JP2014508759A - シアリル化抗体の生産の方法 - Google Patents
シアリル化抗体の生産の方法 Download PDFInfo
- Publication number
- JP2014508759A JP2014508759A JP2013554892A JP2013554892A JP2014508759A JP 2014508759 A JP2014508759 A JP 2014508759A JP 2013554892 A JP2013554892 A JP 2013554892A JP 2013554892 A JP2013554892 A JP 2013554892A JP 2014508759 A JP2014508759 A JP 2014508759A
- Authority
- JP
- Japan
- Prior art keywords
- antibody
- seq
- domain
- sequence
- antiabeta
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 238000000034 method Methods 0.000 title claims abstract description 65
- 230000035772 mutation Effects 0.000 claims abstract description 54
- 108090000141 Sialyltransferases Proteins 0.000 claims abstract description 47
- 102000003838 Sialyltransferases Human genes 0.000 claims abstract description 46
- 125000005629 sialic acid group Chemical group 0.000 claims abstract description 34
- 150000002482 oligosaccharides Chemical class 0.000 claims abstract description 19
- 230000000694 effects Effects 0.000 claims abstract description 18
- 229920001542 oligosaccharide Polymers 0.000 claims abstract description 17
- 102000040430 polynucleotide Human genes 0.000 claims description 53
- 108091033319 polynucleotide Proteins 0.000 claims description 53
- 239000002157 polynucleotide Substances 0.000 claims description 53
- 241000282414 Homo sapiens Species 0.000 claims description 43
- 239000013598 vector Substances 0.000 claims description 41
- 235000001014 amino acid Nutrition 0.000 claims description 32
- 150000001413 amino acids Chemical class 0.000 claims description 27
- 239000000203 mixture Substances 0.000 claims description 25
- 238000006467 substitution reaction Methods 0.000 claims description 17
- 108010046068 N-Acetyllactosamine Synthase Proteins 0.000 claims description 10
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 claims description 8
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 claims description 5
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 claims description 5
- 239000003814 drug Substances 0.000 claims description 5
- 239000008194 pharmaceutical composition Substances 0.000 claims description 5
- 239000004471 Glycine Substances 0.000 claims description 4
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 claims description 4
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 claims description 4
- 239000004472 Lysine Substances 0.000 claims description 4
- 235000004279 alanine Nutrition 0.000 claims description 4
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 claims description 2
- 238000004519 manufacturing process Methods 0.000 abstract description 24
- 108060003306 Galactosyltransferase Proteins 0.000 abstract description 4
- 102000030902 Galactosyltransferase Human genes 0.000 abstract description 4
- 210000004027 cell Anatomy 0.000 description 84
- 125000003275 alpha amino acid group Chemical group 0.000 description 62
- 239000013612 plasmid Substances 0.000 description 47
- 229940027941 immunoglobulin g Drugs 0.000 description 46
- 230000014509 gene expression Effects 0.000 description 45
- 150000007523 nucleic acids Chemical group 0.000 description 35
- 108090000623 proteins and genes Proteins 0.000 description 34
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 31
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 27
- 108091028043 Nucleic acid sequence Proteins 0.000 description 25
- 239000002773 nucleotide Substances 0.000 description 24
- 125000003729 nucleotide group Chemical group 0.000 description 24
- 108090000765 processed proteins & peptides Proteins 0.000 description 24
- 230000009450 sialylation Effects 0.000 description 24
- 239000013613 expression plasmid Substances 0.000 description 23
- 108090001090 Lectins Proteins 0.000 description 22
- 102000004856 Lectins Human genes 0.000 description 22
- 229940024606 amino acid Drugs 0.000 description 22
- 239000002523 lectin Substances 0.000 description 22
- 238000004949 mass spectrometry Methods 0.000 description 19
- 235000018102 proteins Nutrition 0.000 description 19
- 102000004169 proteins and genes Human genes 0.000 description 19
- 230000006870 function Effects 0.000 description 15
- 102000004196 processed proteins & peptides Human genes 0.000 description 15
- 102000004190 Enzymes Human genes 0.000 description 14
- 108090000790 Enzymes Proteins 0.000 description 14
- 108700023372 Glycosyltransferases Proteins 0.000 description 14
- 229940088598 enzyme Drugs 0.000 description 14
- 230000009257 reactivity Effects 0.000 description 14
- 208000024827 Alzheimer disease Diseases 0.000 description 13
- 241001529936 Murinae Species 0.000 description 13
- 125000000539 amino acid group Chemical group 0.000 description 13
- 239000012636 effector Substances 0.000 description 13
- 229930182830 galactose Natural products 0.000 description 13
- 230000001225 therapeutic effect Effects 0.000 description 13
- 208000037259 Amyloid Plaque Diseases 0.000 description 12
- 239000000427 antigen Substances 0.000 description 12
- 102000036639 antigens Human genes 0.000 description 12
- 108091007433 antigens Proteins 0.000 description 12
- 239000013604 expression vector Substances 0.000 description 12
- 108060003951 Immunoglobulin Proteins 0.000 description 11
- 102000018358 immunoglobulin Human genes 0.000 description 11
- 238000011282 treatment Methods 0.000 description 11
- OVRNDRQMDRJTHS-UHFFFAOYSA-N N-acelyl-D-glucosamine Natural products CC(=O)NC1C(O)OC(CO)C(O)C1O OVRNDRQMDRJTHS-UHFFFAOYSA-N 0.000 description 10
- MBLBDJOUHNCFQT-LXGUWJNJSA-N N-acetylglucosamine Natural products CC(=O)N[C@@H](C=O)[C@@H](O)[C@H](O)[C@H](O)CO MBLBDJOUHNCFQT-LXGUWJNJSA-N 0.000 description 10
- 230000003110 anti-inflammatory effect Effects 0.000 description 10
- 229920001184 polypeptide Polymers 0.000 description 10
- 238000001228 spectrum Methods 0.000 description 10
- 108020004414 DNA Proteins 0.000 description 9
- SQVRNKJHWKZAKO-PFQGKNLYSA-N N-acetyl-beta-neuraminic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)O[C@H]1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-PFQGKNLYSA-N 0.000 description 9
- 201000010099 disease Diseases 0.000 description 9
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 9
- 102000051366 Glycosyltransferases Human genes 0.000 description 8
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 8
- 150000001720 carbohydrates Chemical group 0.000 description 8
- 230000004186 co-expression Effects 0.000 description 8
- 239000012634 fragment Substances 0.000 description 8
- 230000013595 glycosylation Effects 0.000 description 8
- 238000006206 glycosylation reaction Methods 0.000 description 8
- 230000007505 plaque formation Effects 0.000 description 8
- 230000002829 reductive effect Effects 0.000 description 8
- 238000001890 transfection Methods 0.000 description 8
- 102100029962 CMP-N-acetylneuraminate-beta-1,4-galactoside alpha-2,3-sialyltransferase Human genes 0.000 description 7
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Natural products CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 7
- SHZGCJCMOBCMKK-DHVFOXMCSA-N L-fucopyranose Chemical compound C[C@@H]1OC(O)[C@@H](O)[C@H](O)[C@@H]1O SHZGCJCMOBCMKK-DHVFOXMCSA-N 0.000 description 7
- OVRNDRQMDRJTHS-RTRLPJTCSA-N N-acetyl-D-glucosamine Chemical compound CC(=O)N[C@H]1C(O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-RTRLPJTCSA-N 0.000 description 7
- WQZGKKKJIJFFOK-PHYPRBDBSA-N alpha-D-galactose Chemical compound OC[C@H]1O[C@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-PHYPRBDBSA-N 0.000 description 7
- 230000001404 mediated effect Effects 0.000 description 7
- 238000012545 processing Methods 0.000 description 7
- 241000894007 species Species 0.000 description 7
- 230000001052 transient effect Effects 0.000 description 7
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 6
- 238000002965 ELISA Methods 0.000 description 6
- PNNNRSAQSRJVSB-SLPGGIOYSA-N Fucose Natural products C[C@H](O)[C@@H](O)[C@H](O)[C@H](O)C=O PNNNRSAQSRJVSB-SLPGGIOYSA-N 0.000 description 6
- 210000004556 brain Anatomy 0.000 description 6
- 238000001514 detection method Methods 0.000 description 6
- 102000045442 glycosyltransferase activity proteins Human genes 0.000 description 6
- 108700014210 glycosyltransferase activity proteins Proteins 0.000 description 6
- 230000004048 modification Effects 0.000 description 6
- 238000012986 modification Methods 0.000 description 6
- 108091026890 Coding region Proteins 0.000 description 5
- 108010037897 DC-specific ICAM-3 grabbing nonintegrin Proteins 0.000 description 5
- 101000863898 Homo sapiens CMP-N-acetylneuraminate-beta-1,4-galactoside alpha-2,3-sialyltransferase Proteins 0.000 description 5
- OVRNDRQMDRJTHS-FMDGEEDCSA-N N-acetyl-beta-D-glucosamine Chemical compound CC(=O)N[C@H]1[C@H](O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-FMDGEEDCSA-N 0.000 description 5
- 241000283984 Rodentia Species 0.000 description 5
- 238000007792 addition Methods 0.000 description 5
- VREFGVBLTWBCJP-UHFFFAOYSA-N alprazolam Chemical compound C12=CC(Cl)=CC=C2N2C(C)=NN=C2CN=C1C1=CC=CC=C1 VREFGVBLTWBCJP-UHFFFAOYSA-N 0.000 description 5
- 239000000872 buffer Substances 0.000 description 5
- 230000001747 exhibiting effect Effects 0.000 description 5
- 210000004962 mammalian cell Anatomy 0.000 description 5
- 150000002772 monosaccharides Chemical class 0.000 description 5
- 229950006780 n-acetylglucosamine Drugs 0.000 description 5
- 102000039446 nucleic acids Human genes 0.000 description 5
- 108020004707 nucleic acids Proteins 0.000 description 5
- 239000000047 product Substances 0.000 description 5
- 102000005962 receptors Human genes 0.000 description 5
- 108020003175 receptors Proteins 0.000 description 5
- 241000282412 Homo Species 0.000 description 4
- 102000009490 IgG Receptors Human genes 0.000 description 4
- 108010073807 IgG Receptors Proteins 0.000 description 4
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 description 4
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 description 4
- 241000699666 Mus <mouse, genus> Species 0.000 description 4
- 240000006028 Sambucus nigra Species 0.000 description 4
- 235000003142 Sambucus nigra Nutrition 0.000 description 4
- 239000002253 acid Substances 0.000 description 4
- 230000002411 adverse Effects 0.000 description 4
- RWZYAGGXGHYGMB-UHFFFAOYSA-N anthranilic acid Chemical compound NC1=CC=CC=C1C(O)=O RWZYAGGXGHYGMB-UHFFFAOYSA-N 0.000 description 4
- 235000014633 carbohydrates Nutrition 0.000 description 4
- 238000000576 coating method Methods 0.000 description 4
- 239000002299 complementary DNA Substances 0.000 description 4
- 239000010432 diamond Substances 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 235000008995 european elder Nutrition 0.000 description 4
- 230000001900 immune effect Effects 0.000 description 4
- 229940072221 immunoglobulins Drugs 0.000 description 4
- 230000003993 interaction Effects 0.000 description 4
- 239000000178 monomer Substances 0.000 description 4
- 239000011780 sodium chloride Substances 0.000 description 4
- 239000002904 solvent Substances 0.000 description 4
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 3
- 108090000288 Glycoproteins Proteins 0.000 description 3
- 102000003886 Glycoproteins Human genes 0.000 description 3
- 102000018071 Immunoglobulin Fc Fragments Human genes 0.000 description 3
- 108010091135 Immunoglobulin Fc Fragments Proteins 0.000 description 3
- 241000124008 Mammalia Species 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- 102000003992 Peroxidases Human genes 0.000 description 3
- 241000700159 Rattus Species 0.000 description 3
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 3
- 101710185494 Zinc finger protein Proteins 0.000 description 3
- 102100023597 Zinc finger protein 816 Human genes 0.000 description 3
- 238000001042 affinity chromatography Methods 0.000 description 3
- 238000004220 aggregation Methods 0.000 description 3
- 238000004458 analytical method Methods 0.000 description 3
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 3
- 238000003556 assay Methods 0.000 description 3
- 210000003719 b-lymphocyte Anatomy 0.000 description 3
- 230000001580 bacterial effect Effects 0.000 description 3
- -1 basic amino acids Chemical class 0.000 description 3
- 230000015572 biosynthetic process Effects 0.000 description 3
- 238000012512 characterization method Methods 0.000 description 3
- 239000003795 chemical substances by application Substances 0.000 description 3
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 3
- 239000011248 coating agent Substances 0.000 description 3
- 230000000295 complement effect Effects 0.000 description 3
- 230000005284 excitation Effects 0.000 description 3
- 238000001917 fluorescence detection Methods 0.000 description 3
- 230000036541 health Effects 0.000 description 3
- 239000008241 heterogeneous mixture Substances 0.000 description 3
- 230000028709 inflammatory response Effects 0.000 description 3
- 238000001990 intravenous administration Methods 0.000 description 3
- 239000003446 ligand Substances 0.000 description 3
- 239000012516 mab select resin Substances 0.000 description 3
- 239000003550 marker Substances 0.000 description 3
- 108040007629 peroxidase activity proteins Proteins 0.000 description 3
- 230000008488 polyadenylation Effects 0.000 description 3
- 238000003259 recombinant expression Methods 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 3
- 235000000346 sugar Nutrition 0.000 description 3
- 239000000725 suspension Substances 0.000 description 3
- 238000013518 transcription Methods 0.000 description 3
- 230000035897 transcription Effects 0.000 description 3
- 230000005030 transcription termination Effects 0.000 description 3
- 238000012546 transfer Methods 0.000 description 3
- 230000009466 transformation Effects 0.000 description 3
- 230000010474 transient expression Effects 0.000 description 3
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 2
- 108010090849 Amyloid beta-Peptides Proteins 0.000 description 2
- 102000013455 Amyloid beta-Peptides Human genes 0.000 description 2
- 101710137189 Amyloid-beta A4 protein Proteins 0.000 description 2
- 101710151993 Amyloid-beta precursor protein Proteins 0.000 description 2
- 102100022704 Amyloid-beta precursor protein Human genes 0.000 description 2
- 102100029963 Beta-galactoside alpha-2,6-sialyltransferase 2 Human genes 0.000 description 2
- 241000282693 Cercopithecidae Species 0.000 description 2
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- 244000290594 Ficus sycomorus Species 0.000 description 2
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 2
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 2
- 241001465754 Metazoa Species 0.000 description 2
- 108020004511 Recombinant DNA Proteins 0.000 description 2
- 239000012505 Superdex™ Substances 0.000 description 2
- 108010017070 Zinc Finger Nucleases Proteins 0.000 description 2
- 238000002835 absorbance Methods 0.000 description 2
- 230000002776 aggregation Effects 0.000 description 2
- 108010064539 amyloid beta-protein (1-42) Proteins 0.000 description 2
- DZHSAHHDTRWUTF-SIQRNXPUSA-N amyloid-beta polypeptide 42 Chemical compound C([C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(=O)NCC(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](C)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C(C)C)C(=O)NCC(=O)NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O)[C@@H](C)CC)C(C)C)NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1N=CNC=1)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC(O)=O)C(C)C)C(C)C)C1=CC=CC=C1 DZHSAHHDTRWUTF-SIQRNXPUSA-N 0.000 description 2
- 230000008827 biological function Effects 0.000 description 2
- OWMVSZAMULFTJU-UHFFFAOYSA-N bis-tris Chemical compound OCCN(CCO)C(CO)(CO)CO OWMVSZAMULFTJU-UHFFFAOYSA-N 0.000 description 2
- 239000001506 calcium phosphate Substances 0.000 description 2
- 229910000389 calcium phosphate Inorganic materials 0.000 description 2
- 235000011010 calcium phosphates Nutrition 0.000 description 2
- 238000004422 calculation algorithm Methods 0.000 description 2
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 2
- 230000003247 decreasing effect Effects 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- 238000012217 deletion Methods 0.000 description 2
- 230000037430 deletion Effects 0.000 description 2
- 230000008021 deposition Effects 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- 239000006185 dispersion Substances 0.000 description 2
- 238000010494 dissociation reaction Methods 0.000 description 2
- 230000005593 dissociations Effects 0.000 description 2
- 239000003937 drug carrier Substances 0.000 description 2
- 238000004520 electroporation Methods 0.000 description 2
- 239000003623 enhancer Substances 0.000 description 2
- 230000004927 fusion Effects 0.000 description 2
- 238000002523 gelfiltration Methods 0.000 description 2
- 230000002068 genetic effect Effects 0.000 description 2
- 150000004676 glycans Chemical class 0.000 description 2
- 239000013628 high molecular weight specie Substances 0.000 description 2
- 238000004128 high performance liquid chromatography Methods 0.000 description 2
- 102000005383 human beta1,4-galactosyltransferase Human genes 0.000 description 2
- 108010045961 human beta1,4-galactosyltransferase Proteins 0.000 description 2
- 230000007062 hydrolysis Effects 0.000 description 2
- 238000006460 hydrolysis reaction Methods 0.000 description 2
- 230000003053 immunization Effects 0.000 description 2
- 230000005847 immunogenicity Effects 0.000 description 2
- 238000009169 immunotherapy Methods 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- 230000002757 inflammatory effect Effects 0.000 description 2
- 238000001802 infusion Methods 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 238000007918 intramuscular administration Methods 0.000 description 2
- 238000007912 intraperitoneal administration Methods 0.000 description 2
- 239000002502 liposome Substances 0.000 description 2
- 210000002540 macrophage Anatomy 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- 238000000520 microinjection Methods 0.000 description 2
- 238000000329 molecular dynamics simulation Methods 0.000 description 2
- 238000013364 oligosaccharide-mapping technique Methods 0.000 description 2
- 230000002018 overexpression Effects 0.000 description 2
- 238000002360 preparation method Methods 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 230000001105 regulatory effect Effects 0.000 description 2
- 238000004007 reversed phase HPLC Methods 0.000 description 2
- 239000000523 sample Substances 0.000 description 2
- 230000028327 secretion Effects 0.000 description 2
- 238000002864 sequence alignment Methods 0.000 description 2
- ALZJERAWTOKHNO-UHFFFAOYSA-M sodium;dodecyl sulfate;3-morpholin-4-ylpropane-1-sulfonic acid Chemical compound [Na+].OS(=O)(=O)CCCN1CCOCC1.CCCCCCCCCCCCOS([O-])(=O)=O ALZJERAWTOKHNO-UHFFFAOYSA-M 0.000 description 2
- 239000000243 solution Substances 0.000 description 2
- 238000007920 subcutaneous administration Methods 0.000 description 2
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 description 2
- 210000001519 tissue Anatomy 0.000 description 2
- 230000000699 topical effect Effects 0.000 description 2
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 2
- 230000003612 virological effect Effects 0.000 description 2
- 210000005253 yeast cell Anatomy 0.000 description 2
- 150000003751 zinc Chemical class 0.000 description 2
- LHZGINQTXBBMKG-PYHIIVFHSA-N (4S,5R,6R)-5-amino-2,4-dihydroxy-5-(2-hydroxyacetyl)-6-[(1R,2R)-1,2,3-trihydroxypropyl]oxane-2-carboxylic acid Chemical compound OCC(=O)[C@@]1(N)[C@@H](O)CC(O)(C(O)=O)O[C@H]1[C@H](O)[C@H](O)CO LHZGINQTXBBMKG-PYHIIVFHSA-N 0.000 description 1
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 1
- GEYOCULIXLDCMW-UHFFFAOYSA-N 1,2-phenylenediamine Chemical compound NC1=CC=CC=C1N GEYOCULIXLDCMW-UHFFFAOYSA-N 0.000 description 1
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 1
- UQBIAGWOJDEOMN-UHFFFAOYSA-N 2-O-(2-O-(alpha-D-mannopyranosyl)-alpha-D-mannopyranosyl)-D-mannopyranose Natural products OC1C(O)C(CO)OC(O)C1OC1C(OC2C(C(O)C(O)C(CO)O2)O)C(O)C(O)C(CO)O1 UQBIAGWOJDEOMN-UHFFFAOYSA-N 0.000 description 1
- 101150094949 APRT gene Proteins 0.000 description 1
- 108010024223 Adenine phosphoribosyltransferase Proteins 0.000 description 1
- 101710186708 Agglutinin Proteins 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 102100026349 Beta-1,4-galactosyltransferase 1 Human genes 0.000 description 1
- 101710120061 Beta-1,4-galactosyltransferase 1 Proteins 0.000 description 1
- 101710136188 Beta-galactoside alpha-2,6-sialyltransferase 2 Proteins 0.000 description 1
- 241001260012 Bursa Species 0.000 description 1
- 101710191381 CD209 antigen-like protein A Proteins 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- 241000557626 Corvus corax Species 0.000 description 1
- 241000699800 Cricetinae Species 0.000 description 1
- 241000699802 Cricetulus griseus Species 0.000 description 1
- 101150074155 DHFR gene Proteins 0.000 description 1
- 102000053602 DNA Human genes 0.000 description 1
- 206010012289 Dementia Diseases 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 206010061818 Disease progression Diseases 0.000 description 1
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 1
- 241000206602 Eukaryota Species 0.000 description 1
- 102000009109 Fc receptors Human genes 0.000 description 1
- 108010087819 Fc receptors Proteins 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 239000004606 Fillers/Extenders Substances 0.000 description 1
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 1
- 108020000311 Glutamate Synthase Proteins 0.000 description 1
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 1
- 102000002068 Glycopeptides Human genes 0.000 description 1
- 108010015899 Glycopeptides Proteins 0.000 description 1
- 208000032843 Hemorrhage Diseases 0.000 description 1
- 229920000209 Hexadimethrine bromide Polymers 0.000 description 1
- 101000766145 Homo sapiens Beta-1,4-galactosyltransferase 1 Proteins 0.000 description 1
- 101710146024 Horcolin Proteins 0.000 description 1
- 241000701024 Human betaherpesvirus 5 Species 0.000 description 1
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 1
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 1
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 1
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 description 1
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- 101710189395 Lectin Proteins 0.000 description 1
- ZTOKCBJDEGPICW-UHFFFAOYSA-N Man3GlcNAc2 Natural products OC1C(NC(=O)C)C(O)OC(CO)C1OC1C(NC(C)=O)C(O)C(OC2C(C(OC3C(C(O)C(O)C(CO)O3)O)C(O)C(COC3C(C(O)C(O)C(CO)O3)O)O2)O)C(CO)O1 ZTOKCBJDEGPICW-UHFFFAOYSA-N 0.000 description 1
- 101710179758 Mannose-specific lectin Proteins 0.000 description 1
- 101710150763 Mannose-specific lectin 1 Proteins 0.000 description 1
- 101710150745 Mannose-specific lectin 2 Proteins 0.000 description 1
- OVRNDRQMDRJTHS-CBQIKETKSA-N N-Acetyl-D-Galactosamine Chemical compound CC(=O)N[C@H]1[C@@H](O)O[C@H](CO)[C@H](O)[C@@H]1O OVRNDRQMDRJTHS-CBQIKETKSA-N 0.000 description 1
- MBLBDJOUHNCFQT-UHFFFAOYSA-N N-acetyl-D-galactosamine Natural products CC(=O)NC(C=O)C(O)C(O)C(O)CO MBLBDJOUHNCFQT-UHFFFAOYSA-N 0.000 description 1
- 108010015197 N-acetyllactosaminide alpha-2,3-sialyltransferase Proteins 0.000 description 1
- 206010030113 Oedema Diseases 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 102000000447 Peptide-N4-(N-acetyl-beta-glucosaminyl) Asparagine Amidase Human genes 0.000 description 1
- 108010055817 Peptide-N4-(N-acetyl-beta-glucosaminyl) Asparagine Amidase Proteins 0.000 description 1
- 206010035226 Plasma cell myeloma Diseases 0.000 description 1
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 1
- 239000004365 Protease Substances 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 241000700584 Simplexvirus Species 0.000 description 1
- 210000001744 T-lymphocyte Anatomy 0.000 description 1
- 108020004440 Thymidine kinase Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- 239000003070 absorption delaying agent Substances 0.000 description 1
- 238000005903 acid hydrolysis reaction Methods 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 230000004913 activation Effects 0.000 description 1
- 239000000910 agglutinin Substances 0.000 description 1
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 1
- ZTOKCBJDEGPICW-GWPISINRSA-N alpha-D-Manp-(1->3)-[alpha-D-Manp-(1->6)]-beta-D-Manp-(1->4)-beta-D-GlcpNAc-(1->4)-beta-D-GlcpNAc Chemical compound O[C@@H]1[C@@H](NC(=O)C)[C@H](O)O[C@H](CO)[C@H]1O[C@H]1[C@H](NC(C)=O)[C@@H](O)[C@H](O[C@H]2[C@H]([C@@H](O[C@@H]3[C@H]([C@@H](O)[C@H](O)[C@@H](CO)O3)O)[C@H](O)[C@@H](CO[C@@H]3[C@H]([C@@H](O)[C@H](O)[C@@H](CO)O3)O)O2)O)[C@@H](CO)O1 ZTOKCBJDEGPICW-GWPISINRSA-N 0.000 description 1
- 230000004075 alteration Effects 0.000 description 1
- 229940126575 aminoglycoside Drugs 0.000 description 1
- 230000003321 amplification Effects 0.000 description 1
- 230000002491 angiogenic effect Effects 0.000 description 1
- 238000005571 anion exchange chromatography Methods 0.000 description 1
- 230000000844 anti-bacterial effect Effects 0.000 description 1
- 230000000843 anti-fungal effect Effects 0.000 description 1
- 230000000340 anti-metabolite Effects 0.000 description 1
- 229940121375 antifungal agent Drugs 0.000 description 1
- 229940100197 antimetabolite Drugs 0.000 description 1
- 239000002256 antimetabolite Substances 0.000 description 1
- 230000006907 apoptotic process Effects 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- 239000003125 aqueous solvent Substances 0.000 description 1
- 125000000613 asparagine group Chemical group N[C@@H](CC(N)=O)C(=O)* 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-N aspartic acid group Chemical group N[C@@H](CC(=O)O)C(=O)O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- 108010064886 beta-D-galactoside alpha 2-6-sialyltransferase Proteins 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 108010057005 beta-galactoside alpha-2,3-sialyltransferase Proteins 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 238000001815 biotherapy Methods 0.000 description 1
- 238000006664 bond formation reaction Methods 0.000 description 1
- 210000005013 brain tissue Anatomy 0.000 description 1
- 238000004364 calculation method Methods 0.000 description 1
- 238000012219 cassette mutagenesis Methods 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 239000006143 cell culture medium Substances 0.000 description 1
- 230000003833 cell viability Effects 0.000 description 1
- 230000007248 cellular mechanism Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 238000006243 chemical reaction Methods 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 210000000349 chromosome Anatomy 0.000 description 1
- 238000012761 co-transfection Methods 0.000 description 1
- 230000003920 cognitive function Effects 0.000 description 1
- 230000024203 complement activation Effects 0.000 description 1
- 239000002131 composite material Substances 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 239000000470 constituent Substances 0.000 description 1
- 239000013078 crystal Substances 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 231100000433 cytotoxic Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- 210000004443 dendritic cell Anatomy 0.000 description 1
- 230000001627 detrimental effect Effects 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 231100000676 disease causative agent Toxicity 0.000 description 1
- 230000005750 disease progression Effects 0.000 description 1
- 239000002612 dispersion medium Substances 0.000 description 1
- 239000002552 dosage form Substances 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 238000005538 encapsulation Methods 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 238000002474 experimental method Methods 0.000 description 1
- 210000003754 fetus Anatomy 0.000 description 1
- 108020001507 fusion proteins Proteins 0.000 description 1
- 102000037865 fusion proteins Human genes 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- 125000000291 glutamic acid group Chemical group N[C@@H](CCC(O)=O)C(=O)* 0.000 description 1
- 150000002337 glycosamines Chemical class 0.000 description 1
- 229930182470 glycoside Natural products 0.000 description 1
- 150000002338 glycosides Chemical class 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 210000005260 human cell Anatomy 0.000 description 1
- 210000004408 hybridoma Anatomy 0.000 description 1
- 239000012642 immune effector Substances 0.000 description 1
- 238000002649 immunization Methods 0.000 description 1
- 229940121354 immunomodulator Drugs 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 208000027866 inflammatory disease Diseases 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 238000002347 injection Methods 0.000 description 1
- 239000007924 injection Substances 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 238000007913 intrathecal administration Methods 0.000 description 1
- 238000005342 ion exchange Methods 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 238000011031 large-scale manufacturing process Methods 0.000 description 1
- 230000000670 limiting effect Effects 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 239000002609 medium Substances 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 230000034217 membrane fusion Effects 0.000 description 1
- 108020004999 messenger RNA Proteins 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- HPNSFSBZBAHARI-UHFFFAOYSA-N micophenolic acid Natural products OC1=C(CC=C(C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-UHFFFAOYSA-N 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 230000007570 microbleeding Effects 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 238000000302 molecular modelling Methods 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- HPNSFSBZBAHARI-RUDMXATFSA-N mycophenolic acid Chemical compound OC1=C(C\C=C(/C)CCC(O)=O)C(OC)=C(C)C2=C1C(=O)OC2 HPNSFSBZBAHARI-RUDMXATFSA-N 0.000 description 1
- 229960000951 mycophenolic acid Drugs 0.000 description 1
- 201000000050 myeloid neoplasm Diseases 0.000 description 1
- AEMBWNDIEFEPTH-UHFFFAOYSA-N n-tert-butyl-n-ethylnitrous amide Chemical compound CCN(N=O)C(C)(C)C AEMBWNDIEFEPTH-UHFFFAOYSA-N 0.000 description 1
- 210000000822 natural killer cell Anatomy 0.000 description 1
- 230000004770 neurodegeneration Effects 0.000 description 1
- 208000015122 neurodegenerative disease Diseases 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- 230000007135 neurotoxicity Effects 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 238000003199 nucleic acid amplification method Methods 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 238000005457 optimization Methods 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 230000002093 peripheral effect Effects 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- 239000000825 pharmaceutical preparation Substances 0.000 description 1
- 229940127557 pharmaceutical product Drugs 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 230000001323 posttranslational effect Effects 0.000 description 1
- 230000003389 potentiating effect Effects 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 230000002265 prevention Effects 0.000 description 1
- 230000000750 progressive effect Effects 0.000 description 1
- 230000000770 proinflammatory effect Effects 0.000 description 1
- 230000002062 proliferating effect Effects 0.000 description 1
- 125000001500 prolyl group Chemical group [H]N1C([H])(C(=O)[*])C([H])([H])C([H])([H])C1([H])[H] 0.000 description 1
- 230000012846 protein folding Effects 0.000 description 1
- 230000009145 protein modification Effects 0.000 description 1
- 238000001742 protein purification Methods 0.000 description 1
- 210000001938 protoplast Anatomy 0.000 description 1
- 230000005180 public health Effects 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 230000009467 reduction Effects 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 239000012266 salt solution Substances 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 239000006152 selective media Substances 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 125000005630 sialyl group Chemical group 0.000 description 1
- 238000004088 simulation Methods 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 238000012244 site-specific gene addition Methods 0.000 description 1
- 238000001542 size-exclusion chromatography Methods 0.000 description 1
- 210000001845 splenic macrophage Anatomy 0.000 description 1
- 239000003381 stabilizer Substances 0.000 description 1
- 230000010473 stable expression Effects 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 239000000126 substance Substances 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 238000004114 suspension culture Methods 0.000 description 1
- 230000008685 targeting Effects 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 102000035160 transmembrane proteins Human genes 0.000 description 1
- 108091005703 transmembrane proteins Proteins 0.000 description 1
- 238000000825 ultraviolet detection Methods 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 239000003981 vehicle Substances 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 238000011179 visual inspection Methods 0.000 description 1
Images


























































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/40—Immunoglobulins specific features characterized by post-translational modification
- C07K2317/41—Glycosylation, sialylation, or fucosylation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- General Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Immunology (AREA)
- Peptides Or Proteins (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
Abstract
Description
従来技術の方法では、医薬製品の開発に見合った量で広範囲にシアリル化された抗体を生産することが可能ではない。βガラクトシルトランスフェラーゼおよび/またはシアリルトランスフェラーゼを過剰発現する細胞株におけるIgG抗体の発現は、きわめて低い生産性の条件においてのみシアリル化抗体をもたらすことが本発明者らによって観察された。同様に、通常の細胞株における、Fcドメイン内で変異された抗体の発現は、きわめて不均一なシアリル化パターンを有する抗体組成物をもたらす。
a)前記抗体の前記Fcドメイン内に変異を導入するステップ、および
b)ステップa)で得られた変異抗体を、β−ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼ活性を発現する細胞株において発現させるステップ
を含む。
a)前記抗体の前記Fcドメイン内に変異を導入するステップ、および
b)ステップa)で得られた変異抗体を、β−ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼ活性を発現する細胞株において発現させるステップ
を含む。

この実施例において、グリコシルトランスフェラーゼの存在下におけるモノクローナル抗体(mAb)の一過的産生は著しく減少することが示されたのに対して、これらのグリコシルトランスフェラーゼをコードするプラスミドの濃度は上昇した。
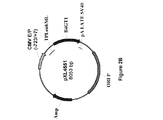
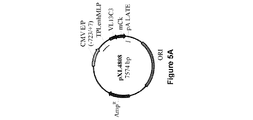
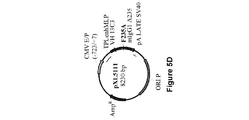
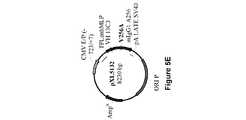
懸濁培養した293−F細胞(ヒト胚性腎HEK293細胞由来であり、Invitrogenで購入した。)において、異なる比率で293fectin(商標)(Invitrogen)と複合体形成させた4種のプラスミドpXL4792、pXL4808、pXL4551およびpXL4555の共トランスフェクションによってAntiAbeta_13C3 mAbの一過的発現を行った。Durocherら、(Nucl.Acids Res.、30:e9、2002)によって報告されているように、EBNAをコードするプラスミドも含めた。細胞培養およびトランスフェクションを、供給業者(Invitrogen)からの推奨に従って振とうフラスコ内で100mL規模で行った。トランスフェクションの8日後、生存可能な細胞を計数して(Vi−CELL XR Cell Viability Analyzer(Beckman Coulter))、280nmにおけるUV検出と連動した分析用HPLC(Poros G/20)によってmAb濃度を決定した。表2に示しているように、mAb産生は、生存可能な細胞が著しく減少したときに回収された細胞と対応していた。
この実施例において、Aβ前原線維に対するantiAbeta_13C3_D257Aの親和性を試験した。この理由は、元のantiAbeta_13C3 mAbがこのリガンドに特異的に結合するからである。
実施例3に記載のα−2,6シアリル化antiAbeta_13C3_D257A mAbは、Fcドメイン内において、広範囲にシアリル化されたN−グリカンの存在によってかなり修飾されている。この修飾は、このドメインにおいて結合することが記載されているFcγ受容体およびC1q成分に対するFc結合に干渉する可能性があった(Shieldsら、J.Biol.Chem.、276:6591−6604、2001;Mershonら、373−382頁、「Therapeutic monoclonal antibodies:from bench to clinic」、編:Zhiqiang An、2009、John Wiley&Sons、Inc.、Hoboken、NJ、USA)。したがって、ネズミタンパク質FcγRsおよびC1qに対して、α−2,6シアリル化antiAbeta_13C3_D257Aの親和性を、Fcによって媒介される強力なエフェクター機能を有するネズミIgG2aモノクローナル抗体(LP09078)と比較して決定した。
α−2,6シアリル化Fcは、エフェクターマクロファージを標的とする抗炎症性の可溶性メディエーターの分泌をもたらす細胞プログラムを誘導したレクチンである、SIGN−R1に関与するという仮説が立てられていた。(Anthonyら、Proc Natl Acad Sci U.S.A.、105:19571−19578、2008)。したがって、この実施例において、SIGN−R1に対するα−2,6シアリル化antiAbeta_13C3_D257Aの親和性を試験した。
この実施例は、ヒトIgG4アイソタイプを有し、Fc内にEU命名法で265位において点変異を含有するα−2,6シアリル化mAbを生産する方法を示している。これは、AntiAbeta_13C3_IgG4における対応している部位では、260位のアスパラギン酸に相当し、ここで、該残基は、分泌されたmAb重鎖の第一番目から番号付けされる。
Claims (21)
- IgG抗体を生産する方法であって、前記抗体のうちの少なくとも80%が、前記抗体の各Fcドメインに結合された2つのシアル酸残基を含有する複合二分岐オリゴ糖を含み、前記方法が、
a)前記抗体の前記Fcドメイン内に変異を導入するステップ、および
b)ステップa)で得られた変異抗体を、β−ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼ活性を発現する細胞株において発現させるステップ
を含む方法。 - β−ガラクトシルトランスフェラーゼがβ−1,4−ガラクトシルトランスフェラーゼであり、シアリルトランスフェラーゼがα−2,6−シアリルトランスフェラーゼである、請求項1に記載の方法。
- β−1,4−ガラクトシルトランスフェラーゼが、配列番号35によって表されるポリヌクレオチド配列によってコードされ、α−2,6−シアリルトランスフェラーゼが、配列番号33によって表されるポリヌクレオチド配列によってコードされる、請求項1または2に記載の方法。
- 前記シアル酸残基が、α−2,6−結合によって抗体に連結されている、請求項1から3のいずれかに記載の方法。
- 抗体がモノクローナル抗体である、請求項1から4のいずれかに記載の方法。
- 抗体がヒト化抗体である、請求項1から5のいずれかに記載の方法。
- 前記変異が、F243、V264およびD265からなる群から選択されるアミノ酸に影響を及ぼす、請求項1から6のいずれかに記載の方法。
- 前記変異が、アラニン(A)、グリシン(G)、ロイシン(L)およびリシン(K)からなる群から選択されるアミノ酸による前記アミノ酸の置換である、請求項1から7のいずれかに記載の方法。
- 前記変異が、D265L、D265KおよびD265Aからなる群から選択される、請求項1から8のいずれかに記載の方法。
- 前記抗体が、ヒトIgG4のFcドメインを含む、請求項1から9のいずれかに記載の方法。
- 前記抗体が、ヒトIgG1のFcドメインを含む、請求項1から10のいずれかに記載の方法。
- β−ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼ活性を発現する前記細胞株が、β−ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼをコードする1種または2種のベクターが安定にトランスフェクトされた細胞株である、請求項1から11のいずれかに記載の方法。
- β−ガラクトシルトランスフェラーゼおよびシアリルトランスフェラーゼ活性を発現する前記細胞株が、前記抗体をコードする1種または2種のベクターが安定にトランスフェクトされた細胞株である、請求項1から12のいずれかに記載の方法。
- 請求項1から13のいずれかに記載の方法によって生産された抗体。
- 請求項14の抗体を含む医薬組成物。
- 医薬として使用するための、請求項14に記載の抗体。
- IgG抗体を含む組成物であって、前記抗体のうちの少なくとも80%が、前記抗体の各Fcドメインに結合された複合二分岐オリゴ糖を含み、前記オリゴ糖が、2つのシアル酸残基を含み、前記Fcドメインが、天然配列のヒトIgGのFcドメインと異なるアミノ配列を含む、組成物。
- 前記シアル酸残基が、α−2,6−結合によって抗体に連結されている、請求項17に記載の組成物。
- 本発明の組成物の抗体が、アミノ酸243、264および265位のうちのいずれか1つ以上におけるアミノ酸置換を含む、請求項17または18に記載の組成物。
- 前記置換が、アラニン(A)、グリシン(G)、ロイシン(L)およびリシン(K)からなる群から選択されるアミノ酸による前記アミノ酸の置換である、請求項19に記載の組成物。
- 前記置換が、D265L、D265KおよびD265Aからなる群から選択される、請求項20に記載の組成物。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP11305200.5 | 2011-02-24 | ||
| EP11305200 | 2011-02-24 | ||
| EP11306090.9 | 2011-09-01 | ||
| EP11306090 | 2011-09-01 | ||
| PCT/EP2012/053065 WO2012113863A1 (en) | 2011-02-24 | 2012-02-23 | Method of production of sialylated antibodies |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2014508759A true JP2014508759A (ja) | 2014-04-10 |
| JP2014508759A5 JP2014508759A5 (ja) | 2015-02-19 |
Family
ID=45774189
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2013554892A Pending JP2014508759A (ja) | 2011-02-24 | 2012-02-23 | シアリル化抗体の生産の方法 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20140046032A1 (ja) |
| EP (1) | EP2678357A1 (ja) |
| JP (1) | JP2014508759A (ja) |
| AR (1) | AR085302A1 (ja) |
| TW (1) | TW201241182A (ja) |
| WO (1) | WO2012113863A1 (ja) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2020515524A (ja) * | 2017-01-31 | 2020-05-28 | サノフイSanofi | ベータアミロイドペプチドのプロトフィブリル形態に特異的な抗体の神経細胞保護作用 |
| JP2021522216A (ja) * | 2018-04-20 | 2021-08-30 | アンスティチュ ナショナル ドゥ ラ サンテ エ ドゥ ラ ルシェルシュ メディカル | 高度にシアル化された自己抗体およびその使用 |
Families Citing this family (26)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US9062106B2 (en) | 2011-04-27 | 2015-06-23 | Abbvie Inc. | Methods for controlling the galactosylation profile of recombinantly-expressed proteins |
| UY34317A (es) | 2011-09-12 | 2013-02-28 | Genzyme Corp | Anticuerpo antireceptor de célula T (alfa)/ß |
| WO2013158279A1 (en) | 2012-04-20 | 2013-10-24 | Abbvie Inc. | Protein purification methods to reduce acidic species |
| WO2013158273A1 (en) | 2012-04-20 | 2013-10-24 | Abbvie Inc. | Methods to modulate c-terminal lysine variant distribution |
| US9067990B2 (en) | 2013-03-14 | 2015-06-30 | Abbvie, Inc. | Protein purification using displacement chromatography |
| US9512214B2 (en) | 2012-09-02 | 2016-12-06 | Abbvie, Inc. | Methods to control protein heterogeneity |
| KR20240023184A (ko) | 2013-03-11 | 2024-02-20 | 젠자임 코포레이션 | 과글리코실화된 결합 폴리펩티드 |
| AU2013381687A1 (en) | 2013-03-12 | 2015-09-24 | Abbvie Inc. | Human antibodies that bind human TNF-alpha and methods of preparing the same |
| US9017687B1 (en) | 2013-10-18 | 2015-04-28 | Abbvie, Inc. | Low acidic species compositions and methods for producing and using the same using displacement chromatography |
| WO2014151878A2 (en) | 2013-03-14 | 2014-09-25 | Abbvie Inc. | Methods for modulating protein glycosylation profiles of recombinant protein therapeutics using monosaccharides and oligosacharides |
| US10450361B2 (en) | 2013-03-15 | 2019-10-22 | Momenta Pharmaceuticals, Inc. | Methods related to CTLA4-Fc fusion proteins |
| WO2014179601A2 (en) * | 2013-05-02 | 2014-11-06 | Momenta Pharmaceuticals, Inc. | Sialylated glycoproteins |
| US10464996B2 (en) | 2013-05-13 | 2019-11-05 | Momenta Pharmaceuticals, Inc. | Methods for the treatment of neurodegeneration |
| JP6594860B2 (ja) * | 2013-05-29 | 2019-10-23 | エフ.ホフマン−ラ ロシュ アーゲー | シアル酸付加の定量的調節 |
| CN105358703A (zh) | 2013-07-05 | 2016-02-24 | 豪夫迈·罗氏有限公司 | 用于使用N-末端截短的β-半乳糖苷-α-2,6-唾液酸转移酶变体的糖蛋白的单-和二唾液酸化的方法 |
| EP2824176A1 (en) * | 2013-07-11 | 2015-01-14 | Siamed'xpress | Methods for producing sialylated therapeutic proteins |
| WO2015051293A2 (en) | 2013-10-04 | 2015-04-09 | Abbvie, Inc. | Use of metal ions for modulation of protein glycosylation profiles of recombinant proteins |
| US20160257754A1 (en) * | 2013-10-16 | 2016-09-08 | Momenta Pharmaceuticals Inc. | Sialylated glycoproteins |
| US9181337B2 (en) | 2013-10-18 | 2015-11-10 | Abbvie, Inc. | Modulated lysine variant species compositions and methods for producing and using the same |
| US9085618B2 (en) | 2013-10-18 | 2015-07-21 | Abbvie, Inc. | Low acidic species compositions and methods for producing and using the same |
| US20150139988A1 (en) | 2013-11-15 | 2015-05-21 | Abbvie, Inc. | Glycoengineered binding protein compositions |
| CN106471010A (zh) | 2014-03-19 | 2017-03-01 | 建新公司 | 靶向模块的位点特异性糖工程化 |
| EP3172333B1 (en) | 2014-07-21 | 2020-05-13 | Glykos Finland Oy | Production of glycoproteins with mammalian-like n-glycans in filamentous fungi |
| EP3037527A1 (en) * | 2014-12-22 | 2016-06-29 | F. Hoffmann-La Roche AG | Sialyltransferase without CMP-dependent sialidase activity |
| EP3237608B1 (en) | 2014-12-22 | 2019-10-09 | F. Hoffmann-La Roche AG | Cmp-dependent sialidase activity |
| WO2023086793A1 (en) * | 2021-11-09 | 2023-05-19 | Amgen Inc. | Production of therapeutic proteins |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2009504569A (ja) * | 2005-06-30 | 2009-02-05 | セントカー・インコーポレーテツド | 向上した治療活性をもつ方法および組成物 |
| JP2010510801A (ja) * | 2006-11-28 | 2010-04-08 | サントリオン | 生物学的活性が向上した修飾可溶性FGF受容体Fc融合物 |
| JP2010512306A (ja) * | 2006-10-27 | 2010-04-22 | ザ ロックフェラー ユニバーシティー | 抗炎症特性が増強され、細胞毒性特性が減少したポリペプチドおよび関連する方法 |
| WO2010130946A1 (fr) * | 2009-05-12 | 2010-11-18 | Sanofi-Aventis | Anticorps humanises spécifiques de la forme protofibrillaire du peptide béta-amyloide |
Family Cites Families (31)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB8607679D0 (en) | 1986-03-27 | 1986-04-30 | Winter G P | Recombinant dna product |
| US5530101A (en) | 1988-12-28 | 1996-06-25 | Protein Design Labs, Inc. | Humanized immunoglobulins |
| GB8928874D0 (en) | 1989-12-21 | 1990-02-28 | Celltech Ltd | Humanised antibodies |
| DE69233482T2 (de) | 1991-05-17 | 2006-01-12 | Merck & Co., Inc. | Verfahren zur Verminderung der Immunogenität der variablen Antikörperdomänen |
| ES2136092T3 (es) | 1991-09-23 | 1999-11-16 | Medical Res Council | Procedimientos para la produccion de anticuerpos humanizados. |
| US5792632A (en) | 1992-05-05 | 1998-08-11 | Institut Pasteur | Nucleotide sequence encoding the enzyme I-SceI and the uses thereof |
| US5474896A (en) | 1992-05-05 | 1995-12-12 | Institut Pasteur | Nucleotide sequence encoding the enzyme I-SceI and the uses thereof |
| US5639641A (en) | 1992-09-09 | 1997-06-17 | Immunogen Inc. | Resurfacing of rodent antibodies |
| US5830729A (en) | 1996-04-18 | 1998-11-03 | Institut Pasteur | I Sce I-induced gene replacement and gene conversion in embryonic stem cells |
| US20020045207A1 (en) * | 1997-10-31 | 2002-04-18 | Lynne A. Krummen | Glycoprotein production process |
| US6761888B1 (en) | 2000-05-26 | 2004-07-13 | Neuralab Limited | Passive immunization treatment of Alzheimer's disease |
| US6750324B1 (en) | 1997-12-02 | 2004-06-15 | Neuralab Limited | Humanized and chimeric N-terminal amyloid beta-antibodies |
| US6737056B1 (en) * | 1999-01-15 | 2004-05-18 | Genentech, Inc. | Polypeptide variants with altered effector function |
| CA2414772C (en) | 2000-07-07 | 2011-06-28 | Jan Naslund | Prevention and treatment of alzheimer's disease |
| ATE466088T1 (de) | 2001-09-14 | 2010-05-15 | Cellectis | Zufällige integration von polynukleotide nach in vivo linearisierung |
| JP4966006B2 (ja) | 2003-01-28 | 2012-07-04 | セレクティス | カスタムメイドメガヌクレアーゼおよびその使用 |
| AR058129A1 (es) | 2005-10-21 | 2008-01-23 | Genzyme Corp | Productos terapeuticos basados en anticuerpos con actividad de adcc mejorada |
| US20080206246A1 (en) * | 2006-04-05 | 2008-08-28 | Ravetch Jeffrey V | Polypeptides with enhanced anti-inflammatory and decreased cytotoxic properties and relating methods |
| CA2632822C (en) | 2005-12-12 | 2018-08-28 | Ruth Greferath | A beta 1-42 specific monoclonal antibodies with therapeutic properties |
| EP3456351A1 (en) | 2006-04-05 | 2019-03-20 | The Rockefeller University | Polypeptides with enhanced anti-inflammatory and decreased cytotoxic properties and relating methods |
| AU2007317755A1 (en) | 2006-10-27 | 2008-05-15 | The Rockefeller University | Polypeptides with enhanced anti-inflammatory and decreased cytotoxic properties and relating methods |
| MX346172B (es) | 2007-08-29 | 2017-03-10 | Sanofi - Aventis | Anticuerpos anti - cxcr5 humanizados, derivados de los mismos y su uso. |
| BRPI0818623A2 (pt) | 2007-10-05 | 2017-05-23 | Ac Immune Sa | composição farmacêutica, e, métodos para reduzir a carga da placa na camada de célula de gânglio retinal de um animal, para reduzir a quantidade de placas na camada de célula de gânglio retinal de um animal, para diminuir a quantidade total de amilóide-beta solúvel na camada de célula de gânglio retinal de um animal, para prevenir, tratar e/ou aliviar os efeitos de uma doença ocular associada com anormalidades patológicas/mudanças no tecido do sistema visual, para monitorar doença ocular residual mínima associada com anormalidades patológicas/mudanças nos tecidos do sistema visual, para predizer responsividade de um paciente, e para reter ou diminuir a pressão ocular nos olhos de um animal |
| WO2009052125A2 (en) | 2007-10-15 | 2009-04-23 | Centocor, Inc. | Human anti-amyloid antibodies, compositions, methods and uses |
| EP2226081A4 (en) | 2007-10-25 | 2011-10-12 | Univ Kagoshima | PEPTIDE VACCINE WITH A MIMIC MOLECULE OF AMYLOID PEPTIDE |
| US8936936B2 (en) | 2007-10-25 | 2015-01-20 | Sangamo Biosciences, Inc. | Methods and compositions for targeted integration |
| US8445649B2 (en) | 2007-10-29 | 2013-05-21 | Tao Health Life Pharma Co., Ltd. | Antibody and use thereof |
| WO2009065054A2 (en) | 2007-11-16 | 2009-05-22 | The Rockefeller University | Antibodies specific for the protofibril form of beta-amyloid protein |
| KR101657637B1 (ko) | 2007-12-11 | 2016-09-19 | 글락소 그룹 리미티드 | 항원 결합성 단백질 |
| EP2233502A1 (en) | 2009-03-27 | 2010-09-29 | Deutsches Rheuma-Forschungszentrum Berlin | Sialylated antigen-specific antibodies for treatment or prophylaxis of unwanted inflammatory immune reactions and methods of producing them |
| US10087236B2 (en) * | 2009-12-02 | 2018-10-02 | Academia Sinica | Methods for modifying human antibodies by glycan engineering |
-
2012
- 2012-02-22 AR ARP120100573A patent/AR085302A1/es unknown
- 2012-02-23 JP JP2013554892A patent/JP2014508759A/ja active Pending
- 2012-02-23 WO PCT/EP2012/053065 patent/WO2012113863A1/en active Application Filing
- 2012-02-23 EP EP12706541.5A patent/EP2678357A1/en not_active Withdrawn
- 2012-02-23 US US14/000,035 patent/US20140046032A1/en not_active Abandoned
- 2012-02-23 TW TW101106076A patent/TW201241182A/zh unknown
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2009504569A (ja) * | 2005-06-30 | 2009-02-05 | セントカー・インコーポレーテツド | 向上した治療活性をもつ方法および組成物 |
| JP2010512306A (ja) * | 2006-10-27 | 2010-04-22 | ザ ロックフェラー ユニバーシティー | 抗炎症特性が増強され、細胞毒性特性が減少したポリペプチドおよび関連する方法 |
| JP2010510801A (ja) * | 2006-11-28 | 2010-04-08 | サントリオン | 生物学的活性が向上した修飾可溶性FGF受容体Fc融合物 |
| WO2010130946A1 (fr) * | 2009-05-12 | 2010-11-18 | Sanofi-Aventis | Anticorps humanises spécifiques de la forme protofibrillaire du peptide béta-amyloide |
Non-Patent Citations (3)
| Title |
|---|
| BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS, vol. 286, JPN6017006279, 2001, pages 243 - 249, ISSN: 0003505805 * |
| SCIENCE, vol. 320, JPN6017006281, 2008, pages 373 - 376, ISSN: 0003505806 * |
| THE JOURNAL OF IMMUNOLOGY, vol. 157, JPN6015007484, 1996, pages 4963 - 4969, ISSN: 0003226087 * |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2020515524A (ja) * | 2017-01-31 | 2020-05-28 | サノフイSanofi | ベータアミロイドペプチドのプロトフィブリル形態に特異的な抗体の神経細胞保護作用 |
| JP2021522216A (ja) * | 2018-04-20 | 2021-08-30 | アンスティチュ ナショナル ドゥ ラ サンテ エ ドゥ ラ ルシェルシュ メディカル | 高度にシアル化された自己抗体およびその使用 |
Also Published As
| Publication number | Publication date |
|---|---|
| AR085302A1 (es) | 2013-09-18 |
| WO2012113863A1 (en) | 2012-08-30 |
| EP2678357A1 (en) | 2014-01-01 |
| TW201241182A (en) | 2012-10-16 |
| US20140046032A1 (en) | 2014-02-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP2014508759A (ja) | シアリル化抗体の生産の方法 | |
| JP7357382B2 (ja) | 抗炎症特性が増強され、細胞毒性特性が減少したポリペプチドおよび関連する方法 | |
| US20220162290A1 (en) | Polypeptides With Enhanced Anti-Inflammatory And Decreased Cytotoxic Properties And Relating Methods | |
| Vattepu et al. | Sialylation as an important regulator of antibody function | |
| DK1896071T3 (en) | Methods and compositions with increased therapeutic activity | |
| CA2674239C (en) | Methods and vectors for generating asialylated immunoglobulins | |
| CA2963760A1 (en) | Anti-myostatin antibodies, polypeptides containing variant fc regions, and methods of use | |
| KR20100074220A (ko) | Cdr의 아미노산 치환에 의해 항체의 등전점을 개변하는 방법 | |
| TWI780097B (zh) | 於活體外進行抗體之糖基化工程 | |
| CN106687481B (zh) | 半乳糖改造的免疫球蛋白1抗体 | |
| CN110100007B (zh) | 用于体外糖工程化抗体的酶的再使用 | |
| MX2013015061A (es) | Anticuerpo anti-cxcr4 con funciones efectoras y su uso para el tratamiento del cancer. | |
| TWI781973B (zh) | 用於以活體外進行抗體糖基化工程之方法 | |
| JP2024518545A (ja) | Trem2のアゴニスト | |
| Krahn et al. | Inhibition of glycosylation on a camelid antibody uniquely affects its FcγRI binding activity | |
| JP2010512306A (ja) | 抗炎症特性が増強され、細胞毒性特性が減少したポリペプチドおよび関連する方法 | |
| NZ597651A (en) | Polypeptides with enhanced anti-inflammatory and decreased cytotoxic properties and relating methods | |
| TW202432574A (zh) | 具有增強的效應子功能之糖基工程化Fc變體多肽 | |
| JP2021006540A (ja) | ガラクトース操作型免疫グロブリン1抗体 | |
| TW201305336A (zh) | 具作用功能的抗cxcr4抗體及其供治療癌症之用途 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20141222 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20141222 |
|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20151207 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20151210 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20160105 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20160330 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20160816 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20161213 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20161220 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20170224 |