KR20220158053A - 중증 급성 호흡기 증후군 코로나바이러스 2(SARS-CoV-2)에 대한 인간 단클론 항체 - Google Patents
중증 급성 호흡기 증후군 코로나바이러스 2(SARS-CoV-2)에 대한 인간 단클론 항체 Download PDFInfo
- Publication number
- KR20220158053A KR20220158053A KR1020227036951A KR20227036951A KR20220158053A KR 20220158053 A KR20220158053 A KR 20220158053A KR 1020227036951 A KR1020227036951 A KR 1020227036951A KR 20227036951 A KR20227036951 A KR 20227036951A KR 20220158053 A KR20220158053 A KR 20220158053A
- Authority
- KR
- South Korea
- Prior art keywords
- antibody
- amino acid
- seq
- acid sequence
- antibody fragment
- Prior art date
Links
- 241001678559 COVID-19 virus Species 0.000 title claims abstract description 190
- 241000282414 Homo sapiens Species 0.000 title claims description 84
- 238000000034 method Methods 0.000 claims abstract description 186
- 230000027455 binding Effects 0.000 claims description 266
- 238000009739 binding Methods 0.000 claims description 262
- 102000008394 Immunoglobulin Fragments Human genes 0.000 claims description 242
- 108010021625 Immunoglobulin Fragments Proteins 0.000 claims description 242
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 170
- 210000004027 cell Anatomy 0.000 claims description 167
- 239000000427 antigen Substances 0.000 claims description 156
- 108091007433 antigens Proteins 0.000 claims description 153
- 102000036639 antigens Human genes 0.000 claims description 153
- 230000035772 mutation Effects 0.000 claims description 100
- 241000700605 Viruses Species 0.000 claims description 73
- 239000012634 fragment Substances 0.000 claims description 73
- 239000000203 mixture Substances 0.000 claims description 71
- 230000003993 interaction Effects 0.000 claims description 64
- 229960005486 vaccine Drugs 0.000 claims description 51
- 230000014509 gene expression Effects 0.000 claims description 49
- 230000000694 effects Effects 0.000 claims description 47
- 238000003556 assay Methods 0.000 claims description 42
- 208000015181 infectious disease Diseases 0.000 claims description 36
- 230000003472 neutralizing effect Effects 0.000 claims description 36
- 238000012360 testing method Methods 0.000 claims description 35
- 230000003612 virological effect Effects 0.000 claims description 35
- 101710198474 Spike protein Proteins 0.000 claims description 33
- 238000006386 neutralization reaction Methods 0.000 claims description 32
- 238000001514 detection method Methods 0.000 claims description 31
- 230000013595 glycosylation Effects 0.000 claims description 31
- 238000006206 glycosylation reaction Methods 0.000 claims description 31
- 238000004519 manufacturing process Methods 0.000 claims description 31
- 229940096437 Protein S Drugs 0.000 claims description 30
- 102000005962 receptors Human genes 0.000 claims description 29
- 108020003175 receptors Proteins 0.000 claims description 29
- 238000012384 transportation and delivery Methods 0.000 claims description 29
- 230000000890 antigenic effect Effects 0.000 claims description 28
- 238000002965 ELISA Methods 0.000 claims description 27
- 210000004408 hybridoma Anatomy 0.000 claims description 26
- 230000001225 therapeutic effect Effects 0.000 claims description 26
- 238000006467 substitution reaction Methods 0.000 claims description 25
- 239000000126 substance Substances 0.000 claims description 23
- 239000013598 vector Substances 0.000 claims description 23
- 238000009472 formulation Methods 0.000 claims description 22
- 208000025721 COVID-19 Diseases 0.000 claims description 19
- 230000002829 reductive effect Effects 0.000 claims description 19
- 108090000975 Angiotensin-converting enzyme 2 Proteins 0.000 claims description 18
- 210000002966 serum Anatomy 0.000 claims description 18
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 claims description 17
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 claims description 17
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 claims description 17
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 claims description 17
- 230000000241 respiratory effect Effects 0.000 claims description 17
- 230000002255 enzymatic effect Effects 0.000 claims description 14
- 150000007523 nucleic acids Chemical group 0.000 claims description 14
- 210000004369 blood Anatomy 0.000 claims description 13
- 239000008280 blood Substances 0.000 claims description 13
- 239000013604 expression vector Substances 0.000 claims description 13
- 230000036541 health Effects 0.000 claims description 13
- 239000007924 injection Substances 0.000 claims description 13
- 238000002347 injection Methods 0.000 claims description 13
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 12
- 239000012530 fluid Substances 0.000 claims description 12
- 238000003127 radioimmunoassay Methods 0.000 claims description 12
- 230000002195 synergetic effect Effects 0.000 claims description 12
- 238000001262 western blot Methods 0.000 claims description 12
- 108020004707 nucleic acids Proteins 0.000 claims description 9
- 102000039446 nucleic acids Human genes 0.000 claims description 9
- 102000040430 polynucleotide Human genes 0.000 claims description 9
- 108091033319 polynucleotide Proteins 0.000 claims description 9
- 239000002157 polynucleotide Substances 0.000 claims description 9
- 238000004113 cell culture Methods 0.000 claims description 8
- 238000010186 staining Methods 0.000 claims description 8
- 208000024172 Cardiovascular disease Diseases 0.000 claims description 7
- 206010061598 Immunodeficiency Diseases 0.000 claims description 7
- 238000012575 bio-layer interferometry Methods 0.000 claims description 7
- 150000004676 glycans Chemical class 0.000 claims description 6
- 208000023504 respiratory system disease Diseases 0.000 claims description 6
- 210000002700 urine Anatomy 0.000 claims description 6
- 101100272788 Arabidopsis thaliana BSL3 gene Proteins 0.000 claims description 5
- 210000001124 body fluid Anatomy 0.000 claims description 5
- 238000004520 electroporation Methods 0.000 claims description 5
- 238000001990 intravenous administration Methods 0.000 claims description 5
- 230000009467 reduction Effects 0.000 claims description 5
- 210000000582 semen Anatomy 0.000 claims description 5
- 238000002198 surface plasmon resonance spectroscopy Methods 0.000 claims description 5
- 206010036790 Productive cough Diseases 0.000 claims description 4
- 241000710960 Sindbis virus Species 0.000 claims description 4
- 210000000416 exudates and transudate Anatomy 0.000 claims description 4
- 210000003608 fece Anatomy 0.000 claims description 4
- 230000002633 protecting effect Effects 0.000 claims description 4
- 210000003296 saliva Anatomy 0.000 claims description 4
- 210000003802 sputum Anatomy 0.000 claims description 4
- 208000024794 sputum Diseases 0.000 claims description 4
- 230000004075 alteration Effects 0.000 claims description 3
- 210000004381 amniotic fluid Anatomy 0.000 claims description 3
- 210000003756 cervix mucus Anatomy 0.000 claims description 3
- 238000012258 culturing Methods 0.000 claims description 3
- 210000003097 mucus Anatomy 0.000 claims description 3
- 210000005059 placental tissue Anatomy 0.000 claims description 3
- 230000029058 respiratory gaseous exchange Effects 0.000 claims description 3
- 210000001138 tear Anatomy 0.000 claims description 3
- 102100025248 C-X-C motif chemokine 10 Human genes 0.000 claims description 2
- 101000897480 Homo sapiens C-C motif chemokine 2 Proteins 0.000 claims description 2
- 101000858088 Homo sapiens C-X-C motif chemokine 10 Proteins 0.000 claims description 2
- 108090001005 Interleukin-6 Proteins 0.000 claims description 2
- 102100035765 Angiotensin-converting enzyme 2 Human genes 0.000 claims 2
- 210000004748 cultured cell Anatomy 0.000 claims 1
- 241000711573 Coronaviridae Species 0.000 abstract description 23
- 108090000623 proteins and genes Proteins 0.000 description 161
- 102000004169 proteins and genes Human genes 0.000 description 114
- 235000018102 proteins Nutrition 0.000 description 111
- 235000001014 amino acid Nutrition 0.000 description 94
- 229940024606 amino acid Drugs 0.000 description 90
- 108090000765 processed proteins & peptides Proteins 0.000 description 74
- 150000001413 amino acids Chemical class 0.000 description 63
- 239000000523 sample Substances 0.000 description 58
- 102000004196 processed proteins & peptides Human genes 0.000 description 53
- 229920001184 polypeptide Polymers 0.000 description 43
- 125000005647 linker group Chemical group 0.000 description 36
- 238000002474 experimental method Methods 0.000 description 34
- 241001465754 Metazoa Species 0.000 description 32
- 210000003719 b-lymphocyte Anatomy 0.000 description 31
- 230000006870 function Effects 0.000 description 28
- 210000001519 tissue Anatomy 0.000 description 28
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 27
- 238000004040 coloring Methods 0.000 description 27
- 201000003176 Severe Acute Respiratory Syndrome Diseases 0.000 description 26
- 239000003814 drug Substances 0.000 description 26
- 101000929928 Homo sapiens Angiotensin-converting enzyme 2 Proteins 0.000 description 24
- 102000048657 human ACE2 Human genes 0.000 description 24
- 229910052739 hydrogen Inorganic materials 0.000 description 24
- 230000004048 modification Effects 0.000 description 24
- 238000012986 modification Methods 0.000 description 24
- 239000002245 particle Substances 0.000 description 24
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 23
- 102100024952 Protein CBFA2T1 Human genes 0.000 description 23
- 238000013459 approach Methods 0.000 description 23
- 230000015572 biosynthetic process Effects 0.000 description 22
- 102000004190 Enzymes Human genes 0.000 description 21
- 108090000790 Enzymes Proteins 0.000 description 21
- 229940088598 enzyme Drugs 0.000 description 21
- 239000003446 ligand Substances 0.000 description 21
- 238000011282 treatment Methods 0.000 description 21
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 20
- 108060003951 Immunoglobulin Proteins 0.000 description 20
- 210000004602 germ cell Anatomy 0.000 description 20
- 239000001257 hydrogen Substances 0.000 description 20
- 102000018358 immunoglobulin Human genes 0.000 description 20
- 239000002773 nucleotide Substances 0.000 description 20
- 125000003729 nucleotide group Chemical group 0.000 description 20
- 239000000243 solution Substances 0.000 description 20
- 239000013078 crystal Substances 0.000 description 19
- 230000004927 fusion Effects 0.000 description 19
- 108020004414 DNA Proteins 0.000 description 18
- 241000699670 Mus sp. Species 0.000 description 18
- 210000001744 T-lymphocyte Anatomy 0.000 description 18
- 238000007792 addition Methods 0.000 description 18
- 239000003795 chemical substances by application Substances 0.000 description 18
- 102000053723 Angiotensin-converting enzyme 2 Human genes 0.000 description 16
- 238000004458 analytical method Methods 0.000 description 16
- 229940079593 drug Drugs 0.000 description 16
- 239000012636 effector Substances 0.000 description 16
- 210000004072 lung Anatomy 0.000 description 16
- 235000018977 lysine Nutrition 0.000 description 16
- 239000004475 Arginine Substances 0.000 description 15
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 15
- 239000004472 Lysine Substances 0.000 description 15
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 15
- 235000009697 arginine Nutrition 0.000 description 15
- 230000001965 increasing effect Effects 0.000 description 15
- 238000011534 incubation Methods 0.000 description 15
- 239000012528 membrane Substances 0.000 description 15
- -1 70% Chemical class 0.000 description 14
- 102100031673 Corneodesmosin Human genes 0.000 description 14
- 101710139375 Corneodesmosin Proteins 0.000 description 14
- 239000011159 matrix material Substances 0.000 description 14
- 230000001404 mediated effect Effects 0.000 description 14
- 210000004379 membrane Anatomy 0.000 description 14
- 229940049595 antibody-drug conjugate Drugs 0.000 description 13
- 239000000562 conjugate Substances 0.000 description 13
- 230000001472 cytotoxic effect Effects 0.000 description 13
- 230000001419 dependent effect Effects 0.000 description 13
- 238000000746 purification Methods 0.000 description 13
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 12
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 12
- 125000003118 aryl group Chemical group 0.000 description 12
- 230000000903 blocking effect Effects 0.000 description 12
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 12
- 238000001727 in vivo Methods 0.000 description 12
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 11
- 108010032595 Antibody Binding Sites Proteins 0.000 description 11
- 206010035226 Plasma cell myeloma Diseases 0.000 description 11
- 230000008859 change Effects 0.000 description 11
- 230000001186 cumulative effect Effects 0.000 description 11
- 231100000433 cytotoxic Toxicity 0.000 description 11
- 201000010099 disease Diseases 0.000 description 11
- 230000002209 hydrophobic effect Effects 0.000 description 11
- 239000002609 medium Substances 0.000 description 11
- 201000000050 myeloid neoplasm Diseases 0.000 description 11
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 11
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 10
- 108010019670 Chimeric Antigen Receptors Proteins 0.000 description 10
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 10
- 101000629318 Severe acute respiratory syndrome coronavirus 2 Spike glycoprotein Proteins 0.000 description 10
- 125000000539 amino acid group Chemical group 0.000 description 10
- 229960002685 biotin Drugs 0.000 description 10
- 235000020958 biotin Nutrition 0.000 description 10
- 239000011616 biotin Substances 0.000 description 10
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 10
- 238000005516 engineering process Methods 0.000 description 10
- 230000028993 immune response Effects 0.000 description 10
- 230000001976 improved effect Effects 0.000 description 10
- 238000000338 in vitro Methods 0.000 description 10
- 239000000463 material Substances 0.000 description 10
- 239000002953 phosphate buffered saline Substances 0.000 description 10
- 238000005406 washing Methods 0.000 description 10
- 239000004971 Cross linker Substances 0.000 description 9
- YZCKVEUIGOORGS-OUBTZVSYSA-N Deuterium Chemical group [2H] YZCKVEUIGOORGS-OUBTZVSYSA-N 0.000 description 9
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 9
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 9
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 9
- 241000315672 SARS coronavirus Species 0.000 description 9
- 150000001720 carbohydrates Chemical class 0.000 description 9
- 230000000875 corresponding effect Effects 0.000 description 9
- 229910052805 deuterium Inorganic materials 0.000 description 9
- 238000011161 development Methods 0.000 description 9
- 230000018109 developmental process Effects 0.000 description 9
- 239000000499 gel Substances 0.000 description 9
- 235000013922 glutamic acid Nutrition 0.000 description 9
- 239000004220 glutamic acid Substances 0.000 description 9
- 230000003053 immunization Effects 0.000 description 9
- 238000002649 immunization Methods 0.000 description 9
- 238000013507 mapping Methods 0.000 description 9
- 230000007246 mechanism Effects 0.000 description 9
- 230000004044 response Effects 0.000 description 9
- 150000003839 salts Chemical class 0.000 description 9
- IJGRMHOSHXDMSA-UHFFFAOYSA-N Atomic nitrogen Chemical compound N#N IJGRMHOSHXDMSA-UHFFFAOYSA-N 0.000 description 8
- 108010073807 IgG Receptors Proteins 0.000 description 8
- 241000699666 Mus <mouse, genus> Species 0.000 description 8
- 208000037847 SARS-CoV-2-infection Diseases 0.000 description 8
- 239000002253 acid Substances 0.000 description 8
- 238000006243 chemical reaction Methods 0.000 description 8
- 230000000295 complement effect Effects 0.000 description 8
- 231100000599 cytotoxic agent Toxicity 0.000 description 8
- 229940127121 immunoconjugate Drugs 0.000 description 8
- 244000052769 pathogen Species 0.000 description 8
- 230000008569 process Effects 0.000 description 8
- 230000001681 protective effect Effects 0.000 description 8
- 230000008685 targeting Effects 0.000 description 8
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 7
- 241000283707 Capra Species 0.000 description 7
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Natural products CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 7
- LFQSCWFLJHTTHZ-UHFFFAOYSA-N Ethanol Chemical compound CCO LFQSCWFLJHTTHZ-UHFFFAOYSA-N 0.000 description 7
- PNNNRSAQSRJVSB-SLPGGIOYSA-N Fucose Natural products C[C@H](O)[C@@H](O)[C@H](O)[C@H](O)C=O PNNNRSAQSRJVSB-SLPGGIOYSA-N 0.000 description 7
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 7
- SHZGCJCMOBCMKK-DHVFOXMCSA-N L-fucopyranose Chemical compound C[C@@H]1OC(O)[C@@H](O)[C@H](O)[C@@H]1O SHZGCJCMOBCMKK-DHVFOXMCSA-N 0.000 description 7
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 7
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 7
- 108020000999 Viral RNA Proteins 0.000 description 7
- 239000012491 analyte Substances 0.000 description 7
- 238000004422 calculation algorithm Methods 0.000 description 7
- 238000003776 cleavage reaction Methods 0.000 description 7
- 239000002254 cytotoxic agent Substances 0.000 description 7
- 230000034994 death Effects 0.000 description 7
- 231100000517 death Toxicity 0.000 description 7
- 230000007423 decrease Effects 0.000 description 7
- 238000012217 deletion Methods 0.000 description 7
- 230000037430 deletion Effects 0.000 description 7
- 230000002068 genetic effect Effects 0.000 description 7
- 230000012010 growth Effects 0.000 description 7
- 238000003018 immunoassay Methods 0.000 description 7
- 229940072221 immunoglobulins Drugs 0.000 description 7
- 230000002458 infectious effect Effects 0.000 description 7
- 230000003389 potentiating effect Effects 0.000 description 7
- 230000002265 prevention Effects 0.000 description 7
- 239000000047 product Substances 0.000 description 7
- 230000006798 recombination Effects 0.000 description 7
- 238000005215 recombination Methods 0.000 description 7
- 238000009738 saturating Methods 0.000 description 7
- 230000007017 scission Effects 0.000 description 7
- 238000000926 separation method Methods 0.000 description 7
- 235000004400 serine Nutrition 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- 208000024891 symptom Diseases 0.000 description 7
- 239000013638 trimer Substances 0.000 description 7
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 6
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 6
- 241000282412 Homo Species 0.000 description 6
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 6
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 6
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 6
- 206010028980 Neoplasm Diseases 0.000 description 6
- MUBZPKHOEPUJKR-UHFFFAOYSA-N Oxalic acid Chemical compound OC(=O)C(O)=O MUBZPKHOEPUJKR-UHFFFAOYSA-N 0.000 description 6
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 6
- 238000011529 RT qPCR Methods 0.000 description 6
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 6
- 108091008874 T cell receptors Proteins 0.000 description 6
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 6
- 239000004473 Threonine Substances 0.000 description 6
- 230000001154 acute effect Effects 0.000 description 6
- 239000002671 adjuvant Substances 0.000 description 6
- 238000000540 analysis of variance Methods 0.000 description 6
- 238000010171 animal model Methods 0.000 description 6
- 229940098773 bovine serum albumin Drugs 0.000 description 6
- 239000000872 buffer Substances 0.000 description 6
- 235000014633 carbohydrates Nutrition 0.000 description 6
- 239000000969 carrier Substances 0.000 description 6
- 230000001413 cellular effect Effects 0.000 description 6
- 239000003153 chemical reaction reagent Substances 0.000 description 6
- 239000002299 complementary DNA Substances 0.000 description 6
- 239000003431 cross linking reagent Substances 0.000 description 6
- 238000010790 dilution Methods 0.000 description 6
- 239000012895 dilution Substances 0.000 description 6
- 231100000673 dose–response relationship Toxicity 0.000 description 6
- 238000002296 dynamic light scattering Methods 0.000 description 6
- 125000000291 glutamic acid group Chemical group N[C@@H](CCC(O)=O)C(=O)* 0.000 description 6
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 6
- 210000000987 immune system Anatomy 0.000 description 6
- 230000002163 immunogen Effects 0.000 description 6
- 230000005847 immunogenicity Effects 0.000 description 6
- 239000013612 plasmid Substances 0.000 description 6
- 238000001556 precipitation Methods 0.000 description 6
- 235000013930 proline Nutrition 0.000 description 6
- 238000000159 protein binding assay Methods 0.000 description 6
- 238000003757 reverse transcription PCR Methods 0.000 description 6
- 238000012163 sequencing technique Methods 0.000 description 6
- 229940125678 AZD7442 Drugs 0.000 description 5
- 108090001008 Avidin Proteins 0.000 description 5
- 102000004127 Cytokines Human genes 0.000 description 5
- 108090000695 Cytokines Proteins 0.000 description 5
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 5
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 5
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 5
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 5
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 5
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 5
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 5
- 239000004365 Protease Substances 0.000 description 5
- 108010076504 Protein Sorting Signals Proteins 0.000 description 5
- 108010001267 Protein Subunits Proteins 0.000 description 5
- 102000002067 Protein Subunits Human genes 0.000 description 5
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 5
- 235000004279 alanine Nutrition 0.000 description 5
- BFNBIHQBYMNNAN-UHFFFAOYSA-N ammonium sulfate Chemical compound N.N.OS(O)(=O)=O BFNBIHQBYMNNAN-UHFFFAOYSA-N 0.000 description 5
- 229910052921 ammonium sulfate Inorganic materials 0.000 description 5
- 235000011130 ammonium sulphate Nutrition 0.000 description 5
- 230000003321 amplification Effects 0.000 description 5
- 235000003704 aspartic acid Nutrition 0.000 description 5
- 239000011324 bead Substances 0.000 description 5
- 230000008901 benefit Effects 0.000 description 5
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 5
- 239000002775 capsule Substances 0.000 description 5
- 239000013553 cell monolayer Substances 0.000 description 5
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 5
- 238000002983 circular dichroism Methods 0.000 description 5
- 238000012875 competitive assay Methods 0.000 description 5
- 229940127089 cytotoxic agent Drugs 0.000 description 5
- 238000006471 dimerization reaction Methods 0.000 description 5
- 238000000684 flow cytometry Methods 0.000 description 5
- 238000003384 imaging method Methods 0.000 description 5
- 239000000543 intermediate Substances 0.000 description 5
- 230000003834 intracellular effect Effects 0.000 description 5
- 238000007918 intramuscular administration Methods 0.000 description 5
- 238000002372 labelling Methods 0.000 description 5
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 5
- 238000005259 measurement Methods 0.000 description 5
- 238000012544 monitoring process Methods 0.000 description 5
- 231100000350 mutagenesis Toxicity 0.000 description 5
- 239000002105 nanoparticle Substances 0.000 description 5
- 238000003199 nucleic acid amplification method Methods 0.000 description 5
- 102000013415 peroxidase activity proteins Human genes 0.000 description 5
- 108040007629 peroxidase activity proteins Proteins 0.000 description 5
- 229920002981 polyvinylidene fluoride Polymers 0.000 description 5
- 238000010149 post-hoc-test Methods 0.000 description 5
- 230000005180 public health Effects 0.000 description 5
- 238000006722 reduction reaction Methods 0.000 description 5
- 230000001105 regulatory effect Effects 0.000 description 5
- 238000011160 research Methods 0.000 description 5
- 239000006152 selective media Substances 0.000 description 5
- 230000035945 sensitivity Effects 0.000 description 5
- 239000011780 sodium chloride Substances 0.000 description 5
- 239000002904 solvent Substances 0.000 description 5
- 229940124597 therapeutic agent Drugs 0.000 description 5
- 235000008521 threonine Nutrition 0.000 description 5
- 230000032258 transport Effects 0.000 description 5
- LPMXVESGRSUGHW-UHFFFAOYSA-N Acolongiflorosid K Natural products OC1C(O)C(O)C(C)OC1OC1CC2(O)CCC3C4(O)CCC(C=5COC(=O)C=5)C4(C)CC(O)C3C2(CO)C(O)C1 LPMXVESGRSUGHW-UHFFFAOYSA-N 0.000 description 4
- 206010069754 Acquired gene mutation Diseases 0.000 description 4
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 4
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 4
- 206010011224 Cough Diseases 0.000 description 4
- 206010012310 Dengue fever Diseases 0.000 description 4
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 4
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 4
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 4
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 4
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 4
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 4
- 230000004988 N-glycosylation Effects 0.000 description 4
- 239000000020 Nitrocellulose Substances 0.000 description 4
- LPMXVESGRSUGHW-GHYGWZAOSA-N Ouabain Natural products O([C@@H]1[C@@H](O)[C@@H](O)[C@@H](O)[C@H](C)O1)[C@H]1C[C@@H](O)[C@@]2(CO)[C@@](O)(C1)CC[C@H]1[C@]3(O)[C@@](C)([C@H](C4=CC(=O)OC4)CC3)C[C@@H](O)[C@H]21 LPMXVESGRSUGHW-GHYGWZAOSA-N 0.000 description 4
- 102000035195 Peptidases Human genes 0.000 description 4
- 108091005804 Peptidases Proteins 0.000 description 4
- NBIIXXVUZAFLBC-UHFFFAOYSA-N Phosphoric acid Chemical compound OP(O)(O)=O NBIIXXVUZAFLBC-UHFFFAOYSA-N 0.000 description 4
- 239000002202 Polyethylene glycol Substances 0.000 description 4
- 108010090804 Streptavidin Chemical class 0.000 description 4
- 244000166550 Strophanthus gratus Species 0.000 description 4
- 108091027544 Subgenomic mRNA Proteins 0.000 description 4
- IEDXPSOJFSVCKU-HOKPPMCLSA-N [4-[[(2S)-5-(carbamoylamino)-2-[[(2S)-2-[6-(2,5-dioxopyrrolidin-1-yl)hexanoylamino]-3-methylbutanoyl]amino]pentanoyl]amino]phenyl]methyl N-[(2S)-1-[[(2S)-1-[[(3R,4S,5S)-1-[(2S)-2-[(1R,2R)-3-[[(1S,2R)-1-hydroxy-1-phenylpropan-2-yl]amino]-1-methoxy-2-methyl-3-oxopropyl]pyrrolidin-1-yl]-3-methoxy-5-methyl-1-oxoheptan-4-yl]-methylamino]-3-methyl-1-oxobutan-2-yl]amino]-3-methyl-1-oxobutan-2-yl]-N-methylcarbamate Chemical compound CC[C@H](C)[C@@H]([C@@H](CC(=O)N1CCC[C@H]1[C@H](OC)[C@@H](C)C(=O)N[C@H](C)[C@@H](O)c1ccccc1)OC)N(C)C(=O)[C@@H](NC(=O)[C@H](C(C)C)N(C)C(=O)OCc1ccc(NC(=O)[C@H](CCCNC(N)=O)NC(=O)[C@@H](NC(=O)CCCCCN2C(=O)CCC2=O)C(C)C)cc1)C(C)C IEDXPSOJFSVCKU-HOKPPMCLSA-N 0.000 description 4
- 150000007513 acids Chemical class 0.000 description 4
- 230000009471 action Effects 0.000 description 4
- 238000001042 affinity chromatography Methods 0.000 description 4
- 230000000840 anti-viral effect Effects 0.000 description 4
- 239000000611 antibody drug conjugate Substances 0.000 description 4
- 230000005888 antibody-dependent cellular phagocytosis Effects 0.000 description 4
- 239000002246 antineoplastic agent Substances 0.000 description 4
- 239000003443 antiviral agent Substances 0.000 description 4
- 235000009582 asparagine Nutrition 0.000 description 4
- 229960001230 asparagine Drugs 0.000 description 4
- 125000004429 atom Chemical group 0.000 description 4
- 230000005540 biological transmission Effects 0.000 description 4
- 201000011510 cancer Diseases 0.000 description 4
- 210000000170 cell membrane Anatomy 0.000 description 4
- 238000005119 centrifugation Methods 0.000 description 4
- 238000012512 characterization method Methods 0.000 description 4
- 238000005520 cutting process Methods 0.000 description 4
- 238000000113 differential scanning calorimetry Methods 0.000 description 4
- 238000001493 electron microscopy Methods 0.000 description 4
- 239000000284 extract Substances 0.000 description 4
- 210000004700 fetal blood Anatomy 0.000 description 4
- 239000012091 fetal bovine serum Substances 0.000 description 4
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 4
- 235000004554 glutamine Nutrition 0.000 description 4
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 4
- 229910052737 gold Inorganic materials 0.000 description 4
- 239000010931 gold Substances 0.000 description 4
- 150000002431 hydrogen Chemical class 0.000 description 4
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 4
- 238000003364 immunohistochemistry Methods 0.000 description 4
- 230000005764 inhibitory process Effects 0.000 description 4
- 238000001155 isoelectric focusing Methods 0.000 description 4
- 108020004999 messenger RNA Proteins 0.000 description 4
- BDAGIHXWWSANSR-UHFFFAOYSA-N methanoic acid Natural products OC=O BDAGIHXWWSANSR-UHFFFAOYSA-N 0.000 description 4
- 229960000485 methotrexate Drugs 0.000 description 4
- 238000004264 monolayer culture Methods 0.000 description 4
- 238000002703 mutagenesis Methods 0.000 description 4
- 229920001220 nitrocellulos Polymers 0.000 description 4
- 229910052757 nitrogen Inorganic materials 0.000 description 4
- LPMXVESGRSUGHW-HBYQJFLCSA-N ouabain Chemical compound O[C@@H]1[C@H](O)[C@@H](O)[C@H](C)O[C@H]1O[C@@H]1C[C@@]2(O)CC[C@H]3[C@@]4(O)CC[C@H](C=5COC(=O)C=5)[C@@]4(C)C[C@@H](O)[C@@H]3[C@@]2(CO)[C@H](O)C1 LPMXVESGRSUGHW-HBYQJFLCSA-N 0.000 description 4
- 229960003343 ouabain Drugs 0.000 description 4
- 230000001717 pathogenic effect Effects 0.000 description 4
- 230000037361 pathway Effects 0.000 description 4
- 229920001223 polyethylene glycol Polymers 0.000 description 4
- 230000036515 potency Effects 0.000 description 4
- 238000002360 preparation method Methods 0.000 description 4
- 235000019419 proteases Nutrition 0.000 description 4
- 238000003908 quality control method Methods 0.000 description 4
- 238000003762 quantitative reverse transcription PCR Methods 0.000 description 4
- 230000002285 radioactive effect Effects 0.000 description 4
- 230000002441 reversible effect Effects 0.000 description 4
- 238000012216 screening Methods 0.000 description 4
- 230000037439 somatic mutation Effects 0.000 description 4
- 238000003786 synthesis reaction Methods 0.000 description 4
- 125000003396 thiol group Chemical group [H]S* 0.000 description 4
- LWIHDJKSTIGBAC-UHFFFAOYSA-K tripotassium phosphate Chemical compound [K+].[K+].[K+].[O-]P([O-])([O-])=O LWIHDJKSTIGBAC-UHFFFAOYSA-K 0.000 description 4
- 229910052720 vanadium Inorganic materials 0.000 description 4
- 210000005253 yeast cell Anatomy 0.000 description 4
- AGNGYMCLFWQVGX-AGFFZDDWSA-N (e)-1-[(2s)-2-amino-2-carboxyethoxy]-2-diazonioethenolate Chemical compound OC(=O)[C@@H](N)CO\C([O-])=C\[N+]#N AGNGYMCLFWQVGX-AGFFZDDWSA-N 0.000 description 3
- TVZGACDUOSZQKY-LBPRGKRZSA-N 4-aminofolic acid Chemical compound C1=NC2=NC(N)=NC(N)=C2N=C1CNC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 TVZGACDUOSZQKY-LBPRGKRZSA-N 0.000 description 3
- 229920001817 Agar Polymers 0.000 description 3
- 241000972773 Aulopiformes Species 0.000 description 3
- 238000011725 BALB/c mouse Methods 0.000 description 3
- 108010012236 Chemokines Proteins 0.000 description 3
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 3
- 208000001490 Dengue Diseases 0.000 description 3
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 3
- 208000000059 Dyspnea Diseases 0.000 description 3
- 206010013975 Dyspnoeas Diseases 0.000 description 3
- 241000588724 Escherichia coli Species 0.000 description 3
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 3
- 108010087819 Fc receptors Proteins 0.000 description 3
- 102000009109 Fc receptors Human genes 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- 101001021527 Homo sapiens Huntingtin-interacting protein 1 Proteins 0.000 description 3
- 101000872458 Homo sapiens Huntingtin-interacting protein 1-related protein Proteins 0.000 description 3
- 101000840258 Homo sapiens Immunoglobulin J chain Proteins 0.000 description 3
- 101001002931 Homo sapiens Immunoglobulin heavy variable 1-58 Proteins 0.000 description 3
- 101001133056 Homo sapiens Mucin-1 Proteins 0.000 description 3
- 101000644689 Homo sapiens Ubiquitin-conjugating enzyme E2 K Proteins 0.000 description 3
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 3
- 102100035957 Huntingtin-interacting protein 1 Human genes 0.000 description 3
- 102000009490 IgG Receptors Human genes 0.000 description 3
- 102100029571 Immunoglobulin J chain Human genes 0.000 description 3
- 102100020774 Immunoglobulin heavy variable 1-58 Human genes 0.000 description 3
- 241000282553 Macaca Species 0.000 description 3
- 241000127282 Middle East respiratory syndrome-related coronavirus Species 0.000 description 3
- 102100034256 Mucin-1 Human genes 0.000 description 3
- QPCDCPDFJACHGM-UHFFFAOYSA-N N,N-bis{2-[bis(carboxymethyl)amino]ethyl}glycine Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(=O)O)CCN(CC(O)=O)CC(O)=O QPCDCPDFJACHGM-UHFFFAOYSA-N 0.000 description 3
- 102100021061 Palmitoyltransferase ZDHHC17 Human genes 0.000 description 3
- 206010057249 Phagocytosis Diseases 0.000 description 3
- 206010035664 Pneumonia Diseases 0.000 description 3
- 108010029485 Protein Isoforms Proteins 0.000 description 3
- 102000001708 Protein Isoforms Human genes 0.000 description 3
- 206010037660 Pyrexia Diseases 0.000 description 3
- 238000012300 Sequence Analysis Methods 0.000 description 3
- ZMANZCXQSJIPKH-UHFFFAOYSA-N Triethylamine Chemical compound CCN(CC)CC ZMANZCXQSJIPKH-UHFFFAOYSA-N 0.000 description 3
- 102100020696 Ubiquitin-conjugating enzyme E2 K Human genes 0.000 description 3
- 241000710959 Venezuelan equine encephalitis virus Species 0.000 description 3
- 230000035508 accumulation Effects 0.000 description 3
- 238000009825 accumulation Methods 0.000 description 3
- 239000013543 active substance Substances 0.000 description 3
- 239000008272 agar Substances 0.000 description 3
- WQZGKKKJIJFFOK-PHYPRBDBSA-N alpha-D-galactose Chemical compound OC[C@H]1O[C@H](O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-PHYPRBDBSA-N 0.000 description 3
- 229960003896 aminopterin Drugs 0.000 description 3
- 239000001166 ammonium sulphate Substances 0.000 description 3
- 102000025171 antigen binding proteins Human genes 0.000 description 3
- 108091000831 antigen binding proteins Proteins 0.000 description 3
- 229950011321 azaserine Drugs 0.000 description 3
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 3
- 239000012472 biological sample Substances 0.000 description 3
- 239000006143 cell culture medium Substances 0.000 description 3
- 239000002738 chelating agent Substances 0.000 description 3
- 238000007385 chemical modification Methods 0.000 description 3
- 239000003593 chromogenic compound Substances 0.000 description 3
- 239000011248 coating agent Substances 0.000 description 3
- 238000000576 coating method Methods 0.000 description 3
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 3
- 150000001875 compounds Chemical class 0.000 description 3
- 230000000139 costimulatory effect Effects 0.000 description 3
- 238000002425 crystallisation Methods 0.000 description 3
- 230000008025 crystallization Effects 0.000 description 3
- 239000012228 culture supernatant Substances 0.000 description 3
- 230000000120 cytopathologic effect Effects 0.000 description 3
- 239000002619 cytotoxin Substances 0.000 description 3
- 230000002950 deficient Effects 0.000 description 3
- 208000025729 dengue disease Diseases 0.000 description 3
- 238000013461 design Methods 0.000 description 3
- 239000000032 diagnostic agent Substances 0.000 description 3
- 229940039227 diagnostic agent Drugs 0.000 description 3
- 230000029087 digestion Effects 0.000 description 3
- 239000000539 dimer Substances 0.000 description 3
- 238000010494 dissociation reaction Methods 0.000 description 3
- 230000005593 dissociations Effects 0.000 description 3
- 238000009826 distribution Methods 0.000 description 3
- 238000001914 filtration Methods 0.000 description 3
- 125000000524 functional group Chemical group 0.000 description 3
- 229930182830 galactose Natural products 0.000 description 3
- 229930195712 glutamate Natural products 0.000 description 3
- 230000036039 immunity Effects 0.000 description 3
- 239000003112 inhibitor Substances 0.000 description 3
- 230000002452 interceptive effect Effects 0.000 description 3
- 150000002500 ions Chemical class 0.000 description 3
- 239000007788 liquid Substances 0.000 description 3
- 238000004949 mass spectrometry Methods 0.000 description 3
- 238000002887 multiple sequence alignment Methods 0.000 description 3
- 239000013642 negative control Substances 0.000 description 3
- 230000007935 neutral effect Effects 0.000 description 3
- 239000003921 oil Substances 0.000 description 3
- 235000019198 oils Nutrition 0.000 description 3
- 230000008520 organization Effects 0.000 description 3
- 238000007500 overflow downdraw method Methods 0.000 description 3
- 230000003647 oxidation Effects 0.000 description 3
- 238000007254 oxidation reaction Methods 0.000 description 3
- 230000005298 paramagnetic effect Effects 0.000 description 3
- 229960003330 pentetic acid Drugs 0.000 description 3
- 238000002823 phage display Methods 0.000 description 3
- 230000008782 phagocytosis Effects 0.000 description 3
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 3
- 210000002381 plasma Anatomy 0.000 description 3
- 239000004033 plastic Substances 0.000 description 3
- 229920003023 plastic Polymers 0.000 description 3
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 3
- 229920000136 polysorbate Polymers 0.000 description 3
- NLKNQRATVPKPDG-UHFFFAOYSA-M potassium iodide Chemical compound [K+].[I-] NLKNQRATVPKPDG-UHFFFAOYSA-M 0.000 description 3
- 239000000843 powder Substances 0.000 description 3
- 150000003141 primary amines Chemical group 0.000 description 3
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 3
- XOJVVFBFDXDTEG-UHFFFAOYSA-N pristane Chemical compound CC(C)CCCC(C)CCCC(C)CCCC(C)C XOJVVFBFDXDTEG-UHFFFAOYSA-N 0.000 description 3
- 230000001737 promoting effect Effects 0.000 description 3
- 238000001742 protein purification Methods 0.000 description 3
- 238000011002 quantification Methods 0.000 description 3
- 230000009257 reactivity Effects 0.000 description 3
- RWWYLEGWBNMMLJ-MEUHYHILSA-N remdesivir Drugs C([C@@H]1[C@H]([C@@H](O)[C@@](C#N)(O1)C=1N2N=CN=C(N)C2=CC=1)O)OP(=O)(N[C@@H](C)C(=O)OCC(CC)CC)OC1=CC=CC=C1 RWWYLEGWBNMMLJ-MEUHYHILSA-N 0.000 description 3
- RWWYLEGWBNMMLJ-YSOARWBDSA-N remdesivir Chemical compound NC1=NC=NN2C1=CC=C2[C@]1([C@@H]([C@@H]([C@H](O1)CO[P@](=O)(OC1=CC=CC=C1)N[C@H](C(=O)OCC(CC)CC)C)O)O)C#N RWWYLEGWBNMMLJ-YSOARWBDSA-N 0.000 description 3
- 230000000717 retained effect Effects 0.000 description 3
- 235000019515 salmon Nutrition 0.000 description 3
- 235000014102 seafood Nutrition 0.000 description 3
- 230000028327 secretion Effects 0.000 description 3
- 208000013220 shortness of breath Diseases 0.000 description 3
- FVAUCKIRQBBSSJ-UHFFFAOYSA-M sodium iodide Chemical compound [Na+].[I-] FVAUCKIRQBBSSJ-UHFFFAOYSA-M 0.000 description 3
- 239000007787 solid Substances 0.000 description 3
- 125000006850 spacer group Chemical group 0.000 description 3
- 241000894007 species Species 0.000 description 3
- 210000000952 spleen Anatomy 0.000 description 3
- 230000000087 stabilizing effect Effects 0.000 description 3
- 238000012916 structural analysis Methods 0.000 description 3
- 238000007920 subcutaneous administration Methods 0.000 description 3
- 239000000758 substrate Substances 0.000 description 3
- 230000009466 transformation Effects 0.000 description 3
- 241000701161 unidentified adenovirus Species 0.000 description 3
- 238000002255 vaccination Methods 0.000 description 3
- 210000003501 vero cell Anatomy 0.000 description 3
- 239000003643 water by type Substances 0.000 description 3
- GKSPIZSKQWTXQG-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 4-[1-(pyridin-2-yldisulfanyl)ethyl]benzoate Chemical group C=1C=C(C(=O)ON2C(CCC2=O)=O)C=CC=1C(C)SSC1=CC=CC=N1 GKSPIZSKQWTXQG-UHFFFAOYSA-N 0.000 description 2
- VBICKXHEKHSIBG-UHFFFAOYSA-N 1-monostearoylglycerol Chemical compound CCCCCCCCCCCCCCCCCC(=O)OCC(O)CO VBICKXHEKHSIBG-UHFFFAOYSA-N 0.000 description 2
- VGIRNWJSIRVFRT-UHFFFAOYSA-N 2',7'-difluorofluorescein Chemical compound OC(=O)C1=CC=CC=C1C1=C2C=C(F)C(=O)C=C2OC2=CC(O)=C(F)C=C21 VGIRNWJSIRVFRT-UHFFFAOYSA-N 0.000 description 2
- OSWFIVFLDKOXQC-UHFFFAOYSA-N 4-(3-methoxyphenyl)aniline Chemical compound COC1=CC=CC(C=2C=CC(N)=CC=2)=C1 OSWFIVFLDKOXQC-UHFFFAOYSA-N 0.000 description 2
- GANZODCWZFAEGN-UHFFFAOYSA-N 5-mercapto-2-nitro-benzoic acid Chemical compound OC(=O)C1=CC(S)=CC=C1[N+]([O-])=O GANZODCWZFAEGN-UHFFFAOYSA-N 0.000 description 2
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 2
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 2
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 2
- 241000710929 Alphavirus Species 0.000 description 2
- QGZKDVFQNNGYKY-UHFFFAOYSA-O Ammonium Chemical compound [NH4+] QGZKDVFQNNGYKY-UHFFFAOYSA-O 0.000 description 2
- 241000112287 Bat coronavirus Species 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- 208000026310 Breast neoplasm Diseases 0.000 description 2
- 108010029697 CD40 Ligand Proteins 0.000 description 2
- 102100032937 CD40 ligand Human genes 0.000 description 2
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 2
- 241000282693 Cercopithecidae Species 0.000 description 2
- 102000019034 Chemokines Human genes 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- 208000035473 Communicable disease Diseases 0.000 description 2
- 208000001528 Coronaviridae Infections Diseases 0.000 description 2
- SRBFZHDQGSBBOR-IOVATXLUSA-N D-xylopyranose Chemical compound O[C@@H]1COC(O)[C@H](O)[C@H]1O SRBFZHDQGSBBOR-IOVATXLUSA-N 0.000 description 2
- 241000702421 Dependoparvovirus Species 0.000 description 2
- FEWJPZIEWOKRBE-JCYAYHJZSA-N Dextrotartaric acid Chemical compound OC(=O)[C@H](O)[C@@H](O)C(O)=O FEWJPZIEWOKRBE-JCYAYHJZSA-N 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 241000196324 Embryophyta Species 0.000 description 2
- 102000004961 Furin Human genes 0.000 description 2
- 108090001126 Furin Proteins 0.000 description 2
- 229910052688 Gadolinium Inorganic materials 0.000 description 2
- 239000004366 Glucose oxidase Substances 0.000 description 2
- 108010015776 Glucose oxidase Proteins 0.000 description 2
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 2
- 108090000288 Glycoproteins Proteins 0.000 description 2
- 102000003886 Glycoproteins Human genes 0.000 description 2
- 102000051366 Glycosyltransferases Human genes 0.000 description 2
- 108700023372 Glycosyltransferases Proteins 0.000 description 2
- 102100026122 High affinity immunoglobulin gamma Fc receptor I Human genes 0.000 description 2
- 101001047619 Homo sapiens Immunoglobulin kappa variable 3-20 Proteins 0.000 description 2
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 2
- 241000711467 Human coronavirus 229E Species 0.000 description 2
- 241001109669 Human coronavirus HKU1 Species 0.000 description 2
- 241000482741 Human coronavirus NL63 Species 0.000 description 2
- 241001428935 Human coronavirus OC43 Species 0.000 description 2
- UFHFLCQGNIYNRP-UHFFFAOYSA-N Hydrogen Chemical compound [H][H] UFHFLCQGNIYNRP-UHFFFAOYSA-N 0.000 description 2
- 206010020751 Hypersensitivity Diseases 0.000 description 2
- 102000018251 Hypoxanthine Phosphoribosyltransferase Human genes 0.000 description 2
- 108010091358 Hypoxanthine Phosphoribosyltransferase Proteins 0.000 description 2
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 2
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 2
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 description 2
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 description 2
- 102000013463 Immunoglobulin Light Chains Human genes 0.000 description 2
- 108010065825 Immunoglobulin Light Chains Proteins 0.000 description 2
- 102100022964 Immunoglobulin kappa variable 3-20 Human genes 0.000 description 2
- 108010076561 Interleukin-23 Subunit p19 Proteins 0.000 description 2
- 102100036705 Interleukin-23 subunit alpha Human genes 0.000 description 2
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 2
- 241000282560 Macaca mulatta Species 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 208000025370 Middle East respiratory syndrome Diseases 0.000 description 2
- OVRNDRQMDRJTHS-RTRLPJTCSA-N N-acetyl-D-glucosamine Chemical compound CC(=O)N[C@H]1C(O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-RTRLPJTCSA-N 0.000 description 2
- 238000005481 NMR spectroscopy Methods 0.000 description 2
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 description 2
- PXHVJJICTQNCMI-UHFFFAOYSA-N Nickel Chemical compound [Ni] PXHVJJICTQNCMI-UHFFFAOYSA-N 0.000 description 2
- 230000004989 O-glycosylation Effects 0.000 description 2
- 102000057297 Pepsin A Human genes 0.000 description 2
- 108090000284 Pepsin A Proteins 0.000 description 2
- ZLMJMSJWJFRBEC-UHFFFAOYSA-N Potassium Chemical compound [K] ZLMJMSJWJFRBEC-UHFFFAOYSA-N 0.000 description 2
- 108010071390 Serum Albumin Proteins 0.000 description 2
- 102000007562 Serum Albumin Human genes 0.000 description 2
- 229920002472 Starch Polymers 0.000 description 2
- 239000004138 Stearyl citrate Substances 0.000 description 2
- NINIDFKCEFEMDL-UHFFFAOYSA-N Sulfur Chemical compound [S] NINIDFKCEFEMDL-UHFFFAOYSA-N 0.000 description 2
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 2
- FEWJPZIEWOKRBE-UHFFFAOYSA-N Tartaric acid Natural products [H+].[H+].[O-]C(=O)C(O)C(O)C([O-])=O FEWJPZIEWOKRBE-UHFFFAOYSA-N 0.000 description 2
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 2
- 108010046334 Urease Proteins 0.000 description 2
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 2
- 238000002835 absorbance Methods 0.000 description 2
- 238000010521 absorption reaction Methods 0.000 description 2
- 235000011054 acetic acid Nutrition 0.000 description 2
- 230000021736 acetylation Effects 0.000 description 2
- 238000006640 acetylation reaction Methods 0.000 description 2
- 230000004913 activation Effects 0.000 description 2
- 230000000996 additive effect Effects 0.000 description 2
- 239000007801 affinity label Substances 0.000 description 2
- 230000002776 aggregation Effects 0.000 description 2
- 238000004220 aggregation Methods 0.000 description 2
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 2
- 238000012867 alanine scanning Methods 0.000 description 2
- 229910000147 aluminium phosphate Inorganic materials 0.000 description 2
- 239000003708 ampul Substances 0.000 description 2
- 230000003042 antagnostic effect Effects 0.000 description 2
- 230000001093 anti-cancer Effects 0.000 description 2
- 230000002223 anti-pathogen Effects 0.000 description 2
- 230000000259 anti-tumor effect Effects 0.000 description 2
- 230000009830 antibody antigen interaction Effects 0.000 description 2
- 229940009098 aspartate Drugs 0.000 description 2
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 2
- 230000001588 bifunctional effect Effects 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- 238000001574 biopsy Methods 0.000 description 2
- 210000004899 c-terminal region Anatomy 0.000 description 2
- 239000011575 calcium Substances 0.000 description 2
- 229910052791 calcium Inorganic materials 0.000 description 2
- 238000005251 capillar electrophoresis Methods 0.000 description 2
- 229910052799 carbon Inorganic materials 0.000 description 2
- 239000005018 casein Substances 0.000 description 2
- BECPQYXYKAMYBN-UHFFFAOYSA-N casein, tech. Chemical compound NCCCCC(C(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(CC(C)C)N=C(O)C(CCC(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(C(C)O)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(COP(O)(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(N)CC1=CC=CC=C1 BECPQYXYKAMYBN-UHFFFAOYSA-N 0.000 description 2
- 235000021240 caseins Nutrition 0.000 description 2
- 230000007541 cellular toxicity Effects 0.000 description 2
- 239000003638 chemical reducing agent Substances 0.000 description 2
- 238000004587 chromatography analysis Methods 0.000 description 2
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Natural products OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 2
- 229910017052 cobalt Inorganic materials 0.000 description 2
- 239000010941 cobalt Substances 0.000 description 2
- GUTLYIVDDKVIGB-UHFFFAOYSA-N cobalt atom Chemical compound [Co] GUTLYIVDDKVIGB-UHFFFAOYSA-N 0.000 description 2
- 230000002860 competitive effect Effects 0.000 description 2
- 230000021615 conjugation Effects 0.000 description 2
- 239000000356 contaminant Substances 0.000 description 2
- 239000007822 coupling agent Substances 0.000 description 2
- 238000004132 cross linking Methods 0.000 description 2
- 238000002447 crystallographic data Methods 0.000 description 2
- 235000018417 cysteine Nutrition 0.000 description 2
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 2
- 230000009089 cytolysis Effects 0.000 description 2
- 230000006240 deamidation Effects 0.000 description 2
- 238000012350 deep sequencing Methods 0.000 description 2
- 230000002939 deleterious effect Effects 0.000 description 2
- 230000008021 deposition Effects 0.000 description 2
- 238000001212 derivatisation Methods 0.000 description 2
- 239000008121 dextrose Substances 0.000 description 2
- 239000003085 diluting agent Substances 0.000 description 2
- 150000002019 disulfides Chemical class 0.000 description 2
- 150000004662 dithiols Chemical class 0.000 description 2
- 239000003937 drug carrier Substances 0.000 description 2
- 229960001484 edetic acid Drugs 0.000 description 2
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 2
- 239000002158 endotoxin Substances 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 230000001747 exhibiting effect Effects 0.000 description 2
- 238000000605 extraction Methods 0.000 description 2
- 229960004887 ferric hydroxide Drugs 0.000 description 2
- 238000005206 flow analysis Methods 0.000 description 2
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 2
- 235000019253 formic acid Nutrition 0.000 description 2
- 108020001507 fusion proteins Proteins 0.000 description 2
- 102000037865 fusion proteins Human genes 0.000 description 2
- UIWYJDYFSGRHKR-UHFFFAOYSA-N gadolinium atom Chemical compound [Gd] UIWYJDYFSGRHKR-UHFFFAOYSA-N 0.000 description 2
- 108010074605 gamma-Globulins Proteins 0.000 description 2
- 238000001502 gel electrophoresis Methods 0.000 description 2
- 230000005182 global health Effects 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- 229940116332 glucose oxidase Drugs 0.000 description 2
- 235000019420 glucose oxidase Nutrition 0.000 description 2
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 2
- 239000001963 growth medium Substances 0.000 description 2
- 125000004435 hydrogen atom Chemical group [H]* 0.000 description 2
- 239000012216 imaging agent Substances 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 239000012642 immune effector Substances 0.000 description 2
- 229940121354 immunomodulator Drugs 0.000 description 2
- 238000011065 in-situ storage Methods 0.000 description 2
- APFVFJFRJDLVQX-AHCXROLUSA-N indium-111 Chemical compound [111In] APFVFJFRJDLVQX-AHCXROLUSA-N 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- 206010022000 influenza Diseases 0.000 description 2
- 238000001802 infusion Methods 0.000 description 2
- 239000004615 ingredient Substances 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 238000011081 inoculation Methods 0.000 description 2
- 238000007689 inspection Methods 0.000 description 2
- 238000007912 intraperitoneal administration Methods 0.000 description 2
- 239000007928 intraperitoneal injection Substances 0.000 description 2
- 229910052742 iron Inorganic materials 0.000 description 2
- IEECXTSVVFWGSE-UHFFFAOYSA-M iron(3+);oxygen(2-);hydroxide Chemical compound [OH-].[O-2].[Fe+3] IEECXTSVVFWGSE-UHFFFAOYSA-M 0.000 description 2
- 238000002955 isolation Methods 0.000 description 2
- 229960000310 isoleucine Drugs 0.000 description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 2
- QWTDNUCVQCZILF-UHFFFAOYSA-N isopentane Chemical compound CCC(C)C QWTDNUCVQCZILF-UHFFFAOYSA-N 0.000 description 2
- JJWLVOIRVHMVIS-UHFFFAOYSA-N isopropylamine Chemical compound CC(C)N JJWLVOIRVHMVIS-UHFFFAOYSA-N 0.000 description 2
- 230000002147 killing effect Effects 0.000 description 2
- 239000008101 lactose Substances 0.000 description 2
- 238000012417 linear regression Methods 0.000 description 2
- 150000002632 lipids Chemical class 0.000 description 2
- 210000001165 lymph node Anatomy 0.000 description 2
- 108010026228 mRNA guanylyltransferase Proteins 0.000 description 2
- HQKMJHAJHXVSDF-UHFFFAOYSA-L magnesium stearate Chemical compound [Mg+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O HQKMJHAJHXVSDF-UHFFFAOYSA-L 0.000 description 2
- 230000003211 malignant effect Effects 0.000 description 2
- 230000035800 maturation Effects 0.000 description 2
- 238000002844 melting Methods 0.000 description 2
- 230000008018 melting Effects 0.000 description 2
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 2
- 230000005012 migration Effects 0.000 description 2
- 238000013508 migration Methods 0.000 description 2
- 235000013336 milk Nutrition 0.000 description 2
- 239000008267 milk Substances 0.000 description 2
- 210000004080 milk Anatomy 0.000 description 2
- 238000002156 mixing Methods 0.000 description 2
- 239000000178 monomer Substances 0.000 description 2
- 230000000869 mutational effect Effects 0.000 description 2
- 108010068617 neonatal Fc receptor Proteins 0.000 description 2
- 210000000440 neutrophil Anatomy 0.000 description 2
- 229920001542 oligosaccharide Polymers 0.000 description 2
- 150000002482 oligosaccharides Chemical class 0.000 description 2
- 235000006408 oxalic acid Nutrition 0.000 description 2
- 239000007800 oxidant agent Substances 0.000 description 2
- 229910052760 oxygen Inorganic materials 0.000 description 2
- 239000001301 oxygen Substances 0.000 description 2
- 239000012188 paraffin wax Substances 0.000 description 2
- 230000036961 partial effect Effects 0.000 description 2
- 229940111202 pepsin Drugs 0.000 description 2
- 230000007030 peptide scission Effects 0.000 description 2
- 239000008194 pharmaceutical composition Substances 0.000 description 2
- 239000000546 pharmaceutical excipient Substances 0.000 description 2
- 239000012071 phase Substances 0.000 description 2
- CTRLRINCMYICJO-UHFFFAOYSA-N phenyl azide Chemical class [N-]=[N+]=NC1=CC=CC=C1 CTRLRINCMYICJO-UHFFFAOYSA-N 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 210000003720 plasmablast Anatomy 0.000 description 2
- 229920000642 polymer Polymers 0.000 description 2
- 239000013641 positive control Substances 0.000 description 2
- 239000011591 potassium Substances 0.000 description 2
- 229910052700 potassium Inorganic materials 0.000 description 2
- 229910000160 potassium phosphate Inorganic materials 0.000 description 2
- 235000011009 potassium phosphates Nutrition 0.000 description 2
- 238000009597 pregnancy test Methods 0.000 description 2
- 230000003449 preventive effect Effects 0.000 description 2
- MFDFERRIHVXMIY-UHFFFAOYSA-N procaine Chemical compound CCN(CC)CCOC(=O)C1=CC=C(N)C=C1 MFDFERRIHVXMIY-UHFFFAOYSA-N 0.000 description 2
- 229960004919 procaine Drugs 0.000 description 2
- 230000000069 prophylactic effect Effects 0.000 description 2
- 238000002818 protein evolution Methods 0.000 description 2
- 230000006337 proteolytic cleavage Effects 0.000 description 2
- 150000003212 purines Chemical class 0.000 description 2
- 238000010791 quenching Methods 0.000 description 2
- 238000003259 recombinant expression Methods 0.000 description 2
- 238000011084 recovery Methods 0.000 description 2
- 230000010076 replication Effects 0.000 description 2
- PYWVYCXTNDRMGF-UHFFFAOYSA-N rhodamine B Chemical compound [Cl-].C=12C=CC(=[N+](CC)CC)C=C2OC2=CC(N(CC)CC)=CC=C2C=1C1=CC=CC=C1C(O)=O PYWVYCXTNDRMGF-UHFFFAOYSA-N 0.000 description 2
- 102220092172 rs753180642 Human genes 0.000 description 2
- 229920006395 saturated elastomer Polymers 0.000 description 2
- 238000013207 serial dilution Methods 0.000 description 2
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 2
- 230000011664 signaling Effects 0.000 description 2
- 239000011734 sodium Substances 0.000 description 2
- 229910052708 sodium Inorganic materials 0.000 description 2
- 210000001082 somatic cell Anatomy 0.000 description 2
- 239000008107 starch Substances 0.000 description 2
- 235000019698 starch Nutrition 0.000 description 2
- 238000003860 storage Methods 0.000 description 2
- 229910052717 sulfur Inorganic materials 0.000 description 2
- 239000011593 sulfur Substances 0.000 description 2
- 239000000725 suspension Substances 0.000 description 2
- 239000011975 tartaric acid Substances 0.000 description 2
- 235000002906 tartaric acid Nutrition 0.000 description 2
- ABZLKHKQJHEPAX-UHFFFAOYSA-N tetramethylrhodamine Chemical compound C=12C=CC(N(C)C)=CC2=[O+]C2=CC(N(C)C)=CC=C2C=1C1=CC=CC=C1C([O-])=O ABZLKHKQJHEPAX-UHFFFAOYSA-N 0.000 description 2
- 238000002560 therapeutic procedure Methods 0.000 description 2
- 230000000699 topical effect Effects 0.000 description 2
- 231100000331 toxic Toxicity 0.000 description 2
- 230000002588 toxic effect Effects 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 230000026683 transduction Effects 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- GETQZCLCWQTVFV-UHFFFAOYSA-N trimethylamine Chemical compound CN(C)C GETQZCLCWQTVFV-UHFFFAOYSA-N 0.000 description 2
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 2
- 238000004704 ultra performance liquid chromatography Methods 0.000 description 2
- 241001529453 unidentified herpesvirus Species 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- 239000003981 vehicle Substances 0.000 description 2
- 239000013603 viral vector Substances 0.000 description 2
- OZFAFGSSMRRTDW-UHFFFAOYSA-N (2,4-dichlorophenyl) benzenesulfonate Chemical compound ClC1=CC(Cl)=CC=C1OS(=O)(=O)C1=CC=CC=C1 OZFAFGSSMRRTDW-UHFFFAOYSA-N 0.000 description 1
- JWDFQMWEFLOOED-UHFFFAOYSA-N (2,5-dioxopyrrolidin-1-yl) 3-(pyridin-2-yldisulfanyl)propanoate Chemical compound O=C1CCC(=O)N1OC(=O)CCSSC1=CC=CC=N1 JWDFQMWEFLOOED-UHFFFAOYSA-N 0.000 description 1
- QBYIENPQHBMVBV-HFEGYEGKSA-N (2R)-2-hydroxy-2-phenylacetic acid Chemical compound O[C@@H](C(O)=O)c1ccccc1.O[C@@H](C(O)=O)c1ccccc1 QBYIENPQHBMVBV-HFEGYEGKSA-N 0.000 description 1
- FFILOTSTFMXQJC-QCFYAKGBSA-N (2r,4r,5s,6s)-2-[3-[(2s,3s,4r,6s)-6-[(2s,3r,4r,5s,6r)-5-[(2s,3r,4r,5r,6r)-3-acetamido-4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-2-[(2r,3s,4r,5r,6r)-4,5-dihydroxy-2-(hydroxymethyl)-6-[(e)-3-hydroxy-2-(octadecanoylamino)octadec-4-enoxy]oxan-3-yl]oxy-3-hy Chemical compound O[C@@H]1[C@@H](O)[C@H](OCC(NC(=O)CCCCCCCCCCCCCCCCC)C(O)\C=C\CCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@@H]([C@@H](N)[C@H](O)C2)C(O)C(O)CO[C@]2(O[C@@H]([C@@H](N)[C@H](O)C2)C(O)C(O)CO)C(O)=O)C(O)=O)[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](CO)O1 FFILOTSTFMXQJC-QCFYAKGBSA-N 0.000 description 1
- KYBXNPIASYUWLN-WUCPZUCCSA-N (2s)-5-hydroxypyrrolidine-2-carboxylic acid Chemical compound OC1CC[C@@H](C(O)=O)N1 KYBXNPIASYUWLN-WUCPZUCCSA-N 0.000 description 1
- WHTVZRBIWZFKQO-AWEZNQCLSA-N (S)-chloroquine Chemical compound ClC1=CC=C2C(N[C@@H](C)CCCN(CC)CC)=CC=NC2=C1 WHTVZRBIWZFKQO-AWEZNQCLSA-N 0.000 description 1
- VYEWZWBILJHHCU-OMQUDAQFSA-N (e)-n-[(2s,3r,4r,5r,6r)-2-[(2r,3r,4s,5s,6s)-3-acetamido-5-amino-4-hydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-6-[2-[(2r,3s,4r,5r)-5-(2,4-dioxopyrimidin-1-yl)-3,4-dihydroxyoxolan-2-yl]-2-hydroxyethyl]-4,5-dihydroxyoxan-3-yl]-5-methylhex-2-enamide Chemical compound N1([C@@H]2O[C@@H]([C@H]([C@H]2O)O)C(O)C[C@@H]2[C@H](O)[C@H](O)[C@H]([C@@H](O2)O[C@@H]2[C@@H]([C@@H](O)[C@H](N)[C@@H](CO)O2)NC(C)=O)NC(=O)/C=C/CC(C)C)C=CC(=O)NC1=O VYEWZWBILJHHCU-OMQUDAQFSA-N 0.000 description 1
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 1
- UVBYMVOUBXYSFV-XUTVFYLZSA-N 1-methylpseudouridine Chemical compound O=C1NC(=O)N(C)C=C1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 UVBYMVOUBXYSFV-XUTVFYLZSA-N 0.000 description 1
- MQLACMBJVPINKE-UHFFFAOYSA-N 10-[(3-hydroxy-4-methoxyphenyl)methylidene]anthracen-9-one Chemical compound C1=C(O)C(OC)=CC=C1C=C1C2=CC=CC=C2C(=O)C2=CC=CC=C21 MQLACMBJVPINKE-UHFFFAOYSA-N 0.000 description 1
- PNDPGZBMCMUPRI-HVTJNCQCSA-N 10043-66-0 Chemical compound [131I][131I] PNDPGZBMCMUPRI-HVTJNCQCSA-N 0.000 description 1
- WUAPFZMCVAUBPE-NJFSPNSNSA-N 188Re Chemical compound [188Re] WUAPFZMCVAUBPE-NJFSPNSNSA-N 0.000 description 1
- 150000003923 2,5-pyrrolediones Chemical class 0.000 description 1
- MIJDSYMOBYNHOT-UHFFFAOYSA-N 2-(ethylamino)ethanol Chemical compound CCNCCO MIJDSYMOBYNHOT-UHFFFAOYSA-N 0.000 description 1
- WFZFMHDDZRBTFH-CZEFNJPISA-N 2-[(e)-2-(5-carbamimidoyl-1-benzofuran-2-yl)ethenyl]-1-benzofuran-5-carboximidamide;dihydrochloride Chemical compound Cl.Cl.NC(=N)C1=CC=C2OC(/C=C/C=3OC4=CC=C(C=C4C=3)C(=N)N)=CC2=C1 WFZFMHDDZRBTFH-CZEFNJPISA-N 0.000 description 1
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 1
- 125000000979 2-amino-2-oxoethyl group Chemical group [H]C([*])([H])C(=O)N([H])[H] 0.000 description 1
- ZOOGRGPOEVQQDX-UUOKFMHZSA-N 3',5'-cyclic GMP Chemical compound C([C@H]1O2)OP(O)(=O)O[C@H]1[C@@H](O)[C@@H]2N1C(N=C(NC2=O)N)=C2N=C1 ZOOGRGPOEVQQDX-UUOKFMHZSA-N 0.000 description 1
- UAIUNKRWKOVEES-UHFFFAOYSA-N 3,3',5,5'-tetramethylbenzidine Chemical compound CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 UAIUNKRWKOVEES-UHFFFAOYSA-N 0.000 description 1
- 101800000504 3C-like protease Proteins 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- PXRKCOCTEMYUEG-UHFFFAOYSA-N 5-aminoisoindole-1,3-dione Chemical compound NC1=CC=C2C(=O)NC(=O)C2=C1 PXRKCOCTEMYUEG-UHFFFAOYSA-N 0.000 description 1
- IDLISIVVYLGCKO-UHFFFAOYSA-N 6-carboxy-4',5'-dichloro-2',7'-dimethoxyfluorescein Chemical compound O1C(=O)C2=CC=C(C(O)=O)C=C2C21C1=CC(OC)=C(O)C(Cl)=C1OC1=C2C=C(OC)C(O)=C1Cl IDLISIVVYLGCKO-UHFFFAOYSA-N 0.000 description 1
- BZTDTCNHAFUJOG-UHFFFAOYSA-N 6-carboxyfluorescein Chemical compound C12=CC=C(O)C=C2OC2=CC(O)=CC=C2C11OC(=O)C2=CC=C(C(=O)O)C=C21 BZTDTCNHAFUJOG-UHFFFAOYSA-N 0.000 description 1
- WEVYAHXRMPXWCK-UHFFFAOYSA-N Acetonitrile Chemical compound CC#N WEVYAHXRMPXWCK-UHFFFAOYSA-N 0.000 description 1
- 101150044980 Akap1 gene Proteins 0.000 description 1
- 102000009027 Albumins Human genes 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 108091093088 Amplicon Proteins 0.000 description 1
- 208000002109 Argyria Diseases 0.000 description 1
- 240000003291 Armoracia rusticana Species 0.000 description 1
- 235000011330 Armoracia rusticana Nutrition 0.000 description 1
- 206010003445 Ascites Diseases 0.000 description 1
- 208000031504 Asymptomatic Infections Diseases 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 108091008875 B cell receptors Proteins 0.000 description 1
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 108700004676 Bence Jones Proteins 0.000 description 1
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 1
- 241000008904 Betacoronavirus Species 0.000 description 1
- 208000031648 Body Weight Changes Diseases 0.000 description 1
- BTBUEUYNUDRHOZ-UHFFFAOYSA-N Borate Chemical compound [O-]B([O-])[O-] BTBUEUYNUDRHOZ-UHFFFAOYSA-N 0.000 description 1
- 206010006187 Breast cancer Diseases 0.000 description 1
- 229940124293 CD30 monoclonal antibody Drugs 0.000 description 1
- 101100220616 Caenorhabditis elegans chk-2 gene Proteins 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 1
- 239000004215 Carbon black (E152) Substances 0.000 description 1
- CURLTUGMZLYLDI-UHFFFAOYSA-N Carbon dioxide Chemical group O=C=O CURLTUGMZLYLDI-UHFFFAOYSA-N 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010078791 Carrier Proteins Proteins 0.000 description 1
- 102000005600 Cathepsins Human genes 0.000 description 1
- 108010084457 Cathepsins Proteins 0.000 description 1
- 102000000844 Cell Surface Receptors Human genes 0.000 description 1
- 108010001857 Cell Surface Receptors Proteins 0.000 description 1
- 229940123150 Chelating agent Drugs 0.000 description 1
- ZAMOUSCENKQFHK-UHFFFAOYSA-N Chlorine atom Chemical compound [Cl] ZAMOUSCENKQFHK-UHFFFAOYSA-N 0.000 description 1
- 241000282552 Chlorocebus aethiops Species 0.000 description 1
- VYZAMTAEIAYCRO-UHFFFAOYSA-N Chromium Chemical compound [Cr] VYZAMTAEIAYCRO-UHFFFAOYSA-N 0.000 description 1
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- JPVYNHNXODAKFH-UHFFFAOYSA-N Cu2+ Chemical compound [Cu+2] JPVYNHNXODAKFH-UHFFFAOYSA-N 0.000 description 1
- 102000008130 Cyclic AMP-Dependent Protein Kinases Human genes 0.000 description 1
- 108010049894 Cyclic AMP-Dependent Protein Kinases Proteins 0.000 description 1
- 102100025698 Cytosolic carboxypeptidase 4 Human genes 0.000 description 1
- 101710112752 Cytotoxin Proteins 0.000 description 1
- 101150097493 D gene Proteins 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 229940021995 DNA vaccine Drugs 0.000 description 1
- 208000034423 Delivery Diseases 0.000 description 1
- 241000725619 Dengue virus Species 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 238000009007 Diagnostic Kit Methods 0.000 description 1
- 206010012735 Diarrhoea Diseases 0.000 description 1
- 101100072149 Drosophila melanogaster eIF2alpha gene Proteins 0.000 description 1
- 101100310856 Drosophila melanogaster spri gene Proteins 0.000 description 1
- 239000012591 Dulbecco’s Phosphate Buffered Saline Substances 0.000 description 1
- 229910052692 Dysprosium Inorganic materials 0.000 description 1
- 101000895909 Elizabethkingia meningoseptica Endo-beta-N-acetylglucosaminidase F1 Proteins 0.000 description 1
- 229910052691 Erbium Inorganic materials 0.000 description 1
- 241000233756 Fabriciana elisa Species 0.000 description 1
- CWYNVVGOOAEACU-UHFFFAOYSA-N Fe2+ Chemical compound [Fe+2] CWYNVVGOOAEACU-UHFFFAOYSA-N 0.000 description 1
- 241000724791 Filamentous phage Species 0.000 description 1
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 1
- 108010001515 Galectin 4 Proteins 0.000 description 1
- 102100039556 Galectin-4 Human genes 0.000 description 1
- GYHNNYVSQQEPJS-OIOBTWANSA-N Gallium-67 Chemical compound [67Ga] GYHNNYVSQQEPJS-OIOBTWANSA-N 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- 108010024636 Glutathione Proteins 0.000 description 1
- 108060005986 Granzyme Proteins 0.000 description 1
- 102000001398 Granzyme Human genes 0.000 description 1
- 208000031886 HIV Infections Diseases 0.000 description 1
- 208000037357 HIV infectious disease Diseases 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 1
- 101000932590 Homo sapiens Cytosolic carboxypeptidase 4 Proteins 0.000 description 1
- 101001055307 Homo sapiens Immunoglobulin heavy constant delta Proteins 0.000 description 1
- 101001037136 Homo sapiens Immunoglobulin heavy variable 3-15 Proteins 0.000 description 1
- 101001091242 Homo sapiens Immunoglobulin kappa joining 1 Proteins 0.000 description 1
- 101000604674 Homo sapiens Immunoglobulin kappa variable 4-1 Proteins 0.000 description 1
- 101000935040 Homo sapiens Integrin beta-2 Proteins 0.000 description 1
- 101000917826 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-a Proteins 0.000 description 1
- 101000917824 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-b Proteins 0.000 description 1
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 description 1
- 101000582320 Homo sapiens Neurogenic differentiation factor 6 Proteins 0.000 description 1
- 101000611183 Homo sapiens Tumor necrosis factor Proteins 0.000 description 1
- 101000851434 Homo sapiens Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 1
- 101000851376 Homo sapiens Tumor necrosis factor receptor superfamily member 8 Proteins 0.000 description 1
- 241000725303 Human immunodeficiency virus Species 0.000 description 1
- 206010020772 Hypertension Diseases 0.000 description 1
- 102100034892 Immunoglobulin kappa joining 1 Human genes 0.000 description 1
- 102100038198 Immunoglobulin kappa variable 4-1 Human genes 0.000 description 1
- 102100022297 Integrin alpha-X Human genes 0.000 description 1
- 102100025390 Integrin beta-2 Human genes 0.000 description 1
- 102000003996 Interferon-beta Human genes 0.000 description 1
- 108090000467 Interferon-beta Proteins 0.000 description 1
- 108010002350 Interleukin-2 Proteins 0.000 description 1
- ZCYVEMRRCGMTRW-AHCXROLUSA-N Iodine-123 Chemical compound [123I] ZCYVEMRRCGMTRW-AHCXROLUSA-N 0.000 description 1
- 101150008942 J gene Proteins 0.000 description 1
- OFFWOVJBSQMVPI-RMLGOCCBSA-N Kaletra Chemical compound N1([C@@H](C(C)C)C(=O)N[C@H](C[C@H](O)[C@H](CC=2C=CC=CC=2)NC(=O)COC=2C(=CC=CC=2C)C)CC=2C=CC=CC=2)CCCNC1=O.N([C@@H](C(C)C)C(=O)N[C@H](C[C@H](O)[C@H](CC=1C=CC=CC=1)NC(=O)OCC=1SC=NC=1)CC=1C=CC=CC=1)C(=O)N(C)CC1=CSC(C(C)C)=N1 OFFWOVJBSQMVPI-RMLGOCCBSA-N 0.000 description 1
- 102100034866 Kallikrein-6 Human genes 0.000 description 1
- YINZYTTZHLPWBO-UHFFFAOYSA-N Kifunensine Natural products COC1C(O)C(O)C(O)C2NC(=O)C(=O)N12 YINZYTTZHLPWBO-UHFFFAOYSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- 102100038609 Lactoperoxidase Human genes 0.000 description 1
- 108010023244 Lactoperoxidase Proteins 0.000 description 1
- 241000713666 Lentivirus Species 0.000 description 1
- 239000000232 Lipid Bilayer Substances 0.000 description 1
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 1
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 1
- 239000012515 MabSelect SuRe Substances 0.000 description 1
- WAEMQWOKJMHJLA-UHFFFAOYSA-N Manganese(2+) Chemical compound [Mn+2] WAEMQWOKJMHJLA-UHFFFAOYSA-N 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- 238000000585 Mann–Whitney U test Methods 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 102000029749 Microtubule Human genes 0.000 description 1
- 108091022875 Microtubule Proteins 0.000 description 1
- 108010006519 Molecular Chaperones Proteins 0.000 description 1
- 102000005431 Molecular Chaperones Human genes 0.000 description 1
- 208000034486 Multi-organ failure Diseases 0.000 description 1
- 241000711408 Murine respirovirus Species 0.000 description 1
- 101001033003 Mus musculus Granzyme F Proteins 0.000 description 1
- 241000282339 Mustela Species 0.000 description 1
- 241000187479 Mycobacterium tuberculosis Species 0.000 description 1
- OVRNDRQMDRJTHS-CBQIKETKSA-N N-Acetyl-D-Galactosamine Chemical compound CC(=O)N[C@H]1[C@@H](O)O[C@H](CO)[C@H](O)[C@@H]1O OVRNDRQMDRJTHS-CBQIKETKSA-N 0.000 description 1
- NQTADLQHYWFPDB-UHFFFAOYSA-N N-Hydroxysuccinimide Chemical compound ON1C(=O)CCC1=O NQTADLQHYWFPDB-UHFFFAOYSA-N 0.000 description 1
- GXCLVBGFBYZDAG-UHFFFAOYSA-N N-[2-(1H-indol-3-yl)ethyl]-N-methylprop-2-en-1-amine Chemical compound CN(CCC1=CNC2=C1C=CC=C2)CC=C GXCLVBGFBYZDAG-UHFFFAOYSA-N 0.000 description 1
- OVRNDRQMDRJTHS-UHFFFAOYSA-N N-acelyl-D-glucosamine Natural products CC(=O)NC1C(O)OC(CO)C(O)C1O OVRNDRQMDRJTHS-UHFFFAOYSA-N 0.000 description 1
- MBLBDJOUHNCFQT-UHFFFAOYSA-N N-acetyl-D-galactosamine Natural products CC(=O)NC(C=O)C(O)C(O)C(O)CO MBLBDJOUHNCFQT-UHFFFAOYSA-N 0.000 description 1
- SQVRNKJHWKZAKO-PFQGKNLYSA-N N-acetyl-beta-neuraminic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)O[C@H]1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-PFQGKNLYSA-N 0.000 description 1
- MBLBDJOUHNCFQT-LXGUWJNJSA-N N-acetylglucosamine Natural products CC(=O)N[C@@H](C=O)[C@@H](O)[C@H](O)[C@H](O)CO MBLBDJOUHNCFQT-LXGUWJNJSA-N 0.000 description 1
- NXTVQNIVUKXOIL-UHFFFAOYSA-N N-chlorotoluene-p-sulfonamide Chemical compound CC1=CC=C(S(=O)(=O)NCl)C=C1 NXTVQNIVUKXOIL-UHFFFAOYSA-N 0.000 description 1
- 206010029260 Neuroblastoma Diseases 0.000 description 1
- 102100030589 Neurogenic differentiation factor 6 Human genes 0.000 description 1
- VEQPNABPJHWNSG-UHFFFAOYSA-N Nickel(2+) Chemical compound [Ni+2] VEQPNABPJHWNSG-UHFFFAOYSA-N 0.000 description 1
- 244000061176 Nicotiana tabacum Species 0.000 description 1
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 1
- 108090001074 Nucleocapsid Proteins Proteins 0.000 description 1
- AWZJFZMWSUBJAJ-UHFFFAOYSA-N OG-514 dye Chemical compound OC(=O)CSC1=C(F)C(F)=C(C(O)=O)C(C2=C3C=C(F)C(=O)C=C3OC3=CC(O)=C(F)C=C32)=C1F AWZJFZMWSUBJAJ-UHFFFAOYSA-N 0.000 description 1
- 241000337007 Oceania Species 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 206010068319 Oropharyngeal pain Diseases 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 240000007594 Oryza sativa Species 0.000 description 1
- 235000007164 Oryza sativa Nutrition 0.000 description 1
- 108010058846 Ovalbumin Proteins 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 241000237988 Patellidae Species 0.000 description 1
- 235000019483 Peanut oil Nutrition 0.000 description 1
- 102100022587 Peroxisomal multifunctional enzyme type 2 Human genes 0.000 description 1
- 101710125609 Peroxisomal multifunctional enzyme type 2 Proteins 0.000 description 1
- 201000007100 Pharyngitis Diseases 0.000 description 1
- 241000283966 Pholidota <mammal> Species 0.000 description 1
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 1
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 1
- OAICVXFJPJFONN-UHFFFAOYSA-N Phosphorus Chemical compound [P] OAICVXFJPJFONN-UHFFFAOYSA-N 0.000 description 1
- 101800001016 Picornain 3C-like protease Proteins 0.000 description 1
- 241000276498 Pollachius virens Species 0.000 description 1
- 239000004698 Polyethylene Substances 0.000 description 1
- 229920002562 Polyethylene Glycol 3350 Polymers 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 239000004793 Polystyrene Substances 0.000 description 1
- 241000288906 Primates Species 0.000 description 1
- 101800000596 Probable picornain 3C-like protease Proteins 0.000 description 1
- XBDQKXXYIPTUBI-UHFFFAOYSA-M Propionate Chemical compound CCC([O-])=O XBDQKXXYIPTUBI-UHFFFAOYSA-M 0.000 description 1
- 102000001253 Protein Kinase Human genes 0.000 description 1
- 241001112090 Pseudovirus Species 0.000 description 1
- IWYDHOAUDWTVEP-UHFFFAOYSA-N R-2-phenyl-2-hydroxyacetic acid Natural products OC(=O)C(O)C1=CC=CC=C1 IWYDHOAUDWTVEP-UHFFFAOYSA-N 0.000 description 1
- 238000002123 RNA extraction Methods 0.000 description 1
- 238000010802 RNA extraction kit Methods 0.000 description 1
- 229940122277 RNA polymerase inhibitor Drugs 0.000 description 1
- 229940022005 RNA vaccine Drugs 0.000 description 1
- 238000003559 RNA-seq method Methods 0.000 description 1
- 239000012980 RPMI-1640 medium Substances 0.000 description 1
- 241000700159 Rattus Species 0.000 description 1
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 1
- 101710100968 Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 101710146873 Receptor-binding protein Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 101710153041 Replicase polyprotein 1a Proteins 0.000 description 1
- 206010057190 Respiratory tract infections Diseases 0.000 description 1
- 241000228636 Rhinolophus Species 0.000 description 1
- 241000608671 Rhinolophus affinis Species 0.000 description 1
- 241000031819 Rhinolophus sinicus Species 0.000 description 1
- 208000036071 Rhinorrhea Diseases 0.000 description 1
- 206010039101 Rhinorrhoea Diseases 0.000 description 1
- 108010039491 Ricin Proteins 0.000 description 1
- 101150010882 S gene Proteins 0.000 description 1
- 108091005634 SARS-CoV-2 receptor-binding domains Proteins 0.000 description 1
- 238000011579 SCID mouse model Methods 0.000 description 1
- 108010084592 Saporins Proteins 0.000 description 1
- 241001678561 Sarbecovirus Species 0.000 description 1
- BUGBHKTXTAQXES-UHFFFAOYSA-N Selenium Chemical compound [Se] BUGBHKTXTAQXES-UHFFFAOYSA-N 0.000 description 1
- 229920005654 Sephadex Polymers 0.000 description 1
- 239000012507 Sephadex™ Substances 0.000 description 1
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 1
- 108010003723 Single-Domain Antibodies Proteins 0.000 description 1
- 239000005708 Sodium hypochlorite Substances 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 108700005078 Synthetic Genes Proteins 0.000 description 1
- PZBFGYYEXUXCOF-UHFFFAOYSA-N TCEP Chemical compound OC(=O)CCP(CCC(O)=O)CCC(O)=O PZBFGYYEXUXCOF-UHFFFAOYSA-N 0.000 description 1
- GKLVYJBZJHMRIY-OUBTZVSYSA-N Technetium-99 Chemical compound [99Tc] GKLVYJBZJHMRIY-OUBTZVSYSA-N 0.000 description 1
- 229910052771 Terbium Inorganic materials 0.000 description 1
- 101710120037 Toxin CcdB Proteins 0.000 description 1
- GYDJEQRTZSCIOI-UHFFFAOYSA-N Tranexamic acid Chemical compound NCC1CCC(C(O)=O)CC1 GYDJEQRTZSCIOI-UHFFFAOYSA-N 0.000 description 1
- 238000010162 Tukey test Methods 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 108060008683 Tumor Necrosis Factor Receptor Proteins 0.000 description 1
- 102100040247 Tumor necrosis factor Human genes 0.000 description 1
- 102100036922 Tumor necrosis factor ligand superfamily member 13B Human genes 0.000 description 1
- 101710181056 Tumor necrosis factor ligand superfamily member 13B Proteins 0.000 description 1
- 102100036857 Tumor necrosis factor receptor superfamily member 8 Human genes 0.000 description 1
- YJQCOFNZVFGCAF-UHFFFAOYSA-N Tunicamycin II Natural products O1C(CC(O)C2C(C(O)C(O2)N2C(NC(=O)C=C2)=O)O)C(O)C(O)C(NC(=O)C=CCCCCCCCCC(C)C)C1OC1OC(CO)C(O)C(O)C1NC(C)=O YJQCOFNZVFGCAF-UHFFFAOYSA-N 0.000 description 1
- 101150117115 V gene Proteins 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 229940122803 Vinca alkaloid Drugs 0.000 description 1
- 108010067390 Viral Proteins Proteins 0.000 description 1
- 230000010530 Virus Neutralization Effects 0.000 description 1
- 229910052769 Ytterbium Inorganic materials 0.000 description 1
- VWQVUPCCIRVNHF-OUBTZVSYSA-N Yttrium-90 Chemical compound [90Y] VWQVUPCCIRVNHF-OUBTZVSYSA-N 0.000 description 1
- PNDPGZBMCMUPRI-XXSWNUTMSA-N [125I][125I] Chemical compound [125I][125I] PNDPGZBMCMUPRI-XXSWNUTMSA-N 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 150000008043 acidic salts Chemical class 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 210000002534 adenoid Anatomy 0.000 description 1
- 230000001464 adherent effect Effects 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 239000000443 aerosol Substances 0.000 description 1
- 238000003314 affinity selection Methods 0.000 description 1
- 238000013019 agitation Methods 0.000 description 1
- 208000026935 allergic disease Diseases 0.000 description 1
- 230000007815 allergy Effects 0.000 description 1
- 229940037003 alum Drugs 0.000 description 1
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 1
- 230000009435 amidation Effects 0.000 description 1
- 238000007112 amidation reaction Methods 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 125000002344 aminooxy group Chemical group [H]N([H])O[*] 0.000 description 1
- 150000008064 anhydrides Chemical class 0.000 description 1
- 150000001450 anions Chemical class 0.000 description 1
- 230000036436 anti-hiv Effects 0.000 description 1
- 230000000852 anti-lentiviral effect Effects 0.000 description 1
- 230000001946 anti-microtubular Effects 0.000 description 1
- 229940125644 antibody drug Drugs 0.000 description 1
- 238000011091 antibody purification Methods 0.000 description 1
- 210000000628 antibody-producing cell Anatomy 0.000 description 1
- 229940041181 antineoplastic drug Drugs 0.000 description 1
- 229940121357 antivirals Drugs 0.000 description 1
- 230000006907 apoptotic process Effects 0.000 description 1
- 239000012736 aqueous medium Substances 0.000 description 1
- 239000008346 aqueous phase Substances 0.000 description 1
- PYMYPHUHKUWMLA-UHFFFAOYSA-N arabinose Natural products OCC(O)C(O)C(O)C=O PYMYPHUHKUWMLA-UHFFFAOYSA-N 0.000 description 1
- 238000003491 array Methods 0.000 description 1
- 125000000613 asparagine group Chemical group N[C@@H](CC(N)=O)C(=O)* 0.000 description 1
- 238000002820 assay format Methods 0.000 description 1
- RYXHOMYVWAEKHL-OUBTZVSYSA-N astatine-211 Chemical compound [211At] RYXHOMYVWAEKHL-OUBTZVSYSA-N 0.000 description 1
- 125000000852 azido group Chemical group *N=[N+]=[N-] 0.000 description 1
- 230000006399 behavior Effects 0.000 description 1
- HFACYLZERDEVSX-UHFFFAOYSA-N benzidine Chemical compound C1=CC(N)=CC=C1C1=CC=C(N)C=C1 HFACYLZERDEVSX-UHFFFAOYSA-N 0.000 description 1
- SRBFZHDQGSBBOR-UHFFFAOYSA-N beta-D-Pyranose-Lyxose Natural products OC1COC(O)C(O)C1O SRBFZHDQGSBBOR-UHFFFAOYSA-N 0.000 description 1
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 1
- 239000011230 binding agent Substances 0.000 description 1
- 230000008275 binding mechanism Effects 0.000 description 1
- 238000005842 biochemical reaction Methods 0.000 description 1
- 239000013060 biological fluid Substances 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 238000005415 bioluminescence Methods 0.000 description 1
- 230000029918 bioluminescence Effects 0.000 description 1
- 229960000074 biopharmaceutical Drugs 0.000 description 1
- JDIBGQFKXXXXPN-UHFFFAOYSA-N bismuth(3+) Chemical compound [Bi+3] JDIBGQFKXXXXPN-UHFFFAOYSA-N 0.000 description 1
- 239000002981 blocking agent Substances 0.000 description 1
- 230000037396 body weight Effects 0.000 description 1
- 230000004579 body weight change Effects 0.000 description 1
- 210000001185 bone marrow Anatomy 0.000 description 1
- 229960000455 brentuximab vedotin Drugs 0.000 description 1
- 239000006227 byproduct Substances 0.000 description 1
- 229940087373 calcium oxide Drugs 0.000 description 1
- 239000004202 carbamide Substances 0.000 description 1
- 150000001718 carbodiimides Chemical class 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 238000000423 cell based assay Methods 0.000 description 1
- 230000011712 cell development Effects 0.000 description 1
- 230000032823 cell division Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 230000003833 cell viability Effects 0.000 description 1
- 230000002032 cellular defenses Effects 0.000 description 1
- 239000001913 cellulose Substances 0.000 description 1
- 229920002678 cellulose Polymers 0.000 description 1
- 239000013522 chelant Substances 0.000 description 1
- 238000012412 chemical coupling Methods 0.000 description 1
- 238000010382 chemical cross-linking Methods 0.000 description 1
- 239000007795 chemical reaction product Substances 0.000 description 1
- 229940044683 chemotherapy drug Drugs 0.000 description 1
- 239000000460 chlorine Substances 0.000 description 1
- 229910052801 chlorine Inorganic materials 0.000 description 1
- 229960003677 chloroquine Drugs 0.000 description 1
- WHTVZRBIWZFKQO-UHFFFAOYSA-N chloroquine Natural products ClC1=CC=C2C(NC(C)CCCN(CC)CC)=CC=NC2=C1 WHTVZRBIWZFKQO-UHFFFAOYSA-N 0.000 description 1
- 229910052804 chromium Inorganic materials 0.000 description 1
- 239000011651 chromium Substances 0.000 description 1
- BFGKITSFLPAWGI-UHFFFAOYSA-N chromium(3+) Chemical compound [Cr+3] BFGKITSFLPAWGI-UHFFFAOYSA-N 0.000 description 1
- 238000000978 circular dichroism spectroscopy Methods 0.000 description 1
- 238000001142 circular dichroism spectrum Methods 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 238000003501 co-culture Methods 0.000 description 1
- 230000004186 co-expression Effects 0.000 description 1
- 239000000701 coagulant Substances 0.000 description 1
- XLJKHNWPARRRJB-UHFFFAOYSA-N cobalt(2+) Chemical compound [Co+2] XLJKHNWPARRRJB-UHFFFAOYSA-N 0.000 description 1
- AGVAZMGAQJOSFJ-WZHZPDAFSA-M cobalt(2+);[(2r,3s,4r,5s)-5-(5,6-dimethylbenzimidazol-1-yl)-4-hydroxy-2-(hydroxymethyl)oxolan-3-yl] [(2r)-1-[3-[(1r,2r,3r,4z,7s,9z,12s,13s,14z,17s,18s,19r)-2,13,18-tris(2-amino-2-oxoethyl)-7,12,17-tris(3-amino-3-oxopropyl)-3,5,8,8,13,15,18,19-octamethyl-2 Chemical compound [Co+2].N#[C-].[N-]([C@@H]1[C@H](CC(N)=O)[C@@]2(C)CCC(=O)NC[C@@H](C)OP(O)(=O)O[C@H]3[C@H]([C@H](O[C@@H]3CO)N3C4=CC(C)=C(C)C=C4N=C3)O)\C2=C(C)/C([C@H](C\2(C)C)CCC(N)=O)=N/C/2=C\C([C@H]([C@@]/2(CC(N)=O)C)CCC(N)=O)=N\C\2=C(C)/C2=N[C@]1(C)[C@@](C)(CC(N)=O)[C@@H]2CCC(N)=O AGVAZMGAQJOSFJ-WZHZPDAFSA-M 0.000 description 1
- 239000003086 colorant Substances 0.000 description 1
- 230000004154 complement system Effects 0.000 description 1
- 239000008139 complexing agent Substances 0.000 description 1
- 230000001010 compromised effect Effects 0.000 description 1
- 238000010205 computational analysis Methods 0.000 description 1
- 238000005094 computer simulation Methods 0.000 description 1
- 239000012141 concentrate Substances 0.000 description 1
- 230000001268 conjugating effect Effects 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 238000001816 cooling Methods 0.000 description 1
- NKLPQNGYXWVELD-UHFFFAOYSA-M coomassie brilliant blue Chemical compound [Na+].C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=2C=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=C1 NKLPQNGYXWVELD-UHFFFAOYSA-M 0.000 description 1
- RYGMFSIKBFXOCR-AKLPVKDBSA-N copper-67 Chemical compound [67Cu] RYGMFSIKBFXOCR-AKLPVKDBSA-N 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000010168 coupling process Methods 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- 230000009260 cross reactivity Effects 0.000 description 1
- 239000008150 cryoprotective solution Substances 0.000 description 1
- 239000011549 crystallization solution Substances 0.000 description 1
- 238000012866 crystallographic experiment Methods 0.000 description 1
- UFULAYFCSOUIOV-UHFFFAOYSA-N cysteamine Chemical compound NCCS UFULAYFCSOUIOV-UHFFFAOYSA-N 0.000 description 1
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 1
- 210000000805 cytoplasm Anatomy 0.000 description 1
- 210000005220 cytoplasmic tail Anatomy 0.000 description 1
- 230000001086 cytosolic effect Effects 0.000 description 1
- 230000003013 cytotoxicity Effects 0.000 description 1
- 231100000135 cytotoxicity Toxicity 0.000 description 1
- 238000002784 cytotoxicity assay Methods 0.000 description 1
- 231100000263 cytotoxicity test Toxicity 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 230000007123 defense Effects 0.000 description 1
- 230000007812 deficiency Effects 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 238000004925 denaturation Methods 0.000 description 1
- 230000036425 denaturation Effects 0.000 description 1
- 230000001627 detrimental effect Effects 0.000 description 1
- 206010012601 diabetes mellitus Diseases 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- 238000009792 diffusion process Methods 0.000 description 1
- 238000007865 diluting Methods 0.000 description 1
- AFABGHUZZDYHJO-UHFFFAOYSA-N dimethyl butane Natural products CCCC(C)C AFABGHUZZDYHJO-UHFFFAOYSA-N 0.000 description 1
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 1
- BNIILDVGGAEEIG-UHFFFAOYSA-L disodium hydrogen phosphate Chemical compound [Na+].[Na+].OP([O-])([O-])=O BNIILDVGGAEEIG-UHFFFAOYSA-L 0.000 description 1
- 229910000397 disodium phosphate Inorganic materials 0.000 description 1
- 235000019800 disodium phosphate Nutrition 0.000 description 1
- 238000013104 docking experiment Methods 0.000 description 1
- 239000002552 dosage form Substances 0.000 description 1
- 238000003110 dot immunobinding assay Methods 0.000 description 1
- 230000003828 downregulation Effects 0.000 description 1
- 208000017574 dry cough Diseases 0.000 description 1
- 239000000975 dye Substances 0.000 description 1
- KBQHZAAAGSGFKK-UHFFFAOYSA-N dysprosium atom Chemical compound [Dy] KBQHZAAAGSGFKK-UHFFFAOYSA-N 0.000 description 1
- 235000013601 eggs Nutrition 0.000 description 1
- 238000002003 electron diffraction Methods 0.000 description 1
- 230000009881 electrostatic interaction Effects 0.000 description 1
- 238000010828 elution Methods 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 239000000839 emulsion Substances 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 230000009088 enzymatic function Effects 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- UYAHIZSMUZPPFV-UHFFFAOYSA-N erbium Chemical compound [Er] UYAHIZSMUZPPFV-UHFFFAOYSA-N 0.000 description 1
- XJRPTMORGOIMMI-UHFFFAOYSA-N ethyl 2-amino-4-(trifluoromethyl)-1,3-thiazole-5-carboxylate Chemical compound CCOC(=O)C=1SC(N)=NC=1C(F)(F)F XJRPTMORGOIMMI-UHFFFAOYSA-N 0.000 description 1
- 238000002270 exclusion chromatography Methods 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 210000003722 extracellular fluid Anatomy 0.000 description 1
- 235000013861 fat-free Nutrition 0.000 description 1
- 235000013312 flour Nutrition 0.000 description 1
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 1
- 238000001917 fluorescence detection Methods 0.000 description 1
- 238000005194 fractionation Methods 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 210000001035 gastrointestinal tract Anatomy 0.000 description 1
- 238000001641 gel filtration chromatography Methods 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 238000007429 general method Methods 0.000 description 1
- 238000010362 genome editing Methods 0.000 description 1
- 238000012268 genome sequencing Methods 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-L glutamate group Chemical group N[C@@H](CCC(=O)[O-])C(=O)[O-] WHUUTDBJXJRKMK-VKHMYHEASA-L 0.000 description 1
- 229960003180 glutathione Drugs 0.000 description 1
- YQEMORVAKMFKLG-UHFFFAOYSA-N glycerine monostearate Natural products CCCCCCCCCCCCCCCCCC(=O)OC(CO)CO YQEMORVAKMFKLG-UHFFFAOYSA-N 0.000 description 1
- SVUQHVRAGMNPLW-UHFFFAOYSA-N glycerol monostearate Natural products CCCCCCCCCCCCCCCCC(=O)OCC(O)CO SVUQHVRAGMNPLW-UHFFFAOYSA-N 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- CBMIPXHVOVTTTL-UHFFFAOYSA-N gold(3+) Chemical compound [Au+3] CBMIPXHVOVTTTL-UHFFFAOYSA-N 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 229910052736 halogen Inorganic materials 0.000 description 1
- 150000002367 halogens Chemical class 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- 230000008821 health effect Effects 0.000 description 1
- 230000005802 health problem Effects 0.000 description 1
- 210000002216 heart Anatomy 0.000 description 1
- 238000003505 heat denaturation Methods 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 229940022353 herceptin Drugs 0.000 description 1
- 238000001239 high-resolution electron microscopy Methods 0.000 description 1
- 239000012145 high-salt buffer Substances 0.000 description 1
- SCKNFLZJSOHWIV-UHFFFAOYSA-N holmium(3+) Chemical compound [Ho+3] SCKNFLZJSOHWIV-UHFFFAOYSA-N 0.000 description 1
- 229920001519 homopolymer Polymers 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 208000033519 human immunodeficiency virus infectious disease Diseases 0.000 description 1
- 150000007857 hydrazones Chemical class 0.000 description 1
- 229930195733 hydrocarbon Natural products 0.000 description 1
- 150000002430 hydrocarbons Chemical class 0.000 description 1
- 235000011167 hydrochloric acid Nutrition 0.000 description 1
- WGCNASOHLSPBMP-UHFFFAOYSA-N hydroxyacetaldehyde Natural products OCC=O WGCNASOHLSPBMP-UHFFFAOYSA-N 0.000 description 1
- 125000002349 hydroxyamino group Chemical group [H]ON([H])[*] 0.000 description 1
- 229910052588 hydroxylapatite Inorganic materials 0.000 description 1
- 238000012872 hydroxylapatite chromatography Methods 0.000 description 1
- 239000005457 ice water Substances 0.000 description 1
- 230000008076 immune mechanism Effects 0.000 description 1
- 238000003119 immunoblot Methods 0.000 description 1
- 238000013115 immunohistochemical detection Methods 0.000 description 1
- 230000003308 immunostimulating effect Effects 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 239000007943 implant Substances 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 230000002757 inflammatory effect Effects 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 150000007529 inorganic bases Chemical class 0.000 description 1
- 102000006495 integrins Human genes 0.000 description 1
- 108010044426 integrins Proteins 0.000 description 1
- 238000005305 interferometry Methods 0.000 description 1
- 229960001388 interferon-beta Drugs 0.000 description 1
- 238000001361 intraarterial administration Methods 0.000 description 1
- 230000004068 intracellular signaling Effects 0.000 description 1
- 238000010255 intramuscular injection Methods 0.000 description 1
- 239000007927 intramuscular injection Substances 0.000 description 1
- 238000010253 intravenous injection Methods 0.000 description 1
- 238000011835 investigation Methods 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 238000005040 ion trap Methods 0.000 description 1
- 238000006317 isomerization reaction Methods 0.000 description 1
- 150000002540 isothiocyanates Chemical class 0.000 description 1
- 210000003292 kidney cell Anatomy 0.000 description 1
- OIURYJWYVIAOCW-VFUOTHLCSA-N kifunensine Chemical compound OC[C@@H]1[C@@H](O)[C@H](O)[C@@H](O)[C@@H]2NC(=O)C(=O)N12 OIURYJWYVIAOCW-VFUOTHLCSA-N 0.000 description 1
- 238000011005 laboratory method Methods 0.000 description 1
- 229940057428 lactoperoxidase Drugs 0.000 description 1
- CZMAIROVPAYCMU-UHFFFAOYSA-N lanthanum(3+) Chemical compound [La+3] CZMAIROVPAYCMU-UHFFFAOYSA-N 0.000 description 1
- 150000002605 large molecules Chemical class 0.000 description 1
- 238000012125 lateral flow test Methods 0.000 description 1
- 231100000518 lethal Toxicity 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 125000001909 leucine group Chemical group [H]N(*)C(C(*)=O)C([H])([H])C(C([H])([H])[H])C([H])([H])[H] 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- 238000011068 loading method Methods 0.000 description 1
- 230000005577 local transmission Effects 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 229940113983 lopinavir / ritonavir Drugs 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 230000002101 lytic effect Effects 0.000 description 1
- 108700021021 mRNA Vaccine Proteins 0.000 description 1
- 229920002521 macromolecule Polymers 0.000 description 1
- ZLNQQNXFFQJAID-UHFFFAOYSA-L magnesium carbonate Chemical compound [Mg+2].[O-]C([O-])=O ZLNQQNXFFQJAID-UHFFFAOYSA-L 0.000 description 1
- 239000001095 magnesium carbonate Substances 0.000 description 1
- 229910000021 magnesium carbonate Inorganic materials 0.000 description 1
- 235000019359 magnesium stearate Nutrition 0.000 description 1
- 230000005291 magnetic effect Effects 0.000 description 1
- 201000004792 malaria Diseases 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 229960002510 mandelic acid Drugs 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 150000002703 mannose derivatives Chemical class 0.000 description 1
- WKPWGQKGSOKKOO-RSFHAFMBSA-N maytansine Chemical class CO[C@@H]([C@@]1(O)C[C@](OC(=O)N1)([C@H]([C@@H]1O[C@@]1(C)[C@@H](OC(=O)[C@H](C)N(C)C(C)=O)CC(=O)N1C)C)[H])\C=C\C=C(C)\CC2=CC(OC)=C(Cl)C1=C2 WKPWGQKGSOKKOO-RSFHAFMBSA-N 0.000 description 1
- 238000010297 mechanical methods and process Methods 0.000 description 1
- 238000005399 mechanical ventilation Methods 0.000 description 1
- 210000001806 memory b lymphocyte Anatomy 0.000 description 1
- 229960003151 mercaptamine Drugs 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- 229910021645 metal ion Inorganic materials 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- DYQNRMCKBFOWKH-UHFFFAOYSA-N methyl 4-hydroxybenzenecarboximidate Chemical group COC(=N)C1=CC=C(O)C=C1 DYQNRMCKBFOWKH-UHFFFAOYSA-N 0.000 description 1
- 229920000609 methyl cellulose Polymers 0.000 description 1
- 239000001923 methylcellulose Substances 0.000 description 1
- 238000000386 microscopy Methods 0.000 description 1
- 210000004688 microtubule Anatomy 0.000 description 1
- 230000003278 mimic effect Effects 0.000 description 1
- 239000002480 mineral oil Substances 0.000 description 1
- 235000010446 mineral oil Nutrition 0.000 description 1
- 150000007522 mineralic acids Chemical class 0.000 description 1
- 210000003470 mitochondria Anatomy 0.000 description 1
- 102000035118 modified proteins Human genes 0.000 description 1
- 108091005573 modified proteins Proteins 0.000 description 1
- 239000003607 modifier Substances 0.000 description 1
- 239000002991 molded plastic Substances 0.000 description 1
- 238000001823 molecular biology technique Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 238000003032 molecular docking Methods 0.000 description 1
- 108010093470 monomethyl auristatin E Proteins 0.000 description 1
- 210000004400 mucous membrane Anatomy 0.000 description 1
- 208000029744 multiple organ dysfunction syndrome Diseases 0.000 description 1
- 210000003205 muscle Anatomy 0.000 description 1
- 239000007923 nasal drop Substances 0.000 description 1
- 229940100662 nasal drops Drugs 0.000 description 1
- 201000009240 nasopharyngitis Diseases 0.000 description 1
- 238000002663 nebulization Methods 0.000 description 1
- 239000006199 nebulizer Substances 0.000 description 1
- QEFYFXOXNSNQGX-UHFFFAOYSA-N neodymium atom Chemical compound [Nd] QEFYFXOXNSNQGX-UHFFFAOYSA-N 0.000 description 1
- PGSADBUBUOPOJS-UHFFFAOYSA-N neutral red Chemical compound Cl.C1=C(C)C(N)=CC2=NC3=CC(N(C)C)=CC=C3N=C21 PGSADBUBUOPOJS-UHFFFAOYSA-N 0.000 description 1
- 229910052759 nickel Inorganic materials 0.000 description 1
- 230000036963 noncompetitive effect Effects 0.000 description 1
- 230000009871 nonspecific binding Effects 0.000 description 1
- 238000010899 nucleation Methods 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 150000003833 nucleoside derivatives Chemical class 0.000 description 1
- 102000026415 nucleotide binding proteins Human genes 0.000 description 1
- 108091014756 nucleotide binding proteins Proteins 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 230000005868 ontogenesis Effects 0.000 description 1
- 210000000056 organ Anatomy 0.000 description 1
- 150000007524 organic acids Chemical class 0.000 description 1
- 235000005985 organic acids Nutrition 0.000 description 1
- 150000007530 organic bases Chemical class 0.000 description 1
- 230000003204 osmotic effect Effects 0.000 description 1
- 229940092253 ovalbumin Drugs 0.000 description 1
- 239000006174 pH buffer Substances 0.000 description 1
- 239000006179 pH buffering agent Substances 0.000 description 1
- VYNDHICBIRRPFP-UHFFFAOYSA-N pacific blue Chemical compound FC1=C(O)C(F)=C2OC(=O)C(C(=O)O)=CC2=C1 VYNDHICBIRRPFP-UHFFFAOYSA-N 0.000 description 1
- 210000002741 palatine tonsil Anatomy 0.000 description 1
- 238000004091 panning Methods 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 244000045947 parasite Species 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 239000000312 peanut oil Substances 0.000 description 1
- 230000006320 pegylation Effects 0.000 description 1
- 230000000149 penetrating effect Effects 0.000 description 1
- XYJRXVWERLGGKC-UHFFFAOYSA-D pentacalcium;hydroxide;triphosphate Chemical compound [OH-].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O XYJRXVWERLGGKC-UHFFFAOYSA-D 0.000 description 1
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 1
- 229930192851 perforin Natural products 0.000 description 1
- KHIWWQKSHDUIBK-UHFFFAOYSA-N periodic acid Chemical compound OI(=O)(=O)=O KHIWWQKSHDUIBK-UHFFFAOYSA-N 0.000 description 1
- 239000003208 petroleum Substances 0.000 description 1
- 239000008177 pharmaceutical agent Substances 0.000 description 1
- 229940124531 pharmaceutical excipient Drugs 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 235000011007 phosphoric acid Nutrition 0.000 description 1
- 229910052698 phosphorus Inorganic materials 0.000 description 1
- 239000011574 phosphorus Substances 0.000 description 1
- 238000006303 photolysis reaction Methods 0.000 description 1
- 230000015843 photosynthesis, light reaction Effects 0.000 description 1
- 238000013081 phylogenetic analysis Methods 0.000 description 1
- 230000035790 physiological processes and functions Effects 0.000 description 1
- 239000006187 pill Substances 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- 238000012123 point-of-care testing Methods 0.000 description 1
- 229920002523 polyethylene Glycol 1000 Polymers 0.000 description 1
- 238000003752 polymerase chain reaction Methods 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 229920002223 polystyrene Polymers 0.000 description 1
- 230000005195 poor health Effects 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- DTBMTXYWRJNBGK-UHFFFAOYSA-L potassium;sodium;phthalate Chemical compound [Na+].[K+].[O-]C(=O)C1=CC=CC=C1C([O-])=O DTBMTXYWRJNBGK-UHFFFAOYSA-L 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 238000011165 process development Methods 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 230000001902 propagating effect Effects 0.000 description 1
- 238000011321 prophylaxis Methods 0.000 description 1
- QQONPFPTGQHPMA-UHFFFAOYSA-N propylene Natural products CC=C QQONPFPTGQHPMA-UHFFFAOYSA-N 0.000 description 1
- 125000004805 propylene group Chemical group [H]C([H])([H])C([H])([*:1])C([H])([H])[*:2] 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 230000006916 protein interaction Effects 0.000 description 1
- 108060006633 protein kinase Proteins 0.000 description 1
- 230000006920 protein precipitation Effects 0.000 description 1
- 229940023143 protein vaccine Drugs 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 239000002213 purine nucleotide Substances 0.000 description 1
- 230000006825 purine synthesis Effects 0.000 description 1
- 150000003230 pyrimidines Chemical class 0.000 description 1
- 238000004445 quantitative analysis Methods 0.000 description 1
- 239000001397 quillaja saponaria molina bark Substances 0.000 description 1
- 238000002708 random mutagenesis Methods 0.000 description 1
- 230000007115 recruitment Effects 0.000 description 1
- 238000004064 recycling Methods 0.000 description 1
- 210000005132 reproductive cell Anatomy 0.000 description 1
- 230000001177 retroviral effect Effects 0.000 description 1
- 238000004366 reverse phase liquid chromatography Methods 0.000 description 1
- WUAPFZMCVAUBPE-IGMARMGPSA-N rhenium-186 Chemical compound [186Re] WUAPFZMCVAUBPE-IGMARMGPSA-N 0.000 description 1
- 235000009566 rice Nutrition 0.000 description 1
- CVHZOJJKTDOEJC-UHFFFAOYSA-N saccharin Chemical compound C1=CC=C2C(=O)NS(=O)(=O)C2=C1 CVHZOJJKTDOEJC-UHFFFAOYSA-N 0.000 description 1
- DOSGOCSVHPUUIA-UHFFFAOYSA-N samarium(3+) Chemical compound [Sm+3] DOSGOCSVHPUUIA-UHFFFAOYSA-N 0.000 description 1
- 238000005070 sampling Methods 0.000 description 1
- 238000003118 sandwich ELISA Methods 0.000 description 1
- 229930182490 saponin Natural products 0.000 description 1
- 150000007949 saponins Chemical class 0.000 description 1
- 238000007790 scraping Methods 0.000 description 1
- 238000010187 selection method Methods 0.000 description 1
- 229910052711 selenium Inorganic materials 0.000 description 1
- 239000011669 selenium Substances 0.000 description 1
- 238000002864 sequence alignment Methods 0.000 description 1
- 206010040400 serum sickness Diseases 0.000 description 1
- 239000004017 serum-free culture medium Substances 0.000 description 1
- 239000008159 sesame oil Substances 0.000 description 1
- 235000011803 sesame oil Nutrition 0.000 description 1
- 208000026425 severe pneumonia Diseases 0.000 description 1
- 230000009450 sialylation Effects 0.000 description 1
- 230000009131 signaling function Effects 0.000 description 1
- 239000000741 silica gel Substances 0.000 description 1
- 229910002027 silica gel Inorganic materials 0.000 description 1
- 239000002356 single layer Substances 0.000 description 1
- 238000009097 single-agent therapy Methods 0.000 description 1
- 238000001542 size-exclusion chromatography Methods 0.000 description 1
- 235000020183 skimmed milk Nutrition 0.000 description 1
- 206010041232 sneezing Diseases 0.000 description 1
- 239000000344 soap Substances 0.000 description 1
- PTLRDCMBXHILCL-UHFFFAOYSA-M sodium arsenite Chemical compound [Na+].[O-][As]=O PTLRDCMBXHILCL-UHFFFAOYSA-M 0.000 description 1
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 1
- SUKJFIGYRHOWBL-UHFFFAOYSA-N sodium hypochlorite Chemical compound [Na+].Cl[O-] SUKJFIGYRHOWBL-UHFFFAOYSA-N 0.000 description 1
- 235000009518 sodium iodide Nutrition 0.000 description 1
- RYYKJJJTJZKILX-UHFFFAOYSA-M sodium octadecanoate Chemical compound [Na+].CCCCCCCCCCCCCCCCCC([O-])=O RYYKJJJTJZKILX-UHFFFAOYSA-M 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 238000005063 solubilization Methods 0.000 description 1
- 230000007928 solubilization Effects 0.000 description 1
- 238000000527 sonication Methods 0.000 description 1
- 238000001179 sorption measurement Methods 0.000 description 1
- 239000003549 soybean oil Substances 0.000 description 1
- 235000012424 soybean oil Nutrition 0.000 description 1
- 238000001228 spectrum Methods 0.000 description 1
- 238000009987 spinning Methods 0.000 description 1
- 239000003381 stabilizer Substances 0.000 description 1
- 239000007858 starting material Substances 0.000 description 1
- 238000010972 statistical evaluation Methods 0.000 description 1
- 239000008227 sterile water for injection Substances 0.000 description 1
- 239000012089 stop solution Substances 0.000 description 1
- 125000001424 substituent group Chemical group 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 230000008093 supporting effect Effects 0.000 description 1
- 239000002344 surface layer Substances 0.000 description 1
- 238000001847 surface plasmon resonance imaging Methods 0.000 description 1
- 239000004094 surface-active agent Substances 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 239000012730 sustained-release form Substances 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 239000003826 tablet Substances 0.000 description 1
- 239000000454 talc Substances 0.000 description 1
- 229910052623 talc Inorganic materials 0.000 description 1
- 238000004885 tandem mass spectrometry Methods 0.000 description 1
- 238000002626 targeted therapy Methods 0.000 description 1
- 229910052713 technetium Inorganic materials 0.000 description 1
- GKLVYJBZJHMRIY-UHFFFAOYSA-N technetium atom Chemical compound [Tc] GKLVYJBZJHMRIY-UHFFFAOYSA-N 0.000 description 1
- GZCRRIHWUXGPOV-UHFFFAOYSA-N terbium atom Chemical compound [Tb] GZCRRIHWUXGPOV-UHFFFAOYSA-N 0.000 description 1
- MPLHNVLQVRSVEE-UHFFFAOYSA-N texas red Chemical compound [O-]S(=O)(=O)C1=CC(S(Cl)(=O)=O)=CC=C1C(C1=CC=2CCCN3CCCC(C=23)=C1O1)=C2C1=C(CCC1)C3=[N+]1CCCC3=C2 MPLHNVLQVRSVEE-UHFFFAOYSA-N 0.000 description 1
- 231100001274 therapeutic index Toxicity 0.000 description 1
- 238000001757 thermogravimetry curve Methods 0.000 description 1
- 150000003568 thioethers Chemical class 0.000 description 1
- CNHYKKNIIGEXAY-UHFFFAOYSA-N thiolan-2-imine Chemical compound N=C1CCCS1 CNHYKKNIIGEXAY-UHFFFAOYSA-N 0.000 description 1
- 150000003573 thiols Chemical group 0.000 description 1
- 125000000341 threoninyl group Chemical group [H]OC([H])(C([H])([H])[H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 229940104230 thymidine Drugs 0.000 description 1
- 239000003104 tissue culture media Substances 0.000 description 1
- 238000003325 tomography Methods 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 238000003146 transient transfection Methods 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- 229960000575 trastuzumab Drugs 0.000 description 1
- 229960001612 trastuzumab emtansine Drugs 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 102000003390 tumor necrosis factor Human genes 0.000 description 1
- 102000003298 tumor necrosis factor receptor Human genes 0.000 description 1
- MEYZYGMYMLNUHJ-UHFFFAOYSA-N tunicamycin Natural products CC(C)CCCCCCCCCC=CC(=O)NC1C(O)C(O)C(CC(O)C2OC(C(O)C2O)N3C=CC(=O)NC3=O)OC1OC4OC(CO)C(O)C(O)C4NC(=O)C MEYZYGMYMLNUHJ-UHFFFAOYSA-N 0.000 description 1
- 238000000108 ultra-filtration Methods 0.000 description 1
- 241000712461 unidentified influenza virus Species 0.000 description 1
- 230000009677 vaginal delivery Effects 0.000 description 1
- LEONUFNNVUYDNQ-UHFFFAOYSA-N vanadium atom Chemical compound [V] LEONUFNNVUYDNQ-UHFFFAOYSA-N 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 230000029812 viral genome replication Effects 0.000 description 1
- 239000005723 virus inoculator Substances 0.000 description 1
- 238000001429 visible spectrum Methods 0.000 description 1
- 239000002699 waste material Substances 0.000 description 1
- 238000009736 wetting Methods 0.000 description 1
- 239000000080 wetting agent Substances 0.000 description 1
- NAWDYIZEMPQZHO-UHFFFAOYSA-N ytterbium Chemical compound [Yb] NAWDYIZEMPQZHO-UHFFFAOYSA-N 0.000 description 1
Images



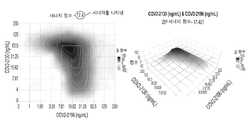




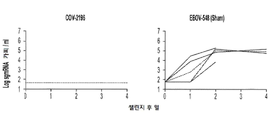










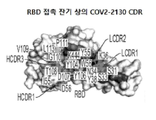

















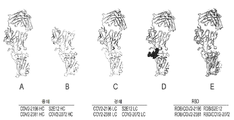













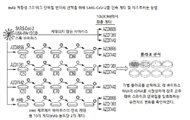


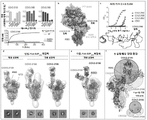


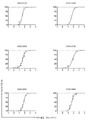

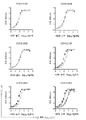

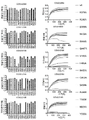
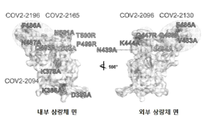
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1002—Coronaviridae
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1002—Coronaviridae
- C07K16/1003—Severe acute respiratory syndrome coronavirus 2 [SARS‐CoV‐2 or Covid-19]
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/569—Immunoassay; Biospecific binding assay; Materials therefor for microorganisms, e.g. protozoa, bacteria, viruses
- G01N33/56983—Viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
- A61K2039/507—Comprising a combination of two or more separate antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/35—Valency
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/55—Fab or Fab'
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/94—Stability, e.g. half-life, pH, temperature or enzyme-resistance
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/165—Coronaviridae, e.g. avian infectious bronchitis virus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2469/00—Immunoassays for the detection of microorganisms
- G01N2469/10—Detection of antigens from microorganism in sample from host
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Virology (AREA)
- Immunology (AREA)
- Molecular Biology (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Biochemistry (AREA)
- Urology & Nephrology (AREA)
- Hematology (AREA)
- Biomedical Technology (AREA)
- Biophysics (AREA)
- Genetics & Genomics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- General Physics & Mathematics (AREA)
- Analytical Chemistry (AREA)
- Physics & Mathematics (AREA)
- Food Science & Technology (AREA)
- Pathology (AREA)
- Microbiology (AREA)
- Cell Biology (AREA)
- Biotechnology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Public Health (AREA)
- Communicable Diseases (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Oncology (AREA)
- Veterinary Medicine (AREA)
- Pulmonology (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
본 개시내용은 SARS-CoV-2로 명명된 코로나바이러스에 결합하고 이를 중화시키는 항체 및 이의 이용 방법에 관한 것이다.
Description
관련 출원에 대한 참조
본 출원은 2020년 3월 26일 출원된 미국 가출원 제63/000,299호, 2020년 3월 31일 출원된 미국 가출원 제63/002,896호, 2020년 4월 1일 출원된 미국 가출원 제63/003,716호, 2020년 5월 12일 출원된 미국 가출원 제63/023,545호, 2020년 5월 13일 출원된 미국 가출원 제63/024,204호, 2020년 5월 13일 출원된 미국 가출원 제63/024,248호, 2020년 5월 19일 출원된 미국 가출원 제63/027,173호, 2020년 6월 11일 출원된 미국 가출원 제63/037,984호, 2020년 6월 17일 출원된 미국 가출원 제63/040,224호, 2020년 6월 17일 출원된 미국 가출원 제63/040,246호, 2021년 1월 27일 출원된 미국 가출원 제63/142,196호 및 2021년 3월 16일 출원된 미국 가출원 제63/161,890호에 대한 우선권을 주장하며, 이들 각각은 전체가 본원에 참조로 포함된다.
연방 지원 자금 공시
본 발명은 미국 국방부 첨단연구 프로젝트국(Defense Advanced Research Projects Agency, DARPA)이 수여한 HR0011-18-2-0001 및 미국 국립알레르기감염병연구소(National Institutes of Allergy and Infection Disease)/국립보건원(National Institutes of Health)이 수여한 HHS 계약 75N93019C00074에 따른 정부 지원으로 이루어졌다. 정부는 본 발명에 특정 권리를 갖는다.
전자 제출된 서열 목록에 대한 참조
전자적으로 제출된 서열 목록의 내용(명칭: 4815-001PC0D_SL_ST25.txt; 크기: 87,769 바이트; 및 생성일: 2021년 3월 24일)은 그 전체가 참조로 본원에 포함된다.
1. 기술분야
본 개시내용은 일반적으로 의학, 감염성 질환 및 면역학 분야에 관한 것이다. 보다 구체적으로, 본 개시내용은 SARS-CoV-2로 명명된 신종 코로나바이러스에 결합하는 인간 항체 및 이의 이용 방법에 관한 것이다.
2. 배경
신종 코로나바이러스(SARS-CoV-2)의 전염병이 중국 본토에 영향을 미치고, 179개의 다른 국가 및 지역에서도 발생했다. 이는 중국 후베이성 성도 우한에서 41명이 명확한 원인 없이 폐렴에 걸린 후 확인됐다. 코로나바이러스 질환 2019(COVID-19)로 명명된 급성 호흡기 질환을 일으키는 바이러스는 사람에서 사람으로 전파될 수 있다. 잠복기(노출부터 증상 발현까지의 시간)는 0~24일이며 평균 3~5일이지만 이 기간과 회복 후에도 전염성이 있을 수 있다. 증상에는 발열, 기침 및 호흡 곤란이 포함된다. 2020년 2월의 사망률 추정치는 확진된 사례의 2%로, 입원이 필요한 사람들에서 더 높았다.
2020년 2월 10일 현재, 중국의 모든 성급 구역을 포함하여 40,627건(6,495건은 심각)이 확인되었다. 더 많은 사람들이 감염되었지만 검출되지 않았을 수 있다(특히 경증의 경우). 2020년 2월 10일 현재, 1월 9일 첫 번째 확인된 사망 이후 910건의 사망이 이 바이러스의 탓으로 돌려졌고, 323명이 회복되었다. 중국 외 지역의 첫 번째 지역 전파는 베트남에서 가족 구성원 간에 발생했으며 가족을 포함하지 않은 첫 번째 국제 전파는 1월 22일 독일에서 발생했다. 중국 이외의 지역에서의 최초 사망은 필리핀에서 발생하였는데, 우한에서 온 한 남성이 2월 1일 사망했다. 2020년 2월 10일 현재, 이 바이러스로 인한 사망자 수는 2003년의 전 세계 SARS 발병을 능가했다.
2020년 2월 초 현재, 허가된 백신이나 특정 치료법은 없으나 여러 백신 접근법과 항바이러스제가 조사되고 있다. 세계보건기구(WHO)는 바이러스가 의료 시스템이 취약한 국가로 확산될 경우 미칠 수 있는 영향을 기반으로 이 발병을 국제적 우려가 있는 공중보건비상사태(Public Health Emergency of International Concern, PHEIC)로 선언하였다. 따라서, SARS-CoV-2의 생물학과 병리학뿐만 아니라, 이 바이러스에 대한 인간 면역 반응의 조사가 시급하다.
따라서, 본 개시내용에 따르면, 대상체에서 SARS-CoV-2에 의한 COVID-19 감염을 검출하는 방법이 제공되며, 이 방법은 (a) 상기 대상체로부터의 샘플을 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편과 접촉시키는 단계; 및 (b) 상기 샘플에서 SARS-CoV-2 항원에 대한 항체 또는 항체 단편의 결합에 의해 상기 샘플에서 SARS-CoV-2를 검출하는 단계를 포함한다. 샘플은 체액, 예컨대, 혈액, 가래, 눈물, 타액, 점액 또는 혈청, 정액, 자궁경부 또는 질 분비물, 양수, 태반 조직, 소변, 삼출액, 누출액, 조직 찰과표본 또는 대변일 수 있다. 검출은 ELISA, RIA, 측면 유동 분석 또는 웨스턴 블롯을 포함할 수 있다. 방법은 단계 (a) 및 (b)를 두 번째로 수행하고 첫 번째 분석과 비교하여 SARS-CoV-2 항원 수준의 변화를 결정하는 단계를 추가로 포함할 수 있다. 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 100%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합할 수 있다. 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다.
또 다른 구현예에서, SARS-CoV-2에 감염된 대상체를 치료하거나 SARS-CoV-2에 감염될 위험이 있는 대상체의 감염 가능성을 감소시키는 방법이 제공되며, 방법은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편을 상기 대상체에게 전달하는 단계를 포함한다. 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 100%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다. 항체는 키메라 항체 또는 이중특이적 항체일 수 있다. 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체 또는 항체 단편은 표면 스파이크 단백질과 같은 SARS-CoV-2 항원에 결합할 수 있다. 항체 또는 항체 단편은 감염 전 또는 감염 후에 투여될 수 있다. 대상체는 60세 이상이거나, 면역저하되거나, 호흡기 및/또는 심혈관 장애를 앓을 수 있다. 전달은 항체 또는 항체 단편 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터를 사용한 유전적 전달을 포함할 수 있다.
또 다른 구현예에서, 단클론 항체가 제공되며, 여기서 항체 또는 항체 단편은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 특징으로 한다. 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 100%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다. 항체는 키메라 항체일 수 있거나, 이중특이적 항체이거나, 인트라바디이다. 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합할 수 있다.
항체 또는 항체 단편을 암호화하는 하이브리도마 또는 조작 세포로서, 항체 또는 항체 단편은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 특징으로 하는 하이브리도마 또는 조작 세포. 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 100%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다. 항체는 키메라 항체, 이중특이적 항체, 또는 인트라바디일 수 있다. 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합할 수 있다.
또 다른 구현예에서, 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 특징으로 하는 하나 이상의 항체 또는 항체 단편을 포함하는 백신 제형이 제공된다. 상기 항체 또는 항체 단편 중 적어도 하나는 표 1의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열, 표 1의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열, 또는 표 1의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 상기 항체 또는 항체 단편 중 적어도 하나는 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 상기 항체 단편 중 적어도 하나는 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. 상기 항체 중 적어도 하나는 키메라 항체, 이중특이적 항체 또는 인트라바디일 수 있다. 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체 또는 항체 단편은 SARS-CoV-2 항원 표면 스파이크 단백질에 결합할 수 있다.
추가 구현예에서, 본원에 기재된 바와 같은 제1 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 포함하는 백신 제형이 제공된다. 발현 벡터(들)는 신드비스(Sindbis) 바이러스 또는 VEE 벡터(들)일 수 있다. 백신은 바늘 주입, 제트 주입 또는 전기천공에 의한 전달을 위해 제형화될 수 있다. 백신 제형은 제26항 내지 제34항의 별개의 항체 또는 항체 단편과 같은 제2 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 추가로 포함할 수 있다.
또 다른 구현예에서, 60세 이상의 대상체, 면역저하 대상체, 또는 SARS-CoV-2에 감염되거나 감염 위험이 있는 호흡기 및/또는 심혈관 장애를 앓는 대상체의 건강을 보호하는 방법이 제공되며, 방법은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편을 상기 대상체에게 전달하는 단계를 포함한다. 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해, 또는 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 100%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다. 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편일 수 있다. 항체는 키메라 항체 또는 이중특이적 항체일 수 있다. 상기 항체 또는 항체 단편은 감염 전 또는 감염 후에 투여될 수 있다. 항체 또는 항체 단편은 표면 스파이크 단백질과 같은 SARS-CoV-2 항원에 결합할 수 있다. 전달은 항체 또는 항체 단편 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터를 사용한 유전적 전달을 포함할 수 있다. 항체 또는 항체 단편은 미처리 대조군과 비교하여 대상체의 호흡을 개선할 수 있고/있거나 미처리 대조군과 비교하여 바이러스 부하를 감소시킬 수 있다.
또 다른 추가 구현예에서, SARS-CoV-2 표면 스파이크 단백질의 항원 무결성, 정확한 형태 및/또는 정확한 서열을 결정하는 방법이 제공되며, 방법은 (a) 상기 항원을 포함하는 샘플을 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 제1 항체 또는 항체 단편과 접촉시키는 단계; 및 (b) 상기 항원에 대한 상기 제1 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 무결성, 정확한 형태 및/또는 정확한 서열을 결정하는 단계를 포함한다. 샘플은 재조합 생산된 항원 또는 백신 제형 또는 백신 생산 배치를 포함할 수 있다. 검출은 ELISA, RIA, 웨스턴 블롯, 표면 플라즈몬 공명 또는 생물층 간섭계를 사용하는 바이오센서, 또는 유세포 분석 염색을 포함할 수 있다. 제1 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열, 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열, 또는 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 제1 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 제1 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다. 방법은 시간 경과에 따른 항원의 항원 안정성을 결정하기 위해 단계 (a) 및 (b)를 두 번째로 수행하는 단계를 추가로 포함할 수 있다.
방법은 (c) 상기 항원을 포함하는 샘플을 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 제2 항체 또는 항체 단편과 접촉시키는 단계; 및 (d) 상기 항원에 대한 상기 제2 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 무결성을 결정하는 단계를 추가로 포함할 수 있다. 제2 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열, 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열, 또는 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화될 수 있다. 제2 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있거나, 또는 표 2의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함할 수 있다. 제2 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편일 수 있다. 방법은 시간 경과에 따른 항원의 항원 안정성을 결정하기 위해 단계 (c) 및 (d)를 두 번째로 수행하는 단계를 추가로 포함할 수 있다.
또한, 인간 단클론 항체 또는 항체 단편, 또는 이를 생산하는 하이브리도마 또는 조작 세포가 제공되며, 상기 항체는 SARS-CoV-2 항원 표면 스파이크 단백질에 결합한다.
본원에 제공된 일 양태(A1)에서, 대상체에서 SARS-CoV-2에 의한 COVID-19 감염을 검출하는 방법은 (a) 상기 대상체로부터의 샘플을 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편과 접촉시키는 단계; 및 (b) 상기 샘플에서 SARS-CoV-2 항원에 대한 항체 또는 항체 단편의 결합에 의해 상기 샘플에서 SARS-CoV-2를 검출하는 단계를 포함한다. A1의 일 양태(A2)에서, 상기 샘플은 체액이다. A1 또는 A2의 일 양태(A3)에서, 상기 샘플은 혈액, 가래, 눈물, 타액, 점액 또는 혈청, 정액, 자궁경부 또는 질 분비물, 양수, 태반 조직, 소변, 삼출액, 누출액, 조직 찰과표본 또는 대변이다. A1 내지 A3 중 어느 하나의 일 양태(A4)에서, 검출은 ELISA, RIA, 측면 유동 분석 또는 웨스턴 블롯을 포함한다. A1 내지 A4 중 어느 하나의 일 양태(A5)에서, 방법은 단계 (a) 및 (b)를 두 번째로 수행하고 첫 번째 분석과 비교하여 SARS-CoV-2 항원 수준의 변화를 결정하는 단계를 추가로 포함한다. A1 내지 A5 중 어느 하나의 일 양태(A6)에서, 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열에 의해 암호화된다. A1 내지 A5 중 어느 하나의 일 양태(A7)에서, 상기 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A1 내지 A5 중 어느 하나의 일 양태(A8)에서, 상기 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 100%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A1 내지 A5 중 어느 하나의 일 양태(A9)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A1 내지 A5 중 어느 하나의 일 양태(A10)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A1 내지 A5 중 어느 하나의 일 양태(A11)에서, 상기 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합한다. A1 내지 A11 중 어느 하나의 일 양태(A12)에서, 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다.
본원에 제공된 일 양태(A13)에서, SARS-CoV-2에 감염된 대상체를 치료하거나 SARS-CoV-2에 감염될 위험이 있는 대상체의 감염 가능성을 감소시키는 방법은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편을 상기 대상체에게 전달하는 단계를 포함한다. A13의 일 양태(A14)에서, 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A13 또는 A14의 일 양태(A15)에서, 상기 항체 또는 항체 단편은 표 1의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A13의 일 양태(A16)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A13의 일 양태(A17)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80% 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A13의 일 양태(A18)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A13 내지 A18 중 어느 하나의 일 양태(A19)에서, 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. A13 내지 A19 중 어느 하나의 일 양태(A20)에서, 상기 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편이다. A13 내지 A18 중 어느 하나의 일 양태(A21)에서, 상기 항체는 키메라 항체 또는 이중특이적 항체이다. A13 내지 A21 중 어느 하나의 일 양태(A22)에서, 상기 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합한다. A13 내지 A22 중 어느 하나의 일 양태(A23)에서, 상기 항체 또는 항체 단편은 감염 전 또는 감염 후에 투여된다. A13 내지 A23 중 어느 하나의 일 양태(A24)에서, 상기 대상체는 60세 이상이거나, 면역저하되었거나, 호흡기 및/또는 심혈관 장애를 앓고 있다. A13 내지 A24 중 어느 하나의 일 양태(A25)에서, 전달은 항체 또는 항체 단편 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터를 사용한 유전적 전달을 포함한다.
일 양태(A26)에서 단클론 항체가 본원에 제공되며, 여기서 항체 또는 항체 단편은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 특징으로 한다. A26의 일 양태(A27)에서, 상기 항체 또는 항체 단편은 표 1의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A26의 일 양태(A28)에서, 상기 항체 또는 항체 단편은 표 1의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A26의 일 양태(A29)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A26의 일 양태(A30)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A26 내지 A30 중 어느 하나의 일 양태(A31)에서, 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. A26 내지 A30 중 어느 하나의 일 양태(A32)에서, 상기 항체는 키메라 항체이거나 이중특이적 항체이다. A26 내지 A32 중 어느 하나의 일 양태(A33)에서, 상기 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편이다. A26 내지 A33 중 어느 하나의 일 양태(A34)에서, 상기 항체 또는 항체 단편은 SARS-CoV-2 항원, 예컨대, 표면 스파이크 단백질에 결합한다. A26 내지 A34 중 어느 하나의 일 양태(A35)에서, 상기 항체는 인트라바디이다.
본원에 제공된 일 양태(A36)에서, 하이브리도마 또는 조작 세포는 항체 또는 항체 단편을 암호화하고, 여기서 항체 또는 항체 단편은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 특징으로 한다. A36의 일 양태(A37)에서, 상기 항체 또는 항체 단편은 표 1의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A36의 일 양태(A38)에서, 상기 항체 또는 항체 단편은 표 1의 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A36의 일 양태(A39)에서, 상기 항체 또는 항체 단편은 표 1의 클론 쌍을 이루는 가변 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A36의 일 양태(A40)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A36의 일 양태(A41)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A36의 일 양태(A42)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A36 내지 A42 중 어느 하나의 일 양태(A43)에서, 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. A36 내지 A43 중 어느 하나의 일 양태(A44)에서, 상기 항체는 키메라 항체, 이중특이적 항체, 또는 인트라바디이다. A36 내지 A43 중 어느 하나의 일 양태(A45)에서, 상기 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편이다. A36 내지 A45 중 어느 하나의 일 양태(A46)에서, 상기 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합한다.
본원에 제공된 일 양태(A47)에서, 백신 제형은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 특징으로 하는 하나 이상의 항체 또는 항체 단편을 포함한다. A47의 일 양태(A48)에서, 상기 항체 또는 항체 단편 중 적어도 하나는 표 1의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A47의 일 양태(A49)에서, 상기 항체 또는 항체 단편 중 적어도 하나는 표 1의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A47의 일 양태(A50)에서, 상기 항체 또는 항체 단편 중 적어도 하나는 표 1의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A47의 일 양태(A51)에서, 상기 항체 또는 항체 단편 중 적어도 하나는 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A47의 일 양태(A52)에서, 상기 항체 또는 항체 단편 중 적어도 하나는 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 90% 또는 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A47 내지 A42 중 어느 하나의 일 양태(A53)에서, 상기 항체 단편 중 적어도 하나는 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. A47 내지 A52 중 어느 하나의 일 양태(A54)에서, 상기 항체 중 적어도 하나는 키메라 항체, 이중특이적 항체 또는 인트라바디이다. A47 내지 A54 중 어느 하나의 일 양태(A55)에서, 상기 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편이다. A47 내지 A55 중 어느 하나의 일 양태(A56)에서, 상기 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합한다.
본원에 제공된 일 양태(A57)에서, 백신 제형은 A26 내지 A34 중 어느 하나에 따른 제1 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 포함한다. A57의 일 양태(A58)에서, 상기 발현 벡터(들)는 신드비스 바이러스 또는 VEE 벡터(들)이다. A57 또는 A58의 일 양태(A59)에서, 백신 제형은 바늘 주입, 제트 주입 또는 전기천공에 의한 전달을 위해 제형화된다. A57의 일 양태(A60)에서, 백신 제형은 A26 내지 A34 중 어느 하나의 별개의 항체 또는 항체 단편과 같은 제2 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 추가로 포함한다.
본원에 제공된 일 양태(A61)에서, 60세 이상의 대상체, 면역저하 대상체, 또는 SARS-CoV-2에 감염되거나 감염 위험이 있는 호흡기 및/또는 심혈관 장애를 앓는 대상체의 건강을 보호하는 방법은 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 항체 또는 항체 단편을 상기 대상체에게 전달하는 단계를 포함한다. A61의 일 양태(A62)에서, 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A61 또는 A62의 일 양태(A63)에서, 항체 또는 항체 단편은 기재된 바와 같이 ___와 적어도 95%의 동일성을 갖는 클론 쌍을 이루는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A61 또는 A62의 일 양태(A64)에서, 상기 항체 또는 항체 단편은 표 1의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A61의 일 양태(A65)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A61의 일 양태(A66)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80% 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A61의 일 양태(A67)에서, 상기 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A61 내지 A67 중 어느 하나의 일 양태(A68)에서, 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. A61 내지 A68 중 어느 하나의 일 양태(A69)에서, 상기 항체는 IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편이다. A61 내지 A67 중 어느 하나의 일 양태(A70)에서, 상기 항체는 키메라 항체 또는 이중특이적 항체이다. A61 내지 A70 중 어느 하나의 일 양태(A71)에서, 상기 항체 또는 항체 단편은 감염 전 또는 감염 후에 투여된다. A61 내지 A71 중 어느 하나의 일 양태(A72)에서, 상기 항체 또는 항체 단편은 SARS-CoV-2 표면 스파이크 단백질에 결합한다. A61 내지 A72 중 어느 하나의 일 양태(A73)에서, 전달은 항체 또는 항체 단편 투여, 또는 항체 또는 항체 단편을 암호화하는 RNA 또는 DNA 서열 또는 벡터를 사용한 유전적 전달을 포함한다. A61의 일 양태(A74)에서, 항체 또는 항체 단편은 미처리 대조군과 비교하여 대상체의 호흡을 개선한다. A61의 일 양태(A75)에서, 항체 또는 항체 단편은 미처리 대조군과 비교하여 바이러스 부하를 감소시킨다.
본원에 제공된 일 양태(A76)에서, SARS-CoV-2 표면 스파이크 단백질의 항원 무결성, 정확한 형태 및/또는 정확한 서열을 결정하는 방법은 (a) 상기 항원을 포함하는 샘플을 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 제1 항체 또는 항체 단편과 접촉시키는 단계; 및 (b) 상기 항원에 대한 상기 제1 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 무결성, 정확한 형태 및/또는 정확한 서열을 결정하는 단계를 포함한다. A76의 일 양태(A77)에서, 상기 샘플은 재조합으로 생산된 항원을 포함한다. A76의 일 양태(A78)에서, 상기 샘플은 백신 제형 또는 백신 생산 배치를 포함한다. A76 내지 A78의 일 양태(A79)에서, 검출은 ELISA, RIA, 웨스턴 블롯, 표면 플라즈몬 공명 또는 생물층 간섭계를 사용하는 바이오센서, 또는 유세포 분석 염색을 포함한다. A76 내지 A79의 일 양태(A80)에서, 제1 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열에 의해 암호화된다. A76 내지 A79의 일 양태(A81)에서, 상기 제1 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A76 내지 A79 중 어느 하나의 일 양태(A82)에서, 상기 제1 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A76 내지 A79 중 어느 하나의 일 양태(A83)에서, 상기 제1 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A76 내지 A79 중 어느 하나의 일 양태(A84)에서, 상기 제1 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A76 내지 A79 중 어느 하나의 일 양태(A85)에서, 상기 제1 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A76 내지 A85 중 어느 하나의 일 양태(A86)에서, 제1 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. A76 내지 A86 중 어느 하나의 일 양태(A87)에서, 방법은 시간 경과에 따른 항원의 항원 안정성을 결정하기 위해 단계 (a) 및 (b)를 두 번째로 수행하는 단계를 추가로 포함한다. A76 내지 A87 중 어느 하나의 일 양태(A88)에서, 방법은 (c) 상기 항원을 포함하는 샘플을 각각 표 3 및 4의 클론 쌍을 이루는 중쇄 및 경쇄 CDR 서열을 갖는 제2 항체 또는 항체 단편과 접촉시키는 단계; 및 (d) 상기 항원에 대한 상기 제2 항체 또는 항체 단편의 검출 가능한 결합에 의해 상기 항원의 항원 무결성을 결정하는 단계를 추가로 포함한다. A88의 일 양태(A89)에서, 제2 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열에 의해 암호화된다. A89의 일 양태(A90)에서, 상기 제2 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 가변 서열과 적어도 70%, 80%, 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A89의 일 양태(A91)에서, 상기 제2 항체 또는 항체 단편은 표 1에 기재된 바와 같이 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열에 의해 암호화된다. A89의 일 양태(A92)에서, 상기 제2 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열에 따른 경쇄 및 중쇄 가변 서열을 포함한다. A89의 일 양태(A93)에서, 상기 제2 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 70%, 80% 또는 90%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A89의 일 양태(A94)에서, 상기 제2 항체 또는 항체 단편은 표 2의 클론 쌍을 이루는 서열과 적어도 95%의 동일성을 갖는 경쇄 및 중쇄 가변 서열을 포함한다. A89의 일 양태(A95)에서, 제2 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편이다. A89의 일 양태(A96)에서, 방법은 시간 경과에 따른 항원의 항원 안정성을 결정하기 위해 단계 (c) 및 (d)를 두 번째로 수행하는 단계를 추가로 포함한다.
일 양태(A97)에서 인간 단클론 항체 또는 항체 단편, 또는 이를 생산하는 하이브리도마 또는 조작 세포가 본원에 제공되며, 상기 항체는 SARS-CoV-2 표면 스파이크 단백질에 결합한다.
청구항 및/또는 명세서에서 "포함하는"이라는 용어와 함께 사용될 때 부정관사("a" 또는 "an")의 사용은 "하나"를 의미할 수 있지만, "하나 이상("one or more" 또는 "one or more than one") 및 "적어도 하나"의 의미와도 일치한다. "약"이라는 단어는 명시된 숫자의 플러스 또는 마이너스 5%를 의미한다.
본원에 기재된 임의의 방법 또는 조성물은 본원에 기재된 임의의 다른 방법 또는 조성물과 관련하여 구현될 수 있음이 고려된다. 본 개시내용의 다른 목적, 특징 및 이점은 다음의 상세한 설명으로부터 명백해질 것이다. 그러나 본 개시내용의 사상 및 범위 내의 다양한 변경 및 수정이 이 상세한 설명으로부터 당업자에게 명백해질 것이므로, 상세한 설명 및 특정 실시예는 본 개시내용의 특정 구현예를 나타내지만, 단지 예시로서만 제공되는 것으로 이해되어야 한다.
본 특허 또는 출원 파일은 컬러 사진의 적어도 하나의 도면을 포함한다. 컬러 도면(들)이 첨부된 본 특허 또는 특허 출원 공보물의 사본은 요청 및 필요한 비용을 지불하면 국제 사무국에서 제공할 것이다.
다음의 도면은 본 명세서의 일부를 형성하고 본 개시내용의 특정 양태를 추가로 증명하기 위해 포함된다. 본 개시내용은 본원에 제시된 특정 구현예의 상세한 설명과 함께 이들 도면 중 하나 이상을 참조하여 더 잘 이해될 수 있다.
도 1. 생 BSL3 SARS-CoV-2 바이러스를 사용하여 COV2-2196 + COV2-2130의 칵테일에 의한 상승적 중화 활성을 평가하기 위한 용량-반응 매트릭스. 정성적으로 개별 mAb(0 ng/mL COV2-2196 + 250 ng/mL COV2-2130 및 250 ng/mL COV2-2196 + 0 ng/mL COV2-2130의 상자)의 최고 테스트 농도(250 ng/mL)에서 작은 분율의 중화되지 않은 바이러스가 있었으나, 콤보에 의한 더 낮은 Ab 농도 범위에서는 완전 중화(100%)가 있었다. 15.6 ng/mL COV2-2196 + 63 ng/mL COV2-2130의 상자는 96%의 바이러스를 중화한 조합에서 15.6 ng/mL의 mAb COV2-2196 및 63 ng/mL의 mAb COV2-2130과 최대 상승효과가 있는 영역을 나타내지만, 개별 Ab는 각각 겨우 6 또는 0%의 중화를 보여주었다(0 ng/mL COV2-2196 + 63 ng/mL COV2-2130 및 15.6 ng/mL COV2-2196 + 0 ng/mL COV2-2130의 타원형). 3중의 기술적 반복실험에 대한 평균 값이 제시되어 있다.
도 2a 내지 2b. BSL3 SARS-CoV-2 생 바이러스를 사용하여 mAb COV2-2196 + mAb COV2-2130의 칵테일에 의한 상승적 중화 활성을 평가하기 위한 용량-반응 매트릭스. 기술적 반복실험에 대한 3중 값의 평균이 제시되어 있다. 데이터를 시각화하고 SynergyFinder 소프트웨어를 사용하여 상승효과를 평가하였다.
도 3. BSL3 SARS-CoV-2 생 바이러스를 사용하여 mAb COV2-2196 + mAb COV2-2130의 칵테일에 의한 상승적 중화 활성을 평가하기 위한 용량-반응 매트릭스. 시너지 점수 해석: <-10: 상호작용이 길항적일 가능성이 높음; -10에서 10: 상호작용이 가산적일 가능성이 높음; >10: 상호작용이 상승적일 가능성이 높음.
도 4a 내지 4c. 확립된 SARS-CoV-2 감염에 대한 중화 인간 mAb의 치료 효능. (도 4a) 105 PFU의 MA-SARS-CoV-2를 비강 경로로 마우스에 접종하고 12시간 후에 복강 내 주사에 의해 표시된 항체 치료를 제공하였다. 플라크 분석을 사용하여 바이러스 챌린지 후 2일에 폐의 바이러스 부하를 측정했다. 개별 마우스로부터의 측정치 및 중앙값 역가가 제시되어 있고, 각 군은 Dunn의 사후 검정과 함께 Kruskal-Wallis ANOVA를 사용하여 이소형 대조군과 비교되었다(* p < 0.05). 데이터는 하나의 실험을 나타낸다. (도 4b) 10 내지 11주령의 BALB/c 마우스(군당 3 내지 9마리의 마우스의 한 실험)를 항-Ifnar1 mAb로 처리하고 하루 후 비강 경로를 통해 AdV-hACE2로 형질도입하였다. 4일 후, 마우스에 105 FFU의 실제(authentic) SARS-CoV-2를 비강 경로를 통해 접종하고, 12시간 후에 복강 내 주사에 의해 표시된 mAb 치료를 제공했다. 플라크 분석을 사용하여 바이러스 챌린지 후 2 dpi에서 폐의 바이러스 부하를 측정했다. 플라크 중화 분석 성능을 위한 2개의 대조군에는 감염되지 않은 개별 또는 mAb COV2-2196 + COV2-2130 칵테일 처리된 마우스의 폐 균질물과 1:1(부피:부피) 혼합된 개별 이소형 처리된 마우스의 폐 균질물이 포함되었다. 개별 마우스로부터의 측정치 및 중앙값 역가가 제시되어 있고, 각 군은 Dunn의 사후 검정과 함께 Kruskal-Wallis ANOVA를 사용하여 이소형 대조군과 비교되었다(** p < 0.01). 데이터는 하나의 실험을 나타낸다. (도 4c) 사이토카인 및 케모카인 유전자 발현은 도 4b에서와 같이 채취한 폐로부터 qPCR 분석에 의해 측정하였다. 개별 마우스로부터의 측정치 및 중앙값이 제시되어 있다. 군은 Mann-Whitney U 검정을 이용하여 비교되었다(* p < 0.05; ** p < 0.01).
도 5. 마카크에 대한 인간 mAb 주입 후의 항체 약동학.
도 6. 야생형 SARS-CoV-2 바이러스를 이용한 비강 내 및 기관 내 챌린지 후 마카크 기관지 폐포 세척액에서의 (새로 제조된 하위 게놈 RNA로 측정한) SARS-CoV-2 바이러스 부하.
도 7. 야생형 SARS-CoV-2 바이러스를 이용한 비강 내 및 기관 내 챌린지 후 마카크 비강 스왑 검체에서의 (새로 제조된 하위 게놈 RNA로 측정한) SARS-CoV-2 바이러스 부하.
도 8a 내지 8e. Fab COV2-2196과 복합된 S 단백질 RBD의 결정 구조. (도 8a) RBD와 복합된 COV2-2196의 카툰 표현. COV2-2196 중쇄는 청록색으로, 경쇄는 자홍색으로, RBD는 녹색으로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8A에서 볼 수 있다. (도 8b) COV2-2196-RBD 복합체의 구조는 RBD 구조를 참조로 사용하여 RBD-인간 ACE2 복합체의 구조(PDB ID: 6M0J)에 중첩된다. COV2-2196-RBD 복합체의 배색은 도 8a와 동일하다. RBD-ACE2 복합체의 RBD는 밝은 파란색으로, 인간 ACE2 펩티다제 도메인은 회색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8B에서 볼 수 있다. (도 8c) COV2-2196-RBD 복합체의 구조는 "위쪽(up)" 형태의 RBD를 참조로 사용하여 "위쪽" 형태의 단일 RBD를 갖는 스파이크 구조(PDB ID: 6XM4)에 중첩된다. COV2-2196-RBD 복합체의 배색은 도 8a와 동일하다. 스파이크의 세 하위 단위는 각각 회색, 노란색 또는 밝은 파란색으로 채색되어 있다.(RBD가 "위쪽" 형태인 하위 단위는 노란색임). 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8C에서 볼 수 있다. (도 8d) COV2-2196에 의해 인식되는 RBD 에피토프의 표면 표현. 에피토프 잔기는 다양한 녹색 음영으로 채색되고 검은색으로 표지된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8D에서 볼 수 있다. (도 8e) COV2-2196과 RBD 사이의 항체-항원 상호작용. RBD는 도 8d에서와 동일한 표면 표현 및 방향으로 제시되어 있다. COV2-2196 파라토프 잔기는 막대 표현으로 제시되어 있다. 중쇄는 청록색으로, 경쇄는 자홍색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8E에서 볼 수 있다.
도 9a 내지 9f. Fab COV2-2196 및 COV2-2130과 복합된 S 단백질 RBD의 결정 구조. (도 9a) COV2-2196 및 COV2-2130 Fab와 복합된 S 단백질 RBD의 결정 구조의 카툰 표현. RBD는 녹색, COV2-2196 중쇄는 청록색, COV2-2196 경쇄는 자홍색, COV2-2130 중쇄는 노란색, COV2-2130 경쇄는 주황색으로 제시되어 있다. COV2-2130의 CDR이 표지되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9A에서 볼 수 있다. (도 9b) COV2-2130-RBD 복합체의 구조는 RBD 구조를 참조로 사용하여 RBD-ACE2 복합체의 구조(PDB ID: 6M0J)에 중첩된다. COV2-2130-RBD 복합체의 배색은 도 9a와 동일하다. RBD-ACE2 복합체의 RBD는 밝은 파란색으로, 인간 ACE2 펩티다제 도메인은 회색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9B에서 볼 수 있다. (도 9c) COV2-2130-RBD 복합체의 구조는 하나의 프로토머의 RBD를 참조로 사용하여 "아래쪽(down)" 형태의 모든 RBD를 갖는 스파이크 구조(PDB ID: 6ZOY)에 중첩된다. COV2-2130-RBD 복합체의 배색은 도 9a와 동일하다. 스파이크의 세 프로토머는 각각 회색, 밝은 파란색, 또는 보라색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9C에서 볼 수 있다. (도 9d) COV2-2196-2130-RBD 복합체의 구조는 "위쪽(up)" 형태의 RBD를 참조로 사용하여 "위쪽" 형태의 하나의 RBD를 갖는 스파이크 구조(PDB ID: 7CAK)에 중첩된다. COV2-2130-RBD 복합체의 배색은 도 9a와 동일하다. 스파이크의 세 프로토머는 각각 회색, 밝은 파란색, 또는 보라색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9D에서 볼 수 있다. (도 9e) COV2-2130에 의해 인식되는 RBD 에피토프의 표면 표현. 에피토프 잔기는 상이한 색으로 표시되고 검은색으로 표지된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9E에서 볼 수 있다. (도 9f) COV2-2130 파라토프 잔기와 에피토프의 상호작용. RBD는 도 9e에서와 동일한 표면 표현 및 방향으로 제시되어 있다. 파라토프 잔기는 막대 표현으로 제시되어 있다. 중쇄는 노란색으로, 경쇄는 주황색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9F에서 볼 수 있다.
도 10a 내지 10b. (도 10a) COV2-2196 중쇄 및 경쇄의 IMGT/DomainGapAlign 결과. 주요 상호작용 잔기 및 이들의 생식계열 유전자의 상응하는 잔기가 상자로 제시되어 있다. 도 10a의 서열에 대한 서열번호는 다음과 같다:

(도 10b) COV2-2196의 점 돌연변이체의 결합 곡선. 위에서 상자로 표시된 중쇄에 대한 점 돌연변이체를 암호화하는 cDNA를 설계하고 DNA로 합성하여 재조합 IgG 단백질을 제조하였고, 스파이크 단백질에 대한 결합 활성을 테스트했다. D108 잔기의 돌연변이체는 위의 좌측 그래프에, 추론된 체세포 돌연변이의 생식계열 서열로의 복귀 돌연변이는 위의 우측 그래프에, P99 돌연변이체는 아래의 좌측 그래프에, HCDR3에서 이황화 결합을 제거하는 돌연변이체는 아래의 우측 그래프에 있다.
도. 11a 내지 11h. 저항성 변이체 선택과 결합된 심층 돌연변이 스캐닝을 통한 COV2-2196 및 COV2-2130에 대한 중요한 잔기 확인. (도 11a) COV2-2196(좌측) 또는 COV2-2130(우측)에 대한 강력한 회피를 나타내는 RBD 부위에서의 모두의 돌연변이 회피 분율의 로고 플롯. 더 큰 문자는 더 큰 항체 결합 회피를 나타낸다. 돌연변이는 인간 ACE2에 대한 RBD 결합을 감소시키는 정도에 기초하여 채색되어 있다. 제시된 데이터는 2개의 독립적인 효모 라이브러리를 사용한 2개의 독립적인 회피 선택 실험의 평균이다; 상관관계는 도 18b 내지 18c에 제시되어 있다. 이 로고 플롯의 대화형 확대/축소 버전은 jbloomlab.github.io/SARS-CoV-2-RBD_MAP_AZ_Abs/에 있다. 본 발명자들은 방법에서 기재된 바와 같이, 0의 값은 변이체가 항상 항체에 의해 결합됨을 의미하고 1의 값은 변이체가 항상 항체 결합을 회피함을 의미하도록, 항체 회피 빈(bin)에 속하는 해당 특정 변이체를 발현하는 세포의 추정된 분율을 나타내는 회피 분율을 결정하였다. (도 11b) 회피 선택에 사용된 Wuhan-Hu-1 참조 균주로부터의 단일 뉴클레오티드 치환에 의해 접근 가능한 COV2-2196 및 COV2-2130에 대한 돌연변이 회피 분율의 로고 플롯(도 11e 내지 11f). ACE2 결합에 대한 각 치환의 효과는 도 11a에 나타낸 바와 같다. (도 11c) 좌측 패널: RBD-COV2-2196 구조의 RBD 표면으로의 COV2-2196에 대한 심층 돌연변이 스캐닝 회피 돌연변이 매핑. COV2-2196 결합을 저해하는 돌연변이를 히트맵을 사용하여 RBD 구조에 표시하였다. 여기서 파란색은 누적 항체 회피가 가장 큰 RBD 부위를 나타내고 흰색은 검출된 회피가 없음을 나타낸다. 회색은 RBD 발현에 대한 해로운 영향이 항체 결합에 대한 돌연변이의 영향 평가를 저해한 잔기를 나타낸다. 우측 패널: RBD의 가장 강력한 회피 부위 주변에서 상호작용하는 잔기를 보여주는 좌측 패널을 확대한 것. COV2-2196 중쇄는 청록색이고 경쇄는 자홍색으로 채색되어 있다. 도 11a에 기재된 바와 같이 독립적인 라이브러리를 이용하여 2회의 반복실험을 수행하였다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 11C에서 볼 수 있다. (도 11d) 우측 패널: RBD-COV2-2130 구조의 RBD 표면으로의 COV2-2130에 대한 심층 돌연변이 스캐닝 회피 돌연변이 매핑. COV2-2130 결합을 저해하는 돌연변이를 도 11c에서와 같이 히트맵을 사용하여 RBD 구조에 표시하였다. 좌측 패널: RBD의 가장 강력한 회피 부위 주변에서 상호작용하는 잔기를 보여주는 좌측 패널을 확대한 것. COV2-2130 중쇄는 노란색이고 경쇄는 연어살색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 11D에서 볼 수 있다. (도 11e) COV2-2196, COV2-2130 및 이들의 조합을 이용한 VSV-SARS-CoV-2 회피 선택 실험의 결과를 보여주는 표. 선택된 회피 돌연변이체의 수 및 수행된 회피 선택 반복실험의 총 횟수 및 회피 돌연변이체 바이러스의 시퀀싱에 의해 확인된 잔기가 기록되어 있다. (도 11f) 하위 중화(sub-neutralizing) 농도의 AZD8895(COV2-2196 기준), AZD1061(COV2-2130 기준) 및 AZD7442(AZD8895 + AZD1061 기준)의 존재 하에서 SARS-CoV-2의 계대 결과를 보여주는 표. 중화 저항성 플라크를 시퀀싱하여 확인된 중화 저항성 관련 바이러스 돌연변이가 기록되어 있다. (도 11g) x축의 돌연변이 회피 분율 및 y축의 ACE2 결합에 대한 효과와 함께, 도 11a의 DMS 데이터를 보여주는 산점도. 십자형은 다중 뉴클레오티드 치환으로만 접근 가능한 돌연변이를 나타내고, 원형은 단일 뉴클레오티드 치환으로 접근 가능한 돌연변이를 나타낸다. VSV-SARS-CoV-2(K444R, K444E) 또는 실제 SARS-CoV-2(R346I)에서 COV2-2130에 의해 선택된 아미노산 치환이 기록되어 있다. (도 11h) 참조 균주 및 관심 SARS-CoV-2 변이체에 대한 FRNT에 의해 측정된 항체 중화. 중화 분석은 2중으로 수행하고 2회 반복했으며, 1회의 반복실험으로부터의 결과가 제시되어 있다. 오차 막대는 각 점의 범위를 나타낸다. WA-1 참조 균주와 비교한 돌연변이가 표시된다. B.1.1.7-OXF는 69-70 및 144-145 결실 및 다음의 치환을 포함한다: N501Y, A570D, D614G, P681H 및 T716I.
도 12. RBD-COV2-2196-2130 복합체 및 RBD-COV2-2196 결정 구조에서 RBD-COV2-2196의 하부 구조 오버레이.
도 13a 내지 13f. RBD-COV2-2196과 스파이크-S2E12 복합체 간에 공유되는 RBD 사이트 F486에서의 유사한 방향족 적층 및 소수성 상호작용 패턴. (도 13a 및 13b) 두 복합체의 구조에서 잔기 F486을 둘러싼 동일한 수소 결합 패턴. (도 13c) COV2-2196과 RBD 사이의 상세한 상호작용. COV2-2196 중쇄는 청록색, 경쇄는 자홍색, RBD는 녹색으로 채색되어 있다. 중요한 상호작용 잔기는 막대 표현으로 제시되어 있다. Ab-Ag 상호작용에 관여하는 물 분자는 분홍색 구체로 표시된다. 직접 수소 결합은 주황색 파선으로 제시되어 있고 물 매개 수소 결합은 노란색 파선으로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2C에서 볼 수 있다. (도 13d) COV2-2196/RBD 결정 구조로의 S2E12/RBD 초저온-EM 구조의 중첩(참조로서 항체의 가변 도메인 포함). COV2-2196 중쇄는 청록색이고, 경쇄는 자홍색이다; S2E12 중쇄는 연청록색이고 경쇄는 연분홍색이다. 두 개의 상응하는 RBD 구조는 각각 녹색 또는 노란색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2D에서 볼 수 있다. (도 13e) COV2-2130 중쇄와 RBD 사이의 상세한 상호작용. 파라토프 잔기는 막대 표현으로 제시되고, 노란색으로 채색되어 있고, 에피토프 잔기는 녹색 막대로 제시되어 있다. 수소 결합 또는 강한 극성 상호작용은 자홍색 파선으로 표시된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2E에서 볼 수 있다. (도 13f) COV2-2130 경쇄와 RBD 사이의 상세한 상호작용. 파라토프 잔기는 막대 표현으로 제시되고, 주황색으로 채색되어 있고, 에피토프 잔기는 녹색 막대로 제시되어 있다. 수소 결합은 자홍색 파선으로 표시된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2F에서 볼 수 있다.
도 14의 A 내지 E. 동일한 결합 메커니즘을 갖는 항-RBD 항체의 공통 클론형. (도 14의 A) COV2-2196/RBD 결정 구조. (도 14의 B) S2E12/RBD 초저온-EM 구조. (도 14의 C) COV2-2381/RBD 상동성 모델. COV2-2072는 회색 구체로 표시된 HCDR3의 N-연결 글리코실화 시퀀(sequon)을 암호화한다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 3D에서 볼 수 있다. (도 14의 D) COV2-2072/RBD 상동성 모델. (도 14의 E) COV2-2196/RBD 결정 구조(도 14의 A) 및 S2E12/RBD 초저온-EM 구조(도 14의 B)의 오버레이.
도 15a 내지 15b. 바이러스 미감염 개체의 항체 가변 유전자 레퍼토리에서 COV2-2196과 유전적으로 유사한 추정상의 공개 클론형 구성원의 확인. COV2-2196 중쇄(도 15a) 및 경쇄(도 15a 및 15b)와 동일한 서열 특징을 갖는 건강한 개체로부터의 항체 가변 유전자 서열이 정렬되어 있다. 3명의 상이한 공여자 및 제대혈로부터의 서열에는 공개 클론형의 특징을 갖는 서열이 포함되었다. COV2-2196에 사용된 서열 특징 및 접촉 잔기는 각각의 다중 서열 정렬 아래에 있는 상자로 강조 표시된다(상자는 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 4A에서 빨간색으로 채색되어 있음). 도 15a의 중쇄 서열에 대한 서열번호는 다음과 같다: HIP1: 서열번호 166, HIP2: 서열번호 167, HIP3: 서열번호 168, 및 CORD: 서열번호 169. 도 15a의 경쇄 서열에 대한 서열번호는 다음과 같다: HIP1: 서열번호 170, HIP2: 서열번호 171, 및 HIP3: 서열번호 172. 도 15b의 HIP1 경쇄 서열에 대한 서열번호는 다음과 같다: 서열번호 173, 서열번호 174, 서열번호 175, 서열번호 176, 및 서열번호 177(위에서 아래로). 도 15b의 HIP2 경쇄 서열에 대한 서열번호는 다음과 같다: 서열번호 178, 서열번호 179, 서열번호 180, 서열번호 181, 및 서열번호 182(위에서 아래로). 도 15b의 HIP3 경쇄 서열에 대한 서열번호는 다음과 같다: 서열번호 183, 서열번호 184, 서열번호 185, 서열번호 186, 및 서열번호 187(위에서 아래로).
도 16의 A 내지 D. (도 16의 A) 상세한 COV2-2130 HCDR3 루프 구조. 루프 형태를 안정화하는 단거리 수소 결합은 파선으로 제시되어 있다(파선은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 5A에서 자홍색으로 채색되어 있음). (도 16의 B) COV2-2130 경쇄의 잔기는 방향족 적층 상호작용 및 HCDR3과의 수소 결합을 형성하여 HCDR3 루프를 추가로 안정화시킨다. (도 16의 C) 긴 LCDR1, HCDR2 및 HCDR3은 RBD 에피토프에 대해 상보적인 결합 표면을 형성한다. RBD는 회색으로 표면 표현으로 제시되어 있다. COV2-2130 중쇄는 노란색으로, HCDR3은 주황색으로, 경쇄는 연어색으로, LCDR1은 자홍색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 3D에서 볼 수 있다. (도 16의 D) 도 16의 C의 180° 회전도.
도 17. COV2-2196 및 COV2-2130과 복합된 RBD의 결정 구조에서 COV2-2196과 COV2-2130 사이의 계면. COV2-2196 중쇄 또는 경쇄는 각각 청록색 또는 자홍색으로 카툰 표현으로 제시되어 있고, COV2-2130 중쇄 또는 경쇄는 각각 노란색 또는 연어색으로 제시되어 있다. RBD는 녹색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 6에서 볼 수 있다. 계면 잔기는 막대 표현으로 제시되어 있다.
도 18a 내지 18i. 항체 결합에 영향을 미치는 돌연변이의 심층 돌연변이 스캐닝에 의한 확인 및 VSV-SARS-CoV-2 바이러스를 이용한 항체 저항성 돌연변이체의 선택 방법. (도 18a) 위: 전면 및 측면 산란(처음 3개의 패널)과 RBD를 발현하는 효모 세포 선택(우측 패널)을 사용하여 단일 효모 세포를 선택하기 위한 대표적인 게이팅 전략을 보여주는 유세포 분석 플롯. 각 플롯은 이전 게이트에서 도출된다. 아래: RBD+, 항체- 효모 세포(즉, RBD를 발현하지만 돌연변이가 항체 결합을 방지하는 세포)에 대한 게이팅을 보여주는 유세포 분석 플롯. COV2-2196 또는 COV2-2130에 대한 선택 실험이 제시되어 있으며, 각각에 대해 2개의 독립 라이브러리가 제시되어 있다. (도 18b) COV2-2196, COV2-2130, 또는 COV2-2196과 COV2-2130의 1:1 혼합물을 사용한 효모 라이브러리 선택 실험 사이의 항체 결합으로부터 관찰된 회피 부위의 상관관계. x축은 라이브러리 1의 각 부위에 대한 누적 회피 분율을 보여주고, y축은 라이브러리 2의 각 부위에 대한 누적 회피 분율을 보여준다. 상관 계수와 n이 각 그래프에 대해 기록되어 있다. (도 18c) COV2-2196, COV2-2130, 또는 COV2-2196과 COV2-2130의 1:1 혼합물을 사용한 효모 라이브러리 선택 실험 사이의 항체 결합을 회피하는 관찰된 돌연변이의 상관관계. x축은 라이브러리 1에 대한 각 아미노산 돌연변이의 회피 분율을 보여주고, y축은 라이브러리 2에 대한 각 아미노산 돌연변이의 회피 분율을 보여준다. 상관 계수와 n이 각 그래프에 대해 기록되어 있다. (도 18d 내지 18f) COV2-2196(도 18d), COV2-2130(도 18e), 또는 COV2-2196과 COV2 2130의 1:1 혼합물(도 18f)에 대한 DMS 결과. 좌측 패널: 전체 RBD에 걸친 회피 부위는 중간 및 우측 패널의 로고 플롯에 상응하는 피크로 표시되어 있다. 중간 패널: 도 11a에서와 같이, COV2-2196, 또는 COV2-2130, 또는 COV2-2196+COV2-2130에 대한 강력한 회피 돌연변이를 갖는 모든 RBD 부위의 누적 회피 돌연변이 분율의 로고 플롯. 돌연변이는 인간 ACE2에 대한 RBD 결합을 저해하는 정도에 기초하여 채색되어 있다. 우측 패널: 다시, 로고 플롯은 누적 회피 분율을 보여주지만, 돌연변이가 효모 디스플레이 시스템에서 RBD 발현에 영향을 미치는 정도에 기초하여 채색되어 있다. 이 로고 플롯의 대화형 확대/축소 버전은 jbloomlab.github.io/SARS-CoV-2-RBD_MAP_AZ_Abs/에 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 7F에서 볼 수 있다. (도 18g) 항체 중화를 회피한 바이러스를 보여주는 대표적인 RTCA 센소그램. 세포변성 효과(CPE)는 포화 농도(5 μg/mL)의 항체 COV2-2130의 존재 하에 바이러스 접종된 Vero E6 세포에서 동역학적으로 모니터링되었다. 회피(자홍색) 또는 검출 가능한 회피 부족(파란색)의 대표적인 사례가 제시되어 있다. 감염되지 않은 세포(녹색) 또는 항체가 없는 바이러스로 접종된 세포(빨간색)가 대조군 역할을 한다. 자홍색 및 파란색 곡선은 단일 대표 우물을 나타낸다. 빨간색과 녹색의 대조군은 2중의 기술적 반복실험의 평균이다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 7G에서 볼 수 있다. (도 18h) 도 18g에서 COV2-2130에 의해 선택된 변이체 바이러스를 검증하는 대표적인 RTCA 센소그램은 실제로 COV2-2130(자홍색)을 회피했지만, COV2-2196(연청색)에 의해 중화되었다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 7H에서 볼 수 있다. (도 18i) COV2-2196, COV2-2130 또는 COV2-2196과 COV2-2130의 1:1 혼합을 사용한 회피 선택 실험에 대한 96웰 E-플레이트 분석의 개별 웰로부터의 예시적인 센소그램. COV2-2130으로부터 회피 사례가 기록된 반면, COV2-2196 또는 COV2-2196+COV2-2130의 존재 하에서는 회피가 검출되지 않았다. 양성 및 음성 대조군은 첫 번째 플레이트에 기록되어 있다.
도 19. 실제 SARS-CoV-2 바이러스를 이용한 항체 저항성 돌연변이체의 선택 방법.
도 20의 a 내지 i. 중화 SARS-CoV-2 mAb의 기능적 특성. (도 20의 a) mAb 중화 활성, hACE2 차단 활성 및 삼량체 S2Pecto 단백질 또는 단량체 SRBD에 대한 결합의 히트맵. mAb는 중화 효능에 따라 정리되어 있다(상단이 가장 높고 하단이 가장 낮음). 파선은 wt 바이러스에 대해 중화 IC50 값이 150 ng/mL 미만인 13개의 항체를 나타낸다. IC50 값은 바이러스 중화 및 hACE2 차단에 대해 시각화된 반면, EC50 값은 결합에 대해 시각화되어 있다. 교차 반응성 SARS-CoV SRBD mAb CR3022의 재조합 형태는 양성 대조군으로 제시된 반면, 항뎅기열 mAb 2D22는 음성 대조군으로 제시되어 있다. 데이터는 각각 2중의 기술적 반복실험으로 수행된 적어도 2개의 독립적인 실험을 나타낸다. 억제 없음은 >10,000 ng/mL의 IC50 값을 나타내는 반면, 결합 없음은 >10,000 ng/mL의 EC50 값을 나타낸다. (도 20의 b 내지 e) mAb의 hACE2 차단, S2Pecto 삼량체 결합, 또는 SRBD 결합과 중화 활성의 상관관계. R2 값은 로그 변환된 값의 선형 회귀 분석에 대해 제시되어 있다. 검은색 원(전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 1B 내지 E에서 보라색으로 제시되어 있음)은 중화 IC50 값이 150 ng/mL 미만인 mAb를 나타낸다. (도 20e) hACE2 차단 및 S2Pecto 삼량체 결합의 상관관계. R2 값은 로그 변환된 값의 선형 회귀 분석에 대해 제시되어 있다. (도 20f) 실제 SARS-CoV-2 바이러스에 대한 중화 분석에서 COV2-2196 및 COV2-2130에 대한 중화 곡선. 계산된 IC50 값이 그래프에 기록되어 있다. 오차 막대는 각 점의 표준 편차를 나타낸다. 데이터는 각각 2중의 기술적 반복실험으로 수행된 적어도 2개의 독립적인 실험을 나타낸다. (도 20g) 유사바이러스(pseudovirus) 중화 분석에서 COV2-2196 및 COV2-2130에 대한 중화 곡선. 오차 막대는 각 점의 표준 편차를 나타낸다. 제시된 값은 단일 실험의 2중의 기술적 반복실험이다. 최소 6회의 실험에서 계산된 IC50 값이 그래프에 기록되어 있다. (도 20h) hACE2 차단 ELISA에서 COV2-2196, COV2-2130, 및 비차단 SARS-CoV mAb rCR3022에 대한 hACE2 차단 곡선. 계산된 IC50 값이 그래프에 기록되어 있다. 오차 막대는 각 점의 표준 편차를 나타낸다. 제시된 값은 2회 반복된 대표적인 실험의 3중의 기술적 반복실험이다. (도 20i) 삼량체 S2Pecto에 대한 COV2-2196, COV2-2130 및 rCR3022의 ELISA 결합. 계산된 EC50 값이 그래프에 기록되어 있다. 오차 막대는 각 점의 표준 편차를 나타낸다. 제시된 값은 2회 반복된 대표적인 실험의 3중의 기술적 반복실험이다.
도 21의 a 내지 d. 경쟁 결합 분석에 의한 mAb의 에피토프 매핑 및 한 쌍의 mAb에 의한 상승 작용적 중화. (도 21의 a) 좌측: 항-마우스 Fc 바이오센서에 로딩된 마우스 Fc (RBD-mFc)에 융합된 RBD에 대한 참조 mAb COV2-2196 및 rCR3022의 결합을 방지하는 mAb의 능력을 측정하는 생물층 간섭계 기반 경쟁 결합 분석. 사각형의 값은 모의 경쟁 대조군 대비 경쟁 mAb의 존재 하의 참조 mAb의 결합의 %이다. 검은색 사각형은 완전한 경쟁을 나타내고(비경쟁 대조군 대비 결합의 <33%), 흰색 사각형은 경쟁 없음을 나타낸다(비경쟁 대조군 대비 결합의 >67%). 우측: hACE2의 결합을 방지하는 mAb의 능력을 측정하는 생물층 간섭계 기반 경쟁 결합 분석. 값은 경쟁이 없는 경우의 hACE2 결합에 대해 정규화된 hACE2의 결합(%)를 나타낸다. 음영은 mAb와 hACE2의 경쟁을 나타낸다. (음영은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 2A에서 빨간색으로 제시되어 있음.) (도 21의 b) 참조 mAb COV2-2130, COV2-2196 또는 rCR3022를 이용한 중화 mAb 패널의 경쟁. 참조 mAb를 비오틴화하고 삼량체 S2Pecto에 대한 참조 mAb의 결합을 경쟁 ELISA에서 포화 양의 각 mAb의 존재 하에 측정했다. 각각의 참조 mAb에 대한 ELISA 신호는 비결합 항-뎅기열 mAb 2D22의 존재 하의 신호에 대해 정규화되었다. 검은색은 완전한 경쟁(참조 mAb의 <25% 결합)을 나타내고, 회색은 부분 경쟁(참조 mAb의 25~60% 결합)을 나타내며, 흰색은 경쟁 없음(참조 mAb의 >60% 결합)을 나타낸다. (도 21의 c) COV2-2196 및 COV2-2130에 의한 야생형 SARS-CoV-2의 상승적 중화. 위: 각 mAb의 연속 희석이 있는 중화 매트릭스. 실험은 3중의 기술적 반복으로 수행되었다. 3중의 기술적 반복으로 수행된 횟수의 대표적인 실험이 제시되어 있다. mAb의 각 조합에 대한 중화(%)가 각 사각형에 제시되어 있다. 흰색에서 검은색으로의 히트맵은 각각 0% 중화에서 100% 중화를 나타낸다. (히트맵은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 2C에서 흰색에서 빨간색까지로 제시되어 있음.) (도 21의 d) 도 21의 c에서 SARS-CoV-2 중화를 기반으로 계산된 상승효과 매트릭스. 더 어두운 색(전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 2D에서 빨간색으로 제시되어 있음)은 상승적 중화가 관찰된 영역을 나타내고, 검은색 상자는 두 mAb 사이의 최대 상승효과 영역을 나타낸다.
도 22의 a 내지 f. mAb의 에피토프 확인 및 구조적 특성화. (도 22의 a) 알라닌 및 아르기닌 돌연변이 유발에 의한 중요한 접촉 잔기의 확인. 위: 생물층 간섭계에 의해 측정된 야생형(wt) 또는 돌연변이 SRBD 구성체에 대한 COV2-2130(금색), COV2-2165(적갈색) 또는 COV2-2196(진보라색)의 결합. y축에는 wt SRBD에 대한 결합에 대해 관찰된 신호에 대해 정규화된 반응이 제시되어 있다. 아래: wt 또는 중요한 접촉 잔기가 돌연변이된 SRBD 구성체에 대한 COV2-2196의 대표적인 결합 곡선. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3A에서 볼 수 있다. (도 22의 b) SARS-CoV-2(파란색) 및 hACE2(녹색)의 결정 구조(PDB(6M0J). hACE2 인식 모티프는 주황색으로 채색되어 있다. COV2-2130에 대한 중요한 접촉 잔기는 금색 구체로 제시되어 있고, COV2-2196에 대한 중요한 접촉 잔기는 보라색 구체로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3B에서 볼 수 있다. (도 22의 c) 60-아미노산 hACE2 인식 모티프에 대한 mAb의 ELISA 결합. 항-뎅기열 mAb인 r2D22는 음성 대조군으로 제시되어 있다. 아래: 보라색으로 제시된 COV2-2196 중요 접촉 잔기가 함께 제시되어 있는 주황색의 hACE2 인식 모티프의 구조. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3C에서 볼 수 있다. (도 22의 d) COV2-2130(금색), COV2-2165(적갈색) 또는 COV2-2196(진보라색)에 대한 음성 착색 전자 현미경으로 시각화된 단일 Fab:S2Pecto 삼량체 복합체. RBD는 파란색으로 제시되어 있고, S N-말단 도메인(NTD)은 빨간색으로 제시되어 있다. 전자 밀도는 회색으로 제시되어 있다. 삼량체 상태(개방 또는 폐쇄)는 각 복합체에 대해 기록되어 있다. 각 복합체에 대한 대표적인 2D 클래스 평균이 하단에 제시되어 있다(상자 크기 128픽셀). 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3D에서 볼 수 있다. (도 22의 e) S2Pecto 삼량체와 복합된 COV2-2130 및 COV2-2196 Fab. S2Pecto 삼량체에 대한 COV2-2130(금색) 및 COV2-2196(보라색) Fab의 동시 결합. 전자 밀도는 회색으로 제시되어 있다. 삼량체 상태(개방 또는 폐쇄)는 기록되어 있다. 복합체에 대한 대표적인 2D 클래스 평균이 하단에 제시되어 있다(상자 크기 128픽셀). 모든 이미지는 Chimera로 제작되었다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3E에서 볼 수 있다. (도 22의 f) S2Pecto 삼량체에서 시각화된 경쟁 결합 분석. CR3022 결정 구조는 이중-Fab:S2Pecto 삼량체 구조에 도킹되었다. CR3022는 청록색으로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3F에서 볼 수 있다. 아래: 정량적 벤 다이어그램은 각 경쟁군의 mAb의 수와 군 간의 중첩을 나타낸다.
도 23의 a 내지 f. SARS-CoV-2 감염에 대한 중화 인간 mAb의 보호 효능. (도 23의 a) SARS-CoV-2 챌린지 모델. 10 내지 11주령의 BALB/c 마우스(군당 4 내지 5마리의 마우스의 두 실험)를 항-Ifnar1 mAb로 처리하고 하루 후 i.n. 경로를 통해 AdV-hACE2로 형질도입하였다. 4일 후, 마우스에 200 μg의 mAb CoV2-2196, CoV2-2130, 또는 조합(1:1 비율) 또는 이소형 대조군 mAb를 i.p. 경로를 통해 처리하였다. 하루 후, SARS-CoV-2를 i.n. 경로를 통해 접종하였다. 분석을 위해 조직을 7 dpi에서 수확하였다(도 23의 c 및 d). (도 23의 b) 패널 a의 마우스의 체중 변화. (Tukey 사후 검정을 사용한 이원 일반 ANOVA: **** P < 0.0001). (도 2의 c) 폐, 비장 및 심장의 바이러스 부하를 RT-qPCR에 의해 측정하였다: Dunn의 사후 검정을 사용한 Kruskal-Wallis ANOVA(*, P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001). 파선은 검출의 분석 한계를 나타낸다. (도 23의 d) 사이토카인 및 케모카인 유전자 발현은 qPCR 분석에 의해 측정하였다. Dunn의 사후 검정을 사용한 Kruskal-Wallis ANOVA(*, P < 0.05, ** P < 0.01, *** P < 0.001). (도 23의 e) MA-SARS-CoV-2 챌린지 모델. 12주령의 BALB/c 마우스(n=10)에 i.n. 경로를 통해 105 PFU의 MA-SARS-CoV-2를 접종하였다. 마우스의 체중 변화가 제시되어 있다. (도 23의 f) (도 23의 e)로부터 RT-qPCR(좌측) 또는 플라크 분석(우측)에 의해 2 dpi에서 폐의 바이러스 부하를 측정하였다: Dunn의 사후 검정을 사용한 Kruskal-Wallis ANOVA(*** P < 0.001, **** P < 0.0001).
도 24. mAb 패널에 대한 SARS-CoV-2 중화 곡선. 인간 mAb에 의한 실제 SARS-CoV-2의 중화. 2중의 기술적 반복실험의 평균 ± SD가 제시되어 있다. 데이터는 둘 이상의 독립적인 실험 중 하나를 나타낸다.
도 25a 내지 25b. hACE2에 대한 S2Pecto 결합의 mAb 억제에 대한 억제 곡선. 항-SARS-CoV-2 중화 인간 mAb에 의한 S2Pecto에 대한 hACE2 결합의 차단. 한 실험의 3중 반복의 평균 ± SD가 제시되어 있다. 항체 CR3022 및 2D22가 대조군으로 사용되었다.
도 26a 내지 26b. 삼량체 SRBD, S2Pecto 또는 SARS-CoV S2Pecto 항원에 대한 항-SARS-CoV-2 중화 인간 mAb의 ELISA 결합. 3중 반복의 평균 ± SD와 두 실험의 대표가 제시되어 있다. 항체 CR3022 및 2D22가 대조군으로 사용되었다.
도 27a 내지 27b. 알라닌 및 아르기닌 돌연변이 유발 및 생물층 간섭계에 의한 mAb의 중요한 접촉 잔기의 매핑. (도 27의 a) 좌측: wt 또는 wt에 대해 정규화된 돌연변이체 SRBD 구성체에 대한 mAb 결합에 대한 반응 값. 별표는 mAb의 해리 증가가 관찰된 잔기를 나타내며, 이는 아마도 잔기가 mAb 에피토프에 근접함을 나타낸다. 우측: wt 또는 돌연변이 SRBD 구성체와의 mAb 결합 및 해리에 대한 전체 반응 곡선. (도 27의 b) 여러 mAb에 대한 중요한 접촉 잔기를 강조하는 RBD의 구조 및 구조에서 이들의 위치.
다음의 도면은 본 명세서의 일부를 형성하고 본 개시내용의 특정 양태를 추가로 증명하기 위해 포함된다. 본 개시내용은 본원에 제시된 특정 구현예의 상세한 설명과 함께 이들 도면 중 하나 이상을 참조하여 더 잘 이해될 수 있다.
도 1. 생 BSL3 SARS-CoV-2 바이러스를 사용하여 COV2-2196 + COV2-2130의 칵테일에 의한 상승적 중화 활성을 평가하기 위한 용량-반응 매트릭스. 정성적으로 개별 mAb(0 ng/mL COV2-2196 + 250 ng/mL COV2-2130 및 250 ng/mL COV2-2196 + 0 ng/mL COV2-2130의 상자)의 최고 테스트 농도(250 ng/mL)에서 작은 분율의 중화되지 않은 바이러스가 있었으나, 콤보에 의한 더 낮은 Ab 농도 범위에서는 완전 중화(100%)가 있었다. 15.6 ng/mL COV2-2196 + 63 ng/mL COV2-2130의 상자는 96%의 바이러스를 중화한 조합에서 15.6 ng/mL의 mAb COV2-2196 및 63 ng/mL의 mAb COV2-2130과 최대 상승효과가 있는 영역을 나타내지만, 개별 Ab는 각각 겨우 6 또는 0%의 중화를 보여주었다(0 ng/mL COV2-2196 + 63 ng/mL COV2-2130 및 15.6 ng/mL COV2-2196 + 0 ng/mL COV2-2130의 타원형). 3중의 기술적 반복실험에 대한 평균 값이 제시되어 있다.
도 2a 내지 2b. BSL3 SARS-CoV-2 생 바이러스를 사용하여 mAb COV2-2196 + mAb COV2-2130의 칵테일에 의한 상승적 중화 활성을 평가하기 위한 용량-반응 매트릭스. 기술적 반복실험에 대한 3중 값의 평균이 제시되어 있다. 데이터를 시각화하고 SynergyFinder 소프트웨어를 사용하여 상승효과를 평가하였다.
도 3. BSL3 SARS-CoV-2 생 바이러스를 사용하여 mAb COV2-2196 + mAb COV2-2130의 칵테일에 의한 상승적 중화 활성을 평가하기 위한 용량-반응 매트릭스. 시너지 점수 해석: <-10: 상호작용이 길항적일 가능성이 높음; -10에서 10: 상호작용이 가산적일 가능성이 높음; >10: 상호작용이 상승적일 가능성이 높음.
도 4a 내지 4c. 확립된 SARS-CoV-2 감염에 대한 중화 인간 mAb의 치료 효능. (도 4a) 105 PFU의 MA-SARS-CoV-2를 비강 경로로 마우스에 접종하고 12시간 후에 복강 내 주사에 의해 표시된 항체 치료를 제공하였다. 플라크 분석을 사용하여 바이러스 챌린지 후 2일에 폐의 바이러스 부하를 측정했다. 개별 마우스로부터의 측정치 및 중앙값 역가가 제시되어 있고, 각 군은 Dunn의 사후 검정과 함께 Kruskal-Wallis ANOVA를 사용하여 이소형 대조군과 비교되었다(* p < 0.05). 데이터는 하나의 실험을 나타낸다. (도 4b) 10 내지 11주령의 BALB/c 마우스(군당 3 내지 9마리의 마우스의 한 실험)를 항-Ifnar1 mAb로 처리하고 하루 후 비강 경로를 통해 AdV-hACE2로 형질도입하였다. 4일 후, 마우스에 105 FFU의 실제(authentic) SARS-CoV-2를 비강 경로를 통해 접종하고, 12시간 후에 복강 내 주사에 의해 표시된 mAb 치료를 제공했다. 플라크 분석을 사용하여 바이러스 챌린지 후 2 dpi에서 폐의 바이러스 부하를 측정했다. 플라크 중화 분석 성능을 위한 2개의 대조군에는 감염되지 않은 개별 또는 mAb COV2-2196 + COV2-2130 칵테일 처리된 마우스의 폐 균질물과 1:1(부피:부피) 혼합된 개별 이소형 처리된 마우스의 폐 균질물이 포함되었다. 개별 마우스로부터의 측정치 및 중앙값 역가가 제시되어 있고, 각 군은 Dunn의 사후 검정과 함께 Kruskal-Wallis ANOVA를 사용하여 이소형 대조군과 비교되었다(** p < 0.01). 데이터는 하나의 실험을 나타낸다. (도 4c) 사이토카인 및 케모카인 유전자 발현은 도 4b에서와 같이 채취한 폐로부터 qPCR 분석에 의해 측정하였다. 개별 마우스로부터의 측정치 및 중앙값이 제시되어 있다. 군은 Mann-Whitney U 검정을 이용하여 비교되었다(* p < 0.05; ** p < 0.01).
도 5. 마카크에 대한 인간 mAb 주입 후의 항체 약동학.
도 6. 야생형 SARS-CoV-2 바이러스를 이용한 비강 내 및 기관 내 챌린지 후 마카크 기관지 폐포 세척액에서의 (새로 제조된 하위 게놈 RNA로 측정한) SARS-CoV-2 바이러스 부하.
도 7. 야생형 SARS-CoV-2 바이러스를 이용한 비강 내 및 기관 내 챌린지 후 마카크 비강 스왑 검체에서의 (새로 제조된 하위 게놈 RNA로 측정한) SARS-CoV-2 바이러스 부하.
도 8a 내지 8e. Fab COV2-2196과 복합된 S 단백질 RBD의 결정 구조. (도 8a) RBD와 복합된 COV2-2196의 카툰 표현. COV2-2196 중쇄는 청록색으로, 경쇄는 자홍색으로, RBD는 녹색으로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8A에서 볼 수 있다. (도 8b) COV2-2196-RBD 복합체의 구조는 RBD 구조를 참조로 사용하여 RBD-인간 ACE2 복합체의 구조(PDB ID: 6M0J)에 중첩된다. COV2-2196-RBD 복합체의 배색은 도 8a와 동일하다. RBD-ACE2 복합체의 RBD는 밝은 파란색으로, 인간 ACE2 펩티다제 도메인은 회색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8B에서 볼 수 있다. (도 8c) COV2-2196-RBD 복합체의 구조는 "위쪽(up)" 형태의 RBD를 참조로 사용하여 "위쪽" 형태의 단일 RBD를 갖는 스파이크 구조(PDB ID: 6XM4)에 중첩된다. COV2-2196-RBD 복합체의 배색은 도 8a와 동일하다. 스파이크의 세 하위 단위는 각각 회색, 노란색 또는 밝은 파란색으로 채색되어 있다.(RBD가 "위쪽" 형태인 하위 단위는 노란색임). 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8C에서 볼 수 있다. (도 8d) COV2-2196에 의해 인식되는 RBD 에피토프의 표면 표현. 에피토프 잔기는 다양한 녹색 음영으로 채색되고 검은색으로 표지된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8D에서 볼 수 있다. (도 8e) COV2-2196과 RBD 사이의 항체-항원 상호작용. RBD는 도 8d에서와 동일한 표면 표현 및 방향으로 제시되어 있다. COV2-2196 파라토프 잔기는 막대 표현으로 제시되어 있다. 중쇄는 청록색으로, 경쇄는 자홍색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 8E에서 볼 수 있다.
도 9a 내지 9f. Fab COV2-2196 및 COV2-2130과 복합된 S 단백질 RBD의 결정 구조. (도 9a) COV2-2196 및 COV2-2130 Fab와 복합된 S 단백질 RBD의 결정 구조의 카툰 표현. RBD는 녹색, COV2-2196 중쇄는 청록색, COV2-2196 경쇄는 자홍색, COV2-2130 중쇄는 노란색, COV2-2130 경쇄는 주황색으로 제시되어 있다. COV2-2130의 CDR이 표지되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9A에서 볼 수 있다. (도 9b) COV2-2130-RBD 복합체의 구조는 RBD 구조를 참조로 사용하여 RBD-ACE2 복합체의 구조(PDB ID: 6M0J)에 중첩된다. COV2-2130-RBD 복합체의 배색은 도 9a와 동일하다. RBD-ACE2 복합체의 RBD는 밝은 파란색으로, 인간 ACE2 펩티다제 도메인은 회색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9B에서 볼 수 있다. (도 9c) COV2-2130-RBD 복합체의 구조는 하나의 프로토머의 RBD를 참조로 사용하여 "아래쪽(down)" 형태의 모든 RBD를 갖는 스파이크 구조(PDB ID: 6ZOY)에 중첩된다. COV2-2130-RBD 복합체의 배색은 도 9a와 동일하다. 스파이크의 세 프로토머는 각각 회색, 밝은 파란색, 또는 보라색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9C에서 볼 수 있다. (도 9d) COV2-2196-2130-RBD 복합체의 구조는 "위쪽(up)" 형태의 RBD를 참조로 사용하여 "위쪽" 형태의 하나의 RBD를 갖는 스파이크 구조(PDB ID: 7CAK)에 중첩된다. COV2-2130-RBD 복합체의 배색은 도 9a와 동일하다. 스파이크의 세 프로토머는 각각 회색, 밝은 파란색, 또는 보라색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9D에서 볼 수 있다. (도 9e) COV2-2130에 의해 인식되는 RBD 에피토프의 표면 표현. 에피토프 잔기는 상이한 색으로 표시되고 검은색으로 표지된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9E에서 볼 수 있다. (도 9f) COV2-2130 파라토프 잔기와 에피토프의 상호작용. RBD는 도 9e에서와 동일한 표면 표현 및 방향으로 제시되어 있다. 파라토프 잔기는 막대 표현으로 제시되어 있다. 중쇄는 노란색으로, 경쇄는 주황색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 9F에서 볼 수 있다.
도 10a 내지 10b. (도 10a) COV2-2196 중쇄 및 경쇄의 IMGT/DomainGapAlign 결과. 주요 상호작용 잔기 및 이들의 생식계열 유전자의 상응하는 잔기가 상자로 제시되어 있다. 도 10a의 서열에 대한 서열번호는 다음과 같다:
(도 10b) COV2-2196의 점 돌연변이체의 결합 곡선. 위에서 상자로 표시된 중쇄에 대한 점 돌연변이체를 암호화하는 cDNA를 설계하고 DNA로 합성하여 재조합 IgG 단백질을 제조하였고, 스파이크 단백질에 대한 결합 활성을 테스트했다. D108 잔기의 돌연변이체는 위의 좌측 그래프에, 추론된 체세포 돌연변이의 생식계열 서열로의 복귀 돌연변이는 위의 우측 그래프에, P99 돌연변이체는 아래의 좌측 그래프에, HCDR3에서 이황화 결합을 제거하는 돌연변이체는 아래의 우측 그래프에 있다.
도. 11a 내지 11h. 저항성 변이체 선택과 결합된 심층 돌연변이 스캐닝을 통한 COV2-2196 및 COV2-2130에 대한 중요한 잔기 확인. (도 11a) COV2-2196(좌측) 또는 COV2-2130(우측)에 대한 강력한 회피를 나타내는 RBD 부위에서의 모두의 돌연변이 회피 분율의 로고 플롯. 더 큰 문자는 더 큰 항체 결합 회피를 나타낸다. 돌연변이는 인간 ACE2에 대한 RBD 결합을 감소시키는 정도에 기초하여 채색되어 있다. 제시된 데이터는 2개의 독립적인 효모 라이브러리를 사용한 2개의 독립적인 회피 선택 실험의 평균이다; 상관관계는 도 18b 내지 18c에 제시되어 있다. 이 로고 플롯의 대화형 확대/축소 버전은 jbloomlab.github.io/SARS-CoV-2-RBD_MAP_AZ_Abs/에 있다. 본 발명자들은 방법에서 기재된 바와 같이, 0의 값은 변이체가 항상 항체에 의해 결합됨을 의미하고 1의 값은 변이체가 항상 항체 결합을 회피함을 의미하도록, 항체 회피 빈(bin)에 속하는 해당 특정 변이체를 발현하는 세포의 추정된 분율을 나타내는 회피 분율을 결정하였다. (도 11b) 회피 선택에 사용된 Wuhan-Hu-1 참조 균주로부터의 단일 뉴클레오티드 치환에 의해 접근 가능한 COV2-2196 및 COV2-2130에 대한 돌연변이 회피 분율의 로고 플롯(도 11e 내지 11f). ACE2 결합에 대한 각 치환의 효과는 도 11a에 나타낸 바와 같다. (도 11c) 좌측 패널: RBD-COV2-2196 구조의 RBD 표면으로의 COV2-2196에 대한 심층 돌연변이 스캐닝 회피 돌연변이 매핑. COV2-2196 결합을 저해하는 돌연변이를 히트맵을 사용하여 RBD 구조에 표시하였다. 여기서 파란색은 누적 항체 회피가 가장 큰 RBD 부위를 나타내고 흰색은 검출된 회피가 없음을 나타낸다. 회색은 RBD 발현에 대한 해로운 영향이 항체 결합에 대한 돌연변이의 영향 평가를 저해한 잔기를 나타낸다. 우측 패널: RBD의 가장 강력한 회피 부위 주변에서 상호작용하는 잔기를 보여주는 좌측 패널을 확대한 것. COV2-2196 중쇄는 청록색이고 경쇄는 자홍색으로 채색되어 있다. 도 11a에 기재된 바와 같이 독립적인 라이브러리를 이용하여 2회의 반복실험을 수행하였다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 11C에서 볼 수 있다. (도 11d) 우측 패널: RBD-COV2-2130 구조의 RBD 표면으로의 COV2-2130에 대한 심층 돌연변이 스캐닝 회피 돌연변이 매핑. COV2-2130 결합을 저해하는 돌연변이를 도 11c에서와 같이 히트맵을 사용하여 RBD 구조에 표시하였다. 좌측 패널: RBD의 가장 강력한 회피 부위 주변에서 상호작용하는 잔기를 보여주는 좌측 패널을 확대한 것. COV2-2130 중쇄는 노란색이고 경쇄는 연어살색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 도 11D에서 볼 수 있다. (도 11e) COV2-2196, COV2-2130 및 이들의 조합을 이용한 VSV-SARS-CoV-2 회피 선택 실험의 결과를 보여주는 표. 선택된 회피 돌연변이체의 수 및 수행된 회피 선택 반복실험의 총 횟수 및 회피 돌연변이체 바이러스의 시퀀싱에 의해 확인된 잔기가 기록되어 있다. (도 11f) 하위 중화(sub-neutralizing) 농도의 AZD8895(COV2-2196 기준), AZD1061(COV2-2130 기준) 및 AZD7442(AZD8895 + AZD1061 기준)의 존재 하에서 SARS-CoV-2의 계대 결과를 보여주는 표. 중화 저항성 플라크를 시퀀싱하여 확인된 중화 저항성 관련 바이러스 돌연변이가 기록되어 있다. (도 11g) x축의 돌연변이 회피 분율 및 y축의 ACE2 결합에 대한 효과와 함께, 도 11a의 DMS 데이터를 보여주는 산점도. 십자형은 다중 뉴클레오티드 치환으로만 접근 가능한 돌연변이를 나타내고, 원형은 단일 뉴클레오티드 치환으로 접근 가능한 돌연변이를 나타낸다. VSV-SARS-CoV-2(K444R, K444E) 또는 실제 SARS-CoV-2(R346I)에서 COV2-2130에 의해 선택된 아미노산 치환이 기록되어 있다. (도 11h) 참조 균주 및 관심 SARS-CoV-2 변이체에 대한 FRNT에 의해 측정된 항체 중화. 중화 분석은 2중으로 수행하고 2회 반복했으며, 1회의 반복실험으로부터의 결과가 제시되어 있다. 오차 막대는 각 점의 범위를 나타낸다. WA-1 참조 균주와 비교한 돌연변이가 표시된다. B.1.1.7-OXF는 69-70 및 144-145 결실 및 다음의 치환을 포함한다: N501Y, A570D, D614G, P681H 및 T716I.
도 12. RBD-COV2-2196-2130 복합체 및 RBD-COV2-2196 결정 구조에서 RBD-COV2-2196의 하부 구조 오버레이.
도 13a 내지 13f. RBD-COV2-2196과 스파이크-S2E12 복합체 간에 공유되는 RBD 사이트 F486에서의 유사한 방향족 적층 및 소수성 상호작용 패턴. (도 13a 및 13b) 두 복합체의 구조에서 잔기 F486을 둘러싼 동일한 수소 결합 패턴. (도 13c) COV2-2196과 RBD 사이의 상세한 상호작용. COV2-2196 중쇄는 청록색, 경쇄는 자홍색, RBD는 녹색으로 채색되어 있다. 중요한 상호작용 잔기는 막대 표현으로 제시되어 있다. Ab-Ag 상호작용에 관여하는 물 분자는 분홍색 구체로 표시된다. 직접 수소 결합은 주황색 파선으로 제시되어 있고 물 매개 수소 결합은 노란색 파선으로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2C에서 볼 수 있다. (도 13d) COV2-2196/RBD 결정 구조로의 S2E12/RBD 초저온-EM 구조의 중첩(참조로서 항체의 가변 도메인 포함). COV2-2196 중쇄는 청록색이고, 경쇄는 자홍색이다; S2E12 중쇄는 연청록색이고 경쇄는 연분홍색이다. 두 개의 상응하는 RBD 구조는 각각 녹색 또는 노란색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2D에서 볼 수 있다. (도 13e) COV2-2130 중쇄와 RBD 사이의 상세한 상호작용. 파라토프 잔기는 막대 표현으로 제시되고, 노란색으로 채색되어 있고, 에피토프 잔기는 녹색 막대로 제시되어 있다. 수소 결합 또는 강한 극성 상호작용은 자홍색 파선으로 표시된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2E에서 볼 수 있다. (도 13f) COV2-2130 경쇄와 RBD 사이의 상세한 상호작용. 파라토프 잔기는 막대 표현으로 제시되고, 주황색으로 채색되어 있고, 에피토프 잔기는 녹색 막대로 제시되어 있다. 수소 결합은 자홍색 파선으로 표시된다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 2F에서 볼 수 있다.
도 14의 A 내지 E. 동일한 결합 메커니즘을 갖는 항-RBD 항체의 공통 클론형. (도 14의 A) COV2-2196/RBD 결정 구조. (도 14의 B) S2E12/RBD 초저온-EM 구조. (도 14의 C) COV2-2381/RBD 상동성 모델. COV2-2072는 회색 구체로 표시된 HCDR3의 N-연결 글리코실화 시퀀(sequon)을 암호화한다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 3D에서 볼 수 있다. (도 14의 D) COV2-2072/RBD 상동성 모델. (도 14의 E) COV2-2196/RBD 결정 구조(도 14의 A) 및 S2E12/RBD 초저온-EM 구조(도 14의 B)의 오버레이.
도 15a 내지 15b. 바이러스 미감염 개체의 항체 가변 유전자 레퍼토리에서 COV2-2196과 유전적으로 유사한 추정상의 공개 클론형 구성원의 확인. COV2-2196 중쇄(도 15a) 및 경쇄(도 15a 및 15b)와 동일한 서열 특징을 갖는 건강한 개체로부터의 항체 가변 유전자 서열이 정렬되어 있다. 3명의 상이한 공여자 및 제대혈로부터의 서열에는 공개 클론형의 특징을 갖는 서열이 포함되었다. COV2-2196에 사용된 서열 특징 및 접촉 잔기는 각각의 다중 서열 정렬 아래에 있는 상자로 강조 표시된다(상자는 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 4A에서 빨간색으로 채색되어 있음). 도 15a의 중쇄 서열에 대한 서열번호는 다음과 같다: HIP1: 서열번호 166, HIP2: 서열번호 167, HIP3: 서열번호 168, 및 CORD: 서열번호 169. 도 15a의 경쇄 서열에 대한 서열번호는 다음과 같다: HIP1: 서열번호 170, HIP2: 서열번호 171, 및 HIP3: 서열번호 172. 도 15b의 HIP1 경쇄 서열에 대한 서열번호는 다음과 같다: 서열번호 173, 서열번호 174, 서열번호 175, 서열번호 176, 및 서열번호 177(위에서 아래로). 도 15b의 HIP2 경쇄 서열에 대한 서열번호는 다음과 같다: 서열번호 178, 서열번호 179, 서열번호 180, 서열번호 181, 및 서열번호 182(위에서 아래로). 도 15b의 HIP3 경쇄 서열에 대한 서열번호는 다음과 같다: 서열번호 183, 서열번호 184, 서열번호 185, 서열번호 186, 및 서열번호 187(위에서 아래로).
도 16의 A 내지 D. (도 16의 A) 상세한 COV2-2130 HCDR3 루프 구조. 루프 형태를 안정화하는 단거리 수소 결합은 파선으로 제시되어 있다(파선은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 5A에서 자홍색으로 채색되어 있음). (도 16의 B) COV2-2130 경쇄의 잔기는 방향족 적층 상호작용 및 HCDR3과의 수소 결합을 형성하여 HCDR3 루프를 추가로 안정화시킨다. (도 16의 C) 긴 LCDR1, HCDR2 및 HCDR3은 RBD 에피토프에 대해 상보적인 결합 표면을 형성한다. RBD는 회색으로 표면 표현으로 제시되어 있다. COV2-2130 중쇄는 노란색으로, HCDR3은 주황색으로, 경쇄는 연어색으로, LCDR1은 자홍색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 3D에서 볼 수 있다. (도 16의 D) 도 16의 C의 180° 회전도.
도 17. COV2-2196 및 COV2-2130과 복합된 RBD의 결정 구조에서 COV2-2196과 COV2-2130 사이의 계면. COV2-2196 중쇄 또는 경쇄는 각각 청록색 또는 자홍색으로 카툰 표현으로 제시되어 있고, COV2-2130 중쇄 또는 경쇄는 각각 노란색 또는 연어색으로 제시되어 있다. RBD는 녹색으로 채색되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 6에서 볼 수 있다. 계면 잔기는 막대 표현으로 제시되어 있다.
도 18a 내지 18i. 항체 결합에 영향을 미치는 돌연변이의 심층 돌연변이 스캐닝에 의한 확인 및 VSV-SARS-CoV-2 바이러스를 이용한 항체 저항성 돌연변이체의 선택 방법. (도 18a) 위: 전면 및 측면 산란(처음 3개의 패널)과 RBD를 발현하는 효모 세포 선택(우측 패널)을 사용하여 단일 효모 세포를 선택하기 위한 대표적인 게이팅 전략을 보여주는 유세포 분석 플롯. 각 플롯은 이전 게이트에서 도출된다. 아래: RBD+, 항체- 효모 세포(즉, RBD를 발현하지만 돌연변이가 항체 결합을 방지하는 세포)에 대한 게이팅을 보여주는 유세포 분석 플롯. COV2-2196 또는 COV2-2130에 대한 선택 실험이 제시되어 있으며, 각각에 대해 2개의 독립 라이브러리가 제시되어 있다. (도 18b) COV2-2196, COV2-2130, 또는 COV2-2196과 COV2-2130의 1:1 혼합물을 사용한 효모 라이브러리 선택 실험 사이의 항체 결합으로부터 관찰된 회피 부위의 상관관계. x축은 라이브러리 1의 각 부위에 대한 누적 회피 분율을 보여주고, y축은 라이브러리 2의 각 부위에 대한 누적 회피 분율을 보여준다. 상관 계수와 n이 각 그래프에 대해 기록되어 있다. (도 18c) COV2-2196, COV2-2130, 또는 COV2-2196과 COV2-2130의 1:1 혼합물을 사용한 효모 라이브러리 선택 실험 사이의 항체 결합을 회피하는 관찰된 돌연변이의 상관관계. x축은 라이브러리 1에 대한 각 아미노산 돌연변이의 회피 분율을 보여주고, y축은 라이브러리 2에 대한 각 아미노산 돌연변이의 회피 분율을 보여준다. 상관 계수와 n이 각 그래프에 대해 기록되어 있다. (도 18d 내지 18f) COV2-2196(도 18d), COV2-2130(도 18e), 또는 COV2-2196과 COV2 2130의 1:1 혼합물(도 18f)에 대한 DMS 결과. 좌측 패널: 전체 RBD에 걸친 회피 부위는 중간 및 우측 패널의 로고 플롯에 상응하는 피크로 표시되어 있다. 중간 패널: 도 11a에서와 같이, COV2-2196, 또는 COV2-2130, 또는 COV2-2196+COV2-2130에 대한 강력한 회피 돌연변이를 갖는 모든 RBD 부위의 누적 회피 돌연변이 분율의 로고 플롯. 돌연변이는 인간 ACE2에 대한 RBD 결합을 저해하는 정도에 기초하여 채색되어 있다. 우측 패널: 다시, 로고 플롯은 누적 회피 분율을 보여주지만, 돌연변이가 효모 디스플레이 시스템에서 RBD 발현에 영향을 미치는 정도에 기초하여 채색되어 있다. 이 로고 플롯의 대화형 확대/축소 버전은 jbloomlab.github.io/SARS-CoV-2-RBD_MAP_AZ_Abs/에 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 7F에서 볼 수 있다. (도 18g) 항체 중화를 회피한 바이러스를 보여주는 대표적인 RTCA 센소그램. 세포변성 효과(CPE)는 포화 농도(5 μg/mL)의 항체 COV2-2130의 존재 하에 바이러스 접종된 Vero E6 세포에서 동역학적으로 모니터링되었다. 회피(자홍색) 또는 검출 가능한 회피 부족(파란색)의 대표적인 사례가 제시되어 있다. 감염되지 않은 세포(녹색) 또는 항체가 없는 바이러스로 접종된 세포(빨간색)가 대조군 역할을 한다. 자홍색 및 파란색 곡선은 단일 대표 우물을 나타낸다. 빨간색과 녹색의 대조군은 2중의 기술적 반복실험의 평균이다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 7G에서 볼 수 있다. (도 18h) 도 18g에서 COV2-2130에 의해 선택된 변이체 바이러스를 검증하는 대표적인 RTCA 센소그램은 실제로 COV2-2130(자홍색)을 회피했지만, COV2-2196(연청색)에 의해 중화되었다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 확장 데이터 도 7H에서 볼 수 있다. (도 18i) COV2-2196, COV2-2130 또는 COV2-2196과 COV2-2130의 1:1 혼합을 사용한 회피 선택 실험에 대한 96웰 E-플레이트 분석의 개별 웰로부터의 예시적인 센소그램. COV2-2130으로부터 회피 사례가 기록된 반면, COV2-2196 또는 COV2-2196+COV2-2130의 존재 하에서는 회피가 검출되지 않았다. 양성 및 음성 대조군은 첫 번째 플레이트에 기록되어 있다.
도 19. 실제 SARS-CoV-2 바이러스를 이용한 항체 저항성 돌연변이체의 선택 방법.
도 20의 a 내지 i. 중화 SARS-CoV-2 mAb의 기능적 특성. (도 20의 a) mAb 중화 활성, hACE2 차단 활성 및 삼량체 S2Pecto 단백질 또는 단량체 SRBD에 대한 결합의 히트맵. mAb는 중화 효능에 따라 정리되어 있다(상단이 가장 높고 하단이 가장 낮음). 파선은 wt 바이러스에 대해 중화 IC50 값이 150 ng/mL 미만인 13개의 항체를 나타낸다. IC50 값은 바이러스 중화 및 hACE2 차단에 대해 시각화된 반면, EC50 값은 결합에 대해 시각화되어 있다. 교차 반응성 SARS-CoV SRBD mAb CR3022의 재조합 형태는 양성 대조군으로 제시된 반면, 항뎅기열 mAb 2D22는 음성 대조군으로 제시되어 있다. 데이터는 각각 2중의 기술적 반복실험으로 수행된 적어도 2개의 독립적인 실험을 나타낸다. 억제 없음은 >10,000 ng/mL의 IC50 값을 나타내는 반면, 결합 없음은 >10,000 ng/mL의 EC50 값을 나타낸다. (도 20의 b 내지 e) mAb의 hACE2 차단, S2Pecto 삼량체 결합, 또는 SRBD 결합과 중화 활성의 상관관계. R2 값은 로그 변환된 값의 선형 회귀 분석에 대해 제시되어 있다. 검은색 원(전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 1B 내지 E에서 보라색으로 제시되어 있음)은 중화 IC50 값이 150 ng/mL 미만인 mAb를 나타낸다. (도 20e) hACE2 차단 및 S2Pecto 삼량체 결합의 상관관계. R2 값은 로그 변환된 값의 선형 회귀 분석에 대해 제시되어 있다. (도 20f) 실제 SARS-CoV-2 바이러스에 대한 중화 분석에서 COV2-2196 및 COV2-2130에 대한 중화 곡선. 계산된 IC50 값이 그래프에 기록되어 있다. 오차 막대는 각 점의 표준 편차를 나타낸다. 데이터는 각각 2중의 기술적 반복실험으로 수행된 적어도 2개의 독립적인 실험을 나타낸다. (도 20g) 유사바이러스(pseudovirus) 중화 분석에서 COV2-2196 및 COV2-2130에 대한 중화 곡선. 오차 막대는 각 점의 표준 편차를 나타낸다. 제시된 값은 단일 실험의 2중의 기술적 반복실험이다. 최소 6회의 실험에서 계산된 IC50 값이 그래프에 기록되어 있다. (도 20h) hACE2 차단 ELISA에서 COV2-2196, COV2-2130, 및 비차단 SARS-CoV mAb rCR3022에 대한 hACE2 차단 곡선. 계산된 IC50 값이 그래프에 기록되어 있다. 오차 막대는 각 점의 표준 편차를 나타낸다. 제시된 값은 2회 반복된 대표적인 실험의 3중의 기술적 반복실험이다. (도 20i) 삼량체 S2Pecto에 대한 COV2-2196, COV2-2130 및 rCR3022의 ELISA 결합. 계산된 EC50 값이 그래프에 기록되어 있다. 오차 막대는 각 점의 표준 편차를 나타낸다. 제시된 값은 2회 반복된 대표적인 실험의 3중의 기술적 반복실험이다.
도 21의 a 내지 d. 경쟁 결합 분석에 의한 mAb의 에피토프 매핑 및 한 쌍의 mAb에 의한 상승 작용적 중화. (도 21의 a) 좌측: 항-마우스 Fc 바이오센서에 로딩된 마우스 Fc (RBD-mFc)에 융합된 RBD에 대한 참조 mAb COV2-2196 및 rCR3022의 결합을 방지하는 mAb의 능력을 측정하는 생물층 간섭계 기반 경쟁 결합 분석. 사각형의 값은 모의 경쟁 대조군 대비 경쟁 mAb의 존재 하의 참조 mAb의 결합의 %이다. 검은색 사각형은 완전한 경쟁을 나타내고(비경쟁 대조군 대비 결합의 <33%), 흰색 사각형은 경쟁 없음을 나타낸다(비경쟁 대조군 대비 결합의 >67%). 우측: hACE2의 결합을 방지하는 mAb의 능력을 측정하는 생물층 간섭계 기반 경쟁 결합 분석. 값은 경쟁이 없는 경우의 hACE2 결합에 대해 정규화된 hACE2의 결합(%)를 나타낸다. 음영은 mAb와 hACE2의 경쟁을 나타낸다. (음영은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 2A에서 빨간색으로 제시되어 있음.) (도 21의 b) 참조 mAb COV2-2130, COV2-2196 또는 rCR3022를 이용한 중화 mAb 패널의 경쟁. 참조 mAb를 비오틴화하고 삼량체 S2Pecto에 대한 참조 mAb의 결합을 경쟁 ELISA에서 포화 양의 각 mAb의 존재 하에 측정했다. 각각의 참조 mAb에 대한 ELISA 신호는 비결합 항-뎅기열 mAb 2D22의 존재 하의 신호에 대해 정규화되었다. 검은색은 완전한 경쟁(참조 mAb의 <25% 결합)을 나타내고, 회색은 부분 경쟁(참조 mAb의 25~60% 결합)을 나타내며, 흰색은 경쟁 없음(참조 mAb의 >60% 결합)을 나타낸다. (도 21의 c) COV2-2196 및 COV2-2130에 의한 야생형 SARS-CoV-2의 상승적 중화. 위: 각 mAb의 연속 희석이 있는 중화 매트릭스. 실험은 3중의 기술적 반복으로 수행되었다. 3중의 기술적 반복으로 수행된 횟수의 대표적인 실험이 제시되어 있다. mAb의 각 조합에 대한 중화(%)가 각 사각형에 제시되어 있다. 흰색에서 검은색으로의 히트맵은 각각 0% 중화에서 100% 중화를 나타낸다. (히트맵은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 2C에서 흰색에서 빨간색까지로 제시되어 있음.) (도 21의 d) 도 21의 c에서 SARS-CoV-2 중화를 기반으로 계산된 상승효과 매트릭스. 더 어두운 색(전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 2D에서 빨간색으로 제시되어 있음)은 상승적 중화가 관찰된 영역을 나타내고, 검은색 상자는 두 mAb 사이의 최대 상승효과 영역을 나타낸다.
도 22의 a 내지 f. mAb의 에피토프 확인 및 구조적 특성화. (도 22의 a) 알라닌 및 아르기닌 돌연변이 유발에 의한 중요한 접촉 잔기의 확인. 위: 생물층 간섭계에 의해 측정된 야생형(wt) 또는 돌연변이 SRBD 구성체에 대한 COV2-2130(금색), COV2-2165(적갈색) 또는 COV2-2196(진보라색)의 결합. y축에는 wt SRBD에 대한 결합에 대해 관찰된 신호에 대해 정규화된 반응이 제시되어 있다. 아래: wt 또는 중요한 접촉 잔기가 돌연변이된 SRBD 구성체에 대한 COV2-2196의 대표적인 결합 곡선. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3A에서 볼 수 있다. (도 22의 b) SARS-CoV-2(파란색) 및 hACE2(녹색)의 결정 구조(PDB(6M0J). hACE2 인식 모티프는 주황색으로 채색되어 있다. COV2-2130에 대한 중요한 접촉 잔기는 금색 구체로 제시되어 있고, COV2-2196에 대한 중요한 접촉 잔기는 보라색 구체로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3B에서 볼 수 있다. (도 22의 c) 60-아미노산 hACE2 인식 모티프에 대한 mAb의 ELISA 결합. 항-뎅기열 mAb인 r2D22는 음성 대조군으로 제시되어 있다. 아래: 보라색으로 제시된 COV2-2196 중요 접촉 잔기가 함께 제시되어 있는 주황색의 hACE2 인식 모티프의 구조. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3C에서 볼 수 있다. (도 22의 d) COV2-2130(금색), COV2-2165(적갈색) 또는 COV2-2196(진보라색)에 대한 음성 착색 전자 현미경으로 시각화된 단일 Fab:S2Pecto 삼량체 복합체. RBD는 파란색으로 제시되어 있고, S N-말단 도메인(NTD)은 빨간색으로 제시되어 있다. 전자 밀도는 회색으로 제시되어 있다. 삼량체 상태(개방 또는 폐쇄)는 각 복합체에 대해 기록되어 있다. 각 복합체에 대한 대표적인 2D 클래스 평균이 하단에 제시되어 있다(상자 크기 128픽셀). 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3D에서 볼 수 있다. (도 22의 e) S2Pecto 삼량체와 복합된 COV2-2130 및 COV2-2196 Fab. S2Pecto 삼량체에 대한 COV2-2130(금색) 및 COV2-2196(보라색) Fab의 동시 결합. 전자 밀도는 회색으로 제시되어 있다. 삼량체 상태(개방 또는 폐쇄)는 기록되어 있다. 복합체에 대한 대표적인 2D 클래스 평균이 하단에 제시되어 있다(상자 크기 128픽셀). 모든 이미지는 Chimera로 제작되었다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3E에서 볼 수 있다. (도 22의 f) S2Pecto 삼량체에서 시각화된 경쟁 결합 분석. CR3022 결정 구조는 이중-Fab:S2Pecto 삼량체 구조에 도킹되었다. CR3022는 청록색으로 제시되어 있다. 채색은 전체가 본원에 참조로 포함되는 미국 특허 제63/161,890호의 실시예 5의 도 3F에서 볼 수 있다. 아래: 정량적 벤 다이어그램은 각 경쟁군의 mAb의 수와 군 간의 중첩을 나타낸다.
도 23의 a 내지 f. SARS-CoV-2 감염에 대한 중화 인간 mAb의 보호 효능. (도 23의 a) SARS-CoV-2 챌린지 모델. 10 내지 11주령의 BALB/c 마우스(군당 4 내지 5마리의 마우스의 두 실험)를 항-Ifnar1 mAb로 처리하고 하루 후 i.n. 경로를 통해 AdV-hACE2로 형질도입하였다. 4일 후, 마우스에 200 μg의 mAb CoV2-2196, CoV2-2130, 또는 조합(1:1 비율) 또는 이소형 대조군 mAb를 i.p. 경로를 통해 처리하였다. 하루 후, SARS-CoV-2를 i.n. 경로를 통해 접종하였다. 분석을 위해 조직을 7 dpi에서 수확하였다(도 23의 c 및 d). (도 23의 b) 패널 a의 마우스의 체중 변화. (Tukey 사후 검정을 사용한 이원 일반 ANOVA: **** P < 0.0001). (도 2의 c) 폐, 비장 및 심장의 바이러스 부하를 RT-qPCR에 의해 측정하였다: Dunn의 사후 검정을 사용한 Kruskal-Wallis ANOVA(*, P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001). 파선은 검출의 분석 한계를 나타낸다. (도 23의 d) 사이토카인 및 케모카인 유전자 발현은 qPCR 분석에 의해 측정하였다. Dunn의 사후 검정을 사용한 Kruskal-Wallis ANOVA(*, P < 0.05, ** P < 0.01, *** P < 0.001). (도 23의 e) MA-SARS-CoV-2 챌린지 모델. 12주령의 BALB/c 마우스(n=10)에 i.n. 경로를 통해 105 PFU의 MA-SARS-CoV-2를 접종하였다. 마우스의 체중 변화가 제시되어 있다. (도 23의 f) (도 23의 e)로부터 RT-qPCR(좌측) 또는 플라크 분석(우측)에 의해 2 dpi에서 폐의 바이러스 부하를 측정하였다: Dunn의 사후 검정을 사용한 Kruskal-Wallis ANOVA(*** P < 0.001, **** P < 0.0001).
도 24. mAb 패널에 대한 SARS-CoV-2 중화 곡선. 인간 mAb에 의한 실제 SARS-CoV-2의 중화. 2중의 기술적 반복실험의 평균 ± SD가 제시되어 있다. 데이터는 둘 이상의 독립적인 실험 중 하나를 나타낸다.
도 25a 내지 25b. hACE2에 대한 S2Pecto 결합의 mAb 억제에 대한 억제 곡선. 항-SARS-CoV-2 중화 인간 mAb에 의한 S2Pecto에 대한 hACE2 결합의 차단. 한 실험의 3중 반복의 평균 ± SD가 제시되어 있다. 항체 CR3022 및 2D22가 대조군으로 사용되었다.
도 26a 내지 26b. 삼량체 SRBD, S2Pecto 또는 SARS-CoV S2Pecto 항원에 대한 항-SARS-CoV-2 중화 인간 mAb의 ELISA 결합. 3중 반복의 평균 ± SD와 두 실험의 대표가 제시되어 있다. 항체 CR3022 및 2D22가 대조군으로 사용되었다.
도 27a 내지 27b. 알라닌 및 아르기닌 돌연변이 유발 및 생물층 간섭계에 의한 mAb의 중요한 접촉 잔기의 매핑. (도 27의 a) 좌측: wt 또는 wt에 대해 정규화된 돌연변이체 SRBD 구성체에 대한 mAb 결합에 대한 반응 값. 별표는 mAb의 해리 증가가 관찰된 잔기를 나타내며, 이는 아마도 잔기가 mAb 에피토프에 근접함을 나타낸다. 우측: wt 또는 돌연변이 SRBD 구성체와의 mAb 결합 및 해리에 대한 전체 반응 곡선. (도 27의 b) 여러 mAb에 대한 중요한 접촉 잔기를 강조하는 RBD의 구조 및 구조에서 이들의 위치.
위에서 논의한 바와 같이, SARS-CoV-2는 활성 사례가 매일 증가하는 주요한 건강 문제이다. 따라서, 이 바이러스의 생물학과 바이러스에 대한 인간 면역 반응의 특성과 정도를 이해하는 것이 가장 중요하다. 본 발명자들은 SARS-CoV-2에 대한 인간 항체의 서열을 확인하였다. 이러한 항체의 서열 및 용도는 본원에 개시되어 있다.
또한, 하나의 항체(COV2-2196)와 RBD의 상호작용을 자세히 연구함으로써, 본 발명자들은 중쇄와 경쇄를 포함하는 복잡한 구조적 배열에 의해 구동되는 SARS-CoV-2에 대한 공개 클론형 선택을 위한 분자적 기초를 확인한다. 이 클론형의 공유된 구조적 특징은 중쇄 및 경쇄에 잔기를 포함하는 파라토프의 형성에 기여하지만, 일반적으로 항원-항체 상호작용을 지배하는 HCDR3과는 현저하게 독립적이다. 본 발명자들은 이 공개 클론형이 SARS-CoV-2 S 단백질 RBD에 대해 인간이 만든 강력한 중화 항체의 더 자주 공유되는 유형 중 하나임을 보여준다. 상세한 구조 연구 덕분에, 일반적으로 형성된 항체 파라토프가 중쇄와 경쇄의 경계면을 둘러싸고 있는 파라토프의 5개의 방향족 잔기에 의해 형성된 "방향족 케이지"에 기여함이 밝혀졌다. 이 케이지 구조는 SARS-CoV-2 S 단백질의 방향족 잔기를 조정하여 이러한 항체의 높은 특이성과 친화성을 설명한다. 놀랍게도, 중쇄와 경쇄가 이 공개 클론형을 형성하는 데 필요하지만(따라서 클론형에서 기본형 IGHV, IGHJ, IGLV 및 IGLJ 유전자를 정의함), HCDR3은 상호작용에 최소한의 영향을 미친다. 이러한 IGHV1-58-IGHJ3 중쇄 및 IGKV3-20-IGKJ1 경쇄 재조합은 면역 전 B 세포 레퍼토리에서 일반적이기 때문에, 많은 개체가 SARS-CoV-2 감염 또는 백신 접종에 대한 반응 중에 이러한 클론을 만들 가능성이 있다. 이 공개 클론형의 복잡한 사전 구성된 구조에 의해 인식되는 항원 부위는 인간 집단에서 B 세포 클론의 빈도와 가변 유전자 세그먼트에 의해 암호화되는 항체의 중화 및 보호 효능 때문에 COVID-19에 대한 보호 백신의 중요한 구성요소일 가능성이 있다.
본 발명의 이들 및 다른 양태는 하기에 상세히 기재되어 있다.
I.
코로나바이러스 2019(SARS-CoV-2)
SARS-CoV-2는 호흡기 감염인 코로나바이러스 질환 2019(COVID-19)로 명명된 급성 호흡기 질환을 일으키는 전염성 바이러스이다. 이는 현재 진행 중인, 세계적인 보건 비상사태인 2019~2020년 코로나바이러스 발병의 원인이다. 게놈 시퀀싱 결과 포지티브 센스(positive-sense)의 단일 가닥 RNA 코로나바이러스임이 밝혀졌다.
진행 중인 발병 동안, 이 바이러스는 흔히 "코로나바이러스", "신종 코로나바이러스" 및 "우한 코로나바이러스"로 통용되어 왔으며, WHO는 "SARS-CoV-2"라는 명칭을 권장한다. 국제바이러스분류위원회(The International Committee on Taxonomy of Viruses, ICTV)는 바이러스의 공식 명칭이 SARS-CoV-2라고 발표했다.
많은 초기 사례가 중국 우한시의 대규모 수산물 및 동물 시장과 관련이 있어, 이 바이러스는 인수공통전염병 기원으로 생각된다. 이 바이러스와 다른 바이러스 샘플의 유전자 서열을 비교한 결과, SARS-CoV(79.5%) 및 박쥐 코로나바이러스(96%)와 유사한 것으로 나타났다. 이 발견은 천산갑과 같은 중간 숙주를 배제할 수는 없지만 궁극적인 기원은 박쥐일 가능성을 제기한다. 바이러스는 2종 이상의 코로나바이러스로 형성된 재조합 바이러스일 수 있다.
바이러스의 사람 간 전파는 확인되었다. 코로나바이러스는 주로 밀접 접촉을 통해, 특히 약 1.8 m(6피트) 범위 내에서 기침과 재채기의 호흡기 비말을 통해 전파된다. 바이러스 RNA는 감염된 환자의 대변 샘플에서도 발견되었다. 잠복기 동안에도 바이러스는 전염성이 있을 수 있다.
식용으로 판매된 동물이 원래 SARS-CoV-2의 저장소 또는 중간 숙주로 의심되었는데, 바이러스에 감염된 것으로 처음 발견된 많은 사람들이 화난(Huanan) 수산물 시장의 노동자였기 때문이다. 2003년 사스(SARS)가 발발했을 때 식용으로 살아있는 동물을 판매하는 시장도 비난을 받았다. 이러한 시장은 신종 병원체의 인큐베이터로 간주된다. 이 발병으로 중국에서 야생 동물의 거래와 소비가 일시적으로 금지되었다. 그러나 일부 연구자들은 화난 수산물 시장이 인간에게 바이러스를 전파하는 근원지가 아닐 수 있다고 제안했다.
충분한 수의 시퀀싱된 게놈으로 바이러스 계열의 돌연변이 이력에 대한 계통수를 재구성하는 것이 가능하다. 2003년 사스 발병의 기원에 대한 연구는 대부분 관박쥐의 리놀로푸스(Rhinolophus) 속에서 기원한, 많은 SARS와 유사한 박쥐 코로나바이러스의 발견으로 이어졌다. SARS-CoV-2는 이 범주의 SARS 관련 코로나바이러스에 속한다. 2015년과 2017년에 발표된 리놀로푸스 시니쿠스(Rhinolophus sinicus)의 두 게놈 서열은 SARS-CoV-2와 80%의 유사성을 보여준다. 윈난(Yunnan) 지방에서 수집된 리놀로푸스 아피니스(Rhinolophus affinis)로부터의 제3 바이러스 게놈인 "RaTG13"은 SARS-CoV-2와 96%의 유사성을 갖는다.[28][29] 비교를 위해, 이러한 바이러스 간의 변이량은 H3N2 인간 인플루엔자 바이러스 균주에서 10년 동안 관찰된 돌연변이의 양과 유사하다.
SARS-CoV-2는 코로나바이러스로 알려진 광범위한 바이러스 계열에 속한다. "nCoV"는 보다 구체적인 명칭이 선택될 때까지 신종 코로나바이러스를 지칭하는 데 사용되는 표준 용어이다. 이는 포지티브 센스의 단일 가닥 RNA(+ssRNA) 바이러스이다. 다른 코로나바이러스는 일반 감기부터 중동 호흡기 증후군(MERS) 및 중증 급성 호흡기 증후군(SARS)과 같은 더욱 중증의 질환에 이르기까지의 질병을 유발할 수 있다. 229E, NL63, OC43, HKU1, MERS-CoV 및 SARS-CoV에 이어 SARS-CoV-2는 사람을 감염시키는 것으로 알려진 일곱 번째 코로나바이러스이다.
SARS-CoV와 마찬가지로, SARS-CoV-2는 사베코바이러스(Sarbecovirus) 아속(베타-CoV 계통 B)의 구성원이다. 이의 RNA 서열은 길이가 약 30,000 염기이다. 1월 12일까지 SARS-CoV-2의 5개 게놈이 우한에서 분리되었으며 중국 질병관리예방센터(Chinese Center for Disease Control and Prevention, CCDC) 및 기타 기관에서 보고했다. 게놈의 수는 1월 26일까지 28개로 증가했다. 가장 초기의 GenBank 게놈을 제외하고, 게놈은 GISAID에서 금수 조치를 받고 있다. 샘플에 대한 계통발생 분석은 Nextstrain을 통해 이용 가능하다.
SARS-CoV-2 게놈의 공개는 바이러스의 스파이크(S) 단백질의 수용체 결합 단백질(RBD)에 대한 여러 단백질 모델링 실험으로 이어졌다. 결과는 S 단백질이 안지오텐신 전환 효소 2(ACE2) 수용체에 대한 충분한 친화성을 유지하여 이를 세포 진입 기전으로 사용할 수 있음을 시사한다. 1월 22일, 완전한 바이러스를 연구하는 중국의 한 집단과 역유전학을 연구하는 미국의 한 집단이 독립적이고 실험적으로 SARS-CoV-2의 수용체로서 인간 ACE2를 설명하였다.
잠재적인 프로테아제 억제제를 찾기 위해, ORF1a 다단백질의 바이러스 3C 유사 프로테아제 M(pro)도 약물 도킹 실험을 위해 모델링되었다. Innophore는 SARS 프로테아제를 기반으로 하는 두 가지 계산 모델을 생성했으며 중국 과학원은 재조합 SARS-CoV-2 프로테아제의 미공개 실험 구조를 생성했다. 또한, 미시간대학교의 연구진은 I-TASSER를 사용하여 SARS-CoV-2 게놈에서 모든 성숙한 펩티드의 구조를 모델링했다.
알려진 최초의 인간 감염은 2019년 12월 초에 발생했다. SARS-CoV-2의 발병은 2019년 12월 중순 중국 우한에서 처음 발견되었으며 감염된 동물 한 마리에서 비롯된 것으로 보인다. 이후, 바이러스는 중국의 모든 지역과 아시아, 유럽, 북아메리카 및 오세아니아의 24개 이상의 다른 국가로 퍼졌다. 이 모든 지역에서 바이러스의 사람 간 전파가 확인되었다. 2020년 1월 30일, SARS-CoV-2는 WHO에 의해 세계적인 보건 비상사태로 지정되었다.
2020년 2월 10일(17:15 UTC) 기준으로 40,645명의 확진된 감염 사례가 있었고, 그 중 40,196명이 중국 본토 내에 있었다. 처음에는 중국 외 지역에서 발생한 거의 모든 사례가 우한에서 이동한 사람이거나 그 지역에서 이동한 사람과 직접 접촉한 사람에게서 발생했다. 나중에는 여행자로부터 다른 나라로 전파되어 전 세계의 많은 나라에서 전파되었다. 확인된 감염을 초래하거나 진단 가능한 SARS-CoV-2 급성 호흡기 질환으로 진행하는 감염의 비율은 아직 명확하지 않지만, 바이러스로 인한 총 사망자 수는 2020년 3월 25일 현재 19,000명 이상이었다.
바이러스의 기초 감염재생산수(R-제로)는 1.4에서 3.9 사이로 추정되었다. 이는 검사하지 않을 경우 바이러스가 일반적으로 확립된 감염당 1.4 내지 3.9건의 새로운 사례를 발생시킴을 의미한다. 바이러스는 적어도 4명의 사슬을 따라 전염될 수 있음이 확립되었다.
2020년 1월, 여러 조직과 기관이 공개된 게놈을 기반으로 SARS-CoV-2에 대한 백신을 만드는 작업을 시작했다. 중국에서는 중국 질병관리예방센터가 신종 코로나바이러스 백신을 개발하고 있다. 홍콩대학교도 백신이 개발 중이라고 발표했다. 상하이 동부병원(Shanghai East Hospital) 또한 생명공학 회사인 Stemirna Therapeutics와 협력하여 백신을 개발하고 있다.
다른 곳에서는, 3개의 백신 프로젝트가 감염병혁신연합(Coalition for Epidemic Preparedness Innovations, CEPI)의 지원을 받고 있으며, 여기에는 생명공학 회사인 모더나(Moderna)와 이노비오 파마슈티컬스(Inovio Pharmaceuticals)의 프로젝트와 퀸즐랜드대학교의 다른 프로젝트가 포함된다. 미국 국립보건원(NIH)은 코로나바이러스 표면의 스파이크와 일치하는 RNA 백신을 만들기 위해 모더나와 협력하고 있다. 2020년 3월 1상 임상 시험이 시작되었다. 이노비오 파마슈티컬스는 2020년 여름에 백신의 인체 시험을 수행하고자 중국 규제 당국의 승인을 가속화하기 위해 DNA 기반 백신을 개발하고 중국 회사와 협력하고 있다. 호주에서는, 퀸즐랜드대학교가 바이러스 단백질을 유전적으로 변형하여 코로나바이러스를 모방하고 면역 반응을 자극하게 하는 분자 클램프 백신의 가능성을 조사하고 있다.
독립적인 프로젝트에서, 캐나다 공중보건국(Public Health Agency of Canada)은 서스캐처원대학교(University of Saskatchewan)의 국제백신센터(International Vaccine Center, VIDO-InterVac)에 백신 연구를 시작하도록 허가했다. VIDO-InterVac은 2020년 3월에 생산 및 동물 테스트를 시작하고 2021년에 인체 테스트를 시작하는 것을 목표로 하고 있다. 런던의 임페리얼대학 의학부는 현재 동물에 대해 백신을 테스트하는 단계에 있다.
COVID-19 급성 호흡기 질환은 SARS-CoV-2에 의한 바이러스성 호흡기 질환이다. 이는 2019~2020년 우한 코로나바이러스 발병 동안 처음 발견되었다. 증상에는 발열, 마른 기침, 숨가쁨이 포함될 수 있다. 2020년 3월 현재 이용 가능한 구체적인 허가된 치료법은 없으며, 증상 완화와 기능 지원에 노력이 집중되고 있다.
감염자는 무증상이거나 발열, 기침, 숨가쁨과 같은 경증에서 중증의 증상을 보일 수 있다. 설사 또는 상부 호흡기 증상(예를 들어, 재채기, 콧물, 인후통)은 덜 빈번하다. 중증 감염의 경우 중증 폐렴, 다기관 부전 및 사망으로 진행될 수 있다. 노출부터 증상 발현까지의 시간은 세계보건기구(WHO)에서는 2~10일로, 미국 질병관리예방센터(CDC)에서는 2~14일로 추정하고 있다.
전세계의 보건 기구는 개인이 SARS-CoV-2 감염 가능성을 줄이기 위해 취할 수 있는 예방 조치를 발표했다. 권장 사항은 이전에 다른 코로나바이러스에 대해 발표된 것과 유사하며 다음을 포함한다: 비누와 물로 손을 자주 씻는다. 씻지 않은 손으로 눈, 코, 입을 만지지 않는다. 좋은 호흡기 위생을 실천한다.
WHO는 SARS-CoV-2에 대한 여러 테스트 프로토콜을 발표했다. 테스트는 실시간 역전사 중합효소 연쇄 반응(rRT-PCR)을 사용한다. 이 테스트는 호흡기 또는 혈액 샘플에서 수행할 수 있다. 결과는 일반적으로 몇 시간에서 며칠 이내에 제공된다.
이 질환에 대한 잠재적 치료법에 대한 연구는 2020년 1월에 시작되었다. 중국 질병관리예방센터는 1월 말부터 코로나바이러스 관련 폐렴에 기존 폐렴 치료법을 테스트하기 시작했다. RNA중합효소 억제제인 렘데시비르와 인터페론 베타에 대한 조사도 있었다. 2020년 1월 말, 중국 의학 연구자들은 탐색 연구에서 세포 수준에서 SARS-CoV-2에 "상당히 좋은 억제 효과"가 있는 것으로 보이는 렘데시비르, 클로로퀸, 로피나비르/리토나비르에 대한 임상 테스트를 시작할 의향을 밝혔다. 2020년 2월 5일 중국은 질환에 대해 렘데시비르 특허 사용을 시작했다.
현재 발병에서 시간이 지남에 따라 치명률이 변할 수 있고 진단 가능한 질환으로 진행되는 감염의 비율이 여전히 불분명하기 때문에 SARS-CoV-2 감염으로 인한 전체 사망률과 이환율은 알려져 있지 않다. 그러나 SARS-CoV-2 급성 호흡기 질환에 대한 예비 연구에서는 치명률이 2%에서 3% 사이로 나타났으며, 2020년 1월 WHO는 치명률이 약 3%임을 시사하였다. 55명의 치명적인 사례에 대한 검토되지 않은 임페리얼대학의 인쇄 전 연구에 따르면, 무증상 감염이 누락됨에 따라 조기 사망률 추정치가 너무 높을 수 있다. 이들은 평균 감염 치사율(감염자 중 사망률)을 무증상 보균자를 포함할 때 0.8%에서 후베이성에서 증상이 있는 사례만 포함할 때 18%까지의 범위로 추정했다.
초기 데이터에 따르면 우한에서 병원에 입원한 처음 41명의 확진 사례 중 13명(32%)이 집중 치료가 필요했고 6명(15%)이 사망했다. 사망자 중 많은 사람들이 처음에는 건강이 좋지 않아, 면역계를 손상시키는 고혈압, 당뇨병 또는 심혈관 질환과 같은 병태를 나타냈다. 사망에 이르게 한 SARS-CoV-2 급성 호흡기 질환의 초기 사례에서 질환의 중앙값은 14일이었고, 총 범위는 6일에서 41일인 것으로 나타났다.
II.
단클론 항체 및 이의 생산
"단리된 항체"는 자연 환경의 구성요소로부터 분리 및/또는 회수된 항체이다. 자연 환경의 오염 성분은 항체의 진단 또는 치료 용도를 방해하는 물질이며, 효소, 호르몬 및 기타 단백질성 또는 비단백질성 용질을 포함할 수 있다. 특정 구현예에서, 항체는 다음과 같이 정제된다: (1) Lowry 방법에 의해 결정 시 95중량% 초과, 가장 특히 99중량% 초과의 항체까지; (2) 스피닝 컵 서열분석기를 사용하여 N 말단 또는 내부 아미노산 서열의 적어도 15개 잔기를 수득하기에 충분한 정도로; 또는 (3) 쿠마시 블루(Coomassie blue) 또는 은 염색을 사용하여 환원 또는 비환원 조건 하에서 SDS-PAGE에 의한 균질성까지. 단리된 항체는 항체의 자연 환경의 적어도 하나의 성분이 존재하지 않을 것이기 때문에 재조합 세포 내의 제자리(in situ) 항체를 포함한다. 그러나 일반적으로 단리된 항체는 적어도 하나의 정제 단계에 의해 제조된다.
기본적인 4쇄 항체 단위는 2개의 동일한 경(L)쇄와 2개의 동일한 중(H)쇄로 구성된 이종사량체 당단백질이다. IgM 항체는 J 쇄라는 추가 폴리펩티드와 함께 5개의 기본 이종사량체 단위로 구성되어 있으므로, 10개의 항원 결합 부위를 포함하는 반면, 분비된 IgA 항체는 중합되어 J 쇄와 함께 기본 4쇄 단위 2~5개를 포함하는 다가 집합체를 형성할 수 있다. IgG의 경우, 4쇄 단위는 일반적으로 약 150,000달톤이다. 각 L 쇄는 하나의 공유 이황화 결합에 의해 H 쇄에 연결되어 있는 반면, 두 개의 H 쇄는 H 쇄 이소형에 따라 하나 이상의 이황화 결합에 의해 서로 연결되어 있다. 각 H 및 L 쇄는 또한 규칙적으로 이격된 쇄 내 이황화 가교를 갖는다. 각 H 쇄는 N 말단에 가변 영역(VH)에 이어, 알파 및 감마 쇄 각각에 대한 3개의 불변 도메인(CH), 그리고 뮤 및 이소형에 대한 4개의 CH 도메인을 갖는다. 각 L 쇄는 N 말단에 가변 영역(VL)에 이어, 다른 쪽 말단에 불변 도메인(CL)을 갖는다. VL은 VH와 정렬되고 CL은 중쇄의 첫 번째 불변 도메인(CH1)과 정렬된다. 특정 아미노산 잔기는 경쇄 및 중쇄 가변 영역 사이의 계면을 형성하는 것으로 여겨진다. VH와 VL의 쌍은 함께 단일 항원 결합 부위를 형성한다. 상이한 부류의 항체의 구조 및 특성에 대해서는, 예를 들어 Basic and Clinical Immunology, 8th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, Conn., 1994, 71페이지 및 제6장 참고.
임의의 척추동물 종의 L 쇄는 불변 도메인(CL)의 아미노산 서열에 따라 카파와 람다라는 두 가지 명확하게 구별되는 유형 중 하나로 지정될 수 있다. 중쇄의 불변 도메인(CH)의 아미노산 서열에 따라 면역글로불린은 상이한 부류 또는 이소형으로 지정될 수 있다. 면역글로불린에는 5가지 부류가 있다: IgA, IgD, IgE, IgG 및 IgM. 이들은 각각 알파, 델타, 엡실론, 감마 및 뮤로 명명된 중쇄를 갖는다. 감마와 알파 부류는 CH 서열과 기능의 상대적으로 사소한 차이에 따라 하위 부류로 더 나뉘며, 인간은 다음의 하위 부류를 발현한다: IgG1, IgG2, IgG3, IgG4, IgA1 및 IgA2.
"가변"이라는 용어는 V 도메인의 특정 세그먼트가 항체 사이에서 서열이 광범위하게 다르다는 사실을 의미한다. V 도메인은 항원 결합을 매개하고 특정 항원에 대한 특정 항체의 특이성을 정의한다. 그러나 가변성은 가변 영역의 110개 아미노산 범위에 걸쳐 고르게 분포되지 않는다. 대신, V 영역은 각각 9~12개 아미노산 길이인 "초가변 영역"이라는 극도의 가변성의 더 짧은 영역으로 분리된 15~30개 아미노산의 프레임워크 영역(FR)이라는 상대적으로 불변의 스트레치로 구성된다. 천연 중쇄 및 경쇄의 가변 영역은 각각 4개의 FR을 포함하는데, 이들은 루프 연결을 형성하고 일부 경우에는 베타 시트 구조의 일부를 형성하는 3개의 초가변 영역에 의해 연결되는 베타 시트 배열을 주로 채택한다. 각 쇄의 초가변 영역은 FR에 의해 밀접하게 함께 유지되고 다른 쇄로부터의 초가변 영역과 함께 항체의 항원 결합 부위 형성에 기여한다(Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991) 참고). 불변 도메인은 항체가 항원에 결합하는 데 직접 관여하지 않지만, 항체 의존성 세포 세포독성(ADCC), 항체 의존성 세포 식세포작용(ADCP), 항체 의존성 호중구 식세포작용(ADNP) 및 항체 의존성 보체 침적(ADCD)의 관여와 같은 다양한 이펙터 기능을 나타낸다.
본원에서 사용되는 "초가변 영역"이라는 용어는 항원 결합을 담당하는 항체의 아미노산 잔기를 지칭한다. 초가변 영역은 일반적으로 "상보성 결정 영역" 또는 "CDR"로부터의 아미노산 잔기(예를 들어, Kabat 넘버링 체계에 따라 넘버링되는 경우 VL에서 약 잔기 24~34(L1), 50~56(L2) 및 89~97(L3) 주변 및 VH에서 약 31~35(H1), 50~65(H2) 및 95~102(H3) 주변; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)); 및/또는 "초가변 루프"로부터의 이러한 잔기(예를 들어, Chothia 넘버링 체계에 따라 넘버링되는 경우 VL에서 잔기 24~34(L1), 50~56(L2) 및 89~97(L3), 및 VH에서 26~32(H1), 52~56(H2) 및 95~101(H3); Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987)); 및/또는 "초가변 루프"/CDR로부터의 이러한 잔기(예를 들어, IMGT 넘버링 체계에 따라 넘버링되는 경우 VL에서 잔기 27~38(L1), 56~65(L2) 및 105~120(L3) 및 VH에서 27~38(H1), 56~65(H2) 및 105~120(H3); Lefranc, M. P. et al. Nucl. Acids Res. 27:209-212 (1999), Ruiz, M. et al. Nucl. Acids Res. 28:219-221 (2000))를 포함한다. 선택적으로 항체는 AHo에 따라 넘버링되는 경우 VL에서 28, 36(L1), 63, 74~75(L2) 및 123(L3) 및 VsubH에서 28, 36(H1), 63, 74~75(H2) 및 123(H3)의 지점 중 하나 이상에서 대칭 삽입을 갖는다;Honneger, A. and Plunkthun, A. J. Mol. Biol. 309:657-670 (2001)).
"생식계열 핵산 잔기"는 불변 또는 가변 영역을 암호화하는 생식계열 유전자에 천연적으로 존재하는 핵산 잔기를 의미한다. "생식계열 유전자"는 생식 세포(즉, 난자 또는 정자가 될 세포)에서 발견되는 DNA이다. "생식계열 돌연변이"는 단일 세포 단계에서 생식 세포 또는 접합체에서 발생한 특정 DNA의 유전 가능한 변화를 지칭하고, 자손에게 전달되는 경우, 이러한 돌연변이는 신체의 모든 세포에 혼입된다. 생식계열 돌연변이는 단일 신체 세포에서 획득된 체세포 돌연변이와는 대조적이다. 일부 경우에, 가변 영역을 암호화하는 생식계열 DNA 서열에서 뉴클레오티드는 돌연변이되고 (즉, 체세포 돌연변이) 상이한 뉴클레오티드로 대체된다.
본원에서 사용되는 "단클론 항체"라는 용어는 실질적으로 균질한 항체 집단으로부터 수득된 항체를 지칭한다(즉, 소량으로 존재할 수 있는 가능한 자연발생적 돌연변이를 제외하고는 집단을 포함하는 개별 항체가 동일함). 단클론 항체는 매우 특이적으로, 단일 항원 부위에 대해 유도된다. 또한, 상이한 결정기(에피토프)에 대해 유도된 상이한 항체를 포함하는 다클론 항체 제제와는 대조적으로, 각각의 단클론 항체는 항원 상의 단일 결정기에 대해 유도된다. 이의 특이성 외에도, 단클론 항체는 다른 항체에 의해 오염되지 않고 합성될 수 있다는 점에서 유리하다. 수식어 "단클론"은 임의의 특정 방법에 의한 항체의 생산을 필요로 하는 것으로 간주되어서는 안 된다. 예를 들어, 본 개시내용에 유용한 단클론 항체는 Kohler et al., Nature, 256:495 (1975)에서 처음 설명된 하이브리도마 방법론에 의해 제조될 수 있거나, 또는 감염 또는 면역화에 반응하는 항원 특이적 B 세포, 항원 특이적 형질모세포의 단일 세포 분류 또는 대량 분류된 항원 특이적 컬렉션의 단일 세포로부터 연결된 중쇄 및 경쇄의 포획 후 세균, 진핵 동물 또는 식물 세포에서 재조합 DNA 방법을 사용하여 제조될 수 있다(예를 들어, 미국 특허 제4,816,567호 참고). "단클론 항체"는 또한 예를 들어, Clackson et al., Nature, 352:624-628 (1991) and Marks et al., J. Mol. Biol., 222:581-597 (1991)에 기재된 기법을 이용하여 파지 항체 라이브러리로부터 단리될 수 있다.
A.
일반 방법
SARS-CoV-2에 결합하는 단클론 항체는 여러 용도가 있음을 이해할 것이다. 여기에는 SARS-CoV-2 감염을 검출, 진단 및 치료하는 데 사용하기 위한 진단 키트의 생산이 포함된다. 이러한 맥락에서, 이러한 항체를 진단제 또는 치료제와 연결하거나, 경쟁 분석에서 포획제 또는 경쟁자로 사용하거나, 부착되는 추가 작용제 없이 개별적으로 사용할 수 있다. 항체는 아래에서 추가로 논의되는 바와 같이 돌연변이되거나 변형될 수 있다. 항체를 제조하고 특성화하는 방법은 당해 분야에 잘 알려져 있다(예를 들어, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988; 미국 특허 제4,196,265호 참고).
단클론 항체(MAb)를 생성하는 방법은 일반적으로 다클론 항체를 제조하기 위한 것과 동일한 맥락에서 시작한다. 이들 방법 둘 다를 위한 제1 단계는 적절한 숙주의 면역화 또는 이전의 천연 감염 또는 허가된 백신 또는 실험적 백신으로 면역성이 있는 대상체의 확인이다. 당해 분야에 널리 주지된 바와 같이, 면역화를 위한 주어진 조성물은 이의 면역원성에서 다양할 수 있다. 따라서, 펩티드 또는 폴리펩티드 면역원을 운반체에 커플링시킴으로써 달성될 수 있는 바와 같이 숙주 면역계를 부스팅하는 것이 자주 필요하다. 예시적이며 바람직한 운반체는 키홀 림펫 호모시아닌(KLH) 및 소 혈청 알부민(BSA)이다. 오브알부민, 마우스 혈청 알부민 또는 토끼 혈청 알부민과 같은 기타 알부민이 또한 운반체로서 사용될 수 있다. 폴리펩티드를 운반체 단백질에 접합시키기 위한 수단은 당해 분야에 널리 공지되어 있고, 글루타르알데히드, m-말레이미도벤코일-N-하이드록시숙신이미드 에스테르, 카보디이미드 및 비스-비아조티화된 벤지딘을 포함한다. 또한, 당해 분야에 널리 공지된 바와 같이, 특정 면역원 조성물의 면역원성은 보조제로서 공지된 면역 반응의 비특이적 자극인자의 사용에 의해 증진될 수 있다. 동물에서 예시적이며 바람직한 보조제는 완전 프로인트 보조제(사멸된 마이코박테리움 튜버큘로시스(Mycobacterium tuberculosis)를 함유하는 면역 반응의 비특이적 자극인자), 불완전 프로인트 보조제 및 수산화알루미늄 보조제를 포함하고, 인간에서는 알룸(alum), CpG, MFP59 및 면역자극 분자의 조합("보조제 시스템", 예를 들어, AS01 또는 AS03)을 포함한다. 물리적 전달 시스템(예를 들어, 지질 나노입자 또는 골드 바이올리스틱 비드)에서 나노입자 백신 또는 DNA 또는 RNA 유전자로서 전달되고 바늘, 유전자총, 경피 전기천공 장치를 사용하여 전달된 유전자 암호화된 항원을 포함하는, SARS-CoV-2 특이적 B 세포를 유도하는 추가의 실험 형태의 접종이 가능하다. 항원 유전자는 또한 아데노바이러스, 아데노 관련 바이러스, 수두바이러스, 헤르페스바이러스, 또는 알파바이러스 레플리콘 또는 대안적으로 바이러스 유사 입자와 같은 복제 적격 또는 결손 바이러스 벡터에 의해 암호화된 바와 같이 운반될 수 있다.
자연 병원체에 대한 인간 항체의 경우, 적절한 접근법은 병원체에 노출된 대상체, 예를 들면, 질환에 걸린 것으로 진단된 대상체 또는 백신 접종을 받아 병원체에 대해 보호 면역을 생성하거나 실험적 백신의 안전성 또는 효능을 테스트한 대상체를 확인하는 것이다. 순환하는 항-병원체 항체를 검출할 수 있고, 이어서 항체 양성 대상체로부터 B 세포를 암호화하거나 생산하는 항체를 수득할 수 있다.
다클론 항체의 생산에 사용되는 면역원 조성물의 양은 면역원의 성질과 면역화에 사용되는 동물에 따라 다르다. 면역원을 투여하기 위해 다양한 경로를 사용할 수 있다(피하, 근육 내, 피내, 정맥 내 및 복강내). 다클론 항체의 생산은 면역화 후의 다양한 시점에서 면역화된 동물의 혈액을 샘플링함으로써 모니터링할 수 있다. 제2의 부스터 주사도 제공될 수 있다. 적합한 역가가 달성될 때까지, 부스팅 및 역가 측정의 과정이 반복된다. 원하는 수준의 면역원성이 획득되면, 면역화된 동물로부터 채혈하고, 혈청을 단리하여 저장하고/하거나, 동물을 사용하여 MAb를 생성할 수 있다.
면역화 후, 항체 생성 가능성이 있는 체세포, 특히 B 림프구(B 세포)가 MAb 생성 프로토콜에 사용하기 위해 선택된다. 이들 세포는 생검된 비장, 림프절, 편도선 또는 아데노이드, 골수 흡인물 또는 생검, 폐 또는 위장관 등의 점막 기관으로부터의 조직 생검 또는 순환 혈액으로부터 수득할 수 있다. 이어서, 면역화된 동물 또는 면역 인간으로부터의 항체 생성 B 림프구를 면역화된 동물 또는 인간 또는 인간/마우스 키메라 세포와 일반적으로 동일한 종의 하나인 불멸성 골수종 세포의 세포와 융합시킨다. 하이브리도마 생성 융합 절차에서 사용하기에 적합한 골수종 세포주는 바람직하게는 항체를 생성하지 않고, 높은 융합 효율을 가지며, 효소 결함에 의해, 원하는 융합 세포(하이브리도마)만의 성장을 지지하는 특정 선택 배지에서 성장할 수 없게 하는 효소 결핍이 있다. 당업자에게 공지된 바와 같이, 다수의 골수종 세포 중 어느 하나를 사용할 수 있다(Goding, pp. 65-66, 1986; Campbell, pp. 75-83, 1984). HMMA2.5 세포 또는 MFP-2 세포는 이러한 세포의 특히 유용한 예이다.
항체 생성 비장 또는 림프절 세포와 골수종 세포의 하이브리드를 생성하는 방법은 일반적으로 체세포와 골수종 세포를 2:1 비율로 혼합하는 단계를 포함하지만, 비율은 세포막의 융합을 촉진하는 작용제(들)(화학적 또는 전기적)의 존재 하에 각각 약 20:1에서 약 1:1까지 달라질 수 있다. 일부 경우, 초기 단계로서 엡스타인 바 바이러스(EBV)로 인간 B 세포를 형질전환하면, B 세포의 크기가 증가하여, 비교적 큰 크기의 골수종 세포와의 융합이 증진된다. EBV에 의한 형질전환 효율은 형질전환 배지에서 CpG 및 Chk2 억제제 약물을 사용함으로써 증진된다. 대안적으로, 인간 B 세포는, IL-21 및 TNF 슈퍼패밀리의 유형 II 구성원인 인간 B 세포 활성화 인자(BAFF)와 같은 추가 가용성 인자를 함유하는 배지에서 CD40 리간드(CD154)를 발현하는 형질감염된 세포주와 공동 배양함으로써 활성화될 수 있다. 센다이(Sendai) 바이러스를 사용한 융합 방법은 Kohler and Milstein(1975; 1976)에 의해 기재되었고, 폴리에틸렌 글리콜(PEG), 예컨대, 37% (v/v) PEG를 이용한 융합 방법은 Gefter et al. (1977)에 의해 기재되었다. 전기적으로 유도된 융합 방법의 사용도 적절하고(Goding, pp. 71-74, 1986), 더 우수한 효율성을 위한 절차가 있다(Yu et al., 2008). 융합 절차는 통상 약 1 × 10-6 내지 1 × 10-8의 낮은 빈도로 생존 가능한 하이브리드를 생산하지만, 최적화된 절차를 사용하면, 200분의 1에 근접하는 융합 효율을 달성할 수 있다(Yu et al., 2008). 그러나 선택 배지에서 배양함으로써, 생존 가능한 융합된 하이브리드가 모체의 주입된 세포(특히 정상적으로 무한정 계속해서 분열하는 주입된 골수종 세포)로부터 분화되기 때문에, 융합 효율이 비교적 낮은 것은 문제를 일으키지 않는다. 선택 배지는 일반적으로 조직 배양 배지에서 뉴클레오티드의 드 노보 합성을 차단하는 작용제를 포함하는 것이다. 예시적이고 바람직한 작용제는 아미노프테린, 메토트렉세이트 및 아자세린이다. 아미노프테린 및 메토트렉세이트는 퓨린과 피리미딘의 드 노보 합성을 차단하는 반면, 아자세린은 퓨린 합성만을 차단한다. 아미노프테린 또는 메토트렉세이트가 사용되는 경우, 배지는 뉴클레오티드의 공급원으로서 하이포잔틴 및 티미딘이 보충된다(HAT 배지). 아자세린이 사용되는 경우, 배지는 하이포잔틴이 보충된다. 골수종에 융합되지 않은 EBV 형질전환 세포주를 제거하기 위해, B 세포 공급원이 EBV 형질전환된 인간 B 세포주인 경우, 우아바인(Ouabain)이 첨가된다.
바람직한 선택 배지는 HAT 또는 우아바인 함유 HAT이다. 뉴클레오티드 구조 경로를 운용할 수 있는 세포만이 HAT 배지에서 생존할 수 있다. 골수종 세포는 구조 경로의 중요한 효소, 예를 들어, 하이포잔틴 포스포리보실 트랜스퍼라제(HPRT)에 결함이 있고, 생존할 수 없다. B 세포는 이 경로를 운용할 수 있지만, 배양 수명이 제한되어 있고, 일반적으로 약 2주 이내에 사망한다. 따라서, 선택 배지에서 생존할 수 있는 유일한 세포는 골수종과 B 세포에서 형성된 하이브리드이다. 융합에 사용되는 B 세포의 공급원이 EBV 형질전환된 B 세포의 계통인 경우, 본원에서와 같이, EBV 형질전환된 B 세포는 약물 사멸에 감수성이 있기 때문에, 우아바인이 하이브리드의 약물 선택에도 사용될 수 있지만, 사용되는 골수종 파트너는 우아바인 저항성이 되도록 선택된다.
배양은 특정 하이브리도마가 선택되는 하이브리도마 집단을 제공한다. 일반적으로, 하이브리도마의 선택은 미세역가 플레이트에서 단일 클론 희석에 의해 세포를 배양하고, 이어서 개별 클론 상청액(약 2~3주 후)을 원하는 반응성에 대해 테스트하여 수행된다. 방사면역분석, 효소 면역분석, 세포독성 분석, 플라크 분석 도트 면역결합 분석 등의 분석은 민감하고 간단하고 신속해야 한다. 이어서, 선택된 하이브리도마는 유세포 분석 분류에 의해 연속 희석하거나 단일 세포로 분류하고, 개별 항체 생산 세포주로 클로닝하고, 이어서 이 클론을 무한정 증식시켜 mAb를 제공할 수 있다. 세포주는 2가지 기본 방식으로 MAb 생산에 이용될 수 있다. 하이브리도마 샘플을 동물(예를 들어, 마우스)에 (종종 복강 내로) 주사할 수 있다. 선택적으로, 동물은 주사 전에 탄화수소, 특히 프리스탄(테트라메틸펜타데칸)과 같은 오일로 프라이밍된다. 인간 하이브리도마를 이와 같이 사용하는 경우, 종양 거부반응을 방지하기 위해 SCID 마우스와 같은 면역저하 마우스를 주사하는 것이 최적이다. 주사된 동물은 융합된 세포 하이브리드에 의해 생성된 특정 단클론 항체를 분비하는 종양을 발달시킨다. 이어서, 혈청 또는 복수액과 같은 동물의 체액을 탭핑하여 고농도의 MAb를 제공할 수 있다. 개별 세포주는 또한 시험관 내에서 배양될 수 있고, 여기서 MAb는 배양 배지로 자연적으로 분비되고, 이로부터 고농도로 용이하게 수득할 있다. 대안적으로, 인간 하이브리도마 세포주는 세포 상청액에서 면역글로불린을 생성하기 위해 시험관 내에서 사용할 수 있다. 세포주는 무혈청 배지에서의 성장에 적합시켜, 고순도의 인간 단클론 면역글로불린을 회수하는 능력을 최적화할 수 있다.
어느 하나의 수단에 의해 생성된 MAb는 원하는 경우 여과, 원심분리 및 FPLC 또는 친화도 크로마토그래피와 같은 다양한 크로마토그래피 방법을 사용하여 추가로 정제할 수 있다. 본 개시내용의 단클론 항체의 단편은 펩신 또는 파파인과 같은 효소에 의한 소화를 포함하는 방법에 의해 및/또는 화학적 환원에 의한 이황화 결합의 절단에 의해 정제된 단클론 항체로부터 수득될 수 있다. 대안적으로, 본 개시내용에 포함되는 단클론 항체 단편은 자동화된 펩티드 합성기를 사용하여 합성될 수 있다.
또한, 단클론 항체를 생성하기 위해 분자 클로닝 접근법을 사용할 수 있는 것으로 고려된다. 형질모세포 또는 활성화된 B 세포 마커로 인해 감염 또는 백신 접종에 반응하는 것으로 확인된 단일 B 세포 또는 관심 항원으로 표지된 기억 B 세포는 상자성 비드 선택 또는 유세포 분석 분류를 사용하여 물리적으로 분류할 수 있으며, 그런 다음 RNA를 단일 세포로부터 단리하고, RT-PCR에 의해 항체 유전자를 증폭시킬 수 있다. 다양한 단일 세포 RNA-seq 방법을 사용하여 단일 세포에서 항체 가변 유전자를 얻을 수 있다. 대안적으로, 항원 특이적 벌크 분류된 세포 집단은 미세소포로 분리되고, 매칭된 중쇄 및 경쇄 가변 유전자는 중쇄 및 경쇄 앰플리콘의 물리적 결합 또는 소포로부터의 중쇄 및 경쇄 유전자의 일반적 바코드를 사용하여 단일 세포에서 회수된다. 단일 세포로부터 매칭된 중쇄 및 경쇄 유전자는 또한 RT-PCR 프라이머, 및 전사체를 세포 당 하나의 바코드로 마킹하기 위한 바코드를 함유하는 세포 침투 나노입자로 처리함으로써 항원 특이적 B 세포 집단으로부터 수득될 수 있다. 항체 가변 유전자는 또한 하이브리도마 계통의 RNA 추출에 의해 단리할 수 있고, 항체 유전자는 RT-PCR에 의해 수득되고, 면역글로불린 발현 벡터에 클로닝될 수 있다. 대안적으로, 조합 면역글로불린 파지미드 라이브러리를 세포주로부터 단리된 RNA로부터 제조하고, 적절한 항체를 발현하는 파지미드를 바이러스 항원을 사용하여 패닝함으로써 선택한다. 통상적인 하이브리도마 기술에 대한 이 접근법의 장점은 단일 라운드에서 약 104배 보다 많은 항체를 생산 및 스크리닝할 수 있고, H 및 L 쇄의 조합에 의해 새로운 특이성이 생성되어 적절한 항체를 발견할 가능성이 보다 높아진다는 점이다.
본 개시내용에 유용한 항체의 생산을 교시하는, 각각이 참조로 본원에 포함되는 기타 미국 특허에는 조합 접근법을 이용한 키메라 항체의 생산을 기재하는 미국 특허 제5,565,332호; 재조합 면역글로불린 제제를 기재하는 미국 특허 제4,816,567호; 및 항체-치료제 접합체를 기재하는 미국 특허 제4,867,973호가 포함된다.
B.
본 개시내용의 항체
본 개시내용에 따른 항체는 우선 이들의 결합 특이성에 의해 정의될 수 있다. 당업자에게 주지된 기술을 사용하여 주어진 항체의 결합 특이성/친화성을 평가함으로써, 당업자는 이러한 항체가 본 청구항의 범위 내에 속하는지를 결정할 수 있다. 예를 들어, 주어진 항체가 결합하는 에피토프는 항원 분자 내에 위치한 3개 이상(예를 들면, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20개)의 아미노산의 단일 인접 서열(예를 들어, 도메인 내의 선형 에피토프)로 구성될 수 있다. 대안적으로, 에피토프는 항원 분자 내에 위치한 복수의 비인접 아미노산(또는 아미노산 서열)으로 구성될 수 있다(예를 들면, 입체형태 에피토프).
SARS-CoV-2 항원의 두 가지 주요 범주는 표면 스파이크(S) 단백질과 내부 단백질, 특히 뉴클레오캡시드(N) 단백질이다. S 단백질에 대한 항체는 예방, 또는 치료, 또는 진단, 또는 백신 특성화에 유용할 것이다. S 단백질 항체는 SARS-CoV-2 S 단백질의 전장 엑토도메인에 결합하는 특정 항체(단량체 또는 올리고머, 예컨대 삼량체로 제시됨; 입체형태 안정화 돌연변이, 예컨대, 중요한 부위에서의 프롤린의 도입("2P 돌연변이") 포함 또는 불포함) 및 (a) 수용체 결합 도메인(RBD)에 결합하는 항-S 단백질 항체, (b) RBD 이외의 도메인에 결합하는 항-S 단백질 항체와 함께, 해당 단백질과의 추가 결합 특이성을 가질 것이다. RBD 이외의 도메인에 결합하는 하위 집합 중 일부는 N 말단 도메인(NTD)에 결합하지만, 다른 일부는 NTD 또는 RBD 이외의 에피토프에 결합하고, (c) S 단백질 항체는 수용체 결합을 중화하지만 차단하지 않는 다른 것들로 SARS-CoV-2 S 단백질의 이의 수용체인 인간 안지오텐신 전환 효소 2(hACE2)에 대한 결합을 차단함으로써, 추가로 SARS-CoV-2를 중화하는 것으로 밝혀질 수 있다. 마지막으로, 항체는 SARS-CoV-2 S 단백질 및 SARS-CoV, MERS-CoV, HCoV-229E, HCoV-OC43, HCoV-NL63 및/또는 HCoV-HKU1과 같은 다른 코로나바이러스의 S 단백질과 교차 반응할 수 있고, SARS-CoV-2와 이러한 다른 코로나바이러스를 교차 중화할 수 있다.
또 다른 특이성은 주로 진단 용도를 가지는 N 항체(또는 기타 내부 표적)에 결합하는 항체일 것이다. 예를 들어, SARS-CoV-2를 특이적으로 인식하거나 SARS-CoV-2 및 SARS-CoV, MERS-CoV, HCoV-229E, HCoV-OC43, HCoV-NL63 및/또는 HCoV-HKU1와 같은 기타 코로나바이러스를 교차 반응하여 인식하는 SARS-CoV-2의 N 또는 기타 내부 단백질에 대한 항체.
항체가 폴리펩티드 또는 단백질 내에서 "하나 이상의 아미노산과 상호작용"하는지를 결정하기 위해 당업자에게 공지된 다양한 기법이 사용될 수 있다. 예시적 기술은, 예를 들어, Antibodies, Harlow and Lane (Cold Spring Harbor Press, Cold Spring Harbor, N.Y.)에 기재된 것과 같은 일상적 교차 차단 분석을 포함한다. 교차 차단은 ELISA, 생물층 간섭계 또는 표면 플라즈몬 공명과 같은 다양한 결합 분석에서 측정할 수 있다. 다른 방법에는 알라닌 스캐닝 돌연변이 분석, 펩티드 블롯 분석(Reineke, Methods Mol. Biol. 248: 443-63, 2004), 펩티드 절단 분석, 단일 입자 재구성을 사용한 고해상도 전자 현미경 기술, 초저온EM 또는 단층 촬영, 결정학 연구 및 NMR 분석이 포함된다. 또한, 에피토프 절제, 에피토프 추출 및 항원의 화학적 변형 등의 방법을 사용할 수 있다(Tomer Prot. Sci. 9: 487-496, 2000). 항체가 상호작용하는 폴리펩티드 내의 아미노산을 확인하기 위해 사용될 수 있는 또 다른 방법은 질량 분광분석에 의해 검출되는 수소/중수소 교환이다. 일반적으로, 수소/중수소 교환 방법은 관심 단백질을 중수소 표지한 후, 항체를 중수소 표지된 단백질에 결합시키는 단계를 포함한다. 이어서, 단백질/항체 복합체는 물로 이동하고, 항체 복합체에 의해 보호되는 아미노산 내의 교환 가능한 양성자는 계면의 일부가 아닌 아미노산 내의 교환 가능한 양성자보다 느린 속도로 중수소로부터 수소로 역교환을 거친다. 그 결과, 단백질/항체 계면의 일부를 형성하는 아미노산은 중수소를 보유할 수 있으므로, 계면에 포함되지 않은 아미노산과 비교하여 상대적으로 더 높은 질량을 나타낼 수 있다. 항체의 해리 후, 표적 단백질을 프로테아제 절단 및 질량 분광분석에 적용함으로써, 항체가 상호작용하는 특정 아미노산에 상응하는 중수소 표지된 잔기를 노출시킨다. 예를 들어, Ehring, Analytical Biochemistry 267: 252-259 (1999); Engen and Smith, Anal. Chem. 73: 256A-265A (2001) 참고. 항체가 SARS-CoV-2를 중화하는 경우, 고농도의 항체 존재 하에 SARS-CoV-2를 시험관 내 또는 동물 모델에서 전파함으로써 항체 회피 돌연변이체 변이체 유기체를 단리할 수 있다. 항체에 의해 표적화된 항원을 암호화하는 SARS-CoV-2 유전자의 서열 분석은 항체 회피를 제공하는 돌연변이(들)를 밝혀, 에피토프 내의 잔기 또는 에피토프의 구조에 알로스테릭하게 영향을 미치는 잔기를 나타난다.
"에피토프"라는 용어는 B 및/또는 T 세포가 반응하는 항원 상의 부위를 지칭한다. B 세포 에피토프는 인접한 아미노산 또는 단백질의 3차 폴딩에 의해 나란히 놓인 인접하지 않은 아미노산 둘 다로부터 형성될 수 있다. 인접한 아미노산으로부터 형성된 에피토프는 변성 용매로의 노출 시 전형적으로 유지되지만, 3차 폴딩에 의해 형성된 에피토프는 변성 용매로의 처리 시 전형적으로 소실된다. 에피토프는 전형적으로 고유한 공간적 입체형태 내에 적어도 3개, 보다 일반적으로 적어도 5개 또는 8~10개의 아미노산을 포함한다.
항원 구조 기반 항체 프로파일링(Antigen Structure-based Antibody Profiling; ASAP)으로도 알려져 있는 변형 지원 프로파일링(Modification-Assisted Profiling; MAP)은 화학적으로 또는 효소적으로 변형된 항원 표면에 대한 각 항체의 결합 프로파일의 유사성에 따라 동일한 항원에 대해 유도된 다수의 단클론 항체(mAb)를 범주화하는 방법이다(본원에서 그 전체가 구체적으로 참조로 포함되는 미국 특허 공개 제2004/0101920호 참고). 각 카테고리는 다른 카테고리로 대표되는 에피토프와는 명백하게 상이하거나, 부분적으로 중첩되는 고유한 에피토프를 반영할 수 있다. 이 기술은 유전적으로 동일한 항체를 신속하게 필터링할 수 있게 하여, 유전적으로 상이한 항체에 초점을 맞추어 특성화를 수행할 수 있다. 하이브리도마 스크리닝에 적용하는 경우, MAP는 원하는 특성을 갖는 mAb를 생성하는 희귀 하이브리도마 클론의 확인을 용이하게 할 수 있다. MAP를 사용하여, 본 개시내용의 항체를 상이한 에피토프에 결합하는 항체군으로 분류할 수 있다.
본 개시내용은 동일한 에피토프, 또는 에피토프의 일부에 결합할 수 있는 항체를 포함한다. 마찬가지로, 본 개시내용은 또한 표적 또는 이의 단편에 대한 결합에 대해 본원에 기재된 임의의 특정 예시적 항체와 경쟁하는 항체를 포함한다. 당해 분야에 공지된 통상의 방법을 사용하여, 항체가 참조 항체와 동일한 에피토프에 결합하는지, 또는 결합에 대해 경쟁하는지를 용이하게 결정할 수 있다. 예를 들어, 시험 항체가 참조와 동일한 에피토프에 결합하는지를 결정하기 위해, 참조 항체를 포화 조건에서 표적에 결합시킨다. 다음으로, 표적 분자에 결합하는 시험 항체의 능력을 평가한다. 시험 항체가 참조 항체와의 포화 결합 후에 표적 분자에 결합할 수 있는 경우, 시험 항체는 참조 항체와는 상이한 에피토프에 결합한다고 결론지을 수 있다. 반면, 참조 항체와의 포화 결합 후에 시험 항체가 표적 분자에 결합할 수 없는 경우, 시험 항체는 참조 항체에 의해 결합된 에피토프와 동일한 에피토프에 결합할 수 있다.
항체가 참조 항-SARS-CoV-2 항체와의 결합에 대해 경쟁하는지를 결정하기 위해, 위에 기재된 결합 방법을 2가지 방향으로 수행한다: 제1 방향에서, 참조 항체를 포화 조건 하에서 SARS-CoV-2 항원에 결합시키고, 이어서 SARS-CoV-2 분자에 대한 시험 항체의 결합을 평가한다. 제2 방향에서, 시험 항체를 포화 조건 하에서 SARS-CoV-2 항원 분자에 결합시키고, 이어서 SARS-CoV-2 분자에 대한 참조 항체의 결합을 평가한다. 양 방향에서, 제1 (포화) 항체만이 SARS-CoV-2에 결합할 수 있는 경우, 시험 항체와 참조 항체는 SARS-CoV-2에 대한 결합에 대해 경쟁하는 것으로 결론지어진다. 당업자가 이해하는 바와 같이, 참조 항체와의 결합을 위해 경쟁하는 항체는 반드시 참조 항체와 동일한 에피토프에 결합할 필요는 없지만, 중첩되거나 인접한 에피토프에 결합함으로써 참조 항체의 결합을 입체적으로 차단할 수 있다.
각각이 항원에 대한 다른 항체의 결합을 경쟁적으로 억제(차단)하는 경우, 2개의 항체는 동일하거나 중첩되는 에피토프에 결합한다. 즉, 1, 5, 10, 20 또는 100배 과량의 하나의 항체는, 경쟁 결합 분석에서 측정 시, 다른 항체의 결합을 적어도 50%, 그러나 바람직하게는 75%, 90% 또는 심지어 99% 억제한다(예를 들어, Junghans et al., Cancer Res. 1990 50:1495-1502 참고). 대안적으로, 하나의 항체의 결합을 감소시키거나 제거하는 항원의 본질적으로 모든 아미노산 돌연변이가 다른 항체의 결합을 감소시키거나 제거하는 경우, 2개의 항체는 동일한 에피토프를 갖는다. 하나의 항체의 결합을 감소시키거나 제거하는 일부 아미노산 돌연변이가 다른 항체의 결합을 감소시키거나 제거하는 경우, 2개의 항체는 중첩되는 에피토프를 갖는다. 일부 양태에서, COV2-2196과 동일하거나 중첩되는 에피토프에 결합하는 항체 또는 항체 단편은 COV2-2130과 동일하거나 중첩되는 에피토프에 결합하는 항체 또는 항체 단편과 조합하여 사용된다.
이어서, 추가의 통상적인 실험(예를 들어, 펩티드 돌연변이 및 결합 분석)을 수행하여, 관찰된 시험 항체의 결합의 부재가 실제로 참조 항체와 동일한 에피토프에 대한 결합에 의한 것인지 또는 관찰된 시험 항체의 결합의 부재의 원인이 입체적 차단(또는 다른 현상)인지를 확인할 수 있다. 이러한 종류의 실험은 ELISA, RIA, 표면 플라즈몬 공명, 유세포 계측 또는 당해 분야에서 이용 가능한 임의의 다른 정량적 또는 정성적 항체 결합 분석법을 사용하여 수행될 수 있다. EM 또는 결정학을 사용한 구조 연구는 결합에 대해 경쟁하는 두 항체가 동일한 에피토프를 인식하는지를 입증할 수도 있다.
또 다른 양태에서, 각각 표 3 및 4에 예시된 바와 같은 중쇄 및 경쇄로부터 클론-쌍을 이루는 CDR을 갖는 단클론 항체가 제공된다. 이러한 항체는 본원에 기재된 방법을 사용하여 아래의 실시예 섹션에서 논의된 클론에 의해 생성될 수 있다.
또 다른 양태에서, 항체는 추가의 "프레임워크" 영역을 포함하는 가변 서열에 의해 정의될 수 있다. 또한, 항체 서열은 선택적으로 아래에서 더 상세히 논의된 방법을 사용하여, 이들 서열과 달라질 수 있다. 예를 들어, 핵산 서열은 (a) 가변 영역이 경쇄 및 중쇄의 불변 도메인으로부터 분리될 수 있거나, (b) 핵산이 이에 의해 암호화된 잔기에 영향을 미치지 않고, 위에 기재된 것들과는 달라질 수 있거나, (c) 핵산이 주어진 백분율, 예를 들어, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 상동성으로 위에 기재된 것들과는 달라질 수 있거나, (d) 핵산이, 약 50℃ 내지 약 70℃의 온도에서 약 0.02 M 내지 약 0.15 M의 NaCl에 의해 제공되는 것과 같은 저염 및/또는 고온 조건 하에서 예시되는 바와 같이, 높은 엄격성 조건 하에서 하이브리드화하는 능력으로 인해 위에 기재된 것들과는 달라질 수 있거나, (e) 아미노산이 주어진 백분율, 예를 들어, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 상동성으로 위에 기재된 것들과는 달라질 수 있거나, 또는 (f) 아미노산이 (아래에 논의된) 보존적 치환을 가능하게 함으로써 위에 기재된 것들과는 달라질 수 있다는 점에서 위에 기재된 것들과는 달라질 수 있다. 위의 각각은 핵산 서열 및 표 2의 아미노산 서열에 적용된다.
폴리뉴클레오티드와 폴리펩티드 서열을 비교하는 경우, 아래에 기재된 바와 같이, 최대 대응을 위해 정렬시킬 때 2개의 서열의 뉴클레오티드 또는 아미노산의 서열이 동일한 경우, 두 서열은 "동일한" 것이라고 한다. 두 서열 간의 비교는 전형적으로 비교 창에서 서열을 비교하여 서열 유사성의 국소 영역을 확인하고 비교함으로써 수행된다. 본원에서 사용되는 "비교 창"은 적어도 약 20개의 인접하는 위치, 통상 30 내지 약 75, 40 내지 약 50개의 세그먼트를 지칭하고, 여기서 서열은, 2개 서열이 최적으로 정렬된 후에 동일한 수의 인접하는 위치의 참조 서열과 비교될 수 있다.
비교를 위한 최적의 서열 정렬은 디폴트 매개변수를 사용하여 생물정보학 소프트웨어의 Lasergene 제품군(DNASTAR, Inc., 미국 위스콘신주 매디슨 소재)에서 Megalign 프로그램을 사용하여 수행될 수 있다. 이 프로그램은 다음의 참고문헌에 기재된 몇몇 정렬 방식을 구현한다: Dayhoff, M. O. (1978) A model of evolutionary change in proteins--Matrices for detecting distant relationships. In Dayhoff, M. O. (ed.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Washington D.C. Vol. 5, Suppl. 3, pp. 345-358; Hein J. (1990) Unified Approach to Alignment and Phylogeny pp. 626-645 Methods in Enzymology vol. 183, Academic Press, Inc., San Diego, Calif.; Higgins, D. G. and Sharp, P. M. (1989) CABIOS 5:151-153; Myers, E. W. and Muller W. (1988) CABIOS 4:11-17; Robinson, E. D. (1971) Comb. Theor 11:105; Santou, N. Nes, M. (1987) Mol. Biol. Evol. 4:406-425; Sneath, P. H. A. and Sokal, R. R. (1973) Numerical Taxonomy--the Principles and Practice of Numerical Taxonomy, Freeman Press, San Francisco, Calif.; Wilbur, W. J. and Lipman, D. J. (1983) Proc. Natl. Acad., Sci. USA 80:726-730.
대안적으로, 비교를 위한 서열의 최적의 정렬은 Smith and Waterman (1981) Add. APL. Math 2:482, 의 국소 동일성 알고리즘에 의해, Needleman and Wunsch (1970) J. Mol. Biol. 48:443의 동일성 정렬 알고리즘에 의해, Pearson and Lipman (1988) Proc. Natl. Acad. Sci. USA 85: 2444의 유사성 방법을 위한 검색에 의해, 이들 알고리즘의 컴퓨터 실행(미국 위스콘신주 메디슨 사이언스 드라이브 575 소재 Genetics Computer Group(GCG)의 위스콘신 제네틱스 소프트웨어 패키지의 GAP, BESTFIT, BLAST, FASTA 및 TFASTA)에 의해, 또는 검사에 의해 수행될 수 있다.
서열 동일성 및 서열 유사성 백분율을 결정하는 데 적합한 알고리즘의 한 특정한 예는 각각 Altschul et al. (1977) Nucl. Acids Res. 25:3389-3402 및 Altschul et al. (1990) J. Mol. Biol. 215:403-410에 기재된 BLAST 및 BLAST 2.0 알고리즘이다. BLAST 및 BLAST 2.0은 본 개시내용의 폴리뉴클레오티드 및 폴리펩티드에 대한 서열 동일성 백분율을 결정하기 위해, 예를 들어, 본원에 기재된 매개변수와 함께 사용될 수 있다. BLAST 분석을 수행하기 위한 소프트웨어는 미국 국립생물공학정보센터(National Center for Biotechnology Information)를 통해 공개적으로 입수 가능하다. 항체 서열의 재배열된 특성 및 각 유전자의 가변 길이는 단일 항체 서열에 대한 여러 라운드의 BLAST 검색을 필요로 한다. 또한, 상이한 유전자의 수동 조립은 어려우며 오류가 발생하기 쉽다. 서열 분석 도구 IgBLAST(ncbi.nlm.nih.gov/igblast/의 월드와이드웹)는 생식세포계열 V, D 및 J 유전자에 대한 매칭, 재배열 접합부의 세부 사항, Ig V 도메인 프레임워크 영역 및 상보성 결정 영역의 설명을 확인한다. IgBLAST는 뉴클레오티드 또는 단백질 서열을 분석할 수 있고, 배치로 서열을 처리할 수 있으며, 생식세포계열 유전자 데이터베이스 및 기타 서열 데이터베이스에 대한 검색을 동시에 허용하여 가장 잘 매칭되는 생식세포계열 V 유전자가 누락될 가능성을 최소화한다.
예시적 일례에서, 누적 스코어는 뉴클레오티드 서열에 대해, 매개변수 M(매칭되는 잔기 쌍에 대한 보상 스코어; 항상 >0) 및 N(미스매칭 잔기에 대한 페널티 스코어; 항상 <0)을 사용하여 계산될 수 있다. 각 방향의 단어 히트의 확장은 다음과 같은 경우에 중지된다: 누적 정렬 스코어가 최대 달성 값으로부터 분량 X만큼 저하되거나; 하나 이상의 음의 스코어의 잔기 정렬의 누적에 의해, 누적 스코어가 0 이하가 되거나; 또는 어느 한 서열의 말단에 도달한 경우. BLAST 알고리즘 매개변수 W, T 및 X는 정렬의 감도 및 속도를 결정한다. BLASTN 프로그램(뉴클레오티드 서열용)은, 디폴트로서, 단어 길이(W) 11, 기대값(E) 10 및 BLOSUM62 스코어링 매트릭스(Henikoff and Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915 참고) 정렬, (B) 50, 기대값(E) 10, M=5, N=-4 및 두 가닥의 비교를 사용한다.
아미노산 서열의 경우, 스코어링 매트릭스를 사용하여 누적 스코어를 계산할 수 있다. 각 방향의 단어 히트의 확장은 다음과 같은 경우에 중지된다: 누적 정렬 스코어가 최대 달성 값으로부터 분량 X만큼 저하되거나; 하나 이상의 음의 스코어의 잔기 정렬의 누적에 의해, 누적 스코어가 0 이하가 되거나; 또는 어느 한 서열의 말단에 도달한 경우. BLAST 알고리즘 매개변수 W, T 및 X는 정렬의 감도 및 속도를 결정한다.
하나의 접근법에서, "서열 동일성의 백분율"은 적어도 20개의 위치의 비교 창에 걸쳐 2개의 최적으로 정렬된 서열을 비교함으로써 결정되고, 여기서 비교 창에서 폴리뉴클레오티드 또는 폴리펩티드 서열의 일부는 2개 서열의 최적 정렬을 위한 참조 서열(이는 부가 또는 결실을 포함하지 않음)과 비교하여 20% 이하, 통상 5 내지 15%, 또는 10 내지 12%의 부가 또는 결실(즉, 갭)을 포함할 수 있다. 백분율은, 양 서열에서 동일한 핵산 염기 또는 아미노산 잔기가 존재하는 위치의 수를 결정하여 매칭된 위치의 수를 산출하고, 매칭된 위치의 수를 참조 서열의 위치의 총 수(즉, 창 크기)로 나누고, 결과에 100을 곱하여 서열 동일성의 백분율을 산출함으로써 계산된다.
항체를 정의하는 또 다른 방법은 아래에 기재된 항체 및 이의 항원 결합 단편의 임의의 "유도체"로 하는 것이다. 용어 "유도체"는 항원에 면역특이적으로 결합하지만, "모"(또는 야생형) 분자에 비하여 1, 2, 3, 4, 5개 이상의 아미노산 치환, 부가, 결실 또는 변형을 포함하는, 항체 또는 이의 항원 결합 단편을 지칭한다. 이러한 아미노산 치환 또는 부가는 자연적으로 발생하는 (즉, DNA 암호화된) 또는 비자연적으로 발생하는 아미노산 잔기를 도입할 수 있다. "유도체"라는 용어는, 예를 들어, 증진된 또는 손상된 이펙터 또는 결합 특성을 나타내는 변이체 Fc 영역을 갖는, 예를 들어, 항체 등을 형성하기 위해 변경된 CH1, 힌지, CH2, CH3 또는 CH4 영역을 갖는 변이체를 포함한다. 용어 "유도체"는 비아미노산 변형, 예를 들어, 글리코실화(예를 들어, 변경된 만노스, 2-N-아세틸글루코사민, 갈락토스, 푸코스, 글루코스, 시알산, 5-N-아세틸뉴라민산, 5-글리콜뉴라민산 등의 함량을 가짐), 아세틸화, 페길화, 인산화, 아미드화, 공지된 보호/차단 기에 의해 유도체화, 단백질 분해 절단, 세포 리간드 또는 기타 단백질 등에 연결될 수 있는 아미노산을 추가로 포함한다. 일부 구현예에서, 변경된 탄수화물 변형은 다음 중 하나 이상을 조절한다: 항체의 가용화, 항체의 세포 이하 수송 및 분비의 촉진, 항체 조립의 촉진, 구조적 무결성 및 항체 매개 이펙터 기능. 특정 구현예에서, 변경된 탄수화물 변형은 탄수화물 변형을 결여하는 항체와 비교하여 항체 매개 이펙터 기능을 증진시킨다. 변경된 항체 매개 이펙터 기능을 유도하는 탄수화물 변형은 당해 분야에 주지되어 있다(예를 들어, Shields, R. L. et al. (2002) "Lack Of Fucose On Human IgG N-Linked Oligosaccharide Improves Binding To Human Fcgamma RIII And Antibody-Dependent Cellular Toxicity," J. Biol. Chem. 277(30): 26733-26740; Davies J. et al. (2001) "Expression Of GnTIII In A Recombinant Anti-CD20 CHO Production Cell Line: Expression Of Antibodies With Altered Glycoforms Leads To An Increase In ADCC Through Higher Affinity For FC Gamma RIII," Biotechnology & Bioengineering 74(4): 288-294 참고). 탄수화물 함량을 변경하는 방법은 당업자에게 공지되어 있다. 예를 들어, Wallick, S. C. et al. (1988) "Glycosylation Of A VH Residue Of A Monoclonal Antibody Against Alpha (1----6) Dextran Increases Its Affinity For Antigen," J. Exp. Med. 168(3): 1099-1109; Tao, M. H. et al. (1989) "Studies Of Aglycosylated Chimeric Mouse-Human IgG. Role of Carbohydrate in The Structure And Effector Functions Mediated By The Human IgG Constant Region," J. Immunol. 143(8): 2595-2601; Routledge, E. G. et al. (1995) "The Effect of Aglycosylation on The Immunogenicity of A Humanized Therapeutic CD3 Monoclonal Antibody," Transplantation 60(8):847-53; Elliott, S. et al. (2003) "Enhancement Of Therapeutic Protein In Vivo Activities Through Glycoengineering," Nature Biotechnol. 21:414-21; Shields, R. L. et al. (2002) "Lack of Fucose on Human IgG N-Linked Oligosaccharide Improves Binding to Human Fcgamma RIII And Antibody-Dependent Cellular Toxicity," J. Biol. Chem. 277(30): 26733-26740 참고).
유도체 항체 또는 항체 단편은, 비드 기반 또는 세포 기반 분석 또는 동물 모델에서 생체 내 연구에 의해 측정 시, 항체 의존성 세포성 세포독성(ADCC), 항체 의존성 세포 식세포작용(ADCP), 항체 의존성 호중구 식세포작용(ADNP) 또는 항체 의존성 보체 침적(ADCD) 기능에서 바람직한 수준의 활성을 부여하기 위해 조작된 서열 또는 글리코실화 상태로 생성될 수 있다.
유도체 항체 또는 항체 단편은, 특이적 화학적 절단, 아세틸화, 제형화, 튜니카마이신의 대사성 합성 등을 포함하나 이에 제한되지는 않는 당업자에게 공지된 기술을 사용한 화학적 변형에 의해 변형될 수 있다. 일 구현예에서, 항체 유도체는 모 항체와 유사하거나 동일한 기능을 가질 것이다. 다른 구현예에서, 항체 유도체는 모 항체에 대해 변경된 활성을 나타낼 것이다. 예를 들어, 유도체 항체(또는 이의 단편)는 그의 에피토프에 모 항체보다 더 단단하게 결합하거나 단백질 분해에 대해 더욱 저항성이 있게 될 수 있다.
C.
항체 서열의 조작
다양한 구현예에서, 개선된 발현, 개선된 교차 반응성 또는 감소된 표적 외 결합 등의 다양한 이유로 확인된 항체의 서열을 조작하도록 선택할 수 있다. 변형된 항체는 표준 분자 생물학 기술을 통한 발현, 또는 폴리펩티드의 화학적 합성을 포함하는, 당업자에게 공지된 임의의 기술에 의해 제조될 수 있다. 재조합 발현 방법은 이 문서의 다른 곳에서 취급한다. 다음은 항체 공학에 대한 관련 목표 기술에 대한 일반 논의이다.
하이브리도마를 배양하고, 이어서 세포를 용해시키고, 총 RNA를 추출할 수 있다. 랜덤 6량체를 RT와 함께 사용하여 RNA의 cDNA 카피를 생성하고, 이어서 모든 인간 가변 유전자 서열을 증폭할 것으로 예상되는 PCR 프라이머의 다중 혼합물을 사용하여 PCR을 수행할 수 있다. PCR 산물은 pGEM-T Easy 벡터에 클로닝하고, 표준 벡터 프라이머를 사용하여 자동화된 DNA 시퀀싱에 의해 서열 분석할 수 있다. 결합 및 중화의 분석은 하이브리도마 상청액으로부터 수집된 항체를 사용하여 수행할 수 있고, 단백질 G 컬럼을 사용하여 FPLC로 정제할 수 있다.
재조합 전장 IgG 항체는 중쇄 및 경쇄 Fv DNA를 클로닝 벡터로부터 IgG 플라스미드 벡터로 서브클로닝하고, 293(예를 들어, Freestyle) 세포 또는 CHO 세포로 형질감염시킴으로써 생성할 수 있고, 항체는 293 또는 CHO 세포 상청액으로부터 수집 및 정제할 수 있다. 다른 적절한 숙주 세포 시스템에는 대장균 등의 세균, 곤충 세포(S2, Sf9, Sf29, 하이 파이브(High Five)), 식물 세포(예를 들어, 담배, 인간 유사 글리칸에 대한 조작의 존재 또는 부재), 조류 또는 마우스, 랫트, 염소 또는 소 등의 다양한 비인간 유전자도입 환경이 포함된다.
후속 항체 정제 및 숙주의 면역화 둘 다의 목적으로, 항체를 암호화하는 핵산의 발현이 또한 고려된다. 항체 암호화 서열은 천연 RNA 또는 변형된 RNA 등의 RNA일 수 있다. 변형된 RNA는 mRNA에 증가된 안정성과 낮은 면역원성을 부여하고, 이에 의해 치료적으로 중요한 단백질의 발현을 촉진하는 특정 화학적 변형을 고려한다. 예를 들어, N1-메틸-슈도우리딘(N1mΨ)은 번역 능력의 관점에서 기타 몇몇 뉴클레오시드 변형 및 이들의 조합을 능가한다. 면역/eIF2α 인산화 의존성의 번역 억제를 오프로 하는 것에 추가하여, 도입된 N1mΨ 뉴클레오티드는 리보솜의 일시 정지 및 mRNA의 밀도를 증가시킴으로써 번역 과정의 역학을 극적으로 변화시킨다. 변형된 mRNA의 리보솜 로딩의 증가는 동일한 mRNA에서 리보솜 재순환 또는 드 노보 리보솜 동원을 도움으로써 이들을 개시에 대해 보다 허용적이 되게 한다. 이러한 변형은 RNA 접종 후에 생체 내에서 항체 발현을 증진시키는 데 사용될 수 있다.
RNA는, 천연 또는 변형된 것이든, 네이키드 RNA로서 또는 지질 나노입자 등의 전달 비히클에서 전달될 수 있다.
대안적으로, 항체를 암호화하는 DNA를 동일한 목적으로 사용할 수 있다. DNA는 그것이 설계된 숙주 세포에서 활성인 프로모터를 포함하는 발현 카세트에 포함된다. 발현 카세트는 유리하게는 종래의 플라스미드 또는 미니벡터 등의 복제 가능한 벡터에 포함된다. 벡터는 수두바이러스, 아데노바이러스, 헤르페스바이러스, 아데노 관련 바이러스 등의 바이러스 벡터를 포함하고, 렌티바이러스가 고려된다. VEE 바이러스 또는 신드비스(Sindbis) 바이러스에 기초한 알파바이러스 레플리콘 등의 항체 유전자를 암호화하는 레플리콘도 또한 고려된다. 이러한 벡터의 전달은 근육 내, 피하 또는 피내 경로를 통한 바늘에 의해, 또는 생체 내 발현이 요구되는 경우는 경피 전기천공에 의해 수행될 수 있다.
최종 cGMP 제조 과정과 동일한 숙주 세포 및 세포 배양 과정에서 생산된 항체의 신속한 가용성은 프로세스 개발 프로그램의 기간을 감소시킬 가능성이 있다. Lonza는 CHO 세포에서 소량(최대 50 g)의 항체를 신속하게 생산하기 위해 CDACF 배지에서 성장한 풀링된(pooled) 형질감염체를 사용하는 일반적 방법을 개발했다. 진정한 일시적 시스템보다 약간 느리긴 하지만, 장점에는 보다 높은 생성물 농도와 생산 세포주와 동일한 숙주 및 과정의 사용이 포함된다. 일회용 생물반응기에서 모델 항체를 발현하는 GS-CHO 풀의 성장 및 생산성의 예를 들어, 유가 배치 모드로 작동되는 일회용 백 생물반응기 배양물(5 L 작업 용적)에서는 2 g/L의 수확 항체 농도가 9주의 형질감염 이내에 달성되었다.
항체 분자는 예를 들어 mAb의 단백질 분해 절단에 의해 생산되는 단편(예컨대, F(ab'), F(ab')2), 또는 예를 들어, 재조합 수단을 통해 생산 가능한 단쇄 면역글로불린을 포함할 것이다. F(ab') 항체 유도체는 1가인 반면, F(ab')2 항체 유도체는 2가이다. 일 구현예에서, 이러한 단편은 서로 조합되거나 다른 항체 단편 또는 수용체 리간드와 조합되어 "키메라" 결합 분자를 형성할 수 있다. 중요하게도, 이러한 키메라 분자는 동일한 분자의 상이한 에피토프에 결합할 수 있는 치환기를 함유할 수 있다.
관련 구현예에서, 항체는 개시된 항체의 유도체, 예를 들어, 개시된 항체의 것과 동일한 CDR 서열을 포함하는 항체(예를 들어, 키메라 또는 CDR 이식된 항체)이다. 대안적으로, 항체 분자에 보존적 변화를 도입하는 것 등의 변형을 수행하는 것을 원할 수 있다. 이러한 변경을 수행할 때, 아미노산의 소수성 지수를 고려할 수 있다. 단백질에 상호작용적 생물학적 기능을 부여하는 데 있어서 수치료학적 아미노산 지수의 중요성은 당해 분야에서 일반적으로 이해된다(Kyte and Doolittle, 1982). 아미노산의 상대적 소수성 특성은 수득되는 단백질의 2차 구조에 기여하는 것으로 인정되고, 이에 의해 단백질과 다른 분자, 예를 들어, 효소, 기질, 수용체, DNA, 항체, 항원 등과의 상호작용이 정의된다.
유사 아미노산의 치환은 친수성에 기초하여 효과적으로 수행될 수 있음이 또한 당해 분야에서 이해된다. 참조로서 본원에 포함되는 미국 특허 제4,554,101호는 인접 아미노산의 친수성에 의해 지배되는 단백질의 최대 국소 평균 친수성이 단백질의 생물학적 특성과 상관관계가 있다고 언급한다. 미국 특허 제4,554,101호에 상세히 기재된 바와 같이, 다음의 친수성 값이 아미노산 잔기에 할당되었다: 염기성 아미노산: 아르기닌(+3.0), 리신(+3.0) 및 히스티딘(-0.5); 산성 아미노산: 아스파르테이트(+3.0 ± 1), 글루타메이트(+3.0 ± 1), 아스파라긴(+0.2) 및 글루타민(+0.2); 친수성, 비이온성 아미노산: 세린(+0.3), 아스파라긴(+0.2), 글루타민(+0.2) 및 트레오닌(-0.4), 황 함유 아미노산: 시스테인(-1.0) 및 메티오닌(-1.3); 소수성, 비방향족 아미노산: 발린(-1.5), 류신(-1.8), 이소류신(-1.8), 프롤린(-0.5 ± 1), 알라닌(-0.5) 및 글리신(0); 소수성, 방향족 아미노산: 트립토판(-3.4), 페닐알라닌(-2.5) 및 티로신(-2.3).
아미노산은 유사한 친수성을 갖는 또 다른 것으로 치환될 수 있고, 생물학적으로 또는 면역학적으로 변형된 단백질을 생성할 수 있는 것으로 이해된다. 이러한 변화에 있어서, 친수성 값이 ±2 이내인 아미노산의 치환이 바람직하고, ±1 이내인 것이 특히 바람직하고, ±0.5 이내인 것이 훨씬 더 특히 바람직하다.
위에서 개설한 바와 같이, 아미노산 치환은 일반적으로 아미노산 측쇄 치환기의 상대적 유사성, 예를 들면, 이들의 소수성, 친수성, 전하, 크기 등을 기반으로 한다. 전술한 다양한 특성을 고려하는 예시적 치환은 당업자에게 주지되어 있고, 다음을 포함한다: 아르기닌 및 리신; 글루타메이트 및 아스파르테이트; 세린 및 트레오닌; 글루타민 및 아스파라긴; 및 발린, 류신 및 이소류신.
본 개시내용은 또한 이소형 변형을 고려한다. 상이한 이소형을 갖도록 Fc 영역을 변형함으로써 상이한 기능을 달성할 수 있다. 예를 들어, IgG1로의 변경은 항체 의존성 세포독성을 증가시킬 수 있고, A 부류로의 스위칭은 조직 분포를 개선할 수 있으며, M 부류로의 스위칭은 원자가를 개선할 수 있다.
대안적으로 또는 추가로, 아미노산 변형을, IL-23p19 결합 분자의 Fc 영역의 C1q 결합 및/또는 보체 의존성 세포독성(CDC) 기능을 변화시키는 하나 이상의 추가 아미노산 변형과 조합하는 것이 유용할 수 있다. 특정한 관심 결합 폴리펩티드는, C1q에 결합하고 보체 의존성 세포독성을 나타내는 것일 수 있다. 기존의 C1q 결합 활성을 갖고 선택적으로 CDC를 매개하는 능력을 추가로 갖는 폴리펩티드는 이들 활성 중 하나 또는 둘 모두가 증진되도록 변형될 수 있다. C1q를 변경하고/하거나 이의 보체 의존적 세포독성 기능을 변경하는 아미노산 변형은, 예를 들어, 본원에서 참조로 포함되는 제WO/0042072호에 기재되어 있다.
예를 들어, C1q 결합 및/또는 FcγR 결합을 변형하고, 이에 의해 CDC 활성 및/또는 ADCC 활성을 변화시킴으로써 이펙터 기능이 변경된 항체의 Fc 영역을 설계할 수 있다. "이펙터 기능"은 생물학적 활성을 활성화 또는 감소시키는 역할을 한다(예를 들어, 대상체에서). 이펙터 기능의 예는 다음을 포함하나 이에 한정되지 않는다: C1q 결합; 보체 의존성 세포독성(CDC); Fc 수용체 결합; 항체 의존성 세포 매개 세포독성(ADCC); 식세포작용; 세포 표면 수용체(예를 들어, B 세포 수용체; BCR) 등의 하향조절. 이러한 이펙터 기능은 Fc 영역을 결합 도메인(예를 들어, 항체 가변 도메인)과 결합시킬 필요가 있고, 다양한 분석(예를 들어, Fc 결합 분석, ADCC 분석, CDC 분석 등)을 사용하여 평가할 수 있다.
예를 들어, 개선된 C1q 결합 및 개선된 FcγRIII 결합(예를 들어, 개선된 ADCC 활성 및 개선된 CDC 활성 둘 모두를 가짐)을 갖는 항체의 변이체 Fc 영역을 생성할 수 있다. 대안적으로, 이펙터 기능을 저하시키거나 제거하는 것이 바람직한 경우, 감소된 CDC 활성 및/또는 감소된 ADCC 활성을 갖는 변이체 Fc 영역이 조작될 수 있다. 다른 구현예에서, 이들 활성 중 하나만을 증가시킬 수 있고, 선택적으로 다른 활성도 감소시킬 수 있다(예를 들어, 개선된 ADCC 활성을 갖지만, 감소된 CDC 활성을 갖는 Fc 영역 변이체를 생성하기 위해 및 그 반대의 경우).
FcRn 결합. Fc 돌연변이는 또한 신생아 Fc 수용체(FcRn)와의 상호작용을 변경하고 약동학적 특성을 개선하기 위해 도입 및 조작할 수 있다. FcRn에 대한 결합이 개선된 인간 Fc 변이체의 콜렉션이 기재되어 있다(Shields et al., (2001). High resolution mapping of the binding site on human IgG1 for FcγRI, FcγRII, FcγRIII, and FcRn and design of IgG1 variants with improved binding to the FcγR, (J. Biol. Chem. 276:6591-6604). 반감기를 증가시킬 수 있는 다수의 방법이 공지되어 있고(Kuo and Aveson, (2011)), 아미노산 변형을 포함하는 것은, 알라닌 스캐닝 돌연변이 유발, 무작위 돌연변이 유발, 및 신생아 Fc 수용체(FcRn)에 대한 결합 및/또는 생체 내 거동을 평가하기 위한 스크리닝을 포함하는 기술을 통해 생성될 수 있다. 컴퓨터 전략에 이은 돌연변이 유발을 사용하여, 돌연변이시킬 아미노산 돌연변이 중의 하나를 선택할 수 있다.
따라서, 본 개시내용은 FcRn에 대한 최적화된 결합을 갖는 항원 결합 단백질의 변이체를 제공한다. 특정 구현예에서, 항원 결합 단백질의 상기 변이체는 상기 항원 결합 단백질의 Fc 영역에 적어도 하나의 아미노산 변형을 포함하고, 여기서 상기 변형은 상기 모 폴리펩티드와 비교하여 Fc 영역의 226, 227, 228, 230, 231, 233, 234, 239, 241, 243, 246, 250, 252, 256, 259, 264, 265, 267, 269, 270, 276, 284, 285, 288, 289, 290, 291, 292, 294, 297, 298, 299, 301, 302, 303, 305, 307, 308, 309, 311, 315, 317, 320, 322, 325, 327, 330, 332, 334, 335, 338, 340, 342, 343, 345, 347, 350, 352, 354, 355, 356, 359, 360, 361, 362, 369, 370, 371, 375, 378, 380, 382, 384, 385, 386, 387, 389, 390, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401 403, 404, 408, 411, 412, 414, 415, 416, 418, 419, 420, 421, 422, 424, 426, 428, 433, 434, 438, 439, 440, 443, 444, 445, 446 및 447로 구성된 군으로부터 선택되고, 여기서 Fc 영역의 아미노산 넘버링은 Kabat의 EU 인덱스의 넘버링이다. 본 개시내용의 추가 양태에서, 변형은 M252Y/S254T/T256E이다.
추가로, 다양한 간행물은, FcRn 결합 폴리펩티드를 분자에 도입하거나, FcRn 결합 친화성이 보존되어 있지만 다른 Fc 수용체에 대한 친화성은 크게 감소되어 있는 항체와 분자를 융합시키거나, 항체의 FcRn 결합 도메인과 융합시킴으로써 반감기가 변형된 생리학적 활성 분자를 수득하는 방법을 기재하고 있다. 예를 들어, Kontermann (2009) 참고.
유도체화 항체는 포유류, 특히 인간에서 모 항체의 반감기(예컨대, 혈청 반감기)를 변경하는 데 이용될 수 있다. 이러한 변경은 15일 초과, 바람직하게는 20일 초과, 25일 초과, 30일 초과, 35일 초과, 40일 초과, 45일 초과, 2개월 초과, 3개월 초과, 4개월 초과, 또는 5개월 초과의 반감기를 초래할 수 있다. 본 개시내용의 항체 또는 이의 단편의 증가된 반감기는 포유동물, 바람직하게는 인간에서, 상기 항체 또는 항체 단편의 포유동물 내 더 높은 혈청 역가를 초래하고, 이에 따라, 상기 항체 또는 항체 단편의 투여 빈도를 낮추고/낮추거나 상기 항체 또는 항체 단편이 투여되는 농도를 낮춘다. 증가된 생체 내 반감기를 갖는 항체 또는 이의 단편은 당업자에게 공지된 기술에 의해 생성될 수 있다. 예를 들어, 증가된 생체 내 반감기를 갖는 항체 또는 이의 단편은 Fc 도메인과 FcRn 수용체 사이의 상호작용에 관여하는 것으로 확인된 아미노산 잔기를 변형(예컨대, 치환, 결실 또는 부가)함으로써 생성될 수 있다.
Beltramello et al. (2010)은, CH2 도메인의 위치 1.3과 1.2의 류신 잔기(C 도메인에 대한 IMGT 고유 넘버링에 따름)가 알라닌 잔기로 치환되어 있는 것을 생성함으로써, 뎅기열 바이러스 감염을 증진시키는 경향이 있기 때문에, 중화 mAb의 변형을 이전에 보고했다. "LALA" 돌연변이로도 공지된 이 변형은 Hessell et al. (2007)에 기재된 바와 같이, FcγRI, FcγRII 및 FcγRIIIa에 대한 항체 결합을 무력화한다. 변이체 및 비변형 재조합 mAb를 4개 뎅기열 바이러스 혈청형에 의한 감염을 중화하고 증진시키는 능력에 대해 비교했다. LALA 변이체는 변형되지 않은 mAb와 동일한 중화 활성을 유지했지만, 증진시키는 활성은 완전히 결여하고 있었다. 따라서, 이러한 성질의 LALA 돌연변이가 본원의 개시된 항체와 관련하여 고려된다.
변경된 글리코실화. 본 개시내용의 특정 구현예는, 시알산, 갈락토스 또는 푸코스를 포함하지 않는 실질적으로 균질한 글리칸을 함유하는, 단리된 단클론 항체 또는 이의 항원 결합 단편이다. 단클론 항체는 중쇄 가변 영역 및 경쇄 가변 영역을 포함하고, 이들 둘 모두는 각각 중쇄 또는 경쇄 불변 영역에 부착될 수 있다. 전술한 실질적으로 균질한 글리칸은 중쇄 불변 영역에 공유 부착될 수 있다.
본 개시내용의 또 다른 구현예는 신규한 Fc 글리코실화 패턴을 갖는 mAb를 포함한다. 단리된 단클론 항체 또는 이의 항원 결합 단편은 GNGN 또는 G1/G2 글리코형에 의해 표시되는 실질적으로 균질한 조성물에 존재한다. Fc 글리코실화는 치료적 mAb의 항바이러스 및 항암 특성에서 중요한 역할을 한다. 본 개시내용은 시험관 내에서 푸코스를 포함하지 않는 항-HIV mAb의 항-렌티바이러스 세포 매개 바이러스 억제의 증가를 나타내는 최근 연구와 일치한다. 코어 푸코스를 결여하는 균질한 글리칸을 사용한 본 개시내용의 이 구현예는 특정 바이러스에 대한 보호가 2배 넘게 증가하는 것으로 나타났다. 코어 푸코스의 제거는 자연 살해(NK) 세포에 의해 매개되는 mAb의 ADCC 활성을 극적으로 개선하지만, 다형핵 세포(PMN)의 ADCC 활성에는 역효과를 갖는 것으로 보인다.
GNGN 또는 G1/G2 글리코형으로 표시되는 실질적으로 균질한 조성물을 포함하는 단리된 단클론 항체 또는 이의 항원 결합 단편은, 실질적으로 균질한 GNGN 글리코형의 부재 및 G0, G1F, G2F, GNF, GNGNF 또는 GNGNFX 함유 글리코형의 존재 하에 동일한 항체와 비교하여 Fc 감마 RI 및 Fc 감마 RIII에 대한 결합 친화성의 증가를 나타낸다. 본 개시내용의 일 구현예에서, 항체는 1 x 10-8 M 이하의 Kd로 Fc 감마 RI로부터 해리되고, 1 x 10-7 M 이하의 Kd로 Fc 감마 RIII로부터 해리된다.
Fc 영역의 글리코실화는 전형적으로 N-연결 또는 O-연결 중 어느 하나이다. N-연결은 아스파라긴 잔기의 측쇄에 대한 탄수화물 모이어티의 부착을 지칭한다. O-연결된 글리코실화는 당 N-아세틸갈락토사민, 갈락토오스 또는 자일로오스 중 하나가 하이드록시아미노산, 가장 일반적으로는 세린 또는 트레오닌에 부착되는 것을 지칭하지만, 5-하이드록시프롤린 또는 5-하이드록시리신도 사용할 수 있다. 아스파라긴 측쇄 펩티드 서열에 대한 탄수화물 모이어티의 효소적 부착에 대한 인식 서열은 아스파라긴-X-세린 및 아스파라긴-X-트레오닌이고, 여기서 X는 프롤린을 제외한 임의의 아미노산이다. 따라서, 폴리펩티드에서 이들 펩티드 서열 중 하나의 존재는 잠재적 글리코실화 부위를 생성한다.
글리코실화 패턴은, 예를 들어, 폴리펩티드에서 발견되는 하나 이상의 글리코실화 부위(들)를 결실시키고/시키거나, 폴리펩티드에 존재하지 않는 하나 이상의 글리코실화 부위(들)를 부가함으로써 변경될 수 있다. 항체의 Fc 영역에 대한 글리코실화 부위의 부가는 전술한 트리펩티드 서열(N-연결된 글리코실화 부위의 경우) 중 하나 이상을 함유하도록 아미노산 서열을 변경함으로써 편리하게 달성된다. 예시적 글리코실화 변이체는 중쇄의 잔기 Asn 297의 아미노산 치환을 갖는다. 변경은 또한 원래의 폴리펩티드의 서열(O-연결된 글리코실화 부위의 경우)에 하나 이상의 세린 또는 트레오닌 잔기를 부가하거나 이들로 치환함으로써 수행할 수 있다. 추가로, Asn 297을 Ala로 변경하면, 글리코실화 부위 중 하나를 제거할 수 있다.
특정 구현예에서, 항체는 베타 (1,4)-N-아세틸글루코사미닐트랜스퍼라제 III(GnT III)를 발현하는 세포에서 발현되고, 그 결과, GnT III는 GlcNAc를 IL-23p19 항체에 부가한다. 이러한 방식으로 항체를 생산하는 방법은 WO/9954342, WO/03011878, 특허 공개 제20030003097A1호 및 Umana et al., Nature Biotechnology, 17:176-180, February 1999에 제공된다. CRISPR(Clustered Regularly Interspaced Short Palindromic Repeats)와 같은 게놈 편집 기술을 사용하여, 글리코실화와 같은 특정 번역 후 변형을 증진시키거나 감소시키거나 제거하도록 세포주를 변경할 수 있다. 예를 들어, CRISPR 기술을 사용하여, 재조합 단클론 항체의 발현에 사용되는 293 또는 CHO 세포에서 글리코실화 효소를 암호화하는 유전자를 제거할 수 있다.
단클론 항체 단백질 서열의 불리한 점의 제거 인간 B 세포에서 수득된 항체 가변 유전자 서열을 조작하여, 이들의 제조 가능성과 안전성을 증진시키는 것이 가능하다. 잠재적 단백질 서열의 불리한 점은 다음을 포함하는 부위와 관련된 서열 모티프를 검색함으로써 확인할 수 있다:
1)
쌍을 이루지 않은 Cys 잔기,
2)
N 연결된 글리코실화,
3)
Asn 탈아미드화,
4)
Asp 이성질체화,
5)
SYE 절단,
6)
Met 산화,
7)
Trp 산화,
8)
N 말단 글루타메이트,
9)
인테그린 결합,
10)
CD11c/CD18 결합, 또는
11)
단편화
이러한 모티프는 재조합 항체를 암호화하는 cDNA에 대한 합성 유전자를 변경하여 제거할 수 있다.
치료적 항체의 개발 분야에서 단백질을 조작하는 노력은 특정 서열 또는 잔기가 용해도 차이와 관련되어 있음을 분명히 나타낸다(Fernandez-Escamilla et al., Nature Biotech., 22 (10), 1302-1306, 2004; Chennamsetty et al., PNAS, 106 (29), 11937-11942, 2009; Voynov et al., Biocon. Chem., 21 (2), 385-392, 2010). 문헌의 용해도 변경 돌연변이로부터의 증거는 아스파르트산, 글루탐산 및 세린과 같은 몇몇 친수성 잔기가 아스파라긴, 글루타민, 트레오닌, 리신 및 아르기닌과 같은 기타 친수성 잔기보다 현저히 유리하게 단백질 용해도에 기여함을 나타낸다.
안정성. 항체는 증진된 생물물리학적 특성을 위해 조작될 수 있다. 평균 겉보기 용융 온도를 사용하여, 고온을 사용하여 항체를 전개하고 상대적 안정성을 결정할 수 있다. 시차 주사 열량계(DSC)는 분자의 열용량(Cp)(1도당 분자의 가온에 필요한 열)을 온도의 함수로서 측정한다. DSC를 사용하여, 항체의 열 안정성을 연구할 수 있다. mAb에 대한 DSC 데이터는, 종종 mAb 구조 내에서 개별 도메인의 전개를 해결하고, 서모그램에서 최대 3개의 피크(Fab, CH2 및 CH3 도메인의 전개로부터)를 생성하기 때문에 특히 흥미롭다. 전형적으로, Fab 도메인의 전개는 가장 강력한 피크를 생성한다. DSC 프로파일 및 Fc 부분의 상대적 안정성은 인간 IgG1, IgG2, IgG3 및 IgG4 하위 부류에 대한 특징적인 차이를 나타낸다(Garber and Demarest, Biochem. Biophys. Res. Commun. 355, 751-757, 2007). CD 분광기로 수행되는 원형 이색성(CD)을 사용하여, 평균 겉보기 용융 온도를 결정할 수도 있다. Far-UV CD 스펙트럼은 0.5 nm의 증분으로 200 내지 260 nm 범위의 항체에 대해 측정된다. 최종 스펙트럼은 20회 누적의 평균으로 결정할 수 있다. 잔류 타원율 값은 배경을 차감한 후에 계산할 수 있다. 항체의 열적 전개(0.1 mg/mL)는 235 nm에서 25~95℃ 및 1℃/min의 가열 속도로 모니터링할 수 있다. 동적 광 산란(DLS)을 사용하여, 응집 경향을 평가할 수 있다. DLS는 단백질을 포함한 다양한 입자의 크기를 특성화하는 데 사용된다. 시스템의 크기가 분산되지 않은 경우, 입자의 평균 유효 직경을 결정할 수 있다. 이 측정은 입자 코어의 크기, 표면 구조의 크기 및 입자 농도에 따라 달라진다. DLS는 본질적으로 입자에 의한 산란 광 강도의 변동을 측정하므로, 입자의 확산 계수를 결정할 수 있다. 상업용 DLA 기기의 DLS 소프트웨어는 다양한 직경의 입자 집단을 표시한다. 안정성 연구는 DLS를 사용하여 편리하게 수행할 수 있다. 샘플의 DLS 측정은 입자의 유체역학적 반경이 증가하는지를 결정함으로써, 입자가 시간의 경과에 따라 응집되는지 또는 온도 변화에 따라 응집되는지를 나타낼 수 있다. 입자가 응집될 경우, 보다 큰 반경의 입자의 더 큰 집단을 볼 수 있다. 온도에 따른 안정성은 그 자리에서 온도를 제어하여 분석할 수 있다. 모세관 전기영동(CE) 기술에는 항체 안정성의 특징을 결정하기 위한 입증된 방법론이 포함된다. iCE 접근법을 사용하여, 탈아미드화, C 말단 리신, 시알릴화, 산화, 글리코실화 및 단백질의 pI의 변화를 초래할 수 있는 단백질에 대한 임의의 기타 변화로 인한 항체 단백질 전하 변이체를 해결할 수 있다. 발현된 각각의 항체 단백질은, Protein Simple Maurice 기기를 사용하여, 모세관 컬럼(cIEF)에서 고처리량, 유리 용액 등전점 포커싱(IEF)에 의해 평가할 수 있다. 등전점(pI)에 초점을 맞춘 분자를 실시간 모니터링하기 위해, 전체 컬럼 UV 흡수 검출을 30초마다 수행할 수 있다. 이 접근법은 종래의 겔 IEF의 고분해능과 컬럼 기반 분리에서 발견되는 정량 및 자동화의 장점을 결합하는 동시에, 동원 단계의 필요성을 제거한다. 이 기술에 의해, 발현된 항체에 대한 동일성, 순도 및 불균일성 프로파일의 재현 가능한 정량 분석이 가능해진다. 이 결과는, 항체의 전하 불균일성 및 분자 크기를 확인하고, 흡광도 및 천연 형광 검출 모드로 0.7 μg/mL까지의 검출 감도를 갖는다.
용해도. 항체 서열의 고유 용해도 스코어를 결정할 수 있다. 고유 용해도 스코어는 CamSol Intrinsic을 사용하여 계산할 수 있다(Sormanni et al., J Mol Biol 427, 478-490, 2015). scFv와 같은 각 항체 단편의 HCDR3에서 잔기 95~102(Kabat 넘버링)에 대한 아미노산 서열은 용해도 스코어를 계산하기 위한 온라인 프로그램을 통해 평가할 수 있다. 또한, 실험실 기술을 사용하여 용해도를 결정할 수 있다. 용액이 포화되어 용해도 한계에 도달할 때까지 동결건조된 단백질을 용액에 첨가하거나, 적절한 분자량 컷오프를 갖는 미세농축기에서 한외여과에 의한 농축을 포함하는 다양한 기술이 존재한다. 가장 간단한 방법은 무정형 침전을 유도하는 것이고, 이는 황산암모늄을 사용하여 단백질 침전을 포함하는 방법을 사용하여 단백질의 용해도를 측정한다(Trevino et al., J Mol Biol, 366: 449-460, 2007). 황산암모늄 침전은 상대적 용해도 값에 대한 신속하고 정확한 정보를 제공한다. 황산암모늄 침전은 명확한 수성 및 고체상을 구비한 침전 용액을 생성하고, 비교적 소량의 단백질을 필요로 한다. 황산암모늄에 의한 무정형 침전의 유도를 사용하여 수행되는 용해도 측정은 다른 pH 값에서도 용이하게 수행할 수 있다. 단백질의 용해도는 pH에 크게 의존하고, pH는 용해도에 영향을 미치는 가장 중요한 외인성 요인으로 간주된다.
자가반응성. 일반적으로, 자가반응 클론은 개체발생 동안 음성 선택에 의해 제거되어야 한다고 생각되지만, 자가반응 특성을 갖는 다수의 인간 천연 유래 항체가 성인의 성숙한 레퍼토리에 존속되고 자가반응성이 병원체에 대한 다수의 항체의 항바이러스 기능을 증진시킬 수 있다는 것이 명백해졌다. 초기 B 세포 발달 동안 항체의 HCDR3 루프는 종종 양전하가 풍부하고 자가반응성 패턴을 나타내는 것으로 주목되었다(Wardemann et al., Science 301, 1374-1377, 2003). 현미경(부착형 HeLa 또는 HEp-2 상피 세포를 사용) 및 유세포 측정 세포 표면 염색(현탁액 Jurkat T 세포 및 293S 인간 배아 신장 세포를 사용)에서 인간 기원 세포에 대한 결합 수준을 평가함으로써, 주어진 항체의 자가반응성을 시험할 수 있다. 자가반응성은 또한 조직 어레이 내의 조직에 대한 결합 평가를 사용하여 조사할 수 있다.
바람직한 잔기("인간 유사성"). 혈액 공여자의 인간 B 세포에 대한 B 세포 레퍼토리 심층 시퀀싱은 다수의 최근 연구에서 광범위하게 수행되고 있다. 인간 항체 레퍼토리의 상당 부분에 대한 서열 정보는 건강한 인간에서 공통된 항체 서열 특징의 통계적 평가를 용이하게 한다. 인간 재조합 항체 가변 유전자 참조 데이터베이스에서 항체 서열의 특징에 대한 지식에 의해, 항체 서열의 "인간 유사성"(HL)의 위치 특이적 정도를 추정할 수 있다. HL은 치료적 항체 또는 백신으로서의 항체 등의 임상 용도에서 항체 개발에 유용한 것으로 나타났다. 목표는 항체의 인간 유사성을 증가시켜, 항체 약물의 효능을 현저히 감소시키거나, 심각한 건강 영향을 유발할 수 있는 잠재적 부작용 및 항-항체 면역 반응을 감소시키는 것이다. 총 약 4억개 서열의 3명의 건강한 인간 혈액 공여자의 조합된 항체 레퍼토리의 항체 특성을 평가하고, 항체의 초가변 영역에 초점을 맞춘 새로운 "상대적 인간 유사성"(rHL) 스코어를 작성할 수 있다. rHL 스코어를 통해, 인간(양성 스코어)과 비인간 서열(음성 스코어)을 용이하게 구별할 수 있다. 항체는 인간 레퍼토리에서 일반적이지 않은 잔기를 제거하도록 조작될 수 있다.
D.
단쇄 항체
단쇄 가변 단편(scFv)은 면역글로불린의 중쇄 및 경쇄 가변 영역의 융합체로, 짧은(보통 세린, 글리신) 링커로 함께 연결된다. 이 키메라 분자는 불변 영역의 제거와 링커 펩티드의 도입에도 불구하고, 원래 면역글로불린의 특이성을 유지한다. 이 변형은 일반적으로 특이성을 변경시키지 않는다. 이러한 분자는 항원 결합 도메인을 단일 펩티드로서 발현시키는 것이 매우 편리한 파지 디스플레이를 용이하게 하기 위해 역사적으로 생성되었다. 대안적으로, scFv는 하이브리도마 또는 B 세포로부터 유래하는 서브클로닝된 중쇄 및 경쇄로부터 직접 생성될 수 있다. 단쇄 가변 단편은 완전한 항체 분자에서 발견되는 불변 Fc 영역을 결여하고 있으므로, 항체의 정제에 사용되는 공통 결합 부위(예를 들어, 단백질 A/G)가 없다. 단백질 L은 카파 경쇄의 가변 영역과 상호작용하기 때문에, 이러한 단편은 종종 단백질 L을 사용하여 정제/고정될 수 있다.
가요성 링커는 일반적으로 알라닌, 세린 및 글리신과 같은 나선 및 턴 촉진 아미노산 잔기로 구성된다. 그러나 다른 잔기도 기능할 수 있다. Tang et al. (1996)은 단백질 링커 라이브러리로부터 단쇄 항체(scFv)를 위해 조정된 링커를 신속하게 선택하는 수단으로서 파지 디스플레이를 사용했다. 중쇄 및 경쇄 가변 도메인에 대한 유전자가 가변 조성의 18-아미노산 폴리펩티드를 암호화하는 세그먼트에 의해 연결된 랜덤 링커 라이브러리를 구축했다. scFv 레퍼토리(약 5 x 106개의 상이한 구성원)를 필라멘트 파지에 표시하고, 합텐과의 친화성 선택을 수행했다. 선택된 변이체의 집단은 결합 활성에서 유의한 증가를 나타냈지만, 상당한 서열 다양성을 유지했다. 이어서, 1054개의 개별 변이체를 스크리닝하자, 가용성 형태로 효율적으로 생성된 촉매 활성 scFv가 수득되었다. 서열 분석은 VH C 말단의 2개 잔기 후의 링커에 보존된 프롤린과, 선택된 테더의 유일한 공통 특징으로서 다른 위치에서 풍부한 아르기닌 및 프롤린을 나타냈다.
본 개시내용의 재조합 항체는 또한 수용체의 이량체화 또는 다량체화를 가능하게 하는 서열 또는 모이어티를 포함할 수 있다. 이러한 서열은 J 쇄와 조합하여 다량체의 형성을 가능하게 하는 IgA로부터 유래된 서열을 포함한다. 또 다른 다량체화 도메인은 Gal4 이량체화 도메인이다. 다른 구현예에서, 쇄는 2개의 항체의 조합을 가능하게 하는 비오틴/아비딘과 같은 작용제로 변형시킬 수 있다.
별도의 구현예에서, 단쇄 항체는 비펩티드 링커 또는 화학 단위를 사용하여 수용체의 경쇄 및 중쇄를 연결함으로써 생성될 수 있다. 일반적으로, 경쇄 및 중쇄는 별개의 세포에서 생성되고, 정제되고, 후속적으로 적절한 방식으로 함께 연결될 것이다(즉, 중쇄의 N 말단이 적절한 화학 가교를 통해 경쇄의 C 말단에 부착됨).
가교 시약은 2개의 상이한 분자의 작용기를 연결하는 분자 가교를 형성하기 위해 사용된다. 예를 들어, 안정화제 및 응고제. 그러나 동일한 유사체의 이량체 또는 다량체 또는 상이한 유사체로 구성된 헤테로머 복합체가 생성될 수 있는 것으로 고려된다. 2개의 상이한 화합물을 단계적으로 연결하기 위해, 원치 않는 단독중합체의 형성을 제거하는 헤테로-이작용성 가교제를 사용할 수 있다.
예시적 헤테로-이작용성 가교제는 2개의 반응성 기를 함유한다: 1급 아민 기와 반응하는 반응성 기(예를 들어, N-하이드록시 석신이미드)와 티올 기와 반응하는 반응성 기(예를 들어, 피리딜 디설파이드, 말레이미드, 할로겐 등). 1급 아민 반응성 기를 통해, 가교제는 하나의 단백질(예를 들어, 선택된 항체 또는 단편)의 리신 잔기(들)와 반응할 수 있고, 티올 반응성 기를 통해, 이미 제1 단백질에 결합된 가교제는 다른 단백질(예를 들어, 선택제)의 시스테인 잔기(유리 설프하이드릴 기)와 반응한다.
혈액 내에서 적당한 안정성을 갖는 가교제를 사용하는 것이 바람직하다. 표적화제 및 치료제/예방제를 접합하기 위해 성공적으로 사용될 수 있는 다수 유형의 이황화 결합 함유 링커가 공지되어 있다. 입체 장애가 있는 이황화 결합을 포함하는 링커는 생체 내에서 보다 큰 안정성을 제공하여, 작용 부위에 도달하기 전에 표적화 펩티드의 방출을 방지하는 것을 증명할 수 있다. 따라서, 이러한 링커는 연결제의 하나의 군이다.
또 다른 가교제는 SMPT로서, 이는 인접한 벤젠 고리 및 메틸 기에 의해 "입체 장애"되는 이황화 결합을 포함하는 이작용성 가교제이다. 이황화 결합의 입체 장애는 조직 및 혈액에 존재할 수 있는 글루타티온과 같은 티올레이트 음이온에 의한 공격으로부터 결합을 보호하는 기능을 하고, 이에 의해 표적 부위로 결합물의 전달 전에 접합체의 디커플링을 방지하는 데 도움을 주는 것으로 여겨진다.
SMPT 가교 시약은, 기타 다수의 공지된 가교 시약과 마찬가지로, 시스테인의 SH 또는 1급 아민(예를 들어, 리신의 엡실론 아미노 기)과 같은 작용기를 가교하는 능력을 제공한다. 다른 가능한 유형의 가교제는 설포석신이미딜-2-(p-아지도 살리실아미도) 에틸-1,3'-디티오프로피오네이트와 같은 절단 가능한 이황화 결합을 함유하는 헤테로-이작용성 광반응성 페닐아지드를 포함한다. N-하이드록시-석신이미딜 기는 1급 아미노 기와 반응하고, 페닐아지드(광분해 시)는 임의의 아미노산 잔기와 비선택적으로 반응한다.
장애된 가교제에 추가하여, 장애되지 않은 링커도 또한 본원에 따라 사용될 수 있다. 보호된 이황화물을 함유하거나 생성하는 것으로 간주되지 않는 기타 유용한 가교제에는 SATA, SPDP 및 2-이미노티올란이 포함된다(Wawrzynczak & Thorpe, 1987). 이러한 가교제의 사용은 당해 분야에서 잘 이해되어 있다. 또 다른 구현예는 가요성 링커의 사용을 포함한다.
미국 특허 제4,680,338호는 아민 함유 중합체 및/또는 단백질과 리간드의 접합체를 생성하기 위해, 특히 킬레이트제, 약물, 효소, 검출 가능한 표지 등과 항체 접합체를 형성하는 데 유용한 이작용성 링커를 기재한다. 미국 특허 제5,141,648호 및 제5,563,250호는 다양한 온화한 조건 하에서 절단 가능한 불안정한 결합을 함유하는 절단 가능한 접합체를 개시하고 있다. 이 링커는 관심 작용제가 링커에 직접 결합될 수 있고, 절단되어 활성제가 방출될 수 있다는 점에서 특히 유용하다. 특정 용도에는 항체 또는 약물과 같은 단백질에 유리 아미노 또는 유리 설프하이드릴 기를 부가하는 것이 포함된다.
미국 특허 제5,856,456호는 폴리펩티드 구성요소를 연결하여 융합 단백질, 예를 들어, 단쇄 항체를 제조하는 데 사용하기 위한 펩티드 링커를 제공한다. 링커는 최대 약 50개 아미노산 길이이고, 하전된 아미노산(바람직하게는 아르기닌 또는 리신)에 이어 프롤린의 적어도 하나의 존재를 포함하고, 보다 큰 안정성과 감소된 응집을 특징으로 한다. 미국 특허 제5,880,270호는 다양한 면역진단 및 분리 기술에 유용한 아미노옥시 함유 링커를 개시하고 있다.
E.
다중특이적 항체
특정 구현예에서, 본 개시내용의 항체는 이중특이적 또는 다중특이적이다. 이중특이적 항체는 적어도 2개의 상이한 에피토프에 대한 결합 특이성을 갖는 항체이다. 예시적인 이중특이적 항체는 단일 항원의 2개의 상이한 에피토프에 결합할 수 있다. 기타 이러한 항체는 제1 항원 결합 부위를 제2 항원에 대한 결합 부위와 조합할 수 있다. 또는, 항-병원체 암(arm)은 T 세포 수용체 분자(예를 들어, CD3)와 같은 백혈구 상의 유발 분자 또는 FcγRI(CD64), FcγRII(CD32) 및 Fc 감마 RIII(CD16)과 같은 IgG의 Fc 수용체(FcγR)에 결합하는 암과 조합시킬 수 있고, 이에 의해 세포 방어 메커니즘을 감염된 세포에 집중시키고 국재화시킨다. 또한, 이중특이적 항체를 사용하여, 세포독성제를 감염된 세포에 국재화시킬 수 있다. 이러한 항체는 병원체 결합 암과 세포독성제(예를 들어, 사포린, 항-인터페론-α, 빈카 알칼로이드, 리신 A 쇄, 메토트렉세이트 또는 방사성 동위원소 합텐)에 결합하는 암을 갖고 있다. 이중특이적 항체는 전장 항체 또는 항체 단편(예를 들어, F(ab')2 이중특이적 항체)으로 제조될 수 있다. 제WO96/16673호는 이중특이적 항-ErbB2/항-Fc 감마 RIII 항체를 기재하고, 미국 특허 제5,837,234호는 이중특이적 항-ErbB2/항-Fc 감마 RI 항체를 개시하고 있다. 이중특이적 항-ErbB2/Fc 알파 항체는 제WO98/02463호에 제시되어 있다. 미국 특허 제5,821,337호는 이중특이적 항-ErbB2/항-CD3 항체를 교시하고 있다.
이중특이적 항체의 제조 방법은 당해 분야에 공지되어 있다. 전장 이중특이적 항체의 종래의 생산은 2개의 면역글로불린 중쇄-경쇄 쌍의 공동 발현을 기반으로 하고, 여기서 2개의 쇄는 상이한 특이성을 갖는다(Millstein et al., Nature, 305:537-539 (1983)). 면역글로불린 중쇄 및 경쇄는 랜덤으로 분류되어 있기 때문에, 이러한 하이브리도마(쿼드로마)는 10개의 상이한 항체 분자의 잠재적 혼합물을 생성하고, 이 중 하나만이 정확한 이중특이적 구조를 갖는다. 일반적으로 친화성 크로마토그래피 단계에 의해 수행되는 정확한 분자의 정제는 다소 번거롭고, 생성물의 수율이 낮다. 유사한 절차는 제WO 93/08829호 및 Traunecker et al., EMBO J., 10:3655-3659 (1991)에 개시되어 있다.
다른 접근법에 따르면, 원하는 결합 특이성을 갖는 항체 가변 영역(항체-항원 결합 부위)을 면역글로불린 불변 도메인 서열에 융합시킨다. 바람직하게는, 융합은 힌지, CH2 및 CH3 영역의 적어도 일부를 포함하는 Ig 중쇄 불변 도메인과의 융합이다. 적어도 하나의 융합에 존재하는, 경쇄 결합에 필요한 부위를 포함하는 제1 중쇄 불변 영역(CH1)을 갖는 것이 바람직하다. 면역글로불린 중쇄 융합물 및 원하는 경우, 면역글로불린 경쇄를 암호화하는 DNA는 개별 발현 벡터에 삽입되고, 적합한 숙주 세포에 동시 형질감염된다. 이것은, 구성에 사용된 3개의 폴리펩티드 쇄의 비균등한 비율이 원하는 이중특이적 항체의 최적 수율을 제공하는 경우, 구현예에서 3개의 폴리펩티드 단편의 상호 비율을 조정하는 데 있어서 보다 큰 유연성을 제공한다. 그러나 동일한 비율의 적어도 2개의 폴리펩티드 쇄의 발현이 높은 수율을 초래하거나 비율이 원하는 쇄 조합의 수율에 유의한 영향을 미치지 않는 경우, 2개 또는 3개의 모든 폴리펩티드 쇄에 대한 암호화 서열을 단일 발현 벡터에 삽입하는 것이 가능하다.
이 접근법의 특정 구현예에서, 이중특이적 항체는 한쪽 암에 제1 결합 특이성을 갖는 하이브리드 면역글로불린 중쇄 및 다른 쪽 암에 하이브리드 면역글로불린 중쇄-경쇄 쌍(제2 결합 특이성을 제공함)으로 구성된다. 이중특이적 분자의 절반에만 면역글로불린 경쇄가 존재함으로써 용이한 분리 방법이 제공되기 때문에, 이러한 비대칭 구조는 원치 않는 면역글로불린 쇄 조합으로부터 원하는 이중특이적 화합물의 분리를 용이하게 하는 것으로 밝혀졌다. 이 접근법은 제WO 94/04690호에 개시되어 있다. 이중특이적 항체의 생성에 대한 추가 상세 내용은 예를 들어, Suresh et al., Methods in Enzymology, 121:210 (1986)을 참고한다.
미국 특허 제5,731,168호에 기재된 또 다른 접근법에 따르면, 항체 분자의 쌍 사이의 계면은 재조합 세포 배양으로부터 회수되는 헤테로이량체의 백분율을 최대화하도록 조작할 수 있다. 바람직한 계면은 CH3 도메인의 적어도 일부를 포함한다. 이 방법에서, 제1 항체 분자의 계면으로부터 하나 이상의 작은 아미노산 측쇄가 보다 큰 측쇄(예를 들어, 티로신 또는 트립토판)로 대체된다. 큰 아미노산 측쇄를 더 작은 아미노산 측쇄(예를 들어, 알라닌 또는 트레오닌)로 대체함으로써 제2 항체 분자의 계면에 큰 측쇄(들)와 동일하거나 유사한 크기의 상보적 "공동(cavities)"이 생성된다. 이것은 호모이량체와 같은 다른 원치 않는 최종 생성물보다 헤테로이량체의 수율을 증가시키는 메커니즘을 제공한다.
이중특이적 항체에는 가교 또는 "헤테로접합체" 항체가 포함된다. 예를 들어, 헤테로접합체의 항체 중 하나는 아비딘에 커플링되고, 다른 하나는 비오틴에 커플링될 수 있다. 이러한 항체는, 예를 들어, 면역계 세포를 원치 않는 세포로 표적화하기 위해(미국 특허 제4,676,980호), 및 HIV 감염의 치료를 위해(제WO 91/00360호, 제WO 92/200373호 및 제EP 03089호) 제안되었다. 헤테로접합체 항체는 임의의 편리한 가교 방법을 사용하여 제조할 수 있다. 적합한 가교제는 당해 분야에 공지되어 있고, 다수의 가교 기술과 함께 미국 특허 제4,676,980호에 개시되어 있다.
항체 단편으로부터 이중특이적 항체를 생성하는 기술도 문헌에 기재되어 있다. 예를 들어, 이중특이적 항체는 화학적 연결을 사용하여 제조할 수 있다. Brennan et al., Science, 229: 81 (1985)은 온전한 항체를 단백질 가수분해로 절단하여 F(ab')2 단편을 생성하는 절차를 기재하고 있다. 이러한 단편은 디티올 착화제인 나트륨 아르세나이트의 존재 하에서 환원되어, 인접 디티올을 안정화하고 분자간 이황화물 형성을 방지한다. 이어서, 생성된 Fab' 단편을 티오니트로벤조에이트(TNB) 유도체로 전환시킨다. 이어서, Fab'-TNB 유도체 중 하나는 메르캅토에틸아민에 의한 환원에 의해 Fab'-티올로 재전환되고, 등몰량의 기타 Fab'-TNB 유도체와 혼합되어 이중특이적 항체를 형성한다. 생성된 이중특이적 항체는 효소의 선택적 고정화를 위한 작용제로 사용될 수 있다.
E. 콜라이로부터 Fab'-SH 단편의 직접적 회수를 촉진하는 기술이 존재하고, 이는 화학적으로 커플링하여 이중특이적 항체를 형성할 수 있다. Shalaby et al., J. Exp. Med., 175: 217-225 (1992)는 인간화 이중특이적 항체 F(ab')2 분자의 생산을 기재하고 있다. 각각의 Fab' 단편은 E. 콜라이로부터 별도로 분비되었고, 시험관 내에서 직접 화학적 커플링을 거쳐 이중특이적 항체를 형성했다. 이렇게 형성된 이중특이적 항체는 ErbB2 수용체를 과발현하는 세포 및 정상 인간 T 세포에 결합할 수 있을 뿐만 아니라, 인간 유방 종양 표적에 대한 인간 세포독성 림프구의 용해 활성을 유발할 수 있다.
재조합 세포 배양물로부터 직접 이중특이적 항체 단편을 제조하고 단리하기 위한 다양한 기술이 또한 기재되어 있다(Merchant et al., Nat. Biotechnol . 16, 677-681 (1998). doi:10.1038/nbt0798-677pmid:9661204). 예를 들어, 이중특이적 항체는 류신 지퍼를 사용하여 생산되었다(Kostelny et al., J. Immunol., 148(5):1547-1553, 1992). Fos 및 Jun 단백질로부터의 류신 지퍼 펩티드는 유전자 융합에 의해 2개의 상이한 항체의 Fab' 부분에 연결되었다. 항체 호모이량체는 힌지 영역에서 환원되어 단량체를 형성하고, 이어서 재산화되어 항체 헤테로이량체를 형성했다. 이 방법은 또한 항체 호모이량체의 생산에도 이용될 수 있다. Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993)에 기재된 "디아바디" 기술은 이중특이적 항체 단편을 제조하기 위한 대체 메커니즘을 제공했다. 단편은, 동일한 쇄의 2개 도메인 사이의 쌍 형성을 가능하게 하기에는 너무 짧은 링커에 의해 VL에 연결된 VH를 포함한다. 따라서, 하나의 단편의 VH 및 VL 도메인은 또 다른 단편의 상보적 VL 및 VH 도메인과 강제적으로 쌍을 형성하게 되고, 이에 의해 2개의 항원 결합 부위가 형성된다. 단쇄 Fv(sFv) 이량체를 사용하여 이중특이적 항체 단편을 제조하는 또 다른 전략도 보고되었다. Gruber et al., J. Immunol., 152:5368 (1994) 참고.
특정 구현예에서, 이중특이적 또는 다중특이적 항체는 DOCK-AND-LOCK™(DNL™) 복합체로서 형성될 수 있다(예를 들어, 미국 특허 제7,521,056호; 제7,527,787호; 제7,534,866호; 제7,550,143호 및 제7,666,400호 참고, 이들 각각의 실시예 섹션은 참조에 의해 본원에 포함됨). 일반적으로, 이 기술은 cAMP 의존성 단백질 키나제(PKA)의 조절(R) 하위 단위의 이량체화 및 도킹 도메인(DDD) 서열과, 임의의 다양한 AKAP 단백질로부터 유래하는 앵커 도메인(AD) 서열 사이에서 발생하는 특이적이고 고친화성의 결합 상호작용을 이용한다(Baillie et al., FEBS Letters. 2005; 579: 3264; Wong and Scott, Nat. Rev. Mol. Cell Biol. 2004; 5: 959). DDD 및 AD 펩티드는 임의의 단백질, 펩티드 또는 기타 분자에 부착될 수 있다. DDD 서열이 자발적으로 이량체화되어 AD 서열에 결합하기 때문에, 이 기술은 DDD 또는 AD 서열에 부착될 수 있는 임의의 선택된 분자 사이의 복합체의 형성을 가능하게 한다.
2가 이상의 원자가를 갖는 항체가 고려된다. 예를 들어, 삼중특이적 항체가 제조될 수 있다(Tutt et al., J. Immunol. 147: 60, 1991; Xu et al., Science, 358(6359):85-90, 2017). 다가 항체는 항체가 결합하는 항원을 발현하는 세포에 의해 2가 항체보다 신속하게 내재화(및/또는 이화)될 수 있다. 본 개시내용의 항체는 3개 이상의 항원 결합 부위를 갖는 다가 항체(예를 들어, 4가 항체)일 수 있고, 이는 항체의 폴리펩티드 쇄를 암호화하는 핵산의 재조합 발현에 의해 용이하게 생성될 수 있다. 다가 항체는 이량체화 도메인 및 3개 이상의 항원 결합 부위를 포함할 수 있다. 바람직한 이량체화 도메인은 Fc 영역 또는 힌지 영역을 포함한다(또는 이로 구성된다). 이 시나리오에서, 항체는 Fc 영역, 및 Fc 영역에 대한 아미노 말단의 3개 이상의 항원 결합 부위를 포함할 것이다. 본원에서 바람직한 다가 항체는 3개 내지 약 8개, 그러나 바람직하게는 4개의 항원 결합 부위를 포함한다(또는 이들로 구성된다). 다가 항체는 적어도 하나의 폴리펩티드 쇄(및 바람직하게는 2개의 폴리펩티드 쇄)를 포함하고, 여기서 폴리펩티드 쇄(들)는 2개 이상의 가변 영역을 포함한다. 예를 들어, 폴리펩티드 쇄(들)는 VD1-(X1)n-VD2-(X2)n-Fc를 포함할 수 있고, 여기서 VD1은 제1 가변 영역이고, VD2는 제2 가변 영역이고, Fc는 Fc 영역의 하나의 폴리펩티드 쇄이고, X1 및 X2는 아미노산 또는 폴리펩티드를 나타내고, n은 0 또는 1이다. 예를 들어, 폴리펩티드 쇄(들)는 다음을 포함할 수 있다: VH-CH1-가요성 링커-VH-CH1-Fc 영역 쇄; 또는 VH-CH1-VH-CH1-Fc 영역 쇄. 본원에서 다가 항체는 바람직하게는 적어도 2개(및 바람직하게는 4개)의 경쇄 가변 영역 폴리펩티드를 추가로 포함한다. 본원에서 다가 항체는 예를 들어, 약 2 내지 약 8개의 경쇄 가변 영역 폴리펩티드를 포함할 수 있다. 본원에서 고려되는 경쇄 가변 영역 폴리펩티드는 경쇄 가변 영역을 포함하고, 선택적으로 CL 도메인을 추가로 포함한다.
전하 변형은 다중특이적 항체의 맥락에서 특히 유용하고, 여기서 Fab 분자의 아미노산 치환에 의해 경쇄와 비매칭 중쇄와의 미스매칭(Bence-Jones 유형 부산물)이 감소하고, 이는 결합 암 중 하나(또는 2개 초과의 항원 결합 Fab 분자를 포함하는 분자의 경우에는 하나 이상)에서 VH/VL 교환을 갖는 Fab 기반 이중/다중특이적 항원 결합 분자의 생성에서 발생할 수 있다(PCT 공개 번호 제WO 2015/150447호 참고, 특히 그 안의 실시예는 참조로서 그 전체가 본원에 포함됨).
따라서, 특정 구현예에서, 치료제에 포함된 항체는
(a) 제1 항원에 특이적으로 결합하는 제1 Fab 분자
(b) 제2 항원에 특이적으로 결합하는 제2 Fab 분자를 포함하고, 여기서 Fab 경쇄 및 Fab 중쇄의 가변 도메인 VL 및 VH는 서로 대체되고,
제1 항원은 활성화 T 세포 항원이고 제2 항원은 표적 세포 항원이거나, 또는 제1 항원은 표적 세포 항원이고 제2 항원은 활성화 T 세포 항원이고;
여기서,
i) a) 하의 제1 Fab 분자의 불변 도메인 CL에서는, 위치 124의 아미노산이 양으로 하전된 아미노산(Kabat에 따른 넘버링)으로 치환되고, a) 하의 제1 Fab 분자의 불변 도메인 CH1에서는, 위치 147의 아미노산 또는 위치 213의 아미노산이 음으로 하전된 아미노산(Kabat EU 인덱스에 따른 넘버링)으로 치환되거나; 또는
ii) b) 하의 제2 Fab 분자의 불변 도메인 CL에서는, 위치 124의 아미노산이 양으로 하전된 아미노산(Kabat에 따른 넘버링)으로 치환되고, b) 하의 제2 Fab 분자의 불변 도메인 CH1에서는, 위치 147의 아미노산 또는 위치 213의 아미노산이 음으로 하전된 아미노산(Kabat EU 인덱스에 따른 넘버링)으로 치환된다.
항체는 i) 및 ii) 하에 언급된 변형을 모두 포함하지 않을 수 있다. 제2 Fab 분자의 불변 도메인 CL 및 CH1은 서로 대체되지 않는다(즉, 교환되지 않은 상태로 유지됨).
항체의 또 다른 구현예에서, a) 하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K), 아르기닌(R) 또는 히스티딘(H)(Kabat에 따른 넘버링)에 의해 독립적으로 (한 가지 바람직한 구현예에서, 리신(K) 또는 아르기닌(R)에 의해 독립적으로) 치환되고, a) 하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산 또는 위치 213의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(Kabat EU 인덱스에 따른 넘버링)에 의해 독립적으로 치환된다.
추가 구현예에서, a) 하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K), 아르기닌(R) 또는 히스티딘(H)(Kabat에 따른 넘버링)에 의해 독립적으로 치환되고, a) 하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(Kabat EU 인덱스에 따른 넘버링)에 의해 독립적으로 치환된다.
특정 구현예에서, a) 하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K), 아르기닌(R) 또는 히스티딘(H)(Kabat에 따른 넘버링)에 의해 독립적으로(한 가지 바람직한 구현예에서, 리신(K) 또는 아르기닌(R)에 의해 독립적으로) 치환되고, 위치 123의 아미노산은 리신(K), 아르기닌(R) 또는 히스티딘(H)(Kabat에 따른 넘버링)에 의해 독립적으로(바람직한 일 구현예에서, 리신(K) 또는 아르기닌(R)에 의해 독립적으로) 치환되고, a) 하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(Kabat EU 인덱스에 따른 넘버링)에 의해 독립적으로 치환되고, 위치 213의 아미노산은 글루탐산(E) 또는 아스파르트산(D)(Kabat EU 인덱스에 따른 넘버링)에 의해 독립적으로 치환된다.
더욱 특별한 구현예에서, a) 하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K)(Kabat에 따른 넘버링)에 의해 치환되고, 위치 123의 아미노산은 리신(K) 또는 아르기닌(R)(Kabat에 따른 넘버링)에 의해 치환되고, a) 하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E)(Kabat EU 인덱스에 따른 넘버링)에 의해 치환되고, 위치 213의 아미노산은 글루탐산(E)(Kabat EU 인덱스에 따른 넘버링)에 의해 치환된다.
훨씬 더 특별한 구현예에서, a) 하의 제1 Fab 분자의 불변 도메인 CL에서, 위치 124의 아미노산은 리신(K)(Kabat에 따른 넘버링)에 의해 치환되고, 위치 123의 아미노산은 아르기닌(R)(Kabat에 따른 넘버링)에 의해 치환되고, a) 하의 제1 Fab 분자의 불변 도메인 CH1에서, 위치 147의 아미노산은 글루탐산(E)(Kabat EU 인덱스에 따른 넘버링)에 의해 치환되고, 위치 213의 아미노산은 글루탐산(E)(Kabat EU 인덱스에 따른 넘버링)에 의해 치환된다.
F.
키메라 항원 수용체
인공 T 세포 수용체(키메라 T 세포 수용체, 키메라 면역수용체, 키메라 항원 수용체(CAR)라고도 알려짐)는 면역 이펙터 세포에 임의의 특이성을 이식하는 조작된 수용체이다. 전형적으로, 이러한 수용체는 단클론 항체의 특이성을 T 세포에 이식하기 위해 사용되고, 레트로바이러스 벡터에 의해 암호화 서열의 전달이 촉진된다. 이러한 방식으로, 입양 세포 전달을 위해, 다수의 표적 특이적 T 세포가 생성될 수 있다. 이 접근법의 1상 임상 연구는 효능을 나타낸다.
이러한 분자의 가장 일반적 형태는 단클론 항체로부터 유래하는 단쇄 가변 단편(scFv)의 융합체이고, CD3-제타 막관통 및 엔도도메인에 융합된다. 이러한 분자는 이의 표적의 scFv에 의한 인식에 응하여 제타 신호의 전달을 초래한다. 이러한 구성체의 예는 하이브리도마 14g2a(디시알로강글리오사이드 GD2를 인식함)로부터 유래하는 scFv의 융합체인 14g2a-제타이다. T 세포가 이 분자를 발현하면(전형적으로 온코레트로바이러스 벡터 형질도입에 의해 달성됨), GD2를 발현하는 표적 세포(예를 들어, 신경모세포종 세포)를 인식하고 사멸시킨다. 악성 B 세포를 표적화하기 위해, 연구자들은 B 계통 분자 CD19에 특이적인 키메라 면역수용체를 사용하여 T 세포의 특이성을 재지시했다.
면역글로불린 중쇄 및 경쇄의 가변 부분은 가요성 링커에 의해 융합되어 scFv를 형성한다. 이 scFv의 전에 신호 펩티드가 있고, 신생 단백질을 소포체로 지시하고, 후속 표면 발현을 지시한다(이는 절단됨). 가요성 스페이서에 의해, scFv를 상이한 방향으로 배향시켜 항원 결합을 가능하게 한다. 막관통 도메인은 세포 내로 돌출되어 원하는 신호를 전달하는 신호전달 엔도도메인의 원래 분자로부터 일반적으로 유래하는 전형적 소수성 알파 나선이다.
유형 I 단백질은 실제로 그 사이에 막관통 알파 나선에 의해 연결된 2개의 단백질 도메인이다. 막관통 도메인이 통과하는 세포막 지질 이중층은 내측 부분(엔도도메인)을 외측 부분(엑토도메인)으로부터 분리하도록 작용한다. 한 단백질의 엑토도메인을 또 다른 단백질의 엔도도메인에 부착하여 전자의 인식을 후자의 신호와 조합시킨 분자를 수득하는 점은 그다지 놀랍지 않다.
엑토도메인. 신호 펩티드는 신생 단백질을 소포체로 지시한다. 이것은 수용체가 글리코실화되고 세포막에 고정되는 경우에 필수적이다. 모든 진핵생물 신호 펩티드 서열은 일반적으로 정상으로 작동한다. 일반적으로, 아미노 말단의 대부분 성분에 자연적으로 부착된 신호 펩티드가 사용된다(예를 들어, 경쇄-링커-중쇄의 방향을 갖는 scFv에서는 경쇄의 천연 신호가 사용된다).
항원 인식 도메인은 일반적으로 scFv이다. 그러나 다수의 대안이 있다. 천연 T 세포 수용체(TCR) 알파 및 베타 단쇄로부터의 항원 인식 도메인은, 단순한 엑토도메인(예를 들어, HIV 감염 세포를 인식하는 CD4 엑토도메인) 및 연결된 사이토카인과 같은 더 색다른 인식 성분(이는 사이토카인 수용체를 갖는 세포의 인식을 유도함)을 갖는 것으로 기재되어 있다. 사실, 주어진 표적에 높은 친화성으로 결합하는 거의 모든 것을 항원 인식 영역으로 사용할 수 있다.
스페이서 영역은 항원 결합 도메인을 막관통 도메인에 연결한다. 이는 항원 인식을 용이하게 하기 위해, 항원 결합 도메인이 상이한 방향으로 배향될 수 있도록 충분히 유연해야 한다. 가장 단순한 형태는 IgG1의 힌지 영역이다. 대안으로는 면역글로불린의 CH2CH3 영역과 CD3의 일부가 포함된다. 대부분의 scFv 기반 구성체에서는 IgG1 힌지로 충분하다. 그러나 최선의 스페이서는 종종 경험적으로 결정되어야 한다.
막관통 도메인. 막관통 도메인은 막에 걸쳐 있는 소수성 알파 나선이다. 일반적으로, 엔도도메인의 가장 막 근위의 구성요소로부터의 막관통 도메인이 사용된다. 흥미롭게도, CD3-제타 막관통 도메인을 사용하면, 인공 TCR을 천연 TCR에 도입할 수 있고, 이는 천연 CD3-제타 막관통 하전 아스파르트산 잔기의 존재에 의존하는 요인이다. 상이한 막관통 도메인은 상이한 수용체의 안정성을 초래한다. CD28 막관통 도메인은 밝게 표시된 안정한 수용체를 초래한다.
엔도도메인. 이것은 수용체의 "가장 말단"이다. 항원 인식 후, 수용체가 클러스터링되고, 신호가 세포로 전달된다. 가장 일반적으로 사용되는 엔도도메인 성분은 3개의 ITAM을 포함하는 CD3-제타이다. 이것은, 항원이 결합된 후, 활성화 신호를 T 세포에 전달한다. CD3-제타는 완전히 유능한 활성화 신호를 제공하지 않을 수 있고, 추가 공동자극 신호가 필요하다.
"1세대" CAR은 전형적으로 내인성 TCR로부터의 신호의 주요 전달물질인 CD3 ξ-쇄로부터의 세포내 도메인을 가졌다. "2세대" CAR은 다양한 공동자극 단백질 수용체(예를 들어, CD28, 41BB, ICOS)로부터의 세포내 신호전달 도메인을 CAR의 세포질 꼬리에 부가하여, T 세포에 추가 신호를 제공한다. 전임상 연구에 따르면, 2세대 CAR 설계가 T 세포의 항종양 활성을 개선하는 것으로 나타났다. 보다 최근에는, "3세대" CAR이 CD3z-CD28-41BB 또는 CD3z-CD28-OX40과 같은 복수의 신호전달 도메인을 조합하여 효능을 추가로 향상시킨다.
G.
ADC
항체 약물 접합체 또는 ADC는 감염성 질환을 앓는 사람들의 치료를 위한 표적 요법으로서 설계된 새로운 부류의 매우 강력한 생물의제이다. ADC는 항체(전체 mAb 또는 항체 단편, 예컨대, 단쇄 가변 단편 또는 scFv)로 구성되는 복잡한 분자이고, 불안정한 결합을 갖는 안정한 화학적 링커를 통해 생물학적 활성 세포독성/항바이러스 페이로드 또는 약물에 연결된다. 항체 약물 접합체는 생체접합체 및 면역접합체의 예이다.
단클론 항체의 독자의 표적화 능력과 세포독성 약물의 암 사멸 능력을 조합함으로써, 항체-약물 접합체는 건강한 조직과 발병 조직을 민감하게 구별할 수 있다. 이는, 종래의 전신 접근법과 대조적으로, 항체-약물 접합체가 감염된 세포를 표적으로 하여 공격하기 때문에, 건강한 세포가 덜 심각하게 영향을 받는다는 것을 의미한다.
ADC 기반 항종양 요법의 개발에서, 항암제(예를 들어, 세포 독소 또는 세포독)를, 특정 세포 마커를 특이적으로 표적화하는 항체(예를 들어, 이상적으로는, 감염된 세포 내에서 또는 감염된 세포 상에서만 발견되는 단백질)에 커플링된다. 항체는 이러한 단백질을 체내에서 추적하여 암 세포의 표면에 달라붙는다. 항체와 표적 단백질(항원) 사이의 생화학적 반응은 종양 세포에서 신호를 촉발하고, 이것이 세포독소와 함께 항체를 흡수하거나 내재화한다. ADC가 내재화된 후, 세포독성 약물이 방출되어 세포를 사멸하거나, 바이러스 복제를 손상시킨다. 이러한 표적화로 인해, 이상적으로는, 이 약물은 다른 작용제보다 부작용이 적고 치료 창이 더 넓어진다.
항체와 세포독성제/항바이러스제 사이의 안정한 연결은 ADC의 중요한 측면이다. 링커는 이황화물, 하이드라존 또는 펩티드(절단 가능) 또는 티오에테르(절단 불가능)를 포함한 화학적 모티프를 기반으로 하고, 세포독성제의 표적 세포로의 분포 및 전달을 제어한다. 절단 가능 및 절단 불가능 유형의 링커는 전임상 및 임상 시험에서 안전한 것으로 입증되었다. 브렌툭시맙 베도틴에는 강력하고 독성이 강한 항미세소관제인 모노메틸 오리스타틴 E 또는 MMAE(합성 항종양제)를 인간 특이적 CD30 양성 악성 세포에 전달하는 효소 민감성 절단 가능한 링커가 포함되어 있다. 튜불린의 중합을 차단하여 세포 분열을 억제하는 MMAE는 독성이 높기 때문에, 단일제 화학요법 약물로서 사용할 수 없다. 그러나 항-CD30 단클론 항체(cAC10, 종양 괴사 인자 또는 TNF 수용체의 세포막 단백질)에 연결된 MMAE의 조합은 세포외 유체에서 안정하고, 카텝신에 의해 절단 가능하고, 치료에 안전한 것으로 입증되었다. 또 다른 승인된 ADC인 트라스투주맙 엠탄신은 마이탄신의 유도체인 미세소관 형성 억제제 메르타신(DM-1)과, 안정한 절단 불가능한 링커에 의해 부착된 항체 트라스투주맙(Herceptin®/Genentech/Roche)의 조합물이다.
보다 우수하고 보다 안정한 링커의 가용성은 화학 결합의 기능을 변경했다. 절단 가능 또는 절단 불가능한 링커의 유형은 세포독성(항암) 약물에 특정한 특성을 부여한다. 예를 들어, 절단 불가능한 링커는 약물을 세포 내에 유지한다. 그 결과, 전체 항체, 링커 및 세포독성제는 표적화된 암 세포로 유입하고, 여기서 항체가 아미노산 수준으로 분해된다. 생성된 복합체-아미노산, 링커 및 세포독성제-는 이제 활성제가 된다. 대조적으로, 절단 가능한 링커는 세포독성제를 방출하는 숙주 세포 내의 효소에 의해 촉매된다.
현재 개발 중의 또 다른 유형의 절단 가능한 링커는 세포독성/항바이러스 약물과 절단 부위 사이에 추가 분자를 추가한다. 이 링커 기술에 의해, 연구자는 절단 속도의 변화를 우려하지 않고 보다 유연성이 있는 ADC를 생성할 수 있다. 연구자들은 또한 펩티드 내의 아미노산을 시퀀싱하는 방법인 에드만(Edman) 분해를 기반으로 하는 새로운 펩티드 절단 방법을 개발하고 있다. ADC의 개발에서 장래의 방향성에는 안정성과 치료 지수를 추가로 개선하기 위한 부위 특이적 접합(TDC)의 개발 및 α 방출 면역접합체 및 항체 접합 나노입자의 개발도 포함된다.
H.
BiTE
이중특이적 T 세포 인게이저(bi-specific t-cell engager, BiTE)는 항암제로 사용하기 위하여 조사되는 인공 이중특이적 단클론 항체의 부류이다. 이들은 감염된 세포에 대하여 숙주의 면역계, 보다 구체적으로 T 세포의 세포독성 활성을 지시한다. BiTE는 Micromet AG의 등록 상표이다.
BiTE는 약 55 킬로달톤의 단일 펩티드 쇄 상의 서로 상이한 항체의 2개 단쇄 가변 단편(scFv) 또는 4개의 상이한 유전자로부터의 아미노산 서열로 구성된 융합 단백질이다. scFv 중 하나는 CD3 수용체를 통해 T 세포에 결합하고, 다른 하나는 특정 분자를 통해 감염된 세포에 결합한다.
다른 이중특이적 항체와 마찬가지로, 그리고 일반적인 단클론 항체와는 달리, BiTE는 T 세포와 표적 세포 사이에 연결을 형성한다. 이것에 의해, T 세포는 MHC I 또는 공동자극 분자의 존재와 무관하게, 퍼포린 및 그랜자임과 같은 단백질을 생성함으로써, 감염된 세포에 대해 세포독성/항바이러스 활성을 발휘한다. 이러한 단백질은 감염된 세포에 유입되어, 세포의 아폽토시스를 개시한다. 이 작용은 감염된 세포에 대한 T 세포 공격 동안 관찰되는 생리학적 과정을 모방한다.
I.
인트라바디
특정 구현예에서, 항체는 세포 내부에서 작용에 적합한 재조합 항체이고, 이러한 항체는 "인트라바디"로서 공지되어 있다. 이러한 항체는 세포 내 단백질 트래피킹(trafficking)의 변화, 효소 기능의 방해, 단백질-단백질 또는 단백질-DNA 상호작용의 차단과 같은 다양한 메커니즘에 의해 표적 기능을 방해할 수 있다. 다수의 점에서, 이들의 구조는 위에서 논의한 단쇄 및 단일 도메인 항체의 구조를 모방하거나 이와 유사하다. 실제로, 단일 전사체/단쇄는 표적 세포에서의 세포내 발현을 가능하게 하고, 또한 세포막을 가로질러 단백질 수송을 보다 실현 가능하게 하는 중요한 특징이다. 그러나 추가 특징이 필요하다.
인트라바디 치료제의 실시에 영향을 미치는 2개의 주요 문제는 세포/조직의 표적화를 포함하는 전달 및 안정성이다. 전달과 관련하여, 조직 지시된 전달, 세포 유형 특이적 프로모터의 사용, 바이러스 기반의 전달 및 세포 투과성/막 전위 펩티드의 사용과 같은 다양한 접근법이 사용되었다. 안정성과 관련하여, 접근법은 일반적으로, 파지 디스플레이를 수반하고 공통 서열의 서열 성숙 또는 발달을 포함할 수 있는 방법을 포함하는 완력에 의한 스크리닝, 또는 삽입 안정화 서열(예를 들어, Fc 영역, 샤페론 단백질 서열, 류신 지퍼) 및 이황화물 치환/변형 등의 더욱 지시된 변형 중의 어느 하나이다.
인트라바디에 필요할 수 있는 추가 특징은 세포내 표적화를 위한 신호이다. 인트라바디(또는 기타 단백질)를 세포질, 핵, 미토콘드리아 및 ER과 같은 세포 이하 영역에 표적화할 수 있는 벡터가 설계되었고, 상업적으로 입수 가능하다(Invitrogen Corp.; Persic et al., 1997).
세포에 유입될 수 있는 능력으로, 인트라바디는 기타 유형의 항체가 달성할 수 없는 추가 용도를 갖는다. 본 발명의 항체의 경우, 살아 있는 세포에서 MUC1 세포질 도메인과 상호작용하는 능력은 신호전달 기능(다른 분자에 결합) 또는 올리고머 형성과 같은 MUC1 CD와 관련된 기능을 방해할 수 있다. 특히, 이러한 항체는 MUC1 이량체 형성을 억제하는 데 사용될 수 있는 것으로 고려된다.
J.
정제
특정 구현예에서, 본 개시내용의 항체는 정제될 수 있다. 본원에서 사용되는 "정제된"이라는 용어는 단백질이 이의 자연적으로 수득할 수 있는 상태에 대하여 임의의 정도까지 정제되는, 다른 성분으로부터 단리 가능한 조성물을 지칭하기 위한 것이다. 따라서, 정제된 단백질은 자연적으로 발생할 수 있는 환경으로부터 유리된 단백질을 지칭한다. "실질적으로 정제된"이라는 용어가 사용되는 경우, 이 명칭은 단백질 또는 펩티드가 조성물의 주요 성분을 형성하는, 예를 들어, 조성물에서 단백질의 약 50%, 약 60%, 약 70%, 약 80%, 약 90%, 약 95% 또는 그 이상을 구성하는 조성물을 지칭한다.
단백질 정제 기술은 당업자에게 잘 알려져 있다. 이러한 기술은 어느 수준에서는 세포 환경의 폴리펩티드 및 비폴리펩티드 분획으로 조 분별하는 것을 포함한다. 폴리펩티드를 다른 단백질로부터 분리한 후, 관심 폴리펩티드는 부분적 또는 완전한 정제(또는 균질성을 위한 정제)를 달성하기 위해 크로마토그래피 및 전기영동 기술을 사용하여 추가로 정제할 수 있다. 순수한 펩티드의 제조에 특히 적합한 분석 방법은 이온 교환 크로마토그래피, 배제 크로마토그래피; 폴리아크릴아미드 겔 전기영동; 등전점 포커싱이다. 단백질 정제를 위한 다른 방법에는 황산암모늄, PEG, 항체 등에 의한 침전 또는 열 변성과 그에 이은 원심분리; 겔 여과, 역상, 하이드록실아파타이트 및 친화성 크로마토그래피; 및 이러한 기술 및 기타 기술의 조합이 포함된다.
본 개시내용의 항체를 정제하는 데 있어서, 원핵생물 또는 진핵생물의 발현 시스템에서 폴리펩티드를 발현시키고, 변성 조건을 사용하여 단백질을 추출하는 것이 바람직할 수 있다. 폴리펩티드는 폴리펩티드의 태깅된 부분에 결합하는 친화성 컬럼을 사용하여 기타 세포 성분으로부터 정제할 수 있다. 당해 분야에 일반적으로 공지된 바와 같이, 다양한 정제 단계를 수행하는 순서를 변경할 수 있거나, 또는 특정 단계를 생략할 수 있으며, 실질적으로 정제된 단백질 또는 펩티드를 제조하기 위한 적합한 방법을 여전히 제공하는 것으로 믿어진다.
일반적으로 완전한 항체는 항체의 Fc 부분에 결합하는 작용제(즉, 단백질 A)를 사용하여 분획화된다. 대안적으로, 항원은 적절한 항체의 정제와 선택을 동시에 수행하기 위해 사용될 수 있다. 이러한 방법은 종종 컬럼, 필터 또는 비드와 같은 지지체에 결합된 선택제를 이용한다. 항체는 지지체에 결합하고, 오염 물질을 제거하고(예를 들어, 세척), 조건(염, 열 등)을 적용함으로써 항체를 방출시킨다.
단백질 또는 펩티드의 정제의 정도를 정량화하는 다양한 방법이 본 개시내용에 비추어 당업자에게 공지될 것이다. 여기에는, 예를 들어, 활성 분획의 특이적 활성을 결정하는 것, 또는 SDS/PAGE 분석에 의해 분획 내의 폴리펩티드의 양을 평가하는 것이 포함된다. 분획의 순도를 평가하는 또 다른 방법은 분획의 특이적 활성을 계산하고, 이를 초기 추출물의 특이적 활성과 비교하여 순도의 정도를 계산하는 것이다. 활성의 양을 나타내기 위해 사용되는 실제 단위는 물론 정제를 추적하기 위해 선택된 특정 분석 기술 및 발현된 단백질 또는 펩티드가 검출 가능한 활성을 나타내는지의 여부에 따라 다를 것이다.
폴리펩티드의 이동은, SDS/PAGE의 조건이 상이하면, 때때로 현저히 변화할 수 있는 것으로 알려져 있다(Capaldi et al., 1977). 따라서, 상이한 전기영동 조건 하에서, 정제 또는 부분적으로 정제된 발현 생성물의 겉보기 분자량이 변화할 수 있음이 이해될 것이다.
III.
SARS-CoV-2 감염의 능동/수동 면역화 및 치료/예방
A.
제형 및 투여
본 개시내용은 항-SARS-CoV-2 바이러스 항체 및 이를 생성하기 위한 항원을 포함하는 제약 조성물을 제공한다. 이러한 조성물은 예방적 또는 치료적 유효량의 항체 또는 이의 단편, 또는 펩티드 면역원, 및 제약상 허용되는 담체를 포함한다. 특정 구현예에서, "제약상 허용되는"이라는 용어는 연방 정부 또는 주 정부의 규제 기관에 의해 승인되었거나 미국 약전 또는 기타 동물용, 더욱 상세하게는 인간용의, 일반적으로 인정되는 약전에 열거되었음을 의미한다. 용어 "담체"는 치료제와 함께 투여되는 희석제, 부형제 또는 비히클을 나타낸다. 이러한 제약 담체는 멸균액, 예컨대, 물 및 오일(석유, 동물유, 식물유 또는 합성유, 예컨대, 땅콩유, 대두유, 광유, 호마유 등 포함)일 수 있다. 제약 조성물이 정맥 내 투여될 때 물이 바람직한 담체이다. 식염수 및 수용성 덱스트로스 및 글리세롤 용액이 또한 특히 주사용액을 위한 액체 담체로서 이용될 수 있다. 적합한 제약 부형제는, 전분, 글루코스, 락토스, 수크로스, 젤라틴, 맥아, 쌀, 밀가루, 백악, 실리카 겔, 스테아르산 나트륨, 글리세롤 모노스테아레이트, 탈크, 염화 나트륨, 건조 탈지유, 글리세롤, 프로필렌, 글리콜, 물, 에탄올 등을 포함한다.
원한다면, 조성물은 또한 적은 양의 습윤제 또는 유화제, 또는 pH 완충제를 함유할 수 있다. 이러한 조성물은 용액, 현탁액, 에멀젼, 정제, 알약, 캡슐, 분말, 서방성 제형 등의 형태를 취할 수 있다. 경구 제형은 제약 등급의 만니톨, 락토스, 전분, 스테아르산 마그네슘, 사카린나트륨, 셀룰로스, 탄산마그네슘 등의 표준 담체를 포함할 수 있다. 적합한 제약 작용제의 예는 "Remington's Pharmaceutical Sciences"에 기재되어 있다. 이러한 조성물은, 예방적 또는 치료적 유효량의 항체 또는 이의 단편을, 바람직하게는 정제된 형태로, 적절한 양의 담체와 함께, 환자에게 적절하게 투여하기 위한 형태를 제공한다. 제형은 경구, 정맥 내, 동맥 내, 협측, 비강 내, 분무, 기관지 흡입, 직장 내, 질, 국소 또는 기계적 인공호흡에 의해 전달될 수 있는 투여 방식에 적합해야 한다.
개시된 것과 같은 항체가 SARS-CoV-2 감염 위험이 있는 대상체에서 생체 내에서 생성되는 경우의 활성 백신도 고려된다. 이러한 백신은 비경구 투여용으로 제형화될 수 있고, 예를 들어, 피내, 정맥 내, 근육 내, 피하, 에어로졸 또는 심지어 복강 내 경로를 통한 주사용으로 제형화될 수 있다. 피내 및 근육 내 경로에 의한 투여가 고려된다. 대안적으로, 백신은 국소 경로에 의해 점막에 직접 투여될 수 있고, 예를 들어, 점비제, 흡입, 분무기에 의해, 또는 직장 내 또는 질 전달을 통해 투여될 수 있다. 제약상 허용되는 염은 산성 염 및, 예를 들어, 염산 또는 인산 등의 무기산, 또는 아세트산, 옥살산, 타르타르산, 만델산 등의 유기산으로 형성된 염을 포함한다. 유리 카르복실 기로 형성된 염은 또한 예를 들어, 나트륨, 칼륨, 암모늄, 칼슘 또는 수산화제2철 등의 무기 염기 및 이소프로필아민, 트리메틸아민, 2-에틸아미노 에탄올, 히스티딘, 프로카인 등과 같은 유기 염기로부터 유래할 수 있다.
인공적으로 획득한 수동 면역으로서 공지된 항체의 수동 전달은 일반적으로 정맥 내 주사 또는 근육 내 주사의 사용을 수반한다. 항체의 형태는, 정맥 내(IVIG) 또는 근육 내(IG) 사용을 위한 풀링된 인간 면역글로불린으로서, 면역화되거나 질환으로부터 회복된 공여자로부터의 고역가 인간 IVIG 또는 IG으로서 및 단클론 항체(MAb)로서, 인간 또는 동물의 혈장 또는 혈청일 수 있다. 이러한 면역은 일반적으로 짧은 기간 동안만 지속되고, 특히 비인간 기원의 감마 글로불린에 의한 과민 반응 및 혈청병의 잠재적 위험이 있다. 그러나 수동 면역은 즉각적 보호를 제공한다. 항체는 주사에 적합한, 즉 멸균되고 주사 가능한 담체에서 제형화될 것이다.
일반적으로, 본 개시내용의 조성물의 성분들은 별도로 또는 단위 투여 형태, 예를 들어, 활성제의 양을 나타낸 앰플 또는 샤세와 같은 기밀하게 밀봉된 용기 내 건조 동결 분말 또는 무수 농축물로 함께 혼합되어 공급된다. 조성물이 주입에 의해 투여되는 경우, 멸균된 제약 등급의 물 또는 염수를 함유하는 주입병으로 분배될 수 있다. 조성물이 주사에 의해 투여되는 경우, 주사용 멸균수 또는 식염수의 앰플이 제공될 수 있으며, 성분들이 투여 전에 혼합될 수 있다.
본 개시내용의 조성물은 중성 또는 염 형태로 제형화될 수 있다. 제약상 허용되는 염은 염산, 인산, 아세트산, 옥살산, 타르타르산 등으로부터 유래된 것 등의 음이온으로 형성된 염 및 나트륨, 칼륨, 암모늄, 칼슘, 수산화제이철, 이소프로필아민, 트리에틸아민, 2-에틸아미노에탄올, 히스티딘, 프로카인 등으로부터 유래된 것과 같은 양이온으로 형성된 염을 포함한다.
2.
ADCC
항체 의존성 세포 매개 세포독성(ADCC)은 면역 이펙터 세포에 의해 항체 코팅된 표적 세포의 용해를 유도하는 면역 메커니즘이다. 표적 세포는 일반적으로 Fc 영역의 N 말단인 단백질 부분을 통해 Fc 영역을 포함하는 항체 또는 이의 단편이 특이적으로 결합하는 세포이다. "항체 의존성 세포 매개 세포독성(ADCC)의 증가/감소를 갖는 항체"는 당업자에게 공지된 임의의 적합한 방법에 의해 결정된 ADCC의 증가/감소를 갖는 항체를 의미한다.
본원에서 사용되는 "증가/감소된 ADCC"라는 용어는, 위에서 정의된 ADCC의 메커니즘에 의해 표적 세포를 둘러싸고 있는 배지 내의 주어진 항체 농도에서 주어진 시간에 용해되는 표적 세포 수의 증가/감소로서 정의되고/되거나, ADCC의 메커니즘에 의해 주어진 시간에 주어진 수의 표적 세포의 용해를 달성하는 데 필요한, 표적 세포를 둘러싸고 있는 배지 내의 항체 농도의 감소/증가로서 정의된다. ADCC의 증가/감소는 동일한 표준 생산, 정제, 제형화 및 저장 방법(이는 당업자에게 공지됨)을 사용하여 동일한 유형의, 그러나 조작되지 않은 숙주 세포에 의해 생성된 동일한 항체에 의해 매개되는 ADCC와 비교된다. 예를 들어, 본원에 기재된 방법에 의해, 변경된 글리코실화 패턴을 갖도록(예를 들어, 글리코실트랜스퍼라제, GnTIII 또는 기타 글리코실트랜스퍼라제를 발현하도록) 조작된 숙주 세포에 의해 생성된 항체에 의해 매개되는 ADCC의 증가는 동일한 유형의 조작되지 않은 숙주 세포에 의해 생성된 동일한 항체에 의해 매개되는 ADCC와 비교된다.
3.
CDC
보체 의존성 세포독성(CDC)은 보체 시스템의 기능이다. 이는 면역계의 항체 또는 세포의 관여 없이 병원체의 막을 손상시킴으로써 병원체를 사멸시키는 면역계의 과정이다. 여기에는 3가지의 주요 과정이 있다. 3가지 모두 치명적 콜로이드 삼투압 팽창, 즉 CDC를 유발하는 병원체에 하나 이상의 막 공격 복합체(MAC)를 삽입한다. 이는 항체 또는 항체 단편이 항바이러스 효과를 발휘하는 메커니즘 중 하나이다.
Ⅳ.
항체 접합체
본 개시내용의 항체는 항체 접합체를 형성하기 위해 적어도 하나의 작용제에 연결될 수 있다. 진단제 또는 치료제로서의 항체 분자의 효능을 증가시키기 위해, 적어도 하나의 원하는 분자 또는 모이어티를 연결하거나, 공유 결합하거나, 또는 복합체를 형성하는 것이 통상적이다. 이러한 분자 또는 모이어티는 적어도 하나의 이펙터 또는 리포터 분자일 수 있지만, 이에 한정되지 않는다. 이펙터 분자는 원하는 활성, 예를 들어, 세포독성 활성을 갖는 분자를 포함한다. 항체에 부착된 이펙터 분자의 비제한적 예에는 독소, 항종양제, 치료적 효소, 방사성핵종, 항바이러스제, 킬레이트제, 사이토카인, 성장 인자 및 올리고뉴클레오티드 또는 폴리뉴클레오티드가 포함된다. 대조적으로, 리포터 분자는 분석을 사용하여 검출될 수 있는 임의의 모이어티로서 정의된다. 항체에 접합된 리포터 분자의 비제한적 예에는 효소, 방사성 표지, 합텐, 형광 표지, 인광 분자, 화학발광 분자, 발색단, 광친화성 분자, 착색 입자 또는 리간드, 예를 들어, 비오틴이 포함된다.
항체 접합체는 일반적으로 진단제로서 사용하기에 바람직하다. 항체 진단제는 일반적으로 2가지 부류, 다양한 면역분석과 같은 시험관 내 진단제에 사용하기 위한 것과, 일반적으로 "항체 지시 이미징"으로 공지된 생체 내 진단 프로토콜에 사용하기 위한 것으로 분류된다. 항체에 이들을 부착하는 방법과 마찬가지로, 다수의 적절한 이미징제가 당해 분야에 공지되어 있다(예를 들어, 미국 특허 제5,021,236호, 제4,938,948호 및 제4,472,509호 참고). 사용된 이미징 모이어티는 상자성 이온, 방사성 동위원소, 형광색소, NMR 검출 가능한 물질 및 X선 이미징제일 수 있다.
상자성 이온의 경우, 예로서, 크롬(III), 망간(II), 철(III), 철(II), 코발트(II), 니켈(II), 구리(II), 네오디뮴(III), 사마륨(III), 이테르븀(III), 가돌리늄(III), 바나듐(II), 테르븀(III), 디스프로슘(III), 홀뮴(III) 및/또는 에르븀(III)과 같은 이온을 들 수 있고, 가돌리늄이 특히 바람직하다. X선 이미징과 같은 기타 맥락에서 유용한 이온에는 란탄(III), 금(III), 납(II), 특히 비스무트(III)가 포함되지만, 이들로 한정되지 않는다.
치료 및/또는 진단용의 방사성 동위원소의 경우, 아스타틴211, 14탄소, 51크롬, 36염소, 57코발트, 58코발트, 구리67, 152Eu, 갈륨67, 3수소, 요오드123, 요오드125, 요오드131, 인듐111, 59철, 32인, 레늄186, 레늄188, 75셀레늄, 35황, 테크니슘99m 및/또는 이트륨90을 들 수 있다. 125I는 특정 구현예에서의 사용에 종종 바람직하고, 테크니시움99m 및/또는 인듐111은 또한 이들의 낮은 에너지 및 장거리 검출 적합성으로 인해 종종 바람직하다. 본 개시내용의 방사성 표지된 단클론 항체는 당해 분야에 주지된 방법에 따라 생성될 수 있다. 예를 들어, 단클론 항체는 요오드화나트륨 및/또는 요오드화칼륨 및 차아염소산나트륨 등의 화학적 산화제 또는 락토퍼옥시다제 등의 효소적 산화제와 접촉시킴으로써 요오드화할 수 있다. 본 개시내용에 따른 단클론 항체는, 예를 들어, 퍼테크네이트를 제1주석 용액으로 환원시키고, 환원된 테크네튬을 Sephadex 컬럼 상에 킬레이트화하고, 항체를 이 컬럼에 적용함으로써 테크네튬99m으로 표지될 수 있다. 대안적으로, 예를 들어, 퍼테크네이트, SNCl2 등의 환원제, 나트륨-칼륨 프탈레이트 용액 등의 완충액 및 항체를 인큐베이션함으로써 직접 표지 기술을 사용할 수 있다. 금속 이온으로서 존재하는 방사성 동위원소를 항체에 결합시키기 위해 자주 사용되는 중간 작용기는 디에틸렌트리아민펜타아세트산(DTPA) 또는 에틸렌 디아민테트라아세트산(EDTA)이다.
접합체로서 사용하기 위해 고려되는 형광성 표지에는 알렉사(Alexa) 350, 알렉사(Alexa) 430, AMCA, BODIPY 630/650, BODIPY 650/665, BODIPY-FL, BODIPY-R6G, BODIPY-TMR, BODIPY-TRX, 카스케이드 블루(Cascade Blue), Cy3, Cy5,6-FAM, 플루오레세인 이소티오시아네이트, HEX, 6-JOE, 오레곤 그린(Oregon Green) 488, 오레곤 그린 500, 오레곤 그린 514, 퍼시픽 블루(Pacific Blue), REG, 로다민 그린(Rhodamine Green), 로다민 레드(Rhodamine Red), 레노그라핀(Renographin), ROX, TAMRA, TET, 테트라메틸로다민(Tetramethylrhodamine) 및/또는 텍사스 레드(Texas Red)가 포함된다.
본 개시내용에서 고려되는 추가 유형의 항체는 주로 시험관 내에서의 사용을 위한 것이고, 여기서 항체는 2차 결합 리간드 및/또는 발색성 기질과의 접촉 시 착색된 생성물을 생성하는 효소(효소 태그)에 연결된다. 적합한 효소의 예는 우레아제, 알칼리성 포스파타제, (서양고추냉이) 수소 퍼옥시다제 또는 글루코스 옥시다제를 포함한다. 바람직한 2차 결합 리간드는 비오틴 및 아비딘 및 스트렙타비딘 화합물이다. 이러한 표지의 사용은 당업자에게 주지되어 있고, 예를 들어, 미국 특허 제3,817,837호, 제3,850,752호, 제3,939,350호, 제3,996,345호, 제4,277,437호, 제4,275,149호 및 제4,366,241호에 기재되어 있다.
항체에 대한 분자의 부위 특이적 부착의 또 다른 공지된 방법은 항체를 합텐 기반 친화성 표지와 반응시키는 것을 포함한다. 기본적으로, 합텐 기반 친화성 표지는 항원 결합 부위의 아미노산과 반응하고, 이에 의해 이 부위를 파괴하고, 특정 항원 반응을 차단한다. 그러나 이는 항체 접합체에 의한 항원 결합의 손실을 초래하기 때문에 유리하지 않을 수 있다.
아지도 기를 포함하는 분자는 또한 낮은 강도의 자외선에 의해 생성되는 반응성 니트렌 중간체를 통해 단백질에 대한 공유 결합을 형성하는 데 사용될 수 있다(Potter and Haley, 1983). 특히, 퓨린 뉴클레오티드의 2-아지도 및 8-아지도 유사체는 조 세포 추출물 중의 뉴클레오티드 결합 단백질을 확인하기 위한 부위 특이적 광프로브로서 사용되었다(Owens & Haley, 1987; Atherton et al., 1985). 2-아지도 및 8-아지도 뉴클레오티드는 또한 정제된 단백질의 뉴클레오티드 결합 도메인을 매핑하기 위해 사용되었고(Khatoon et al., 1989; King et al., 1989; Dholakia et al., 1989), 항체 결합제로서 사용될 수 있다.
접합체 모이어티에 대한 항체의 부착 또는 접합을 위한 몇몇 방법이 당해 분야에 공지되어 있다. 일부 부착 방법은, 예를 들어, 디에틸렌트리아민펜타아세트산 무수물(DTPA) 등의 유기 킬레이트제; 에틸렌트리아민테트라아세트산; N-클로로-p-톨루엔설폰아미드; 및/또는 항체에 부착된 테트라클로로-3α-6α-디페닐글리코우릴-3을 사용하는 금속 킬레이트 착체의 사용을 수반한다(미국 특허 제4,472,509호 및 제4,938,948호). 단클론 항체는 또한 글루타르알데히드 또는 페리오데이트 등의 커플링제의 존재 하에 효소와 반응시킬 수 있다. 플루오레세인 마커와의 접합체는 이러한 커플링제의 존재 하에 또는 이소티오시아네이트와의 반응에 의해 제조된다. 미국 특허 제4,938,948호에서, 유방 종양의 이미징은 단클론 항체를 사용하여 달성되고, 검출 가능한 이미징 모이어티는 메틸-p-하이드록시벤즈이미데이트 또는 N-석신이미딜-3-(4-하이드록시페닐)프로피오네이트 등의 링커를 사용하여 항체에 결합시킨다.
다른 구현예에서, 항체 결합 부위를 변화시키지 않는 반응 조건을 사용하여, 면역글로불린의 Fc 영역에 설프하이드릴 기를 선택적으로 도입함으로써 면역글로불린의 유도체화가 고려된다. 이 방법에 따라 생성된 항체 접합체는 개선된 수명, 특이성 및 감도를 나타내는 것으로 개시되어 있다(미국 특허 제5,196,066호, 참조로서 본원에 포함됨). 리포터 또는 이펙터 분자가 Fc 영역의 탄수화물 잔기에 접합되어 있는, 이펙터 또는 리포터 분자의 부위 특이적 부착도 문헌에 개시되어 있다(O'Shannessy et al., 1987). 이 접근법은 현재 임상 평가 중의 진단 및 치료적으로 유망한 항체를 생산하는 것으로 보고되었다.
V.
면역검출 방법
또 다른 구현예에서, 본 개시내용은 SARS-CoV-2 및 이의 관련 항원을 결합, 정제, 제거, 정량화 및 다르게는 일반적으로 검출하기 위한 면역검출 방법에 관한 것이다. 이러한 방법은 종래의 의미에서 적용될 수 있지만, 백신 및 기타 바이러스 스톡의 품질 관리 및 모니터링에도 사용될 것이고, 여기서 본 개시내용에 따른 항체는 바이러스 중의 항원의 양 또는 무결성(즉, 장기 안정성)을 평가하기 위해 사용될 수 있다. 대안적으로, 이 방법을 사용하여, 적절한/원하는 반응성 프로파일에 대해 다양한 항체를 스크리닝할 수 있다.
다른 면역검출 방법에는 대상체에서 SARS-CoV-2의 존재를 결정하기 위한 특정 분석이 포함된다. 다양한 분석 형식이 고려되지만, 구체적으로는 타액, 혈액, 혈장, 가래, 정액 또는 소변과 같은 대상체로부터 수득된 유체 중의 SARS-CoV-2를 검출하기 위해 사용되는 것들이다. 특히, 정액은 SARS-CoV-2를 검출하기 위한 실행 가능한 샘플로서 입증되었다(Purpura et al., 2016; Mansuy et al., 2016; Barzon et al., 2016; Gornet et al., 2016; Duffy et al., 2009; CDC, 2016; Halfon et al., 2010; Elder et al. 2005). 분석은 가정에서의 임신 검사와 유사한 측면 유동 분석(아래 참고)을 포함하여 비의료(가정)에서의 사용을 위해 유리하게 구성될 수 있다. 이러한 분석은 가족 구성원의 대상체에 의해 사용될 수 있도록 적절한 시약 및 지침을 포함하는 키트 형태로 패키징될 수 있다.
일부 면역검출 방법으로 몇 가지를 나열하자면 효소 결합 면역흡착 분석(ELISA), 방사선면역 분석(RIA), 면역방사성 분석, 형광면역 분석, 화학발광 분석, 생물발광 분석 및 웨스턴 블롯이 포함된다. 특히, 샘플 중의 특정 기생충 에피토프에 대해 지시된 SARS-CoV-2 항체의 검출 및 정량을 위한 경쟁 분석도 제공된다. 다양한 유용한 면역검출 방법의 단계는, 예를 들어, Doolittle and Ben-Zeev (1999), Gulbis and Galand (1993), De Jager et al. (1993) 및 Nakamura et al. (1987)과 같은 과학 문헌에 기재되어 있다. 일반적으로, 면역결합 방법은 SARS-CoV-2를 함유하는 것으로 의심되는 샘플을 수득하고, 경우에 따라, 면역복합체의 형성을 가능하게 하기에 효과적인 조건 하에 본 개시내용에 따라 샘플을 제1 항체와 접촉시키는 단계를 포함한다.
이러한 방법에는 샘플로부터 SARS-CoV-2 또는 관련 항원을 정제하는 방법이 포함된다. 항체는 바람직하게는 컬럼 매트릭스 형태 등의 고체 지지체에 연결되고, SARS-CoV-2 또는 항원 성분을 함유하는 것으로 의심되는 샘플은 고정된 항체에 적용될 것이다. 원치 않는 성분은 컬럼으로부터 세척되어 SARS-CoV-2 항원이 고정된 항체에 면역복합체로 잔류하고, 이어서 고정화된 항체는 컬럼으로부터 유기체 또는 항원을 제거함으로써 수집된다.
면역결합 방법에는 또한 샘플 중의 SARS-CoV-2 또는 관련 성분의 양을 검출 및 정량화하는 방법 및 결합 과정 동안 형성된 임의의 면역 복합체의 검출 및 정량화 방법이 포함된다. 본원에서는, SARS-CoV-2 또는 이의 항원을 함유하는 것으로 의심되는 샘플을 수득하고, 이 샘플을 SARS-CoV-2 또는 이의 성분에 결합하는 항체와 접촉시킨 후, 특정 조건 하에서 형성된 면역 복합체의 양을 검출하고 정량화한다. 항원 검출과 관련하여, 분석된 생물학적 샘플은 조직 절편 또는 표본, 균질화된 조직 추출물, 혈액 및 혈청을 포함한 생물학적 유체, 또는 대변 또는 소변 등의 분비물 등의 SARS-CoV-2 또는 SARS-CoV-2 항원을 함유하는 것으로 의심되는 임의의 샘플일 수 있다.
선택된 생물학적 샘플을 효과적인 조건 하에서 면역 복합체(1차 면역 복합체)의 형성을 가능하게 하기에 충분한 시간 동안 항체와 접촉시키는 것은 일반적으로 항체 조성물을 샘플에 단순히 첨가하고, 항체가 SARS-CoV-2 또는 존재하는 항원과의 면역 복합체를 형성하기에 충분한 시간, 즉 SARS-CoV-2 또는 존재하는 항원에 결합하기에 충분한 시간 동안 혼합물을 인큐베이션하는 것이다. 이 시간 후, 조직 절편, ELISA 플레이트, 도트 블롯 또는 웨스턴 블롯 등의 샘플-항체 조성물을 일반적으로 세척하여 비특이적으로 결합된 항체 종을 제거하여, 1차 면역 복합체 내에서 특이적으로 결합된 항체만이 검출되도록 한다.
일반적으로, 면역복합체 형성의 검출은 당해 분야에 주지되어 있고, 다수의 접근법을 적용함으로써 달성될 수 있다. 이러한 방법은 일반적으로 방사성, 형광성, 생물학적 및 효소적 태그 등의 표지 또는 마커의 검출을 기반으로 한다. 이러한 표지의 사용에 관한 특허에는 미국 특허 제3,817,837호, 제3,850,752호, 제3,939,350호, 제3,996,345호, 제4,277,437호, 제4,275,149호 및 제4,366,241호가 포함된다. 물론, 당해 분야에 공지된 바와 같이, 제2 항체 등의 2차 결합 리간드 및/또는 비오틴/아비딘 리간드 결합 배열을 사용함으로써 추가의 이점을 발견할 수 있다.
검출에 사용된 항체는 그 자체가 검출 가능한 표지에 연결될 수 있고, 이어서 이 표지를 단순히 검출하고, 이에 의해 조성물 중의 1차 면역 복합체의 양을 결정할 수 있다. 대안적으로, 1차 면역 복합체 내에서 결합되는 제1 항체는 항체에 대한 결합 친화성을 갖는 제2 결합 리간드에 의해 검출할 수 있다. 이러한 경우, 제2 결합 리간드는 검출 가능한 표지에 연결될 수 있다. 제2 결합 리간드는 그 자체가 종종 항체이고, 따라서 "제2" 항체로 명명될 수 있다. 1차 면역 복합체는 효과적인 조건 하에 2차 면역 복합체의 형성을 가능하게 하기에 충분한 시간 동안 표지된 제2 결합 리간드 또는 항체와 접촉시킨다. 이어서, 2차 면역 복합체를 일반적으로 세척하여, 비특이적으로 결합된 표지된 제2 항체 또는 리간드를 제거하고, 이어서 2차 면역 복합체의 잔류 표지를 검출한다.
추가 방법에는 2단계 접근법에 의한 1차 면역 복합체의 검출이 포함된다. 전술한 바와 같이, 항체에 대한 결합 친화성을 갖는 항체와 같은 제2 결합 리간드를 사용하여 2차 면역 복합체를 형성한다. 세척 후, 2차 면역 복합체는 다시 효과적인 조건 하에서 면역 복합체(3차 면역 복합체)의 형성을 가능하게 하기에 충분한 기간 동안 제2 항체에 대해 결합 친화성을 갖는 제3 결합 리간드 또는 항체와 접촉시킨다. 제3 리간드 또는 항체는 검출 가능한 표지에 연결되어, 이렇게 형성된 3차 면역 복합체의 검출을 가능하게 한다. 이 시스템은, 이것이 바람직한 경우, 신호 증폭을 제공할 수 있다.
면역검출의 한 가지 방법은 2개의 상이한 항체를 사용한다. 제1 비오틴화된 항체를 사용하여 표적 항원을 검출하고, 이어서 제2 항체를 사용하여 복합체화된 비오틴에 부착된 비오틴을 검출한다. 이 방법에서, 테스트되는 샘플은 먼저 제1 단계의 항체를 포함하는 용액에서 인큐베이션된다. 표적 항원이 존재하는 경우, 항체의 일부는 항원에 결합하여 비오틴화된 항체/항원 복합체를 형성한다. 이어서, 항체/항원 복합체는 스트렙타비딘(또는 아비딘), 비오틴화된 DNA 및/또는 상보적 비오틴화된 DNA의 연속 용액에서 인큐베이션에 의해 증폭되고, 각 단계에서 항체/항원 복합체에 추가 비오틴 부위가 추가된다. 적절한 증폭 수준에 도달할 때까지 증폭 단계가 반복되고, 이 시점에서 샘플은 비오틴에 대한 제2 단계의 항체를 포함하는 용액에서 인큐베이션된다. 이 제2 단계의 항체는, 예를 들어, 발색체 기질을 사용한 조직효소학에 의해 항체/항원 복합체의 존재를 검출하기 위해 사용할 수 있는 효소로 표지된다. 적절한 증폭에 의해, 육안으로 관찰 가능한 접합체를 생성할 수 있다.
면역검출의 또 다른 공지된 방법은 면역-PCR(중합효소 연쇄 반응) 방법론을 이용한다. PCR법은 비오틴화 DNA와의 인큐베이션까지는 Cantor 방법과 유사하지만, 스트렙타비딘과 비오틴화 DNA의 인큐베이션을 복수 회 사용하는 대신에, DNA/비오틴/스트렙타비딘/항체 복합체를 낮은 pH 또는 고염 완충제로 세척하여 항체를 방출시킨다. 이어서, 생성된 세척 용액을 사용하여, 적절한 대조군을 갖는 적절한 프라이머를 사용하여 PCR 반응을 수행한다. 적어도 이론상으로는, PCR의 막대한 증폭 능력과 특이성을 이용하여 단일 항원 분자를 검출할 수 있다.
A.
ELISA
면역분석은 가장 간단하고 직접적인 의미에서 결합 분석이다. 특정한 바람직한 면역분석은 당해 분야에 공지된 다양한 유형의 효소 결합 면역흡착 분석(ELISA) 및 방사선면역분석(RIA)이다. 조직 절편을 사용한 면역조직화학적 검출도 특히 유용하다. 그러나 검출은 이러한 기술에 한정되지 않고, 웨스턴 블롯팅, 도트 블롯팅, FACS 분석 등도 또한 사용될 수 있음을 용이하게 이해할 것이다.
한 가지 예시적 ELISA에서, 본 개시내용의 항체는 폴리스티렌 미세적정 플레이트 내의 웰 등의 단백질 친화성을 나타내는 선택된 표면 상에 고정화된다. 이어서, SARS-CoV-2 또는 SARS-CoV-2 항원을 함유하는 것으로 의심되는 시험 조성물을 웰에 첨가한다. 결합시키고 세척하여 비특이적으로 결합된 면역 복합체를 제거한 후, 결합된 항원을 검출할 수 있다. 검출은 검출 가능한 표지에 연결된 또 다른 항-SARS-CoV-2 항체를 첨가함으로써 달성할 수 있다. 이러한 유형의 ELISA는 단순 "샌드위치 ELISA"이다. 검출은 또한 제2 항-SARS-CoV-2 항체를 첨가하고, 이어서 제2 항체에 대한 결합 친화성을 갖는 제3 항체를 첨가함으로써 달성할 수 있는데, 제3 항체는 검출 가능한 표지에 연결되어 있다.
또 다른 예시적 ELISA에서, SARS-CoV-2 또는 SARS-CoV-2 항원을 함유하는 것으로 의심되는 샘플을 웰 표면에 고정시키고, 이어서 본 개시내용의 항-SARS-CoV-2 항체와 접촉시킨다. 결합시키고 세척하여 비특이적으로 결합된 면역 복합체를 제거한 후, 결합된 항-SARS-CoV-2 항체를 검출한다. 초기 항-SARS-CoV-2 항체가 검출 가능한 표지에 연결된 경우, 면역 복합체를 직접 검출할 수 있다. 다시, 면역 복합체를 제1 항-SARS-CoV-2 항체에 대한 결합 친화성을 갖는 제2 항체를 사용하여 검출할 수 있는데, 제2 항체는 검출 가능한 표지에 연결되어 있다.
사용된 포맷에 관계없이, ELISA는 코팅, 인큐베이션 및 결합, 비특이적으로 결합된 종을 제거하기 위한 세척, 결합된 면역 복합체의 검출 등 공통적인 특정한 특징을 갖는다. 이는 아래에 기재되어 있다.
플레이트를 항원 또는 항체로 코팅하는 경우, 일반적으로, 플레이트의 웰을 항원 또는 항체 용액과 함께 밤새 또는 지정된 시간 동안 인큐베이션한다. 이어서, 플레이트의 웰을 세척하여 불완전하게 흡착된 물질을 제거한다. 이어서, 웰의 나머지 이용 가능한 표면은 시험 항혈청과 관련하여 항원적으로 중성인 비특이적 단백질로 "코팅"된다. 여기에는 소 혈청 알부민(BSA), 카제인 또는 분유 용액이 포함된다. 코팅은 고정화 표면의 비특이적 흡착 부위의 차단을 가능하게 하고, 따라서 표면에 대한 항혈청의 비특이적 결합에 의해 유발된 배경을 감소시킨다.
ELISA에서는, 직접적 절차가 아닌, 제2 또는 제3 검출 수단을 사용하는 것이 보다 일반적일 수 있다. 따라서, 단백질 또는 항체를 웰에 결합시키고, 비반응성 물질로 코팅하여 배경을 감소시키고, 세척하여 비결합된 물질을 제거한 후, 고정화 표면은 면역 복합체(항원/항체)의 형성을 가능하게 하기에 효과적인 조건 하에서 시험되는 생물학적 샘플과 접촉시킨다. 이어서, 면역 복합체의 검출에는, 표지된 제2 결합 리간드 또는 항체, 및 표지된 제3 항체 또는 제3 결합 리간드와 조합한 제2 결합 리간드 또는 항체가 필요하다.
"면역 복합체(항원/항체)의 형성을 가능하게 하기에 효과적인 조건 하에서"는 조건이 바람직하게는 BSA, 소 감마 글로불린(BGG) 또는 인산 완충 식염수(PBS)/트윈 등의 용액으로 항원 및/또는 항체를 희석하는 것을 포함하는 것을 의미한다. 이러한 첨가된 작용제는 또한 비특이적 배경의 감소를 돕는 경향이 있다.
"적합한" 조건은 또한 인큐베이션이 효과적 결합을 가능하게 하기에 충분한 온도 또는 기간 동안 진행되는 것을 의미한다. 인큐베이션 단계는 일반적으로 약 1~2~4시간 정도, 바람직하게는 25℃ 내지 27℃ 정도의 온도에서 또는 약 4℃ 정도에서 밤새 수행할 수 있다.
ELISA의 모든 인큐베이션 단계 후에, 접촉된 표면을 세척하여 복합체를 형성하지 않은 물질을 제거한다. 바람직한 세척 절차에는 PBS/트윈 등의 용액 또는 붕산염 완충액으로 세척하는 것이 포함된다. 시험 샘플과 원래 결합된 물질 사이에 특정의 면역 복합체의 형성 및 후속 세척 후, 미량의 면역 복합체의 존재가 결정될 수 있다.
검출 수단을 제공하기 위해, 제2 또는 제3 항체는 검출을 가능하게 하는 관련 표지를 가질 것이다. 바람직하게는, 이것은 적절한 발색성 기질과의 인큐베이션 시 발색을 생성하는 효소일 것이다. 따라서, 예를 들어, 추가 면역 복합체 형성의 발달을 촉진하는 조건 하에서 일정 기간 동안 우레아제, 글루코스 옥시다제, 알칼리 포스파타제 또는 수소 퍼옥시다제-접합 항체와 1차 및 2차 면역 복합체를 접촉시키거나 인큐베이션하는 것을 원할 것이다(예를 들어, PBS-트윈 등의 PBS 함유 용액에서 실온에서 2시간 동안 인큐베이션).
표지된 항체와 함께 인큐베이션한 후 및 결합되지 않은 물질을 제거하기 위해 세척한 후, 예를 들어, 요소, 브로모크레졸 퍼플 또는 2,2'-아지노-디-(3-에틸-벤즈티아졸린-6-설폰산(ABTS), 또는 효소 표지로서 퍼옥시다제의 경우에 H2O2에 의해 표지의 양을 정량화한다. 이어서, 예를 들어, 가시 스펙트럼 분광 광도계를 사용하여, 생성된 색상 정도를 측정함으로써 정량화를 달성한다.
또 다른 구현예에서, 본 개시내용은 경쟁 포맷의 사용을 고려한다. 이것은 샘플에서 SARS-CoV-2 항체의 검출에 특히 유용하다. 경쟁 기반 분석에서, 미지의 양의 분석물 또는 항체는 공지된 양의 표지된 항체 또는 분석물을 치환하는 능력에 의해 결정된다. 따라서, 신호의 정량화 가능한 손실은 샘플에서 미지의 항체 또는 분석물의 양을 나타낸다.
본원에서, 본 발명자는 샘플 중의 SARS-CoV-2 항체의 양을 결정하기 위해, 표지된 SARS-CoV-2 단클론 항체의 사용을 제안한다. 기본 포맷에는 공지된 양의 SARS-CoV-2 단클론 항체(검출 가능한 표지에 연결됨)를 SARS-CoV-2 항원 또는 입자와 접촉시키는 것이 포함된다. SARS-CoV-2 항원 또는 유기체는 바람직하게는 지지체에 부착된다. 표지된 단클론 항체를 지지체에 결합시킨 후, 샘플을 첨가하고, 샘플 중의 임의의 비표지 항체가 표지된 단클론 항체와 경쟁하고, 따라서 이를 치환할 수 있는 조건 하에 인큐베이션한다. 손실된 표지 또는 잔류하는 표지를 측정(및 결합된 표지의 원래 양으로부터 이를 차감)함으로써, 지지체에 결합된 비표지 항체의 양, 따라서 샘플에 존재하는 항체의 양을 결정할 수 있다.
B.
웨스턴 블롯
웨스턴 블롯(또는 단백질 면역블롯)은 조직 균질물 또는 추출물의 주어진 샘플에서 특정 단백질을 검출하기 위해 사용되는 분석 기술이다. 이는 겔 전기영동을 사용하여 폴리펩티드의 길이(변성 조건) 또는 단백질의 3차원 구조(천연/비변성 조건)에 따라 천연 또는 변성된 단백질을 분리한다. 이어서, 단백질은 막(일반적으로 니트로셀룰로스 또는 PVDF)으로 옮겨지고, 여기서 표적 단백질에 특이적인 항체를 사용하여 프로빙(검출)된다.
샘플은 전체 조직 또는 세포 배양물로부터 채취할 수 있다. 대부분의 경우, 고형 조직은 먼저 블렌더(샘플 용적이 큰 경우), 균질화기(용적이 작은 경우) 또는 초음파 처리를 사용하여 기계적으로 분해된다. 세포는 또한 위의 기계적 방법 중 하나에 의해 파괴되어 개방될 수 있다. 그러나 세균, 바이러스 또는 환경 샘플이 단백질의 공급원으로 될 수 있으므로, 웨스턴 블롯팅은 세포 연구에만 한정되지 않음에 주의해야 한다. 세포의 용해를 촉진하고 단백질을 가용화하기 위해, 각종 계면활성제, 염 및 완충액을 사용할 수 있다. 프로테아제 및 포스파타제 억제제는 그 자체의 효소에 의한 샘플의 소화를 방지하기 위해 종종 첨가된다. 조직의 준비는 종종 단백질의 변성을 회피하기 위해 저온에서 수행된다.
샘플의 단백질은 겔 전기영동을 사용하여 분리된다. 단백질의 분리는 등전점(pI), 분자량, 전하 또는 이러한 요인의 조합에 의해 수행될 수 있다. 분리의 성질은 샘플의 처리와 겔의 성질에 따라 상이하다. 이것은 단백질을 결정하기 위한 매우 유용한 방법이다. 단일 샘플로부터의 단백질을 2차원으로 확산시키는 2차원(2-D) 겔을 사용하는 것도 가능하다. 단백질은 1차원에서는 등전점(중성의 순 전하를 갖는 pH)에 따라, 2차원에서는 분자량에 따라 분리된다.
단백질을 항체 검출에 이용할 수 있도록 하기 위해, 이들은 겔 내로부터 니트로셀룰로스 또는 폴리비닐리덴 디플루오라이드(PVDF)로 제조된 막으로 이동된다. 막을 겔 위에 놓고, 그 위에 여과지 스택을 놓는다. 전체 스택이 완충액에 배치되고, 모세관 작용에 의해 종이 위로 이동하고, 단백질이 함께 운반된다. 단백질을 이동시키는 또 다른 방법은 전기블롯팅(electroblotting)이라고 하고, 전류를 사용하여 단백질을 겔로부터 PVDF 또는 니트로셀룰로스 막으로 끌어당긴다. 단백질은 겔 내의 조직을 유지하면서 겔 내로부터 막으로 이동한다. 이 블롯팅 과정의 결과로서, 단백질은 검출을 위해 얇은 표면층에 노출된다(아래 참고). 두 종류의 막은 비특이적 단백질 결합 특성에 대해 선택된다(즉, 모든 단백질에 동등하게 결합함). 단백질 결합은 소수성 상호작용 및 막과 단백질 사이의 하전 상호작용을 기반으로 한다. 니트로셀룰로스 막은 PVDF보다 저렴하지만, 훨씬 더 손상되기 쉽고, 반복되는 프로빙을 충분히 견디지 못한다. 겔로부터 막으로의 단백질의 이동의 균일성과 전반적인 효과는 쿠마시 브릴리언트 블루(Coomassie Brilliant Blue) 또는 폰소(Ponceau) S 염료로 막을 염색하여 확인할 수 있다. 일단 이동되면, 단백질은 표지된 제1 항체 또는 비표지된 제1 항체를 사용하여 검출하고, 이어서 제1 항체의 Fc 영역에 결합하는 표지된 단백질 A 또는 제2 표지된 항체를 사용하여 간접적으로 검출한다.
C.
측면 유동 분석
측면 유동 면역크로마토그래피 분석으로도 공지된 측면 유동 분석은 특수한 고가의 장비 없이도 샘플(매트릭스) 중의 표적 분석물의 존재(또는 부재)를 검출하는 것을 목적으로 하는 단순한 장치이지만, 판독 장치에 의해 지원되는 다수의 실험실 기반 적용이 존재한다. 전형적으로, 이러한 시험은 가정에서의 시험, 현장 진단(point of care) 시험 또는 실험실 사용을 위해 저 자원의 의료 진단으로 사용된다. 널리 확산되고 주지된 용도는 가정용 임신 검사이다.
이 기술은 다공성 종이 또는 소결 중합체와 같은 일련의 모세관 베드를 기반으로 한다. 이들 요소 각각은 자발적으로 체액(예를 들어, 소변)을 수송하는 능력을 갖는다. 제1 요소(샘플 패드)는 스폰지 역할을 하고, 과량의 샘플 유체를 보유한다. 일단 침지되면, 입자의 표면에 고정된 표적 분자(예를 들어, 항원) 및 이의 화학 파트너(예를 들어, 항체) 사이의 최적화된 화학적 반응을 보증하기 위해 모든 것을 함유하는 염-당 매트릭스에 생리활성 입자의 건조된 포맷(아래 참고), 소위 접합체를 제조업체가 보관한 제2 요소(접합체 패드)로 유체가 이동한다. 샘플 유체는 염-당 매트릭스를 용해하지만, 입자도 용해하고, 하나의 조합된 수송 작용으로, 샘플과 접합체가 다공성 구조를 통해 유동하면서 혼합된다. 이러한 방식으로, 분석물은 제3 모세관 베드를 통해 추가로 이동하면서 입자에 결합한다. 이 물질에는, 제조업체가 제3 분자를 고정시킨 하나 이상의 영역(종종 스트립으로 지칭)이 있다. 샘플-접합체 혼합물이 이러한 스트립에 도달할 때까지, 분석물은 입자에 결합하고, 제3 '포획' 분자는 복합체에 결합한다. 잠시 후, 보다 많은 유체가 스트립을 통과하면, 입자가 축적되고, 스트립 영역의 색상이 변화된다. 전형적으로, 적어도 2개의 스트립이 있다: 하나(대조군)는 임의의 입자를 포획하고, 이에 의해 반응 조건과 기술이 정상으로 기능하는 것을 나타내고, 다른 하나는 특정 포획 분자를 포함하고, 분석물 분자가 고정된 입자만을 포획한다. 이러한 반응 구역을 통과한 후, 유체는 마지막의 다공성 재료 - 심지 -로 유입되고, 이는 단순히 폐기물 용기로서 기능한다. 측면 유동 시험은 경쟁 분석 또는 샌드위치 분석으로 작동할 수 있다. 측면 유동 분석은 미국 특허 제6,485,982호에 개시되어 있다.
D.
면역조직화학
본 개시내용의 항체는 또한 면역조직화학(IHC)에 의한 연구를 위해 제조된, 신선 동결 및/또는 포르말린 고정, 파라핀 포매 조직 블록 둘 모두와 조합하여 사용될 수 있다. 이러한 미립자 표본으로부터 조직 블록을 제조하는 방법은 다양한 예후 인자에 대한 이전의 IHC 연구에서 성공적으로 사용되었고, 당업자에게 주지되어 있다(Brown et al., 1990; Abbondanzo et al., 1990; Allred et al., 1990).
간략하면, 작은 플라스틱 캡슐 내의 인산염 완충 식염수(PBS)에서 실온에서 50 ng의 동결 "분쇄" 조직을 재수화하고; 원심분리에 의해 입자를 펠릿화하고; 점성 포매 매질(OCT)에 이들을 재현탁시키고; 캡슐을 반전시키고/시키거나 원심분리에 의해 다시 펠릿화하고; -70℃ 이소펜탄에서 급속 동결시키고; 플라스틱 캡슐을 절단하고/하거나 동결된 조직 실린더를 제거하고; 크라이오스탯 마이크로톰 척에 조직 실린더를 고정하고/하거나; 캡슐로부터 25~50개의 연속 절편을 절단함으로써 동결 절편을 제조할 수 있다. 대안적으로, 전체 동결 조직 샘플을 연속 절편 절단에 사용할 수 있다.
영구 절편은 플라스틱 마이크로퓨지 튜브에서 50 mg의 샘플을 재수화하고; 펠릿화하고; 10%의 포르말린에 재현탁하여 4시간 동안 고정시키고; 세척/펠릿화하고; 따뜻한 2.5%의 아가에 재현탁시키고; 펠릿화하고; 빙수에서 냉각하여 아가를 고화시키고; 튜브로부터 조직/아가 블록을 제거하고; 파라핀에 블록을 침윤 및/또는 포매하고/하거나; 최대 50개의 연속 영구 절편을 절단하는 것을 수반하는 유사한 방법으로 제조할 수 있다. 또한, 전체 조직 샘플을 대용할 수 있다.
E.
면역검출 키트
또 다른 구현예에서, 본 개시내용은 전술한 면역검출 방법과 함께 사용하기 위한 면역검출 키트에 관한 것이다. 항체는 SARS-CoV-2 또는 SARS-CoV-2 항원을 검출하기 위해 사용될 수 있기 때문에, 항체는 키트에 포함될 수 있다. 따라서, 면역검출 키트는 적절한 용기 수단에서 SARS-CoV-2 또는 SARS-CoV-2 항원에 결합하는 제1차 항체, 및 선택적으로 면역검출 시약을 포함할 것이다.
특정 구현예에서, SARS-CoV-2 항체는 컬럼 매트릭스 및/또는 미세역가 플레이트의 웰 등의 고체 지지체에 사전에 결합시킬 수 있다. 키트의 면역검출 시약은 주어진 항체와 연관되거나 연결된 검출 가능한 표지를 포함하여 다양한 형태 중 어느 하나를 취할 수 있다. 제2 결합 리간드와 연관되거나 부착된 검출 가능한 표지도 또한 고려된다. 예시적인 제2 리간드는 제1 항체에 대해 결합 친화성을 갖는 제2 항체이다.
본 키트에 사용하기 위한 추가의 적합한 면역검출 시약은 제1 항체에 대해 결합 친화성을 갖는 제2 항체 및 제2 항체에 대해 결합 친화성을 갖는 제3 항체를 포함하는 2성분 시약을 포함하고, 제3 항체는 검출 가능한 표지에 연결된다. 위에서 언급된 바와 같이, 다수의 예시적 표지가 당해 분야에 공지되어 있고, 이러한 모든 표지를 본 개시내용과 관련하여 사용할 수 있다.
키트는 검출 분석을 위한 표준 곡선을 작성하기 위해 사용될 수 있는 바와 같이, 표지되든 표지되지 않든, SARS-CoV-2 또는 SARS-CoV-2 항원의 적절하게 분취된 조성물을 추가로 포함할 수 있다. 키트는 완전히 접합된 형태로, 중간체의 형태로, 또는 키트 사용자에 의해 접합되는 별도의 모이어티로서 항체-표지 접합체를 포함할 수 있다. 키트의 구성요소는 수성 매질 또는 동결건조된 형태로 패키징될 수 있다.
키트의 용기 수단은 일반적으로, 항체를 위치시킬 수 있거나, 또는 바람직하게는 적절하게 분취할 수 있는, 적어도 하나의 바이알, 시험관, 플라스크, 병, 주사기 또는 기타 용기 수단을 포함한다. 본 개시내용의 키트는 또한 전형적으로, 상업적 판매를 위해 항체, 항원 및 기타 임의의 시약 용기를 긴밀하게 밀폐하여 수용하기 위한 수단을 포함할 것이다. 이러한 용기는 원하는 바이알이 유지되는 주사 또는 취입 성형 플라스틱 용기를 포함할 수 있다.
F.
백신 및 항원 품질 관리 분석
본 개시내용은 또한 샘플 중의 바이러스 항원의 항원성 무결성을 평가하는 데 사용하기 위한 본원에 기재된 항체 및 항체 단편의 용도를 고려한다. 백신과 같은 생물학적 의약품은 보통 분자적으로 특성화될 수 없다는 점에서 화학 의약품과 상이하다. 항체는 상당히 복잡한 큰 분자이고, 제조마다 광범위하게 달라지는 능력을 갖는다. 이들은 또한 생애 초의 소아를 포함하여 건강한 개체에게 투여되고, 따라서 그 자체가 해를 유발하지 않으면서, 생명을 위협하는 질환의 예방 또는 치료에 효과적임을 가능한 한 보장하기 위해, 이들의 품질에 중점을 두어야 한다.
백신의 생산 및 유통에서 증가하는 세계화는 공중 보건에 대한 우려를 보다 잘 관리하기 위해 새로운 가능성을 열었지만, 다양한 공급원에서 조달된 백신의 동등성과 호환성에 대한 의문도 제기하였다. 따라서, 출발 물질, 생산 및 품질 관리 시험의 국제 표준화 및 이러한 제품의 제조 및 사용 방식에 대한 규제 감독에 대한 높은 기대의 설정은 지속적 성공의 초석으로 되었다. 그러나 이는 끊임없이 변화하는 분야이고, 이 분야에서의 지속적 기술 발전은 가장 오래된 공중 보건의 위협 및 새로운 위협 - 말라리아, 유행성 인플루엔자, HIV 등 - 에 대해 강력한 새로운 무기의 개발 약속을 제공하지만, 또한 제품이 달성 가능한 최고 기준의 품질을 지속적으로 충족하는 것을 보증하기 위해, 제조업체, 규제 당국 및 광범위한 의료계에 큰 압력을 가하고 있다.
따라서, 항원 또는 백신은 임의의 공급원으로부터 또는 제조 과정 중 임의의 시점에서 수득할 수 있다. 따라서, 품질 관리 프로세스는 바이러스 항원에 대한 본원에 개시된 항체 또는 단편의 결합을 확인하는 면역분석을 위한 샘플을 제조하는 것으로부터 개시할 수 있다. 이러한 면역분석은 이 문서의 다른 부분에 개시되어 있고, 이들 중 어느 것을 사용하여, 항원의 구조적/항원 무결성을 평가할 수 있다. 허용 가능한 양의 항원적으로 정확하고 온전한 항원을 포함하는 샘플을 발견하기 위한 기준은 규제 당국에 의해 확립될 수 있다.
항원의 무결성이 평가되는 또 다른 중요한 구현예는 저장 수명 및 저장 안정성을 결정하는 데 있다. 백신을 포함한 대부분의 의약은 시간이 경과됨에 따라 열화될 수 있다. 따라서, 시간 경과에 따라, 백신에서와 같은 항원이 대상체에게 투여될 때 더 이상 항원성이 아니고/아니거나 면역 반응을 생성할 수 없도록, 분해되거나 불안정화하는지 여부, 그 정도를 결정하는 것이 중요하다. 다시, 허용 가능한 양의 항원적으로 온전한 항원을 포함하는 샘플을 발견하기 위한 기준은 규제 당국에 의해 확립될 수 있다.
특정 구현예에서, 바이러스 항원은 한 개 초과의 보호 에피토프를 함유할 수 있다. 이러한 경우, 2, 3, 4, 5 또는 그 이상의 항체와 같은 하나 초과의 항체의 결합을 조사하는 분석을 사용하는 것이 유용할 수 있다. 이러한 항체는 밀접하게 관련된 에피토프에 결합하기 때문에, 서로 인접하거나 심지어 중첩된다. 한편, 이들은 항원의 상이한 부분으로부터 별개의 에피토프를 나타낼 수 있다. 복수의 에피토프의 무결성을 조사함으로써, 항원의 전체 무결성에 대한 보다 완전한 그림을, 따라서 보호 면역 반응을 생성하는 능력을 결정할 수 있다.
본 개시내용에 기재된 바와 같은 항체 및 이의 단편은 또한 보호 SARS-CoV-2 항체의 존재를 검출함으로써 백신접종 절차의 효능을 모니터링하기 위한 키트에 사용될 수 있다. 본 개시내용에 기재된 바와 같은 항체, 항체 단편, 또는 이의 변이체 및 유도체는 또한 원하는 면역원성을 갖는 백신 제조를 모니터링하기 위한 키트에 사용될 수 있다.
G.
실시예
다음의 실시예는 바람직한 구현예를 설명하기 위해 포함된다. 다음의 실시예에 개시된 기술은 구현예의 실시에서 양호하게 기능하기 위해 발명자에 의해 발견된 기술을 나타내고, 따라서 이의 실시를 위한 바람직한 방식을 구성하는 것으로 간주될 수 있음이 당업자에 의해 이해되어야 한다. 그러나 당업자는 본 개시내용에 비추어, 개시되는 특정 구현예에서 다수의 변경이 이루어질 수 있고, 본 개시내용의 사상 및 범위를 벗어나지 않고서 여전히 비슷하거나 유사한 결과를 수득할 수 있음을 인식해야 한다.
실시예 1 - 항체 시너지
시너지는 본원에서 항체의 동일한 총 농도(시험관 내) 또는 용량(생체 내)에서 개별 mAb에 의해 매개되는 것과 비교할 때 2개의 mAb의 칵테일에 의해 매개되는 더 높은 중화 활성으로 정의된다. 2개의 mAb가 SARS-CoV-2를 중화하기 위해 칵테일에서 상승 작용을 하는지 평가하기 위해, 본 발명자는 시너지를 정량화하기 위해 이전에 보고된 접근법을 사용했다(Ianevski A, He L, Aittokallio T, Tang J. Bioinformatics. 33, 2413-2415, 2017). mAb 조합으로 인한 유익한 효과의 중요성을 평가하기 위해, 관찰된 조합 반응(용량-반응 매트릭스)을 시너지 스코어링 모델(Ianevski A, He L, Aittokallio T, Tang J. Bioinformatics. 33, 2413-2415, 2017)에 의해 예상 반응과 비교하였다. 야생형 SARS-CoV-2 및 Vero-E2 세포 배양 단층을 사용하여 기존의 포커스 감소 중화 시험(focus reduction neutralization test, FRNT) 분석에서 바이러스 중화를 측정하였다. 개별 mAb COV2-2196 및 COV2-2130을 상이한 농도로 혼합하여 칵테일에서 상이한 mAb 비율의 중화 활성을 평가하였다. 구체적으로, mAb COV2-2130(500 ng/mL부터 시작)의 7배 희석액 각각을 각 조건당 총 부피 50 μL의 mAb COV2-2196(500 ng/mL부터 시작)의 9개 희석액 각각과 혼합한 다음, 96웰 플레이트에서 성장시킨 합류 Vero-E2 세포에 적용하기 전에 세포 배양 배지(2% FBS가 보충된 RPMI-1640 배지)에서 50 μL의 생 SARS-CoV-2와 함께 인큐베이션하였다. 대조군 값에는 칵테일에서와 동일한 용량으로 평가된 개별 mAb COV2-2196 및 COV2-2130에 대해 개별적으로 측정된 중화 활성의 용량-반응에 대한 값이 포함되었다(도 1 내지 3 참고). 각 실험을 2중 반복실험으로 수행하였다. 다음으로 본 발명자는 각 조건에 대한 바이러스 중화 백분율을 계산한 다음, 시너지 점수 값을 계산하였는데, 이는 칵테일에서 이들 2개의 mAb 사이의 상호작용을 상승 작용적(시너지 점수 = 17.4)으로 정의하였다. 참고로, -10 미만의 시너지 점수는 길항, -10 내지 10의 점수는 부가 효과, 10 초과의 점수는 시너지 효과를 나타낸다. 도 1 내지 3의 예는 용량-반응 매트릭스를 보여주고 칵테일에서 79 ng/mL의 조합된 mAb 용량(16 ng/mL의 COV2-2196 및 63 ng/mL의 COV2-2130)이 250 ng/mL의 각각의 개별 mAb와 동일한 활성을 가짐을 보여준다. 이 발견은 칵테일에서 바이러스 중화에서 동일한 효능을 달성하기 위해 용량을 3배 넘게 줄일 수 있음을 보여준다.
처리의 치료적 효능을 평가하기 위해, 본 발명자들은 먼저 MA-SARS-CoV-2 챌린지 모델을 사용하여 mAb COV2-2196 또는 COV2-2130 또는 이들의 1:1 조합을 테스트하였다. 모든 처리는 바이러스 접종 후 2일째에 폐 조직의 플라크 역가에 의해 측정된 바와 같이 폐에서 감염성 바이러스를 감소시켰다. 400 μg/마우스(약 20 mg/kg)의 용량으로 전달된 칵테일 처리는 폐 부담을 최대 3 x 104배까지 유의하게 감소시켰기 때문에 가장 효율적이었다. 이 처리군의 5마리 중 4마리는 폐에 더 이상 감염성 바이러스가 없었다(도 4a). 유사하게, SARS-CoV-2 수용체 인간 ACE2(AdV-hACE2)를 발현시키기 위해 재조합 아데노바이러스로 형질도입된 폐를 가진 마우스를 실제 SARS-CoV-2 바이러스 챌린지 12시간 후에 400 μg/마우스의 mAb 칵테일로 처리한 결과, 생체 내 폐에서 감염성 바이러스의 완전한 중화가 나타났다(도 4b). 염증 지표인 INF-γ, IL-6, CXCL10 및 CCL2 사이토카인 및 케모카인 유전자의 발현은 또한 이소형 대조군 처리 마우스의 폐와 비교할 때 mAb 칵테일 처리된 마우스의 폐에서 감소되었다(도 4c). 이러한 결과의 종합은 마우스 SARS-CoV-2 챌린지 모델에서 COV2-2196 + COV2-2130의 칵테일에 의해 매개되는 노출 후 치료 효능을 시사하였다.
실시예 2 - 비인간 영장류 챌린지 연구
MAb 생산 및 정제. 합성되고(Twist Bioscience) IgG1 모노시스트론 발현 벡터(pTwist-mCis_G1로 명명됨) 내로 클로닝된 mAb의 서열을 포유동물 세포 배양 mAb 분비에 사용하였다. 이 벡터는 형질감염 시 단일 구성체에서 mAb 중쇄 및 경쇄 유전자의 동시 발현을 허용하는 증진된 2A 서열 및 GSG 링커를 함유한다1. 본 발명자들은 이전에 96웰 플레이트에서 1 mL의 ExpiCHO 배양액에서 mAb의 마이크로규모의 발현을 설명하였다2. 더 큰 규모의 mAb 발현을 위해, 본 발명자들은 공급업체가 설명한 바와 같이 Gibco™ ExpiCHO™ 발현 시스템 및 50 mL의 미니 생물반응기 튜브(Corning)에 대한 프로토콜을 사용하여 CHO 세포 배양물의 형질감염(항체당 1 내지 300 mL)을 수행하였다. 배양 상청액을 24칼럼 병렬 단백질 크로마토그래피 시스템(Protein BioSolutions)에서 HiTrap MabSelect SuRe(Cytiva, 이전 GE Healthcare Life Sciences)를 사용하여 정제하였다. 정제된 mAb를 PBS로 완충액을 교환하고, Amicon® Ultra-4 50KDa 원심분리 필터 유닛(Millipore Sigma)을 사용하여 농축하고 사용할 때까지 4℃에서 보관하였다. 정제된 mAb를 NHP 연구에 대해 <1 EU/mg IgG인 것으로 밝혀진 내독소 수준에 대해 일상적으로 테스트하였다. 내독소 테스트는 감도 범위가 10 내지 0.1 EU/mL인 PTS201F 카트리지(Charles River)와 Endosafe Nexgen-MCS 기기(Charles River)를 사용하여 수행하였다.
본 발명자들은 최근에 설명된 SARS-CoV-2 비인간 영장류(NHP) 챌린지 모델3,4을 사용하여 mAb의 보호 효능을 테스트하였다. 이 모델의 경우, 본 발명자들은 COV2-2196과 동일한 가변 유전자 절편에 의해 암호화되지만 HCDR3 및 LCDR3에서 다수의 주목할만한 아미노산 차이를 사용하는 중화 mAb를 단독요법 COV2-2196으로 테스트하였다. 동물에게 -3일에 하나의 50 mg/kg 용량의 mAb COV2-2196 또는 이소형 대조군 mAb를 정맥 내 투여한 다음, 0일에 1.1 x 104 PFU의 용량의 SARS-CoV-2로 비강 내 및 기관 내 챌린지를 수행하였다. 챌린지 후, 본 발명자들은 기관지 폐포 세척액(BAL) 및 비강 스왑에서 RT-qPCR에 의해 바이러스 RNA를 평가하였다. 비강 스왑에서 7.53(범위 5.37~8.23) 로그10 RNA 카피/스왑의 중앙값 피크와 BAL에서 4.97(범위 3.81~5.24) 로그10 RNA 카피/mL의 중앙값 피크와 함께, 높은 수준의 하위 게놈 바이러스 RNA가 이소형 대조군 mAb 처리 NHP에서 관찰되었다. 하위 게놈 바이러스 RNA는 보호를 나타내는 mAb 처리군(LOD = 50 [1.7 로그10] RNA 카피/스왑 또는 mL 당)의 샘플에서 검출되지 않았다. 약동학 분석은 NHP에서 순환하는 인간 mAb의 안정적인 농도를 보여주었다.
NHP 챌린지 연구. NHP 연구는 실험 동물의 관리 및 사용에 관한 지침서 제8판에 명시된 원칙을 준수하였다. 이 연구가 수행된 시설(Bioqual Inc., 미국 메릴랜드주 록빌)은 국제실험동물관리평가인증협회(AAALAC)에서 완전히 승인을 받았으며 승인된 실험 동물 복지 사무소가 있다. NHP 연구는 모든 관련 지역, 주 및 연방 규정에 따라 수행되었으며, 관련 동물실험윤리위원회(IACUC)의 승인을 받았다.
원주지 인도의 건강한 성체 레서스 마카크원숭이(rhesus macaques, Macaca mulatta) 8마리를 연구하였다. 동물을 항-SARS-CoV-2 mAb 처리군(군당 n = 4)과 하나의 대조(이소형 처리)군(군당 n = 4)에 무작위로 할당하였다. 동물에게 -3일에 하나의 50 mg/kg 용량의 mAb COV2-2196 또는 이소형 대조군 mAb를 정맥 내 투여한 다음, 3일 동안 i.n 경로로 1 mL 및 기관 내 경로로 1 mL로 투여된 1.1 x 104 PFU의 SARS-CoV-2로 챌린지를 수행하였다. 챌린지 후, 바이러스 RNA를 설명된 대로5,6 여러 시점에서 기관지 폐포 세척액 및 비강 스왑에서 RT-qPCR에 의해 평가하였다. 모든 동물은 신체 검사를 받았다. 또한, 모든 동물은 동물실험윤리위원회에서 승인한 내부 스코어링 프로토콜에 따라 매일 모니터링되었다. 이 연구는 맹검 처리되지 않았다.
NHP 혈청에서 순환하는 인간 mAb의 검출. ELISA 플레이트를 1 μg/mL의 염소 항-인간 IgG(H+L) 2차 항체(원숭이 사전 흡착)(Novus Biological)로 4℃에서 밤새 코팅한 다음, 2시간 동안 차단하였다. 혈청 샘플은 PBS(ThermoFisher) 희석제 중 차단제 카제인의 1:3 희석액에서 시작하여 3배 희석으로 분석하였다. 샘플을 주위 온도에서 1시간 동안 인큐베이션한 다음 제거하고,플레이트를 세척하였다. 이어서, 웰을 1:4,000 희석액에서 HRP-접합된 염소 항-인간 IgG(원숭이 사전 흡착)(Southern Biotech)와 함께 1시간 동안 인큐베이션하였다. 웰을 세척한 다음, SureBlue Reserve TMB 마이크로웰 퍼옥시다제 기질(Seracare)(100 μL/웰)과 3분 동안 인큐베이션한 후, TMB 정지 용액(Seracare)과 함께 인큐베이션하여 반응(100 μL/웰)을 정지시켰다. 마이크로플레이트를 450 nm에서 판독하였다. 인간 mAb의 농도를 Prism 소프트웨어 버전 8.0(GraphPad)을 사용하여 정제된 인간 IgG(Sigma) 표준 곡선의 선형 범위에서 보간하였다.
실시예 3 - 실시예 4의 재료 및 방법
SARS-CoV-2 스파이크 단백질의 재조합 수용체 결합 도메인(RBD)의 발현 및 정제. S 단백질 RBD(잔기 319 ~ 528)에 상응하는 DNA 절편을 발현을 위해 서열 최적화하고, 합성하고, IL-2 신호 펩티드(MYRMQLLSCIALSLALVTNS)의 하류에 있는 pTwist-CMV 발현 DNA 플라스미드(Twist Bioscience)로 클로닝하였다. 3개의 아미노산 링커(GSG) 및 His-태그가 단백질 정제를 용이하게 하기 위해 발현 구성체의 C 말단에 통합되었다. Expi293F 세포를 RBD를 암호화하는 플라스미드로 일시적으로 형질감염시키고, 배양 상청액을 5일 후에 수확하였다. RBD를 HisTrap Excel 컬럼(GE Healthcare Life Sciences)을 사용한 니켈 친화성 크로마토그래피에 의해 상청액으로부터 정제하였다. 결정화 시험에 사용되는 단백질 생산을 위해 5 μM의 키푸넨신이 배양 배지에 포함되어 높은 만노스 글리칸이 있는 RBD를 생산하였다. 높은 만노스 당단백질을 이후에 엔도글리코시다제 F1(Millipore)으로 처리하여 균일하게 탈글리코실화된 RBD를 수득하였다.
재조합 COV2-2196 및 COV2-2130 Fab의 발현 및 정제. 인간 IgG1 CH1 도메인을 갖는 COV2-2196 및 COV2-2130 중쇄 가변 도메인 및 인간 카파 쇄 불변 도메인을 갖는 경쇄 가변 도메인에 상응하는 DNA 단편을 합성하고 pTwist 벡터(Twist Bioscience)에 클로닝하였다. 이 벡터는 각 Fab의 중쇄에 이어 GGGGS 링커, 푸린 절단 부위, T2A 리보솜 절단 부위 및 각 Fab의 경쇄를 포함한다. 중쇄 및 경쇄의 발현은 동일한 CMV 프로모터에 의해 유도된다. COV2-2196 및 COV2-2130 Fab는 발현 플라스미드를 사용한 일시적 형질감염에 의해 ExpiCHO 세포에서 발현되었다. 재조합 Fab는 항-CH1 CaptureSelect 컬럼(Thermo Fisher Scientific)을 사용하여 배양 상청액으로부터 정제되었다. RBD/COV2-2196 복합체의 경우, COV2-2196의 wt 서열을 발현에 사용하였다. RBD/COV2-2196/COV2-2130 복합체의 경우, 가변 영역의 처음 2개 아미노산이 QM에서 EV로 돌연변이된 변형된 버전의 COV2-2196 Fab가 사용되었다.
항체-항원 복합체의 결정화 및 구조 결정. 정제된 COV2-2196 Fab를 탈글리코실화된 RBD와 1:1.5의 몰비로 혼합하고 혼합물을 Superdex-200 Increase 컬럼(GE Healthcare Life Sciences)을 이용한 크기 배제 크로마토그래피로 추가 정제하여 항체-항원 복합체를 얻었다. RBD/COV2-2196/COV2-2130 삼중 복합체를 얻기 위해, 정제 및 탈글리코실화된 RBD를 COV2-2196 Fab 및 COV2-2130 Fab와 1:1.5:1.5의 몰비로 혼합하고, 삼중 복합체를 Superdex-200 Increase 컬럼으로 정제하였다. 복합체를 약 10 mg/mL까지 농축하고 결정화 시험을 수행하였다. RBD/COV2-2196 복합체는 16%~18%의 PEG 3350, 0.2 Tris-HCl pH 8.0~8.5에서 결정화되었고, RBD/COV2-2196/COV2-2130 복합체는 5%(w/v) PEG 1000, 100 mM 인산나트륨 이염기성/구연산 pH 4.2, 40%(v/v) 시약 알코올에서 결정화되었다. 동결보호 용액은 두 복합체의 결정에 대해 결정화 용액을 100% 글리세롤과 20:7의 부피비로 혼합하여 제조하였다. 단백질 결정을 동결보호 용액에 신속하게 담근 후 액체 질소에서 급속 동결시켰다. 회절 데이터를 Advanced Photon Source에서 RBD/COV2-2196 복합체의 경우 빔라인 21-ID-F, RBD/COV2-2196/COV2-2130 복합체의 경우 21-ID-G에서 수집하였다. 회절 데이터를 XDS58 및 CCP4 제품군59으로 처리하였다. 프로그램 Phaser60를 사용하여 Fab CC12.1과 복합된 복합체 내의 RBD 구조(PDB ID 6XC2) 및 MR78의 Fab 구조(PDB ID 5JRP)를 사용하여 분자 치환에 의해 결정 구조를 풀었다. 구조를 각각 Phenix61 또는 Coot62를 사용하여 수동으로 정련하고 재건하였다. 모델은 단백질 정보은행(Protein Data Bank)에 기탁되었다. PyMOL 소프트웨어63는 모든 구조 도면을 제작하는 데 사용되었다.
수소 중수소 질량 분석(HDX-MS) 실험. HDX-MS 실험은 M-Class Acquity LC 시스템 및 HDX 매니저(Waters Ltd., 영국 윔슬로우)에 연결된 자동화된 샘플 처리 로봇(LEAP technologies, 미국 플로리다주 포트로더데일)을 사용하여 수행하였다. 7.6 μL의 4.3 μM SARS-CoV-2 S2P 스파이크 삼량체를 단독으로 또는 1:1.12 몰비의 AZD1061 또는 AZD8895와 함께 52.4 μL 표지 완충액(50 mM의 인산칼륨 pD 6.2)에 첨가하고 20℃에서 1분 동안 인큐베이션하였다. 인큐베이션 후, 이 샘플 50 μL를 1℃에서 50 μL의 ?치 용액(50 mM의 인산칼륨, 200 mM의 TCEP, 2 M의 Gdn-HCl, pH 2.3)에 첨가하였다. 50 μL의 ?칭된 샘플을 4분 동안 100 μL min-1(약 4,000 psi)에서 20℃에서 고정된 BEH 펩신 컬럼(Waters Ltd., 영국 윔슬로우)에 통과시킨 후, 생성된 펩티드를 Vanguard 프리컬럼 Acquity UPLC BEH C18 트랩 컬럼(1.7 μm, 2.1 mm x 5 mm, Waters Ltd., 영국 윔슬로우)을 이용하여 포획하였다. 밸브 전환 후, Acquity UPLC BEH C18 분석 컬럼(1.7 μm, 1 mm x 100 mm)을 이용하여 40 μL min-1에서 6분에 걸쳐 H2O(0.2% v/v 포름산) 중 0에서 35%의 MeCN(0.2% v/v 포름산)의 구배 용출에 의해 1℃에서 펩티드를 분리하였다. 120K의 분해능의 오비트랩 검출 모드(중수소화 샘플) 또는 오비트랩-이온트랩 DDA 모드(t=0 샘플)에서 작동하는 오비트랩 융합 질량 분석기(Thermo Fisher, 독일 브레멘)를 사용하여 펩티드를 분석하였다. 펩티드 확인에 사용된 펩티드 MS/MS 데이터는 BioPharma Finder v3.0(Thermo Fisher, 독일 브레멘)을 사용하여 분석하였고, 중수소 흡수는 HDExaminer v2.0(Sierra Analytics)을 사용하여 정량화하였다. PAVED(University of Leeds)64를 사용하여 흡수 플롯 수치와 구조적 히트맵을 생성하기 위해 HDExaminer의 펩티드 풀 결과 및 흡수 요약 테이블 csv 내보내기를 사내 R: 스크립트를 사용하여 다시 포맷팅하였다.
COV2-2196 돌연변이체의 ELISA 결합. 384웰 미세적정 플레이트의 웰을 4℃에서 밤새 정제된 재조합 SARS-CoV-2 S 6P 단백질로 코팅하였다. 플레이트를 1시간 동안 0.05% 트윈-20(DPBS-T)을 함유하는 DPBS에서 2% 무지방 분유 및 2% 정상 염소 혈청으로 차단하였다. 항체를 10 μg/mL까지 희석하고 DPBS-T에서 23회 2배 적정하고, 웰에 첨가하고, 실온에서 1시간 동안 인큐베이션하였다. 결합된 항체는 서양고추냉이 퍼옥시다제와 접합된 염소 항-인간 IgG(Southern Biotech) 및 TMB 기질(Thermo Fischer Scientific)을 사용하여 검출하였다. 반응을 1 N 염산으로 ?칭하고, 분광광도계(Biotek)를 사용하여 450 nm에서 흡광도를 측정하였다.
항체 결합을 회피하는 모든 돌연변이의 매핑. 항체 결합을 회피하는 모든 돌연변이를 DMS 접근법41을 통해 매핑하였다. 본 발명자들은 이전에 설명된 효모 디스플레이 RBD 돌연변이체 라이브러리41,42를 사용하였다. 간략하면, 이중(duplicate) 돌연변이체 라이브러리를 SARS-CoV-2(Wuhan-Hu-1 분리주, Genbank 수탁번호 MN908947, 잔기 N331~T531)의 스파이크 수용체 결합 도메인(RBD)에 구성하였는데, 3,819개의 가능한 아미노산 중 3,804개를 포함하고, >95%가 단일 돌연변이체로 존재한다. 각 RBD 변이체를 하류 시퀀싱을 용이하게 하기 위해 고유한 16개 뉴클레오티드 바코드 서열에 연결하였다. 이전에 설명된 바와 같이, 라이브러리를 RBD 발현 및 ACE2 결합에 대해 분류하여 완전히 미스폴딩되었거나 기능이 없는(즉, 적당한 ACE2 결합 친화력이 없는41) RBD 변이체를 제거하였다.
항체 회피 매핑 실험을 이전에 설명된 대로41, 약간 수정하여, 2개의 독립적인 돌연변이체 RBD 라이브러리를 사용하여 생물학적 2중 반복실험으로 수행하였다. 간략하면, RBD를 발현하도록 유도된 돌연변이체 효모 라이브러리를 세척하고 부드럽게 교반하면서 실온에서 1시간 동안 400 ng/mL의 항체와 함께 인큐베이션하였다. 항체 인큐베이션 후, 라이브러리를 1:100 FITC-접합된 항-MYC 항체(Immunology Consultants Lab, CYMC-45F)로 2차적으로 표지하여 RBD 발현에 대해 표지하고 1:200 PE-접합된 염소 항-인간-IgG(Jackson ImmunoResearch 109-115-098)로 2차적으로 표지하여 결합된 항체에 대한 표지하였다. 라이브러리 샘플의 1% 항체 농도로 표지된 돌연변이되지 않은 SARS-CoV-2 세포를 포획하기 위해 도출된 선택 게이트를 통해 항체 결합이 감소된 RBD 변이체를 발현하는 세포를 농축하기 위해 유세포 분석 분류를 사용하였다. 각 샘플에 대해, 대략 천만 개의 RBD+ 세포가 세포계측기에서 처리되었다. 항체 회피 세포를 SD-CAA(6.7 g/L 효모 질소 염기, 5.0 g/L 카사미노산, 1.065 g/L MES 산 및 2% w/v 덱스트로스)에서 밤새 성장시켜 플라스미드 추출 전에 세포를 확장시켰다.
플라스미드 샘플을 이전에 설명된 대로41 항체 회피 세포의 사전 선택 및 하룻밤 배양물(Zymoprep Yeast Plasmid Miniprep II)에서 준비하였다. 각 RBD 변이체를 확인하는 16개 뉴클레오티드 바코드 서열을 PCR에 의해 증폭시키고, 설명된 대로41,42 50 bp의 단일 말단 리드로 Illumina HiSeq 2500에서 시퀀싱하였다.
회피 분율은 아래에 언급된 대로 약간 수정하여 설명된 대로41 계산하였다. 본 발명자들은 dms_variants 패키지(jbloomlab.github.io/dms_variants/, 버전 0.8.2)를 사용하여 이전에 설명된 바코드/RBD 조회 표65를 사용하여 각 사전 분류 및 항체 회피 집단에서 Illumina 서열을 각각의 바코드화된 RBD 변이체의 카운트로 처리하였다.
각 항체 선택에 대해, 본 발명자들은 원래 및 항체 회피 집단의 각 변이체에 대한 심층 시퀀싱 카운트와 이전에 기재된 식41을 통해 항체 결합을 회피한 라이브러리의 총 분율을 사용하여 각 바코드된 변이체에 대한 "회피 분율"을 계산하였다. 이러한 회피 분율은 0의 값은 변이체가 항상 혈청에 의해 결합됨을 의미하고 1의 값은 변이체가 항상 항체 결합을 회피함을 의미하도록, 항체 회피 빈(bin)에 속하는 해당 특정 변이체를 발현하는 세포의 추정된 분율을 나타낸다. 이어서, 본 발명자들은 효모 세포 표면에서 적절하게 폴딩된 RBD의 낮은 발현을 간단히 유도함으로써 항체 회피를 유발할 수 있는 낮은 시퀀싱 카운트 또는 고도로 해로운 돌연변이를 가진 변이체를 제거하기 위해 계산 필터를 적용하였다41,42. 구체적으로, 그것들은 이전에 설명된 바와 같이42 변이체 수준 및 돌연변이 수준의 심층 돌연변이 스캐닝 스코어를 사용하여, ACE2 결합 스코어 < -2.35 또는 발현 스코어 < -1을 갖는(또는 이러한 돌연변이를 포함하는) 변이체를 제거하였다. 이러한 필터링 기준은 이전에 인간 항체 패널을 매핑하는 데 사용된 기준41보다 약간 더 엄격하지만 mAb65와 다클론 혈청54의 결합에 영향을 미치는 RBD 잔기를 정의하는 최근 연구에서 사용된 기준과 동일하다는 점에 유의한다.
다음으로 본 발명자들은 (jbloomlab.github.io/dms_variants/dms_variants.globalepistasis.html)에서 상술되고 설명된 바와 같이41, dms_variants 패키지에서 구현된 전역 상위성(global epistasis) 모델66을 사용하여 변이체 수준의 회피 스코어를 단일 돌연변이에 대한 회피 분율 추정치로 디콘볼루션하였다. 논문 전체에 걸쳐 보고된 회피 분율은 라이브러리 전체의 평균이다(도 18a 내지 18b에 제시된 상관관계). 이러한 스코어는 표 B에도 있다. 로고 플롯에서 강조 표시하기 위한 각 항체로부터의 강력한 회피 부위를, 합산된 돌연변이 회피 스코어가 선택의 부위별 총합 중앙값의 적어도 10배이고 가장 강력하게 선택된 부위의 부위별 합계의 10배 이내인 부위로 발견적으로 결정하였다. 계산 분석에 대한 전체 문서는 github.com/jbloomlab/SARS-CoV-2-RBD_MAP_AZ_Abs에 있다. 이러한 결과는 또한 https://jbloomlab.github.io/SARS-CoV-2-RBD_MAP_AZ_Abs/에서 대화형으로 이용 가능하다.
VSV-SARS-CoV-2를 이용한 항체 회피 선택 실험. COV2-2196 및 COV2-2130을 사용한 회피 선택 실험을 위해, 본 발명자들은 21개의 아미노산 C 말단이 결실된 SARS-CoV-2의 스파이크 단백질을 암호화하는 복제 가능한 재조합 VSV 바이러스43를 사용하였다. 스파이크 발현 VSV 바이러스를 이전에 설명한 대로43 MA104 세포(아프리카 녹색 원숭이, ATCC CRL-2378.1)에서 증식시켰고, 바이러스 스톡은 Vero E6 세포 단층 배양물에서 적정하였다. 플라크는 중성 적색 염색을 사용하여 시각화하였다. COV2-2196, COV2-2130 또는 COV2-2196과 COV2-2130의 1:1 혼합물로 구성된 칵테일의 존재 하에 선택된 회피 돌연변이를 스크리닝하기 위해, 본 발명자들은 실시간 세포 분석(RTCA) 및 xCELLigence RTCA MP 분석기(ACEA Biosciences Inc.) 및 이전에 설명된 회피 선택 체계41를 사용하였다. 간략하면, 50 μL의 세포 배양 배지(2% FBS가 보충된 DMEM)를 96웰 E-플레이트의 각 웰에 첨가하여 배경 판독값을 얻었다. 50 μL의 세포 배양 배지 중 18,000개의 Vero E6 세포를 웰당 시딩하고, 플레이트를 분석기에 놓았다. 측정은 15분마다 자동으로 이루어졌으며, 센소그램은 RTCA 소프트웨어 버전 2.1.0(ACEA Biosciences Inc)을 사용하여 시각화되었다. VSV-SARS-CoV-2 바이러스(웰당 5e3 플라크 형성 단위(PFU), 약 0.3 MOI)를 총 부피 100 μL 중 포화 중화 농도의 COV2-2196, COV2-2130 또는 COV2-2196과 COV2-2196 항체의 1:1 혼합물(항체의 총 농도 5 μg/mL)과 혼합하고, 37℃에서 1시간 동안 인큐베이션하였다. 세포를 시딩한 후 16~20시간에, 바이러스-항체 혼합물을 세포 단층에 첨가하였다. 항체 부존재 하에 바이러스만 함유하는 웰과 배지에 Vero E6 세포만 함유하는 웰을 각 플레이트에 대조군으로 포함시켰다. 플레이트를 72시간 동안 연속적으로(15분마다) 측정하였다. 회피 돌연변이를 세포 생존력의 저하에 대한 세포 지수를 모니터링하여 확인하였다. 항체 선택으로부터의 회피를 확인하기 위해, COV2-2130의 존재 하에 세포변성 효과가 관찰된 웰을 10 μg/mL의 COV2-2130 또는 COV2-2196의 존재 하에 후속 RTCA 실험에서 평가하였다. COV2-2130에 의한 중화에 대한 선택된 바이러스의 저항성을 확인한 후, 바이러스 분리주를 10 μg/mL의 COV2-2130의 존재 하에 Vero E6 세포에서 확장시켰다. 제조업체 프로토콜에 따라 QiAmp 바이러스 RNA 추출 키트(QIAGEN)를 사용하여 바이러스 RNA를 분리하고, SARS-CoV-2 스파이크 유전자를 역전사시키고, S 유전자의 측면에 있는 프라이머를 이용하여 SuperScript IV 1단계 RT-PCR 키트(ThermoFisher Scientific)를 사용하여 증폭시켰다. 증폭된 PCR 산물을 SPRI 자성 비드(Beckman Coulter)를 1:1 비율로 사용하여 정제하고, RBD의 정방향 및 역방향 리드를 제공하는 프라이머를 사용하여 Sanger 방법으로 시퀀싱하였다.
연속 계대 및 mAb 저항성 돌연변이를 선택하기 위한 SARS-CoV-2의 테스트. SARS-CoV-2 균주 USA-WA1/2020을 각각의 IC50 값에서 시작하여 각 계대에서 IC90 값까지 단계적으로 증가된 농도의 AZD8895, AZD1061 또는 AZD7442를 함유한 Vero 세포 단층 배양물에서 연속적으로 계대하였다. 대조군으로서, 바이러스를 항체의 부재 하에 계대하였다. 최종 계대 후, 플라크 분석에 의해 IC90 농도의 10배의 최종 농도에서 상호 항체에 대한 감수성에 대해 바이러스를 평가하였다. 플라크(n=6)는 AZD1061 배양물에 대해 무작위로 선택되었고, 이들의 바이러스 스파이크 암호화 유전자를 시퀀싱하였다.
변이체 잔기가 있는 바이러스를 포함한, 실제 SARS-CoV-2 바이러스의 단리 또는 생성. UK B.1.1.7-OXF 분리주를 영국 켄트의 감염된 개인의 비인두 스왑에서 얻었다. 서면 동의 후 검체를 얻기 위한 임상 연구는 영국 옥스포드의 존 래드클리프 병원(John Radcliffe Hospital)에서 승인하였다. 샘플을 2% FBS가 포함된 DMEM에 희석하고 0.45 μm 필터를 통과시킨 후, Vero-hACE2-TMPRSS2 세포의 단층에 첨가하였다(A. Creanga와 B. Graham의 기증). 이틀 후, 상청액을 수확하여 계대 제로(p0) 스톡을 확립하였다. SARS-CoV-2의 2019n-CoV/USA_WA1/2019 분리주를 미국 질병관리예방센터(CDC)에서 입수하였고, Vero E6 세포에서 계대하였다. 스파이크 유전자의 개별 점 돌연변이(D614G 및 E484K/D614G)를 이전에 설명한 대로67 2019n-CoV/USA_WA1/2019 균주의 감염성 cDNA 클론에 도입하였다. SARS-CoV-2 야생형 감염성 클론68의 스파이크 유전자를 포함하는 서브클론 puc57-CoV-2-F6에 뉴클레오티드 치환을 도입하였다. 변종 SARS-CoV-2 바이러스의 전장 감염성 cDNA 클론을 이전에 설명된 프로토콜68에 따라 7개의 인접한 cDNA 단편의 시험관 내 결찰에 의해 조립하였다. 이어서, 시험관 내 전사를 수행하여 전장 게놈 RNA를 합성하였다. 돌연변이체 바이러스를 회수하기 위해, RNA 전사체를 Vero E6 세포에 전기천공하였다. 40시간 후에 세포의 상청액으로부터 바이러스를 수집하고 p0 스톡으로 사용하였다. 모든 바이러스 스톡을 시퀀싱에 의해 확인하였다.
포커스 감소 중화 테스트. mAb 또는 혈청의 연속 희석액을 37℃에서 1시간 동안 SARS-CoV-2의 상이한 균주 또는 변이체의 102개의 포커스 형성 단위(FFU)와 함께 인큐베이션하였다. 항체-바이러스 복합체를 96웰 플레이트의 Vero-hACE2-TMPRSS2 세포 단층 배양물에 첨가하고 37℃에서 1시간 동안 인큐베이션하였다. 이어서, 세포를 2% FBS가 보충된 MEM에서 1%(w/v) 메틸셀룰로스로 오버레이하였다. 20시간 후 오버레이를 제거하여 플레이트를 수확하고 PBS 중 4% PFA로 실온에서 20분 동안 고정하였다. 플레이트를 세척하고 0.1% 사포닌 및 0.1% 소 혈청 알부민이 보충된 PBS 중 항-S mAb 및 HRP-접합된 염소 항-인간 IgG의 올리고클론 풀과 함께 순차적으로 인큐베이션하였다. SARS-CoV-2에 감염된 세포 포커스를 TrueBlue 퍼옥시다제 기질(KPL)을 사용하여 시각화하고, ImmunoSpot 미세분석기(Cellular Technologies)에서 정량화하였다.
다중 서열 정렬. 본 발명자들은 COV2-2196을 암호화하는 것과 동일한 특징을 갖는 항체 가변 유전자 서열을 찾고, 조사된 각 개인의 레퍼토리에서 매칭되는 서열을 검색하였다. 이들은 3명의 건강한 개인과 제대혈 레퍼토리(SRP174305에 기탁됨)에 대해 공개적으로 이용 가능한 대규모 항체 서열 레퍼토리에서 유사한 서열을 검색하였다. 중쇄에 대한 검색 매개변수는 P99, D108 및 F110 잔기가 있는 IGHV1-58 및 IGHJ3이 있는 서열이었다. 또한, 경쇄에 대한 검색 매개변수는 Y92 및 W98 잔기가 있는 서열이었다. 각 개체에 속하는 매칭되는 클론형의 서열은 ClustalO(중쇄) 또는 MUSCLE(경쇄)로 정렬하였다. 이어서, WebLogo를 사용하여 정렬된 서열의 LOGO 플롯을 생성하였다.
데이터 및 자료 가용성: 이 논문에서 보고된 결정 구조는 수탁번호 7L7D, 7L7E로 단백질 정보은행에 기탁되었다. 정렬된 인간 항체 유전자 레퍼토리 데이터 세트에 대한 서열 리드 아카이브 기탁물은 NCBI: PRJNA511481에 기탁되어 있다. 다른 모든 데이터는 본문 또는 보충 자료에서 이용할 수 있다.
소프트웨어 가용성. 항체 회피 돌연변이의 심층 돌연변이 스캐닝 분석을 위한 계산 파이프라인은 GitHub: github.com/jbloomlab/SARS-CoV-2-RBD_MAP_AZ_Abs에서 이용할 수 있다. FASTQ 파일은 BioProject PRJNA639956의 일환으로 BioSample SAMN17532001 하에 NCBI 서열 리드 아카이브에서 이용할 수 있다. 돌연변이별 회피 분율은 GitHub (github.com/jbloomlab/SARS-CoV-2-RBD_MAP_AZ_Abs/blob/main/results/supp_data/AZ_cocktail_raw_data.csv) 및 표 B에서 이용할 수 있다.
실시예 4 - 결과 및 논의
COV2-2196 기반 항체는 비경쟁 RBD 특이적 항체 COV2-2130 기반 항체와 조합하여 예방 또는 치료를 위해 조사되고 있다. COV2-2196 및 COV2-2130에 의한 RBD 인식의 분자적 세부 사항과 동물 모델1에서 경쟁하지 않는 두 mAb의 예방적 보호에서 나타난 시너지의 기초가 되는 가능성 있는 구조적 메커니즘을 이해하기 위해, 본 발명자들은 2.50  에서 COV2-2196과 복합된 S 단백질 RBD(도 8a~8e, 표 B) 및 3.00
에서 COV2-2196과 복합된 S 단백질 RBD(도 8a~8e, 표 B) 및 3.00  에서 COV2-2196 및 COV2-2130 둘 다와 복합된 S 단백질 RBD(도 9a~9f, 표 B)의 결정 구조를 결정하였다. RBD-COV2-2196-2130 복합체에서 RBD-COV2-2196의 하부 구조는 RBD-COV2-2196 복합체의 구조와 중첩될 수 있다(도 12). COV2-2196과 RBD 사이의 계면의 매몰 표면적은 두 결정 구조에서 약 650
에서 COV2-2196 및 COV2-2130 둘 다와 복합된 S 단백질 RBD(도 9a~9f, 표 B)의 결정 구조를 결정하였다. RBD-COV2-2196-2130 복합체에서 RBD-COV2-2196의 하부 구조는 RBD-COV2-2196 복합체의 구조와 중첩될 수 있다(도 12). COV2-2196과 RBD 사이의 계면의 매몰 표면적은 두 결정 구조에서 약 650  2이고, COV2-2130과 RBD 사이의 계면의 매몰 표면적은 약 740
2이고, COV2-2130과 RBD 사이의 계면의 매몰 표면적은 약 740  2이다. COV2-2196은 RBD의 수용체 결합 능선에 결합하고, COV2-2130은 잔기 K444 주위의 RBD 가장자리의 일측 및 수용체 결합 모티프(RBM)의 안장 영역에 결합하여, ACE2 결합 부위와 중첩된다(도 8a~8b, 도 9a~9b). 이러한 특징은 본 발명자들의 이전 연구의 RBD 결합에 대한 항체와 ACE2 간의 경쟁을 설명한다. 예를 들어, COV2-2196과 COV2-2130은 인간 수용체 ACE21에 대한 RBD 접근을 차단하여 바이러스를 중화한다. COV2-2196 중쇄 및 경쇄로부터의 방향족 잔기는 RBD 잔기 F486 및 인접한 잔기(G485, N487)를 둘러싸는 소수성 포켓을 형성한다(도 8a, 8d 및 8e; 도 13a 내지 13c). Ab-Ag 상호작용의 이러한 모드는 항체 포켓의 형성이 HCDR3 및 LCDR3의 넓은 공간적 분리에 의해 유발된다는 점에서 이례적이다. 또한, COV2-2196이 인식하는 항원 부위는 S 삼량체 자체의 프로토머 사이의 계면에 매몰되어 있지 않지만, COV2-2196은 인접한 S 프로토머의 RBD와의 입체 충돌로 인해 "아래쪽" 형태의 RBD에 결합할 수 없다. 따라서, COV2-2196은 "위쪽" 형태의 RBD에만 결합한다(도 8c). COV2-2130과 복합된 RBD의 하부 구조(도 9c) 및 COV2-2196 및 COV2-2130과 복합된 RBD의 구조(도 9d)는 COV2-2130이 S 삼량체의 "위쪽" 및 "아래쪽" 형태 둘 모두에서 RBD와 결합할 수 있음을 나타낸다. 이러한 구조적 발견은 음성 염색 전자 현미경을 사용한 복합체에 대한 본 발명자들의 이전의 더 낮은 해상도 결과1와 일치한다.
2이다. COV2-2196은 RBD의 수용체 결합 능선에 결합하고, COV2-2130은 잔기 K444 주위의 RBD 가장자리의 일측 및 수용체 결합 모티프(RBM)의 안장 영역에 결합하여, ACE2 결합 부위와 중첩된다(도 8a~8b, 도 9a~9b). 이러한 특징은 본 발명자들의 이전 연구의 RBD 결합에 대한 항체와 ACE2 간의 경쟁을 설명한다. 예를 들어, COV2-2196과 COV2-2130은 인간 수용체 ACE21에 대한 RBD 접근을 차단하여 바이러스를 중화한다. COV2-2196 중쇄 및 경쇄로부터의 방향족 잔기는 RBD 잔기 F486 및 인접한 잔기(G485, N487)를 둘러싸는 소수성 포켓을 형성한다(도 8a, 8d 및 8e; 도 13a 내지 13c). Ab-Ag 상호작용의 이러한 모드는 항체 포켓의 형성이 HCDR3 및 LCDR3의 넓은 공간적 분리에 의해 유발된다는 점에서 이례적이다. 또한, COV2-2196이 인식하는 항원 부위는 S 삼량체 자체의 프로토머 사이의 계면에 매몰되어 있지 않지만, COV2-2196은 인접한 S 프로토머의 RBD와의 입체 충돌로 인해 "아래쪽" 형태의 RBD에 결합할 수 없다. 따라서, COV2-2196은 "위쪽" 형태의 RBD에만 결합한다(도 8c). COV2-2130과 복합된 RBD의 하부 구조(도 9c) 및 COV2-2196 및 COV2-2130과 복합된 RBD의 구조(도 9d)는 COV2-2130이 S 삼량체의 "위쪽" 및 "아래쪽" 형태 둘 모두에서 RBD와 결합할 수 있음을 나타낸다. 이러한 구조적 발견은 음성 염색 전자 현미경을 사용한 복합체에 대한 본 발명자들의 이전의 더 낮은 해상도 결과1와 일치한다.
RBD와 복합된 COV2-2196의 구조 분석은 COV2-2196이 RBD의 수용체 결합 능선을 인식하는 방법을 보여준다. 주요 접촉 잔기 중 하나인 F486은 결합 부위의 중앙에 위치하여 COV2-2196 중쇄/경쇄 사이의 소수성 포켓(중쇄의 잔기 P99 및 5개의 방향족 측쇄에 의해 형성된 "방향족 케이지")과 소수성 효과 및 반 데르 발스 상호작용을 통해 광범위하게 상호작용한다(도 8d~8e, 도 13a~13b). 4개의 직접적인 Ab-Ag H 결합 및 16개의 물 매개 H 결합으로 구성된 수소 결합(H 결합) 망은 잔기 F486을 둘러싸고 Ab-Ag 상호작용을 강화한다(도 13c). 중요한 것은, 에피토프와 광범위하게 상호작용하는 하나(중쇄의 잔기 P99)를 제외한 모든 잔기의 경우, 생식계열 서열(중쇄의 경우 IGHV1-58*01 및 IGHJ3*02, 경쇄의 경우 IGKV3-20*01 및 IGKJ1*01)에 의해 암호화되거나(도 10A), 또는 중쇄 N107 및 G99 및 경쇄 S94와 같은 이들의 골격 원자만이 Ab-Ag 상호작용에 관여한다는 점이다. 본 발명자들은 COV2-2196과 유사하지만 아마도 상이한 IGHD 유전자를 가진 동일한 IGHV/IGHJ 및 IGKV/IGKJ 재조합에 의해 암호화되는 S2E12라는 문헌의 또 다른 항체에 주목하였다(IGHD2-15 대 IGHD2-2)38. S 단백질과 복합된 S2E12의 초저온-EM 구조(PDB 7K4N)의 비교는, 계면 잔기 측쇄의 입체형태의 변화를 볼 수 있음에도, mAb S2E12가 COV2-2196과 거의 동일한 Ab-Ag 상호작용을 사용할 가능성이 있음을 시사한다(도 13d). 예를 들어, 경쇄 잔기 Y92의 경우, 결정 구조의 페닐 고리는 피팅된 EM 구조의 해당 고리에 수직이다.
본 발명자들은 이러한 구조적 특징이 다른 이들에 의해 분리된 추가 SARS-CoV-2 mAb에 존재하는지 결정하기 위해 유전자 데이터베이스를 검색하고, 클론형의 추가 구성원을 발견하였다(도 10a). 다른 두 연구에서는 여러 COVID-19 회복기 환자로부터 분리된 동일하거나 유사한 클론형의 항체를 보고하였고4,38, 한 연구에서는 D 또는 J 유전자 사용에 대한 정보를 제공하지 않고, 동일한 IGHV1-58 및 IGKV3-20 쌍을 갖는 3개의 항체를 발견하였다39. 이러한 모든 항체는 SARS-CoV-2 RBD에 강력하게 결합하고 높은 효능으로 바이러스를 중화하는 것으로 보고되어 있다1,4,38,39. 현재까지, 이러한 VH(DH)JH 및 VK-JK 재조합에 의해 암호화된 항체의 원자 해상도 구조는 두 가지만 이용 가능한데, 본원에 제시된 COV2-2196의 구조와 S2E12의 구조이다38. 본 발명자들은 COV2-2072 및 COV2-2381로 명명된 항-SARS-CoV-2 항체의 본 발명자들 소유의 패널에서 이 클론형의 2개의 추가 항체에 대한 상동성 모델링을 수행하였다. 예상대로, 이들 항체가 공유된 유전자 클론형의 구성원이라는 점을 감안할 때, COV2-2072/RBD 및 COV2-2381/RBD 복합체의 모델링된 구조는 Ab-Ag 계면에서 COV2-2196/RBD 및 S2E12/RBD의 것과 가상으로 중첩된다(도 14의 A 내지 E). 추가로, CDR의 글리코실화가 항원 인식에 부정적인 영향을 미칠 수 있다는 점을 감안할 때, COV2-2072는 항체의 색다른 특징인 HCDR3(도 14의 D)의 N-연결 글리코실화 시퀀을 암호화한다. 그러나 COV2-2196 구조는 이 클론형에서 이황화물 스테이플링된 HCDR3이 결합 부위에서 떨어져 기울어 있음을 보여주며, 이는 COV2-2072에서 이 특이한 HCDR3 글리코실화가 결합을 손상시키지 않고 어떻게 용인될 수 있는지를 설명한다(도 14의 E).
다음으로 본 발명자들은 SARS-CoV-2를 경험하지 않은 개체의 순환 B 세포의 항체 가변 유전자 레퍼토리에서 이 공개 클론형의 잠재적 전구체를 확인할 수 있는지를 결정하였다. 본 발명자들은 SARS-CoV-2 감염 이력이 없는 3명의 건강한 인간 공여자로부터 유래한 이전에 설명된 종합 레퍼토리 데이터세트와 COVID-19 팬데믹 이전에 수집된 제대혈의 데이터세트40에서 V(D)J 및 VJ 유전자를 검색하였다. 이 SARS-CoV-2 반응성 공개 클론형에 대해 각각 총 386, 193, 47 또는 7개의 중쇄 서열이 각 공여자 또는 제대혈 레퍼토리에서 발견되었다(도 15a). 또한, 본 발명자들은 동일한 경쇄 V-J 재조합(IGKV3-20-IGKJ1*01)을 갖는 516,738개의 인간 항체 서열을 발견하였다. 각각 총 103,534, 191,039 또는 222,165개의 경쇄 서열이 각 공여자에서 이 공개 클론형에 대해 발견되었다. 많은 수의 서열로 인해, 상위 5개의 풍부한 서열을 각 공여자로부터 정렬하였다. ClustalOmega를 사용하여 각 공여자의 서열에 대해 다중 서열 정렬을 생성하고 로고 플롯을 생성하였다. 각 공여자에서 동일한 재조합 이벤트를 갖는 상위 5개 서열은 동일하여 동일한 로고 플롯을 생성하였다(도 15a~15b).
본 발명자들은 항체에서 결합에 중요한 9개의 공통 잔기 중 8개가 생식계열 유전자 서열에 의해 암호화되고, 모두 상이한 4개의 항체-발견 팀으로부터 본원에 나열된 공개 클론형의 14개 구성원 모두에 존재한다는 점에 주목하였다(도 10a). 이들 특징의 중요성을 검증하기 위해, 파라토프에서 점 돌연변이를 갖는 변이체 항체를 발현시켜 보존된 잔기에서 변이의 효과를 결정하였다(도 10b). D108 잔기를 A, N 또는 E로 변경하는 것은 거의 효과가 없었지만, 해당 루프를 경질화하는 HCDR3에서 이황화 결합을 제거하자 결합이 감소하였다. HCDR3 루프를 항원과의 상호작용 부위로부터 멀어지게 하는 P99 잔기도 중요했는데, 이는 P99G 돌연변이체는 감소된 결합을 나타내었고, P99를 함유한 COV2-2196의 생식계열 복귀 돌연변이체 형태는 항원에 결합하였으나, 생식계열 복귀 돌연변이체 배경의 P99G는 그렇지 않았기 때문이다(도 10b).
COV2-2196 가변 영역에 기반한 항체는 임상 시험에서 COV2-2130 가변 영역에 기반한 항체와 조합하여 테스트되고 있다. 22개의 아미노산 잔기를 가진 COV2-2130 HCDR3은 인간 항체에 비해 상대적으로 길며, 추론된 생식계열 IGHD3 유전자에서 고도로 돌연변이된 것이다. HCDR3은 작은 루프로 구성된 길고 구조화된 루프를 형성하고, HCDR3 내에서 단거리 수소 결합 및 소수성 상호작용/방향족 적층에 의해 안정화되며, 경쇄의 잔기들과 상호작용(수소 결합 및 방향족 적층)에 의해 추가로 강화된다(도 16의 A~B). COV2-2130 중쇄 및 경쇄는 각각 생식계열 유전자 IGHV3-15 및 IGKV4-1에 의해 암호화되며, 두 유전자는 COV2-2130에서 사용되는 가장 긴 생식계열 암호화된 HCDR2(10 aa) 및 LCDR1(12 aa) 루프를 암호화한다. 중쇄 V(D)J 재조합, HCDR3 돌연변이, 중쇄와 경쇄의 쌍은, S443 ~ Y449 루프를 중심으로 하는 RBD 영역의 모양과 매칭되는, 중쇄와 경쇄 사이의 결합 틈을 초래한다(도 9a, 도 16의 C) 이러한 구조적 특징과 밀접하게 관련되어, HCDR3, LCDR1, HCDR2 및 LCDR2만이 파라토프의 형성에 관여한다(도 9e~9f, 도 13e~13f). Ab-Ag 계면을 검사하면 상호작용 에너지의 대부분을 차지할 가능성이 있는 영역이 나타난다. RBD 잔기 K444 측쇄는 HCDR3 루프의 하위 루프 Y104 ~ V109로 둘러싸여 있으며, 측쇄 NZ 원자의 양전하는 HCDR3 잔기 D107 측쇄, Y105, D107 및 V109의 3개의 주쇄 카르보닐 산소 원자 및 Y104 페닐 고리의 전자 풍부면(양이온-π 상호작용)에 의해 중화된다(도 13e). 상호작용하는 원자는 용매로부터 완전히 보호되기 때문에 이러한 제한된 공간 내에서 고도로 집중된 상호작용은 에너지 측면에서 유리하다. 나아가, RBD 잔기 R346과 HCDR2 D56 사이의 염교, RBD R346과 HCDR3 Y106의 주쇄 산소 사이의 정전기적 상호작용, RBD N450과 HCDR3 Y105 주쇄 산소 사이의 수소 결합, RBD V445 주쇄 산소와 HCDR3 Y104 측쇄 사이의 수소 결합, V445 측쇄와 HCDR3 L113 및 F118의 측쇄들 사이의 소수성 상호작용을 포함한 Ab-Ag 계면의 이 "핫스팟"은 더 적은 자유 에너지 이득을 갖는 Ab-Ag 상호작용에 의해 용매로 둘러싸여 있거나 용매로부터 보호된다. 또한, HCDR3 잔기 Y105와 LCDR2 잔기 W56 사이의 방향족 적층은 용매로부터 "핫스팟"의 차폐에 참여한다(도 13e). 또한, COV2-2130 경쇄 LCDR1 및 LCDR2는 RBD와 광범위하게 접촉한다. 이 중 LCDR1 S32 측쇄, S33 주쇄 산소, N34 측쇄 및 LCDR2 Y55 측쇄는 RBD E484 측쇄, S494 주쇄 질소, Y449 주쇄 산소 및 G446 주쇄 질소와 수소 결합을 형성한다(도 13f). 잔기 LCDR1 K36, Y38 및 LCDR2 W56은 방향족 적층 및 양이온-π 상호작용을 통해 RBD Y449와 상호작용하여, "상호작용 클러스터"를 형성하지만(도 13f), 이러한 상호작용은 RBD K444의 경우만큼 에너지 측면에서 강하지 않을 가능성이 높다.
COV2-2196 및 COV2-2130과 복합된 RBD의 결정 구조에서, 본 발명자들은 밀접하게 이격된 COV2-2196 및 COV2-2130 Fab 사이의 상호작용에 주목하였다(도 17). RBD 결합 상태에서 두 Fab 간의 상호작용은 이전에 관찰된 SARS-CoV-2의 상승적 중화1에 기여할 수 있는 가능성이 있다. 그러나 얼마나 많은 시너지 효과가 이러한 Fab-Fab 상호작용에서 기인할 수 있는지는 분명하지 않다.
COV2-2196 및 COV2-2130의 결합에 중요한 RBD 부위를 더 잘 이해하기 위해, 본 발명자들은 심층 돌연변이 스캐닝(deep mutational scanning, DMS) 접근법을 사용하여 항체 결합을 회피하는 RBD에 대한 모든 돌연변이를 매핑하였다41; (도 18). 두 항체에 대해, 몇 가지 주요 부위를 확인하였는데, 모두 항체 구조적 발자국에 있었고, RBD 돌연변이가 결합을 강력하게 방해하였다(도 11a 내지 11d). 본 발명자들은 ACE2 결합에 대한 RBD 돌연변이의 효과를 정량화하는 본 발명자들의 이전 작업을 활용하여42 RBD에 대한 항체 결합을 방해한 돌연변이의 ACE2 결합에 대한 효과를 오버레이하였다(도 11a~11b). COV2-2196의 경우, F486 및 N487에 대한 많은 돌연변이는 회피 분율이 1에 근접하여(즉, 해당 RBD 변이체는 테스트한 조건 하에서 항체 결합을 완전히 회피함), 항체 결합에 대한 이들 두 잔기의 기여의 중요성을 강화하였다. 유사하게, COV2-2130의 경우, 위치 K444에서 다른 19개 아미노산 중 임의의 것으로의 돌연변이가 항체 결합을 방해하였으며, 이는 이 위치의 리신이 Ab-Ag 상호작용에 중요함을 나타낸다.
그럼에도, 모든 항체 접촉 잔기가 돌연변이가 결합을 크게 감소시키는 부위로 확인된 것은 아니다. 몇 가지 설명이 가능하다: 1) 일부 결합 부위 잔기는 결합에 중요하지 않을 수 있거나, 2) 일부 잔기는 골격 원자를 사용하여 결합 계면에서 멀어지는 측쇄와 상호 작용할 수 있거나, 3) 일부 부위에 대한 돌연변이가 용인되지 않을 수 있다42. 예를 들어, 잔기 L455, F456 및 Q493은 COV2-2196에 대해 구조적으로 정의된 결합 부위의 일부이지만(도 8d), 이들 부위에 대한 돌연변이는 항체 결합에 검출 가능하게 영향을 미치지 않았으며(도 11a 및 11c), 이는 이러한 잔기들이 중요한 결합 기여를 하지 않음을 시사한다. COV2-2196/RBD 구조를 S2E12/RBD 구조에 중첩시키자 항체가 결합될 때 유지되는 RBD 능선과 나머지 RBD 사이의 유연한 힌지 영역이 명확하게 나타난다(도 13d). 이 발견은 이들 3개 위치에서의 돌연변이가 Ab-Ag 결합에 대해 잘 용인될 수 있고 COV2-2196 또는 S2E12 결합에 대한 이러한 특정 잔기의 비필수적 특성을 뒷받침함을 나타낸다.
중요한 것은, COV2-2196 및 COV2-2130은 RBD에 결합하기 위해 경쟁하지 않으며1, 이는 이들이 예방적 또는 치료적 용도로 회피 저항성 칵테일을 구성할 수 있음을 시사한다는 점이다. 실제로, 이 두 항체에 대한 구조적 결합 부위와 회피 변이체 맵은 중첩되지 않는다. 두 항체의 결합을 회피할 수 있는 단일 돌연변이가 있는지를 테스트하기 위해, 본 발명자들은 COV2-2196 및 COV2-2130 항체의 1:1 혼합물을 이용한 회피 변이체 매핑 실험을 수행했지만, 0.2보다 큰 회피 분율을 갖는 돌연변이는 검출하지 못한 반면, 단일 항체 각각에 대해 가장 큰 효과를 갖는 돌연변이는 대략 1이었다(도 18d).
이러한 실험은 RBD에 대한 항체 결합을 회피하는 모든 돌연변이를 매핑하지만, 본 발명자들은 또한 바이러스 성장 중에 어떤 돌연변이가 발생할 가능성이 있는지를 결정하고자 하였다. 이 질문을 해결하기 위해, 먼저 SARS-CoV-2 S 당단백질을 발현하는 재조합 VSV(VSV-SARS-CoV-2)43를 사용하여 회피 돌연변이를 선택하려고 시도하였다(도 11e). 본 발명자들은 바이러스 성장 중에 선택되는 유일한 아미노산 돌연변이는 1) 단일 뉴클레오티드 RNA 변화에 의해 발생하고, 2) ACE2 결합 및 발현에 대해 최소한의 유해한 영향을 일으키고, 3) 항체 결합에 실질적으로 영향을 미치는 것들뿐이라고 예상하였다41,42. 실제로, 본 발명자들은 단일 뉴클레오티드 접근 가능하고 ACE2 결합에 대한 효과와 관련하여 상대적으로 잘 용인되는 COV2-2196 유도 돌연변이를 검출하지 못했는데(도 11b), 이는 항체의 존재 하에서 재조합 VSV 성장의 88개의 독립적인 반복실험 중 어느 것에서도 회피 돌연변이체가 선택되지 않은 이유를 설명할 수 있다(도 11e, 도 18g). COV2-2130의 경우, 돌연변이에 상대적으로 용인성이 있는 부위인 K444 부위42에 대한 돌연변이는 VSV 키메라 바이러스를 사용한 고처리량 항체 회피 선택에서 항체 결합의 가장 빈번한 회피를 보여주었다. 항체 선택 실험의 40%에서, 항체 결합에 가장 큰 영향을 미치는 단일 뉴클레오티드 돌연변이 중 두 개인 K444R(20개 반복실험 중 6개에서 선택됨)과 K444E(20개 반복실험 중 2개에서 선택됨)가 바이러스 성장 중에 나타났다(도 11e, 도 18g).
실제 감염성 바이러스에 대한 저항성을 조사하기 위해, SARS-CoV-2 균주 USA-WA1/2020을 각각의 IC50 값에서 시작하여 각 계대에서 IC90 값까지 단계적으로 증가된 농도의 COV2-2196(AZD8895), COV2-2130(AZD1061) 또는 이들의 1:1 조합(AZD7442)에 기반한 임상 항체와 함께 Vero 세포 단층 배양물에서 연속적으로 계대하였다(도 19). 대조군으로서, 바이러스를 항체의 부재 하에 계대하였다. 최종 계대 후, 플라크 분석에 의해 IC90 농도의 10배의 최종 농도에서 상호 항체에 대한 감수성에 대해 바이러스를 평가하였다. 본 발명자들은 AZD8895(COV2-2196 기반) 또는 AZD7442 칵테일에 의한 중화 저항성 플라크를 검출하지 못하였다. AZD1061에서 연속적으로 계대된 바이러스는 IC90 값의 10배 농도의 AZD1061의 존재 하에서 1.2 x 107 pfu/mL의 역가까지 플라크를 형성했지만, AZD7442에서는 플라크가 형성되지 않았다. 플라크(n=6)를 무작위로 선택하고, 이들의 바이러스 스파이크 암호화 유전자를 시퀀싱하여, 독립적으로 선택되고 시퀀싱된 플라크 6개 모두에서 동일한 3개의 아미노산 변화를 밝혀냈다: N74K, R346I 및 S686G(도 11f). S686G 변화는 이전에 Vero 세포에서 SARS-CoV-2의 연속 계대와 관련이 있는 것으로 보고되었으며44, 이는 흰 족제비45 또는 NHP46에서의 챌린지 연구에서 분리되었으며 푸린 활성을 감소시킬 것으로 예상된다44. N74K 잔기는 AZD1061 결합 부위 외부의 N 말단 도메인에 위치하고 글리칸의 손실을 초래한다47. R346I 잔기는 AZD1061의 결합 부위에 위치하며 AZD1061 저항성과 연관될 수 있다. S 단백질에 대한 AZD1061(COV2-2130) 결합에 대한 R346I 변화의 영향은 도 11g에 제시되어 있다. VSV-SARS-CoV-2 시스템에서 선택된 K444R 및 K444E 치환과 실제 SARS-CoV-2로 계대하여 선택된 R346I 치환은 단일 뉴클레오티드 치환에 의해 접근 가능하고 ACE2 결합 활성을 보존하며(도 11g), 본 발명자들의 DMS 분석이 COV2-2130 항체의 존재 하에 선택된 돌연변이를 예측함을 나타내었다. 종합하면, 이러한 결과는 COV2-2196 및 COV2-2130의 결합에 대한 모든 아미노산 치환의 효과를 종합적으로 매핑하고 바이러스 진화에 대한 우려 가능성이 있는 부위를 확인시켜 준다. 즉, 잔기 K444 및 R346에 돌연변이를 포함하는 변이체는 GISAID 데이터베이스에 존재하는 모든 시퀀싱된 바이러스 중에서 드물다(12/23/20에 접근할 때 모두 ≤ 0.01%).
최근, 증가된 전염성과 잠재적인 항원 돌연변이를 가진 바이러스 변이체가 임상 분리주에서 보고되었다48~51. 본 발명자들은 신속하게 출현하는 이러한 균주들의 변이체 잔기 중 일부가 강력하게 중화하는 이러한 항체의 활성을 방해하는지를 테스트하였다. 영국 옥스퍼드에서 얻은 비강 샘플에서 바이러스 분리주(UK B.1.1.7-OXF로 명명된 B.1.1.7 바이러스)를 테스트했는데, 여기에는 NTD에서 69~70 및 144~145 결실 및 N501Y, A570D, D614G 및 P681H에서의 치환을 포함한 스파이크 유전자 변화를 정의하는 B.1.1.7 계통이 포함된다49. 본 발명자들은 또한 WA-1 균주 배경(2019n-CoV/USA_WA1/2019, [WA-1])에서 동종 D614G 및 E484K 변이체를 테스트했는데, 모두 실제 SARS-CoV-2 바이러스로서 제조되었고 포커스 감소 중화 테스트43에 사용되었다. E484K 돌연변이가 특별한 관심 대상이었는데, 이 잔기가 Fab 발자국의 바로 주변부에 있지만 Fab 및 RBD의 복합체에서 각 mAb의 4.5  내에 위치하며, 최근 생겨난 계통인 B.1.351(501Y.V2)50 및 P.1(501Y.V3)51에 존재하고, 일부 단클론 항체52,53 및 인간 다클론 혈청 항체54의 결합을 변경하는 것으로 밝혀졌기 때문이다. E484K를 포함하는 변이체는 또한 SARS-CoV-2 생존자의 회복기 혈청 및 혈장에 의해 덜 효율적으로 중화되는 것으로 나타났다55,56. COV2-2196, COV2-2130 및 COV2-2050(이는 잔기 E484와 상호작용하기 때문에 본 발명자들이 비교를 위해 포함시킨 제3 중화 항체임)의 경우, D614G 돌연변이 또는 UK B.1.1.7-OXF 균주에 존재하는 돌연변이 세트의 영향을 사실상 발견하지 못하였다. 오히려, 본 발명자들은 후자의 순환 바이러스에 대해 약간 개선되는 경향(IC50 값의 2배 내지 3배 감소)을 관찰하였다(도 11h). 그러나 D614G/E484K 바이러스의 중화에 대한 효과는 관찰하였다. COV2-2050은 중화 활성을 완전히 상실했으며, 이는 E484K를 COV2-2050 결합을 방해하는 돌연변이로 정의한 본 발명자들의 이전 연구41와 일치한다. 대조적으로, COV2-2196, COV2-2130 및 COV2-2196 + COV2-2130은 단지 약간 더 적은 억제 능력을 나타내었다(IC50 값의 2~5배 증가).
내에 위치하며, 최근 생겨난 계통인 B.1.351(501Y.V2)50 및 P.1(501Y.V3)51에 존재하고, 일부 단클론 항체52,53 및 인간 다클론 혈청 항체54의 결합을 변경하는 것으로 밝혀졌기 때문이다. E484K를 포함하는 변이체는 또한 SARS-CoV-2 생존자의 회복기 혈청 및 혈장에 의해 덜 효율적으로 중화되는 것으로 나타났다55,56. COV2-2196, COV2-2130 및 COV2-2050(이는 잔기 E484와 상호작용하기 때문에 본 발명자들이 비교를 위해 포함시킨 제3 중화 항체임)의 경우, D614G 돌연변이 또는 UK B.1.1.7-OXF 균주에 존재하는 돌연변이 세트의 영향을 사실상 발견하지 못하였다. 오히려, 본 발명자들은 후자의 순환 바이러스에 대해 약간 개선되는 경향(IC50 값의 2배 내지 3배 감소)을 관찰하였다(도 11h). 그러나 D614G/E484K 바이러스의 중화에 대한 효과는 관찰하였다. COV2-2050은 중화 활성을 완전히 상실했으며, 이는 E484K를 COV2-2050 결합을 방해하는 돌연변이로 정의한 본 발명자들의 이전 연구41와 일치한다. 대조적으로, COV2-2196, COV2-2130 및 COV2-2196 + COV2-2130은 단지 약간 더 적은 억제 능력을 나타내었다(IC50 값의 2~5배 증가).
논의. 이러한 구조 분석은 SARS-CoV-2 S RBD의 동일한 영역을 표적으로 하는 중쇄 V-D-J 및 경쇄 V-J 재조합을 공유하는 인간 항체의 공개 클론형의 빈번한 선택을 위한 분자 기초를 정의한다. 중쇄 및 경쇄의 생식계열 항체 유전자 암호화 잔기는 항원 인식에서 중요한 역할을 하며, 이는 체세포 돌연변이가 이 클론형의 항체의 성숙에 거의 요구되지 않음을 시사한다. IGHD2 유전자 암호화된 이황화 결합은 HCDR3이 RBD의 항원 부위에 대한 모양 및 화학적 상보성을 가진 입체형태를 채택하도록 하는 추가 제한을 제공한다. 3개의 서로 다른 IGHD2 유전자(IGHD2-2, IGHD2-8 및 IGHD2-15)가 이 클론형과 관련하여 기능할 수 있는 HCDR3의 부분들을 암호화하는 것으로 보인다. 본 발명자들은 높은 특이성 및 강력한 중화 활성을 가능하게 하는 미리 구성된 구조적 특징을 갖는 공통 생식계열 유전자 암호화 항체의 이러한 발생이 SARS-CoV-2에 대한 1차 인간 면역 반응의 예상치 못한 유익한 특징이라고 제안한다. 이 공개 클론형으로부터 B 세포의 선택은 회피에 저항하는 초강력 중화 항체의 신속한 분리를 가능하게 하였고, 아마도 임상에서 관찰되고 있는 S 단백질 기반 백신의 놀라운 효능을 부분적으로 설명할 수 있다. 이 클론형을 유도하기 위한 인간 면역 반응을 프라이밍하기 위해 이 항원 부위에 대한 도메인 또는 모티프 기반 백신 설계를 사용하여 훨씬 더 높은 중화 효능을 갖는 혈청 중화 항체 역가를 유도할 기회를 구상할 수 있다.
본원에 제시된 RBD-COV2-2196 및 RBD-COV2-2130 복합체의 구조 분석은 두 항체가 계면 표면적 단위당 높은 에너지 밀도로 "리벳과 같은" 상호작용을 형성함으로써 RBD 항원에 결합함을 시사하며, 이러한 발견은 어떻게 그러한 강력한 항체가 Ab-Ag 계면에서 이렇게 상대적으로 작은(< 750  2) 매몰 표면적을 가질 수 있는지를 설명한다. 본원에서 연구된 두 항체에 대한 회피 돌연변이의 매핑은 회피 돌연변이의 유전자좌가 본 발명자들의 Ab-Ag 결정 구조에 의해 결정된 결합 부위와 일치함을 나타내어, DMS 방법론의 정확성을 추가로 확인시켜 준다. 본 발명자들은, 예를 들어 COV2-2130 또는 COV2-2196에 대한 회피 분율이 가장 높은 K444 또는 F486 잔기가 각각 항원-항체 복합체의 구조에서 항체와의 가장 강력한 상호작용을 나타내므로, 회피 분율 데이터가 항원에 대한 항체의 컴퓨터 도킹 또는 보다 일반적으로 단백질에 대한 단백질의 컴퓨터 도킹에 대한 제한으로서 향후 사용될 수 있다고 제안한다.
2) 매몰 표면적을 가질 수 있는지를 설명한다. 본원에서 연구된 두 항체에 대한 회피 돌연변이의 매핑은 회피 돌연변이의 유전자좌가 본 발명자들의 Ab-Ag 결정 구조에 의해 결정된 결합 부위와 일치함을 나타내어, DMS 방법론의 정확성을 추가로 확인시켜 준다. 본 발명자들은, 예를 들어 COV2-2130 또는 COV2-2196에 대한 회피 분율이 가장 높은 K444 또는 F486 잔기가 각각 항원-항체 복합체의 구조에서 항체와의 가장 강력한 상호작용을 나타내므로, 회피 분율 데이터가 항원에 대한 항체의 컴퓨터 도킹 또는 보다 일반적으로 단백질에 대한 단백질의 컴퓨터 도킹에 대한 제한으로서 향후 사용될 수 있다고 제안한다.
알려진 많은 중화 부위에서 증가된 전염성과 변경된 서열을 가진 변종 바이러스 계통의 최근 출현은 개발 및 테스트에서 현재의 항체 대응책을 회피하는 SARS-CoV-2의 능력에 관한 것이다. 본 발명자들은 항체의 활성과 둘 모두의 칵테일을 테스트하고 E484K 돌연변이를 포함하는 바이러스 및 다중 S 유전자 변이를 갖는 B.1.1.7 바이러스를 포함하는 몇 가지 중요한 변이체에 대해 지속적인 활성을 발견하였다. 이러한 광범위한 활성에 대한 유전적 및 구조적 기초는 결정 구조 및 본 발명자들이 본원에서 제시한 DMS 연구에서 밝혀졌다. COV2-2196에서 "방향족 케이지" 도메인에 의한 상대적으로 불변인 F486 잔기의 중심 인식은, 이 잔기의 변이가 감소된 바이러스 적합성과 관련되기 때문에 유익하다. 이 결합 부위를 표적으로 하는 것은 "개방형" 및 "폐쇄형" S 삼량체를 모두 인식하는 mAb COV2-2130과의 조합에서 상승적 중화라는 설정에서 특히 효과적인 것으로 보인다. 따라서, 이 조합은 회피에 저항하는 광범위하고 강력한 억제 메커니즘을 제공하는 것으로 보인다.

* * * * * * * * * * * * * * * * *
실시예 1 및 2에 대한 참고문헌
1.
ter Meulen, J., et al. Human monoclonal antibody as prophylaxis for SARS coronavirus infection in ferrets. Lancet 363, 2139-2141 (2004).
2.
Zhou, P., et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 270-273 (2020).
3.
Robbiani, D.F., et al. Convergent antibody responses to SARS-CoV-2 infection in convalescent individuals. bioRxiv, 2020.2005.2013.092619 (2020).
4.
Brouwer, P.J.M., et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. bioRxiv, 2020.2005.2012.088716 (2020).
5.
Niu, P., et al. Ultrapotent human neutralizing antibody repertoires against Middle East Respiratory Syndrome coronavirus from a recovered patient. J Infect Dis 218, 1249-1260 (2018).
6.
Wang, L., et al. Importance of neutralizing monoclonal antibodies targeting multiple antigenic sites on the Middle East Respiratory Syndrome coronavirus spike glycoprotein to avoid neutralization escape. J Virol 92, e02002-17 (2018).
실시예 3 및 4에 대한 참고문헌
1
Zost, S. J. et al. Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature 584, 443-449, doi:10.1038/s41586-020-2548-6 (2020).
2
Yuan, M. et al. Structural basis of a shared antibody response to SARS-CoV-2. Science 369, 1119-1123, doi:10.1126/science.abd2321 (2020).
3
Nielsen, S. C. A. et al. Human B Cell Clonal Expansion and Convergent Antibody Responses to SARS-CoV-2. Cell Host Microbe 28, 516-525 e515, doi:10.1016/j.chom.2020.09.002 (2020).
4
Robbiani, D. F. et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 584, 437-442, doi:10.1038/s41586-020-2456-9 (2020).
5
Brouwer, P. J. M. et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science 369, 643-650, doi:10.1126/science.abc5902 (2020).
6
Zhou, P. et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 270-273, doi:10.1038/s41586-020-2012-7 (2020).
7
Zhu, N. et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N Engl J Med 382, 727-733, doi:10.1056/NEJMoa2001017 (2020).
8
Hoffmann, M. et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 181, 271-280 e278, doi:10.1016/j.cell.2020.02.052 (2020).
9
Letko, M., Marzi, A. & Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat Microbiol 5, 562-569, doi:10.1038/s41564-020-0688-y (2020).
10
Wahba, L. et al. An Extensive Meta-Metagenomic Search Identifies SARS-CoV-2-Homologous Sequences in Pangolin Lung Viromes. mSphere 5, doi:10.1128/mSphere.00160-20 (2020).
11
Walls, A. C. et al. Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion. Proc Natl Acad Sci U S A 114, 11157-11162, doi:10.1073/pnas.1708727114 (2017).
12
Algaissi, A. et al. SARS-CoV-2 S1 and N-based serological assays reveal rapid seroconversion and induction of specific antibody response in COVID-19 patients. Sci Rep 10, 16561, doi:10.1038/s41598-020-73491-5 (2020).
13
Long, Q. X. et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat Med 26, 845-848, doi:10.1038/s41591-020-0897-1 (2020).
14
Piccoli, L. et al. Mapping Neutralizing and Immunodominant Sites on the SARS-CoV-2 Spike Receptor-Binding Domain by Structure-Guided High-Resolution Serology. Cell, doi:10.1016/j.cell.2020.09.037 (2020).
15
Cao, Y. et al. Potent Neutralizing Antibodies against SARS-CoV-2 Identified by High-Throughput Single-Cell Sequencing of Convalescent Patients' B Cells. Cell 182, 73-84 e16, doi:10.1016/j.cell.2020.05.025 (2020).
16
Hansen, J. et al. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 369, 1010-1014, doi:10.1126/science.abd0827 (2020).
17
Ju, B. et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 584, 115-119, doi:10.1038/s41586-020-2380-z (2020).
18
Liu, L. et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 584, 450-456, doi:10.1038/s41586-020-2571-7 (2020).
19
Rogers, T. F. et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 369, 956-963, doi:10.1126/science.abc7520 (2020).
20
Shi, R. et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 584, 120-124, doi:10.1038/s41586-020-2381-y (2020).
21
Weitkamp, J. H. et al. Infant and adult human B cell responses to rotavirus share common immunodominant variable gene repertoires. J Immunol 171, 4680-4688, doi:10.4049/jimmunol.171.9.4680 (2003).
22
Benton, D. J. et al. Receptor binding and priming of the spike protein of SARS-CoV-2 for membrane fusion. Nature, doi:10.1038/s41586-020-2772-0 (2020).
23
Wrapp, D. et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367, 1260-1263, doi:10.1126/science.abb2507 (2020).
24
Wrobel, A. G. et al. SARS-CoV-2 and bat RaTG13 spike glycoprotein structures inform on virus evolution and furin-cleavage effects. Nat Struct Mol Biol 27, 763-767, doi:10.1038/s41594-020-0468-7 (2020).
25
Zost, S. J. et al. Rapid isolation and profiling of a diverse panel of human monoclonal antibodies targeting the SARS-CoV-2 spike protein. Nat Med 26, 1422-1427, doi:10.1038/s41591-020-0998-x (2020).
26
Tian, C. et al. Immunodominance of the VH1-46 antibody gene segment in the primary repertoire of human rotavirus-specific B cells is reduced in the memory compartment through somatic mutation of nondominant clones. J Immunol 180, 3279-3288, doi:10.4049/jimmunol.180.5.3279 (2008).
27
Wu, X. et al. Focused evolution of HIV-1 neutralizing antibodies revealed by structures and deep sequencing. Science 333, 1593-1602, doi:10.1126/science.1207532 (2011).
28
Zhou, T. et al. Structural Repertoire of HIV-1-Neutralizing Antibodies Targeting the CD4 Supersite in 14 Donors. Cell 161, 1280-1292, doi:10.1016/j.cell.2015.05.007 (2015).
29
Huang, C. C. et al. Structural basis of tyrosine sulfation and VH-gene usage in antibodies that recognize the HIV type 1 coreceptor-binding site on gp120. Proc Natl Acad Sci U S A 101, 2706-2711, doi:10.1073/pnas.0308527100 (2004).
30
Williams, W. B. et al. HIV-1 VACCINES. Diversion of HIV-1 vaccine-induced immunity by gp41-microbiota cross-reactive antibodies. Science 349, aab1253, doi:10.1126/science.aab1253 (2015).
31
Joyce, M. G. et al. Vaccine-Induced Antibodies that Neutralize Group 1 and Group 2 Influenza A Viruses. Cell 166, 609-623, doi:10.1016/j.cell.2016.06.043 (2016).
32
Pappas, L. et al. Rapid development of broadly influenza neutralizing antibodies through redundant mutations. Nature 516, 418-422, doi:10.1038/nature13764 (2014).
33
Sui, J. et al. Structural and functional bases for broad-spectrum neutralization of avian and human influenza A viruses. Nat Struct Mol Biol 16, 265-273, doi:10.1038/nsmb.1566 (2009).
34
Wheatley, A. K. et al. H5N1 Vaccine-Elicited Memory B Cells Are Genetically Constrained by the IGHV Locus in the Recognition of a Neutralizing Epitope in the Hemagglutinin Stem. J Immunol 195, 602-610, doi:10.4049/jimmunol.1402835 (2015).
35
Bailey, J. R. et al. Broadly neutralizing antibodies with few somatic mutations and hepatitis C virus clearance. JCI Insight 2, doi:10.1172/jci.insight.92872 (2017).
36
Giang, E. et al. Human broadly neutralizing antibodies to the envelope glycoprotein complex of hepatitis C virus. Proc Natl Acad Sci U S A 109, 6205-6210, doi:10.1073/pnas.1114927109 (2012).
37
Rappuoli, R., Bottomley, M. J., D'Oro, U., Finco, O. & De Gregorio, E. Reverse vaccinology 2.0: Human immunology instructs vaccine antigen design. J Exp Med 213, 469-481, doi:10.1084/jem.20151960 (2016).
38
Tortorici, M. A. et al. Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms. Science, doi:10.1126/science.abe3354 (2020).
39
Kreer, C. et al. Longitudinal Isolation of Potent Near-Germline SARS-CoV-2-Neutralizing Antibodies from COVID-19 Patients. Cell 182, 843-854 e812, doi:10.1016/j.cell.2020.06.044 (2020).
40
Soto, C. et al. High frequency of shared clonotypes in human B cell receptor repertoires. Nature 566, 398-402, doi:10.1038/s41586-019-0934-8 (2019).
41
Greaney, A. J. et al. Complete mapping of mutations to the SARS-CoV-2 spike receptor-binding domain that escape antibody recognition. Cell Host Microbe, doi:10.1016/j.chom.2020.11.007 (2020).
42
Starr, T. N. et al. Deep mutational scanning of SARS-CoV-2 receptor binding domain reveals constraints on folding and ACE2 binding. Cell 182, 1295-1310 e1220, doi:10.1016/j.cell.2020.08.012 (2020).
43
Case, J. B. et al. Neutralizing antibody and soluble ACE2 inhibition of a replication-competent VSV-SARS-CoV-2 and a clinical isolate of SARS-CoV-2. Cell Host Microbe 28, 475-485 e475, doi:10.1016/j.chom.2020.06.021 (2020).
44
Klimstra, W. B. et al. SARS-CoV-2 growth, furin-cleavage-site adaptation and neutralization using serum from acutely infected hospitalized COVID-19 patients. J Gen Virol 101, 1156-1169, doi:10.1099/jgv.0.001481 (2020).
45
Sawatzki, K. et al. Ferrets not infected by SARS-CoV-2 in a high-exposure domestic setting. bioRxiv, 2020.2008.2021.254995, doi:10.1101/2020.08.21.254995 (2020).
46
Baum, A. et al. REGN-COV2 antibodies prevent and treat SARS-CoV-2 infection in rhesus macaques and hamsters. Science 370, 1110-1115, doi:10.1126/science.abe2402 (2020).
47
Li, Q. et al. The impact of mutations in SARS-CoV-2 spike on viral infectivity and antigenicity. Cell 182, 1284-1294 e1289, doi:10.1016/j.cell.2020.07.012 (2020).
48
Galloway, S. E. et al. Emergence of SARS-CoV-2 B.1.1.7 Lineage - United States, December 29, 2020-January 12, 2021. MMWR Morb Mortal Wkly Rep 70, 95-99, doi:10.15585/mmwr.mm7003e2 (2021).
49
Leung, K., Shum, M. H., Leung, G. M., Lam, T. T. & Wu, J. T. Early transmissibility assessment of the N501Y mutant strains of SARS-CoV-2 in the United Kingdom, October to November 2020. Euro Surveill 26, doi:10.2807/1560-7917.ES.2020.26.1.2002106 (2021).
50
Tegally, H. et al. Emergence and rapid spread of a new severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) lineage with multiple spike mutations in South Africa. medRxiv, 2020.2012.2021.20248640, doi:10.1101/2020.12.21.20248640 (2020).
51
Voloch, C. M. et al. Genomic characterization of a novel SARS-CoV-2 lineage from Rio de Janeiro, Brazil. medRxiv, 2020.2012.2023.20248598, doi:10.1101/2020.12.23.20248598 (2020).
52
Liu, Z. et al. Landscape analysis of escape variants identifies SARS-CoV-2 spike mutations that attenuate monoclonal and serum antibody neutralization. bioRxiv, 2020.2011.2006.372037, doi:10.1101/2020.11.06.372037 (2021).
53
Weisblum, Y. et al. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants. Elife 9, doi:10.7554/eLife.61312 (2020).
54
Greaney, A. J. et al. Comprehensive mapping of mutations to the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human serum antibodies. bioRxiv, 2020.2012.2031.425021, doi:10.1101/2020.12.31.425021 (2021).
55
Wibmer, C. K. et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. bioRxiv, 2021.2001.2018.427166, doi:10.1101/2021.01.18.427166 (2021).
56
Andreano, E. et al. SARS-CoV-2 escape in vitro from a highly neutralizing COVID-19 convalescent plasma. bioRxiv, 2020.2012.2028.424451, doi:10.1101/2020.12.28.424451 (2020).
57
Huo, J. et al. Neutralization of SARS-CoV-2 by destruction of the prefusion spike. Cell Host Microbe 28, 445-454 e446, doi:10.1016/j.chom.2020.06.010 (2020).
58
Kabsch, W. Xds. Acta Crystallogr D Biol Crystallogr 66, 125-132, doi:10.1107/S0907444909047337 (2010).
59
Winn, M. D. et al. Overview of the CCP4 suite and current developments. Acta Crystallogr D Biol Crystallogr 67, 235-242, doi:10.1107/S0907444910045749 (2011).
60
McCoy, A. J. et al. Phaser crystallographic software. J Appl Crystallogr 40, 658-674, doi:10.1107/S0021889807021206 (2007).
61
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D Biol Crystallogr 66, 213-221, doi:10.1107/S0907444909052925 (2010).
62
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr D Biol Crystallogr 60, 2126-2132, doi:10.1107/S0907444904019158 (2004).
63
Schrodinger, LLC. The PyMOL Molecular Graphics System, Version 1.8 (2015).
64
Cornwell, O., Radford, S. E., Ashcroft, A. E. & Ault, J. R. Comparing hydrogen deuterium exchange and fast photochemical oxidation of proteins: a structural characterisation of wild-type and deltaN6 beta2-microglobulin. J Am Soc Mass Spectrom 29, 2413-2426, doi:10.1007/s13361-018-2067-y (2018).
65
Starr, T. N. et al. Prospective mapping of viral mutations that escape antibodies used to treat COVID-19. bioRxiv, doi:10.1101/2020.11.30.405472 (2020).
66
Otwinowski, J., McCandlish, D. M. & Plotkin, J. B. Inferring the shape of global epistasis. Proc Natl Acad Sci U S A 115, E7550-E7558, doi:10.1073/pnas.1804015115 (2018).
67
Plante, J. A. et al. Spike mutation D614G alters SARS-CoV-2 fitness. Nature, doi:10.1038/s41586-020-2895-3 (2020).
68
Xie, X. et al. An infectious cDNA clone of SARS-CoV-2. Cell Host Microbe 27, 841-848 e843, doi:10.1016/j.chom.2020.04.004 (2020).
[표 A]
활성 데이터

실시예 5: SARS-CoV-2 수용체 결합을 차단하고 동물을 보호하는 강력한 중화 인간 항체
저자: Seth J. Zost1*, Pavlo Gilchuk1*, James Brett Case3, Elad Binshtein1, Rita E. Chen2,3, Joseph X. Reidy1, Andrew Trivette1, Rachel S. Nargi1, Rachel E. Sutton1, Naveenchandra Suryadevara1, Lauren E. Williamson4, Elaine C. Chen4, Taylor Jones1, Samuel Day1, Luke Myers1, Ahmed O. Hassan3, Natasha M. Kafai2,3, Emma S. Winkler2,3, Julie M. Fox3, James J. Steinhardt6, Kuishu Ren7, Yueh-Ming Loo7, Nicole L. Kallewaard7, David R. Martinez5, Alexandra Schδfer5, Lisa E. Gralinski5, Ralph S. Baric5, Larissa B. Thackray3, Michael S. Diamond2,3,8,9, Robert H. Carnahan1,10**, James E. Crowe, Jr.1,4,10**
소속:
1미국 37232 테네시주 내쉬빌 반더빌트대학교 의료센터 반더빌트 백신센터
2미국 63110 미주리주 세인트루이스 워싱턴대학교 의과대학 병리학 및 면역학과
3미국 미주리주 63110 세인트루이스 워싱턴대학교 의과대학 의학부4미국 37232 테네시주 내쉬빌 반더빌트대학교 의료센터 병리학, 미생물학 및 면역학과
5미국 27599 노스캐롤라이나주 채플힐 채플힐 소재 노스캐롤라이나대학교 전염병학과
6미국 20878 매릴랜드주 게이더스버그 아스트라제네카 항체 발견 및 단백질 공학, 생물제약 R&D
7미국 20878 매릴랜드주 게이더스버그 아스트라제네카 미생물 과학 및 생물제약 R&D
8미국 63110 미주리주 세인트루이스 워싱턴대학교 의과대학 분자미생물학과
9미국 63110 미주리주 세인트루이스 워싱턴대학교 의과대학 앤드류 M. 및 제인 M. 부르스키 인간면역학 및 면역요법 프로그램 센터(Andrew M. and Jane M. Bursky Center for Human Immunology and Immunotherapy Programs)
10미국 37232 테네시주 내쉬빌 반더빌트대학교 의료센터 소아과
연락처 정보:
제임스 E. 크로, Jr., M.D.
[대표 연락처]
소아학, 병리학, 미생물학 및 면역학과 및 반더빌트 백신센터
우편:
미국 테네시주 37232-0417 내쉬빌 갈런드애비뉴 IV 2213 의학연구빌딩 11475 반더빌트 백신센터
전화
(615) 343-8064
이메일
james.crowe@vumc.org
추가 표제 페이지 각주
* 이 저자들은 동등하게 기여하였다.
**교신저자
키워드: 코로나바이러스; SARS-CoV-2; SARS-CoV; COVID-19; 항체, 단클론; 인간; 적응 면역.
COVID-19 팬데믹은 제한된 의학적 대응책만 존재하는 세계 보건에 대한 큰 위협으로, 체액성 면역의 메커니즘에 대한 완전한 이해가 부족하다
1
,
2
. 감염된 대상체의 B 세포에서 분리된 스파이크(S) 당단백질을 표적으로 하는 단클론 항체(mAb) 패널에서, 유사바이러스 또는 야생형(
wt
) SARS-CoV-2 중화 테스트에서 각각 0.9 또는 15 ng/mL만큼 낮은 IC50 값의 강력한 중화 활성을 나타내는 몇몇 mAb를 확인하였다. 가장 강력한 mAb는 S의 수용체 결합 도메인(SRBD)이 인간 ACE2와 상호작용하는 것을 완전히 차단한다. 경쟁 결합, 구조 및 기능 연구를 통해 SRBD의 주요 항원 부위 내에서 별개의 에피토프를 인식하는 정의된 부류로 mAb를 클러스터링할 수 있었다. 전자 현미경 연구는 이러한 mAb가 삼량체 S 단백질의 뚜렷한 입체형태적 상태를 인식한다는 점을 보여주었다. 고유한 부위를 인식하는 강력한 중화 mAb COV2-2196 및 COV2-2130은 S에 동시에 결합되었고 상승적으로 실제 SARS-CoV-2 바이러스를 중화하였다. SARS-CoV-2 감염의 두 가지 뮤린 모델에서, COV2-2916 또는 COV2-2130 단독 또는 두 mAb의 조합의 수동 전달은 심각한 체중 감소로부터 마우스를 보호하고 폐의 바이러스 부하 및 염증을 감소시켰다. 이러한 결과는 SRBD의 보호 에피토프를 확인하고 합리적인 백신 설계 및 강력한 면역치료 칵테일 선택을 위한 구조 기반 프레임워크를 제공한다.
SARS-CoV-2의 S 단백질은 바이러스 부착, 융합 및 숙주 세포로의 진입의 분자 결정기이다3. SARS-CoV-2에 대한 사전 융합 안정화 삼량체 S 단백질 엑토도메인(S2Pecto)의 초저온-EM 구조는 SARS-CoV S 단백질의 것과 유사한 특징을 나타낸다4. 이 유형 I의 통합 막 단백질 및 클래스 I 융합 단백질은 수용체에 대한 결합을 매개하는 N 말단 하위 단위(S1)와 바이러스-세포막 융합을 매개하는 C 말단 하위 단위(S2)를 가지고 있다. S1 하위 단위는 N 말단 도메인(SNTD)과 수용체 결합 도메인(SRBD)을 포함한다. 게놈에서 약 78%의 서열 동일성을 공유하는 SARS-CoV-2와 SARS-CoV1는 모두 인간 안지오텐신 전환 효소 2(hACE2)를 진입 수용체로 사용한다5-7. SARS-CoV8-12 및 중동호흡기증후군(Middle East respiratory syndrome, MERS)13-22을 포함한 다른 고병원성 인수공통전염병 베타코로나바이러스에 대한 인간 면역에 대한 이전 연구에서는 바이러스 표면 스파이크(S) 당단백질에 대한 Ab가 보호 면역을 매개하는 것으로 나타났다. 가장 강력한 S 단백질 특이적 mAb는 수용체의 결합을 직접 매개하는 SRBD의 영역에 결합함으로써 숙주 세포에 대한 바이러스의 부착을 차단함으로써 베타 코로나바이러스를 중화시키는 것으로 보인다. 인간 Ab는 예방, 노출 후 예방 또는 SARS-CoV-2 감염 치료에 사용될 때 SARS-CoV-2 감염 중에 질환을 변경하는 데 사용할 가능성이 있다23. 회복기 혈장을 평가하는 무작위 대조 시험과 과면역 면역글로불린을 평가하는 1건의 시험을 포함한 많은 연구가 진행 중이지만, 이러한 치료가 이환율이나 사망률을 감소시킬 수 있는지는 아직 명확하지 않다24.
본 발명자들은 중국 우한에서 이전에 SARS-CoV-2에 감염된 2명의 B 세포에서 SARS-CoV-2 S 단백질 반응성 mAb의 대규모 패널을 분리하였다25. 이러한 항체의 하위 집합은 S의 수용체 결합 도메인(SRBD)에 결합하였고 실제 SARS-CoV-2를 사용한 신속 스크리닝 분석에서 중화 활성을 나타냈다25. 여기서, SARS-CoV-2의 항원 환경을 정의하고 SRBD의 어느 부위가 보호 mAb의 표적인지 결정하였다. SARS-CoV-2 균주 WA1/2020을 사용한 정량적 포커스 감소 중화 테스트(FRNT)에서 신속 중화 스크리닝 분석에 의해 이전에 미리 선택한 40개의 항-S 인간 mAb 패널을 테스트하였다. 이러한 분석은 패널이 15 내지 4,000 ng/mL 초과 범위의 절반 최대 억제 농도(IC50) 값의 범위를 나타냄(도 20의 a의 히트맵, 표 B에 제시된 값 및 도 24에 제시된 전체 곡선으로 시각화됨)을 보여주었다. 이러한 SRBD 반응성 mAb 중 다수가 hACE2에 대한 SRBD 결합을 차단함으로써 바이러스 감염을 중화한다고 가정하였다. 실제로, 테스트한 대부분의 중화 mAb는 hACE2와 삼량체 S 단백질의 상호작용을 직접적으로 억제하였다(도 20의 a, 도 25). 이러한 결과와 일치하여, 이들 mAb는 또한 삼량체 S 엑토도메인(S2Pecto) 단백질 또는 단량체 SRBD에 강하게 결합하였다(도 20의 a, 도 26). S2Pecto 또는 SRBD 결합 또는 hACE2-차단 효능이 독립적으로 결합 중화 효능을 예측하는지를 평가했으나, 이러한 측정 중 어느 것도 중화 효능과 상관관계가 없었다(도 20의 b 내지 d). 그러나 가장 높은 중화 효능 단계(IC50<150 ng/mL)의 각 mAb는 또한 hACE2에 대한 가장 강력한 차단 활성(IC50<150 ng/mL) 및 S2Pecto 삼량체 및 SRBD에 대한 탁월한 결합 활성(EC50<2 ng/mL)을 나타냈다(도 20의 e). COV2-2196 및 COV2-2130으로 명명된 2개의 강력하게 중화하는 mAb에 대한 대표적인 중화 곡선이 (도 20의 f)에 제시되어 있다. 강력한 중화는 wt 바이러스보다 훨씬 더 민감한 중화 표현형을 드러내고 mAb 효능 평가를 위한 생 바이러스 분석의 사용 요건을 증명한 유사바이러스 중화 분석을 사용하여 확인하였다(도 20의 g). 이러한 mAb 둘 모두는 S2Pecto 삼량체에 강하게 결합하고 hACE2 결합을 완전히 차단하였다(도 20의 h 내지 i).
다음으로 경쟁 결합 분석에 의해 mAb를 중화하기 위한 SRBD의 주요 항원 부위를 정의하였다. 먼저 강력한 중화 mAb COV2-2196 또는 보존된 암호 에피토프를 인식하는 이전에 설명된 SARS-CoV mAb CR3022의 재조합 버전10,26과의 결합을 위해 경쟁하는 mAb를 스크리닝하기 위해 최소 SRBD 도메인을 이용한 생물층 간섭계 기반 경쟁 분석을 사용하였다. 경쟁하는 mAb의 세 가지 주요 군을 확인하였다(도 21의 a). 가장 큰 mAb 군은 COV2-2196을 차단했지만 rCR3022는 차단하지 않은 반면, 일부 mAb는 rCR3022에 의해 차단되었지만 COV2-2196는 그렇지 않았다. COV2-2130을 포함한 2개의 mAb는 참조 mAb에 의해 차단되지 않았다. 대부분의 mAb는 결합에 대해 hACE2와 경쟁했으며, 이는 SRBD의 hACE2 결합 부위 근처에서 결합했음을 시사한다. 삼량체 S2Pecto 단백질을 사용한 ELISA 기반 경쟁 결합 분석에서 COV2-2196, COV2-2130 및 rCR3022를 사용했으며, 또한 SRBD에 3개의 주요 항원 부위가 포함되어 있고 일부 mAb는 둘 이상의 부위에서 접촉할 가능성이 있음을 발견하였다(도 21의 b). 강력하게 중화하는 mAb의 대부분은 COV2-2196과의 결합에 대해 직접적으로 경쟁하였다.
COV2-2196 및 COV2-2130은 SRBD에 대한 결합에 대해 경쟁하지 않았기 때문에 이러한 mAb가 SARS-CoV mAb에 대해 이전에 관찰된 현상10인 바이러스 중화를 위해 상승 작용을 하는지를 평가하였다. 본 발명자들은 SARS-CoV-2를 사용하여 FRNT에서 조합 반응(용량-반응 중화 매트릭스, 도 21의 c 참고)을 테스트하고, 이러한 실험 값을 시너지 스코어링 모델27에 의해 계산된 예상 반응과 비교하였다. 비교 결과, COV2-2196 + COV2-2130의 조합이 시너지 효과가 있음이 밝혀졌다(17.4의 시너지 점수, 여기서 >10의 점수는 시너지를 나타냄). 도 21의 c의 데이터는 용량-반응 시너지 매트릭스를 보여주고 칵테일에서 79 ng/mL의 조합된 mAb 용량(16 ng/mL의 COV2-2196 및 63 ng/mL의 COV2-2130)이 250 ng/mL의 각각의 개별 mAb와 동일한 활성을 가짐을 보여준다(도 21의 c 참고). 이 발견은 칵테일을 사용하면 시험관 내에서 바이러스 중화의 동일한 효능을 달성하기 위해 각 mAb의 용량이 3배 넘게 감소될 수 있음을 보여준다.
다음으로, 중화를 위해 상승 작용하는 두 가지의 주요 경쟁 결합군에서 대표적인 mAb에 의해 인식되는 에피토프를 정의하였다. 중화 mAb의 결합에 대한 중요한 잔기를 결정하기 위해 알라닌 또는 아르기닌 치환을 사용하여 SRBD의 돌연변이 유발 연구를 수행하였다(도 27). 결합 손실 연구는 COV2-2196에 대한 중요한 잔기로서 F486A 또는 N487A를, COV2-2165에 대한 중요한 잔기로서 N487A를 밝혀냈고, 이들은 결합을 위해 서로 경쟁하며, 마찬가지로 K444A 및 G447R 돌연변이체를 사용하여 COV2-2130에 대한 돌연변이 유발 연구는 이들 잔기가 인식을 위해 중요함을 밝혀냈다(도 22의 a). 이전의 구조 연구는 SRBD와 hACE2 사이의 상호작용을 명확히 밝혀냈다(도 22의 b)28. SRBD에서 상호작용하는 잔기의 대부분은 hACE2 인식 모티프를 정의하는 60개 아미노산 선형 펩티드 내에 함유되어 있다(도 22의 c). 다음으로, 이 최소 펩티드에 대한 인간 mAb의 결합을 테스트하였고, COV2-2196, COV2-2165 및 COV2-2832를 포함한 가장 큰 경쟁 결합군의 강력한 중화 구성원이 이 펩티드를 인식함을 발견했으며(도 22의 c), 이는 이들 mAb가 hACE2 인식 모티브 내에서 중요한 접촉을 수행함을 시사한다.
이러한 mAb에 대한 에피토프를 구조적으로 결정하기 위해 S2Pecto 삼량체/Fab 복합체의 음성 염색 전자 현미경을 사용하였다. 강력하게 중화하는 항체 COV2-2196 및 COV2-2165는 SRBD의 hACE2 인식 모티프에 결합하였고 S2Pecto 삼량체의 '개방' 입체형태 상태를 인식하였다(도 22의 d). 그러나 이러한 두 항체의 결합 방식은, 하나에 대한 스파이크 '몸체'에 대해 결합 자세와 각이 나머지와 비교하여 상이하였기에, 달랐다. 두 번째 경쟁 결합군을 대표하는 COV2-2130은 '폐쇄' 위치에서 S2Pecto 삼량체의 RBD에 결합하였다(도 22의 d). COV2-2196과 COV2-2130은 결합에 대해 경쟁하지 않았기 때문에, 두 Fab의 복합체를 동시에 S2Pecto 삼량체에 결합시키려고 시도하였다. S2Pecto 삼량체가 개방 위치에 있을 때 두 Fab가 동시에 결합하는 것을 발견했으며, 이는 COV2-2130이 두 가지 입체형태에서 SRBD를 인식할 수 있음을 나타낸다(도 22의 e). 2-Fab 복합체를 RBD:CR3022 복합체의 구조26와 중첩하여, 이러한 항체가 경쟁 결합 연구에 기초하여 예측된 바와 같이 SRBD의 3개의 별개의 부위에 결합하는 것을 관찰하였다(도 22의 f).
다음으로, 비강 내 아데노바이러스(AdV-hACE2) 형질도입 후 hACE2가 폐에서 발현되는 BALB/c 마우스에서 새롭게 개발된 SARS-CoV-2 감염 모델에서 COV2-2196 또는 COV2-2130 단독 요법 또는 COV2-2196 + COV2-2130 조합의 예방 효능을 테스트하였다. 이 비교적 엄격한 질환 모델에서, 또한 바이러스 감염 및 발병기전을 강화하기 위해 단일 용량의 항-Ifnar1 항체를 투여했는데, 그 결과 파종성 간질성 폐렴이 발생하였다(A. Hassan 및 M. Diamond, 출판을 위해 제출됨). 4 x 105 PFU의 SARS-CoV-2를 이용한 비강 내 챌린지 1일 전에 AdV-hACE2 형질도입된 마우스에게 단일 용량의 mAb COV2-2196(10 mg/kg), COV2-2130(10 mg/kg), COV2-2196 + COV2-2130의 조합(각각 5 mg/kg) 또는 이소형 대조군 mAb(10 mg/kg)를 수동적으로 전달하였다. COV2-2196 또는 COV2-2130 또는 이들의 조합을 사용한 예방은 감염 첫 주 동안 심각한 SARS-CoV-2 유발 체중 감소를 예방하였다(도 23의 a). 바이러스 RNA 수준은 폐, 및 심장 및 비장을 포함하는 원거리 부위의 7 dpi에서 유의하게 감소하였다(도 23의 b). 염증 지표인 인터페론 감마(INF-g), IL-6, CXCL10 및 CCL2 사이토카인 및 케모카인 유전자의 발현은 또한 처리된 마우스의 폐에서 7 dpi(질환의 정점)에서 감소되었다(도 23의 c).
또한, 마우스 적응(MA) SARS-CoV-2 바이러스29를 사용하여 면역적격 모델에서 예방 효능에 대해 COV2-2196 또는 COV2-2130 또는 이들의 조합을 테스트하였다(도 23의 d). 시험관 내 테스트는 중화에 대한 IC50 값이 이들 mAb에 대한 wt 및 MA SARS-CoV-2 바이러스에 대해 비슷함을 보여주었다(데이터는 제시되지 않음). 200 μg/마우스(약 8 mg/kg)의 용량으로 전달된 각각의 mAb 처리는 이소형 대조군과 비교할 때 폐에서 2 dpi에서 바이러스 RNA 수준을 최대 105배까지 감소시켰다(도 23의 e, 좌측). 이와 일치하여, COV2-2196 및 COV2-2196 + COV2-2130 처리군의 모든 동물 및 COV2-2130 처리의 10마리 중 8마리는 폐 조직의 플라크 역가로 측정 시 폐에서 2 dpi에서 더 이상 감염성 바이러스를 갖고 있지 않았다(도 23의 e, 우측). 종합하면, 마우스에서 이러한 결과는 COV2-2196 또는 COV2-2130 단독 또는 조합이 COVID-19의 치료 또는 예방을 위한 유망한 후보임을 시사하였다.
여기서, SARS-CoV-2에 대한 매우 강력한 mAb의 대규모 패널에 대한 항원 환경을 정의하였다. 수백 개의 더 큰 패널에서 이 mAb 패널을 확인한 이러한 상세한 연구와 스크리닝 연구25는 다양한 인간 중화 항체가 SARS-CoV-2에 의한 자연 감염에 의해 유도되지만 그러한 mAb의 작은 하위 집합만이 높은 효능을 가지고 있으며(생 SARS-CoV-2 바이러스에 대한 IC50<50 ng/mL), 따라서 치료 개발에 적합함을 증명한다. 이러한 강력한 mAb에 대한 생화학적 및 구조적 분석은 SRBD 상의 SARS-CoV 자연 감염에 의해 유발된 인간 mAb에 의한 중화에 취약한 세 가지 주요 항원 부위를 정의하였다. 가장 강력한 2개의 항원 부위로부터의 대표적인 mAb는 시험관 내에서 상승 작용을 일으키고 생체 내 칵테일로서 보호하는 것으로 나타났다. 이 발견은 SARS-CoV-2에 대한 효과적인 체액성 면역의 중요한 특징을 보여주며 다클론 반응에서 상승적 중화 활성의 역할이 더 탐구되어야 함을 시사한다. 또한, SARS-CoV-2가 계속 순환함에 따라, 자연 감염에 의해 유발된 집단 면역이 중화 항체의 선택압에서 회피하는 항원 변이체를 선택하기 시작할 수 있으며, 이는 백신 또는 면역치료제에서 S 단백질의 다중 에피토프를 표적화할 필요성을 강화한다.
현재까지 보고된 전 세계 공통 S 유전자 변이체는 본원에서 연구된 주요 mAb에 대한 돌연변이 연구에서 확인된 잔기 486 또는 487에서의 아미노산 변이체에서 멀리 떨어진 D614G, V483A, L5F, Q675H, H655Y 및 S939F 위치에 있다30. 본원에 설명된 것과 같이 합리적으로 선택된 치료 칵테일은 SARS-CoV-2 회피에 훨씬 더 큰 저항성을 제공할 수 있다. 이 연구는 인간에서 COVID-19 면역치료제로서 사용하기 위한 후보로서 확인된 mAb의 전임상 평가 및 개발을 위한 단계를 설정한다.
데이터 가용성. EM 맵은 수탁 코드 EMBD 21965(S2Pecto apo), EMD-21974(S2Pecto + Fab COVs-2165), EMD-21975(S2Pecto + Fab COVs-2196), EMD-21976(S2Pecto + Fab COVs-2130) 및 EMD-21977(S2Pecto + Fab COV2-2196 + Fab COV2-2130)로 전자현미경 데이터은행에 기탁되었다. 본 연구에서 보고된 자료는 이용이 가능할 것이나, 자료 이전 계약 작성이 필요할 수 있다.
감사의 글. 신속한 시퀀싱을 제공한 반더빌트 VANTAGE 핵심 연구소의 안젤라 존스와 직원, 데이터 관리 및 분석에 도움을 준 로스 트로세스, 유전체학 접근법에 대한 기술 자문을 제공한 VUMC의 로빈 봄바르디와 싱쿠에 소토, 실험 지원 및 주요 시약을 제공한 WUSTL의 아서 킴과 아담 베일리, 로라 밴블라건, 제임스 어니스트, 브록 맥쿤, 스와티 슈리하리, 단백질 및 유사바이러스 시약 및 관련 데이터 생성을 도와 준 아스트라제네카의 케빈 M. 터피와 세메 디알로, 패트릭 M. 맥탐니, 로리 클라크에게 감사드린다. 본 연구는 미국 국방부 첨단연구 프로젝트국(DARPA) 보조금 HR0011-18-2-0001 및 HR00 11-18-3-0001, NIH 계약 75N93019C00074 및 75N93019C00062 및 반더빌트의 돌리 파튼 COVID-19 연구 기금의 지원을 받았다. 본 작업은 Vanderbilt Institute for Clinical and Translational Research에 자리한 차세대 핵산 시퀀서에 대한 NIH 보조금 1S10RR028106-01A1과 NCATS/NIH의 UL1TR002243의 보조금 지원으로 VANTAGE(Vanderbilt Technologies for Advanced Genomics)의 지원을 받았다. S.J.Z.는 NIH T32 AI095202의 지원을 받았다. J.B.C.는 헬렌 헤이 휘트니 재단 박사후 연구원 장학금을 받고 있다. D.R.M.은 NIH T32 AI007151 및 버로우즈 웰컴 기금 박사후 연구원 심화 프로그램 장학금을 받았다. J.E.C.는 연구보조금으로 이 연구를 지원한 독일 다름슈타트의 머크 그룹의 2019 미래 인사이트상 수상자이다. 내용은 전적으로 저자의 책임이며 반드시 미국 정부 또는 기타 후원자의 공식 견해를 나타내는 것은 아니다.
저자 기여. 프로젝트 구상: S.J.Z., P.G., R.H.C., L.B.T., M.S.D., J.E.C.; 자금 확보: J.E.C.와 M.S.D. 실험실 실험 수행: S.J.Z., P.G., J.B.C., E.B., R.E.C., J.X.R., A.T., R.S.N., R.E.S., N.S., L.E.W., A.O.H., N.M.K., E.W., J.M.F., L.B.T., J.J.S., K.R., Y.-M.L., A.S., L.E.G., D.R.M.; 컴퓨터 작업 수행: E.C.C., T.J., S.D., L.M.; 연구 감독: N.L.K, M.S.D., L.B.T., R.S.B., R.H.C., J.E.C. 논문 초고 작성: S.J.Z., P.G., R.H.C., J.E.C.; 모든 저자는 최종 원고를 검토하고 승인하였다.
이익 경쟁. R.S.B.는 백신과 관련된 문제에 대해 다케다(Takeda)와 사노피 파스퇴르의 컨설턴트로 활동하였다. M.S.D.는 이노비스(Inbios), 비어 바이오테크놀로지(Vir Biotechnology), NGM 바이오파마슈티컬스(Biopharmaceuticals), 엘리 릴리(Eli Lilly)의 컨설턴트이고, 모더나(Moderna)의 과학자문위원회에 있으며, 과거에 모더나로부터 관련 없는 연구 보조금을 받았고, 현재는 이머전트 바이오솔루션스(Emergent BioSolutions)에서 관련 없는 연구 보조금을 받고 있다. J.E.C.는 사노피의 컨설턴트로 활동하였고, 컴퓨백스(CompuVax)와 메이사 백신(Meissa Vaccines)의 과학자문위원회에 소속되어 있으며, 모더나와 사노피로부터 이전의 관련 없는 연구 보조금을 받았고, 아이디바이오로직스(IDBiologics, Inc.)의 설립자이다. 반더빌트대학교는 본 작업과 관련이 있는 SARS-CoV-2 항체에 관한 특허를 출원하였다. 아스트라제네카는 본 작업과 관련된 자료/결과에 대해 특허를 출원하였다. J.J.S., K.R., Y.-M.L., N.L.K.는 아스트라제네카의 직원이며 현재 아스트라제네카 주식과 주식매수선택권을 보유하고 있다. 다른 모든 저자는 이익 경쟁을 선언하지 않았다.
추가 정보
본 논문에 대한 보충 정보를 이용할 수 있다.
자료에 대한 서신 및 요청에 대한 수신자는 J.E.C.이다.
참고문헌
1.
Zhou, P., et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 270-273 (2020).
2.
Zhu, N., et al. A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med 382, 727-733 (2020).
3.
Pillay, T.S. Gene of the month: the 2019-nCoV/SARS-CoV-2 novel coronavirus spike protein. J Clin Pathol (2020).
4.
Wrapp, D., et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367, 1260-1263 (2020).
5.
Wan, Y., Shang, J., Graham, R., Baric, R.S. & Li, F. Receptor recognition by the novel Coronavirus from Wuhan: an Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J Virol 94(2020).
6.
Hoffmann, M., et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271-280 e278 (2020).
7.
Li, W., et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426, 450-454 (2003).
8.
Sui, J., et al. Potent neutralization of severe acute respiratory syndrome (SARS) coronavirus by a human mAb to S1 protein that blocks receptor association. Proc Natl Acad Sci U S A 101, 2536-2541 (2004).
9.
ter Meulen, J., et al. Human monoclonal antibody as prophylaxis for SARS coronavirus infection in ferrets. Lancet 363, 2139-2141 (2004).
10.
ter Meulen, J., et al. Human monoclonal antibody combination against SARS coronavirus: synergy and coverage of escape mutants. PLoS Med 3, e237 (2006).
11.
Zhu, Z., et al. Potent cross-reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. Proc Natl Acad Sci U S A 104, 12123-12128 (2007).
12.
Rockx, B., et al. Structural basis for potent cross-neutralizing human monoclonal antibody protection against lethal human and zoonotic severe acute respiratory syndrome coronavirus challenge. J Virol 82, 3220-3235 (2008).
13.
Chen, Z., et al. Human neutralizing monoclonal antibody inhibition of Middle East respiratory syndrome coronavirus replication in the common marmoset. J Infect Dis 215, 1807-1815 (2017).
14.
Choi, J.H., et al. Characterization of a human monoclonal antibody generated from a B- cell specific for a prefusion-stabilized spike protein of Middle East respiratory syndrome coronavirus. PLoS One 15, e0232757 (2020).
15.
Niu, P., et al. Ultrapotent human neutralizing antibody repertoires against Middle East respiratory syndrome coronavirus from a recovered patient. J Infect Dis 218, 1249-1260 (2018).
16.
Wang, L., et al. Importance of neutralizing monoclonal antibodies targeting multiple antigenic sites on the Middle East respiratory syndrome coronavirus spike glycoprotein to avoid neutralization escape. J Virol 92(2018).
17.
Wang, N., et al. Structural definition of a neutralization-sensitive epitope on the MERS- CoV S1-NTD. Cell Rep 28, 3395-3405 e3396 (2019).
18.
Zhang, S., et al. Structural definition of a unique neutralization epitope on the receptor- binding domain of MERS-CoV spike glycoprotein. Cell Rep 24, 441-452 (2018).
19.
Corti, D., et al. Prophylactic and postexposure efficacy of a potent human monoclonal antibody against MERS coronavirus. Proc Natl Acad Sci U S A 112, 10473-10478 (2015).
20.
Jiang, L., et al. Potent neutralization of MERS-CoV by human neutralizing monoclonal antibodies to the viral spike glycoprotein. Sci Transl Med 6, 234ra259 (2014).
21.
Tang, X.C., et al. Identification of human neutralizing antibodies against MERS-CoV and their role in virus adaptive evolution. Proc Natl Acad Sci U S A 111, E2018-2026 (2014).
22.
Ying, T., et al. Exceptionally potent neutralization of Middle East respiratory syndrome coronavirus by human monoclonal antibodies. J Virol 88, 7796-7805 (2014).
23.
Jiang, S., Hillyer, C. & Du, L. Neutralizing antibodies against SARS-CoV-2 and other human coronaviruses. Trends Immunol 41, 355-359 (2020).
24.
Valk, S.J., et al. Convalescent plasma or hyperimmune immunoglobulin for people with COVID-19: a rapid review. Cochrane Database Syst Rev 5, CD013600 (2020).
25.
Zost, S.J., et al. Rapid isolation and profiling of a diverse panel of human monoclonal antibodies targeting the SARS-CoV-2 spike protein. bioRxiv, 2020.2005.2012.091462 (2020).
26.
Yuan, M., et al. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 368, 630-633 (2020).
27.
Ianevski, A., He, L., Aittokallio, T. & Tang, J. SynergyFinder: a web application for analyzing drug combination dose-response matrix data. Bioinformatics 33, 2413-2415 (2017).
28.
Lan, J., et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 581, 215-220 (2020).
29.
Dinnon, K.H., et al. A mouse-adapted SARS-CoV-2 model for the evaluation of COVID- 19 medical countermeasures. bioRxiv, 2020.2005.2006.081497 (2020).
30.
Laha, S., et al. Characterizations of SARS-CoV-2 mutational profile, spike protein stability and viral transmission. bioRxiv, 2020.2005.2003.066266 (2020).


*이전에 보고됨, Zost et al., 2020(참고문헌 25).
방법
항체. 이 논문에서 연구된 인간 항체는 중국에서 획득한 이전에 실험실에서 확인된 증상이 있는 SARS-CoV-2 감염이 있는 북미의 두 대상체의 혈액 샘플에서 분리되었다. 서면 동의 후 검체를 얻기 위한 본래의 임상 연구는 이전에 설명되었으며1 반더빌트대학교 의료센터의 생명윤리위원회와 토론토대학교의 연구윤리위원회의 승인을 받았다. 대상체(56세 남성과 56세 여성)는 부부로, 캐나다 토론토로 여행을 다녀온 중국 우한 거주자이며, 증상 발현 50일 후 백혈구 성분검사를 통해 PBMC를 얻었다. 단일 항원 특이적 B 세포 및 단클론 항체를 암호화하는 항체 가변 유전자의 단리 및 클로닝을 위한 다양한 도구를 사용하여 항체를 단리하였다1.
세포 배양. Vero E6(CRL-1586, ATCC(American Type Culture Collection), Vero CCL81(ATCC), HEK293(ATCC) 및 HEK293T(ATCC)를 10%(vol/vol) 열 불활성화 소 태아 혈청(FBS), 10 mM HEPES pH 7.3, 1 mM 피루브산나트륨, 1x 비필수 아미노산 및 100 U/mL의 페니실린-스트렙토마이신을 포함하는 둘베코 최소 필수 배지(DMEM)에서 5% CO2에서 37℃로 유지시켰다. Vero-퓨린 세포는 T. Pierson(NIH)으로부터 얻었고 이전에 설명되었다2. Expi293F 세포(ThermoFisher Scientific, A1452)를 Expi293F 발현 배지(ThermoFisher Scientific, A1435102)에서 8% CO2에서 37℃로 유지시켰다. ExpiCHO 세포(ThermoFisher Scientific, A29127)를 ExpiCHO 발현 배지(ThermoFisher Scientific, A2910002)에서 8% CO2에서 37℃로 유지시켰다. Expi293F 및 ExpiCHO 배양물의 마이코플라스마 검사를 PCR 기반 마이코플라스마 검출 키트(ATCC, 30-1012K)를 사용하여 매월 수행하였다.
바이러스. SARS-CoV-2 균주 2019 n-CoV/USA_WA1/2020은 질병관리예방센터에서 입수했다(나탈리 손버그의 기증). 바이러스를 Vero CCL81 세포에서 계대하였고, Vero E6 세포에 대한 플라크 분석에 의해 적정하였다. 감염성 SARS-CoV-2에 대한 모든 작업은 워싱턴대학교 의과대학 또는 UNC-채플 힐 기관생물안전성위원회의 승인을 받았으며, 적절한 전동식 공기 정화 호흡기와 개인 보호 장비를 사용하여 승인된 BSL3 시설에서 수행하였다.
재조합 항원 및 단백질. 융합전 입체형태 안정화 SARS-CoV-2 스파이크(S2Pecto) 단백질의 엑토도메인을 암호화하는 유전자를 합성하고 포유동물 세포용 DNA 플라스미드 발현 벡터에 클로닝하였다. 2개의 프롤린이 있고 S의 예비융합 형태의 안정화를 위한 퓨린 절단 부위가 제거된, 유사하게 설계된 S 단백질 항원이 이전에 보고되었다3. 간략하면, 이 유전자는 SARS-CoV-2의 엑토도메인(잔기 1,208까지), T4 피브리틴 삼량체화 도메인, AviTag 부위 특이적 비오틴화 서열 및 C 말단 8x-His 태그를 포함한다. 융합전 입체형태에서 구성체를 안정화하기 위해, 치환 K986P 및 V987P를 포함시키고 잔기 682~685에서 퓨린 절단 부위를 RRAR에서 ASVG로 돌연변이시켰다. 이 재조합 스파이크 2P-안정화 단백질(본원에서 S2Pecto로 명명됨)을 HisTrap Excel 컬럼(GE Healthcare)에서 금속 친화성 크로마토그래피로 단리하고, 단백질 제제를 Superose 6 Increase 10/300 컬럼(GE Healthcare)에서 크기 배제 크로마토그래피로 추가로 정제하였다. 삼량체, 융합 전 입체형태 S 단백질의 존재를 음성 염색 전자 현미경으로 확인하였다1. S 및 Fab를 사용한 전자 현미경을 위하여, 이전에 설명된 것과 유사하게3, AviTag는 결여되어 있지만 C 말단 Twin-Strep-태그를 포함하는 S2Pecto의 변이체를 발현시켰다. 발현된 단백질을 HisTrap Excel 컬럼(GE Healthcare)에서 금속 친화성 크로마토그래피로 단리한 다음, StrepTrap HP 컬럼(GE Healthcare)에서 추가 정제하고, TSKgel G4000SW XL(TOSOH)에서 크기 배제 크로마토그래피를 수행하였다. SARS-CoV-2 S 단백질의 SRBD 하위 도메인을 발현시키기 위해, 잔기 319~541을 IL-2 신호 펩티드의 하류 및 트롬빈 절단 부위, AviTag 및 6x-His 태그의 상류의 포유동물 발현 벡터에 클로닝했다. 마우스 IgG1 Fc 도메인에 융합된 RBD 단백질(RBD-mFc로 명명됨)은 Sino Biological(40592-V05H)에서 구입했다. 알라닌 스캐닝에 의한 에피토프 매핑을 위해 SARS-CoV-2 RBD(잔기 334~526) 또는 RBD 단일 돌연변이 변이체를 N 말단 CD33 리더 서열 및 C 말단 GSSG 링커, AviTag, GSSG 링커 및 8xHis 태그와 함께 클로닝하였다. 스파이크 단백질을 FreeStyle 293 세포(Thermo Fisher)에서 발현시키고 HisTrap 컬럼(GE Healthcare)을 사용한 친화성 크로마토그래피에 이어 Superdex200 컬럼(GE Healthcare)을 사용한 크기 배제 크로마토그래피에 의해 단리하였다. 정제된 단백질을 순도와 적절한 분자량을 보장하기 위해 SDS-PAGE로 분석하였다.
전자 현미경(EM) 염색 그리드 준비, SARS-CoV-2 S2Pecto 단백질 또는 S2Pecto/Fab 복합체의 이미징 및 처리. EM 이미징을 수행하기 위해, 수지에 고정된 시스테인 프로테아제 효소(FabALACTICA, Genovis)를 사용하여 재조합 크로마토그래피 정제된 IgG를 소화시켜 Fab를 생성하였다. 소화는 실온에서 약 16시간 동안 100 mM 인산나트륨, 150 mM NaCl pH 7.2에서 일어났다. 절단된 Fc 및 온전한 IgG를 제거하기 위해, 소화 혼합물을 PBS 완충액에서 RT에서 30분 동안 CaptureSelect Fc 수지(Genovis)와 함께 인큐베이션하였다. 필요한 경우 Fab를 Zeba 스핀 컬럼(Thermo Scientific)을 사용한 원심분리에 의해 Tris 완충액으로 완충액 교환하였다.
인간 Fab와 복합된 음성으로 염색된(NS) SARS-CoV-2 S2Pecto 단백질의 스크리닝 및 이미징을 위해, 단백질을 약 1시간 동안 인큐베이션하고, 약 10~15 μg/mL 농도의 샘플 약 3 μL를 400 평방 메쉬의 구리 EM 그리드(Electron Microscopy Sciences)의 연속 탄소 필름이 있는 글로우 방전 그리드에 적용하였다. 그리드를 0.75%의 우라닐 포르메이트(UF)로 염색하였다4. 200keV에서 작동하고 SerialEM으로 제어하는 FEI TF20(TFS) 투과 전자 현미경을 사용하여 Gatan US4000 4k x 4k CCD 카메라에 이미지를 기록하였다5. 모든 이미지는 1.5~1.8 μm의 디포커스에서 저선량 모드에서 2.18  /pix의 픽셀 크기로 50,000x 배율로 촬영하였다. 현미경 사진의 총 선량은 약 25 내지 38 e-/
/pix의 픽셀 크기로 50,000x 배율로 촬영하였다. 현미경 사진의 총 선량은 약 25 내지 38 e-/ 2였다. 이미지는 cryoSPARC 소프트웨어 패키지를 사용하여 처리하였다6. 이미지를 불러오기 하여 입자를 CTF로 추정하였다. 이어서, 이미지를 노이즈 제거하고 Topaz로 선택하였다7. 입자를 256픽셀의 상자 크기로 추출하였고, 128픽셀로 비닝하였다. 2D 클래스 평균을 수행하고 ab-initio 모델 및 대칭 없는 정제를 위해 양호한 클래스를 선택하였다. SARS-CoV-2 S2Pecto 단백질의 EM 모델 도킹을 위해, EM 맵에 도킹하기 위해 Chimera8에서 폐쇄형 모델(PDB: 6VXX)을 사용하였다(자세한 내용은 표 C 참고). SARS-CoV-2 S2Pecto/Fab COV2-2165 및 SARS-CoV-2 S2Pecto/Fab COV2-2165 복합체의 경우, EM 맵에 도킹하기 위해 Chimera에서 SARS-CoV-2(PDB: 6VYB) 및 Fab(Fab: 12E8)의 개방형 모델을 사용하였다(자세한 내용은 표 C 참조). SARS-Cov-2 S2Pecto/Fab COV2-2130 복합체의 경우, EM 맵에 도킹하기 위해 폐쇄형 모델 및 Fab(PDB: 12E8)를 Chimera에서 사용하였다(자세한 내용은 표 C 참조). 모든 이미지는 Chimera로 제작되었다.
2였다. 이미지는 cryoSPARC 소프트웨어 패키지를 사용하여 처리하였다6. 이미지를 불러오기 하여 입자를 CTF로 추정하였다. 이어서, 이미지를 노이즈 제거하고 Topaz로 선택하였다7. 입자를 256픽셀의 상자 크기로 추출하였고, 128픽셀로 비닝하였다. 2D 클래스 평균을 수행하고 ab-initio 모델 및 대칭 없는 정제를 위해 양호한 클래스를 선택하였다. SARS-CoV-2 S2Pecto 단백질의 EM 모델 도킹을 위해, EM 맵에 도킹하기 위해 Chimera8에서 폐쇄형 모델(PDB: 6VXX)을 사용하였다(자세한 내용은 표 C 참고). SARS-CoV-2 S2Pecto/Fab COV2-2165 및 SARS-CoV-2 S2Pecto/Fab COV2-2165 복합체의 경우, EM 맵에 도킹하기 위해 Chimera에서 SARS-CoV-2(PDB: 6VYB) 및 Fab(Fab: 12E8)의 개방형 모델을 사용하였다(자세한 내용은 표 C 참조). SARS-Cov-2 S2Pecto/Fab COV2-2130 복합체의 경우, EM 맵에 도킹하기 위해 폐쇄형 모델 및 Fab(PDB: 12E8)를 Chimera에서 사용하였다(자세한 내용은 표 C 참조). 모든 이미지는 Chimera로 제작되었다.
MAb 생산 및 정제. 합성되고(Twist Bioscience) IgG1 모노시스트론 발현 벡터(pTwist-mCis_G1로 명명됨) 내로 클로닝된 mAb의 서열을 포유동물 세포 배양 mAb 분비에 사용하였다. 이 벡터는 형질감염 시 단일 구성체에서 mAb 중쇄 및 경쇄 유전자의 동시 발현을 허용하는 증진된 2A 서열 및 GSG 링커를 함유한다9. 본 발명자들은 이전에 96웰 플레이트에서 1 mL의 ExpiCHO 배양액에서 mAb의 마이크로규모의 발현을 설명하였다1. 더 큰 규모의 mAb 발현을 위해, 본 발명자들은 공급업체가 설명한 바와 같이 Gibco™ ExpiCHO™ 발현 시스템 및 50 mL의 미니 생물반응기 튜브(Corning)에 대한 프로토콜을 사용하여 CHO 세포 배양물의 형질감염(항체당 1 내지 300 mL)을 수행하였다. 배양 상청액을 24칼럼 병렬 단백질 크로마토그래피 시스템(Protein BioSolutions)에서 HiTrap MabSelect SuRe(Cytiva, 이전 GE Healthcare Life Sciences)를 사용하여 정제하였다. 정제된 mAb를 PBS로 완충액을 교환하고, Amicon® Ultra- 4 50KDa 원심분리 필터 유닛(Millipore Sigma)을 사용하여 농축하고 사용할 때까지 4℃에서 보관하였다.
ELISA 결합 분석. 96웰 미세적정 플레이트의 웰을 4℃에서 밤새 정제된 재조합 SARS-CoV-2 S 단백질 또는 SARS-CoV-2 SRBD 단백질로 코팅하였다. 플레이트를 1시간 동안 0.05% 트윈-20(DPBS-T)을 함유하는 DPBS에서 2% 무지방 분유 및 2% 정상 염소 혈청으로 차단하였다. 결합된 항체는 HRP(서양고추냉이 퍼옥시다제)와 접합된 염소 항-인간 IgG(Southern Biotech) 및 TMB(3,3',5,5'-테트라메틸벤지미딘) 기질(Thermo Fisher Scientific)을 사용하여 검출하였다. 발색을 모니터링하고, 1N 염산을 첨가하여 반응을 정지시키고, 분광광도계(Biotek)를 사용하여 450 nm에서 흡광도를 측정하였다. 용량-반응 분석을 위해, 정제된 mAb의 연속 희석액을 웰에 3중 반복하여 적용하고, mAb 결합을 위에서 상세히 기재된 바와 같이 검출하였다. S자형 용량-반응 비선형 회귀 분석을 이용하여 mAb 농도의 로그 변환 후 결합에 대한 절반 최대 유효 농도(EC50) 값을 Prism v8.0 소프트웨어(GraphPad)를 사용하여 결정하였다.
RBD 최소 ACE2 결합 모티프 펩티드 결합 ELISA. 384웰 미세적정 플레이트의 웰을 4℃에서 밤새 1 μg/mL의 스트렙타비딘으로 코팅하였다. 플레이트를 1시간 동안 0.05% 트윈-20(DPBS-T)을 함유하는 DPBS에서 0.5% BSA로 차단하였다. 플레이트를 1x PBST로 4x 세척하고, 2 μg/mL의 비오틴화 ACE2 결합 모티프 펩티드(cat. # LT5578, LifeTein, LLC)를 RT에서 1시간 동안 첨가하여 스트렙타디빈을 결합시켰다. 정제된 mAb를 차단 완충액에 희석하고, 웰에 첨가하고, RT에서 1시간 동안 인큐베이션하였다. 결합된 항체는 HRP(서양고추냉이 퍼옥시다제)와 접합된 염소 항-인간 IgG(cat. # 2014-05, Southern Biotech) 및 TMB(3,3',5,5'-테트라메틸벤지미딘) 기질(ThermoFisher Scientific)을 사용하여 검출하였다. 발색을 모니터링하고, 1N 염산을 첨가하여 반응을 정지시키고, 분광광도계(Biotek)를 사용하여 450 nm에서 흡광도를 측정하였다. 용량-반응 분석을 위해, 10 μg/mL 농도에서 시작하는 정제된 mAb의 연속 3배 희석액을 웰에 3중 반복하여 적용하고, mAb 결합을 위에서 상세히 기재된 바와 같이 검출하였다.
알라닌 또는 아르기닌 점 돌연변이가 있는 변이체 RBD 단백질에 대한 항체의 결합 분석. Octet RED96 기기(ForteBio; Pall Life Sciences)와 야생형 RBD 단백질 또는 정의된 위치에서 알라닌 또는 아르기닌으로의 단일 아미노산 변화가 있는 돌연변이체 RBD 단백질을 사용하여 생물층 광 간섭계(BLI)를 수행하였다. RBD 단백질의 결합을 먼저 300초 동안 Penta-His 바이오센서에서 10 μg/mL( 200 nM) 용액에서 옥타-His 태그가 부착된 RBD 야생형 또는 돌연변이체 단백질을 포획하여 확인하였다. 이어서, 바이오센서 팁을 60초간 세척을 위해 결합 완충액(PBS/0.2% Tween 20)에 침지시킨 다음, 150 nM의 mAb를 포함하는 용액에 180초 동안 담그고(결합), 이어서 결합 완충액에 180초 동안 담갔다(해리). 각 RBD 돌연변이체 단백질에 대한 반응을 야생형 RBD 단백질의 반응에 대해 정규화하였다.
200 nM) 용액에서 옥타-His 태그가 부착된 RBD 야생형 또는 돌연변이체 단백질을 포획하여 확인하였다. 이어서, 바이오센서 팁을 60초간 세척을 위해 결합 완충액(PBS/0.2% Tween 20)에 침지시킨 다음, 150 nM의 mAb를 포함하는 용액에 180초 동안 담그고(결합), 이어서 결합 완충액에 180초 동안 담갔다(해리). 각 RBD 돌연변이체 단백질에 대한 반응을 야생형 RBD 단백질의 반응에 대해 정규화하였다.
포커스 감소 중화 테스트(FRNT). mAb의 연속 희석액을 102 FFU의 SARS-CoV-2와 함께 37℃에서 1시간 동안 인큐베이션하였다. mAb-바이러스 복합체를 37℃에서 1시간 동안 96웰 플레이트의 Vero E6 세포 배양 단층에 첨가하였다. 이어서, 세포를 2%의 열 불활성화 FBS를 함유하도록 보충된 최소 필수 배지(MEM)에서 1% (w/v) 메틸셀룰로스로 오버레이하였다. 30시간 후 오버레이를 제거하여 플레이트를 고정하고 PBS 중 4% PFA로 실온에서 20분 동안 고정하였다. 플레이트를 0.1% (w/v) 사포닌(Sigma) 및 0.1% 소 혈청 알부민(BSA)이 보충된 PBS에서 1 μg/mL의 rCR3022 항-S 항체10 및 서양고추냉이 퍼옥시다제(HRP) 접합 염소 항-인간 IgG와 함께 순차적으로 인큐베이션하였다. SARS-CoV-2에 감염된 세포 포커스를 TrueBlue 퍼옥시다제 기질(KPL)을 사용하여 시각화하고, ImmunoSpot 5.0.37 매크로 분석기(Cellular Technologies)에서 정량화하였다. 데이터를 Prism 소프트웨어 버전 8.0(GraphPad)을 사용하여 처리하였다.
S 단백질 유사형 렌티바이러스의 생성 현탁 293 세포를 시딩하고 다음을 암호화하는 패키징 플라스미드와 함께 루시퍼라제를 발현하는 3세대 HIV 기반 렌티바이러스 벡터로 형질감염시켰다: C 말단의 19개 아미노산이 결실된 SARS-CoV-2 스파이크 단백질, Rev 및 Gag-pol. 형질감염 16시간 내지 20시간 후에 배지를 교체하고, 24시간 후에 바이러스를 포함하는 상청액을 수확하였다. 세포 잔해를 저속 원심분리로 제거하고, 상청액을 0.45 μm 필터 유닛에 통과시켰다. 유사바이러스를 초원심분리에 의해 펠릿화하고, 100배 농축 스톡을 위해 PBS에 재현탁시켰다.
유사바이러스 중화 분석. 384-웰 미세역가 플레이트에서 mAb의 연속 희석액을 제조하고, 37℃에서 30분 동안 슈도바이러스와 함께 사전 인큐베이션하고, 여기에 인간 ACE2를 안정적으로 발현하는 293 세포를 첨가하였다. 플레이트를 37℃의 인큐베이터로 되돌린 다음, 48시간 후에 제조업체의 권장사항에 따라 Bright-Glo™ 루시퍼라제 분석 시스템(Promega)을 사용하여 EnVision 2105 다중모드 플레이트 판독기(Perkin Elmer)에서 루시퍼라제 활성을 측정하였다. 억제 백분율을 유사바이러스 단독 대조군과 비교하여 계산하였다. IC50 값을 Prism 소프트웨어 버전 8.1.0(GraphPad)을 사용하여 비선형 회귀에 의해 결정하였다. 각 항체에 대한 평균 IC50 값을 최소 3회의 독립적인 실험에서 결정하였다.
항체 조합에 의한 상승 작용적 중화 측정 시너지는 시험관 내에서 항체의 동일한 총 농도에서 개별 mAb에 의해 매개되는 것과 비교할 때 2개의 mAb의 칵테일에 의해 매개되는 더 높은 중화 활성으로 정의하였다. 2개의 mAb가 SARS-CoV-2를 중화하기 위해 칵테일에서 상승 작용을 하는지 평가하기 위해, 이전에 보고된 접근법을 사용하여 시너지를 정량화하였다11. mAb 조합으로 인한 유익한 효과의 중요성을 평가하기 위해, 관찰된 조합 반응(용량-반응 매트릭스)을 시너지 스코어링 모델에 의해 예상 반응과 비교하였다11. 야생형 SARS-CoV-2 및 Vero-E6 세포 배양 단층을 사용하여 기존의 포커스 감소 중화 시험(focus reduction neutralization test, FRNT) 분석에서 바이러스 중화를 측정하였다. 개별 mAb COV2-2196 및 COV2-2130을 상이한 농도로 혼합하여 칵테일에서 상이한 비율의 mAb의 중화 활성을 평가하였다. 구체적으로, mAb COV2-2130(500 ng/mL부터 시작)의 7배 희석액 각각을 각 조건당 총 부피 50 μL의 mAb COV2-2196(500 ng/mL부터 시작)의 9개 희석액 각각과 혼합한 다음, 96웰 플레이트에서 성장시킨 합류 Vero-E6 세포에 적용하기 전에 세포 배양 배지(2% FBS가 보충된 RPMI-1640 배지)에서 50 μL의 생 SARS-CoV-2와 함께 인큐베이션하였다. 대조군 값에는 칵테일에서와 동일한 용량으로 평가된 개별 mAb COV2-2196 또는 COV2-2130에 대해 개별적으로 측정된 중화 활성의 용량-반응을 결정하기 위한 값이 포함되었다. 각 실험을 2중 반복실험으로 수행하였다. 다음으로 각 조건에 대한 바이러스 중화 백분율을 계산한 다음, 시너지 점수 값을 계산하였는데, 이는 칵테일에서 이들 2개의 mAb 사이의 상호작용을 정의하였다. -10 미만의 시너지 점수는 길항, -10 내지 10의 점수는 부가 효과, 10 초과의 점수는 시너지 효과를 나타낸다11.
MAb 정량화. NanoDrop 분광광도계를 사용하고 인간 IgG의 흡광 계수를 고려하여 UV 분광광도법에 의해 정제된 mAb의 정량화를 수행하였다.
생물층 간섭계를 통한 경쟁 결합 분석. Octet HTX 생물층 간섭계 기기(FortιBio)의 항-마우스 IgG Fc 포획 바이오센서(FortιBio 18-5089)를 1x 동역학 완충액(Molecular Devices 18-1105)에 10분 동안 담그고, 60초 동안 기준선 신호를 측정하였다. 마우스 IgG1에 융합된 재조합 SARS-CoV-2 RBD(RBD-mFc, Sino Biological 40592-V05H)를 바이오센서 팁에 180초 동안 고정시켰다. 30초 동안 1x 동역학 완충액에서의 세척 단계 후, 참조 항체(5 μg/mL)를 항원 함유 바이오센서와 함께 600초 동안 인큐베이션하였다. 참조 항체에는 SARS-CoV 인간 mAb CR3022 및 COV2-2196이 포함되었다. 30초 동안 1x 동역학 완충액에서의 세척 단계 후, 바이오센서 팁을 300초 동안 제2 항체(5 μg/mL)에 담갔다. 각 항체의 최대 결합을 완충액 단독 대조군에 대해 정규화하였다. 자가 대 자가 차단을 차감하였다. 각 항체의 최대 신호 간의 비교를 사용하여 각 항체의 결합 백분율을 결정하였다. 비경쟁 신호의 33% 미만까지의 최대 신호의 감소는, 참조 항체의 존재 하의 제2 항체에 대한 결합의 완전한 경쟁으로 간주하였다. 비경쟁의 33 내지 67%까지의 최대 신호의 감소는, 참조 항체의 존재 하의 제2 항체에 대한 결합의 중간 경쟁으로 간주하였다. 67% 초과의 최대 신호의 결합 백분율은 참조 항체의 존재 하의 제2 항체에 대한 결합의 경쟁 부재로 간주하였다.
고처리량 ACE-2 결합 억제 분석. 384웰 미세적정 플레이트의 웰을 4℃에서 밤새 정제된 재조합 SARS-CoV-2 S2Pecto 단백질로 코팅하였다. 플레이트를 1시간 동안 DPBS-T에서 2% 무지방 분유 및 2% 정상 염소 혈청으로 차단하였다. 마이크로 규모의 발현으로부터 정제된 mAb를 10 μg/mL에서 시작하는 차단 완충액에서 3중 반복으로 2배로 희석하고, 웰에 첨가하고(20 μL/웰), 주위 온도에서 1시간 동안 인큐베이션하였다. C 말단 FLAG 태그 단백질이 있는 재조합 인간 ACE2를 항체 세척 없이 5 μL/웰 부피(ACE2의 최종 0.4 μg/mL 농도)에서 2 μg/mL로 웰에 첨가한 다음, 주위 온도에서 40분 동안 인큐베이션하였다. 플레이트를 세척하고, HRP 접합된 항-FLAG 항체(Sigma) 및 TMB 기질을 사용하여 결합된 ACE2를 검출하였다. 항체가 없는 ACE2 결합은 대조군으로 사용되었다. 테스트한 항체의 각 희석액의 존재 하에 ACE2의 결합에 대해 수득된 신호를, 배경 신호를 뺀 후 항체가 없는 ACE2 결합의 백분율로 표현하였다. S자형 용량-반응 비선형 회귀 분석(Prism 소프트웨어, GraphPad Prism 버전 8.0)을 사용하여 항체 농도의 로그 변환 후 ACE2에 결합하는 S2Pecto 단백질의 mAb에 의한 억제에 대한 절반 최대 억제 농도(IC50) 값을 결정하였다.
생물층 간섭계 바이오센서를 이용한 ACE2 차단 분석. Octet HTX 생물층 간섭계 기기(FortιBio)의 항-마우스 IgG 바이오센서를 1x 동역학 완충액에 10분 동안 담그고, 60초 동안 기준선 신호를 측정하였다. 마우스 IgG1에 융합된 재조합 SARS-CoV-2 RBD(RBD-mFc, Sino Biological 40592-V05H)를 바이오센서 팁에 180초 동안 고정시켰다. 30초 동안 1x 동역학 완충액에서의 세척 단계 후, 항체(5 μg/mL)를 항원이 코팅된 바이오센서와 함께 600초 동안 인큐베이션하였다. 30초 동안 1x 동역학 완충액에서의 세척 단계 후, 바이오센서 팁을 300초 동안 ACE2 수용체(20 μg/mL)(Sigma-Aldrich SAE0064)에 담갔다. ACE2의 최대 결합을 완충액 단독 대조군에 대해 정규화하였다. 항체의 존재 하에 ACE2의 결합 백분율을 ACE2 최대 결합과 비교하였다. 30% 미만까지의 최대 신호의 감소를 ACE2 차단으로 간주하였다.
고처리량 경쟁 결합 분석. 384웰 미세적정 플레이트의 웰을 4℃에서 밤새 정제된 재조합 SARS-CoV-2 S 단백질로 코팅하였다. 플레이트를 1시간 동안 0.05% 트윈-20(DPBS-T)을 함유하는 DPBS에서 2% BSA로 차단하였다. 마이크로 규모의 정제된 비표지 mAb를 차단 완충액에서 10배 희석하고, 웰에 4중 반복하여 첨가하고(20 μL/웰), 주위 온도에서 1시간 동안 인큐베이션하였다. 이전에 설명된 mAb CR302212의 가변 유전자 서열과 SARS-CoV-2 S 단백질의 별개의 항원 영역을 인식하는 새로 확인된 mAb COV2-2096, COV2-2130 및 COV2-2196을 기반으로 하는 재조합 mAb의 비오틴화된 제제를, 표지되지 않은 항체의 세척 없이 5 μL/웰 부피(비오틴화된 mAb의 최종 0.5 μg/mL 농도) 중 2.5 μg/mL의 각각의 mAb가 있는 4개의 웰 각각에 첨가한 다음, 주위 온도에서 1시간 동안 인큐베이션하였다. 플레이트를 세척하고, HRP 접합된 아비딘(Sigma) 및 TMB 기질을 사용하여 결합된 항체를 검출하였다. 표지되지 않은 시험 항체의 존재 하에 비오틴 표지된 참조 항체의 결합에 대해 얻은 신호를, 배경 신호를 뺀 후 참조 항체 단독의 결합의 백분율로 표현하였다. 테스트한 mAb의 존재가 참조 항체 결합을 최대 결합의 41% 미만까지 감소시킨 경우, 테스트한 mAb를 경쟁하는 것으로 간주하였고, 신호가 71%보다 큰 경우 비경쟁으로 간주하였다. 40~70%의 수준을 중간 경쟁으로 간주하였다.
알라닌 또는 아르기닌 RBD 돌연변이체에 대한 mAb의 결합 분석. Octet RED96 기기(ForteBio; Pall Life Sciences)를 사용하여 생물층 광 간섭계를 수행하였다. 결합을 먼저 300초 동안 Penta-His 바이오센서에서 octa-His 태그가 부착된 RBD 돌연변이체 10 μg/mL( 200 nM)을 포획하여 확인하였다. 이어서, 바이오센서를 60초 동안 세척을 위해 결합 완충액(PBS/0.2% TWEEN 20)에 침지시킨 다음, 150 nM의 mAb를 포함하는 용액에 180초 동안 담그고(결합), 이어서 결합 완충액에 180초 동안 담갔다(해리). 각 RBD 돌연변이체에 대한 반응을 야생형 RBD의 반응에 대해 정규화하였다.
200 nM)을 포획하여 확인하였다. 이어서, 바이오센서를 60초 동안 세척을 위해 결합 완충액(PBS/0.2% TWEEN 20)에 침지시킨 다음, 150 nM의 mAb를 포함하는 용액에 180초 동안 담그고(결합), 이어서 결합 완충액에 180초 동안 담갔다(해리). 각 RBD 돌연변이체에 대한 반응을 야생형 RBD의 반응에 대해 정규화하였다.
인간 hACE2 형질도입 마우스를 이용한 마우스 실험. 동물 연구는 실험 동물의 관리 및 사용에 관한 지침서의 권장사항에 따라 수행하였다. 프로토콜은 워싱턴대학교 의과대학의 기관 동물 관리 및 사용 위원회의 승인을 받았다(보장 번호 A3381-01). 염산케타민과 자일라진으로 유도 및 유지된 마취 하에 바이러스 접종을 수행하였고, 동물의 고통을 최소화하기 위해 모든 노력을 기울였다.
BALB/c 마우스는 Jackson Laboratories(균주 000651)에서 구입하였다. 암컷 마우스(10 내지 11주령)에게 2.5 x 108 PFU의 AdV-hACE2를 비강 내 투여하기 하루 전에 2 mg의 항-Ifnar1 mAb(MAR1-5A3 13, Leinco)의 단일 복강 내 주사를 제공하였다. AdV 형질도입 5일 후, 마우스에 비강 경로를 통해 4 x 105 PFU의 SARS-CoV-2를 접종하였다. 항-SARS-CoV-2 인간 mAb 또는 이소형 대조군 mAb를 SARS-CoV-2 접종 24시간 전에 투여하였다. 체중을 매일 모니터링하고 감염 후 5일 또는 7일에 동물을 희생시키고 조직을 수확하였다.
바이러스 부하 측정. 조직을 칭량하고 2% 열 불활성화 FBS가 보충된 DMEM 배지 1 ml에서 MagNA Lyser 기기(Roche Life Science)에서 지르코니아 비드로 균질화하였다. 조직 균질물을 10,000 rpm에서 5분 동안 원심분리하여 정화하고 -80℃에서 보관하였다. MagMax mirVana 총 RNA 분리 키트(Thermo Scientific) 및 Kingfisher Flex 96 웰 추출 기기(Thermo Scientific)를 사용하여 RNA를 추출하였다. TaqMan 프라이머는 SARS-CoV-2(MN908947) 서열을 가이드로 사용하여 N 유전자의 보존 영역을 표적으로 하도록 설계하였다(L 프라이머: ATGCTGCAATCGTGCTACAA; R 프라이머: GACTGCCGCCTCTGCTC; 프로브: /56-FAM/TCAAGGAAC/ZEN/AACATTGCCAA/3IABkFQ/). RNA 표준 곡선을 확립하기 위해, gBlocks 단편(IDT)에서 N 유전자의 연결된 세그먼트를 생성하고 이를 PCR-II topo 벡터(Invitrogen)에 클로닝하였다. 벡터를 선형화하고 시험관 내 T7-DNA 의존성 RNA 전사를 수행하여 정량적 표준 곡선에 대한 물질을 생성하였다.
사이토카인 및 케모카인 mRNA 측정. RNA를 전술한 바와 같이 7 dpi에서 폐 균질물로부터 단리하였다. 제조업체의 프로토콜에 따라 RNase 억제제가 첨가된 고용량 cDNA 역전사 키트(Thermo Scientific)를 이용하여 DNase 처리된 RNA로부터 cDNA를 합성하였다. 사이토카인 및 케모카인 발현을 IFNγ(IDT: Mm.PT.58.41769240), IL-6(Mm.PT.58.10005566), CXCL10(Mm.PT.58.43575827), CCL2(Mm.PT.58.42151692)에 특이적인 상용 프라이머/프로브 세트와 함께 TaqMan Fast Universal PCR master mix(Thermo Scientific)를 이용하여 결정하였고, 결과를 GAPDH(Mm.PT.39a.1) 수준에 대해 정규화하였다. 항-SARS-CoV-2 특이적 또는 이소형 대조군 mAb 처리된 마우스를 미경험 대조군과 비교하는 2-ΔΔCt 방법을 이용하여 배수 변화를 결정하였다.
야생형 마우스를 이용한 마우스 실험. 동물 연구는 실험 동물의 관리 및 사용에 관한 지침서의 권장사항에 따라 수행하였다. 프로토콜은 UNC 채플 힐 의과대학의 기관 동물 관리 및 사용 위원회의 승인을 받았습니다(NIH/PHS 동물 복지 보장 번호는 D16-00256(A3410-01)임). 염산케타민과 자일라진으로 유도 및 유지된 마취 하에 바이러스 접종을 수행하였고, 동물의 고통을 최소화하기 위해 모든 노력을 기울였다.
마우스 적응 SARS-CoV-2(MA-SARS-CoV-2) 바이러스. 바이러스를 이전에 설명한 대로 생성하였다14. 바이러스를 10%의 태아 클론 II 및 1% Pen/Strep을 포함하는 DMEM에서 성장시킨 Vero E6 세포에서 증식시켰다. 바이러스 역가를 플라크 분석에 의해 결정하였다. 간략하면, 바이러스를 연속 희석하고 Vero E6 세포의 컨플루언트 단층에 접종한 다음, 아가로스 오버레이를 수행하였다. 플라크를 중성 적색 염료로 염색한 후 감염 후 2일째에 시각화하였다.
야생형 마우스. Envigo의 12개월령 BALB/c 마우스를 실험에 사용하였다. 실험 시작 전 적어도 72시간 동안 마우스를 BSL3에 적응시켰다. 감염 6시간 전에 복강 내 주사를 통해 200 μg의 인간 단클론 항체로 마우스를 예방적으로 처치하였다. 다음날, 마우스를 케타민과 자일라진의 혼합물로 마취하고, PBS에 희석된 MA-SARS-CoV-2 105 PFU로 비강 내 감염시켰다. 일일 체중 감소를 측정하고, 감염 2일 후 마우스를 조직 수확 전에 이소플루란 과다 투여로 안락사시켰다.
폐 조직 균질물의 플라크 분석. 우측 폐의 하엽을 MagnaLyser(Roche)를 사용하여 1 mL PBS에서 균질화하였다. 바이러스의 연속 희석액을 Vero E6 세포 배양 단층에서 적정하였고, 바이러스 플라크를 접종 후 2일째에 중성 적색 염색에 의해 시각화하였다. 분석에 대한 검출 한계는 폐당 100 PFU이다.
정량화 및 통계 분석. 언급한 바와 같이 기술적 통계 평균 ± SEM 또는 평균 ± SD를 연속 변수에 대해 결정하였다. 기술적 및 생물학적 반복실험은 도면의 범례에 설명되어 있다. 마우스 연구에서, 체중 변화 및 생체 내 바이러스 부하 분석은 각각 2원 ANOVA 및 Mann-Whitney 검정에 의해 결정하였다. 통계 분석은 Prism v8.0(GraphPad)을 사용하여 수행하였다.
온라인 방법에 대한 참고문헌
1
Zost SJ, G. P., Chen RE, Case JB, Reidy JX, Trivette A, Nargi RS, Sutton RE, Suryadevara N, Chen EC, Binshtein E, Shrihari S, Ostrowski M, Chu HY, Didier JE, MacRenaris KW, Jones T, Day S, Myers L, Lee FE-H, Nguyen DC, Sanz I, Martinez DR, Baric RS, Thackray LB,. Diamond MS, Carnahan RH, Crowe JE Jr. . Rapid isolation and profiling of a diverse panel of human monoclonal antibodies targeting the SARS-CoV-2 spike protein. bioRxiv 2020.05.12.091462; doi: https://doi.org/10.1101/2020.05.12.091462 (2020)
2
Mukherjee, S. et al. Enhancing dengue virus maturation using a stable furin over- expressing cell line. Virology 497, 33-40, doi:10.1016/j.virol.2016.06.022 (2016).
3
Wrapp, D. et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367, 1260-1263, doi:10.1126/science.abb2507 (2020).
4
Ohi, M., Li, Y., Cheng, Y. & Walz, T. Negative staining and image classification - Powerful tools in modern electron microscopy. Biol Proced Online 6, 23-34, doi:10.1251/bpo70 (2004).
5
Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. J Struct Biol 152, 36-51, doi:10.1016/j.jsb.2005.07.007 (2005).
6
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat Methods 14, 290-296, doi:10.1038/nmeth.4169 (2017).
7
Bepler, T., Noble, A. J., and Berger, B. Topaz-Denoise: general deep denoising models for cryoEM. bioRxiv. doi:10.1101/838920 (2019).
8
Pettersen, E. F. et al. UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 25, 1605-1612, doi:10.1002/jcc.20084 (2004).
9
Chng, J. et al. Cleavage efficient 2A peptides for high level monoclonal antibody expression in CHO cells. MAbs 7, 403-412, doi:10.1080/19420862.2015.1008351 (2015).
10
Yuan, M. et al. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 368, 630-633, doi:10.1126/science.abb7269 (2020).
11
Ianevski, A., He, L., Aittokallio, T. & Tang, J. SynergyFinder: a web application for analyzing drug combination dose-response matrix data. Bioinformatics 33, 2413-2415, doi:10.1093/bioinformatics/btx162 (2017).
12
ter Meulen, J. et al. Human monoclonal antibody as prophylaxis for SARS coronavirus infection in ferrets. Lancet 363, 2139-2141, doi:10.1016/S0140-6736(04)16506-9 (2004).
13
Sheehan, K. C. et al. Blocking monoclonal antibodies specific for mouse IFN-alpha/beta receptor subunit 1 (IFNAR-1) from mice immunized by in vivo hydrodynamic transfection. J Interferon Cytokine Res 26, 804-819 (2006).
14
Dinnon KH, et al. A mouse-adapted SARS-CoV-2 model for the evaluation of COVID- 19 medical countermeasures. bioRxiv 2020.05.06.081497; doi: https://doi.org/10.1101/2020.05.06.081497 (2020).
* * * * * * * * * * * * * * * * *
[표 1]
항체 가변 영역의 뉴클레오티드 서열



[표 2]
항체 가변 영역에 대한 단백질 서열



[표 3]
중쇄 서열

[표 4]
경쇄 서열

* * * * * * * * * * * * * * * * *
본원에 개시되고 청구된 모든 조성물 및 방법은 본 개시내용에 비추어 과도한 실험 없이 제조 및 실행될 수 있다. 본 개시내용의 조성물 및 방법은 바람직한 구현예의 관점에서 설명되었지만, 당업자에게는 본 개시내용의 개념, 사상 및 범위를 벗어나지 않고도 본원에 기재된 조성물 및 방법, 그리고 방법의 단계 또는 단계의 순서에 변경을 적용할 수 있음이 명백할 것이다. 보다 구체적으로, 화학적으로 및 생리학적으로 관련된 특정 작용제가 본원에 기재된 작용제를 치환할 수 있으며, 동일하거나 유사한 결과가 달성될 것임은 명백할 것이다. 당업자에게 자명한 이러한 모든 유사한 대체물 및 수정은 첨부된 청구범위에 의해 정의된 본 개시내용의 사상, 범위 및 개념의 범위 내에 있는 것으로 간주된다.
VII.
참고문헌
다음의 참고문헌은 이들이 본원에 기재된 것을 보충하는 예시적 절차 또는 기타 상세사항을 제공하는 범위에서 참조로서 본원에 구체적으로 포함된다.
미국 특허 제3,817,837호
미국 특허 제3,850,752호
미국 특허 제3,939,350호
미국 특허 제3,996,345호
미국 특허 제4,196,265호
미국 특허 제4,275,149호
미국 특허 제4,277,437호
미국 특허 제4,366,241호
미국 특허 제4,472,509호
미국 특허 제4,554,101호
미국 특허 제4,680,338호
미국 특허 제4,816,567호
미국 특허 제4,867,973호
미국 특허 제4,938,948호
미국 특허 제5,021,236호
미국 특허 제5,141,648호
미국 특허 제5,196,066호
미국 특허 제5,563,250호
미국 특허 제5,565,332호
미국 특허 제5,856,456호
미국 특허 제5,880,270호
미국 특허 제6,485,982호
"Antibodies: A Laboratory Manual," Cold Spring Harbor Press, Cold Spring Harbor, NY, 1988.
Abbondanzo et al., Am. J. Pediatr. Hematol. Oncol., 12(4), 480-489, 1990.
Allred et al., Arch. Surg., 125(1), 107-113, 1990.
Atherton et al., Biol. of Reproduction, 32, 155-171, 1985.
Barzon et al., Euro Surveill. 2016 Aug 11; 21(32).
Beltramello et al., Cell Host Microbe 8, 271-283, 2010.
Brown et al., J. Immunol. Meth., 12;130(1), :111-121, 1990.
Campbell, In: Monoclonal Antibody Technology, Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 13, Burden and Von Knippenberg, Eds. pp. 75-83, Amsterdam, Elsevier, 1984.
Capaldi et al., Biochem. Biophys. Res. Comm., 74(2):425-433, 1977.
De Jager et al., Semin. Nucl. Med. 23(2), 165-179, 1993.
Dholakia et al., J. Biol. Chem., 264, 20638-20642, 1989.
Diamond et al., J Virol 77, 2578-2586, 2003.
Doolittle and Ben-Zeev, Methods Mol. Biol., 109, :215-237, 1999.
Duffy et al., N. Engl. J. Med. 360, 2536-2543, 2009.
Elder et al. Infections, infertility and assisted reproduction. Part II: Infections in reproductive medicine & Part III: Infections and the assisted reproductive laboratory. Cambridge UK: Cambridge University Press; 2005.
Gefter et al., Somatic Cell Genet., 3:231-236, 1977.
Gornet et al., Semin Reprod Med. 2016 Sep; 34(5):285-292. Epub 2016 Sep 14.
Gulbis and Galand, Hum. Pathol. 24(12), 1271-1285, 1993.
Halfon et al., PLoS ONE 2010; 5 (5) e10569
Hessell et al., Nature 449, 101-4, 2007.
Khatoon et al., Ann. of Neurology, 26, 210-219, 1989.
King et al., J. Biol. Chem., 269, 10210-10218, 1989.
Kohler and Milstein, Eur. J. Immunol., 6, 511-519, 1976.
Kohler and Milstein, Nature, 256, 495-497, 1975.
Kyte and Doolittle, J. Mol. Biol., 157(1):105-132, 1982.
Mansuy et al., Lancet Infect Dis. 2016 Oct; 16(10):1106-7.
Nakamura et al., In: Enzyme Immunoassays: Heterogeneous and Homogeneous Systems, Chapter 27, 1987.
O'Shannessy et al., J. Immun. Meth., 99, 153-161, 1987.
Persic et al., Gene 187:1, 1997
Potter and Haley, Meth. Enzymol., 91, 613-633, 1983.
Purpura et al., Lancet Infect Dis. 2016 Oct; 16(10):1107-8. Epub 2016 Sep 19.
Remington's Pharmaceutical Sciences, 15th Ed., 3:624-652, 1990.
Tang et al., J. Biol. Chem., 271:28324-28330, 1996.
Wawrzynczak & Thorpe, In: Immunoconjugates, Antibody Conuugates In Radioimaging And Therapy Of Cancer, Vogel (Ed.), NY, Oxford University Press, 28, 1987.
Yu et al., J Immunol Methods 336, 142-151, doi:10.1016/j.jim.2008.04.008, 2008.
SEQUENCE LISTING
<110> Vanderbilt University
Crowe, James E Jr.
Zost, Seth
Carnahan, Robert
Gilchuk, Pavlo
<120> Human Monoclonal Antibodies to Severe Acute Respiratory Syndrome
Corona Virus 2 (SARS-CoV-2)
<130> 4815.001PC0D
<150> US 63/000,299
<151> 2020-03-26
<150> US 63/002,896
<151> 2020-03-31
<150> US 63/003,716
<151> 2020-04-01
<150> US 63/023,545
<151> 2020-05-12
<150> US 63/024,204
<151> 2020-05-13
<150> US 63/024,248
<151> 2020-05-13
<150> US 63/027,173
<151> 2020-05-19
<150> US 63/037,984
<151> 2020-06-11
<150> US 63/040,224
<151> 2020-06-17
<150> US 63/040,246
<151> 2020-06-17
<150> US 63/142,196
<151> 2021-01-27
<150> US 63/161,890
<151> 2021-03-16
<160> 187
<170> PatentIn version 3.5
<210> 1
<211> 351
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2165 Heavy
<400> 1
gaggtgcagc tggtggagtc tgggggaggc ttggtccagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggact caccgtccgt agcaactaca tgacctgggt ccgccagact 120
ccagggaagg ggctggaatg ggtgtcagtt atttatagcg gtggtagcac attctacgca 180
gactccgtga agggcagatt caccatctcc agagacaatt ccaagaacac ggtgtatctt 240
caaatgaaca gcctgagagc cgaggacacg gctgtgtatt actgtgcgag agatctcgtg 300
acttacggtt tggacgtctg gggccaaggg accacggtca ccgtctcctc a 351
<210> 2
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2165 Light
<400> 2
gacatccagt tgacccagtc tccatccttc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gggcattagc aattatttag cctggtatca gcaaaaacca 120
gggacagccc ctaacctcct gatctatgct gcatccactt tgcaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagaa ttcactctca caatcagcag cctgcagcct 240
gaagattttg caacttatta ctgtcaacta cttaatagtc accccctcac cttcggccaa 300
gggacacgac tggagattaa a 321
<210> 3
<211> 370
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2838 Heavy
<400> 3
caaatgcagc tggtgcagtc tgggcctgag gtgaagaagc ctgggacctc agtgaaggtc 60
tcctgcaagg cttctggatt cacctttatg agctctgctg tgcagtgggt gcgacaggct 120
cgtggacaac gccttgagtg gataggatgg atcgtcattg gcagtggtaa cacaaactac 180
gcacagaagt tccaggaaag agtcaccatt accagggaca tgtccacaag cacagcctac 240
atggagctga gcagcctgag atccgaggac acggccgtgt attactgtgc ggccccatat 300
tgtagtagta tcagctgcaa tgatggtttt gatatctggg gccaagggac aatggtcacc 360
gtctcttcag 370
<210> 4
<211> 328
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2838 Light
<400> 4
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga gagagccacc 60
ctctcctgca gggccagtca gagtgttagc agcagctact tagcctggta ccagcagaaa 120
cctggccagg ctcccaggct cctcatctat ggtgcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cactatggta gctcacgggg ttggacgttc 300
ggccaaggga ccaaggtgga aatcaaac 328
<210> 5
<211> 351
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2952 heavy
<400> 5
gaggtgcagc tggtggagtc tgggggaggc ttggtccagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggact caccgtccgt agcaactaca tgacctgggt ccgccagact 120
ccagggaagg ggctggaatg ggtgtcagtt atttatagcg gtggtagcac attctacgca 180
gactccgtga agggcagatt caccatctcc agagacaatt ccaagaacac ggtgtatctt 240
caaatgaaca gcctgagagc cgaggacacg gctgtgtatt actgtgcgag agatctcgtg 300
acttacggtt tggacgtctg gggccaaggg accacggtca ccgtctcctc a 351
<210> 6
<211> 322
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2952 light
<400> 6
gacatccagt tgacccagtc tccatccttc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gggcattagc aattatttag cctggtatca gcaaaaacca 120
gggacagccc ctaacctcct gatctatgct gcatccactt tgcaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagaa ttcactctca caatcagcag cctgcagcct 240
gaagattttg caacttatta ctgtcaacta cttaatagtc accccctcac cttcggccaa 300
gggacacgac tggagattaa ac 322
<210> 7
<211> 388
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2514 heavy
<400> 7
gaggtgcagc tggtggagtc tgggggaggt gtggtacggc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt catctttgat gattatgaca tgacctgggt ccgccaagct 120
ccagggaagg ggctggagtg ggtctctggt attaattgga atggtggtag cacaggttat 180
gcagactctg tgaagggccg attcaccatc tccagagaca acgccaagaa ctccctgtat 240
ctgcaaatga acagtctgag agccgaggac acggccttgt atcactgtgc agtgattatg 300
tctccaatcc cccgttatag tggctacgat tgggcgggtg atgcttttga tatctggggc 360
caagggacaa tggtcaccgt ctcttcag 388
<210> 8
<211> 325
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2514 light
<400> 8
tcttctgagc tgactcagga ccctgctgtg tctgtggcct tgggacagac agtcaggatc 60
acatgccaag gagacagcct cagaagctat tatgcaagtt ggtaccagca gaagccagga 120
caggtcccta tacttgtcat ctatgataaa aacaaccggc cctcagggat cccagaccga 180
ttctctggct ccagctcagg aaacacagct tccttgacca tcactggggc tcaggcggaa 240
gatgaggctg actattactg taactcccgg gacagcagtg gtaacgccgt ggtattcggc 300
ggagggacca agctgaccgt cctag 325
<210> 9
<211> 370
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2391 heavy
<400> 9
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggata caccttcggc agctttgata tcaactgggt gcgacaggcc 120
actggacaag ggcttgagtg gatgggacgg atgaactcta acagtgggaa cacagcctat 180
gcacagaagt tccagggcag agtcactatg accagggaca cctccacaaa tacagcctac 240
atggagttga gcagcctgag atctgaggac acggccatgt attactgtgc gagaatgcgc 300
agtggctggc ccacacatgg ccgcccggat gacttctggg gccggggaac cctggtcacc 360
gtctcctcag 370
<210> 10
<211> 331
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2391 light
<400> 10
cagtctgtgc tgactcaggc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcaactc caatatcgga agttatacta taaactggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatttat ggtaatgatc agcggacctc aggggtccct 180
gaccgattct ctggctccaa gtttggcacc tcggcctccc tggccatcag tgggctccag 240
tctgaggatg aaaataatta ttactgtgca gtatgggatg acagcctgaa tggcctggta 300
ttcggcggag ggaccaaact gaccgtccta g 331
<210> 11
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-3025 heavy
<400> 11
gaggtgcagc tggtgcagtc tgggcctgag gtgaagaagc ctgggacctc agtgaaggtc 60
tcctgcaagg cttctggatt cacctttatg agctctgctg tgcagtgggt gcgacaggct 120
cgtggacaac gccttgagtg gataggatgg atcgtcattg gcagtggtaa cacaaactac 180
gcacagaagt tccaggaaag agtcaccatt accagggaca tgtccacaag cacagcctac 240
atggagctga gcagcctgag atccgaggac acggccgtgt attactgtgc ggccccatat 300
tgtagtagta tcagctgcaa tgatggtttt gatatctggg gccaagggac aatggtcacc 360
gtctcttca 369
<210> 12
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-3025 light
<400> 12
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga gagagtcacc 60
ctctcctgca gggccagtca gagtgttagc agcagctact tagcctggta ccagcagaaa 120
cctggccagg ctcccaggct cctcatctat ggtgcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cactatggta gctcacgggg ttggacgttc 300
ggccaaggga ccaaggtgga aatcaaa 327
<210> 13
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2196 heavy
<400> 13
caaatgcagc tggtgcagtc tgggcctgag gtgaagaagc ctgggacctc agtgaaggtc 60
tcctgcaagg cttctggatt cacctttatg agctctgctg tgcagtgggt gcgacaggct 120
cgtggacaac gccttgagtg gataggatgg atcgtcattg gcagtggtaa cacaaactac 180
gcacagaagt tccaggaaag agtcaccatt accagggaca tgtccacaag cacagcctac 240
atggagctga gcagcctgag atccgaggac acggccgtgt attactgtgc ggccccatat 300
tgtagtagta tcagctgcaa tgatggtttt gatatctggg gccaagggac aatggtcacc 360
gtctcttca 369
<210> 14
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2196 light
<400> 14
gaaattgtgt tgacgcagtc tccaggcacc ctgtctttgt ctccagggga gagagccacc 60
ctctcctgca gggccagtca gagtgttagc agcagctact tagcctggta ccagcagaaa 120
cctggccagg ctcccaggct cctcatctat ggtgcatcca gcagggccac tggcatccca 180
gacaggttca gtggcagtgg gtctgggaca gacttcactc tcaccatcag cagactggag 240
cctgaagatt ttgcagtgta ttactgtcag cactatggta gctcacgggg ttggacgttc 300
ggccaaggga ccaaggtgga aatcaaa 327
<210> 15
<211> 387
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2094 heavy
<400> 15
gaggtgcagc tggtggagtc tgggggaggt gtggtacggc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt catctttgat gattatgaca tgacctgggt ccgccaagct 120
ccagggaagg ggctggagtg ggtctctggt attaattgga atggtggtag cacaggttat 180
gcagactctg tgaagggccg attcaccatc tccagagaca acgccaagaa ctccctgtat 240
ctgcaaatga acagtctgag agccgaggac acggccttgt atcactgtgc agtgattatg 300
tctccaatcc cccgttatag tggctacgat tgggcgggtg atgcttttga tatctggggc 360
caagggacaa tggtcaccgt ctcttca 387
<210> 16
<211> 324
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2094 light
<400> 16
tcttctgagc tgactcagga ccctgctgtg tctgtggcct tgggacagac agtcaggatc 60
acatgccaag gagacagcct cagaagctat tatgcaagtt ggtaccagca gaagccagga 120
caggtcccta tacttgtcat ctatgataaa aacaaccggc cctcagggat cccagaccga 180
ttctctggct ccagctcagg aaacacagct tccttgacca tcactggggc tcaggcggaa 240
gatgaggctg actattactg taactcccgg gacagcagtg gtaacgccgt ggtattcggc 300
ggagggacca agctgaccgt ccta 324
<210> 17
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2096 heavy
<400> 17
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggata caccttcggc agctttgata tcaactgggt gcgacaggcc 120
actggacaag ggcttgagtg gatgggacgg atgaactcta acagtgggaa cacagcctat 180
gcacagaagt tccagggcag agtcactatg accagggaca cctccacaaa tacagcctac 240
atggagttga gcagcctgag atctgaggac acggccatgt attactgtgc gagaatgcgc 300
agtggctggc ccacacatgg ccgcccggat gacttctggg gccggggaac cctggtcacc 360
gtctcctca 369
<210> 18
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2096 light
<400> 18
cagtctgtgc tgactcaggc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcaactc caatatcgga agttatacta taaactggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatttat ggtaatgatc agcggacctc aggggtccct 180
gaccgattct ctggctccaa gtttggcacc tcggcctccc tggccatcag tgggctccag 240
tctgaggatg aaaataatta ttactgtgca gtatgggatg acagcctgaa tggcctggta 300
ttcggcggag ggaccaaact gaccgtccta 330
<210> 19
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2130 heavy
<400> 19
gaggtgcagc tggtggagtc tgggggaggc ttggtaaagc ctggggggtc ccttagactc 60
tcctgtgcag cctctggatt cactttcaga gacgtctgga tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggttggccgt attaaaagca aaattgatgg tgggacaaca 180
gactacgctg cacccgtgaa aggcagattc accatctcaa gagatgattc aaaaaacacg 240
ctgtatctgc aaatgaacag cctgaaaacc gaggacacag ccgtgtatta ctgtaccaca 300
gcgggaagct attactatga tactgttggt ccaggcctcc cagagggaaa atttgactac 360
tggggccagg gaaccctggt caccgtctcc tca 393
<210> 20
<211> 336
<212> DNA
<213> Artificial Sequence
<220>
<223> COV2-2130 light
<400> 20
gacatcgtga tgacccagtc tccagactcc ctggctgtgt ctctgggcga gagggccacc 60
atcaactgca agtccagcca gagtgtttta tacagctcca acaataagaa ctacttagct 120
tggtaccagc agaaaccagg acagcctcct aagctgctca tgtactgggc atctacccgg 180
gaatccgggg tccctgaccg attcagtggc agcgggtctg gggcagagtt cactctcacc 240
atcagcagcc tgcaggctga agatgtggca atttattact gtcagcaata ttatagtacc 300
ctcactttcg gcggagggac caaggtggag atcaaa 336
<210> 21
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 heavy
<400> 21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Val Arg Ser Asn
20 25 30
Tyr Met Thr Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Phe Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Leu Val Thr Tyr Gly Leu Asp Val Trp Gly Gln Gly Thr Thr
100 105 110
Val Thr Val Ser Ser
115
<210> 22
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 light
<400> 22
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Thr Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Leu Leu Asn Ser His Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 23
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 heavy
<400> 23
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Met Ser Ser
20 25 30
Ala Val Gln Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Val Ile Gly Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Pro Tyr Cys Ser Ser Ile Ser Cys Asn Asp Gly Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 24
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 light
<400> 24
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Tyr Gly Ser Ser Arg
85 90 95
Gly Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 25
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 heavy
<400> 25
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Val Arg Ser Asn
20 25 30
Tyr Met Thr Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Phe Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Leu Val Thr Tyr Gly Leu Asp Val Trp Gly Gln Gly Thr Thr
100 105 110
Val Thr Val Ser Ser
115
<210> 26
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 light
<400> 26
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Thr Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Leu Leu Asn Ser His Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 27
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 heavy
<400> 27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Asp Asp Tyr
20 25 30
Asp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Asn Trp Asn Gly Gly Ser Thr Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr His Cys
85 90 95
Ala Val Ile Met Ser Pro Ile Pro Arg Tyr Ser Gly Tyr Asp Trp Ala
100 105 110
Gly Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
115 120 125
Ser
<210> 28
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 light
<400> 28
Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Val Arg Ile Thr Cys Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Val Pro Ile Leu Val Ile Tyr
35 40 45
Asp Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Asn Ser Arg Asp Ser Ser Gly Asn Ala
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 29
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 heavy
<400> 29
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Gly Ser Phe
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Met Asn Ser Asn Ser Gly Asn Thr Ala Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Met Arg Ser Gly Trp Pro Thr His Gly Arg Pro Asp Asp Phe
100 105 110
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 30
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 light
<400> 30
Gln Ser Val Leu Thr Gln Ala Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Asn Ser Asn Ile Gly Ser Tyr
20 25 30
Thr Ile Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Asp Gln Arg Thr Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Phe Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Asn Asn Tyr Tyr Cys Ala Val Trp Asp Asp Ser Leu
85 90 95
Asn Gly Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 31
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 heavy
<400> 31
Glu Val Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Met Ser Ser
20 25 30
Ala Val Gln Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Val Ile Gly Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Pro Tyr Cys Ser Ser Ile Ser Cys Asn Asp Gly Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 32
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 light
<400> 32
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Val Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Tyr Gly Ser Ser Arg
85 90 95
Gly Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 33
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 heavy
<400> 33
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Met Ser Ser
20 25 30
Ala Val Gln Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Val Ile Gly Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Pro Tyr Cys Ser Ser Ile Ser Cys Asn Asp Gly Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 34
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 light
<400> 34
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Tyr Gly Ser Ser Arg
85 90 95
Gly Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 35
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 heavy
<400> 35
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Asp Asp Tyr
20 25 30
Asp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Asn Trp Asn Gly Gly Ser Thr Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr His Cys
85 90 95
Ala Val Ile Met Ser Pro Ile Pro Arg Tyr Ser Gly Tyr Asp Trp Ala
100 105 110
Gly Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
115 120 125
Ser
<210> 36
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 light
<400> 36
Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Val Arg Ile Thr Cys Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Val Pro Ile Leu Val Ile Tyr
35 40 45
Asp Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Asn Ser Arg Asp Ser Ser Gly Asn Ala
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 37
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 heavy
<400> 37
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Gly Ser Phe
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Met Asn Ser Asn Ser Gly Asn Thr Ala Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Met Arg Ser Gly Trp Pro Thr His Gly Arg Pro Asp Asp Phe
100 105 110
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 38
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 light
<400> 38
Gln Ser Val Leu Thr Gln Ala Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Asn Ser Asn Ile Gly Ser Tyr
20 25 30
Thr Ile Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Asp Gln Arg Thr Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Phe Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Asn Asn Tyr Tyr Cys Ala Val Trp Asp Asp Ser Leu
85 90 95
Asn Gly Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 39
<211> 131
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 heavy
<400> 39
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Asp Val
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Ile Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr Ala Gly Ser Tyr Tyr Tyr Asp Thr Val Gly Pro Gly
100 105 110
Leu Pro Glu Gly Lys Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
115 120 125
Val Ser Ser
130
<210> 40
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 light
<400> 40
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Met Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Ile Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Thr Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 41
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 CDRH1
<400> 41
Gly Leu Thr Val Arg Ser Asn Tyr
1 5
<210> 42
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 CDRH2
<400> 42
Ile Tyr Ser Gly Gly Ser Thr
1 5
<210> 43
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 CDRH3
<400> 43
Ala Arg Asp Leu Val Thr Tyr Gly Leu Asp Val
1 5 10
<210> 44
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 CDRH1
<400> 44
Gly Phe Thr Phe Met Ser Ser Ala
1 5
<210> 45
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 CDRH2
<400> 45
Ile Val Ile Gly Ser Gly Asn Thr
1 5
<210> 46
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 CDRH3
<400> 46
Ala Ala Pro Tyr Cys Ser Ser Ile Ser Cys Asn Asp Gly Phe Asp Ile
1 5 10 15
<210> 47
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 CDRH1
<400> 47
Gly Leu Thr Val Arg Ser Asn Tyr
1 5
<210> 48
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 CDRH2
<400> 48
Ile Tyr Ser Gly Gly Ser Thr
1 5
<210> 49
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 CDRH3
<400> 49
Ala Arg Asp Leu Val Thr Tyr Gly Leu Asp Val
1 5 10
<210> 50
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 CDRH1
<400> 50
Gly Phe Ile Phe Asp Asp Tyr Asp
1 5
<210> 51
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 CDRH2
<400> 51
Ile Asn Trp Asn Gly Gly Ser Thr
1 5
<210> 52
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 CDRH3
<400> 52
Ala Val Ile Met Ser Pro Ile Pro Arg Tyr Ser Gly Tyr Asp Trp Ala
1 5 10 15
Gly Asp Ala Phe Asp Ile
20
<210> 53
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 CDRH1
<400> 53
Gly Tyr Thr Phe Gly Ser Phe Asp
1 5
<210> 54
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 CDRH2
<400> 54
Met Asn Ser Asn Ser Gly Asn Thr
1 5
<210> 55
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 CDRH3
<400> 55
Ala Arg Met Arg Ser Gly Trp Pro Thr His Gly Arg Pro Asp Asp Phe
1 5 10 15
<210> 56
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 CDRH1
<400> 56
Gly Phe Thr Phe Met Ser Ser Ala
1 5
<210> 57
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 CDRH2
<400> 57
Ile Val Ile Gly Ser Gly Asn Thr
1 5
<210> 58
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 CDRH3
<400> 58
Ala Ala Pro Tyr Cys Ser Ser Ile Ser Cys Asn Asp Gly Phe Asp Ile
1 5 10 15
<210> 59
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 CDRH1
<400> 59
Gly Phe Thr Phe Met Ser Ser Ala
1 5
<210> 60
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 CDRH2
<400> 60
Ile Val Ile Gly Ser Gly Asn Thr
1 5
<210> 61
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 CDRH3
<400> 61
Ala Ala Pro Tyr Cys Ser Ser Ile Ser Cys Asn Asp Gly Phe Asp Ile
1 5 10 15
<210> 62
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 CDRH1
<400> 62
Gly Phe Ile Phe Asp Asp Tyr Asp
1 5
<210> 63
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 CDRH2
<400> 63
Ile Asn Trp Asn Gly Gly Ser Thr
1 5
<210> 64
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 CDRH3
<400> 64
Ala Val Ile Met Ser Pro Ile Pro Arg Tyr Ser Gly Tyr Asp Trp Ala
1 5 10 15
Gly Asp Ala Phe Asp Ile
20
<210> 65
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 CDRH1
<400> 65
Gly Tyr Thr Phe Gly Ser Phe Asp
1 5
<210> 66
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 CDRH2
<400> 66
Met Asn Ser Asn Ser Gly Asn Thr
1 5
<210> 67
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 CDRH3
<400> 67
Ala Arg Met Arg Ser Gly Trp Pro Thr His Gly Arg Pro Asp Asp Phe
1 5 10 15
<210> 68
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 CDRH1
<400> 68
Gly Phe Thr Phe Arg Asp Val Trp
1 5
<210> 69
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 CDRH2
<400> 69
Ile Lys Ser Lys Ile Asp Gly Gly Thr Thr
1 5 10
<210> 70
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 CDRH3
<400> 70
Thr Thr Ala Gly Ser Tyr Tyr Tyr Asp Thr Val Gly Pro Gly Leu Pro
1 5 10 15
Glu Gly Lys Phe Asp Tyr
20
<210> 71
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 CDRL1
<400> 71
Gln Gly Ile Ser Asn Tyr
1 5
<210> 72
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 CDRL2
<400> 72
Ala Ala Ser
1
<210> 73
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2165 CDRL3
<400> 73
Gln Leu Leu Asn Ser His Pro Leu Thr
1 5
<210> 74
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 CDRL1
<400> 74
Gln Ser Val Ser Ser Ser Tyr
1 5
<210> 75
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 CDRL2
<400> 75
Gly Ala Ser
1
<210> 76
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2838 CDRL3
<400> 76
Gln His Tyr Gly Ser Ser Arg Gly Trp Thr
1 5 10
<210> 77
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 CDRL1
<400> 77
Gln Gly Ile Ser Asn Tyr
1 5
<210> 78
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 CDRL2
<400> 78
Ala Ala Ser
1
<210> 79
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2952 CDRL3
<400> 79
Gln Leu Leu Asn Ser His Pro Leu Thr
1 5
<210> 80
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 CDRL1
<400> 80
Ser Leu Arg Ser Tyr Tyr
1 5
<210> 81
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 CDRL2
<400> 81
Asp Lys Asn
1
<210> 82
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2514 CDRL3
<400> 82
Asn Ser Arg Asp Ser Ser Gly Asn Ala Val Val
1 5 10
<210> 83
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 CDRL1
<400> 83
Asn Ser Asn Ile Gly Ser Tyr Thr
1 5
<210> 84
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 CDRL2
<400> 84
Gly Asn Asp
1
<210> 85
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2391 CDRL3
<400> 85
Ala Val Trp Asp Asp Ser Leu Asn Gly Leu Val
1 5 10
<210> 86
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 CDRL1
<400> 86
Gln Ser Val Ser Ser Ser Tyr
1 5
<210> 87
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 CDRL2
<400> 87
Gly Ala Ser
1
<210> 88
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-3025 CDRL3
<400> 88
Gln His Tyr Gly Ser Ser Arg Gly Trp Thr
1 5 10
<210> 89
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 CDRL1
<400> 89
Gln Ser Val Ser Ser Ser Tyr
1 5
<210> 90
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 CDRL2
<400> 90
Gly Ala Ser
1
<210> 91
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 CDRL3
<400> 91
Gln His Tyr Gly Ser Ser Arg Gly Trp Thr
1 5 10
<210> 92
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 CDRL1
<400> 92
Ser Leu Arg Ser Tyr Tyr
1 5
<210> 93
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 CDRL2
<400> 93
Asp Lys Asn
1
<210> 94
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2094 CDRL3
<400> 94
Asn Ser Arg Asp Ser Ser Gly Asn Ala Val Val
1 5 10
<210> 95
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 CDRL1
<400> 95
Asn Ser Asn Ile Gly Ser Tyr Thr
1 5
<210> 96
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 CDRL2
<400> 96
Gly Asn Asp
1
<210> 97
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2096 CDRL3
<400> 97
Ala Val Trp Asp Asp Ser Leu Asn Gly Leu Val
1 5 10
<210> 98
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 CDRL1
<400> 98
Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr
1 5 10
<210> 99
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 CDRL2
<400> 99
Trp Ala Ser
1
<210> 100
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2130 CDRL3
<400> 100
Gln Gln Tyr Tyr Ser Thr Leu Thr
1 5
<210> 101
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> IGHV1-58 FR2-HCDR2
<400> 101
Ile Gly Trp Ile Val
1 5
<210> 102
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> IGHV1-58 HCDR3-FR4
<400> 102
Cys Ala Ala
1
<210> 103
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> IGHD2-2 HCDR3-FR4
<400> 103
Gly Tyr Cys Ser Ser Thr Ser Cys Tyr Ala
1 5 10
<210> 104
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> IGHD2-8 HCDR3-FR4
<400> 104
Gly Tyr Cys Thr Asn Gly Val Cys Tyr Ala
1 5 10
<210> 105
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> IGHD2-15 HCDR3-FR4
<400> 105
Gly Tyr Cys Ser Gly Gly Ser Cys Tyr Ser
1 5 10
<210> 106
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> IGHJ3*02 HCDR3-FR4
<400> 106
Asp Ala Phe Asp Ile Trp
1 5
<210> 107
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> IGKV3-20 LCDR1-FR2
<400> 107
Ser Ser Tyr Leu Ala Trp
1 5
<210> 108
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> IGKV3-20 LCDR3-FR4
<400> 108
Gln Tyr Gly Ser Ser Pro
1 5
<210> 109
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<223> IGKJ1*01 LCDR3-FR4
<400> 109
Trp Thr Phe Gly
1
<210> 110
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 FR2-HCDR2
<400> 110
Ile Gly Trp Ile Val
1 5
<210> 111
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 HCDR3-FR4
<400> 111
Cys Ala Ala Pro Tyr Cys Ser Ser Ile Ser Cys Asn Asp Gly Phe Asp
1 5 10 15
Ile Trp
<210> 112
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 LCDR1-FR2
<400> 112
Ser Ser Tyr Leu Ala Trp
1 5
<210> 113
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2196 LCDR3-FR4
<400> 113
His Tyr Gly Ser Ser Arg Gly Trp Thr Phe Gly
1 5 10
<210> 114
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2381 FR2-HCDR2
<400> 114
Ile Gly Trp Ile Ala
1 5
<210> 115
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2381 HCDR3-FR4
<400> 115
Cys Ala Ala Pro Tyr Cys Ser Arg Thr Ser Cys His Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 116
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2381 LCDR1-FR2
<400> 116
Ser Ser Tyr Leu Ala Trp
1 5
<210> 117
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2381 LCDR3-FR4
<400> 117
His Phe Gly Ser Ser Ser Gln Trp Thr Phe Gly
1 5 10
<210> 118
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2072 FR2-HCDR2
<400> 118
Ile Gly Trp Ile Val
1 5
<210> 119
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2072 HCDR3-FR4
<400> 119
Cys Ala Ala Pro His Cys Asn Arg Thr Ser Cys Tyr Asp Ala Phe Asp
1 5 10 15
Leu Trp
<210> 120
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2072 LCDR1-FR2
<400> 120
Ser Ser Tyr Leu Gly Trp
1 5
<210> 121
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV2-2072 LCDR3-FR4
<400> 121
Gln Tyr Gly Ser Ser Pro Trp Thr Phe Gly
1 5 10
<210> 122
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> MnC5t2p1_G1 FR2-HCDR2
<400> 122
Ile Gly Trp Ile Val
1 5
<210> 123
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> MnC5t2p1_G1 HCDR3-FR4
<400> 123
Cys Ala Ala Pro Arg Cys Ser Gly Gly Ser Cys Tyr Asp Gly Phe Asp
1 5 10 15
Ile Trp
<210> 124
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> MnC5t2p1_G1 LCDR1-FR2
<400> 124
Ser Ser Tyr Leu Ala Trp
1 5
<210> 125
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> MnC5t2p1_G1 LCDR3-FR4
<400> 125
Gln Tyr Gly Ser Ser Pro Trp Thr Phe Gly
1 5 10
<210> 126
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p2_C6 FR2-HCDR2
<400> 126
Ile Gly Trp Ile Val
1 5
<210> 127
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p2_C6 HCDR3-FR4
<400> 127
Cys Ala Ala Pro Tyr Cys Ser Ser Thr Arg Cys Tyr Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 128
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p2_C6 LCDR1-FR2
<400> 128
Ser Ser Tyr Leu Ala Trp
1 5
<210> 129
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p2_C6 LCDR3-FR4
<400> 129
Gln Tyr Gly Arg Ser Pro Trp Thr Phe Gly
1 5 10
<210> 130
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p1_C6 FR2-HCDR2
<400> 130
Ile Gly Trp Ile Val
1 5
<210> 131
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p1_C6 HCDR3-FR4
<400> 131
Cys Ala Ala Pro His Cys Ser Ser Thr Ile Cys Tyr Asp Gly Phe Asp
1 5 10 15
Ile Trp
<210> 132
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p1_C6 LCDR1-FR2
<400> 132
Ser Ser Tyr Leu Ala Trp
1 5
<210> 133
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> HbnC3t1p1_C6 LCDR3-FR4
<400> 133
Gln Tyr Gly Ser Ser Pro Trp Thr Phe Gly
1 5 10
<210> 134
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> S2E12 FR2-HCDR2
<400> 134
Val Gly Trp Ile Val
1 5
<210> 135
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> S2E12 HCDR3-FR4
<400> 135
Cys Ala Ser Pro Tyr Cys Ser Gly Gly Ser Cys Ser Asp Gly Phe Asp
1 5 10 15
Ile Trp
<210> 136
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> S2E12 LCDR1-FR2
<400> 136
Ser Ser Tyr Leu Ala Trp
1 5
<210> 137
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> S2E12 LCDR3-FR4
<400> 137
Gln Tyr Val Gly Leu Thr Gly Trp Thr Phe Gly
1 5 10
<210> 138
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_1 FR2-HCDR2
<400> 138
Ile Gly Trp Ile Val
1 5
<210> 139
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_1 HCDR3-FR4
<400> 139
Cys Ala Ala Pro His Cys Ser Ser Thr Ser Cys Phe Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 140
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_1 LCDR1-FR2
<400> 140
Ser Ser Tyr Leu Ala Trp
1 5
<210> 141
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_1 LCDR3-FR4
<400> 141
Gln Tyr Gly Asn Ser Pro Trp Thr Phe Gly
1 5 10
<210> 142
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_2 FR2-HCDR2
<400> 142
Ile Gly Trp Ile Val
1 5
<210> 143
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_2 HCDR3-FR4
<400> 143
Cys Ala Ala Pro Tyr Cys Ser Gly Gly Ser Cys Ser Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 144
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_2 LCDR1-FR2
<400> 144
Ser Ser Tyr Leu Ala Trp
1 5
<210> 145
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV107_2 LCDR3-FR4
<400> 145
Gln Tyr Gly Ser Ser Pro Trp Thr Phe Gly
1 5 10
<210> 146
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV72 FR2-HCDR2
<400> 146
Ile Gly Trp Ile Val
1 5
<210> 147
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV72 HCDR3-FR4
<400> 147
Cys Ala Ala Val Asp Cys Asn Ser Thr Ser Cys Tyr Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 148
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV72 LCDR1-FR2
<400> 148
Ser Ser Tyr Leu Ala Trp
1 5
<210> 149
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV72 LCDR3-FR4
<400> 149
Gln Tyr Asp Ile Ser Pro Trp Thr Phe Gly
1 5 10
<210> 150
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_1 FR2-HCDR2
<400> 150
Ile Gly Trp Ile Val
1 5
<210> 151
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_1 HCDR3-FR4
<400> 151
Cys Ala Ala Pro His Cys Ser Gly Gly Ser Cys Leu Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 152
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_1 LCDR1-FR2
<400> 152
Ser Ser Tyr Leu Ala Trp
1 5
<210> 153
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_1 LCDR3-FR4
<400> 153
Gln Tyr Gly Ser Ser Pro Trp Thr Phe Gly
1 5 10
<210> 154
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_2 FR2-HCDR2
<400> 154
Ile Gly Trp Ile Val
1 5
<210> 155
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_2 HCDR3-FR4
<400> 155
Cys Ala Ala Pro His Cys Ser Gly Gly Ser Cys Tyr Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 156
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_2 LCDR1-FR2
<400> 156
Ser Ser Tyr Leu Ala Trp
1 5
<210> 157
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV21_2 LCDR3-FR4
<400> 157
Gln Tyr Gly Ser Ser Pro Trp Thr Phe Gly
1 5 10
<210> 158
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_1 FR2-HCDR2
<400> 158
Ile Gly Trp Ile Val
1 5
<210> 159
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_1 HCDR3-FR4
<400> 159
Cys Ala Ala Asn His Cys Ser Gly Gly Ser Cys Tyr Asp Gly Phe Asp
1 5 10 15
Ile Trp
<210> 160
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_1 LCDR1-FR2
<400> 160
Ser Ser Tyr Leu Ala Trp
1 5
<210> 161
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_1 LCDR3-FR4
<400> 161
Gln Tyr Gly Ser Ser Pro Trp Met Phe Gly
1 5 10
<210> 162
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_2 FR2-HCDR2
<400> 162
Ile Gly Trp Ile Val
1 5
<210> 163
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_2 HCDR3-FR4
<400> 163
Cys Ala Ala Pro Tyr Cys Ser Gly Gly Ser Cys Asn Asp Ala Phe Asp
1 5 10 15
Ile Trp
<210> 164
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_2 LCDR1-FR2
<400> 164
Ser Ser Tyr Leu Ala Trp
1 5
<210> 165
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> COV57_2 LCDR3-FR4
<400> 165
Gln Tyr Gly Ser Ser Pro Trp Thr Phe Gly
1 5 10
<210> 166
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP1 HEAVY CHAIN
<220>
<221> MISC_FEATURE
<222> (81)..(81)
<223> Xaa can be Met, Val
<220>
<221> MISC_FEATURE
<222> (100)..(100)
<223> Xaa can be Trp, Ser, Arg
<220>
<221> MISC_FEATURE
<222> (101)..(101)
<223> Xaa can be Asp, Arg, Val
<220>
<221> MISC_FEATURE
<222> (102)..(102)
<223> Xaa can be Leu, Arg, Ala
<220>
<221> MISC_FEATURE
<222> (103)..(103)
<223> Xaa can be Ala, Gly
<220>
<221> MISC_FEATURE
<222> (104)..(104)
<223> Xaa can be Tyr, Ile
<220>
<221> MISC_FEATURE
<222> (107)..(107)
<223> Xaa can be Val, Cys
<220>
<221> MISC_FEATURE
<222> (108)..(108)
<223> Xaa can be Val, Tyr, Thr
<220>
<221> MISC_FEATURE
<222> (109)..(109)
<223> Xaa can be Gly, Val
<220>
<221> MISC_FEATURE
<222> (110)..(110)
<223> Xaa can be Asp, Val
<220>
<221> MISC_FEATURE
<222> (111)..(111)
<223> Xaa can be Cys, Ala
<220>
<221> MISC_FEATURE
<222> (112)..(112)
<223> Xaa can be Ser, Thr
<220>
<221> MISC_FEATURE
<222> (113)..(113)
<223> Xaa can be Arg, His, Ala
<400> 166
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Ser
20 25 30
Ala Val Gln Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Val Val Gly Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Xaa Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Pro Xaa Xaa Xaa Xaa Xaa Cys Gly Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
115 120 125
Ser
<210> 167
<211> 128
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP2 HEAVY CHAIN
<220>
<221> misc_feature
<222> (3)..(3)
<223> Xaa can be Gln, Pro
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa can be Leu, Arg
<220>
<221> misc_feature
<222> (34)..(34)
<223> Xaa can be Met, Val
<220>
<221> misc_feature
<222> (100)..(100)
<223> Xaa can be Gly, Asn, Asp, Gln, Thr, Ser, Leu, Arg
<220>
<221> misc_feature
<222> (101)..(101)
<223> Xaa can be Pro, Gly, Leu, Ser, Val, Lys, Ala
<220>
<221> misc_feature
<222> (102)..(102)
<223> Xaa can be Ser, Gly, Ile, Val, Arg, Pro, Ala, Met
<220>
<221> misc_feature
<222> (103)..(103)
<223> Xaa can be Tyr, Leu, Ile, Arg, Asn, Ala
<220>
<221> misc_feature
<222> (104)..(104)
<223> Xaa can be Cys, Ser, Val, Tyr
<220>
<221> misc_feature
<222> (105)..(105)
<223> Xaa can be Ser, Arg, Val, Tyr, Met
<220>
<221> misc_feature
<222> (106)..(106)
<223> Xaa can be Ser, Gly, Val, Cys
<220>
<221> misc_feature
<222> (107)..(107)
<223> Xaa can be Gly, Thr
<220>
<221> misc_feature
<222> (108)..(108)
<223> Xaa can be Ser, Gly, Val, Thr
<220>
<221> misc_feature
<222> (109)..(109)
<223> Xaa can be Cys, Ala, Thr, Asp
<220>
<221> misc_feature
<222> (110)..(110)
<223> Xaa can be Tyr, Arg, Phe, Pro, Thr
<220>
<221> misc_feature
<222> (111)..(111)
<223> Xaa can be Ala, Pro, Gly, Val, Cys, Arg
<220>
<221> misc_feature
<222> (112)..(112)
<223> Xaa can be Asp, Thr, Ser, Asn, Leu, Ile, Tyr, Pro
<400> 167
Gln Met Xaa Xaa Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Ser
20 25 30
Ala Xaa Gln Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Val Val Gly Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Pro Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120 125
<210> 168
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP3 HEAVY CHAIN
<220>
<221> misc_feature
<222> (3)..(3)
<223> Xaa can be Gln, Pro
<220>
<221> misc_feature
<222> (34)..(34)
<223> Xaa can be Met or Val
<220>
<221> misc_feature
<222> (35)..(35)
<223> Xaa can be Gln, Arg
<220>
<221> misc_feature
<222> (43)..(43)
<223> Xaa can be Gln, Arg
<220>
<221> misc_feature
<222> (100)..(100)
<223> Xaa can be Tyr, Pro, Ala, Arg, Gly
<220>
<221> misc_feature
<222> (101)..(101)
<223> Xaa can be Glu, Leu, Pro, Tyr, Ser
<220>
<221> misc_feature
<222> (102)..(102)
<223> Xaa can be Gly, Val, Ser, Cys, Tyr
<220>
<221> misc_feature
<222> (103)..(103)
<223> Xaa can be Cys, Asp, Val, Gln, Thr
<220>
<221> misc_feature
<222> (104)..(104)
<223> Xaa can be Gly, Asn
<220>
<221> misc_feature
<222> (106)..(106)
<223> Xaa can be Asp, Val
<220>
<221> misc_feature
<222> (108)..(108)
<223> Xaa can be Tyr, Ala, His
<220>
<221> misc_feature
<222> (109)..(109)
<223> Xaa can be Thr, Gln, Ser
<220>
<221> misc_feature
<222> (110)..(110)
<223> Xaa can be Val, His
<220>
<221> misc_feature
<222> (111)..(111)
<223> Xaa can be Thr, Arg
<220>
<221> misc_feature
<222> (112)..(112)
<223> Xaa can be Ala, Pro
<220>
<221> misc_feature
<222> (113)..(113)
<223> Xaa can be Leu, Phe, Ala, Gly, Asn
<400> 168
Gln Met Xaa Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Ser
20 25 30
Ala Xaa Xaa Trp Val Arg Gln Ala Arg Gly Xaa Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Val Val Gly Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Pro Xaa Xaa Xaa Xaa Xaa Gly Xaa Cys Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
115 120 125
Ser
<210> 169
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> CORD HEAVY CHAIN
<220>
<221> misc_feature
<222> (19)..(19)
<223> Xaa can be Arg, Leu
<220>
<221> misc_feature
<222> (78)..(78)
<223> Xaa can be Val, Glu, Gly
<220>
<221> misc_feature
<222> (79)..(79)
<223> Xaa can be Pro, Gly, or His
<220>
<221> misc_feature
<222> (80)..(80)
<223> Xaa can be Cys, Ile, Tyr
<220>
<221> misc_feature
<222> (81)..(81)
<223> Xaa can be Ser, Cys, Val
<220>
<221> misc_feature
<222> (85)..(85)
<223> Xaa can be Cys, Ser, Val
<220>
<221> misc_feature
<222> (86)..(86)
<223> Xaa can be Tyr, Ser, Val
<220>
<221> misc_feature
<222> (87)..(87)
<223> Xaa can be Ser, Thr, Val
<220>
<221> misc_feature
<222> (88)..(88)
<223> Xaa can be Tyr, Ala, Ser
<220>
<221> misc_feature
<222> (89)..(89)
<223> Xaa can be Ser, Ala, Cys
<220>
<221> misc_feature
<222> (90)..(90)
<223> Xaa can be Asp, Tyr
<400> 169
Lys Ala Ser Gly Phe Thr Phe Thr Ser Ser Ala Val Gln Trp Val Arg
1 5 10 15
Gln Ala Xaa Gly Gln Arg Leu Glu Trp Ile Gly Trp Ile Val Val Gly
20 25 30
Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe Gln Glu Arg Val Thr Ile
35 40 45
Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr Met Glu Leu Ser Ser Leu
50 55 60
Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Ala Pro Xaa Xaa Xaa
65 70 75 80
Xaa Gly Gly Ser Xaa Xaa Xaa Xaa Xaa Xaa Asp Ala Phe Asp Ile Trp
85 90 95
Gly Gln Gly Thr Met Val Thr Val Ser Ser
100 105
<210> 170
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP1 LIGHT CHAIN
<220>
<221> misc_feature
<222> (97)..(97)
<223> Xaa can be Pro, Leu, Ser, Thr
<400> 170
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Xaa Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 171
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP2 LIGHT CHAIN
<220>
<221> misc_feature
<222> (97)..(97)
<223> Xaa can be Pro, Leu, Ser, Thr
<400> 171
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Xaa Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 172
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP3 LIGHT CHAIN
<220>
<221> misc_feature
<222> (97)..(97)
<223> Xaa can be Pro, Leu, Ser, Thr
<400> 172
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Xaa Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 173
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP1 Light Chain
<400> 173
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 174
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP1 Light Chain
<400> 174
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 175
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP1 Light Chain
<400> 175
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Leu
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 176
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP1 Light Chain
<400> 176
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Ser
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 177
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP1 Light Chain
<400> 177
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Thr Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 178
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP2 Light Chain
<400> 178
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 179
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP2 Light Chain
<400> 179
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Leu
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 180
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP2 Light Chain
<400> 180
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 181
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP2 Light Chain
<400> 181
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Ser
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 182
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP2 Light Chain
<400> 182
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Thr Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 183
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP3 Light Chain
<400> 183
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 184
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP3 Light Chain
<400> 184
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Leu
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 185
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP3 Light Chain
<400> 185
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 186
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP3 Light Chain
<400> 186
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Ser
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 187
<211> 109
<212> PRT
<213> Artificial Sequence
<220>
<223> HIP3 Light Chain
<400> 187
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Thr Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
Claims (69)
- SARS-CoV-2 표면 스파이크 단백질에 결합하는 단리된 항체 또는 항체 단편으로서,
(a) 서열번호 59의 아미노산 서열을 포함하는 CDRH1, 서열번호 60의 아미노산 서열을 포함하는 CDRH2, 서열번호 61의 아미노산 서열을 포함하는 CDRH3, 서열번호 89의 아미노산 서열을 포함하는 CDRL1, 서열번호 90의 아미노산 서열을 포함하는 CDRH2, 서열번호 91의 아미노산 서열을 포함하는 CDRH3,
(b) 서열번호 68의 아미노산 서열을 포함하는 CDRH1, 서열번호 69의 아미노산 서열을 포함하는 CDRH2, 서열번호 70의 아미노산 서열을 포함하는 CDRH3, 서열번호 98의 아미노산 서열을 포함하는 CDRL1, 서열번호 99의 아미노산 서열을 포함하는 CDRH2, 서열번호 100의 아미노산 서열을 포함하는 CDRH3,
(c) 서열번호 41의 아미노산 서열을 포함하는 CDRH1, 서열번호 42의 아미노산 서열을 포함하는 CDRH2, 서열번호 43의 아미노산 서열을 포함하는 CDRH3, 서열번호 71의 아미노산 서열을 포함하는 CDRL1, 서열번호 72의 아미노산 서열을 포함하는 CDRH2, 서열번호 73의 아미노산 서열을 포함하는 CDRH3,
(d) 서열번호 44의 아미노산 서열을 포함하는 CDRH1, 서열번호 45의 아미노산 서열을 포함하는 CDRH2, 서열번호 46의 아미노산 서열을 포함하는 CDRH3, 서열번호 74의 아미노산 서열을 포함하는 CDRL1, 서열번호 75의 아미노산 서열을 포함하는 CDRH2, 서열번호 76의 아미노산 서열을 포함하는 CDRH3,
(e) 서열번호 47의 아미노산 서열을 포함하는 CDRH1, 서열번호 48의 아미노산 서열을 포함하는 CDRH2, 서열번호 49의 아미노산 서열을 포함하는 CDRH3, 서열번호 77의 아미노산 서열을 포함하는 CDRL1, 서열번호 78의 아미노산 서열을 포함하는 CDRH2, 서열번호 79의 아미노산 서열을 포함하는 CDRH3,
(f) 서열번호 50의 아미노산 서열을 포함하는 CDRH1, 서열번호 51의 아미노산 서열을 포함하는 CDRH2, 서열번호 52의 아미노산 서열을 포함하는 CDRH3, 서열번호 80의 아미노산 서열을 포함하는 CDRL1, 서열번호 81의 아미노산 서열을 포함하는 CDRH2, 서열번호 82의 아미노산 서열을 포함하는 CDRH3,
(g) 서열번호 53의 아미노산 서열을 포함하는 CDRH1, 서열번호 54의 아미노산 서열을 포함하는 CDRH2, 서열번호 55의 아미노산 서열을 포함하는 CDRH3, 서열번호 83의 아미노산 서열을 포함하는 CDRL1, 서열번호 84의 아미노산 서열을 포함하는 CDRH2, 서열번호 85의 아미노산 서열을 포함하는 CDRH3,
(h) 서열번호 56의 아미노산 서열을 포함하는 CDRH1, 서열번호 57의 아미노산 서열을 포함하는 CDRH2, 서열번호 58의 아미노산 서열을 포함하는 CDRH3, 서열번호 86의 아미노산 서열을 포함하는 CDRL1, 서열번호 87의 아미노산 서열을 포함하는 CDRH2, 서열번호 88의 아미노산 서열을 포함하는 CDRH3,
(i) 서열번호 62의 아미노산 서열을 포함하는 CDRH1, 서열번호 63의 아미노산 서열을 포함하는 CDRH2, 서열번호 64의 아미노산 서열을 포함하는 CDRH3, 서열번호 65의 아미노산 서열을 포함하는 CDRL1, 서열번호 66의 아미노산 서열을 포함하는 CDRH2, 서열번호 67의 아미노산 서열을 포함하는 CDRH3,
(j) 서열번호 65의 아미노산 서열을 포함하는 CDRH1, 서열번호 66의 아미노산 서열을 포함하는 CDRH2, 서열번호 67의 아미노산 서열을 포함하는 CDRH3, 서열번호 95의 아미노산 서열을 포함하는 CDRL1, 서열번호 96의 아미노산 서열을 포함하는 CDRH2, 서열번호 97의 아미노산 서열을 포함하는 CDRH3
을 포함하는 항체 또는 항체 단편. - 제1항에 있어서, 다음과 적어도 70%, 80%, 90%, 또는 95%의 동일성을 갖는 중쇄 및 경쇄 가변 서열을 포함하는 항체 또는 항체 단편:
(a) 서열번호 33의 아미노산 서열 및 서열번호 34의 아미노산 서열;
(b) 서열번호 39의 아미노산 서열 및 서열번호 40의 아미노산 서열;
(c) 서열번호 21의 아미노산 서열 및 서열번호 22의 아미노산 서열;
(d) 서열번호 23의 아미노산 서열 및 서열번호 24의 아미노산 서열;
(e) 서열번호 25의 아미노산 서열 및 서열번호 26의 아미노산 서열;
(f) 서열번호 27의 아미노산 서열 및 서열번호 28의 아미노산 서열;
(g) 서열번호 29의 아미노산 서열 및 서열번호 30의 아미노산 서열;
(h) 서열번호 31의 아미노산 서열 및 서열번호 32의 아미노산 서열;
(i) 서열번호 35의 아미노산 서열 및 서열번호 36의 아미노산 서열; 또는
(j) 서열번호 37의 아미노산 서열 및 서열번호 38의 아미노산 서열. - 제1항 또는 제2항에 있어서,
(a) 서열번호 33의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 34의 아미노산 서열을 포함하는 경쇄 가변 서열;
(b) 서열번호 39의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 40의 아미노산 서열을 포함하는 경쇄 가변 서열;
(c) 서열번호 21의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 22의 아미노산 서열을 포함하는 경쇄 가변 서열;
(d) 서열번호 23의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 24의 아미노산 서열을 포함하는 경쇄 가변 서열;
(e) 서열번호 25의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 26의 아미노산 서열을 포함하는 경쇄 가변 서열;
(f) 서열번호 27의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 28의 아미노산 서열을 포함하는 경쇄 가변 서열;
(g) 서열번호 29의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 30의 아미노산 서열을 포함하는 경쇄 가변 서열;
(h) 서열번호 31의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 32의 아미노산 서열을 포함하는 경쇄 가변 서열;
(i) 서열번호 35의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 36의 아미노산 서열을 포함하는 경쇄 가변 서열; 또는
(j) 서열번호 37의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 38의 아미노산 서열을 포함하는 경쇄 가변 서열
을 포함하는 항체 또는 항체 단편. - 제1항 내지 제3항 중 어느 한 항에 있어서, 단클론인 항체 또는 항체 단편.
- 제1항 내지 제4항 중 어느 한 항에 있어서, 항체 단편은 재조합 scFv(단쇄 단편 가변) 항체, Fab 단편, F(ab')2 단편, 또는 Fv 단편인 항체 또는 항체 단편.
- 제1항 내지 제5항 중 어느 한 항에 있어서, YTE 돌연변이를 포함하는 항체 또는 항체 단편.
- 제1항 내지 제6항 중 어느 한 항에 있어서, IgG 또는, 반감기를 증가시키고/시키거나 치료 효능을 증가시키기 위해 FcR 상호작용을 변경(제거 또는 증진)하도록 돌연변이된 Fc 부분, 예를 들어 LALA, LALA PG, N297, GASD/ALIE, DHS, YTE 또는 LS 돌연변이, 또는 글리칸의 효소적 또는 화학적 추가 또는 제거 또는 정의된 글리코실화 패턴으로 조작된 세포주에서의 발현과 같은 FcR 상호작용을 변경(제거 또는 증진)하도록 변형된 글리칸을 포함하는 재조합 IgG 항체 또는 항체 단편인 항체 또는 항체 단편.
- 제1항 내지 제7항 중 어느 한 항에 있어서, Vero-E2 세포 배양 단층을 사용하는 포커스 감소 중화 시험(FRNT) 분석에서 생 BSL3 SARS-CoV-2 바이러스를 중화할 수 있고, 선택적으로 250 ng/mL의 농도에서 생 BSL3 SARS-CoV-2 바이러스의 96%를 중화할 수 있는 항체 또는 항체 단편.
- 제1항 내지 제8항 중 어느 한 항에 있어서, 인간 수용체 안지오텐신 전환 효소 2(ACE2)에 대한 수용체 결합 도메인(RBD) 결합을 차단하고, 선택적으로 hACE2에 대한 활성을 150 ng/mL 미만의 IC50으로 차단하는 항체 또는 항체 단편.
- 제1항 내지 제9항 중 어느 한 항에 있어서, D614G 치환을 포함하는 스파이크 단백질을 포함하는 SARS-CoV-2 바이러스를 중화할 수 있고, 선택적으로 스파이크 단백질은 E484K 치환을 포함하지 않는 것인 항체 또는 항체 단편.
- 제1항 내지 제10항 중 어느 한 항에 있어서, "위쪽" 형태의 RBD에 결합할 수 있는 항체 또는 항체 단편.
- 제1항 내지 제11항 중 어느 한 항에 있어서, "위쪽" 및 "아래쪽" 형태의 RBD에 결합할 수 있는 항체 또는 항체 단편.
- 제1항 내지 제11항 중 어느 한 항에 있어서, "아래쪽" 형태의 RBD에 결합할 수 없는 항체 또는 항체 단편.
- 제1항 내지 제13항 중 어느 한 항에 있어서, 삼량체 스파이크 단백질 엑토도메인에 결합할 수 있고 단량체 스파이크 단백질 RBD에 결합할 수 있으며, 선택적으로 삼량체 스파이크 단백질 엑토도메인에 대한 결합 및/또는 단량체 스파이크 단백질 RBD에 대한 결합은 EC50이 2 ng/mL 미만인 것인 항체 또는 항체 단편.
- 제1항 내지 제14항 중 어느 한 항에 있어서, 검출 가능한 표지를 추가로 포함하는 항체 또는 항체 단편.
- SARS-CoV-2에 감염된 대상체를 치료하거나 SARS-CoV-2에 감염될 위험이 있는 대상체의 감염 가능성을 감소시키는 방법으로서, 제1항 내지 제15항 중 어느 한 항의 제1 항체 또는 항체 단편을 대상체에게 전달하는 단계를 포함하는 방법.
- 60세 이상의 대상체, 면역저하 대상체, 또는 SARS-CoV-2에 감염되거나 감염 위험이 있는 호흡기 및/또는 심혈관 장애를 앓는 대상체의 건강을 보호하는 방법으로서, 제1항 내지 제15항 중 어느 한 항의 제1 항체 또는 항체 단편을 대상체에게 전달하는 단계를 포함하는 방법.
- 제16항 또는 제17항에 있어서, 제2 항체 또는 항체 단편을 대상체에게 전달하는 단계를 추가로 포함하고, 선택적으로 제2 항체 또는 항체 단편은 제1항 내지 제15항 중 어느 한 항의 항체 또는 항체 단편인 방법.
- SARS-CoV-2에 감염된 대상체를 치료하거나 SARS-CoV-2에 감염될 위험이 있는 대상체의 감염 가능성을 감소시키는 방법으로서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편을 대상체에게 전달하는 단계를 포함하고, 제1 및 제2 항체 또는 항체 단편은 SARS-CoV-2를 중화하는 데 상승작용적인 것인 방법.
- 제18항 또는 제19항에 있어서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편은 17.4의 시너지 점수를 갖는 것인 방법.
- 제18항 내지 제20항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편의 용량은, 제1 항체 또는 항체 단편 또는 제2 항체 또는 항체 단편 단독의 용량의 3배를 초과하여 감소되어 바이러스 중화에서 동일한 효능을 달성할 수 있는 것인 방법.
- 제18항 내지 제21항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편은 "위쪽" 형태의 RBD에 결합할 수 있고 "아래쪽" 형태의 RBD에는 결합할 수 없는 것인 방법.
- 제18항 내지 제22항 중 어느 한 항에 있어서, 제2 항체 또는 항체 단편은 "위쪽" 및 "아래쪽" 형태 둘 모두에서 RBD에 결합할 수 있는 것인 방법.
- 제18항 내지 제23항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편은 RBD에 대한 결합에 대해 경쟁하지 않는 것인 방법.
- 제18항 내지 제24항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편은 서열번호 59의 아미노산 서열을 포함하는 CDRH1, 서열번호 60의 아미노산 서열을 포함하는 CDRH2, 서열번호 61의 아미노산 서열을 포함하는 CDRH3, 서열번호 89의 아미노산 서열을 포함하는 CDRL1, 서열번호 90의 아미노산 서열을 포함하는 CDRH2, 서열번호 91의 아미노산 서열을 포함하는 CDRH3을 포함하는 것인 방법.
- 제18항 내지 제25항 중 어느 한 항에 있어서, 제2 항체 또는 항체 단편은 서열번호 68의 아미노산 서열을 포함하는 CDRH1, 서열번호 69의 아미노산 서열을 포함하는 CDRH2, 서열번호 70의 아미노산 서열을 포함하는 CDRH3, 서열번호 98의 아미노산 서열을 포함하는 CDRL1, 서열번호 99의 아미노산 서열을 포함하는 CDRH2, 서열번호 100의 아미노산 서열을 포함하는 CDRH3을 포함하는 것인 방법.
- 제18항 내지 제26항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편은 서열번호 33의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 34의 아미노산 서열을 포함하는 경쇄 가변 서열을 포함하는 것인 방법.
- 제18항 내지 제27항 중 어느 한 항에 있어서, 제2 항체 또는 항체 단편은 서열번호 39의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 40의 아미노산 서열을 포함하는 경쇄 가변 서열을 포함하는 것인 방법.
- 제16항 내지 제28항 중 어느 한 항에 있어서, 전달은 대상체에서 INF-γ, IL-6, CXCL10 및 CCL2의 발현을 감소시키는 것인 방법.
- 제16항 내지 제29항 중 어느 한 항에 있어서, 전달은 정맥 내 전달인 방법.
- 제16항 내지 제30항 중 어느 한 항에 있어서, 대상체는 60세 이상이거나, 면역저하되었거나, 호흡기 및/또는 심혈관 장애를 앓고 있는 대상체인 방법.
- 제16항 내지 제31항 중 어느 한 항에 있어서, 전달은 미처리 대조군과 비교하여 대상체의 호흡을 개선하는 것인 방법.
- 제16항 내지 제32항 중 어느 한 항에 있어서, 전달은 미처리 대조군과 비교하여 바이러스 부하를 감소시키는 것인 방법.
- 제17항 내지 제33항 중 어느 한 항에 있어서, 전달은 감염 전에 이루어지는 것인 방법.
- 제17항 내지 제33항 중 어느 한 항에 있어서, 전달은 감염 후에 이루어지는 것인 방법.
- 하나 이상의, 제1항 내지 제15항 중 어느 한 항의 항체 또는 항체 단편을 포함하는 백신 제형.
- 제36항에 있어서, SARS-CoV-2 표면 스파이크 단백질에 결합하는 제2 항체 또는 항체 단편을 추가로 포함하고, 선택적으로 제2 항체 또는 항체 단편은 제1항 내지 제15항 중 어느 한 항의 항체 또는 항체 단편인 백신 제형.
- 제1항 내지 제15항 중 어느 한 항에 따른 제1 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 포함하는 백신 제형.
- 제38항에 있어서, 발현 벡터(들)는 신드비스 바이러스 또는 VEE 벡터(들)인 백신 제형.
- 제38항 또는 제39항에 있어서, 바늘 주입, 제트 주입 또는 전기천공에 의한 전달을 위해 제형화된 백신 제형.
- 제40항에 있어서, SARS-CoV-2 표면 스파이크 단백질에 결합하는 제2 항체 또는 항체 단편을 암호화하는 하나 이상의 발현 벡터를 추가로 포함하고, 선택적으로 제2 항체 또는 항체 단편은 제1항 내지 제15항 중 어느 한 항의 항체 또는 항체 단편인 백신 제형.
- 대상체에서 SARS-CoV-2에 의한 COVID-19 감염을 검출하는 방법으로서,
(a) 대상체로부터의 샘플을 제1항 내지 제15항 중 어느 한 항의 항체 또는 단편과 접촉시키는 단계; 및
(b) 샘플 내의 SARS-CoV-2 항원에 대한 항체 또는 항체 단편의 결합에 의해 샘플에서 SARS-CoV-2를 검출하는 단계를 포함하는 방법. - 제42항에 있어서, 샘플은 체액인 방법.
- 제42항 또는 제43항에 있어서, 샘플은 혈액, 가래, 눈물, 타액, 점액 또는 혈청, 정액, 자궁경부 또는 질 분비물, 양수, 태반 조직, 소변, 삼출액, 누출액, 조직 찰과표본 또는 대변인 방법.
- 제42항 내지 제44항 중 어느 한 항에 있어서, 검출은 ELISA, RIA, 측면 유동 분석 또는 웨스턴 블롯을 포함하는 것인 방법.
- SARS-CoV-2 표면 스파이크 단백질의 항원 무결성, 정확한 형태 및/또는 정확한 서열을 결정하는 방법으로서,
(a) 항원을 포함하는 샘플을 제1항 내지 제15항 중 어느 한 항의 항체 또는 단편과 접촉시키는 단계; 및
(b) 항원에 대한 항체 또는 항체 단편의 검출 가능한 결합에 의해 항원의 항원 무결성, 정확한 형태 및/또는 정확한 서열을 결정하는 단계를 포함하는 방법. - 제46항에 있어서, 샘플은 재조합으로 생성된 항원을 포함하는 것인 방법.
- 제46항에 있어서, 샘플은 백신 제형 또는 백신 생산 배치를 포함하는 것인 방법.
- 제46항 내지 제48항 중 어느 한 항에 있어서, 검출은 ELISA, RIA, 웨스턴 블롯, 표면 플라즈몬 공명 또는 생물층 간섭계를 사용하는 바이오센서, 또는 유세포 분석 염색을 포함하는 것인 방법.
- 제46항 내지 제49항 중 어느 한 항에 있어서, 시간 경과에 따른 항원의 항원 안정성을 결정하기 위해 단계 (a) 및 (b)를 두 번째로 수행하는 단계를 추가로 포함하는 방법.
- 제1항 내지 제15항 중 어느 한 항의 항체 또는 항체 단편을 암호화하는 폴리뉴클레오티드를 포함하는 하이브리도마 또는 조작 세포.
- 단리된 인간 단클론 항체 또는 항체 단편, 또는 이를 생산하는 하이브리도마 또는 조작 세포로서, 항체는 SARS-CoV-2 표면 스파이크 단백질에 결합하는 것인 단리된 인간 단클론 항체 또는 항체 단편, 또는 이를 생산하는 하이브리도마 또는 조작 세포.
- 제1항 내지 제15항 중 어느 한 항의 항체 또는 항체 단편의 중쇄 가변 영역을 암호화하는 핵산 서열 및/또는 항체 또는 항체 단편의 경쇄 가변 영역을 암호화하는 핵산 서열을 포함하는 단리된 폴리뉴클레오티드.
- 제53항에 있어서, 서열번호 1 내지 20 중 어느 하나의 서열을 포함하는 폴리뉴클레오티드.
- 제54항의 폴리뉴클레오티드를 포함하는 벡터.
- 제53항 또는 제54항의 폴리뉴클레오티드, 제55항의 벡터, 또는 제1항 내지 제15항 중 어느 한 항의 항체 또는 이의 항체 단편의 중쇄 가변 영역을 암호화하는 핵산 분자를 포함하는 제1 벡터 및 경쇄 가변 영역을 암호화하는 핵산 분자를 포함하는 제2 벡터를 포함하는 숙주 세포.
- (a) 제56항의 세포를 배양하는 단계; 및 (b) 배양된 세포로부터 항체 또는 이의 항체 단편을 단리하는 단계를 포함하는 항체 또는 항체 단편의 제조 방법.
- SARS-CoV-2 표면 스파이크 단백질에 결합하는 제1 항체 또는 항체 단편 및 SAR-CoV-2 표면 스파이크 단백질에 결합하는 제2 항체 또는 항체 단편을 포함하는 조성물로서, 선택적으로 제약상 허용되는 조성물인 조성물.
- SARS-CoV-2 표면 스파이크 단백질에 결합하는 제1 항체 또는 항체 단편 및 SAR-CoV-2 표면 스파이크 단백질에 결합하는 제2 항체 또는 항체 단편을 포함하는 키트로서, 선택적으로 SARS-CoV-2에 감염된 대상체를 치료하거나 SARS-CoV-2에 감염될 위험이 있는 대상체의 감염 가능성을 감소시키기 위한 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편의 사용 지침을 추가로 포함하는 키트.
- 제58항 또는 제59항에 있어서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편은 17.4의 시너지 점수를 갖는 것인 조성물 또는 키트.
- 제58항 내지 제60항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편의 용량은, 제1 항체 또는 항체 단편 또는 제2 항체 또는 항체 단편 단독의 용량의 3배를 초과하여 감소되어 바이러스 중화에서 동일한 효능을 달성할 수 있는 것인 조성물 또는 키트.
- 제58항 내지 제61항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편은 "위쪽" 형태의 RBD에 결합할 수 있고 "아래쪽" 형태의 RBD에는 결합할 수 없는 것인 조성물 또는 키트.
- 제58항 내지 제62항 중 어느 한 항에 있어서, 제2 항체 또는 항체 단편은 "위쪽" 및 "아래쪽" 형태 둘 모두에서 RBD에 결합할 수 있는 것인 조성물 또는 키트.
- 제58항 내지 제63항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편은 RBD에 대한 결합에 대해 경쟁하지 않는 것인 조성물 또는 키트.
- 제58항 내지 제64항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편은 서열번호 59의 아미노산 서열을 포함하는 CDRH1, 서열번호 60의 아미노산 서열을 포함하는 CDRH2, 서열번호 61의 아미노산 서열을 포함하는 CDRH3, 서열번호 89의 아미노산 서열을 포함하는 CDRL1, 서열번호 90의 아미노산 서열을 포함하는 CDRH2, 서열번호 91의 아미노산 서열을 포함하는 CDRH3을 포함하는 것인 조성물 또는 키트.
- 제58항 내지 제65항 중 어느 한 항에 있어서, 제2 항체 또는 항체 단편은 서열번호 68의 아미노산 서열을 포함하는 CDRH1, 서열번호 69의 아미노산 서열을 포함하는 CDRH2, 서열번호 70의 아미노산 서열을 포함하는 CDRH3, 서열번호 98의 아미노산 서열을 포함하는 CDRL1, 서열번호 99의 아미노산 서열을 포함하는 CDRH2, 서열번호 100의 아미노산 서열을 포함하는 CDRH3을 포함하는 것인 조성물 또는 키트.
- 제58항 내지 제66항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편은 서열번호 33의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 34의 아미노산 서열을 포함하는 경쇄 가변 서열을 포함하는 것인 조성물 또는 키트.
- 제58항 내지 제67항 중 어느 한 항에 있어서, 제2 항체 또는 항체 단편은 서열번호 39의 아미노산 서열을 포함하는 중쇄 가변 서열 및/또는 서열번호 40의 아미노산 서열을 포함하는 경쇄 가변 서열을 포함하는 것인 조성물 또는 키트.
- 제59항 내지 제68항 중 어느 한 항에 있어서, 제1 항체 또는 항체 단편 및 제2 항체 또는 항체 단편은 개별 용기에 있는 것인 키트.
Applications Claiming Priority (25)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US202063000299P | 2020-03-26 | 2020-03-26 | |
| US63/000,299 | 2020-03-26 | ||
| US202063002896P | 2020-03-31 | 2020-03-31 | |
| US63/002,896 | 2020-03-31 | ||
| US202063003716P | 2020-04-01 | 2020-04-01 | |
| US63/003,716 | 2020-04-01 | ||
| US202063023545P | 2020-05-12 | 2020-05-12 | |
| US63/023,545 | 2020-05-12 | ||
| US202063024204P | 2020-05-13 | 2020-05-13 | |
| US202063024248P | 2020-05-13 | 2020-05-13 | |
| US63/024,248 | 2020-05-13 | ||
| US63/024,204 | 2020-05-13 | ||
| US202063027173P | 2020-05-19 | 2020-05-19 | |
| US63/027,173 | 2020-05-19 | ||
| US202063037984P | 2020-06-11 | 2020-06-11 | |
| US63/037,984 | 2020-06-11 | ||
| US202063040224P | 2020-06-17 | 2020-06-17 | |
| US202063040246P | 2020-06-17 | 2020-06-17 | |
| US63/040,246 | 2020-06-17 | ||
| US63/040,224 | 2020-06-17 | ||
| US202163142196P | 2021-01-27 | 2021-01-27 | |
| US63/142,196 | 2021-01-27 | ||
| US202163161890P | 2021-03-16 | 2021-03-16 | |
| US63/161,890 | 2021-03-16 | ||
| PCT/US2021/024215 WO2021195418A1 (en) | 2020-03-26 | 2021-03-25 | Human monoclonal antibodies to severe acute respiratory syndrome coronavirus 2 (sars-cov-2) |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20220158053A true KR20220158053A (ko) | 2022-11-29 |
Family
ID=75540042
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020227036951A KR20220158053A (ko) | 2020-03-26 | 2021-03-25 | 중증 급성 호흡기 증후군 코로나바이러스 2(SARS-CoV-2)에 대한 인간 단클론 항체 |
Country Status (28)
| Country | Link |
|---|---|
| US (2) | US11345741B2 (ko) |
| EP (2) | EP4356924A2 (ko) |
| JP (1) | JP2023518849A (ko) |
| KR (1) | KR20220158053A (ko) |
| CN (1) | CN115768790A (ko) |
| AU (1) | AU2021242306A1 (ko) |
| BR (1) | BR112022018949A2 (ko) |
| CA (1) | CA3175110A1 (ko) |
| CL (1) | CL2022002594A1 (ko) |
| CO (1) | CO2022014820A2 (ko) |
| CR (1) | CR20220545A (ko) |
| DK (1) | DK4045533T3 (ko) |
| DO (1) | DOP2022000199A (ko) |
| EC (1) | ECSP22083269A (ko) |
| FI (1) | FI4045533T3 (ko) |
| HR (1) | HRP20240182T1 (ko) |
| HU (1) | HUE065039T2 (ko) |
| IL (1) | IL296537A (ko) |
| LT (1) | LT4045533T (ko) |
| MX (1) | MX2022011892A (ko) |
| PE (1) | PE20230415A1 (ko) |
| PL (1) | PL4045533T3 (ko) |
| PT (1) | PT4045533T (ko) |
| RS (1) | RS65199B1 (ko) |
| SI (1) | SI4045533T1 (ko) |
| TW (1) | TW202200610A (ko) |
| UY (1) | UY39135A (ko) |
| WO (1) | WO2021195418A1 (ko) |
Families Citing this family (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US11740240B2 (en) * | 2020-07-20 | 2023-08-29 | Bio-Rad Laboratories, Inc. | Immunoassay for SARS-CoV-2 neutralizing antibodies and materials therefor |
| TW202221025A (zh) * | 2020-08-10 | 2022-06-01 | 英商阿斯特捷利康英國股份有限公司 | 用於治療和預防covid—19的sars—cov—2抗體 |
| EP4228697A1 (en) | 2020-10-16 | 2023-08-23 | Invisishield Technologies Ltd. | Compositions for preventing or treating viral and other microbial infections |
| TW202342095A (zh) * | 2021-11-05 | 2023-11-01 | 英商阿斯特捷利康英國股份有限公司 | 用於治療和預防covid—19之組成物 |
| WO2023108936A1 (zh) * | 2021-12-14 | 2023-06-22 | 杭州安旭生物科技股份有限公司 | 一种可结合SARS-CoV-2病毒的中和抗体及其应用 |
| WO2023209177A1 (en) * | 2022-04-29 | 2023-11-02 | Astrazeneca Uk Limited | Sars-cov-2 antibodies and methods of using the same |
| CN114870061A (zh) * | 2022-05-31 | 2022-08-09 | 康码(上海)生物科技有限公司 | 一种基于病毒阻断剂的空气喷雾剂及其用途 |
| WO2024019110A1 (ja) * | 2022-07-20 | 2024-01-25 | 国立感染症研究所長が代表する日本国 | SARS-CoV-2に対する抗体 |
| CN115925934A (zh) * | 2022-07-26 | 2023-04-07 | 北京昌平实验室 | 一种稳定性提高的人源化单克隆抗体及其应用 |
Family Cites Families (49)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NL154598B (nl) | 1970-11-10 | 1977-09-15 | Organon Nv | Werkwijze voor het aantonen en bepalen van laagmoleculire verbindingen en van eiwitten die deze verbindingen specifiek kunnen binden, alsmede testverpakking. |
| US3817837A (en) | 1971-05-14 | 1974-06-18 | Syva Corp | Enzyme amplification assay |
| US3939350A (en) | 1974-04-29 | 1976-02-17 | Board Of Trustees Of The Leland Stanford Junior University | Fluorescent immunoassay employing total reflection for activation |
| US3996345A (en) | 1974-08-12 | 1976-12-07 | Syva Company | Fluorescence quenching with immunological pairs in immunoassays |
| US4196265A (en) | 1977-06-15 | 1980-04-01 | The Wistar Institute | Method of producing antibodies |
| FR2413974A1 (fr) | 1978-01-06 | 1979-08-03 | David Bernard | Sechoir pour feuilles imprimees par serigraphie |
| US4275149A (en) | 1978-11-24 | 1981-06-23 | Syva Company | Macromolecular environment control in specific receptor assays |
| US4277437A (en) | 1978-04-05 | 1981-07-07 | Syva Company | Kit for carrying out chemically induced fluorescence immunoassay |
| US4366241A (en) | 1980-08-07 | 1982-12-28 | Syva Company | Concentrating zone method in heterogeneous immunoassays |
| US4554101A (en) | 1981-01-09 | 1985-11-19 | New York Blood Center, Inc. | Identification and preparation of epitopes on antigens and allergens on the basis of hydrophilicity |
| US4957939A (en) | 1981-07-24 | 1990-09-18 | Schering Aktiengesellschaft | Sterile pharmaceutical compositions of gadolinium chelates useful enhancing NMR imaging |
| US4867973A (en) | 1984-08-31 | 1989-09-19 | Cytogen Corporation | Antibody-therapeutic agent conjugates |
| US4472509A (en) | 1982-06-07 | 1984-09-18 | Gansow Otto A | Metal chelate conjugated monoclonal antibodies |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| US4676980A (en) | 1985-09-23 | 1987-06-30 | The United States Of America As Represented By The Secretary Of The Department Of Health And Human Services | Target specific cross-linked heteroantibodies |
| US4938948A (en) | 1985-10-07 | 1990-07-03 | Cetus Corporation | Method for imaging breast tumors using labeled monoclonal anti-human breast cancer antibodies |
| US4680338A (en) | 1985-10-17 | 1987-07-14 | Immunomedics, Inc. | Bifunctional linker |
| US5141648A (en) | 1987-12-02 | 1992-08-25 | Neorx Corporation | Methods for isolating compounds using cleavable linker bound matrices |
| US5563250A (en) | 1987-12-02 | 1996-10-08 | Neorx Corporation | Cleavable conjugates for the delivery and release of agents in native form |
| AU2684488A (en) | 1988-06-27 | 1990-01-04 | Carter-Wallace, Inc. | Test device and method for colored particle immunoassay |
| WO1991000360A1 (en) | 1989-06-29 | 1991-01-10 | Medarex, Inc. | Bispecific reagents for aids therapy |
| JPH049249A (ja) | 1990-04-27 | 1992-01-14 | Kusuda:Kk | 塗型剤吹き付け機 |
| JPH06507398A (ja) | 1991-05-14 | 1994-08-25 | リプリジェン コーポレーション | Hiv感染治療のための異種複合抗体 |
| JP4124480B2 (ja) | 1991-06-14 | 2008-07-23 | ジェネンテック・インコーポレーテッド | 免疫グロブリン変異体 |
| US5565332A (en) | 1991-09-23 | 1996-10-15 | Medical Research Council | Production of chimeric antibodies - a combinatorial approach |
| WO1993008829A1 (en) | 1991-11-04 | 1993-05-13 | The Regents Of The University Of California | Compositions that mediate killing of hiv-infected cells |
| EP0656064B1 (en) | 1992-08-17 | 1997-03-05 | Genentech, Inc. | Bispecific immunoadhesins |
| WO1994012520A1 (en) | 1992-11-20 | 1994-06-09 | Enzon, Inc. | Linker for linked fusion polypeptides |
| CA2207869A1 (en) | 1994-12-02 | 1996-06-06 | Chiron Corporation | Method of promoting an immune response with a bispecific antibody |
| US5731168A (en) | 1995-03-01 | 1998-03-24 | Genentech, Inc. | Method for making heteromultimeric polypeptides |
| WO1996040662A2 (en) | 1995-06-07 | 1996-12-19 | Cellpro, Incorporated | Aminooxy-containing linker compounds and their application in conjugates |
| US5837234A (en) | 1995-06-07 | 1998-11-17 | Cytotherapeutics, Inc. | Bioartificial organ containing cells encapsulated in a permselective polyether suflfone membrane |
| US5922845A (en) | 1996-07-11 | 1999-07-13 | Medarex, Inc. | Therapeutic multispecific compounds comprised of anti-Fcα receptor antibodies |
| PT1071700E (pt) | 1998-04-20 | 2010-04-23 | Glycart Biotechnology Ag | Modificação por glicosilação de anticorpos para melhorar a citotoxicidade celular dependente de anticorpos |
| ES2694002T3 (es) | 1999-01-15 | 2018-12-17 | Genentech, Inc. | Polipéptido que comprende una región Fc de IgG1 humana variante |
| US7550143B2 (en) | 2005-04-06 | 2009-06-23 | Ibc Pharmaceuticals, Inc. | Methods for generating stably linked complexes composed of homodimers, homotetramers or dimers of dimers and uses |
| US7534866B2 (en) | 2005-10-19 | 2009-05-19 | Ibc Pharmaceuticals, Inc. | Methods and compositions for generating bioactive assemblies of increased complexity and uses |
| US7527787B2 (en) | 2005-10-19 | 2009-05-05 | Ibc Pharmaceuticals, Inc. | Multivalent immunoglobulin-based bioactive assemblies |
| US7666400B2 (en) | 2005-04-06 | 2010-02-23 | Ibc Pharmaceuticals, Inc. | PEGylation by the dock and lock (DNL) technique |
| WO2002079255A1 (en) | 2001-04-02 | 2002-10-10 | Idec Pharmaceuticals Corporation | RECOMBINANT ANTIBODIES COEXPRESSED WITH GnTIII |
| SE520605C2 (sv) | 2001-06-29 | 2003-07-29 | Flir Systems Ab | Optiskt system innefattande en detektor och en bländare med decentreringsfunktion |
| CN1555411A (zh) | 2001-08-03 | 2004-12-15 | ���迨�����\���ɷݹ�˾ | 抗体-依赖性细胞毒性增大的抗体糖基化变体 |
| US20040101920A1 (en) | 2002-11-01 | 2004-05-27 | Czeslaw Radziejewski | Modification assisted profiling (MAP) methodology |
| EP1874824A4 (en) | 2005-04-06 | 2009-12-30 | Ibc Pharmaceuticals Inc | METHODS OF GENERATING STABLE COMPOUND RELATED COMPLEXES OF HOMODIMERS, HOMOTRATEERS OR DIMER SIZES AND USES THEREOF |
| EP2242768A4 (en) * | 2008-01-17 | 2012-03-14 | Humabs Llc | CROSS-NEUTRALIZATION OF HUMAN MONOCLONAL ANTIBODIES AGAINST SARS-COV AND METHODS FOR THEIR APPLICATION |
| UA117289C2 (uk) | 2014-04-02 | 2018-07-10 | Ф. Хоффманн-Ля Рош Аг | Мультиспецифічне антитіло |
| CN115698057A (zh) * | 2020-02-03 | 2023-02-03 | 维尔生物科技有限公司 | 抗sars-cov-2的抗体和其使用方法 |
| US11168128B2 (en) * | 2020-02-26 | 2021-11-09 | Vir Biotechnology, Inc. | Antibodies against SARS-CoV-2 and methods of using the same |
| US11053304B1 (en) * | 2020-03-23 | 2021-07-06 | Centivax, Inc. | Anti-SARS-Cov-2 antibodies derived from 6nb6 |
-
2021
- 2021-03-25 LT LTEPPCT/US2021/024215T patent/LT4045533T/lt unknown
- 2021-03-25 WO PCT/US2021/024215 patent/WO2021195418A1/en active Application Filing
- 2021-03-25 HR HRP20240182TT patent/HRP20240182T1/hr unknown
- 2021-03-25 AU AU2021242306A patent/AU2021242306A1/en active Pending
- 2021-03-25 SI SI202130109T patent/SI4045533T1/sl unknown
- 2021-03-25 US US17/212,949 patent/US11345741B2/en active Active
- 2021-03-25 CR CR20220545A patent/CR20220545A/es unknown
- 2021-03-25 IL IL296537A patent/IL296537A/en unknown
- 2021-03-25 JP JP2022557735A patent/JP2023518849A/ja active Pending
- 2021-03-25 PT PT217194620T patent/PT4045533T/pt unknown
- 2021-03-25 DK DK21719462.0T patent/DK4045533T3/da active
- 2021-03-25 RS RS20240202A patent/RS65199B1/sr unknown
- 2021-03-25 EP EP23209693.3A patent/EP4356924A2/en active Pending
- 2021-03-25 CN CN202180023056.7A patent/CN115768790A/zh active Pending
- 2021-03-25 CA CA3175110A patent/CA3175110A1/en active Pending
- 2021-03-25 TW TW110110951A patent/TW202200610A/zh unknown
- 2021-03-25 BR BR112022018949A patent/BR112022018949A2/pt unknown
- 2021-03-25 EP EP21719462.0A patent/EP4045533B1/en active Active
- 2021-03-25 PE PE2022002090A patent/PE20230415A1/es unknown
- 2021-03-25 KR KR1020227036951A patent/KR20220158053A/ko active Search and Examination
- 2021-03-25 PL PL21719462.0T patent/PL4045533T3/pl unknown
- 2021-03-25 FI FIEP21719462.0T patent/FI4045533T3/fi active
- 2021-03-25 MX MX2022011892A patent/MX2022011892A/es unknown
- 2021-03-25 UY UY0001039135A patent/UY39135A/es unknown
- 2021-03-25 HU HUE21719462A patent/HUE065039T2/hu unknown
-
2022
- 2022-03-18 US US17/655,503 patent/US20220281958A1/en active Pending
- 2022-09-21 DO DO2022000199A patent/DOP2022000199A/es unknown
- 2022-09-23 CL CL2022002594A patent/CL2022002594A1/es unknown
- 2022-10-19 CO CONC2022/0014820A patent/CO2022014820A2/es unknown
- 2022-10-26 EC ECSENADI202283269A patent/ECSP22083269A/es unknown
Also Published As
| Publication number | Publication date |
|---|---|
| US11345741B2 (en) | 2022-05-31 |
| CO2022014820A2 (es) | 2022-10-21 |
| EP4356924A2 (en) | 2024-04-24 |
| ECSP22083269A (es) | 2022-11-30 |
| EP4045533A1 (en) | 2022-08-24 |
| PL4045533T3 (pl) | 2024-04-15 |
| FI4045533T3 (fi) | 2024-02-02 |
| CN115768790A (zh) | 2023-03-07 |
| WO2021195418A1 (en) | 2021-09-30 |
| LT4045533T (lt) | 2024-02-26 |
| SI4045533T1 (sl) | 2024-03-29 |
| HRP20240182T1 (hr) | 2024-04-26 |
| US20220281958A1 (en) | 2022-09-08 |
| DOP2022000199A (es) | 2022-11-30 |
| BR112022018949A2 (pt) | 2022-11-08 |
| PT4045533T (pt) | 2024-02-14 |
| MX2022011892A (es) | 2022-10-18 |
| RS65199B1 (sr) | 2024-03-29 |
| JP2023518849A (ja) | 2023-05-08 |
| CA3175110A1 (en) | 2021-09-30 |
| CR20220545A (es) | 2023-01-09 |
| HUE065039T2 (hu) | 2024-04-28 |
| TW202200610A (zh) | 2022-01-01 |
| IL296537A (en) | 2022-11-01 |
| EP4045533B1 (en) | 2023-11-15 |
| UY39135A (es) | 2021-10-29 |
| AU2021242306A1 (en) | 2022-11-17 |
| DK4045533T3 (da) | 2024-02-12 |
| CL2022002594A1 (es) | 2023-06-02 |
| PE20230415A1 (es) | 2023-03-07 |
| US20210300999A1 (en) | 2021-09-30 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP4045533B1 (en) | Human monoclonal antibodies to severe acute respiratory syndrome coronavirus 2 (sars-cov-2) | |
| KR102541905B1 (ko) | 치쿤군야 바이러스의 항체-매개 중화 | |
| US20210277092A1 (en) | HUMAN MONOCLONAL ANTIBODIES TO SEVERE ACUTE RESPIRATORY SYNDROME CORONAVIRUS 2 (SARS-CoV-2) | |
| WO2021195326A1 (en) | Human monoclonal antibodies to severe acute respiratory syndrome coronavirus 2 (sars-cov-2) | |
| US20220289828A1 (en) | Human monoclonal antibodies to enterovirus d68 | |
| US20230073075A1 (en) | Human hendra virus and nipah virus antibodies and methods of use therefor | |
| US20230122364A1 (en) | HUMAN MONOCLONAL ANTIBODIES TO SEVERE ACUTE RESPIRATORY SYNDROME CORONAVIRUS 2 (SARS-CoV-2) | |
| WO2021195385A1 (en) | HUMAN MONOCLONAL ANTIBODIES TO SEVERE ACUTE RESPIRATORY SYNDROME CORONAVIRUS 2 (SARS-GoV-2) | |
| US20230065377A1 (en) | Human antibodies to alphaviruses | |
| WO2019210144A1 (en) | Broadly neutralizing antibodies against hepatitis c virus | |
| US20220380442A1 (en) | Human monoclonal antibodies to hantavirus and methods of use therefore | |
| US11939370B2 (en) | Pan-ebola virus neutralizing human antibodies and methods of use therefor | |
| US20230181714A1 (en) | Human monoclonal antibodies to venezuelan equine encephalitis virus and uses therefor | |
| US20230085393A1 (en) | Human antibodies that neutralize zika virus and methods of use therefor | |
| US20230072640A1 (en) | Human monoclonal antibodies against yellow fever virus and uses therefor | |
| AU2020273365A1 (en) | Human antibodies to Ross River virus and methods of use therefor | |
| WO2023187407A1 (en) | Human monoclonal antibodies binding to sars-cov-2 and methods of use thereof | |
| KR20220113346A (ko) | 칸디다에 대한 항체 및 이의 용도 | |
| WO2024015760A2 (en) | Human monoclonal antibodies to omicron variant of severe acute respiratory syndrome coronavirus 2 (sars-cov- 2) | |
| CN117836322A (zh) | 针对SARS-CoV-2的人中和单克隆抗体及其用途 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination |