KR20220108031A - 인공 합성 mRNA 및 그 이용 - Google Patents
인공 합성 mRNA 및 그 이용 Download PDFInfo
- Publication number
- KR20220108031A KR20220108031A KR1020227012511A KR20227012511A KR20220108031A KR 20220108031 A KR20220108031 A KR 20220108031A KR 1020227012511 A KR1020227012511 A KR 1020227012511A KR 20227012511 A KR20227012511 A KR 20227012511A KR 20220108031 A KR20220108031 A KR 20220108031A
- Authority
- KR
- South Korea
- Prior art keywords
- utr
- utr sequence
- sequence
- mrna
- leu
- Prior art date
Links
- 108020004999 messenger RNA Proteins 0.000 title claims abstract description 287
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 73
- 102000004169 proteins and genes Human genes 0.000 claims abstract description 38
- 238000000034 method Methods 0.000 claims abstract description 27
- 108091026898 Leader sequence (mRNA) Proteins 0.000 claims abstract description 21
- 108091036066 Three prime untranslated region Proteins 0.000 claims abstract description 14
- 102000006602 glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 claims description 138
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 claims description 138
- 101001051207 Homo sapiens L-lactate dehydrogenase B chain Proteins 0.000 claims description 53
- 102100024580 L-lactate dehydrogenase B chain Human genes 0.000 claims description 53
- 102100037663 40S ribosomal protein S8 Human genes 0.000 claims description 46
- 101001097439 Homo sapiens 40S ribosomal protein S8 Proteins 0.000 claims description 46
- 102100021519 Hemoglobin subunit beta Human genes 0.000 claims description 42
- 108091005904 Hemoglobin subunit beta Proteins 0.000 claims description 42
- 238000004519 manufacturing process Methods 0.000 claims description 10
- 230000008569 process Effects 0.000 claims description 2
- 238000013519 translation Methods 0.000 abstract description 41
- 108020003589 5' Untranslated Regions Proteins 0.000 description 247
- 108020005345 3' Untranslated Regions Proteins 0.000 description 210
- 210000004027 cell Anatomy 0.000 description 89
- 108020004414 DNA Proteins 0.000 description 85
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 54
- 238000010586 diagram Methods 0.000 description 51
- 241000282414 Homo sapiens Species 0.000 description 40
- 108091034117 Oligonucleotide Proteins 0.000 description 40
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 40
- 238000003780 insertion Methods 0.000 description 39
- 230000037431 insertion Effects 0.000 description 39
- 238000002474 experimental method Methods 0.000 description 37
- 102100028704 Acetyl-CoA acetyltransferase, cytosolic Human genes 0.000 description 24
- 101000837584 Homo sapiens Acetyl-CoA acetyltransferase, cytosolic Proteins 0.000 description 24
- 101000642613 Homo sapiens Sterol O-acyltransferase 2 Proteins 0.000 description 24
- 108700026244 Open Reading Frames Proteins 0.000 description 20
- 238000010362 genome editing Methods 0.000 description 19
- 230000000694 effects Effects 0.000 description 17
- 235000018102 proteins Nutrition 0.000 description 16
- 230000000295 complement effect Effects 0.000 description 13
- 239000013612 plasmid Substances 0.000 description 13
- 238000012217 deletion Methods 0.000 description 12
- 230000037430 deletion Effects 0.000 description 12
- 108010048367 enhanced green fluorescent protein Proteins 0.000 description 8
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 7
- 239000000872 buffer Substances 0.000 description 7
- 108010034529 leucyl-lysine Proteins 0.000 description 7
- 108091033409 CRISPR Proteins 0.000 description 6
- 108010087924 alanylproline Proteins 0.000 description 6
- 108010092854 aspartyllysine Proteins 0.000 description 6
- OSASVXMJTNOKOY-UHFFFAOYSA-N chlorobutanol Chemical compound CC(C)(O)C(Cl)(Cl)Cl OSASVXMJTNOKOY-UHFFFAOYSA-N 0.000 description 6
- 239000002299 complementary DNA Substances 0.000 description 6
- 210000002919 epithelial cell Anatomy 0.000 description 6
- 108010055341 glutamyl-glutamic acid Proteins 0.000 description 6
- 230000006872 improvement Effects 0.000 description 6
- 239000000203 mixture Substances 0.000 description 6
- 239000003755 preservative agent Substances 0.000 description 6
- 238000003757 reverse transcription PCR Methods 0.000 description 6
- 210000000130 stem cell Anatomy 0.000 description 6
- 108091034057 RNA (poly(A)) Proteins 0.000 description 5
- 201000010099 disease Diseases 0.000 description 5
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 5
- 239000003814 drug Substances 0.000 description 5
- 238000009472 formulation Methods 0.000 description 5
- 108010050848 glycylleucine Proteins 0.000 description 5
- 239000012528 membrane Substances 0.000 description 5
- 238000003786 synthesis reaction Methods 0.000 description 5
- JBCLFWXMTIKCCB-UHFFFAOYSA-N H-Gly-Phe-OH Natural products NCC(=O)NC(C(O)=O)CC1=CC=CC=C1 JBCLFWXMTIKCCB-UHFFFAOYSA-N 0.000 description 4
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Chemical compound OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 239000003795 chemical substances by application Substances 0.000 description 4
- 238000005516 engineering process Methods 0.000 description 4
- 108010018006 histidylserine Proteins 0.000 description 4
- 238000000338 in vitro Methods 0.000 description 4
- 238000007852 inverse PCR Methods 0.000 description 4
- 108010057821 leucylproline Proteins 0.000 description 4
- 238000013518 transcription Methods 0.000 description 4
- 230000035897 transcription Effects 0.000 description 4
- 230000014621 translational initiation Effects 0.000 description 4
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 3
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 3
- 241000880493 Leptailurus serval Species 0.000 description 3
- IRMLZWSRWSGTOP-CIUDSAMLSA-N Leu-Ser-Ala Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O IRMLZWSRWSGTOP-CIUDSAMLSA-N 0.000 description 3
- DRRXXZBXDMLGFC-IHRRRGAJSA-N Lys-Val-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCCN DRRXXZBXDMLGFC-IHRRRGAJSA-N 0.000 description 3
- 239000000020 Nitrocellulose Substances 0.000 description 3
- DNIAPMSPPWPWGF-UHFFFAOYSA-N Propylene glycol Chemical compound CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 3
- HEMHJVSKTPXQMS-UHFFFAOYSA-M Sodium hydroxide Chemical compound [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 3
- 238000010459 TALEN Methods 0.000 description 3
- 102000044209 Tumor Suppressor Genes Human genes 0.000 description 3
- 108700025716 Tumor Suppressor Genes Proteins 0.000 description 3
- 241000700605 Viruses Species 0.000 description 3
- 108010017070 Zinc Finger Nucleases Proteins 0.000 description 3
- 108010062796 arginyllysine Proteins 0.000 description 3
- 229960004926 chlorobutanol Drugs 0.000 description 3
- 239000007884 disintegrant Substances 0.000 description 3
- 239000002552 dosage form Substances 0.000 description 3
- 239000003995 emulsifying agent Substances 0.000 description 3
- 108010025306 histidylleucine Proteins 0.000 description 3
- 108010064235 lysylglycine Proteins 0.000 description 3
- 108010054155 lysyllysine Proteins 0.000 description 3
- 230000007246 mechanism Effects 0.000 description 3
- 229920001220 nitrocellulos Polymers 0.000 description 3
- 239000002773 nucleotide Substances 0.000 description 3
- 125000003729 nucleotide group Chemical group 0.000 description 3
- 230000036961 partial effect Effects 0.000 description 3
- 239000000546 pharmaceutical excipient Substances 0.000 description 3
- 238000011160 research Methods 0.000 description 3
- 239000003381 stabilizer Substances 0.000 description 3
- 239000000375 suspending agent Substances 0.000 description 3
- 238000001890 transfection Methods 0.000 description 3
- 108010073969 valyllysine Proteins 0.000 description 3
- 238000012232 AGPC extraction Methods 0.000 description 2
- 229920000936 Agarose Polymers 0.000 description 2
- MDNAVFBZPROEHO-UHFFFAOYSA-N Ala-Lys-Val Natural products CC(C)C(C(O)=O)NC(=O)C(NC(=O)C(C)N)CCCCN MDNAVFBZPROEHO-UHFFFAOYSA-N 0.000 description 2
- IPZQNYYAYVRKKK-FXQIFTODSA-N Ala-Pro-Ala Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O IPZQNYYAYVRKKK-FXQIFTODSA-N 0.000 description 2
- VNFWDYWTSHFRRG-SRVKXCTJSA-N Arg-Gln-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O VNFWDYWTSHFRRG-SRVKXCTJSA-N 0.000 description 2
- OTZMRMHZCMZOJZ-SRVKXCTJSA-N Arg-Leu-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O OTZMRMHZCMZOJZ-SRVKXCTJSA-N 0.000 description 2
- FSNVAJOPUDVQAR-AVGNSLFASA-N Arg-Lys-Arg Chemical compound NC(=N)NCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O FSNVAJOPUDVQAR-AVGNSLFASA-N 0.000 description 2
- PAPSMOYMQDWIOR-AVGNSLFASA-N Arg-Lys-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O PAPSMOYMQDWIOR-AVGNSLFASA-N 0.000 description 2
- ULBHWNVWSCJLCO-NHCYSSNCSA-N Arg-Val-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCN=C(N)N ULBHWNVWSCJLCO-NHCYSSNCSA-N 0.000 description 2
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 2
- BDMIFVIWCNLDCT-CIUDSAMLSA-N Asn-Arg-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=O BDMIFVIWCNLDCT-CIUDSAMLSA-N 0.000 description 2
- OPEPUCYIGFEGSW-WDSKDSINSA-N Asn-Gly-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O OPEPUCYIGFEGSW-WDSKDSINSA-N 0.000 description 2
- NJSNXIOKBHPFMB-GMOBBJLQSA-N Asn-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CC(=O)N)N NJSNXIOKBHPFMB-GMOBBJLQSA-N 0.000 description 2
- UJGRZQYSNYTCAX-SRVKXCTJSA-N Asp-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC(O)=O UJGRZQYSNYTCAX-SRVKXCTJSA-N 0.000 description 2
- LTCKTLYKRMCFOC-KKUMJFAQSA-N Asp-Phe-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O LTCKTLYKRMCFOC-KKUMJFAQSA-N 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- 238000010356 CRISPR-Cas9 genome editing Methods 0.000 description 2
- VTYYLEPIZMXCLO-UHFFFAOYSA-L Calcium carbonate Chemical compound [Ca+2].[O-]C([O-])=O VTYYLEPIZMXCLO-UHFFFAOYSA-L 0.000 description 2
- 229920002134 Carboxymethyl cellulose Polymers 0.000 description 2
- 241000282693 Cercopithecidae Species 0.000 description 2
- HEDRZPFGACZZDS-UHFFFAOYSA-N Chloroform Chemical compound ClC(Cl)Cl HEDRZPFGACZZDS-UHFFFAOYSA-N 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- 241000699800 Cricetinae Species 0.000 description 2
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 2
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 2
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 2
- 108010042407 Endonucleases Proteins 0.000 description 2
- 102000004533 Endonucleases Human genes 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- 108060002716 Exonuclease Proteins 0.000 description 2
- 241000282326 Felis catus Species 0.000 description 2
- MWMJCGBSIORNCD-AVGNSLFASA-N Glu-Leu-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O MWMJCGBSIORNCD-AVGNSLFASA-N 0.000 description 2
- NTBOEZICHOSJEE-YUMQZZPRSA-N Gly-Lys-Ser Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(O)=O NTBOEZICHOSJEE-YUMQZZPRSA-N 0.000 description 2
- 241000282412 Homo Species 0.000 description 2
- BCISUQVFDGYZBO-QSFUFRPTSA-N Ile-Val-Asp Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(O)=O BCISUQVFDGYZBO-QSFUFRPTSA-N 0.000 description 2
- 208000026350 Inborn Genetic disease Diseases 0.000 description 2
- FADYJNXDPBKVCA-UHFFFAOYSA-N L-Phenylalanyl-L-lysin Natural products NCCCCC(C(O)=O)NC(=O)C(N)CC1=CC=CC=C1 FADYJNXDPBKVCA-UHFFFAOYSA-N 0.000 description 2
- YOZCKMXHBYKOMQ-IHRRRGAJSA-N Leu-Arg-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(=O)O)N YOZCKMXHBYKOMQ-IHRRRGAJSA-N 0.000 description 2
- WUFYAPWIHCUMLL-CIUDSAMLSA-N Leu-Asn-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(O)=O WUFYAPWIHCUMLL-CIUDSAMLSA-N 0.000 description 2
- DBSLVQBXKVKDKJ-BJDJZHNGSA-N Leu-Ile-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O DBSLVQBXKVKDKJ-BJDJZHNGSA-N 0.000 description 2
- YRRCOJOXAJNSAX-IHRRRGAJSA-N Leu-Pro-Lys Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)O)N YRRCOJOXAJNSAX-IHRRRGAJSA-N 0.000 description 2
- HQVDJTYKCMIWJP-YUMQZZPRSA-N Lys-Asn-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O HQVDJTYKCMIWJP-YUMQZZPRSA-N 0.000 description 2
- FACUGMGEFUEBTI-SRVKXCTJSA-N Lys-Asn-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CCCCN FACUGMGEFUEBTI-SRVKXCTJSA-N 0.000 description 2
- NNCDAORZCMPZPX-GUBZILKMSA-N Lys-Gln-Ser Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CO)C(=O)O)N NNCDAORZCMPZPX-GUBZILKMSA-N 0.000 description 2
- WVJNGSFKBKOKRV-AJNGGQMLSA-N Lys-Leu-Ile Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WVJNGSFKBKOKRV-AJNGGQMLSA-N 0.000 description 2
- LJADEBULDNKJNK-IHRRRGAJSA-N Lys-Leu-Val Chemical compound CC(C)C[C@H](NC(=O)[C@@H](N)CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O LJADEBULDNKJNK-IHRRRGAJSA-N 0.000 description 2
- HVAUKHLDSDDROB-KKUMJFAQSA-N Lys-Lys-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O HVAUKHLDSDDROB-KKUMJFAQSA-N 0.000 description 2
- JMNRXRPBHFGXQX-GUBZILKMSA-N Lys-Ser-Glu Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O JMNRXRPBHFGXQX-GUBZILKMSA-N 0.000 description 2
- 239000007993 MOPS buffer Substances 0.000 description 2
- 208000024556 Mendelian disease Diseases 0.000 description 2
- RRIHXWPHQSXHAQ-XUXIUFHCSA-N Met-Ile-Lys Chemical compound CSCC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCCN)C(O)=O RRIHXWPHQSXHAQ-XUXIUFHCSA-N 0.000 description 2
- 241000699666 Mus <mouse, genus> Species 0.000 description 2
- YBAFDPFAUTYYRW-UHFFFAOYSA-N N-L-alpha-glutamyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=O YBAFDPFAUTYYRW-UHFFFAOYSA-N 0.000 description 2
- SITLTJHOQZFJGG-UHFFFAOYSA-N N-L-alpha-glutamyl-L-valine Natural products CC(C)C(C(O)=O)NC(=O)C(N)CCC(O)=O SITLTJHOQZFJGG-UHFFFAOYSA-N 0.000 description 2
- 108010047562 NGR peptide Proteins 0.000 description 2
- 101710163270 Nuclease Proteins 0.000 description 2
- 239000004677 Nylon Substances 0.000 description 2
- WEMYTDDMDBLPMI-DKIMLUQUSA-N Phe-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N WEMYTDDMDBLPMI-DKIMLUQUSA-N 0.000 description 2
- IPFXYNKCXYGSSV-KKUMJFAQSA-N Phe-Ser-Lys Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)O)N IPFXYNKCXYGSSV-KKUMJFAQSA-N 0.000 description 2
- DZZCICYRSZASNF-FXQIFTODSA-N Pro-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 DZZCICYRSZASNF-FXQIFTODSA-N 0.000 description 2
- VZKBJNBZMZHKRC-XUXIUFHCSA-N Pro-Ile-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O VZKBJNBZMZHKRC-XUXIUFHCSA-N 0.000 description 2
- LEIKGVHQTKHOLM-IUCAKERBSA-N Pro-Pro-Gly Chemical compound OC(=O)CNC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 LEIKGVHQTKHOLM-IUCAKERBSA-N 0.000 description 2
- 230000006819 RNA synthesis Effects 0.000 description 2
- 241000700159 Rattus Species 0.000 description 2
- MIJWOJAXARLEHA-WDSKDSINSA-N Ser-Gly-Glu Chemical compound OC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(O)=O MIJWOJAXARLEHA-WDSKDSINSA-N 0.000 description 2
- DBMJMQXJHONAFJ-UHFFFAOYSA-M Sodium laurylsulphate Chemical compound [Na+].CCCCCCCCCCCCOS([O-])(=O)=O DBMJMQXJHONAFJ-UHFFFAOYSA-M 0.000 description 2
- 229920002472 Starch Polymers 0.000 description 2
- 101710137500 T7 RNA polymerase Proteins 0.000 description 2
- GULIUBBXCYPDJU-CQDKDKBSSA-N Tyr-Leu-Ala Chemical compound [O-]C(=O)[C@H](C)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]([NH3+])CC1=CC=C(O)C=C1 GULIUBBXCYPDJU-CQDKDKBSSA-N 0.000 description 2
- VBFVQTPETKJCQW-RPTUDFQQSA-N Tyr-Phe-Thr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VBFVQTPETKJCQW-RPTUDFQQSA-N 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 239000004480 active ingredient Substances 0.000 description 2
- 150000001413 amino acids Chemical group 0.000 description 2
- 238000004458 analytical method Methods 0.000 description 2
- 239000000730 antalgic agent Substances 0.000 description 2
- 108010013835 arginine glutamate Proteins 0.000 description 2
- 108010040443 aspartyl-aspartic acid Proteins 0.000 description 2
- 229960000686 benzalkonium chloride Drugs 0.000 description 2
- CADWTSSKOVRVJC-UHFFFAOYSA-N benzyl(dimethyl)azanium;chloride Chemical compound [Cl-].C[NH+](C)CC1=CC=CC=C1 CADWTSSKOVRVJC-UHFFFAOYSA-N 0.000 description 2
- 239000001506 calcium phosphate Substances 0.000 description 2
- 229910000389 calcium phosphate Inorganic materials 0.000 description 2
- 235000011010 calcium phosphates Nutrition 0.000 description 2
- 238000002619 cancer immunotherapy Methods 0.000 description 2
- 239000001768 carboxy methyl cellulose Substances 0.000 description 2
- 235000010948 carboxy methyl cellulose Nutrition 0.000 description 2
- 239000008112 carboxymethyl-cellulose Substances 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- 230000015556 catabolic process Effects 0.000 description 2
- 230000003399 chemotactic effect Effects 0.000 description 2
- 238000006731 degradation reaction Methods 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- 230000002708 enhancing effect Effects 0.000 description 2
- 102000013165 exonuclease Human genes 0.000 description 2
- -1 for example Proteins 0.000 description 2
- 108010063718 gamma-glutamylaspartic acid Proteins 0.000 description 2
- 108010081551 glycylphenylalanine Proteins 0.000 description 2
- 208000002672 hepatitis B Diseases 0.000 description 2
- 238000001727 in vivo Methods 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 239000004615 ingredient Substances 0.000 description 2
- 239000003112 inhibitor Substances 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 230000005764 inhibitory process Effects 0.000 description 2
- 230000003834 intracellular effect Effects 0.000 description 2
- 150000002500 ions Chemical class 0.000 description 2
- 210000003734 kidney Anatomy 0.000 description 2
- 108010017391 lysylvaline Proteins 0.000 description 2
- 210000002901 mesenchymal stem cell Anatomy 0.000 description 2
- LXCFILQKKLGQFO-UHFFFAOYSA-N methylparaben Chemical compound COC(=O)C1=CC=C(O)C=C1 LXCFILQKKLGQFO-UHFFFAOYSA-N 0.000 description 2
- 108020004707 nucleic acids Proteins 0.000 description 2
- 102000039446 nucleic acids Human genes 0.000 description 2
- 150000007523 nucleic acids Chemical class 0.000 description 2
- 229920001778 nylon Polymers 0.000 description 2
- WVDDGKGOMKODPV-ZQBYOMGUSA-N phenyl(114C)methanol Chemical compound O[14CH2]C1=CC=CC=C1 WVDDGKGOMKODPV-ZQBYOMGUSA-N 0.000 description 2
- 108010051242 phenylalanylserine Proteins 0.000 description 2
- 210000001778 pluripotent stem cell Anatomy 0.000 description 2
- 238000002360 preparation method Methods 0.000 description 2
- 230000002335 preservative effect Effects 0.000 description 2
- 239000003531 protein hydrolysate Substances 0.000 description 2
- 238000011002 quantification Methods 0.000 description 2
- 238000004064 recycling Methods 0.000 description 2
- 239000000523 sample Substances 0.000 description 2
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 2
- 235000019333 sodium laurylsulphate Nutrition 0.000 description 2
- 239000000600 sorbitol Substances 0.000 description 2
- 235000010356 sorbitol Nutrition 0.000 description 2
- 239000008107 starch Substances 0.000 description 2
- 235000019698 starch Nutrition 0.000 description 2
- 208000024891 symptom Diseases 0.000 description 2
- 230000002195 synergetic effect Effects 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 2
- 239000013598 vector Substances 0.000 description 2
- 239000013603 viral vector Substances 0.000 description 2
- 230000003612 virological effect Effects 0.000 description 2
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 2
- LNAZSHAWQACDHT-XIYTZBAFSA-N (2r,3r,4s,5r,6s)-4,5-dimethoxy-2-(methoxymethyl)-3-[(2s,3r,4s,5r,6r)-3,4,5-trimethoxy-6-(methoxymethyl)oxan-2-yl]oxy-6-[(2r,3r,4s,5r,6r)-4,5,6-trimethoxy-2-(methoxymethyl)oxan-3-yl]oxyoxane Chemical compound CO[C@@H]1[C@@H](OC)[C@H](OC)[C@@H](COC)O[C@H]1O[C@H]1[C@H](OC)[C@@H](OC)[C@H](O[C@H]2[C@@H]([C@@H](OC)[C@H](OC)O[C@@H]2COC)OC)O[C@@H]1COC LNAZSHAWQACDHT-XIYTZBAFSA-N 0.000 description 1
- BRPMXFSTKXXNHF-IUCAKERBSA-N (2s)-1-[2-[[(2s)-pyrrolidine-2-carbonyl]amino]acetyl]pyrrolidine-2-carboxylic acid Chemical compound OC(=O)[C@@H]1CCCN1C(=O)CNC(=O)[C@H]1NCCC1 BRPMXFSTKXXNHF-IUCAKERBSA-N 0.000 description 1
- IXPNQXFRVYWDDI-UHFFFAOYSA-N 1-methyl-2,4-dioxo-1,3-diazinane-5-carboximidamide Chemical compound CN1CC(C(N)=N)C(=O)NC1=O IXPNQXFRVYWDDI-UHFFFAOYSA-N 0.000 description 1
- VBICKXHEKHSIBG-UHFFFAOYSA-N 1-monostearoylglycerol Chemical compound CCCCCCCCCCCCCCCCCC(=O)OCC(O)CO VBICKXHEKHSIBG-UHFFFAOYSA-N 0.000 description 1
- 108030002617 2'-5' oligoadenylate synthases Proteins 0.000 description 1
- 102100027962 2-5A-dependent ribonuclease Human genes 0.000 description 1
- 108010000834 2-5A-dependent ribonuclease Proteins 0.000 description 1
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- OGHAROSJZRTIOK-KQYNXXCUSA-O 7-methylguanosine Chemical group C1=2N=C(N)NC(=O)C=2[N+](C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OGHAROSJZRTIOK-KQYNXXCUSA-O 0.000 description 1
- 208000030507 AIDS Diseases 0.000 description 1
- 101001082110 Acanthamoeba polyphaga mimivirus Eukaryotic translation initiation factor 4E homolog Proteins 0.000 description 1
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 1
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 1
- 208000009746 Adult T-Cell Leukemia-Lymphoma Diseases 0.000 description 1
- 208000016683 Adult T-cell leukemia/lymphoma Diseases 0.000 description 1
- FJVAQLJNTSUQPY-CIUDSAMLSA-N Ala-Ala-Lys Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN FJVAQLJNTSUQPY-CIUDSAMLSA-N 0.000 description 1
- LWUWMHIOBPTZBA-DCAQKATOSA-N Ala-Arg-Lys Chemical compound NC(=N)NCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CCCCN)C(O)=O LWUWMHIOBPTZBA-DCAQKATOSA-N 0.000 description 1
- XEXJJJRVTFGWIC-FXQIFTODSA-N Ala-Asn-Arg Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N XEXJJJRVTFGWIC-FXQIFTODSA-N 0.000 description 1
- NXSFUECZFORGOG-CIUDSAMLSA-N Ala-Asn-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O NXSFUECZFORGOG-CIUDSAMLSA-N 0.000 description 1
- YSMPVONNIWLJML-FXQIFTODSA-N Ala-Asp-Pro Chemical compound C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N1CCC[C@H]1C(O)=O YSMPVONNIWLJML-FXQIFTODSA-N 0.000 description 1
- VIGKUFXFTPWYER-BIIVOSGPSA-N Ala-Cys-Pro Chemical compound C[C@@H](C(=O)N[C@@H](CS)C(=O)N1CCC[C@@H]1C(=O)O)N VIGKUFXFTPWYER-BIIVOSGPSA-N 0.000 description 1
- LGFCAXJBAZESCF-ACZMJKKPSA-N Ala-Gln-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O LGFCAXJBAZESCF-ACZMJKKPSA-N 0.000 description 1
- FVSOUJZKYWEFOB-KBIXCLLPSA-N Ala-Gln-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](C)N FVSOUJZKYWEFOB-KBIXCLLPSA-N 0.000 description 1
- MVBWLRJESQOQTM-ACZMJKKPSA-N Ala-Gln-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(O)=O MVBWLRJESQOQTM-ACZMJKKPSA-N 0.000 description 1
- YIGLXQRFQVWFEY-NRPADANISA-N Ala-Gln-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O YIGLXQRFQVWFEY-NRPADANISA-N 0.000 description 1
- FUSPCLTUKXQREV-ACZMJKKPSA-N Ala-Glu-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O FUSPCLTUKXQREV-ACZMJKKPSA-N 0.000 description 1
- NWVVKQZOVSTDBQ-CIUDSAMLSA-N Ala-Glu-Arg Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O NWVVKQZOVSTDBQ-CIUDSAMLSA-N 0.000 description 1
- NJPMYXWVWQWCSR-ACZMJKKPSA-N Ala-Glu-Asn Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O NJPMYXWVWQWCSR-ACZMJKKPSA-N 0.000 description 1
- QQACQIHVWCVBBR-GVARAGBVSA-N Ala-Ile-Tyr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O QQACQIHVWCVBBR-GVARAGBVSA-N 0.000 description 1
- HHRAXZAYZFFRAM-CIUDSAMLSA-N Ala-Leu-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O HHRAXZAYZFFRAM-CIUDSAMLSA-N 0.000 description 1
- NINQYGGNRIBFSC-CIUDSAMLSA-N Ala-Lys-Ser Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CO)C(O)=O NINQYGGNRIBFSC-CIUDSAMLSA-N 0.000 description 1
- OINVDEKBKBCPLX-JXUBOQSCSA-N Ala-Lys-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OINVDEKBKBCPLX-JXUBOQSCSA-N 0.000 description 1
- VHEVVUZDDUCAKU-FXQIFTODSA-N Ala-Met-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(O)=O)C(O)=O VHEVVUZDDUCAKU-FXQIFTODSA-N 0.000 description 1
- ZBLQIYPCUWZSRZ-QEJZJMRPSA-N Ala-Phe-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](C)N)CC1=CC=CC=C1 ZBLQIYPCUWZSRZ-QEJZJMRPSA-N 0.000 description 1
- FEGOCLZUJUFCHP-CIUDSAMLSA-N Ala-Pro-Gln Chemical compound [H]N[C@@H](C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O FEGOCLZUJUFCHP-CIUDSAMLSA-N 0.000 description 1
- HOVPGJUNRLMIOZ-CIUDSAMLSA-N Ala-Ser-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@H](C)N HOVPGJUNRLMIOZ-CIUDSAMLSA-N 0.000 description 1
- IOFVWPYSRSCWHI-JXUBOQSCSA-N Ala-Thr-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](C)N IOFVWPYSRSCWHI-JXUBOQSCSA-N 0.000 description 1
- IETUUAHKCHOQHP-KZVJFYERSA-N Ala-Thr-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@H](C)N)[C@@H](C)O)C(O)=O IETUUAHKCHOQHP-KZVJFYERSA-N 0.000 description 1
- QRIYOHQJRDHFKF-UWJYBYFXSA-N Ala-Tyr-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=C(O)C=C1 QRIYOHQJRDHFKF-UWJYBYFXSA-N 0.000 description 1
- VYSRNGOMGHOJCK-GUBZILKMSA-N Arg-Ala-Met Chemical compound C[C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N VYSRNGOMGHOJCK-GUBZILKMSA-N 0.000 description 1
- MUXONAMCEUBVGA-DCAQKATOSA-N Arg-Arg-Gln Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCC(N)=O)C(O)=O MUXONAMCEUBVGA-DCAQKATOSA-N 0.000 description 1
- HJVGMOYJDDXLMI-AVGNSLFASA-N Arg-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H](N)CCCNC(N)=N HJVGMOYJDDXLMI-AVGNSLFASA-N 0.000 description 1
- NABSCJGZKWSNHX-RCWTZXSCSA-N Arg-Arg-Thr Chemical compound NC(N)=NCCC[C@@H](C(=O)N[C@@H]([C@H](O)C)C(O)=O)NC(=O)[C@@H](N)CCCN=C(N)N NABSCJGZKWSNHX-RCWTZXSCSA-N 0.000 description 1
- IIABBYGHLYWVOS-FXQIFTODSA-N Arg-Asn-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O IIABBYGHLYWVOS-FXQIFTODSA-N 0.000 description 1
- ZTKHZAXGTFXUDD-VEVYYDQMSA-N Arg-Asn-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ZTKHZAXGTFXUDD-VEVYYDQMSA-N 0.000 description 1
- RRGPUNYIPJXJBU-GUBZILKMSA-N Arg-Asp-Met Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCSC)C(O)=O RRGPUNYIPJXJBU-GUBZILKMSA-N 0.000 description 1
- TTXYKSADPSNOIF-IHRRRGAJSA-N Arg-Asp-Phe Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O TTXYKSADPSNOIF-IHRRRGAJSA-N 0.000 description 1
- JCAISGGAOQXEHJ-ZPFDUUQYSA-N Arg-Gln-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCCN=C(N)N)N JCAISGGAOQXEHJ-ZPFDUUQYSA-N 0.000 description 1
- QAODJPUKWNNNRP-DCAQKATOSA-N Arg-Glu-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O QAODJPUKWNNNRP-DCAQKATOSA-N 0.000 description 1
- RKRSYHCNPFGMTA-CIUDSAMLSA-N Arg-Glu-Asn Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O RKRSYHCNPFGMTA-CIUDSAMLSA-N 0.000 description 1
- MZRBYBIQTIKERR-GUBZILKMSA-N Arg-Glu-Gln Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O MZRBYBIQTIKERR-GUBZILKMSA-N 0.000 description 1
- GOWZVQXTHUCNSQ-NHCYSSNCSA-N Arg-Glu-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O GOWZVQXTHUCNSQ-NHCYSSNCSA-N 0.000 description 1
- HQIZDMIGUJOSNI-IUCAKERBSA-N Arg-Gly-Arg Chemical compound N[C@@H](CCCNC(N)=N)C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(O)=O HQIZDMIGUJOSNI-IUCAKERBSA-N 0.000 description 1
- RKQRHMKFNBYOTN-IHRRRGAJSA-N Arg-His-Lys Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N RKQRHMKFNBYOTN-IHRRRGAJSA-N 0.000 description 1
- DNUKXVMPARLPFN-XUXIUFHCSA-N Arg-Leu-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O DNUKXVMPARLPFN-XUXIUFHCSA-N 0.000 description 1
- COXMUHNBYCVVRG-DCAQKATOSA-N Arg-Leu-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O COXMUHNBYCVVRG-DCAQKATOSA-N 0.000 description 1
- MJINRRBEMOLJAK-DCAQKATOSA-N Arg-Lys-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCN=C(N)N MJINRRBEMOLJAK-DCAQKATOSA-N 0.000 description 1
- GRRXPUAICOGISM-RWMBFGLXSA-N Arg-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CCCN=C(N)N)N)C(=O)O GRRXPUAICOGISM-RWMBFGLXSA-N 0.000 description 1
- PYZPXCZNQSEHDT-GUBZILKMSA-N Arg-Met-Asn Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N PYZPXCZNQSEHDT-GUBZILKMSA-N 0.000 description 1
- LCBSSOCDWUTQQV-SDDRHHMPSA-N Arg-Met-Pro Chemical compound CSCC[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N LCBSSOCDWUTQQV-SDDRHHMPSA-N 0.000 description 1
- KSUALAGYYLQSHJ-RCWTZXSCSA-N Arg-Met-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O KSUALAGYYLQSHJ-RCWTZXSCSA-N 0.000 description 1
- CZUHPNLXLWMYMG-UBHSHLNASA-N Arg-Phe-Ala Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](C)C(O)=O)CC1=CC=CC=C1 CZUHPNLXLWMYMG-UBHSHLNASA-N 0.000 description 1
- YTMKMRSYXHBGER-IHRRRGAJSA-N Arg-Phe-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N YTMKMRSYXHBGER-IHRRRGAJSA-N 0.000 description 1
- DPLFNLDACGGBAK-KKUMJFAQSA-N Arg-Phe-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N DPLFNLDACGGBAK-KKUMJFAQSA-N 0.000 description 1
- NGYHSXDNNOFHNE-AVGNSLFASA-N Arg-Pro-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O NGYHSXDNNOFHNE-AVGNSLFASA-N 0.000 description 1
- ADPACBMPYWJJCE-FXQIFTODSA-N Arg-Ser-Asp Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O ADPACBMPYWJJCE-FXQIFTODSA-N 0.000 description 1
- FRBAHXABMQXSJQ-FXQIFTODSA-N Arg-Ser-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O FRBAHXABMQXSJQ-FXQIFTODSA-N 0.000 description 1
- AOJYORNRFWWEIV-IHRRRGAJSA-N Arg-Tyr-Asp Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(O)=O)C(O)=O)CC1=CC=C(O)C=C1 AOJYORNRFWWEIV-IHRRRGAJSA-N 0.000 description 1
- IZSMEUDYADKZTJ-KJEVXHAQSA-N Arg-Tyr-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O IZSMEUDYADKZTJ-KJEVXHAQSA-N 0.000 description 1
- FTMRPIVPSDVGCC-GUBZILKMSA-N Arg-Val-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N FTMRPIVPSDVGCC-GUBZILKMSA-N 0.000 description 1
- FMYQECOAIFGQGU-CYDGBPFRSA-N Arg-Val-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O FMYQECOAIFGQGU-CYDGBPFRSA-N 0.000 description 1
- XWGJDUSDTRPQRK-ZLUOBGJFSA-N Asn-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC(N)=O XWGJDUSDTRPQRK-ZLUOBGJFSA-N 0.000 description 1
- GMRGSBAMMMVDGG-GUBZILKMSA-N Asn-Arg-Arg Chemical compound C(C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N)CN=C(N)N GMRGSBAMMMVDGG-GUBZILKMSA-N 0.000 description 1
- MEFGKQUUYZOLHM-GMOBBJLQSA-N Asn-Arg-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O MEFGKQUUYZOLHM-GMOBBJLQSA-N 0.000 description 1
- QHBMKQWOIYJYMI-BYULHYEWSA-N Asn-Asn-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O QHBMKQWOIYJYMI-BYULHYEWSA-N 0.000 description 1
- WPOLSNAQGVHROR-GUBZILKMSA-N Asn-Gln-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CC(=O)N)N WPOLSNAQGVHROR-GUBZILKMSA-N 0.000 description 1
- HCAUEJAQCXVQQM-ACZMJKKPSA-N Asn-Glu-Asp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O HCAUEJAQCXVQQM-ACZMJKKPSA-N 0.000 description 1
- QYXNFROWLZPWPC-FXQIFTODSA-N Asn-Glu-Gln Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O QYXNFROWLZPWPC-FXQIFTODSA-N 0.000 description 1
- BZMWJLLUAKSIMH-FXQIFTODSA-N Asn-Glu-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O BZMWJLLUAKSIMH-FXQIFTODSA-N 0.000 description 1
- FTCGGKNCJZOPNB-WHFBIAKZSA-N Asn-Gly-Ser Chemical compound NC(=O)C[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O FTCGGKNCJZOPNB-WHFBIAKZSA-N 0.000 description 1
- SPCONPVIDFMDJI-QSFUFRPTSA-N Asn-Ile-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O SPCONPVIDFMDJI-QSFUFRPTSA-N 0.000 description 1
- GLWFAWNYGWBMOC-SRVKXCTJSA-N Asn-Leu-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O GLWFAWNYGWBMOC-SRVKXCTJSA-N 0.000 description 1
- DJIMLSXHXKWADV-CIUDSAMLSA-N Asn-Leu-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC(N)=O DJIMLSXHXKWADV-CIUDSAMLSA-N 0.000 description 1
- LSJQOMAZIKQMTJ-SRVKXCTJSA-N Asn-Phe-Asp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O LSJQOMAZIKQMTJ-SRVKXCTJSA-N 0.000 description 1
- RAUPFUCUDBQYHE-AVGNSLFASA-N Asn-Phe-Glu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O RAUPFUCUDBQYHE-AVGNSLFASA-N 0.000 description 1
- HZZIFFOVHLWGCS-KKUMJFAQSA-N Asn-Phe-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O HZZIFFOVHLWGCS-KKUMJFAQSA-N 0.000 description 1
- YUUIAUXBNOHFRJ-IHRRRGAJSA-N Asn-Phe-Met Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCSC)C(O)=O YUUIAUXBNOHFRJ-IHRRRGAJSA-N 0.000 description 1
- NCXTYSVDWLAQGZ-ZKWXMUAHSA-N Asn-Ser-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O NCXTYSVDWLAQGZ-ZKWXMUAHSA-N 0.000 description 1
- AMGQTNHANMRPOE-LKXGYXEUSA-N Asn-Thr-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O AMGQTNHANMRPOE-LKXGYXEUSA-N 0.000 description 1
- KTDWFWNZLLFEFU-KKUMJFAQSA-N Asn-Tyr-His Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CC(=O)N)N)O KTDWFWNZLLFEFU-KKUMJFAQSA-N 0.000 description 1
- CGYKCTPUGXFPMG-IHPCNDPISA-N Asn-Tyr-Trp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O CGYKCTPUGXFPMG-IHPCNDPISA-N 0.000 description 1
- XBQSLMACWDXWLJ-GHCJXIJMSA-N Asp-Ala-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O XBQSLMACWDXWLJ-GHCJXIJMSA-N 0.000 description 1
- BLQBMRNMBAYREH-UWJYBYFXSA-N Asp-Ala-Tyr Chemical compound N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O BLQBMRNMBAYREH-UWJYBYFXSA-N 0.000 description 1
- WSOKZUVWBXVJHX-CIUDSAMLSA-N Asp-Arg-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=O WSOKZUVWBXVJHX-CIUDSAMLSA-N 0.000 description 1
- MFMJRYHVLLEMQM-DCAQKATOSA-N Asp-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(=O)O)N MFMJRYHVLLEMQM-DCAQKATOSA-N 0.000 description 1
- UGKZHCBLMLSANF-CIUDSAMLSA-N Asp-Asn-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O UGKZHCBLMLSANF-CIUDSAMLSA-N 0.000 description 1
- KNMRXHIAVXHCLW-ZLUOBGJFSA-N Asp-Asn-Ser Chemical compound C([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CO)C(=O)O)N)C(=O)O KNMRXHIAVXHCLW-ZLUOBGJFSA-N 0.000 description 1
- RDRMWJBLOSRRAW-BYULHYEWSA-N Asp-Asn-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O RDRMWJBLOSRRAW-BYULHYEWSA-N 0.000 description 1
- SBHUBSDEZQFJHJ-CIUDSAMLSA-N Asp-Asp-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC(O)=O SBHUBSDEZQFJHJ-CIUDSAMLSA-N 0.000 description 1
- CELPEWWLSXMVPH-CIUDSAMLSA-N Asp-Asp-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC(O)=O CELPEWWLSXMVPH-CIUDSAMLSA-N 0.000 description 1
- QXHVOUSPVAWEMX-ZLUOBGJFSA-N Asp-Asp-Ser Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O QXHVOUSPVAWEMX-ZLUOBGJFSA-N 0.000 description 1
- DZQKLNLLWFQONU-LKXGYXEUSA-N Asp-Cys-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC(=O)O)N)O DZQKLNLLWFQONU-LKXGYXEUSA-N 0.000 description 1
- DXQOQMCLWWADMU-ACZMJKKPSA-N Asp-Gln-Ser Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(O)=O DXQOQMCLWWADMU-ACZMJKKPSA-N 0.000 description 1
- VAWNQIGQPUOPQW-ACZMJKKPSA-N Asp-Glu-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O VAWNQIGQPUOPQW-ACZMJKKPSA-N 0.000 description 1
- VILLWIDTHYPSLC-PEFMBERDSA-N Asp-Glu-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O VILLWIDTHYPSLC-PEFMBERDSA-N 0.000 description 1
- RRKCPMGSRIDLNC-AVGNSLFASA-N Asp-Glu-Tyr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O RRKCPMGSRIDLNC-AVGNSLFASA-N 0.000 description 1
- VIRHEUMYXXLCBF-WDSKDSINSA-N Asp-Gly-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O VIRHEUMYXXLCBF-WDSKDSINSA-N 0.000 description 1
- QCVXMEHGFUMKCO-YUMQZZPRSA-N Asp-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(O)=O QCVXMEHGFUMKCO-YUMQZZPRSA-N 0.000 description 1
- TVIZQBFURPLQDV-DJFWLOJKSA-N Asp-His-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CC(=O)O)N TVIZQBFURPLQDV-DJFWLOJKSA-N 0.000 description 1
- CYCKJEFVFNRWEZ-UGYAYLCHSA-N Asp-Ile-Asn Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O CYCKJEFVFNRWEZ-UGYAYLCHSA-N 0.000 description 1
- KQBVNNAPIURMPD-PEFMBERDSA-N Asp-Ile-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(O)=O KQBVNNAPIURMPD-PEFMBERDSA-N 0.000 description 1
- KYQNAIMCTRZLNP-QSFUFRPTSA-N Asp-Ile-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O KYQNAIMCTRZLNP-QSFUFRPTSA-N 0.000 description 1
- JNNVNVRBYUJYGS-CIUDSAMLSA-N Asp-Leu-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O JNNVNVRBYUJYGS-CIUDSAMLSA-N 0.000 description 1
- CLUMZOKVGUWUFD-CIUDSAMLSA-N Asp-Leu-Asn Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O CLUMZOKVGUWUFD-CIUDSAMLSA-N 0.000 description 1
- KFAFUJMGHVVYRC-DCAQKATOSA-N Asp-Leu-Met Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(O)=O KFAFUJMGHVVYRC-DCAQKATOSA-N 0.000 description 1
- LBOVBQONZJRWPV-YUMQZZPRSA-N Asp-Lys-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)NCC(O)=O LBOVBQONZJRWPV-YUMQZZPRSA-N 0.000 description 1
- GKWFMNNNYZHJHV-SRVKXCTJSA-N Asp-Lys-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CC(O)=O GKWFMNNNYZHJHV-SRVKXCTJSA-N 0.000 description 1
- MYLZFUMPZCPJCJ-NHCYSSNCSA-N Asp-Lys-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O MYLZFUMPZCPJCJ-NHCYSSNCSA-N 0.000 description 1
- FAUPLTGRUBTXNU-FXQIFTODSA-N Asp-Pro-Ser Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O FAUPLTGRUBTXNU-FXQIFTODSA-N 0.000 description 1
- CUQDCPXNZPDYFQ-ZLUOBGJFSA-N Asp-Ser-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O CUQDCPXNZPDYFQ-ZLUOBGJFSA-N 0.000 description 1
- QSFHZPQUAAQHAQ-CIUDSAMLSA-N Asp-Ser-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O QSFHZPQUAAQHAQ-CIUDSAMLSA-N 0.000 description 1
- YIDFBWRHIYOYAA-LKXGYXEUSA-N Asp-Ser-Thr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O YIDFBWRHIYOYAA-LKXGYXEUSA-N 0.000 description 1
- JDDYEZGPYBBPBN-JRQIVUDYSA-N Asp-Thr-Tyr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O JDDYEZGPYBBPBN-JRQIVUDYSA-N 0.000 description 1
- BJDHEININLSZOT-KKUMJFAQSA-N Asp-Tyr-Lys Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCCN)C(O)=O BJDHEININLSZOT-KKUMJFAQSA-N 0.000 description 1
- SQIARYGNVQWOSB-BZSNNMDCSA-N Asp-Tyr-Phe Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O SQIARYGNVQWOSB-BZSNNMDCSA-N 0.000 description 1
- WAEDSQFVZJUHLI-BYULHYEWSA-N Asp-Val-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O WAEDSQFVZJUHLI-BYULHYEWSA-N 0.000 description 1
- 210000002237 B-cell of pancreatic islet Anatomy 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 102100021257 Beta-secretase 1 Human genes 0.000 description 1
- 101710150192 Beta-secretase 1 Proteins 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 101100380241 Caenorhabditis elegans arx-2 gene Proteins 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 208000005623 Carcinogenesis Diseases 0.000 description 1
- 241000700198 Cavia Species 0.000 description 1
- 241000700199 Cavia porcellus Species 0.000 description 1
- KRKNYBCHXYNGOX-UHFFFAOYSA-K Citrate Chemical compound [O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O KRKNYBCHXYNGOX-UHFFFAOYSA-K 0.000 description 1
- 108020004705 Codon Proteins 0.000 description 1
- YZKOXEJTLWZOQL-GUBZILKMSA-N Cys-Gln-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CS)N YZKOXEJTLWZOQL-GUBZILKMSA-N 0.000 description 1
- XBELMDARIGXDKY-GUBZILKMSA-N Cys-Pro-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CS)N XBELMDARIGXDKY-GUBZILKMSA-N 0.000 description 1
- 201000003883 Cystic fibrosis Diseases 0.000 description 1
- 102000004127 Cytokines Human genes 0.000 description 1
- 108090000695 Cytokines Proteins 0.000 description 1
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 1
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 1
- 206010013801 Duchenne Muscular Dystrophy Diseases 0.000 description 1
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 1
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 1
- 241000283073 Equus caballus Species 0.000 description 1
- 241000287828 Gallus gallus Species 0.000 description 1
- PHZYLYASFWHLHJ-FXQIFTODSA-N Gln-Asn-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O PHZYLYASFWHLHJ-FXQIFTODSA-N 0.000 description 1
- ZPDVKYLJTOFQJV-WDSKDSINSA-N Gln-Asn-Gly Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O ZPDVKYLJTOFQJV-WDSKDSINSA-N 0.000 description 1
- XEYMBRRKIFYQMF-GUBZILKMSA-N Gln-Asp-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O XEYMBRRKIFYQMF-GUBZILKMSA-N 0.000 description 1
- KVXVVDFOZNYYKZ-DCAQKATOSA-N Gln-Gln-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O KVXVVDFOZNYYKZ-DCAQKATOSA-N 0.000 description 1
- SNLOOPZHAQDMJG-CIUDSAMLSA-N Gln-Glu-Glu Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O SNLOOPZHAQDMJG-CIUDSAMLSA-N 0.000 description 1
- KDXKFBSNIJYNNR-YVNDNENWSA-N Gln-Glu-Ile Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O KDXKFBSNIJYNNR-YVNDNENWSA-N 0.000 description 1
- PNENQZWRFMUZOM-DCAQKATOSA-N Gln-Glu-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O PNENQZWRFMUZOM-DCAQKATOSA-N 0.000 description 1
- GLAPJAHOPFSLKL-SRVKXCTJSA-N Gln-His-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CCC(=O)N)N GLAPJAHOPFSLKL-SRVKXCTJSA-N 0.000 description 1
- GIVHPCWYVWUUSG-HVTMNAMFSA-N Gln-Ile-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCC(=O)N)N GIVHPCWYVWUUSG-HVTMNAMFSA-N 0.000 description 1
- FTIJVMLAGRAYMJ-MNXVOIDGSA-N Gln-Ile-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCC(N)=O FTIJVMLAGRAYMJ-MNXVOIDGSA-N 0.000 description 1
- PSERKXGRRADTKA-MNXVOIDGSA-N Gln-Leu-Ile Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O PSERKXGRRADTKA-MNXVOIDGSA-N 0.000 description 1
- IULKWYSYZSURJK-AVGNSLFASA-N Gln-Leu-Lys Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(O)=O IULKWYSYZSURJK-AVGNSLFASA-N 0.000 description 1
- SHAUZYVSXAMYAZ-JYJNAYRXSA-N Gln-Leu-Phe Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCC(=O)N)N SHAUZYVSXAMYAZ-JYJNAYRXSA-N 0.000 description 1
- ZBKUIQNCRIYVGH-SDDRHHMPSA-N Gln-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)N)N ZBKUIQNCRIYVGH-SDDRHHMPSA-N 0.000 description 1
- CELXWPDNIGWCJN-WDCWCFNPSA-N Gln-Lys-Thr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O CELXWPDNIGWCJN-WDCWCFNPSA-N 0.000 description 1
- MFORDNZDKAVNSR-SRVKXCTJSA-N Gln-Pro-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCC(N)=O MFORDNZDKAVNSR-SRVKXCTJSA-N 0.000 description 1
- BYKZWDGMJLNFJY-XKBZYTNZSA-N Gln-Ser-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(=O)N)N)O BYKZWDGMJLNFJY-XKBZYTNZSA-N 0.000 description 1
- WBBVTGIFQIZBHP-JBACZVJFSA-N Gln-Trp-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CC2=CNC3=CC=CC=C32)NC(=O)[C@H](CCC(=O)N)N WBBVTGIFQIZBHP-JBACZVJFSA-N 0.000 description 1
- WTJIWXMJESRHMM-XDTLVQLUSA-N Gln-Tyr-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C)C(O)=O WTJIWXMJESRHMM-XDTLVQLUSA-N 0.000 description 1
- RUFHOVYUYSNDNY-ACZMJKKPSA-N Glu-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(O)=O RUFHOVYUYSNDNY-ACZMJKKPSA-N 0.000 description 1
- ITYRYNUZHPNCIK-GUBZILKMSA-N Glu-Ala-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O ITYRYNUZHPNCIK-GUBZILKMSA-N 0.000 description 1
- JJKKWYQVHRUSDG-GUBZILKMSA-N Glu-Ala-Lys Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O JJKKWYQVHRUSDG-GUBZILKMSA-N 0.000 description 1
- IYAUFWMUCGBFMQ-CIUDSAMLSA-N Glu-Arg-Cys Chemical compound C(C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)CN=C(N)N IYAUFWMUCGBFMQ-CIUDSAMLSA-N 0.000 description 1
- KKCUFHUTMKQQCF-SRVKXCTJSA-N Glu-Arg-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O KKCUFHUTMKQQCF-SRVKXCTJSA-N 0.000 description 1
- YKLNMGJYMNPBCP-ACZMJKKPSA-N Glu-Asn-Asp Chemical compound C(CC(=O)O)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC(=O)O)C(=O)O)N YKLNMGJYMNPBCP-ACZMJKKPSA-N 0.000 description 1
- CKRUHITYRFNUKW-WDSKDSINSA-N Glu-Asn-Gly Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O CKRUHITYRFNUKW-WDSKDSINSA-N 0.000 description 1
- RDPOETHPAQEGDP-ACZMJKKPSA-N Glu-Asp-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O RDPOETHPAQEGDP-ACZMJKKPSA-N 0.000 description 1
- HJIFPJUEOGZWRI-GUBZILKMSA-N Glu-Asp-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCC(=O)O)N HJIFPJUEOGZWRI-GUBZILKMSA-N 0.000 description 1
- PAQUJCSYVIBPLC-AVGNSLFASA-N Glu-Asp-Phe Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 PAQUJCSYVIBPLC-AVGNSLFASA-N 0.000 description 1
- BUZMZDDKFCSKOT-CIUDSAMLSA-N Glu-Glu-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O BUZMZDDKFCSKOT-CIUDSAMLSA-N 0.000 description 1
- SJPMNHCEWPTRBR-BQBZGAKWSA-N Glu-Glu-Gly Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O SJPMNHCEWPTRBR-BQBZGAKWSA-N 0.000 description 1
- VOORMNJKNBGYGK-YUMQZZPRSA-N Glu-Gly-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CCC(=O)O)N VOORMNJKNBGYGK-YUMQZZPRSA-N 0.000 description 1
- OPAINBJQDQTGJY-JGVFFNPUSA-N Glu-Gly-Pro Chemical compound C1C[C@@H](N(C1)C(=O)CNC(=O)[C@H](CCC(=O)O)N)C(=O)O OPAINBJQDQTGJY-JGVFFNPUSA-N 0.000 description 1
- XOFYVODYSNKPDK-AVGNSLFASA-N Glu-His-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CCC(=O)O)N XOFYVODYSNKPDK-AVGNSLFASA-N 0.000 description 1
- CXRWMMRLEMVSEH-PEFMBERDSA-N Glu-Ile-Asn Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O CXRWMMRLEMVSEH-PEFMBERDSA-N 0.000 description 1
- WTMZXOPHTIVFCP-QEWYBTABSA-N Glu-Ile-Phe Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 WTMZXOPHTIVFCP-QEWYBTABSA-N 0.000 description 1
- ZSWGJYOZWBHROQ-RWRJDSDZSA-N Glu-Ile-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ZSWGJYOZWBHROQ-RWRJDSDZSA-N 0.000 description 1
- HVYWQYLBVXMXSV-GUBZILKMSA-N Glu-Leu-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O HVYWQYLBVXMXSV-GUBZILKMSA-N 0.000 description 1
- VMKCPNBBPGGQBJ-GUBZILKMSA-N Glu-Leu-Asn Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCC(=O)O)N VMKCPNBBPGGQBJ-GUBZILKMSA-N 0.000 description 1
- DNPCBMNFQVTHMA-DCAQKATOSA-N Glu-Leu-Gln Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O DNPCBMNFQVTHMA-DCAQKATOSA-N 0.000 description 1
- IVGJYOOGJLFKQE-AVGNSLFASA-N Glu-Leu-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N IVGJYOOGJLFKQE-AVGNSLFASA-N 0.000 description 1
- NJCALAAIGREHDR-WDCWCFNPSA-N Glu-Leu-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NJCALAAIGREHDR-WDCWCFNPSA-N 0.000 description 1
- ILWHFUZZCFYSKT-AVGNSLFASA-N Glu-Lys-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O ILWHFUZZCFYSKT-AVGNSLFASA-N 0.000 description 1
- ZGEJRLJEAMPEDV-SRVKXCTJSA-N Glu-Lys-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(=O)O)N ZGEJRLJEAMPEDV-SRVKXCTJSA-N 0.000 description 1
- ZQYZDDXTNQXUJH-CIUDSAMLSA-N Glu-Met-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CCC(=O)O)N ZQYZDDXTNQXUJH-CIUDSAMLSA-N 0.000 description 1
- DCBSZJJHOTXMHY-DCAQKATOSA-N Glu-Pro-Pro Chemical compound OC(=O)CC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 DCBSZJJHOTXMHY-DCAQKATOSA-N 0.000 description 1
- SYAYROHMAIHWFB-KBIXCLLPSA-N Glu-Ser-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O SYAYROHMAIHWFB-KBIXCLLPSA-N 0.000 description 1
- BXSZPACYCMNKLS-AVGNSLFASA-N Glu-Ser-Phe Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O BXSZPACYCMNKLS-AVGNSLFASA-N 0.000 description 1
- TWYSSILQABLLME-HJGDQZAQSA-N Glu-Thr-Arg Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O TWYSSILQABLLME-HJGDQZAQSA-N 0.000 description 1
- RGJKYNUINKGPJN-RWRJDSDZSA-N Glu-Thr-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(=O)O)N RGJKYNUINKGPJN-RWRJDSDZSA-N 0.000 description 1
- CQGBSALYGOXQPE-HTUGSXCWSA-N Glu-Thr-Phe Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)O CQGBSALYGOXQPE-HTUGSXCWSA-N 0.000 description 1
- KIEICAOUSNYOLM-NRPADANISA-N Glu-Val-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O KIEICAOUSNYOLM-NRPADANISA-N 0.000 description 1
- YQPFCZVKMUVZIN-AUTRQRHGSA-N Glu-Val-Gln Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O YQPFCZVKMUVZIN-AUTRQRHGSA-N 0.000 description 1
- ZYRXTRTUCAVNBQ-GVXVVHGQSA-N Glu-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N ZYRXTRTUCAVNBQ-GVXVVHGQSA-N 0.000 description 1
- SOYWRINXUSUWEQ-DLOVCJGASA-N Glu-Val-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCC(O)=O SOYWRINXUSUWEQ-DLOVCJGASA-N 0.000 description 1
- VSVZIEVNUYDAFR-YUMQZZPRSA-N Gly-Ala-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)CN VSVZIEVNUYDAFR-YUMQZZPRSA-N 0.000 description 1
- LJPIRKICOISLKN-WHFBIAKZSA-N Gly-Ala-Ser Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O LJPIRKICOISLKN-WHFBIAKZSA-N 0.000 description 1
- XUDLUKYPXQDCRX-BQBZGAKWSA-N Gly-Arg-Asn Chemical compound [H]NCC(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(O)=O XUDLUKYPXQDCRX-BQBZGAKWSA-N 0.000 description 1
- CLODWIOAKCSBAN-BQBZGAKWSA-N Gly-Arg-Asp Chemical compound NC(N)=NCCC[C@H](NC(=O)CN)C(=O)N[C@@H](CC(O)=O)C(O)=O CLODWIOAKCSBAN-BQBZGAKWSA-N 0.000 description 1
- OVSKVOOUFAKODB-UWVGGRQHSA-N Gly-Arg-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCN=C(N)N OVSKVOOUFAKODB-UWVGGRQHSA-N 0.000 description 1
- JVWPPCWUDRJGAE-YUMQZZPRSA-N Gly-Asn-Leu Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O JVWPPCWUDRJGAE-YUMQZZPRSA-N 0.000 description 1
- XCLCVBYNGXEVDU-WHFBIAKZSA-N Gly-Asn-Ser Chemical compound NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O XCLCVBYNGXEVDU-WHFBIAKZSA-N 0.000 description 1
- FMNHBTKMRFVGRO-FOHZUACHSA-N Gly-Asn-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)CN FMNHBTKMRFVGRO-FOHZUACHSA-N 0.000 description 1
- YYPFZVIXAVDHIK-IUCAKERBSA-N Gly-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)CN YYPFZVIXAVDHIK-IUCAKERBSA-N 0.000 description 1
- ZQIMMEYPEXIYBB-IUCAKERBSA-N Gly-Glu-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)CN ZQIMMEYPEXIYBB-IUCAKERBSA-N 0.000 description 1
- BEQGFMIBZFNROK-JGVFFNPUSA-N Gly-Glu-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCC(=O)O)NC(=O)CN)C(=O)O BEQGFMIBZFNROK-JGVFFNPUSA-N 0.000 description 1
- CCQOOWAONKGYKQ-BYPYZUCNSA-N Gly-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)CN CCQOOWAONKGYKQ-BYPYZUCNSA-N 0.000 description 1
- UFPXDFOYHVEIPI-BYPYZUCNSA-N Gly-Gly-Asp Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CC(O)=O UFPXDFOYHVEIPI-BYPYZUCNSA-N 0.000 description 1
- XPJBQTCXPJNIFE-ZETCQYMHSA-N Gly-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)CN XPJBQTCXPJNIFE-ZETCQYMHSA-N 0.000 description 1
- QPCVIQJVRGXUSA-LURJTMIESA-N Gly-Gly-Met Chemical compound CSCC[C@@H](C(O)=O)NC(=O)CNC(=O)CN QPCVIQJVRGXUSA-LURJTMIESA-N 0.000 description 1
- SXJHOPPTOJACOA-QXEWZRGKSA-N Gly-Ile-Arg Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CCCN=C(N)N SXJHOPPTOJACOA-QXEWZRGKSA-N 0.000 description 1
- SCWYHUQOOFRVHP-MBLNEYKQSA-N Gly-Ile-Thr Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SCWYHUQOOFRVHP-MBLNEYKQSA-N 0.000 description 1
- LHYJCVCQPWRMKZ-WEDXCCLWSA-N Gly-Leu-Thr Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LHYJCVCQPWRMKZ-WEDXCCLWSA-N 0.000 description 1
- AFWYPMDMDYCKMD-KBPBESRZSA-N Gly-Leu-Tyr Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 AFWYPMDMDYCKMD-KBPBESRZSA-N 0.000 description 1
- VBOBNHSVQKKTOT-YUMQZZPRSA-N Gly-Lys-Ala Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O VBOBNHSVQKKTOT-YUMQZZPRSA-N 0.000 description 1
- GMTXWRIDLGTVFC-IUCAKERBSA-N Gly-Lys-Glu Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O GMTXWRIDLGTVFC-IUCAKERBSA-N 0.000 description 1
- FXGRXIATVXUAHO-WEDXCCLWSA-N Gly-Lys-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCCN FXGRXIATVXUAHO-WEDXCCLWSA-N 0.000 description 1
- GAFKBWKVXNERFA-QWRGUYRKSA-N Gly-Phe-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CC=CC=C1 GAFKBWKVXNERFA-QWRGUYRKSA-N 0.000 description 1
- DBUNZBWUWCIELX-JHEQGTHGSA-N Gly-Thr-Glu Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O DBUNZBWUWCIELX-JHEQGTHGSA-N 0.000 description 1
- RCHFYMASWAZQQZ-ZANVPECISA-N Gly-Trp-Ala Chemical compound C1=CC=C2C(C[C@@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)CN)=CNC2=C1 RCHFYMASWAZQQZ-ZANVPECISA-N 0.000 description 1
- PNUFMLXHOLFRLD-KBPBESRZSA-N Gly-Tyr-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CC=C(O)C=C1 PNUFMLXHOLFRLD-KBPBESRZSA-N 0.000 description 1
- DNVDEMWIYLVIQU-RCOVLWMOSA-N Gly-Val-Asp Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O DNVDEMWIYLVIQU-RCOVLWMOSA-N 0.000 description 1
- 229920000084 Gum arabic Polymers 0.000 description 1
- 208000028782 Hereditary disease Diseases 0.000 description 1
- IPIVXQQRZXEUGW-UWJYBYFXSA-N His-Ala-His Chemical compound C([C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1NC=NC=1)C(O)=O)C1=CN=CN1 IPIVXQQRZXEUGW-UWJYBYFXSA-N 0.000 description 1
- XINDHUAGVGCNSF-QSFUFRPTSA-N His-Ala-Ile Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O XINDHUAGVGCNSF-QSFUFRPTSA-N 0.000 description 1
- SVHKVHBPTOMLTO-DCAQKATOSA-N His-Arg-Asp Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O SVHKVHBPTOMLTO-DCAQKATOSA-N 0.000 description 1
- MVADCDSCFTXCBT-CIUDSAMLSA-N His-Asp-Asp Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O MVADCDSCFTXCBT-CIUDSAMLSA-N 0.000 description 1
- IMCHNUANCIGUKS-SRVKXCTJSA-N His-Glu-Arg Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O IMCHNUANCIGUKS-SRVKXCTJSA-N 0.000 description 1
- XMENRVZYPBKBIL-AVGNSLFASA-N His-Glu-His Chemical compound N[C@@H](Cc1cnc[nH]1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](Cc1cnc[nH]1)C(O)=O XMENRVZYPBKBIL-AVGNSLFASA-N 0.000 description 1
- FIMNVXRZGUAGBI-AVGNSLFASA-N His-Glu-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O FIMNVXRZGUAGBI-AVGNSLFASA-N 0.000 description 1
- JCOSMKPAOYDKRO-AVGNSLFASA-N His-Glu-Lys Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N JCOSMKPAOYDKRO-AVGNSLFASA-N 0.000 description 1
- BXOLYFJYQQRQDJ-MXAVVETBSA-N His-Leu-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC1=CN=CN1)N BXOLYFJYQQRQDJ-MXAVVETBSA-N 0.000 description 1
- ZSKJIISDJXJQPV-BZSNNMDCSA-N His-Leu-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CN=CN1 ZSKJIISDJXJQPV-BZSNNMDCSA-N 0.000 description 1
- SVVULKPWDBIPCO-BZSNNMDCSA-N His-Phe-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O SVVULKPWDBIPCO-BZSNNMDCSA-N 0.000 description 1
- WCHONUZTYDQMBY-PYJNHQTQSA-N His-Pro-Ile Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WCHONUZTYDQMBY-PYJNHQTQSA-N 0.000 description 1
- VIJMRAIWYWRXSR-CIUDSAMLSA-N His-Ser-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC1=CN=CN1 VIJMRAIWYWRXSR-CIUDSAMLSA-N 0.000 description 1
- DAKSMIWQZPHRIB-BZSNNMDCSA-N His-Tyr-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(O)=O DAKSMIWQZPHRIB-BZSNNMDCSA-N 0.000 description 1
- 101000798114 Homo sapiens Lactotransferrin Proteins 0.000 description 1
- 108091006905 Human Serum Albumin Proteins 0.000 description 1
- 208000015178 Hurler syndrome Diseases 0.000 description 1
- LQSBBHNVAVNZSX-GHCJXIJMSA-N Ile-Ala-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](CC(=O)N)C(=O)O)N LQSBBHNVAVNZSX-GHCJXIJMSA-N 0.000 description 1
- YOTNPRLPIPHQSB-XUXIUFHCSA-N Ile-Arg-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(=O)O)N YOTNPRLPIPHQSB-XUXIUFHCSA-N 0.000 description 1
- NBJAAWYRLGCJOF-UGYAYLCHSA-N Ile-Asp-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N NBJAAWYRLGCJOF-UGYAYLCHSA-N 0.000 description 1
- RGSOCXHDOPQREB-ZPFDUUQYSA-N Ile-Asp-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(C)C)C(=O)O)N RGSOCXHDOPQREB-ZPFDUUQYSA-N 0.000 description 1
- KUHFPGIVBOCRMV-MNXVOIDGSA-N Ile-Gln-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC(C)C)C(=O)O)N KUHFPGIVBOCRMV-MNXVOIDGSA-N 0.000 description 1
- LKACSKJPTFSBHR-MNXVOIDGSA-N Ile-Gln-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N LKACSKJPTFSBHR-MNXVOIDGSA-N 0.000 description 1
- QRTVJGKXFSYJGW-KBIXCLLPSA-N Ile-Glu-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CS)C(=O)O)N QRTVJGKXFSYJGW-KBIXCLLPSA-N 0.000 description 1
- LGMUPVWZEYYUMU-YVNDNENWSA-N Ile-Glu-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N LGMUPVWZEYYUMU-YVNDNENWSA-N 0.000 description 1
- MTFVYKQRLXYAQN-LAEOZQHASA-N Ile-Glu-Gly Chemical compound [H]N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O MTFVYKQRLXYAQN-LAEOZQHASA-N 0.000 description 1
- PNDMHTTXXPUQJH-RWRJDSDZSA-N Ile-Glu-Thr Chemical compound N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H]([C@H](O)C)C(=O)O PNDMHTTXXPUQJH-RWRJDSDZSA-N 0.000 description 1
- KFVUBLZRFSVDGO-BYULHYEWSA-N Ile-Gly-Asp Chemical compound CC[C@H](C)[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC(O)=O KFVUBLZRFSVDGO-BYULHYEWSA-N 0.000 description 1
- DFFTXLCCDFYRKD-MBLNEYKQSA-N Ile-Gly-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(=O)O)N DFFTXLCCDFYRKD-MBLNEYKQSA-N 0.000 description 1
- HYLIOBDWPQNLKI-HVTMNAMFSA-N Ile-His-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N HYLIOBDWPQNLKI-HVTMNAMFSA-N 0.000 description 1
- HUWYGQOISIJNMK-SIGLWIIPSA-N Ile-Ile-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N HUWYGQOISIJNMK-SIGLWIIPSA-N 0.000 description 1
- CSQNHSGHAPRGPQ-YTFOTSKYSA-N Ile-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCCN)C(=O)O)N CSQNHSGHAPRGPQ-YTFOTSKYSA-N 0.000 description 1
- PKGGWLOLRLOPGK-XUXIUFHCSA-N Ile-Leu-Arg Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N PKGGWLOLRLOPGK-XUXIUFHCSA-N 0.000 description 1
- OUUCIIJSBIBCHB-ZPFDUUQYSA-N Ile-Leu-Asp Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O OUUCIIJSBIBCHB-ZPFDUUQYSA-N 0.000 description 1
- YGDWPQCLFJNMOL-MNXVOIDGSA-N Ile-Leu-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N YGDWPQCLFJNMOL-MNXVOIDGSA-N 0.000 description 1
- HUORUFRRJHELPD-MNXVOIDGSA-N Ile-Leu-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N HUORUFRRJHELPD-MNXVOIDGSA-N 0.000 description 1
- ADDYYRVQQZFIMW-MNXVOIDGSA-N Ile-Lys-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N ADDYYRVQQZFIMW-MNXVOIDGSA-N 0.000 description 1
- GVNNAHIRSDRIII-AJNGGQMLSA-N Ile-Lys-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N GVNNAHIRSDRIII-AJNGGQMLSA-N 0.000 description 1
- GLYJPWIRLBAIJH-UHFFFAOYSA-N Ile-Lys-Pro Natural products CCC(C)C(N)C(=O)NC(CCCCN)C(=O)N1CCCC1C(O)=O GLYJPWIRLBAIJH-UHFFFAOYSA-N 0.000 description 1
- RVNOXPZHMUWCLW-GMOBBJLQSA-N Ile-Met-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(=O)N)C(=O)O)N RVNOXPZHMUWCLW-GMOBBJLQSA-N 0.000 description 1
- RCMNUBZKIIJCOI-ZPFDUUQYSA-N Ile-Met-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N RCMNUBZKIIJCOI-ZPFDUUQYSA-N 0.000 description 1
- CIDLJWVDMNDKPT-FIRPJDEBSA-N Ile-Phe-Phe Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)O)N CIDLJWVDMNDKPT-FIRPJDEBSA-N 0.000 description 1
- BJECXJHLUJXPJQ-PYJNHQTQSA-N Ile-Pro-His Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N BJECXJHLUJXPJQ-PYJNHQTQSA-N 0.000 description 1
- ZNOBVZFCHNHKHA-KBIXCLLPSA-N Ile-Ser-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N ZNOBVZFCHNHKHA-KBIXCLLPSA-N 0.000 description 1
- ZLFNNVATRMCAKN-ZKWXMUAHSA-N Ile-Ser-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)NCC(=O)O)N ZLFNNVATRMCAKN-ZKWXMUAHSA-N 0.000 description 1
- JSLIXOUMAOUGBN-JUKXBJQTSA-N Ile-Tyr-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N JSLIXOUMAOUGBN-JUKXBJQTSA-N 0.000 description 1
- DLEBSGAVWRPTIX-PEDHHIEDSA-N Ile-Val-Ile Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)[C@@H](C)CC DLEBSGAVWRPTIX-PEDHHIEDSA-N 0.000 description 1
- SWNRZNLXMXRCJC-VKOGCVSHSA-N Ile-Val-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)[C@@H](C)CC)C(O)=O)=CNC2=C1 SWNRZNLXMXRCJC-VKOGCVSHSA-N 0.000 description 1
- 108010065920 Insulin Lispro Proteins 0.000 description 1
- PWWVAXIEGOYWEE-UHFFFAOYSA-N Isophenergan Chemical compound C1=CC=C2N(CC(C)N(C)C)C3=CC=CC=C3SC2=C1 PWWVAXIEGOYWEE-UHFFFAOYSA-N 0.000 description 1
- LHSGPCFBGJHPCY-UHFFFAOYSA-N L-leucine-L-tyrosine Natural products CC(C)CC(N)C(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 LHSGPCFBGJHPCY-UHFFFAOYSA-N 0.000 description 1
- SENJXOPIZNYLHU-UHFFFAOYSA-N L-leucyl-L-arginine Natural products CC(C)CC(N)C(=O)NC(C(O)=O)CCCN=C(N)N SENJXOPIZNYLHU-UHFFFAOYSA-N 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- LJHGALIOHLRRQN-DCAQKATOSA-N Leu-Ala-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N LJHGALIOHLRRQN-DCAQKATOSA-N 0.000 description 1
- ZRLUISBDKUWAIZ-CIUDSAMLSA-N Leu-Ala-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC(O)=O ZRLUISBDKUWAIZ-CIUDSAMLSA-N 0.000 description 1
- MJOZZTKJZQFKDK-GUBZILKMSA-N Leu-Ala-Gln Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(N)=O MJOZZTKJZQFKDK-GUBZILKMSA-N 0.000 description 1
- WNGVUZWBXZKQES-YUMQZZPRSA-N Leu-Ala-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O WNGVUZWBXZKQES-YUMQZZPRSA-N 0.000 description 1
- PBCHMHROGNUXMK-DLOVCJGASA-N Leu-Ala-His Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CN=CN1 PBCHMHROGNUXMK-DLOVCJGASA-N 0.000 description 1
- WGNOPSQMIQERPK-UHFFFAOYSA-N Leu-Asn-Pro Natural products CC(C)CC(N)C(=O)NC(CC(=O)N)C(=O)N1CCCC1C(=O)O WGNOPSQMIQERPK-UHFFFAOYSA-N 0.000 description 1
- PVMPDMIKUVNOBD-CIUDSAMLSA-N Leu-Asp-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O PVMPDMIKUVNOBD-CIUDSAMLSA-N 0.000 description 1
- RSFGIMMPWAXNML-MNXVOIDGSA-N Leu-Gln-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O RSFGIMMPWAXNML-MNXVOIDGSA-N 0.000 description 1
- HFBCHNRFRYLZNV-GUBZILKMSA-N Leu-Glu-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O HFBCHNRFRYLZNV-GUBZILKMSA-N 0.000 description 1
- OXRLYTYUXAQTHP-YUMQZZPRSA-N Leu-Gly-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](C)C(O)=O OXRLYTYUXAQTHP-YUMQZZPRSA-N 0.000 description 1
- VGPCJSXPPOQPBK-YUMQZZPRSA-N Leu-Gly-Ser Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O VGPCJSXPPOQPBK-YUMQZZPRSA-N 0.000 description 1
- USLNHQZCDQJBOV-ZPFDUUQYSA-N Leu-Ile-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O USLNHQZCDQJBOV-ZPFDUUQYSA-N 0.000 description 1
- XVZCXCTYGHPNEM-UHFFFAOYSA-N Leu-Leu-Pro Natural products CC(C)CC(N)C(=O)NC(CC(C)C)C(=O)N1CCCC1C(O)=O XVZCXCTYGHPNEM-UHFFFAOYSA-N 0.000 description 1
- RXGLHDWAZQECBI-SRVKXCTJSA-N Leu-Leu-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O RXGLHDWAZQECBI-SRVKXCTJSA-N 0.000 description 1
- RZXLZBIUTDQHJQ-SRVKXCTJSA-N Leu-Lys-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(O)=O RZXLZBIUTDQHJQ-SRVKXCTJSA-N 0.000 description 1
- HVHRPWQEQHIQJF-AVGNSLFASA-N Leu-Lys-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O HVHRPWQEQHIQJF-AVGNSLFASA-N 0.000 description 1
- VCHVSKNMTXWIIP-SRVKXCTJSA-N Leu-Lys-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(O)=O VCHVSKNMTXWIIP-SRVKXCTJSA-N 0.000 description 1
- ZAVCJRJOQKIOJW-KKUMJFAQSA-N Leu-Phe-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(O)=O)C(O)=O)CC1=CC=CC=C1 ZAVCJRJOQKIOJW-KKUMJFAQSA-N 0.000 description 1
- AIRUUHAOKGVJAD-JYJNAYRXSA-N Leu-Phe-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O AIRUUHAOKGVJAD-JYJNAYRXSA-N 0.000 description 1
- INCJJHQRZGQLFC-KBPBESRZSA-N Leu-Phe-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)NCC(O)=O INCJJHQRZGQLFC-KBPBESRZSA-N 0.000 description 1
- DRWMRVFCKKXHCH-BZSNNMDCSA-N Leu-Phe-Leu Chemical compound CC(C)C[C@H]([NH3+])C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C([O-])=O)CC1=CC=CC=C1 DRWMRVFCKKXHCH-BZSNNMDCSA-N 0.000 description 1
- PJWOOBTYQNNRBF-BZSNNMDCSA-N Leu-Phe-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCCN)C(=O)O)N PJWOOBTYQNNRBF-BZSNNMDCSA-N 0.000 description 1
- YWKNKRAKOCLOLH-OEAJRASXSA-N Leu-Phe-Thr Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)O)C(O)=O)CC1=CC=CC=C1 YWKNKRAKOCLOLH-OEAJRASXSA-N 0.000 description 1
- RRVCZCNFXIFGRA-DCAQKATOSA-N Leu-Pro-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O RRVCZCNFXIFGRA-DCAQKATOSA-N 0.000 description 1
- VULJUQZPSOASBZ-SRVKXCTJSA-N Leu-Pro-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O VULJUQZPSOASBZ-SRVKXCTJSA-N 0.000 description 1
- CHJKEDSZNSONPS-DCAQKATOSA-N Leu-Pro-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O CHJKEDSZNSONPS-DCAQKATOSA-N 0.000 description 1
- IZPVWNSAVUQBGP-CIUDSAMLSA-N Leu-Ser-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O IZPVWNSAVUQBGP-CIUDSAMLSA-N 0.000 description 1
- XOWMDXHFSBCAKQ-SRVKXCTJSA-N Leu-Ser-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC(C)C XOWMDXHFSBCAKQ-SRVKXCTJSA-N 0.000 description 1
- AMSSKPUHBUQBOQ-SRVKXCTJSA-N Leu-Ser-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)O)N AMSSKPUHBUQBOQ-SRVKXCTJSA-N 0.000 description 1
- GOFJOGXGMPHOGL-DCAQKATOSA-N Leu-Ser-Met Chemical compound CSCC[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(C)C GOFJOGXGMPHOGL-DCAQKATOSA-N 0.000 description 1
- SBANPBVRHYIMRR-GARJFASQSA-N Leu-Ser-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CO)C(=O)N1CCC[C@@H]1C(=O)O)N SBANPBVRHYIMRR-GARJFASQSA-N 0.000 description 1
- SBANPBVRHYIMRR-UHFFFAOYSA-N Leu-Ser-Pro Natural products CC(C)CC(N)C(=O)NC(CO)C(=O)N1CCCC1C(O)=O SBANPBVRHYIMRR-UHFFFAOYSA-N 0.000 description 1
- BRTVHXHCUSXYRI-CIUDSAMLSA-N Leu-Ser-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O BRTVHXHCUSXYRI-CIUDSAMLSA-N 0.000 description 1
- AEDWWMMHUGYIFD-HJGDQZAQSA-N Leu-Thr-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(O)=O AEDWWMMHUGYIFD-HJGDQZAQSA-N 0.000 description 1
- QWWPYKKLXWOITQ-VOAKCMCISA-N Leu-Thr-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC(C)C QWWPYKKLXWOITQ-VOAKCMCISA-N 0.000 description 1
- DAYQSYGBCUKVKT-VOAKCMCISA-N Leu-Thr-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(O)=O DAYQSYGBCUKVKT-VOAKCMCISA-N 0.000 description 1
- HQBOMRTVKVKFMN-WDSOQIARSA-N Leu-Trp-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](C(C)C)C(O)=O HQBOMRTVKVKFMN-WDSOQIARSA-N 0.000 description 1
- VJGQRELPQWNURN-JYJNAYRXSA-N Leu-Tyr-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O VJGQRELPQWNURN-JYJNAYRXSA-N 0.000 description 1
- YIRIDPUGZKHMHT-ACRUOGEOSA-N Leu-Tyr-Tyr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O YIRIDPUGZKHMHT-ACRUOGEOSA-N 0.000 description 1
- FBNPMTNBFFAMMH-AVGNSLFASA-N Leu-Val-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N FBNPMTNBFFAMMH-AVGNSLFASA-N 0.000 description 1
- FBNPMTNBFFAMMH-UHFFFAOYSA-N Leu-Val-Arg Natural products CC(C)CC(N)C(=O)NC(C(C)C)C(=O)NC(C(O)=O)CCCN=C(N)N FBNPMTNBFFAMMH-UHFFFAOYSA-N 0.000 description 1
- AIMGJYMCTAABEN-GVXVVHGQSA-N Leu-Val-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O AIMGJYMCTAABEN-GVXVVHGQSA-N 0.000 description 1
- YQFZRHYZLARWDY-IHRRRGAJSA-N Leu-Val-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCCN YQFZRHYZLARWDY-IHRRRGAJSA-N 0.000 description 1
- 239000012098 Lipofectamine RNAiMAX Substances 0.000 description 1
- MPGHETGWWWUHPY-CIUDSAMLSA-N Lys-Ala-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN MPGHETGWWWUHPY-CIUDSAMLSA-N 0.000 description 1
- XFIHDSBIPWEYJJ-YUMQZZPRSA-N Lys-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN XFIHDSBIPWEYJJ-YUMQZZPRSA-N 0.000 description 1
- NFLFJGGKOHYZJF-BJDJZHNGSA-N Lys-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN NFLFJGGKOHYZJF-BJDJZHNGSA-N 0.000 description 1
- IXHKPDJKKCUKHS-GARJFASQSA-N Lys-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCCN)N IXHKPDJKKCUKHS-GARJFASQSA-N 0.000 description 1
- WXJKFRMKJORORD-DCAQKATOSA-N Lys-Arg-Ala Chemical compound NC(=N)NCCC[C@@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@@H](N)CCCCN WXJKFRMKJORORD-DCAQKATOSA-N 0.000 description 1
- SWWCDAGDQHTKIE-RHYQMDGZSA-N Lys-Arg-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SWWCDAGDQHTKIE-RHYQMDGZSA-N 0.000 description 1
- DNEJSAIMVANNPA-DCAQKATOSA-N Lys-Asn-Arg Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O DNEJSAIMVANNPA-DCAQKATOSA-N 0.000 description 1
- DGWXCIORNLWGGG-CIUDSAMLSA-N Lys-Asn-Ser Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O DGWXCIORNLWGGG-CIUDSAMLSA-N 0.000 description 1
- IWWMPCPLFXFBAF-SRVKXCTJSA-N Lys-Asp-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O IWWMPCPLFXFBAF-SRVKXCTJSA-N 0.000 description 1
- LMVOVCYVZBBWQB-SRVKXCTJSA-N Lys-Asp-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCCCN LMVOVCYVZBBWQB-SRVKXCTJSA-N 0.000 description 1
- PHHYNOUOUWYQRO-XIRDDKMYSA-N Lys-Asp-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCCCN)N PHHYNOUOUWYQRO-XIRDDKMYSA-N 0.000 description 1
- WTZUSCUIVPVCRH-SRVKXCTJSA-N Lys-Gln-Arg Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N WTZUSCUIVPVCRH-SRVKXCTJSA-N 0.000 description 1
- LLSUNJYOSCOOEB-GUBZILKMSA-N Lys-Glu-Asp Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O LLSUNJYOSCOOEB-GUBZILKMSA-N 0.000 description 1
- KZOHPCYVORJBLG-AVGNSLFASA-N Lys-Glu-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CCCCN)N KZOHPCYVORJBLG-AVGNSLFASA-N 0.000 description 1
- QZONCCHVHCOBSK-YUMQZZPRSA-N Lys-Gly-Asn Chemical compound [H]N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O QZONCCHVHCOBSK-YUMQZZPRSA-N 0.000 description 1
- XNKDCYABMBBEKN-IUCAKERBSA-N Lys-Gly-Gln Chemical compound NCCCC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(N)=O XNKDCYABMBBEKN-IUCAKERBSA-N 0.000 description 1
- FHIAJWBDZVHLAH-YUMQZZPRSA-N Lys-Gly-Ser Chemical compound NCCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O FHIAJWBDZVHLAH-YUMQZZPRSA-N 0.000 description 1
- SPCHLZUWJTYZFC-IHRRRGAJSA-N Lys-His-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C(C)C)C(O)=O SPCHLZUWJTYZFC-IHRRRGAJSA-N 0.000 description 1
- QBEPTBMRQALPEV-MNXVOIDGSA-N Lys-Ile-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCCCN QBEPTBMRQALPEV-MNXVOIDGSA-N 0.000 description 1
- JYXBNQOKPRQNQS-YTFOTSKYSA-N Lys-Ile-Ile Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JYXBNQOKPRQNQS-YTFOTSKYSA-N 0.000 description 1
- ZXFRGTAIIZHNHG-AJNGGQMLSA-N Lys-Ile-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)O)NC(=O)[C@H](CCCCN)N ZXFRGTAIIZHNHG-AJNGGQMLSA-N 0.000 description 1
- ONPDTSFZAIWMDI-AVGNSLFASA-N Lys-Leu-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O ONPDTSFZAIWMDI-AVGNSLFASA-N 0.000 description 1
- SKRGVGLIRUGANF-AVGNSLFASA-N Lys-Leu-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O SKRGVGLIRUGANF-AVGNSLFASA-N 0.000 description 1
- XIZQPFCRXLUNMK-BZSNNMDCSA-N Lys-Leu-Phe Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCCCN)N XIZQPFCRXLUNMK-BZSNNMDCSA-N 0.000 description 1
- VUTWYNQUSJWBHO-BZSNNMDCSA-N Lys-Leu-Tyr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O VUTWYNQUSJWBHO-BZSNNMDCSA-N 0.000 description 1
- XOQMURBBIXRRCR-SRVKXCTJSA-N Lys-Lys-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCCN XOQMURBBIXRRCR-SRVKXCTJSA-N 0.000 description 1
- YUAXTFMFMOIMAM-QWRGUYRKSA-N Lys-Lys-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)NCC(O)=O YUAXTFMFMOIMAM-QWRGUYRKSA-N 0.000 description 1
- WBSCNDJQPKSPII-KKUMJFAQSA-N Lys-Lys-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O WBSCNDJQPKSPII-KKUMJFAQSA-N 0.000 description 1
- KJIXWRWPOCKYLD-IHRRRGAJSA-N Lys-Lys-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCCCN)N KJIXWRWPOCKYLD-IHRRRGAJSA-N 0.000 description 1
- PLDJDCJLRCYPJB-VOAKCMCISA-N Lys-Lys-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O PLDJDCJLRCYPJB-VOAKCMCISA-N 0.000 description 1
- BXPHMHQHYHILBB-BZSNNMDCSA-N Lys-Lys-Tyr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O BXPHMHQHYHILBB-BZSNNMDCSA-N 0.000 description 1
- SKUOQDYMJFUMOE-ULQDDVLXSA-N Lys-Met-Phe Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCCCN)N SKUOQDYMJFUMOE-ULQDDVLXSA-N 0.000 description 1
- QBHGXFQJFPWJIH-XUXIUFHCSA-N Lys-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCCN QBHGXFQJFPWJIH-XUXIUFHCSA-N 0.000 description 1
- HYSVGEAWTGPMOA-IHRRRGAJSA-N Lys-Pro-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O HYSVGEAWTGPMOA-IHRRRGAJSA-N 0.000 description 1
- WQDKIVRHTQYJSN-DCAQKATOSA-N Lys-Ser-Arg Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N WQDKIVRHTQYJSN-DCAQKATOSA-N 0.000 description 1
- GHKXHCMRAUYLBS-CIUDSAMLSA-N Lys-Ser-Asn Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O GHKXHCMRAUYLBS-CIUDSAMLSA-N 0.000 description 1
- YFQSSOAGMZGXFT-MEYUZBJRSA-N Lys-Thr-Tyr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O YFQSSOAGMZGXFT-MEYUZBJRSA-N 0.000 description 1
- IEIHKHYMBIYQTH-YESZJQIVSA-N Lys-Tyr-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=C(C=C2)O)NC(=O)[C@H](CCCCN)N)C(=O)O IEIHKHYMBIYQTH-YESZJQIVSA-N 0.000 description 1
- VVURYEVJJTXWNE-ULQDDVLXSA-N Lys-Tyr-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(O)=O VVURYEVJJTXWNE-ULQDDVLXSA-N 0.000 description 1
- RPWQJSBMXJSCPD-XUXIUFHCSA-N Lys-Val-Ile Chemical compound CC[C@H](C)[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CCCCN)C(C)C)C(O)=O RPWQJSBMXJSCPD-XUXIUFHCSA-N 0.000 description 1
- NYTDJEZBAAFLLG-IHRRRGAJSA-N Lys-Val-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(O)=O NYTDJEZBAAFLLG-IHRRRGAJSA-N 0.000 description 1
- OZVXDDFYCQOPFD-XQQFMLRXSA-N Lys-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCCN)N OZVXDDFYCQOPFD-XQQFMLRXSA-N 0.000 description 1
- RIPJMCFGQHGHNP-RHYQMDGZSA-N Lys-Val-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CCCCN)N)O RIPJMCFGQHGHNP-RHYQMDGZSA-N 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- QAHFGYLFLVGBNW-DCAQKATOSA-N Met-Ala-Lys Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN QAHFGYLFLVGBNW-DCAQKATOSA-N 0.000 description 1
- SDTSLIMYROCDNS-FXQIFTODSA-N Met-Cys-Asn Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(O)=O SDTSLIMYROCDNS-FXQIFTODSA-N 0.000 description 1
- GPAHWYRSHCKICP-GUBZILKMSA-N Met-Glu-Glu Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O GPAHWYRSHCKICP-GUBZILKMSA-N 0.000 description 1
- FZUNSVYYPYJYAP-NAKRPEOUSA-N Met-Ile-Ala Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O FZUNSVYYPYJYAP-NAKRPEOUSA-N 0.000 description 1
- QGRJTULYDZUBAY-ZPFDUUQYSA-N Met-Ile-Glu Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(O)=O QGRJTULYDZUBAY-ZPFDUUQYSA-N 0.000 description 1
- AFFKUNVPPLQUGA-DCAQKATOSA-N Met-Leu-Ala Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O AFFKUNVPPLQUGA-DCAQKATOSA-N 0.000 description 1
- BEZJTLKUMFMITF-AVGNSLFASA-N Met-Lys-Arg Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@H](C(O)=O)CCCNC(N)=N BEZJTLKUMFMITF-AVGNSLFASA-N 0.000 description 1
- LNXGEYIEEUZGGH-JYJNAYRXSA-N Met-Phe-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)CCSC)CC1=CC=CC=C1 LNXGEYIEEUZGGH-JYJNAYRXSA-N 0.000 description 1
- KSIPKXNIQOWMIC-RCWTZXSCSA-N Met-Thr-Arg Chemical compound CSCC[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CCCNC(N)=N KSIPKXNIQOWMIC-RCWTZXSCSA-N 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- 206010056886 Mucopolysaccharidosis I Diseases 0.000 description 1
- 101100476480 Mus musculus S100a8 gene Proteins 0.000 description 1
- 241000699670 Mus sp. Species 0.000 description 1
- WUGMRIBZSVSJNP-UHFFFAOYSA-N N-L-alanyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)C)C(O)=O)=CNC2=C1 WUGMRIBZSVSJNP-UHFFFAOYSA-N 0.000 description 1
- XZFYRXDAULDNFX-UHFFFAOYSA-N N-L-cysteinyl-L-phenylalanine Natural products SCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XZFYRXDAULDNFX-UHFFFAOYSA-N 0.000 description 1
- PESQCPHRXOFIPX-UHFFFAOYSA-N N-L-methionyl-L-tyrosine Natural products CSCCC(N)C(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 PESQCPHRXOFIPX-UHFFFAOYSA-N 0.000 description 1
- XMBSYZWANAQXEV-UHFFFAOYSA-N N-alpha-L-glutamyl-L-phenylalanine Natural products OC(=O)CCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XMBSYZWANAQXEV-UHFFFAOYSA-N 0.000 description 1
- BQVUABVGYYSDCJ-UHFFFAOYSA-N Nalpha-L-Leucyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)CC(C)C)C(O)=O)=CNC2=C1 BQVUABVGYYSDCJ-UHFFFAOYSA-N 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 101100068676 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) gln-1 gene Proteins 0.000 description 1
- 239000012124 Opti-MEM Substances 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 241000286209 Phasianidae Species 0.000 description 1
- CGOMLCQJEMWMCE-STQMWFEESA-N Phe-Arg-Gly Chemical compound NC(N)=NCCC[C@@H](C(=O)NCC(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1 CGOMLCQJEMWMCE-STQMWFEESA-N 0.000 description 1
- MQWISMJKHOUEMW-ULQDDVLXSA-N Phe-Arg-His Chemical compound C([C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CC=1NC=NC=1)C(O)=O)C1=CC=CC=C1 MQWISMJKHOUEMW-ULQDDVLXSA-N 0.000 description 1
- AGYXCMYVTBYGCT-ULQDDVLXSA-N Phe-Arg-Leu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O AGYXCMYVTBYGCT-ULQDDVLXSA-N 0.000 description 1
- WMGVYPPIMZPWPN-SRVKXCTJSA-N Phe-Asp-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N WMGVYPPIMZPWPN-SRVKXCTJSA-N 0.000 description 1
- SWZKMTDPQXLQRD-XVSYOHENSA-N Phe-Asp-Thr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SWZKMTDPQXLQRD-XVSYOHENSA-N 0.000 description 1
- GDBOREPXIRKSEQ-FHWLQOOXSA-N Phe-Gln-Phe Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O GDBOREPXIRKSEQ-FHWLQOOXSA-N 0.000 description 1
- KYYMILWEGJYPQZ-IHRRRGAJSA-N Phe-Glu-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1 KYYMILWEGJYPQZ-IHRRRGAJSA-N 0.000 description 1
- KJJROSNFBRWPHS-JYJNAYRXSA-N Phe-Glu-Leu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O KJJROSNFBRWPHS-JYJNAYRXSA-N 0.000 description 1
- PSKRILMFHNIUAO-JYJNAYRXSA-N Phe-Glu-Lys Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N PSKRILMFHNIUAO-JYJNAYRXSA-N 0.000 description 1
- WPTYDQPGBMDUBI-QWRGUYRKSA-N Phe-Gly-Asn Chemical compound N[C@@H](Cc1ccccc1)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O WPTYDQPGBMDUBI-QWRGUYRKSA-N 0.000 description 1
- KRYSMKKRRRWOCZ-QEWYBTABSA-N Phe-Ile-Glu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(O)=O KRYSMKKRRRWOCZ-QEWYBTABSA-N 0.000 description 1
- METZZBCMDXHFMK-BZSNNMDCSA-N Phe-Leu-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CC2=CC=CC=C2)N METZZBCMDXHFMK-BZSNNMDCSA-N 0.000 description 1
- LRBSWBVUCLLRLU-BZSNNMDCSA-N Phe-Leu-Lys Chemical compound CC(C)C[C@H](NC(=O)[C@@H](N)Cc1ccccc1)C(=O)N[C@@H](CCCCN)C(O)=O LRBSWBVUCLLRLU-BZSNNMDCSA-N 0.000 description 1
- XZQYIJALMGEUJD-OEAJRASXSA-N Phe-Lys-Thr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XZQYIJALMGEUJD-OEAJRASXSA-N 0.000 description 1
- BSHMIVKDJQGLNT-ACRUOGEOSA-N Phe-Lys-Tyr Chemical compound C([C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=CC=C1 BSHMIVKDJQGLNT-ACRUOGEOSA-N 0.000 description 1
- TXJJXEXCZBHDNA-ACRUOGEOSA-N Phe-Phe-His Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)N[C@@H](CC3=CN=CN3)C(=O)O)N TXJJXEXCZBHDNA-ACRUOGEOSA-N 0.000 description 1
- RBRNEFJTEHPDSL-ACRUOGEOSA-N Phe-Phe-Lys Chemical compound C([C@@H](C(=O)N[C@@H](CCCCN)C(O)=O)NC(=O)[C@@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 RBRNEFJTEHPDSL-ACRUOGEOSA-N 0.000 description 1
- DBNGDEAQXGFGRA-ACRUOGEOSA-N Phe-Tyr-Lys Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)N[C@@H](CCCCN)C(=O)O)N DBNGDEAQXGFGRA-ACRUOGEOSA-N 0.000 description 1
- 102100026090 Polyadenylate-binding protein 1 Human genes 0.000 description 1
- 101710103012 Polyadenylate-binding protein, cytoplasmic and nuclear Proteins 0.000 description 1
- 239000004698 Polyethylene Substances 0.000 description 1
- 239000002202 Polyethylene glycol Substances 0.000 description 1
- 229920002873 Polyethylenimine Polymers 0.000 description 1
- CGBYDGAJHSOGFQ-LPEHRKFASA-N Pro-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@@H]2CCCN2 CGBYDGAJHSOGFQ-LPEHRKFASA-N 0.000 description 1
- OBVCYFIHIIYIQF-CIUDSAMLSA-N Pro-Asn-Glu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O OBVCYFIHIIYIQF-CIUDSAMLSA-N 0.000 description 1
- VOHFZDSRPZLXLH-IHRRRGAJSA-N Pro-Asn-Phe Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O VOHFZDSRPZLXLH-IHRRRGAJSA-N 0.000 description 1
- DIFXZGPHVCIVSQ-CIUDSAMLSA-N Pro-Gln-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(O)=O DIFXZGPHVCIVSQ-CIUDSAMLSA-N 0.000 description 1
- UAYHMOIGIQZLFR-NHCYSSNCSA-N Pro-Gln-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O UAYHMOIGIQZLFR-NHCYSSNCSA-N 0.000 description 1
- KIPIKSXPPLABPN-CIUDSAMLSA-N Pro-Glu-Asn Chemical compound NC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H]1CCCN1 KIPIKSXPPLABPN-CIUDSAMLSA-N 0.000 description 1
- MGDFPGCFVJFITQ-CIUDSAMLSA-N Pro-Glu-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O MGDFPGCFVJFITQ-CIUDSAMLSA-N 0.000 description 1
- VOZIBWWZSBIXQN-SRVKXCTJSA-N Pro-Glu-Lys Chemical compound NCCCC[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H]1CCCN1)C(O)=O VOZIBWWZSBIXQN-SRVKXCTJSA-N 0.000 description 1
- HAAQQNHQZBOWFO-LURJTMIESA-N Pro-Gly-Gly Chemical compound OC(=O)CNC(=O)CNC(=O)[C@@H]1CCCN1 HAAQQNHQZBOWFO-LURJTMIESA-N 0.000 description 1
- JUJGNDZIKKQMDJ-IHRRRGAJSA-N Pro-His-His Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CNC=N1)C(O)=O JUJGNDZIKKQMDJ-IHRRRGAJSA-N 0.000 description 1
- FKYKZHOKDOPHSA-DCAQKATOSA-N Pro-Leu-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O FKYKZHOKDOPHSA-DCAQKATOSA-N 0.000 description 1
- CDGABSWLRMECHC-IHRRRGAJSA-N Pro-Lys-His Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O CDGABSWLRMECHC-IHRRRGAJSA-N 0.000 description 1
- IWIANZLCJVYEFX-RYUDHWBXSA-N Pro-Phe Chemical compound C([C@@H](C(=O)O)NC(=O)[C@H]1NCCC1)C1=CC=CC=C1 IWIANZLCJVYEFX-RYUDHWBXSA-N 0.000 description 1
- KBUAPZAZPWNYSW-SRVKXCTJSA-N Pro-Pro-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 KBUAPZAZPWNYSW-SRVKXCTJSA-N 0.000 description 1
- SEZGGSHLMROBFX-CIUDSAMLSA-N Pro-Ser-Gln Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(O)=O SEZGGSHLMROBFX-CIUDSAMLSA-N 0.000 description 1
- FNGOXVQBBCMFKV-CIUDSAMLSA-N Pro-Ser-Glu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O FNGOXVQBBCMFKV-CIUDSAMLSA-N 0.000 description 1
- QDDJNKWPTJHROJ-UFYCRDLUSA-N Pro-Tyr-Tyr Chemical compound C([C@@H](C(=O)O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H]1NCCC1)C1=CC=C(O)C=C1 QDDJNKWPTJHROJ-UFYCRDLUSA-N 0.000 description 1
- 108010003201 RGH 0205 Proteins 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 208000007014 Retinitis pigmentosa Diseases 0.000 description 1
- 241000978776 Senegalia senegal Species 0.000 description 1
- BTKUIVBNGBFTTP-WHFBIAKZSA-N Ser-Ala-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)NCC(O)=O BTKUIVBNGBFTTP-WHFBIAKZSA-N 0.000 description 1
- GXXTUIUYTWGPMV-FXQIFTODSA-N Ser-Arg-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(O)=O GXXTUIUYTWGPMV-FXQIFTODSA-N 0.000 description 1
- VQBLHWSPVYYZTB-DCAQKATOSA-N Ser-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CO)N VQBLHWSPVYYZTB-DCAQKATOSA-N 0.000 description 1
- QFBNNYNWKYKVJO-DCAQKATOSA-N Ser-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)CO)CCCN=C(N)N QFBNNYNWKYKVJO-DCAQKATOSA-N 0.000 description 1
- ZXLUWXWISXIFIX-ACZMJKKPSA-N Ser-Asn-Glu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O ZXLUWXWISXIFIX-ACZMJKKPSA-N 0.000 description 1
- BNFVPSRLHHPQKS-WHFBIAKZSA-N Ser-Asp-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O BNFVPSRLHHPQKS-WHFBIAKZSA-N 0.000 description 1
- BGOWRLSWJCVYAQ-CIUDSAMLSA-N Ser-Asp-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O BGOWRLSWJCVYAQ-CIUDSAMLSA-N 0.000 description 1
- OLIJLNWFEQEFDM-SRVKXCTJSA-N Ser-Asp-Phe Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 OLIJLNWFEQEFDM-SRVKXCTJSA-N 0.000 description 1
- MMAPOBOTRUVNKJ-ZLUOBGJFSA-N Ser-Asp-Ser Chemical compound C([C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CO)N)C(=O)O MMAPOBOTRUVNKJ-ZLUOBGJFSA-N 0.000 description 1
- UCOYFSCEIWQYNL-FXQIFTODSA-N Ser-Cys-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCSC)C(O)=O UCOYFSCEIWQYNL-FXQIFTODSA-N 0.000 description 1
- OJPHFSOMBZKQKQ-GUBZILKMSA-N Ser-Gln-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](N)CO OJPHFSOMBZKQKQ-GUBZILKMSA-N 0.000 description 1
- LALNXSXEYFUUDD-GUBZILKMSA-N Ser-Glu-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O LALNXSXEYFUUDD-GUBZILKMSA-N 0.000 description 1
- DSGYZICNAMEJOC-AVGNSLFASA-N Ser-Glu-Phe Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O DSGYZICNAMEJOC-AVGNSLFASA-N 0.000 description 1
- SNVIOQXAHVORQM-WDSKDSINSA-N Ser-Gly-Gln Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(O)=O SNVIOQXAHVORQM-WDSKDSINSA-N 0.000 description 1
- SFTZWNJFZYOLBD-ZDLURKLDSA-N Ser-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CO SFTZWNJFZYOLBD-ZDLURKLDSA-N 0.000 description 1
- WEQAYODCJHZSJZ-KKUMJFAQSA-N Ser-His-Tyr Chemical compound C([C@H](NC(=O)[C@H](CO)N)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CN=CN1 WEQAYODCJHZSJZ-KKUMJFAQSA-N 0.000 description 1
- JIPVNVNKXJLFJF-BJDJZHNGSA-N Ser-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CO)N JIPVNVNKXJLFJF-BJDJZHNGSA-N 0.000 description 1
- ZOPISOXXPQNOCO-SVSWQMSJSA-N Ser-Ile-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CO)N ZOPISOXXPQNOCO-SVSWQMSJSA-N 0.000 description 1
- MUJQWSAWLLRJCE-KATARQTJSA-N Ser-Leu-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O MUJQWSAWLLRJCE-KATARQTJSA-N 0.000 description 1
- CRJZZXMAADSBBQ-SRVKXCTJSA-N Ser-Lys-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CO CRJZZXMAADSBBQ-SRVKXCTJSA-N 0.000 description 1
- BUYHXYIUQUBEQP-AVGNSLFASA-N Ser-Phe-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CO)N BUYHXYIUQUBEQP-AVGNSLFASA-N 0.000 description 1
- UPLYXVPQLJVWMM-KKUMJFAQSA-N Ser-Phe-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O UPLYXVPQLJVWMM-KKUMJFAQSA-N 0.000 description 1
- ADJDNJCSPNFFPI-FXQIFTODSA-N Ser-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CO ADJDNJCSPNFFPI-FXQIFTODSA-N 0.000 description 1
- FKYWFUYPVKLJLP-DCAQKATOSA-N Ser-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CO FKYWFUYPVKLJLP-DCAQKATOSA-N 0.000 description 1
- FLONGDPORFIVQW-XGEHTFHBSA-N Ser-Pro-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CO FLONGDPORFIVQW-XGEHTFHBSA-N 0.000 description 1
- SRSPTFBENMJHMR-WHFBIAKZSA-N Ser-Ser-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O SRSPTFBENMJHMR-WHFBIAKZSA-N 0.000 description 1
- CUXJENOFJXOSOZ-BIIVOSGPSA-N Ser-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)[C@H](CO)N)C(=O)O CUXJENOFJXOSOZ-BIIVOSGPSA-N 0.000 description 1
- WUXCHQZLUHBSDJ-LKXGYXEUSA-N Ser-Thr-Asp Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CC(O)=O)C(O)=O WUXCHQZLUHBSDJ-LKXGYXEUSA-N 0.000 description 1
- YXEYTHXDRDAIOJ-CWRNSKLLSA-N Ser-Trp-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CNC3=CC=CC=C32)NC(=O)[C@H](CO)N)C(=O)O YXEYTHXDRDAIOJ-CWRNSKLLSA-N 0.000 description 1
- FHXGMDRKJHKLKW-QWRGUYRKSA-N Ser-Tyr-Gly Chemical compound OC[C@H](N)C(=O)N[C@H](C(=O)NCC(O)=O)CC1=CC=C(O)C=C1 FHXGMDRKJHKLKW-QWRGUYRKSA-N 0.000 description 1
- BEBVVQPDSHHWQL-NRPADANISA-N Ser-Val-Glu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O BEBVVQPDSHHWQL-NRPADANISA-N 0.000 description 1
- LGIMRDKGABDMBN-DCAQKATOSA-N Ser-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CO)N LGIMRDKGABDMBN-DCAQKATOSA-N 0.000 description 1
- HSWXBJCBYSWBPT-GUBZILKMSA-N Ser-Val-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CO)C(C)C)C(O)=O HSWXBJCBYSWBPT-GUBZILKMSA-N 0.000 description 1
- VMHLLURERBWHNL-UHFFFAOYSA-M Sodium acetate Chemical compound [Na+].CC([O-])=O VMHLLURERBWHNL-UHFFFAOYSA-M 0.000 description 1
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 1
- 108091027544 Subgenomic mRNA Proteins 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- ZUXQFMVPAYGPFJ-JXUBOQSCSA-N Thr-Ala-Lys Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN ZUXQFMVPAYGPFJ-JXUBOQSCSA-N 0.000 description 1
- VFEHSAJCWWHDBH-RHYQMDGZSA-N Thr-Arg-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O VFEHSAJCWWHDBH-RHYQMDGZSA-N 0.000 description 1
- JNQZPAWOPBZGIX-RCWTZXSCSA-N Thr-Arg-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)[C@@H](C)O)CCCN=C(N)N JNQZPAWOPBZGIX-RCWTZXSCSA-N 0.000 description 1
- IRKWVRSEQFTGGV-VEVYYDQMSA-N Thr-Asn-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O IRKWVRSEQFTGGV-VEVYYDQMSA-N 0.000 description 1
- QILPDQCTQZDHFM-HJGDQZAQSA-N Thr-Gln-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O QILPDQCTQZDHFM-HJGDQZAQSA-N 0.000 description 1
- RKDFEMGVMMYYNG-WDCWCFNPSA-N Thr-Gln-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O RKDFEMGVMMYYNG-WDCWCFNPSA-N 0.000 description 1
- LGNBRHZANHMZHK-NUMRIWBASA-N Thr-Glu-Asp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N)O LGNBRHZANHMZHK-NUMRIWBASA-N 0.000 description 1
- JMGJDTNUMAZNLX-RWRJDSDZSA-N Thr-Glu-Ile Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JMGJDTNUMAZNLX-RWRJDSDZSA-N 0.000 description 1
- BNGDYRRHRGOPHX-IFFSRLJSSA-N Thr-Glu-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)[C@@H](C)O)C(O)=O BNGDYRRHRGOPHX-IFFSRLJSSA-N 0.000 description 1
- AQAMPXBRJJWPNI-JHEQGTHGSA-N Thr-Gly-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O AQAMPXBRJJWPNI-JHEQGTHGSA-N 0.000 description 1
- XPNSAQMEAVSQRD-FBCQKBJTSA-N Thr-Gly-Gly Chemical compound C[C@@H](O)[C@H](N)C(=O)NCC(=O)NCC(O)=O XPNSAQMEAVSQRD-FBCQKBJTSA-N 0.000 description 1
- YSXYEJWDHBCTDJ-DVJZZOLTSA-N Thr-Gly-Trp Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N)O YSXYEJWDHBCTDJ-DVJZZOLTSA-N 0.000 description 1
- CRZNCABIJLRFKZ-IUKAMOBKSA-N Thr-Ile-Asp Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N CRZNCABIJLRFKZ-IUKAMOBKSA-N 0.000 description 1
- JRAUIKJSEAKTGD-TUBUOCAGSA-N Thr-Ile-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N JRAUIKJSEAKTGD-TUBUOCAGSA-N 0.000 description 1
- GMXIJHCBTZDAPD-QPHKQPEJSA-N Thr-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N GMXIJHCBTZDAPD-QPHKQPEJSA-N 0.000 description 1
- BVOVIGCHYNFJBZ-JXUBOQSCSA-N Thr-Leu-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O BVOVIGCHYNFJBZ-JXUBOQSCSA-N 0.000 description 1
- MEJHFIOYJHTWMK-VOAKCMCISA-N Thr-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)[C@@H](C)O MEJHFIOYJHTWMK-VOAKCMCISA-N 0.000 description 1
- MECLEFZMPPOEAC-VOAKCMCISA-N Thr-Leu-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)O)N)O MECLEFZMPPOEAC-VOAKCMCISA-N 0.000 description 1
- SCSVNSNWUTYSFO-WDCWCFNPSA-N Thr-Lys-Glu Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O SCSVNSNWUTYSFO-WDCWCFNPSA-N 0.000 description 1
- KKPOGALELPLJTL-MEYUZBJRSA-N Thr-Lys-Tyr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 KKPOGALELPLJTL-MEYUZBJRSA-N 0.000 description 1
- WRQLCVIALDUQEQ-UNQGMJICSA-N Thr-Phe-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O WRQLCVIALDUQEQ-UNQGMJICSA-N 0.000 description 1
- WYLAVUAWOUVUCA-XVSYOHENSA-N Thr-Phe-Asp Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O WYLAVUAWOUVUCA-XVSYOHENSA-N 0.000 description 1
- WTMPKZWHRCMMMT-KZVJFYERSA-N Thr-Pro-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O WTMPKZWHRCMMMT-KZVJFYERSA-N 0.000 description 1
- IWAVRIPRTCJAQO-HSHDSVGOSA-N Thr-Pro-Trp Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O IWAVRIPRTCJAQO-HSHDSVGOSA-N 0.000 description 1
- MFMGPEKYBXFIRF-SUSMZKCASA-N Thr-Thr-Gln Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(O)=O MFMGPEKYBXFIRF-SUSMZKCASA-N 0.000 description 1
- ABCLYRRGTZNIFU-BWAGICSOSA-N Thr-Tyr-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N)O ABCLYRRGTZNIFU-BWAGICSOSA-N 0.000 description 1
- CYCGARJWIQWPQM-YJRXYDGGSA-N Thr-Tyr-Ser Chemical compound C[C@@H](O)[C@H]([NH3+])C(=O)N[C@H](C(=O)N[C@@H](CO)C([O-])=O)CC1=CC=C(O)C=C1 CYCGARJWIQWPQM-YJRXYDGGSA-N 0.000 description 1
- PWONLXBUSVIZPH-RHYQMDGZSA-N Thr-Val-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(=O)O)N)O PWONLXBUSVIZPH-RHYQMDGZSA-N 0.000 description 1
- NWQCKAPDGQMZQN-IHPCNDPISA-N Trp-Lys-Leu Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O NWQCKAPDGQMZQN-IHPCNDPISA-N 0.000 description 1
- DLZKEQQWXODGGZ-KWQFWETISA-N Tyr-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 DLZKEQQWXODGGZ-KWQFWETISA-N 0.000 description 1
- AYPAIRCDLARHLM-KKUMJFAQSA-N Tyr-Asn-Lys Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCCN)C(=O)O)N)O AYPAIRCDLARHLM-KKUMJFAQSA-N 0.000 description 1
- NSTPFWRAIDTNGH-BZSNNMDCSA-N Tyr-Asn-Tyr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O NSTPFWRAIDTNGH-BZSNNMDCSA-N 0.000 description 1
- UABYBEBXFFNCIR-YDHLFZDLSA-N Tyr-Asp-Val Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O UABYBEBXFFNCIR-YDHLFZDLSA-N 0.000 description 1
- RYSNTWVRSLCAJZ-RYUDHWBXSA-N Tyr-Gln-Gly Chemical compound OC(=O)CNC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 RYSNTWVRSLCAJZ-RYUDHWBXSA-N 0.000 description 1
- KIJLSRYAUGGZIN-CFMVVWHZSA-N Tyr-Ile-Asp Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O KIJLSRYAUGGZIN-CFMVVWHZSA-N 0.000 description 1
- BSCBBPKDVOZICB-KKUMJFAQSA-N Tyr-Leu-Asp Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O BSCBBPKDVOZICB-KKUMJFAQSA-N 0.000 description 1
- NKUGCYDFQKFVOJ-JYJNAYRXSA-N Tyr-Leu-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 NKUGCYDFQKFVOJ-JYJNAYRXSA-N 0.000 description 1
- JLKVWTICWVWGSK-JYJNAYRXSA-N Tyr-Lys-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 JLKVWTICWVWGSK-JYJNAYRXSA-N 0.000 description 1
- PMHLLBKTDHQMCY-ULQDDVLXSA-N Tyr-Lys-Val Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O PMHLLBKTDHQMCY-ULQDDVLXSA-N 0.000 description 1
- JXGUUJMPCRXMSO-HJOGWXRNSA-N Tyr-Phe-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=C(O)C=C1 JXGUUJMPCRXMSO-HJOGWXRNSA-N 0.000 description 1
- XGZBEGGGAUQBMB-KJEVXHAQSA-N Tyr-Pro-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CC2=CC=C(C=C2)O)N)O XGZBEGGGAUQBMB-KJEVXHAQSA-N 0.000 description 1
- SOAUMCDLIUGXJJ-SRVKXCTJSA-N Tyr-Ser-Asn Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O SOAUMCDLIUGXJJ-SRVKXCTJSA-N 0.000 description 1
- MQGGXGKQSVEQHR-KKUMJFAQSA-N Tyr-Ser-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 MQGGXGKQSVEQHR-KKUMJFAQSA-N 0.000 description 1
- WQOHKVRQDLNDIL-YJRXYDGGSA-N Tyr-Thr-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O WQOHKVRQDLNDIL-YJRXYDGGSA-N 0.000 description 1
- SQUMHUZLJDUROQ-YDHLFZDLSA-N Tyr-Val-Asp Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O SQUMHUZLJDUROQ-YDHLFZDLSA-N 0.000 description 1
- ABSXSJZNRAQDDI-KJEVXHAQSA-N Tyr-Val-Thr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ABSXSJZNRAQDDI-KJEVXHAQSA-N 0.000 description 1
- 108091023045 Untranslated Region Proteins 0.000 description 1
- 108010064997 VPY tripeptide Proteins 0.000 description 1
- RUCNAYOMFXRIKJ-DCAQKATOSA-N Val-Ala-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN RUCNAYOMFXRIKJ-DCAQKATOSA-N 0.000 description 1
- ZLFHAAGHGQBQQN-GUBZILKMSA-N Val-Ala-Pro Natural products CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O ZLFHAAGHGQBQQN-GUBZILKMSA-N 0.000 description 1
- WGHVMKFREWGCGR-SRVKXCTJSA-N Val-Arg-Arg Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N WGHVMKFREWGCGR-SRVKXCTJSA-N 0.000 description 1
- PAPWZOJOLKZEFR-AVGNSLFASA-N Val-Arg-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(=O)O)N PAPWZOJOLKZEFR-AVGNSLFASA-N 0.000 description 1
- PVPAOIGJYHVWBT-KKHAAJSZSA-N Val-Asn-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](C(C)C)N)O PVPAOIGJYHVWBT-KKHAAJSZSA-N 0.000 description 1
- HZYOWMGWKKRMBZ-BYULHYEWSA-N Val-Asp-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N HZYOWMGWKKRMBZ-BYULHYEWSA-N 0.000 description 1
- VLOYGOZDPGYWFO-LAEOZQHASA-N Val-Asp-Glu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O VLOYGOZDPGYWFO-LAEOZQHASA-N 0.000 description 1
- BMGOFDMKDVVGJG-NHCYSSNCSA-N Val-Asp-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N BMGOFDMKDVVGJG-NHCYSSNCSA-N 0.000 description 1
- NYTKXWLZSNRILS-IFFSRLJSSA-N Val-Gln-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](C(C)C)N)O NYTKXWLZSNRILS-IFFSRLJSSA-N 0.000 description 1
- CVIXTAITYJQMPE-LAEOZQHASA-N Val-Glu-Asn Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O CVIXTAITYJQMPE-LAEOZQHASA-N 0.000 description 1
- GBESYURLQOYWLU-LAEOZQHASA-N Val-Glu-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N GBESYURLQOYWLU-LAEOZQHASA-N 0.000 description 1
- VLDMQVZZWDOKQF-AUTRQRHGSA-N Val-Glu-Gln Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N VLDMQVZZWDOKQF-AUTRQRHGSA-N 0.000 description 1
- ZXAGTABZUOMUDO-GVXVVHGQSA-N Val-Glu-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N ZXAGTABZUOMUDO-GVXVVHGQSA-N 0.000 description 1
- WDIGUPHXPBMODF-UMNHJUIQSA-N Val-Glu-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N WDIGUPHXPBMODF-UMNHJUIQSA-N 0.000 description 1
- XWYUBUYQMOUFRQ-IFFSRLJSSA-N Val-Glu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](C(C)C)N)O XWYUBUYQMOUFRQ-IFFSRLJSSA-N 0.000 description 1
- YTPLVNUZZOBFFC-SCZZXKLOSA-N Val-Gly-Pro Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N1CCC[C@@H]1C(O)=O YTPLVNUZZOBFFC-SCZZXKLOSA-N 0.000 description 1
- LAYSXAOGWHKNED-XPUUQOCRSA-N Val-Gly-Ser Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O LAYSXAOGWHKNED-XPUUQOCRSA-N 0.000 description 1
- KZKMBGXCNLPYKD-YEPSODPASA-N Val-Gly-Thr Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(O)=O KZKMBGXCNLPYKD-YEPSODPASA-N 0.000 description 1
- APQIVBCUIUDSMB-OSUNSFLBSA-N Val-Ile-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](C(C)C)N APQIVBCUIUDSMB-OSUNSFLBSA-N 0.000 description 1
- AGXGCFSECFQMKB-NHCYSSNCSA-N Val-Leu-Asp Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](C(C)C)N AGXGCFSECFQMKB-NHCYSSNCSA-N 0.000 description 1
- WBAJDGWKRIHOAC-GVXVVHGQSA-N Val-Lys-Gln Chemical compound [H]N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(O)=O WBAJDGWKRIHOAC-GVXVVHGQSA-N 0.000 description 1
- ZRSZTKTVPNSUNA-IHRRRGAJSA-N Val-Lys-Leu Chemical compound CC(C)C[C@H](NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)C(C)C)C(O)=O ZRSZTKTVPNSUNA-IHRRRGAJSA-N 0.000 description 1
- JVGHIFMSFBZDHH-WPRPVWTQSA-N Val-Met-Gly Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)NCC(=O)O)N JVGHIFMSFBZDHH-WPRPVWTQSA-N 0.000 description 1
- UEPLNXPLHJUYPT-AVGNSLFASA-N Val-Met-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCCN)C(O)=O UEPLNXPLHJUYPT-AVGNSLFASA-N 0.000 description 1
- GQMNEJMFMCJJTD-NHCYSSNCSA-N Val-Pro-Gln Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O GQMNEJMFMCJJTD-NHCYSSNCSA-N 0.000 description 1
- SSYBNWFXCFNRFN-GUBZILKMSA-N Val-Pro-Ser Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O SSYBNWFXCFNRFN-GUBZILKMSA-N 0.000 description 1
- QWCZXKIFPWPQHR-JYJNAYRXSA-N Val-Pro-Tyr Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 QWCZXKIFPWPQHR-JYJNAYRXSA-N 0.000 description 1
- BZDGLJPROOOUOZ-XGEHTFHBSA-N Val-Thr-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](C(C)C)N)O BZDGLJPROOOUOZ-XGEHTFHBSA-N 0.000 description 1
- MIAZWUMFUURQNP-YDHLFZDLSA-N Val-Tyr-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N MIAZWUMFUURQNP-YDHLFZDLSA-N 0.000 description 1
- VTIAEOKFUJJBTC-YDHLFZDLSA-N Val-Tyr-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N VTIAEOKFUJJBTC-YDHLFZDLSA-N 0.000 description 1
- GUIYPEKUEMQBIK-JSGCOSHPSA-N Val-Tyr-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](Cc1ccc(O)cc1)C(=O)NCC(O)=O GUIYPEKUEMQBIK-JSGCOSHPSA-N 0.000 description 1
- 102100026383 Vasopressin-neurophysin 2-copeptin Human genes 0.000 description 1
- 108020005202 Viral DNA Proteins 0.000 description 1
- 235000010489 acacia gum Nutrition 0.000 description 1
- 239000000205 acacia gum Substances 0.000 description 1
- 108010081404 acein-2 Proteins 0.000 description 1
- 101150092805 actc1 gene Proteins 0.000 description 1
- 101150063416 add gene Proteins 0.000 description 1
- 230000000996 additive effect Effects 0.000 description 1
- 210000001789 adipocyte Anatomy 0.000 description 1
- 201000006966 adult T-cell leukemia Diseases 0.000 description 1
- 238000000246 agarose gel electrophoresis Methods 0.000 description 1
- 108010008685 alanyl-glutamyl-aspartic acid Proteins 0.000 description 1
- 108010024078 alanyl-glycyl-serine Proteins 0.000 description 1
- 108010086434 alanyl-seryl-glycine Proteins 0.000 description 1
- 108010044940 alanylglutamine Proteins 0.000 description 1
- 108010047495 alanylglycine Proteins 0.000 description 1
- 108010070944 alanylhistidine Proteins 0.000 description 1
- 108010028144 alpha-Glucosidases Proteins 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 230000000840 anti-viral effect Effects 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 239000000427 antigen Substances 0.000 description 1
- 210000000612 antigen-presenting cell Anatomy 0.000 description 1
- 102000036639 antigens Human genes 0.000 description 1
- 108091007433 antigens Proteins 0.000 description 1
- 108010008355 arginyl-glutamine Proteins 0.000 description 1
- 108010001271 arginyl-glutamyl-arginine Proteins 0.000 description 1
- 108010043240 arginyl-leucyl-glycine Proteins 0.000 description 1
- 108010059459 arginyl-threonyl-phenylalanine Proteins 0.000 description 1
- 108010084758 arginyl-tyrosyl-aspartic acid Proteins 0.000 description 1
- 239000011668 ascorbic acid Substances 0.000 description 1
- 235000010323 ascorbic acid Nutrition 0.000 description 1
- 229960005070 ascorbic acid Drugs 0.000 description 1
- 108010069205 aspartyl-phenylalanine Proteins 0.000 description 1
- 108010093581 aspartyl-proline Proteins 0.000 description 1
- 108010038633 aspartylglutamate Proteins 0.000 description 1
- 108010047857 aspartylglycine Proteins 0.000 description 1
- 108010068265 aspartyltyrosine Proteins 0.000 description 1
- 238000003556 assay Methods 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 229920001400 block copolymer Polymers 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- UDSAIICHUKSCKT-UHFFFAOYSA-N bromophenol blue Chemical compound C1=C(Br)C(O)=C(Br)C=C1C1(C=2C=C(Br)C(O)=C(Br)C=2)C2=CC=CC=C2S(=O)(=O)O1 UDSAIICHUKSCKT-UHFFFAOYSA-N 0.000 description 1
- 229910000019 calcium carbonate Inorganic materials 0.000 description 1
- 235000010216 calcium carbonate Nutrition 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 230000036952 cancer formation Effects 0.000 description 1
- 229940022399 cancer vaccine Drugs 0.000 description 1
- 238000009566 cancer vaccine Methods 0.000 description 1
- 230000000711 cancerogenic effect Effects 0.000 description 1
- 239000002775 capsule Substances 0.000 description 1
- 231100000504 carcinogenesis Toxicity 0.000 description 1
- 231100000315 carcinogenic Toxicity 0.000 description 1
- 210000004413 cardiac myocyte Anatomy 0.000 description 1
- 125000002091 cationic group Chemical group 0.000 description 1
- 229920006317 cationic polymer Polymers 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 239000003153 chemical reaction reagent Substances 0.000 description 1
- 235000013330 chicken meat Nutrition 0.000 description 1
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 1
- 210000001612 chondrocyte Anatomy 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 238000000975 co-precipitation Methods 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 238000011109 contamination Methods 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 210000000399 corneal endothelial cell Anatomy 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 201000010064 diabetes insipidus Diseases 0.000 description 1
- UGMCXQCYOVCMTB-UHFFFAOYSA-K dihydroxy(stearato)aluminium Chemical compound CCCCCCCCCCCCCCCCCC(=O)O[Al](O)O UGMCXQCYOVCMTB-UHFFFAOYSA-K 0.000 description 1
- FSXRLASFHBWESK-UHFFFAOYSA-N dipeptide phenylalanyl-tyrosine Natural products C=1C=C(O)C=CC=1CC(C(O)=O)NC(=O)C(N)CC1=CC=CC=C1 FSXRLASFHBWESK-UHFFFAOYSA-N 0.000 description 1
- UKWLRLAKGMZXJC-QIECWBMSSA-L disodium;[4-chloro-3-[(3r,5s)-1-chloro-3'-methoxyspiro[adamantane-4,4'-dioxetane]-3'-yl]phenyl] phosphate Chemical compound [Na+].[Na+].O1OC2([C@@H]3CC4C[C@H]2CC(Cl)(C4)C3)C1(OC)C1=CC(OP([O-])([O-])=O)=CC=C1Cl UKWLRLAKGMZXJC-QIECWBMSSA-L 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 210000001671 embryonic stem cell Anatomy 0.000 description 1
- 210000002889 endothelial cell Anatomy 0.000 description 1
- 210000005175 epidermal keratinocyte Anatomy 0.000 description 1
- 210000004920 epithelial cell of skin Anatomy 0.000 description 1
- 210000000981 epithelium Anatomy 0.000 description 1
- ZMMJGEGLRURXTF-UHFFFAOYSA-N ethidium bromide Chemical compound [Br-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CC)=C1C1=CC=CC=C1 ZMMJGEGLRURXTF-UHFFFAOYSA-N 0.000 description 1
- 229960005542 ethidium bromide Drugs 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 239000012091 fetal bovine serum Substances 0.000 description 1
- 210000002950 fibroblast Anatomy 0.000 description 1
- 210000000185 follicular epithelial cell Anatomy 0.000 description 1
- 239000012634 fragment Substances 0.000 description 1
- 239000000499 gel Substances 0.000 description 1
- 238000001415 gene therapy Methods 0.000 description 1
- 210000004602 germ cell Anatomy 0.000 description 1
- 108010013768 glutamyl-aspartyl-proline Proteins 0.000 description 1
- 108010079547 glutamylmethionine Proteins 0.000 description 1
- 235000011187 glycerol Nutrition 0.000 description 1
- HPAIKDPJURGQLN-UHFFFAOYSA-N glycyl-L-histidyl-L-phenylalanine Natural products C=1C=CC=CC=1CC(C(O)=O)NC(=O)C(NC(=O)CN)CC1=CN=CN1 HPAIKDPJURGQLN-UHFFFAOYSA-N 0.000 description 1
- XBGGUPMXALFZOT-UHFFFAOYSA-N glycyl-L-tyrosine hemihydrate Natural products NCC(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 XBGGUPMXALFZOT-UHFFFAOYSA-N 0.000 description 1
- 108010000434 glycyl-alanyl-leucine Proteins 0.000 description 1
- 108010026364 glycyl-glycyl-leucine Proteins 0.000 description 1
- 108010066198 glycyl-leucyl-phenylalanine Proteins 0.000 description 1
- 108010050475 glycyl-leucyl-tyrosine Proteins 0.000 description 1
- 108010048994 glycyl-tyrosyl-alanine Proteins 0.000 description 1
- 108010010147 glycylglutamine Proteins 0.000 description 1
- 108010087823 glycyltyrosine Proteins 0.000 description 1
- 108010037850 glycylvaline Proteins 0.000 description 1
- 239000008187 granular material Substances 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- ZJYYHGLJYGJLLN-UHFFFAOYSA-N guanidinium thiocyanate Chemical compound SC#N.NC(N)=N ZJYYHGLJYGJLLN-UHFFFAOYSA-N 0.000 description 1
- 238000003505 heat denaturation Methods 0.000 description 1
- 108010028295 histidylhistidine Proteins 0.000 description 1
- 108010085325 histidylproline Proteins 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 238000009396 hybridization Methods 0.000 description 1
- 229920001477 hydrophilic polymer Polymers 0.000 description 1
- 229920003063 hydroxymethyl cellulose Polymers 0.000 description 1
- 229940031574 hydroxymethyl cellulose Drugs 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 239000007924 injection Substances 0.000 description 1
- 238000002347 injection Methods 0.000 description 1
- 230000000968 intestinal effect Effects 0.000 description 1
- 238000001361 intraarterial administration Methods 0.000 description 1
- 230000010039 intracellular degradation Effects 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 239000007927 intramuscular injection Substances 0.000 description 1
- 239000007928 intraperitoneal injection Substances 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 238000010253 intravenous injection Methods 0.000 description 1
- 229920000831 ionic polymer Polymers 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 108010027338 isoleucylcysteine Proteins 0.000 description 1
- 210000002510 keratinocyte Anatomy 0.000 description 1
- 238000002372 labelling Methods 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 210000001821 langerhans cell Anatomy 0.000 description 1
- 108010044311 leucyl-glycyl-glycine Proteins 0.000 description 1
- 108010030617 leucyl-phenylalanyl-valine Proteins 0.000 description 1
- 108010073472 leucyl-prolyl-proline Proteins 0.000 description 1
- 108010000761 leucylarginine Proteins 0.000 description 1
- 108010012058 leucyltyrosine Proteins 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- XIXADJRWDQXREU-UHFFFAOYSA-M lithium acetate Chemical compound [Li+].CC([O-])=O XIXADJRWDQXREU-UHFFFAOYSA-M 0.000 description 1
- 210000005229 liver cell Anatomy 0.000 description 1
- 244000144972 livestock Species 0.000 description 1
- HWYHZTIRURJOHG-UHFFFAOYSA-N luminol Chemical compound O=C1NNC(=O)C2=C1C(N)=CC=C2 HWYHZTIRURJOHG-UHFFFAOYSA-N 0.000 description 1
- 108010003700 lysyl aspartic acid Proteins 0.000 description 1
- 108010044348 lysyl-glutamyl-aspartic acid Proteins 0.000 description 1
- 108010045397 lysyl-tyrosyl-lysine Proteins 0.000 description 1
- 108010009298 lysylglutamic acid Proteins 0.000 description 1
- 108010038320 lysylphenylalanine Proteins 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 239000000463 material Substances 0.000 description 1
- 239000002609 medium Substances 0.000 description 1
- 210000003584 mesangial cell Anatomy 0.000 description 1
- 108010005942 methionylglycine Proteins 0.000 description 1
- 229920000609 methyl cellulose Polymers 0.000 description 1
- 239000004292 methyl p-hydroxybenzoate Substances 0.000 description 1
- 235000010270 methyl p-hydroxybenzoate Nutrition 0.000 description 1
- 239000001923 methylcellulose Substances 0.000 description 1
- 235000010981 methylcellulose Nutrition 0.000 description 1
- 229960002216 methylparaben Drugs 0.000 description 1
- 239000000693 micelle Substances 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 239000011859 microparticle Substances 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 1
- 210000000214 mouth Anatomy 0.000 description 1
- 230000035772 mutation Effects 0.000 description 1
- 210000004498 neuroglial cell Anatomy 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- 239000002858 neurotransmitter agent Substances 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 125000003835 nucleoside group Chemical group 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 210000002997 osteoclast Anatomy 0.000 description 1
- 210000004409 osteocyte Anatomy 0.000 description 1
- 210000004738 parenchymal cell Anatomy 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 230000037361 pathway Effects 0.000 description 1
- 108010086507 peptide-chain-release factor 3 Proteins 0.000 description 1
- 102000013415 peroxidase activity proteins Human genes 0.000 description 1
- 108040007629 peroxidase activity proteins Proteins 0.000 description 1
- 229960003742 phenol Drugs 0.000 description 1
- 108010064486 phenylalanyl-leucyl-valine Proteins 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000008055 phosphate buffer solution Substances 0.000 description 1
- 150000003904 phospholipids Chemical class 0.000 description 1
- 239000002504 physiological saline solution Substances 0.000 description 1
- 108010025488 pinealon Proteins 0.000 description 1
- 102000028499 poly(A) binding Human genes 0.000 description 1
- 108091023021 poly(A) binding Proteins 0.000 description 1
- 229920005575 poly(amic acid) Polymers 0.000 description 1
- 229920000573 polyethylene Polymers 0.000 description 1
- 229920001223 polyethylene glycol Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 239000000843 powder Substances 0.000 description 1
- 108010020755 prolyl-glycyl-glycine Proteins 0.000 description 1
- 108010077112 prolyl-proline Proteins 0.000 description 1
- 108010087846 prolyl-prolyl-glycine Proteins 0.000 description 1
- 108010031719 prolyl-serine Proteins 0.000 description 1
- 108010070643 prolylglutamic acid Proteins 0.000 description 1
- 238000012797 qualification Methods 0.000 description 1
- 230000002829 reductive effect Effects 0.000 description 1
- 230000008672 reprogramming Effects 0.000 description 1
- 210000003705 ribosome Anatomy 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 239000012723 sample buffer Substances 0.000 description 1
- 210000002966 serum Anatomy 0.000 description 1
- 108010048818 seryl-histidine Proteins 0.000 description 1
- 108010069117 seryl-lysyl-aspartic acid Proteins 0.000 description 1
- 108010026333 seryl-proline Proteins 0.000 description 1
- 210000000329 smooth muscle myocyte Anatomy 0.000 description 1
- 239000001632 sodium acetate Substances 0.000 description 1
- 235000017281 sodium acetate Nutrition 0.000 description 1
- 235000010413 sodium alginate Nutrition 0.000 description 1
- 239000000661 sodium alginate Substances 0.000 description 1
- 229940005550 sodium alginate Drugs 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- 239000000243 solution Substances 0.000 description 1
- 210000004989 spleen cell Anatomy 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 239000000126 substance Substances 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 239000006188 syrup Substances 0.000 description 1
- 235000020357 syrup Nutrition 0.000 description 1
- 239000003826 tablet Substances 0.000 description 1
- 230000008685 targeting Effects 0.000 description 1
- 230000001225 therapeutic effect Effects 0.000 description 1
- 108010031491 threonyl-lysyl-glutamic acid Proteins 0.000 description 1
- 210000001519 tissue Anatomy 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- 239000001226 triphosphate Substances 0.000 description 1
- 235000011178 triphosphate Nutrition 0.000 description 1
- 108010038745 tryptophylglycine Proteins 0.000 description 1
- 210000004881 tumor cell Anatomy 0.000 description 1
- 108010012567 tyrosyl-glycyl-glycyl-phenylalanyl Proteins 0.000 description 1
- 108010051110 tyrosyl-lysine Proteins 0.000 description 1
- IBIDRSSEHFLGSD-UHFFFAOYSA-N valinyl-arginine Natural products CC(C)C(N)C(=O)NC(C(O)=O)CCCN=C(N)N IBIDRSSEHFLGSD-UHFFFAOYSA-N 0.000 description 1
- 210000003556 vascular endothelial cell Anatomy 0.000 description 1
- 210000003501 vero cell Anatomy 0.000 description 1
- 239000011782 vitamin Substances 0.000 description 1
- 229940088594 vitamin Drugs 0.000 description 1
- 229930003231 vitamin Natural products 0.000 description 1
- 235000013343 vitamin Nutrition 0.000 description 1
- 238000001262 western blot Methods 0.000 description 1
Images







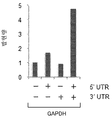







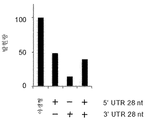












Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/67—General methods for enhancing the expression
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4717—Plasma globulins, lactoglobulin
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0008—Oxidoreductases (1.) acting on the aldehyde or oxo group of donors (1.2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01027—L-Lactate dehydrogenase (1.1.1.27)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y102/00—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2)
- C12Y102/01—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2) with NAD+ or NADP+ as acceptor (1.2.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y102/00—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2)
- C12Y102/01—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2) with NAD+ or NADP+ as acceptor (1.2.1)
- C12Y102/01012—Glyceraldehyde-3-phosphate dehydrogenase (phosphorylating) (1.2.1.12)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y102/00—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2)
- C12Y102/01—Oxidoreductases acting on the aldehyde or oxo group of donors (1.2) with NAD+ or NADP+ as acceptor (1.2.1)
- C12Y102/01009—Glyceraldehyde-3-phosphate dehydrogenase (NADP+) (1.2.1.9)
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Biophysics (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Toxicology (AREA)
- Gastroenterology & Hepatology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
mRNA의 번역 효율이 향상되는 기술을 제공한다. mRNA는 단백질을 코딩하는 mRNA의 5' 비번역 영역과, 상기 5' 비번역 영역에 대한 상보성이 40% 이상 80% 이하인 3' 비번역 영역을 갖춘다.
Description
본 발명은 인공 합성 mRNA 및 그 이용에 관한 것이다. 본 출원은 2019년 10월 17일에 출원된 일본 특허 출원 제2019-189929호에 기초한 우선권을 주장하는 것이며, 당해 특허 출원의 전체 내용은 참고로 원용된다.
지금까지 유전자 치료는 바이러스를 비롯한 DNA를 벡터로 하여 실시되어 왔지만, 게놈에 편입됨으로 인한 발암 등의 위험성이 큰 문제로서 남아 있었다. 한편, mRNA는 DNA와 달리 게놈으로의 삽입 등의 리스크가 없는 안전한 핵산 의약으로서 주목을 받고 있지만, RNA가 본래 가지는 불안정성과 번역 효율의 낮음이 결점으로서 지적되었다(예를 들면, 특허문헌 1 내지 특허문헌 3).
특허문헌 1에는, mRNA의 세포 내에서의 분해 기구를 해명함으로써, 이 기구의 발생을 억제하는 기술로서, 목적 유전자의 mRNA에, 2'-5'-올리고아데닐산 합성효소의 기능 억제 물질을 사용하는 방법이 개시되어 있다.
확실히, 특허문헌 1에 기재된 기술에 의하면, RNA가 본래 가지는 불안정성은 개선된다. 그러나, mRNA의 번역 효율의 향상에 대해서는 개선의 여지가 있었다.
이 때문에, 본 발명자들은 예의 연구를 진행함으로써, mRNA의 번역 효율을 향상시키는 방법을 발명하기에 이르렀다.
본 발명은 상기 과제를 해결하기 위해 이루어진 것으로, 이하의 형태로서 실현하는 것이 가능하다.
(1) 본 발명의 하나의 형태에 의하면, 인공 합성 mRNA가 제공된다. 이 인공 합성 mRNA는 단백질을 코딩하는 mRNA의 5' 비번역 영역과, 상기 5' 비번역 영역에 대한 상보성이 40% 이상 80% 이하인 3' 비번역 영역을 갖춘다.
(2) 상기의 인공 합성 mRNA로서, 상기 단백질은 글리세르알데히드-3-인산 데히드로게나제, β-글로빈과, RPS8와 LDHB로 이루어진 군으로부터 선택되어도 된다.
(3) 상기의 인공 합성 mRNA로서, 상기 3' 비번역 영역은 상기 5' 비번역 영역에 대한 상보성이 50% 이상 75% 이하이어도 된다.
(4) 본 발명의 다른 형태에 의하면, 상기 인공 합성 mRNA를 세포에 도입하는 공정을 포함하는 방법이 제공된다.
(5) 본 발명의 다른 형태에 의하면, 상기 인공 합성 mRNA가 도입된 세포가 제공된다.
(6) 본 발명의 다른 형태에 의하면, 인공 합성 mRNA의 제조 방법이 제공된다. 이 인공 합성 mRNA의 제조 방법은 단백질을 코딩하는 mRNA의 5' 비번역 영역과, 상기 5' 비번역 영역과의 상보성이 40% 이상 80% 이하인 3' 비번역 영역을 갖추는 인공 합성 mRNA를 제조하는 공정을 포함한다.
(7) 본 발명의 다른 형태에 의하면, 인공 합성 mRNA가 제공된다. 이 인공 합성 mRNA는 글리세르알데히드-3-인산 데히드로게나제를 코딩하는 mRNA의 5' 비번역 영역과, 글리세르알데히드-3-인산 데히드로게나제를 코딩하는 mRNA의 3' 비번역 영역을 갖춘다.
(8) 본 발명의 다른 형태에 의하면, 인공 합성 mRNA의 제조 방법이 제공된다. 이 인공 합성 mRNA의 제조 방법은 글리세르알데히드-3-인산 데히드로게나제를 코딩하는 mRNA의 5' 비번역 영역과, 글리세르알데히드-3-인산 데히드로게나제를 코딩하는 mRNA의 3' 비번역 영역을 갖추는 인공 합성 mRNA를 제조하는 공정을 포함한다.
(9) 본 발명의 다른 형태에 의하면, 상기 (7)의 인공 합성 mRNA를 세포에 도입하는 공정을 포함하는 방법이 제공된다.
(10) 본 발명의 다른 형태에 의하면, 상기 (7)의 인공 합성 mRNA가 도입된 세포가 제공된다.
도 1은 제작한 인공 합성 mRNA를 나타내는 도면.
도 2는 인공 합성 mRNA로부터의 단백질 발현량을 측정한 결과를 나타내는 도면.
도 3은 도 1에 나타낸 인공 합성 mRNA의 안정성을 나타내는 도면.
도 4는 제작한 인공 합성 mRNA를 나타내는 도면.
도 5는 인공 합성 mRNA마다의 발현량을 나타내는 도면.
도 6은 인공 합성 mRNA를 신장 유래의 HEK293 세포에 도입한 경우의 인공 합성 mRNA마다의 발현량을 나타내는 도면.
도 7은 제작한 인공 합성 mRNA를 나타내는 도면.
도 8은 GAPDH의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 9는 LDHB의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 10은 ACAT2의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 11은 제작한 인공 합성 mRNA를 나타내는 도면.
도 12는 GAPDH의 5' UTR 서열의 결실의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 13은 제작한 인공 합성 mRNA를 나타내는 도면.
도 14는 GAPDH의 3' UTR 서열의 결실의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 15는 제작한 인공 합성 mRNA를 나타내는 도면.
도 16은 5' UTR 28nt와 3' UTR 28nt의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 17은 제작한 인공 합성 mRNA를 나타내는 도면.
도 18은 상보성에 기인하는 발현량의 차이를 나타내는 도면.
도 19는 제작한 인공 합성 mRNA를 나타내는 도면.
도 20은 상보성에 기인하는 발현량의 차이를 나타내는 도면.
도 21은 제작한 인공 합성 mRNA를 나타내는 도면.
도 22는 암 억제 유전자 p53을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 23은 얻어진 효과의 추정 메커니즘을 설명하기 위한 도면.
도 24는 게놈 편집 유전자 hCas9를 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 25는 게놈 편집 유전자 hCas9를 사용한 경우에 있어서의 게놈 편집량의 차이를 나타내는 도면.
도 26은 β-글로빈의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 27은 RPS8의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 28은 LDHB의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 2는 인공 합성 mRNA로부터의 단백질 발현량을 측정한 결과를 나타내는 도면.
도 3은 도 1에 나타낸 인공 합성 mRNA의 안정성을 나타내는 도면.
도 4는 제작한 인공 합성 mRNA를 나타내는 도면.
도 5는 인공 합성 mRNA마다의 발현량을 나타내는 도면.
도 6은 인공 합성 mRNA를 신장 유래의 HEK293 세포에 도입한 경우의 인공 합성 mRNA마다의 발현량을 나타내는 도면.
도 7은 제작한 인공 합성 mRNA를 나타내는 도면.
도 8은 GAPDH의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 9는 LDHB의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 10은 ACAT2의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 11은 제작한 인공 합성 mRNA를 나타내는 도면.
도 12는 GAPDH의 5' UTR 서열의 결실의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 13은 제작한 인공 합성 mRNA를 나타내는 도면.
도 14는 GAPDH의 3' UTR 서열의 결실의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 15는 제작한 인공 합성 mRNA를 나타내는 도면.
도 16은 5' UTR 28nt와 3' UTR 28nt의 유무에 기인하는 발현량의 차이를 나타내는 도면.
도 17은 제작한 인공 합성 mRNA를 나타내는 도면.
도 18은 상보성에 기인하는 발현량의 차이를 나타내는 도면.
도 19는 제작한 인공 합성 mRNA를 나타내는 도면.
도 20은 상보성에 기인하는 발현량의 차이를 나타내는 도면.
도 21은 제작한 인공 합성 mRNA를 나타내는 도면.
도 22는 암 억제 유전자 p53을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 23은 얻어진 효과의 추정 메커니즘을 설명하기 위한 도면.
도 24는 게놈 편집 유전자 hCas9를 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 25는 게놈 편집 유전자 hCas9를 사용한 경우에 있어서의 게놈 편집량의 차이를 나타내는 도면.
도 26은 β-글로빈의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 27은 RPS8의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
도 28은 LDHB의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면.
(발명을 실시하기 위한 형태)
본 명세서의 개시는 번역 효율이 높은 인공 합성 mRNA 및 그 이용에 관한 것이다. 또한, 인위적인 조작에 의해 조제된다는 특징을 나타냄과 아울러, 세포 내(내재성) mRNA와 구별할 목적으로, 본 발명의 mRNA를 「인공 합성 mRNA」라고 부르는 경우가 있다. 여기에서, 본 명세서에 있어서, 「mRNA」란 단백질에 번역될 수 있는 염기 서열 정보와 구조를 갖는 RNA를 나타낸다. 또한, 2개의 용어 「억제」와 「저해」는 그 의미하는 바가 중복되고, 종종 치환 가능하게 사용된다. 그래서, 본 명세서에서는, 전후의 문맥으로부터 구별이 특별히 필요한 경우를 제외하고, 통일하여 용어 「억제」를 사용한다. 「번역 효율이 높다」는 것은 번역량의 증대를 의미한다.
1. mRNA
본 발명의 mRNA는 목적 유전자의 코드 영역(목적 유전자의 발현 산물인 단백질을 코딩하는 영역)을 갖춘다. 「목적 유전자」란 본 발명의 mRNA를 이용하여 세포 내에서 발현시키는 유전자이다. 다양한 유전자를 목적 유전자로서 채용할 수 있다. 본 발명의 mRNA를 도입함으로써 목적 유전자를 발현시키는 세포를 본원에서 「표적 세포」라고도 부른다. 목적 유전자로서는, 예를 들면, 효소(예를 들면, 뉴클레아제(ZFN(징크핑거 뉴클레아제), TALEN(전사 활성제-유사 이펙터 뉴클레아제), CRISPR-Cas9 등), 사이토카인, 호르몬, 신경전달 물질 등의 유전자, 그 기능 저하(예를 들면, 변이에 의한 것)나 결손 등이 질환의 원인이 되는 유전자, 정상적으로 기능을 하고 있지만 그 발현의 증강이 요망되는 유전자, 표적 세포가 본래 갖지 않는 유전자로서 그것이 발현됨으로써 표적 세포의 생존, 유지 등에 유익한 유전자, 표적 세포에 작용하여, 표적 세포가 본래 가지는 기능을 높이는 단백질 또는 표적 세포가 본래 가지는 기능과는 다른 기능을 발휘시키는 단백질을 코딩하는 유전자, 표적 세포에는 작용하지 않고, 표적 세포로부터 분비되어 주위의 세포에 작용하는 단백질(예를 들면, 세포간 네트워크에 관여하는 단백질)을 코딩하는 유전자 등을 들 수 있다. 표적 세포 및 주위의 세포에 대해서는 실질적으로 작용하지 않는 단백질을 코딩하는 유전자도 목적 유전자가 될 수 있다. 이러한 유전자로서, 예를 들면, 의약품 등에 이용되는 단백질 등을 코딩하는 유전자(예를 들면, 인간 에리트로포이에틴 유전자, 인간 피브리노겐 유전자, 인간 혈청 알부민 유전자, 인간 락토페린 유전자, 인간 α-글루코시다제 유전자)를 들 수 있다. 이러한 유전자를 채용함으로써, 표적 세포 내에서 의약품 등으로서 이용 가능한 재조합 단백질을 생산시키는 것이 가능하다.
표적 세포는 특별히 한정되지 않고, 예를 들면, 각종 진핵 세포를 표적 세포로서 사용할 수 있다. 보다 구체적으로는, 표적 세포로서, 예를 들면, 포유동물(인간, 원숭이, 소, 말, 토끼, 마우스, 래트, 기니피그, 햄스터 등)의 각종 세포, 예를 들면, 심근 세포, 평활근 세포, 지방 세포, 섬유 아세포, 골 세포, 연골 세포, 파골 세포, 실질 세포, 표피각화 세포(케라티노사이트), 상피 세포(피부표피 세포, 각막상피 세포, 결막상피 세포, 구강점막상피, 모포상피 세포, 구강점막상피 세포, 기도점막상피 세포, 장관점막상피 세포 등), 내피 세포(각막내피 세포, 혈관내피 세포 등), 신경 세포, 글리아 세포, 비장 세포, 췌장 β세포, 메산기움 세포, 랑게르한스 세포, 간 세포, 이들의 전구 세포 또는 줄기세포, 혹은 인공 다능성 줄기세포(iPS 세포), 간엽계 줄기세포(MSC), 배성 줄기세포(ES 세포), 배성 생식세포(EG 세포), 배성 종양세포(EC 세포) ) 등을 사용할 수 있습니다. 또한, 표적 세포로서, 예를 들면, 계대 세포, 특정의 세포계보로 분화 유도된 세포, 주화 세포(예를 들면, HeLa 세포, CHO 세포, Vero 세포, HEK293 세포, HepG2 세포, COS-7 세포, NIH3T3 세포, Sf9 세포) 등을 사용할 수 있다.
생체로부터 분리된 상태의 표적 세포(즉, 단리된 표적 세포), 또는 생체를 구성한 상태의 표적 세포에 대하여 본 발명의 mRNA를 도입해도 된다. 따라서, In vitro, In vivo 및 Ex vivo의 임의의 환경하에서도 본 발명을 실시하는 것이 가능하다. 여기서 「단리된」이란 그 본래의 환경(예를 들면, 생체를 구성한 상태)로부터 취출된 상태에 있는 것을 나타낸다. 따라서 통상은 단리된 표적 세포는 배양 용기 내 또는 보존 용기 내에 존재하고, 그것으로의 in vitro에서의 인위적 조작이 가능하다. 구체적으로는, 생체로부터 분리되고, 생체 밖에서 배양 상태에 있는 세포(주화된 세포 포함)는 단리된 표적 세포로서의 적격을 갖는다. 또한, 상기의 의미에 있어서 단리된 상태에 있는 한, 조직체를 형성한 상태이더라도 단리된 세포이다.
단리된 표적 세포는 생체(예를 들면, 환자)로부터 제조할 수 있다. 한편, 독립행정법인 이화학연구소 바이오자원 센터, 독립행정법인 제품평가기술 기반 기구, ATCC(아메리칸 타입 컬쳐 콜렉션), DSMZ(저먼 콜렉션 오브 마이크로오가니즘 앤 셀 컬쳐)) 등으로부터 입수한 세포를 단리된 표적 세포로서 사용해도 된다.
본 발명의 mRNA는 5' 비번역 영역(5' UTR: 5' untranslated region)과 3' 비번역 영역(3' UTR: 3' untranslated region)을 갖춘다.
본 발명의 하나의 실시형태의 mRNA는 단백질을 코딩하는 mRNA의 5' UTR과, 5' UTR과의 상보성이 40% 이상 80% 이하인 3' UTR을 갖춘다. 바꾸어 말하면, 본 발명의 실시형태의 mRNA는 목적 단백질을 코딩하는 번역 영역과, 목적 단백질과는 다른 단백질을 코딩하는 mRNA의 5' UTR과, 5' UTR과의 상보성이 40% 이상 80% 이하인 3' UTR을 갖춘다. 이 형태로 함으로써, 번역 효율을 향상시킬 수 있다. 번역 효율을 보다 향상시키는 관점에서, 3' UTR에 있어서의 5' UTR과의 상보성이 50% 이상 75% 이하인 것이 바람직하다. 번역 효율을 향상시킬 수 있는 결과, 목적 단백질의 발현 효율을 향상시킬 수 있다.
단백질을 코딩하는 mRNA의 5' UTR로서는 특별히 한정되지 않지만, 예를 들면, 세포(예를 들면, HeLa 세포)당의 단백질 발현량이 106 분자 이상의 단백질을 코딩하는 mRNA의 5' UTR이 바람직하다. 이 단백질로서는 글리세르알데히드-3-인산 데히드로게나제와, β-글로빈과, RPS8과, LDHB로 이루어지는 군으로부터 선택되는 단백질이 보다 바람직하다.
또한, 본 발명의 다른 실시형태의 mRNA는 글리세르알데히드-3-인산 데히드로게나제(GAPDH)를 코딩하는 mRNA의 5' UTR과, GAPDH를 코딩하는 mRNA의 3' UTR을 갖춘다. 바꾸어 말하면, 본 발명의 실시형태의 mRNA는 GAPDH를 코딩하는 mRNA의 5' UTR과, GAPDH를 코딩하는 mRNA의 3' UTR과, GAPDH가 아닌 단백질을 코딩하는 ORF(오픈 리딩 프레임)를 갖춘다. 이 형태로 함으로써, 번역 효율을 향상시킬 수 있다. 본 명세서에 있어서, 「~를 코딩하는 mRNA의 UTR」을 단순히 「~의 UTR」라고도 부른다. 예를 들면, 「GAPDH를 코딩하는 mRNA의 5' UTR」을 「GAPDH의 UTR」이라고도 부른다.
본 명세서에 있어서, GAPDH의 UTR로서 인간 유래의 GAPDH의 UTR을 사용하지만, 본 발명은 이것에 한정되지 않는다. GAPDH의 UTR로서, 예를 들면, 다른 생물(예를 들면, 마우스)로부터의 GAPDH의 UTR을 사용해도 된다. 또한, 본 발명의 mRNA가 갖추는 GAPDH의 UTR의 길이는 생물 유래의 GAPDH의 UTR의 7할 이상 13할 이하인 것이 바람직하고, 8할 이상 12할 이하인 것이 보다 바람직하고, 9할 이상 11할 이하인 것이 더욱 바람직하다. 또한, 본 발명의 mRNA가 갖추는 GAPDH의 UTR은 생물 유래의 GAPDH의 UTR과의 일치율이 7할 이상인 것이 바람직하고, 8할 이상인 것이 보다 바람직하고, 9할 이상인 것이 더욱 바람직하다.
본 발명의 다른 실시형태의 mRNA는 GAPDH를 코딩하는 mRNA의 5' UTR과, 5' UTR과의 상보성이 40% 이상 80% 이하인 3' UTR을 갖춘다. 바꾸어 말하면, 본 발명의 실시형태의 mRNA는 GAPDH를 코딩하는 mRNA의 5' UTR과, 5' UTR과의 상보성이 40% 이상 80% 이하인 3' UTR과, GAPDH가 아닌 단백질을 코딩하는 ORF를 포함한다. 이 형태로 함으로써, 번역 효율을 향상시킬 수 있다. 번역 효율을 보다 향상시키는 관점에서, 3' UTR에 있어서의 5' UTR과의 상보성이 50% 이상 75% 이하인 것이 바람직하다.
본 실시형태의 mRNA는 그 번역에 필요한 5' 캡 구조(m7G(7-메틸구아노신)가 5' 말단 뉴클레오시드에 5'-5' 삼인산교를 통해 결합한 구조) 및 폴리 (A)쇄를 갖추어도 된다. 폴리 (A)쇄의 길이는 특별히 한정되지 않지만, 예를 들면, 30 내지 200 염기이다. 5' 캡 구조에는 번역 개시 인자 eIF4E가 결합하고, 폴리 (A)쇄에는 폴리 (A)쇄 결합 단백질 PABP(폴리 (A)-결합 단백질)가 결합함으로써, 양자가 뼈대 단백질인 번역 개시 인자 eIF4G를 통해 복합체를 형성함으로써, mRNA는 환형 구조를 형성한다(Wells SE, et al. Mol Cell. 1998; 2:135-140). 또한, 번역 종결 인자 eRF3은 PABP-eIF4G와 복합체를 형성함으로써 3' UTR을 루프 아웃한다(Uchida N, et al. J Biol Chem. 2002; 277: 50286-50292). 이러한 mRNA의 환상화는 번역 종결 부위와 번역 개시 부위를 물리적으로 가깝게 하여, 번역을 마친 리보솜을, 3' UTR을 거치지 않고 종지 코돈으로부터 다음의 번역 개시로 리사이클함으로써, 번역 개시의 효율에 크게 기여한다. 5' 말단 캡 구조와 폴리 (A)쇄는 이러한 번역의 효율화뿐만 아니라, 엑소뉴클레아제에 의한 말단으로부터의 mRNA 분해를 저해함으로써 mRNA를 안정화하여, 번역의 효율화와 mRNA 안정화 두 과정에서 전사 후의 유전자 발현 조절에 크게 기여한다.
본 발명의 mRNA는, 예를 들면, in vitro 전사계, 화학 합성 등의 방법에 의해 제조할 수 있다. in vitro 전사용 키트(예를 들면, 프로메가사가 제공하는 RiboMAXsystem, 닛폰진사가 제공하는 CUGA7 in vitro 전사 키트, 라이프테크놀로지스사가 제공하는 MEGAscript T7 키트)를 이용함으로써, 간편하게 목적의 mRNA를 조제하는 것이 가능하다. 또한, 5'캡 구조의 부가에 대해서도 공지의 방법으로 행할 수 있고, 예를 들면, New England Biolabs사가 제공하는 3'-O-Me-m7G(5')ppp(5')G RNA Cap Structure Analog를 이용할 수 있다.
본 발명의 mRNA로서, 2종류 이상의 mRNA를 병용하는 것으로 해도 된다. 예를 들면, 특정 유전자의 코드 영역을 갖는 mRNA와, 해당 유전자의 발현 산물과 상호작용하는 발현 산물을 코딩하는 영역을 갖는 mRNA를 병용할 수 있다.
본 발명의 mRNA의 양은, 사용 목적, 사용하는 목적 유전자의 특징, 표적 세포의 종류 등을 고려하면서, 표적 세포 내에서 충분한 양의 발현 산물이 얻어지도록 설정하면 된다. mRNA량의 예를 나타내면, 1회분의 양으로서 3cm 배양 접시당 0.5 내지 1.0㎍의 mRNA를 함유시켜도 된다.
인공 합성 mRNA의 보호를 목적으로, 예를 들면, 엑소뉴클레아제 억제제, 엔도뉴클레아제 억제제, 인지질, 인산칼슘, 폴리에틸렌이민, 나노미셀 형성제인 폴리에틸렌글리콜-폴리 양이온, 완충제, 무기 염류, 2가 이온 등을 사용해도 되고, 세균의 혼입을 저지하는 것을 목적으로 항생 물질 등을 사용해도 되고, 세포의 증식능을 항진시키는 것을 목적으로, 동물 혈청, 성장인자, 당류, 비타민류, 2가 이온 등을 사용해도 된다. 또한, 제제상 허용되는 다른 성분(예를 들면, 담체, 부형제, 붕괴제, 완충제, 유화제, 현탁제, 무통화제, 안정제, 보존제, 방부제, 생리 식염수 등)을 사용해도 된다. 또한, 유효 성분의 세포 도입 효율을 항진시키는 것을 목적으로, 라이프 테크놀로지사가 제공하는 Opti-MEM 등의 특수 합성 배지를 사용해도 된다.
2. 도입 방법
본 발명을 이용하여 표적 세포 내에서 목적 유전자를 발현시키기 위해서는, 표적 세포에 목적 유전자의 mRNA를 도입하는 단계를 행한다.
mRNA의 표적 세포로의 도입은 공지된 방법으로 행할 수 있다. 예를 들면, 인산칼슘 공침강법, 일렉트로포레이션(Potter, H. et al., Proc. Natl. Acad. Sci. USA 81, 7161-7165(1984)), 리포펙션(Felgner, PL et al., Proc. Natl Acad. Sci. USA 84,7413-7417(1987)), 마이크로 인젝션(Graessmann, M. & Graessmann,A., Proc. Natl. Acad. Sci. USA 73,366-370(1976)), Hanahan의 방법(Hanahan, D., J. Mol. Biol. 166, 557-580(1983)), 아세트산 리튬법(Schiestl, R.H. et al., Curr. Genet. 16, 339-346(1989)), 프로토플라스트-폴리에틸렌글리콜법(Yelton, MM et al., Proc. Natl. Acad. Sci. 81, 1470-1474(1984)), 초음파 유전자 도입법, 양이온성 폴리아민산을 이용한 방법(예를 들면, 일본 특개 2011-173802호 공보를 참조), 양이온성 폴리머 세그먼트와 비가전 친수성 폴리머 세그먼트를 갖는 블록 공중합체로 이루어지는 폴리이온 콤플렉스(PIC)형의 고분자 미셀을 이용한 방법(예를 들면, 일본 특개 2004-352972호 공보, 국제공개 제2012/005376호 팜플렛 참조) 등에 의해 실시할 수 있다.
3. 용도
본 발명의 mRNA에 의하면, 표적 세포 내에서 번역 효율이 향상하기 때문에, 목적 단백질이 고발현된다. 따라서, 본 발명은 목적 단백질의 높은 발현이 요망되는 다양한 용도에 적용 가능하다. 본 발명의 용도의 예로서, (A) 각종 바이러스성 질환(예를 들면, B형 간염, 후천성 면역부전 증후군 AIDS, 성인 T세포 백혈병)이나 유전병(예를 들면, 뒤시엔느형 근이영양증, 낭포성 섬유증, β 살라세미아, Hurler 증후군, 망막 색소 변성증, X 연쇄형 신성 요붕증)의 치료, (B) 암면역 요법, (C) iPS 세포의 제작, (D) 줄기세포(예를 들면, iPS 세포나 ES 세포 등의 다분화능 줄기 세포) 또는 전구 세포의 분화 유도 등을 들 수 있다.
상기 (A) 및 (B)는 소위 RNA 의약으로서 본 발명을 이용하는 것이다. (A)의 구체예로서의 B형 간염의 치료에서는, 예를 들면, 게놈에 편입된 바이러스 DNA를 절단·분해하는 뉴클레아제(ZFN, TALEN, 또는 CRISPR-Cas9) 유전자를 목적 유전자로서 편입한 mRNA를 사용함으로써, 종래의 바이러스 벡터를 사용한 방법에서 문제가 되는 발암 리스크를 수반하지 않는 바이러스 치료가 가능하게 된다. 이와 같이, 본 발명은 바이러스 제거제로서도 유용하다. 유전병의 치료에 있어서는, 예를 들면, 질환 원인 유전자(기능 저하 또는 결손에 의해 질환을 일으키는 것)를 목적 유전자로 하고, 본 발명을 적용한다. 상기 (B)의 용도에서는, 본 발명을 이용하여 암 항원의 mRNA를 항원 제시 세포에 도입하고, 암 백신을 체내에서 생산시키게 된다. 상기 (C) 및 (D)의 용도에 본 발명을 적용하면, 바이러스 벡터를 사용하지 않고 초기화 인자를 도입하는 것이 가능하게 되기 때문에, 세포의 암화의 문제를 극복할 수 있다.
본 발명을 RNA 의약으로서 이용하는 경우의 제제화는 통상의 방법에 따라 행할 수 있다. 제형화하는 경우에는, 제제상 허용되는 다른 성분(예를 들면, 완충제, 부형제, 붕괴제, 유화제, 현탁제, 무통화제, 안정제, 보존제, 방부제, 식염수, 담체 등)을 함유시킬 수 있다. 완충제로서는 인산 완충액, 시트르산 완충액 등을 사용할 수 있다. 부형제로서는 유당, 전분, 소르비톨, D-만니톨, 백당 등을 사용할 수 있다. 붕괴제로서는 전분, 카르복시메틸셀룰로오스, 탄산칼슘 등을 사용할 수 있다. 완충제로서는 인산염, 구연산염, 아세트산염 등을 사용할 수 있다. 유화제로서는 아라비아 고무, 알긴산 나트륨, 트라간트 등을 사용할 수 있다. 현탁제로서는 모노스테아르산 글리세린, 모노스테아르산 알루미늄, 메틸셀룰로오스, 카르복시메틸셀룰로오스, 히드록시메틸셀룰로오스, 라우릴황산나트륨 등을 사용할 수 있다. 무통화제로서는 벤질알코올, 클로로부탄올, 소르비톨 등을 사용할 수 있다. 안정제로서는 프로필렌글리콜, 아스코르브산 등을 사용할 수 있다. 보존제로서는 페놀, 염화벤잘코늄, 벤질알코올, 클로로부탄올, 메틸파라벤 등을 사용할 수 있다. 방부제로서는 염화벤잘코늄, 파라옥시벤조산, 클로로부탄올 등을 사용할 수 있다.
제제화하는 경우의 제형도 특별히 한정되지 않는다. 제형의 예는 주사제, 정제, 산말, 미립자, 과립제, 캡슐 및 시럽이다.
본 발명의 RNA 의약은 그 제형에 따라 경구 투여 또는 비경구 투여(정맥내, 동맥내, 피하, 피내, 근육내 또는 복강내 주사, 경피, 경비, 경점막 등)에 의해 대상에 적용된다. 이들의 투여 경로는 서로 배타적이지 않고, 임의로 선택되는 2개 이상을 병용해도 된다(예를 들면, 경구 투여와 동시에 또는 소정 시간 경과 후에 정맥 주사 등을 행하는 등). 여기에서의 「대상」은 특별히 한정되지 않고, 인간 및 인간 이외의 포유동물(애완동물, 가축, 실험동물을 포함한다. 구체적으로는, 예를 들면, 마우스, 래트, 기니피그, 햄스터, 원숭이, 소, 돼지, 염소, 양, 개, 고양이, 닭, 메추라기 등임)를 포함한다. 하나의 바람직한 실시태양에서는 본 발명의 RNA 약제는 인간에 적용된다.
본 발명의 RNA 약제의 투여량은 기대되는 치료 효과가 얻어지도록 설정된다. 치료상 유효 투여량의 설정에 있어서는, 일반적으로 환자의 증상, 연령, 성별 및 체중 등이 고려된다. 또한, 당업자라면 이들 사항을 고려하여 적당한 투여량을 설정하는 것이 가능하다. 투여 스케줄로서는, 예를 들면, 1일 1회 내지 수회, 2일에 1회, 혹은 3일에 1회 등을 채용할 수 있다. 투여 스케줄의 작성에서는, 환자의 증상이나 유효 성분의 효과 지속 시간 등을 고려할 수 있다.
실시예
1. 목적
다양한 임상 응용이 기대되고 있는 인공 합성 mRNA의 표적 세포 내에서의 번역 효율의 향상을 목표로, 이하의 검토를 행했다.
2. 연구 재료 및 방법
(1) 플라스미드
RNA 트랜스펙션용의 벡터로서 pBK-5F-EGFP-pA72를 사용했다. pBK-5F-EGFP-pA72의 제조 방법으로서는 Nogimori et al., "Dom34 mediates targeting of exogenous RNA in the antiviral OAS/RNase L pathway" Nucleic Acids Research, Volume 47, Issue 1, 10 January 2019, Pages 432-449에 기재된 방법을 사용했다.
(2) 비번역 영역의 삽입
·GAPDH의 5' UTR의 삽입
GAPDH의 5' UTR 서열에 관해서는, 서열번호 1과 서열번호 2에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, GAPDH의 5' UTR 서열은 서열번호 27에 나타냈다.
·GAPDH의 3' UTR의 삽입
GAPDH의 3' UTR 서열에 관해서는, HeLa 세포로부터 추출한 총 RNA와, 서열번호 3과 서열번호 4에 나타내는 올리고뉴클레오티드를 사용한 역전사 PCR법에 의해 단리했다. 단리한 서열은 pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, GAPDH의 3' UTR 서열은 서열번호 28에 나타낸다.
·ACAT2의 5' UTR의 삽입
ACAT2의 5' UTR 서열에 관해서는, HeLa 세포로부터 추출한 총 RNA와, 서열번호 5와 서열번호 6에 나타내는 올리고뉴클레오티드를 사용한 역전사 PCR법에 의해 단리했다. 단리한 서열은 pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, ACAT2의 5' UTR 서열은 서열번호 29에 나타낸다.
·ACAT2의 3' UTR의 삽입
ACAT2의 3' UTR 서열에 관해서는, HeLa 세포로부터 추출한 총 RNA와, 서열번호 7과 서열번호 8에 나타내는 올리고뉴클레오티드를 사용한 역전사 PCR법에 의해 단리했다. 단리한 서열은 pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, ACAT2의 3' UTR 서열은 서열번호 30에 나타낸다.
·LDHB의 5' UTR의 삽입
LDHB의 5' UTR 서열에 관해서는, HeLa 세포로부터 추출한 총 RNA와, 서열번호 9와 서열번호 10에 나타내는 올리고뉴클레오티드를 사용한 역전사 PCR법에 의해 단리했다. 단리한 서열은 pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, LDHB의 5' UTR 서열은 서열번호 31에 나타낸다.
·LDHB의 3' UTR의 삽입
LDHB의 3' UTR 서열에 관해서는, HeLa 세포로부터 추출한 총 RNA와, 서열번호 11과 서열번호 12에 나타내는 올리고뉴클레오티드를 사용한 역전사 PCR법에 의해 단리했다. 단리한 서열은 pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, LDHB의 3' UTR 서열은 서열번호 32에 나타낸다.
·GAPDH의 5' UTR 서열과 완전 상보인 3' UTR의 삽입
GAPDH의 5' UTR 서열과 완전 상보인 3' UTR 서열에 관해서는, 서열번호 13과 서열번호 14에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 완전 상보인 3' UTR 서열은 서열번호 33에 나타낸다.
·GAPDH의 5' UTR 서열과 상보성이 94%인 3' UTR의 삽입
GAPDH의 5' UTR 서열과 상보성이 94%인 3' UTR 서열에 관해서는, 서열번호 15와 서열번호 16에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 상보성이 94%인 3' UTR 서열은 서열번호 34에 나타낸다.
·GAPDH의 5' UTR 서열과 상보성이 88%인 3' UTR의 삽입
GAPDH의 5' UTR 서열과 상보성이 88%인 3' UTR 서열에 관해서는, 서열번호 17과 서열번호 18에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 상보성이 88%인 3' UTR 서열은 서열번호 35에 나타낸다.
·GAPDH의 5' UTR 서열과 상보성이 75%인 3' UTR의 삽입
GAPDH의 5' UTR 서열과 상보성이 75%인 3' UTR 서열에 관해서는, 서열번호 19와 서열번호 20에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 상보성이 75%인 3' UTR 서열은 서열번호 36에 나타낸다.
·GAPDH의 5' UTR 서열과 상보성이 50%인 3' UTR의 삽입
GAPDH의 5' UTR 서열과 상보성이 50%인 3' UTR 서열에 관해서는, 서열번호 21과 서열번호 22에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 상보성이 50%인 3' UTR 서열은 서열번호 37에 나타낸다.
·GAPDH의 5' UTR 서열과 상보성이 25%인 3' UTR의 삽입
GAPDH의 5' UTR 서열과 상보성이 25%인 3' UTR 서열에 관해서는, 서열번호 23과 서열번호 24에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 상보성이 25%인 3' UTR 서열은 서열번호 38에 나타낸다.
·베타-글로빈의 5' UTR 삽입
β-글로빈의 5' UTR 서열에 관해서는, 서열번호 41 및 서열번호 42에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, β-글로빈의 5' UTR 서열은 서열번호 94에 나타낸다.
·β-글로빈의 5' UTR 서열과 완전 상보인 3' UTR의 삽입
β-글로빈의 5' UTR 서열과 완전 상보인 3' UTR에 관해서는, 서열번호 43과 서열번호 44에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, β-글로빈의 5' UTR 서열과 완전 상보인 3' UTR 서열은 서열번호 95에 나타낸다.
·β-글로빈의 5' UTR 서열과 상보성이 92%인 3' UTR의 삽입
β-글로빈의 5' UTR 서열과 상보성이 92%인 3' UTR에 관해서는, 서열번호 45와 서열번호 46에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, β-글로빈의 5' UTR 서열과 상보성이 92%인 3' UTR 서열은 서열번호 96에 나타낸다.
·β-글로빈의 5' UTR 서열과 상보성이 88%인 3' UTR의 삽입
β-글로빈의 5' UTR 서열과 상보성이 88%인 3' UTR에 관해서는, 서열번호 47과 서열번호 48에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, β-글로빈의 5' UTR 서열과 상보성이 88%인 3' UTR 서열은 서열번호 97에 나타낸다.
·β-글로빈의 5' UTR 서열과 상보성이 74%인 3' UTR의 삽입
β-글로빈의 5' UTR 서열과 상보성이 74%인 3' UTR에 관해서는, 서열번호 49와 서열번호 50에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, β-글로빈의 5' UTR 서열과 상보성이 74%인 3' UTR 서열은 서열번호 98에 나타낸다.
·β-글로빈의 5' UTR 서열과 상보성이 50%인 3' UTR의 삽입
β-글로빈의 5' UTR 서열과 상보성이 50%인 3' UTR에 관해서는, 서열번호 51과 서열번호 52에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, β-글로빈의 5' UTR 서열과 상보성이 50%인 3' UTR 서열은 서열번호 99에 나타낸다.
·β-글로빈의 5' UTR 서열과 상보성이 24%인 3' UTR의 삽입
β-글로빈의 5' UTR 서열과 상보성이 24%인 3' UTR에 관해서는, 서열번호 53과 서열번호 54에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, β-글로빈의 5' UTR 서열과 상보성이 24%인 3' UTR 서열은 서열번호 100에 나타낸다.
·RPS8의 5' UTR의 삽입
RPS8의 5' UTR 서열에 관해서는, 서열번호 55와 서열번호 56에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 KpnI, XhoI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열은 서열번호 101에 나타낸다.
·RPS8의 5' UTR 서열과 완전 상보인 3' UTR의 삽입
RPS8의 5' UTR 서열과 완전 상보인 3' UTR에 관해서는, 서열번호 57과 서열번호 58에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열과 완전 상보인 3' UTR 서열은 서열번호 102에 나타낸다.
·RPS8의 5' UTR 서열과 상보성이 93%인 3' UTR의 삽입
RPS8의 5' UTR 서열과 상보성이 93%인 3' UTR에 관해서는, 서열번호 59와 서열번호 60에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열과 상보성이 93%인 3' UTR 서열은 서열번호 103에 나타낸다.
·RPS8의 5' UTR 서열과 상보성이 86%인 3' UTR의 삽입
RPS8의 5' UTR 서열과 상보성이 86%인 3' UTR에 관해서는, 서열번호 61과 서열번호 62에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열과 상보성이 86%인 3' UTR 서열은 서열번호 104에 나타낸다.
·RPS8의 5' UTR 서열과 상보성이 71%인 3' UTR의 삽입
RPS8의 5' UTR 서열과 상보성이 71%인 3' UTR에 관해서는, 서열번호 63과 서열번호 64에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열과 상보성이 71%인 3' UTR 서열은 서열번호 105에 나타낸다.
·RPS8의 5' UTR 서열과 상보성이 50%인 3' UTR의 삽입
RPS8의 5' UTR 서열과 상보성이 50%인 3' UTR에 관해서는, 서열번호 65와 서열번호 66에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열과 상보성이 50%인 3' UTR 서열은 서열번호 106에 나타낸다.
·RPS8의 5' UTR 서열과 상보성이 29%인 3' UTR의 삽입
RPS8의 5' UTR 서열과 상보성이 29%인 3' UTR에 관해서는, 서열번호 67과 서열번호 68에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열과 상보성이 29%인 3' UTR 서열은 서열번호 107에 나타낸다.
·RPS8의 5' UTR 서열과 상보성이 14%인 3' UTR의 삽입
RPS8의 5' UTR 서열과 상보성이 14%인 3' UTR에 관해서는, 서열번호 69와 서열번호 70에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, RPS8의 5' UTR 서열과 상보성이 14%인 3' UTR 서열은 서열번호 108에 나타낸다.
·LDHB의 5' UTR 서열과 완전 상보인 3' UTR의 삽입
LDHB의 5' UTR 서열과 완전 상보인 3' UTR에 관해서는, 서열번호 71과 서열번호 72에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, LDHB의 5' UTR 서열과 완전 상보인 3' UTR 서열은 서열번호 109에 나타낸다.
·LDHB의 5' UTR 서열과 상보성이 93%인 3' UTR의 삽입
LDHB의 5' UTR 서열과 상보성이 93%인 3' UTR에 관해서는, 서열번호 73과 서열번호 74에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, LDHB의 5' UTR 서열과 상보성이 93%인 3' UTR 서열은 서열번호 110에 나타낸다.
·LDHB의 5' UTR 서열과 상보성이 87%인 3' UTR의 삽입
LDHB의 5' UTR 서열과 상보성이 87%인 3' UTR에 관해서는, 서열번호 75와 서열번호 76에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, LDHB의 5' UTR 서열과 상보성이 87%인 3' UTR 서열은 서열번호 111에 나타낸다.
·LDHB의 5' UTR 서열과 상보성이 75%인 3' UTR의 삽입
LDHB의 5' UTR 서열과 상보성이 75%인 3' UTR에 관해서는, 서열번호 77과 서열번호 78에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, LDHB의 5' UTR 서열과 상보성이 75%인 3' UTR 서열은 서열번호 112에 나타낸다.
·LDHB의 5' UTR 서열과 상보성이 50%인 3' UTR의 삽입
LDHB의 5' UTR 서열과 상보성이 50%인 3' UTR에 관해서는, 서열번호 79와 서열번호 80에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, LDHB의 5' UTR 서열과 상보성이 50%인 3' UTR 서열은 서열번호 113에 나타낸다.
·LDHB의 5' UTR 서열과 상보성이 25%인 3' UTR의 삽입
LDHB의 5' UTR 서열과 상보성이 25%인 3' UTR에 관해서는, 서열번호 81과 서열번호 82에 나타내는 올리고뉴클레오티드를 혼성화하고, pBK-5F-EGFP-pA72의 EcoRI, XbaI 사이트에 삽입했다. 여기에서, LDHB의 5' UTR 서열과 상보성이 25%인 3' UTR 서열은 서열번호 114에 나타낸다.
·Cas9 mRNA 합성 플라스미드의 제작
Cas9 cDNA 서열은 Mail et al "RNA-guided human genome engineering via Cas9" Science. 2013 Feb 15; 339(6121): 823-6에 기재되어 있는 hCas 플라스미드(Addgene plasmid #41815)와, 서열번호 83과 서열번호 84에 나타내는 올리고뉴클레오티드를 사용한 PCR법에 의해 단리했다. 단리한 서열을 pBK-5F-EGFP-pA72의 HindIII 사이트에 삽입한 플라스미드와, 서열번호 83과 서열번호 85에 나타내는 올리고뉴클레오티드를 사용한 인버스 PCR법에 의해 pBK-F-hCas9-pA72 플라스미드를 얻었다. . 여기에서, hCas9의 cDNA 서열은 서열번호 115에 나타내고, hCas9의 아미노산 서열은 서열번호 116에 나타낸다.
·Cas9 mRNA 합성 플라스미드로의 GAPDH의 5' UTR 삽입
pBK-F-hCas9-pA72 플라스미드와, 서열번호 86과 서열번호 87에 나타낸 올리고 뉴클레오타이드를 사용한 인버스 PCR법에 의해 GAPDH의 5' UTR을 삽입했다. 여기에서, GAPDH의 5' UTR은 서열번호 27에 나타낸다.
·Cas9 mRNA 합성 플라스미드로의 GAPDH의 5' UTR 서열과 상보성이 75%인 3' UTR의 삽입
pBK-F-hCas9-pA72 플라스미드와 서열번호 88과 서열번호 89에 나타내는 올리고뉴클레오티드를 사용한 인버스 PCR법에 의해, GAPDH의 5' UTR 서열과 상보성이 75%인 3' UTR을 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 상보성이 75%인 3' UTR 서열은 서열번호 36에 나타낸다.
·Cas9 mRNA 합성 플라스미드로의 GAPDH의 5' UTR 서열과 상보성이 50%인 3' UTR의 삽입
pBK-F-hCas9-pA72 플라스미드와 서열번호 90과 서열번호 91에 나타내는 올리고뉴클레오티드를 사용한 인버스 PCR법에 의해, GAPDH의 5' UTR 서열과 상보성이 50%인 3' UTR을 삽입했다. 여기에서, GAPDH의 5' UTR 서열과 상보성이 50%인 3' UTR 서열은 서열번호 37에 나타낸다.
·p53의 cDNA의 삽입
p53의 cDNA 서열은 U20S 세포로부터 추출한 총 RNA와, 서열번호 25와 서열번호 26에 나타내는 올리고뉴클레오티드를 사용한 역전사 PCR법에 의해 단리했다. 단리한 p53의 cDNA 서열은 pBK-5F-EGFP-pA72의 HindIII 부위에 삽입했다. 여기에서, p53의 cDNA 서열은 서열번호 39에 나타내고, p53의 아미노산 서열은 서열번호 40에 나타낸다.
(3) RNA 합성
각종 pBK-5F-EGFP-pA72, 각종 pBK-5F-p53-pA72 및 각종 pBK-F-hCas9-pA72 플라스미드를, BsmBI로 처리한 것을 주형으로 하여, 5xFlag-EGFP-pA72, 5xFlag-p53-pA72 및 Flag-hCas9-pA72 mRNA를 합성했다. RNA 합성은 T7 RNA 폴리머라제(타카라 바이오 가부시키가이샤)를 사용하여, 이 T7 RNA 폴리머라제의 취급 설명서에 따라 행했다.
(4) 트랜스펙션
HeLa 세포, 293T 세포, U2OS 세포는 모두 5% 태아 소 혈청을 첨가한 듈베코 변형 이글 배지(닛수세이야쿠 가부시키가이샤)를 사용하고, 5% CO2 존재하 37℃에서 배양했다. HeLa 세포를 약 50% 컨플루언트가 되도록 35mm 접시에 뿌린 후 24시간 동안 배양했다. 그 후, 합성한 각종 RNA를 리포펙타민 RNAiMAX(라이프테크놀로지스 재팬 가부시키가이샤)를 사용하여, 이 취급 설명서에 따라 도입했다.
(5) RNA의 분석
RNA 트랜스펙션 후의 HeLa 세포로부터의 총 RNA의 분리는 구아니딘티오시안산염, 산성 페놀, 클로로포름을 사용한 방법인 AGPC(산 구아니디늄 티오시아네이트-페놀-클로로포름 추출)법에 의해 행했다. 제조한 총 RNA는 아가로스 MOPS 버퍼겔(20mM MOPS(pH 7.0), 5mM 아세트산 나트륨, 1mM EDTA, 2.0% 아가로스, 2.46M 포름알데히드)에 의해 분리한 후, 20xSSC 버퍼 속에서 나일론막 Biodyne-B(니혼폴 가부시키가이샤)에 전사했다. 전사 후의 나일론막은 UV로 고정한 후, DIG 표지 프로브를 사용하여 혼성화했다. DIG 표지 프로브의 제작 및 혼성화는 DIG RNA 라벨링 믹스(로슈 다이아그노스틱스 가부시키가이샤) 및 DIG 세척 및 차단 버퍼 세트(로슈 다이아그노스틱스 가부시키가이샤)를 사용하여 취급 설명서에 따라 행했다. mRNA의 검출은 화학 발광 시약 CDP-Star(로슈 다이아그노스틱스 가부시키가이샤)를 사용하여, LAS3000mini(후지샤신필름 가부시키가이샤)에 의해 검출했다.
(6) 단백질의 분석
단백질의 세포내 발현은 이하에 나타내는 웨스턴 블롯법에 의해 행했다. 도입 후 세포로부터 단백질 라이세이트의 조절은 SDS-PAGE 샘플 버퍼(50mM Tris-HCl(pH 6.8), 4% 글리세롤, 2% SDS, 2% 2-메르캅토에탄올, 0.004% 브로모페놀 블루)를 사용하여 행했다. 단백질 라이세이트는 8, 10, 12 또는 15% 아크릴아미드를 사용하여 SDS-PAGE법으로 분리한 후 니트로셀룰로오스막 BioTrace NC(닛폰 폴 가부시키가이샤)에 전기적으로 전사했다. 전사 후 니트로셀룰로오스막은 항Flag M2 마우스 모노클로날 항체(시그마알드리치 재팬 고도카이샤), 항GAPDH 항체(Saito et al JBC), 항PABPC1 항체(Osawa et al RNA(2012)) 및 퍼옥시다아제 부가 항마우스 IgG(Jackson ImmunoResearch Laboratories, Inc.) 또는 퍼옥시다아제 부가 항토끼 IgG(Jackson ImmunoResearch Laboratories, Inc.)와 배양했다. 니트로셀룰로오스막상의 퍼옥시다아제 효소 활성은 루미놀 화학 발광법을 사용하여 LAS3000mini(후지샤신필름 가부시키가이샤)에 의해 검출했다.
(7) AAVS 영역 게놈 편집 효율의 정량
게놈 편집 효율의 정량은 이하에 나타내는 T7 엔도뉴클레아제 어세이법에 의해 행했다. AAVS 영역을 인식하는 sgRNA(서모피셔사이언티픽 가부시키가이샤)와 합성한 각종 hCas9 mRNA를 HeLa 세포에 도입했다. 도입한 세포를 50mM NaOH로 처리함으로써 게놈 DNA를 조정했다. 조정한 게놈 DNA와, 서열번호 92와 서열번호 93에 나타내는 올리고뉴클레오티드를 사용한 PCR법에 의해 AAVS 게놈 서열을 증폭했다. 증폭한 게놈 서열을 열변성 후 어닐링시킴으로써 미스매치를 포함하는 서열로 했다. 어닐링한 AAVS 게놈 서열을 T7 엔도뉴클레아제 I(뉴잉글랜드 바이오랩 가부시키가이샤)에 의해 처리했다. 처리 후의 AAVS 게놈 서열은 아가로스겔 전기영동에 의해 분리한 후, 에티듐브로마이드로 염색하고, Typhoon 9400(GE Healthcare, Inc.)에 의해 검출했다.
3. 실험 결과
(i) 실험 1
도 1은 제작한 인공 합성 mRNA를 나타내는 도면이다. 도 1에 나타내는 인공 합성 mRNA는 위에서부터 GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열을 갖는 인공 합성 mRNA와, LDHB의 5' UTR 서열 및 LDHB의 3' UTR 서열을 갖는 인공 합성 mRNA와, ACAT2의 5' UTR 서열 및 ACAT2의 3' UTR 서열을 갖는 인공 합성 mRNA를 나타낸다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어 있다. 또한, 이 실험에 사용한 인공 합성 mRNA의 단백질 코드 영역인 ORF에는, 5xFlag 태그를 부가한 EGFP(강화된 녹색 형광 단백질)를 갖는다.
도 2는 인공 합성 mRNA로부터의 단백질 발현량을 측정한 결과를 나타내는 도면이다. 단백질 발현량은 도 1에 나타내는 인공 합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 2는 인공 합성 mRNA마다의 발현량을 나타낸다.
도 2에서는, ACAT2의 UTR 서열을 갖는 인공 합성 mRNA의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다. 도 2에서 참고로, β-글로빈의 3' UTR을 갖는 인공 합성 mRNA의 결과에 대해서도 나타낸다.
도 2의 결과로부터, GAPDH의 UTR 서열을 갖는 인공 합성 mRNA는, ACAT2의 UTR 서열을 갖는 인공 합성 mRNA와 비교하여, 약 10배의 발현량을 나타내는 것을 알 수 있었다. 또한, 발현 효율이 높은 인공 합성 mRNA로서 표준적으로 사용되는 β-글로빈의 3' UTR 서열을 갖는 인공 합성 mRNA와 비교하여, GAPDH의 UTR 서열을 갖는 인공 합성 mRNA는 약 5배의 발현량 보이는 것을 알 수 있었다.
도 3은 도 1에 나타낸 인공 합성 mRNA의 안정성을 나타내는 도면이다. 도 3의 가로축은 세포에 인공 합성 mRNA를 도입한 후의 경과 시간을 나타내고, 도 3의 세로축은 세포에 인공 합성 mRNA를 도입했을 때의 mRNA의 양을 100%로 한 경우의 mRNA의 양을 나타낸다.
도 3의 결과로부터, 어느 인공 합성 mRNA도 HeLa 세포 내에서 동일한 분해 속도를 나타내는 것을 알 수 있었다. 도 2와 도 3에 나타내는 결과로부터, GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열이 발휘하는 효과는 안정성이 향상된다고 하는 효과가 아니고, 번역 효율이 향상되는 효과인 것을 알 수 있었다.
(ii) 실험 2
도 4는 제작한 인공 합성 mRNA를 나타내는 도면이다. 도 4에 나타내는 인공 합성 mRNA는, 위에서부터 차례로, ACAT2의 5' UTR 서열 및 ACAT2의 3' UTR 서열을 가지는 인공 합성 mRNA와, ACAT2의 5' UTR 서열 및 GAPDH의 3' UTR 서열을 가지는 인공 합성 mRNA와, GAPDH의 5' UTR 서열 및 ACAT2의 3' UTR 서열을 가지는 인공 합성 mRNA와, GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열을 가지는 인공 합성 mRNA를 나타낸다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는, 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어 있다. 또, 이 실험에 사용한 인공 합성 mRNA의 단백질 코드 영역인 ORF는 5xFlag 태그를 부가한 EGFP를 가진다.
도 5는 인공 합성 mRNA마다의 발현량을 나타내는 도면이다. 단백질 발현량은 도 4에 나타내는 인공 합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 5에서는, ACAT2의 5' UTR 서열 및 ACAT2의 3' UTR 서열을 갖는 인공 합성 mRNA의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
도 5의 결과로부터, ACAT2의 5' UTR 서열 및 ACAT2의 3' UTR 서열을 갖는 인공 합성 mRNA와 비교하여, GAPDH의 5' UTR 서열 및 ACAT2의 3' UTR 서열을 갖는 인공 합성 mRNA는 약 10배의 발현량을 나타내는 것을 알 수 있었다. 또한, ACAT2의 5' UTR 서열 및 ACAT2의 3' UTR 서열을 갖는 인공 합성 mRNA와 비교하여, GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열을 갖는 인공 합성 mRNA는 약 25배의 발현량을 나타내는 것을 알 수 있었다. 즉, GAPDH의 5' UTR 서열을 가짐으로써 번역 효율의 향상 효과를 얻을 수 있지만, 또한, GAPDH의 3' UTR 서열을 가짐으로써 상승적인 번역 효율의 향상 효과가 얻어지는 것을 알 수 있었다.
도 6은 인공 합성 mRNA를 신장 유래의 HEK293 세포에 도입한 경우의 인공 합성 mRNA마다의 발현량을 나타내는 도면이다. 도 5에 관한 실험과 비교하여, 도입하는 세포를 변경한 것 이외에는, 동일한 방법으로 실험을 행했다. 도 6의 결과로부터도, 도 5의 결과로부터 알 수 있었던 것과 동일한 경향이 보였다. 즉, GAPDH의 5' UTR 서열을 가짐으로써 번역 효율의 향상 효과를 얻을 수 있지만, 또한, GAPDH의 3' UTR 서열을 가짐으로써 상승적인 번역 효율의 향상 효과가 얻어지는 것을 알 수 있었다. 도 5와 도 6에 관한 실험 결과로부터, GAPDH의 UTR 서열을 가짐으로써 얻어지는 번역 효율의 향상 효과는 어느 세포에서도 유효하여, 세포 특이성이 없는 것을 알 수 있었다.
(iii) 실험 3
도 7은 제작한 인공 합성 mRNA를 나타내는 도면이다. 도 7에 나타내는 인공 합성 mRNA는 위에서부터 차례로, UTR 서열을 갖지 않는 인공 합성 mRNA와, 5' UTR 서열을 갖지만 3' UTR 서열을 갖지 않는 인공 합성 mRNA와, 5' UTR 서열을 갖지 않지만 3' UTR 서열을 갖는 인공 합성 mRNA와, 5' UTR 서열 및 3' UTR 서열을 갖는 인공 합성 mRNA를 나타낸다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어있다. 또한, 이 실험에 사용한 인공 합성 mRNA의 단백질 코드 영역인 ORF는 5xFlag 태그를 부가한 EGFP를 갖는다.
도 8은 GAPDH의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면이다. 도 9는 LDHB의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면이다. 도 10은 ACAT2의 UTR 서열의 유무에 기인하는 발현량의 차이를 나타내는 도면이다. 단백질 발현량은 도 7에 나타내는 인공 합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 8, 9, 10에서는 UTR 서열을 갖지 않는 인공 합성 mRNA의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
도 8의 결과로부터, GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열을 갖는 인공 합성 mRNA의 발현량은 5' UTR 서열을 갖지만 3' UTR 서열을 갖지 않는 인공 합성 mRNA의 발현량과, 5' UTR 서열을 갖지 않지만 3' UTR 서열을 갖는 인공 합성 mRNA의 발현량의 합보다 크다. 즉, GAPDH의 5' UTR 서열과 GAPDH의 3' UTR 서열을 가짐으로써, 상승적인 번역 효율의 향상 효과가 얻어지는 것을 알 수 있었다.
반면, 도 9의 결과로부터, LDHB의 5' UTR 서열 및 LDHB의 3' UTR 서열을 갖는 인공 합성 mRNA의 발현량은 5' UTR 서열을 갖지만 3' UTR 서열을 갖지 않는 인공 합성 mRNA의 발현량과, 5' UTR 서열을 갖지 않지만 3' UTR 서열을 갖는 인공 합성 mRNA의 발현량의 합과 동등했다. 즉, LDHB의 5' UTR 서열과 LDHB의 3' UTR 서열을 가짐으로써 얻어지는 효과는 상가적인 효과에 그쳤다.
또한, 도 10의 결과로부터, ACAT2의 UTR 서열을 갖는 인공 합성 mRNA의 발현량은, UTR 서열을 갖지 않는 인공 합성 mRNA의 발현량과 비교하여, 번역 효율이 저하되는 것을 알 수 있었다. 도 8 내지 도 10의 결과로부터, 5' UTR 서열과 3' UTR 서열을 가짐으로써 얻어지는 상승적인 번역 효율의 향상 효과는 GAPDH에 특징적인 효과인 것을 알 수 있었다.
(iv) 실험 4
도 11은 제작한 인공 합성 mRNA를 나타내는 도면이다. 도 11에 나타내는 인공 합성 mRNA는, 위에서부터 차례로, GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열을 갖는 인공 합성 mRNA(이하, 「야생형」이라고도 함)와, 5' UTR 서열을 3분할 한 부분 중 가장 5' 말단측의 부분을 결실한 인공 합성 mRNA(결실 변위체 1)와, 5' UTR 서열을 3분할한 부분 중 중앙 부분을 결실한 인공 합성 mRNA(결실 변위체 2)와, 5' UTR 서열을 3분할한 부분 중 가장 3' 말단측의 부분을 결실한 인공 합성 mRNA(결실 변위체 3)를 나타낸다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어있다. 또한, 이 실험에 사용한 인공 합성 mRNA의 단백질 코드 영역인 ORF는 EGFP를 갖는다.
도 12는 GAPDH의 5' UTR 서열의 결실의 유무에 기인하는 발현량의 차이를 나타내는 도면이다. 단백질 발현량은 도 11에 나타내는 인공 합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 12에서는, 야생형의 발현량을 100으로 한 경우의 발현량의 상대값을 나타낸다.
도 13은 제작한 인공 합성 mRNA를 나타내는 도면이다. 도 13에 나타내는 인공 합성 mRNA는 위에서부터 차례로 GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열을 갖는 인공 합성 mRNA(야생형)와 3' UTR 서열을 3분할한 부분 중 가장 5' 말단측의 부분을 결실한 인공 합성 mRNA(결실 변위체 4)와, 3' UTR 서열을 3 분할한 부분 중 중앙 부분을 결실한 인공 합성 mRNA(결실 변위체 5)와, 3' UTR 서열을 3 분할한 부분 중 가장 3' 말단측의 부분을 결실한 인공 합성 mRNA(결실 변위체 6)를 나타낸다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어있다. 또한, 이 실험에 사용한 인공 합성 mRNA의 단백질 코드 영역인 ORF는 EGFP를 갖는다.
도 14는 GAPDH의 3' UTR 서열의 결실의 유무에 기인하는 발현량의 차이를 나타내는 도면이다. 단백질 발현량은 도 13에 나타내는 인공 합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 14에 있어서, 야생형의 발현량을 100으로 한 경우의 발현량의 상대값을 나타낸다.
도 12, 14의 결과로부터, 어느 결실 변이체에 대해서도, 야생형과 비교하여 발현 효율이 저하된 것을 알 수 있었다. 이것으로부터, GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열은 모두 전체 영역이 존재하는 것이 번역 효율의 향상 효과에 기여하고 있는 것을 알 수 있었다.
(v) 실험 5
도 15는 제작한 인공 합성 mRNA를 나타내는 도면이다. 일반적으로, GAPDH의 5' UTR 서열은 GAPDH의 3' UTR 서열과 전체 영역에 걸쳐 상보성을 가지고 있다. 이 실험에서는 서로의 상보성이 특히 높은 영역인 5' UTR 서열의 일부와 3' UTR 서열의 일부를 갖는 인공 합성 mRNA를 사용했다. 구체적으로는, 5' UTR 서열로서, GAPDH의 5' UTR 서열을 3분할한 부분 중 가장 5' 말단측의 부분인 CU가 풍부한 영역(28뉴클레오티드)(이하,「5' UTR 28nt」라고도 함)를 사용했다. GAPDH의 3' UTR 서열로서, GAPDH의 3' UTR 서열을 3분할한 부분 중 가장 5' 말단측의 부분인 AG가 풍부한 영역(28뉴클레오티드)(이하, 「3' UTR 28nt」라고도 함)을 사용했다.
도 16은 5' UTR 28nt와 3' UTR 28nt의 유무에 기인하는 발현량의 차이를 나타내는 도면이다. 단백질 발현량은 인공합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 16에 있어서, 야생형의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
도 16의 결과로부터, 5' UTR 28nt 및 3' UTR 28nt 중 적어도 하나를 갖는 인공 합성 mRNA는 야생형에 비해 번역 효율이 저하된 것을 알 수 있었다. 이 결과로부터도, GAPDH의 5' UTR 서열 및 GAPDH의 3' UTR 서열은 모두 전체 영역이 존재하는 것이 번역 효율의 향상 효과에 기여하고 있는 것을 알 수 있었다.
(vi) 실험 6
도 17은 제작한 인공 합성 mRNA를 나타내는 도면이다. 이 실험에서는 GAPDH의 5' UTR 서열을 갖고, GAPDH의 5' UTR 서열에 대해 소정의 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA를 사용했다. 구체적으로는, 도 17에 나타내는 인공 합성 mRNA는 위에서부터 차례로 GAPDH의 5' UTR 서열에 94% 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA와, GAPDH의 5' UTR 서열에 대해 88% 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA와, GAPDH의 5' UTR 서열에 대해 75% 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA와, GAPDH 5' UTR 서열에 대해 50% 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA와, GAPDH의 5' UTR 서열에 대해 25% 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA를 나타낸다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어있다. 또한, 이 실험에 사용한 인공 합성 mRNA의 단백질 코드 영역인 ORF는 EGFP를 갖는다.
도 18은 상보성에 기인하는 발현량의 차이를 나타내는 도면이다. 단백질 발현량은 인공합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 18에 있어서, 야생형의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
도 18의 결과로부터, GAPDH의 5' UTR 서열에 대해 50%의 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA와, GAPDH의 5' UTR 서열에 대해 75%의 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA가 야생형의 2.5배 이상 높은 번역 효율을 나타내는 것을 알 수 있었다.
도 19는 제작한 인공 합성 mRNA를 나타내는 도면이다. 이 실험에서는 GAPDH의 5' UTR 서열을 갖고, GAPDH의 5' UTR 서열에 100% 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA를 사용했다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어있다. 또한, 이 실험에 사용한 인공 합성 mRNA의 단백질 코드 영역인 ORF는 EGFP를 갖는다.
도 20은 상보성에 기인하는 발현량의 차이를 나타내는 도면이다. 단백질 발현량은 인공합성 mRNA를 HeLa 세포에 도입함으로써 측정했다. 도 20에서는, 야생형의 발현량을 100으로 한 경우의 발현량의 상대값을 나타낸다.
도 20의 결과로부터, GAPDH의 5' UTR 서열에 대해 100%의 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA는 단백질의 발현이 보이지 않았다. 이 결과로부터, GAPDH의 5' UTR 서열에 부분적인 상보성을 갖는 3' UTR 서열을 갖는 것이 중요하다는 것을 알 수 있었다.
(vii) 실험 7
도 21은 제작한 인공 합성 mRNA를 나타내는 도면이다. 이 실험에 사용한 인공 합성 mRNA의 5' 말단에는 캡 구조가 부가되어 있음과 아울러, 3' 말단에는 폴리 A쇄가 72 염기 부가되어있다. 또한, 이 실험은 실용화를 목적으로 하여, 단백질 코드 영역으로서 암 억제 유전자 p53의 ORF를 사용했다. ORF에는 5xFlag 태그가 추가되었다.
도 22는 암 억제 유전자 p53을 사용한 경우의 발현량의 차이를 나타내는 도면이다. 단백질 발현량은 인공 합성 mRNA를 U2OS 세포에 도입함으로써 측정했다. 도 22에 있어서, GAPDH의 5' UTR 서열에 대하여 75%의 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA의 발현량을 100으로 한 경우의 발현량의 상대값을 나타낸다.
도 22의 결과로부터, 발현 효율이 높은 인공 합성 mRNA로서 표준적으로 사용되는 β-글로빈의 3' UTR 서열을 갖는 인공 합성 mRNA와 비교하여, 야생형은 약 4배 높은 번역 효율을 나타내는 것이 알 수 있고, GAPDH의 5' UTR 서열에 대해 75% 상보성을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA는 약 10배 높은 번역 효율을 나타내는 것을 알 수 있었다.
도 23은 얻어진 효과의 추정 메커니즘을 설명하기 위한 도면이다. 본 실시형태의 인공 합성 mRNA는 GAPDH의 5' 비번역 영역과 GAPDH의 3' 비번역 영역을 갖춘다. 또는, 본 실시형태의 인공 합성 mRNA는 5' 비번역 영역과, 5' 비번역 영역과의 상보성이 40% 이상 80% 이하인 3' 비번역 영역을 갖춘다. 즉, 본 실시형태의 인공 합성 mRNA는 GAPDH의 5' UTR 서열에 대해, 3' UTR 서열은 모두 부분적인 상보성을 갖는다. 이러한 부분적인 상보성이 mRNA의 환상화를 촉진함으로써, 리보솜을 효율적으로 리사이클시키기 때문에, 번역 효율이 향상되는 것으로 생각된다.
(viii) 실험 8
도 24는 게놈 편집 유전자 hCas9를 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면이다. 도면의 세로축은 발현량(발현(배수 증가))을 나타낸다. 이 실험에서는 하기 mRNA를 사용했다. 도면은 5' UTR 서열 및 3' UTR 서열을 갖지 않을 때의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
·5' UTR 서열 및 3' UTR 서열을 갖지 않는 게놈 편집 유전자 hCas9(대조군)
·GAPDH 5' UTR 서열을 갖는 인공 합성 mRNA
·GAPDH 5' UTR 서열을 갖고, GAPDH 5' UTR 서열에 대해 75% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·GAPDH 5' UTR 서열을 갖고, GAPDH 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
도 24의 결과로부터, 5' UTR 서열 및 3' UTR 서열을 갖지 않는 게놈 편집 유전자 hCas9와 비교하여, GAPDH의 5' UTR 서열을 갖는 인공 합성 mRNA는 2배 이상의 번역 효율을 나타내는 것이 알 수 있고, GAPDH의 5' UTR 서열에 대해 75% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA 및 GAPDH의 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA는 5배 이상의 번역 효율을 나타내는 것을 알 수 있었다.
도 25는 게놈 편집 유전자 hCas9를 사용한 경우에 있어서의 게놈 편집량의 차이를 나타내는 도면이다. 도면의 세로축은 게놈 편집량(게놈 편집(배수 증가))을 나타낸다. 이 실험에서는 도 24와 동일한 mRNA를 사용했다. 도면은 5' UTR 서열 및 3' UTR 서열을 갖지 않는 게놈 편집 유전자 hCas9의 게놈 편집량을 1로 한 경우의 게놈 편집량의 상대값을 나타낸다.
도 25의 결과로부터, 5' UTR 서열 및 3' UTR 서열을 갖지 않는 게놈 편집 유전자 hCas9와 비교하여, GAPDH의 5' UTR 서열을 갖는 인공 합성 mRNA는 2배 이상의 게놈 편집량을 나타내는 것 알 수 있고, GAPDH의 5' UTR 서열에 대해 75% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA 및 GAPDH의 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA는 3배 이상의 게놈 편집량을 나타내는 것을 알 수 있었다.
(ix) 실험 9
도 26은 β-글로빈의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면이다. 도면의 세로축은 발현량(발현(배수 증가))을 나타낸다. 이 실험에서는 하기 mRNA를 사용했다. 도면은 5' UTR 서열 및 3' UTR 서열을 갖지 않는 인공 합성 mRNA의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
·β-글로빈의 5' UTR 서열을 갖고, β-글로빈의 5' UTR 서열에 대해 100% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·β-글로빈의 5' UTR 서열을 갖고, β-글로빈의 5' UTR 서열에 대해 92% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·β-글로빈의 5' UTR 서열을 갖고, β-글로빈의 5' UTR 서열에 대해 88% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·β -글로빈의 5' UTR 서열을 갖고, β-글로빈 5' UTR 서열에 대해 74% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·β-글로빈의 5' UTR 서열을 갖고, β-글로빈의 5' UTR 서열에 대해 24% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·β-글로빈의 5' UTR 서열을 갖고, 3' UTR 서열을 갖지 않는 인공 합성 mRNA
·5' UTR 서열 및 3' UTR 서열을 갖지 않는 인공 합성 mRNA
·GAPDH의 5' UTR 서열을 갖고, GAPDH의 3' UTR 서열을 갖는 인공 합성 mRNA
도 26의 결과로부터, β-글로빈의 5' UTR 서열에 대해 74% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA 및 β-글로빈의 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA가 우수한 번역 효율을 나타내는 것을 알 수 있었다.
(x) 실험 10
도 27은 RPS8의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면이다. 도면의 세로축은 발현량(발현(배수 증가))을 나타낸다. 이 실험에서는 이하의 mRNA를 사용했다. 도면은 5' UTR 서열 및 3' UTR 서열을 갖지 않는 인공 합성 mRNA의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
·RPS8의 5' UTR 서열을 갖고, RPS8의 5' UTR 서열에 대해 100% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·RPS8의 5' UTR 서열을 갖고, RPS8의 5' UTR 서열에 대해 93% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·RPS8의 5' UTR 서열을 갖고, RPS8의 5' UTR 서열에 대해 86% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·RPS8의 5' UTR 서열을 갖고, RPS8의 5' UTR 서열에 대하여 71% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·RPS8의 5' UTR 서열을 갖고, RPS8의 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·RPS8의 5' UTR 서열을 갖고, RPS8의 5' UTR 서열에 대해 29% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·RPS8의 5' UTR 서열을 갖고, RPS8의 5' UTR 서열에 대해 14% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·RPS8의 5' UTR 서열을 갖고, 3' UTR 서열을 갖지 않는 인공 합성 mRNA
·5' UTR 서열 및 3' UTR 서열을 갖지 않는 인공 합성 mRNA
·GAPDH의 5' UTR 서열을 갖고, GAPDH의 3' UTR 서열을 갖는 인공 합성 mRNA
도 27의 결과로부터, RPS8의 5' UTR 서열에 대해 71% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA 및 RPS8의 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열 을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA가 우수한 번역 효율을 나타내는 것을 알 수 있었다.
(x) 실험 10
도 28은 LDHB의 5' UTR을 사용한 경우에 있어서의 발현량의 차이를 나타내는 도면이다. 도면의 세로축은 발현량(발현(배수 증가))을 나타낸다. 이 실험에서는 이하의 mRNA를 사용했다. 도면은 LDHB의 5' UTR 서열을 갖고 3' UTR 서열을 갖지 않는 인공 합성 mRNA의 발현량을 1로 한 경우의 발현량의 상대값을 나타낸다.
·LDHB의 5' UTR 서열을 갖고, LDHB의 5' UTR 서열에 대해 100% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·LDHB의 5' UTR 서열을 갖고, LDHB의 5' UTR 서열에 대해 93% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·LDHB의 5' UTR 서열을 갖고, LDHB의 5' UTR 서열에 대해 87% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·LDHB의 5' UTR 서열을 갖고, LDHB의 5' UTR 서열에 대해 75% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·LDHB의 5' UTR 서열을 갖고, LDHB의 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·LDHB의 5' UTR 서열을 갖고, LDHB의 5' UTR 서열에 대해 25% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA
·LDHB의 5' UTR 서열을 갖고, 3' UTR 서열을 갖지 않는 인공 합성 mRNA
도 28의 결과로부터, LDHB의 5' UTR 서열에 대해 75% 상보성의 3' UTR 서열을 갖는 인공 합성 mRNA 및 LDHB의 5' UTR 서열에 대해 50% 상보성의 3' UTR 서열 을 갖는 3' UTR 서열을 갖는 인공 합성 mRNA가 우수한 번역 효율을 나타내는 것을 알 수 있었다.
(산업상 이용가능성)
본 발명에 의하면, 표적 세포에 도입한 인공 합성 mRNA의 번역 효율이 향상되어, 목적 유전자의 고발현이 가능하게 된다. 본 발명의 용도로서, 예를 들면, mRNA 의약(각종 바이러스성 질환의 치료, 암 면역 요법 등), iPS 세포의 제작, 줄기 세포 또는 전구 세포의 분화 유도가 상정된다.
본 발명은 상기 발명의 실시형태 및 실시예의 설명에 전혀 한정되지 않는다. 특허청구범위의 기재를 일탈하지 않고, 당업자가 용이하게 생각할 수 있는 범위에서 다양한 변형 형태도 본 발명에 포함된다. 본 명세서에서 명시한 논문, 공개특허공보 및 특허공보 등의 내용은 그 모든 내용을 원용에 의해 인용하는 것으로 한다.
SEQUENCE LISTING
<110> PUBLIC UNIVERSITY CORPORATION NAGOYA CITY UNIVERSITY
<120> mRNA and use thereof
<130> PA0467WO
<150> JP 2019-189929
<151> 2019-10-17
<160> 116
<170> PatentIn version 3.5
<210> 1
<211> 66
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 1
cgctctctgc tcctcctgtt cgacagtcag ccgcatcttc ttttgcgtcg ccagccgagc 60
cacatc 66
<210> 2
<211> 74
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 2
tcgagatgtg gctcggctgg cgacgcaaaa gaagatgcgg ctgactgtcg aacaggagga 60
gcagagagcg gtac 74
<210> 3
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 3
aaggaattcg acccctggac caccagc 27
<210> 4
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 4
ttttctagaa ctggttgagc acagggta 28
<210> 5
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 5
ttaggtaccg gtgcgcgggg aggtggaggg 30
<210> 6
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 6
tgcctcgagc ttgcttctcc tgccgccgtc 30
<210> 7
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 7
agggaattca ttgcttaaac tttgaacaac 30
<210> 8
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 8
ttttctagaa atgattgatc tgatgttcct 30
<210> 9
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 9
aacggtaccc ccctcctccc tccttgcaga 30
<210> 10
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 10
agtctcgagt ttgcacagga gagagaaggc 30
<210> 11
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 11
gacgaattcc tagtgagctc taggctgtag 30
<210> 12
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 12
ttttctagac acactacaat agttaatttt 30
<210> 13
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 13
aattatgtgg ctcggctggc gacgcaaaag aagatgcggc tgactgtcga acaggaggag 60
cagagagc 68
<210> 14
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 14
ctaggctctc tgctcctcct gttcgacagt cagccgcatc ttcttttgcg tcgccagccg 60
agccacat 68
<210> 15
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 15
aattatgtgg ctcggctggg gacgcaaaag aagatccggc tgactgtcga agaggaggag 60
cagagagg 68
<210> 16
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 16
ctagcctctc tgctcctcct cttcgacagt cagccggatc ttcttttgcg tccccagccg 60
agccacat 68
<210> 17
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 17
aattatgtgg cacggctggg gacgcaacag aagatccggc tgagtgtcga agaggaggaa 60
cagagagg 68
<210> 18
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 18
ctagcctctc tgttcctcct cttcgacact cagccggatc ttctgttgcg tccccagccg 60
tgccacat 68
<210> 19
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 19
aattatgcgg cacggatgga gactcaacag acgatccgga tgagtgtaga agaggcggaa 60
cagtgagg 68
<210> 20
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> CTAGCCTCACTGTTCCGCCTCTTCTACACTCATCCGGATCGTCTGTTGAGTCTCCATCCGTGCCG
CAT
<400> 20
agtctcgagt ttgcacagga gagagaaggc 30
<210> 21
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 21
aattacgcga cacagatcga gtctctacat acgctccaga tcagtctagt agacgcgcaa 60
ctgtgcgg 68
<210> 22
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 22
ctagccgcac agttgcgcgt ctactagact gatctggagc gtatgtagag actcgatctg 60
tgtcgcgt 68
<210> 23
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 23
aattacacga aacacatcaa gtgtctccat tcgccccaaa tctgtcaagt tgacacgcta 60
ctttgcag 68
<210> 24
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 24
ctagctgcaa agtagcgtgt caacttgaca gatttggggc gaatggagac acttgatgtg 60
tttcgtgt 68
<210> 25
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 25
tgcaagcttt cagtctgagt caggccct 28
<210> 26
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 26
gtcaagctta tggaggagcc gcagtcag 28
<210> 27
<211> 64
<212> RNA
<213> Homo sapiens
<400> 27
gcucucugcu ccuccuguuc gacagucagc cgcaucuucu uuugcgucgc cagccgagcc 60
acau 64
<210> 28
<211> 203
<212> RNA
<213> Homo sapiens
<400> 28
gaccccugga ccaccagccc cagcaagagc acaagaggaa gagagagacc cucacugcug 60
gggagucccu gccacacuca gucccccacc acacugaauc uccccuccuc acaguugcca 120
uguagacccc uugaagaggg gaggggccua gggagccgca ccuugucaug uaccaucaau 180
aaaguacccu gugcucaacc agu 203
<210> 29
<211> 131
<212> RNA
<213> Homo sapiens
<400> 29
ggugcgcggg gagguggagg gcgaggggcg gggcuaccuc aggucccgcc cgcggcaggc 60
cugugggcug cgaggaggag cuuugccuag cuugcaggca gcgcagggca gacggcggca 120
ggagaagcaa g 131
<210> 30
<211> 224
<212> RNA
<213> Homo sapiens
<400> 30
auugcuuaaa cuuugaacaa ccucaauuuc uuuuuaaacu aauaaaguac uagguugcaa 60
uaugugaaau cagaggacca aaguacagau ggaaaccauu uccuacauca caaaaaccca 120
aguuuacagc uuguacuuua cuuuaaugug uaauacucaa cucaagguac aagacaauug 180
cauuuaacau uguuauaaau aaaaggaaca ucagaucaau cauu 224
<210> 31
<211> 111
<212> RNA
<213> Homo sapiens
<400> 31
ccccuccucc cuccuugcag agccggcgcc ggaggagacg cacgcagcug acuuugucuu 60
cuccgcacga cuguuacaga ggucuccaga gccuucucuc uccugugcaa a 111
<210> 32
<211> 200
<212> RNA
<213> Homo sapiens
<400> 32
cuagugagcu cuaggcugua gaaauuuaaa aacuacaaug ugauuaacuc gagccuuuag 60
uuuucaucca uguacaugga ucacaguuug cuuugaucuu cuucaauaug ugaauuuggg 120
cucacagaau caaagccuau gcuugguuua augcuugcaa ucugagcucu ugaacaaaua 180
aaauuaacua uuguagugug 200
<210> 33
<211> 64
<212> RNA
<213> Homo sapiens
<400> 33
auguggcucg gcuggcgacg caaaagaaga ugcggcugac ugucgaacag gaggagcaga 60
gagc 64
<210> 34
<211> 64
<212> RNA
<213> Homo sapiens
<400> 34
auguggcucg gcuggggacg caaaagaaga uccggcugac ugucgaagag gaggagcaga 60
gagg 64
<210> 35
<211> 64
<212> RNA
<213> Homo sapiens
<400> 35
auguggcacg gcuggggacg caacagaaga uccggcugag ugucgaagag gaggaacaga 60
gagg 64
<210> 36
<211> 64
<212> RNA
<213> Homo sapiens
<400> 36
augcggcacg gauggagacu caacagacga uccggaugag uguagaagag gcggaacagu 60
gagg 64
<210> 37
<211> 64
<212> RNA
<213> Homo sapiens
<400> 37
acgcgacaca gaucgagucu cuacauacgc uccagaucag ucuaguagac gcgcaacugu 60
gcgg 64
<210> 38
<211> 64
<212> RNA
<213> Homo sapiens
<400> 38
acacgaaaca caucaagugu cuccauucgc cccaaaucug ucaaguugac acgcuacuuu 60
gcag 64
<210> 39
<211> 1182
<212> DNA
<213> Homo sapiens
<400> 39
atggaggagc cgcagtcaga tcctagcgtc gagccccctc tgagtcagga aacattttca 60
gacctatgga aactacttcc tgaaaacaac gttctgtccc ccttgccgtc ccaagcaatg 120
gatgatttga tgctgtcccc ggacgatatt gaacaatggt tcactgaaga cccaggtcca 180
gatgaagctc ccagaatgcc agaggctgct ccccccgtgg cccctgcacc agcagctcct 240
acaccggcgg cccctgcacc agccccctcc tggcccctgt catcttctgt cccttcccag 300
aaaacctacc agggcagcta cggtttccgt ctgggcttct tgcattctgg gacagccaag 360
tctgtgactt gcacgtactc ccctgccctc aacaagatgt tttgccaact ggccaagacc 420
tgccctgtgc agctgtgggt tgattccaca cccccgcccg gcacccgcgt ccgcgccatg 480
gccatctaca agcagtcaca gcacatgacg gaggttgtga ggcgctgccc ccaccatgag 540
cgctgctcag atagcgatgg tctggcccct cctcagcatc ttatccgagt ggaaggaaat 600
ttgcgtgtgg agtatttgga tgacagaaac acttttcgac atagtgtggt ggtgccctat 660
gagccgcctg aggttggctc tgactgtacc accatccact acaactacat gtgtaacagt 720
tcctgcatgg gcggcatgaa ccggaggccc atcctcacca tcatcacact ggaagactcc 780
agtggtaatc tactgggacg gaacagcttt gaggtgcgtg tttgtgcctg tcctgggaga 840
gaccggcgca cagaggaaga gaatctccgc aagaaagggg agcctcacca cgagctgccc 900
ccagggagca ctaagcgagc actgcccaac aacaccagct cctctcccca gccaaagaag 960
aaaccactgg atggagaata tttcaccctt cagatccgtg ggcgtgagcg cttcgagatg 1020
ttccgagagc tgaatgaggc cttggaactc aaggatgccc aggctgggaa ggagccaggg 1080
gggagcaggg ctcactccag ccacctgaag tccaaaaagg gtcagtctac ctcccgccat 1140
aaaaaactca tgttcaagac agaagggcct gactcagact ga 1182
<210> 40
<211> 393
<212> PRT
<213> Homo sapiens
<400> 40
Met Glu Glu Pro Gln Ser Asp Pro Ser Val Glu Pro Pro Leu Ser Gln
1 5 10 15
Glu Thr Phe Ser Asp Leu Trp Lys Leu Leu Pro Glu Asn Asn Val Leu
20 25 30
Ser Pro Leu Pro Ser Gln Ala Met Asp Asp Leu Met Leu Ser Pro Asp
35 40 45
Asp Ile Glu Gln Trp Phe Thr Glu Asp Pro Gly Pro Asp Glu Ala Pro
50 55 60
Arg Met Pro Glu Ala Ala Pro Pro Val Ala Pro Ala Pro Ala Ala Pro
65 70 75 80
Thr Pro Ala Ala Pro Ala Pro Ala Pro Ser Trp Pro Leu Ser Ser Ser
85 90 95
Val Pro Ser Gln Lys Thr Tyr Gln Gly Ser Tyr Gly Phe Arg Leu Gly
100 105 110
Phe Leu His Ser Gly Thr Ala Lys Ser Val Thr Cys Thr Tyr Ser Pro
115 120 125
Ala Leu Asn Lys Met Phe Cys Gln Leu Ala Lys Thr Cys Pro Val Gln
130 135 140
Leu Trp Val Asp Ser Thr Pro Pro Pro Gly Thr Arg Val Arg Ala Met
145 150 155 160
Ala Ile Tyr Lys Gln Ser Gln His Met Thr Glu Val Val Arg Arg Cys
165 170 175
Pro His His Glu Arg Cys Ser Asp Ser Asp Gly Leu Ala Pro Pro Gln
180 185 190
His Leu Ile Arg Val Glu Gly Asn Leu Arg Val Glu Tyr Leu Asp Asp
195 200 205
Arg Asn Thr Phe Arg His Ser Val Val Val Pro Tyr Glu Pro Pro Glu
210 215 220
Val Gly Ser Asp Cys Thr Thr Ile His Tyr Asn Tyr Met Cys Asn Ser
225 230 235 240
Ser Cys Met Gly Gly Met Asn Arg Arg Pro Ile Leu Thr Ile Ile Thr
245 250 255
Leu Glu Asp Ser Ser Gly Asn Leu Leu Gly Arg Asn Ser Phe Glu Val
260 265 270
Arg Val Cys Ala Cys Pro Gly Arg Asp Arg Arg Thr Glu Glu Glu Asn
275 280 285
Leu Arg Lys Lys Gly Glu Pro His His Glu Leu Pro Pro Gly Ser Thr
290 295 300
Lys Arg Ala Leu Pro Asn Asn Thr Ser Ser Ser Pro Gln Pro Lys Lys
305 310 315 320
Lys Pro Leu Asp Gly Glu Tyr Phe Thr Leu Gln Ile Arg Gly Arg Glu
325 330 335
Arg Phe Glu Met Phe Arg Glu Leu Asn Glu Ala Leu Glu Leu Lys Asp
340 345 350
Ala Gln Ala Gly Lys Glu Pro Gly Gly Ser Arg Ala His Ser Ser His
355 360 365
Leu Lys Ser Lys Lys Gly Gln Ser Thr Ser Arg His Lys Lys Leu Met
370 375 380
Phe Lys Thr Glu Gly Pro Asp Ser Asp
385 390
<210> 41
<211> 50
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 41
acatttgctt ctgacacaac tgtgttcact agcaacctca aacagacacc 50
<210> 42
<211> 58
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 42
tcgaggtgtc tgtttgaggt tgctagtgaa cacagttgtg tcagaagcaa atgtgtac 58
<210> 43
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 43
aattggtgtc tgtttgaggt tgctagtgaa cacagttgtg tcagaagcaa atgt 54
<210> 44
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 44
ctagacattt gcttctgaca caactgtgtt cactagcaac ctcaaacaga cacc 54
<210> 45
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 45
aattgctgtc tgtttgaggt tactagtgaa cacagttctg tcagaagcaa atgc 54
<210> 46
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 46
ctaggcattt gcttctgaca gaactgtgtt cactagtaac ctcaaacaga cagc 54
<210> 47
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 47
aattgctgtc tgtgtgaggt tactagtgat cacagttctg tcagacgcaa atgc 54
<210> 48
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 48
ctaggcattt gcgtctgaca gaactgtgat cactagtaac ctcacacaga cagc 54
<210> 49
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 49
aattgctgta tgtgtgatgt tactactgat caccgttctg taagacgcac atgc 54
<210> 50
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 50
ctaggcatgt gcgtcttaca gaacggtgat cagtagtaac atcacacata cagc 54
<210> 51
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 51
aattgctata tctgtcatgc tacgactcat ctccgatctc taacacggac acgc 54
<210> 52
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 52
ctaggcgtgt ccgtgttaga gatcggagat gagtcgtagc atgacagata tagc 54
<210> 53
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 53
aattgctaga tcagtcttgc aacgcctcat ctgcgagctc gaactcggtc actc 54
<210> 54
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 54
ctaggagtga ccgagttcga gctcgcagat gaggcgttgc aagactgatc tagc 54
<210> 55
<211> 57
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 55
gttttacaaa ccgaaccgtg aatctttgcg gtttctcttt ccagccagcg ccgagcg 57
<210> 56
<211> 65
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 56
tcgacgctcg gcgctggctg gaaagagaaa ccgcaaagat tcacggttcg gtttgtaaaa 60
cgtac 65
<210> 57
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 57
aattcgctcg gcgctggctg gaaagagaaa ccgcaaagat tcacggttcg gtttgtaaaa 60
<210> 58
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 58
ctagttttac aaaccgaacc gtgaatcttt gcggtttctc tttccagcca gcgccgagcg 60
<210> 59
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 59
aattagctcg gcgctggcag gaaagagaaa cctcaaagat tcacggctcg gtttgtaaaa 60
<210> 60
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 60
ctagttttac aaaccgagcc gtgaatcttt gaggtttctc tttcctgcca gcgccgagct 60
<210> 61
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 61
aattagctcg gagctggcag gaaagcgaaa cctcaaagac tcacggctcg gttcgtaaaa 60
<210> 62
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 62
ctagttttac gaaccgagcc gtgagtcttt gaggtttcgc tttcctgcca gctccgagct 60
<210> 63
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 63
aattagctgg gagcttgcag gatagcgaat cctcaatgac tcaaggctcg cttcgtacaa 60
<210> 64
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 64
ctagttgtac gaagcgagcc ttgagtcatt gaggattcgc tatcctgcaa gctcccagct 60
<210> 65
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 65
aattagatgt gagattgcag cattgcgcat cctccatcac tgaaggctag cgtcgcacaa 60
<210> 66
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 66
ctagttgtgc gacgctagcc ttcagtgatg gaggatgcgc aatgctgcaa tctcacatct 60
<210> 67
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 67
aattatatgt gatattccac cattgcacat gcttcatcac ggaatgcgag cgtcacacta 60
<210> 68
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 68
ctagtagtgt gacgctcgca ttccgtgatg aagcatgtgc aatggtggaa tatcacatat 60
<210> 69
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 69
aattataggt gatagtccac ccttgcactt gcttcctcac ggcatgcgat cgtcactcta 60
<210> 70
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 70
ctagtagagt gacgatcgca tgccgtgagg aagcaagtgc aagggtggac tatcacctat 60
<210> 71
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 71
aattcagtcg tgcggagaag acaaagtcag ctgcgtgggt ctcctccggc gccggctctg 60
caag 64
<210> 72
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 72
ctagcttgca gagccggcgc cggaggagac ccacgcagct gactttgtct tctccgcacg 60
actg 64
<210> 73
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 73
aattcagtcg tgcggagatg acaaagtcag ctgggtgggt ctcctccgac gccggctctg 60
caac 64
<210> 74
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 74
ctaggttgca gagccggcgt cggaggagac ccacccagct gactttgtca tctccgcacg 60
actg 64
<210> 75
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 75
aattcagacg tgcggtgaag acacagtcag cagcgtgggg ctcctcctgc gccggatctg 60
caac 64
<210> 76
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 76
ctaggttgca gatccggcgc aggaggagcc ccacgctgct gactgtgtct tcaccgcacg 60
tctg 64
<210> 77
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 77
aattcaggcg tacggcgaac acacagtgag cagcgcggga ctcgtcctgc ggcggatcta 60
caac 64
<210> 78
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 78
ctaggttgta gatccgccgc aggacgagtc ccgcgctgct cactgtgtgt tcgccgtacg 60
cctg 64
<210> 79
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 79
aattccggcc taccgcgtac agacagtgac cagagcgcga cgcgtactgg ggcagatgta 60
ctac 64
<210> 80
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 80
ctaggtagta catctgcccc agtacgcgtc gcgctctggt cactgtctgt acgcggtagg 60
ccgg 64
<210> 81
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 81
aattccagcc aaccacgttc agccagggac aagaccgcca cgggtagtgg cgcaaatgga 60
cttc 64
<210> 82
<211> 64
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 82
ctaggaagtc catttgcgcc actacccgtg gcggtcttgt ccctggctga acgtggttgg 60
ctgg 64
<210> 83
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 83
atggacaaga agtactccat tgggctcgat 30
<210> 84
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 84
gattcacacc ttcctcttct tcttggg 27
<210> 85
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 85
ccgggcctcc atggccatca g 21
<210> 86
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 86
catcttcttt tgcgtcgcca gccgagccac atctcgagcc caccatggca tcaatg 56
<210> 87
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 87
cggctgactg tcgaacagga ggagcagaga gcggtaccca attcgcccta tagtga 56
<210> 88
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 88
cggatgagtg tagaagaggc ggaacagtga ggcatttatt ttcattgcaa tgatgtctag 60
<210> 89
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 89
gatcgtctgt tgagtctcca tccgtgccgc attcacacct tcctcttctt cttggg 56
<210> 90
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 90
cagatcagtc tagtagacgc gcaactgtgc ggcatttatt ttcattgcaa tgatgtctag 60
<210> 91
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 91
gagcgtatgt agagactcga tctgtgtcgc gttcacacct tcctcttctt cttggg 56
<210> 92
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 92
ccatttcctg gagccatctc tctcc 25
<210> 93
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> primer
<400> 93
ctctcctggg cttgccaagg actca 25
<210> 94
<211> 50
<212> RNA
<213> Homo sapiens
<400> 94
acauuugcuu cugacacaac uguguucacu agcaaccuca aacagacacc 50
<210> 95
<211> 50
<212> RNA
<213> Homo sapiens
<400> 95
ggugucuguu ugagguugcu agugaacaca guugugucag aagcaaaugu 50
<210> 96
<211> 50
<212> RNA
<213> Homo sapiens
<400> 96
gcugucuguu ugagguuacu agugaacaca guucugucag aagcaaaugc 50
<210> 97
<211> 50
<212> RNA
<213> Homo sapiens
<400> 97
gcugucugug ugagguuacu agugaucaca guucugucag acgcaaaugc 50
<210> 98
<211> 50
<212> RNA
<213> Homo sapiens
<400> 98
gcuguaugug ugauguuacu acugaucacc guucuguaag acgcacaugc 50
<210> 99
<211> 50
<212> RNA
<213> Homo sapiens
<400> 99
gcuauaucug ucaugcuacg acucaucucc gaucucuaac acggacacgc 50
<210> 100
<211> 50
<212> RNA
<213> Homo sapiens
<400> 100
gcuagaucag ucuugcaacg ccucaucugc gagcucgaac ucggucacuc 50
<210> 101
<211> 56
<212> RNA
<213> Homo sapiens
<400> 101
uuuuacaaac cgaaccguga aucuuugcgg uuucucuuuc cagccagcgc cgagcg 56
<210> 102
<211> 56
<212> RNA
<213> Homo sapiens
<400> 102
cgcucggcgc uggcuggaaa gagaaaccgc aaagauucac gguucgguuu guaaaa 56
<210> 103
<211> 56
<212> RNA
<213> Homo sapiens
<400> 103
agcucggcgc uggcaggaaa gagaaaccuc aaagauucac ggcucgguuu guaaaa 56
<210> 104
<211> 56
<212> RNA
<213> Homo sapiens
<400> 104
agcucggagc uggcaggaaa gcgaaaccuc aaagacucac ggcucgguuc guaaaa 56
<210> 105
<211> 56
<212> RNA
<213> Homo sapiens
<400> 105
agcugggagc uugcaggaua gcgaauccuc aaugacucaa ggcucgcuuc guacaa 56
<210> 106
<211> 56
<212> RNA
<213> Homo sapiens
<400> 106
agaugugaga uugcagcauu gcgcauccuc caucacugaa ggcuagcguc gcacaa 56
<210> 107
<211> 56
<212> RNA
<213> Homo sapiens
<400> 107
auaugugaua uuccaccauu gcacaugcuu caucacggaa ugcgagcguc acacua 56
<210> 108
<211> 56
<212> RNA
<213> Homo sapiens
<400> 108
auaggugaua guccacccuu gcacuugcuu ccucacggca ugcgaucguc acucua 56
<210> 109
<211> 60
<212> RNA
<213> Homo sapiens
<400> 109
cagucgugcg gagaagacaa agucagcugc gugggucucc uccggcgccg gcucugcaag 60
<210> 110
<211> 60
<212> RNA
<213> Homo sapiens
<400> 110
cagucgugcg gagaugacaa agucagcugg gugggucucc uccgacgccg gcucugcaac 60
<210> 111
<211> 60
<212> RNA
<213> Homo sapiens
<400> 111
cagacgugcg gugaagacac agucagcagc guggggcucc uccugcgccg gaucugcaac 60
<210> 112
<211> 60
<212> RNA
<213> Homo sapiens
<400> 112
caggcguacg gcgaacacac agugagcagc gcgggacucg uccugcggcg gaucuacaac 60
<210> 113
<211> 60
<212> RNA
<213> Homo sapiens
<400> 113
ccggccuacc gcguacagac agugaccaga gcgcgacgcg uacuggggca gauguacuac 60
<210> 114
<211> 60
<212> RNA
<213> Homo sapiens
<400> 114
ccagccaacc acguucagcc agggacaaga ccgccacggg uaguggcgca aauggacuuc 60
<210> 115
<211> 4140
<212> DNA
<213> Homo sapiens
<400> 115
atggacaaga agtactccat tgggctcgat atcggcacaa acagcgtcgg ctgggccgtc 60
attacggacg agtacaaggt gccgagcaaa aaattcaaag ttctgggcaa taccgatcgc 120
cacagcataa agaagaacct cattggcgcc ctcctgttcg actccgggga gacggccgaa 180
gccacgcggc tcaaaagaac agcacggcgc agatataccc gcagaaagaa tcggatctgc 240
tacctgcagg agatctttag taatgagatg gctaaggtgg atgactcttt cttccatagg 300
ctggaggagt cctttttggt ggaggaggat aaaaagcacg agcgccaccc aatctttggc 360
aatatcgtgg acgaggtggc gtaccatgaa aagtacccaa ccatatatca tctgaggaag 420
aagcttgtag acagtactga taaggctgac ttgcggttga tctatctcgc gctggcgcat 480
atgatcaaat ttcggggaca cttcctcatc gagggggacc tgaacccaga caacagcgat 540
gtcgacaaac tctttatcca actggttcag acttacaatc agcttttcga agagaacccg 600
atcaacgcat ccggagttga cgccaaagca atcctgagcg ctaggctgtc caaatcccgg 660
cggctcgaaa acctcatcgc acagctccct ggggagaaga agaacggcct gtttggtaat 720
cttatcgccc tgtcactcgg gctgaccccc aactttaaat ctaacttcga cctggccgaa 780
gatgccaagc ttcaactgag caaagacacc tacgatgatg atctcgacaa tctgctggcc 840
cagatcggcg accagtacgc agaccttttt ttggcggcaa agaacctgtc agacgccatt 900
ctgctgagtg atattctgcg agtgaacacg gagatcacca aagctccgct gagcgctagt 960
atgatcaagc gctatgatga gcaccaccaa gacttgactt tgctgaaggc ccttgtcaga 1020
cagcaactgc ctgagaagta caaggaaatt ttcttcgatc agtctaaaaa tggctacgcc 1080
ggatacattg acggcggagc aagccaggag gaattttaca aatttattaa gcccatcttg 1140
gaaaaaatgg acggcaccga ggagctgctg gtaaagctta acagagaaga tctgttgcgc 1200
aaacagcgca ctttcgacaa tggaagcatc ccccaccaga ttcacctggg cgaactgcac 1260
gctatcctca ggcggcaaga ggatttctac ccctttttga aagataacag ggaaaagatt 1320
gagaaaatcc tcacatttcg gataccctac tatgtaggcc ccctcgcccg gggaaattcc 1380
agattcgcgt ggatgactcg caaatcagaa gagaccatca ctccctggaa cttcgaggaa 1440
gtcgtggata agggggcctc tgcccagtcc ttcatcgaaa ggatgactaa ctttgataaa 1500
aatctgccta acgaaaaggt gcttcctaaa cactctctgc tgtacgagta cttcacagtt 1560
tataacgagc tcaccaaggt caaatacgtc acagaaggga tgagaaagcc agcattcctg 1620
tctggagagc agaagaaagc tatcgtggac ctcctcttca agacgaaccg gaaagttacc 1680
gtgaaacagc tcaaagaaga ctatttcaaa aagattgaat gtttcgactc tgttgaaatc 1740
agcggagtgg aggatcgctt caacgcatcc ctgggaacgt atcacgatct cctgaaaatc 1800
attaaagaca aggacttcct ggacaatgag gagaacgagg acattcttga ggacattgtc 1860
ctcaccctta cgttgtttga agatagggag atgattgaag aacgcttgaa aacttacgct 1920
catctcttcg acgacaaagt catgaaacag ctcaagaggc gccgatatac aggatggggg 1980
cggctgtcaa gaaaactgat caatgggatc cgagacaagc agagtggaaa gacaatcctg 2040
gattttctta agtccgatgg atttgccaac cggaacttca tgcagttgat ccatgatgac 2100
tctctcacct ttaaggagga catccagaaa gcacaagttt ctggccaggg ggacagtctt 2160
cacgagcaca tcgctaatct tgcaggtagc ccagctatca aaaagggaat actgcagacc 2220
gttaaggtcg tggatgaact cgtcaaagta atgggaaggc ataagcccga gaatatcgtt 2280
atcgagatgg cccgagagaa ccaaactacc cagaagggac agaagaacag tagggaaagg 2340
atgaagagga ttgaagaggg tataaaagaa ctggggtccc aaatccttaa ggaacaccca 2400
gttgaaaaca cccagcttca gaatgagaag ctctacctgt actacctgca gaacggcagg 2460
gacatgtacg tggatcagga actggacatc aatcggctct ccgactacga cgtggatcat 2520
atcgtgcccc agtcttttct caaagatgat tctattgata ataaagtgtt gacaagatcc 2580
gataaaaata gagggaagag tgataacgtc ccctcagaag aagttgtcaa gaaaatgaaa 2640
aattattggc ggcagctgct gaacgccaaa ctgatcacac aacggaagtt cgataatctg 2700
actaaggctg aacgaggtgg cctgtctgag ttggataaag ccggcttcat caaaaggcag 2760
cttgttgaga cacgccagat caccaagcac gtggcccaaa ttctcgattc acgcatgaac 2820
accaagtacg atgaaaatga caaactgatt cgagaggtga aagttattac tctgaagtct 2880
aagctggtct cagatttcag aaaggacttt cagttttata aggtgagaga gatcaacaat 2940
taccaccatg cgcatgatgc ctacctgaat gcagtggtag gcactgcact tatcaaaaaa 3000
tatcccaagc ttgaatctga atttgtttac ggagactata aagtgtacga tgttaggaaa 3060
atgatcgcaa agtctgagca ggaaataggc aaggccaccg ctaagtactt cttttacagc 3120
aatattatga attttttcaa gaccgagatt acactggcca atggagagat tcggaagcga 3180
ccacttatcg aaacaaacgg agaaacagga gaaatcgtgt gggacaaggg tagggatttc 3240
gcgacagtcc ggaaggtcct gtccatgccg caggtgaaca tcgttaaaaa gaccgaagta 3300
cagaccggag gcttctccaa ggaaagtatc ctcccgaaaa ggaacagcga caagctgatc 3360
gcacgcaaaa aagattggga ccccaagaaa tacggcggat tcgattctcc tacagtcgct 3420
tacagtgtac tggttgtggc caaagtggag aaagggaagt ctaaaaaact caaaagcgtc 3480
aaggaactgc tgggcatcac aatcatggag cgatcaagct tcgaaaaaaa ccccatcgac 3540
tttctcgagg cgaaaggata taaagaggtc aaaaaagacc tcatcattaa gcttcccaag 3600
tactctctct ttgagcttga aaacggccgg aaacgaatgc tcgctagtgc gggcgagctg 3660
cagaaaggta acgagctggc actgccctct aaatacgtta atttcttgta tctggccagc 3720
cactatgaaa agctcaaagg gtctcccgaa gataatgagc agaagcagct gttcgtggaa 3780
caacacaaac actaccttga tgagatcatc gagcaaataa gcgaattctc caaaagagtg 3840
atcctcgccg acgctaacct cgataaggtg ctttctgctt acaataagca cagggataag 3900
cccatcaggg agcaggcaga aaacattatc cacttgttta ctctgaccaa cttgggcgcg 3960
cctgcagcct tcaagtactt cgacaccacc atagacagaa agcggtacac ctctacaaag 4020
gaggtcctgg acgccacact gattcatcag tcaattacgg ggctctatga aacaagaatc 4080
gacctctctc agctcggtgg agacagcagg gctgacccca agaagaagag gaaggtgtga 4140
<210> 116
<211> 1379
<212> PRT
<213> Homo sapiens
<400> 116
Met Asp Lys Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val
1 5 10 15
Gly Trp Ala Val Ile Thr Asp Glu Tyr Lys Val Pro Ser Lys Lys Phe
20 25 30
Lys Val Leu Gly Asn Thr Asp Arg His Ser Ile Lys Lys Asn Leu Ile
35 40 45
Gly Ala Leu Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala Thr Arg Leu
50 55 60
Lys Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn Arg Ile Cys
65 70 75 80
Tyr Leu Gln Glu Ile Phe Ser Asn Glu Met Ala Lys Val Asp Asp Ser
85 90 95
Phe Phe His Arg Leu Glu Glu Ser Phe Leu Val Glu Glu Asp Lys Lys
100 105 110
His Glu Arg His Pro Ile Phe Gly Asn Ile Val Asp Glu Val Ala Tyr
115 120 125
His Glu Lys Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys Leu Val Asp
130 135 140
Ser Thr Asp Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala Leu Ala His
145 150 155 160
Met Ile Lys Phe Arg Gly His Phe Leu Ile Glu Gly Asp Leu Asn Pro
165 170 175
Asp Asn Ser Asp Val Asp Lys Leu Phe Ile Gln Leu Val Gln Thr Tyr
180 185 190
Asn Gln Leu Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly Val Asp Ala
195 200 205
Lys Ala Ile Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg Leu Glu Asn
210 215 220
Leu Ile Ala Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu Phe Gly Asn
225 230 235 240
Leu Ile Ala Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys Ser Asn Phe
245 250 255
Asp Leu Ala Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp Thr Tyr Asp
260 265 270
Asp Asp Leu Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln Tyr Ala Asp
275 280 285
Leu Phe Leu Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu Leu Ser Asp
290 295 300
Ile Leu Arg Val Asn Thr Glu Ile Thr Lys Ala Pro Leu Ser Ala Ser
305 310 315 320
Met Ile Lys Arg Tyr Asp Glu His His Gln Asp Leu Thr Leu Leu Lys
325 330 335
Ala Leu Val Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu Ile Phe Phe
340 345 350
Asp Gln Ser Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Gly Ala Ser
355 360 365
Gln Glu Glu Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu Lys Met Asp
370 375 380
Gly Thr Glu Glu Leu Leu Val Lys Leu Asn Arg Glu Asp Leu Leu Arg
385 390 395 400
Lys Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu
405 410 415
Gly Glu Leu His Ala Ile Leu Arg Arg Gln Glu Asp Phe Tyr Pro Phe
420 425 430
Leu Lys Asp Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr Phe Arg Ile
435 440 445
Pro Tyr Tyr Val Gly Pro Leu Ala Arg Gly Asn Ser Arg Phe Ala Trp
450 455 460
Met Thr Arg Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn Phe Glu Glu
465 470 475 480
Val Val Asp Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu Arg Met Thr
485 490 495
Asn Phe Asp Lys Asn Leu Pro Asn Glu Lys Val Leu Pro Lys His Ser
500 505 510
Leu Leu Tyr Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys
515 520 525
Tyr Val Thr Glu Gly Met Arg Lys Pro Ala Phe Leu Ser Gly Glu Gln
530 535 540
Lys Lys Ala Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys Val Thr
545 550 555 560
Val Lys Gln Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys Phe Asp
565 570 575
Ser Val Glu Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser Leu Gly
580 585 590
Thr Tyr His Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe Leu Asp
595 600 605
Asn Glu Glu Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr Leu Thr
610 615 620
Leu Phe Glu Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr Tyr Ala
625 630 635 640
His Leu Phe Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg Arg Tyr
645 650 655
Thr Gly Trp Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile Arg Asp
660 665 670
Lys Gln Ser Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp Gly Phe
675 680 685
Ala Asn Arg Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu Thr Phe
690 695 700
Lys Glu Asp Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp Ser Leu
705 710 715 720
His Glu His Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys Lys Gly
725 730 735
Ile Leu Gln Thr Val Lys Val Val Asp Glu Leu Val Lys Val Met Gly
740 745 750
Arg His Lys Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln
755 760 765
Thr Thr Gln Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys Arg Ile
770 775 780
Glu Glu Gly Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu His Pro
785 790 795 800
Val Glu Asn Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu
805 810 815
Gln Asn Gly Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile Asn Arg
820 825 830
Leu Ser Asp Tyr Asp Val Asp His Ile Val Pro Gln Ser Phe Leu Lys
835 840 845
Asp Asp Ser Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg
850 855 860
Gly Lys Ser Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys
865 870 875 880
Asn Tyr Trp Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys
885 890 895
Phe Asp Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp
900 905 910
Lys Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr
915 920 925
Lys His Val Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr Asp
930 935 940
Glu Asn Asp Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu Lys Ser
945 950 955 960
Lys Leu Val Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys Val Arg
965 970 975
Glu Ile Asn Asn Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val
980 985 990
Val Gly Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser Glu Phe
995 1000 1005
Val Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met Ile Ala
1010 1015 1020
Lys Ser Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr Phe Phe
1025 1030 1035
Tyr Ser Asn Ile Met Asn Phe Phe Lys Thr Glu Ile Thr Leu Ala
1040 1045 1050
Asn Gly Glu Ile Arg Lys Arg Pro Leu Ile Glu Thr Asn Gly Glu
1055 1060 1065
Thr Gly Glu Ile Val Trp Asp Lys Gly Arg Asp Phe Ala Thr Val
1070 1075 1080
Arg Lys Val Leu Ser Met Pro Gln Val Asn Ile Val Lys Lys Thr
1085 1090 1095
Glu Val Gln Thr Gly Gly Phe Ser Lys Glu Ser Ile Leu Pro Lys
1100 1105 1110
Arg Asn Ser Asp Lys Leu Ile Ala Arg Lys Lys Asp Trp Asp Pro
1115 1120 1125
Lys Lys Tyr Gly Gly Phe Asp Ser Pro Thr Val Ala Tyr Ser Val
1130 1135 1140
Leu Val Val Ala Lys Val Glu Lys Gly Lys Ser Lys Lys Leu Lys
1145 1150 1155
Ser Val Lys Glu Leu Leu Gly Ile Thr Ile Met Glu Arg Ser Ser
1160 1165 1170
Phe Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala Lys Gly Tyr Lys
1175 1180 1185
Glu Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys Tyr Ser Leu
1190 1195 1200
Phe Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser Ala Gly
1205 1210 1215
Glu Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys Tyr Val
1220 1225 1230
Asn Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys Gly Ser
1235 1240 1245
Pro Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln His Lys
1250 1255 1260
His Tyr Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe Ser Lys
1265 1270 1275
Arg Val Ile Leu Ala Asp Ala Asn Leu Asp Lys Val Leu Ser Ala
1280 1285 1290
Tyr Asn Lys His Arg Asp Lys Pro Ile Arg Glu Gln Ala Glu Asn
1295 1300 1305
Ile Ile His Leu Phe Thr Leu Thr Asn Leu Gly Ala Pro Ala Ala
1310 1315 1320
Phe Lys Tyr Phe Asp Thr Thr Ile Asp Arg Lys Arg Tyr Thr Ser
1325 1330 1335
Thr Lys Glu Val Leu Asp Ala Thr Leu Ile His Gln Ser Ile Thr
1340 1345 1350
Gly Leu Tyr Glu Thr Arg Ile Asp Leu Ser Gln Leu Gly Gly Asp
1355 1360 1365
Ser Arg Ala Asp Pro Lys Lys Lys Arg Lys Val
1370 1375
Claims (10)
- 단백질을 코딩하는 mRNA의 5' 비번역 영역과, 상기 5' 비번역 영역에 대한 상보성이 40% 이상 80% 이하인 3' 비번역 영역을 갖춘 인공 합성 mRNA.
- 제1항에 있어서, 상기 단백질은 글리세르알데히드-3-인산 데히드로게나제와, β-글로빈과, RPS8과, LDHB로 이루어진 군으로부터 선택되는 인공 합성 mRNA.
- 제1항 또는 제2항에 있어서, 상기 3' 비번역 영역은 상기 5' 비번역 영역에 대한 상보성이 50% 이상 75% 이하인 인공 합성 mRNA.
- 제1항 내지 제3항 중 어느 한 항에 기재된 인공 합성 mRNA를 세포 내로 도입하는 공정을 포함하는 방법.
- 제1항 내지 제3항 중 어느 한 항에 기재된 인공 합성 mRNA가 도입된 세포.
- 단백질을 코딩하는 mRNA의 5' 비번역 영역과, 상기 5' 비번역 영역과의 상보성이 40% 이상 80% 이하인 3' 비번역 영역을 갖춘 인공 합성 mRNA를 제조하는 공정을 포함하는 인공 합성 mRNA의 제조 방법.
- 글리세르알데하이드-3-인산 데히드로게나제를 코딩하는 mRNA의 5' 비번역 영역과, 글리세르알데하이드-3-인산 디하이드로게나제를 코딩하는 mRNA의 3' 비번역 영역을 갖춘 인공 합성 mRNA.
- 글리세르알데히드-3-인산 데히드로게나제를 코딩하는 mRNA의 5' 비번역 영역과, 글리세르알데히드-3-인산 데히드로게나제를 코딩하는 mRNA의 3' 비번역 영역을 갖춘 mRNA를 제조하는 공정을 포함하는 인공 합성 mRNA의 제조 방법.
- 제7항에 기재된 인공 합성 mRNA를 세포에 도입하는 공정을 포함하는 방법.
- 제7항에 기재된 인공 합성 mRNA가 도입된 세포.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JPJP-P-2019-189929 | 2019-10-17 | ||
| JP2019189929 | 2019-10-17 | ||
| PCT/JP2020/039160 WO2021075567A1 (ja) | 2019-10-17 | 2020-10-16 | 人工合成mRNA及びその利用 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20220108031A true KR20220108031A (ko) | 2022-08-02 |
Family
ID=75538171
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020227012511A KR20220108031A (ko) | 2019-10-17 | 2020-10-16 | 인공 합성 mRNA 및 그 이용 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20240026311A1 (ko) |
| EP (1) | EP4047084A4 (ko) |
| JP (1) | JPWO2021075567A1 (ko) |
| KR (1) | KR20220108031A (ko) |
| CN (1) | CN114599789A (ko) |
| WO (1) | WO2021075567A1 (ko) |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2024162360A1 (ja) * | 2023-01-31 | 2024-08-08 | 株式会社シンプロジェン | タンパク質の発現を増強するための人工合成核酸 |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2015221026A (ja) | 2014-05-23 | 2015-12-10 | 公立大学法人名古屋市立大学 | 人工合成mRNAの翻訳効率化方法 |
| JP2015226531A (ja) | 2014-05-09 | 2015-12-17 | 公立大学法人名古屋市立大学 | 人工合成mRNAの安定化方法 |
| JP2018074954A (ja) | 2016-11-09 | 2018-05-17 | 公立大学法人名古屋市立大学 | 人工合成mRNAの発現を効率化する方法 |
Family Cites Families (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP4535229B2 (ja) | 2003-05-08 | 2010-09-01 | 国立大学法人 東京大学 | ポリエチレングリコール−ポリカチオンブロック共重合体 |
| US9017965B2 (en) * | 2005-12-12 | 2015-04-28 | King Faisal Specialist Hospital & Research Centre | Hybrid 3′ untranslated regions suitable for efficient protein expression in mammalian cells |
| JP4655298B1 (ja) | 2010-02-23 | 2011-03-23 | ナノキャリア株式会社 | 短鎖のカチオン性ポリアミノ酸およびその使用 |
| WO2012005376A1 (ja) | 2010-07-09 | 2012-01-12 | 国立大学法人 東京大学 | 核酸送達用組成物及び担体組成物、それを用いた医薬組成物、並びに核酸送達方法 |
| JP5782704B2 (ja) * | 2010-11-11 | 2015-09-24 | 東ソー株式会社 | GAPDHmRNAの測定方法 |
| CA2859452C (en) * | 2012-03-27 | 2021-12-21 | Curevac Gmbh | Artificial nucleic acid molecules for improved protein or peptide expression |
| EP2831240B1 (en) * | 2012-03-27 | 2017-11-15 | CureVac AG | Artificial nucleic acid molecules comprising a 5'top utr |
| SG11201704681QA (en) * | 2014-12-30 | 2017-07-28 | Curevac Ag | Artificial nucleic acid molecules |
| WO2017100551A1 (en) * | 2015-12-09 | 2017-06-15 | Alexion Pharmaceuticals, Inc. | HETEROLOGOUS UTR SEQUENCES FOR ENHANCED mRNA EXPRESSION |
| US10626413B2 (en) * | 2016-01-15 | 2020-04-21 | Enyu Ding | Nucleic acid vector |
| MA46761A (fr) * | 2016-11-10 | 2019-09-18 | Translate Bio Inc | Administration sous-cutanée d'arn messager |
| CN106929513A (zh) * | 2017-04-07 | 2017-07-07 | 东南大学 | mRNA编码的纳米抗体及其应用 |
| JP7172121B2 (ja) | 2018-04-27 | 2022-11-16 | 株式会社豊田中央研究所 | 電気化学反応セル及びそれを用いた人工光合成セル |
-
2020
- 2020-10-16 JP JP2021552479A patent/JPWO2021075567A1/ja active Pending
- 2020-10-16 WO PCT/JP2020/039160 patent/WO2021075567A1/ja active Application Filing
- 2020-10-16 CN CN202080072851.0A patent/CN114599789A/zh active Pending
- 2020-10-16 EP EP20877848.0A patent/EP4047084A4/en active Pending
- 2020-10-16 US US17/769,655 patent/US20240026311A1/en active Pending
- 2020-10-16 KR KR1020227012511A patent/KR20220108031A/ko not_active Application Discontinuation
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2015226531A (ja) | 2014-05-09 | 2015-12-17 | 公立大学法人名古屋市立大学 | 人工合成mRNAの安定化方法 |
| JP2015221026A (ja) | 2014-05-23 | 2015-12-10 | 公立大学法人名古屋市立大学 | 人工合成mRNAの翻訳効率化方法 |
| JP2018074954A (ja) | 2016-11-09 | 2018-05-17 | 公立大学法人名古屋市立大学 | 人工合成mRNAの発現を効率化する方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| US20240026311A1 (en) | 2024-01-25 |
| EP4047084A1 (en) | 2022-08-24 |
| CN114599789A (zh) | 2022-06-07 |
| WO2021075567A1 (ja) | 2021-04-22 |
| JPWO2021075567A1 (ko) | 2021-04-22 |
| EP4047084A4 (en) | 2024-01-17 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7019639B2 (ja) | 修飾型のヌクレオシド、ヌクレオチドおよび核酸、ならびにそれらの使用方法 | |
| KR102666695B1 (ko) | Rna를 편집하기 위한 방법 및 조성물 | |
| US11339409B2 (en) | Methods and products for expressing proteins in cells | |
| JP4804467B2 (ja) | 多重rnaポリメラーゼiiiプロモーター発現構築物 | |
| CN113939591A (zh) | 编辑rna的方法和组合物 | |
| CN118256490A (zh) | 一种环状rna、组合物和治疗疾病的方法 | |
| Kawakubo et al. | Human RNA-specific adenosine deaminase (ADAR1) gene specifies transcripts that initiate from a constitutively active alternative promoter | |
| JP2017119719A (ja) | 非ヒト脊椎動物用の改変核酸及びその使用方法 | |
| JP2015221026A (ja) | 人工合成mRNAの翻訳効率化方法 | |
| JP2022536951A (ja) | 環状ポリリボヌクレオチドを投与する方法 | |
| KR20220108031A (ko) | 인공 합성 mRNA 및 그 이용 | |
| JP7032775B2 (ja) | 人工合成mRNAの発現を効率化する方法 | |
| CN110904046B (zh) | Islr基因在制备治疗肥胖及改善胰岛素抵抗的药物中的应用 | |
| KR101229821B1 (ko) | 그래뉼린-에피테린 전구체 유전자의 발현을 억제하는 안티센스 및 이를 함유하는 약제학적 조성물 | |
| JP2015226531A (ja) | 人工合成mRNAの安定化方法 | |
| EP2843049B1 (en) | Neuronal differentiation promoter | |
| WO2022141406A1 (zh) | 一种能够提供治疗有效量的白介素10的试剂及其抗肿瘤应用 | |
| US20220193110A1 (en) | Nxtar-derived oligonucleotides and uses thereof | |
| JP2023108978A (ja) | HBV由来cccDNAの抑制剤、抑制方法、HBV感染症用医療組成物、およびHBV感染症の治療方法 | |
| CN112618699A (zh) | 一种能够提供治疗有效量的白介素10的试剂及其抗肿瘤应用 | |
| CN113913513A (zh) | 人dsn1基因的用途及相关产品 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| E902 | Notification of reason for refusal |