CN116096732A - 用于预防和治疗冠状病毒感染的组合物和方法-sars-cov-2疫苗 - Google Patents
用于预防和治疗冠状病毒感染的组合物和方法-sars-cov-2疫苗 Download PDFInfo
- Publication number
- CN116096732A CN116096732A CN202180025856.2A CN202180025856A CN116096732A CN 116096732 A CN116096732 A CN 116096732A CN 202180025856 A CN202180025856 A CN 202180025856A CN 116096732 A CN116096732 A CN 116096732A
- Authority
- CN
- China
- Prior art keywords
- protein
- seq
- nucleic acid
- sars
- cov
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 239000000203 mixture Substances 0.000 title claims abstract description 232
- 238000000034 method Methods 0.000 title claims abstract description 103
- 208000001528 Coronaviridae Infections Diseases 0.000 title description 28
- 229940022962 COVID-19 vaccine Drugs 0.000 title description 3
- 241001678559 COVID-19 virus Species 0.000 claims abstract description 271
- 150000007523 nucleic acids Chemical class 0.000 claims abstract description 197
- 102000039446 nucleic acids Human genes 0.000 claims abstract description 180
- 108020004707 nucleic acids Proteins 0.000 claims abstract description 180
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 156
- 102000004169 proteins and genes Human genes 0.000 claims abstract description 122
- 229960005486 vaccine Drugs 0.000 claims abstract description 110
- 101000629318 Severe acute respiratory syndrome coronavirus 2 Spike glycoprotein Proteins 0.000 claims abstract description 97
- 230000002163 immunogen Effects 0.000 claims abstract description 70
- 239000013598 vector Substances 0.000 claims description 195
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 157
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 137
- 229920001184 polypeptide Polymers 0.000 claims description 125
- 241000701161 unidentified adenovirus Species 0.000 claims description 97
- 150000001413 amino acids Chemical class 0.000 claims description 90
- 125000003729 nucleotide group Chemical group 0.000 claims description 70
- 239000002773 nucleotide Substances 0.000 claims description 67
- 208000025721 COVID-19 Diseases 0.000 claims description 64
- 239000012634 fragment Substances 0.000 claims description 64
- 230000035772 mutation Effects 0.000 claims description 53
- 241000701022 Cytomegalovirus Species 0.000 claims description 39
- 230000003053 immunization Effects 0.000 claims description 30
- 241000598171 Human adenovirus sp. Species 0.000 claims description 29
- 238000012217 deletion Methods 0.000 claims description 28
- 230000037430 deletion Effects 0.000 claims description 28
- 238000002649 immunization Methods 0.000 claims description 26
- 108010076504 Protein Sorting Signals Proteins 0.000 claims description 25
- 238000003776 cleavage reaction Methods 0.000 claims description 21
- 230000007017 scission Effects 0.000 claims description 21
- 102000004961 Furin Human genes 0.000 claims description 20
- 108090001126 Furin Proteins 0.000 claims description 20
- 230000004048 modification Effects 0.000 claims description 20
- 238000012986 modification Methods 0.000 claims description 20
- 239000000546 pharmaceutical excipient Substances 0.000 claims description 20
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 claims description 19
- 239000004098 Tetracycline Substances 0.000 claims description 14
- 230000000087 stabilizing effect Effects 0.000 claims description 14
- 229960002180 tetracycline Drugs 0.000 claims description 14
- 230000010076 replication Effects 0.000 claims description 13
- 229930101283 tetracycline Natural products 0.000 claims description 13
- 235000019364 tetracycline Nutrition 0.000 claims description 13
- 150000003522 tetracyclines Chemical class 0.000 claims description 13
- 239000003085 diluting agent Substances 0.000 claims description 12
- 230000000295 complement effect Effects 0.000 claims description 11
- 239000003937 drug carrier Substances 0.000 claims description 10
- 230000002950 deficient Effects 0.000 claims description 7
- 238000004113 cell culture Methods 0.000 claims description 5
- 238000002156 mixing Methods 0.000 claims description 4
- 125000003275 alpha amino acid group Chemical group 0.000 claims 14
- 241000711573 Coronaviridae Species 0.000 abstract description 69
- 230000003472 neutralizing effect Effects 0.000 abstract description 60
- 241000282414 Homo sapiens Species 0.000 abstract description 47
- 102000040430 polynucleotide Human genes 0.000 abstract description 41
- 108091033319 polynucleotide Proteins 0.000 abstract description 41
- 239000002157 polynucleotide Substances 0.000 abstract description 41
- 208000015181 infectious disease Diseases 0.000 abstract description 39
- 102100031673 Corneodesmosin Human genes 0.000 abstract description 37
- 101710139375 Corneodesmosin Proteins 0.000 abstract description 37
- 230000005875 antibody response Effects 0.000 abstract description 25
- 230000001681 protective effect Effects 0.000 abstract description 24
- 229940127002 anti-2019-nCov Drugs 0.000 abstract description 19
- 238000001514 detection method Methods 0.000 abstract description 18
- 239000003814 drug Substances 0.000 abstract description 16
- 238000012544 monitoring process Methods 0.000 abstract description 4
- 229940079593 drug Drugs 0.000 abstract description 3
- 210000004027 cell Anatomy 0.000 description 131
- 230000003612 virological effect Effects 0.000 description 128
- 235000018102 proteins Nutrition 0.000 description 99
- 238000003556 assay Methods 0.000 description 98
- 241001465754 Metazoa Species 0.000 description 93
- 230000004044 response Effects 0.000 description 90
- 108091027544 Subgenomic mRNA Proteins 0.000 description 81
- 210000001519 tissue Anatomy 0.000 description 67
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 63
- 238000002965 ELISA Methods 0.000 description 61
- 241001112090 Pseudovirus Species 0.000 description 57
- 201000010099 disease Diseases 0.000 description 55
- 238000003364 immunohistochemistry Methods 0.000 description 54
- 210000004072 lung Anatomy 0.000 description 54
- 238000009739 binding Methods 0.000 description 51
- 230000027455 binding Effects 0.000 description 50
- 241000700605 Viruses Species 0.000 description 48
- 238000006386 neutralization reaction Methods 0.000 description 45
- 238000012360 testing method Methods 0.000 description 43
- 241000282560 Macaca mulatta Species 0.000 description 42
- 230000014509 gene expression Effects 0.000 description 42
- 238000002255 vaccination Methods 0.000 description 41
- 235000001014 amino acid Nutrition 0.000 description 39
- 108020000999 Viral RNA Proteins 0.000 description 38
- 238000010186 staining Methods 0.000 description 37
- 229940024606 amino acid Drugs 0.000 description 36
- 230000010530 Virus Neutralization Effects 0.000 description 33
- 241000699800 Cricetinae Species 0.000 description 32
- 108020004414 DNA Proteins 0.000 description 32
- 230000028993 immune response Effects 0.000 description 31
- 239000000523 sample Substances 0.000 description 31
- 230000001419 dependent effect Effects 0.000 description 30
- 108010074328 Interferon-gamma Proteins 0.000 description 29
- 229940024452 Janssen COVID-19 vaccine Drugs 0.000 description 29
- 102100037850 Interferon gamma Human genes 0.000 description 28
- 210000001744 T-lymphocyte Anatomy 0.000 description 27
- 239000000427 antigen Substances 0.000 description 27
- 108091007433 antigens Proteins 0.000 description 27
- 102000036639 antigens Human genes 0.000 description 27
- 239000013603 viral vector Substances 0.000 description 27
- 241000282412 Homo Species 0.000 description 26
- 208000024891 symptom Diseases 0.000 description 26
- 102000004127 Cytokines Human genes 0.000 description 24
- 108090000695 Cytokines Proteins 0.000 description 24
- 239000002671 adjuvant Substances 0.000 description 24
- 238000010586 diagram Methods 0.000 description 24
- 238000011282 treatment Methods 0.000 description 24
- 230000028996 humoral immune response Effects 0.000 description 23
- 230000001225 therapeutic effect Effects 0.000 description 23
- 230000000694 effects Effects 0.000 description 22
- 239000002245 particle Substances 0.000 description 22
- 239000008194 pharmaceutical composition Substances 0.000 description 22
- 210000002966 serum Anatomy 0.000 description 21
- 201000003176 Severe Acute Respiratory Syndrome Diseases 0.000 description 20
- 238000004519 manufacturing process Methods 0.000 description 20
- 230000001404 mediated effect Effects 0.000 description 20
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 18
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 18
- 210000000440 neutrophil Anatomy 0.000 description 18
- 229940068196 placebo Drugs 0.000 description 18
- 239000000902 placebo Substances 0.000 description 18
- 229960000187 tissue plasminogen activator Drugs 0.000 description 18
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 17
- 241000699673 Mesocricetus auratus Species 0.000 description 17
- 101001024647 Severe acute respiratory syndrome coronavirus Nucleoprotein Proteins 0.000 description 17
- 230000002146 bilateral effect Effects 0.000 description 17
- 235000013930 proline Nutrition 0.000 description 17
- 238000005829 trimerization reaction Methods 0.000 description 17
- 102000003896 Myeloperoxidases Human genes 0.000 description 16
- 108090000235 Myeloperoxidases Proteins 0.000 description 16
- 230000024932 T cell mediated immunity Effects 0.000 description 16
- 239000013604 expression vector Substances 0.000 description 15
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 14
- 108091028043 Nucleic acid sequence Proteins 0.000 description 14
- 206010057249 Phagocytosis Diseases 0.000 description 14
- 238000009472 formulation Methods 0.000 description 14
- 230000006870 function Effects 0.000 description 14
- 238000007901 in situ hybridization Methods 0.000 description 14
- 230000008782 phagocytosis Effects 0.000 description 14
- 238000011002 quantification Methods 0.000 description 14
- 102000004388 Interleukin-4 Human genes 0.000 description 13
- 108090000978 Interleukin-4 Proteins 0.000 description 13
- 108090001074 Nucleocapsid Proteins Proteins 0.000 description 13
- 210000004369 blood Anatomy 0.000 description 13
- 239000008280 blood Substances 0.000 description 13
- 238000012937 correction Methods 0.000 description 13
- 230000000875 corresponding effect Effects 0.000 description 13
- 229940028885 interleukin-4 Drugs 0.000 description 13
- 239000013612 plasmid Substances 0.000 description 13
- 238000002360 preparation method Methods 0.000 description 13
- 239000000047 product Substances 0.000 description 13
- 230000009870 specific binding Effects 0.000 description 13
- 230000004580 weight loss Effects 0.000 description 13
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 12
- 230000003834 intracellular effect Effects 0.000 description 12
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 12
- 108090000565 Capsid Proteins Proteins 0.000 description 11
- 102100023321 Ceruloplasmin Human genes 0.000 description 11
- 108010002350 Interleukin-2 Proteins 0.000 description 11
- 241000282553 Macaca Species 0.000 description 11
- 241000124008 Mammalia Species 0.000 description 11
- 230000000692 anti-sense effect Effects 0.000 description 11
- 230000008859 change Effects 0.000 description 11
- 238000009826 distribution Methods 0.000 description 11
- 238000011081 inoculation Methods 0.000 description 11
- 229940124597 therapeutic agent Drugs 0.000 description 11
- 238000013518 transcription Methods 0.000 description 11
- 230000035897 transcription Effects 0.000 description 11
- 230000009385 viral infection Effects 0.000 description 11
- 108010041986 DNA Vaccines Proteins 0.000 description 10
- 229940021995 DNA vaccine Drugs 0.000 description 10
- 206010061218 Inflammation Diseases 0.000 description 10
- 238000000585 Mann–Whitney U test Methods 0.000 description 10
- 238000004458 analytical method Methods 0.000 description 10
- 210000000981 epithelium Anatomy 0.000 description 10
- 230000008595 infiltration Effects 0.000 description 10
- 238000001764 infiltration Methods 0.000 description 10
- 230000004054 inflammatory process Effects 0.000 description 10
- 108091026890 Coding region Proteins 0.000 description 9
- 102100039897 Interleukin-5 Human genes 0.000 description 9
- 108010002616 Interleukin-5 Proteins 0.000 description 9
- 108091005461 Nucleic proteins Proteins 0.000 description 9
- 208000037847 SARS-CoV-2-infection Diseases 0.000 description 9
- 230000005888 antibody-dependent cellular phagocytosis Effects 0.000 description 9
- 230000000890 antigenic effect Effects 0.000 description 9
- 239000000306 component Substances 0.000 description 9
- 230000000120 cytopathologic effect Effects 0.000 description 9
- 238000003745 diagnosis Methods 0.000 description 9
- 230000036541 health Effects 0.000 description 9
- 239000012678 infectious agent Substances 0.000 description 9
- 229940100602 interleukin-5 Drugs 0.000 description 9
- 102000005962 receptors Human genes 0.000 description 9
- 108020003175 receptors Proteins 0.000 description 9
- 230000002829 reductive effect Effects 0.000 description 9
- 230000000241 respiratory effect Effects 0.000 description 9
- 238000006467 substitution reaction Methods 0.000 description 9
- 238000012384 transportation and delivery Methods 0.000 description 9
- 239000013638 trimer Substances 0.000 description 9
- 108020004705 Codon Proteins 0.000 description 8
- WZUVPPKBWHMQCE-UHFFFAOYSA-N Haematoxylin Chemical group C12=CC(O)=C(O)C=C2CC2(O)C1C1=CC=C(O)C(O)=C1OC2 WZUVPPKBWHMQCE-UHFFFAOYSA-N 0.000 description 8
- 108010050904 Interferons Proteins 0.000 description 8
- 102000014150 Interferons Human genes 0.000 description 8
- 102000003816 Interleukin-13 Human genes 0.000 description 8
- 108090000176 Interleukin-13 Proteins 0.000 description 8
- 241000699670 Mus sp. Species 0.000 description 8
- 230000006051 NK cell activation Effects 0.000 description 8
- 108010076039 Polyproteins Proteins 0.000 description 8
- 241000315672 SARS coronavirus Species 0.000 description 8
- 230000005867 T cell response Effects 0.000 description 8
- 230000001580 bacterial effect Effects 0.000 description 8
- 230000001086 cytosolic effect Effects 0.000 description 8
- 208000035475 disorder Diseases 0.000 description 8
- 230000003308 immunostimulating effect Effects 0.000 description 8
- 230000001965 increasing effect Effects 0.000 description 8
- 210000002540 macrophage Anatomy 0.000 description 8
- 230000007170 pathology Effects 0.000 description 8
- 238000000513 principal component analysis Methods 0.000 description 8
- 210000001944 turbinate Anatomy 0.000 description 8
- 102000053723 Angiotensin-converting enzyme 2 Human genes 0.000 description 7
- 108090000975 Angiotensin-converting enzyme 2 Proteins 0.000 description 7
- 108060003951 Immunoglobulin Proteins 0.000 description 7
- 206010028851 Necrosis Diseases 0.000 description 7
- 206010037660 Pyrexia Diseases 0.000 description 7
- 208000036142 Viral infection Diseases 0.000 description 7
- 239000004480 active ingredient Substances 0.000 description 7
- 239000000872 buffer Substances 0.000 description 7
- 230000008021 deposition Effects 0.000 description 7
- 238000011161 development Methods 0.000 description 7
- 230000018109 developmental process Effects 0.000 description 7
- 210000002919 epithelial cell Anatomy 0.000 description 7
- 239000012530 fluid Substances 0.000 description 7
- 102000018358 immunoglobulin Human genes 0.000 description 7
- 229960001438 immunostimulant agent Drugs 0.000 description 7
- 239000003022 immunostimulating agent Substances 0.000 description 7
- 238000000338 in vitro Methods 0.000 description 7
- 230000001939 inductive effect Effects 0.000 description 7
- 238000001802 infusion Methods 0.000 description 7
- 238000002347 injection Methods 0.000 description 7
- 239000007924 injection Substances 0.000 description 7
- 238000005259 measurement Methods 0.000 description 7
- 108020004999 messenger RNA Proteins 0.000 description 7
- 210000001616 monocyte Anatomy 0.000 description 7
- 230000017074 necrotic cell death Effects 0.000 description 7
- 230000028327 secretion Effects 0.000 description 7
- 239000011780 sodium chloride Substances 0.000 description 7
- 241001430294 unidentified retrovirus Species 0.000 description 7
- 102000053602 DNA Human genes 0.000 description 6
- 206010015548 Euthanasia Diseases 0.000 description 6
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 6
- 208000000112 Myalgia Diseases 0.000 description 6
- 238000011887 Necropsy Methods 0.000 description 6
- 206010035664 Pneumonia Diseases 0.000 description 6
- 229940022005 RNA vaccine Drugs 0.000 description 6
- 238000010240 RT-PCR analysis Methods 0.000 description 6
- 108091005774 SARS-CoV-2 proteins Proteins 0.000 description 6
- 108700019146 Transgenes Proteins 0.000 description 6
- 238000000540 analysis of variance Methods 0.000 description 6
- 239000003623 enhancer Substances 0.000 description 6
- 230000036039 immunity Effects 0.000 description 6
- 238000003018 immunoassay Methods 0.000 description 6
- 230000005847 immunogenicity Effects 0.000 description 6
- 229940079322 interferon Drugs 0.000 description 6
- 108700021021 mRNA Vaccine Proteins 0.000 description 6
- 239000003550 marker Substances 0.000 description 6
- 238000006384 oligomerization reaction Methods 0.000 description 6
- 238000003753 real-time PCR Methods 0.000 description 6
- 230000008929 regeneration Effects 0.000 description 6
- 238000011069 regeneration method Methods 0.000 description 6
- 239000000243 solution Substances 0.000 description 6
- 230000000638 stimulation Effects 0.000 description 6
- 238000001890 transfection Methods 0.000 description 6
- 230000029812 viral genome replication Effects 0.000 description 6
- 241000008904 Betacoronavirus Species 0.000 description 5
- 241001217856 Chimpanzee adenovirus Species 0.000 description 5
- 206010011224 Cough Diseases 0.000 description 5
- 241000702421 Dependoparvovirus Species 0.000 description 5
- 206010013975 Dyspnoeas Diseases 0.000 description 5
- 108700039887 Essential Genes Proteins 0.000 description 5
- 206010019233 Headaches Diseases 0.000 description 5
- 208000032843 Hemorrhage Diseases 0.000 description 5
- 206010020772 Hypertension Diseases 0.000 description 5
- 102000003814 Interleukin-10 Human genes 0.000 description 5
- 108090000174 Interleukin-10 Proteins 0.000 description 5
- 208000029523 Interstitial Lung disease Diseases 0.000 description 5
- 101100348738 Mus musculus Noc3l gene Proteins 0.000 description 5
- 206010068319 Oropharyngeal pain Diseases 0.000 description 5
- 201000007100 Pharyngitis Diseases 0.000 description 5
- 206010037423 Pulmonary oedema Diseases 0.000 description 5
- 102000009661 Repressor Proteins Human genes 0.000 description 5
- 108010034634 Repressor Proteins Proteins 0.000 description 5
- 230000008901 benefit Effects 0.000 description 5
- 210000001124 body fluid Anatomy 0.000 description 5
- 239000010839 body fluid Substances 0.000 description 5
- 230000037396 body weight Effects 0.000 description 5
- 230000036755 cellular response Effects 0.000 description 5
- 238000007596 consolidation process Methods 0.000 description 5
- 238000013270 controlled release Methods 0.000 description 5
- 230000001186 cumulative effect Effects 0.000 description 5
- 239000012636 effector Substances 0.000 description 5
- 210000001035 gastrointestinal tract Anatomy 0.000 description 5
- 238000001476 gene delivery Methods 0.000 description 5
- 231100000869 headache Toxicity 0.000 description 5
- 229940072221 immunoglobulins Drugs 0.000 description 5
- 238000011532 immunohistochemical staining Methods 0.000 description 5
- 230000002757 inflammatory effect Effects 0.000 description 5
- 229940076144 interleukin-10 Drugs 0.000 description 5
- 150000002632 lipids Chemical class 0.000 description 5
- -1 plasmids Chemical class 0.000 description 5
- 239000003755 preservative agent Substances 0.000 description 5
- 210000002345 respiratory system Anatomy 0.000 description 5
- 239000007787 solid Substances 0.000 description 5
- 238000001228 spectrum Methods 0.000 description 5
- 239000000126 substance Substances 0.000 description 5
- 238000012546 transfer Methods 0.000 description 5
- 230000014616 translation Effects 0.000 description 5
- 238000013519 translation Methods 0.000 description 5
- 230000014599 transmission of virus Effects 0.000 description 5
- 241000990167 unclassified Simian adenoviruses Species 0.000 description 5
- 241000283690 Bos taurus Species 0.000 description 4
- 206010006448 Bronchiolitis Diseases 0.000 description 4
- 208000024172 Cardiovascular disease Diseases 0.000 description 4
- 108010071942 Colony-Stimulating Factors Proteins 0.000 description 4
- 238000011238 DNA vaccination Methods 0.000 description 4
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 4
- 208000000059 Dyspnea Diseases 0.000 description 4
- 238000011510 Elispot assay Methods 0.000 description 4
- 208000010201 Exanthema Diseases 0.000 description 4
- 101001023379 Homo sapiens Lysosome-associated membrane glycoprotein 1 Proteins 0.000 description 4
- 101000934372 Homo sapiens Macrosialin Proteins 0.000 description 4
- 102100035133 Lysosome-associated membrane glycoprotein 1 Human genes 0.000 description 4
- 102100025136 Macrosialin Human genes 0.000 description 4
- 238000011529 RT qPCR Methods 0.000 description 4
- 238000000692 Student's t-test Methods 0.000 description 4
- 108700005077 Viral Genes Proteins 0.000 description 4
- 230000001154 acute effect Effects 0.000 description 4
- 239000000443 aerosol Substances 0.000 description 4
- 238000013357 binding ELISA Methods 0.000 description 4
- 239000002775 capsule Substances 0.000 description 4
- 150000001720 carbohydrates Chemical class 0.000 description 4
- 230000007910 cell fusion Effects 0.000 description 4
- 239000003795 chemical substances by application Substances 0.000 description 4
- 239000003246 corticosteroid Substances 0.000 description 4
- 229960001334 corticosteroids Drugs 0.000 description 4
- 239000006071 cream Substances 0.000 description 4
- 210000005220 cytoplasmic tail Anatomy 0.000 description 4
- 230000034994 death Effects 0.000 description 4
- 231100000517 death Toxicity 0.000 description 4
- 238000013461 design Methods 0.000 description 4
- 229960003957 dexamethasone Drugs 0.000 description 4
- UREBDLICKHMUKA-CXSFZGCWSA-N dexamethasone Chemical compound C1CC2=CC(=O)C=C[C@]2(C)[C@]2(F)[C@@H]1[C@@H]1C[C@@H](C)[C@@](C(=O)CO)(O)[C@@]1(C)C[C@@H]2O UREBDLICKHMUKA-CXSFZGCWSA-N 0.000 description 4
- 206010012601 diabetes mellitus Diseases 0.000 description 4
- 241001493065 dsRNA viruses Species 0.000 description 4
- 239000000839 emulsion Substances 0.000 description 4
- 238000003114 enzyme-linked immunosorbent spot assay Methods 0.000 description 4
- YQGOJNYOYNNSMM-UHFFFAOYSA-N eosin Chemical group [Na+].OC(=O)C1=CC=CC=C1C1=C2C=C(Br)C(=O)C(Br)=C2OC2=C(Br)C(O)=C(Br)C=C21 YQGOJNYOYNNSMM-UHFFFAOYSA-N 0.000 description 4
- 201000005884 exanthem Diseases 0.000 description 4
- 230000002068 genetic effect Effects 0.000 description 4
- 230000002458 infectious effect Effects 0.000 description 4
- 210000004969 inflammatory cell Anatomy 0.000 description 4
- 238000007918 intramuscular administration Methods 0.000 description 4
- 238000004020 luminiscence type Methods 0.000 description 4
- 238000001000 micrograph Methods 0.000 description 4
- 210000005087 mononuclear cell Anatomy 0.000 description 4
- 238000004806 packaging method and process Methods 0.000 description 4
- 230000036961 partial effect Effects 0.000 description 4
- 230000010412 perfusion Effects 0.000 description 4
- 210000003800 pharynx Anatomy 0.000 description 4
- 239000000843 powder Substances 0.000 description 4
- 210000001147 pulmonary artery Anatomy 0.000 description 4
- 238000003762 quantitative reverse transcription PCR Methods 0.000 description 4
- 206010037844 rash Diseases 0.000 description 4
- 230000001105 regulatory effect Effects 0.000 description 4
- 201000004193 respiratory failure Diseases 0.000 description 4
- 230000004043 responsiveness Effects 0.000 description 4
- 230000003248 secreting effect Effects 0.000 description 4
- 239000003381 stabilizer Substances 0.000 description 4
- 238000010561 standard procedure Methods 0.000 description 4
- 238000012353 t test Methods 0.000 description 4
- 241000701447 unidentified baculovirus Species 0.000 description 4
- 230000002792 vascular Effects 0.000 description 4
- 206010067484 Adverse reaction Diseases 0.000 description 3
- 241000004176 Alphacoronavirus Species 0.000 description 3
- 206010001889 Alveolitis Diseases 0.000 description 3
- 241000894006 Bacteria Species 0.000 description 3
- 108010029697 CD40 Ligand Proteins 0.000 description 3
- 102100032937 CD40 ligand Human genes 0.000 description 3
- 241000282472 Canis lupus familiaris Species 0.000 description 3
- 102000001326 Chemokine CCL4 Human genes 0.000 description 3
- 108010055165 Chemokine CCL4 Proteins 0.000 description 3
- 241000288673 Chiroptera Species 0.000 description 3
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 3
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 3
- 206010012735 Diarrhoea Diseases 0.000 description 3
- 102100025137 Early activation antigen CD69 Human genes 0.000 description 3
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- 241000282575 Gorilla Species 0.000 description 3
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 3
- 101000934374 Homo sapiens Early activation antigen CD69 Proteins 0.000 description 3
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 206010028813 Nausea Diseases 0.000 description 3
- 206010030113 Oedema Diseases 0.000 description 3
- 108091034117 Oligonucleotide Proteins 0.000 description 3
- 241000282577 Pan troglodytes Species 0.000 description 3
- 241000709664 Picornaviridae Species 0.000 description 3
- 208000004756 Respiratory Insufficiency Diseases 0.000 description 3
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 3
- 238000007792 addition Methods 0.000 description 3
- 230000006838 adverse reaction Effects 0.000 description 3
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 3
- ILRRQNADMUWWFW-UHFFFAOYSA-K aluminium phosphate Chemical compound O1[Al]2OP1(=O)O2 ILRRQNADMUWWFW-UHFFFAOYSA-K 0.000 description 3
- 230000008382 alveolar damage Effects 0.000 description 3
- 210000002588 alveolar type II cell Anatomy 0.000 description 3
- 229940121363 anti-inflammatory agent Drugs 0.000 description 3
- 239000002260 anti-inflammatory agent Substances 0.000 description 3
- 238000011888 autopsy Methods 0.000 description 3
- 206010006451 bronchitis Diseases 0.000 description 3
- 235000014633 carbohydrates Nutrition 0.000 description 3
- 239000013592 cell lysate Substances 0.000 description 3
- 239000003153 chemical reaction reagent Substances 0.000 description 3
- 238000010367 cloning Methods 0.000 description 3
- 238000000576 coating method Methods 0.000 description 3
- 238000010494 dissociation reaction Methods 0.000 description 3
- 238000002474 experimental method Methods 0.000 description 3
- 201000001155 extrinsic allergic alveolitis Diseases 0.000 description 3
- 206010016256 fatigue Diseases 0.000 description 3
- 238000003304 gavage Methods 0.000 description 3
- 239000000499 gel Substances 0.000 description 3
- 238000001415 gene therapy Methods 0.000 description 3
- 230000008348 humoral response Effects 0.000 description 3
- 206010020718 hyperplasia Diseases 0.000 description 3
- 208000022098 hypersensitivity pneumonitis Diseases 0.000 description 3
- 210000000987 immune system Anatomy 0.000 description 3
- 238000003119 immunoblot Methods 0.000 description 3
- 230000006698 induction Effects 0.000 description 3
- 230000002401 inhibitory effect Effects 0.000 description 3
- 238000003780 insertion Methods 0.000 description 3
- 230000037431 insertion Effects 0.000 description 3
- 239000007927 intramuscular injection Substances 0.000 description 3
- 210000004962 mammalian cell Anatomy 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 229940035032 monophosphoryl lipid a Drugs 0.000 description 3
- 210000000822 natural killer cell Anatomy 0.000 description 3
- 230000008693 nausea Effects 0.000 description 3
- 210000004940 nucleus Anatomy 0.000 description 3
- 238000005457 optimization Methods 0.000 description 3
- 230000001575 pathological effect Effects 0.000 description 3
- 229920000642 polymer Polymers 0.000 description 3
- 125000001500 prolyl group Chemical group [H]N1C([H])(C(=O)[*])C([H])([H])C([H])([H])C1([H])[H] 0.000 description 3
- 239000003380 propellant Substances 0.000 description 3
- 238000012552 review Methods 0.000 description 3
- 206010039083 rhinitis Diseases 0.000 description 3
- 125000006850 spacer group Chemical group 0.000 description 3
- 230000006641 stabilisation Effects 0.000 description 3
- 238000011105 stabilization Methods 0.000 description 3
- 239000006228 supernatant Substances 0.000 description 3
- 239000000725 suspension Substances 0.000 description 3
- 239000003826 tablet Substances 0.000 description 3
- 241001529453 unidentified herpesvirus Species 0.000 description 3
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Chemical compound O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 3
- DIGQNXIGRZPYDK-WKSCXVIASA-N (2R)-6-amino-2-[[2-[[(2S)-2-[[2-[[(2R)-2-[[(2S)-2-[[(2R,3S)-2-[[2-[[(2S)-2-[[2-[[(2S)-2-[[(2S)-2-[[(2R)-2-[[(2S,3S)-2-[[(2R)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[2-[[(2S)-2-[[(2R)-2-[[2-[[2-[[2-[(2-amino-1-hydroxyethylidene)amino]-3-carboxy-1-hydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxypropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1,5-dihydroxy-5-iminopentylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxybutylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1,3-dihydroxypropylidene]amino]-1-hydroxyethylidene]amino]-1-hydroxy-3-sulfanylpropylidene]amino]-1-hydroxyethylidene]amino]hexanoic acid Chemical compound C[C@@H]([C@@H](C(=N[C@@H](CS)C(=N[C@@H](C)C(=N[C@@H](CO)C(=NCC(=N[C@@H](CCC(=N)O)C(=NC(CS)C(=N[C@H]([C@H](C)O)C(=N[C@H](CS)C(=N[C@H](CO)C(=NCC(=N[C@H](CS)C(=NCC(=N[C@H](CCCCN)C(=O)O)O)O)O)O)O)O)O)O)O)O)O)O)O)N=C([C@H](CS)N=C([C@H](CO)N=C([C@H](CO)N=C([C@H](C)N=C(CN=C([C@H](CO)N=C([C@H](CS)N=C(CN=C(C(CS)N=C(C(CC(=O)O)N=C(CN)O)O)O)O)O)O)O)O)O)O)O)O DIGQNXIGRZPYDK-WKSCXVIASA-N 0.000 description 2
- JARGNLJYKBUKSJ-KGZKBUQUSA-N (2r)-2-amino-5-[[(2r)-1-(carboxymethylamino)-3-hydroxy-1-oxopropan-2-yl]amino]-5-oxopentanoic acid;hydrobromide Chemical compound Br.OC(=O)[C@H](N)CCC(=O)N[C@H](CO)C(=O)NCC(O)=O JARGNLJYKBUKSJ-KGZKBUQUSA-N 0.000 description 2
- DRHZYJAUECRAJM-DWSYSWFDSA-N (2s,3s,4s,5r,6r)-6-[[(3s,4s,4ar,6ar,6bs,8r,8ar,12as,14ar,14br)-8a-[(2s,3r,4s,5r,6r)-3-[(2s,3r,4s,5r,6s)-5-[(2s,3r,4s,5r)-4-[(2s,3r,4r)-3,4-dihydroxy-4-(hydroxymethyl)oxolan-2-yl]oxy-3,5-dihydroxyoxan-2-yl]oxy-3,4-dihydroxy-6-methyloxan-2-yl]oxy-5-[(3s,5s, Chemical compound O([C@H]1[C@H](O)[C@H](O[C@H]([C@@H]1O[C@H]1[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O1)O)O[C@H]1CC[C@]2(C)[C@H]3CC=C4[C@@H]5CC(C)(C)CC[C@@]5([C@@H](C[C@@]4(C)[C@]3(C)CC[C@H]2[C@@]1(C=O)C)O)C(=O)O[C@@H]1O[C@H](C)[C@@H]([C@@H]([C@H]1O[C@H]1[C@@H]([C@H](O)[C@@H](O[C@H]2[C@@H]([C@@H](O[C@H]3[C@@H]([C@@](O)(CO)CO3)O)[C@H](O)CO2)O)[C@H](C)O1)O)O)OC(=O)C[C@@H](O)C[C@H](OC(=O)C[C@@H](O)C[C@@H]([C@@H](C)CC)O[C@H]1[C@@H]([C@@H](O)[C@H](CO)O1)O)[C@@H](C)CC)C(O)=O)[C@@H]1OC[C@@H](O)[C@H](O)[C@H]1O DRHZYJAUECRAJM-DWSYSWFDSA-N 0.000 description 2
- CXURGFRDGROIKG-UHFFFAOYSA-N 3,3-bis(chloromethyl)oxetane Chemical compound ClCC1(CCl)COC1 CXURGFRDGROIKG-UHFFFAOYSA-N 0.000 description 2
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 2
- 206010001052 Acute respiratory distress syndrome Diseases 0.000 description 2
- 201000004384 Alopecia Diseases 0.000 description 2
- 241000710929 Alphavirus Species 0.000 description 2
- 208000006820 Arthralgia Diseases 0.000 description 2
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 2
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 2
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 2
- 102000005701 Calcium-Binding Proteins Human genes 0.000 description 2
- 108010045403 Calcium-Binding Proteins Proteins 0.000 description 2
- 102100036956 Chromatin target of PRMT1 protein Human genes 0.000 description 2
- 101710197132 Chromatin target of PRMT1 protein Proteins 0.000 description 2
- 206010010741 Conjunctivitis Diseases 0.000 description 2
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 2
- 241000450599 DNA viruses Species 0.000 description 2
- 241001461743 Deltacoronavirus Species 0.000 description 2
- 101150029662 E1 gene Proteins 0.000 description 2
- 238000012286 ELISA Assay Methods 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- 206010015958 Eye pain Diseases 0.000 description 2
- 241000282326 Felis catus Species 0.000 description 2
- 238000000729 Fisher's exact test Methods 0.000 description 2
- 208000000666 Fowlpox Diseases 0.000 description 2
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 2
- 206010018866 Haematospermia Diseases 0.000 description 2
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 2
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 2
- 208000008982 Hemospermia Diseases 0.000 description 2
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 2
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 2
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 description 2
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 2
- 241000205701 Human adenovirus 26 Species 0.000 description 2
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 2
- 241000701806 Human papillomavirus Species 0.000 description 2
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 2
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 2
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 2
- 102000003815 Interleukin-11 Human genes 0.000 description 2
- 108090000177 Interleukin-11 Proteins 0.000 description 2
- 108010065805 Interleukin-12 Proteins 0.000 description 2
- 102000013462 Interleukin-12 Human genes 0.000 description 2
- 108010002386 Interleukin-3 Proteins 0.000 description 2
- 102000000646 Interleukin-3 Human genes 0.000 description 2
- 102100021592 Interleukin-7 Human genes 0.000 description 2
- 108010002586 Interleukin-7 Proteins 0.000 description 2
- 108090001007 Interleukin-8 Proteins 0.000 description 2
- 102000004890 Interleukin-8 Human genes 0.000 description 2
- 231100000111 LD50 Toxicity 0.000 description 2
- 241000713666 Lentivirus Species 0.000 description 2
- 108090001030 Lipoproteins Proteins 0.000 description 2
- 102000004895 Lipoproteins Human genes 0.000 description 2
- 102100029185 Low affinity immunoglobulin gamma Fc region receptor III-B Human genes 0.000 description 2
- 208000008771 Lymphadenopathy Diseases 0.000 description 2
- 239000004472 Lysine Substances 0.000 description 2
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 2
- TWRXJAOTZQYOKJ-UHFFFAOYSA-L Magnesium chloride Chemical compound [Mg+2].[Cl-].[Cl-] TWRXJAOTZQYOKJ-UHFFFAOYSA-L 0.000 description 2
- 102000018697 Membrane Proteins Human genes 0.000 description 2
- 108010052285 Membrane Proteins Proteins 0.000 description 2
- 102000003792 Metallothionein Human genes 0.000 description 2
- 108090000157 Metallothionein Proteins 0.000 description 2
- NTIZESTWPVYFNL-UHFFFAOYSA-N Methyl isobutyl ketone Chemical compound CC(C)CC(C)=O NTIZESTWPVYFNL-UHFFFAOYSA-N 0.000 description 2
- 241000699666 Mus <mouse, genus> Species 0.000 description 2
- 208000009525 Myocarditis Diseases 0.000 description 2
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 description 2
- 206010053159 Organ failure Diseases 0.000 description 2
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Chemical compound OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 2
- ATUOYWHBWRKTHZ-UHFFFAOYSA-N Propane Chemical compound CCC ATUOYWHBWRKTHZ-UHFFFAOYSA-N 0.000 description 2
- 229940096437 Protein S Drugs 0.000 description 2
- 241000125945 Protoparvovirus Species 0.000 description 2
- 230000004570 RNA-binding Effects 0.000 description 2
- 238000003559 RNA-seq method Methods 0.000 description 2
- 241001068263 Replication competent viruses Species 0.000 description 2
- 206010038687 Respiratory distress Diseases 0.000 description 2
- 108091006197 SARS-CoV-2 Nucleocapsid Protein Proteins 0.000 description 2
- 101710167605 Spike glycoprotein Proteins 0.000 description 2
- 101710198474 Spike protein Proteins 0.000 description 2
- 102220599400 Spindlin-1_D1118H_mutation Human genes 0.000 description 2
- 102220590324 Spindlin-1_D80A_mutation Human genes 0.000 description 2
- 101710137302 Surface antigen S Proteins 0.000 description 2
- 238000011053 TCID50 method Methods 0.000 description 2
- 102000004887 Transforming Growth Factor beta Human genes 0.000 description 2
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 2
- 239000007983 Tris buffer Substances 0.000 description 2
- 206010046865 Vaccinia virus infection Diseases 0.000 description 2
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 2
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 2
- 241000711975 Vesicular stomatitis virus Species 0.000 description 2
- 108010067390 Viral Proteins Proteins 0.000 description 2
- 206010047700 Vomiting Diseases 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 108091005764 adaptor proteins Proteins 0.000 description 2
- 102000035181 adaptor proteins Human genes 0.000 description 2
- 230000002411 adverse Effects 0.000 description 2
- 229910052782 aluminium Inorganic materials 0.000 description 2
- XAGFODPZIPBFFR-UHFFFAOYSA-N aluminium Chemical compound [Al] XAGFODPZIPBFFR-UHFFFAOYSA-N 0.000 description 2
- 230000036528 appetite Effects 0.000 description 2
- 235000019789 appetite Nutrition 0.000 description 2
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- 239000012867 bioactive agent Substances 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 230000015572 biosynthetic process Effects 0.000 description 2
- 210000001185 bone marrow Anatomy 0.000 description 2
- 210000005013 brain tissue Anatomy 0.000 description 2
- 239000001506 calcium phosphate Substances 0.000 description 2
- 229910000389 calcium phosphate Inorganic materials 0.000 description 2
- 235000011010 calcium phosphates Nutrition 0.000 description 2
- 210000000234 capsid Anatomy 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- 230000012292 cell migration Effects 0.000 description 2
- 210000000349 chromosome Anatomy 0.000 description 2
- 238000011260 co-administration Methods 0.000 description 2
- 239000011248 coating agent Substances 0.000 description 2
- 229940001442 combination vaccine Drugs 0.000 description 2
- 238000002790 cross-validation Methods 0.000 description 2
- 230000005593 dissociations Effects 0.000 description 2
- 229940112141 dry powder inhaler Drugs 0.000 description 2
- 238000004520 electroporation Methods 0.000 description 2
- 210000003038 endothelium Anatomy 0.000 description 2
- 239000002158 endotoxin Substances 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 210000003527 eukaryotic cell Anatomy 0.000 description 2
- 238000013265 extended release Methods 0.000 description 2
- 210000001723 extracellular space Anatomy 0.000 description 2
- 238000002618 extracorporeal membrane oxygenation Methods 0.000 description 2
- 239000000835 fiber Substances 0.000 description 2
- 238000000684 flow cytometry Methods 0.000 description 2
- 210000002683 foot Anatomy 0.000 description 2
- 230000002538 fungal effect Effects 0.000 description 2
- 230000004927 fusion Effects 0.000 description 2
- 108010044804 gamma-glutamyl-seryl-glycine Proteins 0.000 description 2
- 230000002496 gastric effect Effects 0.000 description 2
- 230000005182 global health Effects 0.000 description 2
- 208000024963 hair loss Diseases 0.000 description 2
- 230000003676 hair loss Effects 0.000 description 2
- 210000005003 heart tissue Anatomy 0.000 description 2
- 238000007489 histopathology method Methods 0.000 description 2
- 210000005260 human cell Anatomy 0.000 description 2
- 238000009396 hybridization Methods 0.000 description 2
- JYGXADMDTFJGBT-VWUMJDOOSA-N hydrocortisone Chemical compound O=C1CC[C@]2(C)[C@H]3[C@@H](O)C[C@](C)([C@@](CC4)(O)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 JYGXADMDTFJGBT-VWUMJDOOSA-N 0.000 description 2
- 238000010191 image analysis Methods 0.000 description 2
- 238000003317 immunochromatography Methods 0.000 description 2
- 230000002055 immunohistochemical effect Effects 0.000 description 2
- 230000001976 improved effect Effects 0.000 description 2
- CGIGDMFJXJATDK-UHFFFAOYSA-N indomethacin Chemical compound CC1=C(CC(O)=O)C2=CC(OC)=CC=C2N1C(=O)C1=CC=C(Cl)C=C1 CGIGDMFJXJATDK-UHFFFAOYSA-N 0.000 description 2
- 230000000977 initiatory effect Effects 0.000 description 2
- 230000010354 integration Effects 0.000 description 2
- 230000010468 interferon response Effects 0.000 description 2
- 229940047124 interferons Drugs 0.000 description 2
- 229940074383 interleukin-11 Drugs 0.000 description 2
- 229940117681 interleukin-12 Drugs 0.000 description 2
- 229940076264 interleukin-3 Drugs 0.000 description 2
- 229940100994 interleukin-7 Drugs 0.000 description 2
- 229940096397 interleukin-8 Drugs 0.000 description 2
- XKTZWUACRZHVAN-VADRZIEHSA-N interleukin-8 Chemical compound C([C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@@H](NC(C)=O)CCSC)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CCSC)C(=O)N1[C@H](CCC1)C(=O)N1[C@H](CCC1)C(=O)N[C@@H](C)C(=O)N[C@H](CC(O)=O)C(=O)N[C@H](CCC(O)=O)C(=O)N[C@H](CC(O)=O)C(=O)N[C@H](CC=1C=CC(O)=CC=1)C(=O)N[C@H](CO)C(=O)N1[C@H](CCC1)C(N)=O)C1=CC=CC=C1 XKTZWUACRZHVAN-VADRZIEHSA-N 0.000 description 2
- 238000010255 intramuscular injection Methods 0.000 description 2
- 238000001990 intravenous administration Methods 0.000 description 2
- 210000000265 leukocyte Anatomy 0.000 description 2
- 239000002502 liposome Substances 0.000 description 2
- 239000006193 liquid solution Substances 0.000 description 2
- 210000005228 liver tissue Anatomy 0.000 description 2
- 238000007477 logistic regression Methods 0.000 description 2
- 208000018555 lymphatic system disease Diseases 0.000 description 2
- RLSSMJSEOOYNOY-UHFFFAOYSA-N m-cresol Chemical compound CC1=CC=CC(O)=C1 RLSSMJSEOOYNOY-UHFFFAOYSA-N 0.000 description 2
- 238000012423 maintenance Methods 0.000 description 2
- 206010025482 malaise Diseases 0.000 description 2
- 239000002609 medium Substances 0.000 description 2
- 210000004379 membrane Anatomy 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- LXCFILQKKLGQFO-UHFFFAOYSA-N methylparaben Chemical compound COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 2
- 238000010208 microarray analysis Methods 0.000 description 2
- 230000005012 migration Effects 0.000 description 2
- 238000013508 migration Methods 0.000 description 2
- 208000013465 muscle pain Diseases 0.000 description 2
- 238000011587 new zealand white rabbit Methods 0.000 description 2
- 235000015097 nutrients Nutrition 0.000 description 2
- 239000003921 oil Substances 0.000 description 2
- 239000001301 oxygen Substances 0.000 description 2
- 229910052760 oxygen Inorganic materials 0.000 description 2
- 238000010238 partial least squares regression Methods 0.000 description 2
- 244000052769 pathogen Species 0.000 description 2
- 230000001717 pathogenic effect Effects 0.000 description 2
- 230000002688 persistence Effects 0.000 description 2
- 230000036470 plasma concentration Effects 0.000 description 2
- 210000004043 pneumocyte Anatomy 0.000 description 2
- 239000000244 polyoxyethylene sorbitan monooleate Substances 0.000 description 2
- 235000010482 polyoxyethylene sorbitan monooleate Nutrition 0.000 description 2
- 229940068968 polysorbate 80 Drugs 0.000 description 2
- 229920000053 polysorbate 80 Polymers 0.000 description 2
- 230000002064 post-exposure prophylaxis Effects 0.000 description 2
- 238000012809 post-inoculation Methods 0.000 description 2
- 230000002335 preservative effect Effects 0.000 description 2
- 230000001737 promoting effect Effects 0.000 description 2
- 230000000644 propagated effect Effects 0.000 description 2
- 230000001902 propagating effect Effects 0.000 description 2
- QELSKZZBTMNZEB-UHFFFAOYSA-N propylparaben Chemical compound CCCOC(=O)C1=CC=C(O)C=C1 QELSKZZBTMNZEB-UHFFFAOYSA-N 0.000 description 2
- 230000005180 public health Effects 0.000 description 2
- 239000013014 purified material Substances 0.000 description 2
- 238000007637 random forest analysis Methods 0.000 description 2
- 238000003259 recombinant expression Methods 0.000 description 2
- 238000005215 recombination Methods 0.000 description 2
- 230000006798 recombination Effects 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 210000005084 renal tissue Anatomy 0.000 description 2
- 230000003362 replicative effect Effects 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 210000001533 respiratory mucosa Anatomy 0.000 description 2
- 230000036387 respiratory rate Effects 0.000 description 2
- 208000023504 respiratory system disease Diseases 0.000 description 2
- 230000001177 retroviral effect Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 229960000371 rofecoxib Drugs 0.000 description 2
- RZJQGNCSTQAWON-UHFFFAOYSA-N rofecoxib Chemical compound C1=CC(S(=O)(=O)C)=CC=C1C1=C(C=2C=CC=CC=2)C(=O)OC1 RZJQGNCSTQAWON-UHFFFAOYSA-N 0.000 description 2
- 150000003839 salts Chemical class 0.000 description 2
- 230000000405 serological effect Effects 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 210000000952 spleen Anatomy 0.000 description 2
- 210000004988 splenocyte Anatomy 0.000 description 2
- 239000008174 sterile solution Substances 0.000 description 2
- 238000003860 storage Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 238000001356 surgical procedure Methods 0.000 description 2
- 230000004083 survival effect Effects 0.000 description 2
- 238000004114 suspension culture Methods 0.000 description 2
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 description 2
- 231100001274 therapeutic index Toxicity 0.000 description 2
- 210000003371 toe Anatomy 0.000 description 2
- 230000000699 topical effect Effects 0.000 description 2
- 210000005092 tracheal tissue Anatomy 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 230000026683 transduction Effects 0.000 description 2
- 238000012250 transgenic expression Methods 0.000 description 2
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 2
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 2
- 208000007089 vaccinia Diseases 0.000 description 2
- 238000009423 ventilation Methods 0.000 description 2
- 238000012800 visualization Methods 0.000 description 2
- 230000008673 vomiting Effects 0.000 description 2
- 208000016261 weight loss Diseases 0.000 description 2
- RDJGLLICXDHJDY-NSHDSACASA-N (2s)-2-(3-phenoxyphenyl)propanoic acid Chemical compound OC(=O)[C@@H](C)C1=CC=CC(OC=2C=CC=CC=2)=C1 RDJGLLICXDHJDY-NSHDSACASA-N 0.000 description 1
- WRIDQFICGBMAFQ-UHFFFAOYSA-N (E)-8-Octadecenoic acid Natural products CCCCCCCCCC=CCCCCCCC(O)=O WRIDQFICGBMAFQ-UHFFFAOYSA-N 0.000 description 1
- WHTVZRBIWZFKQO-AWEZNQCLSA-N (S)-chloroquine Chemical compound ClC1=CC=C2C(N[C@@H](C)CCCN(CC)CC)=CC=NC2=C1 WHTVZRBIWZFKQO-AWEZNQCLSA-N 0.000 description 1
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 1
- DEIGXXQKDWULML-UHFFFAOYSA-N 1,2,5,6,9,10-hexabromocyclododecane Chemical compound BrC1CCC(Br)C(Br)CCC(Br)C(Br)CCC1Br DEIGXXQKDWULML-UHFFFAOYSA-N 0.000 description 1
- DDMOUSALMHHKOS-UHFFFAOYSA-N 1,2-dichloro-1,1,2,2-tetrafluoroethane Chemical compound FC(F)(Cl)C(F)(F)Cl DDMOUSALMHHKOS-UHFFFAOYSA-N 0.000 description 1
- LQJBNNIYVWPHFW-UHFFFAOYSA-N 20:1omega9c fatty acid Natural products CCCCCCCCCCC=CCCCCCCCC(O)=O LQJBNNIYVWPHFW-UHFFFAOYSA-N 0.000 description 1
- HVCOBJNICQPDBP-UHFFFAOYSA-N 3-[3-[3,5-dihydroxy-6-methyl-4-(3,4,5-trihydroxy-6-methyloxan-2-yl)oxyoxan-2-yl]oxydecanoyloxy]decanoic acid;hydrate Chemical class O.OC1C(OC(CC(=O)OC(CCCCCCC)CC(O)=O)CCCCCCC)OC(C)C(O)C1OC1C(O)C(O)C(O)C(C)O1 HVCOBJNICQPDBP-UHFFFAOYSA-N 0.000 description 1
- FWBHETKCLVMNFS-UHFFFAOYSA-N 4',6-Diamino-2-phenylindol Chemical compound C1=CC(C(=N)N)=CC=C1C1=CC2=CC=C(C(N)=N)C=C2N1 FWBHETKCLVMNFS-UHFFFAOYSA-N 0.000 description 1
- QSBYPNXLFMSGKH-UHFFFAOYSA-N 9-Heptadecensaeure Natural products CCCCCCCC=CCCCCCCCC(O)=O QSBYPNXLFMSGKH-UHFFFAOYSA-N 0.000 description 1
- 108010042708 Acetylmuramyl-Alanyl-Isoglutamine Proteins 0.000 description 1
- 102100022900 Actin, cytoplasmic 1 Human genes 0.000 description 1
- 108010085238 Actins Proteins 0.000 description 1
- 241000701242 Adenoviridae Species 0.000 description 1
- 208000010470 Ageusia Diseases 0.000 description 1
- 102100027211 Albumin Human genes 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 102100022987 Angiogenin Human genes 0.000 description 1
- 102400000068 Angiostatin Human genes 0.000 description 1
- 108010079709 Angiostatins Proteins 0.000 description 1
- 102100030988 Angiotensin-converting enzyme Human genes 0.000 description 1
- 101710185050 Angiotensin-converting enzyme Proteins 0.000 description 1
- 206010002653 Anosmia Diseases 0.000 description 1
- 101100107610 Arabidopsis thaliana ABCF4 gene Proteins 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 102000030907 Aspartate Carbamoyltransferase Human genes 0.000 description 1
- BSYNRYMUTXBXSQ-UHFFFAOYSA-N Aspirin Chemical compound CC(=O)OC1=CC=CC=C1C(O)=O BSYNRYMUTXBXSQ-UHFFFAOYSA-N 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 241000714230 Avian leukemia virus Species 0.000 description 1
- 241000713842 Avian sarcoma virus Species 0.000 description 1
- 102100035526 B melanoma antigen 1 Human genes 0.000 description 1
- 241001485018 Baboon endogenous virus Species 0.000 description 1
- 231100000699 Bacterial toxin Toxicity 0.000 description 1
- 241000112287 Bat coronavirus Species 0.000 description 1
- 241000701106 Bovine adenovirus 3 Species 0.000 description 1
- 241000714266 Bovine leukemia virus Species 0.000 description 1
- 241000701822 Bovine papillomavirus Species 0.000 description 1
- 125000001433 C-terminal amino-acid group Chemical group 0.000 description 1
- 102100037084 C4b-binding protein alpha chain Human genes 0.000 description 1
- 101710159767 C4b-binding protein alpha chain Proteins 0.000 description 1
- 102100027207 CD27 antigen Human genes 0.000 description 1
- 210000001266 CD8-positive T-lymphocyte Anatomy 0.000 description 1
- 102000000905 Cadherin Human genes 0.000 description 1
- 108050007957 Cadherin Proteins 0.000 description 1
- 241000701157 Canine mastadenovirus A Species 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 102100025064 Cellular tumor antigen p53 Human genes 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 206010008469 Chest discomfort Diseases 0.000 description 1
- 102000009016 Cholera Toxin Human genes 0.000 description 1
- 108010049048 Cholera Toxin Proteins 0.000 description 1
- KRKNYBCHXYNGOX-UHFFFAOYSA-K Citrate Chemical compound [O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O KRKNYBCHXYNGOX-UHFFFAOYSA-K 0.000 description 1
- YASYEJJMZJALEJ-UHFFFAOYSA-N Citric acid monohydrate Chemical compound O.OC(=O)CC(O)(C(O)=O)CC(O)=O YASYEJJMZJALEJ-UHFFFAOYSA-N 0.000 description 1
- 238000000959 Cochran–Mantel–Haenszel (CMH) test Methods 0.000 description 1
- 108091033380 Coding strand Proteins 0.000 description 1
- 108700010070 Codon Usage Proteins 0.000 description 1
- 102000007644 Colony-Stimulating Factors Human genes 0.000 description 1
- 108700023317 Coronavirus Receptors Proteins 0.000 description 1
- 108010061994 Coronavirus Spike Glycoprotein Proteins 0.000 description 1
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 1
- 238000000116 DAPI staining Methods 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 239000004375 Dextrin Substances 0.000 description 1
- 229920001353 Dextrin Polymers 0.000 description 1
- 239000004338 Dichlorodifluoromethane Substances 0.000 description 1
- 108091000126 Dihydroorotase Proteins 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 208000032163 Emerging Communicable disease Diseases 0.000 description 1
- 102400001047 Endostatin Human genes 0.000 description 1
- 108010079505 Endostatins Proteins 0.000 description 1
- 241000792859 Enema Species 0.000 description 1
- 241000991587 Enterovirus C Species 0.000 description 1
- 101710091045 Envelope protein Proteins 0.000 description 1
- 102000004190 Enzymes Human genes 0.000 description 1
- 108090000790 Enzymes Proteins 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 101000867232 Escherichia coli Heat-stable enterotoxin II Proteins 0.000 description 1
- 206010015946 Eye irritation Diseases 0.000 description 1
- 108010074860 Factor Xa Proteins 0.000 description 1
- 241000714165 Feline leukemia virus Species 0.000 description 1
- 241000714174 Feline sarcoma virus Species 0.000 description 1
- 101710145505 Fiber protein Proteins 0.000 description 1
- 102000018233 Fibroblast Growth Factor Human genes 0.000 description 1
- 108050007372 Fibroblast Growth Factor Proteins 0.000 description 1
- 241000700662 Fowlpox virus Species 0.000 description 1
- 241000008920 Gammacoronavirus Species 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 101710114810 Glycoprotein Proteins 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 description 1
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 description 1
- 101710121697 Heat-stable enterotoxin Proteins 0.000 description 1
- 241000700721 Hepatitis B virus Species 0.000 description 1
- 241000282375 Herpestidae Species 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 101710094396 Hexon protein Proteins 0.000 description 1
- 101000929928 Homo sapiens Angiotensin-converting enzyme 2 Proteins 0.000 description 1
- 101000874316 Homo sapiens B melanoma antigen 1 Proteins 0.000 description 1
- 101000914511 Homo sapiens CD27 antigen Proteins 0.000 description 1
- 101000654734 Homo sapiens Septin-4 Proteins 0.000 description 1
- 241000700588 Human alphaherpesvirus 1 Species 0.000 description 1
- 241000701074 Human alphaherpesvirus 2 Species 0.000 description 1
- 241001109669 Human coronavirus HKU1 Species 0.000 description 1
- 241000725303 Human immunodeficiency virus Species 0.000 description 1
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 1
- 241000714192 Human spumaretrovirus Species 0.000 description 1
- HEFNNWSXXWATRW-UHFFFAOYSA-N Ibuprofen Chemical compound CC(C)CC1=CC=C(C(C)C(O)=O)C=C1 HEFNNWSXXWATRW-UHFFFAOYSA-N 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 108020005350 Initiator Codon Proteins 0.000 description 1
- 102100034349 Integrase Human genes 0.000 description 1
- 102000002227 Interferon Type I Human genes 0.000 description 1
- 108010014726 Interferon Type I Proteins 0.000 description 1
- 102000006992 Interferon-alpha Human genes 0.000 description 1
- 108010047761 Interferon-alpha Proteins 0.000 description 1
- 102000003996 Interferon-beta Human genes 0.000 description 1
- 108090000467 Interferon-beta Proteins 0.000 description 1
- 102000008070 Interferon-gamma Human genes 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- OFFWOVJBSQMVPI-RMLGOCCBSA-N Kaletra Chemical compound N1([C@@H](C(C)C)C(=O)N[C@H](C[C@H](O)[C@H](CC=2C=CC=CC=2)NC(=O)COC=2C(=CC=CC=2C)C)CC=2C=CC=CC=2)CCCNC1=O.N([C@@H](C(C)C)C(=O)N[C@H](C[C@H](O)[C@H](CC=1C=CC=CC=1)NC(=O)OCC=1SC=NC=1)CC=1C=CC=CC=1)C(=O)N(C)CC1=CSC(C(C)C)=N1 OFFWOVJBSQMVPI-RMLGOCCBSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- 206010025080 Lung consolidation Diseases 0.000 description 1
- 108090000542 Lymphotoxin-alpha Proteins 0.000 description 1
- 102000004083 Lymphotoxin-alpha Human genes 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 241000713821 Mason-Pfizer monkey virus Species 0.000 description 1
- 201000005505 Measles Diseases 0.000 description 1
- SBDNJUWAMKYJOX-UHFFFAOYSA-N Meclofenamic Acid Chemical compound CC1=CC=C(Cl)C(NC=2C(=CC=CC=2)C(O)=O)=C1Cl SBDNJUWAMKYJOX-UHFFFAOYSA-N 0.000 description 1
- 108050008953 Melanoma-associated antigen Proteins 0.000 description 1
- ZRVUJXDFFKFLMG-UHFFFAOYSA-N Meloxicam Chemical compound OC=1C2=CC=CC=C2S(=O)(=O)N(C)C=1C(=O)NC1=NC=C(C)S1 ZRVUJXDFFKFLMG-UHFFFAOYSA-N 0.000 description 1
- 102000006166 Metallocarboxypeptidases Human genes 0.000 description 1
- 108030000089 Metallocarboxypeptidases Proteins 0.000 description 1
- 108700011259 MicroRNAs Proteins 0.000 description 1
- 208000025370 Middle East respiratory syndrome Diseases 0.000 description 1
- 241000127282 Middle East respiratory syndrome-related coronavirus Species 0.000 description 1
- 241001183012 Modified Vaccinia Ankara virus Species 0.000 description 1
- 241000713862 Moloney murine sarcoma virus Species 0.000 description 1
- 241000713333 Mouse mammary tumor virus Species 0.000 description 1
- 241000714177 Murine leukemia virus Species 0.000 description 1
- CMWTZPSULFXXJA-UHFFFAOYSA-N Naproxen Natural products C1=C(C(C)C(O)=O)C=CC2=CC(OC)=CC=C21 CMWTZPSULFXXJA-UHFFFAOYSA-N 0.000 description 1
- 229930193140 Neomycin Natural products 0.000 description 1
- 208000008457 Neurologic Manifestations Diseases 0.000 description 1
- 101100215778 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) ptr-1 gene Proteins 0.000 description 1
- 241000714209 Norwalk virus Species 0.000 description 1
- 108700001237 Nucleic Acid-Based Vaccines Proteins 0.000 description 1
- 108091007494 Nucleic acid- binding domains Proteins 0.000 description 1
- 239000005642 Oleic acid Substances 0.000 description 1
- ZQPPMHVWECSIRJ-UHFFFAOYSA-N Oleic acid Natural products CCCCCCCCC=CCCCCCCCC(O)=O ZQPPMHVWECSIRJ-UHFFFAOYSA-N 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 238000010222 PCR analysis Methods 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 101710173835 Penton protein Proteins 0.000 description 1
- 102000003992 Peroxidases Human genes 0.000 description 1
- 241000283966 Pholidota <mammal> Species 0.000 description 1
- 102100026918 Phospholipase A2 Human genes 0.000 description 1
- 108010058864 Phospholipases A2 Proteins 0.000 description 1
- 229920002565 Polyethylene Glycol 400 Polymers 0.000 description 1
- 241001505332 Polyomavirus sp. Species 0.000 description 1
- 241000188845 Porcine adenovirus Species 0.000 description 1
- 241000288906 Primates Species 0.000 description 1
- 241000282330 Procyon lotor Species 0.000 description 1
- 206010036790 Productive cough Diseases 0.000 description 1
- 101710194807 Protective antigen Proteins 0.000 description 1
- 101710188315 Protein X Proteins 0.000 description 1
- 241000711798 Rabies lyssavirus Species 0.000 description 1
- 241000700159 Rattus Species 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 241000702263 Reovirus sp. Species 0.000 description 1
- 108020005091 Replication Origin Proteins 0.000 description 1
- 208000013616 Respiratory Distress Syndrome Diseases 0.000 description 1
- 241000712907 Retroviridae Species 0.000 description 1
- 208000036071 Rhinorrhea Diseases 0.000 description 1
- 206010039101 Rhinorrhoea Diseases 0.000 description 1
- IWUCXVSUMQZMFG-AFCXAGJDSA-N Ribavirin Chemical compound N1=C(C(=O)N)N=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 IWUCXVSUMQZMFG-AFCXAGJDSA-N 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 241000714474 Rous sarcoma virus Species 0.000 description 1
- 102000044437 S1 domains Human genes 0.000 description 1
- 108700036684 S1 domains Proteins 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 101100068078 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) GCN4 gene Proteins 0.000 description 1
- 206010039491 Sarcoma Diseases 0.000 description 1
- 208000032370 Secondary transmission Diseases 0.000 description 1
- 206010040047 Sepsis Diseases 0.000 description 1
- 206010040070 Septic Shock Diseases 0.000 description 1
- 108010071390 Serum Albumin Proteins 0.000 description 1
- 102000007562 Serum Albumin Human genes 0.000 description 1
- 241000144290 Sigmodon hispidus Species 0.000 description 1
- 241001217859 Simian adenovirus 1 Species 0.000 description 1
- 241001520548 Simian adenovirus 21 Species 0.000 description 1
- 241001492399 Simian adenovirus 23 Species 0.000 description 1
- 241000745901 Simian adenovirus 24 Species 0.000 description 1
- 241000745906 Simian adenovirus 25 Species 0.000 description 1
- 241001618295 Simian adenovirus 26 Species 0.000 description 1
- 241001618250 Simian adenovirus 27.1 Species 0.000 description 1
- 241001618252 Simian adenovirus 28.1 Species 0.000 description 1
- 241001618297 Simian adenovirus 29 Species 0.000 description 1
- 241001618289 Simian adenovirus 30 Species 0.000 description 1
- 241001618244 Simian adenovirus 31.1 Species 0.000 description 1
- 241001618293 Simian adenovirus 32 Species 0.000 description 1
- 241001618291 Simian adenovirus 33 Species 0.000 description 1
- 241001618283 Simian adenovirus 34 Species 0.000 description 1
- 241001618247 Simian adenovirus 35.1 Species 0.000 description 1
- 241001618278 Simian adenovirus 36 Species 0.000 description 1
- 241001618234 Simian adenovirus 37.2 Species 0.000 description 1
- 241001618277 Simian adenovirus 39 Species 0.000 description 1
- 241001618230 Simian adenovirus 40.1 Species 0.000 description 1
- 241001618232 Simian adenovirus 41.1 Species 0.000 description 1
- 241001618225 Simian adenovirus 42.1 Species 0.000 description 1
- 241001618271 Simian adenovirus 43 Species 0.000 description 1
- 241001618269 Simian adenovirus 44 Species 0.000 description 1
- 241001618261 Simian adenovirus 45 Species 0.000 description 1
- 241001618255 Simian adenovirus 46 Species 0.000 description 1
- 241000813758 Simian adenovirus 48 Species 0.000 description 1
- 241000813756 Simian adenovirus 49 Species 0.000 description 1
- 241000813754 Simian adenovirus 50 Species 0.000 description 1
- 241000701155 Simian adenovirus 7 Species 0.000 description 1
- 241000991300 Simian adenovirus 8 Species 0.000 description 1
- 241000745902 Simian adenovirus E22 Species 0.000 description 1
- 241000713311 Simian immunodeficiency virus Species 0.000 description 1
- 239000004147 Sorbitan trioleate Substances 0.000 description 1
- PRXRUNOAOLTIEF-ADSICKODSA-N Sorbitan trioleate Chemical compound CCCCCCCC\C=C/CCCCCCCC(=O)OC[C@@H](OC(=O)CCCCCCC\C=C/CCCCCCCC)[C@H]1OC[C@H](O)[C@H]1OC(=O)CCCCCCC\C=C/CCCCCCCC PRXRUNOAOLTIEF-ADSICKODSA-N 0.000 description 1
- 101710172711 Structural protein Proteins 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- 208000000389 T-cell leukemia Diseases 0.000 description 1
- 208000028530 T-cell lymphoblastic leukemia/lymphoma Diseases 0.000 description 1
- 108091023040 Transcription factor Proteins 0.000 description 1
- 102000040945 Transcription factor Human genes 0.000 description 1
- 102000009618 Transforming Growth Factors Human genes 0.000 description 1
- 108010009583 Transforming Growth Factors Proteins 0.000 description 1
- 102400001320 Transforming growth factor alpha Human genes 0.000 description 1
- 101800004564 Transforming growth factor alpha Proteins 0.000 description 1
- 102100039094 Tyrosinase Human genes 0.000 description 1
- 108060008724 Tyrosinase Proteins 0.000 description 1
- 241000700618 Vaccinia virus Species 0.000 description 1
- 241000710959 Venezuelan equine encephalitis virus Species 0.000 description 1
- 241000714205 Woolly monkey sarcoma virus Species 0.000 description 1
- 241001115400 Zaire ebolavirus Species 0.000 description 1
- UZQJVUCHXGYFLQ-AYDHOLPZSA-N [(2s,3r,4s,5r,6r)-4-[(2s,3r,4s,5r,6r)-4-[(2r,3r,4s,5r,6r)-4-[(2s,3r,4s,5r,6r)-3,5-dihydroxy-6-(hydroxymethyl)-4-[(2s,3r,4s,5s,6r)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyoxan-2-yl]oxy-3,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-3,5-dihydroxy-6-(hy Chemical compound O([C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O)O[C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O)O[C@H]1CC[C@]2(C)[C@H]3CC=C4[C@@]([C@@]3(CC[C@H]2[C@@]1(C=O)C)C)(C)CC(O)[C@]1(CCC(CC14)(C)C)C(=O)O[C@H]1[C@@H]([C@@H](O[C@H]2[C@@H]([C@@H](O[C@H]3[C@@H]([C@@H](O[C@H]4[C@@H]([C@@H](O[C@H]5[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O5)O)[C@H](O)[C@@H](CO)O4)O)[C@H](O)[C@@H](CO)O3)O)[C@H](O)[C@@H](CO)O2)O)[C@H](O)[C@@H](CO)O1)O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O UZQJVUCHXGYFLQ-AYDHOLPZSA-N 0.000 description 1
- HIHOWBSBBDRPDW-PTHRTHQKSA-N [(3s,8s,9s,10r,13r,14s,17r)-10,13-dimethyl-17-[(2r)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1h-cyclopenta[a]phenanthren-3-yl] n-[2-(dimethylamino)ethyl]carbamate Chemical compound C1C=C2C[C@@H](OC(=O)NCCN(C)C)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HIHOWBSBBDRPDW-PTHRTHQKSA-N 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 230000002159 abnormal effect Effects 0.000 description 1
- 238000010521 absorption reaction Methods 0.000 description 1
- 150000001242 acetic acid derivatives Chemical class 0.000 description 1
- 229960001138 acetylsalicylic acid Drugs 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 206010001053 acute respiratory failure Diseases 0.000 description 1
- 108700010877 adenoviridae proteins Proteins 0.000 description 1
- 238000004115 adherent culture Methods 0.000 description 1
- 230000001464 adherent effect Effects 0.000 description 1
- 201000000028 adult respiratory distress syndrome Diseases 0.000 description 1
- 235000019666 ageusia Nutrition 0.000 description 1
- AZDRQVAHHNSJOQ-UHFFFAOYSA-N alumane Chemical class [AlH3] AZDRQVAHHNSJOQ-UHFFFAOYSA-N 0.000 description 1
- NTGGOTYRTOXKMQ-UHFFFAOYSA-K aluminum;potassium;phosphate Chemical compound [Al+3].[K+].[O-]P([O-])([O-])=O NTGGOTYRTOXKMQ-UHFFFAOYSA-K 0.000 description 1
- 210000002821 alveolar epithelial cell Anatomy 0.000 description 1
- 230000006229 amino acid addition Effects 0.000 description 1
- 210000004381 amniotic fluid Anatomy 0.000 description 1
- 229960000723 ampicillin Drugs 0.000 description 1
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 1
- 238000004873 anchoring Methods 0.000 description 1
- 108010072788 angiogenin Proteins 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 239000003146 anticoagulant agent Substances 0.000 description 1
- 229940127219 anticoagulant drug Drugs 0.000 description 1
- 229940111131 antiinflammatory and antirheumatic product propionic acid derivative Drugs 0.000 description 1
- 239000002246 antineoplastic agent Substances 0.000 description 1
- 239000003963 antioxidant agent Substances 0.000 description 1
- 235000006708 antioxidants Nutrition 0.000 description 1
- 230000007416 antiviral immune response Effects 0.000 description 1
- 230000004596 appetite loss Effects 0.000 description 1
- 238000013459 approach Methods 0.000 description 1
- 239000008365 aqueous carrier Substances 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 210000004507 artificial chromosome Anatomy 0.000 description 1
- 210000003567 ascitic fluid Anatomy 0.000 description 1
- 235000010323 ascorbic acid Nutrition 0.000 description 1
- 229960005070 ascorbic acid Drugs 0.000 description 1
- 239000011668 ascorbic acid Substances 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- FZCSTZYAHCUGEM-UHFFFAOYSA-N aspergillomarasmine B Natural products OC(=O)CNC(C(O)=O)CNC(C(O)=O)CC(O)=O FZCSTZYAHCUGEM-UHFFFAOYSA-N 0.000 description 1
- 208000006673 asthma Diseases 0.000 description 1
- 208000004668 avian leukosis Diseases 0.000 description 1
- 210000003719 b-lymphocyte Anatomy 0.000 description 1
- 239000000688 bacterial toxin Substances 0.000 description 1
- XUZMWHLSFXCVMG-UHFFFAOYSA-N baricitinib Chemical compound C1N(S(=O)(=O)CC)CC1(CC#N)N1N=CC(C=2C=3C=CNC=3N=CN=2)=C1 XUZMWHLSFXCVMG-UHFFFAOYSA-N 0.000 description 1
- 229950000971 baricitinib Drugs 0.000 description 1
- 238000010923 batch production Methods 0.000 description 1
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 238000010364 biochemical engineering Methods 0.000 description 1
- 229920000249 biocompatible polymer Polymers 0.000 description 1
- 238000001574 biopsy Methods 0.000 description 1
- 239000012503 blood component Substances 0.000 description 1
- 210000000133 brain stem Anatomy 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 229910052793 cadmium Inorganic materials 0.000 description 1
- BDOSMKKIYDKNTQ-UHFFFAOYSA-N cadmium atom Chemical compound [Cd] BDOSMKKIYDKNTQ-UHFFFAOYSA-N 0.000 description 1
- AXCZMVOFGPJBDE-UHFFFAOYSA-L calcium dihydroxide Chemical compound [OH-].[OH-].[Ca+2] AXCZMVOFGPJBDE-UHFFFAOYSA-L 0.000 description 1
- 239000000920 calcium hydroxide Substances 0.000 description 1
- 229910001861 calcium hydroxide Inorganic materials 0.000 description 1
- 239000007963 capsule composition Substances 0.000 description 1
- 230000003197 catalytic effect Effects 0.000 description 1
- 229960000590 celecoxib Drugs 0.000 description 1
- RZEKVGVHFLEQIL-UHFFFAOYSA-N celecoxib Chemical compound C1=CC(C)=CC=C1C1=CC(C(F)(F)F)=NN1C1=CC=C(S(N)(=O)=O)C=C1 RZEKVGVHFLEQIL-UHFFFAOYSA-N 0.000 description 1
- 210000000170 cell membrane Anatomy 0.000 description 1
- 210000001175 cerebrospinal fluid Anatomy 0.000 description 1
- 239000002738 chelating agent Substances 0.000 description 1
- 238000006243 chemical reaction Methods 0.000 description 1
- 229960003677 chloroquine Drugs 0.000 description 1
- WHTVZRBIWZFKQO-UHFFFAOYSA-N chloroquine Natural products ClC1=CC=C2C(NC(C)CCCN(CC)CC)=CC=NC2=C1 WHTVZRBIWZFKQO-UHFFFAOYSA-N 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 229960002303 citric acid monohydrate Drugs 0.000 description 1
- 229940047120 colony stimulating factors Drugs 0.000 description 1
- 239000013066 combination product Substances 0.000 description 1
- 229940127555 combination product Drugs 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 238000010924 continuous production Methods 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 230000002596 correlated effect Effects 0.000 description 1
- 229940111134 coxibs Drugs 0.000 description 1
- 230000009260 cross reactivity Effects 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 125000004122 cyclic group Chemical group 0.000 description 1
- 239000003255 cyclooxygenase 2 inhibitor Substances 0.000 description 1
- 229940127089 cytotoxic agent Drugs 0.000 description 1
- 230000002354 daily effect Effects 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 230000003111 delayed effect Effects 0.000 description 1
- 210000000852 deltoid muscle Anatomy 0.000 description 1
- KXGVEGMKQFWNSR-LLQZFEROSA-N deoxycholic acid Chemical compound C([C@H]1CC2)[C@H](O)CC[C@]1(C)[C@@H]1[C@@H]2[C@@H]2CC[C@H]([C@@H](CCC(O)=O)C)[C@@]2(C)[C@@H](O)C1 KXGVEGMKQFWNSR-LLQZFEROSA-N 0.000 description 1
- 229960003964 deoxycholic acid Drugs 0.000 description 1
- KXGVEGMKQFWNSR-UHFFFAOYSA-N deoxycholic acid Natural products C1CC2CC(O)CCC2(C)C2C1C1CCC(C(CCC(O)=O)C)C1(C)C(O)C2 KXGVEGMKQFWNSR-UHFFFAOYSA-N 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 239000003599 detergent Substances 0.000 description 1
- 235000019425 dextrin Nutrition 0.000 description 1
- 238000002405 diagnostic procedure Methods 0.000 description 1
- 230000035487 diastolic blood pressure Effects 0.000 description 1
- PXBRQCKWGAHEHS-UHFFFAOYSA-N dichlorodifluoromethane Chemical compound FC(F)(Cl)Cl PXBRQCKWGAHEHS-UHFFFAOYSA-N 0.000 description 1
- 235000019404 dichlorodifluoromethane Nutrition 0.000 description 1
- UMNKXPULIDJLSU-UHFFFAOYSA-N dichlorofluoromethane Chemical compound FC(Cl)Cl UMNKXPULIDJLSU-UHFFFAOYSA-N 0.000 description 1
- 229940099364 dichlorofluoromethane Drugs 0.000 description 1
- 229940087091 dichlorotetrafluoroethane Drugs 0.000 description 1
- 229960001259 diclofenac Drugs 0.000 description 1
- DCOPUUMXTXDBNB-UHFFFAOYSA-N diclofenac Chemical compound OC(=O)CC1=CC=CC=C1NC1=C(Cl)C=CC=C1Cl DCOPUUMXTXDBNB-UHFFFAOYSA-N 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 150000002016 disaccharides Chemical class 0.000 description 1
- 230000008034 disappearance Effects 0.000 description 1
- 208000028659 discharge Diseases 0.000 description 1
- 208000018459 dissociative disease Diseases 0.000 description 1
- 238000012137 double-staining Methods 0.000 description 1
- 229960001850 droxicam Drugs 0.000 description 1
- OEHFRZLKGRKFAS-UHFFFAOYSA-N droxicam Chemical compound C12=CC=CC=C2S(=O)(=O)N(C)C(C2=O)=C1OC(=O)N2C1=CC=CC=N1 OEHFRZLKGRKFAS-UHFFFAOYSA-N 0.000 description 1
- 238000001647 drug administration Methods 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 1
- 208000025887 endotheliitis Diseases 0.000 description 1
- 239000007920 enema Substances 0.000 description 1
- 229940079360 enema for constipation Drugs 0.000 description 1
- 231100000655 enterotoxin Toxicity 0.000 description 1
- 244000309457 enveloped RNA virus Species 0.000 description 1
- 230000008472 epithelial growth Effects 0.000 description 1
- 229960005293 etodolac Drugs 0.000 description 1
- XFBVBWWRPKNWHW-UHFFFAOYSA-N etodolac Chemical compound C1COC(CC)(CC(O)=O)C2=N[C]3C(CC)=CC=CC3=C21 XFBVBWWRPKNWHW-UHFFFAOYSA-N 0.000 description 1
- 229960004945 etoricoxib Drugs 0.000 description 1
- MNJVRJDLRVPLFE-UHFFFAOYSA-N etoricoxib Chemical compound C1=NC(C)=CC=C1C1=NC=C(Cl)C=C1C1=CC=C(S(C)(=O)=O)C=C1 MNJVRJDLRVPLFE-UHFFFAOYSA-N 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 230000003203 everyday effect Effects 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 210000003722 extracellular fluid Anatomy 0.000 description 1
- 239000003889 eye drop Substances 0.000 description 1
- 229940012356 eye drops Drugs 0.000 description 1
- 231100000013 eye irritation Toxicity 0.000 description 1
- ZCGNOVWYSGBHAU-UHFFFAOYSA-N favipiravir Chemical compound NC(=O)C1=NC(F)=CNC1=O ZCGNOVWYSGBHAU-UHFFFAOYSA-N 0.000 description 1
- 229950008454 favipiravir Drugs 0.000 description 1
- ZWJINEZUASEZBH-UHFFFAOYSA-N fenamic acid Chemical class OC(=O)C1=CC=CC=C1NC1=CC=CC=C1 ZWJINEZUASEZBH-UHFFFAOYSA-N 0.000 description 1
- 229960001419 fenoprofen Drugs 0.000 description 1
- 229940126864 fibroblast growth factor Drugs 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 239000000834 fixative Substances 0.000 description 1
- 229960004369 flufenamic acid Drugs 0.000 description 1
- LPEPZBJOKDYZAD-UHFFFAOYSA-N flufenamic acid Chemical compound OC(=O)C1=CC=CC=C1NC1=CC=CC(C(F)(F)F)=C1 LPEPZBJOKDYZAD-UHFFFAOYSA-N 0.000 description 1
- 238000000198 fluorescence anisotropy Methods 0.000 description 1
- 229960002390 flurbiprofen Drugs 0.000 description 1
- SYTBZMRGLBWNTM-UHFFFAOYSA-N flurbiprofen Chemical compound FC1=CC(C(C(O)=O)C)=CC=C1C1=CC=CC=C1 SYTBZMRGLBWNTM-UHFFFAOYSA-N 0.000 description 1
- 239000012537 formulation buffer Substances 0.000 description 1
- 238000012395 formulation development Methods 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 239000003862 glucocorticoid Substances 0.000 description 1
- 239000008103 glucose Substances 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 150000004676 glycans Chemical class 0.000 description 1
- 210000002288 golgi apparatus Anatomy 0.000 description 1
- 239000008187 granular material Substances 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 229940093915 gynecological organic acid Drugs 0.000 description 1
- 230000009931 harmful effect Effects 0.000 description 1
- 229910001385 heavy metal Inorganic materials 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 208000027700 hepatic dysfunction Diseases 0.000 description 1
- 208000006454 hepatitis Diseases 0.000 description 1
- 231100000283 hepatitis Toxicity 0.000 description 1
- 208000021760 high fever Diseases 0.000 description 1
- 102000048657 human ACE2 Human genes 0.000 description 1
- 229960000890 hydrocortisone Drugs 0.000 description 1
- 239000000017 hydrogel Substances 0.000 description 1
- 229920001477 hydrophilic polymer Polymers 0.000 description 1
- XXSMGPRMXLTPCZ-UHFFFAOYSA-N hydroxychloroquine Chemical compound ClC1=CC=C2C(NC(C)CCCN(CCO)CC)=CC=NC2=C1 XXSMGPRMXLTPCZ-UHFFFAOYSA-N 0.000 description 1
- 229960004171 hydroxychloroquine Drugs 0.000 description 1
- 229960001680 ibuprofen Drugs 0.000 description 1
- 230000014726 immortalization of host cell Effects 0.000 description 1
- 210000002865 immune cell Anatomy 0.000 description 1
- 238000010166 immunofluorescence Methods 0.000 description 1
- 238000003125 immunofluorescent labeling Methods 0.000 description 1
- 239000000677 immunologic agent Substances 0.000 description 1
- 229940124541 immunological agent Drugs 0.000 description 1
- 239000002955 immunomodulating agent Substances 0.000 description 1
- 229940121354 immunomodulator Drugs 0.000 description 1
- 239000007943 implant Substances 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 230000000415 inactivating effect Effects 0.000 description 1
- 239000005414 inactive ingredient Substances 0.000 description 1
- 229960000905 indomethacin Drugs 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- 230000002452 interceptive effect Effects 0.000 description 1
- 229960001388 interferon-beta Drugs 0.000 description 1
- 239000000543 intermediate Substances 0.000 description 1
- 210000002570 interstitial cell Anatomy 0.000 description 1
- 208000028774 intestinal disease Diseases 0.000 description 1
- 238000001361 intraarterial administration Methods 0.000 description 1
- 238000010212 intracellular staining Methods 0.000 description 1
- 238000007917 intracranial administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 230000002601 intratumoral effect Effects 0.000 description 1
- QXJSBBXBKPUZAA-UHFFFAOYSA-N isooleic acid Natural products CCCCCCCC=CCCCCCCCCC(O)=O QXJSBBXBKPUZAA-UHFFFAOYSA-N 0.000 description 1
- 238000000111 isothermal titration calorimetry Methods 0.000 description 1
- 229950002252 isoxicam Drugs 0.000 description 1
- YYUAYBYLJSNDCX-UHFFFAOYSA-N isoxicam Chemical compound OC=1C2=CC=CC=C2S(=O)(=O)N(C)C=1C(=O)NC=1C=C(C)ON=1 YYUAYBYLJSNDCX-UHFFFAOYSA-N 0.000 description 1
- 229960000318 kanamycin Drugs 0.000 description 1
- 229930027917 kanamycin Natural products 0.000 description 1
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 1
- 229930182823 kanamycin A Natural products 0.000 description 1
- DKYWVDODHFEZIM-UHFFFAOYSA-N ketoprofen Chemical compound OC(=O)C(C)C1=CC=CC(C(=O)C=2C=CC=CC=2)=C1 DKYWVDODHFEZIM-UHFFFAOYSA-N 0.000 description 1
- 229960000991 ketoprofen Drugs 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 230000002045 lasting effect Effects 0.000 description 1
- 239000010410 layer Substances 0.000 description 1
- 230000003902 lesion Effects 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- GZQKNULLWNGMCW-PWQABINMSA-N lipid A (E. coli) Chemical compound O1[C@H](CO)[C@@H](OP(O)(O)=O)[C@H](OC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCCCC)[C@@H](NC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCC)[C@@H]1OC[C@@H]1[C@@H](O)[C@H](OC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](NC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](OP(O)(O)=O)O1 GZQKNULLWNGMCW-PWQABINMSA-N 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 229920006008 lipopolysaccharide Polymers 0.000 description 1
- 239000007788 liquid Substances 0.000 description 1
- 239000006194 liquid suspension Substances 0.000 description 1
- 229940113983 lopinavir / ritonavir Drugs 0.000 description 1
- 229960002202 lornoxicam Drugs 0.000 description 1
- OXROWJKCGCOJDO-JLHYYAGUSA-N lornoxicam Chemical compound O=C1C=2SC(Cl)=CC=2S(=O)(=O)N(C)\C1=C(\O)NC1=CC=CC=N1 OXROWJKCGCOJDO-JLHYYAGUSA-N 0.000 description 1
- 208000019017 loss of appetite Diseases 0.000 description 1
- 235000021266 loss of appetite Nutrition 0.000 description 1
- 208000030500 lower respiratory tract disease Diseases 0.000 description 1
- 238000003670 luciferase enzyme activity assay Methods 0.000 description 1
- 229960000994 lumiracoxib Drugs 0.000 description 1
- KHPKQFYUPIUARC-UHFFFAOYSA-N lumiracoxib Chemical compound OC(=O)CC1=CC(C)=CC=C1NC1=C(F)C=CC=C1Cl KHPKQFYUPIUARC-UHFFFAOYSA-N 0.000 description 1
- 210000002751 lymph Anatomy 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 230000000527 lymphocytic effect Effects 0.000 description 1
- 210000003563 lymphoid tissue Anatomy 0.000 description 1
- 229910001629 magnesium chloride Inorganic materials 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 239000011159 matrix material Substances 0.000 description 1
- 238000005399 mechanical ventilation Methods 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 229960003803 meclofenamic acid Drugs 0.000 description 1
- 229960003464 mefenamic acid Drugs 0.000 description 1
- HYYBABOKPJLUIN-UHFFFAOYSA-N mefenamic acid Chemical compound CC1=CC=CC(NC=2C(=CC=CC=2)C(O)=O)=C1C HYYBABOKPJLUIN-UHFFFAOYSA-N 0.000 description 1
- 229960001929 meloxicam Drugs 0.000 description 1
- 210000004779 membrane envelope Anatomy 0.000 description 1
- 230000034217 membrane fusion Effects 0.000 description 1
- 230000037323 metabolic rate Effects 0.000 description 1
- 239000002923 metal particle Substances 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- 239000002679 microRNA Substances 0.000 description 1
- 238000012737 microarray-based gene expression Methods 0.000 description 1
- 230000000813 microbial effect Effects 0.000 description 1
- 239000003094 microcapsule Substances 0.000 description 1
- 239000004005 microsphere Substances 0.000 description 1
- 230000003278 mimic effect Effects 0.000 description 1
- 230000002438 mitochondrial effect Effects 0.000 description 1
- 238000009126 molecular therapy Methods 0.000 description 1
- 150000002772 monosaccharides Chemical class 0.000 description 1
- 210000004400 mucous membrane Anatomy 0.000 description 1
- 238000012243 multiplex automated genomic engineering Methods 0.000 description 1
- 229940031348 multivalent vaccine Drugs 0.000 description 1
- BSOQXXWZTUDTEL-ZUYCGGNHSA-N muramyl dipeptide Chemical compound OC(=O)CC[C@H](C(N)=O)NC(=O)[C@H](C)NC(=O)[C@@H](C)O[C@H]1[C@H](O)[C@@H](CO)O[C@@H](O)[C@@H]1NC(C)=O BSOQXXWZTUDTEL-ZUYCGGNHSA-N 0.000 description 1
- 210000003205 muscle Anatomy 0.000 description 1
- 210000004165 myocardium Anatomy 0.000 description 1
- OHDXDNUPVVYWOV-UHFFFAOYSA-N n-methyl-1-(2-naphthalen-1-ylsulfanylphenyl)methanamine Chemical compound CNCC1=CC=CC=C1SC1=CC=CC2=CC=CC=C12 OHDXDNUPVVYWOV-UHFFFAOYSA-N 0.000 description 1
- 239000002105 nanoparticle Substances 0.000 description 1
- 229960002009 naproxen Drugs 0.000 description 1
- CMWTZPSULFXXJA-VIFPVBQESA-N naproxen Chemical compound C1=C([C@H](C)C(O)=O)C=CC2=CC(OC)=CC=C21 CMWTZPSULFXXJA-VIFPVBQESA-N 0.000 description 1
- 229920005615 natural polymer Polymers 0.000 description 1
- 239000013642 negative control Substances 0.000 description 1
- 229960004927 neomycin Drugs 0.000 description 1
- 230000009251 neurologic dysfunction Effects 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- 229910052757 nitrogen Inorganic materials 0.000 description 1
- 229940021182 non-steroidal anti-inflammatory drug Drugs 0.000 description 1
- 239000002736 nonionic surfactant Substances 0.000 description 1
- 229940023146 nucleic acid vaccine Drugs 0.000 description 1
- 239000007764 o/w emulsion Substances 0.000 description 1
- 239000002674 ointment Substances 0.000 description 1
- ZQPPMHVWECSIRJ-KTKRTIGZSA-N oleic acid Chemical compound CCCCCCCC\C=C/CCCCCCCC(O)=O ZQPPMHVWECSIRJ-KTKRTIGZSA-N 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 210000003463 organelle Anatomy 0.000 description 1
- 150000007524 organic acids Chemical class 0.000 description 1
- 235000005985 organic acids Nutrition 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 229960002739 oxaprozin Drugs 0.000 description 1
- OFPXSFXSNFPTHF-UHFFFAOYSA-N oxaprozin Chemical compound O1C(CCC(=O)O)=NC(C=2C=CC=CC=2)=C1C1=CC=CC=C1 OFPXSFXSNFPTHF-UHFFFAOYSA-N 0.000 description 1
- 229960004662 parecoxib Drugs 0.000 description 1
- TZRHLKRLEZJVIJ-UHFFFAOYSA-N parecoxib Chemical compound C1=CC(S(=O)(=O)NC(=O)CC)=CC=C1C1=C(C)ON=C1C1=CC=CC=C1 TZRHLKRLEZJVIJ-UHFFFAOYSA-N 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 239000006072 paste Substances 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 230000002085 persistent effect Effects 0.000 description 1
- 229940124531 pharmaceutical excipient Drugs 0.000 description 1
- 239000008180 pharmaceutical surfactant Substances 0.000 description 1
- 239000002831 pharmacologic agent Substances 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- WVDDGKGOMKODPV-ZQBYOMGUSA-N phenyl(114C)methanol Chemical compound O[14CH2]C1=CC=CC=C1 WVDDGKGOMKODPV-ZQBYOMGUSA-N 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 239000006187 pill Substances 0.000 description 1
- 229960002702 piroxicam Drugs 0.000 description 1
- QYSPLQLAKJAUJT-UHFFFAOYSA-N piroxicam Chemical compound OC=1C2=CC=CC=C2S(=O)(=O)N(C)C=1C(=O)NC1=CC=CC=N1 QYSPLQLAKJAUJT-UHFFFAOYSA-N 0.000 description 1
- 229920001983 poloxamer Polymers 0.000 description 1
- 229920002627 poly(phosphazenes) Polymers 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920001282 polysaccharide Polymers 0.000 description 1
- 239000005017 polysaccharide Substances 0.000 description 1
- 229920000136 polysorbate Polymers 0.000 description 1
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 1
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 1
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 1
- 239000013641 positive control Substances 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 229960004618 prednisone Drugs 0.000 description 1
- XOFYZVNMUHMLCC-ZPOLXVRWSA-N prednisone Chemical compound O=C1C=C[C@]2(C)[C@H]3C(=O)C[C@](C)([C@@](CC4)(O)C(=O)CO)[C@@H]4[C@@H]3CCC2=C1 XOFYZVNMUHMLCC-ZPOLXVRWSA-N 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 230000000770 proinflammatory effect Effects 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000002062 proliferating effect Effects 0.000 description 1
- 239000001294 propane Substances 0.000 description 1
- 238000011321 prophylaxis Methods 0.000 description 1
- 150000005599 propionic acid derivatives Chemical class 0.000 description 1
- 239000004405 propyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010232 propyl p-hydroxybenzoate Nutrition 0.000 description 1
- 229960003415 propylparaben Drugs 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 208000010707 pulmonary consolidation Diseases 0.000 description 1
- 208000005333 pulmonary edema Diseases 0.000 description 1
- 230000002685 pulmonary effect Effects 0.000 description 1
- 210000003314 quadriceps muscle Anatomy 0.000 description 1
- 238000004445 quantitative analysis Methods 0.000 description 1
- 239000001397 quillaja saponaria molina bark Substances 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- RWWYLEGWBNMMLJ-MEUHYHILSA-N remdesivir Drugs C([C@@H]1[C@H]([C@@H](O)[C@@](C#N)(O1)C=1N2N=CN=C(N)C2=CC=1)O)OP(=O)(N[C@@H](C)C(=O)OCC(CC)CC)OC1=CC=CC=C1 RWWYLEGWBNMMLJ-MEUHYHILSA-N 0.000 description 1
- RWWYLEGWBNMMLJ-YSOARWBDSA-N remdesivir Chemical compound NC1=NC=NN2C1=CC=C2[C@]1([C@@H]([C@@H]([C@H](O1)CO[P@](=O)(OC1=CC=CC=C1)N[C@H](C(=O)OCC(CC)CC)C)O)O)C#N RWWYLEGWBNMMLJ-YSOARWBDSA-N 0.000 description 1
- 230000008085 renal dysfunction Effects 0.000 description 1
- 230000036454 renin-angiotensin system Effects 0.000 description 1
- 230000000754 repressing effect Effects 0.000 description 1
- 230000000284 resting effect Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 230000002207 retinal effect Effects 0.000 description 1
- 229960000329 ribavirin Drugs 0.000 description 1
- HZCAHMRRMINHDJ-DBRKOABJSA-N ribavirin Natural products O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1N=CN=C1 HZCAHMRRMINHDJ-DBRKOABJSA-N 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 231100000279 safety data Toxicity 0.000 description 1
- 210000003296 saliva Anatomy 0.000 description 1
- 239000012898 sample dilution Substances 0.000 description 1
- 229930182490 saponin Natural products 0.000 description 1
- 150000007949 saponins Chemical class 0.000 description 1
- 238000013341 scale-up Methods 0.000 description 1
- 230000036303 septic shock Effects 0.000 description 1
- 230000035939 shock Effects 0.000 description 1
- 208000013220 shortness of breath Diseases 0.000 description 1
- 210000000813 small intestine Anatomy 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- NLJMYIDDQXHKNR-UHFFFAOYSA-K sodium citrate Chemical compound O.O.[Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O NLJMYIDDQXHKNR-UHFFFAOYSA-K 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 235000019337 sorbitan trioleate Nutrition 0.000 description 1
- 229960000391 sorbitan trioleate Drugs 0.000 description 1
- 239000000600 sorbitol Substances 0.000 description 1
- 230000002269 spontaneous effect Effects 0.000 description 1
- 239000007921 spray Substances 0.000 description 1
- 210000003802 sputum Anatomy 0.000 description 1
- 208000024794 sputum Diseases 0.000 description 1
- 238000012289 standard assay Methods 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 230000001954 sterilising effect Effects 0.000 description 1
- 238000004659 sterilization and disinfection Methods 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 229940031626 subunit vaccine Drugs 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 150000005846 sugar alcohols Chemical class 0.000 description 1
- 229960000894 sulindac Drugs 0.000 description 1
- MLKXDPUZXIRXEP-MFOYZWKCSA-N sulindac Chemical compound CC1=C(CC(O)=O)C2=CC(F)=CC=C2\C1=C/C1=CC=C(S(C)=O)C=C1 MLKXDPUZXIRXEP-MFOYZWKCSA-N 0.000 description 1
- 239000000829 suppository Substances 0.000 description 1
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 239000012730 sustained-release form Substances 0.000 description 1
- 238000003786 synthesis reaction Methods 0.000 description 1
- 229920001059 synthetic polymer Polymers 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 229940037128 systemic glucocorticoids Drugs 0.000 description 1
- 230000035488 systolic blood pressure Effects 0.000 description 1
- 239000007916 tablet composition Substances 0.000 description 1
- 230000008685 targeting Effects 0.000 description 1
- 229960002871 tenoxicam Drugs 0.000 description 1
- WZWYJBNHTWCXIM-UHFFFAOYSA-N tenoxicam Chemical compound O=C1C=2SC=CC=2S(=O)(=O)N(C)C1=C(O)NC1=CC=CC=N1 WZWYJBNHTWCXIM-UHFFFAOYSA-N 0.000 description 1
- 210000001550 testis Anatomy 0.000 description 1
- 238000004448 titration Methods 0.000 description 1
- 229960003989 tocilizumab Drugs 0.000 description 1
- 229960002905 tolfenamic acid Drugs 0.000 description 1
- YEZNLOUZAIOMLT-UHFFFAOYSA-N tolfenamic acid Chemical compound CC1=C(Cl)C=CC=C1NC1=CC=CC=C1C(O)=O YEZNLOUZAIOMLT-UHFFFAOYSA-N 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 210000003437 trachea Anatomy 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- ODLHGICHYURWBS-LKONHMLTSA-N trappsol cyclo Chemical compound CC(O)COC[C@H]([C@H]([C@@H]([C@H]1O)O)O[C@H]2O[C@@H]([C@@H](O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O3)[C@H](O)[C@H]2O)COCC(O)C)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O)[C@@H]3O[C@@H]1COCC(C)O ODLHGICHYURWBS-LKONHMLTSA-N 0.000 description 1
- CYRMSUTZVYGINF-UHFFFAOYSA-N trichlorofluoromethane Chemical compound FC(Cl)(Cl)Cl CYRMSUTZVYGINF-UHFFFAOYSA-N 0.000 description 1
- 229940029284 trichlorofluoromethane Drugs 0.000 description 1
- 230000001960 triggered effect Effects 0.000 description 1
- 230000010415 tropism Effects 0.000 description 1
- KCFYEAOKVJSACF-UHFFFAOYSA-N umifenovir Chemical compound CN1C2=CC(Br)=C(O)C(CN(C)C)=C2C(C(=O)OCC)=C1CSC1=CC=CC=C1 KCFYEAOKVJSACF-UHFFFAOYSA-N 0.000 description 1
- 229960004626 umifenovir Drugs 0.000 description 1
- 241000712461 unidentified influenza virus Species 0.000 description 1
- 210000000689 upper leg Anatomy 0.000 description 1
- 229960002004 valdecoxib Drugs 0.000 description 1
- LNPDTQAFDNKSHK-UHFFFAOYSA-N valdecoxib Chemical compound CC=1ON=C(C=2C=CC=CC=2)C=1C1=CC=C(S(N)(=O)=O)C=C1 LNPDTQAFDNKSHK-UHFFFAOYSA-N 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 210000003556 vascular endothelial cell Anatomy 0.000 description 1
- 229940126580 vector vaccine Drugs 0.000 description 1
- 230000017613 viral reproduction Effects 0.000 description 1
- 239000005723 virus inoculator Substances 0.000 description 1
- 239000008215 water for injection Substances 0.000 description 1
- 239000001993 wax Substances 0.000 description 1
- 238000001262 western blot Methods 0.000 description 1
- UGZADUVQMDAIAO-UHFFFAOYSA-L zinc hydroxide Chemical compound [OH-].[OH-].[Zn+2] UGZADUVQMDAIAO-UHFFFAOYSA-L 0.000 description 1
- 229910021511 zinc hydroxide Inorganic materials 0.000 description 1
- 229940007718 zinc hydroxide Drugs 0.000 description 1
Images















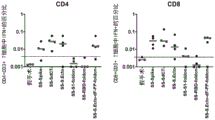

































































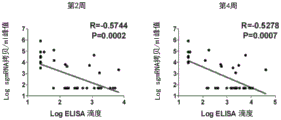










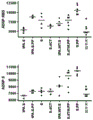





































































































































































































































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/215—Coronaviridae, e.g. avian infectious bronchitis virus
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1002—Coronaviridae
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1002—Coronaviridae
- C07K16/1003—Severe acute respiratory syndrome coronavirus 2 [SARS‐CoV‐2 or Covid-19]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N7/00—Viruses; Bacteriophages; Compositions thereof; Preparation or purification thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5256—Virus expressing foreign proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/53—DNA (RNA) vaccination
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/545—Medicinal preparations containing antigens or antibodies characterised by the dose, timing or administration schedule
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55505—Inorganic adjuvants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/10011—Adenoviridae
- C12N2710/10311—Mastadenovirus, e.g. human or simian adenoviruses
- C12N2710/10341—Use of virus, viral particle or viral elements as a vector
- C12N2710/10343—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14111—Dependovirus, e.g. adenoassociated viruses
- C12N2750/14141—Use of virus, viral particle or viral elements as a vector
- C12N2750/14143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14111—Dependovirus, e.g. adenoassociated viruses
- C12N2750/14151—Methods of production or purification of viral material
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14111—Dependovirus, e.g. adenoassociated viruses
- C12N2750/14171—Demonstrated in vivo effect
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/20011—Coronaviridae
- C12N2770/20022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/20011—Coronaviridae
- C12N2770/20034—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/20011—Coronaviridae
- C12N2770/20071—Demonstrated in vivo effect
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2800/00—Nucleic acids vectors
- C12N2800/22—Vectors comprising a coding region that has been codon optimised for expression in a respective host
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Virology (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Engineering & Computer Science (AREA)
- Immunology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Microbiology (AREA)
- Pharmacology & Pharmacy (AREA)
- Veterinary Medicine (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- General Engineering & Computer Science (AREA)
- Communicable Diseases (AREA)
- Pulmonology (AREA)
- Oncology (AREA)
- Epidemiology (AREA)
- Mycology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Gastroenterology & Hepatology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Abstract
本发明涉及含有冠状病毒(例如,2019‑nCoV;也称为SARS‑CoV‑2)蛋白或编码冠状病毒(例如,2019‑nCoV;SARS‑CoV‑2)蛋白的多核苷酸的免疫原性组合物和疫苗及其用途。本发明还提供了通过向受试者(例如,人)给药免疫原性组合物或疫苗来治疗和/或预防冠状病毒(例如2019‑nCoV;SARS‑CoV‑2)感染的方法。本发明还提供了检测和/或监测保护性抗冠状病毒(例如2019‑nCoV;SARS‑CoV‑2)抗体应答(例如,抗冠状病毒抗体应答,例如抗2019‑nCoV抗体应答,例如抗刺突抗体应答,例如抗刺突中和抗体应答)。本发明涉及编码冠状病毒S蛋白,特别是SARS‑CoV‑2S蛋白的分离的核酸和/或重组的核酸,还涉及冠状病毒S蛋白,以及涉及所述核酸和/或其蛋白在疫苗中的用途。
Description
技术领域
本发明涉及病毒学和医学领域。具体地,本发明涉及用于预防由SARS-CoV-2引起的疾病的疫苗。
相关申请的交叉引用
本申请要求以下优先权:2020年1月31日提交的美国临时申请号62/969,008;2020年3月25日提交的美国临时申请号62/994,630;2020年4月23日提交的美国临时申请号63/014,467;2020年5月15日提交的美国临时申请号63/025,782;2020年6月15日提交的美国临时申请号62/705,187;2020年6月21日提交的美国临时申请号62/705,308;2020年6月23日提交的美国临时申请号63/043,090;2020年8月12日提交的美国临时申请号62/706,366;2020年8月14日提交的美国临时申请号63/066,147;2020年9月2日提交的美国临时申请号62/706,676;2020年9月18日提交的美国临时申请号62/706,937;2020年9月21日提交的美国临时申请号62/706,958;2020年9月28日提交的美国临时申请号63/198,089;2020年10月9日提交的美国临时申请号63/198,306;2020年11月12日提交的美国临时申请号63/112,900;2020年11月28日提交的加拿大专利申请号3,101,131;2020年12月4日提交的美国临时申请号63/121,482;2021年1月5日提交的美国临时申请号63/133,969;2021年1月8日提交的美国临时申请号63/135,182;2021年1月26日提交的美国临时申请号63/141,913;2021年1月28日提交的美国临时申请号63/142,977。每个公开内容整体援引加入本文。
序列表
本申请含有已以ASCII格式电子提交的序列表,并且在此通过援引整体加入本文。ASCII副本创建于2021年1月29日,名为CRU6043WOPCT,大小为1.36MB。
关于联邦资助研究的声明
本发明是根据HHSO100201700018C协议在政府支持下完成的,由HHS资助。政府对本发明有一定的权利。
背景技术
SARS-CoV-2是一种β-冠状病毒,与MERS-CoV和SARS-CoV一样,它们都起源于蝙蝠。目前有来自多个国家的数名患者的数个序列,表明这种病毒可能是单一的、最近从动物储存宿主(reservoir)中出现的。由病毒导致的这种疾病的名称是2019冠状病毒病,缩写为COVID-19。对于确诊的COVID-19病例,COVID-19的症状范围从轻度症状到严重疾病和死亡。
如上所述,SARS-CoV-2与蝙蝠冠状病毒具有强的遗传相似性,它可能起源于蝙蝠冠状病毒,尽管认为也涉及诸如穿山甲的中间储存宿主。从分类学的角度来看,将SARS-CoV-2归类为与严重急性呼吸道综合症(SARS)相关的冠状病毒种的一株。
冠状病毒是有包膜的RNA病毒。主要表面蛋白是大的三聚体刺突糖蛋白(S),其介导与宿主细胞受体的结合以及病毒和宿主细胞膜的融合。S蛋白由分别负责受体结合和膜融合的N端S1亚基和C端S2亚基构成。最近对alpha-、beta-和delta冠状病毒的CoV三聚体S结构的冷冻电镜重构揭示,S1亚基包含两个不同的结构域:N末端结构域(S1 NTD)和受体结合结构域(S1 RBD)。SARS-CoV-2利用其S1 RBD与人血管紧张素转换酶2(ACE2)结合。
COVID-19大流行的迅速扩大使开发SARS-CoV-2疫苗成为全球卫生优先事项。自2019年底在人中首次观察到新型SARS-CoV-2病毒以来,已感染超过800万人,数十万人因COVID-19而死亡。SARS-CoV-2,和更概括地,冠状病毒,缺乏有效的治疗,导致大量未满足的医疗需求。此外,目前没有可用于预防冠状病毒引起的疾病(COVID-19)的疫苗。目前预防疾病的最优方法是避免暴露于这种病毒。由于新出现的传染病,例如COVID-19对公共卫生构成主要威胁,因此迫切需要可用于预防冠状病毒引起的呼吸道疾病的新型疫苗。
已知2019-nCoV会导致可能造成死亡的呼吸道症状和发烧。
世界卫生组织于2020年1月30日宣布2019-nCoV爆发为国际关注的突发公共卫生事件,并已在16个国家确诊超过11,000例病例。虽然快速开发安全有效的2019-nCoV疫苗是全球卫生优先事项,但目前对2019-nCoV免疫学和免疫保护机制知之甚少。
因此,2019-nCoV治疗领域有未满足的需求。
发明内容
在导致本发明的研究中,构建了某些稳定化的SARS-CoV-2 S蛋白,证明其可用作用于诱导针对SARS-CoV-2的保护性免疫应答的免疫原。
本发明提供了一种分离的核酸分子,其包含编码2019-NCOV刺突(S)蛋白(在本文中也称为SARS-CoV-2 S蛋白)的核苷酸序列,所述2019-NCOV刺突(S)蛋白包含对SEQ IDNO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
在一些实施方案中,这些是对SEQ ID NO:29的序列进行的唯一修饰。在其他实施方案中,所述分离的核酸分子编码2019-NCOV刺突(S)蛋白,所述2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
c.信号序列的缺失。
在一些实施方案中,编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(SEQ IDNO:219)启动子,优选CMV立即早期启动子可操作地连接。在一些实施方案中,编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(CMV)启动子可操作地连接,所述CMV启动子包含至少一个四环素操纵子(TetO)基序。在具体实施方案中,所述包含至少一个TetO基序的CMV启动子包含SEQ ID NO:219的核苷酸序列。在一些实施方案中,CMV启动子由SEQ ID NO:219的核苷酸序列组成。这些核酸通常形成载体的部分。
在一方面,本发明因此涉及分离的和/或重组的核酸,其编码稳定化的冠状病毒S蛋白,特别是SARS-CoV-2 S蛋白,或其片段,所述核酸包含选自SEQ ID NO:211-218的核苷酸序列。
在一个优选的实施方案中,本发明涉及一种分离的和/或重组的核酸,其编码稳定化的冠状病毒S蛋白,特别是SARS-CoV-2 S蛋白,或其片段,所述核酸包含SEQ ID NO:211的核苷酸。
本发明提供了一种分离的2019-NCOV刺突(S)蛋白(本文也称为SARS-CoV-2 S蛋白),其包含对SEQ ID NO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
在一些实施方案中,这些是对SEQ ID NO:29的序列进行的唯一修饰。在其他实施方案中,所述分离的2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
c.信号序列的缺失。
在另一方面,本发明涉及分离的和/或重组的冠状病毒S蛋白,其包含选自SEQ IDNO:205-210的氨基酸序列或其片段;以及涉及编码这种冠状病毒S蛋白或其片段的核酸。
在一个优选的实施方案中,本发明涉及一种分离的和/或重组的冠状病毒S蛋白,所述冠状病毒S蛋白包含SEQ ID NO:205的氨基酸序列,或其片段;以及涉及编码这种冠状病毒S蛋白或其片段的核酸。
在又一方面,本发明涉及包含这种核酸的载体。在某些实施方案中,所述载体是血清型26的重组人腺病毒。
在另一方面,本发明涉及包含这种核酸、蛋白和/或载体的组合物和疫苗。
在另一方面,本发明涉及向受试者接种针对COVID-19的疫苗的方法,所述方法包括向所述受试者给药本发明的疫苗或组合物。
在另一方面,本发明涉及一种分离的宿主细胞,其包含血清型26的重组人腺病毒,所述重组人腺病毒包含编码SARS-CoV-2 S蛋白或其片段的核酸。
在另一方面,本发明涉及制备针对COVID-19的疫苗的方法,所述方法包括提供血清型26的重组人腺病毒,所述重组人腺病毒包含编码SARS-CoV-2 S蛋白或其片段的核酸;使所述重组腺病毒在宿主细胞培养物中增殖;分离和纯化所述重组腺病毒;以及将所述重组腺病毒配制于药学上可接受的组合物中。这个方面的重组人腺病毒可以是本文所述的任何腺病毒。
在另一方面,本发明涉及一种分离的重组核酸,其形成血清型26的重组人腺病毒的基因组,所述重组人腺病毒包含编码SARS-CoV-2 S蛋白或其片段的核酸。腺病毒也可以是上述实施方案中描述的任何腺病毒。
本发明还涉及一种组合物,其用于预防有需要的受试者的分子确诊的中度至严重/危重COVID-19,包括向所述受试者给药如本文所述的本发明的组合物或免疫原性组合物,其中所述组合物在一剂方案(即单剂)中以5x1010vp/剂量的剂量给药。
本发明的特征在于优化的和/或非天然存在的冠状病毒(例如,2019-nCoV)核酸分子和多肽,用于产生DNA或RNA疫苗、抗体和免疫原性组合物,以及它们在预防、减少和/或治疗受试者(例如,哺乳动物受试者(例如,人))的冠状病毒(例如,2019-nCoV)感染的方法中的用途。
本发明的一方面的特征在于一种分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:1-84中任一项的氨基酸序列具有至少85%序列相同性的多肽。在一些实施方案中,a)所述多肽能够在受试者中引发免疫应答;或b)所述多肽与SEQ ID NO:1-84中任一项具有至少86、87、88、89、90、91、92、93、94、95、96、97、98或99%的序列相同性,或者具有SEQ ID NO:1-84中任一项的多肽序列。在一些实施方案中,所述多肽具有SEQ IDNO:56的氨基酸序列。在一些实施方案中,所述多肽具有SEQ ID NO:51的氨基酸序列。
另一方面的特征在于一种分离的核酸分子,其包含核苷酸序列,所述核苷酸序列与SEQ ID NO:93-181、190-195和199-204或其互补序列中任一项的全部或部分具有至少85%序列相同性。在一些实施方案中,所述核酸分子与SEQ ID NO:93-181、190-195和199-204中任一项具有至少86、87、88、89、90、91、92、93、94、95、96、97、98或99%的序列相同性,或者具有SEQ ID NO:93-181、190-195和199-204中任一项的核苷酸序列。在一些实施方案中,所述核酸分子或其部分能够在受试者中引发免疫应答。在一些实施方案中,所述核酸分子具有SEQ ID NO:195的核苷酸序列。在一些实施方案中,所述核酸分子具有SEQ ID NO:143的核苷酸序列。在一些实施方案中,所述核酸分子具有SEQ ID NO:204的核苷酸序列。在一些实施方案中,所述核酸分子具有SEQ ID NO:204的核苷酸19-3837的核苷酸序列。
另一方面的特征在于一种分离的多肽,其包含氨基酸序列,所述氨基酸序列与SEQID NO:1-84中任一项的全部或部分具有至少85%序列相同性。在一些实施方案中,所述多肽与SEQ ID NO:1-84中任一项具有至少86、87、88、89、90、91、92、93、94、95、96、97、98或99%的序列相同性,或者具有SEQ ID NO:1-84中任一项的氨基酸序列。在一些实施方案中,所述多肽或其部分能够在受试者中引发免疫应答。在一些实施方案中,所述多肽具有SEQID NO:28的氨基酸序列。在一些实施方案中,所述多肽具有SEQ ID NO:51的氨基酸序列。
另一方面的特征在于一种分离的载体,其包含一种或多种上述核酸分子。在一些实施方案中,所述载体是复制缺陷的(例如,缺乏E1、E3和/或E4区)。在一些实施方案中,所述载体是哺乳动物、细菌或病毒载体。在一些实施方案中,所述载体是表达载体。在一些实施方案中,所述病毒载体是选自逆转录病毒、腺病毒、腺相关病毒、细小病毒、冠状病毒、负链RNA病毒、正粘病毒、弹状病毒、副粘病毒、正链RNA病毒、小核糖核酸病毒(picornavirus)、甲病毒、双链DNA病毒、疱疹病毒、爱泼斯坦-巴尔病毒(Epstein-Barrvirus)、巨细胞病毒、鸡痘(fowlpox)和金丝雀痘的病毒。在一些实施方案中,所述载体是腺病毒。在一些实施方案中,所述腺病毒选自Ad2、Ad5、Ad11、Ad12、Ad24、Ad26、Ad34、Ad35、Ad40、Ad48、Ad49、Ad50、Ad52、Ad59和Pan9。在一些实施方案中,所述Ad52是恒河猴Ad52或者所述Ad59是恒河猴Ad59。在一些实施方案中,所述腺病毒是Ad26。在其他实施方案中,所述腺病毒是Ad26载体,其包含核酸分子,所述核酸分子具有SEQ ID NO:204的核苷酸19-3837或SEQ ID NO:204的全部核苷酸。在另一个实施方案中,所述腺病毒是Ad26载体,其包含核酸分子,所述核酸分子编码与SEQ ID NO:51的多肽具有至少85%或更多(例如,90%、95%、99%或100%)序列相同性的多肽。
另一方面的特征在于一种分离的抗体,其特异性结合任何上述多肽。在一些实施方案中,通过用核酸、多肽或载体免疫哺乳动物来产生所述抗体。在一些实施方案中,哺乳动物是人、牛、羊、小鼠或兔。在一些实施方案中,所述抗体是人源化的。在一些实施方案中,所述抗体是IgG。在一些实施方案中,所述抗体是bis-Fab、Fv、Fab、Fab′-SH、F(ab′)2、双抗体、线性抗体或scFV。
另一方面的特征在于一种产生抗2019-nCoV抗体的方法,所述方法包括向受试者给药一定量的核酸分子、多肽和/或载体,在向所述受试者给药后,所述量的核酸分子、多肽和/或载体足以引发中和的抗2019-nCoV抗血清的产生。
另一方面的特征在于通过任何上述方法产生的分离的抗2019-nCoV抗体。在一些实施方案中,抗体结合SEQ ID NO:1-84中任一项中的表位。
另一方面的特征在于一种组合物,其包含所述核酸分子、多肽、载体或抗体。在一些实施方案中,所述组合物进一步包含药学上可接受的载体、赋形剂或稀释剂。在一些实施方案中,所述组合物进一步包含佐剂或免疫刺激剂。
另一方面的特征在于一种免疫原性组合物,其包含所述核酸分子、多肽、载体或抗体。在一些实施方案中,所述免疫原性组合物是疫苗。在一些实施方案中,所述免疫原性组合物能够在有需要的受试者中治疗或降低冠状病毒感染(例如,2019-nCoV感染)的风险。在一些实施方案中,免疫原性组合物在向所述受试者给药后引发中和抗2019-nCoV抗血清的产生。在一些实施方案中,受试者是哺乳动物。在一些实施方案中,哺乳动物是人。在一些实施方案中,人具有潜在的健康状况。在一些实施方案中,潜在的健康状况是高血压、糖尿病或心血管疾病。
另一方面的特征在于一种鉴定、诊断和/或预测受试者对冠状病毒感染的易感性的方法,所述方法包括在来自受试者的样品中确定所述受试者是否具有保护水平的抗冠状病毒抗体(例如抗刺突抗体),其中优选地,保护水平是:(i)使用假病毒中和测定确定的等于或高于至少约70滴度的水平;(ii)使用活病毒中和测定确定的等于或高于至少约25滴度的水平;(iii)通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列中抗冠状病毒抗体中值水平的至少80%的水平。在一些实施方案中,抗冠状病毒抗体的保护水平是足以预防或减少严重疾病发展的水平。在一些实施方案中,所述方法进一步包括向具有低于保护水平的抗冠状病毒抗体的受试者给药有效量的所述组合物或免疫原性组合物。在一些实施方案中,所述方法进一步包括鉴定所述抗冠状病毒抗体(例如,抗刺突抗体)的亚类和/或效应功能。在一些实施方案中,所述亚类是IgM、IgA、IgG1、IgG2、IgG3或FcgR2A。在一些实施方案中,效应功能是抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性补体沉积(ADCD)、抗体依赖性单核细胞吞噬作用(ADCP)或抗体依赖性NK细胞激活。在一些实施方案中,所述样品是来自受试者的体液,其中优选地,体液是血液。在一些实施方案中,所述冠状病毒是2019-nCoV。
另一方面的特征在于一种治疗有需要的受试者的冠状病毒感染或降低其风险的方法,所述方法包括向所述受试者给药治疗有效量的所述组合物或免疫原性组合物。在一些实施方案中,所述方法进一步包括测量受试者中的抗冠状病毒抗体(例如,抗刺突抗体)的水平。在一些实施方案中,在给药之前和/或之后测量受试者中的抗冠状病毒抗体水平。在一些实施方案中,在给药后约1、2、3、4、5或6天,1、2、3、4、5、6或7周,2、3、4、5或6个月,1、2、3、4或5年内测量受试者中的抗冠状病毒抗体水平一次或多次。在一些实施方案中,受试者的抗冠状病毒抗体水平低于保护水平,并且其中所述方法进一步包括向受试者重新给药所述组合物或免疫原性组合物或者向受试者给药不同的抗冠状病毒组合物。在一些实施方案中,保护水平是足以减少冠状病毒介导的疾病的症状或持续时间的水平(例如,其特征在于体重减轻(例如,至少约5%(例如,至少约7.5%、至少约10%、至少约12.5%、至少约15%、至少约20%、至少约25%或更多)的体重减轻),肺炎的发展和/或呼吸衰竭,和/或死亡风险增加)。在-些实施方案中,保护水平是:(i)使用假病毒中和测定确定的等于或高于至少约70滴度的水平;(ii)使用活病毒中和测定确定的等于或高于至少约25滴度的水平;或(iii)通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列中的抗冠状病毒抗体中值水平的至少80%的水平。在一些实施方案中,冠状病毒是2019-nCoV。在一些实施方案中,所述方法进一步包括测量来自受试者的样品中的冠状病毒(例如,2019-nCoV)病毒载量。在一些实施方案中,样品是支气管肺泡灌洗液(BAL)或鼻拭子(NS)。在一些实施方案中,样品是来自受试者的体液(例如,血液,例如,全血或血浆)。在一些实施方案中,样品是来自受试者的组织样品(例如,呼吸道组织样品)。在一些实施方案中,病毒载量是可检测的核酸(例如,亚基因组mRNA)水平或可检测的蛋白(例如,核衣壳蛋白(N))水平。在一些实施方案中,可检测的核酸(例如,亚基因组mRNA)通过RNA-seq、RT-qPCR、qPCR、多重qPCR或RT-qPCR、LAMP、微阵列分析或杂交(例如如,ISH(例如,FISH))确定。在一些实施方案中,可检测的蛋白(例如,核衣壳蛋白(N))通过免疫测定(例如,免疫组织化学(IHC)测定或侧流免疫层析测定)确定。在一些实施方案中,可检测的病毒载量表明受试者对疾病(例如,2019-nCoV介导的疾病,例如,COVID-19,例如,严重的COVID-19疾病)易感。在一些实施方案中,大于至少约3.5log10 sgmRNA拷贝/mL的病毒载量。在一些实施方案中,BAL中大于3.85log10sgmRNA拷贝/mL或NS中大于3.78log10 sgmRNA拷贝/mL的病毒载量表明受试者对疾病(例如,2019-nCoV介导的疾病,例如,COVID-19,例如,严重的COVID-19疾病)易感。在一些实施方案中,BAL中大于3.85log10 sgmRNA拷贝/mL或NS中大于3.78log10 sgmRNA拷贝/mL的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,大于约2.0log10 sgmRNA拷贝/g组织的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,肺组织中大于约8.0log10sgmRNA拷贝/g、鼻孔组织中大于约7.0log10 sgmRNA拷贝/g、气管组织中大于约6.0log10sgmRNA拷贝/g、心脏组织中大于约5.5log10 sgmRNA拷贝/g,或者胃肠道(GI)、脾脏、肝脏、肾脏或脑组织中大于约2.0log10 sgmRNA拷贝/g的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,通过ISH染色的大于约3%的SARS-CoV-2vRNA的病毒载量表明受试者对疾病易感。在一些实施方案中,通过ISH染色的大于约5%的SARS-CoV-2vRNA的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,在感染后约1、2、3、4、5或6天或者1、2、3、4、5、6或7周内测量冠状病毒(例如2019-nCoV)病毒载量一次或多次。在一些实施方案中,向已确定为对疾病(例如,2019-nCoV介导的疾病,例如,COVID-19,例如,严重的COVID-19疾病)易感的受试者单独或者与其他治疗剂组合给药本公开的组合物或免疫原性组合物。
另一方面的特征在于降低感染有2019-nCoV的受试者中的冠状病毒介导的活性(例如如,2019-nCoV介导的活性)的方法,其包括向所述受试者给药治疗有效量的组合物或免疫原性组合物。在一些实施方案中,治疗有效量足以在受试者中产生大于2的log血清抗刺突抗体滴度,如通过ELISA测定测量的。在一些实施方案中,治疗有效量为15μg-300μg的组合物或免疫原性组合物。在一些实施方案中,所述活性是病毒滴度、病毒传播、感染或细胞融合。在一些实施方案中,在给药组合物或免疫原性组合物后病毒滴度降低。在一些实施方案中,病毒滴度降低25%或更多。在一些实施方案中,病毒滴度降低50%或更多。在一些实施方案中,病毒滴度降低75%或更多。在一些实施方案中,冠状病毒在所述给药后是不可检测的。在一些实施方案中,给药发生在暴露于冠状病毒之前。在一些实施方案中,给药发生在暴露于所述冠状病毒之前至少1小时。在一些实施方案中,给药发生在暴露于所述冠状病毒之前至少1周、1个月或一年。在一些实施方案中,给药发生在暴露于冠状病毒之后。在一些实施方案中,给药发生在暴露于所述冠状病毒后至少15分钟。在一些实施方案中,给药发生在暴露于所述冠状病毒后至少1小时、1天、1周。在一些实施方案中,向受试者给药至少一个剂量的所述核酸分子、多肽、载体、组合物、免疫原性组合物和抗体。在一些实施方案中,向受试者给药至少两个剂量。在一些实施方案中,将所述核酸分子、多肽、载体、组合物或免疫原性组合物作为初免、加强或作为初免加强向所述受试者给药。在一些实施方案中,所述核酸分子、多肽、载体、组合物、免疫原性组合物或抗体经肌内、静脉内、皮内、经皮、动脉内、腹膜内、病灶内、颅内、关节内、前列腺内、胸膜内、气管内、鼻内、玻璃体内、阴道内、直肠内、局部(topically)、肿瘤内、腹膜、皮下、结膜下、血管内(intravesicularlly)、黏膜内、心包内、脐内、眼内、口服、外用(topically)、局部(locallly)、通过吸入、通过注射、通过输注、通过连续输注、通过直接对靶细胞局部灌注、通过导管、通过灌洗、通过灌胃、在乳膏中或在脂质组合物中给药。在一些实施方案中,肌内给药所述核酸分子、多肽、载体、组合物、免疫原性组合物或抗体。在一些实施方案中,受试者是哺乳动物。在一些实施方案中,哺乳动物是人。在一些实施方案中,人具有潜在的健康状况。在一些实施方案中,潜在的健康状况是高血压、糖尿病或心血管疾病。在一些实施方案中,所述方法促进所述受试者中的免疫应答。在一些实施方案中,免疫应答是体液免疫应答。在一些实施方案中,体液免疫应答是IgG应答。
另一方面的特征在于一种组合物,其用于治疗有需要的受试者的冠状病毒感染(例如2019-nCoV感染)或降低其风险,其包括治疗有效量的组合物或免疫原性组合物。
另一方面的特征在于一种组合物,其用于降低感染有2019-nCoV的受试者中冠状病毒介导的活性(例如,2019-nCoV介导的活性),其包括治疗有效量的组合物或免疫原性组合物。
另一方面的特征在于一种制备免疫原性组合物的方法,所述免疫原性组合物用于治疗有需要的受试者的冠状病毒(例如,2019-nCoV)感染或降低其风险,所述方法包括以下步骤:(a)将所述核酸分子、多肽、载体、组合物和抗体中的至少一种与药学上可接受的载剂、赋形剂或稀释剂混合以形成所述免疫原性组合物;和(b)将所述免疫原性组合物置于容器中。
另一方面的特征在于一种试剂盒,其包含:(a)第一容器,其包含所述核酸分子、多肽、载体、组合物、免疫原性组合物和抗体中的至少一种;(b)其使用说明;以及任选存在的(c)第二容器,其包含药学上可接受的载剂、赋形剂或稀释剂。在一些实施方案中,第一容器进一步包含药学上可接受的载剂、赋形剂或稀释剂。试剂盒任选地包括佐剂和/或免疫刺激剂。
另一方面的特征在于一种试剂盒,其包含:一种或多种用于确定来自受试者的样品(例如,血液样品)中抗冠状病毒抗体(例如抗刺突抗体)的存在的试剂和用于鉴定、诊断和/或预测受试者对冠状病毒感染的易感性的说明。在一些实施方案中,试剂盒进一步包含用于鉴定抗冠状病毒抗体的亚类和/或效应功能的试剂。在一些实施方案中,试剂盒进一步包含用于比较的标准品或样品。
附图说明
包括附图以说明本发明的实施方案并进一步理解其实施方式。当结合附图阅读时,将更好地理解本发明的上述概述以及以下详细描述。应当理解,本发明不限于附图中所示的具体实施方案。
专利或申请文件含有至少一幅彩色附图。专利局将根据请求和必要费用的支付提供本专利或专利申请出版物的具有彩色附图的副本。
图1是示出刺突蛋白免疫原的图。2019-nCoV刺突(SEQ ID NO:1)的注释的结构域包括S1(SEQ ID NO:4)、S2(SEQ ID NO:1的氨基酸665-1191)、TM(SEQ ID NO:86)和CT(SEQID NO:85)结构域。标记了全长刺突(SEQ ID NO:1)、SdCT(SEQ ID NO:2)、S.Ecto(SEQ IDNO:3)、S1-foldon(SEQ ID NO:9)、RBD-foldon(SEQ ID NO:10)和S.Ecto-PP-foldon(SEQID NO:22)蛋白免疫原。白框表示折叠子(foldon)结构域,S.Ecto-PP-foldon中的双相交线表示两个稳定化突变的大致位置(对应于SEQ ID NO:1的氨基酸K969和V970的脯氨酸取代)。
图2是示出通过多克隆抗SARS抗血清识别重组2019-nCoV蛋白的免疫印迹(western blot)。使用多克隆抗SARS抗血清检测用编码SS-Spike(泳道1)、SS-SdCT(泳道2)、SS-S.Ecto(泳道3)和SS-S.Ecto-dF-PP-foldon (泳道4)的DNA转染的细胞的细胞裂解物(左图)和上清液(右图)。每个印迹左侧的数字标记的黑线表示以kDa为单位的大约分子量,并且每个印迹顶部的数字表示泳道编号。
图3是示出在2019-nCoV疫苗接种的小鼠中产生的抗体识别全长刺突的图。从用编码SS-Spike(泳道1)、SS-SdCT(泳道2)、SS-S.Ecto(泳道3)、SS-S1-foldon(泳道4)、SS-RBD-foldon (泳道5)和SS-S.Ecto-dF-PP-foldon(泳道6)的DNA疫苗接种后4周的小鼠收集血清并且用于使用全长刺突(SEQ ID NO:1)的ELISA。灰色条代表平均ELISA滴度。
图4是示出在2019-nCoV疫苗接种的小鼠中产生的抗体识别S.dTM.PP的图。从用编码SS-Spike(泳道1)、SS-SdCT(泳道2)、SS-S.Ecto(泳道3)、SS-S1-foldon(泳道4)、SS-RBD-foldon(泳道5)和SS-S.Ecto-dF-PP-foldon(泳道6)的DNA疫苗接种后4周的小鼠收集血清并用于使用全长胞外域S.Ecto-PP(SEQ ID NO:19)的ELISA。灰色条代表平均ELISA滴度。
图5是示出在2019-nCoV疫苗接种的小鼠中产生的抗体的中和活性的图。从用编码SS-Spike(泳道1)、SS-SdCT(泳道2)、SS-S.Ecto(泳道3)、SS-S1-foldon(泳道4)、SS-RBD-foldon(泳道5)和SS-S.Ecto-dF-PP-foldon(泳道6)的DNA疫苗接种后4周的小鼠收集血清并且用于体外2019-nCoV刺突假病毒中和测定。灰色条代表平均IC50滴度。
图6A-6E是示出在免疫接种的恒河猴中的体液免疫应答的图。免疫后通过(图6A)结合抗体ELISA、(图6B)假病毒中和测定和(图6C)活病毒中和测定评估体液免疫应答。(图6D)疫苗接种的猕猴(所有动物和SS-Spike/SS-SdCT组)、已从2019-nCoV感染恢复的9只恢复的猕猴的队列(cohort)和27个来自波士顿的恢复的人的队列中的假病毒中和滴度比较。(图6E)示出了S-和RBD-特异性抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性补体沉积(ADCD)、抗体依赖性单核细胞吞噬作用(ADCP)和抗体依赖性NK细胞激活(IFN-γ分泌、CD107a脱粒和MIP-1β表达)。雷达图示出了疫苗组中抗体特征的分布。楔形的大小和颜色强度表示相应组的特征的中值(蓝色表示抗体功能,红色表示抗体同种型/亚类/FcγR结合)。主成分分析(PCA)图示出了跨组的多变量抗体谱。每个点代表一种动物,点的颜色表示组,椭圆示出组的分布,置信水平为70%,假设为多变量正态分布,红色条反映应答中值。虚线反映了检测的测定极限。
图7A-7B是示出在疫苗接种的恒河猴中的细胞免疫应答的图。免疫后通过应答于合并的S肽的(图7A)IFN-γELISPOT测定和(图7B)多参数细胞内细胞因子染色测定评估细胞免疫应答。红色条反映平均应答。
图8A-8D是示出2019-nCoV攻击的恒河猴中病毒载量的图。通过鼻内和气管内途径用1.2x1012VP(1.1x104PFU)2019-nCoV攻击恒河猴。(图8A)在攻击后的多个时间点,在假对照中的支气管肺泡灌洗液(BAL)和鼻拭子(NS)中评估Log10 sgmRNA拷贝/mL或拷贝/拭子(限度50拷贝/mL)。(图8B)BAL中log10 sgmRNA拷贝/mL和(图8C)疫苗接种的动物NS中log10sgmRNA拷贝/拭子。(图8D)攻击后BAL和NS中的病毒载量峰值。红线反映病毒载量中值。P值表示双边Mann-Whitney检验。
图9A-9C是示出保护的免疫相关因素的图。攻击前(图9A)假病毒NAb滴度和(图9B)活性NAb滴度与BAL中log sgmRNA拷贝/mL峰值或攻击后鼻拭子中log sgmRNA拷贝/拭子峰值的相关性。红线反映了这些变量之间的最优拟合关系。P和R值反映了双边Spearman等级相关性检验。(图9C)热图(上图)示出了BAL中抗体特征和logsgmRNA拷贝/mL峰值之间的Spearman和Pearson相关性(*q<0.05,**q<0.01,***q<0.001,其中通过多重检验的Benjamini-Hochberg校正获得q值)。条形图(左下图)示出了由偏最小二乘回归(PLSR)和随机森林(RF)回归的递归特征消除定义的最具预测性组合或单个抗体特征的Pearson相关性等级。相关性热图(右下图)表示所有动物特征之间的成对Pearson相关性。
图10是示出在疫苗接种的猕猴中假病毒和活病毒NAb测定的相关性的图。红线反映了这些变量之间的最优拟合关系。P和R值反映了双边Spearman等级相关性检验。
图11是示出在假对照中BAL和鼻拭子中2019-nCoV攻击后病毒RNA的图。红线反映病毒载量的中值。
图12是示出在疫苗接种的动物中BAL中2019-nCoV攻击后病毒RNA的图。红线反映病毒载量的中值。
图13是示出在疫苗接种的动物中鼻拭子中2019-nCoV攻击后病毒RNA的图。红线反映病毒载量的中值。
图14是示出在疫苗接种的动物中BAL和鼻拭子中2019-nCoV攻击后的病毒RNA峰值的图。红线反映病毒载量的中值。P值表示双边Wilcoxon秩和检验。
图15是示出攻击前log ELISA滴度与攻击后BAL和鼻拭子中log sgmRNA的相关性的图。红线反映了这些变量之间的最优拟合关系。P和R值反映了双边Spearman等级相关性检验。
图16是示出攻击前log ELISPOT应答与攻击后BAL和鼻拭子中log sgmRNA的相关性的图。红线反映了这些变量之间的最优拟合关系。P和R值反映了双边Spearman等级相关性检验。
图17是示出攻击前log CD4+ICS应答与攻击后BAL和鼻拭子中log sgmRNA的相关性的图。红线反映了这些变量之间的最优拟合关系。P和R值反映了双边Spearman等级相关性检验。
图18是示出攻击前log CD8+ICS应答与攻击后BAL中log sgmRNA拷贝/mL和鼻拭子中log sgmRNA拷贝/拭子的相关性的图。红线反映了这些变量之间的最优拟合关系。P和R值反映了双边Spearman等级相关性检验。
图19是示出攻击后抗体再生的ELISA应答的图。示出了攻击后第0天和第14天的应答。红线反映应答中值。
图20是示出攻击后抗体再生的假病毒NAb应答的图。示出了攻击后第0天和第14天的应答。红线反映应答中值。
图21是示出攻击后抗体再生的活病毒NAb应答的图。示出了攻击后第0天和第14天的应答。红线反映应答中值。
图22是示出攻击后的抗体再生ELISPOT应答的图。示出了攻击后第0天和第14天的应答。红线反映应答中值。
图23A-23C是示出2019-nCoV攻击的恒河猴中病毒载量的图。通过鼻内和气管内途径向恒河猴接种1.1x106PFU(第1组;N=3)、1.1x105PFU(第2组;N=3)或1.1x104PFU(第3组;N=3)2019-nCoV。(图23A)在攻击后的多个时间点评估在支气管肺泡灌洗液(BAL)中的Log10病毒RNA拷贝/mL(限度50拷贝/mL)。在攻击后的多个时间点评估在鼻拭子(NS)中的(图23B)log10病毒RNA拷贝/拭子和(图23C)log10 sgmRNA拷贝/拭子(限度50拷贝/拭子)。红色水平条反映病毒载量的中值。
图24A-24F是示出2019-nCoV攻击的恒河猴中的免疫应答的图。在第35天通过(图24A)结合抗体ELISA、(图24B)假病毒中和测定、(图24C)活病毒中和测定和(图24D)包括抗体亚类和对受体结合结构域(RBD)、可溶性刺突(S)胞外域和核衣壳(N)蛋白的效应功能的系统血清学谱在攻击后评估体液免疫应答。示出了抗体依赖性补体沉积(ADCD)、抗体依赖性细胞吞噬作用(ADCP)、抗体依赖性中性粒细胞吞噬作用(ADNP),以及抗体依赖性NK细胞脱粒(NK CD107a)和细胞因子分泌(NK MIP1β,NK IFNγ)。在攻击后还通过应答于合并的S肽的(图24E)IFN-γELISPOT测定和(图24F)多参数细胞内细胞因子染色测定来评估细胞免疫应答。红色水平条反映平均应答。
图25A-25L是一组显微照片,示出2019-nCoV诱导急性病毒性间质性肺炎。(图25A-25F)来自感染2019-nCoV的恒河猴攻击后2天的固定的肺组织的H&E切片示出(图25A)间质水肿和局部肺实变,(图25B)肺泡内水肿和中性粒细胞浸润,(图25C-25D)细支气管上皮脱落和坏死,(图25E)细支气管上皮合胞细胞形成,和(图25F)肺泡间隔内的透明膜。(图25G-25H)SARS核衣壳的IHC示出间质空间内的病毒感染细胞,包括(图25G)管腔内的病毒合胞细胞和(图25H)病毒感染的肺泡衬细胞。(图25I)通过 原位杂交示出含有2019-nCoV RNA的多个细胞的炎症浸润。(图25J-25L)支气管呼吸道上皮示出(图25J)粘膜下层内的炎症和炎症细胞迁移到支气管纤毛柱状呼吸道上皮中,(图25K)2019-nCoV RNA,和(图25L)SARS核衣壳。比例尺(图25A)=200微米;(25C、25I、25K-25L)=100微米;(图25G)=50微米;(图25B、25D-25F、25J)=20微米,并且(图25H)=10微米。H&E=苏木精和伊红;IHC=免疫组化;RNAscope=2019-nCoV RNA染色。
原位杂交示出含有2019-nCoV RNA的多个细胞的炎症浸润。(图25J-25L)支气管呼吸道上皮示出(图25J)粘膜下层内的炎症和炎症细胞迁移到支气管纤毛柱状呼吸道上皮中,(图25K)2019-nCoV RNA,和(图25L)SARS核衣壳。比例尺(图25A)=200微米;(25C、25I、25K-25L)=100微米;(图25G)=50微米;(图25B、25D-25F、25J)=20微米,并且(图25H)=10微米。H&E=苏木精和伊红;IHC=免疫组化;RNAscope=2019-nCoV RNA染色。
图26A-26K是一组显微照片,示出在恒河猴中2019-nCoV感染肺泡上皮细胞。感染2019-nCoV的恒河猴的固定的肺组织在攻击后2天进行循环免疫荧光(CyCIF)染色。(图26A)肺的全视野数字切片,具有细胞核的DAPI染色、核实变区域(箭头)和病毒复制病灶(方框)。(图26B)示出(图26C)SARS核衣壳阳性细胞(绿色)和细胞核的DAPI(蓝色)的插入框的高倍放大图像。(图26D)来自(图26B和图26C)中所示相应肺区域的SARS核衣壳的明场IHC。(图26E-26K)SARS-N与CD206(图26E和图26K)、泛细胞角蛋白(pan-CK)(图26G和图26H)、CD68(图26I)和Iba-1(图26J)共染,示出病毒感染的上皮细胞和感染的上皮细胞附近的巨噬细胞。比例尺(图26F-26K)=50微米。
图27A-27F是示出2019-nCoV再攻击后恒河猴中病毒载量的图。在初始感染后的第35天(参见图23A-23C和24A-24F),通过鼻内和气管内途径用1.1x106PFU(第1组;N=3),1.1x105PFU(第2组;N=3)或1.1x104PFU(第3组;N=3)的2019-nCoV再次攻击恒河猴。将三只幼稚 的动物包括在内作为再攻击实验中的阳性对照。(图27A)在再次攻击后的多个时间点在支气管肺泡灌洗液(BAL)中评估Log10病毒RNA拷贝/mL(限度50拷贝/mL)。无法灌洗其中一只幼稚动物。(图27B)初次攻击和再攻击后BAL中病毒RNA的比较。在再次攻击后的多个时间点在鼻拭子(NS)中评估(图27C)log10病毒RNA拷贝/mL和(图27E)log10 sgmRNA拷贝/拭子(限度50拷贝/mL)。初次攻击和再攻击后NS中(图27D)病毒RNA和(图27F)sgmRNA的比较。红色水平条反映病毒载量的中值。P值反映了双边Mann-Whitney检验。
的动物包括在内作为再攻击实验中的阳性对照。(图27A)在再次攻击后的多个时间点在支气管肺泡灌洗液(BAL)中评估Log10病毒RNA拷贝/mL(限度50拷贝/mL)。无法灌洗其中一只幼稚动物。(图27B)初次攻击和再攻击后BAL中病毒RNA的比较。在再次攻击后的多个时间点在鼻拭子(NS)中评估(图27C)log10病毒RNA拷贝/mL和(图27E)log10 sgmRNA拷贝/拭子(限度50拷贝/mL)。初次攻击和再攻击后NS中(图27D)病毒RNA和(图27F)sgmRNA的比较。红色水平条反映病毒载量的中值。P值反映了双边Mann-Whitney检验。
图28是示出了2019-nCoV再攻击后在恒河猴中的抗体再生的免疫应答的一系列图。在2019-nCoV再攻击之前和之后7天进行病毒特异性结合抗体ELISA、假病毒中和测定、活病毒中和测定和IFN-γELISPOT测定。红线反映平均应答。P值反映了双边Mann-Whitney检验。
图29是示出2019-nCoV攻击的恒河猴中血浆病毒载量的一系列图。在图23中描述了第1-3组在攻击后的多个时间点在血浆中评估Log10病毒RNA拷贝/mL(限度为50拷贝/mL)。红色水平条反映病毒载量的中值。
图30是示出2019-nCoV攻击的恒河猴临床评分的一系列图。图23中描述了第1-3组。基于外观、呼吸困难、躺卧、食欲和应答能力的动物半定量临床评分。攻击后多个时间点的呼吸窘迫。红色水平条反映临床评分的中值。
图31是示出2019-nCoV攻击的恒河猴中血液学的图。示出了中性粒细胞和淋巴细胞计数(K/μL)。红色水平条反映中值。
图32是示出2019-nCoV攻击的恒河猴中组织病毒载量的图。向恒河猴接种2019-nCoV,并在攻击后的第2天(N=2)和第4天(N=2)进行尸检。在多个组织中评估了Log10病毒RNA拷贝/g组织(限度200拷贝/g)。红色水平条反映病毒载量的中值。
图33A-33J是显微照片,示出2019-nCoV复制诱导与1型干扰素应答相关的多形核和单个核炎症浸润。在(图33A-33E)攻击后第2天2019-nCoV感染的恒河猴和(图33F-33G)未感染的恒河猴中,(图33A和图33F)检测2019-nCoV RNA,(图33B和图33G)髓过氧化物酶(MPO)的IHC,(图33C和图33H)CD4(棕色)和巨噬细胞的双染色(CD68和CD163,红色),(图33D和图33I)CD8 T淋巴细胞,和(图33E和图33J)MX1(1型干扰素应答基因)。比例尺=100微米。
图34A-34B是示出肺组织中炎症浸润的定量分析的图。在2019-nCoV感染与未感染恒河猴中(图34A)肺泡多形核(PMN)细胞浸润和(图34B)MX1染色程度。P值反映了双边Mann-Whitney检验。
图35A-35J是显微照片,示出了2019-nCoV急性感染中肺实变区域内的炎症浸润。(图35A)感染后2天的一个肺切片,其含有三个感兴趣区域(ROI),显示组织实变(ROI 1-3);比例尺=5mm。(图35B)支气管相关淋巴组织(BALT)和(图35C和图35D)ROI 1内的细支气管上皮显示细支气管坏死,与(图35C)CD3+T淋巴细胞迁移到细支气管腔和(图35D)有大量髓过氧化物酶(MPO)表达的中性粒细胞和CD16+巨噬细胞的细支气管周围炎症;CD3(灰色)、CD20(蓝色)、CD68(黄色)、HLADR(青色)、MPO(洋红色)、E-Cadherin(绿色);比例尺=100μm。(图35E和图35F)在实变组织(DNA,灰色)中的CD16阳性(红色)和(图35E、图35G和图35I)在ROI 1中低2019-nCoV阳性与(图35E、图35G和图35J)在ROI 3中的高2019-nCoV阳性相比。(图35G)ROI 1与(图35H)ROI3中的SARS-N(绿色)和pan-CK(上皮,红色)染色对比。在ROI 1(低SARS CoV2阳性)(图35I)与ROI 3(高SARS CoV2阳性)(图35J)中的CD3(灰色)、CD20(红色)和CD68(绿色)阳性对比;比例尺=0.5mm。
图36是示出在2019-nCoV初次攻击和再攻击后BAL和鼻拭子的噬斑测定的图。示出了在初次攻击或再攻击后1-7天,BAL每毫升或鼻拭子每拭子的噬斑形成单位(PFU)峰值。红色水平条反映PFU滴度的中值。
图37是示出2019-nCoV再攻击的恒河猴的临床评分的图。图23中描述了第1-3组。基于外观、呼吸困难、躺卧、食欲和应答能力的动物半定量临床评分。攻击后多个时间点的呼吸窘迫。
图38A是示出Ad26载体的构建的图。产生了七种表达SARS-CoV-2 S蛋白变体的Ad26载体:(i)tPA前导序列与全长S(tPA.S(SEQ ID NO:1,tPA前导序列与N末端融合)),(ii)tPA前导序列与具有弗林蛋白酶切割位点的突变和两个脯氨酸稳定化突变的全长S,(tPA.S.PP(SEQ ID NO:23,tPA前导序列与N末端融合)),(iii)野生型前导序列与天然全长S(S(SEQ ID NO:29)),(iv)野生型前导序列与细胞质尾缺失的S(S.dCT(SS-SdCT)(SEQ IDNO:30)),(v)串联tPA和野生型前导序列与全长S(tPA.WT.S(SEQ ID NO:29,tPA前导序列与N-末端融合)),(vi)野生型前导序列与跨膜区和细胞质尾缺失的S,反映可溶性胞外域,弗林蛋白酶切割位点突变、脯氨酸稳定化突变和折叠子三聚化结构域(S.dTM.PP(SS-S.Ecto-dF-PP-foldon)(SEQ ID NO:56)),和(vii)野生型前导序列与具有弗林蛋白酶切割位点的突变和脯氨酸稳定化突变的全长S,(S.PP(SS-Spike-dF-PP)(SEQ ID:51))。红色三角形表示tPA前导序列,黑色三角形表示野生型前导序列,红色X表示弗林蛋白酶切割位点突变,红色垂直线表示脯氨酸突变,空心正方形表示折叠子三聚化结构域。
图38B是示出通过多克隆抗SARS抗体识别SARS-CoV-2 S变体的免疫印迹。使用抗SARS多克隆抗体免疫印迹分析在细胞裂解液中,来自编码tPA.S(泳道1)、tPA.S.PP(泳道2)、S(泳道3)、S.dCT(SS-SdCT)(泳道4)、tPA.WT S(泳道5)、S.dTM.PP(SS-S.Ecto-dF-PP-foldon)(泳道6)和S.PP(SS-Spike-dF-PP)(泳道7)的Ad26载体的表达。
图39A-39D是示出在疫苗接种的恒河猴中的体液免疫应答的图。通过(图39A)RBD特异性结合抗体ELISA、(图39B)假病毒中和测定和(图39C)活病毒中和测定在第0、2和4周评估体液免疫应答。红色条反映应答的中值。虚线反映定量的测定限。(图39D)在第4周的S-和RBD-特异性抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性单核细胞吞噬作用(ADCP)、抗体依赖性补体沉积(ADCD)和抗体依赖性NK细胞激活(ADNKA)以雷达图示出。楔形的大小和颜色强度表示相应组的特征的中值(蓝色表示抗体功能,红色表示抗体同种型/亚类/FcγR结合)。主成分分析(PCA)图示出了跨组的多变量抗体谱。每个点代表一种动物,点的颜色表示组,椭圆示出组的分布,假设为多变量正态分布,置信水平为70%。
图40A-40B是示出在疫苗接种的恒河猴中的细胞免疫应答的图。在免疫后第4周,通过(图40A)IFN-γELISPOT测定和(图40B)应答于合并的S肽的IFN-γ+CD4+和IFN-γ+CD8+T细胞细胞内细胞因子染色测定细胞免疫应答。红色条反映应答中值。虚线反映定量的测定限。
图41A-41D是示出SARS-CoV-2攻击后恒河猴中病毒载量的图。通过鼻内和气管内途径用1.2x108VP(1.1x104 PFU)SARS-CoV-2攻击恒河猴。(图41A-41B)攻击后评估在假对照和在疫苗接种的动物的支气管肺泡灌洗液(BAL)中的Log10 sgmRNA拷贝/mL(定量限50拷贝/mL)。(图41C-41D)攻击后评估在假对照和疫苗接种的动物的鼻拭子(NS)中的Log10sgmRNA拷贝/拭子(定量限50拷贝/拭子)。S.dTM.PP(SS-S.Ecto-dF-PP-foldon)组中的一只动物在攻击后没有获得BAL样品峰值。红线反映中值。
图42是示出SARS-CoV-2攻击后病毒载量峰值总结的图。攻击后BAL和NS中的病毒载量峰值。在攻击后的1-4天出现不同的病毒载量峰值。红线反映病毒载量的中值。P值表示双边Mann-Whitney检验(*P<0.05,**P<0.001,***P<0.0001)。
图43A-43D是示出保护的抗体相关因素的图。攻击后在第2周和第4周的(图43A)结合ELISA滴度、(图43B)假病毒NAb滴度和(图43C)活病毒NAb滴度与BAL中log sgmRNA拷贝/mL峰值的相关性。红线反映这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。(图43D)热图示出完全受保护(N=17)和部分受保护和未受保护(N=22)动物之间z分数特征的平均值的差异。两组通过双边Mann-Whitney检验进行比较,星号表示Benjamini-Hochberg校正的q值(*q<0.05)。点图示出了最能区分完全受保护和部分受保护动物的特征的差异,包括NAb滴度、S特异性抗体依赖性NK细胞激活(ADNKA)和抗体依赖性单核细胞吞噬作用(ADCP)。P值表示双边Mann-Whitney检验。条形图使用逻辑回归模型中x轴上指示的特征示出接收者操作特征曲线下的交叉验证区域。示出了前三个1-特征和2-特征模型。误差线表示100次重复10倍交叉验证的平均值和标准偏差。
图44A-44B是示出SARS-CoV-2攻击后的免疫应答的图。(图44A)攻击前和攻击后第14天的假病毒NAb滴度和(图44B)通过细胞内细胞因子染色测定攻击后第14天在假对照组和疫苗接种Ad26-S.PP(Ad26 SS-Spike-dF-PP)的动物中应答于合并的刺突(S1,S2)、核衣壳(NCAP)和非结构蛋白(N6、N7a、N8)肽的IFN-γ+CD8+和IFN-γ+CD4+T细胞应答。红色条反映应答的中值。虚线反映定量的测定限。
图45是示出在疫苗接种的猕猴中假病毒NAb滴度和ELISA或活病毒NAb测定的相关性的一对图。红线反映了这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。
图46是示出在疫苗接种的猕猴和恢复期的猕猴和人中的假病毒NAb滴度的比较图。将疫苗接种Ad26-S.PP(Ad26 SS-Spike-dF-PP)的猕猴中的假病毒NAb与之前报道的从SARS-CoV-2感染中恢复的恢复期猕猴和恢复期人的队列的假病毒NAb进行比较。
图47是示出在疫苗接种的恒河猴中BAL中的体液免疫应答的一对图。通过ELISA在第4周在假对照和疫苗接种Ad26-S.PP(Ad26 SS-Spike-dF-PP)动物中的BAL中的S特异性IgG和IgA。红色条反映应答的中值。虚线反映定量的测定限。
图48是示出在疫苗接种的恒河猴中的体液免疫应答的一系列图。示出了S和RBD特异性抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性单核细胞吞噬作用(ADCP)、抗体依赖性补体沉积(ADCD)和抗体依赖性NK细胞激活(ADNKA)。红色条反映应答的中值。
图49是示出在疫苗接种的恒河猴中的细胞免疫应答的一对图。在疫苗接种Ad26-S.dTM.PP(Ad26 SS-S.Ecto-dF-PP-foldon)和Ad26-S.PP(Ad26 SS-Spike-dF-PP)的动物中应答于合并的S肽的IFN-γ+、IL-2+、IL-4+和IL-10+CD4+T细胞细胞内细胞因子染色测定。红色条反映应答中值。虚线反映定量的测定限。
图50是示出保护的ELISA相关因素的一对图。攻击后第2周和第4周结合ELISA滴度与NS中log sgmRNA拷贝/拭子峰值的相关性。红线反映了这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。
图51是示出保护的假病毒NAb相关因素的一对图。攻击后第2周和第4周的假病毒NAb滴度与NS中log sgmRNA拷贝/拭子峰值的相关性。红线反映了这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。
图52是示出保护的活病毒NAb相关因素的一对图。攻击后第2周和第4周的活病毒NAb滴度与NS中log sgmRNA拷贝/拭子峰值的相关性。红线反映了这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。
图53是示出保护的抗体相关因素的一系列图。点图示出完全受保护和部分受保护动物之间的特征差异。P值表示双边Mann-Whitney检验。
图54是示出SARS-CoV-2攻击后的NAb滴度一系列图。在疫苗接种的动物中攻击前和攻击后第14天的假病毒NAb滴度。红色条反映应答中值。虚线反映定量的测定限。
图55A-55C是示出仓鼠中感染SARS-CoV-2后临床疾病的图。通过鼻内途径用5x104TCID50(低剂量;N=4)或5x105TCID50(高剂量;N=16)的SARS-CoV-2感染叙利亚金仓鼠(10-12周龄;雄性和雌性;N=20)。(图55A)攻击后重量变化的百分比中值。这些数字反映了每个时间点的动物数量。在高剂量组中,4只动物在第2天进行尸检,4只动物在第4天进行尸检,4只动物在第6天满足安乐死标准,2只动物在第7天满足安乐死标准。(图55B)单个动物在攻击之后的体重变化百分比。体重减轻的中值用红色表示。星号表示死亡率。灰线表示在第2天和第4天进行安排的尸检的动物。(图55C)在第2天和第4天的安排的尸检和在第6-7天满足安乐死标准的6只动物中的2-5只中通过log10RNA拷贝/克组织测量的组织病毒载量(定量限100拷贝/g)。第6天未收获扩展的组织。
图56A-56L是示出在仓鼠中高剂量SARS-CoV-2感染的病理特征一系列图。(图56A)鼻甲中的坏死和炎症(箭头),H&E(d2)。(图56B)细支气管上皮坏死,管腔内有细胞碎片和变性中性粒细胞(箭头),血管壁内有炎症细胞迁移(箭头),H&E(d2)。(图56C)间质性肺炎、出血和肺实质实变,H&E(d2)。(图56D)鼻甲上皮通过IHC(d2)示出对SARS-CoV-N的强阳性。(图56E)通过IHC(d2)示出细支气管上皮和管腔细胞碎片对SARS-CoV-N的强阳性。(图56F)通过IHC(d2)示出肺细胞和肺泡间隔对SARS-CoV-N的多灶性强阳性。(图56G)在肺间质(箭头,间质性肺炎)和细支气管上皮(箭头;d2)内通过 进行的扩散vRNA染色。(图56H)在肺间质(d4)内通过
进行的扩散vRNA染色。(图56H)在肺间质(d4)内通过 进行的扩散vRNA染色。(图56I)肺间质(d7)内的Iba-1IHC(巨噬细胞)。(图56J)肺间质内的CD3+T淋巴细胞,CD3 IHC(d4)。(图56K)MPO(中性粒细胞髓过氧化物酶)IHC指示间质中性粒细胞的存在(d7)。(图56L)干扰素诱导基因MX1、IHC在整个肺(d4)中示出强且弥漫的阳性。H&E,苏木精和伊红;IHC,免疫组化;Iba1,离子钙结合衔接蛋白1。示出了代表性切片。比例尺=20μm(图56B、56D);50μm(图56A、56E、56F);100μm(图56C、56G-56L)。
进行的扩散vRNA染色。(图56I)肺间质(d7)内的Iba-1IHC(巨噬细胞)。(图56J)肺间质内的CD3+T淋巴细胞,CD3 IHC(d4)。(图56K)MPO(中性粒细胞髓过氧化物酶)IHC指示间质中性粒细胞的存在(d7)。(图56L)干扰素诱导基因MX1、IHC在整个肺(d4)中示出强且弥漫的阳性。H&E,苏木精和伊红;IHC,免疫组化;Iba1,离子钙结合衔接蛋白1。示出了代表性切片。比例尺=20μm(图56B、56D);50μm(图56A、56E、56F);100μm(图56C、56G-56L)。
图57A-57F是示出肺中病毒复制和相关炎症的纵向定量图像分析的一系列图。(图57A)通过 检测的反义SARS-CoV-2病毒RNA(vRNA)的阳性肺面积百分比。(图57B)IHC检测的SARS-CoV-N蛋白(核或细胞质)阳性的总细胞百分比。(图57C)IHC每单位面积的Iba-1阳性细胞。(图57D)每单位面积的CD3阳性细胞。(图57E)每单位面积的MPO阳性细胞。(图57F)MX1阳性肺组织占肺总面积的百分比。ISH,原位杂交;IHC,免疫组化;SARS-N,SARS-CoV核衣壳;MPO,髓过氧化物酶;MX1,粘病毒蛋白1(一种1型干扰素诱导基因)。每个点代表一只动物。
检测的反义SARS-CoV-2病毒RNA(vRNA)的阳性肺面积百分比。(图57B)IHC检测的SARS-CoV-N蛋白(核或细胞质)阳性的总细胞百分比。(图57C)IHC每单位面积的Iba-1阳性细胞。(图57D)每单位面积的CD3阳性细胞。(图57E)每单位面积的MPO阳性细胞。(图57F)MX1阳性肺组织占肺总面积的百分比。ISH,原位杂交;IHC,免疫组化;SARS-N,SARS-CoV核衣壳;MPO,髓过氧化物酶;MX1,粘病毒蛋白1(一种1型干扰素诱导基因)。每个点代表一只动物。
图58A-58O是示出肺病毒动力学和ACE2受体表达模式的一系列图。在攻击之前(SARS-CoV-2阴性)或在高剂量的SARS-CoV-2攻击之后的第2天(D2)、第4天(D4)、第7天(D7)和第14天(D14)对仓鼠进行尸检。对肺组织的连续切片进行对于vRNA反义 (图58A-58E)、vRNA正义
(图58A-58E)、vRNA正义 (图58F-58J)和ACE2 IHC (图58K-58O)的染色。反义
(图58F-58J)和ACE2 IHC (图58K-58O)的染色。反义 使用正义探针;正义
使用正义探针;正义 使用反义探针。IHC,免疫组化。示出了代表性切片。比例尺=100μm。
使用反义探针。IHC,免疫组化。示出了代表性切片。比例尺=100μm。
图59A-59L是示出肺外病理学的一系列图。(图59A)攻击后第2天脑干中的反义SARS-CoV-2病毒RNA(vRNA)。(图59B)更高放大倍数示出在没有炎症和病理的情况下神经元中的细胞质vRNA染色。(图59C)攻击后第2天小肠绒毛固有层中的反义SARS-CoV-2vRNA染色。(图59D)更高放大倍数示出在没有炎症和组织病状的情况下单个核细胞中的细胞质和核vRNA染色。(图59E)攻击后第4天,在心肌内和沿着心脏的心外膜表面进行反义SARS-CoV-2vRNA染色。(图59F)更高放大倍数示出与局灶性心肌炎一致的炎性单个核细胞浸润染色。(图59G)攻击后第4天(d4)肺血管示出内皮炎。(图59H)通过IHC示出CD3+T淋巴细胞染色的肺血管粘附于内皮和血管壁内,攻击后d4。(图59I)通过IHC肺血管示出沿内皮和血管周围的Iba-1+染色的巨噬细胞,d4。(图59J)通过IHC肺血管示出对SARS-CoV-N的最小血管染色,d4。(图59K)来自(图59E、59F)的心脏示出如通过IHC对细胞的CD3+T淋巴细胞染色(图59L)所确认的局灶性淋巴细胞性心肌炎,d4。比例尺=500μm(图59A、59C、59E);100μm(图59B、59D、59F、59G-59L)。
图60A-60F是示出血液单个核细胞和骨髓中的SARS-CoV-2的一系列图。(图60A-60C)攻击后第2天肺血栓内单个核细胞内的SARS-CoV-2反义vRNA染色。(图60D)攻击后4天来自鼻甲骨的骨髓示出(图60E)造血细胞(H&E),其示出(图60F)SARS-CoV-N IHC阳性染色。vRNA,病毒RNA;H&E,苏木精和伊红;IHC,免疫组化。比例=500μm(图60A);200μm(图60D);100μm(图60B、60C、60E、60F)。
图61A-61F是示出疫苗接种的仓鼠的体液免疫应答的图。(图61A)SARS-CoV-2刺突(S)免疫原,具有(i)跨膜区和细胞质尾缺失,反映具有折叠子三聚化结构域(S.dTM.PP)的可溶性胞外域或(ii)全长S(S.PP),都具有弗林蛋白酶切割位点的突变和两个脯氨酸稳定化突变。红色X表示弗林蛋白酶切割位点突变,红色垂直线表示脯氨酸突变,空心方块表示折叠子三聚化结构域。S1和S2代表S蛋白的第一个和第二个结构域,TM表示跨膜区,以及CT表示细胞质结构域。用1010vp或109vp的Ad26-S.dTM.PP或Ad26-S.PP或假对照(N=10/组)对仓鼠进行疫苗接种。通过(图61B)RBD特异性结合抗体ELISA和(图61C)假病毒中和测定在第0、2和4周评估体液免疫应答。红色条反映应答的中值。虚线反映定量的测定限。(图61D)第4周的S-和RBD-特异性IgG亚类、FcγR和ADCD应答以雷达图示出。楔形的大小和颜色强度表示相应组的特征的中值(抗体亚类,红色;FcγR结合,蓝色;ADCD,绿色)。(图61E)主成分分析(PCA)图,示出了跨疫苗接种组的多变量抗体谱。每个点代表一只动物,点的颜色表示组,椭圆示出组的分布,假设为多变量正态分布,置信水平为70%。(图61F)热图示出疫苗组S.PP和S.dTM.PP之间z分数特征的平均值的差异。两组通过双边Mann-Whitney检验进行比较,星号表示Benjamini-Hochberg校正的q值(*q<0.05,**q<0.01,***q<0.001)。
图62A-62C是示出抗体滴度和存活曲线的相关性的图。(图62A)在第2周和第4周结合ELISA滴度和假病毒NAb滴度的相关性。红线反映了这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。(图62B)疫苗研究的存活曲线。P值表示双边Fisher精确检验。N表示每组中的动物数量。(图62C)涉及接受5x105TCID50攻击剂量并纵向追踪的所有动物的两项仓鼠研究的组合分析。P值表示双边Fisher精确检验。N表示每组中的动物数量。
图63A-63D是示出在仓鼠中高剂量SARS-CoV-2攻击后临床疾病的图。(图63A)攻击后重量变化的百分比中值。(图63B)单个动物攻击后体重变化百分比。体重减轻的中值用红色表示。星号表示死亡率。灰线表示在第4天进行安排的尸检的动物。(图63C)Ad26-S.dTM.PP(N=14)、Ad26-S.PP(N=14)和假对照(N=7)组中的最大体重减轻,不包括在第4天进行尸检的动物。P值表示双边Mann-Whitney检验。(图63D)在攻击后第4天,与对照仓鼠相比,来自Ad26-S.dTM.PP和Ad26-S.PP疫苗接种仓鼠的组织切片中对反义vRNA阳性的肺面积百分比的定量。P值代表双边Mann-Whitney检验。
图64A-64B是示出临床保护的抗体相关因素的图。(图64A)在第2周和第4周的结合ELISA滴度和(图64B)假病毒NAb滴度与攻击后最大体重减轻百分比的相关性。红线反映了这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。
图65A-65B是示出第4天和第14天组织病毒载量的图。在攻击后(图65A)第4天(N=6反映了每种疫苗的两个剂量组)和(图65B)第14天(N=14反映每种疫苗的两个剂量组)。红线反映中值。每个点代表一只动物。
图66A-66D是示出保护的抗体相关因素的图。第2周和第4周的(图66A,66C)结合ELISA滴度和(图66B,66D)假病毒NAb滴度与第4天进行尸检的动物中的log10RNA拷贝/g(图66A,66B)肺和(图66C,66D)鼻甲组织的相关性。红线反映了这些变量之间的最优线性拟合关系。P和R值反映了双边Spearman等级相关性检验。
图67A-67B是示出保护和抗体再生应答的抗体相关因素的图。(图67A)热图示出抗体特征与体重减轻(N=35)、肺病毒载量(N=12)和鼻甲病毒载量(N=12)之间的Spearman等级相关性。在使用Benjamini-Hochberg程序(*q<0.05,**q<0.01,***q<0.001)进行多重检验校正后,星号表示显著相关性。(图67B)SARS-CoV-2攻击后第14天存活仓鼠的ELISA和NAb应答。
图68A-68S是示出Ad26疫苗接种预防SARS-CoV-2病理的一系列图。(图68A-68E、68K-68N)假对照和(图68F-J、68O-R)Ad26-S.PP疫苗接种仓鼠的组织病理学H&E评价在假对照(图68A)中示出严重的肺实质实变和炎症细胞浸润,(图68B)细支气管上皮合胞和坏死,(图68C)SARS-CoV-N阳性(IHC)细支气管上皮细胞,(图68D)细支气管周CD3+T淋巴细胞浸润,和(图68E)细支气管周围巨噬细胞浸润(Iba-1;IHC),和(图68F-J)在疫苗接种Ad26-S.PP的动物中最小至没有相应的病理学。(图68O-68R)在攻击后第4天,(图68N)假对照中的SARS-CoV-2反义 ISH(图68K)、间质CD3+T淋巴细胞(图68L)通过IHC染色的MPO和通过IHC染色的MX1与疫苗接种Ad26-S.PP的动物相似区域的比较。在攻击后4天,与对照仓鼠相比,对来自疫苗接种Ad26-S.dTM.PP和Ad26-S.PP的仓鼠的组织切片中的反义vRNA(图68S)阳性肺面积百分比进行定量。示出了代表性切片。P值代表双边Mann-Whitney检验。比例尺=20μm(图68B、68C、68G、68H、68I);50μm(图68A、68D、68E、68J);100μm(图68F、68K-68R)。
ISH(图68K)、间质CD3+T淋巴细胞(图68L)通过IHC染色的MPO和通过IHC染色的MX1与疫苗接种Ad26-S.PP的动物相似区域的比较。在攻击后4天,与对照仓鼠相比,对来自疫苗接种Ad26-S.dTM.PP和Ad26-S.PP的仓鼠的组织切片中的反义vRNA(图68S)阳性肺面积百分比进行定量。示出了代表性切片。P值代表双边Mann-Whitney检验。比例尺=20μm(图68B、68C、68G、68H、68I);50μm(图68A、68D、68E、68J);100μm(图68F、68K-68R)。
图69:几种SARS-CoV-2 S构建体设计的示意图(S1浅灰色,S2深灰色)。在底层设计中,野生型(wt)信号肽变为tPA信号肽。S1和S2之间的垂直线表示弗林蛋白酶切割位点的突变。虚线表示位置986和987处的双脯氨酸突变。Delta ERRS表示含有内质网驻留信号的C末端残基的缺失。
图70:在用几种不同的S变体转染后,使用基于细胞的ELISA,基于发光强度,用几种SARS-CoV-1特异性抗体在细胞表面检测SARS-CoV-2 S的表达。数据表示为平均值±SEM。
图71:在转染几种不同的S变体后,使用流式细胞仪,用几种SARS-CoV-1特异性抗体和一种对HIV gp120(2G12)上的高甘露斑(high-mannose patch)特异性的抗体,检测细胞表面SARS-CoV-2 S的表达。数据表示为平均值±SEM。
图72:第19天通过ELISA测定的血清SARS-CoV-2刺突特异性抗体滴度。ULOQ,检测上限;LLOD,检测下限。统计分析:ANOVA和Bonferroni多重比较校正;ns,不显著(p>0.05)。
图73:不同信号肽对蛋白表达的影响。使用基于细胞的ELISA,基于发光强度,用ACE2-抗体构建体和SARS-CoV-1特异性抗体(CR3022)检测具有不同信号肽的SARS-CoV-2 S构建体在细胞表面的表达。数据表示为平均值±SEM。
图74:在线性化的目标载体中克隆SARS-CoV-2编码序列。将编码SARS-CoV-2刺突蛋白部分的两个DNA片段组装至它们的目标载体中。
图75:将编码SARS-CoV-2序列的表达盒引入Ad26载体的E1区。
图76A-76B:通过ELISA测量的S蛋白结合抗体滴度。(图76A)第2周;(图76B)第4周。每组的应答中值用水平线表示。虚线表示LLOD。Ad26NCOV030(Ad26COVS1)、Ad26NCOV006和Ad26NCOV028(数据未示出)之间的跨剂量比较是通过ANOVA的t检验和Bonferroni多重比较校正进行的。*:p<0.05。LLOD=检测下限。
图77A-77B:通过wtVNA测量的中和抗体滴度确定病毒分离物Leiden1(L-001)对Vero E6细胞的细胞病变效应(CPE)。(图77A)第2周;(图77B)第4周。每组的应答中值用水平线表示。虚线表示LLOD。应答等于或低于LLOD的动物以空心符号示出。wtVNA=野生型病毒中和测定。Ad26NCOV030(Ad26COVS1)、Ad26NCOV006和Ad26NCOV028(数据未示出)之间的跨剂量比较是用Tobit ANOVA的z检验和Bonferroni多重检验校正进行的。**:p<0.01。
图78:SARS-CoV-2 S蛋白在小鼠中的特异性细胞应答;示出了来自用肽池1和2刺激的SFU的总和。在肽刺激18小时后,通过ELISpot测量脾细胞产生的IFN-γ。虚线表示用培养基和DMSO孵育的200个IFN-γSFU/106未刺激脾细胞的LLOD。应答等于或低于LLOD的动物以空心符号示出。
图79A-79C:通过基于多重ELISA的分析确定细胞上清液中的细胞因子浓度,并计算比例。(图79A)IFNγ/IL-4比例;(图79B)IFNγ/IL-5比例;(图79C)IFNγ/IL-10比例。每组的比例中值用水平线表示。Ad26COVS1(Ad26NCOV030)和用佐剂的S蛋白之间的统计差异通过Mann-Whitney U检验和2倍Bonferroni校正计算。***:p<0.001。
图80A-80C:通过ELISA测量S蛋白结合IgG1和IgG2a滴度。(图80A)IgGl;(图80B)IgG2a。每组的应答中值用水平线表示。虚线表示测定的LLOD和ULOD。应答等于或低于LLOD或等于或高于ULOD的动物以空心符号示出。(图80C)计算IgG2a/IgG1滴度的比例。每组的应答中值用水平线表示。虚线表示IgG2a/IgG1比例为1。在IgG2a ELISA中应答等于或低于LLOD的动物以空心符号示出。Ad26COVS1(Ad26NCOV030)与用佐剂的S蛋白的统计比较通过Mann-Whitney U-test检验和2倍Bonferroni校正分析。**:p<0.01。
图81A-81B:(图81A)通过ELISA测量的S蛋白结合抗体滴度。(图81B)通过wtVNA测量中和抗体滴度测定病毒分离物Leiden1(L-001)对Vero E6细胞的细胞病变效应(CPE)。每组的应答中值用水平线表示。下面的虚线表示ELISA的1.699log10和wtVNA的5.7的LLOD。应答等于或低于LLOD的动物以空心符号示出。使用来自ANOVA(ELISA)的t检验和Cochran-Mantel-Haenszel检验(wtVNA)进行跨剂量比较,并对多重比较进行Bonferroni校正。
图82:通过ELISpot测量肽刺激18小时后的PBMC产生的IFN-γ。示出了来自用肽池1和2刺激的SFU的总和。虚线表示4.5IFN-γSFU/106PBMC的LOB,以来自用培养基和DMSO孵育的未刺激细胞的SFU的第95个百分位数计算。应答等于或低于LOB的动物以空心符号示出。使用来自ANOVA的t检验进行跨剂量比较,并对多重比较进行Bonferroni校正。
图83A-83B:(图83A)通过ELISA测量的S蛋白结合抗体滴度。Ad26.空样品N=18。(图83B)通过wtVNA测量的中和抗体滴度测定病毒分离物Leiden1(L-001)对Vero E6细胞的细胞病变效应(CPE)。Ad26.Empty组以来自第7组(N=6)和第8组(N=6)的2个合并的血清组示出。每组的应答中值用水平线表示。虚线表示LLOD。应答等于或低于LLOD的动物以空心符号示出。通过Mann-Whitney U-检验在Ad26COVS1(Ad26NCOV030)和Ad26.Empty之间进行每个剂量的ELISA数据比较。没有对多重比较进行校正。***:p<0.001。
图84:通过噬斑测定测量的具有复制能力的病毒(TCID50/克肺)。每组的中值病毒载量用水平线表示。应答等于或低于LLOD的动物以空心符号示出。通过Mann-Whitney U检验在Ad26.Empty组与Ad26COVS1(Ad26NCOV030)和Ad26NCOV006组之间进行比较。没有对多重比较进行校正。**:p<0.01。
图85:在攻击前在相对于攻击第[C]0天和攻击后4天在第C4天测量的体重。体重的相对变化表示为相对于第C0天的体重%。每组体重的变化中值用水平线表示。通过ANOVA的t检验在Ad26.Empty组与Ad26COVS1(Ad26NCOV030)和Ad26NCOV006组之间进行比较。没有对多重比较进行校正。***:p<0.001;**:p<0.01;*:p<0.05。
图86A-86D:(图86A)通过ELISA测量的S蛋白结合抗体滴度。(图86B)通过ppVNA测量的中和抗体滴度。(图86C)通过wtVNA使用Seattle Washington分离物(设计用于表达荧光素酶和GFP,在Vero E6细胞上孵育)测量的中和抗体滴度。通过Nano-Glo荧光素酶测定系统测量滴度。(图86D)通过ELISpot测量SARS-CoV-2肽刺激的PBMC的IFN-γ产生。示出了用肽池1和2刺激的SFU的总和。虚线表示50IFN-γSFU/106PBMC的LOB。应答等于或低于LOB的动物以空心符号示出。每组的应答中值用水平线表示。虚线表示ELISA的25的LLOD,ppVNA的20的LLOD,wtVNA的4的LLOD。应答等于或低于LLOD的动物以空心符号示出。ppVNA=假型病毒中和测定;SFU=斑点形成单位;vp=病毒颗粒;wtVNA=野生型病毒中和测定。
图87:使用抗SARS多克隆抗体免疫印迹分析在细胞裂解物中来自编码tPA.S(泳道1)、tPA.S.PP(泳道2)、S(泳道3)、S.dCT(泳道4)、tPA.WT.S(泳道5)、S.dTM.PP(泳道6)和S.PP(泳道7)的Ad26载体的表达。
图88A-88B:疫苗接种的恒河猴中的体液免疫应答。通过(图88A)RBD特异性结合抗体ELISA和(图88B)假病毒中和测定在第0、2和4周评估体液免疫应答。红色(实心)条反映应答中值。
图89A-89B:疫苗接种的恒河猴中的细胞免疫应答。在免疫后第4周,通过(图89A)IFN-γELISPOT测定和(图89B)应答于合并的S肽的IFN-γ+CD4+和IFN-γ+CD8+T细胞细胞内细胞因子染色测定来评估细胞免疫应答。红色(实心)条反映应答中值。
图90A-90D:SARS-CoV-2攻击后恒河猴中的病毒载量。通过鼻内和气管内途径用1.2x108VP(1.1x104PFU)的SARS-CoV-2攻击恒河猴。(图90A、图90B)在攻击后,评估在假对照和疫苗接种的动物的支气管肺泡灌洗液(BAL)中的Log10 sgmRNA拷贝/ml(限度50拷贝/ml)。(图90C,图90D)在攻击后,评估在假对照和疫苗接种的动物的鼻拭子(NS)中的Log10 sgmRNA拷贝/拭子(限度50拷贝/拭子)。红线反映中值。
图91:SARS-CoV-2攻击后的病毒载量峰值总结。攻击后BAL和NS中的病毒载量峰值。在攻击后的第1-4天出现不同的病毒载量峰值。红线反映病毒载量中值。P值表示双边Mann-Whitney检验(*P<0.05,**P<0.001,***P<0.0001)。
图92:SARS-CoV-2攻击后的免疫应答。攻击前和攻击后第14天的假病毒NAb滴度。红色条反映应答的中值。
图93:在疫苗接种的猕猴和恢复期猕猴和人中的假病毒NAb滴度的比较。将疫苗接种Ad26.S.PP的猕猴中的假病毒NAb与之前报道的从SARS-CoV-2感染中恢复的恢复期猕猴和恢复期人的队列进行比较。
图94:在疫苗接种的恒河猴中BAL中的体液免疫应答。通过ELISA在第4周在假对照和疫苗接种Ad26.S.PP动物的BAL中的S特异性IgG和IgA。红色条反映应答的中值。
图95:疫苗接种的恒河猴中的细胞免疫应答。在疫苗接种Ad26.S.dTM.PP和Ad26.S.PP的动物中应答于合并的S肽的IFN-γ+、IL-2+、IL-4+和IL-10+CD4+T细胞细胞内细胞因子染色测定。红色条反映应答的中值。
图96A-96C:(图96A)攻击后重量变化的百分比中值。这些数字反映了每个时间点的动物数量。在高剂量组中,在第2天对4只动物进行尸检,在第4天对4只动物进行尸检,在第6天4只动物满足安乐死标准,在第7天2只动物满足安乐死标准。(图96B)在单个动物中攻击后的体重变化百分比。体重减轻的中值用红色表示。星号表示死亡率。灰线表示在第2天和第4天进行安排的尸检的动物。(图96C)在第2天和第4天的安排的尸检和在第6-7天满足安乐死标准的6只动物中的2-5只中通过log10RNA拷贝/克组织测量的组织病毒载量(定量限100拷贝/g)。第6天未收获扩展组织。
图97A-97L:仓鼠中高剂量SARS-CoV-2感染的病理特征。(图97A)鼻甲中的坏死和炎症(箭头),H&E(d2)。(图97B)细支气管上皮坏死,管腔中有细胞碎片和退化的中性粒细胞(箭头),血管壁有炎症细胞迁移(箭头),H&E(d2)。(图97C)间质性肺炎、出血和肺实质实变,H&E(d2)。(图97D)鼻甲上皮通过IHC(d2)示出对SARS-CoV-N的强阳性。(图97E)通过IHC(d2)示出细支气管上皮和管腔细胞碎片对SARS-CoV-N的强阳性。(图97F)通过IHC(d2)示出肺细胞和肺泡间隔对SARS-CoV-N的多灶性强阳性。(图97G)通过RNAscope在肺间质内(箭头,间质性肺炎)和细支气管上皮内(箭头;d2)的扩散vRNA染色。(图97H)通过RNAscope在肺间质(d4)内的扩散vRNA染色。(图97I)肺间质(d7)内的Iba-1 IHC(巨噬细胞)。(图97J)肺间质内的CD3+T淋巴细胞,CD3 IHC(d4)。(图97K)MPO(中性粒细胞髓过氧化物酶)IHC指示间质中性粒细胞(d7)的存在。(图97L)干扰素诱导基因,MX1,IHC在整个肺(d4)中示出强且弥漫的阳性。H&E,苏木精和伊红;IHC,免疫组化;Iba1,离子钙结合衔接蛋白1。示出了代表性部分。实验至少重复3次,有着相似的结果。比例尺=20μm(b,d);50μtm(a、e、f);100μm(c,g-1)。
图98A-98F:疫苗接种的仓鼠的体液免疫应答。(图98A)SARS-CoV-2刺突(S)免疫原,具有(i)跨膜区和细胞质尾缺失,反映具有折叠子三聚化结构域(S.dTM.PP)的可溶性胞外域或(ii)全长S(S.PP),都具有弗林蛋白酶切割位点的突变和两个脯氨酸稳定化突变。红色X表示弗林蛋白酶切割位点突变,红色垂直线表示脯氨酸突变,空心方块表示折叠子三聚化结构域。S1和S2代表S蛋白的第一个和第二个结构域,TM表示跨膜区,以及CT表示细胞质结构域。用1010vp或109vp的Ad26-S.dTM.PP或Ad26-S.PP或假对照(N=10/组)对仓鼠进行疫苗接种。通过(图98B)RBD-特异性结合抗体ELISA和(图98C)假病毒中和测定在第0、2和4周评估体液免疫应答。红色条反映应答的中值。虚线反映定量的测定限。(图98D)第4周的S-和RBD特异性IgG亚类、FcγR和ADCD应答以雷达图示出。楔形的大小和颜色强度表示相应组的特征的中值(抗体亚类,红色;FcγR结合,蓝色;ADCD,绿色)。(图98E)主成分分析(PCA)图示出跨疫苗接种组的多变量抗体谱。每个点代表一只动物,点的颜色表示组,椭圆示出组的分布,假设为多变量正态分布,置信水平为70%。(图98F)热图示出疫苗组S.PP和S.dTM.PP之间z分数特征的平均值的差异。两组通过双边Mann-Whitney检验进行比较,星号表示Benjamini-Hochberg校正的q值(*q<0.05,**q<0.01,***q<0.001)。
图99A-99C:高剂量SARS-CoV-2攻击后仓鼠中的临床疾病。(图99A)攻击后重量变化的百分比中值。(图99B)单个动物在攻击后的体重变化百分比。体重减轻的中值用红色表示。星号表示死亡率。灰线表示在第4天进行安排的尸检的动物。(图99C)Ad26-S.dTM.PP(N=14)、Ad26-S.PP(N=14)和假对照(N=7)组的合并的最大体重减轻,不包括在第4天进行尸检的动物。P值表示双边Mann Whitney检验。N反映了所有在第4天体重减轻且未进行尸检的动物。
图100A-100F:病毒复制和肺相关炎症的纵向定量图像分析。(图100A)通过RNAscope ISH检测的反义SARS-CoV-2病毒RNA(vRNA)阳性肺面积百分比。(图100B)通过IHC检测的SARS-CoV-N蛋白(核或细胞质)阳性的总细胞百分比。(图100C)通过IHC检测的每单位面积的Iba-1阳性细胞。(图100D)每单位面积的CD3阳性细胞。(图100E)每单位面积的MPO阳性细胞。(图100F)MX1阳性肺组织占肺总面积的百分比。ISH,原位杂交;IHC,免疫组化;SARS-N,SARS-CoV核衣壳;MPO,髓过氧化物酶;MX1,粘病毒蛋白1(一种1型干扰素诱导基因)。每个点代表一种动物。
图101:同时在比利时和美国的站点招募参与者。参与者以1∶1∶1∶1∶1的比例平行随机分配到五个疫苗接种组之一,接受一次或两次IM注射的5x1010vp或1x1011vp两种剂量水平的Ad26.COV2.S,或安慰剂。对于每个队列,在给5名哨兵参与者(每个剂量水平一个和安慰剂一个)第一次疫苗接种后24小时没有临床显著发现的情况下,给所有组中的另外10名参与者疫苗接种。直到28天的安全性数据随后由内部数据审查委员会审查,然后将其余参与者随机分配。
图102A-102C:队列1a(图102A)、队列1b(图102B)和队列3(图102C)的流程图。
图103A-103D:体液和细胞免疫应答。(图103A)在基线和疫苗接种后29天,在所有参与者中,根据队列1a和3中的计划,通过ELISA(每毫升的ELISA单位[EU/mL])测量的血清SARS-CoV-2结合抗体的log几何平均滴度(GMT-如水平条和每个时间点下方的数字所示)。虚线表示测定的定量下限(LLOQ)和定量上限(ULOQ),误差条表示95%置信区间(CI)。对于低于LLOQ的值,绘制LLOQ/2的值。(图103B)在基线和疫苗接种后29天,在一组参与者中,根据在队列1a和3的计划,通过50%微量中和测定测量的血清SARS-CoV-2中和抗体的Log GMT(ID50 Log GMT-如水平条和每个时间点下方的数字所示)。虚线表示测定的LLOQ和ULOQ,误差条表示95%CI。对于低于LLOQ的值,绘制LLOQ/2的值。(图103C)通过细胞内细胞因子染色(ICS)测量CD4 T细胞Th1(IFNγ和/或IL-2,不是IL-4、IL-5和IL-13)和Th2(IL-4和/或IL-5和/或IL-13和CD40L))细胞因子的表达。在基线和疫苗接种后15天,在来自队列1a和3的一部分参与者中,按计划进行,中值(如水平条和每个时间点下方的数字所示)以及对外周血单个核细胞中对SARS-CoV-2 S蛋白肽池的应答的个体ICS。百分比表示Th1或Th2细胞因子阳性T细胞的百分比。虚线表示LLOQ。(图103D)通过ICS测量CD8T细胞的IFNγ和/或IL-2细胞因子的表达。在基线和疫苗接种后15天,在来自队列1a和3中的一部分参与者,按计划进行,中值(如水平条和每个时间点下方的数字所示)和外周血单个核细胞中对SARS-CoV-2 S蛋白肽池的应答的个体ICS。百分比表示对IFNγ和/或IL-2细胞因子呈阳性的CD8 T细胞的百分比。虚线表示LLOQ。
图104:随时间变化的针对SARS-CoV-2(几何平均滴度[GMTs]与相应的95%CI)的VNA应答的图示。
图105:描述了年龄对到第29天的中和抗体应答的影响。
图106:描述了研究队列中的中和抗体应答和应答率。
图107:描述了5×1010vp疫苗组和1×1011vp疫苗组的反向累积分布曲线。
图108:通过ELISA测量的SARS-CoV-2 S蛋白结合抗体应答的图示。
图109:描述了在第29天年龄对结合抗体应答的影响。
图110:通过ELISA测量的SARS-CoV-2 S蛋白结合抗体应答的图示。
图111:描述了5×1010vp疫苗组和1×1011vp疫苗组的反向累积分布曲线。
图112:示出在第15天和第29天,wtVNA滴度与ELISA滴度高度相关,Spearman相关系数分别为0.734和0.72。
图113:示出了表达IFNγ和/或IL-2(Th1)而不是Th2细胞因子并且表达IL-4和/或IL-5/IL-13和CD40L(Th2)的CD4+T细胞的百分比。
图114:示出组合方案谱。
图115:示出应答于SARS-CoV-2 S肽刺激产生IFNγ和/或IL-2的CD8+T细胞的描述性统计。
图116:示出组合方案谱。
图117A-117C:在成年恒河猴中对1剂和2剂Ad26.COV2.S疫苗方案的SARS-CoV-2特异性体液免疫应答。(图117A)使用哺乳动物细胞中产生的三聚体、可溶性稳定S蛋白作为包被抗原,用合格的ELISA对人样品随时间测量刺突(S)蛋白结合抗体水平。单个动物水平用灰点表示,成对测量用灰线连接。每组结合抗体应答的几何平均滴度(GMT)用红线表示。虚线表示检测下限(LLOD)和定量下限(LLOQ)。(图117B)使用合格的psVNA对人样品随时间测量S蛋白中和抗体水平,使用由修饰的水泡性口炎病毒(VSVΔG)骨架制成并带有SARS-CoV-2的S糖蛋白的假病毒颗粒。中和抗体应答测量为能达到50%的中和(IC50)的样品稀释度的倒数。单个动物水平用灰点表示,成对测量用灰线连接。每组中和抗体应答的GMT用红线表示。虚线表示LLOD和LLOQ。(图117C)除了假对照组和第0周(基线)之外的所有组和时间点的每只动物的S-特异性结合抗体水平和中和抗体滴度之间的相关性。虚线表示每个测定的LLOD。
图118A-118C:老年恒河猴疫苗接种后的SARS-CoV-2特异性体液和细胞免疫应答。(图118A)使用哺乳动物细胞中产生的三聚体、可溶性稳定S蛋白作为包被抗原,用合格的ELISA对人样品随时间测量刺突(S)蛋白结合抗体水平。单个动物水平用灰点表示,成对测量用灰线连接。每组结合抗体应答的几何平均滴度(GMT)用红线表示。虚线表示检测下限(LLOD)和定量下限(LLOQ)。(图118B)通过wtVNA测量的SARS-Cov-2中和抗体滴度随时间的变化。单个动物水平用灰点表示,成对测量用灰线连接。每组的GMT用红线表示。虚线表示LLOD。(图118C)除假对照组和第0周外,所有组和时间点的每只动物的S特异性结合抗体水平和中和抗体滴度之间的相关性。虚线表示每个测定的LLOD。
图119A-119C:老年恒河猴疫苗接种后的SARS-CoV-2特异性细胞免疫应答。(图119A)用IFN-γ/IL-4双色ELISpot在指定时间点测量的刺突(S)蛋白特异性T细胞应答。每组的几何平均滴度(GMT)应答用水平线表示。出于可视化目的,将背景减去计数低于或等于0的样品设置为10,由虚线表示。(图119B)在指定时间点通过细胞内细胞因子染色测量的刺突(S)蛋白特异性T细胞应答。CD4+CD69+T细胞表达细胞因子的频率。每组的几何平均滴度(GMT)应答用水平线表示。虚线表示技术阈值。空心符号表示等于技术阈值的样品。(图119C)表达Th1(IFN-γ或IL-2)或Th2(IL-4、IL-5、IL-13)细胞因子的CD4+CD69+T细胞的比例。每组的几何平均滴度(GMT)应答用水平线表示。空心符号表示值是表达Th1、Th2或任何细胞因子的细胞等于技术阈值。出于可视化目的,将水平虚线设置为比例为1。
图120:在指定时间点通过细胞内细胞因子染色测量的刺突(S)蛋白特异性T细胞应答。CD8+CD69+T细胞表达细胞因子的频率。每组的几何平均滴度(GMT)应答用水平线表示。虚线表示技术阈值。空心符号表示等于技术阈值的样品。
图121A-121D:在基线和疫苗接种后第29天,在所有参与者中以及在第57天和第71天对队列1a中可用的,根据队列1a(18-55岁)的计划(图121A)和队列3(>65岁)的计划(图121B),通过ELISA(每毫升的ELISA单位[EU/mL])测量的血清中SARS-CoV-2刺突蛋白结合抗体的log几何平均滴度(GMT-如水平条和每个时间点下方的数字所示)。虚线表示测定的定量下限(LLOQ)和定量上限(ULOQ),误差条表示95%置信区间(CI)。对于低于LLOQ的值,绘制LLOQ/2值。
在基线和疫苗接种后第29天、第57天和第71天,在一部分参与者中,根据计划,队列1a(18-55岁)(图121C)和队列3(>65岁)(图121D),通过50%中和测定(IC50 Log GMT-如水平条和每个时间点下方的数字所示)测量的血清SARS-CoV-2中和抗体(wtVNA)的LogGMT。虚线表示使用当前用于疫苗样品的预稀释测定的LLOQ和ULOQ,误差条表示95%CI。对于低于LLOQ的值,绘制LLOQ/2值。
图122A-122F:通过80%中和测定测量的血清SARS-CoV-2中和抗体(wtVNA)的LogGMT。
(IC80 Log GMT-如黑点和每个时间点下方的数字所示),在基线和疫苗接种后第29天、第57天和第71天,在一部分参与者中,根据计划,队列1a(18-55岁)和疫苗接种后第1天、第15天和第29天队列3(>65岁)。
图123:Ad26和SARS-CoV-2中和抗体滴度之间的相关性图:基线Ad26 VNA基线与第29天的wtVNA,以及第57天的Ad26 VNA与第71天的wtVNA。
图124A-124C:CD4+T细胞表达的Th1(IFN-γ和/或IL-2,而不是IL-4、IL-5和IL-13)(图124A)和Th2(IL-4和/或IL-5和/或IL-13和CD40L)(图124B)通过细胞内细胞因子染色(ICS)测量。给出中值(如水平条和每个时间点下方的数字所示)和个体在外周血单个核细胞中对SARS-CoV-2 S蛋白肽池的ICS应答,在基线和疫苗接种后15天,在来自队列1a(18-55岁)和队列3(>65岁)的一部分参与者,根据计划。Y轴表示Th1或Th2细胞因子阳性的T细胞百分比。虚线表示LLOQ。低于LLOQ的值绘制为LLOQ/2。
(图124C)通过ICS测量CD8+T细胞的IFN-γ和/或IL-2细胞因子的表达。给出中值(如水平条和每个时间点下方的数字所示)和在外周血单个核细胞中对SARS-CoV-2 S蛋白肽池应答的个体ICS,在基线和疫苗接种后15天,在来自队列1a(18-55岁)和队列3(>65岁)的一部分参与者中,根据计划。Y轴表示IFN-γ和/或IL-2细胞因子阳性的CD8+T细胞的百分比。虚线表示LLOQ。低于LLOQ的值绘制为LLOQ/2。
图125A-125B:在sPER.C6和PER.C6 TetR细胞中Ad26NCOV030(JNJ 78436735)和Ad26NCOV028纯化材料的相对产率。
A)sPER.C6细胞B)sPER.C6 TetR细胞。在摇瓶中用Ad26载体的纯化材料(70或300VP/细胞)转导细胞。在感染后第0、1、2、3和4天采集样品,并通过VP-qPCR测量载体颗粒浓度。标准对照(Ad26.ZEBOV)和低对照(26ZIK001)。VP,病毒颗粒;qPCR,定量聚合酶链反应;TetR,四环素阻遏物。
图126A-126E:SARS-CoV-2攻击剂量的滴定和叙利亚仓鼠的组织病理学特征。用102、103.3、104.6或105.9的TCID50SARS-CoV-2 BetaCoV/Munich/BavPat1/2020鼻内接种叙利亚仓鼠(每组N=12),或用Vero E6细胞-上清液模拟接种。每天采集咽拭子,在感染后第2、3、4和7天,每组牺牲3只仓鼠,收集鼻和肺组织用于病毒学分析和组织病理学。(图126A)肺组织、(图126B)鼻组织和(图126C)咽拭子中具有复制能力的病毒载量通过对Vero E6细胞的TCID50测定来测定。每只动物每克或每毫升组织计算LLOD,应答等于或低于LLOD的动物以空心符号示出。(图126D)分析肺组织并对肺泡炎、肺泡损伤、肺泡水肿、肺泡出血、II型肺细胞增生、支气管炎、细支气管炎、支气管周围和血管套状白细胞的存在和严重度进行评分。分数总和表示为LRT疾病参数的总和(潜在范围:0-24)。(图126E)分析鼻组织并在0到3的范围内对鼻炎的严重度进行评分。虚线表示组织病理学的最小和最大评分。每组的应答中值用水平线表示,图c中的误差条表示范围。p.i.=接种后;LLOD=检测下限;LRT=下呼吸道;N=动物数量;TCID50/g=每克组织50%组织培养物感染剂量;TCID50/ml=每毫升样品50%组织培养物感染剂量;vp=病毒颗粒。
图127A-127C:在叙利亚仓鼠中由1剂和2剂Ad26.COV2.S疫苗方案引发的SARS-CoV-2中和抗体应答。(图127A)用109或1010VP(N=12/剂量水平)的基于Ad26的候选疫苗,或用1010vp的没有基因插入的Ad26载体作为对照(Ad26.empty,N=6)免疫叙利亚仓鼠。免疫后4周,每组一半的仓鼠用相同的基于Ad26的候选疫苗进行第二次免疫(每组N=6)。(图127B)在剂量1后4周和(图127C)在剂量2后4周通过野生型VNA测量SARS-CoV-2中和滴度以测定SARS-CoV-2对Vero E6细胞的细胞病变效应的抑制作用。将来自Ad26.Empty免疫的叙利亚仓鼠的血清分2组作为阴性对照样品。每组的应答中值用水平线表示。虚线表示LLOD。应答等于或低于LLOD的动物在LLOD上以空心符号示出。CPE=细胞病变效应;LLOD=检测下限;p.i.=接种后;VNA=病毒中和测定;VP=病毒颗粒。
图128A-128C:在叙利亚仓鼠中由1剂和2剂Ad26疫苗方案引发的SARS-CoV-2刺突蛋白结合抗体应答,以及与中和抗体应答的比例。用109或1010VP的基于Ad26的候选疫苗Ad26.S、Ad26.dTM.PP或Ad26.COV2.S免疫叙利亚仓鼠,或用1010VP的没有基因插入的Ad26载体免疫作为对照(Ad26.empty)。免疫后4周,用相同的基于Ad26的候选疫苗第二次免疫每组一半的仓鼠(每组N=6)。一次免疫后2周和4周,以及第二次免疫后1、2和4周(分别为第5、6和8周),收集血样并分离血清用于血清学分析。(图128A)通过ELISA测量接受两次免疫的仓鼠(每组N=6)的SARS-CoV-2刺突蛋白特异性抗体结合滴度。(图128B)在剂量1后4周(仓鼠接受一次免疫,每组N=6)和(图128C)在剂量2后4周(接受两次免疫的仓鼠,每组N=6)测量中和滴度。SARS-CoV-2中和抗体与结合抗体的比例是通过用wtVNA测量的抗体滴度除以ELISA测量的抗体滴度来计算的。中和滴度等于或低于LLOD的动物以空心符号示出。每组的应答中值用水平线表示。LLOD=检测下限;ELISA=酶联免疫吸附测定;wtVNA=野生型病毒中和测定;VP=病毒颗粒。
图129A-129C:2剂Ad26疫苗方案在新西兰白兔中引发的SARS-CoV-2刺突蛋白结合抗体应答和中和抗体应答。用5x109vp或5x1010vp的Ad26.S、Ad26.dTM.PP、Ad26.COV2.S或盐水的2剂方案肌内免疫新西兰白兔(图129A)。在免疫前(第-3天)和第14、21、35、42、56、63、70、84天和牺牲时(第99-101天,在图中表示为第100天)对血清进行取样。(图129B)通过ELISA测量SARS-CoV-2刺突蛋白特异性抗体结合滴度。(图129C)在第14、35、56、70天和牺牲时测量中和滴度。每组的应答中值用水平线表示,垂直线表示组范围。ELISA=酶联免疫吸附测定;wtVNA=野生型病毒中和试验;LLOQ=定量下限;ULOQ=定量上限;LLOD=检测下限;VP=病毒颗粒。
图130A-130F:在用基于Ad26的候选疫苗免疫的叙利亚仓鼠中针对SARS-CoV-2病毒复制的保护。用Ad26.S、Ad26.dTM.PP、Ad26.COV2.S或Ad26.empty(Ad26载体不编码任何SARS-CoV-2抗原)的1剂方案和2剂方案肌内免疫叙利亚仓鼠。仓鼠在剂量1后4周(第4周)或剂量2后4周(第8周)接受了102TCID50 SARS-CoV-2株BetaCoV/Munich/BavPat1/2020的鼻内接种。在4天接种阶段结束时收集(图130A、130B)右肺组织和(图130C、130D)右鼻甲用于病毒载量分析。通过TCID50测定测量具有复制能力的病毒。(图130E、130F)在接种后每天采集咽拭子样品,并计算四天追踪期间曲线下的病毒载量面积为TCID50/ml x天。每组的病毒载量中值用水平线表示。计算每只动物的LLOD,应答等于或低于LLOD的动物在LLOD上以空心符号示出。在Ad26.S、Ad26.dTM.PP和Ad26.COV2.S组之间进行跨剂量水平的比较,通过Mann-Whitney U检验与Ad26.empty组进行比较。星号表示统计差异:*:p<0.05;**:p<0.01。LLOD=检测下限;TCID50/g=每克组织50%组织培养物感染剂量;TCID50/ml=每毫升样品50%组织培养物感染剂量;VP=病毒颗粒。
图131A-131D:用基于Ad26的候选疫苗免疫的叙利亚仓鼠肺组织中针对SARS-CoV-2IHC和组织病理的保护。用Ad26.S、Ad26.dTM.PP、Ad26.COV2.S或Ad26.empty(Ad26载体不编码任何SARS-CoV-2抗原)的1剂方案和2剂方案肌内免疫叙利亚仓鼠。在剂量1后4周(第4周)或剂量2后4周(第8周),仓鼠接受了102TCID50 SARS-CoV-2株的BetaCoV/Munich/BavPat1/2020的鼻内接种。接种后4天分离左肺组织用于免疫组织化学SARS-CoV-NP染色和组织病理学分析。由于技术故障,只可以分析用一剂Ad26.Emtpy免疫的8只仓鼠中的2只的组织。(图131A和131B)SARS-CoV-2NP的存在通过免疫组织化学染色确定。(图131C和131D)对肺组织的肺泡炎、肺泡损伤、肺泡水肿、肺泡出血、II型肺细胞增生、支气管炎、细支气管炎、支气管周围和血管周围套状白细胞的存在和严重度进行评分。分数总和表示为LRT疾病参数的总和。每组的中值分数用水平线表示。虚线表示LLOD。在疫苗组之间进行跨剂量水平的比较,通过Mann-Whitney U检验与Ad26.Empty组进行比较,除了图c。星号表示统计差异:*:p<0.05;**:p<0.01。LLOD=检测下限;VP=病毒颗粒;NP=核衣壳蛋白;IHC=免疫组织化学;LRT=下呼吸道。
图132A-132D:用基于Ad26的候选疫苗免疫的叙利亚仓鼠鼻组织中针对SARS-CoV-2IHC和组织病理的保护。用Ad26.S、Ad26.dTM.PP、Ad26.COV2.S或Ad26.empty(Ad26载体不编码任何SARS-CoV-2抗原)的1剂方案和2剂方案肌内免疫叙利亚仓鼠。在第1剂给药后4周(第4周)或第2剂给药后4周(第8周),仓鼠接受了102TCID50 SARS-CoV-2株BetaCoV/Munich/BavPat1/2020的鼻内接种。接种后4天分离左鼻组织用于免疫组织化学SARS-CoV-NP染色和组织病理学分析。由于技术故障,只可以分析用一剂Ad26.Emtpy免疫的8只仓鼠中的5只的组织。(图132A和132B)SARS-CoV-NP的存在通过免疫组织化学染色确定。(图132C和132D)对鼻组织的鼻炎严重度进行评分。
每组的中值分数用水平线表示。虚线表示LLOD。通过Mann-Whitney U-检验在疫苗组和Ad26.Empty组之间进行跨剂量水平的比较。星号表示统计差异:*:p<0.05;**:p<0.01 LLOD=检测下限;VP=病毒颗粒;NP=核衣壳蛋白;IHC=免疫组织化学。
图133A-133D:仓鼠中Ad26.COV2.S对免疫原性和肺病毒载量的剂量应答性。用每组107、108、109或1010VP的Ad26.COV2.SN=8或1010VP的Ad26.Irr(不编码任何SARS-CoV-2抗原的Ad26载体,N=8)免疫叙利亚仓鼠。一次免疫后4周,确定SARS-CoV-2刺突蛋白特异性抗体结合滴度(图133A)和SARS-CoV-2中和抗体(图133B)。每组的中值抗体应答用水平线表示。虚线表示LLOD。将应答等于或低于LLOD的动物置于LLOD并以空心符号示出。仓鼠在免疫后4周(第4周)接受鼻内接种102TCID50 SARS-CoV-2株的BetaCoV/Munich/BavPat1/2020。接种后4天分离右肺组织用于病毒学分析和免疫组织化学。(图133C)通过对Vero E6细胞的TCID50测定确定肺病毒载量。每组的中值病毒载量用水平线表示。每只动物计算LLOD,应答等于或低于LLOD的动物以空心符号示出。(图133D)SARS-CoV-2NP的存在通过免疫组织化学染色确定。通过Mann-Whitney U检验在Ad26.COV2.S剂量水平组与Ad26.Irr组之间进行比较。星号表示统计差异:*:p<0.05;**:p<0.01;***:p<0.001。Ad26.Irr=不编码任何SARS-CoV-2抗原的Ad26载体;LLOD=检测下限;N=动物数量;TCID 50/g=50%组织培养物感染剂量/克组织;VP=病毒颗粒;NP=核衣壳蛋白。
图134:在接种SARS-CoV-2的Ad26免疫的叙利亚仓鼠中没有出现VAERD的迹象,在IN接种102TCID50SARS-CoV-2后四天(每组N=8),a)分离肺组织并评分肺泡炎的严重度、肺泡损伤、肺泡水肿、肺泡出血、II型肺细胞增生、支气管炎、细支气管炎、支气管周围和血管周围套状白细胞(cuffing)的存在和严重度。分数总和表示为LRT疾病参数的总和。b)接种后四天,分离鼻组织并对炎症(鼻炎)的严重度进行评分。
水平线表示病理学评分为0,表示没有组织病理。红色符号表示来自具有突破性肺病毒载量(>102TCID50/g)的仓鼠样品。通过Mann-Whitney U检验在Ad26.COV2.S剂量水平组与Ad26.Irr组之间进行比较。星号表示统计差异:*:p<0.05;**:p<0.01;***:p<0.001。Ad26.Irr=不编码任何SARS-CoV-2抗原的Ad26载体;LRT=下呼吸道;N=动物数量;VP=病毒颗粒。
图135A-135D:结合和中和抗体与保护相关。每个疫苗构建体的保护定义为肺组织中低于102TCID50/g的病毒载量,与疫苗方案和剂量水平无关(参见图129A和B,以及图133C)。用109或1010VP的Ad26.S或Ad26.dTM.PP(N=24每个构建体)或107、108、109、1010VP的Ad26.CoV.S对叙利亚仓鼠进行一次或两次免疫(N=56)。给仓鼠接种102TCID50SARS-CoV-2,四天后牺牲以进行肺组织的病毒学分析。在病毒接种之前分析血清样品的(图135A)抗体结合滴度和(图135B)病毒中和抗体。每组的抗体应答的中值用水平线表示。虚线表示LLOD。(图135C)用来自合并的方案和Ad26.COV2.S剂量水平的结合和中和抗体滴度的Firth校正建立保护概率逻辑回归模型。虚线表示95%置信区间。LLOD=检测下限;N=动物数量;TCID50/g=每克组织50%组织培养物感染剂量;VP=病毒颗粒。
图136A-136C:Ad26.COV2.S疫苗接种后快速诱导结合和中和抗体。高剂量(HD)和低剂量(LD)Ad26.COV2.S或安慰剂(PL)的接受者在第1天和第8天的(图136A)通过ELISA的S特异性结合抗体、(图136B)通过ELISA的RBD特异性结合抗体和(图136C)假病毒中和抗体(psVNA)。红色条反映几何平均滴度(GMT)。P值反映了双边Mann-Whitney检验。
图137A-137B:Ad26.COV2.S疫苗接种后结合和中和抗体的动力学和量级。Ad26.COV2.S疫苗接种后的(A)通过ELISA的S和RBD特异性结合抗体和(B)SARS-CoV-2假病毒中和抗体(psVNA)和Ad26病毒中和抗体(Ad26 VNA)应答。LD,低剂量;HD,高剂量;PL,安慰剂。红色条反映几何平均滴度(GMT)。虚线反映了定量的下限。
图138:体液免疫应答的相关性。第29天log10 S特异性ELISA滴度、log10 RBD特异性ELISA滴度和log10中和抗体(NAb)滴度的相关性。P和R值反映双边Spearman等级相关性检验。
图139A-139B:Ad26.COV2.S疫苗接种后的抗体交叉应答性。电化学发光测定(MesoScale Discovery SARS-CoV-2 IgG Panel 2;K15369U-2)评估在Ad26.COV2.S疫苗接种后结合抗体对来自(图139A)SARS-CoV-2和SARS-CoV-1以及(图139B)CoV-229E、CoV-HKU1、CoV-NL63和CoV-OC43的S蛋白的应答。LD,低剂量;HD,高剂量;PL,安慰剂。红色条反映几何平均应答。虚线反映了定量的下限。
图140A-140B:Ad26.COV2.S疫苗接种后细胞免疫应答的动力学和量级。在Ad26.COV2.S疫苗接种后(图140A)IFN-γ和IL-4ELISPOT应答和(图140B)通过ICS测定的IFN-γ中央记忆CD27+CD45RA-CD4+和CD8+T细胞应答。在第71/85天,对具有足够外周血单个核细胞(PBMC)的一部分参与者进行了ICS测定。SFC,斑点形成细胞;LD,低剂量;HD,高剂量;PL,安慰剂。红色条反映几何平均应答。虚线反映了定量的下限。
图141:细胞和体液免疫应答的相关性。第29天log10 ELISPOT应答与log10 S特异性ELISA滴度、log10 RBD特异性ELISA滴度和log10中和抗体(NAb)滴度的相关性。P和R值反映双边Spearman等级相关性检验。
图142:分子确诊的在疫苗接种后至少1天发病的中度至严重/危重COVID-19病例的累积发病率,完整分析集。
图143:分子确诊的在疫苗接种后至少1天发病直至第29天的中度至严重/危重COVID-19病例的血清状态的累积发病率;完整分析集。
图144:分子确诊的在疫苗接种后至少1天发病的严重/危重COVID-19病例的累积发病率,完整分析集。
图145:感染力。不同国家的安慰剂COVID-19发病率。
具体实施方式
定义
如本文所用,术语“约”是指所述值的+/-10%。
术语“腺病毒载体(adenovirus vector)”和“腺病毒载体(adenoviral vector)”可互换使用,其是指经基因工程化改造的腺病毒,设计所述腺病毒以将感兴趣的多核苷酸(例如,编码2019-nCoV免疫原的多核苷酸)插入真核细胞,从而随后表达所述多核苷酸。可用作病毒载体的腺病毒的实例包括具有或衍生自血清型Ad2、Ad5、Ad11、Ad12、Ad24、Ad26、Ad34、Ad35、Ad40、Ad48、Ad49、Ad50、Ad52(例如,RhAd52)、Ad59(例如,RhAd59)和Pan9(也称为AdC68)的那些;这些载体可以衍生自例如人、黑猩猩或恒河猴腺病毒。在一些实施方案中,腺病毒是Ad26。
术语“佐剂”是指药理学或免疫学药剂,其改变其他药剂(例如疫苗)的作用,而当其自身给予时几乎没有直接作用。它们通常包括在疫苗中,以增强接受者对所提供抗原的免疫应答,同时将注射的外来物质保持在最低。
如本文所用,“给药”是指将一定剂量的药物组合物(例如,免疫原性组合物(例如,疫苗(例如,2019-nCoV疫苗)))给予受试者的方法。在本文所述的方法中使用的组合物可以例如肌内、静脉内、皮内、经皮、动脉内、腹膜内、病灶内、颅内、关节内、前列腺内、胸膜内、气管内、鼻内、玻璃体内、阴道内、直肠内、局部、肿瘤内、腹膜、皮下、结膜下、血管内、黏膜内、心包内、脐内、眼内、口服、外用、局部、通过吸入、通过注射、通过输注、通过连续输注、通过直接对靶细胞局部灌注、通过导管、通过灌洗、通过灌胃、在乳膏中或在脂质组合物中给药。优选的给药方法可以根据各种要素(例如,所给药的组合物的成分和所治疗的病况的严重度)而变化。
术语“抗体”和“免疫球蛋白(Ig)”在最广义上可互换使用,并且包括单克隆抗体(例如,全长或完整的单克隆抗体)、多克隆抗体、多价抗体、多特异性抗体(例如,双特异性抗体,只要它们表现出所期望的生物活性)并且还可以包括某些抗体片段。抗体通常包含“轻链”和“重链”。基于其恒定结构域的氨基酸序列,可以将来自任何脊椎动物物种的抗体(免疫球蛋白)的轻链分成两种明显不同的类型中的一种,称为kappa(κ)和lambda(λ)。根据其重链恒定结构域的氨基酸序列,免疫球蛋白可分为不同的类别。有五种主要的免疫球蛋白类别:IgA、IgD、IgE、IgG和IgM,并且这些中的一些可以进一步分为亚类(同种型),例如IgG1、IgG2、IgG3、IgG4、IgA1和IgA2。对应于免疫球蛋白不同类别的重链恒定结构域分别称为α、δ、ε、γ和μ。不同类别的免疫球蛋白的亚基结构和三维构型是熟知的。
如本文所用的术语“密码子”指给定信使RNA分子或DNA编码链中三个连续核苷酸碱基的任何组,其指定特定氨基酸或翻译的起始或终止信号。术语密码子也指DNA链中的碱基三联体。
在整个说明书和权利要求书中,词语“包含(comprise)”或诸如“包含(comprises)”或“包含(comprising)”的变化将理解为暗示包括所陈述的整数或整数组,但不排除任何其他整数或整数组。
如本文所用的术语“恢复期”是指已经恢复或正在从冠状病毒感染(例如,2019-nCoV)中恢复的受试者。“恢复期人的队列”是指具有共同特征(例如,性别、年龄、体重、病史、种族、民族或环境)并且已经或正在从冠状病毒感染(例如,2019-nCoV)中恢复的人的队列。优选地,恢复期人的队列将与具有2019-nCoV感染风险或怀疑对冠状病毒感染易感的受试者具有共同特征。优选地,来自恢复期人的样品会在记录恢复(例如,用阴性鼻拭子确定的)后至少7天获得。
术语“胞外域(ectodomain)”和“胞外结构域(extracellular domain)”是指冠状病毒刺突多肽中延伸超出跨膜结构域进入胞外空间的部分。胞外域介导刺突多肽与一种或多种冠状病毒受体(例如,ACE2)的结合。例如,胞外域包括刺突多肽的S1结构域(例如,SEQID NO:4)和RBD(例如,SEQ ID NO:5)。
“基因递送载体”定义为任何可以携带插入的多核苷酸进入宿主细胞的分子。基因递送载体的实例是脂质体、生物相容性聚合物,包括天然聚合物和合成聚合物;脂蛋白;多肽;多糖;脂多糖;人工病毒包膜;金属颗粒;和细菌,或病毒,例如杆状病毒、腺病毒和逆转录病毒、噬菌体、粘粒、质粒、真菌载体和本领域通常使用的已将其描述用于在多种真核和原核宿主中表达,并且可以用于基因治疗以及简单的蛋白表达的其他重组载体。
如本文所用的“基因递送”、“基因转移”等是指将外源多核苷酸(有时称为“转基因”)引入宿主细胞的术语,与用于引入的方法无关。这种方法包括多种技术,例如载体介导的基因转移(例如,病毒感染/转染,或各种其他基于蛋白或基于脂质的基因递送复合物)以及促进“裸”多核苷酸递送的技术(例如如电穿孔、“基因枪”递送和用于引入多核苷酸的各种其他技术)。引入的多核苷酸可以稳定地或瞬时地维持在宿主细胞中。稳定维持通常需要引入的多核苷酸含有与宿主细胞相兼容的复制起点或整合至宿主细胞的复制子中,例如染色体外复制子(例如质粒)或核或线粒体染色体。许多载体能够介导基因向哺乳动物细胞的转移。
“基因产物”意在包括从基因转录的mRNA或其他核酸(例如,microRNA),以及从那些mRNA翻译的多肽。在一些实施方案中,基因产物来自病毒(例如,2019-nCoV)并且可以包括例如本文所述的任何一种或多种病毒蛋白或其片段。
“异源核酸分子”是指可以编码衍生自或获得自病原生物如病毒的蛋白的核苷酸序列,其可以掺入多核苷酸或载体中。异源核酸也可以编码合成的或人工的蛋白,例如免疫原性表位,构建以诱导免疫。异源核酸分子的一个实例是编码一种或多种衍生自冠状病毒(例如,2019-nCoV)的免疫原性肽或多肽的核酸分子。异源核酸分子是通常不与掺入异源核酸分子的多核苷酸或载体中发现的其他核酸分子相关联的核酸分子。
术语“宿主细胞”是指已引入外源核酸的细胞,包括这种细胞的后代。宿主细胞包括“转化体”和“转化细胞”,其包括初级转化细胞和由其衍生的后代,不考虑传代次数。宿主细胞包括受试者(例如哺乳动物受试者(例如人))体内已引入外源核酸的细胞。
“免疫原”是指在给药后可以在受试者中诱导免疫应答的任何多肽。在一些实施方案中,免疫原由核酸分子编码,该核酸分子可以掺入例如多核苷酸或载体中,用于随后表达免疫原(例如,感兴趣的基因产物,或其片段(例如,多肽)))。
如本文所用的术语“免疫原性组合物”定义为用于引发免疫应答并且可以在向受试者给药免疫原性组合物后赋予免疫性的材料。
术语“免疫刺激剂”是指通过诱导其任何成分的激活或增加其活性来刺激免疫系统的物质(例如,药物和营养物)。免疫刺激剂包括细胞因子(例如,粒细胞巨噬细胞集落刺激因子)和干扰素(例如,IFN-α和/或IFN-γ)。
“分离的”是指从其天然环境的组分中分离、回收或纯化。例如,核酸分子或多肽可以从其天然环境的组分中分离出1%(2%、3%、4%、5%、6%、7%、8%、9%、10%、20%、30%、40%、50%、60%、70%、80%或90%)或更多。
“药物组合物”是指含有治疗活性剂或生物活性剂的任何组合物,例如免疫原性组合物或疫苗(例如,2019-nCoV核酸分子、载体和/或多肽),优选包括编码感兴趣的抗原基因产物或其片段的核苷酸序列,适用于向受试者给药并治疗或预防疾病(例如,2019-nCoV感染)或减轻或改善疾病的一种或多种症状(例如,2019-nCoV病毒滴度、病毒传播、感染和/或细胞融合)。为了本发明的目的,药物组合物包括疫苗,并且适用于递送治疗剂或生物活性剂的药物组合物可以包括例如片剂、囊形片、胶囊剂、丸剂、粉剂、颗粒剂、悬浮剂、乳剂、溶液剂、凝胶剂、水凝胶、口服凝胶、糊剂、滴眼剂、软膏、乳膏、膏药、灌药器(drenches)、递送装置、栓剂、灌肠剂、注射剂、植入物、喷雾剂或气雾剂。这些制剂中的任何都可以通过本领域公知和公认的方法制备。例如,参见Remington:The Science and Practice ofPharmacy(21sted.),ed.A.R.Gennaro,Lippincott Williams&Wilkins,2005;以及Encyclopedia of Pharmaceutical Technology,ed.J.Swarbrick,Informa Healthcare,2006,每个援引加入本文。
如本文所用的术语“连接(linked)”或“连接(links)”或“连接(link)”是指分别通过肽键或磷酸二酯键将两个氨基酸序列或两个核酸序列共价连接在一起,这种连接可以包括在连接的两个氨基酸序列或核酸序列之间的任何数量的附加氨基酸或核酸序列。
“核酸分子”或“多核苷酸”,如本文中互换使用的,其是指任何长度的核苷酸的聚合物,并且包括DNA和RNA。核苷酸可以是脱氧核糖核苷酸、核糖核苷酸、修饰的核苷酸或碱基和/或它们的类似物,或者可以通过DNA或RNA聚合酶或通过合成反应掺入聚合物中的任何底物。多核苷酸可以包含修饰的核苷酸,例如甲基化核苷酸和它们的类似物。如果存在,可以在聚合物组装之前或之后对核苷酸结构进行修饰。可以用非核苷酸成分打断核苷酸序列。多核苷酸可在合成后进一步修饰,例如通过与标记缀合。
“核酸疫苗”是指疫苗,其包括用于在受试者中表达,在启动子控制下的异源核酸分子。可以将异源核酸分子掺入表达载体,如质粒。“DNA疫苗”是指其中核酸为DNA的疫苗。“RNA疫苗”是指其中核酸为RNA(例如,mRNA)的疫苗。
当核酸处于与另一个核酸序列的结构或功能关系中时,它是“可操作地连接”的。例如,如果一个DNA区段和另一个DNA区段彼此相对定位在同一个连续的DNA分子上并具有结构或功能上的关系,则一个DNA区段可能与另一个DNA区段可操作地连接,如启动子或增强子相对于编码序列定位以促进编码序列的转录;核糖体结合位点相对于编码序列定位以促进翻译;或者前序列或分泌前导序列相对于编码序列定位以促进前蛋白(例如,参与编码的多肽的分泌的前蛋白)的表达。在其他实例中,可操作地连接的核酸序列不是连续的,而是以使它们作为核酸或作为由它们表达的蛋白彼此具有功能关系的方式定位。例如,增强子不一定是连续的。连接可以通过在方便的限度性位点处连接(ligation)或通过使用合成的寡核苷酸接头(adaptor)或连接子(linker)来完成。
“优化的”是指非天然存在的肽、多肽或蛋白的免疫原性多肽,如非天然存在的病毒多肽(例如刺突多肽)。优化的病毒多肽序列最初是通过修饰一种或多种天然存在的病毒基因产物(例如肽、多肽和蛋白)的氨基酸序列来产生的,以增加在对受试者(例如人)进行免疫(例如,当掺入组合物,例如疫苗)时产生的抗病毒免疫应答(例如,细胞或体液免疫应答)的广度、强度、深度或持续时间(longevity)。因此,优化的病毒多肽可能对应于“亲本”病毒基因序列;或者,优化的病毒多肽可能不对应于特定的“亲本”病毒基因序列,但可能对应于来自病毒的各种毒株或准种(quasi-species)的类似序列。可以包括在优化的病毒多肽中的对病毒基因序列的修饰包括氨基酸添加、取代和缺失。在一个实施方案中,优化的多肽是刺突多肽,已将其进一步改变为包括用于最大蛋白表达的前导/信号序列(例如刺突信号序列或tPA信号序列)、Xa因子位点、折叠子三聚化结构域(参见,例如,SEQ ID NO:87)和/或连接子或间隔区(例如,SEQ ID NO:88或89)序列。优化的多肽可以但不必还包括切割位点突变(例如弗林蛋白酶切割位点突变(例如SEQ ID NO:91))。产生优化的病毒多肽的方法描述于例如Fisher et al.“Polyvalent Vaccine for Optimal Coverage of PotentialT-Cell Epitopes in Global HIV-1 Variants,”Nat.Med.13(1):100-106(2007)和国际专利申请公开WO 2007/024941,援引加入本文。一旦产生了优化的病毒多肽序列,就可以通过标准技术(例如重组病毒载体,如国际专利申请公开WO 2006/040330和WO 2007/104792中公开的腺病毒载体,援引加入本文)产生或给药以及任选地组装以形成稳定化的多肽三聚体。
如本文所用的术语“优化的密码子”和“密码子优化”是指将密码子序列修饰以匹配靶标(例如受试者)或宿主生物体中的密码子频率,但不改变原始翻译的蛋白的氨基酸序列。
“药学上可接受的稀释剂、赋形剂、载剂或佐剂”是指受试者生理上可接受的稀释剂、赋形剂、载剂或佐剂,同时保留与其一起给药的药物组合物的治疗特性。一种示例性的药学上可接受的载剂是生理盐水。其他生理学上可接受的稀释剂、赋形剂、载剂或佐剂及其制剂是本领域技术人员已知的(参见例如美国公布号2012/0076812)。
“部分”或“片段”是指整体的一部分。一部分可包含多核苷酸或多肽序列区域的全长的至少10%、20%、30%、40%、50%、60%、70%、80%、90%或95%。例如,对于多核苷酸,一部分可以包括参考多核苷酸分子的至少5、6、7、8、9、10、20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700、1800或更多个连续核苷酸。例如,对于多肽,一部分可以包括参考多肽分子的至少5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500、525、550、575或600个或者更多个连续氨基酸。
在一些情况下,核酸分子的片段可以包括多核苷酸SS-Spike(SEQ ID NO:121)的至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-Spike-dF-PP(SEQ ID NO:143和204)的至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-SdCT(SEQID NO:122)的至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-S.Ecto(SEQ ID NO:123)的至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-S1-foldon(SEQ ID NO:129)的至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-RBD-foldon(SEQ ID NO:130)的至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-S.Ecto-dF-foldon(SEQ ID NO:136或193)的至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-S.Ecto-PP-foldon(SEQ IDNO:142)的至少5、6、7、8、9、10、20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700、1800、1900、2000或更多个连续核苷酸。在一些情况下,核酸分子的片段可以包括多核苷酸SS-S.Ecto-dF-PP-foldon(SEQ ID NO:195)的至少5、6、7、8、9、10、20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700、1800、1900、2000或更多个连续核苷酸。
在一些情况下,多肽的片段可以包括多肽SS-Spike(SEQ ID NO:29)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500、525、550或更多个连续氨基酸。在一些情况下,多肽的片段可以包括多肽SS-SdCT(SEQ IDNO:30)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500或更多个连续氨基酸。在一些情况下,多肽的片段可以包括多肽SS-Spike-dF-PP(SEQ ID NO:51)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500或更多个连续氨基酸。在一些情况下,多肽的片段可以包括SS-S.Ecto(SEQ ID NO:31)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475或更多个连续氨基酸。在一些情况下,多肽的片段可以包括多肽SS-S1-foldon(SEQ ID NO:37)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475或更多个连续氨基酸。在一些情况下,多肽的片段可以包括多肽SS-RBD-foldon(SEQ ID NO:38)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500或更多个连续氨基酸。在一些情况下,多肽的片段可以包括多肽SS-S.Ecto-dF-foldon(SEQ IDNO:44)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425或更多个连续氨基酸。在一些情况下,多肽的片段可以包括多肽S.Ecto-PP-foldon(SEQ ID NO:50)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400或更多个连续氨基酸。在一些情况下,多肽的片段可以包括多肽SS-S.Ecto-dF-PP-foldon(SEQ ID NO:56)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500、525、550、575、600、625、650、675或更多个连续氨基酸。在一些情况下,多肽的片段可以包括具有JEV Stem/TM的多肽prM-Env(SEQ IDNO:27)的至少20、25、50、75、90、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500、525、550、575、600、625、650、675或更多个连续氨基酸。
在一些情况下,向受试者给药多核苷酸(例如,SEQ ID NO:93-181、190-195和199-204)和/或多肽(例如,SEQ ID NO:1-84,例如,SEQ ID NO:51)的片段可以在受试者中引发免疫应答。
“启动子”是能够启动信使RNA中的基因序列转录的核酸序列,这种转录是通过RNA聚合酶在启动子上或启动子附近的结合而启动的。
“促进免疫应答”是指引发针对例如,一种或多种感染性物质(例如,病毒(例如,2019-nCoV))或已给药药物组合物(例如,免疫原性组合物或疫苗)的受试者中的蛋白靶标的体液应答(例如,抗体的产生)或细胞应答(例如,T细胞、巨噬细胞、中性粒细胞和/或自然杀伤细胞的激活)。
术语“2019-nCoV介导的疾病”在本文中与“2019冠状病毒病(COVID-19)”及其语法变体互换使用,其是指本领域已知由(单独或与其他中介物相关联)2019-nCoV(SARS-CoV-2)感染或暴露于患有该疾病的受试者中引起、加重或与之相关联的任何病理或后遗症。疾病可以是急性的(例如,发烧)或慢性的(例如,慢性疲劳)、轻度的(例如,脱发)或严重的(例如,器官衰竭)和早发的(例如,感染后2-14天)或迟发性(例如,感染后2周)。严重疾病的非限度性实例包括肺炎、急性呼吸窘迫综合征(ARDS)、急性呼吸衰竭、肺水肿、器官衰竭或死亡。症状的非限制性实例包括体重减轻、发烧、咳嗽、呼吸困难、疲劳、头痛、味觉或嗅觉丧失、脱发、皮疹、喉咙痛、恶心和腹泻。症状可以是轻度的或严重的(例如,一周内体重减轻超过约5%和高烧)和暂时性的或永久性的。
“保护水平”是指标志物(例如抗体、细胞(例如免疫细胞,例如T细胞、B细胞、NK细胞或中性粒细胞))的量或水平,其是表示部分或完全保护免受冠状病毒感染或疾病。高于保护水平的标志物的量或水平表明保护免受冠状病毒感染(例如,2019-nCoV感染)或疾病(例如,2019-nCoV介导的疾病,例如,COVID-19,例如,严重的COVID-19疾病)。低于保护水平的标志物的量或水平表明对冠状病毒感染或疾病(例如,2019-nCoV介导的疾病,例如,COVID-19,例如严重的临床疾病)的易感性。标志物可以是单一测量值(例如,中和抗体水平)或标志物可以是多个测量值的组合(例如,中和抗体水平和RBD特异性IgG2水平)。在一些情况下,保护水平是使用本文所述的假病毒中和测定测量的至少约70的抗冠状病毒抗体滴度,使用本文所述的活病毒中和测定测量的至少约25的抗冠状病毒抗体滴度,或如通过本文所述的假病毒中和测定或活病毒中和测定确定的高于恢复期人的队列的中值或平均水平的至少约80%的水平的抗冠状病毒抗体滴度。在一些情况下,保护水平是使用本文所述的假病毒中和测定测量的至少约100的抗冠状病毒抗体滴度。
如本文所用,术语“样品”是获得自或衍生自受试者的组合物,其含有待表征和/或鉴定的细胞和/或其他分子实体,例如基于物理、生化、化学和/或生理特征。样品可以是来自新鲜、冷冻和/或保存的器官、组织样品、活检和/或抽出物的实体组织;血液或任何血液成分,如血浆;体液,如脑脊髓液、羊水、腹膜液或间质液。样品也可以是原代或培养的细胞或细胞系。样品可能含有在自然界中不与组织自然混合的化合物,如防腐剂、抗凝剂、缓冲剂、固定剂、蜡、营养物、抗生素等。
“序列相同性”或“序列相似性”是指两个或更多个氨基酸序列或者两个或更多个核苷酸序列之间的相同性或相似性,分别以序列之间的相同性或相似性表示。序列相同性可以用“百分比(%)相同性”来测量,其中较高的百分比表明序列之间共享的相同性较大。序列相似性可以用百分比相似性来测量(考虑到保守氨基酸取代);百分比越高,序列之间共享的相似性越多。当使用标准方法比对时,核酸或氨基酸序列的同源物或直系同源物具有相对高度的序列相同性/相似性。序列相同性可以在默认设置下使用序列分析软件测量(例如,遗传计算机组的序列分析软件包,威斯康星大学生物技术中心,1710 UniversityAvenue,Madison,WI 53705)。这样的软件可以通过将同源度分配给各种取代、缺失和其他修饰来匹配相似的序列。序列相同性/相似性可以在所比较的两个或更多个序列的全部或定义部分中测定。
“信号肽”是指在多肽的N-末端处的短肽(例如,长度为5-30个氨基酸,如长度为17个氨基酸,例如,SEQ ID NO:92),其引导多肽向分泌途径(例如,细胞外空间)。通常在多肽分泌期间切割信号肽。信号序列可以将多肽引导至细胞内区室或细胞器,例如高尔基体。信号序列可以通过与肽的同源性或生物活性来鉴定,该肽具有将多肽靶向细胞的特定区域的已知功能。本领域普通技术人员可以通过使用容易获得的软件(例如,遗传计算机组的序列分析软件包,威斯康星大学生物技术中心,1710 University Avenue,Madison,Wis.53705,BLAST或PILEUP/PRETTYBOX程序)鉴定信号肽。信号肽可以是例如与SEQ ID NO:92的氨基酸序列基本上相同的肽。
如本文所用,短语“特异性结合”是指结合反应,其对于例如通过抗体或其抗原结合片段特异性识别的抗原在蛋白和其他生物分子的异质群体中的存在是决定的。特异性结合抗原的抗体或其抗原结合片段会以小于100nM的KD结合抗原。例如,特异性结合抗原的抗体或其抗原结合片段会以达100nM(例如,l pM-100nM)的KD结合抗原。对特定抗原或其表位没有表现出特异性结合的抗体或其抗原结合片段会对那个特定抗原或其表位表现出大于100nM(例如,大于500nm、1μM、100μM、500μM或1mM)的KD。多种免疫测定形式可用于选择与特定蛋白或碳水化合物特异性免疫反应的抗体。例如,固相ELISA免疫测定常规地用于选择与蛋白或碳水化合物特异性免疫反应的抗体。对于可用于确定特定免疫反应性的免疫测定形式和条件的描述,参见Harlow&Lane,Antibodies,A Laboratory Manual,Cold SpringHarbor Press,New York(1988)和Harlow&Lane,Using Antibodies,A LaboratoryManual,Cold Spring Harbor Press,New York(1999)。
如本文所用,术语“稳定化的多肽三聚体”或“稳定化的三聚体”是指但不限于包括增加三聚体结构稳定性的蛋白和/或多肽序列的寡聚体(例如,通过一种或多种寡聚化结构域的存在)(例如,减少三聚体解离成单体单元)。例如,稳定化的多肽三聚体可以是同源三聚体。“寡聚化结构域”是指但不限于可用于增加寡聚包膜蛋白的稳定性的多肽序列,例如增加刺突三聚体的稳定性。寡聚化结构域可用于增加同源寡聚多肽以及异源寡聚多肽的稳定性。寡聚化结构域是本领域中是熟知的并且包括“三聚化结构域”。三聚化结构域是指稳定三聚多肽(例如,由一种或多种刺突多肽组成的三聚体)的寡聚化结构域。三聚化结构域的实例包括但不限于T4-纤蛋白“折叠子”三聚化结构域;衍生自GCN4的卷曲螺旋三聚化结构域(Yang et al.(2002)J.Virol.76:4634);和大肠杆菌天冬氨酸转氨甲酰酶的催化亚基作为三聚体标签(Chen et al.(2004)J.Virol.78:4508)。
“受试者”是脊椎动物,例如哺乳动物(例如,灵长类动物和人,特别是具有潜在健康状况(例如,高血压、糖尿病或心血管疾病)的人)。哺乳动物还包括但不限于农场动物(如牛)、运动动物(例如马)、宠物(如猫和狗)、小鼠、大鼠、蝙蝠、果子狸和貉。待根据本文所述方法治疗的受试者(例如,需要保护免受2019-nCoV感染或患有2019-nCoV感染的受试者可以是已由医生诊断为具有这种需要或感染的受试者。可以通过任何合适的方式进行诊断。其中正在预防感染发展的受试者可能已经或可能没有接受这种诊断。本领域技术人员会理解待根据本发明治疗的受试者可能已经经过标准测试,或者可能未经检查就已将其鉴定为疑似感染或由于存在一种或多种风险要素(例如,暴露于2019-nCoV、例如,由于前往2019-nCoV感染流行的地区旅行)而处于感染的高风险中。此外,将具有潜在健康状况(例如,高血压、糖尿病或心血管疾病)的人鉴定为处于感染冠状病毒(例如,2019-nCoV)的高风险中的受试者。因此,用组合物治疗人受试者的方法在治疗、减少和/或预防具有潜在健康状况的人的2019-nCoV感染方面特别有用。
如本文所用,术语“转染”是指常见地用于将外源核酸分子(例如,DNA,如表达载体)引入原核或真核宿主细胞的多种技术中的任一种,例如,电穿孔、脂质转染、磷酸钙沉淀、DEAE-葡聚糖转染等。
如本文所用并且如本领域所熟知的,“治疗”是用于获得有益或期望结果,如临床结果的方法。有益或期望的结果可以包括但不限于减轻或改善一种或多种症状(例如发热、关节痛、皮疹、结膜炎、肌肉痛、头痛、眶后痛、水肿、淋巴结病、不适、虚弱、喉咙痛、咳嗽、恶心、呕吐、腹泻和血精症(hematospermia))或病况(Zammarchi et al.,J.Clin.Virol.63:32-5,2015;Waddell et al.,PLoS One 11(5):e0156376,2016);减轻疾病、病症或病况的程度;稳定疾病、病症或病况的状态(例如,不恶化);预防疾病、病症或病况的传播;延迟或减缓疾病、病症或病况的进展;疾病、病症或病况的改善或缓和;和缓解(无论是部分的还是全部的),无论是可检测的还是不可检测的。“缓和”疾病、病症或病况是指与没有治疗的程度或时间过程相比,疾病、病症或病况的程度和/或不期望的临床表现减轻,和/或进展的时间过程减慢或延长。治疗可以包括一种或多种治疗剂,如本文所述的一种或多种组合物和/或一种或多种其他治疗剂。其他治疗剂可以包括刺激(例如干扰素)或抑制(例如抗炎剂,如皮质类固醇,例如地塞米松)免疫应答的试剂。治疗可以包括一种或多种治疗干预,例如手术或俯卧位通气(prone positioning)。
如本文所用的术语“疫苗”定义为用于引发免疫应答并在向受试者给药疫苗后的一段时间内赋予免疫力的材料。
“载体”是指包括一个或多个多核苷酸或其片段的DNA构建体,例如来自病毒种,如2019-nCoV种。载体可用于感染受试者的细胞,其导致载体的多核苷酸翻译成蛋白产物。一种类型的载体是“质粒”,其是指其中可以连接其他DNA片段的环状双链DNA环。某些载体能够在它们所引入的宿主细胞中自主复制(例如,具有细菌复制起点的细菌载体和游离型哺乳动物载体)。其他载体(例如,非游离型哺乳动物载体)可以在引入宿主细胞后整合到宿主细胞的基因组中,从而与宿主基因组一起复制。此外,某些载体能够指导与其可操作地连接的基因的表达。这类载体在本文中称为“重组表达载体”(或简称为“重组载体”)。一般而言,在重组DNA技术中有用的表达载体经常是质粒的形式。在本说明书中,“质粒”和“载体”有时可以互换使用,因为质粒是最常用的载体形式。其他载体包括例如病毒载体,如腺病毒载体(例如Ad26载体),特别是本文所述的那些。
如本文所用,术语“病毒”定义为不能在宿主细胞外生长或增殖并且感染哺乳动物(例如人)的感染剂。
“病毒载体”定义为重组产生的病毒或病毒颗粒,其包含待递送至宿主细胞中的多核苷酸。病毒载体的实例包括逆转录病毒载体、腺病毒载体、腺相关病毒载体(例如参见PCT公开号WO 2006/002203)、甲病毒载体等。
在基因转移由DNA病毒载体如腺病毒(Ad(例如,Ad26))或腺相关病毒(AAV)介导的方面,载体构建体是指包含病毒基因组或其部分的多核苷酸和转基因。Ad是相对较好表征的同源病毒组,包括超过50种血清型(WO 95/27071)。Ad易于生长并且不需要整合至宿主细胞基因组中。还构建了重组Ad衍生的载体,特别是那些降低重组和产生野生型病毒的潜能的载体(WO 95/00655和WO 95/11984)。含有启动子和克隆位点二者的载体是本领域已知的,在克隆位点中多核苷酸可以可操作地连接。这类载体能够在体外或体内转录RNA。为了优化表达和/或体外转录,可能需要去除、添加或改变克隆的5′和/或3′非翻译部分,以消除额外的、潜在的不适当的可选的翻译起始密码子或在转录或翻译水平上可能干扰或减少表达的其他序列。
从以下详细描述、附图和权利要求书中,可以看出其他特征和优点。
发明详述
本发明提供一种分离的核酸分子,其包含编码2019-NCOV刺突蛋白(S)蛋白(在本文中也称为SARS-CoV-2 S蛋白)的核苷酸序列,所述2019-NCOV刺突蛋白(S)蛋白包含对SEQID NO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
SEQ ID NO:29提供了全长2019-NCOV刺突(S)蛋白的氨基酸序列;也可参见NCBI参考序列:YP_009724390.1。稳定化突变是从这个序列中的原始氨基酸到脯氨酸。如SEQ IDNO:29所示,第986位的原始氨基酸是赖氨酸(lys,K),第987位的原始氨基酸是缬氨酸(val,V)。
2019-NCOV刺突(S)蛋白内的弗林蛋白酶切割位点包含或具有氨基酸序列RARR(SEQ ID NO:90)。合适的突变可以包含突变为SRAG(SEQ ID NO:225)或GGSG(SEQ ID NO:91)。SRAG突变是通过将R682S和R685G突变引入氨基酸序列来实现的。GGSG突变是通过将R682G、R683G、A684S和R685G突变引入氨基酸序列来实现的。
在一些实施方案中,这些是对SEQ ID NO:29的序列进行的唯一修饰。因此,一个优选的核酸分子编码全长2019-NCOV刺突(S)蛋白,其具有稳定化突变和对弗林蛋白酶切割位点的突变作为唯一的修饰。在其他实施方案中,分离的核酸分子编码2019-NCOV刺突(S)蛋白,所述2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
c.信号序列的缺失。
在一些实施方案中,编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(CMV)启动子,优选CMV立即早期启动子可操作地连接。在一些实施方案中,编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(CMV)启动子可操作地连接,所述CMV启动子包含至少一个四环素操纵子(TetO)基序。在具体实施方案中,所述包含至少一个TetO基序的CMV启动子包含SEQ IDNO:219的核苷酸序列。在一些实施方案中,CMV启动子由SEQ ID NO:219的核苷酸序列组成。这些核酸通常形成载体的部分。载体在本文中更详细地描述。
本发明还提供一种分离的2019-NCOV刺突(S)蛋白(本文也称为SARS-CoV-2 S蛋白),其包含对SEQ ID NO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
在一些实施方案中,这些是对SEQ ID NO:29的序列进行的唯一修饰。在其他实施方案中,分离的2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
c.信号序列的缺失。
如本文所用,用于核酸、蛋白和/或腺病毒的术语“重组”表示已将它通过人工修饰,例如在腺载体的情况下,它具有主动克隆在其中的改变的末端和/或它包含异源基因,即它不是天然存在的野生型腺病毒。
如本领域习惯,本文提供的核苷酸序列是从5′到3′的方向。
冠状病毒科含有Alphacoronavirus、Betacoronavirus、Gammacoronavirus和Deltacoronavirus属。所有这些属都含有致病病毒,其可以感染多种动物,包括鸟、猫、狗、牛、蝙蝠和人。这些病毒引起一系列疾病,包括肠道和呼吸道疾病。宿主范围主要由介导病毒进入宿主细胞的病毒刺突蛋白(S蛋白)决定。可以感染人的冠状病毒在Alphacoronavirus属和Betacoronavirus属中都有发现。已知会在人中导致呼吸道疾病的冠状病毒是Betacoronavirus属的成员。这些包括SARS-CoV-1、SARS-CoV-2和MERS、OC43和HKU1。
如上所述,SARS-CoV-2可在人中引起严重的呼吸道疾病。为了结束COVID-19大流行可能需要安全有效的SARS-CoV-2疫苗。
SARS CoV-2病毒刺突(S)蛋白与血管紧张素转换酶2(ACE2)结合,所述ACE2是SARS-CoV-2使用的进入受体。ACE2是一种I型跨膜金属羧肽酶,其与ACE具有同源性,ACE是一种长期已知的肾素-血管紧张素系统(RAS)的关键参与者和治疗高血压的靶点的酶。它特别是在血管内皮细胞、肾小管上皮和睾丸的间质细胞中表达。PCR分析揭示ACE-2也在肺、肾脏和胃肠道中表达,已证明这些组织含有SARS-CoV-2。冠状病毒的刺突(S)蛋白是一种主要表面蛋白,是感染的患者中的中和抗体的靶标(Lester et al.,Access Microbiology2019;1),因此认为其是疫苗设计的潜在保护性抗原。在导致本发明的研究中,设计了几种基于SARS-CoV-2病毒S蛋白的抗原构建体。令人惊讶地发现,本发明的核酸(即SEQ ID NO:211)在表达时具有优异的免疫原性,并且可以以高产率制备含有这种核酸的腺载体。含有编码SEQ ID NO:205的SARS CoV-2 S蛋白的核酸的Ad26载体可诱导稳健的中和抗体应答,并在SARS-CoV-2攻击后在支气管肺泡灌洗液中提供完全保护和/或在鼻拭子中提供近乎完全的保护。此外,如实施例中所示,它显示出在非人灵长类动物中针对SARS-CoV-2的稳健的单次疫苗保护作用。
在一方面,本发明因此提供分离的和/或重组的核酸,其编码稳定化的冠状病毒S蛋白,特别是SARS-CoV-2 S蛋白,或其片段,所述核酸包含选自SEQ ID NO:211-218的核苷酸序列。
在一个优选的实施方案中,本发明提供一种分离的和/或重组的核酸,其编码稳定化的冠状病毒S蛋白,特别是SARS-CoV-2 S蛋白,或其片段,所述核酸包含SEQ ID NO:211的核苷酸序列,或者由SEQ ID NO:211的核苷酸序列或其片段组成。
本发明还提供分离的和/或重组的冠状病毒S蛋白,其包含选自SEQ ID NO:205-210或其片段的氨基酸序列,以及编码这种冠状病毒S蛋白或其片段的核酸。
在一个优选的实施方案中,本发明提供一种分离的和/或重组的冠状病毒S蛋白,其包含SEQ ID NO:205的氨基酸序列或其片段,以及编码这种冠状病毒S蛋白或其片段的核酸。S蛋白可以包含或不包含信号肽(或前导序列)。信号肽可以包含SEQ ID NO:205的氨基酸1-13。在某些实施方案中,冠状病毒S蛋白由SEQ ID NO:205的氨基酸序列组成。在某些实施方案中,冠状病毒S蛋白由不具有信号肽的SEQ ID NO:205的氨基酸序列组成。
技术人员应理解,由于遗传密码的简并性,许多不同的核酸可以编码相同的多肽或蛋白。还应理解,技术人员可以使用常规技术进行不影响由核酸编码的氨基酸序列的核苷酸取代,以反映要在其中表达多肽的任何特定宿主生物的密码子使用。因此,除非另有说明,“编码氨基酸序列的核苷酸序列”包括彼此为简并形式的并且编码相同氨基酸序列的所有核苷酸序列。编码蛋白和RNA的核苷酸序列可以包括内含子。
因此,本发明还提供编码冠状病毒S蛋白,特别是SEQ ID NO:205的SARS-CoV-2 S蛋白或其片段的核酸。在某些实施方案中,优化核酸密码子以在人细胞中表达。
如本文所用的术语“片段”是指具有氨基末端的和/或羧基末端的和/或内部的缺失的蛋白或(多)肽,但其中保留的氨基酸序列与SARS-CoV-2 S蛋白,例如SARS-CoV-2 S蛋白的全长序列的序列中的相应位置相同。应当理解,为了诱导免疫应答和通常为了疫苗接种目的,蛋白不需要是全长的,也不需要具有所有的其野生型的功能,并且蛋白的片段(即不具有信号肽)同样有用。
根据本发明的片段是免疫学活性片段,并且通常包含SARS-CoV-2 S蛋白的至少15个氨基酸,或至少30个氨基酸。在某些实施方案中,它包含SARS-CoV-2 S蛋白的至少50、75、100、150、200、250、300、350、400、450、500或550个氨基酸。
本领域技术人员还会理解可以对蛋白进行改变,例如通过氨基酸取代、缺失、添加等,例如使用常规分子生物学程序。通常,可以应用保守氨基酸取代而不丧失多肽的功能或免疫原性。这可以根据本领域技术人员熟知的常规程序容易地检查。
本发明进一步提供一种载体,其包含本发明的核酸序列。
在本发明的某些实施方案中,载体是腺病毒载体,如重组人腺病毒载体。本发明的腺病毒属于腺病毒科,优选属于哺乳动物腺病毒属。它可以是人腺病毒,也可以是感染其他物种的腺病毒,包括但不限于牛腺病毒(例如,牛腺病毒3,BAdV3)、犬腺病毒(例如,CAdV2)、猪腺病毒(例如,PAdV3或5),或猿猴腺病毒(其包括猴腺病毒和猿腺病毒,如黑猩猩腺病毒或大猩猩腺病毒)。优选地,腺病毒是人腺病毒(HAdV或AdHu),或猿猴腺病毒,如黑猩猩或大猩猩腺病毒(ChAd、AdCh或SAdV),或恒河猴腺病毒(RhAd)。在本发明中,如果在不指明物种的情况下称为Ad,则表示人腺病毒,例如,简短的符号“Ad26”表示与HAdV26相同,HAdV26是人腺病毒血清型26。同样如本文所用,符号“rAd”表示重组腺病毒,例如“rAd26”是指重组人腺病毒26。
已经使用人腺病毒进行了最先进的研究,并且根据本发明的某些方面,人腺病毒是优选的。在某些优选实施方案中,本发明的重组腺病毒基于人腺病毒。在优选实施方案中,重组腺病毒基于人腺病毒血清型5、11、26、34、35、48、49、50、52等。根据本发明特别优选的实施方案,腺病毒是血清型26的人腺病毒。这些血清型的优点包括人群中的低血清流行率和/或低预先存在的中和抗体滴度,以及在临床试验中在人受试者中使用的经验。
猿猴腺病毒通常在人群中也具有低血清流行率和/或低预先存在的中和抗体滴度,并且已经报道了使用黑猩猩腺病毒载体的大量工作(例如,US6083716;WO 2005/071093;WO 2010/086189;WO 2010/085984;Farina et al,2001,J Virol 75:11603-13;Cohen et al,2002,J GenVirol 83:151-55;Kobinger et al,2006,Virology 346:394-401;Tatsis et al.,2007,Molecular Therapy 15:608-17;还参见综述Bangari andMittal,2006,Vaccine 24:849-62;和综述Lasaro and Ertl,2009,Mol Ther 17:1333-39)。因此,在其他实施方案中,本发明的重组腺病毒基于猿猴腺病毒,例如黑猩猩腺病毒。在某些实施方案中,重组腺病毒基于猿猴腺病毒1、7、8、21、22、23、24、25、26、27.1、28.1、29、30、31.1、32、33、34、35.1、36、37.2、39、40.1、41.1、42.1、43、44、45、46、48、49、50或SA7P型。在某些实施方案中,重组腺病毒基于黑猩猩腺病毒,如ChAdOx 1(参见例如WO 2012/172277)或ChAdOx 2(参见例如WO 2018/215766)。在某些实施方案中,重组腺病毒基于诸如BZ28的黑猩猩腺病毒(参见例如WO 2019/086466)。在某些实施方案中,重组腺病毒基于诸如BLY6(参见例如WO 2019/086456)或BZ1(参见例如WO2019/086466)的大猩猩腺病毒。
在本发明的一个优选实施方案中,腺病毒载体包含来自稀有血清型例如包括Ad26的衣壳蛋白。在典型的实施方案中,载体是rAd26病毒。“腺病毒衣壳蛋白”是指腺病毒(例如,Ad26、Ad35、rAd48、rAd5HVR48载体)衣壳上的蛋白,其参与确定特定腺病毒的血清型和/或嗜性。腺病毒衣壳蛋白通常包括纤维、五邻体和/或六邻体蛋白。如本文所用,特定腺病毒的“衣壳蛋白”,如“Ad26衣壳蛋白”可以是,例如,包括Ad26衣壳蛋白的至少一部分的嵌合衣壳蛋白。在某些实施方案中,衣壳蛋白是Ad26的完整衣壳蛋白。在某些实施方案中,六邻体、五邻体和纤维是Ad26的。
本领域普通技术人员会认识到衍生自多种血清型的元件可以组合在单个重组腺病毒载体中。因此,可以产生组合了来自不同血清型的期望特性的嵌合腺病毒。因此,在一些实施方案中,本发明的嵌合腺病毒可以将第一血清型的预先存在的免疫性的缺失与诸如温度稳定性、组装、锚定、产量、重定向或改进的感染、DNA在靶标细胞中的稳定性等特征组合起来。参见例如关于嵌合腺病毒Ad5HVR48的WO 2006/040330,其包括具有来自Ad48的部分衣壳的Ad5骨架,以及也关于嵌合腺病毒Ad26HVRPtr1、Ad26HVRPtr12和Ad26HVRPtr13的例如WO 2019/086461,其包括分别具有Ptr1、Ptr12和Ptr13的部分衣壳蛋白的Ad26病毒骨架)。
在某些优选实施方案中,用于本发明的重组腺病毒载体主要或完全衍生自Ad26(即,载体是rAd26)。在一些实施方案中,腺病毒是复制缺陷的,例如,因为它在基因组的E1区中含有缺失。对于衍生自非C组的腺病毒,例如Ad26或Ad35,通常将腺病毒的E4-orf6编码序列与诸如Ad5的人C亚组的腺病毒的E4-orf6交换。这允许这类腺病毒在表达Ad5的E1基因的熟知的互补细胞系中增殖,例如293细胞、PER.C6细胞等(参见,例如,Havenga,et al.,2006,J Gen Virol 87:2135-43;WO 03/104467)。然而,这类腺病毒将不能在不表达Ad5的E1基因的非互补细胞中复制。
重组腺病毒载体的制备是本领域熟知的。rAd26载体的制备描述于例如WO2007/104792和Abbink et al.,(2007)Virol 81(9):4654-63中。Ad26的示例性基因组序列见于GenBank登录号EF 153474和WO 2007/104792的SEQ ID NO:1。可用于本发明的载体的实例例如包括在WO2012/082918中描述的那些,其公开内容整体援引加入本文。
通常,用于本发明的载体是使用包含整个重组腺病毒基因组的核酸(例如,质粒、粘粒或杆状病毒载体)产生的。因此,本发明还提供分离的核酸分子,其编码本发明的腺病毒载体。本发明的核酸分子可以是通过克隆获得或合成产生的RNA形式或DNA形式。DNA可以是双链的或单链的。
用于本发明的腺病毒载体通常是复制缺陷的。在这些实施方案中,通过缺失或失活对病毒复制至关重要的区域,如E1区域,使病毒复制缺陷。可以通过例如在该区域内插入感兴趣的基因,如编码合成SARS CoV2 S蛋白的基因(通常与启动子连接)或编码SARS CoV2S抗原性多肽的基因(通常与启动子连接)基本上缺失或失活该区域。在一些实施方案中,本发明的载体可以含有在其他区域如E2、E3或E4区中的缺失,或者在这些区域的一个或多个内与启动子连接的异源基因的插入。对于E2和/或E4突变的腺病毒,通常使用E2和/或E4互补细胞系来产生重组腺病毒。腺病毒的E3区中的突变不需要由细胞系补充,因为复制不需要E3。
在某些实施方案中,重组人腺病毒具有腺病毒基因组的E1区中的缺失、E3区中的缺失或E1和E3区两者中的缺失。因此,在某些实施方案中,本发明的腺病毒载体缺乏病毒复制所需的腺病毒基因组的E1区(例如E1a区和/或E1b区)的至少一种必需基因功能。在某些实施方案中,本发明的腺病毒载体缺乏非必需E3区的至少部分。在某些实施方案中,载体缺乏E1区的至少一种必需基因功能和非必需E3区的至少一部分。腺病毒载体可以是“多重缺陷的”,意味着腺病毒载体在腺病毒基因组的两个或更多个区域的每一个中都缺乏一种或多种必需基因功能。例如,上述E1-缺陷或E1-、E3-缺陷的腺病毒载体可以进一步缺乏E4区的至少一种必需基因和/或E2区(例如,E2A区和/或E2B区)的至少一种必需基因。
在某些实施方案中,载体是血清型26的重组人腺病毒(rAd26载体)。这种血清型通常在人群中具有低血清流行率和/或低预先存在的中和抗体滴度。rAd26载体的制备描述于例如WO 2007/104792和Abbink et al.,(2007)Virol 81(9):4654-63中。Ad26的示例性基因组序列见于GenBank登录号EF 153474和WO 2007/104792的SEQ ID NO:1。
在优选的实施方案中,腺病毒是复制缺陷的,例如因为它在基因组的E1区中含有缺失。如本领域技术人员所知,在从腺病毒基因组缺失必需区域的情况下,这些区域编码的功能必须以反式提供,优选由生产细胞提供,即当E1、E2和/或E4区的部分或全部从腺病毒中缺失时,这些必须存在于生产细胞中,例如整合到其基因组中,或以所谓的辅助腺病毒或辅助质粒的形式存在。腺病毒也可以在E3区中具有缺失,其对于复制是可有可无的,因此不必补充这种缺失。
在一个优选的实施方案中,将重组的、无复制能力的人26型腺病毒(Ad26)载体构建为编码稳定在其融合前构象的SARS-CoV-2刺突(S)蛋白。优选地,腺病毒包含SEQ ID NO:205的核酸。
包装细胞系通常用于产生足够量的用于本发明的腺病毒载体。包装细胞是这样的细胞,其包含在复制缺陷载体中已缺失或失活的那些基因,从而允许病毒在细胞中复制。在E1区中具有缺失的腺病毒的合适包装细胞系包括例如PER.C6、911、293和E1A549。
在本发明的一个优选实施方案中,所述载体是腺病毒载体,更优选rAd26载体,最优选在腺病毒基因组的E1区中具有至少一个缺失的rAd26载体,例如在Abbink,J Virol,2007.81(9):p.4654-63中的描述的那个,其援引加入本文。通常,将编码合成的SARS CoV-2S抗原的核酸序列克隆至腺病毒基因组的E1和/或E3区中。
在某些实施方案中,编码冠状病毒S蛋白的核酸与巨细胞病毒(CMV)启动子可操作地连接,所述CMV启动子包含至少一个四环素操纵子(TetO)基序。这允许具有成本效益、大规模制造包含SARS CoV-2 S蛋白插入物的腺病毒颗粒。不打算受理论限制,据信SARS CoV-2 S蛋白导致较低水平的腺病毒颗粒生产。将TetO基序添加到CMV启动子允许较高水平的腺病毒颗粒生产。
如本文所用,“启动子”是能够启动信使RNA中的基因序列转录的核酸序列,这种转录由RNA聚合酶在启动子上或附近的结合启动。
如上文所定义,在某些实施方案中,启动子是包含至少一个四环素操纵子(TetO)基序的巨细胞病毒启动子。TetO基序可以指“调控序列”或“调控元件”,如本文所用,其指通常但不限于DNA的核酸区段,其调节与它可操作地连接的核酸序列的转录,并且因此充当转录调谐子(modulator)。调控序列经常包含作为转录结合结构域的核酸序列,其由转录蛋白和/或转录因子、增强子或阻遏物等的核酸结合结构域所识别。例如,可操作地将阻遏物序列偶联至启动子是可能的,阻遏蛋白可以结合该阻遏序列,所述阻遏蛋白可以减少或阻止转基因在表达阻遏蛋白的生产细胞系中的表达。这可以改善传代后和/或在生产细胞系中大量生产时核酸分子的遗传稳定性和/或表达水平。这类系统在本领域中已有所描述。调节序列可以包括一个或多个四环素操纵子(TetO)基序/序列,使得在四环素阻遏蛋白(TetR)存在的情况下抑制表达。在没有四环素的情况下,TetR蛋白能够与TetO位点结合并抑制与TetO基序/序列可操作地连接的转基因(例如,SARS CoV-2 S抗原)的转录。在某些实施方案中,编码SARS-CoV-2 S蛋白的核酸,当存在于腺病毒载体中时,与包含至少一个四环素操纵子(TetO)基序的巨细胞病毒(CMV)启动子可操作地连接,从而使SARS CoV-2 S蛋白在表达TetR蛋白的生产细胞系中产生的重组腺病毒中受到抑制。当将重组腺病毒载体引入受试者或不表达TetR蛋白的细胞时,表达不会受到抑制。然而,本发明不限于使用包含至少一个四环素操纵子(TetO)基序的巨细胞病毒启动子。
如本文所用,术语“阻遏物”是指具有抑制、干扰、延缓和/或阻抑重组表达载体(例如如腺病毒载体)的异源蛋白产物产生的能力的分子(例如蛋白)。阻遏物可以通过干扰沿表达载体如在表达盒中的适当位置处的结合位点(例如TetR可以结合CMV启动子中的TetO基序)抑制表达。在载体增殖期间阻遏载体转基因表达可以防止转基因不稳定性并且可以增加在生产期间具有转基因的载体的产量。
当核酸处于与另一个核酸序列的结构或功能关系中时,它是“可操作地连接”的。例如,如果一个DNA区段和另一个DNA区段彼此相对定位在同一个连续的DNA分子上并具有结构或功能上的关系,则一个DNA区段可能与另一个DNA区段可操作地连接,如启动子或增强子相对于编码序列定位以促进编码序列的转录;核糖体结合位点相对于编码序列定位以促进翻译;或者前序列或分泌前导序列相对于编码序列定位以促进前蛋白(例如,参与编码的多肽的分泌的前蛋白)的表达。在其他实例中,可操作地连接的核酸序列不是连续的,而是以使它们作为核酸或作为由它们表达的蛋白彼此具有功能关系的方式定位。例如,增强子不一定是连续的。连接可以通过在方便的限度性位点处连接或通过使用合成的寡核苷酸接头或连接子来完成。
在某些实施方案中,所述包含至少一个TetO基序的CMV启动子包含SEQ ID NO:219的核苷酸序列,优选地,CMV启动子由SEQ ID NO:219组成。
在一个优选的实施方案中,腺病毒载体在PER.C6-四环素阻遏物(TetR)细胞中产生。
本发明进一步提供组合物,其包含本发明的核酸、蛋白和/或载体。为了向人给药,本发明可以使用包含核酸、蛋白和/或载体以及药学上可接受的载剂或赋形剂的药物组合物。在本文中,术语“药学上可接受的”是指载剂或赋形剂,在所使用的剂量和浓度下不会对给药的受试者造成任何不希望的或有害的影响。这类药学上可接受的载剂和赋形剂是本领域熟知的(参见Remington′s Pharmaceutical Sciences,18th edition,A.R.Gennaro,Ed.,Mack Publishing Company[1990];Pharmaceutical Formulation Development ofPeptides and Proteins,S.Frokjaer and L.Hovgaard,Eds.,Taylor&Francis[2000];和Handbook of Pharmaceutical Excipients,3rd edition,A.Kibbe,Ed.,PharmaceuticalPress[2000])。纯化的核酸、蛋白和/或载体优选以无菌溶液配制和给药,尽管也可以使用冻干制剂。通过无菌过滤或通过本领域本身已知的其他方法制备无菌溶液。然后将溶液冻干或填充到药物剂量容器中。溶液的pH通常在pH 3.0-9.5的范围内,优选在pH 5.0-7.5的范围内。核酸、蛋白和/或载体通常在具有合适的药学上可接受的缓冲液的溶液中,并且所述溶液还可以含有盐。可任选存在稳定剂,如白蛋白。在某些实施方案中,添加去污剂。在某些实施方案中,核酸、蛋白和/或载体可以配制成可注射制剂。这些含有有效量的核酸、蛋白和/或载体的制剂可以是无菌液体溶液、液体悬浮液或冻干形式,并且任选地含有稳定剂或赋形剂。
例如,腺病毒可以储存在也用于腺病毒世界标准的缓冲液中(Hoganson et al.,Development of a stable adenoviral vector formulation,Bioprocessing March2002,p.43-48):20mM Tris pH 8、25mM NaCl、2.5%甘油。适合向人给药的另一种有用的制剂缓冲液是20mM Tris、2mM MgCl2、25mM NaCl、蔗糖10%w/v、聚山梨醇酯800.02%w/v。显然,可以使用许多其他缓冲液,在欧洲专利号0853660、美国专利6,225,289和国际专利申请WO 99/41416、WO 99/12568、WO 00/29024、WO 01/66137、WO 03/049763、WO 03/078592、WO03/061708中可以找到用于储存和用于纯化的(腺)病毒制剂的药物给药的几个合适制剂的实例。
在某些实施方案中,腺病毒组合物是用于肌内注射的无菌悬浮液,其含有以下非活性成分:
·一水柠檬酸
·柠檬酸三钠二水合物
·乙醇
·2-羟丙基-β-环糊精(HBCD)
·聚山梨醇酯80
·氯化钠
·氢氧化钠
·注射用水
和任选地盐酸,
并且其中产品不含防腐剂。”
在一个优选的实施方案中,当在-25℃--15℃(-13°F-5°F)的推荐储存条件下冷冻储存时,腺病毒载体疫苗的初始保质期为24个月,并且在这24个月内,在2℃-8℃(36°F-46°F)下储存时,可保存3个月。在某些实施方案中,本发明的组合物包含本发明的(腺)载体,与进一步的活性成分组合。这种进一步的活性成分可以包含一种或多种SARS-CoV-2蛋白抗原,例如本发明的SARS-CoV-2蛋白,或任何其他SARS-CoV-2蛋白抗原,或包含编码相似或替代的SARS-CoV-2抗原的核酸的其他载体。这类载体也可以是非腺病毒或腺病毒的,其中后者可以是任何血清型。
在某些实施方案中,包含腺病毒的组合物进一步包含一种或多种佐剂。本领域已知佐剂进一步增加对所应用的抗原决定簇的免疫应答,并且包含腺病毒和合适佐剂的药物组合物例如在WO 2007/110409中公开,其援引加入本文。术语“佐剂”和“免疫刺激剂”互换使用,定义为引起免疫系统刺激的一种或多种物质。在这个上下文中,佐剂用于增强对本发明的腺病毒载体的免疫应答。合适的佐剂的实例包括铝盐,如氢氧化铝和/或磷酸铝;油乳液组合物(或水包油组合物),包括角鲨烯-水乳液,如MF59(参见例如WO 90/14837);皂苷制剂,如QS21和免疫刺激复合物(ISCOMS)(参见例如US 5,057,540;WO 90/03184、WO 96/11711、WO 2004/004762、WO 2005/002620);细菌或微生物衍生物,其实例为单磷酰脂质A(MPL)、3-O-脱酰基MPL(3dMPL)、含有CpG基序的寡核苷酸、ADP-核糖基化细菌毒素或其突变体,如大肠杆菌耐热肠毒素LT、霍乱毒素CT等。使用载体编码的佐剂也是可能的,例如通过使用编码C4结合蛋白(C4bp)的寡聚化结构域与感兴趣抗原的融合物(例如,Solabomi etal,2008,Infect Immun 76:3817-23)的异源核酸。在某些实施方案中,本发明的组合物包含铝作为佐剂,例如以氢氧化铝、磷酸铝、磷酸铝钾或其组合的形式,每剂量的铝含量浓度为0.05-5mg,例如0.075-1.0mg。
在其他实施方案中,组合物不包含佐剂。
本发明进一步提供针对COVID-19的疫苗,其包含本发明的核酸、蛋白和/或载体。术语“疫苗”是指含有活性成分的药剂或组合物,所述活性成分有效地在受试者中诱导针对某种病原体或疾病的治疗程度的免疫。根据本发明,疫苗可包含有效量的血清型26的重组腺病毒,其编码SARS CoV-2 S蛋白,特别是包含SEQ ID NO:1的氨基酸序列的SARS CoV-2蛋白或其抗原片段,其导致针对SARS CoV-2的S蛋白的免疫应答,优选保护性免疫应答。本发明的疫苗可用于预防导致住院的严重下呼吸道疾病和降低因在受试者中的SARS-CoV-2感染和复制引起的如肺炎和细支气管炎的并发症的频率的方法。疫苗也可用于所谓的暴露后预防(PEP),即用于在潜在或记录在案的暴露于冠状病毒后预防疾病和/或降低感染的二次传播的风险。本发明的“疫苗”通常包括药学上可接受的稀释剂、载剂或赋形剂。它可以包含或不包含进一步的活性成分。
在某些实施方案中,它可以是组合疫苗,其进一步包含诱导免疫应答的其他成分,例如针对SARS.CoV2的其他蛋白和/或针对其他传染原的免疫应答。
在某些实施方案中,疫苗是组合疫苗,其包含本发明的载体和SARS CoV-2 S蛋白以及任选存在的佐剂,其中所述载体和SARS CoV-2蛋白用于同时给药。SARS CoV-2蛋白可以是如本文所述的蛋白或本领域已知的任何其他合适的SARS CoV-2 S蛋白。
根据本发明,载体和蛋白优选用于同时给药(即同时地给药)。“同时给药(concurrent administration)或共同给药(co-administration)”在向受试者给药载体和蛋白的上下文中是指组合使用载体和蛋白,其中在24小时的时间段内向给受试者给药所述载体和蛋白。
在某些实施方案中,载体和蛋白例如与药学上可接受的缓冲液、载剂、赋形剂和/或佐剂共同配制在用于给药的单一组合物中,例如混合,并同时一起向受试者给药。在其他实施方案中,载体和蛋白例如与药学上可接受的缓冲液、载剂、赋形剂和/或佐剂配制在分开的组合物中,并在24小时内,如12小时内、10小时内、8小时内、6小时内、4小时内、2小时内或在1小时内或更短时间内,例如大约同时在受试者的同一臂或两个不同臂中,以分开的组合物向受试者给药。
本发明提供用于主动免疫的方法,以预防18岁以上的个体的严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)引起的2019冠状病毒病(COVID-19)。
本发明提供用于主动免疫的方法,以预防16岁以上的个体的严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)引起的2019冠状病毒病(COVID-19)。
本发明提供用于主动免疫的方法,以预防12岁以上的个体的严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)引起的2019冠状病毒病(COVID-19)。
本发明提供用于主动免疫的方法,以预防2个月以上的个体的严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)引起的2019冠状病毒病(COVID-19)。
本发明还提供用于主动免疫的方法,以预防严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)变体引起的2019冠状病毒病(COVID-19)。
SARS CoV-2变体优选选自英国变体(SARS-CoV-2谱系(lin.)B.1.1.7)、南非(SARS-CoV-2501Y.V2)和丹麦变体(Cluster 5)。
SARS CoV-2变体可以包含一个或多个突变,所述突变选自de169-70、del145、A570D、D614G、P681H、T716I、S982A、D1118H、D80A、D215G、L242H、R246I、K417N、E484K、N501Y、Y435F、I1692V、M1229I和A701V。
SARS CoV-2变体可具有E484K突变和一个或多个选自del69-70、del145、A570D、D614G、P681H、T716I、S982A、D1118H、D80A、D215G、L242H、R246I、K417N、E484K、N501Y、Y435F、I1692V、M1229I和A701V的突变。这类SARS CoV-2变体的实例是B.1.351、B.1.1.7、P.1和CAL20C。
本发明还提供一种用于在有需要的受试者中诱导SARS-CoV-2结合抗体(如通过例如ELISA测量的)的方法,其包括向所述受试者给药如本文所述的组合物或疫苗。
本发明还提供一种用于在有需要的受试者中诱导SARS-CoV-2中和抗体(如通过例如VNA测量的)的方法,其包括向所述受试者给药如本文所述的组合物或疫苗。
本发明还提供一种用于在有需要的受试者中诱导SARS-CoV-2特异性T细胞应答(如例如在SARS-CoV-2 S蛋白肽刺激外周血单个核细胞(PBMC)并细胞内染色后通过流式细胞术评估的)的方法,其包括向所述受试者给药如本文所述的组合物或疫苗。
本发明还提供一种用于减少SARS-CoV-2或其变体在例如受试者的鼻道和肺中的感染和/或复制的方法,其包括向所述受试者给药如本文所述的组合物或疫苗。这会减少受试者中的SARS-CoV-2感染引起的不良反应,从而有助于保护受试者免受这类不良反应。在某些实施方案中,可以基本上预防SARS-COV-2感染的不良反应,即降低到低水平以致它们在临床上不相关。重组腺病毒可以是本发明的疫苗的形式,包括上述实施方案。进一步的活性成分的给药例如可以通过单独给药或通过给药本发明疫苗的组合产品来完成。
本发明还提供一种用于预防分子确诊的中度至严重/危重COVID-19的方法,其包括当以一剂或两剂疫苗给予时,向受试者给药如本文所述的组合物或疫苗。在某些实施方案中,本发明提供一种方法,用于预防SARS-CoV-2血清阴性的成人的分子确诊的中度至严重/危重COVID-19,与安慰剂相比,所述方法包括当以一剂或两剂疫苗给予时,向受试者给药如本文所述的组合物或疫苗。
本发明还提供一种用于预防分子确诊的严重/危重COVID-19的方法,其包括当以一剂或两剂疫苗给予时,向受试者给药如本文所述的组合物或疫苗。在某些实施方案中,本发明提供一种用于预防SARS-CoV-2血清阴性的成人的分子确诊的严重/危重COVID-19的方法,与安慰剂相比,所述方法包括当以一剂或两剂疫苗给予时,向受试者给药如本文所述的组合物或疫苗。
根据本发明,中度COVID-19定义为:在中心实验室分子确诊的来自任何可用呼吸道样品或其他样品的SARS-CoV-2阳性RT-PCR或分子检测结果,并且
在观察期间的任何时间,直到体征和症状消失:
以下新的或恶化的体征或症状中任何一种:
呼吸频率≥20次呼吸/分钟;
氧饱和度(SpO2)异常,但在海平面的室内空气中仍>93%;
肺炎的临床或放射学证据;
DVT的放射学证据;
呼吸急促或呼吸困难;
或者
以下新的或恶化的体征或症状中任何2种:
发烧(≥38.0℃或≥100.4°F);
心率≥90次心跳/分;
寒战或僵硬;
新的或不断变化的嗅觉或味觉障碍;
咽喉痛;
不适;
头痛;
咳嗽;
肌肉疼痛(肌痛);
胃肠道症状;
脚或脚趾看起来发红或有淤青。
根据本发明,严重/危重COVID-19定义为:在中心实验室分子确诊的来自任何可用呼吸道样品或其他样品的SARS-CoV-2阳性RT-PCR或分子检测结果;和
表明严重全身性疾病的静息临床体征(呼吸频率≥30次呼吸/分钟,心率≥125次心跳/分钟,SpO2在海平面的室内空气中≤93%,或PaO2/FiO2<300mmHg);
呼吸衰竭(定义为需要高流量氧气、无创通气、机械通气或ECMO[体外膜肺氧合])
休克的证据(定义为收缩压<90mmHg,舒张压<60mmHg,或需要血管加压药);
严重的急性肾脏、肝脏或神经功能障碍;
入住重症监护室;
死亡。
在某些实施方案中,严重COVID-19由FDA指南定义。
根据本发明,轻度COVID-19定义为:在中心实验室分子确诊的来自任何可用呼吸道样品或其他样品的SARS-CoV-2阳性RT-PCR或分子检测结果,和
以下症状之一:
发烧(≥38.0℃或≥100.4°F)
咽喉痛,
头痛,
肌肉疼痛(肌痛),
胃肠道症状,
咳嗽,
胸闷,
流鼻涕,
气喘,
皮疹
眼睛刺激或分泌物,
发冷,
新的或不断变化的嗅觉或味觉障碍,
脚或脚趾看起来发红或有淤青,
寒战或僵硬,
不适(食欲不振、身体不适、疲劳、身体虚弱。
将符合上述病例定义但不符合中度至严重/危重定义的病例视为轻度。
无症状或未检测到的SARS CoV-2感染定义为:
基于体征和症状,参与者不符合疑似COVID-19的标准;和
具有来自任何可用的呼吸道样品(例如,鼻拭子样品、痰样品、咽喉拭子样品、唾液样品)或其他样品的SARS-CoV-2阳性RT-PCR或分子检测结果;
或者
出现血清学(非S蛋白)检测阳性(血清转化)。
本发明还提供一种方法,用于减少SARS-CoV-2病毒载量,如通过定量逆转录酶聚合酶链式反应(RT-PCR)在具有分子确诊的中度至严重/危重COVID-19的参与者中评估的。
本发明还提供一种方法,用于预防或减少与任何分子确诊的COVID-19相关的肺炎发生,当与安慰剂相比时。
本发明还提供一种用于减少由SARS CoV-2感染引起的症状的方法。
本发明还提供一种方法,用于预防或减少与任何分子确诊的COVID-19相关的住院,当与安慰剂相比时。
本发明还提供一种方法,用于预防或减少与任何分子确诊的COVID-19相关的急性呼吸窘迫综合征发生,当与安慰剂相比时。
本发明还提供一种方法,用于预防或减少与任何分子确诊的COVID-19相关的败血症发生,当与安慰剂相比时。
本发明还提供一种方法,用于预防或减少与任何分子确诊的COVID-19相关的感染性休克发生,当与安慰剂相比时。
本发明还提供一种方法,用于预防或降低与任何分子确诊的COVID-19相关的死亡率。
在一个优选的实施方案中,所述方法包括将组合物或疫苗以0.5mL的单剂给药,其包含5×1010vp的本发明的疫苗。
在某些实施方案中,疫苗的效果(即诱导SARS-CoV-2结合和/或中和抗体和/或诱导SARS-CoV-2特异性T细胞应答)在疫苗给药后8天已经发生。
在某些实施方案中,取决于方案,疫苗的效果在第一剂研究疫苗后至少14天发生。在某些实施方案中,取决于方案,疫苗的效果在第一剂研究疫苗后至少28天发生。
在优选的实施方案中,本发明提供一种用于预防分子确诊的中度至严重COVID-19的方法。
本发明的组合物或疫苗优选地针对分子确诊的中度至严重COVID-19具有至少50、55、60、65或70%的疫苗效力。优选地,本发明的组合物或疫苗针对在疫苗接种后至少14天发病的分子确诊的中度至严重COVID-19具有至少60%、优选至少65%的疫苗效力。
根据本发明,组合物或疫苗有效对抗由SARS CoV-2引起的COVID-10,以及至少一些流通的SARS-CoV-2变体,所述流通的SARS-CoV-2变体与快速增加的病例数相关联并且在英国(B1.1.7/501Y.V1)、南非212(501Y.V2)和巴西(B1.1.28/501.V3)具有特别的流行率。
根据本发明,已经显示单剂疫苗在预防中度至严重/危重COVID-19方面是有效的,疫苗接种后第14天和第28天后的疫苗效力(VE)均为66%。
第28天后,针对中度/严重疾病的疫苗效力在美国为71%,在巴西为66%,在南非为57%,其中大多数毒株(90%)是变体501Y.V2(南非变体)。注意到针对严重/危重COVID-19的高疫苗效力(总体为85%),在18-59岁人群中的效力超过90%。这一发现在各个国家和地区(北美和南美、南非)是一致的,包括几乎所有病例都感染了SARS-CoV-2的新变体的南非。
估计在第14天效力开始,直到第56天效力增加,特别是针对严重疾病,与从第14天起检测到的中和抗体滴度的发现一致,继续增加直到第56天,直至第85天没有减弱的迹象。
可以向受试者例如人受试者给药本发明的组合物或疫苗。在一次给药期间提供给受试者的腺病毒的总剂量通常在1x107病毒颗粒(vp)和1x1012vp之间,优选在1x108vp和1x1011vp之间,例如在3x108和5x1010vp之间,例如在109和3x1010vp之间。
在一个优选的实施方案中,在一剂或两剂方案中以1.25x1010、2.5x1010、5x1010或1x1011vp/剂的剂量向人受试者给药本发明的疫苗,其中所述剂量间隔约1、2或3个月给药。
在某些实施方案中,在一剂方案中以1x1011vp/剂的剂量向人受试者给药本发明的疫苗,并在6、12或24个月时以相同剂量进行第二次疫苗接种。
在一个优选的实施方案中,在一剂方案中以5x1010vp/剂的剂量向人受试者给药本发明的疫苗。
在另一个优选的实施方案中,在一剂方案中以1x1011vp/剂的剂量向人受试者给药本发明的疫苗。
在另一个优选的实施方案中,以两剂方案向人受试者给药疫苗,所述方案包括第一次给药5×1010vp/剂的剂量和间隔约2个月(8周或56天)给药5×1010vp/剂的第二剂。
在某些实施方案中,在间隔约2个月(8周)给药的两剂方案中以5x1010vp/剂的剂量向人受试者给药疫苗,然后在8个月、14个月和26个月(即完成2剂方案后6个月、12个月或24个月)以相同剂量进一步疫苗接种。
在一个优选的实施方案中,在一剂方案中以5x1010vp/剂的剂量给药组合物。
可以使用标准给药途径进行腺病毒组合物的给药。非限制性实施方案包括肠胃外给药,如通过注射,例如肌内、皮内等,或皮下、经皮或粘膜给药,例如鼻内、口服等。然而,本发明特别优选肌内给药疫苗。肌内给药的优点是简单且完善,并且不存在用于6个月以下婴儿鼻内应用的安全问题。在一个实施方案中,通过肌内注射将组合物给药至例如手臂的三角肌或大腿的股外侧肌中。
如本文所用的受试者优选地是哺乳动物,例如啮齿动物,例如小鼠、棉鼠、或非人灵长类动物或人。优选地,受试者是人受试者。受试者可以是任何年龄,例如约1个月-100岁、例如约2个月-约80岁、约6个月-约3岁、约3岁-约18岁、约12岁-约18岁、约18岁-约55岁、约50岁-约75岁等。在某些优选实施方案中,受试者是2岁以上的人。在其他优选的实施方案中,人受试者是18岁以上的人,优选60岁以上的人,或65岁以上的人。
在某些实施方案中,多次向受试者给药组合物或疫苗,例如一年一次。在某些实施方案中,疫苗接种的方法包括向受试者单次给药组合物或疫苗。提供本发明的疫苗的一次或多次第二次(或加强)给药也是可能的。如果进行第二次疫苗接种,通常,在第一次向受试者给药组合物(在这种情况下,有时称为“初次疫苗接种”)后的一周至一年之间,优选两周至四个月之间的时刻向同一受试者施用这种第二次疫苗接种。
本发明进一步提供分离的宿主细胞,其包含血清型26的重组人腺病毒,所述重组人腺病毒包含编码SARS-CoV-2 S蛋白或其片段的核酸。
可以使用的宿主细胞(有时在本领域和本文中也称为“包装细胞”或“互补细胞”或“生产细胞”)可以是其中可以增殖的期望的腺病毒的任何宿主细胞。宿主细胞系通常用于产生足量的本发明腺病毒载体。宿主细胞是包含在复制缺陷载体中已缺失或失活的那些基因的细胞,从而允许病毒在细胞中复制。合适的细胞系包括例如PER.C6、911、293和E1A549。
在某些实施方案中,宿主细胞进一步包含编码四环素阻遏物(TetR)蛋白的核苷酸序列。例如,编码TetR蛋白的核苷酸序列可以整合到宿主细胞的基因组中。例如,编码TetR蛋白的核苷酸序列可以整合到1号染色体中。宿主细胞系可以是例如PER.C6细胞。
在一个优选的实施方案中,宿主细胞是PER.C6细胞,其包含编码四环素阻遏物(TetR)蛋白的核苷酸序列。这类“PER.C6 TetR”细胞是这样的PER.C6细胞(通过E1永生化的人视网膜细胞),已向其引入编码TetR的核苷酸,如PCT/EP2018/053201(援引加入本文)中所述。在这类实施方案中,优选地,腺病毒包含编码SARS-CoV-2 S蛋白或其片段的核酸,其与包含一个或多个Tet操纵子(TetO)基序的启动子可操作地连接。在优选的实施方案中,启动子是包含一个或多个TetO基序的CMV启动子。优选地,启动子是包含两个TetO基序的CMV启动子。优选地,启动子是CMV启动子并且包含SEQ ID NO:219的核苷酸序列。在一些实施方案中,启动子由SEQ ID NO:219的核苷酸序列组成。
本发明进一步提供用于制备针对SARS冠状病毒(SARS-COV-2)的疫苗的方法,其包括提供血清型26的重组人腺病毒,所述重组人腺病毒包含编码如本文所述的SARS-COV-2 S蛋白或其片段的核酸;在宿主细胞的培养物中增殖所述重组腺病毒;分离和纯化所述重组腺病毒;并将所述重组腺病毒置于药学上可接受的组合物中。在某些实施方案中,本文提供生产包含SARS-CoV-2抗原的腺病毒颗粒的方法。所述方法包括(a)使本发明的宿主细胞与本发明的腺病毒载体接触;和(b)在产生包含所述SARS-CoV-2抗原的腺病毒颗粒的条件下培养所述宿主细胞。根据熟知的方法,可以制备重组腺病毒并在宿主细胞中增殖,其需要(entail)感染有腺病毒的宿主细胞的细胞培养物细胞培养物可以是任何类型的细胞培养物,包括贴壁细胞培养物,例如附着在培养容器表面或微载体上的细胞,以及悬浮培养物。
大多数大规模悬浮培养都以分批或补料分批的过程进行操作,因为它们最容易操作和扩大规模。如今,基于灌注原理的连续过程变得越来越普遍并且也是合适的(参见例如WO 2010/060719和WO 201I/098592,均援引加入本文,其描述了用于获得和纯化大量重组腺病毒的合适方法)。
本发明进一步提供一种分离的重组核酸,其形成血清型26的重组人腺病毒的基因组,所述重组人腺病毒包含编码SARS-CoV2 S蛋白或其片段的核酸。
当向感染有或暴露于冠状病毒(例如2019-nCoV)的受试者(例如,人受试者)给药时,2019-nCoV多肽可以用于引发针对冠状病毒感染(例如,2019-nCoV感染)的保护性和治疗性免疫应答(例如,体液应答或细胞应答)。可以制备用于向受试者给药的组合物包括2019-nCoV蛋白(例如,刺突(S)蛋白或其部分(例如,具有SEQ ID NO:1-84中任一项序列的多肽,例如SEQ ID NO:51,或与具有SEQ ID NO:1-84中任一项序列的多肽具有至少85%(例如,至少90%、95%、99%或更多)序列相同性的多肽)或载体(例如,表达载体,如质粒或病毒载体,如腺病毒(例如,Ad26)、痘病毒、腺相关病毒、逆转录病毒或其他病毒载体,或裸露的或包裹的DNA),所述载体含有编码2019-nCoV蛋白的核酸序列(例如,具有SEQ ID NO:93-181、190-195和199-204中任一项序列的核酸分子;或者与具有SEQ ID NO:93-181、190-195和199-204中任一项序列的核酸分子具有至少85%(例如,至少90%、95%、99%或更多)序列相同性的核酸分子)。
描述了表达2019-nCoV刺突(S)蛋白的DNA疫苗的产生。2019-nCoV DNA疫苗可以通过将编码S或其部分(例如,SEQ ID NO:1-84,例如,SEQ ID NO:51,或与其具有达85%或更多序列相同性的变体)的多核苷酸(例如,SEQ ID NO:93-181、190-195和199-204或与其具有达85%或更多序列相同性的变体)掺入哺乳动物表达载体(例如,pcDNA3.1+;Invitrogen,CA,USA)来产生疫苗。
还描述了表达2019-nCoV刺突(S)蛋白的重组病毒载体(例如Ad26病毒载体)的产生。2019-nCoV病毒载体可以通过将编码S或将其部分(例如,SEQ ID NO:1-84,或与其具有达85%或更多序列相同性的变体,例如,SEQ ID NO:51)掺入病毒载体(例如,Ad26病毒载体)中来产生。
来自受试者(例如,人受试者)的样品中存在的抗冠状病毒抗体(例如,抗2019-nCoV抗体,例如抗刺突抗体,例如抗刺突中和抗体)可用于检测和/或监测保护性抗体应答。抗冠状病毒抗体(例如,抗刺突抗体,例如,抗刺突中和抗体)可以在给药治疗性组合物(例如,本文所述的任何组合物或免疫原性组合物)后的短时间范围内(例如,在给药后1天和给药后8周之间)或更长的时间范围内(例如,在给药后2个月和给药后15年之间)检测。
本文描述了治疗和预防2019-nCoV感染的核酸分子、多肽、载体、疫苗、组合物、抗体和方法。
I.组合物和方法
核酸分子
基于2019-nCoV设计核酸分子(例如,SEQ ID NO:93-181、190-195和199-204或与其具有达85%或更多序列相同性的变体)。核酸分子编码2019-nCoV刺突(S)蛋白的区域,例如,全长(SEQ ID NO:121)、具有细胞质区域的缺失的刺突(SEQ ID NO:94)、胞外域(SEQ IDNO:95)、S1(SEQ ID NO:96)和受体结合结构域(SEQ ID NO:97)。本发明的特征还在于对上述S区域的额外修饰,包括缺失或包含信号序列(例如,SEQ ID NO:189)、稳定化突变(例如,对应于SEQ ID NO:1的氨基酸K969和V970的脯氨酸取代)、对弗林蛋白酶切割位点的突变(例如,SEQ ID NO:188)、引入三聚化结构域(例如,折叠子三聚化结构域,例如,SEQ ID NO:184)、引入接头或间隔序列(例如,SEQ ID NO:185和186),以及它们的组合。核酸分子已相对于野生型2019-nCoV刺突核苷酸序列进行了优化,以改善在宿主细胞(例如哺乳动物(例如人)宿主细胞)中的表达。优化可以包括添加前导序列、限制性位点和/或Kozak序列。
核酸分子具有与SEQ ID NO:93-181、190-195和199-204中任一项的全部或部分或其互补序列具有至少85%(例如,至少86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)的序列相同性的核苷酸序列。例如,核酸分子可以具有SEQ ID NO:195的核苷酸序列。或者,分离的核酸分子具有编码2019-nCoV多肽的核苷酸序列,所述多肽与SEQ ID NO:1-84中任一项的氨基酸序列具有至少85%(例如,至少86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列相同性。例如,分离的核酸分子可以具有编码具有SEQ ID NO:56的氨基酸序列的多肽的核苷酸序列。
核酸分子可以进一步优化,如通过密码子优化,以在靶向的哺乳动物受试者(例如,人或用于疫苗生产的非人动物)中表达。
核酸分子也可以插入表达载体中,如质粒或病毒载体,例如腺病毒、痘病毒、腺相关病毒、逆转录病毒或其他病毒载体,或制备为裸露的或包裹的DNA并掺入组合物。
多肽
多肽是冠状病毒多肽(例如,2019-nCoV多肽),其对应于例如2019-nCoV刺突(S)蛋白的区域(SEQ ID NO:1-84),例如,全长(SEQ ID NO:29)、具有细胞质区域的缺失的刺突(SEQ ID NO:2)、胞外域(SEQ ID NO:3)、S1(SEQ ID NO:4)、受体结合结构域(SEQ ID NO:5)和与SEQ ID NO:1-84中任一项的全部或部分具有至少85%(例如,至少86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列相同性的变体。多肽可以包括SEQ ID NO:1-84中任一项的至少5、6、7、8、9、10、20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900、1000、1100、1200、1300、1400、1500、1600、1700或1800或更多个连续或非连续氨基酸。多肽还可以包括信号序列(例如,SEQ ID NO:92)的缺失或包含、稳定化突变(例如,对应于SEQ ID NO:1的氨基酸K969和V970的脯氨酸取代)、对弗林蛋白酶切割位点的突变(例如,SEQ ID NO:91)、引入三聚化结构域(例如,折叠子三聚化结构域,例如,SEQ ID NO:87)、引入接头或间隔序列(例如,SEQ ID NO:88和89),以及它们的组合。例如,多肽可以具有SEQ ID NO:28的氨基酸序列。多肽也可以从其他组分(例如,多肽与其天然相关联的组分)中分离并掺入组合物中。
载体
本发明的特征还在于包括上述多核苷酸中的任何一种或多种的重组载体(例如Ad26病毒载体)。载体可用于递送表达免疫原(例如,SEQ ID NO:1-84中的一个或多个或者与其具有至少85-99%序列相同性,例如至少大于90%序列相同性的变体)的核酸,并包括哺乳动物、病毒和细菌表达载体。例如,载体可用于递送核酸(例如,含有SEQ ID NO:104或204的核苷酸序列的核酸),所述核酸表达具有SEQ ID NO:51的氨基酸序列的免疫原。例如,载体可用于递送核酸(例如,含有SEQ ID NO:195的核苷酸序列的核酸),所述核酸表达具有SEQ ID NO:56的氨基酸序列的免疫原。可以遗传修饰哺乳动物、病毒和细菌载体以含有一种或多种如SEQ ID NO:93-181、190-195和199-204中所示的核酸序列或者与其具有至少85-99%序列相同性,例如至少大于90%序列相同性的其变体及其互补物。
载体可以是例如质粒、人工染色体(例如BAG、PAC、YAC)和病毒或噬菌体载体,并且可以任选地包括用于多核苷酸表达的启动子、增强子或调节子。载体还可以含有一种或多种选择性标志基因,例如在细菌质粒的情况下的氨苄青霉素、新霉素和/或卡那霉素抗性基因或真菌载体的抗性基因。载体可用于体外,例如产生DNA或RNA,或用于转染或转化宿主细胞,例如哺乳动物宿主细胞,例如用于产生由载体编码的蛋白。载体也可适用于体内,例如DNA疫苗接种、RNA疫苗接种或基因治疗的方法。
可以选择启动子和其他表达调控信号以与为其设计表达的宿主细胞相容。例如,哺乳动物启动子包括金属硫蛋白启动子和β-肌动蛋白启动子,金属硫蛋白启动子可以受诱导应答于诸如镉的重金属。也可以使用病毒启动子,其可以从病毒(例如多瘤病毒、鸡痘病毒、腺病毒(A)、牛乳头状瘤病毒、禽肉瘤病毒、巨细胞病毒(CMV)、逆转录病毒、肝炎-B病毒,和猿猴病毒40(SV40)和人乳头状瘤病毒(HPV))的基因组中获得。这些启动子在本领域中是熟知的并且容易获得。
优选的启动子元件是CMV立即早期启动子。在一些实施方案中,表达质粒是pcDNA3.1+(Invitrogen,CA,USA)。在一些实施方案中,表达载体是病毒载体,如衍生自腺病毒或痘病毒的载体。
病毒基因组提供载体的丰富来源,可用于将外源基因有效递送到细胞(例如,真核或原核细胞)的基因组中。病毒基因组是用于基因递送的特别有用的载体,因为包含在这类基因组中的多核苷酸通常通过一般的或特殊化的转导掺入靶细胞的基因组中。这些过程作为自然病毒复制周期的一部分发生,不需要添加蛋白或试剂来诱导基因整合。可用于递送表达免疫原(例如,SEQ ID NO:1-84中的一个或多个或者与其具有至少85-99%序列相同性,例如至少大于90%序列相同性的变体)的核酸的病毒载体的实例包括逆转录病毒、腺病毒(例如,Ad2、Ad5、Ad11、Ad12、Ad24、Ad26、Ad34、Ad35、Ad40、Ad48、Ad49、Ad50、Ad52(例如,RhAd52)、Ad59(例如,RhAd59)和Pan9(也称为AdC68))、细小病毒(例如腺相关病毒)、冠状病毒、负链RNA病毒如正粘病毒(例如流感病毒)、弹状病毒(例如狂犬病毒和水疱性口炎病毒)、副粘病毒(例如麻疹和仙台)、正链RNA病毒,如小核糖核酸病毒和甲病毒,以及双链DNA病毒,包括腺病毒、疱疹病毒(例如1型和2型单纯疱疹病毒、爱泼斯坦-巴尔病毒、巨细胞病毒)和痘病毒(例如,痘苗、改良的安卡拉痘苗(MVA)、鸡痘和金丝雀痘)。可用于递送编码免疫原(例如多肽)的多核苷酸的其他病毒包括例如,诺瓦克(Norwalk)病毒、披盖(togavirus)病毒、冠状病毒、呼肠病毒、乳多空病毒、嗜肝DNA病毒和肝炎病毒。逆转录病毒的实例包括:禽白血病肉瘤(leukosis-sarcoma)、哺乳动物C型、B型病毒、D型病毒、HTLV-BLV组、慢病毒、泡沫病毒(Coffin,J.M.,Retroviridae:The viruses and theirreplication,In Fundamental Virology,Third Edition,B.N.Fields,et al.,Eds.,Lippincott-Raven Publishers,Philadelphia,1996)。例如,载体可以是Ad26。这些腺病毒载体可以衍生自例如人、黑猩猩或恒河猴腺病毒。其他实例包括鼠白血病病毒、鼠肉瘤病毒、小鼠乳腺肿瘤病毒、牛白血病病毒、猫白血病病毒、猫肉瘤病毒、禽白血病病毒(leukemia virus)、人T细胞白血病病毒、狒狒内源性病毒、长臂类人猿白血病病毒、梅森辉瑞(Mason Pfizer)猴病毒、猿猴免疫缺陷病毒、猿猴肉瘤病毒、劳氏肉瘤病毒和慢病毒。载体的其他实例描述于例如,McVey等人(美国专利号5,801,030);其整体援引加入本文。可以将病毒载体的核酸材料(例如,包括核酸分子)包裹在例如脂质膜中或由结构蛋白(例如,衣壳蛋白)包裹,其可以包括一种或多种病毒多肽(例如,糖蛋白)。病毒载体可用于感染受试者的细胞,其转而促进病毒载体的异源基因翻译成免疫原。例如,可以将病毒载体遗传修饰以含有一种或多种SEQ ID NO:93-181、190-195和199-204示出的核酸序列或者与其具有至少85-99%序列相同性,例如至少大于90%序列相同性的其变体,及其互补物。
国际专利申请公布WO 2006/040330和WO 2007/104792中公开的腺病毒载体,每篇援引加入本文,作为载体特别有用。这些腺病毒载体可以编码和/或递送免疫原(例如,2019-nCoV多肽)的一种或多种以治疗具有与病毒感染(例如,2019-nCoV感染)相关的病理病况的受试者。在一些实施方案中,可以向受试者给药一种或多种重组腺病毒载体以表达多于一种类型的免疫原(例如,2019-nCoV多肽)。在一些实施方案中,可以修饰重组腺病毒载体以改变六邻体HVR结构域(例如,用不同血清型的那些替代一个或多个HVR)。除了腺病毒载体之外,本领域已知可用于促进免疫原的一种或多种在受试者(例如人)中的递送和/或表达的其他病毒载体和技术。这些病毒包括痘病毒(例如,痘苗病毒和改良的痘苗病毒安卡拉(MVA);参见,例如,美国专利号4,603,112和5,762,938,每个援引加入本文)、疱疹病毒、披盖病毒(例如,委内瑞拉马脑炎病毒;参见,例如,美国专利号5,643,576,援引加入本文)、小核糖核酸病毒(例如脊髓灰质炎病毒;参见例如美国专利号5,639,649,援引加入本文)、杆状病毒和由Wattanapitayakul和Bauer(Biomed.Pharmacother.54:487(2000),援引加入本文)描述的其他。
使用这些病毒的基因转移技术是本领域技术人员已知的。例如,逆转录病毒载体可用于将多核苷酸稳定整合到宿主基因组中,尽管这种重组不是优选的。相比之下,复制缺陷的腺病毒载体保持游离,因此允许瞬时表达。
可以使用能够在昆虫细胞(例如杆状病毒载体)、人细胞、酵母或细菌中驱动表达的载体,以产生大量由本发明的多核苷酸编码的2019-nCoV蛋白,例如,用作亚单位疫苗或用于免疫测定。
抗体
抗2019-nCoV抗体能够特异性结合2019-nCoV多肽,并且能够抑制受试者(例如,人)中2019-nCoV介导的活性(例如,病毒传播、感染和/或细胞融合)。这种结合的结果可以是,例如,在向感染有2019-nCoV的受试者给药抗体后病毒滴度(例如,病毒载量)降低约1%(例如,2%、3%、4%、5%、6%、7%、8%、9%、10%、20%、30%、40%、50%、60%、70%、80%或90%)或更多。抗2019-nCoV抗体可以选择性地结合包含2019-nCoV多聚蛋白的Env区域的全部或部分的表位。特别地,抗2019-nCoV抗体可以选择性地结合包含SEQ ID NO:1-84中任一项的全部或部分的表位。例如,抗体可以结合包含SEQ ID NO:28的全部或部分的表位。因此,抗体可用于预防或治疗2019-nCoV感染。
抗体或抗体片段与2019-nCoV多聚蛋白的特异性结合可以通过多种建立的方法中的任何一种来测定。亲和力可以通过各种测量来定量表示,包括实现体外病毒传播(例如病毒滴度)的半数最大抑制所需的抗体浓度(IC50)和抗体-2019-nCoV多聚蛋白复合物解离的平衡常数(KD)。描述2019-nCoV多聚蛋白与抗体相互作用的平衡常数KD是2019-nCoV多聚蛋白-抗体复合物解离反应成不互相作用的溶剂分离的2019-nCoV多聚蛋白和抗体分子的化学平衡常数。
抗体是以小于1μM(例如,900nM、800nM、700nM、600nM、500nM、400nM、300nM、200nM、100nM、95nM、90nM、85nM、80nM、75nM、70nM、65nM、60nM、55nM、50nM、45nM、40nM、35nM、30nM、25nM、20nM、15nM、10nM、5nM、4nM、3nM、2nM或1nM)的KD值特异性结合2019-nCoV多聚蛋白(例如,2019-nCoV的Env区域)的那些。在某些情况下,抗体是以小于1nM(例如,990pM、980pM、970pM、960pM、950pM、940pM、930pM、920pM、910pM、900pM、890pM、880pM、870pM、860pM、850pM、840pM、830pM、820pM、810pM、800pM、790pM、780pM、770pM、760pM、750pM、740pM、730pM、720pM、710pM、700pM、690pM、680pM、670pM、660pM、650pM、640pM、630pM、620pM、610pM、600pM、590pM、580pM、570pM、560pM、550pM、540pM、530pM、520pM、510pM、500pM、490pM、480pM、470pM、460pM、450pM、440pM、430pM、420pM、410pM、400pM、390pM、380pM、370pM、360pM、350pM、340pM、330pM、320pM、310pM、300pM、290pM、280pM、270pM、260pM、250pM、240pM、230pM、220pM、210pM、200pM、190pM、180pM、170pM、160pM、150pM、140pM、130pM、120pM、110pM、100pM、90pM、80pM、70pM、60pM、50pM、40pM、30pM、20pM、10pM、5pM或1pM)的KD值特异性结合2019-nCoV多聚蛋白的那些。
抗体也可以通过多种体外结合测定来表征。可用于确定2019-nCoV抗体的KD或IC50的实验实例包括例如表面等离子体共振、等温滴定量热法、荧光各向异性和基于ELISA的测定等。ELISA代表了一种特别有用的分析抗体活性的方法,因为这种测定通常需要最低浓度的抗体。在典型的ELISA测定中分析的一个常见信号是发光,其通常是与特异性结合一抗(例如2019-nCoV抗体)的二抗缀合的过氧化物酶的活性的结果。抗体能够结合2019-nCoV及其衍生的表位,如含有SEQ ID NO:1-84中任一项的一个或多个残基的表位,以及衍生自2019-nCoV的分离的肽,所述分离的肽在结构上预先组织各种残基以模拟天然蛋白中这些氨基酸的构象。例如,抗体可以结合含有SEQ ID NO:1-84中任一项的氨基酸序列的肽,或含有SEQ ID NO:1-84中任一项的约10-约30个连续或不连续氨基酸的肽。在直接ELISA实验中,可以定量这种结合,例如,通过分析在HRP底物(例如,2,2′-叠氮基-二-3-乙基苯并噻唑啉磺酸盐)与结合HRP缀合的二抗的抗原-抗体复合物一起孵育时发生的发光。
抗体包括通过用SEQ ID NO:1-84的多肽免疫宿主(例如哺乳动物宿主,例如人)而产生的那些。如果在其中产生抗2019-nCoV抗体的宿主不是人,则可以重组制备抗体,并在必要时进行人源化,以便随后向人受者给药。
组合物
组合物包括含有异源核酸分子的DNA或RNA载体,所述异源核酸分子编码来自2019-nCoV的抗原性或治疗性基因产物或其片段(例如,SEQ ID NO:93-181、190-195和199-204中任一项的核酸分子的全部或部分,或者与SEQ ID NO:93-181、190-195和199-204中任一项具有至少85%序列相同性(例如,86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)的变体,及其互补物)。例如,组合物可以含有这样的DNA载体,所述DNA载体含有SEQ ID NO:143或SEQ ID NO:204的核酸分子。例如,组合物可以含有这样的DNA载体,所述DNA载体含有SEQ ID NO:195的核酸分子。其他组合物包括来自2019-nCoV多聚蛋白的免疫原性多肽或其片段(例如,SEQ ID NO:1-84的多肽的全部或部分,或者与SEQ ID NO:1-84具有至少85%(例如,86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%)序列相同性的其变体)。例如,组合物可以包括具有SEQ ID NO:51的氨基酸序列的免疫原性多肽。例如,组合物可以包括具有SEQ IDNO:28的氨基酸序列的免疫原性多肽。组合物还可以包括能够结合2019-nCoV及其衍生的表位的2019-nCoV抗体(例如,抗刺突抗体),如含有SEQ ID NO:1-84中任一项的一个或多个残基的表位。例如,组合物可以包括能够结合含有SEQ ID NO:28的一个或多个残基的表位的抗体。所述抗体可以通过用SEQ ID NO:1-84中任一项的多肽免疫宿主来产生。例如,可以通过用具有SEQ ID NO:28的氨基酸序列的多肽免疫宿主来产生抗体。
任选地,可以配制组合物,例如,用于通过病毒载体(例如,腺病毒载体或痘病毒载体)给药。重组腺病毒在用于表达例如一种或多种免疫原(例如,2019-nCoV多肽)的载体上具有几个显著的优势。病毒可以制备成高滴度,可以感染非复制细胞,并且可以在与靶细胞群接触后赋予靶细胞离体的高效转导。此外,腺病毒不将它们的DNA整合到宿主基因组中。因此,它们作为表达载体的使用降低诱发自发性增殖性病症的风险。在动物模型中,通常发现腺病毒载体介导大约一周的高水平表达。转基因表达(核酸分子的表达)的持续时间可以通过使用细胞或组织特异性启动子来延长。腺病毒载体本身的分子工程化的其他改进产生了较持久的转基因表达和较少的炎症。这可以通过在额外的早期腺病毒基因中包含特定突变的所谓“第二代”载体和利用Cre-Lox策略缺失几乎所有病毒基因的“无胆”载体看到(Engelhardt et al.,Proc.Natl.Acad.Sci.USA91:6196(1994)和Kochanek et al.,Proc.Natl.Acad.Sci.USA 93:5731(1996),each herein incorporated by reference,每个援引加入本文)。
使用本领域已知的标准方法,通过将具有所期望纯度的活性成分与任选的生理学上可接受的载剂、赋形剂或稳定剂混合(Remington’s Pharmaceutical Sciences(20thedition),ed.A.Gennaro,2000,Lippincott,Williams&Wilkins,Philadelphia,PA)制备组合物的治疗性制剂用于向受试者(例如人)给药。使用本领域已知的标准方法,通过将具有所期望纯度的活性成分与任选的生理学上可接受的载剂、赋形剂或稳定剂混合(Remington’s Pharmaceutical Sciences(20th edition),ed.A.Gennaro,2000,Lippincott,Williams&Wilkins,Philadelphia,PA)制备组合物的治疗性制剂。可接受的载剂,包括盐水或缓冲液,如磷酸盐、柠檬酸盐和其他有机酸;抗氧化剂,包括抗坏血酸;低分子量(少于约10个残基)的多肽;蛋白,如血清白蛋白、明胶或免疫球蛋白;亲水性聚合物如聚乙烯吡咯烷酮,氨基酸如甘氨酸、谷氨酰胺、天冬酰胺、精氨酸或赖氨酸;单糖、二糖和其他碳水化合物,包括葡萄糖、甘露糖或糊精;螯合剂,如EDTA;糖醇,如甘露醇或山梨糖醇;成盐的平衡离子(counterion)如钠;和/或非离子表面活性剂,如TWEENTM、PLURONICSTM或PEG。
任选地,但优选地,制剂含有药学上可接受的盐,优选氯化钠,并且优选为约生理浓度。任选地,制剂可以含有药学上可接受的防腐剂。防腐剂浓度可以在约0.1-约2.0%的范围内,通常为v/v。合适的防腐剂包括制药领域已知的那些,如苯甲醇、苯酚、间甲酚、对羟基苯甲酸甲酯和对羟基苯甲酸丙酯。任选地,制剂可以包括浓度为约0.005-约0.02%的药学上可接受的表面活性剂。
任选地,可以配制组合物以包括用于共同给药或与顺序给药的佐剂和/或免疫刺激剂,(例如蛋白),如受体分子、核酸、免疫原性蛋白、药物、化疗剂和辅助细胞因子。例如,白细胞介素-3(IL-3)、白细胞介素-4(IL-4)、白细胞介素-5(IL-5)、白细胞介素-7(IL-7)、白细胞介素-8(IL-8)、白细胞介素-10(IL-10)、白细胞介素-11(IL-11)、白细胞介素-12(IL-12)、白细胞介素-13(IL-13)、脂质A、磷脂酶A2、内毒素、葡萄球菌肠毒素B、I型干扰素、II型干扰素、转化生长因子-β(TGF-β)、淋巴毒素迁移抑制因子、粒细胞-巨噬细胞集落刺激因子(CSF)、单核细胞-巨噬细胞CSF、粒细胞CSF、血管上皮生长因子(VEGF)、血管生成素、转化生长因子(TGF-α)、热休克蛋白(HSP)、血型碳水化合物部分(carbohydrate moietiesofblood groups)、Rh因子、成纤维细胞生长因子、核苷酸、DNA、RNA、mRNA、MART、MAGE、BAGE、突变型p53、酪氨酸酶、AZT、血管抑素、内皮抑素或其组合可包括在组合物的制剂中或与组合物共同给药。
可以以治疗有效量给药药物组合物,所述治疗有效量提供针对感染剂(例如,2019-nCoV)的免疫原性和/或保护性作用。在一些实施方案中,可以配制包含核酸分子、多肽、载体和/或抗体的组合物以用于以至少1-1,000μg(例如,至少15、16、17、18、19、20、30、40、50、60、70、80、90、100、125、150、175、200、225、250、275或300μg或更多)的剂量给药。剂量可以是0.2mL-1.0mL或达1L的体积(例如,如果制备为输液)。在一些实施方案中,包含核酸分子、载体和/或疫苗的组合物以50μg的剂量给药。
可配制用于本文所述方法的组合物,例如用于肌内、静脉内、皮内、经皮、动脉内、腹膜内、病灶内、颅内、关节内、前列腺内、胸膜内、气管内、鼻内、玻璃体内、阴道内、直肠内、局部、肿瘤内、腹膜、皮下、结膜下、血管内、黏膜内、心包内、脐内、眼内、口服、外用、局部、通过吸入、通过注射、通过输注、通过连续输注、通过直接对靶细胞进行局部灌注、通过导管、通过灌洗、通过灌胃、在乳膏中或在脂质组合物中给药。
可以配制本文所述的本发明的药物组合物以在给药后(例如,靶向递送)立即或者在给药后的任何预定时间段使用控释或缓释制剂(extended release)释放组合物。以控释或缓释制剂给药药物组合物是有用的,其中组合物单独或组合具有(i)窄治疗指数(例如,导致有害副作用或毒性反应的血浆浓度与导致治效力的血浆浓度的差异小;通常,治疗指数TI定义为致死剂量(LD50)的中值与有效剂量(ED50)的中值之比;(ii)释放部位(如胃肠道)的窄吸收窗;或(iii)短生物半衰期,因此需要在一天中频繁给药以维持治疗水平。
可以采用许多策略来获得控释或缓释,其中释放速率超过药物组合物的代谢速率。例如,通过适当选择制剂参数和成分,包括例如适当的控释组合物和包衣,可以获得控释。合适的制剂是本领域技术人员已知的。实例包括单或多单元片剂或胶囊组合物、油溶液、悬浮液、乳剂、微胶囊、微球、纳米颗粒、贴剂和脂质体。
组合物可通过常规灭菌技术灭菌或可无菌过滤。得到的水溶液可以原样包装使用,或冻干,冻干制剂可以粉末形式给药或在给药前与无菌水性载剂组合。制剂的pH通常为3-11,更优选5-9或6-8,最优选7-8,如7-7.5。所得固体形式的组合物可以包装在多个单剂单位中,每个单位含有固定量的免疫原性组合物(例如,疫苗或抗2019-nCoV抗体)以及,如果需要,一种或多种免疫调节剂,如在片剂或胶囊的密封包装中,或在能够给药一剂或多剂的合适的干粉吸入器(DPI)中。
使用组合物的治疗方法
药物组合物(例如,免疫原性组合物和抗2019-nCoV抗体)可用于治疗处于冠状病毒(例如,2019-nCoV)暴露风险中(例如,由于前往冠状病毒(例如,2019-nCoV)感染流行的地区)的受试者(例如,人)、对冠状病毒(例如,2019-nCoV)感染易感的受试者,或治疗具有冠状病毒(例如,2019-nCoV)感染的受试者。特别地,组合物可用于治疗(暴露前或暴露后)2019-nCoV感染。在一些实施方案中,治疗可以诱导保护性水平的抗冠状病毒抗体(例如抗2019-nCoV抗体,例如抗刺突抗体,例如抗刺突中和抗体)。在一些实施方案中,保护水平是使用本文所述的假病毒中和测定测量的至少约70的滴度、使用本文所述的活病毒中和测定测量的至少约25的滴度,或高于如本文所述,通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值或平均水平的至少约80%的水平。在一些实施方案中,用组合物治疗可降低受试者中2019-nCoV介导的活性,如病毒滴度、病毒传播、感染和/或细胞融合。在一些实施方案中,2019-nCoV介导的活性是呼吸道(例如,上呼吸道和/或下呼吸道)中的病毒载量。在一些实施方案中,2019-nCoV介导的活性是肺、鼻孔和/或气管中的病毒载量。在一些实施方案中,2019-nCoV病毒载量减少约1%或更多(例如,2%、3%、4%、5%、6%、7%、8%、9%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、99%、99.9%、99.99%或更多)。在一些实施方案中,在向受试者给药组合物(例如疫苗)后,感染有2019-nCoV的治疗的受试者中的2019-nCoV滴度降低至少约1%或更多(例如,2%、3%、4%、5%、6%、7%、8%、9%、10%、20%、30%、40%、50%、60%、70%、80%或90%、95%、99%、99.9%、99.99%或更多)。
组合物(例如本文所述的任何组合物)可用于在受试者(例如人受试者)中诱导免疫应答(例如体液和/或细胞免疫应答)。取决于所给药的组合物或组合物的组合,诱导的免疫应答可能不同(例如,特异性、稳健性或持久性不同)。例如,组合物可以诱导具有不同抗体类型(例如,不同比例的IgM、IgA、IgG1、IgG2、IgG3或FcgR2A.1)或不同功能特征(例如,诱导抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性补体沉积(ADCD)、抗体依赖性单核细胞吞噬作用(ADCP)或抗体依赖性NK细胞激活(IFN-γ分泌、CD107a脱粒和MIP-1β表达)的能力)。本文所述的组合物(例如,SS-Spike和SS-SdCT)可以诱导可以监测的ADCD应答(例如,以评估治疗效力)。本文所述的组合物(例如,SS-RBD-foldon和SS-S.Ecto-dF-PP-foldon)可以诱导可以监测的抗体依赖性NK细胞激活应答(例如,以评估治疗效力)。组合物还可以诱导具有不同特征的细胞应答(例如,Th1、Th2或Th17应答)。本文所述的组合物(例如,SS-Spike、SS-SdCT和SS-S.Ecto-dF-PP-foldon)可以诱导可以监测的S特异性CD4+或CD8+T细胞应答(例如,以评估治疗效力)。
在预防和/或治疗2019-nCoV感染的方法中,可以用载体(例如,哺乳动物、细菌或病毒(例如,Ad26)衍生的表达载体)向受试者递送表达免疫原(例如,SEQ ID NO:1-84中的一个或多个或者与其具有至少85-99%序列相同性,例如至少大于90%序列相同性的其变体)的核酸。例如,载体可用于递送表达具有SEQ ID NO:51的氨基酸序列的免疫原的核酸(例如,含有SEQ ID NO:104或204的核苷酸序列的核酸)。例如,载体可用于递送表达具有SEQ ID NO:56的氨基酸序列的免疫原的核酸(例如,含有SEQ ID NO:195的核苷酸序列的核酸)。可以遗传修饰载体(例如,哺乳动物、细菌或病毒衍生的表达载体)以含有如SEQ IDNO:93-181、190-195和199-204中所示的一种或多种核酸序列或与其具有至少85-99%序列相同性,例如至少大于90%序列相同性的其变体,及其互补物。特别地,在国际专利申请公布WO 2006/040330和WO 2007/104792中公开的腺病毒载体(例如,衍生自Ad2、Ad5、Ad11、Ad12、Ad24、Ad26、Ad34、Ad35、Ad40、Ad48、Ad49、Ad50、Ad52(RhAd52)、Ad59(RhAd59)和Pan9(也称为AdC68)),每个援引加入本文,在将免疫原递送至受试者的方法中作为载体特别有用。例如,载体可以是Ad26。载体的其他实例描述于例如McVey等人(美国专利号5,801,030);整体援引加入本文。
用于将免疫原递送至有需要的受试者的有用基因治疗方法包括在PCT公开号WO2006/060641、美国专利号US 7,179,903和PCT公开号WO 2001/036620中描述的那些,其描述了例如腺病毒载体(例如衍生自Ad2、Ad5、Ad11、Ad12、Ad24、Ad26、Ad34、Ad35、Ad40、Ad48、Ad49、Ad50、Ad52(RhAd52)、Ad59(RhAd59)和Pan9(也称为AdC68))用于治疗性蛋白递送的用途。
本文所述的任何组合物(例如药物组合物(例如免疫原性组合物和抗2019-nCoV抗体))中的一种或多种可用于治疗。治疗可以包括一种或多种其他治疗剂(例如促炎剂(l如干扰素)或抗炎剂(例如皮质类固醇,例如地塞米松))和/或一种或多种治疗干预(例如手术和俯卧位通气)。治疗剂和/或干预可以顺序给药(例如,在疾病之前或在疾病的早期(例如,在症状发作的一周内)给药本文所述的任何组合物中的一种或多种,然后给药其他治疗剂(例如,在疾病的后期(例如,症状发作一周后)的抗炎剂(例如,皮质类固醇,例如,地塞米松)))或同时(例如,给药本文所述的任何组合物中的一种或多种和/或一种或多种其他治疗剂)。其他治疗剂可以包括皮质类固醇(例如糖皮质激素(例如地塞米松、泼尼松和氢化可的松))、干扰素(例如干扰素β)、脱氧胆酸、集落刺激因子(例如G-CSF和GM-CSF)和非-甾体抗炎药(例如,阿司匹林、丙酸衍生物如布洛芬(ibuprofen)、非诺洛芬(fenoprofen)、酮洛芬(ketoprofen)、氟比洛芬(flurbiprofen)、奥沙普秦(oxaprozin)和萘普生(naproxen)、乙酸衍生物如舒林酸(sulindac)、吲哚美辛(indomethacin)、依托度酸(etodolac)、双氯芬酸(diclofenac)、烯醇酸衍生物如吡罗昔康(piroxicam)、美洛昔康(meloxicam)、替诺昔康(tenoxicam)、屈昔康(droxicam)、氯诺昔康(lornoxicam)和异昔康(isoxicam),芬那酸衍生物如甲芬那酸(mefenamic acid)、甲氯芬那酸(meclofenamic acid)、氟芬那酸(flufenamic acid)、托芬那酸(tolfenamic acid)和COX-2抑制剂如塞来昔布(celecoxib)、依托昔布(etoricoxib)、罗美昔布(lumiracoxib)、帕瑞昔布(parecoxib)、罗非昔布(rofecoxib)、罗非昔布和伐地昔布(valdecoxib)。可以与本文所述的组合物组合给药的其他药剂包括瑞德西韦(remdesivir)、氯喹(chloroquine)、羟氯喹(hydroxychloroquine)、巴瑞克替尼(baricitinib)、洛匹那韦(lopinavir)/利托那韦(ritonavir)、阿比朵尔(umifenovir)、法匹拉韦(favipiravir)、托珠单抗(tocilizumab)和利巴韦林(ribavirin)。
给药
药物组合物可以在暴露于感染剂(例如冠状病毒,如2019-nCoV)之前或之后向受试者(例如人)给药,以治疗、预防、改善、抑制感染的进展,或减轻感染(例如,冠状病毒感染,例如2019-nCoV感染)的一种或多种症状的严重度。例如,可以向具有2019-nCoV感染的受试者给药组合物。可以使用组合物治疗的由病毒感染(如2019-nCoV)引起的疾病的症状的实例包括例如发烧、肺炎、呼吸衰竭、体重减轻、关节痛、皮疹、结膜炎、肌肉疼痛、头痛、眼眶后疼痛、水肿、淋巴结病、不适、虚弱、喉咙痛、咳嗽、恶心、呕吐、腹泻和血精症。这些症状及其在治疗期间的消退可以由例如医生在体格检查期间或通过本领域已知的其他测试和方法来测量。本文所述的药物组合物可以在暴露于感染剂(例如冠状病毒,如2019-nCoV)之前或之后向受试者(例如人)给药,以降低或预防由感染剂引起的死亡的风险。
优选的给药方法可以取决于各种要素(例如,所给药的组合物的组分,以及所治疗病况的严重度)而变化。适用于口服或鼻腔给药的制剂可以由液体溶液组成,如溶解在稀释剂(例如水、盐水或PEG-400)中的有效量的组合物、胶囊、散剂、片剂或凝胶,每一种含有预先指定量的嵌合Ad5载体组合物。药物组合物还可以是用于例如吸入支气管通道的气雾剂制剂。气雾剂制剂可以与加压的药学上可接受的抛射剂(例如二氯二氟甲烷、丙烷或氮气)混合。特别地,吸入给药可以通过使用例如含有脱水山梨糖醇三油酸酯或油酸的气雾剂与例如三氯氟甲烷、二氯氟甲烷、二氯四氟乙烷或任何其他生物相容的抛射剂气体一起来完成。
如果组合物与免疫刺激剂和/或佐剂共同给药,则可以显著改善组合物的免疫原性。本领域技术人员熟知的合适佐剂包括,例如,磷酸铝、氢氧化铝、QS21、Quil A(及其衍生物和其组分)、磷酸钙、氢氧化钙、氢氧化锌、糖脂类似物、十八烷基酯氨基酸(octodecylesters of an amino acid)、胞壁酰二肽、聚磷腈、脂蛋白、ISCOM基质、DC-Chol、DDA、细胞因子和其他佐剂及其衍生物。
可以给药组合物以提供暴露前预防或在受试者诊断为具有病毒感染(例如,2019-nCoV感染)或受试者暴露于诸如病毒(例如,冠状病毒感染,如2019-nCoV)的感染剂之后给药组合物。组合物可以在暴露于2019-nCoV之前,例如,1、2、3、4、5、6、7、8、9、10、15、20、30、35、40、45、50、55或60分钟,2、4、6、10、15或24小时,2、3、5或7天,2、4、6或8周,或甚至3、4或6个月前给药,或可在暴露于冠状病毒(例如,2019-nCoV)之后15-30分钟或1、2、3、4、5、6、7、8、9、10、12、20、24、48或72小时,2、3、5、或7天,2、4、6或8周,3、4、6或9个月,1、2、3、4、5、6、7、8、9、10、15或20年向受试者给药。
当治疗病毒感染(例如,2019-nCoV感染)时,可以在出现症状或明确诊断之前或者在诊断或症状变得明显之后向受试者给药组合物。例如,可以例如在诊断或临床识别症状后立即或者在诊断或检测到症状后2、4、6、10、15或24小时,2、3、5或7天后给药组合物。
可向有需要的受试者给药一剂或多剂(例如,1、2、3、4、5、6、7、8、9或10剂)免疫原性组合物或者含有抗2019-nCoV抗体的组合物。在一些实施方案中,向受试者给药至少一剂。在一些实施方案中,向受试者给药至少两剂。在一些实施方案中,在同一天给药剂量。在一些实施方案中,在不同的日子给药剂量。在一些实施方案中,将免疫原性组合物作为初免、加强或作为初免-加强向有需要的受试者给药。在一些实施方案中,在初免-加强方案的初免之后的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27或28天,5、6、7、8、9、10、11或12周,4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22或23个月,2、3、4、5、6、7、8、9、10、15或20年后加强给药。在其他实施方案中,给药多个加强剂量,其中每个加强剂量至少间隔1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27或28天,5、6、7、8、9、10、11或12周,4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22或23个月,2、3、4、5、6、7、8、9、10、15或20年给药。
本文所述的任何组合物(例如本文所述的任何免疫原性组合物)的一剂或多剂可与一种或多种其他治疗剂依次或同时给药。
剂量
组合物的剂量或使用组合物的治疗数目可以基于受试者中疾病的严重度、发生或进展(例如,基于例如病毒感染的一种或多种症状的严重度)增加或减少。
可以以治疗有效量给药药物组合物,所述治疗有效量提供针对感染剂(例如,2019-nCoV)的免疫原性和/或保护性作用。在一些实施方案中,包含核酸分子、多肽、载体和/或抗体的组合物可以以至少1μg-100mg(例如,至少10μg、20μg、30μg、40μg、50μg、60μg、70μg、80μg、90μg、100μg、125μg、150μg、175μg、200μg、225μg、250μg、275μg、300μg、325μg、350μg、375μg、400μg、425μg、450μg、475μg、500μg、525μg、550μg、575μg、600μg、625μg、650μg、675μg、700μg、725μg、750μg、775μg、800μg、825μg、850μg、875μg、900μg、925μg、950μg、975μg、1mg、2mg、3mg、4mg、5mg、6mg、7mg、8mg、9mg、10mg、11mg、12mg、13mg、14mg、15mg、16mg、17mg、18mg、19mg、20mg、25mg、30mg、40mg、50mg、60mg、70mg、80mg、90mg或更多)的剂量给药。在一些实施方案中,包含核酸分子、载体和/或抗体的组合物以约50μg的剂量(例如,约25μg-约75μg的剂量)给药。在一些实施方案中,包含核酸分子、载体和/或抗体的组合物以约5mg的剂量(例如,约1mg-约10mg的剂量)给药。
在一些情况下,给药有效量的组合物(例如,免疫原,如SEQ ID NO:1-84)诱导抗冠状病毒抗体(例如抗2019-nCoV抗体,例如抗刺突抗体,例如抗刺突中和抗体)的保护水平(例如,高于如使用本文所述的假病毒中和测定测量的至少约70的滴度,高于如使用本文所述的活病毒中和测定测量的至少约25的滴度,或高于如通过本文所述的假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值或平均水平的至少80%的水平)。在一些情况下,保护水平是如使用本文所述的假病毒中和测定测量的至少约70(例如,至少约80、至少约100或至少约120)的滴度。在一些情况下,保护水平是如使用本文所述的假病毒中和测定测量的至少约100的滴度。在一些情况下,给药有效量的组合物导致抗冠状病毒抗体(例如抗2019-nCoV抗体,例如抗刺突抗体,例如抗刺突中和抗体)的保护水平维持至少约1、2、3、4、5、6、7、8、9、10、11或12个月或者1、2、3、4、5、6、7、8、9、10、11、12、13、14、15年或更长时间。
在一些情况下,与在给药有效量的组合物之前从患者确定的病毒载量相比,给药有效量的组合物(例如,免疫原,如SEQ ID NO:1-84)将从具有2019-nCoV感染的受试者确定的2019-nCoV血清病毒载量降低至少约5%、10%、20%、30%、40%、50%、60%、70%、80%、90%或更多。在一些情况下,与在给药有效量的组合物之前从患者确定的病毒载量相比,给药有效量的组合物将血清病毒载量降低至不可检测的水平。在一些情况下,给药有效量的组合物导致降低的和/或不可检测的血清病毒载量,这可以维持至少约1、2、3、4、5、6、7天;1、2、3、4周;1、2、3、4、5、6、7、8、9、10、11或12个月;或者1年或更长时间。
给药的剂量取决于待治疗的受试者(例如,年龄、体重、免疫系统的能力和接受治疗的受试者的一般健康状况)、给药的形式(例如,作为固体或液体)、给药的方式(例如,通过注射、吸入或干粉抛射剂)和靶向的细胞(例如,上皮细胞,如血管上皮细胞、鼻上皮细胞或肺上皮细胞)。组合物优选以提供足够水平的抗原性或治疗性基因产物或其片段的量给药(例如引起免疫应答,而没有由抗原性基因产物在宿主中引起的不适当的不良生理影响的抗原性基因产物的水平)。
递送方法,例如递送DNA或RNA疫苗的方法,也可以决定剂量。在一些情况下,通过静脉内(iv)或肌内(im)途径注射给药的剂量可能需要不同量的DNA或RNA疫苗,例如10μg-1mg。然而,使用基因枪给药可能需要0.2μg-20μg(例如,0.2、0.1、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20μg)的DNA或RNA疫苗的剂量。在一些情况下,使用基因枪递送一剂DNA或RNA疫苗可能仅需要ng量的DNA或RNA,例如10ng-200ng (例如,10、12、13、14、15、16、17、18、19、20、30、40、50、60、70、80、90、100、125、150、175或200ng)。
在其中递送载体是病毒(例如,Ad26病毒)的其他实施方案中,可以以至少约1x103病毒颗粒(VP)/剂或1x101-1x1020VP/剂(例如,1x101、1x102、1x103、1x104、1x105、1x106、1x107、1x108、1x109、1x1010、1x1011、1x1012、1x1013、1x1014、1x1015、1x1016、1x1017、1x1018、1x1019、或1x1020VP/剂)向受试者给药。例如,可以以约1x106-约1x1014VP/剂(例如,约1x107、约1x108、约1x109、约1x1010、约1x1011、约1x1012、约1x1013、约1x1014,或约1x1015VP/剂)向受试者给药。例如,可以以约1x1011、约1x1012、约1x1013或约1x1014VP/剂向受试者给药。
此外,可以向受试者单次或多次(例如,一次给药或给药两次或更多次)给药本发明的组合物(暴露前或暴露后和/或诊断前或诊断后)。例如,对感染例如病毒感染(例如,2019-nCoV感染)特别易感的受试者可能需要多次治疗来建立和/或维持针对病毒的保护。本文所述的药物组合物提供的诱导的免疫的水平可以通过例如测量中和的分泌抗体和血清抗体的量来监测。然后可以根据需要调整或重复剂量以触发所期望水平的免疫应答。例如,由组合物的单次给药(初免)触发的免疫应答可能不足以有效和/或持久以提供有效的保护。因此,在一些实施方案中,重复给药(加强),从而建立初免加强方案,可以显著增强对组合物的抗原的体液和细胞应答。
或者,可以通过监测组合物给药后受试者(例如人)中表达的抗原性或治疗性基因产物或其片段的水平来确定治疗的效力。例如,可以使用例如本领域已知的标准测定来检测受试者的血液或淋巴液的抗原性或治疗性基因产物或其片段。
在一些情况下,可以通过监测在给药有效量的组合物(例如,免疫原,例如SEQ IDNO:1-84中的任一项)之前和之后从受试者获得的样品中的血清病毒载量的变化来确定治疗的效力。与在给药有效量的组合物之前从受试者确定的病毒载量相比,血清病毒载量减少至少约5%、10%、20%、30%、40%、50%、60%、70%、80%、90%或更多,可以表明受试者正在从治疗中受益。如果在给药组合物后病毒载量没有降低至少约10%、20%、30%或更多,则可以增加要给药的组合物的剂量。例如,通过增加向受试者给药的DNA疫苗(例如,含有SEQ ID NO:93-181、190-195和199-204中的一个或多个的DNA疫苗)的μg或mg量,或通过增加基于腺病毒载体的疫苗(例如,含有SEQ ID NO:93-181、190-195和199-204中的一个或多个的基于腺病毒载体的疫苗)的病毒颗粒(VP)的数目。
单剂的组合物可以实现保护,暴露前或诊断前。此外,暴露后或诊断后给药的单个剂量可用作本发明的治疗。
单剂的组合物也可用于在接受治疗感染(例如冠状病毒感染,如2019-nCoV感染)的受试者中实现治疗。必要时,也可以向这些受试者给药多个剂量(例如,2、3、4、5、6、7、8、9、10、15、20或更多剂量)。
诊断和预测对冠状病毒感染的易感性的方法
诊断方法
本文提供用于识别、诊断和/或预测受试者对冠状病毒感染的易感性的方法。所述方法包括测量来自受试者的样品(例如全血样品,例如血清或血浆样品)中的抗冠状病毒抗体(例如抗刺突抗体)的水平或量。在一些实施方案中,冠状病毒是2019-nCoV。在一些实施方案中,抗冠状病毒抗体(例如,抗刺突抗体)是中和抗体。在一些实施方案中,如果抗冠状病毒抗体(例如,抗刺突抗体)量或水平低于保护水平(例如,低于如使用本文所述的假病毒中和测定测量的至少约70的滴度,低于如使用本文所述的活病毒中和测定测量的至少约25的滴度,或低于如通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值水平的80%(例如,已恢复或正在从冠状病毒感染(例如2019-nCoV)中恢复的人的队列),则确定受试者对冠状病毒感染易感,并且如果抗冠状病毒抗体(例如,抗刺突抗体)水平高于保护水平,则确定受试者对冠状病毒感染不易感。在一些实施方案中,保护水平是如在假病毒中和测定中确定的至少约70(例如,约70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、110、115、120、125、130、140、150、175、200、225、250、275、300、325、350或更多)的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。在一些实施方案中,保护水平是如在假病毒中和测定中确定的至少约83的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。在一些实施方案中,保护水平是如在活病毒中和测定中确定的至少约25(例如约25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、110、115、120、125、130、140、150、175、200或更多)的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。在一些实施方案中,保护水平是如在活病毒中和测定中确定的至少约35的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。在一些实施方案中,保护水平是如通过本文所述的假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值或平均水平的至少约60%(例如约61%、约62%、约63%、约64%、约65%、约66%、约67%、约68%、约69%、约70%、约71%、约72%、约73%、约74%、约75%、约76%、约77%、约78%、约79%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、约110%、约120%)的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。在一些实施方案中,保护水平是如通过本文所述的假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值或平均水平的至少约80%的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。可以向确定为对冠状病毒感染易感的受试者(具有低于保护水平(例如,低于如使用本文所述的假病毒中和测定测量的至少约70的滴度,低于如使用本文所述的活病毒中和测定测量的至少约25的滴度,或低于如通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列(例如,已恢复或正在从冠状病毒感染(例如,2019-nCoV)中恢复的人的队列)的中值水平的80%)的抗冠状病毒抗体(例如,抗刺突抗体)的量或水平的受试者)给予治疗(例如给药本文所述的任何组合物),如有效量的本文所述的一种或多种药物组合物(例如,免疫原性组合物和抗2019-nCoV抗体)。可以重新向受试者给予治疗,直到确定受试者对冠状病毒感染不易感(例如,直到受试者具有抗冠状病毒抗体(例如,抗刺突抗体)水平高于保护水平(例如,高于如使用本文所述的假病毒中和测定测量的至少约70的滴度的水平,高于如使用本文所述的活病毒中和测定测量的至少约25的滴度的水平,或等于如通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列(例如,已经恢复或正在从冠状病毒感染中恢复的人的队列(例如,2019-nCoV))的抗冠状病毒抗体的中值水平的至少80%的水平(并且优选地等于或高于中值水平)。所述方法还可以涉及确定抗刺突抗体是否是RBD特异性抗体。所述方法还可以涉及确定抗刺突抗体是否是S1特异性抗体。所述方法还可以涉及确定抗刺突抗体是否是S2特异性抗体。所述方法还可以涉及识别抗冠状病毒抗体的亚类(例如如,IgM、IgA、IgG1、IgG2、IgG3或FcgR2A.1)和/或效应功能(例如,抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性补体沉积(ADCD)、抗体依赖性单核细胞吞噬作用(ADCP)或抗体依赖性NK细胞激活(IFN-γ分泌、CD107a脱粒和MIP-1β表达))。所述方法可以进一步包括向确定的需要进一步治疗的受试者给药本文所述的一种或多种药物组合物(例如,免疫原性组合物和抗2019-nCoV抗体)。
所述方法可以包括测量来自受试者的样品中的冠状病毒(例如,2019-nCoV)的病毒载量。在一些实施方案中,样品是支气管肺泡灌洗液(BAL)或鼻拭子(NS)。在一些实施方案中,样品是来自受试者的体液(例如,血液,例如,全血或血浆)。在一些实施方案中,样品是来自受试者的组织样品(例如,呼吸道组织样品)。在一些实施方案中,病毒载量是可检测的核酸(例如,亚基因组mRNA)水平或可检测的蛋白(例如,核衣壳蛋白(N))水平。在一些实施方案中,可检测的核酸(例如,亚基因组mRNA)通过RNA-seq、RT-qPCR、qPCR、多重qPCR或RT-qPCR、LAMP、微阵列分析或杂交(例如,ISH(例如,FISH))确定。在一些实施方案中,可检测的蛋白(例如,核衣壳蛋白(N))通过免疫测定(例如,免疫组织化学(IHC)测定或侧流免疫层析测定)确定。在一些实施方案中,可检测的病毒载量表明受试者对疾病(例如,2019-nCoV介导的疾病,例如,COVID-19,例如,严重的COVID-19疾病)易感。在一些实施方案中,大于至少约3.5log10 sgmRNA拷贝/mL(例如,约3.75log10 sgmRNA拷贝/mL、约3.8log10 sgmRNA拷贝/mL、约3.9log10 sgmRNA拷贝/mL、约4.0log10sgmRNA拷贝/mL、约4.25log10 sgmRNA拷贝/mL、约4.5log10sgmRNA拷贝/mL、约4.75log10 sgmRNA拷贝/mL、约5.0log10sgmRNA拷贝/mL、约5.5log10sgmRNA拷贝/mL、约6.0log10 sgmRNA拷贝/mL、约6.5log10 sgmRNA拷贝/mL、约7.0log10 sgmRNA拷贝/mL、约7.5log10 sgmRNA拷贝/mL、约8.0log10 sgmRNA拷贝/mL、约8.5log10 sgmRNA拷贝/mL、约9log10 sgmRNA拷贝/mL、约10log10 sgmRNA拷贝/mL、约11log10sgmRNA拷贝/mL、约12log10 sgmRNA拷贝/mL、约13log10sgmRNA拷贝/mL或更多)的病毒载量。在一些实施方案中,BAL中大于3.85log10sgmRNA拷贝/mL或NS中大于3.78log10 sgmRNA拷贝/mL的病毒载量表明受试者对疾病(例如,2019-nCoV介导的疾病,例如,COVID-19,例如严重的COVID-19疾病)易感。在一些实施方案中,BAL中大于3.85log10 sgmRNA拷贝/mL或NS中大于3.78log10 sgmRNA拷贝/mL的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,大于约2.0log10 sgmRNA拷贝/g(例如,约2.0log10 sgmRNA拷贝/g、约2.5log10sgmRNA拷贝/g、约3.0log10 sgmRNA拷贝/g、约3.5log10sgmRNA拷贝/g、约4.0log10sgmRNA拷贝/g、约4.25log10 sgmRNA拷贝/g、约4.5log10sgmRNA拷贝/g、约4.75log10sgmRNA拷贝/g、约5.0log10sgmRNA拷贝/g、约5.5log10sgmRNA拷贝/g、约6.0log10sgmRNA拷贝/g、约6.5log10sgmRNA拷贝/g、约7.0log10sgmRNA拷贝/g、约7.5log10sgmRNA拷贝/g、约8.0log10sgmRNA拷贝/g、约8.5log10sgmRNA拷贝/g、约9log10sgmRNA拷贝/g、约10log10sgmRNA拷贝/g、约11log10 sgmRNA拷贝/g、约12log10 sgmRNA拷贝/g、约13log10 sgmRNA拷贝/g或更多)的组织的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,肺组织中大于约8.0log10 sgmRNA拷贝/g、鼻孔组织中大于约7.0log10 sgmRNA拷贝/g、气管组织中大于约6.0log10 sgmRNA拷贝/g、心脏组织中大于约5.5log10 sgmRNA拷贝/g,或胃肠道、脾脏、肝脏、肾脏或脑组织中约2.0log10 sgmRNA拷贝/g的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,通过ISH染色的大于约3%(例如,约4%、约5%、约6%、约7%、约8%、约9%、约10%)的SARS-CoV-2vRNA的病毒载量表明受试者对疾病(例如,2019-nCoV介导的疾病,例如COVID-19,例如严重的COVID-19疾病)易感。在一些实施方案中,通过ISH染色的大于约5%(例如,约5%、约6%、约7%、约8%、约9%、约10%)的SARS-CoV-2vRNA的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,通过ISH染色的大于约5%(例如,约5%、约6%、约7%、约8%、约9%、约10%)的SARS-CoV-2vRNA的病毒载量表明受试者对严重的COVID-19疾病易感。在一些实施方案中,在感染后约1、2、3、4、5或6天或者1、2、3、4、5、6或7周内测量冠状病毒(例如2019-nCoV)病毒载量一次或多次。
监控应答性
本文提供用于监测受试者对用于治疗冠状病毒感染或降低冠状病毒感染风险的治疗性组合物(例如,本文所述或本领域已知的任何组合物或免疫原性组合物)的抗冠状病毒免疫应答的方法。所述方法包括测量受试者中抗冠状病毒抗体(例如,抗刺突抗体)的水平或量。在一些实施方案中,冠状病毒是2019-nCoV。在一些实施方案中,抗冠状病毒抗体(例如,抗刺突抗体)是中和抗体。抗冠状病毒抗体(例如,抗刺突抗体,例如抗刺突中和抗体)可以在给药治疗性组合物(例如,本文所述的任何组合物或免疫原性组合物)后的短时间范围内(例如,以测量抗体应答的稳健性)或较长时间范围内(例如如,以测量抗体应答的持久性)测量。在一些实施方案中,在向受试者给药治疗性组合物(例如,本文所述的任何组合物或免疫原性组合物)后的约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27或28天,约5、6、7、8、9、10、11或12周,约4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22或23个月,或者约2、3、4、5、6、7、8、9、10、15或20年测量抗冠状病毒抗体(例如,抗刺突抗体)。
如果在受试者中(例如,在受试者的血液中)检测到的抗冠状病毒抗体(例如,抗刺突抗体)高于保护水平(例如,高于如使用本文所述的假病毒中和测定测量的至少约70的滴度,高于如使用本文所述的活病毒中和测定测量的至少约25的滴度,或等于如通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列(例如,已经恢复或正在从冠状病毒感染(例如,2019-nCoV)中恢复的人的队列)的抗冠状病毒抗体的中值水平的至少80%的水平(并且优选地等于或高于中值水平)),则确定受试者对治疗性组合物有应答。或者,如果在受试者中检测到的抗冠状病毒抗体(例如,抗刺突抗体)低于保护水平(例如,低于如使用本文所述的假病毒中和测定测量的至少约70的滴度,低于如使用本文所述的活病毒中和测定测量的至少约25的滴度,或等于如通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值水平的至少80%的水平),则确定受试者对治疗性组合物没有应答。抗冠状病毒抗体(例如,抗刺突中和抗体)的保护水平对应于如在假病毒中和测定(例如本文所述的假病毒中和测定)中确定的至少约70(例如,约70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、110、115、120、125、130、140、150、175、200、225、250、275、300、325、350或更多)的滴度。在一些实施方案中,保护水平是如在活病毒中和测定(例如,本文所述的假病毒中和测定)中确定的至少约25(例如,约25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、110、115、120、125、130、140、150、175、200或更多)的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。在一些实施方案中,保护水平是如通过本文所述的假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值或平均水平的至少约60%(例如,约61%、约62%、约63%、约64%、约65%、约66%、约67%、约68%、约69%、约70%、约71%、约72%、约73%、约74%、约75%、约76%、约77%、约78%、约79%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、约110%、约120%)的抗冠状病毒抗体滴度(例如,抗刺突中和抗体滴度)。
如果随着时间,受试者(例如,受试者血液中)的抗冠状病毒抗体(例如,抗刺突抗体)滴度降至低于或未能达到保护水平(例如,低于如使用本文所述的假病毒中和测定测量的至少约70的滴度,低于如使用本文所述的活病毒中和测定测量的至少约25的滴度,或低于如通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列的中值水平的80%),可以向受试者单独或与其他治疗剂(例如本文所述的一种或多种其他治疗剂)组合给药或者单独或与其他治疗剂组合重新给药冠状病毒疫苗组合物(例如,本文所述的一种或多种治疗性或免疫原性组合物)。向有需要的受试者给药本公开的组合物可以在一天或多天(例如,1、2、3、4、5、6或7天)、一周或多周(例如,1、2、3、4、5、6、7或8周)、一个月或多个月(例如,2、3、4、5、6、7、8、9、10、11或12个月)或者一年或多年(例如,1、2、3、4、5、6、7、8、9、10或更多年),或者在受试者的整个生命中根据需要进行一次或多次(例如,1、2、3、4、5、6、7、8、9、10、11、12、13、14、15或更多次)以在受试者中维持抗冠状病毒抗体的保护水平,从而保护受试者免受冠状病毒感染(例如,受2019-nCoV感染)。
所述方法可以包括测量来自受试者的样品中的冠状病毒(例如,2019-nCoV)的病毒载量。在一些实施方案中,冠状病毒是2019-nCoV。在一些实施方案中,病毒载量在感染后约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、30、36、42或48小时或者约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27或28天测量。在一些实施方案中,样品是支气管肺泡灌洗液(BAL)或鼻拭子(NS)。在一些实施方案中,样品是来自受试者的体液(例如,血液,例如,全血或血浆)。在一些实施方案中,样品是来自受试者的组织样品(例如,呼吸道组织样品)。在一些实施方案中,病毒载量是可检测的核酸(例如,亚基因组mRNA)水平或可检测的蛋白(例如,核衣壳蛋白(N))水平。在一些实施方案中,可检测的核酸(例如,亚基因组mRNA)通过RNA-seq、RT-qPCR、qPCR、多重qPCR或RT-qPCR、LAMP、微阵列分析或杂交(例如,ISH(例如,FISH))确定。在一些实施方案中,可检测的蛋白(例如,核衣壳蛋白(N))通过免疫测定(例如,免疫组织化学(IHC)测定或侧流免疫层析测定)确定。如果病毒载量低于预先指定的水平,则确定受试者对治疗性组合物有应答。在一些实施方案中,预先指定的水平为少于约3.5log10sgmRNA拷贝/mL BAL或NS或者少于约5.0log10sgmRNA拷贝/g组织(例如,肺、鼻孔、气管、心脏、胃肠道、脾脏、肝脏、肾脏或脑组织)。在一些实施方案中,如果受试者中的病毒载量降低,则确定受试者对治疗性组合物有应答。
如果根据病毒载量确定受试者没有应答,则可以向受试者单独或与其他治疗剂(例如本文所述的一种或多种其他治疗剂)组合给药或者单独或与其他治疗剂组合重新给药冠状病毒疫苗组合物(例如,本文所述的一种或多种治疗性或免疫原性组合物)。
向有需要的受试者给药本公开的组合物可以在一天或多天(例如,1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27或28天)根据需要进行一次或多次(例如,1、2、3、4、5、6、7、8、9、10、11、12、13、14、15或更多次)以减少病毒载量。
其他实施方案
本说明书中提及的所有出版物、专利和专利申请都援引加入本文,其程度与具体地和单独地将每个独立出版物或专利申请援引加入的程度相同。
虽然本发明已结合其特定实施方案进行了描述,但应当理解,它能够进行进一步修改,并且本申请旨在涵盖遵循一般原则以及包括与本发明所属领域内的已知或常规技术的偏离并且可以应用于上文所述的基本特征,在权利要求的范围内的任何变化、使用或改造。
本发明还提供:
实施方案1是一种分离的核酸分子,其包含编码2019-NCOV刺突(S)蛋白的核苷酸序列,所述2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
实施方案2是实施方案1的分离的核酸分子,其包含SEQ ID NO:211的核苷酸序列。
实施方案3是实施方案1或2的分离的核酸分子,其包含编码2019-NCOV刺突(S)蛋白的核苷酸序列,所述2019-NCOV刺突(S)蛋白包含SEQ ID NO:205的氨基酸序列。
实施方案4是实施方案1的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:51的氨基酸序列具有至少85%序列相同性的多肽。
实施方案5是实施方案1或4的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:51的氨基酸序列具有至少99%序列相同性的多肽。
实施方案6是实施方案1的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:54的氨基酸序列具有至少85%序列相同性的多肽。
实施方案7是实施方案1的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:56的氨基酸序列具有至少85%序列相同性的多肽。
实施方案8是实施方案1的分离的核酸分子,其包含与SEQ ID NO:121或其互补序列具有至少85%序列相同性的核苷酸序列。
实施方案9是实施方案4或5的分离的核酸分子,其包含与SEQ ID NO:143或其互补序列具有至少85%序列相同性的核苷酸序列。
实施方案10是实施方案6或7的分离的核酸分子,其包含与SEQ ID NO:146或其互补序列具有至少85%序列相同性的核苷酸序列。
实施方案11是实施方案6或7的分离的核酸分子,其包含与SEQ ID NO:148或其互补序列具有至少85%序列相同性的核苷酸序列。
实施方案12是实施方案1的分离的核酸分子,其编码2019-NCOV刺突(S)蛋白,所述2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
C.信号序列的缺失。
实施方案13是实施方案1或12的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:23的氨基酸序列具有至少85%序列相同性的多肽。
实施方案14是实施方案1、12或13的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:23的氨基酸序列具有至少99%序列相同性的多肽。
实施方案15是实施方案1或12的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:26的氨基酸序列具有至少85%序列相同性的多肽。
实施方案16是实施方案1、12或13的分离的核酸分子,其包含与SEQ IDNO:115或其互补序列具有至少85%序列相同性的核苷酸序列。
实施方案17是实施方案15的分离的核酸分子,其包含与SEQ ID NO:118或其互补序列具有至少85%序列相同性的核苷酸序列。
实施方案18是前述实施方案中任一项的分离的核酸分子,其中编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(CMV)启动子,优选CMV立即早期启动子可操作地连接。
实施方案19是前述实施方案中任一项的分离的核酸分子,其中编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(CMV)启动子可操作地连接,所述CMV启动子包含至少一个四环素操纵子(TetO)基序。
实施方案20是实施方案19的分离的核酸分子,其中包含至少一个TetO基序的CMV启动子包含SEQ ID NO:219的核苷酸序列。
实施方案21是实施方案19或20的分离的核酸分子,其中所述CMV启动子由SEQ IDNO:219的核苷酸序列组成。
实施方案22是一种分离的2019-NCOV刺突(S)蛋白,其包含对SEQ ID NO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
实施方案23是实施方案22的分离的2019-NCOV刺突(S)蛋白,其包含SEQ ID NO:205的氨基酸序列。
实施方案24是实施方案22的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ ID NO:51的氨基酸序列具有至少85%序列相同性的氨基酸序列。
实施方案25是实施方案22或23的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ IDNO:51的氨基酸序列具有至少99%序列相同性的氨基酸序列。
实施方案26是实施方案22的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ ID NO:54的氨基酸序列具有至少85%序列相同性的氨基酸序列。
实施方案27是实施方案22的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ ID NO:56的氨基酸序列具有至少85%序列相同性的氨基酸序列。
实施方案28是实施方案22的分离的2019-NCOV刺突(S)蛋白,其包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
c.信号序列的缺失。
实施方案29是实施方案22或28的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ IDNO:23的氨基酸序列具有至少85%序列相同性的氨基酸序列。
实施方案30是实施方案22、28或29的分离的2019-NCOV刺突(S)蛋白,其包含与SEQID NO:23的氨基酸序列具有至少99%序列相同性的氨基酸序列。
实施方案31是实施方案22或28的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ IDNO:26的氨基酸序列具有至少85%序列相同性的氨基酸序列。
实施方案32是一种分离的载体,其包含实施方案1-21中任一项的核酸分子中的一种或多种。
实施方案33是实施方案32的载体,其中所述载体是复制缺陷的。
实施方案34是实施方案32的载体,其中所述载体是哺乳动物、细菌或病毒载体。
实施方案35是实施方案32的载体,其中所述载体是表达载体。
实施方案36是实施方案32的载体,其中所述病毒载体是选自以下的病毒:逆转录病毒、腺病毒、腺相关病毒、细小病毒、冠状病毒、负链RNA病毒、正粘病毒、弹状病毒、副粘病毒、正链RNA病毒、小核糖核酸病毒、甲病毒、双链DNA病毒、疱疹病毒、爱泼斯坦-巴尔(Epstein-Barr)病毒、巨细胞病毒、鸡痘和金丝雀痘。
实施方案37是实施方案36的载体,其中所述载体是腺病毒。
实施方案38是实施方案37的载体,其中所述腺病毒选自Ad26、Ad52、Ad59、Ad2、Ad5、Ad11、Ad12、Ad24、Ad34、Ad35、Ad40、Ad48、Ad49、Ad50和Pan9,特别是Ad26。
实施方案39是一种组合物,其包含实施方案1-21中任一项所述的核酸分子、实施方案22-31中任一项所述的多肽或实施方案32-38中任一项所述的载体。
实施方案40是实施方案39的组合物,其进一步包含药学上可接受的载剂、赋形剂或稀释剂。
实施方案41是实施方案39或40的组合物,其还包含佐剂或免疫刺激剂。
实施方案42是一种免疫原性组合物,其包含实施方案1-21中任一项的核酸分子、实施方案22-31中任一项的多肽或实施方案32-38中任一项的载体。
实施方案43是实施方案42的免疫原性组合物,其中所述免疫原性组合物是疫苗。
实施方案44是一种组合物,其用于治疗有需要的受试者的冠状病毒感染例如2019-nCoV感染或降低其风险,所述组合物包含治疗有效量的实施方案39-41中任一项的组合物或者实施方案42或43的免疫原性组合物。
实施方案45是一种组合物,其用于降低感染有2019-nCoV的受试者中的冠状病毒介导的活性(例如2019-nCoV介导的活性),所述组合物包含治疗有效量的实施方案39-41中任一项的组合物或者实施方案42或43的免疫原性组合物。
实施方案46一种组合物,其用于预防有需要的受试者的分子确诊的中度至严重/危重COVID-19,其包括向所述受试者给药实施方案39-41中任一项的组合物或者实施方案的免疫原性组合物42或43,其中所述组合物用于在一剂方案中以5x1010vp/剂的剂量给药。
实施方案47是一种制造免疫原性组合物的方法,所述免疫原性组合物用于治疗有需要的受试者的冠状病毒(例如,2019-NCOV)感染或降低其风险,所述方法包括以下步骤:
(a)将实施方案1-21中任一项的核酸分子、实施方案22-31中任一项的多肽、实施方案32-38中任一项的载体、实施方案39-41中任一项的组合物中的至少一种与药学上可接受的载剂、赋形剂或稀释剂混合以形成所述免疫原性组合物;和
(b)将所述免疫原性组合物置于容器中。
实施方案48是一种试剂盒,其包含:
(a)第一容器,其包含实施方案1-12中任一项的核酸分子、实施方案22-31中任一项的多肽、实施方案32-38中任一项的载体、实施方案39-41中任一项的组合物,和实施方案42或43的免疫原性组合物中的至少一种;
(b)其使用说明;以及
任选存在的(c)第二容器,其包含药学上可接受的载剂、赋形剂或稀释剂。
实施方案49是实施方案48的试剂盒,其中所述第一容器还包含药学上可接受的载剂、赋形剂或稀释剂。
实施方案50是实施方案48或49的试剂盒,其中试剂盒任选地包含佐剂和/或免疫刺激剂。
实施方案51是一种编码冠状病毒S蛋白或其片段的分离的和/或重组的核酸,其包含SEQ ID NO:211的核苷酸序列。
实施方案52是一种分离的和/或重组的冠状病毒S蛋白,其包含SEQ ID NO:205的氨基酸序列,或其片段。
实施方案53是一种核酸,其编码实施方案52的冠状病毒S蛋白。
实施方案54是实施方案51或53的核酸,其为在人细胞中表达而进行密码子优化。
实施方案55是一种载体,其包含实施方案51、53或54的核酸。
实施方案56是一种载体,其包含编码实施方案52的蛋白的核酸。
实施方案57是实施方案55或56的载体,其中所述载体是重组人腺病毒载体。
实施方案58是实施方案57的载体,其中编码冠状病毒S蛋白的核酸与巨细胞病毒(CMV)启动子可操作地连接,所述CMV启动子包含至少一个四环素操纵子(TetO)基序。
实施方案59是实施方案58的载体,其中包含至少一个TetO基序的CMV启动子包含SEQ ID NO:219的核苷酸序列。
实施方案60为实施方案57、58或59的载体,其中所述重组人腺病毒具有腺病毒基因组的E1区缺失、E3区缺失或E1和E3区均缺失。
实施方案61是实施方案58-60中任一项的载体,其中所述载体是血清型26的重组人腺病毒。
实施方案62是一种组合物,其包含实施方案51、53或54的核酸、实施方案52的蛋白和/或实施方案55-61中任一项的载体。
实施方案63是针对COVID-19的疫苗,其包含实施方案51、53或54中任一项的核酸、实施方案52的蛋白和/或实施方案55-61中任一项的载体。
实施方案64是实施方案63的疫苗,其包含血清型26的重组人腺病毒,所述重组人腺病毒包含编码SARS-CoV-2 S蛋白的核酸,所述SARS-CoV-2 S蛋白包含SEQ IDNO:205的氨基酸序列,或其片段。
实施方案65是一种用于对受试者进行针对COVID-19的疫苗接种的方法,其包括向所述受试者给药实施方案63或64的疫苗。
实施方案66是一种用于减少受试者中SARS-CoV-2的感染和/或复制的方法,其包括向所述受试者给药实施方案59的组合物或者实施方案63或64的疫苗。
实施方案67是一种用于预防受试者的分子确诊的中度至严重/危重COVID-19的方法,其包括向所述受试者给药实施方案60或61的疫苗,所述疫苗以一剂或两剂疫苗方案给予。
实施方案68是一种与安慰剂相比,预防SARS-CoV-2血清反应阴性的成年受试者的分子确诊的中度至严重/危重COVID-19的方法,其包括向所述受试者给药实施方案63或64的疫苗,所述疫苗以一剂或两剂疫苗方案给予。
实施方案69是一种用于在受试者中降低通过定量逆转录酶聚合酶链反应(RT-PCR)评估的参与者的SARS-CoV-2病毒载量的方法,所述参与者具有分子确诊的中度至严重/危重COVID-19,所述方法包括向所述受试者给药实施方案63或64的疫苗,所述疫苗以一剂或两剂疫苗方案给予。
实施方案70为实施方案65-69中任一项的方法,其中所述受试者疑似患有SARS-CoV-2感染或诊断其患有SARS-CoV-2感染。
实施方案71是实施方案65-70中任一项的方法,其中肌内给药所述疫苗。
实施方案72是实施方案65-71中任一项的方法,其中所述疫苗以两剂疫苗方案给药,所述疫苗方案包括间隔约8周给予5×1010vp/剂或1×1011vp/剂的剂量。
实施方案73是实施方案65-72中任一项的方法,其由向所述受试者单次给药5×1010vp/剂或1×1011vp/剂的剂量的疫苗组成。
实施方案74是一种分离的宿主细胞,其包含血清型26的重组人腺病毒,所述重组人腺病毒包含编码SARS-CoV-2 S蛋白或其片段的核酸。
实施方案75是一种制备针对COVID-19的疫苗的方法,其包括提供血清型26的重组人腺病毒,其包含编码SARS-CoV-2 S蛋白或其片段的核酸,使所述重组腺病毒在宿主细胞培养物中增殖,分离和纯化所述重组腺病毒,以及将所述重组腺病毒配制于药学上可接受的组合物中。
实施方案76是一种分离的重组核酸,其形成血清型26的重组人腺病毒的基因组,所述重组人腺病毒包含编码SARS-CoV-2 S蛋白或其片段的核酸。
实施方案77是一种分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ ID NO:1-84中任一项的氨基酸序列具有至少85%序列相同性的多肽。
实施方案78是实施方案77的核酸分子,其中
a)所述多肽能够在受试者中引发免疫应答;或者
b)所述多肽与SEQ ID NO:1-84中任一项具有至少86、87、88、89、90、91、92、93、94、95、96、97、98或99%的序列相同性,或者所述多肽具有SEQ ID NO:1-84中任一项的多肽序列。
实施方案79是实施方案77或78的核酸分子,其中所述多肽具有SEQ ID NO:56的氨基酸序列。
实施方案80是一种分离的核酸分子,其包含与SEQ ID NO:93-181、190-195和199-204或其互补序列中任一项的全部或部分具有至少85%序列相同性的核苷酸序列。
实施方案81是实施方案80的分离的核酸分子,其中所述核酸分子与SEQ ID NO:93-181、190-195和199-204中任一项具有至少86、87、88、89、90、91、92、93、94、95、96、97、98或99%的序列相同性,或者具有SEQ ID NO:93-181、190-195和199-204中任一项的核苷酸序列。
实施方案82是实施方案80或81的分离的核酸分子,其中所述核酸分子或其部分能够在受试者中引发免疫应答。
实施方案83是实施方案80-82中任一项的分离的核酸分子,其中所述核酸分子具有SEQ ID NO:195的核苷酸序列。
实施方案84是一种分离的多肽,其包含与SEQ ID NO:1-84中任一项的全部或部分具有至少85%序列相同性的氨基酸序列。
实施方案85是实施方案84的分离的多肽,其中所述多肽与SEQ ID NO:1-84中任一项具有至少86、87、88、89、90、91、92、93、94、95、96、97、98或99%的序列相同性,或者具有SEQ ID NO:1-84中任一项的氨基酸序列。
实施方案86是实施方案84或85的分离的多肽,其中所述多肽或其部分能够在受试者中引发免疫应答。
实施方案87是实施方案84-86中任一项的分离的多肽,其中所述多肽具有SEQ IDNO:28的氨基酸序列。
实施方案88是一种分离的载体,其包含实施方案77-83中任一项的核酸分子中的一种或多种。
实施方案89是实施方案88的载体,其中所述载体是复制缺陷的。
实施方案90是实施方案88或89的载体,其中所述载体是哺乳动物、细菌或病毒载体。
实施方案91是实施方案90的载体,其中所述载体是表达载体。
实施方案92是实施方案90的载体,其中所述病毒载体是选自以下的病毒:逆转录病毒、腺病毒、腺相关病毒、细小病毒、冠状病毒、负链RNA病毒、正粘病毒、弹状病毒、副粘病毒、正链RNA病毒、小核糖核酸病毒、甲病毒、双链DNA病毒、疱疹病毒、爱泼斯坦-巴尔病毒、巨细胞病毒、鸡痘和金丝雀痘。
实施方案93是实施方案92的载体,其中所述载体是腺病毒。
实施方案94是实施方案93的载体,其中所述腺病毒选自Ad26、Ad52、Ad59、Ad2、Ad5、Ad11、Ad12、Ad24、Ad34、Ad35、Ad40、Ad48、Ad49、Ad50和Pan9。
实施方案95是实施方案94的载体,其中所述Ad52是恒河猴Ad52或者所述Ad26是恒河猴Ad26。
实施方案96是一种分离的抗体,其与实施方案84-87中任一项的多肽特异性结合。
实施方案97是实施方案96的抗体,其中所述抗体是通过用实施方案77-82中任一项的核酸、实施方案84-87中任一项的多肽或实施方案88-95中任一项的载体免疫哺乳动物产生的。
实施方案98是实施方案97的抗体,其中所述哺乳动物是人、牛、羊、小鼠或兔。
实施方案99是实施方案96或97的抗体,其中所述抗体是人源化的。
实施方案100是实施方案96-99中任一项的抗体,其中所述抗体是IgG。
实施方案101是实施方案96-100中任一项的抗体,其中所述抗体是双Fab、Fv、Fab、Fab′-SH、F(ab′)2、双抗体、线性抗体或scFV。
实施方案102是一种产生抗2019-nCoV抗体的方法,其包括向受试者给药一定量的实施方案77-83中任一项的核酸分子、实施方案84-87中任一项的多肽,和/或实施方案88-95中任一项的载体,所述量在向所述受试者给药后足以引发中和的抗2019-nCoV抗血清的产生。
实施方案103是一种分离的抗2019-nCoV抗体,其通过实施方案102的方法产生。
实施方案104是实施方案103的抗体,其中所述抗体结合SEQ ID NO:1-84中任一项内的表位。
实施方案105是一种组合物,其包含实施方案77-83中任一项的核酸分子、实施方案84-87中任一项的多肽、实施方案88-95中任一项的载体或者实施方案96-101或103-104中任一项的抗体。
实施方案106是实施方案105的组合物,其还包含药学上可接受的载剂、赋形剂或稀释剂。
实施方案107是实施方案105或106的组合物,其还包含佐剂或免疫刺激剂。
实施方案108是一种免疫原性组合物,其包含实施方案77-82中任一项的核酸分子、实施方案84-87中任一项的多肽、实施方案88-95中任一项的载体或者实施方案96-101或103-104中任一项的抗体。
实施方案109是实施方案108的免疫原性组合物,其中所述免疫原性组合物是疫苗。
实施方案110是实施方案108的免疫原性组合物,其中所述免疫原性组合物能够治疗有需要的受试者的冠状病毒感染(例如,2019-nCoV感染)或降低其风险。
实施方案111是实施方案108-110中任一项的免疫原性组合物,其中在向所述受试者给药所述免疫原性组合物后引发中和的抗2019-nCoV抗血清的产生。
实施方案112是实施方案108-111中任一项的免疫原性组合物,其中所述受试者是哺乳动物。
实施方案113是实施方案112的免疫原性组合物,其中所述哺乳动物是人。
实施方案114是实施方案113的免疫原性组合物,其中所述人具有潜在的健康状况。
实施方案115是实施方案114的免疫原性组合物,其中所述潜在的健康状况是高血压、糖尿病或心血管疾病。
实施方案116是一种鉴定、诊断和/或预测受试者对冠状病毒感染的易感性的方法,其包括测定所述受试者在来自受试者的样品中是否具有保护水平的抗冠状病毒抗体(例如抗刺突抗体),其中优选保护水平是:
(i)等于或高于使用假病毒中和测定确定的至少约70的滴度的水平;或者
(ii)等于或高于使用活病毒中和测定确定的至少约25的滴度的水平;或者
(iii)通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列中抗冠状病毒抗体中值水平的至少80%的水平。
实施方案117是实施方案116的方法,其中所述方法还包括向具有低于保护水平的抗冠状病毒抗体的受试者给药有效量的实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物。
实施方案118是实施方案116或117的方法,其中所述方法还包括鉴定抗冠状病毒抗体(例如,抗刺突抗体)的类。
实施方案119是实施方案118的方法,其中所述类是IgG。
实施方案120是实施方案116-119中任一项的方法,其中所述样品是来自受试者的体液,其中优选地所述体液是血液。
实施方案121是实施方案116-120中任一项的方法,其中所述冠状病毒是2019-nCoV。
实施方案122是一种治疗有需要的受试者的冠状病毒感染或降低其风险的方法,其包括向所述受试者给药治疗有效量的实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物。
实施方案123是实施方案122的方法,其还包括测量所述受试者中的抗冠状病毒抗体(例如,抗刺突抗体)水平。
实施方案124是实施方案123的方法,其中在给药之前和/或之后测量所述受试者中的抗冠状病毒抗体水平。
实施方案125是实施方案124的方法,其中在给药后约1、2、3、4、5或6天,1、2、3、4、5、6或7周,2、3、4、5或6个月,1、2、3、4或5年内测量所述受试者中的抗冠状病毒抗体水平一次或多次。
实施方案126是实施方案122-125中任一项的方法,其中所述受试者的抗冠状病毒抗体水平低于保护水平,并且其中所述方法还包括向所述受试者再给药实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物,或者向所述受试者给药不同的抗冠状病毒组合物。
实施方案127是实施方案126的方法,其中所述保护水平是足以减少冠状病毒介导的疾病的症状或持续时间的水平。
实施方案128是实施方案126或127的方法,其中所述保护水平是:
(i)等于或高于使用假病毒中和测定确定的至少约70的滴度的水平;或者
(ii)等于或高于使用活病毒中和测定确定的至少约25的滴度的水平;或者
(iii)通过假病毒中和测定或活病毒中和测定确定的恢复期人的队列中抗冠状病毒抗体中值水平的至少80%。
实施方案129是实施方案122-128中任一项的方法,其中所述冠状病毒是2019-nCoV。
实施方案130是一种降低感染有2019-nCoV的受试者中冠状病毒介导的活性(例如,2019-nCoV介导的活性)的方法,其包括向所述受试者给药治疗有效量的实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物。
实施方案131是实施方案130的方法,其中所述治疗有效量是通过ELISA测定测量的足以在所述受试者中产生大于2的log血清抗刺突抗体滴度。
实施方案132是实施方案131的方法,其中所述治疗有效量是15μg-300μg的实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物。
实施方案133是实施方案130-132中任一项的方法,其中所述活性是病毒滴度、病毒传播、感染或细胞融合。
实施方案134是实施方案133的方法,其中所述病毒滴度在给药实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物后降低。
实施方案135是实施方案134的方法,其中所述病毒滴度降低25%或更多。
实施方案136是实施方案135的方法,其中所述病毒滴度降低50%或更多。
实施方案137是实施方案136的方法,其中所述病毒滴度降低75%或更多。
实施方案138是实施方案137的方法,其中所述冠状病毒在所述给药后是不可检测的。
实施方案139是实施方案130-138中任一项的方法,其中所述给药发生在暴露于冠状病毒之前。
实施方案140是实施方案139的方法,其中所述给药发生在暴露于冠状病毒之前至少1小时。
实施方案141是实施方案140的方法,其中所述给药发生在暴露于冠状病毒之前至少1周、1个月或1年。
实施方案142是实施方案130-138中任一项的方法,其中所述给药发生在暴露于冠状病毒后。
实施方案143是实施方案142的方法,其中所述给药发生在暴露于冠状病毒后至少15分钟。
实施方案144是实施方案143的方法,其中所述给药发生在暴露于冠状病毒后至少1小时、1天、1周。
实施方案145是实施方案130-144中任一项的方法,其中向所述受试者给药至少一剂的所述核酸分子、多肽、载体、组合物、免疫原性组合物和抗体。
实施方案146是实施方案145的方法,其中向所述受试者给药至少两个剂量。
实施方案147是实施方案146的方法,其中将所述核酸分子、多肽、载体、组合物或免疫原性组合物作为初免、加强或初免-加强向所述受试者给药。
实施方案148是实施方案130-147中任一项的方法,其中所述核酸分子、多肽、载体、组合物、免疫原性组合物或抗体通过肌内、静脉内、皮内、经皮、动脉内、腹膜内、病灶内、颅内、关节内、前列腺内、胸膜内、气管内、鼻内、玻璃体内、阴道内、直肠内、局部、肿瘤内、腹膜、皮下、结膜下、血管内、黏膜内、心包内、脐内、眼内、口服、外用、局部、通过吸入、通过注射、通过输注、通过连续输注、通过直接对靶细胞进行局部灌注、通过导管、通过灌洗、通过灌胃、在乳膏中或在脂质组合物中给药。
实施方案149是实施方案130-148中任一项的方法,其中所述受试者是哺乳动物。
实施方案150是实施方案149的方法,其中所述哺乳动物是人。
实施方案151是实施方案150的方法,其中所述人具有潜在的健康状况。
实施方案152是实施方案151的方法,其中所述潜在健康状况是高血压、糖尿病或心血管疾病。
实施方案153是实施方案130-152中任一项的方法,其中所述方法促进所述受试者的免疫应答。
实施方案154是实施方案153的方法,其中所述免疫应答是体液免疫应答。
实施方案155是实施方案154的方法,其中所述体液免疫应答是IgG应答。
实施方案156是一种组合物,其用于治疗有需要的受试者的冠状病毒感染例如2019-nCoV感染或降低其风险,所述组合物包含治疗有效量的实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物。
实施方案157是一种组合物,其用于降低感染有2019-nCoV的受试者中的冠状病毒介导的活性(例如,2019-nCoV介导的活性),所述组合物包含治疗有效量的实施方案105-107中任一项的组合物或实施方案108-115中任一项的免疫原性组合物。
实施方案158是一种制备免疫原性组合物的方法,所述免疫原性组合物用于治疗有需要的受试者的冠状病毒(例如,2019-nCoV)感染或降低其风险,所述方法包括以下步骤:
(a)将实施方案77-83中任一项的核酸分子、实施方案84-87中任一项的多肽、实施方案88-95中任一项的载体、实施方案105-107中任一项的组合物,以及实施方案96-101或102-104中任一项的抗体中的至少一种与药学上可接受的载体、赋形剂或稀释剂混合以形成所述免疫原性组合物;和
(b)将所述免疫原性组合物置于容器中。
实施方案159是一种试剂盒,其包含:
(a)第一容器,其包含实施方案77-83中任一项的核酸分子、实施方案84-87中任一项的多肽、实施方案88-95中任一项的载体、实施方案105-107中任一项的组合物、实施方案108-115任一项的免疫原性组合物、以及实施方案96-101或103-104中任一项的抗体中的至少一种;
(b)其使用说明;以及
任选存在的(c)第二容器,其包含药学上可接受的载剂、赋形剂或稀释剂。
实施方案160是实施方案159的试剂盒,其中所述第一容器还包含药学上可接受的载剂、赋形剂或稀释剂。
实施方案161是实施方案159或160的试剂盒,其中所述试剂盒任选地包含佐剂和/或免疫刺激剂。
实施方案162是实施方案80-82中任一项的分离的核酸分子,其中所述核酸分子具有SEQ ID NO:143的核苷酸序列。
实施方案163是实施方案80-82中任一项的分离的核酸分子,其中所述核酸分子具有SEQ ID NO∶204的核苷酸序列。
实施方案164是实施方案80-82中任一项的分离的核酸分子,其中所述核酸分子具有SEQ ID NO:204的核苷酸19-3837的核苷酸序列。
实施方案165是实施方案84-86中任一项的分离的多肽,其中所述多肽具有SEQ IDNO:51的氨基酸序列。
实施方案166是实施方案88的载体,其中载体是Ad26。
实施方案167是实施方案93的抗体,其中所述抗体通过用包含SEQ ID NO:143、SEQID NO:204的核苷酸19-3837或SEQ ID NO:204的核酸、包含SEQ ID NO:51的氨基酸序列的多肽、或者包含核酸的Ad26载体(所述核酸包含SEQ ID NO:143、SEQ ID NO:204的核苷酸19-3837或SEQ ID NO:204)免疫哺乳动物产生。
实施方案168是实施方案102的方法,其包括给药一定量的包含SEQ ID NO:143、SEQ ID NO:204的核苷酸19-3837或SEQ ID NO:204的核酸分子、包含SEQ ID NO:51的氨基酸序列的多肽,或者包含核酸的Ad26载体,所述核酸包含SEQ ID NO:143、SEQ ID NO:204的核苷酸19-3837或SEQ ID NO:204,所述量足以在向所述受试者给药后引发中和的抗2019-nCoV抗血清的产生。
实施方案169是一种组合物,其包含实施方案162-164中任一项的核酸分子、实施方案165的多肽、实施方案166的载体或者实施方案167或168的抗体。
实施方案170是一种免疫原性组合物,其包含实施方案162-164中任一项的核酸分子、实施方案165的多肽、实施方案166的载体或者实施方案167或168的抗体。
实施方案171是一种治疗有需要的受试者的冠状病毒感染或降低其风险的方法,其包括向所述受试者给药治疗有效量的实施方案169的组合物或实施方案170的免疫原性组合物。
实施方案172是一种降低感染有2019-nCoV的受试者中的冠状病毒介导的活性(例如2019-nCoV介导的活性)的方法,其包括向所述受试者给药治疗有效量的实施方案169的组合物或实施方案170的免疫原性组合物。
实施方案173是一种组合物,其用于治疗有需要的受试者的冠状病毒感染例如2019-nCoV感染或降低其风险,其包含治疗有效量的实施方案169的组合物或实施方案170的免疫原性组合物。
实施方案174是一种组合物,其用于降低感染有2019-nCoV的受试者中的冠状病毒介导的活性(例如,2019-nCoV介导的活性),所述组合物包含治疗有效量的实施方案169的组合物或实施方案170的免疫原性组合物。
实施方案175是实施方案1-21中任一项的核酸分子、实施方案22-31中任一项的多肽、实施方案32-38中任一项的载体、实施方案39-41中任一项的组合物和实施方案42或43的免疫原性组合物中的至少一种在制备用于治疗有需要的受试者的冠状病毒感染例如2019-nCoV感染或降低其风险的药物中的用途。
实施方案176是实施方案1-21中任一项的核酸分子、实施方案22-31中任一项的多肽、实施方案32-38中任一项的载体、实施方案39-41中任一项的组合物和实施方案42或43的免疫原性组合物中的至少一种在制备用于降低感染有2019-nCoV的受试者的冠状病毒介导的活性(例如2019-nCoV介导的活性)的药物中的用途。
实施方案177是实施方案1-21中任一项的核酸分子、实施方案22-31中任一项的多肽、实施方案32-38中任一项的载体、实施方案39-41中任一项的组合物和实施方案42或43的免疫原性组合物中的至少一种在制备用于预防有需要的受试者的分子确诊的中度至严重/危重COVID-19的药物中的用途,其中所述组合物在单剂方案中,以5x1010vp/剂的剂量给药。
实施方案178是实施方案63或64的疫苗,其用于为受试者接种针对COVID-19的疫苗。
实施方案179是实施方案63或64的疫苗或实施方案62的组合物,用于减少受试者中SARS-CoV-2的感染和/或复制。
实施方案180是实施方案64或64的疫苗,其用于在一剂或两剂疫苗方案中预防受试者的分子确诊的中度至严重/危重COVID-19。
实施方案181是实施方案63或64的疫苗,其用于在SARS-CoV-2血清阴性的成人受试者中,与安慰剂相比,以一剂或两剂疫苗方案,预防分子确诊的中度至严重/危重COVID-19。
实施方案182是实施方案63或64的疫苗,其用于以一剂或两剂疫苗方案减少在受试者中通过定量逆转录酶聚合酶链反应(RT-PCR)评估的参与者中的SARS-CoV-2病毒载量,所述参与者具有分子确诊的中度至严重/危重COVID-19。
实施方案183是实施方案178-182中任一项的使用的疫苗,其中所述受试者疑似患有SARS-CoV2感染或诊断其患有SARS-CoV2感染。
实施方案184是实施方案178-183中任一项使用的疫苗,其中所述疫苗配制为用于肌内给药。
实施方案185为实施方案181-184中任一项使用的疫苗,其中所述疫苗用于以两剂疫苗方案给药,所述疫苗方案包括间隔约8周给予5x1010vp或1x1011vp/剂的剂量。
实施方案186是实施方案178-185中任一项使用的疫苗,其包括向所述受试者单次给药5×1010vp或1×1011vp/剂的剂量的疫苗。
实施方案187是实施方案51、53或54中任一项的核酸、实施方案52的蛋白和/或实施方案55-61中任一项的载体在制备用于向受试者接种针对COVID-19的疫苗的药物中的用途。
实施方案188是实施方案51、53或54中任一项的核酸、实施方案52的蛋白和/或实施方案55-61中任一项的载体在制备用于减少受试者中SARS-CoV-2的感染和/或复制的药物中的用途。
实施方案189是实施方案51、53或54中任一项的核酸、实施方案52的蛋白和/或实施方案55-61中任一项的载体在制备用于以一剂或两剂疫苗方案预防受试者的分子确诊的中度至严重/危重COVID-19的药物中的用途。
实施方案190是实施方案51、53或54中任一项的核酸、实施方案52的蛋白和/或实施方案55-61中任一项的载体在制备用于在SARS-CoV-2血清反应阴性的成人受试者中,与安慰剂相比,以一剂或两剂疫苗方案预防分子确诊的中度至严重/危重COVID-19的药物中的用途。
实施方案191是实施方案51、53或54中任一项的核酸、实施方案52的蛋白和/或实施方案55-61中任一项的载体在制备用于以一剂或两剂疫苗方案减少受试者中通过定量逆转录酶聚合酶链反应(RT-PCR)评估的参与者中的SARS-CoV-2病毒载量的药物中的用途,所述参与者具有分子确诊、中度至严重/危重COVID-19。
实施方案192是实施方案187-191中任一项的用途,其中所述受试者疑似患有SARS-CoV2感染或诊断其患有SARS-CoV2感染。
实施方案193是实施方案187-192中任一项的用途,其中所述疫苗配制为用于肌内给药。
实施方案194是实施方案187-193中任一项的用途,其中所述疫苗用于以两剂疫苗方案给药,所述疫苗方案包括间隔约8周给予5x1010vp或1x1011vp/剂的剂量。
实施方案195是实施方案187-194中任一项的用途,用于以5x1010vp或1x1011vp/剂的剂量单次向受试者给药。
实施方案196是实施方案77-83中任一项的核酸分子、实施方案84-87中任一项的多肽、实施方案88-95中任一项的载体或者实施方案96-101或103-104中任一项的抗体在制备用于治疗有需要的受试者的冠状病毒感染(例如2019-nCoV感染)或降低其风险的药物中的用途。
实施方案197是实施方案77-83中任一项的核酸分子、实施方案84-87中任一项的多肽、实施方案88-95中任一项的载体或者实施方案99-101或103-104中任一项的抗体在制备用于降低感染有2019-nCoV的受试者中的冠状病毒介导的活性(例如,2019-nCoV介导的活性)的药物中的用途。
实施方案198是实施方案162-164中任一项的核酸分子、实施方案165的多肽、实施方案166的载体或者实施方案167或168的抗体在制备用于治疗有需要的受试者的冠状病毒感染,例如2019-nCoV感染或降低其风险的药物中的用途。
实施方案199是实施方案162-164中任一项的核酸分子、实施方案165的多肽、实施方案166的载体或者实施方案167或168的抗体在制备用于减少感染有2019-nCoV的受试者中的冠状病毒介导的活性(例如,2019-nCoV介导的活动)的药物中的用途。
具体实施方案在所述实施方案之内。
本发明在以下实施例中进一步解释。实施例不以任何方式限制本发明。它们仅用于阐明本发明。
实施例
以下实施例用于说明本发明。它们不意味着以任何方式限制本发明。
实施例1. 2019-nCoV DNA疫苗的开发
2019-nCoV免疫原和2019-nCoV DNA疫苗的设计
用2019-nCoV刺突蛋白(SEQ ID NO:29)设计用于合成产生的核酸分子SEQ ID NO:93-181、190一195和199-204)。对核酸分子进行了一些优化以增强转基因表达。可以通过将SEQ ID NO:93-181、190-195和199-204的核酸分子引入哺乳动物表达载体(例如pcDNA3.1+;Invitrogen,CA,USA)中来产生DNA疫苗。缺失的突变体可以缺少信号序列、细胞质区域、跨膜区、S2或其组合。
实施例2.向人受试者给药核酸疫苗
根据这些方法,可以在暴露于2019-nCoV之前或之后向人受试者给药组合物。人受试者可以是被鉴定为具有高感染风险的人,例如已经或将要前往2019-nCoV感染流行地区(例如湖北省)的个人,或被鉴定为出现与2019-nCoV感染症状一致的人。
例如,可以向具有潜在健康状况(例如,高血压、糖尿病或心血管疾病),并被鉴定为具有2019-nCoV感染风险的人给药含有编码2019-nCoV核酸的核酸分子的核酸疫苗(例如,SEQ ID NO:93-181、190-195和199-204中的任一项),例如在腺病毒载体中,剂量为10μg-10mg。优选地,核酸疫苗(例如,DNA疫苗或RNA疫苗)可以包含SEQ ID NO:136、193、142、148、195、121、122、123、129和/或130。然后可以监测患者的2019-nCoV感染症状的表现、症状的消退和/或2019-nCoV抗体的产生。如果需要,可以给药第二剂或额外剂量的核酸疫苗。
实施例3.向人受试者给药免疫原性2019-nCoV多肽
根据这些方法,可以在暴露于2019-nCoV之前或之后向人受试者给药组合物。人受试者可以是被鉴定为具有高感染风险的人,例如已经或将要前往2019-nCoV感染流行地区(例如湖北省)的个人,或被鉴定为出现与2019-nCoV感染症状一致的人。
例如,可以向具有潜在健康状况(例如,高血压、糖尿病或心血管疾病),并被鉴定为具有2019-nCoV感染风险的人给药2019-nCoV免疫原(例如,SEQ ID NO:1-84中的任一项),例如,在腺病毒载体中,剂量为10μg-10mg。优选地,免疫原是由SEQ ID NO:44、50、56、29、30、31、37或38编码的一种或多种多肽。然后可以监测患者2019-nCoV感染症状的表现、症状的消退和/或2019-nCoV抗体的产生。如果需要,可以给药第二剂或额外剂量的免疫原。
实施例4.向有2019-nCoV感染风险的人受试者给药抗2019-nCoV抗体
可以向被鉴定为具有2019-nCoV感染风险的人受试者(例如,由于前往2019-nCoV感染流行地区(例如,湖北省),或者是具有潜在健康状况(例如,高血压、糖尿病或心血管疾病))的人受试者给药与刺突(SEQ ID NO:29)多肽内的表位结合的抗2019-nCoV抗体(例如,该抗体可以是针对SEQ ID NO:1-84中任一项的多肽产生的),剂量在1-1,000mg之间,其作为预防性治疗。可以在暴露于2019-nCoV之前或之后向受试者给药抗2019-nCoV抗体作为预防性治疗。然后可以监测患者2019-nCoV感染症状的表现或症状的消退。如果需要,可以给药第二剂或额外剂量的抗2019-nCoV抗体。
实施例5.向出现2019-nCoV感染症状的人受试者给药抗2019-nCoV抗体
可以向被鉴定为出现2019-nCoV症状的人受试者给药抗2019-nCoV抗体,所述抗体与刺突(SEQ ID NO:29)多肽内的表位结合(例如,该抗体可以是针对SEQ ID NO:1-84中任一项的多肽产生的),剂量在1-1,000mg之间。受试者(例如,人,特别是具有潜在健康状况(例如,高血压、糖尿病或心血管疾病)的人)最近可能去过2019-nCoV感染流行地区(例如,湖北省)。在医生诊断出2019-nCoV感染后,可以向受试者给药一剂抗2019-nCoV抗体。然后可以监测患者症状的消退。如果需要,可以给药第二剂或额外剂量的抗2019-nCoV抗体。
实施例6.用多克隆抗SARS抗血清检测2019-nCoV DNA疫苗编码的免疫原的反应性
为了用抗SARS抗体评估2019-nCoV DNA疫苗编码的免疫原的反应性,用含有编码2019-nCoV免疫原(分别为SEQ ID NO:29、30、31和56)的SS-Spike(SEQ ID NO:121)、SS-SdCT(SEQ ID NO:122)、SS-S.Ecto(SEQ ID NO:123)和SS-S.Ecto-dF-PP-foldon(SEQ IDNO:195)的质粒转染细胞。从转染的细胞中收集细胞裂解物和上清液,并在还原和变性条件下跑胶。随后使用从豚鼠分离的多克隆抗SARS抗血清(BEI Resources,NIAID,NIH;货号:NR-10361)通过免疫印迹分析样品。所有测试的DNA疫苗都能够产生与抗血清中存在的抗SARS抗体交叉反应的蛋白(图2)。刺突蛋白显示为一条大小约为200kDa的条带。
实施例7. 2019-nCoV DNA疫苗能够引发中和抗刺突抗体应答
检测含有编码2019-nCoV免疫原(分别为SEQ ID NO:29、30、31、37、38和56)的SS-Spike(SEQ ID NO:121)、SS-SdCT(SEQ ID NO:122)、SS-S.Ecto(SEQ ID NO:123)、SS-S1-foldon(SEQ ID NO:129)、SS-RBD-folden(SEQ ID NO:130)和SS-S.Ecto-dF-PP-foldon(SEQ ID NO:195)的DNA疫苗产生中和抗体应答的能力。向雌性Balb/c小鼠(8-12周龄)肌内(IM)注射50μg的DNA疫苗的一种,并在4周后肌内注射第二剂。疫苗接种后4周收集来自处理的小鼠的血清样品,并通过ELISA分析与全长刺突(SEQ ID NO:1)(图3)和S.Ecto-PP(SEQID NO:19)的结合(图4)。所有测试的DNA疫苗都能够在处理的小鼠中引发识别全长刺突(SEQ ID NO:1)和S.Ecto-PP (SEQ ID NO:19)的抗体应答。与检测的其他DNA疫苗相比,编码SS-S.Ecto-dF-PP-foldon的DNA疫苗产生了更好的抗体应答。
进一步分析来自疫苗接种后4周小鼠的血清的中和活性。使用体外基于荧光素酶的假病毒中和测定评估中和活性。中和测定使用了2019-nCoV刺突蛋白(SEQ ID NO:1)的假逆转录病毒核心。在用2019-nCoV受体人血管紧张素转换酶2(ACE2)转导的293细胞中测试了感染性,以支持假病毒的进入。在用DNA疫苗SS-S.Ecto、SS-RBD-foldon和SS-S.Ecto-dF-PP-foldon处理的小鼠中观察到了显著的中和能力。用SS-S.Ecto-dF-PP-foldon DNA疫苗处理的小鼠表现出最强的中和能力(图5)。
实施例8. 2019-nCoV DNA疫苗能够引发针对冠状病毒感染的保护性免疫应答
迅速扩大的COVID-19大流行使开发安全、有效和可部署的疫苗成为全球的关键优先事项(1-8)。然而,目前对2019-nCoV保护的免疫相关因素的理解非常有限。这些知识对于开发2019-nCoV疫苗以及其他免疫治疗干预措施至关重要。在这项研究中,构建了一组原型DNA疫苗,它们表达各种形式的2019-nCoV刺突(S)蛋白,并评估了它们在恒河猴中对2019-nCoV攻击的免疫原性和保护效力。
方法
动物和研究设计.将35只6-12岁的远交系的印度裔成年雄性和雌性恒河猴(Macaca mulatta)随机分组。所有动物都饲养在Bioqual,Inc.(Rockville,MD)。动物接受了表达SS-Spike(N=4)、SS-SdCT(N=4)、SS-S.Ecto(N=4)、Sl(N=4)、SS-RBD-FOLDON(N=4)、SS-S.Ecto-dF-PP-foldon(N=5)和假对照(N=10)的DNA疫苗。动物在第0周和第3周接受5mg DNA疫苗。在第6周,用1.2x108VP(1.1x104)PFU 2019-nCoV攻击所有动物。通过鼻内(IN)途径给药1ml病毒(每个鼻孔0.5ml)和通过气管内(IT)途径给药1ml。所有免疫学和病毒学测定均在盲法下进行。所有动物研究均按照所有相关的地方、州和联邦法规进行,并得到生物质量机构动物管理和使用委员会(IACUC)的批准。
人样品.来自马萨诸塞州波士顿的2019-nCoV恢复期个体的27份去识别的人血清样品是在鼻拭子为阴性,记录为恢复后至少7天后从个体获得的。所有人研究均按照所有相关的地方、州和联邦法规进行,并得到合作伙伴机构审查委员会(Partners InstitutionalReview Board)(IRB)的批准。
DNA疫苗.DNA疫苗是基于2019-nCoV刺突(S)蛋白序列(Wuhan/WIV04/2019)设计的。序列经过密码子优化和商业合成(Integrated DNA Technologies,NJ,USA)。产生了刺突的六种形式(version)(全长SS-Spike;细胞质结构域缺失SS-SdCT;可溶性胞外域SS-S.Ecto;带有折叠子三聚化标签的S1结构域SS-S1-foldon;带有折叠子三聚化标签的受体结合结构域SS-RBD-foldon;带有弗林蛋白酶切割位点缺失,PP稳定化突变和折叠子三聚化标签的可溶性胞外域SS-S.Ecto-dF-PP-foldon)。将合成基因克隆至哺乳动物表达质粒pcDNA3.1+(Invitrogen,CA,USA)中,并用无内毒素gigaprep试剂盒(Machery-Nagel,Düüren,Germany)进行扩增。所有DNA疫苗序列均由Sanger DNA测序确认。
免疫印迹.使用Lipofectamine 2000(Invitrogen)用2019-nCoV DNA表达质粒(10μg DNA/构建体)瞬时转染接种有70-80%汇合度的293T细胞的T-25培养瓶,将转染后48小时收获的上清液和细胞裂解物分别与还原样品缓冲液(Pierce)混合,在95℃加热5分钟,并且在预制的4一15%SDS-PAGE凝胶(Bio-Rad)上跑胶。使用iBlot干式印迹系统(Invitrogen)将蛋白转移到聚偏二氟乙烯(PVDF)膜上,并在4℃下在含有0.2%Tween20(Sigma)(V/V)和5%(W/V)脱脂奶粉的Dulbecco磷酸盐缓冲盐水T(D-PBST)中进行膜封闭过夜。过夜封闭后,将PVDF膜在含有1∶10,000稀释的(交叉反应性)多克隆豚鼠抗SARS冠状病毒抗血清(BEI resources)的3%牛奶DPBS-T中孵育1小时。孵育后,PVDF膜用5%牛奶DPBS-T洗涤5次,并且随后与3%牛奶DPBS-T中的1∶30,000的抗豚鼠辣根过氧化物酶(HRP)缀合的二抗(Jackson Immunoresearch)一起孵育。最后,将PVDF膜用5%牛奶DPBS-T再次洗涤5次,并使用Amersham ECL Plus免疫印迹检测系统(GE Healthcare)显影。
病毒RNA测定.用RT-PCR测定监测病毒载量,基本上如前所述(16)。简而言之,使用QIAcube HT(Qiagen,Germany)和Cador病原体HT试剂盒从支气管肺泡灌洗液(BAL)上清液和鼻拭子中提取RNA。使用superscript VILO(Invitrogen)对RNA进行逆转录,并根据制造商的说明使用QuantStudio 6和7Flex实时PCR系统(Applied Biosystems)一式两份运行。病毒载量计算为每毫升或每拭子的病毒RNA拷贝,测定灵敏度为50拷贝。扩增的靶标是2019-nCoV N(核衣壳)基因。靶标的引物和探针是:
2019-nCoV_N1-F:5′-GACCCCAAAATCAGCGAAAT-3′(SEQ ID NO:196)
2019-nCoV_N1-R:5′-TCTGGTTACTGCCAGTTGAATCTG-3′(SEQ ID NO:197)
2019-nCoV_N1-P:5′-FAM-ACCCCGCATTACGTTTGGTGGACC-BHQ1-3′(SEQ ID NO:198)
亚基因组mRNA测定.通过RT-PCR使用与先前描述的方法相似的方法评估2019-nCoV E基因亚基因组mRNA(sgmRNA)(17)。为了生成标准曲线,将2019-nCoV E基因sgmRNA克隆至pcDNA3.1表达质粒中;使用AmpliCap-Max T7 High Yield Message Maker Kit(Cellscript)转录此插入物,以获得标准的RNA。在RT-PCR之前,根据制造商的说明,使用Superscript III VILO(Invitrogen)将从受攻击的动物或标准中收集的样品进行逆转录。使用靶向E基因sgmRNA(17)的序列设计了Taqman定制基因表达测定(ThermoFisherScientific)。根据制造商的说明,在QuantStudio 6和7Flex实时PCR系统(AppliedBiosystems)上进行反应。标准曲线用于计算每毫升或每拭子sgmRNA的拷贝数;定量测定的灵敏度为每毫升或每拭子50个拷贝。
PFU测定.对于噬斑测定,在6孔板中制备Vero E6细胞的汇合单层。将从受攻击的动物中收集的指定样品连续稀释,添加到孔中,并在37℃下孵育1小时。孵育后,将1.5mL0.5%甲基纤维素培养基添加至每个孔中,并将板在37℃和5%CO2下孵育2天。通过每孔加入400pL冰冷的甲醇并在-20℃下孵育30分钟来固定板。固定后弃去甲醇,用每孔600μL的0.23%结晶紫染色细胞单层30分钟。染色后,弃去结晶紫,用600μL水洗板一次以可视化和计数噬斑。
ELISA.简而言之,用1xDPBS中的1μg/mL 2019-nCoV刺突(S)蛋白(SinoBiological)包被96孔板,并在4℃下孵育过夜。孵育后,用洗涤缓冲液(1X DPBS中的0.05%Tween20)洗涤板一次,并在室温下用350μL酪蛋白封闭液/孔封闭2-3小时。孵育后,弃去封闭溶液并将板吸干。将在酪蛋白封闭液中稀释的热灭活血清的连续稀释液添加至孔中,并将板在室温下孵育1小时,然后再洗涤3次,随后与1∶1000稀释的抗猕猴IgG HRP(NIH NHPReagent Program)在室温下避光孵育1小时。然后用洗涤缓冲液洗涤板3次,并且每孔加入100μL KPL TMB SureBlue Start溶液;通过每孔添加100μL
KPL TMB SureBlue Start溶液;通过每孔添加100μL KPLTMB Stop溶液来停止板的显色。使用VERSAMAXTM或
KPLTMB Stop溶液来停止板的显色。使用VERSAMAXTM或 微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
假病毒中和测定.通过与之前描述的方法相似的方法产生表达荧光素酶报告基因的2019-nCoV假病毒(9)。简而言之,通过磷酸钙,将包装构建体psPAX2(AIDS Resource andReagent Program)、荧光素酶报告质粒pLenti-CMV Puro-Luc(Addgene)和表达刺突蛋白的pcDNA3.1-SARS CoV-2 SΔCT共转染到HEK293T细胞中。转染后48小时收集含有假病毒的上清液;通过用0.45μm过滤器过滤来纯化假病毒。为了确定来自疫苗接种动物的抗血清的中和活性,将HEK293T-hACE2细胞以1.75×104个细胞/孔的密度接种在96孔组织培养板中过夜。制备热灭活血清样品的两倍连续稀释液,并与50μL假病毒混合。在加入HEK293T-hACE2细胞之前,将混合物在37℃下孵育1小时。感染后48小时,根据制造商的说明,在 Luciferase Assay(Promega)中裂解细胞。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
Luciferase Assay(Promega)中裂解细胞。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
活病毒中和测定.设计基于Seattle Washington分离株的全长2019-nCoV病毒用于表达荧光素酶和GFP,并通过反向遗传学回收,如先前所描述(13,14)。病毒在VeroE6USAMRID细胞中滴定,以获得仅细胞对照背景的至少10倍的相对光单位(RLU)信号。VeroE6 USAMRID细胞在前一天以每孔20,000个细胞接种在透明底黑壁96孔板中。中和抗体血清样品以1∶40的起始稀释度进行检测,并连续稀释4倍,最多8个稀释度。抗体-病毒复合物在37℃和5%CO2下孵育1小时。孵育后,去除生长培养基并将病毒-抗体稀释复合物一式两份地添加到细胞中。每个中和测定板中包括仅病毒对照和仅细胞对照。感染后,将平板在37℃和5%CO2下孵育48小时。孵育48小时后,裂解细胞并根据制造商说明,通过Nano-GloLuciferase Assay System(Promega)测量荧光素酶活性。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
系统血清学.对于血清样品的功能分析,使用基于珠的测定来定量抗体依赖性细胞吞噬作用(ADCP)、抗体依赖性中性粒细胞吞噬作用(ADNP)和抗体依赖性补体沉积(ADCD),如前所述(15)。蛋白抗原包括受体结合结构域(RBD;感谢Aaron Schmidt、RagonInstitute和MassCPR提供)、融合前稳定化的刺突胞外域(S;感谢Bing Chen、儿童医院和MassCPR提供)和核衣壳(N;Sino Biological)。荧光链霉亲和素珠(Thermo Fisher)与生物素化的RBD、N和S偶联,并与稀释的血清(ADCP和ADNP 1∶100、ADCD 1∶10)一起孵育。对于ADCP,将THP添加到免疫复合物中并在37℃下孵育16小时。对于ADNP,使用氯化铵钾(ACK)裂解缓冲液从全血中分离原代中性粒细胞。在37℃孵育1小时后,中性粒细胞用抗CD66bPacBlue检测抗体(Biolegend)染色。对于ADCD测定,根据制造商的说明,将冻干的豚鼠补体(Cedarlane)重悬,并稀释在有钙和镁的明胶弗洛拿(veronal)缓冲液(BostonBioProducts)中。孵育后,用荧光素缀合的山羊IgG豚鼠补体C3组分(Fluorescein-Conjugated Goat IgG Fraction to Guinea Pig Complement C3)(Mpbio)检测C3。为了检测抗体依赖性NK细胞活性,使用基于ELISA的方法。简而言之,用3ug/mL抗原(如上所述)包被板,并以1∶50稀释度添加样品,并在37℃下孵育2小时。前一天通过RosetteSep(StemCell Technologies)从健康的血沉棕黄层中分离出NK细胞,并在1ng/mL IL-15(Stemcell)中过夜。NK细胞与免疫复合物在37℃下用含有CD107a PE-Cy5(BD)、Golgi stop(BD)和Brefeldin A(BFA,Sigma Aldrich)的染色混合物孵育5小时。NK细胞孵育后,固定细胞(Perm A,Life Tech)并在固定时使用抗CD16 APC-Cy7(BD)、抗CD56 PE-Cy7(BD)和抗CD3PacBlue(BD)对表面标志物进行染色。使用Perm B(Life Tech)、抗IFN-γFITC(BD)和抗MIP-1βPE(BD)抗体进行透化后的细胞内染色。所有测定均通过流式细胞仪iQue(Intellicyt)和S-Lab机器人(PAA)获得。对于ADCP,以珠阳性细胞门控事件,其中将中性粒细胞定义为CD66b阳性,然后以珠阳性中性粒细胞进行门控。ADCP和ADNP的吞噬作用评分计算为(珠阳性细胞的百分比)x(珠阳性细胞的MFI)除以10000。ADCD报告为C3沉积的MFI。将NK细胞定义为CD3-、CD16+和CD56+。数据报告为对CD107a、MIP-1-α或IFN-γ呈阳性的细胞的百分比。
Pearson相关性和Spearman相关性分别用于探索抗体特征与BAL中log10sgmRNA拷贝/mL峰值之间的线性和非线性关系。用Benjamini-Hochberg校正来校正多重比较。此外,用Pearson相关性测试抗体特征的所有配对相关性。为了确定与保护相关的最优特征,使用递归特征消除进行了偏最小二乘法回归(PLSR)和随机森林回归(RFR)。首先,所有的同种型/亚类和Fc受体结合数据都进行了log10转换。使用R包“ropls”构建PCA,以比较多变量谱。使用R包“ropls”执行PLSR,并使用R包“randomForest”执行随机森林。每个模型(即每组特征)都进行了10次重复的5倍交叉验证。在特征消除的每一步中,将对于PLSR来说,具有最低平均(跨折叠)变量投影重要性(VIP)得分的特征和对于RFR来说,具有最低平均(跨折叠)重要性的特征测量为节点杂质并去除。对两个特征的所有可能组合进行了测试。
ELISPOT测定.用来自BD Pharmingen的小鼠抗人IFN-γ单克隆抗体以5μg/孔的浓度包被ELISPOT板,在4℃下过夜。用含有0.25%Tween20的DPBS洗涤板,并在37℃下用R10培养基(含有11%FBS和1.1%青霉素-链霉素的RPMI)封闭1小时。以2μg/孔的浓度制备刺突蛋白1和刺突蛋白2肽池,并添加200,000个细胞/孔。将肽和细胞在37℃下孵育18-24小时。此孵育后的所有步骤均在室温下进行。用库尔特(coulter)缓冲液洗涤板并与来自U-Cytech的兔多克隆抗人IFN-γ生物素(1μg/mL)一起孵育2小时。将板第二次洗涤并与来自Southern Biotechnology的链霉亲和素-碱性磷酸酶抗体(1μg/mL)一起孵育2小时。最后一次洗涤后添加氯化硝基四氮唑蓝/5-溴-4-氯-3′-吲哚基磷酸对甲苯胺盐(NBT/BCIP显色液)底物溶液,保持7分钟。弃去显色液(chromagen),将板用水洗涤并在暗处干燥24小时。在Cellular Technologies Limited Immunospot Analyzer上扫描和计数板。
细胞内细胞因子染色测定.将106PBMC/孔重悬在100μL的补充有CD49d单克隆抗体(1μg/mL)的R10培养基中。使用模拟(100μL的R10加0.5%DMSO;背景对照)、刺突1肽池和刺突2肽池(2μg/mL)或10pg/mL豆蔻酰佛波醇乙酯(phorbol myristate acetate)(PMA)和1μg/mL离子霉素(Sigma-Aldrich)(100μL;阳性对照)对每个样品进行评估并在37℃下孵育1小时。孵育后,将50μL R10中的0.25μL GolgiStop和0.25μL GolgiPlug添加到每个孔中,并在37℃下孵育8小时,然后在4℃下保持过夜。第二天,细胞用DPBS洗涤两次,用近红外活/死染料染色10分钟,然后用针对CD279(克隆EH12.1、BB700)、CD38(克隆OKT10、PE)、CD28(克隆28.2,PE CY5)、CD4(克隆L200,BV510)、CD45(克隆D058-1283,BUV615)、CD95(克隆DX2,BUV737)、CD8(克隆SK1,BUV805)的预先指定滴度的单克隆抗体染色,持续30分钟。然后用2%FBS/DPBS缓冲液洗涤细胞两次,并用200μL BD CytoFix/CytoPerm Fixation/Permeabilization溶液孵育15分钟。用1xPerm洗涤缓冲液(将CytoFix/CytoPermFixation/Permeabilization试剂盒中的BD Perm/WashTM Buffer 10X用Milli Q水稀释并通过0.22μm过滤器)洗涤细胞两次,并用针对Ki67(克隆B56,FITC)、CD69(克隆TP1.55.3,ECD)、IL10(克隆JES3-9D7,PE CY7)、IL13(克隆JES10-5A2,BV421)、TNF-α(克隆Mab11,BV650)、IL4(克隆MP4-25D2,BV711)、IFN-γ(克隆B27;BUV395)、IL2(克隆MQ1-17H12、APC)、CD3(克隆SP34.2、Alexa 700)的单克隆抗体进行细胞内染色30分钟。细胞用1X Perm洗涤缓冲液洗涤两次,并用250μL新鲜制备的1.5%甲醛固定。将固定的细胞转移至96孔圆底板中并通过BD FACSymphonyTM系统进行分析。
统计分析.使用GraphPad Prism 8.4.2(GraphPad软件)对病毒学和免疫学数据进行分析。使用双边Mann-Whitney检验进行组间数据比较。通过双边Spearman等级相关性检验评估相关性。认为P值小于0.05是显著的。
结果
DNA疫苗的构建和免疫原性
产生了表达2019-nCoV S蛋白六种变体的DNA疫苗:全长(SS-Spike)、细胞质尾缺失(SS-SdCT)(9)、跨膜结构域和反映可溶性胞外域的细胞质尾缺失(SS-S.Ecto)(9)、带有折叠子三聚化标签的S1结构域(SS-S1-foldon)、带有折叠子三聚化标签的受体结合结构域(SS-RBD-foldon),以及带有弗林蛋白酶切割位点缺失、两个脯氨酸突变和折叠子三聚化标签的融合前稳定化的可溶性胞外域(SS-S.Ecto-dF-PP-foldon)(10-12)(图1)。免疫印迹分析确认了S、SS-SdCT、SS-S.Ecto和SS-S.Ecto-dF-PP-foldon构建体在细胞裂解物中的表达,以及可溶性SS-S.Ecto和SS-S.Ecto-dF-PP-foldon构建体在培养上清液中的表达(图6)。对于SS-S.Ecto而不是SS-S.Ecto-dF-PP-foldon注意到分泌蛋白的蛋白水解切割,可能是由于SS-S.Ecto-dF-PP-foldon中弗林蛋白酶切割位点的突变。
用DNA疫苗对在以下组中的35只6-12岁的成年恒河猴进行免疫:SS-Spike(N=4)、SS-SdCT(N=4)、SS-S.Ecto(N=4)、S1(N=4)、SS-RBD-FOLDON(N=4)、SS-S.Ecto-dF-PP-foldon(N=5)和假对照(N=10)。动物在第0周和第3周接受5mg DNA疫苗。在第5周加强免疫后,通过ELISA(图6A)观到S特异性结合抗体和使用假病毒中和测定(9)(图6B)和活病毒中和测定(13,14)(图6C)观察中和抗体(NAb)。通过ELISA,两只动物在基线时具有结合抗体,这可能反映了其他天然灵长类冠状病毒的交叉反应性。通过假病毒中和测定测量的NAb滴度与通过活病毒中和测定测量的NAb滴度相关(P<0.0001,R=0.8052,双边Spearman等级相关性检验;图10)。此外,疫苗接种的猕猴的NAb滴度(滴度中值74;S和SS-SdCT组的滴度中值170)在量上与从2019-nCoV感染中恢复的9只恢复期猕猴的队列(滴度中值106)和27个恢复期人的队列(滴度中值93)的NAb滴度相当(图6D)。
疫苗接种后的猕猴中的S特异性和RBD特异性抗体包括不同的亚类和效应功能,包括抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性补体沉积(ADCD)、抗体依赖性单核细胞吞噬作用(ADCP)和抗体NK细胞激活(IFN- 分泌、CD107a脱粒和MIP-1β表达)(15)(图6E)。在S和SS-SdCT组中观察到更高的ADCD应答趋势,而在SS-RBD-FOLDON和SS-S.Ecto-dF-PP-foldon组中观察到更高的NK细胞激活。功能和生物物理抗体特征的主成分分析显示不同疫苗组的重叠,并且在S和RBD组中具有更不同的谱(图6E)。
分泌、CD107a脱粒和MIP-1β表达)(15)(图6E)。在S和SS-SdCT组中观察到更高的ADCD应答趋势,而在SS-RBD-FOLDON和SS-S.Ecto-dF-PP-foldon组中观察到更高的NK细胞激活。功能和生物物理抗体特征的主成分分析显示不同疫苗组的重叠,并且在S和RBD组中具有更不同的谱(图6E)。
通过IFN-γELISPOT测定在大多数疫苗接种的动物中也观察到了对合并的S肽的细胞免疫应答(图7A)。细胞内细胞因子染色测定证明了诱导S特异性CD4+和CD8+T细胞应答,以及较短的SS-S1-foldon和SS-RBD-Foldon免疫原诱导的应答较低(图7B)。
针对2019-nCoV攻击的保护效力
在第6周,即加强免疫后3周,用1.2x108VP(1.1x104PFU)的2019-nCoV攻击所有动物,通过鼻内(IN)途径给药1ml,通过气管内途径(IT)给药1ml。攻击后,通过RT-PCR(16)在支气管肺泡灌洗液(BAL)和鼻拭子(NS)中评估病毒RNA水平。血浆中病毒RNA呈阴性,动物仅表现出轻度的临床症状。在假对照中观察到高水平的病毒RNA,在BAL中的峰值中值为6.46(范围4.81-7.99)log10RNA拷贝/mL,在NS中的峰值中值为6.82(范围5.96-7.96)log10RNA拷贝/拭子(图11)。在疫苗组中观察到较低水平的病毒RNA(图12、图13),其包括在疫苗接种SS-Spike的动物中,与假对照相比,BAL和NS中的峰值中值病毒RNA分别降低了1.92和2.16log10(P=0.02和P=0.04,双边Mann-Whitney检验)(图14)。病毒RNA测定通过PFU测定确认。
攻击后BAL和NS中的大部分病毒RNA被认为代表输入攻击病毒,因此还评估了亚基因组mRNA(sgmRNA)的水平,其被认为反映了病毒复制的细胞中间体,这些中间体没有被包装到毒粒中,因此假定在细胞中复制病毒(17)。在假对照中观察到高水平的sgmRNA,其在BAL中的峰值中值为5.35(范围3.97-6.95)log10 sgmRNA拷贝/mL,NS中的峰值中值为6.40(范围4.91-7.01)log10 sgmRNA拷贝/拭子(图8A)。与假对照相比,在疫苗接种SS-Spike的动物中,在疫苗组中观察到显著降低的sgmRNA水平(图8B、8C),包括在BAL和NS中的峰值中值sgmRNA分别降低>3.1和>3.7log10(P=0.03和P=0.01,双边Mann-Whitney检验)(图8D)。尽管在SS-S.Ecto组中观察到较低的保护作用,但在其他疫苗组中也观察到sgmRNA水平降低,这包括SS-SdCT、SS-S1一foldon、SS-RBD-Foldon和SS-S.Ecto-dF-PP-foldon,其确认了融合前胞外域稳定化的重要性,如先前报道(12)。在攻击后的任何时间点,25只疫苗接种的动物中共有8只在BAL和NS中未表现出可检测到的sgmRNA。
保护的免疫相关因素
本研究中保护效力的可变性促进了对保护的免疫相关因素的分析。第5周的log10假病毒NAb滴度与BAL(P<0.0001,R=-0.6877,双边Spearman等级相关性检验)和NS(P=0.0199,R=-0.4162)中的log10 sgmRNA峰值负相关(图9A)。类似地,第5周的log10活病毒NAb滴度与BAL(P<0.0001,R=-0.7702)和NS(P=0.1006,R=-0.3360)中的log10 sgmRNA峰值水平负相关(图9B)。这些数据表明,疫苗引发的血清NAb滴度可能是防止2019-nCoV攻击的保护的强免疫相关因素。由于收集的拭子的内在可变性,与NS中的病毒载量相比,推测与BAL中的病毒载量的相关性更强。第5周的log10ELISA滴度也与BAL(P=0.0041,R=-0.4733)中的log10 sgmRNA峰值水平负相关(图15)。疫苗引发的ELISPOT应答(图16)、CD4+ICS应答(图17)和CD8+ICS应答(图18)与保护不相关。
接下来探讨了其他抗体效应功能对保护的免疫相关因素的潜在贡献。除了NAb滴度之外,S-和RBD-特异性ADCD应答与BAL中的log10 sgmRNA峰值水平负相关(图9C,上图)。然后使用两种正交无偏机器学习方法来定义保护的最小组合相关因素。非线性随机森林分析和线性偏最小二乘回归分析表明,利用两个特征提高了与保护的相关性,例如RBD特异性Fc R2a-1结合与ADCD应答,或NAb滴度与RBD特异性IgG2应答(图9C,左下图)。此外,NAb滴度与大多数抗体效应功能相关,除了抗体介导的NK细胞激活(图9C,右下图)。总之,这些数据表明NAb在防止2019-nCoV中的主要作用是由诸如ADCD的先天免疫效应功能支持的。
R2a-1结合与ADCD应答,或NAb滴度与RBD特异性IgG2应答(图9C,左下图)。此外,NAb滴度与大多数抗体效应功能相关,除了抗体介导的NK细胞激活(图9C,右下图)。总之,这些数据表明NAb在防止2019-nCoV中的主要作用是由诸如ADCD的先天免疫效应功能支持的。
攻击后的抗体再生免疫应答
所有动物在攻击后都表现出抗体再生体液和细胞免疫应答,包括在攻击后的第14天增加的ELISA滴度(图19)、假病毒NAb滴度(图20)、活病毒NAb滴度(图21)和IFN-γELISPOT应答(图22)。这些数据表明疫苗保护不是消除病毒(sterilizing),而是可能由攻击后的快速病毒学控制介导,所述数据包括在25只动物中的8只中,在攻击后的任何时间点在BAL和NS中未检测到sgmRNA。
讨论
结束全球COVID-19大流行可能需要一种安全有效的2019-nCoV疫苗。一些候选疫苗已开始临床试验,还有许多其他候选疫苗处于临床前开发阶段。然而,目前对动物模型中候选2019-nCoV疫苗的保护和保护效力的免疫相关因素知之甚少。在这项研究中,产生了一系列表达各种S免疫原的原型DNA疫苗,并评估了在恒河猴中对鼻内和气管内2019-nCoV攻击的保护效力。我们证实了强的疫苗保护作用,与假对照相比,S免疫动物的BAL和NS的中值病毒载量分别大幅降低>3.1和>3.7log10。保护不是消除病毒,而是可能由攻击后的快速免疫控制介导。
我们的数据扩展了之前对小鼠、雪貂和猕猴的SARS和MERS疫苗保护的研究(9,18-21)。还进行了针对SARS和MERS候选疫苗的1期临床研究(22),但尚未对这些疫苗在人中的效力进行测试。我们的数据表明,在猕猴中防止2019-nCoV的保护是可行的。在上呼吸道和下呼吸道中观察到病毒复制的显著减少。使用全长S免疫原实现了最优保护,而使用可溶性构建体和较小的片段则观察到了降低的保护作用。然而,较小的免疫原可能在某些情况下有用(例如,不同的患者群体)。
进一步的研究需要解决保护性免疫的持久性和人2019-nCoV疫苗的最优疫苗平台等重要问题。尽管数据仅限于DNA疫苗,但这些发现也可以推广到其他基于基因的疫苗,包括RNA疫苗和基于重组载体的疫苗。进一步的研究还需要解决可能由抗体依赖性增强引起的呼吸道疾病加重的问题(23,24)。未能保护的次优疫苗构建体未观察到增强的临床疾病。
在本研究中,确定通过两种独立测定(假病毒中和和活病毒中和)测量的血清NAb滴度为保护的强相关因素。如果证明这种相关因素在NHP和人的多项疫苗研究中是可推广的,那么这将是2019-nCoV疫苗临床开发的有用基准。诸如ADCD的先天免疫效应功能也可能有助于保护效力。
总之,已经用一系列原型DNA疫苗证明了在恒河猴中防止2019-nCoV攻击的疫苗保护,并且已确定血清NAb滴度为这个模型中保护作用的相关因素。这种相关因素应在多种环境中进一步研究,但如果得到确认,可能证明它可以用作在人中测试2019-nCoV疫苗的基准。
参考文献
1.F.Wu et al.,A new coronavirus associated with human respiratorydisease in China.Nature 579,265-269(2020).
2.P.Zhou et al.,A pneumonia outbreak associated with a newcoronavirus of probable bat origin.Nature 579,270-273(2020).
3.M.L.Holshue et al.,First Case of 2019Novel Coronavirus in theUnited States.N Engl JMed 382,929-936(2020).
4.Q.Li et al.,Early Transmission Dynamics in Wuhan,China,of NovelCoronavirus-Infected Pneumonia.NEngl JMed,(2020).
5.N.Zhu et al.,A Novel Coronavirus from Patients with Pneumonia inChina,2019.N Engl JMed 382,727-733(2020).
6.N.Chen et al.,Epidemiological and clinical characteristics of99cases of 2019novel coronavirus pneumonia in Wuhan,China:a descriptivestudy.Lancet 395,507-513(2020).
7.C.Huang et al.,Clinical features of patients infected with2019novel coronavirus in Wuhan,China.Lancet 395,497-506(2020).
8.J.F.Chan et al.,A familial cluster of pneumonia associated with the2019novel coronavirus indicating person-to-person transmission:a study of afamily cluster.Lancet 395,514-523(2020).
9.Z.Y.Yang et al.,A DNA vaccine induces SARS coronavirusneutralization and protective immunity in mice.Nature 428,561-564(2004).
10.R.N.Kirchdoerfer et al.,Pre-fusion structure of a humancoronavirus spike protein.Nature 531,118-121(2016).
11.J.Pallesen et al.,Immunogenicity and structures of a rationallydesigned prefusion MERS-CoV spike antigen.ProcNatl Acad Sci USA 114,E7348-E7357(2017).
12.D.Wrapp et al.,Cryo-EM structure of the 2019-nCoV spike in theprefusion conformation.Science 367,1260-1263(2020).
13.T.Scobey et al.,Reverse genetics with a full-length infectiouscDNA of the Middle East respiratory syndrome coronavirus.Proc NatlAcad SciUSA 110,16157-16162(2013).
14.B.Yount et al.,Reverse genetics with a full-length infectious cDNAof severe acute respiratory syndrome coronavirus.Proc Natl Acad Sci USA 100,12995-13000(2003).
15.A.W.Chung et al.,Dissecting Polyclonal Vaccine-Induced HumoralImmunity against HIV Using Systems Serology.Cell 163,988-998(2015).
16.P.Abbink et al.,Lack of therapeutic efficacy of an antibody toalpha4beta7 in SIVmac251-infected rhesus macaques.Sciencc 365,1029-1033(2019).
17.R.Wolfel et al.,Virological assessment of hospitalized patientswith COVID-2019.Nature,(2020).
18.R.Liu et al.,A recombinant VSV-vectored MERS-CoV vaccine inducesneutralizing antibody and T cell responses in rhesus monkeys after singledose immunization.Antiviral research150,30-38(2018).
19.K.Muthumani et al.,A synthetic consensus anti-spike protein DNAvaccine induces protective immunity against Middle East respiratory syndromecoronavirus in nonhuman primates.Sci TranslMed 7,301ra132(2015).
20.G.PKobinger et al.,Adenovirus-based vaccine prevents pneumonia inferrets challenged with the SARS coronavirus and stimulates robust immuneresponses in macaques.Vaccine 25,5220-5231(2007).
21.J.Zhou et al.,Immunogenicity,safety,and protective efficacy of aninactivated SARS-associated coronavirus vaccine in rhesus monkeys.Vaccine23,3202-3209(2005).
22.J.E.Martin et al.,A SARS DNA vaccine induces neutralizing antibodyand cellular immune responses in healthy adults in a Phase I clinicaltrial.Vaccine 26,6338-6343(2008).
23.C.T.Tseng et al.,Immunization with SARS coronavirus vaccines leadsto pulmonary immunopathology on challenge with the SARS virus.PLoS ONE7,e35421(2012).
24.L.Liu et al.,Anti-spike IgG causes severe acute lung injury byskewing macrophage responses during acute SARS-CoV infection.JCI insight 4,(2019).
实施例9.针对冠状病毒感染的2019-nCoV自然免疫应答
COVID-19大流行的爆炸性传播使开发应对措施成为全球紧迫的优先事项(1-8)。然而,目前对2019-nCoV的免疫发病机制的了解非常有限。特别是,目前尚不清楚是否2019-nCoV感染会诱导自然免疫,从而防止人再次暴露。这些信息对于疫苗策略、流行病学建模和公共卫生方法至关重要。为了探索这个问题,我们开发了2019-nCoV感染的恒河猴模型,并评估了感染的病毒学、免疫学和病理学特征,以及对再次攻击的保护性免疫。
方法
动物和研究设计.将13只6-12岁的远交系的印度裔成年雄性和雌性恒河猴(Macaca mulatta)随机分组。所有动物都饲养在Bioqual,Inc.(Rockville,MD)。在第一项研究中,9只动物接种了2019-nCoV,总剂量为1.1x106噬斑形成单位(PFU)(第1组;N=3)、1.1x105PFU(第2组;N=3)、或1.1x104PFU(第3组;N=3)。这些剂量通过鼻内(IN)途径给药1ml(每个鼻孔0.5ml),通过气管内(IT)途径给药1ml。在攻击后的第35天,用与初始攻击中使用的剂量相匹配的剂量的2019-nCoV再次攻击动物。半定量临床评分涉及由兽医人员对临床表现、呼吸困难、躺卧和应答能力进行盲法评估(0-40范围内每个类别10分;轻度疾病<12;中度疾病13-20;严重疾病>20)。在第二项研究中,4只恒河猴接种了总剂量为1.1x105PFU的2019-nCoV,再次通过IN和IT途径给药。在攻击后第2天(N=2)或第4天(N=2)对猕猴进行尸检。所有免疫学和病毒学测定均在盲法下进行。所有动物研究均按照所有相关的地方、州和联邦法规进行,并得到生物质量机构动物管理和使用委员会(IACUC)的批准。
2019-nCoV原种.2019-nCoV USA-WA1/2020原种来自于BEI Resource(NR-52281;货号#70033175;感谢疾病控制和预防中心的Natalie Thornburg提供),在Vero E6细胞中扩增,并在感染后第5天,在90%的细胞病理学效应(CPE)下收获病毒攻击原种。全基因组测序显示与亲本病毒序列100%相同性(GenBank MN985325.1;感谢威斯康星大学David O’Connor,Shelby O’Connor提供)。
病毒RNA测定.使用靶向2019-nCoV核衣壳的RT-PCR测定对病毒RNA进行定量。简而言之,使用Qiagen MinElute virus spin kit或RNA-STAT 60(Tel-test B)从攻击前或攻击后指定时间点从猕猴收集的样品(例如,支气管肺泡灌洗液(BAL)上清液)中分离RNA并重悬于无RNA酶的水中。将分离的RNA一式两份加样并在Applied Biosystems 7500序列检测器中使用以下程序进行扩增:i)48℃30分钟,ii)95℃10分钟,和iii)95℃15秒的40个循环和55℃1分钟。该测定表现每毫升液体或每克组织5x101-5x108RNA拷贝的实际范围。正向(F)和反向(R)引物序列和探针序列(P)是:
2019-nCoV_N1-F:5′-GACCCCAAAATCAGCGAAAT-3′(SEQ ID NO:196)
2019-nCoV_N1-R:5′-TCTGGTTACTGCCAGTTGAATCTG-3′(SEQ ID NO:197)
2019-nCoV_N1-P:5′-FAM-ACCCCGCATTACGTTTGGTGGACC-BHQ1-3′(SEQ ID NO:198)
亚基因组mRNA测定.通过RT-PCR使用与先前描述的方法相似的方法评估2019-nCoVE基因亚基因组mRNA(sgmRNA)(9)。为了生成标准曲线,将2019-nCoV E基因sgmRNA克隆至pcDNA3.1表达质粒中;使用AmpliCap-Max T7 High Yield Message Maker Kit(Cellscript)转录此插入物,以获得标准的RNA。在RT-PCR之前,根据制造商的说明,使用Superscript III VILO(Invitrogen)将从受攻击的动物或标准中收集的样品进行逆转录。使用靶向E基因sgmRNA(9)的序列设计了Taqman定制基因表达测定(ThermoFisherScientific)。根据制造商的说明,在QuantStudio 6和7Flex实时PCR系统(AppliedBiosystems)上进行反应。标准曲线用于计算每毫升或每拭子sgmRNA的拷贝数;定量测定的灵敏度为每毫升或每拭子50个拷贝。
PFU测定.对于噬斑测定,在6孔板中制备Vero E6细胞的汇合单层。将从受攻击的动物中收集的指定样品连续稀释,添加到孔中,并在37℃下孵育1小时。孵育后,将1.5mL0.5%甲基纤维素培养基添加至每个孔中,并将板在37℃和5%CO2下孵育2天。通过每孔加入400μL冰冷的甲醇并在-20℃下孵育30分钟来固定板。固定后弃去甲醇,用每孔600μL的0.23%结晶紫染色细胞单层30分钟。染色后,弃去结晶紫,用600μL水洗板一次以可视化和计数噬斑。
ELISA.简而言之,用1xDPBS中的1μg/mL 2019-nCoV刺突(S)蛋白(SinoBiological)包被96孔板,并在4℃下孵育过夜。孵育后,用洗涤缓冲液(1X DPBS中的0.05%Tween20)洗涤板一次,并在室温下用350μL酪蛋白封闭液/孔封闭2-3小时。孵育后,弃去封闭溶液并将板吸干。将在酪蛋白封闭液中稀释的热灭活血清的连续稀释液添加至孔中,并将板在室温下孵育1小时,然后再洗涤3次,随后用1∶1000稀释的抗猕猴IgG HRP(NIH NHPReagent Program)在室温下避光孵育1小时。然后用洗涤缓冲液洗涤板3次,并且每孔加入100μL KPL TMB SureBlue Start溶液;通过每孔添加100μL
KPL TMB SureBlue Start溶液;通过每孔添加100μL KPLTMB Stop溶液来停止板的显色。使用VERSAMAXTM或
KPLTMB Stop溶液来停止板的显色。使用VERSAMAXTM或 微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
假病毒中和测定.通过与之前描述的方法相似的方法产生表达荧光素酶报告基因的2019-nCoV假病毒(10)。简而言之,通过磷酸钙,将包装构建体psPAX2(AIDS Resourceand Reagent Program)、荧光素酶报告质粒pLenti-CMV Puro-Luc(Addgene)和表达刺突蛋白的pcDNA3.1-SARS CoV-2 SACT共转染到HEK293T细胞中。转染后48小时收集含有假病毒的上清液;通过用0.45μm过滤器过滤来纯化假病毒。为了确定来自疫苗接种动物的抗血清的中和活性,将HEK293T-hACE2细胞以1.75×104个细胞/孔的密度接种在96孔组织培养板中过夜。制备热灭活血清样品的两倍连续稀释液,并与50μL假病毒混合。在加入HEK293T-hACE2细胞之前,将混合物在37℃下孵育1小时。感染后48小时,根据制造商的说明,在 Luciferase Assay(Promega)中裂解细胞。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
Luciferase Assay(Promega)中裂解细胞。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
活病毒中和测定.设计基于Seattle Washington分离株的全长2019-nCoV病毒用于表达荧光素酶和GFP,并通过反向遗传学回收,如先前所描述(11,12)。病毒在VeroE6USAMRID细胞中滴定,以获得仅细胞对照背景的至少10倍的相对光单位(RLU)信号。VeroE6 USAMRID细胞在前一天以每孔20,000个细胞接种在透明底黑壁96孔板中。中和抗体血清样品以1∶40的起始稀释度进行检测,并连续稀释4倍,最多8个稀释度。抗体-病毒复合物在37℃和5%CO2下孵育1小时。孵育后,去除生长培养基并将病毒-抗体稀释复合物一式两份地添加到细胞中。每个中和测定板中包括仅病毒对照和仅细胞对照。感染后,将平板在37℃和5%CO2下孵育48小时。孵育48小时后,裂解细胞并根据制造商说明,通过Nano-GloLuciferase Assay System(Promega)测量荧光素酶活性。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
系统血清学.对于血浆样品的功能分析,使用基于珠的测定来定量抗体依赖性细胞吞噬作用(ADCP)、抗体依赖性中性粒细胞吞噬作用(ADNP)和抗体依赖性补体沉积(ADCD),如前所述(13)。蛋白抗原包括受体结合结构域(RBD;感谢Aaron Schmidt、RagonInstitute和MassCPR提供)、融合前稳定化的刺突胞外域(S;感谢Bing Chen、儿童医院和MassCPR提供)和核衣壳(N;Sino Biological)。荧光链霉亲和素珠(Thermo Fisher)与生物素化的RBD、N和S偶联,并与稀释的血浆(ADCP和ADNP 1∶100、ADCD 1∶10)一起孵育。对于ADCP,将THP添加到免疫复合物中并在37℃下孵育16小时。对于ADNP,使用氯化铵钾(ACK)裂解缓冲液从全血中分离原代中性粒细胞。在37℃孵育1小时后,中性粒细胞用抗CD66bPacBlue检测抗体(Biolegend)染色。对于ADCD测定,根据制造商的说明,将冻干的豚鼠补体(Cedarlane)重悬,并稀释在有钙和镁的明胶弗洛拿缓冲液(Boston BioProducts)中。孵育后,用荧光素缀合的山羊IgG豚鼠补体C3组分(Mpbio)检测C3。为了检测抗体依赖性NK细胞活性,使用基于ELISA的方法。简而言之,用3ug/mL抗原(如上所述)包被板,并以1∶50稀释度添加样品,并在37℃下孵育2小时。前一天通过RosetteSep(Stem Cell Technologies)从健康的血沉棕黄层中分离出NK细胞,并在1ng/mL IL-15(Stemcell)中过夜。NK细胞与免疫复合物在37℃下用含有CD107a PE-Cy5(BD)、Golgi stop(BD)和Brefeldin A(BFA,SigmaAldrich)的染色混合物孵育5小时。NK细胞孵育后,固定细胞(Perm A,Life Tech)并在固定时使用抗CD16 APC-Cy7 (BD)、抗CD56 PE-Cy7 (BD)和抗CD3 PacBlue (BD)对表面标志物进行染色。使用Perm B(Life Tech)、抗IFN-γFITC(BD)和抗MIP-1βPE(BD)抗体进行透化后的细胞内染色。所有测定均通过流式细胞仪和iQue (Intellicyt)和S-Lab机器人(PAA)获得。对于ADCP,以珠阳性细胞门控事件,其中将中性粒细胞定义为CD66b阳性,然后以珠阳性中性粒细胞进行门控。ADCP和ADNP的吞噬作用评分计算为(珠阳性细胞的百分比)x(珠阳性细胞的MFI)除以10000。ADCD报告为C3沉积的MFI。将NK细胞定义为CD3-、CD16+和CD56+。数据报告为对CD107a、MIP-1-α或IFN-γ呈阳性的细胞的百分比。
ELISPOT测定.用来自BD Pharmingen的小鼠抗人IFN-γ单克隆抗体以5μg/孔的浓度包被ELISPOT板,在4℃下过夜。用含有0.25%Tween20的DPBS洗涤板,并在37℃下用R10培养基(含有11%FBS和1.1%青霉素-链霉素的RPMI)封闭1小时。以2μg/孔的浓度制备刺突蛋白1和刺突蛋白2肽池,并添加200,000个细胞/孔。将肽和细胞在37℃下孵育18-24小时。此孵育后的所有步骤均在室温下进行。用库尔特(coulter)缓冲液洗涤板并与来自U-Cytech的兔多克隆抗人IFN-γ生物素(1μg/mL)一起孵育2小时。将板第二次洗涤并与来自Southern Biotechnology的链霉亲和素-碱性磷酸酶抗体(1μg/mL)一起孵育2小时。最后一次洗涤后添加氯化硝基四氮唑蓝/5-溴-4-氯-3′-吲哚基磷酸对甲苯胺盐(NBT/BCIP显色液)底物溶液,保持7分钟。弃去显色液(chromagen),将板用水洗涤并在暗处干燥24小时。在Cellular Technologies Limited Immunospot Analyzer上扫描和计数板。
细胞内细胞因子染色测定.将106PBMC/孔重悬在100μL的补充有CD49d单克隆抗体(1μg/mL)的R10培养基中。使用模拟(100μL的R10加0.5%DMSO;背景对照)、刺突1肽池和刺突2肽池(2μg/mL)或10pg/mL豆蔻酰佛波醇乙酯(phorbol myristate acetate)(PMA)和1μg/mL离子霉素(Sigma-Aldrich)(100μL;阳性对照)对每个样品进行评估并在37℃下孵育1小时。孵育后,将50μL R10中的0.25μL GolgiStop和0.25μL GolgiPlug添加到每个孔中,并在37℃下孵育8小时,然后在4℃下保持过夜。第二天,细胞用DPBS洗涤两次,用近红外活/死染料染色10分钟,然后用针对CD279(克隆EH12.1、BB700)、CD38(克隆OKT10、PE)、CD28(克隆28.2,PE CY5)、CD4(克隆L200,BV510)、CD45(克隆D058-1283,BUV615)、CD95(克隆DX2,BUV737)、CD8(克隆SK1,BUV805)的预先指定滴度的单克隆抗体染色,持续30分钟。然后用2%FBS/DPBS缓冲液洗涤细胞两次,并用200μL BD CytoFix/CytoPerm Fixation/Permeabilization溶液孵育15分钟。用1X Perm洗涤缓冲液(将CytoFix/CytoPermFixation/Permeabilization试剂盒中的BD Perm/WashTM Buffer 10X用Milli Q水稀释并通过0.22μm过滤器)洗涤细胞两次,并用针对Ki67(克隆B56,FITC)、CD69(克隆TP1.55.3,ECD)、IL10(克隆JES3-9D7,PE CY7)、IL13(克隆JES10-5A2,BV421)、TNF-α(克隆Mab11,BV650)、IL4(克隆MP4-25D2,BV711)、IFN-γ(克隆B27;BUV395)、IL2(克隆MQ1-17H12、APC)、CD3(克隆SP34.2、Alexa 700)的单克隆抗体进行细胞内染色30分钟。细胞用1X Perm洗涤缓冲液洗涤两次,并用250μL新鲜制备的1.5%甲醛固定。将固定的细胞转移至96孔圆底板中并通过BD FACSymphonyTM系统进行分析。
组织病理学和免疫组织化学.组织在新鲜制备的4%多聚甲醛中固定24小时,转移到70%乙醇中,并在7天内包埋石蜡,并以5μm切片。载玻片在65℃下烘烤30-60分钟,然后在二甲苯中脱蜡并且通过一系列分级乙醇至蒸馏水进行复水。对于SARS-N和Iba-1的免疫组织化学,使用压力锅在蒸汽设置下在柠檬酸盐缓冲液(Thermo;AP-9003-500)中进行25分钟的热诱导表位修复(HIER),然后用3%过氧化氢处理。然后在蒸馏水中冲洗载玻片并封闭蛋白(BioCare,BE965H)15分钟,然后在1x磷酸盐缓冲盐水中冲洗。使用1∶250的兔抗SARS-核蛋白一抗(Novus;NB100-56576)和兔抗Iba-1抗体(Wako;019-19741),然后使用兔Mach-2HRP-Polymer(BioCare;RHRP520L)30分钟,再用苏木精复染,然后用0.25%氨水染成蓝色。SARS-N和Iba-1的标记是在Biogenex i6000全自动染色机(v3.02)上进行的。CD4+T细胞、巨噬细胞(CD68/CD163)、中性粒细胞(MPO)和1型IFN应答(Mx1)如前所述使用BiocareintelliPATH全自动染色机(14)进行IHC,并且所有抗体在室温下孵育1小时。使用1∶200的CD4(abcam货号ab133616;克隆EPR6855),并使用兔Polink-1 HRP(GBI Labs货号D13-110)检测,使用1∶400的CD68(Biocare货号CM033C;克隆KP1),并使用小鼠Polink-2 AP(GBILabs货号D69-110)检测,使用1∶400的CD163(Thermo货号MA5-11458;克隆10D6),并使用小鼠Polink-2 AP(GBI Labs货号D69-110)检测,使用1∶1000的MPO(Dako货号A0398;多克隆),并使用兔Polink-1 HRP(GBI Labs货号D13-110)检测,使用1∶1000的Mx1(EMD Millipore货号MABF938;克隆M143/CL143),并使用小鼠Polink-2 HRP (GBI Labs货号D37-110)检测。组织病理学由两名专业认证的兽医病理学家(AJM和ADM)独立评估。使用HALO软件(v2.3.2089.27,Indica Labs)进行定量图像分析。对于SARS CoV-2感染的动物(n=4),分析了左右肺各四个区域,对于未感染的对照(n=4),分析了一个区域。对于Mx1定量,AreaQuantification v2模块用于确定MX1蛋白占总组织面积的百分比。对于MPO(中性粒细胞)定量,首先通过排除血管(>5mm2)、支气管、细支气管、软骨和结缔组织训练HALO AI软件来检测肺的肺泡部分;随后,CytoNuclear v1.6模块用于检测MPO+细胞,并表示为占总肺泡组织的比率(PMNs/mm2)。在所有情况下,对每个样本都进行了手动处理,以确保注释准确并校正假阳性/假阴性。



循环免疫荧光.如前所述进行循环免疫荧光(CyCIF)(16)。简而言之,将载玻片上的多聚甲醛固定的样品脱蜡并在Leica Bond RX上进行抗原修复。然后使用以下抗体对载玻片进行循环免疫荧光(CyCIF)(表1)。使用配备20x、0.8NA物镜的
 扫描仪扫描载玻片,然后使用BaSiC工具校正不均匀照明的图像。使用ASHLAR软件处理平场校正后的图像,以在所有循环中将图像和通道相互对准(align)。
扫描仪扫描载玻片,然后使用BaSiC工具校正不均匀照明的图像。使用ASHLAR软件处理平场校正后的图像,以在所有循环中将图像和通道相互对准(align)。
表1:组织循环免疫荧光抗体

统计分析.使用GraphPad Prism 8.4.2(GraphPad软件)对病毒学和免疫学数据进行分析。使用双边Mann-Whitney检验进行组间数据比较。认为P值小于0.05是显著的。
结果
恒河猴中的2019-nCoV感染的病毒学和免疫学
9只成年恒河猴(6-12岁)共接种1.1x106PFU(第1组;N=3)、1.1x105PFU(第2组;N=3)或1.1x104PFU(第3组;N=3)的2019-nCoV,通过鼻内(IN)途径给药1ml,并且通过气管内(IT)途径给药1ml。病毒攻击后,通过RT-PCR在多个解剖区室中评估病毒RNA水平。在支气管肺泡灌洗液(BAL)(图23A)和鼻拭子(NS)(图23B)中观察到高水平的病毒RNA,BAL中的峰值中值为6.56(范围5.32-8.97)log10 RNA拷贝/mL以及NS中的峰值中值为7.00(范围5.06-8.55)log10 RNA拷贝/拭子。从第1天到第2天,所有动物的NS中的病毒RNA增加,这表明病毒复制。病毒RNA在第2天达到峰值,并且通常到第10-14天在BAL中消退和到第21-28天在NS中消退。第2天后,无论剂量如何,BAL和NS中的病毒载量在所有组中都具有可比性。在血浆中检测不到病毒RNA(图29)。在高剂量组中动物表现出适度降低的食欲和反应性,提示轻度临床疾病(图30)以及轻度短暂的中性粒细胞减少和淋巴细胞减少(图31),但未观察到发热、体重减轻、呼吸窘迫和死亡。
为了帮助区分输入攻击病毒和新复制病毒,开发了一种RT-PCR测定来评估E基因亚基因组mRNA(sgmRNA)。E基因sgmRNA反映了病毒复制的细胞中间体,这些中间体没有被包装到毒粒中,因此代表了细胞中假定的复制病毒(9)。与总病毒RNA(图23B)相比,NS中的sgmRNA水平在第1天较低,中值为5.11(范围<1.70-5.94)log10 sgmRNA拷贝/拭子,但随后在第2天增加至6.50的中值(范围4.16-7.81)log10 sgmRNA拷贝/拭子(图23C)。
接下来评估了这些动物的2019-nCoV特异性体液和细胞免疫应答。所有9只猕猴都通过ELISA(图24A)产生了对2019-nCoV刺突(S)蛋白的结合抗体应答,并使用假病毒中和测定(10)(图24B)和活病毒中和测定(11、12)(图24C)产生了中和抗体(NAb)应答。在第35天,无论剂量组如何,在所有动物中观察到大约100的NAb滴度(通过假病毒中和测定的范围为83-197,通过活病毒中和测定的范围为35-326)。观察到多个亚类对受体结合结构域(RBD)、融合前S胞外域(S)和核衣壳(N)的抗体应答,并且抗体表现出多种效应功能,包括抗体依赖性补体沉积(ADCD)、抗体-依赖性细胞吞噬作用(ADCP)、抗体依赖性中性粒细胞吞噬作用(ADNP)和抗体依赖性NK细胞脱粒(NK CD107a)和细胞因子分泌(NK MIP1β,NK IFNγ)(13)(图24D)。在第35天,通过IFN-γELISPOT测定在大多数动物中观察到对合并的S肽的细胞免疫应答,在较低剂量组中具有较低应答的趋势(图24E)。细胞内细胞因子染色测定证明了诱导S特异性CD8+和CD4+T细胞应答(图24F)。
恒河猴中SARS CoV-2感染诱导急性病毒性间质性肺炎
目前只能获得来自2019-nCoV感染的人的有限病理学数据。为了评估在恒河猴中感染2019-nCoV的病理特征,4只动物通过上述IN和IT途径接种1.1x105PFU的病毒,并在攻击后第2天(N=2)和第4天(N=2)尸检。收获上呼吸道、下呼吸道、胃肠道、淋巴结和其他器官的多个区域进行病毒学和病理学分析。在所有鼻粘膜、咽部、气管和肺组织中观察到高水平的病毒RNA,在胃肠道、肝脏和肾脏中发现较低水平的病毒(图32)。在气管旁淋巴结中很容易检测到病毒RNA,但在远端淋巴结和脾脏中仅偶尔发现(图32)。
将上呼吸道黏膜、气管和肺多聚甲醛固定、石蜡包埋,并通过组织病理学评估。在攻击后的第2天,两只尸检的动物都表现出多灶性炎症区域和病毒性肺炎的迹象,包括肺泡间隔扩张,以及单个核细胞浸润、实变和水肿(图25A、25B)。水肿区域还含有大量多形核细胞,主要是中性粒细胞。末端细支气管上皮坏死并脱落,在气道内和远端肺泡空间内检测到上皮细胞团块(图25C、25D),偶尔形成细支气管上皮合胞细胞(图25E)。在肺泡间隔内偶尔观察到透明膜,这与对I型和II型肺细胞的损伤一致(图25F)。扩散的反应性肺泡巨噬细胞充满肺泡,并且一些是多核的并且通过免疫组织化学标记为核衣壳阳性(图25G)。肺泡衬细胞(肺细胞)也显著标记为核衣壳阳性(图25H)。
通过免疫组织化学和原位RNA杂交 (14,15)(图25I)检测,整个肺实质中存在多灶性病毒感染细胞簇。
(14,15)(图25I)检测,整个肺实质中存在多灶性病毒感染细胞簇。 观察到正义和反义病毒RNA,表明在肺组织中的病毒复制。致密的炎症浸润包括通过内源性髓过氧化物酶染色(MPO)检测的多形核细胞、CD68和CD163阳性巨噬细胞、CD4+和CD8+T淋巴细胞,以及1型干扰素基因MX1的扩散上调(图33)。与未感染的动物相比,2019-nCoV感染导致肺泡的多形核细胞浸润显著增加(P=0.0286),并且在大约30%的总肺组织中出现广泛的MX1染色(P=0.0286)(图34)。在较大呼吸道的呼吸道上皮粘膜下层中也检测到炎症浸润,伴有炎症细胞迁移到细支气管腔中(图25J)。纤毛上皮细胞也对SARS CoV-2RNA (图25K)和SARS核衣壳(图25L)呈染色阳性。到感染后第4天,炎症和病毒性肺炎的程度已经减轻,但在肺实质和中性粒细胞浸润中仍检测到病毒,并且1型干扰素应答持续存在(图34)。
观察到正义和反义病毒RNA,表明在肺组织中的病毒复制。致密的炎症浸润包括通过内源性髓过氧化物酶染色(MPO)检测的多形核细胞、CD68和CD163阳性巨噬细胞、CD4+和CD8+T淋巴细胞,以及1型干扰素基因MX1的扩散上调(图33)。与未感染的动物相比,2019-nCoV感染导致肺泡的多形核细胞浸润显著增加(P=0.0286),并且在大约30%的总肺组织中出现广泛的MX1染色(P=0.0286)(图34)。在较大呼吸道的呼吸道上皮粘膜下层中也检测到炎症浸润,伴有炎症细胞迁移到细支气管腔中(图25J)。纤毛上皮细胞也对SARS CoV-2RNA (图25K)和SARS核衣壳(图25L)呈染色阳性。到感染后第4天,炎症和病毒性肺炎的程度已经减轻,但在肺实质和中性粒细胞浸润中仍检测到病毒,并且1型干扰素应答持续存在(图34)。
为了进一步表征受感染的组织,进行了循环免疫荧光(CyCIF)成像,这是一种对多聚甲醛固定的组织样品(16)进行多重免疫表型分析的方法。对组织进行核衣壳(SARS-N)、泛细胞角蛋白(识别上皮细胞)、Iba-1(离子钙结合适配分子作为泛巨噬细胞标志物)、CD68(单核细胞/巨噬细胞标志物)和CD206(巨噬细胞标志物)染色,此外还有一组标志物用于识别其他免疫细胞和解剖结构,以及对DNA进行复染以标记所有细胞核。病毒感染细胞的病灶随机分散在整个肺中,并且与炎症浸润可变地相关(图26A-D)。一些实质性巩固(parenchymal consolidation)和炎症区域几乎不含病毒或不含病毒(图26A,箭头;图35)。病毒感染的细胞经常与泛细胞角蛋白共染色(图26E-H),表明它们是肺泡上皮细胞(肺细胞)。在病毒感染的上皮细胞附近也经常检测到未感染的Iba-1+CD68+CD206+激活的巨噬细胞(图26E,I-K)。这些数据表明,2019-nCoV诱导多灶性急性炎症和病毒性肺炎,涉及感染的肺细胞、纤毛支气管上皮细胞和可能的其他细胞类型。
在恒河猴中2019-nCoV对再次攻击的保护效力
在初始病毒感染后的第35天(图23-24),用与初次感染使用的相同剂量的2019-nCoV,即1.1x106PFU(第1组;N=3),1.1x105PFU(第2组;N=3),或1.1x104PFU(第3组;N=3)再次攻击所有9只恒河猴。重新攻击实验中包括3只幼稚(naive)动物作为阳性对照。在两只第1组动物和一只第2组动物再次攻击后的第1天,在BAL中观察到非常有限的病毒RNA,在随后的时间点没有检测到病毒RNA(图27A)。相反,正如预期的那样,在同时受到攻击的幼稚 动物中观察到高水平的病毒RNA(图27A)。与初次攻击相比,再攻击后BAL中的病毒载量峰值的中值降低>5.1log10(P<0.0001,双边Mann-Whitney检验;图27B)。与BAL相比,再次攻击后NS中的病毒RNA较高,但表现出剂量依赖性和快速下降(图27C),再次攻击后NS中的病毒载量峰值的中值仍比初次攻击低>1.7log10(P=0.0011,双边Mann-Whitney检验;图27D)。
动物中观察到高水平的病毒RNA(图27A)。与初次攻击相比,再攻击后BAL中的病毒载量峰值的中值降低>5.1log10(P<0.0001,双边Mann-Whitney检验;图27B)。与BAL相比,再次攻击后NS中的病毒RNA较高,但表现出剂量依赖性和快速下降(图27C),再次攻击后NS中的病毒载量峰值的中值仍比初次攻击低>1.7log10(P=0.0011,双边Mann-Whitney检验;图27D)。
再次攻击后在NS中检测到的大多数病毒推测为输入攻击病毒,因此在再次攻击后在NS中评估sgmRNA水平。在再次攻击后的第1天,在9只动物中的4只,在NS中仍观察到低但可检测水平的sgmRNA,但sgmRNA水平迅速下降(图27E),并且在再次攻击后NS中的sgmRNA峰值水平的中值相比初次攻击降低>4.8log10(P=0.0003,双边Mann-Whitney检验;图27F)。与这些数据一致,再攻击后BAL和NS样品中的噬斑测定显示没有可恢复的病毒并且低于初次感染后的水平(分别为P=0.009和0.002,双边Mann-Whitney检验;图36)。此外,在再次攻击后的动物中几乎没有观察到临床疾病或没有观察到临床疾病(图37)。
在2019-nCoV再次攻击后,动物表现出快速的抗体再生免疫应答,包括在再次攻击后第7天时,增加的病毒特异性ELISA滴度(P=0.0034,双边Mann-Whitney检验)、增加的假病毒NAb滴度(P=0.0003)和增加的活病毒NAb滴度(P=0.0003),以及IFN-γELISPOT应答增加的趋势(P=0.1837)(图28)。无论在BAL或NS中是否存在残留的病毒RNA或sgmRNA,所有动物在再次攻击后都产生了抗体再生抗体应答,因此推测对再次攻击的保护效力是由快速免疫控制介导的。
讨论
从某些病毒感染中恢复的个体通常会产生病毒特异性抗体应答,以提供强的保护性免疫以防止再次暴露,但一些病毒不会产生保护性天然免疫,例如HIV-1(17)。对普通感冒冠状病毒229E的人攻击研究表明,可能存在部分天然免疫(18)。然而,目前尚无数据表明从2019-nCoV感染中恢复的人是否可以避免再次暴露(世界卫生组织,Scientific Brief,2020年4月24日)。这是一个关键问题,对疫苗开发、公共卫生策略、基于抗体的疗法和群体免疫的流行病学模型具有深远的影响。在这项研究中,证明在恒河猴中感染2019-nCoV对2019-nCoV再次攻击具有显著的保护效力。
开发了2019-nCoV感染的恒河猴模型,再现了人2019-nCoV感染的许多方面,包括上呼吸道和下呼吸道的高水平病毒复制(图23)和病毒性肺炎的明确病理证据(图25、26)。组织病理学、免疫组织化学、 和CyCIF成像证明在急性炎症区域存在多灶性病毒感染细胞簇,有证据表明肺泡肺细胞和纤毛支气管上皮细胞的病毒感染。这些数据表明,恒河猴可用作2019-nCoV感染模型,用于测试疫苗和治疗方法以及研究免疫发病机制,这些发现补充和扩展了最近发表的食蟹猴数据(19)。然而,这两种非人灵长类动物模型都不会导致呼吸衰竭或死亡,因此需要进一步研究来开发一种严重的COVID-19疾病的非人灵长类动物模型。
和CyCIF成像证明在急性炎症区域存在多灶性病毒感染细胞簇,有证据表明肺泡肺细胞和纤毛支气管上皮细胞的病毒感染。这些数据表明,恒河猴可用作2019-nCoV感染模型,用于测试疫苗和治疗方法以及研究免疫发病机制,这些发现补充和扩展了最近发表的食蟹猴数据(19)。然而,这两种非人灵长类动物模型都不会导致呼吸衰竭或死亡,因此需要进一步研究来开发一种严重的COVID-19疾病的非人灵长类动物模型。
恒河猴中的2019-nCoV感染导致强的体液和细胞免疫应答(图2),并提供防止再次攻击的强的保护(图27)。2019-nCoV再攻击后,一部分动物的鼻拭子中残留的低水平亚基因组mRNA(图27)和所有动物的抗体再生免疫应答(图28)表明保护是由快速免疫控制介导的,可能不是消除病毒。
恒河猴中的2019-nCoV感染导致通过假病毒中和测定和活病毒中和测定诱导的中和抗体滴度约为100,但中和抗体、其他功能性抗体、细胞免疫和先天免疫对防止2019-nCoV的保护效力的相对重要性仍有待确定。此外,还需要其他的研究来确定自然免疫的持久性。
总之,恒河猴中的2019-nCoV感染诱导了体液和细胞免疫应答,并提供了防止2019-nCoV再次攻击的强保护效力。这些数据提出了一种可能性,即预防和治疗2019-nCoV感染的免疫方法实际上是可能的。
参考文献
1.F.Wu et al.,A new coronavirus associated with human respiratorydisease in China.Nature579,265-269(2020).
2.P.Zhou et al.,A pneumonia outbreak associated with a newcoronavirus of probable bat origin.Nature 579,270-273(2020).
3.M.L.Holshue et al.,First Case of 2019Novel Coronavirus in theUnited States.N Engl JMed 382,929-936(2020).
4.Q.Li et al.,Early Transmission Dynamics in Wuhan,China,of NovelCoronavirus-Infected Pneumonia.NEngl JMed,(2020).
5.N.Zhu et al.,A Novel Coronavirus from Patients with Pneumonia inChina,2019.N Engl JMed 382,727-733(2020).
6.N.Chen et al.,Epidemiological and clinical characteristics of99cases of 2019 novel coronavirus pneumonia in Wuhan,China:a descriptivestudy.Lancet 395,507-513(2020).
7.C.Huang et al.,Clinical features of patients infected with 2019novel coronavirus in Wuhan,China.Lancet 395,497-506(2020).
8.J.F.Chan et al.,Afamilial cluster of pneumonia associated with the2019 novel coronavirus indicating person-to-person transmission:a study of afamily cluster.Lancet 395,514-523(2020).
9.R.Wolfel et al.,Virological assessment of hospitalized patientswith COVID-2019.Nature,(2020).
10.Z.Y.Yang et al.,A DNA vac cine induces SARS coronavirusneutralization and protective immunity in mice.Nature 428,561-564(2004).
11.T.Scobey et al.,Reverse genetics with a full-length infectiouscDNA of the Middle East respiratory syndrome coronavirus.Proc Natl Acad SciUSA 110,16157-16162(2013).
12.B.Yount et al.,Reverse genetics with a full-length infectious cDNAof severe acute respiratory syndrome coronavirus.Proc NatlAcad Sci USA 100,12995-13000(2003).
13.A.W.Chung et al.,Dissecting Polyclonal Vaccine-Induced HumoralImmunity against HIV Using Systems Serology.Cell 163,988-998(2015).
14.C.Deleage et al.,Impact of early cART in the gut during acute HIVinfection.JCI insight1,(2016).
15.C.Deleage et al.,Defining HIV and SIV Reservoirs in LymphoidTissues.Pathog Immun 1,68-106(2016).
16.J.R.Lin et al.,Highly multiplexed immunofluorescence imaging ofhuman tissues and tumors using t-CyCIF and conventional opticalmicroscopes.eLife 7,(2018).
17.M.Altfeld et al.,HIV-1 superinfection despite broad CD8+ T-cellresponses containing replication of the primary virus.Nature420,434-439(2002).
18.K.A.Callow,H.F.Parry,M.Sergeant,D.A.Tyrrell,The time course oftheimmune response to experimental coronavirus infection of man.Epidemiol Infect105,435-446(1990).
19.B.Rockx et al.,Comparative pathogenesis of COVID-19,MERS,and SARSin a nonhuman primate model.Science,(2020).
实施例10.向人受试者给药或再给药抗冠状病毒组合物
基于不存在抗冠状病毒抗体水平(例如,在受试者的血液中)或抗冠状病毒抗体水平低于如本文所定义的保护水平,可鉴定人受试者为对冠状病毒感染易感。根据本文所述的方法,可以向受试者给药抗冠状病毒组合物(例如本文所述的任何组合物或免疫原性组合物)作为第一次给药(初免)或作为再给药(加强)。治疗后,可以通过测量人受试者(例如,受试者血液中)的抗冠状病毒抗体水平来测试人受试者的治疗有效性。如果该水平低于如本文所定义的保护水平,则可以随后给予人受试者抗冠状病毒组合物的加强或可以用与先前给药的抗冠状病毒组合物不同的抗冠状病毒组合物治疗人受试者。
可以在治疗后不久(例如,在给药后1天和给药后8周之间)测量抗冠状病毒抗体水平,以确定是否需要额外剂量(例如,加强)来达到抗冠状病毒抗体的保护水平。还可以在更长的时间范围内(例如,在给药后2个月和给药后15年之间)监测受试者中的抗冠状病毒抗体水平,并且如果抗冠状病毒抗体低于如本文所定义的保护水平可以重新给药抗冠状病毒组合物。
实施例11.单剂疫苗可防止恒河猴中的2019-nCoV
要结束COVID-19大流行,可能需要一种安全有效的2019-nCoV疫苗1-8。对于全球部署和大流行控制,只需要单次免疫的疫苗将是最优的。本文显示了单剂的基于腺病毒血清型26(Ad26)载体的疫苗的免疫原性和保护效力,所述疫苗在非人灵长类动物中表达2019-nCoV刺突(S)蛋白。用表达一组S变体之一或假对照的Ad26载体免疫52只恒河猴,并通过鼻内和气管内途径9,10用2019-nCoV攻击恒河猴。最优Ad26疫苗可诱导强的中和抗体应答,并在2019-nCoV攻击后在支气管肺泡灌洗液和鼻拭子提供完全或接近完全的保护。疫苗引发的中和抗体滴度与保护效力相关,表明保护的免疫相关因素。这些数据证明了在非人灵长类动物中防止2019-nCoV的强的单次疫苗保护,这是一种本领域接受的人疾病治疗动物模型。针对2019-nCoV的最优的基于Ad26载体的疫苗,称为Ad26.COV2.S,计划将其用于即将进行的临床试验。
COVID-19大流行的迅速扩大使开发2019-nCoV疫苗成为全球卫生优先事项。已显示腺病毒血清型26(Ad26)载体11在非人灵长类动物和人中诱导对多种病原体的强体液和细胞免疫应答。开发了一系列表达2019-nCoV刺突(S)蛋白不同变体的Ad26载体,并评估了它们在恒河猴中防止2019-nCoV攻击的免疫原性和保护效力。
材料和方法
动物和研究设计.将52只6-12岁的远交系的印度裔成年雄性和雌性恒河猴(Macaca mulatta)随机分组。所有动物都饲养在Bioqual,Inc.(Rockville,MD)。动物接受表达tPA.S(N=4)、tPA.S.PP(N=4)、S(N=4)、S.dCT(SS-SdCT)(N=4)、tPA.WT.S(N=4)、S.dTM.PP(SS-S.Ecto-dF-PP-foldon)(N=6)、S.PP (SS-Spike-dF-PP)(N=6)和假对照(N=20)的Ad26载体。通过无佐剂的肌内途径,动物在第0周接受了1011病毒颗粒(vp)Ad26载体的单次免疫。在第6周,用1.2x108VP(1.1x104PFU)的2019-nCoV攻击所有动物,所述2019-nCoV来源于USA-WA1/2020(NR-52281,BEI Resources)9。通过鼻内(IN)途径给药1ml病毒(每个鼻孔0.5ml)和通过气管内(IT)途径给药1ml。所有免疫学和病毒学测定均在盲法下进行。所有动物研究均按照所有相关的地方、州和联邦法规进行,并得到生物质量机构动物管理和使用委员会(IACUC)的批准。
Ad26载体.Ad26载体由2019-nCoV刺突(S)蛋白序列(Wuhan/WIV04/2019;GenbankMN996528.1)的七个变体构建而成。序列经过密码子优化和合成。使用含有完整Ad26载体基因组和转基因表达盒的质粒在PER.C6.TetR细胞中产生无复制能力、E1/E3缺失的Ad26载11。在使用前对载体进行测序和表达测试。
免疫印迹.用2019-nCoV DNA表达质粒瞬时转染接种有70-80%汇合度的293T细胞的T-25培养瓶,在转染后48小时收获的细胞裂解物,并分别与还原样品缓冲液(Pierce)混合,在95℃加热5分钟,并且在预制的4-15%SDS-PAGE凝胶(Bio-Rad)上跑胶。使用iBlot干式印迹系统(Invitrogen)将蛋白转移到聚偏二氟乙烯(PVDF)膜上,并在4℃下在含有0.2%Tween 20(Sigma)(V/V)和5%(W/V)脱脂奶粉的Dulbecco磷酸盐缓冲盐水T(D-PBST)中进行膜封闭过夜。过夜封闭后,将PVDF膜在含有1∶10,000稀释的多克隆豚鼠抗SARS抗体(BEIresources)的3%牛奶DPBS-T中孵育1小时。孵育后,PVDF膜用5%牛奶DPBS-T洗涤5次,并且随后与3%牛奶DPBS-T中的1∶30,000抗豚鼠或抗兔辣根过氧化物酶(HRP)缀合的二抗(Jackson Immunoresearch)一起孵育。最后,将PVDF膜用5%牛奶DPBS-T再次洗涤5次,并使用Amersham ECL Plus免疫印迹检测系统(GE Healthcare)显影。
亚基因组mRNA测定.是通过RT-PCR使用与先前描述的方法相似的方法评估2019-nCoV E基因亚基因组mRNA(sgmRNA)9,10,20。简而言之,为了生成标准曲线,将2019-nCoV E基因sgmRNA克隆至pcDNA3.1表达质粒中;使用AmpliCap-Max T7High Yield Message MakerKit (Cellscript)转录此插入物,以获得标准的RNA。在RT-PCR之前,根据制造商的说明,使用Superscript III VILO(Invitrogen)将从受攻击的动物或标准中收集的样品进行逆转录。使用靶向E基因sgmRNA20的序列设计了Taqman定制基因表达测定(ThermoFisherScientific)。根据制造商的说明,在QuantStudio 6和7Flex实时PCR系统(AppliedBiosystems)上进行反应。标准曲线用于计算每毫升或每拭子sgmRNA的拷贝数;定量测定的灵敏度为每毫升或每拭子50个拷贝。
PFU测定.对于噬斑测定,在6孔板中制备Vero E6细胞的汇合单层。将从受攻击的动物中收集的指定样品连续稀释,添加到孔中,并在37℃下孵育1小时。孵育后,将1.5mL0.5%甲基纤维素培养基添加至每个孔中,并将板在37℃和5%CO2下孵育2天。通过每孔加入400μL冰冷的甲醇并在-20℃下孵育30分钟来固定板。固定后弃去甲醇,用每孔600μL的0.23%结晶紫染色细胞单层30分钟。染色后,弃去结晶紫,用600μL水洗板一次以可视化和计数噬斑。
ELISA.基本上如所述9,10,通过ELISA评估RBD特异性结合抗体。简而言之,用1xDPBS中的1μg/mL 2019-nCoV RBD蛋白(Aaron Schmidt,MassCPR)包被96孔板,并在4℃下孵育过夜。孵育后,用洗涤缓冲液(1X DPBS中的0.05%Tween20)洗涤板一次,并在室温下用350μL酪蛋白封闭液/孔封闭2-3小时。孵育后,弃去封闭溶液并将板吸干。将在酪蛋白封闭液中稀释的热灭活血清的连续稀释液添加至孔中,并将板在室温下孵育1小时,然后再洗涤3次,随后用1∶1000稀释的抗猕猴IgG HRP(NIH NHP Reagent Program)在室温下避光孵育1小时。然后洗涤板3次,并且每孔加入100μL KPL TMB SureBlue Start溶液;通过每孔添加100μL
KPL TMB SureBlue Start溶液;通过每孔添加100μL KPL TMB Stop溶液来停止板的显色。使用VERSAMAXTM或
KPL TMB Stop溶液来停止板的显色。使用VERSAMAXTM或 微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
假病毒中和测定.通过与之前描述的方法相似的方法产生表达荧光素酶报告基因的2019-nCoV假病毒9,10,16。简而言之,通过磷酸钙,将包装构建体psPAX2(AIDS Resourceand Reagent Program)、荧光素酶报告质粒pLenti-CMV Puro-Luc(Addgene)和表达刺突蛋白的pcDNA3.1-SARS CoV-2 SΔCT共转染到HEK293T细胞中。转染后48小时收集含有假病毒的上清液;通过用0.45μm过滤器过滤来纯化假病毒。为了确定来自疫苗接种动物的抗血清的中和活性,将HEK293T-hACE2细胞以1.75×104个细胞/孔的密度接种在96孔组织培养板中过夜。制备热灭活血清样品的两倍连续稀释液,并与50μL假病毒混合。在加入HEK293T-hACE2细胞之前,将混合物在37℃下孵育1小时。48小时后,根据制造商的说明,在 Luciferase Assay(Promega)中裂解细胞。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
Luciferase Assay(Promega)中裂解细胞。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
活病毒中和测定.设计基于Seattle Washington分离株的全长2019-nCoV病毒用于表达荧光素酶和GFP,并通过反向遗传学回收,如先前所描述17,18。病毒在Vero E6USAMRID细胞中滴定,以获得仅细胞对照背景的至少10倍的相对光单位(RLU)信号。Vero E6USAMRID细胞在前一天以每孔20,000个细胞接种在透明底黑壁96孔板中。中和抗体血清样品以1∶4的起始稀释度进行检测,并连续稀释4倍,最多8个稀释度。抗体-病毒复合物在37℃和5%CO2下孵育1小时。孵育后,去除生长培养基并将病毒-抗体稀释复合物一式两份地添加到细胞中。每个中和测定板中包括仅病毒对照和仅细胞对照。感染后,将平板在37℃和5%CO2下孵育48小时。然后,裂解细胞并根据制造商说明,通过Nano-Glo LuciferaseAssay System(Promega)测量荧光素酶活性。将2019-nCoV中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
系统血清学.
Luminex
如前所述29,使用了一种定制的多样的方法来定量相对抗原特异性抗体滴度。因此,具有独特荧光的微球(Luminex)通过EDC(Thermo Scientific)和Sulfo-NHS(ThermoScientific)通过共价N-羟基琥珀酰亚胺(NHS)-酯键与2019-nCoV抗原(包括刺突蛋白(S)和受体结合结构域(RBD))偶联。在384孔板(Greiner)的每孔中,添加1.2×103珠每个区域/抗原,并与稀释的血清样品一起孵育(所有同种型/亚类以1∶100,除了IgG1以及Fc受体结合以1∶250稀释)在4℃下以900rmp振动16小时。在形成免疫复合物后,用自动洗板机(Tecan)在0.1%BSA和0.05%Tween-20(Luminex测定缓冲液)中洗涤微球3次。将抗恒河猴IgG1、IgG2、IgG3、IgA(NIH NHP Reagent Program)和IgM(Life Diagnostic)检测抗体在Luminex测定缓冲液中稀释至0.65ug/mL,并在室温下与珠一起孵育1小时,同时以900rpm振动。洗涤染色的免疫复合物后,将三级山羊抗小鼠IgG-PE抗体(Southern Biotech)以0.5ug/mL添加到每个孔中,并在室温下在振荡器上孵育1小时。类似地,对于Fc受体结合谱,生物素化(Thermo Scientific)重组恒河猴FcγR2A-1、FcγR2A-2、FcγR2A-3、FcγR2A-4、FcγR3A和人FcγR2B(Duke Protein Production facility)并与链霉亲和素-PE偶联10分钟(Southern Biotech)。然后洗涤包被的珠并在连接有机械臂(PAA)的流式细胞仪iQue(Intellicyt)上读数。在每个珠区域对事件进行门控,报告珠阳性事件的PE中值荧光。每个二级检测试剂均一式两份运行样品。
基于珠的ADNP、ADCP、ADCD
如前所述30-32进行抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性细胞吞噬作用(ADCP)和抗体依赖性补体沉积(ADCD)。简而言之,生物素化(Thermo Fisher)2019-nCoV S和RBD并在37℃下与1μm黄色(ADCP、ADNP)和红色(ADCD)荧光珠偶联2小时。通过用PBS中的0.1%BSA洗涤两次来去除多余的抗原。随后,将1.82×108个抗原包被的珠添加到96孔板的每个孔中,并与稀释的样品(ADCP和ADNP 1∶100,ADCD 1∶10)在37℃下孵育2小时以促进免疫复合物的形成。孵育后,洗涤复合珠,对于ADCP,每孔加入2.5×104THP-1细胞(美国典型培养物保藏中心)并在37℃孵育16小时。对于ADNP,通过添加氯化铵钾(Ammonium-Chloride-Potassium,ACK)裂解(Thermo fisher)通过裂解红细胞从健康供体中分离出来外周血单个核细胞(PBMC),每孔添加5×104个细胞并在37℃下孵育1小时。随后,用抗Cd66b Pac blue检测抗体(BioLegend)对原代血细胞进行染色。对于ADCD,根据制造商的说明(Cedarlane)用水重构冻干豚鼠补体,并在含有Mg2+和Ca2+(GVB++,BostonBioProducts)的明胶弗洛拿缓冲液中每孔加入4μL,在37℃下持续20分钟。用PBS中的15mM EDTA洗涤两次后,用荧光素缀合的山羊IgG豚鼠补体C3组分(Mpbio)染色免疫复合物。对于ADCD和ADNP,在与THP-s孵育和细胞染色后,用4%多聚甲醛(PFA)固定细胞样品,并使用机械臂(PAA)通过流式细胞仪(Intellicyt,iQue Screener plus)进行样品采集。以单细胞和珠阳性事件门控所有事件,对于ADCP和ADNP,吞噬作用评分计算为珠阳性细胞×GMFI/1,000的百分比。对于ADCD,报告了C3阳性事件的中值。所有样品在不同的天一式两份进行。
ADNKA
为了分析NK细胞相关应答,使用了基于ELISA的测定。因此,在37℃下用2019-nCoVS包被96孔ELISA板(Thermo Fisher)2小时。然后洗涤板并且用PBS中的5%BSA将板在4℃下封闭过夜。使用RosetteSep分离试剂盒(Stem Cell Technologies)从健康供体(MGH献血中心)的血沉棕黄层中分离出NK细胞,并在补充IL-15(Stemcell)的情况下将NK细胞静置过夜。将血清样品以1∶50稀释并在ELISA板上于37℃孵育2小时。将抗CD107a-PE-Cy5染色剂(BD)、布雷菲德菌素A(brefeldin A)(Sigma)和GolgiStop(BD)的染色混合物添加到NK细胞中,每孔添加5×104个NK细胞并在37℃下孵育5小时。使用Perm A和B(Thermo Fisher)固定和透化NK细胞,并用抗CD16APC-Cy7(BD)、抗CD56 PE-Cy7(BD)和抗CD3 AlexaFluor 700抗体(BD)对表面标志物进行染色。细胞内染色包括抗IFNγAPC(BD)和抗MIP-1βPE(BD)。通过配备机械臂(PAA)的流式细胞仪iQue(Intellicyt)进行采集。定义NK细胞为CD3-、CD16+和CD56+。ADNKA测定在两个献血者中一式两份进行。
分析
所有同种型/亚类、Fc受体结合和ADCD数据都进行了log10转换。对于雷达图,在使用组内的中值之前,对每个抗体特征进行标准化,使其最小值为0,组间的最大值为1。使用R包“ropls”构建主成分分析(PCA)以比较多变量谱。将完全受保护的动物定义为在BAL和NS中没有可检测的sgmRNA拷贝/mL。使用双边Mann-Whitney检验比较完全受保护与部分受保护和未受保护的动物。对于热图中的可视化,显示了完全和部分受保护的组的z分数特征的平均值的差异。为了表明热图中的显著性,使用Benjamini-Hochberg校正来校正多重比较。为了评估特征及其组合预测保护的能力,在10倍交叉验证框架中对逻辑回归模型进行了100次重复训练,并计算了接收者操作特征(AUROC)曲线下的面积。测试了两个特征的所有潜在组合。为此,使用了R包“glm”和“pROC”。
ELISPOT测定.用来自BD Pharmingen的小鼠抗人IFN-γ单克隆抗体以5μg/孔的浓度包被ELISPOT板,在4℃下过夜。用含有0.25%Tween20的DPBS洗涤板,并在37℃下用R10培养基(含有11%FBS和1.1%青霉素-链霉素的RPMI)封闭1小时。刺突1和刺突2肽池含有15个氨基酸肽,这些肽有11个氨基酸重叠,它们涵盖蛋白序列并分别反映蛋白的N端和C端半部分。以2μg/孔的浓度制备刺突蛋白1和刺突蛋白2肽池,并添加200,000个细胞/孔。将肽和细胞在37℃下孵育18-24小时。此孵育后的所有步骤均在室温下进行。用库尔特(coulter)缓冲液洗涤板并与来自U-Cytech的兔多克隆抗人IFN-γ生物素(1μg/mL)一起孵育2小时。将板第二次洗涤并与来自Southern Biotechnology的链霉亲和素-碱性磷酸酶抗体(1μg/mL)一起孵育2小时。最后一次洗涤后添加氯化硝基四氮唑蓝/5-溴-4-氯-3′-吲哚基磷酸对甲苯胺盐(NBT/BCIP显色液)底物溶液,保持7分钟。弃去显色液(chromagen),将板用水洗涤并在暗处干燥24小时。在Cellular Technologies Limited Immunospot Analyzer上扫描和计数板。
细胞内细胞因子染色测定.将106PBMC/孔重悬在100μL的补充有CD49d单克隆抗体(1μg/mL)的R10培养基中。使用模拟(100μL的R10加0.5%DMSO;背景对照)、刺突1肽池和刺突2肽池(2μg/mL)或10pg/mL豆蔻酰佛波醇乙酯(phorbol myristate acetate)(PMA)和1μg/mL离子霉素(Sigma-Aldrich)(100μL;阳性对照)对每个样品进行评估并在37℃下孵育1小时。孵育后,将50μL R10中的0.25μL GolgiStop和0.25μL GolgiPlug添加到每个孔中,并在37℃下孵育8小时,然后在4℃下保持过夜。第二天,细胞用DPBS洗涤两次,用近红外活/死染料染色10分钟,然后用针对CD279(克隆EH12.1、BB700)、CD38(克隆OKT10、PE)、CD28(克隆28.2,PE CY5)、CD4(克隆L200,BV510)、CD45(克隆D058-1283,BUV615)、CD95(克隆DX2,BUV737)、CD8(克隆SK1,BUV805)的预先指定滴度的单克隆抗体染色,持续30分钟。然后用2%FBS/DPBS缓冲液洗涤细胞两次,并用200μL BD CytoFix/CytoPerm Fixation/Permeabilization溶液孵育15分钟。用1X Perm洗涤缓冲液(将CytoFix/CytoPermFixation/Permeabilization试剂盒中的BD Perm/WashTM Buffer 10X用Milli Q水稀释并通过0.22μm过滤器)洗涤细胞两次,并用针对Ki67(克隆B56,FITC)、CD69(克隆TP1.55.3,ECD)、IL10(克隆JES3-9D7,PE CY7)、IL13(克隆JES10-5A2,BV421)、TNF-α(克隆Mab11,BV650)、IL4(克隆MP4-25D2,BV711)、IFN-γ(克隆B27;BUV395)、IL2(克隆MQ1-17H12、APC)、CD3(克隆SP34.2、Alexa 700)的单克隆抗体进行细胞内染色30分钟。细胞用1x Perm洗涤缓冲液洗涤两次,并用250μL新鲜制备的1.5%甲醛固定。将固定的细胞转移至96孔圆底板中并通过BD FACSymphonyTM系统进行分析。
统计分析.使用GraphPad Prism 8.4.2(GraphPad软件)对病毒学和免疫学数据进行分析。使用双边Mann-Whitney检验进行组间数据比较。通过双边Spearman等级相关性检验评估相关性。认为P值小于0.05是显著的。
结果
Ad26候选疫苗的产生和免疫原性
产生了七种表达2019-nCoV S变体的Ad26载体,这些变体反映了不同的启动子、抗原形式和稳定化突变:(i)组织纤溶酶原激活物(tPA)前导序列和全长S(tPA.S(SEQ ID NO:1,tPA前导序列与N端融合))12,(ii)tPA前导序列和具有弗林蛋白酶切割位点突变和两个脯氨酸稳定化突变的全长S(tPA.S.PP(SEQ ID NO:23),tPA前导序列与N端融合))13-15,(iii)野生型前导序列和天然全长S(S(SEQ ID NO:29)),(iv)野生型前导序列和细胞质尾缺失的S(S.dCT(SS-SdCT)(SEQ ID NO:30))16,(v)串联tPA和野生型前导序列以及全长S(tPA.WT.S(SEQ ID NO:29,tPA前导序列与N端融合))12,(vi)野生型前导序列和S,所述S具有反映可溶性胞外域的跨膜区和细胞质尾的缺失,弗林蛋白酶切割位点的突变,脯氨酸稳定化突变以及折叠子三聚化结构域(S.dTM.PP(SS-S.Ecto-dF-PP-foldon)(SEQ ID NO:56))15和(vii)野生型前导序列和具有弗林蛋白酶切割位点突变和脯氨酸稳定化突变的全长S序列(S.PP(SS-Spike-dF-PP)(SEQ ID NO:51))(图38a)。免疫印迹分析确认了在细胞裂解物中来自所有载体的S表达(图38b)。
用表达tPA.S(N=4)、tPA.S.PP(N=4)、S(N=4)、S.dCT(SS-SdCT)(N=4)、tPA.WT.S(N=4)、S.dTM.PP(SS-S.Ecto-dF-PP-foldon)(N=6)、S.PP(SS-Spike-dF-PP)(N=6)和假对照(N=20)的Ad26载体免疫52只6-12岁的成年恒河猴。动物在第0周通过无佐剂的肌内途径接受了1011病毒颗粒(vp)Ad26载体的单次免疫。通过ELISA,到第2周,在32只疫苗接种动物中的31只和到第4周,在所有疫苗接种动物中观察到RBD特异性结合抗体(图39a)。使用假病毒中和测定9,10,16(图39b)和活病毒中和测定9,10,17,18(图39c)评估中和抗体(NAb)应答。在第2周在大多数疫苗接种的动物中观察到通过两种测定测量的NAb滴度,并且通常在第4周增加。在第4周,Ad26-S.PP(Ad26SS-Spike-dF-PP)疫苗引发最高的假病毒NAb滴度(中值为408;范围208-643)和活病毒NAb滴度(中值为113;范围53-233)。假病毒NAb滴度与ELISA滴度和活病毒NAb滴度相关(分别为P<0.0001,R=0.8314和P<0.0001,R=0.8427,双边Spearman等级相关性检验;图45)。疫苗接种Ad26-S.PP的猕猴的NAb滴度的中值是先前报道从2019-nCoV感染恢复后的9只恢复期猕猴9和27个恢复期人10的队列中的NAb滴度的中值的4倍(图46)。Ad26-S.PP疫苗还在支气管肺泡灌洗液(BAL)中诱导低但可检测到的S特异性IgG和IgA应答(图47)。
疫苗接种动物中的S特异性和RBD特异性抗体应答通过系统血清学19进一步表征。观察到多种Fc效应功能,其包括抗体依赖性中性粒细胞吞噬作用(ADNP)、抗体依赖性单核细胞吞噬作用(ADCP)、抗体依赖性补体沉积(ADCD)和抗体依赖性NK细胞激活(ADNKA),以及多个Ig亚类和FcR结合(图39d,图48)。用Ad26-tPA.S.PP和Ad26-S.PP疫苗观察到最高的结合抗体应答,并且用Ad26-S.PP疫苗观察到最高的效应功能应答。主成分分析显示疫苗组有大量重叠,尽管Ad26-S.PP是最不同的组(图39d)。
在第4周,通过IFN-γELISPOT测定,使用合并的S肽在32只接种动物中的30只中诱导了细胞免疫应答(图40a)。使用多参数细胞内细胞因子染色测定来评估IFN-γ+CD4+和CD8+T细胞应答(图40b)。大多数疫苗组的应答相当,尽管用Ad26-S.PP疫苗有降低细胞免疫应答的趋势。对于Ad26-S.dTM.PP(Ad26 SS-S.Ecto-dF-PP-foldon)和Ad26-S.PP疫苗,IFN-γ+和IL-2+CD4+T细胞应答高于IL-4+和IL-10+CD4+T细胞应答(图49),表明诱导Th1偏向应答。
Ad26候选疫苗的保护效力
在第6周,通过鼻内(IN)和气管内(IT)途径9,10,用1.2x108VP(1.1x104PFU)的2019-nCoV攻击所有动物。通过RT-PCR测定亚基因组mRNA(sgmRNA)评估支气管肺泡灌洗液(BAL)和鼻拭子(NS)中的病毒载量,其被认为可以测量复制的病毒9,20。所有20个假对照被感染并且在BAL中示出4.89(范围3.85-6.51)log10sgmRNA拷贝/mL峰值的中值(图41a)。相比之下,接受Ad26-S.PP(Ad26 SS-Spike-dF-PP)的动物在BAL中未显示出可检测的病毒(定量限为1.69log10 sgmRNA拷贝/mL)。尽管偶尔动物在BAL中表现出低水平的sgmRNA(图41b),但使用其他疫苗也观察到了显著的保护作用。类似地,假对照在NS中示出5.59(范围3.78-8.01)log10 sgmRNA峰值的中值(图41c)。只有一只接受Ad26-S.PP疫苗的动物在NS中示出少量病毒。与对照相比,尽管Ad26-S.PP的保护是最优的,接受其他疫苗的动物通常在NS中表现出降低的病毒载量(图41d)。
疫苗接种动物的病毒载量峰值的比较表明,BAL中的保护通常比NS中更强(图42)。Ad26-S.PP疫苗在下呼吸道和上呼吸道都提供了完全的保护,除了一只动物在NS中示出少量病毒,并且与假对照相比,在BAL和NS中,Ad26-S.PP疫苗分别导致sgmRNA峰值的中值降低>3.2和>3.9log10(分别为P<0.0001和P<0.0001,双边Mann-Whitney检验)(图42)。在32只疫苗接种的猕猴中,17只动物受到完全保护并且在攻击后在BAL或NS中未检测到sgmRNA,另外5只动物在BAL中没有sgmRNA但在NS中示出一些病毒。
保护的免疫相关因素
这项研究的规模和不同疫苗构建体的结果的差异促进了免疫相关因素的分析。第2周和第4周的log10 ELISA滴度、假病毒NAb滴度和活病毒NAb滴度与在BAL(图43)和NS(图50-52)中的log10 sgmRNA峰值负相关。一般而言,第4周的滴度相关性好于第2周的滴度,NAb滴度的相关性优于ELISA滴度。第4周的log10假病毒NAb滴度和活病毒NAb滴度与BAL中(分别为P<0.0001,R=-0.6880和P<0.0001,R=-0.6562,双边Spearman等级-相关性检验)(图43b-c)和NS中(分别为P<0.0001,R=-0.5839和P<0.0001,R=-0.5714,双边Spearman等级相关性检验)(图51和52)的log10sgmRNA峰值负相关。连同之前公布的数据10,这些发现表明血清抗体滴度可能证明是2019-nCoV疫苗保护的有用免疫相关因素。相比之下,疫苗引发的ELISPOT应答、CD4+ICS应答和CD8+ICS应答与保护无关。
为了进一步了解保护的抗体相关因素,定义了区分完全受保护动物(定义为在攻击后在BAL或NS中未检测到sgmRNA的动物)和部分受保护或未受保护动物的抗体参数。与部分受保护或未受保护动物相比,NAb滴度是完全受保护动物中最丰富的参数(P=0.0009,双边Mann-Whitney检验),其次是ADNKA(P=0.0044)和ADCP(P=0.0092)应答(图43d,图53)。此外,逻辑回归分析显示利用两个特征,例如NAb滴度和FcγR2A-3、IgM或ADCD应答,改善了与保护的相关性(图43d)。这些数据表明,NAb主要负责防止2019-nCoV的保护,但其他结合和功能性抗体也可能发挥作用。
攻击后疫苗接种动物的免疫应答
到2019-nCoV攻击后的第14天,假对照和大多数疫苗接种的动物发展出显著更高的假病毒NAb应答(图44a、图54)以及CD8+和CD4+T细胞应答(图44b)。在这些动物中,CD8+和CD4+T细胞应答针对多种2019-nCoV蛋白,其包括刺突蛋白(S1、S2)、核衣壳蛋白(NCAP)和非结构蛋白(NS6、NS7a、NS8)(图44b)。相比之下,接受Ad26-S.PP(Ad26 SS-Spike-dF-PP)疫苗的动物在攻击之后没有表现出抗体再生NAb应答(图44a),仅表现出针对是疫苗抗原的刺突(S1,S2)的低T细胞应答(图44b)。这些发现与疫苗接种Ad26-S.PP的动物(图41和42)中基本上检测不到的病毒载量一致,并且表明这些动物中病毒复制水平极低,如果有的话,是在攻击之后。
讨论
开发安全有效的2019-nCoV疫苗是一项重要的全球优先事项。这些数据表明,用表达融合前稳定S免疫原的Ad26载体进行单次免疫可诱导强的NAb应答,并在恒河猴中提供防止2019-nCoV攻击的完全或接近完全的保护。本研究评估的最优疫苗是Ad26-S.PP(Ad26SS-Spike-dF-PP),它含有野生型前导序列、全长膜结合S、弗林蛋白酶切割位点突变和两个脯氨酸稳定化突变15。
这些数据扩展了最近在非人灵长类动物中对2019-nCoV灭活病毒疫苗和DNA疫苗的临床前研究10,21。灭活病毒疫苗、核酸疫苗和蛋白亚单位疫苗通常需要两次或多次免疫,而腺病毒载体可以在单次免疫后诱导强和持久的NAb应答22-24。与用于大规模疫苗接种活动和大流行控制的两剂疫苗相比,单次2019-nCoV疫苗将具有重要的统筹(logistic)和实际优势。然而,Ad26-HIV载体的同源加强可以将非人灵长类动物和人的抗体滴度提高至10倍以上25-27,这表明Ad26-S.PP疫苗的单剂和两剂方案都可以成为有效的方法。
Ad26-S.PP在单次免疫后诱导了强的NAb应答,并提供了防止2019-nCoV攻击的完全保护,除了一只动物在NS中具有低水平的病毒。此外,疫苗接种Ad26-S.PP的动物中的NAb滴度和T细胞应答在攻击后没有扩大,T细胞应答也没有扩大到非疫苗抗原,例如核衣壳和非结构蛋白。相比之下,假对照和其他疫苗在攻击后对多种2019-nCoV蛋白产生更高的NAb滴度和T细胞应答,这与之前对DNA疫苗的观察结果一致10。这些数据表明,在2019-nCoV攻击后,疫苗接种Ad26-S.PP的动物中病毒复制很少甚至没有,这可能与这些动物中较高的NAb应答有关。
攻击前疫苗引发的NAb滴度与攻击后BAL和NS中的保护相关,这与先前的研究结果10一致。这些数据表明,血清NAb滴度可能是疫苗保护的潜在生物标志物,尽管这需要在非人灵长类动物和人的其他2019-nCoV疫苗效力研究中得到确认。此外,其他功能性抗体应答也可能有助于保护,例如ADNKA、ADCP和ADCD应答。T细胞应答在疫苗保护中的作用仍有待确定,与其他载体相比,使用Ad26-S.PP的较低的细胞免疫应答的趋势需要进一步研究。
Ad26-S.dTM.PP(Ad26 SS-S.Ecto-dF-PP-foldon)和Ad26-S.PP疫苗引发偏向Th1而不是偏向Th2的CD4+T细胞应答,以及具有亚保护性的NAb滴度的动物没有表现出增强的病毒复制或临床疾病。这是反对感染的抗体依赖性增强的可能性的证据28。
总之,这些数据表明,针对2019-nCoV的基于Ad26载体的疫苗的单次免疫引发了强的NAb滴度,并在恒河猴中提供了防止2019-nCoV攻击的完全或接近完全的保护。保护的NAb相关因素的鉴定确认了这些2019-nCoV疫苗在临床上的有效性。本研究中的一种最优Ad26-S.PP疫苗,称为Ad26.COV2.S,可能对人有效。
参考文献
1 Wu,F.et al.A new coronavirus associated with human respiratorydisease in China.Nature 579,265-269,doi:10.1038/s41586-020-2008-3(2020).
2 Zhou,P.et al.A pneumonia outbreak associated with a new coronavirusof probable bat origin.Nature 579,270-273,doi:10.1038/s41586-020-2012-7(2020).
3 Holshue,M.L.et al.First Case of 2019Novel Coronavirus in the UnitedStates.N Engl J Med 382,929-936,doi:10.1056/NEJMoa2001191(2020).
4 Li,Q.et al.Early Transmission Dynamics in Wuhan,China,of NovelCoronavirus-Infected Pneumonia.N Engl J Med,doi:10.1056/NEJMoa2001316(2020).
5 Zhu,N.et al.A Novel Coronavirusfrom Patients with Pneumonia inChina,2019.N Engl J Med 382,727-733,doi:10.1056/NEJMoa2001017(2020).
6 Chen,N.et al.Epidemiological and clinical characteristics of 99cases of 2019 novel coronavirus pneumonia in Wuhan,China:a descriptiVestudy.Lancet 395,507-513,doi:10.1016/S0140-6736(20)30211-7(2020).
7 Huang,C.et al.Clinical features of patients infected with 2019novelcoronavirus in Wuhan,China.Lancet 395,497-506,doi:10.1016/S0140-6736(20)30183-5(2020).
8 Chan,J.F.et al.A familial cluster of pneumonia associated with the2019 novel coronavirus indicating person-to-person transmission:a study of afamily cluster.Lancet 395,514-523,doi:10.1016/S0140-6736(20)30154-9(2020).
9 Chandrashekar,A.et al.SARS-CoV-2 infection protects againstrechallenge in rhesus macaques.Science,doi:10.1126/science.abc4776(2020).
10 Yu,J.et al.DNA vaccine protection against SARS-CoV-2 in rhesusmacaques.Science,doi:10.1126/science.abc6284(2020).
11 Abbink,P.et al.Comparative seroprevalence and immunogenicity ofsix rare serotype recombinant adenovirus vaccine vectors from subgroups B andD.J Virol 81,4654-4663,doi:JVI.02696-06[pii]10.1128/JVI.02696-06(2007).
12 Alharbi,N.K.et al.ChAdOx1 and MVA based vaccine candidates againstMERS-CoV elicit neutralising antibodies and cellular immune responses inmice.Vaccine 35,3780-3788,doi:10.1016/j.vaccine.2017.05.032(2017).
13 Kirchdocrfer,R.N.et al.Pre-fusion structure of a human coronavirusspike protein.Nature 531,118-121,doi:10.1038/nature17200(2016).
14 Pallesen,J.et al.Immunogenicity and structures of a rationallydesigned prefusion MERS-CoV spike antigen.Proc Natl Acad Sci U S A 114,E7348-E7357,doi:10.1073/pnas.1707304114(2017).
15 Wrapp,D.et al.Cryo-EM structure of the 2019-nCoV spike in theprefusion conformation.Science 367,1260-1263,doi:10.1126/science.abb2507(2020).
16 Yang,Z.Y.et al.A DNA vaccine induces SARS coronavirusneutralization and protective immunity in mice.Nature 428,561-564,doi:10.1038/nature02463(2004).
17 Scobey,T.et al.Reverse genetics with a full-length infectious cDNAof the Middle East respiratory syndrome coronavirus.Proc Natl Acad Sci U S A110,16157-16162,doi:10.1073/pnas.1311542110(2013).
18 Yount,B.et al.Reverse genetics with a full-length infectious cDNAof severe acute respiratory syndrome coronavirus.Proc Natl Acad Sci U S A100,12995-13000,doi:10.1073/pnas.1735582100(2003).
19 Chung,A.W.et al.Dissecting Polyclonal Vaccine-Induced HumoralImmunity against HIV Using Systems Serology.Cell 163,988-998,doi:10.1016/j.cell.2015.10.027(2015).
20 Wolfel,R.et al.Virological assessment of hospitalized patientswith COVID-2019.Nature,doi:10.1038/s41586-020-2196-x(2020).
21 Gao,Q.et al.Rapid development of an inactivated vaccine candidatefor SARS-CoV-2.Science,doi:10.1126/science.abc1932(2020).
22 Abbink,P.et al.Durability and correlates of vaccine protectionagainst Zika virus in rhesus monkeys.Sci Transl Med 9,doi:10.1126/scitranslmed.aao4163(2017).
23 Abbink,P.et al.Protective efficacy of multiple vaccine platformsagainst Zika virus challenge in rhesus monkeys.Science 353,1129-1132,doi:10.1126/science.aah6157(2016).
24 Cox,F.et al.Adenoviral vector type 26 encoding Zika virus(ZIKV)M-Env antigen induces humoral and cellular immune responses and protects miceand nonhuman primates against ZIKV challenge.PLoS ONE 13,e0202820,doi:10.1371/journal.pone.0202820(2018).
25 Barouch,D.H.et al.Evaluation of a mosaic HIV-1 vaccine in amulticentre,randomised,double-blind,placebo-controlled,phase 1/2a c(inicaltrial(APPROACH)and in rhesus monkeys(NHP 13-19).Lancet 392,232-243,doi:10.1016/S0140-6736(18)31364-3(2018).
26 Barouch,D.H.et al.Protective efricacy of adenovirus/proteinvaccines against SIV challenges in rhesus monkeys.Science 349,320-324,doi:10.1126/science.aab3886(2015).
27 Baden,L.R.et al.First-in-human evaluation of the safety andimmunogenicity of a recombinant adenovirus serotype 26 HIV-1 Env vaccine(IPCAVD 001).J Infect Dis 207,240-247,doi:10.1093/infdis/jis670(2013).
28 Graham,B.S.Rapid COVID-19 vaccine development.Science,doi:10.1126/science.abb8923(2020).
29 Brown,E.P.et al.Multiplexed Fc array for evaluation of antigen-specific antibody effector profiles.J Immunol Methods 443,33-44,doi:10.1016/j.jim.2017.01.010(2017).
30 Ackerman,M.E.et al.A robust,high-throughput assay to determine thephagocytic activity of clinical antibody samples.J Immunol Methods 366,8-19,doi:10.1016/j.jim.2010.12.016(2011).
31 Lu,L..L.et al.A Functional Role for Antibodies inTuberculosis.Cell 167,433-443e414,doi:10.1016/j.cell.2016.08.072(2016).
32 Fischinger,S.et al.A high-throughput,bead-based,antigen-specificassay to assess the ability of antibodies to induce complement activation.JImmunol Methods 473,112630,doi:10.1016/j.jim.2019.07.002(2019).
实施例12.单剂疫苗可防止仓鼠中的2019-nCoV严重临床疾病
人中的COVID-19疾病在临床上通常是一种轻度疾病,但有些个体会发展为严重的肺炎、呼吸衰竭和死亡1-4。在仓鼠5-7和非人灵长类动物8-10中的SARS-CoV-2(2019-nCoV)感染的研究普遍报告了轻度的临床疾病,临床前SARS-CoV-2疫苗研究表明在非人灵长类动物的上呼吸道和下呼吸道中的病毒复制减少11-13。
如下文所述,在仓鼠中的高剂量鼻内SARS-CoV-2感染会导致严重的临床疾病,其包括组织中高水平的病毒复制、广泛性肺炎、体重减轻和部分动物死亡。使用基于腺病毒血清型26(Ad26)载体的疫苗进行单次免疫能引发结合抗体应答和中和抗体应答,并防止SARS-CoV-2引起的体重减轻、肺炎和死亡,所述疫苗表达编码稳定化的SARS-CoV-2刺突(S)蛋白(SS-Spike-dF-PP,SEQ ID NO:51)的核苷酸(SS-Spike-dF-PP,SEQ ID NO:204)。这些数据证明了防止SARS-CoV-2临床疾病的疫苗保护。这个模型应该证明了对SARS-CoV-2疫苗、治疗和发病机制的临床前研究有用。
材料和方法
动物和研究设计.将70只10-12周龄,雄性和雌性叙利亚金仓鼠(Envigo)随机分组。所有动物都饲养在Bioqual,Inc.(Rockville,MD)。动物接受表达编码S.dTM.PP(SS-S.Ecto-dF-PP-foldon,SEQ ID NO:56)的核苷酸(SS-S.Ecto-dF-PP-foldon,SEQ ID NO:195)或编码S.PP(SS-Spike-dF-PP,SEQ ID NO:51)的核苷酸(SS-Spike-dF-PP,SEQ ID NO:204)的Ad26载体或假对照(N=10/组)。通过无佐剂的肌内途径,动物在第0周接受了1010或109病毒颗粒(vp)Ad26载体的单次免疫。在第4周,用5.0x105TCID50(6x108VP,5.5x104PFU)或5.0x104TCID50(6x107VP,5.5x103PFU)的SARS-CoV-2攻击所有动物,所述SARS-CoV-2来源于USA-WA1/2020(NR-52281;BEI Resources)10的1代。病毒通过鼻内(IN)途径给药100μL(每个鼻孔50μL)。每天评估体重。所有免疫学和病毒学测定均在盲法下进行。在第4天,因组织病毒载量和病理学将一部分动物安乐死。所有动物研究均按照所有相关的地方、州和联邦法规进行,并得到生物质量机构动物管理和使用委员会(IACUC)的批准。
Ad26载体.Ad26载体由2019-nCoV刺突(S)蛋白序列(Wuhan/WIV04/2019;GenbankMN996528.1)的两个变体构建而成。序列经过密码子优化和合成。使用含有完整Ad26载体基因组和转基因表达盒的质粒在PER.C6.TetR细胞中产生无复制能力、E1/E3缺失的Ad26载体19。在使用前对载体进行测序和表达测试。
组织病理学和免疫组织化学.组织在新鲜制备的4%多聚甲醛中固定24小时,转移到70%乙醇中,并在7天内包埋石蜡,并以5μm切片。载玻片在65℃下烘烤30-60分钟,然后在二甲苯中脱蜡并且通过一系列分级乙醇至蒸馏水进行复水。对于SARS-CoV-N、Iba-1和CD3的免疫组织化学,使用压力锅在蒸汽设置下在柠檬酸盐缓冲液(Thermo;AP-9003-500)中进行25分钟的热诱导表位修复(HIER),然后用3%过氧化氢处理。然后在蒸馏水中冲洗载玻片并封闭蛋白(BioCare,BE965H)15分钟,然后在1x磷酸盐缓冲盐水中冲洗。使用兔抗SARS-CoV-核蛋白一抗(Novus;NB100-56576,1∶500或1∶1000)、兔抗Iba-1抗体(Wako;019-19741,1∶500)或兔抗CD3(Dako,A0452,1∶300)30分钟,然后使用兔Mach-2 HRP-Polymer(BioCare;RHRP520L)30分钟,再用苏木精复染,然后用0.25%氨水染成蓝色。SARS-N、Iba-1和CD3的标记是在Biogenex i6000全自动染色机(v3.02)上进行的。在一些情况下,使用1∶400的CD3(Thermo货号RM-9107-S;克隆SP7)、1∶500的Iba-1(BioCare货号CP290A;多克隆)或ACE-2(Abcam;ab108252)染色CD3、Iba-1和ACE-2,所有这些均使用兔Polink-1 HRP(GBI Labs货号D13-110)检测。使用1∶1000的MPO(Dako货号A0398;多克隆),并使用兔Polink-1 HRP检测中性粒细胞(MPO),以及使用1∶1000的Mx1(EMD Millipore货号MABF938;克隆M143/CL143),并使用小鼠Polink-2 HRP(GBI Labs货号D37-110)检测1型IFN应答。CD3、Iba-1、MPO和Mx1的免疫组织化学的染色如前所述使用Biocare intelliPATH全自动染色机进行,所有抗体在室温下孵育1小时。组织病理学由两名兽医病理学家独立评估。



定量图像分析.使用HALO软件(v2.3.2089.27或v3.0.311.405;Indica Labs)对每只动物的至少一个肺叶横截面进行定量图像分析。在>1个横截面可用的情况下,将每个肺叶定量为一个单独的数据点。对于SARS-CoV-N,Multiplex IHC v2.3.4算法与酸性血红素的排除筛选一起使用,以确定SAR-N蛋白阳性细胞占细胞总数的百分比。对于Iba-1,使用Multiplex IHC v2.3.4算法进行定量。对于SARS-CoV-2 ISH和Mx1定量,使用Area Quantification v2.1.3模块来确定总SARS-CoV-2反义或正义探针的百分比,或Mx1蛋白占总组织面积的百分比。对于MPO(中性粒细胞)和CD3+细胞定量,对载玻片进行注释以排除血管(>5mm2)、支气管、细支气管、软骨和结缔组织;随后,使用Cytonuclear v1.6模块检测MPO+或CD3+细胞并计算为占总肺泡组织的比率(PMNs/mm2),该比率通过运行AreaQuantification v2.1.3模块确定。在所有情况下,对每个样本都进行了所有图像的手动检查,以确保注释准确。
ISH和Mx1定量,使用Area Quantification v2.1.3模块来确定总SARS-CoV-2反义或正义探针的百分比,或Mx1蛋白占总组织面积的百分比。对于MPO(中性粒细胞)和CD3+细胞定量,对载玻片进行注释以排除血管(>5mm2)、支气管、细支气管、软骨和结缔组织;随后,使用Cytonuclear v1.6模块检测MPO+或CD3+细胞并计算为占总肺泡组织的比率(PMNs/mm2),该比率通过运行AreaQuantification v2.1.3模块确定。在所有情况下,对每个样本都进行了所有图像的手动检查,以确保注释准确。
亚基因组mRNA测定.通过RT-PCR使用先前描述的引物和探针评估SARS-CoV-2E基因亚基因组mRNA(sgmRNA)10,11,23。简而言之,使用QIAcube HT(Qiagen)和RNeasy 96QIAcube HTKit(Qiagen)从几个解剖部位的组织匀浆中提取总RNA。为了生成标准曲线,将SARS-CoV-2 E基因sgmRNA克隆至pcDNA3.1表达质粒中;使用AmpliCap-Max T7 High YieldMessage MakerKit(Cellscript)转录此插入物。在RT-PCR之前,根据制造商的说明,使用Superscript III VILO(Invitrogen)将从受攻击的动物或标准中收集的样品进行逆转录。使用靶向E基因sgmRNA的序列设计了Taqman定制基因表达测定(ThermoFisherScientific)。根据制造商的说明,在QuantStudio 6和7Flex实时PCR系统(AppliedBiosystems)上进行反应。标准曲线用于计算每克组织的sgmRNA拷贝数;定量测定的灵敏度为100个拷贝。
ELISA.通过ELISA评估RBD特异性结合抗体,基本上如10,11所述。简而言之,用1xDPBS中的1μg/mL SARS-CoV-2RBD蛋白(Aaron Schmidt,MassCPR)或1μg/mL SARS-CoV-2刺突(S)蛋白(Sino Biological)包被96孔板,并在4℃下孵育过夜。孵育后,用洗涤缓冲液(1X DPBS中的0.05%Tween20)洗涤板一次,并在室温下用350μL酪蛋白封闭液/孔封闭2-3小时。孵育后,弃去封闭溶液并将板吸干。将在酪蛋白封闭液中稀释的热灭活血清的三倍连续稀释液添加至孔中,并将板在室温下孵育1小时,然后再洗涤3次,随后与在酪蛋白封闭液中的0.1μg/mL的抗仓鼠IgG HRP(Southern Biotech)在室温下避光孵育1小时。洗涤板3次,然后每孔加入100μL KPL TMB
KPL TMB Start溶液;通过每孔添加100μL
Start溶液;通过每孔添加100μL KPL TMB Stop溶液来停止板的显色。使用VERSAMAXTM或
KPL TMB Stop溶液来停止板的显色。使用VERSAMAXTM或 微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生是背景2倍的最高血清稀释度的倒数。
微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生是背景2倍的最高血清稀释度的倒数。
假病毒中和测定.通过与之前描述的方法相似的方法产生表达荧光素酶报告基因的SARS-CoV-2假病毒10,11,21。简而言之,通过lipofectamine 2000(ThermoFisher)将包装构建体psPAX2(AIDS Resource and Reagent Program)、荧光素酶报告质粒pLenti-CMVPuro-Luc(Addgene)和表达刺突蛋白的pcDNA3.1-SARS CoV-2 SΔCT共转染到HEK293T细胞中。转染后48小时收集含有假病毒的上清液;通过用0.45μm过滤器过滤来纯化假病毒。为了确定来自疫苗接种动物的抗血清的中和活性,将HEK293T-hACE2细胞以1.75×104个细胞/孔的密度接种在96孔组织培养板中过夜。制备热灭活血清样品的三倍连续稀释液,并与50μL假病毒混合。在加入HEK293T-hACE2细胞之前,将混合物在37℃下孵育1小时。感染后48小时,根据制造商的说明,在 Luciferase Assay(Promega)中裂解细胞。将SARS-CoV-2中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
Luciferase Assay(Promega)中裂解细胞。将SARS-CoV-2中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
Luminex.为了检测抗原特异性抗体滴度的相对量,如前所述25进行了定制的Luminex测定。因此,荧光标记的微球(Luminex)通过EDC(Thermo Scientific)和Sulfo-NHS(Thermo Scientific)通过共价N-羟基琥珀酰亚胺(NHS)-酯键与包括刺突蛋白(S)(EricFischer,Dana Farber Cancer Institute)、S1和S2(Sino Biological)以及受体结合结构域(RBD)(Aaron Schmidt,Ragon Institute)在内的SARS-CoV-2抗原偶联。将1.2×103珠每个区域和抗原添加至384孔板(Greiner)的每孔中,并与稀释的血清样品一起孵育(对于IgG2a、IgG3和IgM,1∶90;对于总IgG和Fc受体结合测定,1∶500)在4℃下以900rmp振动16小时。在形成免疫复合物后,用自动洗板机(Tecan)在0.1%BSA和0.05%Tween-20(Luminex测定缓冲液)中洗涤微球3次。将PE标记的山羊抗小鼠IgG、IgG2a、IgG3和IgM检测抗体(southern biotech)在Luminex测定缓冲液中稀释至0.65ug/mL,并在室温下与珠一起孵育1小时,同时以900rpm振动。类似地,对于Fc受体结合谱,生物素化(Thermo Scientific)重组小鼠FcγR2、FcγR3和FcγR4(Duke Protein Production facility)并与链霉亲和素-PE偶联10分钟(Southern Biotech),然后添加至样品。这些小鼠抗体和蛋白与仓鼠有交叉反应。然后洗涤包被的珠并在连接有机械臂(PAA)的流式细胞仪iQue(Intellicyt)上读数。在每个珠区域对事件进行门控,报告珠阳性事件的PE荧光中值。每个二级检测试剂均一式两份运行样品。
抗体依赖性补体沉积(ADCD).如前所述26进行ADCD测定。简而言之,生物素化(Thermo Fisher)SARS-CoV-2 S和RBD并在37℃下与1μm红色荧光中性亲和素-珠偶联2小时,然后洗去多余的抗原。为了形成免疫复合物,将1.82x108个抗原包被的珠添加到96孔板圆底板的每个孔中,并与1∶10稀释的样品在37℃下孵育2小时。根据制造商的说明(Cedarlane)用水重构冻干豚鼠补体,并在含有Mg2+和Ca2+(GVB++,BostonBioProducts)的明胶弗洛拿缓冲液中每孔向免疫组合物加入4μL,在37℃下持续20分钟。用PBS中的15mM EDTA洗涤免疫组合物,并且添加荧光素缀合的山羊IgG豚鼠补体C3组分(MpBio)。染色后,用4%多聚甲醛(PFA)固定细胞样品,并使用机械臂(PAA)通过流式细胞仪(Intellicyt,iQueScreener plus)进行样品采集。以单细胞和珠阳性事件门控所有事件,报告C3阳性事件的中值。所有样品在不同的天一式两份进行。
统计分析.使用GraphPad Prism 8.4.2(GraphPad软件)对免疫学、病毒学和体重数据进行分析。使用双边Mann-Whitney检验进行组间数据比较。通过双边Fisher’s精确检验评估死亡率。通过双边Spearman等级相关性检验评估相关性。将P值小于0.05认为是显著的。所有系统血清学数据都经过log10转换。对于雷达图,在使用组内的中值之前,对每个抗体特征进行标准化,使其最小值为0,跨组的最大值为1。使用R包“ropls”构建主成分分析(PCA)以比较多变量谱。对于热图中的可视化,显示了S.dTM.PP和S.PP组Z分数特征的平均值的差异。为了表明热图中的显著性,使用Benjamini-Hochberg校正来校正一行内的多重比较。
结果与讨论
SARS-CoV-2可以感染非人灵长类动物8-10、仓鼠5-7、雪貂14-16、hACE2转基因小鼠17,18和其他物种16,但这些模型中的临床疾病通常是轻度的。重症肺炎模型可用于SARS-CoV-2疫苗和其他应对措施的临床前评估,因为人感染SARS-CoV-2可导致严重的临床疾病、呼吸衰竭和死亡1-4。评估了仓鼠中高剂量SARS-CoV-2感染的临床和病毒学特征,并在这个严格的模型中评估了基于Ad26载体的疫苗19的保护效力,此疫苗编码稳定化的SARS-CoV-2刺突(S)。
20只叙利亚金仓鼠(10-12周龄)通过鼻内途径接种5x104TCID50(N=4;低剂量)或5x105TCID50(N=16;高剂量)的SARS-CoV-2。在高剂量组中,在第2天对4只动物尸检,在第4天对4只动物进行尸检,以检测组织病毒载量和组织病理学,其余8只动物进行纵向随访。在第14天对所有剩余的动物进行尸检。在低剂量组中,仓鼠到第6天时体重的中值下降了14.7%,但到第14天完全恢复(图55A-B),这与之前的研究5-7一致。在高剂量组中,仓鼠到第6天时体重的中值下降了19.9%。在这个组中纵向追踪的8只动物中,有4只在第6天时满足IACUC人道安乐死标准,即体重减轻>20%和呼吸窘迫,另外2只动物在第7天时满足这些标准。其余2只动物在第14天恢复。这些数据表明,仓鼠中的高剂量SARS-CoV-2感染导致严重的体重减轻和部分死亡。
对接受高剂量SARS-CoV-2并在第2天进行尸检的4只动物、在第4天进行尸检的4只动物以及在第6-7天满足安乐死标准的6只动物中的5只动物评估组织病毒载量(图55C)。在第2天观察到,肺组织中1012RNA拷贝/g以及鼻孔和气管中108-109RNA拷贝/g的高组织病毒载量中值,以及心脏、胃肠道、脑、脾脏、肝脏和肾脏中105-108RNA拷贝/g的中值,这表明播散性感染。到第6-7天,组织病毒载量降低了大约2log,尽管体重持续下降。
在第2天(N=4)、第4天(N=4)、第6-7天(N=6)和第14天(N=2)通过组织病理学评估感染了高剂量SARS-CoV-2的仓鼠。感染与鼻甲(图56A)和细支气管上皮的显著炎症浸润和多灶性上皮坏死相关,其导致管腔中的退化的中性粒细胞和细胞碎片(图56B)。附近血管的内皮与单个核细胞与内皮的粘附反应并在血管壁内迁移,表明内皮炎(图56B)。早在SARS-CoV-2感染后第2天,就有中度至严重多灶性间质性肺炎,其特征是影响了30-60%的肺实质的肺实变(图56C)。炎症浸润由大量巨噬细胞和中性粒细胞组成,淋巴细胞较少。通过免疫组织化学(IHC)在鼻甲上皮(图56D)和细支气管上皮细胞(图56E)的炎症和坏死区域对SARS核衣壳蛋白(SARS-CoV-N)呈强阳性。SARS-CoV-N的免疫组织化学还显示肺泡间隔和间质单个核细胞的局部广泛染色,其形态与巨噬细胞一致(图56F)。类似地,在细支气管上皮和肺间质中的炎症区域中观察到大量SARS-CoV-2病毒RNA(vRNA)(图56G-H)。
肺中SARS-CoV-2vRNA和SARS-CoV-N蛋白的表达水平在第2天最高,到第4天下降,到第7天检测到最少的vRNA和SARS-CoV-N蛋白(图57A-57B)。肺炎的特征在于肺间质中Iba-1+巨噬细胞的大量炎性浸润以及CD3+T淋巴细胞(图56I-56J)。在整个肺,特别是在坏死区域中检测到大量存活和退化的中性粒细胞,其中中性粒细胞髓过氧化物酶(MPO)在整个肺中高表达(图56K)。还在肺中检测到干扰素诱导基因产物MX1的扩散表达(图56L)。与在第2天达到峰值的SARS-CoV-2vRNA和SARS-CoV-N检测的动力学相反,这些炎症标志物在第7天达到峰值(图57C-57F),与最大的体重减轻和死亡率一致(图55A-55B)。通过 检测肺中的vRNA并不仅仅反映病毒接种物,因为从攻击后的第2天到第7天不仅检测到反义vRNA(图58A-58E),而且还检测到正义vRNA(图58F-58J),其在位置和模式上重叠。SARS-CoV-2 vRNA表达(反义和正义)存在于肺中,具有强ACE2受体表达(图58K-58O)。
检测肺中的vRNA并不仅仅反映病毒接种物,因为从攻击后的第2天到第7天不仅检测到反义vRNA(图58A-58E),而且还检测到正义vRNA(图58F-58J),其在位置和模式上重叠。SARS-CoV-2 vRNA表达(反义和正义)存在于肺中,具有强ACE2受体表达(图58K-58O)。
在包括脑干、胃肠道和心肌的远端组织中也检测到全身性vRNA(图59A-59F)。尽管SARS-CoV-N染色很少(图59G-59J),但在这些组织中观察到显著的内皮炎和巨噬细胞以及淋巴细胞的血管周围炎症。在一只动物中观察到局灶性淋巴细胞性心肌炎并且对应于vRNA的存在(图59K-59L)。病毒检测的其他部位包括肺血栓中的外周血单个核细胞(图60A-60C)和鼻甲骨骨髓(图60D-60F)。
产生了重组的、无复制能力的Ad26载体,其编码(i)SARS-CoV-2刺突(S),其具有缺失的跨膜区和细胞质尾,反映具有折叠子三聚化结构域的可溶性胞外域(S.dTM.PP(SS-S.Ecto-dF-PP-foldon)或(ii)全长S(S.PP(Ad26 SS-Spike-dF-PP)),两者都具有弗林蛋白酶切割位点的突变和两个脯氨酸稳定化突变20(图61A)。最近报道了这些疫苗在恒河猴中对SARS-CoV-2攻击的免疫原性和保护效力13。
用1010或109个病毒颗粒(vp)Ad26载体免疫50只叙利亚金仓鼠,此载体编码S.dTM.PP或S.PP(N=10/组)或假对照(N=10)。通过肌内途径动物在第0周接受单次疫苗接种。在第2周和第4周,在所有动物中通过ELISA10,11观察受体结合结构域(RBD)特异性结合抗体(图61B),并通过假病毒中和测定10,11,21观察中和抗体(NAb)(图61C)。在第4周,与Ad26-S.dTM.PP(Ad26 SS-S.Ecto-dF-PP-foldon)(1014,1185)相比,Ad26-S.PP引发的ELISA滴度中值(4470、4757)是4.0-4.7倍(图61B;P<0.0001,双边Mann-Whitney检验)。类似地,与Ad26-S.dTM.PP(139,211)相比,Ad26-S.PP引发的NAb IC50滴度中值(359,375)是1.8-2.6倍(P<0.05,双边Mann-Whitney检验)。对于每个载体,测试的两个剂量似乎是可比的。ELISA和NAb数据在第2周和第4周都相关(分别为R=0.7074,P<0.0001和R=0.7849,P<0.0001,双边Spearman等级相关性检验;图62A)。
通过系统血清学22,在第4周对疫苗接种动物的S特异性和RBD特异性抗体应答进行了表征。评估了IgG、IgG2a、IgG3、IgM、Fc受体FcRγ2、FcRγ3和FcRγ4,以及抗体依赖性补体沉积(ADCD)应答(图61D-F)。与Ad26.S.dTM.PP相比,使用Ad26-S.PP观察到更高和更一致的应答(图61D、61F),并且这些抗体特征的主成分分析确认这两种疫苗具有不同的谱(图61E)。
在第4周,通过鼻内途径用5x105TCIDs0 SARS-CoV-2攻击所有动物。在第4天对每组中的3只动物进行尸检,以检测组织病毒载量和组织病理学,并对每组中剩余的7只动物进行追踪直至第14天。在假对照中,到第7天,仓鼠体重的中值下降了19.6%,在第6-7天纵向追踪的动物中有43%(7只中的3只)满足安乐死标准(图63A-63B)。接种Ad26-S.dTM.PP的动物体重中值下降8.7%,接种Ad26-S.PP的动物体重中值下降4.0%(图63A-63B)。与假对照相比,两个疫苗接种组的最大体重减轻百分比均显著降低(P<0.0001,双边Mann-Whitney检验;图63C),接受Ad26-S.PP的动物比接受Ad26.S.dTM.PP的动物表现出更少的体重减轻(P<0.0001,双边Mann-Whitney检验;图63C)。与假对照相比,两种疫苗都防止死亡,将死亡定义为满足人道安乐死标准(P=0.02,双边Fisher精确检验;图62B)。两个仓鼠实验的组合分析确认两种疫苗都有效防止死亡(P=0.007,双边Fisher精确检验;图62C)。第2周(R=-0.8992,P<0.0001)和第4周(R=-0.9344,P<0.0001)的ELISA应答与最大体重减轻百分比负相关(图64A)。第2周(R=-0.7380,P<0.0001)和第4周(R=-0.8075,P<0.0001)的NAb应答也与最大体重减轻百分比负相关(图64B)。
在第4天尸检的一部分动物和第14天剩余的存活动物中评估组织病毒载量。在高剂量SARS-CoV-2攻击后的第4天,通过亚基因组RNA RT-PCR在所有动物的组织中检测病毒,亚基因组RNA RT-PCR被认为能测量复制的病毒10,23(图65A)。肺组织中的病毒载量中值在假对照中约为1012RNA拷贝/g,而在Ad26-S.dTM.PP疫苗接种动物中为108RNA拷贝/g,在Ad26-S.PP疫苗接种的动物中为106RNA拷贝/g。到第14天,仍然在存活的假对照的肺和鼻孔中检测到病毒,但仅在少数Ad26-S.dTM.PP疫苗接种动物中观察到,而在Ad26-S.dTM.PP疫苗接种动物中均未观察到(图65B)。
第2周(R=-0.8133,P=0.0004)和第4周(R=-0.9288,P<0.0001)的ELISA应答与第4天的肺病毒载量负相关(图66A),以及第2周(R=-0.7469,P=0.0020)和第4周(R=-0.6004,P=0.0199)的NAb应答与第4天的肺病毒载量负相关(图66B)。ELISA和NAb应答也与鼻甲中的病毒载量负相关(图66c、d)。对免疫相关因素的更深入分析揭示了多种抗体特征与体重减轻和组织病毒载量负相关(图67A)。
在攻击后第4天,与假对照相比,疫苗接种的动物也表现出减少的病理学(图68)。疫苗接种Ad26-S.PP的动物表现出极少甚至没有病毒性间质性肺炎、细支气管上皮破坏或CD3+T淋巴细胞和巨噬细胞的细支气管周围聚集体的证据。与假对照相比,疫苗接种Ad26-S.dTM.PP和Ad26-S.PP的仓鼠的所有肺叶中的组织细胞性和嗜中性炎症浸润显著减少,SARS-CoV-2 vRNA显著减少(分别为P=0.004和P=0.004,双边Mann-Whitney检验;图63D)。
在攻击后的第14天,存活的假对照产生了有效的结合抗体应答和中和抗体应答(图67B)。疫苗接种的动物在攻击后也表现出更高的ELISA和NAb应答(图67B),这与组织病毒载量一致,在高剂量SARS-CoV-2攻击后在这些动物中显示出低和短暂的病毒复制水平。
在这项研究中,证明编码全长融合前稳定化的S免疫原(S.PP)的Ad26载体的单次免疫可以在仓鼠接受高剂量SARS-CoV-2攻击后防止严重的临床疾病。假对照表现出明显的体重减轻、严重肺炎和部分死亡。相反地,疫苗接种的动物显示很少的体重减轻和肺炎,并且没有死亡。疫苗引发的结合抗体应答和中和抗体应答与防止临床疾病以及减少上呼吸道和下呼吸道中的病毒复制相关。
这个模型中这种临床疾病的严重度与先前涉及仓鼠5-7和其他物种8-10,14-18的SARS-CoV-2感染的研究形成鲜明对比。仓鼠是SARS-CoV-2的允许模型,因为它们与人ACE2受体同源5,并且据报道SARS-CoV-2在仓鼠之间传播6。在本研究中,高攻击剂量导致广泛的临床疾病,尽管可能影响临床疾病程度的生物学要素,例如动物年龄、动物来源和攻击原种,仍有待严格研究。
迄今为止,在非人灵长类动物身上进行的SARS-CoV-2疫苗研究已证明可以防止感染或减少上呼吸道和下呼吸道中的病毒复制11,12。最近有报道称,Ad26-S.PP的单次免疫可在恒河猴中提供防止SARS-CoV-2攻击的完全或接近完全的保护13。然而,非人灵长类动物中的SARS-CoV-2感染不会导致严重的临床疾病或死亡8-10。严重疾病模型将有助于补充当前的非人灵长类动物攻击模型,因为防止病毒复制的保护并不一定意味着防止严重疾病的保护。事实上,在本研究中在仓鼠的组织病理学分析中,肺中的病毒载量从第2天到第7天下降,而炎症标志物在此期间持续升高并与持续的体重减轻相关。这些数据表明,仓鼠的进行性临床疾病主要是一种炎症过程,由感染引发,即使当病毒复制减少,炎症也会继续增加。
由于人的COVID-19可发展为严重的临床疾病,因此在再现严重临床疾病(包括暴发性肺炎和死亡)的临床前模型中测试SARS-CoV-2候选疫苗非常重要。本文所述的高剂量仓鼠模型满足许多这些标准,因此代表了人疾病和治疗的有用模型,特别是关于严重疾病的发病机制。这个模型中临床疾病的主要表现是严重肺炎,而不是某些hACE2转基因小鼠模型中报道的脑炎24。此外,结合抗体应答和中和抗体应答与保护相关。
总之,这些数据表明,在与人中的疾病发展和进展非常相似的动物模型中,Ad26-S.PP的单次免疫提供了防止高剂量SARS-CoV-2感染后严重临床疾病的强的保护。这种防止严重SARS-CoV-2肺炎和死亡的疫苗保护以前没有报道。Ad26-S.PP,也称为Ad26.COV2.S,目前正在临床试验中进行评估。这种仓鼠严重疾病模型还可用于测试SARS-CoV-2疫苗、治疗和其他应对措施。
参考文献
1 Li,Q.et al.Early Transmission Dynamics in Wuhan,China,ofNovelCoronavirus-Infected Pneumonia.NEnglJMed,doi:10.1056/NEJMoa2001316(2020).
2 Zhu,N.et al.A Novel Coronavirus from Patients with Pneumonia inChina,2019.N Engl J Med 382,727-733,doi:10.1056/NEJMoa2001017(2020).
3 Chen,N.et al.Epidemiological and clinical characteristics of99cases of 2019 novel coronavirus pneumonia in Wuhan,China:a descriptivestudy.Lancet 395,507-513,doi:10.1016/S0140-6736(20)30211-7(2020).
4 Huang,C.et al.Clinical features of patients infected with 2019novel coronavirus in Wuhan,China.Lancet 395,497-506,doi:10.1016/S0140-6736(20)30183-5(2020).
5 Chan,J.F.et al.Simulation of the clinical and pathologicalmanifestations of Coronavirus Disease 2019(COVID-19)in golden Syrian hamstermodel:implications for disease pathogenesis and transmissibility.Clin InfectDis,doi:10.1093/cid/ciaa325(2020).
6 Sia,S.F.et al.Pathogenesis and transmission of SARS-CoV-2 in goldenhamsters.Nature,doi:10.1038/s41586-020-2342-5(2020).
7 Imai,M.et al.Syrian hamsters as a small animal model for SARS-CoV-2infection and countermeasure development.Proc Natl Acad Sci U S A 117,16587-16595,doi:10.1073/pnas.2009799117(2020).
8 Munster,V.J.et al.Respiratory disease in rhesus macaques inoculatedwith SARS-CoV-2.Nature,doi:10.1038/s41586-020-2324-7(2020).
9 Rockx,B.et al.Comparative pathogenesis of COVID-19,MERS,and SARS ina nonhuman primate model.Science,doi:10.1126/science.abb7314(2020).
10 Chandrashekar,A.et al.SARS-CoV-2 infection protects againstrechallenge in rhesus macaques.Science,doi:10.1126/science.abc4776(2020).
11 Yu,J.et al.DNA vaccine protection against SARS-CoV-2 in rhesusmacaques.Science,doi:10.1126/science.abc6284(2020).
12 Gao,Q.et al.Rapid development of an inactivated vaccine candidatefor SARS-CoV-2.Science,doi:10.1126/science.abc1932(2020).
13 Mercado,N.B.et al.Single-shot Ad26 vaccine protects against SARS-CoV-2 in rhesus macaques.Nature,doi:10.1038/s41586-020-2607-z(2020).
14 Blanco-Melo,D.et al.Imbalanced Host Response to SARS-CoV-2 DrivesDevelopment of COVID-19.Cell 181,1036-1045 e1039,doi:10.1016/j.cell.2020.04.026(2020).
15 Kim,Y.I.et al.Infection and Rapid Transmission of SARS-CoV-2 inFerrets.Cell Host Microbe 27,704-709 e702,doi:10.1016/j.chom.2020.03.023(2020).
16 Shi,J.et al.Susceptibility of ferrets,cats,dogs,and otherdomesticated animals to SARS-coronavirus 2.Science 368,1016-1020,doi:10.1126/science.abb7015(2020).
17 Bao,L.et al.The pathogenicity of SARS-CoV-2in hACE2 transgenicmice.Nature,doi:10.1038/s41586-020-2312-y(2020).
18 Sun,S.H.et al.A Mouse Model of SARS-CoV-2 Infection andPathogenesis.Cell Host Microbe 28,124-133e124,doi:10.1016/j.chom.2020.05.020(2020).
19 Abbink,P.et al.Comparative seroprevalence and immunogenicity ofsix rare serotype recombinant adenovirus vaccine vectorsfrom subgroups B andD.J Virol 81,4654-4663,doi:JVI.02696-06[pii]10.1128/JVI.02696-06(2007).
20 Wrapp,D.et al.Cryo-EM structure of the 2019-nCoV spike in theprefusion conformation.Science 367,1260-1263,doi:10.1126/science.abb2507(2020).
21 Yang,Z.Y.et al.A DNA vaccine induces SARS coronavirusneutralization and protectiVe immunity in mice.Nature 428,561-564,doi:10.1038/nature02463(2004).
22 Chung,A.W.et al.Dissecting Polyclonal Vaccine-Induced HumoralImmunity against HIV Using Systems Serology.Cell163,988-998,doi:10.1016/j.cell.2015.10.027(2015).
23 Wolfel,R.et al.Virological assessment of hospitalized patientswith COVID-2019.Nature,doi:10.1038/s41586-020-2196-x(2020).
24 Netland,J.,Meyerholz,D.K.,Moore,S.,Cassell,M.&Perlman,S.Severeacute respiratory syndrome coronavirus infection causes neuronal death in theabsence of encephalitis in mice trahsgenic for human ACE2.J Virol82,7264-7275,doi:10.1128/JVI.00737-08(2008).
25 Brown,E.P.et al.High-throughput,multiplexed IgG subclassing ofantigen-specific antibodies from clinical samples.J Immunol Methods 386,117-123,doi:10.1016/j.jim.2012.09.007(2012).
26 Fischinger,S.et al.A high-throughput,bead-based,antigen-specificassay to assess the ability of antibodies to induce complement activation.JImmunol Methods 473,112630,doi:10.1016/j.jim.2019.07.002(2019).
实施例13.抗原设计
设计了几种基于全长Wuhan-CoV S蛋白序列的抗原,其中一些在图69中示意性地示出。所有序列均基于SARS-CoV-2刺突全长蛋白(YP_009724390.1-SEQ ID NO:29)。
对于不同的抗原,使用不同的信号肽/前导序列,例如COR200006、200007和200008)中的天然野生型信号肽、tPA信号肽(COR200009、200010和200011)或嵌合前导序列(COR200018)。
此外,一些构建体含有野生型弗林蛋白酶切割位点(wt)(即,200006、200009和200018),并且在一些构建体(即,200007、200008、200010和200011)中,通过改变弗林蛋白酶位点氨基酸序列RRAR(wt)(SEQ ID NO:90和224)至SRAG(dFur)(SEQ ID NO:225)而去除弗林蛋白酶切割位点,即通过引入R682 S和R685G突变(其中氨基酸位置的编号是根据氨基酸序列YP_009724390-SEQ ID NO:29中的编号),以优化稳定性和表达。
在一些构建体中,在铰链环位置986和987处引入稳定(脯氨酸)突变以优化稳定性和表达,特别是COR20007、20008、200010和200011包含K986P和V987P突变(其中氨基酸位置的编号是根据氨基酸序列YP_009724390-SEQ ID NO:29中的编号)。
最后,在构建体200008和200011中,缺失ER驻留信号以优化运输和表达,即通过去除最后5个氨基酸KLHYT(SEQ ID NO:226)。
在基于细胞的Elisa(CBE)和荧光激活细胞分选(FACS)实验中测试了一些SARS-CoV-2免疫原设计的表达和抗原性,包括COR200010、200011和200018。
对于CBE,第1天以100%的汇合度将HEK293细胞接种到黑壁聚-D-赖氨酸包被的微孔板上。在第2天使用lipofectamine用质粒转染细胞,并且在第4天在4℃下进行基于细胞的ELISA。没有使用固定步骤。用BM化学发光ELISA底物(Roche)检测二抗。Ensight机器用于测量细胞汇合度和发光强度。
使用了一些与SARS-CoV-2 S蛋白交叉反应的SARS-CoV抗体。已知抗体CR3022(在WO06/051091中公开)以低效力中和SARS-CoV(Ter Meulen et al.(2006),PLOSMedicine)。它不会中和SARS-CoV-2。它仅在至少两个受体结合区(RBD)处于向上位置时才结合(Yuan et al.,Science.2020Apr 3.pii:eabb7269.doi:10.1126/science.abb7269.[Epub ahead of print];Joyce et al.doi:https://doi.org/10.1101/2020.03.15.992883)。CR3015(在WO2005/012360中公开)已知是非中和SARS-CoV。也认为CR3023、CR3046、CR3050、CR3054和CR3055是非中和抗体。结果示于图70。
如图所示,COR200010具有最高的中和:非中和Ab结合比,这反映了S蛋白的有利抗原性和正确的天然折叠,因此可能表明该蛋白主要处于融合前状态。基于这些数据和FACS数据(参见图71)取消选择COR200011并且没有进一步测试。此外,取消选择COR200008(数据未示出)。
此外,用100mcg的相应DNA构建体或作为对照的磷酸盐缓冲盐水肌内免疫6-8周龄的Balb/C小鼠。在免疫后第19天使用重组可溶性稳定刺突靶抗原通过ELISA测定血清SARS-CoV-2刺突特异性抗体滴度。弗林蛋白酶位点敲除(KO)和脯氨酸突变(PP)增加了免疫原性(对Furin KO+PP-S蛋白的ELISA,参见图72)。
此外,ER驻留信号(dERRS)的去除降低了CBE中CR3022的结合并降低了免疫原性。
基于CBE中CR3022:CR3015结合比(连同FACS中的COR200011数据)、WB上的表达水平(数据未示出)、小鼠DNA免疫后的ELISA滴度(与COR200009、COR200010和COR200011相比)(数据未示出),以及用COR200010DNA观察到的中和作用,COR2000010似乎是最好的抗原构建体,并选其为腺载体构建的抗原。
因为,对于膜结合的S蛋白,tPA信号肽(ST)当与非稳定形式中的wt SP相比时似乎没有有益效果(基于CR3022结合),所以也选择了COR200007用于腺载体构建。
图73示出COR200007与ACE2的结合比COR200010更好。
实施例14:将SARS-CoV-2载体克隆到Ad26中
如实施例13所述,为基于SARS-CoV-2序列的S蛋白(全基因组序列NC 045512)的Ad26设计了一些不同的插入物。插入物是膜锚定形式COR20007、COR20008、COR20009、COR200010、COR200011和COR200018。
SARS-CoV2刺突蛋白的克隆包括两个不同的步骤:
1:SARS-CoV2刺突蛋白的设计和克隆至穿梭表达载体中
这个载体包含人巨细胞病毒(hCMV)启动子,该启动子在hCMV启动子内携带两个四环素操纵子(tetO)序列和SV40多腺苷酸化信号。将表达载体用于体外和体内实验,并用于产生携带SARS-CoV2刺突蛋白的Ad26病毒。
2:将表达载体克隆至Ad26病毒中
设计并且克隆SARS-CoV2刺突蛋白的设计至穿梭表达载体中
如实施例13所述,基于野生型SARS-CoV2刺突蛋白(参考,YP_009724390-SEQ IDNO:29),设计了一些基于全长SARS-CoV-刺突蛋白的构建体(命名为COR20007、COR20008、COR20009、COR 200010、COR200011和COR200018),其中引入了不同的信号肽,弗林蛋白酶切割位点的去除,稳定化突变和跨膜区变异(见表)

*注:nt序列只编码编码序列;即不包括下面描述的额外添加的5′-和3′-序列。
这些蛋白序列使用最常见的人密码子使用(CLC-main workbench;Qiagen)进行反向翻译,使用开源生物信息学工具(GeneOptimizer;GeneArt,剪接位点和启动子预测;fruitfly.org)对GC含量、隐性剪接位点、SD序列、TATA盒和终止信号进行校正。接下来,在不同设计之间协调密码子使用,并将核苷酸添加至5′-末端,其包含与穿梭载体的重叠、HindIII限制性位点和Kozak序列(CGGCCGGGAACGGTGCATTGGAAGCTTGCCACC(SEQ ID NO:220)并且添加了一个双终止密码子、XbaI限制性位点和与穿梭表达载体的重叠序列至3′-末端(TGATAATCTAGACGAGATCCGAACTTGTTTATTG(SEQ ID NO:221)。然后通过Gibson组装( HiFi DNA Assembly Master Mix,NEB,Cat E2621)将两个编码SARS-CoV2刺突蛋白序列的部分的DNA片段(gBlocks由Integrated DNA Technologies,IDT提供)组装到它们的HindIII+XbaI线性化靶载体中(ID 7346_pUC_E1-CMVdel134TO-stuffer,SEQ ID NO:13),如图74示意性地描述的那样。这产生了编码抗原构建体COR20009、COR200010、COR200011和COR200018的表达载体。
HiFi DNA Assembly Master Mix,NEB,Cat E2621)将两个编码SARS-CoV2刺突蛋白序列的部分的DNA片段(gBlocks由Integrated DNA Technologies,IDT提供)组装到它们的HindIII+XbaI线性化靶载体中(ID 7346_pUC_E1-CMVdel134TO-stuffer,SEQ ID NO:13),如图74示意性地描述的那样。这产生了编码抗原构建体COR20009、COR200010、COR200011和COR200018的表达载体。
图74示出了功能性表达SARS-CoV-2刺突蛋白的克隆的示意图。编码SARS-CoV-2刺突蛋白的两个大约2kb的DNA片段(gBlocks;IDT)相互重叠,并在5′-和3′-末端重叠,这允许两个DNA片段体外组装(IVA)至线性化载体中(如Gibson et al.;(2009)Nat Methods 6(5):345-345所述)。
为了构建COR20007和COR20008表达载体,分别将载体pUC-CMVdel134TO_COR200010和CMVdel134_COR200011用作定点诱变方法的模板,以将tPA前导替换为野生型刺突前导。至此,根据制造商的方案,使用Q5-ste定向诱变试剂盒(NEB;E0554S)进行定点PCR。使用引物7_fw(gctccccctcgtctccagtCAATGCGTGAACCTGACC(SEQ ID NO:222)和8_Rv(agtaccagaaacacgaacatGGTGGCAAGCTTCCAATG(SEQ ID NO:223))和PCR程序在98℃下变性30秒,然后在98℃下20秒,在64℃解链20秒并在72℃下延伸2分钟,进行25个循环。根据制造商的方案,从凝胶中分离得到的扩增子并用KLD混合物处理,得到COR20007和COR20008表达载体。
将穿梭表达构建体用于体外和体内测定(例如,如实施例13中所述)和用于产生携带SARS-CoV-2刺突序列的Ad26载体基因组质粒,如下所述。
Ad26载体基因组质粒pAd26.dE1.dE3.5orf6的产生
pAd26.dE1.dE3.5orf6是一种质粒,其含有一个全长Ad26载体基因组,该基因组携带一个E1缺失、一个E3缺失,并用Ad5替代Ad26 E4 orf6,如先前生成的基于Ad26的载体所述(Abbink et al.,(2007);J Virol,81(9),4654-4663)。pAd26.dE1.dE3.5orr6的质粒骨架包含pMB1复制起点和氨苄青霉素抗性基因,并且其来源于pBR322(GenBankd登录号J01749.1)。pAd26.dE1.dE3.5orf6还含有一个独特的AsiSI限制性位点来代替载体基因组的E1缺失。这些位点可用于在相应位置插入来自穿梭载体(pShuttle_E1.CMVdel134TO;SEQID NO:217)的转基因盒。最后,pAd26.dE1.dE3.5orf6内的腺病毒载体基因组在其每个末端的侧翼有一个SwaI限制性位点,允许其通过SwaI消化从质粒骨架中释放。
携带SARS-CoV-2刺突序列的Ad26载体基因组质粒的产生
质粒pShuttle_E1.CMVdel134TO表达载体带有一个人巨细胞病毒(hCMV)启动子,该启动子在hCMV启动子内带有两个四环素操纵子(tetO)序列和一个SV40多腺苷酸化信号。此外,这个载体在转基因盒的上游和下游都含有一个SapI限制性位点,其侧翼为与接收的Ad26载体(ID6598_pAd26.dE1.dE3.5orf6;SEQ ID NO:218)同源的20nt,这允许将转基因盒体外组装(Gibson;2009,supra)插入Ad26载体中。
图75示出了将表达SARS-CoV-2刺突蛋白的穿梭载体克隆到Ad26载体的E1区的示意图。Ad26载体用限制酶AsiSI线性化。穿梭载体用SapI消化,以将表达盒与其骨架分开。表达盒含有与Ad26骨架中AsiSI位点侧翼的序列同源的序列,这允许两个DNA片段的体外组装(IVA)(如Gibson et al.;2009所述)。
为此,用SapI消化携带SARS-CoV-2刺突序列的pShuttle_E1.CMVdel134TO表达载体,并且将表达盒从凝胶中分离出来,体外组装( HiFi DNA Assembly MasterMix,NEB,货号E2621)到AsiSI线性化的Ad26载体(ID6598_pAd26.dE1.dE3.5orf6;SEQ IDNO:218)。这产生携带SARS-Cov-2刺突变体的Ad26载体;pAd26.E1.CMVdel134-TO.COR20007、pAd26.E1.CMVdel134-TO.COR20008、pAd26.E1.CMVdel134-TO.COR20009、pAd26.E1.CMVdel134-TO.COR200010、pAd26.E1.CMVdel134-TO.COR200011和pAd26.E1.CMVdel134-TO.COR200018。
HiFi DNA Assembly MasterMix,NEB,货号E2621)到AsiSI线性化的Ad26载体(ID6598_pAd26.dE1.dE3.5orf6;SEQ IDNO:218)。这产生携带SARS-Cov-2刺突变体的Ad26载体;pAd26.E1.CMVdel134-TO.COR20007、pAd26.E1.CMVdel134-TO.COR20008、pAd26.E1.CMVdel134-TO.COR20009、pAd26.E1.CMVdel134-TO.COR200010、pAd26.E1.CMVdel134-TO.COR200011和pAd26.E1.CMVdel134-TO.COR200018。
重组Ad26载体的产生
使用标准操作程序,通过将全基因组质粒转染至E1互补悬浮PER.C6 TetR(sPER.C6 TetR)细胞中,生成含有SARS-CoV2插入物的Ad26载体。在转染到sPER.C6 TetR中之前,质粒在补充了10%胎牛血清(FBS)和10mM MgCl2的Dulbecco改良Eagle培养基(DMEM)中在PLL包被板上生长,Ad载体质粒用SwaI消化以释放来自质粒的腺病毒载体基因组。根据标准程序使用Lipofectamine转染试剂(Invitrogen:Carlsbad,CA)进行转染。在收获病毒拯救转染后,通过在Permexcis培养基中培养的sPER.C6 TetR细胞上的几轮连续感染进一步扩增病毒。如前所述,使用两步氯化铯(CsCl)密度梯度超速离心程序从粗病毒收获物中纯化病毒(Havenga et al.,“Novel replication-incompetent adenoviral B-groupvectors:high vector stability and yield in PER.6cells,”J.Gen.Virol.87(8):2135-43(2006))。通过先前描述的基于分光光度法的程序测量病毒颗粒(VP)滴度(Maizelet al.,“The polypeptides of adenovirus:I.Evidence for multiple proteincomponents in the virion and a comparison of types 2,7A,and 12”Virology,36(1):115-25(1968))。
实施例15:小鼠中的体液免疫原性
用单剂的108、109或1010vp的Ad26COVS1(N=10/剂量水平)肌内(IM)免疫BALB/c小鼠。注意,名称Ad26NCOV030和Ad26COVS1用于相同的构建体并且可以互换使用。阴性对照组接受没有转基因的1010vp Ad26载体(Ad26.Empty,N=5)。Ad26NCOV006载体编码没有稳定化突变的天然全长S蛋白,可以与Ad26COVS1进行比较,以评估稳定化突变的益处。一组(第1组,N=5)接受107vp Ad26NCOV006以产生血清,这组的结果未呈现。在免疫后第2周和第4周采集血清以测量S蛋白结合抗体应答和SARS-CoV-2中和抗体应答。通过ELISA测量S蛋白结合抗体滴度。
S蛋白结合抗体应答
用Ad26COVS1单次免疫导致S蛋白结合抗体的剂量水平依赖性诱导,免疫后2周就具有高抗体滴度(图76A)。与编码无稳定化突变的S蛋白的Ad26NCOV006载体相比,Ad26COVS1在第2周和第4周时间点诱导显著更高的结合抗体滴度。每组的应答中值用水平线表示。虚线表示LLOD。
中和抗体应答
用接受1010vp Ad26COVS1的小鼠的血清样品确定中和抗体的存在。通过wtVNA测量中和抗体滴度,确定病毒分离株Leiden1(L-001)对Vero E6细胞的细胞病变效应(CPE)。每组的应答中值用水平线表示。虚线表示LLOD。应答等于或低于LLOD的动物显示为空心符号。早在免疫后2周,用Ad26COVS1进行单次免疫即可诱导SARS-CoV-2中和抗体(图77)。从第2周到第4周,由Ad26COVS1诱导的wtVNA中和抗体滴度的中值增加至4倍,达到64(滴度范围:22.6-90.5)。与Ad26NCOV006相比,Ad26COVS1在第4周诱导显著更高的中和抗体滴度。
实施例16:小鼠中免疫应答的极化
这个实验旨在确定由2个基于Ad26的SARS-CoV-2载体(包括Ad26COVS1)诱导的Th1/Th2平衡。这是对疫苗相关疾病增强的理论风险的间接测量。用单剂的1010vpAd26COVS1(N=6)IM免疫BALB/c小鼠(倾向于产生Th2型应答)。对照组接受50μg重组稳定S蛋白,以100μg磷酸铝(AlPO4; )为佐剂,通常已知磷酸铝诱导Th2偏向的免疫应答(N=6)。阴性对照组仅接受100μg AlPO4佐剂(N=3)。在免疫后第2周采集脾脏和血清以测量细胞和体液免疫应答。
)为佐剂,通常已知磷酸铝诱导Th2偏向的免疫应答(N=6)。阴性对照组仅接受100μg AlPO4佐剂(N=3)。在免疫后第2周采集脾脏和血清以测量细胞和体液免疫应答。
S蛋白特异性细胞因子应答
为了测量细胞因子的产生,用野生型S蛋白肽池离体刺激脾细胞。在免疫后第13天,牺牲小鼠,将脾脏加工成单细胞悬液,并用2个覆盖完整的野生型SARS-CoV-2S蛋白序列的有11个氨基酸重叠的15元肽的肽池离体刺激脾细胞。示出了来自用肽池1和2刺激的SFU的总和。在肽刺激18小时后,通过ELISpot测量脾细胞产生的IFN-γ。通过IFN-γELISpot、ICS和多重ELISA(使用10-plex meso scale discovery[MSD]pro-inflammatory panel[mouse]kit)确定细胞免疫应答。
单剂的Ad26COVS1导致通过ELISpot测量的大量IFN-γ分泌,而以A1PO4为佐剂的重组S蛋白诱导不可检测或低的细胞免疫应答(图78)。这些结果由CD8+T细胞的ICS确认(数据未示出)。
免疫后第13天牺牲小鼠,将脾脏处理成单细胞悬液。用2个覆盖完整的野生型SARS-CoV-2 S蛋白序列的有11个氨基酸重叠的15元肽地肽池离体刺激脾细胞48小时。示出的结果是池1刺激和池2刺激之和的比例。通过多重ELISA测量细胞培养上清液中的细胞因子浓度。在用Ad26COVS1免疫后观察到大量分泌的IFN-γ,其被认为是Th1极化的标志,但与Th2型免疫应答相关的典型细胞因子IL-4、IL-5和IL-10的分泌较低(数据未示出)。认为IFN-γ与IL-4、IL-5或IL-10的细胞因子浓度的比例是免疫应答的Th1极化的指示,并在图79中示出。用Ad26COVS1免疫使脾细胞应答偏向于IFN-γ分泌。相比之下,用以A1PO4为佐剂的S蛋白免疫没有偏向于IFN-γ分泌,并且这组的总体细胞因子分泌较低。
S蛋白特异性IgG亚型应答
在小鼠中,IgG1在任何类型的免疫应答期间产生,而IgG2a主要在Th1极化免疫应答期间产生,因此IgG2a/IgG1比例的增加表明免疫应答的Th1偏向。免疫后的IgG1和IgG2a滴度和IgG2a/IgG1比例在图80中示出。免疫后13天对血清进行取样。通过ELISA测量S蛋白结合IgG1和IgG2a滴度。Ad26COVS1在所有动物中诱导S蛋白结合IgG1和IgG2a抗体。相反,在所有动物中,以AlPO4为佐剂的S蛋白诱导IgG1,但IgG2a滴度接近或等于检测下限。这导致在Ad26COVS1免疫后的IgG2a/IgG1比例主要偏向Th1,并且在统计学上不同于用以A1PO4为佐剂的S蛋白免疫后观察到的比例,A1PO4与Th2偏向的应答相关。
实施例17:新西兰白兔的免疫原性
用单剂的5x109vp或临床剂量水平5x1010vp的Ad26COVS1 IM免疫新西兰白兔(N=5/剂量水平)。阴性对照组接受生理盐水(N=5)。Ad26NCOV006载体编码没有稳定化突变的天然全长S蛋白,可以与Ad26COVS1进行比较,以评估稳定化突变的益处。在剂量1后第2周采集的血清中测量S蛋白结合抗体应答和SARS-CoV-2中和抗体应答。在剂量1后第3周采集的外周血单个核细胞(PBMC)中测量S蛋白特异性细胞应答。
S蛋白结合抗体应答
用Ad26COVS1单次免疫导致S蛋白结合抗体的剂量水平依赖性诱导(图81A)。与编码无稳定化突变的S蛋白的Ad26NCOV006载体相比,Ad26COVS 1诱导显著更高的结合抗体滴度。
中和抗体应答
用Ad26COVS1单次免疫导致SARS-CoV-2中和抗体的剂量水平依赖性诱导,所有接受5x1010vp更高剂量水平的动物都有应答(图81B)。中和滴度低且多变,这可能是由于早期测定时间点(第2周)。与编码无稳定化突变的S蛋白的Ad26NCOV006载体相比,Ad26COVS1诱导的中和抗体滴度显著升高。
S蛋白特异性细胞因子应答
为了测量细胞免疫原性,用野生型S蛋白肽池离体刺激PBMC,并通过IFN-γELISpot测定细胞免疫应答。在免疫后第21天,分离出PBMC并用2个覆盖完整的野生型SARS-CoV-2 S蛋白序列的有11个氨基酸重叠的15元肽肽池离体刺激PBMC。在肽刺激18小时后通过ELISpot测量PBMC产生的IFN-γ。当在剂量1后3周测量时,Ad26COVS1在所有动物中诱导了与剂量水平无关的细胞应答,尽管该应答低于用编码天然S蛋白,即没有稳定化突变的Ad26NCOV006载体免疫后的应答(图82)。这确认了在用Ad26COVS1进行的鼠类研究中已经观察到的Th1相关细胞因子IFN-γ的诱导。
实施例18:叙利亚仓鼠中的免疫原性和效力
用109或1010vp Ad26COVS1(每组N=6)在1剂方案(第5组和第6组)或间隔4周的2剂方案(第13组和第14组)中IM免疫叙利亚仓鼠。Ad26NCOV006编码没有稳定化突变的天然全长S蛋白,其以与Ad26COVS1相同的剂量水平和方案给药(每组N=6,1剂第1组和第2组,2剂第9组和第10组)并且可以进行比较以评估Ad26COVS1稳定化突变的益处。阴性对照组在1剂和2剂方案(分别为第7组和第8组以及第15组)中接受Ad26.Empty(每组N=6)。在剂量1后第4周采集的血清中测量了S蛋白结合抗体应答和SARS-CoV-2中和抗体应答。在第4周用SARS-CoV-2攻击接受1剂方案的动物,追踪期为4天。
S蛋白结合抗体应答和中和抗体应答
免疫后28天对血清进行取样。通过ELISA测量S蛋白结合抗体滴度。Ad26.Empty N=18(图83A)。通过wtVNA测量中和抗体滴度以确定病毒分离株Leiden1(L-001)对Vero E6细胞的细胞病变效应(CPE)(图83B)。用Ad26COVS1单次免疫导致S蛋白结合抗体和SARS-CoV-2中和抗体的诱导,在剂量1后第4周测量,即在接受1剂方案的动物的攻击时(图83A和83B)。
病毒载量
叙利亚仓鼠用1×109或1×1010vp Ad26COVS1(Ad26NCOV030)或Ad26NCVO006(编码天然S蛋白的Ad26载体)(N=6/剂量水平)或Ad26.Empty(Ad26载体不编码任何COVID-19抗原,N=6)肌内免疫。仓鼠在免疫后4周(第28天)接受102TCID50 SARS-CoV-2株BetaCoV/Munich/BavPat1/2020的鼻内攻击。在攻击阶段(第C4天)结束时分离肺组织用于病毒载量分析。通过噬斑测定测量复制能力病毒(TCID50/克肺)。攻击后4天通过噬斑测定确定肺组织中的复制能力病毒。
接受Ad26.Empty的模拟免疫对照组在攻击后4天具有高肺病毒载量。与Ad26.Empty模拟免疫动物相比,单剂的Ad26COVS1显著降低了病毒载量。与编码没有稳定化突变的S蛋白的Ad26NCOV006载体相比,用Ad26COVS1免疫导致病毒载量显著降低(图84)。大多数用Ad26COVS1免疫的动物的病毒载量低于检测限。
攻击后体重减轻
在用SARS-CoV-2攻击后,确定体重减轻为疾病严重度的衡量。与在攻击后4天测量的Ad26.Empty对照免疫的动物相比,单剂的Ad26COVS1显著减少了体重减轻(图85)。在Ad26COVS1和Ad26NCOV006免疫组之间,不同剂量水平的体重减轻没有显著差异(p=0.091来自方差分析[ANOVA]的t检验以及3倍Bonferroni校正)。
实施例19:非人灵长类动物(NHP)中的免疫原性
用单剂的1011vp Ad26COVS1(N=6)IM免疫NHP,这对应于用于临床使用的高疫苗剂量。阴性对照组仅接受生理盐水(N=4)。在免疫前(第0周,基线)和免疫后第2周以及第4周采集用于评估体液免疫原性的血清。在第0周和第4周分离用于基于细胞的测定的PBMC。
S蛋白结合抗体应答和中和抗体应答
在NHP中,早在免疫后第2周,用Ad26COVS1进行的单次免疫诱导S蛋白结合抗体(图86A)和通过ppVNA(图86B)和wtVNA(图86C)测量的中和抗体。所有测定都在Chandrashekaret al.Science.2020;eabc4776.doi:10.1126/science.abc4776中进行了描述。
与第2周相比,第4周结合抗体滴度中值增加至3.3倍,达到16,497(滴度范围:4,450-57,790)。从第2周到第4周,ppVNA中和抗体滴度的中值增加至5.8倍,达到408(滴度范围:208-643)。野生型VNA中和抗体滴度通常较低,但随着时间的推移显示出相当的增加;第4周中和抗体滴度的中值增加至6.2倍,达到113(滴度范围53-234)。
S蛋白特异性细胞因子应答
为了测量细胞免疫原性,用野生型S蛋白肽池离体刺激PBMC,并通过IFN-γELISpot测定细胞免疫应答。当免疫后4周测量时,Ad26COVS1在6只动物中的5只中诱导了细胞应答(图86D)。这确认了已经在具有Ad26COVS1的小鼠和兔中观察到的Th1相关细胞因子IFN-γ的诱导。
实施例20:单次Ad26疫苗在恒河猴中防止SARS-CoV-2攻击
Ad26候选疫苗的产生和免疫原性
如上所述,产生了一些表达SARS-CoV-2 S蛋白变体的Ad26载体:(i)使用组织纤溶酶原激活物(tPA)前导序列(LS)代替野生型(wt)LS(tPA.S;COR200009:SEQ ID NO:207),(ii)tPA前导LS代替野生型LS和具有弗林蛋白酶切割位点突变和两个脯氨酸稳定化突变的全长S(tPA.S.PP;COR200010:SEQ ID NO:208)(iii)天然全长S(S;Wuhan/WIV04/2019),(iv)tPA和野生型前导序列以及全长S(tPA.WT.S;COR200018:SEQ ID NO:210)),和(v)具有弗林蛋白酶切割位点突变和脯氨酸稳定化突变的全长S(S.PP;COR200007:SEQ ID NO:205)。免疫印迹分析确认了在已经接种了所有载体中的每一种的细胞的细胞裂解物中刺突蛋白的表达,并且在具有突变的弗林蛋白酶切割位点的刺突变体的细胞裂解物中不存在弗林蛋白酶切割产物(图87)。
用表达tPA.S(N=4)、tPA.S.PP(N=4)、S(N=4)、tPA.WT.S(N=4)、S.PP(N=6)的Ad26载体和假对照(N=20)免疫52只6-12岁的混合性别的成年恒河猴。动物在第0周接受了无佐剂的1011病毒颗粒(vp)Ad26载体的肌内单次免疫。到第2周,通过ELISA在几乎所有疫苗接种的动物中观察到RBD特异性结合抗体,并且到第4周在所有疫苗接种的动物中观察到RBD特异性结合抗体(图88A)。使用假病毒中和测定(Chandrashekar,A.et al.Science,doi:10.1126/science.abc4776(2020);Yu,J.et al.Science,doi:10.1126/science.abc6284(2020);Yang,ZY et al.Nature 428,561-564,doi:10.1038/nature02463(2004))(图88B)和活病毒中和测定(数据未示出)评估中和抗体(NAb)应答。在第2周,在大多数疫苗接种的动物中观察到通过两种中和测定测量的NAb滴度,并在第4周增加。Ad26-S.PP(Ad26COVS1)疫苗引发第4周时最高的假病毒NAb滴度(中值408;范围208-643)和活病毒NAb滴度(中值113;范围53-233)。疫苗接种Ad26-S.PP的猕猴的NAb滴度的中值是先前报道的从SARS-CoV-2感染10恢复的9只恢复期猕猴和27个恢复期人的队列的NAb滴度的中值的4倍(图93)。Ad26-S.PP疫苗还在支气管肺泡灌洗液(BAL)中诱导了低但可检测到的S特异性IgG和IgA应答(图94)。
在第4周,使用合并的S肽通过IFN-γELISPOT测定在大多数疫苗接种的动物中诱导细胞免疫应答(图89A)。使用多参数细胞内细胞因子染色测定来评估IFN-γ+CD4+和CD8+T细胞应答(图89B)。尽管Ad26-S.PP疫苗的应答有降低的趋势,但在各个疫苗组的应答相当。IFN-γ+和IL-2+CD4+T细胞应答高于IL-4+和IL-10+CD4+T细胞应答(图95),表明诱导了Th1偏向的细胞免疫应答。
Ad26候选疫苗的保护效力
在第6周,通过鼻内(IN)和气管内(IT)途径用1.2x108VP(1.1x104PFU)SARS-CoV-2攻击所有动物(Chandrashekar,A.et al.Science,doi:10.1126/science.abc4776(2020);Yu,J.et al.Science,doi:10.1126/science.abc6284(2020))。通过RT-PCR检测亚基因mRNA(sgmRNA)评估支气管肺泡灌洗液(BAL)和鼻拭子(NS)中的病毒载量,认为其代表复制的病毒(Chandrashekar,A.et al.同上;Wolfel,R.et al.Nature,doi:10.1038/s41586-020-2196-x(2020)。
所有20个假对照被感染并且在BAL中示出4.89(范围3.85-6.51)log10 sgmRNA拷贝/ml峰值的中值(图90A)。相比之下,接受Ad26-S.PP的动物在BAL中没有示出可检测的病毒(检测限为1.69log10 sgmRNA拷贝/ml)。用其他疫苗也观察到了显著的保护作用,动物偶尔在BAL中表现出低水平的sgmRNA(图90B)。类似地,假对照在NS中示出5.59(范围3.78-8.01)log10 sgmRN A峰值的中值(图90C)。一只接受Ad26.S.PP的动物在NS中的病毒量很低。与对照相比,尽管用Ad26.S.PP观察到最好的保护作用,其他疫苗通常表现出在NS中的病毒载量降低(图90D)。
疫苗接种动物的病毒载量峰值的比较表明,BAL中的保护通常比NS中更强(图91)。Ad26-S.PP疫苗在下呼吸道提供完全保护,在上呼吸道保护5/6只动物,与假对照相比,BAL和NS中的sgmRNA峰值的中值分别降低>3.2和>3.9log10(分别为P<0.0001和P<0.0001,双边Mann-Whitney检验)(图91)。这些数据支持S免疫原融合前稳定化的相关性。在32名疫苗接种者中,17只动物受到完全保护,在攻击后BAL或NS中未检测到sgmRNA,另外5只动物在BAL中未检测到sgmRNA,但在NS中显示出一些病毒。
攻击后Ad26.S.PP疫苗接种动物的免疫应答
在SARS-CoV-2攻击后的第14天,假对照产生了强的假病毒NAb应答(图92)和CD8+和CD4+T细胞应答(数据未示出)。这些动物中的CD8+和CD4+T细胞应答针对多种SARS-CoV-2蛋白,包括刺突蛋白(S1、S2)、核衣壳(NCAP)和非结构蛋白(NS6、NS71、NS8)。相比之下,接受Ad26.S.PP疫苗的动物没有表现出抗体再生NAb应答(图92),并且在攻击后仅表现出对刺突(S1、S2)的低T细胞应答。这些发现与来自这些动物的几乎不存在的病毒载量数据一致(图90-91)并且表明在这些动物中病毒复制水平极低,如果有的话,在攻击之后。
开发安全有效的SARS-CoV-2疫苗是一项重要的全球优先事项。本发明证明,用表达工程化改造的S免疫原的Ad26载体进行单次免疫可诱导强的NAb应答并在恒河猴中防止SARS-CoV-2攻击。本研究中最好的疫苗是Ad26-S.PP(Ad26COVS1或Ad26NCOV030),它含有野生型前导序列、全长膜结合S、弗林蛋白酶位点突变和两个脯氨酸稳定化突变。
这些数据扩展了最近对非人灵长类动物中SARS-CoV-2灭活病毒和DNA疫苗的临床前研究(Yu,J.et al.Science,doi:10.1126/science.abc6284(2020);Gao,Q.etal.Science,doi:10.1126/science.abc1932(2020))。灭活病毒疫苗和核酸疫苗通常需要至少2次免疫,而基于腺病毒载体的疫苗可以在单次免疫后诱导强而持久的抗体应答。与用于大规模疫苗接种活动和大流行控制的两剂疫苗相比,单次SARS-CoV-2疫苗将具有重要的统筹和实际优势。然而,之前也显示,Ad26-HIV载体的同源加强可以在非人灵长类动物和人中增加至10倍以上的抗体滴度,这表明Ad26-S.PP疫苗的单剂和两剂方案都可以在临床试验中进行评估。在本研究中,Ad26.S.PP在单次免疫后诱导NAb应答,并提供防止SARS-CoV-2攻击的完全或接近完全的保护。此外,这些动物中的NAb滴度和T细胞应答在攻击后没有显著扩大,并且T细胞应答没有扩大到非疫苗抗原,例如核衣壳和非结构蛋白,而假对照在攻击后对多种SARS-CoV-2蛋白产生了强的NAb滴度和T细胞应答。这些数据表明,在攻击后用Ad26COVS1疫苗接种的动物中病毒复制很少甚至没有。
此外,值得注意的是,Ad26疫苗引发了Th1偏向而不是Th2偏向的CD4+T细胞应答,并且具有亚保护NAb滴度的动物没有表现出增强的病毒复制。
材料和方法
动物和研究设计.将52只远交系的6-12岁的印度裔成年雄性和雌性恒河猴(Macaca mulatta)随机分组。所有动物都饲养在Bioqual,Inc.(Rockville,MD)。动物接受表达tPA.S(N=4)、tPA.S.PP(N=4)、S(N=4)、tPA.WT.S(N=4)、S.PP(N=6)的Ad26载体和假对照(N=20)。通过无佐剂的肌内途径,动物在第0周接受了1011病毒颗粒(vp)Ad26载体的单次免疫。在第6周,用1.2x108VP(1.1x104PFU)SARS-CoV-2(USA-WA1/2020;NR-52281原种;BEI Resources)攻击所有动物。通过鼻内(IN)途径给药1ml病毒(每个鼻孔0.5ml)和通过气管内(IT)途径给药1 ml。所有免疫学和病毒学测定均在盲法下进行。所有动物研究均按照所有相关的地方、州和联邦法规进行,并得到生物质量机构动物管理和使用委员会(IACUC)的批准。
Ad26载体由多个形式的SARS-CoV-2刺突(S)蛋白序列(Wuhan/WIV04/2019)构建而成。序列经过密码子优化和合成。使用含有完整Ad26载体基因组和转基因表达盒的质粒在PER.C6.TetR细胞中产生无复制能力的、E1/E3缺失的Ad26载体(Abbink,P.et al.J Virol81,4654-4663,doi:JVI.02696-06[pii]10.1128/JVI.02696-06(2007))。在使用前对载体进行测序和表达测试。
免疫印迹.用SARS-Cov-2 Ad26载体瞬时转染接种有70-80%汇合度的293T细胞的T-25培养瓶,并且在转染后48小时,收获细胞裂解物,并分别与还原样品缓冲液(Pierce)混合,在95℃加热5分钟,并且在预制的4-15%SDS-PAGE凝胶(Bio-Rad)上跑胶。使用iBlot干式印迹系统(Invitrogen)将蛋白转移到聚偏二氟乙烯(PVDF)膜上,并在4℃下在含有0.2%Tween 20(V/V)(Sigma)和5%(W/V)脱脂奶粉的Dulbecco磷酸盐缓冲盐水T(D-PBST)中进行膜封闭过夜。过夜封闭后,将PVDF膜在含有1∶10,000稀释的多克隆豚鼠抗SARS抗体(BEIresources)的3%牛奶DPBS-T中孵育1小时。孵育后,PVDF膜用5%牛奶DPBS-T洗涤5次,并且随后与在3%牛奶DPBS-T中的1∶30,000抗豚鼠或抗兔辣根过氧化物酶(HRP)缀合的二抗(Jackson Immunoresearch)一起孵育。最后,将PVDF膜用5%牛奶DPBS-T再次洗涤5次,并使用Amersham ECL Plus免疫印迹检测系统(GE Healthcare)显影。
亚基因组mRNA测定.通过RT-PCR使用先前描述的相同的方法评估SARS-CoV-2E基因亚基因组mRNA(sgmRNA)9,10,20。为了生成标准曲线,将SARS-CoV-2E基因sgmRNA克隆至pcDNA3.1表达质粒中;使用AmpliCap-Max T7 High Yield Message Maker Kit(Cellscript)转录此插入物,以获得标准的RNA。在RT-PCR之前,根据制造商的说明,使用Superscript III VILO(Invitrogen)将从受攻击的动物或标准中收集的样品进行逆转录。使用靶向E基因sgmRNA的序列设计了Taqman定制基因表达测定(ThermoFisherScientific)。根据制造商的说明,在QuantStudio 6和7Flex实时PCR系统(AppliedBiosystems)上进行反应。标准曲线用于计算每毫升或每拭子sgmRNA的拷贝数;定量测定的灵敏度为每毫升或每拭子50个拷贝。
PFU测定.对于噬斑测定,在6孔板中制备Vero E6细胞的汇合单层。将从受攻击的动物中收集的指定样品连续稀释,添加到孔中,并在37℃下孵育1小时。孵育后,将1.5mL0.5%甲基纤维素培养基添加至每个孔中,并将板在37℃和5%CO2下孵育2天。通过每孔加入400μL冰冷的甲醇并在-20℃下孵育30分钟来固定板。固定后弃去甲醇,用每孔600μL的0.23%结晶紫染色细胞单层30分钟。染色后,弃去结晶紫,用600μL水洗板一次以可视化和计数噬斑。
ELISA.简而言之,用1xDPBS中的1μg/mL SARS-CoV-2RBD蛋白(Aaron Schmidt,MassCPR)包被96孔板,并在4℃下孵育过夜。孵育后,用洗涤缓冲液(1X DPBS中的0.05%Tween20)洗涤板一次,并在室温下用350μL酪蛋白封闭液/孔封闭2-3小时。孵育后,弃去封闭溶液并将板吸干。将在酪蛋白封闭液中稀释的热灭活血清的连续稀释液添加至孔中,并将板在室温下孵育1小时,然后再洗涤3次,随后用1∶1000稀释的抗猕猴IgG HRP(NIH NHPReagent Program)在室温下避光孵育1小时。然后用洗涤缓冲液洗涤板3次,并且每孔加入100μL SeraCare KPL TMB SureBlue Start溶液;通过每孔添加100μL SeraCare KPL TMBStop溶液来停止板的显色。使用VERSAMAXTM或 微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生吸光度>0.2的最高血清稀释度的倒数。报告为Log10终点滴度。
假病毒中和测定.通过与之前描述的方法相似的方法产生表达荧光素酶报告基因的SARS-CoV-2假病毒9,10,16。简而言之,通过磷酸钙,将包装构建体psPAX2(AIDS Resourceand Reagent Program)、荧光素酶报告质粒pLenti-CMV Puro-Luc(Addgene)和表达刺突蛋白的pcDNA3.1-SARS CoV-2 SΔCT共转染到HEK293T细胞中。转染后48小时收集含有假病毒的上清液;通过用0.45μm过滤器过滤来纯化假病毒。为了确定来自疫苗接种动物的抗血清的中和活性,将HEK293T-hACE2细胞以1.75×104个细胞/孔的密度接种在96孔组织培养板中过夜。制备热灭活血清样品的两倍连续稀释液,并与50μL假病毒混合。在加入HEK293T-hACE2细胞之前,将混合物在37℃下孵育1小时。感染后48小时,根据制造商的说明,在Steady-Glo Luciferase Assay(Promega)中裂解细胞。将SARS-CoV-2中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
ELISPOT检测.用来自BD Pharmingen的小鼠抗人IFN-γ单克隆抗体以5μg/孔的浓度包被ELISPOT板,在4℃下过夜。用含有0.25%Tween20的DPBS洗涤板,并在37℃下用R10培养基(含有11%FBS和1.1%青霉素-链霉素的RPMI)封闭1小时。刺突1和刺突2肽池含有15个氨基酸肽,这些肽有11个氨基酸重叠,它们涵盖蛋白序列并分别反映蛋白的N端和C端半部分。以2μg/孔的浓度制备刺突蛋白1和刺突蛋白2肽池,并添加200,000个细胞/孔。将肽和细胞在37℃下孵育18-24小时。此孵育后的所有步骤均在室温下进行。用库尔特(coulter)缓冲液洗涤板并与来自U-Cytech的兔多克隆抗人IFN-γ生物素(1μg/mL)一起孵育2小时。将板第二次洗涤并与来自Southern Biotechnology的链霉亲和素-碱性磷酸酶抗体(1μg/mL)一起孵育2小时。最后一次洗涤后添加氯化硝基四氮唑蓝/5-溴-4-氯-3′-吲哚基磷酸对甲苯胺盐(NBT/BCIP显色液)底物溶液,保持7分钟。弃去显色液(chromagen),将板用水洗涤并在暗处干燥24小时。在Cellular Technologies Limited Immunospot Analyzer上扫描和计数板。
细胞内细胞因子染色测定.将106PBMC/孔重悬在100μL的补充有CD49d单克隆抗体(1μg/mL)的R10培养基中。使用模拟(100μL的R10加0.5%DMSO;背景对照)、肽池(2μg/mL)或10pg/mL豆蔻酰佛波醇乙酯(phorbol myristate acetate)(PMA)和1μg/mL离子霉素(Sigma-Aldrich)(100μL;阳性对照)对每个样品进行评估并在37℃下孵育1小时。孵育后,将50μL R10中的0.25μL GolgiStop和0.25μL GolgiPlug添加到每个孔中,并在37℃下孵育8小时,然后在4℃下保持过夜。第二天,细胞用DPBS洗涤两次,用近红外活/死染料染色10分钟,然后用针对CD279(克隆EH12.1、BB700)、CD38(克隆OKT10、PE)、CD28(克隆28.2,PECY5)、CD4(克隆L200,BV510)、CD45(克隆D058-1283,BUV615)、CD95(克隆DX2,BUV737)、CD8(克隆SK1,BUV805)的预先指定滴度的单克隆抗体染色,持续30分钟。然后用2%FBS/DPBS缓冲液洗涤细胞两次,并用200μL BD CytoFix/CytoPerm Fixation/Permeabilization溶液孵育15分钟。用1X Perm洗涤缓冲液(将CytoFix/CytoPerm Fixation/Permeabilization试剂盒中的BD Perm/WashTM Buffer 10X用Milli Q水稀释并通过0.22μm过滤器)洗涤细胞两次,并用针对Ki67(克隆B56,FITC)、CD69(克隆TP1.55.3,ECD)、IL10(克隆JES3-9D7,PECY7)、IL13(克隆JES10-5A2,BV421)、TNF-α(克隆Mab11,BV650)、IL4(克隆MP4-25D2,BV711)、IFN-γ(克隆B27;BUV395)、IL2(克隆MQ1-17H12、APC)、CD3(克隆SP34.2、Alexa700)的单克隆抗体进行细胞内染色30分钟。细胞用1x Perm洗涤缓冲液洗涤两次,并用250μL新鲜制备的1.5%甲醛固定。将固定的细胞转移至96孔圆底板中并通过BD FACSymphonyTM系统进行分析。
统计分析.使用GraphPad Prism 8.4.2(GraphPad软件)对病毒学和免疫学数据进行分析。使用双边Mann-Whitney检验进行组间数据比较。通过双边Spearman等级相关性检验评估相关性。认为P值小于0.05是显著的。
实施例21:Ad26疫苗在仓鼠中防止SARS-CoV-2严重临床疾病
评估了仓鼠中高剂量SARS-CoV-2感染的临床和病毒学特征,并评估了严格的模型中根据本发明的基于Ad26载体的疫苗的保护效力,所述基于Ad26载体的疫苗编码稳定化的SARS-CoV-2刺突(S)。
通过鼻内途径用5x104TCID50(N=4;低剂量)或5x105TCID50(N=16;高剂量)SARS-CoV-2接种20只叙利亚金仓鼠。在高剂量组中,在第2天对4只动物进行尸检,在第4天对4只动物进行尸检,以检测组织病毒载量和组织病理学,并且其余8只动物进行纵向追踪。
在第14天对所有剩余的动物进行尸检。在低剂量组中,仓鼠到第6天时体重的中值下降了14.7%,但在第14天完全恢复(图96A、96B),这与之前的研究一致(Chan et al.ClinInfect Dis,doi:10.1093/cid/ciaa325,(2020);Sia et al.,Nature,doi:10.1038/s41586-020-2342-5(2020);Imai et al.,Procc Natl Acad Sci USA 117,16587-16595,(2020))。在高剂量组中,仓鼠到第6天时体重的中值下降了19.9%。在该组纵向追踪的8只动物中,4只在第6天满足IACUC人道安乐死标准,体重减轻>20%并且呼吸窘迫,另外2只动物在第7天满足这些标准。其余2只动物在第14天恢复。
这些数据表明,仓鼠中的高剂量SARS-CoV-2感染会导致严重的体重减轻和部分死亡。
对接受高剂量SARS-CoV-2并在第2天进行尸检的4只动物、在第4天进行尸检的4只动物以及在第6-7天满足安乐死标准的6只动物中的5只动物评估组织病毒载量(图96C)。在第2天观察到,肺组织中1012RNA拷贝/g以及鼻孔和气管中108-109RNA拷贝/g的高组织病毒载量中值,以及心脏、胃肠道、脑、脾脏、肝脏和肾脏中105-108RNA拷贝/g的中值,这表明播散性感染。到第6-7天,组织病毒载量降低了大约2log,尽管体重持续下降。
在第2天(N=4)、第4天(N=4)、第6-7天(N=6)和第14天(N=2)通过组织病理学评估感染了高剂量SARS-CoV-2的仓鼠。感染与鼻甲(图97a)和细支气管上皮的显著炎症浸润和多灶性上皮坏死相关,其导致管腔中的退化的中性粒细胞和细胞碎片(图97b)。附近血管的内皮与单个核细胞与内皮的粘附反应并在血管壁内迁移,表明内皮炎(图97b)。早在SARS-CoV-2感染后第2天,就有中度至严重多灶性间质性肺炎,其特征是影响了30-60%的肺实质的肺实变(图97c)。炎症浸润由大量巨噬细胞和中性粒细胞组成,淋巴细胞较少。通过免疫组织化学(IHC)在鼻甲上皮(图97d)和细支气管上皮细胞(图97e)的炎症和坏死区域对SARS核衣壳蛋白(SARS-CoV-N)呈强阳性。SARS-CoV-N的免疫组织化学还显示肺泡间隔和间质单个核细胞的局部广泛染色,其形态与巨噬细胞一致(图97f)。类似地,在细支气管上皮和肺间质中的炎症区域中观察到大量SARS-CoV-2病毒RNA(vRNA)(图97g,97h)。
肺中SARS-CoV-2vRNA和SARS-CoV-N蛋白的表达水平在第2天最高,到第4天下降,到第7天检测到最少的vRNA和SARS-CoV-N蛋白(图100)。肺炎的特征在于肺间质中Iba-1+巨噬细胞的大量炎性浸润以及CD3+T淋巴细胞(图97i,97j)。在整个肺,特别是在坏死区域中检测到大量存活和退化的中性粒细胞,其中中性粒细胞髓过氧化物酶(MPO)在整个肺中高表达(图97k)。还在肺中检测到干扰素诱导基因产物MX1的扩散表达(图971)。与在第2天达到峰值的SARS-CoV-2vRNA和SARS-CoV-N检测的动力学相反,这些炎症标志物在第7天达到峰值(图100),与最大的体重减轻和死亡率一致(图96A,96B)。通过 检测肺中的vRNA并不仅仅反映病毒接种物,因为从攻击后的第2天到第7天我们不仅检测到反义vRNA,而且还检测到正义vRNA,其在位置和模式上重叠。SARS-CoV-2vRNA表达(反义和正义)存在于肺中,具有强ACE2受体表达(数据未示出)。
检测肺中的vRNA并不仅仅反映病毒接种物,因为从攻击后的第2天到第7天我们不仅检测到反义vRNA,而且还检测到正义vRNA,其在位置和模式上重叠。SARS-CoV-2vRNA表达(反义和正义)存在于肺中,具有强ACE2受体表达(数据未示出)。
我们产生了重组的、无复制能力的Ad26载体,其编码(i)SARS CoV-2刺突(S),其跨膜区和细胞质尾缺失,反映了具有折叠子三聚化结构域的可溶性胞外域(S.dTM.PP)或(ii)全长S(S.PP),都具有弗林蛋白酶切割位点的突变和两个脯氨酸稳定化突变(图98A)。
在实施例20中,描述了S.PP疫苗在恒河猴中防止SARS-CoV-2攻击的免疫原性和保护效力。
在本实施例中,用1010或109个病毒颗粒(vp)的编码S.dTM.PP或S.PP的Ad26载体(N=10/组)或假对照(N=10)免疫50只叙利亚金仓鼠。动物在第0周通过肌内途径接受单次疫苗接种。在第2周和第4周,在所有动物中,我们通过ELISA观察受体结合结构域(RBD)特异性结合抗体(图98B),并通过假病毒中和测定观察到中和抗体(NAb)(图98C)。在第4周,与Ad26-S.dTM.PP(1014,1185)相比,Ad26-S.PP引发的ELISA滴度的中值(4470,4757)是4.0-4.7倍(图98B;P<0.0001,双边Mann-Whitney检验)。类似地,与Ad26-S.dTM.PP(139,211)相比,Ad26-S.PP引发的NAb IC50滴度的中值(359,375)是1.8-2.6倍(P<0.05,双边Mann-Whitney检验)。对于每个载体,测试的两个剂量似乎是可比的。ELISA和NAb数据在第2周和第4周均相关(分别为R=0.7074,P<0.0001和R=0.7849,P<0.0001,双边Spearman等级相关性检验)。
我们通过系统血清学进一步表征了疫苗接种动物中在第4周时的S特异性和RBD特异性抗体应答(Chung,et al.,Cell 163,988-998,doi:10.1016/j.cell.2015.10.027(2015))。评估了IgG、IgG2a、IgG3、IgM、Fc受体FcRγ2、FcRγ3和FcRγ4,以及抗体依赖性补体沉积(ADCD)应答(图98D-98F)。与Ad26.S.dTM.PP相比,Ad26-S.PP观察到了更高和更一致的应答(图98D,98F),并且这些抗体特征的主成分分析确认这两种疫苗具有不同的谱(图98E)。
在第4周通过鼻内途径用5x105TCID50 SARS-CoV-2攻击所有动物。在第4天,对每组中的3只动物进行尸检,以检测组织病毒载量和组织病理学,并对每组中剩余的7只动物进行追踪直至第14天。在假对照中,到第7天,仓鼠体重的中值下降了19.6%,在第6-7天纵向追踪的动物中有43%(7只中的3只)满足安乐死标准(图99A、99B)。疫苗接种Ad26-S.dTM.PP的动物体重的中值下降8.7%,而疫苗接种Ad26-S.PP的动物体重的中值下降了4.0%(图99A、99B)。与假对照相比,两个疫苗接种组的最大体重减轻百分比均显著降低(P<0.0001,双边Mann-Whitney检验;图99C),并且接受Ad26-S.PP的动物比接受Ad26.S.dTM.PP的动物表现出更少的体重减轻(P<0.0001,双边Mann-Whitney检验;图99C)。与假对照相比,两种疫苗都防止死亡,将死亡定义为满足人道安乐死标准(P=0.02,双边Fisher精确检验)。对两个仓鼠实验的组合分析确认两种疫苗都能有效防止死亡(P=0.007,双边Fisher精确检验)。第2周(R=-0.8992,P<0.0001)和第4周(R=-0.9344,P<0.0001)的ELISA应答与最大体重减轻百分比负相关。第2周(R=-0.7380,P<0.0001)和第4周(R=-0.8075,P<0.0001)的NAb应答也与最大体重减轻百分比负相关。
与假对照相比,疫苗接种Ad26-S.dTM.PP和Ad26-S.PP的仓鼠的所有肺叶中的组织细胞性和嗜中性炎症浸润显著减少,SARS-CoV-2vRNA显著减少(P=0.004和P=0.004,分别为双边Mann-Whitney检验)。
在本实施例中,证明了编码全长融合前稳定S免疫原(S.PP)的Ad26载体的单次免疫可在仓鼠中防止高剂量SARS-CoV-2攻击后的严重的临床疾病。假对照表现出明显的体重减轻、严重肺炎和部分死亡。相比之下,疫苗接种的动物显示出较少的体重减轻和肺炎,而且没有死亡。疫苗引发的结合抗体应答和中和抗体应答与防止临床疾病以及减少上呼吸道和下呼吸道中的病毒复制相关。
这个模型中这种临床疾病的严重度与先前涉及仓鼠和其他物种的SARS-CoV-2感染的研究形成鲜明对比。仓鼠是SARS-CoV-2的允许模型,因为它们与人ACE2受体同源,并且据报道SARS-CoV-2在仓鼠之间传播(Sia,et al.,Nature,doi:10.1038/s41586-020-2342-5(2020))。在本研究中,高攻击剂量导致广泛的临床疾病,尽管可能影响临床疾病程度的生物学要素,例如动物年龄、动物来源和病毒攻击原种,仍有待完全确定。
迄今为止,在非人灵长类动物身上进行的SARS-CoV-2疫苗研究已证明可以防止感染或减少上呼吸道和下呼吸道中的病毒复制。在实施例20中,它报道了Ad26-S.PP的单次免疫可在恒河猴中提供防止SARS-CoV-2攻击的完全或接近完全的保护。然而,非人灵长类动物中的SARS-CoV-2感染不会导致严重的临床疾病或死亡。严重疾病模型将有助于补充当前的非人灵长类动物攻击模型,因为防止病毒复制的保护并不一定意味着防止严重疾病的保护。事实上,在本研究中在仓鼠的组织病理学分析中,肺中的病毒载量从第2天到第7天下降,而炎症标志物在此期间持续升高并与持续的体重减轻相关。这些数据表明,仓鼠的进行性临床疾病主要是一种炎症过程,由感染引发,即使当病毒复制减少,炎症也会继续增加。
由于人的COVID-19可发展为严重的临床疾病,因此在再现严重临床疾病(包括暴发性肺炎和死亡)的临床前模型中测试SARS-CoV-2候选疫苗非常重要。在这个实施例所述的高剂量仓鼠模型满足许多这些标准,因此可用于研究严重疾病的发病机制以及测试应对措施。这个模型中临床疾病的主要表现是严重肺炎,而不是某些hACE2转基因小鼠模型中报道的脑炎。此外,结合抗体应答和中和抗体应答与保护相关。
总之,这些数据表明,Ad26-S.PP的单次免疫提供了防止高剂量SARS-CoV-2感染后的严重临床疾病的强的保护。据我们所知,这种防止严重SARS-CoV-2肺炎和死亡的疫苗保护以前没有报道。Ad26-S.PP,也称为Ad26.COV2.S,目前正在临床试验中进行评估。这种仓鼠严重疾病模型还可用于测试SARS-CoV-2疫苗、治疗和其他应对措施。
方法
动物和研究设计.将70只10-12周龄的雄性和雌性叙利亚金仓鼠(Envigo)随机分组。所有动物都饲养在Bioqual,Inc.(Rockville,MD)。动物接受表达S.dTM.PP或S.PP的Ad26载体或假对照(N=10/组)。通过无佐剂的肌内途径,动物在第0周接受了1010或109病毒颗粒(vp)的Ad26载体的单次免疫接种。在第4周,用5.0x105TCID50(6x108VP,5.5x104PFU)或5.0x104TCID50(6x107VP,5.5x103PFU)的SARS-CoV-2攻击所有动物,所述SARS-CoV-2来源于USA-WA1/2020(NR52281;BEI Resources)的1代。通过鼻内(IN)途径给药100μL的病毒(每个鼻孔50μL)。每天评估体重。所有免疫学和病毒学测定均在盲法下进行。在第4天,因组织病毒载量和病理学将一部分动物安乐死。所有动物研究均按照所有相关的地方、州和联邦法规进行,并得到生物质量机构动物管理和使用委员会(IACUC)的批准。
Ad26载体.Ad26载体由SARS-CoV-2刺突(S)蛋白序列(Wuhan/WIV04/2019;GenbankMN996528.1)的两个变体构建而成。序列经过密码子优化和合成。使用含有完整Ad26载体基因组和转基因表达盒的质粒在PER.C6.TetR细胞中产生无复制能力、E1/E3缺失的Ad26载体。假对照包括Ad26-Empty载体。在使用前对载体进行测序和表达测试。
组织病理学和免疫组织化学.组织在新鲜制备的4%多聚甲醛中固定24小时,转移到70%乙醇中,并在7-10天内包埋石蜡,并以5μm切片。载玻片在65℃下烘烤30-60分钟,然后在二甲苯中脱蜡并且通过一系列分级乙醇至蒸馏水进行复水。对于SARS-CoV-N、Iba-1和CD3的免疫组织化学,使用压力锅在蒸汽设置下在柠檬酸盐缓冲液(Thermo;AP-9003-500)中进行25分钟的热诱导表位修复(HIER),然后用3%过氧化氢处理。然后在蒸馏水中冲洗载玻片并封闭蛋白(BioCare,BE965H)15分钟,然后在1x磷酸盐缓冲盐水中冲洗。
应用兔抗SARS-CoV-核蛋白一抗(Novus;NB100-56576,1∶500或1∶1000)、兔抗Iba-1抗体(Wako;019-19741,1∶500)或兔抗CD3(Dako,A0452,1∶300)30分钟,然后使用兔Mach-2HRP-Polymer(BioCare;RHRP520L)30分钟,再用苏木精复染,然后用0.25%氨水染成蓝色。SARS-N、Iba-1和CD3的标记是在Biogenex i6000全自动染色机(v3.02)上进行的。在一些情况下,使用1∶400的CD3(Thermo货号RM-9107-S;克隆SP7)、1∶500的Iba-1(BioCare货号CP290A;多克隆)或ACE-2(Abcam;ab108252)染色CD3、Iba-1和ACE-2,所有这些均使用兔Polink-1 HRP(GBI Labs货号D13-110)检测。使用1∶1000的MPO(Dako货号A0398;多克隆),并使用兔Polink-1 HRP检测中性粒细胞(MPO),以及使用1∶1000的Mx1(EMD Millipore货号MABF938;克隆M143/CL143),并使用小鼠Polink-2HRP(GBI Labs货号D37-110)检测1型IFN应答。CD3、Iba-1、MPO和Mx1的免疫组织化学的染色如前所述使用Biocare intelliPATH全自动染色机进行,所有抗体在室温下孵育1小时。组织病理学由两名兽医病理学家独立评估。
RNAscope原位杂交.RNAscope原位杂交如前所述进行(Chandrashekar,A.et al.,Science,doi:10.1126/science.abc4776(2020)),使用SARS-CoV2反义特异性探针v-nCoV2019-S(ACD货号848561)靶向正义病毒RNA,以及SARS-CoV2正义特异性探针vnCoV2019-orf1ab-sense(ACD货号859151)靶向反义基因组病毒RNA。简而言之,载玻片在二甲苯中脱蜡并通过一系列分级乙醇至蒸馏水进行复水,在95-98℃下,在ACD P2修复缓冲液(ACD货号322000)中进行修复30分钟,然后用PBS中1∶10稀释的蛋白酶III(ACD货号322337)进行处理,在40℃下孵育20分钟。然后将载玻片与PBS中的3%H2O2在室温下孵育10分钟。在杂交之前,使用微型离心机以13,000rpm的速度将探针原液离心10分钟,然后在探针稀释剂(ACD货号300041)中以1∶2的比例稀释以减少探针聚集组织伪影。载玻片使用 2.5 HD Detection Reagents-RED(ACD货号322360)显色。
2.5 HD Detection Reagents-RED(ACD货号322360)显色。
定量图像分析.使用HALO软件(v2.3.2089.27或v3.0.311.405;Indica Labs)对每只动物的至少一个肺叶横截面进行定量图像分析。在>1个横截面可用的情况下,将每个肺叶定量为一个单独的数据点。对于SARS-CoV-N,Multiplex IHC v2.3.4算法与酸性血红素的排除筛选一起使用,以确定SAR-N蛋白阳性细胞占细胞总数的百分比。对于Iba-1,使用Multiplex IHC v2.3.4算法进行定量。对于SARS-CoV-2 RNAscope ISH和Mx1定量,使用Area Quantification v2.1.3模块来确定总SARS-CoV-2反义或正义探针的百分比,或Mx1蛋白占总组织面积的百分比。对于MPO(中性粒细胞)和CD3+细胞定量,对载玻片进行注释以排除血管(>5mm2)、支气管、细支气管、软骨和结缔组织;随后,使用Cytonuclear v1.6模块检测MPO+或CD3+细胞并计算为占总肺泡组织的比率(PMNs/mm2),该比率通过运行AreaQuantification v2.1.3模块确定。在所有情况下,对每个样本都进行了所有图像的手动检查,以确保注释准确。
亚基因组mRNA测定.通过RT-PCR使用先前描述的引物和探针评估SARS-CoV-2E基因亚基因组mRNA(sgmRNA)10,11,23。简而言之,使用QIAcube HT(Qiagen)和RNeasy 96QIAcube HTKit(Qiagen)从几个解剖部位的组织匀浆中提取总RNA。为了生成标准曲线,将SARS-CoV-2E基因sgmRNA克隆至pcDNA3.1表达质粒中;使用AmpliCap-Max T7 High YieldMessage MakerKit(Cellscript)转录此插入物。在RT-PCR之前,根据制造商的说明,使用Superscript III VILO(Invitrogen)将从受攻击的动物或标准中收集的样品进行逆转录。使用靶向E基因sgmRNA的序列设计了Taqman定制基因表达测定(ThermoFisherScientific)。根据制造商的说明,在QuantStudio 6和7Flex实时PCR系统(AppliedBiosystems)上进行反应。标准曲线用于计算每克组织sgmRNA的拷贝数;定量测定的灵敏度为100个拷贝。
ELISA.通过ELISA评估RBD特异性结合抗体,基本上如(Chandrashekar,et al.,Science,doi:10.1126/science.abc4776(2020);Yu,et al.,Science,doi:10.1126/science.abc6284(2020))10,11所述。简而言之,用1xDPBS中的1μg/mL SARS-CoV-2 RBD(Aaron Schmidt,MassCPR)或1μg/mL SARS-CoV-2刺突(S)蛋白(Sino Biological)包被96孔板,并在4℃下孵育过夜。孵育后,用洗涤缓冲液(1X DPBS中的0.05%Tween20)洗涤板一次,并在室温下用350μL酪蛋白封闭液/孔封闭2-3小时。孵育后,弃去封闭溶液并将板吸干。
将在酪蛋白封闭液中三倍稀释的热灭活血清的连续稀释液添加至孔中,并将板在室温下孵育1小时,然后再洗涤3次,随后与在酪蛋白封闭液中的0.1μg/mL的抗仓鼠IgG HRP(Southern Biotech)在室温下避光孵育1小时。然后用洗涤缓冲液洗涤板3次,并且每孔加入100μL SeraCare KPL TMB SureBlue Start溶液;通过每孔添加100μL SeraCare KPLTMB Stop溶液来停止板的显色。使用VERSAMAX或OMEGA微孔板读数仪记录450nm处的吸光度。将ELISA终点滴度定义为产生是背景2倍的吸光度的最高血清稀释度倒数。
假病毒中和测定.通过与之前描述的方法相似的方法产生表达荧光素酶报告基因的SARS-CoV-2假病毒10,11,21。
简而言之,通过lipofectamine 2000(ThermoFisher),将包装构建体psPAX2(AIDSResource and Reagent Program)、荧光素酶报告质粒pLenti-CMV Puro-Luc(Addgene)和表达刺突蛋白的pcDNA3.1-SARS CoV-2 SACT的共转染到HEK293T细胞中。转染后48小时收集含有假病毒的上清液;通过用0.45μm过滤器过滤来纯化假病毒。为了确定来自疫苗接种动物的抗血清的中和活性,将HEK293T-hACE2细胞以1.75×104个293细胞/孔的密度接种在96孔组织培养板中过夜。制备热灭活血清样品的三倍连续稀释液,并与50μL假病毒混合。在加入HEK293T-hACE2细胞之前,将混合物在37℃下孵育1小时。感染后48小时,根据制造商的说明,在Steady-Glo Luciferase Assay (Promega)中裂解细胞。将SARS-CoV-2中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。
Luminex.为了检测抗原特异性抗体滴度的相对量,如前所述25进行了定制的Luminex测定。因此,荧光标记的微球(Luminex)通过EDC(Thermo Scientific)和Sulfo-NHS(Thermo Scientific)通过共价N505羟基琥珀酰亚胺(NHS)-酯键与包括刺突蛋白(S)(EricFischer,Dana Farber Cancer Institute)、S1和S2(Sino Biological)以及受体结合结构域(RBD)(Aaron Schmidt,Ragon Institute)在内的SARS-CoV-2抗原偶联。将1.2×103506珠每个区域和抗原添加至384孔板(Greiner)中,并与稀释的血清一起孵育(对于IgG2a、IgG3和IgM,1∶90,对于总IgG和Fc受体结合测定,1∶500)在4℃下以900rmp振动16小时。在形成免疫复合物后,用自动洗板机(Tecan)在0.1%BSA和0.05%Tween-20(Luminex测定缓冲液)中洗涤微球3次。将PE标记的山羊抗小鼠IgG、IgG2a、IgG3和IgM检测抗体(southernbiotech)在Luminex测定缓冲液中稀释至0.65ug/mL,并在室温下与珠一起孵育1小时,同时以900rpm振动。类似地,对于Fc受体结合谱,生物素化(Thermo Scientific)重组小鼠FcγR2、FcγR3和FcγR4(Duke Protein Production facility)并与链霉亲和素-PE偶联10分钟(Southern Biotech),然后添加至样品。这些小鼠抗体和蛋白与仓鼠有交叉反应。然后洗涤包被的珠并在连接有机械臂(PAA)的流式细胞仪iQue(Intellicyt)上读数。在每个珠区域对事件进行门控,报告珠阳性事件的PE荧光中值。每个二级检测试剂均一式两份运行样品。
抗体依赖性补体沉积(ADCD).如前所述26进行ADCD测定。简而言之,生物素化(Thermo Scientific)SARS-CoV-2 S和RBD并在37℃下与1μm红色荧光中性亲和素-珠偶联2小时,然后洗去多余的抗原。为了形成免疫复合物,将1.82x108个抗原包被的珠添加到96孔板圆底板的每个孔中,并与1∶10稀释的样品在37℃下孵育2小时。根据制造商的说明(Cedarlane)用水重构冻干豚鼠补体,并在含有Mg2+和Ca2+(GVB++,BostonBioProducts)的明胶弗洛拿缓冲液中每孔向免疫组合物加入4μL,在37℃下持续20分钟。用PBS中的15mM EDTA洗涤免疫组合物,并且添加荧光素缀合的山羊IgG豚鼠补体C3组分(MpBio)。
染色后,用4%多聚甲醛(PFA)固定细胞样品,并使用机械臂(PAA)通过流式细胞仪(Intellicyt,iQue Screener plus)进行样品采集。以单细胞和珠阳性事件门控所有事件,报告C3阳性事件的中值。所有样品在不同的天一式两份进行。
统计分析.使用GraphPad Prism 8.4.2(GraphPad软件)对免疫学、病毒学和体重数据进行分析。使用双边Mann-Whitney检验进行组间数据比较。通过双边Fisher’s精确检验评估死亡率。通过双边Spearman等级相关性检验评估相关性。将P值小于0.05认为是显著的。所有系统血清学数据都经过log10转换。对于雷达图,在使用组内的中值之前,对每个抗体特征进行标准化,使其最小值为0,跨组的最大值为1。使用版本为3.6.1的R包“ropls”构建主成分分析(PCA)以比较多变量谱。对于热图中的可视化,显示了S.dTM.PP和S.PP组Z分数特征的平均值的差异。为了表明热图中的显著性,使用Benjamini-Hochberg校正来校正一行内的多重比较。
实施例22:在18岁以上成人中评估Ad26.COV2.S预防SARS-CoV-2介导的COVID-19的效力和安全性的随机、双盲、安慰剂为对照3期研究
进行了一项随机、双盲、安慰剂为对照的3期研究,以评估Ad26.COV2.S在18岁以上成人中预防SARS-CoV-2介导的COVID-19的效力和安全性。
Ad26.COV2.S是一种单价疫苗,其由重组的、无复制能力的腺病毒26型(Ad26)载体组成,如本文描述将其构建为编码严重急性呼吸道综合征冠状病毒-2(SARS-CoV-2)刺突(S)蛋白。
这是一项在18岁以上成人中的多中心、随机、双盲、安慰剂为对照、3期关键效力和安全性研究。在居住或前往有高SARS-CoV-2活性地区的COVID-19风险较高的参与者(基于年龄、性别、种族/民族、职业、合并症)中,在给药2剂研究疫苗后,评估Ad26.COV2.S的效力、安全性和免疫原性。
参与者将按1∶1的比例平行随机接受Ad26.COV2.S或安慰剂的肌内(IM)注射,如下表所示。Ad26.COV2.S将以1×1011病毒颗粒(vp)的剂量水平给药,或以1x1010的剂量水平在一剂或两剂给药方案中给药。
预计至少约25%的招募的参与者将≥60岁,最多约25%的招募的参与者将≥18至<40岁。
将使用交错招募策略:
·第1阶段:最初,基于可接受的第29天安全性和足够的包括T辅助细胞1/T辅助细胞2(Th1/Th2)的免疫原性数据,将1,000名没有与进展为严重COVID-19的风险增加相关的合并症的参与者(包括500名Ad26.COV2.S接受者和500名安慰剂接受者;在年龄组之间平均分配)招募入2个年龄依赖性的亚群(阶段1a为≥18岁至<60岁,阶段1b为≥60岁),其来自首次人(FIH)研究VAC31518COV1001的相应年龄组(关于更多详细信息,请参阅本文档的正文)。
·第2阶段:在第1阶段暂停疫苗接种后,允许独立数据监测委员会(IDMC,也称为数据安全监测委员会[DSMB])检查为期3天的安全数据(包括所有4级AE和所有SAE),如果没有发现安全问题,将继续进行招募,扩大招募范围,以包括在2个年龄依赖性亚组(阶段2a为≥18岁至<60岁,阶段2b为≥60岁)中存在与进展为严重COVID-19的风险增加相关的合并症的参与者。
与进展为严重COVID-19的风险增加相关或可能相关的合并症(或风险要素)包括:中度至严重哮喘;慢性肺病,例如慢性阻塞性肺病(COPD)(包括肺气肿和慢性支气管炎)、特发性肺纤维化和囊性纤维化;糖尿病(包括1型、2型或妊娠);严重的心脏病,包括心力衰竭、冠状动脉疾病、先天性心脏病、心肌病和(肺)高血压或高血压;肥胖(体重指数[BMI]≥30kg/m2);慢性肝病,包括肝硬化;镰状细胞疾病;地中海贫血;脑血管疾病;神经系统疾病(痴呆);晚期肾脏疾病;器官移植;癌症;人类免疫缺陷病毒(HIV)感染和其他免疫缺陷乙型肝炎感染;睡眠呼吸暂停;帕金森病;癫痫;缺血性中风;颅内出血;吉兰-巴雷综合征;脑病;脑膜脑炎;以及住在疗养院或长期护理机构的参与者。关于详细信息和例外情况,请参阅本文档中的排除标准。
个人参与(包括筛选)的持续时间最长为2年3个月。如果参与者无法完成研究,但没有撤回同意,将进行提前退出访视。将研究中最后一位参与者的最后一次访视的完成视为研究结束。
主要效力评估包括对COVID-19样体征和症状的监测、与COVID-19相关的住院和并发症的记录,以及通过分子测定(基于RT-PCR)和通过抗SARS-CoV-2血清学对SARS-CoV-2感染的实验室确诊。还将进行免疫原性评估,尤其是对体液免疫应答的评估,重点是中和抗体和结合抗体。关键安全性评估将包括对征求的(solicited)和非征求的(unsolicited)AE的监测(仅在安全子集中),以及在所有参与者中SAE和MAAE的收集。将在确诊的COVID-19病例中评估SARS-CoV-2的病毒载量。还将研究与SARS-CoV-2感染和COVID-19严重度相关的生物标志物。将对所有经分子确诊的、有症状的COVID-19的参与者记录疫苗接种后的医疗资源利用情况(MRU)。在选定的站点,如果当地法规允许,可以收集与当前工作状况、生活状况和社区互动有关的其他基线特征,用于协变量分析。
在第2次疫苗接种后1年内,将通过电子临床结果评估(eCOA)每周至少两次询问每位参与者是否出现任何可能与感染SARS-CoV-2相关的新症状或健康问题。从第2次疫苗接种后1年起,直至2年随访期结束,通过eCOA将该监测间题的频率降至每2周一次。在最后一次研究访视之前,所有参与者仍需接受疾病增强监测。对于所有通过eCOA追踪失访且未记录住院的参与者,将尽一切努力记录他们的状态。
在COVID-19第1-2天和第3-5天,所有具有COVID-19样体征或症状满足疑似COVID-19预设标准的参与者都应进行COVID-19程序(如本方案正文中所述),直到症状发作后14天(COVID-19第15天)或直到COVID-19发作消退,以最后发生者为准,除非确认在COVID-19第1-2天和第3-5天采集的鼻拭子对于SARS-CoV-2均为阴性。将COVID-19发作的消退定义为连续2次SARS-CoV-2阴性鼻拭子和连续2天没有与COVID-19相关的体征或症状。在COVID-19发作消退时,将根据临床病例定义应用收集到的信息。
应采取一切必要的预防措施(根据当地法规)保护医务人员和参与者的其他接触者,所述参与者疑似患有COVID-19,直到通过分子技术证明为阴性,或者是阳性并且在COVID-19第1-2天和第3-5天满足预先指定的疑似COVID-19标准,直到他们不再是阳性。如果确认感染SARS-CoV-2,将通知参与者和参与者的医疗服务提供者和/或当地卫生局(如果需要),并要求参与者根据当地法规遵守规定的适当措施和限制。
中度至严重COVID-19的病例定义
对于主要终点,将考虑所有中度和严重/危重COVID-19病例。
中度COVID-19病例定义
来自任何可用呼吸道样品(例如,鼻拭子样品、痰样品、咽喉拭子样品、唾液样品)或其他样品的SARS-CoV-2阳性RT-PCR或分子检测结果。
以及在观察过程中的任何时间:
呼吸急促(呼吸困难)
或者
以下任何一种新的或恶化的体征和以下任何一种新的或恶化的症状:

*SpO2标准将根据海拔进行调整。
或者
以下任何2种新的或恶化的症状:


*具有症状的2种或更多要素(例如,呕吐和腹泻或疲劳和食欲不振)仅计为病例定义的1种症状。为了符合病例定义,参与者需要至少有2种不同的症状。
严重/危重COVID-19的病例定义
来自任何可用呼吸道样品(例如,鼻拭子样品、痰样品、咽喉拭子样品、唾液样品)或其他样品的SARS-CoV-2阳性RT-PCR或分子检测结果。
以及在观察过程中的任何时间的以下任何一项:
静息时表明严重全身性疾病的临床体征(呼吸率≥30次呼吸/分钟,心率≥125次心跳/分钟,在海平面的室内空气中氧饱和度(SpO2)≤93%*,或氧分压/吸入氧气分率(PaO2/FiO2)<300mmHg)
*SpO2标准将根据海拔进行调整。
呼吸衰竭(定义为需要高流量氧气、无创通气、机械通气或体外膜氧合[ECMO])
休克的证据(定义为收缩压<90mmHg,舒张压<60mmHg,或需要血管加压药)
严重的急性肾脏、肝脏或神经功能障碍
入住ICU
死亡
轻度COVID-19病例定义
来自任何可用呼吸道样品(例如,鼻拭子样品、痰样品、咽喉拭子样品、唾液样品)或其他样品的SARS-CoV-2阳性RT-PCR或分子检测结果;
以及在观察过程中的任何时间:
以下症状之一:发烧(≥38.0℃或≥100.4°F)、喉咙痛、不适(食欲不振、一般不适、疲劳、身体虚弱)、头痛、肌肉疼痛(肌痛)、胃肠道症状、咳嗽、胸闷、流鼻涕、气喘、皮疹、眼睛刺激或分泌物、发冷或新的味觉或嗅觉丧失。
将符合上述病例定义但不符合中度至严重/危重定义的病例视为临床轻度。
实施例23.在18岁至≤55岁和年龄≥65岁的成年人中评估Ad26.COV2.S的安全性、反应原性和免疫原性的随机、双盲、安慰剂为对照、首次人(FIH)1/2a期多中心研究
设计了1/2a期随机、双盲、安慰剂为对照的临床研究,评估Ad26.COV2.S的安全性、反应原性和免疫原性,Ad26.COV2.S是一种基于非复制型腺病毒26的载体,其表达稳定化的SARS-CoV-2蛋白的融合前刺突(S),根据本发明,以每次疫苗接种5x1010或1x1011个病毒颗粒(vp)的剂量水平,以单剂或以间隔56天的两剂方案向健康成人(18-55岁;队列1a;n=377)和健康老年人(>65岁;队列3;n=394)给药。通过ELISA测量疫苗引发的S特异性抗体水平,在野生型病毒中和测定(wtVNA)中测量中和滴度。通过细胞内细胞因子染色(ICS)评估Th1和Th2。
方法
研究设计和参与者
该研究是一项多中心、随机、双盲、安慰剂为对照的试验,旨在评估在18-55岁和>65岁的健康成年人中,以单剂或间隔8周的两剂方案,肌内给药5x1010或1x1011vpAd26.COV2.S的安全性、反应原性和免疫原性。在图101中给出临床试验的概述。
Ad26.COV2.S是一种重组的、无复制能力的Ad26载体,其编码来源于Wuhan毒株的第一个临床分离株(全基因组序列NC_045512)的工程化改造的形式的SARS-CoV-2S蛋白,如本文所述。该研究在比利时和美国的多个临床站点进行。通过收集鼻腔样品进行PCR筛查所有受试者的COVID-19,如果阳性将排除他们,并通过当地可用的血清学测定来检测以前的SARS-CoV-2感染,队列1a和队列3之间最多允许25名血清阳性参与者。该研究由机构审查委员会审查和批准。所有参与者在招募前都提供了书面知情同意书。
随机化和盲法
将402名18-55岁的合格参与者和405名>65岁的参与者随机分配接受一或两次5x1010或1x1011vp剂量的疫苗或安慰剂(PL)的疫苗接种(1∶1∶1∶1∶1每个年龄组):5x1010vp/5x1010vp;5x1010vp/PL;1x1011vp/1x1011vp;1x1011vp/PL;或PL/PL(图101)。随机化由交互式网络应答系统(IWRS)完成,并使用随机排列的区块按站点进行分层。参与者和研究者将在整个研究过程中保持盲态。在掩蔽的相同注射器中提供疫苗和安慰剂。当所有参与者完成第29天访视或提前停止时,申办者和统计学家对中期分析进行分组揭盲。
终点
支持每个方案的安全性和反应原性的主要目标的终点是每次疫苗接种后28天的不良事件(AE)、每次疫苗接种后7天的局部和全身反应原性以及整个研究期间的严重不良事件(SAE)。根据FDA指导文件“Toxicity Grading Scale for Healthy Adult andAdolescent Volunteers Enrolled in Preventive Vaccine Clinical Trials”对AE进行分级。次要终点是对SARS-CoV-2的S蛋白的体液免疫应答,如通过S特异性ELISA和通过wtVNA中针对野生型SARS-CoV-2的中和滴度,以及通过ICS测量的细胞免疫应答证明。
程序
在第一天参与者接受在1mL体积中5x1010vp或1x1011vp Ad26.COV2.S或安慰剂(0.9%盐水)的在三角肌的肌内(IM)注射。在日记卡上收集疫苗接种后7天的征求的AE、疫苗接种后28天的非征求的AE和整个研究过程中的SAE。用于血清化学、血液学和确定免疫读数的血液样品已经并将在整个研究的几个时间点收集;在疫苗接种之前收集用于妊娠测试的尿液样品。
在基线和第29天,通过ELISA测量刺突(S)特异性结合抗体。将血清阳性定义为滴度>50.3EU/mL。使用英国公共卫生部(PHE)的Victoria/1/2020SARS-CoV-2毒株在wtVNA中测量SARS-CoV-2血清中和抗体滴度。wtVNA中的血清阳性定义为IC50滴度>58。在基线和第15天,使用两个通过11个重叠的15元(mer)的S肽池,通过细胞内细胞因子染色(ICS)测量SARS-CoV-2 S特异性T细胞应答。Th1应答的特征在于表达IFNg和/或IL-2而不是IL-4、IL-5和/或IL13的CD4+T细胞,并且Th2应答的特征在于CD4+T细胞表达IL-4、IL-5和/或IL-13以及CD40L。所有测定均以盲法进行,如下所述。
SARS-CoV-2野生型病毒中和测定:
使用由英国公共卫生部(PHE)开发和认证的野生型病毒微量中和测定(MNA)对能够抑制野生型病毒感染的中和抗体进行定量。使用的病毒原种来自Victoria/1/2020毒株。
简而言之,在96孔转移板中制备了6个热灭活人血清样品的两倍连续稀释液。将SARS-CoV-2野生型病毒以目标工作浓度(约100个噬斑形成单位[PFU]/孔)依次添加到血清稀释液中,并在37℃和5%CO2下孵育60-90分钟。然后将血清-病毒混合物转移到测定板上,预先用Vero E6非洲绿猴肾细胞接种过夜,并在37℃和5%CO2下孵育60-90分钟,然后添加羧甲基纤维素(CMC)覆盖培养基并进一步孵育24小时。孵育后,固定细胞并且使用对SARS-CoV-2 RBD S蛋白双特异性的抗体染色细胞,并使用TrueBlueTM底物可视化免疫斑块。使用来自CTL的免疫斑点分析仪计数免疫斑块。将免疫斑块计数输出到SoftMaxpro,并将血清样品的中和滴度计算为对应于该样品的50%中和抗体滴度(ID50)的血清稀释度倒数。
刺突酶联免疫吸附测定(ELISA)
使用人SARS-CoV-2 Pre-Spike IgG ELISA测定SARS-CoV-2 Pre-Spike特异性结合抗体浓度,这是一种基于抗体/抗原相互作用的间接ELISA。使用的SARS-CoV-2抗原是在ES-293细胞中产生的稳定化的融合前刺突蛋白((2P)、Δfurin、T4 foldon、His-Tag)。ELISA是在加拿大Laval的Nexelis开发并用于人血清鉴定。
简而言之,将纯化的SARS-CoV-2 Pre-SpikeAntigen吸附到微孔板的孔中,并添加稀释的血清样品(测试样品、标准品和质量控制)。洗去未结合的样品,并添加酶缀合的抗人IgG。洗去多余的缀合物后,加入3,3′,5,5′-四甲基联苯胺(TMB)比色底物。在确定的时间段之后,停止反应。用每个测试板上的参考标准,根据标准指定的单位(每毫升ELISA实验室单位:ELU/mL)定量样品中针对SARS-CoV-2 Pre-Spike的抗体量。
细胞内细胞因子染色(ICS)
对SARSCoV2的S蛋白的T细胞应答由HIV Vaccine Trials NetworkLaboratories,Fred Hutchinson Cancer Research Center(HVTN,FHCRC),Seattle,WA,United States的27色ICS表征。
将冷冻保存的外周血单个核细胞(PBMC)解冻并静置过夜,然后在37℃下用两个覆盖SARSCoV2 S蛋白全长的共有肽池、二甲基亚砜(DMSO)(阴性对照)或葡萄球菌肠毒素B(SEB)(阳性对照)刺激6小时。在刺激过程中加入Brefeldin A以预防细胞因子释放。首先用活性染料对细胞进行染色,然后用细胞外抗体混合物对细胞进行染色,然后固定和透化并且用细胞内标志物(包括Th1和Th2细胞因子)的抗体混合物对细胞进行染色。使用5-激光30-参数Becton-Dickinson FACSymphony细胞仪获得细胞荧光,并使用FlowJo软件进行分析。
统计方法
本研究旨在评估安全性和免疫原性。在完整分析集中对安全性数据进行了描述性分析,并在符合方案(per protocol)的免疫原性群体中分析了免疫原性数据。SARS-CoV-2刺突(S)结合抗体滴度表示为每毫升的ELISA单位(EU/mL)和wtVNA中的中和抗体滴度,表示为中和50%测试病毒剂量(50%抑制浓度[IC50])的血清稀释度倒数),以log10比例显示,并使用GMT和95%置信区间(CI)进行描述。对于这两种测定,将血清转化定义为如果基线滴度低于LLOQ,则疫苗接种后抗体滴度高于定量下限(LLOQ),或者如果基线滴度高于LLOQ,则疫苗接种后抗体滴度是基线的4倍。将ICS应答描述为亲本群体的百分比。通过比较未刺激与S肽刺激孔的单边Fisher精确检验确定样品阳性。LLOQ为0.22%,估算不可定量的值为LLOQ/2。如果Th1和/或Th2应答为阳性且高于2xLLOQ,则计算Th1/Th2比例。如果来自参与者的Th1或Th应答不满足这些标准,如果没有Th2应答,则认为Th1/Th2比例高于1,如果没有Th1应答,则认为Th1/Th2比例低于1。
结果
研究参与者
自2020年7月13日对参与者进行筛查,队列1a和1b参与者(年龄18-55岁)的疫苗接种起始于2020年7月22日(队列1a)、2020年7月22日(队列1b),以及队列1a的第一次疫苗接种完成于2020年8月4日,队列1b完成于2020年8月7日。筛选了575名志愿者,其中队列1a招募了380人(377人疫苗接种),筛选了28人,队列1b招募了25人(25人疫苗接种)(分别为图102A和102B)。7名(1.7%)参与者在筛选时对SARS-CoV-2呈血清阳性,Ad26的血清阳性率将在稍后报告。队列3的参与者(年龄>65岁)的疫苗接种于起始2020年8月3日,第一次疫苗接种完成于2020年8月24日。筛选了660名志愿者,其中招募了405人(394人疫苗接种)(图102C)。队列3只有前15名参与者的免疫原性结果可用。基线特征在各组之间具有广泛的可比性。
疫苗安全性和反应原性
在疫苗接种后7天在日记卡上收集征求的AE,在疫苗接种后28天收集非征求的AE,并在整个研究过程中收集SAE。为了让评估这两项研究的参与者和研究者保持盲态,在本中期报告中,安全性数据在没有分组揭盲的情况下呈现。
在队列1a和1b(年龄18-≤55岁)中,研究者对402名参与者的反应原性评估可用,其中288名(72%)参与者报告了征求的AE。235名(58%)参与者报告了征求的局部AE——主要是1级/2级。三名参与者报告了3级疼痛/压痛。最常见的AE是注射部位疼痛。258名(64%)参与者报告了征求的全身性AE,主要是1级/2级,46名(11%)参与者报告了3级全身性AE。最常见的AE是疲劳、头痛和肌痛。76名(19%)参与者报告了发烧,22名(5%)参与者报告了3级发烧。总体而言,98名参与者报告了178起非征求的AE,其中12名起报告了3级AE。
在队列3(≥65岁)总体中,研究者对394名参与者的反应原性评估可用,其中183名(46%)参与者报告了征求的AE。108名(27%)参与者报告了征求的局部AE,其中大多数为1级/2级,1名参与者报告3级肿胀和红斑。最常见的AE是注射部位疼痛。140名(36%)参与者报告了征求的全身性AE,其中大多数为1级/2级,其中3名参与者报告了3级AE。最常见的AE是头痛、疲劳和肌痛。14名(4%)参与者报告了1级或2级轻度或中度发热,其中只有1名是2级。队列3中没有报告限制日常生活活动(3级)的高烧或其他发热。总体而言,46参与者报告了77起非征求的AE。4名参与者报告了非征求的3级AE。在任何队列中均未报告任何4级AE,无论是征求的还是非征求的。
没有参与者因AE终止研究。有两个SAE:因为过去有反复低血压的病史,调查人员判断一个低血压与疫苗无关,另一个发烧的参与者因为怀疑COVID-19而住院过夜,并在12小时内恢复。
Ad26.COV2.S的免疫原性
通过ELISA测量针对稳定化的SARS-CoV-2全长刺突蛋白的抗体。在队列1a中,在基线时94%和98%的参与者对SARS-CoV-2 S蛋白的几何平均滴度(GMT)低于LLOQ,在疫苗接种后第29天,对于5x1010和1x1011vp剂量组,该滴度分别增加到528(95%CI:442;630)和695(95%CI:596;810),每个剂量组有99%的血清转化(图103A)。对于5x1010和1x1011vp剂量组,分别为88%(7/8)和67%(2/3)的队列1a参与者在基线时血清阳性表明了将结合抗体滴度增加至4倍的预设标准视为疫苗应答者。队列3中所有15名首次招募的参与者在基线时均为血清阴性,并在疫苗接种后29天发生血清转化,对于5x1010和1x1011的剂量,GMT分别为507(95%CI:181;1418)和248(95%CI:122;506)。在本研究中使用的人恢复期血清(HCS)组中观察到899的GMT,每个队列中两个剂量的GMT的95%CI有重叠,表明与疫苗引发的滴度没有重大差异。
在一部分参与者中(5x1010vp和1x1011vp剂量组中的每一个,n=50),通过wtVNA测量了SARS-CoV-2中和抗体水平。在队列1a中,在第29天,对于5x1010和1x1011vp组,两个剂量组在基线时的GMT<LLOQ分别增加到214(95%CI:177;259)和243(95%CI:200;295)的IC50GMT。在队列3的哨兵组中观察到类似的结果,在第29天,对于5x1010和1x1011vp组,基线时的GMT<LLOQ分别增加到196(95%CI:69;560)和127(95%CI:<LLOQ;327)的GMT(图103B)。为了比较,在这个分析中使用的HCS组中测量了522的GMT,HCS组与剂量组和队列之间的GMT的95%CI有重叠。几个疫苗研究样品达到了疫苗样品分析运行的定量上限(ULOQ=640),并且正在重新分析。测试更高稀释度的样品将允许确定具有更高ULOQ的更高滴度。HCS样品在比疫苗研究样品更高的稀释度下进行测试,因此其中一些样品的滴度高于此处提供的ULOQ。在疫苗接种后第29天,队列1a中98%的参与者对SARS-CoV-2的中和抗体呈阳性,与所给予的疫苗剂量无关。队列1a中5x1010和1x1011vp组在第29天的血清转化率都为92%,并且队列3中5x1010和1x1011vp组为100%(6/6)和83%(5/6)。在队列1a中,在5x1010和1x1011vp组中,总共分别有82%和94%的参与者达到了大于100的滴度,这表明大多数参与者在用Ad26.COV2.S单次接种疫苗后诱导了强的应答。wtVNA和S-ELISA队列1a滴度高度相关,Spearman相关性为0.86。来自表达SARS-CoV-2 S蛋白中和测定的假病毒的数据正在等待中,并将包括在我们的剂量1后中期分析的完整公开中。
在基线和疫苗接种后15天,在一部分研究参与者中,对SARS-CoV-2 S特异性CD4+和CD8+T细胞应答进行表征。用覆盖整个S蛋白的肽进行刺激后,在队列1a参与者中,在疫苗接种15天后,对于5x1010和1x1011vp组,CD4+Th1应答分别从基线的不可检测到0.08%(95%CI:0.05;0.16)和0.11(95%CI:0.07;0.16)以及在队列3的哨兵组中,对于5x1010和1x1011vp组,分别从基线的不可检测到0.36%(95%CI:0.15;0.89)和0.13(95%CI:0.04;0.50)组(图103C)。在队列1a中,5x1010和1x1011vp剂量的接受者分别观察到76%(95%CI:65;86)和83%(95%CI:73;91)Th1阳性应答。在队列3的前15名参与者中,在5x1010和1x1011vp剂量的接受者中分别观察到100%(95%CI:54;100)和67%(95%CI:22;96)Th1阳性应答。除了队列1a中5x1010vp组中的一名参与者之外,在任一队列中都没有可定量的Th2应答(值推算为LLOQ/2)(图103C)。然而,这个参与者的Th1/Th2比例为28.9,表明存在Th1偏向表型。在所有其他具有可测量的Th1和/或Th2应答的参与者中,Th1/Th2比例范围为1.0-68.5。总体而言,这些数据表明Ad26.COV2.S在两个年龄组中都诱导了Th1偏向的应答。
通过在S肽刺激后IFNg和/或IL-2细胞因子的表达来鉴定S特异性CD8+T细胞应答(图103D)。接种疫苗后15天,在队列1a中,对于5x1010和1x1011vp组,S特异性CD8+T细胞应答量分别为0.07%(95%CI:0.03;0.19)和0.09%(95%CI:0.05;0.19),以及在队列3中,对于5x1010和1x1011vp组分别为0.05%(0.02;0.24)和0.07%(0.02;0.14)。在队列1a中,对于5x1010和1x1011vp组,分别有51%(95%CI:39;63)和64%(95%CI:52;75)的参与者对S肽刺激具有阳性CD8+T细胞应答,并且在队列3中,两个剂量组的33%(95%CI:4%;78%)的参与者显示出强的疫苗诱导的CD8+T细胞应答。
讨论
我们对1/2a期研究的中期分析显示,候选疫苗Ad26.COV2.S具有可接受的安全性和反应原性谱,并且在5x1010vp或1x1011vp剂量下均具有免疫原性。
单剂的Ad26.COV2.S在绝大多数疫苗接受者中引起强烈的体液应答。在队列1a中99%的参与者和在队列3中100%的第一批参与者中通过ELISA测量的S结合抗体水平从基线到疫苗接种后第29天增加,与给予的疫苗剂量水平无关。类似地,在wtVNA中观察到高应答率。疫苗接种后29天,98%的参与者具有可检测的中和抗体。92%的队列1a参与者和在队列3中接受5x1010vp和1x1011vp剂量的分别为6名接受者中的6名和6名接受者中的5名对SARS-CoV-2中和抗体进行了血清转化。在队列1a中,84%-92%的参与者的中和抗体滴度高于100。这里报告的几乎所有参与者的中和抗体应答都是在单剂Ad26.COV2.S后获得的。
这些临床数据表明,两种剂量水平的Ad26.COV2.S在绝大多数健康状况稳定的健康年轻和年长参与者中都诱导了强烈的中和抗体应答。尽管到目前为止,只有将少数队列3的参与者包括在分析中,但Ad26.COV2.S在18-55岁和>65岁的成年人中在这里观察到的相似免疫原性是令人鼓舞的,因为对有极高风险发展为严重COVID-19的老年人的保护极其重要,尤其是要在这次大流行期间减轻医疗保健系统的压力。与COVID-19人恢复期血清相比,Ad26.COV2.S诱导的结合抗体和中和抗体的水平似乎较低,尽管对于大多数参与者来说在相同的范围内。然而,由于人血清组组成的可变性以及恢复期血清抗体测定的非标准化,这种比较的意义尚未确定。使用不同的HCS组,与疫苗样品滴度的比较(比例)会有所不同。例如年龄、疾病严重度和疾病发作后的时间以及小组中的样本数量的人口统计数据可能会对测量的抗体水平产生影响,从而使得与疫苗样品的适当比较变得随意(arbitrary)。
之前对SARS-CoV和MERS-CoV疫苗动物模型的经验引起了对疫苗相关增强型呼吸道疾病(VAERD)的担忧。有人提出了这种安全间题与体液免疫的中和效力差以及Th2偏向之间的关联。这里显示,Ad26.COV2.S引发的T细胞应答绝大多数是Th1偏向,没有或有非常低的Th2应答。事实上,在所有应答者中,Th1/Th2比例均高于1,这与我们之前使用基于Ad26的疫苗平台的经验一致。这种强烈的Th1应答伴随着疫苗接种后强烈的CD8+T细胞应答。除了由Ad26.COV2.S引发的强烈体液应答外,CD4+Th1和CD8+T细胞应答的总体强的诱导将VAERD的理论风险降至最低。
显然,与用于当前大流行的两剂疫苗方案相比,有效的单剂COVID-19疫苗将具有巨大优势,并显著增加对可以控制SARS-CoV-2大流行的时间范围产生直接影响的可能性。通过单剂Ad26.COV2.S引发的免疫应答可以防止SARS.CoV.2感染或COVID-19,一个重要的未知数是这种免疫应答的持久性。这项正在进行的ph1/2a研究将提供有关单剂Ad26.COV2.S引发的免疫应答持久性以及第二剂Ad26.COV2.S后免疫应答的数据。
总之,单次疫苗接种后的安全性谱和免疫原性支持Ad26.COV2.S作为针对COVID-19的潜在保护性疫苗的进一步临床开发。这个中期分析表明,单剂的Ad26.COV2.S,无论是剂量为5x1010vp还是1x1011vp,都是安全的、耐受性良好且具有高度免疫原性的。基于两种剂量水平的相似免疫原性,已选择5x1010vp剂量进行进一步评估。在第一个3期研究中,将评估单剂5x1010vp的Ad26.COV2.S候选疫苗的效力。另一项3期研究正在准备中,以评估使用5x1010vp Ad26.COV2.S疫苗的两剂方案的免疫效力和持久性。
实施例24.在年龄≥18至≤55岁和年龄≥65岁的成年人中评估Ad26.COV2.S的安全性、反应原性和免疫原性的随机、双盲、安慰剂为对照、首次人(FIH)1/2a期多中心研究-队列3中期分析免疫原性报告
这个队列3中期分析包括来自野生型病毒中和测定(wtVNA)和酶联免疫吸附测定(ELISA)的分组揭盲体液免疫原性数据,收集直到(包括)第一剂后28天的访视(第29天)。此外,还描述了对于一部分参与者的来自细胞内细胞因子染色(ICS)的细胞免疫原性数据,收集直到(包括)第一剂后14天(第15天)。
目标和终点
本研究的目标/终点,适用于队列3的中期分析,提供如下:

方法
研究设计概述
在实施例23描述了研究设计。
免疫原性
对于体液免疫原性(wtVNA和ELISA),如果该值严格大于LLOQ(>LLOQ),则认为样品为阳性。体液免疫原性的应答者定义为:
-基线样本值小于或等于LLOQ(≤LLOQ),并且基线后样本严格大于LLOQ(>LLOQ)
-基线样本值严格大于LLOQ(>LLOQ),并且基线后样本值表示从基线样本值增加至至少4倍(≥4倍)。
对于细胞免疫原性(ICS),样本的阳性由Fisher精确检验定义,其比较模拟和刺激条件。
Th1/Th2应答比例的评估
基于组合的SARS-Cov2-S肽池,并仅使用基线后时间点,将为满足以下两个条件中的至少一个的样本计算Th1/Th2应答比例:
-Th1应答(“IFN-g或IL2不是TH2”)均为阳性且≥2 x LLOQ,
或者
-Th2应答(“IL4或IL5或IL13和CD40L”)均为阳性且≥2 x LLOQ
出于Thl/Th2比例分析的目的,两个细胞群(Th1和Th2)的LLOQ均为0.022%。
如果两个细胞群(Th1和Th2)均为阳性且≥2xLLOQ,则Th1/Th2的比例将计算为数值结果。
如果只有一个细胞群(Th1或Th2)呈阳性且≥2xLLOQ,则将使用以下规则来确定Th1/Th2比例的定性评估:
-如果一个细胞群为阳性而另一个为阴性,则阳性细胞群大于阴性细胞群:如果Th1应答为阳性而Th2应答为阴性,则Th1/Th2比例将设置为“>1”。如果Th1应答为阴性而Th2应答为阳性,则Th1/Th2比例将设置为“<1”
-如果两个细胞群均为阳性,则≥2 x LLOQ的细胞群大于<2 x LLOQ的细胞群:如果Th1应答≥2 x LLOQ且Th2应答<2 x LLOQ,则Th1/Th2比例将设置为“>1”。如果Th1应答<2 x LLOQ且Th2应答≥2 x LLOQ,则Th1/Th2比例将设置为“<1”。
结果
本实施例描述了在队列3中,直到(和包括)剂量1后28天的访视1(第29天)收集的体液免疫应答数据(wtVNA和ELISA)的分析。此外,本实施例描述了在队列3中,对于一部分参与者,直到(和包括)剂量1后14天的访视1(第15天)收集的细胞免疫应答数据(ICS)。下文提供了免疫原性数据的总结。
人口统计和基线特征
参与者的人口统计和基线特征总结在表2中。大多数参与者是白人(09.5%)。总体而言,50.1%的参与者是女性,49.9%是男性。年龄中值为69.00岁(范围:65.0-88.0岁),体重指数(BMI)中值为25.700kg/m2(范围:16.60-29.90kg/m2)。疫苗接种组之间的人口统计和基线特征通常很好地平衡。


免疫原性结果
分析的数据集
免疫原性结果的分析基于符合方案的免疫原性群体,该群体由可获得免疫原性数据的所有随机和疫苗接种的参与者组成。将第1组和第2组(5x1010vp)以及第3组和第4组(1x1011vp)的疫苗接种后1免疫原性结果汇总。
在完整分析集中,5×1010vp疫苗组的1名参与者、1×1011vp疫苗组的2名参与者和安慰剂组的1名参与者在基线时对SARS-COV-2呈血清阳性(表2)。
体液免疫应答
在野生型病毒中和测定中测量了队列3 COV1001参与者的中和抗体。由非盲的独立外部统计学家根据预定义的子集选择策略(按疫苗分配分层的前瞻性随机抽样)选择125名研究参与者(每组25名)的随机子集。
针对SARS-CoV-2的中和抗体应答
野生型病毒中和测定
对于这个测定,由非盲的独立外部统计学家根据预定义的子集选择策略(按疫苗分配分层的前瞻性随机抽样)选择125名研究参与者(每组25名)的随机子集。
通过wtVNA测量针对SARS-CoV-2的中和抗体滴度,其表示为50%抑制浓度(IC50)单位。在图106中呈现VNA对SARS-CoV-2的应答随时间变化的图形表示(几何平均滴度[GMT]与相应的95%CI)。
在基线时,在5×1010vp疫苗组中的47名参与者、在1×1011vp疫苗组中的45名参与者和安慰剂组中的24名参与者不具有可检测的SARS-CoV-2 IC50滴度(该测定的定量下限[LLOQ]为58的IC50)。在5x1010vp疫苗组的2名参与者、1×1011vp疫苗组中的5名参与者以及安慰剂组中无参与者中观察到SARS-CoV-2 IC50滴度的可检测的基线水平,这可能表明之前的SARS-CoV-2暴露(图106)。
在第15天,两个活性疫苗组中75%的参与者达到了大于100的IC50滴度,这表明在大多数参与者中诱导了快速、强的应答。在第15天,5×1010vp疫苗组的应答率为87%,并且1×1011vp疫苗组的应答率为83%。在第15天,5×1010vp和1×1011vp疫苗组的GMT分别为197(95%CI:137;284)和180(95%CI:117;276)。两个疫苗组在第15天的标准偏差(SD)相似。
第29天的中和抗体应答与第15天的应答相当。在第29天,5×1010vp疫苗组中总共81%的参与者和1×1011vp疫苗组中88%的参与者达到大于100的IC50滴度。在第29天5×1010vp疫苗组的应答率为90%和1×1011vp疫苗组的应答率为91%。在第29天,5×100vp和1×1011vp疫苗组的GMT分别为221(95%CI:160;307)和210(95%CI:155;285)。两个疫苗组的标准偏差(SD)相似。
目前的分析可能低估了中和抗体应答的量以及应答率,因为一些参与者的疫苗接种后滴度达到了测定的定量上限(ULOQ)。这些样品将在稍后阶段以更高的稀释度重新分析,以允许确定个体滴度。在第15天或第29天,安慰剂组未观察到增加的wtVNA滴度。
三名参与者的应答状态缺失,显示基线时可检测到SARS-CoV-2中和抗体水平。其余四名参与者都在积极的疫苗方案中,在第15天和第29天显示GMT增加。但是,并非所有这些参与者都符合预先指定的应答者标准。
没有观察到中和抗体应答随年龄增加而下降的趋势。在比较第29天两个疫苗组的队列3和队列1a中和抗体数据时,队列3中抗体应答的量(GMT,队列3与队列1a:5x1010vp疫苗组的221与214和1x1011vp疫苗组的210与243)和应答率与队列1a在同一范围内。
5x1010vp组中在疫苗接种后使用(185GMT,78%应答者)和未使用(238GMT,95%应答者)退热药/镇痛药的参与者在第29天的抗体水平相似。在第29天,1×1011vp组中在疫苗接种后使用(229GMT,90%应答者)和未使用(202GMT,91%应答者)退热药/镇痛药的参与者的抗体水平也相似,表明使用这些药物似乎不会影响通过wtVNA测量的中和抗体应答。
5x1010vp疫苗组和1×1011vp疫苗组在第15天和第29天的反向累积分布曲线相似(图107)。
针对SARS-CoV-2 S蛋白的结合抗体应答(ELISA)
通过ELISA测量的Ad26.COV2.S诱导的SARS-CoV-2 S蛋白结合抗体应答的描述性统计、实际值和相对于基线的倍数增加(相应的95%CI)在图108中以图形表示。
5×1010vp疫苗组的2名参与者、1×1011vp疫苗组的3名参与者,以及安慰剂组的3名参与者中观察到可检测的SARS-CoV-2 S ELISA滴度基线水平,这可能表明之前的SARS-CoV-2暴露。其中一名在1×1011vp疫苗组的参与者在筛查时表现出SARS-COV-2血清阳性状态。
在第15天5x1010vp和1×1011vp疫苗组的应答率分别为75%和77%。5×1010vp和1×1011vp组的相应的GMT分别为122(95%CI:97;152)和141(95%CI:114;175)(图108)。在安慰剂组中未观察到蛋白结合抗体的增加。
第29天的结合抗体应答高于第15天的应答。在第29天,两个活性疫苗组的应答率为96%。在第29天,5x1010vp和1×1011vp组的GMT分别为312(95%CI:246;396)和350(95%CI:284;431)。两个疫苗组的SD相似。在两个时间点,安慰剂组均未观察到疫苗接种后蛋白结合抗体的增加,无论是在GMT和应答率方面。
没有观察到在队列3内随着年龄的增加结合抗体应答下降的趋势。在第29天比较两个疫苗组的队列3和队列1a结合抗体数据时,队列3的抗体应答的量(GMT,队列3与队列1a:5x1010vp疫苗组的312与528和1x1011vp疫苗组的350与695)较低,但在与队列1a相同的范围内(图110)。队列1a和队列3的结合抗体应答率相当,分别为99%和96%(图108和图110)。
疫苗接种后使用退热药/镇痛药的参与者在第15天和第29天的结合抗体水平与未使用的参与者相似。5×1010vp组使用退热药/镇痛药的在第29天的GMT和应答率分别为357和94%。5×1010vp组未使用退热药/镇痛药的在第29天的GMT和应答率分别为302和97%。
1×1011vp组使用退热药/镇痛药的在第29天的GMT和应答率分别为357和94%。5×1010vp组未使用退热药/镇痛药的在第29天的GMT和应答率分别为302和97%。
5x1010vp疫苗组和1×1011vp疫苗组的反向累积分布曲线相似(图111)。
wtVNA与ELISA
对体液测定相关性的进一步检查表明,在第15天和第29天,wtVNA滴度与ELISA滴度高度相关,Spearman相关性系数分别为0.734和0.72(图112)。在两种测定的低和高范围内,相关性是强的,这表明ELISA可用作未来分析中wtVNA的替代物。
针对SARS-CoV-2 S肽的细胞免疫应答
通过ICS在第1天(疫苗接种前)和剂量1后第15天评估Th1和Th2应答。PBMC在参与者的子集中收集。
CD4+/CD8+T细胞应答(ICS)
用SARS-CoV-2 S肽(肽池S1和S2)刺激PBMC后,通过ICS确定CD4+和CD8+T细胞应答的诱导。通过产生IFNγ和/或IL-2而不是IL-4、IL-5和/或IL-13的CD4+T细胞的百分比表征T-辅助(Th)1应答的测量。确定Th2应答为表达IL-4和/或IL5/IL13和CD40L的CD4+T细胞的百分比。这个数据用于确定Th1/Th2比例。通过产生IFNγ和/或IL-2的CD8+T细胞的百分比表征CD8+T细胞应答的检测。在以下CD4+和CD8+T细胞应答的总结中,介绍并讨论了S1和S2肽池的组合值。
CD4+T细胞应答
表达IFNγ和/或IL-2(Th1)而不是Th2细胞因子的CD4+T细胞以及表达IL-4和/或IL-5/IL-13和CD40L(Th2)的CD4+T细胞的百分比在图113中呈现。图114中提供了组合方案谱。
Th1应答
对于活性疫苗组和安慰剂,基线Th1应答在第1天低于LLOQ(<0.022%)。疫苗接种后14天,在5x1010vp疫苗接种组的参与者的Th1应答的中值增加至0.09%(Q1;Q3:0.04;0.17),以及在1x1011vp剂量组的参与者则增加至0.11%(Q1;Q3:0.04;0.15)(图45)。第一次疫苗接种后14天,安慰剂组中的Th1应答没有增加。这些应答与队列1a中的年轻成年人相当,在第15天队列1a中的年轻成年人显示在5x1010vp疫苗接种组中Th1应答的中值为0.08%(Q1;Q3:0.05;0.16),在1x1011vp剂量组中Th1应答的中值为0.11%(Q1;Q3:0.07;0.16)。
5x1010vp疫苗组和1×1011vp疫苗组的SD分别为0.41和0.36。
在疫苗接种后14天,在5x1010vp疫苗组中总共60%(95%CI:46%;74%)的参与者和在1x1011vp疫苗组中总共67%(95%CI:53%;79%)的参与者显示出CD4+T细胞应答。这略低于队列1a中所见的情况,其中在5x1010vp疫苗组中76%(95%CI:65%;86%)的参与者和在1x1011vp疫苗组中83%(95%CI:73%;91%)的参与者显示出可检测的CD4+T细胞应答。
Th2应答
在活性疫苗组和安慰剂组中,在基线和第15天检测不到Th2应答(图113)。这些结果与队列1a中的年轻成年人相当。只有一名参与者有Th2应答。这名包括在1x1011vp疫苗组中的参与者也有Th1应答,其高于Th2应答,Th1/Th2比例为20.12,表明Th1偏向表型。
Th1/Th2比例计算
在疫苗接种Ad26.COV2.S后,计算Th1/Th2比例以评估T细胞应答阳性的参与者的T细胞表型偏向。Ad26.COV2.S疫苗接种组中的所有参与者的Th1/Th2比例均高于1,表明总体而言,Ad26.COV2.S诱导Th1偏向应答。这与队列1a中的结果相当,队列1a中的结也显示100%的用Ad26.COV2.S疫苗接种的参与者的Th1/Th2比例高于1。
CD8+T细胞应答
对SARS-CoV-2 S肽刺激应答并产生IFNγ和/或IL-2的CD8+T细胞的描述性统计数据在图115中显示。图116中提供了组合方案谱。
在疫苗组和安慰剂组中,疫苗接种前的基线CD8+T细胞应答均低于LLOQ (<0.022%)。疫苗接种后14天,5x1010vp组的CD8+T细胞应答的中值水平增加到0.06%(Q1;Q3:0.02;0.12),1x1011vp组增加到0.02%(Q1;Q3:0.01;0.08)。疫苗接种后安慰剂组中的CD8+T细胞应答没有增加。5x1010vp组中的这些CD8+T细胞应答与队列1a中年轻成年人中所见的应答相当(0.7%,Q1;Q3:0.02;0.12)。1x1011vp组中的CD8+T细胞应答有低于年轻成年人中所见的CD8+T细胞应答的趋势(0.09%,Q1;Q3:0.05;0.19)。
两个疫苗组的SD为5×1010vp组的0.40和1×1011vp组的0.41。
疫苗接种后14天,在5x1010vp组中总共36%(95%CI:23%;51%)的参与者和1x1011vp组中24%(95%CI:13%;37%)的参与者显示出CD8+T细胞应答。这低于在第15天,队列1a中年轻成年人显示的CD8+应答的百分比(5×1010vp组=51%和1×1011vp组=64%)。
结论与讨论
先前报道了来自队列3的15名哨兵参与者的免疫学分析。完整的队列3中期分析与哨兵报告一致,显示Ad26.COV2.S在65岁及以上的参与者中以5x1010vp或1x1011vp的剂量水平单次疫苗接种后具有高度免疫原性。
在Ad26.COV2.S疫苗组疫苗接种后14天和28天,观察到SARS-CoV-2中和和S结合抗体应答相较基线的增加。对于wtVNA和ELISA,5x1010vp疫苗组的应答率分别为90%和96%。对于wtVNA和ELISA,1x1011vp疫苗组的应答率分别为91%和96%。
经ICS评估,Ad26.COV2.S还诱导细胞免疫应答。在用5x1010vp或1x1011vp进行单次疫苗接种后14天观察到CD4+和CD8+T细胞应答的增加。疫苗接种后,CD4+T细胞具有Th1-偏向表型,并且没有观察到或非常有限的Th2应答。在所有具有T细胞应答的参与者中,Th1/Th2比例>1。
如先前报道的,队列3中体液应答的总体量和应答率与队列1a中观察到的应答相当。细胞CD4+T细胞应答与队列1a中的年轻成年人相似,CD8+T细胞应答通常低于队列1a中年轻成年人中所见的CD8+T细胞应答。
总体而言,在65岁的参与者中,单次接种5x1010vp或1x1011vp Ad26.CoV.2会诱导强的中和抗体应答和结合抗体应答、Th1偏向表型以及升高的CD4+和CD8+T细胞应答。
实施例25:在18岁及以上成人中评估Ad26.COV2.S预防SARS-CoV-2介导的COVID-19的效力和安全性的随机、双盲、安慰剂为对照的3期研究
该研究将招募多达30,000名参与者,以评估Ad26.COV2.S在成人参与者中与安慰剂相比在预防分子确诊的中度至严重/危重2019冠状病毒病(COVID-19)的效力。
详细描述COVID-19疫苗临床开发计划的目的是开发一种安全有效的疫苗来预防COVID-19。目前,没有可用于预防COVID-19的疫苗。正在开发Ad26.COV2.S中,其是一种基于编码SARS-CoV-2 S蛋白的人无复制能力的Ad26载体的COVID-19疫苗,构建Ad26.COV2.S以编码严重急性呼吸道综合征冠状病毒-2(SARS-CoV-2)病毒刺突(S)蛋白。该研究将包括:筛选阶段(最多28天)、双盲研究期(60周)和长期随访期(额外一年)。参与者的总研究时间最长为2年3个月。将在整个研究过程中进行效力(COVID-19样体征和症状等)、免疫原性(例如体液免疫应答)和安全性(例如AE监测)等评估。
参与者将在第1天和第57天接受肌内(IM)注射Ad26.COV2.S疫苗或安慰剂。
实施例26:Ad26.COV2.S COVID-19候选疫苗的一剂和两剂方案在成年和老年恒河猴中的免疫原性
如上文实施例20和21所示,在非临床效力研究中,单剂Ad26.COV2.S在恒河猴的上气道和下气道中提供了针对SARS-CoV-2攻击的强的保护,并且保护叙利亚金仓鼠免受严重临床疾病。保护效力与NHP血清中病毒中和活性的存在密切相关。这些数据确认了先前报道的关于SARS-CoV的发现,该发现显示针对SARS-CoV刺突(S)蛋白的中和抗体应答与在非临床模型中对SARS-CoV的保护有关,所述刺突(S)蛋白与SARS-CoV-2结合相同的细胞受体以进入细胞。
已显示根据本发明的Ad26.COV2.S候选疫苗具有可接受的安全性和反应原性谱,并且在中期分析中,在单次剂量后在成人和老年人中引发迅速和强烈的免疫应答,在1/2a期研究中测量到免疫后第29天。免疫应答满足预先指定的最低标准,可以在3期研究中继续测试Ad26.COV2.S。
在本实施例中,报告了在成年和老年NHP中单剂和两剂Ad26.COV2.S方案后的在第一次疫苗接种后追踪期长达14周的免疫原性数据。
方法
动物
成年NHP-包括成年动物的NHP研究在Charles River Laboratories(CRL)Montreal ULC,Laval Site(CA)进行。动物获自中国昆明生物医学国际有限公司。在从实验设施地(colony)转移之前,对所有动物进行健康评估,并通过皮内注射结核菌素对其进行至少一次结核病测试。通过皮下注射对每只动物进行驱虫处理。评估由技术人员按照标准操作程序进行。动物实验批准由CRL Montreal ULC,Laval Site(CA)的机构动物管理和使用委员会(IACUC)提供。动物实验是按照加拿大动物管理委员会发布的指南和加拿大国家研究委员会发布的实验动物管理和使用指南进行的。实验设施已获得加拿大动物管理委员会(CCAC)和美国实验动物管理认证协会(AAALAC)的认可。此外,该研究是根据EMA指南ICHM3(R2)进行:人临床试验和药品上市许可的非临床安全性研究指南和FDA指南,红皮书2000:设计和进行毒性研究的一般指南。
老年NHP-使用老年NHP的研究在荷兰Rijswijk的生物医学灵长类动物研究中心(AAALAC认可的机构)进行。为了研究目的,人工增殖并成群饲养动物。动物饲养室符合非人灵长类动物管理和使用的国际指南(欧洲理事会指令86/609/EEC和ETS 123公约,包括修订的附录A以及“外国机构人道管理和实验动物使用标准”识别号A5539-01,由美国国立卫生研究院(NIH)卫生与公众服务部提供)。根据荷兰法律,所有动物处理均在动物科学系(ASD)内进行。有大量经验丰富的工作人员可参与,包括全职兽医和病理学家。ASD由负责机构(Voedsel en Waren Autoriteit,VWA)和独立的动物福利官员定期检查。一些动物对猿猴疱疹B病毒抗体呈血清阳性,而模拟免疫对照组的一些动物对猿猴T细胞白血病病毒和猿猴逆转录病毒抗体呈阳性。根据体检和全血细胞计数和血清化学的评估,将所有动物归类为健康。生物医学灵长类动物研究中心(dierexperimentencommissie,DEC-BPRC)的机构动物管理和使用委员会批准了根据严格的国际伦理和科学标准和指南制定的研究方案。关于动物实验的荷兰法律要求这个委员会成员的资格,包括他们独立于研究所的资格。
疫苗
Ad26.COV2.S疫苗已如本文所述产生。Ad26载体的制造在互补细胞系PER.C6TetR(Wunderlich et al.,2018)(Zahn et al.,2012)中进行。阴性对照载体Ad26.RSV.gLuc编码RSV F蛋白,RSV F蛋白与Gaussia萤火虫荧光素酶融合作为由2A肽序列分隔的单个转基因,从而导致两种单个蛋白的表达。载体的制造在PER.C6中进行。
研究设计动物实验
成年NHP-通过基于体重的随机分层系统,分配60只(57只雌性和3只雄性。分配3只雄性到第3、4和5测试组,每组1只雄性)来自中国的3.3-5.0岁的恒河猴(Macaca Mulatta)到五个组,每个疫苗组包括14只动物,以及假对照组包括4只动物。第1组(n=4)是假对照组,在第0周和第8周接受生理盐水注射,第2组和第3组(每组n=14)分别在第0周接受1x1011病毒颗粒(vp)和5x1010vp Ad26.COV2.S的一次免疫,第4组和第5组(每组n=14)接受两次5x1010vp Ad26.COV.2的免疫,分别间隔4周(第0周和第4周)和8周(第0周和第8周)。所有免疫均通过左后腿股四头肌的肌内途径进行。在第一剂疫苗之前和随后每2周采集血液以获得血清,直至研究的第14周。
老年NHP-将20只年龄在13.75-21.9岁的雌性恒河猴(Macaca Mulatta)分布在4个实验处理组中,并饲养在ABSL-III设施中,与社会相容的动物成对饲养。第1组(n=6)在第0周接受1x1011病毒颗粒(vp)的Ad26.COV2.S。第2组(n=6)在第0周和第8周接受5x1010vp的Ad26.COV2.S。第3组(n=4)在第0周和第8周接受100μg S蛋白,以500μg氢氧化铝(Al(OH)3;2%Alhydrogel,InvivoGen)为佐剂。用1x1011vp Ad26.RSV.gLuc,一种表达无关抗原的Ad26载体免疫假对照组(第4组,n=4)。所有免疫均在左后腿的股四头肌内进行。如文中所述获得用于血清和外周血单个核细胞(PBMC)分离的血液。
酶联免疫吸附测定(ELISA)
使用基于Wuhan-Hu-1 SARS-CoV-2株(MN908947)的重组S蛋白抗原,通过ELISA测量IgG与SARS-CoV-2 S蛋白的结合。在4℃下,将SARS-CoV-2 S蛋白抗原吸附在96孔微孔板上至少16小时。孵育后,将板在PBS/0.05%Tween-20中洗涤并在室温下用在PBS/0.05%Tween-20中的5%脱脂牛奶封闭1小时。稀释血清标准品、对照和NHP血清样品并在平板上室温孵育1小时。洗涤板,然后在室温下用过氧化物酶缀合的山羊抗人IgG孵育1小时,洗涤,并在室温下用四甲基联苯胺(TMB)底物避光显色30分钟,然后用H2SO4终止。在450/620nm处读取光密度。根据标准反算抗体浓度,并基于所有稀释度生成可报告值,以ELISA单位[EU]/mL表示。检测下限(LLOD)为3.4EU/mL,基于标准最低插值范围浓度乘以每稀释系数,并用作信息性LLOD。定量下限(LLOQ)基于对人样品进行的定性,已设置为50.3EU/mL。
假病毒中和测定(psVNA)
通过假病毒中和测定测量SARS-CoV-2 S中和抗体滴度。假病毒颗粒由改良的水泡性口炎病毒(VSVΔG)骨架制成,并带有SARS-CoV-2的S糖蛋白。假颗粒含有用于检测的荧光素酶报告基因。在96孔转移板中制备热灭活NHP血清样品的连续稀释液。将SARS-CoV-2假病毒依次添加到血清稀释液中,并在37℃以及补充有5%CO2中孵育60±5分钟。然后将血清-病毒复合物转移到预先用Vero E6细胞接种过夜的板上,并在37℃和5%CO2下孵育20±2小时。在此孵育之后,将荧光素酶底物添加到细胞中以评估每孔的发光水平。然后在发光读板器上读板。发光强度以相对发光单位(RLU)进行定量。血清样品的中和滴度计算为对应于该样品的50%中和抗体滴度(IC50)的血清稀释度倒数。LLOD为10,这是用作信息性LLOD的第一个样品稀释度(1∶10)。LLOQ基于对人样品的定性,已设置为33IC50。
野生型中和测定(wtVNA)
使用如前所述的微中和测定进行针对活SARS-CoV-2的中和测定(Bos et al.,2020),并使用了不同株的修饰。从咽拭子中分离出临床分离株SARS-CoV-2/human/NLD/Leiden-0008/2020(Leiden-0008),并在Vero E6细胞中传代两次。这个病毒分离株的源自NGS的完整基因组可在GenBank登录号MT705206.1下获得。分离株Leiden-0008在Vero E6细胞中增殖和滴定。
ELISpot
对新鲜分离的PBMC进行IFN-γ/IL-4双染色。使用Ficoll梯度离心(10ml在1∶4DPBS稀释的血液中的92%Ficoll-Paque(GE Healthcare)Plus)从乙二胺四乙酸(EDTA)全血中分离PBMC。根据制造商方案,使用ImmunoSpot人IFN-γ/IL-4双染色酶ELISpot测定试剂盒(Cellular Technology Limited)进行ELISpot。用抗人IFN-γ和IL-4捕获抗体包被乙醇激活的96孔ELISpot板过夜。细胞以每孔250,000个细胞的浓度铺板,并且在DMSO存在下用任一细胞培养基刺激,用2个肽浓度为2μg/mL的具有11个氨基酸重叠,涵盖SARS-CoV-2 S蛋白的整个长度的连续15元肽池(JPT)刺激,或用1μg/mL作为阳性对照的PHA刺激22小时。使用CTL ImmunoSpot Analyzer和ImmunoSpot软件(Cellular Technology)进行分析。每1.0×106PBMC的斑点形成单位是通过减去每只动物的单个肽池的培养基刺激计数并在2个肽池中求和来计算的。
细胞内细胞因子染色(ICS)
为了分析细胞内细胞因子的产生,将1x106新鲜分离的PBMC在37℃下,在存在GolgiStop(BD Biosciences)的情况下,用任一细胞培养基、2μg/mL SARS-CoV-2 S蛋白肽池(如ELISpot所述)、培养基或用1μg/mL PHA刺激过夜(约15小时)。首先用LIVE/DEAD Aqua活性染料(Invitrogen)孵育受刺激的细胞,然后用抗人单克隆抗体CD3-PerCP-Cy5.5、CD4-APC H7、CD8-BV650、CD14-BV605、CD69-BV786(BD Biosciences)和CD20-BV605(Biolegend)进行表面染色。随后用Cytofix/Cytoperm缓冲液(BD Biosciences)固定细胞并用抗人IL-2-PE、IFN-g-APC(BD Biosciences)、IL-5-Vi0515(Miltenyi Biotec)、IL-4-PE Dazzle594和IL-13-BV421(Biolegend)进行细胞内染色。在LSR Fortessa(BD Biosciences)上进行样品采集,并在FlowJo V10(TreeStar)中分析数据。通过对单细胞(FSC-H与FSC-A)、活细胞、大小(淋巴细胞)(FSC-A与SSC-A)、CD3+、CD4+或CD8+细胞和CD69+加细胞因子-阳性进行连续门控来鉴定抗原特异性T细胞(门控策略在补充图2中显示)。细胞因子阳性应答在减去在每只单个动物的相应培养基刺激样品中检测到的背景应答后呈现。应答者由技术阈值定义(Bowyer et al.,2018),即在细胞因子门中检测至少1个事件的理论能力,并且此处定义为每次测定运行培养基和肽池刺激的样品的CD4或CD8 T细胞平均数量的倒数。Th1与Th2细胞因子的比例通过Th1(表达IFN-γ或IL-2的CD4+CD69+T细胞)和Th2(表达IL-4或IL-5或IL-13的CD4+CD69+T细胞)亚群的布尔门控(Boolean gating)计算。表达IL-4或IL-5或IL-13)的T细胞亚群。
统计分析
ELISA和psVNA
对于结合和psVNA中和抗体数据,使用方差分析(ANOVA)中的双样本t检验对特定疫苗组进行比较。用每个疫苗组的成对t检验比较连续时间点。p值是根据log10转换值的平均值计算的。
wtVNA、ELISpot和ICS
用Mann-Whitney U检验将疫苗组与阴性对照组进行比较。如果少于50%的滴度等于LLOD,则使用以疫苗为因素的Tobit ANOVA进行疫苗组之间的成对比较。疫苗之间的成对比较是用z检验完成的。如果对于测定任何疫苗组有50%或更多的删失,则使用Mann-Whitney U检验进行成对比较。
对于每次测定,每只动物计算连续时间点之间的滴度差异。
根据删失测量的数量,将差异与Tobit ANOVA进行比较,然后进行事后z检验或符号检验。
对于所有统计检验,显著性水平为5%。未应用多重比较校正。所有统计计算均在SAS 9.4中完成。(SAS Institute Inc.,Cary,NC,US)。
相关性分析
通过双边Spearman等级相关性检验计算结合抗体浓度和测量的中和抗体滴度之间的相关性分析。
结果
一剂和两剂Ad26.COV2.S疫苗方案在成年恒河猴中的免疫原性
在第0周用单剂1x1011vp或5x1010vp Ad26.COV2.S(每组n=14)或者用间隔4或8周(每组n=14)的两剂的5x1010vp Ad26.COV2.S免疫60只成年恒河猴的队列(3.3-5.0岁,57只雌性和3只雄性)。假对照组(n=4)在第0周和第8周接受生理盐水注射。通过酶联免疫吸附测定(ELISA)和假病毒中和测定在第一次免疫后每两周至14周追踪S特异性抗体应答(psVNA)并早在免疫后两周在所有接种疫苗的动物中检测到S特异性抗体应答,并且S特异性抗体应答在免疫后4周显著增加(p≤0.030,ANOVA t检验)(图1A和B)。相对于用5x1010vpAd26.CoV2.S免疫(组合第3、4和5组)的动物,接受1x1011vp Ad26.CoV2.S(第2组)的动物具有1.6倍的结合抗体和2倍的中和抗体(分别为p=0.008和p=0.004,ANOVA t检验)。在整个观察期间,应答水平的类似差异保持不变。S-结合抗体水平比中和抗体水平下降得更快,与动物接受的疫苗剂量无关。
相对于剂量2前时间点,在第一次接种后4或8周给予的第二次疫苗剂量引起了S特异性抗体应答的显著增加(p≤0.001,ANOVA t检验)(图117A和117B)。与用5x1010vpAd26.COV2.S的单次免疫相比,在第一剂后4和8周给予的第二次免疫(在第2剂后2周测量)分别引起结合抗体浓度的至5.7和11.8倍的增加(p<0.001,ANOVA t检验),以及中和抗体滴度的至7.6倍和15.2倍的增加(p<0.001,ANOVA t检验)。当比较由两剂疫苗方案引发的抗体应答与由单次1x1011vp疫苗剂量引发的抗体应答时,观察到了类似的差异。
虽然具有4周和8周间隔的两剂疫苗方案在第二次免疫后两周引发相当的S特异性结合抗体浓度,但与4周方案相比,8周的中和抗体滴度的几何平均值是2.2倍(p=0.005,ANOVA t检验)。在第二次免疫后第4周和第6周,两个剂量组的结合和中和抗体水平均下降,具有相似的动力学,保持中和抗体量的相对差异(对于8周方案,在4周和6周分别是2.1和2.4倍,分别为p=0.021和p=0.001,ANOVAt检验)。
尽管在接受1剂方案的动物中,结合抗体滴度相对于中和抗体滴度下降得更快,但我们观察到所有测试方案的不同时间点的结合抗体水平和中和抗体水平之间存在良好的总体相关性(R=0.7875,p<0.001,Spearman等级相关性检验)(图117C)。
一剂和两剂Ad26.COV2.S疫苗方案在老年恒河猴中的免疫原性。
随着老年人中COVID-19严重度和死亡率的增加,我们还分析了Ad26.COV2.S在老年恒河猴(20只雌性,13.75-21.9岁)中的免疫原性。包括以氢氧化铝(Al(OH)3)为佐剂的在其融合前构象中稳定化的可溶性三聚化刺突蛋白作为T辅助2(Th2)偏向对照疫苗。在第0周,用1x1011vp Ad26.COV2.S的单剂方案(n=6),用间隔8周的5x1010vp Ad26.COV2.S的两剂方案(n=6)或用间隔8周的以Al(OH)3为佐剂的100μg S蛋白的两剂方案(n=4)免疫各组以促进Th2型应答。假对照组在第0周和第8周接受编码无关抗原(Ad26.RSV.gLuc;假对照;n=4)的Ad26载体。在第一次免疫后每隔两周直至10周测量SARS-CoV-2 S蛋白特异性结合抗体水平和中和抗体水平,在第4周和第10周测量S蛋白特异性细胞应答。
从第2周开始,每种疫苗接种方案的S蛋白特异性结合抗体浓度显著增加(p≤0.034,ANOVA与事后成对t检验比较第0周与第2周)。在第6周和第8周时,与以Al(OH)3为佐剂的S蛋白诱导的浓度相比,Ad26.COV2.S诱导的抗体浓度显著增加(p≤0.036,ANOVA与事后t检验)。直至第8周,在通过两Ad26.COV2.S剂量水平引发的抗体应答中未检测到统计学上的显著差异。在第10周,即第二剂后2周,与单剂1x1011vp Ad26.COV2.S的接受者相比,接受第二剂5x1010vp Ad26 COV2.S的组具有显著更高的抗体浓度(5x1010vp Ad26.COV2.S组的4.4倍,p=0.002),而S抗体浓度在以Al(OH)3为佐剂的S蛋白的接收者中相似(p=0.482)(图118A)。
通过野生型病毒中和测定确定中和抗体滴度的动力学。在第2周用单剂的1x1011vpAd26.COV2.S诱导中和抗体滴度,与之前的时间点相比,在第4周时所有动物的中和抗体滴度均显著增加(p=0.031,符号检验),并且此后一直保持稳定直至第10周。类似地,两剂5x1010vp Ad26.COV2.S方案在所有动物中诱导中和抗体滴度,相比之前的时间点,中和抗体滴度在第4周(p=0.031,符号检验)和第6周(p=0.008,to bit方差分析与事后z检验)显著增加。在第10周,即第二剂后2周,与第8周相比,抗体滴度增加至3倍(p<0.001,to bit方差分析与事后z检验)。以Al(OH)3为佐剂的S蛋白在第一剂后仅在4只动物中的2只中诱导低水平和瞬时水平的中和抗体。然而,在第10周,即第二剂后2周,所有4只动物都具有与Ad26.COV2.S组相同范围的中和抗体(由于用佐剂的蛋白组的组规模小,无法进行统计分析)。第10周疫苗组的成对比较表明,与单剂1x1011vp Ad26.COV2.S组相比,两剂5x1010vpAd26.COV2.S方案诱导显著更高的中和抗体滴度(10倍,p<0.00,to bit方差分析和事后z检验),而接受S蛋白组的NHP中的滴度相似(p=0.303)(图118B)。用ELISA测量的S-特异性结合IgG水平与野生型中和抗体滴度高度相关(R=0.92,p=<0.001,Spearman等级相关性),显示出更高的ELISA灵敏度(图118C)。
使用通过涵盖完整的S蛋白的有11个重叠的15元肽,用ELISpot和细胞内细胞因子染色(ICS)测量S蛋白特异性T细胞应答。在第一次免疫后4周,Ad26.COV2.S方案以及以Al(OH)3为佐剂的刺突蛋白均诱导了IFN-γ应答(图119A和119B)。在第二剂5x1010vpAd26.COV2.S和用佐剂的刺突蛋白组后两周(第10周),5x1010vp Ad26.COV2.S组的IFN-γ应答与第4周时间点相当,但低于1x1011vp Ad26.COV2.S和用佐剂的S蛋白组,这表明第二剂Ad26.COV2.S提供了更好的T细胞应答持久性。通过ELISpot在第4周和第10周仅观察到以Al(OH)3为佐剂的刺突蛋白组的大量IL-4应答(图119A)。也通过ICS分析了CD4+和CD8+T细胞细胞因子应答。S蛋白特异性CD4+T细胞IFN-γ和IL-4表达模式确认了ELISpot应答,在第4周所有疫苗组诱导的IFN-γ水平显著高于假对照组(p≤0.029,Mann-Whitney-U检验)并在第10周下降,除了两剂5x1010vp Ad26.COV2.S组与假组相比仍然显著更高(p=0.010)。与假对照组相比,仅以Al(OH)3为佐剂的刺突蛋白组在两个时间点的IL-4表达显著更高(p=0.029)(图119B)。测量了其他的细胞因子IL-2或IL-5和IL-13,并分别反映了IFN-γ或IL-4所见的模式,对于接受第二剂的组来说,应答的提升很小或没有,且仅接受以Al(OH)3为佐剂的刺突蛋白的组诱导强水平的Th2细胞因子IL-4、IL-5和IL-13(图119C)。这通过计算Th1与Th2比例得到确认,在Ad26.COV2.S免疫后观察到明显的Th1偏向,与方案无关(图119C)。Ad26.COV2.S诱导的刺突蛋白特异性CD8+T细胞主要产生IFN-γ和IL-2,而以Al(OH)3为佐剂的刺突蛋白仅产生IL-2。没有一种免疫方案诱导CD8+T细胞产生显著量的IL-4、IL-5和IL-13(图120)。
在本实施例中,评估了单剂和两剂Ad26.COV2.S方案在成年和老年恒河猴中首次给药后直至14周的免疫原性,以深入了解单次给药候选疫苗后的免疫持久性以及第二剂对S蛋白特异性免疫应答的量的影响。
在成年猕猴和老年猕猴中,早在第一次免疫后两周就检测到S结合和SARS-CoV-2中和抗体应答,到第4周显著增加。在所有测试的Ad26.COV2.S疫苗方案的成年和老年恒河猴中的抗体应答的动力学和量似乎相似。这些观察结果与用Ad26.COV2.S单次免疫后第4周在成人和老年人中的观察结果一致。
需要2剂以Al(OH)3为佐剂的S蛋白才能引发显著水平的中和抗体,这在3期临床试验中基于蛋白或mRNA平台的其他COVID-19疫苗中也观察到,这些疫苗在NHP和人中单次给药后没有足够的免疫原性(Corbett et al.,The New England Journal of Medicine,383(16),1544-1555,2020)(Guebre-Xabier et al.,BioRxiv,1-16,2020)。在成年和老年NHP研究中,发现结合抗体应答和中和抗体应答之间存在高度相关性,尽管在这两项研究中使用了不同的中和测定。这表明S蛋白结合抗体ELISA可用作中和抗体应答的替代读数,我们早期的观察也支持这一点(Mercado et al.,Nature.https://doi.org/10.1038/s41586-020-2607,2020),
在成年猕猴中,单剂Ad26.COV2.S引发的体液免疫应答至少维持到免疫后第14周。随着时间的推移,结合抗体应答确实显示出一些下降,而中和抗体应答保持更稳定,提供了我们的候选疫苗引发的免疫持久性的早期迹象。与接受5x1010vp疫苗剂量的NHP相比,接受1x1011vp剂量的NHP中的体液免疫应答略高,但差异太小,不足以保证在进一步的临床开发中使用更高剂量。
与单剂方案相比,在第一次免疫后8周给予的第二剂Ad26.COV2.S在成年和老年NHP中导致刺突蛋白特异性结合显著增加,更重要的是中和抗体应答。这与其他基于Ad26的疫苗的观察结果一致,其中第二剂总是在动物模型和人中引发更高和更持久的免疫应答。NHP中两剂Ad26.COV2.S方案的发现表明,与我们的单剂方案相比,两剂Ad26.COV2.S疫苗方案也有可能使人中产生更高和更持久的免疫应答。
成年NHP中两次疫苗剂量之间间隔长度的潜在影响的评估表明,虽然在4周或8周间隔的两次剂量后引发的结合抗体浓度水平相似,但以8周的间隔接受两剂的动物中的中和抗体应答更高。因此,在2剂之间间隔8周的疫苗方案优于间隔4周的方案。
开发COVID-19疫苗时要评估的一个重要方面是疫苗相关增强型呼吸道疾病(VAERD)的潜在风险和理论风险,这通常被认为与非中和抗体应答和Th2偏向细胞免疫应答有关。在这项研究中显示,Ad26.COV2.S在老年NHP中引发了Th1偏向的CD4+T细胞应答,如先前在成年NHP以及成人和老年人中所显示的那样,并且与其他在NHP和人中使用S蛋白抗原的基因疫苗平台所发现的相似(van Doremalen et al.,Nature,May.https://doi.org/10.1038/s41586-020-2608-y,2020)(Yu et al.,Science(New York,NY),369(6505),806-811.https://doi.org/10.1126/science.abc6284,2020)(Vogel et al.,BioRxiv,2020.09.08.280818.https://doi.org/10.1101/2020.09.08.280818,2020)(Corbett etal.,同上,2020)。相比之下,以Al(OH)3为佐剂的S蛋白诱导了更多的Th2偏向免疫应答,正如这种佐剂所预期的那样,并确认在这种NHP动物模型中可以引发Th2偏向应答。NHP中的Th1偏向应答以及Ad26.COV2.S诱导的强的和持久的中和抗体应答一起降低了这种疫苗的VAERD的可能性。
总之,这些数据表明,一剂和两剂Ad26.COV2.S疫苗方案在成年和老年NHP中引发相似的抗体应答。重要的是,与单剂疫苗相比,在第一次免疫后8周给药第二剂疫苗,在成年和老年动物中均诱导了(中和)抗体应答的显著增加。此外,对成年猕猴免疫原性的追踪表明,抗体应答在第一次免疫后可维持长达14周,这提供了单剂Ad26.COV2.S引发的持久免疫应答的早期迹象。
实施例27:使用Ad26.COV2.S Covid-19疫苗进行的1/2a期试验的中期结果
如上所述,在以单剂或两剂方案(56天间隔)接受5x1010或1x1011病毒颗粒剂量的Ad26.COV2.S的健康成人(年龄18-55岁;队列1,n=405或>65岁;队列3;n=405)中设计多中心1/2a期随机,双盲,安慰剂为对照的试验。
在本实施例中,报告了第一次(队列1和3)和第二次(队列1)疫苗接种后的结果。
方法
研究设计和参与者
该研究是一项多中心、随机、双盲、安慰剂为对照的1/2a期试验,旨在评估在18-55岁的健康成人中Ad26.COV2.S在1ml体积中5x1010(低剂量,LD)或1x1011病毒颗粒(vp)(更高剂量,HD)的安全性、反应原性和免疫原性,以单剂或间隔56天的两剂方案肌内(IM,三角肌)给药(队列1a:目标n=375;队列1b(深入免疫原性的探索性队列):目标n=25;和>65岁(队列3:目标n=375)。
Ad26.COV2.S是一种重组的、无复制能力的Ad26载体,其编码全长和稳定化的SARS-CoV-2 S蛋白,SARS-CoV-2 S蛋白源自Wuhan株的第一个临床分离株(全基因组序列NC_045512),如本专利申请中所述。该研究在比利时和美国进行,并得到了当地伦理委员会和机构审查委员会(IRB)的审查和批准。所有参与者在招募前都提供了书面知情同意书。
程序
将队列1和队列3中的405名合格参与者随机分配(1∶1∶1∶1∶1)在以下治疗组之一中:第一剂和第二剂为LD/LD;LD/安慰剂;HD/HD;HD/安慰剂;安慰剂/安慰剂。随机化由交互式网络应答系统(IWRS)完成,并使用随机排列的区块按站点进行分层。参与者和研究者将在整个研究过程中保持盲态。为了满足盲法研究的标准,只有当所有参与者都达到剂量2后第8天时,申办者和统计学家才对队列1和队列3的主要分析进行分组揭盲分析。
主要终点是每个剂量方案的安全性和反应原性。疫苗接种后7天在日记卡上收集征求的不良事件(AE),疫苗接种后28天的非征求的不良事件和整个研究过程中的SAE。根据FDA指导文件“Toxicity Grading Scale for Healthy Adult and AdolescentVolunteers Enrolled in Preventive Vaccine Clinical Trials”对AE进行分级。队列1和队列3的安全数据截至到截止日期2020年10月30日。
次要终点是对SARS-CoV-2 S蛋白的体液和细胞免疫。在基线和第15、29、57和71天,通过ELISA测量SARS-CoV-2 S特异性结合抗体。血清阳性定义为高于测定定量下限(定量下限[LLOQ]=50.3EU/mL)的滴度。使用英国公共卫生部的Victoria/1/2020SARS-CoV-2株在微量中和wtVNA中的随机样本子集中测量SARS-CoV-2血清中和抗体滴度,血清阳性定义为IC50滴度>58(LLOQ)。还在这些时间点评估了IC80 wtVNA滴度。在基线和第15天,通过ICS,使用两个有11个氨基酸重叠的15元的S肽池的池测量S特异性T细胞应答。Th1应答的特征在于CD4+T细胞表达IFN- 和/或IL-2而不是IL-4、IL-5和/或IL13,而Th2应答的特征在于表达IL-4、IL-5和/或IL-13和CD40L。所有测定均以盲法进行,并在补充材料中进行了详细描述。
和/或IL-2而不是IL-4、IL-5和/或IL13,而Th2应答的特征在于表达IL-4、IL-5和/或IL-13和CD40L。所有测定均以盲法进行,并在补充材料中进行了详细描述。
统计方法
在完整分析集中对安全性数据进行了描述性分析,并在符合方案的免疫原性群体中分析了免疫原性数据。详见补充材料和统计分析计划。
结果
研究参与者
队列1参与者(年龄18-55岁)的第一次疫苗接种起始于2020年7月22日。到2020年8月7日,筛选了593名参与者,招募了405人,并且用第一剂免疫了402人,到2020年11月7日队列1中用第二剂免疫(1a和1b组合。队列3参与者(年龄>65岁)的免疫起始于2020年8月3日,到2020年8月24日,已筛选660人,招募了405人,用第一剂免疫了403人。队列1a和队列3中对SARS-CoV-2 S特异性抗体的基线血清阳性率分别为1.9%和1.0%。各组的基线特征大致相似。
Ad26.COV2.S的疫苗安全性和反应原性
收集和分析了两个队列的分组水平的揭盲安全数据。在疫苗接种后7天在日记卡上收集征求的AE,在疫苗接种后28天收集非征求的AE,并在整个研究过程中收集SAE。
研究者对剂量1后反应原性的评估可用于队列1中的402名参与者和队列3中的403名参与者。在这两个队列中,征求的局部AE大多为1级或2级,最常见的是注射部位疼痛。在队列1中,在103名LD接受者(64%)、123名HD接受者(78%)和7名(9%)给药安慰剂的接受者中发现了征求的局部AE。在队列3中,66名LD接受者(41%)、68名HD接受者(42%)和11名给药安慰剂的安慰剂接受者(14%)报告了征求的局部AE。
在这两个队列中,最多的征求的全身性AE为1级或2级,最常见的是疲劳、头痛和肌痛。在队列1中,105名LD接受者(65%)、133名HD接受者(84%)和21名安慰剂接受者(26%)报告了征求的全身性AE。在队列3中,74名LD接受者(46%)、88名HD接受者(55%)和19名安慰剂接受者(23%)报告了征求的全身性AE。在队列1中,15名LD接受者(9%)和32名HD接受者(20%)报告了3级全身性AE,而安慰剂组中没有。比较队列1a中具有一个或多个征求的3级AE的参与者显示,在18-30岁年龄组中,LD、HD和安慰剂组分别为24%、26%和0%,在这些组中31-45岁年龄组分别为3%、14%和0%以及在这些组中46-55岁年龄组分别为3%、11%和0%。在队列3中,1名给予LD的参与者(0.6%)报告了3级全身性AE,4名给予HD的参与者(2.5%)报告了3级全身性AE,没有给予安慰剂的参与者报告了3级全身性AE。在队列1中,25名LD接受者(15%)和62名HD接受者(39%)报告了发烧,而安慰剂组没有报告,8名LD接受者(5%)和15名HD接受者(9%)报告了3级发热。在队列3中,7名LD接受者(4%)和14名HD接受者(9%)报告了发热,而接受LD或安慰剂的参与者不具有3级发热以及2名HD接受者(1%)具有3级发热。所有发烧都在免疫后两天内发生,并在一到两天内消退,超过80%的人在症状出现时使用退烧药。
在队列1中,LD、HD和安慰剂组分别报告了34例(21%)、56例(35%)和14例(17%)的非征求的AE。在队列3中,LD、HD和安慰剂组27名(17%)、38名(24%)和13名(16%)参与者分别报告了非征求的AE。在任何队列中均未报告任何4级AE,无论是征求的还是非征求的。
在队列1a中,给药后,363名参与者可获得两项安全性数据。LD组和HD组分别有77.0%和79.7%的参与者出现一个或多个征求的AE,而33.8%和31.3%的在第一剂疫苗后接受安慰剂作为第二剂的参与者出现一个或多个征求的AE以及21.6%的仅接受安慰剂的参与者出现一个或多个征求的AE。与剂量1后相比,在LD和HD组参与者中分别有1.4%和6.8%的参与者出现了3级或更高级别的征求的AE,而此前接受过第一剂疫苗的对照组分别为1.4%和1.5%以及从未接种过疫苗的对照组的0%的参与者出现了3级或更高级别的征求的AE。接种第二剂疫苗后,任何组均未出现3级发热。
没有参与者因AE终止研究。有5起SAE:1起因既往有复发性低血压病史而研究者判断其为与疫苗无关的低血压,1起有肾结石病史(不相关)的患者中的双侧肾结石,1起与军团菌肺炎的无关病例,1起恶化的多发性硬化症,基于MRI发现,其大约存在8-10年,但仅在纳入研究后才诊断出来(不相关),以及一名发热的参与者,其因怀疑Covid-19而住院过夜,在12小时内恢复,随后研究者判断发烧与疫苗有关。详情见补充材料。
Ad26.COV2.S的免疫原性
这个中期分析的免疫原性数据通过剂量水平揭盲。在队列1a的所有5组中,针对稳定化的SARS-CoV-2全长刺突蛋白测量结合抗体浓度(EU/mL GMC)。基线GMC小于LLOQ,并且在疫苗接种后第29天LD/安慰剂、LD/LD、HD/安慰剂和HD/HD组分别增加到478(95%置信区间:379;603)、586(445;771)、625(505;773)和788(628;988),在所有组中具有>99%的血清转化(图121A)。到第57天,GMC分别进一步增加到660(513;849)、754(592;961)、873(701;1087)和1100(908;1332)。剂量1后血清转化率除HD/安慰剂组(97%)外全部为100%。第二剂后14天,LD/LD和HD/HD组的GMC分别达到1,677(1,334;2,109)和2,292(1,846;2,845),每组的血清转化率为100%。在LD/安慰剂组和HD/安慰剂组中,第71天的GMC(分别为600(443;814)和951(696;1,300))与第57天相似。在队列3中,基线时GMT<LLOQ。到用LD或HD疫苗接种后第15天,GMT分别增加到122(97;152)和141(114;175),两个剂量组的血清转化率均为77%(图121B)。到第29天,GMT为312(246;396)和350(281;429),血清转化率为96%和96%。
通过wtVNA在队列1a和3的随机参与者子集中测量SARS-CoV-2中和抗体滴度(IC50)。基线时几何平均滴度(GMT)小于LLOQ,并在疫苗接种后第29天队列1a中的LD/安慰剂、LD/LD、HD/安慰剂和HD/HD组分别增加到224(158;318)、224(168;298)、215(16;273)和354(220;571),具有96%、88%、96%和92%的血清转化率(图121C)。LD/安慰剂、LD/LD、HD/安慰剂和HD/HD组的GMT在第57天进一步分别增加至310(228;422)、288(221;376)、370(268;511)和488(334;714),LD/安慰剂组的血清转化率为100%,其他组的血清转化率为96%。在队列1a中第二剂后14天,LD/LD和HD/HD组的GMT分别达到775(508;1183)和1272(746;2169),两个剂量组的血清转化率均为100%(图121B,左图)。在第71天,LD/安慰剂和HD/安慰剂组中的GMT分别为315(227;438)和388(290;509),与第57天的GMT相似。两组都有100%的应答者。在队列3中,基线时GMT<LLOQ,在第15天LD和HD组分别增加到212(137;284)和171(119;269),在第29天增加到277(193;307)和212(163;266)(图121D)。在第15天,每个剂量水平组的血清转化率为92%,在第29天,在队列3中的LD和HD组血清转化率为98%(83;99)和94%(76;99)。这些数据得到IC80 wtVNA的确认(图122)。wtVNA和ELISA抗体水平在两个队列中强烈相关(图122A),尽管相关性在队列3中具有更宽的椭圆形状,这表明老年人中的中和抗体滴度和结合抗体滴度之间的关系具有更大的可变性。
不同实验室进行的体液免疫评估分析中包括不同人恢复期血清(HCS)组中的抗体水平以及疫苗接受者的血清中的抗体水平在相同范围内。
基线或剂量1后的Ad26中和抗体分别对在第29天和第71天(剂量2后)的第一次和第二次免疫诱导SARS-CoV-2中和抗体的能力没有任何影响(图123)。
Ad26.COV2.S引发了S特异性CD4+Th1和Th2应答,CD8+T细胞应答在基线和剂量1后15天的一部分参与者中得到表征。在队列1a中,76%(95%CI:65;86)和83%(73;91)以及在队列3中60%(46;74)和67%(53;79)的LD和HD接受者分别有对S肽的可检测的Th1应答。队列1a中LD和HD接受者对S肽的CD4+Th1应答中值分别从基线时不可检测到在疫苗接种后15天增加至0.08%(Q1;Q3:0.05;0.16)和0.11%(Q1;Q3:0.07;0.16)的中值以及队列3中从基线不可检测到0.09%(Q1;Q3:0.04;0.17)和0.11%(Q1;Q3:0.04;0.15)的中值(图124A)。队列1a的一名参与者(LD接受者)和队列3中的一名参与者(HD接受者)具有可测量的Th2应答(图124B),但所有具有可测量的Th1和/或Th2应答的参与者的Th1/Th2比例均远高于1。
S特异性CD8+T细胞应答,如通过S肽刺激后IFN-γ和/或IL-2细胞因子的表达所确定(图124C),在两个队列的基线中均不存在。在疫苗接种后第15天,51%(95%CI:39;63)和64%(95%CI:52;75)的队列1a参与者具有CD8+T细胞应答,对于LD和HD组,S特异性CD8+T细胞应答量的中值分别为0.07%(Q1;Q3:0.03;0.19)和0.09%(Q1;Q3:0.05;0.19)。在队列3中,CD8+T细胞应答较低,36%(95%CI:23;51)和24%(95%CI:13;37)的应答者,对于LD和HD组,量的中值分别为0.06%(Q1;Q3:0.02;0.12)和0.02%(Q1;Q3:0.01;0.08)。在两个队列中CD4+Th1和CD8+T细胞应答之间的相关性都很差(图122B)。
结论
这项1/2a期研究的中期分析表明,候选疫苗Ad26.COV2.S具有可接受的安全性和反应原性谱,并且在用5x1010或1x1011vp剂量单次疫苗接种后具有免疫原性。在剂量1后,随着疫苗剂量水平的提高,征求的全身性AE的发生率呈上升趋势,随着年龄的增长,3级AE的下降趋势明显,这是有效Covid-19疫苗的预期目标人群。局部和全身性反应发生在免疫当天或次日,并且一般在24小时内消退。全身性反应对退热药响应非常灵敏,无需预防性使用。在18-55岁的第二次接种后,对于较低和较高的疫苗剂量,3级征求的全身性AE的发生率远低于第一次免疫后,这与基于mRNA的疫苗的观察结果形成对比,后者似乎是第二剂与增加的反应原性有关。
虽然目前正在开发的所有其他Covid-19疫苗都需要两剂,但单剂Ad26.COV2.S在绝大多数疫苗接种者中引起了强的体液应答,超过90%的参与者中具有S结合和中和抗体,与年龄组和疫苗剂量无关。此外,在目前可用的剂量1后71天的随访时间内,抗体滴度进一步增加和稳定,这表明Ad26.COV2.S引发的免疫应答的持久性。
与两剂疫苗相比,有效的单剂Covid-19疫苗具有明显的统筹,特别是在大流行环境中使用。在此观察到,在第57天在18-55岁成年人中的第二次疫苗剂量进一步增加了抗体滴度,这再次与NHP中最近的观察结果一致。由第一次疫苗接种引发的针对Ad26载体的中和抗体滴度与剂量2后应答的量不相关。目前正在一项3期临床试验中研究是否需要第二剂来提高人中的效力和/或持久性。
缺乏标准和使用不同的检测方法使得比较正在开发的不同Covid-19疫苗的性能变得复杂。此外,与HCS组的比较相当随意,因为它们具有不同的GMT,可能与组的组成有关(即Covid-19供体的严重度、自疾病发作以来的采样时间等)。
疫苗相关增强型呼吸道疾病(VAERD)的理论风险与中和性差的体液免疫和Th2偏向细胞免疫应答相关。在这里,所有Ad26.COV2.S引发的CD4+T细胞应答都是Th1偏向的,这与之前使用基于Ad26的疫苗平台的经验一致。伴随的一致的CD8+T细胞应答(尽管在老年人中较低)和强烈的体液应答进一步降低了VAERD的理论风险。
总之,我们的中期分析表明Ad26.COV2.S是安全的、耐受性良好且具有免疫原性。两项正在进行的3期研究正在评估单剂或两剂方案的5x1010vp Ad26.COV2.S的效力。
实施例28:Ad26.COV2.S的产率
Ad26.COV2.S(也称为Ad26NCOV030、Ad26COVS1)的产率,如通过每毫升病毒颗粒的滴度(VP/mL)所定义,对于在生物反应器中扩大载体生产规模以提供足够的用于不同的下游处理步骤的材料至关重要。因此,通过将疫苗候选载体与各自的基准对照进行比较,在小规模实验中评估产率,其中在10-L过程强化(PIN)-生物反应器过程中的性能是已知的。
Ad26NCOV030候选疫苗的产率与Ad26NCOV028一起在一个实验中进行了测试,因为最初考虑这两种构建体作为先导候选的选择。简而言之,用70和300VP/细胞的纯化Ad26NCOV030和Ad26NCOV028材料以及两个Ad26基准对照Ad26.ZEBOV和26ZIK001载体(分别为标准和低对照)转导sPER.C6和sPER.C6 TetR细胞。
在感染后第0、1、2、3和4天采集样品,并通过病毒颗粒-定量聚合酶链反应(VP-qPCR)确定Ad26载体滴度。如图125A所示,Ad26NCOV030在sPER.C6细胞中的产率介于低和标准对照之间,并且高于Ad26NCOV028,这与低对照相似。Ad26NCOV030和Ad26NCOV028(均与标准对照相似)在sPER.C6 TetR细胞(图125B)中的产率最大化。使用PER.C6和sPER.C6 TetR细胞运行2L生物反应器,加强了小规模产率评估的结果(下表)。这些数据表明,Ad26NCOV028和Ad26NCOV030在2LPIN平台上的PER.C6 TetR细胞上都是高生产者,而Ad26NCOV028和Ad26NCOV030在PER.C6细胞上则分别低于平均,和平均生产者。
表(Ad26NCOV030)和Ad26NCOV028 2L生物反应器运行。

VP,病毒颗粒;qPCR,定量聚合酶链反应;TetR,四环素阻遏物;
实施例29:Ad26.COV2.S引发的免疫可在叙利亚仓鼠中防止614G刺突变体SARS-CoV-2感染,并且具有在次优疫苗给药后出现的突破性感染的受攻击的动物的呼吸道疾病不会加重
在本实施例中,描述了另一种SARS-CoV-2仓鼠感染模型,该模型用于确认Ad26.COV2.S的免疫原性并确定其对SARS-CoV-2的614G刺突变体(BetaCoV/Munich/BavPat1/2020)鼻内感染的保护效力。基于组织病理学发现,在未疫苗接种的动物中,感染这种病毒会导致上呼吸道和下呼吸道感染,从而导致轻度至中度疾病。Ad26.COVS.2在单剂后引发结合和中和抗体,在4周间隔后使用第二剂疫苗可进一步增加。与已评估的另外两种基于Ad26的COVID-19原型疫苗相比,Ad26.COV2.S引发了最高水平的抗体和保护。Ad26.COV2.S的次优剂量引起了体液应答,该应答在攻击后不能防止肺中的病毒复制,但通过监测呼吸道病毒复制和详细的组织病理学分析没有发现VAERD的证据。
材料和方法
疫苗
基于Ad26的疫苗是如本文中描述的产生的。简而言之,它们基于编码融合前稳定化的SARS-COV-2刺突蛋白序列(Wuhan Hu1;GenBank登录号:MN908947)的无复制能力的腺病毒血清型26(Ad26)载体。使用AdVac系统(Abbink et al.,J.Virol.8 1,4654-4663(2007)),使用含有包括转基因表达盒的Ad26载体基因组的单质粒技术,对无复制能力、E1/E3缺失的Ad26载体进行工程化改造。将密码子优化、融合前稳定化的SARS-COV-2刺突蛋白编码基因插入Ad26载体基因组的E1位。Ad26载体的制造在互补细胞系PER.C6 TetR中进行(Wunderlich et al.,Potent and short promoter for expression ofheterologousgenes.(2018))(Zahn et al.,PLoS One 7,1-13(2012))。阴性对照载体Ad26.Irr(RSV-FA2-2A-GLuc)编码RSV F蛋白,RSV F蛋白与Gaussia萤火虫荧光素酶融合成为由2A肽序列分隔的单个转基因,从而导致两种单个蛋白的表达。载体的制造在PER.C6中进行。在使用前对腺病毒载体的生物载量和内毒素水平进行了测试。
研究设计动物实验
仓鼠研究
动物实验得到了动物科学程序中央管理局(Centrale Commissie Dierproeven)的批准,并按照欧洲指南(欧盟动物试验指令86/609/EEC)和荷兰当地的动物实验立法进行。生存阶段在Viroclinics Biosciences BV进行。所有参与进行临床观察和需要解释数据的实验室分析的Viroclinics人员在研究完成前的任何时候都不知道治疗分配(Treatment Allocation Key),并通过为每个收集和分析的样品分配一个独特的样本号来进行盲法。
雄性叙利亚(金色)仓鼠(Mesocricetus auratus),品系HsdHan:AURA,在研究开始时年龄为9-11周,购自Envigo++++。在异氟醚麻醉下,通过肌内途径用100μl疫苗(每条后腿50μl)对仓鼠进行免疫。用含有102TCID50的SARS-CoV-2(BetaCoV/Munich/BavPat1/2020,在S1片段中含有D614G取代)的100μl对仓鼠进行鼻内接种。已对攻击原种的序列进行了表征,并已显示与亲本株一致。在感染当天、接种前和每天直到感染后四天,在异氟醚麻醉下收集咽拭子。将咽拭子收集在病毒转运培养基中,等分并储存至分析时间。在异氟醚麻醉下经由眶后出血途径收集中间血样。处理血液用于血清分离。实验结束时,在麻醉下,颈椎脱臼牺牲动物并进行尸检。分析尸检后收集的呼吸道组织的病毒载量和组织病理学变化。
兔研究
兔实验得到了当地动物福利机构的批准,并按照欧洲指南(欧盟关于保护用于科学目的的动物的指令2010/63/EU)和比利时当地的动物实验立法进行。生命阶段在AAALAC批准的实验室Janssen Research and Development的非临床安全Beerse站点进行。研究开始时大约4个月大的雌性新西兰白兔购自法国Charles River Laboratories。在研究的第0周和第8周,经由肌内途径用0.5mL体积的5x109或5x1010vp Ad26.S、Ad26.dTM.PP或Ad26.COV2.S对兔进行免疫。作为对照组,用盐水免疫五只兔。经由侧耳收集用于血清处理的中间血样。在实验结束时,通过静脉内注射戊巴比妥钠牺牲动物,然后通过股动脉放血。
组织病理学
组织病理学由Viroclinics Biosciences BV的病理学家和Janssen Non-Clinical Safety (Beerse,Belgium)的病理学家评估。
术后四天,通过打开胸腔和腹腔并检查所有主要器官对所有动物进行尸检。基于对受影响肺组织百分比的目测估计来评估肺实变的程度。收集左鼻甲、气管和左肺进行组织病理学检查和IHC分析。轻轻灌输,和/或浸入10%中性缓冲福尔马林中对所有组织进行固定。对肺和气管进行常规处理,石蜡包埋,在载玻片上显微切片至3μm,并用苏木精和伊红(H&E)染色以进行组织病理学评估。对取样固定的鼻甲进行脱钙处理并包埋在石蜡块中,同样进行切割和染色。通过光学显微镜检查H&E染色的组织切片,使用放大倍率为25x、100x、200x和400x的Zeiss Axioplan或Leica DM2500光学显微镜进行组织病理学评分以及检查任何其他病变的存在。针对鼻炎和气管炎对鼻甲和气管中炎性细胞浸润的严重度进行评分:0=无炎性细胞,1=炎性细胞很少,2=炎性细胞数量适中,3=炎性细胞很多。
对于肺组织,检查每张整个载玻片,并对是否存在肺泡水肿、肺泡出血和II型肺细胞增生进行评分(0=否,1=是)。针对肺泡炎和支气管炎/细支气管炎对肺泡、支气管/细支气管中炎性细胞浸润和损伤的程度和严重度进行评分:0=无炎性细胞,1=炎性细胞很少,2=炎性细胞数量适中,3=炎性细胞很多。支气管周围/血管周围套状白细胞的程度:0=无,1=1-2细胞厚度,2=3-10细胞厚度,3=超过10细胞厚度。此外,对每张载玻片的肺泡炎/肺泡损伤程度进行评分:0=0%、1=<25%、2=25-50%、3=>50%。
肺组织炎症程度和严重度的累积评分(总和)提供了总下呼吸道(LRT)评分,可能的最高评分为24。以下组织病理学参数包括在下呼吸道疾病参数的总和中:肺泡炎、肺泡损伤、肺泡水肿、肺泡出血、II型肺细胞增生、支气管炎、细支气管炎、支气管周围和血管周围套状白细胞。
免疫组织化学
对肺、鼻子和气管组织样品进行取样,在10%福尔马林(肺灌注)中固定14天,并通过Viroclinics Biosciences BV包埋在石蜡中。组织块由来自Janssen Non-clinicalSafety(Beerse,比利时)的病理学家交付和评估。
使用对SARS-CoV-2NP有交叉反应的兔多克隆抗SARS-CoV核衣壳蛋白抗体(NP,Novus NB100-56576,1/300)对所有动物的肺、气管和鼻子切片的石蜡切片进行自动染色(Ventana Discovery Ultra,Roche,France)。对这些切片的免疫反应细胞数进行半定量评分,并分级为0:无阳性免疫反应细胞,1:最小(少数/局部)阳性细胞数,2中等(局部/多局部)阳性细胞数和3:许多/高(局部广泛/多灶性)免疫反应细胞数量。
病毒中和测定
使用Algaissi和Hashem先前描述的微量中和测定对活SARS-CoV-2进行中和测定(Algaissi和Hashem,Evaluation of MERS-CoV Neutralizing Antibodies in SeraUsing Live Virus Microneutralization Assay in 107-116(2020)。doi:10.1007/978-1-0716-0211-9_9)。Vero E6细胞[CRL-1580,美国典型培养物保藏中心(ATCC)]在补充有8%胎牛血清(FCS;Bodinco BV)、1%青霉素-链霉素(Sigma-Aldrich,P4458)和2mM L-谷氨酰胺(PAA)的Eagle最低基础培养基(EMEM;Lonza)中生长。细胞在含有5%CO2的湿润空气中保持在37℃。从鼻咽样品中分离出临床分离株SARS-CoV-2/human/NLD/Leiden-0008/2020(Leiden L-0008),其特征将在别处描述(准备中的手稿)。这个病毒分离株的下一代测序衍生序列可在GenBank登录号MT705206下获得,与Wuhan序列相比,Leiden-0008病毒有1个突变,导致刺突蛋白第614位处的Asp>Gly(D614G)。使用TCID50终点稀释法在Vero E6细胞中增殖和滴定分离株Leiden-0008。TCID50由Spearman- 算法计算,如所述(Hierholzer&Killington,Virology Methods Manual-Virus isolation and quantitation(1996).doi:10.1016/B978-0-12-465330-6.X5000-3)。有活的SARS-CoV-2的所有工作在LeidenUniversity Medical Center的生物安全3级设施中进行。
算法计算,如所述(Hierholzer&Killington,Virology Methods Manual-Virus isolation and quantitation(1996).doi:10.1016/B978-0-12-465330-6.X5000-3)。有活的SARS-CoV-2的所有工作在LeidenUniversity Medical Center的生物安全3级设施中进行。
在感染前1天,将Vero-E6细胞以12,000个细胞/孔接种在96孔组织培养板中。一式两份分析热灭活(56℃下30分钟)血清样品。将血清组一式两份连续稀释2倍,在补充有青霉素、链霉素、2mM L-谷氨酰胺和2%FCS的60μL EMEM培养基中初始稀释度为1∶10,最终稀释度为1∶1280。将稀释的血清与等体积的120TCID50/60μL Leiden-0001病毒混合,并在37℃下孵育1小时。然后将病毒-血清混合物添加到Vero-E6细胞单层上,并在37℃和5%CO2的湿润空气中孵育。未暴露于病毒或与120TCID50/60μL SARS-CoV-2混合的细胞分别用作阴性(未感染)和阳性(感染)对照。在感染后3天,用40μL 37%甲醛/PBS溶液/孔在4℃下固定和灭活细胞过夜。从细胞中去除固定剂,并用50μL/孔结晶紫溶液对簇进行染色,孵育10分钟并用水冲洗。评估干板的病毒细胞病变效应。中和滴度的计算方法是用完全抑制病毒诱导的细胞病变效应的阳性孔数除以重复数,并加2.5以稳定计算的比例。中和抗体滴度定义为该值的log2倒数。每次测定运行都包括SARS-CoV-2反滴定,以确认所用接种物的剂量在30-300TCID50的可接受范围内。
ELISA
通过ELISA测量IgG与SARS-CoV-2刺突抗原的结合,并且用内部产生的全长刺突蛋白COR200099(仓鼠研究)或COR200153(兔研究)。COR200099是一种内部产生的SARS-CoV-2刺突蛋白,通过S1/S2连接处的敲除弗林蛋白酶切割位点的两个点突变和S2铰链区中两个新引入的脯氨酸来稳定。此外,已用折叠结构域取代跨膜和细胞质区域以进行三聚化突变,从而使蛋白以可溶性蛋白的形式产生。COR200153是一种内部产生的SARS-CoV-2刺突蛋白,基于Wuhan-Hu-1 SARS-CoV-2株(MN908947),并通过S1/S2连接处的敲除弗林蛋白酶切割位点的两个点突变(R682A、R685G),以及S2铰链区中的两个连续脯氨酸(K986P、V987P)和A942P来稳定。此外,已用纤维蛋白折叠结构域取代跨膜和细胞质区域以进行三聚化。该蛋白通过分选酶A反应在C末端进行生物素化。
对于仓鼠,用蛋白包被96孔Perkin Elmer白色1/2面积板过夜。对于兔,在37℃下孵育2小时以包被板。孵育后,洗涤板,封闭1小时,随后用封闭缓冲液中3倍连续稀释的血清样品孵育1小时,一式两份。洗涤后,将板与在封闭缓冲液中的兔抗仓鼠IgG HRP(Invitrogen,货号A18895)或抗兔IgG-HRP(Jackson ImmunoResearch)孵育1小时,再次洗涤并使用ECL底物显色。使用BioTek Synergy Neo仪器(仓鼠样品)或Envision Multimode读板器(兔样品)进行发光读数。仓鼠抗体滴度报告为Log10终点,兔滴度报告为与参考标准相比的Log10相对效力。
统计分析
对于ELISA测量的S特异性结合抗体、病毒中和测定(VNA)测量的中和滴度、TCID50测量的病毒载量、组织病理学和IHC评分,对免疫方案之间不同剂量水平的统计学差异进行了双边评估。Ad26.S、Ad26.dTM.PP和Ad26.COV2.S组之间的跨剂量水平比较是使用来自ANOVA的t检验进行的并且疫苗和剂量作为测量因素,在LLOD或LLOQ不删失,或者使用来自Tobit ANOVA测量和z检验,最多删失50%,或者用Cochran-Mantel-Haenszel检验测量,删失至少50%。通过3倍Bonferroni校正对多重比较结果进行校正。使用上述方法或Mann-Whitney U检验,对每个剂量水平进行探索性比较以及对Ad26.S、Ad26.dTM.PP和Ad26.COV2.S组与用无关抗原Ad26.Empty和Ad26.Irr免疫的组的跨剂量水平进行探索性比较。由于这些分析的探索性,结果未针对多重比较进行校正。
使用SAS 9.4版(SAS Institute Inc.Cary,NC,US)和R3.6.1版(2019-07-05)进行统计分析。统计检验是在总体显著性水平α=0.05的情况下双边进行的。
相关性分析
将仓鼠归类为感染或免受SARS-CoV-2感染,将其定义为肺病毒载量分别高于或低于102TCID50/g。从Ad26.COV2.S不同方案和剂量水平汇总的结合和中和抗体数据,采用Firth校正建立逻辑回归模型,以保护结果作为因变量,并且将接种前wtVNA和Log10转换的ELISA数据作为自变量。
结果
建立另一种SARS-CoV-2叙利亚仓鼠攻击模型。
为了评估疫苗在仓鼠中的免疫原性、效力和VAERD,我们建立了基于SARS-CoV-2株的另一种叙利亚仓鼠攻击模型,其中刺突蛋白中主要是D614G取代。雄性动物(每个接种剂量水平n=12)接种SARS-CoV-2BetaCoV/Munich/BavPat1/2020(在S1片段中含有D614G取代),剂量水平为102、103.3、104.6和105。通过鼻内途径给药950%组织培养感染剂量(TCID50)。每天采集咽拭子并在接种后2、3、4和7天(dpi)进行尸检(每个时间点n=3),以监测咽拭子、肺和鼻组织中的病毒载量,并研究呼吸道病理。如图126A和B所示,肺和鼻组织病毒载量评估显示在2dpi时在所有接种动物中通过TCID50测量的有复制能力的病毒的高滴度,与接种物的大小无关。2dpi后观察到的肺和鼻病毒载量动力学在所有测试的接种量中具有可比性。用103.3、104.6和105.9TCID50接种的仓鼠在1dpi时咽拭子中的病毒载量最高(图126C),然后在接种后最多5天(dpi)病毒载量降至检测限以下。相比之下,接种102TCID50的最低SARS-CoV-2剂量第会导致传染性病毒载量从1dpi增加至2dpi,这表明肺复制,之后在接种后4天病毒载量下降到检测限以下。
用102TCID50攻击后的组织学分析显示,通过免疫组织化学,在严重炎症区域大量存在SARS核衣壳蛋白抗原(SARS-CoV-2 NP),其特征是上、下呼吸道上皮细胞多灶性中度至严重变性和坏死(数据未示出)。与较高的攻击剂量相比,102TCID50在整个呼吸道中诱导了相当程度和严重度的炎症和损伤,这由盲法半定量评分确定(图126D和E),在较低剂量水平下肺组织病理学评分略低。总之,这些结果表明,与较高的病毒剂量攻击相比,低剂量攻击接种物诱导了相当的病毒载量和疾病病理学。对于随后的实验,我们基于组织病理学结果选择了与轻度至中度疾病相关的102TCID50攻击剂量,从而可以在该模型中检测到更严重疾病的发生,并解决了VAERD的理论风险。选择攻击后4天的追踪时间作为同时评估肺组织病毒载量和组织病理学的最优时间点。
在叙利亚仓鼠中Ad26.COV2.A的免疫原性。
在上述新建立的攻击模型中评估了我们的Ad26.COV2.S候选疫苗的免疫原性和保护效力。为了比较,使用了两种基于Ad26的原型疫苗来表达膜结合的全长野生型刺突蛋白(Ad26.S)或具有C末端折叠子取代跨膜结构域的可溶性融合前稳定化的刺突蛋白(Ad26.dTM.PP)。用109或1010病毒颗粒(VP)的Ad26.COV2.S和两种原型疫苗对仓鼠进行免疫。对于每个剂量水平,在单次给药后的不同时间点以及在第4周给予第二次同源剂量后评估免疫原性。在最后一次疫苗剂量后4周攻击动物(图127A)。在第4周,与Ad26.S(滴度中值109VP 8.5、1010VP 9.7;8/12应答者,p<0.001)和Ad26.dTM.PP(滴度中值109VP 8.5、1010VP16.0;8/12应答者,p<0.001)相比,Ad26.COV2.S在不同的剂量水平上引发了最高的中和抗体滴度和响应动物的频率(滴度中值109VP 22.6,1010VP 38.6;12/12应答者)。(图127B)。无论使用何种疫苗,第二剂均增加了中和滴度(第8周;图127C)。Ad26.COV2.S疫苗在两剂后也具有最强的免疫原性,两剂109VP后的中和滴度中值为128,以及两剂1010VP后的中和滴度中值为219,而Ad26.S分别为32和55(p=0.003),A26.dTM.PP为16和61(p=0.002)。
通过ELISA测量的结合抗体显示了Ad26.COV2.S和两种原型疫苗之间的相同差异(图128A),这通过可比较的中和与结合抗体滴度比例反映出来(图128B和C)。然而,第4周的第二剂仅在第5周短暂地增加了结合抗体滴度中值(图128A)。抗体滴度随后下降,并且在第8周时与第4周剂量2之前观察到的水平相当或更低。
我们确认了Ad26.COV2.S和两种原型疫苗的免疫原性,以及使用8周间隔的2剂方案中第二剂对兔的益处,该间隔也在临床研究中进行评估。疫苗在5x109和5x1010VP的剂量水平上进行了测试,其中后者代表3期临床试验中使用的人剂量。所有测试的疫苗早在第1剂后2周就引发了结合和中和抗体滴度,与Ad26.S和Ad26.dTM.PP相比,Ad26.COV2.S再次诱导更高的抗体滴度(补充图129A和B)。当跨剂量与第二剂前水平相比时,在第8周第二个同源剂量显著提高了第10周(第二剂后2周)的结合和中和抗体滴度(p<0.001)。
在叙利亚仓鼠中Ad26.COV2.S防止SARS-CoV-2攻击的保护效力。
接下来,我们研究了Ad26.COV2.S和两种原型疫苗作为1剂或2剂方案给药的保护效力,然后在最后一次给药4周后鼻内接种102TCID50的SARS-CoV-2D614G病毒,即在叙利亚仓鼠中,1剂方案在第4周给药,2剂方案在第8周给药。在4dpi,牺牲动物,并分析肺、鼻甲和咽拭子的病毒载量和病理变化。为了增加统计分析的能力,并且由于我们没有观察到病毒学读数的明显剂量反应性,我们合并了病毒载量读数以与Ad26.Empty对照组进行比较。跨剂量水平比较三种不同疫苗(Ad26.COV2.S、Ad26.S、Ad26.dTM.PP)。
在单次疫苗接种和随后的攻击之后,所有疫苗组中的肺病毒载量中值与Ad26.Empty对照组相比显著降低(病毒载量中值为107.3TCID50/g)(图130A)。在用单剂Ad26.S免疫的12只动物中有11只的肺中检测到病毒(病毒载量中值109VP 105.6TCID50/g,1010VP 105TCID50/g),在用单剂A26.dTM.PP免疫的12只动物中有10只检测到病毒(病毒载量中值109VP 105.3TCID50/g,1010VP 105.7TCID50/g),以及未观察到疫苗剂量的明显影响。相比之下,用单剂Ad26.COV2.S免疫的12只动物中只有3只在肺中有可检测的病毒(合并剂量水平的滴度中值为101.2TCID50/g)。在再次疫苗接种第二剂Ad26.COV2.S的仓鼠中,12只动物中有9只在两种疫苗剂量水平下都检测不到病毒载量,这表明第二剂没有附加价值。突破性感染动物中的病毒载量也与一次疫苗接种后突破性感染动物中的病毒载量相似。第二剂109VP Ad26.S或Ad26.dTM.PP对第8周攻击后的肺感染率也几乎没有影响(6只动物中有5只显示每种疫苗可检测的病毒载量),但这些原型疫苗的第二剂1010VP与1剂方案相比,与较低的肺病毒载量中值相关(滴度中值分别为102TCID50/g和104TCID50/g),表明2剂方案的益处(图130B)。
为了确定疫苗对上呼吸道病毒载量的影响,在4dpi牺牲后测定鼻甲病毒载量,以及感染后每天测定咽拭子病毒载量,并分析为每只动物直到感染后4天的曲线下面积(AUC)。在接受单次疫苗剂量的动物中,与对照组相比,观察到Ad26.dTM.PP在攻击后在鼻甲的病毒载量有限但具有统计学显著性的降低,但Ad26.COV2.S和Ad26.S未观察到。在2疫苗剂量后,与Ad26.Empty组相比,所有三种疫苗在攻击后均诱导鼻甲中的病毒载量显著降低。相比之下,咽拭子病毒载量数据显示,在单次免疫和随后接种SARS-CoV-2后,没有一种疫苗能降低咽喉中的病毒载量(图130C和E)并且仅有用两剂Ad26.COV2.S免疫的动物与对照相比,其咽喉病毒载量显著降低(图130E和F)。
在肺和鼻组织中SARS-CoV-2NP的免疫组织化学(IHC)染色和组织病理学进一步支持了观察到的保护效力结果(图131和132)。肺和鼻IHC和组织病理学评分总体上与病毒载量数据一致,与Ad26.Emtpy组相比,免疫组的肺中值评分较低(图131),并且与独立于疫苗方案的Ad26.empty对照组相比,疫苗接种的动物鼻组织中的病毒感染量和后续病理学没有减少(图132)。
用次优剂量水平A26.COV2.S免疫的叙利亚仓鼠在接种SARS-CoV-2和突破性感染后未出现VAERD迹象。
基于观察到的免疫原性和效力,选择Ad26.COV2.S在剂量滴定研究中进行进一步评估,以解决在SARS-CoV-2攻击后允许突破性感染的次优免疫应答条件下VAERD的理论风险。用107、108、109或1010VP的单剂Ad26.COV.S对仓鼠组进行免疫(4组;n=8/组)。对照组接受1010VP的编码无关抗原(Ad26.Irr)的对照疫苗。在免疫后4周,我们观察到结合抗体和中和抗体的明显剂量应答,无论是应答动物的数量还是抗体滴度,在107VP的最低剂量水平下,8只动物中有3只没有检测到中和抗体滴度(图133和B)。对于109和1010VP剂量,结合抗体的中值水平(分别为103.7和103.9的中值终点滴度)和中和抗体应答的中值(分别为45和91的中值中和滴度)与先前研究所观察到的一致。在用102TCID50SARS-CoV-2攻击后,给药109和1010VP Ad26.COV2.S的仓鼠显示出与先前研究中的可比组相似的突破性肺感染频率(图130A),其中分别8只动物中有3只以及8只动物中有2只在剂量滴定研究中具有可检测的肺病毒载量。尽管在较低的Ad26.COV2.S剂量水平下发生突破性感染的动物数量增加(接受108VP的8只动物中有6只,接受107VP的8只动物中有8只),肺病毒载量滴度中值(108VP时的中值为104.8TCID50/g,107VP时的中值为105TCID50/g)和这些组中SARS-CoV-2NP的IHC染色(评分中值1)低于对照组(肺病毒载量中值106.8TCID50/g和IHC评分中值2)(图133C和D)。与肺病毒载量和IHC染色结果一致,与模拟免疫仓鼠相比,用108、109和1010VP免疫显著降低了下呼吸道的组织病理学(图134A)。用109和1010VP免疫分别导致8只仓鼠中有4只和8只仓鼠中有3只没有任何下呼吸道组织病理学迹象。值得注意的是,尽管在所有给药107VP的仓鼠和大多数用108VP免疫的仓鼠中都可检测到突破性肺感染,但与模拟免疫组相比,下呼吸道组织病理学评分中值较低。鼻组织(鼻炎)的炎症评分在疫苗接种组和对照组之间没有显示出显著差异(图134B)。总的来说,这些数据表明,与模拟疫苗相比,次优Ad26.COV2.S疫苗引发的低水平中和抗体的存在不会加重受攻击叙利亚仓鼠的肺疾病。
结合抗体和中和抗体与保护相关。
为了确定保护的推定相关因素,将对于Ad26.COV2.S来自不同方案和剂量水平的结合抗体和中和抗体滴度合并(N=56),并在受保护动物和未受保护动物之间进行比较(图135)。将防止SARS-CoV-2感染的保护定义为肺病毒载量低于102TCID/g,基于观察到仅少数动物的可检测的病毒载量低于此界限,这可能与每只动物可用样本量的变化相关(图129A和B,以及图133C)。用Ad26.COV2.S给药的受保护动物与未受保护动物相比,具有显著(2.3倍)更高的结合抗体滴度中值(p<0.001,两个样本t检验)(图135A。对中和抗体滴度中值的类似分析观察到相似的结果,与未受保护的动物相比,在肺病毒载量不可检测的Ad26.COV2.S免疫动物中中和抗体滴度中值也显著增加(4倍)(p<0.001,两个样本t检验)(图135B)。
为了更定量地了解免疫应答水平和保护结果之间的关系,我们使用Firth校正建立了逻辑回归模型(图135C)。将仓鼠归类为感染或防止SARS-CoV-2感染,其定义为肺病毒载量分别高于或低于102TCID50/g。结合抗体和中和抗体都显示出与保护概率的强的相关性。结合抗体滴度与保护概率显著相关(p=0.0004),终点滴度高于103.6似乎与防止肺感染的保护有关。用中和抗体滴度观察到相当的显著斜率(p=0.0002),其中超过32的滴度似乎与保护结果相关。
结论
在使用原型早期SARS-CoV-2分离株的先前研究(实施例21)中,其证明单剂Ad26.COV2.S引发的免疫应答可以降低病毒载量并保护仓鼠免受严重的临床疾病。然而,在持续的SARS-CoV-2大流行期间,出现了一种在刺突蛋白中具有D614G取代的病毒变体。这种突变与增加的病毒适应性、增强的传染性有关,现在已成为世界大部分地区的主要变体,尽管随着时间的推移,不断出现的新变体可能会取代它。在叙利亚仓鼠中,已确认D614G变体与上呼吸道中较高的传染性病毒滴度相关,但与肺无关。在这里,我们描述了使用含有流行的D614G取代的SARS-CoV-2株建立的轻度至中度疾病的另一种仓鼠攻击模型。我们使用这个模型来测试我们的COVID-19候选疫苗Ad26.COV2.S和两种编码不同SARS-CoV-2刺突变体的基于Ad26的原型疫苗所引发的免疫应答的保护效力。我们的研究表明,这种仓鼠攻击模型中的疾病进展显示出患有明显组织病理学肺疾病的人中度疾病过程的特征,这种疾病仅通过接种更高剂量的病毒而略微恶化。肺病毒载量峰值不受鼻内接种剂量水平的影响,表明大多数原发性易感肺细胞的感染导致在接种后第2天出现病毒血症峰值。与我们之前对NHP和仓鼠的研究一致,Ad26.COV2.S疫苗接种将病毒复制比在对照动物中观察到的水平减少了6 log10,其中许多接受更高疫苗剂量的动物显示检测不到病毒复制。Ad26.COV2.S在免疫原性和保护效力方面均显著优于两种原型疫苗。与本研究相结合的早期数据表明,Ad26.COV2.S引发的免疫应答对带有和不带有D614G刺突突变的SARS-CoV-2变体提供了足够的保护。
我们通过评估第二次同源剂量的Ad26.COV2.S和原型疫苗扩展了先前的研究,仅适度提高结合抗体水平,而中和抗体滴度显著提高。这与NHP中的观察结果形成鲜明对比,其中第二疫苗剂量显著提高了中和抗体和结合抗体的水平。可能的解释包括仓鼠和NHP之间剂量水平的有限可翻译性,第一剂引起的抗Ad26载体应答对物种之间第二剂同源剂量的免疫原性的不同影响,以及较短的免疫间隔可以减少第二剂的影响,如先前在NHP中观察到的。此外,仓鼠单次免疫诱导的高结合抗体水平可能代表饱和水平,而中和抗体在第二剂后仍可能增加,这可能反映了亲和力成熟的延长。在用相同的基于Ad26的疫苗免疫的兔中也观察到第二次同源疫苗剂量对体液SARS-CoV-2 S特异性免疫应答的优势,并确认了我们的临床数据(Sadoff,le Gars,NEJM in press)。2剂方案是否也是提高疫苗效力的首选方案还有待观察。
有趣的是,与用这些原型疫苗单次疫苗接种仓鼠所达到的低保护相比,第二剂Ad26.S或Ad26.dTM.PP在攻击后增加了防止肺病毒载量的保护。相比之下,第二剂Ad26.COV2.S并没有进一步提高单剂所建立的已经很高的效力水平。我们的相关分析支持这一点,其中保护概率随着抗体滴度的升高而增加,并且如果达到某个抗体滴度,保护概率仅适度增加。抗体和肺保护的这种负相关确认了我们之前在NHP在这种SARS-CoV-2-D614G叙利亚仓鼠模型中的研究,并且与疫苗、剂量水平或方案无关。一旦获得效力数据,需要进一步研究S特异性体液免疫与人保护的相关性,并可能为下一代COVID-19疫苗设计和其他正在开发的疫苗提供信息。
除了防止COVID-19的保护之外,理想情况下,疫苗引发的免疫还可以通过减少上呼吸道的病毒载量来防止无症状感染以及防止病毒传播。正如在其他SARS-CoV-2动物模型和其他呼吸道病毒感染中观察到的那样,上呼吸道似乎更难以防止病毒感染和通过亲代免疫而脱落。
在这个仓鼠攻击模型中,尽管使用了低病毒接种剂量,但我们观察到鼻甲中具有高复制能力的病毒水平,这与刺突D614G取代增加SARS-CoV-2在攻击的仓鼠中上呼吸道的感染性的观察结果一致。由于攻击接种物的剂量很低,因此4天后检测到的鼻甲中的病毒载量不太可能来自原始接种物。无论使用何种疫苗,鼻组织中病毒载量的减少都需要两剂疫苗。两剂Ad26.COV2.S是唯一也降低咽拭子病毒滴度的方案。与下呼吸道相比,上呼吸道病毒血症的减少是有限的,这与我们NHP研究中观察到的鼻病毒载量几乎完全减少形成对比。这可能是由于鼻上皮对病毒复制的易感性不同,或者仓鼠和灵长类动物呼吸道中存在的免疫细胞组成可能不同,以及Ad26对局部上呼吸道免疫的诱导不同。Ad26.COV2.S引发的免疫是否可以防止无症状感染和SARS-CoV-2传播仍有待在未来的动物研究和人数据中确定。
先前针对SARS-CoV和MERS的候选冠状病毒疫苗的研究表明,某些疫苗引发的免疫应答会加剧疾病。然而,在临床前动物模型或正在进行的临床研究中(包括基于mRNA和其他腺病毒载体或灭活疫苗的3期研究的效力报告)用SARS-CoV-2候选疫苗疫苗接种后,均未报告VAERD或抗体介导的疾病增强。然而,迄今为止,临床研究中的疫苗效力很高,VAERD的理论可能需要进一步研究,特别是在次优免疫诱导的环境中。因此,我们评估了当动物只接受了低剂量的Ad26.COV2.S,Ad26.COV2.S在疫苗诱导的抗体水平不足以阻止病毒在肺中复制的情况下对VAERD易感的可能性。重要的是,即使在免疫不足不能阻止肺病毒复制的情况下,与对照组相比,免疫动物的下呼吸道组织病理学评分也没有显示出VAERD的迹象。相反,与对照动物相比,大多数具有突破性感染的疫苗接种的动物仍然表现出组织病理学降低。这些结果意味着Ad26.COV2.S使对VAERD易感的理论风险很小。
我们的研究确认了Ad26.COV2.S候选疫苗具有高度免疫原性,可以保护仓鼠免受SARS-CoV-2病毒614G变体的攻击。Ad26.COV2.S的卓越效力和缺乏对VAERD易感的数据支持其在正在进行的3期临床试验中在单剂和两剂方案中继续评估。
实施例30:Ad26.COV2.S COVID-19疫苗在人中的免疫原性
作为Ad26.COV2.S的随机、双盲、安慰剂为对照的1期临床试验的一部分(另见实施例23、24和27),在单个临床站点招募25名参与者以深度评估这个疫苗的免疫原性。在本实施例中,报告了Ad26.COV2.S的单次和两次方案引发的体液和细胞免疫应答的动力学和量。
材料和方法
研究设计.
这项研究是作为多中心、随机、双盲、安慰剂为对照的1/2a期试验的一部分(队列1b)进行的,以评估在健康成人中5x1010或1x1011vp Ad26.COV2.S在以单次或相隔56天两次疫苗接种方案肌内(IM)给药时的安全性、反应原性和免疫原性。队列1a招募了18-55岁的成年人(n=375),并专注于收集主要安全性和免疫原性终点。队列1b(在此报告)也招募了18-55岁的成年人(n=25),但包括两次额外的访视和增加的血清、血浆和外周血单个核细胞采样,以便研究探索性终点。队列3(n=375)招募了年龄大于65岁的参与者,并测试了与队列1中评估的相同的方案。在马萨诸塞州波士顿的Beth Israel Deaconess Medical Center的单个站点招募队列1b的参与者。该方案得到了BIDMC机构审查委员会的批准,并在ClinicalTrials.gov(NCT04436276)上注册。
研究参与者.
合格的参与者为18-55岁(包含端点),通过鼻咽PCR和血清免疫球蛋白检测筛查SARS-CoV-2感染呈阴性。合格的参与者的体重指数(BMI)也为≤30.0kg/m2,并且在研究者的临床判断中是健康的,这由病史、体检、临床实验室评估和筛查时进行的生命体征所确认,并且没有患有与严重COVID-19风险增加相关的合并症。要求有生育能力的参与者使用高效避孕措施,并且不能怀孕或哺乳。如果参与者目前从事的职业暴露于SARS-CoV-2的风险很高,或者调查人员认为他们因任何其他原因感染COVID-19的风险增加,则排除他们在外。所有参与者都签署了书面知情同意书,并在研究程序开始前成功完成了理解评估。研究方案中描述了完整的资格标准。
随机化和掩蔽.
随机化由交互式网络应答系统(IWRS)使用随机排列的块完成。申办者、临床工作人员、研究者、参与者和实验室工作人员对分配不知情。当所有参与者完成第29天的访视或提前终止时,申办者和统计学家对中期分析进行分组揭盲。主要负责研究产品制备和配药的药剂师对分组分配不知情。为了保持盲法,他/她在注射器上放置了一个覆盖层。
研究产品
Ad26.COV2.S是一种重组的、无复制能力的Ad26载体,其编码全长和稳定化的SARS-CoV-2 S蛋白,SARS-CoV-2 S蛋白源自Wuhan株的第一个临床分离株(全基因组序列NC_045512),如本文所述。Ad26.COV2.S以1x1011vp/mL的浓度作为悬浮液提供在一次性小瓶中,可提取体积为0.5mL。制剂缓冲液以15mM柠檬酸盐、5%(w/w)羟丙基-β-环糊精、0.4%(w/w)乙醇、0.03%(w/w)聚山梨醇酯80、75mM NaCl、pH 6.2提供,以及安慰剂为0.9%氯化钠。出于盲法的目的,对所有参与者进行了相同的体积(1mL)给药。研究疫苗通过肌内注射给药入三角肌,优选非优势臂。安慰剂由1mL的0.9%NaCl溶液组成。
安全性评估.
从疫苗接种时间到疫苗接种后7天,通过日记收集对每次疫苗接种记录的征求AE。从疫苗接种时间到疫苗接种后28天,每次疫苗接种都报告了所有其他非征求的AE和特殊报告情况,无论是严重的还是非严重的(此处未报告第二次疫苗接种后的数据)。将记录发病日期在上述时间范围之外(先前研究疫苗接种后>28天)且在随后疫苗接种当天正在进行的非征求的AE。从第一次疫苗接种的那一刻起,直到参与者的最后一次研究相关程序的完成,其中可能包括联系以进行安全随访,所有参与者的所有导致终止研究/疫苗接种的SAE和AE(无论因果关系)都要报告。将追踪所有AE,直到消退或临床稳定。内部数据审查委员会(DRC)由不直接参与研究实施、数据管理或统计分析的成员组成,并将监测数据,以确保参与本研究的参与者的持续安全。
样本量和统计分析.
没有测试安全性或免疫原性的正式统计假设。为这项研究选择了参与者的数量,以提供初步的安全性和免疫原性评估。出于盲法和安全性目的,包括安慰剂接受者在内,并为免疫原性测定提供额外的对照样本。完整分析集(FAS)包括记录了至少一次疫苗接种的所有参与者。符合方案的免疫原性(PPI)人群包括所有可获得免疫原性数据的随机和疫苗接种的参与者,不包括预期会影响免疫原性结果的主要方案偏差的参与者。此外,将在错过疫苗接种后获得的样本或在筛选后发生自然感染的参与者(如果适用)排除在分析集之外。没有计划对安全性数据进行正式的统计测试。安全性数据按疫苗组进行描述性分析。没有测试关于免疫原性的正式假设。在所有可用时间点计算连续免疫学参数的描述性统计数据。对PPI群体进行免疫原性分析。使用GraphPad Prism8.4.2(GraphPad软件)进行免疫数据分析。使用双边Mann-Whitney检验进行组间数据比较。通过双边Spearman等级相关性检验评估相关性。认为P值小于0.05是显著的。
酶联免疫吸附测定(ELISA).
通过ELISA评估S-特异性结合抗体和RBD-特异性结合抗体,基本上如(Chandrashekar,et al.,Science 369,812-817(2020);Yu et al.,Science 369,806-811(2020))所述。简而言之,用1xDPBS中的1μg/mL SARS-CoV-2 S或2μg/mL SARS-CoV-2 RBD包被96孔板,并在4℃下孵育过夜。孵育后,用洗涤缓冲液(1X DPBS中的0.05%Tween20)洗涤板一次,并在室温下用350μL酪蛋白封闭液/孔封闭2-3小时。孵育后,弃去封闭溶液并将板吸干。将在酪蛋白封闭液中稀释的热灭活血清的连续稀释液添加至孔中,并将板在室温下孵育1小时,然后再洗涤3次,随后与1∶4000稀释的抗人IgG HRP(Invitrogen)在室温下避光孵育1小时。然后洗涤板3次,并且每孔加入100μL SeraCare KPL TMB SureBlue Start溶液;通过每孔添加100μL SeraCare KPL TMB Stop溶液来停止板的显色。使用VERSAMAX微孔板读数仪记录450nm处的吸光度,参考为650nm处。对于每个样品,在Graphpad Prism软件中计算ELISA终点滴度,使用四参数逻辑曲线拟合计算血清稀释度的倒数,产生0.2的校正吸光度值(450nm-650nm)。报告为Log10端点滴度。在Virology and Vaccine Research atBeth Israel Deaconess Medical Center对人血清进行ELISA测定并鉴定。
假病毒中和测定(psVNA).
通过与之前描述的方法相似的方法产生表达荧光素酶报告基因的SARS-CoV-2假病毒(Chandrashekar et al.,Science 369,812-817(2020);Yu et al.,Science 369,806-811(2020))。简而言之,用lipofactamine 2,000(ThermoFisher Scientific)将包装构建体psPAX2(AIDS Resource and Reagent Program)、荧光素酶报告质粒pLenti-CMVPuro-Luc(Addgene)和表达刺突蛋白的pcDNA3.1-SARS CoV-2 SΔCT共转染到HEK293T细胞中。转染后48小时收集含有假病毒的上清液;通过用0.45μm过滤器过滤来纯化假病毒。为了确定来自疫苗接种动物的抗血清的中和活性,将HEK293T-hACE2细胞以1.75×104个细胞/孔的密度接种在96孔组织培养板中过夜。制备热灭活血清样品的三倍连续稀释液,并与50μL假病毒混合。在加入HEK293T-hACE2细胞之前,将混合物在37℃下孵育1小时。48小时后,根据制造商的说明,在Steady-Glo Luciferase Assay (Promega)中裂解细胞。将SARS-CoV-2中和滴度定义为相对于病毒对照孔的平均值所观察到的RLU降低50%的样品稀释度。在Center for Virology and Vaccine Research at Beth Israel Deaconess MedicalCenter.对人血清进行psVNA测定并鉴定。
Ad26中和测定.
A549人肺癌细胞在感染前一天在96孔板中以每孔1x104个细胞的密度接种。E1/E3缺失、无复制能力的rAd26.Luc报告病毒与3倍连续稀释的热灭活人血清混合,然后以500的感染复数(MOI)应用于A549细胞。感染后24小时,使用Steady-Glo Luciferase ReagentSystem(Promega,Madison,WI)测量细胞中的荧光素酶活性。中和滴度定义为中和90%荧光素酶活性的最大血清稀释度。Ad26中和测定是在Center for Virology and VaccineResearch at Beth Israel Deaconess Medical Center针对人血清进行的。
电化学发光测定(ECLA).
设计和产生ECLA板(MesoScale Discovery SARS-CoV-2 IgG Panel2;K15369U-2),在每个孔中具有9个抗原点。包括的抗原是SARS-CoV-2 S、SARS-CoV-1 S、hCoV-229E S、hCoV-NL63 S、hCoV-OC43 S和hCoV-HKU1 S。用50uL Blocker A溶液封闭板,在室温下用数字微孔板振荡器以700rpm的速度振动至少30m。在封闭期间,将血清在稀释剂100中以1∶5,000稀释。然后用150μL MSD试剂盒洗涤缓冲液将板洗涤3次,吸干,然后将50μL稀释的样品一式两份加入板中并设置为在室温下以700rpm的速度振动至少2小时。将板再次洗涤3次,将50μL在稀释剂100中稀释至1x的SULFO-标签的抗人IgG检测抗体添加到每个孔中,并在室温下以700rpm的速度振荡孵育至少1小时。然后将板洗涤3次并将150μL MSD GOLD ReadBuffer B添加到每个孔中,然后立即在MESO QuickPlex SQ 120机器上读取板。每个样品的MSD滴度报告为相对光单位(RLU),其计算为每个样品的每个点的样品RLU减去空白RLU。每次测定的检测限定义为1000RLU。
IFN-γ酶联免疫斑点(ELISPOT)测定.
用来自MabTech的小鼠抗人IFN-γ单克隆抗体以1μg/孔的浓度包被ELISPOT板在4℃下过夜。用DPBS洗涤板,并用R10培养基(RPMI和10%热灭活FBS和1%的100x青霉素-链霉素、1M HEPES、100mM丙酮酸钠、200mM L-谷氨酰胺和0.1%55mM 2-巯基乙醇)在37℃下封闭2-4小时。刺突1和刺突2肽池含有15个氨基酸肽,这些肽有11个氨基酸重叠,它们涵盖蛋白序列并分别反映蛋白的N端和C端半部分。以2μg/孔的浓度制备刺突1和刺突2肽池,并添加200,000个细胞/孔到板中。将肽和细胞在37℃下孵育15-20小时。此孵育后的所有步骤均在室温下进行。将板用ELISPOT洗涤缓冲液洗涤,并与来自MabTech的生物素化小鼠抗人IFN-γ单克隆抗体(1μg/mL)一起孵育2-4小时。将板第二次洗涤并与来自Vector Laboratories的碱性磷酸酶缀合的抗生物素单克隆抗体(1.33μg/mL)一起孵育2-3小时。最后一次洗涤后,添加氯化硝基四氮唑蓝/5-溴-4-氯-3′-吲哚基磷酸对甲苯胺盐(NBT/BCIP显色液)底物溶液,保持7分钟。弃去显色液,将板用水洗涤并在暗处干燥24小时。在CellularTechnologies Limited Immunospot Analyzer上扫描和计数板。IFN-γELISPOT测定是在Center for Virology and Vaccine Research at Beth Israel Deaconess MedicalCenter针对人血清进行和鉴定的。
IL-4 ELISPOT测定.
用来自MabTech的IL4-I单克隆抗体以1.5μg/孔的浓度包被ELISPOT板,在4℃下过夜。用含有1x PBS洗涤板,并用R10培养基(RPMI和10%热灭活FBS和1%的100x青霉素-链霉素、1M HEPES、100mM丙酮酸钠、200mM L-谷氨酰胺和0.1%的55mM 2-巯基乙醇)封闭30分钟-4小时。以2μg/孔的浓度制备刺突蛋白1和刺突蛋白2肽池,并添加200,000个细胞/孔。将肽和细胞在37℃下孵育18-48小时。此孵育后的所有步骤均在室温下进行。将板用1xPBS洗涤并与来自MabTech的生物素化单克隆抗体IL4-II(1μg/孔)孵育2小时。将板第二次洗涤并与来自MabTech(1∶1000)的链霉亲和素-碱性磷酸酶一起孵育1小时。最后一次洗涤后添加氯化硝基四氮唑蓝/5-溴-4-氯-3′-吲哚基磷酸对甲苯胺盐(NBT/BCIP显色液)底物溶液,保持12-25分钟。弃去显色液(chromagen),将板用水洗涤并在暗处干燥24小时。在CellularTechnologies Limited Immunospot Analyzer上扫描和计数板。IL-4ELISPOT测定是在Center for Virology and Vaccine Research at Beth Israel Deaconess MedicalCenter针对人血清进行和鉴定的。
细胞内细胞因子染色(ICS)测定.
将106PBMC/孔重悬在100μL的补充有抗CD28,CD49d的单克隆抗体(1μg/mL)的R10培养基中。使用模拟(100μL的R10加0.5%DMSO;背景对照)、肽池(2μg/mL)或10pg/mL豆蔻酰佛波醇乙酯(phorbol myristate acetate)(PMA)和1μg/mL离子霉素(Sigma-Aldrich)(100μL;阳性对照)对每个样品进行评估并在37℃下孵育1小时。孵育后,将50μL R10中的0.25μLGolgiStop和0.25μL GolgiPlug添加到每个孔中,并在37℃下孵育8小时,然后在4℃下保持过夜。第二天,细胞用DPBS洗涤两次,用Aqua活/死染料(LifeTechnologies)染色10分钟,然后用针对CD279(克隆EH12.1,BB700,BD Pharmingen)、CD4(克隆L200,BV510,BD)、CD27(克隆M-T271,BUV563,BD Pharmingen)、CD8(克隆SK1,BUV805,BD)、CD45RA(克隆5H9,APC H7,BD)的预先指定滴度的单克隆抗体染色,持续30分钟。然后用2%FBS/DPBS缓冲液洗涤细胞两次,并用200μL BD CytoFix/CytoPerm Fixation/Permeabilization溶液孵育15分钟。用1X Perm洗涤缓冲液(将CytoFix/CytoPerm Fixation/Permeabilization试剂盒中的BDPerm/WashTM Buffer 10X用Milli Q水稀释并通过0.22μm过滤器)洗涤细胞两次,并用针对Ki67(克隆B56,FITC,BD)、IL21(克隆3A3-N2.1,PE,BD Pharmingen)、CD69(克隆TP1.55.3,ECD,Beckman Coulter)、IL10(克隆JES3-9D7,PE CY7,Biolegend)、IL13(克隆JES10-5A2,BV421,BD Pharmingen)、IL4(克隆MP4-25D2.BV605,Biolegend)、TNF-α(克隆Mab11,BV650,BD)、IL17(克隆N49-653,BV750,BD)、IFN-γ(克隆B27,BUV395,BD)、IL2(克隆MQ1-17H12,BUV737,BD Pharmingen)、IL6(克隆MQ2-13A5,APC,BD)、CD3(克隆SP34.2,Alexa 700,BD)的单克隆抗体进行细胞内染色30分钟。细胞用1X Perm洗涤缓冲液洗涤两次,并用250μL新鲜制备的1.5%甲醛固定。将固定的细胞转移至96孔圆底板中并通过BD FACSymphonyTM系统进行分析。ICS测定是在Center for Virology and Vaccine Research at Beth IsraelDeaconess Medical Center针对人血清进行的。
结果
研究设计
2020年7月27日至2020年8月7日期间,在马萨诸塞州波士顿的Beth IsraelDeaconess Medical Center对28名参与者进行了筛查。两名参与者因筛查时对SARS-CoV-2的阳性诊断测试不合格,一名参与者因体重指数>30kg/m2而不合格。将25名参与者随机分配并接受至少一剂疫苗或安慰剂。将参与者随机分配到五个实验组之一(N=5/组):(i)第1天和第57天(LD/LD)5x1010vp Ad26.COV2.S(低剂量;LD);(ii)第1天LD和第57天安慰剂作为单次注射疫苗(LD/PL);(iii)第1天和第57天(HD/HD)1x1011vp Ad26.COV2.S(高剂量;HD);(iv)第1天的HD和第57天的安慰剂作为单次注射疫苗(HD/PL);或(v)第1天和第57天安慰剂(PL/PL)。已分别报告这25名参与者的安全性数据(Sadoff et al.,New Engl.J.Med.,印刷中)。
结合和中和抗体应答的动力学和量
到免疫后第8天,通过ELISA,在65%(20名中的13名)疫苗接受者中观察到针对全长刺突(S)的结合抗体(图136A;P=0.0194,双边Mann-Whitney检验)以及在90%(20名中的18名)疫苗接受者中观察到针对受体结合结构域(RBD)的结合抗体(图136B;P=0.0033,双边Mann-Whitney检验)。在第8天,通过基于荧光素酶的假病毒中和抗体(psVNA)测定(5-7),在25%(20名中的5名)疫苗接受者中也观察到病毒中和抗体的趋势(图136C)。这些数据表明在人疫苗接种Ad26.COV2.S后快速诱导结合和中和抗体应答。
到第15天,在100%(20名中的20名)疫苗接受者中观察到S特异性和RBD特异性结合抗体(图137A),在85%(20名中的17名)疫苗接受者中观察到中和抗体(图137B)。在第29、57和71天,结合抗体和中和抗体继续增加。到第57天和第71天,100%(20名中的20名)疫苗接受者显示出中和抗体以及S特异性结合抗体和RBD特异性结合抗体。第71天,在LD/LD、LD/PL、HD/HD、HD/PL和PL/PL组中S特异性结合抗体的几何平均滴度(GMT)分别为2432、3249、5729、2852和20,RBD特异性结合抗体的GMT分别为1018、2023、3666、2372和21,以及中和抗体的GMT分别为242、375、449、387和13(图137A、B)。总之,这些数据证明了Ad26.COV2.S的单次注射方案和两次注射方案后结合抗体应答和中和抗体应答的快速动力学和强的量。在单次注射组中,通过第71天抗体应答的持续性和增加表明这些应答的持久性。
第57天的第二次免疫使在LD/LD和HD/HD组中的结合抗体滴度平均增加至2.56倍(范围1.58-3.04)和中和抗体滴度平均增加至4.62倍(范围3.56-5.68),在第71天,类似于之前的结果(参见实施例27)。然而,在第71天,两次注射疫苗组的总体GMT与单次注射疫苗组的GMT相当,这可能反映了每组参与者人数较少以及在加强之前LD/LD和HD/HD组中有两个低应答者的事实。与LD/LD组相比,在HD/HD组中观察到更高的结合抗体应答和中和抗体应答的趋势。在第71天,LD/LD和HD/HD组中Ad26载体特异性中和抗体的增加也表明了对加强的免疫应答(图127B)。
第29天的S特异性结合抗体滴度和RBD特异性结合抗体滴度彼此强烈相关(P<0.0001,R=0.9838,双边Spearman等级相关性检验;图138),并且都与第29天的中和抗体滴度相关(P<0.0001,R=0.8809和P<0.0001,R=0.8604,分别为双边Spearman等级相关性检验;图138)。这些数据表明,疫苗引发的抗体通常结合S和RBD并表现出强的病毒中和能力。
为了评估抗体对SARS-CoV-2应答的特异性,我们使用了电化学发光测定(ECLA;Meso Scale Discovery SARS-CoV-2 IgG Panel 2;K15369U-2)。我们评估了对来自SARS-CoV-2、SARS-CoV-1、CoV-229E、CoV-HKU1、CoV-NL63和CoV-OC43的全长S蛋白的反应性。通过ECLA(图139A)Ad26.COV2.S疫苗接种诱导SARS-CoV-2 S特异性抗体,与ELISA(图137A)相当。还观察到对来自SARS-CoV-1的S蛋白的交叉反应,其量大约是SARS-CoV-2的十分之一(图139A)。来自普通感冒冠状病毒CoV-229E、CoV-HKU1、CoV-NL63和CoV-OC43的S蛋白在基线时观察到高背景水平的抗体,而疫苗接种后这些应答没有明显增加(图139B)。这些数据表明,Ad26.CoV.S引发了低水平的针对SARS-CoV-1的交叉反应性抗体,但针对关系较远的普通感冒CoV的抗体很少甚至没有。
细胞免疫应答
我们在所有参与者的外周血单个核细胞(PBMC)中通过IFN-γ和IL-4ELISPOT测定评估S特异性细胞免疫应答,以及在有足够的PBMC的一部分参与者中进行多参数细胞内细胞因子染色(ICS)测定。到第15天,在65%(20名中的13名)疫苗接种者中观察到IFN-γELISPOT应答,到第71天在84%(19名中的16名)疫苗接种者中观察到IFN-γELISPOT应答,各组之间的细胞免疫应答没有明显差异(图140A)。没有观察到IL-4反应,表明强烈的TH1偏向的细胞免疫应答。多参数ICS测定确认了在除了一名研究的疫苗接受者之外的所有疫苗接受者中都诱导了中央记忆CD27+CD45RA-CD4+和CD8+T细胞应答(图140B)。在第29天,IFN-γELISPOT应答与S特异性结合抗体滴度、RBD特异性结合抗体滴度和中和抗体滴度相关(分别为P=0.0045,R=0.5492;P=0.0056,R=0.5378;和P=0.0031,R=0.5676,双边Spearman等级相关性检验;图141)。
讨论和结论
我们在此报告了在1期临床试验中接受Ad26.COV2.S或安慰剂的各种方案的25名个体的详细免疫分析。总体而言,免疫原性与亲本研究中的810个个体相当(参见实施例27和Sadoffet al.,New Engl.J.Med.,印刷中)。我们通过在疫苗接种后一周显示抗体的快速诱导来扩展这些先前的结果,到第8天在90%的疫苗接受者中检测到RBD特异性结合抗体。这些数据表明Ad26.COV2.S诱导快速和复杂的抗体应答,具有可能的持久性以及单次免疫后的细胞免疫应答。
当前大多数COVID-19疫苗计划的一个中心假设是,针对SARS-CoV-2 S蛋白的抗体具有保护作用。因此,大多数项目都专注于诱导病毒特异性中和抗体的策略,但其他抗体功能也可能有助于保护。结合、中和和功能性非中和抗体都与恒河猴中DNA和Ad26疫苗接种后的保护效力相关(Mercado et al.,Nature 586,583-588(2020):Yu et al.,Science 369,806-811(2020))。我们最近表明,从恢复期猕猴中过继转移纯化的IgG以剂量依赖性方式保护幼稚(naive)猕猴免受SARS-CoV-2攻击(Chandrashekar et al.,Science 369,812-817(2020);McMahan et al.,Correlates of protection against SARS-CoV-2in rhesusmacaques.Nature(2020))。400的S特异性结合抗体滴度、100的RBD特异性结合抗体滴度和50的中和抗体滴度代表了这个临床前模型中的保护阈值,但尚未确定保护的相关因素以防止COVID-19感染或人疾病。然而,值得注意的是,Ad26.COV2.S(图137)在人中诱导的抗体滴度大大超过了这些先前定义的阈值,这些先前定义的阈值使用基本相同的测定。T细胞应答也可能有助于保护,特别是在抗体应答减弱或临界的情况下。
快速、稳健地和持久地诱导多种功能性抗体应答可能对COVID-19疫苗至关重要。Ad26.COV2.S在单次注射免疫后具有高度免疫原性,并且不需要零度以下的冷冻冷链,这两者都可以为大规模疫苗接种活动提供实质性的统筹和实际优势。
到初次免疫后第8天,结合抗体迅速出现在大多数疫苗接种者中,并且在一部分个体中观察到中和抗体。单次免疫后第57天,100%的疫苗接受者检测到结合抗体和中和抗体。这些数据表明,Ad26.COV2.S可诱导具有快速动力学的抗体应答以及细胞免疫应答。目前正在进行两项3期临床试验,以确定Ad26.COV2.S疫苗在人中的效力。
实施例31:在18岁及以上成人中评估Ad26.COV2.S预防SARS-CoV-2介导的COVID-19的效力和安全性的随机、双盲、安慰剂为对照的3期研究(ENSEMBLE)
本研究是一项正在进行的、随机、双盲、安慰剂为对照的3期关键效力、安全性和免疫原性研究,在北美、中美洲、南美洲和南非进行,在有或没有与进展为严重COVID-19的风险增加相关的合并症的18岁至<60岁(阶段1)和≥60岁(阶段2)的成年人中进行。单剂Ad26.COV2.S的效力、安全性和免疫原性正在参与者中进行评估,这些参与者居住在获得SARS CoV 2感染的高风险地区或前往获得SARS CoV 2感染的高风险地区。通过随机分层实现了研究群体中年龄和存在/不存在合并症的良好代表性。还努力确保在种族、民族和性别方面有良好的代表性。共同主要目标是证明单剂5×1010vp Ad26.COV2.S与安慰剂相比,在SARS-CoV-2血清阴性的成年人中,在预防疫苗接种后至少14天和至少28天发病的分子确诊、中度至严重/危重COVID-19的效力。其他关键终点包括预防严重/危重COVID-19的效力(包括死亡和医疗干预),预防所有有症状的COVID-19的效力(包括通过严重度校正加权BOD分析后测量的轻度、中度和严重/危重疾病),以及对60岁以上成人的效力。
本实施例描述了在这个3期研究中评估的Ad26.COV2.S的效力。在这项大型、多国关键效力研究中,显示该疫苗可有效预防种族和地理多样化的成年人群中的中度至严重/危重COVID-19。
最初计划的60,000名参与者的样本量是在研究开始前的操作截止日期计算的,其基于流行病学情况的不确定性以及在假设60%VE的疫苗的研究的8个月内提供高概率(约90%)到达信号()的时间的能力。一旦发现COVID-19的发病率明显高于研究开始时最初假设的发病率,样本量就会减少到大约40,000名参与者。
如临床研究方案中所述,在整个研究过程对SARS-CoV-2感染和有症状的COVID-19进行了鉴定和分子确诊。中心实验室(华盛顿大学[UW病毒学实验室])对SARS-CoV-2感染的分子确诊(使用Abbott Real Time SARS-CoV-2 RT-PCR测定[Abbott2020])用于分析案例定义。
术语严重/危重COVID-19病例是指由临床严重度判定委员会进行临床评估并判定为重度/危重的病例。严重度的分类是基于观察期间的最高严重度。除非另有说明,包括严重/危重病例在内的所有分析均基于判定。
在整个研究过程中监测COVID-19相关住院和COVID-19相关并发症(例如但不限于高炎症综合征、肺炎、神经或血管并发症、重症肺炎、严重神经或血管事件、急性呼吸窘迫综合征、肾脏并发症、脓毒症、感染性休克、死亡[WHO 2020d])的发生。
COV3001研究的主要目的是证明Ad26.COV2.S在SARS-CoV-2血清阴性成人中,与安慰剂相比,在预防分子确诊的(定义为中央实验室使用基于PCR或其他分子诊断测试的SARS-CoV-2病毒RNA结果呈阳性)、中度至严重/危重COVID-19中的效力。
完整分析集(FAS)包含所有接受至少一次疫苗接种的随机参与者。
符合方案(PP)集包含FAS的子集,不包括(i)具有影响效力的主要方案偏差的受试者;(ii)基线血清阳性(第1天);(iii)基线PCR+,如果有的话*(第1天)(*所有病例均针对第1天PCR进行分析)。
除非另有说明,否则所有效力分析均在PP(符合方案)集上进行。PP集包括接受研究疫苗以及在接种疫苗时呈血清阴性且没有其他认为可能影响疫苗效力的主要方案偏差的参与者。
主要分析补充了针对年龄组(18至<60岁,≥60岁)和合并症(是/否)存在的亚组分析,采用描述性总结,包括(未校正)95%CI来使用相同方法描述每个亚群中的VE。
在主要分析时,测试了以下次要终点:
·针对所有在疫苗接种后至少28天和至少14天发病的有症状的COVID-19的VE,通过(疾病负担(BOD)终点进行评估。BOD评估针对预防有症状的COVID-19的严重度校正的VE。为严重感染分配更高的权重(严重病例得分为2,非严重病例得分为1),因此,BOD终点旨在提供更高的统计能力,以区分安慰剂疫苗对严重感染的增强的保护。
·针对在疫苗接种后至少28天和/或至少14天发作的严重/危重COVID-19的VE。
还对以下终点进行了描述性分析(包括95%CI):
·针对在疫苗接种后至少28天和至少14天发病(分别从第29天和第15天发病的23例病例必须可用)的需要医疗干预的COVID-19的VE。需要医疗干预的COVID-19(例如住院、ICU入院、机械通气和ECMO的复合终点,与例如氧合减少、X射线或CT发现的客观措施相关)是基于医疗资源利用数据确定的,这些数据是通过就医COVID-19表格收集。
·针对从第29天开始发病的所有SARS-CoV-2感染的VE(>15,000名参与者必须获得第71天的样本)。
·针对从第29天发病的无症状/或未检测到的病例(所有参与者必须进行至少6个月的随访)的VE。
描述性描述的其他次要终点包括针对分子确诊的轻度COVID-19的VE和针对符合美国FDA统一的COVID-19病例定义的COVID-19的VE,通过年龄、是否存在合并症、性别、地区、国家、种族和HIV状态以及感染后的病毒载量进行亚组分析。为了评估VE的潜在时间效应,使用Kaplan-Meier方法随时间累积发生率图。
由于研究进行期间COVID-19的高发病率,在初步分析时,并非所有病例都由中心实验室(UoW)确认。有大量病例的症状符合COVID-19病例定义,并且至少有1例来自其他来源(本地或中央)的PCR结果呈阳性。因此,主要分析补充了敏感性分析,其包括至少有一个阳性PCR结果的病例,无论该测试的来源如何。尽管分析是非推断性的,但使用与各自推断性分析相同的α水平。
参与者信息:
将总共43,783名参与者随机分组(Ad26.COV2.S 5x1010组21,895人,安慰剂组21,888人),他们都接受了研究疫苗接种,并且包括39,321人(Ad26.COV2.S 5x1010组19,630人,安慰剂组19,691人)在PP集中。本节中提供的数据是PP集的数据。
在PP集的参与者中,没有参与者已经完成了研究,130名(0.3%;(Ad26.COV2.S5x1010组中有41名[0.2%]参与者以及安慰剂组有89名[0.5%]参与者)提前终止研究,主要是由于撤回同意(分别为30[0.2%]名参与者和62名[0.3%]名参与者)。在初步分析时,疫苗接种后的随访中值为58天并且PP集中的21,491名参与者具有至少2个月的随访。
有资格获得授权/许可的COVID-19疫苗的研究参与者可以根据方案要求揭盲。截至截止日期2021年1月22日,在本研究过程中,揭盲2,257名(5.2%)参与者以接受授权/许可的COVID-19疫苗。
在Ad26.COV2.S组和安慰剂组之间没有观察到人口统计学和基线特征的相关差异。随机化按站点、年龄和合并症的存在/不存在进行分层,并努力确保在种族、民族和性别方面有良好的代表性:
研究群体人口统计的主要发现:
年龄分布>=60(33,5%),>=65(19,6%),>=75(3,5%),>=80(1%)
性别:45%女性,55%男性
地区:44%美国,41%中美洲和南美洲,15%南非
种族:19.5%是黑人或非裔美国人(美国13%)
合并症(总体41%、肥胖症(28.5%)、2型糖尿病(7.3%)、高血压(10.3%)、HIV(2.8%)以及其他免疫功能低下的参与者也在研究中
在PP集中,大部分参与者来自北美(46.7%)和中美洲和南美洲(40.6%);其他参与者来自南非(12.7%)。总体而言,62.1%的参与者是白人,17.2%是黑人或非裔美国人;8.3%是美洲印第安人/阿拉斯加原住民,3.5%是亚洲人;45.1%的参与者是西班牙裔或拉丁裔。
在PP集中,44.5%的参与者是女性,55.5%是男性。年龄中值为53.0岁(范围:18-100)。至少约30%≥60岁的参与者和约20%≥18至<40岁的参与者满足预期的招募目标,其中34.6%的参与者≥60岁,22.2%的参与者≥18至<40岁。总共有20.4%和3.7%的参与者分别≥65岁和≥75岁。体重指数(BMI)中值为26.90kg/m2(范围:2.3;82.6kg/m2)。
在完整的分析集中(FAS,即有记录的研究疫苗给药的所有随机参与者,无论是否发生方案偏差和招募时的血清状况),9.6%的参与者在基线时为SARS-CoV-2血清阳性。排除这些参与者在PP集之外。
总体而言,在PP集中,39.9%的参与者在基线时患有一种或多种与进展为严重COVID-19的风险增加相关的合并症。最常见的合并症是肥胖(27.5%)、高血压(10.3%)、2型糖尿病(7.2%)、严重心脏病(2.4%)和哮喘(1.3%)。<1%的参与者存在其他合并症。2.5%的参与者在基线时报告了HIV感染。
效力分析
基于疫苗接种后至少14天和至少28天发生的COVID-19病例数的效力分析总结分别见表1和表2。
在以下部分中,除非另有明确说明,否则将描述血清阴性参与者中确认的COVID-19病例的疫苗效力。如果一个终点观察到<6例,则不显示VE。
在评估统计假设时(主要终点、针对任何有症状的感染[BOD]终点的VE和针对严重/危重COVID-19的VE和针对需要医疗干预的COVID-19的VE),文本、表格和图显示了假阳性对照的95%置信区间(CI)(进一步称为“校正后的CI”),否则使用95%CI对数据进行描述性总结。
对于分子确诊的COVID-19的参与者,随访时间定义为从疫苗接种到病例发病时间之间的时间。对于所有未确诊COVID-19的受试者,随访时间定义为从疫苗接种到最后一次可用测量(对于正处于研究中的受试者)或研究中止/完成的时间。请注意,在所有分析的随访计算中,除了包括确诊病例和非确诊病例的敏感性分析之外,非确诊病例均计为“无病例”;在这些分析中,对未确诊病例的随访定义为从疫苗接种到任何来源的PCR检测呈阳性的时间之间的时间。
值得注意的是,该试验是在8个国家和三个地区的COVID-19大流行高峰期进行的,在此期间,疾病在全球范围内的传播速度加快,导致人们对病毒的暴露增加,而且正处于新出现冠状病毒株的时期,包括出现一些更具传染性的冠状病毒株(参见图145)。
表1:

表2:

结果:
在SARS-CoV-2的发病率非常高且新的病毒谱系出现时,该研究在多个地区(北美洲和南美洲、非洲)产生了高质量的数据。
该研究满足它的共同主要目标:
单剂Ad26.COV2.S疫苗可有效预防中度至严重/危重COVID-19,疫苗接种后第14天和第28天后VE为67%和66%。
在美国,单剂Ad26.COV2.S疫苗可有效预防中度至严重/危重COVID-19,疫苗接种后第14天和第28天后VE为77%和72%。
自第14天起效力开始明显,至第56天效力增加,尤其是针对严重疾病。这一发现与现有的免疫原性结果一致,从第14天开始检测到中和抗体滴度和结合抗体滴度,直到第56天持续增加,直到第84天没有减弱的迹象。
疫苗组的中度COVID-19病例报告的症状少于安慰剂组的中度COVID-19病例。
当仅使用中央实验室确认的PCR+病例或当使用所有PCR+病例而不考虑中央实验室确认时的结果是一致的。
疫苗效力随严重度而增加:
观察到对严重/危重COVID-19的高疫苗效力(第28天后总体为85%),早在第7天就观察到了效力(自第14天为77%)。
这种高疫苗效力在包括南非在内的国家和地区是一致的,几乎所有病例都是由于SARS-CoV-2的新变体(B.1.351)引起的(基于部分可用的测序数据:45/48有B.1.351)。
对于需要医疗干预(定义为住院、ICU、ECMO或机械通气)的COVID-19发作的参与者,第14天后疫苗接种组报告了2例,安慰剂组报告了8例以及第28天后疫苗接种组没有病例,安慰剂组报告了5例。对可能的低报(under-reporting)的调查正在进行中。
所有COVID-19确诊死亡(5)均发生在安慰剂组。
共同主要终点是在血清阴性参与者中出现确诊的中度至严重/危重COVID-19病例,与安慰剂相比,在疫苗接种Ad26.COV2.S后至少28天和至少14天发病。成功的主要终点结论要求VE大于30%,点估计值≥50%,并且Ad26.COV2.S和安慰剂组之间有良好的分割,对于严重病例,在安慰剂组中至少有5例病例。在主要分析时,在PP集中,有259例中度至严重/危重COVID-19病例发生在至少28天,464例发生在疫苗接种后至少14天。两个共同主要终点均满足成功主要分析的预先指定的标准(见上文),因此,Ad26.COV2.S对中度和严重/危重COVID-19的效力已在疫苗接种后14天后确定。
疫苗接种后至少28天发生的针对中度至严重危重COVID-19的VE(校正的95%CI)分别为66.1%(55.01;74.80)和66.9%(59.03;73.40),在疫苗接种后至少14天进行评估。在Ad26.COV2.S组与安慰剂组中,疫苗接种后至少28天发生的病例的病例分割分别为66与193。在疫苗接种后至少14天发生的病例,分别有116和348例。疫苗接种后至少28天和至少14天发生的严重/危重病例的病例分割分别为5与34和14与60。
补充分析按年龄类别(≥18至<60岁,≥60岁)评估了VE对中度至严重/危重COVID-19的影响。≥60岁参与者的VE(CI)与<60岁的成年参与者相比至少同样高:在≥60岁的参与者中分别为66.2%(36.74;82.99)和76.3%(61.58;86.04)在疫苗接种后至少28天和至少14天发病,以及<60岁的参与者分别为66.1%(53.30;75.77)和63.7%(53.87;71.58)。下面讨论了通过基线SARS-CoV-2血清状况以及人口统计学和基线特征进行的其他亚组分析。
图142和图143显示了对确诊的中度至严重/危重COVID-19的早期保护,其中展现了首次出现确诊的中度至严重/危重COVID-19并在疫苗接种后至少1天发病的累积发病率。Ad26.COV2.S和安慰剂组在疫苗接种后14天左右的累积分布曲线开始分离,并且VE继续增加直到第56天,直到第85天没有减弱的迹象,这与在1期试验的发现一致,1期试验中从第15天开始观察到中和抗体应答,直到第57天,GMT和应答者百分比都在增加。
对严重疾病的效力随着时间的推移而增加,疫苗接种的参与者在第49天后没有报告病例。
在疫苗接种后至少28天和至少14天分别发生的259和464例中度至严重/危重COVID-19病例中,39和74例为严重/危重,220和390例为中度。此外,截至分析截止日期,报告了2例和4例轻度COVID-19病例,分别在接种疫苗后至少28天和14天发病。
针对严重/危重COVID-19的疫苗效力
针对VE<0%的无效假设对严重/危重COVID-19疫苗效力进行了统计测试。基于疫苗接种后至少28天发病的病例以及至少14天发病的病例(校正的95%CI大于0),满足预先指定的成功标准,因此,对严重/危重COVID-19的效力从疫苗接种后14天开始。疫苗接种后至少28天和14天发生的针对严重/危重COVID-19的VE(CI)分别为85.4%(54.15;96.90)和76.7%(54.56;89.09)。
图144显示了在疫苗接种后至少1天发病,首次出现严重/危重COVID-19的累积发病率,并证明了防止严重/危重COVID-19的早期保护。在疫苗接种后7天左右,Ad26.COV2.S和安慰剂组之间的累积分布曲线开始分离,并且VE增加至第56天。
在ENSEMBLE中需要医疗干预(住院、ICU入院、机械通气、ECMO)的或在疫苗接种后至少28天导致的原发性死亡的所有确诊的COVID-19病例都发生在接受安慰剂的参与者中。
针对中度和轻度COVID-19的疫苗效力
疫苗接种后至少28天和至少14天发生的针对中度COVID19的VE(CI)分别为62.0%(48.68;72.21)和64.8%(55.75;72.21)。对于轻度COVID-19病例(<6例),Ad26.COV2.S和安慰剂组之间存在良好的分割,在疫苗接种后至少28天分别发生0例和2例,以及在疫苗接种后14天分别发生1和3例。
表3:针对疫苗接种后至少14天发病的中度COVID-19的疫苗效力总结;根据中度症状数量的符合方案集:

表3:针对疫苗接种后至少14天发病的中度COVID-19的疫苗效力总结;根据中度症状数量的符合方案集:

表4:针对疫苗接种后至少28天发病的中度COVID-19的疫苗效力总结;根据中度症状数量的符合方案集(研究VAC31518COV3001):
疫苗组的中度COVID-19病例报告的症状少于安慰剂组的中度COVID-19病例,这表明在发生突破性感染(在“中度疾病”类别中)的情况下,疫苗接种的人的症状明显少于接受安慰剂的人。
针对任何有症状的COVID-19的疫苗效力
考虑到所有有症状的COVID-19病例,无论严重度如何,报告了总共有285和468例有症状的COVID-19病例,在疫苗接种后至少28天和疫苗接种后至少14天发病。通过BOD终点的平均值评估针对所有有症状COVID-19的VE,该终点评估针对预防有症状COVID-19的严重度校正的VE。为严重感染分配更高的权重(严重病例得分为2分,非严重病例得分为1分),因此,BOD终点旨在提供更高的统计能力来区分安慰剂疫苗对严重感染的增强的保护。根据BOD终点测量,针对疫苗接种后至少28天和至少14天发生的所有有症状的COVID-19的VE分别为69.0%(56.68、77.64)和68.1%(60.26、74.32)。由于两个终点的校正的95%CI的下限>0,因此满足了预先指定的成功标准,因此,Ad26.COV2.S对所有有症状的COVID-19的效力已在疫苗接种后14天确定。
由于针对严重疾病的VE高于针对中度和轻度疾病的VE,因此针对所有有症状的COVID-19的VE(所有严重度权重均等)与严重度校正的VE相比略低:基于疫苗接种后至少28天和至少14天发生的病例分别为66.5%(55.50;75.05)和66.9%(59.07;73.37)。
根据美国FDA统一定义,针对COVID-19的VE与针对所有有症状的COVID-19观察到的VE一致:疫苗接种后至少28天和至少14天分别为66.7%(55.63;75.23)和67.2%(59.32;73.67)。
所有SARS-CoV-2感染,包括无症状或未检测到的病例
使用依赖于SARS-CoV-2N蛋白的ELISA和/或SARS-CoV-2免疫球蛋白测定评估Ad26.COV2.S对确诊的无症状或未检测到的SARS-CoV-2感染发生的影响。在第1天(疫苗接种前)和疫苗接种后第71天、6个月和1年之间评估血清转化。此外,在第1天(疫苗接种前)和疫苗接种后第29天之间评估血清转化。在中期分析时,可获得第71天和第29天的有限数据。针对疫苗接种后至少28天发生的无症状COVID-19感染的疫苗效力(CI)为74.0%(27.89;92.40)(参见表5)。
表5:

Ad26.COV2.S在预防疫苗接种后至少28天发病的所有SARS-CoV-2感染(血清学和/或分子确诊的有症状和无症状感染的组合)中的疫苗效力(CI)为67.2%(56.86;75.26)(参见上表2)。
亚组分析
通过包括年龄(18-59岁和≥60岁)、存在/不存在合并症(包括哮喘、癌症、脑血管疾病、囊性纤维化、慢性肾病、COPD、严重心脏病、高血压、肝病、神经系统疾病、肥胖、肺纤维化、镰状细胞性贫血、1型或2型糖尿病或地中海贫血),年龄和合并症,性别(男性/女性)、国家(阿根廷、巴西、智利、哥伦比亚、墨西哥、秘鲁、美国或南非)、种族(美洲印第安人/阿拉斯加、亚洲人、黑人或非裔美国人、夏威夷原住民/其他、白人,多种族),民族(西班牙裔/拉丁裔,非西班牙裔/拉丁裔),地区(拉丁美洲,北美洲和南部非洲)和艾滋病毒状况的人口统计和基线特征针对在疫苗接种后至少28天和至少14天发病的分子确诊的中度至严重/危重COVID-19的效力分析正在进行中。初步结果如下所示:
针对中度/严重疾病和亚组中严重疾病的疫苗效力(VE):
疫苗在各个年龄段都有效,估计疫苗对中度/严重疾病的效力:第14天后至少64%,以及第28天后至少66%。
疫苗在有和没有合并症的参与者中是有效的,在有合并症的组中,对中度/严重疾病的估计的疫苗效力在第14天后至少为63%,在第28天后至少为49%。无论是否经中心实验室确认,包括所有PCR+的补充分析显示,第14天后至少为64%,第28天后至少为59%。需要更多的随访数据来了解有和没有合并症亚组之间观察到的潜在差异。
黑人/非洲裔美国人与白人以及西班牙裔与非西班牙裔美国人的疫苗效力是一致的。
自第14天起,针对严重疾病的疫苗效力很明显,以及到第28天,在有足够可用数据的国家(南非、哥伦比亚)中,疫苗效力约为85%或更多。
疫苗接种后28天,在美国对中度和严重COVID-19感染的保护水平为72%,拉丁美洲为66%,南非为57%。
针对不同病毒株的疫苗效力
在研究期间,已在研究中包括的国家发现了不同的突变变体。例如,在南非,导致COVID-19感染的绝大多数毒株是变体501Y.V2(RSA变体),特别是在毒株(45/48测序)中,94%是B.1.351(501Y.V2)SARS-CoV-2谱系。
针对B1.351(携带501Y突变)变体和携带E484K突变的变体,针对在疫苗接种后至少28天发病的首次出现的分子确诊的中度至严重/危重COVID-19的疫苗效力为67.0(-8.87-92.24)。
单次Ad26.COV2.S 5x1010vp剂量直至第29天后的体液免疫原性
鉴于SARS-CoV-2病毒形式的增多,在病毒S蛋白中已知为中和抗体靶标的区域出现突变,因此在疫苗接种后第1天和第29天通过S-ELISA评估了Ad26.COV2.S对疫苗插入物的免疫原性,使用来自两个随机选择的感染率高的巴西站点以及其他随机巴西、南非和美国站点的样本。
总体而言,在巴西、南非和美国参与者之间,Ad26.COV2.S诱导的结合抗体水平没有观察到差异。巴西参与者(第29天的GMC)的结合抗体浓度范围从在巴西站点BR10003的317(95%CI 227;444)到巴西站点BR10004的473(95%CI 268;836)。在第29天,南非和美国参与者的结合抗体浓度与GMC相似,分别为388(95%CI 297;506)和412(95%CI 306;554)。
来自所有3个国家的站点的应答率相似,活性疫苗组的应答率>93%,代表几何平均数从基线增加至6倍至9.3倍以上。
巴西站点的GMR(与美国参与者相比)为0.77(95%CI0.49;1.20)至1.15(95%CI0.64;2.07),南非站点为0.94(95%CI0.63;1.42)。
与COV1001数据相比,各国观察到了类似的GMC。
总之,在初步分析中表明:
单剂Ad26.COV2.S可在18岁以上成人(包括≥60岁的成人)中防止COVID 19。基于这项关键研究的主要效力分析,包括接受Ad26.COV2.S的19;630名参与者和接受安慰剂的19;691名参与者,当考虑疫苗接种后至少14天的病例时,在疫苗接种时血清阴性的参与者中针对分子确诊的中度至严重/危重COVID-19的共同主要终点的疫苗效力(校正的95%CI)为66.9%(59.03;73.40),以及当考虑疫苗接种后至少28天的病例时,疫苗效力为66.1%(55.01;74.80)。
针对在单次Ad26.COV2.S剂量后至少28天和14天发生的严重/危重COVID-19的VE分别为85.4%(54.15;96.90)和76.7%(54.56;89.09),在第28天后的18-59岁人群中为90%效力。这一发现在包括南非的国家和地区(北美和南美、南非)之间是一致的,在南非几乎所有病例都感染了新的SARS-CoV-2变体(501Y.V2)。
在第14天估计效力开始,效力增加到第56天,尤其是针对严重疾病。这一发现与从第14天起检测到的中和抗体滴度和结合抗体滴度的免疫学发现一致,直到第56天持续增加,直到第85天没有减弱的迹象。
针对所有有症状的疾病的疫苗效力与主要终点一致。针对在单次Ad26.COV2.S剂量后至少28天和至少14天发生的所有有症状的COVID-19的疫苗效力,通过严重度校正加权分析(BOD)测量,包括轻度、中度和严重/危重VE,分别为69.0%和68.1%。对于在疫苗接种后至少28天和至少14天的发生,针对严重/危重COVID-19(85.4%和76.7%)的效力高于针对中度COVID-19(62.0%和64.8%)的效力。非加权分析与这些结果一致。
在COVID-19疫苗接种的主要目标人群(例如老年人和患有合并症的(老年)成人)中的疫苗效力与在总体人群中观察到的效力一致。一般来说,描述性亚组分析显示,在疫苗接种时,Ad26.COV2.S在不同年龄组、性别、种族/民族、地理、合并症、年龄和合并症组合的疫苗效力没有差异。
初步数据表明疫苗对无症状感染有作用,通过无症状PCR+结果和血清转化进行评估。针对疫苗接种后至少28天发生的无症状COVID-19感染的疫苗效力(CI)为74.0%(27.89;92.40)。
此外,Ad26.COV2.S的安全性和反应原性谱与其他基于Ad26的疫苗一致。尽管1期和2期Ad26.COV2.S研究中的发热频率高于之前观察到的频率,尤其是对于年轻人来说,但这一发现并未在更大的3期Ad26.COV2.S研究中重现。Ad26.COV2.S的反应原性明显是短暂的,并在1-2天内消退。疫苗接种后可根据需要使用退热药缓解症状。
序列
>COR200007_SEQ ID NO:205

>COR200008_SEQ ID NO:206


>COR200009_SEQ ID NO:207

>COR200010_SEQ ID NO:208


>COR200011_SEQ ID NO:209

>COR200018_SEQ ID NO:210


*粗体和下划线:理论信号肽序列
>COR200007_SEQ ID NO:211


>COR200008_SEQ ID NO:212



>COR200009_SEQ ID NO:213


>COR200010_SEQ ID NO:214


>COR200011_SEQ ID NO:215



>COR200018_SEQ ID NO:216


>pShuttle_E1.CMVdel134TO;SEQ ID NO:217


注:粗体:CMVdel134启动子
>ID_6598_pAd26.dE1.dE3.5orf6;SEQ ID NO:218












>CMVdel134包括2x TO;SEQ ID NO:219

Claims (33)
1.一种分离的核酸分子,其包含编码2019-NCOV刺突(S)蛋白的核苷酸序列,所述2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
2.权利要求1的分离的核酸分子,其包含SEQ ID NO:211的核苷酸序列。
3.权利要求1或2的分离的核酸分子,其包含编码2019-NCOV刺突(S)蛋白的核苷酸序列,所述2019-NCOV刺突(S)蛋白包含SEQ ID NO:205的氨基酸序列。
4.权利要求1的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ IDNO:51的氨基酸序列具有至少85%序列相同性的多肽。
5.权利要求1或4的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQID NO:51的氨基酸序列具有至少99%序列相同性的多肽。
6.权利要求1的分离的核酸分子,其包含核苷酸序列,所述核苷酸序列编码与SEQ IDNO:56具有至少85%序列相同性的多肽。
7.权利要求4或5的分离的核酸分子,其包含与SEQ ID NO:143或其互补序列具有至少85%序列相同性的核苷酸序列。
8.权利要求6的分离的核酸分子,其包含与SEQ ID NO:148或其互补序列具有至少85%序列相同性的核苷酸序列。
9.实施方案权利要求1-8中任一项的分离的核酸分子,其编码2019-NCOV刺突(S)蛋白,所述2019-NCOV刺突(S)蛋白包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
c.信号序列的缺失。
10.前述权利要求中任一项的分离的核酸分子,其中编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(CMV)启动子,优选CMV立即早期启动子可操作地连接。
11.前述权利要求中任一项的分离的核酸分子,其中编码2019-NCOV刺突(S)蛋白的核酸与巨细胞病毒(CMV)启动子可操作地连接,所述CMV启动子包含至少一个四环素操纵子(TetO)基序。
12.权利要求11的分离的核酸分子,其中所述包含至少一个TetO基序的CMV启动子包含SEQ ID NO:219的核苷酸序列。
13.一种分离的2019-NCOV刺突(S)蛋白,其包含对SEQ ID NO:29的全长氨基酸序列的以下修饰:
a.在氨基酸986和987处突变为脯氨酸的稳定化突变;和
b.对弗林蛋白酶切割位点(SEQ ID NO:90)的突变。
14.权利要求13的分离的2019-NCOV刺突(S)蛋白,其包含SEQ ID NO:205的氨基酸序列。
15.权利要求13的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ ID NO:51的氨基酸序列具有至少85%序列相同性的氨基酸序列。
16.权利要求13或15的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ ID NO:51的氨基酸序列具有至少99%序列相同性的氨基酸序列。
17.权利要求13的分离的2019-NCOV刺突(S)蛋白,其包含与SEQ ID NO:56的氨基酸序列具有至少85%序列相同性的氨基酸序列。
18.权利要求13的分离的2019-NCOV刺突(S)蛋白,其包含对SEQ ID NO:29的全长氨基酸序列的以下进一步修饰:
c.信号序列的缺失。
19.一种分离的载体,其包含权利要求1-12中任一项的核酸分子中的一种或多种。
20.权利要求19的载体,其中所述载体是复制缺陷的。
21.权利要求19或20的载体,其中所述载体是血清型26的腺病毒。
22.一种组合物,其包含权利要求1-12中任一项的核酸分子、权利要求13-18中任一项的蛋白或者权利要求19-21中任一项的载体。
23.权利要求22的组合物,其进一步包含药学上可接受的载剂、赋形剂或稀释剂。
24.权利要求22或23的组合物,其用作疫苗。
25.权利要求22-24中任一项的组合物,其用于主动免疫以预防18岁以上的成人的2019冠状病毒病(COVID-19)。
26.一种用于预防18岁以上的个体的严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)引起的2019冠状病毒病(COVID-19)的主动免疫方法,其包括向受试者给药权利要求22-25中任一项的组合物。
27.权利要求26的方法,其包括以一剂方案给药实施方案22-25中任一项的组合物。
28.权利要求26的方法,其包括以两剂方案给药权利要求22-25中任一项的组合物,其中所述剂量间隔约1、2或3个月给药。
29.权利要求26-28中任一项的方法,其中以5x1010vp/剂的剂量给药所述组合物。
30.一种制备用于预防18岁以上的个体的严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)引起的2019冠状病毒病(COVID-19)的免疫原性组合物的方法,所述方法包括以下步骤:
(a)将权利要求1-12中任一项的核酸分子、权利要求13-18中任一项的蛋白、权利要求19-21中任一项的载体、权利要求22-25中任一项的组合物中的至少一种与药学上可接受的载剂、赋形剂或稀释剂混合以形成所述免疫原性组合物;和
(b)将所述免疫原性组合物置于容器中。
31.一种试剂盒,其包含:
(a)第一容器,其包含权利要求1-12中任一项的核酸分子、权利要求13-18中任一项的蛋白、权利要求16-21中任一项的载体、权利要求22-25中任一项的组合物中的至少一种;
(b)其使用说明;以及任选存在的
(c)第二容器,其包含药学上可接受的载剂、赋形剂或稀释剂。
32.一种制备针对COVID-19以预防18岁以上的个体的严重急性呼吸道综合征冠状病毒2(SARS-CoV-2)引起的2019冠状病毒病(COVID-19)的疫苗的方法,其包括
提供血清型26的重组人腺病毒,其包含编码SARS-CoV-2S蛋白或其片段的核酸,
使所述重组腺病毒在宿主细胞培养物中增殖,
分离和纯化所述重组腺病毒,以及
将所述重组腺病毒配制于药学上可接受的组合物中。
33.一种分离的重组核酸,其形成血清型26的重组人腺病毒的基因组,所述重组人腺病毒包含编码SARS-CoV-2S蛋白或其片段的核酸。
Applications Claiming Priority (43)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US202062969008P | 2020-01-31 | 2020-01-31 | |
| US62/969,008 | 2020-01-31 | ||
| US202062994630P | 2020-03-25 | 2020-03-25 | |
| US62/994,630 | 2020-03-25 | ||
| US202063014467P | 2020-04-23 | 2020-04-23 | |
| US63/014,467 | 2020-04-23 | ||
| US202063025782P | 2020-05-15 | 2020-05-15 | |
| US63/025,782 | 2020-05-15 | ||
| US202062705187P | 2020-06-15 | 2020-06-15 | |
| US62/705,187 | 2020-06-15 | ||
| US202062705308P | 2020-06-21 | 2020-06-21 | |
| US62/705,308 | 2020-06-21 | ||
| US202063043090P | 2020-06-23 | 2020-06-23 | |
| US63/043,090 | 2020-06-23 | ||
| US202062706366P | 2020-08-12 | 2020-08-12 | |
| US62/706,366 | 2020-08-12 | ||
| US202063066147P | 2020-08-14 | 2020-08-14 | |
| US63/066,147 | 2020-08-14 | ||
| US202062706676P | 2020-09-02 | 2020-09-02 | |
| US62/706,676 | 2020-09-02 | ||
| US202062706937P | 2020-09-18 | 2020-09-18 | |
| US62/706,937 | 2020-09-18 | ||
| US202062706958P | 2020-09-21 | 2020-09-21 | |
| US62/706,958 | 2020-09-21 | ||
| US202063198089P | 2020-09-28 | 2020-09-28 | |
| US63/198,089 | 2020-09-28 | ||
| US202063198306P | 2020-10-09 | 2020-10-09 | |
| US63/198,306 | 2020-10-09 | ||
| US202063112900P | 2020-11-12 | 2020-11-12 | |
| US63/112,900 | 2020-11-12 | ||
| CA3101131A CA3101131A1 (en) | 2020-01-31 | 2020-11-28 | Compositions and methods for preventing and treating coronavirus infection - sarscov-2 vaccines |
| CA3101131 | 2020-11-28 | ||
| US202063121482P | 2020-12-04 | 2020-12-04 | |
| US63/121,482 | 2020-12-04 | ||
| US202163133969P | 2021-01-05 | 2021-01-05 | |
| US63/133,969 | 2021-01-05 | ||
| US202163135182P | 2021-01-08 | 2021-01-08 | |
| US63/135,182 | 2021-01-08 | ||
| US202163141913P | 2021-01-26 | 2021-01-26 | |
| US63/141,913 | 2021-01-26 | ||
| US202163142977P | 2021-01-28 | 2021-01-28 | |
| US63/142,977 | 2021-01-28 | ||
| PCT/US2021/015946 WO2021155323A1 (en) | 2020-01-31 | 2021-01-30 | Compositions and methods for preventing and treating coronavirus infection-sars-cov-2 vaccines |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN116096732A true CN116096732A (zh) | 2023-05-09 |
Family
ID=77079265
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN202180025856.2A Pending CN116096732A (zh) | 2020-01-31 | 2021-01-30 | 用于预防和治疗冠状病毒感染的组合物和方法-sars-cov-2疫苗 |
Country Status (9)
| Country | Link |
|---|---|
| US (3) | US11498944B2 (zh) |
| EP (1) | EP4097122A4 (zh) |
| JP (1) | JP2023512519A (zh) |
| KR (1) | KR20220134622A (zh) |
| CN (1) | CN116096732A (zh) |
| BR (1) | BR112022014808A2 (zh) |
| TW (1) | TW202204380A (zh) |
| UY (1) | UY39060A (zh) |
| WO (1) | WO2021155323A1 (zh) |
Families Citing this family (54)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20210260181A1 (en) * | 2014-09-16 | 2021-08-26 | Altimmune, Inc | Coronavirus immunogenic compositions and uses thereof |
| US10953089B1 (en) | 2020-01-27 | 2021-03-23 | Novavax, Inc. | Coronavirus vaccine formulations |
| KR20220134622A (ko) | 2020-01-31 | 2022-10-05 | 베쓰 이스라엘 디코니스 메디칼 센터 인크 | 코로나바이러스 감염 예방 및 치료용 조성물 및 방법 - sars-cov-2 백신 |
| US11241493B2 (en) | 2020-02-04 | 2022-02-08 | Curevac Ag | Coronavirus vaccine |
| US12194089B2 (en) | 2020-02-04 | 2025-01-14 | CureVac SE | Coronavirus vaccine |
| KR20220143684A (ko) | 2020-02-07 | 2022-10-25 | 알엔에이이뮨, 인코포레이티드 | 신종 코로나바이러스 감염에 대한 mRNA 백신의 조성물 및 방법 |
| CA3167611A1 (en) * | 2020-02-13 | 2021-08-19 | Etienne Simon-Loriere | Nucleic acid vaccine against the sars-cov-2 coronavirus |
| AU2021220971A1 (en) * | 2020-02-14 | 2022-10-06 | Altimmune, Inc. | Coronavirus immunogenic compositions and uses thereof |
| IL295697A (en) | 2020-02-25 | 2022-10-01 | Inovio Pharmaceuticals Inc | Vaccines against coronavirus and methods of use |
| US11213482B1 (en) * | 2020-03-05 | 2022-01-04 | University of Pittsburgh—Of the Commonwealth System of Higher Educat | SARS-CoV-2 subunit vaccine and microneedle array delivery system |
| GB2594364A (en) | 2020-04-22 | 2021-10-27 | Biontech Rna Pharmaceuticals Gmbh | Coronavirus vaccine |
| US20210341480A1 (en) * | 2020-04-29 | 2021-11-04 | Shi-long LU | Viral testing in saliva |
| US12226431B2 (en) | 2021-09-11 | 2025-02-18 | Porosome Therapeutics, Inc. | Aerosol and topical administration of a formulation containing cyclodextrin, quercetin and zinc, in combination or separately, to mitigate infection by enveloped viruses |
| CN116322758A (zh) * | 2020-05-29 | 2023-06-23 | 库尔维科欧洲股份公司 | 基于核酸的组合疫苗 |
| WO2021249116A1 (en) | 2020-06-10 | 2021-12-16 | Sichuan Clover Biopharmaceuticals, Inc. | Coronavirus vaccine compositions, methods, and uses thereof |
| US20230314432A1 (en) * | 2020-07-10 | 2023-10-05 | Covid Diagnostics Ltd. | Compositions, methods, and systems for detecting immune response |
| EP4182687A4 (en) * | 2020-07-20 | 2024-09-11 | Bio-Rad Laboratories, Inc. | IMMUNOASSAY FOR SARS-COV-2 NEUTRALISING ANTIBODIES AND MATERIALS THEREFOR |
| US20220193225A1 (en) * | 2020-08-31 | 2022-06-23 | Bruce Lyday | Compositions and methods for sars-2 vaccine with virus replicative particles and recombinant glycoproteins |
| MX2023007574A (es) | 2020-12-22 | 2023-09-29 | CureVac SE | "vacuna de arn contra variantes de sars-cov-2. |
| US20220226489A1 (en) * | 2021-01-20 | 2022-07-21 | Singh Biotechnology, Llc | Therapeutics directed against coronavirus |
| JP7598538B2 (ja) * | 2021-01-29 | 2024-12-12 | 東亞合成株式会社 | SARS-CoV-2由来のアミノ酸配列およびその利用 |
| EP4313143A4 (en) * | 2021-03-22 | 2025-06-18 | Novavax Inc | CORONAVIRUS VACCINE FORMULATIONS |
| WO2022204219A1 (en) | 2021-03-22 | 2022-09-29 | Recode Therapeutics, Inc. | Compositions and methods for targeted delivery to cells |
| MX2023011243A (es) | 2021-03-23 | 2023-12-14 | Recode Therapeutics Inc | Composiciones de polinucleotidos, formulaciones relacionadas y metodos de uso de los mismos. |
| EP4333887A4 (en) * | 2021-05-05 | 2025-05-21 | Inovio Pharmaceuticals Inc | CORONAVIRUS VACCINES AND METHODS OF USE |
| EP4384214A2 (en) | 2021-08-11 | 2024-06-19 | Rnaimmune, Inc. | Composition and methods of mrna vaccines against novel coronavirus infection |
| CN113832168B (zh) * | 2021-08-15 | 2023-08-04 | 北京科兴中维生物技术有限公司 | mRNA疫苗及其制备方法和应用 |
| WO2023020939A1 (en) * | 2021-08-17 | 2023-02-23 | Janssen Pharmaceuticals, Inc. | Sars-cov-2 vaccines |
| WO2023026182A1 (en) | 2021-08-24 | 2023-03-02 | Janssen Pharmaceuticals, Inc. | Sars-cov-2 vaccines |
| WO2023023940A1 (zh) * | 2021-08-24 | 2023-03-02 | 复旦大学 | 一种诱导广谱抗冠状病毒的t细胞疫苗免疫原及其应用 |
| WO2023025287A1 (zh) * | 2021-08-26 | 2023-03-02 | 江苏瑞科生物技术股份有限公司 | 新型冠状病毒免疫原性物质、其制备方法和应用 |
| CN113913464B (zh) * | 2021-09-28 | 2022-11-22 | 武汉枢密脑科学技术有限公司 | 一种表达载体、重组腺相关病毒及其在制备2019新型冠状病毒疫苗中的应用 |
| WO2023092100A2 (en) * | 2021-11-19 | 2023-05-25 | Meissa Vaccines, Inc. | Methods of administering chimeric vaccines |
| CN116178527A (zh) * | 2021-11-26 | 2023-05-30 | 深圳先进技术研究院 | 抗sar-cov-2全人源单克隆抗体6g18及其制法与应用 |
| US12186387B2 (en) | 2021-11-29 | 2025-01-07 | BioNTech SE | Coronavirus vaccine |
| AU2022399829A1 (en) * | 2021-11-30 | 2024-06-27 | Novavax, Inc. | Coronavirus vaccine formulations |
| TWI843471B (zh) * | 2022-03-11 | 2024-05-21 | 財團法人國家衛生研究院 | 含抗原和dna之組成物及其用途 |
| CN114574502B (zh) * | 2022-04-11 | 2023-07-14 | 四川大学 | 一种以复制缺陷腺相关病毒为载体的新型冠状病毒疫苗 |
| WO2023198815A1 (en) | 2022-04-14 | 2023-10-19 | Janssen Vaccines & Prevention B.V. | Sequential administration of adenoviruses |
| WO2023205589A2 (en) * | 2022-04-20 | 2023-10-26 | Inovio Pharmaceuticals, Inc. | Immunogenic compositions against sars-cov-2 variants and their methods of use |
| WO2023212446A1 (en) * | 2022-04-29 | 2023-11-02 | Xequel Bio, Inc. | Compositions and methods for treating complications of viral infections and other respiratory disorders |
| WO2023240278A2 (en) * | 2022-06-10 | 2023-12-14 | The Trustees Of Columbia University In The City Of New York | Uses of glycolipids as a vaccine adjuvant and methods thereof |
| US11654121B1 (en) | 2022-06-22 | 2023-05-23 | Flagship Pioneering Innovations Vi, Llc | Combination therapies for the treatment of viral infections |
| WO2023250111A1 (en) | 2022-06-22 | 2023-12-28 | Flagship Pioneering Innovations Vi, Llc | Combination therapies for the treatment of viral infections |
| WO2024002985A1 (en) | 2022-06-26 | 2024-01-04 | BioNTech SE | Coronavirus vaccine |
| EP4549452A1 (en) * | 2022-07-07 | 2025-05-07 | WestVac Biopharma Co., Ltd. | Pharmaceutical composition for resisting infection with sars-cov-2 or mutant thereof, and combined drug thereof |
| CN118119646A (zh) * | 2022-08-08 | 2024-05-31 | 神州细胞工程有限公司 | 一种可诱导广谱中和活性重组五组分新冠病毒三聚体蛋白疫苗的制备及应用 |
| WO2024076805A2 (en) * | 2022-10-07 | 2024-04-11 | Zeptometrix Llc | Control for protein-based assay |
| DE202023106198U1 (de) | 2022-10-28 | 2024-03-21 | CureVac SE | Impfstoff auf Nukleinsäurebasis |
| US12280104B2 (en) | 2022-11-01 | 2025-04-22 | RNAimmune, Inc. | Composition and methods for MRNA vaccines against novel omicron coronavirus infections |
| CN116024269B (zh) * | 2022-11-18 | 2024-02-02 | 复百澳(苏州)生物医药科技有限公司 | 一种冠状病毒假病毒颗粒的制备方法 |
| CN116077635A (zh) * | 2022-11-23 | 2023-05-09 | 珠海市丽珠单抗生物技术有限公司 | 新型冠状病毒融合蛋白疫苗其应用 |
| WO2024229199A2 (en) * | 2023-05-03 | 2024-11-07 | Partner Therapeutics, Inc. | Humoral modulation in vaccinated and virally infected subjects with granulocyte-macrophage colony-stimulating factor (gm-csf) |
| WO2024229195A1 (en) * | 2023-05-03 | 2024-11-07 | Partner Therapeutics, Inc. | Viral infection modulation in vaccinated subjects treated with granulocyte-macrophage colony-stimulating factor (gm-csf) |
Family Cites Families (52)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4603112A (en) | 1981-12-24 | 1986-07-29 | Health Research, Incorporated | Modified vaccinia virus |
| US5057540A (en) | 1987-05-29 | 1991-10-15 | Cambridge Biotech Corporation | Saponin adjuvant |
| NZ230747A (en) | 1988-09-30 | 1992-05-26 | Bror Morein | Immunomodulating matrix comprising a complex of at least one lipid and at least one saponin; certain glycosylated triterpenoid saponins derived from quillaja saponaria molina |
| US5298416A (en) | 1989-01-18 | 1994-03-29 | British Technology Group Ltd. | Attenuated polioviruses |
| JPH0832638B2 (ja) | 1989-05-25 | 1996-03-29 | カイロン コーポレイション | サブミクロン油滴乳剤を含んで成るアジュバント製剤 |
| DE69233158T2 (de) | 1991-03-07 | 2004-05-13 | Connaught Technology Corp., Greenville | Gentechnologisch hergestellter stamm für impfstoffe |
| US5505947A (en) | 1994-05-27 | 1996-04-09 | The University Of North Carolina At Chapel Hill | Attenuating mutations in Venezuelan Equine Encephalitis virus |
| AUPM873294A0 (en) | 1994-10-12 | 1994-11-03 | Csl Limited | Saponin preparations and use thereof in iscoms |
| US5801030A (en) | 1995-09-01 | 1998-09-01 | Genvec, Inc. | Methods and vectors for site-specific recombination |
| FR2751343B1 (fr) | 1996-07-16 | 1998-12-18 | Transgene Sa | Procede de conservation de virus recombinants infectieux, suspension aqueuse virale et utilisation comme medicament |
| WO1998010087A1 (en) | 1996-09-06 | 1998-03-12 | Trustees Of The University Of Pennsylvania | Chimpanzee adenovirus vectors |
| US6210683B1 (en) | 1997-09-05 | 2001-04-03 | Merck & Co., Inc. | Stabilizers containing recombinant human serum albumin for live virus vaccines |
| DE69942708D1 (de) | 1998-02-17 | 2010-10-07 | Schering Corp | Virus enthaltende Zusammensetzungen und Methoden zur Konzentration von Viruspräparaten |
| DK1133316T3 (da) | 1998-11-16 | 2009-05-25 | Introgen Therapeutics Inc | Adenovirus-formulering til genterapi |
| US6225289B1 (en) | 1998-12-10 | 2001-05-01 | Genvec, Inc. | Methods and compositions for preserving adenoviral vectors |
| ATE360082T1 (de) | 1999-11-16 | 2007-05-15 | Genzyme Corp | Vektoren und transgene mit regulatorischen elementen zur genverabreichung in leber |
| EP1263461A4 (en) | 2000-03-07 | 2009-08-12 | Merck & Co Inc | ADENOVIRUS FORMULATIONS |
| DE60233047D1 (de) | 2001-05-14 | 2009-09-03 | Gbp Ip Llc | Lentivirale vektoren kodierend für gerinnungsfaktoren für die gentherapie |
| JP2005517394A (ja) | 2001-12-12 | 2005-06-16 | エフ エイチ フォールディング アンド カンパニー リミテッド | ウイルス保存のための組成物 |
| IL162404A0 (en) | 2002-01-18 | 2005-11-20 | Schering Ag | Stabilized formulations of adenovirus |
| US20030180936A1 (en) | 2002-03-15 | 2003-09-25 | Memarzadeh Bahram Eric | Method for the purification, production and formulation of oncolytic adenoviruses |
| SE0202110D0 (sv) | 2002-07-05 | 2002-07-05 | Isconova Ab | Iscom preparation and use thereof |
| EP1649053A4 (en) | 2003-06-18 | 2006-10-18 | Chinese Nat Human Genome Ct Sh | CHARACTERIZATION OF THE EARLY LEVELS OF THE VIRUS OF HEAVY ACUTE RESPIRATORY SYNDROME (SARS) AND USES THEREOF |
| SE0301998D0 (sv) | 2003-07-07 | 2003-07-07 | Isconova Ab | Quil A fraction with low toxicity and use thereof |
| KR101206206B1 (ko) | 2003-07-22 | 2012-11-29 | 크루셀 홀란드 비.브이. | 사스-코로나바이러스에 대한 결합분자 및 그것의 용도 |
| WO2005013904A2 (en) | 2003-08-04 | 2005-02-17 | University Of Massachusetts | Sars nucleic acids, proteins, vaccines, and uses thereof |
| US7129042B2 (en) * | 2003-11-03 | 2006-10-31 | Diagnostic Hybrids, Inc. | Compositions and methods for detecting severe acute respiratory syndrome coronavirus |
| CA2880061C (en) | 2004-01-23 | 2018-03-13 | Agostino Cirillo | Chimpanzee adenovirus vaccine carriers |
| EA010433B1 (ru) | 2004-10-13 | 2008-08-29 | Круселл Холланд Б.В. | Усовершенствованные аденовирусные векторы и их применение |
| BRPI0515819A (pt) | 2004-12-01 | 2008-08-05 | Genzyme Corp | métodos para o fornecimento direcionado de material genético ao fìgado |
| WO2007024941A2 (en) | 2005-08-23 | 2007-03-01 | The Regents Of The University Of California | Polyvalent vaccine |
| WO2007104792A2 (en) | 2006-03-16 | 2007-09-20 | Crucell Holland B.V. | Recombinant adenoviruses based on serotype 26 and 48, and use thereof |
| EP1998804B1 (en) | 2006-03-27 | 2014-04-16 | Crucell Holland B.V. | Compositions comprising a recombinant adenovirus and an adjuvant |
| CN102203242B (zh) | 2008-11-03 | 2013-06-12 | 克鲁塞尔荷兰公司 | 产生腺病毒载体的方法 |
| US9017691B2 (en) | 2008-11-18 | 2015-04-28 | Beth Israel Deaconess Medical Center | Antiviral vaccines with improved cellular immunogenicity |
| KR101763093B1 (ko) | 2009-02-02 | 2017-07-28 | 글락소스미스클라인 바이오로지칼즈 에스.에이. | 시미안 아데노바이러스 핵산- 및 아미노산-서열, 이를 포함하는 벡터 및 이의 용도 |
| WO2010085984A1 (en) | 2009-02-02 | 2010-08-05 | Okairos Ag | Simian adenovirus nucleic acid- and amino acid-sequences, vectors containing same, and uses thereof |
| AU2011214262B2 (en) | 2010-02-15 | 2015-05-21 | Crucell Holland B.V. | Method for the production of Ad26 adenoviral vectors |
| EA029504B1 (ru) | 2010-12-14 | 2018-04-30 | Дзе Гавермент Оф Дзе Юнайтед Стейтс Оф Америка Эз Репрезентед Бай Дзе Секретэри Оф Дзе Департмент Оф Хелт Энд Хьюмэн Сервисиз | Способ индуцирования иммунного ответа против филовирусного антигена у пациента с использованием аденовирусов серотипа 26 и серотипа 35 |
| GB201108879D0 (en) | 2011-05-25 | 2011-07-06 | Isis Innovation | Vector |
| EP3532095A1 (en) * | 2016-10-25 | 2019-09-04 | The U.S.A. as represented by the Secretary, Department of Health and Human Services | Prefusion coronavirus spike proteins and their use |
| MX2019009316A (es) | 2017-02-09 | 2019-09-19 | Janssen Vaccines & Prevention Bv | Promotor corto y potente para la expresion de genes heterologos. |
| MA47790A (fr) * | 2017-03-17 | 2021-05-05 | Modernatx Inc | Vaccins à base d'arn contre des maladies zoonotiques |
| GB201708444D0 (en) | 2017-05-26 | 2017-07-12 | Univ Oxford Innovation Ltd | Compositions and methods for inducing an immune response |
| WO2019086456A1 (en) | 2017-10-31 | 2019-05-09 | Janssen Vaccines & Prevention B.V. | Adenovirus and uses thereof |
| AU2018360253B2 (en) | 2017-10-31 | 2023-11-09 | Janssen Vaccines & Prevention B.V. | Adenovirus and uses thereof |
| MX2020004490A (es) | 2017-10-31 | 2020-08-13 | Janssen Vaccines & Prevention Bv | Vectores adenovirales y usos de estos. |
| US10953089B1 (en) * | 2020-01-27 | 2021-03-23 | Novavax, Inc. | Coronavirus vaccine formulations |
| CA3168902A1 (en) * | 2020-01-28 | 2021-08-05 | Modernatx, Inc. | Coronavirus rna vaccines |
| KR20220134622A (ko) | 2020-01-31 | 2022-10-05 | 베쓰 이스라엘 디코니스 메디칼 센터 인크 | 코로나바이러스 감염 예방 및 치료용 조성물 및 방법 - sars-cov-2 백신 |
| JP2023524860A (ja) * | 2020-05-11 | 2023-06-13 | ヤンセン ファーマシューティカルズ,インコーポレーテッド | SARS-CoV-2ワクチン |
| US10906944B2 (en) | 2020-06-29 | 2021-02-02 | The Scripps Research Institute | Stabilized coronavirus spike (S) protein immunogens and related vaccines |
-
2021
- 2021-01-30 KR KR1020227030114A patent/KR20220134622A/ko active Pending
- 2021-01-30 EP EP21747489.9A patent/EP4097122A4/en not_active Withdrawn
- 2021-01-30 JP JP2022546341A patent/JP2023512519A/ja not_active Withdrawn
- 2021-01-30 WO PCT/US2021/015946 patent/WO2021155323A1/en active Application Filing
- 2021-01-30 TW TW110103610A patent/TW202204380A/zh unknown
- 2021-01-30 BR BR112022014808A patent/BR112022014808A2/pt unknown
- 2021-01-30 US US17/163,357 patent/US11498944B2/en active Active
- 2021-01-30 CN CN202180025856.2A patent/CN116096732A/zh active Pending
- 2021-01-30 US US17/759,803 patent/US20230075527A1/en active Pending
- 2021-02-01 UY UY0001039060A patent/UY39060A/es unknown
- 2021-07-28 US US17/387,105 patent/US11384122B2/en active Active
Also Published As
| Publication number | Publication date |
|---|---|
| EP4097122A4 (en) | 2024-07-17 |
| JP2023512519A (ja) | 2023-03-27 |
| TW202204380A (zh) | 2022-02-01 |
| US20230075527A1 (en) | 2023-03-09 |
| WO2021155323A1 (en) | 2021-08-05 |
| EP4097122A1 (en) | 2022-12-07 |
| US20210246170A1 (en) | 2021-08-12 |
| KR20220134622A (ko) | 2022-10-05 |
| US11384122B2 (en) | 2022-07-12 |
| US20210388032A1 (en) | 2021-12-16 |
| BR112022014808A2 (pt) | 2022-09-20 |
| UY39060A (es) | 2021-10-29 |
| US11498944B2 (en) | 2022-11-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US11498944B2 (en) | Compositions and methods for preventing and treating coronavirus infection—SARS-CoV-2 vaccines | |
| CA3101131A1 (en) | Compositions and methods for preventing and treating coronavirus infection - sarscov-2 vaccines | |
| Dangi et al. | Improved control of SARS-CoV-2 by treatment with a nucleocapsid-specific monoclonal antibody | |
| Chmielewska et al. | Combined adenovirus vector and hepatitis C virus envelope protein prime-boost regimen elicits T cell and neutralizing antibody immune responses | |
| US10898566B2 (en) | Zika virus vaccines | |
| JP7544399B2 (ja) | Covid-19に対する保護のための弱毒化されたポックスウイルスベクターをベースとするワクチン | |
| Zu Natrup et al. | Stabilized recombinant SARS-CoV-2 spike antigen enhances vaccine immunogenicity and protective capacity | |
| Liu et al. | Heterologous prime-boost immunizations with chimpanzee adenoviral vectors elicit potent and protective immunity against SARS-CoV-2 infection | |
| Hayashi et al. | Preclinical study of a DNA vaccine targeting SARS-CoV-2 | |
| Kurup et al. | Measles-based Zika vaccine induces long-term immunity and requires NS1 antibodies to protect the female reproductive tract | |
| WO2018152526A1 (en) | Zika virus vaccines | |
| JP2024539609A (ja) | コロナウイルスワクチン | |
| JP2023529124A (ja) | コロナウイルスワクチンコンストラクトおよびこれを作製し使用する方法 | |
| US20230242940A1 (en) | Methods of making and using a vaccine against coronavirus | |
| Wang et al. | The pentameric complex is not required for vertical transmission of cytomegalovirus in seronegative pregnant rhesus macaques | |
| Hao et al. | An adenovirus-vectored RVF vaccine confers complete protection against lethal RVFV challenge in A129 mice | |
| KR20240137110A (ko) | 오미크론 코로나바이러스 백신 구조체 및 이를 제조하고 사용하는 방법 | |
| JP2025518717A (ja) | 弱毒化生SARS-CoV-2ワクチン | |
| WO2022136921A1 (en) | A new hace2 transgenic animal with remarkable permissiveness of lung and central nervous system to replication of viruses targeting hace2 - an experimental model for vaccine, drug and neuro/immune/physio-pathology of covid-19 and other pathologies linked to viruses or coronaviruses using hace2 as a cellular receptor | |
| How et al. | As investigative journalist Jon Rappoport put it:“BY THEIR OWN STANDARDS, the FDA should never have allowed the Pfizer COVID vaccine to be shot into a single arm. The Agencyʼs Emergency Use Authorization was a crime—according to their own data.” | |
| Salvati | Active monitoring of long-term immunity to SARS-CoV-2 infection and vaccination | |
| Class | Vaccines Against SARS-CoV-2 and Intrahost Viral Adaptations | |
| Choudhary et al. | Broad-Spectrum Heavily Mutated Monoclonal Antibody Isolated from COVID-19 Convalescent Vaccinee with Capacity to Neutralize SARS-CoV2 Variants Ranging from B. 1 to BQ. 1.1. | |
| Malhi | Designing a Next-generation Epstein-Barr Virus Vaccine | |
| Schnell et al. | Measles-based Zika vaccine induces long-term immunity and requires NS1 antibodies to protect the female reproductive tract. |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |