JP4383855B2 - パイエル板および/またはm細胞を標的とするリガンド - Google Patents
パイエル板および/またはm細胞を標的とするリガンド Download PDFInfo
- Publication number
- JP4383855B2 JP4383855B2 JP2003510683A JP2003510683A JP4383855B2 JP 4383855 B2 JP4383855 B2 JP 4383855B2 JP 2003510683 A JP2003510683 A JP 2003510683A JP 2003510683 A JP2003510683 A JP 2003510683A JP 4383855 B2 JP4383855 B2 JP 4383855B2
- Authority
- JP
- Japan
- Prior art keywords
- seq
- peptide
- synthetic polypeptide
- amino acid
- purified synthetic
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
- 239000003446 ligand Substances 0.000 title abstract description 73
- 210000001986 peyer's patch Anatomy 0.000 title abstract description 39
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 252
- 102000004196 processed proteins & peptides Human genes 0.000 claims abstract description 130
- 229920001184 polypeptide Polymers 0.000 claims abstract description 102
- 238000000034 method Methods 0.000 claims abstract description 32
- 210000000981 epithelium Anatomy 0.000 claims abstract description 12
- 239000003795 chemical substances by application Substances 0.000 claims abstract description 11
- 150000001413 amino acids Chemical class 0.000 claims description 87
- 241000700159 Rattus Species 0.000 claims description 62
- 239000003814 drug Substances 0.000 claims description 36
- 229940079593 drug Drugs 0.000 claims description 29
- 239000000427 antigen Substances 0.000 claims description 27
- 108091007433 antigens Proteins 0.000 claims description 27
- 102000036639 antigens Human genes 0.000 claims description 27
- 108090000623 proteins and genes Proteins 0.000 claims description 25
- 241000894006 Bacteria Species 0.000 claims description 19
- 241000699666 Mus <mouse, genus> Species 0.000 claims description 18
- 210000004347 intestinal mucosa Anatomy 0.000 claims description 17
- 210000004379 membrane Anatomy 0.000 claims description 17
- 239000012528 membrane Substances 0.000 claims description 17
- 239000002105 nanoparticle Substances 0.000 claims description 17
- 241000700605 Viruses Species 0.000 claims description 16
- 239000011859 microparticle Substances 0.000 claims description 15
- 102000004169 proteins and genes Human genes 0.000 claims description 15
- 239000002502 liposome Substances 0.000 claims description 14
- 241000282472 Canis lupus familiaris Species 0.000 claims description 12
- 241001524679 Escherichia virus M13 Species 0.000 claims description 12
- 230000004927 fusion Effects 0.000 claims description 11
- 239000002671 adjuvant Substances 0.000 claims description 9
- 108020004414 DNA Proteins 0.000 claims description 7
- 230000001717 pathogenic effect Effects 0.000 claims description 7
- 230000004572 zinc-binding Effects 0.000 claims description 7
- 150000007523 nucleic acids Chemical group 0.000 claims description 6
- 244000052769 pathogen Species 0.000 claims description 6
- 229940124597 therapeutic agent Drugs 0.000 claims description 6
- 238000013519 translation Methods 0.000 claims description 6
- 241000699670 Mus sp. Species 0.000 claims description 5
- 241000282898 Sus scrofa Species 0.000 claims description 5
- 230000015572 biosynthetic process Effects 0.000 claims description 5
- 229960005486 vaccine Drugs 0.000 claims description 5
- 241000124008 Mammalia Species 0.000 claims description 4
- 241000282887 Suidae Species 0.000 claims description 3
- 239000000969 carrier Substances 0.000 claims description 3
- 239000002773 nucleotide Substances 0.000 claims description 3
- 125000003729 nucleotide group Chemical group 0.000 claims description 3
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 2
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 claims description 2
- 238000007918 intramuscular administration Methods 0.000 claims description 2
- 238000001990 intravenous administration Methods 0.000 claims description 2
- 238000007920 subcutaneous administration Methods 0.000 claims description 2
- 230000003612 virological effect Effects 0.000 claims description 2
- 239000011701 zinc Substances 0.000 claims description 2
- 229910052725 zinc Inorganic materials 0.000 claims description 2
- 238000003786 synthesis reaction Methods 0.000 claims 3
- 125000003275 alpha amino acid group Chemical group 0.000 claims 1
- 238000004519 manufacturing process Methods 0.000 claims 1
- 210000001519 tissue Anatomy 0.000 abstract description 28
- 230000000968 intestinal effect Effects 0.000 abstract description 19
- 230000008685 targeting Effects 0.000 abstract description 9
- 239000003937 drug carrier Substances 0.000 abstract 1
- 210000003126 m-cell Anatomy 0.000 abstract 1
- 239000008177 pharmaceutical agent Substances 0.000 abstract 1
- 230000027455 binding Effects 0.000 description 134
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 82
- 238000002835 absorbance Methods 0.000 description 62
- 210000004027 cell Anatomy 0.000 description 62
- 235000001014 amino acid Nutrition 0.000 description 59
- 229960002685 biotin Drugs 0.000 description 41
- 235000020958 biotin Nutrition 0.000 description 41
- 239000011616 biotin Substances 0.000 description 41
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 39
- 238000002965 ELISA Methods 0.000 description 26
- 239000002245 particle Substances 0.000 description 22
- 238000003556 assay Methods 0.000 description 21
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 19
- 238000001514 detection method Methods 0.000 description 18
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 15
- 235000018102 proteins Nutrition 0.000 description 13
- 210000000813 small intestine Anatomy 0.000 description 11
- 230000032258 transport Effects 0.000 description 11
- 239000012634 fragment Substances 0.000 description 10
- 241000282465 Canis Species 0.000 description 8
- 239000004201 L-cysteine Substances 0.000 description 7
- 235000013878 L-cysteine Nutrition 0.000 description 7
- 241000894007 species Species 0.000 description 7
- 102000000905 Cadherin Human genes 0.000 description 6
- 108050007957 Cadherin Proteins 0.000 description 6
- 102000010834 Extracellular Matrix Proteins Human genes 0.000 description 6
- 108010037362 Extracellular Matrix Proteins Proteins 0.000 description 6
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 6
- 210000004369 blood Anatomy 0.000 description 6
- 239000008280 blood Substances 0.000 description 6
- 239000000872 buffer Substances 0.000 description 6
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 6
- 238000012377 drug delivery Methods 0.000 description 6
- 210000002919 epithelial cell Anatomy 0.000 description 6
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical compound FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 6
- 238000012216 screening Methods 0.000 description 6
- 241000222722 Leishmania <genus> Species 0.000 description 5
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 5
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 5
- 238000004458 analytical method Methods 0.000 description 5
- 210000002744 extracellular matrix Anatomy 0.000 description 5
- 210000000936 intestine Anatomy 0.000 description 5
- 230000037361 pathway Effects 0.000 description 5
- 239000006228 supernatant Substances 0.000 description 5
- 241000588724 Escherichia coli Species 0.000 description 4
- 102000002274 Matrix Metalloproteinases Human genes 0.000 description 4
- 108010000684 Matrix Metalloproteinases Proteins 0.000 description 4
- 108091034117 Oligonucleotide Proteins 0.000 description 4
- 241000223960 Plasmodium falciparum Species 0.000 description 4
- 239000004793 Polystyrene Substances 0.000 description 4
- 108010090804 Streptavidin Proteins 0.000 description 4
- 241000224526 Trichomonas Species 0.000 description 4
- 102000003990 Urokinase-type plasminogen activator Human genes 0.000 description 4
- 108090000435 Urokinase-type plasminogen activator Proteins 0.000 description 4
- 239000002246 antineoplastic agent Substances 0.000 description 4
- 210000001035 gastrointestinal tract Anatomy 0.000 description 4
- 238000001727 in vivo Methods 0.000 description 4
- 210000002490 intestinal epithelial cell Anatomy 0.000 description 4
- 239000013642 negative control Substances 0.000 description 4
- 229920002223 polystyrene Polymers 0.000 description 4
- 238000012360 testing method Methods 0.000 description 4
- XZKIHKMTEMTJQX-UHFFFAOYSA-N 4-Nitrophenyl Phosphate Chemical compound OP(O)(=O)OC1=CC=C([N+]([O-])=O)C=C1 XZKIHKMTEMTJQX-UHFFFAOYSA-N 0.000 description 3
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 3
- 102000011632 Caseins Human genes 0.000 description 3
- 108010076119 Caseins Proteins 0.000 description 3
- 241000242711 Fasciola hepatica Species 0.000 description 3
- 241000233866 Fungi Species 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- 102000009438 IgE Receptors Human genes 0.000 description 3
- 108010073816 IgE Receptors Proteins 0.000 description 3
- 102100037852 Insulin-like growth factor I Human genes 0.000 description 3
- 241001465754 Metazoa Species 0.000 description 3
- 229920001213 Polysorbate 20 Polymers 0.000 description 3
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 3
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 3
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 3
- 108010042352 Urokinase Plasminogen Activator Receptors Proteins 0.000 description 3
- 102000004504 Urokinase Plasminogen Activator Receptors Human genes 0.000 description 3
- 210000000170 cell membrane Anatomy 0.000 description 3
- 239000000032 diagnostic agent Substances 0.000 description 3
- 229940039227 diagnostic agent Drugs 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- 208000006275 fascioliasis Diseases 0.000 description 3
- 238000011990 functional testing Methods 0.000 description 3
- 238000001476 gene delivery Methods 0.000 description 3
- 210000004072 lung Anatomy 0.000 description 3
- 108020004707 nucleic acids Proteins 0.000 description 3
- 102000039446 nucleic acids Human genes 0.000 description 3
- 244000045947 parasite Species 0.000 description 3
- 239000008188 pellet Substances 0.000 description 3
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 3
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 3
- 229920000136 polysorbate Polymers 0.000 description 3
- 238000012163 sequencing technique Methods 0.000 description 3
- 239000000758 substrate Substances 0.000 description 3
- 239000013598 vector Substances 0.000 description 3
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 3
- JKMHFZQWWAIEOD-UHFFFAOYSA-N 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid Chemical compound OCC[NH+]1CCN(CCS([O-])(=O)=O)CC1 JKMHFZQWWAIEOD-UHFFFAOYSA-N 0.000 description 2
- 229920001817 Agar Polymers 0.000 description 2
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 2
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 2
- 241000224489 Amoeba Species 0.000 description 2
- 102000055006 Calcitonin Human genes 0.000 description 2
- 108060001064 Calcitonin Proteins 0.000 description 2
- 102000016362 Catenins Human genes 0.000 description 2
- 108010067316 Catenins Proteins 0.000 description 2
- 241000242722 Cestoda Species 0.000 description 2
- 108010014258 Elastin Proteins 0.000 description 2
- 102000016942 Elastin Human genes 0.000 description 2
- 102000003951 Erythropoietin Human genes 0.000 description 2
- 108090000394 Erythropoietin Proteins 0.000 description 2
- 102000018233 Fibroblast Growth Factor Human genes 0.000 description 2
- 108050007372 Fibroblast Growth Factor Proteins 0.000 description 2
- 102100023600 Fibroblast growth factor receptor 2 Human genes 0.000 description 2
- 108700012941 GNRH1 Proteins 0.000 description 2
- 239000000579 Gonadotropin-Releasing Hormone Substances 0.000 description 2
- 102000018997 Growth Hormone Human genes 0.000 description 2
- 108010051696 Growth Hormone Proteins 0.000 description 2
- 239000000095 Growth Hormone-Releasing Hormone Substances 0.000 description 2
- 102000038461 Growth Hormone-Releasing Hormone Human genes 0.000 description 2
- 239000007995 HEPES buffer Substances 0.000 description 2
- WZUVPPKBWHMQCE-UHFFFAOYSA-N Haematoxylin Chemical compound C12=CC(O)=C(O)C=C2CC2(O)C1C1=CC=C(O)C(O)=C1OC2 WZUVPPKBWHMQCE-UHFFFAOYSA-N 0.000 description 2
- 241000238631 Hexapoda Species 0.000 description 2
- MHAJPDPJQMAIIY-UHFFFAOYSA-N Hydrogen peroxide Chemical compound OO MHAJPDPJQMAIIY-UHFFFAOYSA-N 0.000 description 2
- 102000004157 Hydrolases Human genes 0.000 description 2
- 108090000604 Hydrolases Proteins 0.000 description 2
- 108090000723 Insulin-Like Growth Factor I Proteins 0.000 description 2
- 108090001090 Lectins Proteins 0.000 description 2
- 102000004856 Lectins Human genes 0.000 description 2
- 102000000422 Matrix Metalloproteinase 3 Human genes 0.000 description 2
- 241001529936 Murinae Species 0.000 description 2
- VQPRNSWQIAHPMS-HNNXBMFYSA-N N(6)-dansyl-L-lysine Chemical group C1=CC=C2C(N(C)C)=CC=CC2=C1S(=O)(=O)NCCCC[C@H](N)C(O)=O VQPRNSWQIAHPMS-HNNXBMFYSA-N 0.000 description 2
- 239000002202 Polyethylene glycol Substances 0.000 description 2
- 238000012228 RNA interference-mediated gene silencing Methods 0.000 description 2
- 241000242680 Schistosoma mansoni Species 0.000 description 2
- 241000607768 Shigella Species 0.000 description 2
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 2
- 101710142969 Somatoliberin Proteins 0.000 description 2
- 102000013275 Somatomedins Human genes 0.000 description 2
- 102000000591 Tight Junction Proteins Human genes 0.000 description 2
- 108010002321 Tight Junction Proteins Proteins 0.000 description 2
- 239000013543 active substance Substances 0.000 description 2
- 239000000853 adhesive Substances 0.000 description 2
- 230000001070 adhesive effect Effects 0.000 description 2
- 239000008272 agar Substances 0.000 description 2
- 239000013566 allergen Substances 0.000 description 2
- 229940035676 analgesics Drugs 0.000 description 2
- 239000000730 antalgic agent Substances 0.000 description 2
- 239000004004 anti-anginal agent Substances 0.000 description 2
- 230000002686 anti-diuretic effect Effects 0.000 description 2
- 230000003474 anti-emetic effect Effects 0.000 description 2
- -1 anti-migraine drugs Substances 0.000 description 2
- 239000003146 anticoagulant agent Substances 0.000 description 2
- 229940127219 anticoagulant drug Drugs 0.000 description 2
- 229940124538 antidiuretic agent Drugs 0.000 description 2
- 229940125683 antiemetic agent Drugs 0.000 description 2
- 239000002111 antiemetic agent Substances 0.000 description 2
- 239000002220 antihypertensive agent Substances 0.000 description 2
- 229940124433 antimigraine drug Drugs 0.000 description 2
- 229940034982 antineoplastic agent Drugs 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 229960004015 calcitonin Drugs 0.000 description 2
- BBBFJLBPOGFECG-VJVYQDLKSA-N calcitonin Chemical compound N([C@H](C(=O)N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N1[C@@H](CCC1)C(N)=O)C(C)C)C(=O)[C@@H]1CSSC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N1 BBBFJLBPOGFECG-VJVYQDLKSA-N 0.000 description 2
- 239000002327 cardiovascular agent Substances 0.000 description 2
- 229940125692 cardiovascular agent Drugs 0.000 description 2
- 239000005018 casein Substances 0.000 description 2
- BECPQYXYKAMYBN-UHFFFAOYSA-N casein, tech. Chemical compound NCCCCC(C(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(CC(C)C)N=C(O)C(CCC(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(C(C)O)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(COP(O)(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(N)CC1=CC=CC=C1 BECPQYXYKAMYBN-UHFFFAOYSA-N 0.000 description 2
- 235000021240 caseins Nutrition 0.000 description 2
- 239000013553 cell monolayer Substances 0.000 description 2
- 239000002738 chelating agent Substances 0.000 description 2
- 238000006243 chemical reaction Methods 0.000 description 2
- 238000000576 coating method Methods 0.000 description 2
- 235000018417 cysteine Nutrition 0.000 description 2
- 230000001086 cytosolic effect Effects 0.000 description 2
- 230000001419 dependent effect Effects 0.000 description 2
- 201000010099 disease Diseases 0.000 description 2
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 2
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 2
- 230000000694 effects Effects 0.000 description 2
- 229920002549 elastin Polymers 0.000 description 2
- 229940105423 erythropoietin Drugs 0.000 description 2
- DEFVIWRASFVYLL-UHFFFAOYSA-N ethylene glycol bis(2-aminoethyl)tetraacetic acid Chemical compound OC(=O)CN(CC(O)=O)CCOCCOCCN(CC(O)=O)CC(O)=O DEFVIWRASFVYLL-UHFFFAOYSA-N 0.000 description 2
- 229940126864 fibroblast growth factor Drugs 0.000 description 2
- 230000009368 gene silencing by RNA Effects 0.000 description 2
- 210000004837 gut-associated lymphoid tissue Anatomy 0.000 description 2
- 210000003630 histaminocyte Anatomy 0.000 description 2
- 239000012216 imaging agent Substances 0.000 description 2
- 238000000338 in vitro Methods 0.000 description 2
- NOESYZHRGYRDHS-UHFFFAOYSA-N insulin Chemical compound N1C(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(NC(=O)CN)C(C)CC)CSSCC(C(NC(CO)C(=O)NC(CC(C)C)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CCC(N)=O)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(N)=O)C(=O)NC(CC=2C=CC(O)=CC=2)C(=O)NC(CSSCC(NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2C=CC(O)=CC=2)NC(=O)C(CC(C)C)NC(=O)C(C)NC(=O)C(CCC(O)=O)NC(=O)C(C(C)C)NC(=O)C(CC(C)C)NC(=O)C(CC=2NC=NC=2)NC(=O)C(CO)NC(=O)CNC2=O)C(=O)NCC(=O)NC(CCC(O)=O)C(=O)NC(CCCNC(N)=N)C(=O)NCC(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC=CC=3)C(=O)NC(CC=3C=CC(O)=CC=3)C(=O)NC(C(C)O)C(=O)N3C(CCC3)C(=O)NC(CCCCN)C(=O)NC(C)C(O)=O)C(=O)NC(CC(N)=O)C(O)=O)=O)NC(=O)C(C(C)CC)NC(=O)C(CO)NC(=O)C(C(C)O)NC(=O)C1CSSCC2NC(=O)C(CC(C)C)NC(=O)C(NC(=O)C(CCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(NC(=O)C(N)CC=1C=CC=CC=1)C(C)C)CC1=CN=CN1 NOESYZHRGYRDHS-UHFFFAOYSA-N 0.000 description 2
- 230000003993 interaction Effects 0.000 description 2
- 108010029560 keratinocyte growth factor receptor Proteins 0.000 description 2
- 239000002523 lectin Substances 0.000 description 2
- 210000001165 lymph node Anatomy 0.000 description 2
- 238000005259 measurement Methods 0.000 description 2
- 230000001404 mediated effect Effects 0.000 description 2
- 210000000110 microvilli Anatomy 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- BQJCRHHNABKAKU-KBQPJGBKSA-N morphine Chemical compound O([C@H]1[C@H](C=C[C@H]23)O)C4=C5[C@@]12CCN(C)[C@@H]3CC5=CC=C4O BQJCRHHNABKAKU-KBQPJGBKSA-N 0.000 description 2
- 230000035772 mutation Effects 0.000 description 2
- 239000003887 narcotic antagonist Substances 0.000 description 2
- 229940094443 oxytocics prostaglandins Drugs 0.000 description 2
- 238000002823 phage display Methods 0.000 description 2
- 229920001223 polyethylene glycol Polymers 0.000 description 2
- OXCMYAYHXIHQOA-UHFFFAOYSA-N potassium;[2-butyl-5-chloro-3-[[4-[2-(1,2,4-triaza-3-azanidacyclopenta-1,4-dien-5-yl)phenyl]phenyl]methyl]imidazol-4-yl]methanol Chemical compound [K+].CCCCC1=NC(Cl)=C(CO)N1CC1=CC=C(C=2C(=CC=CC=2)C2=N[N-]N=N2)C=C1 OXCMYAYHXIHQOA-UHFFFAOYSA-N 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 239000000047 product Substances 0.000 description 2
- 150000003180 prostaglandins Chemical class 0.000 description 2
- 238000000159 protein binding assay Methods 0.000 description 2
- 102000005962 receptors Human genes 0.000 description 2
- 108020003175 receptors Proteins 0.000 description 2
- 210000005084 renal tissue Anatomy 0.000 description 2
- 230000008439 repair process Effects 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 229940125723 sedative agent Drugs 0.000 description 2
- 239000000932 sedative agent Substances 0.000 description 2
- 239000002924 silencing RNA Substances 0.000 description 2
- 239000000243 solution Substances 0.000 description 2
- 238000001179 sorption measurement Methods 0.000 description 2
- 210000000952 spleen Anatomy 0.000 description 2
- 108091007196 stromelysin Proteins 0.000 description 2
- 210000001578 tight junction Anatomy 0.000 description 2
- 229960000187 tissue plasminogen activator Drugs 0.000 description 2
- 241001515965 unidentified phage Species 0.000 description 2
- 239000013603 viral vector Substances 0.000 description 2
- FELGMEQIXOGIFQ-CYBMUJFWSA-N (3r)-9-methyl-3-[(2-methylimidazol-1-yl)methyl]-2,3-dihydro-1h-carbazol-4-one Chemical compound CC1=NC=CN1C[C@@H]1C(=O)C(C=2C(=CC=CC=2)N2C)=C2CC1 FELGMEQIXOGIFQ-CYBMUJFWSA-N 0.000 description 1
- RMTXUPIIESNLPW-UHFFFAOYSA-N 1,2-dihydroxy-3-(pentadeca-8,11-dienyl)benzene Natural products CCCC=CCC=CCCCCCCCC1=CC=CC(O)=C1O RMTXUPIIESNLPW-UHFFFAOYSA-N 0.000 description 1
- LEBVLXFERQHONN-UHFFFAOYSA-N 1-butyl-N-(2,6-dimethylphenyl)piperidine-2-carboxamide Chemical compound CCCCN1CCCCC1C(=O)NC1=C(C)C=CC=C1C LEBVLXFERQHONN-UHFFFAOYSA-N 0.000 description 1
- VOXZDWNPVJITMN-ZBRFXRBCSA-N 17β-estradiol Chemical compound OC1=CC=C2[C@H]3CC[C@](C)([C@H](CC4)O)[C@@H]4[C@@H]3CCC2=C1 VOXZDWNPVJITMN-ZBRFXRBCSA-N 0.000 description 1
- SGTNSNPWRIOYBX-UHFFFAOYSA-N 2-(3,4-dimethoxyphenyl)-5-{[2-(3,4-dimethoxyphenyl)ethyl](methyl)amino}-2-(propan-2-yl)pentanenitrile Chemical compound C1=C(OC)C(OC)=CC=C1CCN(C)CCCC(C#N)(C(C)C)C1=CC=C(OC)C(OC)=C1 SGTNSNPWRIOYBX-UHFFFAOYSA-N 0.000 description 1
- VKUYLANQOAKALN-UHFFFAOYSA-N 2-[benzyl-(4-methoxyphenyl)sulfonylamino]-n-hydroxy-4-methylpentanamide Chemical compound C1=CC(OC)=CC=C1S(=O)(=O)N(C(CC(C)C)C(=O)NO)CC1=CC=CC=C1 VKUYLANQOAKALN-UHFFFAOYSA-N 0.000 description 1
- QARRXYBJLBIVAK-UEMSJJPVSA-N 3-[(8e,11e)-pentadeca-8,11-dienyl]benzene-1,2-diol;3-[(8e,11e)-pentadeca-8,11,14-trienyl]benzene-1,2-diol;3-[(8e,11e,13e)-pentadeca-8,11,13-trienyl]benzene-1,2-diol;3-[(e)-pentadec-8-enyl]benzene-1,2-diol;3-pentadecylbenzene-1,2-diol Chemical compound CCCCCCCCCCCCCCCC1=CC=CC(O)=C1O.CCCCCC\C=C\CCCCCCCC1=CC=CC(O)=C1O.CCC\C=C\C\C=C\CCCCCCCC1=CC=CC(O)=C1O.C\C=C\C=C\C\C=C\CCCCCCCC1=CC=CC(O)=C1O.OC1=CC=CC(CCCCCCC\C=C\C\C=C\CC=C)=C1O QARRXYBJLBIVAK-UEMSJJPVSA-N 0.000 description 1
- IYROWZYPEIMDDN-UHFFFAOYSA-N 3-n-pentadec-8,11,13-trienyl catechol Natural products CC=CC=CCC=CCCCCCCCC1=CC=CC(O)=C1O IYROWZYPEIMDDN-UHFFFAOYSA-N 0.000 description 1
- USSIQXCVUWKGNF-UHFFFAOYSA-N 6-(dimethylamino)-4,4-diphenylheptan-3-one Chemical compound C=1C=CC=CC=1C(CC(C)N(C)C)(C(=O)CC)C1=CC=CC=C1 USSIQXCVUWKGNF-UHFFFAOYSA-N 0.000 description 1
- 229930000680 A04AD01 - Scopolamine Natural products 0.000 description 1
- 241000235389 Absidia Species 0.000 description 1
- 206010063409 Acarodermatitis Diseases 0.000 description 1
- 241000589291 Acinetobacter Species 0.000 description 1
- 241000186046 Actinomyces Species 0.000 description 1
- 102000007469 Actins Human genes 0.000 description 1
- 108010085238 Actins Proteins 0.000 description 1
- 241000701242 Adenoviridae Species 0.000 description 1
- 244000058084 Aegle marmelos Species 0.000 description 1
- 235000003930 Aegle marmelos Nutrition 0.000 description 1
- 238000008940 Alkaline Phosphatase assay kit Methods 0.000 description 1
- 108020000948 Antisense Oligonucleotides Proteins 0.000 description 1
- 241000293035 Apophysomyces Species 0.000 description 1
- 108091023037 Aptamer Proteins 0.000 description 1
- 241000712892 Arenaviridae Species 0.000 description 1
- 241000228212 Aspergillus Species 0.000 description 1
- 241000228197 Aspergillus flavus Species 0.000 description 1
- 241001225321 Aspergillus fumigatus Species 0.000 description 1
- 241000228245 Aspergillus niger Species 0.000 description 1
- 241000223838 Babesia bovis Species 0.000 description 1
- 241000223848 Babesia microti Species 0.000 description 1
- 241000606125 Bacteroides Species 0.000 description 1
- 241000335423 Blastomyces Species 0.000 description 1
- 108010006654 Bleomycin Proteins 0.000 description 1
- 241000589968 Borrelia Species 0.000 description 1
- 241000244203 Caenorhabditis elegans Species 0.000 description 1
- 241000714198 Caliciviridae Species 0.000 description 1
- 241000222122 Candida albicans Species 0.000 description 1
- 241000222173 Candida parapsilosis Species 0.000 description 1
- 241000222178 Candida tropicalis Species 0.000 description 1
- 101710132601 Capsid protein Proteins 0.000 description 1
- BVKZGUZCCUSVTD-UHFFFAOYSA-L Carbonate Chemical compound [O-]C([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-L 0.000 description 1
- 102000053642 Catalytic RNA Human genes 0.000 description 1
- 108090000994 Catalytic RNA Proteins 0.000 description 1
- 241000606161 Chlamydia Species 0.000 description 1
- 241000588923 Citrobacter Species 0.000 description 1
- GJSURZIOUXUGAL-UHFFFAOYSA-N Clonidine Chemical compound ClC1=CC=CC(Cl)=C1NC1=NCCN1 GJSURZIOUXUGAL-UHFFFAOYSA-N 0.000 description 1
- 241000193163 Clostridioides difficile Species 0.000 description 1
- 241000193403 Clostridium Species 0.000 description 1
- 101710094648 Coat protein Proteins 0.000 description 1
- 108091026890 Coding region Proteins 0.000 description 1
- 102000008186 Collagen Human genes 0.000 description 1
- 108010035532 Collagen Proteins 0.000 description 1
- 108010071942 Colony-Stimulating Factors Proteins 0.000 description 1
- 102000007644 Colony-Stimulating Factors Human genes 0.000 description 1
- 241000207892 Convolvulus Species 0.000 description 1
- 241000711573 Coronaviridae Species 0.000 description 1
- 201000007336 Cryptococcosis Diseases 0.000 description 1
- 241000221204 Cryptococcus neoformans Species 0.000 description 1
- 241000223936 Cryptosporidium parvum Species 0.000 description 1
- 102000004127 Cytokines Human genes 0.000 description 1
- 108090000695 Cytokines Proteins 0.000 description 1
- 108010041986 DNA Vaccines Proteins 0.000 description 1
- 229940021995 DNA vaccine Drugs 0.000 description 1
- 108010000437 Deamino Arginine Vasopressin Proteins 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- 239000012591 Dulbecco’s Phosphate Buffered Saline Substances 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 102000002322 Egg Proteins Human genes 0.000 description 1
- 108010000912 Egg Proteins Proteins 0.000 description 1
- 102400001368 Epidermal growth factor Human genes 0.000 description 1
- 101800003838 Epidermal growth factor Proteins 0.000 description 1
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 1
- 108010054218 Factor VIII Proteins 0.000 description 1
- 102000001690 Factor VIII Human genes 0.000 description 1
- 241000282324 Felis Species 0.000 description 1
- 108090000385 Fibroblast growth factor 7 Proteins 0.000 description 1
- 241000711950 Filoviridae Species 0.000 description 1
- 206010016717 Fistula Diseases 0.000 description 1
- 241000710781 Flaviviridae Species 0.000 description 1
- GHASVSINZRGABV-UHFFFAOYSA-N Fluorouracil Chemical compound FC1=CNC(=O)NC1=O GHASVSINZRGABV-UHFFFAOYSA-N 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 1
- 241000606790 Haemophilus Species 0.000 description 1
- 241000258937 Hemiptera Species 0.000 description 1
- 241000700739 Hepadnaviridae Species 0.000 description 1
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 1
- 241000700586 Herpesviridae Species 0.000 description 1
- 102000007625 Hirudins Human genes 0.000 description 1
- 108010007267 Hirudins Proteins 0.000 description 1
- 241000228402 Histoplasma Species 0.000 description 1
- 241000282412 Homo Species 0.000 description 1
- 102000002265 Human Growth Hormone Human genes 0.000 description 1
- 108010000521 Human Growth Hormone Proteins 0.000 description 1
- 239000000854 Human Growth Hormone Substances 0.000 description 1
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 1
- STECJAGHUSJQJN-GAUPFVANSA-N Hyoscine Natural products C1([C@H](CO)C(=O)OC2C[C@@H]3N([C@H](C2)[C@@H]2[C@H]3O2)C)=CC=CC=C1 STECJAGHUSJQJN-GAUPFVANSA-N 0.000 description 1
- 102000038455 IGF Type 1 Receptor Human genes 0.000 description 1
- 108010031794 IGF Type 1 Receptor Proteins 0.000 description 1
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 1
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 1
- 102000004877 Insulin Human genes 0.000 description 1
- 108090001061 Insulin Proteins 0.000 description 1
- 108010005714 Interferon beta-1b Proteins 0.000 description 1
- 102000006992 Interferon-alpha Human genes 0.000 description 1
- 108010047761 Interferon-alpha Proteins 0.000 description 1
- 108090000467 Interferon-beta Proteins 0.000 description 1
- 102000003996 Interferon-beta Human genes 0.000 description 1
- 102000008070 Interferon-gamma Human genes 0.000 description 1
- 108010074328 Interferon-gamma Proteins 0.000 description 1
- 108010050904 Interferons Proteins 0.000 description 1
- 102000014150 Interferons Human genes 0.000 description 1
- 102000000588 Interleukin-2 Human genes 0.000 description 1
- 108010002350 Interleukin-2 Proteins 0.000 description 1
- 102000015696 Interleukins Human genes 0.000 description 1
- 108010063738 Interleukins Proteins 0.000 description 1
- 241000588748 Klebsiella Species 0.000 description 1
- 241000968140 Lactobacillus hominis Species 0.000 description 1
- 101710143474 Lectin-related protein Proteins 0.000 description 1
- 241000589248 Legionella Species 0.000 description 1
- 108010000817 Leuprolide Proteins 0.000 description 1
- JAQUASYNZVUNQP-USXIJHARSA-N Levorphanol Chemical compound C1C2=CC=C(O)C=C2[C@]23CCN(C)[C@H]1[C@@H]2CCCC3 JAQUASYNZVUNQP-USXIJHARSA-N 0.000 description 1
- NNJVILVZKWQKPM-UHFFFAOYSA-N Lidocaine Chemical compound CCN(CC)CC(=O)NC1=C(C)C=CC=C1C NNJVILVZKWQKPM-UHFFFAOYSA-N 0.000 description 1
- 239000006142 Luria-Bertani Agar Substances 0.000 description 1
- 208000016604 Lyme disease Diseases 0.000 description 1
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- 102000004318 Matrilysin Human genes 0.000 description 1
- 108090000855 Matrilysin Proteins 0.000 description 1
- 102000005741 Metalloproteases Human genes 0.000 description 1
- 108010006035 Metalloproteases Proteins 0.000 description 1
- 102000014171 Milk Proteins Human genes 0.000 description 1
- 108010011756 Milk Proteins Proteins 0.000 description 1
- 241000219470 Mirabilis Species 0.000 description 1
- 241000235395 Mucor Species 0.000 description 1
- 101000851053 Mus musculus Elastin Proteins 0.000 description 1
- 101001027408 Mus musculus Fibroblast growth factor 7 Proteins 0.000 description 1
- 241000204031 Mycoplasma Species 0.000 description 1
- STECJAGHUSJQJN-UHFFFAOYSA-N N-Methyl-scopolamin Natural products C1C(C2C3O2)N(C)C3CC1OC(=O)C(CO)C1=CC=CC=C1 STECJAGHUSJQJN-UHFFFAOYSA-N 0.000 description 1
- UBQYURCVBFRUQT-UHFFFAOYSA-N N-benzoyl-Ferrioxamine B Chemical compound CC(=O)N(O)CCCCCNC(=O)CCC(=O)N(O)CCCCCNC(=O)CCC(=O)N(O)CCCCCN UBQYURCVBFRUQT-UHFFFAOYSA-N 0.000 description 1
- CMWTZPSULFXXJA-UHFFFAOYSA-N Naproxen Natural products C1=C(C(C)C(O)=O)C=CC2=CC(OC)=CC=C21 CMWTZPSULFXXJA-UHFFFAOYSA-N 0.000 description 1
- 241000588653 Neisseria Species 0.000 description 1
- 241000244206 Nematoda Species 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 239000000020 Nitrocellulose Substances 0.000 description 1
- SNIOPGDIGTZGOP-UHFFFAOYSA-N Nitroglycerin Chemical compound [O-][N+](=O)OCC(O[N+]([O-])=O)CO[N+]([O-])=O SNIOPGDIGTZGOP-UHFFFAOYSA-N 0.000 description 1
- 239000000006 Nitroglycerin Substances 0.000 description 1
- 101710141454 Nucleoprotein Proteins 0.000 description 1
- 108010026867 Oligo-1,6-Glucosidase Proteins 0.000 description 1
- 102000003840 Opioid Receptors Human genes 0.000 description 1
- 108090000137 Opioid Receptors Proteins 0.000 description 1
- 241000713112 Orthobunyavirus Species 0.000 description 1
- 241000712464 Orthomyxoviridae Species 0.000 description 1
- UQCNKQCJZOAFTQ-ISWURRPUSA-N Oxymorphone Chemical compound O([C@H]1C(CC[C@]23O)=O)C4=C5[C@@]12CCN(C)[C@@H]3CC5=CC=C4O UQCNKQCJZOAFTQ-ISWURRPUSA-N 0.000 description 1
- 102400000050 Oxytocin Human genes 0.000 description 1
- 101800000989 Oxytocin Proteins 0.000 description 1
- XNOPRXBHLZRZKH-UHFFFAOYSA-N Oxytocin Natural products N1C(=O)C(N)CSSCC(C(=O)N2C(CCC2)C(=O)NC(CC(C)C)C(=O)NCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(CCC(N)=O)NC(=O)C(C(C)CC)NC(=O)C1CC1=CC=C(O)C=C1 XNOPRXBHLZRZKH-UHFFFAOYSA-N 0.000 description 1
- 241001537205 Paracoccidioides Species 0.000 description 1
- 241000935974 Paralichthys dentatus Species 0.000 description 1
- 241000711504 Paramyxoviridae Species 0.000 description 1
- 208000002606 Paramyxoviridae Infections Diseases 0.000 description 1
- 208000030852 Parasitic disease Diseases 0.000 description 1
- 241000701945 Parvoviridae Species 0.000 description 1
- 241000606856 Pasteurella multocida Species 0.000 description 1
- 241000517307 Pediculus humanus Species 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 108010067902 Peptide Library Proteins 0.000 description 1
- 229940083963 Peptide antagonist Drugs 0.000 description 1
- 102000003992 Peroxidases Human genes 0.000 description 1
- 102000013566 Plasminogen Human genes 0.000 description 1
- 108010051456 Plasminogen Proteins 0.000 description 1
- 241000233870 Pneumocystis Species 0.000 description 1
- 241000700625 Poxviridae Species 0.000 description 1
- 101710083689 Probable capsid protein Proteins 0.000 description 1
- 229940124158 Protease/peptidase inhibitor Drugs 0.000 description 1
- 108010067787 Proteoglycans Proteins 0.000 description 1
- 102000016611 Proteoglycans Human genes 0.000 description 1
- 241000588769 Proteus <enterobacteria> Species 0.000 description 1
- 241000589516 Pseudomonas Species 0.000 description 1
- 241000711798 Rabies lyssavirus Species 0.000 description 1
- 241000702247 Reoviridae Species 0.000 description 1
- 241000712907 Retroviridae Species 0.000 description 1
- 241000711931 Rhabdoviridae Species 0.000 description 1
- 241000235402 Rhizomucor Species 0.000 description 1
- 241000952054 Rhizopus sp. Species 0.000 description 1
- 241000606701 Rickettsia Species 0.000 description 1
- 241000702670 Rotavirus Species 0.000 description 1
- 241000607142 Salmonella Species 0.000 description 1
- 241000293871 Salmonella enterica subsp. enterica serovar Typhi Species 0.000 description 1
- 241000447727 Scabies Species 0.000 description 1
- 241000242677 Schistosoma japonicum Species 0.000 description 1
- 241000356562 Selepa celtis Species 0.000 description 1
- 241000607720 Serratia Species 0.000 description 1
- 241000700584 Simplexvirus Species 0.000 description 1
- 241000710960 Sindbis virus Species 0.000 description 1
- 108010056088 Somatostatin Proteins 0.000 description 1
- 102000005157 Somatostatin Human genes 0.000 description 1
- 241000191967 Staphylococcus aureus Species 0.000 description 1
- 241000191963 Staphylococcus epidermidis Species 0.000 description 1
- 241001478878 Streptobacillus Species 0.000 description 1
- 241000194019 Streptococcus mutans Species 0.000 description 1
- 241000187747 Streptomyces Species 0.000 description 1
- 102100030416 Stromelysin-1 Human genes 0.000 description 1
- 101710108790 Stromelysin-1 Proteins 0.000 description 1
- 101710172711 Structural protein Proteins 0.000 description 1
- 102100027918 Sucrase-isomaltase, intestinal Human genes 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 241000710924 Togaviridae Species 0.000 description 1
- 241000159241 Toxicodendron Species 0.000 description 1
- 241000223997 Toxoplasma gondii Species 0.000 description 1
- 108010033576 Transferrin Receptors Proteins 0.000 description 1
- 102000007238 Transferrin Receptors Human genes 0.000 description 1
- 102100023935 Transmembrane glycoprotein NMB Human genes 0.000 description 1
- 241000589886 Treponema Species 0.000 description 1
- 241000243774 Trichinella Species 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- 241000223104 Trypanosoma Species 0.000 description 1
- 102100022563 Tubulin polymerization-promoting protein Human genes 0.000 description 1
- 101710158555 Tubulin polymerization-promoting protein Proteins 0.000 description 1
- 108091005906 Type I transmembrane proteins Proteins 0.000 description 1
- 241000202898 Ureaplasma Species 0.000 description 1
- GXBMIBRIOWHPDT-UHFFFAOYSA-N Vasopressin Natural products N1C(=O)C(CC=2C=C(O)C=CC=2)NC(=O)C(N)CSSCC(C(=O)N2C(CCC2)C(=O)NC(CCCN=C(N)N)C(=O)NCC(N)=O)NC(=O)C(CC(N)=O)NC(=O)C(CCC(N)=O)NC(=O)C1CC1=CC=CC=C1 GXBMIBRIOWHPDT-UHFFFAOYSA-N 0.000 description 1
- 108010004977 Vasopressins Proteins 0.000 description 1
- 102000002852 Vasopressins Human genes 0.000 description 1
- 241000607626 Vibrio cholerae Species 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000004913 activation Effects 0.000 description 1
- 208000009956 adenocarcinoma Diseases 0.000 description 1
- 108700010877 adenoviridae proteins Proteins 0.000 description 1
- 208000026935 allergic disease Diseases 0.000 description 1
- AZDRQVAHHNSJOQ-UHFFFAOYSA-N alumane Chemical class [AlH3] AZDRQVAHHNSJOQ-UHFFFAOYSA-N 0.000 description 1
- 230000003321 amplification Effects 0.000 description 1
- 230000000118 anti-neoplastic effect Effects 0.000 description 1
- 230000030741 antigen processing and presentation Effects 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 229940030600 antihypertensive agent Drugs 0.000 description 1
- 229940127088 antihypertensive drug Drugs 0.000 description 1
- 239000000074 antisense oligonucleotide Substances 0.000 description 1
- 238000012230 antisense oligonucleotides Methods 0.000 description 1
- 238000013459 approach Methods 0.000 description 1
- KBZOIRJILGZLEJ-LGYYRGKSSA-N argipressin Chemical compound C([C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CSSC[C@@H](C(N[C@@H](CC=2C=CC(O)=CC=2)C(=O)N1)=O)N)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCN=C(N)N)C(=O)NCC(N)=O)C1=CC=CC=C1 KBZOIRJILGZLEJ-LGYYRGKSSA-N 0.000 description 1
- 230000001746 atrial effect Effects 0.000 description 1
- 230000002238 attenuated effect Effects 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 210000003651 basophil Anatomy 0.000 description 1
- 229940049706 benzodiazepine Drugs 0.000 description 1
- 150000001557 benzodiazepines Chemical class 0.000 description 1
- 229940021459 betaseron Drugs 0.000 description 1
- 238000007413 biotinylation Methods 0.000 description 1
- 230000006287 biotinylation Effects 0.000 description 1
- 229960001561 bleomycin Drugs 0.000 description 1
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 229960003150 bupivacaine Drugs 0.000 description 1
- RMRJXGBAOAMLHD-IHFGGWKQSA-N buprenorphine Chemical compound C([C@]12[C@H]3OC=4C(O)=CC=C(C2=4)C[C@@H]2[C@]11CC[C@]3([C@H](C1)[C@](C)(O)C(C)(C)C)OC)CN2CC1CC1 RMRJXGBAOAMLHD-IHFGGWKQSA-N 0.000 description 1
- 229960001736 buprenorphine Drugs 0.000 description 1
- IFKLAQQSCNILHL-QHAWAJNXSA-N butorphanol Chemical compound N1([C@@H]2CC3=CC=C(C=C3[C@@]3([C@]2(CCCC3)O)CC1)O)CC1CCC1 IFKLAQQSCNILHL-QHAWAJNXSA-N 0.000 description 1
- 229960001113 butorphanol Drugs 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 230000024245 cell differentiation Effects 0.000 description 1
- 230000009087 cell motility Effects 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 229940044683 chemotherapy drug Drugs 0.000 description 1
- 108091006116 chimeric peptides Proteins 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 229960002896 clonidine Drugs 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- 238000003501 co-culture Methods 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 229920001436 collagen Polymers 0.000 description 1
- 230000011382 collagen catabolic process Effects 0.000 description 1
- 210000001072 colon Anatomy 0.000 description 1
- 230000000295 complement effect Effects 0.000 description 1
- 238000009833 condensation Methods 0.000 description 1
- 230000005494 condensation Effects 0.000 description 1
- 238000004624 confocal microscopy Methods 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 239000006059 cover glass Substances 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 150000001945 cysteines Chemical class 0.000 description 1
- 210000004292 cytoskeleton Anatomy 0.000 description 1
- 229940127089 cytotoxic agent Drugs 0.000 description 1
- 230000007123 defense Effects 0.000 description 1
- 229960000958 deferoxamine Drugs 0.000 description 1
- 230000002939 deleterious effect Effects 0.000 description 1
- 239000000412 dendrimer Substances 0.000 description 1
- 210000004443 dendritic cell Anatomy 0.000 description 1
- 229920000736 dendritic polymer Polymers 0.000 description 1
- 229960004281 desmopressin Drugs 0.000 description 1
- NFLWUMRGJYTJIN-NXBWRCJVSA-N desmopressin Chemical compound C([C@H]1C(=O)N[C@H](C(N[C@@H](CC(N)=O)C(=O)N[C@@H](CSSCCC(=O)N[C@@H](CC=2C=CC(O)=CC=2)C(=O)N1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(N)=O)=O)CCC(=O)N)C1=CC=CC=C1 NFLWUMRGJYTJIN-NXBWRCJVSA-N 0.000 description 1
- 238000002405 diagnostic procedure Methods 0.000 description 1
- DCOPUUMXTXDBNB-UHFFFAOYSA-N diclofenac Chemical compound OC(=O)CC1=CC=CC=C1NC1=C(Cl)C=CC=C1Cl DCOPUUMXTXDBNB-UHFFFAOYSA-N 0.000 description 1
- 229960001259 diclofenac Drugs 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- XYYVYLMBEZUESM-UHFFFAOYSA-N dihydrocodeine Natural products C1C(N(CCC234)C)C2C=CC(=O)C3OC2=C4C1=CC=C2OC XYYVYLMBEZUESM-UHFFFAOYSA-N 0.000 description 1
- 229960004166 diltiazem Drugs 0.000 description 1
- HSUGRBWQSSZJOP-RTWAWAEBSA-N diltiazem Chemical compound C1=CC(OC)=CC=C1[C@H]1[C@@H](OC(C)=O)C(=O)N(CCN(C)C)C2=CC=CC=C2S1 HSUGRBWQSSZJOP-RTWAWAEBSA-N 0.000 description 1
- 239000002934 diuretic Substances 0.000 description 1
- FGXWKSZFVQUSTL-UHFFFAOYSA-N domperidone Chemical compound C12=CC=CC=C2NC(=O)N1CCCN(CC1)CCC1N1C2=CC=C(Cl)C=C2NC1=O FGXWKSZFVQUSTL-UHFFFAOYSA-N 0.000 description 1
- 229960001253 domperidone Drugs 0.000 description 1
- 239000000428 dust Substances 0.000 description 1
- 208000001848 dysentery Diseases 0.000 description 1
- 239000008393 encapsulating agent Substances 0.000 description 1
- 239000003623 enhancer Substances 0.000 description 1
- 210000001842 enterocyte Anatomy 0.000 description 1
- 238000006911 enzymatic reaction Methods 0.000 description 1
- 210000003979 eosinophil Anatomy 0.000 description 1
- 229940116977 epidermal growth factor Drugs 0.000 description 1
- 210000005081 epithelial layer Anatomy 0.000 description 1
- 210000004783 epithelial tight junction Anatomy 0.000 description 1
- 229960003133 ergot alkaloid Drugs 0.000 description 1
- 229960005309 estradiol Drugs 0.000 description 1
- 229930182833 estradiol Natural products 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 229960000301 factor viii Drugs 0.000 description 1
- 229960002428 fentanyl Drugs 0.000 description 1
- PJMPHNIQZUBGLI-UHFFFAOYSA-N fentanyl Chemical compound C=1C=CC=CC=1N(C(=O)CC)C(CC1)CCN1CCC1=CC=CC=C1 PJMPHNIQZUBGLI-UHFFFAOYSA-N 0.000 description 1
- 239000010419 fine particle Substances 0.000 description 1
- 230000003890 fistula Effects 0.000 description 1
- 229960002949 fluorouracil Drugs 0.000 description 1
- 235000013305 food Nutrition 0.000 description 1
- 229940044627 gamma-interferon Drugs 0.000 description 1
- 102000034356 gene-regulatory proteins Human genes 0.000 description 1
- 108091006104 gene-regulatory proteins Proteins 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 229960003711 glyceryl trinitrate Drugs 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 239000000122 growth hormone Substances 0.000 description 1
- 208000019622 heart disease Diseases 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 230000002440 hepatic effect Effects 0.000 description 1
- 238000013537 high throughput screening Methods 0.000 description 1
- WQPDUTSPKFMPDP-OUMQNGNKSA-N hirudin Chemical compound C([C@@H](C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC=1C=CC(OS(O)(=O)=O)=CC=1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CCCCN)NC(=O)[C@H]1N(CCC1)C(=O)[C@@H](NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)CNC(=O)[C@@H](NC(=O)[C@@H](NC(=O)[C@H]1NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(O)=O)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CO)NC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H]2CSSC[C@@H](C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(=O)N[C@H](C(NCC(=O)N[C@@H](CCC(N)=O)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N2)=O)CSSC1)C(C)C)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]1NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)CNC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=2C=CC(O)=CC=2)NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)C(C)C)[C@@H](C)O)CSSC1)C(C)C)[C@@H](C)O)[C@@H](C)O)C1=CC=CC=C1 WQPDUTSPKFMPDP-OUMQNGNKSA-N 0.000 description 1
- 229940006607 hirudin Drugs 0.000 description 1
- 125000001183 hydrocarbyl group Chemical group 0.000 description 1
- LLPOLZWFYMWNKH-CMKMFDCUSA-N hydrocodone Chemical compound C([C@H]1[C@H](N(CC[C@@]112)C)C3)CC(=O)[C@@H]1OC1=C2C3=CC=C1OC LLPOLZWFYMWNKH-CMKMFDCUSA-N 0.000 description 1
- 229960000240 hydrocodone Drugs 0.000 description 1
- OROGSEYTTFOCAN-UHFFFAOYSA-N hydrocodone Natural products C1C(N(CCC234)C)C2C=CC(O)C3OC2=C4C1=CC=C2OC OROGSEYTTFOCAN-UHFFFAOYSA-N 0.000 description 1
- WVLOADHCBXTIJK-YNHQPCIGSA-N hydromorphone Chemical compound O([C@H]1C(CC[C@H]23)=O)C4=C5[C@@]12CCN(C)[C@@H]3CC5=CC=C4O WVLOADHCBXTIJK-YNHQPCIGSA-N 0.000 description 1
- 229960001410 hydromorphone Drugs 0.000 description 1
- 230000036039 immunity Effects 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 229940125396 insulin Drugs 0.000 description 1
- 229940079322 interferon Drugs 0.000 description 1
- 229940047122 interleukins Drugs 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 1
- YWXYYJSYQOXTPL-SLPGGIOYSA-N isosorbide mononitrate Chemical compound [O-][N+](=O)O[C@@H]1CO[C@@H]2[C@@H](O)CO[C@@H]21 YWXYYJSYQOXTPL-SLPGGIOYSA-N 0.000 description 1
- 229960003827 isosorbide mononitrate Drugs 0.000 description 1
- 150000002596 lactones Chemical class 0.000 description 1
- 210000001821 langerhans cell Anatomy 0.000 description 1
- GFIJNRVAKGFPGQ-LIJARHBVSA-N leuprolide Chemical compound CCNC(=O)[C@@H]1CCCN1C(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](CC(C)C)NC(=O)[C@@H](NC(=O)[C@H](CO)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H]1NC(=O)CC1)CC1=CC=C(O)C=C1 GFIJNRVAKGFPGQ-LIJARHBVSA-N 0.000 description 1
- 229960004338 leuprorelin Drugs 0.000 description 1
- 229960003406 levorphanol Drugs 0.000 description 1
- 229960004194 lidocaine Drugs 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 210000005228 liver tissue Anatomy 0.000 description 1
- 230000033001 locomotion Effects 0.000 description 1
- 229920002521 macromolecule Polymers 0.000 description 1
- 238000012423 maintenance Methods 0.000 description 1
- 201000004792 malaria Diseases 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 239000002609 medium Substances 0.000 description 1
- 229960001797 methadone Drugs 0.000 description 1
- TTWJBBZEZQICBI-UHFFFAOYSA-N metoclopramide Chemical compound CCN(CC)CCNC(=O)C1=CC(Cl)=C(N)C=C1OC TTWJBBZEZQICBI-UHFFFAOYSA-N 0.000 description 1
- 229960004503 metoclopramide Drugs 0.000 description 1
- 235000021239 milk protein Nutrition 0.000 description 1
- 239000003226 mitogen Substances 0.000 description 1
- 210000001616 monocyte Anatomy 0.000 description 1
- 229960005181 morphine Drugs 0.000 description 1
- UZHSEJADLWPNLE-GRGSLBFTSA-N naloxone Chemical compound O=C([C@@H]1O2)CC[C@@]3(O)[C@H]4CC5=CC=C(O)C2=C5[C@@]13CCN4CC=C UZHSEJADLWPNLE-GRGSLBFTSA-N 0.000 description 1
- 229960004127 naloxone Drugs 0.000 description 1
- DQCKKXVULJGBQN-XFWGSAIBSA-N naltrexone Chemical compound N1([C@@H]2CC3=CC=C(C=4O[C@@H]5[C@](C3=4)([C@]2(CCC5=O)O)CC1)O)CC1CC1 DQCKKXVULJGBQN-XFWGSAIBSA-N 0.000 description 1
- 229960003086 naltrexone Drugs 0.000 description 1
- 229960002009 naproxen Drugs 0.000 description 1
- CMWTZPSULFXXJA-VIFPVBQESA-N naproxen Chemical compound C1=C([C@H](C)C(O)=O)C=CC2=CC(OC)=CC=C21 CMWTZPSULFXXJA-VIFPVBQESA-N 0.000 description 1
- 230000001452 natriuretic effect Effects 0.000 description 1
- 229960001597 nifedipine Drugs 0.000 description 1
- HYIMSNHJOBLJNT-UHFFFAOYSA-N nifedipine Chemical compound COC(=O)C1=C(C)NC(C)=C(C(=O)OC)C1C1=CC=CC=C1[N+]([O-])=O HYIMSNHJOBLJNT-UHFFFAOYSA-N 0.000 description 1
- 229920001220 nitrocellulos Polymers 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 238000003199 nucleic acid amplification method Methods 0.000 description 1
- 229960005343 ondansetron Drugs 0.000 description 1
- 239000003399 opiate peptide Substances 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 229940082615 organic nitrates used in cardiac disease Drugs 0.000 description 1
- 210000004681 ovum Anatomy 0.000 description 1
- 229960005118 oxymorphone Drugs 0.000 description 1
- XNOPRXBHLZRZKH-DSZYJQQASA-N oxytocin Chemical compound C([C@H]1C(=O)N[C@H](C(N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CSSC[C@H](N)C(=O)N1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CC(C)C)C(=O)NCC(N)=O)=O)[C@@H](C)CC)C1=CC=C(O)C=C1 XNOPRXBHLZRZKH-DSZYJQQASA-N 0.000 description 1
- 229960001723 oxytocin Drugs 0.000 description 1
- 239000012188 paraffin wax Substances 0.000 description 1
- 235000016236 parenteral nutrition Nutrition 0.000 description 1
- 239000011236 particulate material Substances 0.000 description 1
- 229940051027 pasteurella multocida Drugs 0.000 description 1
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 1
- 125000001151 peptidyl group Chemical group 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 108700010839 phage proteins Proteins 0.000 description 1
- 150000002990 phenothiazines Chemical class 0.000 description 1
- 229940012957 plasmin Drugs 0.000 description 1
- 230000033885 plasminogen activation Effects 0.000 description 1
- 239000002797 plasminogen activator inhibitor Substances 0.000 description 1
- 239000004033 plastic Substances 0.000 description 1
- 229920003023 plastic Polymers 0.000 description 1
- 201000000317 pneumocystosis Diseases 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 230000002265 prevention Effects 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 230000035755 proliferation Effects 0.000 description 1
- 235000019833 protease Nutrition 0.000 description 1
- 238000002731 protein assay Methods 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 210000003689 pubic bone Anatomy 0.000 description 1
- 238000011552 rat model Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 230000008929 regeneration Effects 0.000 description 1
- 238000011069 regeneration method Methods 0.000 description 1
- 108091092562 ribozyme Proteins 0.000 description 1
- 108010038196 saccharide-binding proteins Proteins 0.000 description 1
- 208000005687 scabies Diseases 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- STECJAGHUSJQJN-FWXGHANASA-N scopolamine Chemical compound C1([C@@H](CO)C(=O)O[C@H]2C[C@@H]3N([C@H](C2)[C@@H]2[C@H]3O2)C)=CC=CC=C1 STECJAGHUSJQJN-FWXGHANASA-N 0.000 description 1
- 229960002646 scopolamine Drugs 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- NHXLMOGPVYXJNR-ATOGVRKGSA-N somatostatin Chemical compound C([C@H]1C(=O)N[C@H](C(N[C@@H](CO)C(=O)N[C@@H](CSSC[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@@H](CC=2C3=CC=CC=C3NC=2)C(=O)N[C@@H](CCCCN)C(=O)N[C@H](C(=O)N1)[C@@H](C)O)NC(=O)CNC(=O)[C@H](C)N)C(O)=O)=O)[C@H](O)C)C1=CC=CC=C1 NHXLMOGPVYXJNR-ATOGVRKGSA-N 0.000 description 1
- 229960000553 somatostatin Drugs 0.000 description 1
- 229960004532 somatropin Drugs 0.000 description 1
- 230000009870 specific binding Effects 0.000 description 1
- 239000012798 spherical particle Substances 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 210000002536 stromal cell Anatomy 0.000 description 1
- 238000006467 substitution reaction Methods 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- GGCSSNBKKAUURC-UHFFFAOYSA-N sufentanil Chemical compound C1CN(CCC=2SC=CC=2)CCC1(COC)N(C(=O)CC)C1=CC=CC=C1 GGCSSNBKKAUURC-UHFFFAOYSA-N 0.000 description 1
- 229960004739 sufentanil Drugs 0.000 description 1
- 238000009495 sugar coating Methods 0.000 description 1
- KQKPFRSPSRPDEB-UHFFFAOYSA-N sumatriptan Chemical compound CNS(=O)(=O)CC1=CC=C2NC=C(CCN(C)C)C2=C1 KQKPFRSPSRPDEB-UHFFFAOYSA-N 0.000 description 1
- 229960003708 sumatriptan Drugs 0.000 description 1
- 239000000725 suspension Substances 0.000 description 1
- 230000001225 therapeutic effect Effects 0.000 description 1
- 230000017423 tissue regeneration Effects 0.000 description 1
- 230000007838 tissue remodeling Effects 0.000 description 1
- 231100000033 toxigenic Toxicity 0.000 description 1
- 230000001551 toxigenic effect Effects 0.000 description 1
- LLPOLZWFYMWNKH-UHFFFAOYSA-N trans-dihydrocodeinone Natural products C1C(N(CCC234)C)C2CCC(=O)C3OC2=C4C1=CC=C2OC LLPOLZWFYMWNKH-UHFFFAOYSA-N 0.000 description 1
- 230000018889 transepithelial transport Effects 0.000 description 1
- 108091007466 transmembrane glycoproteins Proteins 0.000 description 1
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 1
- 241000701161 unidentified adenovirus Species 0.000 description 1
- 241000712461 unidentified influenza virus Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 description 1
- DQTMTQZSOJMZSF-UHFFFAOYSA-N urushiol Natural products CCCCCCCCCCCCCCCC1=CC=CC(O)=C1O DQTMTQZSOJMZSF-UHFFFAOYSA-N 0.000 description 1
- 210000004291 uterus Anatomy 0.000 description 1
- 238000002255 vaccination Methods 0.000 description 1
- 210000001177 vas deferen Anatomy 0.000 description 1
- 229940124549 vasodilator Drugs 0.000 description 1
- 239000003071 vasodilator agent Substances 0.000 description 1
- 229960003726 vasopressin Drugs 0.000 description 1
- 229960001722 verapamil Drugs 0.000 description 1
- 229940118696 vibrio cholerae Drugs 0.000 description 1
- OGWKCGZFUXNPDA-XQKSVPLYSA-N vincristine Chemical compound C([N@]1C[C@@H](C[C@]2(C(=O)OC)C=3C(=CC4=C([C@]56[C@H]([C@@]([C@H](OC(C)=O)[C@]7(CC)C=CCN([C@H]67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)C[C@@](C1)(O)CC)CC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-XQKSVPLYSA-N 0.000 description 1
- 229960004528 vincristine Drugs 0.000 description 1
- OGWKCGZFUXNPDA-UHFFFAOYSA-N vincristine Natural products C1C(CC)(O)CC(CC2(C(=O)OC)C=3C(=CC4=C(C56C(C(C(OC(C)=O)C7(CC)C=CCN(C67)CC5)(O)C(=O)OC)N4C=O)C=3)OC)CN1CCC1=C2NC2=CC=CC=C12 OGWKCGZFUXNPDA-UHFFFAOYSA-N 0.000 description 1
- 108010047303 von Willebrand Factor Proteins 0.000 description 1
- 102100036537 von Willebrand factor Human genes 0.000 description 1
- 229960001134 von willebrand factor Drugs 0.000 description 1
- 239000011534 wash buffer Substances 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
Images























































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/08—Linear peptides containing only normal peptide links having 12 to 20 amino acids
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/001—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof by chemical synthesis
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4725—Proteoglycans, e.g. aggreccan
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/02—Linear peptides containing at least one abnormal peptide link
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/06—Linear peptides containing only normal peptide links having 5 to 11 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- General Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Gastroenterology & Hepatology (AREA)
- Zoology (AREA)
- Toxicology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicinal Preparation (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Compounds Of Unknown Constitution (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Description
プチドによって与えられる能力と少なくとも同じであることを特徴とする、精製合成ポリペプチドリガンドである。
る群から選択される12アミノ酸長L‐ペプチドのD型である12アミノ酸長D‐ペプチドまたはその相同体を有する精製合成ポリペプチドリガンドであって、前記相同体は前記群から選択された12アミノ酸長D‐ペプチドに対して少なくとも9/12の相同性を有し、前記12アミノ酸長D‐ペプチドまたはその相同体はM13ファージのN末端PIII融合ペプチドとして組み込まれると、該ファージにCaco‐2細胞、IEC‐6細胞、ラット、マウス、ブタまたはイヌのいずれかのホモジネート膜画分に結合する能力を与え、前記能力は、同様に組み込まれた配列番号67の翻訳である12アミノ酸長ペプチドによって与えられる能力と少なくとも同じであることを特徴とする、精製合成ポリペプチドリガンドである。
特定の12アミノ酸長L‐ペプチドもしくは天然に生じる12アミノ酸長相同体、それらのD型およびそれらの反転型に関する本発明に関して、相同体は少なくとも10/12の相同性を有することが好ましく、11/12の相同性を有することがより好ましい。同様に、12アミノ酸長の断片は長さが少なくとも8アミノ酸であることが好ましい。相同体または断片よりも、完全な特定のL‐ペプチド、D型、または反転型の存在が最も好ましい。
前述の精製合成ポリペプチドリガンドをコードする核酸分子もまた本発明の態様である。長さが600ヌクレオチド以下であるものが好ましい。特定の12アミノ酸長ペプチド、モチーフ、または天然に生じる相同体のうち1つからなる精製合成ポリペプチドをコードするものが非常に好ましい。
本発明の特定の実施形態では、前述の精製合成ポリペプチドリガンドのうち1つが薬剤を含む担体実体に共有結合または非共有結合している。たとえば、該担体実体はナノ粒子、微粒子、リポソーム、細菌、ファージ(バクテリオファージ)およびウイルス(好ましくは哺乳類ウイルス、最も好ましくはヒトウイルス、特に組換えまたはその他の技術によって作製された非病原性型)からなる群から選択される。本明細書の他所で詳細に述べるように、ナノ粒子、微粒子またはリポソームの大きさは最大の寸法が10nm〜500μmの範囲にあることが好ましい。本発明の特定の実施形態では、薬剤は薬物または治療薬である。その他の特定の実施形態では、薬剤は病原体抗原である。
一態様では、本発明は腸上皮を有する生物に薬剤を投与する方法であって、前記方法は担体実体に共有結合、または非共有結合した前述の精製合成ポリペプチドリガンドの1つと前記腸上皮とを接触させることからなる方法である。好ましい実施形態では、この生物は哺乳類である。最も好ましくは、この哺乳類はヒトである。
溶液中のポリペプチドのほとんどまたは全てが特定の合成ポリペプチドリガンドである溶液中の合成ポリペプチドリガンド集団は、精製合成ポリペプチドリガンドの一例である。

12アミノ酸長ペプチド配列の分析によって、いくつかのペプチドは共通モチーフを含有することが明らかになった。したがって、本発明は、これらのモチーフおよびこれらのモチーフを有するポリペプチドリガンドをも含んでおり、このポリペプチドリガンドによって薬剤を腸上皮、M細胞またはパイエル板組織に侵入または横断して輸送することが容易になる。
薬剤を搭載した、またはカプセル化したナノ粒子および微粒子を、腸上皮組織、たとえばM細胞またはパイエル板組織を標的とするポリペプチドリガンド、たとえば本発明のポリペプチドリガンドで被覆することが可能である。被覆は共有結合または非共有結合によって実施することが可能である。共有結合は、吸着または任意のその他の被覆工程によって実施することが可能である。いずれの場合においても、この結合の形成により完成した粒子とするか、またはその後粒子の一部を形成する粒子成分とすることが可能である。
あるいは、薬剤を直接ポリペプチドリガンドに結合することが可能である。薬剤自身がポリペプチドまたはペプチドの場合、生成物は薬剤とターゲティング(標的を目標とする)部分の両方からなるキメラポリペプチドである。細菌ベクターはその表面上にターゲティングリガンドを発現することが可能であり、同時に表面上に抗原を発現するか、または抗原をコードする遺伝子を有することも可能である。ウイルスベクターはその表面上にタ
ーゲティングリガンドを発現することが可能であり、表面上に抗原を発現するか、または抗原をコードする遺伝子を有することもまた可能である。
薬物には、たとえば、鎮痛剤、抗偏頭痛薬、抗血液凝固剤、制吐薬、心臓血管薬、抗高血圧症薬、麻薬拮抗薬、キレート剤、抗狭心症薬、化学療法剤、鎮静剤、抗新生物薬、プロスタグランジンおよび抗利尿薬、アンチセンスオリゴヌクレオチド、遺伝子修正(gene‐correcting)ハイブリッドオリゴヌクレオチド、リボザイム、RNA干渉(RNAi)オリゴヌクレオチド、サイレンシングRNA(siRNA)オリゴヌクレオチド、アプタマー・オリゴヌクレオチドおよび三重鎖形成オリゴヌクレオチドが含まれる。
球菌、狂犬病ウイルスおよびエプスタイン‐バーウイルスが含まれる。
ル属の菌、クンニングアメラ属の菌、アポフィソミセス(Apophysomyces)属の菌(A.サクセナエ(saksenaea)、A.ムコールおよびA.アブシジアなど)、スポロトリクス・シェンキィ、パラコクシジオイデス・ブラジリエンシス、シューダレシェリア・ボイディ(Pseudallescheria boydii)、トルロプシス・グラブラタおよび皮膚糸状菌(Dermatophyres)が含まれるが、これらだけに限定されない。
腸上皮を有する生物に担体実体を投与するための本発明の好ましい方法には、担体実体が腸上皮に進入もしくは横断して、またはM細胞もしくはパイエル板などの腸の好ましい領域を進入もしくは横断して輸送されるように、担体実体の存在下で腸上皮を本発明のポリペプチドリガンドと接触させることが含まれる。
ファージディスプレイペプチドライブラリ(1.5×1011pfuの12アミノ酸長)をラットループモデル(n=5)に十二指腸内接種した。血液試料を0、30、60、90および120分の時点でラットループから採取した。次に動物を殺処分して、ループを切除した。これらのループから、パイエル板および非パイエル板組織を単離して、洗浄してホモジナイズした。これらの組織試料に存在するバクテリオファージを大腸菌内で増幅して、ポリエチレングリコール(PEG)沈殿によって単離した。パイエル板特異的ファージの力価を測定して、その後のスクリーニングサイクルで使用するために選択した。
実施例1で得られたファージプールをトップアガーとともにLB寒天プレート上に広げ、Caco‐2およびIEC‐6細胞モデルのほか様々な種から得られたパイエル板組織に対する結合をELISA分析で評価するために、ファージクローンを選択した。
ファージクローンのハイスループットスクリーニングのために、ビオチン‐ExtrAvidin(登録商標)アルカリホスファターゼアッセイを確立した。最初のスクリーニングで、高結合クローンとして500個のクローンから55個を同定した(吸光度の読取り値>0.75)。
さらに吸光度が0.5〜0.75のクローンを50個使用したとき、これらの結果には再現性があった。
100個の高結合クローンについて、パイエル板組織ホモジネートを含む、および含まない、ブタ、イヌおよびマウスの小腸ホモジネートへの結合特性をアッセイした(図8、9、10参照)。これらのホモジネートは、前述のラットホモジネートを得る方法と同様の方法で調製した。ラット組織で認められたように、クローン全てについてパイエル板組織および非パイエル板組織への結合の差は無視できるものであり、これらのクローンが両方の組織のタイプに特異的な部位に結合していることが示唆された。ラットモデルにおいて最も高い結合を示した数個のクローンは、イヌおよびブタのモデル系においても高結合クローンとして分類された。1.002、1.009、1.016、1.038、1.083、2.078、2.080、3.084、3.087および5.074と番号づけられたこれらのクローンを、高、中程度および低結合性を示す100個のクローンと一緒に、結合機能に関連し得るペプチドインサートの性質を決定するための配列決定用に選択した。
70個のクローンについて、細胞ホモジネート(IEC‐6およびCaco‐2ホモジネート)への結合をアッセイした。IEC‐6細胞はラット正常小腸上皮細胞株、Caco‐2細胞はヒトの小腸上皮細胞の特性を示すと考えられるヒト結腸上皮腺癌細胞株である。
ELISAによるファージクローンのCaco‐2細胞膜画分への結合分析を以下の通り実施した。96ウェルELISAプレートを一晩4℃でCaco‐2細胞膜画分(0.05M炭酸緩衝液(pH9.6)中に10μg/ml、100μl/ウェル)で被覆した。次に、このプレートを1.5%BSA‐TBS(100μl/ウェル)で室温で1時間ブロッキングしてからTBS/Tween20(0.05%)で3回洗浄した。ファージクローン(1.5%BSA‐TBSで1:2に希釈)を連続希釈してプレートに入れ、室温で1〜2時間インキュベートした。TBS/Tween20で3回洗浄後、ファージをビオチン化マウス抗M13MAb(1.5% BSA‐TBSで1:5000に希釈、RDI社、100μl/ウェル)とともに室温で1時間インキュベートした。該プレートを3回洗浄してからExtrAvidin(登録商標)AP(1.5% BSA‐TBSで1:5000に希釈、シグマ(Sigma)、100μl/ウェル)とともに室温で1時間インキュベートした。このプレートを再度TBS/Tween20で3回洗浄した。アルカリホスファターゼ活性を基質p‐NPP(p‐ニトロフェニルホスフェート)を使用して検出した。30分後、酵素反応の進行を3M NaOH(100μl/ウェル)の添加により停止させた。ELISAプレートリーダーを使用して、プレートを405nmで読み取った。
高結合クローン全てと、選択した中程度および低結合クローンを含む実施例3のファージクローン100個の配列を決定して、ペプチドインサートの性質を測定した。
細胞の増殖をもたらす、マイトジェンである繊維芽細胞増殖因子(FGF)ファミリーの一種、KGFと相互作用する。KGF‐KGFR相互作用は、上皮修復過程において役割を担っているものと考えられる。非特許文献3、4を参照のこと。
進行に重要である。
水した。この切片をPBSで洗浄した。組織上の抗原決定基を、2.1g/Lの酢酸中で5分間マイクロ波を当てて露出させ、プラスティックラップで覆って室温で20分間冷却させた。20分後、この切片をPBSで洗浄して、次にメタノールに溶かした1%過酸化水素中において内在性ペルオキシダーゼで10分間ブロッキングした。すすぎを繰り返し、この切片をPBSに溶かした2%BSAで室温で20分間ブロッキングした。この切片をPBS中の2%BSAに溶かしたペプチド50μg/mlとともに室温で1時間インキュベートした。対照組織はBSA単独で処理した。切片をPBSに溶かした0.05%Tweenですすいだ。2%BSAで1/500にしたストレプトアビジン‐HRPを添加し室温で30〜60分間とした。再度、この切片をPBS/Tweenですすいだ。DAB基質を添加して最大5分とし、スライドを水に浸漬することによって反応を停止させた。この切片をヘマトキシリンで50秒間対比染色して、次に水ですすいだ。このスライドを1%酸アルコール中で5〜10秒間識別化(differentiate)して、次に水ですすいだ。このスライドを、水性の封入剤およびカバーガラスを使用して封入した。陰性対照ペプチドは結合を示さなかった。配列番号14(ELISAで測定したところ低から中程度の結合であった)も、この実験では結合は陰性であった。ヒトパイエル板に対する結合は、配列番号8および25で陽性であった。いずれのペプチドもヒト腸細胞の先端側に陽性染色をもたらした。
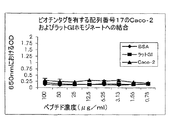
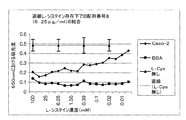
濃度6.25μg/ml、12.5μg/mlおよび25μg/mlのペプチド配列番号8を濃度範囲100mMから0.003mMにわたるL‐システインと一緒にインキュベートした。濃度6.25μg/mlの配列番号25を濃度範囲100mMから0.003mMにわたるL‐システインと一緒にインキュベートした。遊離L‐システインの存在により結合が妨害され、したがってこれらのペプチドの結合にはシステイン基も関与することが示された(図16Aから16D)。
配列番号8の腸上皮組織に対する結合は中程度である。2個のシステイン残基のうちいずれかをアラニン残基で置換した場合(配列番号31および32)、結合は依然として維持される。両方のシステインをアラニン残基で置換すると(配列番号33)、上皮への結合が消失する。ビオチンタグをアミノ末端またはカルボキシル末端のいずれかに添加しても、結合親和性の差は認められなかった(図17A参照)。
Caco‐2ホモジネートへの結合を、ELISAをベースとしたアッセイで試験した。試験した全例において結合が維持されていた。さらに、2重変異ペプチドリガンド(配列番号33)をポリスチレン粒子に吸着させると、元のペプチドの約60%の結合が認められる(図19A〜19B参照)。
高結合ファージクローン番号1.009、5.074、2.078および4.009を前述のようにラット腸ループに注射した。1群当たりマウスは3匹とした。0、30、60、90および120分に血液試料を採取した。次に、動物を殺処分してループを切除した。各血液試料100μlをLBで連続希釈して、IPTG/Xgalを含有するトップアガープレートに広げた。37℃で一晩インキュベートした後、青いプラークを計数した。PBS対照およびm13mp19対照を置いた。
基をアラニン残基で置換すると、結合が消失する。配列番号25の安定化されたD型および反転D型は強い結合性を維持していた。
Claims (46)
- 配列番号1〜34、配列番号38〜39および配列番号42からなる群から選択される12アミノ酸長ペプチドを有する精製合成ポリペプチドであって、前記12アミノ酸長ペプチドはM13ファージのN末端PIII融合ペプチドとして組み込まれると、該ファージにCaco‐2細胞、IEC‐6細胞、ラット、マウス、ブタまたはイヌのいずれかのホモジネート膜画分に結合する能力を与え、前記能力は、同様に組み込まれた配列番号67の翻訳である12アミノ酸長ペプチドによって与えられる能力と少なくとも同じであることを特徴とする、精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドは、12アミノ酸L−ペプチド、該12アミノ酸L−ペプチドのD型、および該12アミノ酸L−ペプチドの反転型から選択される請求項1に記載の精製合成ポリペプチド。
- 亜鉛結合ドメインを含む請求項1または2に記載の精製合成ポリペプチド。
- 配列番号1〜34、配列番号38、39、および配列番号42からなる群から選択されたアミノ酸配列からなる請求項1に記載の精製合成ポリペプチド。
- 配列番号1〜34、配列番号38、39、および配列番号42からなる群から選択された12アミノ酸長ペプチドまたはその反転型からなる請求項1に記載の精製合成ポリペプチド。
- 長さが多くとも200アミノ酸である請求項1〜3のいずれかに記載の精製合成ポリペプチド。
- 長さが少なくとも30アミノ酸である請求項1〜3のいずれかに記載の精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドがLETTCASLCYPS(配列番号8)、LETTAASLCYPS(配列番号31)、LETTCASLAYPS(配列番号32)、LET
TAASLAYPS(配列番号34)からなる群から選択される請求項1〜7のいずれかに記載の精製合成ポリペプチド。 - 前記12アミノ酸長ペプチドがLETTCASLCYPS(配列番号8)である請求項1〜7のいずれかに記載の精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドがVPPHPMTYSCQY(配列番号25)、VPPHPMTYSSQY(配列番号39)およびVPPHPMTYSAQY(配列番号38)からなる群から選択される12アミノ酸長L‐ペプチドである請求項1〜7のいずれかに記載の精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドがVPPHPMTYSCQY(配列番号25)である請求項1〜7のいずれかに記載の精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドがVCSNMYFSCRLS(配列番号24)およびVSSNMYFSSRLS(配列番号42)からなる群から選択される請求項1〜7のいずれかに記載の精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドがVCSNMYFSCRLS(配列番号24)である請求項1〜7のいずれかに記載の精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドが配列番号8、配列番号24、および配列番号25の12アミノ酸長L−ペプチドのD型から選択される請求項1〜7のいずれかに記載の精製合成ポリペプチド。
- 前記12アミノ酸長ペプチドが配列番号8および配列番号25の12アミノ酸長L−ペプチドの反転型から選択される請求項1〜7のいずれかに記載の精製合成ポリペプチド。
- HESSH(配列番号97)およびNVYTXXXXSPXP(配列番号98)からなる群から選択されるL‐ペプチド、そのD‐ペプチド型、またはその反転型からなる精製合成ポリペプチドであって、前記L‐ペプチド、そのD‐ペプチド型、またはその反転型はM13ファージのN末端PIII融合ペプチドとして組み込まれると、該ファージにCaco‐2細胞、IEC‐6細胞、ラット、マウス、ブタまたはイヌのいずれかのホモジネート膜画分に結合する能力を与え、前記能力は、同様に組み込まれた配列番号67の12アミノ酸長ペプチドによって与えられる能力と少なくとも同じであることを特徴とする、精製合成ポリペプチド。
- 長さが200アミノ酸以下である請求項16に記載の精製合成ポリペプチド。
- 長さが50アミノ酸以下である請求項16に記載の精製合成ポリペプチド。
- 長さ200アミノ酸以下の精製合成ポリペプチドであって、配列番号74〜配列番号96からなる群から選択されたペプチドを有し、前記ペプチドはM13ファージのN末端PIII融合ペプチドとして組み込まれると、該ファージにCaco‐2細胞、IEC‐6細胞、ラット、マウス、ブタまたはイヌのいずれかのホモジネート膜画分に結合する能力を与え、前記能力は、同様に組み込まれた配列番号67の翻訳である12アミノ酸長ペプチドによって与えられる能力と少なくとも同じであることを特徴とする、精製合成ポリペプチド。
- 前記ペプチドは、配列番号74〜配列番号96からなる群から選択されたペプチドのL型、D型、または反転型である請求項19に記載の精製合成ポリペプチド。
- 長さが多くとも50アミノ酸である請求項19または20に記載の精製合成ポリペプチド。
- ファージのタンパク質に組み込まれることを特徴とする、請求項1〜21のいずれか一項に記載の精製合成ポリペプチド。
- ファージの表面上で発現されるポリペプチドであって、該ファージが同様に表面上に発現される抗原および同抗原をコードする遺伝子のうち少なくとも一方をさらに含むことを特徴とする、請求項22に記載のポリペプチド。
- 細菌の表面上で発現されるポリペプチドであって、該細菌が同様に表面上に発現される抗原および同抗原をコードする遺伝子のうち少なくとも一方をさらに含むことを特徴とする、請求項22に記載のポリペプチド。
- 薬剤を含む担体実体に共有結合または非共有結合される請求項1〜21のいずれか一項に記載の精製合成ポリペプチド。
- 前記担体実体がナノ粒子、微粒子、リポソーム、細菌、ファージおよびウイルスからなる群から選択される請求項25に記載の精製合成ポリペプチド。
- 前記担体実体の最大の寸法が10nm〜500μmの範囲にある請求項26に記載の精製合成ポリペプチド。
- 前記薬剤が薬物または治療薬である請求項25に記載のポリペプチド。
- 前記薬剤が病原体抗原である請求項25に記載のポリペプチド。
- 前記薬剤がアジュバントである請求項25に記載のポリペプチド。
- 前記担体実体がファージおよびウイルスからなる群から選択される請求項25に記載の精製合成ポリペプチド。
- 請求項1に記載の精製合成ポリペプチドをコードする、ヌクレオチド配列の長さが600ヌクレオチド以下である精製核酸配列。
- 腸上皮を有する生物に投与される薬剤の製造における、担体実体に共有結合または非共有結合された請求項1〜31に記載の精製合成ポリペプチドの使用方法。
- 前記生物が哺乳類である請求項33に記載の使用方法。
- 前記哺乳類がヒトである請求項34に記載の使用方法。
- 前記担体実体がナノ粒子、微粒子リポソーム、細菌、ファージ、およびウイルス担体からなる群から選択される請求項34に記載の使用方法。
- 前記担体実体がナノ粒子、微粒子およびリポソーム担体実体の主要な寸法が10nm〜500μmの範囲にある請求項36に記載の使用方法。
- 前記ナノ粒子、微粒子、またはリポソームに薬剤を搭載するか、あるいは前記ナノ粒子、微粒子、またはリポソームを薬剤とともにカプセル化する請求項36または37に記載の使用方法。
- 前記投与が経口経路によって実施される請求項33〜38のいずれかに記載の使用方法。
- 前記投与が直腸、皮下、筋肉内、鼻腔内または静脈内のいずれかの経路によって実施される請求項33〜38のいずれかに記載の使用方法。
- 前記薬剤がワクチンである請求項33〜40のいずれかに記載の使用方法。
- 前記精製合成ポリペプチドが、ナノ粒子または微粒子のいずれかである担体の表面に被覆、吸着または共有結合される、ファージのタンパク質に組み込まれたペプチドである、請求項33〜41のいずれかに記載の使用方法。
- 前記ファージが抗原をコードするDNAを含有するように修飾される請求項42に記載の使用方法。
- 前記ナノ粒子または微粒子に薬剤を搭載するか、あるいは前記ナノ粒子または微粒子を薬剤とともにカプセル化する請求項42に記載の使用方法。
- 前記精製合成ポリペプチドが亜鉛結合モチーフを含み、該精製合成ポリペプチドを亜鉛の存在下で前記上皮に接触させることを特徴とする請求項42に記載の使用方法。
- 前記担体実体がファージおよびウイルス担体からなる群に由来する請求項42に記載の使用方法。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US30259101P | 2001-07-02 | 2001-07-02 | |
| PCT/IB2002/003401 WO2003004517A2 (en) | 2001-07-02 | 2002-06-28 | Peyers's patch and/or m-celle targeting ligands |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2005503782A JP2005503782A (ja) | 2005-02-10 |
| JP2005503782A5 JP2005503782A5 (ja) | 2008-12-18 |
| JP4383855B2 true JP4383855B2 (ja) | 2009-12-16 |
Family
ID=23168408
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2003510683A Expired - Fee Related JP4383855B2 (ja) | 2001-07-02 | 2002-06-28 | パイエル板および/またはm細胞を標的とするリガンド |
Country Status (8)
| Country | Link |
|---|---|
| US (3) | US6916789B2 (ja) |
| EP (1) | EP1432729B1 (ja) |
| JP (1) | JP4383855B2 (ja) |
| AT (1) | ATE414100T1 (ja) |
| AU (1) | AU2002326070A1 (ja) |
| CA (1) | CA2451741C (ja) |
| DE (1) | DE60229867D1 (ja) |
| WO (1) | WO2003004517A2 (ja) |
Families Citing this family (11)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6521737B1 (en) * | 1995-11-10 | 2003-02-18 | Elan Corporation, Plc | Peptides which enhance transport across tissues and methods of identifying and using the same |
| JP4383855B2 (ja) * | 2001-07-02 | 2009-12-16 | メリオン リサーチ スリー リミテッド | パイエル板および/またはm細胞を標的とするリガンド |
| EP1631582A2 (en) * | 2003-06-10 | 2006-03-08 | University of Saskatchewan | Chimeric adenovirus capsid proteins |
| AU2012201102B2 (en) * | 2004-06-23 | 2014-05-08 | Ferguson, Ian Andrew | Agents and methods for early diagnosis and monitoring of Alzheimer's disease and other neurological disorders |
| DE102005031755B9 (de) * | 2005-07-01 | 2010-04-08 | Technische Universität Dresden | Salmonella spp. bindende Peptide, dafür kodierende Nukleinsäuren, deren Verwendungen und Verfahren und Kits zur Anreicherung, Immobilisierung und zum Nachweis von Salmonella spp. |
| WO2007137174A2 (en) * | 2006-05-18 | 2007-11-29 | The Ohio State University Research Foundation | Compositions and methods for the detection of vulnerable plaque |
| CN104884065B (zh) | 2012-09-21 | 2019-01-01 | 强烈治疗剂公司 | 治疗癌症的方法 |
| EP2989122B1 (en) | 2013-04-23 | 2019-04-03 | President and Fellows of Harvard College | Genetic reprogramming of bacterial biofilms |
| US20180258435A1 (en) * | 2015-04-06 | 2018-09-13 | President And Fellows Of Harvard College | Biosynthetic amyloid-based materials displaying functional protein sequences |
| US11098133B2 (en) | 2016-05-19 | 2021-08-24 | President And Fellows Of Harvard College | Methods of making gels and films using curli nanofibers |
| US10400013B2 (en) | 2016-10-26 | 2019-09-03 | Chao-Wei Liao | Fusion polypeptide for immuno-enhancement and method for enhancing stimulation of immune response using the same |
Family Cites Families (21)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ATE89005T1 (de) * | 1985-02-05 | 1993-05-15 | Synergen Biolog Inc | Rekombinantes vektorsystem fuer metalloproteinase- inhibitorsequenz und dessen verwendung und herstellung nach dem rekombinant-dns-verfahren. |
| IL84167A (en) * | 1986-10-24 | 1991-04-15 | Southern Res Inst | Oral delivery of bioactive agents to and through the peyer's patch by use of microencapsulation |
| US5254342A (en) | 1991-09-30 | 1993-10-19 | University Of Southern California | Compositions and methods for enhanced transepithelial and transendothelial transport or active agents |
| GB9201139D0 (en) * | 1992-01-20 | 1992-03-11 | Connaught Lab | Synthetic peptides for a rubella vaccine |
| AU3937693A (en) | 1992-04-10 | 1993-11-18 | Brigham And Women's Hospital | Methods and compositions for oral delivery of therapeutic agents |
| US5534496A (en) | 1992-07-07 | 1996-07-09 | University Of Southern California | Methods and compositions to enhance epithelial drug transport |
| EP0750630A4 (en) * | 1994-03-11 | 1998-01-07 | Ariad Pharma Inc | METHOD AND MATERIALS FOR IDENTIFYING INHIBITORS OF MOLECULAR INTERACTIONS, WHICH ARE BROUGHT BY THE SH3 DOMAINS |
| US5831008A (en) * | 1994-08-18 | 1998-11-03 | La Jolla Cancer Research Foundation | Retinoblastoma protein-interacting zinc finger proteins |
| US6086875A (en) | 1995-01-17 | 2000-07-11 | The Brigham And Women's Hospital, Inc. | Receptor specific transepithelial transport of immunogens |
| US6030613A (en) | 1995-01-17 | 2000-02-29 | The Brigham And Women's Hospital, Inc. | Receptor specific transepithelial transport of therapeutics |
| IE80466B1 (en) | 1995-11-10 | 1998-07-29 | Elan Corp Plc | Peptides which enhance transport across tissues and methods of identifying and using the same |
| CA2234685A1 (en) * | 1995-11-10 | 1997-05-15 | Elan Corporation, Plc | Peptides which enhance transport across tissues and methods of identifying and using the same |
| US6261787B1 (en) | 1996-06-03 | 2001-07-17 | Case Western Reserve University | Bifunctional molecules for delivery of therapeutics |
| US6060082A (en) | 1997-04-18 | 2000-05-09 | Massachusetts Institute Of Technology | Polymerized liposomes targeted to M cells and useful for oral or mucosal drug delivery |
| JP4129298B2 (ja) * | 1997-05-15 | 2008-08-06 | サイトジェン コーポレーション | 胃腸管(git)輸送受容体に結合するランダムペプチド、及び関連した方法 |
| CA2386231C (en) * | 1999-09-27 | 2010-09-14 | Daniel J. O'mahony | Membrane translocating peptide drug delivery system |
| JP2001245675A (ja) * | 1999-12-28 | 2001-09-11 | Kyogo Ito | 腫瘍抗原 |
| WO2002072015A2 (en) * | 2001-03-12 | 2002-09-19 | Montana State University-Bozeman | M cell directed vaccines |
| CN1323825A (zh) * | 2000-05-16 | 2001-11-28 | 上海博德基因开发有限公司 | 一种新的多肽——人网格蛋白轻链9和编码这种多肽的多核苷酸 |
| CA2443644A1 (en) * | 2001-04-04 | 2003-01-16 | Elan Corporation, Plc | Genetic analysis of peyers's patches and m cells and methods and compositions targeting peyer's patches and m cell receptors |
| JP4383855B2 (ja) * | 2001-07-02 | 2009-12-16 | メリオン リサーチ スリー リミテッド | パイエル板および/またはm細胞を標的とするリガンド |
-
2002
- 2002-06-28 JP JP2003510683A patent/JP4383855B2/ja not_active Expired - Fee Related
- 2002-06-28 CA CA2451741A patent/CA2451741C/en not_active Expired - Fee Related
- 2002-06-28 DE DE60229867T patent/DE60229867D1/de not_active Expired - Lifetime
- 2002-06-28 EP EP02760458A patent/EP1432729B1/en not_active Expired - Lifetime
- 2002-06-28 US US10/185,815 patent/US6916789B2/en not_active Expired - Fee Related
- 2002-06-28 AU AU2002326070A patent/AU2002326070A1/en not_active Abandoned
- 2002-06-28 WO PCT/IB2002/003401 patent/WO2003004517A2/en active Application Filing
- 2002-06-28 AT AT02760458T patent/ATE414100T1/de not_active IP Right Cessation
-
2005
- 2005-06-07 US US11/147,083 patent/US7485321B2/en not_active Expired - Fee Related
-
2008
- 2008-12-19 US US12/340,019 patent/US20100016549A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| AU2002326070A1 (en) | 2003-01-21 |
| US6916789B2 (en) | 2005-07-12 |
| US20100016549A1 (en) | 2010-01-21 |
| EP1432729A2 (en) | 2004-06-30 |
| WO2003004517A3 (en) | 2004-04-29 |
| JP2005503782A (ja) | 2005-02-10 |
| DE60229867D1 (de) | 2008-12-24 |
| US7485321B2 (en) | 2009-02-03 |
| US20050227924A1 (en) | 2005-10-13 |
| EP1432729B1 (en) | 2008-11-12 |
| CA2451741C (en) | 2014-02-18 |
| ATE414100T1 (de) | 2008-11-15 |
| CA2451741A1 (en) | 2003-01-16 |
| WO2003004517A2 (en) | 2003-01-16 |
| US20030096354A1 (en) | 2003-05-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20100016549A1 (en) | Peyer's patch and/or m-cell targeting ligands | |
| US7563866B2 (en) | Modular antigen transporter molecules (MAT molecules) for modulating immune reactions, associated constructs, methods and uses | |
| Rafferty et al. | Peptide therapeutics and the pharmaceutical industry: barriers encountered translating from the laboratory to patients | |
| JP3830525B2 (ja) | 組織横断輸送を強化するペプチドとその同定法および使用法 | |
| KR101216008B1 (ko) | 바이포달 펩타이드 바인더 | |
| Siebert et al. | Tuftsin-properties and analogs | |
| Higgins et al. | In vivo phage display to identify M cell-targeting ligands | |
| CN103533949A (zh) | 作为缺血性脑损伤的有效神经保护剂并用于治疗疼痛的psd-95的高亲和力的二聚抑制剂 | |
| WO2011071280A2 (ko) | 세포내 타겟 결합용 바이포달 펩타이드 바인더 | |
| CN116801908A (zh) | 用于降解病毒和宿主蛋白的嵌合缀合物和使用方法 | |
| JP2013535954A (ja) | ペプチド、構造体およびその使用 | |
| JP6218175B2 (ja) | ヘルパーt細胞誘導性ポリペプチドの改変 | |
| US20130109616A1 (en) | Antimicrobial peptides | |
| JP2005503782A5 (ja) | ||
| US10073109B2 (en) | Phages of biomarker capture and methods of use | |
| CN104177476A (zh) | 一种靶向人癌细胞的多肽及其应用 | |
| US20130059793A1 (en) | Egf receptor mimicking peptides | |
| CN109310729A (zh) | 用于改善转染效率的合成化合物 | |
| JP2023533168A (ja) | タイトジャンクション透過性のモジュレーター | |
| CN101684159B (zh) | 人颗粒酶b蛋白衍生物及其在靶向治疗腺癌中的用途 | |
| WO2013165262A1 (en) | Peptides, constructs and uses therefor | |
| WO2018144817A1 (en) | Modified cxcl12 polypeptides and uses thereof | |
| US20050277575A1 (en) | Therapeutic compositions and methods for treating diseases that involve angiogenesis | |
| JP4238557B2 (ja) | ハイモビリティーグループタンパクと親和性を有するペプチド | |
| WO2013149237A1 (en) | Targeting intracellular organelle zip codes with functional homing ligands from cell-internalizing phage combinatorial libraries |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20050624 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20050624 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20061212 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20080603 |
|
| A524 | Written submission of copy of amendment under article 19 pct |
Free format text: JAPANESE INTERMEDIATE CODE: A524 Effective date: 20080822 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20081024 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20081031 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20081216 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20090316 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20090324 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090605 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20090804 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090812 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20090915 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20090924 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121002 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131002 Year of fee payment: 4 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| LAPS | Cancellation because of no payment of annual fees |