JP2005287403A - 脂質生産菌の育種方法およびその利用 - Google Patents
脂質生産菌の育種方法およびその利用 Download PDFInfo
- Publication number
- JP2005287403A JP2005287403A JP2004107512A JP2004107512A JP2005287403A JP 2005287403 A JP2005287403 A JP 2005287403A JP 2004107512 A JP2004107512 A JP 2004107512A JP 2004107512 A JP2004107512 A JP 2004107512A JP 2005287403 A JP2005287403 A JP 2005287403A
- Authority
- JP
- Japan
- Prior art keywords
- lipid
- breeding
- gene
- fatty acid
- producing bacterium
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
- 238000000034 method Methods 0.000 title claims abstract description 150
- 150000002632 lipids Chemical class 0.000 title claims abstract description 148
- 241000894006 Bacteria Species 0.000 title claims abstract description 97
- 238000009395 breeding Methods 0.000 title claims abstract description 81
- 230000001488 breeding effect Effects 0.000 title claims abstract description 64
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 148
- 230000014509 gene expression Effects 0.000 claims abstract description 64
- 241000235575 Mortierella Species 0.000 claims abstract description 51
- 239000013604 expression vector Substances 0.000 claims description 45
- 108091030071 RNAI Proteins 0.000 claims description 44
- 230000009368 gene silencing by RNA Effects 0.000 claims description 44
- 108091032973 (ribonucleotides)n+m Proteins 0.000 claims description 43
- 238000003259 recombinant expression Methods 0.000 claims description 33
- 102000040650 (ribonucleotides)n+m Human genes 0.000 claims description 29
- 238000004519 manufacturing process Methods 0.000 claims description 29
- 235000020777 polyunsaturated fatty acids Nutrition 0.000 claims description 28
- 241000907999 Mortierella alpina Species 0.000 claims description 26
- 230000009466 transformation Effects 0.000 claims description 24
- 150000004665 fatty acids Chemical group 0.000 claims description 21
- 239000003153 chemical reaction reagent Substances 0.000 claims description 16
- 102000004190 Enzymes Human genes 0.000 claims description 13
- 108090000790 Enzymes Proteins 0.000 claims description 13
- 108010087894 Fatty acid desaturases Proteins 0.000 claims description 12
- 102000009114 Fatty acid desaturases Human genes 0.000 claims description 11
- 238000010276 construction Methods 0.000 claims description 10
- 108010005155 delta-12 fatty acid desaturase Proteins 0.000 claims description 10
- 230000037356 lipid metabolism Effects 0.000 claims description 10
- 239000002245 particle Substances 0.000 claims description 9
- 238000002716 delivery method Methods 0.000 claims description 8
- 230000001629 suppression Effects 0.000 claims description 8
- 238000004520 electroporation Methods 0.000 claims description 6
- 230000004129 fatty acid metabolism Effects 0.000 claims description 5
- 108010073542 Delta-5 Fatty Acid Desaturase Proteins 0.000 claims description 4
- 238000012258 culturing Methods 0.000 claims description 4
- 108010037138 Linoleoyl-CoA Desaturase Proteins 0.000 claims description 3
- 102100020754 Putative fatty acid desaturase 2-like protein FADS2B Human genes 0.000 claims description 3
- 108010022240 delta-8 fatty acid desaturase Proteins 0.000 claims description 3
- 108010086283 delta-9 fatty acid desaturase Proteins 0.000 claims description 3
- 108010033653 omega-3 fatty acid desaturase Proteins 0.000 claims description 3
- 239000000126 substance Substances 0.000 claims description 3
- 238000009738 saturating Methods 0.000 claims 1
- 230000008569 process Effects 0.000 abstract description 16
- 239000013612 plasmid Substances 0.000 description 44
- 108020004414 DNA Proteins 0.000 description 43
- 102000053602 DNA Human genes 0.000 description 43
- 239000012634 fragment Substances 0.000 description 31
- YZXBAPSDXZZRGB-DOFZRALJSA-N arachidonic acid Chemical compound CCCCC\C=C/C\C=C/C\C=C/C\C=C/CCCC(O)=O YZXBAPSDXZZRGB-DOFZRALJSA-N 0.000 description 22
- 210000004027 cell Anatomy 0.000 description 22
- 108091008146 restriction endonucleases Proteins 0.000 description 20
- 241000233866 Fungi Species 0.000 description 18
- 150000004671 saturated fatty acids Chemical class 0.000 description 17
- 239000013598 vector Substances 0.000 description 17
- 238000006243 chemical reaction Methods 0.000 description 14
- 235000014113 dietary fatty acids Nutrition 0.000 description 13
- 229930195729 fatty acid Natural products 0.000 description 13
- 239000000194 fatty acid Substances 0.000 description 13
- 230000035772 mutation Effects 0.000 description 12
- VLKZOEOYAKHREP-UHFFFAOYSA-N n-Hexane Chemical compound CCCCCC VLKZOEOYAKHREP-UHFFFAOYSA-N 0.000 description 12
- 235000021342 arachidonic acid Nutrition 0.000 description 11
- 229940114079 arachidonic acid Drugs 0.000 description 11
- 239000002609 medium Substances 0.000 description 11
- 238000002844 melting Methods 0.000 description 10
- 230000008018 melting Effects 0.000 description 10
- 230000000295 complement effect Effects 0.000 description 8
- 239000003550 marker Substances 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 7
- 230000000694 effects Effects 0.000 description 7
- 239000003623 enhancer Substances 0.000 description 7
- 235000003441 saturated fatty acids Nutrition 0.000 description 7
- UNSRRHDPHVZAHH-YOILPLPUSA-N (5Z,8Z,11Z)-icosatrienoic acid Chemical compound CCCCCCCC\C=C/C\C=C/C\C=C/CCCC(O)=O UNSRRHDPHVZAHH-YOILPLPUSA-N 0.000 description 6
- UNSRRHDPHVZAHH-UHFFFAOYSA-N 6beta,11alpha-Dihydroxy-3alpha,5alpha-cyclopregnan-20-on Natural products CCCCCCCCC=CCC=CCC=CCCCC(O)=O UNSRRHDPHVZAHH-UHFFFAOYSA-N 0.000 description 6
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 6
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 6
- 238000000855 fermentation Methods 0.000 description 6
- 230000004151 fermentation Effects 0.000 description 6
- 239000000463 material Substances 0.000 description 6
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 5
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 5
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 5
- 235000011130 ammonium sulphate Nutrition 0.000 description 5
- 125000004432 carbon atom Chemical group C* 0.000 description 5
- 239000007788 liquid Substances 0.000 description 5
- 244000005700 microbiome Species 0.000 description 5
- 239000003921 oil Substances 0.000 description 5
- 238000011426 transformation method Methods 0.000 description 5
- 229920001817 Agar Polymers 0.000 description 4
- 206010059866 Drug resistance Diseases 0.000 description 4
- 241000588724 Escherichia coli Species 0.000 description 4
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 4
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 4
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 4
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 4
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 4
- 239000008272 agar Substances 0.000 description 4
- 229940024606 amino acid Drugs 0.000 description 4
- 150000001413 amino acids Chemical class 0.000 description 4
- 230000000692 anti-sense effect Effects 0.000 description 4
- 230000006696 biosynthetic metabolic pathway Effects 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 210000000349 chromosome Anatomy 0.000 description 4
- 230000029087 digestion Effects 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 230000001747 exhibiting effect Effects 0.000 description 4
- 239000003925 fat Substances 0.000 description 4
- 235000019197 fats Nutrition 0.000 description 4
- 239000008103 glucose Substances 0.000 description 4
- XMHIUKTWLZUKEX-UHFFFAOYSA-N hexacosanoic acid Chemical compound CCCCCCCCCCCCCCCCCCCCCCCCCC(O)=O XMHIUKTWLZUKEX-UHFFFAOYSA-N 0.000 description 4
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 4
- VKOBVWXKNCXXDE-UHFFFAOYSA-N icosanoic acid Chemical compound CCCCCCCCCCCCCCCCCCCC(O)=O VKOBVWXKNCXXDE-UHFFFAOYSA-N 0.000 description 4
- 238000001727 in vivo Methods 0.000 description 4
- 108020004999 messenger RNA Proteins 0.000 description 4
- 239000000203 mixture Substances 0.000 description 4
- 235000020665 omega-6 fatty acid Nutrition 0.000 description 4
- 239000000047 product Substances 0.000 description 4
- 238000012216 screening Methods 0.000 description 4
- 239000007787 solid Substances 0.000 description 4
- HOBAELRKJCKHQD-UHFFFAOYSA-N (8Z,11Z,14Z)-8,11,14-eicosatrienoic acid Natural products CCCCCC=CCC=CCC=CCCCCCCC(O)=O HOBAELRKJCKHQD-UHFFFAOYSA-N 0.000 description 3
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 3
- 235000021355 Stearic acid Nutrition 0.000 description 3
- 239000002253 acid Substances 0.000 description 3
- 230000003321 amplification Effects 0.000 description 3
- 229910052799 carbon Inorganic materials 0.000 description 3
- 150000001875 compounds Chemical class 0.000 description 3
- HOBAELRKJCKHQD-QNEBEIHSSA-N dihomo-γ-linolenic acid Chemical compound CCCCC\C=C/C\C=C/C\C=C/CCCCCCC(O)=O HOBAELRKJCKHQD-QNEBEIHSSA-N 0.000 description 3
- 238000005194 fractionation Methods 0.000 description 3
- 238000010230 functional analysis Methods 0.000 description 3
- VZCCETWTMQHEPK-QNEBEIHSSA-N gamma-linolenic acid Chemical compound CCCCC\C=C/C\C=C/C\C=C/CCCCC(O)=O VZCCETWTMQHEPK-QNEBEIHSSA-N 0.000 description 3
- 235000020664 gamma-linolenic acid Nutrition 0.000 description 3
- 230000002068 genetic effect Effects 0.000 description 3
- 239000006455 gy-medium Substances 0.000 description 3
- 230000006801 homologous recombination Effects 0.000 description 3
- 238000002744 homologous recombination Methods 0.000 description 3
- 229910052757 nitrogen Inorganic materials 0.000 description 3
- 238000003199 nucleic acid amplification method Methods 0.000 description 3
- 235000015097 nutrients Nutrition 0.000 description 3
- QIQXTHQIDYTFRH-UHFFFAOYSA-N octadecanoic acid Chemical compound CCCCCCCCCCCCCCCCCC(O)=O QIQXTHQIDYTFRH-UHFFFAOYSA-N 0.000 description 3
- OQCDKBAXFALNLD-UHFFFAOYSA-N octadecanoic acid Natural products CCCCCCCC(C)CCCCCCCCC(O)=O OQCDKBAXFALNLD-UHFFFAOYSA-N 0.000 description 3
- 239000008117 stearic acid Substances 0.000 description 3
- 230000001131 transforming effect Effects 0.000 description 3
- 229940035893 uracil Drugs 0.000 description 3
- OYHQOLUKZRVURQ-NTGFUMLPSA-N (9Z,12Z)-9,10,12,13-tetratritiooctadeca-9,12-dienoic acid Chemical compound C(CCCCCCC\C(=C(/C\C(=C(/CCCCC)\[3H])\[3H])\[3H])\[3H])(=O)O OYHQOLUKZRVURQ-NTGFUMLPSA-N 0.000 description 2
- WRIDQFICGBMAFQ-UHFFFAOYSA-N (E)-8-Octadecenoic acid Natural products CCCCCCCCCC=CCCCCCCC(O)=O WRIDQFICGBMAFQ-UHFFFAOYSA-N 0.000 description 2
- 108020004463 18S ribosomal RNA Proteins 0.000 description 2
- LQJBNNIYVWPHFW-UHFFFAOYSA-N 20:1omega9c fatty acid Natural products CCCCCCCCCCC=CCCCCCCCC(O)=O LQJBNNIYVWPHFW-UHFFFAOYSA-N 0.000 description 2
- QSBYPNXLFMSGKH-UHFFFAOYSA-N 9-Heptadecensaeure Natural products CCCCCCCC=CCCCCCCCC(O)=O QSBYPNXLFMSGKH-UHFFFAOYSA-N 0.000 description 2
- 229930024421 Adenine Natural products 0.000 description 2
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 2
- 239000004475 Arginine Substances 0.000 description 2
- 235000021298 Dihomo-γ-linolenic acid Nutrition 0.000 description 2
- 241000196324 Embryophyta Species 0.000 description 2
- YQYJSBFKSSDGFO-UHFFFAOYSA-N Epihygromycin Natural products OC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1O YQYJSBFKSSDGFO-UHFFFAOYSA-N 0.000 description 2
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 2
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 2
- 239000004472 Lysine Substances 0.000 description 2
- ZQPPMHVWECSIRJ-UHFFFAOYSA-N Oleic acid Natural products CCCCCCCCC=CCCCCCCCC(O)=O ZQPPMHVWECSIRJ-UHFFFAOYSA-N 0.000 description 2
- 239000005642 Oleic acid Substances 0.000 description 2
- 108091081021 Sense strand Proteins 0.000 description 2
- 101150044776 URA5 gene Proteins 0.000 description 2
- 229960000643 adenine Drugs 0.000 description 2
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 2
- 229940041514 candida albicans extract Drugs 0.000 description 2
- 230000003247 decreasing effect Effects 0.000 description 2
- AGDANEVFLMAYGL-UHFFFAOYSA-N docosanoic acid Chemical compound CCCCCCCCCCCCCCCCCCCCCC(O)=O.CCCCCCCCCCCCCCCCCCCCCC(O)=O AGDANEVFLMAYGL-UHFFFAOYSA-N 0.000 description 2
- 238000004945 emulsification Methods 0.000 description 2
- 235000019387 fatty acid methyl ester Nutrition 0.000 description 2
- 235000013305 food Nutrition 0.000 description 2
- VZCCETWTMQHEPK-UHFFFAOYSA-N gamma-Linolensaeure Natural products CCCCCC=CCC=CCC=CCCCCC(O)=O VZCCETWTMQHEPK-UHFFFAOYSA-N 0.000 description 2
- 229960002733 gamolenic acid Drugs 0.000 description 2
- FUKUFMFMCZIRNT-UHFFFAOYSA-N hydron;methanol;chloride Chemical compound Cl.OC FUKUFMFMCZIRNT-UHFFFAOYSA-N 0.000 description 2
- QXJSBBXBKPUZAA-UHFFFAOYSA-N isooleic acid Natural products CCCCCCCC=CCCCCCCCCC(O)=O QXJSBBXBKPUZAA-UHFFFAOYSA-N 0.000 description 2
- 229930182817 methionine Natural products 0.000 description 2
- 150000004702 methyl esters Chemical class 0.000 description 2
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 2
- 230000000813 microbial effect Effects 0.000 description 2
- 238000000520 microinjection Methods 0.000 description 2
- 235000021281 monounsaturated fatty acids Nutrition 0.000 description 2
- ZQPPMHVWECSIRJ-KTKRTIGZSA-N oleic acid Chemical compound CCCCCCCC\C=C/CCCCCCCC(O)=O ZQPPMHVWECSIRJ-KTKRTIGZSA-N 0.000 description 2
- BASFCYQUMIYNBI-UHFFFAOYSA-N platinum Chemical compound [Pt] BASFCYQUMIYNBI-UHFFFAOYSA-N 0.000 description 2
- 230000006798 recombination Effects 0.000 description 2
- 238000005215 recombination Methods 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 238000010839 reverse transcription Methods 0.000 description 2
- 238000003757 reverse transcription PCR Methods 0.000 description 2
- 238000010187 selection method Methods 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- CBYCSRICVDBHMZ-UHFFFAOYSA-N tetracosanoic acid Chemical compound CCCCCCCCCCCCCCCCCCCCCCCC(O)=O.CCCCCCCCCCCCCCCCCCCCCCCC(O)=O CBYCSRICVDBHMZ-UHFFFAOYSA-N 0.000 description 2
- 235000021122 unsaturated fatty acids Nutrition 0.000 description 2
- 150000004670 unsaturated fatty acids Chemical class 0.000 description 2
- 238000011144 upstream manufacturing Methods 0.000 description 2
- 239000012138 yeast extract Substances 0.000 description 2
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- 102100034542 Acyl-CoA (8-3)-desaturase Human genes 0.000 description 1
- 241000351920 Aspergillus nidulans Species 0.000 description 1
- 108010006654 Bleomycin Proteins 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 239000004970 Chain extender Substances 0.000 description 1
- 101150094690 GAL1 gene Proteins 0.000 description 1
- 102100028501 Galanin peptides Human genes 0.000 description 1
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- 101100121078 Homo sapiens GAL gene Proteins 0.000 description 1
- GRRNUXAQVGOGFE-UHFFFAOYSA-N Hygromycin-B Natural products OC1C(NC)CC(N)C(O)C1OC1C2OC3(C(C(O)C(O)C(C(N)CO)O3)O)OC2C(O)C(CO)O1 GRRNUXAQVGOGFE-UHFFFAOYSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 1
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 1
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- OYHQOLUKZRVURQ-HZJYTTRNSA-N Linoleic acid Chemical compound CCCCC\C=C/C\C=C/CCCCCCCC(O)=O OYHQOLUKZRVURQ-HZJYTTRNSA-N 0.000 description 1
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 1
- 241000907973 Mortierella capitata Species 0.000 description 1
- 241000133028 Mortierella cogitans Species 0.000 description 1
- 241001219224 Mortierella elongata Species 0.000 description 1
- 241000048020 Mortierella exigua Species 0.000 description 1
- 241000907975 Mortierella gamsii Species 0.000 description 1
- 241000133355 Mortierella hygrophila Species 0.000 description 1
- 241001238138 Mortierella turficola Species 0.000 description 1
- 241000221961 Neurospora crassa Species 0.000 description 1
- MUPFEKGTMRGPLJ-RMMQSMQOSA-N Raffinose Natural products O(C[C@H]1[C@@H](O)[C@H](O)[C@@H](O)[C@@H](O[C@@]2(CO)[C@H](O)[C@@H](O)[C@@H](CO)O2)O1)[C@@H]1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 MUPFEKGTMRGPLJ-RMMQSMQOSA-N 0.000 description 1
- 108020005091 Replication Origin Proteins 0.000 description 1
- 101100370749 Streptomyces coelicolor (strain ATCC BAA-471 / A3(2) / M145) trpC1 gene Proteins 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- MUPFEKGTMRGPLJ-UHFFFAOYSA-N UNPD196149 Natural products OC1C(O)C(CO)OC1(CO)OC1C(O)C(O)C(O)C(COC2C(C(O)C(O)C(CO)O2)O)O1 MUPFEKGTMRGPLJ-UHFFFAOYSA-N 0.000 description 1
- 241000306282 Umbelopsis isabellina Species 0.000 description 1
- 241000180122 Umbelopsis vinacea Species 0.000 description 1
- 235000020661 alpha-linolenic acid Nutrition 0.000 description 1
- DTOSIQBPPRVQHS-PDBXOOCHSA-N alpha-linolenic acid Chemical compound CC\C=C/C\C=C/C\C=C/CCCCCCCC(O)=O DTOSIQBPPRVQHS-PDBXOOCHSA-N 0.000 description 1
- 229960000723 ampicillin Drugs 0.000 description 1
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 1
- 238000004458 analytical method Methods 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 230000001851 biosynthetic effect Effects 0.000 description 1
- 229960001561 bleomycin Drugs 0.000 description 1
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 1
- 239000000872 buffer Substances 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 229960005091 chloramphenicol Drugs 0.000 description 1
- WIIZWVCIJKGZOK-RKDXNWHRSA-N chloramphenicol Chemical compound ClC(Cl)C(=O)N[C@H](CO)[C@H](O)C1=CC=C([N+]([O-])=O)C=C1 WIIZWVCIJKGZOK-RKDXNWHRSA-N 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 239000000470 constituent Substances 0.000 description 1
- PYRZPBDTPRQYKG-UHFFFAOYSA-N cyclopentene-1-carboxylic acid Chemical compound OC(=O)C1=CCCC1 PYRZPBDTPRQYKG-UHFFFAOYSA-N 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- 239000000839 emulsion Substances 0.000 description 1
- 235000004626 essential fatty acids Nutrition 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 238000004108 freeze drying Methods 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 238000004817 gas chromatography Methods 0.000 description 1
- 230000007614 genetic variation Effects 0.000 description 1
- 229960002518 gentamicin Drugs 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- GRRNUXAQVGOGFE-NZSRVPFOSA-N hygromycin B Chemical compound O[C@@H]1[C@@H](NC)C[C@@H](N)[C@H](O)[C@H]1O[C@H]1[C@H]2O[C@@]3([C@@H]([C@@H](O)[C@@H](O)[C@@H](C(N)CO)O3)O)O[C@H]2[C@@H](O)[C@@H](CO)O1 GRRNUXAQVGOGFE-NZSRVPFOSA-N 0.000 description 1
- 229940097277 hygromycin b Drugs 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000009776 industrial production Methods 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 235000020778 linoleic acid Nutrition 0.000 description 1
- OYHQOLUKZRVURQ-IXWMQOLASA-N linoleic acid Natural products CCCCC\C=C/C\C=C\CCCCCCCC(O)=O OYHQOLUKZRVURQ-IXWMQOLASA-N 0.000 description 1
- 229960004488 linolenic acid Drugs 0.000 description 1
- KQQKGWQCNNTQJW-UHFFFAOYSA-N linolenic acid Natural products CC=CCCC=CCC=CCCCCCCCC(O)=O KQQKGWQCNNTQJW-UHFFFAOYSA-N 0.000 description 1
- -1 lipid A lipid Chemical class 0.000 description 1
- 239000002502 liposome Substances 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 230000004060 metabolic process Effects 0.000 description 1
- 230000007269 microbial metabolism Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 235000014593 oils and fats Nutrition 0.000 description 1
- 235000021315 omega 9 monounsaturated fatty acids Nutrition 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 230000002018 overexpression Effects 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 230000035790 physiological processes and functions Effects 0.000 description 1
- 229910052697 platinum Inorganic materials 0.000 description 1
- 230000032361 posttranscriptional gene silencing Effects 0.000 description 1
- 239000002244 precipitate Substances 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- MUPFEKGTMRGPLJ-ZQSKZDJDSA-N raffinose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO[C@@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)O)O1 MUPFEKGTMRGPLJ-ZQSKZDJDSA-N 0.000 description 1
- 230000003252 repetitive effect Effects 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 230000028070 sporulation Effects 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 150000003626 triacylglycerols Chemical class 0.000 description 1
- 101150016309 trpC gene Proteins 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 108700026220 vif Genes Proteins 0.000 description 1
- 235000013343 vitamin Nutrition 0.000 description 1
- 239000011782 vitamin Substances 0.000 description 1
- 229940088594 vitamin Drugs 0.000 description 1
- 229930003231 vitamin Natural products 0.000 description 1
- 150000003722 vitamin derivatives Chemical class 0.000 description 1
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/64—Fats; Fatty oils; Ester-type waxes; Higher fatty acids, i.e. having at least seven carbon atoms in an unbroken chain bound to a carboxyl group; Oxidised oils or fats
- C12P7/6436—Fatty acid esters
- C12P7/6445—Glycerides
- C12P7/6472—Glycerides containing polyunsaturated fatty acid [PUFA] residues, i.e. having two or more double bonds in their backbone
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/14—Fungi; Culture media therefor
- C12N1/145—Fungal isolates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/88—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation using microencapsulation, e.g. using amphiphile liposome vesicle
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/64—Fats; Fatty oils; Ester-type waxes; Higher fatty acids, i.e. having at least seven carbon atoms in an unbroken chain bound to a carboxyl group; Oxidised oils or fats
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/64—Fats; Fatty oils; Ester-type waxes; Higher fatty acids, i.e. having at least seven carbon atoms in an unbroken chain bound to a carboxyl group; Oxidised oils or fats
- C12P7/6436—Fatty acid esters
- C12P7/6445—Glycerides
- C12P7/6463—Glycerides obtained from glyceride producing microorganisms, e.g. single cell oil
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/645—Fungi ; Processes using fungi
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Biotechnology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Oil, Petroleum & Natural Gas (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Cell Biology (AREA)
- Botany (AREA)
- Mycology (AREA)
- Biophysics (AREA)
- Tropical Medicine & Parasitology (AREA)
- Virology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Farming Of Fish And Shellfish (AREA)
- Fodder In General (AREA)
Abstract
【解決手段】 モルティエレラ(Mortierella)属に属する脂質生産菌の育種方法であって、上記脂質生産菌における特定の遺伝子の発現を抑制する発現抑制工程を含む脂質生産菌の育種方法によれば、モルティエレラ属に属する脂質生産菌を効率的かつ有効に育種することができる。
【選択図】 なし
Description
変異処理により集団作出工程を行う場合、様々な手法で微生物に突然変異を生じさせて集団を作出するが、突然変異そのものは無作為に多くの種類が生じることになる。そのため、その後の選抜工程にて、目的の形質を示す品種(株)を取得できたとしても、目的の形質に関わる遺伝子以外の他の遺伝子に予想外の損傷が生じている可能性が否めない。例えば、上記脂質生産菌の場合では、生産される脂質の種類に変化が生じても、増殖や胞子形成能などが悪くなること等があり得る。したがって、変異処理による集団作出工程では、必ずしも生産性のよい株を得ることができるとは限らない。
これに対して、形質転換により集団作出工程を行う場合、育種しようとする有用生物を宿主として、目的の性質を獲得するために必要なDNA断片を導入(形質転換)することにより形質転換体の集団を得る。すなわち、目的に合わせた特定の遺伝子のみを発現制御した集団を作出することになる。そのため、その後の選抜工程では、得られた形質転換体の中からより望ましい品種(株)のみを選抜すればよいことになるので、スクリーニングが容易になるだけでなく、上記他の遺伝子に予想外の損傷が生じることも回避できる。したがって、育種にかける労力を著しく低減することができる。
(a)上記特定の遺伝子が有する塩基配列の全体または一部に相当する2本鎖RNAを発現する組換え発現ベクター。
(b)(a)の組換え発現ベクターを構築するための試薬類。
(c)(a)の組換え発現ベクターを脂質生産菌に導入するための試薬類。
(d)脂質生産菌及び/又は(a)の組換え発現ベクターが導入された形質転換株を培養するための試薬類。
本発明に係る育種方法の対象となるモルティエレラ属の菌類としては、特に限定されるものではなく、モルティエレラ属に分類される各種糸状菌を挙げることができる。モルティエレラ属は、MortierellaとMicromucorとの2亜属に分けられる。Mortierella亜属の菌は全て、アラキドン酸等の炭素数20の脂肪酸を生産するが、Micromucor亜属は炭素数18以下の脂肪酸しか生産しない。本発明の対象となるモルティエレラ属の菌類は上記2亜属の何れであってもよい。
本発明に係る育種方法は、上記発現抑制工程を含むものであればよく、その他の工程、材料、条件等は、特に限定されるものではない。ここでいう「発現抑制工程」とは、脂質生産菌における特定の遺伝子の発現を抑制する工程であればよく、その具体的な手法等は従来公知の方法を利用でき、特に限定されるものではない。例えば、相同組換えによる染色体上の遺伝子を欠失させる方法、コサプレッション法、アンチセンス法、RNAi法などを用いることができよう。これらの中でも、RNAi法がその手法の簡便性や優れた効果等の理由から特に好ましい。
本発明において行われるRNAi工程は、脂質生産菌における特定の遺伝子の発現を、RNAi法により抑制する工程であればよく、その他の具体的な方法、条件、材料等は特に限定されるものではない。
本発明において行われる発現ベクター構築工程は、モルティエレラ属の菌類(脂質生産菌)において、発現を抑制したい所定の遺伝子が有する塩基配列の全体または一部に相当する2本鎖RNAを発現する組換え発現ベクターを構築する工程であればよい。すなわち、モルティエレラ属の菌類において、RNAiを引き起こす2本鎖RNAが発現されるように、プロモーターの支配下に相当な配列のDNAを連結した組換え発現ベクターを構築する工程であれば特に限定されるものではない。つまり、プロモーターによって上記2本鎖RNAをコードする遺伝子が発現するように組換え発現ベクターを構築する工程ともいえる。
本発明において行われる形質転換工程は、上記脂質生産菌に、上記特定の遺伝子が有する塩基配列の全体または一部に相当する2本鎖RNAを発現する組換え発現ベクターを導入する工程であればよく、その他の具体的な工程、条件、材料等は特に限定されるものではない。つまり、上記発現ベクター構築工程において構築した組換え発現ベクターを、上記脂質生産菌に導入(形質転換)する工程であればよい。
本発明に係る育種方法においては、上記形質転換工程や、さらに上記組換え発現ベクター構築工程が含まれていてもよいが、さらに他の工程が含まれていてもよい。具体的には、形質転換後の脂質生産菌群から適切な形質転換株を選抜する選抜工程等を挙げることができる。
〔3−1〕脂質生産菌(新品種)
本発明に係る脂質生産菌の育種方法は、上記のように、特定の遺伝子の発現を抑制して、より望ましい性質(形質)を示す品種を取得するようになっている。それゆえ、モルティエレラ属に属する脂質生産菌を原種として、新規品種(新規株)を効率的かつ有効に生産することが可能となる。モルティエレラ属の菌は脂質生産菌として良く知られており、M. alpina のように信頼性の高いものも含まれているため、例えば、脂質生産性をより一層高めた株を容易かつ効率的に生産することが可能となる。
本発明に係る育種キットは、上記〔2〕欄で説明した育種方法を実施するためのものであればよく、これに含まれる具体的な構成、材料、機器等は、特に限定されるものではない。具体的には、上記育種方法の各工程を実施するための物が含まれていればよい。
本発明に係る脂質の生産方法は、上述の〔3−1〕欄で説明した脂質生産菌からPUFA含有脂質を生産する方法であればよい。例えば、上記脂質生産菌を培養することにより、PUFA含有脂質を簡便に生産することができる。
・上記の脂質の生産方法によって生産された脂質であって、該脂質を構成する全脂肪酸に占めるω9系PUFAの割合が8%以上である脂質。
・上記の脂質の生産方法によって生産された脂質であって、該脂質を構成する全脂肪酸に占めるミード酸の割合が1.3%以上である脂質。
・上記の脂質の生産方法によって生産された脂質であって、該脂質を構成する全脂肪酸に占めるアラキドン酸の割合が10%以上で、かつ該脂質を構成する全脂肪酸に占める超長鎖飽和脂肪酸の割合が0.1%以下である脂質。
MAELO遺伝子を酵母で発現させるためのプラスミドを以下のとおり作製した。
MAELO-S1:5’-gcactagtttagatgtgcttgctgttggag-3’(配列番号16)
プラスミドpY2MELにより、S.cerevisiae INVSc1株を形質転換し、ウラシルを含まないプレート(グルコース2%、Yeast Nitrogen Base w/o Amino Acids and Ammonium Sulfate (Difco社製)0.17%、硫酸アンモニウム0.5%、ヒスチジン 20mg/l、ロイシン60mg/l、トリプトファン40mg/l、Bacto agar2%)上で生育してきた株を形質転換株として選択した。形質転換株を、ラフィノース2%、Yeast Nitrogen Base w/o Amino Acids and Ammonium Sulfate (Difco社製)0.17%、硫酸アンモニウム0.5%、タージトール タイプNP-40 1%、ヒスチジン 20mg/l、ロイシン60mg/l、トリプトファン40mg/l、ステアリン酸(18:0) 0.05%を含む培地に1白金時植菌し、6時間振とう培養した。つづいてプラスミドpY2MEL上でGAL1プロモーター下流に連結されたMAELO遺伝子の発現を誘導するために2%(W/V)がラクトースを加えて、さらに28℃にて42時間振とう培養した。遠心分離により菌体を集め、凍結乾燥した後、塩酸メタノール法により菌体内の脂肪酸残基をメチルエステルに誘導した後、ヘキサンで抽出し、ヘキサンを留去したのち得られる脂肪酸メチルエステルをガスクロマトグラフィーで分析した。培養液あたりの超長鎖飽和脂肪酸の生成量を表に示す。
MAELO遺伝子の一部分に対応する2本鎖RNAを過剰発現させるためのプラスミドを以下のとおり構築した。
TrpCRX;5’-gaagaattccctctaaacaagtgtacctgt-3’(配列番号2)
M. alpina をGY液体培地(2%グルコース、1%酵母エキス、pH6.0に調整)にて、28℃で5日間培養し、得られた菌体から文献(E. Sakuradani et al. Eur J. Biochem., 260, 208-216, 1999)の記載の方法に従い、ゲノムDNAを調整した。
MAELORNAi3-1;5’-aaccatggtcatccctaggtggaagtaatg-3’(配列番号4)
M. alpina のゲノムDNAを鋳型として、プライマーMAELORNAi1とプライマーMAELORNAi5とで、LA Taq(タカラバイオ)を用いてPCRを行った。反応条件は、94℃ 1分、55℃ 1分、72℃ 1分を1サイクルとして、30サイクル行った。こうして増幅された約0.7kbのDNA断片をベクターpTBlueT-Vector(タカラバイオ)にTAクローニングした。塩基配列を確認した後、制限酵素NcoIとBlnIとで消化し、得られた約0.7kbのDNA断片をプラスミドpBlueMEi3のNcoI−BlnIサイトに挿入し、プラスミドpBlueMEi5を構築した。
MAERORNAi5;5’-tgatctcctaggtggaacactgatagccac-3’(配列番号6)
プラスミドpBlueMEi5を制限酵素EcoRIで消化して得られた約3.3kbの断片をプラスミドpDuraのEcoRIサイトに挿入し、プラスミドpDura5MEi51を構築した。
次に、M.alpinaのゲノムDNAを鋳型として、プライマーMAELORNAi1とプライマーMAELORNAi4で、LA Taq(タカラバイオ)を用いてPCRを行った。PCRは、94℃ 1分、55℃ 1分、72℃ 1分を1サイクルとして、30サイクルの反応を行った。こうして増幅された約0.5kbのDNA断片をベクターpT7Blue T-Vector(タカラバイオ)にTAクローニングした。塩基配列を確認したあと、制限酵素NcoIとBlnIで消化し、得られた約0.7kbのDNA断片をプラスミドpBlueMEi2のNcoI−BlnIサイトに挿入し、プラスミドpBlueMEi4を構築した。
プラスミドpBlueMEi4を制限酵素EcoRIで消化して得られた約2.9kbの断片をプラスミドpDura5のEcoRIサイトに挿入し、プラスミドpDura5Mei41を構築した。プラスミドpDura5Mei51では約700bp、プラスミドpDura5Mei41では約500bpのMAELO遺伝子に対応する2本鎖RNAを過剰発現させることが可能である。
プライマーRDNA2;5’-cgctgcgttcttcatcgatg-3’(配列番号8)
上記形質転換株を、GY液体培地10mlを入れた試験管に植菌し、28℃で12日間振とう培養し、ろ過により集菌した。
プライマーMAELO-2;5’-cggtgtcagccaactcccagtactt-3’(配列番号10)
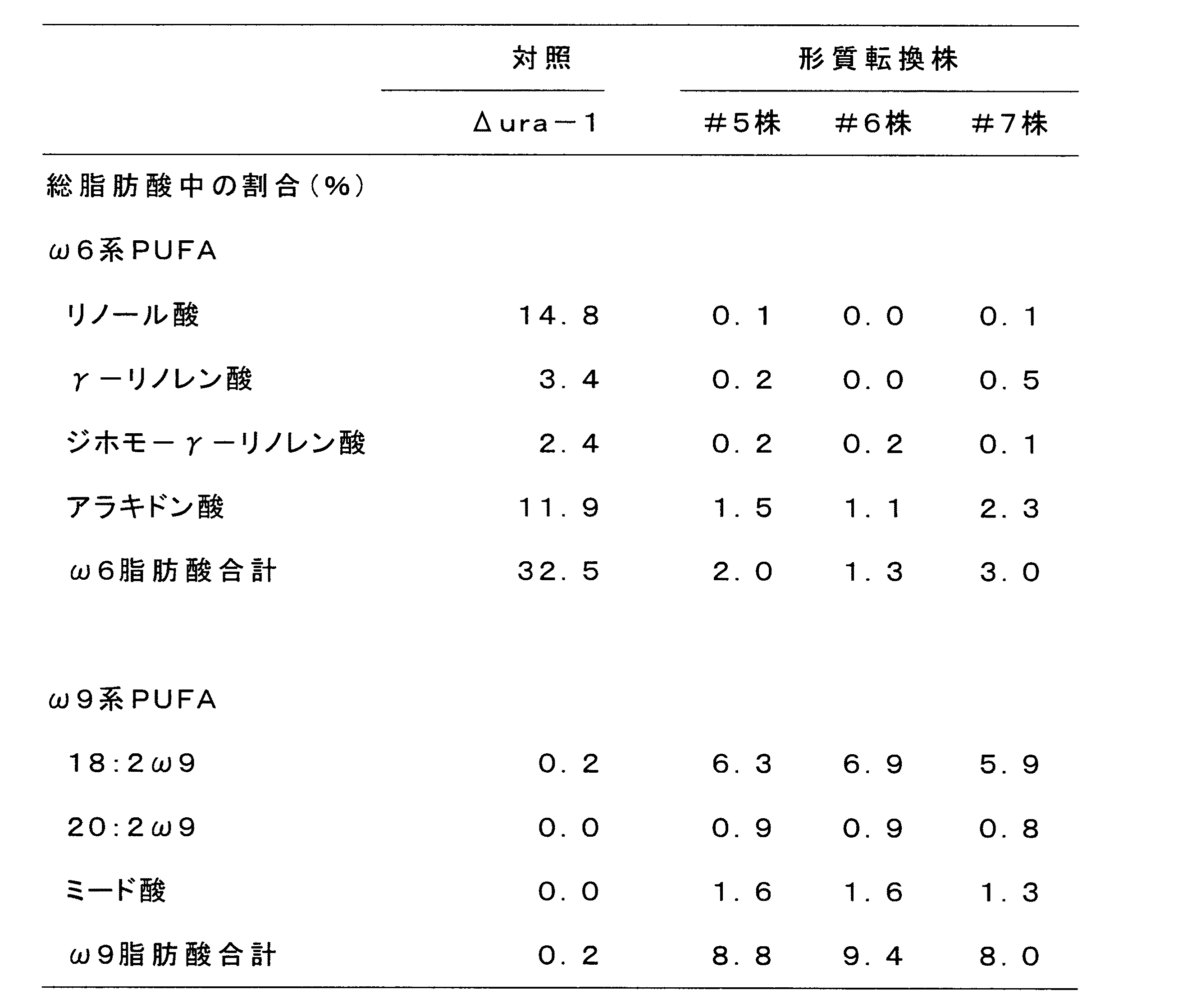
得られたPCR産物を電気泳動し、約340bpのフラグメントのバンドについて蛍光強度を比較した。その結果、宿主としたΔura-1株では明確なバンドが確認できたのに対して、形質転換株#1〜#4ではいずれもバンドを確認することができなかった。このことから、形質転換株#1〜#4ではMAELO遺伝子のmRNAの発現が抑制されていることが確認できた。残りの菌体を乾燥し、塩酸メタノール法により菌体内の脂肪酸残基をメチルエステルに誘導した後、ヘキサンで抽出し、ヘキサンを留去したのち得られる脂肪酸メチルエステルをガスクロマトグラフィーで分析した。その結果を下記表2に示す。
Δ12脂肪酸不飽和化酵素遺伝子の一部分に対応する2本鎖RNAを過剰発現させるために以下のベクターを構築した。
プライマーΔ12−2;5’-agaggccttcataataaggtacgcaggc-3’(配列番号12)
増幅されたDNA断片を制限酵素BamHIおよびStuIにて消化し、プラスミドpMOD10をBamHIおよびMscIで消化して得られた約3.7kbの断片とligation high(東洋紡)を用いて連結し、プラスミドpBΔ12RNAiを得た。続いて、プラスミドpBΔ12RNAiを制限酵素EcoRIで消化し、DNA blunting kit(タカラバイオ)を用いて末端を平滑化した後、制限酵素BamHIで消化して約1.1kbの断片を得た。一方、プラスミドpBlueHptを制限酵素NcoIで消化し、DNA blunting kit(タカラバイオ)を用いて末端を平滑化した後、制限酵素BamHIで消化して約4.7kbのDNA断片を得た。これら2つの断片をligation high(東洋紡)を用いて連結し、プラスミドpBlueΔ12RNAiを得た。これを制限酵素EcoRIで消化して得られた2.8kbのDNA断片をプラスミドpDura5のEcoRIサイトに挿入することによってプラスミドpDura5Δ12RNAiを得た。
プライマーΔ12−4;5’-tgggaacaaagacctggtccttgg-3’(配列番号14)
得られたPCR産物を電気泳動し、約310bpのフラグメントのばんどについて蛍光強度を比較した。その結果、宿主としてΔura-1株では明確なバンドが確認できたのに対して、形質転換株#5〜#7ではいずれもバンドを確認することができなかった。このことから、形質転換株#5〜#7では、Δ12脂肪酸不飽和化酵素遺伝子のmRNAの発現が抑制されていることが確認できた。
Claims (15)
- モルティエレラ(Mortierella)属に属する脂質生産菌の育種方法であって、
上記脂質生産菌における特定の遺伝子の発現を抑制する発現抑制工程を含むことを特徴とする脂質生産菌の育種方法。 - 上記発現抑制工程は、特定の遺伝子の発現をRNAi法により抑制するRNAi工程を含むことを特徴とする請求項1に記載の脂質生産菌の育種方法。
- 上記RNAi工程は、上記脂質生産菌に、上記特定の遺伝子が有する塩基配列の全体または一部に対応する2本鎖RNAを発現する組換え発現ベクターを導入する形質転換工程を含むことを特徴とする請求項2に記載の脂質生産菌の育種方法。
- 上記RNAi工程は、さらに、上記組換え発現ベクターを構築する発現ベクター構築工程を含むことを特徴とする請求項3に記載の脂質生産菌の育種方法。
- 上記形質転換工程では、エレクトロポレーション法、またはパーティクルデリバリー法が用いられることを特徴とする請求項3または4に記載の脂質生産菌の育種方法。
- 上記脂質生産菌は、モルティエレラ アルピナ(Mortierella alpina)であることを特徴とする請求項1〜5のいずれか1項に記載の脂質生産菌の育種方法。
- 上記特定の遺伝子は、脂質代謝遺伝子であることを特徴とする請求項1〜6のいずれか1項に記載の脂質生産菌の育種方法。
- 上記脂質代謝遺伝子は、脂肪酸代謝遺伝子であることを特徴とする請求項7に記載の脂質生産菌の育種方法。
- 上記脂肪酸代謝遺伝子は、脂肪酸鎖長延長酵素または脂肪酸不飽和化酵素をコードする遺伝子であることを特徴とする請求項8に記載の脂質生産菌の育種方法。
- 上記脂肪酸鎖長延長酵素をコードする遺伝子は、GLELO遺伝子またはMAELO遺伝子であることを特徴とする請求項9に記載の脂質生産菌の育種方法。
- 上記脂肪酸不飽和化酵素をコードする遺伝子は、Δ5脂肪酸不飽和化酵素、Δ6脂肪酸不飽和化酵素、Δ8脂肪酸不飽和化酵素、Δ9脂肪酸不飽和化酵素、Δ12脂肪酸不飽和化酵素、Δ15脂肪酸不飽和化酵素、Δ17脂肪酸不飽和化酵素、ω3脂肪酸不飽和化酵素から選ばれる1つの酵素をコード遺伝子であることを特徴とする請求項9に記載の脂質生産菌の育種方法。
- 請求項1〜11のいずれか1項に記載の脂質生産菌の育種方法を実施するための育種キット。
- 上記育種キットは、以下の(a)〜(d)のうち、少なくとも1つの物質を含むことを特徴とする請求項12に記載の育種キット。
(a)上記特定の遺伝子が有する塩基配列の全体または一部に相当する2本鎖RNAを発現する組換え発現ベクター。
(b)(a)の組換え発現ベクターを構築するための試薬類。
(c)(a)の組換え発現ベクターを脂質生産菌に導入するための試薬類。
(d)脂質生産菌及び/又は(a)の組換え発現ベクターが導入された形質転換株を培養するための試薬類。 - 請求項1〜13のいずれか1項に記載の脂質生産菌の育種方法または育種キットによって得られることを特徴とする脂質生産菌。
- 請求項14に記載の脂質生産菌からPUFA含有脂質を生産することを特徴とする脂質の生産方法。
Priority Applications (14)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2004107512A JP4481702B2 (ja) | 2004-03-31 | 2004-03-31 | 脂質生産菌の育種方法およびその利用 |
| MYPI20051356A MY142164A (en) | 2004-03-31 | 2005-03-28 | Method of breeding lipid-producing strain and utilization of the same |
| PCT/JP2005/005786 WO2005095597A1 (ja) | 2004-03-31 | 2005-03-28 | 脂質生産菌の育種方法およびその利用 |
| CN2005800106128A CN1946843B (zh) | 2004-03-31 | 2005-03-28 | 脂质生产菌的育种方法及其利用 |
| US10/594,772 US20080138874A1 (en) | 2004-03-31 | 2005-03-28 | Breeding Method of Lipid Producing Fungi and Use of Such a Method |
| SG200901710-4A SG151257A1 (en) | 2004-03-31 | 2005-03-28 | Method of breeding lipid-producing strain and utilization of the same |
| DK05727395.5T DK1748068T3 (da) | 2004-03-31 | 2005-03-28 | Fremgangsmåde til dyrkning af en lipidproducerende stamme og udnyttelse deraf |
| AT05727395T ATE459713T1 (de) | 2004-03-31 | 2005-03-28 | Verfahren zur züchtung eines lipidproduzierenden stamms und nutzung davon |
| DE602005019717T DE602005019717D1 (de) | 2004-03-31 | 2005-03-28 | Verfahren zur züchtung eines lipidproduzierenden stamms und nutzung davon |
| KR1020067022753A KR101158591B1 (ko) | 2004-03-31 | 2005-03-28 | 지질 생산균의 육종 방법 및 그 이용 |
| EP05727395A EP1748068B1 (en) | 2004-03-31 | 2005-03-28 | Method of breeding lipid-producing strain and utilization of the same |
| AU2005227827A AU2005227827B2 (en) | 2004-03-31 | 2005-03-28 | Method of breeding lipid-producing strain and utilization of the same |
| CA2560952A CA2560952C (en) | 2004-03-31 | 2005-03-28 | Method for the production of lipid-producing fungi |
| TW094110401A TWI349703B (en) | 2004-03-31 | 2005-03-31 | Breeding method of lipid producing fungi and use of such a method |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2004107512A JP4481702B2 (ja) | 2004-03-31 | 2004-03-31 | 脂質生産菌の育種方法およびその利用 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2005287403A true JP2005287403A (ja) | 2005-10-20 |
| JP4481702B2 JP4481702B2 (ja) | 2010-06-16 |
Family
ID=35063777
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2004107512A Expired - Lifetime JP4481702B2 (ja) | 2004-03-31 | 2004-03-31 | 脂質生産菌の育種方法およびその利用 |
Country Status (14)
| Country | Link |
|---|---|
| US (1) | US20080138874A1 (ja) |
| EP (1) | EP1748068B1 (ja) |
| JP (1) | JP4481702B2 (ja) |
| KR (1) | KR101158591B1 (ja) |
| CN (1) | CN1946843B (ja) |
| AT (1) | ATE459713T1 (ja) |
| AU (1) | AU2005227827B2 (ja) |
| CA (1) | CA2560952C (ja) |
| DE (1) | DE602005019717D1 (ja) |
| DK (1) | DK1748068T3 (ja) |
| MY (1) | MY142164A (ja) |
| SG (1) | SG151257A1 (ja) |
| TW (1) | TWI349703B (ja) |
| WO (1) | WO2005095597A1 (ja) |
Cited By (22)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2007145943A (ja) * | 2005-11-25 | 2007-06-14 | Mitsubishi Paper Mills Ltd | 蓄熱材マイクロカプセル、蓄熱材マイクロカプセル分散液および蓄熱材マイクロカプセル固形物 |
| WO2009008466A1 (ja) * | 2007-07-11 | 2009-01-15 | Suntory Holdings Limited | ホスファチジン酸ホスファターゼホモログとその利用 |
| WO2009047992A1 (ja) | 2007-10-12 | 2009-04-16 | Suntory Holdings Limited | Udp-グルクロン酸転移酵素およびそれをコードするポリヌクレオチド |
| WO2010084879A1 (ja) | 2009-01-21 | 2010-07-29 | サントリーホールディングス株式会社 | フラボノイド3位グルクロン酸転移酵素、及びそれをコードするポリヌクレオチド |
| WO2010134519A1 (ja) | 2009-05-19 | 2010-11-25 | サントリーホールディングス株式会社 | フロフラン型リグナン4位グルコース転移酵素、及びそれをコードするポリヌクレオチド |
| WO2011034199A1 (ja) | 2009-09-18 | 2011-03-24 | サントリーホールディングス株式会社 | グリセロール-3-リン酸アシル基転移酵素 |
| US7927845B2 (en) | 2006-05-08 | 2011-04-19 | Suntory Holdings Limited | Fatty acid synthetase, polynucleotide encoding the same, and uses thereof |
| WO2011078134A1 (ja) | 2009-12-21 | 2011-06-30 | サントリーホールディングス株式会社 | ジアシルグリセロールアシル基転移酵素遺伝子及びその用途 |
| WO2011093509A1 (ja) | 2010-02-01 | 2011-08-04 | サントリーホールディングス株式会社 | アシル-CoAシンセターゼホモログをコードするポリヌクレオチド及びその用途 |
| WO2012023583A1 (ja) | 2010-08-19 | 2012-02-23 | サントリーホールディングス株式会社 | フラボノール8-水酸化活性を有するフラビン酵素及びその用途 |
| WO2012091165A1 (ja) | 2010-12-28 | 2012-07-05 | サントリーホールディングス株式会社 | モノテルペン配糖体化酵素の利用方法 |
| WO2013018879A1 (ja) | 2011-08-04 | 2013-02-07 | サントリーホールディングス株式会社 | 脂肪酸鎖長延長促進活性を有するタンパク質、これをコードする遺伝子及びその用途 |
| WO2013118906A1 (ja) | 2012-02-06 | 2013-08-15 | サントリーホールディングス株式会社 | ホップ由来モノテルペン配糖体化酵素及びその利用方法 |
| WO2013137487A1 (ja) | 2012-03-16 | 2013-09-19 | サントリーホールディングス株式会社 | ステビオール配糖体化酵素およびそれをコードする遺伝子 |
| WO2013146555A1 (ja) | 2012-03-27 | 2013-10-03 | サントリーホールディングス株式会社 | ステビオール配糖体の製造方法 |
| WO2013180306A1 (ja) | 2012-05-30 | 2013-12-05 | サントリーホールディングス株式会社 | ステビオール配糖体化酵素およびそれをコードする遺伝子 |
| KR20140032939A (ko) * | 2010-10-01 | 2014-03-17 | 고쿠리쓰다이가쿠호진 규슈다이가쿠 | 스트라메노파일의 형질 전환 방법 |
| WO2014157736A1 (ja) | 2013-03-27 | 2014-10-02 | サントリーホールディングス株式会社 | モルティエレラ属微生物内で高発現活性を示すプロモーター |
| WO2015016393A1 (ja) | 2013-08-02 | 2015-02-05 | サントリーホールディングス株式会社 | ヘキセノール配糖体化酵素の利用方法 |
| WO2018124143A1 (ja) | 2016-12-27 | 2018-07-05 | サントリーホールディングス株式会社 | 新規ステビオール配糖体およびその製造方法、ならびにそれを含む甘味料組成物 |
| WO2018181515A1 (ja) | 2017-03-31 | 2018-10-04 | サントリーホールディングス株式会社 | 新規ステビオール配糖体およびその製造方法、ならびにそれを含む甘味料組成物 |
| WO2021020516A1 (ja) | 2019-07-31 | 2021-02-04 | サントリーホールディングス株式会社 | 新規ステビオール配糖体およびその製造方法、ならびにそれを含む甘味料組成物 |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20090169893A1 (en) * | 2005-11-17 | 2009-07-02 | Koshiro Ikegami | Thermal Storage Material Microcapsules, Thermal Storage Material Microcapsule Dispersion and Thermal Storage Material Microcapsule Solid |
| CN110747133B (zh) * | 2019-11-18 | 2021-03-02 | 河南农业大学 | 一株头孢被孢霉、包括头孢被孢霉的菌剂及其制备方法和应用 |
Family Cites Families (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5204250A (en) * | 1986-03-31 | 1993-04-20 | Suntory Limited | Process for production of arachidonic acid |
| JP3995290B2 (ja) * | 1996-08-23 | 2007-10-24 | サントリー株式会社 | オメガ9系高度不飽和脂肪酸及びそれを含有する脂質の製造方法 |
| US6015703A (en) * | 1998-03-10 | 2000-01-18 | Iogen Corporation | Genetic constructs and genetically modified microbes for enhanced production of beta-glucosidase |
| US6403349B1 (en) * | 1998-09-02 | 2002-06-11 | Abbott Laboratories | Elongase gene and uses thereof |
| CN1362522A (zh) * | 2001-12-26 | 2002-08-07 | 武汉烯王生物工程有限公司 | 用离子束生物工程诱变菌生产含花生四烯酸油脂的方法 |
-
2004
- 2004-03-31 JP JP2004107512A patent/JP4481702B2/ja not_active Expired - Lifetime
-
2005
- 2005-03-28 WO PCT/JP2005/005786 patent/WO2005095597A1/ja active Application Filing
- 2005-03-28 AT AT05727395T patent/ATE459713T1/de not_active IP Right Cessation
- 2005-03-28 EP EP05727395A patent/EP1748068B1/en not_active Not-in-force
- 2005-03-28 DE DE602005019717T patent/DE602005019717D1/de active Active
- 2005-03-28 SG SG200901710-4A patent/SG151257A1/en unknown
- 2005-03-28 CN CN2005800106128A patent/CN1946843B/zh not_active Expired - Fee Related
- 2005-03-28 AU AU2005227827A patent/AU2005227827B2/en not_active Ceased
- 2005-03-28 US US10/594,772 patent/US20080138874A1/en not_active Abandoned
- 2005-03-28 CA CA2560952A patent/CA2560952C/en not_active Expired - Fee Related
- 2005-03-28 MY MYPI20051356A patent/MY142164A/en unknown
- 2005-03-28 KR KR1020067022753A patent/KR101158591B1/ko active IP Right Grant
- 2005-03-28 DK DK05727395.5T patent/DK1748068T3/da active
- 2005-03-31 TW TW094110401A patent/TWI349703B/zh not_active IP Right Cessation
Cited By (27)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2007145943A (ja) * | 2005-11-25 | 2007-06-14 | Mitsubishi Paper Mills Ltd | 蓄熱材マイクロカプセル、蓄熱材マイクロカプセル分散液および蓄熱材マイクロカプセル固形物 |
| US7927845B2 (en) | 2006-05-08 | 2011-04-19 | Suntory Holdings Limited | Fatty acid synthetase, polynucleotide encoding the same, and uses thereof |
| JP5180960B2 (ja) * | 2007-07-11 | 2013-04-10 | サントリーホールディングス株式会社 | ホスファチジン酸ホスファターゼホモログとその利用 |
| WO2009008466A1 (ja) * | 2007-07-11 | 2009-01-15 | Suntory Holdings Limited | ホスファチジン酸ホスファターゼホモログとその利用 |
| WO2009047992A1 (ja) | 2007-10-12 | 2009-04-16 | Suntory Holdings Limited | Udp-グルクロン酸転移酵素およびそれをコードするポリヌクレオチド |
| WO2010084879A1 (ja) | 2009-01-21 | 2010-07-29 | サントリーホールディングス株式会社 | フラボノイド3位グルクロン酸転移酵素、及びそれをコードするポリヌクレオチド |
| WO2010134519A1 (ja) | 2009-05-19 | 2010-11-25 | サントリーホールディングス株式会社 | フロフラン型リグナン4位グルコース転移酵素、及びそれをコードするポリヌクレオチド |
| WO2011034199A1 (ja) | 2009-09-18 | 2011-03-24 | サントリーホールディングス株式会社 | グリセロール-3-リン酸アシル基転移酵素 |
| WO2011078134A1 (ja) | 2009-12-21 | 2011-06-30 | サントリーホールディングス株式会社 | ジアシルグリセロールアシル基転移酵素遺伝子及びその用途 |
| EP2930236A1 (en) | 2010-02-01 | 2015-10-14 | Suntory Holdings Limited | Polynucleotide encoding acyl-CoA synthetase homolog and method for using the same |
| WO2011093509A1 (ja) | 2010-02-01 | 2011-08-04 | サントリーホールディングス株式会社 | アシル-CoAシンセターゼホモログをコードするポリヌクレオチド及びその用途 |
| EP2772538A1 (en) | 2010-02-01 | 2014-09-03 | Suntory Holdings Limited | Polynucleotide encoding acyl-CoA synthetase homolog and method for using the same |
| WO2012023583A1 (ja) | 2010-08-19 | 2012-02-23 | サントリーホールディングス株式会社 | フラボノール8-水酸化活性を有するフラビン酵素及びその用途 |
| KR20140032939A (ko) * | 2010-10-01 | 2014-03-17 | 고쿠리쓰다이가쿠호진 규슈다이가쿠 | 스트라메노파일의 형질 전환 방법 |
| KR101964168B1 (ko) | 2010-10-01 | 2019-04-01 | 고쿠리쓰다이가쿠호진 규슈다이가쿠 | 스트라메노파일의 형질 전환 방법 |
| WO2012091165A1 (ja) | 2010-12-28 | 2012-07-05 | サントリーホールディングス株式会社 | モノテルペン配糖体化酵素の利用方法 |
| US8980591B2 (en) | 2011-08-04 | 2015-03-17 | Suntory Holdings Limited | Protein having activity to promote fatty acid chain elongation, gene encoding same and use thereof |
| WO2013018879A1 (ja) | 2011-08-04 | 2013-02-07 | サントリーホールディングス株式会社 | 脂肪酸鎖長延長促進活性を有するタンパク質、これをコードする遺伝子及びその用途 |
| WO2013118906A1 (ja) | 2012-02-06 | 2013-08-15 | サントリーホールディングス株式会社 | ホップ由来モノテルペン配糖体化酵素及びその利用方法 |
| WO2013137487A1 (ja) | 2012-03-16 | 2013-09-19 | サントリーホールディングス株式会社 | ステビオール配糖体化酵素およびそれをコードする遺伝子 |
| WO2013146555A1 (ja) | 2012-03-27 | 2013-10-03 | サントリーホールディングス株式会社 | ステビオール配糖体の製造方法 |
| WO2013180306A1 (ja) | 2012-05-30 | 2013-12-05 | サントリーホールディングス株式会社 | ステビオール配糖体化酵素およびそれをコードする遺伝子 |
| WO2014157736A1 (ja) | 2013-03-27 | 2014-10-02 | サントリーホールディングス株式会社 | モルティエレラ属微生物内で高発現活性を示すプロモーター |
| WO2015016393A1 (ja) | 2013-08-02 | 2015-02-05 | サントリーホールディングス株式会社 | ヘキセノール配糖体化酵素の利用方法 |
| WO2018124143A1 (ja) | 2016-12-27 | 2018-07-05 | サントリーホールディングス株式会社 | 新規ステビオール配糖体およびその製造方法、ならびにそれを含む甘味料組成物 |
| WO2018181515A1 (ja) | 2017-03-31 | 2018-10-04 | サントリーホールディングス株式会社 | 新規ステビオール配糖体およびその製造方法、ならびにそれを含む甘味料組成物 |
| WO2021020516A1 (ja) | 2019-07-31 | 2021-02-04 | サントリーホールディングス株式会社 | 新規ステビオール配糖体およびその製造方法、ならびにそれを含む甘味料組成物 |
Also Published As
| Publication number | Publication date |
|---|---|
| US20080138874A1 (en) | 2008-06-12 |
| DK1748068T3 (da) | 2010-06-07 |
| EP1748068A4 (en) | 2007-08-08 |
| TW200533747A (en) | 2005-10-16 |
| CN1946843B (zh) | 2011-10-12 |
| KR101158591B1 (ko) | 2012-06-22 |
| WO2005095597A1 (ja) | 2005-10-13 |
| AU2005227827B2 (en) | 2009-10-01 |
| AU2005227827A1 (en) | 2005-10-13 |
| SG151257A1 (en) | 2009-04-30 |
| EP1748068A1 (en) | 2007-01-31 |
| TWI349703B (en) | 2011-10-01 |
| CA2560952C (en) | 2013-04-30 |
| KR20070058377A (ko) | 2007-06-08 |
| EP1748068B1 (en) | 2010-03-03 |
| CN1946843A (zh) | 2007-04-11 |
| MY142164A (en) | 2010-10-15 |
| JP4481702B2 (ja) | 2010-06-16 |
| CA2560952A1 (en) | 2005-10-13 |
| DE602005019717D1 (de) | 2010-04-15 |
| ATE459713T1 (de) | 2010-03-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP4481702B2 (ja) | 脂質生産菌の育種方法およびその利用 | |
| US11203763B2 (en) | Microbial oil containing fatty acids obtained from stramenopile and method of producing the same | |
| US10815505B2 (en) | Microbial oil extracted from stramenopiles | |
| Sakuradani et al. | Single cell oil production by Mortierella alpina | |
| JP4537319B2 (ja) | 脂質生産菌の育種方法 | |
| Kikukawa et al. | Microbial production of dihomo-γ-linolenic acid by Δ5-desaturase gene-disruptants of Mortierella alpina 1S-4 | |
| JP6723002B2 (ja) | 油脂の製造方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20061218 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20071205 |
|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20071213 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A712 Effective date: 20090422 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20090908 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20091104 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20091201 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20100118 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20100309 |
|
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20100318 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130326 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20130326 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20140326 Year of fee payment: 4 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |