BRPI1008267B1 - Processo para a produção fermentativa de ácido succínico - Google Patents
Processo para a produção fermentativa de ácido succínico Download PDFInfo
- Publication number
- BRPI1008267B1 BRPI1008267B1 BRPI1008267-0A BRPI1008267A BRPI1008267B1 BR PI1008267 B1 BRPI1008267 B1 BR PI1008267B1 BR PI1008267 A BRPI1008267 A BR PI1008267A BR PI1008267 B1 BRPI1008267 B1 BR PI1008267B1
- Authority
- BR
- Brazil
- Prior art keywords
- glycerol
- succinic acid
- process according
- acid
- strain
- Prior art date
Links
Images


Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/40—Preparation of oxygen-containing organic compounds containing a carboxyl group including Peroxycarboxylic acids
- C12P7/44—Polycarboxylic acids
- C12P7/46—Dicarboxylic acids having four or less carbon atoms, e.g. fumaric acid, maleic acid
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
- C12N1/205—Bacterial isolates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/32—Processes using, or culture media containing, lower alkanols, i.e. C1 to C6
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
- C12N15/746—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora for lactic acid bacteria (Streptococcus; Lactococcus; Lactobacillus; Pediococcus; Enterococcus; Leuconostoc; Propionibacterium; Bifidobacterium; Sporolactobacillus)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
- C12N15/75—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora for Bacillus
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
- C12N15/76—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora for Actinomyces; for Streptomyces
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1025—Acyltransferases (2.3)
- C12N9/1029—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y203/00—Acyltransferases (2.3)
- C12Y203/01—Acyltransferases (2.3) transferring groups other than amino-acyl groups (2.3.1)
- C12Y203/01054—Formate C-acetyltransferase (2.3.1.54), i.e. pyruvate formate-lyase or PFL
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/01—Bacteria or Actinomycetales ; using bacteria or Actinomycetales
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02P—CLIMATE CHANGE MITIGATION TECHNOLOGIES IN THE PRODUCTION OR PROCESSING OF GOODS
- Y02P20/00—Technologies relating to chemical industry
- Y02P20/50—Improvements relating to the production of bulk chemicals
- Y02P20/52—Improvements relating to the production of bulk chemicals using catalysts, e.g. selective catalysts
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Medicinal Chemistry (AREA)
- Virology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Molecular Biology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Furan Compounds (AREA)
- Pyrrole Compounds (AREA)
- Organic Low-Molecular-Weight Compounds And Preparation Thereof (AREA)
Abstract
cepa bacteriana, processos para a produção fermentativa de um ácido orgânico e de ácido succínico, e para produção de ácido succínico, de tetraidrofurano (thf) e/ou 1,4-butanediol (bdo) e/ou gama-butirolactona (gbl), e de pirrolidonas, e, uso de uma cepa bacteriana a presente invenção diz repeito a cepas bacterianas capazes de utilizar glicerol como uma fonte de carbono para a produção fermentativa de ácido succínico, em que as ditas cepas são geneticamente modificadas, de maneira tal que compreendam uma desregulação de sua atividade da enzima piruvato-formato-líase endógena, bem como a métodos para produzir ácidos orgânicos, em particular, ácido succínico, fazendo uso de tal microrganismo. a presente invenção também diz respeito ao processamento à jusante dos ácidos orgânicos produzidos por cromatografia de troca catiônica.
Description
[0001] A presente invenção diz respeito a cepas bacterianas capazes de utilizar glicerol como uma fonte de carbono para a produção fermentativa de ácido succínico, em que as ditas cepas são geneticamente modificadas, de maneira tal que compreendam uma desregulação de sua atividade da enzima piruvato-formato-liase endógena, bem como a métodos de produzir ácidos orgânicos, em particular, ácido succínico, usando tal microrganismo.
[0002] A produção fermentativa de ácido succínico (SA) a partir da biomassa já atraiu muita atenção em decorrência de o dito ácido representar um importante constituinte de resinas sintéticas ou ser uma fonte de compostos químicos de baixo peso molecular muito valiosos, em particular tetraidrofurano (THF), 1,4-butanediol (BDO), gama-butirolactona (GBL) e pirrolidonas (WO-A-2006/066839).
[0003] Uma bactéria que produz SA isolada de rúmen bovino foi descrita por Lee et al (2002a). A bactéria é um cocobacilo ou bastonete gram- negativo não móvel, não formador de esporo, mesófila e capnofílica. A análise filogenética com base na sequência de RNAr 16S e a análise fisiológica indicaram que a cepa pertence ao gênero Mannheimia, como uma espécie inédita, e foi denominada Mannheimia succiniciproducens MBEL55E. Em condições de 100 % de CO2, cresce bem na faixa de pH de 6,0-7,5 e produz SA, ácido acético e ácido fórmico em uma razão constante de 2:1:1. Quando M. succiniciproducens MBEL55E foi cultivada anaerobicamente em saturação de CO2 com glicose como fonte de carbono, 19,8 g/L de glicose foram consumidos e 13,3 g/L de SA foram produzidos em 7,5 horas de incubação. Além do mais, neste microrganismo, a produção de SA melhorou com a mutação/deleção de genes metabólicos. A mutação/deleção combinada dos genes lactato desidrogenase ldhA, piruvato- formato-liase pflB, fosfotransacetilase pta e acetato quinase ackA resultou em uma cepa que converte carbono em SA com um rendimento (YP/S) de 0,6 g de SA por g de fonte de carbono fonte adicionada. Observou-se que o rendimento de espaço-tempo para a produção de SA é 1,8 g/litro/hora. (Lee 2006)
[0004] Lin et al 2005 descrevem uma cepa mutante de E. coli que carrega mutações no gene ldh, bem como no pfl, descrita como SB202. Entretanto, esta cepa foi caracterizada por crescimento de lento e pela incapacidade de fermentar um sacarídeo por completo em condições anaeróbicas. O ldh e pfl inativos não fazem com que o fluxo de carbono seja contido na protuberância de piruvato, fazendo com que o piruvato se acumule como o principal produto. Com relação a isto, observou-se que o rendimento de carbono (YP/S) de succinato na fonte de carbono é menor que 0,15g /g de SA/Carbono.
[0005] Sanchez et al. 2005 descrevem cepas de E. coli que carregam mutações nos genes ldh, adhE, ack-pta e iclR. Nestes experimentos, as células foram crescidas aerobicamente no meio complexo, coletadas, concentradas e incubadas com fontes de carbono em condições anaeróbicas. Nestas condições específicas para a conversão direta de um carboidrato e, SA, foram observados os rendimentos de carbono YP/S de 0,98 a 1,13 g de SA por g de fonte de carbono, com um rendimento espaço-tempo de 0,79 g/1 hora de SA. A utilização de carbono para a geração da biomassa antes da fase de conversão anaeróbica não foi explicitamente incluída neste cálculo e é não é descrita adicionalmente.
[0006] Hong e Lee (2001) descrevem cepas de E. coli que carregam mutações nos genes ldh e pfl. Estas cepas produzem SA a partir da fermentação de carboidrato, entretanto, com utilização lenta do carboidrato e pouco espaço-tempo e rendimentos de carbono (YP/S) de SA a partir da glicose na fonte de carbono e carboidrato. Além do mais, os ácidos succínico, acético e lático foram produzidos em uma razão de 1:0,034:1,6. Nesta análise, o crescimento da cepa que carrega mutações nos genes ldh e pfl foi mais lento se comparado à cepa mãe não mutada.
[0007] Zhu et al. 2005 descrevem uma cepa de E. coli mutada no gene pfl que não produz ácido succínico, mas lactato, e exibe pouco crescimento quando crescida em glicose como o único substrato.
[0008] Um obstáculo significativo do organismo Mannheimia succiniciproducens é, entretanto, sua incapacidade de metabolizar glicerol que, como um constituinte de triacil gliceróis (TAGs), se torna facilmente disponível, por exemplo, como subproduto na reação de transesterificação da produção de biodiesel (Dharmadi et al., 2006).
[0009] A produção fermentativa de SA a partir do glicerol descrita na literatura científica (Lee et al., 2001; Dharmadi et al., 2006) e com maiores rendimentos de glicerol [massa de SA produzido/massa de material bruto consumido] do que com açúcares comuns como a glicose foi atingida (Lee et al., 2001). Entretanto, o rendimento espaço-tempo obtido com glicerol foi substancialmente menor que com a glicose (0,14 vs. 1,0 g de SA/[L h]).
[00010] Apenas em poucos casos a metabolização anaeróbica do glicerol em produtos de fermentação foi descrita. A E. coli é capaz de fermentar glicerol em condições muito específicas, tal como pH acídico que evita o acúmulo do gás hidrogênio na fermentação, e composição de meio apropriado (Dharmadi et al 2006, Yazdani e Gonzalez 2007). Muitos microrganismos são capazes de metabolizar glicerol na presença de aceptores de elétron externos (metabolismo respiratório), poucos são capazes de realizar tão fermentativamente (isto é, na ausência de aceptores de elétrons). O metabolismo fermentativo de glicerol foi estudado com mais detalhes em várias espécies da família Enterobacteriaceae, tais como Citrobacter freundii e Klebsiella pneumoniae. A dissimilação de glicerol nestes organismos é estritamente ligada a sua capacidade de sintetizar o produto altamente reduzido 1,3-propanediol (1,3-PDO) (Dharmadi et al 2006). A conversão de glicerol em SA usando Anaerobiospirillum succiniciproducens foi relatada (Lee et al. 2001). Este estudo demonstrou que SA pode ser produzido com pouca formação de subproduto de ácido acético usando glicerol como uma fonte de carbono, facilitando assim a purificação de SA. O maior rendimento foi obtido alimentando intermitentemente glicerol e extrato de levedura, uma estratégia que resultou na produção de cerca de 19 g/L de SA. Entretanto, nota-se que os componentes nutricionais não identificados presentes no extrato de levedura foram necessários para que a fermentação de glicerol ocorresse. Os sacarídeos, entretanto, podem ser teoricamente convertidos em SA com um rendimento significativamente menor que o glicerol em virtude do menor estado de redução dos sacarídeos comparado ao poliol glicerol. Observou-se que a combinação de sacarídeos com glicerol funciona em organismos anaeróbicos que produzem SA (Lee et al. 2001), entretanto sem atingir s titulações de SA além de 29 g/L. Além disso, observou-se que o rendimento de carbono YP/S é apenas 92 % e observou-se que a relação SA/AA é 4,9:1. Apenas 4 g/L de glicerol foram convertidos em ácido succínico no máximo.
[00011] As reações de carboxilação de oxaloacetato catalisadas pelas enzimas fosfoenolpiruvato carboxilase (PEPC), fosfoenolpiruvato carboxiquinase (PEPCK) e piruvato carboxilase (PycA) utilizam HCO3- como uma fonte de CO2 (Peters-Wendisch, PG et al 1996, 1998). Portanto, as fontes de hidrogenocarbonato, tais como NaHCO3, KHCO3, NH4HCO3 e assim por diante, podem ser aplicadas nos meios de fermentação e cultivo para melhorar a disponibilidade de HCO3- na metabolização de substratos em SA. Não observou-se até o momento que a produção de SA a partir da glicose é dependente da adição de HCO3- na técnica anterior.
[00012] A produção de biomassa por organismos anaeróbicos é limitada pela quantidade de ATP produzido a partir das vias fermentativas. O rendimento da biomassa de glicerol em organismos anaeróbicos é menor que o de sacarídeos, como hexoses, tal como glicose, frutose, pentoses tal como xilose, arabinose, ou dissacarídeos tal como sacarose ou maltose (Lee et al. 2001, Dharmadi 2007).
[00013] O pedido de patente anterior PCT/EP2008/006714, cujo conteúdo é aqui incorporado pela referência, revela uma cepa bacteriana, sendo um elemento da família Pasteurellaceae, originalmente isolado do rúmem e capaz de utilizar glicerol como uma fonte de carbono e variantes e cepas mutantes derivadas destes que mantêm a dita capacidade, em particular, uma cepa bacteriana determinada DD1, da maneira depositada em DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Inhoffenstr. 7B, D-38124 Braunschweig, Alemanha) com o número de depósito DSM 18541 (ID 06-614), e com a capacidade de produzir ácido succínico e variante ou cepas mutantes derivadas destes que mantêm pelo menos a dita capacidade de produzir o ácido succínico. A cepa DD1 pertence à espécie Basfia succiniciproducens e à família Pasteurellaceae, da maneira classificada por Kuhnert et al., 2010.
[00014] Portanto, existe uma necessidade de cepas bacterianas inéditas, que têm a capacidade de produzir ácidos orgânicos, em particular SA, a partir de glicerol. Em particular, tais cepas podem produzir os ditos ácidos com alta produtividade a partir do glicerol, especialmente se o glicerol bruto, por exemplo, a partir da produção de biodiesel, puder ser usado sem purificação prévia. É um objetivo da presente invenção fornecer tais cepas inéditas e processos de produção.
[00015] Os presentes inventores, que isolaram uma cepa bacteriana determinada DD1, resolveram de maneira surpreendente o dito objetivo mutando a dita cepa, de maneira tal que a atividade da proteína PFL diminuiu, de maneira tal que a dita cepa apresente as características metabólicas desejadas. Assim, forneceram um novo tipo de cepa bacteriana, capaz de utilizar glicerol como uma fonte de carbono para a produção fermentativa de ácido succínico, em que a dita cepa é geneticamente modificada, de maneira tal que compreenda uma desregulação de sua atividade de enzima PFL endógena.
[00016] Os presentes inventores observaram surpreendentemente que uma cepa bacteriana mutada como esta, com as características metabólicas desejadas, mostrou comportamento técnico muito melhor na fermentação de SA.
[00017] A figura 1 representa um mapa esquemático de plasmídeo pSacB (SEQ ID NO:3).
[00018] A figura 2 representa um mapa esquemático de plasmídeo pSacB (delta pfl) (SEQ ID NO:4).
[00019] A figura 3 representa um mapa esquemático de plasmídeo pSacB (delta ldh) (SEQ ID NO:5).
[00020] a) Definição geral de termos particulares: O termo “célula bacteriana”, da maneira aqui usada, refere-se a um organismo procarioto, isto é, uma bactéria. As bactérias podem ser classificadas com base em suas propriedades bioquímicas e microbiológicas, bem como sua morfologia. Estes critérios de classificação são bem conhecidos na técnica.
[00021] A palavra “ácido” (no contexto de ácidos orgânicos mono ou dicarboxílicos da maneira aqui referida, isto é, acético, lático e SA) é entendida em seu sentido mais amplo e também inclui sais deste, por exemplo, como sais de metal alcalino, como sais de Na e K, ou sais alcalinos terrosos, como sais de Mg e Ca, ou sais de amônio, ou anidretos dos ditos ácidos.
[00022] A “identidade” ou “homologia” entre duas sequências de nucleotídeo significa a identidade dos resíduos com relação ao tamanho completo das sequências alinhadas tal como, por exemplo, a identidade calculada (preferivelmente para sequências similares) com o auxílio do programa needle a partir do pacote de software de bioinformática EMBOSS (Versão 5.0.0, http://emboss.sourceforge.net/what/) com os parâmetros padrões que são: -gapopen (penalidade de abertura de intervalo): 10,0 -gapextend (penalidade de extensão de intervalo): 0.5 -datafile (arquivo de matriz de classificação incluído no pacote): EDNAFUL
[00023] O termo “cepa bacteriana que contém um gene mutado que codifica uma enzima piruvato-formato-liase com menor atividade” inclui uma célula bacteriana modificada que apresenta uma menor atividade, ou mesmo nenhuma atividade detectável de PFL. Os métodos para a detecção e determinação da atividade de PFL podem ser encontrados em Knappe et al. 1990 e Knappe 1993 e suas referências. Além do mais, o termo inclui uma célula bacteriana que apresenta uma atividade de PFL significativamente reduzida quando comparada a uma célula bacteriana que exibe níveis de atividade fisiológica de piruvato-formato-liase. Se uma redução for significativa, ela pode ser determinada por métodos estatísticos bem conhecidos pelos versados na técnica. As células bacterianas que são deficientes na atividade de PFL podem ocorrer naturalmente, isto é, em virtude de mutações espontâneas. Uma célula bacteriana pode ser modificada para precisar ou ter a atividade de PFL significativamente reduzida por várias técnicas. Preferivelmente, tais células bacterianas são obtidas por tratamento ou radiação química. Para esta finalidade, as células bacterianas serão tratadas, por exemplo, por um agente químico mutagênico, raios-X ou luz UV. Em uma etapa subsequente, aquelas células bacterianas que precisam de PFL, ou que pelo menos apresentam uma menor atividade de PFL, serão selecionadas. As células bacterianas também são obtidas por técnicas de recombinação homóloga, que ajudam a mutar, interromper ou excisar o PFL no genoma da célula bacteriana, ou introduzir mutações que levarão a um gene mutado que codifica uma proteína com menor atividade. Uma técnica preferida para recombinação, em particular, para introduzir mutações ou para deletar sequências, é descrita a seguir.
[00024] A definição anterior também se aplica a outros genes que codificam uma outra enzima aqui mencionada para ser modulada, em particular, cuja atividade é para ser finalizada, diminuída ou interrompida.
[00025] O termo “menor atividade” inclui, por exemplo, a expressão de um produto de gene (por exemplo, piruvato-formato-liase (pfl), lactato desidrogenase (ldh) ou outros) pelo dito microrganismo geneticamente manipulado (por exemplo, geneticamente modificado) em um nível menor que aquele expresso antes da manipulação do microrganismo. A manipulação genética pode incluir, mas sem limitação, alterar ou modificar as sequências regulatórias ou sítios associados à expressão de um gene particular (por exemplo, removendo promotores fortes, promotores indutíveis ou promotores múltiplos), modificar o local cromossômico de um gene particular, alterar as sequências de ácido nucléico adjacentes a um gene particular tal como uma sequência na região promotora, incluindo sequências regulatórias importantes para a atividade promotora de um sítio de ligação de ribossomo ou finalizador de transcrição, diminuir o número de cópias de um gene particular, modificar proteínas (por exemplo, proteínas regulatórias, supressores, melhoradores, ativadores transcricionais e similares) envolvidas na transcrição de um gene particular e/ou tradução de um produto particular de gene, ou qualquer outro meio convencional de diminuir a expressão de um gene particular comum na técnica (incluindo, mas sem limitação, o uso de moléculas de ácido nucléico anti-sentido ou outros métodos para neutralizar ou bloquear a expressão da proteína alvo).
[00026] Em particular, o gene pode ser manipulado aquele um ou mais nucleotídeos são deletados do cromossomo do organismo hospedeiro. A menor atividade do produto de gene, por exemplo, de uma molécula de piruvato-formato-liase, também pode ser obtida introduzindo uma ou mais mutações de genes que levam a uma menor atividade do produto de gene. A menor atividade pode ser uma redução da atividade enzimática em >50 % da atividade enzimática não mutada ou não alterada, ou redução da atividade enzimática em >90 %, ou mais preferivelmente uma redução da atividade enzimática em >95 %, ou mais preferivelmente uma redução da atividade enzimática em >98 %, ou ainda mais preferivelmente uma redução da atividade enzimática em >99 % ou ainda mais preferivelmente uma redução da atividade enzimática em >99,9 %.
[00027] O termo microrganismo “recombinante” inclui um microrganismo (por exemplo, bactérias, célula de levedura, célula fúngica, etc.) que foi geneticamente alterado, modificado ou modificado por engenharia genética (por exemplo, geneticamente modificado), de maneira tal que exibe um genótipo e/ou fenótipo alterado, modificado ou diferente (por exemplo, quando a modificação genética afeta as sequências codificantes de ácido nucléico do microrganismo) comparado ao microrganismo de ocorrência natural a partir do qual foi derivado. O termo “promotor” refere-se a uma sequência de DNA, que direciona a transcrição de um gene estrutural para produzir RNAm. Tipicamente, um promotor está localizado na região 5’ de um gene, próximo ao códon inicial de um gene estrutural. Se um promotor for um promotor indutível, então a taxa de transcrição aumenta em resposta a um agente de indução. Ao contrário, a taxa de transcrição não é regulada por um agente de indução se o promotor for um promotor constitutivo.
[00028] O termo “melhorador” refere-se a um elemento promotor. Um melhorador pode aumentar a eficiência com a qual um gene particular é transcrito em RNAm independente da distância ou orientação do melhorador com relação ao sítio inicial de transcrição.
[00029] O termo “vetor de clonagem” refere-se a uma molécula de DNA, tal como um plasmídeo, cosmídeo, fagomídeo ou bacteriófago, que apresenta a capacidade de replicar autonomamente em uma célula hospedeira e que é usada para transformar células para a manipulação de gene. Os vetores de clonagem contêm tipicamente um ou um número pequeno de sítios de reconhecimento de endonuclease de restrição, em que as sequências de DNA estrangeiro podem ser inseridas de uma maneira determinada sem perda de uma função biológica essencial do vetor, bem como um gene marcador, que é adequado para uso na identificação e seleção de células transformadas com o vetor de clonagem. Os genes marcadores incluem tipicamente genes que fornecem resistência à tetraciclina ou resistência à ampicilina.
[00030] A palavra “vetor” refere-se a uma molécula de DNA que compreende um gene estrutural clonado que codifica uma proteína estrangeira, que fornece um gene em um hospedeiro recombinante. Tipicamente, no caso de um vetor destinado para a integração no genoma do hospedeiro, o gene clonado é colocado ou operavelmente ligado a certas sequências à montante ou à jusante homólogas ou idênticas à sequência genética do hospedeiro.
[00031] O termo “hospedeiro recombinante” refere-se a um hospedeiro que pode ser qualquer célula de procarioto ou de eucarioto, que contém tanto um vetor de clonagem quanto um vetor de expressão. Este termo também significa incluir aquelas células de procarioto ou eucarioto que foram geneticamente modificadas para conter o(s) gene(s) clonado(s) no cromossomo ou genoma da célula hospedeira. Para exemplos de hospedeiros adequados, ver Sambrook et al., 1989.
[00032] As palavras “expressa”, “expressando”, “expressou” e “expressão” referem-se à expressão de um produto de gene (por exemplo, uma enzima biossintética de um gene a partir de uma via ou reação definida e descrita neste pedido) em um nível que a atividade enzimática resultante desta proteína codificada, ou a via ou reação na qual ela se refere, permite o fluxo metabólico por meio desta via ou reação no organismo no qual este gene/via é expresso. A expressão pode ser realizada por alteração genética do microrganismo que é usado como um organismo inicial. Em algumas modalidades, um microrganismo pode ser geneticamente alterado (por exemplo, geneticamente modificado) para expressar um produto de gene em um nível maior com relação àquele produzido pelo microrganismo inicial ou em um microrganismo comparável que não foi alterado. A alteração genética inclui, mas sem limitação, alterar ou modificar as sequências regulatórias ou sítios associados com expressão de um gene particular (por exemplo, adicionando promotores fortes, promotores indutíveis ou promotores múltiplos ou removendo sequências regulatórias, de maneira tal que expressão seja constitutiva), modificar o local cromossômico de um gene particular, alterar as sequências de ácido nucléico adjacentes a um gene particular, tal como um sítio de ligação de ribossomo ou finalizador de transcrição, aumentar o número de cópia de um gene particular, modificar proteínas (por exemplo, proteínas regulatórias, supressores, melhoradores, ativadores transcricionais e similares) envolvidas na transcrição de um gene particular e/ou tradução de um produto de gene particular, ou qualquer outro meio convencional de desregular a expressão de um gene particular usando técnicas comuns (incluindo, mas sem limitação, o uso de moléculas de ácido nucléico anti-sentido, por exemplo, para bloquear a expressão de proteínas repressoras).
[00033] Um microrganismo pode ser física ou ambientalmente “alterado” ou “modificado” para expressar um produto de gene em um maior ou menor nível com relação ao nível de expressão do produto de gene pelo microrganismo inicial. Por exemplo, um microrganismo pode ser tratado ou cultivado na presença de um agente (químico ou genético) conhecido ou suspeito de aumentar ou diminuir a transcrição e/ou tradução de um gene particular e/ou tradução de um produto de gene particular, de maneira tal que a transcrição e/ou tradução sejam aumentadas ou diminuídas. Alternativamente, um microrganismo pode ser cultivado em uma temperatura selecionada para aumentar ou diminuir a transcrição e/ou tradução de um gene particular e/ou tradução de um produto de gene particular, de maneira tal que a transcrição e/ou tradução seja maior ou menor. “Geneticamente modificado” refere-se a um microrganismo alterado no sentido anterior por meio de técnicas de engenharia genética disponíveis na técnica, por exemplo, como transformação, mutação, recombinação homóloga.
[00034] As palavras “desregula”, “desregulado” e “desregulação” referem-se à alteração ou modificação de pelo menos um gene em um microrganismo, em que a alteração ou modificação resulta em maior eficiência de SA no microrganismo com relação à produção de SA na ausência da alteração ou modificação. Em algumas modalidades, um gene que é alterado ou modificado codifica uma enzima em uma via biossintética ou uma proteína de transporte, de maneira tal que o nível ou atividade da enzima biossintética no microrganismo seja alterada ou modificada, ou que a especificidade ou eficiência do transporte seja alterada ou modificada. Em algumas modalidades, pelo menos um gene que codifica uma enzima em uma via biossintética é alterado ou modificado de maneira tal que o nível ou atividade da enzima seja melhor ou maior com relação ao nível na presença do gene inalterado ou tipo selvagem. A desregulação também inclui alterar a região codificante de um ou mais genes para render, por exemplo, uma enzima que é resistente à retroalimentação ou apresenta uma maior ou menor atividade específica. Igualmente, a desregulação inclui adicionalmente alteração genética de genes que codificam fatores transcricionais (por exemplo, ativadores, repressores) que regulam a expressão de genes que codificam enzimas ou proteínas de transporte. Mais especificamente, a desregulação pode resultar em “menor” atividade enzimática, em que a atividade enzimática resultante que é menos de 100 % da atividade enzimática observada no estado não desregulado é “interrompida”, isto é reversível ou irreversivelmente, não mais presente ou pelo menos não mais detectável por uma ferramenta analítica convencional, como um ensaio de atividade enzimática.
[00035] O termo “capaz de utilizar” refere-se à capacidade de um microrganismo da invenção converter um substrato, por exemplo, como glicerol, em pelo menos um produto químico estruturalmente e/ou estericamente diferente.
[00036] Uma “atividade enzimática envolvida ou associada com a conversão fermentativa de glicerol em succinato” significa qualquer atividade catalítica ou regulatória de uma enzima que influencia a conversão de glicerol em succinato e ou subprodutos, como pode ser determinada por qualquer um dos conjuntos de parâmetros da maneira aqui definida a seguir.
[00037] Os diferentes parâmetros de rendimento da maneira aqui descrita (“Rendimento” ou YP/S; “rendimento de produtividade específica” ou rendimento espaço-tempo (STY)) são bem conhecidos na técnica e são determinados da maneira descrita, por exemplo, por Song e Lee, 2006.
[00038] “Rendimento” e “YP/S” (cada qual expresso em massa de produto sintetizado/massa de material consumido) são aqui usados como sinônimos.
[00039] O rendimento de produtividade específica descreve a quantidade de um produto, como SA, que é produzido por hora e litro de caldo de fermentação por grama de biomassa seca. A quantidade de peso seco de célula declarada como DCW descreve a quantidade de microrganismo biologicamente ativo em uma reação bioquímica. O valor é fornecido como grama de produto por grama de DCW por hora (isto é g/gDCW-1 h-1).
[00040] O termo “produção fermentativa” ou “fermentação” refere-se à capacidade de um microrganismo (auxiliado pela atividade enzimática contida ou gerada pelo dito microrganismo) produzir um composto químico em cultura de célula utilizando pelo menos uma fonte de carbono adicionada na incubação.
[00041] Entende-se que o termo "caldo de fermentação" significa uma solução aquosa que baseia-se em um processo fermentativo e que foi ou não desenvolvida, por exemplo, da maneira aqui descrita. b) Definição geral para microrganismos diferentes
[00042] A “célula bacteriana” ou “cepa bacteriana”, referida de acordo com a presente invenção, é selecionada da família de Enterobacteriaceae, Pasteurellaceae, Bacilli ou Actinobacteria.
[00043] “Enterobacteriaceae” representa uma grande família de bactérias, incluindo muitas das bactérias mais familiares, tais como Salmonella e Escherichia coli. Elas pertencem à Proteobacteria e são fornecidas em sua própria ordem (Enterobacteriales). Os elementos da família Enterobacteriaceae são em forma de bastonete. Como outras que pertencem ao filo Proteobacteria, apresentam coloração Gram-negativa, e são anaeróbias facultativas que fermentam açúcares para produzir ácido lático e vários outros produtos finais, tal como ácido succínico. A maioria também reduz nitrato em nitrito. Diferente da maioria das bactérias similares, as Enterobacteriaceae precisam em geral de citocromo C oxidase. A maioria apresenta flagelos que são usados para locomoção, mas muitos gêneros não são móveis. Não formam esporos e são, principalmente, catalase positivas. Muitos elementos desta família são uma parte normal da flora do aparelho digestivo, encontrados nos intestinos de humanos e outros animais, embora outros sejam encontrados na água ou solo, ou sejam parasitas em uma variedade de diferentes animais e plantas. A Escherichia coli, mais conhecida como E. coli, é um dos organismos modelo mais importantes, e sua genética e bioquímica foram muito estudadas. Muitos elementos de Enterobacteriaceae apresentam fímbrias tipo I peritríquias envolvidas na adesão das células bacterianas em seus hospedeiros. Exemplos das Enterobacteriaceae são E. coli, Proteus, Salmonella, Klebsiella.
[00044] “Pasteurellaceae” compreende uma grande e diversificada família de Proteobacteria gram negativa com elementos que variam de bactérias tal como Haemophilus influenzae a comensais da mucosa humana ou animal. Muitos elementos vivem como comensais nas superfícies mucosas de pássaros e mamíferos, especialmente no trato respiratório superior. As bactérias da família Pasteurellaceae são tipicamente em forma de bastonete e são um grupo notável de anaeróbios facultativos. Elas podem ser distinguidas das Enterobacteriaceae relacionadas pela presença de oxidase, e de muitas outras bactérias similares pela ausência de flagelos. As bactérias da família Pasteurellaceae foram classificadas em inúmeros gêneros com base nas propriedades metabólicas e suas sequências do RNA 16S e 23S. Uma definição mais precisa de Pasteurellacea pode ser encontrada em Dousse et al. 2008 e Kuhnert, P. 2008 e em suas referências. Muitas das bactérias da família Pasteurellaceae contêm os genes piruvato-formato-liase e são capazes de fermentar anaerobicamente fontes de carbono em ácidos orgânicos.
[00045] A palavra “Bacilli” refere-se a uma classe taxonômica de bactérias. Inclui duas ordens, Bacillales e Lactobacillales, a espécie Bacillus representa grandes bactérias cilíndricas (~4-8 x1,5μm) que podem crescer em condições aeróbicas a 37 °C. São tipicamente não patogênicas. O gênero Bacillales contém as espécies Alicyclobacillaceae, Bacillaceae, Caryophanaceae, Listeriaceae, Paenibacillaceae, Planococcaceae, Sporolactobacillaceae, Staphylococcaceae, Thermoactinomycetaceae, Turicibacteraceae. Muitas das bactérias do gênero Bacilli contêm genes piruvato-formato-liase e são capazes de fermentar anaerobicamente fontes de carbono em ácidos orgânicos.
[00046] A palavra “Actinobacteria” ou ”Actinomycetes” refere-se a um grupo de bactérias gram positivas com alta razão G+C. Elas incluem alguns dos seres vivos mais comuns do solo, desempenhando um importante papel na decomposição de materiais orgânicos. Outras Actinobacteria habitam plantas e animais, e são exemplos Mycobacterium, Corynebacterium, Nocardia, Rhodococcus e Streptomyces. Algumas Actinobacteria formam filamentos de ramificação que, em alguns casos, se assemelham ao micélio dos fungos não relacionados, entre os quais eles foram originalmente classificados com o nome mais antigo, Actinomycetes. A maioria dos elementos é aeróbia, mas alguns podem crescer em condições anaeróbicas. Ao contrário das Firmicutes, o outro principal grupo de bactérias gram positivas, apresentam DNA com um alto teor de GC.
[00047] As cepas bacterianas preferidas são do gênero “Basfia”. As bactérias do gênero Basfia são gram negativas e anaeróbias facultativas. As espécies de Basfia não são móveis, são pleomórficas e a maioria é frequentemente catalase e oxidase positiva (Kuhnert e Christensen, 2008, ISBN 978-1-904455-34-9). Preferivelmente, a célula bacteriana é uma célula bacteriana Basfia e, mais preferivelmente, uma célula da cepa Basfia DD1.
[00048] Acima de tudo preferivelmente, a cepa DD1 de Basfia é a cepa bacteriana depositada no Budapest Treaty com DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen, GmbH), Alemanha, com o número de depósito DSM 18541. Esta cepa foi originalmente isolada do rúmen de uma vaca de origem alemã.
[00049] As bactérias Basfia podem ser isoladas do trato gastrointestinal de animais e, preferivelmente, de mamíferos. A cepa bacteriana DD1, em particular, pode ser isolada de rúmen bovino e é capaz de utilizar glicerol (incluindo glicerol bruto) como uma fonte de carbono. Preferivelmente, a dita cepa apresenta a capacidade de produzir SA a partir do glicerol (incluindo glicerol bruto), em particular, em condições anaeróbicas. Além do mais, a cepa DD1 Basfia exibe pelo menos uma das características metabólicas adicionais a seguir: a) produção de SA a partir da sacarose; em particular, em condições anaeróbicas; b) produção de ácido succínico a partir da maltose; em particular, em condições anaeróbicas; c) produção de SA a partir da D-frutose; em particular, em condições anaeróbicas; d) produção de SA a partir de D-galactose; em particular, em condições anaeróbicas; e) produção de SA a partir de D-manose; em particular, em condições anaeróbicas; f) produção de SA a partir de D-glicose; em particular, em condições anaeróbicas; g) produção de SA a partir de D-xilose; em particular, em condições anaeróbicas; h) produção de SA a partir de L-arabinose; em particular, em condições anaeróbicas; i) nenhuma utilização de xilitol, inositol e sorbitol; j) crescimento tanto em condições aeróbicas quanto anaeróbicas; k) crescimento em concentrações iniciais de glicose de 75 g/L ou mais; l) tolerância a amônia.
[00050] Em particular, a dita cepa mostra pelo menos 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 ou todas as ditas características metabólicas.
[00051] A cepa DD1 foi analisada com relação à capacidade de co- metabolizar um sacarídeo e o poliol glicerol (PCT/EP2008/006714). Observa- se que DD1 é capaz de co-metabolizar maltose e glicerol resultando na formação da biomassa, formação de SA e simultânea utilização de maltose e glicerol. c) Modalidades preferidas
[00052] Uma primeira modalidade da invenção diz respeito a cepa bacteriana capaz de utilizar glicerol como uma fonte de carbono para a produção fermentativa de SA, em que a dita cepa é geneticamente modificada de maneira tal que compreenda uma desregulação de sua atividade da enzima piruvato-formato-liase endógena. Em particular, a dita atividade da enzima piruvato-formato-liase é menor ou interrompida.
[00053] A dita bactéria mutada, que contém uma piruvato-formato- liase com uma menor atividade, pode ser construída por meio da genética, bem como por indução de mutações aplicando métodos para mutação bem conhecidos na literatura da técnica anterior (exemplos e descrições para a modificação de genomas bacterianos podem ser encontrados em Saier, Milton H Jr 2008, Foster, Patricia L, 2007, Witkin, E M 1969, Eisenstark, A 1971, Walker, G C et al. 1983 e 1984, Botstein, D, e Shortle, D 1985 e suas referências), é capaz de utilizar misturas de diferentes fontes de carbono, tais como sacarídeos e glicerol, ou utilizar apenas glicerol. Os métodos para isolar cepas com mutações no gene pfl podem ser encontrados em Varenne S et al.1975., e em Pascal, M et al. 1981.
[00054] Preferivelmente a dita cepa apresenta a capacidade de produzir SA a partir de diferentes fontes de carbono (incluindo glicerol), em particular, em condições anaeróbicas.
[00055] Em uma outra modalidade da dita cepa, pelos menos uma atividade enzimática adicional envolvida ou associada com a conversão fermentativa de glicerol em succinato é desregulada.
[00056] Em particular, a dita cepa é derivada de um microrganismo selecionado de um microrganismo da família de Enterobacteriaceae, Pasteurellaceae, Bacilli ou Actinobacteria.
[00057] Em particular, a dita cepa é derivada de um microrganismo da família Pasteurellaceae com um DNAr 16S de SEQ ID NO: 1; ou uma sequência que mostra uma homologia de sequência de pelo menos 96, 97, 98, 99 ou 99,9 %, e/ ou com um DNAr 23S de SEQ ID NO: 2; ou uma sequência que mostra uma homologia de sequência de pelo menos 95, 96, 97, 98, 99 ou 99.9 %. Em uma modalidade da presente invenção, a cepa bacteriana é derivada de um microrganismo da família Pasteurellaceae e pertence à espécie Basfia succiniciproducens. A espécie Basfia succiniciproducens é definida por Kuhnert et al., 2010, aqui incorporada pela referência.
[00058] A cepa bacteriana da presente invenção mostra adicionalmente pelo menos uma das características metabólicas adicionais a seguir: a) produção de ácido succínico a partir de sacarose; b) produção de ácido succínico a partir de maltose; c) produção de ácido succínico a partir de maltodextrina; d) produção de ácido succínico a partir de D-frutose; e) produção de ácido succínico a partir de D-galactose; f) produção de ácido succínico a partir de D-manose; g) produção de ácido succínico a partir de D-glicose; h) produção de ácido succínico a partir de D-xilose; i) produção de ácido succínico a partir de L-arabinose; j) produção de ácido succínico a partir de lactose; k) produção de ácido succínico a partir de rafinose; l) produção de ácido succínico a partir de glicerol; m) crescimento em concentrações iniciais de glicose de 75 g/l ou mais; n) crescimento em concentrações iniciais de glicerol de 70 g/l ou mais. por exemplo, como uma combinação de 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 ou todas as ditas características, com a característica l (glicerol- >SA) como um constituinte obrigatório de cada uma das ditas combinações.
[00059] Em uma modalidade adicional, a cepa da invenção converte sacarose, maltose, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, lactose, D-manose, rafinose e/ou glicerol em ácido succínico, com um coeficiente de rendimento YP/S de pelo menos 0,5 g/g, preferivelmente até cerca de 1,28 g/g; por exemplo, como um coeficiente de rendimento YP/S de pelo menos 0,6 g/g, de pelo menos 0,7 g/g, de pelo menos 0,75 g/g, de pelo menos 0,8 g/g, de pelo menos 0,85 g/g, de pelo menos 0,9 g/g, de pelo menos 0,95 g/g, de pelo menos 1,0 g/g, de pelo menos 1,05 g/g, de pelo menos 1,07 g/g, de pelo menos 1,09 g/g de pelo menos 1,10 g/g, de pelo menos 1,11 g/g, de pelo menos 1,22 g/g, ou de pelo menos 1,24 g/g
[00060] Em uma modalidade adicional, a cepa da invenção mostra pelo menos uma das características a seguir: a) converter pelo menos 25 g/L de glicerol em pelo menos 25,1 g/L de ácido succínico, com um coeficiente de rendimento YP/S de pelo menos 1,01 g/g; b) converter pelo menos uma fonte de carbono selecionada de sacarose, maltose, maltodextrina, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, lactose, D-manose, rafinose e/ou glicerol em ácido succínico com um rendimento de produtividade específica de pelo menos 0,58 g DCW-1 h-1 de ácido succínico; c) converter pelo menos uma fonte de carbono selecionada de sacarose, maltose, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, lactose, D-manose e/ou glicerol em ácido succínico com um rendimento espaço-tempo para ácido succínico de pelo menos 2,2 g/(L h) de ácido succínico; d) converter pelo menos 25 g/L de pelo menos uma fonte de carbono selecionada de sacarose, maltose, D-frutose, D-glicose, D-xilose, L- arabinose, D-galactose, lactose, D-manose e/ou glicerol em ácido succínico com um rendimento espaço-tempo para ácido succínico de pelo menos 2,2 g/(L h); e) converter pelo menos uma fonte de carbono selecionada de sacarose, maltose, maltodextrina, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, lactose, D-manose, rafinose e/ou glicerol em ácido succínico com um rendimento de produtividade específica de pelo menos 0,58 g g DCW-1 h-1 de ácido succínico e um rendimento espaço-tempo para ácido succínico de pelo menos 2,2 g/(L h).
[00061] Ainda de acordo com uma outra modalidade, a cepa bacteriana da invenção converte pelo menos 28 g/L de glicerol em pelo menos 28.1 g/L de SA, com um coeficiente de rendimento YP/S de pelo menos 1,0 g/g, ou de >1,0 g/g, ou de > 1,05 g/g, ou de >1,1 g/g, ou de >1,15 g/g, ou de >1,20 g/g, ou de >1,22 g/g, ou de >1,24 g/g, até cerca de 1,28 g/g. Por exemplo, 28 g/L de glicerol podem ser convertidos em até cerca de 40 ou até cerca de 35 g/L de SA.
[00062] Ainda de acordo com uma outra modalidade, a cepa bacteriana da invenção converte pelo menos uma fonte de carbono selecionada de sacarose, maltose, rafinose, maltodextrina, D-frutose, D-glicose, D-xilose, L- arabinose, D-galactose, D-manose e/ou glicerol to SA com um rendimento de produtividade específica de pelo menos 0.6 g g DCW-1 h-1 de SA, ou de pelo menos 0,65, de pelo menos 0,7 g g DCW-1 h-1, de pelo menos 0,75 g g DCW-1 h-1, ou de pelo menos 0,77 g g DCW-1 h-1 de SA.
[00063] Ainda de acordo com uma outra modalidade, a cepa bacteriana da invenção converte pelo menos uma fonte de carbono selecionada de sacarose, maltose, rafinose, maltodextrina, D-frutose, D-glicose, D-xilose, L- arabinose, D-galactose, D-manose e/ou glicerol to SA com um rendimento espaço-tempo para SA de pelo menos 2,2 g/(L h) ou de pelo menos 2,5, pelo menos 2,75, pelo menos 3, pelo menos 3,25, pelo menos 3,5 ou pelo menos 3,7 g/(L*h) de SA.
[00064] Ainda de acordo com uma outra modalidade, a cepa bacteriana da invenção converte pelo menos 28 g/L de pelo menos uma fonte de carbono selecionada de sacarose, maltose, rafinose, maltodextrina, D-frutose, D- glicose, D-xilose, L-arabinose, D-galactose, D-manose e/ou glicerol em SA com um rendimento espaço-tempo para SA de pelo menos 2,2 g/(L h), ou de pelo menos 2,5, pelo menos 2,75, pelo menos 3, pelo menos 3,25, pelo menos 3,5 ou pelo menos 3,7 g/(L*h).
[00065] De acordo com uma outra modalidade, a cepa bacteriana da invenção converte pelo menos uma fonte de carbono selecionada de sacarose, maltose, rafinose, maltodextrina, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, D-manose e/ou glicerol em SA com um rendimento de produtividade específica de pelo menos 0,6 g gDCW-1 h-1, ou de pelo menos 0,65, ou de pelo menos 0,7 g gDCW-1 h-1 de SA, ou de pelo menos 0,77 g gDCW-1 h-1 de SA, e um rendimento espaço-tempo para SA de pelo menos 2,2 g/(L h), ou de pelo menos 2,5, pelo menos 2,75, pelo menos 3, pelo menos 3,25, pelo menos 3,5 ou pelo menos 3,7 g/(L*h).
[00066] Preferivelmente, a dita cepa da invenção pode ser derivada da cepa DD1, da maneira depositada com DSMZ com o número de depósito DSM 18541, ou pode ser derivada de um variante ou cepa de DD1 mutante com a capacidade de produzir ácido succínico.
[00067] As cepas particulares da invenção são produtoras de ácido succínico (SA) e subprodutos (SSP) em uma proporção SA/SSP de >10:1, ou > 12,5:1, ou > 15:1, ou > 17,5.1, ou > 20:1, ou > 25:1, ou > 30:1, ou > 33:1, em que SSP representa o soma de subprodutos de ácido lático (LA), ácido fórmico (FA), ácido acético (AA) e ácido málico (MA), cada qual em uma quantidade que é expressa em g/L.
[00068] As cepas particulares adicionais são produtoras de ácido succínico (SA) e subproduto de ácido acético (AA) em uma proporção SA/AA de >10:1, ou > 12,5:1, ou > 15:1, ou > 17,5:1, ou > 20:1, ou > 25:1, ou > 30:1, ou > 40:1 ou > 50:1, ou > 75:1, ou > 90:1, cada qual em uma quantidade que é expressa em g/L.
[00069] As cepas particulares adicionais são produtoras de ácido succínico (SA) e subprodutos de ácido fórmico (FA) em uma proporção SA/FA de > 90:1, ou > 100:1, cada quantidade sendo expressa em g/L.
[00070] Uma outra modalidade da invenção diz respeito a um processo para a produção fermentativa de um ácido orgânico ou um sal ou derivado deste, cujo processo compreende as etapas de: a) incubar uma cepa bacteriana da maneira definida em uma das reivindicações precedentes em um meio que contém uma fonte de carbono assimilável e que cultiva a dita cepa em condições que favorecem a formação do ácido orgânico desejado; e b) obter o dito ácido orgânico, em particular SA, ou sal ou derivado deste a partir do meio.
[00071] De acordo com um processo particular, a fermentação é realizada em uma temperatura na faixa de cerca de 10 a 60 °C, por exemplo, como 20 a 50 °C, 30 a 45 °C, ou 25 a 35 °C, e em um pH de 5,0 a 9,0, por exemplo, como 5,5 a 8, ou 6 a 7, e na presença de dióxido de carbono. O pH pode ser controlado pela adição de NH4HCO3, (NH4)2CO3, NaOH, Na2CO3, NaHCO3, KOH, K2CO3, KHCO3, Mg(OH)2, MgCO3, MgH(CO3)2, Ca(OH)2, CaCO3, Ca(HCO3)2, CaO, CH6N2O2, C2H7N e/ou misturas destes.
[00072] Em particular, a dita fonte de carbono assimilável é selecionada de glicerol, sacarose, maltose, maltodextrina, D-frutose, D- galactose, D-manose, lactose, D-glicose, D-xilose, L-arabinose, rafinose, produtos de decomposição de amido, celulose, hemiceluloses e lignocelulose; e misturas destes.
[00073] Em particular, a dita fonte de carbono é glicerol ou uma mistura de glicerol e pelo menos uma fonte de carbono adicional selecionada de sacarose, maltose, D-frutose, D-galactose, lactose, D-manose, D-glicose, D-xilose, rafinose e L-arabinose.
[00074] De acordo com uma modalidade particular do dito processo, a concentração da fonte de carbono assimilável é ajustada para um valor em uma faixa de 5 a 80 g/l, por exemplo, como 10 a 60.
[00075] A presente invenção fornece adicionalmente um processo para a produção fermentativa de ácido succínico, ou um sal, ou derivado deste, cujo processo compreende as etapas de: a) incubar uma cepa bacteriana em um meio que contém pelo menos uma fonte de carbono assimilável e cultivar a dita cepa em condições que favorecem a formação do ácido orgânico desejado; b) obter o dito ácido orgânico ou sal ou derivado deste do meio; caracterizado adicionalmente em pelo menos uma das características a seguir: c) conversão de pelo menos 25 g/L de glicerol em pelo menos 25,1 g/L de ácido succínico, com um coeficiente de rendimento YP/S de pelo menos 1,0 g/g; d) conversão de pelo menos uma fonte de carbono selecionada de sacarose, maltose, D-frutose, D-glicose, D-xilose, L-arabinose, D- galactose, D-manose, rafinose e/ou glicerol em ácido succínico com um rendimento de produtividade específica de pelo menos 0,58 g gDCW-1 h-1 de ácido succínico; e) conversão de pelo menos uma fonte de carbono selecionada de sacarose, maltose, D-frutose, D-glicose, D-xilose, L-arabinose, D- galactose, lactose, D-manose, rafinose e/ou glicerol em ácido succínico com um rendimento espaço-tempo para ácido succínico de pelo menos 2,2 g/(L h) de ácido succínico; f) conversão de pelo menos 25 g/L de pelo menos uma fonte de carbono selecionada de sacarose, maltose, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, lactose, D-manose, rafinose e/ou glicerol em ácido succínico com um rendimento espaço-tempo para ácido succínico de pelo menos 2,2 g/(L h); g) conversão de pelo menos uma fonte de carbono selecionada de sacarose, maltose, D-frutose, D-glicose, D-xilose, L-arabinose, D- galactose, lactose, D-manose, rafinose e/ou glicerol em ácido succínico com um rendimento de produtividade específica de pelo menos 0,6 g gDCW-1 h-1 ácido succínico e um rendimento espaço-tempo para ácido succínico de pelo menos 2,2 g/(L h); h) produção de ácido succínico (SA) e subprodutos (SSP) em uma proporção SA/SSP de >10:1, ou > 12,5:1, ou > 15:1, ou > 17,5.1, ou > 20:1, ou > 25:1, ou > 30:1, ou > 33:1, em que SSP representa a soma de subprodutos de ácido lático (LA), ácido fórmico (FA), ácido acético (AA) e ácido málico (MA), cada quantidade sendo expressa em g/L; i) produção de ácido succínico (SA) e do subproduto de ácido acético (AA) em uma proporção de SA/AA de >10:1, ou > 12,5:1, ou > 15:1, ou > 17,5:1, ou > 20:1, ou > 25:1, ou > 30:1, ou > 50:1, ou > 75:1, ou > 90:1, cada quantidade sendo expressa em g/L.
[00076] De acordo com uma modalidade particular do dito processo, a dita cepa bacteriana é uma cepa geneticamente modificada da maneira definida anteriormente.
[00077] Os processos da invenção podem ser realizados descontinua ou continuamente. O curso da produção do ácido pode ser monitorado por meios convencionais, por exemplo, como análise HPLC ou GC.
[00078] Preferivelmente, SA é produzido em condições anaeróbicas. As condições anaeróbicas podem ser estabelecidas por meio de técnicas convencionais, por exemplo, desgaseificando os constituintes do meio de reação e mantendo as condições anaeróbicas pela introdução de dióxido de carbono ou nitrogênio, ou misturas destes, e opcionalmente hidrogênio em uma taxa de fluxo de, por exemplo, 0,1 a 1 ou 0,2 a 0,5 vvm.
[00079] As condições aeróbicas podem ser estabelecidas por meio de técnicas convencionais, por exemplo, introduzindo ar ou oxigênio em uma taxa de fluxo de, por exemplo, 0,1 a 1 ou 0,2 a 0,5 vvm.
[00080] Se apropriado, uma pressão muito leve de 0,01 MPa a 0,15 MPa (0,1 a 1,5 bar) pode ser aplicada de acordo com a invenção.
[00081] Em uma outra modalidade, a invenção fornece um processo para a produção de ácido succínico e/ou sais de amônio e ácido succínico, cujo método compreende a produção fermentativa de ácido succínico da maneira definida anteriormente e, adicionalmente, controlar o pH com uma base de amônia, ou uma solução aquosa desta, ou com NH4HCO3, (NH4)2CO3, NaOH, Na2CO3, NaHCO3, KOH, K2CO3, KHCO3, Mg(OH)2, MgCO3, MgH(CO3)2, Ca(OH)2, CaCO3, Ca(HCO3)2, CaO, CH6N2O2, C2H7N e misturas destes. Em geral, a condição física da pode ser qualquer de uma solução aquosa, suspensão aquosa, gasosa ou sólida.
[00082] Em uma modalidade o ácido orgânico, em particular ácido succínico, e/ou sais deste são produzidos por um dos métodos mencionados anteriormente ou a seguir e são isolados e/ou purificados adicionalmente pelas etapas a seguir: filtração e/ou centrifugação, cromatografia de troca iônica e/ou cristalização.
[00083] Preferivelmente, o ácido orgânico e/ou sais deste são isolados e/ou purificados adicionalmente pelas etapas a seguir: filtração, seguido por cromatografia de troca iônica, seguido por cristalização.
[00084] A filtração pode ser usada para separar as células bacterianas do líquido contendo ácido succínico. A filtração pode ser uma diafiltração, filtração de fluxo cruzado e/ou ultrafiltração.
[00085] O material usado para a cromatografia de troca iônica pode ser uma resina de troca catiônica ácida forte. Uma resina de troca catiônica ácida forte carrega, por exemplo, grupos de ácido sulfônico. Em particular, o material usado para a cromatografia de troca iônica pode ser grupos de ácido sulfônico que carregam estirol-divinil benzol-copolimerisado na forma de H+. A forma de H+ significa que os grupos de ácido sulfônico estão presentes na forma de ácido. Preferivelmente, o tamanho médio da partícula da resina de cromatografia de troca iônica é 0,3 a 1,5, mais preferivelmente 0,55 a 0,75 mm e/ou a densidade aparente é 700 a 800 g/L. A resina de cromatografia de troca iônica pode ser macroporosa. Macroporosa significa que preferivelmente o diâmetro médio do poro da resina de troca catiônica é de 20 a 120 nm, preferivelmente de 20 a 100 nm e, mais preferivelmente, de 20 a 40 nm. A distribuição de partícula é preferivelmente monodispera. Preferivelmente, a capacidade total do material de cromatografia de troca iônica é 0,5 a 2,0, mais preferivelmente 0,8 a 1,7, mais preferivelmente 1,0 a 1,5, mais preferivelmente 1,4 a 1,9 min eq./L. x eq./L significa que 1 L de resina de troca catiônica carrega x mol de grupos de ácido sulfônico. Dessa maneira, eq./L é calculado com relação a uma molécula de carga única. O sal de ácido succínico a ser purificado pode ser um sal de Na, K, Ca, Mg e/ou amônio. Por exemplo, a resina de troca catiônica ácida forte pode ser Type Lewatit Monoplus SP 112 da Lanxess.
[00086] Preferivelmente, a cromatografia de troca iônica é realizada em uma temperatura de 20 a 60 °C, mais preferivelmente de 45 a 60°C.
[00087] Os métodos de produzir SA adicionalmente preferidos são descritos a seguir: Método 1:
[00088] Em uma outra modalidade, a presente invenção fornece um processo para a produção fermentativa de SA ou um sal ou derivado deste, cujo processo compreende as etapas de: a) incubar uma cepa bacteriana em um meio que contém pelo menos uma fonte de carbono assimilável e cultivar a dita cepa em condições que favorecem a formação do ácido orgânico desejado; b) obter o dito ácido orgânico, ou sal ou derivado deste, do meio; e cujo processo é adicionalmente caracterizado pela conversão de pelo menos 50 g/L de glicerol em pelo menos 50 g/L de SA, com um coeficiente de rendimento YP/S de pelo menos 1,0 g/g, ou de >1,0 g/g, ou de > 1,05 g/g, ou de >1,1 g/g, ou de >1,15 g/g, ou de >1,20 g/g, ou de >1,22 g/g, ou de >1,24 g/g; até cerca de 1,28 g/g; por exemplo, como um coeficiente de rendimento YP/S de pelo menos 0,6 g/g, de pelo menos 0,7 g/g, de pelo menos 0,75 g/g, de pelo menos 0,8 g/g, de pelo menos 0,85 g/g, de pelo menos 0,9 g/g, de pelo menos 0,95 g/g, de pelo menos 1,0 g/g, de pelo menos 1,05 g/g, de pelo menos 1,1 g/g, de pelo menos 1,15 g/g, de pelo menos 1,20 g/g, de pelo menos 1,22 g/g, ou de pelo menos 1,24 g/g. Por exemplo, 50 g/L de glicerol podem ser convertidos em até cerca de 65 ou até 62,5 g/L de SA ou até 60 g/L de SA. Método 2:
[00089] Em uma outra modalidade, a presente invenção fornece um processo para a produção fermentativa de SA ou um sal ou derivado deste, cujo processo compreende as etapas de: a) incubar uma cepa bacteriana com uma enzima piruvato- formato-liase com menor atividade em um meio que contém pelo menos uma fonte de carbono assimilável, e cultivar a dita cepa em condições que favorecem a formação do ácido orgânico desejado; b) obter o dito ácido orgânico, ou sal ou derivado deste, do meio; e cujo processo é adicionalmente caracterizado pela conversão de uma fonte de carbono selecionada de sacarose, maltose, maltodextrina, rafinose, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, D-manose e/ou glicerol em SA com um rendimento de produtividade específica de pelo menos 0,42 g gDCW-1 h-1 de AS, ou de pelo menos 0,45 ou de pelo menos 0,47 g g DCW-1 h-1 de SA, ou de pelo menos 0,49 g gDCW-1 h-1 de SA. Método 3:
[00090] Em uma outra modalidade, a presente invenção fornece um processo para a produção fermentativa de SA ou um sal ou derivado deste, cujo processo compreende as etapas de: a) incubar uma cepa bacteriana com uma enzima piruvato- formato-liase com menor atividade em um meio que contém pelo menos uma fonte de carbono assimilável, e cultivar a dita cepa em condições que favorecem a formação do ácido orgânico desejado; b) obter o dito ácido orgânico, ou sal ou derivado deste, do meio; e cujo processo é adicionalmente caracterizado pela conversão de uma fonte de carbono selecionada de sacarose, maltose, maltodextrina, rafinose, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, D-manose e/ou glicerol em SA com um rendimento espaço-tempo para SA de pelo menos 2,22 g/(L h), ou de pelo menos 2,5, pelo menos 2,75, pelo menos 2,9, g/(L*h) de SA. Método 4:
[00091] Em uma outra modalidade, a presente invenção fornece um processo para a produção fermentativa de SA ou um sal ou derivado deste, cujo processo compreende as etapas de: a) incubar uma cepa bacteriana em um meio que contém pelo menos uma fonte de carbono assimilável e cultivar a dita cepa em condições que favorecem a formação do ácido orgânico desejado; b) obter o dito ácido orgânico, ou sal ou derivado deste, do meio; e cujo processo é adicionalmente caracterizado pela conversão de pelo menos 50 g/L de uma fonte selecionada de sacarose, maltose, maltodextrina, rafinose, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, D-manose e/ou glicerol em SA com um rendimento espaço-tempo para SA de pelo menos 2,2 g/(L h), ou de pelo menos 2.5, pelo menos 2,75, pelo menos 3, pelo menos 3,25, pelo menos 3,5 ou pelo menos 3,7 g/(L*h). Método 5:
[00092] Em uma outra modalidade, a presente invenção fornece um processo para a produção fermentativa de SA ou um sal ou derivado deste, cujo processo compreende as etapas de: a) incubar uma cepa bacteriana em um meio que contém pelo menos uma fonte de carbono assimilável e cultivar a dita cepa em condições que favorecem a formação do ácido orgânico desejado; b) obter o dito ácido orgânico, ou sal ou derivado deste, do meio; e cujo processo é adicionalmente caracterizado pela conversão de uma fonte de carbono selecionada de sacarose, maltose, maltodextrina, rafinose, D-frutose, D-glicose, D-xilose, L-arabinose, D-galactose, D-manose e/ou glicerol em SA com um rendimento de produtividade específica de pelo menos 0,6 g gDCW-1 h-1 de SA, ou de pelo menos 0,65, ou de pelo menos 0,7 g gDCW-1 h-1 de SA, ou de pelo menos 0,75 g gDCW-1 h-1 de SA, ou de pelo menos 0,77 g gDCW-1 h-1 de SA e um rendimento espaço-tempo para SA de pelo menos 2,2 g/(L h), ou de pelo menos 2,5, pelo menos 2,75, pelo menos 3, pelo menos 3,25, pelo menos 3,5 ou pelo menos 3,7 g/(L*h).
[00093] Em uma outra modalidade dos métodos 1 a 5 de produzir SA identificados anteriormente, a fonte de carbono é glicerol ou uma mistura de glicerol e pelo menos uma fonte de carbono adicional selecionada de sacarose, maltose, rafinose, maltodextrina, D-frutose, D-galactose, D-manose, D-glicose, D-xilose e L-arabinose.
[00094] As condições particularmente adequadas para produzir SA são: Fonte de carbono: Glicose, xilose, maltose ou maltodextrina, rafinose e/ou glicerol (incluindo glicerol bruto); Temperatura: 30 a 45 °C; pH: 5,5 a 7,0, controlado por uma base da maneira descrita anteriormente, preferivelmente por uma fonte HCO3-, tal como Na2CO3, NaHCO3, Mg(HCO3)2, Ca(HCO3)2 ou, Mg(OH)2, MgCO3,, Ca(OH)2, CaCO3; gás fornecido: CO2
[00095] O SA e/ou sais de SA produzidos podem ser isolados de maneira convencional por métodos conhecidos na técnica, por exemplo, como cristalização, filtração, eletrodiálise, cromatografia. Por exemplo, podem ser isolados precipitando um produto de succinato de cálcio no fermentador durante a fermentação usando hidróxido, óxido, carbonato ou hidrogenocarbonato de cálcio para neutralização e filtração do precipitado. O produto de SA desejado é recuperado do succinato de cálcio precipitado por acidificação do succinato com ácido sulfúrico, seguido por filtração, para remover o sulfato de cálcio (gesso) ou que precipita. A solução resultante pode ser purificada adicionalmente por meio de cromatografia de troca iônica, a fim de remover íons residuais indesejados.
[00096] Em uma outra modalidade, a presente invenção fornece um processo para a produção de tetraidrofurano (THF) e/ou 1,4-butanediol (BDO) e/ou gama-butirolactona (GBL), que compreende: b) a produção fermentativa de ácido succínico e/ou sais de ácido succínico, da maneira definida anteriormente, e b1) tanto a hidrogenação catalítica direta do ácido livre obtido em THF e/ou BDO e/ou GBL quanto b2) a esterificação química de ácido succínico livre obtido e/ou sais de ácido succínico em seu éster de di-alquila inferior correspondente e subsequente hidrogenação catalítica do dito éster em THF e/ou BDO e/ou GBL.
[00097] Em uma outra modalidade, a presente invenção fornece um processo para a produção de pirrolidonas que compreende: a) a produção fermentativa de sais de amônio e ácido succínico da maneira definida anteriormente, e b) a conversão química de sais de amônio e ácido succínico em pirrolidonas de uma maneira conhecida por si.
[00098] Em uma modalidade particular dos processos, o dito glicerol, que é usado como fonte de carbono assimilável, é glicerol bruto, em particular obtido por clivagem de éster de triacilglicerídeos.Por exemplo, o glicerol é um produto residual obtido a partir da fabricação de biodiesel.
[00099] A presente invenção também diz respeito ao uso de uma cepa bacteriana da maneira definida anteriormente para a produção fermentativa de um produto químico orgânico refinado, por exemplo, como ácido succínico, ou um sal ou derivado deste. d) Modalidades particulares adicionais d1) Manipulações genéticas
[000100] Ainda de acordo com uma outra modalidade, a cepa bacteriana da invenção contém um gene que codifica uma enzima mutada da enzima piruvato-formato-liase (pfl), da maneira que sua atividade enzimática é definida pelo numero EC EC 2.3.1.54. Por exemplo, a atividade enzimática de pfl é influenciada negativamente pelas mutações no gene pflA ou afetando a regulação expressional do gene pflA. A sequência do gene pflA e do produto de gene pflA pode ser encontrada nos números de acesso a seguir GeneID:6268899, YP_001880903. Os homólogos deste gene são conhecidos nos números de acesso: NCBI-GeneID 945514, 945444, 947623, 948454, 3075405, as respectivas proteínas nos números de acesso: UniProt: P09373, P75793, P42632, P32674, Q65VK2.
[000101] Estão também no escopo desta invenção os genes que codificam as enzimas de ativação de piruvato-formato-liase que são definidas pelo número EC EC 1.97.1.4 e são descritos em Knappe et al. 1990 e 1993 com atividade menor ou desregulada. Isto pode ser realizado por meio da introdução de mutações ou deleções de gene por métodos descritos nesta invenção. Exemplos para esta enzima cuja atividade pode ser menor ou cujo gene codificante pode ser mutado ou desregulado são codificados pelo pfl que ativa o gene da enzima pflA e o gene yfiD, o gene de K12 E. coli conhecido no acesso GeneID: 947068,o gene ybiY, com o número de acesso NCBI- GeneID: 945445 e a respectiva proteína NP_415345, o gene de Mannheimia succiniproducens conhecido no acesso GeneID: AAU37008, as respectivas proteínas no acesso YP_087593, NP_417074 e YP_087564, bem como os homólogos deste gene. São descritos os número de acessos das sequências de gene não mutado que são submetidas a mutações ou deleções descritas nesta invenção.
[000102] Estão também no escopo desta invenção as cepas que mostram uma atividade reduzida da proteína arcA, por exemplo, acesso: ECK4393 (também conhecida nas descrições a seguir: cpxC, fexA, sfrA, msp), ou fnr carregando mutações genéticas nos respectivos genes, conhecido no número de acesso NCBI-GeneID: 948874 para arcA ou NCBI-GeneID: 945908 para fnr. As sequências da respectiva proteína podem ser encontradas no acesso P0A9E5. Os genes similares são conhecidos por outros organismos, tal como, a saber, Mannheimia succinicproducens. NCBI-GeneID: 3076294 e a respectiva proteína YP_088696 para arcA e para fnr NCBI-GeneID:3075449 e UniProt: Q65TM6.
[000103] Estão também no escopo desta invenção as cepas que mostram uma atividade reduzida da lactato desidrogenase definida pelos números EC EC 1.1.1.27 e EC 1.1.1.28 que codificam enzimas com uma especificidade de produzir ácido D-lático ou L-lático, ou ambos. Os exemplos são os genes de E. coli NCBI-GeneID: 946315 e as respectivas proteína NP_415898 ou o gene de M succiniproducens NCBI-GeneID: 3075603 e a respectiva proteína YP_089271.
[000104] Ainda de acordo com uma outra modalidade, a cepa bacteriana da invenção contém: (1) um gene mutado que codifica uma enzima piruvato- formato-liase definida pela nomenclatura como EC 2.3.1.54, com menor atividade; e/ou (2) um gene mutado que codifica a enzima que ativa piruvato- formato-liase definida pela nomenclatura as EC 1.97.1.4 com menor atividade; e /ou(3) um gene mutado que codifica a proteína arcA e/ou (4) um gene mutado que codifica uma lactato desidrogenase definida pela nomenclatura como EC 1,1.1,27 ou EC 1,1.1,28 com menor atividade.
[000105] Um método particular para preparar cepas bacterianas geneticamente modificadas da invenção é uma técnica que também é algumas vezes aqui referida como a “recombinação do tipo Campbell” (Leenhouts et al., 1989, Appl Env Microbiol 55, 394-400). “Em Campbell”, da maneira aqui usada, refere-se à preparação de um transformante de uma célula hospedeira original, em que uma molécula de DNA de fita dupla circular inteira (por exemplo, um plasmídeo) integrou-se em um cromossomo por um único evento de recombinação homóloga (um cruzamento em evento), e que resulta eficientemente na inserção de uma versão linearizada da dita molécula de DNA circular em uma primeira sequência de DNA do cromossomo que é homóloga a uma primeira sequência de DNA da dita molécula de DNA circular. “Realizada em Campbell” refere-se à sequência linearizada de DNA que foi integrada no cromossomo de um transformante A de “em Campbell”. “Em Campbell” contém uma duplicação da primeira sequência homóloga de DNA, em que cada cópia inclui e circunda uma cópia do ponto de cruzamento de recombinação homóloga.
[000106] “Fora de Campbell”, da maneira aqui usada, refere-se a uma célula descendente de um transformante “em Campbell”, em que um segundo evento de recombinação homóloga (um cruzamento fora do evento) ocorreu entre uma segunda sequência de DNA que está contida no DNA linearizado inserido do DNA “realizado em Campbell”, e uma segunda sequência de DNA de origem cromossômica, que é homóloga à segunda sequência de DNA da dita inserção linearizada. O segundo evento de recombinação resulta na deleção (descarte) de uma porção da sequência de DNA integrada, mas de maneira importante, também resulta em uma porção (esta pode ser tão pequena quanto uma única base) do DNA integrado “realizado em Campbell” que permanece no cromossomo, de maneira tal que comparada à célula hospedeira original, a célula “fora de Campbell” contém uma ou mais mudanças intencional no cromossomo (por exemplo, uma deleção da sequência de DNA, uma única substituição de base, múltiplas substituições de base, inserção de um gene ou sequência de DNA heterólogos, inserção de uma cópia ou cópias adicionais de um gene homólogo ou um gene homólogo modificado, ou inserção de uma sequência de DNA que compreende mais de um destes exemplos anteriormente mencionados listados anteriormente).
[000107] Uma célula “fora de Campbell” é, preferivelmente, obtida por uma contra-seleção contra um gene que está contido em uma porção (a porção que é desejada para ser descartada) da sequência de DNA “realizada em Campbell”, por exemplo, o gene sacB de Bacillus subtilis, que é letal quando expresso em uma célula que cresce na presença de cerca de 5 % a 10 % de sacarose. Tanto com quanto sem uma contra-seleção, uma célula “fora de Campbell” desejada pode ser obtida ou identificada selecionando a célula desejada pelo uso de qualquer fenótipo selecionável tais como, mas sem limitação, morfologia de colônia, cor da colônia, presença ou ausência de resistência a antibióticos, presença ou ausência de uma dada sequência de DNA por reação em cadeia da polimerase, presença ou ausência de um auxotrófico, presença ou ausência de uma enzima, presença ou ausência de uma atividade enzimática tal como uma atividade de piruvato formato liase ou uma atividade de lactato desidrogenase, hibridização de ácido nucléico da colônia, seleção de anticorpo, etc. O termo “em Campbell” e “fora de Campbell” também pode ser usado como verbos em vários tempos verbais para se referir ao método ou processo descrito anteriormente.
[000108] Entende-se que o evento de recombinação homóloga que leva a um “em Campbell” ou “fora de Campbell” pode ocorrer em uma faixa de bases de DNA na sequência homóloga de DNA e, uma vez que as sequências homólogas serão idênticas umas as outras em pelo menos parte desta faixa, em geral não é possível especificar exatamente onde o evento do cruzamento ocorreu. Em outras palavras, não é possível especificar precisamente qual sequência era originalmente do DNA inserido, e qual era originalmente do DNA cromossômico. Além do mais, a primeira sequência homóloga de DNA e a segunda sequência homóloga de DNA são em geral separadas por uma região de não homologia parcial, e é esta região de não homologia que permanece depositada em um cromossomo da célula “fora de Campbell”.
[000109] Preferivelmente, a primeira e segunda sequência homóloga de DNA têm pelo menos cerca de 200 pares de base de tamanho, e podem ter até milhares de pares de base de tamanho. Entretanto, o procedimento pode ser realizado para funcionar com sequências mais curtas ou mais longas. Por exemplo, um tamanho para a primeira e segunda sequências homólogas pode variar de cerca de 500 a 2.000 bases, e a obtenção de um “fora de Campbell” a partir de um “em Campbell” é facilitada arranjando a primeira e segunda sequências homólogas para terem aproximadamente o mesmo tamanho, preferivelmente com uma diferença de menos que 200 pares de base, e acima de tudo preferivelmente com a menor das duas tendo pelo menos 70 % do tamanho da maior em pares de base.
[000110] Aplicando o método de modificação genética anterior, as cepas mutantes de uma cepa particular produtora de SA (isto é, DD1) foram preparadas deletando os genes da enzima endógena piruvato-formato-liase e/ou enzima lactato desidrogenase da maneira descrita com mais detalhes nos exemplos a seguir. d2) Etapas de fermentação:
[000111] Uma fermentação usada de acordo com a presente invenção, por exemplo, pode ser realizada em fermentadores agitados, colunas de bolha e reatores de circulação. Uma perspectiva compreensiva dos possíveis tipos de método, incluindo tipos de agitador e desenhos geométricos, pode ser encontrada em "Chmiel: Bioprozesstechnik: Einführung in die Bioverfahrenstechnik, Band 1". No processo da invenção, os variantes típicos disponíveis são os variantes a seguir conhecidos pelos versados na técnica ou explicados, por exemplo, em "Chmiel, Hammes and Bailey: Biochemical Engineering", tais como em lotes, em lote alimentado, em lote alimentado repetido ou ainda fermentação contínua com e sem reciclagem da biomassa. Dependendo da cepa produtora, a dispersão pelo ar, oxigênio, dióxido de carbono, hidrogênio, nitrogênio ou mistura gasosa apropriada pode ser realizada a fim de atingir bom rendimento (YP/S).
[000112] Antes da conversão química pretendida em um caldo de fermentação ser realizada no processo de acordo com a invenção, o caldo de fermentação pode ser pré-tratado; por exemplo, a biomassa do caldo pode ser removida. Os processos para remover a biomassa são conhecidos pelos versados na técnica, por exemplo, a filtração, sedimentação e flotação. Consequentemente, a biomassa pode ser removida, por exemplo, com centrífugas, separadores, decantadores, filtros ou em aparelhos de flotação. Para recuperação máxima do produto de valor, a lavagem da biomassa é frequentemente aconselhável, por exemplo, na forma de uma diafiltração. A seleção do método é dependente do conteúdo da biomassa no caldo fermentador e das propriedades da biomassa, e também da interação da biomassa com o produto de valor. Em uma modalidade, o caldo de fermentação pode ser esterilizado ou pasteurizado.
[000113] Em uma modalidade adicional, o caldo de fermentação é concentrado. Dependendo da exigência, esta concentração pode ser realizada em lotes ou continuamente. A faixa de pressão e temperatura pode ser selecionada de maneira tal que em primeiro lugar nenhum dano do produto ocorra, e em segundo lugar que seja necessário o uso mínimo do aparelho e da energia. A seleção especializada de níveis de pressão e temperatura para uma evaporação em multiestágios, em particular, possibilita a economia de energia.
[000114] Tanques com agitação, evaporadores de filme descendente, evaporadores de filme fino, evaporadores de circulação rápida forçada e outros tipos de evaporadores podem ser utilizados no modo de circulação natural ou forçada. d3) Esterificação de SA e hidrogenação:
[000115] As condições experimentais adequadas para realizar a esterificação química, seguido por hidrogenação catalítica direta, são bem conhecidas e, por exemplo, descritas no pedido de patente Européia 06007118.0 aqui incorporado pela referência. a) Processo de esterificação:
[000116] O processo de esterificação, que pode compreender uma destilação reativa pode ser realizado usando um aparelho conhecido por si em vários modelos.
[000117] Por exemplo, uma aparelhagem de esterificação, que é operada no modo contínuo, pode ser usada compreendendo uma coluna de retificação com um numero apropriado de estágios teórico atingidos por instalação de bandejas ou embalagens. A carga aquosa que compreende o sal de amônio de SA é alimentada na parte superior da coluna de um vaso reservatório assim que um perfil de temperatura em estado de equilíbrio se forma na coluna, em função da alimentação em alcanol que é evaporado no evaporador de circulação aderente ao recipiente da coluna. A reação forma um fluxo contracorrente de líquido e condensado contendo sal de amônio descendente e fase de vapor contendo alcanol ascendente. Para catalisar a reação de esterificação, um catalisador homogêneo pode ser adicionado à carga inicial do sal de amônio. Alternativamente, o catalisador heterogêneo pode ser fornecido nas partes internas da coluna. O éster carboxílico formado é líquido nas condições do processo e passa por meio da menor extremidade da coluna no recipiente da coluna de destilação, e é continuamente retirado do recipiente. Os componentes gasosos, por exemplo, as misturas azeotrópicas, compreendem alcanol-água e/ou amônia, são removidos da coluna de reação e consequentemente da reação em equilíbrio na parte superior da coluna.
[000118] As modificações adicionais das modalidades específicas descritas anteriormente podem ser implementadas pelos versados na técnica sem esforço excessivo.
[000119] As faixas de parâmetro do processo adequadas para o processo de esterificação, de acordo com a invenção, podem ser determinadas facilmente pelos versados na técnica dependendo da configuração do aparelho usado, por exemplo, tipo de partes internas da coluna usadas, tipo e quantidade dos reagentes, tipo e quantidade do catalisador usado, se apropriado. Por exemplo, sem ser restritivo a estes, os parâmetros individuais podem ser ajustados nas faixas de parâmetro a seguir: Temperatura da coluna: 0-300 °C, em particular 40-250 °C, ou 70-200 °C; Pressão: de 0,01 MPa a 0,6 MPa (0,1 a 6 bar), em particular pressão padrão;
[000120] Tempo de permanência: poucos segundos (por exemplo, de 1 a 60) até dias (por exemplo, de 1 a 5), em particular de poucos minutos (por exemplo, de 1 a 60) a poucas horas (por exemplo, de 1 a 15), mais preferivelmente de poucos minutos (por exemplo, de 5 a 20) a 2 horas. b) Processo de hidrogenação
[000121] Os ésteres de SA ou SA preparados de acordo com a invenção por si, são hidrogenados de uma maneira conhecida por si usando os processos, aparelhos e auxiliares, tais como catalisadores, familiares aos versados na técnica.
[000122] Em particular, uma fase de hidrogenação gasosa contínua ou em lotes é realizada na presença de um catalisador heterogêneo adequado para a hidrogenação do éster. Os parâmetros ideais do processo podem ser estabelecidos pelos versados na técnica para o éster particular sem maior esforço. Por exemplo, a temperatura da reação está na faixa de cerca de 100 a cerca de 300 °C, preferivelmente na faixa de cerca de 200 a 280 °C, e a pressão é de cerca de 0,5 MPa a 10 MPa (5 a 100 bar), por exemplo, de 1 MPa a 5 MPa (10 a 50 bar). A razão molar do reagente para hidrogênio é ajustada na faixa de cerca de 1:100 a cerca de 1:2000, por exemplo, de 1:800 a 1:1500.
[000123] Os catalisadores usados para a reação de hidrogenação são conhecidos pelos versados na técnica. Por exemplo, vários catalisadores de cobre podem ser usados. A técnica anterior descreve, por exemplo, o uso de catalisadores cromito de cobre reduzidos, que são obtidos com o nome 85/1 da Davy Process Technology Ltd., Inglaterra. Entretanto, os catalisadores particularmente adequados de acordo com a invenção são catalisadores de cobre suportados em óxido, o óxido de cobre sendo aplicado em materiais auxiliares de alumínio ou sílica. Os exemplos da hidrogenação de ésteres succínicos em BDO (1,4-Butanediol)/GBL (gama-butirlactona)/THF com catalisadores de cobre também são descritos nas tesis a seguir: Schlander, Jan., Feb. 2000, University of Karlsruhe, "Gasphasenhydrierung von Maleinsauredimethylester zu 1,4-Butandiol, gamma-Butyrolacton und Tetrahydrofuran an Kupfer-Katalysatoren".
[000124] A presente invenção será descrita com mais detalhes por meio dos exemplos a seguir. Os exemplos a seguir são para propósitos ilustrativos e não pretende-se que limitem o escopo da invenção. EXEMPLOS - Modificação genética e cultivo Exemplo 1: Método geral para a transformação de DD1 Tabela 1: Nomenclatura da DD1 tipo selvagem e mutantes referidas nos exemplos.
[000125] A cepa LU13843 de Basfia (DD1 tipo selvagem) foi transformada com DNA por eletroporação usando o protocolo a seguir:
[000126] Para preparar uma pré-cultura, LU 13843 foi inoculada a partir de um crescimento recente em placa de ágar BHI em 40 mL de BHI (infusão de cérebro e coração, Difco) em 100 mL de frasco com agitação. A incubação foi realizada por toda a noite a 30 °C; 200 rpm.
[000127] Para preparar a cultura principal, 50 mL de BHI foram colocados em um frasco de agitação de 100 mL e inoculado em uma OD final (610nm) de 0,4 com a pré-cultura. A incubação foi realizada por aproximadamente 1,5 hora a 30 °C, 200 rpm. As células foram coletadas em uma OD de aproximadamente 1,3, os precipitados foram lavados uma vez com glicerol gelado 10 % a 4 °C e ressuspensos em 1,7 mL de glicerol 10 % (4 °C).
[000128] 100 μL de células competentes foram misturados com 5-10 μg de DNA (10-20 μL) e mantidos no gelo por 2 minutos em uma cubeta de eletroporação com uma espessura de 0,2 cm. A eletroporação ocorreu nas seguintes condições: 800 Q; 25 μF; 2 kV (Gene Pulser, Bio-Rad). 1 mL de BHI foi adicionado imediatamente após a eletroporação e uma incubação foi realizada por 2 horas a 30 °C.
[000129] As células foram plaqueadas em BHI com 5 mg/L de cloranfenicol e incubadas por 2-5 dias a 30 °C até que as colônias dos transformantes ficassem visíveis. Os clones foram isolados e re-estriados em BHI com 5 mg/L de cloranfenicol até que a pureza dos clones fosse obtida. Exemplo 2: Produção de construtos de seleção
[000130] Os plasmídeos de mutação/deleção foram construídos com base no vetor pSacB (SEQ ID NO 3). A figura 1 mostra um mapa esquemático de plasmídeo pSacB. As regiões 5’ e 3’que flanqueiam do fragmento cromossômico, que podem ser deletadas, foram amplificadas por PCR a partir do DNA cromossômico de LU 13843 e introduzidas no dito vetor usando técnicas padrões. Em geral, pelo menos 80 % do ORF foram alvejados para uma deleção. Desta maneira, foram construídos os plasmídeos de deleção para a piruvato-formato-liase pfl, pSacB (Δpfl) (SEQ ID NO 4) e a lactato desidrogenase ldhA, pSacB (ΔldhA) (SEQ ID NO 5). As figuras 2 e 3 mostram mapas esquemáticos do plasmídeo pSacB (Δpfl) e pSacB (ΔldhA).
[000131] Na sequência do plasmídeo pSacB (SEQ ID NO:3) o gene sacB está contido nas 5169-6590. O gene do cloranfenicol está contido nas bases 526-984. O promotor sacB está contido nas bases 3802-4264. O gene do cloranfenicol está contido nas bases 526-984. A origem de replicação para E. coli (ori EC) está contido nas bases 1477-2337.
[000132] Na sequência do plasmídeo pSacB delta pfl (SEQ ID NO:4) a região flanqueante 3’ do gene pfl, que é homóloga ao genoma de DD1, está contida nas bases 65-1533, enquanto a região flanqueante 5’ do gene PFL, que é homóloga ao genoma de DD1, está contida nas bases 1534-2956. O gene sacB está contido nas bases 5256-6677. O promotor sacB está contido nas bases 6678-7140. O gene cloranfenicol está contido nas bases 3402-3860. A origem de replicação para E. coli (ori EC) está contida nas bases 43535213.
[000133] No plasmídeo pSacB delta ldh (SEQ ID NO:5) a região flanqueante 5’ do gene ldh, que é homóloga ao genoma de DD1, está contida nas bases 2850-1519, enquanto a região flanqueante 3’ do gene ldh, que é homóloga ao genoma de DD1, está contida nas bases 1518-63. O gene sacB está contido nas bases 5169-6590. O promotor sacB está contido nas bases 6591-7053. O gene de cloranfenicol está contido nas bases 3315-3773. A origem de replicação para E. coli (ori EC) está contida nas bases 4266-5126.
[000134] Exemplo 3: Produção de cepas que produzem succinato melhor
[000135] a) LU 13843 foi transformada da maneira descrita anteriormente com o pSacB (Δpfl) e “realizada em Campbell” para render uma cepa “em Campbell”. A transformação e integração no genoma de LU 13843 foi confirmada por PCR que rende bandas para o evento integracional do plasmídeo no genoma de LU 13843.
[000136] A cepa “em Campbell” foi então “realizada fora de Campbell” usando placas de ágar contendo sacarose como um meio de contra seleção, selecionado a perda (de função) do gene sacB. Portanto, as cepas “em Campbell” foram incubadas em 25-35 mL de meio não seletivo (BHI sem atibiótico) a 37 °C, 220 rpm por toda a noite. As culturas crescidas por toda a noite foram então estriadas em BHI recentemente preparado contendo placas de sacarose (10 %, sem antibióticos) e incubadas por toda a noite a 37 °C (“primeira transferência para a sacarose”). Colônias individuais obtidas da primeira transferência foram novamente estriadas em BHI recentemente preparado contendo placas de sacarose (10 %) e incubadas por toda a noite a 37 °C (“segunda transferência para a sacarose”). Este procedimento foi repetido até uma mínima finalização de cinco transferências (“terceira, quarta, quinta transferência para a sacarose”) em sacarose. O termo “primeira a quinta transferência para a sacarose” refere-se à transferência de uma cepa após integração cromossômica de um vetor contendo um gene levansucrase sacB em sacarose e meio de crescimento que contém placas com ágar com o propósito de selecionar cepas com a perda do gene sacB e as sequências de plasmídeo ao redor. Colônias isoladas de placas da quinta transferência foram inoculadas em 25-35 mL de meio não seletivo (BHI sem antibiótico) e incubadas a 37 °C, 220 rpm por toda a noite. A cultura crescida por toda a noite foi diluída em série e plaqueada em placas de BHI para obter colônias isoladas.
[000137] As cepas “realizadas fora de Campbell” contendo a mutação/deleção do gene pfl foram confirmadas por sensibilidade ao cloranfenicol. Os mutantes de mutação/deleção entre estas cepas foram identificados e confirmados por análise de PCR. Isto leva ao mutante de mutação/deleção de pfl DD1 delta pfl LU 15348.
[000138] b) LU15348 foi transformada com pSacB (Δldh) da maneira descrita anteriormente e “realizada em Campbell” para render uma cepa “em Campbell”. A transformação e integração foram confirmadas por PCR. A cepa “em Campbell” foi então “realizada fora de Campbell” da maneira descrita anteriormente. Os mutantes de deleção entre estas cepas foram identificados e confirmados por análise de PCR. Isto levou ao mutante de delação dupla de pfl ldhA LU15224.
[000139] c) LU13843 foi transformada com pSacB (Δldh) da maneira descrita anteriormente e “realizada em Campbell” para render uma cepa “em Campbell”. A transformação e integração foram confirmadas por PCR. A cepa “em Campbell” foi então “realizada fora de Campbell” da maneira descrita anteriormente. Os mutantes de deleção entre estas cepas foram identificados e confirmados por análise de PCR. Isto levou ao mutante de deleção de ldhA LU15050. Exemplo 4: Preparação do banco de célula 1. Preparação dos meios A composição dos meios de cultivo é descrita na tabela 2.
 a As concentrações de glicose foram 15 g/L (em placas) e 20 ou 50 g/L (em meios líquidos). b MgCO3 (Riedel-de Haen, número do produto: 13117 da Sigma-Aldrich Laborchemikalien GmbH) as concentrações foram 5 g/L (em placas) e 0 ou 30 g/L (em meios líquidos).
a As concentrações de glicose foram 15 g/L (em placas) e 20 ou 50 g/L (em meios líquidos). b MgCO3 (Riedel-de Haen, número do produto: 13117 da Sigma-Aldrich Laborchemikalien GmbH) as concentrações foram 5 g/L (em placas) e 0 ou 30 g/L (em meios líquidos).
[000140] 5 g de extrato de levedura, 5 g de peptona, MgCO3 e (para meios sólidos) 12 g de Bacto-Agar foram misturados em 900 mL de água destilada e autoclavados (20 minutos). Após resfriamento a cerca de 65°C, os componentes que faltam foram adicionados como soluções estoque estéreis. Glicose, sulfato de amônio e K2HPO4 foram todos autoclavados separadamente. Cloretos de Ca, Mg e Na foram autoclavados juntos. 2. Preparação de MCB
[000141] O banco de célula principal (MCB) para a inoculação dos experimentos individuais foi realizado da maneira a seguir. Duas placas de ágar foram inoculadas recentemente com a cepa desejada e incubadas a 37 °C em uma jarra anaeróbica (Anaerocult A, Merck) por toda a noite. A biomassa foi retirada das placas e ressuspensas no meio líquido sem MgCO3 com 20 g/L de glicose para ajustar a OD6OO~1,O. A inoculação foi realizada com 0,5 mL desta suspensão celular. Os cultivos foram realizados em garrafas de soro de 100 mL com tampas de borracha butílica a prova e gás (Ochs GmbH, Bovenden/Lenglern, Alemanha) contendo 50 mL do meio líquido com 20 g/L de glicose e 30 g/L deMgCO3 e uma atmosfera de CO2 com 0,08 MPa (0,8 bar) de pressão excessiva. As garrafas de soro (em total 10) foram incubadas a 37 °C, uma velocidade rotatória de 160 rpm e um diâmetro de agitação de 2,5 cm.
[000142] Para monitorar o consume de glicose, o cultivo de uma garrafa parou e foram realizadas amostragem e análise HPLC após 0, 3, 4, 5, 7, 8 e 8,5 horas. Após 8,5 h (a concentração de glicose foi 3,4 g/L) o cultivo parou. Alíquotas de 0,5 mL de suspensão celular e 0,5 mL de glicerol estéril foram preenchidas em criofrascos, misturadas e armazenadas por 13 horas a -20 e em seguida a -80 °C como MCB. O MCB foi testado com relação à pureza estriando uma alça do último criofrasco em placas de ágar, para controle da contaminação e conferir em cultura líquida (meios descritos na tabela 8) o espectro do produto, e com relação à contaminação (por microscopia).
[000143] O consumo de glicose e a formação de SA e subprodutos foram quantificados por meio da análise HPLC dos sobrenadantes sem célula não diluídos do frasco de cultivo usando detecção RI. As amostras de caldo foram obtidas com uma seringa estéril por meio do plugue de borracha butílica, a separação celular foi realizada por filtração (0,22 μm). Uma coluna Aminex HPX-87 H (Biorad) 300 x 7,8 mm I. D. e H2SO4 5 mm foram usadas como fase estacionária e móvel, respectivamente. A temperatura da coluna foi 30 °C, a vazão foi 0,5 mL min-1.
[000144] Um frasco do MCB foi usado para inocular uma garrafa de soro de 100 mL com tampa de borracha butílica a prova de gás (ver anteriormente) contendo 50 mL do meio líquido com 50 g/L de glicose. A incubação foi realizada por 10 horas a 37 °C em um incubador com agitação (velocidade de rotação: 180 rpm, diâmetro da agitação: 2,5 cm). No final do cultivo, a concentração de glicose foi 20 g/L e o pH em torno de 6,5. Alíquotas de 0,5 mL de suspensão celular e 0,5 mL de glicerol estéril foram preenchidas em criofrascos, misturadas e armazenadas a -80 °C como WCB. A pureza conferida foi a mesma do MCB. As condições de HPLC foram as mesmas daquelas descritas anteriormente. Exemplo 5: Cultivo de várias cepas DD1 em glicerol ou glicerol e maltose
[000145] A produtividade da cepa mutante DD1Δpfl (LU15348) e DD1Δpfl Δldh (LU15224) na presença de glicerol ou glicerol e maltose como uma fonte de carbono foi analisada adicionalmente utilizando o meio e as condições de incubação a seguir. 1. Meio preparação
[000146] A composição e preparação do meio de cultivo é da maneira descrita na tabela 3 a seguir. Tabela 3: Composição do meio para cultivo de DD1 nos substratos glicerol ou glicerol e maltose. Meio de crescimento sintético alternativo
Meio de crescimento sintético alternativo
[000147] É favorável usar um meio de crescimento sintético sem ingredientes complexos para a fermentação a fim de melhorar o processamento à jusante e projetar um meio de crescimento sintético para fermentação mais barata. Preparação do meio
[000148] O meio de crescimento sintético foi desenvolvido com relação a outros meios de crescimento sintéticos para bactérias do rúmen (Nili e Brooker, 1995, McKinlay et al, 2005), experiência interna prévia com outras bactérias e realizando experimentos de omissão simples. Finalmente, o meio continha 50 g/L de glicose, 1 g/L de (NH4)2SO4, 0,2 g/L CaCl2*2H2O, 0,2 g/L de MgCl2*6H2O, 1 g/L de NaCl, 3 g/L de K2HPO4, 1 mg/L de ácido nicotínico, 1,5 mg/L de ácido pantotênico, 5 mg/L de piridoxina, 5 mg/L de riboflavina, 5 mg/L de biotina, 1,5 mg/L de tiamina HCl, 0,26 g/L de lisina, 0,15 g/L de treonina, 0,05 g/L de metionina, 0,71 g/L de ácido glutâmico, 0,06 g/L de histidina, 0,07 g/L de triptofano, 0,13 g/L de fenilalanina, 0,06 g/L de tirosina, 0,5 g/L de serina, 0,5 g/L de glicina, 0,5 g/L de cisteína, 0,1 g/L de β-alanina, 0,27 g/L de alanina, 0,19 g/L de valina, 0,23 g/L de leucina, 0,16 g/L de isoleucina, 0,33 g/L de ácido aspártico, 0,1 g/L de asparagina, 0,13 g/L de prolina, 0,15 g/L de arginina e/ou 0,1 g/L de glutamina.
[000149] As garrafas de soro contendo 50 mL de meio de crescimento sintético foram autoclavadas com água e 30g/L de MgCO3 como o sistema tampão. Glicose, sulfato de amônio e fosfato de potássio foram esterilizados separadamente. Cloretos de Ca, Mg e Na foram esterilizados juntos. As vitaminas e aminoácidos foram montadas em várias soluções estoques e esterilizadas por filtro. Após resfriamento das garrafas de soro, os componentes foram adicionados como soluções estoque estéreis. 2. Cultivos e análises
[000150] Para crescer a cultura semente, um frasco do WCB foi usado para inocular uma garrafa de soro de 100 mL com tampa de borracha butílica a prova de gás (ver anteriormente) contendo 50 mL do meio líquido descrito na tabela 2, mas com 20 g/L de glicose e uma atmosfera de CO2 com 0,08 MPa (0,8 bar) de pressão excessiva. A incubação foi realizada por várias horas específicas de mutante (tabela 4) a 37 °C e 160 rpm (diâmetro de agitação: 2,5 cm). A suspensão celular foi centrifugada (Biofuge primo R, Heraeus,) com 5.000 g por 5 minutos e o precipitado celular foi lavado, e a seguir ressuspenso em 50 mL meio sem uma fonte de carbono e sem MgCO3 para gerar um inoculo sem glicose (todas as etapas em temperatura ambiente e na câmara anaeróbica). Tabela 4: Tempo de incubação de várias culturas sementes de mutante DD1
[000151] As principais culturas foram crescidas em garrafas de soro de 100 mL contendo 10 mL de meio líquido tanto com 50g/L de glicerol quanto com 50g/L de glicerol e 10 g/L de D-maltose, e em ambos os casos, uma atmosfera de CO2 com 0,08 MPa (0,8 bar) de pressão excessiva. A qualidade “glicerol com 99 % de pureza” (Riedel-de Haen, número do produto: 15523- 1L-R por Sigma-Aldrich Laborchemikalien GmbH, Seelze, Alemanha) foi usada para todos os experimentos. A inoculação foi realizada com 1,5 mL do inoculo sem glicose. As garrafas foram incubadas a 37 °C e 160 rpm (diâmetro da agitação: 2,5 cm).
[000152] O consumo das fontes de C e a produção de ácidos carboxílicos foram quantificados por meio de HPLC, da maneira descrita no exemplo 4, após 24 horas. A medida que o glicerol foi medido, a temperatura da coluna foi ajustada para 50 °C para atingir uma separação suficiente de SA, ácido lático e glicerol, que apresentaram períodos de retenção similares.
[000153] O crescimento celular foi medido avaliando a absorbância a 660nm (OD600) usando um espectrofotômetro (Ultrospec3000, Amersham Biosciences, Uppsala, Suécia). A concentração celular definida como peso celular seco em grama (DCW) por litro foi calculada a partir da curva padrão pré-determinada relacionando a OD600 com DCW (1 OD600 = 0,27g g DCW l-1). 3. Resultados
[000154] Os resultados dos experimentos de cultivo com as diferentes cepas DD1 cepas são mostrados na tabela 5 com relação ao substrato glicerol e tabela 6 com relação ao substrato mistura de glicerol e maltose. Tabela 5: Cultivo de várias cepas DD1 em glicerol a tempo de cultivo. b consumo de substrato (glicerol, maltose). 4. formação de ácido succínico, lático, fórmico, acético, pirúvico e málico. 5. soma de subprodutos de ácido lático, fórmico, acético, pirúvico e málico. 6. razão de SA por soma de subprodutos. 7. razão de SA por subproduto (FA = ácido fórmico; AA = ácido acético). 8. rendimento espaço-tempo e rendimento (YP/S) para SA. 9. Observou-se que os limites de detecção para ácido acético, ácido lático, ácido málico e ácido fórmico são menores que 0,01g/L no dado método HPLC.
a tempo de cultivo. b consumo de substrato (glicerol, maltose). 4. formação de ácido succínico, lático, fórmico, acético, pirúvico e málico. 5. soma de subprodutos de ácido lático, fórmico, acético, pirúvico e málico. 6. razão de SA por soma de subprodutos. 7. razão de SA por subproduto (FA = ácido fórmico; AA = ácido acético). 8. rendimento espaço-tempo e rendimento (YP/S) para SA. 9. Observou-se que os limites de detecção para ácido acético, ácido lático, ácido málico e ácido fórmico são menores que 0,01g/L no dado método HPLC.
[000155] No experimento de cultivo em glicerol mostra-se que a neutralização do gene piruvato formato liase pfl em um organismo produtor de SA, por exemplo, como DD1, leva ao rendimento de carbono significativamente maior (YP/S) e STY para SA quanto à tipo selvagem quando crescida em glicerol como um substrato. O rendimento de carbono (YP/S) é aumentado de 1,12 g/g para a cepa DD1 para 1,15 g/g para a cepa mutante Δpfl LU15348.
[000156] Sem ser relatado por Lee et al, 2006 ou Lin et al 2005 com relação à bactérias produtoras de SA em glicose, a neutralização de apenas o gene lactato desidrogenase ldhA em DD1 (LU15050) não mostra mostra nenhuma melhoria das características relevantes técnicas desta fermentação, tal como STY de SA, e apenas pouco aumento no rendimento de carbono (YP/S). A quantidade de ácido acético, entretanto, ainda é maior se a atividade enzimática de lactato desidrogenase for menor. Surpreendente e inesperadamente, a partir das análises do comportamento de cepas com as mutações únicas, observa-se que na cepa mutante, que carrega a combinação das mutações de gene no gene pfl e no gene ldh, a fermentação de glicerol em SA mostra melhorias não aditivas ainda maiores com relação ao rendimento de carbono alcançado (YP/S), da maneira esperada para as mutações simples de LU 15050 e LU15348. O rendimento de carbono (YP/S) de 1,26 g/g observado é próximo do rendimento de carbono teórico potencial (YP/S) de 1,28 g/g para a conversão de 1 Mol de glicerol + 1 Mol de CO2 a 1 Mol de SA.
[000157] Igualmente, a soma de subprodutos (SSP) gerada em LU15348 é significativamente menor, crescendo ao mesmo tempo no glicerol, já que nenhum ácido fórmico e menos ácido acético são produzidos. Da maneira mencionada anteriormente, a concentração de SA é significativamente maior comparada ao tipo selvagem ou LU15050. Esta observação é expressa nas razões de SA com relação à soma aritmética de subprodutos (SSP) SA:SSP g/g, que excede 40 para LU15348 e excede 100 para LU15224 comparada a um nível de 10 para LU13843 e LU15050.
[000158] No experimento anteriormente mencionado, observa-se que o STY da fermentação de glicerol em SA não excedeu 1,5g/(l*h) para as cepas que carregam mutações nos genes pfl e ldh. Portanto, um processo melhor que foi desenvolvido mostrou melhores valores de STY para a produção de SA em uma fermentação anaeróbica. Este processo é descrito no pedido PCT/EP2008/006714 nas páginas 44-46. Este processo foi adaptado para a produção de succinato utilizando cepas que carregam mutações nos genes. Tabela 6: Cultivo de várias cepas DD1 em glicerol e maltose a tempo de cultivo. b consumo de substrato (glicerol, maltose). c formação de ácido succínico, lático, fórmico, acético, pirúvico e málico. d soma de subprodutos de ácido lático, fórmico, acético, pirúvico e málico. e razão de SA por soma de subprodutos. f razão de SA por subproduto (FA = ácido fórmico; AA = ácido acético). g rendimento espaço-tempo e rendimento (YP/S) para SA. h Densidade ótica a 600nm, diluindo a amostra 1:20 com HCl 1M antes de medir em um Ultrospec2000, Amersham Biosciences, Uppsala, Suécia. i g Biomassa como peso celular seco (DCW) h Produtividade específica: g de SA por g de biomassa (peso celular seco) por hora i Observou-se que os limites de detecção para ácido acético, ácido lático, ácido málico e ácido fórmico são menores que 0,01g/L no dado método HPLC.
a tempo de cultivo. b consumo de substrato (glicerol, maltose). c formação de ácido succínico, lático, fórmico, acético, pirúvico e málico. d soma de subprodutos de ácido lático, fórmico, acético, pirúvico e málico. e razão de SA por soma de subprodutos. f razão de SA por subproduto (FA = ácido fórmico; AA = ácido acético). g rendimento espaço-tempo e rendimento (YP/S) para SA. h Densidade ótica a 600nm, diluindo a amostra 1:20 com HCl 1M antes de medir em um Ultrospec2000, Amersham Biosciences, Uppsala, Suécia. i g Biomassa como peso celular seco (DCW) h Produtividade específica: g de SA por g de biomassa (peso celular seco) por hora i Observou-se que os limites de detecção para ácido acético, ácido lático, ácido málico e ácido fórmico são menores que 0,01g/L no dado método HPLC.
[000159] Crescer DD1 em glicerol simultaneamente com um outro sacarídeo maltose mostrou permitir um maior STY e rendimento de SA (YP/S) e uma maior concentração de subprodutos, comparados usando glicerol como o único substrato (PCT/EP2008/006714 nas páginas 44-46).
[000160] A comparação de LU15348 à DD1 tipo selvagem mostra maior quantidade de SA, STY e rendimento de carbono (YP/S) se a atividade da enzima Pfl for menor. Surpreendentemente, ao contrário de outros exemplos descritos na técnica declarada (Lee et al 2006, Lin et al 2005), nenhuma deficiência no crescimento foi observada nas cepas mutantes com relação à cepa DD1 não mutada. Esta observação é de grande relevância técnica, uma vez que o bom crescimento de uma cepa é essencial para um processo técnico de produção. O crescimento da célula é maior em todos os mutantes comparado ao tipo selvagem. A neutralização tanto de pfl quanto de ldh apresenta um efeito positivo no crescimento da cepa bacteriana mutada.
[000161] Em virtude da necessidade de ácido fórmico detectável, menor quantidade de ácido acético e maior concentração de SA, a razão SA/SSP é maior em cepas mutantes contendo menor atividade enzimática de Pfl deduzida por mutações genéticas. Entretanto, o subproduto do ácido lático aumentou comparado ao tipo selvagem. O LU15224 neutralizado duas vezes apresentou maior rendimento adicional (YP/S) e STY, enquanto LU15050 não mostrou nenhuma melhoria no rendimento de carbono (YP/S), STY ou o SSP observada.
[000162] Nota-se que a pfl mutação é necessária e suficiente para melhorar a fermentação de glicerol com e sem um segundo substrato de sacarídeo com relação ao desempenho de uma cepa tipo selvagem, em um processo de AS, com base na metabolização de glicerol. Ao contrário destes achados, uma mutação pfl na técnica anterior em uma cepa tipo selvagem derivada não mostrou induzir a fermentação de SA (Zhu 2005). Apenas as combinações de várias mutações incluindo pfl e ldh não resultaram em uma produção mensurável de ácido succínico, embora em menor crescimento e pouco desempenho de STY (Lin 2005, Lee 2006). Os resultados deste trabalho preceituam a construção de um processo melhor para a produção fermentativa de AS, que consiste em uma cepa mutada junto com um processo específico para render um processo com desempenho superior com relação à técnica anterior.
[000163] Surpreendentemente, a produtividade específica de SA é superior para a cepa mutante pfl LU15348 com relação a LU15050 e a LU15224 que carregam mutações no gene ldh e tanto no gene ldh quanto pfl. Os versados na técnica sabem que, dependendo do processo, uma atividade específica elevada de formação de produto é uma característica elevada de um processo técnico. Obviamente, o efeito negativo da neutralização de lactato desidrogenase diminui a produtividade específica de LU15348 abaixo do valor do tipo selvagem. Exemplo 6: Cultivo de LU15348 em misturas de glicerol com vários carboidratos
[000164] A produtividade da cepa mutante LU15348 na presença de glicerol e vários carboidratos como uma fonte de carbono foi analisada utilizando o meio e as condições de incubação a seguir. 1. Preparação do meio, cultivo e análises
[000165] A composição e preparação do meio de cultivo é da maneira descrita na tabela 3 do exemplo 5. O cultivo e análises ocorrem da maneira descrita no exemplo 5.
[000166] A qualidade “maltodextrina” (Maldex150, Cat.no.: 50499 by Boom, 7942 JE Meppel, Holanda) foi usada neste experimento. Em virtude da mistura não definida de cadeias de sacarídeo com vários tamanhos, a concentração de maltodextrina não foi analisada em análises HPLC de extensão total. Portanto, o teor de maltodextrina foi determinado gravimetricamente de maneira precisa antes de ser adicionada ao meio de cultivo. A fim de calcular o menor limite do rendimento teórico atingido(YP/S), assume-se que toda a maltodextrina adicionada na fermentação foi consumida, estando ciente de que isto permitirá apenas o cálculo do menor limite de rendimento carbono (YP/S) após a fermentação de SA. Mais provavelmente, os valores mais exatos do rendimento de carbono (YP/S) serão maiores que os valores descritos em virtude do consumo potencialmente incompleto do substrato maltodextrina que não é detectado. 2. Resultados
[000167] Os resultados dos experimentos de cultivo para LU15348 são mostrados na tabela 7 para o substrato glicerol em co-fermentação com vários carboidratos, por exemplo, maltose, maltodextrina ou rafinose. Tabela 7: Cultivo de LU15348 em glicerol em co-fermentação com vários carboidratos a tempo de cultivo. b consumo de substrato (glicerol, maltose). c formação de ácido succínico, lático, fórmico, acético, pirúvico e málico. d soma de subprodutos de ácido lático, fórmico, acético, pirúvico e málico. e razão de SA por soma de subprodutos. f razão de SA por subproduto (FA = ácido fórmico; AA = ácido acético). g rendimento espaço-tempo e rendimento (YP/S) para SA. i Observa-se que os limites de detecção para ácido acético, ácido lático, ácido málico e ácido são menores que 0,01g/L no dado método HPLC. - menor limite de rendimento (YP/S) em virtude de concentração residual de maltodextrina desconhecida cuja não foi analisada.
a tempo de cultivo. b consumo de substrato (glicerol, maltose). c formação de ácido succínico, lático, fórmico, acético, pirúvico e málico. d soma de subprodutos de ácido lático, fórmico, acético, pirúvico e málico. e razão de SA por soma de subprodutos. f razão de SA por subproduto (FA = ácido fórmico; AA = ácido acético). g rendimento espaço-tempo e rendimento (YP/S) para SA. i Observa-se que os limites de detecção para ácido acético, ácido lático, ácido málico e ácido são menores que 0,01g/L no dado método HPLC. - menor limite de rendimento (YP/S) em virtude de concentração residual de maltodextrina desconhecida cuja não foi analisada.
[000168] O cultivo da cepa LU15348 em glicerol em co-fermentação com diversos sacarídeos, tal como maltose, uma mistura de sacarídeo de alto peso molecular, tal como maltodextrina, ou ainda um outro sacarídeo rafinose, leva a resultados similares que mostram que a fermentação simultânea de diversas fontes de carbono, incluindo glicerol como uma fonte de carbono, leva a inúmeras melhorias tecnicamente relevantes da produção de SA com relação à técnica declarada anteriormente descrita. Os exemplos são maiores taxas e melhores quantidades totais de glicerol consumido pelo processo, o que leva a maior titulação de SA comparada ao da técnica declarada. Adicionalmente, o STY é maior com relação ao controle que não contém um sacarídeo. A concentração do subproduto é em geral menor, exceto no caso da maltose como co-substrato, onde o ácido lático é o maior subproduto comparado com o cultivo apenas com glicerol. O rendimento de SA (YP/S) é similar ou apenas levemente diminuído comparado ao glicerol como o único substrato.
[000169] Sumário dos experimentos - microrganismos que diminuíram atividade de pfl mostram mais fermentação no glicerol com relação à técnica declarada; - combinação de diferentes fontes de carbono são convertidas de maneira eficiente em ácido succínico; - microrganismos que diminuíram a atividade de pfl mostram mais fermentação em uma mistura de glicerol e um sacarídeo com relação à técnica declarada.
[000170] Conclusão: O novo processo para a produção de ácido succínico (SA) apresenta um excelente potencial para a produção de SA e/ou sais de SA, com alto rendimento de carbono (YP/S) e rendimento espaço- tempo, bem como com poucos subprodutos.
[000171] No contexto da presente invenção, uma cepa bacteriana DD1 (ID 06-614) foi depositada com DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Inhoffenstr. 7B, D-38124 Braunschweig, Alemanha) em 11 de agosto de 2006, com o número de depósito DSM 18541. Neste context, é feita referência à prioridade dos pedidos de patente Européia EP 09152959.4 e EP 09171250,5, em que o depósito foi mencionado em um primeiro momento no contexto da presente invenção. Além do mais, é feita referência à WO 2009/024294, em que a cepa DD1 é descrita pela primeira vez, incluindo o depósito em DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Inhoffenstr. 7B, D-38124 Braunschweig, Alemanha) em 11 de agosto de 2006.
[000172] O teor dos documentos aqui citados é incorporado pela referência. EXEMPLOS - Processamento à jusante Exemplo 7: Método para isolamento de ácido succínico a partir dos caldos de fermentação por resina de troca catiônica
[000173] Um caldo de fermentação neutralizado por NH4OH (25 % em peso, calculado no peso total da solução NH4OH) durante o processo de fermentação foi filtrado. O caldo de fermentação aquoso sem célula contendo 15 % (p/p) de ácido succínico (neutralizado como sal) foi usado para o processo à jusante.
[000174] Uma resina de troca catiônica (tipo Lewatit Monoplus SP 112 da Lanxess; 471 mL) foi carregada em uma coluna de vidro com temperatura controlada (50 °C) (elevação do leito: 24 cm) como fase estacionária e lavada com água. Após esta etapa de lavagem, a resina transbordou em uma maneira de cima para baixo com a solução aquosa de ácido succínico (156 mL, contendo 25g de ácido succínico, densidade: 1,069 kg/L).
[000175] A velocidade do fluxo da solução foi em média 33 mL / min, correspondendo a uma velocidade de 4,2 volumes do leito (BV) por hora.
[000176] Assim uma solução limpa e sem cor contendo aproximadamente 24,9 g de ácido succínico livre foi obtida na primeira fração (453 mL). A concentração de ácido succínico desta fração foi em média 5,38 % em peso.
[000177] Além do índice de refração, o valor do pH e o adsorção (350 nm) da solução que sai da coluna foram medidos.
[000178] A capacidade de ligação resultante da resina de traça catiônica ácida forte usada foi em média aproximadamente 0,89 equivalentes (eq) por litro de resina.
[000179] Após o processo de ligação, a resina foi lavada com água (546 mL) e finalmente regenerada na forma catiônica com 5 % de ácido clorídrico (919 mL; velocidade: 66 mL/min) que transbordou a resina de maneira ascendente. Como uma última etapa, a resina foi lavada novamente com água (889 mL). Além do fato de que a resina liberou o ácido succínico, a resina descoloriu o caldo e uma solução muito pura e sem cor de ácido succínico foi obtida. Exemplo 8: Método para medir uma curva por meio de intervalo com uma resina de troca iônica ácida forte
[000180] O caldo de fermentação aquoso sem célula contendo uma quantidade de 15 % (p/o) de ácido succínico (neutralizado como sal) foi usado para o processo à jusante após a filtração.
[000181] Uma resina de troca iônica ácida forte (tipo Lewatit MonoPlus SP 112 da Lanxess; 689 mL) foi carregada em uma coluna de vidro com temperatura controlada (50 °C) (elevação do leito: 97,5 cm) e lavada com água. Após esta etapa de lavagem, a resina transbordou de uma maneira de cima para baixo com a sulução aquosa de ácido succínico (468 mL, contendo 75g de ácido succínico; densidade: 1,069 kg/L).
[000182] A velocidade do fluxo da solução foi em média 24 mL / min, o que correposnde a uma velocidade de 2,1 volumes do leito (BV) por hora.
[000183] Assim como no exemplo 7, uma solução clara (457 mL) contendo aproximadamente 53,8 g de ácido succínico livre foi obtida na primeira fração. A concentração de ácido succínico desta fração variou em média 11,53 % em peso.
[000184] Diferente do ensaio no exemplo 7, neste caso a amostragem da primeira fração parou no momento que os cátions diminuíram. Este momento foi detectado com o valor medido do pH que aumentou repentinamente (de um valor de aproximadamente 1,4) em virtude do rompimento por meio do sal de ácido succínico.
[000185] A fração amostrada após esta solução clara continha sal de ácido succínico e apresentou uma cor marrom similar ao caldo de fermentação original.
[000186] Neste ensaio, a capacidade de ligação resultante da resina de troca catiônica ácida forte usada variou em média aproximadamente 1,32 equivalentes (eq) por litro resina.
[000187] Após o processo de ligação, a resina foi lavada com água (678 mL) e regenerada na forma catiônica com 5 % de ácido clorídrico (2.034 mL; velocidade: 92 mL/min). Finalmente, a resina foi lavada novamente com água (824 mL).
[000188] Além do fato de que a resina liberou o ácido succínico, o resina descoloriu o caldo e uma solução muito pura e sem cor de ácido succínico foi obtida.
[000189] Exemplo 9: Método para purificar ácido succínico a partir dos caldos de fermentação seguido por concentração e cristalização da solução dessalinizada resultante
[000190] Duas amostras foram usadas para a etapa de cristalização e foram obtidas da mesma maneira descrita no exemplo 7.
[000191] Em cada caso, caldo de fermentação aquoso sem célula contendo uma quantidade de 15 % (p/p) de ácido succínico (neutralizado como sal) foi usado para o processo à jusante. As duas amostras que foram purificadas e dessalinizadas usando uma resina de troca catiônica (tipo Lewatit MonoPlus SP 112 da Lanxess) foram usadas para a etapa de cristalização.
[000192] As tabelas a seguir mostram o volume de caldo de fermentação, resina e produtos químicos e as quantidades e capacidades obtidas nestes experimentos.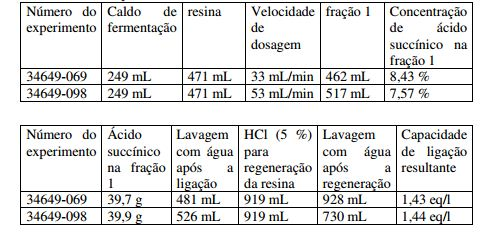
[000193] Assim, duas soluções claras e sem cor foram obtidas como primeiras frações e combinadas para obter uma solução succínica (com aproximadamente 76 g de ácido succínico livre; com diferença em virtude da amostragem).
[000194] Esta solução foi concentrada destilando água para obter 380,2 g de solução com uma concentração de ácido succínico de 20 % em peso. Após a etapa de concentração, a solução foi agitada e resfriada. Uma vez que a solução apresentou uma temperatura de 50 °C, ela foi semeada. Logo após, a cristalização começou e os cristais de ácido succínico precipitaram.
[000195] A suspensão de ácido succínico foi agitada por toda a noite em temperatura ambiente e resfriada em gelo/banho maria por 1 hora.
[000196] Os cristais forma filtrados no fio e a torta foi lavada duas vezes com 20 mL de água gelada cada. Após a etapa de lavagem, os cristais foram secos em um fluxo de gás nitrogênio. Assim, 65,2 g de cristais sem cor de ácido succínico com uma pureza de 99,8 % foram obtidos.
[000197] Subsequentemente, os cristais de ácido succínico foram secos em um fluido para secar. Referências Botstein, D, Shortle, D (1985) Strategies and applications of in vitro mutagenesis. Science 229 1193-201. Dharmadi Y, Murarka A, Gonzalez R (2006) Anaerobic fermentation of glycerol by Escherichia coli: A new platform for metabolic engineering. Biotech Bioeng 94: 821-829. Dousse et al. (2008) Routine phenotypic identification of bacterial species of the family Pasteurellaceae isolated from animals Dousse, Fjournal of veterinary diagnostic investigation 20 716-724. Eisenstark, A (1971) Mutagenic and lethal effects of visible and near-ultraviolet light on bacterial cells. Adv Genet 16 167-98. Hong SH, Lee SY (2001) Metabolic Flux analysis for SA production by recombinant Escherichia coli with amplified malic enzyme activity. Biotechnology and Bioengineering 74/2: 89-95. Foster, Patricia L, (2007) Stress-induced mutagenesis in bacteria. Crit Rev Biochem Mol Biol 42 373-97. Knappe, J, et al. (1993) Pyruvate formate-lyase mechanism involving the protein-based glycyl radical. Biochem Soc Trans 21 731-4. Knappe, J, Sawers, G (1990) A radical-chemical route to acetyl-CoA: the anaerobically induced pyruvate formate-lyase system of Escherichia coli. FEMS Microbiol Rev 6 383-98. Kuhnert, P. and Christensen, H. 2008 "Pasteurellaceae: Biology, Genomics and Molecular Aspects."; ISBN 978-1-904455-34-9. Kuhnert P, Scholten E, Haefner S, Mayor D and Frey J (2010), Basfia succiniproducens gen. nov., sp. nov., A new member of the family Pasteurellaceae isolated from bovine rumen (2010) International Journal of Systematic and Evolutionary Microbiology, 60, 44-50. Lee PC, Lee SY, Hong SA, Chang HN (2002a) Isolation and characterization of a new SA-producing bacterium, Mannheimia succiniciproducens MBEL 55E, from bovine rumen. Appl Microbiol Biotechnol 58: 663-668. Lee SJ, Song H, Lee SY (2006) Genome-Based Metabolic engineering of Mannheimia succiniciproducens for SA production. Appl Environ Biotechnol 72/3: 1939-1948. Leenhouts et al., 1989, Appl Env Microbiol 55, 394-400 Lin H, San K-Y, Bennett GN (2005) Effect of Sorghum vulgare phosphoenolpyruvate carboxylase and Lactococcus lactis pyruvate carboxylase coexpression on succinate production in mutant strains of Escherichia coli. Appl Microbiol Biotechnol 67: 515-523. Pascal, M et al. (1981) Mutants of escherichia-coli-k12 with defects in anaerobic pyruvate metabolism Journal of General Microbiology 124 35-42. Peters-Wendisch, PG et al. (1996) Archives of Microbiology 165 387-396. Peters-Wendisch PG, (1998). Microbiology. 144, 915-27. Saier, Milton H Jr (2008) The bacterial chromosome Crit Rev Biochem Mol Biol 43 89-134. Sambrook et al., (1989) MOLECULAR CLONING: A LABORATORY MANUAL, Second Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. ["Sambrook"]. Sanchez AM, Bennet GN, San K-Y (2005) Novel pathway engineering design of the anaerobic central metabolic pathway in Escherichia coli to increase succinate yield and productivity. Metabolic Engineering 7: 229-239. Sawers, G (1993): specific transcriptional requirements for positive regulation of the anaerobically inducible pfl operon by arca and fnr molecular microbiology 10 737-747. Song H and Lee S (2006) Production of SA by bacterial fermentation. Enz Microb Tech 39: 352-361. Varenne s et al. (1975) Mutant of escherichia-coli deficient in pyruvate formate lyase, molecular & general genetics 141 181-184. Walker, G C (1984) Mutagenesis and inducible responses to deoxyribonucleic acid damage in Escherichia coli. Microbiol Rev 48 60-93. Walker, G C et al. (1983) Regulation and function of cellular gene products involved in UV and chemical mutagenesis in E. coli. Basic Life Sci 23 181-202; Witkin, E M (1969) Ultraviolet-induced mutation and DNA repair. Annu Rev Microbiol 23 487-514. Yazdani S, Gonzalez R (2007) Anaerobic fermentation of glycerol: a path to economic viability for the biofuels industry. Curr Opinion Biotechnol 18: 213-219. Zhu, et al. (2005) Effect of a single-gene knockout on the metabolic regulation in Escherichia coli for D-lactate production under microaerobic condition Metab Eng 7 104-15.
Claims (16)
1. Processo para a produção fermentativa de ácido succínico ou um sal ou derivado do mesmo, caracterizado pelo fato de que processo compreende as etapas de: a) incubar uma cepa bacteriana em um meio contendo pelo menos glicerol como fonte de carbono assimilável e cultivar a dita cepa sob condições que favorecem a formação do ácido succínico desejado; e b) obter o dito ácido succínico, ou sal ou derivado do mesmo, do meio, em que a referida cepa bacteriana é da família Pasteurellaceae pertencente à espécie Basfia succiniciproducens, capaz de utilizar glicerol como fonte de carbono para a produção fermentativa de ácido succínico, em que a referida cepa contém um gene mutado codificando uma enzima piruvato-formato-liase com atividade diminuída ou indetectável, método que compreende a produção fermentativa de ácido succínico ou sal ou derivado do mesmo e controlar o pH com NH4HCO3, (NH4)2CO3, NaOH, Na2CO3, NaHCO3, KOH, K2CO3, KHCO3, CaCO3, CaO, CH6N2O2, C2H7N e/ou misturas dos mesmos.
2. Processo, de acordo com a reivindicação 1, caracterizado pelo fato de que a fermentação é realizada em uma temperatura na faixa de 10 a 60 °C em um pH de 5,0 a 9,0, na presença de dióxido de carbono.
3. Processo, de acordo com a reivindicação 1 ou 2, caracterizado pelo fato de que a fonte de carbono assimilável é selecionada de glicerol, sacarose, maltose, maltodextrina, D-frutose, D-galactose, D-manose, lactose, D-glicose, D-xilose, L-arabinose, rafinose, produtos de decomposição de amido, celulose, hemiceluloses e lignocelulose e misturas dos mesmos.
4. Processo, de acordo com a reivindicação 3, caracterizado pelo fato de que a fonte de carbono é glicerol ou uma mistura de glicerol e pelo menos uma fonte de carbono adicional selecionada de sacarose, maltose, D-frutose, D-galactose, lactose, D-manose, D-glicose, D-xilose, rafinose e L- arabinose.
5. Processo, de acordo com qualquer uma das reivindicações 1 a 4, caracterizado pelo fato de que a concentração da fonte de carbono assimilável é ajustada para um valor em uma faixa de 5 a 80 g/L.
6. Processo, de acordo com qualquer uma das reivindicações 1 a 5, caracterizado pelo fato de que é realizado descontinuamente ou continuamente.
7. Processo, de acordo com qualquer uma das reivindicações 1 a 6, caracterizado pelo fato de que o ácido succínico e/ou sais dos mesmos são isolados e/ou purificados adicionalmente por filtração, cristalização, eletrodiálise e/ou cromatografia de troca catiônica.
8. Processo, de acordo com qualquer uma das reivindicações 1 a 7, caracterizado pelo fato de que o ácido succínico e/ou sais dos mesmos são isolados e/ou purificados adicionalmente pelas etapas a seguir: filtração seguido por cromatografia de troca catiônica seguido por cristalização.
9. Processo, de acordo com a reivindicação 7 ou 8, caracterizado pelo fato de que o material usado para a cromatografia de troca catiônica é uma resina de troca catiônica na forma de H+ que carrega grupos de ácido sulfônico com uma capacidade total de 0,5 a 2,0 equivalentes/L de resina de troca catiônica.
10. Processo, de acordo com qualquer uma das reivindicações 7 a 9, caracterizado pelo fato de que a cromatografia de troca catiônica é realizada em uma temperatura de 45 a 60 °C.
11. Processo, de acordo com qualquer uma das reivindicações 1 a 10, caracterizado pelo fato de que o dito glicerol, que é usado como fonte de carbono assimilável, é obtido por clivagem do éster de triacilglicerídeos.
12. Processo, de acordo com a reivindicação 11, caracterizado pelo fato de que o glicerol é um produto residual obtido a partir da fabricação do biodiesel.
13. Processo de acordo com qualquer uma das reivindicações 1 a 12, caracterizado pelo fato de que a cepa bacteriana da família Pasteurellaceae possui um rDNA 16S de SEQ ID NO: 1.
14. Processo de acordo com qualquer uma das reivindicações 1 a 13, caracterizado pelo fato de que a cepa bacteriana da família Pasteurellaceae possui um rDNA 23S de SEQ ID NO: 2.
15. Processo de acordo com qualquer uma das reivindicações 1 a 14, caracterizado pelo fato de que a cepa bacteriana possui ao menos uma das seguintes características metabólicas adicionais: a) produção de ácido succínico a partir de D-glicose; b) produção de ácido succínico a partir de D-xilose; c) crescimento em concentrações iniciais de glicose de 75 g/L ou mais; d) crescimento em concentrações iniciais de glicerol de 70 g/L ou mais.
16. Processo de acordo com qualquer uma das reivindicações 1 a 15, caracterizado pelo fato de que a cepa bacteriana está depositada com o número de depósito DSM 18541.
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP09152959 | 2009-02-16 | ||
| EP09152959.4 | 2009-02-16 | ||
| US24530609P | 2009-09-24 | 2009-09-24 | |
| EP0917250.5 | 2009-09-24 | ||
| EP09171250 | 2009-09-24 | ||
| US61/245,306 | 2009-09-24 | ||
| PCT/EP2010/051798 WO2010092155A1 (en) | 2009-02-16 | 2010-02-12 | Novel microbial succinic acid producers and purification of succinic acid |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| BRPI1008267A2 BRPI1008267A2 (pt) | 2015-08-25 |
| BRPI1008267B1 true BRPI1008267B1 (pt) | 2021-06-22 |
Family
ID=42125502
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| BRPI1008267-0A BRPI1008267B1 (pt) | 2009-02-16 | 2010-02-12 | Processo para a produção fermentativa de ácido succínico |
Country Status (9)
| Country | Link |
|---|---|
| US (3) | US8673598B2 (pt) |
| EP (3) | EP3345997A1 (pt) |
| JP (1) | JP5826634B2 (pt) |
| KR (1) | KR101823695B1 (pt) |
| CN (2) | CN102317438A (pt) |
| BR (1) | BRPI1008267B1 (pt) |
| CA (1) | CA2751280C (pt) |
| MY (1) | MY161185A (pt) |
| WO (1) | WO2010092155A1 (pt) |
Families Citing this family (31)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2005051170A2 (en) | 2003-11-19 | 2005-06-09 | Dexcom, Inc. | Integrated receiver for continuous analyte sensor |
| ES2587402T3 (es) | 2007-08-17 | 2016-10-24 | Basf Se | Miembro productor de ácido carboxílico de la familia Pasteurellaceae |
| ES2560534T3 (es) * | 2008-12-23 | 2016-02-19 | Basf Se | Células bacterianas que muestran actividad de formato deshidrogenasa para la fabricación de ácido succínico |
| EP2202294B1 (en) * | 2008-12-23 | 2015-10-21 | Basf Se | Bacterial cells having a glyoxylate shunt for the manufacture of succinic acid |
| EA201290533A1 (ru) | 2009-12-17 | 2013-01-30 | Винтерсхол Хольдинг Гмбх | Способ получения гомополисахаридов |
| CA2788596A1 (en) | 2010-02-11 | 2011-08-18 | Metabolix, Inc. | Process for producing a monomer component from a genetically modified polyhydroxyalkanoate biomass |
| EP2360137B1 (en) * | 2010-02-12 | 2013-08-07 | PURAC Biochem BV | Process for manufacturing succinic acid |
| KR101221557B1 (ko) * | 2010-08-30 | 2013-01-14 | 한국과학기술원 | 수크로오즈와 글리세롤을 동시에 이용하는 신규 숙신산 생성 변이 미생물 및 이를 이용한 숙신산 제조방법 |
| GB201106686D0 (en) * | 2011-04-20 | 2011-06-01 | Univ Manchester | Production of succinic acid |
| WO2012170793A1 (en) * | 2011-06-08 | 2012-12-13 | Metabolix, Inc. | Biorefinery process for thf production |
| BR112014001382B1 (pt) * | 2011-07-21 | 2020-11-10 | Archer Daniels Midland Company | método de produção de composto derivado do ácido c4 |
| CN103842495B (zh) | 2011-07-22 | 2018-04-13 | 麦兰特公司 | 甘油发酵为有机酸 |
| ES2817793T3 (es) * | 2012-05-07 | 2021-04-08 | Archer Daniels Midland Co | Purificación de ácido succínico |
| CN104350034B (zh) | 2012-06-08 | 2018-07-31 | Cj 第一制糖株式会社 | 可再生丙烯酸生产和自其制备的产物 |
| US9657316B2 (en) | 2012-08-27 | 2017-05-23 | Genomatica, Inc. | Microorganisms and methods for enhancing the availability of reducing equivalents in the presence of methanol, and for producing 1,4-butanediol related thereto |
| US9932611B2 (en) | 2012-10-22 | 2018-04-03 | Genomatica, Inc. | Microorganisms and methods for enhancing the availability of reducing equivalents in the presence of methanol, and for producing succinate related thereto |
| US10150976B2 (en) | 2012-12-17 | 2018-12-11 | Genomatica, Inc. | Microorganisms and methods for enhancing the availability of reducing equivalents in the presence of methanol, and for producing adipate, 6-aminocaproate, hexamethylenediamine or caprolactam related thereto |
| JP2014150747A (ja) * | 2013-02-06 | 2014-08-25 | Sekisui Chem Co Ltd | 変異微生物、並びに、コハク酸の生産方法 |
| EP3039121A4 (en) * | 2013-08-30 | 2017-05-03 | Basf Se | Modified microorganism for improved production of alanine |
| ES2731335T3 (es) * | 2014-02-07 | 2019-11-15 | Basf Se | Microorganismos mejorados para la producción de ácido succínico |
| WO2015118051A1 (en) * | 2014-02-07 | 2015-08-13 | Basf Se | Modified microorganism for improved production of fine chemicals on sucrose |
| KR102304838B1 (ko) * | 2014-02-07 | 2021-09-28 | 바스프 에스이 | 개선된 바이오매스 분리 거동을 갖는 변형된 미생물 |
| JP6672162B2 (ja) * | 2014-03-19 | 2020-03-25 | ビーエーエスエフ ソシエタス・ヨーロピアBasf Se | 発酵のための炭水化物の制限的供給を伴うグリセロールの使用 |
| MY193587A (en) | 2014-05-08 | 2022-10-19 | Basf Se | Genetically modified microorganism for improved production of fine chemicals on sucrose |
| WO2016030373A1 (en) * | 2014-08-28 | 2016-03-03 | Basf Se | Modified microorganism for improved production of fine chemicals on sucrose |
| WO2016110520A1 (de) * | 2015-01-09 | 2016-07-14 | Basf Se | Verfahren zur herstellung von bernsteinsäureanhydrid und hochreiner bernsteinsäure oder ihren estern oder amiden |
| WO2016110556A1 (de) * | 2015-01-09 | 2016-07-14 | Basf Se | Verfahren zur herstellung von tetrahydrofuran, 1,4-butandiol oder gamma-butyrolacton |
| CN105779513B (zh) * | 2016-05-10 | 2019-12-06 | 华东理工大学 | 利用重组大肠杆菌以甘油为碳源发酵生产琥珀酸的方法 |
| EP3625337B1 (en) | 2017-05-19 | 2021-02-24 | Basf Se | Process for producing an organic compound |
| EP3502241A1 (en) | 2017-12-21 | 2019-06-26 | Basf Se | Modified microorganism for improved production of succinate |
| CN112111418B (zh) * | 2019-06-20 | 2022-09-20 | 华东师范大学 | 琥珀酰多糖高产菌株及琥珀酰多糖的制备和应用 |
Family Cites Families (32)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4550185A (en) | 1983-12-22 | 1985-10-29 | E. I. Du Pont De Nemours And Company | Process for making tetrahydrofuran and 1,4-butanediol using Pd/Re hydrogenation catalyst |
| US5341206A (en) | 1992-12-03 | 1994-08-23 | Hewlett-Packard Company | Method for calibrating a spectrograph for gaseous samples |
| US5504004A (en) | 1994-12-20 | 1996-04-02 | Michigan Biotechnology Institute | Process for making succinic acid, microorganisms for use in the process and methods of obtaining the microorganisms |
| US5573931A (en) | 1995-08-28 | 1996-11-12 | Michigan Biotechnology Institute | Method for making succinic acid, bacterial variants for use in the process, and methods for obtaining variants |
| EP0805208B1 (en) | 1995-09-12 | 2003-03-05 | Kirin Beer Kabushiki Kaisha | Promoter/terminator of formate dehydrogenase gene of candida boidinii |
| DE19748570A1 (de) | 1997-11-04 | 1999-05-06 | Miele & Cie | Gasbeheiztes Gerät mit einer Einrichtung zur Regelung des Heizbetriebes |
| EP0917125A1 (en) | 1997-11-11 | 1999-05-19 | SYNCO ITALIANA S.r.l. | Visual communication system with LCD pricetags updatable by remote control |
| KR100329019B1 (ko) | 1999-04-13 | 2002-03-18 | 윤덕용 | 유기산의 고효율 생산방법 |
| KR100372218B1 (ko) | 2000-06-29 | 2003-02-14 | 바이오인포메틱스 주식회사 | 유기산을 생산하는 균주 및 이를 이용한 유기산의 생산방법 |
| US6743610B2 (en) * | 2001-03-30 | 2004-06-01 | The University Of Chicago | Method to produce succinic acid from raw hydrolysates |
| US6951643B2 (en) | 2001-07-24 | 2005-10-04 | Oklahoma State University | Direct-fed microbial |
| US7256016B2 (en) | 2001-11-02 | 2007-08-14 | Rice University | Recycling system for manipulation of intracellular NADH availability |
| US7163812B2 (en) | 2003-08-06 | 2007-01-16 | Board Of Trustees Of Michigan State University | Actinobacillus succinogenes shuttle vector and methods of use |
| KR100556099B1 (ko) * | 2003-11-27 | 2006-03-03 | 한국과학기술원 | 루멘 박테리아 변이균주 및 이를 이용한 숙신산의 제조방법 |
| DE602004022584D1 (de) * | 2003-11-27 | 2009-09-24 | Korea Advanced Inst Sci & Tech | Neue kaumagenbakterienvarianten sowie verfahren zur herstellung von bernsteinsäure unter verwendung davon |
| CA2565727C (en) * | 2004-05-03 | 2012-02-21 | Ut-Battelle, Llc | A method to produce succinic acid from raw hydrolysates |
| US7262046B2 (en) | 2004-08-09 | 2007-08-28 | Rice University | Aerobic succinate production in bacteria |
| WO2006031424A2 (en) * | 2004-08-27 | 2006-03-23 | Rice University | Mutant e. coli strain with increased succinic acid production |
| WO2006034156A2 (en) | 2004-09-17 | 2006-03-30 | Rice University | High succinate producing bacteria |
| US8017790B2 (en) | 2004-12-21 | 2011-09-13 | Basf Se | Method for producing pyrrolidones from succinates from fermentation broths |
| KR100679638B1 (ko) | 2005-08-19 | 2007-02-06 | 한국과학기술원 | 포메이트 디하이드로게나제 d 또는 e를 코딩하는 유전자로 형질전환된 미생물 및 이를 이용한 숙신산의 제조방법 |
| EP1842843A1 (de) | 2006-04-04 | 2007-10-10 | Basf Aktiengesellschaft | Verfahren zur Herstellung eines Carbonsäurealkylesters |
| JP5602982B2 (ja) | 2006-07-03 | 2014-10-08 | 三菱化学株式会社 | コハク酸の製造方法 |
| US20080293101A1 (en) | 2006-07-27 | 2008-11-27 | Peters Matthew W | Engineered microorganisms for increasing product yield in biotransformations, related methods and systems |
| KR100780324B1 (ko) | 2006-07-28 | 2007-11-29 | 한국과학기술원 | 신규 순수 숙신산 생성 변이 미생물 및 이를 이용한 숙신산제조방법 |
| CA2715093A1 (en) | 2006-12-01 | 2008-11-27 | Gevo, Inc. | Engineered microorganisms for producing n-butanol and related methods |
| CA2688938C (en) * | 2007-03-20 | 2018-01-16 | University Of Florida Research Foundation, Inc. | Materials and methods for efficient succinate and malate production |
| ES2587402T3 (es) | 2007-08-17 | 2016-10-24 | Basf Se | Miembro productor de ácido carboxílico de la familia Pasteurellaceae |
| CN101240259B (zh) * | 2008-01-16 | 2010-12-08 | 南京工业大学 | 新构建的高产富马酸基因工程菌及其生产富马酸的方法 |
| CN101945997B (zh) | 2008-02-21 | 2014-12-03 | 巴斯夫欧洲公司 | 产生γ-氨基丁酸的方法 |
| EP2202294B1 (en) | 2008-12-23 | 2015-10-21 | Basf Se | Bacterial cells having a glyoxylate shunt for the manufacture of succinic acid |
| ES2560534T3 (es) | 2008-12-23 | 2016-02-19 | Basf Se | Células bacterianas que muestran actividad de formato deshidrogenasa para la fabricación de ácido succínico |
-
2010
- 2010-02-12 WO PCT/EP2010/051798 patent/WO2010092155A1/en active Application Filing
- 2010-02-12 CA CA2751280A patent/CA2751280C/en active Active
- 2010-02-12 EP EP18151548.7A patent/EP3345997A1/en not_active Withdrawn
- 2010-02-12 MY MYPI2011003541A patent/MY161185A/en unknown
- 2010-02-12 EP EP10708150.7A patent/EP2396401B1/en active Active
- 2010-02-12 KR KR1020117021474A patent/KR101823695B1/ko active IP Right Grant
- 2010-02-12 JP JP2011549580A patent/JP5826634B2/ja not_active Expired - Fee Related
- 2010-02-12 EP EP19150461.2A patent/EP3505619A1/en not_active Withdrawn
- 2010-02-12 CN CN2010800079634A patent/CN102317438A/zh active Pending
- 2010-02-12 BR BRPI1008267-0A patent/BRPI1008267B1/pt active IP Right Grant
- 2010-02-12 CN CN201510246300.2A patent/CN104877935B/zh active Active
- 2010-02-12 US US13/201,547 patent/US8673598B2/en active Active
-
2014
- 2014-01-08 US US14/150,206 patent/US9023632B2/en active Active
-
2015
- 2015-04-01 US US14/676,317 patent/US9932612B2/en active Active
Also Published As
| Publication number | Publication date |
|---|---|
| CA2751280A1 (en) | 2010-08-19 |
| CA2751280C (en) | 2019-03-12 |
| CN104877935A (zh) | 2015-09-02 |
| EP2396401B1 (en) | 2018-12-19 |
| KR20110127690A (ko) | 2011-11-25 |
| US9932612B2 (en) | 2018-04-03 |
| US8673598B2 (en) | 2014-03-18 |
| CN102317438A (zh) | 2012-01-11 |
| KR101823695B1 (ko) | 2018-03-14 |
| EP3345997A1 (en) | 2018-07-11 |
| US20140127764A1 (en) | 2014-05-08 |
| US20110300589A1 (en) | 2011-12-08 |
| US9023632B2 (en) | 2015-05-05 |
| BRPI1008267A2 (pt) | 2015-08-25 |
| EP3505619A1 (en) | 2019-07-03 |
| CN104877935B (zh) | 2021-10-01 |
| JP5826634B2 (ja) | 2015-12-02 |
| US20150197776A1 (en) | 2015-07-16 |
| MY161185A (en) | 2017-04-14 |
| EP2396401A1 (en) | 2011-12-21 |
| WO2010092155A1 (en) | 2010-08-19 |
| JP2012521190A (ja) | 2012-09-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US9932612B2 (en) | Microbial succinic acid producers and purification of succinic acid | |
| US8877466B2 (en) | Bacterial cells having a glyoxylate shunt for the manufacture of succinic acid | |
| EP3006556B1 (en) | Recombinant escherichia coli for producing succinic acid and application thereof | |
| US8883466B2 (en) | Bacterial cells exhibiting formate dehydrogenase activity for the manufacture of succinic acid | |
| RU2422526C2 (ru) | СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ, ПРИНАДЛЕЖАЩИХ К РОДУ Yarrowia | |
| JP6608392B2 (ja) | ショ糖におけるファインケミカルの改善された生産のための遺伝的改変微生物 | |
| US10287558B2 (en) | Microorganisms for succinic acid production | |
| US9850506B2 (en) | Modified microorganism for improved production of fine chemicals on sucrose | |
| WO2016030373A1 (en) | Modified microorganism for improved production of fine chemicals on sucrose | |
| ES2715930T3 (es) | Productores microbianos de ácido succínico novedosos y purificación del ácido succínico | |
| KR20070096347A (ko) | 4―hydroxybutyrate(4HB)생성능을가지는 변이체 및 이를 이용한 4HB의 제조방법 | |
| EP3625337A1 (en) | Process for producing an organic compound |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| B06F | Objections, documents and/or translations needed after an examination request according [chapter 6.6 patent gazette] | ||
| B07A | Application suspended after technical examination (opinion) [chapter 7.1 patent gazette] | ||
| B06A | Patent application procedure suspended [chapter 6.1 patent gazette] | ||
| B09A | Decision: intention to grant [chapter 9.1 patent gazette] | ||
| B09W | Decision of grant: rectification |
Free format text: A NOTIFICACAO FOI EFETUADA COM INCORRECAO NO QUADRO 1 DO PARECER DE DEFERIMENTO. |
|
| B16A | Patent or certificate of addition of invention granted [chapter 16.1 patent gazette] |
Free format text: PRAZO DE VALIDADE: 20 (VINTE) ANOS CONTADOS A PARTIR DE 12/02/2010, OBSERVADAS AS CONDICOES LEGAIS. PATENTE CONCEDIDA CONFORME ADI 5.529/DF, , QUE DETERMINA A ALTERACAO DO PRAZO DE CONCESSAO. |