JP3987104B2 - シャペロニン10のアンタゴニスト - Google Patents
シャペロニン10のアンタゴニスト Download PDFInfo
- Publication number
- JP3987104B2 JP3987104B2 JP51529695A JP51529695A JP3987104B2 JP 3987104 B2 JP3987104 B2 JP 3987104B2 JP 51529695 A JP51529695 A JP 51529695A JP 51529695 A JP51529695 A JP 51529695A JP 3987104 B2 JP3987104 B2 JP 3987104B2
- Authority
- JP
- Japan
- Prior art keywords
- cpn10
- epf
- antibody
- cells
- peptide
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images



















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4715—Pregnancy proteins, e.g. placenta proteins, alpha-feto-protein, pregnancy specific beta glycoprotein
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P15/00—Drugs for genital or sexual disorders; Contraceptives
- A61P15/06—Antiabortive agents; Labour repressants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P15/00—Drugs for genital or sexual disorders; Contraceptives
- A61P15/08—Drugs for genital or sexual disorders; Contraceptives for gonadal disorders or for enhancing fertility, e.g. inducers of ovulation or of spermatogenesis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P15/00—Drugs for genital or sexual disorders; Contraceptives
- A61P15/18—Feminine contraceptives
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
- A61P17/02—Drugs for dermatological disorders for treating wounds, ulcers, burns, scars, keloids, or the like
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/20—Hypnotics; Sedatives
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
- A61P35/02—Antineoplastic agents specific for leukemia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/06—Immunosuppressants, e.g. drugs for graft rejection
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/08—Antiallergic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P5/00—Drugs for disorders of the endocrine system
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Engineering & Computer Science (AREA)
- Immunology (AREA)
- Reproductive Health (AREA)
- Gynecology & Obstetrics (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Endocrinology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Pregnancy & Childbirth (AREA)
- Zoology (AREA)
- Dermatology (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Biomedical Technology (AREA)
- Hematology (AREA)
- Neurology (AREA)
- Neurosurgery (AREA)
- Anesthesiology (AREA)
- Rheumatology (AREA)
- Oncology (AREA)
- Pain & Pain Management (AREA)
- Diabetes (AREA)
- Transplantation (AREA)
- Pulmonology (AREA)
Description
この発明は「cpn10」として知られるシャペロニン(Chaperonin)10のアンタゴニストに関する。
先行技術
シャペロニン類は、より広いクラスの分子シャペロン類に属し、翻訳後のひだ形成、ターゲッティングおよび他のタンパク質の集合に含まれる分子であるが、Ellisら,1991,Annu.Rev.Biochem.60,321−347に論じられているように、それ自体は最終的な集合構造の部分を形成しない。大部分のシャペロン類は“熱ショック”または“ストレス”蛋白質(hsp)であり、すなわち、その生成は、種々の細胞傷害(例えば代謝性破壊、酸素ラジカル、炎症、感染および変形)により誘導または増大される。Lindquistら,1988,Annu.Rev.Genet.22,631-677で評されているように、熱がよりよい研究ストレスの唯一のものである。特定のタンパク質レベルにおけるこれらの量的変化と同様に、上記のLindquistらに言及されているように、ストレスは本質的に生成したストレスタンパク質の異なる細胞区分への移動を誘導し得る。熱ショック反応は最も高く維持されている遺伝系の一つであり、そして種々の熱ショックタンパク質が最も進化論的に安定なタンパク質の中に存在している。逆境条件の下で細胞が切り抜けられるように、これらの類のあるものは正常細胞内で基本的な機能を果す。
2つの型のcpn分子、すなわちcpn60(単量体Mr〜60000)とcpn10(単量体Mr〜10000)がある。cpn60はよく研究されてきた。これはすべての細菌、ミトコンドリア、色素体で認められ、そして細胞質型、TCP−1は真核細胞中に認められる。いくつかの細胞の表面上でのその存在が報告されている。しかし上記のEllis文献やvan Eden, 1991, Immunol.Reviews 121,5−28では疑問視されて来た。非常に最近までcpn10は細菌中でのみ認められていたが、構造的および機能的に同等なものが葉緑体中(Bertschら,1992,Proceedings of the National Academy of Sciences USA 89,8696-8700)、ラット中(Hartmanら,1992,Proceedings of the National Academy of Sciences USA 89,3394−3398)および牛肝ミトコンドリア中(Lubbenら,1990,Proceedings of the National Academy of Sciences USA 87,7683−7687)で見い出された。
ATPの存在下でcpn60とcpn10は機能的に相互作用をおこし、タンパク質の集合を仲介する。cpn60から独立して働くcpn10の例は報告されていないが、cpn60は単独で働き、本質的に相違する事象に関係している。例えば、それは細菌感染における抗体とT細胞反応の免疫支配標的であり、しかし、タンパク質が高く保持されているので自己反応性が生成される。van Eden,1991,Immunol.Reviews 121,5−28で論じられているように、健常人は変形および感染自己細胞を排除するためにこの自己認識を用いることができ、かかる認識の調節を欠いている者は自己免疫疾患になり得る。当然のことであるが、cpn60はリウマチ性関節炎などの状態に関係している。分子シャペロン類は、多様なポリペプチド類の構造と結合したりあるいは変化する能力があり、タンパク質生物発生以外の細胞機能において最も重要な役割をなし得る。Hartmanら,1993,Proceedings of the National Academy of Sciences USA 90,2276−2280を参照、これはcpn10およびcpn60を用いたタンパク質分子の安定化を記載している。
哺乳動物のcpn10については、非常に近似した配列相同性であることが確証できた。例えば、ラットcpn10分子(Hartmanら,1992)は、J.E.Walker,MRC Lab.of Molecular Biology,Hills Road,Cambridge,UK提供のMT BTC PN10においてEMBL Data Base Directryに報告されている中のcpn10の構造と99%以上の相同姓を有している。このことは、細菌cpn10がラットシャペロニン10とわずか34%平均相同性しか有していないことと対称的である。
初期妊娠因子(EPF)
EPFは最初、妊娠関連物質として言及され(Mortonら,1976,Proc.R.Soc.B.193,413-419)、この発見は受精から6−24時間以内に妊娠の有無を検査できるので非常に興味を呼んだ。最初にEPFは、リンパ球から免疫抑制因子を放出する能力により、免疫抑制剤としての役割が考えられた(Rolfeら,1988,Clin.exp.Immunol 73,219-225)。これらの抑制因子はマウスにおいて遅延型の過敏症を低下せしめ、よって、抗原的に異質の胎児に対する母体の免疫反応を抑制するのであろう。さらに最近の研究によると、EPFの生成は妊娠に限られていない。これは初期および新生細胞増殖の産物であり、これらの状態において成長因子として働く(Quinnら,1990,Clin.exp.Immunol.80,100-108;Cancer Immunol.Immunother.1992,34,265−271)。EPFは血小板活性化の産物でもあり、したがって、創傷治癒および皮膚再生に関係することが言われている(Cavanaghら,1991,Journal Reproduction and Fertility 93,355−365)。
EPFは、Mortonら、Early Pregnancy Factor, Seminars Reproduction Endocrinology 10,72-82 1992で詳細に再検討された。EPF生成の場所および調節が記載され、さらに血小板からのEPFの精製および精製物と他の源から導かれたEPFとの関係についても述べている。この再検討は、精製工程のモニタリングおよび生成源の研究を含み、EPFの生物検定法(すなわち、ロゼット阻害試験)についても論じている。EPFの生物活性が論じられ、EPFとそのアンタゴニストの可能な臨床適用が記載されている。
Mortonら,1992,Repord.Fertil.Dev.4,411−422は、EPFの免疫抑制および成長因子の性質に関して以前なされた発表を再検討している。前胚保持におけるEPFの役割についても、この文献で論じられている。
今まで、ロゼット阻害試験が複合生物学的混合物中のEPFを検出する唯一の手段であった(Mortonら,1976,Proc.R.Soc.B. 413−419)。この検定法は、免疫抑制の抗リンパ球血清(ALS)がリンパ球と非定型赤血球とのインビトロ自発性ロゼット形成を阻止し得るとのBachおよびAntoine,1968,Nature(Lond)217,658−659の最初の知見に基づいている。EPFを検出するために修正法が導入された。まず、EPF中でプレインキュベーションしたリンパ球が同じ提供者のEPF未処理のリンパ球に比してALSで有意に高いロゼット阻害価(RIT)を有していることが上記の1976文献で示された。この試験は、詳細に、上記の1976文献およびMortonら,1987,“In Current Topics in Developmental Biology”Vol.23,73−92,Academic Press,San Dirgoに記載されており、簡単には、EPFがリンパ球に結合し、続いて抑制因子の放出を誘導する一連の流れに関している(Rolfeら,1988,Clin.exp.Immunol. 73,219-225)(Rolfeら,1989,Immunol.Cell Biol. 67,205−208)。
Athanasas-Platsisら,1989,Jornal Reproduction and Fertility 87,495−502およびAthanasas-Platsisら,1991,Jornal Reproduction and Fertility 92,443−451に、EPFに対するモノクローナルおよびポリクローナル抗体の生成および胚生存の欠如をもたらすこれらの抗体による妊娠ラットの受動免疫法について記載されている。これらの研究で妊娠がうまく成立するためにEPFが必要であることがはっきりした。
Quinnら,1990,Clin.exp. Immunol. 80,100−108に、EPFが培養腫瘍および変形細胞の成長調節物であることが示されている。これらの細胞は継続成長のためにEPFに依存しており、すなわち、EPFがオートクリンモードで働く。
腫瘍細胞の成長が抗EPFmAbsでかなり遅延されることから、EPFが腫瘍の成長促進に役割を演じていることが確証されている。さらに、高い親和力の抗EPF抗体を生成するハイブリドーマは本来、不安定であろうことをこの文献は示している。
Quinnら,1992,Cancer Immunol. Immunother,34 265−271に、移植可能マウス腫瘍のインビボ成長に対する抗EPFのモノクローナル抗体(mAbs)の作用が記載されている。この文献の要点は、EPFの中和がインビボ腫瘍の成長を遅らすことにある。
癌が非常に初期段階にあるとき、抗EPFmAbによるEPFの中和が、癌の拡大を止めることはQuinnら、1992の文献により明らかである。しかし、癌が大きくなってしまうと、これらのmAbsでの処置は癌の成長を止めるが完全に破壊しない。
腫瘍成長におけるEPFの役割に関する他の文献として、Quinn,1991,Immunol.Cell.Biol.69,1−6およびQuinn.K.A.の1991年オーストラリアのクイーンズランド大学での“初期妊娠因子:細胞増殖に関する新因子”なる博士論文がある。
上記のことから、EPFは、胚の存在の指標でありまた必要であり、免疫抑制作用を有していると評価される。最近の研究は、変形、新生および正常細胞の成長調節剤としてのEPFの重要性を示唆しており、血小板中でのその存在は炎症および創傷治癒において役割を演じる。
この特異な結合作用を保持する分子は大きい意義を有しているが、今まで、この分子の構造は明らかでなかった。
意外にも、哺乳動物のシャペロン10が初期妊娠因子(EPF)と同じアミノ酸配列を有することが確証された。
発明の要約
本発明は、一面において、cpn10、cpn10誘導体またはcpn10から誘導されたペプチドの生産を提供するものである。
他の一面において、本発明は、抗cpn10抗体又は他のアンタゴニストの使用に関する。抗cpn10抗体又は他のアンタゴニストの使用は、生長抑制作用又は免疫上昇作用を含むことができる。
本発明は、その範囲に、(i)原核性、真核性、組換え又は合成cpn10或いはそれらの修飾体又は断片に対するモノクローナル又はポリクローナル抗体若しくはそれら抗体の活性部位又は中心に基づく人為的加工構成体、又は(ii)その分子又は断片の構造の修飾に基づくcpn10のアンタゴニストの使用を含む。
実験
A. cpn10の精製と抗体の生産
(i)ヒト血小板からのヒトEPFの精製(図1a、1b、1c、1d)
抽 出
臨床的に7日まで経過の濃縮血小板(血液銀行より)をチロード(Tyrodes)緩衝液で洗い、Method in Enzymology, 1989, 167, 7−11に記載の技法を用い、液体N2で凍らせて−70℃で保存した。
精製の直前に、約100洗浄血小板単位を水浴で溶かし、75−85℃で15分間断続的に緩やかに撹拌した。氷冷した後、細胞片を遠心分離(8000g、20分、4℃)で除き、ペレットを0.05M酢酸/0.1M NaCl/0.1mg/mlナトリウムアジド pH3.0で均等化により2度抽出し、次いで遠心分離した(8000g、15分、4℃)。この3上清液をためて、全抽出量が500−600mlとなった。
イオン交換クロマトグラフィー
100血小板単位からの抽出液を濃塩酸でpH3.0に調整し、0.05M酢酸/0.1M NaCl pH3.0であらかじめ平衡化した250ml SP−Sephadex C−25(Pharmacia−LKB)と共に緩やかに一晩、4℃で撹拌した。ゲルを同じ緩衝液20volで洗ったカラムに入れ、0.5Mリン酸ナトリウム緩衝液/0.05M NaCl pH7.5で溶出した。次いで、ゲルを、捨てた。
親和性クロマトグラフィー
SP−Sephadex溶出物を、濃塩酸でpH6.3−6.4に調整し、0.05Mリン酸ナトリウム緩衝液/0.05M NaCl pH6.3であらかじめ平衡化したHeparin−Sepharose CL−6Bのカラム(2.5×7.5cm;Pharmacia−LKB)にかけた。カラムを同じ緩衝液5volで洗い、0.05Mトリス−HCl/5mM CaCl2/0.2M NaCl pH7.5の5volで溶出し、サンプル適用のために用いるカラムに逆方向でかけた。
高性能疎水性相互作用クロマトグラフィー(HIC−h.p.l.c.)
固体の(NH4)2SO4をHeparin−Sepharose溶出液に最終濃度が2Mになるまで加え、0.45μmフィルターを通して後、0.1Mトリス−HCl pH7.0/5mM CaCl2/2M(NH4)2SO4であらかじめ平衡化したTSK Phenyl 5PWカラム(7.5×75mm、Pharmacia−LKB)上の専用溶媒ラインを通してサンプルをポンプで送った。このカラムを同じ緩衝液10volで洗い、この緩衝液から0.1M トリス−HCl pH7.5/5mM CaCl2/10%アセトニトリルの50分直線勾配溶出を行った(図1a)。
RP−h.p.l.c.−1
活性HIC−h.p.l.c.フラクションをプールし、A、0.04M トリス/HCl pH7.0/5mM−CaCl2およびB、0.04M トリス/HCl pH7.0/5mM CaCl2/80%(v/v)アセトニトリルからなる溶媒を用いてC3カラム(Ultrapore RPSC,Beckman Institute)上で分画した。カラムをサンプル適用する前に溶媒Aで平衡化し、その後溶媒A5volで洗い、この溶媒から30分75%溶媒Bまでの直線勾配で溶出を行った(図1b)。
RP−h.p.l.c.−2
いくつかの100単位血小板のRP−h.p.l.c.−1からの活性フラクションをプールし、EDTAとDTTをそれぞれの最終濃度が20mMと1mMになるように加え、この混合物を0.5−1時間4℃で放置した。溶媒Aの2volで希釈した後、C3カラムにかけ、この工程および次の工程を行い、RP−h.p.l.c.−1に記載されたように、しかしCaCl2を除き、分画した(図1c)。
RP−h.p.l.c.−3
RP−h.p.l.c.−2からの活性フラクションをプールし、トリフルオロ酢酸(TFA)を最終濃度が0.1%になるように加え、0.1%TFA2volで希釈し、0.1%TFAであらかじめ平衡化したC3カラムに混合物をかけた。この溶媒と60%(v/v)アセトニトリル/0.1%TFAの30分直線勾配溶出を、次いで90%(v/v)アセトニトリル/0.1%TFAの3分直線勾配溶出を行った。活性フラクションをプールした(図1d)。
1単位は約500mlである単一の血液提供からの血小板を表す。“活性フラクション”はロゼット阻害試験における活性フラクションであった。
他の供給源からのEPFの精製
Cavanagh & Morton,1994,Eur.J.Biochem.222,551−560に論じられたように、種々の供給源からEPFを精製した。
すべての例において、生物活性はヒト血小板について述べられている複合精製スキームを通して同じパターンであった。さらに、すべての供給源からの最終活性フラクションは固定化モノクローナル−抗EPFにより特に結合され、定量的に回収し得る(図1e)。
これらの研究は次の理由により重要である:
A.すべての材料で観察された生化学的および免疫的類似性は、生物検定法が種々の生物的状況において作用する単一の物質または近接する類の物質を検出することの証拠である。
B.これらの材料から精製された活性体は、EPFであるとして上記Mortonら、1992参照おいて以前に報告されたすべての物質よりも多くの程度より強力である。これは、上に論じたEPFの生物検定法の詳細な分析を基に、恐らくEPFに関している活性が非常に少量の不純物の存在を反映しているに違いないとの推測を確認している。
C.(活性の妨害として)タンパク質レベルで研究するのに充分なEPFを提供する唯一の供給源材料は血小板と再生肝であり、量はそれぞれ平均100単位当たり15μg(〜50リットル血に相当)および40g組織当たり5μg(6匹ラットよりの摘出肝)。EPFは細胞外に表れるよりも細胞中に多く存在している。しかし、蓄積されたEPF生物学的知識(上記のMortonら、1992参照)はこの細胞外での出現が偶然のものでないことを示している。
ヒト血小板誘導EPFは、最も豊富であり、かなり詳細に研究されてきた。SDS−PAGEにおいて、Mrの単一バンド約8500として表れ、生物活性に一致した(図2a);再生ラット肝臓由来のEPFは同様の行動を示した。質量分析計により血小板資料について分子量10843.5±2Daが明確かつ精密に決定され、この高いレベルでの同質性の明白な証拠であった(図2b)。エドマン分解法の後、分子がN−ブロックされていることを示すが、約4nmol EPFのタンパク質分解分裂が行われた。得たペプチドフラグメントを逆相HPLCで分離し、エドマン分解法で配列を決めた。12(フラグメント1)、27(フラグメント2)および33(フラグメント3)の残基を含む3配列領域は、ラットのミトコンドリアcpn10における残基7−18、27−53および69−101(C末端)に対応することが判明した。フラグメント2の残基52は異なっていた(cpn10中のS、ラットcpn10中のG;この相違のみでラットcpn10よりcpn10が30Da大きいことが説明できよう)。他のすべての残基は同一で、シャペロニン類の高く保持された本質に一致した(図2c参照)。
EPFとcpn10の配列の同一性が確認されて以来、ラットミトコンドリアcpn10、E.coli cpn10(groESとして知られる)およびE.coli cpn60(groEL)を用いて、機能的な関係についていくつかの研究がなされてきた。最初に、cpn10がEPFとして働くことが証明されている。ラットcpn10がEPF生物検定法で検討され、予測された希釈範囲以上にポジティブであることが見いだされた。この活性はEPFに対するモノクローナル抗体で中和され得た(表1)。興味あることに、ラットcpn10と〜40%の相同性を有するE.coli cpn10は生物検定で活性を示さなかった。これは、すべての哺乳動物の細胞系および酵母細胞が活性であるのに対し、E.coliの培地は活性ではないとの知見に一致している。cpn60は生物検定で不活性であり、EPF活性に作用しなかった。EPFはcpn10のように働き得ることを示していた。EPFをATPの存在下または非存在下でcpn60と混合し、この混合物をTSK G3000SWゲル浸透カラムで分析した。得られたフラクションをSDS−PAGEで分析した。cpn60はデカテトラマー(decatetramer)であり、このカラムの排除体積中に溶出する(排除限界 300000)。ATPの存在下では、EPFもこの分画に存在し、cpn60と安定な複合物の形成を示す。ATPの非存在下ではそうでない。このフラクションは生物検定において活性であったがATP非存在下の実験での同様のフラクション(EPFがcpn60と結合していなかった)は活性でない(図3a参照)。このようにEPFおよびcpn10活性は同じ分子に属する。
これらの研究者は、血小板誘導EPFがcpn10と構造的および機能的に相同体であるとの明白な証拠を提供する。cpn10とロゼット阻害試験における活性との関係が試験された(図3b)。ATPの存在下において、ATPの非存在下ではそうでないのであるが、固定化cpn60は原型源資料、妊娠血清からすべての活性を除去し得た。そして固定化複合物からATPを取り除くと活性は回復し得た。図3aに記載した実験のように、このATPの必要性はcpn60と活性分子との相互関係の特殊性を示している。cpn10はEPF生物検定法において反応を司る分子単位である。
cpn10とのEPFの同一性の確認は本主題についての研究を進める主段階であり、今日までなされた多くの知見を説明する助けとなった。EPF生成がこのように多様な生物的状態、例えば、着床前−後妊娠初期および主要細胞増殖および血小板活性において生じるとの主張に対して批判があった。hsp(heat stress protein:熱ストレスタンパク質)としてのその役割において、EPF生成の速い開始が期待されるすべての条件がある。細胞の生存に必須であるhspの機能は、上記のLinquistらに示されているように細胞内である。逆に、今日まで記載のEPF活性は細胞外である。例えば、Mortonら,1987,Current Topics in Development Biology,Vol.23,73−92に論じられているように、交配後4−6時間のマウス血清およびQuinn博士論文(1991)に示されているようにラットの部分的肝切除後4−8時間にその活性が表れる。上述のQuinnら1990文献に論じられたようにオートクリン相または上述のRolfeら1988で論じられた外分泌相において、EPFが働き得ることを我々は示して来た。これらはhspについて以前に記された役割ではない。
EPFの構造が知られたので、組換えDNA技法または化学合成などの適当な技術によって商業的な量でEPFを製造し得ることが評価される。
(ii)ANTI−CPN10誘導ペプチドの生産
抗cpn10誘導ペプチドの生産に使用された方法と得られた結果について記載する。cpn10のペプチドは、次のアミノ酸配列を含むことができる:
(i)AGQAFRKFLPL;
(ii)Ac−AGQAFRKFLPL;
(iii)EKSQGKVLQAT
(iv)A1AGQAFRKFLPLA2;
(v)AGQAFRKFLPLA2;
(vi)A1AGQAFRKFLPL;
(vii)Ac−A1AGQAFRKFLPLA2;
(viii)Ac−AGQAFRKFLPLA2;
(ix)Ac−A1AGQAFRKFLPL;
(x)A1EKSQGKVLQATA2;
(xi)EKSQGKVLQATA2;
(xii)A1EKSQGKVLQAT;
これらの式中、A1とA2は分子(i)〜(xii)の一つ又は各の末端に追加されてもよいアミノ酸配列であり、Acはアセチルである。
抗cpn10誘導ペプチド抗体は、上記アミノ酸配列(i)〜(xii)のいずれか一つに対する抗体を含むことが出来る。例えば、抗体は、N末端断片(Ac−AGQAFRKFLPLC)と内部断片(EKSQGKVLQATC)に対して産生された。
上記ペプチドについて、それが一個のアミノ酸の付加、欠失又は置換を含むことが出来、本発明にはそのようなペプチドに対して産生された抗体も含まれることが理解されてよい。
方 法
cpn10誘導ペプチドの合成
ペプチドは、cpn10のN末端フラグメント(N−ペプチド、即ちAc−AGQAFRKFLPLC)および内部フラグメント(I−ペプチド、即ちEKSQGKVLQATC)と一致するように合成した。
卵白アルブミンに対するペプチドの結合
ペプチドを、異種2機能性試薬SPDPにより、製造者(Pharmacia LKB.Biotechnolkogy,Uppsala,Sweden)の指示に従って、卵白アルブミンに結合された。
免疫化スケジュール
成熟非近交系ニュージーランドウサギを、接合体の一つを週4回注射し、続いて数カ月追加免疫することにより免疫化した。
注射のために、抗原を0.9%食塩水(Mr12−15000除去透析管、Visking,Union Carbide,IL,USA)で透析し、等量のフロインドアジュバント(一回目は完全、それ以降不完全)に乳濁化した。免疫化はs.c.経路経由であった。注射された抗原の量を第1表に示す。
抗血清のスクリーニング
抗血清を、適切な抗原(即ち、I−ペプチドまたはN−ペプチド;卵白アルブミン)(5mg/ml)に対してELISAで試験した。結合IgGは、o−フェニレンジアミンを基質として、ビオチン−ストレプトアビジン系(Amersham)で検出した。吸光度は492nmで読んだ。
抗N−ペプチドAbsもまた、血小板誘導EPF(1mg/ml)(Cavanagh et al.,1994, Eur.J.Biochem., 222, 551-560)とN−ペプチド(5mg/ml)に対して、抗EPFAbs♯810及び♯816(Athanasis-Platsis et al., 1989, J.Reprod.Fert.87, 495-502)と平行して試験された。
抗体の精製
IgGを親和性クロマトグラフィーにより血清から精製した。製造者の指示に従って、N及びIペプチドと卵白アルブミンを別々にHiTrap(商標)親和性カラム(HiTrap NHS−活性化1ml, Pharmacia−LKB)にかけた。各カラムを0.05NaPi−0.5M NaCl(pH7.4)で平衡化し、適切な抗血清を製造者の指示に従って適用した。平衡バッファーでよく洗浄した後、結合ウサギIgGを0.2Mグリシン(pH2.5)で溶出した。溶出液のpHは、2Mトリスで約7.4に調節した。
溶出液中のAbs純度は、SDS−PAGEで決定し、最強のフラクションをプールした。
タンパク質の評価
タンパク質(IgG)は、商業用に調整されたFolin−Ciocalteu試薬(Stansens,Qld,Australia)を使用して、Lowryの方法(Lowry et al., 1951, J.Biol.Chem., 193, 265-275)により決定した。標準曲線は、精製ウサギIgG製剤(20mg/ml;Silenus, Hawthorne, Australia)で作成した。
結 果
抗体のELISAスクリーニングは、若干の興味ある結果を与えた。
抗ペプチドAbsタイターは、追加免疫(boosting)を行っても低下したが(図4a,4b)、抗卵白アルブミンコントロールAbsの産生は正規のレスポンスを示した(図4c)。ペプチド結合体で免疫したウサギにおける抗卵白アルブミンAbsのタイターも同様に減少した(図4a,4b)ことが注目される。
交差反応性の検討は、図5に示される。
親和性精製Absのタイターは、ウシ血清アルブミン(BSA)に結合した免疫ペプチドに対するELISAによって決定された。この試験は、また、方法の効率をも証明した。結果は、第2表に示される。
これらのAb製剤は、SDSポリアクリルアミドゲル電気泳動で約95%の純度であることが示された。
結 論
免疫スケジュールの間のAbsの減少タイターは、B細胞クローンの増殖におけるcpn10に対する役割を示唆する。抗EPF抗体を生産する抗体産生B細胞クローンの不安定性は、先に記載された。抗EPF抗体産生のパターンは、上記したように、最初の免疫後5週間で最高のタイターとなり、その後、追加免疫を繰り返しても低下する。イン・ビトロにおいて、抗EPF抗体を産生するハイブリドーマは、本来不安定であった(Quinn et al., 1990, Clin.Exp.Immunol.,80, 100-108)。抗EPF/cpn10抗体を生産するハイブリドーマの安定なセルラインを作り出すことの困難性は、細胞増殖におけるEPF/cpn10のオートクリン作用、すなわち、抗体が増殖細胞によってそれら自身の利益のための生産されるEPF/cpn10を中和することによるものであろう。
(iii)組換えCPN10に対する抗体の生産
cpn10をコードするヒトcDNAのクローニングおよびcpn10の製造
商業的使用のための製造は、哺乳動物cpn10遺伝子、望ましくはヒドcDNAcpn10遺伝子をpGEX系、pET系からのプラスミドのような適当なベクターに挿入し、コードされた哺乳動物cpn10を発現し、組換えcpn10を精製することにより達成され得る。
略号:
ANGIS オーストラリア国立遺伝情報サービス
bp 塩基対
BSA 牛血清アルブミン
cDNA 相補的DNA
cpn10 シャペロニン10
DNA デオキシリボ核酸
E.coli 大腸菌
GSH グルタチオン(還元型)
GST グルタチオン−S−トランスフェラーゼ
LB ルリア−ベルタニブロス
M モル
ORF オープンリーディングフレーム(翻訳領域)
PCR ポリメラーゼ連鎖反応
rEPF 組換え初期妊娠因子
RSP 逆配列プライマー
SDS ドデシル硫酸ナトリウム
SDS−PAGE ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動
Tris トリ(ヒドロキシメチル)アミノメタン
USP 正配列プライマー
材料および方法
ヒトcpn10オープンリーディングフレームのクローニング
メラノーマ細胞系A2058cDNAラムダライブラリー(Stratagene)からヒトcpn10cDNAのORF(274bp)部分を最初に増幅するためにポリメラーゼ連鎖反応(PCR)を用いた。変性cpn10アンプリマー(P1)は、ヒトcpn10のアミノ酸残基83−91に対応するアミノ酸配列VLDDKDYFLからつくられた。プライマーP1は配列5’ARRAARTARTCYTTRTCRTC3’を有し、RはAまたはG、YはCまたはTである。逆配列プライマーは、PCR増幅(非特異的プライマー)および配列DNA構築に用いられ、配列5’CAGGAAACAGCTATGAC3’を有する。正配列プライマーは配列5’GTAAAACGACGGCCAGT3’を有している。ファージ・ライブラリーのPCR増幅は、非特異的上流アンプリマー(RSP)およびP1を、それぞれ0.5μM最終濃度、1.5mM MgCl2(Pharmacia Biotech)、1Xポリメラーゼ緩衝液(Boehringer Mannheim)、テルムスアクアティカス(Thermus aquaticus)DNAポリメラーゼ(Boehringer Mannheim)5単位および最終容量の50μL中で用いて、達成された。30サイクルについてのパラメーターは、94℃、1分間での変性、40℃、30秒間のアニーリングおよび72℃、3分間のエクステンションであった。72℃で7分間の最終エクステンションが4℃10分間のソークサイクルに続いて、なされた。1μLのアリコートがコピー数を増すために同じ条件で増幅された。
オープンリーディングフレイムを有する2つのcpn10特異アンプリマーが作られた。上流プライマーP2、5’−GCGCGGATCCATGGCAGGACAAGCGTTTAG−3’が最初のPCRフラグメントの配列から作られた。下流プライマーP3、5’−ATATGAATTCAGTCTACGTACTTTCC−3’は、Expressed Sequence Tag database via ANGIS(Accession No.HUM00TB037)から得られる配列から作られた。アニーリング温度を50℃にした以外は上述と同じ反応およびサイクリングの条件を用いてファージ・ライブラリーからA319bpフラグメントが増幅された。
DNA構築および分析
Boehringer Mannheimから得た制限酵素およびバッファーを用いて、Sambrookら(Sambrookら,1989,Molecular Cloning:A Laboratory Manual. 2nd Ed. Cold Spring Harbor Press, Cold Spring Harbor, NY)に従って、PCR生成物およびベクターのすべての制限酵素による切断を行った。最初PCRフラグメントをEcoR1で消化し、プラスミドpRM1を形成するpBluescript KS(+)(Stratagene)のEcoR1とSma I位でライゲートした(Sambrook et al., 1989, Molecular Cloning:A Laboratory Manual. 2nd Ed.Cold Spring Harbor Press, Cold Spring Harbor, NY)(図6a;部分cpn10挿入274bp)。319bp生成物をBamHIおよびEcoR1で消化し、プラスミドpRM2を形成する発現プラスミドpGEX−2T(Pharmacia Biotech)に最初にクローン化した(図6b)。この同一性を確認するために、BamHI−EcoR1フラグメントをpBluescript(SK+)にサブクローン化し(pRM3;図6c)、配列決定した。DNAを、エチディウムブロマイドを含む0.8−1.0%(w/v)アガローズゲルで分析し、電気泳動の後、UVイルミネーションで観測した。
E.coliの形質転換
適格なE.coli DH5αの細胞(100μL)をプラスミドで熱律動法(Sambrookら,1989,Molecular Cloning:A Laboratory Manual. 2nd Ed. Cold Spring Harbor Press, Cold Spring Harbor, NY)により形質転換した。細胞とDNAの混合物(10−100ng)を30分間氷の上におき、正確に2分間42℃で熱律動せしめ、2分間氷の上にもどした。LBの0.9mLを加えた後1時間後振り、37℃で細胞を回収した。100μLの部分標本を最終濃度100μg/mLでアンピシリンを補ってLB寒天プレート上に置いた。37℃で一夜、培養した後、ランダムコロニーを次の実験に選択した。
DNA配列決定
PCR生成物の制限フラグメントをpBluescript中にクローンし、製造業者(Pharmacia Biotech)の指示に従ってT7ポリメラーゼキットを用いてジデオキシチェインターミネーション法によって両定位に配列した。約2μgのプラスミドDNAを変性し、エタノール沈殿し、USP、RSPまたはP3のいずれかにアニーリングした。配列反応を8%アクリルアミド/46%ウレア ゲル上で電気泳動した。固定し乾燥した後、X−線フィルムをゲルに一夜さらし、そして現像した。
E.coliにおける組換えcpn10の発現および精製
Smithら(Smithら,1988,Gene 67(1)31−40)に記載された方法により小培地スケール(2ml)でグルタチオン−S−トランスフェラーゼ融合タンパク質の発現のためにpRM2で組換えたクローンをスクリーニングした。一夜置いた培地を希釈し、0.1mM IPTGを加え、37℃で数時間おいて融合タンパク質を発現せしめた。細胞をペレット化し、PBS/0.1%Triton X−100に溶解し、溶解質を50%グルタチオン−アガローゼ ビーズ(Sigma Chemical Company)と混合した。組換え融合タンパク質をSDSバッファー中で煮沸することにより親和性ビーズから溶出した。サンプルのアリコートを10%SDS−PAGEゲルにかけた。ゲルを固定し、コーマッシーブルー(Coomassie blue)で着色した。融合タンパク質の発現を確認した後GST分子からのrcpn10の精製がより大きい規模で行われた。
上述のように細胞を生長、誘導せしめて、細胞ペレットをPBS中に懸濁し、超音波処理し(output level 4,50% duty cycle,2×30 sec)そして細胞溶解質を−30℃で保存した。10リットル細胞培地からの溶媒質を解凍し、rLIFの分離のためにGearingら(Gearingら., 1989, Biotechnology 7, 1157−1161)が用いたと同じ方法によりrcpn10を分離した。直ちに、TritonX−100を最終濃度0.1%になるまで加え、そして細胞切片を遠心分離で除いた(15分、15000rpm、4℃)。10mlのグルタチオン−セファローズ4Bゲル(Pharmacia−LKB Biotechnology)を上清に加え、スラリーを4℃で2時間混合した。ゲルをペレットし、50mlのPBS/0.1% TritonX−100で5回、50mlの0.05M Tris−HCl pH8.0/0.15M NaClで1回および0.05M Tris−HCl pH8.0/0.15M NaCl/2.5mM CaCl2で1回洗った。ゲルを4mlの0.05M トリス−HCl pH8.0/0.15M NaCl /2.5mM CaCl2バッファー中に懸濁し、1000単位のトロンビン(SigmaT6884)を加え、スラリーを振動水浴中、37℃で1時間混合した。ゲルをペレットし、上清を保持し、ゲルを3回4ml0.05M トリス−HCl pH8.0/0.15M NaClで洗った。rcpn10を包有するものである、これらの洗液と上清をプールすると4−5mgの組換えタンパク質が得られた。ゲルと非特異的に結合している追加のrcpn10は次のように回収された。4mlの0.05M Tris−HCl pH8.0/2M NaClを加え、そしてスラリーを4℃で2時間混合した。
ペレットした後、ゲルを3回、2mlのこの0.05M Tris−HCl pH8.0/2M NaCl bufferで洗い、洗液を最初の上清をプールして、さらに約1mgのrcpn10が得られた。タンパク質濃度はLawryらの方法(Lawryら., 1951, J.Biol.Chem.193, 265−275)によって測定された。15%Tris−Tricineゲルを用いてSDS−PAGEによってタンパク質が分析された(Schagger et al., 1987, Anal.Biochem.166, 368−379)。
組換えcpn10は、そのN末端に二つの追加のアミノ酸を有している。組換えタンパク質のN末端は、Gly−Ser−Alaであるのに対し、天然タンパク質のN末端はAc−Alaである。組換えcpn10のアミノ酸配列は、次の通りである:

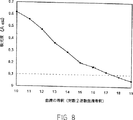
組換えタンパク質に対する抗体は、抗ペプチド抗体を生産する為に記載されたのと同じスケジュールを使用し、ウサギで産生された。各注射に対し、約10μgのタンパク質が使用された。ウサギ血清は、プレートを最初cpn10(5μg/ml)で被覆した以外は、抗ペプチド抗体をスクリーニングするために記載された方法を使用し、ELISAによって抗cpn抗体に対してスクリーニングされた。cpn10に対する及びウサギ♯42の血清中の全融合タンパク質に対する(この場合は、GST:rcpn10,5μg/mlをプレートに結合させた。)抗体(Ab)タイターは、図7に示される。第4回のブースター後にこのウサギから採取されたcpn10に対する血清試料の滴定を図8に説明する。
B. 妊娠の終結
本発明の他の一面において、妊娠患者に対してcpn10特異性抗体を投与することにより、妊娠を終わらせることが出来る。抗体は、cpn10又はその誘導体に対して産生させ得る。これらの抗体の投与は、好ましくは、前着床段階(1〜2細胞段階)又は近着床段階において行われる。抗cpn10抗体による妊娠の終結は、マウスモデル系を例として以下のように証明される。このマウスモデル系は、単なる例示であって、この方法がマウスに限定されるものではない。この方法は、ヒトを含むいかなる適当な哺乳類種に対しても適用されてよい。
(i)CPN10ペプチドに対して産生された抗体による前着床段階での妊娠の終結
抗cpn10抗体
これらの抗体の製法及び特性は、既に記載されたとおりである。これらの実験において、使用された抗体は、N−末端ペプチド(cpnN)及び内部ペプチド(cpnI)に対して製造されたものである;cpnNとcpnIはロゼット阻止試験において活性である。IgGは、抗血清から、45%硫酸アンモニウムによって沈殿したものであり、濃度はLowryとゲル電気泳動によって決定された。IgG製剤は、ウシ血清アルブミンに結合した免疫ペプチドに対するELISAで試験された。製剤はまた、マウス妊娠血清において活性を中和するそれらの活性について試験された。種々の濃度の抗体を等容量のマウス血清と共に培養し、次いでその混合物をロゼット阻止試験における活性について試験した。EPF活性を安全に中和できた抗体の最低濃度を決定した(Cavanagh et al., 1994, Eur.J.Biochem., 222, 551-560参照)。10pgの抗N−ペプチドAbは、妊娠血清1mlの活性を中和したが、4ngの抗I−ペプチドが完全な中和に必要であった。
受動免疫化
成熟非近交系雄および雌カッケンブッシュマウスを、7:30a.m.に対でケージに入れ、8:30p.m.に離した。膣栓のある雌マウスに、妊娠1日目(交配の日)および2日目の9:00a.m.および5:00p.m.に、抗−N−ペプチド/卵白アルブミン、抗−I−ペプチド/卵白アルブミンまたは抗−卵白アルブミンIgG製剤を注射した。2用量レジメにおける特異的IgGの注射量は、約1mg/マウス/日と概算した。7日目に、マウスをCO2で安楽死させ、着床胚および計数した黄体(CL)について子宮を試験した。それぞれの群において、試験IgGで処理したマウスの胚/CLの数を、同量の対照IgGを投与したものの数と比較した(χ2試験)。
結 果
第3表に示した結果は、妊娠血清における活性の中和が妊娠の早期段階において、胎児の生存能力に逆に影響することを明らかに証明する。ロゼット阻止試験においてcpn10活性を中和する抗体の能力は、妊娠に逆に影響するそれらのイン・ビボ能力のイン・ビトロモニターである。
C. ガンと腫瘍
本発明の更に他の局面は、患者に対するcpn10のアンタゴニストの投与による異常細胞の生長の抑制である。上記の異常細胞や正常細胞の異常生長は腫瘍やガンを含むものである;正常細胞の異常生長は、乾癬やレイター氏症候群のような疾病を含む。腫瘍細胞には、良性の生長を示すものと悪性の生長を示すものの両方が含まれる。固形腫瘍や血液ガンのような悪性疾病の細胞も含まれる。腫瘍細胞成長の抑制効果の例は、ネズミB16メラノーマやMCA−2繊維芽肉腫細胞を使用する実験によって証明される。
(i)抗CPN10誘導ペプチド抗体(Abs)のイン・ビトロ腫瘍細胞の成長に対する効果
導 入
次の研究は、イン・ビトロで腫瘍細胞によって生産されたcpn10が当該細胞の継続生長にも必要である可能性を調査したものである。
方 法
細胞培養
セルラインを、標準条件下、基礎培地としてのダルベッコ修飾イーグル培地(DMEM;IC Biochemicals Australia Pty. Ltd., Australia)に10%ウシ胎児血清(FCN,ICN)、20mMグルタミン(ICN)及び抗生物質[100μg/mlストレプトマイシン(ICN)、100U/mlペニシリン(CSL,Melbourne, Australia)]を補充した培地中、37℃において、空気中に5%CO2を含む湿気含有雰囲気下に培養した。
細胞は、成長の対数期に維持した。単層を、血清フリー培地中で洗浄してから、37℃で0.1%w/vトリプシンと0.02%w/vバーセンのカルシウムとマグネシウムフリーの平衡塩溶液に短時間暴露することにより、解離された。トリプシンの作用を2%v/vFCS含有培地の添加によって中和し、細胞を200gに5分間遠心分離して回収し、血清フリー培地で2回洗浄し、その後、培養皿又は96ウエルプレート(NUNC)に接種した。
セルラインのストックは、常に液安中で凍結して保持した。
共培養実験用抗ペプチドAbsの調整
親和性精製抗NAbs、抗IAbs及び抗卵白アルブミン抗体(コントロール抗体)をDMEMに交換し、最終濃度1mg/mlに調節した。この製剤を、0.2μMカットオフフィルター(Minisart, Sartorius Gmbh, Gottingen, Germany)を通過させて、殺菌した。コントロール培地として、DEME単独を同様に処理した。
腫瘍細胞と抗ペプチドAbsとの共培養
ネズミB16メラノーマ及びMCA−2繊維芽肉腫セルラインについて研究した。細胞(103)を、抗体ペプチドAbs又はコントロールAbの62.5〜500μgAb/ml(最終濃度)用量を含む0.2ml培養培地(DMEM+10%FCS(熱不活化))中に、3回接種した。細胞は、同様に、抗体を含まない濾過培地に接種した。培地を96時間培養後、試験した。生存度をトリパンブルー排除で評価し、メチル−[3H]チミジン5’−トリホスフェート([3H]チミジン;Amersham International, Amersham, UK)の取り込みを細胞分割速度をモニターするために使用した。各抗体用量に対する相対的[3H]チミジンの取り込みは、配合した平均cpm(3倍のウエルから)を抗体不含有ウエル中に配合された平均cpmの百分率として表すことにより計算した。
細胞生存能の決定
トリプシンによって解離された細胞を等容量の0.1%w/vトリパンブルーPBS溶液と混合し、ヘキサイトメーター(haemocytometer)上に広げた。細胞生存能は、染料除外細胞の百分率として計算した。
[3H]チミジン取り込みの決定
80時間培養後、細胞をさらに16時間、ウエル当たり0.5μCi[3H]チミジンと共に培養した。培養後、粘着性細胞の上澄培地を除去し、各ウエルを温DMEMで2回にわたり洗浄した。酸沈殿性物質を、各ウエルに対して250μl氷冷5%w/v三塩化酢酸(TCA,BDH Chemicals, Australia Pty Led. Kilsyth, Victoria, Australia)を添加することにより、分離した(Plate, 1974, J.Exp.Med., 139, 851−861)。沈殿物をTCAで2回にわたり洗浄し、0.3ml 0.25N NaOH中で溶解化し、この調製物250μlを2のmlシンチレーションカクテル(Emulsifier safe, Packard Instruments Co., Meriden, CT, USA)と混合し、酸沈殿性物質中に配合されたcpmを各ウエルにつきベータ計数によって決定した。
免疫細胞化学
ヒトT細胞白血病細胞Molt4(ATCC CRL1582)を、RPMI+10%FCS中で対数期に維持した。細胞をRPMI+10%FCS中で3回にわたって洗浄し、10μg(0.1ml中)の親和性精製抗NペプチドAb、抗IペプチドAbまたはコントロール抗体(抗卵白アルブミンAb)と共に培養した(106細胞)。コントロール試験は、106正常脾臓細胞を含んでいた。結合抗体を抗ウサギ、ビオチニル化IgG、F(ab’2)断片(Armsham)、次いで製造業者の指示書に従ってストレプタビジン−フルオレスセイン処理を行い、検出した。結合は、UV顕微鏡により可視化された。
結果
腫瘍細胞成長は、抗cpn10誘導ペプチドAbsとの共培養によって行う。
抗ペプチドAbsの濃度を上昇させて行ったB16メラノーマとMCA−2繊維芽肉腫細胞の培養は、96時間後、細胞分割の顕著な減少と細胞死滅の増加をもたらした(図9a、9b、10a、10b)。コントロールAbの類似の濃度における細胞培養は、効果がなかった(図9c、10c)。
抗Iペプチド抗体は、Molt4細胞の表面中でcpn10に結合した。Molt4細胞上において、抗Nペプチドまたは抗卵白アルブミン抗体により、又は正常な脾臓細胞上において、上記抗体のいずれかにより、結合は検出されなかった。これは、細胞外cpn10の最初の可視図である(図11)。
結論
ここに記載された研究により、抗cpn10誘導ペプチドAbsは腫瘍細胞の成長を阻止することが確立された。抗ペプチドAbsの用量を増加された場合の培養B16およびMCA−2セルラインの抗増殖効果は、それらの細胞の成長がcpn10の継続的存在に依存するものであることの証明である。これらの研究は、腫瘍細胞がイン・ビトロ増殖のためにcpn10を必要とすることを確証した。
発明の他の局面
上記N末端フラグメントおよび内部フラグメントは、ロゼット阻止試験において活性であり、従って活性中心として機能する分子の領域である。
薬理学的アンタゴニストは、常套手段により、それら活性中心の構造を修飾し、標的部位、たとえば腫瘍細胞に対する結合が標的を賦活させることなく生ずるように、構成することができる。腫瘍細胞上の全体のcpn10分子の作用を干渉することによって、これらのアンタゴニストは抗cpn10抗体に対して記述した抗増殖効果と同様な効果を示す。
本発明は、またその範囲に、次の工程を含むことを特徴とする、試料中の抗cpn10抗体を測定する方法を有する:(1)本質的に精製したcpn10を当該試料と反応させること、(2)抗体とcpn10の間の結合を決定することによって当該試料中の抗cpn10抗体の量を決定すること。
本発明は、また上記のことから、抗EPF抗体がマウスモデルの腫瘍の成長を抑制するのに有用であると言うデータが存在することを承知するであろう(たとえば、Quinn et al., 1992, Cancer Immunol. Immunother. 34, 265-271)。このようなデータは、抗EPF抗体がイン・ビボまたはイン・ビトロにおける腫瘍の成長を抑制するであろうと言う主張を指示するものである。
アンタゴニストまたは抗体の投与において利用される投与量は、アンタゴニストに対しては体重のkg当たり1〜1,000(好ましくは50〜200)μgの範囲であり、抗体に対しては体重のkg当たり1〜1,000(好ましくは50〜200)mgの範囲である。これらの投与量はcpn10と同じ分子量を有する分子に基づくものであり、従って用量を適宜に調節すべきである。
C. ガンと腫瘍
本発明の更に他の局面は、患者に対するcpn10のアンタゴニストの投与による異常細胞の生長の抑制である。上記の異常細胞や正常細胞の異常生長は腫瘍やガンを含むものである;正常細胞の異常生長は、乾癬やレイター氏症候群のような疾病を含む。腫瘍細胞には、良性の生長を示すものと悪性の生長を示すものの両方が含まれる。固形腫瘍や血液ガンのような悪性疾病の細胞も含まれる。腫瘍細胞成長の抑制効果の例は、ネズミB16メラノーマやMCA−2繊維芽肉腫細胞を使用する実験によって証明される。
(i)抗CPN10誘導ペプチド抗体(Abs)のイン・ビトロ腫瘍細胞の成長に対する効果
導 入
次の研究は、イン・ビトロで腫瘍細胞によって生産されたcpn10が当該細胞の継続生長にも必要である可能性を調査したものである。
方 法
細胞培養
セルラインを、標準条件下、基礎培地としてのダルベッコ修飾イーグル培地(DMEM;IC Biochemicals Australia Pty. Ltd., Australia)に10%ウシ胎児血清(FCN,ICN)、20mMグルタミン(ICN)及び抗生物質[100μg/mlストレプトマイシン(ICN)、100U/mlペニシリン(CSL,Melbourne, Australia)]を補充した培地中、37℃において、空気中に5%CO2を含む湿気含有雰囲気下に培養した。
細胞は、成長の対数期に維持した。単層を、血清フリー培地中で洗浄してから、37℃で0.1%w/vトリプシンと0.02%w/vバーセンのカルシウムとマグネシウムフリーの平衡塩溶液に短時間暴露することにより、解離させた。トリプシンの作用を2%v/vFCS含有培地の添加によって中和し、細胞を200gに5分間遠心分離して回収し、血清フリー培地で2回洗浄し、その後、培養皿又は96ウエルプレート(NUNC)に接種した。
セルラインのストックは、常に液安中で凍結して保持した。
共培養実験用抗ペプチドAbsの調製
親和性精製抗NAbs、抗IAbs及び抗卵白アルキレン抗体(コントロール抗体)をDMEMに交換し、最終濃度1mg/mlに調節した。この製剤を、0.2μMカットオフフィルター(Minisart, Sartorius Gmbh, Gottingen, Germany)を通過させて、殺菌した。コントロール培地として、DEME単独を同様に処理した。
腫瘍細胞と抗ペプチドAbsとの共培養
ネズミB16メラノーマ及びMCA−2繊維芽肉腫セルラインについて研究した。細胞(103)を、抗体ペプチドAbs又はコントロールAbの62.5〜500μgAb/ml(最終濃度)用量を含む0.2ml培養培地(DMRM+10%FCS(熱不活化))中に、3回接種した。細胞は、同様に、抗体を含まない濾過培地に接種した。培地を96時間培養後、試験した。生存度をトリパンブルー排除で評価し、メチル−[3H]チミジン5’−トリホスフェート([3H]チミジン;Amersham International, Amersham, UK)の取り込みを細胞分割速度をモニターするために使用した。各抗体用量に対する相対的[3H]チミジンの取り込みは、配合した平均cpm(3倍のウエルから)を抗体不含有ウエル中に配合された平均cpmの百分率として表すことにより計算した。
細胞生存能の決定
トリプシンによって解離された細胞を等容量の0.1%w/vトリパンブルーPBS溶液と混合し、ヘモサイトメーター(haemocytometer)上に広げた。細胞生存能は、染料除外細胞の百分率として計算した。
[3H]チミジン取り込みの決定
80時間培養後、細胞をさらに16時間、ウエル当たり0.5μCi[3H]チミジンと共に培養した。培養後、粘着性細胞の上澄培地を除去し、各ウエルを温DMEMで2回にわたり洗浄した。酸沈殿性物質を、各ウエルに対して250μl氷冷5%w/v三塩化酢酸(TCA,BDH Chemicals, Australia Pty Ltd. Kilsyth, Victoria, Australia)を添加することにより、分離した(Plate, 1974, J.Exp.Med., 139, 851−861)。沈殿物をTCAで2回にわたり洗浄し、0.3ml 0.25N NaOH中で溶解化し、この調製物250μlを2のmlシンチレーションカクテル(Emulsifier safe, Packard Instruments Co., Meriden, CT, USA)と混合し、酸沈殿性物質中に配合されたcpmを各ウエルにつきベータ計数によって決定した。
免疫細胞化学
ヒトT細胞白血病細胞Molt4(ATCC CRL1582)を、RPMI+10%FCS中で対数期に維持した。細胞をRPMI+10%FCS中で3回にわたって洗浄し、10μg(0.1ml中)の親和性精製抗NペプチドAb、抗IペプチドAbまたはコントロール抗体(抗卵白アルブミンAb)と共に培養した(106細胞)。コントロール試験は、106正常脾臓細胞を含んでいた。結合抗体を抗ウサギ、ビオチニル化IgG、F(ab’2)断片(Armsham)、次いで製造業者の指示書に従ってストレプタビジンーフルオレスセイン処理を行い、検出した。結合は、UV顕微鏡により可視化された。
結果
腫瘍細胞成長は、抗cpn10誘導ペプチドAbsとの共培養によって行う。
抗ペプチドAbsの濃度を上昇させて行ったB16メラノーマとMCA−2繊維芽肉腫細胞の培養は、96時間後、細胞分割の顕著な減少と細胞死滅の増加をもたらした(図9a、9b、10a、10b)。コントロールAbの類似の濃度における細胞培養は、効果がなかった(図9c、10c)。
抗Iペプチド抗体は、Molt4細胞の表面中でcpn10に結合した。Molt4細胞上において、抗Nペプチドまたは抗卵白アルブミン抗体により、又は正常な脾臓細胞上において、上記抗体のいずれかにより、結合は検出されなかった。これは、細胞外cpn10の最初の可視図である(図11)。
結論
ここに記載された研究により、抗cpn10誘導ペプチドAbsは腫瘍細胞の成長を阻止することが確立された。抗ペプチドAbsの用量を増加させた場合の培養B16およびMCA−2セルラインの抗増殖効果は、それらの細胞の成長がcpn10の継続的存在に依存するものであることの証明である。これらの研究は、腫瘍細胞がイン・ビトロ増殖のためにcpn10を必要とすることを確証した。
発明の他の局面
上記N末端フラグメントおよび内部フラグメントは、ロゼット阻止試験において活性であり、従って活性中心として機能する分子の領域である。
薬理学的アンタゴニストは、常套手段により、それら活性中心の構造を修飾し、標的部位、たとえば腫瘍細胞に対する結合が標的を賦活させることなく生ずるように、構成することができる。腫瘍細胞上の全体のcpn10分子の作用を干渉することによって、これらのアンタゴニストは抗cpn10抗体に対して記述した抗増殖効果と同様な効果を示す。
本発明は、またその範囲に、次に工程を含むことを特徴とする、試料中の抗cpn10抗体を測定する方法を有する:(1)本質的に精製したcpn10を当該試料と反応させること、(2)抗体とcpn10の間の結合を決定することによって当該試料中の抗cpn10抗体の量を決定すること。
本発明は、また上記のことから、抗EPF抗体がマウスモデルの腫瘍の成長を抑制するのに有用であると言うデータが存在することを承知するであろう(たとえば、Quinn et al., 1992, Cancer Immunol. Immunother. 34, 265-271)。このようなデータは、抗EPF抗体がイン・ビボまたはイン・ビトロにおける腫瘍の成長を抑制するであろうと言う主張を指示するものである。
アンタゴニストまたは抗体の投与において利用される投与量は、アンタゴニストに対しては体重のkg当たり1〜1,000(好ましくは50〜200)μgの範囲であり、抗体に対しては体重のkg当たり1〜1,000(好ましくは50〜200)mgの範囲である。これらの投与量はcpn10と同じ分子量を有する分子に基づくものであり、従って用量を適宜に調節すべきである。
表1
ウサギに投与した抗体調製物の注射当たりの抗原の量
表2
親和性精製抗−cpn10ペプチド抗体および対照抗−卵白アルブミン抗体のタイター。
表3
確認交配マウスに、cpn10由来ペプチドに対するを抗体1日および2日目のp.c.で受動免疫化した、p.c.7日目の着床胚および黄体の数における効果。
*(ヘテロセダスティックt−検定)
図面の説明
図1a
EPFの精製。熱抽出ヒト血小板(100単位)をSP−セファデックスおよびヘパリンセファロースで分画し、次いで、TSK−フェニル5PWカラムにかけ、逆塩勾配で溶出した。フラクションはロゼット阻害試験で試験した(免疫抑制抗リンパ球血清のロゼット阻害活性を増大させるEPFの能力を基本にして)。
図1b
(a)の活性フラクション
図1c
(b)の活性フラクション
図1d
(c)の活性フラクション
図1e
(d)由来の活性フラクションを伴う固定化モノクローナル抗EPF抗体5/341とヒト妊娠血清、妊娠6日(10ml);ヒト妊娠尿、妊娠1カ月まで(10リットル);プロラクチンで刺激した発情マウス卵巣(100)により条件付した培地とマウス胚条件付培地(卵巣CM);ウシ腎臓細胞系MDBK(MDBK−CM;ATCC CCL22、10リットル)により条件付した無血清培地;部分的肝切除後24時間に得たラット血清(ポスト−pH、10ml);ポスト−pH24時間のラット肝臓(40g);(a)から(d)の全フラクションの等価フラクションの相互作用。抗EPF結合および非結合フラクションは、ロゼット阻害試験で試験し、活性が結合しない無関係抗体を使用した平衡実験と比較することにより特異性を証明した。
図2a
図1Aのように300単位ヒト血小板から精製したEPFの分析。モノマーサイズの決定。ヨウ素化EPFをSDS−PAGE29で分画し、ゲルスライス(2mm広さのスライス)し、放射活性および生物活性の分布を比較した。(挿入図)同様の調製物における直接クマーシーブルー染色。
図2b
EPFのイオン噴霧マススペクトル、多重プロトン化分子イオンとして表示。(挿入図)分子質量のコンピューター復元。
図2c
ラットcpn10(下線)と比較した、ヒトEPF由来のペプチドのアミノ酸配列(1文字標記)。EPFをエンドプロテイナーゼlys Cおよびエンドプロテイナーゼglu Cで消化し、残ったペプチドをRP−HPLCにより分離し、配列決定した。この個々のフラグメントの配列を示す;74−101以外は、lys消化由来である。
図3
EPFとcpn60(groEL)の相互作用。
図3a
cpn60−EPF混合物+Mg2+ATPをかけた後のTSK G3000SWゲル透過カラムの除去量におけるピークフラクションをSDS−PAGE(Schaggerら, 1987)で分析し、銀染色した(Morrissey, 1981)。左レーン +ATP、右レーン −ATP。(cpn60は14量体、M 840000、カラム溶出限界>300000。SDSゲルの高い方のMrバンドはgroELの多量体形である)。
図3b
固定化cpn60をヒト妊娠血清(妊娠6日目)とMg2+ATP存在下または非存在下で混合した。非結合および結合フラクション(後者はゲルから、EDTAによるATPの除去により回収した)を、次いで、ロゼット阻害試験で試験した。結果は限界量、ロゼット阻害試験で陽性結果となるサンプルの最も高い希釈として示す。
図4a,4b、4c
cpn10−ペプチド/卵白アルブミン結合体に対するウサギ抗体。抗体を免疫抗原に対するELISAで試験した。
図5
抗N−ペプチド、抗EPF♯816、抗EPF♯810及びコントロール抗卵白アルブミン抗体(100ng/ml)を□N−ペプチド(5μg/ml)及び EPF/cpn10(1μg/ml)に対するELISAで試験した。結合IgGは、基質としてo−フェニレンジアミンを使用するビオチンーストレプトビジン系(Amersham)により、検出した。吸光度は492nmで読み取った。
図6a
pRM1
図6b
pRM2
図6c
pRM3
図7
cpn10に対する抗体の調製。融合蛋白質(GST:rcpn10)
図8
ELISAによるウサギ血清中の抗cpn10抗体の検出。血清は抗原の第4回の追加免疫後採取された。
図9
抗cpn10誘導ペプチド抗体と共に96時間培養後、B16メラノーマ細胞の相対的[3H]チミジン取り込み(−■−)及び生存能(--□--)。増殖は、抗体と共に培養した細胞中への抗[3H]チミジンの取り込みによって評価し、抗体なしで培養された[3H]チミジンの百分率として表した。
*p<0.05,**p<0.01,***p<0.001,(学生のtテスト)n=3
図10
抗cpn10誘導ペプチド抗体を共に96時間培養後、MCA−2繊維芽肉腫の相対的[3H]チミジン取り込み(−■−)及び生存能(--□--)。増殖は、抗体と共に培養した細胞中への抗[3H]チミジンの取り込みによって評価し、抗体なしで培養された[3H]チミジンの百分率として表した。
*p<0.05,**p<0.01,***p<0.001,(学生のtテスト)n=3
図11
抗cpn10 I−ペプチドAbsは、ヒトMolt4白血病細胞の表面上のcpn10を検出する。
Claims (8)
- EPF/cpn10を認識する抗体の製造方法であって、下記の工程:
−ペプチドAc−AGQAFRKFLPLC、AGQAFRKFLPLCまたはEKSQGKVLQATCで、ヒト以外の宿主を免疫すること、および
−得られる抗体を取得すること、
を含む方法。 - EPF/cpn10を認識する抗体の製造方法であって、下記の工程:
−次の(i)−(iii)よりなる群から選ばれるペプチドで、ヒト以外の宿主を免疫すること
(i)AGQAFRKFLPL
(ii)Ac−AGQAFRKFLPL
(iii)EKSQGKVLQAT、および
−得られる抗体を取得すること、
を含む方法。 - 次のいずれかのアミノ酸配列で表されるペプチドに対して産生された抗体:
Ac−AGQAFRKFLPLC、AGQAFRKFLPLCまたはEKSQGKVLQATC。 - 次のいずれかのペプチドに対して産生された抗体:
(i)AGQAFRKFLPL
(ii)Ac−AGQAFRKFLPL
(iii)EKSQGKVLQAT。 - 細胞成長を抑制するか、または免疫学的活性を増強するための、下記からなる群から選ばれる抗体:
(a)ペプチドAc−AGQAFRKFLPLC、AGQAFRKFLPLCまたはEKSQGKVLQATCに対して産生された抗体、
(b)下記のペプチドに対して産生された抗体:
(i)AGQAFRKFLPL
(ii)Ac−AGQAFRKFLPL
(iii)EKSQGKVLQAT。 - 妊娠を終了させるための、請求項5記載の抗体。
- 腫瘍細胞の成長を抑制させるための、請求項5記載の抗体。
- 細胞が白血球である、請求項7記載の抗体。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AUPM270593 | 1993-11-30 | ||
| AU8234 | 1994-09-16 | ||
| AU2705 | 1994-09-16 | ||
| AUPM8234A AUPM823494A0 (en) | 1994-09-16 | 1994-09-16 | Diagnostic and therapeutic uses of chaperonin 10 |
| PCT/AU1994/000742 WO1995015339A1 (en) | 1993-11-30 | 1994-11-30 | Antagonists to chaperonin 10 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPH09505811A JPH09505811A (ja) | 1997-06-10 |
| JP3987104B2 true JP3987104B2 (ja) | 2007-10-03 |
Family
ID=25644587
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP51529495A Expired - Fee Related JP3545408B2 (ja) | 1993-11-30 | 1994-11-30 | シャペロニン10 |
| JP51529695A Expired - Fee Related JP3987104B2 (ja) | 1993-11-30 | 1994-11-30 | シャペロニン10のアンタゴニスト |
| JP2003319869A Expired - Fee Related JP3986481B2 (ja) | 1993-11-30 | 2003-09-11 | シャペロニン10 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP51529495A Expired - Fee Related JP3545408B2 (ja) | 1993-11-30 | 1994-11-30 | シャペロニン10 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2003319869A Expired - Fee Related JP3986481B2 (ja) | 1993-11-30 | 2003-09-11 | シャペロニン10 |
Country Status (11)
| Country | Link |
|---|---|
| US (3) | US6113899A (ja) |
| EP (2) | EP0731811B1 (ja) |
| JP (3) | JP3545408B2 (ja) |
| AT (1) | ATE282634T1 (ja) |
| AU (2) | AU684577B2 (ja) |
| CA (1) | CA2177179C (ja) |
| DE (1) | DE69434137T2 (ja) |
| ES (1) | ES2233938T3 (ja) |
| NZ (2) | NZ276670A (ja) |
| PT (1) | PT731811E (ja) |
| WO (2) | WO1995015338A1 (ja) |
Families Citing this family (17)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1995015338A1 (en) * | 1993-11-30 | 1995-06-08 | The University Of Queensland | Chaperonin 10 |
| JP2001509396A (ja) * | 1997-07-10 | 2001-07-24 | メディカル リサーチ カウンシル | シャペロン断片 |
| US6007821A (en) * | 1997-10-16 | 1999-12-28 | Fordham University | Method and compositions for the treatment of autoimmune disease using heat shock proteins |
| US6475490B1 (en) | 1998-10-19 | 2002-11-05 | Fordham University | Compositions and methods for promoting tissue repair using heat shock proteins |
| AUPP823999A0 (en) * | 1999-01-20 | 1999-02-11 | University Of Queensland, The | A treatment |
| GB0028122D0 (en) * | 2000-11-17 | 2001-01-03 | St Georges Entpr Ltd | Method |
| JP4326326B2 (ja) * | 2001-06-27 | 2009-09-02 | ジヤンセン・フアーマシユーチカ・ナームローゼ・フエンノートシヤツプ | Epf受容体アッセイ、化合物および治療用組成物 |
| AU2002952492A0 (en) | 2002-11-06 | 2002-11-21 | Cbio Limited | Chaperonin 10 immunosuppression |
| GB0226105D0 (en) * | 2002-11-08 | 2002-12-18 | St Georges S Entpr Ltd | Pain relief agents |
| CA2553325A1 (en) | 2004-01-16 | 2005-07-28 | Cbio Limited | Chaperonin 10 modulation of toll-like receptor-inducible cytokine and chemokine secretion |
| US7977458B2 (en) * | 2005-08-31 | 2011-07-12 | Cbio Limited | Modified chaperonin 10 |
| AU2006303826B2 (en) * | 2005-10-20 | 2012-07-19 | Cbio Limited | Treatment of hypersensitivity |
| BRPI0617681A2 (pt) * | 2005-10-20 | 2011-08-02 | Cbio Ltd | tratamento de hipersensibilidade |
| EP2007409A4 (en) * | 2006-03-02 | 2012-04-25 | Cbio Ltd | REGULATION OF IMMUNE RESPONSES BY MODULATING THE FUNCTION OF CELLS HAVING ANTIGENS |
| CA2720617A1 (en) | 2008-04-11 | 2009-10-15 | Cbio Limited | Modified cpn10 and prr signalling |
| SG10201505023TA (en) * | 2009-10-09 | 2015-07-30 | Cbio Ltd | Chaperonin 10 Variants |
| US20110243921A1 (en) * | 2010-03-30 | 2011-10-06 | Cbio Limited | Prevention and treatment of cutaneous lupus erythematosus |
Family Cites Families (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0262119A1 (en) * | 1985-03-12 | 1988-04-06 | University Of Queensland | Method for detecting early pregnancy factor (epf) in mammals, purifying epf and method for producing a monoclonal antibody |
| AU599021B2 (en) * | 1985-03-12 | 1990-07-12 | University Of Queensland, The | Method for detecting early pregnancy factor (epf) in mammals,purifying epf and method for producing a monoclonal antibody |
| JPH02501160A (ja) * | 1986-12-22 | 1990-04-19 | ユニバーシティー オブ クイーンスランド | 早期妊娠因子(epf)に対する抗体を使用する哺乳動物の治療方法 |
| AU1086988A (en) * | 1986-12-22 | 1988-07-15 | University Of Queensland, The | Method of treatment of mammals using antibodies to early pregnancy factor (epf) |
| WO1995015338A1 (en) * | 1993-11-30 | 1995-06-08 | The University Of Queensland | Chaperonin 10 |
-
1994
- 1994-11-30 WO PCT/AU1994/000740 patent/WO1995015338A1/en active IP Right Grant
- 1994-11-30 EP EP95902003A patent/EP0731811B1/en not_active Expired - Lifetime
- 1994-11-30 JP JP51529495A patent/JP3545408B2/ja not_active Expired - Fee Related
- 1994-11-30 DE DE69434137T patent/DE69434137T2/de not_active Expired - Lifetime
- 1994-11-30 ES ES95902003T patent/ES2233938T3/es not_active Expired - Lifetime
- 1994-11-30 JP JP51529695A patent/JP3987104B2/ja not_active Expired - Fee Related
- 1994-11-30 AT AT95902003T patent/ATE282634T1/de not_active IP Right Cessation
- 1994-11-30 NZ NZ276670A patent/NZ276670A/en not_active IP Right Cessation
- 1994-11-30 PT PT95902003T patent/PT731811E/pt unknown
- 1994-11-30 CA CA002177179A patent/CA2177179C/en not_active Expired - Fee Related
- 1994-11-30 EP EP95902005A patent/EP0733071A4/en not_active Withdrawn
- 1994-11-30 AU AU11034/95A patent/AU684577B2/en not_active Ceased
- 1994-11-30 AU AU11036/95A patent/AU684578B2/en not_active Ceased
- 1994-11-30 WO PCT/AU1994/000742 patent/WO1995015339A1/en not_active Application Discontinuation
- 1994-11-30 NZ NZ276672A patent/NZ276672A/en not_active IP Right Cessation
-
1996
- 1996-05-29 US US08/654,618 patent/US6113899A/en not_active Expired - Lifetime
-
1999
- 1999-02-23 US US09/255,703 patent/US6417334B1/en not_active Expired - Fee Related
-
2002
- 2002-05-21 US US10/151,006 patent/US7358329B2/en not_active Expired - Fee Related
-
2003
- 2003-09-11 JP JP2003319869A patent/JP3986481B2/ja not_active Expired - Fee Related
Also Published As
| Publication number | Publication date |
|---|---|
| EP0733071A4 (en) | 1997-02-26 |
| US20030175280A1 (en) | 2003-09-18 |
| JP2004067698A (ja) | 2004-03-04 |
| US6417334B1 (en) | 2002-07-09 |
| JP3545408B2 (ja) | 2004-07-21 |
| NZ276670A (en) | 1998-01-26 |
| DE69434137D1 (en) | 2004-12-23 |
| US6113899A (en) | 2000-09-05 |
| JPH09506431A (ja) | 1997-06-24 |
| AU684578B2 (en) | 1997-12-18 |
| CA2177179C (en) | 2006-10-17 |
| ES2233938T3 (es) | 2005-06-16 |
| US7358329B2 (en) | 2008-04-15 |
| EP0733071A1 (en) | 1996-09-25 |
| WO1995015339A1 (en) | 1995-06-08 |
| AU1103495A (en) | 1995-06-19 |
| JP3986481B2 (ja) | 2007-10-03 |
| CA2177179A1 (en) | 1995-06-08 |
| AU1103695A (en) | 1995-06-19 |
| ATE282634T1 (de) | 2004-12-15 |
| DE69434137T2 (de) | 2005-12-01 |
| WO1995015338A1 (en) | 1995-06-08 |
| EP0731811A1 (en) | 1996-09-18 |
| AU684577B2 (en) | 1997-12-18 |
| EP0731811A4 (en) | 2003-05-28 |
| PT731811E (pt) | 2005-04-29 |
| NZ276672A (en) | 1998-01-26 |
| EP0731811B1 (en) | 2004-11-17 |
| JPH09505811A (ja) | 1997-06-10 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP3987104B2 (ja) | シャペロニン10のアンタゴニスト | |
| Auron et al. | Studies on the molecular nature of human interleukin 1. | |
| Husby et al. | Amyloid fibril protein subunit,‘protein AS’: distribution in tissue and serum in different clinical types of amyloidosis including that associated with myelomatosis and Waldenström's macroglobulinaemia | |
| US5731167A (en) | Autotaxin: motility stimulating protein useful in cancer diagnosis and therapy | |
| JPS63502271A (ja) | 天然のコロニ−促進因子‐1の精製 | |
| IBRAHIM et al. | Reproductive tract secretions and bull spermatozoa contain different clusterin isoforms that cluster cells and inhibit complement‐induced cytolysis | |
| Koshland et al. | Differences in amino acid composition related to allotypic and antibody specificity of rabbit IgG heavy chains | |
| CZ6997A3 (en) | Proteins, extractable from mammal livers | |
| CA2176948C (en) | Chaperonin 10 | |
| US5434247A (en) | Peptides for inducing monocyte cytotoxicity in diagnostics | |
| JP2000515001A (ja) | ダウンレギュレーション耐性c3コンバターゼ | |
| US7939635B2 (en) | Autotaxin: motility stimulating protein useful in cancer diagnosis and therapy | |
| Sarfati et al. | T-cell-derived IgE-binding factors. II. Purification and characterization of IgE-binding factors produced by human T cell leukemia/lymphoma virus-1-transformed T lymphocytes. | |
| JPH04505614A (ja) | タンパク質複合体 | |
| JPH03504248A (ja) | 外来抗原に対する寛容の誘発 | |
| Teuscher et al. | Experimental allergic orchitis: the isolation and partial characterization of an aspermatogenic polypeptide (AP3) with an apparent sequential disease-inducing determinant (s). | |
| EP0515494A1 (en) | Use of inhibitor of cytokine activity | |
| US6387875B1 (en) | Method of use for murine leukaemia inhibitory factor-binding protein (mLBP) | |
| Percy et al. | Functional Differentiation of B Lymphocytes in Congenital Agammaglobulinemia: II. Immunochemical Analysis of the in Vitro Primary Immune Response | |
| JPH03502446A (ja) | 動物およびヒトのレクチン類の少なくとも一部分の配列を再現するアミノ酸配列,それを得る方法,およびその診断および治療への用途 | |
| WO1993016715A1 (en) | Fsf-1 and the early detection of fibrosis | |
| JP2608023B2 (ja) | インターロイキン−1β誘導体及び医薬 | |
| MAIRE et al. | FUNCTIONAL DIFFERENTIATION OF B LYMPHOCYTES IN CONGENITAL AGAMMAGLOBULINEMIA | |
| JPH08253422A (ja) | インターロイキン−1β誘導体及び医薬 | |
| JPH08225596A (ja) | インターロイキン−1β誘導体及び医薬 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20040210 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20040329 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20040416 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20040615 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20040914 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20050111 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20050311 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20060131 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20060428 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20060619 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20060726 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20070612 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20070712 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20100720 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20100720 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110720 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20110720 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20120720 Year of fee payment: 5 |
|
| LAPS | Cancellation because of no payment of annual fees |