JP4255209B2 - 結核ワクチン - Google Patents
結核ワクチン Download PDFInfo
- Publication number
- JP4255209B2 JP4255209B2 JP2000507804A JP2000507804A JP4255209B2 JP 4255209 B2 JP4255209 B2 JP 4255209B2 JP 2000507804 A JP2000507804 A JP 2000507804A JP 2000507804 A JP2000507804 A JP 2000507804A JP 4255209 B2 JP4255209 B2 JP 4255209B2
- Authority
- JP
- Japan
- Prior art keywords
- nucleic acid
- domain
- recombinant
- cell
- polypeptide
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
- 229960002109 tuberculosis vaccine Drugs 0.000 title 1
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 70
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 58
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 57
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 57
- 102000004196 processed proteins & peptides Human genes 0.000 claims abstract description 46
- 229920001184 polypeptide Polymers 0.000 claims abstract description 44
- 201000008827 tuberculosis Diseases 0.000 claims abstract description 42
- 239000013598 vector Substances 0.000 claims abstract description 28
- 210000004027 cell Anatomy 0.000 claims description 69
- 108091007433 antigens Proteins 0.000 claims description 57
- 102000036639 antigens Human genes 0.000 claims description 57
- 239000000427 antigen Substances 0.000 claims description 56
- 230000028993 immune response Effects 0.000 claims description 26
- 230000004927 fusion Effects 0.000 claims description 25
- 239000002773 nucleotide Substances 0.000 claims description 23
- 125000003729 nucleotide group Chemical group 0.000 claims description 23
- 241000186366 Mycobacterium bovis Species 0.000 claims description 21
- 230000003834 intracellular effect Effects 0.000 claims description 19
- 238000000034 method Methods 0.000 claims description 18
- 210000002540 macrophage Anatomy 0.000 claims description 15
- 241000124008 Mammalia Species 0.000 claims description 14
- 101710088334 Diacylglycerol acyltransferase/mycolyltransferase Ag85B Proteins 0.000 claims description 13
- 230000001580 bacterial effect Effects 0.000 claims description 12
- 102000043129 MHC class I family Human genes 0.000 claims description 11
- 108091054437 MHC class I family Proteins 0.000 claims description 11
- 108010076504 Protein Sorting Signals Proteins 0.000 claims description 11
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 10
- 210000000680 phagosome Anatomy 0.000 claims description 10
- 206010028980 Neoplasm Diseases 0.000 claims description 9
- 230000002688 persistence Effects 0.000 claims description 8
- 241000186359 Mycobacterium Species 0.000 claims description 7
- 238000012258 culturing Methods 0.000 claims description 6
- 230000003612 virological effect Effects 0.000 claims description 6
- 230000005867 T cell response Effects 0.000 claims description 4
- 244000045947 parasite Species 0.000 claims description 4
- 101710166488 6 kDa early secretory antigenic target Proteins 0.000 claims description 3
- 101710088335 Diacylglycerol acyltransferase/mycolyltransferase Ag85A Proteins 0.000 claims description 3
- 241000186781 Listeria Species 0.000 claims description 3
- 230000002759 chromosomal effect Effects 0.000 claims description 2
- 230000003248 secreting effect Effects 0.000 claims description 2
- 230000001939 inductive effect Effects 0.000 claims 6
- 230000000295 complement effect Effects 0.000 claims 1
- 230000036039 immunity Effects 0.000 abstract description 9
- 230000001681 protective effect Effects 0.000 abstract description 7
- 108010008038 Synthetic Vaccines Proteins 0.000 abstract description 2
- 229940124551 recombinant vaccine Drugs 0.000 abstract description 2
- 108090000623 proteins and genes Proteins 0.000 description 22
- 239000013612 plasmid Substances 0.000 description 18
- 230000002163 immunogen Effects 0.000 description 17
- 239000006228 supernatant Substances 0.000 description 17
- 208000015181 infectious disease Diseases 0.000 description 14
- 241000699670 Mus sp. Species 0.000 description 12
- 235000018102 proteins Nutrition 0.000 description 12
- 102000004169 proteins and genes Human genes 0.000 description 12
- 102000003855 L-lactate dehydrogenase Human genes 0.000 description 11
- 108700023483 L-lactate dehydrogenases Proteins 0.000 description 11
- 210000001744 T-lymphocyte Anatomy 0.000 description 11
- 229960005486 vaccine Drugs 0.000 description 11
- 239000012634 fragment Substances 0.000 description 10
- 230000002949 hemolytic effect Effects 0.000 description 10
- 241000186779 Listeria monocytogenes Species 0.000 description 9
- 241000187479 Mycobacterium tuberculosis Species 0.000 description 9
- 238000002255 vaccination Methods 0.000 description 9
- 241000092431 Listeria monocytogenes EGD Species 0.000 description 8
- 230000003013 cytotoxicity Effects 0.000 description 8
- 231100000135 cytotoxicity Toxicity 0.000 description 8
- 108020001507 fusion proteins Proteins 0.000 description 8
- 102000037865 fusion proteins Human genes 0.000 description 8
- 241001467552 Mycobacterium bovis BCG Species 0.000 description 7
- 230000000694 effects Effects 0.000 description 7
- 241000894006 Bacteria Species 0.000 description 6
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 6
- 229940024606 amino acid Drugs 0.000 description 6
- 235000001014 amino acid Nutrition 0.000 description 6
- 150000001413 amino acids Chemical class 0.000 description 6
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 6
- 238000003780 insertion Methods 0.000 description 6
- 230000037431 insertion Effects 0.000 description 6
- 229930027917 kanamycin Natural products 0.000 description 6
- 229960000318 kanamycin Drugs 0.000 description 6
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 6
- 229930182823 kanamycin A Natural products 0.000 description 6
- 239000002953 phosphate buffered saline Substances 0.000 description 6
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 6
- 241000588724 Escherichia coli Species 0.000 description 5
- 241000699666 Mus <mouse, genus> Species 0.000 description 5
- 108091028043 Nucleic acid sequence Proteins 0.000 description 5
- 230000007969 cellular immunity Effects 0.000 description 5
- 230000002950 deficient Effects 0.000 description 5
- 230000006870 function Effects 0.000 description 5
- 230000012010 growth Effects 0.000 description 5
- 244000005700 microbiome Species 0.000 description 5
- 230000028327 secretion Effects 0.000 description 5
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 4
- 102000006303 Chaperonin 60 Human genes 0.000 description 4
- 108010058432 Chaperonin 60 Proteins 0.000 description 4
- 244000026610 Cynodon dactylon var. affinis Species 0.000 description 4
- 229930182566 Gentamicin Natural products 0.000 description 4
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 4
- 108700027766 Listeria monocytogenes hlyA Proteins 0.000 description 4
- 102100028389 Melanoma antigen recognized by T-cells 1 Human genes 0.000 description 4
- 238000004458 analytical method Methods 0.000 description 4
- 239000013592 cell lysate Substances 0.000 description 4
- 210000000349 chromosome Anatomy 0.000 description 4
- 229960002518 gentamicin Drugs 0.000 description 4
- 238000012744 immunostaining Methods 0.000 description 4
- 239000006166 lysate Substances 0.000 description 4
- 239000002609 medium Substances 0.000 description 4
- 239000012528 membrane Substances 0.000 description 4
- 244000052769 pathogen Species 0.000 description 4
- 230000004083 survival effect Effects 0.000 description 4
- 238000005406 washing Methods 0.000 description 4
- 235000014469 Bacillus subtilis Nutrition 0.000 description 3
- 101710132601 Capsid protein Proteins 0.000 description 3
- 108020004414 DNA Proteins 0.000 description 3
- 102400001302 Gasdermin-B, N-terminal Human genes 0.000 description 3
- 241000589248 Legionella Species 0.000 description 3
- 208000007764 Legionnaires' Disease Diseases 0.000 description 3
- 241001429274 Mycobacterium virus L5 Species 0.000 description 3
- 101710189818 Non-structural protein 2a Proteins 0.000 description 3
- 108700026244 Open Reading Frames Proteins 0.000 description 3
- 101710151911 Phosphoprotein p30 Proteins 0.000 description 3
- 241000607142 Salmonella Species 0.000 description 3
- 238000012512 characterization method Methods 0.000 description 3
- 210000000172 cytosol Anatomy 0.000 description 3
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 3
- 239000013613 expression plasmid Substances 0.000 description 3
- 230000002068 genetic effect Effects 0.000 description 3
- 238000009396 hybridization Methods 0.000 description 3
- 230000010354 integration Effects 0.000 description 3
- 238000010369 molecular cloning Methods 0.000 description 3
- PHEDXBVPIONUQT-RGYGYFBISA-N phorbol 13-acetate 12-myristate Chemical compound C([C@]1(O)C(=O)C(C)=C[C@H]1[C@@]1(O)[C@H](C)[C@H]2OC(=O)CCCCCCCCCCCCC)C(CO)=C[C@H]1[C@H]1[C@]2(OC(C)=O)C1(C)C PHEDXBVPIONUQT-RGYGYFBISA-N 0.000 description 3
- 230000002829 reductive effect Effects 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 230000004044 response Effects 0.000 description 3
- 238000012546 transfer Methods 0.000 description 3
- 241000712461 unidentified influenza virus Species 0.000 description 3
- 108010088751 Albumins Proteins 0.000 description 2
- 102000009027 Albumins Human genes 0.000 description 2
- 244000063299 Bacillus subtilis Species 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- 108050001049 Extracellular proteins Proteins 0.000 description 2
- 206010018910 Haemolysis Diseases 0.000 description 2
- 108010006464 Hemolysin Proteins Proteins 0.000 description 2
- 101000578784 Homo sapiens Melanoma antigen recognized by T-cells 1 Proteins 0.000 description 2
- 241000701806 Human papillomavirus Species 0.000 description 2
- 241000589242 Legionella pneumophila Species 0.000 description 2
- 108010010995 MART-1 Antigen Proteins 0.000 description 2
- 102000011931 Nucleoproteins Human genes 0.000 description 2
- 108010061100 Nucleoproteins Proteins 0.000 description 2
- 101710105714 Outer surface protein A Proteins 0.000 description 2
- 239000002033 PVDF binder Substances 0.000 description 2
- 241001494479 Pecora Species 0.000 description 2
- 201000005485 Toxoplasmosis Diseases 0.000 description 2
- 239000002671 adjuvant Substances 0.000 description 2
- 210000000612 antigen-presenting cell Anatomy 0.000 description 2
- 201000011510 cancer Diseases 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- 238000010367 cloning Methods 0.000 description 2
- 238000010276 construction Methods 0.000 description 2
- 230000001461 cytolytic effect Effects 0.000 description 2
- 210000000805 cytoplasm Anatomy 0.000 description 2
- 239000003085 diluting agent Substances 0.000 description 2
- 238000010790 dilution Methods 0.000 description 2
- 239000012895 dilution Substances 0.000 description 2
- 210000003743 erythrocyte Anatomy 0.000 description 2
- 239000000499 gel Substances 0.000 description 2
- 230000008588 hemolysis Effects 0.000 description 2
- 238000004191 hydrophobic interaction chromatography Methods 0.000 description 2
- 230000003053 immunization Effects 0.000 description 2
- 238000002649 immunization Methods 0.000 description 2
- 238000001727 in vivo Methods 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 229940115932 legionella pneumophila Drugs 0.000 description 2
- 231100000518 lethal Toxicity 0.000 description 2
- 230000001665 lethal effect Effects 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- 239000000203 mixture Substances 0.000 description 2
- 210000001616 monocyte Anatomy 0.000 description 2
- 238000006386 neutralization reaction Methods 0.000 description 2
- 230000001717 pathogenic effect Effects 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 229920002401 polyacrylamide Polymers 0.000 description 2
- 229920002981 polyvinylidene fluoride Polymers 0.000 description 2
- 210000001236 prokaryotic cell Anatomy 0.000 description 2
- 230000035755 proliferation Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 239000000523 sample Substances 0.000 description 2
- 238000013207 serial dilution Methods 0.000 description 2
- 239000013605 shuttle vector Substances 0.000 description 2
- 239000000725 suspension Substances 0.000 description 2
- 230000001988 toxicity Effects 0.000 description 2
- 231100000419 toxicity Toxicity 0.000 description 2
- 210000003934 vacuole Anatomy 0.000 description 2
- 108091006112 ATPases Proteins 0.000 description 1
- 102000057290 Adenosine Triphosphatases Human genes 0.000 description 1
- 229920001817 Agar Polymers 0.000 description 1
- 101710197318 Asparagine-rich protein Proteins 0.000 description 1
- 231100000699 Bacterial toxin Toxicity 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 239000012619 Butyl Sepharose® Substances 0.000 description 1
- 241000700198 Cavia Species 0.000 description 1
- 101710117490 Circumsporozoite protein Proteins 0.000 description 1
- 208000003322 Coinfection Diseases 0.000 description 1
- 102000012410 DNA Ligases Human genes 0.000 description 1
- 108010061982 DNA Ligases Proteins 0.000 description 1
- 108010041986 DNA Vaccines Proteins 0.000 description 1
- 230000007023 DNA restriction-modification system Effects 0.000 description 1
- 229940021995 DNA vaccine Drugs 0.000 description 1
- 101100408262 Danio rerio pi15a gene Proteins 0.000 description 1
- 206010059866 Drug resistance Diseases 0.000 description 1
- 102000004190 Enzymes Human genes 0.000 description 1
- 108090000790 Enzymes Proteins 0.000 description 1
- 241000701533 Escherichia virus T4 Species 0.000 description 1
- 241000192125 Firmicutes Species 0.000 description 1
- 102100028972 HLA class I histocompatibility antigen, A alpha chain Human genes 0.000 description 1
- 102100036242 HLA class II histocompatibility antigen, DQ alpha 2 chain Human genes 0.000 description 1
- 108010075704 HLA-A Antigens Proteins 0.000 description 1
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 1
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 1
- 101710147195 Hemolysin A Proteins 0.000 description 1
- 241000282412 Homo Species 0.000 description 1
- 101000930801 Homo sapiens HLA class II histocompatibility antigen, DQ alpha 2 chain Proteins 0.000 description 1
- 101000605534 Homo sapiens Prostate-specific antigen Proteins 0.000 description 1
- 241000725303 Human immunodeficiency virus Species 0.000 description 1
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 1
- 102100034343 Integrase Human genes 0.000 description 1
- 108010061833 Integrases Proteins 0.000 description 1
- 102100037850 Interferon gamma Human genes 0.000 description 1
- 108010074328 Interferon-gamma Proteins 0.000 description 1
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 1
- 241000222722 Leishmania <genus> Species 0.000 description 1
- 241000985284 Leuciscus idus Species 0.000 description 1
- 206010024305 Leukaemia monocytic Diseases 0.000 description 1
- 108090001030 Lipoproteins Proteins 0.000 description 1
- 102000004895 Lipoproteins Human genes 0.000 description 1
- 241000617782 Listeria monocytogenes serotype 1/2a Species 0.000 description 1
- 101710164436 Listeriolysin O Proteins 0.000 description 1
- 206010024641 Listeriosis Diseases 0.000 description 1
- 239000001971 Middlebrook 7H10 Agar Substances 0.000 description 1
- 239000000020 Nitrocellulose Substances 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 108010058846 Ovalbumin Proteins 0.000 description 1
- 108010056995 Perforin Proteins 0.000 description 1
- 102000004503 Perforin Human genes 0.000 description 1
- 208000037581 Persistent Infection Diseases 0.000 description 1
- 206010057249 Phagocytosis Diseases 0.000 description 1
- 101000777480 Phyllodiscus semoni DELTA-alicitoxin-Pse1b Proteins 0.000 description 1
- 241000224016 Plasmodium Species 0.000 description 1
- 241000223960 Plasmodium falciparum Species 0.000 description 1
- 208000035109 Pneumococcal Infections Diseases 0.000 description 1
- 206010036790 Productive cough Diseases 0.000 description 1
- 102100038358 Prostate-specific antigen Human genes 0.000 description 1
- 239000012083 RIPA buffer Substances 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 108020005091 Replication Origin Proteins 0.000 description 1
- 229940124842 Salmonella vaccine Drugs 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- 101710172711 Structural protein Proteins 0.000 description 1
- 230000024932 T cell mediated immunity Effects 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 101710131114 Threonine-rich protein Proteins 0.000 description 1
- 208000007536 Thrombosis Diseases 0.000 description 1
- 102000002938 Thrombospondin Human genes 0.000 description 1
- 108060008245 Thrombospondin Proteins 0.000 description 1
- 241000223996 Toxoplasma Species 0.000 description 1
- 241000223997 Toxoplasma gondii Species 0.000 description 1
- 102000004338 Transferrin Human genes 0.000 description 1
- 108090000901 Transferrin Proteins 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 description 1
- 108010078814 Tumor Suppressor Protein p53 Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 238000005273 aeration Methods 0.000 description 1
- 239000008272 agar Substances 0.000 description 1
- 230000001355 anti-mycobacterial effect Effects 0.000 description 1
- 230000030741 antigen processing and presentation Effects 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 239000003926 antimycobacterial agent Substances 0.000 description 1
- 230000002238 attenuated effect Effects 0.000 description 1
- 210000003719 b-lymphocyte Anatomy 0.000 description 1
- 229960000190 bacillus calmette–guérin vaccine Drugs 0.000 description 1
- 244000052616 bacterial pathogen Species 0.000 description 1
- 239000000688 bacterial toxin Substances 0.000 description 1
- 102000015736 beta 2-Microglobulin Human genes 0.000 description 1
- 108010081355 beta 2-Microglobulin Proteins 0.000 description 1
- 229940098773 bovine serum albumin Drugs 0.000 description 1
- 210000004556 brain Anatomy 0.000 description 1
- 239000000872 buffer Substances 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 230000030833 cell death Effects 0.000 description 1
- 239000006285 cell suspension Substances 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000011217 control strategy Methods 0.000 description 1
- 230000009260 cross reactivity Effects 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 230000001186 cumulative effect Effects 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 210000004443 dendritic cell Anatomy 0.000 description 1
- 229960003964 deoxycholic acid Drugs 0.000 description 1
- KXGVEGMKQFWNSR-LLQZFEROSA-N deoxycholic acid Chemical compound C([C@H]1CC2)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(O)=O)C)[C@@]2(C)[C@@H](O)C1 KXGVEGMKQFWNSR-LLQZFEROSA-N 0.000 description 1
- KXGVEGMKQFWNSR-UHFFFAOYSA-N deoxycholic acid Natural products C1CC2CC(O)CCC2(C)C2C1C1CCC(C(CCC(O)=O)C)C1(C)C(O)C2 KXGVEGMKQFWNSR-UHFFFAOYSA-N 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 239000012470 diluted sample Substances 0.000 description 1
- 201000010099 disease Diseases 0.000 description 1
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 239000003814 drug Substances 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 210000001163 endosome Anatomy 0.000 description 1
- 238000002474 experimental method Methods 0.000 description 1
- 239000013604 expression vector Substances 0.000 description 1
- 210000000416 exudates and transudate Anatomy 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 230000009036 growth inhibition Effects 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 239000003228 hemolysin Substances 0.000 description 1
- 230000028996 humoral immune response Effects 0.000 description 1
- 210000002865 immune cell Anatomy 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 238000010874 in vitro model Methods 0.000 description 1
- 230000002458 infectious effect Effects 0.000 description 1
- 208000037797 influenza A Diseases 0.000 description 1
- 101150062334 int gene Proteins 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- 210000000936 intestine Anatomy 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 230000009545 invasion Effects 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 238000011813 knockout mouse model Methods 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 239000007788 liquid Substances 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 229940042470 lyme disease vaccine Drugs 0.000 description 1
- 230000002132 lysosomal effect Effects 0.000 description 1
- 210000003712 lysosome Anatomy 0.000 description 1
- 230000001868 lysosomic effect Effects 0.000 description 1
- 201000004792 malaria Diseases 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000001404 mediated effect Effects 0.000 description 1
- 210000002752 melanocyte Anatomy 0.000 description 1
- 201000006894 monocytic leukemia Diseases 0.000 description 1
- 230000009670 mycobacterial growth Effects 0.000 description 1
- 230000017066 negative regulation of growth Effects 0.000 description 1
- 229920001220 nitrocellulos Polymers 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 229940092253 ovalbumin Drugs 0.000 description 1
- 230000003647 oxidation Effects 0.000 description 1
- 238000007254 oxidation reaction Methods 0.000 description 1
- 230000003071 parasitic effect Effects 0.000 description 1
- 239000008188 pellet Substances 0.000 description 1
- 102000013415 peroxidase activity proteins Human genes 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 230000008782 phagocytosis Effects 0.000 description 1
- 239000008194 pharmaceutical composition Substances 0.000 description 1
- 239000002644 phorbol ester Substances 0.000 description 1
- 238000007747 plating Methods 0.000 description 1
- 108010040473 pneumococcal surface protein A Proteins 0.000 description 1
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 1
- 235000010482 polyoxyethylene sorbitan monooleate Nutrition 0.000 description 1
- 229920000053 polysorbate 80 Polymers 0.000 description 1
- 239000013641 positive control Substances 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 239000000047 product Substances 0.000 description 1
- 230000002035 prolonged effect Effects 0.000 description 1
- 238000011002 quantification Methods 0.000 description 1
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 1
- 230000000284 resting effect Effects 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 230000001177 retroviral effect Effects 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 230000000392 somatic effect Effects 0.000 description 1
- 241000894007 species Species 0.000 description 1
- 210000003046 sporozoite Anatomy 0.000 description 1
- 210000003802 sputum Anatomy 0.000 description 1
- 208000024794 sputum Diseases 0.000 description 1
- 230000000638 stimulation Effects 0.000 description 1
- 210000002784 stomach Anatomy 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 238000006467 substitution reaction Methods 0.000 description 1
- 125000003396 thiol group Chemical group [H]S* 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- 239000012581 transferrin Substances 0.000 description 1
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 1
- 230000002476 tumorcidal effect Effects 0.000 description 1
- 230000005760 tumorsuppression Effects 0.000 description 1
- 210000000626 ureter Anatomy 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- 238000001262 western blot Methods 0.000 description 1
Images






Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/35—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Mycobacteriaceae (F)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/62—DNA sequences coding for fusion proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/01—Fusion polypeptide containing a localisation/targetting motif
- C07K2319/02—Fusion polypeptide containing a localisation/targetting motif containing a signal sequence
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/40—Fusion polypeptide containing a tag for immunodetection, or an epitope for immunisation
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Medicinal Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Zoology (AREA)
- Gastroenterology & Hepatology (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Pharmacology & Pharmacy (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Communicable Diseases (AREA)
- General Chemical & Material Sciences (AREA)
- Oncology (AREA)
- Veterinary Medicine (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
Description
本発明は特に結核に対する防御免疫を提供する新規な組換えワクチンに関する。さらに、本発明は新規な組換え核酸分子、この核酸分子を含有するベクター、この核酸分子で形質転換した細胞、およびこの核酸分子によってコードされるポリペプチドに関する。
【0002】
Mycobacterium tuberculosisによって引き起こされる結核(TB)は重要でグローバルな問題のままである。世界の人口の三分の一が M.tuberculosisに感染し ているものと推定される(Kochi,1991)。多くの国で、TBの制御のための唯一の手段は M.bovis bacille Calmette-Guerin(BCG)でのワクチン接種であった。しかし、TBに対するBCGの総合的なワクチン効率は約50%であり、各種の実用試験間で0%〜80%の範囲の極端な変動がある(Rocheら、1995)。したがって、よりよいTB防御用のワクチンを提供するために、例えば遺伝子工学によってBCGを改良すべきである(Murrayら、1996;Hess and Kaufmann,1993)。多数の薬剤耐性 M.tuberculosis株の広範な出現によって、新規なTBワクチンへの緊急の要請がさらに強調されている(Grange,1996)。
【0003】
M.tuberculosisは休止マクロファージのファゴソーム液胞内で複製される細胞内細菌の群に属するので、TBに対する防御はT細胞性免疫に依存する(Kaufmann,1993)。しかし、マウスおよびヒトにおけるいくつかの研究では、マイコバクテリアは抗原特異的な主要組織適合性遺伝子複合体(MHC)クラスIIまたはクラスI拘束CD4およびCD8T細胞をそれぞれ刺激することが示された(Kaufmann,1993 )。
【0004】
MHCクラスI拘束CD8T細胞の重要な役割は、β2-ミクログロブリン(β2m) 欠失マウスが実験的な M.tuberculosisの感染を防御することができないことに よって、説得力のある証明がなされた(Flynnら、1993)。これらの突然変異マ ウスはMHCクラスIを欠損しているので、機能性CD8T細胞が発生することができ ない。M.tuberculosisの感染とは対照的に、β2m-欠失マウスはBCGワクチン株の一定の感染性用量を制御することができる(Flynnら、1993;Ladelら、1995)。さらに、BCGで免疫したC57BL/6がTBに耐性であるところから、β2m-欠失マウス のBCGワクチン接種によって、その後の M.tuberculosisの感染の後の生存が延長された(Flynnら、1993)。M.tuberculosisおよびBCG間のこの差異のある CD8T細胞依存性は以下のように説明することができる:M.tuberculosis抗原はBCGか らの抗原よりも細胞質へのより良い接近手段を獲得して、より明白なMHCクラスIの提示が導かれる(Hess and Kaufmann、1993)。その結果、M.tuberculosisによ ってより効果的なCD8T細胞応答が産生される。この見解は、BCGによるよりも M.tuberculosisでの抗原提示細胞(APC)の同時感染によって、無関係な抗原、オボアルブミンのMHCクラスIの提示が増加することから、最近支持された(Mazzaccaroら、1996)。
【0005】
M.tuberculosisの分泌タンパク質はMHCクラスI提示にとって有効な抗原の起源を含んでいる。最近、この分泌された抗原 Ag85AをコードするDNAワクチンがマ ウスにおいてMHCクラスI拘束CD8T細胞応答を誘発したので、これがTBに対す る防御に寄与するのかも知れない(Huygenら、1996)。一般的に、分泌されたM.tuberculosisのタンパク質抗原による免疫化がモルモットおよびマウスにおいてTBに対するなんらかの防御を誘導するという証拠が蓄積されてきている(Horwitz ら、1995;Andersen,1994)。したがって、BCGに基づく改良されたTBワクチンの開発の重要な目標は、分泌されたBCG特異的抗原の感染したAPCの細胞質への接近能力を増強することである。これらの分泌されたタンパク質から誘導されるペプチドのその後のMHCクラスI提示経路中への送達は、TB防御のためにすでに存在するBCG特異的免疫応答を増強すると考えられる。
【0006】
L.monocytogenesのファゴリソソーム脱出(escape)はリステリア抗原のMHCクラスI抗原提示を助長するための独特の機構を説明するものである(Bercheら、1987;Portnoyら、1988)。細孔形成性のスルフヒドリルで活性化される細胞溶解素であるリステリオリシン(lysteriolysin)(Hly)は宿主細胞のファゴリソソーム液胞から細胞質ゾルへの L.monocytogenes微生物体の放出に必須である(Gaillardら、1987;Portnoyら、1988)。最近、この脱出機能が Bacillus subtilis および弱毒化 Salmonella ssp.株に移された(Bieleckiら、1991;Gentschevら、1995;Hess and Kaufmann,1997)。無胞子性 B.subtilis突然変異株によるかまたはSalmonella ssp.中でのHlyの発現の結果、J774マクロファージ様細胞のファゴリソソームから細胞質ゾル中に細菌が脱出する(Bieleckiら、1991;Gentschev ら、1995;Hess and Kaufmann,1997)。
【0007】
このように、リソソーム脱出機能の異種微生物への転移は生成する組換え微生物の毒性の上昇の原因となることがある。この理由によって、組換え生ワクチンの調製のためのこれらのリソソーム脱出機能の使用はすぐには考慮に入れられなかった。
【0008】
本発明において、溶血活性があるHlyを分泌する組換えBCG株を構築した。これは改良されたMHCクラスI拘束免疫応答を示し、そして驚くべきことに、改変しない天然のBCG株に比較して等しいかむしろ低い細胞傷害性を示す。すなわち、 これらの組換え生物はTBに対するワクチンの確実な候補である。
【0009】
本発明の第1の態様は、哺乳類中で免疫応答を誘発する能力があるマイコバクテリウム・ポリペプチド由来の少なくとも1つのドメイン、および(b) ファゴリソソーム脱出ドメイン1種、を含む融合ポリペプチドをコードする組換え核酸分子である。
【0010】
この第1の態様の特定の1実施形態は配列番号1中の核酸分子である。この核酸分子はシグナルペプチド1種をコードする配列(ヌクレオチド1-120)、免疫 原性ドメインをコードする配列(ヌクレオチド121-153)、ペプチドリンカーを コードする配列(ヌクレオチド154-210)、ファゴリソソームドメインをコード する配列(ヌクレオチド211-1722)、別のペプチドリンカーをコードする配列(ヌクレオチド1723-1800)およびランダムペプチドをコードする配列(ヌクレオチド1801-1870)を含む。対応するアミノ酸配列を配列番号2に示す。
【0011】
本発明の核酸はマイコバクテリウム属の生物、好ましくは Mycobacterium tuberculosisまたは Mycobacterium bovis由来のポリペプチド由来の少なくとも1つの免疫原性ドメインを含有する。このドメインは長さ6以上、好ましくは8以上のアミノ酸を有する。この免疫原性ドメインは好ましくは天然の マイコバクテリウム・ポリペプチドの一部分である。しかし、天然の免疫原性ドメインから1または数個のアミノ酸の置換、欠失および/または添加によって誘導される、修飾された免疫原性ドメインもまた本発明の範囲内である。
【0012】
この免疫原性ドメインは哺乳類中に免疫応答を誘発することができる。この免疫応答はB細胞性免疫応答も可能である。しかし、好ましくはこの免疫原性ドメインはT細胞性免疫応答、より好ましくはMHCクラスI拘束CD8T細胞応答を誘 発する能力があるものである。
【0013】
免疫応答を誘発することができるドメインは好ましくは M.bovisもしくは M.tuberculosisからの免疫原性ペプチドもしくはポリペプチド、またはこれらの免 疫原性フラグメントから選択される。好適な抗原の特定の例は、M.tuberculosis由来の Ag85B(p30)(Harthら、1996)、M.bovis BCG由来の Ag85B(α抗原) (Matsuoら、1988)、M.tuberculosis由来のAg85A(Huygenら、1996)および M.tuberculosi由来の ESAT-6(Sorensenら、1996、Harboeら、1996 および Andersenら、1995)である。さらに好ましくは、免疫原性ドメインは抗原 Ag85Bから誘導される。最も好ましくは免疫原性ドメインは配列番号2中のaa.41からaa.51の配列を含む。
【0014】
本発明に従う組換え核酸分子はさらに、ファゴリソソーム脱出ドメイン、すなわち融合ポリペプチドを哺乳類細胞のファゴリソソームから細胞質ゾル中へ脱出させる手段を与えるポリペプチドドメインを含む。好ましくはこのファゴリソソーム脱出ドメインは リステリア属生物由来である。さらに好ましくは、ファゴリ ソソーム脱出ドメインは L.monocytogenes生物由来である。最も好ましくは、ファゴリソソーム脱出ドメインは、(a) 配列番号1中に示されるヌクレオチド211-1722からのヌクレオチド配列、(b) (a)からの配列と同一のアミノ酸配列をコードするヌクレオチド配列、および(c) (a)または(b)からの配列とストリンジェント条件下でハイブリダイズするヌクレオチド配列、から選択される核酸分子によってコードされる。
【0015】
配列番号1に図示するヌクレオチド配列とは別に、本発明はこれらとハイブリダイズする核酸配列をも含む。本発明において、用語「ハイブリダイゼーション」は Sambrookら(Molecular Cloning.A Laboratory manual,Cold Spring Harbor Laboratory Press(1989),1.101-1.104)の定義にしたがって使用する。本発明において、用語「ハイブリダイゼーション」は、55℃、好ましくは62℃、さらに好ましくは68℃で1時間、 1×SSCおよび0.1% SDS で、特に55℃、好ましくは62℃、さらに好ましくは68℃で1時間、 0.2×SSCおよび0.1% SDS での洗浄後もなお陽性ハイブリダイゼーションシグナルを観察することができる場合に、使用する。こうした洗浄条件下で配列番号1によるヌクレオチド配列とハイブリダイズする配列は、本発明にとって好ましい、ファゴリソソーム脱出ドメインをコードするヌクレオチド配列である。
【0016】
好ましくは、融合ポリペプチドをコードする組換え核酸分子はシグナルペプチドをコードする配列を含んでいる。さらに好ましくは、このシグナル配列はマイコバクテリア、好ましくは M.bovis中で活性なシグナル配列であり、例えば天然の M.bovisシグナル配列である。好適なシグナル配列の好ましい一例は、配列番号1中のヌクレオチド1-120で示される、Ag85Bシグナルペプチドをコードするヌクレオチド配列である。
【0017】
さらに、免疫原性ドメインとファゴリソソーム脱出ドメインとの間にペプチドリンカーを配置するのが好ましい。好ましくは、このペプチドリンカーは長さ5 から50のアミノ酸である。さらに好ましくは、配列番号1中のヌクレオチド154-210に示すリンカーをコードする配列、または遺伝コードの縮重性に関してこの 配列に相当する配列である。
【0018】
本発明の別の主題は、上に定義した核酸分子の少なくとも1つのコピーを含む組換えベクターに関する。好ましくは、この組換えベクターは原核ベクター、すなわち原核細胞中での複製または/およびゲノムへの組み込みのためのエレメントを含有するベクターである。好ましくは、この組換えベクターは発現制御配列に機能し得るように連結した本発明の核酸分子を保有する。この発現制御配列は好ましくはマイコバクテリア、特に M.bovis中で活性な発現制御配列である。このベクターは染色体外ベクターまたは染色体中に組み込むのに好適なベクターとすることができる。こうしたベクターの例は当分野の技術者に知られており、例えば Sambrookら、上記、に記載されている。
【0019】
本発明のさらに別の主題は、上に定義した組換え核酸またはベクターを含む細胞である。好ましくは、この細胞は原核細胞、特に マイコバクテリウムの細胞である。さらに、この細胞が本発明の核酸分子を発現することができることが好ましい。
【0020】
本発明の第2の態様において、(a) 哺乳類中で免疫応答を誘発する能力がある少なくとも1つのドメイン、および(b) ファゴリソソーム脱出ドメインを含む融合ポリペプチドをコードする少なくとも1つの組換え核酸分子を含む、組換えMycobacterium bovis 細胞が提供される。この態様によれば、免疫原性ドメインはマイコバクテリウムの抗原に限定されるものではなく、自己抗原、腫瘍抗原、および一般的なウイルス抗原、寄生体抗原、細菌抗原などの病原体抗原、ならびにそれらの免疫原性フラグメントから選択することができる。好適な腫瘍抗原の特定の例は、p53腫 瘍サプレッサー遺伝子産物などのヒト腫瘍抗原(Houbiersら、1993)およびメラノサイト分化抗原、例えば Melan-A/MART-1および gp100である(van Elsasら、1996)。好適なウイルス抗原の特定の例は、ヒト乳頭腫ウイルス抗原、例えば抗原 E6およびE7などのヒト腫瘍ウイルス抗原(Boschら、1991)、インフルエンザウイルス抗原、例えばインフルエンザウイルス核タンパク質(Matsuiら、1995;Fuら、1997)またはHIV抗原、例えばHIV-1抗原p17、p24、RTおよび Envなどのレトロウイルス抗原(Harrerら、1996;Haasら、1996)である。好適な寄生体抗原の特定の例は、肝期(liver stage)抗原(LSA-1)などのマラリア原虫(Plasmodium)抗原、サーカムスポロゾイト(circumsporozoite)タンパク質(CSまたは対立変異体 cp26若しくは cp29)、トロンボスポンジン(thrombospondin)関連アモニマス(amonymous)タンパク質(TRAP)、Plasmodium falciparumからのスポロゾイトトレオニンおよびアスパラギンに富むタンパク質(STARP)(Aidooら、1995)ならびに Toxoplasma gondiiからのp30などの Toxoplasma抗原(Khanら、1991;Bulow and Boothroyd,1991)である。好適な細菌抗原の特定の例は Legionella pneumophilaからの主要分泌タンパク質(Blander and Horwitz,1991)などの レジオネラの抗原である。
【0021】
本発明に従う細胞は好ましくは本発明の核酸分子によってコードされる融合タンパク質を分泌することができ、そしてMHCクラスI拘束抗原認識に好適な形態でそれを提供することができる。
本発明の第3の態様においては、ファゴリソソーム脱出ペプチドまたはポリペプチドをコードする少なくとも1つの核酸分子を含む、組換えMycobacterium bovis 細胞が提供される。ファゴリソソーム脱出ペプチドまたはポリペプチドが抗原と融合しなくても、免疫原特性の驚くベき改善が見られる。
【0022】
本発明にしたがって提供される組換えMycobacterium bovis細胞は、哺乳類中 で免疫応答を誘発する能力があるペプチドまたはポリペプチドをコードする少なくとも1つのさらに別の組換え体、例えば異種核酸分子を含んでいてもよい。このさらに別の免疫原性ペプチドまたはポリペプチドはマイコバクテリウムの抗原から、またはさらに広い意味で、自己抗原、腫瘍抗原、病原体抗原、およびそれらの免疫原性フラグメントから選択することができる。このさらに別のペプチドまたはポリペプチドをコードする核酸分子は融合遺伝子と同じベクター上に位置させてもよい。しかし、例えば融合遺伝子とは無関係に、別のプラスミド上に位置させてもよく、または染色体として統合してもよい。
【0023】
驚くべきことに、本発明に従うマイコバクテリウムの細胞は感染した細胞、例えばマクロファージ中で細胞内持続性を有し、これは組換え核酸分子を含有しない、対応する天然の マイコバクテリウムの細胞の細胞内持続性に等しいかまたはこれより低いことがわかった。
【0024】
本発明のさらに別の主題は、上の定義の核酸分子によってコードされた組換え融合ポリペプチドである。本発明に従うこの融合ポリペプチドは細胞に改善されたMHCクラスI拘束抗原認識の能力を与える。
【0025】
本発明は活性成分として上の定義の細胞または融合ポリペプチドを、任意に薬学的に許容される希釈剤、担体およびアジュバントとともに含む医薬組成物にも関する。好ましくは、この組成物は哺乳類、好ましくはヒトに投与するのに好適な生ワクチンである。実際に選択されるワクチン接種経路はワクチン投与するベクターの選択に依存する。投与は一回の用量でまたは間隔をとって反復して実施してもよい。適切な投与量はワクチン用ベクター自体または投与経路などの各種のパラメーターに依存する。粘膜表面(例えば眼球、鼻腔内、経口、胃、腸、肛門、腟若しくは尿管)または非経口(例えば皮下、皮内、筋内、静脈内または腹腔内)投与を選択することができる。
【0026】
さらに、本発明は上記定義の組換え細菌細胞の調製方法に関する。第1の態様によれば、この方法は、(i) (a)哺乳類中で免疫応答を誘発する能力があるマイコバクテリウム・ポリペプチド由来の少なくとも1つのドメイン、および(b) ファゴリソソーム脱出ドメインを含む融合ポリペプチドをコードする組換え核酸分子を細菌細胞中に挿入し、そして(ii) ステップ(i)によって取得した細胞を好適な条件下で培養する、ステップを含む。好ましくはこの核酸分子を発現することができる細胞を取得する。好ましくは、その細胞は M.bovis細胞である。
【0027】
第2の態様によれば、この方法は、(i) (a)哺乳類中で免疫応答を誘発する能力があるポリペプチド由来の少なくとも1つのドメイン、および(b)ファゴリソソーム脱出ドメイン、を含む融合ポリペプチドをコードする組換え核酸分子を Mycobacterium bovis 細胞中に挿入し、そして(ii) (i)によって取得した細胞を好適な条件下で培養する、ステップを含む。
【0028】
第3の態様によれば、この方法は、(i) ファゴリソソーム脱出ペプチドまたはポリペプチドをコードする組換え核酸分子を Mycobacterium bovis 細胞中に挿入し、そして(ii) (i)によって取得した細胞を好適な条件下で 培養する、ステップを含む。
【0029】
所望ならば、本発明の方法に、哺乳類中で免疫応答を誘発する能力があるペプチドまたはポリペプチドをコードする、少なくとも1つの別の組換え核酸分子をさらに挿入することを含ませる。
【0030】
最後に、本発明は、薬学的に有効な量の医薬として許容される希釈剤、担体および/またはアジュバント中の上記組換え細胞を製剤することを含む、生ワクチンの調製方法に関する。
【0031】
本発明を以下の図面および配列表によってさらに説明する。
配列番号1: 本発明に従う核酸分子のヌクレオチド配列を示す。
配列番号2: 配列番号1の核酸分子に対応するアミノ酸配列を示す。
【0032】
(実施例)
1.実験操作
1.1 細菌株およびプラスミド
Dubos培地用アルブミン(Difco)を補充したDubosブロスベース(Difco)中、37℃で、M.bovis BCG株 Chicago(ATCC 27289)を培養した。中期対数期の培養 物を分注し、使用時まで-70℃で保存した。G.B.Mackanessから最初に入手した L.monocytogenes EGD Sv 1/2a(Domann and Chakraborty,1989)を、脳心臓浸出 物(BHI)ブロス(Difco)中、37℃でエアレーションしながら増殖させた。プラスミド plLH-1は Dr.I.Gentschev および Dr.W.Goebel(W(rzburg大学、ドイツ )の寛大な寄贈によった。マイコバクテリア-大腸菌シャトルベクター、pAT261 およびpMV306は MedImmune(Gaithersburg,U.S.A.)から入手した。
【0033】
1.2 酵素および一般的な遺伝子技法
制限酵素(Boehringer Mannheim)およびT4 DNAリガーゼ(Pharmacia)を製造者の指示にしたがって使用した。分子クローニングおよび組換えDNA技法を標準 プロトコル(Sambrookら、1989)にしたがって実施した。
【0034】
1.3 DNA操作およびシークエンシング
染色体外 pAT261(親ベクター pAB261;Stoverら、1993)および組み込み用(integrative) pMV306(親ベクター pMV361;Stoverら、1991)発現プラスミドをHly分泌のために使用した。プラスミド pAT261および pMV306は発現カセット1個、選択マーカーとしてカナマイシン耐性を与えるTn903由来の aph遺伝子、およびpUC19由来の大腸菌複製起源を含む、共通のエレメントを共有している。これらはマイコバクテリアプラスミドの複製起源(pAT261)かまたはマイコバクテリオファージL5の attPおよび int遺伝子(pMV306)の挿入について異なっている 。プラスミド構築物 pAT261中に挿入された M.bovis BCG特異的Ag85B遺伝子のDNA断片はBCG hsp60プロモーターの制御下にある。成熟Ag85Bタンパク質のための コード配列の下流の Pst I制限部位(位置4404、MedImmune)を使用して、L.monocytogenes EGDからの天然のHlyの溶血活性が維持され、N-末端 Ag85B特異的シ グナルペプチドによって移送される、Hly由来の融合物を構築した。pAT261:Hly の構築のために、プラスミド plLH-1(Gentschevら、1995;Hessら、1996)によってコードされた当初の遺伝子融合物 hly-hlyAの 1.7 kb Pst-I断片(当初の位置 1357および4277;Hessら、1986)を使用した。コードされている Ag85B-Hly ハイブリッドタンパク質のためのプラスミド pAT261:Hly中の hsp60プロモータ ーを含む完全 Xba I-Sal I DNA発現カセットをプラスミド pMV306中に導入した。この結果生成した構築物を pMV306:Hlyと命名した。以下のオリゴヌクレオチド、 BCG-Hly5-GCTTTGTCCTGCTGおよびBCG-Hly3-GGAAGTCAGGGTGA (Sequiserve,Vaterstetten,Germany)を使用して、これらのプラスミドの断片挿入部位の正しいDNA配列を決定した。DNA配列分析から、hly-hlyA遺伝子融合物の3'末端に11 aaをコードする短い Pst I-DNA断片のランダム挿入がわかった。
【0035】
1.4 組換えM.bovis BCG株の特性決定
標準的なエレクトロポレーションプロトコル(Langermannら、1994)によって、プラスミド pAT261:Hlyまたは pMV306:Hlyを M.bovis BCG株 Chicago中に導入し、次にカナマイシン(15μg/ml)を補充した Middlebrook 7H10アガー上で組換え体コロニーを選択した。10% Dubos培地用アルブミン(Difco)および15μg/mlカナマイシンを含有する Dubos液体培地(Difco)中で3週間、カナマイシン耐性コロニーを中期対数期まで増殖させた。Tween 80を含むリン酸塩緩衝化生理食塩水(PBS)中で細胞を洗浄した後、RIPAバッファー(1% NP-40、0.5% デオキシコール酸、0.1% SDS、50 mM Tris、pH 8.0)中で細胞懸濁液を20倍に濃縮して、細胞を溶解した。これらの培養物の細菌を含まない上清(1 ml)を0.2μm 膜フィルターでろ過した。この上清中のHly融合タンパク質を、回転装置中、室温で30分、ブチルセファロース(Pharmacia)100μlとともにインキュベートすることによって、富化した(Schoelら、1994)。遠心分離(3000 rpm)後、ペレットを Laemmliバッファー(Laemmli,1970)中に溶解した。次に、前記(Laemmli,1970)にしたがい、10% SDS/ポリアクリルアミドゲル電気泳動によって、タンパク質を分離し、Hybond-PVDF膜(Amersham Life Science)に移した。抗Hly mAbH14-3(Natoら、1991)およびペルオキシダーゼと複合体化した2次抗体(Boehringer Mannheim)によって免疫染色を実施した。製造者[BM Western Blotting Kit(Mouse/Rabbit)(Boehringer Mannheim)]の記載にしたがって、洗浄操作およびケミルミネセンス免疫検出を実施した。X線フィルム(Kodak,XOMAT-AR)上での シグナルの現像を1分間実施した。
【0036】
0.1%ウシ血清アルブミンを含有するリン酸塩緩衝化生理食塩水(PBS)でサンプルを連続希釈することによって、BCG pAT261:Hly、BCG pMV306:Hly、BCGおよ び L.monocytogenesの上清および全細菌懸濁液の溶血活性を判定した。次に、希釈したサンプル(100μl)をシステイン(最終濃度 20 mM)の添加によって活性化し、96ウェルプレート中、37℃で45分間、ヒツジ赤血球(PBS中 6×108細胞/ml、pH6.0) 50μlとともにインキュベートした。溶血活性は、完全溶血を検出することができた最高希釈率の逆数と定義される、完全CHUである(Gentschevら、1995)。
【0037】
1.5 マイコバクテリアの増殖のin vitro分析
それぞれヒトおよびマウスのマクロファージ様細胞である THP-1(ATCC TIB-202)および J774A.1(ATCC TIB-67)を24ウェルプレートに付着させた(1ウェ ルについて106)。THP-1の場合、感染の48時間前に 10 nM PMA(Sigma)で刺激 することによって、付着を成功させた。1細胞につき10マイコバクテリア(BCG 、BCG pAT261:HlyまたはBCG pMV306:Hly)の感染多重度(moi)で3時間、細胞に感染させた。感染直後に、Bacto Middlebrook OADC(Difco)および適切な 15μg/mlカナマイシンで富化した7H10アガー上に、上清および細胞溶解物の連続希釈物をプレーティングすることによって、CFUを測定した。マクロファージによるマイコバクテリアの取り込みは同程度だった。感染したマクロファージの残ったサンプルをPBSで洗浄し、200μg/mlゲンタマイシンの存在下でさらに14日間インキュベートした。感染後(p.i.)1、8または15日目に、CFU分析によって、組換えBCG株の細胞内増殖を判定した。
【0038】
1.6 LDH放出
感染したJ774A.1マクロファージによるLDH放出を測定することによって、組換えBCG株および陽性対照としての L.Monocytogenes EGDの細胞傷害性を判定した。Promegaより入手した定量用キットを使用して、BCG、BCG pAT261:Hly、BCG pMV306:Hlyまたは L.monocytogenes EGDを感染させた J774A.1マクロファージの培養上清および細胞溶解物のLDH活性をアッセイした。J774A.1細胞(1ウェルについて104)を96ウェルプレートに播種し、10 moiで感染させた。感染後1時間目 に、サンプルにゲンタマイシン(最終濃度 200μg/ml)を添加した。製造者の指示にしたがって、p.i.3、4、5または24時間目にLDH活性を定量分析した。細 胞傷害性のパーセンテージを以下のようにして算出した:細胞傷害性%=(感染J774A.1 - 自発的J774A.1)/(最大J774A.1 - 自発的J774A.1×100)。
【0039】
2. 結果
2.1 マイコバクテリア-大腸菌シャトル発現ベクター pAT261:HlyおよびpMV306:Hlyの構築
[L.monocytogenes EGD Sv 1/2a(Domann and Chakraborty,1989)のHlyによって仲介される]ファゴリソソーム脱出機能をBCG Chicagoに転移させるため、2つの別の大腸菌-マイコバクテリアシャトルベクター、pAT261および pMV306を使用した。pMV261の誘導体である第2世代ベクター pAT261(Stoverら、1991)は1つのBCGゲノムについて約5プラスミドコピーの染色体外Hly発現を指令し、pMV361の誘導体 である組み込み用(integrative)プラスミド pMV306はHlyの安定な染色体内発現 を可能にする(図1)(Stoverら、1991)。
【0040】
hly-hlyA(大腸菌 pHly152特異的溶血素A)オープンリーディングフレーム(ORF)をコードする plLH-1由来の 1.7 kb Pst I-DNA断片をプラスミド pAT261のPst I部位に挿入した(Gentschevら、1995;Stoverら、1993)。これによって生成する遺伝子融合物は BCG特異的Ag85Bシグナルペプチド(Matsuoら、1990)によって上清に分泌するように指令されるタンパク質の発現をコードする。この構築物を pAT261:Hlyと命名し、次に hsp60マイコバクテリアプロモーターの転写制御下にあるそのXba I-Sal I DNA発現カセットを使用して、親pMV306ベクター中に挿入し、構築物 pMV306:Hlyを生成させた(図1)。BCG特異的Ag85Bシグナルペ プチド(Matsuoら、1990)のためのコード配列を含む、両マイコバクテリア発現プラスミド中の hly特異的挿入部位のDNA配列を分析した。誘導された完全Hly融合タンパク質のアミノ酸配列を図2に示す。成熟Hly融合タンパク質は、融合体のN-末端に30アミノ酸(aa)、およびC-末端部分に52 aaの、元来大腸菌のHlyAに属するアミノ酸で構成される(Gentschevら、1995)。
【0041】
続いて、各プラスミド構築物 pAT261:Hlyまたは pMV306:Hlyを BCG Chicago株中にエレクトロポレートし、それぞれプラスミドまたは染色体 Hly発現をする BCG pAT261:Hlyまたは BCG pMV306:Hlyを生成させた。
【0042】
2.2 BCG pAT261:Hlyおよび BCG pMV306:Hly中のHly発現の分析
BCG pAT261:Hlyまたは BCG pMV306:Hly株による Hly分泌を特性付けるため、Stoverら(1993)にしたがって、中期対数期の増殖培養物の適切な上清およびマイコバクテリア溶菌液を調製した。免疫染色のために利用可能な抗Hlyモノクローナル抗体(mAb)に観察される交差反応性を克服するために、Hly融合体を疎水性相互作用クロマトグラフィーによって富化した(Schoelら、1994;Natoら、1991)。両マイコバクテリア株、BCG pAT261:Hlyおよび BCG pMV306:Hlyの溶菌液 および上清において、Hly融合タンパク質が検出可能である(図3)。Hly由来のポリペプチドの推定サイズ、62 kDaは元来の L.monocytogenesの 58 kDa Hlyタ ンパク質よりもわずかに大きい。
【0043】
BCG pAT261:Hlyおよび BCG pMV306:Hlyによって分泌されるHly融合タンパク質の細孔形成能力を特性付けるため、全細菌懸濁液および上清の溶血活性を判定した。BCG pAT261:Hlyおよび BCG pMV306:Hlyのサンプルはヒツジ赤血球に対して溶血活性を表し(表1)、これはマイコバクテリア種への細胞溶解性のHlyの機能の転移が成功したことを明白に証明するものである。
【0044】
【表1】

で与えられる。
b 細胞外および膜結合溶血活性。
c ND、検出されない。
【0045】
2.3 組換えBCG株のマクロファージ中の増殖
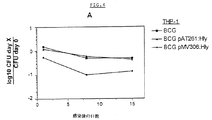
感染後(p.i.)1、8および15日目に、感染させたマクロファージのマイコバクテリア CFUによって、宿主細胞中の BCG pAT261:Hlyまたは BCG pMV306:Hly微生物の生存をモニターした。マイコバクテリアの標的細胞として、ヒト単球細胞系 THP-1(ATCC TIB-202)およびマウスマクロファージ様細胞系 J774A.1(ATCC TIB-67)を使用した。ホルボールミリステートアセテート(PMA)で刺激したTHP-1細胞は天然のヒト単球由来のマクロファージに類似している(Tsuchiyaら、1982)。THP-1または J774A.1細胞の感染後3時間目に、マイコバクテリア の食作用の効力を判定した。続いて200μg/mlゲンタマイシンの存在下で長期間の培養を実施して、上清中の放出されたかまたは食作用を受けなかったマイコバクテリアを死滅させた。図4に示すように、各BCG株、BCG pAT261:Hlyおよび BCG pMV306:Hlyはいずれの宿主細胞中でも増殖することができなかった。さらに、BCG pMV306:Hly細菌は親BCG株に比較して、THP-1および J774A.1宿主細胞中での細胞内持続性の減退を示した。THP-1マクロファージ中の BCG pMV306:Hly細菌の細胞内生存率は BCGまたは BCG pAT261:Hly感染サンプルの数値に関して p.i.1日目にすでに低下したことは注目に値する。
【0046】
対照的に、BCG pMV306:Hlyの細胞内持続性は、THP-1中でのBCGに匹敵した(図4)。興味深いことに、p.i.15日目に可視 BCG pAT261:Hly細菌は感染した J774A.1細胞中で検出されず、少なくともゲンタマイシン存在下でのこれらのマイコバクテリア構築物の完全増殖阻害が示唆された。
【0047】
BCG pAT261:Hlyおよび BCG pMV306:Hly株の減退した細胞内持続性の本質を究明するため、短期培養におけるこれらの組換えBCG株の J774A.1マクロファージに対する細胞傷害性を判定した。BCG;BCG pAT261:Hly;BCG pMV306:Hly;または L.monocytogenes EGDで感染させた宿主細胞の p.i.3、4、5および24時間目の上清中のラクテートデヒドロゲナーゼ(LDH)活性を測定することによって、細胞傷害性を分析した。p.i.24時間目、上清中に放出されたLDHの量は、親BCG、BCG pAT261:Hly若しくは BCG pMV306:Hlyで感染させるかまたは感染させない宿主細胞間で有意な差異はなかった(図4)。対照的に、増殖が早い溶血性 L.monocytogenes EGD株はp.i.24時間以内に上清中に顕著なLDH放出を引き起こした 。これらのデータは、組換えBCG株による溶血性Hlyの分泌は親BCG株の細胞傷害性を変性させなかったことを示唆している。むしろ、BCG pAT261:Hlyおよび BCG pMV306:Hly株の両者ともに非組換えBCG保有体に比較して、マウスマクロファージ中で減退した持続性を示した。
【0048】
(引用文献)
Aidoo,M.,Lalvani,A.,Allsopp,C.E.M.ら(1995)、マラリアに対する細胞傷害性Tリンパ球を誘導するワクチンのための保存された抗原性成分の同定、The Lancet 345:1003。
【0049】
Andersen,P.(1994)、分泌されたマイコバクテリアタンパク質の可溶性混合物 による、Mycobacterium tuberculosis感染に対するマウスの有効なワクチン接種、Infect.Immun.62:2536-2544。
【0050】
Andersen,P.,Andersen,A.B.,Sorensen,A.L.and Nagai,S.(1995)、マウスにお ける Mycobacterium tuberculosis感染に対する長期存続免疫のリコール(recall)、J.Immunol.154:3359。
【0051】
Berche,P.,Gaillard,J.L.,and Sansonetti,P.J.(1987)、T細胞性免疫のin vivo誘発のための必須要件としての L.monocytogenesの細胞内増殖、J.Immunol.138:2266-2276。
【0052】
Bielecki,J.Youngman,P.,Connelly,P.,and Portnoy,D.A.(1990)、Listeria monocytogenesからの溶血素遺伝子を発現する Bacillus subtilisは哺乳類細胞中 で増殖することができる、Nature 354:175-176。
【0053】
Blander,S.J.and Horwitz,M.A.(1991)、レジオネラの主要分泌タンパク質によるワクチン接種は、Legionella pneumophilaの別種のセログループ(serogroup) および別種のレジオネラ間にまたがる体液性および細胞性免疫応答ならびに防御免疫を誘発する、J.Immunol.147:285。
【0054】
Bosch,F.X.,Durst,M.,Schwarz,E.,Boukamp,P.,Fusenig,N.E.and zur Hausen,H.(1991)、腫瘍抑制のターゲットとしての癌関連ヒト乳頭腫ウイルスの初期遺伝子E6およびE7?、Behring Inst.Mitt.108。
【0055】
Bulow,R.and Boothroyd,J.C.(1991)、リポソーム中のp30抗原での免疫化によ る致死性Toxoplasma gondii感染からのマウスの防御、J.Immunol.147:3496。
【0056】
Clemens,D.L.,と Horwitz,M.A.(1996)、Mycobacterium tuberculosisファゴ ソームは初期エンドソームと相互作用し、外部から投与したトランスフェリンに接近することができる、J.Exp.Med.184:1349-1355。
【0057】
Darji,A.,Chakraborty,T.,Wehland,J.,and Weiss,S.(1996)、リステリオリシ ンは主要組織適合性遺伝子複合体クラス Iによる外来抗原の提示のための経路を作成する、Eur.J.Immunol.25:2967-2971。
【0058】
Domann,E.,and Chakraborty,T.(1989)、Listeria monocytogenesセロタイプ 1/2a株からのリステリオリシン遺伝子のヌクレオチド配列、Nucleic Acids Res.17:6406。
【0059】
Flesch,I.,Hess,J.H.,Oswald,I.P.,and Kaufmann,S.H.E.(1994)、IFN-γで刺 激されたマクロファージによる Mycobacterium bovisの増殖阻害:内在性腫瘍壊死因子αおよびIL-10による調節、Int.Immunol.6:693-700。
【0060】
Flynn,J.L.,Goldstein,M.M.,Triebold,K.J.,Koller,B.,and Bloom,B.R.(1992)、Mycobacterium tuberculosis感染に対する耐性のためには、主要組織適合性遺伝子複合体クラス I拘束T細胞が必要とされる、Proc.Natl.Acad.Sci.USA 89:12013-12017。
【0061】
Fu,T.M.,Friedman,A.,Ulmer,J.B.,Liu,M.A.and Donnelly,J.J.(1997)、防御性細胞免疫:細胞傷害性Tリンパ球はインフルエンザウイルス核タンパク質の優性および劣性エピトープを誘発するDNAでの免疫に対して応答する、J.Virol.71:2715。
【0062】
Gaillard,J.L.,Berche,P.,Mounier,J.,Richard,S.,and Sansonetti,P.J.(1987)、ヒトエンテロサイト(enterocyte)様細胞系 Caco-2中の Listeria monocytogenesの侵入および細胞内増殖の in vitroモデル、Infect.Immun.55:2822-2829。
【0063】
Gentschev,I.,Sokolovic,Z.,Mollenkopf,H.-J,Hess,J.,Kaufmann,S.H.E.,Kuhn,M.,Krohne,G.F.,and Goebel,W.(1995)、サルモネラが分泌する活性なリステリオリシンはその細胞内局在位置を変更する、Infect.Immun.63:4202-4205 。
【0064】
Grange,J.M.(1996)、マイコバクテリアおよびヒト疾病における薬剤耐性の疫 学的様相、Arnold,London,pp.124-125。
【0065】
Haas,G.,Plikat,U.,Debre,P.,Lucchiari,M.,Katlama,C.,Dudoit,Y.,Bonduelle,O.,Bauer,M.,Ihlenfeldt,H.G.,Jung,G.,Maier,B.,Meyerhans,A.and Autran,B.(1996)、HIV-1 Nefおよび特異的細胞毒性Tリンパ球中のウイルス変異体の in vivo 動態、J.Immunol.157:4212。
【0066】
Harboe,M.,Oettinger,T.,Wiker,H.G.ら(1996)、Mycobacterium tuberculosis および毒性Mycobacterium bovis中の ESAT-6タンパク質の出現、ならびに Mycobacterium bovis BCG中のその不在の証拠、Infect.Immun.64:16。
【0067】
Harrer,T.,Harrer,E.,Kalams,S.A.,Barbosa,P.,Trocha,A.,Johnson,R.P.,Elbeik,T.,Feinberg,M.B.,Buchbinder,S.P.and Walker,B.D.(1996)、無症候性長期非進行性 HIV-1感染症中の細胞毒性Tリンパ球。長期感染および低ウイルス負荷患者における応答の範囲および特異性ならびに in vivoウイルス準種との関連、J.Immunol.156:2616。
【0068】
Harth,G.,Lee,B.-Y.,Wang.J.,Clemens,D.L.,and Horwitz,M.A.(1996)、Mycobacterium tuberculosisの 30キロダルトンの主要細胞外タンパク質の遺伝学、生 化学および免疫細胞化学に対する新規な洞察、Infect.Immun.64:3038-3047。
【0069】
Hess,J.,Wels,W.,Vogel,M.,and Goebel,W.(1986)、プラスミドにコードされた溶血素決定基のヌクレオチド配列および対応する染色体溶血素配列との比較、FEMS Lett.34:1-11。
【0070】
Hess,J.and Kaufmann,S.H.E.(1993)、細胞内微生物に対するワクチン接種戦略、FEMS Microbiol.Immunol.7:95-103。
【0071】
Hess,J.,Gentschev,I.,Miko,D.,Welzel,M.,Ladel,C.,Goebel,W.and Kaufmann,S.H.E.(1996)、組換えサルモネラワクチンが誘発するリステリア症に対する防御における、体性 p60またはリステリオリシン抗原表示に対する分泌の優れた効力、Proc.Natl.Acad.Sci.USA 93:1458-1463。
【0072】
Hess,J.and Kaufmann,S.H.E.(1997)、細胞内病原体に対する宿主応答における、細胞内病原体に対するワクチン接種戦略の基礎となる細胞性免疫の原理、S.H.E.Kaufmann(ed),R.G.Landes Co.,Austin,pp.75-90。
【0073】
Horwitz,M.A.,Lee,B.-W.E.,Dillon,B.J.,and Harth,G.(1995)、Mycobacterium tuberculosisの主要細胞外タンパク質でのワクチン接種によって誘発される、 結核に対する防御免疫、Proc.Natl.Acad.Sci.USA 92:1530-1534。
【0074】
Houbiers,J.G.A.,Nijman,H.W.,van der Burg,S.H.,Drijfhout,J.W.,Kenemans,P.,van de Velde,C.J.H.,Brand,A.,Momburg,F.,Kast,W.M.and Melief,C.J.M.(1993)、変異型および野生型 p53のペプチドに対するヒト細胞傷害性Tリンパ球応 答の in vitro誘発、Eur.J.Immunol.23:2072。
【0075】
Huygen,K.,Content,J.,Denis,O.,Montgomery,D.L.,Yawman,A.M.,Deck,R.R.,DeWitt,C.M.,Orme,I.M.,Baldwin,S.,D'Souza,C.,Drowart,A.,Lozes,E.,Vandenbussche,P.,Van Vooren,J.-P.,Liu,M.A.,and Ulmer,J.B.(1996)、結核DNAワクチンの免疫原性および防御効力、Nat.Med.2:893-898。
【0076】
Kaufmann,S.H.E.(1993)、細胞内細菌に対する免疫、Annu.Rev.Immunol.11:129-163。
【0077】
Khan,I.A.,Ely,K.H.and Kasper,L.H.(1991)、精製された寄生体抗原(p30)がマウスにおける致死性 Toxoplasma gondii感染に対する CD8T細胞免疫を仲介する、J.Immunol.147:3501。
【0078】
King,C.H.,Mundayoor,S.,Crawford,J.T.and Schinnik,T.M.(1993)、Mycobacterium tuberculosisによる接触依存性細胞傷害活性の発現およびその活性をコー ドするゲノム遺伝子座の単離、Infect.Immun.61:2708-2712。
【0079】
Kochi,A.(1991)、世界の結核の現況および 世界保健機構の新しい制御戦略、Tubercle 72:1-6。
【0080】
Ladel,C.H.,Daugelat,S.,and Kaufmann,S.H.E.(1995)、主要組織適合性遺伝子複合体クラス I-および II-欠失ノックアウトマウスにおける Mycobacterium bovis bacille Calmette Gu(rin感染に対する免疫応答:獲得される耐性に対するCD4およびCD8T細胞の寄与、Eur.J.Immunol.25:377-384。
【0081】
Laemmli,U.K.(1970)、バクテリオファージ T4の頭部の組み立て中の構造タン パク質の開裂、Nature 227:680-685。
【0082】
Langermann,S.,Palaszynski,S.R.,Burlein,J.E.,Koenig,S.,Hanson,M.S.,Briles,D.E.,and Stover,C.K.(1994)、肺炎球菌表面タンパク質Aを発現する組換えBacille Calmette Gu(rinワクチンによって誘発されるマウスの肺炎球菌感染に 対する防御性体液応答、J.Exp.Med.180:2277-2286。
【0083】
Matsui,M.,Moots,R.J.,Warburton,R.J.,Peace-Brewer,A.,Tussey,L.G.,Quinn,D.G.,McMichael,A.J.and J.A.Frelinger(1995)、インフルエンザAマトリックスペプチドに特異的な CTL認識の細胞内ペプチド間の差異に関する遺伝子的証拠、J.Immunol.154:1088。
【0084】
Matsuo,K.,Yamaguchi,R.,Yamazaki,A.,Tasaka,H.,Terasaka,K.,and Yamada,T.(1990)、細胞外アルファ抗原のための Mycobacterium bovis BCG遺伝子のクローニングおよび発現、J.Bacteriol.170:3847-3854。
【0085】
Mazzaccaro,R.Z.,Gedde,M.,Jensen,E.R.,Van Santen,H.M.,Ploegh H.L.,Rock,K.L.,and Bloom,B.R.(1996)、Mycobacterium tuberculosis感染によって助長さ れる可溶性抗原の主要組織適合性クラス Iの提示、Proc.Natl.Acad.Sci.USA 93:11786-11791。
【0086】
McDonough,K.A.,Kress,Y.,and Bloom,B.R.(1993)、結核の病因論:Mycobacterium tuberculosisとマクロファージの相互作用、Infect.Immun.61:2763-2773。
【0087】
Murray,P.J.,Aldovini,A.,and Young,R.A.(1996)、サイトカインを分泌する組換え Bacille Calmette-Gu(rin株を使用する抗マイコバクテリア免疫の操作および有力化、Proc.Natl.Acad.Sci.USA 93:934-939。
【0088】
Nato,F.,Reich,K.,Lhopital,S.,Rouye,S.,Geoffroy,C.,Mazie,J.C.,and Cossart,P.(1991)、リステリオリシンOに対する無力化および非無力化モノクローナ ル抗体の製造および特性決定、Infect.Immun.59:4641-4646。
【0089】
Portnoy,D.A.,Jacks,P.S.,and Hinrichs,D.J.(1988)、Listeria monocytogenesの細胞内増殖のための溶血素の役割、J.Exp.Med.167:1459-1471。
【0090】
Roche,P.W.,Triccas,J.A.,and Winter,N.(1995)、結核に対するBCG接種:過去の失望および将来の希望、Trends Microbiol.3:397-401。
【0091】
Russell,D.G.(1995)、Mycobacteriumおよび Leishmania:エンドソームネットワーク内の隠れ場所、Trends in Cell Biology 5:125-128。
【0092】
Sambrook,J.,Fritsch,E.F.,and Maniatis,T.(1989)、Molecular cloning:a laboratory manual,2nd edition,Cold Spring Harbor Laboratory Press,New York。
【0093】
Schoel,B.,Welzel,M.,and Kaufmann,S.H.E.(1994)、細胞溶解性細菌毒素の精 製のための疎水性相互作用クロマトグラフィー、J.Chromatography A 667:131-139。
【0094】
Sorensen,A.L.,Nagai,S.,Houen,G.,Andersen,P.and Andersen,A.B.(1995)、Mycobacterium tuberculosisによって分泌される低分子量T細胞抗原の精製および特性決定、Infect.Immun.63:1710。
【0095】
Stover,C.K.,Bansal,G.P.,Hanson,M.S.,Burlein,J.E.,Palaszynski,S.R.,Young,J.F.,Koenig,S.,Young,D.B.,Sadziene,A.,Barbour,A.G.(1993)、組換えBacille Calmette Gu(rin(BCG)が発現する外表面タンパク質A(OspA)リポタンパク 質によって誘発される防御免疫:ライム(lyme)病ワクチンの候補、J.Exp.Med.178:197-209。
【0096】
Stover,C.K.,de la Cruz,V.F.,Fuerst,T.R.,Burlein,J.E.,Benson,L.A.,Bennett,L.T.,Bansal,G.P.,Young,J.F.,Lee,M.H.,Hatfull,G.F.,Snapper,S.B.,Barletta,R.G.,Jacobs,W.R.,Jr.,and Bloom,B.R.(1991)、組換えワクチンのためのBCG の新規な利用、Nature 351:456-460。
【0097】
Sturgill-Koszycki,S.,Schlesinger,P.H.,Chakrasborty,P.,Haddix,P.L.,Collins,H.L.,Fok,A.K.,Allen,R.D.,Gluck,S.L.,Heuser,J.and Russell,D.G.(1994) 、小胞状プロトンATPアーゼの排除によって産生される Mycobacterium ファゴソームの酸化作用の欠如、Science 263:678-681。
【0098】
Towbin,H.,Staehelin,T.,and Gordon,J.(1979)、ポリアクリルアミドゲルからニトロセルロースシートへのタンパク質の電気泳動転移:操作法および適用例、Proc.Natl.Acad.Sci.USA 76:4350-4354。
【0099】
Tsuchiya,S.,Kobayashi,Y.,Goto,Y.,Okumura,H.,Nakae,S.,Konno,T.,and Tada,K.(1982)、ホルボールジエステルによる培養ヒト単球性白血病細胞の成熟の誘 導、Cancer Res.42:1530-1536。
【0100】
Tweten,R.K.(1995)、グラム陽性細菌の細孔形成性毒素、細菌性病原体の毒性機構、J.A.Rothら(ed),American Society for Microbiology,Washington,D.C.,pp.207-228。
【0101】
van Elsas,A.,van der Burg,S.H.,Van der Minne,C.E.,Borghi,M.,Mourer,J.S.Melief,C.J.M.and Schrier,P.I.(1996)、ペプチドパルス(pulse)した樹状細胞 は健康なドナーから、Melan-A/MART-1自己抗原からの安定な HLA-A'0201-結合性ペプチドに対する腫瘍撲滅性(tumoricidal)細胞傷害性Tリンパ球を誘発する、Eur.J.Immunol.26:1683。
【図面の簡単な説明】
【図1】 組換えBCG株によるHly分泌のためのプラスミドマップを示す。
A.大腸菌-マイコバクテリアシャトルプラスミド pAT261:Hlyによる染色体外Hly発現。成熟Hlyタンパク質のDNA配列をコードするplLH-1由来の1.7 kb Pst I 断片の挿入。省略語:Mrep、マイコバクテリアレプリコン;Erep、大腸菌複製起源;kan、カナマイシン耐性遺伝子;hsp、熱ショックタンパク質プロモーター。 B.マイコバクテリアによるHly発現のための染色体組み込み用シャトルベク ター pMV306:Hly。挿入されるhsp60プロモーターを含むDNA制限断片(Xba I-Sal I)はプラスミド pAT261:Hly由来である。省略語:attP、マイコバクテリオフ ァージL5の付着部位;MCS、多重クローニング部位;int、マイコバクテリオファージL5のインテグラーゼ。
【図2】 BCG pAT261:Hlyまたは BCG pMV306:Hlyによって発現されるHly融合体のアミノ酸配列を示す。hly遺伝子特異的オープンリーディングフレームに対応するアミ ノ酸配列はマイコバクテリア発現プラスミド pAT261:Hlyまたは pMV306:HlyのDNA配列由来である。Hly融合タンパク質は以下の異なるポリペプチド配列で構成される:シグナルペプチドを含むBCG特異的 Ag85B、下線を付けた1文字コードの アミノ酸配列(以前はα抗原と称された;Matsuoら、1988);大腸菌 pHly152特異的HlyA、イタリック文字、(Hessら、1986);成熟Hly、太字、(Domann andChakraborty,1989);ランダムアミノ酸配列、通常文字。対応する遺伝子融合体について使用する制限部位(Pst IおよびNsi I)をそのアミノ酸配列の下に示す。
【図3】 組換えBCGによるHly発現の分析を示す。BCG、BCG pAT261:Hlyまたは BCG pMV306:Hly株の溶菌液(L)または上清(S)中のHly融合タンパク質の免疫染色によ る検出。各種マイコバクテリア株の培養物溶菌液および富化した上清を SDS/10 %ポリアクリルアミドゲル上で分離し、Hybond-PVDF膜に移した。62 kDa Hlyハ イブリッドタンパク質のケミルミネセンス免疫染色のために使用した一次抗体は抗Hly mAb H14-3(Natoら、1991)である。
【図4】 組換えBCG株の細胞内増殖および細胞傷害性を示す。
A.ヒトマクロファージ様細胞 THP-1中の野生型BCG(■)、BCG pAT261:Hly (△)およびBCG pMV306:Hly(◆)株の生存。
B.マウス J774A.1マクロファージ様細胞中の野生型BCG(■)、BCG pAT261:Hly(△)およびBCG pMV306:Hly(◆)株の生存。感染3時間後に感染細胞溶解 液からrBCG特異的CFUを判定し、0日から15日目までモニターした。データを平 均±SD(n=3)で示す。
C.BCGまたはrBCG感染後のLDH活性について、J774A.1の上清および細胞溶解 液をアッセイした。J774A.1(□)、BCG(◇)、BCG pMV306:Hly(◆)、BCG pAT261:Hly(△)または L.monocytogenes EGD(■)。上清中で検出した総LDH活 性の累積パーセンテージで示す(平均±SD)。これは3つの実験の代表例である。上清中に放出されたLDHのパーセンテージを測定して細胞死の尺度とした。
【配列表】
配列番号1:
(i)配列の特色:
(A)配列の長さ:1881塩基対
(B)配列の型: 核酸
(C)鎖の数: 両形態
(D)トポロジー:直鎖状
(ix)配列の特徴:
(A)特徴を表す記号:CDS
(B)存在位置: 1..1878
(xi)配列:
(i)配列の特色:
(A)配列の長さ:626アミノ酸
(B)配列の型:アミノ酸
(D)トポロジー:直鎖状
(ii)配列の種類:タンパク質
(xi) 配列:
Claims (28)
- (a) 哺乳類中で免疫応答を誘発する能力があるマイコバクテリウム・ポリペプチド由来の少なくとも1つのドメイン、および(b) ファゴリソソーム脱出ドメイン、を含む融合ポリペプチドをコードする組換え核酸分子。
- ファゴリソソーム脱出ドメインがリステリア属の生物に由来する、請求項1に記載の核酸。
- ファゴリソソーム脱出ドメインが、
(a) 配列番号1に示すヌクレオチド211-1722のヌクレオチド配列、
(b) (a)からの配列と同一のアミノ酸配列をコードするヌクレオチド配列、および、
(c) (a)または(b)からの配列の相補鎖配列とストリンジェント条件下でハイブリダイズするヌクレオチド配列、
から選択される核酸分子によってコードされる、請求項1または2に記載の核酸。 - 免疫応答を誘発する能力があるドメインがMHCクラスI拘束CD8T細胞応答を誘発する能力があるペプチドまたはポリペプチドである、請求項1〜3のいずれか1項に記載の核酸。
- 免疫応答を誘発する能力があるドメインがマイコバクテリウム抗原Ag85B(M.tuberculosis)、Ag85B(M.bovis)、Ag85A(M.tuberculosis)およびESAT-6(M.tuberculosis)から選択される、請求項1〜4のいずれか1項に記載の核酸。
- 免疫応答を誘発する能力があるドメインが抗原 Ag85B である、請求項5に記載の核酸。
- 融合ポリペプチドがシグナルペプチド配列によって先行される、請求項1〜6のいずれか1項に記載の核酸。
- 免疫応答誘発ドメインとファゴリソソーム脱出ドメインの間にペプチドリンカーが位置する、請求項1〜7のいずれか1項に記載の核酸。
- 請求項1〜8のいずれか1項に記載の核酸分子の少なくとも1コピーを含む組換えベクター。
- 核酸分子が発現制御配列に機能し得る形で連結されている、請求項9に記載のベクター。
- 発現制御配列がマイコバクテリア中で活性である、請求項10に記載のベクター。
- 染色体外ベクターである、請求項9、10または11に記載のベクター。
- 染色体ベクターである、請求項9、10または11に記載のベクター。
- 請求項1〜8のいずれか1項に記載の組換え核酸分子または請求項9〜13のいずれか1項に記載のベクターを含む細胞。
- (a) 哺乳類中で免疫応答を誘発する能力がある少なくとも1つのドメイン、および(b) ファゴリソソーム脱出ドメイン、を含む融合ポリペプチドをコードする少なくとも1つの組換え核酸分子を含む、組換えMycobacterium bovis細胞。
- ファゴリソソーム脱出ペプチドまたはファゴリソソーム脱出ポリペプチドをコードする組換え核酸分子を少なくとも1つ含む、組換えMycobacterium bovis細胞。
- 哺乳類中で免疫応答を誘発する能力があるペプチドまたはポリペプチドをコードする別の組換え核酸分子を少なくとも1つ含む、請求項16に記載の細胞。
- 免疫応答を誘発する能力があるドメインまたはペプチド若しくはポリペプチドが自己抗原、腫瘍抗原、ウイルス抗原、寄生体抗原、および細菌抗原から選択される、請求項15または17に記載の細胞。
- 少なくとも1つの前記組換え核酸分子を発現する能力がある、請求項14〜18のいずれか1項に記載の細胞。
- 少なくとも1つの前記核酸分子によってコードされたポリペプチドを分泌する能力がある、請求項14または19に記載の細胞。
- 天然のマイコバクテリウム細胞の細胞内持続性に等しいかまたはそれよりも低い、感染マクロファージ中の細胞内持続性を有する、請求項14〜20のいずれか1項に記載の細胞。
- 請求項1〜8のいずれか1項に記載の核酸分子によってコードされた組換え融合ポリペプチド。
- 請求項14に記載の組換え細菌細胞の調製方法であって、
(i) (a)哺乳類中で免疫応答を誘発する能力があるマイコバクテリウム・ポリペプチド由来の少なくとも1つのドメイン、および(b) ファゴリソソーム脱出ドメイン、を含む融合ポリペプチドをコードする組換え核酸分子を細菌細胞中に挿入し、そして、
(ii) (i)によって取得した細胞を好適な条件下で培養する、
各ステップを含む方法。 - 細胞がM.bovis細胞である、請求項23に記載の方法。
- 請求項15に記載の組換え細菌細胞の調製方法であって、
(i) (a)哺乳類中で免疫応答を誘発する能力があるポリペプチド由来の少なくとも1つのドメイン、および(b) ファゴリソソーム脱出ドメイン、を含む融合ポリペプチドをコードする組換え核酸分子をMycobacterium bovis 細胞中に挿入し、そして、
(ii) (i)によって取得した細胞を好適な条件下で培養する、
各ステップを含む方法。 - 請求項16に記載の組換え細菌細胞の調製方法であって、
(i) ファゴリソソーム脱出ペプチドまたはファゴリソソーム脱出ポリペプチドをコードする組換え核酸分子を Mycobacterium bovis 細胞中に挿入し、そして、
(ii) (i)によって取得した細胞を好適な条件下で培養する、
各ステップを含む方法。 - 哺乳類中で免疫応答を誘発する能力があるペプチドまたはポリペプチドをコードする別の組換え核酸分子をMycobacterium bovis細胞に少なくとも1つ挿入することを含む、請求項25または26に記載の方法。
- 免疫応答を誘発する能力があるドメインまたはペプチド若しくはポリペプチドが自己抗原、腫瘍抗原、ウイルス抗原、寄生体抗原、および細菌抗原から選択される、請求項25または27に記載の方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP97114614A EP0902086A1 (en) | 1997-08-22 | 1997-08-22 | Tuberculosis vaccine |
| EP97114614.7 | 1997-08-22 | ||
| PCT/EP1998/005109 WO1999010496A1 (en) | 1997-08-22 | 1998-08-12 | Tuberculosis vaccine |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2001514000A JP2001514000A (ja) | 2001-09-11 |
| JP2001514000A5 JP2001514000A5 (ja) | 2006-01-05 |
| JP4255209B2 true JP4255209B2 (ja) | 2009-04-15 |
Family
ID=8227261
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2000507804A Expired - Lifetime JP4255209B2 (ja) | 1997-08-22 | 1998-08-12 | 結核ワクチン |
Country Status (13)
| Country | Link |
|---|---|
| US (1) | US6673353B1 (ja) |
| EP (2) | EP0902086A1 (ja) |
| JP (1) | JP4255209B2 (ja) |
| AT (1) | ATE347599T1 (ja) |
| AU (1) | AU9435098A (ja) |
| CA (1) | CA2301879C (ja) |
| CY (1) | CY1106350T1 (ja) |
| DE (1) | DE69836588T2 (ja) |
| DK (1) | DK1007686T3 (ja) |
| ES (1) | ES2274580T3 (ja) |
| PT (1) | PT1007686E (ja) |
| SI (1) | SI1007686T1 (ja) |
| WO (1) | WO1999010496A1 (ja) |
Families Citing this family (33)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US8791237B2 (en) | 1994-11-08 | 2014-07-29 | The Trustees Of The University Of Pennsylvania | Compositions and methods for treatment of non-hodgkins lymphoma |
| US7662396B2 (en) | 2001-03-26 | 2010-02-16 | The Trustees Of The University Of Pennsylvania | Compositions and methods for enhancing the immunogenicity of antigens |
| US8956621B2 (en) | 1994-11-08 | 2015-02-17 | The Trustees Of The University Of Pennsylvania | Compositions and methods for treatment of cervical dysplasia |
| US6051237A (en) | 1994-11-08 | 2000-04-18 | The Trustees Of The University Of Pennsylvania | Specific immunotherapy of cancer using a live recombinant bacterial vaccine vector |
| US7794729B2 (en) | 1994-11-08 | 2010-09-14 | The Trustees Of The University Of Pennsylvania | Methods and compositions for immunotherapy of cancer |
| US7635479B2 (en) | 2000-03-29 | 2009-12-22 | The Trustees Of The University Of Pennsylvania | Composition and methods for enhancing immunogenecity of antigens |
| US7820180B2 (en) * | 2004-09-24 | 2010-10-26 | The Trustees Of The University Of Pennsylvania | Listeria-based and LLO-based vaccines |
| US8114414B2 (en) | 1994-11-08 | 2012-02-14 | The Trustees Of The University Of Pennsylvania | Compositions and methods for treatment of cervical cancer |
| ID29858A (id) | 1998-11-04 | 2001-10-18 | Isis Innovation | Uji diagnostik tuberkulosis |
| GB0006693D0 (en) * | 2000-03-20 | 2000-05-10 | Glaxo Group Ltd | Vaccine |
| US9012141B2 (en) | 2000-03-27 | 2015-04-21 | Advaxis, Inc. | Compositions and methods comprising KLK3 of FOLH1 antigen |
| US6855320B2 (en) | 2000-03-29 | 2005-02-15 | The Trustees Of The University Of Pennsylvania | Fusion of non-hemolytic, truncated form of listeriolysin O to antigens to enhance immunogenicity |
| US7700344B2 (en) | 2001-03-26 | 2010-04-20 | The Trustees Of The University Of Pennsylvania | Compositions and methods for enhancing the immunogenicity of antigens |
| US8771702B2 (en) | 2001-03-26 | 2014-07-08 | The Trustees Of The University Of Pennsylvania | Non-hemolytic LLO fusion proteins and methods of utilizing same |
| ATE552843T1 (de) * | 2003-02-06 | 2012-04-15 | Aduro Biotech | Listerien deren eindringen in nicht-phagozytische zellen abgeschwächt ist, impfstoffe, die diese listerien enthalten und deren verwendungen |
| CA2523084C (en) | 2003-04-23 | 2012-09-04 | Max-Planck-Gesellschaft Zur Foerderung Der Wissenschaften E.V. | Tuberculosis vaccine with improved efficacy |
| EP1649869A1 (en) * | 2004-10-21 | 2006-04-26 | Vakzine Projekt Management GmbH | Combination of a recombinant mycobacterium and a biologically active agent as a vaccine |
| DK1827504T3 (da) | 2004-12-01 | 2011-08-29 | Aeras Global Tb Vaccine Foundation | Rekombinante BCG-stammer med forøget evne til at undslippe fra endosomet |
| DK2977456T3 (en) | 2006-08-15 | 2018-01-22 | Univ Pennsylvania | Compositions comprising HMW-MAA and fragments thereof for the treatment of cancer |
| US8268326B2 (en) | 2006-08-15 | 2012-09-18 | The Trustees Of The University Of Pennsylvania | Compositions comprising HMW-MAA and fragments thereof, and methods of use thereof |
| ES2741730T3 (es) | 2008-05-19 | 2020-02-12 | Advaxis Inc | Sistema de administración doble para antígenos heterólogos que comprende una cepa de Listeria recombinante atenuada por la mutación de dal/dat y la deleción de ActA que comprende una molécula de ácido nucleico que codifica una proteína de fusión de listeriolisina O - antígeno prostático específico |
| US9650639B2 (en) | 2008-05-19 | 2017-05-16 | Advaxis, Inc. | Dual delivery system for heterologous antigens |
| US9017660B2 (en) | 2009-11-11 | 2015-04-28 | Advaxis, Inc. | Compositions and methods for prevention of escape mutation in the treatment of Her2/neu over-expressing tumors |
| EP2403935B1 (en) | 2009-03-04 | 2017-05-10 | The Trustees Of The University Of Pennsylvania | Compositions comprising angiogenic factors and methods of use thereof |
| US10016617B2 (en) | 2009-11-11 | 2018-07-10 | The Trustees Of The University Of Pennsylvania | Combination immuno therapy and radiotherapy for the treatment of Her-2-positive cancers |
| ES2784451T3 (es) * | 2010-09-20 | 2020-09-25 | Vakzine Projekt Man Gmbh | Mycobacterium recombinante como vacuna para uso en seres humanos |
| CN107412756A (zh) | 2010-10-01 | 2017-12-01 | 宾夕法尼亚大学理事会 | 李斯特菌疫苗载体用于在寄生虫感染的个体中扭转免疫无应答的用途 |
| WO2012085099A1 (en) * | 2010-12-21 | 2012-06-28 | MAX-PLANCK-Gesellschaft zur Förderung der Wissenschaften e.V. | Determination of the efficacy of an anti-mycobacterial vaccination |
| CN108434448A (zh) * | 2010-12-21 | 2018-08-24 | 马克斯·普朗克科学促进学会 | 作为疫苗的重组分枝杆菌 |
| CA2829960A1 (en) | 2011-03-11 | 2012-09-20 | John Rothman | Listeria-based adjuvants |
| BR102012003790A2 (pt) | 2012-02-17 | 2014-04-22 | Fundacao Butantan | Cepa de mycobacterium recombinante, composição imunogênica e uso |
| JP2015511602A (ja) | 2012-03-12 | 2015-04-20 | アドバクシス, インコーポレイテッド | リステリアワクチン処理後のサプレッサー細胞機能抑制 |
| AU2013243922A1 (en) * | 2012-04-02 | 2014-10-02 | The Arizona Board Of Regents For And On Behalf Of Arizona State University | Recombinant bacterium for induction of cellular immune response |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5504005A (en) * | 1987-03-02 | 1996-04-02 | Albert Einstein College Of Medicine Of Yeshiva University | Recombinant mycobacterial vaccine |
-
1997
- 1997-08-22 EP EP97114614A patent/EP0902086A1/en not_active Withdrawn
-
1998
- 1998-08-12 US US09/485,717 patent/US6673353B1/en not_active Expired - Lifetime
- 1998-08-12 PT PT98947427T patent/PT1007686E/pt unknown
- 1998-08-12 AT AT98947427T patent/ATE347599T1/de active
- 1998-08-12 SI SI9830871T patent/SI1007686T1/sl unknown
- 1998-08-12 AU AU94350/98A patent/AU9435098A/en not_active Abandoned
- 1998-08-12 DE DE69836588T patent/DE69836588T2/de not_active Expired - Lifetime
- 1998-08-12 WO PCT/EP1998/005109 patent/WO1999010496A1/en active IP Right Grant
- 1998-08-12 ES ES98947427T patent/ES2274580T3/es not_active Expired - Lifetime
- 1998-08-12 CA CA2301879A patent/CA2301879C/en not_active Expired - Lifetime
- 1998-08-12 JP JP2000507804A patent/JP4255209B2/ja not_active Expired - Lifetime
- 1998-08-12 DK DK98947427T patent/DK1007686T3/da active
- 1998-08-12 EP EP98947427A patent/EP1007686B1/en not_active Expired - Lifetime
-
2007
- 2007-03-01 CY CY20071100297T patent/CY1106350T1/el unknown
Also Published As
| Publication number | Publication date |
|---|---|
| CA2301879C (en) | 2014-02-04 |
| DE69836588D1 (de) | 2007-01-18 |
| ES2274580T3 (es) | 2007-05-16 |
| AU9435098A (en) | 1999-03-16 |
| ATE347599T1 (de) | 2006-12-15 |
| DK1007686T3 (da) | 2007-04-10 |
| WO1999010496A1 (en) | 1999-03-04 |
| CA2301879A1 (en) | 1999-03-04 |
| US6673353B1 (en) | 2004-01-06 |
| DE69836588T2 (de) | 2007-04-05 |
| PT1007686E (pt) | 2007-03-30 |
| CY1106350T1 (el) | 2011-10-12 |
| EP1007686B1 (en) | 2006-12-06 |
| EP1007686A1 (en) | 2000-06-14 |
| SI1007686T1 (sl) | 2007-04-30 |
| JP2001514000A (ja) | 2001-09-11 |
| EP0902086A1 (en) | 1999-03-17 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4255209B2 (ja) | 結核ワクチン | |
| US8545854B2 (en) | Tuberculosis vaccine with improved efficacy | |
| Skeiky et al. | T cell expression cloning of a Mycobacterium tuberculosis gene encoding a protective antigen associated with the early control of infection | |
| JP4940479B2 (ja) | マイコバクテリウム・ツベルクローシス融合蛋白質及びその応用 | |
| KR101329323B1 (ko) | 엔도솜을 이탈하는 능력이 보강된 재조합 비씨지 균주 | |
| KR20100122130A (ko) | 감염 잠복기 동안 발현되는 항원을 포함하는 결핵 백신 | |
| Bai et al. | Expression and purification of Mycobacterium tuberculosis ESAT-6 and MPT64 fusion protein and its immunoprophylactic potential in mouse model | |
| US6776993B2 (en) | Tuberculosis vaccine | |
| AU2002222098A1 (en) | Projection against mycobacterial infections | |
| Hess et al. | Vaccination strategies against intracellular microbes | |
| Dow et al. | Immunization with f-Met peptides induces immune reactivity against Mycobacterium tuberculosis | |
| Martin et al. | Comparative protective effects of recombinant DNA and Mycobacterium bovis bacille Calmette–Guérin vaccines against M. avium infection | |
| EP2303316A1 (en) | Constructing a dna chimera for vaccine development against leishmaniasis and tuberculosis | |
| Rezwan | Lipoprotein maturation is required for virulence of Mycobacterium tuberculosis and is essential for correct lipoprotein localization | |
| Al-Zarouni | Expression of Recombinant Antigen in BCG | |
| Kamanová | Bordetella Adenylate Cyclase: Molecular Mechanism of Action and Its Use for Antigen Delivery |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20050711 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20050711 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20080415 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20080617 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20080624 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20080813 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20080909 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20081209 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20090113 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20090127 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120206 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120206 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130206 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130206 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20140206 Year of fee payment: 5 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |