ES2747363T3 - Estructuras inmunorreguladoras de proteínas que se encuentran normalmente - Google Patents
Estructuras inmunorreguladoras de proteínas que se encuentran normalmente Download PDFInfo
- Publication number
- ES2747363T3 ES2747363T3 ES16189090T ES16189090T ES2747363T3 ES 2747363 T3 ES2747363 T3 ES 2747363T3 ES 16189090 T ES16189090 T ES 16189090T ES 16189090 T ES16189090 T ES 16189090T ES 2747363 T3 ES2747363 T3 ES 2747363T3
- Authority
- ES
- Spain
- Prior art keywords
- peptide
- peptides
- seq
- hsa
- albumin
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/06—Immunosuppressants, e.g. drugs for graft rejection
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/76—Albumins
- C07K14/765—Serum albumin, e.g. HSA
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5044—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics involving specific cell types
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/574—Immunoassay; Biospecific binding assay; Materials therefor for cancer
- G01N33/57484—Immunoassay; Biospecific binding assay; Materials therefor for cancer involving compounds serving as markers for tumor, cancer, neoplasia, e.g. cellular determinants, receptors, heat shock/stress proteins, A-protein, oligosaccharides, metabolites
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/34—Identification of a linear epitope shorter than 20 amino acid residues or of a conformational epitope defined by amino acid residues
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/435—Assays involving biological materials from specific organisms or of a specific nature from animals; from humans
- G01N2333/52—Assays involving cytokines
- G01N2333/54—Interleukins [IL]
- G01N2333/55—IL-2
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/435—Assays involving biological materials from specific organisms or of a specific nature from animals; from humans
- G01N2333/76—Assays involving albumins other than in routine use for blocking surfaces or for anchoring haptens during immunisation
- G01N2333/765—Serum albumin, e.g. HSA
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2500/00—Screening for compounds of potential therapeutic value
- G01N2500/10—Screening for compounds of potential therapeutic value involving cells
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Biomedical Technology (AREA)
- Cell Biology (AREA)
- Urology & Nephrology (AREA)
- Hematology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biophysics (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Toxicology (AREA)
- Pharmacology & Pharmacy (AREA)
- Public Health (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Veterinary Medicine (AREA)
- General Chemical & Material Sciences (AREA)
- Animal Behavior & Ethology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Physics & Mathematics (AREA)
- Pathology (AREA)
- Biotechnology (AREA)
- General Physics & Mathematics (AREA)
- Microbiology (AREA)
- Analytical Chemistry (AREA)
- Food Science & Technology (AREA)
- Zoology (AREA)
- Gastroenterology & Hepatology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Hospice & Palliative Care (AREA)
- Oncology (AREA)
- Transplantation (AREA)
- Peptides Or Proteins (AREA)
Abstract
Un fragmento de seroalbúmina humana (HSA) que comprende la secuencia de aminoácidos SEQ ID NO: 81 para su uso en el tratamiento de una afección inflamatoria, en donde dicho fragmento de HSA tiene 10-50 aminoácidos de longitud.
Description
DESCRIPCIÓN
Estructuras inmunorreguladoras de proteínas que se encuentran normalmente
Campo técnico
La presente invención se refiere a ciertas secuencias de proteína identificadas en la seroalbúmina humana (HAS) para su uso en el tratamiento de una afección inflamatoria.
Antecedentes de la invención
Aunque los datos indican que el sistema inmunitario tiene la mayor importancia en el control del cáncer, los tumores malignos continúan creciendo y la eficacia de la inmunoterapia es bastante pobre con una tasa de remisión objetivo del 15-30 %. Puede haber varias razones de esta aparente paradoja:
• Los tumores evitan el reconocimiento por el sistema inmunitario no expresando antígenos asociados al tumor apropiadamente.
• Los antígenos asociados al tumor (a menudo autoantígenos), que son demasiado débiles para dar lugar a una respuesta inmunitaria adecuada.
• Inducción de tolerancia
• Inmunosupresión relativa al cáncer, que evita una respuesta inmunitaria adecuada.
Estas alternativas necesitan estrategias terapéuticas completamente diferentes, sea para la estimulación apropiada del sistema inmunitario o el control de los mecanismos inmunosupresores relacionados con el cáncer.
La inmunosupresión del cáncer se caracteriza principalmente por: Disminución de la capacidad proliferativa y citotóxica de los linfocitos, en particular los linfocitos que infiltran el tumor, la pobre migración de las células inflamatorias, disminución de la producción y respuesta a la IL-2, dificultad para dar lugar a una respuesta inmunitaria mediante vacunación, también contra otros distintos a los antígenos relacionados con el tumor y la producción patológica de citocinas. Esta mala regulación del sistema inmunitario da como resultado un control del cáncer mediado de manera inmunitaria débil y del síndrome paraneoplásico (subfiebre, fatiga, anorexia, pérdida de peso y deterioro de los parámetros de laboratorio).
Las estrategias terapéuticas inmunoestimulantes que utilizan citocinas (por ejemplo, interferones, interleucinas) o la vacunación, con el objetivo de aumentar la reactividad mediada inmunitariamente contra el tumor, se ha intentado durante varias décadas, pero hasta ahora solo han tenido un éxito muy limitado. Esto indica que la inmunoestimulación con el fin de superar la pobre respuesta inmunitaria en los pacientes con cáncer se puede suprimir por otros mecanismos no identificados hasta ahora.
El documento WO 2004/082617 A2 se refiere a marcadores biológicos para la artritis reumatoide (RA). El documento WO 2004/082617 A2 también desvela los usos de dichos marcadores para diagnosticar y tratar la RA, controlar la progresión de la enfermedad, evaluar las intervenciones terapéuticas, y explorar fármacos candidatos en un ensayo clínico o preclínicos.
El documento WO 2006/043891 A desvela un anticuerpo monoclonal A capaz de unirse y bloquear la actividad inmunosupresora de estructuras relacionadas con la HSA en pacientes de cáncer i.a.
Los inventores describieron en dos solicitudes de patente anteriores dos mecanismos inmunorreguladores fundamentales de relevancia para todos los tipos de tumores malignos. En la primera de estas solicitudes se demostró la importancia de la modulación de receptor de Fc y las maneras para superar esta inmunosupresión relacionada con el cáncer modulando el entrecruzamiento del receptor Fc. En esta solicitud de patente también se demostró que los fragmentos proteolíticos de proteínas de origen normal inducen la producción patológica de monocinas. En la segunda solicitud de patente se descubrió que estas neo-estructuras se unían/bloqueaban la integrina y su existencia y actividad inmunorreguladora se analizó adicionalmente utilizando anticuerpos monoclonales dirigidos contra las neo-estructuras derivadas de la albúmina. En la última solicitud de patente también se describe la existencia y la importancia de los autoanticuerpos para estas neo-estructuras.
Sumario de la presente invención
Se describen varias secuencias proteicas, tal como péptidos, fragmentos de péptido, neo-estructuras y/o neo-epítopos, de una proteína de origen normal en el suero, que se unen a células inmunitarias. Algunas de sus actividades inmunorreguladoras se desvelan en el presente documento.
Se desvela adicionalmente que se ha identificado la estructura de varios de estos péptidos, fragmentos de péptidos, neo-estructuras y/o neo-epítopos, que tienen actividad inmunorreguladora mediante la unión a receptores de células inmunitarias.
En un aspecto, la presente invención proporciona un fragmento de seroalbúmina humana (HSA) que comprende la secuencia de aminoácidos SEQ ID NO: 81 para su uso en el tratamiento de una afección inflamatoria, en el que dicho fragmento de HSA tiene 10-50 aminoácidos de longitud.
En otro aspecto, la presente invención proporciona un péptido que consiste en la secuencia de aminoácidos SEQ ID NO: 80 para su uso en el tratamiento de una afección inflamatoria.
En un aspecto adicional, la presente invención proporciona un péptido que consiste en la secuencia de aminoácidos SEQ ID NO: 81 para su uso en el tratamiento de una afección inflamatoria.
En un aspecto adicional más, la presente invención proporciona el uso de un fragmento de seroalbúmina humana (HSA) que comprende la secuencia de aminoácidos S e Q ID NO: 81 para la fabricación de un medicamento para el tratamiento de una afección inflamatoria, en el que dicho fragmento de HSA tiene 10-50 aminoácidos de longitud.
En un aspecto adicional más, la presente invención proporciona el uso de un péptido que consiste en la secuencia de aminoácidos SEQ ID NO: 80 para la fabricación de un medicamento para el tratamiento de una afección inflamatoria.
En un aspecto adicional más, la presente invención proporciona el uso de un péptido que consiste en la secuencia de aminoácidos SEQ ID NO: 81 para la fabricación de un medicamento para el tratamiento de una afección inflamatoria.
Descripción detallada de la presente invención
Se desvela una secuencia proteica de unión a una célula inmunitaria de una proteína que existe normalmente en el suero, tal como un péptido de unión celular aislado, un fragmento peptídico, una neo-estructura y/o un neo-epítopo de una proteína que existe normalmente en el suero, que está presente en un tejido humano, en el que dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo tienen actividad inmunorreguladora y dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo son el resultado de un aumento de la actividad proteolítica y/o desnaturalización en un tejido inflamatorio y/o un tumor maligno. Ejemplos específicos de dichas secuencias proteicas de unión a una célula inmunitaria se seleccionan de entre las secuencias de aminoácidos enumeradas, por ejemplo, como las SEQ ID NO: 1-81, tal como en particular las secuencias que se corresponden con las SEQ ID NO: 26, 80, y 81.
Se desvela el uso de una secuencia proteica de unión a una célula inmunitaria de una proteína que existe normalmente en el suero, tal como un péptido de unión a una célula aislado, un fragmento peptídico, una neo-estructura y/o un neoepítopo, que están presentes en un tejido humano, en el que dicho péptido, fragmento de péptido, neo-estructura y/o neo-epítopo tienen una actividad inmunorreguladora y dicho péptido, fragmento de péptido, neo-estructura y/o neoepítopo es el resultado de un aumento de la actividad proteolítica y/o desnaturalización en un tejido inflamatorio y/o un tumor maligno, que se seleccionan de entre las secuencias de aminoácidos enumeradas como SEQ ID NO: 1-81, tal como en particular las seleccionadas de entre las secuencias que se corresponden con SEQ ID NO: 26, 80, y 81.
Se desvela el uso de una secuencia proteica de unión a una célula inmunitaria de una proteína que existe normalmente en el suero, de acuerdo con la presente invención, seleccionada de entre las secuencias de aminoácidos enumeradas como SEQ ID NO: 1-81, tal como en particular las seleccionadas de entre las secuencias que se corresponden con SEQ ID NO: 26, 80, y 81, para el diagnóstico, tratamiento y/o prevención del cáncer en un paciente que necesita la misma.
Se desvela un anticuerpo monoclonal dirigido contra una secuencia proteica de unión a una célula inmunitaria de una proteína que existe normalmente en el suero, tal como un péptido de unión a una célula aislado, un fragmento peptídico, una neo-estructura y/o un neo-epítopo, que están presentes en un tejido humano, en el que dicho péptido, fragmento de péptido, neo-estructura y/o neo-epítopo tienen una actividad inmunorreguladora y dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo son el resultado de un aumento de la actividad proteolítica y/o desnaturalización en un tejido inflamatorio y/o un tumor maligno. En una realización actualmente preferida, dicho anticuerpo monoclonal se dirige contra al menos una de las secuencias proteicas correspondientes a una secuencia seleccionada de entre las secuencias de aminoácidos enumeradas como SEQ ID NO: 1-81, tal como en particular las seleccionadas de entre las secuencias que se corresponden con SEQ ID NO: 26, 80, y 81.
Se desvela adicionalmente un anticuerpo anti-3028 de conejo, que se ha demostrado en el presente documento, así como un anticuerpo policlonal anti-3218 o anti-3315 de conejo que se dirige contra al menos una de las secuencias proteicas correspondientes a las SEQ ID NO: 26, 80, o 81. Dicho anticuerpo se utiliza normalmente para diferentes métodos de diagnóstico y/o investigación.
Se desvela en el presente documento un método para el diagnóstico de la presencia de un tumor maligno determinando la respuesta de un anticuerpo como se ha descrito anteriormente.
Se desvela un compuesto que inhibe la actividad de una secuencia proteica de unión a una célula inmunitaria de una
proteína que existe normalmente en el suero, tal como un péptido de unión a una célula aislado, un fragmento peptídico, una neo-estructura y/o un neo-epítopo, que están presentes en un tejido humano, en el que dicho péptido, fragmento de péptido, neo-estructura y/o neo-epítopo tienen una actividad inmunorreguladora y dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo son el resultado de un aumento de la actividad proteolítica y/o desnaturalización en un tejido inflamatorio y/o un tumor maligno.
Se desvela adicionalmente un método para el tratamiento de cualquier tumor maligno administrando un compuesto que inhibe la existencia de una secuencia proteica de unión a una célula inmunitaria de una proteína que se encuentra normalmente en el suero, tal como un péptido de unión celular aislado, un fragmento peptídico, una neo-estructura y/o un neo-epítopo, de acuerdo con la presente invención, en el que dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo tienen una actividad inmunorreguladora y son el resultado de un cáncer o tumor maligno.
Se desvela un anticuerpo que se genera contra dicho péptido de unión celular, fragmento peptídico, neo-estructura y/o neo-epítopo para la administración en una cantidad suficiente para dar lugar a una respuesta inmunitaria contra cualquier tumor maligno.
Se desvela adicionalmente un método para el tratamiento de un tumor maligno inhibiendo la actividad de dicho péptido inmunorregulador, fragmento peptídico, neo-estructura y/o neo-epítopo utilizando principios farmacológicos convencionales de desarrollo de fármacos que producen fármacos bloqueantes de receptor o fármacos que inhiben la transducción de la señal de los receptores de dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo.
Se desvela en el presente documento por primera vez que una secuencia proteica de unión a una célula inmunitaria de una proteína que se encuentra normalmente en el suero, de acuerdo con la presente invención, tal como un péptido de unión celular aislado, fragmento peptídico, neo-estructura y/o neo-epítopo, que es el resultado de un aumento de la actividad proteolítica y/o la desnaturalización en un tejido inflamatorio y/o un tumor maligno, tienen una actividad inmunorreguladora inhibidora, es decir, que sea un inmunoinhibidor fisiológico. Se desvela adicionalmente el uso de un péptido de unión celular aislado, un fragmento peptídico, neo-estructura y/o neo-epítopo de una proteína que se encuentra normalmente en el suero, de acuerdo con la presente invención, para la inmunorregulación no solo del cáncer, sino también de afecciones inflamatorias y/o dependientes de la interleucina-2 y/o enfermedades tales como la psoriasis, linfoma de células T, rechazo de aloinjerto, GVH, lesión por isquemia-reperfusión, enfermedades inflamatorias crónicas y/o enfermedades autoinmunitarias.
En una realización, el fragmento de seroalbúmina humana (HSA) para su uso como se define en las reivindicaciones es para su uso en el tratamiento de una afección inflamatoria seleccionada de entre el grupo que consiste en psoriasis, rechazo de aloinjerto, enfermedad del injerto contra el huésped (GVH), lesión de isquemia-reperfusión, una enfermedad inflamatoria crónica, y una enfermedad autoinmunitaria.
Se desvela adicionalmente un método para el tratamiento de afecciones y/o enfermedades inflamatorias y/o dependientes de la interleucina-2 administrando una cantidad terapéuticamente eficaz del péptido inmunosupresor, fragmento peptídico, neo-estructura y/o neo-epítopo de una proteína que se encuentra normalmente en el suero, de acuerdo con la presente divulgación. Se desvela en el presente documento una secuencia proteica, tal como un péptido, fragmento peptídico, neo-estructura y/o neo-epítopo de seroalbúmina normal que tiene un primer ácido glutámico a una distancia de 3 a 7 aminoácidos desde cualquier lisina presente en dicha secuencia, preferentemente 4 a 6 aminoácidos desde cualquier lisina presente en dicha secuencia, más preferentemente 5 a 6 aminoácidos desde cualquier lisina presente en dicha secuencia, y que tiene una actividad inmunorreguladora.
Preferentemente, dicha secuencia desvelada contiene un ácido glutámico adicional a una distancia de desde 2 a 3 aminoácidos desde dicho primer ácido glutámico.
Preferentemente, el péptido, fragmento peptídico, neo-estructura y/o neo-epítopo de seroalbúmina normal desvelados en el presente documento tienen una secuencia peptídica seleccionada de entre las secuencias de aminoácido enumeradas como SEQ ID NO: 1-81.
En una realización, dicha secuencia contiene adicionalmente un aminoácido ácido a una distancia de -12±1 aminoácidos desde el primer ácido glutámico, y a una distancia de 3±1 aminoácidos desde la lisina.
Se desvela adicionalmente un anticuerpo monoclonal dirigido contra una o más secuencias proteicas, tal como un péptido, fragmento peptídico, neo-estructura y/o neo-epítopo de seroalbúmina normal que tiene un primer ácido glutámico a una distancia de 3 a 7 aminoácidos desde cualquier lisina presente en dicha secuencia, preferentemente 4 a 6 aminoácidos desde cualquier lisina presente en dicha secuencia, más preferentemente 5 a 6 aminoácidos desde cualquier lisina presente en dicha secuencia, y que tiene una actividad inmunorreguladora.
El anticuerpo desvelado puede dirigirse contra un péptido, fragmento peptídico, neo-estructura y/o neo-epítopo de seroalbúmina normal correspondientes con una o más secuencias peptídicas seleccionadas de entre las secuencias de aminoácido enumeradas como SEQ ID NO: 1-81. Se desvela un método para el diagnóstico de la presencia opcional de un cáncer o tumor maligno inmunosupresor, determinando la presencia de un péptido, fragmento
peptídico, neo-estructura y/o neo epítopo de seroalbúmina humana normal que tienen un primer ácido glutámico a una distancia de 3 a 7 aminoácidos desde cualquier lisina presente en dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo, preferentemente 4 a 6 aminoácidos desde cualquier lisina presente en dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo, más preferentemente 5 a 6 aminoácidos desde cualquier lisina presente en dicho péptido, fragmento peptídico, neo-estructura y/o neo-epítopo, y tienen actividad inmunorreguladora, como se muestra en uno o más ensayos inmunitarios convencionales / ensayos convencionales sobre la función inmunitaria, tal como la producción de citocinas, proliferación de linfocitos, bloqueo de la unión del anticuerpo anti-integrina a su receptor.
Las secuencias actualmente preferidas de un péptido, fragmento peptídico, neo-estructura y/o neo-epítopo desveladas en el presente documento se enumeran de la siguiente manera:
EENFK SEQ ID NO 1
EDHVK SEQ ID NO 2
ENCDK SEQ ID NO 3
ETFLK SEQ ID NO 4
ERAFK SEQ ID NO 5
ECCEK SEQ ID NO 6
ECYAK SEQ ID NO 7
ERQIK SEQ ID NO 8
EKCCK SEQ ID NO 9
EEGKK SEQ ID NO: 10
EETFLK SEQ ID NO: 11
ETFLKK SEQ ID NO: 12
ETTLEK SEQ ID NO: 13
ETYVPK SEQ ID NO: 14
ERQIKK SEQ ID NO: 15
ELVKHK SEQ ID NO: 16
EVAHRFK SEQ ID NO: 17
EVTEFAK SEQ ID NO: 18
ECFLQHK SEQ ID NO: 19
EETFLKK SEQ ID NO: 20
ELLFFAK SEQ ID NO: 21
ELRDEGK SEQ ID NO: 22
EFAEVSK SEQ ID NO: 23
EKPLLEK SEQ ID NO: 24
ESKDVCK SEQ ID NO: 25
EPQNLIK SEQ ID NO: 26
EQLGEYK SEQ ID NO: 27
EKERQIK SEQ ID NO: 28
ESAENCDK SEQ ID NO: 29
EMADCCAK SEQ ID NO: 30
ECCQAADK SEQ ID NO: 31
EGKASSAK SEQ ID NO: 32
EEPQNLIK SEQ ID NO: 33
EVSRNLGK SEQ ID NO: 34
EKERQIKK SEQ ID NO: 35
ELVKHKPK SEQ ID NO: 36
ENQDSISSK SEQ ID NO: 37
EKCCKADDK SEQ ID NO: 38
ETCFAEEGK SEQ ID NO: 39
KDLGE SEQ ID NO: 40
KLVNE SEQ ID NO: 41
KQEPE SEQ ID NO: 42
KYLYE SEQ ID NO: 43
KVHTE SEQ ID NO: 44
KYICE SEQ ID NO: 45
KECCE SEQ ID NO: 46
KPLLE SEQ ID NO: 47
KNYAE SEQ ID NO: 48
KVFDE SEQ ID NO: 49
KPLVE SEQ ID NO: 50
KQNCE SEQ ID NO: 51
KCCTE SEQ ID NO: 52
(continuación)
KATKE SEQ ID NO: 53
KDLGEE SEQ ID NO: 54
KKYLYE SEQ ID NO: 55
KAAFTE SEQ ID NO: 56
KAEFAE SEQ ID NO: 57
KPLVEE SEQ ID NO: 58
KEFNAE SEQ ID NO: 59
KADDKE SEQ ID NO: 60
KTCVADE SEQ ID NO: 61
KLKECCE SEQ ID NO: 62
KSHCIAE SEQ ID NO: 63
KCCKHPE SEQ ID NO: 64
KRMPCAE SEQ ID NO: 65
KQTALVE SEQ ID NO: 66
KPKATKE SEQ ID NO: 67
KETCFAE SEQ ID NO: 68
KLVNEVTE SEQ ID NO: 69
KQEPERNE SEQ ID NO: 70
KLDELRDE SEQ ID NO: 71
KTYETTLE SEQ ID NO: 72
KQNCELFE SEQ ID NO: 73
KKQTALVE SEQ ID NO: 74
KETCFAEE SEQ ID NO: 75
KRYKAAFTE SEQ ID NO: 76
KDVCKNYAE SEQ ID NO: 77
KHKPKATKE SEQ ID NO: 78
EKDDAKCCK SEQ ID NO: 79
VFDEFKPLVEEPQNLIK SEQ ID NO: 80
VFDEFKPLVE SEQ ID NO: 81
A continuación, el término "tejido" como se utiliza en el presente documento significará la sangre completa, suero, plasma, fluido linfático, saliva, orina, heces, líquido ascítico, derrame pleural, pus, así como cualquier tejido, incluyendo, músculo, grasa, y tejido conjuntivo, incluyendo células inflamatorias.
En el presente contexto, la expresión "secuencia proteica" se utiliza para describir una o más proteínas, polipéptidos, péptidos, fragmentos peptídicos, neo-estructuras y/o neo-epítopos que se generan como resultado de la fragmentación proteolítica, desnaturalización y/o cambios conformacionales de una proteína que se encuentra normalmente en el suero. Como entiende fácilmente un experto en la técnica, un cambio conformacional de una proteína por supuesto no dará lugar necesariamente siempre a su fragmentación, sino que también puede resultar simplemente en la formación y/o presentación de una nueva estructura y/o epítopo. En el presente contexto se desvelan varias nuevas estructuras y/o epítopos que siguen unidas y presentados por la proteína original que se encuentra normalmente en el suero.
Un fragmento de una proteína que se encuentra normalmente en el suero se define en el presente contexto como que incluyen fragmentos de proteínas, polipéptidos y/o péptidos, sin referencia a una longitud específica de dicha secuencia proteica.
En el presente contexto, "desnaturalización" significa cualquier cambio de la estructura de una proteína a partir de la estructura normal, natural, tal como, por ejemplo, debido a estrés oxidativo.
Las proteínas son macromoléculas biológicas constituidas por restos de aminoácido unidos por enlaces peptídicos. Las proteínas, como polímeros lineales de aminoácidos, también se llaman polipéptidos. Normalmente, las proteínas tienen de 50-800 restos de aminoácido y por tanto tienen pesos moleculares en el intervalo de desde aproximadamente 6.000 a aproximadamente varios cientos de miles de Daltons o más. Las proteínas pequeñas se llaman péptidos, oligopéptidos o polipéptidos. En el contexto de la presente invención, un "péptido" o "fragmento peptídico" para su uso de acuerdo con la presente invención, se refiere a un polipéptido que puede tener, aunque no se limita a, 5-50 aminoácidos de longitud, tal como 5, 10, 15, 20, 25, 30, 35, 40, 41,42, 43, 44, 45, 46, 47, 47, 48, 49 o 50 aminoácidos. Dichos péptidos pueden ser también más largos de 50 aminoácidos.
Se desvelan en el presente documento secuencias de aminoácidos que son al menos un 70 % idénticas, tal como las que son al menos un 72 %, 75 %, 77 %, 80 %, 82 %, 85 %, 87 %, 90 %, 91 %, 92 %, 93 %, 94 %, 95 %, 96 %, 97 %, 98 %, o 99 % idénticas a la secuencia de aminoácidos de un péptido y/o fragmento peptídico de una secuencia como se enumera en SEQ ID NO: 1-81.
Por un péptido, fragmento peptídico, neo-estructura y/o neo-epítopo que tiene una secuencia de aminoácidos al
menos, por ejemplo, un 95 % idéntica a una secuencia de aminoácidos de referencia se entiende que la secuencia de aminoácidos de por ejemplo el péptido es idéntica a la secuencia de referencia, excepto en que la secuencia de aminoácidos puede incluir hasta 5 mutaciones puntuales por cada 100 aminoácidos de la secuencia de aminoácidos de referencia. En otras palabras, para obtener un péptido que tiene una secuencia de aminoácidos al menos un 95 % idéntica a una secuencia de aminoácidos de referencia: hasta un 5 % de los aminoácidos de la secuencia de referencia pueden eliminarse o sustituirse con otro aminoácido, o se pueden insertar en la secuencia de referencia varios aminoácidos hasta el 5 % del total de aminoácidos de la secuencia de referencia. Estas mutaciones de la secuencia de referencia pueden producirse en posiciones del extremo carboxilo y/o amino de la secuencia de aminoácidos de referencia o en cualquiera entre esas posiciones terminales, intercaladas individualmente entre los aminoácidos de la secuencia de referencia o en uno o más grupos contiguos dentro de la secuencia de referencia.
En el contexto de la presente invención, lo que se ajusta más adecuadamente es un programa de algoritmo local para determinar la identidad. Los programas de algoritmo local (tal como el de Smith Waterman) comparan una subsecuencia en una secuencia con una subsecuencia en una segunda secuencia y encuentra la combinación de subsecuencias y el alineamiento de esas subsecuencias que da lugar a la mayor puntuación de similitud total. Los huecos internos, si se permiten, están penalizados. Los algoritmos locales trabajan bien para comparar dos proteínas multidominio, que tienen un dominio único o solo un sitio de unión en común.
Los métodos para determinar la identidad y similitud se codifican en programas disponibles públicamente. Los métodos de programas de computadora preferidos para determinar la identidad y similitud entre dos secuencias incluyen, pero no se limitan al paquete de programas GCG (Devereux, J et al (1994)) BLASTP, BLASTN, and FASTA (Altschul, S.F. et al (1990)). El programa BLASTX está disponible públicamente en el NCBI y otras fuentes (BLAST Manual, Altschul, S.F. et al, Altschul S.F. et al (1990)). Cada programa de análisis de secuencia tiene una matriz de valoración por defecto y penalizaciones de huecos por defecto. En general, un biólogo molecular esperaría utilizar los ajustes por defecto establecidos por el programa de software que se utilice.
Resultados
Mapeo con espectrometría de masas del epítopo de un anticuerpo monoclonal de ratón específico para seroalbúmina humana desnaturalizada (dHSA)
Dos anticuerpos monoclonales dirigidos contra la HSA desnaturalizada demostraron tener una actividad inmunomoduladora. La estructura del epítopo de uno de estos mAb se investigó adicionalmente.
Se utilizaron dos estrategias similares para el mapeo del epítopo con una espectrometría de masas de desorción/ionización láser asistida por matriz con tiempo de vuelo (MALDI-TOF ms)
con el fin de definir el sitio o sitios posibles de la seroalbúmina humana en el que se une un anticuerpo monoclonal de ratón específico de la albúmina desnaturalizada. Una estrategia aprovecha el hecho de que los péptidos trípticos a los que se une el anticuerpo no generarán espectros de masas característicos en MALDI ya que se ocultan del análisis (3). Otra estrategia aprovecha el hecho de que los sitios en la proteína en los que el anticuerpo se ha unido están protegidos de la proteólisis (1, 2).
Unión de péptidos generados por tripsinización de dHSA por el anticuerpo monoclonal A (mAb A)
La seroalbúmina humana purificada (HSA) se desnaturalizó con urea, se redujo con DTT y se alquiló como se ha descrito (4). La HSA desnaturalizada se sometió entonces a un tratamiento con tripsina a baja concentración (0,02-2 ng/ml) de tripsina. Sin embargo, los espectros obtenidos con MALDI no eran satisfactorios ya que las masas de péptido típicas para la albúmina no se encontraban. Basándose en electroforesis en gel, se descubrió que esta preparación (digerida con 0,02 ng/ml de tripsina) contenía cantidades sustanciales de albúmina sin digerir. Por lo tanto, se continuó con la digestión con tripsina, a una concentración más alta (5 pg/ml) con el fin de obtener los espectros de masas utilizados habitualmente para la identificación de proteínas por MALDI.
Se incubó algo de la solución de albúmina ahora completamente escindida con el mAb A. Se llevó a cabo la MALDI-TOF ms y se compararon los espectros obtenidos de la albúmina desnaturalizada tratada con la enzima en presencia y ausencia del mAb A. Catorce de las albúminas que se sometieron a la espectrometría de masas estaban ausentes o disminuyeron después de la incubación con el mAb A (Tabla 1 A, Columna D). La secuencia de aminoácidos de estos péptidos se muestra en la Tabla 1B. Los espectros representan múltiples áreas que engloban los restos 66 a 508 de la molécula de albúmina.
Tabla 1 A. Restos peptídicos de HSA unidos al mAb A. Columna C: Área del pico de péptidos antes de la adsorción con el mAb A. Columna D: Área del pico de péptidos antes de la adsorción con el mAb A. Columna E: Área del pico de péptidos cuando la digestión de la dHSA estaba protegida por la unión al mAb
A

Tabla 1B n i min i l r í i H A ni l mAb A.

Con el fin de confirmar estos resultados, se permitió que el anticuerpo monoclonal mAb A se uniera a la albúmina desnaturalizada (previamente digerida con tripsina a una concentración de 0,02 ng/ml) con el fin de proteger las secuencias peptídicas del epítopo. El complejo se trató entonces de nuevo con tripsina. Se llevó a cabo entonces la MALDI-TOF ms y se compararon los espectros de masas generados de la albúmina con los espectros generados de la albúmina desnaturalizada tratada con tripsina en ausencia de anticuerpo. Los mismos catorce espectros de masas de los 39 espectros de masas de albúmina desaparecían completamente o estaban reducidos significativamente en la muestra cuando estaba presente el mAb durante el tratamiento con tripsina (Tabla 1 A, Columna E). Se tomaron múltiples lecturas para verificar los resultados.
No se pudieron identificar importantes fragmentos peptídicos debido a la posibilidad de que el epítopo de unión al mAb de la albúmina se hubieran escindido por la tripsina, dando como resultado a fragmentos del epítopo con una afinidad
de unión demasiado baja para unirse al mAb. Por lo tanto, también se utilizó un método alternativo. Se repitió el mapeo del epítopo por MALDI del mAb A basándose en la protección de la proteólisis por el mAb. Esta vez se utilizó una estrategia ligeramente diferente. Se incubó la HSA desnaturalizada con el mAb A. La albúmina no unida por el anticuerpo se retiró de la muestra mediante exclusión por tamaño en un ultrafiltrador. Los mAb libres resultantes y los complejos de mAb-albúmina se digirieron entonces con tripsina (las secuencias de la molécula de albúmina a las que se unía el mAb deberían resistir la digestión con tripsina). Los fragmentos pequeños de mAb escindidos y la albúmina no protegida se retiraron entonces de la muestra mediante ultrafiltración (30 kD). Los complejos de mAb y los fragmentos de albúmina unidos se disociaron mediante disminución del pH a 2,7. De nuevo, se llevó a cabo una ultrafiltración a 30 kD para separar los mAb completos de los fragmentos de albúmina menores de 30 kD. El análisis MALDI-TOF de estos fragmentos no identificaba los espectros típicos de la albúmina. Razonablemente, debido a que los fragmentos que contienen el epítopo de mAb A seguían siendo demasiado grandes. Este filtrado (< 30 kD) se digirió adicionalmente con tripsina (para la escisión de los sitios previamente protegidos por el mAb) con el fin de generar masas peptídicas adecuadas para el análisis por MALDI-TOF ms.
Después de este segundo tratamiento con tripsina, ocho de los 32 espectros de masas detectados por MALDI TOF ms coincidían con la albúmina (Tabla 2). Por lo tanto, estas secuencias de aminoácidos identificadas ahora comprenden una parte del epítopo, que también contiene secuencias del otro lado del punto de escisión de la tripsina.
Tabl 2 P i l min n r r ri iniz i n fr m n m r n l í l mAb A.

Seis de las ocho masas peptídicas (marcadas con * en la Tabla 2) eran masas peptídicas que también desaparecían cuando se analizaron previamente cuando la albúmina escindida completamente se incubaba con el mAb A antes del análisis MALDI-TOF (Tablas 1A y B).
El epítopo de este anticuerpo se estableció de esta manera. Es importante señalar que muchas de dichas estructuras están presentes en la molécula de albumina, que pueden producir entonces un entrecruzamiento de los receptores a los que se unen. Un estudio previo de la antigenicidad de la albúmina, basado en 13 anticuerpos monoclonales diferentes, ha demostrado que existe una reactividad intramolecular entre diferentes dominios en la albúmina humana (5), por lo tanto, se puede esperar que haya múltiples sitios de epítopo en el mAb A o la albúmina.
Basándose en estos resultados consistentes, se encontró un patrón común. Se encontró un ácido glutámico a una distancia de 5 o 6 aminoácidos de la lisina, tanto en las secuencias identificadas mediante MALDI-TOF como en la secuencia adyacente a la secuencia peptídica identificada por esta técnica (que están en el otro lado del punto de escisión de la tripsina, en K (lisina), (Tabla 3)). Es interesante señalar que en 4 de estas secuencias se encontraba un ácido glutámico adicional a una distancia de 2 o 3 aminoácidos del primer resto de ácido glutámico. Estos restos de ácido glutámico adicionales pueden tener importancia en la afinidad o la transducción de la señal de estos péptidos. La actividad biológica de estos péptidos también puede estar influenciada por la existencia de aminoácidos ácidos a una distancia de 12±1 aminoácidos (posición -12) del primer resto de ácido glutámico E (en la estructura E5K) y a una distancia de 3±1 aminoácidos (posición 3) del resto de lisina K (en la estructura E5K). Debido a la longitud de los dos aminoácidos importantes, el ácido glutámico (E) y la lisina (K), en el epítopo del mAb A, la distancia exacta fijada entre estos aminoácidos no es necesaria para la actividad inmunorreguladora de estos fragmentos. Por lo tanto, la secuencia de E3-7K puede tener una actividad inmunorreguladora similar a la de la secuencia E5K (Tabla 4A y B).
Tabla 3. Secuencias peptídicas que rodean las estructuras E5K y E6K seleccionadas para la síntesis de péptidos para el ensayo de la actividad inmunológica. Se incluye un péptido con la estructura E6K en la tabla secuencia 2.

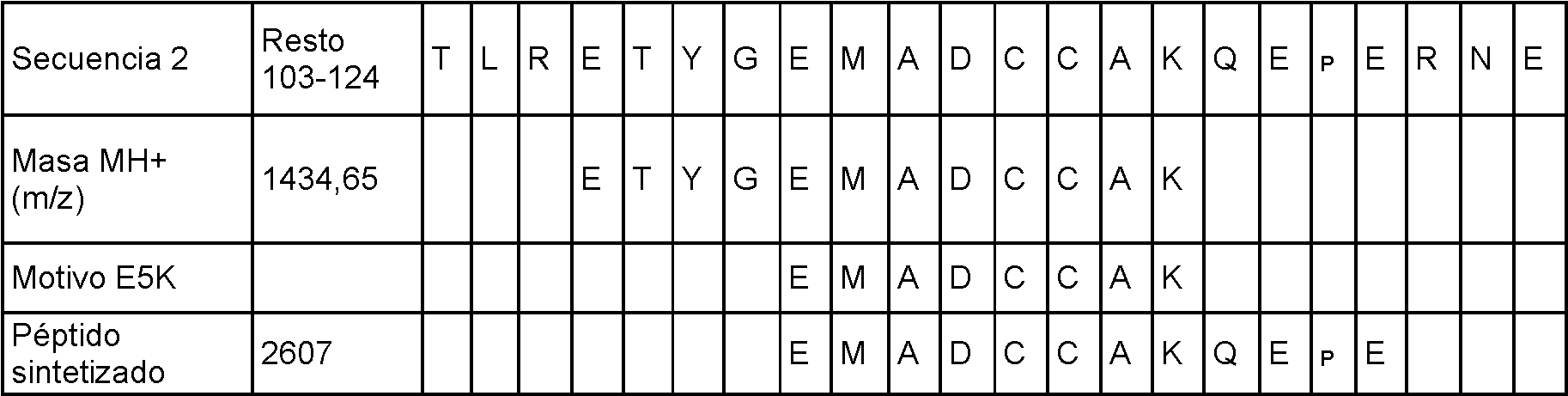



Se sintetizaron cinco de estos péptidos (Tabla 3) y se ha investigado sus funciones inmunorreguladoras. Basándose en estos estudios se postuló que ambas secuencias peptídicas estimulante e inhibidora están presentes en la seroalbúmina.
Conclusión- mapeo de epítopo por MALDI-TOF MS
El epítopo del mAb A se ha identificado como la estructura E5-6K
La estructura biológicamente relevante es por tanto E3-7K, posiblemente con restos de aminoácido ácidos en las posiciones -12 y 3. En conjunto, estos resultados indican que el mAb A se puede unir a múltiples regiones de la molécula de albúmina. Aunque los experimentos se llevaron a cabo con albúmina desnaturalizada, estos epítopos probablemente no son sitios generados por la combinación de restos cuando la molécula está plegada.
Actividad de unión de péptidos E5K de la unión del mAb A a la dHSA
Con el fin de ensayar la especificidad de los péptidos sintetizados, se ensayaron en un ELISA en el que se analizó la inhibición de la unión del mAb A a placas revestidas con dHSA. Una alta unión del anticuerpo a la placa es por tanto consistente con una actividad no inhibidora y esta unión se reduce cuando se añade una sustancia inhibidora al sistema. Como se muestra en la Fig. 1, cuatro de los cinco péptidos presentaban una inhibición dependiente de la dosis del anticuerpo contra las placas revestidas de dHSA, confirmando que contienen una estructura que reacciona con el anticuerpo.
Expresión del epítopo E5K en células tumorales - correlación con la supervivencia
Se había demostrado previamente, utilizando la tinción inmunohistoquímica con mAb A, que el epítopo/estructura de E5K se expresa mediante varios tipos de células cancerosas (documento WO 06/043891). Una serie de 20 biopsias de pacientes con melanoma se tiñeron utilizando esta técnica y se valoró la intensidad de tinción desde a ++ utilizando un microscopio óptico. Se observó una variación de intensidad de tinción considerable. Basándose en las áreas teñidas más intensamente de las secciones se clasificaron los pacientes desde altos a bajos expresadores de E5K. El número de pacientes se dividió entonces en dos grupos iguales de altos y bajos expresadores, y se analizó una posible diferencia de supervivencia entre estos grupos de acuerdo con los análisis de Kaplan-Meier y rango logarítmico. Como se muestra en la Fig. 2 se encontró una gran diferencia estadísticamente significativa para los altos y bajos expresadores. Actividad inmunomoduladora de los péptidos E5K.
Efecto de los péptidos sobre la proliferación de PBMC inducida por PHA
Se ensayó el efecto de dos péptidos de albúmina, 2605 y 2608, sobre la proliferación de PBMC inducida por PHA de una control sano y dos pacientes de cáncer. Como se muestra en la Figura 3A y B, el patrón de respuesta es bastante diferente entre los individuos, presumiblemente debido al grado de estimulación inmunitaria de las PBMC in vivo y posiblemente debido también a la existencia de autoanticuerpos de las neo-estructuras representadas por los péptidos. La importancia del grado de estimulación inmunitaria se demuestra comparando el efecto de los péptidos cuando se estimulan las PBMC con 5 o 10 pg/ml de PHA (compárese La Fig. 3 A y B). Además de sus diferencias interindividuales, también se descubrió un patrón de respuesta bifásica, por ejemplo, utilizando el péptido 2605 con las PBMC de K92, la menor concentración era inhibidora, la concentración media era estimuladora y de nuevo a la concentración más alta, se inhibía la respuesta proliferativa. También es interesante señalar que se descubrió una actividad estimuladora clara con la concentración más baja de ambos péptidos en el paciente P46. También hay algunas diferencias en la actividad de los dos péptidos ensayados especialmente cuando se estimulan las PBMC con PHA a una concentración de 5 pg/ml. Cuando se utilizaba una concentración de PHA de 10 pg/ml la actividad de los dos péptidos era similar. El modelo de cultivo con el grado meno de estimulación menor es por supuesto más sensible a la variación en la estructura de unión del receptor. Este ejemplo, demuestra así que una vez que se ha demostrado una secuencia peptídica biológicamente activa, los cambios en la secuencia de aminoácidos puede modular su actividad biológica.
Para analizar adicionalmente las diferencias interindividuales en el efecto de estos péptidos, se analizaron las PBMC de 5 controles sanos y 4 pacientes (Fig. 4). Se utilizó PHA a una concentración de 5 pg/ml y los péptidos a una concentración de 10 pg/ml. De nuevo se demostró una clara diferencia entre los individuos. El péptido 2605 tenía un efecto inhibidor o estimulador en un control de cada y un paciente de cada. El péptido 2608 tenía un efecto estimulador en 1/4 de los controles mientras que 3/4 de los pacientes se estimulaban.
Efecto de la dHSA sobre la proliferación de células mononucleares de sangre periférica (PBMC) inducida por PHA El efecto de la dHSA sobre la proliferación de PBMC inducida por PHA en controles sanos y en pacientes de cáncer es bastante variable (Fig. 5). De nuevo esto puede ser debido al grado de estimulación de las PBMC in vivo y posiblemente a la presencia de autoanticuerpos contra la dHSA. La adición de dHSA a estos cultivos puede dar como resultado tanto la estimulación como la inhibición de la tasa de proliferación, pero frecuentemente da como resultado la estimulación de la tasa proliferativa. Es extraordinario que un control no respondiera para nada y que esta persona no respondiera tampoco a una concentración más alta de PHA. En un paciente (P41) la adición de dHSA inhibía la proliferación, especialmente a una concentración de PHA de 10 pg/ml. La variación en la respuesta entre los pacientes demuestra la necesidad del diagnóstico del estado inmunitario individual de los pacientes de cáncer.
Efecto de los péptidos sobre la proliferación de PBMC inducida por PHA modulada por dHSA
A continuación, se analizó el efecto de los péptidos 2605 y 2608, sobre la proliferación inducida por PHA aumentada por dHSA. Como se muestra en la Figura 6 A y B la adición de dHSA a una concentración de 8 pg/ml aumentaba significativamente la tasa proliferativa de las PBMC estimuladas por PHA en dos controles sanos diferentes. A concentraciones menores de dHSA, la actividad estimuladora decaía. De manera interesante, la adición de los péptidos a una concentración de 10 pg/ml inhibía significativamente la actividad estimuladora de las dos concentraciones de dHSA más altas, mientras que a concentraciones más bajas los péptidos, por el contrario, estimulaban la tasa proliferativa. Por lo tanto, el efecto estimulador de la dHSA a 8^pg/ml se inhibía mediante la adición de los péptidos, pero la misma concentración de péptidos estimulaba la tasa proliferativa a una concentración menor de dHSA. Razonablemente, el entrecruzamiento del receptor de E5K estaba implicado en la actividad estimuladora de la dHSA ya que la unión monomérica de los péptidos a este receptor inhibe el efecto estimulador de la dHSA. La
misma concentración de los péptidos tiene entonces una actividad bastante diferente en presencia de diferentes concentraciones de dHSA, a 0,8 |jg/ml sigue habiendo una actividad ligeramente inhibidora mientras que a concentraciones menores de dHSA la tasa proliferativa aumenta significativamente. Una explicación razonable para esto es que la actividad estimuladora de los péptidos se bloquea por una neo-estructura inhibidora de albúmina a la concentración de 8 jg/m l de dHSA.
Efecto de los péptidos sobre la producción de monocinas por las PBMC
El efecto de los péptidos de albúmina, 2604-2608 sobre la producción de IL-6 inducida por LPS se muestra en la Figura 7 A-C. De nuevo se observaron diferencias considerables interindividuales en la actividad de los péptidos. Analizando las PBMC de un control sano, el péptido 2604 era estimulador a la concentración más baja, mientras que los péptidos 2606 y 2608 a esta concentración eran inhibidores (Fig. 7A). La actividad también variaba entre los dos pacientes de melanoma, pero en uno de estos pacientes todos los péptidos tenían actividad estimuladora (Fig. 7C).
Por lo tanto, los cinco péptidos tienen actividad inmunomoduladora, pero el efecto varía dependiendo del estado inmunitario del individuo investigado.
Efecto de los péptidos de albúmina sobre la tinción inmunohistoquímica de las PBMC utilizando un anticuerpo antiintegrina
La importancia inmunobiológica de la secuencia de aminoácidos de la albúmina se estudió adicionalmente analizando su influencia sobre la unión de un anticuerpo monoclonal a la integrina P2 LFA-1 (CD11) sobre las células inmunitarias (Fig. 8 A-F). Esta molécula se escogió para estos experimentos, ya que se sabe que la unión de ciertos mAb a esta molécula podía modular/inhibir seriamente funciones del sistema inmunitario. Se ha demostrado que el anticuerpo particular escogido para estos experimentos inhibe la unión de LFA-1 a ICAM-1, AND, ICAM-3.
Las preparaciones en cytospin de las células mononucleares de la sangre de controles sanos, pacientes de cáncer y líneas celulares monocíticas, THP-1, se prepararon (como se describe en materiales y métodos), se secaron y se secaron a -70 °C. En la tinción inmunocitológica, se bloqueó la tinción inespecífica mediante la incubación con un 10 % de suero AB humano. Algunos de los portaobjetos se preincubaron durante 60 minutos con péptidos de albúmina a una concentración de 40 jg/ml, al que se añadía un 10 % de suero AB, como se indica en las Figuras 8 A-F. El procedimiento de tinción se continuó entonces como se describe en los materiales y métodos. La intensidad de la tinción de los portaobjetos teñidos con y sin preincubación con los péptidos se registró semicuantitativamente utilizando un microscopio óptico convencional.
Como se muestra en las Figuras 8 A-F, se puede inhibir la unión del mAb al LFA-1 mediante la preincubación con los péptidos. Como se ha mencionado en otras secciones del presente documento, el estado inmunitario del donante de las células sanguíneas puede tener influencia en el resultado de los análisis inmunológicos. En consecuencia, la tinción de las PBMC de algunos donantes parece que no está influenciada por la preincubación con los péptidos en algunos casos con una baja tinción inicial incluso se observó un aumento de la tinción. La unión del mAb al LFA-1 en la línea celular monocítica THP-1 estaba claramente aumentada por la preincubación con el péptido 2606 (Fig. 8 A, B). Estos resultados claramente muestran que la estructura de E5K interactúa con la integrina p2 de una manera, que tiene importancia para la función del sistema inmunitario.
Unión de los péptidos generados por tripsinización de la dHSA por los receptores de superficie celular
Basándose en la observación de que las secuencias de péptido inmunorreguladoras están presentes en la seroalbúmina, existe la posibilidad de que otras secuencias distintas al epítopo del mAb tengan una función inmunorreguladora. Por lo tanto, se preparó una superficie celular artificial (ACS) como se describe en materiales y métodos. La mezcla de péptidos obtenida después de la tripsinización se adsorbió por la ACS y se identificó la unión de los péptidos comparando las soluciones de péptidos adsorbidos y no adsorbidos utilizado una técnica MALDI TOF ms. Estos péptidos se muestran en la Tabla 5 A.
Tabla 5 A. Péptidos generados por degradación con tripsina de dHSA y grado de adsorción de los receptores de ACS. Los aminoácidos entre paréntesis muestran el punto de escisión de la proteasa y no están incluidos en las masas identificadas.

continuación

Basándose en su grado de unión y su relación espacial con las estructuras E5K de la albúmina, se seleccionaron cuatro nuevos péptidos para sintetizarse e investigarse en cuanto a su actividad inmunorreguladora (Tabla 5 B).

Actividad inmunomoduladora de péptidos generados por tripsina
Efecto de los péptidos sobre la proliferación de PBMC inducida por PHA modulada por dHSA Dos de los péptidos en la nueva serie 3026 y 3028, se ensayaron y compararon con el péptido 2605 en el análisis en cuanto a su efecto sobre la proliferación estimulada por PHA modulada por dHSA (Fig. 9). La proliferación de PBMC inducida por PHA de dos controles sanos se estimularon adicionalmente mediante dHSA. Como se muestra en la Figura 9 todos los péptidos inhibían la actividad estimuladora de la dHSA. También, en este experimento, el grado de estimulación de las PBMC tiene un impacto sobre los resultados.
Efecto de los péptidos sobre la proliferación de PBMC inducida por interleucina-2
Los péptidos de esta nueva serie, 3026-3029, también se ensayaron en cuanto a su efecto sobre la proliferación inducida por IL-2. Como se muestra en la Figura 10 A, 3/4 péptidos, 3026, 3027 y 3029, no tenían una actividad estadísticamente significativa. Por el contrario, el péptido 3028 era altamente inhibidor (p = 0,005). Es interesante señalar que este efecto inhibidor se invertía completamente mediante la modulación del entrecruzamiento con el receptor de Fc (Figura 10 B) de manera similar a la situación previamente descrita para la inmunosupresión relacionada con IL-2 en el carcinoma de células renales (documento WO 03/099312 A1).
Efecto de la nueva serie de péptidos sobre la producción de monocinas por las PBMC
El efecto de la nueva serie de péptidos presentaba una diferencia considerable en su efecto incluso entre individuos de control sanos (Figura 11). El péptido 3026 no tenía cierto efecto en uno de los controles (PBMC 2), pero tenía un efecto bifásico claro en el otro (PBMC 1). En el último caso, se estimuló la producción de IL-6 con las tres concentraciones más altas y se inhibía claramente a la concentración más baja. El péptido 3027 era ligeramente estimulador en uno de los controles y tenía un efecto inhibidor en el otro. Se encontraron resultados similares como el péptido 3028. El péptido 3029 tenía un efecto ligeramente estimulador solo en uno de los controles, a las dos concentraciones más altas. Es interesante señalar que todos los péptidos excepto el 3029 tenían un efecto inmunomodulador con una concentración tan baja como de 10 ng/ml. Por lo tanto, todos los péptidos tenían un efecto en al menos uno de los controles analizados.
De manera similar al efecto de la nueva serie de péptidos sobre las PBMC de los controles sanos, también las PBMC de los pacientes de cáncer presentaban diferencias interindividuales considerables (Figura 12). Los péptidos 3026 y 3027 tenían un efecto estimulador en el paciente de carcinoma de células renales y además el péptido 3027 también estimulaba uno de los pacientes de melanoma. Los otros dos péptidos, 3028 y 3029, esencialmente no tenían efectos en estos ensayos. Al contrario que en la situación de los controles, no se veían efectos inhibidores.
Unión de los péptidos generados por degradación de asparaginasa de la dHSA por los receptores de superficie celular
La secuencia peptídica completa de la albúmina no se recuperó utilizando la técnica MALDI-TOF después de la degradación con tripsina. Además, algunas secuencias con la capacidad para unirse a los receptores de superficie celulares de las células inmunitarias podían haberse degradado por el tratamiento con tripsina. Por lo tanto, se utilizó el mismo procedimiento experimental que se ha descrito anteriormente para una mezcla peptídica obtenida por la degradación utilizando asparaginasa. Los péptidos unidos a los ACS resultantes se muestran en la Tabla 6 A y B.
Además de los péptidos generados por la degradación con tripsina, se encontraron otros seis péptidos con un peso molecular de 700 - 3600 Da que se adsorbían eficazmente (> 65 %) por las estructuras de superficie celular en la columna ACS (Tabla 6A).
Tabla 6 A. DHSA-ASP adsorbida en ACS

Se descubrió que siete péptidos de un peso molecular entre 3200 y 9000 Da se adsorbían completamente por la ACS y uno de los péptidos de este grupo se unía al 37 %. En este análisis otros 9 péptidos no eran adsorbidos por la ACS.
Tabla 6 B. DHSA-ASP adsorbida en ACS

Dos péptidos de este grupo no se unían y se obtenían resultados contradictorios para uno de los péptidos (SISSKLKECCEKPLLEK SHCIAEVEN DEMPA) con respecto a la adsorción por la ACS.
Por lo tanto, el tratamiento con asparaginasa genera secuencias peptídicas diferentes a las generadas por la tripsina, que se unen eficazmente a estructuras de superficie celular de las células inmunitarias. Basándose en los resultados descritos anteriormente estas estructuras serán las que más probablemente tengan actividad inmunomoduladora.
Existencia de fragmentos de unión a la ACS de IgG en pacientes con cáncer
Con el fin de identificar adicionalmente la existencia de estructuras de unión a células inmunitarias, se preparó plasma sanguíneo, y se llevó a cabo una cromatografía de afinidad como se ha descrito anteriormente. Las sustancias unidas por la columna de ACS se eluyeron, fraccionaron con una electroforesis en gel 2D y se identificaron utilizando la técnica MALDI-TOF. Como se esperaba, se identificaron las áreas correspondientes a la albúmina y las inmunoglobulinas en el gel 2D. Además, también se identificaron otras sustancias de unión a las células inmunitarias (Fig. 13). La unión de algunas variedades de albúmina, presumiblemente de albúmina dañada que tenía cambios conformacionales, a las células inmunitarias se había descrito previamente por varios grupos. Las nuevas estructuras de unión a células inmunitarias que se encontraron en esta investigación se resumen en la Tabla 7.
T l Pr ín i n ifi r MALDI-T F m

Por lo tanto, además de la seroalbúmina, hay también otras proteínas de origen natural que son sustratos para la generación de fragmentos inmunorreguladores.
Por lo tanto, se puede concluir y se demuestra claramente en la presente solicitud de patente que secuencias de proteínas de origen natural tal como la seroalbúmina e IgG se unen a los receptores de superficie celular de células inmunitarias y tienen actividad inmunorreguladora. Se han identificado secuencias tanto estimuladoras como inhibidoras. Además, se descubrió que el entrecruzamiento de receptores de células inmunitarias era un mecanismo por el que la función de estas células puede modularse.
Modelo humano ex vivo para la evaluación de la inmunosupresión en pacientes con cáncer
La IL-2 tiene una importancia fundamental en el inicio y estimulación de la respuesta inmunitaria y la actividad de esta citocina a menudo está inhibida en la inmunosupresión relacionada con el cáncer. Por lo tanto, se construyó un modelo humano ex vivo para la inmunosupresión en pacientes con cáncer (Fig. 14 y 15) para la evaluación de péptidos inmunorreguladores posiblemente inhibidores.
Se demostró que la respuesta a IL-2 en este modelo se correlacionaba con la supervivencia total de los pacientes (Fig. 15). La inmunosupresión en este modelo humano ex vivo está mediada por factores del suero, ya que se inhibe significativamente la capacidad proliferativa de las PBMC de los controles sanos si estas células se cultivan con suero de pacientes con cáncer en el medio (Fig. 16).
Identificación de péptidos inmunorreguladores adicionales
Se utilizaron las columnas de superficie celular artificial (ACS) con el fin de identificar secuencias peptídicas de la albúmina sérica que se unan a la superficie de células inmunitarias. Después de la biotinilación, se descubrió que dichos receptores se unían a perlas de estreptavidina. Los péptidos que se unen a dichas columnas se identificaron tras la elución mediante la técnica MALDI-TOF. Basándose en estos resultados y su relación con péptidos de albúmina identificados previamente, se sintetizaron cinco péptidos. Su actividad inmunorreguladora se ensayó primariamente sobre la respuesta a la IL-2.
El efecto de los diferentes péptidos sobre la proliferación inducida por IL-2 se analizó en el modelo humano ex vivo. El péptido 3028 inhibe regularmente la proliferación inducida por IL-2 pero ninguno de los otros péptidos identificados por su unión a la superficie celular artificial tenía actividad inhibidora (Fig. 17). Como la parte del extremo C del péptido 3028 contiene una estructura inmunorreguladora, E5K, previamente identificada, el efecto de los cinco péptidos que contenían esta estructura también se ensayó sobre la proliferación inducida por IL-2, pero estos solo demostraron ninguna o una mínima actividad inhibidora.
La actividad inhibidora del péptido 3028 sobre la proliferación inducida por IL-2 también se puede demostrar en cultivos con PBMC de pacientes con cáncer, incluso si la respuesta a la IL-2 ya se había suprimido (Fig. 18). Como la inmunosupresión en el cáncer se caracteriza por una pobre respuesta a la IL-2, la inhibición de la actividad de esta neo-estructura de albúmina en los pacientes con cáncer tiene una gran capacidad para superar la inmunosupresión relacionada con el cáncer. Este péptido inhibe uno de los mecanismos fundamentales en el inicio y regulación positiva de una respuesta inmunitaria, por lo tanto, será más probable que sea más útil en la regulación negativa de la reactividad inmunitaria en enfermedades inflamatorias crónicas y autoinmunitarias.
Caracterización adicional del efecto del péptido 3028 sobre la proliferación inducida por IL
Como se había descubierto previamente que ciertas neo-estructuras de albúmina tienen actividad inmunomoduladora y la parte del extremo C del péptido 3028 tenía una estructura similar, se sintetizaron las partes del extremo C y el extremo N del péptido 3028 y se analizaron por separado y en combinación. Obviamente, la actividad inhibidora de las dos partes del péptido 3028 es mucho más débil (Fig. 19).
Caracterización de un antisuero de conejo y anticuerpos de conejo purificados por afinidad dirigidos contra el péptido 3028
El antisuero de conejo dirigido contra el péptido 3028 de albúmina se une a dHSA y en menor extensión a la kHSA. Se ensayaron dos antisueros, R y L, de dos conejos diferentes. Estos anticuerpos del suero se unen preferentemente al fragmento 3325 pero no al 3218 de 3028. También se obtenían resultados similares con los anticuerpos purificados por afinidad (véase la figura 21).
Efecto inmunomodulador de los anticuerpos de conejo purificados por afinidad dirigidos contra el péptido 3028
Como se muestra en la Figura 22, la inhibición de la respuesta proliferativa a la IL-2 estaba superada en los pacientes con cáncer inmunosuprimidos (Fig. 22A) y en los controles normales con regulación negativa de la reactividad inmunitaria (Fig. 22 B) que tienen una tasa proliferativa de menos de 100000 dpm en el modelo humano ex vivo. Los anticuerpos anti-3028 no tenían efecto cuando la tasa proliferativa está en el intervalo normal.
La IgG policlonal de conejo se añadió a los cultivos de control con el fin de estar seguros de que el efecto de los anticuerpos purificados por afinidad no era debido a una actividad inespecífica de la IgG de conejo en este modelo. La IgG de conejo solo tenía una actividad mínima. La especificidad de los anticuerpos anti-3028 se demostró adicionalmente ya que el efecto estimulador de estos anticuerpos se neutralizaba mediante una pequeña cantidad de péptido 3028 que no tenía actividad inhibidora per se. Además, la adsorción del suero inhibidor mediante un gel al que se unían los anticuerpos anti-3028 reducía la actividad inhibidora de dicho suero.
De manera similar que los resultados en el modelo ex vivo autólogo, la actividad inmunosupresora de los sueros de personas con una baja respuesta proliferativa a IL-2 se superaba por la adición de los anticuerpos anti-3028 a los cultivos.
Unión de anticuerpos anti-3028 en / Expresión del epítopo 3028 en / tumores malignos
Las estructuras a las que se unían los anticuerpos anti-3028 se expresan ampliamente en los tumores malignos humanos, por ejemplo, en melanoma maligno, carcinoma de células renales y cáncer colorrectal (véase la figura 23).
El receptor del péptido 3028:
Unión del 3028 a LFA-1
De manera similar a los resultados descritos anteriormente para los sueros de los pacientes de cáncer y los péptidos inmunorreguladores identificados previamente el péptido 3028 tiene la capacidad de modular la unión del anticuerpo LFA-1 (HI 111) al LFA-1 de las células mononucleares sanguíneas. Tanto la inhibición (Fig. 24) como el aumento de la unión se habían demostrado, dependiendo razonablemente de la estructura del LFA-1 (en forma activada o inactivada) cuando se prepararon las preparaciones cytospin de las células. También. Se ha demostrado que las partes del extremo C y N de este péptido tenían alguna actividad inhibidora (Fig. 24).
De acuerdo con estos resultados y el efecto del péptido 3028 sobre la proliferación inducida por IL-2, es interesante señalar que el anticuerpo anti-LFA1 utilizado en estos experimentos es un potente inhibidor de la proliferación inducida por IL-2. Resultados similares se habían publicado previamente por Vyth-Dreese et al. (1993).
Unión de 3028 a la cadena a (CD25) del receptor de IL-2
Como el péptido 3028 inhibe significativamente la respuesta proliferativa a IL-2, se comparó la secuencia de aminoácidos de este péptido a la de la IL-2 y se encontraron ciertas similitudes en el sitio de unión del receptor de IL-2 (Tabla 8).
Tabla 8. Homologías en la secuencia de aminoácidos del péptido de albúmina 3028 y un segmento de la interleucina-2 humana, que participa en la interacción entre la interleucina-2 con el receptor alfa de la interleucina-2 (CD25).
Péptido 3028: V  K
K
IL-2 humana: E L K P L E E
(a.a. 61-72)
Basándose en esta observación, se estudió el efecto del péptido 3028 sobre la unión de IL-2 a CD25. La proteína de fusión de CD25 y la parte Fc de la IgG se unieron en microplacas revestidas de proteína G /placas de ELISA y se incubaron las placas con IL-2 biotinilada con o sin la presencia del péptido 3028. Sorprendentemente, la unión de la IL-2 al CD25 aumentaba con el péptido 3028, indicando una interacción de tres partes entre IL-2, CD25 y 3028. Incluso si la unión de la IL-2 al CD25 aumentaba el ensamblaje apropiado del receptor de alta afinidad y/o la transducción de la señal se bloquean ya que el péptido 3028 es un potente inhibidor de la proliferación inducida por IL-2 (véase anteriormente).
A continuación, se demostró utilizando un modelo molecular asistido por computadora que el péptido 3028 se une a CD25 en el sitio de unión de IL-2 (Fig. 25). Se puede concluir, por tanto, que el péptido 3028 tiene una capacidad inmunorreguladora dual uniéndose tanto a LFA-1 como al receptor de la IL-2.
Péptido 3028, estructura inmunosupresora óptima:
El péptido inhibidor fisiológico
Basándose en los resultados descritos anteriormente (diferencia entre la actividad antiproliferativa de los péptidos 3216 y 3325, especificidad de la afinidad de los anticuerpos purificados dirigidos contra el péptido 3325 y no al péptido 3218, la actividad inmunomoduladora de estos anticuerpos, y el efecto de estos péptidos sobre la unión del mAb anti-LFA-1 contra las células inmunitarias) se puede concluir que ninguno de los péptidos menores, 3218 o 3325 son tan eficaces como el péptido completo 3028 (Fig. 26). Sin embargo, ambos péptidos inhiben la unión del mAb HI 111 al LFA-1. Una explicación razonable a esto es que ambos péptidos menores contribuyen a la actividad completa del efecto inhibidor del péptido 3028. Por lo tanto, es lógico extender el péptido 3325 con los aminoácidos del extremo N del péptido 3218. Como la extensión del extremo C del péptido 3325 es una lisina sería interesante producir péptidos más largos con el fin de ensayar la posibilidad de que péptidos más largos sean incluso más eficaces que el péptido 3028.
Con el fin de mantener la naturaleza fisiológica de este péptido inhibidor, las únicas modificaciones relevantes de su estructura son cambiar su longitud como se ha expuesto anteriormente.
Este programa clarificará de esta manera la estructura óptima del péptido 3028 que se va a utilizar como fármaco inmunosupresor para el tratamiento de afecciones patológicas/enfermedades dependientes/relacionadas con la IL-2 tales como enfermedades malignas de células T, rechazo de injertos o trasplantes de órganos, enfermedad del injerto contra el huésped (GVH), enfermedades inflamatorias crónicas tales como psoriasis y algunas enfermedades autoinmunitarias. La racional para el uso terapéutico del péptido inmunoinhibidor 3028 en estas condiciones se demuestra por la actividad terapéutica de los anticuerpos monoclonales dirigidos contra CD25 (el receptor Tac).
Tabla 9

Comentarios sobre el presente mecanismo inmunorregulador
Como la inmunosupresión del cáncer se caracteriza por una pobre respuesta a la inhibición de IL-2 de la actividad de esta neo-estructura de albúmina en los pacientes de cáncer tiene una gran capacidad para superar la inmunosupresión relacionada con el cáncer. Este péptido inhibe uno de los mecanismos fundamentales en el inicio y regulación positiva de una respuesta inmunitaria, por lo tanto, será más probable que sea más útil en la regulación negativa de la reactividad inmunitaria en enfermedades inflamatorias crónicas y autoinmunitarias.
La estructura 3028 inmunorreguladora descrita en la presente solicitud se genera por un mecanismo fisiológico presente en la inflamación y el cáncer. Las estrategias terapéuticas basadas en estas dianas serán por lo tanto aplicables en general.
Basándose en los datos actuales el mecanismo de acción es específico de la especie y por tanto no son aplicables los modelos análogos en animales. La prueba del concepto se obtiene en un modelo humano ex vivo en el que los resultados se correlacionan con la supervivencia total en pacientes con cáncer.
Anticuerpos específicos para el péptido 3028 de albúmina para su uso terapéutico
Los anticuerpos, de longitud completa o fragmentos, con especificidad para 3028, así como para cualquiera de los fragmentos desvelados en la s Eq ID NO: 1-81, preferentemente deberían ser humanizados o completamente humanos para las aplicaciones terapéuticas. Dichos anticuerpos se pueden producir utilizando varias de las tecnologías establecidas.
Para humanizar un anticuerpo monoclonal animal (por ejemplo, de ratón), se utilizan estrategias recombinantes para injertar las regiones determinantes de complementariedad (CDR) desde el ADNc de inmunoglobulina de un animal derivado de hibridoma en las regiones correspondientes de una ADNc de inmunoglobulina humana coincidente. El anticuerpo recombinante resultante se puede expresar entonces y producirse en varios organismos, por ejemplo, bacterias o líneas celulares de mamífero.
Los anticuerpos completamente humanos se pueden obtener primariamente mediante tres estrategias diferentes; 1) por rescate de los anticuerpos de origen natural a partir de donantes inmunitarios humanos mediante una transformación con virus de Epstein Barr (EBV) de células B o mediante PCR de clonación y fago de presentación. 2) inmunizando y produciendo hibridomas a partir de ratones transgénicos, que se han creado con un repertorio de secuencias genéticas de la línea germinal humana. 3) explorando bibliotecas de fagos sintéticos que contienen genes de la región variable (V) de un anticuerpo humano y seleccionando las regiones V de unión al antígeno mediante fagos de presentación. El anticuerpo seleccionado se clona entonces.
Hay ahora múltiples compañías comerciales que desarrollan anticuerpos humanos contra una proteína/péptido definido por encargo. Además, se desarrollan rápidamente nuevas moléculas "tipo anticuerpos" (por ejemplo, anticalinas, afilina, aficuerpos) y se producen como potenciales candidatos a fármacos. (Para una revisión, véase, por ejemplo: Peterson NC. Advances in monoclonal antibody technology: Genetic engineering of mice, cells and immunoglobulins. ILAR Journal, 2005, 46:314-9.)
Efecto de los péptidos de albúmina sobre la actividad citotóxica cito lítica natural (NK) de donantes de sangre sanos
Resultados
Se ensayó la actividad citotóxica NK de las células mononucleares de la sangre de cuatro donantes sanos. Como se ve en la figura XX la presencia del péptido 3028 y, en un menor grado, el péptido 3026 reducían el porcentaje de lisis específica de células diana K562 por los cuatro donantes. No se veía inhibición en presencia del péptido 3027, sin embargo.
Materiales y métodos
Preparación de seroalbúmina humana desnaturalizada (dHSA)
Se desnaturalizó una solución en infusión de seroalbúmina humana (HSA) (Pharmacia, Uppsala, Suecia) y se reduje resuspendiéndola a una concentración final de 10 mg/ml en 8 M de urea y 10 mM de ditiotreitol (ambos de Sigma Chemical Co, St. Louis, MO) en 50 mM de Tris-HCl (pH 7,9) durante 2 h a 25 °C. La HSA se alquiló entonces mediante la adición de 60 mM de yodoacetamida (Sigma) y se incubó adicionalmente durante 2 h a 25 °C en oscuridad. La solución de HSA se diluyó a una concentración de 100 pg/ml con solución salina tampón de fosfato (PBS, Gibco BRL y se dializó extensamente contra el PBS utilizando entubado de diálisis Spectrapore 4 con un corte de pm de 12000 (Spectrum Europe, Breda, Países Bajos). Se preparó una HSA de control en paralelo incubando la HSA a 10 mg/ml en Tris-HCl (pH 7,9) seguido por la diálisis. Antes de su uso en los experimentos de cultivo tisular la dHSA se filtró a través de un filtro de jeringa de 0,22 Dm (Millipore Co, MA, USA). La dHSA se almacenó a 4 °C o se secó por congelación a -20 °C.
Escisión enzimática de dHSA con una dosis baja de tripsina
Se llevó a cabo un intercambio de tampón a 25 mM de NH4HCO3 , pH 8, en la HSA desnaturalizada con filtración en gel Sephadex-G25 (PD-10, columnas de desalación, Amersham Biosciences Europe, Uppsala, Suecia). Se determinó el intercambio proteico con un ensayo proteico Bio-Rad basado en el procedimiento de unión al colorante de Bradford siguiendo las recomendaciones del fabricante (Bio-Rad Laboratories AB, Sundbyberg, Suecia). Se añadió tripsina modificada de calidad de secuenciación (Promega, Madison, concentración tras el tampón WI) a una concentración final de 2, 0,2 o 0,02 ng/ml a la HSA desnaturalizada (49 pg/ml). De manera alternativa, como control, se añadió una cantidad equivalente de tampón de dilución de tripsina (50 mM de mM C2H4O2). La mezcla se incubó a 37 °C durante 18 horas. La actividad de la tripsina se detuvo por pasaje de la muestra sobre una columna de inhibidor de tripsina de soja entre cruzada con agarosa activada CNBr (Sigma).
Escisión enzimática completa de dHSA con tripsina a altas dosis seguido por la incubación con mAb para el mapeo de epítopos
Se secaron por congelación ocho pg de dHSA tratada con tripsina a dosis bajas y se disolvió entonces en 16 □ de tripsina modificada de calidad de secuenciación (a 5 pg/ml) (Promega) y se incubaron a 37 °C durante 18 horas. Una parte (10 pl) de los péptidos trípticos digeridos se hicieron reaccionar con el anticuerpo monoclonal (mAb A) a una concentración final de 0,2 mg/ml durante 2 horas a temperatura ambiente. Las muestras se almacenaron a 4 °C durante una noche y después se analizaron mediante MALDI-TOF MS (véase posteriormente).
Incubación de dHSA con el mAb seguido por la escisión completa con tripsina para el mapeo del epítopo
Se incubó la HSA tratada con tripsina a bajas dosis, desnaturalizada (8 pg) en 25 mM de NH4HCO3 , pH 8, con 8 pg del anticuerpo monoclonal (mAb A) o con PBS de control durante 2 horas a 4 °C. También se incubó en paralelo un control por separado que consistía en 8 pg de anticuerpo monoclonal en 25 mM de NH4HCO3 solo. Las muestras se removieron brevemente durante 10 min. Las muestras se secaron inmediatamente después durante una noche en un concentrador al vacío SpeedVac (Savant, Farmingdale, NY). Las muestras se disolvieron entonces en 16 pl de tripsina modificada de calidad de secuenciación a 5 pg/ml (Promega) y se incubaron a 37 °C durante 18 horas. Las muestras se almacenaron a 4 °C durante una noche y después se analizaron mediante MALDI-TOF MS (véase posteriormente).
Incubación de dHSA con el mAb seguido por escisión enzimática con tripsina con ultrafiltración en condiciones ácidas para el mapeo del epítopo
La HSA desnaturalizada (80 pg) se incubó con el mAb A (10 pg) en PBS durante 18 h a temperatura ambiente. Para retirar la dHSA libre, la mezcla de reacción dHSA-mAb se centrifugó durante 5 min a 3000 rpm en un ultrafiltrador Amicon Ultra-15 con un corte de peso molecular de 100.000 Da (Millipore Co., Billerica, MA). El retenido se diluyó en 25 mM de NH4HCO3 y se centrifugó de nuevo como se ha descrito anteriormente. El retenido se transfirió a un tubo de microcentrífuga Eppendorf en 0,4 ml de NH4HCO325 mM y se añadieron 0,4 pg de tripsina modificada de calidad de secuenciación (Promega). Se llevó a cabo la digestión a 37 °C durante una noche con agitación suave. La tripsina y los fragmentos de albúmina libres (no unidos al anticuerpo) se retiraron mediante ultrafiltración en un filtro Amicon Ultra-4 (corte de pm de 30.000 Da, Millipore Co.) durante 5 min a 3000 rpm. Esto se repitió tres veces. El retenido se transfirió a un nuevo ultra filtro donde se disoció el mAb A de su unión con la albúmina mediante la adición de 600 pl de glicina-HCl 0,1 M, pH 2,7 durante 30 min a temperatura ambiente tras lo cual se centrifugó el ultra filtro durante 10 min a 3000 rpm. El filtrado se transfirió a un tubo de microcentrífuga estéril y se neutralizó con Tris-HCl, pH 9. La muestra se secó inmediatamente durante una noche en un concentrador al vacío SpeedVac. Las muestras se
disolvieron entonces en 16 |jl de tripsina modificada (a 5 |jg/ml) (Promega) y se incubaron a 37 °C durante 18 horas. Se utilizaron puntas de pipeta Zip Tip (Millipore) que contenían medio de cromatografía ds para la desalación antes de analizar la muestra por MALDI-TOF ms (véase posteriormente).
Espectrometría de masas por MALDI-TOF
Se mezcló 1 j l de cada muestra de la digestión tríptica con 1 j l de una solución saturada de ácido a-ciano-4-hidroxicinámico (0,02 mg/ml) en un 70 % de acetonitrilo/0,3 % de ácido trifluoroacético. 1 j l de esa muestra se distribuyó puntualmente sobre una placa diana de acero inoxidable y se analizó utilizando la MALDI-TOF ms (Voyager-DE PRO, Applied Biosystems, CA, US) equipado con un láser N2 de 337 nm. Las búsquedas en la base de datos correspondientes con la seroalbúmina humana en los espectros resultantes se llevaron a cabo en el NCBI o SwissProt con el MS-Fit como motor de búsqueda.
Péptidos de albúmina
Todos los péptidos de albúmina sintéticos que se utilizaron en el presente documento se prepararon a medida en CSBio Co, Menlo Park, CA. Los péptidos eran > 95 % puros como se confirmó por HPLC. Los péptidos se mantuvieron secados por congelación a menos 20 °C. Los péptidos se reconstituyeron en H2O estéril (Sigma) para su uso en un ELISA o en RPMI 1640 (GIBCO) para su uso en los experimentos de cultivo celular. Los péptidos se esterilizaron por filtración a través de un filtro de jeringa de 0,22 jm (Millipore Co) antes de su uso en experimentos de cultivo celular.
ELISA anti-dHSA, co-incubación de mAb A anti dHSA con los péptidos de albúmina sintéticos Se revistieron por duplicado los pocillos de placas de microtitulación unidos a Hi (Costar 2592, Corning Inc., NY, USA) con 100 j l de dHSA diluida en PBS a 4,5 jg/m l y se incubaron a temperatura ambiente durante una noche. Los pocillos se lavaron entonces con tampón de lavado que consistía en un 0,05 % de Tween-20 en PBS (Sigma) seguido por bloqueo durante 1 h a 25 °C con 200 j l de gelatina al 0,5 % preparada a partir de piel bovina (Sigma) en PBS seguido por lavado en tampón de lavado. Se preincubó el anticuerpo monoclonal mAb A, diluido en el diluyente del reactivo de ELISA (gelatina al 0,01 % y un 0,05 % de Tween-20 en 20 mM de solución salina tampón Tris (TBS Sigma)) a 4 jg/m l durante 1 h a temperatura ambiente con las concentraciones indicadas de péptido. Se añadieron entonces 100 j l del anticuerpo monoclonal solo, o de manera alternativa, el anticuerpo monoclonal mezclado con los péptidos, a cada pocillo y se incubaron durante 1,5 h a 25 °C seguido por un lavado. Se añadió Envision-HRP (DakoCytomation Norden A/S, Glostrup, Dinamarca) diluido 1/10 en el diluyente del reactivo de ELISA y se incubaron las placas durante 15 min a 25 °C seguido por un lavado. Finalmente, se añadió una solución de sustrato que consistía en H2O2 y tetrametilbencidina (R&D Systems Europe, Ltd, Abingdon, UK). La reacción se detuvo con 1 M de H2SO4 y se midió la densidad óptica como la absorbancia (A) a las longitudes de onda duales, 450 nm y 570 nm, con un lector de microplacas Multiscan EX (Labsystems).
Correlación de la tinción inmunohistológica de las biopsias con un mAb anti-dHSA (mAb A) y supervivencia en pacientes con melanoma maligno
Las biopsias de tumores obtenidos de veinte pacientes diagnosticados con un melanoma maligno metastático se congelaron instantáneamente en nitrógeno líquido y se almacenaron a -70 °C hasta su uso. Se cortaron secciones de tejido congelado de 6-7 jm de grosor, se descongelaron y se fijaron con acetona durante 5 min a temperatura ambiente. Las secciones se bloquearon primero con un 10% de suero AB humano normal durante 1 h antes de la tinción. Entonces se añadió el anticuerpo primario que consistía en un anticuerpo de ratón anti-albúmina humana desnaturalizada (mAb-A) diluido en solución salina tampón Tris (TBS, pH 7,6) a 10 jg/ml, y se incubaron los portaobjetos durante 30 min. Los portaobjetos se lavaron en TBS seguido por Envision-Alkaline Phosphatase (DakoCytomation) durante 30 min. Después de un lavado adicional en TBS, se incubaron los portaobjetos en un sustrato de fosfatasa alcalina que consistía en una sal Fast Red TR (Sigma) naftol AS-MX (Sigma) y 5 mM de levamisol (Sigma) para bloquear la actividad de la fosfatasa alcalina endógena, durante 20 min seguido por un lavado en TBS. Se contra tiñeron en hematoxilina de Mayer durante 1 minuto y se montaron en Glycergel (Dakopatts). La IgG1 de ratón monoclonal contra un antígeno irrelevante (glucosa oxidasa de Aspergillus niger, DakoCytomation), se utilizó como control negativo. Todas las incubaciones se llevaron a cabo a temperatura ambiente en una cámara de humedad. La intensidad de la tinción se evaluó en un microscopio óptico y se clasificó como baja, media o alta. Se analizó la supervivencia entre los grupos de acuerdo con Kaplan Meier y un análisis de rango logarítmico.
Aislamiento de células mononucleares de sangre periférica (PBMC)
Se extrajo sangre venosa de voluntarios sanos o de pacientes de cáncer en tubos de cristal al vacío con una solución A de citrato de dextrosa ácido como anticoagulante (Vacutainer, Becton Dickinson, Franklin Lakes, NJ). Se retiraron los eritrocitos mediante sedimentación en una solución de dextrano T500 al 2 % (Amersham Pharmacia Biotech AB, Uppsala, Suecia) en un 0.9% de NaCI (esta etapa se omitió en los cultivos con estimulación con PHA, véase posteriormente). Entonces se aislaron las PBMC mediante centrifugación en un gradiente de densidad Ficoll-paque Plus (GE Healthcare Bio-Sciences AB, Uppsala, Suecia) tras lo cual, las células se lavaron dos veces en RPMI 1640 con modificación de Dutch (Gibco, Invitrogen AB, Stockholm, Suecia) con un 2 % de seroalbúmina humana (HSA) (Pharmacia & Upjohn, Stockholm, Suecia) (RPMI/2 % HSA). Para los cultivos celulares con estimulación con PHA, se
lavaron las PBMC en solución salina equilibrada de Hank (HBSS) con un 10% de plasma autólogo en vez del RPMI/2 % HSA. Se evaluó la viabilidad celular mediante exclusión en un 0,05 % de azul tripano y era siempre por encima del 95 %. La suspensión celular se tiñó con solución de Turk y se hizo el recuento del número de linfocitos y monocitos en la preparación de PBMC en un hemocitómetro. Las PBMC se suspendieron en RPMI/2 % HSA y se ajustó la concentración celular a 5 x 105 linfocitos/ml.
Suero
Se recolectó suero humano en tubos de recolección de suero sin aditivos (Vacutainer, Becton Dickinson, Franklin Lakes, NJ) al mismo tiempo que las muestras de sangre para el aislamiento de las PBMC. Los sueros se inactivaron por calor a 56 °C durante 30 minutos.
Proliferación de PBMC inducida por PHA en presencia de péptidos de albúmina y/o dHSA
Se resuspendieron las PBMC de voluntarios sanos en RPMI 1640 con o sin la adición de dHSA y/o péptidos de albúmina, como se indica, a una concentración celular de 5 x 105 linfocitos/ml. Se sembraron 100 pl de la suspensión celular en placas de microtitulación de fondo redondo (Corning NY, USA) seguido por 100 pl de medio de cultivo que consistía en RPMI 1640 (Flow Laboratories, Irvine, Scotland) suplementado con 200 Ul/ml de Penicilina, 200 pg/ml de Estreptomicina (Flow laboratories) y un 20 % de suero autólogo inactivado por calor. Entonces se añadió Fitohematoaglutinina (PHA-P, Sigma Chemical Co, St. Louis, MO) a una concentración final de 5 o 10 pg/ml. Todas las condiciones del cultivo se prepararon en pocillos por triplicado. Las células se cultivaron durante 3 días en una atmósfera humidificada con un 5% de CO2 a 37 °C. Se ensayó la proliferación incorporando 1,6 pCi/pocillo de [3H]timidina (Amersham International, U K) durante las últimas 18 h. Se utilizaron los valores medios de las desintegraciones por minuto (dpm) de los triplicados para los cálculos.
Efecto de los péptidos sobre la producción de IL-6 inducida por LPS
Se añadieron 100 pl de medio de cultivo que consistía en RPMI 1640 suplementado con 200 Ul/ml de penicilina, 200 pg/ml de estreptomicina, 4 mM de L-glutamina (Sigma Chemical, MO, US) y un 20 % de suero autólogo inactivado por calor a las placas de microtitulación no revestidas o pre-revestidas con 100 pl de suspensión de PBMC (5 x 104 linfocitos) en RPMI/2 % HSA con los péptidos a las concentraciones indicadas. Se añadieron los liposacáridos (LPS, Sigma Chemical Co, MO, US) a una concentración final de 0,05 ng/ml. Las células se cultivaron en una atmósfera humidificada, con un 5 % de CO2 a 37 °C. Todas las condiciones del ensayo se prepararon en pocillos por triplicado. Se recolectaron los sobrenadantes (SN) tras 24 h y se retiraron las celas restantes mediante centrifugación en una centrífuga refrigerada (Beckman) a 2600 x g durante 5 minutos. Los SN se congelaron y almacenaron a -70 °C. La IL-6 de los SN del cultivo se midió mediante un ELISA utilizando el kit de desarrollo de ELISA DuoSet© para la IL-6 humana (R&D Systems Europe, Ltd., Abingdon, UK) siguiendo los procedimientos recomendados por el fabricante. El límite menor de detección era de 3,1 pg/ml. Las muestras se analizaron como medias de los pocillos triplicados.
Proliferación de PBMC inducida por interleucina-2 (IL-2) en presencia de péptidos de albúmina en placas de cultivo tisular revestidas o sin revestimiento
Se pre-revistieron placas de cultivo tisular de 96 pocillos de fondo redondo (Costar, Corning Inc. NY, US) solo con HSA o con HSA y un agrupamiento de IgG humanas para inyección intravenosa (Gammagard, Baxter AS, DK) de la siguiente manera; se diluyó la HSA en RPMI 1640 sin suplementos a una concentración de 10 mg/ml. También se preparó una mezcla de 1 mg/ml de IgG en una solución de 9 mg/ml de HSA en RPMI (HSA/IgG). Se añadieron entonces 200 pl de HSA o HSA/IgG a cada pocillo de la placa. Las placas se incubaron a 4 °C durante 30 minutos tras lo cual se lavaron los pocillos dos veces con 200 pl de RPMI 1640. Las placas revestidas se utilizaron inmediatamente. Se añadieron 100 pl de RPMI 1640 suplementado con 200 UI/ml de penicilina, 200 pl/ml de estreptomicina, 4 mM de L-glutamina (todos de Sigma Chemical Co. MO, US) y un 20 % de suero humano (autólogo) inactivado por calor a los pocillos de microtitulación de cultivo tisular revestidos con HSA o HSA/IgG. Las PBMC, aisladas de individuos sanos, se diluyeron en RPMI/2 % de HSA y se añadieron los péptidos directamente a la suspensión celular a una concentración de 10 pg/ml. Cien pl de esta suspensión celular (5 x 104 linfocitos) se añadieron entonces por pocillo dando una concentración final de 5 pg/ml de péptido por pocillo. Se añadió a los pocillos la IL-2 (Proleukin, Chiron, NL), a una concentración final de 120 UI/pocillo. Las células se cultivaron durante 7 días en una atmósfera humidificada con un 5 % de CO2 a 37 °C. Se ensayó la proliferación mediante la incorporación de 1,6 pCi/pocillo de [3H]-timidina (Amersham Int., UK) durante las últimas 18 h. Se utilizaron las medias de los valores de dpm (desintegraciones por minuto) de los triplicados para los cálculos.
Tinción inmunohistoquímica de PBMC y una línea celular monocítica humana con un anticuerpo anti-integrina (CD11a) en presencia o ausencia de péptidos de albúmina
Las PBMC se prepararon como se describió anteriormente. Las células THP-1 cultivadas (obtenidas en la colección americana de cultivos tipo a través de LGL Nordic AB, Suecia) se lavaron cuidadosamente y se suspendieron en RPMI 1640. Las PBMC y THP-1 se aglomeraron por centrifugación inmediatamente en portaobjetos de microscopio pre limpiados en un Cytospin Shandon (Shandon Scientific Ltd, UK) a 1000 rpm durante 7 min a 2,5 o 5 x 104 células por
portaobjetos. Los portaobjetos se dejaron secar a temperatura ambiente durante una noche, tras lo cual se envolvieron en parafilm y se almacenaron a -70 °C. Inmediatamente antes de su uso, los cytospin se descongelaron y se fijaron con acetona durante 5 min a temperatura ambiente. Los cytospin se bloquearon primero con un 10 % de suero AB humano normal con y sin los péptidos de albúmina (40 □g/ml) durante 1 h antes de la tinción. Se añadió el anticuerpo primario, que consistía en un anticuerpo monoclonal de ratón anti-CD11a humano (clon HI111, BD Biosciences) diluido en solución salina tampón Tris (TBS, pH 7,6) a 5 pg/ml (de THP-1) o 1 □g/ml (PBMC). Los portaobjetos se incubaron durante 30 min y entonces se lavaron en TBS seguido por Envision-Alkaline Phosphatase (Dako Norden A/S, Dinamarca) durante 30 min. después de un lavado adicional en TBS, se incubaron los portaobjetos en un sustrato de fosfatasa alcalina que consistía en sal TR Fast Red (Sigma, naftol AS-MX (Sigma) y 5 mM de levamisol (Sigma) para bloquear la actividad de la fosfatasa alcalina endógena, durante 20 min seguido por lavado en TBS. Se contra tiñeron entonces en hematoxilina de Mayer durante 1 minuto y se montaron en Glycergel (Dako Norden A/S). La IgG1 de ratón monoclonal contra un antígeno irrelevante (glucosa oxidasa de Aspergillus niger, Dako Norden A/S), se utilizó como control negativo. Todas las incubaciones se llevaron a cabo a temperatura ambiente en una cámara de humedad.
Fragmentación proteolítica de la seroalbúmina humana desnaturalizada (dHSA) con tripsina o endoproteinasa ASP-N
La dHSA secada por congelación (0,5 mg) se reconstituyó en 25 mM de NH4HCO3, pH 8, que contenía 10 mg de tripsina modificada de calidad de secuenciación (Promega Corporation, Wl) o 2 mg de Endoproteinasa ASP-N (Sigma) y se incubó a 37 °C durante una noche. Para retirar la albúmina no fragmentada y la enzima, la muestra se ultrafiltró a través de un filtro de centrífuga Amicon Ultra 4 (corte de pm de 5000) o a Centriplus (corte de pm de 10000) (Millipore AB, Solna, Suecia). El filtrado que contenía la dHSA fragmentada sin enzimas, se recolectó y se diluyó con PBS con Ca y Mg (GIBCO).
Preparación del lisado celular de PBMC con proteínas de superficie celular biotiniladas (ACS)
Las capas leucocitarias generadas de 450 ml de sangre se recolectaron cada una de 4 donantes sanos. Se retiraron los eritrocitos por sedimentación en una solución de un 2 % de dextrano T500 (Amersham Pharmacia Biotech AB, Uppsala Suecia) en un 0,9 % NaCI. Las células mononucleares (PBMC) se aislaron entonces por centrifugación en gradiente de densidad Ficoll-Paque Plus (GE Healthcare Bioscience AB Suecia). Las PBMC se suspendieron entonces en solución salina tampón fosfato (PBS) que contenía Ca y Mg (GIBCO) a una concentración de 10 x 106 /ml. Se añadió EZ Link Sulfo-NHS-biotin (Pierce USA) a una concentración final de 0,2 mg/ml y se incubó la mezcla en un agitador a temperatura ambiente durante 10 min. Se retiró entonces el exceso de biotina lavando las PBMC en PBS. Las PBMC biotiniladas se lisaron entonces añadiendo 1,0 ml de tampón de lisis enfriado en hielo (50 mM de Tris-HCl, pH 7,5, con 0,15 M de NaCl, 5 mM de MgCl2 que contenía 100 mM de octil glucósido y 1 mM de fenilmetilsulfonil fluoruro) por cada 2 x 107 de células aglomeradas con agitado suave, se incubaron después durante 30 min. sobre hielo. Los desechos se retiraron mediante centrifugación a 5000 x g a 4 °C durante 10 min y se recolectaron y agruparon los sobrenadantes de los cuatro donantes. El lisado se almacenó entonces a -70 °C en tubos de plástico de polipropileno.
Preparación de la columna de afinidad con proteínas de superficie celular biotiniladas de las células mononucleadas acopladas a estreptavidina-sepharose (utilizadas para la adsorción de la dHSA fragmentada con tripsina) 18 ml de lisado celular biotinilado en tampón de lisado se diluyeron 1/10 en tampón de unión (20 mM de NaH2PO4, 0,15 M de NaCI, pH 7,5). Esta cantidad de lisado se corresponde con 36 x 107 células mononucleares. Se añadió a una columna de afinidad Hitrap Streptavidin HP de 1 ml (Amersham Biosciences) para bloquear la posible biotina libre restante, se añadieron 5 ml de glicina 0,1 M (Sigma) a la columna. La estreptavidina no saturada de la columna se hizo reaccionar entonces con 150 pg de biotina (Sigma) en el tampón de unión. La columna se lavó cuidadosamente con PBS y se almacenó en PBS con un 0,1 % de NaN3 a 4 °C hasta su uso.
Preparación de la columna de afinidad con proteínas de superficie celular biotiniladas de las células mononucleadas acopladas a estreptavidina-sepharose (utilizadas para la adsorción de la dHSA fragmentada con ASP-N) El lisado celular biotinilado en tampón de lisado se sometió a un intercambio de tampón mediante diálisis en tubo con Spectrapore 4 (Spectrum Europe, Breda, Países Bajos) en tampón de unión (20 mM de NaH2PO4, 0,15 M de NaCI, pH 7,5). 27 ml de lisado celular biotinilado en tampón de unión (correspondiente a 54 x 107 células mononucleares) se añadieron a 1,5 ml de Sepharose Streptavidin HP lavada (Amersham Biosciences). Para bloquear la posible biotina libre restante, se añadieron 25 ml de glicina 0,1 M (Sigma) a la Sepharose Streptavidin. La estreptavidina no saturada se hizo reaccionar entonces con 225 pg de biotina (Sigma) en el tampón de unión. La Sepharose Streptavidin se lavó cuidadosamente con PBS. Un ml del lisado celular biotinilado acoplado a estreptavidina Sepharose se empaquetó entonces en una columna vacía (Tricorn Empty High Performance Column, Amersham Bioscience) y se lavó con solución salina tampón de fosfato (PBS) que contiene Ca y Mg (GIBCO).
Adsorción de la dHSA fragmentada con enzima utilizando una columna de afinidad con proteínas de superficie celular (ACS) biotiniladas
Dos ml de dHSA fragmentada por enzimas en PBS correspondientes a un total de 0,2 mg de proteína, se pasó sobre la columna de ACS preparada como se ha descrito anteriormente. El flujo continuo se recolectó teniendo en consideración la escasez de volumen y la dilución de la muestra adsorbida recolectando en pequeñas porciones de
0,2 ml. Se secaron treinta |jl de cada muestra, incluyendo la muestra de control que no se había adsorbido en una centrífuga Speed-Vac.
Espectrometría de masas
Las mezclas secadas se reconstituyeron en 10 j l de un 0,1 % de TFA. Se utilizaron puntas de pipeta Zip Tip (Millipore, USA) que contiene medio de C18 en fase inversa para desalar las muestras reconstituidas. Para el análisis de las muestras en el intervalo de peso de 700 - 3600 Da, un j l de cada muestra eluída Zip Tip se mezclaron con 1 j l de una solución saturada de ácido a-ciano-4-hidroxicinámico (0,02 mg/ml) en acetonitrilo al 70 % / ácido trifluoroacético al 0,3 %. Para el análisis de las muestras en el intervalo de peso de 1500 - 9000 Da, un j l de cada muestra eluída Zip Tip se mezcló con 1 j l de ácido sinapínico (ácido 3-metoxi-4-hidroxicinámico). 1 j l de la muestra se distribuyó puntualmente sobre la placa MALDI y se analizó utilizando la MALDI-TOF MS (Voyager-DE PRO, Applied Biosystems, CA, US). La búsqueda de identidad de los espectros resultantes se llevó a cabo en las bases de datos de SwissProt o NCBI utilizando MS-Fit.
Identificación de proteínas en el plasma humano que se une a ACS
Cromatografía de afinidad del plasma con ACS
Se obtuvo el plasma mediante plasmaféresis de un paciente diagnosticado de melanoma maligno. El plasma se congeló a -20 °C. Al descongelar el plasma se coaguló inmediatamente mediante la adición de CaCl2 hasta una concentración final de 13 mM. El coágulo de plasma gelificado se retiró mediante centrifugación a 3500 rpm durante 7 min a 4 °C. El plasma se dializó entonces extensamente contra PBS utilizando la diálisis en tubo Spectrapore 4 (Spectrum Europe, Breda, Países Bajos). Se preparó una columna de afinidad con proteínas de superficie celular biotiniladas de células mononucleares acopladas a estreptavidina-sepharose (ACS-sepharose) como se ha descrito anteriormente. Se incubaron 45 ml de plasma con la ACS-sepharose durante una noche a 4 °C con agitado suave. El plasma-ACS-sepharose se lavó extensamente con PBS. La ACS-sepharose lavada se almacenó durante cinco días a 4 °C en PBS con un 0,1 % de NaN3. La ACS-sepharose se empaquetó en una columna vacía (Tricorn Empty High Performance Column, Amersham Bioscience) y las proteínas unidas se eluyeron con 2 ml de Glicina-HCl 0,1 M, pH 2,7. La fracción eluída se neutralizó inmediatamente con 1 M de tampón Tris a un pH 9 y se secaron por congelación.
Electroforesis en gel 2-D
Las muestras secadas por congelación se desalaron reconstituyéndolas con H2O y un 10 % de ácido tricloroacético (TCA) con 20 mM de DTT en acetona. Las muestras se centrifugaron a 13000 RPM durante 5 min. El aglomerado resultante se lavó dos veces con 20 mM de DTT en acetona para retirar el TCA y finalmente se disolvieron en tampón de rehidratación (8 M de urea, un 4 % de CHAPS, 10 mM de DTT, un 0,5 % de tampón IPG y una traza de naranja G). Se llevó a cabo la electroforesis en gel bidimensional en una construcción 2-D horizontal (Multiphore/IPGphore, Pharmacia Biotech, SE) basándose en un enfoque isoeléctrico (IEF) en la primera dimensión y el peso molecular en la segunda dimensión. En resumen, las muestras se aplicaron en geles IPG (Immobiline™ Dry strip, pH 3-10 NL, (GE Healthcare)) y se enfocan durante una noche a 38060 Vh. Entonces se llevó a cabo la s DS-PAGe con geles de poliacrilamida Exelgel XL SDS 12-14 con plancha prefundida (Amersham Biosciences). Se incluyeron referencias de pesos moleculares en cada ejecución. Las proteínas separadas se detectaron mediante tinción con plata de acuerdo con el método de Shevenco. Las manchas de proteína digeridas con tripsina se escindieron del gel y se analizaron con el MALDI-TOF ms como se ha descrito anteriormente.
Pacientes de cáncer
Los pacientes incluidos en el análisis de supervivencia total de acuerdo con la respuesta proliferativa de las células mononucleares de sangre periférica (PBMC) a la interleucina-2, se habían diagnosticado de carcinoma de células renales metastático sistémico. No se habían tratado previamente y se había programado un tratamiento con interleucina-2 (Proleukin, Chiron, NL). Se tomaron muestras de sangre antes del inicio del tratamiento. Los pacientes incluidos en otros estudios en la presente solicitud de patente se describen brevemente según sea adecuado en la sección de resultados.
Modelo ex vivo de inmunosupresión relacionada con IL-2; proliferación de PBMC inducida por interleucina (IL-2)
Se aislaron las PBMC a partir de muestras de sangre venosa de donantes de sangre sanos (controles) o pacientes de cáncer. Se añadieron cien j l de medio de cultivo (RPMI 1640 con la modificación de Dutch (Gibco, Invitrogen AB, Stockholm, Suecia) suplementado con 200 UI/ml de penicilina, 200 jl/m l de estreptomicina, 4 mM de L-glutamina (todos de Sigma Chemical Co. MO, US) y un 20 % de suero humano inactivado por calor) a placas de cultivo tisular de 96 pocillos con el fondo redondeado (Costar, Corning Inc. NY, US). Se añadieron entonces cien j l de PBMC en RPMI/2 % de HSA (5 x 104 linfocitos) por pocillo seguido por IL-2 (Proleukin, Chiron, NL) a una concentración final de 120 UI/pocillo. Los pocillos de control sin IL-2 se construyeron en paralelo. Las células se cultivaron durante 7 días en una atmósfera humidificada con un 5 % de CO2 a 37 °C. Se ensayó la proliferación mediante la incorporación de 1,6 jCi/pocillo de [3H]-timidina (Amersham Int., UK) durante las últimas 18-24 h. Se utilizaron las medias de los valores
de dpm (desintegraciones por minuto) de los pocilios triplicados para los cálculos. En los cultivos en los que se recolectó el suero de pacientes de cáncer se utilizaron en vez del suero autólogo, las PBMC eran de donantes compatibles con el grupo sanguíneo.
La interleucina-2 (IL-2) inducía la proliferación de PBMC en presencia de péptidos de albúmina Los cultivos de proliferación inducida por IL-2 se construyeron con PBMC de donantes sanos y suero autólogo como se ha descrito anteriormente con la excepción de que las PBMC se preincubaron primero durante 30 min a temperatura ambiente con los péptidos de albúmina indicados a una concentración de 10 pg/ml.
Generación de antisuero de conejo específico para el péptido de albúmina 3028
Se sintetizó el péptido 3028 con una cisteína añadida en el extremo N y luego se conjugó con hemocianina de lapa californiana (k Lh ) como proteína portadora. Se generaron los sueros policlonales mediante inmunizaciones repetidas de conejos con el péptido 3028 conjugado con KLH y adyuvantes de Freund. En algunos experimentos, los antisueros estaban purificados por afinidad mediante cromatografía sobre geles de yodoacetilo Ultralink conjugados con péptido 3028 (Pierce Biotechnology Inc.). Para los experimentos de cultivo celular, se llevó a cabo el intercambio de tampón de RPMI 1640 con modificación de Dutch (Gibco, InVitrogen AB, Stockholm, Suecia) por el pasaje sobre columnas de sephadex PD-10 (Amersham Biosciences, Uppsala, Suecia) seguido por esterilización por filtrado sobre filtros de jeringa Millex de 0,22 pm (Millipore Co., MA, USA). Las inmunizaciones de los conejos y purificación de los antisueros se llevaron a cabo en Agrisera AB, Suecia.
ELISA con el antisuero anti-3028 de conejo
Se revistieron pocillos por duplicado en placas de microtitulación Hi-binding (Costar 2592, Corning Inc, NY, USA) con 100 pl de péptido 3028 (10 pg/ml), HSA desnaturalizada (dHSA, 4,5 pg/ml) o HSA de control (4,5 pg/ml). Todos los reactivos de revestimiento se diluyeron en PBS y se incubaron a temperatura ambiente durante una noche. Los pocillos se lavaron entonces con tampón de lavado que consistía en un 0,05 % de Tween-20 en PBS (Sigma) seguido por bloqueo durante 1 h a 25 °C con 200 pl de gelatina al 0,5 % preparada a partir de piel bovina (Sigma) en PBS seguido por lavado en tampón de lavado. Los sueros preinmunes o suero anti-3028, diluido 1/1000000 en diluyente de reactivo de ELISA (un 0,01 % de gelatina y un 0,05 % de Tween-20 en PBS), se añadieron e incubaron durante 1 h a 25 °C seguido por un lavado. Se añadió entonces un anticuerpo de caballo anti-IgG de conejo/ratón (Vectastain ELITE, Vetor Laboratories lnc; CA, USA) diluido 1/5 en diluyente de reactivo ELISA y se incubaron las placas durante 1 h a 25 °C seguido por un lavado. A continuación, se añadió estreptavidina conjugada con HRP (R&D systems Europe, Ltd, UK). Finalmente, después del lavado en tampón de lavado, se añadió una solución de sustrato que consistía en H2O2 y tetrametilbencidina (R&D Systems). La reacción se detuvo con 1 M de H2SO4 y se midió la densidad óptica como la absorbancia (A) a las longitudes de onda duales, 450 nm y 570 nm, con un lector de microplacas Multiscan EX (Labsystems).
ELISA de inhibición de anti-3028 de conejo por péptidos de albúmina
Para ensayar si los péptidos de albúmina inhibían la unión del anti-3028 de conejo a los pocillos revestidos de 3028, el antisuero de conejo diluido 1/1000000 en diluyente de reactivo de ELISA, se preincubó durante 1 h a temperatura ambiente con las concentraciones indicadas de los péptidos. Se añadieron entonces 100 pl del anticuerpo monoclonal solo, o de manera alternativa, el anticuerpo monoclonal mezclado con los péptidos, a los pocillos revestidos con 3028 y se llevó a cabo el ELISA como se ha descrito.
Proliferación de PBMC inducida por interleucina-2 (IL-2) en presencia de anti-3028 de conejo
Los cultivos para ensayar el efecto de inmunomodulación de anticuerpos de conejo purificados por afinidad específicos de 3028 se llevaron a cabo como se ha descrito anteriormente para la proliferación inducida por IL-2 con las siguientes excepciones; se omitió el 2 % de HSA del medio de lavado y del medio de suspensión de PBMC. El medio de cultivo contenía suero (100 pl/pocillo) se preincubó con 20 pg/ml de anticuerpos de conejo durante 30 min a temperatura ambiente antes de la adición de 100 pl de suspensión de PBMC a los pocillos de cultivo.
Tinción inmunohistoquímica de biopsias tumorales con anti-3028 de conejo
Se prepararon las secciones de tejido a partir de biopsias fijadas en formalina de pacientes de cáncer. Se desparafinaron las secciones y se bloquearon con un 10 % de suero AB humano normal en solución salina equilibrada de Hank suplementada con 0,01 M de Hepes (BSS, GIBCO BRL) durante una h antes de la tinción. Se tiñeron entonces las secciones con 10 pg/ml de anti-3028 de conejo purificado diluido en BSS con un 2 % de suero AB y 0,1 g/ml de saponina durante 30 min. Después del lavado en b Ss con 0,1 g/ml de saponina, se añadió un polímero de fosfatasa alcalina Ultravison One específico para Ig de ratón y conejo (Lab Vision Co., CA, USA). Se lavó entonces el exceso de polímero de las secciones con BSS con 0,1 g/ml de saponina. La unión del complejo de polímero se detectó mediante un sustrato de naftol fosfato y cromógeno líquido Fast Red (Lab Vision Corp.). Las secciones se contra tiñeron con hematoxilina de Mayer y se montaron en Glycergel.
Efecto de los péptidos de albúmina sobre la actividad citotóxica de células cito líticas naturales (NK) de donantes de sangre sanos
Se separaron las células mononucleares mediante centrifugación en gradiente de densidad Ficoll-paque Plus (Pharmacia AB, Suecia) convencional a partir de la sangre heparinizada obtenida de donantes sanos. Se ensayó entonces la actividad citotóxica de células NK de las células mononucleares utilizando un kit comercial (NKTEST, Orpegen Pharma GmbH, Heidelberg, Alemania) siguiendo el protocolo de los fabricantes. En resumen, el kit contiene células diana sensibles a NK crioconservadas (K562) marcadas con un colorante lipófilo de membrana fluorescente verde, que hace posible la discriminación entre células efectoras y diana. Después de la incubación con las células efectoras, las células diana destruidas se identifican mediante una tinción de ADN, que penetra y tiñe específicamente los núcleos de las células diana muertas. De esta manera se puede determinar el porcentaje de dianas destruidas mediante citometría de flujo. Las células mononucleares se preincubaron durante 30 min a 37 °C con los péptidos indicados (los péptidos se han descrito previamente) a 10 pg/ml. Las células diana se añadieron entonces, dando una relación de efector:diana de 40:1, y se incubó la mezcla celular a 37 °C durante 3-4 horas. Se analizaron las muestras en un FACSCalibur (BD Biosciences, San Jose, CA).
LEYENDAS DE LAS FIGURAS
Figura 1. Este diagrama muestra que algunos péptidos que contienen la secuencia E5K inhiben la unión del mAb A a placas de ELISA revestidas con dHSA. Obviamente, hay algunas diferencias en la actividad inhibidora de los péptidos ensayados.
Figura 2. Efecto de la expresión de E5K detectado por mAb A sobre la supervivencia de pacientes con melanoma maligno metastático (p = 0,009).
Baja expresión WWziWí x - - x
Alta expresión RHBfl o - - o
Figura 3 A. Se ensayó el efecto de dos péptidos de albúmina, 2605 y 2608, sobre la proliferación de PBMC inducida por PHA (5 pg/ml) de un control sano y dos pacientes de cáncer. Las tres diferentes concentraciones de los péptidos (pg/ml) se indican en la figura. Los pacientes P45 y 46 sufrían carcinoma de células renales y melanoma maligno Figura 3 B. Se ensayó el efecto de dos péptidos de albúmina, 2605 y 2608, sobre la proliferación de PBMC inducida por PHA (10 pg/ml) de un control sano y dos pacientes de cáncer. Las tres concentraciones de los péptidos de (pg/ml) que se utilizan se indican en la figura.
Figura 4. Variación interindividual en el efecto de los péptidos de albúmina sobre la proliferación de PBMC inducida por PHA de controles sanos y pacientes de cáncer. Se utilizó PHA a una concentración de 5 Cpg/ml y los péptidos a una concentración de 10 pg/ml. Los pacientes tenían los siguientes diagnósticos: Melanoma maligno (P46) y carcinoma de células renales (P39, P41, y P45).
Figura 5. Efecto de dHSA, 8 pg/ml, sobre la proliferación de PBMC inducida por PHA de 5 controles y 4 pacientes de cáncer.
Figura 6 A. Efecto del péptido 2605 sobre la proliferación inducida por PHA aumentada por dHSA. Los detalles se describen en el texto.
Figura 6 B. Efecto del péptido 2608 sobre la proliferación inducida por PHA aumentada por dHSA. Los detalles se describen en el texto.
Figura 7. Efecto de los péptidos de albúmina sobre la producción de IL-6 inducida por LPS por las PBMC de A) un donante sano, B) y C) pacientes de cáncer con melanoma maligno. Las PBMC se estimularon con LPS durante 24 h en presencia de los péptidos indicados a 0,01 pg/ml (barras negras), 0,1 pg/ml (barras grises), 1 pg/ml (barras blancas) o 10 pg/ml (barras sombreadas). La cantidad de IL-6 liberada en el medio de cultivo se midió por ELISA. Los resultados se expresan como el porcentaje de IL-6 liberado en los cultivos de control sin la adición de péptidos. Esto era 1072 pg/ml, 997 pg/ml y 902 pg/ml en A), B), y C), respectivamente.
Figura 8 A-F. Efecto de los péptidos de albúmina sobre la unión de anticuerpos a LFA-1 en células inmunitarias. La línea celular monocítica THP-1 (paneles A y B), PBMC de dos controles sanos (paneles C, D y E, F, respectivamente). El péptido 2606 se utilizó en los experimentos que se muestran en B y D y el péptido se utilizó en 2605 en F.
Figura 9. Efecto de los péptidos sobre la proliferación de PBMC inducida por p Ha modulada por dHSA.
Figura 10. Efecto de la nueva serie de péptidos sobre la proliferación inducida por IL-2 (A) y la inversa de la actividad inhibidora del péptido 3028 por la modulación del receptor Fc (B).
Figura 11. Efecto de la nueva serie de péptidos sobre la producción de IL-6 inducida por LPS de las PBMC de dos controles sanos.
Figura 12. Efecto de la nueva serie de péptidos sobre la producción de IL-6 inducida por LPS por las PBMC de tres pacientes de cáncer con melanoma maligno (MM) y con carcinoma de células renales (RCC).
Figura 13. Electroforesis en gel de 2-D de proteínas del plasma después de la cromatografía de afinidad en ACS. El plasma se obtuvo de un paciente con melanoma maligno.
Figura 14. La proliferación de PBMC inducida por IL-2 de controles sanos y PBMC de pacientes de carcinoma de células renales (RCC) cultivadas en un 10 % de suero autólogo.
Figura 15. Un análisis de Kaplan-Meier de pacientes con carcinoma de células renales de acuerdo con la respuesta proliferativa a IL-2. Una tasa proliferativa baja indica una pobre supervivencia.
Figura 16. Cultivo de los PBMc de controles sanos en los sueros de pacientes de cáncer que se sabe que tienen una
Claims (14)
1. Un fragmento de seroalbúmina humana (HSA) que comprende la secuencia de aminoácidos SEQ ID NO: 81 para su uso en el tratamiento de una afección inflamatoria, en donde dicho fragmento de HSA tiene 10-50 aminoácidos de longitud.
2. El fragmento de seroalbúmina humana (HSA) para su uso de acuerdo con la reivindicación 1, en donde la afección inflamatoria se selecciona de entre el grupo que consiste en psoriasis, rechazo de aloinjerto, enfermedad del injerto contra el huésped (GVH), lesión de isquemia-reperfusión, una enfermedad inflamatoria crónica y una enfermedad autoinmunitaria.
3. El fragmento de seroalbúmina humana (HSA) para su uso de acuerdo con la reivindicación 1, en donde dicho fragmento de HSA comprende la SEQ ID n O: 80.
4. El fragmento de seroalbúmina humana (HSA) para su uso de acuerdo con la reivindicación 3, en donde la afección inflamatoria se selecciona de entre el grupo que consiste en psoriasis, rechazo de aloinjerto, enfermedad del injerto contra el huésped (GVH), lesión de isquemia-reperfusión, una enfermedad inflamatoria crónica y una enfermedad autoinmunitaria.
5. Un péptido que consiste en la secuencia de aminoácidos de SEQ ID NO: 80 para su uso en el tratamiento de una afección inflamatoria.
6. El péptido para su uso de acuerdo con la reivindicación 5, en donde la afección inflamatoria se selecciona de entre el grupo que consiste en psoriasis, rechazo de aloinjerto, enfermedad del injerto contra el huésped (GVH), lesión de isquemia-reperfusión, una enfermedad inflamatoria crónica y una enfermedad autoinmunitaria.
7. Un péptido que consiste en la secuencia de aminoácidos de SEQ ID NO: 81 para su uso en el tratamiento de una afección inflamatoria.
8. El péptido para su uso de acuerdo con la reivindicación 7, en donde la afección inflamatoria se selecciona de entre el grupo que consiste en psoriasis, rechazo de aloinjerto, enfermedad del injerto contra el huésped (GVH), lesión de isquemia-reperfusión, una enfermedad inflamatoria crónica y una enfermedad autoinmunitaria.
9. El uso de un fragmento de seroalbúmina humana (HSA) que comprende la secuencia de aminoácidos SEQ ID NO: 81 para la fabricación de un medicamento para el tratamiento de una afección inflamatoria, en donde dicho fragmento de HSA tiene 10-50 aminoácidos de longitud.
10. El uso del fragmento de seroalbúmina humana (HSA) de acuerdo con la reivindicación 9, en donde dicho fragmento de HSA comprende la SEQ ID NO: 80.
11. El uso de un péptido que consiste en la secuencia de aminoácidos de SEQ ID NO: 80 para la fabricación de un medicamento para el tratamiento de una afección inflamatoria.
12. El uso del péptido de acuerdo con una cualquiera de las reivindicaciones 9 -11, en donde la afección inflamatoria se selecciona de entre el grupo que consiste en psoriasis, rechazo de aloinjerto, enfermedad del injerto contra el huésped (GVH), lesión de isquemia-reperfusión, una enfermedad inflamatoria crónica y una enfermedad autoinmunitaria.
13. El uso de un péptido que consiste en la secuencia de aminoácidos de SEQ ID NO: 81 para la fabricación de un medicamento para el tratamiento de una afección inflamatoria.
14. El uso del péptido de acuerdo con la reivindicación 13, en donde la afección inflamatoria se selecciona de entre el grupo que consiste en psoriasis, rechazo de aloinjerto, enfermedad del injerto contra el huésped (GVH), lesión de isquemia-reperfusión, una enfermedad inflamatoria crónica y una enfermedad autoinmunitaria.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| SE0701099 | 2007-05-08 | ||
| SE0701100 | 2007-05-08 | ||
| SE0702520 | 2007-11-15 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2747363T3 true ES2747363T3 (es) | 2020-03-10 |
Family
ID=39709034
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES16189090T Active ES2747363T3 (es) | 2007-05-08 | 2008-05-08 | Estructuras inmunorreguladoras de proteínas que se encuentran normalmente |
| ES12164229.2T Active ES2608803T3 (es) | 2007-05-08 | 2008-05-08 | Estructuras inmunorreguladoras procedentes de proteínas naturales |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES12164229.2T Active ES2608803T3 (es) | 2007-05-08 | 2008-05-08 | Estructuras inmunorreguladoras procedentes de proteínas naturales |
Country Status (5)
| Country | Link |
|---|---|
| US (4) | US9120874B2 (es) |
| EP (4) | EP3636667A3 (es) |
| DK (2) | DK2487186T3 (es) |
| ES (2) | ES2747363T3 (es) |
| WO (1) | WO2008136736A2 (es) |
Families Citing this family (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP1507548B1 (en) * | 2002-05-27 | 2014-10-01 | CanImGuide Therapeutics AB | Method for determining immune system affecting compounds |
| SE0402536D0 (sv) | 2004-10-20 | 2004-10-20 | Therim Diagnostica Ab | Immunoregulation in cancer, chronic inflammatory and autoimmune diseases |
| EP1875246B1 (en) | 2005-04-15 | 2016-10-05 | CanImGuide Therapeutics AB | Diagnostic method for detecting cancer by measuring amount of a cytokine like il-6 |
| WO2008136736A2 (en) | 2007-05-08 | 2008-11-13 | Canimguide Therapeutics Ab | Immunoregulatory structures from normally occurring proteins |
| US8455624B2 (en) * | 2011-06-03 | 2013-06-04 | University Of South Alabama | Methods and compositions for detecting endometrial or ovarian cancer |
| CN111732652A (zh) * | 2013-09-09 | 2020-10-02 | 卡尼姆盖德治疗学公司 | 免疫系统调节器 |
| CN103694342B (zh) * | 2013-11-12 | 2015-11-25 | 北京理工大学 | 检测人老龄化的多肽标志物 |
| WO2016131962A1 (en) * | 2015-02-20 | 2016-08-25 | Pharis Biotec Gmbh | Polypeptide with activity against diphtheria toxin |
| CA2978617A1 (en) * | 2015-03-06 | 2016-09-15 | Canimguide Therapeutics Ab | Immune system modulators and compositions |
| CN112789030A (zh) | 2018-09-28 | 2021-05-11 | 卡尼姆盖德治疗学公司 | 肽抑制剂的药物制剂 |
| CN109294998B (zh) * | 2018-09-30 | 2020-11-06 | 北京鼎成肽源生物技术有限公司 | 一种rff1细胞 |
| CN109294982B (zh) * | 2018-09-30 | 2020-11-06 | 北京鼎成肽源生物技术有限公司 | 一种rff2细胞 |
| CN111499746B (zh) * | 2020-04-28 | 2020-11-24 | 优睿赛思(武汉)生物科技有限公司 | 一种针对人白介素-2的高亲和力兔单克隆抗体及其应用 |
Family Cites Families (21)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS6216430A (ja) | 1985-07-12 | 1987-01-24 | Ajinomoto Co Inc | インタ−ロイキン2レセプタ−発現誘導物質a |
| DE378732T1 (de) | 1989-01-20 | 1991-05-02 | Aziende Chimiche Riunite Angelini Francesco A.C.R.A.F. S.P.A., Rom/Roma, It | Serum-proteine in bezug auf autoimmunerkrankungen. |
| CA2015515C (en) | 1990-01-03 | 1999-12-07 | Jean-Marie Saint-Remy | Pharmaceutical compositions containing antigen-antibody complexes and uses therefor |
| US6737057B1 (en) | 1997-01-07 | 2004-05-18 | The University Of Tennessee Research Corporation | Compounds, compositions and methods for the endocytic presentation of immunosuppressive factors |
| US5948609A (en) * | 1997-12-03 | 1999-09-07 | Carter; Daniel C. | Oxygen-transporting albumin-based blood replacement composition and blood volume expander |
| AU1243500A (en) | 1998-11-06 | 2000-05-29 | Emory University | Biomarkers for oxidative stress |
| EP1326892A2 (en) | 2000-10-12 | 2003-07-16 | University of Rochester | Compositions that inhibit proliferation of cancer cells |
| AU2002312410A1 (en) | 2001-06-08 | 2002-12-23 | Target Protein Technologies, Inc. | Tissue-specific endothelial membrane proteins |
| EP1507548B1 (en) | 2002-05-27 | 2014-10-01 | CanImGuide Therapeutics AB | Method for determining immune system affecting compounds |
| AU2003298689A1 (en) | 2002-11-21 | 2004-06-18 | Wyeth | Methods for diagnosing rcc and other solid tumors |
| WO2004082617A2 (en) * | 2003-03-14 | 2004-09-30 | Ppd Biomarker Discovery Sciences, Llc | Biological markers for diagnosing rheumatoid arthritis |
| JP4068538B2 (ja) | 2003-09-12 | 2008-03-26 | 株式会社サン・クロレラ | サイトカイン遊離阻害剤 |
| SE0402536D0 (sv) | 2004-10-20 | 2004-10-20 | Therim Diagnostica Ab | Immunoregulation in cancer, chronic inflammatory and autoimmune diseases |
| KR100664587B1 (ko) | 2004-12-22 | 2007-01-04 | 김현기 | 인간 암 억제 유전자, 이에 의해 코딩되는 단백질, 이를포함하는 발현 벡터 및 이 벡터로 형질전환된 세포 |
| EP1875246B1 (en) | 2005-04-15 | 2016-10-05 | CanImGuide Therapeutics AB | Diagnostic method for detecting cancer by measuring amount of a cytokine like il-6 |
| US8232066B2 (en) | 2006-07-03 | 2012-07-31 | The Johns Hopkins University | Peptide antibody depletion and its application to mass spectrometry sample preparation |
| WO2008136736A2 (en) | 2007-05-08 | 2008-11-13 | Canimguide Therapeutics Ab | Immunoregulatory structures from normally occurring proteins |
| RU2360924C1 (ru) | 2007-12-21 | 2009-07-10 | Институт биохимии им. А.А. Баха РАН | Маркер рака предстательной железы |
| US8455624B2 (en) | 2011-06-03 | 2013-06-04 | University Of South Alabama | Methods and compositions for detecting endometrial or ovarian cancer |
| CN111732652A (zh) | 2013-09-09 | 2020-10-02 | 卡尼姆盖德治疗学公司 | 免疫系统调节器 |
| CN103694342B (zh) | 2013-11-12 | 2015-11-25 | 北京理工大学 | 检测人老龄化的多肽标志物 |
-
2008
- 2008-05-08 WO PCT/SE2008/000314 patent/WO2008136736A2/en active Application Filing
- 2008-05-08 EP EP19190281.6A patent/EP3636667A3/en not_active Withdrawn
- 2008-05-08 EP EP16189090.0A patent/EP3130602B1/en active Active
- 2008-05-08 ES ES16189090T patent/ES2747363T3/es active Active
- 2008-05-08 US US12/599,484 patent/US9120874B2/en not_active Expired - Fee Related
- 2008-05-08 DK DK12164229.2T patent/DK2487186T3/da active
- 2008-05-08 EP EP12164229.2A patent/EP2487186B1/en not_active Not-in-force
- 2008-05-08 EP EP09178421A patent/EP2226334A3/en not_active Withdrawn
- 2008-05-08 ES ES12164229.2T patent/ES2608803T3/es active Active
- 2008-05-08 DK DK16189090T patent/DK3130602T3/da active
-
2015
- 2015-07-31 US US14/815,471 patent/US9796777B2/en active Active
-
2017
- 2017-08-02 US US15/667,058 patent/US20180030126A1/en not_active Abandoned
-
2019
- 2019-02-11 US US16/272,876 patent/US20200048331A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| US9120874B2 (en) | 2015-09-01 |
| WO2008136736A8 (en) | 2009-07-23 |
| ES2608803T3 (es) | 2017-04-17 |
| DK3130602T3 (da) | 2019-11-04 |
| WO2008136736A2 (en) | 2008-11-13 |
| DK2487186T3 (da) | 2017-01-16 |
| EP2226334A3 (en) | 2010-12-29 |
| EP2226334A2 (en) | 2010-09-08 |
| EP3636667A2 (en) | 2020-04-15 |
| US9796777B2 (en) | 2017-10-24 |
| US20160046702A1 (en) | 2016-02-18 |
| EP3130602B1 (en) | 2019-08-07 |
| EP3130602A1 (en) | 2017-02-15 |
| EP2487186A1 (en) | 2012-08-15 |
| US20110262470A1 (en) | 2011-10-27 |
| US20180030126A1 (en) | 2018-02-01 |
| EP2487186B1 (en) | 2016-09-28 |
| EP3636667A3 (en) | 2020-07-29 |
| US20200048331A1 (en) | 2020-02-13 |
| WO2008136736A3 (en) | 2009-02-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2747363T3 (es) | Estructuras inmunorreguladoras de proteínas que se encuentran normalmente | |
| ES2358917T3 (es) | Anticuerpo nonoclonal ykl-40. | |
| JP6886816B2 (ja) | 免疫系調節薬 | |
| AU2014317884A1 (en) | Immune system modulators | |
| US5681700A (en) | Assay for pathogenicity of anti-DNA antibodies | |
| EP0563627A2 (en) | Anti-tumor method and anti-tumor agent | |
| Brandt et al. | Functional and molecular characterization of a monoclonal antibody against human interleukin 2 | |
| ES2928791T3 (es) | Anticuerpos contra Mac-1 | |
| KR100190910B1 (ko) | C형 간염 바이러스에 대한 인체 면역조절기능을 가진펩타이드 | |
| JP2024508834A (ja) | 好中球細胞外トラップを特異的に認識する抗切断ヒストンh3モノクローナル抗体 | |
| KR20190042281A (ko) | 돼지 조직인자 세포외 도메인 단백질을 특이적으로 인식하는 단일클론항체 및 이의 용도 | |
| JP2002543091A (ja) | インターロイキン12に由来するペプチドホモダイマーおよびペプチドヘテロダイマー | |
| JPH08511941A (ja) | ヒト細胞接着性タンパク質aamp‐1及びその利用 | |
| JP2002543090A (ja) | インターフェロンα2に由来するペプチドホモダイマーおよびペプチドヘテロダイマー | |
| JP2016008194A (ja) | 抗ヒト心筋トロポニンtモノクローナル抗体、その製法、ハイブリドーマ、ヒト心筋トロポニンtの測定方法及びヒト心筋トロポニンtの測定キット |