ES2923953T3 - Mejoramiento de patatas de semilla híbridas - Google Patents
Mejoramiento de patatas de semilla híbridas Download PDFInfo
- Publication number
- ES2923953T3 ES2923953T3 ES19152162T ES19152162T ES2923953T3 ES 2923953 T3 ES2923953 T3 ES 2923953T3 ES 19152162 T ES19152162 T ES 19152162T ES 19152162 T ES19152162 T ES 19152162T ES 2923953 T3 ES2923953 T3 ES 2923953T3
- Authority
- ES
- Spain
- Prior art keywords
- potato
- plants
- plant
- diploid
- homozygous
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01H—NEW PLANTS OR NON-TRANSGENIC PROCESSES FOR OBTAINING THEM; PLANT REPRODUCTION BY TISSUE CULTURE TECHNIQUES
- A01H6/00—Angiosperms, i.e. flowering plants, characterised by their botanic taxonomy
- A01H6/82—Solanaceae, e.g. pepper, tobacco, potato, tomato or eggplant
- A01H6/827—Solanum tuberosum [potato]
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01H—NEW PLANTS OR NON-TRANSGENIC PROCESSES FOR OBTAINING THEM; PLANT REPRODUCTION BY TISSUE CULTURE TECHNIQUES
- A01H5/00—Angiosperms, i.e. flowering plants, characterised by their plant parts; Angiosperms characterised otherwise than by their botanic taxonomy
- A01H5/04—Stems
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Botany (AREA)
- Developmental Biology & Embryology (AREA)
- Environmental Sciences (AREA)
- Physiology (AREA)
- Natural Medicines & Medicinal Plants (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- General Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Pretreatment Of Seeds And Plants (AREA)
- Medicines Containing Plant Substances (AREA)
Abstract
La presente invención se refiere a líneas de patata diploides, fértiles, autocompatibles y esencialmente homocigóticas, en las que dichas líneas comprenden un rasgo agronómicamente deseable como el vigor. La invención se refiere además a métodos para producir tales plantas ya semillas híbridas obtenidas cruzando tales líneas de patata homocigóticas ya plantas de patata cultivadas a partir de dichas semillas. (Traducción automática con Google Translate, sin valor legal)
Description
DESCRIPCIÓN
Mejoramiento de patatas de semilla híbridas
Campo de la invención
La invención se refiere al campo de la agricultura, en particular a las nuevas plantas de patata. La invención también se refiere a plantas de patata obtenidas por métodos para mejorar la constitución genética de dichas plantas. La invención se refiere además a semillas híbridas y a plantas de patata obtenidas a partir de dichas semillas híbridas.
Antecedentes de la invención
La patata (Solanum tuberosum L.) es una parte integral del sistema alimentario mundial. Es el producto alimenticio no cereal número uno del mundo, con una producción que alcanzó un récord de 325 millones de toneladas en 2007.
A diferencia de otros cultivos de campo importantes, las patatas se reproducen vegetativamente a partir de otras patatas. Por lo tanto, una parte de la cosecha de cada año, que oscila entre el 5 y el 15 por ciento, según la calidad de los tubérculos cosechados, se reserva para su reutilización en la próxima temporada de siembra. La mayoría de los agricultores de los países en desarrollo seleccionan y almacenan sus propios tubérculos-semilla. En los países desarrollados, es más probable que los agricultores compren "semillas certificadas" libres de enfermedades de proveedores dedicados.
Las patatas de semilla son más difíciles de producir y suministrar que las semillas de cereales o legumbres. Una proporción semilla:cosecha de 1:20 para patatas se considera buena, en comparación con 1:400 para maíz o 1:10.000 para tomate. Por lo tanto, una hectárea puede requerir dos toneladas de material de semilla para maximizar el rendimiento de los productos cosechables, en comparación con los 18 kg del maíz. Para romper la latencia, las patatas de semilla deben almacenarse durante varias semanas antes de que puedan plantarse. Las condiciones adecuadas durante el almacenamiento, tal como la cantidad de luz, la temperatura y la humedad, son cruciales para garantizar una buena calidad de "semilla".
Además de la mala proporción semilla:cosecha, las patatas de semilla atraen y transportan plagas y enfermedades. Estos incluyen (entre otros) el mildiú de la patata, los gorgojos andinos de la patata, los nematodos, las polillas y los virus de los tubérculos. Estos últimos son transmitidos en el campo por pulgones y luego llevados de generación en generación en la semilla. Dicha infección de virus puede disminuir los rendimientos hasta en un 20 por ciento.
Las patatas de semilla tienen altos costes de transporte debido a las grandes distancias entre las principales áreas de producción de semillas y las principales áreas de producción de consumo y el peso relativamente alto de los tubérculos individuales de patata de semilla.
Para alimentar a la creciente población mundial ahora y en el futuro, la industria de la patata debe seguir creciendo para satisfacer las necesidades del público consumidor. Se dedican importantes esfuerzos de investigación y desarrollo a la modernización de la siembra y la cosecha de campos y el procesamiento de patatas, y al desarrollo de variedades de patata económicamente ventajosas. A través del cruzamiento de patatas, los investigadores esperan obtener patatas con las características deseables de buen procesamiento, tanto para consumo en fresco como para fines industriales, alto contenido de sólidos solubles, alto rendimiento, resistencia a enfermedades y plagas y adaptabilidad a diversas áreas y condiciones de cultivo.
La investigación que conduce a variedades de patata que combinan las características ventajosas mencionadas anteriormente es en gran parte empírica. Esta investigación requiere grandes inversiones de tiempo, mano de obra y dinero. El desarrollo de un cultivo de patata a menudo puede llevar hasta ocho años o más, seguidos de al menos cinco años de propagación para obtener cantidades suficientes para uso comercial. El mejoramiento comienza con una cuidadosa selección de progenitores superiores para incorporar las características más importantes a la progenie. Dado que todos los rasgos deseados por lo general no aparecen en una progenie, la mejoramiento es un procedimiento continuo de selección de los mejores recombinantes, combinando rasgos favorables de los ancestros.
La ardua tarea de producir una nueva variedad de patata se comprende mejor cuando se comprende la genética de la patata. La patata comercial tiene un genoma tetraploide. Los tubérculos diploides son generalmente demasiado pequeños para aplicaciones comerciales importantes. Además, el genoma tetraploide es extremadamente heterocigótico y, a menudo, alberga múltiples alelos por locus. Se cree que la autoincompatibilidad, que se experimenta principalmente a nivel diploide, y la depresión endogámica son responsables del mantenimiento de la alta variabilidad genética que se encuentra en la patata y que la sobredominancia de los alelos heterocigotos (heterosis) da como resultado plantas vigorosas. En una descendencia de patata típica obtenida de un cruce entre dos líneas parentales no relacionadas, los alelos deletéreos pueden, por lo tanto, contribuir a una aptitud reducida en caso de tendencia homocigótica o a un aumento del vigor en caso de tendencia heterocigota. Está claro que un seleccionador necesita grandes poblaciones para maximizar la posibilidad de encontrar individuos que porten un número relativamente alto de loci heterocigotos y un número bajo de loci homocigotos, al mismo tiempo que exhiben combinaciones beneficiosas de características agronómicamente deseables.
Las técnicas actuales de mejoramiento de la patata se basan en el cruzamiento controlado de clones parentales que a su vez son el resultado de un desarrollo integral previo a la mejoramiento durante el cual, entre otras, se aplican técnicas especiales tales como la duplicación de cromosomas, el rescate de embriones y la fusión somática para introducir las características beneficiosas de por ejemplo especies salvaje y primitiva de Solanum en estos clones. El material parental que se considera apropiado para la mejoramiento posterior después de un procedimiento de selección fenotípica se cruza mutuamente y las semillas híbridas no uniformes resultantes se siembran en grandes cantidades en invernaderos. De decenas de miles de plántulas F1 individuales se cosechan tubérculos y se conservan para la siembra del próximo año. Al año siguiente, se planta en el campo un solo tubérculo "semilla" de cada plántula resultante. Se debe tener mucho cuidado para evitar la introducción de virus y enfermedades, ya que el material solo se expande clonalmente (vegetativamente) antes de venderse a clientes individuales años después. Después del segundo año, se toman muestras de tubérculos para mediciones de densidad y pruebas preliminares de alevines para determinar la idoneidad de los tubérculos para uso comercial. Una multitud de tubérculos de plantas que han sobrevivido al procedimiento de selección hasta este punto se plantan luego en el tercer año para una serie más completa de pruebas de alevines y determinaciones de densidad. En la etapa de desarrollo del cuarto año, se cultiva un número decreciente de selecciones supervivientes en cantidades cada vez mayores y las plantas de las mismas se someten a pruebas de campo en varias fases para determinar su adaptabilidad a diferentes condiciones de crecimiento. Eventualmente, las variedades que tienen cualidades agronómicas superiores se transfieren a otras granjas y la "semilla" (en forma de tubérculos) se aumenta a escala comercial. Dado que un tubérculo "semilla" puede generar entre 6 y 20 tubérculos cosechados, este procedimiento de incremento de escala puede tardar años antes de que se produzca suficiente "semilla". Generalmente, para este momento, se han invertido ocho o más años de siembra, cosecha y pruebas para intentar desarrollar una variedad de patata nueva y mejorada.
Para reducir la depresión por endogamia, un seleccionador puede introducir nuevos genes de un progenitor genéticamente más remoto, tal como de especies salvajes y primitivas con niveles de ploidía que van desde diploide a hexaploide. Sin embargo, cuando se cruzan dos plantas de patata no relacionadas genéticamente, el nivel de tendencia heterocigota puede aumentar pero, al mismo tiempo, también se introducen más genes nocivos. Como consecuencia, un seleccionador por lo general hará cruces adicionales con más germoplasma comercial para enriquecer la población en alelos favorables. En total, dicho programa de mejoramiento de cruzamiento múltiple puede llevar decenas de años, ya que la selección de los genotipos favorables en cada generación ya puede llevar cinco años. Por lo tanto, el mejoramiento de la patata es actualmente un ejercicio predominantemente empírico, fuertemente caracterizado por prueba y error.
Las patatas y sus especies salvajes afines (especies Solanum productoras de tubérculos) son en su mayoría exogamia porque la autofecundación se ve obstaculizada por un sistema de autoincompatibilidad gametofítica. La autoincompatibilidad (SI) es un nombre general para varios mecanismos genéticos en las angiospermas, que previenen la autofertilización y la endogamia. En plantas con SI, cuando un grano de polen producido en una planta alcanza un estigma de la misma planta o de otra planta con un genotipo similar, el procedimiento de germinación del polen, crecimiento del tubo polínico, fertilización del óvulo y desarrollo del embrión se detiene en una de sus fases, y en consecuencia no se producen semillas. La autoincompatibilidad no se encuentra en las patatas tetraploides.
La provisión de tales clones autocompatibles puede facilitar la generación de progenies de patata autofecundadas y, por consiguiente, la producción de líneas de patata (altamente) homocigotas. Esto podría brindar una gran oportunidad para el desarrollo de líneas de mejoramiento homocigóticas élite en patata. Sin embargo, hasta la fecha, el desarrollo de líneas élite homocigóticas con rasgos agronómicamente deseables genéticamente fijadas, que permitirían la producción de semilla de patata híbrida genéticamente uniforme, no ha tenido éxito.
La provisión de líneas élite homocigóticas se ve obstaculizada por causas desconocidas. La autofecundación de un clon autocompatible ocasionalmente encontrado da como resultado una disminución más lenta de la tendencia heterocigota de lo esperado teóricamente. La disminución más lenta de la tendencia heterocigota puede ser el resultado de una selección involuntaria pero inevitable durante la autofecundación de las plantas heterocigotas en la descendencia que exhiben mayor vigor, fertilidad y germinación de semillas. Se da a entender que, a medida que se reduce la tendencia heterocigota por la autofecundación, también se reducen la fertilidad y el vigor y las plantas pueden volverse débiles y completamente estériles. La situación quizás empeore por el establecimiento de configuraciones homocigotas de genes recesivos nocivos. Este fenómeno, al que generalmente se hace referencia como depresión endogámica, ha dificultado en gran medida el desarrollo de líneas de patata homocigóticas y, por consiguiente, la producción de semilla de patata híbrida uniforme.
Existe un gran prejuicio contra la producción de líneas de mejoramiento homocigóticas en patata debido a la depresión endogámica. Uijtewaal et al. (Euphytica 36 (1987) 745-753) indicaron que debido a problemas de esterilidad, los clones homocigotos de patata serían de poca importancia para el mejoramiento práctico. Se consideró el desarrollo de líneas puras homocigóticas, pero se hizo imposible en patata (Umaerus, 1987, Proceeding of the 10th Triennial Conference of the European Association of Potato Research, Aalborg, Denmark, pp 72-103 as cited in Almekinders et al. 2009 Potato Research 52:275-293). Durante mucho tiempo se ha presumido que las rutas que implican la duplicación de haploides son prometedoras. Sin embargo, hasta el día de hoy la opinión predominante es que la depresión endogámica en la patata diploide es demasiado fuerte para dar lugar a plantas homocigóticas vigorosas.
Birhman and Hosaka (Genome 43: 495-502 (2000)) han sugerido la posibilidad de usar el gen Sli derivado de S. chacoense para el desarrollo de líneas de semilla de patata verdadera altamente homocigóticas (TPS) y mejoramiento por heterosis de la patata. Sin embargo, hasta la fecha, nunca se han informado líneas homocigotas con buenas características agronómicas, como un buen rendimiento de tubérculos, a partir de esta línea de investigación propuesta. Más bien, los homocigotos que se han producido no exhiben ningún rendimiento de tubérculos agronómicamente relevante.
Rommens en 2010 (Genetic modification of Plants, Kempen & Jung, eds, In: Biotechnology in Agriculture and Forestry 64(1): 61-77 (2010)), aboga por la vía de la transformación genética, debido a que los esfuerzos por mejorar el rendimiento y la calidad de este cultivo se ven obstaculizados por la depresión endogámica.
En resumen, la producción de líneas puras de cultivo de patata se considera imposible. Debido a esto, el mejoramiento de la patata no puede escapar a los esquemas tradicionales basados en cruces de heterocigotos tetraploides. Como resultado, el desafío se considera formidable para combinar diversos rasgos de uso (relacionados con usos frescos y de procesamiento), resistencias a patógenos y plagas, y muchos otros rasgos agronómicos relevantes con mejoras en el rendimiento en un variedad comercialmente aceptable (Douches et al. 1999, Crop Science 36(6):1544-1552).
Sumario de la invención
Los presentes inventores han intentado conseguir líneas de mejoramiento parentales homocigóticas para la propagación generativa de patatas a través de un número de rutas, ninguna de las cuales tuvo éxito. Estas rutas fallidas incluyeron androgénesis, ginogénesis, duplicación de haploides y autofecundaciones continuas. Sólo mediante la autofecundación repetida de una planta autocompatible que comprenda el gen Sli derivado de S. chacoense llegaron los inventores al producto de la presente invención. La presente invención proporciona ahora en un primer aspecto una línea de patata diploide, fértil, autocompatible y esencialmente homocigótica Solanum tuberosum que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, en la que las plantas de dicha línea tienen un nivel de tendencia homocigótica superior al 75 % cuando se prueban 100 loci, obtenible mediante un método que comprende (a) proporcionar una primera planta de patata, cuya primera planta de patata es una planta de una primera línea de patata S. tuberosum diploide, autocompatible, fértil y esencialmente homocigótica seleccionada del número de acceso de NCIMB 41663, número de acceso de NCIMB 41664, número de acceso de NCIMB 41665, o número de acceso de NCIMB 41765, las semillas representativas de dichas líneas se han depositado en NCIMB, Aberdeen, Escocia, con las referencias de los obtenedores AGVD1, AGVD2, AGVD3 y AGVD17, respectivamente, en las que dicho rasgo de autocompatibilidad se debe a la presencia de un gen inhibidor del locus S (Sli); (b) proporcionar una segunda planta de patata S. tuberosum, en la que dicha segunda planta de patata es una planta de una línea de patata diploide que puede tener cualquier nivel de tendencia homocigótica, comprendiendo dicha línea de dicha segunda planta de patata plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta; (c) polinizar de forma cruzada dicha primera y segunda planta de patata para proporcionar semillas y recolectar dichas semillas para proporcionar así una generación descendiente en forma de una semilla descendiente híbrida; (d) hacer crecer dicha semilla descendiente híbrida en una población de plantas de patata descendientes híbridas y seleccionar de dicha población plantas que produzcan tubérculos con un rendimiento expresado en gramos de peso fresco de al menos 200 gramos por planta, y (e) autofecundar dicha planta de patata descendiente híbrida para entre 1 y 8 autofecundaciones, seleccionando así una descendencia fértil para proporcionar así una línea de mejoramiento de patata diploide, autocompatible y esencialmente homocigótica que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, en la que las plantas de dicha línea tienen un nivel de tendencia homocigótica que es superior al 75 %, y en la que dicho rasgo de autocompatibilidad se debe a la presencia de dicho gen inhibidor del locus S (Sli). Las plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, en términos generales, también se denominan en este documento como vigorosas.
Dichas etapas a-e preferiblemente se repiten, en las que la etapa b comprende la provisión de una planta de patata adicional, en las que dicha planta de patata adicional es un donante de un rasgo agronómicamente deseable adicional.
Dicho rasgo adicional deseable agronómicamente se selecciona preferiblemente de tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades (incluyendo, pero no limitando a, resistencia a la sarna causada por Streptomyces spp, sarna pulverulenta, Rhizoctonia, sarna plateada, Phytophthora infestans), tolerancia a herbicidas, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad y resistencia al endulzamiento por frío.
Las plantas de dicha línea cuando maduran alcanzan preferiblemente un peso fresco promedio del follaje y los brotes de al menos 500 gramos por planta. Las plantas que alcanzan un peso fresco promedio del follaje y los brotes de al menos 500 gramos por planta, en términos generales, también se denominan vigorosas en este documento.
En este documento se describe una planta de una línea según la invención como se describe anteriormente. Preferentemente, dicha planta es diploide, fértil, autocompatible y esencialmente homocigótica y tiene un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta.
En otro aspecto, la presente invención proporciona una semilla de patata híbrida S. tuberosum, cuya semilla, cuando se siembra, produce una planta de patata diploide, fértil, autocompatible y esencialmente heterocigota que tiene un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta y en la que dicho rasgo de autocompatibilidad se debe a la presencia de un gen inhibidor del locus S (Sli), pudiendo obtenerse dicha semilla cruzando plantas de una primera y segunda línea de mejoramiento de patatas diploides según una cualquiera de las reivindicaciones 1-5, en la que dichas plantas de dicha primer y segunda línea de mejoramiento de patatas diploides contienen al menos un 20 % de loci homocigotos contrastantes como se puede determinar por un análisis de marcador molecular, al probar 100 loci. Dicha semilla puede ser homocigota (es decir, representar una semilla autofecundada), pero también puede ser heterocigota para los alelos en los que difieren los progenitores. Dicha semilla es preferiblemente una semilla de patata híbrida uniforme. Tal semilla de patata híbrida uniforme es el resultado de un cruce entre dos plantas homocigóticas, autocompatibles, fértiles y vigorosas de una línea según la invención como se ha descrito anteriormente, preferiblemente de dos líneas diferentes, en la que las líneas contienen al menos un 20 % de loci homocigotos contrastantes como se puede determinar por un análisis de marcador molecular, al probar 100 loci
En este documento se describe adicionalmente un método de producción de una línea de patata diploide, fértil, autocompatible y esencialmente homocigótica S. tuberosum que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, que comprende:
(a) proporcionar una primera planta de patata, cuya primera planta de patata es una planta de una primera línea de patata diploide, autocompatible, fértil y esencialmente homocigótica;
(b) proporcionar una segunda planta de patata, en la que dicha segunda planta de patata es una planta de una línea de patata diploide o tetraploide que puede tener cualquier nivel de tendencia homocigótica, comprendiendo dicha línea de dicha segunda planta de patata plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta;
(c) polinizar de forma cruzada dicha primera y segunda planta de patata para proporcionar semillas y recolectar dichas semillas para proporcionar así una generación descendiente en forma de una semilla descendiente híbrida;
(d) hacer crecer dicha semilla descendiente híbrida en una población de plantas de patata descendientes híbridas y seleccionar de dicha población plantas que produzcan tubérculos con un rendimiento expresado en gramos de peso fresco de al menos 200 gramos por planta, y
(e) autofecundar dicha planta de patata descendiente híbrida o retrocruzamiento de dicha planta de patata descendiente híbrida con una planta de patata de dicha línea de dicha primera planta de patata, entre 1 y 8 autofecundaciones o retrocruces, para proporcionar así una línea de mejoramiento de patata diploide, autocompatible y esencialmente homocigótica que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta.
Una primera planta de patata en un método como el descrito en este documento es preferiblemente una planta de una primera línea de patata diploide, autocompatible, fértil y esencialmente homocigótica, en la que la autocompatibilidad está condicionada por la presencia de un alelo dominante de un gen inhibidor de locus S (locus de autoincompatibilidad) (Sli) que no es alelo del locus S como se describe en Hosaka and Hanneman 1998 Euphytica, 99: 191-197.
El método anterior, cuando se realiza a una escala suficientemente grande (al menos más de 10.000 plantas) y se realiza con el uso de una segunda planta de patata apropiada, que es preferiblemente un clon de patata diploide que esencialmente no es homocigoto, dicha línea de dicha segunda patata planta que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, preferiblemente una línea tal como IVPAA-096-18 (D2) como se describe en los ejemplos a continuación, en este documento, da como resultado la producción de una línea de mejoramiento esencialmente no homocigótica, fértil, vigorosa y autocompatible, en la que dicho vigor está representado en particular por un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta. La provisión de dicha línea de mejoramiento es una etapa crucial en la provisión de semilla de patata verdadera genéticamente homocigota (TPS).
La primera y la segunda línea de mejoramiento se pueden usar posteriormente en la producción de TPS cuando tanto la primera como la segunda línea de mejoramiento comprenden características agronómicamente deseables, tienen buenas habilidades generales de combinación y preferiblemente tienen al menos un 20 % de loci homocigotos que contrastan como se puede determinar por un análisis de marcador molecular.
En un método preferido como se describe en este documento para producir una línea de patata diploide, fértil, autocompatible y esencialmente homocigótica, dichas etapas a-e se repiten en el que la etapa b comprende la provisión de una tercera (o más) planta de patata en la que dicha tercera planta de patata es una planta de una línea de patata diploide o tetraploide que puede tener cualquier nivel de tendencia homocigótica, en la que dicha tercera planta de patata es donante de un rasgo agronómicamente deseable adicional. Preferiblemente, dicho rasgo se selecciona del grupo que consiste en tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades, que incluye, pero no se limita a, resistencia a la sarna causada por Streptomyces spp, sarna pulverulenta, Rhizoctonia,
sarna plateada, Phytophthora infestans, tolerancia a herbicidas, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad y resistencia al endulzamiento por frío, o cualquier otro rasgo agronómicamente beneficioso o característica deseable como se define en este documento.
La segunda o tercera planta de patata como se ha descrito anteriormente puede ser adecuadamente un clon de patata diploide seleccionado de las plantas D1-D21 que se enumeran en la tabla 6 del ejemplo 3 a continuación. El experto en la técnica comprenderá que también se pueden usar otros clones diploides de patata. Muy preferido es el uso del clon de patata IVPAA-096-18 (D2) que está disponible en el Laboratory of Plant Breeding (IVP) (Dr. Ir Ronald B.C. Hutten), Wageningen University, Droevendaalsesteeg 1,6708 PB Wageningen, Países Bajos.
Las fuentes apropiadas (comerciales) de variedades tetraploides se proporcionan a través de la base de datos European Cultivated Potato (ECPD) en http://www.europotato.org/menu.php y/o la base de datos Potato Pedigree del expertise group Plant Breeding (a cooperation between the Laboratory of Plant breeding of Wageningen UR, the Netherlands) y the business unit Biodiversity and breeding of Plant Research International (PRI), Wageningen, Países Bajos en http://www.plantbreeding.wur.nl/potatopedigree/.
Dicha primera planta de patata es preferiblemente una planta de la línea de patata NCIMB 41663, NCIMB 41664, NCIMB 41665 o NCIMB 41765, habiéndose depositado semillas representativas de dicha línea en NCIMB, Aberdeen, Escocia.
Dicha segunda planta de patata es preferiblemente una planta de una segunda línea de patata diploide, fértil, autocompatible y esencialmente homocigótica.
Dicha segunda planta de patata es preferiblemente una planta de una segunda línea de patata diploide, fértil, autoincompatible.
Una línea de mejoramiento de patatas diploides puede obtenerse además por métodos que comprenden las etapas de potenciar el valor de mejoramiento de la línea de mejoramiento de patatas diploides mediante cruzamiento, autofecundación y retrocruzamiento mientras se selecciona continuamente para rasgos agronómicamente deseables. Los cruces experimentales se realizan para evaluar las habilidades generales de combinación (capacidad de entrecruzamiento).
En otro aspecto, la presente invención proporciona un semilla de patata híbrida S. tuberosum, cuya semilla, cuando se siembra, produce una planta de patata diploide, fértil, autocompatible y esencialmente heterocigota que tiene un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta y en la que dicho rasgo de autocompatibilidad se debe a la presencia de un gen inhibidor del locus S (Sli), pudiendo obtenerse dicha semilla cruzando plantas de una primera y segunda línea de mejoramiento de patatas diploides según la presente invención como se ha descrito anteriormente, en el que dichas plantas de dicha primera y segunda línea de mejoramiento de patatas diploides contienen al menos un 20 % de loci homocigotos contrastantes como se puede determinar por un análisis de marcador molecular, al probar 100 loci
Dicha primera y segunda planta de patata son preferiblemente de líneas de patata que comprenden plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, preferiblemente en la que dicha primera y segunda planta de patata son plantas que tienen un rendimiento de tubérculo de al menos 200 gramos de peso fresco por planta.
Dicha al menos una de dichas primera y segunda plantas de patata comprende preferiblemente al menos un rasgo agronómicamente deseable, preferiblemente seleccionado del grupo que consiste en tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades (incluyendo, pero sin limitarse a, resistencia a la sarna causada por Streptomyces spp, sarna pulverulenta, Rhizoctonia, sarna plateada, Phytophthora infestans), tolerancia a herbicidas, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad y resistencia al endulzamiento por frío.
Dicha primera línea de patata es preferiblemente una planta de patata descendiente diploide, autocompatible y esencialmente homocigótica que comprende el gen Sli, preferiblemente en el que dicha primera planta de patata es una planta de las líneas de patata NCIMB 41663, NCIMB 41664, NCIMB 41665 o NCIMB 41765, habiéndose depositado semillas representativas de dichas líneas en el NCIMB, Aberdeen, Escocia, o una planta de patata descendiente diploide, autocompatible y esencialmente homocigótica de la misma.
Dicha segunda planta de patata es preferiblemente una planta de una segunda línea de patata diploide, autocompatible y esencialmente homocigótica que comprende preferiblemente al menos dos rasgos agronómicamente deseables seleccionadas del grupo que consiste en tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades (incluyendo, pero sin limitarse a, resistencia a la sarna causada por Streptomyces spp, sarna pulverulenta, Rhizoctonia, sarna plateada, Phytophthora infestans), tolerancia a herbicidas, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad y resistencia al endulzamiento por frío.
En este documento se describe adicionalmente una semilla de las líneas de patata NCIMB 41663, NCIMB 41664, NCIMB 41665 o NCIMB 41765, habiéndose depositado semillas representativas de dicha línea en NCIMB, Aberdeen, Escocia.
En otro aspecto, la presente invención proporciona una planta de patata híbrida, o parte de esta, producida cultivando la semilla de patata híbrida S. tuberosum de la presente invención.
En otro aspecto, la presente invención proporciona un tubérculo de la planta de patata híbrida S. tuberosum de la presente invención.
En otro aspecto, la presente invención proporciona un cultivo de tejido de las plantas de patata híbridas S. tuberosum de la presente invención.
En otro aspecto, la presente invención proporciona un conjunto de al menos 2 plantas de patata intercruzables de diferentes líneas de patatas diploides, fértiles, autocompatibles y esencialmente homocigótica S. tuberosum según la invención, que comprenden plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, en la que cada una de dichas líneas se diferencia de otra línea en dicho conjunto por tener al menos un 20 % de loci homocigotos contrastantes como se puede determinar por un análisis de marcador molecular, al probar 100 loci, y en la que dicho rasgo de autocompatibilidad un gen inhibidor del locus S (Sli).
Además, en este documento se describen métodos que comprenden las etapas de:
(a) proporcionar una primera planta de patata, cuya primera planta de patata es una planta de una primera línea de patata diploide, autocompatible y esencialmente homocigótica que lleva cromosomas con alelos para rasgos agronómicamente deseables que se van a expresar en generaciones descendientes,
(b) proporcionar una segunda planta de patata, en la que dicha segunda planta de patata es una planta de una segunda línea de patata diploide, autocompatible y esencialmente homocigótica que lleva cromosomas con alelos para rasgos agronómicamente deseables que se van a expresar en generaciones descendientes y que pueden s ser diferentes de los rasgos en dicha primera planta, y
(c) polinizar de forma cruzada dicha primera y segunda planta de patata de manera que una de dichas plantas progenitoras produzca bayas con semillas y recolectar dichas semillas de dichas bayas para proporcionar una generación descendiente en forma de una descendencia híbrida portadora de combinaciones de cromosomas con dichos alelos.
Por lo tanto, la planta de la primera línea de patata puede portar cromosomas con alelos para rasgos agronómicamente deseables que se expresarán en generaciones descendientes, en cuyo caso el rasgo agronómicamente deseable en la planta de la segunda línea de patata puede ser diferente de los rasgos en dicha primera planta.
En este documento se describe adicionalmente un método de prevención de la transmisión de infecciones virales entre generaciones de patatas, que comprende:
(a) proporcionar una primera planta de patata, cuya primera planta de patata es una planta de una primera línea de patata diploide, autocompatible y esencialmente homocigótica,
(b) proporcionar una segunda planta de patata, en la que dicha segunda planta de patata es una planta de una línea de patata que puede tener cualquier nivel de ploidía y/o cualquier nivel de tendencia homocigótica, llevando dicha segunda planta de patata un cromosoma con un alelo para un rasgo agronómicamente deseable que se va a expresar en una generación descendiente;
(c) polinizar de forma cruzada dicha primera y segunda planta de patata de manera que una de dichas plantas progenitoras produzca bayas con semillas y recolectar dichas semillas de dichas bayas para proporcionar una generación descendiente en forma de una semilla descendiente híbrida que lleva un cromosoma con dicho alelo;
(d) hacer crecer dicha semilla descendiente híbrida en una planta de patata descendiente híbrida que lleva un cromosoma con dicho alelo, y
(e1) cosechar tubérculos de dicha planta de patata híbrida para proporcionar tubérculos que expresen dicho rasgo, o
(e2) autofecundar dicha planta de patata descendiente híbrida o retrocruzar dicha planta de patata descendiente híbrida con una planta de patata de dicha línea de dicha primera planta de patata, proporcionando así más plantas descendientes que llevan un cromosoma con dicho alelo y donde dichas plantas descendientes son cada vez más homocigotas después de cada etapa de retrocruzamiento o autofecundación.
En este documento se describe además una línea de patata diploide, fértil, autocompatible y esencialmente homocigótica que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, siendo dicha planta obtenible mediante:
(a) proporcionar una primera planta de patata, cuya primera planta de patata es una planta de una primera línea de patata diploide, fértil, autocompatible y esencialmente homocigótica,
(b) proporcionar una segunda planta de patata, en la que dicha segunda planta de patata es una planta de una línea de patata diploide que tiene un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta,
(c) polinizar de forma cruzada dicha primera y segunda planta de patata y permitir que una de dichas plantas progenitoras produzca bayas con semillas y recolectar dichas semillas de dichas bayas para proporcionar una generación descendiente en forma de una semilla descendiente híbrida,
(d) hacer crecer dicha semilla descendiente híbrida en una planta de patata descendiente híbrida,
(e) retrocruzar dicha planta de patata descendiente híbrida con una planta de patata de dicha línea de dicha primera planta de patata, y seleccionar de las plantas descendientes generadas así una planta que es diploide, autocompatible y vigorosa porque produce tubérculos con un rendimiento expresado en gramos de peso fresco de al menos 200 gramos por planta, y
(f) autofecundar dicha planta diploide, autocompatible y vigorosa entre 1 y 8 generaciones mientras se selecciona por autocompatibilidad y vigor como se define en (e).
Los etapas de seleccionar o proporcionar plantas que son homocigóticas, o la etapa de seleccionar o proporcionar plantas que tienen al menos un 20 % de loci contrastantes como se define en este documento anteriormente, pueden comprender selección asistida por marcadores usando los marcadores genéticos moleculares como se proporciona en este documento.
Descripción de los dibujos
La figura 1 muestra una presentación esquemática de un esquema de desarrollo para el desarrollo de variedades híbridas de patata usando los métodos y plantas como se describe en este documento, en el que se indican las diversas etapas para obtener híbridos comerciales de patata:
0, germoplasma de patata: El origen comprende en principio todos los genotipos de patata dentro de las especies de patata cultivadas (Solanum tuberosum) y especies que se pueden cruzar relacionadas;
1, patata diploide autocompatible: patata cultivada (Solanum tuberosum) suele ser incompatible consigo misma. La autocompatibilidad se introduce a partir de especies relacionadas (véase, por ejemplo, Hermsen, J.G.Th. 1978, Euphytica 27, 1-11);
2, patata diploide autocompatible homocigótica fértil: Los diploides homocigotos se pueden obtener duplicando los haploides (Uijtewaal et al., 1987, Theor. Appl. Genet. 73, 751-78) por cultivo de anteras (Jacobsen and Sopory, 1977, Theor. Appl. Genet. 52, 119-123), por cultivo de óvulos o por autofecundaciones repetidas (Phumichai & Hosaka, 2006, Euphytica 149, 251-258). El ejemplo de esto último se ilustra en este documento;
3, patata diploide autocompatible homocigota fértil vigorosa: El vigor se mejora cruzando con otras patatas diploides, autofecundadas y retrocruzadas mientras se selecciona para mayor vigor;
4, líneas de mejoramiento de patatas diploides: Los rasgos agronómicamente deseables se introducen mediante el cruzamiento con otras patatas diploides, la autofecundación y el retrocruzamiento mientras se seleccionan estos rasgos;
5, líneas de mejoramiento de patatas diploides de élite: El valor de mejoramiento se mejora aún más mediante el entrecruzamiento, la autofecundación y el retrocruzamiento, mientras se selecciona continuamente para obtener rasgos agronómicamente deseables. Se realizan cruces experimentales para evaluar las habilidades combinatorias generales;
6, variedades híbridas de patata: Se cruzan buenas líneas de mejoramiento combinadas. Las semillas obtenidas son semillas de patata verdaderas híbridas que son uniformes.
La figura 2 muestra una presentación esquemática de las genealogías de las poblaciones, depósitos y plantas como se describe en los ejemplos, a partir de un cruce entre una planta de patata diploide, esencialmente homocigótica y autocompatible ("H") y tres clones diploides diferentes (D1, D2 y D3) que son esencialmente no homocigotos y autoincompatibles y que actúan como líneas donantes de uno o más rasgos agronómicos deseables. La producción de una matriz de líneas esencialmente homocigóticas desde líneas retrocruzadas hasta clones completamente autofecundados, cada una con uno o más rasgos deseables derivados de las líneas donantes, proporciona un conjunto de líneas de mejoramiento élite para la producción de una matriz de variedades híbridas de patata en forma de semilla híbrida. ® Se refiere a las autofecundaciones. Como puede verse, y como se detalla en los ejemplos, sólo la línea donante D2 dio como resultado la producción de un gran número de plantas autocompatibles que mostrarían vigor
como se define en este documento. También se podrían obtener plantas vigorosas de los otros cruces, aunque con una frecuencia mucho menor (0.1-1 %).
La figura 3 muestra el mapa de ligamiento genético ultradenso del genoma del clon de mejoramiento diploide RH89-039-16. El número a la izquierda del grupo de ligamiento indica el número acumulativo de eventos de recombinación contados desde arriba. El número de marcadores AFLP en cada compartimiento está representado por tonos de gris según la leyenda de la figura. Las posiciones putativas del centrómero se indican con "Cen" junto al cromosoma (van Os et al., 2006 Genetics 173(2):1075-1087). Los números a la derecha del grupo de ligamiento indican la ubicación aproximada del marcador PotSNP correspondiente como se usa en este documento, en los que se hace referencia a la tabla 5B en el ejemplo 3, a continuación, para las anotaciones del marcador. La figura indica claramente la distribución de marcadores informativos sobre el genoma de patata de este clon. Se observó una distribución similar para estos marcadores con otro clon usado en este documento, el clon SH83-92-488.
La figura 4 muestra datos de marcadores de 100 marcadores sobre 20 genotipos diploides de patata diferentes, que representan el germoplasma de cultivo de patata diploide (Ejemplo 2).
La figura 5 muestra el resultado del análisis del marcador SNP como se describe en el ejemplo 3 en un conjunto de seis poblaciones F3 y sus respectivos progenitores.
Descripción detallada de la invención
Definiciones
El término "patata" se usa en este documento para referirse a material que es esencialmente de tuberosum, pero puede incluir segmentos de introgresión de otros especies Solanum de tubérculos tales como Solanum chacoense, Solanum phureja, Solanum andigena, Solanum demissum.
El término "planta de patata" se usa en este documento para referirse a una plántula o planta madura que crece a partir de un cultivo celular o una semilla. Los expertos en la técnica reconocerán que cuando se usa el término "planta de patata" en el contexto de la presente invención, esto también incluye variedades derivadas que conservan las características distintivas esenciales de, por ejemplo, las variedades de patata que son objeto de la presente invención, tales como una planta convertida de un solo gen de esa variedad o un derivado transgénico que tenga incorporados uno o más genes de valor agregado (tales como resistencia a herbicidas o plagas). Los métodos de retrocruzamiento se pueden usar para mejorar o introducir una característica en la variedad.
El término "cruzamiento" como se usa en este documento se refiere a la fertilización de plantas (o gametos) femeninas por plantas (o gametos) masculinos. El término "gameto" se refiere a la célula reproductiva haploide (óvulo o espermatozoide) producida en plantas por mitosis a partir de un gametofito e implicada en la reproducción sexual, durante la cual dos gametos de sexo opuesto se fusionan para formar un cigoto diploide. El término generalmente incluye referencias a un polen (incluido el espermatozoide) y un óvulo (incluido el óvulo). Por lo tanto, "cruzamiento" se refiere generalmente a la fertilización de óvulos de un individuo con polen de otro individuo, mientras que "autofecundación" se refiere a la fertilización de óvulos de un individuo con polen del mismo individuo. El cruzamiento se usa ampliamente en el mejoramiento de plantas y da como resultado una mezcla de información genómica entre las dos plantas cruzadas, un cromosoma de la madre y un cromosoma del progenitor. Esto dará como resultado una nueva combinación de rasgos heredados genéticamente. En general, la progenie de un cruce se designa como: "F1". Si la F1 no es uniforme (segrega), generalmente se designa como "población F1". La "autofecundación" de una planta homocigota generalmente dará como resultado una planta genéticamente idéntica ya que no hay variación genética. La "autofecundación" de una F1 dará como resultado una descendencia que se segrega para todos los rasgos que tienen locus heterocigotos en la F1. Tal descendencia se designa: "F2" o "población F2"
Al referirse a "cruzamiento" en el contexto de lograr la introgresión de una región o segmento genómico, el experto en la técnica entenderá que para lograr la introgresión de solo una parte de un cromosoma de una planta en el cromosoma de otra planta, se requiere que porciones aleatorias de los genomas de ambas líneas parentales se recombinen durante el cruzamiento debido a la ocurrencia de eventos de entrecruzamiento en la producción de los gametos en las líneas parentales. Por lo tanto, los genomas de ambos progenitores deben combinarse en una sola célula mediante un cruce, donde tras la producción de gametos a partir de dicha célula y su fusión en la fecundación dará lugar a un evento de introgresión.
El término "intercruzable", como se usa en este documento, se refiere a la capacidad de producir plantas progenies después de realizar cruces entre plantas progenitoras.
Como se usan en este documento, los términos "introgresión", "introgresar" e "introgresión" se refieren tanto a un procedimiento natural como artificial mediante el cual genes individuales o cromosomas completos se trasladan de un individuo, especie, variedad o variedad cultivada al genoma de otro individuo, especie, variedad o variedad cultivada, cruzando esos individuos, especies, variedades o variedades cultivadas. En el mejoramiento de la planta, el procedimiento suele implicar la autofecundación o el retrocruzamiento con el progenitor recurrente para proporcionar una planta cada vez más homocigótica que tenga esencialmente las características del progenitor recurrente además del gen o rasgo que ha sido sometido ha introgresión.
El término "introgresión" se refiere al resultado de un evento de introgresión.
El término "retrocruzada" se refiere al resultado de un procedimiento de "retrocruzamiento" en el que la planta resultante de un cruce entre dos líneas parentales se cruza (repetidamente) con una de sus líneas parentales, en el que la línea parental usada en el retrocruzamiento se denomina el progenitor recurrente. El retrocruzamiento repetido da como resultado la sustitución de fragmentos del genoma del progenitor donante por los del recurrente. La descendencia de un retrocruzamiento se denomina "BCx" o "población BCx", donde "x" representa el número de retrocruzamientos.
El término "retrocruzamiento", como se usa en este documento, se refiere al cruce repetido de una progenie híbrida con los progenitores recurrentes. La planta de patata parental que aporta el gen para la característica deseada se denomina progenitor no recurrente o donante. Esta terminología se refiere al hecho de que el progenitor no recurrente se usa una vez en el protocolo de retrocruzamiento y, por lo tanto, no se repite. La planta de patata parental a la que se transfiere el gen o los genes del progenitor no recurrente se conoce como progenitor recurrente, ya que se usa durante varias rondas en el protocolo de retrocruzamiento. En un protocolo típico de retrocruzamiento, la variedad original de interés (progenitor recurrente) se cruza con una segunda variedad (progenitor no recurrente) que lleva el único gen de interés que se va a transferir. La progenie resultante de este cruce se vuelve a cruzar con el progenitor recurrente y el procedimiento se repite hasta que se obtiene una planta de patata en la que esencialmente todas las características morfológicas y fisiológicas deseadas del progenitor recurrente se recuperan en la planta convertida, además de la gen único o un número limitado de genes transferidos del progenitor no recurrente.
La selección de un progenitor recurrente apropiado es una etapa importante para un procedimiento de retrocruzamiento exitoso. El objetivo de un protocolo de retrocruzamiento es alterar o sustituir un solo rasgo o característica en la variedad original. Para lograr esto, se modifica, sustituye o complementa un solo gen de la variedad recurrente con el gen deseado del progenitor no recurrente, conservando esencialmente todo el resto de los genes deseados y, por lo tanto, la constitución fisiológica y morfológica deseada de la variedad original. La elección del progenitor particular no recurrente dependerá del propósito del retrocruzamiento. Uno de los propósitos principales es agregar algunas características comercialmente deseables y agronómicamente importantes al progenitor recurrente. El protocolo de retrocruzamiento exacto dependerá de la característica o rasgo que se modifique o agregue para determinar un protocolo de prueba apropiado. Aunque los métodos de retrocruzamiento se simplifican cuando la característica que se transfiere es un alelo dominante, también se puede transferir un alelo recesivo. En este caso, puede ser necesario introducir una prueba de la progenie para determinar si la característica deseada se ha transferido con éxito. Preferiblemente, tales genes se controlan mediante marcadores moleculares de diagnóstico.
Asimismo, los transgenes se pueden introducir en la planta usando cualquiera de una variedad de métodos recombinantes establecidos bien conocidos por los expertos en la técnica.
Se han identificado muchos rasgos de un solo gen que no se seleccionan regularmente para el desarrollo de una nueva variedad, pero que se pueden mejorar mediante retrocruzamiento y técnicas de ingeniería genética. Los rasgos de un solo gen pueden o no ser transgénicos, los ejemplos de estos rasgos incluyen pero no se limitan a: resistencia a herbicidas; resistencia a enfermedades bacterianas, fúngicas o virales; resistencia a insectos; uniformidad o aumento en la concentración de almidón y otros carbohidratos; calidad nutricional potenciada; disminución en la tendencia del tubérculo a magullarse; y disminución en la tasa de conversión de almidón a azúcares.
El término "autofecundación" se refiere al procedimiento de autofertilización en el que un individuo es polinizado o fertilizado con su propio polen. La autofecundación repetida finalmente da como resultado una descendencia homocigota.
Una "línea", como se usa en este documento, se refiere a una población de plantas derivada de un solo cruce, retrocruzamiento o autofecundación. Las plantas descendientes individuales no son necesariamente idénticas entre sí. Es posible que las plantas descendientes individuales no sean vigorosas, fértiles o autocompatibles debido a la variabilidad natural. Sin embargo, está previsto que las plantas apropiadas que sean vigorosas, fértiles y autocompatibles puedan identificarse fácilmente en una línea y usarse para fines de mejoramiento adicionales.
Como se usa en este documento, el término "alelo(s)" significa cualquiera de una o más formas alternativas de un gen, todos los cuales alelos se relacionan con al menos un rasgo o característica. En una célula u organismo diploide, las dos copias de un gen ocupan loci correspondientes en un par de cromosomas homólogos. Cada copia puede ser un alelo distinto.
Un "gen" se define en este documento como una unidad hereditaria (a menudo indicada por una secuencia de ADN) que ocupa una ubicación específica en un cromosoma y que contiene la instrucción genética para una contribución a las características o rasgos fenotípicos potenciales en una planta.
Un "locus" se define en este documento como la posición que ocupa un gen dado en un cromosoma de una especie de planta dada.
Como se usa en este documento, el término "homocigoto" significa una condición genética que existe cuando alelos idénticos residen en loci correspondientes en cromosomas homólogos.
El término "esencialmente homocigótica" se refiere a un nivel de tendencia homocigótica de al menos el 25 %, preferiblemente al menos el 50 %, más preferiblemente al menos el 75 %, aún más preferiblemente al menos 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 o 100 % de tendencia homocigótica cuando se prueba 100, preferiblemente 1000, más preferiblemente al menos 10.000 loci
Los niveles de tendencia homocigótica son valores promedio para la población y se refieren preferiblemente a aquellos loci en los que difieren los progenitores.
Como se usa en este documento, el término "heterocigoto" significa una condición genética que existe cuando diferentes alelos residen en loci correspondientes en cromosomas homólogos.
El término "recombinación" o "recombinar" se refiere al intercambio de información entre dos cromosomas homólogos durante la meiosis. En una planta "recombinante", el ADN que originalmente está presente en una ubicación específica dentro del cromosoma, por ejemplo, ligado a un gen/locus, se intercambia por ADN de otra planta (es decir, materno por paterno o viceversa). Para intercambiar sólo el material requerido y mantener la valiosa información original en el cromosoma tanto como sea posible, generalmente se requerirán dos eventos de recombinación o cruce flanqueante. En un recombinante doble, este intercambio ha tenido lugar en ambos lados de un gen/locus, una forma de encontrar dicho recombinante doble es seleccionar una población de plantas F2. Esta población tiene que ser grande, ya que la doble recombinación se produce sólo con una frecuencia limitada. Alternativamente, los recombinantes dobles dentro de una unidad genética pueden ser el resultado de un retrocruzamiento posterior. La frecuencia de la doble recombinación es el producto de las frecuencias de los recombinantes solos. (Por ejemplo, se puede encontrar un recombinante en un área de 10 cM con una frecuencia del 10 %, se encuentran recombinantes dobles con una frecuencia del 10 % x 10 % = 1 %).
Como se usa en este documento, el término " progenie " significa (a) descendencia o descendiente(s) genético (s).
Como se usa en este documento, el término "población" significa una colección genéticamente heterogénea de plantas que comparten una derivación genética común.
Un "evento de recombinación" se refiere a un evento de cruce mitótico o meiótico, incluido un evento de GMO.
Como se usa en este documento, el término "híbrido" significa cualquier descendiente de un cruce entre dos individuos genéticamente diferentes, más preferiblemente el término se refiere al cruce entre dos líneas de mejoramiento (élite o endogámicas) que no se reproducirán fielmente al progenitor a partir de la semilla.
El término "segregar", como se usa en este documento, se refiere a la separación de alelos emparejados durante la meiosis de modo que los miembros de cada par de alelos aparezcan en diferentes gametos. El término incluye la referencia al resultado de este fenómeno genético en el que la población descendiente de un cruce en el que al menos uno de los progenitores es heterocigoto para un gen alelo no es uniforme con respecto al rasgo fenotípico conferido por dicho gen.
El término "línea de mejoramiento", como se usa en este documento, se refiere a una línea de patata cultivada que tiene características comercialmente valiosas o agronómicamente deseables, en contraposición a las variedades salvajes o variedades locales. El término incluye la referencia a una línea de mejoramiento de élite o línea de élite, que representa una línea de plantas en esencia homocigóticas, generalmente endogámicas, usadas para producir híbridos F1 comerciales. Una línea de mejoramiento de élite se obtiene mediante el mejoramiento y la selección para obtener un rendimiento agronómico superior que comprenda una multitud de rasgos agronómicamente deseables. Una planta élite es cualquier planta de una línea élite. Rendimiento agronómico superior se refiere a una combinación deseada de características agronómicamente deseables como se define en este documento, en el que es deseable que la mayoría, preferiblemente todos los rasgos agronómicamente deseables se mejoren en la línea de mejoramiento de élite en comparación con una línea de mejoramiento que no es de élite. Las líneas de mejoramiento de élite son esencialmente homocigóticas y son preferiblemente líneas endogámicas.
El término "línea de élite", como se usa en este documento, se refiere a cualquier línea que haya resultado del mejoramiento y la selección para obtener un rendimiento agronómico superior. Una línea élite preferiblemente es una línea que tiene múltiples, preferiblemente al menos 3, 4, 5, 6 o más (genes para) rasgos agronómicos deseables como se define en este documento.
Los términos "variedad cultivada" y "variedad" se usan indistintamente en este documento y denotan una planta que se ha desarrollado deliberadamente mediante mejoramiento, cruzamiento cq y selección, con el propósito de ser comercializada, usada cq por agricultores y cultivadores, para producir productos agrícolas para autoconsumo o para comercialización (consumo en fresco, procesamiento, pienso, etc). El término "germoplasma de mejoramiento" denota una planta que tiene un estado biológico distinto del estado "salvaje", estado "salvaje" que indica el estado original no cultivado o natural de una planta o accesión.
El término "germoplasma de mejoramiento" incluye, pero no se limita a, seminatural, semisalvaje, maleza, cultivo tradicional, variedad local, material de mejoramiento, material de investigación, línea de mejoramiento, población sintética, híbrido, población fundadora/población base, línea endogámica (progenitor de la variedad híbrida), población
segregante, stock genético/mutante, clase de mercado y variedad avanzada/mejorada. Los ejemplos de variedades incluyen variedades cultivadas como Bintje, Russet Burbank, Figenheimer y Nicola.
Como se usa en este documento, los términos "de raza pura", "endogámica pura" o "endogámica" son intercambiables y se refieren a una planta o línea de planta sustancialmente homocigótica obtenida por autofecundación y/o retrocruzamiento repetidos.
Como se usa en este documento, el término "marcador genético molecular" o "marcador" abreviado se refiere a un indicador que se usa en métodos para visualizar diferencias en las características de las secuencias de ácidos nucleicos. Ejemplos de tales indicadores son marcadores de polimorfismo de longitud de fragmentos de restricción (RFLP), marcadores de polimorfismo de longitud de fragmento amplificado (AFLP), polimorfismos de un solo nucleótido (SNP), mutaciones de inserción/deleción (INDEL), marcadores de microsatélites (SSR), regiones amplificadas caracterizadas por secuencia (SCAR), marcadores de secuencia polimórfica amplificada escindida (CAPS) o marcadores de isoenzimas o combinaciones de los marcadores descritos en este documento que definen una ubicación genética y cromosómica específica.
Como se usa en este documento, el término "parte de la planta" indica una parte de la planta de patata, incluidos organelos, células solas y tejidos celulares tales como células vegetales que están intactas en las plantas, grupos de células y cultivos de tejidos a partir de los cuales se pueden regenerar plantas de patata. Los ejemplos de partes de plantas incluyen, pero no se limitan a, células solas y tejidos de polen, óvulos, hojas, embriones, raíces, puntas de raíces, tubérculos, anteras, flores, frutos, tallos y semillas; así como polen, óvulos, hojas, embriones, raíces, puntas de raíces, anteras, flores, frutos, tallos, brotes, vástagos, portainjertos, semillas, tubérculos, protoplastos, callos y similares.
A menos que se indique expresamente lo contrario, el término "semilla", como se usa en esta memoria descriptiva, se refiere al cuerpo a partir del cual se desarrolla una nueva planta (o núcleo en algunas plantas) que contiene la pequeña planta embrionaria encerrada en una cubierta de semilla, generalmente junto con algunos alimentos almacenados. Esta semilla, denominada semilla botánica o "verdadera", es el producto del óvulo maduro de plantas gimnospermas y angiospermas que se produce después de la fertilización y cierto crecimiento dentro de la planta madre.
Como se usan en este documento, los términos "vigoro" y "vigoroso" se refieren a la cantidad relativa de tejidos superficiales o subterráneos de una planta, cantidad relativa que puede ser más o menos independiente entre sí.
La cantidad relativa de tejidos aéreos se puede expresar como observaciones visuales de la cantidad de follaje, en términos de altura de la planta, número de tallos, número de hojas evaluado en escalas ordinales y/o en términos de peso fresco o materia seca. Este rasgo tiene relevancia durante el primer período del ciclo de crecimiento de la planta con una duración promedio de 100 días de cultivo (intervalo 75 - 130 días) en condiciones normales de crecimiento que prevalecen en el noroeste de Europa durante la temporada de mayo a agosto. En estas condiciones, las plantas alcanzan un peso del follaje y de los brotes (planta aérea) expresado en gramos de peso fresco o seco que es de al menos el 50 %, más preferiblemente al menos el 60 %, 70 %, 80 %, 90 % o más del clon de mejoramiento diploide (esencialmente heterocigoto) RH89-039-16, que, a su vez, tiene el 68 % (intervalo 44 - 86 %) del rendimiento de tubérculos de las variedades de patata tetraploides en base a nueve años de prueba. El clon RH89-039-16 está disponible como referencia en Dr. Ir Ronald B.C. Hutten, Laboratory of Plant Breeding, Wageningen University, Droevendaalsesteeg 1,6708 PB Wageningen, Países Bajos.
Alternativamente, o además, el término vigor puede usarse en relación con un cierto rendimiento mínimo de tubérculos. El término "rendimiento de tubérculos", como se usa en este documento, se referirá generalmente a un rendimiento de tubérculos expresado en gramos de peso fresco que en la cosecha en las plantas es de al menos 30-50 %, más preferiblemente de al menos 60 %, 70 %, 80 %, 90 % o más del clon de mejoramiento diploide (esencialmente heterocigoto) RH89-039-16, que, a su vez, tiene el 68 % (intervalo 44 - 86 %) del rendimiento de tubérculos de las variedades de patata tetraploides en base a nueve años de pruebas de campo bajo condiciones de un período de cultivo de 100 días con condiciones de crecimiento normales que prevalecen en el noroeste de Europa durante la temporada de mayo a agosto. Como se indicó anteriormente, el clon RH89-039-16 está disponible con fines de referencia del Dr. Ir Ronald B.C. Hutten, Laboratory of Plant Breeding, Wageningen University
El rendimiento de los tubérculos, a menos que se indique expresamente lo contrario, se basa en los tubérculos formados a partir de una planta de plántulas, en contraste con los tubérculos formados a partir de una planta cultivada en tubérculo, cuyo rendimiento puede ser un 50-200 % mayor que el rendimiento de una plántula.
Alternativamente, o además, el término vigor puede referirse a un rendimiento de tubérculo expresado en gramos de peso fresco o seco que en la cosecha es de al menos 30-50 %, más preferiblemente al menos 60 %, 70 %, 80 %, 90 % o más del clon reproductor diploide RH89-039-16. Esto significa que el rendimiento del tubérculo expresado en gramos de peso fresco puede alcanzar niveles de al menos 200, más preferiblemente al menos 300, 400 o 500 gramos por planta cultivada a partir de semilla en suelo arcilloso en condiciones normales de verano en el noroeste de Europa, e incluso 400, 500, 600, 700, 800 o más, tal como 900 o 1000 gramos de peso de tubérculo fresco o más por planta cuando dicha planta se cultiva a partir de tubérculos y se cultiva en suelo arcilloso en condiciones normales de verano en el noroeste de Europa.
El término "una línea que comprende plantas", en la que se indica que dichas plantas tienen una determinada característica, tal como el rendimiento de tubérculos, debe entenderse preferiblemente como una línea que consiste esencialmente en plantas que tienen dicho rasgo, al tiempo que permite alguna variación biológica.
El término "diploide", como se usa en este documento, se refiere a una planta en la que cada célula vegetativa contiene dos juegos de cromosomas (2x = 2n, donde n es el número de cromosomas). Cada progenitor dona un conjunto de cromosomas.
El término "tetraploide", como se usa en este documento, se refiere a una planta en la que cada célula vegetativa contiene cuatro conjuntos de cromosomas (2x = 4n).
El término "resistencia a nematodos", como se usa en este documento, se refiere a una planta en la que se ha introgresado un gen de resistencia funcional que impide la multiplicación de al menos una población o aislado de nematodos.
El término "resistencia a enfermedades", como se usa en este documento, se refiere a la capacidad de mostrar una reducción de más del 50 % de la cantidad de superficie foliar enferma o del volumen del tubérculo, o la cantidad de multiplicación de un insecto o microorganismo patógeno, incluyendo, pero no limitando a, Streptomyces sp, rizoctonia, sarna plateada y, Phytophthora infestans,
El término "tolerancia a herbicidas", como se usa en este documento, se refiere a una planta que muestra menos del 50 % de daño en el follaje que el daño habitual tras la aplicación de una dosis específica de herbicida.
El término "tolerancia al frío", como se usa en este documento, se refiere a la cantidad promedio de superficie foliar de una planta que muestra daño por heladas, daño que en plantas tolerantes al frío es menor que el daño promedio observado a una temperatura de menos 3 °C para variedades de referencia relativamente sensibles al frío, tales como, por ejemplo, Caribe y/o Kennebec.
El término "tolerancia a la sequía", como se usa en este documento, se refiere a una planta que muestra un daño inferior al promedio en condiciones de agua limitantes en comparación con variedades de referencia relativamente sensibles a la sequía tales como Caribe y/o Carlton.
El término "tolerancia a la inundación", como se usa en este documento, se refiere a una proporción 50 % menor de tubérculos afectados (degradación anaeróbica) en relación con una planta que muestra un daño promedio a la inundación.
El término "tolerancia a la pudrición húmeda" como se usa en este documento se refiere a una planta con el más mínimo nivel de resistencia a la especie Erwinia (actualmente denominada pectobacteria spp.).
El término "tolerancia a la pudrición seca" como se usa en este documento se refiere a una planta con el más mínimo nivel de resistencia a la especie fusarium.
El término "tolerancia a la salinidad", como se usa en este documento, se refiere a una planta que muestra menos daño que el promedio en condiciones de salinidad.
El término "tasa de crecimiento", como se usa en este documento, se refiere al aumento de la biomasa vegetal por unidad de tiempo.
El término "defectos en el desarrollo de los tubérculos" (por ejemplo, tubérculos deformados o dañados)" como se usa en este documento se refiere a una planta que muestra malformaciones tanto visuales como internas de los tubérculos durante el desarrollo de la planta y/o la cosecha del tubérculo.
El término "rendimiento de tubérculos", como se usa en este documento, se refiere al peso total de los tubérculos de una planta o de una población de plantas generalmente expresado en gramos de peso fresco.
El término "tamaño del tubérculo", como se usa en este documento, se refiere a la anchura, altura y longitud absolutas de un tubérculo. Los tamaños de tubérculo beneficiosos incluyen tamaños de tubérculo promedio de planta de alrededor de 80-160 gramos por tubérculo. El número medio de tubérculos para plantas comercialmente valiosas es de aproximadamente 8 -12 tubérculos por planta.
El término "color de la piel del tubérculo", como se usa en este documento, se refiere al color de la piel de un tubérculo después de la cosecha como resultado de la acumulación de antocianina en los tejidos de la piel del tubérculo.
El término "profundidad del ojo" como se usa en este documento se refiere a la distancia relativa entre la superficie de la piel y el primordio del brote de un tubérculo.
El término "forma de tubérculo", como se usa en este documento, se refiere a la proporción largo/ancho, para indicar la variación continua de tubérculos redondos, ovalados a largos, así como la proporción alto/ancho, para indicar la variación continua de cilíndrico a la cantidad de planitud de un tubérculo.
El término "color de la pulpa del tubérculo" como se usa en este documento se refiere al color del interior de la pulpa del tubérculo después de la cosecha como resultado de la ausencia o presencia de compuestos carotenoides que causan el color de la pulpa blanca o amarilla, respectivamente, así como la ausencia de compuestos de antocianinas que causan tonos rojos, azules, púrpuras del color de la carne, independientemente de que la presencia de color en los patrones sea parcial o total.
El término "sabor a tubérculo", como se usa en este documento, se refiere a la apreciación de los consumidores al consumir un tubérculo de patata cocida.
Los términos "vida útil del tubérculo" y "capacidad de almacenamiento del tubérculo" como se usan en este documento son sinónimos y se refieren a la falta de cambios en la apariencia de un tubérculo durante el almacenamiento.
El término "período de latencia del tubérculo", como se usa en este documento, se refiere al período de tiempo entre la siembra y la brotación de un tubérculo, mientras se siembra en condiciones normales.
El término "resistencia a la deshidratación del tubérculo", como se usa en este documento, se refiere a una planta que muestra menos deshidratación del tubérculo que el promedio en condiciones de baja humedad (véase, vida útil).
El término "contenido de almidón de tubérculo" como se usa en este documento se refiere al peso de almidón sobre el peso fresco total de un tubérculo.
El término "contenido de materia seca del tubérculo", como se usa en este documento, se refiere al peso de los componentes secos de una patata dividido entre el peso fresco total.
El término "calidad de cocción del tubérculo", como se usa en este documento, se refiere a la variación continua de firme a harinoso de un tubérculo después de la cocción.
El término "calidad para freír los tubérculos", como se usa en este documento, se refiere a la apreciación de los consumidores por un tubérculo después de freírlo.
El término "calidad de fabricación de chips de tubérculos", como se usa en este documento, se refiere a la apreciación de los consumidores de un tubérculo después de hacer chips.
El término "uniformidad del tubérculo", como se usa en este documento, se refiere a la desviación estándar de los rasgos del tubérculo, por lo cual una variación estándar baja se refiere a una uniformidad alta y viceversa.
El término "resistencia al edulcorante en frío", como se usa en este documento, se refiere a la resistencia de un tubérculo para producir glucosa y/o fructosa mientras se almacena a temperaturas inferiores a 8 °C.
El término "tetraploide", como se usa en este documento, se refiere a una planta en la que cada célula vegetativa contiene cuatro conjuntos de cromosomas (4n). Los términos "fértil" y "fertilidad" se refieren en términos generales a la capacidad de reproducirse, es decir, de concebir y producir descendencia, preferiblemente descendencia fértil. De este modo, el término fertilidad se refiere a los individuos que se pueden cruzar para producir descendencia fértil en el sentido de que las flores, cuando se polinizan, producen bayas que contienen semillas que, cuando se siembran, dan como resultado una planta que lleva flores fértiles, etc.
El término "descendencia fértil" o "semilla fértil" se define en este documento como semilla capaz de crecer en una planta de patata productora de flores en la que las flores son fértiles masculinas y femeninas. De este modo, el término se refiere preferiblemente a una planta o semilla que, cuando se convierte en una planta, es capaz de producir descendencia como progenitor masculino y femenino en virtud de la presencia de óvulos fértiles y polen fértil (es decir, tanto las flores masculinas como las femeninas son fértiles).
El término "planta fértil" se define en este documento como una planta capaz de producir bayas portadoras de semillas fértiles. Preferiblemente, dichas bayas llevan cada una al menos 5, más preferiblemente al menos 15-20, aún más preferiblemente al menos 50 semillas fértiles, aún más preferiblemente entre 50 y 500 semillas. Alternativamente, se dice que una planta es fértil cuando el número de semillas producidas por baya es al menos el 10 %, más preferiblemente al menos el 30 %, 40 %, 50 %, 60 %, 70 %, 80 %, 90 %, 100 %, 150 % 200 % o más del número encontrado en las bayas del clon de mejoramiento diploide RH89-039-16. El término también, o alternativamente, se refiere a líneas de plantas que tienen más del 25 %, preferiblemente más del 90 % de plantas fértiles.
El término "autocompatible" se refiere a la capacidad de desarrollar semillas en bayas que son el resultado de la autopolinización, la autofertilización y la producción de una progenie fértil.
El término "capaz de realizar retrocruzamiento/autofecundación/cruzamiento y dar como resultado una descendencia (auto)fértil y autocompatible" se refiere a la capacidad de producir una o más bayas con semilla como resultado de un retrocruzamiento, autofecundación o cruzamiento y en el que más del 60 %, preferiblemente más del 70 %, 80 %, 85 %, 90 % o más de dicha semilla en dicha una o más bayas es capaz de convertirse en plantas que son (auto)fértiles y autocompatibles.
Para otros términos como se usan en este documento, se hace referencia a Allard, R. W. Principies of Plant Breeding, 2nd Edition, Wiley Nueva York, 1999, y específicamente al Glosario del mismo.
Descripción general
Los presentes inventores descubrieron que se pueden producir con éxito líneas de mejoramiento de élite para el mejoramiento de patatas. El cruce de dos líneas de mejoramiento de élite proporciona semillas híbridas f 1, tales semillas híbridas F1, cuando se convierten en plantas, dan como resultado plantas de rendimiento agronómico superior. Más importante aún, todas las semillas son genéticamente esencialmente idénticas, lo que significa que, excepto por el cruce ocasional o el evento de mutación, todas las semillas son uniformes.
En este documento se describe un método que permite la provisión de líneas de mejoramiento de patata de élite que, cuando se cruzan entre sí, proporcionan semillas de patata híbridas F1 uniformes que, cuando se convierten en plantas, dan como resultado plantas que tienen un rendimiento agronómico superior. Las líneas de mejoramiento de patata de élite pueden tener características complementarias y, como resultado de su tendencia homocigótica, las plantas F1 que crecen a partir de semillas híbridas F1 son uniformes, pero cuando estas plantas F1 se realiza la autofecundación o polinización cruzada posteriormente, las plantas F2 resultantes se segregarán para el diversos rasgos
De este modo, en contraste con las verdaderas semillas de patata (TPS) actualmente disponibles generadas de las formas convencionales de la técnica anterior y cosechadas a partir de bayas en las que cada semilla individual es genéticamente diferente de cualquier otra semilla, las semillas obtenidas del cruce de las líneas de patata y las líneas de mejoramiento de patata de élite son genéticamente esencialmente idénticas.
Las variedades de TPS de la técnica anterior se derivan de cruces entre progenitores seleccionados pero heterocigotos y, por consiguiente, se espera variabilidad en plantas y tubérculos. El mejoramiento híbrido de patata, como se propone en este documento, difiere de la producción convencional de TPS en que los TPS actuales son el resultado del cruce de dos líneas puras de mejoramiento, lo que resulta en variedades uniformes, mientras que las variedades TPS de la técnica anterior son el resultado de la polinización abierta o el cruce entre dos líneas genéticamente no homocigotas y a menudo tetraploides que dan como resultado variedades no uniformes.
Las plantas proporcionadas actualmente no sólo son novedosas porque combinan un número de propiedades, nunca antes alcanzadas juntas en una sola línea de patata, sino que también permiten métodos de mejoramiento novedosos y muy ventajosos. Por consiguiente, las plantas son un intermediario en la producción de nuevas variedades de patata, variedades que pueden producirse mediante métodos como los descritos en este documento. Los métodos de mejoramiento ventajosos permiten la eliminación de muchos de los inconvenientes del mejoramiento convencional de patata como se indicó anteriormente, incluida la eliminación de la selección de clones que requiere mucho tiempo para llegar a una nueva variedad de patata y la posibilidad de proporcionar semilla de patata híbrida uniforme. Además, las nuevas plantas y métodos también permiten eliminar los inconvenientes relacionados con la propagación y producción clonal, incluida la posibilidad de eliminar los altos costes de transporte y la persistencia clonal de plagas y enfermedades.
Nuevas plantas de patata
Líneas de mejoramiento parentales
En este documento se describen plantas de patata Solanum tuberosum esencialmente homocigóticas, diploides, vigorosas, fértiles y autocompatibles.
Preferiblemente, las plantas tienen fijadas dentro de su genoma secuencias de ácido nucleico que codifican genes que confieren a la planta un rasgo agronómicamente deseable. Tales secuencias de ácido nucleico pueden ser introgresiones o transgenes derivados de germoplasma de patata con rasgos agronómicamente deseables, preferiblemente de germoplasma diploide.
- Esencialmente homocigoto
En el mejoramiento, las plantas se propagan por reproducción sexual, por lo general mediante el cruce de progenitores seleccionados con rasgos deseables para producir una progenie mejorada. La progenie hereda genes para rasgos deseables e indeseables de ambos progenitores. La selección repetida de individuos meritorios de cada generación que servirán como progenitores de la siguiente conservará las características deseadas y eliminará las indeseables. El progenitor definitivo en el mejoramiento de la planta es una raza pura, una línea altamente homocigota que, cuando se usa en cruces con otras razas puras, da como resultado una descendencia idéntica, tanto fenotípica como genotípicamente, dentro de una sola generación y entre generaciones consecutivas obtenidas de otros cruces de los mismos progenitores. Las razas puras, o endogámicas puras, se pueden obtener mediante varias generaciones de autofecundación o endogamia (cruzamiento de parientes cercanos, tales como plantas hermano con hermana o descendiente con el progenitor (retrocruzamiento)), mostrando cada generación sucesiva un mayor grado de uniformidad genética u tendencia homocigótica. El uso de homocigotos endogámicos como progenitores de mejoramiento resulta en una descendencia que es idéntica, tanto fenotípica como genotípicamente, dentro de una
sola generación y entre generaciones consecutivas obtenidas de otros cruces de esos progenitores. Por lo tanto, los endógamos puros se pueden usar repetidamente para producir las mismas plantas descendientes.
En la patata, la autofecundación y la endogamia dan como resultado una severa depresión endogámica, o la expresión de genes recesivos nocivos, cuya expresión se revela por la pérdida de vigor y la pérdida de fertilidad. Por consiguiente, hasta ahora ha sido imposible producir homocigotos puros endogámicos en patata ya que hasta la presente invención no se podían alcanzar altos niveles de tendencia homocigótica en patata con suficiente vigor. La presente invención ahora, por primera vez, proporciona plantas de patata diploides Solanum tuberosum que tienen un nivel de tendencia homocigótica que es superior al 75 % y que se combine con la fertilidad, la autocompatibilidad y el vigor de la planta.
- Diploide
Las plantas de patata Solanum tuberosum son plantas diploides, a diferencia de las variedades comerciales de patata, que son tetraploides. Cabe señalar que en dichas plantas están previstos expresamente mayores niveles de ploidía, ya que la etapa diploide se usa principalmente durante el desarrollo de las líneas de mejoramiento. Una vez que se han establecido las razas puras, las líneas pueden hacerse tetraploides. Por consiguiente, las plantas también pueden ser tetraploides. Las plantas tetraploides se pueden producir a partir de células tetraploides o cultivos de tejidos, que a su vez se pueden producir duplicando el genoma de células diploides o cultivos de tejidos mediante métodos conocidos per se, por ejemplo, usando oryzalin como describe Barandalla et al. (Barandalla et al. Potato Research (2006) 49:143-154). Las plantas tetraploides pueden tener la ventaja de que proporcionan mayores rendimientos.
- Vigorosa
Aunque las variedades comerciales de patata tetraploide son fértiles, autocompatibles y vigorosas porque crecen bien, no producen plantas o frutos deformados, sino que se manifiestan como plantas sanas con un alto rendimiento de tubérculos, su progenie autofecundada muestra una grave depresión endogámica, es decir, dan como resultado plantas de bajo crecimiento y bajo rendimiento con tubérculos deformados. Las líneas diploides y esencialmente homocigóticas de la técnica anterior, tales como las descritas por Phumichai et al. como se hace referencia en este documento, también crecen muy mal y se manifiestan como plantas muy débiles y pequeñas, que tienen una mala formación de semillas. De hecho, las plantas de patata diploides resultantes de la endogamia y que alcanzan una tendencia homocigótica de al menos un 25 % o un 50 % en los alelos son invariablemente de crecimiento débil.
Las plantas de patata Solanum tuberosum no son débiles ni frágiles, y aunque no tienen por qué ser necesariamente tan vigorosas como las variedades tetraploides comerciales actuales, no muestran depresión por endogamia. Por consiguiente, son vigorosas. La formación de tubérculos ejemplifica el vigor de las plantas de las líneas de patata Solanum tuberosum de la presente invención. Aunque todas las variedades de patata comerciales (tetraploides y derivadas de tubérculos-semillas) producen tubérculos grandes, o al menos tubérculos de tamaño suficiente, las líneas diploides de la técnica anterior y las plantas esencialmente homocigóticas producen sólo tubérculos pequeños, que, por ejemplo, no sirven para la producción de chips de patatas o patatas fritas. Las líneas diploides de la técnica anterior y las plantas esencialmente homocigóticas producen también muy pocos tubérculos, de modo que el rendimiento de tubérculos por planta es al menos diez veces menor que el de las plantas diploides heterocigotas y las variedades comerciales de patata tetraploide. En cambio, las plantas de las líneas de patata Solanum tuberosum de la presente invención exhiben características beneficiosas de formación de tubérculos, incluidos tubérculos de perfil de tamaño beneficioso, y un rendimiento de tubérculo que puede ser de al menos un 50 % en comparación con las plantas heterocigotas diploides usadas como donantes en los ejemplos descritos en este documento (D1, D2, y/o D3).
Como se encontró en este documento, el rendimiento de tubérculo de los diploides F1 fue en promedio de 443 gramos por planta, con valores tan altos como 600 y 634 gramos.
El retrocruzamiento con la F1: "IVP08-06" produjo en promedio 155 y 209 gramos, al realizar el retrocruzamiento con la F1: "IVP08-07" produjo en promedio 209 y 259 gramos.
Las "pseudo F2" produjeron en promedio 172 y 332 gramos por planta, y ocasionalmente produjeron más que los diploides F1.
Las F2 produjeron en promedio 69 y 252 gramos por planta. Varias plantas se superponen con los valores de rendimiento de las diploides F1.
- fértil
Las plantas de patata Solanum tuberosum, aunque esencialmente homocigóticas, también son fértiles. Eso significa que pueden usarse en programas de mejoramiento porque su descendencia también será fértil.
- autocompatible
Las plantas de patata Solanum tuberosum son autocompatibles. Esta característica se introduce para permitir la autofecundación de las plantas y la producción de líneas homocigóticas. La base preferida de la autocompatibilidad se deriva del gen inhibidor del locus S dominante (Sli) (Hosaka and Hanneman, 1998, Euphytica, 99: 191-197).
Las plantas de patata Solanum tuberosum esencialmente homocigóticas tienen una ventaja de que pueden cruzarse con otras plantas de patata esencialmente homocigóticas por lo que la descendencia híbrida de dicho cruce es esencialmente genéticamente uniforme y no se segregará.
En este documento se describen plantas de patata Solanum tuberosum esencialmente homocigóticas, diploides, vigorosos, fértiles y autocompatibles que tienen características agronómicamente deseables.
Estas nuevas plantas de patata son capaces de:
• ser retrocruzadas con un progenitor recurrente esencialmente homocigótica y dar como resultado una descendencia (auto) fértil y autocompatible;
• ser autofecundadas (autofertilización de 1 descendiente individual) y dar como resultado descendencia (auto)fértil y autocompatible;
• ser cruzadas mutuamente (fertilización entre 2 descendientes individuales) y dar como resultado una descendencia (auto) fértil y autocompatible.
Dichas plantas de patata Solanum tuberosum incluyen la línea con la referencia de los obtentores AGVD1, semillas representativas de dicha línea que se depositaron el 23 de octubre de 2009 en NCIMB, Aberdeen, Escocia, con el número de acceso de NCIMB 41663 y la referencia de los obtentores AGVD1, cuya planta representa una F2 obtenida cruzando una planta de progenitor IVP07-1001/4 (véase, el ejemplo 1), que es una línea de patata esencialmente homocigótica, diploide, fértil y autocompatible pero no vigorosa con una planta del progenitor D1, que es un clon de mejoramiento diploide élite que es esencialmente no homocigoto, y no es autocompatible, pero que tiene el rasgo agronómico deseable de color de pulpa amarilla y exhibe una buena calidad de cocción en el sentido de que la pulpa no se cambia a gris después de la cocción y autofecundación de las plantas descendientes F1 resultantes. Se estima que AGVD1 es aproximadamente homocigoto en un 50 %, lo que significa que aproximadamente el 50 % de los loci en los que difieren los progenitores son homocigotos.
Dichas plantas de patata Solanum tuberosum también incluyen la línea con la referencia de obtentor AGVD2, semillas representativas de dicha línea que se depositaron el 23 de octubre de 2009 en NCIMB, Aberdeen, Escocia, con el número de acceso de NCIMB 41664 y con la referencia de obtentor AGVD2, cuya línea representa una línea (F1 x F1) (o pseudo F2) obtenida cruzando una planta del progenitor IVP07-1001/4 como se indicó anteriormente con una planta del progenitor D1 como se indicó anteriormente, y cruzando mutuamente plantas descendientes F1 individuales, que comparten el progenitor IVP07-1001/4 pero tienen diferentes individuos del progenitor D1. Se estima que AGVD2 es aproximadamente un 25 % homocigoto para los loci en los que difieren los progenitores.
Dichas plantas de patata Solanum tuberosum también incluyen la línea con la referencia de los obtenedores AGVD3, semillas representativas de dicha línea que se depositaron el 23 de octubre de 2009 en NCIMB, Aberdeen, Escocia, con el número de acceso de NCIMB 41665 y con la referencia de los obtenedores AGVD3, línea que representa una población BC1 obtenida por cruzamiento una planta del progenitor IVP07-1001/4 como se indicó anteriormente con una planta del progenitor D1 como se indicó anteriormente, y retrocruzamiento de las plantas descendientes F1 resultantes con el progenitor IVP07-1001/4. Se estima que AGVD3 es aproximadamente homocigoto en un 75 % para los loci en los que difieren los progenitores.
Dichas plantas de patata Solanum tuberosum también incluyen la línea con la referencia de obtentor AGVD17, semillas representativas de dicha línea que se depositaron el 5 de octubre de 2010 en NCIMB, Aberdeen, Escocia, con el número de acceso de NCIMB 41765 y con la referencia de obtentor AGVD17, línea que representa una población F3 obtenida por cruce una planta del progenitor IVP07-1001/4 como se indicó anteriormente con una planta del progenitor D2 como se indicó anteriormente, y retrocruzamiento de las plantas descendientes F1 resultantes con el progenitor IVP07-1001/4, seguido de dos etapas de autofecundación. Se estima que AGVD17 es > 80 % homocigoto para los loci en los que difieren los progenitores.
Una planta de patata Solanum tuberosum tal como se describe se puede usar de forma muy apropiada como progenitor recurrente en el que se incorporan rasgos agronómicamente deseables adicionales mediante cruzamiento, o se introducen por otros medios tales como por medios transgénicos. Una vez que un número de rasgos agronómicamente deseables se han acumulado en una planta esencialmente homocigótica, vigorosa, diploide, fértil y autocompatible, esa planta se puede usar como línea de mejoramiento de élite para la producción de semilla de patata híbrida F1 comercial. Asimismo, se puede proporcionar una segunda línea de mejoramiento de élite que pueda servir como progenitor del sexo opuesto para que ambas plantas puedan cruzarse para producir la semilla F1 comercial de una nueva variedad de patata.
El mejoramiento de semillas híbridas uniformes depende de un número de condiciones técnicas. Los requisitos previos para lograr una semilla híbrida uniforme son que i) se pueda realizar un cruce de introgresión inicial entre una planta
esencialmente homocigótica, diploide, fértil y autocompatible y una planta que actúe como donante de un rasgo agronómicamente deseable, que ii) se pueda realizar un retrocruzamiento producido entre la descendencia de ese cruce inicial y la planta esencialmente homocigótica, diploide, fértil y autocompatible, y que iii) la descendencia de la misma puede ser autofecundada. Las líneas de patata AGVD1, AGVD2 y AGVD3 exhiben estas características y las plantas individuales de estas líneas se pueden usar en cruces de introgresión adicionales, retrocruzamientos y autofecundaciones para llegar a las líneas de mejoramiento de élite apropiadas para usar como progenitores para semillas híbridas F1 comerciales que se pueden cultivar en planta de patata híbrida de una nueva variedad de patata.
Una planta muy preferida es una planta de la línea AGVD2 o una planta descendiente de la misma que tenga todas las características fisiológicas y morfológicas de la planta de la línea AGVD2.
Otra línea depositada como ejemplo de realizaciones de la presente invención es AGVD17 (NCIMB 41765, depositada el 5 de octubre de 2010). La ventaja de AGVD17 es que es una línea altamente homocigota (> 80 % homocigota) que combina vigor y fertilidad con un rendimiento de tubérculo de 300 gramos (peso fresco) en promedio por planta. Las plantas individuales de poblaciones relacionadas incluso mostraron rendimientos de tubérculo de 500 gramos por planta. Estos datos se generaron a partir de plantas cultivadas en el campo que se criaron a partir de plántulas. Las plantas de patata cultivadas a partir de clones generalmente producen tubérculos de un 50 a un 100 % más altos que las plantas cultivadas a partir de plántulas. Por consiguiente, se espera que la producción de tubérculos expresada en gramos de peso fresco de plantas como se reivindica en este documento, cuando se propagan por clonación y se crían a partir de tubérculos y se cultivan en suelo arcilloso en condiciones normales de verano en el noroeste de Europa, alcance niveles de al menos alrededor de 1000 gramos de peso fresco por planta.
De este modo, la línea AGVD17 demuestra en particular la posibilidad de entrecruzar líneas élite individuales de la presente invención (es decir, que son autocompatibles, fértiles y esencialmente homocigóticas) para llegar a la semilla híbrida que es el objetivo de la presente invención. Por consiguiente, una planta muy preferida es una planta de la línea AGVD17 o una planta descendiente de la misma que tenga todas las características fisiológicas y morfológicas de la planta de la línea AGVD17.
Híbridos
Las plantas híbridas se pueden producir mediante un método como se ejemplifica en la figura 1. Las plantas híbridas se producen mediante el cruce de dos líneas de mejoramiento parentales de patata de élite esencialmente homocigóticas que comprenden entre ellas un número de rasgos agronómicamente deseables seleccionadas del grupo que consta de tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades (incluyendo, pero no limitando a, resistencia a la sarna causada por Streptomyces spp, sarna pulverulenta, Rhizoctonia, sarna plateada, Phytophthora infestans), tolerancia a herbicidas, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad, resistencia al endulzamiento por frío y resistencia a la deshidratación del tubérculo. Además de estos rasgos agronómicamente deseables, las líneas de mejoramiento parental pueden exhibir las características deseadas en mercados de uso específico, tal como el color deseable de la piel del tubérculo, la forma deseable del tubérculo, el color deseable de la pulpa del tubérculo, el sabor deseable del tubérculo, el contenido deseable de almidón en el tubérculo, el contenido de materia seca deseable del tubérculo, calidad de cocción deseable de los tubérculos o calidad deseable para freír los tubérculos. Además de estas características específicas del mercado, las líneas de mejoramiento parentales, cuando se cruzan, preferiblemente dan como resultado híbridos que exhiben rasgos beneficiosos como tasa de crecimiento beneficiosa, rendimiento de tubérculo beneficioso, tamaño de tubérculo beneficioso, capacidad de almacenamiento de tubérculo beneficioso, período de latencia de tubérculo beneficioso, vida útil del tubérculo, calidad beneficiosa para hacer chips de tubérculo y uniformidad beneficiosa del tubérculo.
En este documento se describe un conjunto de líneas de plantas de patata, cuyas plantas o líneas de plantas son: diploides (2n); esencialmente homocigóticas (preferiblemente más del 60 % en todos los alelos); autocompatible; fértil; y vigoroso. Hasta ahora ha resultado imposible proporcionar la combinación de todas estas características en una sola planta (línea) de patata. El conjunto de plantas de patata de la invención se puede usar para producir nuevas variedades o variedades cultivadas.
En particular, se describe en este documento un conjunto de al menos 2 plantas diploides (2n), esencialmente homocigóticas (preferiblemente más del 60 % en todos los alelos), autocompatibles, fértiles y vigorosas que se pueden cruzar para producir plantas de patata híbridos Solanum tuberosum.
Nuevo método de mejoramiento de semillas de patata
Los presentes inventores han generado plantas de patata que son esencialmente homocigóticas, diploides, vigorosas, fértiles y autocompatibles y que tienen fijadas dentro de su genoma secuencias de ácidos nucleicos que codifican genes que confieren a la planta un rasgo agronómicamente deseable derivado de líneas donantes diploides apropiadas. Las dichas secuencias de ácido nucleico son el resultado de introgresiones de germoplasma de patata que tiene rasgos agronómicamente deseables. Convencionalmente, al menos en la mayoría de las especies distintas de la patata, las plantas homocigóticas se pueden obtener en principio en tan solo dos generaciones mediante la duplicación de monohaploides generados a partir de células generativas no fertilizadas, tales como óvulos o células
de polen, mediante métodos conocidos como tecnología de doble haploide (DH). Esto no es posible en la patata, ya que las plantas resultantes han demostrado hasta ahora ser infértiles, debido a lo que se considera, la expresión de genes perjudiciales recesivos. El método DH fue ampliamente probado por los presentes inventores, pero resultó infructuoso.
Los presentes inventores usaron la estrategia de autofecundación, retrocruzamiento y cruce repetidos de germoplasma de patata de una sola línea de patata autocompatible y una o más líneas de patata autoincompatibles que albergan rasgos agronómicos. Las plantas homocigóticas se obtuvieron por autofecundación continua de plantas de patata diploides y selección de descendencia fértil (además de la selección inherente por autocompatibilidad). Este es un enfoque completamente nuevo, ya que en los métodos de la técnica anterior, las plantas se obtienen por selección de rasgos agronómicos, tales como formación de tubérculos o características generales de crecimiento de las plantas descendientes, no por selección de descendencia fértil de autofecundaciones. Además, y a diferencia de los métodos de la técnica anterior, los procedimientos se realizan usando líneas de plantas diploides, en lugar de plantas tetraploides. Inesperadamente, este enfoque radicalmente nuevo resultó exitoso para obtener plantas descendientes diploides cada vez más homocigóticas, que eran autocompatibles y fértiles. Además, cuando se cruzan (exteriormente) con plantas de líneas diploides que albergan rasgos beneficiosas, tales como la resistencia o el color blanco del tubérculo, estos rasgos podrían introducirse en el germoplasma esencialmente homocigótico sin pérdida de autocompatibilidad y fertilidad.
En esencia, el método para obtener las plantas es el siguiente:
El punto de partida es una línea de patata autocompatible diploide (se han depositado ejemplos de líneas apropiadas bajo las referencias de los depositantes AGVD 1, AGDV 2, AGDV 3 y AGVD17). En general, una línea que es autocompatible debido a la presencia del gen Sli se puede usar como punto de partida. Las plantas de esta línea autocompatible diploide se autofecundan y las semillas obtenidas de bayas portadoras de semillas se siembran y se cultivan en plantas descendientes. Las plantas descendientes que llevan flores se autofecundan nuevamente y las semillas obtenidas de las bayas portadoras de semillas se siembran nuevamente y se cultivan en plantas descendientes. No se presta atención, o al menos se presta poca, a la apariencia agronómica de las plantas en esta etapa. Pueden verse marchitas y frágiles, con sólo unas pocas flores. No obstante, la autofecundación se repite al menos tres veces, pero preferiblemente 4 o 5 o incluso 6-8 veces. De esta manera se obtiene una línea de plantas de patata esencialmente homocigóticas, fértiles, autocompatibles y diploides, esencialmente por autofecundación y selección de plantas que producen descendencia fértil.
A continuación, se agrega un rasgo agronómicamente deseable al genotipo diploide de patata esencialmente homocigótico, fértil, autocompatible, cruzando una planta de esa línea con una planta de una línea de patata diploide esencialmente heterocigota no relacionada, preferiblemente una línea que contiene un rasgo agronómicamente deseable. Esta línea esencialmente heterocigota (cuyo término simplemente refleja que la línea no es una línea esencialmente homocigótica como se usa en este documento) funciona como donante de un rasgo deseable agronómicamente, en este caso el vigor.
El híbrido resultante se autofecunda y/o se retrocruza con la línea autocompatible mientras se selecciona para mayor vigor. De esta manera, se potencia el vigor de la línea de plantas de patata dipolides esencialmente homocigóticas, fértiles, autocompatibles y, al mismo tiempo, se mejora agronómicamente la línea. Estos etapas darán como resultado una patata diploide autocompatible homocigota fértil vigorosa: El vigor puede potenciarse aún más cruzando con otras líneas de patata diploides esencialmente heterocigóticas, cuyas líneas adicionales sirven como donantes de características agronómicamente deseables adicionales, seguido de autofecundación y/o retrocruzamiento con la línea autocompatible mientras se seleccionan estos rasgos adicionales, que las etapas darán como resultado la provisión de líneas de mejoramiento de patata diploides.
Con el fin de producir líneas de mejoramiento de patatas diploides de élite, el valor de mejoramiento de las líneas de mejoramiento se potencia aún más mediante el entrecruzamiento, la autofecundación y el retrocruzamiento mientras se seleccionan continuamente los rasgos agronómicamente deseables proporcionados a la línea de mejoramiento, y mediante el establecimiento de líneas que exhiben buenas habilidades de combinación con otras lineas Tales habilidades de combinación se evalúan mediante el uso de cruces experimentales.
Finalmente, las variedades híbridas de patata se pueden producir cruzando líneas individuales de mejoramiento de patatas diploides de élite que muestren buenas habilidades de combinación. Las semillas resultantes de este cruce de progenitores homocigotos son semillas híbridas de patata verdaderas que son uniformes.
Usando los métodos que se describen en este documento, generalmente, alrededor de 7 a 8 ciclos de autofecundación darán como resultado plantas esencialmente homocigóticas, pero el experto en la técnica comprenderá que también se pueden usar más generaciones de autofecundación. También se pueden usar menos generaciones, como seis generaciones, para proporcionar plantas de patata Solanum tuberosum esencialmente homocigóticas. El número de generaciones de retrocruzamiento puede incluso reducirse a tres para alcanzar más del 95 % de tendencia homocigótica al aplicar marcadores genéticos moleculares.
Esencialmente, el método novedoso de producción de variedades híbridas de patata proporciona la posibilidad de producir una nueva variedad de patata dentro de aproximadamente 5 años. En el ejemplo 5 se proporciona un procedimiento detallado para la producción de patatas híbridas a partir de AGVD17 (línea F3). Esta línea es >80 % homocigota, de modo que se necesitan muy pocas autofecundaciones para obtener una planta que sea aproximadamente 95 %-100 % homocigota. Además, AGVD17 ya es vigorosa como se define en este documento.
En este documento se describe un método de producción de una semilla de patata verdadera (TPS) genéticamente homogénea o uniforme. Este método es entre otros útil en un método de prevención de la transmisión de infecciones víricas entre generaciones de patata, pero también para superar muchos de los otros problemas asociados con el mejoramiento clásico de patata, producción y transporte y cultivo de patata de “semilla". Además, el material parental actual para la producción de semilla híbrida puede mantenerse en forma de semilla y no en forma de material vegetativo como requiere la producción tradicional de TPS.
En este documento se describe un método de producción de una variedad de patata híbrida que comprende el uso del material vegetal depositado proporcionado en este documento como material de partida.
Alternativamente, se pueden desarrollar otras líneas de plantas esencialmente homocigóticas, diploides, fértiles, vigorosas y autocompatibles diferentes de los depósitos a los que se hace referencia en este documento y la producción de una variedad de patata híbrida puede comenzar desde ese nivel de fase temprana usando la enseñanza proporcionada en este documento ya que los presentes inventores han demostrado que esta vía, aunque laboriosa, tiene éxito. Dicho material de mejoramiento alternativo también es vigoroso, esencialmente homocigótico, diploide, fértil y autocompatible.
Como una primera etapa, dicho desarrollo en fase temprana puede comprender la provisión de una planta de una primera especie de patata cultivada (Solanum tuberosum) o una especie cruzada relacionada (generalmente un tubérculo que porta especie Solanum).
A continuación, dicha planta se desarrolla en una línea de plantas de patata diploides autocompatibles, preferiblemente una Solanum tuberosum. La patata diploide suele ser autoincompatible. La autocompatibilidad puede introducirse adecuadamente a partir de especies relacionadas (véase, por ejemplo, Hermsen, J.G.Th. 1978, Euphytica 27, 1-11).
La línea resultante de plantas de patata diploides autocompatibles se desarrolla luego en una línea de patatas diploides autocompatibles, fértiles, homocigóticas. Los diploides homocigotos se pueden obtener duplicando los haploides (Uijtewaal et al., 1987, Theor. Appl. Genet. 73, 751-78) por cultivo de anteras (Jacobsen and Sopory, 1977, Theor. Appl. Genet. 52, 119-123), por cultivo de óvulos o por autofecundaciones repetidas (Phumichai & Hosaka, 2006, Euphytica 149, 251-258). El último método se usó para producir la línea de patata diploide homocigota fértil autocompatible que formó la base de las plantas vigorosas como se describe en este documento.
A continuación, la línea de patata diploide homocigótica, fértil, autocompatible se desarrolla en una línea de patata diploide homocigótica, autocompatible, fértil, vigorosa. El vigor se puede potenciar cruzando con otras plantas de patata diploides, autofecundadas y retrocruzadas mientras se selecciona para mayor vigor. Debe señalarse que se necesitan poblaciones altas y que incluso para algunas líneas de patata diploides esencialmente heterocigotas no relacionadas que se usan como donantes del rasgo agronómicamente deseable, se pueden encontrar muy pocas plantas autocompatibles en una población de 1000 plantas. En general, tales intentos se terminan, y sólo cuando aproximadamente el 10-50 % de las plantas descendientes son autocompatibles (tal como en el caso de D2 en el ejemplo 2 a continuación), la línea donante exhibe una capacidad de combinación general tan buena que está previsto una introducción exitosa del rasgo agronómico en el linaje autocompatible.
A continuación, la línea de patata diploide, vigorosa, fértil, homocigota, autocompatible, se desarrolla en una línea de mejoramiento de patata diploide. Lo más preferiblemente, se producen al menos dos líneas de patata diploides autocompatibles homocigotas fértiles vigorosas separadas y se desarrollan al menos dos líneas de mejoramiento de patatas diploides distintas. Tales líneas de mejoramiento comprenden un alto número de rasgos agronómicamente deseables. Los rasgos agronómicamente deseables se introducen mediante el cruzamiento con otras plantas de patata diploides, la autofecundación y el retrocruzamiento mientras se seleccionan estos rasgos. Este programa de mejoramiento puede comprender pruebas de enfermedades, tales como si los brotes o tubérculos están libres de enfermedades, o si son resistentes a las enfermedades.
A continuación, las líneas de mejoramiento de patatas diploides se desarrollan en líneas de mejoramiento de patatas diploides de élite. Esto se logra potenciando el valor de mejoramiento por medio de cruces, autofecundaciones y retrocruzamientos mientras se seleccionan continuamente los rasgos agronómicamente deseables. En esta fase se realizan cruces experimentales para evaluar las habilidades combinatorias generales. Las líneas de élite diploides ahora también pueden convertirse en líneas de élite tetraploides (4x) mediante métodos conocidos per sé, tales como por tratamiento químico con, por ejemplo, colchicina, o por duplicación "espontánea" del genoma mediante cultivo de tejidos.
Finalmente, las variedades híbridas de patata se producen a partir de estas líneas de mejoramiento de patatas diploides o tetraploides de élite mediante el cruce de líneas de mejoramiento de buena combinación. Las semillas obtenidas son semillas de patata verdaderas híbridas que son uniformes.
Un método de producción de un variedad híbrida de patata comprende las etapas de producir una línea de mejoramiento diploide de patata. Dicho método comprende los etapas de:
(a) proporcionar una primera planta de patata, cuya primera planta de patata es una planta de una línea de patata diploide, autocompatible y esencialmente homocigótica, preferiblemente una planta de una línea de patata diploide homocigótica, autocompatible, vigorosa, fértil, más preferiblemente una línea de mejoramiento de patata diploide;
(b) proporcionar una segunda planta de patata de una línea de patata que puede tener cualquier nivel de ploidía, preferiblemente diploide, y/o cualquier nivel de tendencia homocigótica en la que dicha segunda planta de patata sirve como donante para un rasgo agronómicamente deseable cuyo rasgo se va a introgresar en dicha línea esencialmente homocigótica de dicha primera planta de patata o al menos en una planta descendiente resultante de un cruce entre dicha primera y segunda planta, en la que dicho rasgo se selecciona del grupo que consiste en tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades (incluyendo pero no limitado a la resistencia a la sarna causada por Streptomyces spp, sarna pulverulenta, Rhizoctonia, sarna plateada, Phytophthora infestans), tolerancia a herbicidas, capacidad de enraizamiento, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad, tasa de crecimiento, defectos de desarrollo del tubérculo (por ejemplo, tubérculos deformados o dañados), rendimiento del tubérculo, tamaño del tubérculo, color de la piel del tubérculo, profundidad del ojo, forma del tubérculo, color de la pulpa del tubérculo, sabor del tubérculo, capacidad de almacenamiento del tubérculo, período de latencia del tubérculo, vida útil del tubérculo, resistencia a la deshidratación del tubérculo, contenido de almidón del tubérculo, contenido de materia seca del tubérculo, calidad de cocción del tubérculo, calidad para freír el tubérculo, calidad para hacer chips de tubérculo, uniformidad del tubérculo y resistencia al endulzamiento por frío.
Tras la provisión de estas dos plantas, el método comprende entonces la etapa de:
(c) polinizar de forma cruzada dicha primera y segunda planta de patata de manera que una de dichas plantas progenitoras produzca frutos portadores de semillas (bayas) y recoger dichas semillas de dichos frutos para proporcionar una semilla descendiente híbrida.
A continuación, las semillas se cultivan hasta convertirse en plantas y las plantas que comprenden el rasgo agronómicamente deseable de la planta donante se seleccionan, ya sea directamente o siguiendo etapas adicionales de autofecundación y retrocruzamiento, como es bien conocido en la técnica del mejoramiento de la planta. En estos métodos, se pueden usar marcadores para ayudar en el procedimiento de mejoramiento.
La semilla descendiente híbrida se cultiva preferiblemente en una planta que posteriormente se retrocruza con una planta a la línea parental recurrente
Las siguientes etapas no son esenciales para ninguno de los métodos anteriores. Sin embargo, estas etapas adicionales pueden proveer la producción de tubérculos de consumo mediante el cultivo de las semillas híbridas de la invención y/o pueden proveer la producción de nuevas líneas de mejoramiento diploides homocigóticas, autocompatibles y de élite que pueden servir como progenitores para la inclusión de aún más rasgos en una generación descendiente. Las plantas son fértiles y vigorosas. Estos beneficios adicionales se logran mediante las siguientes etapas adicionales de:
(d) hacer crecer dicha semilla descendiente en una planta de patata descendiente híbrida, y
(e1) cosechar tubérculos de dicha planta de patata híbrida para proporcionar tubérculos que porten dicho rasgo, o
(e2) autofecundación de dicha planta de patata descendiente híbrida o retrocruzamiento de dicha planta de patata descendiente híbrida con una planta de patata de dicha línea de dicha primera planta de patata para proporcionar más plantas descendientes que lleven dicho rasgo y en el que dichas plantas descendientes son cada vez más homocigóticas después de cada etapa de retrocruzamiento o autofecundación.
Preferiblemente, dicha segunda planta de patata usada en el método descrito anteriormente es una planta de patata descendiente híbrida como se define en la etapa (d) anterior y obtenida mediante el método descrito anteriormente. Más preferiblemente, dicha segunda planta de patata usada en el método descrito anteriormente es una planta de patata descendiente híbrida como se define en la etapa (e2) anterior y obtenida mediante el método descrito anteriormente.
Preferiblemente, dicha etapa (e2) comprende de 2 a 7 retrocruzamientos usando una planta de la línea AGVD1, AGVD2, AGVD3 o AGVD17 como progenitor recurrente y probando plantas descendientes para detectar la presencia del rasgo (por ejemplo, en forma de presencia homocigota o heterocigota de alelos responsables del rasgo), seguido de uno o dos (o más) autofecundaciones, para proporcionar una planta esencialmente homocigótica que es genéticamente igual a una planta de población a Gv D1, AGVD2, AGVD3 o AGVD17, pero en la que los alelos responsables del rasgo están presentes de forma heterocigótica, preferiblemente homocigótica. Esta planta tiene al menos un alelo adicional no disponible en la población AGVD1, a Gv D2, AGVD3 o AGVD17.
El método anterior proporciona ahora un procedimiento de mejoramiento para la producción de líneas de mejoramiento homocigotas o líneas de mejoramiento de élite en base al antecedente de la población AGVD1, AGVD2, AGVD3 o AGVD17, al cual se le agregan sucesivamente alelos adicionales de antecedente, proporcionando así líneas de mejoramiento de élite en las que los rasgos deseables están acumulados.
Los métodos descritos en este documento proporcionan ahora por primera vez un método de prevención de la transmisión de infecciones virales entre generaciones de patatas y, al mismo tiempo, resuelven muchos de los otros problemas de la técnica anterior relacionados con el mejoramiento de nuevas variedades de patatas y la producción comercial de patatas para el consumo. Esto ha sido posible gracias al suministro de semilla de patata verdadera, es decir, semilla botánica, cuya semilla puede convertirse en una planta de patata que produce tubérculos para el consumo, y cuyas semillas proceden de un cruce en el que al menos uno de los progenitores es una planta de una línea de patata diploide, autocompatible y esencialmente homocigótica, preferiblemente dichas semillas proceden de un cruce en el que ambos progenitores son plantas de una línea de mejoramiento diploide, autocompatible y esencialmente homocigótica.
Una ventaja importante de este nuevo método es que, de esta forma, se puede prevenir eficazmente la transmisión de una infección por un virus que no se transmite a través de los gametos y las semillas.
En este documento se describe adicionalmente un método para el mejoramiento adicional de líneas homocigotas que son fáciles de manejar y que exhiben una fertilidad y un vigor de la planta mejorados en comparación con las líneas endogámicas de la técnica anterior, mientras se mantiene la autocompatibilidad y un alto nivel de tendencia homocigótica . Se apreciará que, al igual que en el tomate, no siempre se necesita una tendencia homocigótica del 100 % para proporcionar un programa de mejoramiento eficaz. Esto no es un problema siempre que se mantenga la autocompatibilidad.
La transferencia genética de rasgos deseables o beneficiosas agronómicamente en una línea de patata homocigótica ahora se ha demostrado con éxito por primera vez.
Basándose en los medios y métodos proporcionados en este documento, el experto en la técnica podrá producir con éxito líneas de mejoramiento de patata homocigóticas basadas en su propio germoplasma. Además, el experto en la técnica apreciará que se proporcionan medios y métodos mediante los cuales puede, por ejemplo, acumular rasgos adicionales en las líneas autocompatibles y en esencia homocigóticas proporcionadas actualmente para establecer una población de líneas que se enriquecen para rasgos agronómicamente deseables y que pueden servir como germoplasma para el mejoramiento comercial por derecho propio.
Ejemplos
Ejemplo 1. Descripción general del procedimiento de mejoramiento y genealogías.
Se hicieron muchos intentos para generar genotipos diploides de patata homocigotos, fértiles y autocompatibles sin éxito. Estos intentos incluyeron la formación de haploides dobles y la posterior cribado de flores fértiles. Se emplearon varios métodos, como el cultivo de anteras para regenerar plantas homocigóticas haploides y eventualmente dobles haploides, autofecundaciones repetidas para aumentar gradualmente el nivel de tendencia homocigótica en el germoplasma diploide existente y polinización por espinas mediante el uso de S. phureja como donante de polen para inducir la formación de embriones a partir de óvulos no fertilizados. Especialmente el último método se realizó a gran escala, analizando más de dos millones de descendientes pero sin el éxito deseado. Ninguna de estas pruebas resultó en plantas autocompatibles vigorosas.
Se obtuvo una línea de patata esencialmente homocigótica mediante múltiples generaciones de autofecundación de plantas de una línea derivada de germoplasma público cuyas plantas tienen la autocompatibilidad controlando el gen Sli (Phumichai et al. Euphytica (2006) 148: 227-234). Estas plantas, denominadas "H" o IVP07-1001/4 en este documento, crecieron mal, tenían muy pocas flores y cada baya contenía sólo unas pocas semillas. Además, las plantas eran muy pequeñas (máx. 60 cm). Las plantas casi no produjeron tubérculos y los que se formaron fueron muy pequeños (peso promedio de tubérculo de unos 5 gramos/tubérculo). En total, las plantas no tuvieron ningún valor para el mejoramiento comercial de la patata, ya que exhibieron un peso de la planta y un rendimiento de tubérculo inferior al 20 % del del clon de mejoramiento diploide RH89-039-16. Las plantas simplemente contenían alelos homocigotos en muchos de los loci de su genoma, eran autocompatibles y diploides.
En el verano de 2008, las plantas de esta línea IVP07-1001/4 se cruzaron con diferentes líneas donantes diploides (D1, D2 y D3, véase la genealogía completa en la figura 2). D1 es temprano, tiene tubérculos ovalados de tamaño largo, pulpa de color amarillo claro y buena calidad para cocinar patatas fritas y buena calidad para cocinar y se usó como progenitor masculino (Tabla 6). La progenie resultante se denomina F1(D1) en este ejemplo. Se obtuvieron otras plantas descendientes F1 de manera similar pero usando el donante D2 como progenitor femenino. D2 es un clon de mejoramiento diploide de élite que es esencialmente no homocigoto y no es autocompatible, pero que tiene el rasgo agronómico deseable de cocción temprana, tubérculos ovalados, pulpa amarilla y buena calidad para cocinar chips. Esto resultó en líneas designadas como F1 (D2).
Los F1 anteriores fueron (D1) se autofecundaron para producir una F2 (D1). Esta línea fue depositada bajo la referencia de obtenedores AGVD1. Se estima que AGVDl es aproximadamente homocigota en un 50 %, lo que significa que aproximadamente el 50 % de los loci en los que difieren los progenitores son homocigotos.
F1 (D1) se cruzó con F1 (D2) para producir un pseudo F2. Esta línea (F2(D1/2)) fue depositada bajo la referencia de obtenedores AGVD2. Se estima que AGVD2 contiene aproximadamente un 25 % de alelos homocigotos que se originan en IVP07-1001/4.
Las F1 fueron(D1) también se retrocruzaron con IVP07-1001/4 para producir una línea de BC1. Esta línea (BC1(D1) y BC1(D2)) fue depositada bajo la referencia de obtenedores AGVD3. Se estima que AGVD3 es aproximadamente homocigoto en un 75 % para los loci en los que difieren los progenitores.
Se obtuvieron otras plantas descendientes de F1 de manera similar usando el donante D3 como progenitor femenino. D3 es un clon de mejoramiento diploide de pulpa amarilla y con los genes de resistencia R3, H1, Gpa2, RXadg. Esto resultó en una línea designada F1 (D3).
De manera similar, como se describió anteriormente, F1 (D3) se autofecundó para generar una F2 (D3) y se retrocruzó con el donante del gen Sli para generar la descendencia BC1 (D3).
F1 (D1) se cruzó con F1 (D2) para producir una pseudo F2. Esta línea (F2(D1/2)) fue depositada bajo la referencia de obtenedores AGVD2. Se estima que AGVD2 es aproximadamente un 25 % homocigoto para los loci en los que difieren los progenitores.
F1(D1) también se retrocruzó con IVP07-1001/4 para producir una línea BC1. Esta línea (BC1(D1)) fue depositada bajo la referencia de criadores AGVD3. Se estima que AGVD3 es aproximadamente homocigoto en un 75 % para los loci en los que difieren los progenitores.
Los BC1 se autofecundaron en un vivero de invierno dando como resultado poblaciones de BC1S1. El próximo verano (2010) las F2, las pseudo F2 y las BC1S1 se sembraron en semilleros en un vivero y se trasplantaron en pequeñas macetas de turba. En junio de 2010, las plántulas se trasplantaron al campo en un suelo arcilloso y arenoso en los Países Bajos. En total, se cultivaron más de 15000 plantas a partir de plántulas y se seleccionaron por su autocompatibilidad y por su buena calidad y rendimiento de los tubérculos. La mayoría de las poblaciones F2 y BC1S1 mostraron una frecuencia muy baja de autocompatibilidad. Se seleccionaron plantas raras con un desarrollo razonablemente bueno de bayas y semillas para pruebas de progenie sin más evaluaciones agronómicas. Sólo dos poblaciones F2, que se originaron en el cruce D2 x H, mostraron aproximadamente un 25 % de autocompatibilidad. Para cada una de las poblaciones más autocompatibles, se seleccionaron como máximo quince plantas con la mejor calidad y cantidad de tubérculos para las pruebas de progenie.
Las plantas de los pseudo F2 mostraron mucho más vigor que las otras poblaciones, probablemente porque estas plantas no albergan ningún locus con alelos homocigotos de los progenitores donantes. Una de estas poblaciones mostró más del 50 % de autocompatibilidad. De manera similar a lo descrito anteriormente, también se seleccionaron un máximo de quince plantas de las poblaciones más autocompatibles para la prueba de progenie.
Además, se realizaron cruces con plantas autocompatibles del campo como donantes de polen sobre dos a doce plantas de los donantes D1-D19 (Ejemplo 3). Después de la evaluación de la calidad y cantidad de los tubérculos, sólo se seleccionaron cruces con plantas seleccionadas o sus hermanos completos, en caso de falta de un número suficiente de cruces, para la prueba de progenie.
De esta forma, en cada generación, las plantas son seleccionadas por su autocompatibilidad y buen desempeño agronómico. Las semillas de las bayas autofecundadas de estas plantas seleccionadas se recolectan para realizar más pruebas con el fin de generar líneas homocigóticas. Además, se realizan cruces de las plantas seleccionadas con otros donantes diploides u otras líneas de mejoramiento para potenciar la variación genética. Estas semillas F1 son heterocigotas y se necesitan varias rondas de endogamia para generar descendencia homocigota con los rasgos deseados.
De esta forma se pueden hacer dos generaciones de selecciones, autofecundaciones y cruces al año. Este procedimiento se mejora aún más al explotar el mejoramiento asistido por marcadores mediante el uso de marcadores de diagnóstico para rasgos específicos y marcadores aleatorios para seleccionar el genoma parental recurrente. Cuando se han obtenido progenitores de élite, se pueden obtener derivados de estas líneas mediante dos o tres generaciones de retrocruzamiento y dos generaciones de autofecundación.
Ejemplo 2 Mejoramiento de patata diploide autocompatible.
1. Introducción
Una característica común de la patata diploide es la autoincompatibilidad (SI); tras la autofertilización no se genera progenie vital. El gen inhibidor del locus S (Sli) procedente de Solanum chacoense ha sido descrito (Euphytica 99,191 197, 1998) para inhibir esta autoincompatibilidad y hacer que la patata diploide sea autocompatible (SC). Tras
repetidas autofecundaciones, se han generado clones de patata diploides autocompatibles casi homocigotos (Genoma 48, 977-984, 2005). La calidad agrícola de estos clones fue muy pobre, lo que refleja una germinación, un crecimiento de la planta, una calidad de los tubérculos y un rendimiento de los tubérculos deficientes.
2. Experimentos
2.1 Materiales
Con el fin de desarrollar clones de patata autocompatibles con un nivel mucho más alto de rasgos agrícolas, se cruzó el donante de genes Sli con diversos clones de patata, designado: IVP97-079-9, IVPAA-096-18 y SH83-92-488. Estos clones están disponibles en Dr. R.C.B. Hutten, Laboratory of Plant Breeding, Wageningen University, Países Bajos. Como se usa en este documento, estos clones se designan como clones D1, D2 y D3 resp. Las progenies (F1) fueron autofecundadas y retrocruzadas con el donante de genes Sli que genera poblaciones F2 y BC1, respectivamente. La BC1 se autofecundó para generar poblaciones BC1S1. Todos estos cruces y autofertilizaciones se realizaron sin otra selección que la autocompatibilidad de las plantas.
2.2 Evaluación de progenies en una prueba de invierno
En un experimento piloto, se cultivaron varias progenies aleatorias de BC1 y F2 en un invernadero climatizado con 16 horas de luz artificial mientras las temperaturas exteriores estaban bajo cero durante casi tres meses. El % autocompatible se evaluó como el número de plantas con bayas después de la polinización manual y el rendimiento como el peso de tubérculos por planta (Tabla 2).
Resultados
El donante Sli es autocompatible (SC) y la mayoría de las plantas F1 y BC1 también eran autocompatibles, mientras que una minoría de las plantas F2 eran autocompatibles. El donante Sli mostró un rendimiento de tubérculo extremadamente bajo, mientras que la BC1 y una F2 fueron aproximadamente el 50 % del rendimiento de tubérculo F1. El rendimiento de tubérculos de algunas plantas BC1 fue similar al de las plantas F1 y algunas plantas F2 (no seleccionadas) fueron mejores que las plantas F1 más débiles.
Tabla 2. Algunas características de las plantas cultivadas en invierno de un programa de mejoramiento de patata diploide para introducir el gen Sli.

2.3 Evaluaciones de progenies en campo
En el verano de 2010, las plantas descendientes de las poblaciones F2 y BC1S1 se probaron en el campo para seleccionar plantas descendientes autocompatibles con buen rendimiento agrícola (Tabla 3).
Tabla 3. Esquema de mejoramiento para introducir autocompatibilidad (SC) en patata diploide. En negrita se indican los materiales más prometedores.

* Conjunto de semillas suficiente
Resultados
Se probaron 52 poblaciones F2 que comprenden de 30 a 840 plantas y 62 poblaciones BC1S1 que comprenden de 1 a 390 plantas, en total 9660 plantas F2 y 3901 plantas BC1S1 resp. La mayoría de las poblaciones mostraron un crecimiento vegetal débil y una baja frecuencia de autocompatibilidad. Sólo dos poblaciones F2 derivadas del progenitor D2 mostraron aproximadamente un 25 % de plantas autocompatibles. Las mejores plantas de estas poblaciones fueron seleccionadas sobre la base del rendimiento y la calidad de los tubérculos.
3. Conclusión
Sólo dos de las 52 poblaciones F2 y ninguna de las poblaciones BC1S1 mostraron un nivel razonable de autocompatibilidad. Sin embargo, inesperadamente, se pudieron obtener las plantas deseadas, a partir de material que de otro modo habría sido descuidado por los obtenedores. Las principales diferencias de las plantas descritas en este documento y las de la técnica anterior se dan en la tabla 4 a continuación.
Tabla 4. Características de las líneas parentales de patata de la presente invención propuestas para la producción de semilla de patata híbrida (verdadera).

Ejemplo 3 Desarrollo de una plataforma de marcadores moleculares
Introducción
Numerosos sistemas de marcadores están disponibles para apoyar la investigación genética. Para la presente invención se desarrolló una plataforma de marcadores moleculares en patata y se usó para evaluar el nivel de tendencia homocigótica versus tendencia heterocigota en patata diploide.
Selección de las secuencias más informativas para ensayos de SNP
Para analizar SNPs en patata, el protocolo GoldenGate estándar de Illumina (Shen et al. 2005. Mutation Research 573:70-82) se ha usado previamente (Anithakumari et al. 2010. Mol. Breeding, 26:65-75). Esta plataforma comprende de 384 SNP y posteriormente se amplió con otros 768 SNP. Estos SNP 384 y 768 se han desarrollado usando secuencias e St , que representan el ARNm sin secuencias de intrones. Dado que los presentes inventores tienen acceso al primer borrador de secuencia del genoma de la patata (www.potatogenome.net), fue posible comparar el ARNm con la secuencia de ADN genómico, incluidas las secuencias intermedias. Esta comparación con la secuencia del genoma de la patata se realizó con the Basic Local Alignment Search Tool (BLAST; Altschul et al. 1990. J. Mol. Biol. 215: 403-410). Esto dio como resultado 538 y 1681 aciertos, respectivamente. Se consideró que la aparición de múltiples aciertos de BLAST reflejaba parálogos o secuencias que abarcan intrones. Estos se eliminaron de la colección inicial de SNP que se desarrollaron por primera vez para el análisis GoldenGate (Anithakumari et al. 2010). Esto dio como resultado 279 y 453 aciertos BLAST únicos que representan la aparición de una sóla copia en la secuencia del genoma de la patata de Solanum tuberosum Group Phureja DM1-3 516R44 (CIP801092) como se conoce hasta ahora (Potato Genome Sequencing Consortium, www.potatogemome.net).
Del total de 732 SNP con aciertos BLAST únicos, sólo se omitieron unos pocos (31 casos, distribuidos equitativamente en el conjunto 384 y 768) debido al alto nivel de desajustes de secuencia (3-6 SNP dentro de 101 pb). Debe esperarse una falta de coincidencia y se toleró la inclusión de un segundo SNP en las secuencias que abarcan SNP seleccionadas.
Para el presente diseño de SNP, la longitud de secuencia mínima se fijó en 101 pb. Por lo tanto, se eliminaron 81 secuencias porque eran más cortas. Las 553 secuencias resultantes tienen una longitud de al menos 101 pb. Luego se eliminó un total de 100 secuencias por tener coincidencias incompletas de sólo 35 hasta 94 nucleótidos dentro de un andamio de un monoploide doble (DM-3516R44; www.potatogenome.net). También se eliminaron las secuencias con brechas de alineación introducidos y las secuencias que tenían otros posibles problemas de desajuste.
Aunque veinticinco secuencias no cumplían los criterios descritos anteriormente, se mantuvieron porque se sabía que eran muy informativas en los ensayos GoldenGate SNP. Estos SNP permitieron incluso discriminar entre las cinco clases de genotipos que se dan en los tetraploides (Voorrips et al, EAPR-EUCARPIA congress "Potato Breeding after completion of the DNA Sequence of the Potato Genome", 27-30 June 2010, Wageningen, the Netherlands, epublication at http://edepot.wur.nl/143559, pg 42).
Finalmente, se seleccionaron 515 secuencias. La selección final de 100 secuencias se basó en la posición conocida en el mapa de 237 marcadores (de los 515) en el mapa genético de la patata (Anithakumari et al, 2010 and Van Os et al. Genetics 173: 1075-1087, 2006). Los marcadores seleccionados se distribuyeron uniformemente en los 12 cromosomas de patata, incluidos los marcadores teloméricos y centroméricos más distales y con preferencia por los SNP que eran heterocigotos en el clon RH (RH89-039-16; www.potatogenome.net), porque el clon Rh es un progenitor del material usado en la presente investigación. O en otras palabras, los alelos del clon RH también están presentes en el material usado en la presente investigación (véase, la tabla 2 y la figura 3).
Prueba de marcadores
Las 100 secuencias de SNP seleccionadas como se describe anteriormente se usaron en un sistema de genotipado de SNP KASPar (KBioscience Ltd. Hoddesdon, UK; http://www.kbioscience.co.uk/lab%20services/SNP%20Genotyping/genotyping_chemistry.html). Los marcadores se probaron en un conjunto de 20 genotipos diploides de patata (véase la figura 4), incluidos los tres progenitores diploides usados en el presente estudio y, además, en el donante del gen Sli, clon 07-1004-1 (véase también el ejemplo 2 anterior).
Resultados
Todas las secuencias de SNP generaron polimorfismos en el sistema de genotipificación KASPar SNP (véase, la figura 4), lo que corresponde a una tasa de éxito del 100 %. En total, se probaron 100 marcadores x 20 genotipos, que comprenden 2000 puntos de datos. Se observaron 29 datos faltantes, que es menos del 2 %. Como control, se probó un genotipo dos veces, usando muestras de plantas de diferentes lugares (Figura 4, columnas 4 y 7). Todos los datos eran idénticos. Estos resultados ilustran que el sistema de genotipado KASPar SNP es una plataforma de marcadores eficiente, confiable y reproducible para la patata diploide. Esto indicó que el sistema de genotipado KASPar SNP es eficiente, confiable y reproducible para la patata diploide.
Todos los marcadores mostraron una alta frecuencia de ocurrencia en el germoplasma diploide de patata usado en este experimento (Apéndice III). La frecuencia de heterocigotos en el germoplasma al demandar estos 100 marcadores osciló entre 26 % y 77 %, mientras que el donante del gen Sli era 100 % homocigoto de acuerdo con las expectativas (Phumichai et al. 2005 Genome 48: 977-984).
El número de marcadores informativos entre el donante Sli y los tres progenitores usados en el presente estudio fue de 54, 52 y 62 resp. Esto indica que el sistema de genotipado KASPar SNP también es informativo para la patata diploide.
Con base en lo anterior, se concluyó que el sistema de genotipado KASPar SNP como se describe en este documento y basado en los marcadores desarrollados en este documento es eficiente, confiable y reproducible y muy informativo para describir el nivel de tendencia homocigótica/tendencia heterocigota en patata diploide.
Tabla 5A. Secuencias de 100 marcadores SNP en patata





Tabla 5B: Lista de marcadores PotSNP y su anotación específica para la posición en el mapa SH y RH de la figura 3.


Tabla 6. Genotipos diploides de patata usados para el desarrollo y prueba de marcadores SNP


Ejemplo 4. Desarrollo de plantas de patata diploides homocigotas y generación de progenie híbrida F1 heterocigota Introducción
Los objetivos de este ejemplo fueron desarrollar líneas de patata diploides homocigóticas que sean autocompatibles (SC) y tengan un buen rendimiento agronómico. Un programa de mejoramiento para combinar la autocompatibilidad con un buen rendimiento agronómico se describe anteriormente en el ejemplo 2 anterior. El objetivo del experimento actual es demostrar que los genotipos de patata autocompatibles diploides homocigotos pueden generarse mediante endogamia y que las plantas de patata diploides que contienen loci homocigotos contrastantes dispersos por el genoma de la patata pueden cruzarse (son intercruzables) para generar una progenie híbrida F1 heterocigótica. Materiales y métodos
Un genotipo de patata homocigoto diploide autocompatible, que alberga el gen Sli como se describe en los ejemplos anteriores se cruzó con dos clones de patata, designados D1 y D2 (véanse los ejemplos 1 y 2). Las progenies F1 se autofecundaron y se generaron dos progenies F2 a partir de dos plantas SC F1 aleatorias. Las plantas aleatorias autocompatibles de estas dos poblaciones F2 se autofecundaron y por población F2 se probaron tres poblaciones F3 para determinar el nivel de tendencia homocigótica y la segregación de marcadores SNP.
Se probó el nivel de ploidía de las 6 plantas F2 (mediante cuantificación fluorescente de ADN) y se encontró que todas eran diploides.
Se usó un conjunto de 36 marcadores en 265 plantas F3 individuales aplicando el sistema de genotipado KASPar SNP (véase, el ejemplo 3) que cubría los 24 brazos cromosómicos de la patata (véase, la figura 5).
Resultados
Se probaron subconjuntos de 35 a 56 plantas de las seis poblaciones F3, en total 265 plantas, con al menos 24 marcadores informativos por par de progenitores. En total se usaron 36 marcadores (véase, la figura 5). Para la posición de los marcadores en el genoma de la patata, véase el ejemplo 3 anterior. los Sli el donante era 100 % homocigoto para estos marcadores probados, mientras que los progenitores D1 y D2 contenían 20 y 19 loci resp. que eran heterocigotos de los 24 marcadores informativos probados por par de progenitores, mientras que los otros cinco y seis loci resp. contenía alelos homocigotos que se originaban en el progenitor donante (D1 o D2).
Seis plantas F3 homocigotas
Como era de esperar, las poblaciones F3 segregaron para marcadores que eran heterocigotos en F2 y no para marcadores que estaban fijos en F2 (Figura 5). El nivel medio de heterocigotos osciló entre 84 y 95 %, mientras que el de las plantas individuales osciló entre 71 y 100 % (Tabla 7).
Tabla 7. Frecuencias de homocigotos (%) de seis poblaciones F3
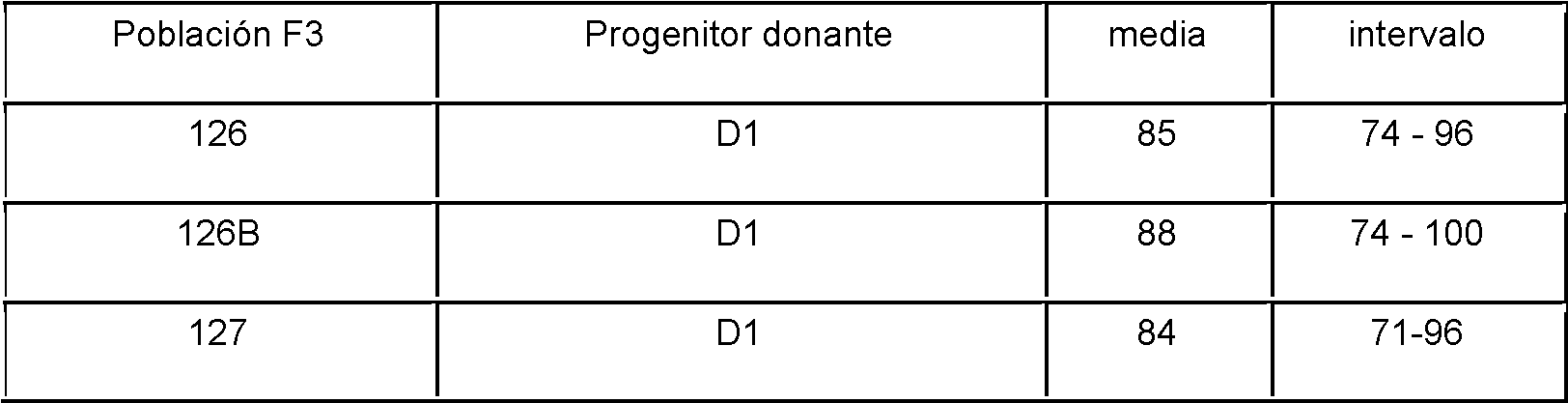

Tres poblaciones F3 de los cruces D1 x H y de los cruces H x D2 se analizaron para determinar la frecuencia de loci homocigotos basándose en los datos de la figura 5 como se muestra en la tabla 9.
Se espera que estas frecuencias proporcionen las frecuencias alélicas de los progenitores. Había seis plantas de tres poblaciones que ya eran 100 % homocigóticas. Estos albergaban principalmente loci homocigotos con alelos del donante Sli. Esto se espera como el donante Sli alberga alelos homocigotos, mientras que los otros progenitores contenían un alelo común con el donante Sli por marcador heterocigoto. Así que, los alelos derivados del donante Sli fueron abundantes en todas las poblaciones descendientes. Estas seis plantas F3 homocigóticas, denominadas 122 34, 123-6, 123-20, 123-23, 123-24 y 126B-17, contenían dos, cinco, cuatro, tres, cinco y siete loci resp. con alelos homocigotos del progenitor donante. Así, ya en dos generaciones autofecundadas se alcanzó el 100 % de tendencia homocigótica para 24 loci marcadores que representan 24 brazos cromosómicos.
Sin alelo letal absoluto
A menudo se afirma que las autofecundaciones repetidas en patatas diploides sólo darán como resultado plantas débiles debido a la depresión endogámica. Esto debería deberse a numerosos alelos nocivos que reducen la aptitud de la planta, cuando estos alelos están presentes de forma homocigótica. Por lo tanto, se probaron la segregación de 36 marcadores en las seis poblaciones F3 mencionadas (véase, la figura 5).
Estos marcadores, que se eligieron porque son informativos para los pares de progenitores usados en este estudio, también se probaron para las frecuencias en 19 genotipos diploides de patata, que se eligieron como donantes para el programa de mejoramiento (véase el ejemplo 3, la figura 4 y la tabla 9). Si hay alelos letales presentes, entones los loci con alelos homocigotos deberían estar ausentes. De hecho, siete marcadores (designados como PotSNP165, PotSNP205, PotSNP753, PotSNP 238, PotSNP 700, PotSNP 100 y PotSNP 607) no mostraron alelos homocigotos que fueran diferentes de los alelos donantes Sli en este germoplasma diploide. La mayoría de estos marcadores ya estaban fijados en el F2, la mayoría de ellos para alelos donantes Sli ya que estos eran más abundantes en los progenitores, pero también se observaron ejemplos en los que la población F3 se fijó para el alelo del progenitor no Sli (PotSNP753 en las poblaciones 126B y 127). Tres de estos siete marcadores (PotSNP205, PotSNP753 y PotSNP100) segregaron en las poblaciones F3, por lo que PotSNP753 segregaba normalmente y PotSNP205 y PotSNP100 mostraban una segregación distorsionada. Estos pueden representar alelos nocivos. Sólo se identificó una planta con alelos donantes PotSNP100 homocigotos. Sin desear ceñirse a ninguna teoría, se cree que esto puede representar un alelo muy perjudicial. Como se ha identificado una sola planta con un alelo homocigoto del progenitor donante, esto no proporciona una prueba completa de un alelo letal absoluto.
Cruces F3 x F3
Las 265 plantas F3 probadas se criaron en un invernadero sin calefacción en el verano de 2010. Además, se cultivaron en el campo con suelo arcilloso aproximadamente 600 plantas F3 adicionales (véase también el ejemplo 2). Todas estas plantas procedían de plantas F2 no seleccionadas cultivadas en un vivero de invierno en los Países Bajos (véase el ejemplo 2). Los cruces se realizaron aplicando polen de plantas autocompatibles cultivadas en invernadero y en campo sobre el estigma de plantas F3 autocompatibles cultivadas en invernadero. Sólo se realizaron cruces entre plantas F3 con diferentes bisabuelos (plantas F3 de las poblaciones 121 -123 cruzadas con 126 - 127). Las plantas de invernadero designadas 126-24, 126-31, 126B-2 y 127-21 se cruzaron con éxito como progenitor femenino autocompatible con una planta F3 autocompatible de la población 123 como progenitor masculino. Las plantas F3 femeninas eran más del 80 % homocigotas, mientras que la planta F3 en el campo no se probó, pero el nivel medio de tendencia homocigótica de la población fue del 94 % con un intervalo desde 88 a 100 %.
Los progenitores de los cruces F3 x F3 se compararon en busca de loci que ya estaban fijados en las poblaciones F3 (véase, la tabla 8). Se identificaron diez loci homocigotos en nueve cromosomas que al menos contrastaban entre uno de los cuatro progenitores femeninos y uno masculino usados para los cruces F3 x F3. Como consecuencia, la progenie híbrida F1 será al menos heterocigota para esos loci contrastantes. Como cada par de progenitores individuales contenía al menos cinco loci contrastantes, cada planta descendiente individual albergará al menos cinco regiones cromosómicas contrastantes.
Tabla 8. Contraste de loci fijos entre los progenitores de los cruces F3 x F3.

Los códigos 126-24, 126-31, 126B-2 y 127-21 se refieren a plantas F3 individuales que se usaron como hembras en cruces con una planta cultivada en el campo de la población F3 denominada "123" (véase, arriba). El nivel de tendencia homocigótica se calculó como el porcentaje de loci homocigotos sobre todos los loci probados (Figura 5). La "x" se refiere a marcadores homocigotos que tienen alelos contrastantes entre las cuatro plantas hembra y la planta de la población 123.
Conclusiones
Se encontró que después de dos generaciones de autofecundaciones se obtuvieron plantas de patata diploides homocigóticas que comprendían alelos de ambos progenitores. Ni un solo marcador de los 36 marcadores probados proporcionó pruebas de un alelo letal. Además, las plantas con más del 84 % de loci homocigotos se pueden cruzar con éxito para obtener plantas híbridas con al menos cinco regiones cromosómicas heterocigóticas. Esto significa que ahora se han producido semillas híbridas que son esencialmente uniformes y que representan verdaderas semillas de patata.
Tabla 9. Frecuencias aléllcas de marcadores SNP de todo el genoma en germoplasma
diplolde y poblaciones F3.

#: número de plantas: FIFI: locus donante SU homoclgoto: locus donante homoclgoto:
FDc locus heteroclgoto: % es porcentaje de plantas heteroclgotas, números bajos Indican
nivel alto de tendencia homoclgútlca en F3.
Ejemplo 5. Esquema de mejoramiento para una variedad comercial de patata
Partiendo de materiales F3 obtenidos de cruces entre un donante Sli y otra patata diploide como se describe en los ejemplos 2 y 4 anteriores, a continuación en este documento se expone un esquema de mejoramiento para producir variedades comerciales de patata. Este esquema consta de un número de etapas consecutivos que, en combinación
con la enseñanza que se describe en este documento, permitirán al experto llegar a una variedad de patata híbrida diploide o tetraploide que es tan buena o preferible mejor que los híbridos comerciales existentes y que se puede multiplicar mucho más rápido que las variedades mejoradas convencionales en cinco años.
1. Definición de la variedad del producto.
Como se describe en este documento, la producción de una nueva variedad de patata basada en la producción de híbridos comerciales no es una cuestión de azar, sino un procedimiento de diseño dirigido, que comienza con la articulación del perfil de producto de la variedad. En este documento, se debe definir el perfil de producto deseado (perfil de producto diana) para un mercado específico (patatas fritas, industria del almidón, patata de consumo). Además, se deben definir métodos de evaluación para los rasgos relevantes en el perfil del producto. Preferiblemente, la presencia de rasgos relevantes se evalúa mediante marcadores de diagnóstico. Tales marcadores son bien conocidos en la técnica de mejoramiento convencional de patatas.
2. Pruebas de campo
Dependiendo de la temporada en la que se inicie el procedimiento de desarrollo, se selecciona un campo de prueba apropiado. Al comenzar en septiembre u octubre, se prefiere seleccionar un campo de prueba apropiado ubicado en el hemisferio sur. Los ensayos de campo pueden comenzar con al menos 2, preferiblemente más poblaciones F3, seleccionadas de al menos uno, pero preferiblemente más cruces de un diploide. El donante Sli y otro genotipo de patata diploide que sirve como donante para un rasgo agronómico, preferiblemente el rasgo de vigor, cuyas poblaciones F3 representan las líneas de patata diploides, fértiles, autocompatibles, esencialmente homocigóticas y vigorosas de la invención como se ejemplifica en el ejemplo 2. Además de estas poblaciones F3, las progenies F1 se toman de al menos dos, preferiblemente más plantas F2 seleccionadas (como se ejemplifica en el ejemplo 2 anteriormente en este documento) cruzadas con al menos dos, preferiblemente más genotipos de patata diploides que son diferentes de los abuelos de las plantas F2 seleccionadas. Al menos 50, preferiblemente más plantas individuales de estas poblaciones F3 y F1 se cultivan luego en el campo de patata seleccionado y se autofecundan.
3. Evaluaciones y selección
Durante el crecimiento en campo se evalúa la frecuencia de plantas autocompatibles que lleven bayas en un periodo de 2-6 semanas desde el inicio de la floración, y al menos dos, preferiblemente más poblaciones F3 y F1 con al menos un 10 %, preferiblemente se seleccionan plantas más autocompatibles. Además, durante la temporada de crecimiento, se evalúan las características agronómicas definidas en el perfil del producto de las plantas autocompatibles en las poblaciones seleccionadas. Se recolectan muestras de hojas de todas las plantas autocompatibles de las poblaciones seleccionadas para la prueba de marcadores de ADN.
Al final de la temporada de crecimiento, se evalúa la calidad y cantidad de tubérculos de las plantas autocompatibles en las poblaciones seleccionadas, y los genotipos marcadores de ADN se evalúan mediante el uso de marcadores de diagnóstico y aleatorios. Con base en esta evaluación y valoración, se seleccionan al menos las cinco, preferiblemente más, plantas autocompatibles de cada población en función de: (i) la combinación de rasgos más cercana al perfil del producto diana; (ii) el nivel más alto de tendencia homocigótica, y (iii) con el número máximo de loci homocigotos contrastantes. Las plantas F3 y F1 seleccionadas y autofecundadas proporcionan progenies F4 y F2, respectivamente, en forma de semillas.
4. Cosecha y envío de tubérculos y semillas
Los tubérculos de las plantas F3 y F1 seleccionadas así como las semillas F2 y F4 en las bayas de estas plantas seleccionadas se cosechan en un período de al menos seis semanas, preferiblemente más, después de la floración. Las semillas F4 se recolectan para ensayos de campo adicionales que implican la prueba de las capacidades de combinación generales entre las plantas F4 cultivadas de esa forma, y las semillas F2 proporcionan bancos de genes adicionales para el mejoramiento y se recolectan para ensayos de campo adicionales. Dependiendo de la temporada y con el fin de ahorrar tiempo, las semillas pueden enviarse adecuadamente al hemisferio norte para crecer en un invernadero, siempre que se cumplan todas las regulaciones fitosanitarias de los países iimplicados.
5. Almacenamiento y evaluación de tubérculos
Los tubérculos de las plantas F1 y F3 seleccionadas se almacenan preferiblemente en la región de cultivo para evitar la propagación no deseada de patógenos y plagas, y estos tubérculos almacenados se evalúan en busca de algunas características intrínsecas relevantes de los tubérculos almacenados, tales como se define en el perfil del producto.
6. Propagación de tubérculos y evaluaciones después de 1 año
Los tubérculos de las plantas F1 y F3 seleccionadas se cultivan la próxima temporada, preferiblemente en la misma región que el campo de patata original para evitar la propagación de patógenos o plagas no deseadas. Se usan al menos dos, preferiblemente más tubérculos de patata por parcela y preferiblemente se usan una preferiblemente más repeticiones por clon. Se incluye como control al menos una variante comercial, preferiblemente más relevante. Preferiblemente se incluyen hileras laterales con variedades comerciales para evitar efectos secundarios. Se evalúan
las características relevantes de la planta como se definen en el perfil del producto (por ejemplo, cocinar, freír, hacer chips, contenido de almidón, etc.). Además, se evalúan el rendimiento de tubérculos por parcela, así como las características intrínsecas relevantes de los tubérculos, como se definen en el perfil del producto. Tales evaluaciones se usan para la caracterización familiar y la selección de progenie. Estos ensayos se repiten cada año, preferiblemente en lugares de ambos hemisferios para seleccionar continuamente las propiedades ancestrales de las plantas cultivadas en el campo. En fases posteriores, cuando se obtienen híbridos que se prueban para introducciones comerciales, los ensayos de evaluación se extienden a por lo menos dos, preferiblemente más, repeticiones en tamaños de parcela de al menos una, preferiblemente más m2 y en dos, preferiblemente más, ubicaciones preferiblemente en pero no restringidas a los mercados diana.
7. Cruces de prueba con semillas F4 y pruebas de campo
Poco después de la recolección de las semillas de la progenie F4 de las plantas autofecundadas F3 seleccionadas, se siembran al menos cincuenta, preferiblemente más, de estas semillas. Si es necesario, esta siembra puede ocurrir en un invernadero climatizado. Las plántulas se trasplantan preferiblemente en macetas pequeñas aproximadamente 1 mes después de la siembra y luego se transfieren al campo en un lugar apropiado algunas semanas después. Las evaluaciones y selecciones como se describió anteriormente continúan y al menos dos, preferiblemente más plantas F4 seleccionadas por población, se cruzan de prueba con al menos dos, preferiblemente más, otras plantas de progenie F4 seleccionadas de dos, preferiblemente más plantas progenitoras seleccionadas con el máximo número de loci contrastantes para producir híbridos de prueba (F4 x F4 -> híbrido). Adicionalmente, al menos dos, preferiblemente más plantas F4 seleccionadas por población, pueden cruzarse con al menos un, preferiblemente más genotipos diploides con características o rasgos deseables que faltan en las plantas seleccionadas para potenciar la diversidad genética del germoplasma de mejoramiento. Además, al menos dos, preferiblemente más plantas F4 seleccionadas por población, pueden autofecundarse para producir semillas F5. Aún más, pueden esterilizarse partes de plantas de al menos dos, preferiblemente más plantas F4 seleccionadas por población, in vitro se propagan y se tratan con productos químicos apropiados (colchicina/orizalina) para producir genotipos tetraploides F4, y se realizan cruces de prueba con plantas F4 tetraploides entre los mismos progenitores, como se describió anteriormente. Además, las semillas de progenie F2 similares se siembran, trasplantan y evalúan, se cruzan y se autofecundan, pero no se prueban/cruzan.
El esquema de mejoramiento anterior se repite cada año mediante el cual se logra un aumento incremental en los rasgos de la planta como se define en el perfil del producto. Las evaluaciones de los tubérculos de cruces de prueba son fundamentales para determinar las habilidades generales de combinación de los progenitores. La capacidad de combinación general se define en este documento como el rendimiento promedio de una línea en un número de cruces. Para cada mercado se logran al menos dos, preferiblemente más líneas parentales de élite que producirán semillas híbridas comerciales. Se espera que en cinco años se genere una variedad híbrida diploide o tetraploide que sea tan buena o preferible que los híbridos comerciales existentes y que se pueda multiplicar mucho más rápido que las variedades mejoradas convencionales.
Claims (11)
1. Una línea de mejoramiento de patata Solanum tuberosum diploide, fértil, autocompatible y esencialmente homocigótica que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, en la que las plantas de dicha línea tienen un nivel de tendencia homocigótica que es superior al 75 % cuando se prueban 100 loci, obtenible por un método que comprende:
(a) proporcionar una primera planta de patata, cuya primera planta de patata es una planta de una primera línea de patata diploide, autocompatible, fértil y esencialmente homocigótica S. tuberosum seleccionada del número de acceso de NCIMB 41663, número de acceso de NCIMB 41664, número de acceso de NCIMB 41665 o número de acceso de NCIMB 41765, semillas representativas de dichas líneas han sido depositadas en NCIMB, Aberdeen, Escocia, con las referencias de los obtentores AGVD1, AGVD2, AGVD3, y AGVD17 respectivamente, donde dicho rasgo de autocompatibilidad se debe a la presencia de un gen inhibidor del locus S (Sli);
(b) proporcionar una segunda planta de patata S. tuberosum, en la que dicha segunda planta de patata es una planta de una línea de patata diploide que puede tener cualquier nivel de tendencia homocigótica, comprendiendo dicha línea de dicha segunda planta de patata plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta;
(c) polinizar de forma cruzada dicha primera y segunda planta de patata para proporcionar semillas y recolectar dichas semillas para proporcionar así una generación descendiente en forma de una semilla descendiente híbrida;
(d) hacer crecer dicha semilla descendiente híbrida en una población de plantas de patata descendientes híbridas y seleccionar de dicha población plantas que produzcan tubérculos con un rendimiento expresado en gramos de peso fresco de al menos 200 gramos por planta, y
(e) autofecundar dicha planta de patata descendiente híbrida durante entre 1 y 8 autofecundaciones, seleccionando así una descendencia fértil para proporcionar así una línea de mejoramiento de patata diploide, autocompatible y esencialmente homocigótica que comprende plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, en la que las plantas de dicha línea tienen un nivel de tendencia homocigótica que es superior al 75 %, y en la que dicho rasgo de autocompatibilidad se debe a la presencia de dicho gen inhibidor del locus S (Sli).
2. La línea de mejoramiento de patata diploide S. tuberosum de la reivindicación 1, en la que dichas etapas a-e se repiten y en la que la etapa b comprende la provisión de una planta de patata adicional, en la que dicha planta de patata adicional es un donante de un rasgo agronómicamente deseable adicional.
3. La línea de mejoramiento de patata diploide S. tuberosum según la reivindicación 2, en la que dicho rasgo se selecciona del grupo que consiste en tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades, tolerancia a herbicidas, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad y resistencia al endulzamiento por frío.
4. La línea de mejoramiento de patata diploide S. tuberosum según la reivindicación 3, en la que la resistencia a enfermedades se selecciona de la resistencia a la sarna provocada por Streptomyces spp, resistencia a sarna pulverulenta, resistencia a Rhizoctonia, resistencia a sarna plateada, o resistencia a Phytophthora infestans.
5. La línea de mejoramiento de patata diploide S. tuberosum de una cualquiera de las reivindicaciones 1-4, en la que dicha segunda planta de patata es una planta de una segunda línea de patata diploide, fértil, autocompatible y esencialmente homocigótica, o en la que dicha segunda planta de patata es una planta de una segunda línea de patata diploide, fértil y autocompatible.
6. Una semilla de patata híbrida S. tuberosum, cuya semilla, cuando se siembra, produce una planta de patata diploide, fértil, autocompatible y esencialmente heterocigota que tiene un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta y en la que dicho rasgo de autocompatibilidad se debe a la presencia de un gen inhibidor del locus S (Sli), pudiendo obtenerse dicha semilla cruzando plantas de una primera y segunda línea de mejoramiento de patatas diploides según una cualquiera de las reivindicaciones 1-5, en la que dichas plantas de dicha primera y segunda líneas de mejoramiento de patatas diploides contienen al menos un 20 % de loci homocigoto que contrastan como se puede determinar por un análisis de marcador molecular, al probar 100 loci.
7. La semilla de patata híbrida S. tuberosum de la reivindicación 6, en la que al menos una de dichas primera y segunda plantas de patata comprende al menos un rasgo agronómicamente deseable seleccionado del grupo que consiste en tolerancia a insectos, resistencia a nematodos, resistencia a enfermedades, tolerancia a herbicidas, tolerancia al frío, tolerancia a la sequía, tolerancia a la humedad, tolerancia a la pudrición seca y húmeda, tolerancia a la salinidad y resistencia al endulzamiento por frío.
8. La semilla de patata híbrida S. tuberosum de la reivindicación 6 o 7, en la que dicha resistencia a enfermedades se selecciona de la resistencia a la sarna provocada por Streptomyces spp, resistencia a sarna pulverulenta, resistencia a Rhizoctonia, resistencia a sarna plateada, y resistencia a Phytophthora infestans.
9. Una planta de patata híbrida S. tuberosum, producida cultivando la semilla de una cualquiera de las reivindicaciones 6-8.
10. Un tubérculo o un cultivo de tejido de la planta de patata híbrida S. tuberosum de la reivindicación 9.
11. Un conjunto de al menos 2 plantas de patata intercruzables de diferentes líneas de patatas diploides, fértiles, autocompatibles y esencialmente homocigótica S. tuberosum según una cualquiera de las reivindicaciones 1-5, comprendiendo dichas líneas plantas que tienen un rendimiento de tubérculo promedio expresado en gramos de peso fresco de al menos 200 gramos por planta, en el que cada una de dichas líneas se diferencia de otra línea en dicho conjunto por tener al menos 20 % de loci homocigoto que contrastan como se puede determinar por un análisis de marcador genético molecular, al probar 100 loci, y en el que dicho rasgo de autocompatibilidad se debe a la presencia del gen inhibidor del locus S (Sli).
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US25498009P | 2009-10-26 | 2009-10-26 | |
| EP09013474 | 2009-10-26 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2923953T3 true ES2923953T3 (es) | 2022-10-03 |
Family
ID=42035614
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES19152162T Active ES2923953T3 (es) | 2009-10-26 | 2010-10-26 | Mejoramiento de patatas de semilla híbridas |
Country Status (14)
| Country | Link |
|---|---|
| US (3) | US10524436B2 (es) |
| EP (4) | EP4215040A1 (es) |
| CN (2) | CN105766658B (es) |
| AU (1) | AU2010313913B2 (es) |
| CA (1) | CA2778779C (es) |
| DE (1) | DE202010018273U1 (es) |
| DK (1) | DK3536146T3 (es) |
| EA (1) | EA032461B9 (es) |
| ES (1) | ES2923953T3 (es) |
| IL (2) | IL219460A (es) |
| MX (1) | MX337427B (es) |
| PL (1) | PL3536146T3 (es) |
| WO (1) | WO2011053135A2 (es) |
| ZA (1) | ZA201203153B (es) |
Families Citing this family (21)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US10524436B2 (en) * | 2009-10-26 | 2020-01-07 | Agventure B.V. | Hybrid seed potato breeding |
| EP2514303A1 (en) * | 2011-04-21 | 2012-10-24 | Agventure B.V. | Hybrid seed potato breeding |
| BR112015006119B1 (pt) | 2012-09-19 | 2022-05-03 | Biodiscovery New Zealand Limited | Método iterativo de triagem de traço fenotípico ou genotípico de planta para selecionar um ou mais microorganismos capazes de transmitir pelo menos um traço fenotípico ou genotípico benéfico à uma planta, método para a produção de uma composição, métodos para a seleção de composições e métodos para a seleção de um ou mais micro-organismos. |
| US9732335B2 (en) | 2012-09-19 | 2017-08-15 | Biodiscovery New Zealand Limited | Methods of screening for microorganisms that impart beneficial properties to plants |
| US9777267B2 (en) | 2012-09-19 | 2017-10-03 | Biodiscovery New Zealand Limited | Methods of screening for microorganisms that impart beneficial properties to plants |
| CN103477876B (zh) * | 2013-09-23 | 2016-01-13 | 青海省农林科学院 | 一种马铃薯杂交育种的方法 |
| EP3994978A1 (en) * | 2014-05-23 | 2022-05-11 | BioConsortia, Inc. | Integrated plant breeding methods for complementary pairings of plants and microbial consortia |
| CN105052724A (zh) * | 2015-07-24 | 2015-11-18 | 中国农业科学院蔬菜花卉研究所 | 一种提高二倍体马铃薯自交结实率的方法及其应用 |
| US11058090B2 (en) | 2016-07-25 | 2021-07-13 | Bejo Zaden B.V. | Hybrid true potato seed of tetraploid hybrid 1490185 |
| CN108849471A (zh) * | 2018-06-14 | 2018-11-23 | 中国农业科学院农业基因组研究所 | 一种使马铃薯自交亲和的方法 |
| US20220267386A1 (en) | 2019-05-07 | 2022-08-25 | Agventure B.V. | Self-compatibility in cultivated potato |
| CN110622851A (zh) * | 2019-09-03 | 2019-12-31 | 内蒙古坤元太和农业科技有限公司 | 一种马铃薯选育方法 |
| US20210137041A1 (en) * | 2019-11-08 | 2021-05-13 | Agventure B.V. | Processes for production of large quantities of uniform potato tubers from true seeds |
| CN110938120B (zh) * | 2019-12-04 | 2021-12-03 | 中国农业科学院农业基因组研究所 | 改变二倍体马铃薯材料自交不亲和性的StSCI蛋白 |
| CN110894539B (zh) * | 2019-12-04 | 2022-01-25 | 云南师范大学 | 鉴定二倍体马铃薯是否自交亲和的方法 |
| CN111011217B (zh) * | 2019-12-31 | 2023-04-28 | 青海省农林科学院 | 杂合二倍体马铃薯rh的高效再生方法 |
| EP4438285A2 (en) | 2020-02-07 | 2024-10-02 | Sasib S.p.A. | Manufacturing machine and manufacturing method for the production of a tubular element, in particular for a smoking article |
| CN116803248B (zh) * | 2022-03-16 | 2024-10-18 | 中国农业科学院深圳农业基因组研究所(岭南现代农业科学与技术广东省实验室深圳分中心) | 自交不亲和的双单倍体马铃薯品系及其选育方法和杂交马铃薯及其制种方法 |
| CN114946647B (zh) * | 2022-05-31 | 2023-08-22 | 吉林省蔬菜花卉科学研究院 | 一种马铃薯生态穿梭育种方法 |
| CN116200518B (zh) * | 2022-09-20 | 2023-10-27 | 宁夏农林科学院农业生物技术研究中心(宁夏农业生物技术重点实验室) | 一种与马铃薯淀粉含量相关的kasp标记的开发及应用 |
| CN118318728A (zh) * | 2024-06-11 | 2024-07-12 | 云南师范大学 | 一种使用桥梁种S. verrucosum进行马铃薯间接杂交的方法 |
Family Cites Families (14)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA1289094C (en) | 1986-01-27 | 1991-09-17 | Robert D. Reichert | Method of solubilizing potato starch |
| CA2275885C (en) * | 1997-02-10 | 2007-01-09 | Her Majesty The Queen In Right Of Canada As Represented By The Minister Of Agriculture And Agri-Food Canada | Transgenic potatoes having reduced levels of alpha glucan l- or h-type tuber phosphorylase activity with reduced cold-sweetening |
| CR5828A (es) * | 1997-07-30 | 1999-10-25 | Wisconsin Alumni Res Found | Plasma de germen y marcadores moleculares para resistenciaen enfermedades de papas |
| JPH11127712A (ja) * | 1997-10-31 | 1999-05-18 | Kobe University | ミニポテト |
| AU2000251248B2 (en) * | 2000-04-27 | 2005-11-17 | J.R. Simplot Company | Method for in vitro selection of potato clones resistant to blackspot bruising and the potatoes produced therefrom |
| EP1425958A1 (en) * | 2002-12-04 | 2004-06-09 | Coöperatieve Verkoop- en Productievereniging van Aardappelmeel en Derivaten 'AVEBE' B.A. | Potatoes with increased protein content |
| JP2005130770A (ja) * | 2003-10-30 | 2005-05-26 | Ajinomoto Co Inc | 植物体あたり得られるデンプン量が増加したバレイショおよびその作出法 |
| US10524436B2 (en) | 2009-10-26 | 2020-01-07 | Agventure B.V. | Hybrid seed potato breeding |
| EP2514303A1 (en) | 2011-04-21 | 2012-10-24 | Agventure B.V. | Hybrid seed potato breeding |
| US20220267386A1 (en) | 2019-05-07 | 2022-08-25 | Agventure B.V. | Self-compatibility in cultivated potato |
| US20210029956A1 (en) | 2019-07-29 | 2021-02-04 | Agventure B.V. | Diploid, fertile and highly homozygous potato line solyntus |
| US20210137041A1 (en) | 2019-11-08 | 2021-05-13 | Agventure B.V. | Processes for production of large quantities of uniform potato tubers from true seeds |
| CN110938120B (zh) | 2019-12-04 | 2021-12-03 | 中国农业科学院农业基因组研究所 | 改变二倍体马铃薯材料自交不亲和性的StSCI蛋白 |
| CN110894539B (zh) | 2019-12-04 | 2022-01-25 | 云南师范大学 | 鉴定二倍体马铃薯是否自交亲和的方法 |
-
2010
- 2010-10-26 US US13/504,156 patent/US10524436B2/en active Active
- 2010-10-26 EP EP22210656.9A patent/EP4215040A1/en active Pending
- 2010-10-26 AU AU2010313913A patent/AU2010313913B2/en active Active
- 2010-10-26 EA EA201290178A patent/EA032461B9/ru not_active IP Right Cessation
- 2010-10-26 PL PL19152162.4T patent/PL3536146T3/pl unknown
- 2010-10-26 EP EP19152162.4A patent/EP3536146B1/en active Active
- 2010-10-26 MX MX2012004920A patent/MX337427B/es active IP Right Grant
- 2010-10-26 DE DE201020018273 patent/DE202010018273U1/de not_active Expired - Lifetime
- 2010-10-26 EP EP22210666.8A patent/EP4215041A1/en active Pending
- 2010-10-26 WO PCT/NL2010/050716 patent/WO2011053135A2/en active Application Filing
- 2010-10-26 ES ES19152162T patent/ES2923953T3/es active Active
- 2010-10-26 CN CN201510920986.9A patent/CN105766658B/zh active Active
- 2010-10-26 CN CN201080059512.5A patent/CN102770017B/zh active Active
- 2010-10-26 EP EP10770643A patent/EP2493286A2/en active Pending
- 2010-10-26 CA CA2778779A patent/CA2778779C/en active Active
- 2010-10-26 DK DK19152162.4T patent/DK3536146T3/da active
-
2012
- 2012-04-29 IL IL219460A patent/IL219460A/en active IP Right Grant
- 2012-05-02 ZA ZA2012/03153A patent/ZA201203153B/en unknown
-
2016
- 2016-04-20 IL IL245246A patent/IL245246A0/en active IP Right Grant
-
2019
- 2019-08-14 US US16/541,071 patent/US11140841B2/en active Active
-
2021
- 2021-08-30 US US17/461,445 patent/US20220095576A1/en active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| DK3536146T3 (da) | 2022-07-18 |
| MX2012004920A (es) | 2012-08-23 |
| ZA201203153B (en) | 2014-10-29 |
| DE202010018273U1 (de) | 2015-03-26 |
| IL219460A0 (en) | 2012-06-28 |
| EP4215040A1 (en) | 2023-07-26 |
| CA2778779C (en) | 2023-10-17 |
| US20200029520A1 (en) | 2020-01-30 |
| US20120284852A1 (en) | 2012-11-08 |
| CN102770017B (zh) | 2015-12-16 |
| US11140841B2 (en) | 2021-10-12 |
| US10524436B2 (en) | 2020-01-07 |
| EA032461B9 (ru) | 2019-09-30 |
| CN105766658B (zh) | 2018-06-26 |
| EA032461B1 (ru) | 2019-05-31 |
| CN105766658A (zh) | 2016-07-20 |
| MX337427B (es) | 2016-03-04 |
| AU2010313913B2 (en) | 2014-03-20 |
| EP4215041A1 (en) | 2023-07-26 |
| AU2010313913A1 (en) | 2012-05-24 |
| CA2778779A1 (en) | 2011-05-05 |
| EA201290178A1 (ru) | 2013-07-30 |
| IL245246A0 (en) | 2016-06-30 |
| IL219460A (en) | 2016-05-31 |
| PL3536146T3 (pl) | 2022-08-08 |
| EP2493286A2 (en) | 2012-09-05 |
| EP3536146A1 (en) | 2019-09-11 |
| US20220095576A1 (en) | 2022-03-31 |
| EP3536146B1 (en) | 2022-04-27 |
| WO2011053135A2 (en) | 2011-05-05 |
| WO2011053135A3 (en) | 2011-07-28 |
| CN102770017A (zh) | 2012-11-07 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US11140841B2 (en) | Hybrid seed potato breeding | |
| AU2017239475B2 (en) | Hybrid seed potato breeding | |
| US20130198884A1 (en) | Spinach hybrid variety nun 01084 | |
| US20150173318A1 (en) | Novel genetic factor capable of increasing yield in maize and method thereof | |
| US20130254914A1 (en) | Spinach hybrid variety callisto | |
| BR112012003139B1 (pt) | Método para produzir uma planta híbrida, planta, material de planta ou semente | |
| US20210137041A1 (en) | Processes for production of large quantities of uniform potato tubers from true seeds | |
| US20210029956A1 (en) | Diploid, fertile and highly homozygous potato line solyntus | |
| US10813319B2 (en) | Spinach variety NUN 06258 SPS | |
| US9012724B2 (en) | Hybrid carrot variety PURPLE SNAX | |
| US9131650B2 (en) | Hybrid carrot variety trooper | |
| US9000266B2 (en) | Hybrid carrot variety purple elite | |
| US9012725B2 (en) | Hybrid carrot variety slendercut | |
| US10785934B2 (en) | Spinach variety nun 06303 SPS | |
| US8952220B2 (en) | Hybrid carrot variety rebel | |
| US8962923B2 (en) | Hybrid carrot variety NUN 89141 CAC | |
| Gupta | Plant Breeding Plant Propagation and Biotechnology |