짧은 바이오패닝된(biopanned) 펩티드의 이용이 기재되어 있다(미국 특허 제7,220,405호; 제7,309,482호; 제7,285,264호 및 제7,807,141호; 미국 특허 출원 공개 제2005-0226839 A1호; 제2007-0196305 A1호; 제2006-0199206 A1호; 제2007-0065387 A1호; 제2008-0107614 A1호; 제2007-0110686 A1호; 제2006-0073111 A1호; 제2010-0158846호; 제2010-0158847호; 및 제2010-0247589호; 및 PCT 출원 공개 WO2008/054746호; WO2004/048399호 및 WO2008/073368호). 미국 특허 제7,807,141호(후앙(Huang) 등)에는 치아 표면에 구강 관리 유익제를 결합시키기에 적절한 펩티드계 구강 관리 표면 시약이 개시된다. 과산 유익제의 생성을 위해 활성 CE-7 과가수분해효소(즉, "표적화된 과가수분해효소")를 결합시키기 위한 구강 표면에 대하여 친화성을 갖는 펩티드 물질을 이용하는 것이 기재된 적이 없다.
이와 같이, 해결해야 할 추가의 문제는 효소에 의한 과산 생성을 구강 표면에 표적화시키기에 적절한 조성물 및 방법을 제공하는 것이다.
과산계 유익제를 효소에 의해 생성하고, 이를 구강 표면에 전달하기 위한 성분을 포함하는 조성물 및 방법이 제공된다.
일 실시형태에서, 구강 관리 응용, 예를 들어, 구강 표면 탈색, 치아 미백, 소독, 탈염, 탈취, 치아 우식증의 치료, 치아 우식증의 예방, 치아 우식증과 관련된 구강 박테리아의 감소 및 구강 생물막(예를 들어, 치아 플라크)의 치료 또는 제거에서 사용하기 위한 과산 유익제를 효소에 의해 생성하기 위하여, CE-7 과가수분해효소를 사용하는 구강 관리 조성물 및 방법이 제공된다.
일 실시형태에서,
1) a) i) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티(moiety)이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
ii) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
iii) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
iv) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
b) 과산소원; 및
c) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유(CLUSTALW)를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
i) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
ii) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
iii) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - 를 포함하는 반응 성분의 세트를 제공하는 단계;
2) (1)의 반응 성분을 적절한 반응 조건 하에서 배합하는 단계 - 이에 의해, 적어도 하나의 과산이 효소에 의해 생성된다 - ; 및
3) 구강 표면을 적어도 하나의 과산과 접촉시키는 단계 - 이에 의해, 구강 표면에 탈색, 치아 미백, 소독, 탈염, 탈취, 생물막의 감소 또는 제거 및 그들의 조합으로 이루어진 군으로부터 선택되는 과산 기반의 이익이 제공된다 - 를 포함하는 방법이 제공된다.
일 실시형태에서, 구강 표면은 치아 에나멜(enamel), 치아 균막(pellicle), 구강 내의 연조직(예를 들어, 잇몸, 혀) 또는 구강 생물막(예를 들어, 구강 플라크)이다.
다른 실시형태에서, 과가수분해 효소 및 구강 표면에 대하여 친화성을 갖는 펩티드 성분을 포함하며, 2가지 성분이 임의로 펩티드 스페이서(spacer)에 의해 분리될 수 있는 융합 단백질(즉, "표적화된 과가수분해효소")의 사용을 포함하는 조성물 및 방법이 제공된다.
일 실시형태에서,
1) a) i) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
ii) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
iii) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
iv) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
b) 과산소원; 및
c) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 일반 구조
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
(여기서,
PAH는 과가수분해 활성을 갖는 효소이며;
OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이고;
L은 길이가 1 내지 100개 아미노산 범위인 임의의 펩티드 링커이며;
y는 0 또는 1이다)을 갖는 융합 단백질을 포함한다 - 를 포함하는 반응 성분의 세트를 제공하는 단계;
2) (1)의 반응 성분을 적절한 반응 조건 하에서 배합하는 단계 - 이에 의해, 적어도 하나의 과산이 효소에 의해 생성된다 - ; 및
3) 구강 표면을 적어도 하나의 과산과 접촉시키는 단계 - 이에 의해, 구강 표면에 탈색, 치아 미백, 소독, 탈염, 탈취, 생물막의 감소 또는 제거 및 그들의 조합으로 이루어진 군으로부터 선택되는 과산 기반의 이익이 제공된다 - 를 포함하는 방법이 제공된다.
융합 단백질은 리파제, 프로테아제, 에스테라제, 아실 트랜스퍼라제, 아릴 에스테라제, 탄수화물 에스테라제 및 그들의 조합으로 이루어진 군으로부터 선택되는 과가수분해 효소를 포함할 수 있다.
일 실시형태에서, 융합 단백질은 마이코박테리움 스메그마티스로부터의 과가수분해 아릴 에스테라제(ArE)를 포함한다. 다른 실시형태에서, 융합 단백질은 서열 번호 460으로 제공된 S54V 마이코박테리움 스메그마티스 아릴 에스테라제와 적어도 95% 동일성을 갖는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
일 실시형태에서, 융합 단백질은 슈도모나스 플루오레슨스(Pseudomonas fluorescens)로부터의 과가수분해 에스테라제를 포함한다. 다른 실시형태에서, 융합 단백질은 서열 번호 477로 제공된 슈도모나스 플루오레슨스 에스테라제와 적어도 95% 동일성을 갖는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
다른 실시형태에서, 융합 단백질은 서열 번호 424, 425, 426, 427, 428, 429, 430, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 476, 477, 478 및 479로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
다른 실시형태에서, 융합 단백질은 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 CE-7 과가수분해효소를 포함하며, 상기 시그니처 모티프는
i) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
ii) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
iii) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다.
다른 실시형태에서, 구강 표면에 대하여 친화성을 갖는 펩티드 성분은 바람직하게는 적어도 하나의 구강 표면-결합 펩티드를 포함하는 단쇄 펩티드이다. 추가의 실시형태에서, 구강 표면-결합 펩티드는 치아 에나멜, 치아 균막 또는 치아 에나멜 및 치아 균막 둘 모두에 대하여 친화성을 갖는 펩티드이다.
다른 실시형태에서, 하기의 일반 구조를 포함하는 융합 단백질이 제공된다:
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
상기 식에서,
1) PAH는 과가수분해 활성을 갖는 효소이고;
2) OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이며;
3) L은 길이가 1 내지 100개 아미노산 범위인 임의의 펩티드 링커이며;
4) y는 0 또는 1이다.
다른 실시형태에서, 하기의 일반 구조를 포함하는 융합 단백질이 제공된다:
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
상기 식에서,
a) PAH는 과가수분해 활성을 갖는 CE-7 탄수화물 에스테라제이며, PAH는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖고, 상기 시그니처 모티프는
i) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
ii) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
iii) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함하고;
b) OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이며;
c) L은 길이가 1 내지 100개 아미노산 범위인 임의의 펩티드 링커이고;
d) y는 0 또는 1이다.
다른 실시형태에서,
1) 상기 임의의 과가수분해 융합 단백질을 포함하는 효소 촉매;
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
d) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
3) 과산소원; 및
4) 구강에 허용가능한 담체 매질을 포함하는 구강 관리 제품이 제공된다.
다른 실시형태에서,
1) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
a) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
b) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
c) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - ;
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
d) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
3) 과산소원; 및
4) 구강에 허용가능한 담체 매질을 포함하는 구강 관리 제품이 제공된다.
다른 실시형태에서, 구강 표면에 대하여 친화성을 갖는 분리된 폴리펩티드가 제공되며, 상기 폴리펩티드는 서열 번호 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421 및 422로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는다.
다른 실시형태에서, 구강 관리 제품에서 구강 물질/표면의 탈색, 미백, 소독, 탈염, 탈취 또는 생물막의 제거를 위한 유효 농도의 적어도 하나의 과산을 생성하기 위한 과가수분해 활성을 갖는 CE-7 탄수화물 에스테라제의 용도도 또한 제공된다.
다른 실시형태에서,
a) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
i) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
ii) 서열ㅌ번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
iii) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - ;
b) 1) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
2) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
3) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
d) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질; 및
c) 과산소원을 포함하는 - 이에 의해, (a), (b) 및 (c)의 동시의 또는 단계적 방식(그러나, 특정 순서 없이)의 혼합 시에 과산이 생성된다 - 치아 우식증, 치은염, 구강 칸디다증 또는 치근막염의 치료 또는 예방을 위한 과산 생성 조성물의 용도가 제공된다.
다른 실시형태에서, 구강 제품에서 하기의 일반 구조를 포함하는 융합 단백질의 용도가 제공된다:
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
상기 식에서,
1) PAH는 서열 번호 460과 적어도 95% 아미노산 동일성을 갖는 아미노산 서열을 갖는 과가수분해 활성을 갖는 효소이고;
2) OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이며;
3) L은 길이가 1 내지 100개 아미노산 범위인 펩티드 링커이며;
4) y는 0 또는 1이다.
본 명세서에 기재된 일부 에스테르 기질(표 20)은 특히 과산화수소와 반응 시에 화학적으로 과가수분해되어, 과아세트산을 생성하기 쉽다. 다른 실시형태에서, 1,2,3,5-테트라-O-아세틸-리보푸라노스; 1,2,3,4-테트라-O-아세틸-리보피라노스; 2-아세트아미도-2-데옥시-1,3,4,6-테트라아세틸-β-D-글루코피라노스; β-D-글루코피라노스, 1,2,3,4-테트라아세테이트; 2,3,4,6-테트라아세틸-β-D-글루코피라노스; 1,3,4,6-테트라-O-아세틸-만노피라노스; 및 α-D-만노피라노스 펜타아세테이트로 이루어진 군으로부터 선택되는 과산 전구체를 포함하는 개인 관리 제품이 제공된다. 바람직한 실시형태에서, 개인 관리 제품은 구강 관리 제품이다.
다른 실시형태에서,
a) i) 1,2,3,5-테트라-O-아세틸-리보푸라노스; 1,2,3,4-테트라-O-아세틸-리보피라노스; 2-아세트아미도-2-데옥시-1,3,4,6-테트라아세틸-β-D-글루코피라노스; β-D-글루코피라노스, 1,2,3,4-테트라아세테이트; 2,3,4,6-테트라아세틸-β-D-글루코피라노스; 1,3,4,6-테트라-O-아세틸-만노피라노스; 및 α-D-만노피라노스 펜타아세테이트로 이루어진 군으로부터 선택되는 과산 전구체; 및
ii) 과산소원을 포함하는 반응 성분의 세트를 제공하는 단계;
b) 체표면을 물의 존재 하에 반응 성분의 세트를 배합하여 생성된 유효량의 과아세트산과 접촉시키는 단계 - 이에 의해, 과아세트산은 체표면에 이익을 제공한다 - 를 포함하는 방법도 또한 제공된다. 바람직한 태양에서, 상기 방법에서 체표면은 구강 조직, 예를 들어, 치아 및/또는 잇몸이다.
생물학적 서열의 간단한 설명
하기의 서열들은 37 C.F.R. §§ 1.821-1.825("뉴클레오티드 서열 및/또는 아미노산 서열 개시를 포함하는 특허출원에 관한 요건 - 서열 규정")를 따르며 세계지적재산권기구(World Intellectual Property Organization, WIPO) 표준 ST.25(2009), 및 유럽 특허 조약(EPC) 및 특허 협력 조약(PCT)의 서열 목록 요건(규정 5.2 및 49.5(a-bis)) 및 시행세칙의 섹션 208 및 부칙 C에 부합한다. 뉴클레오티드 및 아미노산 서열 데이터에 사용된 기호 및 체제는 37 C.F.R. §1.822에 제시된 규정을 따른다.
서열 번호 1은 바실러스 서브틸리스(Bacillus subtilis) ATCC(등록 상표) 31954(상표명)로부터의 세팔로스포린 C 데아세틸라제를 암호화하는 핵산 서열이다.
서열 번호 2는 바실러스 서브틸리스 ATCC(등록 상표) 31954(상표명)로부터의 세팔로스포린 C 데아세틸라제의 아미노산 서열이다.
서열 번호 3은 바실러스 서브틸리스 아종 서브틸리스 균주 168로부터의 세팔로스포린 C 데아세틸라제를 암호화하는 핵산 서열이다.
서열 번호 4는 바실러스 서브틸리스 아종 서브틸리스 균주 168로부터의 세팔로스포린 C 데아세틸라제의 아미노산 서열이다.
서열 번호 5는 바실러스 서브틸리스 ATCC(등록 상표) 6633(상표명)으로부터의 세팔로스포린 C 데아세틸라제를 암호화하는 핵산 서열이다.
서열 번호 6은 바실러스 서브틸리스 ATCC(등록 상표) 6633(상표명)으로부터의 세팔로스포린 C 데아세틸라제의 아미노산 서열이다.
서열 번호 7은 바실러스 리케니포르미스(B. licheniformis) ATCC(등록 상표) 14580(상표명)으로부터의 세팔로스포린 C 데아세틸라제를 암호화하는 핵산 서열이다.
서열 번호 8은 바실러스 리케니포르미스 ATCC(등록 상표) 14580(상표명)으로부터의 세팔로스포린 C 데아세틸라제의 추정된 아미노산 서열이다.
서열 번호 9는 바실러스 푸밀러스(B. pumilus) PS213으로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 10은 바실러스 푸밀러스 PS213으로부터의 아세틸 자일란 에스테라제의 추정된 아미노산 서열이다.
서열 번호 11은 클로스트리듐 써모셀룸(Clostridium thermocellum) ATCC(등록 상표) 27405(상표명)로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 12는 클로스트리듐 써모셀룸 ATCC(등록 상표) 27405(상표명)로부터의 아세틸 자일란 에스테라제의 추정된 아미노산 서열이다.
서열 번호 13은 써모토가 네아폴리타나(Thermotoga neapolitana)로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 14는 써모토가 네아폴리타나로부터의 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 15는 써모토가 마리티마(Thermotoga maritima) MSB8로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 16은 써모토가 마리티마 MSB8로부터의 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 17은 써모아나에로박테리움(Thermoanaerobacterium) 종 JW/SL YS485로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 18은 써모아나에로박테리움 종 JW/SL YS485로부터의 아세틸 자일란 에스테라제의 추정된 아미노산 서열이다.
서열 번호 19는 바실러스 종 NRRL B-14911로부터의 세팔로스포린 C 데아세틸라제의 핵산 서열이다. 진뱅크(GENBANK)(등록 상표) 수탁 번호 ZP_01168674에 보고된 바와 같이, 바실러스 종 NRRL B-14911로부터의 세팔로스포린 C 데아세틸라제를 암호화하는 핵산 서열이 다른 세팔로스포린 C 데아세틸라제와의 서열 정렬, 및 보고된 길이(340개 아미노산) 대 다른 CAH 효소의 관찰된 길이(전형적으로, 길이가 318 내지 325개 아미노산; 본 명세서에 참조로 포함되는 미국 특허 출원 공개 제US-2010-0087528-A1호 참조)의 비교에 기초하여, 아마도 부정확할 것 같은 15개 아미노산 N-말단 부가물을 암호화하는 것으로 보이는 것에 주의해야 한다. 이와 같이, 본 명세서에 보고된 바와 같은 핵산 서열은 진뱅크(등록 상표) 수탁 번호 ZP_01168674 하에 보고된 N-말단 15개 아미노산이 없는 바실러스 종 NRRL B-14911로부터의 세팔로스포린 C 데아세틸라제 서열을 암호화한다.
서열 번호 20은 서열 번호 19의 핵산 서열에 의해 암호화되는 바실러스 종 NRRL B-14911로부터의 세팔로스포린 C 데아세틸라제의 추정된 아미노산 서열이다.
서열 번호 21은 바실러스 할로두란스(Bacillus halodurans) C-125로부터의 세팔로스포린 C 데아세틸라제를 암호화하는 핵산 서열이다.
서열 번호 22는 바실러스 할로두란스 C-125로부터의 세팔로스포린 C 데아세틸라제의 추정된 아미노산 서열이다.
서열 번호 23은 바실러스 클라우시이(Bacillus clausii) KSM-K16으로부터의 세팔로스포린 C 데아세틸라제를 암호화하는 핵산 서열이다.
서열 번호 24는 바실러스 클라우시이 KSM-K16으로부터의 세팔로스포린 C 데아세틸라제의 추정된 아미노산 서열이다.
서열 번호 25는 바실러스 서브틸리스 ATCC(등록 상표) 29233(상표명) 세팔로스포린 C 데아세틸라제(CAH)를 암호화하는 핵산 서열이다.
서열 번호 26은 바실러스 서브틸리스 ATCC(등록 상표) 29233(상표명) 세팔로스포린 C 데아세틸라제(CAH)의 추정된 아미노산 서열이다.
서열 번호 27은 미국 특허 출원 공개 제2010-0087529호(본 명세서에 그 전문이 참고로 포함)로부터의 써모토가 네아폴리타나 아세틸 자일란 에스테라제 변이체의 추정된 아미노산 서열(여기서, 위치 277에서의 Xaa 잔기는 Ala, Val, Ser 또는 Thr이다)이다.
서열 번호 28은 미국 특허 출원 공개 제2010-0087529호로부터의 써모토가 마리티마 MSB8 아세틸 자일란 에스테라제 변이체의 추정된 아미노산 서열(여기서, 위치 277에서의 Xaa 잔기는 Ala, Val, Ser 또는 Thr이다)이다.
서열 번호 29는 미국 특허 출원 공개 제2010-0087529호로부터의 써모토가 레틴가에(Thermotoga lettingae) 아세틸 자일란 에스테라제 변이체의 추정된 아미노산 서열(여기서, 위치 277에서의 Xaa 잔기는 Ala, Val, Ser 또는 Thr이다)이다.
서열 번호 30은 미국 특허 출원 공개 제2010-0087529호로부터의 써모토가 페트로필라(Thermotoga petrophila) 아세틸 자일란 에스테라제 변이체의 추정된 아미노산 서열(여기서, 위치 277에서의 Xaa 잔기는 Ala, Val, Ser 또는 Thr이다)이다.
서열 번호 31은 미국 특허 출원 공개 제2010-0087529호로부터의 "RQ2(a)" 유래의 써모토가 종 RQ2 아세틸 자일란 에스테라제 변이체의 추정된 아미노산 서열(여기서, 위치 277에서의 Xaa 잔기는 Ala, Val, Ser 또는 Thr이다)이다.
서열 번호 32는 미국 특허 출원 공개 제2010-0087529호로부터의 "RQ2(b)" 유래의 써모토가 종 RQ2 아세틸 자일란 에스테라제 변이체의 추정된 아미노산 서열(여기서, 위치 278에서의 Xaa 잔기는 Ala, Val, Ser 또는 Thr이다)이다.
서열 번호 33은 써모토가 레틴가에 아세틸 자일란 에스테라제의 추정된 아미노산 서열이다.
서열 번호 34는 써모토가 페트로필라 아세틸 자일란 에스테라제의 추정된 아미노산 서열이다.
서열 번호 35는 본 명세서에서 "RQ2(a)"로 기재된 써모토가 종 RQ2로부터의 제1 아세틸 자일란 에스테라제의 추정된 아미노산 서열이다.
서열 번호 36은 본 명세서에서 "RQ2(b)"로 기재된 써모토가 종 RQ2로부터의 제2 아세틸 자일란 에스테라제의 추정된 아미노산 서열이다.
서열 번호 37은 써모아네아로박테리움 사카롤리티쿰(Thermoanearobacterium saccharolyticum) 세팔로스포린 C 데아세틸라제를 암호화하는 코돈 최적화된 핵산 서열이다.
서열 번호 38은 써모아네아로박테리움 사카롤리티쿰 세팔로스포린 C 데아세틸라제의 추정된 아미노산 서열이다.
서열 번호 39는 락토코커스 락티스(Lactococcus lactis)로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열(진뱅크(등록 상표) 수탁 번호 EU255910)이다.
서열 번호 40은 락토코커스 락티스로부터의 아세틸 자일란 에스테라제의 아미노산 서열(진뱅크(등록 상표) 수탁 번호 ABX75634.1)이다.
서열 번호 41은 메소리조비움 로티(Mesorhizobium loti)로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열(진뱅크(등록 상표) 수탁 번호 NC_002678.2)이다.
서열 번호 42는 메소리조비움 로티로부터의 아세틸 자일란 에스테라제의 아미노산 서열(진뱅크(등록 상표) 수탁 번호 BAB53179.1)이다.
서열 번호 43은 게오바실러스 스테아로써모필러스(Geobacillus stearothermophilus)로부터의 아세틸 자일란 에스테라제를 암호화하는 핵산 서열(진뱅크(등록 상표) 수탁 번호 AF038547.2)이다.
서열 번호 44는 게오바실러스 스테아로써모필러스로부터의 아세틸 자일란 에스테라제의 아미노산 서열(진뱅크(등록 상표) 수탁 번호 AAF70202.1)이다.
서열 번호 45는 야생형 써모토가 마리티마 아세틸 자일란 에스테라제 아미노산 서열에 대하여 하기의 치환을 갖는 변이체 아세틸 자일란 에스테라제(변이체 "A3")를 암호화하는 핵산 서열이다: (F24I/S35T/Q179L/N275D/C277S/S308G/F317S).
서열 번호 46은 "A3" 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 47은 N275D/C277S 변이체 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 48은 N275D/C277S 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 49는 C277S/F317S 변이체 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 50은 C277S/F317S 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 51은 S35T/C277S 변이체 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 52는 S35T/C277S 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 53은 Q179L/C277S 변이체 아세틸 자일란 에스테라제를 암호화하는 핵산 서열이다.
서열 번호 54는 Q179L/C277S 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 55는 야생형 써모토가 마리티마 아세틸 자일란 에스테라제 아미노산 서열에 대하여 하기의 치환을 갖는 변이체 아세틸 자일란 에스테라제 843H9를 암호화하는 핵산 서열이다: (L8R/L125Q/Q176L/V183D/F247I/C277S/P292L).
서열 번호 56은 843H9 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 57은 야생형 써모토가 마리티마 아세틸 자일란 에스테라제 아미노산 서열에 대하여 하기의 치환을 갖는 변이체 아세틸 자일란 에스테라제 843F12를 암호화하는 핵산 서열이다: K77E/A266E/C277S.
서열 번호 58은 843F12 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 59는 야생형 써모토가 마리티마 아세틸 자일란 에스테라제 아미노산 서열에 대하여 하기의 치환을 갖는 변이체 아세틸 자일란 에스테라제 843C12를 암호화하는 핵산 서열이다: F27Y/I149V/A266V/C277S/I295T/N302S.
서열 번호 60은 843C12 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 61은 야생형 써모토가 마리티마 아세틸 자일란 에스테라제 아미노산 서열에 대하여 하기의 치환을 갖는 변이체 아세틸 자일란 에스테라제 842H3을 암호화하는 핵산 서열이다: L195Q/C277S.
서열 번호 62는 842H3 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 63은 야생형 써모토가 마리티마 아세틸 자일란 에스테라제 아미노산 서열에 대하여 하기의 치환을 갖는 변이체 아세틸 자일란 에스테라제 841A7을 암호화하는 핵산 서열이다: Y110F/C277S.
서열 번호 64는 841A7 변이체 아세틸 자일란 에스테라제의 아미노산 서열이다.
서열 번호 65 내지 221, 271 및 368은 모발에 대하여 친화성을 갖는 펩티드의 아미노산 서열의 비제한적인 목록이다.
서열 번호 217 내지 269는 피부에 대하여 친화성을 갖는 펩티드의 아미노산 서열이다.
서열 번호 270 및 271은 네일(nail)에 대하여 친화성을 갖는 펩티드의 아미노산 서열이다.
서열 번호 272 내지 382는 구강 표면에 대하여 친화성을 갖는 펩티드의 아미노산 서열이다. 서열 번호 272 내지 291 및 312 내지 382는 치아 균막에 대하여 친화성을 갖는다. 서열 번호 292 내지 311은 치아 에나멜에 대하여 친화성을 갖는다.
서열 번호 383 내지 396은 펩티드 링커/스페이서의 아미노산 서열이다.
서열 번호 397은 발현 플라스미드 pLD001의 핵산 서열이다.
서열 번호 398은 시퀀싱(sequencing) 프라이머의 핵산 서열이다.
서열 번호 399 내지 410은 실시예 2로부터의 치아 에나멜-결합 및 치아 균막-결합 펩티드의 아미노산 서열이다.
서열 번호 411은 표 4에 나타낸 바와 같이 C-말단 라이신이 있는 치아-결합 펩티드 DenP03의 아미노산 서열이다.
서열 번호 412 내지 422는 표 4에 나타낸 바와 같이 C-말단 라이신을 갖는 치아 에나멜-결합 펩티드 및 치아 균막-결합 펩티드의 아미노산 서열이다.
서열 번호 423은 펩티드 HC263의 아미노산 서열이다.
서열 번호 424는 본 출원에서 효소 "EZ-1"로도 지칭되는 써모토가 마리티마 변이체 C277S의 아미노산 서열이다.
서열 번호 425 내지 430 및 437 내지 467 및 479는 표 5 및/또는 표 6에 개시된 바와 같은 다양한 과가수분해효소 구축물의 아미노산 서열이다.
서열 번호 431 내지 436 및 468 내지 475는 실시예 4에 개시된 다양한 표적화 서열의 아미노산 서열이다.
서열 번호 476은 하기의 치환을 갖는 써모토가 마리티마 변이체 HTS-007-D5의 아미노산 서열이다: C277T/R296P.
서열 번호 477은 과가수분해 활성을 갖는 슈도모나스 플루오레슨스 에스테라제의 아미노산 서열이다(미국 특허 제7,384,787호; "L29P" 변이체). 치환의 넘버링이 처음의 메티오닌 잔기를 포함하지 않는 개시된 특허로부터 이어짐을 주의한다. 서열 번호 477은 처음의 메티오닌이 본 서열에 포함되기 때문에, 잔기 위치 번호 30에서 L29P 치환을 포함한다).
서열 번호 478은 야생형 마이코박테리움 스메그마티스 아릴 에스테라제의 아미노산 서열이다(미국 특허 제7,754,460호).
본 개시내용에서, 다수의 용어 및 약어가 사용된다. 하기의 정의는 특별히 다르게 언급되지 않는 한 적용된다.
본 명세서에서 사용되는 바와 같이, 본 발명의 요소 또는 성분 앞의 부정 관사("a", "an") 및 정관사("the")는 요소 또는 성분의 경우(즉, 발생)의 수에 관해서는 비제한적인 것으로 의도된다. 따라서, 부정 관사("a", "an") 및 정관사("the")는 하나 또는 적어도 하나를 포함하는 것으로 파악되어야 하며, 요소 또는 성분의 단수형은 그 수가 명백하게 단수임을 의미하는 것이 아니라면 복수형도 포함한다.
본 명세서에 사용되는 바와 같이, 용어 "포함하는"은 특허청구범위에서 인용된 바와 같은 언급된 특징, 정수, 단계, 또는 성분의 존재를 의미하지만, 하나 이상의 다른 특징, 정수, 단계, 성분 또는 그의 그룹의 존재 또는 부가를 배제하지 않는다. 용어 "포함하는"은 용어 "본질적으로 이루어진" 및 "이루어진"에 의해 포함되는 실시형태를 포함하고자 한다. 유사하게, 용어 "본질적으로 이루어진"은 용어 "이루어진"에 의해 포함되는 실시형태를 포함하고자 한다.
본 명세서에 사용되는 바와 같이, 사용되는 성분 또는 반응물질의 양을 수식하는 용어 "약"은 예를 들어, 현장에서 농축물 또는 사용 용액을 제조하는 데 사용되는 전형적인 측정 및 액체 취급 절차를 통해; 이들 과정에서의 우발적인 오차를 통해; 조성물을 제조하거나 방법을 실행하기 위해 적용된 성분의 제조, 공급원 또는 순도의 차이 등을 통해 발생할 수 있는 수치 분량의 변이를 말한다. 용어 "약"은 또한 특정 초기 혼합물로부터 유발되는 조성물에 대한 상이한 평형 조건으로 인해 달라지는 양을 포함한다. 용어 "약"에 의한 수식 여부를 불문하고, 특허청구범위는 분량의 균등물을 포함한다.
모든 범위는 존재하는 경우 포괄적이며 조합가능하다. 예를 들어, "1 내지 5"의 범위가 기재되는 경우, 기재된 범위는 범위 "1 내지 4", "1 내지 3", "1 내지 2", "1 내지 2 및 4 내지 5", "1 내지 3 및 5" 등을 포함하는 것으로 간주되어야 한다.
본 명세서에서 사용되는 "접촉"은 원하는 결과(표적 표면 결합, 과산 기반의 효과 등)를 달성하는데 충분한 기간 동안 조성물을 표적 체표면과 접촉하게 배치하는 것을 말한다 일 실시형태에서, "접촉"은 원하는 결과를 달성하는데 충분한 기간 동안 유효 농도의 과산을 포함하는(또는 이를 생성할 수 있는) 조성물을 표적 체표면과 접촉하게 배치하는 것을 말할 수 있다. 다른 실시형태에서, "접촉"은 또한 개인 관리 조성물의 적어도 하나의 성분, 예를 들어, 효소에 의한 과가수분해에 사용되는 하나 이상의 반응 성분을 표적 체표면과 접촉하게 배치하는 것을 말할 수 있다. 접촉은 유효 농도의 과산을 포함하는 과산 용액 또는 조성물, 유효 농도의 과산을 형성하는 용액 또는 조성물, 또는 유효 농도의 과산을 형성하는 조성물의 한 성분의 체표면으로의 분무, 처리, 침지, 플러싱(flushing), 그의 위 또는 안으로의 주입, 혼합, 조합, 페인팅, 코팅, 도포, 부착 및 다르게는 전달을 포함한다.
본 명세서에서 사용되는 용어 "기질", "적절한 기질" 및 "카르복실산 에스테르 기질"은 상호교환적으로, 구체적으로
(a) 구조
[X]mR5
(여기서,
X는 화학식 R6C(O)O의 에스테르기이며;
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 하나 이상의 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ; 또는
(b) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 하나 이상의 글리세리드; 또는
(c) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 또는
(d) 하나 이상의 아세틸화 단당류, 아세틸화 이당류 또는 아세틸화 다당류; 또는
(e) (a) 내지 (d)의 임의의 조합을 말한다.
본 명세서에서 사용되는 용어 "과산"은 퍼옥시산, 퍼옥시카르복실산, 퍼옥시산, 퍼카르복실산 및 퍼옥소산과 같은 의미이다.
본 명세서에서 사용되는 용어 "과아세트산"은 "PAA"로 약칭되며, 퍼옥시아세트산, 에탄퍼옥소산 및 CAS 등록 번호 79-21-0의 모든 다른 유의어와 같은 의미이다.
본 명세서에서 사용되는 용어 "모노아세틴"은 글리세롤 모노아세테이트, 글리세린 모노아세테이트 및 글리세릴 모노아세테이트와 같은 의미이다.
본 명세서에서 사용되는 용어 "다이아세틴"은 글리세롤 다이아세테이트; 글리세린 다이아세테이트, 글리세릴 다이아세테이트 및 CAS 등록 번호 25395-31-7의 모든 다른 유의어와 같은 의미이다.
본 명세서에서 사용되는 용어 "트라이아세틴"은 글리세린 트라이아세테이트; 글리세롤 트라이아세테이트; 글리세릴 트라이아세테이트, 1,2,3-트라이아세톡시프로판; 1,2,3-프로판트라이올 트라이아세테이트 및 CAS 등록 번호 102-76-1의 모든 다른 유의어와 같은 의미이다.
본 명세서에서 사용되는 용어 "모노부티린"은 글리세롤 모노부티레이트, 글리세린 모노부티레이트 및 글리세릴 모노부티레이트와 같은 의미이다.
본 명세서에서 사용되는 용어 "다이부티린"은 글리세롤 다이부티레이트 및 글리세릴 다이부티레이트와 같은 의미이다.
본 명세서에서 사용되는 용어 "트라이부티린"은 글리세롤 트라이부티레이트, 1,2,3-트라이부티릴글리세롤 및 CAS 등록 번호 60-01-5의 모든 다른 유의어와 같은 의미이다.
본 명세서에서 사용되는 용어 "모노프로피오닌"은 글리세롤 모노프로피오네이트, 글리세린 모노프로피오네이트 및 글리세릴 모노프로피오네이트와 같은 의미이다.
본 명세서에 사용되는 용어 "다이프로피오닌"은 글리세롤 다이프로피오네이트 및 글리세릴 다이프로피오네이트와 같은 의미이다.
본 명세서에서 사용되는 용어 "트라이프로피오닌"은 글리세릴 트라이프로피오네이트, 글리세롤 트라이프로피오네이트, 1,2,3-트라이프로피오닐글리세롤 및 CAS 등록 번호 139-45-7의 모든 다른 유의어와 같은 의미이다.
본 명세서에서 사용되는 용어 "아세틸화 당" 및 "아세틸화 당류"는 적어도 하나의 아세틸기를 포함하는 단당류, 이당류 및 다당류를 말한다. 예에는, 글루코스 펜타아세테이트; 자일로스 테트라아세테이트; 아세틸화 자일란; 아세틸화 자일란 단편; β-D-리보푸라노스-1,2,3,5-테트라아세테이트; 트라이-O-아세틸-D-갈락탈; 및 트라이-O-아세틸-글루칼이 포함되나 이들에 한정되지 않는다.
본 명세서에서 사용되는 용어 "하이드로카르빌", "하이드로카르빌기" 및 "하이드로카르빌 모이어티"는 단일, 이중 또는 삼중 탄소-탄소 결합 및/또는 에테르 결합으로 연결되고, 이에 따라 수소 원자로 치환된 직쇄, 분지형 또는 환형 배열의 탄소 원자를 의미한다. 이러한 하이드로카르빌기는 지방족 및/또는 방향족일 수 있다. 하이드로카르빌기의 예에는 메틸, 에틸, 프로필, 아이소프로필, 부틸, 아이소부틸, t-부틸, 사이클로프로필, 사이클로부틸, 펜틸, 사이클로펜틸, 메틸사이클로펜틸, 헥실, 사이클로헥실, 벤질 및 페닐이 포함된다. 바람직한 실시형태에서, 하이드로카르빌 모이어티는 단일 탄소-탄소 결합 및/또는 에테르 결합으로 연결되고 이에 따라 수소 원자로 치환된 직쇄, 분지형 또는 환형 배열의 탄소 원자이다.
본 명세서에서 사용되는 용어 1,2-에탄다이올; 1,2-프로판다이올; 1,3-프로판다이올; 1,2-부탄다이올; 1,3-부탄다이올; 2,3-부탄다이올; 1,4-부탄다이올; 1,2-펜탄다이올; 2,5-펜탄다이올; 1,5-펜탄다이올; 1,6-펜탄다이올; 1,2-헥산다이올; 2,5-헥산다이올; 1,6-헥산다이올의 "모노에스테르" 및 "다이에스테르" 및 그들의 혼합물은 화학식 RC(O)O(여기서, R은 C1 내지 C7 선형 하이드로카르빌 모이어티이다)의 적어도 하나의 에스테르기를 포함하는 상기 화합물을 말한다. 일 실시형태에서, 카르복실산 에스테르 기질은 프로필렌 글리콜 다이아세테이트(PGDA), 에틸렌 글리콜 다이아세테이트(EDGA) 및 그들의 혼합물로 이루어진 군으로부터 선택된다.
본 명세서에서 사용되는 용어 "프로필렌 글리콜 다이아세테이트"는 1,2-다이아세톡시프로판, 프로필렌 다이아세테이트, 1,2-프로판다이올 다이아세테이트 및 CAS 등록 번호 623-84-7의 모든 다른 유의어와 같은 의미이다.
본 명세서에서 사용되는 용어 "에틸렌 글리콜 다이아세테이트"는 1,2-다이아세톡시에탄, 에틸렌 다이아세테이트, 글리콜 다이아세테이트 및 CAS 등록 번호 111-55-7의 모든 다른 유의어와 같은 의미이다.
본 명세서에서 사용되는 용어 "적절한 효소적 반응 혼합물", "과산의 동소 생성에 적절한 성분", "적절한 반응 성분", "적절한 수성 반응 혼합물", "반응 혼합물" 및 "과산-생성 성분"은 반응물질 및 과가수분해 효소 촉매가 접촉되는 물질 및 물을 말한다. 과산-생성 성분은 적어도 과가수분해 활성을 갖는 효소를 포함하는 것이며, 바람직하게는 과가수분해 효소는 적어도 하나의 CE-7 과가수분해효소(임의로, 체표면에 표적화된 융합 단백질 형태), 적어도 하나의 적절한 카르복실산 에스테르 기질, 과산소원 및 물(과산소원, 예를 들어 과산화수소를 포함하는 수용액)이다. 일 실시형태에서, CE-7 탄수화물 에스테라제의 분류에 속하지 않는 임의의 과가수분해 효소는 조건부로, 표적 표면, 바람직하게는 구강 표면에 대하여 친화성을 갖는 적어도 하나의 펩티드 성분을 갖는 융합 단백질의 형태로 사용될 것이다.
본 명세서에서 사용되는 용어 "과가수분해"는 과산을 형성하기 위한 선택된 기질과 과산화물의 반응으로 정의된다. 전형적으로, 무기 과산화물은 촉매의 존재 하에 선택된 기질과 반응하여, 퍼옥시카르복실산을 생성한다. 본 명세서에 사용되는 용어 "화학적 과가수분해"는 기질(퍼옥시카르복실산 전구체)을 과산화수소원과 배합하는 과가수분해 반응을 포함하며, 퍼옥시카르복실산은 효소 촉매의 부재 하에 형성된다. 본 명세서에서 사용되는 용어 "효소에 의한 과가수분해"는 카르복실산 에스테르 기질(과산 전구체)을 과산화수소원 및 물과 배합하여, 효소 촉매가 과산의 형성을 촉매작용시키는 과가수분해 반응을 포함한다.
본 명세서에서 사용되는 용어 "과가수분해효소 활성"은 단백질, 건조 세포 중량 또는 고정화된 촉매 중량의 단위 질량(예를 들어, 밀리그램)당 촉매 활성을 말한다.
본 명세서에서 사용되는 "효소 활성 1 유닛" 또는 "활성 1 유닛" 또는 "U"는 특정 온도에서 분당 1 μmol의 퍼옥시카르복실산 생성물의 생성에 필요한 과가수분해효소 활성의 양으로 정의된다.
본 명세서에서 사용되는 용어 "효소 촉매" 및 "과가수분해효소 촉매"는 과가수분해 활성을 갖는 효소를 포함하는 촉매를 말하며, 온전한 미생물 세포, 투과화된(permeabilized) 미생물 세포(들), 미생물 세포 추출물의 하나 이상의 세포 성분, 부분 정제된 효소 또는 정제된 효소의 형태일 수 있다. 또한, 효소 촉매는 화학적으로 변형될 수 있다(예를 들어, 페길화(pegylation)에 의하여 또는 가교 결합 시약과의 반응에 의하여). 또한, 과가수분해효소 촉매는 당업자에게 널리 공지되어 있는 방법을 사용하여 용해성 또는 불용성 지지체 상에 고정화될 수 있으며; 예를 들어, 문헌[Immobilization of Enzymes and Cells; Gordon F. Bickerstaff, Editor; Humana Press, Totowa, NJ, USA; 1997]을 참조한다. 일 실시형태에서, 과가수분해효소 촉매는 구강 관리 스트립(예를 들어, 화이트닝 스트립(whitening strip)) 또는 덴탈 트레이(dental tray) 내에 또는 그 상에 비-공유적으로 또는 공유적으로 고정화될 수 있다. 고정화된 효소는 구강 관리 스트립 또는 덴탈 트레이(예를 들어, 이산화티탄, 수산화인회석, 구강에 허용가능한 접착제, 폴리에틸렌, 폴리프로필렌 등) 내에서 폴리머 지지체 및/또는 성분에 직접 결합될 수 있다. 추가의 실시형태에서, 스트립 또는 덴탈 트레이에 대한 비-공유적 고정화는 스트립 또는 트레이 내의 또는 그 상의 물질에 대하여 강력한 친화성을 갖는 펩티드 결합 도메인(예를 들어, 임의의 펩티드 스페이서를 통해 펩티드 결합 도메인에 결합된 과가수분해 효소를 포함하는 융합 단백질)의 사용을 통해 이루어질 수 있다. 다른 실시형태에서, 덴탈 트레이는 변형가능한 트레이이다. 추가의 실시형태에서, 과가수분해효소 촉매는 치과용 인상(dental impression)의 형성 후에 변형가능한 트레이 내에서 또는 그 상에서 고정화된다.
본 명세서에 사용되는 "아세틸 자일란 에스테라제"는 아세틸화 자일란 및 기타 아세틸화 당류의 탈아세틸화를 촉매작용시키는 효소(E.C. 3.1.1.72; AXE)를 말한다. 본 명세서에 예시된 바와 같이, 유의미한 과가수분해 활성을 갖는 아세틸 자일란 에스테라제로 분류되는 몇몇 효소가 제공된다.
본 명세서에 사용되는 용어 "세팔로스포린 C 데아세틸라제" 및 "세팔로스포린 C 아세틸 가수분해효소"는 세팔로스포린, 예를 들어, 세팔로스포린 C 및 7-아미노세팔로스포란산의 탈아세틸화를 촉매작용시키는 효소(E.C. 3.1.1.41)를 말한다(문헌[Mitsushima et al., (1995) Appl. Env. Microbiol. 61(6):2224-2229]). 유의미한 과가수분해 활성을 갖는 몇몇 세팔로스포린 C 데아세틸라제의 아미노산 서열이 본 명세서에 제공된다.
본 명세서에서 사용되는 용어 "바실러스 서브틸리스 ATCC(등록 상표) 31954(상표명)"는 국제 기탁소 수탁 번호가 ATCC(등록 상표) 31954(상표명)인 아메리칸 타입 컬쳐 콜렉션(American Type Culture Collection)(ATCC)에 기탁된 박테리아 세포를 말한다. 본 명세서에 기재된 바와 같이, 바실러스 서브틸리스 ATCC(등록 상표) 31954(상표명)로부터의 유의미한 과가수분해효소 활성을 갖는 효소는 서열 번호 2로 제공된다(미국 특허 출원 공개 제2010-0041752호 참조).
본 명세서에 사용되는 용어 "써모토가 마리티마 MSB8"은 아세틸 자일란 에스테라제 활성을 갖는 것으로 보고된 박테리아 세포이다(진뱅크(등록 상표) NP_227893.1; 미국 특허 출원 공개 제2008-0176299호 참조). 써모토가 마리티마 MSB8로부터의 과가수분해효소 활성을 갖는 효소의 아미노산 서열은 서열 번호 16으로 제공되어 있다.
본 명세서에서 사용되는 바와 같은 "분리된 핵산 분자", "분리된 폴리뉴클레오티드" 및 "분리된 핵산 단편"은 상호호환적으로 사용될 것이며, 임의로 합성, 비-천연 또는 변경된 뉴클레오티드 염기를 함유하는 단일 가닥 또는 이중 가닥의 RNA 또는 DNA의 폴리머를 지칭할 것이다. DNA의 폴리머 형태인 분리된 핵산 분자는 cDNA, 게놈 DNA 또는 합성 DNA의 하나 이상의 세그먼트로 이루어질 수 있다.
용어 "아미노산"은 단백질 또는 폴리펩티드의 기본 화학 구조 단위를 지칭한다. 하기의 약어는 특정 아미노산을 확인하기 위해 본 명세서에서 사용된다.

예를 들어, 주어진 부위에 화학적으로 균등한 아미노산의 생성을 유발하면서 암호화된 단백질의 기능적 특성에 영향을 미치지 않는 유전자의 변경이 흔하다는 것은 당업계에 주지되어 있다. 본 발명의 목적상, 치환은 하기 5 개 그룹 중 하나의 그룹 내에서의 교환으로 정의된다:
1. 비극성 또는 약간 극성인 작은 지방족 잔기: Ala, Ser, Thr (Pro, Gly);
2. 음으로 하전된 극성 잔기 및 그의 아미드: Asp, Asn, Glu, Gln;
3. 양으로 하전된 극성 잔기: His, Arg, Lys;
4. 비극성인 큰 지방족 잔기: Met, Leu, Ile, Val (Cys); 및
5. 큰 방향족 잔기: Phe, Tyr 및 Trp.
따라서, 소수성 아미노산인 아미노산 알라닌에 대한 코돈은 소수성이 보다 낮은 다른 잔기(예를 들어, 글리신) 또는 소수성이 보다 높은 잔기(예를 들어, 발린, 류신 또는 아이소류신)를 암호화하는 코돈으로 치환될 수 있다. 마찬가지로, 음으로 하전된 하나의 잔기로 다른 하나를 치환하거나(예를 들어 글루탐산을 아스파르트산으로) 양으로 하전된 하나의 잔기로 다른 하나를 치환하는(예를 들어, 아르기닌을 라이신으로) 변화도 기능적으로 균등한 생성물을 생성시키는 것으로 예상될 수 있다. 많은 경우에, 단백질 분자의 N-말단 및 C-말단 부분의 변경을 유발하는 뉴클레오티드 변화도 단백질의 활성을 변경하지 않을 것으로 예상된다. 각각의 제안된 변형은 당업계의 통상적 기술로 충분히 가능하며, 암호화된 생성물의 생물학적 활성의 보유를 결정하는 것도 그러하다.
본 명세서에서 사용되는 용어 "시그니처 모티프" 및 "진단 모티프"는 정의된 활성을 갖는 효소의 패밀리 중에 공유되는 보존된 구조를 지칭한다. 시그니처 모티프를 사용하여, 정의된 기질의 패밀리에 대하여 유사한 효소 활성을 갖는 구조적으로 관련된 효소의 패밀리를 정의하고/거나 동정할 수 있다. 시그니처 모티브는 단일의 연속 아미노산 서열 또는 함께 시그니처 모티프를 형성하는 보존된 불연속 모티프의 집합일 수 있다. 전형적으로, 보존된 모티프(들)는 아미노산 서열로 나타낸다. 일 실시형태에서, 과가수분해 효소는 CE-7 탄수화물 에스테라제 시그니처 모티프를 포함한다.
본 명세서에서 사용되는 용어 "코돈 최적화된"은 그것이 다양한 숙주의 형질전환을 위한 핵산 분자의 유전자 또는 암호화 영역을 지칭하기 때문에, DNA가 암호화하는 폴리펩티드를 변경시키지 않고, 숙주 유기체의 전형적인 코돈 사용을 반영하기 위한 핵산 분자의 유전자 또는 암호화 영역에서의 코돈의 변경을 지칭한다.
본 명세서에서 사용되는 "합성 유전자"는 당업자에게 공지되어 있는 절차를 사용하여 화학적으로 합성된 올리고뉴클레오티드 빌딩 블록(building block)으로부터 조립될 수 있다. 이들 빌딩 블록을 라이게이션시키고 어닐링시켜, 유전자 세그먼트를 형성하고, 이어서, 이를 효소에 의해 조립하여 전체 유전자를 구축한다. DNA 서열에 관계된 바와 같은 "화학적으로 합성된"은 뉴클레오티드 성분이 시험관 내에서 조립되는 것을 의미한다. DNA의 수동의 화학적 합성은 널리 확립되어 있는 절차를 사용하여 달성될 수 있거나, 자동화된 화학적 합성은 다수의 상업적으로 입수할 수 있는 장치 중 하나를 사용하여 수행할 수 있다. 따라서, 유전자는 숙주 세포의 코돈 편향을 반영하기 위한 뉴클레오티드 서열의 최적화에 기초하여 최적의 유전자 발현을 위해 맞춤화될 수 있다. 당업자는 코돈 사용이 숙주가 선호하는 코돈에 편향된다면, 성공적인 유전자 발현이 가능함을 인식한다. 바람직한 코돈의 결정은 서열 정보를 이용할 수 있는 숙주 세포 유래의 유전자의 조사에 기초할 수 있다.
본 명세서에서 사용되는 "유전자"는 암호화 서열에 앞서는 조절 서열(5' 비암호화 서열) 및 암호화 서열에 뒤따르는 조절 서열(3' 비암호화 서열)을 포함하는, 특정 단백질을 발현하는 핵산 단편을 말한다. "천연 유전자"는 그 자체의 조절 서열과 함께 자연에서 발견되는 유전자를 지칭한다. "키메라 유전자"는 자연에서 함께 발견되지 않는 조절 및 암호화 서열을 포함하는, 천연 유전자가 아닌 임의의 유전자를 지칭한다. 따라서, 키메라 유전자는 상이한 공급원 유래의 조절 서열과 암호화 서열, 또는 동일한 공급원 유래이지만, 자연에서 발견되는 것과 상이한 방식으로 배열된 조절 서열과 암호화 서열을 포함할 수 있다. "내인성 유전자"는 유기체의 게놈에서 그의 자연적 위치에 있는 천연 유전자를 지칭한다. "외래" 유전자는 숙주 유기체 내에서 정상적으로 발견되는 것이 아니라 유전자 전달에 의해 숙주 유기체 내로 도입된 유전자를 지칭한다. 외래 유전자는 비-천연 유기체 내로 삽입된 천연 유전자, 또는 키메라 유전자를 포함할 수 있다. "트랜스유전자(transgene)"는 형질전환 절차에 의해 게놈 내로 도입된 유전자이다.
본 명세서에 사용되는 "암호화 서열"은 특정 아미노산 서열을 암호화하는 DNA 서열을 말한다. "적절한 조절 서열"은 암호화 서열의 상류(5' 비-암호화 서열), 암호화 서열의 내부 또는 암호화 서열의 하류(3' 비-암호화 서열)에 위치하고, 전사, RNA 프로세싱 또는 안정성 또는 연계된 암호화 서열의 번역에 영향을 미치는 뉴클레오티드 서열을 지칭한다. 조절 서열은 프로모터, 번역 리더 서열(translation leader sequence), RNA 프로세싱 부위, 이펙터(effector) 결합 부위 및 스템-루프 구조(stem-loop structure)를 포함할 수 있다.
본 명세서에 사용되는 용어 "작동가능하게 연결된"은 하나의 기능이 다른 것에 의해 영향을 받게 하는 단일의 핵산 분자 상의 핵산 서열의 회합을 말한다. 예를 들어, 프로모터는 그것이 암호화 서열의 발현에 영향을 미칠 수 있는 경우, 다시 말하면, 암호화 서열이 프로모터의 전사적 제어 하에 있는 경우, 그 암호화 서열과 작동가능하게 연결된다. 암호화 서열은 센스 또는 안티센스 배향으로 조절 서열에 작동가능하게 연결될 수 있다.
본 명세서에 사용되는 용어 "발현"은 본 발명의 핵산 분자로부터 유래된 센스(mRNA) 또는 안티센스 RNA의 전사 및 안정한 축적을 말한다. 또한, 발현은 mRNA의 폴리펩티드로의 번역을 말할 수 있다.
본 명세서에서 사용되는 "형질전환"은 핵산 분자가 숙주 유기체의 게놈으로 전달되어, 유전적으로 안정한 유전성이 야기되는 것을 말한다. 본 발명에서, 숙주 세포의 게놈은 염색체 및 염색체외(예를 들어, 플라스미드) 유전자를 포함한다. 형질전환된 핵산 분자를 포함하는 숙주 유기체는 "트랜스제닉", "재조합" 또는 "형질전환된" 유기체로 지칭된다.
본 명세서에서 사용되는 용어 "서열 분석 소프트웨어"는 뉴클레오티드 또는 아미노산 서열을 분석하는데 유용한 임의의 컴퓨터 알고리듬 또는 소프트웨어 프로그램을 지칭한다. "서열 분석 소프트웨어"는 상업적으로 입수할 수 있거나 독립적으로 개발할 수 있다. 통상적인 서열 분석 소프트웨어는 GCG 프로그램 모음(위스콘신 패키지 버전 9.0, 미국 캘리포니아주 샌 디에고 소재의 악셀리즈 소프트웨어 코포레이션(Accelrys Software Corp.)), BLASTP, BLASTN, BLASTX(문헌[Altschul et al., J. Mol Biol. 215:403-410 (1990)]) 및 DNASTAR(미국 위스콘신주 53715 파크 스트리트 매디슨 1228 S 소재의 디엔에이스타, 인포레이티드(DNASTAR, Inc.)), 클러스털더블유(예를 들어, 버전 1.83; 문헌[Thompson et al ., Nucleic Acids Research, 22(22):4673-4680 (1994)]) 및 스미스 워터만 알고리듬(Smith-Waterman algorithm)이 통합된 FASTA 프로그램(문헌[W. R. Pearson, Comput. Methods Genome Res., [Proc. Int. Symp.] (1994), Meeting Date 1992, 111-20. Editor(s): Suhai, Sandor. Publisher: Plenum, New York, NY]), 벡터(Vector) NTI (미국 메릴랜드주 베데스다 소재의 인포맥스(Informax)) 및 시퀀처(Sequencher) v. 4.05를 포함할 것이나 이들에 한정되지 않는다. 본 출원과 관련하여, 서열 분석 소프트웨어가 분석을 위해 사용된 경우에는, 그 분석 결과가 달리 명시되지 않는 한, 참조한 프로그램의 "디폴트 값"에 기초할 것임을 이해할 것이다. 본 명세서에서 사용된 "디폴트 값"은 최초 초기화할 때에 소프트웨어에 원래 로딩된 소프트웨어 제조처에 의해 설정된 임의의 수치 또는 파라미터의 세트를 의미할 것이다.
용어 "체표면"은 유익제, 예를 들어 과산 유익제에 대한 표적으로 삼을 수 있는 임의의 인간 체표면을 말한다. 본 발명의 방법 및 조성물은 구강 관리 응용 및 제품에 관한 것이다. 이와 같이, 체표면은 구강 물질/표면을 포함한다. 일 실시형태에서, 구강 물질은 치아 에나멜, 치아 균막, 연조직, 예를 들어, 볼, 혀 및 잇몸 및 구강 생물막(예를 들어, 구강 플라크)을 포함한다.
본 명세서에서 사용되는 용어 "생물학적 오염물질"은 미생물, 포자, 바이러스, 프리온(prion) 및 그들의 혼합물을 포함하나 이들에 한정되지 않는 하나 이상의 원치않는 및/또는 병원성 생물학적 엔티티(entity)를 지칭한다. 일 실시형태에서, 생물학적 오염물질을 줄이고/거나 그의 존재를 제거하는데 유용한 유효 농도의 적어도 하나의 과산을 효소에 의해 생성하는 방법이 제공된다.
본 명세서에서 사용되는 용어 "소독한다"는 생물학적 오염물질의 파괴 또는 그의 성장의 방지 과정을 말한다. 본 명세서에 사용되는 용어 "소독제"는 인간 구강 내의 생물학적 오염 물질, 예를 들어, 치아 우식증, 치은염, 구강 칸디다증 또는 치근막염과 관련된 미생물을 포함할 수 있는 생물학적 오염물질을 파괴하거나, 중화시키거나 그의 성장을 억제함으로써 소독하는 제제를 말한다. 본 명세서에 사용되는 용어 "소독"은 소독 행위 또는 과정을 말한다. 본 명세서에 사용되는 용어 "보존제"는 질환-운반형(disease-carrying) 미생물의 성장을 억제하는 화학적 제제를 말한다. 일 태양에서, 생물학적 오염물질은 병원성 미생물이다.
본 명세서에 사용되는 용어 "위생적인"은 전형적으로 건강에 해로울 수 있는 제제를 제거하거나, 방지하거나 조절함에 의한, 건강의 회복 또는 유지를 의미하거나 그와 관련이 있다. 본 명세서에 사용되는 용어 "살균하다"는 위생적이게 만드는 것을 의미한다. 본 명세서에 사용되는 용어 "살균제"는 살균시키는 제제를 말한다. 본 명세서에 사용되는 용어 "살균"은 살균 행위 또는 과정을 말한다.
본 명세서에 사용되는 용어 "살생물제"는 미생물을 불활성화시키거나 파괴시키는 전형적으로 광범위의 화학적 제제를 말한다. 미생물을 불활성화시키거나 파괴하는 능력을 나타내는 화학적 제제는 "살생물" 활성을 갖는 것으로 기재된다. 과산은 살생물 활성을 가질 수 있다. 대안의 전형적인 살생물제는 예를 들어, 염소, 이산화염소, 클로로아이소시아누레이트, 차아염소산염, 오존, 아크롤레인, 아민, 염화 페놀 화합물, 구리 염, 유기-황 화합물 및 4차 암모늄 염을 포함할 수 있으나, 이들에 한정되지 않는다.
본 명세서에 사용되는 어구 "최소 살생물 농도"는 생존가능한 표적화된 미생물의 집단에서 소정의 접촉 시간 동안 원하는 치사의 비가역적인 감소가 생기게 할 살생물제의 최소 농도를 말한다. 유효성은 처리 후에 생존가능한 미생물의 log10 감소로 측정될 수 있다. 일 태양에서, 처리 후의 생존가능한 미생물의 표적화된 감소는 적어도 3-log10 감소, 더욱 바람직하게는 적어도 4-log10 감소, 가장 바람직하게는 적어도 5-log10 감소이다. 다른 태양에서, 최소의 살생물 농도는 생존가능한 미생물 세포의 적어도 6-log10 감소이다.
본 명세서에 사용되는 "세정 조성물" 및 "세정 제형"은 치아로부터 원치않는 화합물을 제거하는데 사용되는 조성물(구강 청결제, 치약 등)을 말한다. 상기 용어는 조성물이 과가수분해효소 및 조성물에 사용되는 다른 효소(들)와 상용성인 한, 원하는 특정 유형의 세정 조성물 및 제품의 형태(예를 들어, 액체, 페이스트, 겔, 에멀젼, 과립 또는 분무 조성물)를 위해 선택되는 임의의 물질/화합물을 포함한다.
본 명세서에 사용되는 "구강 세정 조성물"은 치분(dentifrice), 치약, 투스겔(toothgel), 투스파우더(toothpowder), 구강세정제, 구강 스프레이, 구강 젤, 츄잉 검(chewing gum), 로젠지(lozenge), 샤세(sachet), 정제, 바이오겔(biogel), 예방 페이스트(prophylaxis paste), 치아 처리 용액 등을 말한다. 본 발명의 과가수분해효소와 함께 사용되는 구강 관리 조성물은 당업계에 널리 알려져 있다(예를 들어, 모두의 전문이 본 명세서에 참고로 포함되는 미국 특허 제5,601,750호; 제6,379,653호; 및 제5,989,526호 참조).
본 명세서에서 사용되는 "약제학적으로 허용가능한"은 그 용어가 기재되는 약물, 약제 및/또는 불활성 성분이 합리적인 이익/위험 비와 상응하는 과도한 독성, 배합금기, 불안정성, 자극, 알러지 반응 등 없이 인간 및 다른 동물의 조직과 접촉하여 사용하기에 적절한 것을 의미한다.
본 명세서에서 사용되는 "개인 관리 제품"은 샴푸, 바디 로션, 샤워젤, 국소 모이스처라이저, 치약, 투스겔, 구강청결제, 구강헹굼제(mouthrinse), 플라그 방지 헹굼제(anti-plaque rinse) 및/또는 기타 국소 세정제(cleanser)를 포함하나 이들에 한정되지 않는 모발, 피부, 두피 및 치아의 세정, 탈색 및/또는 소독에 사용되는 제품을 의미한다. 특히 바람직한 일부 실시형태에서, 이들 제품은 인간에서 사용되는 한편, 다른 실시형태에서, 이들 제품은 비-인간 동물(예를 들어, 수의학 응용에서)에서 이용된다.
본 명세서에 사용되는 용어 "치아 미백" 및 "치아 탈색"은 치아(들)의 밝기를 향상시키는(예를 들어, 미백하는) 것을 지칭하는 것으로 상호교환가능하게 사용된다. 상기 용어는 본 발명을 포함하는 치아를 미백시키기에 적절한 임의의 방법뿐 아니라 화학적 처리, 약산 처리, 연마제 치아 미백 및 레이저 치아 미백을 포함하고자 한다. 특히 바람직한 실시형태에서, 본 발명은 치아를 미백시키기에 적절한 과가수분해효소 및 과가수분해효소-함유 조성물을 제공한다.
본 명세서에 사용되는 치아의 "고유 착색(stain)"은 에나멜 및 아래의 상아질 내의 색소원(chromogen)으로부터 생성된 색을 말한다. 인간 치아의 고유색은 에나멜의 세선화(thinning) 및 아래의 황색 상아질의 암화(darkening)에 기인하여 노화와 함께 더욱 황색이 되는 경향이 있다. 고유 착색의 제거는 통상 에나멜을 침투하고 내부 색소원을 탈색시키는 과산화물 또는 다른 산화 화학물질의 사용을 필요로 한다.
고유 착색과 대조적으로, "외적 착색"은 통상 천연적으로 치아를 코팅하는 균막 내에서, 외인성 색소원 물질이 에나멜에 결합되는 경우 치아의 표면 상에 형성된다. 대부분의 사람들은 시간이 지남에 따라, 그들의 치아 상에 보기 흉한 어느 정도의 외적 착색을 축적한다. 이러한 착색 과정은 (1) 탄닌-함유 식품 및 음료, 예를 들어, 커피, 차 또는 레드 와인의 섭취; (2) 담배 제품의 사용; 및/또는 (3) 특정 양이온성 물질(예를 들어, 주석, 철 및 클로르헥시딘)에 대한 노출과 같은 인자에 의해 증진된다. 이들 물질은 에나멜의 수산화인회석 구조에 부착되는 경향이 있으며, 이는 치아 변색 및 동시의 치아 백색도의 감소를 야기한다. 수년의 기간에 걸쳐, 외적 착색은 에나멜 층을 침투하고, 고유 착색을 야기할 수 있다.
본 명세서에 사용되는 용어 "탈취"는 불쾌한 냄새를 제거하거나 예방하는 것을 의미한다.
본 명세서에 사용되는 용어 "탈염시키다" 또는 "탈염"은 구강 표면으로부터 착색을 제거하는 과정을 말한다. 착색(들)은 고유 착색, 외적 착색 또는 그들의 조합일 수 있다.
본 명세서에 사용되는 과가수분해효소-함유 조성물에서의 "향상된 성능"은 치과 분야에서의 표준 방법을 사용하여 결정시, 다른 조성물에 비하여 탈색-감수성 착색의 세정을 증가시키는 것으로 정의된다. 특정 실시형태에서, 본 발명의 과가수분해효소는 산화 성능의 증가 및 착색의 제거를 제공한다. 추가의 실시형태에서, 본 발명의 과가수분해효소는 착색의 제거 및/또는 탈색의 성능 증가를 제공한다.
본 명세서에서 사용되는 "유효량의 과가수분해효소"는 특정 응용에 필요한 효소적 양을 달성하는데 필요한 양의 과가수분해효소를 말한다. 이러한 유효량은 당업자에 의해 용이하게 인식되며, 많은 인자, 예를 들어, 사용되는 특정 효소 변이체, 세정 응용, 세정 조성물의 특정 조성 및 액체 조성물이 필요한지 또는 비-액체(예를 들어, 에멀젼) 조성물이 필요한지의 여부 등에 좌우된다.
본 명세서에 사용되는 용어 "과산소원" 및 "과산소의 공급원"은 과산화수소, 과산화수소 부가물(예를 들어, 우레아-과산화수소 부가물(카르바미드 퍼옥시드)), 과붕산염 및 과탄산염을 포함하나 이들에 한정되지 않는, 수용액에 존재하는 경우 약 1 mM 이상의 농도로 과산화수소를 제공할 수 있는 화합물을 말한다. 본 명세서에 기재된 바와 같이, 수성 반응 제형 중의 과산소 화합물에 의해 제공되는 과산화수소의 농도는 반응 성분의 배합시에 초기에는 적어도 0.1 mM 또는 그 이상이다. 일 실시형태에서, 수성 반응 제형 중의 과산화수소 농도는 적어도 0.5 mM이다. 일 실시형태에서, 수성 반응 제형 중의 과산화수소 농도는 적어도 1 mM이다. 다른 실시형태에서, 수성 반응 제형 중의 과산화수소 농도는 적어도 10 mM이다. 다른 실시형태에서, 수성 반응 제형 중의 과산화수소 농도는 적어도 100 mM이다. 다른 실시형태에서, 수성 반응 제형 중의 과산화수소 농도는 적어도 200 mM이다. 다른 실시형태에서, 수성 반응 제형 중의 과산화수소 농도는 500 mM 이상이다. 또 다른 실시형태에서, 수성 반응 제형 중의 과산화수소 농도는 1000 mM 이상이다. 수성 반응 제형 중의 과산화수소 대 효소 기질, 예를 들어, 트라이글리세리드, (H2O2:기질)의 몰비는 약 0.002 내지 20, 바람직하게는 약 0.1 내지 10, 가장 바람직하게는 약 0.5 내지 5일 수 있다.
본 명세서에 사용되는 용어 "올리고당"은 글리코시드 결합에 의해 결합되는 2 내지 적어도 24개의 단당류 단위를 함유하는 화합물을 말한다. 용어 "단당류"는 실험식 (CH2O)n(여기서, n은 3 이상이고, 탄소 골격은 비분지형이며, 하나의 탄소 원자를 제외한 각 탄소 원자는 하이드록실기를 함유하고, 나머지 탄소 원자는 탄소 원자 1에서 알데히드 또는 케톤이다)의 화합물을 지칭한다. 또한, 용어 "단당류"는 세포내 환형 헤미아세탈 또는 헤미케탈 형태를 지칭한다.
본 명세서에 사용되는 용어 "부형제"는 제형 중의 활성 성분에 대한 담체로 사용되는 불활성 물질을 말한다. 부형제를 사용하여 활성 성분의 저장 안정성과 같이, 제형 중의 활성 성분을 안정화시킬 수 있다. 또한, 부형제는 때때로 활성 성분을 함유하는 제형의 부피를 늘리기 위해 사용된다. 본 명세서에 기재되는 "활성 성분"은 과가수분해 활성을 갖는 효소, 적절한 반응 조건 하에서 과가수분해 효소에 의해 생성되는 과산 또는 그들의 조합일 수 있다.
용어 "실질적으로 물이 없는"은 카르복실산 에스테르에 존재하는 경우 효소 또는 효소 분말의 저장 안정성에 해로운 영향을 주지 않는 제형 중의 물의 농도를 지칭할 것이다. 카르복실산 에스테르는 매우 낮은 농도의 물을 함유할 수 있으며, 예를 들어, 트라이아세틴은 전형적으로 180 ppm 내지 300 ppm의 물을 갖는다. 일 실시형태에서, 과가수분해 효소는 실질적으로 물이 없는 카르복실산 에스테르 기질 중에 저장된다. 추가의 실시형태에서, "실질적으로 물이 없는"은 효소(또는 효소 분말) 및 카르복실산 에스테르를 포함하는 제형 중의 2000 ppm 미만, 바람직하게는 1000 ppm 미만, 더욱 바람직하게는 500 ppm 미만, 더더욱 바람직하게는 250 ppm 미만의 물을 의미할 수 있다. 일 실시형태에서, 과가수분해 효소는 효소가 수용액(예를 들어, 저장 동안 효소에 의해 가수분해될 수 있는 상당한 농도의 카르복실산 에스테르 기질을 함유하지 않는 용액) 중에 안정적이도록 생성 시스템이 고안된다면, 수용액 중에 저장될 수 있다. 일 실시형태에서, 과가수분해 효소는 물 및 하나 이상의 완충제(예를 들어, 바이카르보네이트, 시트레이트, 아세테이트, 포스페이트, 피로포스페이트, 메틸포스포네이트, 석시네이트, 말레이트, 푸마레이트, 타르트레이트 및 말레에이트의 나트륨 및/또는 칼륨 염)가 실질적으로 없는 카르복실산 에스테르 기질을 포함하는 혼합물 중에 저장될 수 있다.
과가수분해 활성을 갖는 효소
과가수분해 활성을 갖는 효소는 효소가 본 발명의 하나 이상의 기질에 대하여 과가수분해 활성을 갖는 한, 리파제, 프로테아제, 에스테라제, 아실 트랜스퍼라제, 아릴 에스테라제, 탄수화물 에스테라제 및 조합으로 분류된 일부 효소를 포함할 수 있다. 예에는 과가수분해 프로테아제(서브틸리신 칼스버그(subtilisin Carlsberg) 변이체; 미국 특허 제7,510,859호), 과가수분해 아릴 에스테라제(슈도모나스 플루오레슨스; 서열 번호 477; 미국 특허 제7,384,787호), 마이코박테리움 스메그마티스로부터의 과가수분해 아릴 에스테라제/아실 트랜스퍼라제(서열 번호 460 및 478; 미국 특허 제7,754,460호; WO2005/056782호; 및 제EP1689859 B1호) 및 과가수분해효소 탄수화물 에스테라제가 포함될 수 있으나 이들에 한정되지 않는다. 바람직한 태양에서, 과가수분해 탄수화물 에스테라제는 CE-7 탄수화물 에스테라제이다.
일 실시형태에서, 적절한 과가수분해효소는 비-CE-7 과가수분해효소가 표적화된 과가수분해효소 응용에 제한되지 않는다면(다시 말하면, CE-7 탄수화물 에스테라제 패밀리에 속하지 않는 과가수분해 효소는 적어도 하나의 펩티드 표적화 도메인을 포함하는 융합 단백질의 형태로 사용된다), 본 명세서에 보고된 임의의 아미노산 서열에 대하여 적어도 30%, 33%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 아미노산 동일성을 갖는 아미노산 서열을 포함하는 효소를 포함할 수 있다.
다른 실시형태에서, 적절한 과가수분해효소는 비-CE-7 과가수분해효소가 표적화된 과가수분해효소 응용에 제한되지 않는다면(다시 말하면, CE-7 탄수화물 에스테라제 패밀리에 속하지 않는 과가수분해 효소는 적어도 하나의 펩티드 표적화 도메인을 포함하는 융합 단백질의 형태로 사용된다), 서열 번호 460, 477 및 478에 대하여 적어도 30%, 33%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 아미노산 동일성을 갖는 아미노산 서열을 포함하는 효소를 포함할 수 있다. 실질적으로 유사한 과가수분해 효소를 동정하기 위해 사용되는 동일성 백분율 비교 및 서열 정렬이 과가수분해 효소를 포함하는 융합 단백질의 부분(다시 말하면, 표적화 도메인 및 링커는 포함되지 않는다)에 대하여 행해지는 것이 이해된다.
다른 실시형태에서, 융합 단백질은 서열 번호 460으로 제공된 S54V 마이코박테리움 스메그마티스 아릴 에스테라제와 적어도 95% 동일성을 갖는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
일 실시형태에서, 융합 단백질은 슈도모나스 플루오레슨스로부터의 과가수분해 에스테라제를 포함한다. 다른 실시형태에서, 융합 단백질은 서열 번호 477로 제공된 슈도모나스 플루오레슨스 에스테라제와 적어도 95% 동일성을 갖는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
다른 실시형태에서, 융합 단백질은 서열 번호 424, 425, 426, 427, 428, 429, 430, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 476, 477, 478 및 479로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
다른 실시형태에서, 실질적으로 유사한 과가수분해 효소는 비-CE-7 과가수분해효소가 표적화된 과가수분해효소 응용에 제한되지 않는다면(다시 말하면, CE-7 탄수화물 에스테라제 패밀리에 속하지 않는 과가수분해 효소는 적어도 하나의 펩티드 표적화 도메인을 포함하는 융합 단백질의 형태로 사용된다), 매우 엄격한 혼성화 조건(0.1X SSC, 0.1% SDS, 65℃, 및 2X SSC, 0.1% SDS로의 세정에 이은, 0.1X SSC, 0.1% SDS, 65℃의 최종 세정) 하에서 본 발명의 임의의 과가수분해 효소를 암호화하는 폴리뉴클레오티드 서열에 혼성화되는 폴리뉴클레오티드 서열에 의해 암호화되는 것을 포함할 수 있다.
CE-7 과가수분해효소
바람직한 실시형태에서, 구강 관리 조성물 및 방법은 구조적으로 효소의 탄수화물 패밀리 에스테라제 패밀리 7(CE-7 패밀리)의 구성원으로 분류되는 과가수분해 활성을 갖는 효소를 포함한다(문헌[Coutinho, P.M., Henrissat, B. "Carbohydrate-active enzymes: an integrated database approach" in Recent Advances in Carbohydrate Bioengineering, H.J. Gilbert, G. Davies, B. Henrissat and B. Svensson eds., (1999) The Royal Society of Chemistry, Cambridge, pp. 3-12] 참조). 효소의 CE-7 패밀리는 과산소원과 배합되는 경우 다양한 카르복실산 에스테르 기질로부터 퍼옥시카르복실산을 생성하는데 특히 효과적인 것으로 증명되었다(미국 특허 제7,794,378호; 제7,951,566호; 제7,723,083호; 및 제7,964,378호 및 미국 특허 출원 공개 제2008-0176299호, 제2010-0087529호, 제2011-0081693호 및 제2011-0236335호(DiCosimo et al.); 각각은 본 명세서에 참고로 포함됨)
CE-7 패밀리의 구성원은 세팔로스포린 C 데아세틸라제(CAH; E.C. 3.1.1.41) 및 아세틸 자일란 에스테라제(AXE; E.C. 3.1.1.72)를 포함한다. CE-7 에스테라제 패밀리의 구성원은 보존된 시그니처 모티프를 공유한다(문헌[incent et al., J. Mol. Biol., 330:593-606(2003)]). CE-7 시그니처 모티프("CE-7 과가수분해효소") 및/또는 실질적으로 유사한 구조를 포함하는 과가수분해효소는 본 명세서에 기재된 조성물과 방법에 사용하기에 적절하다. 실질적으로 유사한 생물학적 분자를 동정하는 수단은 당업계에 널리 알려져 있다(예를 들어, 서열 정렬 프로토콜, 핵산 혼성화 및/또는 보존된 시그니처 모티프의 존재). 일 태양에서, 과가수분해효소는 CE-7 시그니처 모티프, 및 본 명세서에 기재된 서열 중 하나에 대하여 적어도 20%, 바람직하게는 적어도 30%, 더욱 바람직하게는 적어도 33%, 더욱 바람직하게는 적어도 40%, 더욱 바람직하게는 적어도 42%, 더욱 바람직하게는 적어도 50%, 더욱 바람직하게는 적어도 60%, 더욱 바람직하게는 적어도 70%, 더욱 바람직하게는 적어도 80%, 더욱 바람직하게는 적어도 90%, 가장 바람직하게는 적어도 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 아미노산 동일성을 포함하는 효소를 포함한다.
본 명세서에 사용되는 어구 "효소는 구조적으로 CE-7 효소로 분류된다", "CE-7 과가수분해효소" 또는 "구조적으로 탄수화물 에스테라제 패밀리 7 효소로 분류된"은 구조적으로 CE-7 탄수화물 에스테라제로 분류된 과가수분해 활성을 갖는 효소를 지칭하기 위해 사용될 것이다. 이러한 효소의 패밀리는 시그니처 모티프의 존재에 의해 정의될 수 있다(상기 문헌[Vincent et al.]). CE-7 에스테라제에 대한 시그니처 모티프는 3가지 보존된 모티프를 포함한다(서열 번호 2의 참조 서열에 대한 잔기 위치 넘버링; 바실러스 서브틸리스 ATCC(등록 상표) 31954(상표명)로부터의 CE-7 과가수분해효소):
a) Arg118-Gly119-Gln120;
b) Gly179-Xaa180-Ser181-Gln182-Gly183; 및
c) His298-Glu299.
전형적으로, 아미노산 잔기 위치 180에서의 Xaa는 글리신, 알라닌, 프롤린, 트립토판 또는 트레오닌이다. 촉매 트라이아드에 속하는 3개의 아미노산 잔기 중 2개를 볼드체로 나타내었다. 일 실시형태에서, 아미노산 잔기 위치 180에서의 Xaa는 글리신, 알라닌, 프롤린, 트립토판 및 트레오닌으로 이루어진 군으로부터 선택된다.
CE-7 탄수화물 에스테라제 패밀리 내의 보존된 모티프의 추가의 분석에 의해, CE-7 탄수화물 에스테라제 패밀리에 속하는 과가수분해효소를 추가로 정의하기 위해 사용될 수 있는 추가의 보존된 모티프(서열 번호 2의 아미노산 위치 267 내지 269에서의 LXD)의 존재가 나타난다. 추가의 실시형태에서, 상기 정의된 시그니처 모티프는 하기로 정의된 추가의(제4) 보존된 모티프를 포함할 수 있다:
Leu267-Xaa268-Asp269.
아미노산 잔기 위치 268에서의 Xaa는 전형적으로, 아이소류신, 발린 또는 메티오닌이다. 제4 모티프는 촉매 트라이아드(Ser181-Asp269-His298)에 속하는 아스파르트산 잔기(볼드체)를 포함한다.
CE-7 과가수분해효소는 적어도 하나의 체표면에 대하여 친화성을 갖는 적어도 하나의 펩티드 성분을 갖는 융합 단백질의 형태일 수 있다. 일 실시형태에서, 표적화된 과가수분해효소(융합 단백질)가 CE-7 시그니처 모티프를 포함하는지를 결정하기 위해 사용되는 모든 정렬은 체표면에 대하여 친화성을 갖는 펩티드 성분이 없는 과가수분해 효소의 아미노산 서열에 기초할 것이다.
많은 널리 공지된 전반적인 정렬 알고리듬(즉, 서열 분석 소프트웨어)을 사용하여 과가수분해효소 활성을 갖는 효소를 대표하는 둘 이상의 아미노산 서열을 정렬하여, 효소가 본 발명의 시그니처 모티프로 이루어지는지 결정할 수 있다. 정렬된 서열(들)을 참조 서열(서열 번호 2)과 비교하여, 시그니처 모티프의 존재를 결정한다. 일 실시형태에서, 참조 아미노산 서열을 사용하는 클러스털(CLUSTAL) 정렬(예를 들어, 클러스털더블유)(본 명세서에 사용되는 바와 같이, 바실러스 서브틸리스 ATCC(등록 상표) 31954(상표명)로부터의 과가수분해효소 서열(서열 번호 2))을 사용하여 CE-7 에스테라제 패밀리에 속하는 과가수분해효소를 동정한다. 보존된 아미노산 잔기의 상대적 넘버링은 참조 아미노산 서열의 잔기 넘버링에 기초하여, 정렬된 서열 내의 작은 삽입 또는 결실(예를 들어, 전형적으로 5개 이하의 아미노산)을 설명한다.
(참조 서열에 비하여) 본 발명의 시그니처 모티프를 포함하는 서열을 동정하기 위해 사용될 수 있는 다른 적절한 알고리듬의 예에는 니들만(Needleman) 및 분쉬(Wunsch)(문헌[J. Mol. Biol. 48, 443-453 (1970)]; 전체 정렬 도구) 및 스미스-워터만(Smith-Waterman)(문헌[J. Mol. Biol. 147:195-197 (1981)]; 국소 정렬 도구)아 포함되나 이들에 한정되지 않는다. 일 실시형태에서, 스미스-워터만 정렬은 디폴트 파라미터를 사용하여 이행된다. 적절한 디폴트 파라미터의 예에는 블로섬(BLOSUM)62 스코어링 매트릭스(scoring matrix)의 사용이 포함되며, 갭 개방 페널티(GAP open penalty)는 10이고, 갭 연장 페널티(GAP extension penalty)는 0.5이다.
과가수분해효소 중의 전체 동일성 백분율의 비교에 의해, (시그니처 모티프를 보유하면서) 서열 번호 2에 대하여 33%만큼 적은 아미노산 동일성을 갖는 효소가 유의미한 과가수분해효소 활성을 나타내는 것으로 표현되며, 구조적으로 CE-7 탄수화물 에스테라제로 분류된다. 일 실시형태에서, 적절한 과가수분해효소는 CE-7 시그니처 모티프, 및 서열 번호 2에 대하여 적어도 20%, 바람직하게는 적어도 30%, 33%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 아미노산 동일성을 포함하는 효소를 포함한다.
과가수분해 활성을 갖는 적절한 CE-7 탄수화물 에스테라제의 예에는 아미노산 서열, 예를 들어, 서열 번호 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476을 갖는 효소가 포함되나 이들에 한정되지 않는다. 일 실시형태에서, 효소는 14, 16, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476으로 이루어진 군으로부터 선택되는 아미노산 서열을 포함한다.
본 명세서에서 사용되는 용어 "CE-7 변이체", "변이체 과가수분해효소" 또는 "변이체"는 CE-7 시그니처 모티프 및 관련 과가수분해 활성이 유지되는 한, 변이체가 유래되는 해당하는 효소(전형적으로 야생형 효소)에 비하여, 적어도 하나의 아미노산 부가, 결실 및/또는 치환을 야기하는 유전자 변형을 갖는 CE-7 과가수분해효소를 지칭할 것이다. 또한, CE-7 변이체 과가수분해효소는 본 발명의 조성물 및 방법에서 사용될 수 있다. CE-7 변이체의 예는 서열 번호 27, 28, 29, 30, 31, 32, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476으로 제공된다. 일 실시형태에서, 변이체는 서열 번호 27, 28, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476을 포함할 수 있다.
당업자는 실질적으로 유사한 CE-7 과가수분해효소 서열(시그니처 모티프 보유)이 또한 본 발명의 조성물 및 방법에 사용될 수 있음을 인식한다. 일 실시형태에서, 실질적으로 유사한 서열은 매우 엄격한 조건 하에서 본 명세서에 예시된 서열과 관련된 핵산 분자와 혼성화하는 그들의 능력에 의해 정의된다. 다른 실시형태에서, 서열 정렬 알고리듬을 사용하여, 본 명세서에 제공되는 DNA 또는 아미노산 서열에 대한 동일성 백분율에 기초하여 실질적으로 유사한 효소를 정의할 수 있다.
본 명세서에서 사용되는 핵산 분자는 제1 분자의 단일의 가닥이 적절한 온도 및 용액 이온 세기의 조건 하에서 다른 분자에 어닐링될 수 있는 경우, 다른 핵산 분자, 예를 들어, cDNA, 게놈 DNA 또는 RNA에 "혼성화가능하다". 혼성화 및 세정 조건은 널리 공지되어 있으며, 문헌[Sambrook, J. and Russell, D., T. Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor (2001)]에 예시되어 있다. 온도 및 이온 세기 조건은 혼성화의 "엄격성(stringency)"을 결정한다. 엄격성 조건을 조정하여, 관계가 먼 유기체로부터의 상동 서열과 같은 중등으로 유사한 분자 내지 밀접하게 관련된 유기체로부터의 기능성 효소를 중복시키는 유전자와 같은 고도로 유사한 분자를 스크리닝할 수 있다. 혼성화 후 세정이 전형적으로 엄격성 조건을 결정한다. 한 세트의 바람직한 조건은 실온에서 15분 동안 6X SSC, 0.5% SDS로 시작한 다음, 45℃에서 30분 동안 2X SSC, 0.5% SDS로 반복한 다음, 50℃에서 30분 동안 0.2X SSC, 0.5% SDS로 2회 반복한 일련의 세정을 사용한다. 더욱 바람직한 세트의 조건은 더 높은 온도를 사용하며, 여기서, 세정은 0.2X SSC, 0.5% SDS에서의 최종 2회의 30분 세정의 온도가 60℃로 증가된 것을 제외하고 상기와 동일하다. 다른 바람직한 세트의 고도로 엄격한 혼성화 조건은 0.1X SSC, 0.1% SDS, 65℃이며, 2X SSC, 0.1% SDS로 세정하고, 0.1X SSC, 0.1% SDS, 65℃의 최종 세정으로 이어진다.
혼성화의 엄격성에 따라 염기 간의 미스매치가 가능하지만, 혼성화를 위해서는 2개의 핵산이 상보적 서열을 함유하는 것이 요구된다. 혼성화되는 핵산에 대해 적절한 엄격성은, 당업계에 주지된 변수인 핵산의 길이 및 상보성의 정도에 의존한다. 2개 뉴클레오티드 서열 사이의 유사성 또는 상동성의 정도가 클수록, 이들 서열을 가진 핵산의 하이브리드에 대한 Tm의 값이 커진다. 핵산 혼성화의 상대적 안정성(높은 Tm에 상응함)은 하기의 순서로 감소한다: RNA:RNA, DNA:RNA, DNA:DNA. 100개 뉴클레오티드 길이를 초과하는 하이브리드에 대하여, Tm을 계산하는 수학식이 유도되었다(상기 문헌[Sambrook and Russell]). 더 짧은 핵산, 즉, 올리고뉴클레오티드의 혼성화에 있어서는 미스매치의 위치가 더욱 중요해지며 올리고뉴클레오티드의 길이가 그의 특이성을 결정한다(상기 문헌[Sambrook and Russell]). 일 태양에서, 혼성화가능한 핵산의 길이는 적어도 약 10개 뉴클레오티드이다. 바람직하게는, 혼성화가능한 핵산의 최소의 길이는 적어도 약 15개 뉴클레오티드 길이, 더욱 바람직하게는 적어도 약 20개 뉴클레오티드 길이, 더더욱 바람직하게는 적어도 30개 뉴클레오티드 길이, 더더욱 바람직하게는 적어도 300개 뉴클레오티드 길이, 가장 바람직하게는 적어도 800개 뉴클레오티드 길이이다. 또한, 프로브의 길이와 같은 인자에 따라 필요한 경우에 온도 및 세정 용액 염 농도를 조정할 수 있음을 당업자는 인식할 것이다.
본 명세서에서 사용되는 용어 "동일성 백분율"은 2개 이상의 폴리펩티드 서열 또는 2개 이상의 폴리뉴클레오티드 서열 사이의 관계로서, 서열을 비교함으로써 결정된다. 당업계에서, "동일성"은 또한 경우에 따라서, 서열의 스트링(string) 간의 매치에 의해 결정되는, 폴리펩티드 또는 폴리뉴클레오티드 서열 간의 서열 관련성의 정도를 의미한다. "동일성" 및 "유사성"은 문헌[Computational Molecular Biology (Lesk, A. M., ed.) Oxford University Press, NY (1988)]; 문헌[Biocomputing: Informatics and Genome Projects (Smith, D. W., ed.) Academic Press, NY (1993)]; 문헌[Computer Analysis of Sequence Data, Part I (Griffin, A. M., and Griffin, H. G., eds.) Humana Press, NJ (1994)]; 문헌[Sequence Analysis in Molecular Biology (von Heinje, G., ed.) Academic Press (1987)]; 및 문헌[Sequence Analysis Primer (Gribskov, M. and Devereux, J., eds.) Stockton Press, NY (1991)]에 기재된 것을 포함하나 이들에 한정되지 않는 공지되어 있는 방법에 의해 용이하게 계산할 수 있다. 동일성 및 유사성을 결정하는 방법은 공중에게 이용가능한 컴퓨터 프로그램에 명시되어 있다. 레이저진(LASERGENE) 바이오인포매틱스 컴퓨팅 스위트(bioinformatics computing suite)(미국 위스콘신주 매디슨 소재의 디엔에이스타 인코포레이티드)의 멕얼라인(Megalign) 프로그램, 벡터 NTI v. 7.0(미국 메릴랜드주 베데스다 소재의 인포맥스, 인코포레이티드)의 얼라인엑스(AlignX) 프로그램, 또는 엠보스 오픈 소프트웨어 스위트(EMBOSS Open Software Suite)(EMBL-EBI; 문헌[Rice et al., Trends in Genetics 16, (6):276-277 (2000)])를 사용하여 서열 정렬 및 동일성 백분율 계산을 수행할 수 있다. 서열의 다중의 정렬은 디폴트 파라미터와 함께 클러스털 정렬 방법(예를 들어, 클러스털더블유; 예를 들어, 버전 1.83)(문헌[Higgins and Sharp, CABIOS, 5:151-153 (1989)]; 문헌[Higgins et al., Nucleic Acids Res. 22:4673-4680 (1994)]; 및 문헌[Chenna et al., Nucleic Acids Res 31 (13):3497-500 (2003)], 유럽 바이오인포매틱스 연구소(European Bioinformatics Institute)를 통하여 유럽 분자 생물학 실험실로부터 입수가능함)을 사용하여 수행할 수 있다. 클러스털더블유 단백질 정렬에 적절한 파라미터는 갭 존재 페널티(GAP Existence penalty)=15, 갭 연장=0.2, 매트릭스=고넷(Gonnet)(예를 들어, 고넷250), 단백질 엔드갭(ENDGAP)=-1, 단백질 갭디스트(GAPDIST)=4, 및 KTUPLE=1을 포함한다. 일 실시형태에서, 고속 또는 저속 정렬은 디폴트 설정으로 사용되며, 저속 정렬이 바람직하다. 대안적으로, 클러스털더블유 방법(예를 들어, 버전 1.83)을 사용하는 파라미터는 또한 KTUPLE=1, 갭 페널티=10, 갭 연장=1, 매트릭스=블로섬(예를 들어, 블로섬64), 윈도우(WINDOW)=5 및 탑 디아고날 세이브드(TOP DIAGONALS SAVED)=5를 사용하여 변경될 수 있다.
일 태양에서, 적절한 분리된 핵산 분자는 본 명세서에 기재된 아미노산 서열에 대하여 적어도 약 20%, 바람직하게는 적어도 30%, 33%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 동일한 아미노산 서열을 갖는 폴리펩티드를 암호화한다. 다른 태양에서, 적절한 분리된 핵산 분자는 본 명세서에 기재된 아미노산 서열에 대하여 적어도 약 20%, 바람직하게는 적어도 30%, 33%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 동일한 아미노산 서열을 갖는 폴리펩티드를 암호화하되; 단, 폴리펩티드는 CE-7 시그니처 모티프를 보유한다. 적절한 핵산 분자는 상기 상동성을 가질 뿐 아니라, 또한 전형적으로 약 210 내지 340개 아미노산 길이, 약 300 내지 약 340개 아미노산, 바람직하게는 약 310 내지 약 330개 아미노산, 가장 바람직하게는 약 318 내지 약 325개 아미노산 길이를 갖는 폴리펩티드를 암호화하며, 각 폴리펩티드는 과가수분해 활성을 갖는 것을 특징으로 한다.
표적화된 과가수분해효소
본 명세서에 사용되는 용어 "표적화된 과가수분해효소" 및 "과가수분해 활성을 갖는 표적화된 효소"는 표적 표면, 바람직하게는 표적화된 체표면에 대하여 친화성을 갖는 적어도 하나의 펩티드 성분에 융합/결합된 적어도 하나의 과가수분해 효소(야생형 또는 그의 변이체)를 포함하는 융합 단백질을 지칭할 것이다. 표적화된 과가수분해효소 내의 과가수분해 효소는 임의의 과가수분해 효소일 수 있으며, 효소가 본 발명의 하나 이상의 기질에 대하여 과가수분해 활성을 갖는 한, 리파제, 프로테아제, 에스테라제, 아실 트랜스퍼라제, 아릴 에스테라제, 탄수화물 에스테라제 및 조합물을 포함할 수 있다. 예에는 과가수분해 프로테아제(예를 들어, 서브틸리신 변이체; 미국 특허 제7,510,859호), 과가수분해 에스테라제(예를 들어, 슈도모나스 플루오레슨스; 미국 특허 제7,384,787호; 서열 번호 477) 및 과가수분해 아릴 에스테라제(예를 들어, 마이코박테리움 스메그마티스; 미국 특허 제7,754,460호; WO2005/056782호; 및 제EP1689859 B1호; 서열 번호 460[S54V 변이체] 및 478[야생형])가 포함될 수 있으나, 이들에 한정되지 않는다.
일 실시형태에서, 융합 단백질은 서열 번호 460으로 제공된 S54V 마이코박테리움 스메그마티스 아릴 에스테라제와 적어도 95% 동일성을 갖는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
일 실시형태에서, 융합 단백질은 슈도모나스 플루오레슨스(Pseudomonas fluorescens)로부터의 과가수분해 에스테라제를 포함한다. 다른 실시형태에서, 융합 단백질은 서열 번호 477로 제공된 슈도모나스 플루오레슨스 에스테라제와 적어도 95% 동일성을 갖는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
다른 실시형태에서, 융합 단백질은 서열 번호 424, 425, 426, 427, 428, 429, 430, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 476, 477, 478 및 479로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 과가수분해 효소를 포함한다.
본 명세서에서 사용되는 용어 "펩티드 성분", "구강 표면에 대하여 친화성을 갖는 펩티드 성분" 및 "OCBD"는 펩티드 결합에 의해 결합된 둘 이상의 아미노산의 적어도 하나의 폴리머를 포함하는 과가수분해 효소의 부분이 아닌 융합 단백질의 성분을 지칭할 것이며; 성분은 표적 구강 표면에 대하여 친화성을 갖는다.
일 실시형태에서, 체표면에 대하여 친화성을 갖는 펩티드 성분은 항체, Fab 항체 단편, 단쇄 가변 단편(scFv) 항체, 카멜리대 항체(문헌[Muyldermans, S., Rev. Mol. Biotechnol. (2001) 74:277-302]), 비-항체 스캐폴드 디스플레이 단백질(다양한 스캐폴드-보조 방법의 검토를 위한 문헌[Hosse et al., Prot. Sci. (2006) 15(1): 14-27] 및 문헌[Binz, H. et al . (2005) Nature Biotechnology 23, 1257-1268]) 또는 면역글로불린 폴드(immunoglobulin fold)가 결여된 단쇄 폴리펩티드일 수 있다. 다른 태양에서, 체표면에 대하여 친화성을 갖는 펩티드 성분은 면역글로불린 폴드가 결여된 단쇄 펩티드이다(즉, 구강 표면에 대하여 친화성을 갖는 체표면-결합 펩티드 또는 적어도 하나의 체표면-결합 펩티드를 포함하는 체표면-결합 도메인). 바람직한 실시형태에서, 펩티드 성분은 구강 표면에 대하여 친화성을 갖는 하나 이상의 체표면-결합 펩티드를 포함하는 단쇄 펩티드이다.
구강 표면에 대하여 친화성을 갖는 펩티드 성분은 임의의 펩티드 링커에 의해 과가수분해효소로부터 분리될 수 있다. 소정의 펩티드 링커/스페이서는 길이가 1 내지 100개 또는 1 내지 50개 아미노산이다. 일부 실시형태에서, 펩티드 스페이서는 길이가 약 1 내지 약 25개, 3 내지 약 40개, 또는 3 내지 약 30개 아미노산이다. 다른 실시형태에서, 스페이서는 길이가 약 5 내지 약 20개 아미노산이다. 다수의 펩티드 링커가 사용될 수 있다. 일 실시형태에서, 적어도 하나의 펩티드 링커가 존재하며, 최대 10번 반복될 수 있다.
이전에 하나의 체표면에 대하여 친화성을 갖는 것으로 동정된 펩티드는 구강 표면에 대한 친화성도 또한 가질 수 있다. 이와 같이, 융합 펩티드는 다른 체표면, 예를 들어, 모발(서열 번호 65 내지 221, 271 및 368); 피부(서열 번호 217 내지 269); 또는 네일(서열 번호 270 및 271)에 대하여 친화성을 갖는 것으로 이전에 보고된 적어도 하나의 것을 포함할 수 있다. 일 실시형태에서, 융합 펩티드는 서열 번호 272 내지 382 및 399 내지 422를 포함하는 군으로부터의 적어도 하나의 구강 표면-결합 펩티드를 포함한다. 일 실시형태에서, 융합 펩티드는 서열 번호 272 내지 382, 399 내지 410 및 412 내지 422로 이루어진 군으로부터 선택되는 적어도 하나의 구강 표면-결합 펩티드를 포함하며; 서열 번호 272 내지 291 및 312 내지 382는 치아 균막에 대하여 친화성을 가지며; 서열 번호 292 내지 311은 치아 에나멜에 대하여 친화성을 갖고; 서열 번호 399 내지 410 및 412 내지 422는 치아 에나멜 또는 균막에 대하여 친화성을 갖는다. 일부 체표면-결합 펩티드는 1개 초과의 체표면에 대하여 강력한 친화성을 가질 수 있으며, 이와 같이 상이한 체표면에 대하여 과가수분해 효소를 표적화하기 위해 사용될 수 있다. 다른 실시형태에서, 융합 펩티드는 표적 체표면에 정전기 인력을 갖도록 고안된 임의의 체표면-결합 펩티드(예를 들어, 표적 체표면에 정전기적으로 결합되도록 엔지니어링된 체표면-결합 펩티드)를 포함할 수 있다.
다른 실시형태에서, 표적 표면은 패키징 및/또는 구강으로의 전달 방법의 일부의 물질이다. 펩티드 성분은 폴리머, 플라스틱 및 막과 같은 이용에서 물질 또는 물질들에 대한 친화성에 대해 선택된다. 표적화된 과가수분해효소 융합 단백질 고안은 비제한적으로 마우쓰(mouth) 트레이 또는 스트립과 같은 제거가능한 장치에서 이를 유지함으로써 사용자로부터의 과가수분해효소의 전달의 조절 및 제거를 가능하게 한다.
표적화된 CE-7 과가수분해효소
바람직한 실시형태에서, "표적화된 과가수분해효소"는 과가수분해 활성을 갖는 표적화된 CE-7 탄수화물 에스테라제이다. 본 명세서에 사용되는 용어 "표적화된 CE-7 과가수분해효소" 및 "표적화된 CE-7 탄수화물 에스테라제"는 표적화된 표면, 바람직하게는 표적화된 체표면에 대하여 친화성을 갖는 적어도 하나의 펩티드 성분에 융합/결합된 적어도 하나의 CE-7 과가수분해효소(야생형 또는 변이체 과가수분해효소)를 포함하는 융합 단백질을 지칭할 것이다. 체표면에 대하여 친화성을 갖는 펩티드 성분은 상기 기재된 임의의 것일 수 있다. 바람직한 태양에서, 표적화된 CE-7 과가수분해효소의 펩티드 성분은 면역글로불린 폴드가 결여된 단쇄 펩티드이다(즉, 구강 표면에 대하여 친화성을 갖는 체표면-결합 펩티드 또는 적어도 하나의 체표면-결합 펩티드를 포함하는 체표면-결합 도메인). 바람직한 실시형태에서, 펩티드 성분은 구강 표면에 대하여 친화성을 갖는 하나 이상의 체표면-결합 펩티드를 포함하는 단쇄 펩티드이다.
구강 표면에 대하여 친화성을 갖는 펩티드 성분은 임의의 펩티드 링커에 의해 CE-7 과가수분해효소로부터 분리될 수 있다. 소정의 펩티드 링커/스페이서는 길이가 1 내지 100개 또는 1 내지 50개 아미노산이다. 일부 실시형태에서, 펩티드 스페이서는 길이가 약 1 내지 약 25개, 3 내지 약 40개, 또는 3 내지 약 30개 아미노산이다. 다른 실시형태에서, 스페이서는 길이가 약 5 내지 약 20개 아미노산이다. 다수의 펩티드 링커가 사용될 수 있다.
이와 같이, 표적화된 CE-7 과가수분해효소의 예에는 구강 표면에 대하여 친화성을 갖는 펩티드 성분에 결합된 서열 번호 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476으로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 임의의 CE-7 과가수분해효소가 포함될 수 있으나, 이들에 한정되지 않는다. 바람직한 실시형태에서, 표적화된 과가수분해효소의 예에는 구강 표면에 대하여 친화성을 갖는 하나 이상의 체표면-결합 펩티드에 (임의로 펩티드 스페이서를 통해) 결합된 서열 번호 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476으로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 임의의 CE-7 과가수분해효소가 포함될 수 있으나, 이들에 한정되지 않는다.
다른 실시형태에서, 표적화된 CE-7 과가수분해효소는 또한 구강 관리 표면에 대하여 친화성을 가질 수 있는, 하나의 체표면에 대하여 친화성을 갖는 것으로 이전에 동정된 펩티드를 포함할 수 있다. 이와 같이, 융합 펩티드는 다른 체표면, 예를 들어, 모발(서열 번호 65 내지 221, 271 및 368); 피부(서열 번호 217 내지 269); 또는 네일(서열 번호 270 및 271)에 대하여 친화성을 갖는 것으로 이전에 보고된 적어도 하나의 것을 포함할 수 있다. 일 실시형태에서, 융합 펩티드는 서열 번호 272 내지 382 및 399 내지 422를 포함하는 군으로부터의 적어도 하나의 구강 표면-결합 펩티드를 포함한다. 일 실시형태에서, CE-7 과가수분해효소 융합 펩티드는 서열 번호 272 내지 382, 399 내지 410 및 412 내지 422로 이루어진 군으로부터 선택되는 적어도 하나의 구강 표면-결합 펩티드를 포함하며; 서열 번호 272 내지 291 및 312 내지 382는 치아 균막에 대하여 친화성을 가지며; 서열 번호 292 내지 311은 치아 에나멜에 대하여 친화성을 갖고; 서열 번호 399 내지 410 및 412 내지 422는 치아 에나멜 또는 균막에 대하여 친화성을 갖는다. 일부 체표면-결합 펩티드는 1개 초과의 체표면에 대하여 강력한 친화성을 가질 수 있으며, 이와 같이 상이한 체표면에 대하여 과가수분해 효소를 표적화하기 위해 사용될 수 있다. 다른 실시형태에서, CE-7 과가수분해효소 융합 펩티드는 표적 체표면에 정전기 인력을 갖도록 고안된 임의의 체표면-결합 펩티드(예를 들어, 표적 체표면에 정전기적으로 결합되도록 엔지니어링된 체표면-결합 펩티드)를 포함할 수 있다.
다른 실시형태에서, 표적 표면은 패키징 및/또는 구강으로의 전달 방법의 일부인 물질이다. 펩티드 성분은 폴리머, 플라스틱 및 막과 같은 이용에서 물질 또는 물질들에 대한 친화성에 대해 선택된다. 표적화된 CE-7 과가수분해효소 융합 단백질 고안은 마우쓰 트레이 또는 스트립과 같은 제거가능한 장치에서 이를 유지함으로써 사용자로부터의 과가수분해효소의 전달의 조절 및 제거를 가능하게 한다.
체표면에 대하여 친화성을 갖는 펩티드
구강 표면에 결합할 수 있는 면역글로불린 폴드가 결여된 단쇄 펩티드는 "구강 표면-결합 펩티드"(OCBP)로 지칭되며, 예를 들어, 치아 표면에 결합하는 펩티드(치아-결합 펩티드), 연조직, 예를 들어, 잇몸에 대하여 친화성을 갖는 펩티드 또는 구강에서 사용하기에 안전한 구강에 허용가능한 물질에 대하여 친화성을 갖는 펩티드를 포함할 수 있다. 치아-결합 펩티드는 치아 에나멜에 대하여 친화성을 갖는 펩티드("치아 에나멜-결합 펩티드") 및 치아 균막에 대하여 친화성을 갖는 펩티드("치아 균막-결합 펩티드")를 포함할 수 있다.
모발(서열 번호 65 내지 221, 271 및 368로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 모발-결합 펩티드), 피부(피부-결합 펩티드는 서열 번호 217 내지 269로 이루어진 군으로부터 선택되는 아미노산 서열을 포함한다) 및 네일(네일-결합 펩티드는 서열 번호 270 및 271로 이루어진 군으로부터 선택되는 아미노산 서열을 포함한다)에 대하여 친화성을 갖는 것을 포함하는 적어도 하나의 체표면에 대하여 친화성을 갖는 비제한적인 펩티드의 목록이 본 명세서에 제공된다. 구강 표면에 대하여 친화성을 갖는 펩티드(구강-결합 펩티드)의 예에는 서열 번호 272 내지 382 및 399 내지 422로 이루어진 군으로부터 선택되는 아미노산 서열이 포함된다. 바람직한 태양에서, 구강 표면에 대하여 친화성을 갖는 펩티드는 서열 번호 272 내지 382, 399 내지 410 및 412 내지 422로 이루어진 군으로부터 선택되며; 서열 번호 272 내지 291 및 312 내지 382는 치아 균막에 대하여 친화성을 갖고; 서열 번호 292 내지 311은 치아 에나멜에 대하여 친화성을 가지며; 서열 번호 399 내지 410 및 412 내지 422는 치아 에나멜 또는 균막에 대하여 친화성을 갖는다.
일 실시형태에서, 구강 표면에 대하여 친화성을 가질 수 있는 펩티드는 또한 서열 번호 65 내지 382, 399 내지 410 및 412 내지 422 중 하나 이상을 포함할 수 있다. 바람직하게는 본 발명의 조성물 및 방법에 사용되는 펩티드는 서열 번호 272 내지 382, 399 내지 410 및 412 내지 422로 이루어진 군으로부터 선택된다. 다른 실시형태에서, 구강 표면-결합 펩티드는 일부 구강 표면(예를 들어, 잇몸)에 대한 피부-결합 펩티드를 포함할 수 있다. 다른 실시형태에서, 융합 펩티드는 표적 체표면에 정전기 인력을 갖도록 고안된 임의의 체표면-결합 펩티드(예를 들어, 표적 체표면에 정전기적으로 결합되도록 엔지니어링된 체표면-결합 펩티드)를 포함할 수 있다.
다른 실시형태에서, 본 발명의 조성물 및 방법은 서열 번호 399 내지 410 및 412 내지 422로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 적어도 하나의 구강 표면-결합 펩티드를 포함한다.
일부 실시형태에서, 구강 표면-결합 도메인은 길이가 최대 약 60개 아미노산인 구강 표면-결합 펩티드로 이루어진다. 일 실시형태에서 구강 표면-결합 펩티드는 길이가 5 내지 60개 아미노산이다. 다른 실시형태에서, 표면-결합 펩티드는 길이가 7 내지 50개 아미노산이거나, 길이가 7 내지 30개 아미노산이다. 또 다른 실시형태는 길이가 7 내지 27개 아미노산인 구강 표면-결합 펩티드이다.
구강 표면-결합 펩티드를 포함하는 융합 펩티드가 본 발명의 특정 실시형태이지만, 본 발명의 다른 실시형태에서 다수의 구강 표면결합 펩티드를 사용하는 것이 유리할 수 있다. 다수, 즉, 둘 이상의 구강 표면-결합 펩티드의 포함은 예를 들어, 심지어 단일 구강 표면-결합 펩티드를 포함하는 결합 요소보다 더 내구성 있는 펩티드 성분을 제공할 수 있다. 일부 실시형태에서, 구강 표면-결합 도메인(다시 말하면, 다수, 즉, 2개 이상의 구강 표면-결합 펩티드)은 2 내지 약 50개 또는 2 내지 약 25개 구강 표면-결합 펩티드를 포함한다. 다른 실시형태는 2 내지 약 10개 또는 2 내지 5개 구강 표면-결합 펩티드를 포함하는 구강 표면-결합 도메인을 포함한다.
다수의 결합 요소(즉, 구강 표면-결합 펩티드 또는 구강 표면-결합 도메인)는 서로 직접 결합될 수 있거나, 이들은 펩티드 스페이서를 사용하여 함께 결합될 수 있다. 소정의 펩티드 스페이서/링커는 길이가 1 내지 100개 또는 1 내지 50개 아미노산이다. 일부 실시형태에서, 펩티드 스페이서는 길이가 약 1 내지 약 25개, 3 내지 약 40개, 또는 3 내지 약 30개 아미노산이다. 다른 실시형태에서, 스페이서는 길이가 약 1 내지 약 20개 또는 약 5 내지 약 20개 아미노산이다.
구강 표면-결합 도메인 및 그들을 구성하는 더 짧은 구강 표면-결합 펩티드는 예를 들어, 임의의 공지되어 있는 바이오패닝 기술, 예를 들어, 파지 디스플레이, 박테리아 디스플레이, 효모 디스플레이, 리보솜 디스플레이, mRNA 디스플레이 및 그들의 조합을 비롯한 당업계에 공지되어 있는 많은 방법을 사용하여 동정할 수 있다. 전형적으로, 펩티드의 무작위 또는 실질적인 무작위(사건에서 편향이 존재) 라이브러리를 표적 체표면에 대하여 바이오패닝하여, 표적 체표면에 대하여 친화성을 갖는 라이브러리 내의 펩티드를 동정한다.
펩티드의 무작위 라이브러리의 생성은 널리 공지되어 있으며, 박테리아 디스플레이(문헌[Kemp, D.J.; Proc. Natl. Acad. Sci. USA 78(7):4520-4524 (1981)] 및 문헌[Helfman et al., Proc. Natl. Acad. Sci. USA 80(1):31-35, (1983)]), 효모 디스플레이(문헌[Chien et al., Proc Natl Acad Sci USA 88(21):9578-82 (1991)]), 조합 고체상 펩티드 합성(미국 특허 제5,449,754호, 미국 특허 제5,480,971호, 미국 특허 제5,585,275호, 미국 특허 제5,639,603호) 및 파지 디스플레이 기술(미국 특허 제5,223,409호, 미국 특허 제5,403,484호, 미국 특허 제5,571,698호, 미국 특허 제5,837,500호); 리보솜 디스플레이(미국 특허 제5,643,768호; 미국 특허 제5,658,754호; 및 미국 특허 제7,074,557호),및 mRNA 디스플레이 기술(프로퓨젼(상표명); 미국 특허 제6,258,558호; 제6,518,018호; 제6,281,344호; 제6,214,553호; 제6,261,804호; 제6,207,446호; 제6,846,655호; 제6,312,927호; 제6,602,685호; 제6,416,950호; 제6,429,300호; 제7,078,197호; 및 제6,436,665호 참조)을 포함하는 다양한 기술에 의해 달성될 수 있다.
결합 친화성
구강 표면에 대하여 친화성을 갖는 펩티드 성분은 10-5 몰(M) 이하의 구강 표면에 대한 결합 친화성을 포함한다. 특정 실시형태에서, 펩티드 성분은 10-5 몰(M) 이하의 인간 모발, 피부, 네일 또는 구강에 대한 결합 친화성을 갖는 하나 이상의 구강 표면-결합 펩티드 및/또는 결합 도메인(들)이다. 일부 실시형태에서, 결합 펩티드 또는 도메인은 적어도 약 50 내지 500 mM 염의 존재 하에 10-5 M 이하의 결합 친화성 값을 가질 것이다. 용어 "결합 친화성"은 결합 펩티드와 그의 각 기질, 이 경우에는 인간 구강 표면(잇몸, 치아 등)의 상호작용의 세기를 말한다. 결합 친화성은 결합 펩티드의 해리 상수("KD") 또는 "MB50"에 관하여 정의되거나 측정될 수 있다.
"KD"는 표적 상의 결합 부위가 절반 점유된, 다시 말하면, 펩티드가 결합된 표적(결합 표적 물질)의 농도가 펩티드가 결합되지 않은 표적의 농도와 같은 경우의 펩티드의 농도에 해당한다. 해리 상수가 더 작을수록, 펩티드가 더욱 단단히 결합된다. 예를 들어, 나노몰(nM) 해리 상수를 갖는 펩티드는 마이크로몰(μM) 해리 상수를 갖는 펩티드보다 더욱 단단히 결합한다. 본 발명의 특정 실시형태는 KD 값이 10-5 이하일 것이다.
"MB50"은 ELISA-기반의 결합 검정법에서 수득되는 최대 신호의 50%의 신호를 제공하는 결합 펩티드의 농도를 말한다. 예를 들어, 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2005/022683호의 실시예 3을 참고한다. MB50은 복합체의 성분의 결합 상호작용 또는 친화성의 세기의 표시를 제공한다. MB50의 값이 더 낮을수록, 펩티드와 그의 해당하는 기질의 상호작용이 더욱 강력하며, 다시 말하면, 더 낫다. 예를 들어, 나노몰(nM) MB50을 갖는 펩티드는 마이크로몰(μM) MB50을 갖는 펩티드보다 더욱 단단하게 결합한다. 본 발명의 특정 실시형태는 MB50 값이 10-5 M 이하일 것이다.
일부 실시형태에서, 구강 표면에 대하여 친화성을 갖는 펩티드 성분은 KD 또는 MB50 값으로 측정시, 결합 친화성이 약 10-5 M 이하, 약 10-6 M 이하, 약 10-7 M 이하, 약 10-8 M 이하, 약 10-9 M 이하, 또는 약 10-10 M 이하일 수 있다.
일부 실시형태에서, 구강 표면-결합 펩티드 및/또는 구강 표면-결합 도메인은 KD 또는 MB50 값으로 측정시, 결합 친화성이 약 10-5 M 이하, 약 10-6 M 이하, 약 10-7 M 이하, 약 10-8 M 이하, 약 10-9 M 이하, 또는 약 10-10 M 이하일 수 있다.
본 명세서에서 사용되는 용어 "강력한 친화성"은 KD 또는 MB50 값이 약 10-5 M 이하, 바람직하게는 약 10-6 M 이하, 더욱 바람직하게는 약 10-7 M 이하, 더욱 바람직하게는 약 10-8 M 이하, 약 10-9 M 이하, 또는 가장 바람직하게는 약 10-10 M 이하인 결합 친화성을 지칭할 것이다.
다성분
퍼옥시카르복실산
생성 시스템
다수의 활성 성분을 분리하고 배합하는 시스템 및 수단의 고안은 당업계에 알려져 있으며, 일반적으로 개별 반응 성분의 물리적 형태에 좌우될 것이다. 예를 들어, 다수의 활성 유체(액체-액체) 시스템은 전형적으로 멀티-챔버 디스펜서(multi-chamber dispenser) 병 또는 2상 시스템(예를 들어, 미국 특허 출원 공개 제2005/0139608호; 미국 특허 제5,398,846호; 미국 특허 제5,624,634호; 미국 특허 제6,391,840호; 유럽 특허 제0807156B1호; 미국 특허 출원 공개 제2005/0008526호; 및 PCT 공개 WO 00/61713호), 예컨대 원하는 탈색제가 반응성 유체의 혼합시에 생성되는 일부 탈색 응용에서 관찰되는 것을 사용한다. 퍼옥시카르복실산을 생성하기 위해 사용되는 다른 형태의 다성분 시스템은 하나 이상의 고체 성분 또는 고체-액체 성분의 조합, 예를 들어, 분말(예를 들어, 미국 특허 제5,116,575호), 다층 정제(예를 들어, 미국 특허 제6,210,639호), 다수의 구획을 갖는 수용해성 패킷(packet)(예를 들어, 미국 특허 제6,995,125호) 및 물의 첨가 시에 반응하는 고체 응집체(예를 들어, 미국 특허 제6,319,888호)를 위해 고안된 것들을 포함할 수 있으나, 이들에 한정되지 않는다.
다른 실시형태에서, 제1 성분 중의 카르복실산 에스테르는 모노아세틴, 다이아세틴, 트라이아세틴 및 그들의 조합으로 이루어진 군으로부터 선택된다. 다른 실시형태에서, 제1 성분 중의 카르복실산 에스테르는 아세틸화 당류이다. 다른 실시형태에서, 제1 성분 중의 효소 촉매는 미립자 고체, 액체 또는 겔일 수 있다. 다른 실시형태에서, 제1 반응 성분은 고체 정제 또는 분말일 수 있다.
퍼옥시카르복실산은 매우 반응성이 있으며, 일반적으로 시간이 지남에 따라 농도가 감소된다. 이는 특히 종종 장기간 안정성이 결여된 시판용 사전-형성된 퍼옥시카르복실산 조성물에 대하여 그러하다. 또한, 특히 큰 용기 및/또는 매우 농축된 퍼옥시카르복실산 용액을 더 긴 거리로 운송하는 경우, 사전-형성된 퍼옥시카르복실산 수용액은 취급 및/또는 운송의 어려움을 나타낼 수 있다. 추가로, 사전-형성된 퍼옥시카르복실산 용액은 특정 표적 응용을 위해 원하는 농도의 퍼옥시카르복실산을 제공할 수 없을 수 있다. 이와 같이, 특히 액체 제형을 위해 다양한 반응 성분을 분리되게 유지하는 것이 매우 바람직하다.
원하는 퍼옥시카르복실산을 생성하기 위해 배합되는 둘 이상의 성분을 포함하는 다성분 퍼옥시카르복실산 생성 시스템의 이용이 보고되어 있다. 개별 성분은 연장된 기간 동안 안정적이며, 취급하기에 안전해야 한다(다시 말하면, 혼합 시에 생성되는 퍼옥시카르복실산의 농도로 측정시). 일 실시형태에서, 다성분 효소적 퍼옥시카르복실산 생성 시스템의 저장 안정성은 효소 촉매 안정성에 관하여 측정될 수 있다.
원하는 퍼옥시카르복실산 농도를 갖는 과산 수용액을 빠르게 생성하기 위해 효소 촉매를 사용하는, 다성분 퍼옥시카르복실산 생성 제형을 포함하는 개인 관리 제품이 본 명세서에 제공된다. 혼합은 사용 직전에 및/또는 적용 부위(동소)에서 발생할 수 있다. 일 실시형태에서, 개인 관리 제품 제형은 사용될 때까지 분리되어 유지되는 적어도 2개의 성분으로 이루어질 것이다. 성분의 혼합에 의해, 과산 수용액이 신속하게 형성된다. 생성된 과산 수용액이 의도된 최종 용도에 적절한 유효 농도의 과산을 포함하도록 각 성분이 고안된다. 개별 성분의 조성은 (1) 연장된 저장 안정성을 제공하고/거나 (2) 퍼옥시카르복실산으로 이루어진 적절한 수성 반응 제형의 형성을 증진시키는 능력을 제공하도록 고안되어야 한다.
다성분 제형은 적어도 2개의 실질적으로 액체인 성분으로 이루어질 수 있다. 일 실시형태에서, 다성분 제형은 제1 액체 성분 및 제2 액체 성분을 포함하는 2성분 제형일 수 있다. 용어 "제1" 또는 "제2" 액체 성분의 이용은 상대적이되, 단 특정 성분을 포함하는 2가지 상이한 액체 성분은 사용할 때까지 분리되어 유지된다. 최소한도로, 다성분 퍼옥시카르복실산 제형은 (1) 과가수분해 활성을 가지며, 바람직하게는 CE-7 에스테라제로 분류된 적어도 하나의 효소 촉매, (2) 카르복실산 에스테르 기질 및 (3) 과산소원 및 물을 포함하며, 제형은 성분의 혼합시에 원하는 과산을 효소에 의해 생성한다.
2성분 제형에 사용되는 다양한 성분의 유형 및 양은 (1) 각 성분의 저장 안정성, 특히 효소 촉매의 과가수분해 활성 및 (2) 용해성 및/또는 원하는 퍼옥시카르복실산 수용액을 효율적으로 형성하는 능력을 증진시키는 물리적 특성을 제공하도록 조심히 선택되고, 균형을 이루어야 한다(예를 들어, 수성 반응 혼합물 중의 에스테르 기질의 용해성을 증진시키는 성분 및/또는 적어도 하나의 액체 성분의 점도 및/또는 농도를 변경하는 성분[다시 말하면, 효소에 의한 과가수분해 활성에 대한 상당한 역효과를 갖지 않는 적어도 하나의 공용매]).
효소에 의한 과산 생성 시스템의 성능 및/또는 촉매 안정성을 향상시키는 다양한 방법이 개시된다. 미국 특허 출원 공개 제2010-0048448 A1호에는 용해성 및/또는 특정 에스테르 기질의 혼합 특징을 증진시키는 적어도 하나의 공용매의 이용이 기재된다. 또한, 본 발명의 개인 관리 조성물 및 방법은 공용매를 사용할 수 있다. 일 실시형태에서, 카르복실산 에스테르 기질 및 과가수분해효소 촉매를 포함하는 성분은 Log P 값이 약 2 미만인 유기 용매를 포함하며, Log P는 P = [용질]옥탄올/[용질]물로 표현되는 옥탄올과 물 사이의 기질의 분배 계수의 로그로 정의된다. 효소 활성에 대한 유의미한 역효과를 갖지 않는 log P 값이 2 이하인 몇몇 공용매가 기재된다. 다른 실시형태에서, 공용매는 카르복실산 에스테르 기질 및 효소를 포함하는 반응 성분 내에서 약 20 wt% 내지 약 70 wt%이다. 카르복실산 기질 및 효소를 포함하는 반응 성분은 임의로, 하나 이상의 완충제(예를 들어, 바이카르보네이트, 시트레이트, 아세테이트, 포스페이트, 피로포스페이트, 메틸포스포네이트, 석시네이트, 말레이트, 푸마레이트, 타르트레이트 및 말레에이트의 나트륨 및/또는 칼륨 염)를 포함할 수 있다.
미국 특허 출원 공개 제2010-0086534 A1호에는 2성분 시스템의 용도가 기재되어 있으며, 여기서, 제1 성분은 액체 카르복실산 에스테르와 고체 효소 분말의 제형을 포함하며, 상기 효소 분말은 (a) 과가수분해 활성을 갖는 적어도 하나의 CE-7 에스테라제 및 (b) 적어도 하나의 올리고당 부형제의 제형을 포함하며; 제2 성분은 과산소원 및 과산화수소 안정화제를 갖는 물을 포함한다. 본 발명의 개인 관리 조성물 및 방법은 미국 특허 출원 공개 US 2010-0086534 A1호에 기재된 시스템과 유사한 2성분 제형을 사용할 수 있다. 이와 같이, 올리고당 부형제를 사용하여 효소 활성의 안정화를 도울 수 있다. 일 실시형태에서, 올리고당 부형제는 수평균 분자량이 적어도 약 1250일 수 있고, 중량평균 분자량이 적어도 약 9000일 수 있다. 다른 실시형태에서, 올리고당 부형제는 수평균 분자량이 적어도 약 1700이고, 중량평균 분자량이 적어도 약 15000이다. 다른 실시형태에서, 올리고당은 말토덱스트린이다.
또한, 미국 특허 출원 공개 제2010-0086535-A1호에는 2성분 시스템이 기재되어 있으며, 여기서, 제1 성분은 액체 카르복실산 에스테르와 고체 효소 분말의 제형을 포함하며, 상기 제형은 (a) 과가수분해 활성을 갖는 적어도 하나의 CE-7 에스테라제 및 적어도 하나의 올리고당 부형제 및 적어도 하나의 계면활성제를 포함하는 효소 분말; 및 (b) 적어도 하나의 완충제를 포함하고, 바람직한 실시형태에서, 완충제는 분리된(다시 말하면, 효소 분말과 분리된) 불용성 성분으로서 카르복실산 에스테르 기질에 첨가되고; 제2 성분은 과산소원 및 과산화수소 안정화제를 갖는 물을 포함한다. 본 발명의 개인 관리 조성물 및 방법은 미국 특허 출원 공개 제2010-0086535 A1호에 기재된 시스템과 유사한 2성분 제형을 사용할 수 있다. 일 실시형태에서, 부형제는 수평균 분자량이 적어도 약 1250이고, 중량평균 분자량이 적어도 약 9000인 올리고당 부형제일 수 있다. 다른 실시형태에서, 올리고당 부형제는 수평균 분자량이 적어도 약 1700일 수 있고, 중량평균 분자량이 적어도 약 15000일 수 있다. 다른 실시형태에서, 올리고당은 말토덱스트린이다. 추가의 실시형태에서, 임의의 pH 완충제는 바이카르보네이트 완충제이다. 추가의 실시형태에서, 과산화수소 안정화제는 터피날(TURPINAL)(등록 상표) SL이다.
효소 분말
일부 실시형태에서, 개인 관리 조성물은 안정화된 효소 분말 형태의 효소 촉매를 사용할 수 있다. 효소 분말을 포함하는 제형을 제조하고 안정화시키는 방법은 미국 특허 출원 공개 제2010-0086534호 및 제2010-0086535호에 기재되어 있다.
일 실시형태에서, 효소는 효소 분말의 건조 중량에 기초하여, 약 5 중량 퍼센트(wt%) 내지 약 75 wt% 범위의 양으로 효소 분말에 존재할 수 있다. 효소 분말/분무-건조된 혼합물 중의 효소의 바람직한 중량% 범위는 약 10 wt% 내지 50 wt%이며, 효소 분말/분무-건조된 혼합물 중의 효소의 더욱 바람직한 중량% 범위는 약 20 wt% 내지 33 wt%이다.
일 실시형태에서, 효소 분말은 부형제를 추가로 포함할 수 있다. 일 태양에서, 부형제는 효소 분말의 건조 중량에 기초하여 약 95 wt% 내지 약 25 wt%의 범위의 양으로 제공된다. 효소 분말 중의 부형제의 바람직한 wt% 범위는 약 90 wt% 내지 50 wt%이며, 효소 분말 중의 부형제의 더욱 바람직한 wt% 범위는 약 80 wt% 내지 67 wt%이다.
일 실시형태에서, 효소 분말을 제조하기 위해 사용되는 부형제는 올리고당 부형제일 수 있다. 일 실시형태에서, 올리고당 부형제는 수평균 분자량이 적어도 약 1250이고, 중량평균 분자량이 적어도 약 9000이다. 일부 실시형태에서, 올리고당 부형제는 수평균 분자량이 적어도 약 1700이고, 중량평균 분자량이 적어도 약 15000이다. 특정 올리고당은 말토덱스트린, 자일란, 만난, 후코이단, 갈락토만난, 키토산, 라피노스, 스타키오스, 펙틴, 인슐린, 레반, 그라미난, 아밀로펙틴, 수크로스, 락툴로스, 락토스, 말토스, 트레할로스, 셀로비오스, 니제로트라이오스(nigerotriose), 말토트라이오스, 멜레지토스, 말토트라이울로스(maltotriulose), 라피노스, 케스토스(kestose) 및 그들의 혼합물을 포함할 수 있으나, 이들에 한정되지 않는다. 바람직한 실시형태에서, 올리고당 부형제는 말토덱스트린이다. 또한, 올리고당계 부형제는 수용성 비이온성 셀룰로스 에테르, 예를 들어, 하이드록시메틸-셀룰로스 및 하이드록시프로필메틸셀룰로스 및 그들의 혼합물을 포함할 수 있지만, 이들에 한정되지 않는다. 추가의 실시형태에서, 부형제는 하기의 화합물 중 하나 이상으로부터 선택될 수 있으나 이들에 한정되지 않는다: 트레할로스, 락토스, 수크로스, 만니톨, 소르비톨, 글루코스, 셀로비오스, α-사이클로덱스트린 및 카르복시메틸셀룰로스.
제형은 적어도 하나의 임의의 계면활성제를 포함할 수 있으며, 여기서, 적어도 하나의 계면활성제의 존재가 바람직하다. 계면활성제는 이온성 및 비이온성 계면활성제 또는 습윤제, 예를 들어, 에톡실화 피마자유, 폴리글리콜화 글리세리드, 아세틸화 모노글리세리드, 소르비탄 지방산 에스테르, 폴록사머, 폴리옥시에틸렌 소르비탄 지방산 에스테르, 폴리옥시에틸렌 유도체, 모노글리세리드 또는 그의 에톡실화 유도체, 다이글리세리드 또는 그의 폴리옥시에틸렌 유도체, 소듐 도쿠세이트, 소듐 라우릴설페이트, 콜린산 또는 그의 유도체, 레시틴, 인지질, 에틸렌 글리콜 및 프로필렌 글리콜의 블록 코폴리머 및 비이온성 유기규소를 포함할 수 있으나, 이들에 한정되지 않는다. 바람직하게는, 계면활성제는 폴리옥시에틸렌 소르비탄 지방산 에스테르이며, 폴리소르베이트 80이 더욱 바람직하다.
제형이 효소 분말을 포함하는 경우, 분말을 제조하는데 사용되는 계면활성제는 효소 분말에 존재하는 단백질의 중량에 기초하여, 약 5 wt% 내지 0.1 wt%, 바람직하게는 효소 분말에 존재하는 단백질의 중량에 기초하여, 약 2 wt% 내지 0.5 wt% 범위의 양으로 존재할 수 있다.
효소 분말은 추가로 하나 이상의 완충제(예를 들어, 바이카르보네이트, 시트레이트, 아세테이트, 포스페이트, 피로포스페이트, 메틸포스포네이트, 석시네이트, 말레이트, 푸마레이트, 타르트레이트 및 말레에이트의 나트륨 및/또는 칼륨 염), 및 효소 안정화제(예를 들어, 에틸렌다이아민테트라아세트산, (1-하이드록시에틸리덴)비스포스폰산)를 포함할 수 있다.
효소 분말을 형성하기 위한 제형의 분무 건조는 예를 들어, 문헌[Spray Drying Handbook, 5th ed., K. Masters, John Wiley & Sons, Inc., NY, N.Y. (1991)] 및 PCT 특허 출원 공개 WO 97/41833호 및 WO 96/32149호(Platz, R. et al.)에 일반적으로 기재된 바와 같이 수행된다.
일반적으로, 분무 건조는 고도로 분산된 액체와 충분한 양의 고온 공기가 함께 있게 하여 증발을 초래하는 하는 단계 및 액적을 건조시키는 단계로 구성된다. 전형적으로, 공급물은 용매를 증발시키고 건조된 생성물을 수집기로 이동시키는 따뜻한 여과된 공기의 흐름으로 분무된다. 소모된 공기는 이어서 용매와 함께 배기된다. 당업자는 몇몇 상이한 유형의 장치를 사용하여 원하는 제품을 제공할 수 있음을 인식할 것이다. 예를 들어, 부치 리미티드(Buchi Ltd.)(스위스 포스트파치 소재) 또는 지이에이 니로 코포레이션(GEA Niro Corp.)(덴마크 코펜하겐 소재)에 의해 제조되는 시판용 분무 건조기는 원하는 크기의 입자를 효율적으로 생성할 것이다. 특히 이중 노즐 기술을 사용하는 2가지 용액의 동시의 분무와 같은 특정 응용을 위해 이들 분무 건조기 및 그들의 아토마이저(atomizer)가 변경되거나 맞춤화될 수 있음을 추가로 인식할 것이다. 더욱 구체적으로, 유중수형 에멀젼은 하나의 노즐로부터 무화될 수 있으며, 만니톨과 같은 부착-방지제(anti-adherent)를 함유하는 용액을 제2 노즐로부터 공동-무화할 수 있다. 다른 경우에, 고압 액체 크로마토그래피(HPLC) 펌프를 사용하여 맞춤 설계된 노즐을 통해 공급 용액을 밀어내는 것이 바람직할 수 있다. 정확한 형태 및/또는 조성을 포함하는 미세구조체가 생성된다면, 장치의 선택은 중요하지 않으며, 본 명세서의 교시에 비추어 당업자에게 명백할 것이다.
분무된 재료를 건조시키는데 사용되는 기체의 양쪽 입구 및 출구의 온도는 분무된 재료에서 효소의 분해를 야기하지 않는 정도이다. 이러한 온도는 전형적으로 실험에 의해 측정되지만, 일반적으로, 입구 온도는 약 50℃ 내지 약 225℃ 범위이면서 출구 온도는 약 30℃ 내지 약 150℃ 범위일 것이다. 바람직한 파라미터는 약 0.14 ㎫ 내지 1.03 ㎫(20 내지 150 psi), 바람직하게는 약 0.21-0.28 ㎫ 내지 0.69 ㎫(30-40 내지 100 psi) 범위의 무화 압력을 포함한다. 전형적으로, 사용되는 무화 압력은 다음 중 하나일 것이다(㎫): 0.14, 0.21, 0.28, 0.34, 0.41, 0.48, 0.55, 0.62, 0.69, 0.76, 0.83 또는 그 이상.
일 실시형태에서, "실질적으로 그의 효소적 활성을 유지한다"는 카르복실산 에스테르 중의 효소 분말 또는 효소 분말 제형이 카르복실산 에스테르 및 효소 분말로 이루어진 제형의 제조 전의 효소 분말의 초기 효소 활성에 비하여, 주위 온도에서의 연장된 저장 기간 후에 및/또는 고온(주위 온도 초과)에서의 짧은 저장 기간 후에, 카르복실산 에스테르 및 효소 분말로 이루어진 제형에서의 효소 분말 또는 효소 분말 제형 중의 효소의 효소 활성의 적어도 약 75%를 유지하는 것을 의미한다. 연장된 저장 기간은 주위 온도에서의 약 1년 내지 약 2년의 기간이다. 일 실시형태에서, 고온에서 짧은 저장 기간은 카르복실산 에스테르 및 효소 분말로 이루어진 제형이 40℃에서 생성되는 때로부터 40℃에서 약 8주까지의 기간이다. 다른 실시형태에서, 고온은 약 30℃ 내지 약 52℃의 범위이다. 바람직한 실시형태에서, 고온은 약 30℃ 내지 약 40℃ 범위이다.
일부 실시형태에서, 효소 분말은 40℃에서 카르복실산 에스테르 및 효소 분말로 이루어진 제형의 제조 전의 효소 분말의 초기 효소 활성에 비하여, 40℃에서 8주의 저장 후에 카르복실산 에스테르 및 효소 분말로 이루어진 제형 중에, 효소 활성의 적어도 75%를 유지한다. 다른 실시형태에서, 효소 분말은 40℃에서 카르복실산 에스테르 및 효소 분말로 이루어진 제형의 제조 전의 효소 분말의 초기 효소 활성에 비하여, 40℃에서의 8주의 저장 후에 카르복실산 에스테르 및 효소 분말로 이루어진 제형에서 적어도 하나의 효소의 효소 활성의 적어도 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 또는 100%를 유지한다. 바람직하게는, 과가수분해 활성은 미국 특허 출원 공개 제2010-0086510호의 실시예 8 내지 13에 기재된 바와 같이 측정되나; 과가수분해 활성의 임의의 측정 방법이 사용될 수 있다.
언급된 기간에 걸친 효소 활성의 추가의 향상은 미국 특허 출원 공개 제2010-0086534호에 기재된 바와 같이, 약 5.5 내지 약 9.5의 pH 범위의 완충 능력을 갖는 완충제를 카르복실산 에스테르 및 분무-건조된 효소 분말로 이루어진 제형에 첨가함으로써 달성될 수 있다. 적절한 완충제는 바이카르보네이트, 피로포스페이트, 포스페이트, 메틸포스포네이트, 시트레이트, 아세테이트, 말레이트, 푸마레이트, 타르트레이트, 말레에이트 또는 석시네이트의 나트륨 염, 칼륨 염 또는 나트륨 또는 칼륨 염의 혼합물을 포함할 수 있으나 이들에 한정되지 않는다. 카르복실산 에스테르 및 분무-건조된 효소 분말로 이루어진 제형에서 사용하기에 바람직한 완충제는 바이카르보네이트, 피로포스페이트, 포스페이트, 메틸포스포네이트, 시트레이트, 아세테이트, 말레이트, 푸마레이트, 타르트레이트, 말레에이트, 또는 석시네이트의 나트륨 염, 칼륨 염, 또는 나트륨 또는 칼륨 염의 혼합물을 포함한다.
완충제가 카르복실산 에스테르 및 효소 분말 제형에 존재할 수 있는 실시형태에서, 완충제는 카르복실산 에스테르 및 효소 분말로 이루어진 제형 중의 카르복실산 에스테르의 중량에 기초하여, 약 0.01 wt% 내지 약 50 wt% 범위의 양으로 존재할 수 있다. 완충제는 카르복실산 에스테르 및 효소 분말로 이루어진 제형 중의 카르복실산 에스테르의 중량에 기초하여, 약 0.10% 내지 약 10%의 더욱 바람직한 범위로 존재할 수 있다. 추가로, 이들 실시형태에서, 효소의 과가수분해 활성 간의 비교는 카르복실산 에스테르, 약 5.5 내지 약 9.5의 pH 범위에서 완충 능력을 갖는 완충제 및 효소 분말로 이루어진 제형의 제조 전의 효소 분말의 초기 과가수분해 활성에 비한, 40℃에서의 8주의 저장 후에 카르복실산 에스테르, 약 5.5 내지 약 9.5의 pH 범위에서 완충 능력을 갖는 완충제 및 효소 분말로 이루어진 제형에서 적어도 하나의 효소의 과가수분해 활성의 적어도 75%를 유지하는 효소 분말 간의 비교로 결정된다.
건조 효소 분말이 적어도 하나의 효소에 대한 기질인 유기 화합물, 예를 들어, 트라이아세틴 중의 제형으로 저장되는 것으로 의도된다. 첨가되는 과산화수소의 부재 하에, 트라이아세틴은 보통 CE-7 탄수화물 에스테라제에 의해 수용액 중에서 가수분해되어, 다이아세틴 및 아세트산을 생성하며, 아세트산의 생성에 의해, 반응 혼합물의 pH의 감소가 야기된다. 트라이아세틴 중의 효소의 장기간 저장 안정성을 위한 하나의 요건은 트라이아세틴 중에 존재할 수 있는 임의의 물과 트라이아세틴의 유의미한 반응이 존재하지 않는 것이며; 하나의 시판용 트라이아세틴(벨기에 브뤼셀 소재의 테센델로 그룹(Tessenderlo Group)에 의해 공급) 중의 수분 함량에 대한 명시는 0.03 wt%의 물(300 ppm)이다. 트라이아세틴 중의 효소의 저장 동안 발생하는 트라이아세틴의 임의의 가수분해에 의해, 아세트산이 생성될 것이며, 이는 CE-7 과가수분해효소의 활성의 감소 또는 CE-7 과가수분해효소의 불활성화를 야기할 수 있고; 과가수분해효소는 전형적으로 pH 5.0 이하에서 불활성화된다(미국 특허 출원 공개 제2009-0005590호(DiCosimo, R., et al) 참조). 본 출원서에서 사용하기 위해 선택된 부형제는 아세트산이 제형 중의 낮은 농도의 물의 존재에 기인하여 생성될 수 있는 조건 하의 효소를 위하여, 유기 기질 중에서의 효소의 안정성을 제공해야 한다. 건조 효소 분말은 적어도 하나의 효소에 대한 기질인 유기 화합물 중의 제형으로 저장될 수 있으며, 여기서, 제형은 추가로 부형제 및 하나 이상의 완충제(예를 들어, 바이카르보네이트, 시트레이트, 아세테이트, 포스페이트, 피로포스페이트, 메틸포스포네이트, 석시네이트, 말레이트, 푸마레이트, 타르트레이트 및 말레에이트의 나트륨 및/또는 칼륨 염)를 포함한다.
효소에 의해 촉매작용되는,
카르복실산
에스테르 및 과산화수소로부터의
과산의
제조에 적절한 반응 조건
본 발명의 개인 관리 조성물 및 방법에서, 과가수분해 활성을 갖는 하나 이상의 효소를 사용하여 유효 농도의 원하는 과산(들)을 생성할 수 있다. 원하는 퍼옥시카르복실산은 카르복실산 에스테르를 과가수분해 활성을 갖는 효소 촉매의 존재 하에 과산화수소, 과산화아연, 과산화나트륨, 우레아 퍼옥시드, 과산화칼슘, 과붕산나트륨, 과탄산나트륨, 또는 과산화수소의 복합체를 포함하나 이들에 한정되지 않는 과산소원과 반응시킴으로써 제조될 수 있다.
표적화된 과가수분해효소 내의 과가수분해 효소는 임의의 과가수분해 효소일 수 있으며, 효소가 본 발명의 하나 이상의 기질에 대하여 과가수분해 활성을 갖는 한, 리파제, 프로테아제, 에스테라제, 아실 트랜스퍼라제, 아릴 에스테라제, 탄수화물 에스테라제 및 조합물을 포함할 수 있다. 예에는 과가수분해 프로테아제(서브틸리신 변이체; 미국 특허 제7,510,859호), 과가수분해 에스테라제(슈도모나스 플루오레슨스; 미국 특허 제7,384,787호; "L29P" 변이체 서열 번호 477) 및 과가수분해 아릴 에스테라제(마이코박테리움 스메그마티스; 미국 특허 제7,754,460호; WO2005/056782호; 및 EP1689859 B1호; 서열 번호 460[S54V 변이체] 및 478[야생형])가 포함될 수 있으나 이들에 한정되지 않는다.
일 실시형태에서, 효소 촉매는 과가수분해효소 활성을 갖는 적어도 하나의 효소를 포함하며, 상기 효소는 구조적으로, CE-7 탄수화물 에스테라제 패밀리(CE-7; 상기 문헌[Coutinho, P.M., and Henrissat, B.] 참조)의 구성원으로 분류된다. 다른 실시형태에서, 과가수분해효소 촉매는 구조적으로, 세팔로스포린 C 데아세틸라제로 분류된다. 다른 실시형태에서, 과가수분해효소 촉매는 구조적으로, 아세틸 자일란 에스테라제로 분류된다.
일 실시형태에서, 과가수분해효소 촉매는 과가수분해 활성, 및
a) 서열 번호 2의 아미노산 잔기 118 내지 120과 정렬되는 RGQ 모티프;
b) 서열 번호 2의 아미노산 잔기 179 내지 183과 정렬되는 GXSQG 모티프; 및
c) 서열 번호 2의 아미노산 잔기 298 및 299와 정렬되는 HE 모티프를 포함하는 시그니처 모티프를 갖는 효소를 포함한다.
바람직한 실시형태에서, 서열 번호 2의 참조 서열에 대한 정렬은 클러스털더블유를 사용하여 수행된다.
추가의 실시형태에서, 추가의 CE-7 시그니처 모티프가 포함될 수 있으며, 추가의(즉, 제4) 모티프는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬하는 경우 아미노산 잔기 267 내지 269에서의 LXD 모티프로 정의된다.
다른 실시형태에서, 과가수분해효소 촉매는 과가수분해효소 활성을 갖는 효소를 포함하며, 상기 효소는 서열 번호 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476으로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는다.
다른 실시형태에서, 과가수분해효소 촉매는 과가수분해효소 활성을 갖는 효소를 포함하며, 상기 효소는 서열 번호 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 424, 437 및 476으로 이루어진 군으로부터 선택되는 아미노산 서열을 가지며, 상기 효소는 시그니처 모티프가 보존되고, 과가수분해효소 활성이 유지되는 한 하나 이상의 부가, 결실 또는 치환을 가질 수 있다.
상술된 바와 같이, CE-7 과가수분해효소는 CE-7 과가수분해효소를 포함하는 제1 부분, 및 과가수분해효소가 원하는 체표면에 "표적화"되도록 표적 체표면에 대하여 친화성을 갖는 펩티드 성분을 포함하는 제2 부분을 갖는 융합 단백질일 수 있다. 일 실시형태에서, 임의의 CE-7 과가수분해효소(CE-7 시그니처 모티프의 존재로 정의)는 효소를 체표면에 표적화시킬 수 있는 임의의 펩티드 성분/결합 요소에 융합될 수 있다. 일 태양에서, 구강 표면에 대하여 친화성을 갖는 펩티드 성분은 항체, 항체 단편(Fab)뿐 아니라 단쇄 가변 단편(scFv; 면역글로불린의 중쇄(VH) 및 경쇄(VL)의 가변 영역의 융합), 단일 도메인 카멜리드 항체, 스캐폴드 디스플레이 단백질 및 면역글로불린 폴드가 결여된 단쇄 친화성 펩티드를 포함할 수 있다. 항체, 항체 단편 및 다른 면역글로불린-유래의 결합 요소뿐 아니라 라지 스캐폴드 디스플레이 단백질을 포함하는 조성물은 종종 경제적으로 실행가능하지 않다. 이와 같이, 그리고 바람직한 태양에서, 펩티드 성분/결합 요소는 면역글로불린 폴드 및/또는 면역글로불린 도메인이 결여된 단쇄 친화성 펩티드이다. 짧은 단쇄 체표면-결합 펩티드는 실험에 의해 생성하거나(예를 들어, 음으로 하전된 표면에 표적화된 양으로 하전된 폴리펩티드) 또는 표적 체표면에 대한 바이오패닝을 사용하여 생성할 수 있다. 많은 디스플레이 기술(예를 들어, 파지 디스플레이, 효모 디스플레이, 박테리아 디스플레이, 리보솜 디스플레이 및 mRNA 디스플레이)을 사용하여 친화성 펩티드를 확인/수득하는 방법은 당업계에 널리 공지되어 있다. 개별 구강 표면-결합 펩티드는 임의의 스페이서/링커를 통해 함께 결합되어 더 큰 결합 "도메인"(본 명세서에서 결합 "손(hand)"으로도 지칭)을 형성하여, 과가수분해 효소의 표적 구강 표면으로의 부착/국소화를 증진시킬 수 있다.
또한, 융합 단백질은 CE-7 과가수분해효소 및 구강 표면-결합 도메인 및/또는 상이한 구강 표면-결합 펩티드(예를 들어, 복수의 구강 표면 결합 펩티드가 함께 결합되어 더 큰 표적 구강 표면 결합 도메인을 형성하는 경우) 사이를 분리하는 하나 이상의 펩티드 링커/스페이서를 포함할 수 있다. 다수의 펩티드 링커/스페이서가 존재할 수 있으며, 링커의 수는 최대 10회 반복될 수 있다. 예시적인 펩티드 스페이서의 비제한적인 목록은 서열 번호 383 내지 396의 아미노산 서열 및 표 5에 예시된 것으로 제공된다.
구강 표면에 대하여 친화성을 갖는 적절한 펩티드는 본 명세서에서 상기에 기재되어 있다. 상기 "디스플레이" 기술 중 임의의 것을 사용하는 추가의 구강 표면-결합 펩티드의 확인 방법은 당업계에 널리 알려져 있으며, 이를 사용하여 추가의 구강 표면-결합 펩티드를 확인할 수 있다.
적절한 카르복실산 에스테르 기질은 하기의 화학식을 갖는 에스테르를 포함할 수 있다:
(a) 구조
[X]mR5
(여기서,
X는 화학식 R6C(O)O의 에스테르기이며;
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 하나 이상의 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ; 또는
(b) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 하나 이상의 글리세리드; 또는
(c) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 또는
(d) 하나 이상의 아세틸화 단당류, 아세틸화 이당류 또는 아세틸화 다당류; 또는
(e) (a) 내지 (d)의 임의의 조합.
또한, 적절한 기질은 아실화 단당류, 이당류 및 다당류로 이루어진 군으로부터 선택되는 하나 이상의 아실화 당류를 포함할 수 있다. 다른 실시형태에서, 아실화 당류는 아세틸화 자일란; 아세틸화 자일란의 단편; 아세틸화 자일로스(예컨대, 자일로스 테트라아세테이트); 아세틸화 글루코스(예컨대, α-D-글루코스 펜타아세테이트; β-D-글루코스 펜타아세테이트; 1-티오-β-D-글루코스-2,3,4,6-테트라아세테이트); β-D-갈락토스 펜타아세테이트;소르비톨 헥사아세테이트; 수크로스 옥타아세테이트; β-D-리보푸라노스-1,2,3,5-테트라아세테이트; β-D-리보푸라노스-1,2,3,4-테트라아세테이트; 트라이-O-아세틸-D-갈락탈; 트라이-O-아세틸-D-글루칼; β-D-자일로푸라노스 테트라아세테이트, α-D-글루코피라노스 펜타아세테이트; β-D-글루코피라노스-1,2,3,4-테트라아세테이트; β-D-글루코피라노스-2,3,4, 6-테트라아세테이트; 2-아세트아미도-2-데옥시-1,3,4,6-테트라아세틸-β-D-글루코피라노스; 2-아세트아미도-2-데옥시-3,4,6-트라이아세틸-1-클로라이드-α-D-글루코피라노스; α-D-만노피라노스 펜타아세테이트 및 아세틸화 셀룰로스로 이루어진 군으부터 선택된다. 바람직한 실시형태에서, 아세틸화 당류는 β-D-리보푸라노스-1,2,3,5-테트라아세테이트; 트라이-O-아세틸-D-갈락탈; 트라이-O-아세틸-D-글루칼; 수크로스 옥타아세테이트; 및 아세틸화 셀룰로스로 이루어진 군으로부터 선택된다.
다른 실시형태에서, 추가의 적절한 기질은 또한 5-아세톡시메틸-2-푸르알데히드; 3,4-다이아세톡시-1-부텐; 4-아세톡시벤조산; 바닐린 아세테이트; 프로필렌 글리콜 메틸 에테르 아세테이트; 메틸 락테이트; 에틸 락테이트; 메틸 글리콜레이트; 에틸 글리콜레이트; 메틸 메톡시아세테이트; 에틸 메톡시아세테이트; 메틸 3-하이드록시부티레이트; 에틸 3-하이드록시부티레이트; 및 트라이에틸 2-아세틸 시트레이트를 포함할 수 있다.
다른 실시형태에서, 적절한 기질은 모노아세틴; 다이아세틴; 트라이아세틴; 모노프로피오닌; 다이프로피오닌; 트라이프로피오닌; 모노부티린; 다이부티린; 트라이부티린; 글루코스 펜타아세테이트; 자일로스 테트라아세테이트; 아세틸화 자일란; 아세틸화 자일란 단편; β-D-리보푸라노스-1,2,3,5-테트라아세테이트; 트라이-O-아세틸-D-갈락탈; 트라이-O-아세틸-D-글루칼; 1,2-에탄다이올; 1,2-프로판다이올; 1,3-프로판다이올; 1,2-부탄다이올; 1,3-부탄다이올; 2,3-부탄다이올; 1,4-부탄다이올; 1,2-펜탄다이올; 2,5-펜탄다이올; 1,5-펜탄다이올; 1,6-펜탄다이올; 1,2-헥산다이올; 2,5-헥산다이올; 1,6-헥산다이올의 모노에스테르 또는 다이에스테르; 및 그들의 혼합물로 이루어진 군으로부터 선택된다. 다른 실시형태에서, 기질은 하나 이상의 에스테르기를 포함하는 C1 내지 C6 폴리올이다. 바람직한 실시형태에서, C1 내지 C6 폴리올 상의 하나 이상의 하이드록실기는 하나 이상의 아세톡시기(예를 들어, 1,3-프로판다이올 다이아세테이트; 1,2-프로판다이올 다이아세테이트; 1,4-부탄다이올 다이아세테이트; 1,5-펜탄다이올 다이아세테이트 등)로 치환된다. 추가의 실시형태에서, 기질은 프로필렌 글리콜 다이아세테이트(PGDA), 에틸렌 글리콜 다이아세테이트(EGDA) 또는 그들의 혼합물이다.
추가의 실시형태에서, 적절한 기질은 모노아세틴, 다이아세틴, 트라이아세틴, 모노프로피오닌, 다이프로피오닌, 트라이프로피오닌, 모노부티린, 다이부티린 및 트라이부티린으로 이루어진 군으로부터 선택된다. 또 다른 태양에서, 기질은 다이아세틴 및 트라이아세틴으로 이루어진 군으로부터 선택된다. 가장 바람직한 실시형태에서, 적절한 기질은 트라이아세틴을 포함한다.
바람직한 실시형태에서, 카르복실산 에스테르는 모노아세틴, 다이아세틴, 트라이아세틴 및 그들의 조합(즉, 혼합물)으로 이루어진 군으로부터 선택되는 액체 기질이다. 카르복실산 에스테르는 효소-촉매작용되는 과가수분해시에 원하는 농도의 퍼옥시카르복실산을 생성하기에 충분한 농도로 반응 제형에 존재한다. 카르복실산 에스테르는 반응 제형 중에서 완전히 용해성일 필요는 없으나, 과가수분해효소 촉매에 의한 에스테르의 상응하는 퍼옥시카르복실산으로의 전환을 가능하게 하기에 충분한 용해성을 갖는다. 카르복실산 에스테르는 반응 제형의 0.05 wt% 내지 40 wt%의 농도, 바람직하게는 반응 제형의 0.1 wt% 내지 20 wt%의 농도, 더욱 바람직하게는 반응 제형의 0.5 wt% 내지 10 wt%의 농도로 반응 제형 중에 존재한다.
과산소원은 과산화수소, 과산화수소 부가물(예를 들어, 우레아-과산화수소 부가물(카르바미드 퍼옥시드)), 과붕산염, 과탄산염 및 과산화염을 포함할 수 있으나 이들에 한정되지 않는다. 반응 제형 중의 과산소 화합물의 농도는 0.0033 wt% 내지 약 50 wt%, 바람직하게는 0.033 wt% 내지 약 40 wt%, 더욱 바람직하게는 0.1 wt% 내지 약 30 wt%의 범위일 수 있다.
또한, 과산소원(즉, 과산화수소)은 유효량의 과산화수소를 생성할 수 있는 효소를 사용하여 효소에 의해 생성될 수 있다. 예를 들어, 글루코스 산화효소, 락토스 산화효소, 탄수화물 산화효소, 알코올 산화효소, 에틸렌 글리콜 산화효소, 글리세롤 산화효소 및 아미노산 산화효소를 포함하나 이들에 한정되지 않는 다양한 산화효소가 본 발명의 조성물 및 방법에서 사용되어, 유효량의 과산화수소를 생성할 수 있다.
많은 과가수분해효소 촉매(전체 세포, 투과화된 전체 세포 및 부분 정제된 전체 세포 추출물)는 카탈라아제 활성(EC 1.11.1.6)을 갖는 것으로 보고되었다. 카탈라아제는 과산화수소의 산소 및 물로의 전환을 촉매작용시킨다. 일 태양에서, 과가수분해 촉매에는 카탈라아제 활성이 결여되어 있다. 다른 태양에서, 카탈라아제 억제제는 반응 제형에 첨가될 수 있다. 당업자는 필요에 따라 카탈라아제 억제제의 농도를 조절할 수 있다. 카탈라제 억제제의 농도는 통상 0.1 mM 내지 약 1 M; 바람직하게는 약 1 mM 내지 약 50 mM; 더욱 바람직하게는 약 1 mM 내지 약 20 mM의 범위이다.
다른 실시형태에서, 효소 촉매에는 상당한 카탈라아제 활성이 결여되거나, 카탈라아제 활성을 감소시키거나 제거하도록 엔지니어링될 수 있다. 숙주 세포 내의 카탈라아제 활성은 트랜스포손(transposon) 돌연변이유발, RNA 안티센스 발현, 표적화된 돌연변이유발 및 무작위 돌연변이유발을 포함하나 이에 한정되지 않는 널리 공지되어 있는 기술을 사용하여 카탈라아제 활성을 담당하는 유전자(들)의 발현을 파괴시킴으로써 하향 조절되거나 제거될 수 있다. 바람직한 실시형태에서, 내인성 카탈라아제 활성을 암호화하는 유전자(들)는 하향 조절되거나 파괴된다(즉, 낙-아웃). 본 명세서에 사용되는 "파괴된" 유전자는 변형된 유전자에 의해 암호화된 단백질의 활성 및/또는 기능이 더이상 존재하지 않는 것이다. 유전자를 파괴시키는 수단은 해당 분야에 널리 공지되어 있으며, 해당하는 단백질의 활성 및/또는 기능이 더이상 존재하지 않는 한, 유전자에 대한 삽입, 결실 또는 돌연변이를 포함할 수 있으나, 이들에 한정되지 않는다. 추가의 바람직한 실시형태에서, 생성 숙주는 katG 및 katE로 이루어진 군으로부터 선택되는 파괴된 카탈라아제 유전자를 포함하는 에스케리키아 콜라이(E. coli) 생성 숙주이다(미국 특허 출원 공개 제2008-0176299호 참조). 다른 실시형태에서, 생성 숙주는 katG 및 katE 카탈라아제 유전자 둘 모두에서의 하향 조절 및/또는 파괴를 포함하는 에스케리키아 콜라이 균주이다.
수성 반응 제형 중의 촉매의 농도는 촉매의 특정 촉매 활성에 좌우되며, 원하는 반응 속도를 수득하도록 선택된다. 과가수분해 반응물 중의 촉매의 중량은 전형적으로 총 반응 부피 ㎖당 0.0001 ㎎ 내지 10 ㎎, 바람직하게는 ㎖당 0.001 ㎎ 내지 2.0 ㎎ 범위이다. 또한, 촉매는 당업자에게 널리 공지되어 있는 방법을 사용하여 용해성 또는 불용성 지지체 상에 고정화될 수 있으며; 예를 들어, 문헌[Immobilization of Enzymes and Cells; Gordon F. Bickerstaff, Editor; Humana Press, Totowa, NJ, USA; 1997]을 참조한다. 고정화된 촉매의 사용에 의해, 이후의 반응에서 촉매의 회수 및 재사용이 가능하게 된다. 효소 촉매는 전체 미생물 세포, 투과화된 미생물 세포, 미생물 세포 추출물, 부분 정제되거나 정제된 효소 및 그들의 혼합물의 형태일 수 있다.
일 태양에서, 카르복실산 에스테르의 화학적 과가수분해 및 효소에 의한 과가수분해의 조합에 의해 생성되는 퍼옥시카르복실산의 농도는 선택된 개인 관리 응용을 위해 유효 농도의 퍼옥시카르복실산을 제공하기에 충분하다. 다른 태양에서, 본 발명은 효소 및 효소 기질의 조합을 제공하여, 원하는 유효 농도의 퍼옥시카르복실산을 생성하며, 여기서, 첨가되는 효소의 부재 하에서, 유의미하게 더 낮은 농도의 퍼옥시카르복실산의 생성이 존재한다. 일부 경우에, 무기 과산화물과 효소 기질의 직접적인 화학적 반응에 의한 효소 기질의 상당한 화학적 과가수분해가 존재할 수 있지만, 원하는 응용에서 유효 농도의 퍼옥시카르복실산을 제공하기 위해 생성되는 충분한 농도의 퍼옥시카르복실산이 존재하지 않을 수 있으며, 총 퍼옥시카르복실산 농도의 유의미한 증가는 적절한 과가수분해효소 촉매를 반응 제형에 첨가함으로써 달성된다.
적어도 하나의 카르복실산 에스테르의 과가수분해에 의해 생성되는 퍼옥시카르복실산(예를 들어, 과아세트산)의 농도는 과가수분해 반응의 개시 10분 내, 바람직하게는 5분 내에 적어도 약 0.1 ppm, 바람직하게는 적어도 0.5 ppm, 1 ppm, 5 ppm, 10 ppm, 20 ppm, 100 ppm, 200 ppm, 300 ppm, 500 ppm, 700 ppm, 1000 ppm, 2000 ppm, 5000 ppm 또는 10,000 ppm의 과산이다. 퍼옥시카르복실산을 포함하는 제품 제형은 임의로, 표적 적용에 대해 원하는 더 낮은 농도의 퍼옥시카르복실산 베이스를 갖는 제형을 생성하도록 물 또는 주로 물로 이루어진 용액으로 희석될 수 있다. 당업자가 반응 성분 및/또는 희석량을 조절하여, 선택된 개인 관리 제품을 위한 원하는 과산 농도를 달성할 수 있는 것이 명백하다.
일 태양에서, 원하는 농도의 과산을 생성하는데 필요한 반응 시간은 약 2시간 이하, 바람직하게는 약 30분 이하, 더욱 바람직하게는 약 10분 이하, 가장 바람직하게는 약 5분 이하이다. 다른 태양에서, 구강 표면은 반응 성분의 배합 5분 내에 본 명세서에 기재된 방법에 따라 형성되는 퍼옥시카르복실산과 접촉된다. 일 실시형태에서, 표적 구강 표면은 상기 반응 성분의 배합 약 5분 내지 약 168시간 내에, 또는 상기 반응 성분의 배합 약 5분 내지 약 48시간 내에, 또는 약 5분 내지 2시간 내에, 또는 그 안의 임의의 이러한 시간 간격으로 본 명세서에 기재된 방법에 따라 생성되는 퍼옥시카르복실산과 접촉된다.
본 명세서에 기재된 방법에 따라 형성된 과산은 개인 관리 제품/응용에서 사용되며, 과산은 표적 구강 표면과 접촉하여, 과산 기반의 이익을 구강에 제공한다. 일 실시형태에서, 표적 체표면을 위한 과산의 생성 방법은 동소에서 행해진다.
반응의 온도를 선택하여, 반응 속도와 효소 촉매 활성의 안정성 둘 모두를 조절할 수 있다. 명백하게, 특정 개인 관리 응용을 위하여, 표적 체표면의 온도(예를 들어, 구강 내의 37℃)는 반응 온도일 수 있다. 반응 온도는 반응 제형의 동결점 바로 위의 온도(대략 0℃) 내지 약 95℃ 범위일 수 있으며, 바람직한 범위는 5℃ 내지 약 75℃이고, 더욱 바람직한 반응 온도 범위는 약 5℃ 내지 약 55℃이다.
퍼옥시카르복실산을 함유하는 최종 반응 제형의 pH는 약 2 내지 약 9, 바람직하게는 약 3 내지 약 8, 더욱 바람직하게는 약 5 내지 약 8, 더더욱 바람직하게는 약 5.5 내지 약 8, 보다 바람직하게는 약 6.0 내지 약 7.5이다. 반응물 및 최종 반응 제형의 pH는 임의로 포스페이트, 피로포스페이트, 바이카르보네이트, 아세테이트 또는 시트레이트를 포함하나 이들에 한정되지 않는 적절한 완충제의 첨가에 의해 조절될 수 있다. 완충제가 사용되는 경우, 완충제의 농도는 전형적으로 0.1 mM 내지 1.0 M, 바람직하게는 1 mM 내지 300 mM, 가장 바람직하게는 10 mM 내지 100 mM이다.
다른 태양에서, 효소에 의한 과가수분해 반응 제형은 분산제로 작용하는 유기 용매를 함유하여, 반응 제형 중의 카르복실산 에스테르의 용해 속도를 증진시킬 수 있다. 이러한 용매는 프로필렌 글리콜 메틸 에테르, 아세톤, 사이클로헥산온, 다이에틸렌 글리콜 부틸 에테르, 트라이프로필렌 글리콜 메틸 에테르, 다이에틸렌 글리콜 메틸 에테르, 프로필렌 글리콜 부틸 에테르, 다이프로필렌 글리콜 메틸 에테르, 사이클로헥산올, 벤질 알코올, 아이소프로판올, 에탄올, 프로필렌 글리콜 및 그들의 혼합물을 포함하나 이들에 한정되지 않는다.
단일 단계 대 다단계 적용 방법
전형적으로, 과산 유익제를 효소에 의해 생성하기 위한 최소 반응 성분 세트는 (1) 본 명세서에 기재된 과가수분해 활성을 갖는 적어도 하나의 효소, 예를 들어, CE-7 과가수분해효소(임의로, 표적화된 융합 단백질 형태), (2) 적어도 하나의 적절한 카르복실산 에스테르 기질 및 (3) 과산소원을 포함할 것이다.
개인 관리 조성물의 과산 생성 반응 성분은 사용할 때까지 분리되어 유지될 수 있다. 일 실시형태에서, 과산-생성 성분을 배합한 다음, 표적 체표면과 접촉시켜, 이에 의해, 생성되는 과산계 유익제가 체표면에 이익을 제공한다. 성분을 배합한 다음 표적 체표면과 접촉시키거나, 또는 표적 체표면 상에서 배합할 수 있다. 일 실시형태에서, 과산-생성 성분은 과산이 동소에서 생성되도록 배합한다.
또한, 다단계 적용이 사용될 수 있다. 효소에 의한 과산 생성에 필요한 나머지 성분을 적용하기 전에, 과산 생성 시스템(즉, 체표면 상에서의, 3가지 기본 반응 성분 중 적어도 하나의 순차적 적용) 조성물의 1가지 또는 2가지의 개별 성분을 구강 표면과 접촉시킬 수 있다. 일 실시형태에서, 과가수분해 효소는 구강 표면을 카르복실산 에스테르 기질 및/또는 과산소원과 접촉시키기 전에 구강 표면과 접촉시킨다(즉, "2-단계 적용"). 일 실시형태에서, 과가수분해 활성을 갖는 효소는 효소에 의한 과산 생성에 필요한 나머지 성분을 배합하기 전에 구강 표면에 적용되는 표적화된 과가수분해효소이다.
바람직한 실시형태에서, 과가수분해 활성을 갖는 효소는 효소에 의한 과산 생성에 필요한 나머지 성분을 배합하기 전에 구강 표면에 적용되는(즉, 2-단계 적용 방법) "표적화된 CE-7 과가수분해효소"(즉, CE-7 융합 단백질)이다. 표적화된 과가수분해효소는 구강 표면에 대한 융합 단백질의 비공유 결합을 촉진시키는 적절한 조건 하에서 구강 표면과 접촉한다. 나머지 반응 성분을 배합하기 전에, 임의의 헹굼 단계를 사용하여 과잉의 및/또는 미결합 융합 단백질을 제거할 수 있다.
추가의 실시형태에서, 과가수분해 효소(임의로, 구강 표면에 표적화된 융합 단백질 형태) 및 카르복실산 에스테르를 과산소원의 첨가 전에 표적 구강 표면에 적용한다.
추가의 실시형태에서, 과가수분해 효소(임의로 구강 표면에 표적화된 융합 단백질 형태) 및 과산소원(예를 들어, 과산화수소를 포함하는 수용액)을 카르복실산 에스테르 기질의 첨가 전에 구강 표면에 적용한다.
추가의 실시형태에서, 카르복실산 에스테르 기질 및 과산소원(예를 들어, 과산화수소를 포함하는 수용액)을 과가수분해 효소(임의로 구강 표면에 표적화된 융합 단백질 형태)의 첨가 전에 구강 표면에 적용한다.
또 다른 실시형태에서, 본 명세서에 기재된 조성물 또는 방법 중 임의의 것이 본 발명을 실시하기 위한 키트에 통합될 수 있다. 키트는 효소에 의한 과산의 생성을 용이하게 하는 재료 및 시약을 포함할 수 있다. 예시적인 키트에는 기질, 과산소원 및 과가수분해 활성을 갖는 효소 촉매가 포함되며, 여기서, 효소 촉매는 임의로 구강 표면에 표적화될 수 있다. 기타 키트 성분은 제한 없이, 다음의 것 중 하나 이상을 포함할 수 있다: 시료 튜브, 고체 지지체, 지시 자료 및 과산을 효소에 의해 생성하는데 유용한 기타 용액 또는 기타 화학 시약, 예를 들어, 허용가능한 성분 또는 담체.
구강 관리 조성물
구강에 허용가능한 성분/담체/매질
또한, 본 발명의 조성물 및 방법은 구강에 허용가능한 담체뿐 아니라 추가의(즉, 과산계 유익제에 더하여) 구강 관리 유익제를 포함할 수 있다. 본 명세서에서 사용되는 용어 "구강 관리 유익제"는 원하는/유익한 효과를 제공하거나 구강 표면에 기여하는 화합물 또는 물질에 적용되는 일반 용어이다. 일 실시형태에서, 구강 표면에 대한 유익제는 백색 안료, 예를 들어, 이산화티탄 및 백색 미네랄(white mineral), 예를 들어, 수산화인회석 또는 지르콘을 포함하나 이들에 한정되지 않는 착색제를 (과산계 유익제에 더하여) 포함할 수 있다. 다른 실시형태에서, 구강 관리 유익제는 또한 미백제 및 추가의 효소, 예를 들어, 옥시다제, 퍼옥시다제, 프로테아제, 리파제, 글리코시다제, 에스테라제 및 다당류 가수분해효소를 포함할 수 있다. 다른 태양에서, 유익제는 항-플라크제, 항-착색제 및 항미생물제를 포함할 수 있다. 항미생물제에는 항미생물 펩티드, 마가이닌(magainin), 세크로핀(cecropin), 살미생물제, 트리클로산(triclosan), 클로르헥시딘, 세틸피리디늄 클로라이드, 4차 암모늄 화합물, 클로록실레놀, 클로록시에탄올, 프탈산 및 그의 염, 티몰 및 그들의 조합이 포함될 수 있으나, 이들에 한정되지 않는다. 또한, 구강 관리 유익제는 항충치제, 예를 들어, 플루오르화나트륨 또는 나트륨 모노플루오로포스페이트 및 향미제, 예를 들어, 동록유(wintergreen), 페퍼민트 오일, 스피어민트 오일, 또는 메틸 살리실레이트, 유칼립톨 또는 바닐린을 포함할 수 있다. 또한, 구강 관리 유익제는 몇가지만 말하자면, 청량제(coolant), 예를 들어, 석시네이트계 청량제 화합물 및 타액분비제(salivating agent)를 포함할 수 있다. 본 명세서에 사용되는 용어 "타액분비제"는 구강 관리 조성물에 존재하는 경우, 사용자의 타액분비를 더 크게 증진시키는 물질을 말한다. 일 실시형태에서, 유익제는 구강 관리 제품에 사용하는 것으로 승인된 구강에 허용가능한 물질이다. 다른 실시형태에서, 구강에 허용가능한 유익제를 사용하여, 치아의 미용적 외관을 개선시킨다.
구강에 허용가능한 담체 매질에 종종 사용되는 성분의 비-제한적인 목록은 본 명세서에 그들의 전문이 각각 참고로 포함되는 미국 특허 제6,740,311호(White et al.); 미국 특허 제6,706,256호(Lawler et al.); 미국 특허 제6,264,925호(Fuglsang et al.); 및 미국 특허 출원 공개 제2005-0069501호(Ibrahim et al.)에 기재되어 있다. 예를 들어, 구강 관리 조성물은 연마제, 계면활성제, 산화방지제, 킬레이트제, 플루오르화물 공급원, 증점제, 완충제, 용매, 보습제, 담체, 증량제(bulking agent), 항-플라크제, 항-착색제, 항미생물제, 항충치제, 항-염증제, 탈감작화제(desensitizing agent), 감미제, 착향제, 호흡-청량(breath-freshening)제, 청량제, 영양제 및 타액분비제 중 하나 이상을 포함할 수 있다.
혼합물 중의 성분은 구강 관리 조성물이 원하는 과산 유익제를 효소에 의해 생성하는 능력을 유지하도록 선택되는 것이 인식될 것이다. 본 명세서에 개시된 구강 관리 시스템의 적절한 혼합물은 통상의 실험을 사용하여 당업자에 의해 결정될 수 있다. 구강 관리 제형이 있는 구강 관리 유익제의 총 농도는 구강 관리 조성물의 총 중량에 비하여 약 0.001 중량% 내지 약 90 중량%일 수 있다.
구강 관리 조성물은 치약, 덴탈 크림(dental cream), 투스젤, 또는 투스 파우더, 구강 세정제, 호흡 청량제 및 치실(dental floss)을 포함할 수 있으나, 이들에 한정되지 않는다. 추가의 실시형태는 마우스 트레이를 통하여 구강 환경에 적용되는 페이스트 또는 겔의 반응 성분의 적용을 포함한다. 또한, 하나 이상의 반응 성분은 먼저, 에나멜에 부착되는 플라스틱 스트립 상에 침착되어, 하나 이상의 반응 성분을 전달하여, 과산 유익제를 생성할 수 있다. 전달 장치, 예를 들어, 스트립 상의 과가수분해효소 융합의 침착의 경우에, 과가수분해효소 융합은 스트립의 물질에 대하여 친화성을 갖는 결합 요소를 포함하여, 사용과 사용 후의 장치의 제거 동안 과가수분해효소의 스트립으로의 침착 및 보유를 지원하도록 고안될 수 있다.
구강 질환과 관련된 미생물을 감소시키거나 원치않는 생물막을 제거하기 위한 과산계 구강 관리 제품
과산계 구강 관리 제품을 사용하여 치아 우식증(예를 들어, 스트렙토코커스 뮤탄스(Streptococcus mutans)), 치은염, 구강 칸디다증 또는 치근막염과 관련된 구강 박테리아를 줄일 수 있다. 과산계 구강 관리 제품을 사용하여, 구강 생물막(들)을 감소시키거나 제거할 수 있다.
일 실시형태에서, 유효 농도의 적어도 하나의 과산을 생성하기 위한 구강 관리 제품에서, 구강 표면을 탈색시키거나, 미백하거나, 소독하거나, 탈염시키거나, 탈취시키거나 또는 그로부터 생물막을 제거하기 위한 과가수분해 활성을 갖는 효소의 용도가 제공된다.
일 실시형태에서, 과가수분해 활성을 갖는 효소는 표적화된 과가수분해효소이며, 효소가 본 발명의 하나 이상의 기질에 대하여 과가수분해 활성을 갖는 한, 리파제, 프로테아제, 에스테라제, 아실 트랜스퍼라제, 아릴 에스테라제, 탄수화물 에스테라제 및 조합을 포함할 수 있다. 예에는 과가수분해 프로테아제(예를 들어, 서브틸리신 변이체; 미국 특허 제7,510,859호), 과가수분해 에스테라제(예를 들어, 슈도모나스 플루오레슨스; 미국 특허 제7,384,787호; 서열 번호 477) 및 과가수분해 아릴 에스테라제(예를 들어, 마이코박테리움 스메그마티스; 미국 특허 제7,754,460호; WO2005/056782호; 및 제EP1689859 B1호; 서열 번호 460[S54V 변이체] 및 478[야생형])가 포함될 수 있으나, 이들에 한정되지 않는다.
다른 실시형태에서, 구강 관리 제품에서 구강 표면의 탈색, 미백, 소독, 탈염, 탈취 또는 생물막의 제거를 위한 유효 농도의 적어도 하나의 과산을 생성하기 위한 과가수분해 활성을 갖는 CE-7 탄수화물 에스테라제의 용도가 제공된다.
다른 실시형태에서,
a) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
i) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
ii) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
iii) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - ;
b) 1) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티, 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
M은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
2) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
3) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
4) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질; 및
c) 과산소원을 포함하는 - 이에 의해, (a), (b) 및 (c)의 혼합시에 과산이 형성되며, 과산 생성 제형은 치아 우식증, 치은염, 구강 칸디다증 또는 치근막염의 치료 또는 예방을 위해 사용된다 - 과산 생성 조성물의 용도도 또한 제공된다.
퍼옥시카르복실산 및 과산화수소의 농도를 결정하기 위한 HPLC 검정 방법.
적정, 고성능 액체 크로마토그래피(HPLC), 기체 크로마토그래피(GC), 질량 분석기(MS), 모세관 전기영동(CE), 문헌[U. Pinkernell et al., (Anal . Chem ., 69(17):3623-3627 (1997))]에 기재된 분석 절차, 및 본 실시예에 기재된 바와 같은 2,2'-아지노-비스 (3-에틸벤조타졸린)-6-설포네이트(ABTS) 검정법(문헌[U. Pinkernell et. al. Analyst, 122: 567-571 (1997)] 및 문헌[Dinu et. al. Adv. Funct. Mater ., 20: 392-398 (2010)])을 포함하나 이에 한정되지 않는 다양한 분석 방법을 본 발명의 방법에 사용하여 반응물 및 생성물을 분석할 수 있다.
퍼옥시카르복실산의 최소 살생 농도의 결정
특정 개인 관리 응용, 예를 들어, 몇가지만 말하자면, 체취 및 진균 감염, 및 치아 우식증의 발생과 관련된 것과 같은 원치않는 미생물의 제거와 관련이 있을 수 있다. 이와 같이, 표적 개인 관리 응용을 위한 최소 살생 농도를 측정하고자 할 수 있다. 문헌[J. Gabrielson, et al. (J. Microbiol . Methods 50: 63-73 (2002))]에 기재되어 있는 방법은 퍼옥시카르복실산 또는 과산화수소 및 효소 기질의 최소 살생 농도(MBC)의 결정을 위해 사용할 수 있다. 검정 방법은 XTT 환원 억제를 기반으로 하며, 여기서, XTT((2,3-비스[2-메톡시-4-니트로-5-설포페닐]-5-[(페닐아미노)카르보닐]-2H-테트라졸륨, 내염, 모노나트륨 염)는 490 ㎚ 또는 450 ㎚에서 측정한 광학 밀도(OD)의 변화에 의해 미생물 호흡 활성을 나타내는 산화환원 염료이다. 그러나, 플레이트 생존 계수, 직접적인 현미경 계수, 건조 중량, 탁도 측정, 흡광도 및 생물발광을 포함하나 이들에 한정되지 않는 소독제 및 보존제의 활성의 시험에 이용할 수 있는 다양한 다른 방법이 존재한다(예를 들어, 문헌[Brock, Semour S., Disinfection, Sterilization, and Preservation, 5th edition, Lippincott Williams & Wilkins, Philadelphia, PA, USA; 2001] 참조).
재배합 미생물 발현
본 발명의 서열의 유전자 및 유전자 산물은 이종 숙주 세포, 특히 미생물 숙주 세포에서 생성될 수 있다. 본 발명의 유전자 및 핵산 분자의 발현에 바람직한 이종 숙주 세포는 진균 또는 박테리아 패밀리에서 관찰될 수 있고, 넓은 범위의 온도, pH 값 및 용매 용인성에 걸쳐 성장하는 미생물 숙주이다. 예를 들어, 박테리아, 효모 및 사상 진균 중 임의의 것이 본 발명의 핵산 분자의 발현에 적절한 숙주일 수 있는 것으로 고려된다. 과가수분해효소는 세포내, 세포외 또는 세포내와 세포외 둘 모두의 조합에서 발현될 수 있으며, 여기서, 세포외 발현은 발효 생성물로부터의 원하는 단백질의 회수를 세포내 발현에 의해 생성되는 단백질의 회수 방법보다 더 용이하게 한다. 전사, 번역 및 단백질 생합성 장치는 세포 바이오매스를 생성하는데 사용되는 세포 공급원료에 비하여 변함 없이 유지되며; 기능적 유전자는 관련 없이 발현될 것이다. 숙주 균주의 예에는 박테리아, 진균 또는 효모 종, 예를 들어, 아스페르길루스(Aspergillus), 트리코더마(Trichoderma), 사카로마이세스(Saccharomyces), 피키아(Pichia), 파피아(Phaffia), 클루이베로마이세스(Kluyveromyces), 칸디다(Candida), 한세눌라(Hansenula), 야로위아(Yarrowia), 살모넬라(Salmonella), 바실러스(Bacillus), 아키네토박터(Acinetobacter), 자이모모나스(Zymomonas), 아그로박테리움(Agrobacterium), 에리트로박터(Erythrobacter), 클로로븀(Chlorobium), 크로마튬(Chromatium), 플라보박테리움(Flavobacterium), 사이토파가(Cytophaga), 로도박터(Rhodobacter), 로도코커스(Rhodococcus), 스트렙토마이세스(Streptomyces), 브레비박테리움(Brevibacterium), 코리네박테리아(Corynebacteria), 마이코박테리움(Mycobacterium), 데이노코커스(Deinococcus), 에스케리키아(Escherichia), 에르위니아(Erwinia), 판토에아(Pantoea), 슈도모나스(Pseudomonas), 스핑고모나스(Sphingomonas), 메틸로모나스(Methylomonas), 메틸로박터(Methylobacter), 메틸로코커스(Methylococcus), 메틸로시너스(Methylosinus), 메틸로마이크로븀(Methylomicrobium), 메틸로시스티스(Methylocystis), 알칼리게네스(Alcaligenes), 시네코시스티스(Synechocystis), 시네코코커스(Synechococcus), 아나바에나(Anabaena), 티오바실러스(Thiobacillus), 메타노박테리움(Methanobacterium), 클레브시엘라(Klebsiella) 및 마익소코커스(Myxococcus)가 포함되나 이들에 한정되지 않는다. 일 실시형태에서, 박테리아 숙주 균주에는 에스케리키아, 바실러스, 클루이베로마이세스 및 슈도모나스가 포함된다. 바람직한 실시형태에서, 박테리아 숙주 세포는 바실러스 서브틸리스 또는 에스케리키아 콜라이이다.
대규모 미생물 성장 및 기능적 유전자 발현은 매우 다양한 단순 또는 복합 탄수화물, 유기산 및 알코올 또는 포화 탄화수소, 예를 들어, 메탄 또는 광합성 숙주 또는 화학자가영양(chemoautotrophic) 숙주의 경우에 이산화탄소, 소정의 형태 및 양의 질소, 인, 황, 산소, 탄소, 또는 작은 무기 이온을 포함하는 임의의 미량의 미량영양소를 사용할 수 있다. 성장 속도의 조절은 특정 조절 분자의 배양물로의 첨가 또는 무첨가에 의해 영향을 받을 수 있으며, 통상 영양분 또는 에너지원을 고려하지 않는다.
적절한 숙주 세포의 형질전환에 유용한 벡터 또는 카세트가 당업계에 널리 알려져 있다. 전형적으로, 벡터 또는 카세트는 관련 유전자의 전사 및 번역을 유도하는 서열, 선별가능한 마커, 및 자가 복제 또는 염색체 통합을 허용하는 서열을 포함한다. 적절한 벡터는 전사 개시 제어부를 품고 있는 유전자의 5'의 영역 및 전사 종결을 제어하는 DNA 단편의 3'의 영역을 포함한다. 둘 모두의 제어 영역이 형질전환된 숙주 세포에 대하여 동종의 및/또는 생성 숙주에 대해 천연의 유전자로부터 유래되는 경우가 가장 바람직하지만, 그러한 제어 영역이 그렇게 유래될 필요는 없다.
바람직한 숙주 세포에서 본 발명의 세팔로스포린 C 데아세틸라제 암호화 영역의 발현을 작동시키기에 유용한 개시 제어 영역 또는 프로모터는 많으며, 당업자에게 잘 알려져 있다. 사실상, CYC1, HIS3, GAL1, GAL10, ADH1, PGK, PHO5, GAPDH, ADC1, TRP1, URA3, LEU2, ENO, TPI (사카로마이세스에서의 발현에 유용함); AOX1 (피키아에서의 발현에 유용함); 및 lac, araB, tet, trp, lPL, lPR, T7, tac 및 trc (에스케리키아 콜라이에서의 발현에 유용함)뿐 아니라 amy, apr, npr 프로모터 및 바실러스에서의 발현에 유용한 다양한 파지 프로모터를 포함하나 이들에 한정되지 않는 이들 유전자를 작동시킬 수 있는 임의의 프로모터가 본 발명에 적절하다.
종결 제어 영역은 또한 바람직한 숙주 세포의 다양한 천연의 유전자로부터 유래될 수 있다. 일 실시형태에서, 종결 제어 영역의 포함은 선택적이다. 다른 실시형태에서, 키메라 유전자는 바람직한 숙주 세포 유래의 종결 제어 영역을 포함한다.
산업적 생성
다양한 배양 방법을 적용하여, 과가수분해효소 촉매를 생성할 수 있다. 예를 들어, 재조합 미생물 숙주로부터 과발현되는 특정 유전자 산물의 대규모 생성은 배치식(batch), 유가식(fed-batch) 및 연속식(continuous) 배양 방법에 의해 생성될 수 있다. 배치식 및 유가식 배양 방법은 통상적이며, 해당 분야에 널리 알려져 있으며, 예는 문헌[Thomas D. Brock in Biotechnology: A Textbook of Industrial Microbiology, Second Edition, Sinauer Associates, Inc., Sunderland, MA (1989)] 및 문헌[Deshpande, Mukund V., Appl. Biochem. Biotechnol., 36:227-234 (1992)]에서 찾을 수 있다.
또한, 목적 과가수분해효소 촉매의 상업적 생성은 연속식 배양으로 달성될 수 있다. 연속식 배양은 정의된 배양 배지를 생물반응기에 연속 첨가함과 동시에, 동량의 조절된 배지를 처리를 위해 제거하는 개방형 시스템이다. 연속식 배양은 일반적으로 세포를 고정된 높은 액상 밀도로 유지시키며, 여기서, 세포는 주로 대수기 성장으로 존재한다. 대안적으로, 연속식 배양은 고정화된 세포로 실시될 수 있고, 여기서, 탄소 및 영양분을 지속적으로 공급하고, 가치있는 생성물, 부산물 또는 폐기물을 세포 매스로부터 지속적으로 제거한다. 세포 고정화는 천연 및/또는 합성 재료로 이루어진 매우 다양한 고체 지지체를 사용하여 수행할 수 있다.
배치식 발효, 유가식 발효 또는 연속식 배양으로부터의 목적 과가수분해효소 촉매의 회수는 당업계에 공지되어 있는 임의의 방법에 의해 달성될 수 있다. 예를 들어, 효소 촉매가 세포 내에서 생성되는 경우, 세포 페이스트(paste)를 원심분리 또는 막 여과에 의해 배양 배지로부터 분리하고, 임의로, 물 또는 원하는 pH의 수성 완충제로 세정한 다음, 원하는 pH에서의 수성 완충제 중의 세포 페이스트의 현탁액을 균질화시켜, 원하는 효소 촉매를 함유하는 세포 추출물을 생성한다. 효소 촉매 용액으로부터 원치않는 단백질을 침전시키기 위한 열-처리 단계 전에, 세포 추출물을 임의로, 적절한 여과 조제, 예를 들어 셀라이트 또는 실리카를 통해 여과하여, 세포 데브리스를 제거할 수 있다. 이어서, 원하는 효소 촉매를 함유하는 용액을 막 여과 또는 원심분리에 의하여 침전된 세포 데브리스 및 단백질로부터 분리할 수 있으며, 얻은 부분-정제된 효소 촉매 용액을 추가의 막 여과에 의해 농축시킨 다음, 임의로 적절한 담체(예를 들어, 말토덱스트린, 포스페이트 완충제, 시트레이트 완충제 또는 그들의 혼합물)와 혼합하고, 분무 건조시켜, 원하는 효소 촉매를 함유하는 고체 분말을 생성할 수 있다.
양, 농도, 또는 다른 값 또는 파라미터가 범위, 바람직한 범위 또는 바람직한 상한값 및 바람직한 하한값의 목록으로서 주어지는 경우, 범위가 별도로 개시되는 지에 상관없이 임의의 상한 범위 또는 바람직한 값 및 임의의 하한 범위 또는 바람직한 값의 임의의 쌍으로부터 형성된 모든 범위를 구체적으로 개시하는 것으로 이해되어야 한다. 수치 값의 범위가 본 명세서에서 언급될 경우, 달리 기술되지 않는다면, 그 범위는 그 종점 및 그 범위 내의 모든 정수와 분수를 포함하고자 하는 것이다. 범위를 정의할 때 열거된 특정 값으로 본 발명의 범주를 한정하고자 하는 것은 아니다.
일반 방법
본 발명의 바람직한 태양을 보여주기 위해 하기 실시예가 제공된다. 실시예에 개시된 기술은 본 발명의 실시에서 잘 기능하는 기술을 따르며, 이에 따라, 그의 바람직한 실시 방식을 구성하는 것으로 간주될 수 있음이 당업자에 의해 인식되어야 한다. 그러나, 본 개시내용의 견지에서, 당업자는 개시된 특정 실시형태에서 많은 변경이 이루어질 수 있으며, 여전히 본 명세서에 개시된 방법 및 실시예의 목적 및 범주로부터 벗어남 없이, 비슷하거나 유사한 결과가 수득되는 것을 인식해야 한다.
모든 시약 및 재료는 달리 특정하지 않는 한, 디프코 래보러터리즈(DIFCO Laboratories)(미국 미시간주 디트로이트 소재), GIBCO/BRL(미국 메릴랜드주 개더스버그 소재), 티씨아이 아메리카(TCI America)(미국 오리건주 포틀랜드 소재), 로슈 디아그노스틱스 코포레이션(Roche Diagnostics Corporation)(미국 인디애나주 인디애나폴리스), 써모 사이언티픽(미국 일리노이주 록포드 소재의 피어스 프로틴 리서츠 프로덕츠(Pierce Protein Research Products)) 또는 시그마/알드리치 케미컬 컴퍼니(Sigma/Aldrich Chemical Company)(미국 미주리주 세인트 루이스 소재)로부터 수득하였다.
설명에서 하기의 약어는 후술되는 바와 같은 측정 유닛, 기술, 특성 또는 화합물에 해당한다: "sec" 또는 "s"는 초를 의미하고, "min"은 분을 의미하며, "h" 또는 "hr"은 시간을 의미하고, "㎕"은 마이크로리터를 의미하며, "㎖"은 밀리리터를 의미하고, "L"은 리터를 의미하며, "mM"은 밀리몰을 의미하고, "M"은 몰을 의미하며, "mmol"은 밀리몰을 의미하고, "ppm"은 백만분율을 의미하며, "wt"는 중량을 의미하고, "wt%"는 중량 백분율을 의미하며, "g"는 그램을 의미하고, "㎎"는 밀리그램을 의미하며, "㎍"는 마이크로그램을 의미하고, "ng"는 나노그램을 의미하며, "g"는 중력을 의미하고, "HPLC"는 고성능 액체 크로마토그래피를 의미하며, "dd H2O"는 증류수 및 탈이온수를 의미하고, "dcw"는 건조 세포 중량을 의미하며, "ATCC" 또는 "ATCC(등록 상표)"는 아메리칸 타입 컬쳐 콜렉션(미국 버지니아주 머내서스 소재)을 의미하고, "U"는 과가수분해효소 활성의 단위를 의미하며, "rpm"은 분당 회전수를 의미하고, "Tg"는 유리 전이 온도를 의미하며, "EDTA"는 에틸렌다이아민테트라아세트산을 의미한다.
발현 벡터 pLD001
플라스미드 pLD001(서열 번호 397)은 이전에 에스케리키아 콜라이에 대한 적절한 발현 벡터로 보고된 적이 있다(본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2010-0158823 A1호(Wang et al.) 참조).
벡터 pLD001은 상업적으로 입수할 수 있는 벡터 pDEST17(미국 캘리포니아주 칼스배드 소재의 인비트로겐(Invitrogen)) 유래의 것이었다. 이는 상업적으로 입수할 수 있는 벡터 pET31b(미국 위스콘신주 매디슨 소재의 노바겐(Novagen)) 유래의 서열을 포함하며, 효소 케토스테로이드 아이소머라제(KSI)의 단편을 암호화한다. KSI 단편은 에스케리키아 콜라이에서 펩티드의 불용성 봉입체로의 분할을 증진시키기 위한 융합 파트너로 포함시켰다. pET31b로부터의 KSI-암호화 서열을 표준 돌연변이유발 절차(퀵체인지(QuickChange) II, 미국 캘리포니아주 라 졸라 소재의 스트라타진(Stratagene))를 사용하여 야생형 KSI 서열에서 관찰되는 하나의 Cys 코돈에 더하여 3개의 추가의 Cys 코돈을 포함하도록 변형시켰다. 또한, 암호화 서열 내의 모든 Asp 코돈을 Glu 코돈으로 대체하였다. 서열 번호 397로 제공되는 플라스미드 pLD001을 당업자에게 널리 공지되어 있는 표준 재조합 DNA 방법을 사용하여 구축하였다.
실시예 1
치아 미백제로서의 과아세트산의 유효성
본 실시예에는 에나멜 표면이 착색된 모델에서의 탈색 효과를 달성하기 위한 과아세트산의 이용이 기재되어 있다. 소 에나멜 절치(incisor)를 SE 덴탈(Dental)(미국 루이지애나주 배턴 루지 소재)로부터 수득하였다. 치아를 섹션화하고(sectioned), 다이아몬드 칼날과 함께 드레멜(Dremel)(등록 상표) 회전톱(미국 일리노이주 시카고 소재의 로버트 보슈 파워 툴 코포레이션(Robert Bosch Power Tool Corporation))을 사용하여 각 측 상에 대략 7 ㎜의 에나멜 슬라브(slab)로 절단하였다. 에나멜 슬라브를 세정하고, 부드럽게 닦아 표면 데브리스를 제거하였다. 에나멜을 1 내지 5일 동안 커피 및 차의 혼합물로 사전처리하여, 인간 착색된 치아와 유사한 색상으로 착색시켰다.
각 에나멜 블록(block)을 사용 전에 적어도 1시간 동안 수 중에서 수화시켰다. 기질에 대한 색상 측정을 시험 용액에 대한 노출 전에 수득하였다. 과아세트산의 용액을 500 mM 인산나트륨 완충제, pH 7.2 중에 32% 스톡(stock)으로부터 제조하였다. 또한, 2.5% H2O2의 용액을 동일한 완충제에서 제조하였다. 다수의 에나멜 블록을 각 용액에 1분 동안 노출시키고, 5분, 10분, 15분 및 30분의 추가의 노출로 이어졌다. 각 처리를 위하여, 새로운 과아세트산 및 과산화수소 용액을 스톡 용액으로부터 제조하였다. 각 처리 후에, 에나멜 블록을 물로 헹구고, 코니카-미놀타(Konica-Minolta) 2600d 분광광도계를 사용하여 측정하였다. 백색도를 표 1 및 2에 열거된 각 시료에 대하여 결정하였다.
백색도(WI)는 국제 조명 위원회(International Commission on Illumination)(CIE)에 의해 정의되고, ASTM 방법 E313-05에 기재되어 있으며, 하기와 같이 D65/10 입사광에 대하여 계산하였다:
WI = Y + 800*(0.3138-x) + 1700*(0.3310-y)
상기 식에서, Y, x 및 y는 각 에나멜 기질의 휘도율 및 색도 좌표이다.


보다 양의 값으로의 백색도의 변화는 미백 효과를 나타낸다. 또한, 시료의 육안의 검사에 의해, 완충제 및 과산화수소 대조군에 비하여 과아세트산 처리된 시료에 대한 감지할 수 있는 미백 효과가 나타났다. 이러한 데이터에 의해, 과아세트산이 효과적인 탈색제이며, 보다 낮은 농도에서 과산화수소에 대하여 뛰어난 성능을 제공하는 것이 입증된다.
실시예 2
표준
바이오패닝을
사용한 치아 에나멜 및
균막
결합 펩티드의 선택
본 실시예의 목적은 표준 파지 디스플레이 바이오패닝을 사용하여 치아 에나멜 및 균막에 결합하는 파지 펩티드를 동정하려는 것이다.
소 에나멜 절치를 SE 덴탈(미국 루이지애나주 배턴 루지 소재)로부터 수득하였다. 치아를 대략 5 ㎣로 절단하고, 닦아서 표면 데브리스를 제거하였다. 에나멜 블록을 사용 전에 살균시켰다. 에나멜 블록을 몰딩 재료를 함유하는 웰 플레이트에 포매시켜, 웰 내의 에나멜 표면 만을 노출시켰다. 30분 동안 입 안에서의 인큐베이션을 위하여 고정되는 왁스 상에 블록을 고정시켜, 균막 코팅된 표면을 형성함으로써, 균막을 추가의 에나멜 블록 상에 형성하였다. 균막 코팅된 에나멜 기질에 콜게이트(Colgate)(등록 상표) 맥스프레시(MaxFresh)(등록 상표) 치약(미국 뉴욕주 뉴욕 소재의 콜게이트-팔몰리브(Colgate-Palmolive))의 1:2 슬러리로 솔질하고, 추가 30분 동안 다시 인큐베이션시켰다. 블록의 일부를 왁스로부터 제거하고, 웰 플레이트 내에 포매하는 한편, 다른 것들은 웰 플레이트 내의 포매 전에 다시 솔질하였다. 포매 과정은 오직 에나멜 및 균막-코팅된 에나멜 표면과의 용액 접촉을 가능하게 하였다.
이어서, 기질을 실온(약 22℃)에서, 0.1% 트윈(등록 상표)20이 있는 블로킹 완충제(포스페이트 완충된 염수 pH 7.2 중의 1 ㎎/㎖ 소 혈청 알부민(피어스(Pierce) BupH(상표명) #28372))(PBST)에서 1시간 동안 인큐베이션시키고, PBST를 사용한 2회의 세정으로 이어졌다. 길이가 15 내지 20개 아미노산인 무작위 펩티드 삽입물을 함유하는 파지의 라이브러리(1011 pfu)를 각 웰에 첨가하였다. 최종 결합 용액에는 1011 pfu 파지, 10% UV 처리된 전 타액(whole saliva) 및 0.1% 트윈(등록 상표)20 (PBST) 중의 1 ㎎/㎖ BSA가 함유되어 있다. 50 rpm에서의 쉐이킹과 함께 37℃에서 30분의 인큐베이션 후에, 각 웰 밖으로 액체를 흡입시킴으로써 미결합 파지를 제거하고, 1.0 ㎖ PBST로의 6회의 세정으로 이어졌다.
이어서, 에나멜 블록을 깨끗한 튜브로 옮기고, 0.2 M 글리신-HCl, pH 2.2 중의 1 ㎎/㎖ BSA로 이루어진 용리 완충제 1 ㎖을 각 웰에 첨가하고, 10분 동안 인큐베이션시켜 결합 파지를 용리시켰다. 이어서, 1 M Tris-HCl, pH 9.1로 이루어진 중화 완충제 167 ㎕를 각 웰에 첨가하였다. 용리 완충제 중에 있는 파지 입자뿐 아니라 에나멜 블록 상에 있는 파지 입자는 37℃에서 4.5시간 동안 LB 배지에서 1:100 희석한 밤샘 배양물로부터의 20 ㎖의 희석된 에스케리키아 콜라이 ER2738 세포와 함께 인큐베이션시킴으로써 증식시켰다. 이후에, 세포 배양물을 2분 동안 원심분리시키고, 상측의 15 ㎖의 상층액을 새로운 튜브로 옮기고, 2.5 ㎖의 PEG/NaCl(20% 폴리에틸렌 글리콜-800, 2.5 M 염화나트륨)을 첨가하고, 파지가 4℃에서 하룻밤 침전되게 하였다. 침전물을 4℃에서 10,000 x g에서의 원심분리에 의해 수집하고, 생성된 펠렛을 1 ㎖의 PBS 중에 재현탁화시켰다. 이는 1차 증폭된 스톡이었다. 이어서, 증식된 1차 파지 스톡을 표준 프로토콜에 따라 적정하였다. 이후의 회차의 바이오패닝을 위하여, 이전의 회차로부터의 2 x 1011 초과의 pfu의 파지 스톡을 사용하였다. 1차 후의 각 추가의 회차는 또한 인간 전체 타액(실온에서 2시간 동안 UV 처리)으로의 추가의 세정, 탄산염 완충제 pH 9.4(피어스 BupH(상표명) 탄산염-중탄산염 완충제 #28382)로의 2회의 세정, 50 mM 포스페이트 완충제, pH 2.5로의 2회의 세정에 이은 보통의 PBST로의 2회의 세정을 포함하였다.
3차의 바이오패닝 및 이후의 각 회차 후에, 95개의 무작위 단일 파지 플라크를 분리하고, 단일 가닥의 파지 게놈 DNA를 일러스타 템플리피(Illustra Templiphi) 500 증폭 키트(미국 뉴저지주 피스카타웨이 소재의 지이 헬쓰케어(GE Healthcare))를 사용하여 제조하고, -96 gIII 시퀀싱 프라이머(5'-CCCTCATAGTTAGCGTAACG-3'; 서열 번호 398)를 사용하여 듀폰 시퀀싱 시설(DuPont Sequencing Facility)에서 시퀀싱하였다. 디스플레이된 펩티드는 유전자 III의 신호 펩티드 직후에 위치한다. 펩티드 서열에 기초하여, 12개의 파지 후보물질을 표 3에 나타낸 바와 같이 추가의 결합 분석을 위해 동정하였다.

실시예 3
에나멜 상의 치아-결합 후보물질의 특성화
본 실시예의 목적은 합성에 의해 생성된 펩티드를 사용하여 에나멜 표면 상의 펩티드 조성물의 결합을 확인하려는 것이다.
총 11개의 합성 펩티드를 표 3으로부터 수득된 서열을 사용하여 제작하였다. 펩티드를 C-말단에서 비오틴 표지된 라이신을 사용하여 신바이오사이 코포레이션(SynBioSci Corp.)(미국 캘리포니아주 리버모어 소재)로부터 수득하였다.
에나멜 기질을 실시예 2에 기재된 바와 같이 제조하였다. 각 기질을 PBST(0.1% 트윈(등록 상표) 20이 있는 피어스 BupH(상표명) #28372) 중의 1 ㎎/㎖ BSA로 이루어진 블로킹 완충제 1 ㎖과 함께 실온(약 22℃)에서 1시간 동안 인큐베이션시켰다. 블로킹 완충제를 각 웰 밖으로 액체를 흡입시킴으로써 제거하였다. 튜브를 PBST로 이루어진 세정 완충제로 2회 헹구었다. 웰을 블로킹 완충제 중에 희석시킴으로써 제조된 20 μM 펩티드 용액 500 ㎕로 채웠다. 시료를 37℃에서 저속으로 쉐이킹시키면서 30분 동안 인큐베이션시켰다. 비-결합 펩티드를 PBST로의 6회의 세정에 의해 제거하였다. 이어서, PBST 중에 1:1000 희석된 호스래디쉬 퍼옥시다제(horseradish peroxidase)/스트렙트아비딘 컨쥬게이트(피어스 #22127) 500 ㎕를 첨가하고, 실온(약 22℃)에서 1시간 동안 인큐베이션시켰다. 컨쥬게이트 용액을 제거하고, 에나멜 블록을 PBST로 4회 세정하였다.
각 에나멜 기질을 웰로부터 제거하고, 10 ㎖의 PBST가 있는 15-㎖ 시험 튜브에서 다시 세정하였다. 이어서, 각 에나멜 기질을 에나멜 표면만이 노출되게 깨끗한 웰 플레이트에 고정시켰다. 200 ㎕의 쿠안타블루(QuantaBlu)(상표명) 기질 용액(미국 일리노이주 록포드 소재의 써모-피셔(Thermo-Fisher); #1856187)을 각 에나멜 블록에 직접 첨가하였다. 용액을 실온에서 20분 동안 인큐베이션시켰다. 200 ㎕의 쿠안타블루(상표명) 스탑(Stop) 용액(써모 피셔)을 첨가하였다. 혼합 후에, 200 ㎕의 용액을 깨끗한 96-웰 블랙(black) 원심분리용 플레이트로 옮겼다. 플레이트의 형광을 마이크로플레이트 분광광도계(미국 캘리포니아주 서니베일 소재의 몰레큘라 디바이스즈(Molecular Devices))를 사용하여 컷 오프 파장 없이 325 ㎚ 여기 및 420 ㎚ 방출을 사용하여 측정하였다. 생성된 형광 값은 표 4에 제공되어 있다. 11개의 균막/에나멜 결합 후보물질의 분석을 공지되어 있는 결합 펩티드, DenP03과 비교하였다. 각 서열을 3개의 반복검증 에나멜 기질을 사용하여 시험하였다.

실시예 4
과가수분해효소 및 과가수분해효소 융합의 구축
이러한 실시예에는 에나멜-결합 서열을 통하여 에나멜에 표적화된 과가수분해효소의 생성을 위한 발현 시스템의 고안이 기재되어 있다.
에나멜-결합 도메인에 대한 과가수분해 활성을 갖는 효소("과가수분해효소")의 융합을 암호화하는 유전자는 3'-말단에서 다양한 아미노산 가요성 링커를 암호화하는 뉴클레오티드 서열에 융합된 표 5에 열거된 다양한 효소의 폴리뉴클레오티드 서열을 갖도록 고안하였으며; 각 링커는 표 6에 기재된 바와 같은 에나멜-결합 도메인 또는 비-결합 서열 대조군에 추가로 융합된다. 유전자를 에스케리키아 콜라이에서의 발현을 위해 코돈 최적화시키고, DNA2.0(미국 캘리포니아주 멘로 파크 소재)에 의해 합성하였다. 암호화 서열을 플라스미드 내에서, T7 프로모터(발현 벡터 pLD001(서열 번호 397)) 또는 pBAD 프로모터 뒤에, NdeI과 AscI 제한 부위 사이에 클로닝하여, 플라스미드를 생성하였다. 융합 단백질을 발현시키기 위하여, 플라스미드를 적절한 발현 숙주에 전달하였다: T7 프로모터 하의 구축물을 위한 에스케리키아 콜라이 균주 BL21AI(미국 캘리포니아주 칼스배드 소재의 인비트로겐) 또는 pBAD 프로모터 하의 구축물을 위한 에스케리키아 콜라이 MG1655의 AraBAD 유도균주.
표 5에 열거된 비-표적화된 과가수분해효소 변이체를 유사하게 클로닝하였다. 써모토가 마리티마 변이체의 제조 및 재조합 발현은 이전에 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2010-0087529호(DiCosimo et al.)에서 기재되었다.
락토코커스 락티스(아세틸 자일란 에스테라제; 서열 번호 40), 메소리조비움 로티(아세틸 자일란 에스테라제; 서열 번호 42) 및 바실러스 푸밀러스(아세틸 자일란 에스테라제; 서열 번호 10)로부터의 추가의 CE-7 과가수분해효소를 유사한 방식으로 클로닝하였다. 락토코커스 락티스, 메소리조비움 로티 및 바실러스 푸밀러스로부터의 CE-7 과가수분해효소의 클로닝 및 발현은 이전에, 각각이 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2011-0081693호 및 미국 특허 제7,951,566호(DiCosimo et al.)에 기재되었다.
과가수분해효소의 CE-7 패밀리에 속하지 않는 과가수분해 효소도 또한 유사한 방식으로 클로닝하였다. 과가수분해 활성을 갖는 마이코박테리움 스메그마티스 아릴 에스테라제("ArE"; 야생형 서열은 서열 번호 478이며; S54V 변이체는 서열 번호 460으로 제공된다)는 미국 특허 제7,754,460호에 기재되어 있다. 과가수분해 활성을 갖는 슈도모나스 플루오레슨스 에스테라제 변이체 L29P("Pfl"; 서열 번호 477)는 미국 특허 제7,384,787호에 기재되어 있다.





실시예
5
융합 단백질의 생성
이러한 실시예에는 구강 표면에 대한 결합을 위한 표적화 서열이 있거나 없는 과가수분해효소의 발현 및 정제가 기재되어 있다.
균주를 50 ㎎/L의 스펙티노마이신을 함유하는 1 리터의 자가유도(autoinduction) 배지(10 g/L의 트립톤, 5 g/L의 효모 추출물, 5 g/L의 NaCl, 50 mM Na2HPO4, 50 mM KH2PO4, 25 mM (NH4)2SO4, 3 mM MgSO4, 0.75% 글리세롤, 0.075% 글루코스 및 0.05% 아라비노스)에서 200 rpm 진탕 하에 37℃에서 20시간 동안 성장시켰다. 비표적화된 과가수분해효소의 생성은 이전에 미국 특허 출원 공개 제2010-0087529호(DiCosimo et al.)에서 기재되었다. 표적화된 과가수분해효소의 생성은 유사한 프로토콜을 따랐다. 세포를 8000 rpm에서의 원심분리에 의해 수집하고, 세포 펠렛을 1 mM DTT를 함유하는 20 ㎖의 50 mM 인산칼륨 완충제, pH 7.1에 재현탁화시킴으로써 세정하였다. 용액을 8000 rpm에서 다시 원심분리하고, 상층액을 제거하고, 펠렛을 DTT를 함유하는 포스페이트 완충제에 다시 재분산시켰다. 이어서, 용액을 30초 동안 균질화시켜, 펠렛을 분산시켰다(브린크만 호모지나이저 모델(Brinkman Homogenizer model) PCU11). 이어서, 용액을 89.6 ㎫(약 13,000 psi)에서 프렌치 프레스(French Press)(SLM 인스트루먼츠(Instruments))를 통과하게 처리함으로써 세포를 용해시켰다. 용액을 추가 2회 프레스를 통과하게 처리하여, 완전한 용해를 달성하였다. 이어서, 세포 용액을 코니컬 튜브(conical tube)로 옮기고, 8500 rpm에서 5분 동안 원심분리하였다. 써모토가 마리티마 구축물을 위하여, 상층액을 제거하고, 80℃에서 30분 동안 가열하였다. 용액을 다시 원심분리하고, 상층액을 깨끗한 바이얼로 옮겼다.
비-호열성 효소에 대하여, 오염시키는 세포 성분으로부터 효소를 정제하기 위하여 열 처리를 사용하지 않았다. 대신에, 시료를 Co-NTA 아가로스(히스푸르 코발트 레진(HisPur Cobalt Resin), 써모 사이언티픽(Thermo Scientific), 제품 번호: 89965)를 사용하는 금속 킬레이트화 크로마토그래피에 의해 효소의 C-종말단에 융합된 His6 태그를 사용하여 정제하였다. 전형적으로, 세포 추출물을 4 부피의 평형화 완충제(10 mM Tris HCl pH 7.5, 10% 글리세롤, 1 mM 이미다졸 및 150 mM NaCl)로 평형화된 Co-NTA 아가로스의 5 내지 10 ㎖ 컬럼에 로딩하였다. 컬럼 상에 로딩된 각 추출물의 양을 Co-NTA 아가로스 비드 ㎖당 5 내지 10 ㎎의 과가수분해효소 융합을 함유하도록 조정하였다. 레진을 2 충진 부피의 평형화 완충제로 세정하고, 2 부피의 용리 완충제(10 mM Tris HCl pH 7.5, 10% 글리세롤, 150 mM 이미다졸, 500 mM NaCl)로 용리시켰다. 분획을 수집하고, 전장의 정제된 단백질의 존재를 PAGE에 의해 확인하였다.
세포 생성 후에 구축물 EZ-19- 내지 EZ-26의 생성을 위하여, 세포를 8000 rpm에서의 원심분리에 의해 수집하고, 세포 펠렛을 20 ㎖의 50 mM 인산칼륨 완충제, pH 7.2에 재현탁화시킴으로써 세정하였다. 용액을 8000 rpm에서 다시 원심분리하고, 상층액을 제거하고, 펠렛을 포스페이트 완충제에 다시 재분산시켰다. 이어서, 용액을 30초 동안 균질화시켜, 펠렛을 분산시켰다(브린크만 호모지나이저 모델 PCU11). 이어서, 용액을 89.6 ㎫(약 13,000 psi)에서 프렌치 프레스(SLM 인스트루먼츠)를 통과하게 처리함으로써 세포를 용해시켰다. 용액을 추가 2회 프레스를 통과하게 처리하여, 완전한 용해를 달성하였다. 이어서, 세포 용액을 코니컬 튜브로 옮기고, 8500 rpm에서 5분 동안 원심분리하였다. 불용성 용해물 펠렛을 50 ㎖의 세포 용해물 펠렛당 3 ㎖의 완충제로, 사르코실(sarkosyl) 완충제(50 mM 포스페이트 완충제 pH 7.2, 2% 트리톤(등록 상표)-X100 및 1.5% 사르코실) 중에 용해시켰다. 용액을 원심분리하고, 상층액을 새로운 튜브로 옮겼다. 융합 단백질을 써모 사이언티픽(미국 일리노이주 록포드 소재)으로부터의 히스푸르(상표명) 코발트 레진 키트를 사용함으로써 정제하였다.
이러한 생성 및 정제 프로토콜의 산출량은 통상 ㎖당 2 내지 10 ㎎의 단백질을 제공하며, 융합 과가수분해효소의 순도는 폴리아크릴아미드 겔 전기영동(PAGE) 분석에 의해 추정시 90% 내지 75%의 단백질이었다. 표준물질로서 소 혈청 알부민의 용액을 사용하여 총 단백질을 BCA(비신코닌산(bicinchoninic acid)) 검정법(써모 사이언티픽)에 의해 정량화하였다.
실시예 6
에나멜-표적화된 과가수분해효소 융합의 수산화인회석으로의 결합
이러한 실시예에는 과가수분해효소의 수산화인회석 입자로의 결합이 기재된다. 수산화인회석은 에나멜에 대한 효과적인 모방체(mimic)이다.
표 6에 열거된 과가수분해효소는 수산화인회석으로의 결합에 대해 평가하였다. 수산화인회석 나노입자(알드리치 677418)의 분산액을 10 mM 포스페이트 완충제 pH 7.2에 0.5% 고형물로 제조하였다. 효소 스톡 용액을 수산화인회석 분산액에 10 μM의 최종 농도로 첨가하고, 온건하게 진탕시키면서 원심분리용 튜브에서 30분 동안 인큐베이션시켰다. 각 시료를 10000 rpm에서 5분 동안 원심분리하였다. 상층액을 제거하고, 추가의 완충제를 첨가하였다. 입자를 재현탁화시키고, 새로운 튜브에 옮겼다. 상기 과정을 추가 2회 반복하였다. 원래 효소 투입 시료, 각 세정 단계를 사용하여 입자로부터 제거된 상층액 및 최종 입자를 SDS-PAGE 분석을 위해 60 ㎕의 시료, 20 ㎕의 LDS 완충제 및 8 ㎕의 환원제(Nu-PAGE)를 배합함으로써 제조하였다. 시료를 85℃에서 10분 동안 가열하였다. 25 ㎕의 각 시료를 4-12% 비스트리스(BisTris) 겔(인비트로겐(Invitrogen)) 상에 로딩하고, 1.5시간 동안 115 볼트에서 전개시켰다. 겔을 제거하고, 심플리 블루 스테인(Simply Blue stain)(인비트로겐)의 1:1 희석액으로 하룻밤 염색하고, 탈이온수 중에서 4시간 동안 탈염시켰다. 겔을 분석하고, 각 분획 중의 효소의 검출을 밴드의 존재에 기초하여 추정하였다. 분석의 결과는 수산화인회석 표면과 각 효소 간의 결합의 세기를 결정하기 위하여, 효소 투입(투입 = 1)에 비한 겔에서의 각 밴드의 상대 세기의 표시와 함께 표 7에 제공되어 있다.
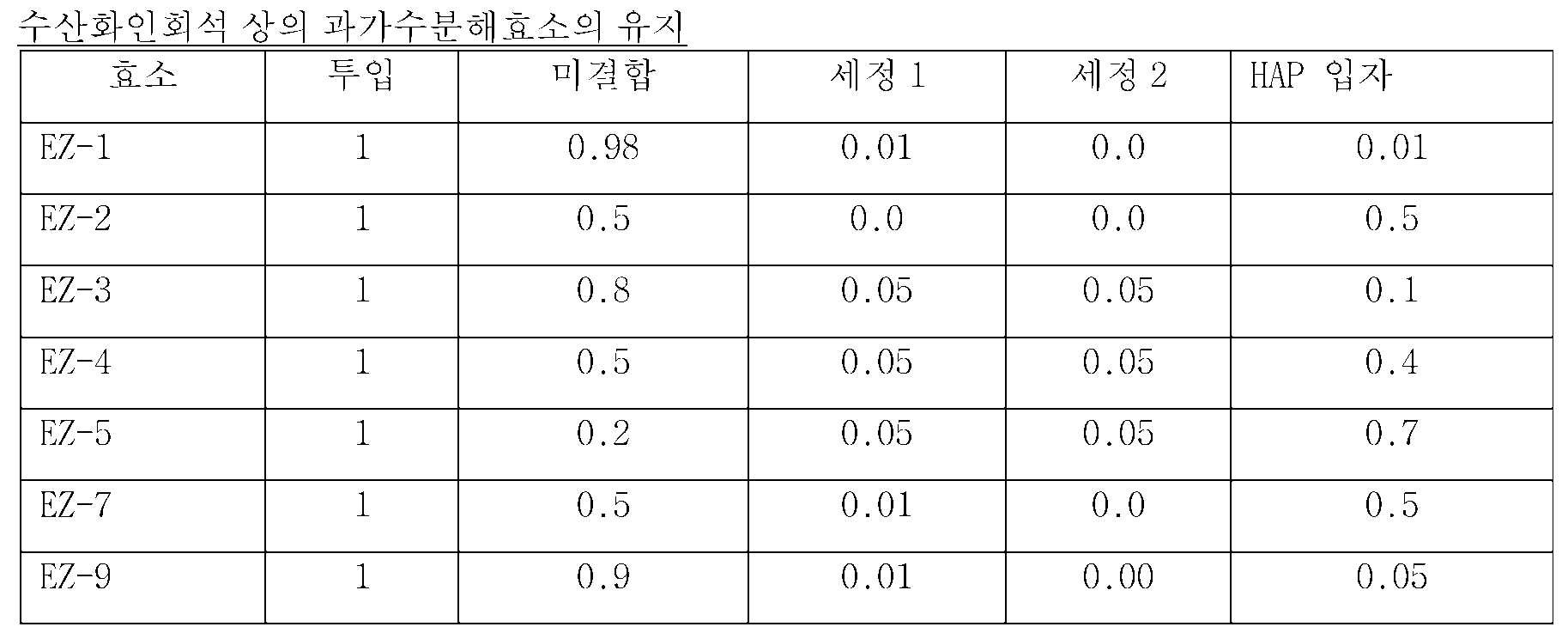
표 7의 데이터에 의해, 표적화 서열과의 과가수분해효소 융합이 세정 후에 수산화인회석에 남아 있는 한편, 비표적화 과가수분해효소는 그렇지 않았다.
실시예 7
수산화인회석에 대한 결합 및 용액 중의 과가수분해효소 활성의 정량화
이러한 실시예에는 과아세트산을 생성하기 위해 트라이아세틴 및 과산화수소를 사용하는 과가수분해효소의 과가수분해효소 활성을 통한 과가수분해효소의 검출 및 정량화 방법이 기재되어 있다.
과아세트산의 검출은 과아세트산에 의한 2,2'-아지노-비스(3-에틸벤조티아졸린)-6-설포네이트(ABTS) 산화의 비색 검출을 사용하여 문헌[Pinkernell et. al. (Analyst, 1997, 122, 567)]에 기재된 방법을 따랐다. 트라이아세틴 및 과산화수소를 첨가하여 과아세트산을 형성한 후에, 90 ㎕의 용액을 웰 플레이트 내의 10 ㎕의 0.1 M H3PO4에 첨가하였다. 50 ㎕의 1 M 아세트산, 50 ㎕의 0.5 g/L ABTS 및 50 ㎕의 0.002 g/L KI를 첨가하였다. 용액을 5분 동안 발색(develop)되게 하였다. 용액의 흡광도를 마이크로플레이트 판독기를 사용하여 405 ㎚에서 측정하였다. 과아세트산 농도를 동시에 과아세트산 시약 용액을 사용하여 만든 표준 곡선에 기초하여 계산하였다.
용액 중의 효소 활성을 50 mM 포스페이트 완충제, pH 7.2 중의 0.625 ㎍/㎖의 각 효소의 용액을 제조함으로써 측정하였다. 10 ㎕의 효소 용액을 90 ㎕의 완충제, 30 ㎕의 수 중의 3% 트라이아세틴, 30 ㎕의 30 mM H2O2와 혼합하였다. 용액을 5분 동안 인큐베이션시켰다. 90 ㎕의 분취액을 상기 기재된 바와 같이 ABTS 산화를 통한 검출을 위해 제거하였다. 결과는 표 8에 열거되어 있다.

HAP 디스크(disc) 표면에 1회 결합된 융합의 과가수분해효소 활성을 동일한 ABTS 방법을 사용하여 결정하였다. 수산화인회석 디스크(미국 뉴욕주 올드 베쓰페이지 소재의 하이메드 인코포레이티드(HiMed Inc); 5 ㎜ 직경 X 1.8㎜ 두께)를 20 μM 효소 용액(50 mM 인산칼륨 완충제 pH 7.2) 중에 60분 동안 인큐베이션시키고, 6회의 세정(50 mM 인산칼륨 완충제 pH 7.2)으로 이어졌다. 효소가 흡수된 디스크를 새로운 웰로 옮기고, 200 ㎕의 포스페이트 완충제(10 mM pH 7.2), 30 ㎕의 3% 트라이아세틴(0.346%의 최종 농도) 및 30 ㎕의 30 mM H2O2(3.46 mM의 최종 농도)를 첨가하였다. 용액을 실온에서 5분 동안 인큐베이션되게 하였다. 90 ㎕의 용액을 10 ㎕의 100 mM H3PO4를 함유하는 새로운 웰로 피펫팅하였다. 50 ㎕의 아세트산, 50 ㎕의 KI, 50 ㎕의 ABTS를 상기 기재된 바와 같이 첨가하였다. 용액을 실온에서 5분 동안 발색시키고, A405㎚에서 판독하였다. 결과는 표 9에 열거되어 있다.

이러한 실험에는 EZ-5 및 EZ-7이 수산화인회석에 결합되는 경우 활성 효소이며, 트라이아세틴 및 과산화수소의 첨가와 함께 과아세트산이 생성되는 것이 입증된다. EZ-1과 함께 검출된 낮은 과아세트산의 값은 EZ-1이 수산화인회석(실시예 6)에 결합되지 않으며, 과아세트산을 생성하기 위하여 표면 상에 존재하지 않는다는 다른 관찰과 일치하는 것이다.
실시예 8
추가의
CE
-7
과가수분해효소
구축물을 위한 용액 중의
과가수분해효소
및
결합된
과가수분해효소 활성의 정량화
이러한 실시예에는 써모토가 마리티마, 바실러스 푸밀러스(Bpu), 메소리조비움 로티(Mlo) 및 락토바실러스 락티스(Lla)로부터의 수산화인회석에 결합하는 경우, 그리고 용액 중의 CE-7 과가수분해효소 및 각 융합 단백질의 트라이아세틴 및 과산화수소를 사용하여 과아세트산을 생성하는 과가수분해 활성이 기재되어 있다.
표 6에 열거된 과가수분해효소를 용액 활성에 대하여 평가하였다. 100 mM 포스페이트 완충제, pH 7.2 중의 0.5 μM의 효소, 100 mM의 트라이아세틴, 100 mM의 H2O2를 함유하는 500 ㎕ 용액을 제조함에 의한 용액 중의 과가수분해효소 활성의 측정 방법은 실시예 7에 기재되어 있다. 용액을 혼합하고, 37℃에서 10분 동안 인큐베이션시켰다. 10 ㎕의 분취액을 제거하고, 실시예 7에 기재된 바와 같이 검출을 위해 희석하였다. 과아세트산 농도는 과아세트산의 스톡 용액(알드리치)을 사용하여 생성된 표준 곡선을 사용하여 결정하였다. 효소 부재를 대조군으로 제시하는 실험도 또한 수행하였다.

이들 구축물의 수산화인회석으로의 결합을 평가하기 위하여, 33 ㎎의 수산화인회석 입자(마크로-프렙 세라믹 하이드로아파타이트 타입(Macro-prep Ceramic Hydroxyapatite TYPE) I, 80 ㎛ 사이즈, 미국 캘리포니아주 헤르큘레스 소재의 바이오래드(BioRad))를 10 mM 포스페이트 완충제 pH 7.2로 세정하였다. 상층액을 제거한 후에, 효소 스톡 용액을 수산화인회석 분산액에 10 μM의 최종 농도로 첨가하고, 온건하게 진탕시키면서 원심분리용 튜브에서 30분 동안 인큐베이션시켰다. 각 시료를 10000 rpm에서 1분 동안 원심분리하였다. 상층액을 제거하고, 추가의 완충제를 첨가하였다. 입자를 재현탁화시키고, 새로운 튜브에 옮겼다. 상기 과정을 추가 2회 반복하였다. 수산화인회석 결합된 효소 활성을 100 mM 포스페이트 완충제, pH 7.2 중의 100 mM 트라이아세틴, 100 mM H2O2를 함유하는 500 ㎕ 용액을 첨가함으로써 측정하였다. 용액을 37℃에서 30분 동안 인큐베이션시켰다. 분취액을 제거하고, H3PO4와 혼합하고, 실시예 7에 기재된 바와 같은 ABTS 산화를 통한 검출을 위해 적절하게 희석하였다. 실험을 상이한 날에 수행하였으며, 각 실험에 대하여 효소 부재를 대조군으로서 사용하였다. 결과는 표 11에 열거되어 있다.

이러한 실험에서는 Bpu, Mlo 및 Llo 효소로부터의 모든 CE-7 과가수분해효소 융합이 효소 부재 대조군에 비하여, 용액 중에서 뿐만 아니라 수산화인회석 표면에 대한 결합 후에 유의미한 과가수분해효소 활성이 나타난 것이 입증된다. 바실러스 푸밀러스로부터의 모든 3가지 CE-7 과가수분해효소 융합은 마이코박테리움 로티 또는 락토코커스 락티스로부터의 융합 효소에 비하여 효소 활성이 더 높았다. 바실러스 푸밀러스로부터의 표적화된 EZ-29는 비표적화된 EZ-27에 비하여 더 높은 결합 활성을 나타냈다.
실시예 9
CXH
펩티드가 있는
표적화된
C277S
과가수분해효소
융합에 대한 용액 중의, 그리고 수산화인회석에
결합된
효소 활성의 정량화
이러한 실시예에는 CE-7 C277S 및 CXH 펩티드와의 융합, 및 용액 중에서, 그리고 수산화인회석에 결합되는 경우에, 트라이아세틴 및 과산화수소를 사용하여 과아세트산을 생성하는 그들의 과가수분해효소 활성이 기재되어 있다. 수산화인회석은 에나멜에 대한 효과적인 모방체이다.
표 6에 열거된 과가수분해효소 EZ-19 내지 EZ-26을 용액 중의 효소 활성에 대하여 평가하였다. 용액 중의 과가수분해효소 활성을 측정하는 방법은 실시예 7에 기재되어 있다. 용액 중의 효소 활성을 100 mM 트라이아세틴, 100 mM H2O2 및 10 mM 포스페이트 완충제, pH 7.2 중의 0.5 μM의 각 효소의 500 ㎕ 사르코실 완충제 용액을 제조함으로써 측정하였다. 용액을 37℃에서 10분 동안 인큐베이션시켰다. 반응을 H3PO4와 혼합함으로써 중단시킨 후에, 10 ㎕ 분취액을 적절한 희석을 위해 제거한 다음, 실시예 7에 기재된 바와 같이 ABTS 산화를 통해 검출하였다. 결과는 표 12에 열거되어 있다.
또한, 동일한 과가수분해효소 융합 효소를 수산화인회석에 대한 결합에 대해 평가하였다. 수화된 수산화인회석 디스크(5 ㎜ 직경 × 1.8 ㎜ 두께, 하이메드 인코포레이티드로부터의)를 사용하였다. 디스크를 10 mM 포스페이트 완충제 pH 7.2에서 10분 동안 평형화시켰다. 200㎕의 효소 용액을 10 mM 포스페이트 완충제 중의 10 μM의 최종 농도로 수산화인회석 디스크에 첨가하고, 온건하게 진탕시키면서 원심분리용 튜브에서 30분 동안 인큐베이션시켰다. 상층액을 제거하고, 미결합 효소 활성 검정법을 위해 사용하였다. 디스크를 새로운 튜브에 옮기고, 포스페이트 완충제로 헹구었다. 상기 과정을 추가 2회 반복하였다. 결합된 효소가 있는 수산화인회석 디스크의 효소 활성을 100 mM 트라이아세틴, 100 mM H2O2 및 100 mM 포스페이트 완충제, pH 7.2를 함유하는 500 ㎕ 용액을 디스크에 첨가하고, 37℃에서 30분 동안 인큐베이션시킴으로써 측정하였다. 반응을 H3PO4와 혼합함으로써 중단시킨 후에, 10 ㎕ 분취액을 적절한 희석을 위해 제거한 다음, 실시예 7에 기재된 바와 같이 ABTS 산화를 통해 검출하였다. 실험을 몇일에 걸쳐 수행하고, 효소 부재 용액을 각 날에 대조군으로 사용하였다. 효소 부재 대조군을 제한 후의 결과는 표 12에 열거되어 있다.

실험에 의해, CXH 표적화 서열이 있는 모든 C277S 변이체가 용액 중에서, 그리고 수산화인회석에 결합되는 경우에, 활성이 있으며 충분한 양의 과아세트산을 생성하는 것이 입증되었다.
실시예 10
비 CE-7 과가수분해효소 구축물에 대한, 용액 중의, 그리고 수산화인회석에 결합되는 과가수분해효소 활성의 정량화
본 실시예의 목적은 용액 중에서, 그리고 수산화인회석에 결합되는 경우에 과아세트산을 생성하기 위한, 마이코박테리움 스메그마티스로부터의 표적화 또는 비표적화 아릴 에스테라제 효소 변이체 및 표 6에 열거된 슈도모나스 플루오레슨스로부터의 과가수분해효소 변이체의 용도를 증명하려는 것이다.
아릴 에스테라제 구축물을 위하여, 용액 중의 효소 활성은 100 mM 포스페이트 완충제, pH 7.2 중의 0.5 μM의 각 효소, 100 mM 트라이아세틴 및 100 mM H2O2의 1 ㎖ 용액을 제조함으로써 측정하였다. 용액을 혼합하고, 37℃에서 30분 동안 인큐베이션시켰다. 실시예 7에 기재된 바와 같이, 반응은 일부를 H3PO4로 옮김으로써 중단시키고, 10 ㎕ 분취액을 제거하고, ABTS 산화를 통한 검출을 위해 희석하였다.
수산화인회석 상의 결합 및 활성의 평가를 위하여, 아릴 에스테라제 효소를 10mM 포스페이트 완충제 중의 10 μM 용액을 사용하여, 완충제 세정된 HAP 입자(마크로-프렙 세라믹 하이드로아파타이트(Macro-prep Ceramic Hydroxyapatite) 타입 I, 80 ㎛ 사이즈, 미국 캘리포니아주 헤르큘레스 소재의 바이오래드) 33 ㎎을 사용하여 실시예 8에 기재된 바와 같이 수산화인회석 입자에 노출시켰다. 원심분리 및 효소 용액의 제거 후에, 원심분리 및 상층액의 제거에 의하여 입자를 포스페이트 완충제로 헹구었다. 100 mM 트라이아세틴, 100 mM H2O2 및 100 mM 포스페이트 완충제, pH 7.2를 함유하는 200 ㎕ 용액을 입자에 첨가하고, 37℃에서 30분 동안 인큐베이션시켰다. 실시예 7에 기재된 바와 같이, 반응은 일부를 H3PO4로 옮김으로써 중단시키고, 10 ㎕ 분취액을 제거하고, ABTS 산화를 통한 검출을 위해 희석하였다. 용액 및 표면 결합 검정법 둘 모두에 대한 결과는 표 13에 열거되어 있다.

슈도모나스 플루오레슨스 구축물을 위하여, 용액 활성을 1 M 아세트산나트륨 완충제, pH 5.5 중의 2 μM 효소, 100 mM H2O2를 함유하는 1 ㎖ 용액을 혼합함으로써 측정하였다. 용액을 37℃에서 30분 동안 인큐베이션시켰다. 실시예 7에 기재된 바와 같이, 반응은 일부를 H3PO4로 옮김으로써 중단시키고, 10 ㎕ 분취액을 제거하고, ABTS 산화를 통한 검출을 위해 희석하였다. 수산화인회석 상의 결합 및 활성의 평가를 위하여, 슈도모나스 플루오레슨스 효소를 완충제 세정된 HAP 입자(마크로-프렙 세라믹 하이드로아파타이트 타입 I, 80 ㎛ 사이즈, 미국 캘리포니아주 헤르큘레스 소재의 바이오래드) 100 ㎎을 사용하여 실시예 8에 기재된 바와 같이 10 mM 포스페이트 완충제 중에 20 μM로 수산화인회석 입자에 노출시켰다. 원심분리 및 효소 용액의 제거 후에, 원심분리 및 상층액의 제거에 의하여 입자를 포스페이트 완충제로 헹구었다. 1 M 아세트산나트륨 완충제, pH 5.5 중의 300 mM H2O2를 함유하는 200 ㎕ 용액을 입자에 첨가하고, 37℃에서 10분 동안 인큐베이션시켰다. 실시예 7에 기재된 바와 같이, 반응은 일부를 H3PO4로 옮김으로써 중단시키고, 10 ㎕ 분취액을 제거하고, ABTS 산화를 통한 검출을 위해 희석하였다. 용액 및 표면 결합 검정법 둘 모두에 대한 결과는 표 14에 열거되어 있다.

이들 실험에 의해, CE-7 패밀리를 벗어난 패밀리로부터의 과가수분해효소가 표적화 서열을 포함하는 구축물에 대하여 용액 중에서, 그리고 수산화인회석에 결합되는 경우에 활성이 있음이 입증된다.
실시예 11
용액 중의 및 소 에나멜에 결합된 에나멜-표적화 C277S 및 C277T 변이체 과가수분해효소 융합의 과가수분해 활성의 정량화
이러한 실시예에는 과가수분해효소 융합 단백질의 소 에나멜으로의 결합, 및 용액 중의 그리고 소 에나멜에 결합되는 경우의 효소 활성의 측정이 기재된다.
표 5에 열거된 과가수분해효소를 실시예 7에 기재된 바와 같이 용액 중의 효소 활성에 대하여 평가하였다. 용액 중의 효소 활성을 100 mM 트라이아세틴, 100 mM H2O2 및 10 mM 포스페이트 완충제, pH 7.2 중의 0.5 μM의 각 효소의 500 ㎕ 용액을 제조함으로써 측정하였다. 용액을 37℃에서 10분 동안 인큐베이션시켰다. 반응을 분취액을 H3PO4와 혼합함으로써 중단시킨 후에, 10 ㎕ 분취액을 적절한 희석을 위해 제거한 다음, 실시예 7에 기재된 바와 같이 ABTS 산화를 통해 검출하였다. 결과는 표 15에 열거되어 있다.
또한, 과가수분해효소를 소 에나멜 기질로의 결합에 대해 평가하였다. 에나멜 기질을 실시예 2 및 3에 기재된 바와 같이 제조하였다. 각 에나멜 블록을 실온(약 22℃)에서 하룻밤 동안 수 중에 수화시켰다. 이어서, 에나멜 블록을 10분 동안 10 mM 인산칼륨 완충제, pH7.2로 평형화시켰다. 에나멜 기질을 완충제로 3회 헹구었다. 에나멜 웰을 10 mM 포스페이트 완충제 중에서 희석시킴으로써 제조된 10 μM 효소 용액 500 ㎕로 채웠다. 시료를 37℃에서 저속으로 쉐이킹시키면서 30분 동안 인큐베이션시켰다. 비-결합 효소를 포스페이트 완충제로의 4회의 세정에 의해 제거하였다. 이어서, 각 에나멜 블록을 퍼티(putty)가 채워진 24-웰 플레이트 내측에 포매시키고, 오직 에나멜 상측 표면 만을 노출시켰다. 에나멜 결합 효소로부터의 과가수분해효소 활성을 실시예 8에 기재된 바와 같이 측정하였다. 100 ㎕의 반응 혼합물(100 mM 포스페이트 완충제, pH7.2 및 100 mM H2O2 및 100 mM 트라이아세틴)을 에나멜의 상측에 첨가하고, 37℃에서 30분 동안 인큐베이션시켰다. 90 ㎕의 분취액을 실시예 8에 기재된 바와 같이 ABTS 산화를 통한 검출을 위해 제거하였다. 결과는 표 15에 열거되어 있다. 각 시료 데이터점은 3가지 독립적인 에나멜 블록의 평균을 나타낸다.

이러한 실험에 의해, 써모토가 마리티마 서열 변이체로부터의 C227S 및 C277T를 사용하는 융합이 표적 서열을 포함하는 구축물에 대하여, 용액 중에서, 그리고 소 에나멜에 결합되는 경우에 활성이 있으며, 치아를 미백시키기에 충분한 수준의 과아세트산을 생성함이 입증된다.
실시예 12
1-단계 적용에서 과가수분해효소를 사용한 치아 탈색 효능
이러한 실시예의 목적은 1-단계 적용에서 효소에 의해 생성된 과아세트산의 치아 탈색 효과를 보여주고 화학적으로 유도된 과아세트산으로 달성되는 효능과 비교하려는 것이다. 표적 치아 탈색 수준을 달성하기 위하여 과가수분해 효소(CE-7 과가수분해효소) 시스템을 사용하는 2가지 방법을 개발하였다. 제1 방법(본 명세서에서 "1-단계 방법"으로 지칭)은 에나멜 착색 모델을 사용하여 상이한 양의 적어도 하나의 CE-7 과가수분해효소를 트라이아세틴(일 예의 적절한 에스테르 기질) 및 과산화수소와 배합하여, 과아세트산을 생성하는 것을 포함한다. 소 에나멜 절치를 실시예 1에 기재된 바와 같이 제조하였다. 착색된 소 에나멜 블록을 기질의 색상을 안정화시키기 위하여 사용하기 적어도 1시간 전에 수 중에 수화시켰다. 각 에나멜 블록에 대한 색상을 수화 후에, 실험의 시작 전에 측정하였다. 3가지 에나멜 시료를 각 용액 유형에 대하여 처리하였다. 사용된 용액은 완충제만의 대조군, 2.5% H2O2, 1% 과아세트산, 10 μM EZ-1(C277S; 서열 번호 424), 100 mM 트라이아세틴 및 250 mM H2O2를 함유하는 과가수분해효소 조성물 및 100 mM 트라이아세틴 및 250 mM H2O2의 효소 부재 대조군이었다. 모든 용액을 500 mM 인산나트륨 완충제, pH 7.2 중에서 제조하였다. 중성의 pH에서 1% 과아세트산 용액을 유지하는데 높은 완충제 세기가 필요하였다. 모든 용액을 각 처리점을 위해 새로이 제조하였다. 효소-트라이아세틴-H2O2 조합물 및 트라이아세틴- H2O2 조합물을 에나멜 노출 직전에 혼합하였다. 각 처리 단계 후에, 각 에나멜 블록을 물로 헹구고, 색상 측정치를 수득하였다. 총 61분의 처리 시간을 1분, 5분, 10분, 15분 및 30분의 노출 기간과 함께 사용하였다. 1분, 10분 및 30분 처리 후의 용액 중의 과아세트산의 농도를 ABTS 산화의 비색 검출에 의해 평가하였다. 결과는 표 16 및 17에 제공되어 있다.

비색 방법에 의한 반응 혼합물 중의 과아세트산(PAA) 농도의 결정을 상기 문헌[Dinu et. al.]에 기재된 방법에 따라 수행하였다. 125 mM 시트르산칼륨 완충제, pH 5.0 중의 1 mM 2,2'-아지노-비스(3-에틸벤조티아졸린)-6-설포네이트(ABTS), 50 μM 요오드화칼륨의 시약 용액을 제조하였다. 25 ㎕의 시료를 975 ㎕의 이러한 검출 시약과 혼합하고, 5분 동안 인큐베이션되게 하였다. 용액을 마이크로플레이트 판독기를 사용하여 405 ㎚에서 흡광도에 대하여 분석하였다. 용액 중의 과아세트산의 상대량의 비교를 흡광도 값을 직접 비교함으로써 결정하였다.

표 16의 데이터에 의해, 효소-에스테르-과산화물 조성물이 치아의 미백에 효과적인 것이 확인된다. 표 17의 데이터에 의해, 과아세트산이 EZ-1(C277S; 서열 번호 424) 효소, 트라이아세틴 및 H2O2의 조합으로부터 생성되는 것이 입증된다. 또한, 화학적 과아세트산에 대한 흡광도 데이터의 비교에 의해, 효소적 탈색 시스템이 놀랍게도 더 나은 미백 성능과 함께 더 낮은 검출가능한 과아세트산을 보이는 것으로 나타났다. 또한, 트라이아세틴 및 H2O2를 함유하는 효소-부재에 대하여 낮은 수준의 과아세트산이 시간이 지남에 따라 생성되며, 이는 ABTS 시약으로의 1:10 희석으로 검출되었다(표에는 포함되지 않음). 이는 효소 촉매작용된 과아세트산의 높은 수준의 생성에 비하여 훨씬 더 느린 속도로 검출가능한 탈색 성능으로 이어졌다.
실시예 13
1-단계 적용에서 에나멜
표적화된
과가수분해
효소를 사용한 치아 탈색 효능
이러한 실시예의 목적은 표적화된 CE-7 과가수분해효소를 사용하는 1-단계 적용에서 효소에 의해 생성된 과아세트산의 치아 탈색 효과를 보여주고 화학적으로 유도된 과아세트산으로 달성되는 효능과 비교하려는 것이다.
소 에나멜 절치를 실시예 1에 기재된 바와 같이 제조하였다. 착색된 소 에나멜 블록을 기질의 색상을 안정화시키기 위하여 사용하기 적어도 1시간 전에 수 중에 수화시켰다. 각 에나멜 블록에 대한 색상을 수화 후에, 실험의 시작 전에 측정하였다. 2가지 에나멜 시료를 각 용액 유형에 대하여 처리하였다. 사용된 용액은 완충제만의 대조군, 0.1% 과아세트산, 0.52 μM EZ-7(에나멜 결합 도메인이 있는 C277S; 서열 번호 429), 100 mM 트라이아세틴 및 32.6 mM H2O2를 포함하는 과가수분해효소 조성물이었다. 모든 용액을 100 mM 인산나트륨 완충제, pH 7.2 중에서 제조하였다. 모든 용액을 각 처리점을 위해 새로이 제조하였다. 효소-트라이아세틴-H2O2 조합물을 에나멜 노출 직전에 혼합하였다. 각 처리 단계 후에, 각 에나멜 블록을 물로 헹구고, 색상 측정치를 수득하였다. 30분의 처리 시간을 각 에나멜 기질의 세트에 대하여 4회 반복하였다. 30분 처리 후의 용액 중의 과아세트산의 농도를 ABTS 산화의 비색 검출에 의해 평가하였다. 결과는 표 18 및 19에 제공되어 있다.


표 18의 데이터에 의해, 표적화된 효소-에스테르-과산화물 조성물이 1-단계 과정에서 치아를 미백시키는데 효과적인 것이 확인된다. 표 19의 데이터에 의해, 과아세트산이 EZ-7 효소, 트라이아세틴 및 H2O2의 조합으로부터 생성되는 것이 입증된다.
실시예 14
다양한 에스테르 기질에 대한 CE-7 과가수분해효소를 사용하는 과아세트산 생성
이러한 실시예의 목적은 에나멜 표적화 과가수분해효소를 포함하는 매우 다양한 CE-7 과가수분해효소가 구강 관리 특이적 조건 하에서 매우 다양한 에스테르로부터의 과아세트산의 형성을 촉매작용시키는 것을 입증하려는 것이었다.
써모토가 마리티마 과가수분해효소의 변이체를 클로닝하고, 재조합으로 발현하고, 실시예 4에 기재된 것과 유사한 방식으로 정제하였다. 이들 과가수분해효소 변이체의 서열 변화가 표 5에 열거되어 있다.
다양한 에스테르 기질을 표 5에 열거된 적어도 2개 이상의 과가수분해효소 변이체에 대하여 시험하였다. 맞춤 합성한 3가지 에스테르를 제외하고, 모든 다른 에스테르를 시그마 알드리치(미국 미주리주 세인트 루이스 소재), 티씨아이 아메리카(미국 오리건주 포틀랜드 소재), 알파 아에사르(Alpha Aesar)(미국 매사추세츠주 워드 힐 소재) 또는 테센들로 컴퍼니(Tessendlo Company)(미국 애리조나주 포에닉스 소재)로부터 구하였다.
에스테르 기질에 대한 다양한 효소를 시험하기 위하여, 각 개별 반응의 시작을 0.5 내지 1분의 시차를 둠으로써 최대 4개의 반응을 동시에 수행하는 것이 편리하였다. 각 개별 반응을 자성 교반 막대가 구비된 유리관(높이: 76 ㎜; 외경: 33 ㎜; 내경: 24 ㎜)에서 수행하였다. 4개의 반응관을 함께 묶고, 재킷형(jacketed) 1 리터, 스테인리스 강, 템퍼링 비커(tempering beaker)(케이지더블유 아이소썸(KGW IsoTherm), # TSS-G 1000W)에서 불변의 온도로 유지하고, 순환수를 써모 네스랩(Thermo NesLab) 재순환 배쓰(모델 # RTE-7 디지털 원(Digital One))에 의해 조절하였다. 완충제(6 ㎖, 100 mM 인산나트륨 완충제, pH 7.2)를 각 반응관에 첨가하고, 37℃에서 평형화되게 하였다. 대상 에스테르 기질을 첨가하여 100 mM 농도를 달성하였다. 반응을 40 ppm 효소 및 60 mM H2O2 (37 ㎕, 30% 과산화수소 용액)의 동시의 첨가에 의해 개시하였다. 과아세트산의 생성을 추적하기 위하여, 각 반응으로부터의 80 ㎕ 시료를 1 내지 15분의 특정 간격으로 제거하였다. 시료를 즉시, 효소적 반응을 중단시키고(pH를 2 내지 3으로 낮춤으로써) 편리한 HPLC 분석을 위해 시료를 희석하기에 충분한 소정의 부피 및 농도의 인산 용액을 함유하는 원심분리기 필터 튜브(나노세프(NanoSep) 30K VWR 카탈로그 번호 82031-354)에서 켄칭하였다. 산-켄칭된 시료를 5분 동안 원심분리하여, 임의의 미립자를 제거하였다. 일단 여과되면, 과아세트산에 대한 카르스트(Karst) 반응 및 HPLC 분석을 즉시 미국 특허 제7,829,315호(DiCosimo et al.)에 이전에 기재된 방법을 사용하여 각 시료 세트 상에서 수행하였다. 각 에스테르 기질에 의해 생성되는 최대의 과아세트산이 표 20에 열거되어 있다.


이들 에스테르 기질을 표 20에 대한 데이터를 생성하기 위하여 사용된 동일한 반응 조건 하에서, 표적화된 C277S 과가수분해효소 변이체, EZ-7을 사용하여 과아세트산 생성에 대하여 시험하였다. EZ-7 과가수분해효소 시스템을 사용한 프로필렌 글리콜 다이아세테이트 또는 수크로스 옥타아세테이트를 사용한 과아세트산 생성이 표 21에 나타나 있다. 나타낸 데이터에 의해, 대안적인 에스테르가 실시예 5에 기재된 바와 같은 표적화된 과가수분해효소의 변이체에 대한 효율적인 기질임이 입증된다.

실시예 15
다양한 에스테르 기질에 대한 CE-7 과가수분해효소를 사용하는 치아 탈색
이러한 실시예의 목적은 구강 관리 관련 조건 하에서 4가지 에스테르로부터의 과아세트산의 형성을 촉매작용시키는 CE-7 과가수분해효소를 사용하는 치아 탈색 효능을 입증하려는 것이었다.
이러한 실시예를 위하여, 써모토가 마리티마 과가수분해효소의 C277S(EZ-1) 변이체를 실시예 4에서와 같이 클로닝하고, 재조합에 의해 발현하고, 정제하였다. 트라이아세틴(테센들로 컴퍼니(미국 애리조나주 포에닉스 소재))을 제외하고, 모든 기질을 시그마-알드리치(미국 미주리주 세인트 루이스 소재)로부터 구매하였다.
기질 트라이아세틴(TA), α-D-글루코스 펜타아세테이트(GPA), 수크로스 옥타아세테이트(SOC) 및 프로필렌 글리콜 다이아세테이트(PGDA)를 사용하여 EZ-1(C277S; 서열 번호 424)의 효소 매개의 치아 미백을 입증하였다. 각 기질과 함께 과아세트산의 생성에 사용되는 최적의 조건은 실시예 14에서의 연구로부터 유래된 것이며, 다음과 같이 아래에 기재되어 있다:
40 ppm EZ-1, 100 mM 기질, X mM H2O2, 95 mM 포스페이트, pH 7.2(여기서, X는 기질의 아이덴티티에 따라 달라진다); 100 mM H2O2 (TA), 360 mM H2O2 (PGDA 및 SOC), 60 mM H2O2 (GPA).
소 에나멜 절치를 실시예 1에 기재된 바와 같이 제조하였다. 착색된 에나멜 블록을 상아질 측을 아래로 향하게 하여, 24웰 플레이트에 배치하고, 포스페이트 완충제, pH 7.2에서 하룻밤 수화시켰다. 처리 전에, 각 치아의 색상 측정을 코니카-미놀타(Konica-Minolta) 2600d 분광광도계를 사용하여 수행하고, 백색도를 실시예 1에 상세화된 바와 같이 계산하였다.
각 효소 용액을 1.5-㎖ 원심분리기용 튜브(에펜도르프(Eppendorf), #2243102-1)에서 제조하고, 1 ㎖의 용액을 즉시, 사전-수화된 치아를 함유하는 24 웰 플레이트로 옮겼다. 대조군 시료는 100 mM 포스페이트 완충제, pH 7.2 및 100 mM 시트레이트/포스페이트 완충제, pH 5.3 중의 9% H2O2로 이루어진다. 치아를 실온에서 30분 동안 인큐베이션시키고, 제거하고, 100 mM 포스페이트, pH 7.2로 헹구고, 색상 측정을 취하였다. 치아를 다시 24 웰 플레이트에 배치하고, 이 때 새로운 효소 용액을 제조하고, 각 웰에 첨가하였다. 이러한 과정을 총 3번의 미백 처리(처리당 30분)에 대하여 반복하였다(표 22).

표 22에서 관찰된 바와 같이, TA, PGDA, SOC 및 GPA를 사용하여, 효소에 의해 생성된 모든 과아세트산 시료는 각 연속적인 처리와 함께, 보다 양의 값으로의 이동에 의해 나타나는 바와 같은 백색도의 유의미한 변화를 나타내었다. 완충제 대조군에 대한 백색도의 변화는 미미하며, ΔWI는 11.4이다. 또한, 이들 측정은 각 30분 처리 후에 치아의 육안의 검사와 일치한다. 이러한 데이터에 의해, EZ-1과 함께 다양한 상이한 기질을 사용하여, 효소에 의해 생성된 과아세트산이 커피-차로 착색된 소 치아의 미백에 효과적임이 입증되었다.
실시예 16
2-단계 적용에서 과가수분해효소를 사용한 치아 탈색 효능
본 실시예의 목적은 2-단계 적용에서의 과가수분해효소 시스템의 치아 탈색 효능을 증명하려는 것이다.
소 에나멜 절치를 실시예 1에 기재된 바와 같이 제조하였다. 착색된 에나멜 블록을 48 웰 플레이트 내에 포매시켜, 상아질 후면을 용액 노출로부터 보호하였다. 10 mM 포스페이트 완충제, pH 7.2 중의 10 μM의 각 효소의 용액을 제조하고, 500 ㎕의 용액을 각 에나멜 기질과 함께 60분 동안 인큐베이션시켰다. 효소 용액을 제거하고, 각 웰을 추가 500 ㎕의 완충제로 3회 헹구었다. 에나멜 블록을 미백 과정을 위해 새로운 웰에 배치하였다. 50 mM 포스페이트 pH 7.2 완충제 중의 40 mM 트라이아세틴, 100 mM H2O2의 최종 농도가 있는 용액을 새로이 제조하고, 500 ㎕의 용액을 각 에나멜 블록에 첨가하였다. 1시간 경과에 걸쳐, 에나멜 블록을 제거하고, 색상 측정을 수득한 다음, 각각을 웰 플레이트 내의 용액으로 복귀시켰다. 각 시료에 대한 백색도를 모니터링하였다. 결과는 표 23에 열거되어 있다.

이러한 실시예에는 표면 결합 과가수분해효소가 에나멜의 표면에서 과아세트산의 형성을 촉매작용시킴으로써 치아를 미백시키는데 사용될 수 있음이 입증된다. 이러한 실시예에서 탈색 성능은 수산화인회석 상에서의 각 효소의 유지의 관찰과 관련이 있다. 비표적화 EZ-1(C277S; 서열 번호 424)은 수산화인회석 및 에나멜 상에서 불량한 유지를 보였으며, 이에 따라 2-단계 과정에서 적절한 미백을 달성하는 능력이 낮았다. 에나멜 상에서 효소를 유지하기 위한 효과적인 표적화 서열의 첨가에 의해, 2-단계 적용 과정에서 과아세트산의 생성이 가능하게 된다.
실시예 17
2-단계 적용에서 과가수분해효소를 사용한 치아 탈색 효능
본 실시예의 목적은 2-단계 적용에서의 표적화 및 비표적화 과가수분해효소 시스템의 치아 탈색 효능을 증명하려는 것이다.
소 에나멜 절치를 실시예 1에 기재된 바와 같이 제조하였다. 착색된 에나멜 블록을 실리 퍼티(silly putty)(등록 상표)(미국 펜실베이니아주 이스턴 소재의 크래욜라 엘엘씨(Crayola LLC))가 채워진 웰, 24-웰 플레이트에 포매시켜, 상아질 후면을 용액 노출로부터 보호하였다. 10 mM 포스페이트 완충제, pH 7.2 중의 20 μM의 각 효소의 용액을 제조하고, 500 ㎕의 효소 용액을 각 에나멜 블록에 첨가하고, 37℃에서 10분 동안 인큐베이션시켰다. 이어서, 에나멜 블록을 매번 추가 500 ㎕의 완충제로 3회 헹구었다. 미백 과정을 위하여, 에나멜 블록을 옮기고, 새로운 퍼티 웰에 포매시켰다. 100 mM 포스페이트 완충제, 100 mM 트라이아세틴, 100 mM H2O2, pH 7.2를 함유하는 용액(200 ㎕)을 에나멜에 첨가하고, 37℃에서 10분 동안 인큐베이션시켰다. 에나멜 블록을 겸자(forcep)를 사용하여 웰로부터 제거한 다음, 헹구고, 색상 측정을 위하여 1.5 ㎖의 물로 채운 웰에 저장하였다. 2-단계 과정을 5회 반복하였다(총 50분). 2-단계 과정 후의 커피-차로 착색된 소 에나멜의 색상 측정의 결과는 표 24에 나타나 있다. 각 시료 데이터점은 독립적인 에나멜 블록의 2번의 반복을 나타낸다.
시스템에서 생성된 과아세트산의 수준을 측정하기 위하여, ABTS 방법을 사용하고, 90 ㎕의 반응 혼합물을 10 ㎕의 정지 완충제(1.33 M H3PO4)를 함유하는 새로운 웰로 옮겼다. 시료를 100 mM 포스페이트 완충제를 사용하여 1:100으로 희석하고, 실시예 8에 나타낸 바와 같이 ABTS 검출 시약에 첨가하였다. 결과는 표 25에 열거되어 있다. 각 시료 데이터점은 독립적인 에나멜 블록의 2번의 반복을 나타낸다.


이러한 실시예에는 표면 결합 과가수분해효소가 에나멜의 표면에서 과아세트산의 형성을 촉매작용시킴으로써 치아를 미백시키는데 사용될 수 있음이 입증된다. 소 에나멜을 사용하는 이러한 실시예에서 탈색 성능은 수산화인회석 상에서의 각 효소의 유지의 관찰과 관련이 있다. 비표적화 EZ-1(C277S)은 수산화인회석 및 에나멜 상에서 불량한 유지를 보였으며, 이에 따라 2-단계 과정에서 적절한 미백을 달성하는 능력이 낮았다. 에나멜 상에서 효소를 유지하기 위한 효과적인 표적화 서열의 첨가에 의해, 2-단계 적용 과정에서 과아세트산의 생성이 가능하게 된다.
SEQUENCE LISTING
<110> E.I. duPont de Nemours and Company Inc.
Di Cosimo, Robert
Cunningham, Scott
Butterick, Lisa
Fosser, Kari
Gruber, Tanja
Wang, Hong
Rouviere, Pierre
Payne, Mark
<120> ENZYMATICA PERACID GNERATION FOR USE IN ORAL CARE PRODUCTS
<130> CL5256 PCT
<150> US 61/424,903
<151> 2010-12-20
<160> 479
<170> PatentIn version 3.5
<210> 1
<211> 960
<212> DNA
<213> Bacillus subtilis
<220>
<221> CDS
<222> (1)..(960)
<400> 1
atg caa cta ttc gat ctg ccg ctc gac caa ttg caa aca tat aag cct 48
Met Gln Leu Phe Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
gaa aaa aca gca ccg aaa gat ttt tct gag ttt tgg aaa ttg tct ttg 96
Glu Lys Thr Ala Pro Lys Asp Phe Ser Glu Phe Trp Lys Leu Ser Leu
20 25 30
gag gaa ctt gca aaa gtc caa gca gaa cct gat tta cag ccg gtt gac 144
Glu Glu Leu Ala Lys Val Gln Ala Glu Pro Asp Leu Gln Pro Val Asp
35 40 45
tat cct gct gac gga gta aaa gtg tac cgt ctc aca tat aaa agc ttc 192
Tyr Pro Ala Asp Gly Val Lys Val Tyr Arg Leu Thr Tyr Lys Ser Phe
50 55 60
gga aac gcc cgc att acc gga tgg tac gcg gtg cct gac aag caa ggc 240
Gly Asn Ala Arg Ile Thr Gly Trp Tyr Ala Val Pro Asp Lys Gln Gly
65 70 75 80
ccg cat ccg gcg atc gtg aaa tat cat ggc tac aat gca agc tat gat 288
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
ggt gag att cat gaa atg gta aac tgg gca ctc cat ggc tac gcc gca 336
Gly Glu Ile His Glu Met Val Asn Trp Ala Leu His Gly Tyr Ala Ala
100 105 110
ttc ggc atg ctt gtc cgc ggc cag cag agc agc gag gat acg agt att 384
Phe Gly Met Leu Val Arg Gly Gln Gln Ser Ser Glu Asp Thr Ser Ile
115 120 125
tca ctg cac ggt cac gct ttg ggc tgg atg acg aaa gga att ctt gat 432
Ser Leu His Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Asp
130 135 140
aaa gat aca tac tat tac cgc ggt gtt tat ttg gac gcc gtc cgc gcg 480
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
ctt gag gtc atc agc agc ttc gac gag gtt gac gaa aca agg atc ggt 528
Leu Glu Val Ile Ser Ser Phe Asp Glu Val Asp Glu Thr Arg Ile Gly
165 170 175
gtg aca gga gga agc caa ggc gga ggt tta acc att gcc gca gca gcg 576
Val Thr Gly Gly Ser Gln Gly Gly Gly Leu Thr Ile Ala Ala Ala Ala
180 185 190
ctg tca gac att cca aaa gcc gcg gtt gcc gat tat cct tat tta agc 624
Leu Ser Asp Ile Pro Lys Ala Ala Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
aac ttc gaa cgg gcc att gat gtg gcg ctt gaa cag ccg tac ctt gaa 672
Asn Phe Glu Arg Ala Ile Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
atc aat tcc ttc ttc aga aga aat ggc agc ccg gaa aca gaa gtg cag 720
Ile Asn Ser Phe Phe Arg Arg Asn Gly Ser Pro Glu Thr Glu Val Gln
225 230 235 240
gcg atg aag aca ctt tca tat ttc gat att atg aat ctc gct gac cga 768
Ala Met Lys Thr Leu Ser Tyr Phe Asp Ile Met Asn Leu Ala Asp Arg
245 250 255
gtg aag gtg cct gtc ctg atg tca atc ggc ctg att gac aag gtc acg 816
Val Lys Val Pro Val Leu Met Ser Ile Gly Leu Ile Asp Lys Val Thr
260 265 270
ccg ccg tcc acc gtg ttt gcc gcc tac aat cat ttg gaa aca gag aaa 864
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Glu Lys
275 280 285
gag ctg aag gtg tac cgc tac ttc gga cat gag tat atc cct gct ttt 912
Glu Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Ala Phe
290 295 300
caa acg gaa aaa ctt gct ttc ttt aag cag cat ctt aaa ggc tga taa 960
Gln Thr Glu Lys Leu Ala Phe Phe Lys Gln His Leu Lys Gly
305 310 315
<210> 2
<211> 318
<212> PRT
<213> Bacillus subtilis
<400> 2
Met Gln Leu Phe Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
Glu Lys Thr Ala Pro Lys Asp Phe Ser Glu Phe Trp Lys Leu Ser Leu
20 25 30
Glu Glu Leu Ala Lys Val Gln Ala Glu Pro Asp Leu Gln Pro Val Asp
35 40 45
Tyr Pro Ala Asp Gly Val Lys Val Tyr Arg Leu Thr Tyr Lys Ser Phe
50 55 60
Gly Asn Ala Arg Ile Thr Gly Trp Tyr Ala Val Pro Asp Lys Gln Gly
65 70 75 80
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Glu Ile His Glu Met Val Asn Trp Ala Leu His Gly Tyr Ala Ala
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gln Ser Ser Glu Asp Thr Ser Ile
115 120 125
Ser Leu His Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Asp
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Ser Ser Phe Asp Glu Val Asp Glu Thr Arg Ile Gly
165 170 175
Val Thr Gly Gly Ser Gln Gly Gly Gly Leu Thr Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Ala Ala Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Ile Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Phe Phe Arg Arg Asn Gly Ser Pro Glu Thr Glu Val Gln
225 230 235 240
Ala Met Lys Thr Leu Ser Tyr Phe Asp Ile Met Asn Leu Ala Asp Arg
245 250 255
Val Lys Val Pro Val Leu Met Ser Ile Gly Leu Ile Asp Lys Val Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Glu Lys
275 280 285
Glu Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ala Phe Phe Lys Gln His Leu Lys Gly
305 310 315
<210> 3
<211> 957
<212> DNA
<213> Bacillus subtilis
<400> 3
atgcaactat tcgatctgcc gctcgaccaa ttgcaaacat ataagcctga aaaaacagca 60
ccgaaagatt tttctgagtt ttggaaattg tctttggagg aacttgcaaa agtccaagca 120
gaacctgatt tacagccggt tgactatcct gctgacggag taaaagtgta ccgtctcaca 180
tataaaagct tcggaaacgc ccgcattacc ggatggtacg cggtgcctga caaggaaggc 240
ccgcatccgg cgatcgtgaa atatcatggc tacaatgcaa gctatgatgg tgagattcat 300
gaaatggtaa actgggcact ccatggctac gccacattcg gcatgcttgt ccgcggccag 360
cagagcagcg aggatacgag tatttcaccg cacggtcacg ctttgggctg gatgacgaaa 420
ggaattcttg ataaagatac atactattac cgcggtgttt atttggacgc cgtccgcgcg 480
cttgaggtca tcagcagctt cgacgaggtt gacgaaacaa ggatcggtgt gacaggagga 540
agccaaggcg gaggtttaac cattgccgca gcagcgctgt cagacattcc aaaagccgcg 600
gttgccgatt atccttattt aagcaacttc gaacgggcca ttgatgtggc gcttgaacag 660
ccgtaccttg aaatcaattc cttcttcaga agaaatggca gcccggaaac agaagtgcag 720
gcgatgaaga cactttcata tttcgatatt atgaatctcg ctgaccgagt gaaggtgcct 780
gtcctgatgt caatcggcct gattgacaag gtcacgccgc cgtccaccgt gtttgccgcc 840
tacaatcatt tggaaacaaa gaaagagctg aaggtgtacc gctacttcgg acatgagtat 900
atccctgctt ttcaaactga aaaacttgct ttctttaagc agcatcttaa aggctga 957
<210> 4
<211> 318
<212> PRT
<213> Bacillus subtilis s
<400> 4
Met Gln Leu Phe Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
Glu Lys Thr Ala Pro Lys Asp Phe Ser Glu Phe Trp Lys Leu Ser Leu
20 25 30
Glu Glu Leu Ala Lys Val Gln Ala Glu Pro Asp Leu Gln Pro Val Asp
35 40 45
Tyr Pro Ala Asp Gly Val Lys Val Tyr Arg Leu Thr Tyr Lys Ser Phe
50 55 60
Gly Asn Ala Arg Ile Thr Gly Trp Tyr Ala Val Pro Asp Lys Glu Gly
65 70 75 80
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Glu Ile His Glu Met Val Asn Trp Ala Leu His Gly Tyr Ala Thr
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gln Ser Ser Glu Asp Thr Ser Ile
115 120 125
Ser Pro His Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Asp
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Ser Ser Phe Asp Glu Val Asp Glu Thr Arg Ile Gly
165 170 175
Val Thr Gly Gly Ser Gln Gly Gly Gly Leu Thr Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Ala Ala Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Ile Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Phe Phe Arg Arg Asn Gly Ser Pro Glu Thr Glu Val Gln
225 230 235 240
Ala Met Lys Thr Leu Ser Tyr Phe Asp Ile Met Asn Leu Ala Asp Arg
245 250 255
Val Lys Val Pro Val Leu Met Ser Ile Gly Leu Ile Asp Lys Val Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Lys Lys
275 280 285
Glu Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ala Phe Phe Lys Gln His Leu Lys Gly
305 310 315
<210> 5
<211> 957
<212> DNA
<213> Bacillus subtilis
<400> 5
atgcaactat tcgatctgcc gctcgaccaa ttgcaaacgt ataagcctga aaaaacaaca 60
ccgaacgatt tttctgagtt ttggaaatcg tctttggacg aacttgcgaa agtcaaagca 120
gcacctgatt tacagctggt tgattatcct gctgatggag tcaaggtgta ccgcctcaca 180
tataaaagct tcggaaacgc ccgcattacc ggatggtacg cagtgcctga caaggaagga 240
ccgcatccgg cgatcgtcaa atatcatggc tacaacgcta gctatgacgg tgagattcat 300
gaaatggtaa actgggcgct ccacggttac gccgcattcg gcatgctagt ccgcggccag 360
cagagcagcg aggatacgag tatttctcca catggccatg ctttgggctg gatgacgaaa 420
ggaatccttg ataaagatac atactattac cggggcgttt atttggacgc tgtccgcgcg 480
cttgaggtca tcagcagctt tgacgaagtt gacgaaacaa gaatcggtgt gacaggcgga 540
agccaaggag gcggcttaac cattgccgca gccgctctgt cagacattcc aaaagccgcg 600
gttgccgatt atccttattt aagcaacttt gaacgggcca ttgatgtggc gcttgaacag 660
ccgtaccttg aaatcaattc cttctttaga agaaatggaa gcccggaaac ggaagagaag 720
gcgatgaaga cactttcata tttcgatatt atgaatctcg ctgaccgagt gaaggtccct 780
gtcctgatgt cgatcggtct gattgacaag gtcacgccgc cgtccaccgt gtttgccgca 840
tacaaccact tggagacaga gaaagagctc aaagtgtacc gctacttcgg gcatgagtat 900
atccctgcct ttcaaacaga aaaacttgct ttctttaagc agcatcttaa aggctga 957
<210> 6
<211> 318
<212> PRT
<213> Bacillus subtilis
<400> 6
Met Gln Leu Phe Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
Glu Lys Thr Thr Pro Asn Asp Phe Ser Glu Phe Trp Lys Ser Ser Leu
20 25 30
Asp Glu Leu Ala Lys Val Lys Ala Ala Pro Asp Leu Gln Leu Val Asp
35 40 45
Tyr Pro Ala Asp Gly Val Lys Val Tyr Arg Leu Thr Tyr Lys Ser Phe
50 55 60
Gly Asn Ala Arg Ile Thr Gly Trp Tyr Ala Val Pro Asp Lys Glu Gly
65 70 75 80
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Glu Ile His Glu Met Val Asn Trp Ala Leu His Gly Tyr Ala Ala
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gln Ser Ser Glu Asp Thr Ser Ile
115 120 125
Ser Pro His Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Asp
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Ser Ser Phe Asp Glu Val Asp Glu Thr Arg Ile Gly
165 170 175
Val Thr Gly Gly Ser Gln Gly Gly Gly Leu Thr Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Ala Ala Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Ile Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Phe Phe Arg Arg Asn Gly Ser Pro Glu Thr Glu Glu Lys
225 230 235 240
Ala Met Lys Thr Leu Ser Tyr Phe Asp Ile Met Asn Leu Ala Asp Arg
245 250 255
Val Lys Val Pro Val Leu Met Ser Ile Gly Leu Ile Asp Lys Val Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Glu Lys
275 280 285
Glu Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ala Phe Phe Lys Gln His Leu Lys Gly
305 310 315
<210> 7
<211> 957
<212> DNA
<213> Bacillus licheniformis
<400> 7
atgcagcagc cttatgatat gccgcttgaa cagctttatc agtataaacc tgaacggacg 60
gcaccggccg attttaaaga gttctggaag ggttcattgg aggaattggc aaatgaaaaa 120
gcgggaccgc agcttgaacc gcatgaatat ccggctgacg gggtaaaagt ctactggctt 180
acatacagaa gcatcggggg agcgcgaatt aaaggctggt acgcagtacc cgaccgccaa 240
gggcctcatc ctgcgatcgt caaataccac ggctataacg caagctatga cggagacatt 300
cacgatattg tcaattgggc tcttcacggc tatgcggcat tcggtatgct ggtccgcgga 360
cagaacagca gtgaagatac agagatctct catcacggac atgtacccgg ctggatgaca 420
aaaggaatcc tcgatccgaa aacatattac tacagagggg tctatttaga tgccgtacga 480
gcagtcgaag tggtcagcgg ttttgctgaa gtcgatgaaa agcggatcgg ggtgatcggg 540
gcaagccaag gaggcgggct ggccgtcgcg gtttcggcgc tgtccgatat tccaaaagca 600
gccgtgtcag aataccctta tttaagcaat tttcaacgag cgatcgatac agcgatcgac 660
cagccatatc tcgaaatcaa ctcctttttc agaagaaaca ccagtccgga tattgagcag 720
gcggccatgc ataccctgtc ttatttcgat gtcatgaacc ttgcccaatt ggtcaaagcg 780
accgtactca tgtcgatcgg actggttgac accatcactc cgccatccac cgtctttgcg 840
gcttacaatc acttggaaac ggataaagaa ataaaagtgt accgttattt tggacacgaa 900
tacatcccgc cgttccaaac cgaaaagctg gcgtttctga gaaagcatct gaaataa 957
<210> 8
<211> 318
<212> PRT
<213> Bacillus licheniformis
<400> 8
Met Gln Gln Pro Tyr Asp Met Pro Leu Glu Gln Leu Tyr Gln Tyr Lys
1 5 10 15
Pro Glu Arg Thr Ala Pro Ala Asp Phe Lys Glu Phe Trp Lys Gly Ser
20 25 30
Leu Glu Glu Leu Ala Asn Glu Lys Ala Gly Pro Gln Leu Glu Pro His
35 40 45
Glu Tyr Pro Ala Asp Gly Val Lys Val Tyr Trp Leu Thr Tyr Arg Ser
50 55 60
Ile Gly Gly Ala Arg Ile Lys Gly Trp Tyr Ala Val Pro Asp Arg Gln
65 70 75 80
Gly Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr
85 90 95
Asp Gly Asp Ile His Asp Ile Val Asn Trp Ala Leu His Gly Tyr Ala
100 105 110
Ala Phe Gly Met Leu Val Arg Gly Gln Asn Ser Ser Glu Asp Thr Glu
115 120 125
Ile Ser His His Gly His Val Pro Gly Trp Met Thr Lys Gly Ile Leu
130 135 140
Asp Pro Lys Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg
145 150 155 160
Ala Val Glu Val Val Ser Gly Phe Ala Glu Val Asp Glu Lys Arg Ile
165 170 175
Gly Val Ile Gly Ala Ser Gln Gly Gly Gly Leu Ala Val Ala Val Ser
180 185 190
Ala Leu Ser Asp Ile Pro Lys Ala Ala Val Ser Glu Tyr Pro Tyr Leu
195 200 205
Ser Asn Phe Gln Arg Ala Ile Asp Thr Ala Ile Asp Gln Pro Tyr Leu
210 215 220
Glu Ile Asn Ser Phe Phe Arg Arg Asn Thr Ser Pro Asp Ile Glu Gln
225 230 235 240
Ala Ala Met His Thr Leu Ser Tyr Phe Asp Val Met Asn Leu Ala Gln
245 250 255
Leu Val Lys Ala Thr Val Leu Met Ser Ile Gly Leu Val Asp Thr Ile
260 265 270
Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Asp
275 280 285
Lys Glu Ile Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Pro
290 295 300
Phe Gln Thr Glu Lys Leu Ala Phe Leu Arg Lys His Leu Lys
305 310 315
<210> 9
<211> 963
<212> DNA
<213> Bacillus pumilus
<400> 9
atgcaattgt tcgatttatc actagaagag ctaaaaaaat ataaaccaaa gaaaacagca 60
cgtcctgatt tctcagactt ttggaagaaa tcgctcgaag aactgcgcca agtggaggca 120
gagccaacac ttgaatctta tgactatcca gtgaaaggcg tcaaggtgta ccgcctgacg 180
tatcaaagct ttggacattc taaaattgaa ggcttttatg ctgtgcctga tcaaactggt 240
ccgcatccag cgctcgttcg ttttcatggc tataatgcca gctatgacgg cggcattcac 300
gacatcgtca actgggcgct gcacggctat gcaacatttg gtatgctcgt ccgcggtcaa 360
ggtggcagtg aagacacatc agtgacacca ggcgggcatg cattagggtg gatgacaaaa 420
ggcattttat cgaaagatac gtactattat cgaggcgttt atctagatgc tgttcgtgca 480
cttgaagtca ttcagtcttt ccccgaagta gatgaacacc gtatcggcgt gatcggtgga 540
agtcaggggg gtgcgttagc gattgcggcc gcagcccttt cagacattcc aaaagtcgtt 600
gtggcagact atccttactt atcaaatttt gagcgtgcag ttgatgttgc cttggagcag 660
ccttatttag aaatcaattc atactttcgc agaaacagtg atccgaaagt ggaggaaaag 720
gcatttgaga cattaagcta ttttgattta atcaatttag ctggatgggt gaaacagcca 780
acattgatgg cgatcggtct gattgacaaa ataaccccac catctactgt gtttgcggca 840
tacaaccatt tagaaacaga taaagacctg aaagtatatc gctattttgg acacgagttt 900
atccctgctt ttcaaacaga gaagctgtcc tttttacaaa agcatttgct tctatcaaca 960
taa 963
<210> 10
<211> 320
<212> PRT
<213> Bacillus pumilus
<400> 10
Met Gln Leu Phe Asp Leu Ser Leu Glu Glu Leu Lys Lys Tyr Lys Pro
1 5 10 15
Lys Lys Thr Ala Arg Pro Asp Phe Ser Asp Phe Trp Lys Lys Ser Leu
20 25 30
Glu Glu Leu Arg Gln Val Glu Ala Glu Pro Thr Leu Glu Ser Tyr Asp
35 40 45
Tyr Pro Val Lys Gly Val Lys Val Tyr Arg Leu Thr Tyr Gln Ser Phe
50 55 60
Gly His Ser Lys Ile Glu Gly Phe Tyr Ala Val Pro Asp Gln Thr Gly
65 70 75 80
Pro His Pro Ala Leu Val Arg Phe His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Gly Ile His Asp Ile Val Asn Trp Ala Leu His Gly Tyr Ala Thr
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gly Gly Ser Glu Asp Thr Ser Val
115 120 125
Thr Pro Gly Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Ser
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Gln Ser Phe Pro Glu Val Asp Glu His Arg Ile Gly
165 170 175
Val Ile Gly Gly Ser Gln Gly Gly Ala Leu Ala Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Val Val Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Val Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Tyr Phe Arg Arg Asn Ser Asp Pro Lys Val Glu Glu Lys
225 230 235 240
Ala Phe Glu Thr Leu Ser Tyr Phe Asp Leu Ile Asn Leu Ala Gly Trp
245 250 255
Val Lys Gln Pro Thr Leu Met Ala Ile Gly Leu Ile Asp Lys Ile Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Asp Lys
275 280 285
Asp Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Phe Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ser Phe Leu Gln Lys His Leu Leu Leu Ser Thr
305 310 315 320
<210> 11
<211> 963
<212> DNA
<213> Clostridium thermocellum
<400> 11
atggcacaat tatatgatat gcctttggag gaattaaaaa aatataagcc tgcgcttaca 60
aaacagaaag attttgatga gttttgggaa aaaagcctta aagagctggc tgaaattcct 120
ttaaaatatc aacttatacc ttatgatttt ccggcccgga gggtaaaagt tttcagagtt 180
gaatatcttg gttttaaagg tgcaaatatt gaagggtggc ttgccgttcc cgagggagaa 240
gggttgtatc ccgggcttgt acagtttcac ggatacaact gggcgatgga tggatgtgtt 300
cccgatgtgg taaattgggc tttgaatgga tatgccgcat ttcttatgct tgttcgggga 360
cagcagggaa gaagcgtgga caatattgtg cccggcagcg gtcatgcttt gggatggatg 420
tcgaaaggta ttttgtcacc ggaggaatat tattatagag gagtatatat ggatgcggtt 480
cgtgctgttg aaattttggc ttcgcttcct tgtgtggatg aatcgagaat aggagtgaca 540
gggggcagcc agggtggagg acttgcactg gcggtggctg ctctgtccgg cataccgaaa 600
gttgcagccg tgcattatcc gtttctggca cattttgagc gtgccattga cgttgcgccg 660
gacggccctt atcttgaaat taacgaatat ttaagaagaa acagcggtga agaaatagaa 720
agacaggtaa agaaaaccct ttcctatttt gatatcatga atcttgctcc ccgtataaaa 780
tgccgtactt ggatttgcac tggtcttgtg gatgagatta ctcctccgtc aacggttttt 840
gcagtgtaca atcacctcaa atgcccaaag gaaatttcgg tattcagata ttttgggcat 900
gaacatatgc caggaagcgt tgaaatcaag ctgaggatac ttatggatga gctgaatccg 960
taa 963
<210> 12
<211> 320
<212> PRT
<213> Clostridium thermocellum
<400> 12
Met Ala Gln Leu Tyr Asp Met Pro Leu Glu Glu Leu Lys Lys Tyr Lys
1 5 10 15
Pro Ala Leu Thr Lys Gln Lys Asp Phe Asp Glu Phe Trp Glu Lys Ser
20 25 30
Leu Lys Glu Leu Ala Glu Ile Pro Leu Lys Tyr Gln Leu Ile Pro Tyr
35 40 45
Asp Phe Pro Ala Arg Arg Val Lys Val Phe Arg Val Glu Tyr Leu Gly
50 55 60
Phe Lys Gly Ala Asn Ile Glu Gly Trp Leu Ala Val Pro Glu Gly Glu
65 70 75 80
Gly Leu Tyr Pro Gly Leu Val Gln Phe His Gly Tyr Asn Trp Ala Met
85 90 95
Asp Gly Cys Val Pro Asp Val Val Asn Trp Ala Leu Asn Gly Tyr Ala
100 105 110
Ala Phe Leu Met Leu Val Arg Gly Gln Gln Gly Arg Ser Val Asp Asn
115 120 125
Ile Val Pro Gly Ser Gly His Ala Leu Gly Trp Met Ser Lys Gly Ile
130 135 140
Leu Ser Pro Glu Glu Tyr Tyr Tyr Arg Gly Val Tyr Met Asp Ala Val
145 150 155 160
Arg Ala Val Glu Ile Leu Ala Ser Leu Pro Cys Val Asp Glu Ser Arg
165 170 175
Ile Gly Val Thr Gly Gly Ser Gln Gly Gly Gly Leu Ala Leu Ala Val
180 185 190
Ala Ala Leu Ser Gly Ile Pro Lys Val Ala Ala Val His Tyr Pro Phe
195 200 205
Leu Ala His Phe Glu Arg Ala Ile Asp Val Ala Pro Asp Gly Pro Tyr
210 215 220
Leu Glu Ile Asn Glu Tyr Leu Arg Arg Asn Ser Gly Glu Glu Ile Glu
225 230 235 240
Arg Gln Val Lys Lys Thr Leu Ser Tyr Phe Asp Ile Met Asn Leu Ala
245 250 255
Pro Arg Ile Lys Cys Arg Thr Trp Ile Cys Thr Gly Leu Val Asp Glu
260 265 270
Ile Thr Pro Pro Ser Thr Val Phe Ala Val Tyr Asn His Leu Lys Cys
275 280 285
Pro Lys Glu Ile Ser Val Phe Arg Tyr Phe Gly His Glu His Met Pro
290 295 300
Gly Ser Val Glu Ile Lys Leu Arg Ile Leu Met Asp Glu Leu Asn Pro
305 310 315 320
<210> 13
<211> 978
<212> DNA
<213> Thermotoga neapolitana
<400> 13
atggccttct tcgatatgcc ccttgaggaa ctgaaaaagt accggcctga aaggtacgag 60
gagaaagatt tcgatgagtt ctggagggaa acacttaaag aaagcgaagg attccctctg 120
gatcccgtct ttgaaaaggt ggactttcat ctcaaaacgg ttgaaacgta cgatgttact 180
ttctctggat acagggggca gagaataaag ggctggcttc ttgttccgaa gttggcggaa 240
gaaaagcttc catgcgtcgt gcagtacata ggttacaatg gtggaagggg ttttccacac 300
gactggctgt tctggccgtc aatgggttac atctgttttg tcatggacac cagggggcag 360
ggaagcggct ggatgaaggg agacacaccg gattaccctg agggtccagt cgatccacag 420
taccccggat tcatgacgag gggcattctg gatccgggaa cctattacta caggcgagtc 480
ttcgtggatg cggtcagggc ggtggaagca gccatttcct tcccgagagt ggattccagg 540
aaggtggtgg tggccggagg cagtcagggt gggggaatcg cccttgcggt gagtgccctg 600
tcgaacaggg tgaaggctct gctctgcgat gtgccgtttc tgtgccactt cagaagggcc 660
gtgcaacttg tcgacacaca cccatacgtg gagatcacca acttcctcaa aacccacagg 720
gacaaagagg agattgtttt cagaacactt tcctacttcg atggtgtgaa ctttgcagca 780
agggcaaagg tgcccgccct gttttccgtt gggctcatgg acaccatctg tcctccctcg 840
acggtcttcg ccgcttacaa ccactacgcc ggtccaaagg agatcagaat ctatccgtac 900
aacaaccacg aaggtggagg ttctttccag gcaattgagc aggtgaaatt cttgaagaga 960
ctatttgagg aaggctag 978
<210> 14
<211> 325
<212> PRT
<213> Thermotoga neapolitana
<400> 14
Met Ala Phe Phe Asp Met Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Arg Glu Thr Leu
20 25 30
Lys Glu Ser Glu Gly Phe Pro Leu Asp Pro Val Phe Glu Lys Val Asp
35 40 45
Phe His Leu Lys Thr Val Glu Thr Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Ala Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Met Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Gly Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Val Asp Ala Val Arg Ala Val Glu Ala Ala Ile Ser Phe Pro Arg
165 170 175
Val Asp Ser Arg Lys Val Val Val Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Asn Arg Val Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Val Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Val Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Thr Ile Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Ile Glu Gln Val Lys Phe Leu Lys Arg
305 310 315 320
Leu Phe Glu Glu Gly
325
<210> 15
<211> 978
<212> DNA
<213> Thermotoga maritima
<400> 15
atggccttct tcgatttacc actcgaagaa ctgaagaaat atcgtccaga gcggtacgaa 60
gagaaagact tcgatgagtt ctgggaagag acactcgcag agagcgaaaa gttcccctta 120
gaccccgtct tcgagaggat ggagtctcac ctcaaaacag tcgaagcgta cgatgtcacc 180
ttctccggat acaggggaca gaggatcaaa gggtggctcc ttgttccaaa actggaagaa 240
gaaaaacttc cctgcgttgt gcagtacata ggatacaacg gtggaagagg attccctcac 300
gactggctgt tctggccttc tatgggttac atatgtttcg tcatggatac tcgaggtcag 360
ggaagcggct ggctgaaagg agacacaccg gattaccctg agggtcccgt tgaccctcag 420
tatccaggat tcatgacaag aggaatactg gatcccagaa cttactacta cagacgagtc 480
ttcacggacg ctgtcagagc cgttgaagct gctgcttctt ttcctcaggt agatcaagaa 540
agaatcgtga tagctggagg cagtcagggt ggcggaatag cccttgcggt gagcgctctc 600
tcaaagaaag caaaggctct tctgtgcgat gtgccgtttc tgtgtcactt cagaagagca 660
gtacagcttg tggatacgca tccatacgcg gagatcacga actttctaaa gacccacaga 720
gacaaggaag aaatcgtgtt caggactctt tcctatttcg atggagtgaa cttcgcagcc 780
agagcgaaga tccctgcgct gttttctgtg ggtctcatgg acaacatttg tcctccttca 840
acggttttcg ctgcctacaa ttactacgct ggaccgaagg aaatcagaat ctatccgtac 900
aacaaccacg agggaggagg ctctttccaa gcggttgaac aggtgaaatt cttgaaaaaa 960
ctatttgaga aaggctaa 978
<210> 16
<211> 325
<212> PRT
<213> Thermotoga maritima
<400> 16
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 17
<211> 963
<212> DNA
<213> Thermoanaerobacterium sp.
<400> 17
atgggacttt tcgacatgcc attacaaaaa cttagagaat acactggtac aaatccatgc 60
cctgaagatt tcgatgagta ttggaatagg gctttagatg agatgaggtc agttgatcct 120
aaaattgaat tgaaagaaag tagctttcaa gtatcctttg cagaatgcta tgacttgtac 180
tttacaggtg ttcgtggtgc cagaattcat gcaaagtata taaaacctaa gacagaaggg 240
aaacatccag cgttgataag atttcatgga tattcgtcaa attcaggcga ctggaacgac 300
aaattaaatt acgtggcggc aggcttcacc gttgtggcta tggatgtaag aggtcaagga 360
gggcagtctc aagatgttgg cggtgtaact gggaatactt taaatgggca tattataaga 420
gggctagacg atgatgctga taatatgctt ttcaggcata ttttcttaga cactgcccaa 480
ttggctggaa tagttatgaa catgccagaa gttgatgaag atagagtggg agtcatggga 540
ccttctcaag gcggagggct gtcgttggcg tgtgctgcat tggagccaag ggtacgcaaa 600
gtagtatctg aatatccttt tttatctgac tacaagagag tttgggactt agaccttgca 660
aaaaacgcct atcaagagat tacggactat ttcaggcttt ttgacccaag gcatgaaagg 720
gagaatgagg tatttacaaa gcttggatat atagacgtta aaaaccttgc gaaaaggata 780
aaaggcgatg tcttaatgtg cgttgggctt atggaccaag tatgtccgcc atcaactgtt 840
tttgcagcct acaacaacat acagtcaaaa aaagatataa aagtgtatcc tgattatgga 900
catgaaccta tgagaggatt tggagattta gcgatgcagt ttatgttgga actatattca 960
taa 963
<210> 18
<211> 320
<212> PRT
<213> Thermoanaerobacterium sp.
<400> 18
Met Gly Leu Phe Asp Met Pro Leu Gln Lys Leu Arg Glu Tyr Thr Gly
1 5 10 15
Thr Asn Pro Cys Pro Glu Asp Phe Asp Glu Tyr Trp Asn Arg Ala Leu
20 25 30
Asp Glu Met Arg Ser Val Asp Pro Lys Ile Glu Leu Lys Glu Ser Ser
35 40 45
Phe Gln Val Ser Phe Ala Glu Cys Tyr Asp Leu Tyr Phe Thr Gly Val
50 55 60
Arg Gly Ala Arg Ile His Ala Lys Tyr Ile Lys Pro Lys Thr Glu Gly
65 70 75 80
Lys His Pro Ala Leu Ile Arg Phe His Gly Tyr Ser Ser Asn Ser Gly
85 90 95
Asp Trp Asn Asp Lys Leu Asn Tyr Val Ala Ala Gly Phe Thr Val Val
100 105 110
Ala Met Asp Val Arg Gly Gln Gly Gly Gln Ser Gln Asp Val Gly Gly
115 120 125
Val Thr Gly Asn Thr Leu Asn Gly His Ile Ile Arg Gly Leu Asp Asp
130 135 140
Asp Ala Asp Asn Met Leu Phe Arg His Ile Phe Leu Asp Thr Ala Gln
145 150 155 160
Leu Ala Gly Ile Val Met Asn Met Pro Glu Val Asp Glu Asp Arg Val
165 170 175
Gly Val Met Gly Pro Ser Gln Gly Gly Gly Leu Ser Leu Ala Cys Ala
180 185 190
Ala Leu Glu Pro Arg Val Arg Lys Val Val Ser Glu Tyr Pro Phe Leu
195 200 205
Ser Asp Tyr Lys Arg Val Trp Asp Leu Asp Leu Ala Lys Asn Ala Tyr
210 215 220
Gln Glu Ile Thr Asp Tyr Phe Arg Leu Phe Asp Pro Arg His Glu Arg
225 230 235 240
Glu Asn Glu Val Phe Thr Lys Leu Gly Tyr Ile Asp Val Lys Asn Leu
245 250 255
Ala Lys Arg Ile Lys Gly Asp Val Leu Met Cys Val Gly Leu Met Asp
260 265 270
Gln Val Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Asn Ile Gln
275 280 285
Ser Lys Lys Asp Ile Lys Val Tyr Pro Asp Tyr Gly His Glu Pro Met
290 295 300
Arg Gly Phe Gly Asp Leu Ala Met Gln Phe Met Leu Glu Leu Tyr Ser
305 310 315 320
<210> 19
<211> 978
<212> DNA
<213> Bacillus sp.
<220>
<221> CDS
<222> (1)..(978)
<400> 19
atg aac ctt ttt gat atg ccc ctt gag gag ctg cag cat tac aag cct 48
Met Asn Leu Phe Asp Met Pro Leu Glu Glu Leu Gln His Tyr Lys Pro
1 5 10 15
gcc cag acc agg cag gat gat ttt gag tca ttc tgg aaa aag cgg att 96
Ala Gln Thr Arg Gln Asp Asp Phe Glu Ser Phe Trp Lys Lys Arg Ile
20 25 30
gag gag aac agt caa tat ccg ctg aat ata gaa gta atg gag cgg gtt 144
Glu Glu Asn Ser Gln Tyr Pro Leu Asn Ile Glu Val Met Glu Arg Val
35 40 45
tat ccg gtt ccg gga gtg aga gta tat gat att tat ttt gac ggg ttc 192
Tyr Pro Val Pro Gly Val Arg Val Tyr Asp Ile Tyr Phe Asp Gly Phe
50 55 60
cgg aat tcc cgc atc cat ggg gtg tat gtt act cca gaa act ccg gga 240
Arg Asn Ser Arg Ile His Gly Val Tyr Val Thr Pro Glu Thr Pro Gly
65 70 75 80
gcg gac act cct gcg gca gtg att ttt cac ggc tat aac tgg aac acg 288
Ala Asp Thr Pro Ala Ala Val Ile Phe His Gly Tyr Asn Trp Asn Thr
85 90 95
ctg cag ccg cat tac agc ttc aag cac gtg att cag ggg att cct gta 336
Leu Gln Pro His Tyr Ser Phe Lys His Val Ile Gln Gly Ile Pro Val
100 105 110
ctg atg gtg gag gtg cgg gga caa aat ctc ttg tct cca gat aga aat 384
Leu Met Val Glu Val Arg Gly Gln Asn Leu Leu Ser Pro Asp Arg Asn
115 120 125
cat tat ggg aat gga ggt ccg gga ggc tgg atg aca ctc ggc gtg atg 432
His Tyr Gly Asn Gly Gly Pro Gly Gly Trp Met Thr Leu Gly Val Met
130 135 140
gat ccc gat caa tat tat tac agc ctg gta tat atg gac tgc ttc cgc 480
Asp Pro Asp Gln Tyr Tyr Tyr Ser Leu Val Tyr Met Asp Cys Phe Arg
145 150 155 160
agc att gat gct gtc agg gaa ctg tcg agg aag aga agt gtg ttt gtg 528
Ser Ile Asp Ala Val Arg Glu Leu Ser Arg Lys Arg Ser Val Phe Val
165 170 175
gaa ggc gga agc cag gga ggt gca ctg gcg att gcc gca gcc gcc ctg 576
Glu Gly Gly Ser Gln Gly Gly Ala Leu Ala Ile Ala Ala Ala Ala Leu
180 185 190
cag gat gac atc ctg ctt gca ctc gcc gac atc cct ttt ctc acc cat 624
Gln Asp Asp Ile Leu Leu Ala Leu Ala Asp Ile Pro Phe Leu Thr His
195 200 205
ttc aag cgt tcc gtg gag ctt tcc tcg gat gga ccg tat cag gag att 672
Phe Lys Arg Ser Val Glu Leu Ser Ser Asp Gly Pro Tyr Gln Glu Ile
210 215 220
tcc cac tac ttc aaa gtt cat gat cct ctt cat caa acg gaa gag cag 720
Ser His Tyr Phe Lys Val His Asp Pro Leu His Gln Thr Glu Glu Gln
225 230 235 240
gta tat cag acg ctc agc tat gtg gac tgc atg aac atg gcc agc atg 768
Val Tyr Gln Thr Leu Ser Tyr Val Asp Cys Met Asn Met Ala Ser Met
245 250 255
gtt gaa tgt cca gtc ctt ctt tca gcc ggt ctg gaa gac atc gtt tgt 816
Val Glu Cys Pro Val Leu Leu Ser Ala Gly Leu Glu Asp Ile Val Cys
260 265 270
ccc ccg tcc agt gca ttt gca ctg ttc aac cat ctc ggc ggg cca aaa 864
Pro Pro Ser Ser Ala Phe Ala Leu Phe Asn His Leu Gly Gly Pro Lys
275 280 285
gaa ata cgg gcc tat ccg gaa tac gcc cat gaa gta ccg gct gtc cat 912
Glu Ile Arg Ala Tyr Pro Glu Tyr Ala His Glu Val Pro Ala Val His
290 295 300
gaa gag gaa aag ctg aag ttt ata tct tca agg cta aaa aat aga gaa 960
Glu Glu Glu Lys Leu Lys Phe Ile Ser Ser Arg Leu Lys Asn Arg Glu
305 310 315 320
aag agg tgc cgg cca tga 978
Lys Arg Cys Arg Pro
325
<210> 20
<211> 325
<212> PRT
<213> Bacillus sp.
<400> 20
Met Asn Leu Phe Asp Met Pro Leu Glu Glu Leu Gln His Tyr Lys Pro
1 5 10 15
Ala Gln Thr Arg Gln Asp Asp Phe Glu Ser Phe Trp Lys Lys Arg Ile
20 25 30
Glu Glu Asn Ser Gln Tyr Pro Leu Asn Ile Glu Val Met Glu Arg Val
35 40 45
Tyr Pro Val Pro Gly Val Arg Val Tyr Asp Ile Tyr Phe Asp Gly Phe
50 55 60
Arg Asn Ser Arg Ile His Gly Val Tyr Val Thr Pro Glu Thr Pro Gly
65 70 75 80
Ala Asp Thr Pro Ala Ala Val Ile Phe His Gly Tyr Asn Trp Asn Thr
85 90 95
Leu Gln Pro His Tyr Ser Phe Lys His Val Ile Gln Gly Ile Pro Val
100 105 110
Leu Met Val Glu Val Arg Gly Gln Asn Leu Leu Ser Pro Asp Arg Asn
115 120 125
His Tyr Gly Asn Gly Gly Pro Gly Gly Trp Met Thr Leu Gly Val Met
130 135 140
Asp Pro Asp Gln Tyr Tyr Tyr Ser Leu Val Tyr Met Asp Cys Phe Arg
145 150 155 160
Ser Ile Asp Ala Val Arg Glu Leu Ser Arg Lys Arg Ser Val Phe Val
165 170 175
Glu Gly Gly Ser Gln Gly Gly Ala Leu Ala Ile Ala Ala Ala Ala Leu
180 185 190
Gln Asp Asp Ile Leu Leu Ala Leu Ala Asp Ile Pro Phe Leu Thr His
195 200 205
Phe Lys Arg Ser Val Glu Leu Ser Ser Asp Gly Pro Tyr Gln Glu Ile
210 215 220
Ser His Tyr Phe Lys Val His Asp Pro Leu His Gln Thr Glu Glu Gln
225 230 235 240
Val Tyr Gln Thr Leu Ser Tyr Val Asp Cys Met Asn Met Ala Ser Met
245 250 255
Val Glu Cys Pro Val Leu Leu Ser Ala Gly Leu Glu Asp Ile Val Cys
260 265 270
Pro Pro Ser Ser Ala Phe Ala Leu Phe Asn His Leu Gly Gly Pro Lys
275 280 285
Glu Ile Arg Ala Tyr Pro Glu Tyr Ala His Glu Val Pro Ala Val His
290 295 300
Glu Glu Glu Lys Leu Lys Phe Ile Ser Ser Arg Leu Lys Asn Arg Glu
305 310 315 320
Lys Arg Cys Arg Pro
325
<210> 21
<211> 960
<212> DNA
<213> Bacillus halodurans
<400> 21
ttagagatca gataaaaatt gaaaaatccg atcacgatgg cctggcaaat cttcgtgagc 60
aaagtctgga tataactcga tactttttgt cgtcgtgagt ttgttataca tggcaaattg 120
tgtagacggc gggcaaaccg tatccattaa cccaacagca agtaagactt ctccctttac 180
gagtggagca agatgctgaa tatcaatata gcctagcttc gtaaagattt cagcctcacg 240
tcggtgctgt ggatcaaagc gacgaaaata cgtttgcaat tcgtcataag ctttctcggc 300
taaatccatc tcccatacgc gttggtaatc gctaaggaaa ggataaacag gagctacctt 360
tttaattttc ggttccaaag ccgcacaagc aatcgctaag gcccctcctt gtgaccaacc 420
tgtcactgcc acgcgctctt catcgacttc aggaaggttc atcacaatgt tggcaagctg 480
agccgtatca agaaacacat gacggaacaa taattgatca gcattatcat cgagtccgcg 540
tattatatga ccggaatgag tattcccctt cacgcctcct gtgtcttcag acaagcctcc 600
ttgcccgcga acgtccattg caagaacaga atatccgagg gctgcgtaat gaagtaaacc 660
cgtccattcc cccgcattca tcgtatatcc gtgaaaatga ataaccgccg ggtgtgtccc 720
gctcgtgtgt cttgggcgca cgtattttgc gtgaattcta gcacccctaa cccctgtaaa 780
atataggtgg aagcattctg catacgtggt ttgaaaatca ctcggtatga gctctacgtt 840
tggatttacc tttctcatct cttgtaaagc acgatcccaa tactcagtaa agtcatctgg 900
ctttggatta cgtcccatgt actcttttaa ttcggttaac ggcatgtcta ttagtggcat 960
<210> 22
<211> 319
<212> PRT
<213> Bacillus halodurans
<400> 22
Met Pro Leu Ile Asp Met Pro Leu Thr Glu Leu Lys Glu Tyr Met Gly
1 5 10 15
Arg Asn Pro Lys Pro Asp Asp Phe Thr Glu Tyr Trp Asp Arg Ala Leu
20 25 30
Gln Glu Met Arg Lys Val Asn Pro Asn Val Glu Leu Ile Pro Ser Asp
35 40 45
Phe Gln Thr Thr Tyr Ala Glu Cys Phe His Leu Tyr Phe Thr Gly Val
50 55 60
Arg Gly Ala Arg Ile His Ala Lys Tyr Val Arg Pro Arg His Thr Ser
65 70 75 80
Gly Thr His Pro Ala Val Ile His Phe His Gly Tyr Thr Met Asn Ala
85 90 95
Gly Glu Trp Thr Gly Leu Leu His Tyr Ala Ala Leu Gly Tyr Ser Val
100 105 110
Leu Ala Met Asp Val Arg Gly Gln Gly Gly Leu Ser Glu Asp Thr Gly
115 120 125
Gly Val Lys Gly Asn Thr His Ser Gly His Ile Ile Arg Gly Leu Asp
130 135 140
Asp Asn Ala Asp Gln Leu Leu Phe Arg His Val Phe Leu Asp Thr Ala
145 150 155 160
Gln Leu Ala Asn Ile Val Met Asn Leu Pro Glu Val Asp Glu Glu Arg
165 170 175
Val Ala Val Thr Gly Trp Ser Gln Gly Gly Ala Leu Ala Ile Ala Cys
180 185 190
Ala Ala Leu Glu Pro Lys Ile Lys Lys Val Ala Pro Val Tyr Pro Phe
195 200 205
Leu Ser Asp Tyr Gln Arg Val Trp Glu Met Asp Leu Ala Glu Lys Ala
210 215 220
Tyr Asp Glu Leu Gln Thr Tyr Phe Arg Arg Phe Asp Pro Gln His Arg
225 230 235 240
Arg Glu Ala Glu Ile Phe Thr Lys Leu Gly Tyr Ile Asp Ile Gln His
245 250 255
Leu Ala Pro Leu Val Lys Gly Glu Val Leu Leu Ala Val Gly Leu Met
260 265 270
Asp Thr Val Cys Pro Pro Ser Thr Gln Phe Ala Met Tyr Asn Lys Leu
275 280 285
Thr Thr Thr Lys Ser Ile Glu Leu Tyr Pro Asp Phe Ala His Glu Asp
290 295 300
Leu Pro Gly His Arg Asp Arg Ile Phe Gln Phe Leu Ser Asp Leu
305 310 315
<210> 23
<211> 954
<212> DNA
<213> Bacillus clausii
<400> 23
atgccattag tcgatatgcc gttgcgcgag ttgttagctt atgaaggaat aaaccctaaa 60
ccagcagatt ttgaccaata ctggaaccgg gccaaaacgg aaattgaagc gattgatccc 120
gaagtcactc tagtcgaatc ttctttccag tgttcgtttg caaactgtta ccatttctat 180
tatcgaagcg ctggaaatgc aaaaatccat gcgaaatacg tacagccaaa agcaggggag 240
aagacgccag cagtttttat gttccatggg tatggggggc gttcagccga atggagcagc 300
ttgttaaatt atgtagcggc gggtttttct gttttctata tggacgtgcg tggacaaggt 360
ggaacttcag aggatcctgg gggcgtaagg gggaatacat ataggggcca cattattcgc 420
ggcctcgatg ccgggccaga cgcacttttt taccgcagcg ttttcttgga caccgtccaa 480
ttggttcgtg ctgctaaaac attgcctcac atcgataaaa cacggcttat ggccacaggg 540
tggtcgcaag ggggcgcctt aacgcttgcc tgtgctgccc ttgttcctga aatcaagcgt 600
cttgctccag tatacccgtt tttaagcgat tacaagcgag tgtggcaaat ggatttagcg 660
gttcgttcgt ataaagaatt ggctgattat ttccgttcat acgatccgca acataaacgc 720
catggcgaaa tttttgaacg ccttggctac atcgatgtcc agcatcttgc tgaccggatt 780
caaggagatg tcctaatggg agttggttta atggatacag aatgcccgcc gtctacccaa 840
tttgctgctt ataataaaat aaaggctaaa aaatcgtatg agctctatcc tgattttggc 900
catgagcacc ttccaggaat gaacgatcat atttttcgct ttttcactag ttga 954
<210> 24
<211> 317
<212> PRT
<213> Bacillus clausii
<400> 24
Met Pro Leu Val Asp Met Pro Leu Arg Glu Leu Leu Ala Tyr Glu Gly
1 5 10 15
Ile Asn Pro Lys Pro Ala Asp Phe Asp Gln Tyr Trp Asn Arg Ala Lys
20 25 30
Thr Glu Ile Glu Ala Ile Asp Pro Glu Val Thr Leu Val Glu Ser Ser
35 40 45
Phe Gln Cys Ser Phe Ala Asn Cys Tyr His Phe Tyr Tyr Arg Ser Ala
50 55 60
Gly Asn Ala Lys Ile His Ala Lys Tyr Val Gln Pro Lys Ala Gly Glu
65 70 75 80
Lys Thr Pro Ala Val Phe Met Phe His Gly Tyr Gly Gly Arg Ser Ala
85 90 95
Glu Trp Ser Ser Leu Leu Asn Tyr Val Ala Ala Gly Phe Ser Val Phe
100 105 110
Tyr Met Asp Val Arg Gly Gln Gly Gly Thr Ser Glu Asp Pro Gly Gly
115 120 125
Val Arg Gly Asn Thr Tyr Arg Gly His Ile Ile Arg Gly Leu Asp Ala
130 135 140
Gly Pro Asp Ala Leu Phe Tyr Arg Ser Val Phe Leu Asp Thr Val Gln
145 150 155 160
Leu Val Arg Ala Ala Lys Thr Leu Pro His Ile Asp Lys Thr Arg Leu
165 170 175
Met Ala Thr Gly Trp Ser Gln Gly Gly Ala Leu Thr Leu Ala Cys Ala
180 185 190
Ala Leu Val Pro Glu Ile Lys Arg Leu Ala Pro Val Tyr Pro Phe Leu
195 200 205
Ser Asp Tyr Lys Arg Val Trp Gln Met Asp Leu Ala Val Arg Ser Tyr
210 215 220
Lys Glu Leu Ala Asp Tyr Phe Arg Ser Tyr Asp Pro Gln His Lys Arg
225 230 235 240
His Gly Glu Ile Phe Glu Arg Leu Gly Tyr Ile Asp Val Gln His Leu
245 250 255
Ala Asp Arg Ile Gln Gly Asp Val Leu Met Gly Val Gly Leu Met Asp
260 265 270
Thr Glu Cys Pro Pro Ser Thr Gln Phe Ala Ala Tyr Asn Lys Ile Lys
275 280 285
Ala Lys Lys Ser Tyr Glu Leu Tyr Pro Asp Phe Gly His Glu His Leu
290 295 300
Pro Gly Met Asn Asp His Ile Phe Arg Phe Phe Thr Ser
305 310 315
<210> 25
<211> 960
<212> DNA
<213> Bacillus subtilis
<220>
<221> CDS
<222> (1)..(960)
<400> 25
atg caa cta ttc gat ctg ccg ctc gac caa ttg caa aca tat aag cct 48
Met Gln Leu Phe Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
gaa aaa aca gca ccg aaa gat ttt tct gag ttt tgg aaa ttg tct ttg 96
Glu Lys Thr Ala Pro Lys Asp Phe Ser Glu Phe Trp Lys Leu Ser Leu
20 25 30
gag gaa ctt gca aaa gtc caa gca gaa cct gat cta cag ccg gtt gac 144
Glu Glu Leu Ala Lys Val Gln Ala Glu Pro Asp Leu Gln Pro Val Asp
35 40 45
tat cct gct gac gga gta aaa gtg tac cgt ctc aca tat aaa agc ttc 192
Tyr Pro Ala Asp Gly Val Lys Val Tyr Arg Leu Thr Tyr Lys Ser Phe
50 55 60
gga aac gcc cgc att acc gga tgg tac gcg gtg cct gac aag caa ggc 240
Gly Asn Ala Arg Ile Thr Gly Trp Tyr Ala Val Pro Asp Lys Gln Gly
65 70 75 80
ccg cat ccg gcg atc gtg aaa tat cat ggc tac aat gca agc tat gat 288
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
ggt gag att cat gaa atg gta aac tgg gca ctc cat ggc tac gcc gca 336
Gly Glu Ile His Glu Met Val Asn Trp Ala Leu His Gly Tyr Ala Ala
100 105 110
ttc ggc atg ctt gtc cgc ggc cag cag agc agc gag gat acg agt att 384
Phe Gly Met Leu Val Arg Gly Gln Gln Ser Ser Glu Asp Thr Ser Ile
115 120 125
tca ccg cac ggt cac gct ttg ggc tgg atg acg aaa gga att ctt gat 432
Ser Pro His Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Asp
130 135 140
aaa gat aca tac tat tac cgc ggt gtt tat ttg gac gcc gtc cgc gcg 480
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
ctt gag gtc atc agc agc ttc gac gag gtt gac gaa aca agg atc ggt 528
Leu Glu Val Ile Ser Ser Phe Asp Glu Val Asp Glu Thr Arg Ile Gly
165 170 175
gtg aca gga gga agc caa ggc gga ggt tta acc att gcc gca gca gcg 576
Val Thr Gly Gly Ser Gln Gly Gly Gly Leu Thr Ile Ala Ala Ala Ala
180 185 190
ctg tca gac att cca aaa gcc gcg gtt gcc gat tat cct tat tta agc 624
Leu Ser Asp Ile Pro Lys Ala Ala Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
aac ttc gaa cgg gcc att gat gtg gcg ctt gaa cag ccg tac ctt gaa 672
Asn Phe Glu Arg Ala Ile Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
atc aat tcc ttc ttc aga aga aat ggc agc ccg gaa aca gaa gtg cag 720
Ile Asn Ser Phe Phe Arg Arg Asn Gly Ser Pro Glu Thr Glu Val Gln
225 230 235 240
gcg atg aag aca ctt tca tat ttc gat att atg aat ctc gct gac cga 768
Ala Met Lys Thr Leu Ser Tyr Phe Asp Ile Met Asn Leu Ala Asp Arg
245 250 255
gtg aag gtg cct gtc ctg atg tca atc ggc ctg att gac aag gtc acg 816
Val Lys Val Pro Val Leu Met Ser Ile Gly Leu Ile Asp Lys Val Thr
260 265 270
ccg cca tcc acc gtg ttt gcc gcc tac aat cat ttg gaa aca gag aaa 864
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Glu Lys
275 280 285
gag ctg aag gtg tac cgc tac ttc gga cat gag tat atc cct gct ttt 912
Glu Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Ala Phe
290 295 300
caa acg gaa aaa ctt gct ttc ttt aag cag cat ctt aaa ggc tga taa 960
Gln Thr Glu Lys Leu Ala Phe Phe Lys Gln His Leu Lys Gly
305 310 315
<210> 26
<211> 318
<212> PRT
<213> Bacillus subtilis
<400> 26
Met Gln Leu Phe Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
Glu Lys Thr Ala Pro Lys Asp Phe Ser Glu Phe Trp Lys Leu Ser Leu
20 25 30
Glu Glu Leu Ala Lys Val Gln Ala Glu Pro Asp Leu Gln Pro Val Asp
35 40 45
Tyr Pro Ala Asp Gly Val Lys Val Tyr Arg Leu Thr Tyr Lys Ser Phe
50 55 60
Gly Asn Ala Arg Ile Thr Gly Trp Tyr Ala Val Pro Asp Lys Gln Gly
65 70 75 80
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Glu Ile His Glu Met Val Asn Trp Ala Leu His Gly Tyr Ala Ala
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gln Ser Ser Glu Asp Thr Ser Ile
115 120 125
Ser Pro His Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Asp
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Ser Ser Phe Asp Glu Val Asp Glu Thr Arg Ile Gly
165 170 175
Val Thr Gly Gly Ser Gln Gly Gly Gly Leu Thr Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Ala Ala Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Ile Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Phe Phe Arg Arg Asn Gly Ser Pro Glu Thr Glu Val Gln
225 230 235 240
Ala Met Lys Thr Leu Ser Tyr Phe Asp Ile Met Asn Leu Ala Asp Arg
245 250 255
Val Lys Val Pro Val Leu Met Ser Ile Gly Leu Ile Asp Lys Val Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Glu Lys
275 280 285
Glu Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ala Phe Phe Lys Gln His Leu Lys Gly
305 310 315
<210> 27
<211> 325
<212> PRT
<213> Thermotoga neapolitana
<220>
<221> MISC_FEATURE
<222> (277)..(277)
<223> Xaa is Ala, Val, Ser, or Thr.
<400> 27
Met Ala Phe Phe Asp Met Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Arg Glu Thr Leu
20 25 30
Lys Glu Ser Glu Gly Phe Pro Leu Asp Pro Val Phe Glu Lys Val Asp
35 40 45
Phe His Leu Lys Thr Val Glu Thr Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Ala Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Met Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Gly Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Val Asp Ala Val Arg Ala Val Glu Ala Ala Ile Ser Phe Pro Arg
165 170 175
Val Asp Ser Arg Lys Val Val Val Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Asn Arg Val Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Val Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Val Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Thr Ile Xaa Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Ile Glu Gln Val Lys Phe Leu Lys Arg
305 310 315 320
Leu Phe Glu Glu Gly
325
<210> 28
<211> 325
<212> PRT
<213> Thermotoga maritima
<220>
<221> MISC_FEATURE
<222> (277)..(277)
<223> Xaa is Ala, Val, Ser, or Thr.
<400> 28
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Xaa Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 29
<211> 326
<212> PRT
<213> Thermotoga lettingae
<220>
<221> MISC_FEATURE
<222> (277)..(277)
<223> Xaa is Ala, Val, Ser, or Thr.
<400> 29
Met Val Tyr Phe Asp Met Pro Leu Glu Asp Leu Arg Lys Tyr Leu Pro
1 5 10 15
Gln Arg Tyr Glu Glu Lys Asp Phe Asp Asp Phe Trp Lys Gln Thr Ile
20 25 30
His Glu Thr Arg Gly Tyr Phe Gln Glu Pro Ile Leu Lys Lys Val Asp
35 40 45
Phe Tyr Leu Gln Asn Val Glu Thr Phe Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Lys Ile Lys Gly Trp Leu Ile Leu Pro Lys Phe Arg Asn
65 70 75 80
Gly Lys Leu Pro Cys Val Val Glu Phe Val Gly Tyr Gly Gly Gly Arg
85 90 95
Gly Phe Pro Tyr Asp Trp Leu Leu Trp Ser Ala Ala Gly Tyr Ala His
100 105 110
Phe Ile Met Asp Thr Arg Gly Gln Gly Ser Asn Trp Met Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Glu Asp Asn Pro Ser Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Leu Thr Lys Gly Val Leu Asn Pro Glu Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Met Asp Ala Phe Met Ala Val Glu Thr Ile Ser Gln Leu Glu Gln
165 170 175
Ile Asp Ser Gln Thr Ile Ile Leu Ser Gly Ala Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Ser Lys Val Met Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Tyr Lys Arg Ala Val Gln Ile Thr
210 215 220
Asp Ser Met Pro Tyr Ala Glu Ile Thr Arg Tyr Cys Lys Thr His Ile
225 230 235 240
Asp Lys Ile Gln Thr Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Cys Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asp Ile Xaa Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Glu Lys Asp Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe His Thr Leu Glu Lys Leu Lys Phe Val Lys Lys
305 310 315 320
Thr Ile Ser Met Arg Glu
325
<210> 30
<211> 325
<212> PRT
<213> Thermotoga petrophilia
<220>
<221> MISC_FEATURE
<222> (277)..(277)
<223> Xaa is Ala, Val, Ser, or Thr.
<400> 30
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Gly Thr Leu
20 25 30
Ala Glu Asn Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Met Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Met Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Asp Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Arg
165 170 175
Val Asp His Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Val Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Xaa Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Ile Glu Gln Val Lys Phe Leu Lys Arg
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 31
<211> 325
<212> PRT
<213> Thermotoga sp. RQ2a
<220>
<221> MISC_FEATURE
<222> (277)..(277)
<223> Xaa is Ala, Val, Ser, or Thr.
<400> 31
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Lys Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Val Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Asp Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Arg
165 170 175
Val Asp His Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Val Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Xaa Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Ile Glu Gln Val Lys Phe Leu Lys Arg
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 32
<211> 329
<212> PRT
<213> Thermotoga sp. RQ2b
<220>
<221> MISC_FEATURE
<222> (278)..(278)
<223> Xaa is Ala, Val, Ser, or Thr.
<400> 32
Met Ala Leu Phe Asp Met Pro Leu Glu Lys Leu Arg Ser Tyr Leu Pro
1 5 10 15
Asp Arg Tyr Glu Glu Glu Asp Phe Asp Leu Phe Trp Lys Glu Thr Leu
20 25 30
Glu Glu Ser Arg Lys Phe Pro Leu Asp Pro Ile Phe Glu Arg Val Asp
35 40 45
Tyr Leu Leu Glu Asn Val Glu Val Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Ala Trp Leu Ile Leu Pro Val Val Lys Lys
65 70 75 80
Glu Glu Arg Leu Pro Cys Ile Val Glu Phe Ile Gly Tyr Arg Gly Gly
85 90 95
Arg Gly Phe Pro Phe Asp Trp Leu Phe Trp Ser Ser Ala Gly Tyr Ala
100 105 110
His Phe Val Met Asp Thr Arg Gly Gln Gly Thr Ser Arg Val Lys Gly
115 120 125
Asp Thr Pro Asp Tyr Cys Asp Glu Pro Ile Asn Pro Gln Phe Pro Gly
130 135 140
Phe Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg
145 150 155 160
Val Phe Thr Asp Ala Val Arg Ala Val Glu Thr Ala Ser Ser Phe Pro
165 170 175
Gly Ile Asp Pro Glu Arg Ile Ala Val Val Gly Thr Ser Gln Gly Gly
180 185 190
Gly Ile Ala Leu Ala Val Ala Ala Leu Ser Glu Ile Pro Lys Ala Leu
195 200 205
Val Ser Asn Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Ile
210 215 220
Thr Asp Asn Ala Pro Tyr Ser Glu Ile Val Asn Tyr Leu Lys Val His
225 230 235 240
Arg Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly
245 250 255
Val Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Ala
260 265 270
Leu Met Asp Lys Thr Xaa Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn
275 280 285
His Tyr Ala Gly Pro Lys Glu Ile Lys Val Tyr Pro Phe Asn Glu His
290 295 300
Glu Gly Gly Glu Ser Phe Gln Arg Met Glu Glu Leu Arg Phe Met Lys
305 310 315 320
Arg Ile Leu Lys Gly Glu Phe Lys Ala
325
<210> 33
<211> 326
<212> PRT
<213> Thermotoga lettingae
<400> 33
Met Val Tyr Phe Asp Met Pro Leu Glu Asp Leu Arg Lys Tyr Leu Pro
1 5 10 15
Gln Arg Tyr Glu Glu Lys Asp Phe Asp Asp Phe Trp Lys Gln Thr Ile
20 25 30
His Glu Thr Arg Gly Tyr Phe Gln Glu Pro Ile Leu Lys Lys Val Asp
35 40 45
Phe Tyr Leu Gln Asn Val Glu Thr Phe Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Lys Ile Lys Gly Trp Leu Ile Leu Pro Lys Phe Arg Asn
65 70 75 80
Gly Lys Leu Pro Cys Val Val Glu Phe Val Gly Tyr Gly Gly Gly Arg
85 90 95
Gly Phe Pro Tyr Asp Trp Leu Leu Trp Ser Ala Ala Gly Tyr Ala His
100 105 110
Phe Ile Met Asp Thr Arg Gly Gln Gly Ser Asn Trp Met Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Glu Asp Asn Pro Ser Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Leu Thr Lys Gly Val Leu Asn Pro Glu Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Met Asp Ala Phe Met Ala Val Glu Thr Ile Ser Gln Leu Glu Gln
165 170 175
Ile Asp Ser Gln Thr Ile Ile Leu Ser Gly Ala Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Ser Lys Val Met Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Tyr Lys Arg Ala Val Gln Ile Thr
210 215 220
Asp Ser Met Pro Tyr Ala Glu Ile Thr Arg Tyr Cys Lys Thr His Ile
225 230 235 240
Asp Lys Ile Gln Thr Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Cys Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asp Ile Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Glu Lys Asp Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe His Thr Leu Glu Lys Leu Lys Phe Val Lys Lys
305 310 315 320
Thr Ile Ser Met Arg Glu
325
<210> 34
<211> 325
<212> PRT
<213> Thermotoga petrophilia
<400> 34
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Gly Thr Leu
20 25 30
Ala Glu Asn Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Met Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Met Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Asp Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Arg
165 170 175
Val Asp His Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Val Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Ile Glu Gln Val Lys Phe Leu Lys Arg
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 35
<211> 325
<212> PRT
<213> Thermotoga sp. RQ2
<400> 35
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Lys Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Val Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Asp Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Arg
165 170 175
Val Asp His Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Val Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Ile Glu Gln Val Lys Phe Leu Lys Arg
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 36
<211> 329
<212> PRT
<213> Thermotoga sp. RQ2
<400> 36
Met Ala Leu Phe Asp Met Pro Leu Glu Lys Leu Arg Ser Tyr Leu Pro
1 5 10 15
Asp Arg Tyr Glu Glu Glu Asp Phe Asp Leu Phe Trp Lys Glu Thr Leu
20 25 30
Glu Glu Ser Arg Lys Phe Pro Leu Asp Pro Ile Phe Glu Arg Val Asp
35 40 45
Tyr Leu Leu Glu Asn Val Glu Val Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Ala Trp Leu Ile Leu Pro Val Val Lys Lys
65 70 75 80
Glu Glu Arg Leu Pro Cys Ile Val Glu Phe Ile Gly Tyr Arg Gly Gly
85 90 95
Arg Gly Phe Pro Phe Asp Trp Leu Phe Trp Ser Ser Ala Gly Tyr Ala
100 105 110
His Phe Val Met Asp Thr Arg Gly Gln Gly Thr Ser Arg Val Lys Gly
115 120 125
Asp Thr Pro Asp Tyr Cys Asp Glu Pro Ile Asn Pro Gln Phe Pro Gly
130 135 140
Phe Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg
145 150 155 160
Val Phe Thr Asp Ala Val Arg Ala Val Glu Thr Ala Ser Ser Phe Pro
165 170 175
Gly Ile Asp Pro Glu Arg Ile Ala Val Val Gly Thr Ser Gln Gly Gly
180 185 190
Gly Ile Ala Leu Ala Val Ala Ala Leu Ser Glu Ile Pro Lys Ala Leu
195 200 205
Val Ser Asn Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Ile
210 215 220
Thr Asp Asn Ala Pro Tyr Ser Glu Ile Val Asn Tyr Leu Lys Val His
225 230 235 240
Arg Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly
245 250 255
Val Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Ala
260 265 270
Leu Met Asp Lys Thr Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn
275 280 285
His Tyr Ala Gly Pro Lys Glu Ile Lys Val Tyr Pro Phe Asn Glu His
290 295 300
Glu Gly Gly Glu Ser Phe Gln Arg Met Glu Glu Leu Arg Phe Met Lys
305 310 315 320
Arg Ile Leu Lys Gly Glu Phe Lys Ala
325
<210> 37
<211> 960
<212> DNA
<213> artificial sequence
<220>
<223> Synthetic gene - codon optimized
<400> 37
atgggtctgt tcgatatgcc actgcaaaaa ctgcgtgaat ataccggtac caacccatgt 60
cctgaggatt tcgatgaata ctgggatcgc gcactggacg aaatgcgtag cgttgatcct 120
aaaatcaaga tgaagaagag ctcctttcaa gttccgttcg cggaatgtta cgatctgtat 180
tttaccggcg ttcgtggtgc ccgcattcac gcgaaataca ttcgtccgaa aaccgaaggc 240
aaacacccgg cgctgattcg cttccatggt tactccagca actctggtga ttggaacgac 300
aagctgaact acgttgcggc tggttttacc gtagtagcga tggacgctcg tggccagggt 360
ggccaatctc aggacgtcgg cggtgttaat ggcaacaccc tgaacggtca catcatccgt 420
ggcctggacg atgatgcaga taacatgctg ttccgtcata ttttcctgga caccgcgcag 480
ctggctggta tcgttatgaa catgccggaa atcgatgagg accgcgtagc tgttatgggt 540
ccgtcccagg gcggcggtct gtccctggcg tgtgcggctc tggaacctaa aatccgtaaa 600
gtagtgtccg aatatccgtt cctgagcgac tacaagcgtg tgtgggatct ggatctggcc 660
aaaaatgcgt accaagaaat cactgactat ttccgtctgt tcgacccacg ccacgaacgt 720
gagaacgagg tttttactaa actgggttac attgacgtaa agaacctggc gaaacgtatc 780
aaaggtgatg ttctgatgtg cgtgggcctg atggatcagg tctgcccgcc gagcaccgta 840
tttgcagcat acaacaacat ccagtccaag aaggacatca aagtctaccc ggactatggt 900
cacgaaccga tgcgtggctt cggtgacctg gctatgcagt tcatgctgga actgtattct 960
<210> 38
<211> 320
<212> PRT
<213> Thermoanaerobacterium saccharolyticum
<400> 38
Met Gly Leu Phe Asp Met Pro Leu Gln Lys Leu Arg Glu Tyr Thr Gly
1 5 10 15
Thr Asn Pro Cys Pro Glu Asp Phe Asp Glu Tyr Trp Asp Arg Ala Leu
20 25 30
Asp Glu Met Arg Ser Val Asp Pro Lys Ile Lys Met Lys Lys Ser Ser
35 40 45
Phe Gln Val Pro Phe Ala Glu Cys Tyr Asp Leu Tyr Phe Thr Gly Val
50 55 60
Arg Gly Ala Arg Ile His Ala Lys Tyr Ile Arg Pro Lys Thr Glu Gly
65 70 75 80
Lys His Pro Ala Leu Ile Arg Phe His Gly Tyr Ser Ser Asn Ser Gly
85 90 95
Asp Trp Asn Asp Lys Leu Asn Tyr Val Ala Ala Gly Phe Thr Val Val
100 105 110
Ala Met Asp Ala Arg Gly Gln Gly Gly Gln Ser Gln Asp Val Gly Gly
115 120 125
Val Asn Gly Asn Thr Leu Asn Gly His Ile Ile Arg Gly Leu Asp Asp
130 135 140
Asp Ala Asp Asn Met Leu Phe Arg His Ile Phe Leu Asp Thr Ala Gln
145 150 155 160
Leu Ala Gly Ile Val Met Asn Met Pro Glu Ile Asp Glu Asp Arg Val
165 170 175
Ala Val Met Gly Pro Ser Gln Gly Gly Gly Leu Ser Leu Ala Cys Ala
180 185 190
Ala Leu Glu Pro Lys Ile Arg Lys Val Val Ser Glu Tyr Pro Phe Leu
195 200 205
Ser Asp Tyr Lys Arg Val Trp Asp Leu Asp Leu Ala Lys Asn Ala Tyr
210 215 220
Gln Glu Ile Thr Asp Tyr Phe Arg Leu Phe Asp Pro Arg His Glu Arg
225 230 235 240
Glu Asn Glu Val Phe Thr Lys Leu Gly Tyr Ile Asp Val Lys Asn Leu
245 250 255
Ala Lys Arg Ile Lys Gly Asp Val Leu Met Cys Val Gly Leu Met Asp
260 265 270
Gln Val Cys Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Asn Ile Gln
275 280 285
Ser Lys Lys Asp Ile Lys Val Tyr Pro Asp Tyr Gly His Glu Pro Met
290 295 300
Arg Gly Phe Gly Asp Leu Ala Met Gln Phe Met Leu Glu Leu Tyr Ser
305 310 315 320
<210> 39
<211> 939
<212> DNA
<213> Lactococcus lactis
<400> 39
atgacaaaaa taaacaattg gcaagattat caaggaagtt cacttaaacc agaggatttt 60
gataaatttt gggatgaaaa aattaatttg gtttcaaatc atcaatttga atttgaatta 120
atagaaaaaa atctttcctc taaggtagtt aacttttatc atttgtggtt tacagctatt 180
gatggagcta aaattcatgc tcagttaatt gttcccaaga atttgaaaga gaaataccca 240
gccatcttac aatttcatgg ttatcattgc gatagtgggg attgggtcga taaaataggg 300
atagttgccg aagggaatgt agttcttgcg cttgattgtc gaggacaagg tggtttaagt 360
caagataata ttcaaactat ggggatgaca atgaagggac tcattgttcg aggaattgat 420
gaagggtatg aaaatctcta ttacgttcgc caatttatgg acttaataac tgcaaccaaa 480
attttatccg agtttgattt tgttgatgaa acaaatataa gtgcacaagg tgcttctcaa 540
ggtggagcgc ttgccgttgc ttgcgccgca ctttctcctc ttataaaaaa ggtgactgcc 600
acttacccct ttctttcaga ttatcgcaaa gcttatgagc ttggtgccga ggaatctgct 660
ttcgaagaac ttccatattg gtttcagttt aaagatccac ttcatctaag agaagactgg 720
ttttttaatc agttggaata cattgatatt caaaatttag caccaagaat taaggctgag 780
gtcatttgga tcctaggcgg caaagatact gttgttcctc cgattacgca aatggcggct 840
tacaataaaa tacaaagtaa aaaatctctc tatgtcttac ctgaatacgg ccatgaatat 900
cttcctaaaa ttagcgactg gttaagagag aatcaataa 939
<210> 40
<211> 312
<212> PRT
<213> Lactococcus lactis
<400> 40
Met Thr Lys Ile Asn Asn Trp Gln Asp Tyr Gln Gly Ser Ser Leu Lys
1 5 10 15
Pro Glu Asp Phe Asp Lys Phe Trp Asp Glu Lys Ile Asn Leu Val Ser
20 25 30
Asn His Gln Phe Glu Phe Glu Leu Ile Glu Lys Asn Leu Ser Ser Lys
35 40 45
Val Val Asn Phe Tyr His Leu Trp Phe Thr Ala Ile Asp Gly Ala Lys
50 55 60
Ile His Ala Gln Leu Ile Val Pro Lys Asn Leu Lys Glu Lys Tyr Pro
65 70 75 80
Ala Ile Leu Gln Phe His Gly Tyr His Cys Asp Ser Gly Asp Trp Val
85 90 95
Asp Lys Ile Gly Ile Val Ala Glu Gly Asn Val Val Leu Ala Leu Asp
100 105 110
Cys Arg Gly Gln Gly Gly Leu Ser Gln Asp Asn Ile Gln Thr Met Gly
115 120 125
Met Thr Met Lys Gly Leu Ile Val Arg Gly Ile Asp Glu Gly Tyr Glu
130 135 140
Asn Leu Tyr Tyr Val Arg Gln Phe Met Asp Leu Ile Thr Ala Thr Lys
145 150 155 160
Ile Leu Ser Glu Phe Asp Phe Val Asp Glu Thr Asn Ile Ser Ala Gln
165 170 175
Gly Ala Ser Gln Gly Gly Ala Leu Ala Val Ala Cys Ala Ala Leu Ser
180 185 190
Pro Leu Ile Lys Lys Val Thr Ala Thr Tyr Pro Phe Leu Ser Asp Tyr
195 200 205
Arg Lys Ala Tyr Glu Leu Gly Ala Glu Glu Ser Ala Phe Glu Glu Leu
210 215 220
Pro Tyr Trp Phe Gln Phe Lys Asp Pro Leu His Leu Arg Glu Asp Trp
225 230 235 240
Phe Phe Asn Gln Leu Glu Tyr Ile Asp Ile Gln Asn Leu Ala Pro Arg
245 250 255
Ile Lys Ala Glu Val Ile Trp Ile Leu Gly Gly Lys Asp Thr Val Val
260 265 270
Pro Pro Ile Thr Gln Met Ala Ala Tyr Asn Lys Ile Gln Ser Lys Lys
275 280 285
Ser Leu Tyr Val Leu Pro Glu Tyr Gly His Glu Tyr Leu Pro Lys Ile
290 295 300
Ser Asp Trp Leu Arg Glu Asn Gln
305 310
<210> 41
<211> 972
<212> DNA
<213> Mesorhizobium loti
<400> 41
atgccgttcc cggatctgat ccagcccgaa ctgggcgctt atgtcagcag tgtcggcatg 60
ccggacgact ttgcccaatt ctggacgtcg accatcgccg aggctcgcca ggccggcggt 120
gaggtcagta tcgtgcaggc gcagacgaca ctgaaggcgg tccagtcctt cgatgtcacg 180
tttccaggat acggcggtca tccaatcaaa ggatggctga tcttgccgac gcaccacaag 240
gggcggcttc ccctcgtcgt gcagtatatc ggctatggcg gcggccgcgg cttggcgcat 300
gagcaactgc attgggcggc gtcaggcttt gcctatttcc gaatggatac acgcgggcag 360
ggaagcgact ggagcgtcgg tgagaccgcc gatcccgtcg gctcgacctc gtccattccc 420
ggctttatga cgcgtggcgt gctggacaag aatgactact attaccggcg cctgttcacc 480
gatgccgtga gggcgataga tgctctgctc ggactggact tcgtcgatcc cgaacgcatc 540
gcggtttgcg gtgacagtca gggaggcggt atttcgctcg ccgttggcgg catcgacccg 600
cgcgtcaagg ccgtaatgcc cgacgttcca tttctgtgcg actttccgcg cgctgtgcag 660
actgccgtgc gcgatcccta tttggaaatc gttcgctttc tggcccagca tcgcgaaaag 720
aaggcggcag tctttgaaac gctcaactat ttcgactgcg tcaacttcgc ccggcggtcc 780
aaggcgccgg cgctgttttc ggtggccctg atggacgaag tctgcccgcc ctctaccgtg 840
tatggcgcat tcaatgccta tgcaggcgaa aagaccatca cagagtacga attcaacaat 900
catgaaggcg ggcaaggcta tcaagagcgc caacagatga cgtggctcag caggctgttc 960
ggtgtcggct ga 972
<210> 42
<211> 323
<212> PRT
<213> Mesorhizobium loit
<400> 42
Met Pro Phe Pro Asp Leu Ile Gln Pro Glu Leu Gly Ala Tyr Val Ser
1 5 10 15
Ser Val Gly Met Pro Asp Asp Phe Ala Gln Phe Trp Thr Ser Thr Ile
20 25 30
Ala Glu Ala Arg Gln Ala Gly Gly Glu Val Ser Ile Val Gln Ala Gln
35 40 45
Thr Thr Leu Lys Ala Val Gln Ser Phe Asp Val Thr Phe Pro Gly Tyr
50 55 60
Gly Gly His Pro Ile Lys Gly Trp Leu Ile Leu Pro Thr His His Lys
65 70 75 80
Gly Arg Leu Pro Leu Val Val Gln Tyr Ile Gly Tyr Gly Gly Gly Arg
85 90 95
Gly Leu Ala His Glu Gln Leu His Trp Ala Ala Ser Gly Phe Ala Tyr
100 105 110
Phe Arg Met Asp Thr Arg Gly Gln Gly Ser Asp Trp Ser Val Gly Glu
115 120 125
Thr Ala Asp Pro Val Gly Ser Thr Ser Ser Ile Pro Gly Phe Met Thr
130 135 140
Arg Gly Val Leu Asp Lys Asn Asp Tyr Tyr Tyr Arg Arg Leu Phe Thr
145 150 155 160
Asp Ala Val Arg Ala Ile Asp Ala Leu Leu Gly Leu Asp Phe Val Asp
165 170 175
Pro Glu Arg Ile Ala Val Cys Gly Asp Ser Gln Gly Gly Gly Ile Ser
180 185 190
Leu Ala Val Gly Gly Ile Asp Pro Arg Val Lys Ala Val Met Pro Asp
195 200 205
Val Pro Phe Leu Cys Asp Phe Pro Arg Ala Val Gln Thr Ala Val Arg
210 215 220
Asp Pro Tyr Leu Glu Ile Val Arg Phe Leu Ala Gln His Arg Glu Lys
225 230 235 240
Lys Ala Ala Val Phe Glu Thr Leu Asn Tyr Phe Asp Cys Val Asn Phe
245 250 255
Ala Arg Arg Ser Lys Ala Pro Ala Leu Phe Ser Val Ala Leu Met Asp
260 265 270
Glu Val Cys Pro Pro Ser Thr Val Tyr Gly Ala Phe Asn Ala Tyr Ala
275 280 285
Gly Glu Lys Thr Ile Thr Glu Tyr Glu Phe Asn Asn His Glu Gly Gly
290 295 300
Gln Gly Tyr Gln Glu Arg Gln Gln Met Thr Trp Leu Ser Arg Leu Phe
305 310 315 320
Gly Val Gly
<210> 43
<211> 990
<212> DNA
<213> Geobacillus stearothermophilus
<400> 43
atgttcgata tgccgttagc acaattacag aaatacatgg ggacaaatcc gaagccggct 60
gattttgctg acttttggag tcgagcgttg gaggaattat ctgcccaatc gttgcattat 120
gagctgattc cggcaacatt tcaaacgaca gtggcgagtt gctaccattt gtatttcacg 180
ggagtcggcg gggctagagt ccattgtcag ttagtaaaac cgagagagca gaagcagaaa 240
ggcccggggt tggtatggtt tcatggctac catacgaata gcggcgattg ggtcgataaa 300
ctggcatatg ctgcggcagg ttttactgta ttggcgatgg attgccgcgg ccaaggagga 360
aaatcagagg ataatttgca agtgaaaggc ccaacattga agggccatat tattcgcgga 420
attgaggatc caaatcctca tcatctttat tatcgaaatg tttttttaga tacagttcag 480
gcggtaagaa ttttatgctc tatggatcat attgatcgtg aacgaattgg tgtatatggc 540
gcttcccaag gaggagcgtt ggcattagcg tgtgctgctc tggaaccatc ggtggtgaaa 600
aaagcggttg tgctctatcc atttttatcg gattataagc gggcgcaaga gttggatatg 660
aaaaataccg cgtatgagga aattcattat tattttcgat ttttagatcc cacacatgag 720
cgggaagaag aagtatttta caaactaggc tatattgata ttcaactctt agccgatcgg 780
atttgtgccg atgttttatg ggctgttgcg ctagaagacc atatttgtcc cccgtccaca 840
caatttgctg tttataataa aattaagtca aaaaaagaca tggttttgtt ttacgagtat 900
ggtcatgagt atttaccgac tatgggagac cgtgcttatc tgtttttttg cccgatcttc 960
tttccaatcc aaaagagaaa cgttaagtaa 990
<210> 44
<211> 329
<212> PRT
<213> Geobacillus stearothermophilus
<400> 44
Met Phe Asp Met Pro Leu Ala Gln Leu Gln Lys Tyr Met Gly Thr Asn
1 5 10 15
Pro Lys Pro Ala Asp Phe Ala Asp Phe Trp Ser Arg Ala Leu Glu Glu
20 25 30
Leu Ser Ala Gln Ser Leu His Tyr Glu Leu Ile Pro Ala Thr Phe Gln
35 40 45
Thr Thr Val Ala Ser Cys Tyr His Leu Tyr Phe Thr Gly Val Gly Gly
50 55 60
Ala Arg Val His Cys Gln Leu Val Lys Pro Arg Glu Gln Lys Gln Lys
65 70 75 80
Gly Pro Gly Leu Val Trp Phe His Gly Tyr His Thr Asn Ser Gly Asp
85 90 95
Trp Val Asp Lys Leu Ala Tyr Ala Ala Ala Gly Phe Thr Val Leu Ala
100 105 110
Met Asp Cys Arg Gly Gln Gly Gly Lys Ser Glu Asp Asn Leu Gln Val
115 120 125
Lys Gly Pro Thr Leu Lys Gly His Ile Ile Arg Gly Ile Glu Asp Pro
130 135 140
Asn Pro His His Leu Tyr Tyr Arg Asn Val Phe Leu Asp Thr Val Gln
145 150 155 160
Ala Val Arg Ile Leu Cys Ser Met Asp His Ile Asp Arg Glu Arg Ile
165 170 175
Gly Val Tyr Gly Ala Ser Gln Gly Gly Ala Leu Ala Leu Ala Cys Ala
180 185 190
Ala Leu Glu Pro Ser Val Val Lys Lys Ala Val Val Leu Tyr Pro Phe
195 200 205
Leu Ser Asp Tyr Lys Arg Ala Gln Glu Leu Asp Met Lys Asn Thr Ala
210 215 220
Tyr Glu Glu Ile His Tyr Tyr Phe Arg Phe Leu Asp Pro Thr His Glu
225 230 235 240
Arg Glu Glu Glu Val Phe Tyr Lys Leu Gly Tyr Ile Asp Ile Gln Leu
245 250 255
Leu Ala Asp Arg Ile Cys Ala Asp Val Leu Trp Ala Val Ala Leu Glu
260 265 270
Asp His Ile Cys Pro Pro Ser Thr Gln Phe Ala Val Tyr Asn Lys Ile
275 280 285
Lys Ser Lys Lys Asp Met Val Leu Phe Tyr Glu Tyr Gly His Glu Tyr
290 295 300
Leu Pro Thr Met Gly Asp Arg Ala Tyr Leu Phe Phe Cys Pro Ile Phe
305 310 315 320
Phe Pro Ile Gln Lys Arg Asn Val Lys
325
<210> 45
<211> 978
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<400> 45
atggcgttct tcgacctgcc tctggaagaa ctgaagaaat accgtccaga gcgttacgaa 60
gagaaggaca tcgacgagtt ctgggaggaa actctggcgg agaccgaaaa gtttccgctg 120
gacccagtgt tcgagcgtat ggaatctcac ctgaaaaccg tggaggcata tgacgttact 180
ttttctggtt accgtggcca gcgtatcaaa ggctggctgc tggttccgaa actggaggaa 240
gaaaaactgc cgtgcgtagt tcagtacatc ggttacaacg gtggccgtgg ctttccgcac 300
gattggctgt tctggccgtc tatgggctac atttgcttcg tcatggatac tcgtggtcag 360
ggttccggct ggctgaaagg cgatactccg gattatccgg agggcccggt agacccgcag 420
taccctggct tcatgacgcg tggtattctg gatccgcgta cctattacta tcgccgcgtt 480
tttaccgatg cagttcgtgc cgtagaggcc gcggcttctt tccctcaggt tgacctggag 540
cgtattgtta tcgctggtgg ctcccagggt ggcggcatcg ccctggcggt atctgcgctg 600
agcaagaaag ctaaggcact gctgtgtgac gtcccgttcc tgtgtcactt ccgtcgcgct 660
gttcagctgg tagataccca tccgtacgcg gagattacta acttcctgaa aactcaccgc 720
gacaaagaag aaatcgtttt ccgcaccctg tcctatttcg acggcgttaa cttcgcggct 780
cgtgcaaaaa ttccggcact gttctctgtt ggtctgatgg acgacatcag ccctccttct 840
accgttttcg cggcatataa ctattatgcg ggtccgaaag aaatccgtat ctatccgtac 900
aacaaccacg aaggcggtgg tggctttcag gctgttgaac aagtgaaatc cctgaagaaa 960
ctgtttgaga agggctaa 978
<210> 46
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 46
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Ile Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Thr Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Leu Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asp Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Gly Phe Gln Ala Val Glu Gln Val Lys Ser Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 47
<211> 978
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<400> 47
atggcgttct tcgacctgcc tctggaagaa ctgaagaaat accgtccaga gcgttacgaa 60
gagaaggact tcgacgagtt ctgggaggaa actctggcgg agagcgaaaa gtttccgctg 120
gacccagtgt tcgagcgtat ggaatctcac ctgaaaaccg tggaggcata tgacgttact 180
ttttctggtt accgtggcca gcgtatcaaa ggctggctgc tggttccgaa actggaggaa 240
gaaaaactgc cgtgcgtagt tcagtacatc ggttacaacg gtggccgtgg ctttccgcac 300
gattggctgt tctggccgtc tatgggctac atttgcttcg tcatggatac tcgtggtcag 360
ggttccggct ggctgaaagg cgatactccg gattatccgg agggcccggt agacccgcag 420
taccctggct tcatgacgcg tggtattctg gatccgcgta cctattacta tcgccgcgtt 480
tttaccgatg cagttcgtgc cgtagaggcc gcggcttctt tccctcaggt tgaccaggag 540
cgtattgtta tcgctggtgg ctcccagggt ggcggcatcg ccctggcggt atctgcgctg 600
agcaagaaag ctaaggcact gctgtgtgac gtcccgttcc tgtgtcactt ccgtcgcgct 660
gttcagctgg tagataccca tccgtacgcg gagattacta acttcctgaa aactcaccgc 720
gacaaagaag aaatcgtttt ccgcaccctg tcctatttcg acggcgttaa cttcgcggct 780
cgtgcaaaaa ttccggcact gttctctgtt ggtctgatgg acgacatcag ccctccttct 840
accgttttcg cggcatataa ctattatgcg ggtccgaaag aaatccgtat ctatccgtac 900
aacaaccacg aaggcggtgg tagctttcag gctgttgaac aagtgaaatt cctgaagaaa 960
ctgtttgaga agggctaa 978
<210> 48
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 48
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asp Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 49
<211> 978
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic construct
<400> 49
atggcgttct tcgacctgcc tctggaagaa ctgaagaaat accgtccaga gcgttacgaa 60
gagaaggact tcgacgagtt ctgggaggaa actctggcgg agagcgaaaa gtttccgctg 120
gacccagtgt tcgagcgtat ggaatctcac ctgaaaaccg tggaggcata tgacgttact 180
ttttctggtt accgtggcca gcgtatcaaa ggctggctgc tggttccgaa actggaggaa 240
gaaaaactgc cgtgcgtagt tcagtacatc ggttacaacg gtggccgtgg ctttccgcac 300
gattggctgt tctggccgtc tatgggctac atttgcttcg tcatggatac tcgtggtcag 360
ggttccggct ggctgaaagg cgatactccg gattatccgg agggcccggt agacccgcag 420
taccctggct tcatgacgcg tggtattctg gatccgcgta cctattacta tcgccgcgtt 480
tttaccgatg cagttcgtgc cgtagaggcc gcggcttctt tccctcaggt tgaccaggag 540
cgtattgtta tcgctggtgg ctcccagggt ggcggcatcg ccctggcggt atctgcgctg 600
agcaagaaag ctaaggcact gctgtgtgac gtcccgttcc tgtgtcactt ccgtcgcgct 660
gttcagctgg tagataccca tccgtacgcg gagattacta acttcctgaa aactcaccgc 720
gacaaagaag aaatcgtttt ccgcaccctg tcctatttcg acggcgttaa cttcgcggct 780
cgtgcaaaaa ttccggcact gttctctgtt ggtctgatgg acaacatcag ccctccttct 840
accgttttcg cggcatataa ctattatgcg ggtccgaaag aaatccgtat ctatccgtac 900
aacaaccacg aaggcggtgg tagctttcag gctgttgaac aagtgaaatc cctgaagaaa 960
ctgtttgaga agggctaa 978
<210> 50
<211> 325
<212> PRT
<213> Artificial sequence
<220>
<223> synthetic construct
<400> 50
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Ser Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 51
<211> 978
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic construct
<400> 51
atggcgttct tcgacctgcc tctggaagaa ctgaagaaat accgtccaga gcgttacgaa 60
gagaaggact tcgacgagtt ctgggaggaa actctggcgg agaccgaaaa gtttccgctg 120
gacccagtgt tcgagcgtat ggaatctcac ctgaaaaccg tggaggcata tgacgttact 180
ttttctggtt accgtggcca gcgtatcaaa ggctggctgc tggttccgaa actggaggaa 240
gaaaaactgc cgtgcgtagt tcagtacatc ggttacaacg gtggccgtgg ctttccgcac 300
gattggctgt tctggccgtc tatgggctac atttgcttcg tcatggatac tcgtggtcag 360
ggttccggct ggctgaaagg cgatactccg gattatccgg agggcccggt agacccgcag 420
taccctggct tcatgacgcg tggtattctg gatccgcgta cctattacta tcgccgcgtt 480
tttaccgatg cagttcgtgc cgtagaggcc gcggcttctt tccctcaggt tgaccaggag 540
cgtattgtta tcgctggtgg ctcccagggt ggcggcatcg ccctggcggt atctgcgctg 600
agcaagaaag ctaaggcact gctgtgtgac gtcccgttcc tgtgtcactt ccgtcgcgct 660
gttcagctgg tagataccca tccgtacgcg gagattacta acttcctgaa aactcaccgc 720
gacaaagaag aaatcgtttt ccgcaccctg tcctatttcg acggcgttaa cttcgcggct 780
cgtgcaaaaa ttccggcact gttctctgtt ggtctgatgg acaacatcag ccctccttct 840
accgttttcg cggcatataa ctattatgcg ggtccgaaag aaatccgtat ctatccgtac 900
aacaaccacg aaggcggtgg tagctttcag gctgttgaac aagtgaaatt cctgaagaaa 960
ctgtttgaga agggctaa 978
<210> 52
<211> 325
<212> PRT
<213> Artificial sequence
<220>
<223> synthetic construct
<400> 52
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Thr Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 53
<211> 978
<212> DNA
<213> Artificial sequence
<220>
<223> synthetic construct
<400> 53
atggcgttct tcgacctgcc tctggaagaa ctgaagaaat accgtccaga gcgttacgaa 60
gagaaggact tcgacgagtt ctgggaggaa actctggcgg agagcgaaaa gtttccgctg 120
gacccagtgt tcgagcgtat ggaatctcac ctgaaaaccg tggaggcata tgacgttact 180
ttttctggtt accgtggcca gcgtatcaaa ggctggctgc tggttccgaa actggaggaa 240
gaaaaactgc cgtgcgtagt tcagtacatc ggttacaacg gtggccgtgg ctttccgcac 300
gattggctgt tctggccgtc tatgggctac atttgcttcg tcatggatac tcgtggtcag 360
ggttccggct ggctgaaagg cgatactccg gattatccgg agggcccggt agacccgcag 420
taccctggct tcatgacgcg tggtattctg gatccgcgta cctattacta tcgccgcgtt 480
tttaccgatg cagttcgtgc cgtagaggcc gcggcttctt tccctcaggt tgacctggag 540
cgtattgtta tcgctggtgg ctcccagggt ggcggcatcg ccctggcggt atctgcgctg 600
agcaagaaag ctaaggcact gctgtgtgac gtcccgttcc tgtgtcactt ccgtcgcgct 660
gttcagctgg tagataccca tccgtacgcg gagattacta acttcctgaa aactcaccgc 720
gacaaagaag aaatcgtttt ccgcaccctg tcctatttcg acggcgttaa cttcgcggct 780
cgtgcaaaaa ttccggcact gttctctgtt ggtctgatgg acaacatcag ccctccttct 840
accgttttcg cggcatataa ctattatgcg ggtccgaaag aaatccgtat ctatccgtac 900
aacaaccacg aaggcggtgg tagctttcag gctgttgaac aagtgaaatt cctgaagaaa 960
ctgtttgaga agggctaa 978
<210> 54
<211> 325
<212> PRT
<213> Artificial sequence
<220>
<223> synthetic construct
<400> 54
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Leu Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 55
<211> 978
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<220>
<221> CDS
<222> (1)..(978)
<400> 55
atg gcg ttc ttc gac ctg cct cgg gaa gaa ctg aag aaa tac cgt cca 48
Met Ala Phe Phe Asp Leu Pro Arg Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
gag cgt tac gaa gag aag gac ttc gac gag ttc tgg gag gaa act ctg 96
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
gcg gag agc gaa aag ttt ccg ctg gac cca gtg ttc gag cgt atg gaa 144
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
tct cac ctg aaa acc gtg gag gca tat gac gtt act ttt tct ggt tac 192
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
cgt ggc cag cgt atc aaa ggc tgg ctg ctg gtt ccg aaa ctg gag gaa 240
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
gaa aaa ctg ccg tgc gta gtt cag tac atc ggt tac aac ggt ggc cgt 288
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
ggc ttt ccg cac gat tgg ctg ttc tgg ccg tct atg ggc tac att tgc 336
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
ttc gtc atg gat act cgt ggt cag ggt tcc ggc tgg cag aaa ggc gat 384
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Gln Lys Gly Asp
115 120 125
act ccg gat tat ccg gag ggc ccg gta gac ccg cag tac cct ggc ttc 432
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
atg acg cgt ggt att ctg gat ccg cgt acc tat tac tat cgc cgc gtt 480
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
ttt acc gat gca gtt cgt gcc gta gag gcc gcg gct tct ttc cct ctg 528
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Leu
165 170 175
gtt gac cag gag cgt att gat atc gct ggt ggc tcc cag ggt ggc ggc 576
Val Asp Gln Glu Arg Ile Asp Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
atc gcc ctg gcg gta tct gcg ctg agc aag aaa gct aag gca ctg ctg 624
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
tgt gac gtc ccg ttc ctg tgt cac ttc cgt cgc gct gtt cag ctg gta 672
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
gat acc cat ccg tac gcg gag att act aac ttc ctg aaa act cac cgc 720
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
gac aaa gaa gaa atc gtt atc cgc acc ctg tcc tat ttc gac ggc gtt 768
Asp Lys Glu Glu Ile Val Ile Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
aac ttc gcg gct cgt gca aaa att ccg gca ctg ttc tct gtt ggt ctg 816
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
atg gac aac atc agc cct cct tct acc gtt ttc gcg gca tat aac tat 864
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
tat gcg ggt ctg aaa gaa atc cgt atc tat ccg tac aac aac cac gaa 912
Tyr Ala Gly Leu Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
ggc ggt ggt agc ttt cag gct gtt gaa caa gtg aaa ttc ctg aag aaa 960
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
ctg ttt gag aag ggc taa 978
Leu Phe Glu Lys Gly
325
<210> 56
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 56
Met Ala Phe Phe Asp Leu Pro Arg Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Gln Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Leu
165 170 175
Val Asp Gln Glu Arg Ile Asp Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Ile Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Leu Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 57
<211> 978
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<220>
<221> CDS
<222> (1)..(978)
<400> 57
atg gcg ttc ttc gac ctg cct ctg gaa gaa ctg aag aaa tac cgt cca 48
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
gag cgt tac gaa gag aag gac ttc gac gag ttc tgg gag gaa act ctg 96
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
gcg gag agc gaa aag ttt ccg ctg gac cca gtg ttc gag cgt atg gaa 144
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
tct cac ctg aaa acc gtg gag gca tat gac gtt act ttt tct ggt tac 192
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
cgt ggc cag cgt atc aaa ggc tgg ctg ctg gtt ccg gaa ctg gag gaa 240
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Glu Leu Glu Glu
65 70 75 80
gaa aaa ctg ccg tgc gta gtt cag tac atc ggt tac aac ggt ggc cgt 288
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
ggc ttt ccg cac gat tgg ctg ttc tgg ccg tct atg ggc tac att tgc 336
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
ttc gtc atg gat act cgt ggt cag ggt tcc ggc tgg ctg aaa ggc gat 384
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
act ccg gat tat ccg gag ggc ccg gta gac ccg cag tac cct ggc ttc 432
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
atg acg cgt ggt att ctg gat ccg cgt acc tat tac tat cgc cgc gtt 480
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
ttt acc gat gca gtt cgt gcc gta gag gcc gcg gct tct ttc cct cag 528
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
gtt gac cag gag cgt att gtt atc gct ggt ggc tcc cag ggt ggc ggc 576
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
atc gcc ctg gcg gta tct gcg ctg agc aag aaa gct aag gca ctg ctg 624
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
tgt gac gtc ccg ttc ctg tgt cac ttc cgt cgc gct gtt cag ctg gta 672
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
gat acc cat ccg tac gcg gag att act aac ttc ctg aaa act cac cgc 720
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
gac aaa gaa gaa atc gtt ttc cgc acc ctg tcc tat ttc gac ggc gtt 768
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
aac ttc gcg gct cgt gca aaa att ccg gaa ctg ttc tct gtt ggt ctg 816
Asn Phe Ala Ala Arg Ala Lys Ile Pro Glu Leu Phe Ser Val Gly Leu
260 265 270
atg gac aac atc agc cct cct tct acc gtt ttc gcg gca tat aac tat 864
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
tat gcg ggt ccg aaa gaa atc cgt atc tat ccg tac aac aac cac gaa 912
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
ggc ggt ggt agc ttt cag gct gtt gaa caa gtg aaa ttc ctg aag aaa 960
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
ctg ttt gag aag ggc taa 978
Leu Phe Glu Lys Gly
325
<210> 58
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 58
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Glu Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Glu Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 59
<211> 978
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<220>
<221> CDS
<222> (1)..(978)
<400> 59
atg gcg ttc ttc gac ctg cct ctg gaa gaa ctg aag aaa tac cgt cca 48
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
gag cgt tac gaa gag aag gac ttc gac gag tac tgg gag gaa act ctg 96
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Tyr Trp Glu Glu Thr Leu
20 25 30
gcg gag agc gaa aag ttt ccg ctg gac cca gtg ttc gag cgt atg gaa 144
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
tct cac ctg aaa acc gtg gag gca tat gac gtt act ttt tct ggt tac 192
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
cgt ggc cag cgt atc aaa ggc tgg ctg ctg gtt ccg aaa ctg gag gaa 240
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
gaa aaa ctg ccg tgc gta gtt cag tac atc ggt tac aac ggt ggc cgt 288
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
ggc ttt ccg cac gat tgg ctg ttc tgg ccg tct atg ggc tac att tgc 336
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
ttc gtc atg gat act cgt ggt cag ggt tcc ggc tgg ctg aaa ggc gat 384
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
act ccg gat tat ccg gag ggc ccg gta gac ccg cag tac cct ggc ttc 432
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
atg acg cgt ggt gtt ctg gat ccg cgt acc tat tac tat cgc cgc gtt 480
Met Thr Arg Gly Val Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
ttt acc gat gca gtt cgt gcc gta gag gcc gcg gct tct ttc cct cag 528
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
gtt gac cag gag cgt att gtt atc gct ggt ggc tcc cag ggt ggc ggc 576
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
atc gcc ctg gcg gta tct gcg ctg agc aag aaa gct aag gca ctg ctg 624
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
tgt gac gtc ccg ttc ctg tgt cac ttc cgt cgc gct gtt cag ctg gta 672
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
gat acc cat ccg tac gcg gag att act aac ttc ctg aaa act cac cgc 720
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
gac aaa gaa gaa atc gtt ttc cgc acc ctg tcc tat ttc gac ggc gtt 768
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
aac ttc gcg gct cgt gca aaa att ccg gta ctg ttc tct gtt ggt ctg 816
Asn Phe Ala Ala Arg Ala Lys Ile Pro Val Leu Phe Ser Val Gly Leu
260 265 270
atg gac aac atc agc cct cct tct acc gtt ttc gcg gca tat aac tat 864
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
tat gcg ggt ccg aaa gaa acc cgt atc tat ccg tac aac agc cac gaa 912
Tyr Ala Gly Pro Lys Glu Thr Arg Ile Tyr Pro Tyr Asn Ser His Glu
290 295 300
ggc ggt ggt agc ttt cag gct gtt gaa caa gtg aaa ttc ctg aag aaa 960
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
ctg ttt gag aag ggc taa 978
Leu Phe Glu Lys Gly
325
<210> 60
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 60
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Tyr Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Val Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Val Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Thr Arg Ile Tyr Pro Tyr Asn Ser His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 61
<211> 978
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<220>
<221> CDS
<222> (1)..(978)
<400> 61
atg gcg ttc ttc gac ctg cct ctg gaa gaa ctg aag aaa tac cgt cca 48
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
gag cgt tac gaa gag aag gac ttc gac gag ttc tgg gag gaa act ctg 96
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
gcg gag agc gaa aag ttt ccg ctg gac cca gtg ttc gag cgt atg gaa 144
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
tct cac ctg aaa acc gtg gag gca tat gac gtt act ttt tct ggt tac 192
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
cgt ggc cag cgt atc aaa ggc tgg ctg ctg gtt ccg aaa ctg gag gaa 240
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
gaa aaa ctg ccg tgc gta gtt cag tac atc ggt tac aac ggt ggc cgt 288
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
ggc ttt ccg cac gat tgg ctg ttc tgg ccg tct atg ggc tac att tgc 336
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
ttc gtc atg gat act cgt ggt cag ggt tcc ggc tgg ctg aaa ggc gat 384
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
act ccg gat tat ccg gag ggc ccg gta gac ccg cag tac cct ggc ttc 432
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
atg acg cgt ggt att ctg gat ccg cgt acc tat tac tat cgc cgc gtt 480
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
ttt acc gat gca gtt cgt gcc gta gag gcc gcg gct tct ttc cct cag 528
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
gtt gac cag gag cgt att gtt atc gct ggt ggc tcc cag ggt ggc ggc 576
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
atc gcc cag gcg gta tct gcg ctg agc aag aaa gct aag gca ctg ctg 624
Ile Ala Gln Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
tgt gac gtc ccg ttc ctg tgt cac ttc cgt cgc gct gtt cag ctg gta 672
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
gat acc cat ccg tac gcg gag att act aac ttc ctg aaa act cac cgc 720
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
gac aaa gaa gaa atc gtt ttc cgc acc ctg tcc tat ttc gac ggc gtt 768
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
aac ttc gcg gct cgt gca aaa att ccg gca ctg ttc tct gtt ggt ctg 816
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
atg gac aac atc agc cct cct tct acc gtt ttc gcg gca tat aac tat 864
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
tat gcg ggt ccg aaa gaa atc cgt atc tat ccg tac aac aac cac gaa 912
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
ggc ggt ggt agc ttt cag gct gtt gaa caa gtg aaa ttc ctg aag aaa 960
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
ctg ttt gag aag ggc taa 978
Leu Phe Glu Lys Gly
325
<210> 62
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 62
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Gln Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 63
<211> 978
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<220>
<221> CDS
<222> (1)..(978)
<400> 63
atg gcg ttc ttc gac ctg cct ctg gaa gaa ctg aag aaa tac cgt cca 48
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
gag cgt tac gaa gag aag gac ttc gac gag ttc tgg gag gaa act ctg 96
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
gcg gag agc gaa aag ttt ccg ctg gac cca gtg ttc gag cgt atg gaa 144
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
tct cac ctg aaa acc gtg gag gca tat gac gtt act ttt tct ggt tac 192
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
cgt ggc cag cgt atc aaa ggc tgg ctg ctg gtt ccg aaa ctg gag gaa 240
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
gaa aaa ctg ccg tgc gta gtt cag tac atc ggt tac aac ggt ggc cgt 288
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
ggc ttt ccg cac gat tgg ctg ttc tgg ccg tct atg ggc ttc att tgc 336
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Phe Ile Cys
100 105 110
ttc gtc atg gat act cgt ggt cag ggt tcc ggc tgg ctg aaa ggc gat 384
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
act ccg gat tat ccg gag ggc ccg gta gac ccg cag tac cct ggc ttc 432
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
atg acg cgt ggt att ctg gat ccg cgt acc tat tac tat cgc cgc gtt 480
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
ttt acc gat gca gtt cgt gcc gta gag gcc gcg gct tct ttc cct cag 528
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
gtt gac cag gag cgt att gtt atc gct ggt ggc tcc cag ggt ggc ggc 576
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
atc gcc ctg gcg gta tct gcg ctg agc aag aaa gct aag gca ctg ctg 624
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
tgt gac gtc ccg ttc ctg tgt cac ttc cgt cgc gct gtt cag ctg gta 672
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
gat acc cat ccg tac gcg gag att act aac ttc ctg aaa act cac cgc 720
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
gac aaa gaa gaa atc gtt ttc cgc acc ctg tcc tat ttc gac ggc gtt 768
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
aac ttc gcg gct cgt gca aaa att ccg gca ctg ttc tct gtt ggt ctg 816
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
atg gac aac atc agc cct cct tct acc gtt ttc gcg gca tat aac tat 864
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
tat gcg ggt ccg aaa gaa atc cgt atc tat ccg tac aac aac cac gaa 912
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
ggc ggt ggt agc ttt cag gct gtt gaa caa gtg aaa ttc ctg aag aaa 960
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
ctg ttt gag aag ggc taa 978
Leu Phe Glu Lys Gly
325
<210> 64
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic Construct
<400> 64
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Phe Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 65
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 65
Arg Val Pro Asn Lys Thr Val Thr Val Asp Gly Ala
1 5 10
<210> 66
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 66
Asp Arg His Lys Ser Lys Tyr Ser Ser Thr Lys Ser
1 5 10
<210> 67
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 67
Lys Asn Phe Pro Gln Gln Lys Glu Phe Pro Leu Ser
1 5 10
<210> 68
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 68
Gln Arg Asn Ser Pro Pro Ala Met Ser Arg Arg Asp
1 5 10
<210> 69
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 69
Thr Arg Lys Pro Asn Met Pro His Gly Gln Tyr Leu
1 5 10
<210> 70
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 70
Lys Pro Pro His Leu Ala Lys Leu Pro Phe Thr Thr
1 5 10
<210> 71
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 71
Asn Lys Arg Pro Pro Thr Ser His Arg Ile His Ala
1 5 10
<210> 72
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 72
Asn Leu Pro Arg Tyr Gln Pro Pro Cys Lys Pro Leu
1 5 10
<210> 73
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 73
Arg Pro Pro Trp Lys Lys Pro Ile Pro Pro Ser Glu
1 5 10
<210> 74
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 74
Arg Gln Arg Pro Lys Asp His Phe Phe Ser Arg Pro
1 5 10
<210> 75
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa = Thr or Pro
<220>
<221> MISC_FEATURE
<222> (12)..(12)
<223> Xaa = Thr or Pro
<400> 75
Ser Val Pro Asn Lys Xaa Val Thr Val Asp Gly Xaa
1 5 10
<210> 76
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 76
Thr Thr Lys Trp Arg His Arg Ala Pro Val Ser Pro
1 5 10
<210> 77
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 77
Trp Leu Gly Lys Asn Arg Ile Lys Pro Arg Ala Ser
1 5 10
<210> 78
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 78
Ser Asn Phe Lys Thr Pro Leu Pro Leu Thr Gln Ser
1 5 10
<210> 79
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 79
Ser Val Ser Val Gly Met Lys Pro Ser Pro Arg Pro
1 5 10
<210> 80
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 80
Asp Leu His Thr Val Tyr His
1 5
<210> 81
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 81
His Ile Lys Pro Pro Thr Arg
1 5
<210> 82
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 82
His Pro Val Trp Pro Ala Ile
1 5
<210> 83
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 83
Met Pro Leu Tyr Tyr Leu Gln
1 5
<210> 84
<211> 26
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 84
His Leu Thr Val Pro Trp Arg Gly Gly Gly Ser Ala Val Pro Phe Tyr
1 5 10 15
Ser His Ser Gln Ile Thr Leu Pro Asn His
20 25
<210> 85
<211> 41
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 85
Gly Pro His Asp Thr Ser Ser Gly Gly Val Arg Pro Asn Leu His His
1 5 10 15
Thr Ser Lys Lys Glu Lys Arg Glu Asn Arg Lys Val Pro Phe Tyr Ser
20 25 30
His Ser Val Thr Ser Arg Gly Asn Val
35 40
<210> 86
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 86
Lys His Pro Thr Tyr Arg Gln
1 5
<210> 87
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 87
His Pro Met Ser Ala Pro Arg
1 5
<210> 88
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 88
Met Pro Lys Tyr Tyr Leu Gln
1 5
<210> 89
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 89
Met His Ala His Ser Ile Ala
1 5
<210> 90
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 90
Ala Lys Pro Ile Ser Gln His Leu Gln Arg Gly Ser
1 5 10
<210> 91
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 91
Ala Pro Pro Thr Pro Ala Ala Ala Ser Ala Thr Thr
1 5 10
<210> 92
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 92
Asp Pro Thr Glu Gly Ala Arg Arg Thr Ile Met Thr
1 5 10
<210> 93
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 93
Leu Asp Thr Ser Phe Pro Pro Val Pro Phe His Ala
1 5 10
<210> 94
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 94
Leu Asp Thr Ser Phe His Gln Val Pro Phe His Gln
1 5 10
<210> 95
<211> 11
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 95
Leu Pro Arg Ile Ala Asn Thr Trp Ser Pro Ser
1 5 10
<210> 96
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 96
Arg Thr Asn Ala Ala Asp His Pro Ala Ala Val Thr
1 5 10
<210> 97
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 97
Ser Leu Asn Trp Val Thr Ile Pro Gly Pro Lys Ile
1 5 10
<210> 98
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 98
Thr Asp Met Gln Ala Pro Thr Lys Ser Tyr Ser Asn
1 5 10
<210> 99
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 99
Thr Ile Met Thr Lys Ser Pro Ser Leu Ser Cys Gly
1 5 10
<210> 100
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 100
Thr Pro Ala Leu Asp Gly Leu Arg Gln Pro Leu Arg
1 5 10
<210> 101
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 101
Thr Tyr Pro Ala Ser Arg Leu Pro Leu Leu Ala Pro
1 5 10
<210> 102
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 102
Ala Lys Thr His Lys His Pro Ala Pro Ser Tyr Ser
1 5 10
<210> 103
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 103
Thr Asp Pro Thr Pro Phe Ser Ile Ser Pro Glu Arg
1 5 10
<210> 104
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 104
Ser Gln Asn Trp Gln Asp Ser Thr Ser Tyr Ser Asn
1 5 10
<210> 105
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 105
Trp His Asp Lys Pro Gln Asn Ser Ser Lys Ser Thr
1 5 10
<210> 106
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 106
Leu Asp Val Glu Ser Tyr Lys Gly Thr Ser Met Pro
1 5 10
<210> 107
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 107
Asn Thr Pro Lys Glu Asn Trp
1 5
<210> 108
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 108
Asn Thr Pro Ala Ser Asn Arg
1 5
<210> 109
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 109
Pro Arg Gly Met Leu Ser Thr
1 5
<210> 110
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 110
Pro Pro Thr Tyr Leu Ser Thr
1 5
<210> 111
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 111
Thr Ile Pro Thr His Arg Gln His Asp Tyr Arg Ser
1 5 10
<210> 112
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 112
Thr Pro Pro Thr His Arg Leu
1 5
<210> 113
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 113
Leu Pro Thr Met Ser Thr Pro
1 5
<210> 114
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 114
Leu Gly Thr Asn Ser Thr Pro
1 5
<210> 115
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 115
Thr Pro Leu Thr Gly Ser Thr Asn Leu Leu Ser Ser
1 5 10
<210> 116
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 116
Thr Pro Leu Thr Lys Glu Thr
1 5
<210> 117
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 117
Lys Gln Ser His Asn Pro Pro
1 5
<210> 118
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 118
Gln Gln Ser His Asn Pro Pro
1 5
<210> 119
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 119
Thr Gln Pro His Asn Pro Pro
1 5
<210> 120
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 120
Ser Thr Asn Leu Leu Arg Thr Ser Thr Val His Pro
1 5 10
<210> 121
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 121
His Thr Gln Pro Ser Tyr Ser Ser Thr Asn Leu Phe
1 5 10
<210> 122
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 122
Ser Leu Leu Ser Ser His Ala
1 5
<210> 123
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 123
Gln Gln Ser Ser Ile Ser Leu Ser Ser His Ala Val
1 5 10
<210> 124
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 124
Asn Ala Ser Pro Ser Ser Leu
1 5
<210> 125
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 125
His Ser Pro Ser Ser Leu Arg
1 5
<210> 126
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> X= H, R or N
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> X= His, Arg or Asn
<400> 126
Lys Xaa Ser His His Thr His
1 5
<210> 127
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> X = H, R or N
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> X = His, Arg or Asn
<400> 127
Glu Xaa Ser His His Thr His
1 5
<210> 128
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 128
Ser His His Thr His Tyr Gly Gln Pro Gly Pro Val
1 5 10
<210> 129
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 129
Leu Glu Ser Thr Ser Leu Leu
1 5
<210> 130
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 130
Asp Leu Thr Leu Pro Phe His
1 5
<210> 131
<211> 8
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 131
Arg Thr Asn Ala Ala Asp His Pro
1 5
<210> 132
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 132
Ile Pro Trp Trp Asn Ile Arg Ala Pro Leu Asn Ala
1 5 10
<210> 133
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 133
Glu Gln Ile Ser Gly Ser Leu Val Ala Ala Pro Trp Glu Gly Glu Gly
1 5 10 15
Glu Arg
<210> 134
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 134
Thr Pro Pro Glu Leu Leu His Gly Ala Pro Arg Ser
1 5 10
<210> 135
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 135
Leu Asp Thr Ser Phe His Gln Val Pro Phe His Gln Lys Arg Lys Arg
1 5 10 15
Lys Asp
<210> 136
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 136
Glu Gln Ile Ser Gly Ser Leu Val Ala Ala Pro Trp Lys Arg Lys Arg
1 5 10 15
Lys Asp
<210> 137
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 137
Thr Pro Pro Glu Leu Leu His Gly Asp Pro Arg Ser Lys Arg Lys Arg
1 5 10 15
Lys Asp
<210> 138
<211> 13
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 138
Asn Thr Ser Gln Leu Ser Thr Glu Gly Glu Gly Glu Asp
1 5 10
<210> 139
<211> 13
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 139
Thr Pro Pro Glu Leu Leu His Gly Asp Pro Arg Ser Cys
1 5 10
<210> 140
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 140
His Ile Asn Lys Thr Asn Pro His Gln Gly Asn His His Ser Glu Lys
1 5 10 15
Thr Gln Arg Gln
20
<210> 141
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 141
His Ala His Lys Asn Gln Lys Glu Thr His Gln Arg His Ala Ala
1 5 10 15
<210> 142
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 142
His Glu His Lys Asn Gln Lys Glu Thr His Gln Arg His Ala Ala
1 5 10 15
<210> 143
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 143
His Asn His Met Gln Glu Arg Tyr Thr Glu Pro Gln His Ser Pro Ser
1 5 10 15
Val Asn Gly Leu
20
<210> 144
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 144
Thr His Ser Thr His Asn His Gly Ser Pro Arg His Thr Asn Ala Asp
1 5 10 15
Ala
<210> 145
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 145
Gly Ser Cys Val Asp Thr His Lys Ala Asp Ser Cys Val Ala Asn Asn
1 5 10 15
Gly Pro Ala Thr
20
<210> 146
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 146
Ala Gln Ser Gln Leu Pro Asp Lys His Ser Gly Leu His Glu Arg Ala
1 5 10 15
Pro Gln Arg Tyr
20
<210> 147
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 147
Ala Gln Ser Gln Leu Pro Ala Lys His Ser Gly Leu His Glu Arg Ala
1 5 10 15
Pro Gln Arg Tyr
20
<210> 148
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 148
Ala Gln Ser Gln Leu Pro Glu Lys His Ser Gly Leu His Glu Arg Ala
1 5 10 15
Pro Gln Arg Tyr
20
<210> 149
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 149
Thr Asp Met Met His Asn His Ser Asp Asn Ser Pro Pro His Arg Arg
1 5 10 15
Ser Pro Arg Asn
20
<210> 150
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 150
Thr Pro Pro Glu Leu Ala His Thr Pro His His Leu Ala Gln Thr Arg
1 5 10 15
Leu Thr Asp Arg
20
<210> 151
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 151
Arg Leu Leu Arg Leu Leu Arg Leu Leu Arg Leu Leu
1 5 10
<210> 152
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 152
Thr Pro Pro Glu Leu Leu His Gly Glu Pro Arg Ser
1 5 10
<210> 153
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 153
Thr Pro Pro Glu Leu Leu His Gly Ala Pro Arg Ser
1 5 10
<210> 154
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 154
Glu Gln Ile Ser Gly Ser Leu Val Ala Ala Pro Trp
1 5 10
<210> 155
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 155
Asn Glu Val Pro Ala Arg Asn Ala Pro Trp Leu Val
1 5 10
<210> 156
<211> 13
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 156
Asn Ser Pro Gly Tyr Gln Ala Asp Ser Val Ala Ile Gly
1 5 10
<210> 157
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 157
Ala Lys Pro Ile Ser Gln His Leu Gln Arg Gly Ser
1 5 10
<210> 158
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 158
Leu Asp Thr Ser Phe Pro Pro Val Pro Phe His Ala
1 5 10
<210> 159
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 159
Ser Leu Asn Trp Val Thr Ile Pro Gly Pro Lys Ile
1 5 10
<210> 160
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 160
Thr Gln Asp Ser Ala Gln Lys Ser Pro Ser Pro Leu
1 5 10
<210> 161
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 161
Lys Glu Leu Gln Thr Arg Asn Val Val Gln Arg Glu
1 5 10
<210> 162
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 162
Gln Arg Asn Ser Pro Pro Ala Met Ser Arg Arg Asp
1 5 10
<210> 163
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 163
Thr Pro Thr Ala Asn Gln Phe Thr Gln Ser Val Pro
1 5 10
<210> 164
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 164
Ala Ala Gly Leu Ser Gln Lys His Glu Arg Asn Arg
1 5 10
<210> 165
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 165
Glu Thr Val His Gln Thr Pro Leu Ser Asp Arg Pro
1 5 10
<210> 166
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 166
Lys Asn Phe Pro Gln Gln Lys Glu Phe Pro Leu Ser
1 5 10
<210> 167
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 167
Leu Pro Ala Leu His Ile Gln Arg His Pro Arg Met
1 5 10
<210> 168
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 168
Gln Pro Ser His Ser Gln Ser His Asn Leu Arg Ser
1 5 10
<210> 169
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 169
Arg Gly Ser Gln Lys Ser Lys Pro Pro Arg Pro Pro
1 5 10
<210> 170
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 170
Thr His Thr Gln Lys Thr Pro Leu Leu Tyr Tyr His
1 5 10
<210> 171
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 171
Thr Lys Gly Ser Ser Gln Ala Ile Leu Lys Ser Thr
1 5 10
<210> 172
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 172
Thr Ala Ala Thr Thr Ser Pro
1 5
<210> 173
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 173
Leu Gly Ile Pro Gln Asn Leu
1 5
<210> 174
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 174
Thr His Ser Thr His Asn His Gly Ser Pro Arg His Thr Asn Ala Asp
1 5 10 15
Ala Gly Asn Pro
20
<210> 175
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 175
Gln Gln His Lys Val His His Gln Asn Pro Asp Arg Ser Thr Gln Asp
1 5 10 15
Ala His His Ser
20
<210> 176
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 176
His His Gly Thr His His Asn Ala Thr Lys Gln Lys Asn His Val
1 5 10 15
<210> 177
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 177
Ser Thr Leu His Lys Tyr Lys Ser Gln Asp Pro Thr Pro His His
1 5 10 15
<210> 178
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 178
Ser Val Ser Val Gly Met Lys Pro Ser Pro Arg Pro
1 5 10
<210> 179
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 179
Thr Pro Pro Thr Asn Val Leu Met Leu Ala Thr Lys
1 5 10
<210> 180
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 180
Thr Pro Pro Glu Leu Leu His Gly Asp Pro Arg Ser
1 5 10
<210> 181
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 181
Asn Thr Ser Gln Leu Ser Thr
1 5
<210> 182
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 182
Ser Thr Leu His Lys Tyr Lys Ser Gln Asp Pro Thr Pro His His
1 5 10 15
<210> 183
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 183
Gly Met Pro Ala Met His Trp Ile His Pro Phe Ala
1 5 10
<210> 184
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 184
His Asp His Lys Asn Gln Lys Glu Thr His Gln Arg His Ala Ala
1 5 10 15
<210> 185
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 185
His Asn His Met Gln Glu Arg Tyr Thr Asp Pro Gln His Ser Pro Ser
1 5 10 15
Val Asn Gly Leu
20
<210> 186
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic hair-binding peptide
<400> 186
Thr Ala Glu Ile Gln Ser Ser Lys Asn Pro Asn Pro His Pro Gln Arg
1 5 10 15
Ser Trp Thr Asn
20
<210> 187
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 187
Ser Ser Ala Asp Phe Ala Ser Phe Gly Phe Phe Gly Phe Ser Ala Ala
1 5 10 15
Ser Ala Asp Ser Arg
20
<210> 188
<211> 23
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 188
Ser Ser Phe Ala Glu Ala Trp Ser Arg Ala Trp Pro Arg Ala Glu Val
1 5 10 15
Phe Phe Pro Ser Arg Gly Tyr
20
<210> 189
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 189
Ser Ser Phe Ser Val Asn Glu Pro His Ala Trp Met Ala Pro Leu Ser
1 5 10 15
Arg
<210> 190
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 190
Ser Ser Phe Ser Trp Val Tyr Gly His Gly Gly Leu Gly Phe Ala Ser
1 5 10 15
Arg
<210> 191
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 191
Ser Ser Phe Val Ser Trp Ser Pro Tyr Lys Ser Pro Pro Glu Leu Ser
1 5 10 15
Arg
<210> 192
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 192
Ser Ser Phe Tyr Gly Ser Ser Ala Phe Val Ser Ser Gly Val Ser Val
1 5 10 15
Ala Tyr Gly Ser Arg
20
<210> 193
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 193
Ser Ser Gly Ser Val Ala Val Ser Ala Glu Ala Ser Trp Phe Ser Gly
1 5 10 15
Val Ala Ala Ser Arg
20
<210> 194
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 194
Ser Ser His Asp Glu His Tyr Gln Tyr His Tyr Tyr Ser Ser Arg
1 5 10 15
<210> 195
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 195
Ser Ser His Tyr Tyr Tyr Asn Asp Tyr Asp His Gln Ser Ser Arg
1 5 10 15
<210> 196
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 196
Ser Ser Leu Phe Asn Met Tyr Gly His Gln Ser Val Leu Gly Pro Ser
1 5 10 15
Arg
<210> 197
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 197
Ser Ser Leu Phe Ser Asp Val His Tyr Gly Ser Asn Lys Ala Leu Ser
1 5 10 15
Arg
<210> 198
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 198
Ser Ser Leu Leu Ser Asp Phe His Tyr Gly Asp Met Trp Asp Ala Ser
1 5 10 15
Arg
<210> 199
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 199
Ser Ser Asn Tyr Asn Tyr Asn Tyr Asn Tyr Gln Tyr Ser Ser Arg
1 5 10 15
<210> 200
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> synethetic construct
<400> 200
Ser Ser Asn Tyr Asn Tyr Asn Tyr Asn Tyr Gln Tyr Ser Ser Arg Glu
1 5 10 15
Gly Glu Gly Glu Arg
20
<210> 201
<211> 21
<212> PRT
<213> ARTIFICIAL SEQUENCE
<220>
<223> Synthetic construct
<400> 201
Ser Ser Asn Tyr Asn Tyr Asn Tyr Asn Tyr Gln Tyr Ser Ser Arg Lys
1 5 10 15
Arg Lys Arg Lys Asp
20
<210> 202
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 202
Ser Ser Gln Tyr Tyr Gln Asp Tyr Gln Tyr Tyr His Ser Ser Arg
1 5 10 15
<210> 203
<211> 23
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 203
Ser Ser Ser Cys Met Gly Ser His Asn Pro Arg Met Ser Val Glu Glu
1 5 10 15
Ser Thr Arg Asn Cys Ser Arg
20
<210> 204
<211> 23
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 204
Ser Ser Ser Cys Asn Asn Asn Trp Phe Tyr Ser Ser Thr Leu Pro Gly
1 5 10 15
Gly Asp His Ala Cys Ser Arg
20
<210> 205
<211> 23
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 205
Ser Ser Ser Cys Tyr Asp Val Glu Cys Ser Ser Phe Val Ala Trp Met
1 5 10 15
Arg Gly Pro Ser Ser Ser Arg
20
<210> 206
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 206
Ser Ser Ser Phe Ala Ala Ser Ser Ala Phe Ser Phe Leu Val Asp Ala
1 5 10 15
Val Ala Trp Ser Arg
20
<210> 207
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 207
Ser Ser Ser Phe Ala Tyr Leu Val Pro Asp Asp Gly Trp Leu Ser Ser
1 5 10 15
Arg
<210> 208
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 208
Ser Ser Ser Gly Ala Val Phe Ser Ser Gly Gly Ala Asp Ala Gly Trp
1 5 10 15
Gly Val Trp Ser Arg
20
<210> 209
<211> 23
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 209
Ser Ser Ser Ser Ala Asp Ala Ala Tyr Gly His Cys Cys Gly Ala Gly
1 5 10 15
Phe Ser Thr Phe Ser Ser Arg
20
<210> 210
<211> 23
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 210
Ser Ser Ser Ser Asp Val His Asn Ser Ile Ile Gly Trp Asp Phe Tyr
1 5 10 15
His Ser Arg Gly Ser Ser Arg
20
<210> 211
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 211
Ser Ser Ser Ser Leu Asp Phe Phe Ser Tyr Ser Ala Phe Ser Gly Gly
1 5 10 15
Val Ala Glu Ser Arg
20
<210> 212
<211> 23
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 212
Ser Ser Ser Ser Asn Asp Ser Asn Val Ser Trp Phe His Tyr Tyr Ala
1 5 10 15
Ser Gly Leu Thr Ser Ser Arg
20
<210> 213
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 213
Ser Ser Val Asp Tyr Glu Val Pro Leu Ala Val Ala Ala Glu Trp Gly
1 5 10 15
Phe Ser Val Ser Arg
20
<210> 214
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 214
Ser Ser Tyr His Tyr Asp Tyr Asp His Tyr Tyr Glu Ser Ser Arg
1 5 10 15
<210> 215
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 215
Ser Ser Tyr Tyr Asn Tyr His Tyr Gln Tyr Gln Asp Ser Ser Arg
1 5 10 15
<210> 216
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Dyed-hair-bindg peptide
<400> 216
Ser Ser Tyr Tyr Tyr Asp Tyr Tyr Gln Gln Asp Tyr Ser Ser Arg
1 5 10 15
<210> 217
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 217
Lys Arg Gly Arg His Lys Arg Pro Lys Arg His Lys
1 5 10
<210> 218
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 218
Arg Leu Leu Arg Leu Leu Arg
1 5
<210> 219
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 219
His Lys Pro Arg Gly Gly Arg Lys Lys Ala Leu His
1 5 10
<210> 220
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 220
Lys Pro Arg Pro Pro His Gly Lys Lys His Arg Pro Lys His Arg Pro
1 5 10 15
Lys Lys
<210> 221
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 221
Arg Gly Arg Pro Lys Lys Gly His Gly Lys Arg Pro Gly His Arg Ala
1 5 10 15
Arg Lys
<210> 222
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 222
Thr Pro Phe His Ser Pro Glu Asn Ala Pro Gly Ser
1 5 10
<210> 223
<211> 13
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 223
Thr Pro Phe His Ser Pro Glu Asn Ala Pro Gly Ser Lys
1 5 10
<210> 224
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 224
Thr Pro Phe His Ser Pro Glu Asn Ala Pro Gly Ser Gly Gly Gly Ser
1 5 10 15
<210> 225
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 225
Thr Pro Phe His Ser Pro Glu Asn Ala Pro Gly Ser Gly Gly Gly Ser
1 5 10 15
Ser
<210> 226
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 226
Thr Pro Phe His Ser Pro Glu Asn Ala Pro Gly Ser Gly Gly Gly
1 5 10 15
<210> 227
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 227
Phe Thr Gln Ser Leu Pro Arg
1 5
<210> 228
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 228
Lys Gln Ala Thr Phe Pro Pro Asn Pro Thr Ala Tyr
1 5 10
<210> 229
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 229
His Gly His Met Val Ser Thr Ser Gln Leu Ser Ile
1 5 10
<210> 230
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 230
Leu Ser Pro Ser Arg Met Lys
1 5
<210> 231
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 231
Leu Pro Ile Pro Arg Met Lys
1 5
<210> 232
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 232
His Gln Arg Pro Tyr Leu Thr
1 5
<210> 233
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 233
Phe Pro Pro Leu Leu Arg Leu
1 5
<210> 234
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 234
Gln Ala Thr Phe Met Tyr Asn
1 5
<210> 235
<211> 11
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 235
Val Leu Thr Ser Gln Leu Pro Asn His Ser Met
1 5 10
<210> 236
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 236
His Ser Thr Ala Tyr Leu Thr
1 5
<210> 237
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 237
Ala Pro Gln Gln Arg Pro Met Lys Thr Phe Asn Thr
1 5 10
<210> 238
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 238
Ala Pro Gln Gln Arg Pro Met Lys Thr Val Gln Tyr
1 5 10
<210> 239
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 239
Pro Pro Trp Leu Asp Leu Leu
1 5
<210> 240
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 240
Pro Pro Trp Thr Phe Pro Leu
1 5
<210> 241
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 241
Ser Val Thr His Leu Thr Ser
1 5
<210> 242
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 242
Val Ile Thr Arg Leu Thr Ser
1 5
<210> 243
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 243
Asp Leu Lys Pro Pro Leu Leu Ala Leu Ser Lys Val
1 5 10
<210> 244
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 244
Ser His Pro Ser Gly Ala Leu Gln Glu Gly Thr Phe
1 5 10
<210> 245
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 245
Phe Pro Leu Thr Ser Lys Pro Ser Gly Ala Cys Thr
1 5 10
<210> 246
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 246
Asp Leu Lys Pro Pro Leu Leu Ala Leu Ser Lys Val
1 5 10
<210> 247
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 247
Pro Leu Leu Ala Leu His Ser
1 5
<210> 248
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 248
Val Pro Ile Ser Thr Gln Ile
1 5
<210> 249
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 249
Tyr Ala Lys Gln His Tyr Pro Ile Ser Thr Phe Lys
1 5 10
<210> 250
<211> 7
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 250
His Ser Thr Ala Tyr Leu Thr
1 5
<210> 251
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 251
Ser Thr Ala Tyr Leu Val Ala Met Ser Ala Ala Pro
1 5 10
<210> 252
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 252
Ser Val Ser Val Gly Met Lys Pro Ser Pro Arg Pro
1 5 10
<210> 253
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 253
Thr Met Gly Phe Thr Ala Pro Arg Phe Pro His Tyr
1 5 10
<210> 254
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 254
Asn Leu Gln His Ser Val Gly Thr Ser Pro Val Trp
1 5 10
<210> 255
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 255
Gln Leu Ser Tyr His Ala Tyr Pro Gln Ala Asn His His Ala Pro
1 5 10 15
<210> 256
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 256
Asn Gln Ala Ala Ser Ile Thr Lys Arg Val Pro Tyr
1 5 10
<210> 257
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 257
Ser Gly Cys His Leu Val Tyr Asp Asn Gly Phe Cys Asp His
1 5 10
<210> 258
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 258
Ala Ser Cys Pro Ser Ala Ser His Ala Asp Pro Cys Ala His
1 5 10
<210> 259
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 259
Asn Leu Cys Asp Ser Ala Arg Asp Ser Pro Arg Cys Lys Val
1 5 10
<210> 260
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 260
Asn His Ser Asn Trp Lys Thr Ala Ala Asp Phe Leu
1 5 10
<210> 261
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 261
Gly Ser Ser Thr Val Gly Arg Pro Leu Ser Tyr Glu
1 5 10
<210> 262
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 262
Ser Asp Thr Ile Ser Arg Leu His Val Ser Met Thr
1 5 10
<210> 263
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 263
Ser Pro Leu Thr Val Pro Tyr Glu Arg Lys Leu Leu
1 5 10
<210> 264
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 264
Ser Pro Tyr Pro Ser Trp Ser Thr Pro Ala Gly Arg
1 5 10
<210> 265
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 265
Val Gln Pro Ile Thr Asn Thr Arg Tyr Glu Gly Gly
1 5 10
<210> 266
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 266
Trp Pro Met His Pro Glu Lys Gly Ser Arg Trp Ser
1 5 10
<210> 267
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 267
Asp Ala Cys Ser Gly Asn Gly His Pro Asn Asn Cys Asp Arg
1 5 10
<210> 268
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 268
Asp His Cys Leu Gly Arg Gln Leu Gln Pro Val Cys Tyr Pro
1 5 10
<210> 269
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 269
Asp Trp Cys Asp Thr Ile Ile Pro Gly Arg Thr Cys His Gly
1 5 10
<210> 270
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 270
Ala Leu Pro Arg Ile Ala Asn Thr Trp Ser Pro Ser
1 5 10
<210> 271
<211> 12
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 271
Tyr Pro Ser Phe Ser Pro Thr Tyr Arg Pro Ala Phe
1 5 10
<210> 272
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 272
Ala His Pro Glu Ser Leu Gly Ile Lys Tyr Ala Leu Asp Gly Asn Ser
1 5 10 15
Asp Pro His Ala
20
<210> 273
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 273
Ala Ser Val Ser Asn Tyr Pro Pro Ile His His Leu Ala Thr Ser Asn
1 5 10 15
Thr Thr Val Asn
20
<210> 274
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 274
Asp Glu Cys Met Glu Pro Leu Asn Ala Ala His Cys Trp Arg
1 5 10
<210> 275
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 275
Asp Glu Cys Met His Gly Ser Asp Val Glu Phe Cys Thr Ser
1 5 10
<210> 276
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 276
Asp Leu Cys Ser Met Gln Met Met Asn Thr Gly Cys His Tyr
1 5 10
<210> 277
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 277
Asp Leu Cys Ser Ser Pro Ser Thr Trp Gly Ser Cys Ile Arg
1 5 10
<210> 278
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 278
Asp Pro Asn Glu Ser Asn Tyr Glu Asn Ala Thr Thr Val Ser Gln Pro
1 5 10 15
Thr Arg His Leu
20
<210> 279
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 279
Glu Pro Thr His Pro Thr Met Arg Ala Gln Met His Gln Ser Leu Arg
1 5 10 15
Ser Ser Ser Pro
20
<210> 280
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 280
Gly Asn Thr Asp Thr Thr Pro Pro Asn Ala Val Met Glu Pro Thr Val
1 5 10 15
Gln His Lys Trp
20
<210> 281
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 281
Asn Gly Pro Asp Met Val Gln Ser Val Gly Lys His Lys Asn Ser
1 5 10 15
<210> 282
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 282
Asn Gly Pro Glu Val Arg Gln Ile Pro Ala Asn Phe Glu Lys Leu
1 5 10 15
<210> 283
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 283
Asn Asn Thr Ser Ala Asp Asn Pro Pro Glu Thr Asp Ser Lys His His
1 5 10 15
Leu Ser Met Ser
20
<210> 284
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 284
Asn Asn Thr Trp Pro Glu Gly Ala Gly His Thr Met Pro Ser Thr Asn
1 5 10 15
Ile Arg Gln Ala
20
<210> 285
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 285
Asn Pro Thr Ala Thr Pro His Met Lys Asp Pro Met His Ser Asn Ala
1 5 10 15
His Ser Ser Ala
20
<210> 286
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 286
Asn Pro Thr Asp His Ile Pro Ala Asn Ser Thr Asn Ser Arg Val Ser
1 5 10 15
Lys Gly Asn Thr
20
<210> 287
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 287
Asn Pro Thr Asp Ser Thr His Met Met His Ala Arg Asn His Glu
1 5 10 15
<210> 288
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 288
Gln His Cys Ile Thr Glu Arg Leu His Pro Pro Cys Thr Lys
1 5 10
<210> 289
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 289
Thr Pro Cys Ala Pro Ala Ser Phe Asn Pro His Cys Ser Arg
1 5 10
<210> 290
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 290
Thr Pro Cys Ala Thr Tyr Pro His Phe Ser Gly Cys Arg Ala
1 5 10
<210> 291
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 291
Trp Cys Thr Asp Phe Cys Thr Arg Ser Thr Pro Thr Ser Thr Ser Arg
1 5 10 15
Ser Thr Thr Ser
20
<210> 292
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 292
Ala Pro Pro Leu Lys Thr Tyr Met Gln Glu Arg Glu Leu Thr Met Ser
1 5 10 15
Gln Asn Lys Asp
20
<210> 293
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 293
Glu Pro Pro Thr Arg Thr Arg Val Asn Asn His Thr Val Thr Val Gln
1 5 10 15
Ala Gln Gln His
20
<210> 294
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 294
Gly Tyr Cys Leu Arg Gly Asp Glu Pro Ala Val Cys Ser Gly
1 5 10
<210> 295
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 295
Leu Ser Ser Lys Asp Phe Gly Val Thr Asn Thr Asp Gln Arg Thr Tyr
1 5 10 15
Asp Tyr Thr Thr
20
<210> 296
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 296
Asn Phe Cys Glu Thr Gln Leu Asp Leu Ser Val Cys Thr Val
1 5 10
<210> 297
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 297
Asn Thr Cys Gln Pro Thr Lys Asn Ala Thr Pro Cys Ser Ala
1 5 10
<210> 298
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 298
Pro Ser Glu Pro Glu Arg Arg Asp Arg Asn Ile Ala Ala Asn Ala Gly
1 5 10 15
Arg Phe Asn Thr
20
<210> 299
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 299
Thr His Asn Met Ser His Phe Pro Pro Ser Gly His Pro Lys Arg Thr
1 5 10 15
Ala Thr
<210> 300
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 300
Thr Thr Cys Pro Thr Met Gly Thr Tyr His Val Cys Trp Leu
1 5 10
<210> 301
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 301
Tyr Cys Ala Asp His Thr Pro Asp Pro Ala Asn Pro Asn Lys Ile Cys
1 5 10 15
Gly Tyr Ser His
20
<210> 302
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 302
Ala Ala Asn Pro His Thr Glu Trp Asp Arg Asp Ala Phe Gln Leu Ala
1 5 10 15
Met Pro Pro Lys
20
<210> 303
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 303
Asp Leu His Pro Met Asp Pro Ser Asn Lys Arg Pro Asp Asn Pro Ser
1 5 10 15
Asp Leu His Thr
20
<210> 304
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 304
Glu Ser Cys Val Ser Asn Ala Leu Met Asn Gln Cys Ile Tyr
1 5 10
<210> 305
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 305
His Asn Lys Ala Asp Ser Trp Asp Pro Asp Leu Pro Pro His Ala Gly
1 5 10 15
Met Ser Leu Gly
20
<210> 306
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 306
Leu Asn Asp Gln Arg Lys Pro Gly Pro Pro Thr Met Pro Thr His Ser
1 5 10 15
Pro Ala Val Gly
20
<210> 307
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 307
Asn Thr Cys Ala Thr Ser Pro Asn Ser Tyr Thr Cys Ser Asn
1 5 10
<210> 308
<211> 14
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 308
Ser Asp Cys Thr Ala Gly Leu Val Pro Pro Leu Cys Ala Thr
1 5 10
<210> 309
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 309
Thr Ile Glu Ser Ser Gln His Ser Arg Thr His Gln Gln Asn Tyr Gly
1 5 10 15
Ser Thr Lys Thr
20
<210> 310
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 310
Val Gly Thr Met Lys Gln His Pro Thr Thr Thr Gln Pro Pro Arg Val
1 5 10 15
Ser Ala Thr Asn
20
<210> 311
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> Synthetic construct
<400> 311
Tyr Ser Glu Thr Pro Asn Asp Gln Lys Pro Asn Pro His Tyr Lys Val
1 5 10 15
Ser Gly Thr Lys
20
<210> 312
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 312
Asn Gly Asn Asn His Thr Asp Ile Pro Asn Arg Ser Ser Tyr Thr Gly
1 5 10 15
Gly Ser Phe Ala
20
<210> 313
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 313
Thr Met Thr Asn His Val Tyr Asn Ser Tyr Thr Glu Lys His Ser Ser
1 5 10 15
Thr His Arg Ser
20
<210> 314
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 314
Thr Thr Tyr His Tyr Lys Asn Ile Tyr Gln Glu Ser Tyr Gln Gln Arg
1 5 10 15
Asn Pro Ala Val
20
<210> 315
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 315
Val Glu Pro Ala Thr Lys Asn Met Arg Glu Ala Arg Ser Ser Thr Gln
1 5 10 15
Met Arg Arg Ile
20
<210> 316
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 316
Tyr Leu Leu Pro Lys Asp Gln Thr Thr Ala Pro Gln Val Thr Pro Ile
1 5 10 15
Val Gln His Lys
20
<210> 317
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 317
Ala Ser Asn Leu Asp Ser Thr Phe Thr Ala Ile Asn Thr Pro Ala Cys
1 5 10 15
Cys Thr
<210> 318
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 318
Glu Phe Pro Tyr Tyr Asn Asp Asn Pro Pro Asn Pro Glu Arg His Thr
1 5 10 15
Leu Arg
<210> 319
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 319
Gly Met Pro Thr Arg Tyr Tyr His Asn Thr Pro Pro His Leu Thr Pro
1 5 10 15
Lys Phe
<210> 320
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 320
His Lys Asn Ala Ile Gln Pro Val Asn Asp Ala Thr Thr Leu Asp Thr
1 5 10 15
Thr Met
<210> 321
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 321
Ala Val Val Pro Ala Asp Leu Asn Asp His Ala Asn His Leu Ser
1 5 10 15
<210> 322
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 322
Asp Leu Gly Thr Phe Pro Asn Arg Thr Leu Lys Met Ala Ala His
1 5 10 15
<210> 323
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 323
Phe Asp Gly Ile Gly Leu Gly Thr Ala Thr Arg His Gln Asn Arg
1 5 10 15
<210> 324
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 324
Gln Ala Ala Gln Val His Met Met Gln His Ser Arg Pro Thr Thr
1 5 10 15
<210> 325
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 325
Ser Glu Ala Arg Ala Arg Thr Phe Asn Asp His Thr Thr Pro Met Pro
1 5 10 15
Ile Ile
<210> 326
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 326
Glu Leu Asp His Asp Ser Arg His Tyr Met Asn Gly Leu Gln Arg Lys
1 5 10 15
Val Thr
<210> 327
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 327
Gly Pro Gln His Val Leu Met Gln Asp Thr His Gln Gly Tyr Ala Phe
1 5 10 15
Asp Asn
<210> 328
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 328
Thr Thr Gly Ser Ser Ser Gln Ala Asp Thr Ser Ala Ser Met Ser Ile
1 5 10 15
Val Pro Ala His
20
<210> 329
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 329
Lys Ala Pro Ile Ala Asn Met Leu Gln Pro His Ser Tyr Gln Tyr Ser
1 5 10 15
Val Ala
<210> 330
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 330
Thr Tyr Gln Gly Val Pro Ser Trp Pro Ala Val Ile Asp Asp Ala Ile
1 5 10 15
Arg Arg
<210> 331
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 331
Val Asn Pro Asn Trp Val Glu Thr Gln Ala Leu His Gln Pro Pro Gly
1 5 10 15
Asn Thr
<210> 332
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 332
Asp His Asn Asn Arg Gln His Ala Val Glu Val Arg Glu Asn Lys Thr
1 5 10 15
His Thr Ala Arg
20
<210> 333
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 333
Ile Tyr Pro Asn Glu Ser Met Ser Thr Ser Asn Val Arg Gly Pro Tyr
1 5 10 15
His Pro
<210> 334
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 334
His Asp Pro Asn His Leu Thr His Gln Ala Arg Thr Ile Tyr Arg Asn
1 5 10 15
Ala Asn His Thr
20
<210> 335
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 335
Ser Asn Ala Thr Met Tyr Asn Ile Gln Ser His Ser His His Gln
1 5 10 15
<210> 336
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 336
Ala Asn Glu Leu Ser Thr Tyr Ala Gln Thr Asn Pro Gly Ser Gly
1 5 10 15
<210> 337
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 337
Asp Thr Ile His Pro Asn Lys Met Lys Ser Pro Ser Ser Pro Leu
1 5 10 15
<210> 338
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 338
Ala Pro Pro Thr Tyr Gln Thr Ala Ser Tyr Pro His Asn Leu Pro Ser
1 5 10 15
Lys Arg Lys Met
20
<210> 339
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 339
Gln Val Pro Asp Tyr Leu Ser Pro Thr His Gln Lys Lys Ala Phe Leu
1 5 10 15
Glu Ile Pro Thr
20
<210> 340
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 340
Thr Asn Asp Leu His Ala Asn Pro Phe Thr Gly Thr Tyr Ile Ala Pro
1 5 10 15
Asp Pro Thr Ser
20
<210> 341
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 341
His Lys Asn Glu Asn Ile Met Gln Tyr Asn Val Asn Asp Arg Trp His
1 5 10 15
Ile Thr Pro Ala
20
<210> 342
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 342
Ile Asp Gly Pro His His Ser Pro Val His Arg Tyr His Thr Pro Ser
1 5 10 15
Ile Thr
<210> 343
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 343
Ala Ile Glu Tyr Gln His Ser Ala Thr Thr Pro Trp Thr Met Arg Thr
1 5 10 15
Arg Leu Pro Pro
20
<210> 344
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 344
Glu Phe Tyr Pro Phe Ala Glu Val Pro Pro Glu Lys Ser Gly Ile Gly
1 5 10 15
Arg Gln Val Phe
20
<210> 345
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 345
Gly Val His Gln Tyr Ser Arg Pro Thr Val Pro Ser Tyr Leu Trp Thr
1 5 10 15
Ser Gly Gln His
20
<210> 346
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 346
Gly Tyr Gln Pro His Tyr Val Asp His Thr Ile Gly Trp Gln Pro Met
1 5 10 15
Ile Arg Pro Asn
20
<210> 347
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 347
Gln Phe Asn Gln Thr Ser His Ser Phe Met His Gly Thr Ser Gly Tyr
1 5 10 15
Val Pro Gly Lys
20
<210> 348
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 348
Ser Phe Ser Trp His Arg Gly Asp Trp Glu Leu Gly His Gln Ser Lys
1 5 10 15
Thr Met Gly Met
20
<210> 349
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 349
Ser Met Trp His Asp Ile Thr Lys Arg Tyr Arg Asn Pro Ser Glu Met
1 5 10 15
Val Ser Ala Tyr
20
<210> 350
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 350
Thr His Gly Asn Lys His Gln Ser Trp Thr Tyr Pro Ser Glu Ile Asn
1 5 10 15
His Lys Asn Tyr
20
<210> 351
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 351
Trp His Glu Pro His Gln Phe Ser Gly Glu Asn Thr Asp Tyr Ser Ser
1 5 10 15
Ser Met Gly Thr
20
<210> 352
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 352
Thr His Gly Asn Lys His Gln Ser Trp Thr Tyr Pro Ser Glu Ile Asn
1 5 10 15
His Lys Asn Tyr
20
<210> 353
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 353
Asp Gly Tyr Lys Leu Gln Thr Ser Leu Asp Trp Gln Met Trp Asn Pro
1 5 10 15
<210> 354
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 354
Phe Pro Ser Lys Trp Tyr Asn His His Arg His Ile Thr Gly His Val
1 5 10 15
<210> 355
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 355
Gly Gly Met Gly Ala Leu Glu Ser Tyr Arg Gln Trp Asn His Leu Ala
1 5 10 15
<210> 356
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 356
Gly Ile Asn Lys Gly Gln Arg Pro Pro Trp Glu Ser Trp His Glu Asn
1 5 10 15
<210> 357
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 357
Gly Tyr Gly Gln Tyr Val Ser Gln Gln Thr Trp Ala His Ser Asn Lys
1 5 10 15
<210> 358
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 358
His Asp His Leu Ser Trp Trp Gly Gln Phe Asp Arg Gln Asn Leu Leu
1 5 10 15
<210> 359
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 359
Met Pro Gly His Gln Glu Ser Ile Lys Val Gln Asn Trp Asn Arg Val
1 5 10 15
<210> 360
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 360
Asn Leu His Ser Pro Trp Pro Ser His Ala Ala His His Trp Ser Thr
1 5 10 15
<210> 361
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 361
Asn Gln Gln Met Lys Leu Val Pro Gln His Trp His Arg Ala Gln Pro
1 5 10 15
<210> 362
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic oral-surface binding peptide
<400> 362
Ser Glu Lys Trp Phe Asn Pro Gly Pro Trp Pro Lys Leu Ala Thr Gln
1 5 10 15
<210> 363
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 363
Ser Ser Arg Pro Asn Gly Asn Asn His Thr Asp Ile Pro Asn Arg Ser
1 5 10 15
Ser Tyr Thr Gly Gly Ser Phe Ala Lys
20 25
<210> 364
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 364
Ser Ser Arg Pro Thr Met Thr Asn His Val Tyr Asn Ser Tyr Thr Glu
1 5 10 15
Lys His Ser Ser Thr His Arg Ser Lys
20 25
<210> 365
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 365
Ser Ser Arg Pro Thr Thr Tyr His Tyr Lys Asn Ile Tyr Gln Glu Ser
1 5 10 15
Tyr Gln Gln Arg Asn Pro Ala Val Lys
20 25
<210> 366
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 366
Ser Ser Arg Pro Val Glu Pro Ala Thr Lys Asn Met Arg Glu Ala Arg
1 5 10 15
Ser Ser Thr Gln Met Arg Arg Ile Lys
20 25
<210> 367
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 367
Ser Ser Arg Pro Tyr Leu Leu Pro Lys Asp Gln Thr Thr Ala Pro Gln
1 5 10 15
Val Thr Pro Ile Val Gln His Lys Lys
20 25
<210> 368
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 368
Ser Ser Arg Pro Glu Phe Pro Tyr Tyr Asn Asp Asn Pro Pro Asn Pro
1 5 10 15
Glu Arg His Thr Leu Arg Lys
20
<210> 369
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 369
Ser Ser Arg Pro Asp Leu Gly Thr Phe Pro Asn Arg Thr Leu Lys Met
1 5 10 15
Ala Ala His Lys
20
<210> 370
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 370
Ser Ser Arg Pro Phe Asp Gly Ile Gly Leu Gly Thr Ala Thr Arg His
1 5 10 15
Gln Asn Arg Lys
20
<210> 371
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 371
Ser Ser Arg Pro Gln Ala Ala Gln Val His Met Met Gln His Ser Arg
1 5 10 15
Pro Thr Thr Lys
20
<210> 372
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 372
Ser Ser Arg Pro Ser Glu Ala Arg Ala Arg Thr Phe Asn Asp His Thr
1 5 10 15
Thr Pro Met Pro Ile Ile Lys
20
<210> 373
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 373
Ser Ser Arg Pro Glu Leu Asp His Asp Ser Arg His Tyr Met Asn Gly
1 5 10 15
Leu Gln Arg Lys Val Thr Lys
20
<210> 374
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 374
Ser Ser Arg Pro Gly Pro Gln His Val Leu Met Gln Asp Thr His Gln
1 5 10 15
Gly Tyr Ala Phe Asp Asn Lys
20
<210> 375
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 375
Ser Ser Arg Pro Thr Thr Gly Ser Ser Ser Gln Ala Asp Thr Ser Ala
1 5 10 15
Ser Met Ser Ile Val Pro Ala His Lys
20 25
<210> 376
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 376
Ser Ser Arg Pro Thr Tyr Gln Gly Val Pro Ser Trp Pro Ala Val Ile
1 5 10 15
Asp Asp Ala Ile Arg Arg Lys
20
<210> 377
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 377
Ser Ser Arg Pro Val Asn Pro Asn Trp Val Glu Thr Gln Ala Leu His
1 5 10 15
Gln Pro Pro Gly Asn Thr Lys
20
<210> 378
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 378
Ser Ser Arg Pro Ile Tyr Pro Asn Glu Ser Met Ser Thr Ser Asn Val
1 5 10 15
Arg Gly Pro Tyr His Pro Lys
20
<210> 379
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 379
Ser Ser Arg Pro His Asp Pro Asn His Leu Thr His Gln Ala Arg Thr
1 5 10 15
Ile Tyr Arg Asn Ala Asn His Thr Lys
20 25
<210> 380
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 380
Ser Ser Arg Pro Ala Pro Pro Thr Tyr Gln Thr Ala Ser Tyr Pro His
1 5 10 15
Asn Leu Pro Ser Lys Arg Lys Met Lys
20 25
<210> 381
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 381
Ser Ser Arg Pro Gln Val Pro Asp Tyr Leu Ser Pro Thr His Gln Lys
1 5 10 15
Lys Ala Phe Leu Glu Ile Pro Thr Lys
20 25
<210> 382
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 382
Ser Ser Arg Pro His Lys Asn Glu Asn Ile Met Gln Tyr Asn Val Asn
1 5 10 15
Asp Arg Trp His Ile Thr Pro Ala Lys
20 25
<210> 383
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic construct - caspace 3 cleavable linker
<400> 383
Leu Glu Ser Gly Asp Glu Val Asp
1 5
<210> 384
<211> 37
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 384
Thr Ser Thr Ser Lys Ala Ser Thr Thr Thr Thr Ser Ser Lys Thr Thr
1 5 10 15
Thr Thr Ser Ser Lys Thr Thr Thr Thr Thr Ser Lys Thr Ser Thr Thr
20 25 30
Ser Ser Ser Ser Thr
35
<210> 385
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 385
Gly Gln Gly Gly Tyr Gly Gly Leu Gly Ser Gln Gly Ala Gly Arg Gly
1 5 10 15
Gly Leu Gly Gly Gln Gly
20
<210> 386
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 386
Gly Pro Gly Gly Tyr Gly Pro Gly Gln Gln
1 5 10
<210> 387
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 387
Gly Gly Ser Gly Pro Gly Ser Gly Gly
1 5
<210> 388
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 388
Gly Gly Pro Lys Lys
1 5
<210> 389
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 389
Gly Pro Gly Val Gly
1 5
<210> 390
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 390
Gly Gly Gly Cys Gly Gly Gly
1 5
<210> 391
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 391
Gly Gly Gly Cys
1
<210> 392
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 392
Pro His Met Ala Ser Met Thr Gly Gly Gln Gln Met Gly Ser
1 5 10
<210> 393
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 393
Gly Pro Glu Glu Ala Ala Lys Lys Glu Glu Ala Ala Lys Lys Pro Ala
1 5 10 15
<210> 394
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 394
Gly Ser Gly Gly Gly Gly Ser Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 395
<211> 37
<212> PRT
<213> Artificial Sequence
<220>
<223> synthetic construct
<400> 395
Gly Pro Glu Pro Glu Pro Glu Pro Glu Pro Ile Pro Glu Pro Pro Lys
1 5 10 15
Glu Ala Pro Val Val Ile Glu Lys Pro Lys Pro Lys Pro Lys Pro Lys
20 25 30
Pro Lys Pro Pro Ala
35
<210> 396
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 396
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
1 5 10 15
Gly Ser
<210> 397
<211> 6368
<212> DNA
<213> artificial sequence
<220>
<223> synthetic construct
<400> 397
agatctcgat cccgcgaaat taatacgact cactataggg agaccacaac ggtttccctc 60
tagaaataat tttgtttaac tttaagaagg agatatacat atgcacactc cagaacatat 120
caccgcagta gtacagcgtt ttgtggcagc tctgaacgcg ggcgagctgg aaggtattgt 180
ggcgctgttc gcggaagaag ccaccgtgga agaaccggtg ggttctgaac cgcgttccgg 240
caccgcagcc tgccgtgaat tttacgcaaa cagcctgaag ctgccgctgg cggttgaact 300
gacccaagaa tgtcgtgcgg tggctaacga agccgctttc gcgttcaccg tgtccttcga 360
ataccagggt cgtaagaccg ttgtggcgcc atgcgaacac tttcgtttca acggcgcagg 420
caaagtggtt tccatccgcg cactgttcgg tgaaaagaac atccatgctt gtcagggatc 480
cgatccgact ccgccgacga atgtactgat gctggcaacc aaaggcggtg gtacgcattc 540
cacgcacaac catggcagcc cgcgccacac gaatgctgac gcaggcaatc cgggcggcgg 600
caccccacca accaatgtcc tgatgctggc tactaaaggc ggcggcacgc attctaccca 660
caaccatggt agcccgcgcc atactaatgc agatgccggc aacccgggcg gtggtacccc 720
gccaaccaac gttctgatgc tggcgacgaa aggtggcggt acccattcca cgcataatca 780
tggcagccct cgccacacca acgctgatgc tggtaatcct ggtggcggta agaagaaata 840
ataaggcgcg ccgacccagc tttcttgtac aaagtggttg attcgaggct gctaacaaag 900
cccgaaagga agctgagttg gctgctgcca ccgctgagca ataactagca taaccccttg 960
gggcctctaa acgggtcttg aggggttttt tgctgaaagg aggaactata tccggatatc 1020
cacaggacgg gtgtggtcgc catgatcgcg tagtcgatag tggctccaag tagcgaagcg 1080
agcaggactg ggcggcggcc aaagcggtcg gacagtgctc cgagaacggg tgcgcataga 1140
aattgcatca acgcatatag cgctagcagc acgccatagt gactggcgat gctgtcggaa 1200
tggacgatat cccgcaagag gcccggcagt accggcataa ccaagcctat gcctacagca 1260
tccagggtga cggtgccgag gatgacgatg agcgcattgt tagatttcat acacggtgcc 1320
tgactgcgtt agcaatttaa ctgtgataaa ctaccgcatt aaagcttgca gtggcggttt 1380
tcatggcttg ttatgactgt ttttttgggg tacagtctat gcctcgggca tccaagcagc 1440
aagcgcgtta cgccgtgggt cgatgtttga tgttatggag cagcaacgat gttacgcagc 1500
agggcagtcg ccctaaaaca aagttaaaca tcatgaggga agcggtgatc gccgaagtat 1560
cgactcaact atcagaggta gttggcgtca tcgagcgcca tctcgaaccg acgttgctgg 1620
ccgtacattt gtacggctcc gcagtggatg gcggcctgaa gccacacagt gatattgatt 1680
tgctggttac ggtgaccgta aggcttgatg aaacaacgcg gcgagctttg atcaacgacc 1740
ttttggaaac ttcggcttcc cctggagaga gcgagattct ccgcgctgta gaagtcacca 1800
ttgttgtgca cgacgacatc attccgtggc gttatccagc taagcgcgaa ctgcaatttg 1860
gagaatggca gcgcaatgac attcttgcag gtatcttcga gccagccacg atcgacattg 1920
atctggctat cttgctgaca aaagcaagag aacatagcgt tgccttggta ggtccagcgg 1980
cggaggaact ctttgatccg gttcctgaac aggatctatt tgaggcgcta aatgaaacct 2040
taacgctatg gaactcgccg cccgactggg ctggcgatga gcgaaatgta gtgcttacgt 2100
tgtcccgcat ttggtacagc gcagtaaccg gcaaaatcgc gccgaaggat gtcgctgccg 2160
actgggcaat ggagcgcctg ccggcccagt atcagcccgt catacttgaa gctagacagg 2220
cttatcttgg acaagaagaa gatcgcttgg cctcgcgcgc agatcagttg gaagaatttg 2280
tccactacgt gaaaggcgag atcaccaagg tagtcggcaa ataatgtcta acaattcgtt 2340
caagcttatc gatgataagc tgtcaaacat gagaattctt gaagacgaaa gggcctcgtg 2400
atacgcctat ttttataggt taatgtcatg ataataatgg tttcttagac gtcaggtggc 2460
acttttcggg gaaatgtgcg cggaacccct atttgtttat ttttctaaat acattcaaat 2520
atgtatccgc tcatgagaca ataaccctga taaatgcttc aataatattg aaaaaggaag 2580
agtatgagta ttcaacattt ccgtgtcgcc cttattccct tttttgcggc attttgcctt 2640
cctgtttttg ctcacccaga aacgctggtg aaagtaaaag atgctgaaga tcagttgggt 2700
gcacgagtgg gttacatcga actggatctc aacagcggta agatccttga gagttttcgc 2760
cccgaagaac gttttccaat gatgagcact tttaaagttc tgctatgtgg cgcggtatta 2820
tcccgtgttg acgccgggca agagcaactc ggtcgccgca tacactattc tcagaatgac 2880
ttggttgagt actcaccagt cacagaaaag catcttacgg atggcatgac agtaagagaa 2940
ttatgcagtg ctgccataac catgagtgat aacactgcgg ccaacttact tctgacaacg 3000
atcggaggac cgaaggagct aaccgctttt ttgcacaaca tgggggatca tgtaactcgc 3060
cttgatcgtt gggaaccgga gctgaatgaa gccataccaa acgacgagcg tgacaccacg 3120
atgcctgcag caatggcaac aacgttgcgc aaactattaa ctggcgaact acttactcta 3180
gcttcccggc aacaattaat agactggatg gaggcggata aagttgcagg accacttctg 3240
cgctcggccc ttccggctgg ctggtttatt gctgataaat ctggagccgg tgagcgtggg 3300
tctcgcggta tcattgcagc actggggcca gatggtaagc cctcccgtat cgtagttatc 3360
tacacgacgg ggagtcaggc aactatggat gaacgaaata gacagatcgc tgagataggt 3420
gcctcactga ttaagcattg gtaactgtca gaccaagttt actcatatat actttagatt 3480
gatttaaaac ttcattttta atttaaaagg atctaggtga agatcctttt tgataatctc 3540
atgaccaaaa tcccttaacg tgagttttcg ttccactgag cgtcagaccc cgtagaaaag 3600
atcaaaggat cttcttgaga tccttttttt ctgcgcgtaa tctgctgctt gcaaacaaaa 3660
aaaccaccgc taccagcggt ggtttgtttg ccggatcaag agctaccaac tctttttccg 3720
aaggtaactg gcttcagcag agcgcagata ccaaatactg tccttctagt gtagccgtag 3780
ttaggccacc acttcaagaa ctctgtagca ccgcctacat acctcgctct gctaatcctg 3840
ttaccagtgg ctgctgccag tggcgataag tcgtgtctta ccgggttgga ctcaagacga 3900
tagttaccgg ataaggcgca gcggtcgggc tgaacggggg gttcgtgcac acagcccagc 3960
ttggagcgaa cgacctacac cgaactgaga tacctacagc gtgagctatg agaaagcgcc 4020
acgcttcccg aagggagaaa ggcggacagg tatccggtaa gcggcagggt cggaacagga 4080
gagcgcacga gggagcttcc agggggaaac gcctggtatc tttatagtcc tgtcgggttt 4140
cgccacctct gacttgagcg tcgatttttg tgatgctcgt caggggggcg gagcctatgg 4200
aaaaacgcca gcaacgcggc ctttttacgg ttcctggcct tttgctggcc ttttgctcac 4260
atgttctttc ctgcgttatc ccctgattct gtggataacc gtattaccgc ctttgagtga 4320
gctgataccg ctcgccgcag ccgaacgacc gagcgcagcg agtcagtgag cgaggaagcg 4380
gaagagcgcc tgatgcggta ttttctcctt acgcatctgt gcggtatttc acaccgcata 4440
tatggtgcac tctcagtaca atctgctctg atgccgcata gttaagccag tatacactcc 4500
gctatcgcta cgtgactggg tcatggctgc gccccgacac ccgccaacac ccgctgacgc 4560
gccctgacgg gcttgtctgc tcccggcatc cgcttacaga caagctgtga ccgtctccgg 4620
gagctgcatg tgtcagaggt tttcaccgtc atcaccgaaa cgcgcgaggc agctgcggta 4680
aagctcatca gcgtggtcgt gaagcgattc acagatgtct gcctgttcat ccgcgtccag 4740
ctcgttgagt ttctccagaa gcgttaatgt ctggcttctg ataaagcggg ccatgttaag 4800
ggcggttttt tcctgtttgg tcactgatgc ctccgtgtaa gggggatttc tgttcatggg 4860
ggtaatgata ccgatgaaac gagagaggat gctcacgata cgggttactg atgatgaaca 4920
tgcccggtta ctggaacgtt gtgagggtaa acaactggcg gtatggatgc ggcgggacca 4980
gagaaaaatc actcagggtc aatgccagcg cttcgttaat acagatgtag gtgttccaca 5040
gggtagccag cagcatcctg cgatgcagat ccggaacata atggtgcagg gcgctgactt 5100
ccgcgtttcc agactttacg aaacacggaa accgaagacc attcatgttg ttgctcaggt 5160
cgcagacgtt ttgcagcagc agtcgcttca cgttcgctcg cgtatcggtg attcattctg 5220
ctaaccagta aggcaacccc gccagcctag ccgggtcctc aacgacagga gcacgatcat 5280
gcgcacccgt ggccaggacc caacgctgcc cgagatgcgc cgcgtgcggc tgctggagat 5340
ggcggacgcg atggatatgt tctgccaagg gttggtttgc gcattcacag ttctccgcaa 5400
gaattgattg gctccaattc ttggagtggt gaatccgtta gcgaggtgcc gccggcttcc 5460
attcaggtcg aggtggcccg gctccatgca ccgcgacgca acgcggggag gcagacaagg 5520
tatagggcgg cgcctacaat ccatgccaac ccgttccatg tgctcgccga ggcggcataa 5580
atcgccgtga cgatcagcgg tccagtgatc gaagttaggc tggtaagagc cgcgagcgat 5640
ccttgaagct gtccctgatg gtcgtcatct acctgcctgg acagcatggc ctgcaacgcg 5700
ggcatcccga tgccgccgga agcgagaaga atcataatgg ggaaggccat ccagcctcgc 5760
gtcgcgaacg ccagcaagac gtagcccagc gcgtcggccg ccatgccggc gataatggcc 5820
tgcttctcgc cgaaacgttt ggtggcggga ccagtgacga aggcttgagc gagggcgtgc 5880
aagattccga ataccgcaag cgacaggccg atcatcgtcg cgctccagcg aaagcggtcc 5940
tcgccgaaaa tgacccagag cgctgccggc acctgtccta cgagttgcat gataaagaag 6000
acagtcataa gtgcggcgac gatagtcatg ccccgcgccc accggaagga gctgactggg 6060
ttgaaggctc tcaagggcat cggtcgatcg acgctctccc ttatgcgact cctgcattag 6120
gaagcagccc agtagtaggt tgaggccgtt gagcaccgcc gccgcaagga atggtgcatg 6180
caaggagatg gcgcccaaca gtcccccggc cacggggcct gccaccatac ccacgccgaa 6240
acaagcgctc atgagcccga agtggcgagc ccgatcttcc ccatcggtga tgtcggcgat 6300
ataggcgcca gcaaccgcac ctgtggcgcc ggtgatgccg gccacgatgc gtccggcgta 6360
gaggatcg 6368
<210> 398
<211> 20
<212> DNA
<213> artificial sequence
<220>
<223> primer
<400> 398
ccctcatagt tagcgtaacg 20
<210> 399
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 399
Ser Asn Ala Thr Met Tyr Asn Ile Gln Ser His Ser His His Gln
1 5 10 15
<210> 400
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 400
Gln Ala Ala Gln Val His Met Met Gln His Ser Arg Pro Thr Thr
1 5 10 15
<210> 401
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 401
His Asp Pro Tyr Thr Met Lys Ser Ala Leu Arg Gln Ser Thr Ser
1 5 10 15
<210> 402
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 402
Asp Leu Gly Thr Phe Pro Asn Arg Thr Leu Lys Met Ala Ala His
1 5 10 15
<210> 403
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 403
Asp Thr Ile His Pro Asn Lys Met Lys Ser Pro Ser Ser Pro Leu
1 5 10 15
<210> 404
<211> 15
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 404
Gly Ser Asn Asn His Leu Pro Ser Thr Val Pro Arg Leu Thr Val
1 5 10 15
<210> 405
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 405
Ser Asn Pro Ile Pro Asn Phe Ala His Asp Leu Arg His Ser Lys Tyr
1 5 10 15
Asn Ser
<210> 406
<211> 18
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 406
Thr Lys Pro Pro Arg Thr Pro Thr Ala Asn Thr Ser Arg Pro His His
1 5 10 15
Asn Phe
<210> 407
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 407
Ala Asn Ser Gly Phe Pro Ile Trp Leu Gln Lys Tyr Pro Trp Ser Glu
1 5 10 15
Val Gln Gln Glu
20
<210> 408
<211> 19
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 408
Ala Thr Pro Arg Leu Thr Pro Glu Ala His His Lys Ala Gly Asn Trp
1 5 10 15
Tyr Ala Ser
<210> 409
<211> 20
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 409
Ala Thr Pro Ser Gln His Arg Tyr Gly Leu Met Gln Asn His Ala Pro
1 5 10 15
Asn Gly Ile Glu
20
<210> 410
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 410
Gly Met Gly Ser Glu Val Leu Ser Gln Tyr Pro Gln Ala Pro Val Gly
1 5 10 15
<210> 411
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 411
Thr Thr Tyr His Tyr Lys Asn Ile Tyr Gln Glu Ser Tyr Gln Gln Arg
1 5 10 15
Asn Pro Ala Val Lys
20
<210> 412
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 412
Ser Asn Ala Thr Met Tyr Asn Ile Gln Ser His Ser His His Gln Lys
1 5 10 15
<210> 413
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 413
Gln Ala Ala Gln Val His Met Met Gln His Ser Arg Pro Thr Thr Lys
1 5 10 15
<210> 414
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 414
His Asp Pro Tyr Thr Met Lys Ser Ala Leu Arg Gln Ser Thr Ser Lys
1 5 10 15
<210> 415
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 415
Asp Leu Gly Thr Phe Pro Asn Arg Thr Leu Lys Met Ala Ala His Lys
1 5 10 15
<210> 416
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 416
Asp Thr Ile His Pro Asn Lys Met Lys Ser Pro Ser Ser Pro Leu Lys
1 5 10 15
<210> 417
<211> 16
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 417
Gly Ser Asn Asn His Leu Pro Ser Thr Val Pro Arg Leu Thr Val Lys
1 5 10 15
<210> 418
<211> 19
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 418
Ser Asn Pro Ile Pro Asn Phe Ala His Asp Leu Arg His Ser Lys Tyr
1 5 10 15
Asn Ser Lys
<210> 419
<211> 19
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 419
Thr Lys Pro Pro Arg Thr Pro Thr Ala Asn Thr Ser Arg Pro His His
1 5 10 15
Asn Phe Lys
<210> 420
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 420
Ala Asn Ser Gly Phe Pro Ile Trp Leu Gln Lys Tyr Pro Trp Ser Glu
1 5 10 15
Val Gln Gln Glu Lys
20
<210> 421
<211> 21
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 421
Ala Thr Pro Ser Gln His Arg Tyr Gly Leu Met Gln Asn His Ala Pro
1 5 10 15
Asn Gly Ile Glu Lys
20
<210> 422
<211> 17
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 422
Gly Met Gly Ser Glu Val Leu Ser Gln Tyr Pro Gln Ala Pro Val Gly
1 5 10 15
Lys
<210> 423
<211> 88
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 423
Pro Ser Ala Gln Ser Gln Leu Pro Asp Lys His Ser Gly Leu His Glu
1 5 10 15
Arg Ala Pro Gln Arg Tyr Gly Pro Glu Pro Glu Pro Glu Pro Glu Pro
20 25 30
Ile Pro Glu Pro Pro Lys Glu Ala Pro Val Val Ile Glu Lys Pro Lys
35 40 45
Pro Lys Pro Lys Pro Lys Pro Lys Pro Pro Ala His Asp His Lys Asn
50 55 60
Gln Lys Glu Thr His Gln Arg His Ala Ala Gly Ser Gly Gly Gly Gly
65 70 75 80
Ser Pro His His His His His His
85
<210> 424
<211> 325
<212> PRT
<213> Thermotoga maritima
<400> 424
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 425
<211> 431
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 425
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser Pro Ser Ala Gln Ser Gln Leu Pro Asp
340 345 350
Lys His Ser Gly Leu His Glu Arg Ala Pro Gln Arg Tyr Gly Pro Glu
355 360 365
Pro Glu Pro Glu Pro Glu Pro Ile Pro Glu Pro Pro Lys Glu Ala Pro
370 375 380
Val Val Ile Glu Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro
385 390 395 400
Pro Ala His Asp His Lys Asn Gln Lys Glu Thr His Gln Arg His Ala
405 410 415
Ala Gly Ser Gly Gly Gly Gly Ser Pro His His His His His His
420 425 430
<210> 426
<211> 333
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 426
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Ser His His His His His His
325 330
<210> 427
<211> 353
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 427
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser Gly Lys Gly Lys Gly Lys Gly Lys Gly
340 345 350
Lys
<210> 428
<211> 359
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 428
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser Gly Lys Gly Lys Gly Lys Gly Lys Gly
340 345 350
Lys His His His His His His
355
<210> 429
<211> 375
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 429
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser Thr Lys Pro Pro Arg Thr Pro Thr Ala
340 345 350
Asn Thr Ser Arg Pro His His Asn Phe Gly Ser Gly Gly Gly Gly Ser
355 360 365
Pro His His His His His His
370 375
<210> 430
<211> 349
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 430
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser His His His His His His
340 345
<210> 431
<211> 106
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 431
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
1 5 10 15
Gly Ser Pro Ser Ala Gln Ser Gln Leu Pro Asp Lys His Ser Gly Leu
20 25 30
His Glu Arg Ala Pro Gln Arg Tyr Gly Pro Glu Pro Glu Pro Glu Pro
35 40 45
Glu Pro Ile Pro Glu Pro Pro Lys Glu Ala Pro Val Val Ile Glu Lys
50 55 60
Pro Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro Pro Ala His Asp His
65 70 75 80
Lys Asn Gln Lys Glu Thr His Gln Arg His Ala Ala Gly Ser Gly Gly
85 90 95
Gly Gly Ser Pro His His His His His His
100 105
<210> 432
<211> 8
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 432
Gly Ser His His His His His His
1 5
<210> 433
<211> 28
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 433
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
1 5 10 15
Gly Ser Gly Lys Gly Lys Gly Lys Gly Lys Gly Lys
20 25
<210> 434
<211> 34
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 434
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
1 5 10 15
Gly Ser Gly Lys Gly Lys Gly Lys Gly Lys Gly Lys His His His His
20 25 30
His His
<210> 435
<211> 50
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 435
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
1 5 10 15
Gly Ser Thr Lys Pro Pro Arg Thr Pro Thr Ala Asn Thr Ser Arg Pro
20 25 30
His His Asn Phe Gly Ser Gly Gly Gly Gly Ser Pro His His His His
35 40 45
His His
50
<210> 436
<211> 24
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 436
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
1 5 10 15
Gly Ser His His His His His His
20
<210> 437
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 437
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 438
<211> 375
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 438
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser Thr Lys Pro Pro Arg Thr Pro Thr Ala
340 345 350
Asn Thr Ser Arg Pro His His Asn Phe Gly Ser Gly Gly Gly Gly Ser
355 360 365
Pro His His His His His His
370 375
<210> 439
<211> 349
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 439
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser His His His His His His
340 345
<210> 440
<211> 431
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 440
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser Pro Ser Ala Gln Ser Gln Leu Pro Asp
340 345 350
Lys His Ser Gly Leu His Glu Arg Ala Pro Gln Arg Tyr Gly Pro Glu
355 360 365
Pro Glu Pro Glu Pro Glu Pro Ile Pro Glu Pro Pro Lys Glu Ala Pro
370 375 380
Val Val Ile Glu Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro
385 390 395 400
Pro Ala His Asp His Lys Asn Gln Lys Glu Thr His Gln Arg His Ala
405 410 415
Ala Gly Ser Gly Gly Gly Gly Ser Pro His His His His His His
420 425 430
<210> 441
<211> 333
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 441
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Ser His His His His His His
325 330
<210> 442
<211> 359
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 442
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
325 330 335
Ser Ala Gly Gly Pro Gly Ser Gly Lys Gly Lys Gly Lys Gly Lys Gly
340 345 350
Lys His His His His His His
355
<210> 443
<211> 386
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 443
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly Gly
325 330 335
Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Glu Pro Glu Pro
340 345 350
Glu Trp Lys Thr Lys Lys Ile Leu Leu Ser Arg Thr Arg Arg Ile Met
355 360 365
Arg Gln Val Val Arg Ser Val Met His Lys Ile Trp His His His His
370 375 380
His His
385
<210> 444
<211> 387
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 444
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu
325 330 335
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
340 345 350
Gly Ser Trp Lys Thr Lys Lys Ile Leu Leu Ser Arg Thr Arg Arg Ile
355 360 365
Met Arg Gln Val Val Arg Ser Val Met His Lys Ile Trp His His His
370 375 380
His His His
385
<210> 445
<211> 386
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 445
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly Gly
325 330 335
Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Glu Pro Glu Pro
340 345 350
Glu Pro Leu Trp Arg Arg Ile Thr Lys Arg Lys Leu Val Arg Pro Val
355 360 365
Ala Thr Leu Met Trp Tyr Trp Phe Thr Ser Lys Arg His His His His
370 375 380
His His
385
<210> 446
<211> 387
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 446
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu
325 330 335
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
340 345 350
Gly Ser Pro Leu Trp Arg Arg Ile Thr Lys Arg Lys Leu Val Arg Pro
355 360 365
Val Ala Thr Leu Met Trp Tyr Trp Phe Thr Ser Lys Arg His His His
370 375 380
His His His
385
<210> 447
<211> 382
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 447
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Gly Pro Gly Ser Gly Gly Ala Gly
325 330 335
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Glu Pro Glu Arg Met Leu
340 345 350
Ser Arg Ile Leu Arg Met Phe Val Arg Ile Leu Lys Arg Glu Arg Leu
355 360 365
Ser Gln Val Arg Gly Leu Phe Val His His His His His His
370 375 380
<210> 448
<211> 383
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 448
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Pro Glu Pro Glu Gly Pro Gly Ser
325 330 335
Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Arg Met
340 345 350
Leu Ser Arg Ile Leu Arg Met Phe Val Arg Ile Leu Lys Arg Glu Arg
355 360 365
Leu Ser Gln Val Arg Gly Leu Phe Val His His His His His His
370 375 380
<210> 449
<211> 390
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 449
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Pro Glu Pro Glu Gly Pro Gly Ser
325 330 335
Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Glu Pro
340 345 350
Glu Pro Glu Pro Glu Leu Arg Phe Leu Ala Arg Arg Phe Leu Lys Leu
355 360 365
Arg Arg Ala Arg Lys Trp Trp Asn Ala Trp Lys Val Trp Val Thr Arg
370 375 380
His His His His His His
385 390
<210> 450
<211> 391
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 450
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Ser Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Arg Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu
325 330 335
Pro Glu Pro Glu Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser
340 345 350
Ala Gly Gly Pro Gly Ser Leu Arg Phe Leu Ala Arg Arg Phe Leu Lys
355 360 365
Leu Arg Arg Ala Arg Lys Trp Trp Asn Ala Trp Lys Val Trp Val Thr
370 375 380
Arg His His His His His His
385 390
<210> 451
<211> 344
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 451
Met Gln Leu Phe Asp Leu Ser Leu Glu Glu Leu Lys Lys Tyr Lys Pro
1 5 10 15
Lys Lys Thr Ala Arg Pro Asp Phe Ser Asp Phe Trp Lys Lys Ser Leu
20 25 30
Glu Glu Leu Arg Gln Val Glu Ala Glu Pro Thr Leu Glu Ser Tyr Asp
35 40 45
Tyr Pro Val Lys Gly Val Lys Val Tyr Arg Leu Thr Tyr Gln Ser Phe
50 55 60
Gly His Ser Lys Ile Glu Gly Phe Tyr Ala Val Pro Asp Gln Thr Gly
65 70 75 80
Pro His Pro Ala Leu Val Arg Phe His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Gly Ile His Asp Ile Val Asn Trp Ala Leu His Gly Tyr Ala Thr
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gly Gly Ser Glu Asp Thr Ser Val
115 120 125
Thr Pro Gly Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Ser
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Gln Ser Phe Pro Glu Val Asp Glu His Arg Ile Gly
165 170 175
Val Ile Gly Gly Ser Gln Gly Gly Ala Leu Ala Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Val Val Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Val Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Tyr Phe Arg Arg Asn Ser Asp Pro Lys Val Glu Glu Lys
225 230 235 240
Ala Phe Glu Thr Leu Ser Tyr Phe Asp Leu Ile Asn Leu Ala Gly Trp
245 250 255
Val Lys Gln Pro Thr Leu Met Ala Ile Gly Leu Ile Asp Lys Ile Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Asp Lys
275 280 285
Asp Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Phe Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ser Phe Leu Gln Lys His Leu Leu Leu Ser Thr
305 310 315 320
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
325 330 335
Gly Ser His His His His His His
340
<210> 452
<211> 426
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 452
Met Gln Leu Phe Asp Leu Ser Leu Glu Glu Leu Lys Lys Tyr Lys Pro
1 5 10 15
Lys Lys Thr Ala Arg Pro Asp Phe Ser Asp Phe Trp Lys Lys Ser Leu
20 25 30
Glu Glu Leu Arg Gln Val Glu Ala Glu Pro Thr Leu Glu Ser Tyr Asp
35 40 45
Tyr Pro Val Lys Gly Val Lys Val Tyr Arg Leu Thr Tyr Gln Ser Phe
50 55 60
Gly His Ser Lys Ile Glu Gly Phe Tyr Ala Val Pro Asp Gln Thr Gly
65 70 75 80
Pro His Pro Ala Leu Val Arg Phe His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Gly Ile His Asp Ile Val Asn Trp Ala Leu His Gly Tyr Ala Thr
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gly Gly Ser Glu Asp Thr Ser Val
115 120 125
Thr Pro Gly Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Ser
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Gln Ser Phe Pro Glu Val Asp Glu His Arg Ile Gly
165 170 175
Val Ile Gly Gly Ser Gln Gly Gly Ala Leu Ala Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Val Val Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Val Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Tyr Phe Arg Arg Asn Ser Asp Pro Lys Val Glu Glu Lys
225 230 235 240
Ala Phe Glu Thr Leu Ser Tyr Phe Asp Leu Ile Asn Leu Ala Gly Trp
245 250 255
Val Lys Gln Pro Thr Leu Met Ala Ile Gly Leu Ile Asp Lys Ile Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Asp Lys
275 280 285
Asp Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Phe Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ser Phe Leu Gln Lys His Leu Leu Leu Ser Thr
305 310 315 320
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
325 330 335
Gly Ser Pro Ser Ala Gln Ser Gln Leu Pro Asp Lys His Ser Gly Leu
340 345 350
His Glu Arg Ala Pro Gln Arg Tyr Gly Pro Glu Pro Glu Pro Glu Pro
355 360 365
Glu Pro Ile Pro Glu Pro Pro Lys Glu Ala Pro Val Val Ile Glu Lys
370 375 380
Pro Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro Pro Ala His Asp His
385 390 395 400
Lys Asn Gln Lys Glu Thr His Gln Arg His Ala Ala Gly Ser Gly Gly
405 410 415
Gly Gly Ser Pro His His His His His His
420 425
<210> 453
<211> 372
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 453
Met Gln Leu Phe Asp Leu Ser Leu Glu Glu Leu Lys Lys Tyr Lys Pro
1 5 10 15
Lys Lys Thr Ala Arg Pro Asp Phe Ser Asp Phe Trp Lys Lys Ser Leu
20 25 30
Glu Glu Leu Arg Gln Val Glu Ala Glu Pro Thr Leu Glu Ser Tyr Asp
35 40 45
Tyr Pro Val Lys Gly Val Lys Val Tyr Arg Leu Thr Tyr Gln Ser Phe
50 55 60
Gly His Ser Lys Ile Glu Gly Phe Tyr Ala Val Pro Asp Gln Thr Gly
65 70 75 80
Pro His Pro Ala Leu Val Arg Phe His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Gly Ile His Asp Ile Val Asn Trp Ala Leu His Gly Tyr Ala Thr
100 105 110
Phe Gly Met Leu Val Arg Gly Gln Gly Gly Ser Glu Asp Thr Ser Val
115 120 125
Thr Pro Gly Gly His Ala Leu Gly Trp Met Thr Lys Gly Ile Leu Ser
130 135 140
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
Leu Glu Val Ile Gln Ser Phe Pro Glu Val Asp Glu His Arg Ile Gly
165 170 175
Val Ile Gly Gly Ser Gln Gly Gly Ala Leu Ala Ile Ala Ala Ala Ala
180 185 190
Leu Ser Asp Ile Pro Lys Val Val Val Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
Asn Phe Glu Arg Ala Val Asp Val Ala Leu Glu Gln Pro Tyr Leu Glu
210 215 220
Ile Asn Ser Tyr Phe Arg Arg Asn Ser Asp Pro Lys Val Glu Glu Lys
225 230 235 240
Ala Phe Glu Thr Leu Ser Tyr Phe Asp Leu Ile Asn Leu Ala Gly Trp
245 250 255
Val Lys Gln Pro Thr Leu Met Ala Ile Gly Leu Ile Asp Lys Ile Thr
260 265 270
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Asp Lys
275 280 285
Asp Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Phe Ile Pro Ala Phe
290 295 300
Gln Thr Glu Lys Leu Ser Phe Leu Gln Lys His Leu Leu Leu Ser Thr
305 310 315 320
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
325 330 335
Gly Ser Asp Pro Thr Lys Pro Pro Arg Thr Pro Thr Ala Asn Thr Ser
340 345 350
Arg Pro His His Asn Phe Gly Ser Gly Gly Gly Gly Ser Pro His His
355 360 365
His His His His
370
<210> 454
<211> 347
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 454
Met Pro Phe Pro Asp Leu Ile Gln Pro Glu Leu Gly Ala Tyr Val Ser
1 5 10 15
Ser Val Gly Met Pro Asp Asp Phe Ala Gln Phe Trp Thr Ser Thr Ile
20 25 30
Ala Glu Ala Arg Gln Ala Gly Gly Glu Val Ser Ile Val Gln Ala Gln
35 40 45
Thr Thr Leu Lys Ala Val Gln Ser Phe Asp Val Thr Phe Pro Gly Tyr
50 55 60
Gly Gly His Pro Ile Lys Gly Trp Leu Ile Leu Pro Thr His His Lys
65 70 75 80
Gly Arg Leu Pro Leu Val Val Gln Tyr Ile Gly Tyr Gly Gly Gly Arg
85 90 95
Gly Leu Ala His Glu Gln Leu His Trp Ala Ala Ser Gly Phe Ala Tyr
100 105 110
Phe Arg Met Asp Thr Arg Gly Gln Gly Ser Asp Trp Ser Val Gly Glu
115 120 125
Thr Ala Asp Pro Val Gly Ser Thr Ser Ser Ile Pro Gly Phe Met Thr
130 135 140
Arg Gly Val Leu Asp Lys Asn Asp Tyr Tyr Tyr Arg Arg Leu Phe Thr
145 150 155 160
Asp Ala Val Arg Ala Ile Asp Ala Leu Leu Gly Leu Asp Phe Val Asp
165 170 175
Pro Glu Arg Ile Ala Val Cys Gly Asp Ser Gln Gly Gly Gly Ile Ser
180 185 190
Leu Ala Val Gly Gly Ile Asp Pro Arg Val Lys Ala Val Met Pro Asp
195 200 205
Val Pro Phe Leu Cys Asp Phe Pro Arg Ala Val Gln Thr Ala Val Arg
210 215 220
Asp Pro Tyr Leu Glu Ile Val Arg Phe Leu Ala Gln His Arg Glu Lys
225 230 235 240
Lys Ala Ala Val Phe Glu Thr Leu Asn Tyr Phe Asp Cys Val Asn Phe
245 250 255
Ala Arg Arg Ser Lys Ala Pro Ala Leu Phe Ser Val Ala Leu Met Asp
260 265 270
Glu Val Cys Pro Pro Ser Thr Val Tyr Gly Ala Phe Asn Ala Tyr Ala
275 280 285
Gly Glu Lys Thr Ile Thr Glu Tyr Glu Phe Asn Asn His Glu Gly Gly
290 295 300
Gln Gly Tyr Gln Glu Arg Gln Gln Met Thr Trp Leu Ser Arg Leu Phe
305 310 315 320
Gly Val Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala
325 330 335
Gly Gly Pro Gly Ser His His His His His His
340 345
<210> 455
<211> 429
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 455
Met Pro Phe Pro Asp Leu Ile Gln Pro Glu Leu Gly Ala Tyr Val Ser
1 5 10 15
Ser Val Gly Met Pro Asp Asp Phe Ala Gln Phe Trp Thr Ser Thr Ile
20 25 30
Ala Glu Ala Arg Gln Ala Gly Gly Glu Val Ser Ile Val Gln Ala Gln
35 40 45
Thr Thr Leu Lys Ala Val Gln Ser Phe Asp Val Thr Phe Pro Gly Tyr
50 55 60
Gly Gly His Pro Ile Lys Gly Trp Leu Ile Leu Pro Thr His His Lys
65 70 75 80
Gly Arg Leu Pro Leu Val Val Gln Tyr Ile Gly Tyr Gly Gly Gly Arg
85 90 95
Gly Leu Ala His Glu Gln Leu His Trp Ala Ala Ser Gly Phe Ala Tyr
100 105 110
Phe Arg Met Asp Thr Arg Gly Gln Gly Ser Asp Trp Ser Val Gly Glu
115 120 125
Thr Ala Asp Pro Val Gly Ser Thr Ser Ser Ile Pro Gly Phe Met Thr
130 135 140
Arg Gly Val Leu Asp Lys Asn Asp Tyr Tyr Tyr Arg Arg Leu Phe Thr
145 150 155 160
Asp Ala Val Arg Ala Ile Asp Ala Leu Leu Gly Leu Asp Phe Val Asp
165 170 175
Pro Glu Arg Ile Ala Val Cys Gly Asp Ser Gln Gly Gly Gly Ile Ser
180 185 190
Leu Ala Val Gly Gly Ile Asp Pro Arg Val Lys Ala Val Met Pro Asp
195 200 205
Val Pro Phe Leu Cys Asp Phe Pro Arg Ala Val Gln Thr Ala Val Arg
210 215 220
Asp Pro Tyr Leu Glu Ile Val Arg Phe Leu Ala Gln His Arg Glu Lys
225 230 235 240
Lys Ala Ala Val Phe Glu Thr Leu Asn Tyr Phe Asp Cys Val Asn Phe
245 250 255
Ala Arg Arg Ser Lys Ala Pro Ala Leu Phe Ser Val Ala Leu Met Asp
260 265 270
Glu Val Cys Pro Pro Ser Thr Val Tyr Gly Ala Phe Asn Ala Tyr Ala
275 280 285
Gly Glu Lys Thr Ile Thr Glu Tyr Glu Phe Asn Asn His Glu Gly Gly
290 295 300
Gln Gly Tyr Gln Glu Arg Gln Gln Met Thr Trp Leu Ser Arg Leu Phe
305 310 315 320
Gly Val Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala
325 330 335
Gly Gly Pro Gly Ser Pro Ser Ala Gln Ser Gln Leu Pro Asp Lys His
340 345 350
Ser Gly Leu His Glu Arg Ala Pro Gln Arg Tyr Gly Pro Glu Pro Glu
355 360 365
Pro Glu Pro Glu Pro Ile Pro Glu Pro Pro Lys Glu Ala Pro Val Val
370 375 380
Ile Glu Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro Pro Ala
385 390 395 400
His Asp His Lys Asn Gln Lys Glu Thr His Gln Arg His Ala Ala Gly
405 410 415
Ser Gly Gly Gly Gly Ser Pro His His His His His His
420 425
<210> 456
<211> 375
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 456
Met Pro Phe Pro Asp Leu Ile Gln Pro Glu Leu Gly Ala Tyr Val Ser
1 5 10 15
Ser Val Gly Met Pro Asp Asp Phe Ala Gln Phe Trp Thr Ser Thr Ile
20 25 30
Ala Glu Ala Arg Gln Ala Gly Gly Glu Val Ser Ile Val Gln Ala Gln
35 40 45
Thr Thr Leu Lys Ala Val Gln Ser Phe Asp Val Thr Phe Pro Gly Tyr
50 55 60
Gly Gly His Pro Ile Lys Gly Trp Leu Ile Leu Pro Thr His His Lys
65 70 75 80
Gly Arg Leu Pro Leu Val Val Gln Tyr Ile Gly Tyr Gly Gly Gly Arg
85 90 95
Gly Leu Ala His Glu Gln Leu His Trp Ala Ala Ser Gly Phe Ala Tyr
100 105 110
Phe Arg Met Asp Thr Arg Gly Gln Gly Ser Asp Trp Ser Val Gly Glu
115 120 125
Thr Ala Asp Pro Val Gly Ser Thr Ser Ser Ile Pro Gly Phe Met Thr
130 135 140
Arg Gly Val Leu Asp Lys Asn Asp Tyr Tyr Tyr Arg Arg Leu Phe Thr
145 150 155 160
Asp Ala Val Arg Ala Ile Asp Ala Leu Leu Gly Leu Asp Phe Val Asp
165 170 175
Pro Glu Arg Ile Ala Val Cys Gly Asp Ser Gln Gly Gly Gly Ile Ser
180 185 190
Leu Ala Val Gly Gly Ile Asp Pro Arg Val Lys Ala Val Met Pro Asp
195 200 205
Val Pro Phe Leu Cys Asp Phe Pro Arg Ala Val Gln Thr Ala Val Arg
210 215 220
Asp Pro Tyr Leu Glu Ile Val Arg Phe Leu Ala Gln His Arg Glu Lys
225 230 235 240
Lys Ala Ala Val Phe Glu Thr Leu Asn Tyr Phe Asp Cys Val Asn Phe
245 250 255
Ala Arg Arg Ser Lys Ala Pro Ala Leu Phe Ser Val Ala Leu Met Asp
260 265 270
Glu Val Cys Pro Pro Ser Thr Val Tyr Gly Ala Phe Asn Ala Tyr Ala
275 280 285
Gly Glu Lys Thr Ile Thr Glu Tyr Glu Phe Asn Asn His Glu Gly Gly
290 295 300
Gln Gly Tyr Gln Glu Arg Gln Gln Met Thr Trp Leu Ser Arg Leu Phe
305 310 315 320
Gly Val Gly Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala
325 330 335
Gly Gly Pro Gly Ser Asp Pro Thr Lys Pro Pro Arg Thr Pro Thr Ala
340 345 350
Asn Thr Ser Arg Pro His His Asn Phe Gly Ser Gly Gly Gly Gly Ser
355 360 365
Pro His His His His His His
370 375
<210> 457
<211> 336
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 457
Met Thr Lys Ile Asn Asn Trp Gln Asp Tyr Gln Gly Ser Ser Leu Lys
1 5 10 15
Pro Glu Asp Phe Asp Lys Phe Trp Asp Glu Lys Ile Asn Leu Val Ser
20 25 30
Asn His Gln Phe Glu Phe Glu Leu Ile Glu Lys Asn Leu Ser Ser Lys
35 40 45
Val Val Asn Phe Tyr His Leu Trp Phe Thr Ala Ile Asp Gly Ala Lys
50 55 60
Ile His Ala Gln Leu Ile Val Pro Lys Asn Leu Lys Glu Lys Tyr Pro
65 70 75 80
Ala Ile Leu Gln Phe His Gly Tyr His Cys Asp Ser Gly Asp Trp Val
85 90 95
Asp Lys Ile Gly Ile Val Ala Glu Gly Asn Val Val Leu Ala Leu Asp
100 105 110
Cys Arg Gly Gln Gly Gly Leu Ser Gln Asp Asn Ile Gln Thr Met Gly
115 120 125
Met Thr Met Lys Gly Leu Ile Val Arg Gly Ile Asp Glu Gly Tyr Glu
130 135 140
Asn Leu Tyr Tyr Val Arg Gln Phe Met Asp Leu Ile Thr Ala Thr Lys
145 150 155 160
Ile Leu Ser Glu Phe Asp Phe Val Asp Glu Thr Asn Ile Ser Ala Gln
165 170 175
Gly Ala Ser Gln Gly Gly Ala Leu Ala Val Ala Cys Ala Ala Leu Ser
180 185 190
Pro Leu Ile Lys Lys Val Thr Ala Thr Tyr Pro Phe Leu Ser Asp Tyr
195 200 205
Arg Lys Ala Tyr Glu Leu Gly Ala Glu Glu Ser Ala Phe Glu Glu Leu
210 215 220
Pro Tyr Trp Phe Gln Phe Lys Asp Pro Leu His Leu Arg Glu Asp Trp
225 230 235 240
Phe Phe Asn Gln Leu Glu Tyr Ile Asp Ile Gln Asn Leu Ala Pro Arg
245 250 255
Ile Lys Ala Glu Val Ile Trp Ile Leu Gly Gly Lys Asp Thr Val Val
260 265 270
Pro Pro Ile Thr Gln Met Ala Ala Tyr Asn Lys Ile Gln Ser Lys Lys
275 280 285
Ser Leu Tyr Val Leu Pro Glu Tyr Gly His Glu Tyr Leu Pro Lys Ile
290 295 300
Ser Asp Trp Leu Arg Glu Asn Gln Gly Pro Gly Ser Gly Gly Ala Gly
305 310 315 320
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser His His His His His His
325 330 335
<210> 458
<211> 418
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 458
Met Thr Lys Ile Asn Asn Trp Gln Asp Tyr Gln Gly Ser Ser Leu Lys
1 5 10 15
Pro Glu Asp Phe Asp Lys Phe Trp Asp Glu Lys Ile Asn Leu Val Ser
20 25 30
Asn His Gln Phe Glu Phe Glu Leu Ile Glu Lys Asn Leu Ser Ser Lys
35 40 45
Val Val Asn Phe Tyr His Leu Trp Phe Thr Ala Ile Asp Gly Ala Lys
50 55 60
Ile His Ala Gln Leu Ile Val Pro Lys Asn Leu Lys Glu Lys Tyr Pro
65 70 75 80
Ala Ile Leu Gln Phe His Gly Tyr His Cys Asp Ser Gly Asp Trp Val
85 90 95
Asp Lys Ile Gly Ile Val Ala Glu Gly Asn Val Val Leu Ala Leu Asp
100 105 110
Cys Arg Gly Gln Gly Gly Leu Ser Gln Asp Asn Ile Gln Thr Met Gly
115 120 125
Met Thr Met Lys Gly Leu Ile Val Arg Gly Ile Asp Glu Gly Tyr Glu
130 135 140
Asn Leu Tyr Tyr Val Arg Gln Phe Met Asp Leu Ile Thr Ala Thr Lys
145 150 155 160
Ile Leu Ser Glu Phe Asp Phe Val Asp Glu Thr Asn Ile Ser Ala Gln
165 170 175
Gly Ala Ser Gln Gly Gly Ala Leu Ala Val Ala Cys Ala Ala Leu Ser
180 185 190
Pro Leu Ile Lys Lys Val Thr Ala Thr Tyr Pro Phe Leu Ser Asp Tyr
195 200 205
Arg Lys Ala Tyr Glu Leu Gly Ala Glu Glu Ser Ala Phe Glu Glu Leu
210 215 220
Pro Tyr Trp Phe Gln Phe Lys Asp Pro Leu His Leu Arg Glu Asp Trp
225 230 235 240
Phe Phe Asn Gln Leu Glu Tyr Ile Asp Ile Gln Asn Leu Ala Pro Arg
245 250 255
Ile Lys Ala Glu Val Ile Trp Ile Leu Gly Gly Lys Asp Thr Val Val
260 265 270
Pro Pro Ile Thr Gln Met Ala Ala Tyr Asn Lys Ile Gln Ser Lys Lys
275 280 285
Ser Leu Tyr Val Leu Pro Glu Tyr Gly His Glu Tyr Leu Pro Lys Ile
290 295 300
Ser Asp Trp Leu Arg Glu Asn Gln Gly Pro Gly Ser Gly Gly Ala Gly
305 310 315 320
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Pro Ser Ala Gln Ser Gln
325 330 335
Leu Pro Asp Lys His Ser Gly Leu His Glu Arg Ala Pro Gln Arg Tyr
340 345 350
Gly Pro Glu Pro Glu Pro Glu Pro Glu Pro Ile Pro Glu Pro Pro Lys
355 360 365
Glu Ala Pro Val Val Ile Glu Lys Pro Lys Pro Lys Pro Lys Pro Lys
370 375 380
Pro Lys Pro Pro Ala His Asp His Lys Asn Gln Lys Glu Thr His Gln
385 390 395 400
Arg His Ala Ala Gly Ser Gly Gly Gly Gly Ser Pro His His His His
405 410 415
His His
<210> 459
<211> 363
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 459
Met Thr Lys Ile Asn Asn Trp Gln Asp Tyr Gln Gly Ser Ser Leu Lys
1 5 10 15
Pro Glu Asp Phe Asp Lys Phe Trp Asp Glu Lys Ile Asn Leu Val Ser
20 25 30
Asn His Gln Phe Glu Phe Glu Leu Ile Glu Lys Asn Leu Ser Ser Lys
35 40 45
Val Val Asn Phe Tyr His Leu Trp Phe Thr Ala Ile Asp Gly Ala Lys
50 55 60
Ile His Ala Gln Leu Ile Val Pro Lys Asn Leu Lys Glu Lys Tyr Pro
65 70 75 80
Ala Ile Leu Gln Phe His Gly Tyr His Cys Asp Ser Gly Asp Trp Val
85 90 95
Asp Lys Ile Gly Ile Val Ala Glu Gly Asn Val Val Leu Ala Leu Asp
100 105 110
Cys Arg Gly Gln Gly Gly Leu Ser Gln Asp Asn Ile Gln Thr Met Gly
115 120 125
Met Thr Met Lys Gly Leu Ile Val Arg Gly Ile Asp Glu Gly Tyr Glu
130 135 140
Asn Leu Tyr Tyr Val Arg Gln Phe Met Asp Leu Ile Thr Ala Thr Lys
145 150 155 160
Ile Leu Ser Glu Phe Asp Phe Val Asp Glu Thr Asn Ile Ser Ala Gln
165 170 175
Gly Ala Ser Gln Gly Gly Ala Leu Ala Val Ala Cys Ala Ala Leu Ser
180 185 190
Pro Leu Ile Lys Lys Val Thr Ala Thr Tyr Pro Phe Leu Ser Asp Tyr
195 200 205
Arg Lys Ala Tyr Glu Leu Gly Ala Glu Glu Ser Ala Phe Glu Glu Leu
210 215 220
Pro Tyr Trp Phe Gln Phe Lys Asp Pro Leu His Leu Arg Glu Asp Trp
225 230 235 240
Phe Phe Asn Gln Leu Glu Tyr Ile Asp Ile Gln Asn Leu Ala Pro Arg
245 250 255
Ile Lys Ala Glu Val Ile Trp Ile Leu Gly Gly Lys Asp Thr Val Val
260 265 270
Pro Pro Ile Thr Gln Met Ala Ala Tyr Asn Lys Ile Gln Ser Lys Lys
275 280 285
Ser Leu Tyr Val Leu Pro Glu Tyr Gly His Glu Tyr Leu Pro Lys Ile
290 295 300
Ser Asp Trp Leu Arg Glu Asn Gln Gly Pro Gly Ser Gly Gly Ala Gly
305 310 315 320
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Asp Pro Thr Lys Pro Pro
325 330 335
Arg Thr Pro Thr Ala Asn Thr Ser Arg Pro His His Asn Phe Gly Ser
340 345 350
Gly Gly Gly Gly Ser Pro His His His His His
355 360
<210> 460
<211> 216
<212> PRT
<213> Mycobacterium smegmatis
<400> 460
Met Ala Lys Arg Ile Leu Cys Phe Gly Asp Ser Leu Thr Trp Gly Trp
1 5 10 15
Val Pro Val Glu Asp Gly Ala Pro Thr Glu Arg Phe Ala Pro Asp Val
20 25 30
Arg Trp Thr Gly Val Leu Ala Gln Gln Leu Gly Ala Asp Phe Glu Val
35 40 45
Ile Glu Glu Gly Leu Val Ala Arg Thr Thr Asn Ile Asp Asp Pro Thr
50 55 60
Asp Pro Arg Leu Asn Gly Ala Ser Tyr Leu Pro Ser Cys Leu Ala Thr
65 70 75 80
His Leu Pro Leu Asp Leu Val Ile Ile Met Leu Gly Thr Asn Asp Thr
85 90 95
Lys Ala Tyr Phe Arg Arg Thr Pro Leu Asp Ile Ala Leu Gly Met Ser
100 105 110
Val Leu Val Thr Gln Val Leu Thr Ser Ala Gly Gly Val Gly Thr Thr
115 120 125
Tyr Pro Ala Pro Lys Val Leu Val Val Ser Pro Pro Pro Leu Ala Pro
130 135 140
Met Pro His Pro Trp Phe Gln Leu Ile Phe Glu Gly Gly Glu Gln Lys
145 150 155 160
Thr Thr Glu Leu Ala Arg Val Tyr Ser Ala Leu Ala Ser Phe Met Lys
165 170 175
Val Pro Phe Phe Asp Ala Gly Ser Val Ile Ser Thr Asp Gly Val Asp
180 185 190
Gly Ile His Phe Thr Glu Ala Asn Asn Arg Asp Leu Gly Val Ala Leu
195 200 205
Ala Glu Gln Val Arg Ser Leu Leu
210 215
<210> 461
<211> 240
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 461
Met Ala Lys Arg Ile Leu Cys Phe Gly Asp Ser Leu Thr Trp Gly Trp
1 5 10 15
Val Pro Val Glu Asp Gly Ala Pro Thr Glu Arg Phe Ala Pro Asp Val
20 25 30
Arg Trp Thr Gly Val Leu Ala Gln Gln Leu Gly Ala Asp Phe Glu Val
35 40 45
Ile Glu Glu Gly Leu Val Ala Arg Thr Thr Asn Ile Asp Asp Pro Thr
50 55 60
Asp Pro Arg Leu Asn Gly Ala Ser Tyr Leu Pro Ser Cys Leu Ala Thr
65 70 75 80
His Leu Pro Leu Asp Leu Val Ile Ile Met Leu Gly Thr Asn Asp Thr
85 90 95
Lys Ala Tyr Phe Arg Arg Thr Pro Leu Asp Ile Ala Leu Gly Met Ser
100 105 110
Val Leu Val Thr Gln Val Leu Thr Ser Ala Gly Gly Val Gly Thr Thr
115 120 125
Tyr Pro Ala Pro Lys Val Leu Val Val Ser Pro Pro Pro Leu Ala Pro
130 135 140
Met Pro His Pro Trp Phe Gln Leu Ile Phe Glu Gly Gly Glu Gln Lys
145 150 155 160
Thr Thr Glu Leu Ala Arg Val Tyr Ser Ala Leu Ala Ser Phe Met Lys
165 170 175
Val Pro Phe Phe Asp Ala Gly Ser Val Ile Ser Thr Asp Gly Val Asp
180 185 190
Gly Ile His Phe Thr Glu Ala Asn Asn Arg Asp Leu Gly Val Ala Leu
195 200 205
Ala Glu Gln Val Arg Ser Leu Leu Gly Pro Gly Ser Gly Gly Ala Gly
210 215 220
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser His His His His His His
225 230 235 240
<210> 462
<211> 322
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 462
Met Ala Lys Arg Ile Leu Cys Phe Gly Asp Ser Leu Thr Trp Gly Trp
1 5 10 15
Val Pro Val Glu Asp Gly Ala Pro Thr Glu Arg Phe Ala Pro Asp Val
20 25 30
Arg Trp Thr Gly Val Leu Ala Gln Gln Leu Gly Ala Asp Phe Glu Val
35 40 45
Ile Glu Glu Gly Leu Val Ala Arg Thr Thr Asn Ile Asp Asp Pro Thr
50 55 60
Asp Pro Arg Leu Asn Gly Ala Ser Tyr Leu Pro Ser Cys Leu Ala Thr
65 70 75 80
His Leu Pro Leu Asp Leu Val Ile Ile Met Leu Gly Thr Asn Asp Thr
85 90 95
Lys Ala Tyr Phe Arg Arg Thr Pro Leu Asp Ile Ala Leu Gly Met Ser
100 105 110
Val Leu Val Thr Gln Val Leu Thr Ser Ala Gly Gly Val Gly Thr Thr
115 120 125
Tyr Pro Ala Pro Lys Val Leu Val Val Ser Pro Pro Pro Leu Ala Pro
130 135 140
Met Pro His Pro Trp Phe Gln Leu Ile Phe Glu Gly Gly Glu Gln Lys
145 150 155 160
Thr Thr Glu Leu Ala Arg Val Tyr Ser Ala Leu Ala Ser Phe Met Lys
165 170 175
Val Pro Phe Phe Asp Ala Gly Ser Val Ile Ser Thr Asp Gly Val Asp
180 185 190
Gly Ile His Phe Thr Glu Ala Asn Asn Arg Asp Leu Gly Val Ala Leu
195 200 205
Ala Glu Gln Val Arg Ser Leu Leu Gly Pro Gly Ser Gly Gly Ala Gly
210 215 220
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Pro Ser Ala Gln Ser Gln
225 230 235 240
Leu Pro Asp Lys His Ser Gly Leu His Glu Arg Ala Pro Gln Arg Tyr
245 250 255
Gly Pro Glu Pro Glu Pro Glu Pro Glu Pro Ile Pro Glu Pro Pro Lys
260 265 270
Glu Ala Pro Val Val Ile Glu Lys Pro Lys Pro Lys Pro Lys Pro Lys
275 280 285
Pro Lys Pro Pro Ala His Asp His Lys Asn Gln Lys Glu Thr His Gln
290 295 300
Arg His Ala Ala Gly Ser Gly Gly Gly Gly Ser Pro His His His His
305 310 315 320
His His
<210> 463
<211> 250
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 463
Met Ala Lys Arg Ile Leu Cys Phe Gly Asp Ser Leu Thr Trp Gly Trp
1 5 10 15
Val Pro Val Glu Asp Gly Ala Pro Thr Glu Arg Phe Ala Pro Asp Val
20 25 30
Arg Trp Thr Gly Val Leu Ala Gln Gln Leu Gly Ala Asp Phe Glu Val
35 40 45
Ile Glu Glu Gly Leu Val Ala Arg Thr Thr Asn Ile Asp Asp Pro Thr
50 55 60
Asp Pro Arg Leu Asn Gly Ala Ser Tyr Leu Pro Ser Cys Leu Ala Thr
65 70 75 80
His Leu Pro Leu Asp Leu Val Ile Ile Met Leu Gly Thr Asn Asp Thr
85 90 95
Lys Ala Tyr Phe Arg Arg Thr Pro Leu Asp Ile Ala Leu Gly Met Ser
100 105 110
Val Leu Val Thr Gln Val Leu Thr Ser Ala Gly Gly Val Gly Thr Thr
115 120 125
Tyr Pro Ala Pro Lys Val Leu Val Val Ser Pro Pro Pro Leu Ala Pro
130 135 140
Met Pro His Pro Trp Phe Gln Leu Ile Phe Glu Gly Gly Glu Gln Lys
145 150 155 160
Thr Thr Glu Leu Ala Arg Val Tyr Ser Ala Leu Ala Ser Phe Met Lys
165 170 175
Val Pro Phe Phe Asp Ala Gly Ser Val Ile Ser Thr Asp Gly Val Asp
180 185 190
Gly Ile His Phe Thr Glu Ala Asn Asn Arg Asp Leu Gly Val Ala Leu
195 200 205
Ala Glu Gln Val Arg Ser Leu Leu Gly Pro Gly Ser Gly Gly Ala Gly
210 215 220
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Gly Lys Gly Lys Gly Lys
225 230 235 240
Gly Lys Gly Lys His His His His His His
245 250
<210> 464
<211> 268
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 464
Met Ala Lys Arg Ile Leu Cys Phe Gly Asp Ser Leu Thr Trp Gly Trp
1 5 10 15
Val Pro Val Glu Asp Gly Ala Pro Thr Glu Arg Phe Ala Pro Asp Val
20 25 30
Arg Trp Thr Gly Val Leu Ala Gln Gln Leu Gly Ala Asp Phe Glu Val
35 40 45
Ile Glu Glu Gly Leu Val Ala Arg Thr Thr Asn Ile Asp Asp Pro Thr
50 55 60
Asp Pro Arg Leu Asn Gly Ala Ser Tyr Leu Pro Ser Cys Leu Ala Thr
65 70 75 80
His Leu Pro Leu Asp Leu Val Ile Ile Met Leu Gly Thr Asn Asp Thr
85 90 95
Lys Ala Tyr Phe Arg Arg Thr Pro Leu Asp Ile Ala Leu Gly Met Ser
100 105 110
Val Leu Val Thr Gln Val Leu Thr Ser Ala Gly Gly Val Gly Thr Thr
115 120 125
Tyr Pro Ala Pro Lys Val Leu Val Val Ser Pro Pro Pro Leu Ala Pro
130 135 140
Met Pro His Pro Trp Phe Gln Leu Ile Phe Glu Gly Gly Glu Gln Lys
145 150 155 160
Thr Thr Glu Leu Ala Arg Val Tyr Ser Ala Leu Ala Ser Phe Met Lys
165 170 175
Val Pro Phe Phe Asp Ala Gly Ser Val Ile Ser Thr Asp Gly Val Asp
180 185 190
Gly Ile His Phe Thr Glu Ala Asn Asn Arg Asp Leu Gly Val Ala Leu
195 200 205
Ala Glu Gln Val Arg Ser Leu Leu Gly Pro Gly Ser Gly Gly Ala Gly
210 215 220
Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Asp Pro Thr Lys Pro Pro
225 230 235 240
Arg Thr Pro Thr Ala Asn Thr Ser Arg Pro His His Asn Phe Gly Ser
245 250 255
Gly Gly Gly Gly Ser Pro His His His His His His
260 265
<210> 465
<211> 296
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 465
Met Ser Thr Phe Val Ala Lys Asp Gly Thr Gln Ile Tyr Phe Lys Asp
1 5 10 15
Trp Gly Ser Gly Lys Pro Val Leu Phe Ser His Gly Trp Pro Leu Asp
20 25 30
Ala Asp Met Trp Glu Tyr Gln Met Glu Tyr Leu Ser Ser Arg Gly Tyr
35 40 45
Arg Thr Ile Ala Phe Asp Arg Arg Gly Phe Gly Arg Ser Asp Gln Pro
50 55 60
Trp Thr Gly Asn Asp Tyr Asp Thr Phe Ala Asp Asp Ile Ala Gln Leu
65 70 75 80
Ile Glu His Leu Asp Leu Lys Glu Val Thr Leu Val Gly Phe Ser Met
85 90 95
Gly Gly Gly Asp Val Ala Arg Tyr Ile Ala Arg His Gly Ser Ala Arg
100 105 110
Val Ala Gly Leu Val Leu Leu Gly Ala Val Thr Pro Leu Phe Gly Gln
115 120 125
Lys Pro Asp Tyr Pro Gln Gly Val Pro Leu Asp Val Phe Ala Arg Phe
130 135 140
Lys Thr Glu Leu Leu Lys Asp Arg Ala Gln Phe Ile Ser Asp Phe Asn
145 150 155 160
Ala Pro Phe Tyr Gly Ile Asn Lys Gly Gln Val Val Ser Gln Gly Val
165 170 175
Gln Thr Gln Thr Leu Gln Ile Ala Leu Leu Ala Ser Leu Lys Ala Thr
180 185 190
Val Asp Cys Val Thr Ala Phe Ala Glu Thr Asp Phe Arg Pro Asp Met
195 200 205
Ala Lys Ile Asp Val Pro Thr Leu Val Ile His Gly Asp Gly Asp Gln
210 215 220
Ile Val Pro Phe Glu Thr Thr Gly Lys Val Ala Ala Glu Leu Ile Lys
225 230 235 240
Gly Ala Glu Leu Lys Val Tyr Lys Asp Ala Pro His Gly Phe Ala Val
245 250 255
Thr His Ala Gln Gln Leu Asn Glu Asp Leu Leu Ala Phe Leu Lys Arg
260 265 270
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
275 280 285
Gly Ser His His His His His His
290 295
<210> 466
<211> 306
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 466
Met Ser Thr Phe Val Ala Lys Asp Gly Thr Gln Ile Tyr Phe Lys Asp
1 5 10 15
Trp Gly Ser Gly Lys Pro Val Leu Phe Ser His Gly Trp Pro Leu Asp
20 25 30
Ala Asp Met Trp Glu Tyr Gln Met Glu Tyr Leu Ser Ser Arg Gly Tyr
35 40 45
Arg Thr Ile Ala Phe Asp Arg Arg Gly Phe Gly Arg Ser Asp Gln Pro
50 55 60
Trp Thr Gly Asn Asp Tyr Asp Thr Phe Ala Asp Asp Ile Ala Gln Leu
65 70 75 80
Ile Glu His Leu Asp Leu Lys Glu Val Thr Leu Val Gly Phe Ser Met
85 90 95
Gly Gly Gly Asp Val Ala Arg Tyr Ile Ala Arg His Gly Ser Ala Arg
100 105 110
Val Ala Gly Leu Val Leu Leu Gly Ala Val Thr Pro Leu Phe Gly Gln
115 120 125
Lys Pro Asp Tyr Pro Gln Gly Val Pro Leu Asp Val Phe Ala Arg Phe
130 135 140
Lys Thr Glu Leu Leu Lys Asp Arg Ala Gln Phe Ile Ser Asp Phe Asn
145 150 155 160
Ala Pro Phe Tyr Gly Ile Asn Lys Gly Gln Val Val Ser Gln Gly Val
165 170 175
Gln Thr Gln Thr Leu Gln Ile Ala Leu Leu Ala Ser Leu Lys Ala Thr
180 185 190
Val Asp Cys Val Thr Ala Phe Ala Glu Thr Asp Phe Arg Pro Asp Met
195 200 205
Ala Lys Ile Asp Val Pro Thr Leu Val Ile His Gly Asp Gly Asp Gln
210 215 220
Ile Val Pro Phe Glu Thr Thr Gly Lys Val Ala Ala Glu Leu Ile Lys
225 230 235 240
Gly Ala Glu Leu Lys Val Tyr Lys Asp Ala Pro His Gly Phe Ala Val
245 250 255
Thr His Ala Gln Gln Leu Asn Glu Asp Leu Leu Ala Phe Leu Lys Arg
260 265 270
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
275 280 285
Gly Ser Gly Lys Gly Lys Gly Lys Gly Lys Gly Lys His His His His
290 295 300
His His
305
<210> 467
<211> 324
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 467
Met Ser Thr Phe Val Ala Lys Asp Gly Thr Gln Ile Tyr Phe Lys Asp
1 5 10 15
Trp Gly Ser Gly Lys Pro Val Leu Phe Ser His Gly Trp Pro Leu Asp
20 25 30
Ala Asp Met Trp Glu Tyr Gln Met Glu Tyr Leu Ser Ser Arg Gly Tyr
35 40 45
Arg Thr Ile Ala Phe Asp Arg Arg Gly Phe Gly Arg Ser Asp Gln Pro
50 55 60
Trp Thr Gly Asn Asp Tyr Asp Thr Phe Ala Asp Asp Ile Ala Gln Leu
65 70 75 80
Ile Glu His Leu Asp Leu Lys Glu Val Thr Leu Val Gly Phe Ser Met
85 90 95
Gly Gly Gly Asp Val Ala Arg Tyr Ile Ala Arg His Gly Ser Ala Arg
100 105 110
Val Ala Gly Leu Val Leu Leu Gly Ala Val Thr Pro Leu Phe Gly Gln
115 120 125
Lys Pro Asp Tyr Pro Gln Gly Val Pro Leu Asp Val Phe Ala Arg Phe
130 135 140
Lys Thr Glu Leu Leu Lys Asp Arg Ala Gln Phe Ile Ser Asp Phe Asn
145 150 155 160
Ala Pro Phe Tyr Gly Ile Asn Lys Gly Gln Val Val Ser Gln Gly Val
165 170 175
Gln Thr Gln Thr Leu Gln Ile Ala Leu Leu Ala Ser Leu Lys Ala Thr
180 185 190
Val Asp Cys Val Thr Ala Phe Ala Glu Thr Asp Phe Arg Pro Asp Met
195 200 205
Ala Lys Ile Asp Val Pro Thr Leu Val Ile His Gly Asp Gly Asp Gln
210 215 220
Ile Val Pro Phe Glu Thr Thr Gly Lys Val Ala Ala Glu Leu Ile Lys
225 230 235 240
Gly Ala Glu Leu Lys Val Tyr Lys Asp Ala Pro His Gly Phe Ala Val
245 250 255
Thr His Ala Gln Gln Leu Asn Glu Asp Leu Leu Ala Phe Leu Lys Arg
260 265 270
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
275 280 285
Gly Ser Asp Pro Thr Lys Pro Pro Arg Thr Pro Thr Ala Asn Thr Ser
290 295 300
Arg Pro His His Asn Phe Gly Ser Gly Gly Gly Gly Ser Pro His His
305 310 315 320
His His His His
<210> 468
<211> 61
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 468
Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
1 5 10 15
Ser Ala Gly Gly Pro Gly Ser Glu Pro Glu Pro Glu Trp Lys Thr Lys
20 25 30
Lys Ile Leu Leu Ser Arg Thr Arg Arg Ile Met Arg Gln Val Val Arg
35 40 45
Ser Val Met His Lys Ile Trp His His His His His His
50 55 60
<210> 469
<211> 62
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 469
Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly
1 5 10 15
Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Trp Lys Thr
20 25 30
Lys Lys Ile Leu Leu Ser Arg Thr Arg Arg Ile Met Arg Gln Val Val
35 40 45
Arg Ser Val Met His Lys Ile Trp His His His His His His
50 55 60
<210> 470
<211> 61
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 470
Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly
1 5 10 15
Ser Ala Gly Gly Pro Gly Ser Glu Pro Glu Pro Glu Pro Leu Trp Arg
20 25 30
Arg Ile Thr Lys Arg Lys Leu Val Arg Pro Val Ala Thr Leu Met Trp
35 40 45
Tyr Trp Phe Thr Ser Lys Arg His His His His His His
50 55 60
<210> 471
<211> 62
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 471
Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly
1 5 10 15
Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro Gly Ser Pro Leu Trp
20 25 30
Arg Arg Ile Thr Lys Arg Lys Leu Val Arg Pro Val Ala Thr Leu Met
35 40 45
Trp Tyr Trp Phe Thr Ser Lys Arg His His His His His His
50 55 60
<210> 472
<211> 57
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 472
Glu Pro Glu Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala
1 5 10 15
Gly Gly Pro Gly Ser Glu Pro Glu Arg Met Leu Ser Arg Ile Leu Arg
20 25 30
Met Phe Val Arg Ile Leu Lys Arg Glu Arg Leu Ser Gln Val Arg Gly
35 40 45
Leu Phe Val His His His His His His
50 55
<210> 473
<211> 58
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 473
Glu Pro Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly Gly Ala Gly Ser
1 5 10 15
Pro Gly Ser Ala Gly Gly Pro Gly Ser Arg Met Leu Ser Arg Ile Leu
20 25 30
Arg Met Phe Val Arg Ile Leu Lys Arg Glu Arg Leu Ser Gln Val Arg
35 40 45
Gly Leu Phe Val His His His His His His
50 55
<210> 474
<211> 65
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 474
Glu Pro Glu Pro Glu Pro Glu Gly Pro Gly Ser Gly Gly Ala Gly Ser
1 5 10 15
Pro Gly Ser Ala Gly Gly Pro Gly Ser Glu Pro Glu Pro Glu Pro Glu
20 25 30
Leu Arg Phe Leu Ala Arg Arg Phe Leu Lys Leu Arg Arg Ala Arg Lys
35 40 45
Trp Trp Asn Ala Trp Lys Val Trp Val Thr Arg His His His His His
50 55 60
His
65
<210> 475
<211> 66
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 475
Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu Gly
1 5 10 15
Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro Gly
20 25 30
Ser Leu Arg Phe Leu Ala Arg Arg Phe Leu Lys Leu Arg Arg Ala Arg
35 40 45
Lys Trp Trp Asn Ala Trp Lys Val Trp Val Thr Arg His His His His
50 55 60
His His
65
<210> 476
<211> 325
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 476
Met Ala Phe Phe Asp Leu Pro Leu Glu Glu Leu Lys Lys Tyr Arg Pro
1 5 10 15
Glu Arg Tyr Glu Glu Lys Asp Phe Asp Glu Phe Trp Glu Glu Thr Leu
20 25 30
Ala Glu Ser Glu Lys Phe Pro Leu Asp Pro Val Phe Glu Arg Met Glu
35 40 45
Ser His Leu Lys Thr Val Glu Ala Tyr Asp Val Thr Phe Ser Gly Tyr
50 55 60
Arg Gly Gln Arg Ile Lys Gly Trp Leu Leu Val Pro Lys Leu Glu Glu
65 70 75 80
Glu Lys Leu Pro Cys Val Val Gln Tyr Ile Gly Tyr Asn Gly Gly Arg
85 90 95
Gly Phe Pro His Asp Trp Leu Phe Trp Pro Ser Met Gly Tyr Ile Cys
100 105 110
Phe Val Met Asp Thr Arg Gly Gln Gly Ser Gly Trp Leu Lys Gly Asp
115 120 125
Thr Pro Asp Tyr Pro Glu Gly Pro Val Asp Pro Gln Tyr Pro Gly Phe
130 135 140
Met Thr Arg Gly Ile Leu Asp Pro Arg Thr Tyr Tyr Tyr Arg Arg Val
145 150 155 160
Phe Thr Asp Ala Val Arg Ala Val Glu Ala Ala Ala Ser Phe Pro Gln
165 170 175
Val Asp Gln Glu Arg Ile Val Ile Ala Gly Gly Ser Gln Gly Gly Gly
180 185 190
Ile Ala Leu Ala Val Ser Ala Leu Ser Lys Lys Ala Lys Ala Leu Leu
195 200 205
Cys Asp Val Pro Phe Leu Cys His Phe Arg Arg Ala Val Gln Leu Val
210 215 220
Asp Thr His Pro Tyr Ala Glu Ile Thr Asn Phe Leu Lys Thr His Arg
225 230 235 240
Asp Lys Glu Glu Ile Val Phe Arg Thr Leu Ser Tyr Phe Asp Gly Val
245 250 255
Asn Phe Ala Ala Arg Ala Lys Ile Pro Ala Leu Phe Ser Val Gly Leu
260 265 270
Met Asp Asn Ile Thr Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn Tyr
275 280 285
Tyr Ala Gly Pro Lys Glu Ile Pro Ile Tyr Pro Tyr Asn Asn His Glu
290 295 300
Gly Gly Gly Ser Phe Gln Ala Val Glu Gln Val Lys Phe Leu Lys Lys
305 310 315 320
Leu Phe Glu Lys Gly
325
<210> 477
<211> 272
<212> PRT
<213> Pseudomonas fluorescens
<400> 477
Met Ser Thr Phe Val Ala Lys Asp Gly Thr Gln Ile Tyr Phe Lys Asp
1 5 10 15
Trp Gly Ser Gly Lys Pro Val Leu Phe Ser His Gly Trp Pro Leu Asp
20 25 30
Ala Asp Met Trp Glu Tyr Gln Met Glu Tyr Leu Ser Ser Arg Gly Tyr
35 40 45
Arg Thr Ile Ala Phe Asp Arg Arg Gly Phe Gly Arg Ser Asp Gln Pro
50 55 60
Trp Thr Gly Asn Asp Tyr Asp Thr Phe Ala Asp Asp Ile Ala Gln Leu
65 70 75 80
Ile Glu His Leu Asp Leu Lys Glu Val Thr Leu Val Gly Phe Ser Met
85 90 95
Gly Gly Gly Asp Val Ala Arg Tyr Ile Ala Arg His Gly Ser Ala Arg
100 105 110
Val Ala Gly Leu Val Leu Leu Gly Ala Val Thr Pro Leu Phe Gly Gln
115 120 125
Lys Pro Asp Tyr Pro Gln Gly Val Pro Leu Asp Val Phe Ala Arg Phe
130 135 140
Lys Thr Glu Leu Leu Lys Asp Arg Ala Gln Phe Ile Ser Asp Phe Asn
145 150 155 160
Ala Pro Phe Tyr Gly Ile Asn Lys Gly Gln Val Val Ser Gln Gly Val
165 170 175
Gln Thr Gln Thr Leu Gln Ile Ala Leu Leu Ala Ser Leu Lys Ala Thr
180 185 190
Val Asp Cys Val Thr Ala Phe Ala Glu Thr Asp Phe Arg Pro Asp Met
195 200 205
Ala Lys Ile Asp Val Pro Thr Leu Val Ile His Gly Asp Gly Asp Gln
210 215 220
Ile Val Pro Phe Glu Thr Thr Gly Lys Val Ala Ala Glu Leu Ile Lys
225 230 235 240
Gly Ala Glu Leu Lys Val Tyr Lys Asp Ala Pro His Gly Phe Ala Val
245 250 255
Thr His Ala Gln Gln Leu Asn Glu Asp Leu Leu Ala Phe Leu Lys Arg
260 265 270
<210> 478
<211> 216
<212> PRT
<213> Mycobacterium smegmatis
<400> 478
Met Ala Lys Arg Ile Leu Cys Phe Gly Asp Ser Leu Thr Trp Gly Trp
1 5 10 15
Val Pro Val Glu Asp Gly Ala Pro Thr Glu Arg Phe Ala Pro Asp Val
20 25 30
Arg Trp Thr Gly Val Leu Ala Gln Gln Leu Gly Ala Asp Phe Glu Val
35 40 45
Ile Glu Glu Gly Leu Ser Ala Arg Thr Thr Asn Ile Asp Asp Pro Thr
50 55 60
Asp Pro Arg Leu Asn Gly Ala Ser Tyr Leu Pro Ser Cys Leu Ala Thr
65 70 75 80
His Leu Pro Leu Asp Leu Val Ile Ile Met Leu Gly Thr Asn Asp Thr
85 90 95
Lys Ala Tyr Phe Arg Arg Thr Pro Leu Asp Ile Ala Leu Gly Met Ser
100 105 110
Val Leu Val Thr Gln Val Leu Thr Ser Ala Gly Gly Val Gly Thr Thr
115 120 125
Tyr Pro Ala Pro Lys Val Leu Val Val Ser Pro Pro Pro Leu Ala Pro
130 135 140
Met Pro His Pro Trp Phe Gln Leu Ile Phe Glu Gly Gly Glu Gln Lys
145 150 155 160
Thr Thr Glu Leu Ala Arg Val Tyr Ser Ala Leu Ala Ser Phe Met Lys
165 170 175
Val Pro Phe Phe Asp Ala Gly Ser Val Ile Ser Thr Asp Gly Val Asp
180 185 190
Gly Ile His Phe Thr Glu Ala Asn Asn Arg Asp Leu Gly Val Ala Leu
195 200 205
Ala Glu Gln Val Arg Ser Leu Leu
210 215
<210> 479
<211> 376
<212> PRT
<213> artificial sequence
<220>
<223> synthetic construct
<400> 479
Met Ser Thr Phe Val Ala Lys Asp Gly Thr Gln Ile Tyr Phe Lys Asp
1 5 10 15
Trp Gly Ser Gly Lys Pro Val Leu Phe Ser His Gly Trp Pro Leu Asp
20 25 30
Ala Asp Met Trp Glu Tyr Gln Met Glu Tyr Leu Ser Ser Arg Gly Tyr
35 40 45
Arg Thr Ile Ala Phe Asp Arg Arg Gly Phe Gly Arg Ser Asp Gln Pro
50 55 60
Trp Thr Gly Asn Asp Tyr Asp Thr Phe Ala Asp Asp Ile Ala Gln Leu
65 70 75 80
Ile Glu His Leu Asp Leu Lys Glu Val Thr Leu Val Gly Phe Ser Met
85 90 95
Gly Gly Gly Asp Val Ala Arg Tyr Ile Ala Arg His Gly Ser Ala Arg
100 105 110
Val Ala Gly Leu Val Leu Leu Gly Ala Val Thr Pro Leu Phe Gly Gln
115 120 125
Lys Pro Asp Tyr Pro Gln Gly Val Pro Leu Asp Val Phe Ala Arg Phe
130 135 140
Lys Thr Glu Leu Leu Lys Asp Arg Ala Gln Phe Ile Ser Asp Phe Asn
145 150 155 160
Ala Pro Phe Tyr Gly Ile Asn Lys Gly Gln Val Val Ser Gln Gly Val
165 170 175
Gln Thr Gln Thr Leu Gln Ile Ala Leu Leu Ala Ser Leu Lys Ala Thr
180 185 190
Val Asp Cys Val Thr Ala Phe Ala Glu Thr Asp Phe Arg Pro Asp Met
195 200 205
Ala Lys Ile Asp Val Pro Thr Leu Val Ile His Gly Asp Gly Asp Gln
210 215 220
Ile Val Pro Phe Glu Thr Thr Gly Lys Val Ala Ala Glu Leu Ile Lys
225 230 235 240
Gly Ala Glu Leu Lys Val Tyr Lys Asp Ala Pro His Gly Phe Ala Val
245 250 255
Thr His Ala Gln Gln Leu Asn Glu Asp Leu Leu Ala Phe Leu Lys Arg
260 265 270
Gly Pro Gly Ser Gly Gly Ala Gly Ser Pro Gly Ser Ala Gly Gly Pro
275 280 285
Gly Ser Pro Ser Ala Gln Ser Gln Leu Pro Asp Lys His Ser Gly Leu
290 295 300
His Glu Arg Ala Pro Gln Arg Tyr Gly Pro Glu Pro Glu Pro Glu Pro
305 310 315 320
Glu Pro Ile Pro Glu Pro Pro Lys Glu Ala Pro Val Val Ile Glu Lys
325 330 335
Pro Lys Pro Lys Pro Lys Pro Lys Pro Lys Pro Pro Ala His Asp His
340 345 350
Lys Asn Gln Lys Glu Thr His Gln Arg His Ala Ala Gly Ser Gly Gly
355 360 365
Gly Gly Ser Pro His His His His
370 375
Claims (44)
1) a) i) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티(moiety)이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
ii) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
iii) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
iv) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
b) 과산소원; 및
c) 과가수분해(perhydrolytic) 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유(CLUSTALW)를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처(signature) 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
i) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
ii) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
iii) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - 를 포함하는 반응 성분의 세트를 제공하는 단계;
2) (1)의 반응 성분을 적절한 반응 조건 하에서 배합하는 단계 - 이에 의해, 적어도 하나의 과산이 효소에 의해 생성된다 - ; 및
3) 구강 표면을 적어도 하나의 과산과 접촉시키는 단계 - 이에 의해, 구강 표면에 탈색(bleaching), 치아 미백, 소독, 탈염(destaining), 탈취, 생물막(biofilm)의 감소 또는 제거 및 그들의 조합으로 이루어진 군으로부터 선택되는 과산 기반의 이익이 제공된다 - 를 포함하는 방법.
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티(moiety)이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
ii) 구조
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
iii) 화학식
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
iv) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
b) 과산소원; 및
c) 과가수분해(perhydrolytic) 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유(CLUSTALW)를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처(signature) 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
i) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
ii) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
iii) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - 를 포함하는 반응 성분의 세트를 제공하는 단계;
2) (1)의 반응 성분을 적절한 반응 조건 하에서 배합하는 단계 - 이에 의해, 적어도 하나의 과산이 효소에 의해 생성된다 - ; 및
3) 구강 표면을 적어도 하나의 과산과 접촉시키는 단계 - 이에 의해, 구강 표면에 탈색(bleaching), 치아 미백, 소독, 탈염(destaining), 탈취, 생물막(biofilm)의 감소 또는 제거 및 그들의 조합으로 이루어진 군으로부터 선택되는 과산 기반의 이익이 제공된다 - 를 포함하는 방법.
제1항에 있어서, 반응 성분이 구강 내에서 배합되는 방법.
제1항에 있어서, 반응 성분이 구강 표면과의 접촉 전에 구강의 외측에서 배합되는 방법.
제1항에 있어서, 과가수분해 활성을 갖는 효소가 과산 유익제(benefit agent)의 생성 전에 구강 내에 존재하는 방법.
1) a) i) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
ii) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
iii) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
iv) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
b) 과산소원; 및
c) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 일반 구조
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
(여기서,
PAH는 과가수분해 활성을 갖는 효소이며;
OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이고;
L은 길이가 1 내지 100개 아미노산 범위인 임의의 펩티드 링커이며;
y는 0 또는 1이다)를 갖는 융합 단백질을 포함한다 - 를 포함하는 반응 성분의 세트를 제공하는 단계;
2) (1)의 반응 성분을 적절한 반응 조건 하에서 배합하는 단계 - 이에 의해, 적어도 하나의 과산이 효소에 의해 생성된다 - ; 및
3) 구강 표면을 적어도 하나의 과산과 접촉시키는 단계 - 이에 의해, 구강 표면에 탈색, 치아 미백, 소독, 탈염, 탈취, 생물막의 감소 또는 제거 및 그들의 조합으로 이루어진 군으로부터 선택되는 과산 기반의 이익이 제공된다 - 를 포함하는 방법.
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
m은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
ii) 구조
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
iii) 화학식
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
iv) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
b) 과산소원; 및
c) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 일반 구조
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
(여기서,
PAH는 과가수분해 활성을 갖는 효소이며;
OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이고;
L은 길이가 1 내지 100개 아미노산 범위인 임의의 펩티드 링커이며;
y는 0 또는 1이다)를 갖는 융합 단백질을 포함한다 - 를 포함하는 반응 성분의 세트를 제공하는 단계;
2) (1)의 반응 성분을 적절한 반응 조건 하에서 배합하는 단계 - 이에 의해, 적어도 하나의 과산이 효소에 의해 생성된다 - ; 및
3) 구강 표면을 적어도 하나의 과산과 접촉시키는 단계 - 이에 의해, 구강 표면에 탈색, 치아 미백, 소독, 탈염, 탈취, 생물막의 감소 또는 제거 및 그들의 조합으로 이루어진 군으로부터 선택되는 과산 기반의 이익이 제공된다 - 를 포함하는 방법.
제5항에 있어서, 과가수분해 활성을 갖는 효소가 리파제, 프로테아제, 에스테라제, 아실 트랜스퍼라제, 아릴 에스테라제, 탄수화물 에스테라제 및 그들의 조합의 군으로부터 선택되는 방법.
제6항에 있어서, 탄수화물 에스테라제가 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 CE-7 탄수화물 에스테라제이며, 상기 시그니처 모티프가
1) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
2) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
3) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함하는 방법.
1) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
2) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
3) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함하는 방법.
제5항, 제6항 또는 제7항에 있어서, 구강 표면에 대하여 친화성을 갖는 펩티드 성분이 항체, Fab 항체 단편, 단쇄 가변 단편(scFv) 항체, 카멜리대(Camelidae) 항체, 스캐폴드(scaffold) 디스플레이 단백질 또는 면역글로불린 폴드(fold)가 결여된 단쇄 폴리펩티드인 방법.
제6항에 있어서, 구강 표면에 대하여 친화성을 갖는 펩티드 성분이 면역글로불린 폴드가 결여된 단쇄 폴리펩티드인 방법.
제9항에 있어서, 단쇄 폴리펩티드가 구강 표면에 대한 KD 값 또는 MB50 값이 10-5 M 이하인, 길이가 5 내지 60개 아미노산 범위인 적어도 하나의 구강 표면-결합 펩티드를 포함하는 방법.
제10항에 있어서, 단쇄 폴리펩티드가 2 내지 50개 구강 표면-결합 펩티드를 포함하며, 구강 표면-결합 펩티드는 독립적으로 그리고 임의로, 길이가 1 내지 100개 아미노산 범위인 폴리펩티드 스페이서에 의해 분리되는 방법.
제1항 또는 제5항에 있어서, 구강 표면이 치아 표면인 방법.
제12항에 있어서, 치아 표면이 치아 에나멜(enamel), 치아 균막(pellicle) 또는 그들의 조합을 포함하는 방법.
제10항에 있어서, 구강 표면-결합 펩티드가 치아 에나멜-결합 펩티드인 방법.
제10항에 있어서, 구강 표면-결합 펩티드가 치아 균막-결합 펩티드인 방법.
제10항에 있어서, 구강 표면-결합 펩티드가 피부-결합 펩티드인 방법.
제1항 또는 제5항에 있어서, 과산이 반응 성분의 세트의 배합 5분 내에 500 ppb 내지 10,000 ppm의 농도로 생성되는 방법.
제17항에 있어서, 과산이 1시간 미만 동안 구강 관리 표면과 접촉되는 방법.
제1항 또는 제5항에 있어서, 과산이 과아세트산인 방법.
제19항에 있어서, 반응 성분의 배합 5분 내에, 유효 농도의 과아세트산이 효소에 의해 생성되고, 구강 표면과 접촉되는 방법.
제1항 또는 제5항에 있어서, 기질이 트라이아세틴을 포함하는 방법.
하기의 일반 구조를 포함하는 융합 단백질:
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
상기 식에서,
1) PAH는 과가수분해 활성을 갖는 효소이고;
2) OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이며;
3) L은 길이가 1 내지 100개 아미노산 범위인 펩티드 링커이고;
4) y는 0 또는 1이다.
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
상기 식에서,
1) PAH는 과가수분해 활성을 갖는 효소이고;
2) OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이며;
3) L은 길이가 1 내지 100개 아미노산 범위인 펩티드 링커이고;
4) y는 0 또는 1이다.
제22항에 있어서, 과가수분해 활성을 갖는 효소가 리파제, 프로테아제, 에스테라제, 아실 트랜스퍼라제, 아릴 에스테라제, 탄수화물 에스테라제 및 그들의 조합의 군으로부터 선택되는 융합 단백질.
제23항에 있어서, 효소가 과가수분해 활성을 갖는 CE-7 탄수화물 에스테라제이며; PAH는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖고, 상기 시그니처 모티프는
1) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
2) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
3) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함하는 융합 단백질.
1) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
2) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
3) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함하는 융합 단백질.
제23항에 있어서, 과가수분해 활성을 갖는 효소가 마이코박테리움 스메그마티스(Mycobacterium smegmatis)로부터의 아릴 에스테라제인 융합 단백질.
제23항, 제24항 또는 제25항에 있어서, 구강 표면에 대하여 친화성을 갖는 펩티드 성분이 면역글로불린 폴드가 결여된 단쇄 폴리펩티드인 융합 단백질.
제26항에 있어서, 단쇄 폴리펩티드는 길이가 5 내지 60개 아미노산 범위인 적어도 하나의 구강 표면-결합 펩티드를 포함하는 융합 단백질.
제27항에 있어서, 상기 적어도 하나의 구강 표면-결합 펩티드는 KD 값 또는 MB50 값이 10-5 M 이하인 융합 단백질.
제26항에 있어서, 단쇄 폴리펩티드가 2 내지 50개 구강 표면-결합 펩티드를 포함하며, 구강 표면-결합 펩티드는 독립적으로 그리고 임의로, 길이가 1 내지 100개 아미노산 범위인 폴리펩티드 스페이서에 의해 분리되는 융합 단백질.
제22항, 제23항, 제24항 및 제25항 중 어느 한 항에 있어서, 구강 표면에 대하여 친화성을 갖는 펩티드 성분이 200개 이하의 길이의 아미노산을 포함하는 융합 단백질.
1) 제23항의 융합 단백질을 포함하는 효소 촉매;
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
M은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
c) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
3) 과산소원; 및
4) 구강에 허용가능한 담체 매질을 포함하는 구강 관리 제품.
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
M은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
c) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
3) 과산소원; 및
4) 구강에 허용가능한 담체 매질을 포함하는 구강 관리 제품.
1) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
a) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
b) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
c) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - ;
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
M은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
d) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
3) 과산소원; 및
4) 구강에 허용가능한 담체 매질을 포함하는 구강 관리 제품.
a) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
b) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
c) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - ;
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
M은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
d) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질;
3) 과산소원; 및
4) 구강에 허용가능한 담체 매질을 포함하는 구강 관리 제품.
제32항에 있어서, 과가수분해 활성을 갖는 효소가 제22항의 융합 단백질인 구강 관리 제품.
제33항에 있어서, 제2 부분이 적어도 하나의 치아 에나멜-결합 또는 치아 균막-결합 펩티드를 포함하는 단쇄 펩티드인 구강 관리 제품.
제34항에 있어서, 적어도 하나의 치아 에나멜-결합 또는 치아 균막-결합 펩티드는 길이가 5 내지 60개 아미노산 범위인 구강 관리 제품.
제31항, 제32항, 제33항, 제34항 또는 제35항에 있어서, 구강 관리 제품이 분말, 페이스트, 겔, 액체, 연고, 정제, 헹굼제(rinse) 또는 그들의 임의의 조합의 형태인 구강 관리 제품.
제36항에 있어서, 구강 관리 제품이 치약, 치과용 크림(dental cream), 투스 겔(tooth gel), 투스 파우더(tooth powder), 구강 세정제(mouth wash), 호흡 청량제(breath freshener), 스트립(strip) 또는 치실(dental floss)인 구강 관리 제품.
제36항에 있어서, 구강 관리 제품이 미백 스트립(whitening strip) 또는 치과용 트레이(dental tray)의 형태인 구강 관리 제품.
제31항 또는 제32항에 있어서, 효소 촉매가 구강 관리 제품의 사용 전에, 기질, 과산소원, 또는 기질 및 과산소원 둘 모두로부터 분리되어 유지되는 구강 관리 제품.
서열 번호 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421 및 422로 이루어진 군으로부터 선택되는 아미노산 서열을 갖는 구강 표면에 대하여 친화성을 갖는 분리된 폴리펩티드.
구강 관리 제품에서 구강 표면의 탈색, 미백, 소독, 탈염, 탈취 또는 생물막의 제거를 위한 유효 농도의 적어도 하나의 과산을 생성하기 위한 과가수분해 활성을 갖는 CE-7 탄수화물 에스테라제의 용도.
1) 과가수분해 활성을 갖는 효소 촉매 - 상기 효소 촉매는 클러스털더블유를 사용하여 서열 번호 2의 참조 서열과 정렬되는 CE-7 시그니처 모티프를 갖는 효소를 포함하며, 상기 시그니처 모티프는
a) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
b) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
c) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - ;
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
M은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식

(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
d) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질; 및
3) 과산소원을 포함하는 - 이에 의해, (1), (2) 및 (3)의 혼합 시에 과산이 생성된다 - 치아 우식증, 치은염, 구강 칸디다증 또는 치근막염의 치료 또는 예방을 위한 과산 생성 조성물의 용도.
a) 서열 번호 2의 위치 118 내지 120에 해당하는 위치에서 RGQ 모티프;
b) 서열 번호 2의 위치 179 내지 183에 해당하는 위치에서 GXSQG 모티프; 및
c) 서열 번호 2의 위치 298 및 299에 해당하는 위치에서 HE 모티프를 포함한다 - ;
2) a) 구조
[X]mR5
(여기서, X는 화학식 R6C(O)O의 에스테르기이며,
R6은 하이드록실기 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 선형, 분지형 또는 환형 하이드로카르빌 모이어티이고, R6은 C2 내지 C7인 R6에 대하여, 하나 이상의 에테르 결합을 임의로 포함하고;
R5는 하이드록실기로 임의로 치환된 C1 내지 C6 선형, 분지형 또는 환형 하이드로카르빌 모이어티 또는 5-원 환형 헤테로방향족 모이어티 또는 6-원 환형 방향족 또는 헤테로방향족 모이어티이며; R5에서 각 탄소 원자는 각각 1개 이하의 하이드록실기 또는 1개 이하의 에스테르기 또는 카르복실산기를 포함하고; R5는 임의로 하나 이상의 에테르 결합을 포함하며;
M은 1 내지 R5에서의 탄소 원자수 범위의 정수이다)를 갖는 에스테르 - 상기 에스테르는 25℃에서 적어도 5 ppm의 수 용해도를 갖는다 - ;
b) 구조
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R3 및 R4는 각각 H 또는 R1C(O)이다)를 갖는 글리세리드;
c) 화학식
(여기서, R1은 하이드록실 또는 C1 내지 C4 알콕시기로 임의로 치환된 C1 내지 C7 직쇄 또는 분지쇄 알킬이며, R2는 C1 내지 C10 직쇄 또는 분지쇄 알킬, 알케닐, 알키닐, 아릴, 알킬아릴, 알킬헤테로아릴, 헤테로아릴, (CH2CH2O)n 또는 (CH2CH(CH3)-O)nH이고, n은 1 내지 10이다)의 하나 이상의 에스테르; 및
d) 아세틸화 단당류, 아세틸화 이당류 및 아세틸화 다당류로 이루어진 군으로부터 선택되는 아세틸화 당류로 이루어진 군으로부터 선택되는 적어도 하나의 기질; 및
3) 과산소원을 포함하는 - 이에 의해, (1), (2) 및 (3)의 혼합 시에 과산이 생성된다 - 치아 우식증, 치은염, 구강 칸디다증 또는 치근막염의 치료 또는 예방을 위한 과산 생성 조성물의 용도.
제41항에 있어서, 과가수분해 활성을 갖는 효소가 구강 표면에 대하여 친화성을 갖는 적어도 하나의 부분을 포함하는 융합 단백질의 형태인 용도.
구강 제품에서의 하기의 일반 구조를 포함하는 융합 단백질의 용도:
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
상기 식에서,
1) PAH는 서열 번호 460과 적어도 95% 아미노산 동일성을 갖는 아미노산 서열을 갖는 과가수분해 활성을 갖는 효소이고;
2) OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이며;
3) L은 길이가 1 내지 100개 아미노산 범위인 펩티드 링커이고;
4) y는 0 또는 1이다.
PAH-[L]y-OCBD
또는
OCBD-[L]y-PAH
상기 식에서,
1) PAH는 서열 번호 460과 적어도 95% 아미노산 동일성을 갖는 아미노산 서열을 갖는 과가수분해 활성을 갖는 효소이고;
2) OCBD는 구강 표면에 대하여 친화성을 갖는 펩티드 성분이며;
3) L은 길이가 1 내지 100개 아미노산 범위인 펩티드 링커이고;
4) y는 0 또는 1이다.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201061424903P | 2010-12-20 | 2010-12-20 | |
| US61/424,903 | 2010-12-20 | ||
| PCT/US2011/065912 WO2012087970A2 (en) | 2010-12-20 | 2011-12-19 | Enzymatic peracid generation for use in oral care products |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20130132936A true KR20130132936A (ko) | 2013-12-05 |
Family
ID=46314812
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR20137019091A KR20130132936A (ko) | 2010-12-20 | 2011-12-19 | 구강 관리 제품에 사용하기 위한 효소에 의한 과산 생성 |
Country Status (13)
| Country | Link |
|---|---|
| US (1) | US8663616B2 (ko) |
| EP (1) | EP2654695B8 (ko) |
| JP (2) | JP6359831B2 (ko) |
| KR (1) | KR20130132936A (ko) |
| CN (1) | CN103269679B (ko) |
| AU (1) | AU2011349451A1 (ko) |
| BR (1) | BR112013015459B8 (ko) |
| CA (1) | CA2822422C (ko) |
| ES (1) | ES2784495T3 (ko) |
| MX (1) | MX2013006994A (ko) |
| RU (1) | RU2660373C2 (ko) |
| WO (1) | WO2012087970A2 (ko) |
| ZA (1) | ZA201303372B (ko) |
Families Citing this family (16)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101801521B (zh) | 2007-05-14 | 2015-06-17 | 纽约州立大学研究基金会 | 生物膜中细菌细胞内的生理学分散响应诱导 |
| MX349237B (es) | 2011-09-14 | 2017-07-18 | Colgate Palmolive Co | Tira blanqueadora de dientes. |
| AU2015201460B2 (en) * | 2011-12-19 | 2017-03-16 | Colgate-Palmolive Company | System providing enzyme-catalyzed reaction |
| BR112014014771B1 (pt) * | 2011-12-19 | 2021-05-25 | E.I. Du Pont De Nemours And Company | tira de clareamento de dente e método de clareamento dos dentes utilizando a referida tira |
| CA2859793C (en) * | 2011-12-19 | 2019-10-01 | Colgate-Palmolive Company | Oral care system providing perhydrolase-catalyzed peracid formation |
| BR112014014626A2 (pt) * | 2011-12-19 | 2017-06-13 | Colgate Palmolive Co | sistema que fornece reação catalisada por enzima |
| JP6574376B2 (ja) | 2012-03-30 | 2019-09-11 | イー・アイ・デュポン・ドウ・ヌムール・アンド・カンパニーE.I.Du Pont De Nemours And Company | 過酸生成に有用な酵素 |
| BR112014024424B1 (pt) | 2012-03-30 | 2022-04-05 | E.I. Du Pont De Nemours And Company | Processo para a produção de um ácido peroxicarboxílico, composição, sistema de geração e fornecimento de perácido e polipeptídeo isolado |
| BR112014024217B1 (pt) | 2012-03-30 | 2022-04-05 | E.I. Du Pont De Nemours And Company | Processo de produção de um ácido peroxicarboxílico, composição e sistema de geração e fornecimento de perácido |
| BR112014024207B1 (pt) | 2012-03-30 | 2022-04-05 | E.I. Du Pont De Nemours And Company | Processo para a produção de um ácido peroxicarboxílico, composição e sistema de geração e fornecimento de perácido |
| EP2831252B1 (en) | 2012-03-30 | 2021-11-24 | DuPont US Holding, LLC | Enzymes useful for peracid production |
| TWI561248B (en) * | 2012-12-22 | 2016-12-11 | Colgate Palmolive Co | Peracid-generating compositions |
| AU2017382664B2 (en) * | 2016-12-20 | 2020-05-14 | Colgate-Palmolive Company | Oral care compositions and methods for whitening teeth |
| CA3044407A1 (en) * | 2016-12-20 | 2018-06-28 | Colgate-Palmolive Company | Two-phase oral care whitening compositions having enzyme catalyzing peracetic acid |
| MX2019006904A (es) * | 2016-12-20 | 2019-08-22 | Colgate Palmolive Co | Composicion para el cuidado bucal y metodos para blanquear los dientes. |
| BR112021012152A2 (pt) | 2018-12-27 | 2021-12-14 | Colgate Palmolive Co | Composições para higiene bucal |
Family Cites Families (93)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3974082A (en) | 1972-08-21 | 1976-08-10 | Colgate-Palmolive Company | Bleaching compositions |
| JPS526867B2 (ko) | 1972-09-14 | 1977-02-25 | ||
| US4138476A (en) | 1977-08-03 | 1979-02-06 | The United States Of America As Represented By The Secretary Of The Navy | Plaque dispersing enzymes as oral therapeutic agents by molecular alteration |
| US4444886A (en) | 1982-08-11 | 1984-04-24 | Eastman Kodak Company | Diacetinase from Bacillus subtilis |
| US5116575A (en) | 1986-02-06 | 1992-05-26 | Steris Corporation | Powdered anti-microbial composition |
| US5296161A (en) | 1986-06-09 | 1994-03-22 | The Clorox Company | Enzymatic perhydrolysis system and method of use for bleaching |
| US5364554A (en) | 1986-06-09 | 1994-11-15 | The Clorox Company | Proteolytic perhydrolysis system and method of use for bleaching |
| DE69005309T2 (de) | 1989-09-12 | 1994-05-05 | Novonordisk As | Verfahren zur herstellung von peroxykarbonsäuren unter verwendung eines enzymkatalysators. |
| GB9021671D0 (en) | 1990-10-05 | 1990-11-21 | Unilever Plc | Delivery of agents |
| AU641736B2 (en) | 1990-03-21 | 1993-09-30 | Quest International B.V. | Delivery of agents |
| AU642979B2 (en) | 1990-03-21 | 1993-11-04 | Quest International B.V. | Utilization and delivery of enzymes |
| AU642980B2 (en) | 1990-03-21 | 1993-11-04 | Quest International B.V. | Ultilization of enzymes |
| GB9122048D0 (en) | 1991-10-17 | 1991-11-27 | Interox Chemicals Ltd | Compositions and uses thereof |
| US5279816A (en) | 1991-11-22 | 1994-01-18 | Colgate-Palmolive Co. | Oral composition having improved tooth whitening effect |
| BE1006057A3 (fr) | 1992-07-06 | 1994-05-03 | Solvay Interox | Procede pour la delignification d'une pate a papier chimique. |
| US5302375A (en) | 1992-11-19 | 1994-04-12 | Colgate-Palmolive Company | Oral composition having improved tooth whitening effect |
| US5683724A (en) | 1993-03-17 | 1997-11-04 | Ecolab Inc. | Automated process for inhibition of microbial growth in aqueous food transport or process streams |
| US5398846A (en) | 1993-08-20 | 1995-03-21 | S. C. Johnson & Son, Inc. | Assembly for simultaneous dispensing of multiple fluids |
| GB2297976A (en) | 1995-02-01 | 1996-08-21 | Reckitt & Colmann Prod Ltd | Improvements in or relating to a bleaching process |
| EP1011620A1 (en) | 1996-10-11 | 2000-06-28 | Novo Nordisk A/S | CELLULOSE BINDING DOMAINS (CBDs) FOR ORAL CARE PRODUCTS |
| US6830745B1 (en) | 1997-10-16 | 2004-12-14 | Pharmacal Biotechnologies, Llc | Compositions for treating biofilm |
| US5871714A (en) | 1997-10-16 | 1999-02-16 | Pharmacal Biotechnologies, Inc. | Compositions for controlling bacterial colonization |
| US6221341B1 (en) | 1997-11-19 | 2001-04-24 | Oraceutical Llc | Tooth whitening compositions |
| FR2774588B1 (fr) | 1998-02-11 | 2000-05-05 | Oreal | Composition cosmetique ou dermatologique contenant au moins une proteine de soie d'arachnides naturelle, recombinante ou un analogue |
| AU758625B2 (en) | 1998-08-20 | 2003-03-27 | Ecolab Inc. | The treatment of meat products |
| US6010729A (en) | 1998-08-20 | 2000-01-04 | Ecolab Inc. | Treatment of animal carcasses |
| US6210639B1 (en) | 1998-10-26 | 2001-04-03 | Novartis Ag | Apparatus, method and composition for cleaning and disinfecting |
| CA2364560A1 (en) | 1999-04-12 | 2000-10-19 | Unilever Plc | Multiple component bleaching compositions |
| AU6373000A (en) | 1999-07-22 | 2001-02-13 | Pericor Science, Inc. | Lysine oxidase linkage of agents to tissue |
| US6245729B1 (en) | 1999-07-27 | 2001-06-12 | Ecolab, Inc. | Peracid forming system, peracid forming composition, and methods for making and using |
| EP1360500B1 (en) | 2000-04-14 | 2010-01-13 | Genencor International, Inc. | Methods for selective targeting |
| AU2002311922A1 (en) | 2001-05-15 | 2002-11-25 | The Procter And Gamble Company | Oral care compositions |
| RU2269337C2 (ru) | 2001-06-25 | 2006-02-10 | Дзе Проктер Энд Гэмбл Компани | Абразивная композиция для чистки зубов и способ чистки и полировки зубов |
| US6635286B2 (en) | 2001-06-29 | 2003-10-21 | Ecolab Inc. | Peroxy acid treatment to control pathogenic organisms on growing plants |
| US7448556B2 (en) | 2002-08-16 | 2008-11-11 | Henkel Kgaa | Dispenser bottle for at least two active fluids |
| US20060171885A1 (en) | 2002-11-25 | 2006-08-03 | Giselle Janssen | Skin or hair binding peptides |
| DE10260903A1 (de) * | 2002-12-20 | 2004-07-08 | Henkel Kgaa | Neue Perhydrolasen |
| US20060161121A1 (en) | 2003-02-06 | 2006-07-20 | Dehli-Cleve Collection As | Peroxide compositions |
| DE60320778D1 (de) | 2003-07-08 | 2008-06-19 | Procter & Gamble | Flüssige Bleichaktivatorzusamensetzung |
| US20050069501A1 (en) | 2003-08-15 | 2005-03-31 | Sayed Ibrahim | Rapid temporary tooth whitening composition |
| US20050226839A1 (en) | 2003-09-08 | 2005-10-13 | Xueying Huang | Pepetide-based body surface reagents for personal care |
| US7285264B2 (en) | 2003-09-08 | 2007-10-23 | E.I. Du Pont De Nemours And Company | Peptide-based body surface coloring reagents |
| US20050054752A1 (en) | 2003-09-08 | 2005-03-10 | O'brien John P. | Peptide-based diblock and triblock dispersants and diblock polymers |
| US7807141B2 (en) | 2003-09-08 | 2010-10-05 | E.I. Du Pont De Nemours And Company | Peptide-based oral care surface reagents for personal care |
| US7220405B2 (en) | 2003-09-08 | 2007-05-22 | E. I. Du Pont De Nemours And Company | Peptide-based conditioners and colorants for hair, skin, and nails |
| US7754460B2 (en) | 2003-12-03 | 2010-07-13 | Danisco Us Inc. | Enzyme for the production of long chain peracid |
| MXPA06005652A (es) | 2003-12-03 | 2006-08-17 | Genencor Int | Perhidrolasa. |
| US20070196305A1 (en) | 2005-03-01 | 2007-08-23 | Hong Wang | Method for identifying hair conditioner-resistant hair-binding peptides and hair benefit agents therefrom |
| EP1877566B1 (en) | 2005-04-29 | 2009-02-18 | E.I. Du Pont De Nemours And Company | Enzymatic production of peracids using perhydrolytic enzymes |
| US7550420B2 (en) | 2005-04-29 | 2009-06-23 | E. I. Dupont De Nemours And Company | Enzymatic production of peracids using perhydrolytic enzymes |
| US7384787B2 (en) | 2005-06-06 | 2008-06-10 | Regents Of The University Of Minnesota | Increasing perhydrolase activity in esterases and/or lipases |
| US20070065387A1 (en) | 2005-09-16 | 2007-03-22 | Beck William A | Method for enhancing the effect of particulate benefit agents |
| US20070082832A1 (en) | 2005-10-06 | 2007-04-12 | Dicosimo Robert | Enzymatic production of peracids from carboxylic acid ester substrates using non-heme haloperoxidases |
| US7563758B2 (en) | 2005-10-06 | 2009-07-21 | E. I. Du Pont De Nemours And Company | Enzymatic production of peracids using Lactobacilli having perhydrolysis activity |
| US7951566B2 (en) * | 2005-12-13 | 2011-05-31 | E.I. Du Pont De Nemours And Company | Production of peracids using an enzyme having perhydrolysis activity |
| US8518675B2 (en) | 2005-12-13 | 2013-08-27 | E. I. Du Pont De Nemours And Company | Production of peracids using an enzyme having perhydrolysis activity |
| US7723083B2 (en) | 2005-12-13 | 2010-05-25 | E.I. Du Pont De Nemours And Company | Production of peracids using an enzyme having perhydrolysis activity |
| US7964378B2 (en) * | 2005-12-13 | 2011-06-21 | E.I. Du Pont De Nemours And Company | Production of peracids using an enzyme having perhydrolysis activity |
| US7928076B2 (en) | 2005-12-15 | 2011-04-19 | E. I. Du Pont De Nemours And Company | Polypropylene binding peptides and methods of use |
| US7700716B2 (en) | 2005-12-15 | 2010-04-20 | E. I. Du Pont De Nemours And Company | Polytetrafluoroethylene binding peptides and methods of use |
| US7906617B2 (en) | 2005-12-15 | 2011-03-15 | E. I. Du Pont De Nemours And Company | Polyethylene binding peptides and methods of use |
| US7858581B2 (en) | 2005-12-15 | 2010-12-28 | E. I. Du Pont De Nemours And Company | PMMA binding peptides and methods of use |
| US7632919B2 (en) | 2005-12-15 | 2009-12-15 | E.I. Du Pont De Nemours And Company | Polystyrene binding peptides and methods of use |
| US7709601B2 (en) | 2005-12-15 | 2010-05-04 | E. I. Du Pont De Nemours And Company | Nylon binding peptides and methods of use |
| WO2007106293A1 (en) * | 2006-03-02 | 2007-09-20 | Genencor International, Inc. | Surface active bleach and dynamic ph |
| CN101394832A (zh) * | 2006-03-03 | 2009-03-25 | 金克克国际有限公司 | 用于牙齿美白的过水解酶 |
| US20080280810A1 (en) * | 2006-10-30 | 2008-11-13 | O'brien John P | Peptides having affinity for body surfaces |
| DE102007036392A1 (de) * | 2007-07-31 | 2009-02-05 | Henkel Ag & Co. Kgaa | Zusammensetzungen enthaltend Perhydrolasen und Alkylenglykoldiacetate |
| ATE557085T1 (de) | 2008-08-13 | 2012-05-15 | Du Pont | Steuerung der enzymatischen herstellung von persäuren |
| ES2735230T3 (es) | 2008-10-03 | 2019-12-17 | Du Pont | Estabilización de perhidrolasas con excipientes |
| US8697654B2 (en) | 2008-12-18 | 2014-04-15 | E I Du Pont De Nemours And Company | Peptide linkers for effective multivalent peptide binding |
| US8287845B2 (en) | 2008-12-18 | 2012-10-16 | E I Du Pont De Nemours And Company | Hair-binding peptides |
| US20100158846A1 (en) | 2008-12-18 | 2010-06-24 | E. I. Du Pont De Nemours And Company | Hair-binding peptides |
| US7662587B1 (en) | 2009-03-05 | 2010-02-16 | E. I. Du Pont De Nemours And Company | Gene knockout mutations that increase peptide production |
| WO2010114638A1 (en) * | 2009-03-30 | 2010-10-07 | E. I. Du Pont De Nemours And Company | Peptide-based tooth whitening reagents |
| US8389675B2 (en) | 2009-05-20 | 2013-03-05 | E I Du Pont De Nemours And Company | PMMA binding peptides |
| US8206693B2 (en) | 2009-05-20 | 2012-06-26 | E I Du Pont De Nemours And Company | PMMA binding peptides |
| US8263737B2 (en) | 2009-05-20 | 2012-09-11 | E I Du Pont De Nemours And Company | PMMA binding peptides |
| US20100298535A1 (en) | 2009-05-20 | 2010-11-25 | E. I. Du Pont De Nemours And Company | Pmma binding peptides |
| US8455614B2 (en) | 2009-05-20 | 2013-06-04 | E.I. Du Pont De Nemours And Company | PMMA binding peptides |
| US20100298531A1 (en) | 2009-05-20 | 2010-11-25 | E.I. Du Pont De Nemours And Company | Pmma binding peptides |
| US8378065B2 (en) | 2009-05-20 | 2013-02-19 | E I Du Pont De Nemours And Company | PMMA binding peptides |
| US8404214B2 (en) | 2009-05-20 | 2013-03-26 | E I Du Pont De Nemours And Company | PMMA binding peptides |
| US20100310495A1 (en) | 2009-06-08 | 2010-12-09 | E. I. Du Pont De Nemours And Company | Peptides having affinity for poly (benzyl methacrylate-co-methacrylic acid) potassium salt copolymers and methods of use |
| US8222012B2 (en) | 2009-10-01 | 2012-07-17 | E. I. Du Pont De Nemours And Company | Perhydrolase for enzymatic peracid production |
| US7932072B1 (en) | 2009-12-07 | 2011-04-26 | E. I. Du Pont De Nemours And Company | Perhydrolase providing improved peracid stability |
| US7923233B1 (en) | 2009-12-07 | 2011-04-12 | E. I. Du Pont De Nemours And Company | Perhydrolase providing improved peracid stability |
| US7910347B1 (en) | 2009-12-07 | 2011-03-22 | E. I. Du Pont De Nemours And Company | Perhydrolase providing improved peracid stability |
| US7927854B1 (en) | 2009-12-07 | 2011-04-19 | E. I. Du Pont De Nemours And Company | Perhydrolase providing improved peracid stability |
| US7960528B1 (en) | 2009-12-07 | 2011-06-14 | E. I. Du Pont De Nemours And Company | Perhydrolase providing improved peracid stability |
| WO2011119714A1 (en) * | 2010-03-26 | 2011-09-29 | E.I. Dupont De Nemours And Company | Perhydrolase providing improved specific activity |
| WO2011119703A1 (en) | 2010-03-26 | 2011-09-29 | E.I. Dupont De Nemours And Company | Process for the purification of proteins |
| US8389254B2 (en) | 2010-03-26 | 2013-03-05 | E.I. Du Pont De Nemours And Company | Perhydrolase providing improved specific activity |
-
2011
- 2011-12-19 AU AU2011349451A patent/AU2011349451A1/en not_active Abandoned
- 2011-12-19 CN CN201180061614.5A patent/CN103269679B/zh active Active
- 2011-12-19 CA CA2822422A patent/CA2822422C/en active Active
- 2011-12-19 RU RU2013133831A patent/RU2660373C2/ru active
- 2011-12-19 US US13/330,261 patent/US8663616B2/en active Active
- 2011-12-19 EP EP11850480.2A patent/EP2654695B8/en active Active
- 2011-12-19 BR BR112013015459A patent/BR112013015459B8/pt active IP Right Grant
- 2011-12-19 KR KR20137019091A patent/KR20130132936A/ko not_active Application Discontinuation
- 2011-12-19 WO PCT/US2011/065912 patent/WO2012087970A2/en active Application Filing
- 2011-12-19 ES ES11850480T patent/ES2784495T3/es active Active
- 2011-12-19 MX MX2013006994A patent/MX2013006994A/es unknown
- 2011-12-19 JP JP2013546292A patent/JP6359831B2/ja active Active
-
2013
- 2013-05-09 ZA ZA2013/03372A patent/ZA201303372B/en unknown
-
2016
- 2016-12-22 JP JP2016248580A patent/JP6410786B2/ja active Active
Also Published As
| Publication number | Publication date |
|---|---|
| RU2660373C2 (ru) | 2018-07-05 |
| CA2822422A1 (en) | 2012-06-28 |
| AU2011349451A1 (en) | 2013-05-30 |
| JP6410786B2 (ja) | 2018-10-24 |
| MX2013006994A (es) | 2013-07-29 |
| BR112013015459B1 (pt) | 2018-04-10 |
| RU2018123151A3 (ko) | 2022-02-08 |
| CN103269679B (zh) | 2019-04-12 |
| JP2017081958A (ja) | 2017-05-18 |
| EP2654695A4 (en) | 2015-05-27 |
| ES2784495T3 (es) | 2020-09-28 |
| JP6359831B2 (ja) | 2018-07-18 |
| EP2654695A2 (en) | 2013-10-30 |
| EP2654695B1 (en) | 2020-01-22 |
| US8663616B2 (en) | 2014-03-04 |
| WO2012087970A3 (en) | 2012-11-22 |
| ZA201303372B (en) | 2014-08-27 |
| RU2013133831A (ru) | 2015-01-27 |
| BR112013015459B8 (pt) | 2018-06-12 |
| CA2822422C (en) | 2021-02-02 |
| CN103269679A (zh) | 2013-08-28 |
| US20120328534A1 (en) | 2012-12-27 |
| RU2018123151A (ru) | 2019-03-06 |
| JP2014506250A (ja) | 2014-03-13 |
| EP2654695B8 (en) | 2020-03-04 |
| WO2012087970A2 (en) | 2012-06-28 |
| BR112013015459A2 (pt) | 2017-06-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR20130132936A (ko) | 구강 관리 제품에 사용하기 위한 효소에 의한 과산 생성 | |
| KR20130128442A (ko) | 모발 관리 제품에 사용하기 위한 효소에 의한 과산 생성 | |
| KR20140114269A (ko) | 표적화된 과가수분해효소 | |
| JP6235482B2 (ja) | ペルヒドロラーゼ触媒反応を与えるシステム | |
| JP6181666B2 (ja) | 過酸−生成組成物 | |
| WO2004058955A2 (de) | Cholinoxidasen | |
| CN101394832A (zh) | 用于牙齿美白的过水解酶 | |
| RU2793140C2 (ru) | Ферментативное образование перкислоты для применения в продуктах для ухода за полостью рта |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| WITN | Application deemed withdrawn, e.g. because no request for examination was filed or no examination fee was paid |