KR20170105079A - 천연 또는 가공된 진핵 세포의 생리에 긍정적으로 영향을 미치는 박테리아 샤페론의 조합물 - Google Patents
천연 또는 가공된 진핵 세포의 생리에 긍정적으로 영향을 미치는 박테리아 샤페론의 조합물 Download PDFInfo
- Publication number
- KR20170105079A KR20170105079A KR1020177022784A KR20177022784A KR20170105079A KR 20170105079 A KR20170105079 A KR 20170105079A KR 1020177022784 A KR1020177022784 A KR 1020177022784A KR 20177022784 A KR20177022784 A KR 20177022784A KR 20170105079 A KR20170105079 A KR 20170105079A
- Authority
- KR
- South Korea
- Prior art keywords
- chaperone
- expression
- protein
- groel
- groes
- Prior art date
Links
Images

Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/80—Vectors or expression systems specially adapted for eukaryotic hosts for fungi
- C12N15/81—Vectors or expression systems specially adapted for eukaryotic hosts for fungi for yeasts
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/24—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Enterobacteriaceae (F), e.g. Citrobacter, Serratia, Proteus, Providencia, Morganella, Yersinia
- C07K14/245—Escherichia (G)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/37—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from fungi
- C07K14/39—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from fungi from yeasts
- C07K14/395—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from fungi from yeasts from Saccharomyces
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/02—Preparation of peptides or proteins having a known sequence of two or more amino acids, e.g. glutathione
Abstract
본 발명은 진핵 세포 내에서 3개의 박테리아 샤페론의 특정 조합의 발현에 관한 것이며, 이는 이의 성장 특성 및 특히 상기 세포가 적어도 하나의 비-천연 유전자를 사용한 추가의 가공을 포함하는 경우 물리화학적 스트레스에 대한 내성과 관련된 이의 특성을 유의적으로 증진시킨다. 샤페론의 바람직한 조합은 이.콜라이(E. coli)의 샤페론 GroES 및 GroEL 및 시네초코쿠스 엘롱가투스(Synechococcus elongatus)의 샤페론 RbcX를 포함한다.
Description
본 발명은 세포 가공, 특히 진핵 세포의 분야에 관한 것이다. 보다 상세하게는, 본 발명은 개선된 성장 및/또는 대사 특성을 갖는 진핵 세포, 및 목적한 화합물의 생산을 위한 이의 용도에 관한 것이다. 본 발명은 특히 샤페론(chaperon)의 특수한 조합물을 발현하는 진핵 세포에 관한 것이다. 본 발명은 특히 재조합체 단백질의 생산 분야에서 응용된다.
생명공학, 식품 가공, 의약 산업 또는 진단 연구, 기본 및/또는 응용 분야와 같은 다양한 분야에서 사용되는 목적한 단백질에 대해 많은 발현 시스템이 현재 존재한다. 이들 발현 시스템 중에서, 진핵 세포, 및 효모는 특히 산업 조건에서 이들의 글리코실화 능력 및 이들의 배양 용이성으로 인하여 특히 중요한 역할을 한다. 그러나, 이들 세포는 예를 들면, 목적한 생성물의 축적(예를 들면, 고 농도의 에탄올 또는 다른 알코올)과 관련된 또는 발현된 단백질의 직접적인 또는 간접적인 독성 또는 발현 플라스미드의 존재를 필요로 하는 에너지 부하(energy load)와 관련된 독성 제한으로 인하여 한계를 가지고 있다.
발명의 요약
본 발명은 최적화된 발현 시스템의 개발을 가능하도록 하는, 개선된 수행능력을 갖는 진핵 세포를 제공한다.
효모에서 합성 캘빈 주기(Calvin cycle)를 도입하는 것과 관련된 계획안과 관련하여, 본 발명자들은 놀랍게도, 효모에서 3개의 박테리아 샤페론(RbcX, GroES 및 GroEL)의 세트의 동시-발현이 특히 이종 또는 내인성 단백질의 발현 및 기능적 폴딩(functional folding) 측면에서 현저한 대사 특성을 진핵 세포에게 부여한다는 것을 관찰하였다. 이들의 시험을 지속하기 위해, 본 발명자들은 또한 샤페론의 이러한 조합이 효모의 성장을 가속화시키고 예를 들면, 키나제 PRK가 세포내에서 동시-발현되는 경우 다른 가공의 독성 효과를 제거하도록 할 수 있음을 입증하였다. 본 발명자들은 또한 다양한 진핵 세포에서의 수행능에 있어 동일한 효과를 확인하고 수득하여, 이들 예측하지 못한 결과의 이점 및 큰 잠재능을 확인하였다.
따라서, 본 발명의 목적은 샤페론 RbcX, GroES 및 GroEL을 발현하는 것을 특징으로 하는, 진핵 세포에 관한 것이다.
특수한 구현예에서, 본 발명은 다음을 함유함을 특징으로 하는, 형질전환된 진핵 세포에 관한 것이다:
(i) 적합한 프로모터의 전사 조절 하에서 박테리아의 제I형 RuBisCO 효소의 폴딩(folding)에 관여하는 샤페론 RbcX를 암호화하는 서열을 함유하는 발현 카세트(expression cassette);
(ii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroES를 암호화하는 서열을 함유하는 발현 카세트; 및
(iii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroEL을 암호화하는 서열을 함유하는 발현 카세트.
본 발명은 또한:
(i) 적합한 프로모터의 전사 조절 하에서 샤페론 RbcX를 암호화하는 서열;
(ii) 적합한 프로모터의 전사 조절 하에서 샤페론 GroES를 암호화하는 서열; 및
(iii) 적합한 프로모터의 전사 조절 하에서 샤페론 GroEL을 암호화하는 서열을 함유하는 진핵 세포에 관한 것이다.
특수한 구현예에 따라서, 본 발명의 진핵 세포는 박테리아의 제I형 RuBisCO 효소의 RbcL 및/또는 RbcS 소단위를 암호화하는 서열을 함유하지 않는다.
본 발명은 특히 상기 나타낸 바와 같은 형질전환된 효모를 제안한다.
본 발명은 또한 목적으로서 진핵 세포의 생리를 증진시키기 위한, 특히 상기 진핵 세포의 성장율을 증가시키고/시키거나 환경 스트레스에 대한 상기 진핵 세포의 내성을 증가시키고/시키거나 진핵 세포에 의해 합성된 화합물의 독성에 대해 상기 세포의 내성을 증가시키고/시키거나 재조합체 단백질을 생산하기 위한, 샤페론 RbcX 및 샤페론 GroES 및 GroEL의 발현을 가능하게 하는 발현 카세트의 조합물의 용도를 갖는다.
본 발명은 또한 목적으로서 진핵 세포에 의한 화학적 분자 및 단백질로부터 선택된 적어도 하나의 화합물의 합성을 가능하도록 하는 조건하에서 본 발명에 따른 진핵 세포를 배양하는 단계, 및 상기 화합물을 수집하는 단계를 포함함을 특징으로 하는, 상기 화학적 분자 및 단백질로부터 선택된 적어도 하나의 화합물 생화학적 생산 방법을 갖는다.
본 발명은 보다 상세하게는 (i) RbcX, GroES 및 GroEL을 발현하는 진핵 세포내로 재조합체 단백질을 암호화하는 서열을 삽입하는 단계, (ii) 상기 세포를 상기 서열의 발현을 가능하도록 하는 조건 하에 배양하는 단계 및 임의로 (iii) 상기 단백질을 수집하거나 정제하는 단계를 포함하는, 재조합체 단백질의 생산 방법을 제안한다.
도 1: 균주 1b, 18b, 102, 15, 14b의 에탄올 생산의 역학을 나타낸다. 오차 바아(error bar)는 3개의 독립된 배양물의 표준 편차를 나타낸다.
상세한 설명
진핵 세포에서 특정의 박테리아 샤페론 단백질의 발현을 연구하는 동안에, 본 발명자들은, 매우 놀랄만한 방식으로, 박테리아 샤페론이 진핵 세포의 세포질에서 발현될 수 있으며 상기 진핵 세포내에서 이들의 샤페론 기능을 보유함을 발견하였다. 박테리아 샤페론 기능은 진핵 세포내에 이미 존재하는 세포질 샤페론 기능에 부가된다. 본 발명자들은 또한, 특수한 3개의 샤페론, 즉, 박테리아 샤페론 GroEL 및 GroES 및 샤페론 RbcX의 발현이 이들을 발현하는 형질전환된 진핵 세포에서 성장, 발현 및 기능성 단백질 폴딩의 측면에서 특히 유리한 특성을 부여함을 발견하였다.
필수적으로 관찰된 효과는 적어도 2개의 상이한 미생물로부터 우선적으로 오는, 인용된 3개의 샤페론의 동시 존재를 필요로 한다. 샤페론의 동시-발현이 박테리아 시스템내에서 이종 단백질의 폴딩을 증진시킴으로써 잠재적으로 이에 의존한 생물공정의 수행능을 증진시키는 경향이 있음이 알려져 있는 경우, 3개의 박테리아 샤페론 단백질의 특수한 유기체-간 조합이 진핵세포 측면에서 발현됨에 의해 특히 효과적임이 입증됨은 예측할 수 없었다. 본 발명의 효과를 생성하는 분자 메카니즘은, 이들이 아마도 적어도 부분적으로 단백질 폴딩 또는 이들의 세포내 운명(destiny)의 조절과 관련되어 있는 경우에도 현재 알려져 있지 않다. 놀라운 일반적인 역할은 상보적인 역할을 하는 샤페론 GroES 및 GroES의 존재하에서 RbcX 계열(선행 기술에서 광합성 유기체의 RuBisCO 복합체의 기능적 연합에 대해 구체적으로 기술되어 있다)의 단백질에 의해 수행된다. 본 발명의 이러한 특징은, 관찰된 효과가 촉진된 폴딩의 역할에 전체적으로 기여할 수 없으며 특수한 단백질의 수명, 복합체의 조립 또는 이의 특성의 변형, 또는 규정될 특성의 구체적인 효과의 조정과 같은 다른 메카니즘을 포함할 수 있음을 제안한다.
이와 관련하여, 본 발명은 따라서 진핵 세포를 형질전환시켜 이들이 특수한 3중 샤페론, 즉 박테리아 샤페론 GroES 및 GroEL, 및 샤페론 RbcX을 발현하도록 하는 것을 제안한다. 이러한 형질전환된 세포는 특히 재조합체 단백질의 생산과 관련하여, 많은 응용을 발견할 수 있다.
따라서, 본 발명은 샤페론 RbcX, GroES 및 GroEL을 발현함을 특징으로 하는, 진핵 세포를 목적으로 한다.
샤페론 GroEL 및 GroES는 열-쇼크 단백질(HSP)의 계열에 속한다. 이들 샤페론은 많은 박테리아 속에 존재한다. 본 발명과 관련하여, 이들 샤페론은, 이들이 동시-작용하여 매우 많은 수의 단백질의 효과적인 폴딩을 가능하도록 함이 공지되어 있다는 의미에서 "일반 샤페론"으로 언급된다(M. Mayhew etal. 1996, "Protein folding in the central cavity of the GroEL-GroES chaperonin complex" Nature 1996 Feb 1;379(6564):420-6). 본 발명에 따라서, 샤페론 GroEL 및 GroES는 이들을 발현하는 어떠한 박테리아, 및 특히 예를 들면, 이. 콜라이 (E. coli)(유전자 확인 번호: 948655 및 948665), 에스. 엘롱가투스(S. elongatus)(유전자 확인 번호: 3199735, 3199535 및 3198035), 에스. 뉴모니아(S. pneumonia)(진뱅크(GenBank) 수탁 번호: AF117741), 에스. 피오게네스(S. pyogenes)(진뱅크 수탁 번호: SPGROELGN), 에스. 아우레우스(S. aureus)(진뱅크 수탁 번호: STAHSP) 또는 피. 아에루기노사(P. aeruginosa)(진뱅크 수탁 번호: ATCC9027)로부터 올 수 있다. 당해 분야의 숙련가는 박테리아 속에서 이들 샤페론중의 하나 또는 다른 하나를 암호화하는 경향이 있는 핵산 서열을 완벽하게 확인할 수 있다. 정보 목적을 위하여, 샤페론의 서열 유사성(정렬에서 동일한 아미노산의 %)은 에스. 엘롱가투스로부터의 GroEL1와 이.콜라이로부터의 GroEL 사이에서 61%; 에스. 엘롱가투스로부터의 GroEL2와 E. coli로부터의 GroEL 사이에서 56%; 에스. 엘롱가투스로부터의 GroEL1과 GroEL2 사이에서 63%임이 주목될 것이다.
본 발명에 따른 세포는 또한 시아노박테리아 및 식물에서 공지된 샤페론 RbcX를 발현하여 Rubisco의 RbcL 및 RbcS 소단위의 정확한 조립에 관여한다. 본 발명의 내용에서, 당해 샤페론은, 당해 단백질이 Rubisco를 사용한 경우에 주목되는 바와 같이, 단백질 복합체의 기능적 연합에 있어서 역할을 담당하는 것으로 알려져 있다는 의미에서 "특수한 샤페론(specific chaperone)"으로 언급된다(S. Saschenbrecker et al. 2007, "Structure and function of RbcX, an assembly chaperone for hexadecameric Rubisco", Cell. 2007 Jun 15; 129(6):1189-200). 본 발명에 따라서, 샤페론 RbcX는 이를 발현하는 어떠한 시아노박테리아, 및 특히 예를 들면, 에스. 엘롱가투스(서열 번호: 3), 시네코시스티스 아종(Synechocystis sp.)(Kaneko et al., "Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-codingregions." DNA Res. 3(3), 109-136 (1996)), 아나바에나 아종(Anabaena sp.)(Li et al. J. Bacteriol. (1997), 179(11), 3793-3796), 마이크로시스티스 아종(Microcystis sp.), 티코네마 아종(Tychonema sp.), 플랑크토트릭스 아종(Planktothrix sp.) 또는 노스톡 아종(Nostoc sp.)(Rudi et al., J. Bacteriol. (1998), 180(13), 3453-3461)으로부터 기원될 수 있다.
본 발명의 내용에서, "샤페론 활성"은 단백질 폴딩 및/또는 단백질 복합체의 기능적 연합에 있어서의 작용을 말한다.
본 발명의 특수한 구현예에서, 사용된 샤페론은 우선적으로 2개의 상이한 유기체로부터 기원된다. 본 발명에 따라서, 샤페론은 하나 이상의 상이한 박테리아으로부터 올 수 있다. 바람직하게는, 3개의 샤페론은, 이의 적어도 하나가 시아노박테리아인 적어도 2개의 명백한 그람-음성 박테리아 종으로부터 온다.
본 발명의 다른 특수한 시행에 따라서, 일반 샤페론 GroES 및 GroEL 중의 적어도 하나는 시아노박테리아 또는 RuBisCO 복합체를 발현하는 다른 박테리아으로부터도 기원되지 않는다. 본 발명에 따라서, 샤페론 GroES 및 GroEL은 동일한 박테리아 또는 2개의 상이한 박테리아으로부터 기원될 수 있다. 특수한 예시적인 구현예에서, 샤페론 GroES 및 GroEL은 E. coli로부터 기원한다.
특수한 예시적인 구현예에서, 샤페론 GroES 및 GroEL은 E. coli로부터 기원되고 샤페론 RbcX는 시네초코쿠스 엘롱가투스(Synechococcus elongatus)로부터 온다.
다른 예시적인 구현예에서, 3개의 샤페론 GroES, GroEL 및 RbcX는 시네초코쿠스 엘롱가투스로부터 기원된다. 이러한 경우에, 형질전환된 세포는 샤페론 GroEL의 하나 또는 다른 이소형(isoform) 또는 이소형 둘 다(GroEL1 및 GroEL2), 바람직하게는 이소형 둘 다를 발현할 수 있다.
본 발명과 관련하여, 단백질은 95% 이상의 아미노산 서열 동질성(amino acid sequence identity)을 갖고 유기체로부터 고려된 단백질과 동일한 기능을 가진 경우 제공된 상기 유기체로부터 "기원되는(come)" 것으로 고려된다.
본 내용에서, "GroES" 및 "GroEL"은 E. coli K12로부터의 GroES 및 GroEL 각각과 65% 내지 100% 아미노산 동질성을 갖고 샤페론 활성을 갖는 특정 단백질을 말한다. 또한, "GroES" 및 "GroEL"은 에스. 엘롱가투스로부터의 일반 샤페론과 같이, 보다 낮은 동질성 퍼센트, 및 특히 55% 내지 65%, 및 보다 상세하게는 56% 내지 63%의 동질성 퍼센트를 갖는 일반 샤페론을 말한다. E. coli로부터의 일반 샤페론 GroES 및 GroEL의 변이체의 샤페론 활성은 예를 들면, 하기 기술된 다양한 실시예에서 E. coli로부터의 음성 GroES 또는 GroEL을 암호화하는 발현 카세트를 평가할 샤페론의 변이체로 치환함으로써 확인할 수 있다.
샤페론 RbcX는 GroEL 및 GroES와는 매우 상이하며 이의 서열은 이들 2개의 샤페론의 서열과 정렬될 수 없다.
본 내용에서, "RbcX"는 샤페론 활성을 갖고 서열 번호:3의 서열에 의해 암호화된 샤페론 RbcX와 50% 이상의 아미노산 서열 동질성을 가지며 당해 단백질의 특수한 샤페론 활성을 보유하는, 특히 시아노박테리아로부터의 특정 단백질을 말한다. 특수한 샤페론 활성은 서열 번호: 3을 포함하는 발현 카세트를 평가할 다른 서열로 치환하여 세포 추출물 상에서 이렇게 수득된 RuBisCO 활성을 시험관내 시험(in vitro test)에 의해 측정함으로써, 에스. 엘롱가투스로부터의 RuBisCO의 RbcL 및 RbcS 소단위를 발현하는 효모에서 확인할 수 있다.
바람직하게는, 본 발명은 서열 번호: 3에 의해 암호화된 샤페론 RbcX와 아미노산 서열 동질성이 80% 이상, 바람직하게는 90% 이상, 보다 바람직하게는 95% 이상, 심지어 보다 바람직하게는 99% 이상인 샤페론 RbcX를 사용하여 시행한다.
본 발명은 단세포 또는 다세포 유기체로부터의 어떠한 유형의 진핵 세포로 시행할 수 있다. 특히, 본 발명에 따른 샤페론의 조합은 효모 세포, 진균 세포, 식물 세포, 포유동물 세포 등과 같은 동물 세포에서 발현시킬 수 있다.
특히, 본 발명은 특수한 샤페론 RbcX, 박테리아의 일반 샤페론 GroES, 및 박테리아의 일반 샤페론 GroEL을 발현하는 형질전환된 효모에 관한 것이다.
보다 상세하게는, 본 발명은:
(i) 적합한 프로모터의 전사 조절 하에서 박테리아의 제I형 RuBisCO 효소의 폴딩에 관여하는 샤페론 RbcX를 암호화하는 서열을 함유하는 발현 카세트;
(ii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroES를 암호화하는 서열을 함유하는 발현 카세트; 및
(iii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroEL을 암호화하는 서열을 함유하는 발현 카세트를 함유함을 특징으로 하는, 형질전환된 효모에 관한 것이다.
본 발명은 목적한 임의의 효모로 시행할 수 있다. 유리하게는, 효모는 사카로마이세스(Saccharomyces), 야로위아(Yarrowia) 및 피치아(Pichia)로부터 선택된다. 예를 들면, 본 발명에 따르는 형질전환된 효모는 사카로마이세스 세레비지아에(Saccharomyces cerevisiae) 세포이다. 다른 예에서, 본 발명에 따른 형질전환된 효모는 야로위아 리폴리티카(Yarrowia lipolytica) 세포, 또는 피치아 파스토리스(Pichia pastoris) 세포이다.
피치아 파스토리스의 사용은 재조합체 단백질의 생산을 위해 특히 유리하다. 실제로, 피치아는 분비 및 세포내 발현의 측면 둘 다에서 고-성능의 진핵세포 단백질 발현 시스템을 갖는다. 이는 재조합체 진핵세포 단백질의 대규모 생산에 특히 적합하다. 특히, 피치아는 포유동물 세포 발현 시스템과 관련된 것들과 비교하여 생산과 비용 시간을 줄이기 위하여, 분비된 단백질을 고 수율로 생산하는데 사용할 수 있다.
야로위아 리폴리티카(Yarrowia lipolytica)가 또한 재조합체 단백질의 생산에 사용하기에 적합하다. 실제로, 야로위아는 (i) 고-밀도 성장, (ii) 고 분비율, (iii) 알칼리성 프로테아제 AEP의 부재 및 (iv) 탄소원으로서 사카로즈의 사용을 허용하는 에스. 세레비지아에(S. cerevisiae)를 생산하는 능력(Nicaudet al., 1989)을 갖는다. 후자의 특성은, 당해 균주가 당밀(molasses)과 같은 저렴한 기질 상에서 효율적으로 성장할 수 있으므로, 산업 생산의 경우에 특히 유리하다.
본 발명은 또한 특수한 샤페론 RbcX, 박테리아의 일반 샤페론 GroES 및 박테리아의 일반 샤페론 GroEL을 발현하도록 형질전환된 동물 세포 및 특히 포유동물 세포와 같은 다세포 유기체로부터의 진핵 세포에 관한 것이다.
예시적인 구현예에서, 이러한 진핵 세포는 형질전환되어 다음을 함유한다:
(i) 적합한 프로모터의 전사 조절 하에서, 박테리아의 제I형 RuBisCO 효소의 폴딩에 관여하는 샤페론 RbcX를 암호화는 서열을 함유하는 발현 카세트;
(ii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroES를 암호화하는 서열을 함유하는 발현 카세트; 및
(iii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroEL을 암호화하는 서열을 함유하는 발현 카세트.
본 발명은 특히 본 발명에 따른 3중의 샤페론을 발현하는 형질전환된 CHO 세포에 관한 것이다.
본 발명에 따라서, 샤페론 GroEL, GroES 및 RbcX를 암호화하는 유전자는 상기 세포내에서 이들의 발현을 가능하도록 하는 형태로 진핵 세포내에 도입된다. 따라서, 샤페론을 암호화하는 서열은 이들의 전사를 가능하도록 하는 프로모터 서열과 관련된다. 일 구현예에서, 동일한 프로모터 서열이 3중의 샤페론을 암호화하는 서열과 관련된다. 다른 구현예에서, 샤페론 RbcX는 샤페론 GroEL 및 GroES와 관련된 프로모터(들)과는 상이한 특수한 프로모터와 관련된다. 다른 구현예에서, 각각의 샤페론은 상이한 특수한 프로모터와 관련된다.
본 발명의 내용에서 이용가능한 프로모터는 구성적 프로모터(constitutive promoter), 즉 대부분의 세포 상태 및 환경 조건 하에서 활성인 프로모터, 및 외인성의 물리적 또는 화학적 자극에 의해 활성화되거나 저해되어 이들 자극의 존재 또는 부재의 기능으로서 다양한 수준의 발현을 유도하는 유도성 프로모터를 포함한다.
효모내의 발현 카세트의 경우, 구성적 프로모터의 예는 유전자 TEF1, TDH3, PGI1, PGK, ADH1의 것이다. 유도성 프로모터의 예는 프로모터 tetO-2, GAL10, GAL10 - CYC1, PHO5이다. 바람직하게는, 사용된 프로모터는 하나의 카세트로부터 다른 것까지 상이할 것이다. 본 발명의 발현 카세트는 또한 전사 터미네이터와 같은 일반적인 서열, 및 필요에 따라 다른 전사 조절 성분을 추가로 포함한다. 본 발명에 따라서 발현 카세트는 숙주 세포의 염색체 DNA내로 삽입될 수 있고/있거나 하나 이상의 염색체외 레플리콘(들)에 의해 수반된다. 이들 단백질의 상대적인 입체화학은 본 발명의 최적의 시행에 중요한 역할을 담당하는 경향이 있다. 하기 실험 단락에 기술된 동시-발현은 이러한 국면에서 특히 관련되어 있다. 그러나, 본 발명은 이들 시스템의 용도에 한정되지 않으며, 이는 적어도 동일한 효과를 갖는 것으로 언급된 성분의 발현의 어떠한 변이체로도 시행될 수 있으므로, 이들은 예를 들면, 상기 세포에 대한 표준 배지 속에서 형질전환된 세포의 성장을 측정함으로써 측정할 수 있다.
유리한 구현예에 따라서, 3개의 발현 카세트는 유전 정보의 연속적인 블록을 형성한다. 이는 또한 3개의 샤페론의 발현 카세트가 단일의 에피솜 유전 성분에 의해 수반되는 것이 또한 유리할 수 있다. 따라서, 본 발명의 특히 유리한 국면은 복제의 중심 오리진(origin)을 갖는 에피소옴 성분에 의해 수반된 단일의 "유전 플러그 인(genetic plug-in)"(연속된 DNA 서열)이다. 당해 성분에 의한 형질전환은 야생형 효모에서 목적한 특성 또는 어떠한 가공도 수반하는 것들을 도입하는데 충분하다.
본 발명에 따라서, 각각의 샤페론을 암호화하는 유전자는 하나 이상의 카피로 세포내에 도입될 수 있다. 특히, GroES를 암호화하는 서열을 함유하는 1, 2, 3개 이상의 카세트, GroEL을 암호화하는 서열을 함유하는 1, 2, 3개 이상의 카세트, 및 RbcX를 암호화하는 서열을 함유하는 1, 2, 3개 이상의 카세트를 도입하는 것이 가능하다. 유사하게, 카세트는 GroES, GroEL 또는 RbcX를 암호화하는 서열의 수개의 카피를 함유할 수 있다. 샤페론을 암호화하는 유전자의 수개의 카피가 세포내로 도입되는 경우에, 즉, 동일한 박테리아으로부터 오는 동일한 서열이 매번 바람직하게 사용된다. 상이한 박테리아으로부터 오는 서열을 사용하는 것도 또한 가능하다.
위에서 언급한 바와 같이, 본 발명에 따른 세포는 개선된 특성(성장율, 내성, 생산능 등)을 갖는다. 이들 세포는 따라서 단백질 또는 다른 화합물을 생산하거나, 발효 공정을 개선시키는데 유용하다.
단백질의 생산 (production of proteins)
본 발명은 또한 위에서 기술한 바와 같은 샤페론의 조합을 발현하도록 형질전환되고 상기 샤페론 이외의 이종 단백질에 대한 적어도 하나의 발현 카세트를 추가로 포함하고/하거나 내인성 단백질의 발현 수준 및/또는 서열을 개질시키는 서열 가공(sequence engineering)을 거친 진핵 세포에 관한 것이다.
본 발명의 바람직한 시행에 따라서, 형질전환된 진핵 세포는 효모이다. 본 발명의 다른 바람직한 시행에 따라서, 형질전환된 진핵 세포는 CHO 세포이다.
따라서, 본 발명은 목적으로서 상기 기술된 바와 같은 샤페론의 조합을 발현하도록 형질전환되고, 및:
(iv) 상기 샤페론 이외의 이종 단백질에 대한 발현 카세트를 갖고/갖거나
(v) 내인성 단백질의 발현 수준 및/또는 서열을 개질시키는 서열 가공을 거친 효모 또는 CHO 세포를 갖는다.
본 발명에 따라서, 하나 이상의 목적한 단백질을 생산하도록 형질전환된 세포를 사용하는 것이 가능하다.
유리하게는, 목적한 단백질은 Rubisco가 아니다. 유사하게, 우선적으로 목적한 단백질은 PKR 이외의 단백질이다. 또한, 형질전환된 진핵 세포가 Rubisco 및/또는 PKR를 발현하는 경우, 이는 유리하게는 적어도 하나의 다른 이종 단백질을 발현한다.
특수한 구현예에서, 3중의 샤페론 GroES, GroEL 및 RbcX를 발현하는 형질전환된 세포는 재조합체 단백질을 발현하여 분비하도록 추가로 변형된다.
본 발명은 또한, 화학 분자, 효소, 호르몬, 항체 및 단백질로부터 선택된 적어도 하나의 화합물을 생산하기 위한 생명공학 공정에 관한 것이며, 이것이 상기 기술한 바와 같은 형질전환된 세포를 당해 세포에 의해 상기 화합물의 합성을 가능하도록 하는 조건 하에서 배양하는 단계, 및 임의로 상기 화합물을 수집하고/하거나 정제하는 단계를 포함함을 특징으로 한다.
본 발명은 또한 (i) 3중의 샤페론 RbcX, GroES 및 GroEL을 발현하는 진핵 세포내로 재조합체 단백질을 암호화하는 서열을 삽입하는 단계, (ii) 상기 서열의 발현을 가능하도록 하는 조건 하에서 상기 세포를 배양하는 단계 및 임의로 (iii) 상기 단백질을 수집 및/또는 정제하는 단계를 포함하여, 상기 재조합체 단백질을 생산하는 방법에 관한 것이다. 유리하게는, 상기 단백질은 효소 또는 호르몬이다.
특수 구현예에서, 본 발명에 따른 세포는 적어도 하나의 이종 효소를 생산하도록 형질전환된다. 예를 들면, 세포는 바실러스 투링지엔시스(Bacillus thuringiensis) 내독소, 리파제, 서브틸리신, 셀룰라제 및 루시퍼라제와 같은 내독소로부터 선택된 효소를 생산하도록 형질전환된다.
다른 특수한 구현예에서, 본 발명에 따른 세포는 의학 목적의 적어도 하나의 분자를 생산하도록 형질전환된다. 예를 들면, 세포는 호르몬, 성장 인자, 항체 등을 생산하도록 형질전환된다. 바람직하게는, 의학 목적의 분자는 에리트로포이에틴, 제I형 및/또는 제II형 알파-인터페론, 과립구 콜로니-자극 인자, 인슐린, 성장 호르몬, 조직 플라스미노겐 활성인자로부터 선택된다.
유리하게는, 본 발명에 다른 형질전환된 세포는 본 발명의 샤페론의 조합물을 발현하지 않는 재조합체 세포와 비교하여, 목적한 단백질(들)의 생산이 증진되어 있다. 본 발명의 내용에서, "증진된" 생산은 양 및/또는 품질의 측면을 의미한다. 특히, 본 발명에 따라 형질전환된 세포는 보다 활성이고/이거나 보다 안정하여서, 거의 분해되지 않는 경향이 있어, 본 발명의 샤페론의 조합물을 발현하지 않는 재조합체 세포에 의해 생산된 이종 단백질과 비교하여, 상기 단백질의 보다 큰 축적을 가능하도록 하는 목적한 단백질을 생산할 수 있다. 따라서, 본 발명에 따른 형질전환된 진핵 세포에 의한 재조합체 단백질의 발현 수준에서의 증가는 보다 큰 안정성 및 따라서 세포 및/또는 배양 배지 속에서 상기 단백질의 보다 큰 축적에 의해 설명된다. 더욱이, 형질전환된 세포에 의한 샤페론의 조합물의 발현은 재조합체 세포가 발현하는 재조합체 단백질에 대한 상기 세포의 내성을 유리하게 증가시킬 수 있으므로, 또한 생산 수율을 증가시키는데 관여한다.
본 발명은 또한 진핵 세포내에서 특이적인 샤페론 RbcX 및 일반 샤페론 GroES 및 GroEL의 발현을 가능하도록 하는 발현 카세트의 조합을 사용하여 상기 진핵 세포의 생리 및/또는 수행능(특히 성장율)을 증진시키는 것에 관한 것이다. 본 발명의 당해 국면의 바람직한 시행에 따라서, 이의 증진된 생리이 추구되는 진핵 세포는 효모이다. 다른 바람직한 시행에 따라서, 상기 진핵 세포는 박테리아의 제I형 RuBisCO 효소의 RbcL 소단위를 암호화하는 서열, 및/또는 상기 RuBisCO 효소의 RbcS 소단위를 암호화하는 서열을 발현하도록 형질던환되지 않는다.
상기 언급한 바와 같이, 일반 샤페론 GroES 및 GroEL 및 특이적인 샤페론 RbcX의 발현을 가능하도록 하는 발현 카세트의 조합물이 발현의 수준을 변형시키도록 가공되는 서열 및/또는 예를 들면, 에피소옴 유전 성분의 형태로, 상기 샤페론 이외의 다른 이종 단백질에 대한 적어도 하나의 발현 카세트를 포함하는 서열 가공을 겪은 진핵 세포의 생리적 증진시키는데 특히 유용하다.
특히, 세포내에서, 일반 샤페론 GroES 및 GroEL 및 특이적인 샤페론 RbcX의 동시 발현은 상기 세포의 성장율을 증가시키고/시키거나 환경 스트레스, 특히 세포의 배양 배지 속에 존재하는 성분의 독성으로 인한 스트레스에 대한 상기 세포의 내성을 증가시키는 것을 가능하도록 한다. 다른 유리한 응용은 이에 의해 합성된 화합물의 독성에 대한 세포의 내성을 증가시킴으로써 목적한 화합물의 생산을 증가시키는 것이다.
본 발명의 매우 많은 응용, 특히 형질전환된 진핵 세포에 대해 동종이거나 이종인 단백질의 폴딩 또는 안정성(화학제 또는 열 제제에 대한 내성, 고유의 수명)을 증진시키는 것과 관련된 모든 응용이 고려될 수 있다. 이들은 효소 촉매작용에서 목적한, 또는 이들의 고유의 특성(항체, 치료학적 단백질, 복합체내 구조 단백질 등)으로 인하여 단백질 자체일 수 있으며; 적용은 합성 또는 반-합성 대사 쇄(형질전환된 진핵 세포의 단백질을 포함함)의 개선된 성능과 관련되고; 이 경우에, 장점은 주로 목적한 생성물(화학 분자)의 생산에 있어서 이들 작용의 결과의 증진이다. 당해 효과는 폴딩 또는 안정성에 있어서의 개선 뿐만 아니라, 다른 현상, 예를 들면, 아세포 수송(subcellular transport), 복합체의 촉진되거나 변형된 형성, 커플링 메카니즘의 조정 등; 성장, 생존능, 적응성, 스트레스에 대한 내성 또는 이로부터의 회복, 공지되거나 설명가능한 메카니즘의 부재하의 목적한 생성물의 형성의 측면에서 유기체에서 긍정적인 전체적 효과를 생성하는 응용이다.
본 발명의 산업적 응용의 예는 다음을 포함하나, 이에 한정되지 않는다:
단백질
폴딩
/
안정성에 있어서의
효과
가용성 및 기능성 형태를 생산하는 것이 어렵거나 심지어 불가능한 것으로 고려되는 목적한 특정의 재조합체 단백질의 생산은, 본 발명의 시행에 의해 증진될 수 있다. 이는 특히 본 발명에 따른 형질전환된 진핵 세포에서 상기 단백질의 폴딩 결함의 교정에 의한다. 이는 특히 건강, 에너지, 화학, 식품 가공 등의 분야에서 유용할 수 있다.
목적한 가공 생성물 또는 당해 가공의 중간체에 의해 유도된
독성에 대한 보호
숙주 세포에 대해 독성인 분자의 생물생산 또는 이의 생산 공정이 독성 중간체, 예를 들면, 의약(하이드로코르티손, 아르테미신산 또는 트릭토시나이드, 개발된 공정을 취할 특정의 플라보노이드) 또는 포화되지 않은 케톤, 알데하이드(예를 들면, 바닐린) 등과 같은 반응성 분자를 포함하는 경우 문제가 될 수 있다. 다른 예는 고농도에서 독성이 되는 분자, 예를 들면, 에탄올 또는 다른 알코올을 생산하는 공정에 상응한다. 예를 들면, 에탄올 생산은 알코올 고 농도에 대한 효모 균주의 내성에 의해 제한된다. 다른 예는 매우 다양한 산업적 화학 분자, 현재 생성물의 전구체 또는 화학 중간체 및 물론 생물연료의 생산에 관한 것이다. 이들은 또한 재조합체 세포에 의한 이의 생산이 물질대사를 불균형화시켜 세포 사멸을 유도하는 경향이 있는 재조합체 단백질일 수 있다. 본 발명에 따른 샤페론의 조합물을 발현하는 진핵 세포의 사용은, 이것이 발현하는 재조합체 단백질에 의해 유도된 형질전환된 세포 세포독성의 내성을 증가시키는 것이 유리하게는 가능하도록 한다.
열 내성
특정 효소 또는, 효모와 같은 유기체의 열 내성을 증진시키는 것은 예를 들면, 많은 난용성 분자의 생명공학적 생산성에 있어 중요한 인자이다.
고정된 양의
탄소원으로부터
생산된 미생물의
생물량(biomass)의
증가
이것은 분자 수준에서 이해되지 않지만 하기 설명된 제1 실험에서 관찰된 효과이다.
상기 언급한 바와 같이, 본 발명은 또한:
(i) 적합한 프로모터의 전사 조절 하에서, 박테리아의 제I형 RuBisCO 효소의 폴딩에 관여된 샤페론 RbcX를 암호화하는 서열을 함유하는 발현 카세트;
(ii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroES를 암호화하는 서열을 함유하는 발현 카세트; 및
(iii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroEL을 암호화하는 서열을 함유하는 발현 카세트를 포함하는 핵산 분자에 관한 것이다.
이러한 "유전적 플러그-인(genetic plug-in)"의 예는 복제의 동원체 기원(centromeric origin)을 갖는 에피좀 성분에 의해 운반된, 위에서 언급한 3개의 카세트(임의의 순서)를 포함하는 연속된 DNA 서열이다.
하기 실험 단락은 다음을 나타냄으로써 본 발명을 예증하나, 이에 한정되지는 않는다:
- 생성된 작제물 및 시행된 방법(실시예 1);
- 2개의 상이한 유형에 속하는 16개의 펩타이드 쇄의 조립으로 이루어진 제I형 RuBisCO의 활성의 재구성(reconstruction)에 있어서 다양한 샤페론의 조합의 효과를 비교하는 제1의 일련의 시험들. 샤페론의 다양한 연합의 발현은 "제I형 RuBisCO"의 RbcL 및 RbcS 폴리펩타이드의 발현과 조합되었다. 이는 효모 세포 추출물에서 시험관내 측정된 RuBisCO 활성에 있어서 샤페론의 조합물의 역할을 분석하는 것을 가능하도록 하였다(실시예 2);
- 에스. 엘롱가투스의 포스포리불로키나제(PRK)를 조건적으로 발현하는 효모에서 샤페론 시스템의 동시-발현을 포괄하는 제2의 일련의 시험. 본 발명의 부재하에서, PRK의 유도는 에스. 세레비지아에 균주 및 배양 조건(배지, 산소 수준, pH)에 의존하여 매우 느린 성장으로부터 총 치사도(>99.99%) 범위의 주요 독성 효과를 효모내에서 유도한다. 예시화는 본 발명이 완전한 활성을 유지하면서, PRK 효소의 활성을 방해하지 않고 당해 병적 상태를 "치유함"을 예증한다(실시예 3);
- 본 발명이 심지어 중성의 플라스미드(자체는 이들 자체의 복제이외의 작용의 발현을 유도하지 않는다)에 의해 보완된 독립영양을 수반하는 균주 속에서 야생 수준의 성장을 회복함을 나타내는 제3의 일련의 시험. 당해 상황은 에피소옴 유전 성분을 포괄하는 대표적인 물질대사 가공이다. 실시예에서, 본 발명은 전체적으로 에피소옴 유전 구조의 존재와 관련된 성장에 있어서 부정적인 영향을 총체적으로 정정한다(실시예 4); 및
- 본 발명의 세포가 화합물 및 특히 재조합체 단백질의 생산에 특히 유용함을 예증하는 다른 일련의 시험(실시예 5).
실시예:
하기 실시예에서, 및 달리 규정하지 않는 한, 동일한 접두어가 표마다 및 실험마다 사용되어 동일한 성분을 나타낸다.
실시예
1: 물질 및 방법 - "
샤페론
플러그-인" 및 벡터의
작제
- 다양한 균주의
작제
- 배양 및 측정 방법
1.1. 에스 . 세레비지아에에 대한 " 샤페론 플러그-인" 및 벡터의 작제
하기 기술된 특정의 작제물은 RuBisCO(pFPP45)의 2개의 RbcS 및 RbcL 소단위의 발현 및 시네초코쿠스 엘롱가투스 pCC6301으로부터 포스포리불로키나제(PRK)(pFPP20)의 발현을 가능하도록 한다. 하기 기술된 다른 작제물은 단일 발현 벡터로부터 시네초코쿠스 엘롱가투스로부터의 특이적인 샤페론 RbcX 및 E. coli로부터의 일반 샤페론 GroES(유전자 확인 번호: 948655), GroEL(유전자 확인 번호: 948665) 또는 시네초코쿠스 엘롱가투스로부터의 이들의 동족체 GroES(유전자 확인 번호: 3199735), GroEL1(유전자 확인 번호: 3199535) 및 GroEL2(유전자 확인 번호: 3198035)의 발현의 다양한 조합을 작업하는 것을 가능하도록 하기 위해 생성시켰다.
RbcS(유전자 확인 번호: 3200023) 및 RbcL(유전자 확인 번호: 3200134) 소단위 및 시네초코쿠스 엘롱가투스 pCC6301로부터의 RuBisCO의 특이적인 샤페론 RbcX (유전자 확인 번호: 3199060)를 암호화하고, 효모내에서의 발현을 위해 최적화된 합성 유전자를 제조하여 플라스미드 pBSII(Genecust)내로 클로닝하였다. HA 태그(tag)가 암호화 서열의 3' 말단에 첨가된, 효모내에서 발현을 위해 최적화된 변이체를 또한 작제하였다. 이들 합성 유전자의 서열(HA 태그 없음)은 각각 서열 번호: 1 내지 서열 번호: 3 하에 부록에서의 서열 목록에 나타낸다.
유사하게, 시네초코쿠스 엘롱가투스 pCC6301로부터의 포스포리불로키나제(PRK)(서열 번호: 4)(pFPP20), 및 일반 샤페론 GroES(서열 번호: 5), GroEL1(서열 번호: 6) 및 GroEL2(서열 번호: 7)을 암호화하고 효모내에서의 발현을 위해 최적화된 합성 유전자를 작제하여 클로닝하였다.
E. coli로부터의 샤페론 GroES 및 GroEL을 암호화하는 서열을 E. coli 배양물로부터 증폭시키고 플라스미드 pSC-B-amp/kan(Stratagene)내로 클로닝하였다.
클로닝 벡터로부터 회수된 서열을 효모 발현 벡터내로 도입하였다. 이들 숙주 벡터는 하기 표 I에 나열한다.
[표 I] 사용된 벡터의 목록
| 명칭 | 효모 복제 오리진 | 선택 마커 | 전사 카세트(프로모터-터미네이터) | E. coli 레플리콘 |
| pFPP5 | 2μ | URA3 | pGAL10-CYC1-tPGK | 예 (AmpR) |
| pFPP10 | 2μ | URA3 | pTDH3- -tADH | 예(AmpR) |
| pFPP11 | 2μ | URA3 | pTDH3- -tCYC1 | 예(AmpR) |
| pFPP12 | 2μ | URA3 | pTGI1- -tCYC1 | 예(AmpR) |
| pFPP13 | ARS-CEN6 | LEU2 | pTEF1-tPGK | 예(AmpR) |
참고: pGAL10-CYC1: GAL10 유전자의 UAS 및 CYC1 유전자의 전사 개시로 구성된 합성 프로모터(Pompon et al., Methods Enzymol, 272,51-64, 1996).
이렇게 수득된 발현 카세트는 하기 표 II에 나열한다.
[표 II] 발현 카세트
| 명칭 | 프로모터 | 개방 판독 프레임(Open reading frame) | 태그 | 터미네이터 |
| CAS6 | TDH3p | RbcL | 없음 | ADH1 |
| CAS7 | TetO7p | PRK | 없음 | CYC1t |
| CAS16 | TEF1 | RbcS | 없음 | PGK |
| CAS19 | TEF1p | RbcX | 없음 | PGK |
| CAS21 | PGI1p | GroES E. coli | 없음 | CYC1 |
| CAS22 | TDH3 | GroEL E. coli | 없음 | ADH |
| CAS23 | PGl1p | GroES S. elongatus | 없음 | CYC1t |
| CAS25 | TDH3p | GroEL2 S. elongatus | 없음 | ADH1t |
| CAS28 | PGI1p | 폴리링커 | 없음 | CYC1t |
| CAS33 | TEF1p | 폴리링커 | 없음 | PGKt |
특정의 벡터에서, 2개 또는 3개의 카세트가 삽입되었다. 이러한 목적으로, 플라스미드를 박테리아 에스케리키아 콜라이(Escherichia coli) DH5α내에서 증폭시키고 막시프렙(maxiprep)으로 제조한 후, 적합한 제한 효소로 분해하였다. 마지막으로, 단편을 T4 리가제(FERMENTAS) 또는 동종 재조합에 의해 효모내에서 직접 숙주 벡터내로 통합시켰다. 작제된 벡터의 목록을 하기 표 III에 나타낸다.
[표 III] 발현 벡터
| 명칭 | 오리진 유형 | 카세트 1 | 카세트 2 | 카세트 3 | 마커 | 숙주 벡터 |
| pFPP13 | ARS415-CEN6 | CAS33 | 없음 | 없음 | LEU2 | pFL36 |
| pFPP20 | ARS416-CEN4 | CAS7 | 없음 | 없음 | TRP | pCM185 |
| pFPP45 | 2μ | CAS6 | CAS16 | 없음 | URA3 | pFPP5/pFPP10 |
| pFFP53 | ARS415-CEN6 | CAS19 | CAS28 | 없음 | LEU2 | pFL36 |
| pFPP56 | ARS415-CEN6 | CAS19 | CAS21 | CAS22* | LEU2 | pFPP13 |
| pFB05 | ARS415-CEN6 | CAS19 | CAS25* | CAS21 | LEU2 | pFFP56 |
| pFB07 | ARS415-CEN6 | CAS23 | CAS22* | CAS19 | LEU2 | pFFP56 |
| pFB08 | ARS415-CEN6 | CAS23 | CAS25* | CAS19 | LEU2 | pFFP56 |
| pFB09 | ARS415-CEN6 | CAS21 | CAS22* | 없음 | LEU2 | pFFP56 |
1.2. 다양한 에스. 세레비지아에 균주의 작제
상기 다양한 플라스미드를 작제하여 효모 균주내에서 개개 성분의 의도된 발현 또는 이들 성분의 연합을 가능하도록 하였다. 따라서 다양한 벡터 또는 벡터의 조합물을 사용하여 효모 에스. 세레비지아에(W303.1B, FY1679 및 CEN.PK 1605)의 7개 균주를 형질전환시켰다. CEN.PK 1605는 균주 1605의 독립영양 버젼이다. 따라서, 이는 "생리적 거동"을 위한 양성 대조군이다. 하기 표 IV의 첫번째 컬럼에 편집된 각각의 번호는 동일한 표의 상응하는 라인에 기술된 벡터의 연합에 상응한다. 사용된 각각의 균주의 경우, 따라서, 연합된 참고 번호는 재작제된 가공에 대한 정보를 제공한다. CEN.PK 1605 균주만이 명확성의 목적으로 예시(표 IV)되었지만, 명명법은 다른 2개의 균주에 대해 동일하다.
[표 IV] 플라스미드 및 균주의 조합(표 III 참고)
| 조합물 번호 | 모 균주 | 벡터 1 | 벡터 2 | 벡터 3 | 발현된 단백질 | |||||
| RbcS | RbcL | RbcX | PRK syn | GroES | GroEL | |||||
| 1b | CEN.PK 1605 | PYEDP51 | pCM185 | pFPP13 | ||||||
| 2 | CEN.PK 1605 | pFPP45 | pCM185 | pFPP56 | X | X | X | 이.콜라이 | 이.콜라이 | |
| 3 | CEN.PK 1605 | pFPP45 | pFPP20 | pFPP56 | X | X | X | syn | 이.콜라이 | 이.콜라이 |
| 4 | CEN.PK 1605 | pFPP45 | pFPP20 | pFPP53 | X | X | X | syn | ||
| 5 | CEN.PK 1605 | pFPP45 | pCM185 | pFPP53 | X | X | X | |||
| 12 | CEN.PK 1605 | PYEDP51 | pCM185 | pFPP53 | X | |||||
| 13b | CEN.PK 1605 | PYEDP51 | pCM185 | pFPP56 | X | 이.콜라이 | 이.콜라이 | |||
| 14b | CEN.PK 1605 | PYEDP51 | pFPP20 | pFPP53 | X | Syn | ||||
| 15 | CEN.PK 1605 | PYEDP51 | pFPP20 | pFPP56 | X | syn | 이.콜라이 | 이.콜라이 | ||
| 16b | CEN.PK 1605 | pFPP45 | pCM185 | pFPP13 | X | X | ||||
| 17b | CEN.PK 1605 | pFPP45 | pFPP20 | pFPP13 | X | X | syn | |||
| 18b | CEN.PK 1605 | PYEDP51 | pFPP20 | pFPP13 | syn | |||||
| 101 | CEN.PK 1605 | pFPP45 | pFPP20 | pFB08 | X | X | X | Syn | syn | L2 syn |
| 102 | CEN.PK 1605 | PYEDP51 | pFPP20 | pFB09 | Syn | 이.콜라이 | 이.콜라이 | |||
| 102b | CEN.PK 1605 | pFPP45 | pFPP20 | pFB09 | X | X | syn | 이.콜라이 | 이.콜라이 | |
| 103 | CEN.PK 1605 | PYEDP51 | pCM185 | pFB09 | 이.콜라이 | 이.콜라이 | ||||
(Syn: 시네초코쿠스 엘롱가투스; L2 syn: GroEL2 시네초코쿠스 엘롱가투스, L1 syn: GroEL1 시네초코쿠스 엘롱가투스)
특정 표들과 관련된 주석:
1. pCM185: 시판 플라스미드(ATCC 87659)
2. pFL36: 시판 플라스미드(ATCC 77202)
3. PYeDP51: 다음 논문에 기술된 "빈(Empty)" 플라스미드: Urban P, Mignotte C, Kazmaier M, Delorme F, Pompon D. Cloning, yeast expression, and characterization of the coupling of two distantly related Arabidopsis thaliana NADPH - cytochrome P450 reductases with P450 CYP73A5. J Biol Chem. 1997 Aug 1; 272(31):19176-86.
4. GroES E. coli 유전자 확인 번호: 6061370; GroEL E. coli 유전자 확인 번호: 6061450
5. 에스. 세레비지아에 균주 CEN.PK 113-7D: Mat 독립영양성
6. 에스. 세레비지아에 균주 CEN.PK 1605: CEN.PK 113-7D로부터 생성된 Mat HIS3leu2-3.112trp1-289 ura3-52 MAL.28c. 균주.
7. 다른 약어는 데이타 뱅크에 기술된 에스. 세레비지아에를 말한다.
8. 합성 유전자: RuBisCO 소단위, RuBisCO 조립체에 특이적인 샤페론(RbcX), PRK 및 일반 샤페론 GroES, GroEL1 및 GroEL2를 암호화하는 시네초코쿠스 엘롱가투스 유전자를 불균질 코돈 바이아스(inhomogeneous codon bias)를 시행하는 효모에 대해 재-암호화한 후 합성하여 pCC6301(시판됨)내로 클로닝하였다. 이들 단백질(재-암호화한 후)의 암호화 서열은 별첨에 나타낸다 (서열 번호: 1: RbcS 암호화 서열; 서열 번호: 2: RbcL 암호화 서열; 서열 번호: 3: RbcX 암호화 서열; 서열 번호: 4: PRK 암호화 서열; 서열 번호: 5: GroES 암호화 서열; 서열 번호: 6: GroEL1 암호화 서열; 서열 번호: 7: GroEL2 암호화 서열).
9. E. coli 샤페론 GroES 및 GroEL의 암호화 서열을 박테리아로부터 증폭시켜, pSC-B-amp/kan(Stratagene) 내로 클로닝하고 발현 벡터내에서 재-암호화없이 조립하였다(상기 참조).
10. 시네초코쿠스 엘롱가투스 샤페론을 암호화하는 cDNA의 재-암호화된 서열을 효모내에서 2개의 분자를 동시-형질전환시킴으로써 앞서 선형화된 벡터 pUC57내로 동종 재조합체 의해 삽입하였다. 유사하게, ORF를 앞서의 작제물로부터 PCR에 의해 증폭시켜 벡터 pFPP56에 의해 수반된 프로모터 및 터미네이터에 대해 동종인 플랭킹 영역(flanking region)을 생성시켰다. 이는 이미 선형화된 벡터 pFPP56으로 효모 균주내 당해 PCR 생성물을 동시-형질감염시킴으로써 동종 재조합체 의한 클로닝을 허용하여, 표 II에 기술된 카세트에 따라 표 III에 기술된 각종 발현 벡터를 생성하였다.
1.3. 상이한 오리진의 샤페론을 통합시키는 다양한 사카로마이세스 세레비지아에 균주의 작제
다양한 박테리아으로부터의 샤페론을 평가하였다. 따라서, 동일한 종(에스. 엘롱가투스)으로부터의 단백질을 포함하는 조합물을 또한 에스. 엘롱가투스로부터의 RbcX를 에스. 엘롱가투스로부터의 GroES 및 GroEL1 및/또는 GroEL2와 조합하여 시험하였다.
[표 V] 발현 카세트
| 명칭 | 프로모터 | 개방 판독 프레임 | 태그 | 터미네이터 |
| CAS19 | TEF1p | RbcX 최적화된 에스. 엘롱가투스 | 없음 | PGKt |
| CAS21 | PGI1p | GroES E. coli | 없음 | CYC1t |
| CAS22 | TDH3p | GroEL E. coli | 없음 | ADH1t |
| CAS23 | PGl1p | GroES 최적화된 에스. 엘롱가투스 | 없음 | CYC1t |
| CAS24 | TEF2p | GroEL1 최적화된 에스. 엘롱가투스 | 없음 | TEF1t |
| CAS25 | TDH3p | GroEL2 최적화된 에스. 엘롱가투스 | 없음 | ADH1t |
[표 VI] 발현 벡터
| 명칭 | 오리진 유형 | 카세트 1 | 카세트 2 | 카세트 3 | 카세트 4 | 독립영양 마커 | 숙주 벡터 |
| pFFP56 | ARS415-CEN6 | CAS19 | CAS21 | CAS22* | LEU2 | pFL36 | |
| pFB08 | ARS415-CEN6 | CAS23 | CAS25* | CAS19 | LEU2 | pFFP56 | |
| pCB02 | ARS415-CEN6 | CAS23 | CAS25* | CAS19 | CAS24 | LEU2 | pFB08 |
[표 VII] 플라스미드 및 균주의 조합
| 조합 | 모 균주 | 벡터 1 | 벡터 2 | 벡터 3 | 발현된 단백질 | ||
| RbcX | GroES | GroEL | |||||
| 1b | CEN.PK 1605 | V51TEF | pCM185 | pFL36 | |||
| 13b | CEN.PK 1605 | V51TEF | pCM185 | pFPP56 | syn | E. coli | E. coli |
| 116 | CEN.PK 1605 | V51TE | pCM185 | pFB08 | Syn | syn | L2 syn |
| 111 | CEN.PK 1605 | V51TEF | pCM185 | pCB02 | Syn | syn | L2 syn L1 syn |
(Syn: 시네초코쿠스 엘롱가투스; L2 syn: GroEL2; L1 syn: GroEL1 시네초코쿠스 엘롱가투스)
1.4. 피키아 파스토리스( Pichia pastoris ) 균주의 작제
플러그-인에 함유된 유전자의 기능적 발현을 유지하기 위하여, 프로모터 및 터미네이터를 피키아 파스토리스 GS115에서 기능성인 프로모터 및 터미네이터로 대체하였다(Thermo Fisher Scientific C181-00).
플라스미드 pFPP56에서(표 III), CAS19, CAS20 및 CAS21(표II)로부터 RbcX, GroES 및 GroEL 유전자의 발현을 조절하는 각각의 프로모터를 문헌에 따라 혼화성 프로모터(하기 표 VIII)로 대체하였다.
[표 VIII] 발현 카세트
| 명칭 | 프로모터 | 개방 판독 프레임 | 터미네이터 |
| CAS50 | PEX8 | RbcX | TEF1 |
| CAS51 | AOX1 | GroES | PGK |
| CAS52 | FLD1 | GroEL | ADH1 |
하기 3개의 발현 카세트(표 IX)를 포함하는 영역을 PCR로 증폭시키고 시판되는 플라스미드 pPIC3.5(Thermo Fisher K1710-01)내로 클로닝하였다.
[표 IX] 발현 벡터
| 명칭 | 오리진 유형 | 카세트 1 | 카세트 2 | 카세트 3 | 마커 | 숙주 벡터 |
| pCB05 | ARS415-CEN6 | CAS50 | CAS51 | CAS52 | LEU2 | pFL36 |
| pCB06 | 통합성 | CAS50 | CAS51 | CAS52 | HIS4 | pPIC3.5 |
이들 플라스미드를 선형화하고 히스티딘에 대해 자가영양성인 피키아 파스토리스 균주 GS115(Thermo Fisher C181-00)에서 이지셀렉트 피키아 발현 키트 프로토콜(EasySelect Pichia Expression Kit protocol)(Thermo Fisher)에 따라 개별적으로 형질전환하여 최소 배지 및 글루코즈 상에 30℃에서 선택하였다.
[표 X] 플라스미드 및 균주
| 명칭 | 모 균주 | 벡터 1 | 발현된 단백질 | ||
| RbcX | GroES | GroEL | |||
| PPGC115_01 | 피키아 파스토리스 GS115 | pIC3.5 | - | - | - |
| PPGC115_02 | 피키아 파스토리스 GS115 | pCB06 | 에스. 엘롱가투스 | E. coli | E. coli |
1.5. 야로위아 리폴리티카( Yarrowia lipolytica ) 균주의 작제
폐수로부터 분리한 야생형 W29(ATCC 20460, MatA)를 사용하였다.
플라스미드 pFPP56(표 III)에, CAS19, CAS20 및 CAS21(표 II)로부터 RbcX, GroES 및 GroEL 유전자의 발현을 조절하는 각각의 프로모터를 문헌에 따라 혼화성 프로모터(하기 표 XI)로 대체하였다.
[표 XI] 발현 카세트
| 명칭 | 프로모터 | 개방 판독 프레임 | 터미네이터 |
| CAS55 | TEF | RbcX | TEF1 |
| CAS56 | EXP | GroES | PGK |
| CAS57 | GDP | GroEL | ADH1 |
상기 3개의 발현 카세트(표 XI)를 포함하는 영역을 PCR로 증폭시키고 시판되는 플라스미드 pYLEX1내로 클로닝하였다.
[표 XII] 발현 벡터
| 명칭 | 오리진 유형 | 카세트 1 | 카세트 2 | 카세트 3 | 마커 | 숙주 벡터 |
| pCB07 | 통합성 | CAS19 | CAS20 | CAS21 | LEU2 | pYLEX1 |
이들 플라스미드를 선형화하고 루이신에 대해 독립영양성인 야로위아 리폴리티카 균주 속에서 YLOS 형질전환 키틈 프로토콜(Yeastearn Biotech)에 따라 개별적으로 형질감염시키고 최소 배지 및 글루코즈에서 28℃에서 선택하였다(YLEX 발현 키트, Yeastearn Biotech, 제품 번호: FYY201-1KT).
[표 XIII] 플라스미드 및 균주
| 명칭 | 모 균주 | 벡터 1 | 발현된 단백질 | ||
| RbcX | GroES | GroEL | |||
| PO1f_01 | 야로위아 리폴리티카 | YLEX1 | - | - | - |
| PO1f_02 | 야로위아 리폴리티카 | CB07 | 에스. 엘롱가투스 | E. coli | E. coli |
야로위아 리폴리티카 PO1f (ATCC®MYA2613TM)
유전형: MATA ura3 -302 leu2 -270 xpr2 -322 axp2 - deltaNU49 XPR2 :: SUC2
샤페론의 영향의 평가를 28℃에서 72시간 동안 루이신이 들어있지 않은 합성 배지 속에서 균주 PO1f_01 및 PO1f_02의 배양물에서 수행하였다.
1.6. CHO 세포의 작제
고등 진핵세포의 세포의 세트에서 "샤페론" 플러그-인의 용이하고 다양한 취급 및 고-수행능 전달을 보증하기 위하여, 4세대 렌티바이러스 형질도입 시스템을 선택하였다. 이들 렌티바이러스 입자는 플러그-인을 예를 들면, 사람 또는 쥐 세포와 같은 고등 진핵세포의 다양한 종으로부터 원시의, 무한증식되거나 형질전환된 세포에서 동일하게 전달할 수 있도록 한다.
플라스미드 pFPP56(표 III)에서, CAS20 및 CAS21(표 II)로부터의 GroES 및 GroEL 유전자의 발현을 조절하는 프로모터를 문헌에 따라 혼화성 프로모터(표 XIV)로 대체하였다.
[표 XIV] 발현 카세트
| 명칭 | 프로모터 | 개방 판독 프레임 | 터미네이터 |
| CAS54 | CMV | RbcX | h베타 글로빈 |
| CAS55 | hEF-1a | GroES | hPKG1 |
| CAS56 | hUBC | GroEL | hGAPDH |
RbcX의 개방 판독 프레임을 포함하는 영역 및 하기 2개의 발현 카세트 CAS55 및 CAS56을 PCR로 증폭시키고 XhoI-KpnI을 시판되는 플라스미드 pLVX-Puro(Clontech, 제품 번호 632164)내에 클로닝하였다.
[표 XV] 발현 벡터
| 명칭 | 오리진 유형 | 카세트 1 | 카세트 2 | 카세트 3 | 마커 | 숙주 벡터 |
| pCB10 | 통합성 | CAS54 | CAS55 | CAS56 | Puro | pLVX-Puro |
플라스미드 pLVX-Puro 또는 pCB10를 렌티-X 293T 세포(Clontech) 중 렌티-X 패키징 시스템(Lenti-X Packaging System)(Clontech)을 사용하여 키트 프로토콜에 따라 형질전환시켰다. 바이러스 입자를 함유하는 상층액을 여과하고 1/5 또는 1/2 희석에서 5ml의 배지의 최종 용적에 대해 10cm의 페트리디쉬(Petridish) 속에 배양된 CHO 세포에 가하였다.
형질도입 24시간 후, 세포를 PBS로 세척하고 2μg/ml의 푸로마이신이 보충된 새로운 배양 배지를 선택을 위해 37℃에서 48시간 동안 가한다.
이렇게 확립된 세포주를 배양 배지 속에서 0.5μg/ml의 푸로마이신의 농도하에 유지시킨다.
[표 XVI] 플라스미드 및 균주
| 명칭 | 모 균주 | 벡터 1 | 발현된 단백질 | ||
| RbcX | GroES | GroEL | |||
| CHO-01 | CHO | pLVX-Puro | - | - | - |
| CHO-02 | CHO | pCB10 | 에스. 엘롱가투스 | E. coli | E. coli |
1.7. 방법
배양 방법 1:
글루코즈에서의
성장
형질전환된 세포를 30℃에서 주위 공기하에 6.7g/l의 황산암모늄, 20g/l의 글루코즈, 형질전환에 사용된 플라스미드의 선택 마커(-ura, -leu, -trp)에 적합한 시판되는 CSM 배지(MP Biomedicals)가 보충된 한천에 대한 20g/l의 한천이 보충된 YNB 배지(질소 기재가 없는 효모) 상에서 및 2μ/ml의 독시사이클린의 존재하에서 성장시킨다. 배양을 4℃에서 대수상의 종결전 세대를 냉각시켜 정지시킨다. 배양물을 원심분리한 후, 원형질체를 고장성(hypertonic) 소르비톨 배지(1.2M 소르비톨) 속에서 자이몰리아-사이토헬리카제 혼합물로 세포 벽을 효소 분해하여 제조한다. 원형질체를 포화 농도의 PMSF 및 EDTA(프로테아제 억제제)의 존재하에서 고장성 소르비톨 배지 속에서 세척한 후, 반복된 피펫팅 및 온화한 초음파에 의해 등장성 소르비톨 배지(0.6 M) 속에서 파괴하였다. 저속(1500rpm)에서 원심분리하여 거대한 부스러기를 온화한 속도(4000rpm)로 제거하여 중간 크기의 부스러기 및 미토콘드리아를 수집한 후에, 상층액을 수집하고 단백질 농도를 브래드포드법(Bradford method)으로 정량화한다.
시험관내에서
RuBisCO
활성에 대한 시험
시험관내에서, 당해 세포 분해물로부터의 15μg의 단백질 샘플을 200μl의 반응 용적에 대해, 적합한 완충액(50 mM 트리스/HCl pH7.4, 10 mM MgCl2 +, 60 mM 중탄산나트륨) 속에서 합성 분자 RuBP(최종 2mM)에 가하여, RuBisCO 복합체가 효모 분해물 속에서 발현되도록 하고, 포스포글리세르산 분자의 형성을 촉매한다. 다양한 시점에서, 반응을 HCl(12.1M)을 가하여 정지시키고 반응 생성물을 HPLC/MS으로 분석하여 시간에 따라 생산된 포스포글리세르산을 평가함으로써 단백질 샘플의 카복실라제 활성을 평가하였다.
조절된 배지 속의 배양물
예비배양물을 화학적으로 정의된 배지에서 제조하였다. 해동 후, 1ml의 스톡 튜브(-80℃)를 취하여 10ml의 배양 배지를 함유하는 페니실린 병(100 ml)에 접종하고 18시간 동안 30℃ 및 120rpm에서 항온처리하였다. 예비배양물을 혐기생활(질소로 이미 플러싱한 병)에서 및 독시사이클린의 존재하에서 제조하여 PRK 유전자의 존재하에 관찰된 독성 문제를 피하였다. 이후에, 예비배양물을 생리적 염수(NaCl, 9g/l)로 3회(원심분리, 재현탁, 15초 동안 와동) 세척한 후, 세포를 독시사이클린이 없는 배양 배지 속에서 재현탁시켰다.
예비배양물로부터의 줄기생성하는(stemming) 이들 세포를 이후 접종하여 초기 광학 밀도가 0.05(또는 0.1g/l)가 되도록 한다. 출발 배양물 용적은 혐기생활(250ml의 배플화된 에를렌마이어 플라스크(baffled Erlenmeyer flask)) 속에서 50ml 또는 혐기생활(100ml의 페니실린 병)에서 35ml였다.
모든 글루코즈가 소비되거나 에탄올 생산이 중지된 후에 배양을 정지시켰다. 각각의 배양물을 3개 제조하였다.
분석: 세포외 대사물질의 특성화
글루코즈, 포름산 및 기본 대사물질(에탄올; 글리세롤; 아세트산, 석신산 및 피루브산)의 농도를 고-성능 액체 크로마토그래피(HPLC)로 측정하였다. 사용된 장치는 Aminex HPX 87-H+(300 mm x 7.8mm) 컬럼이 장착된 크로마토그래프(Waters, Alliance 2690)이었다. 분자의 검출은 굴절률 검출기(Waters 2414 굴절기)로 제공하였다. 용출물은 0.5ml/min의 유동 속도에서 8mM의 H2SO4이었고 컬럼 온도는 50℃로 설정하였다. 혐기생활에서, 당해 분석을 각각의 균주의 1개 병에서 수행하였다. 이 경우에, 표준 편차의 계산은 질량의 손실로 수행한 후, 대사물질에 적용하였다.
실시예
2: 효모에서
제I형
RuBisCO의
카복실라제
활성의 재구성에 대한
샤페론
조합물의 효과
캘빈 사이클(Calvin cycle)은, 식물 및 시아노박테리아가 이산화탄소로부터 글루코즈를 생산하도록 한다. 주요 단계는 5개의 탄소를 갖는 분자인, 리보즈-1,5-비스포스페이트(RuBP)에서 CO2의 고정이다. 당해 단계는 RuBisCO(리불로즈-1,5-비스포스페이트카복실레이트/옥시게나제에 대해)라고 불리는 효소를 필요로 한다. 당해 효소는 2개의 3개-탄소 3-포스포글리세레이트 분자를 신속하게 제공하는 불안정한 6개-탄소 분자의 형성을 가능하도록 한다. 수개 형태의 RuBisCO가 존재한다. 제I형은 2개의 소단위 유형으로 이루어진다: 거대한 소단위(RbcL) 및 작은 소단위(RbcS), 이의 정확한 조립은 또한 적어도 하나의 특이적인 샤페론: RbcX의 개입을 필요로 한다. RuBisCO의 기질인, RuBP는 리불로즈-5-포스페이트와 ATP의 반응에 의해 형성되며; 당해 반응은 포스포리불로키나제(PRK)에 의해 촉매된다.
당해 실시예에서, 인공적인 캘빈 주기는 효모 균주 CEN의 동시-형질전환에 의해 재구성된다. 상기 표 IV의 3 및 4번 벡터의 조합에 의한 PK 1605는 재조합 번호에 따라 E. coli 또는 시네초코쿠스로부터의 일반 샤페론 GroEL 및 GroES의 존재(조합 3 및 101) 또는 부재(조합 4)하에, RuBisCO의 RbcS 및 RbcL 소단위, 시네초코쿠스 엘롱가투스로부터의 특이적인 샤페론 RbcX 및 PRK 효소의 동시 발현을 가능하도록 한다.
따라서, 3개의 별도의 실험(A, B 및 C)에서 RuBisCO 활성에 대한 시험을 글루코즈 상의 효모 배양물로부터의 단백질 추출물로부터 수행하며(상기 1.3 지점에서 상세히 설명된 프로노콜); 이들의 수율은 시간의 함수로서 포스포글리세르산 생산을 측정함으로써 평가한다. 결과는 하기 표 XVII에 나타낸다.
표 XVII: 시험관내 RuBisCO 활성에 대한 시험은 글루코즈에서 성장되고 제1 컬럼에 나타낸 가공물을 함유하는 CEN-PK 균주의 추출물로부터 수행하였다. 당해 시험은 2mM 리불로즈 디포스페이트를 함유하는 200μl의 반응 용적 속에서 효모의 가용성 추출물의 단백질 0.01 내지 0.02mg을 사용하여 80분의 항온처리 동안 실온에서 수행한다. 활성은 추출물 속의 nmol의 형성된 3-포스포글리세레이트의 mg으로 제공된다.
[표 17]

실험 A는 샤페론 RbcX 단독이, RuBisCO 복합체에 특이적임에 불구하고 활성 효소 복합체의 발현을 가능하도록 하는데 충분하지 않음을 나타낸다. RbcX의 존재에 적합한 화학양론으로 연합된, E. coli로부터의 일반 샤페론 GroES 및 GroEL 둘 다의 조합만이 시간에 따른 포스포글리세르산 생산의 증가를 검출할 수 있도록 한다.
더욱이, 실험 C는 RuBisCO 활성이 시네초코쿠스 엘롱가투스로부터의 RbcL/RbcS 소단위가 동일한 유기체로부터의 동종 샤페론 RbcX, GroES, GroEL2와 연합된 경우 90% 이상까지 현저하게 강하됨을 나타낸다. 이는 이종 샤페로닌(chaperonin)의 연합의 장점을 명확하게 나타낸다.
본 실시예는 이종 박테리아의 일반 샤페론과 박테리아의 특수한 샤페론 RbcX의 연합이 효모에서 합성 RuBisCO 복합체의 활성을 최적화하는데 필수적임을 명확하게 측정할 수 있도록 한다.
실시예 3: 재조합체 단백질의 독성에 대한 샤페론의 조합의 보호 효과
RuBisCO와는
독립적으로,
생체내(in vivo)에서
리불로키나제
발현과 관련된 독성을
제거하는데 있어서의
효과
시행된 방법 및 분석은 상기 실시예 1에 기술한다.
효모(균주 18b)에서 리불로키나제만의 발현은 야생형 균주(WT)와 비교하여 장기간의 잠재 상(latency phase)(50시간 이상) 및 이의 최대 성장율에 있어서 현저한 강하(호기 생활에서 70% 및 혐기 생활에서 82%)를 포함한다(표 XVIII).
표 XVIII: 균주 WT, 1b, 18b, 102, 15, 14b의 유전형, 최대 성장율 및 성장 지연
[표 18]

PRK에 의해 유도된 당해 독성은, E. coli로부터의 샤페론 GroES/GroEL의 균주 102 내 동시-발현에 의해서(혐기생활에서 26% 및 호기생활에서 42%의 성장율에서 독성의 제거) 또는 균주 14b 내 시네초코쿠스 엘롱가투스로부터의 샤페론 RbcX 의 동시-발현에 의해 (혐기생활에서 34% 및 호기생활에서 10%의 성장율에서 독성의 제거) 부분적으로 제거할 수 있다.
E. coli로부터의 샤페론 GroES/GroEL 및 시네초코쿠스 엘롱가투스로부터의 RbcX의 균주 15에서 동시-발현은 전체 성장 근처까지 회복(혐기생활에서 78% 및 호기생활에서 63%의 성장율에서 독성의 제거)을 가능하도록 한다. 당해 동시-발현은 또한 장기간의 잠재 상을 완전히 제거하는 것을 가능하도록 한다.
리불로키나제(PRK)의 발현과 관련된 독성은 잠재 상(latency phase)의 존재 및 성장율의 저하와 직접적으로 관련된 에탄올 생산성(균주 18b)(도 1)에 있어서의 강하를 특징으로 하는 알코올 발효에 영향을 미친다. "샤페론" 가공체(engineering)(E. coli로부터의 GroES+ GroEL + 시네초코쿠스 엘롱가투스로부터의 RbcX)의 발현은 리불로키나제(PRK)의 독성, 보다 상세하게는 PRK-촉매된 반응의 생성물의 축적을 완전히 제거할 뿐만 아니라, 결과적으로 에탄올 생산성(균주 15)의 증가가 가능하도록 하는 반면, 일반적인 샤페론 쌍 GroES/GroEL은 부분적인 효과(균주 102)만을 가지며 RbcX 만의 발현은 효과가 없었다(균주 14b).
실시예
4: 형질전환된 세포의
성장에 있어서
일반적인 효과의 예시
4.1. 발효시 사카로마이세스 세레비지아에(Saccharomyces cerevisiae)의 성장에 있어서 일반적인 효과
시행된 방법 및 분석은 상기 실시예 1에 기술되어 있다. 결과는 하기 표 XIX 에 나타낸다.
[표 XIX] 균주 WT, 1b, 103, 13b의 유전형, 최대 성장율 및 성장 지연.
[표 19]
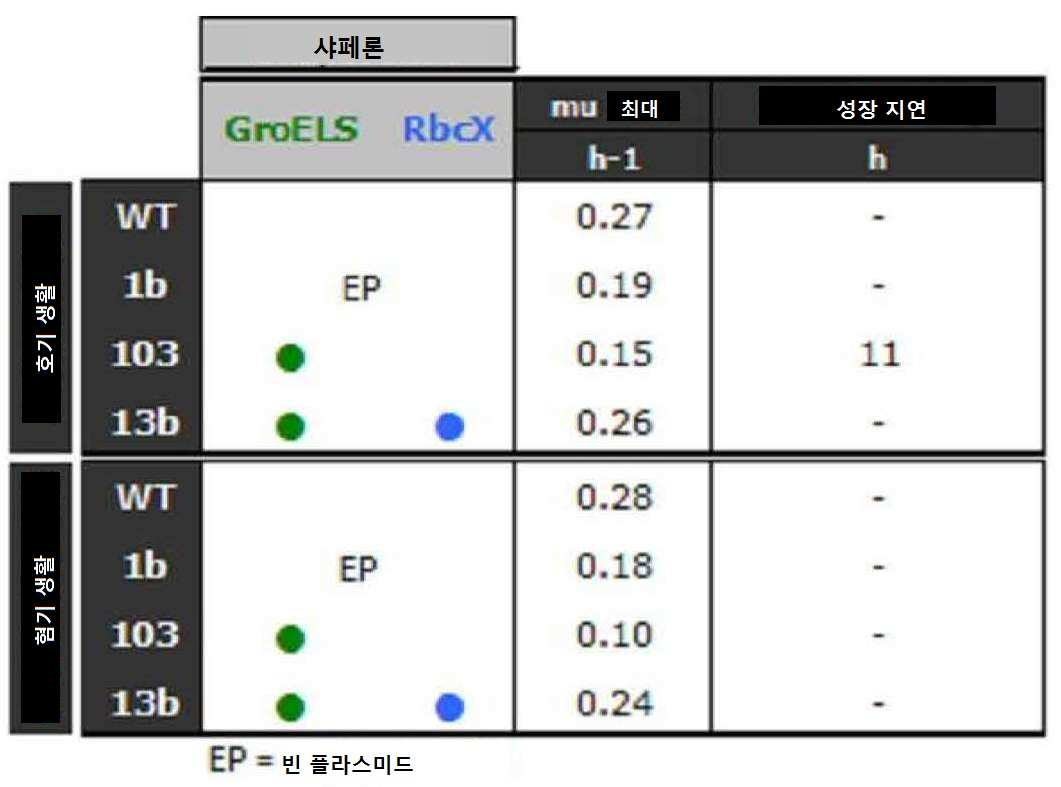
유리하게는, "샤페론" 가공의 발현은 3개의 "빈" 플라스미드를 함유하는 대조군 균주(균주 1b), 또는 E. coli의 일반 샤페론 GroES/GroEL 만을 발현하는 균주 103과 비교하여 균주 13b(RbcX+(Gros/GroEL) E. coli)에 대해 30%의 증식성 장점(proliferative advantage)을 제공한다.
더욱이, 균주 13b의 성장율은 형질전환되지 않아서 호기생활 및 혐기생활 각각에서 스트레스받지 않는 야생형(WT) CEN.PK 113-7D의 성장율의 96% 및 86%와 동일하다(표 XVIII).
4.2. 발효시 피키아 파스토리스(Pichia pastoris)의 성장에 있어서의 일반적인 효과
샤페론의 영향의 평가는 글리세롤이 들어있는 최소 배지상에서 30℃에서 14시간 동안 균주 PPGC115_01 및 PPGC115_02의 배양물 및 F. Wang etal. 2015 (PLoS One. 2015 Mar 17;10(3):e0120458. "High-level expression of endo-β-N-acetylglucosaminidase H from Streptomyces plicatus in Pichia pastoris and its application for the deglycosylation of glycoproteins.")에 기술된 프로토콜에 따라서 48시간 내지 100시간 동안 1% 메탄올을 사용한 유도로 수행하였다.
2개 균주를 100시간의 발효에서 평가된 동일한 세포 농도에서 접종하였고, 성장 곡선의 대수상에서 게산된 균주의 최대 mu는 균주 PPGC115_02의 경우 대조군 균주 PPGC115_01의 것에 대해 30% 정도의 증식성 장점을 갖는다.
4.3. 발효시 야로위아 리폴리티카의 성장에 있어서의 일반적인 효과
균주 PO1f_01 및 PO1f_02를 JM Nicaud et al. 2002(Protein expression and secretion in the yeast Yarrowia lipolytica. FEMS Yeast Res. 2002 Aug;2(3):371-9)에 기술된 프로토콜에 따라서 평가하였다. 표현형은 샤페론의 조합을 발현하는 균주에 대한 증가된 성장을 나타낸다. 증식성 장점은 35% 이상에서 평가된다.
4.4. CHO 세포의 성장에 있어서의 일반적인 효과
세포주(lines) CHO-01 및 CHO-02를 동일한 밀도(10cm 디쉬(dish)당 2x106개의 세포)로 접종하고 성장을 4일에 걸쳐 평가한다. 세포를 탈착시키고 트립신으로 처리하여 개별화하고 각각의 날에 자동 계수기를 사용하여 계수한다. 세포주 CHO-02의 세포의 성장율은 대조군 세포주 CHO-01의 것보다 25% 정도로 더 높다. 샤페론의 조합은 세포가 2배로 되는 시간에 있어서의 효과를 갖는다.
이들 연구는, 이들 "샤페론" 가공이 (i) 다른 가공체를 함유하거나 함유하지 지만 물리화학적 스트레스가 적용되는 진핵 세포의 정상적인 성장을 회복시키고 (ii) 동일한 조건하에서 사용된 균주/세포와 관련하여 증식성 장점을 제공하는 것이 가능하도록 함을 나타낸다.
실시예
5:
재조합체
단백질의 생산에 대한 효과의 예시
5.1. 사카로마이세스 세레비지아에에서의 인간 성장 호르몬의 생산
인간 성장 호르몬 유전자(진뱅크(GenBank): K02382.1)를 합성하고 하기 카세트에 따라서 구성적 프로모터 TEF1의 하부에 클로닝하였다.
[표 XX] 발현 카세트
| 명칭 | 프로모터 | 개방 판독 프레임 | 터미네이터 |
| CAS54 | TEF1p | hGH | PGK |
[표 XXI] 발현 벡터
| 명칭 | 오리진 유형 | 카세트 1 | 카세트 2 | 카세트 3 | 마커 | 숙주 벡터 |
| pCB09 | 2μ | CAS54 | URA3 | PYeDP51 | ||
| pFPP56 | ARS415-CEN6 | CAS19 | CAS21 | CAS22 | LEU2 | pFL36 |
이들 플라스미드를 앞서 기술한 형질전환 프로토콜에 따라 균주 CEN.PK 1605에서 공동으로 형질전환하였다. 균주를 합성 배지(-leu-ura)에서 선택하였다.
[표 XXII] 균주
| 명칭 | 모 균주 | 벡터 1 | 벡터 2 | 발현된 단백질 | |||
| RbcX | GroES | GroEL | hGH | ||||
| 200 | CEN.PK1605 | PYeDP51 | pFPP56 | 에스. 엘롱가투스 | E. coli | E. coli | |
| 230 | CEN.PK1605 | pCB09 | pFPP56 | 에스. 엘롱가투스 | E. coli | E. coli | hGH |
| 231 | CEN.PK1605 | pCB09 | pFL36 | hGH | |||
균주 200, 230 및 231을 OD 600nm가 0.7이 될 때까지 합성 배지(-leu-ura)에서 성장시켰다. 세포를 수집하고 1ml의 냉 분해 완충액(1X PBS pH 7.4, 1 mM PMSF)으로 1회 세척한 후, 0.3ml의 램믈리 완충액(Laemmli buffer) 속에 재현탁시키고 5분 동안 98℃에서 항온처리하였다. 일련의 희석물(1:10)을 제조하고 각각의 샘플의 동일한 용적을 SDS-PAGEgel(구배: 4% 내지 20%)에 침착시키고, 니트로셀룰로즈 막으로 이전시켰다. hGH 발현을 항체 [GH-1](ab9821, Abcam)로 평가하고 편재한 유전자(ubiquitous gene) GAPDH(ab9485, Abcam)의 발현과 관련하여 표준화하였다. 또한, hGH 발현을 성장 호르몬 ELISA 키트, 사람(Thermo Scientific, 제품 번호: EHGH1)의 프로토콜을 사용하여 및 이에 따라 ELISA로 정량화한다.
균주 230에서 평가된 생산된 hGH 단백질의 양은 균주 211에서 수득된 것보다 40% 더 높다.
5.2.
사카로마이세스
세레비지아에에서의
내인성
루시퍼라제
활성의 평가
개똥벌레 루시퍼라제 유전자를 벡터 pGL4(Promega)로부터 증폭시키고 하기 카세트에 따라 구성적 프로모터 TEF1의 하부에 클로닝하였다.
[표 XXIII] 발현 카세트
| 명칭 | 프로모터 | 개방 판독 프레임 | 터미네이터 |
| CAS53 | TEF1p | fLuc | PGK |
[표 XXIV] 발현 벡터
| 명칭 | 오리진 유형 | 카세트 1 | 카세트 2 | 카세트 3 | 마커 | 숙주 벡터 |
| pCB08 | 2μ | CAS53 | URA3 | PYeDP51 | ||
| pFPP56 | ARS415-CEN6 | CAS19 | CAS21 | CAS22 | LEU2 | pFL36 |
이들 플라스미드를 앞서 기술한 형질전환 프로토콜에 따라서 균주 CEN.PK 1605에서 공동으로 형질전환시켰다. 균주를 합성 배지(-leu-ura)에서 선택하였다.
[표 XXV] 플라스미드 및 균주
| 명칭 | 모 균주 | 벡터 1 | 벡터 2 | 발현된 단백질 | |||
| RbcX | GroES | GroEL | fLUC | ||||
| 200 | CEN.PK1605 | PYeDP51 | pFPP56 | 에스. 엘롱가투스 | E. coli | E. coli | |
| 210 | CEN.PK1605 | pCB08 | pFPP56 | 에스. 엘롱가투스 | E. coli | E. coli | fLuc |
| 211 | CEN.PK1605 | pCB08 | pFL36 | fLuc | |||
균주 200, 210 및 211를 합성 배지(-leu-ura) 속에서 OD 600nm가 0.7이 될때까지 성장시켰다. 세포를 수집하고 1ml의 냉 분해 완충액(1X PBS pH 7.4, 1mM PMSF)으로 1회 세척한 후, 0.3ml의 동일한 완충액 속에 재현탁시켰다. 현탁된 세포를 유리 비드(Fast Prep)로 분해하였다.
조 분해물의 농도를 브래드포드법(BioRad)으로 측정하고 0.5mg/ml로 희석시키고, 루시퍼라제 활성을 샘플당 5μl의 분해물을 사용하여 루시퍼라제 검정 시스템(Promega)으로 측정하고 발광성을 광도계로 평가하였다.
활성을 총 단백질의 양에 대해 표준화한다.
균주 210에서 평가한 루시퍼라제 활성은 균주 211에서 평가된 것보다 60% 더 높다.
5.3. 사카로마이세스 세레비지아에에서의 재조합체 셀룰라제의 활성의 평가
샤페론 가공은 프로모터 TEF2하에서 탈라로마이세스 에메르소니이(Talaromyces emersonii)(진뱅크 수탁 번호 AAL89553)로부터의 셀룰라제인 셀로바이오하이드롤라제 1(CBH1)의 발현에 대한 가공과 관련된다. 셀룰라제 활성의 분석은 Y. Ito et al. 2015 (Combinatorial Screening for Transgenic Yeasts with High Cellulase Activities in Combination with a Tunable Expression System. PLoS One. 2015 Dec21;10(12))에 기술된 바와 같이 수행하였다.
사페론의 존재하에 셀룰라제 가공물을 동시-발현하는 균주에 대해 기록한 활성은 셀룰라제 가공만을 발현하는 균주보다 활성 수율이 37% 더 높다.
5.4. 단백질 생산의 증진
사카로마이세스에 대해 앞서 기술된 단백질 생산을 개선시키는 샤페론의 용도는 목적한 진핵 세포 및 특히 피키아 및 야로위아에서 용이하게 시행함으로써 내인성 또는 외인성 단백질의 수율 및/또는 활성을 최적화할 수 있다.
당해 분야의 숙련가는 특히 본 발명에 따른 샤페론의 조합물을 발현하는 효모가 식품-가공 산업, 약제 분야, 생물량 가수분해, 에너지 등을 위한 목적한 다양한 단백질의 생산을 가능하도록 하는 하기 문헌을 특히 언급할 수 있다:
* Spohner SC, M?ller H, Quitmann H, Czermak P. Expression of enzymes for the usage in food and feed industry with Pichia pastoris. J Biotechnol. 2015 May 20;202:118-34.
* Kim H, Yoo SJ, Kang HA. Yeast synthetic biology for the production of recombinant therapeutic proteins. FEMS Yeast Res. 2014 Aug 12.
* Ahmad M, Hirz M, Pichler H, Schwab H. Protein expression in Pichia pastoris: recent achievements and perspectives for heterologous protein production. Appl Microbiol Biotechnol. 2014 Jun;98(12):5301-17. doi: 10.1007/s00253-014-5732-5. Epub 2014 Apr 18. Review.
* Weinacker D, Rabert C, Zepeda AB, Figueroa CA, Pessoa A, Far?as JG. Applications of recombinant Pichia pastoris in the healthcare industry. Braz J Microbiol. 2014 Mar 10;44(4):1043-8
* Rabert C, Weinacker D, Pessoa A Jr, Far?as JG. Recombinants proteins for industrial uses: utilization of Pichia pastoris expression system. Braz J Microbiol. 2013 Oct 30;44(2):351-6.
* Spadiut O, Capone S, Krainer F, Glieder A, Herwig C. Microbials for the production of monoclonal antibodies and antibody fragments. Trends Biotechnol. 2014 Jan;32(1):54-60.
* Espejo-Mojica AJ, Almeciga-D?az CJ, Rodriguez A, Mosquera A, Diaz D, Beltran L, Diaz S, Pimentel N, Moreno J, Sanchez J, Sanchez OF, Cordoba H, Poutou-Pinales RA, Barrera LA. Human recombinant lysosomal enzymes produced in microorganisms. Mol Genet Metab. 2015 Sep-Oct;116(1-2):13-23.
* Gunduz Ergun B, Calik P. Lignocellulose degrading extremozymes produced by Pichia pastoris: current status and future prospects. Bioprocess Biosyst Eng. 2015 Oct 23.
* Ledesma-Amaro R, Dulermo T, Nicaud JM. Engineering Yarrowia lipolytica to produce biodiesel from raw starch. Biotechnol Biofuels. 2015 Sep 15;8:148.
* Kalyani D, Tiwari MK, Li J, Kim SC, Kalia VC, Kang YC, Lee JK. A highly efficient recombinant laccase from the yeast Yarrowia lipolytica and its application in the hydrolysis of biomass. PLoS One. 2015 Mar 17;10(3):e0120156.
* Zinjarde SS. Food-related applications of Yarrowia lipolytica. Food Chem. 2014;152:1-10.
* Hughes SR, Lopez-Nunez JC, Jones MA, Moser BR, Cox EJ, Lindquist M, Galindo-Leva LA, Riano-Herrera NM, Rodriguez-Valencia N, Gast F, Cedeno DL, Tasaki K, Brown RC, Darzins A, Brunner L. Sustainable conversion of coffee and other crop wastes to biofuels and bioproducts using coupled biochemical and thermochemical processes in a multi-stage biorefinery concept. Appl Microbiol Biotechnol. 2014 Oct;98(20):8413-31.
* Groenewald M, Boekhout T, Neuveglise C, Gaillardin C, van Dijck PW, Wyss M. Yarrowia lipolytica: safety assessment of an oleaginous yeast with a great industrial potential. Crit Rev Microbiol. 2014 Aug;40(3):187-206.
* Celik E, Calik P. Production of recombinant proteins by yeast cells. Biotechnol Adv. 2012 Sep-Oct;30(5):1108-18.
* Sabirova JS, Haddouche R, Van Bogaert I, Mulaa F, Verstraete W, Timmis K, Schmidt-Dannert C, Nicaud J, Soetaert W. The 'LipoYeasts' project: using the oleaginous yeast Yarrowia lipolytica in combination with specific bacterial genes for the bioconversion of lipids, fats and oils into high-value products. Microb Biotechnol. 2011 Jan;4(1):47-54.
SEQUENCE LISTING
<110> INSTITUT NATIONAL DE LA RECHERCHE AGRONOMIQUE
INSTITUT NATIONAL DES SCIENCES APPLIQUEES DE TOULOUSE
CENTRE NATIONAL DE LA RECHERCHE SCIENTIFIQUE
<120> COMBINAISON DE CHAPERONNES BACTERIENNES AFFECTANT POSITIVEMENT LA PHYSIOLOGIE D?NE CELLULE EUCARYOTE NATIVE OU PORTANT UNE INGENIERIE
<130> IP20173217/FR
<160> 7
<170> KopatentIn 2.0
<210> 1
<211> 336
<212> DNA
<213> Synechococcus elongatus
<400> 1
atgtcaatga agacgctccc taaagaaagg agatttgaaa cgttttcata tctgccccct 60
ctctctgata gacaaatcgc tgctcaaatc gaatatatga tcgaacaagg ttttcatcca 120
ttaatcgaat ttaatgaaca ttctaatcca gaagaatttt attggactat gtggaagctc 180
cctctttttg attgtaaatc tcctcaacag gttttagatg aagtgagaga gtgtagatct 240
gaatatggtg attgttatat cagagttgct ggttttgata atatcaaaca atgtcaaact 300
gtttctttta tcgttcatag acctggaaga tactaa 336
<210> 2
<211> 1419
<212> DNA
<213> Synechococcus elongatus
<400> 2
atgcctaaga ctcaatcggc tgccggttac aaggcaggtg taaaagatta caaactaact 60
tattatactc cagattatac acccaaagac actgacttac tagccgcctt tcgcttttcg 120
ccccagccag gtgttccagc tgatgaagct ggtgctgcta tcgctgctga atcttctact 180
ggtacttgga ctactgtttg gactgattta ttaactgata tggacagata caaaggcaaa 240
tgttaccata ttgaaccggt tcaaggtgag gaaaattctt attttgcttt tatcgcatac 300
cctctagatc tttttgaaga gggttctgtt actaatatct taacttctat cgtcggtaat 360
gtctttggct ttaaggccat tcgtagccta cgtcttgaag acatcaggtt tccagttgct 420
ttagttaaaa cttttcaagg tccaccacat ggtatccaag tagaacggga tcttttaaat 480
aaatatggca gaccgatgct cgggtgcacg attaagccga agctcgggct ttctgctaaa 540
aattatggta gagctgttta tgaatgttta cgtggtggtt tagattttac taaagatgat 600
gaaaatatca attctcaacc gttccagcgt tggcgggacc gattcctctt tgtggccgac 660
gcgatccata aatctcaagc tgaaactggt gaaatcaaag gtcattattt aaatgtaacg 720
gcgcctacat gtgaagaaat gatgaagcga gcagaatttg ctaaggaact aggtatgcct 780
atcatcatgc atgatttttt aactgctggt tttactgcta atactacttt agctaaatgg 840
tgccgggaca atggagtcct attacatatc catcgtgcca tgcacgcggt cattgatcgt 900
caaaggaatc atggtatcca ttttagagtt ttagctaaat gtttaagatt atctggtggt 960
gatcatttac attctggtac tgtcgtggga aagcttgagg gtgacaaggc atctacatta 1020
ggttttgttg atttaatgag agaagatcat atcgaagctg atagatctag aggtgttttt 1080
tttactcaag actgggcgtc gatgccgggg gtgctcccag ttgcttctgg tggtatccat 1140
gtttggcaca tgccggcgtt agttgaaatc tttggtgatg attctgtttt acaatttggt 1200
ggtggtactt taggtcatcc atggggtaat gcaccaggtg ctactgctaa tagagttgct 1260
ttagaagctt gtgttcaagc tagaaatgaa ggtagagatt tatatagaga gggtggtgat 1320
attttaaggg aagcaggtaa atggtcgcct gaactggcag ccgccctcga tttatggaaa 1380
gaaatcaaat ttgaatttga aactatggat aaattataa 1419
<210> 3
<211> 486
<212> DNA
<213> Synechococcus elongatus
<400> 3
atgcaattta tgggtactgc ttctaggatg gcgtcgacgc aacgggccaa gcctatggag 60
atgccgagga ttagccgtga tactgctaga atgttagtta attatttaac ttatcaagct 120
gtttgtgtta tcagagatca attagctgaa actaatccag ctggtgccta tagattacaa 180
gttttttctg ctgaattttc ttttcaagat ggtgaagctt atttagctgc tttattaaat 240
catgatagag aattaggact aagggtgatg acggtaaggg aacatttagc tgaacatatt 300
ctagattatc ttccagaaat gacgattgcc caaattcaag aggccaacat taaccataga 360
agagcacttt tagaaaggtt aacaggcctt ggggctgagc catctttacc ggaaacggag 420
gtctcagaca gaccctcaga ttctgctact ccagatgatg cttctaatgc ttctcatgct 480
gattaa 486
<210> 4
<211> 1002
<212> DNA
<213> Synechococcus elongatus
<400> 4
atgtctaaac cagatagagt tgttttaatc ggtgttgctg gtgattctgg ttgtggtaaa 60
tctacatttc ttaacaggtt agctgattta tttggtactg aattaatgac tgttatttgt 120
ttagatgatt atcattcgtt agatcgtaaa ggcagaaagg aagcgggtgt aactgcttta 180
gatcctagag ctaataattt tgatttaatg tatgaacaag ttaaagcttt aaaaaatggt 240
gaaactatca tgaaaccaat ctataatcat gaaactggtt taatcgatcc acctgaaaag 300
atcgagccaa acagaattat tgtaattgaa gggttacacc cactttatga cgaacgagtt 360
cgcgaacttt tagatttttc tgtttattta gatatcgatg atgaagttaa aatcgcttgg 420
aaaatccaaa gagatatggc cgaaagaggt cattcttatg aagatgtttt agcctcaatt 480
gaggctagaa ggccagattt taaagcatat attgaaccgc aacggggaca cgctgatatc 540
gttattcgtg taatgcccac tcaacttatc ccgaatgaca ctgagaggaa agtcctaagg 600
gtacaattaa tccagagaga aggaagggat ggatttgaac cagcttattt atttgatgaa 660
ggttctacaa ttcaatggac gccttgtggc agaaagttaa catgtagcta tcctggcatt 720
cgcttagctt atggtccaga tacttattat ggtcatgaag tttctgtcct tgaagtggat 780
ggacaatttg aaaatttaga agaaatgatt tacgttgaag gtcatttatc taaaactgat 840
actcaatatt atggtgaatt aactcatcta cttttacaac acaaagatta tccaggttct 900
aataatggta ctggtttatt ccaagtgcta acgggtctca agatgcgggc cgcctatgaa 960
aggttaactt ctcaagctgc tccagttgct gcttctgttt aa 1002
<210> 5
<211> 312
<212> DNA
<213> Synechococcus elongatus
<400> 5
atggctgccg tctcattatc tgtttctact gttactccat taggtgatag agtttttgtt 60
aaagttgctg aagctgaaga aaaaactgct ggtggtatca tcttaccaga taatgctaaa 120
gaaaaaccac aagtcggtga aattgtcgct gttggtccag gtaaaagaaa tgatgatggt 180
tcaagacaag ctccagaagt taaaatcggt gataaagttt tatattctaa atatgctggt 240
actgatatta aattaggtaa tgatgattat gttcttttat ctgaaaaaga tatcttagct 300
gttgtcgctt aa 312
<210> 6
<211> 1668
<212> DNA
<213> Synechococcus elongatus
<400> 6
atggctaaat taatcttatt tcatgaagat tcaagacaag cattagaaag gggtgttaat 60
gctttagcta atgctgttaa agttacttta ggtccaagag gtagaaatgt tttattagaa 120
aaaaaatttg gtgctccaga aatcatcaat gatggtgttt ctatcgctaa agaaatcgaa 180
ttagaagatc cacatgaaaa tgcaggtgca agactagttc aagaagttgc tgctaaaact 240
aaagaaatcg ctggtgatgg tactactact gctactgttt tagctcaagc tatcgttaga 300
gaaggtttaa ctaatgttgc tgctggtgct aatccaatcg ttttaagaag aggtatcgaa 360
aaagctgttg ctactttagt tgaagctatc gctgctaaag ctcaaccagt tgctgatgaa 420
gctgctatca gatctatcgc tgctgtttct gctggtaatg atgatgaagt tggtcaaatg 480
atcgctgatg ctgttgctaa agttactaaa gatggtgtta tcacagttga agaatctaaa 540
tctttagcta ctgaattaga agtcgttgaa ggtatgcaat ttgatagagg ttatttatct 600
ccatattttg ttactgatca agatagacaa gtagttgaat atgataatcc attaatctta 660
ttaactgata aaaaaatcgc ttctatccaa gatttagttc cagttttaga agatgttgct 720
agagctggta gaccattatt aatcatcgct gaagatatcg aaggtgaagc tttagctact 780
ttagttgtta ataaagctag aggtgtttta aatactgttg ctgttaaagc tccagctttt 840
ggtgatagaa gaaaagctat cttacaagat atcgctgttt taactggtgg tcaagttatc 900
tctgaagaag ttggtttatc tttagctgat gctaattctt ctgttttagg taaagctcaa 960
aaaatcacta tctctaaaga tactactatc atcgttgctg gtgatgaaaa taaagctgat 1020
gttgctgcta gaatcgctca aatcagaaga tctttagaag aaactgattc tgattatgat 1080
agagaaaaat tacaagaaag aatcgctaaa ttagctggtg gtgttgctgt tatcaaagtt 1140
ggtgctccaa ctgaaactga attaaaaaat agaaaattaa gaatcgaaga tgctttaaat 1200
gctactagag ctgctatcga agaaggagtt gttccaggtg gtggtactac tttattacat 1260
ttagcttctg ctttaacttc tttacaagct tctttaactg ttgctgatga aaaattaggt 1320
gttgaaatcg ttgctagagc tttagaagct ccattaagac aaattgctga taatgctggt 1380
gcagaaggtt ctgttgttgt cgaaaaatta agagataaag attttaattt tggttataat 1440
gctttaactg gtcaatatga agatttagtt gcttctggta tcttagatcc agctaaagtt 1500
gttagatctg ctttacaaga tgctgcttct gttgcttctt taatcttaac tactgaagtt 1560
ttagttgttg atcaacctga accagaacca gctatgcctg ctggtggtga tatgggtggt 1620
atgggtggta tgggtatgcc tggtatgggt ggtatgggta tgatgtaa 1668
<210> 7
<211> 1635
<212> DNA
<213> Synechococcus elongatus
<400> 7
atggctaaaa gaatcatcta taatgaaaat gctagaagag ctttagaaaa aggtatcgat 60
atcttagctg aagctgttgc tgttacttta ggtccaaaag gtagaaatgt cgtcttagaa 120
aagaaatttg gtgcaccaca aattatcaat gatggtgtta ctatcgctaa agaaatcgaa 180
ttagaagatc atatcgaaaa tactggtgtt gctttaatca gacaagcagc ttcaaaaaca 240
aatgatgctg ctggtgatgg tactactact gctactgttt tagctcatgc tgttgtcaaa 300
gaaggtttaa gaaatgttgc tgctggtgct aatgctatct tattaaaaag aggtatcgat 360
aaagctacaa attttcttgt cgaacaaatt aaatcacatg ctcgtccagt cgaagattct 420
aaatctatcg cacaagttgg tgcaatctct gctggtaatg attttgaagt tggtcaaatg 480
atcgctgatg ctatggataa agttggtaaa gaaggtgtta tctctttaga agaaggtaaa 540
tctatgacta ctgaattaga agttactgaa ggtatgcgtt ttgataaagg ttatatctct 600
ccatattttg ctactgatac tgaaagaatg gaagccgtct ttgatgaacc atttatctta 660
atcactgata aaaaaatcgg attagttcaa gatcttgtcc cagttttaga acaagttgct 720
agagctggta gaccattagt tattatcgca gaagatatcg aaaaagaagc tttagctact 780
ttagttgtta atagattaag aggtgtctta aatgttgcag ctgtcaaagc tccaggtttt 840
ggtgatagaa gaaaagctat gttagaagat atcgctgttc ttacaggtgg tcaacttatc 900
acagaagatg ctggtttaaa attagatact actaaattag atcaattagg taaagctaga 960
agaatcacta tcactaaaga taatactact atcgttgctg aaggtaatga agctgctgtt 1020
aaagctagag tcgatcaaat tagaaggcaa attgaagaaa cagaaagctc ttatgataaa 1080
gaaaagttac aagaaagatt agctaaatta tctggtggtg tcgcagttgt caaagttggt 1140
gctgctactg aaactgaaat gaaagataga aaattaagat tagaagatgc tatcaatgct 1200
actaaagctg ctgttgaaga aggtatcgtt ccaggtggtg gtactacttt agctcattta 1260
gctccacaat tagaagaatg ggcaactgct aatttatctg gtgaagaatt aactggtgct 1320
caaatcgttg ctagagcttt aactgctcca ttaaaaagaa tcgctgaaaa tgctggttta 1380
aatggtgctg ttatctctga aagagtcaaa gaattaccat ttgatgaagg ttatgatgca 1440
tcaaataatc aatttgttaa tatgtttact gctggtattg ttgatccagc taaagttaca 1500
agatcagctt tacaaaatgc tgcttctatc gctgctatgg ttttaactac tgaatgtatc 1560
gttgttgata aaccagaacc aaaagaaaaa gctccagctg gtgctggtgg tggtatgggt 1620
gattttgatt attaa 1635
Claims (20)
- 하기를 함유하는 것을 특징으로 하는, 형질전환된 진핵 세포:
(i) 적합한 프로모터의 전사 조절 하에서 박테리아의 제I형 Rubisco 효소의 폴딩(folding)에 관여하는 샤페론 RbcX를 암호화하는 서열을 함유하는 발현 카세트;
(ii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroES를 암호화하는 서열을 함유하는 발현 카세트; 및
(iii) 적합한 프로모터의 전사 조절 하에서 박테리아의 일반 샤페론 GroEL을 암호화하는 서열을 함유하는 발현 카세트. - 청구항 1에 있어서,
상기 샤페론 RbcX가 시아노박테리아 샤페론임을 특징으로 하는 진핵 세포. - 청구항 1 또는 청구항 2에 있어서,
상기 일반 샤페론 GroES 및 GroEL 중의 적어도 하나는 시아노박테리아 또는 RuBisCO 복합체를 발현하는 다른 박테리아으로부터 기원되지 않는 것을 특징으로 하는 진핵 세포. - 청구항 1 내지 청구항 3 중의 어느 한 항에 있어서,
상기 3개의 발현 카세트가 유전 정보의 연속된 블록(continuous block)을 형성하는 것을 특징으로 하는 진핵 세포. - 청구항 1 내지 청구항 4 중의 어느 한 항에 있어서,
상기 3개의 샤페론의 발현 카세트가 단일의 에피소옴 유전 성분(episomal genetic element)에 의해 수반되는 것을 특징으로 하는 진핵 세포. - 청구항 1 내지 청구항 5 중의 어느 한 항에 있어서,
상기 샤페론 이외의 이종 단백질에 대한 적어도 하나의 발현 카세트를 추가로 포함하거나, 내인성 단백질의 발현 수준 및/또는 서열을 개질시키는 서열 가공(sequence engineering)을 거치는 것을 특징으로 하는 진핵 세포. - 청구항 1 내지 청구항 6 중의 어느 한 항에 있어서,
박테리아의 제I형 RuBisCO 효소의 RbcL 또는 RbcS 소단위(subunit)를 암호화하는 서열을 함유하지 않음을 특징으로 하는 진핵 세포. - 청구항 1 내지 청구항 7 중의 어느 한 항에 있어서,
효모, 바람직하게는 사카로마이세스 세레비지아에(Saccharomyces cerevisiae), 야로위아 리폴리티카(Yarrowia lipolytica) 및 피키아 파스토리스(Pichia pastoris)로부터 선택되는 것임을 특징으로 하는 진핵 세포. - 진핵 세포의 생리(physiology)를 증진시키기 위한, 샤페론 RbcX 및 샤페론 GroES 및 GroEL을 발현할 수 있는 발현 카세트의 조합물의 용도.
- 청구항 9에 있어서,
효모 균주의 생리를 증진시키기 위한, 용도. - 청구항 9 또는 청구항 10에 있어서,
상기 샤페론 이외의 이종 단백질에 대한 적어도 하나의 발현 카세트를 포함하거나, 또는 내인성 단백질의 발현 수준 및/또는 서열을 개질시키는 서열 가공을 겪은 진핵 세포의 생리를 증진시키기 위한, 용도. - 청구항 8 내지 청구항 11 중의 어느 한 항에 있어서,
상기 진핵 세포의 성장율을 증가시키기 위한, 용도. - 청구항 8 내지 청구항 12 중의 어느 한 항에 있어서,
환경 스트레스에 대한 상기 진핵 세포의 내성을 증가시키기 위한, 용도. - 청구항 13에 있어서,
상기 환경 스트레스가 진핵 세포의 배양 배지 속에 존재하는 성분의 독성에 기인되는, 용도. - 청구항 8 내지 청구항 14 중의 어느 한 항에 있어서,
진핵 세포에 의해 합성된 화합물의 독성에 대한 상기 세포의 내성을 증가시키기 위한 용도. - 재조합체 단백질의 생산을 위한 청구항 1 내지 청구항 8 중의 어느 한 항에 따른 진핵 세포의 용도.
- 청구항 1 내지 청구항 8 중의 어느 한 항에 따른 진핵 세포에 의한, 화학 분자 및 단백질로부터 선택된 적어도 하나의 화합물의 합성을 가능하게 하는 조건 하에서 상기 진핵 세포를 배양하는 단계, 및 임의로 상기 화합물을 수집하는 단계를 포함함을 특징으로 하는, 상기 화학 분자 및 단백질로부터 선택된 적어도 하나의 화합물의 생명공학적 생산 방법.
- (i) RbcX, GroES 및 GroEL을 발현하는 진핵 세포 내로 재조합체 단백질을 암호화하는 서열을 삽입하는 단계, (ii) 상기 서열의 발현을 가능하게 하는 조건 하에서 상기 세포를 배양하는 단계 및 임의로 (iii) 상기 단백질을 수집하거나 정제하는 단계를 포함하는, 재조합체 단백질의 생산 방법.
- 청구항 17 또는 청구항 18에 있어서,
상기 단백질이 효소임을 특징으로 하는 방법. - 청구항 17 또는 청구항 18에 있어서,
상기 단백질이 호르몬임을 특징으로 하는 방법.
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP15305048.9 | 2015-01-16 | ||
| EP15305049.7 | 2015-01-16 | ||
| EP15305049 | 2015-01-16 | ||
| EP15305048 | 2015-01-16 | ||
| PCT/EP2016/050832 WO2016113418A1 (fr) | 2015-01-16 | 2016-01-15 | Combinaison de chaperonnes bacteriennes affectant positivement la physiologie d'une cellule eucaryote native ou portant une ingenierie |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20170105079A true KR20170105079A (ko) | 2017-09-18 |
Family
ID=55168274
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020177022784A KR20170105079A (ko) | 2015-01-16 | 2016-01-15 | 천연 또는 가공된 진핵 세포의 생리에 긍정적으로 영향을 미치는 박테리아 샤페론의 조합물 |
Country Status (9)
| Country | Link |
|---|---|
| US (1) | US20180187204A1 (ko) |
| EP (1) | EP3245285A1 (ko) |
| JP (1) | JP2018501810A (ko) |
| KR (1) | KR20170105079A (ko) |
| CN (1) | CN107257851A (ko) |
| AU (1) | AU2016207978A1 (ko) |
| BR (1) | BR112017015201A2 (ko) |
| CA (1) | CA2973912A1 (ko) |
| WO (1) | WO2016113418A1 (ko) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN109536398B (zh) * | 2013-02-22 | 2023-08-04 | 帝斯曼知识产权资产管理有限公司 | 用于产量增加的方法中的重组体微生物 |
| CN110563825B (zh) * | 2019-07-19 | 2023-09-05 | 中国科学院武汉病毒研究所 | 细菌内co2固定类细胞器的合成及应用 |
| CN110734480B (zh) * | 2019-11-07 | 2021-08-24 | 中国科学院遗传与发育生物学研究所 | 大肠杆菌分子伴侣GroEL/ES在协助合成植物Rubisco中的应用 |
| CN113201552B (zh) * | 2021-04-29 | 2024-03-22 | 江南大学 | 一种分子伴侣质粒系统及其应用 |
| CN113755518B (zh) * | 2021-08-13 | 2023-10-20 | 中国科学院天津工业生物技术研究所 | 一种构建重组解脂耶氏酵母的方法与应用 |
Family Cites Families (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN103882014A (zh) * | 2012-12-21 | 2014-06-25 | 中国科学院青岛生物能源与过程研究所 | 用于固定二氧化碳的构建体、菌株及其制备方法 |
| CN109536398B (zh) * | 2013-02-22 | 2023-08-04 | 帝斯曼知识产权资产管理有限公司 | 用于产量增加的方法中的重组体微生物 |
| FR3016371B1 (fr) * | 2014-01-16 | 2018-02-02 | Institut National De La Recherche Agronomique | Levures modifiees pour utiliser le dioxyde de carbone |
-
2016
- 2016-01-15 WO PCT/EP2016/050832 patent/WO2016113418A1/fr active Application Filing
- 2016-01-15 US US15/542,682 patent/US20180187204A1/en not_active Abandoned
- 2016-01-15 CN CN201680005924.8A patent/CN107257851A/zh active Pending
- 2016-01-15 AU AU2016207978A patent/AU2016207978A1/en not_active Abandoned
- 2016-01-15 JP JP2017537495A patent/JP2018501810A/ja active Pending
- 2016-01-15 BR BR112017015201-0A patent/BR112017015201A2/pt not_active Application Discontinuation
- 2016-01-15 EP EP16700765.7A patent/EP3245285A1/fr not_active Withdrawn
- 2016-01-15 CA CA2973912A patent/CA2973912A1/fr not_active Abandoned
- 2016-01-15 KR KR1020177022784A patent/KR20170105079A/ko unknown
Also Published As
| Publication number | Publication date |
|---|---|
| WO2016113418A1 (fr) | 2016-07-21 |
| JP2018501810A (ja) | 2018-01-25 |
| AU2016207978A1 (en) | 2017-07-13 |
| EP3245285A1 (fr) | 2017-11-22 |
| US20180187204A1 (en) | 2018-07-05 |
| CN107257851A (zh) | 2017-10-17 |
| CA2973912A1 (fr) | 2016-07-21 |
| BR112017015201A2 (pt) | 2018-03-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US11976284B2 (en) | Promoter variants for protein production | |
| US11168117B2 (en) | Constitutive promoter | |
| JP6982607B2 (ja) | 目的タンパク質を発現させるための組換え宿主細胞 | |
| Rantasalo et al. | Novel genetic tools that enable highly pure protein production in Trichoderma reesei | |
| CN106661541B (zh) | 被工程化为过表达辅助蛋白的重组宿主细胞 | |
| JP5107910B2 (ja) | 遺伝子発現技術 | |
| KR20170105079A (ko) | 천연 또는 가공된 진핵 세포의 생리에 긍정적으로 영향을 미치는 박테리아 샤페론의 조합물 | |
| Terfrüchte et al. | Tackling destructive proteolysis of unconventionally secreted heterologous proteins in Ustilago maydis | |
| Zhang et al. | Efficient secretory expression and purification of food-grade porcine myoglobin in Komagataella phaffii | |
| Seppälä et al. | Heterologous transporters from anaerobic fungi bolster fluoride tolerance in Saccharomyces cerevisiae | |
| KR20130000883A (ko) | 클루이베로마이세스 마르시아누스 내에서의 향상된 단백질 생산 | |
| US10428123B2 (en) | Constitiutive promoter | |
| Allgaier et al. | Expression of ribonuclease A and ribonuclease N 1 in the filamentous fungus Neurospora crassa | |
| Bulakhov et al. | Properties of Chimeric Polysaccharide Monooxygenase with an Attached Cellulose Binding Module and Its Use in the Hydrolysis of Cellulose-Containing Materials in the Composition of Cellulase Complexes | |
| US20150037842A1 (en) | Modified bacterial cell |