1995年にGuoとKemphuesは、C.エレガンス(C.elegans)でアンチセンスを用いた遺伝子発現を抑制する上で、アンチセンスRNAと同様にセンスRNAも有効であることを発見し、その原因を糾明するための研究を行い始め、1998年にFire等が初めて、二重鎖RNA(double stranded RNA,dsRNA)を注入してそれに対応するmRNAが特異的に分解されて遺伝子の発現が抑制される現象を確認し、これをRNA干渉(RNA interference,RNAi)と名付けた。RNAiは遺伝子発現を抑制するために用いられる方法で、簡便でありながら少ない費用で遺伝子抑制効果を明確に得ることができるので、その技術の応用分野が多様化しつつある。
このような遺伝子の発現を抑制する技術は、特定遺伝子の発現を調節できるので、特定遺伝子の過剰発現が原因とされる癌、遺伝疾患などの標的遺伝子をmRNAレベルで除去でき、疾病治療のための治療剤開発及び標的検証の重要なツールとして活用可能である。標的遺伝子の発現を抑制する従来の技術としては、標的遺伝子に対する転移遺伝子を導入する技術が開示されているが、プロモーターを基準に逆方向(アンチセンス)に転移遺伝子を導入する方法と、プロモーター基準に順方向(sense)に移植遺伝子を導入する方法がある。
このようなRNAを標的にするRNA治療法は、標的RNAに対するオリゴヌクレオチドを用いて当該遺伝子の機能を除去する方法で、既存の抗体と化合物(small molecule)のような従来の治療剤が主にタンパク質を標的にすることとは異なる方法であると言える。RNAを標的にする接近法には大きく2種類があるが、一つは、二重螺旋RNAが媒介されたRNAiであり、他の一つは、アンチセンスオリゴヌクレオチド(ASO)である。現在、様々な疾病においてRNAを標的にして臨床が試みられている。
アンチセンスオリゴヌクレオチド(ASO、以下、‘ASO’という。)は、ワトソン-クリック塩基反応によって目的遺伝子に結合するようにデザインされた短い長さの合成DNAであり、遺伝子の特定塩基配列の発現を特異的に抑制できる点で、遺伝子の機能を研究して癌のような疾患を分子レベルで治療できる治療剤を開発するために用いられてきた。このようなASOは、遺伝子発現を抑制する目標を様々に設定し、容易に作製可能である長所から、発癌遺伝子の発現と癌細胞の成長を抑制することに活用しようとする研究が行われてきた。ASOが特定遺伝子の発現を抑制する過程は、相補的なmRNA配列と結合してRNase H活性を誘導してmRNAを除去したり、或いはタンパク質翻訳のためのリボソーム複合体の形成及び進行を妨害することによってなされる。また、ASOはゲノムDNAと結合してトリプルヘリックス(Triple helix)構造を形成することによって遺伝子の転写を抑制することも報告されている。ASOは上記のような潜在的可能性があるが、これを臨床に活用するためには、ヌクレアーゼ(nuclease)に対する安定性が向上するとともに、目的遺伝子の塩基配列に特異的に結合するように標的組織や細胞内に効率的に伝達される必要がある。また、遺伝子mRNAの2次及び3次構造はASOの特異的結合に重要な要素であり、mRNAの2次構造が少ない部分が、ASOが接近するのに非常に有利なので、ASOを合成する前に、mRNAの2次構造が少なく生成される領域を体系的に分析し、生体外だけでなく生体内でも遺伝子特異的抑制を効果的に達成するために努力してきた。このようなASOは、RNAの種類であるsiRNAに比べて非常に安定的であり、水と生理食塩水などによく溶解される長所があり、現在、3つのASOがFDA(Federal Drug Administration)に承認されている(Jessica,C.,J Postdoc Res,4:35-50,2016)。
干渉RNA(RNA interference、以下、‘RNAi’という。)は、その役目が発見されて以来、種々の哺乳動物細胞(mammalian cell)において配列特異的mRNAに作用するという事実が明らかにされた(Barik,S.,J Mol.Med.(2005)83:764-773)。長いRNA二重鎖が細胞に伝達されると、伝達されたRNA二重鎖は、ダイサー(Dicer)というエンドヌクレアーゼ(endonuclease)によって21~23個の二重鎖(base pair,bp)にプロセシングされた短い干渉RNA(small interfering RNA、以下、‘siRNA’という。)に変換され、siRNAは、RISC(RNA-induced silencing complex)に結合し、ガイド(アンチセンス)鎖がターゲットmRNAを認識して分解する過程によって、ターゲット遺伝子の発現を配列特異的に阻害する。SiRNAを用いた遺伝子の発現抑制技術は、標的細胞で標的遺伝子の発現を抑制させ、これによる変化を観察することであり、標的細胞における標的遺伝子の機能を糾明する研究に有用に用いられる。特に、感染性ウイルス又は癌細胞などで標的遺伝子の機能を抑制することは、当該疾病の治療方法を開発するのに有用であろうが、生体外(in vitro)における研究及び実験動物を用いた生体内(in vivo)研究を行った結果、siRNAによる標的遺伝子の発現抑制が可能であると報告されたことがある。
ベルトラン(Bertrand)研究陣によれば、同じターゲット遺伝子に対するsiRNAが、アンチセンスオリゴヌクレオチド(Antisense oligonucleotide,ASO)に比べて、生体内/外(in vitro及びin vivo)においてmRNA発現の阻害効果に優れ、当該効果が長く持続する効果を有することが明らかにされた。また、siRNAの作用機序は、ターゲットmRNAと相補的に結合して配列特異的にターゲット遺伝子の発現を調節するので、既存の抗体基盤医薬品や化学物質医薬品(small molecule drug)に比べて適用可能な対象を画期的に拡大できるという長所がある(MA Behlke,MOLECULAR THERAPY.2006 13(4):664-670)。
siRNAの優れた効果及び様々な使用範囲にもかかわらず、siRNAが治療剤として開発されるためには、体内におけるsiRNAの安定性(stability)の改善と細胞伝達効率の改善によってsiRNAがターゲット細胞に効果的に伝達されるようにしなければならない(FY Xie,Drug Discov.Today.2006 Jan;11(1-2):67-73)。体内安定性の向上及びsiRNAの非特異的な細胞免疫反応(innate immune stimulation)の問題を解決するために、siRNAの一部ヌクレオチド又は骨格(backbone)を、核酸分解酵素抵抗性を有するように修飾(modification)したり、或いはウイルス性ベクター(viral vector)、リポソーム又はナノ粒子(nanoparticle)などの伝達体を用いるなど、これに対する研究が活発に試みられている。
アデノウイルスやレトロウイルスなどのウイルス性ベクターを用いた伝達システムは、形質注入効率(transfection efficacy)が高いが、免疫原性(immunogenicity)及び発癌性(oncogenicity)が高い。一方、ナノ粒子を含む非ウイルス性(non-viral)伝達システムは、ウイルス性伝達システムに比べて細胞伝達効率は低いが、生体内(in vivo)における安定性(stability)が高く、ターゲット特異的に伝達が可能であり、内包されているRNAiオリゴヌクレオチドを細胞又は組織に吸収(uptake)及び内在化(internalization)し、細胞毒性及び免疫誘発(immune stimulation)がほとんどないという長所から、今のところウイルス性伝達システムに比べて有力な伝達方法として評価されている(Akhtar S,J Clin Invest.2007 December 3;117(12):3623-3632)。
非ウイルス性伝達システムのうち、ナノ伝達体(nanocarrier)を用いる方法は、リポソーム、陽イオン高分子複合体などの様々な高分子を使用することによってナノ粒子を形成し、siRNAをこのようなナノ粒子(nanoparticle)、すなわちナノ伝達体(nanocarrier)に担持して細胞に伝達する方法である。ナノ伝達体を用いる方法において主に活用される方法には、高分子ナノ粒子(polymeric nanoparticle)、高分子ミセル(polymer micelle)、リポプレックス(lipoplex)などがあるが、なかでもリポプレックスを用いた方法は、陽イオン性脂質で構成され、細胞のエンドソーム(endosome)の陰イオン性脂質と相互作用することで、エンドソームの脱安定化効果を誘発して細胞内に伝達する役割を担う。
また、siRNAパッセンジャー(passenger;センス(sense))鎖の末端部位に化学物質などを連結し、増進した薬物動態学(pharmacokinetics)的特徴を有させ、生体内(in vivo)で高い効率を誘導できるということが知られている(J Soutschek,Nature 11;432(7014):173-8,2004)。このとき、siRNAセンス(sense;パッセンジャー(passenger))又はアンチセンス(antisence;ガイド(guide))鎖の末端に結合した化学物質の性質によってsiRNAの安定性が変わる。例えば、ポリエチレングリコール(polyethylene glycol,PEG)のような高分子化合物が接合された形態のsiRNAは、陽イオン性物質が存在する条件で、siRNAの陰イオン性リン酸基と相互作用して複合体を形成することによって、改善されたsiRNA安定性を持つ伝達体となる(SH Kim,J Control Release 129(2):107-16,2008)。特に、高分子複合体で構成されたミセル(micelle)は、薬物伝達運搬体として用いられる他のシステムである、微小球体(microsphere)又はナノ粒子(nanoparticle)などに比べて、そのサイズが極小さいながらも分布が非常に一定であり、且つ自発的に形成される構造であるので、製剤の品質管理及び再現性の確保がしやすいという長所がある。
siRNAの細胞内伝達効率性を向上させるために、siRNAに生体適合性高分子である親水性物質(例えば、ポリエチレングリコール(polyethylene glycol,PEG))を単純共有結合又はリンカー媒介(linker-mediated)共有結合で接合させたsiRNA接合体を用いて、siRNAの安定性確保及び効率的な細胞膜透過性のための技術が開発された(大韓民国登録特許第883471号)。しかし、siRNAの化学的修飾及びポリエチレングリコール(polyethylene glycol,PEG)を接合させること(PEGylation)だけでは、生体内における低い安定性と標的臓器への円滑な伝達の課題が依然として残る。このような課題を解決するために、オリゴヌクレオチド、特にsiRNAのような二重鎖オリゴRNAに親水性及び疎水性物質が結合した二重鎖オリゴRNA構造体が開発されたが、該構造体は、疎水性物質の疎水性相互作用によってSAMiRNATM(self assembled micelle inhibitory RNA)と名付けられた自己組立ナノ粒子を形成するようになるが(大韓民国登録特許第1224828号)、SAMiRNATM技術は、既存の伝達技術に比べて非常にサイズが小さいながらも均一な(homogenous)ナノ粒子が得られるという長所がある。
SAMiRNATM技術の具体的な例では、親水性物質としてPEG(polyethyleneglycol)又はHEG(Hexaethylenglycol)が用いられるが、PEGは、合成ポリマー(synthetic polymer)でしばしば医薬品、特にタンパク質の水溶性(solubility)増加及び薬物動態学の調節のために用いられる。PEGは多分散系(polydisperse)物質であり、1つのバッチ(batch)のポリマーは、異なる個数の単量体(monomer)の総和からなり、分子量がガウス曲線の形態を示し、多分散指数(polydisperse value,Mw/Mn)で物質の同質性の程度を表現する。すなわち、PEGが低い分子量(3~5kDa)であるとき、約1.01の多分散指数を示し、高い分子量(20kDa)のとき、約1.2という高い多分散指数を示し、高い分子量であるほど、物質の同質性が相対的に低い特徴を示す。したがって、PEGを医薬品に結合させた場合、接合体にPEG多分散的特徴が反映され、単一物質の検証がし難いという短所があり、PEGの合成及び精製過程の改善によって低い多分散指数を持つ物質を生産している趨勢であるが、特に、分子量の小さい物質にPEGを結合させた場合、容易に結合されたかどうか確認し難い不都合があるなど、物質の多分散性特徴に伴う問題点がある。(Francesco M.VDRUG DISCOVERY TODAY(2005)10(21):1451-1458)。
これによって、最近では、既存の自己組立ナノ粒子であるSAMiRNATM技術の改良された形態であり、SAMiRNATMを構成する二重鎖オリゴヌクレオチド構造体の親水性物質を、一定の分子量を持つ均一な1~15個の単量体(monomer)、及び必要によってリンカー(linker)を含む基本単位をブロック(block)化し、これを必要に応じて適切な個数を使用することによって、既存のSAMiRNATMに比べてより小さいサイズを有するとともに多分散性が画期的に改善された新しい形態の伝達体技術が開発されている。また、体内にsiRNAを注入した場合、血液中に存在する様々な酵素によってsiRNAが急速に分解され、標的細胞又は組織などへの伝達効率がよくないことが既に知られているが、改良されたSAMiRNATMにおいても標的遺伝子によって安定性及び発現阻害率にバラツキがあった。したがって、本発明者らは、改良された自己組立ナノ粒子であるSAMiRNATMを用いて標的遺伝子をより安定的で効果的に発現阻害するために、ガイドであるセンス鎖はASOであるDNA配列を、パッセンジャーであるアンチセンス鎖はRNA配列を用いて、DNA-RNAハイブリッド形態の二重鎖オリゴヌクレオチドを適用することによって、標的遺伝子に対する発現阻害効果と安定性を増進させようとした。
線維症(Fibrosis)の一種である特発性肺線維化症(Idiopathic Pulmonary Fibrosis、以下、‘IPF’という。)は、肺胞(肺の袋)の壁に慢性炎症細胞が侵入して肺を硬くする様々な変化が発生しながら肺組織の大きな構造的変化を引き起こし、肺機能が漸次低下して結局としては死亡に至る疾患であり、今のところ効果的な治療方法がなく、通常、症状を感じて診断をすると、平均生存期間が約3~5年しかならない、極めて予後の悪い疾病である。発生頻度は、海外の場合、人口10万名につき約3~5名程度と報告されており、通常、50代以後に発病率が高く、男子が女子に比べて2倍程度発病率が高いと知られている。
IPFの発病原因はまだ明確にされていないが、喫煙者において頻度が高く、抗うつ剤、胃食道逆流による慢性的肺吸入、金属粉塵、木材粉塵、又は溶媒剤吸入などがIPFの発生と関連している危険因子として報告されたことがあるが、大部分の患者において確実な因果関係を持つ因子が報告されたことはない。最も言及されている因子としては、いかなる原因であれ、Th1/Th2反応、凝固カスケード(coagulation cascades)などが活性化すると、これらによって線維化性サイトカイン(cytokine)が分泌され、活性化されたサイトカインは線維芽細胞を刺激してECM(Extracellular Matrix)を増加させ、これで肺線維化が起きることが知られている。勿論、この過程で肺の炎症を伴うことによって肺の線維化が起きることがあるが、最近では、肺の炎症に関係なく直接的に肺線維化を起こし得るという意見がより支配的である。最近の仮設は、上皮-間葉細胞相互関係(Epithelial-mesenchymal interaction)の異常信号伝達体系によって傷治癒過程で病理的な肺線維化が起きるということである。上皮細胞が損傷を受けると、上皮細胞のアポトーシスが増加し、移動が制限され、分化調節がされず、増殖が抑制されて液性因子(soluble factors)(TGF、HGF、KGF、angiotensin II、ROSなど)が分泌され、ECMと共に間葉細胞のアポトーシスが抑制されることで、筋線維芽細胞の分化増加、ECM浸潤に関して肺線維化を誘発したり或いは上皮細胞を再び刺激することになる。すなわち、肺の炎症が肺線維化に直接つながるとはいえないが、ただし、まず肺の炎症が起き、続いて正常組織として治癒される過程で、IPF患者と正常人の差によって肺線維化が起きるということを意味する。また、IPFはTh1/Th2サイトカインの不調和によって誘発されることがあるが、Th1サイトカイン反応は細胞媒介性免疫に関係するもので、組織回復に関する損傷部位を正常に回復させるのに対し、Th2サイトカインは、線維芽細胞の活性化と増殖によってECMの浸潤及び線維化を誘発する。ブレオマイシンを用いた肺線維化モデルにおいて、IFN-γを投与すると、TGF-βとプロコラーゲンのmRNAを減少させ、肺線維化を防ぐことができるという報告があるが、病因については正確に知られておらず、将来、線維化を起こす初期の原因因子を糾明し、IPFと関連した遺伝子及びTGF-β信号伝達体系を抑制できる物質の開発などが必要である。
IPFは、治療しないと悪化し続き、患者の約50%以上が3~5年内に死亡すると知られており、しかも、一応症状が進行して完全に線維化すると、いかなる治療をしても好転しないため、治療する場合には、早期治療が有効である可能性が高いと予測している。現在用いられている治療剤には、ステロイド(steroid)とアザチオプリン(azathioprine)、又はシクロホスファミド(cyclophosphamide)の併合療法を用いる方法が知られてはいるが、特に効果があるとはいえず、様々な線維化抑制剤が動物実験及び小規模の患者に対して試みられたが、明確な効果が立証されたことがない現状である。特に、末期IPF患者にとっては肺移植の他には有効な治療方法がない実情である。このため、より効率的なIPFの治療剤の開発が切実な状況である。
線維症(Fibrosis)とは、ある理由によって、組織や臓器が、結合組織の過剰線維化によって硬くなる病症を総称するが、線維化が起きる全ての過程は、部位を問わず、傷跡が治療される過程と同じ経路を経る。線維化症状は現在まで完治方法がないとされており、治療方法を開発及び研究中である。効果的な線維化治療剤は、代表的な線維症である肝硬変(cirrhosis)、肝線維症(liver fibrosis)、骨髄線維症(myelofibrosis)、心筋線維症(myocardial fibrosis)、腎臓線維症(renal fibrosis)、肺線維症(pulmonary fibrosis)だけでなく、線維症を伴う様々な疾病に適用可能な点から、効率的な線維症の治療剤の開発が切実な状況である。
一方、アンフィレグリンは、上皮細胞成長因子受容体に結合して上皮細胞受容体経路(EGFR pathway)を活性化させ、細胞増殖に関与するという事実が知られており、アンフィレグリン特異的siRNAによってアンフィレグリンの発現を阻害させることができ、これは特定タイプの乳癌に治療効果を示すことが開示されている。また、アンフィレグリンに対するshRNAを用いて炎症性乳癌における細胞侵入を抑制でき(Andrea Baillo,J Cell Physiol.2011 226(10):2691-2701)、アンフィレグリン特異的shRNAを用いてアンフィレグリン発現を抑制すると、タバコ煙に露出されたマウスにおいて肺動脈再形成(pulmonary artery remodeling)が抑制されるという事実が開示されている。気道平滑筋(airway smooth muscle;ASM)過増殖(hyperplasia)と血管新生にアンフィレグリンが関連しており、特に、喘息患者の気道再形成(airway remodeling)を促進するということと、急性喘息による組織再形成において過剰分泌される上皮細胞成長因子(EGF)及びアンフィレグリンが関与すること、が開示されている。
以上述べたように、アンフィレグリンの呼吸器疾患及び線維症、特にCOPD及びIPF治療剤ターゲットとしての可能性が提示されている状況であるが、まだアンフィレグリンに対するRNAi治療剤及びその伝達技術に関する技術開発はわずかな状況であり、特異的且つ高効率にアンフィレグリン発現を阻害できる二重鎖オリゴヌクレオチド治療剤及びその伝達技術に対する市場の需要は非常に高い状況である。
そこで、本発明者らは、IPFをはじめとする線維化症に関連した遺伝子としてアンフィレグリンを選定し、アンフィレグリンを標的にする二重鎖オリゴヌクレオチドを選別してアンフィレグリン発現を阻害できるRNAi治療剤及びその伝達体を確認し、本発明を完成するに至った。
本発明の目的は、アンフィレグリンの発現を非常に特異的で高効率に阻害できる二重鎖オリゴヌクレオチド、好ましくは、RNA/RNA、DNA/DNA、又はDNA/RNAハイブリッド形態の配列を含む二重鎖オリゴヌクレオチド、該二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体、及び該二重鎖オリゴヌクレオチド構造体を含むナノ粒子を提供することにある。
本発明の他の目的は、前記二重鎖オリゴヌクレオチド、これを含む二重鎖オリゴヌクレオチド構造体及び/又は前記二重鎖オリゴヌクレオチド構造体を含むナノ粒子を有効成分として含有する線維症又は呼吸器疾患の予防又は治療用医薬組成物を提供することにある。
本発明のさらに他の目的は、前記線維症又は呼吸器疾患の予防又は治療用医薬組成物を、線維症又は呼吸器疾患の予防又は治療を必要とする個体に投与する段階を含む、線維症又は呼吸器疾患の予防又は治療方法を提供することにある。
本発明のさらに他の目的は、線維症又は呼吸器疾患の予防又は治療に用いるための前記二重鎖オリゴヌクレオチド、これを含む二重鎖オリゴヌクレオチド構造体及び前記二重鎖オリゴヌクレオチド構造体を含むナノ粒子を提供することにある。
本発明のさらに他の目的は、線維症又は呼吸器疾患の予防又は治療に用いるための前記薬学的組成物を提供することにある。
本発明のさらに他の目的は、線維症又は呼吸器疾患の予防又は治療のための薬物の製造のための前記二重鎖オリゴヌクレオチド、これを含む二重鎖オリゴヌクレオチド構造体及び前記二重鎖オリゴヌクレオチド構造体を含むナノ粒子の用途を提供することにある。
前記目的を達成するために、本発明は、配列番号1~14からなる群、より好ましくは配列番号10、11及び12からなる群から選ばれるいずれか一つの配列を含むセンス鎖(sense strand)とこれに相補的な配列を含むアンチセンス鎖(anti-sense strand)を含む二重鎖オリゴヌクレオチドを提供する。
本発明はまた、前記二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体及び前記二重鎖オリゴヌクレオチド構造体を含むナノ粒子を提供する。
本発明はまた、配列番号1~14からなる群、より好ましくは配列番号10、11及び12からなる群から選ばれるいずれか一つの配列を含むセンス鎖(sense strand)とこれに相補的な配列を含むアンチセンス鎖(anti-sense strand)を含む二重鎖オリゴヌクレオチド、前記二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体、又は前記二重鎖オリゴヌクレオチド構造体を含むナノ粒子を含む線維症又は呼吸器疾患の予防又は治療用医薬組成物を提供する。
本発明はまた、前記線維症又は呼吸器疾患の予防又は治療用医薬組成物を、線維症又は呼吸器疾患の予防又は治療を必要とする個体に投与する段階を含む、線維症又は呼吸器疾患の予防又は治療方法を提供する。
本発明に係る配列番号1~14からなる群、より好ましくは配列番号10、11及び12からなる群から選ばれるいずれか一つの配列を含むセンス鎖(sense strand)とこれに相補的な配列を含むアンチセンス鎖(anti-sense strand)を含む二重鎖オリゴヌクレオチド、これを含む二重鎖オリゴヌクレオチド構造体、これを含むナノ粒子は、アンフィレグリンの発現を非常に効率的に阻害できるので、本発明に係る二重鎖オリゴヌクレオチド、これを含む二重鎖オリゴヌクレオチド構造体及びナノ粒子のそれぞれを線維症又は呼吸器疾患の予防又は治療に有用に用いることができる。
前記目的を達成するために、提供される好ましい二重鎖オリゴヌクレオチドに含まれる配列番号10、11及び12の配列は、次の通りである。
5’-CACCTACTCTGGGAAGCGT-3’(配列番号10)
5’-ACCTACTCTGGGAAGCGTG-3’(配列番号11)
5’-CTGGGAAGCGTGAACCATT-3’(配列番号12)
本発明に係る二重鎖オリゴヌクレオチドは、一般のRNAi(RNA interference)作用を持つ全ての物質を含む概念であり、前記アンフィレグリンタンパク質をエンコードするmRNA特異的二重鎖オリゴヌクレオチドにはアンフィレグリン特異的shRNAなども含まれるということが、本発明の属する技術の分野における通常の知識を有する者には明らかであろう。すなわち、前記オリゴヌクレオチドは、siRNA、shRNA又はmiRNAであることを特徴とし得る。
また、アンフィレグリンに対する特異性が維持される限り、前記配列番号10、11及び12からなる群から選ばれるいずれか一つの配列を含むセンス鎖又はこれに相補的なアンチセンス鎖において、一つ以上の塩基が置換、欠失、又は挿入された配列を含むセンス鎖及びアンチセンス鎖を含むアンフィレグリン特異的siRNA、アンチセンスオリゴヌクレオチドも本発明の権利範囲に含まれるということは、本発明の属する技術の分野における通常の知識を有する者には明らかであろう。
本発明において、前記センス又はアンチセンス鎖は独立してDNA又はRNAであることを特徴とし、また、センス鎖はDNA、アンチセンス鎖はRNAであるか、センス鎖はRNA、アンチセンス鎖はDNAである、ハイブリッド(hybrid)形態が用いられてもよい。
本発明において、前記配列番号10、11及び12はDNA形態であると記載されているが、RNA形態が用いられる場合、配列番号10、11及び12の配列は、それに対応するRNA配列、すなわち、TがUに変更された配列を用いることができる。
また、本発明に係る二重鎖オリゴヌクレオチドは、アンフィレグリンに対する特異性が維持される限り、配列のセンス鎖がアンフィレグリン遺伝子の結合部位と100%相補的な塩基配列である場合、すなわち、完全一致(perfect match)する場合だけでなく、一部の塩基配列が一致しない場合、すなわち不完全一致(mismatch)がある場合も含む。
本発明に係る二重鎖オリゴヌクレオチドは、一側又は両側の鎖の3’末端に1つ又はそれ以上の非結合した(unpaired)ヌクレオチドを含む構造であるオーバーハング(overhang)を含むことができる。
本発明において、前記センス鎖又はアンチセンス鎖は、好ましくは、19~31個のヌクレオチドで構成されることを特徴とし得るが、これに限定されるものではない。
本発明において、配列番号10、11及び12からなる群から選ばれるいずれか一つの配列を含むセンス鎖とこれに相補的な配列を含むアンチセンス鎖を含む二重鎖オリゴヌクレオチドは、アンフィレグリン(amphiregulin)に特異的であることを特徴とし得るが、これに限定されるものではない。
本発明において、前記二重鎖オリゴヌクレオチドのセンス鎖又はアンチセンス鎖は、生体内安定性の向上のために、又は核酸分解酵素抵抗性の付与及び非特異的免疫反応の減少のために、様々な化学的修飾(chemical modification)を含むことを特徴とし、化学的修飾は、ヌクレオチド内糖(sugar)構造の2’炭素位置で水酸化基(-OH)がメチル基(-CH3)、メトキシ基(-OCH3)、アミン基(-NH2)、フッ素(-F)、O-2-メトキシエチル基、O-プロピル基、O-2-メチルチオエチル基、O-3-アミノプロピル基、O-3-ジメチルアミノプロピル基、O-N-メチルアセトアミド基及びO-ジメチルアミドオキシエチルからなる群から選ばれるいずれか一つに置換される修飾;ヌクレオチド内糖構造の酸素が硫黄に置換される修飾;ヌクレオチド結合がホスホロチオエート(phosphorothioate)結合、ボラノホスフェート(boranophosphate)結合及びメチルホスホネート(methyl phosphonate)結合からなる群から選ばれるいずれか一つの結合となる修飾;及びPNA(peptide nucleic acid)、LNA(locked nucleic acid)及びUNA(unlocked nucleic acid)形態への修飾;DNA-RNAハイブリッド形態への修飾;からなる群から選ばれるいずれか一つ以上であることを特徴とし得るが(Ann.Rev.Med.55,61-65 2004;US5,660,985;US5,958,691;US6,531,584;US5,808,023;US6,326,358;US6,175,001;Bioorg.Med.Chem.Lett.14:1139-1143,2003;RNA,9:1034-1048,2003;Nucleic Acid Res.31:589-595,2003;Nucleic Acids Research,38(17)5761-773,2010;Nucleic Acids Research,39(5):1823-1832,2011)、これに限定されるものではない。
本発明において、二重鎖オリゴヌクレオチドのアンチセンス鎖の5’末端に1つ以上のリン酸基(Phosphate group)が結合していることを特徴とし、好ましくは1~3個のリン酸基が結合していることを特徴とし得る。
本発明は、他の観点において、下記構造式(1)の構造を含む二重鎖オリゴヌクレオチド構造体に関し、下記構造式(1)において、Aは親水性物質、Bは疎水性物質、X及びYはそれぞれ独立して単純共有結合又はリンカーが媒介された共有結合を意味し、Rは二重鎖オリゴヌクレオチドを意味する。
一つの好ましい具体例として、本発明に係るアンフィレグリン特異的配列を含む二重鎖オリゴヌクレオチド構造体は、構造式(1)のような構造を有することが好ましい。
前記構造式(1)において、Aは親水性物質、Bは疎水性物質、X及びYはそれぞれ独立して単純共有結合又はリンカーが媒介された共有結合を意味し、Rはアンフィレグリン特異的二重鎖オリゴヌクレオチドを意味する。
本発明に係る二重鎖オリゴヌクレオチドの形態としては、DNA-RNAハイブリッド、siRNA(short interfering RNA)、shRNA(short hairpin RNA)及びmiRNA(microRNA)などが好ましいが、これに限定されず、miRNAに対する拮抗剤(antagonist)の役割ができる単一鎖のmiRNA阻害剤も可能である。
以下、本発明に係る二重鎖オリゴヌクレオチドはRNAを中心に説明するが、本発明の二重鎖オリゴヌクレオチドと同じ特性を有する他の二重鎖オリゴヌクレオチドにも適用可能であることは、当業界における通常の技術者にとって明らかであろう。
より好ましくは、本発明に係るアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体は、下記構造式(2)の構造を有する。
前記構造式(2)において、A、B、X及びYは、前記構造式(1)における定義と同一であり、Sはアンフィレグリン特異的二重鎖オリゴヌクレオチドのセンス鎖、ASはアンフィレグリン特異的二重鎖オリゴヌクレオチドのアンチセンス鎖を意味する。
より好ましくは、アンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体は、下記構造式(3)又は(4)の構造を有する。
前記構造式(3)及び構造式(4)において、A、B、S、AS、X及びYは、前記構造式(2)における定義と同一であり、5’及び3’は、アンフィレグリン特異的二重鎖オリゴヌクレオチドセンス鎖の5’末端及び3’末端を意味する。
前記親水性物質は、ポリエチレングリコール(PEG)、ポリビニルピロリドン及びポリオキサゾリンからなる群から選ばれることを特徴とし得るが、これに限定されない。
上記の、構造式(1)~構造式(4)における前記アンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体は、アンチセンス鎖の5’末端にリン酸基(phosphate group)が1~3個結合でき、RNAに代えてshRNAが用いられてもよいことは、本発明の属する技術分野における通常の知識を有する者には明らかであろう。
前記構造式(1)~構造式(4)における親水性物質は、分子量が200~10,000である高分子物質であることが好ましく、より好ましくは、1,000~2,000である高分子物質である。例えば、親水性高分子物質には、ポリエチレングリコール、ポリビニルピロリドン、ポリオキサゾリンなどの非イオン性親水性高分子化合物を用いることが好ましいが、必ずしもこれに限定されない。
特に、構造式(1)~構造式(4)における親水性物質(A)は、下記構造式(5)又は構造式(6)のような形態の親水性物質ブロック(block)の形態で用いることができるが、このような親水性物質ブロックを必要によって適切な個数(構造式(5)又は構造式(6)におけるn)を使用することによって、一般合成高分子物質などを使用する場合に発生し得る多分散性による問題点を大きく改善することができる。
前記構造式(5)又は構造式(6)において、A’は親水性物質単量体(monom er)、Jは、m個の親水性物質単量体同士又はm個の親水性物質単量体と二重鎖オリゴヌクレオチドとを連結するリンカー、mは1~15の整数、nは1~10の整数を意味し、(A’
m-J)又は(J-A’
m)で表示される反復単位が親水性物質ブロックの基本単位に該当する。
前記構造式(5)又は構造式(6)のような親水性物質ブロックを有する場合、本発明に係るアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体は、下記構造式(7)又は構造式(8)のような構造を有することができる。
前記構造式(7)及び構造式(8)において、X、R、Y及びBは、構造式(1)における定義と同一であり、A’、J、m及びnは、構造式(5)及び構造式(6)における定義と同一である。
前記構造式(5)及び構造式(6)において、親水性物質単量体(A’)は、非イオン性親水性高分子の単量体のうち、本発明の目的に符合するものであればいずれも用いることができ、好ましくは、表1に記載された化合物(1)~化合物(3)から選択された単量体、より好ましくは、化合物(1)の単量体を用いることができ、化合物(1)においてGは、好ましくはO、S及びNHから選択可能である。
特に、親水性物質単量体の中でも特に化合物(1)で表示される単量体は、様々な官能基を導入でき、生体内親和性が良く、免疫反応を少なく誘導するなど、生体適合性(bio-compatibility)に優れるだけでなく、構造式(7)又は構造式(8)による構造体内に含まれた二重鎖オリゴヌクレオチドの生体内安定性を増加させ、伝達効率を増加させることができるという長所から、本発明に係る構造体の製造に非常に適している。
前記構造式(5)~構造式(8)における親水性物質は、総分子量が1,000~2,000の範囲に含まれることが特に好ましい。したがって、例えば、構造式(7)及び構造式(8)において、化合物(1)によるヘキサエチレングリコール(Hexaethylene glycol)、すなわち、GがOであり、mが6である物質が用いられる場合、ヘキサエチレングリコールスぺーサ(spacer)の分子量が344であるので、反復回数(n)は3~5であることが好ましい。特に、本発明は、必要によって、前記構造式(5)及び構造式(6)において(A’m-J)n又は(J-A’m)nで表示される親水性グループの反復単位、すなわち、親水性物質ブロック(block)として、nで表示される適切な個数を使用できることを特徴とする。前記各親水性物質ブロック内に含まれる親水性物質単量体であるA及びリンカーであるJは、独立して、各親水性物質ブロック間において同一でもよく、互いに異なってもよい。すなわち、親水性物質ブロックが3個用いられる場合(n=3)、一番目のブロックには化合物(1)による親水性物質単量体が、二番目のブロックには化合物(2)による親水性物質単量体が、三番目のブロックには化合物(3)による親水性物質単量体、が用いられるなど、全ての親水性物質ブロック別に異なる親水性物質単量体が用いられもよく、全ての親水性物質ブロックに、化合物(1)~化合物(3)による親水性物質単量体から選択されたいずれか一つの親水性物質単量体が同一に用いられてもよい。同様に、親水性物質単量体の結合を媒介するリンカーも、各親水性物質ブロック別に全て同じリンカーが用いられてもよく、各親水性物質ブロック別に異なるリンカーが用いられてもよい。また、親水性物質単量体の個数であるmも、各親水性物質ブロックにおいて同一でも異なってもよい。すなわち、一番目の親水性物質ブロックでは親水性物質単量体が3個連結(m=3)され、二番目の親水性物質ブロックでは親水性物質単量体が5個(m=5)、三番目の親水性物質ブロックでは親水性物質単量体が4個連結(m=4)されるなど、異なる個数の親水性物質単量体が用いられてもよく、全ての親水性物質ブロックにおいて同じ個数の親水性物質単量体が用いられてもよい。
また、本発明において、前記リンカー(J)は、-PO3
--、-SO3-及び-CO2-からなる群から選ばれることが好ましいが、これに制限されず、用いられる親水性物質の単量体などに応じて、発明の目的に符合するものであれば如何なるリンカーも使用可能であることは、通常の技術者には明らかであろう。
前記構造式(1)~構造式(4)、構造式(7)及び構造式(8)における疎水性物質(B)は、疎水性相互作用によって、構造式(1)~構造式(4)、構造式(7)及び構造式(8)による二重鎖オリゴヌクレオチド構造体で構成されたナノ粒子を形成する役割を担う。前記疎水性物質は、分子量が250~1,000が好ましく、ステロイド(steroid)誘導体、グリセリド(glyceride)誘導体、グリセロールエーテル(glycerol ether)、ポリプロピレングリコール(polypropylene glycol)、C12~C50の不飽和又は飽和炭化水素(hydrocarbon)、ジアシルホスファチジルコリン(diacylphosphatidylcholine)、脂肪酸(fatty acid)、リン脂質(phospholipid)、リポポリアミン(lipopolyamine)、脂質(lipid)、トコフェロール(tocopherol)及びトコトリエノール(tocotrienol)からなる群から選ばれるいずれか一つを用いることができるが、これに制限されず、本発明の目的に符合するものであればいかなる疎水性物質も用いることができるという点は、本発明の属する技術の分野における通常の知識を有する者には明らかであろう。
前記ステロイド(steroid)誘導体は、コレステロール、コレスタノール、コール酸、コレステリルホルメート、コテスタニルホルメート及びコレステリルアミンからなる群から選択可能であり、前記グリセリド誘導体は、モノ-、ジ-及びトリ-グリセリドなどから選択可能であるが、このとき、グリセリドの脂肪酸は、C12~C50の不飽和又は飽和脂肪酸が好ましい。
特に、前記疎水性物質の中でも飽和又は不飽和炭化水素又はコレステロールが、本発明に係る二重鎖オリゴヌクレオチド構造体の合成段階において容易に結合させることができるので好ましく、C24炭化水素、特にジスルフィド結合を含む形態が最も好ましい。
前記疎水性物質は、親水性物質の反対側の末端(distal end)に結合し、二重鎖オリゴヌクレオチドのセンス鎖又はアンチセンス鎖のいずれの位置に結合しても構わない。
本発明に係る構造式(1)~構造式(4)、構造式(7)及び構造式(8)における親水性物質又は疎水性物質とアンフィレグリン特異的二重鎖オリゴヌクレオチドは、単純共有結合又はリンカーが媒介された共有結合(X又はY)によって結合する。前記共有結合を媒介するリンカーは、親水性物質、又は疎水性物質とアンフィレグリン特異的二重鎖オリゴヌクレオチドの末端で共有結合し、必要によって特定環境で分解可能な結合を提供する限り、特に限定されるものではない。したがって、前記リンカーは、本発明に係る二重鎖オリゴヌクレオチド構造体の製造過程においてアンフィレグリン特異的二重鎖オリゴヌクレオチド及び/又は親水性物質(又は疎水性物質)を活性化するために結合させるいかなる化合物も用いることができる。前記共有結合は、非分解性結合又は分解性結合のいずれであってもよい。このとき、非分解性結合にはアミド結合又はリン酸化結合があり、分解性結合には、二硫化結合、酸分解性結合、エステル結合、アンヒドリド結合、生分解性結合又は酵素分解性結合などがあるが、これに限定されるものではない。
また、前記構造式(1)~構造式(4)、構造式(7)及び構造式(8)におけるR(又は、S及びAS)で表示されるアンフィレグリン特異的二重鎖オリゴヌクレオチドは、アンフィレグリンのmRNAと特異的に結合できる特性を持つ二重鎖オリゴヌクレオチドであればいずれも制限なく使用可能であり、好ましくは、本発明では、配列番号10、11及び12番から選択されたいずれか一つの配列を含むセンス鎖とそれに相補的配列を含むアンチセンス鎖で構成される。
本発明はまた、本発明に係るアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体において、前記構造体内の親水性物質のオリゴヌクレオチドと結合した反対側の末端部位に、アミン基又はポリヒスチジン(polyhistidine)基がさらに導入され得る。
これは、本発明に係るアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体の伝達体の細胞内導入とエンドソーム脱出を容易にするためのものであり、既に量子ドット(Quantum dot)、デンドリマー(Dendrimer)、リポソーム(liposome)などの伝達体の細胞内導入とエンドソーム脱出を容易にするために、アミン基の導入とポリヒスチジン基を用いることができる点及びその効果が報告されたことがある。
具体的に、伝達体の末端或いは外側に修飾された一次アミン基は、生体内pHで陽性子化されながら陰電荷を帯びる遺伝子と静電気的相互作用によって結合体を形成し、細胞内流入後にエンドソームの低いpHで緩衝効果を持つ内部3次アミンによってエンドソームの脱出が容易になることから、リソソームの分解から伝達体を保護できると知られており(高分子基盤ハイブリッド物質を用いた遺伝子伝達及び発現抑制。Polymer Sci.Technol.,Vol.23,No.3,pp254-259)、
非必須アミノ酸の一つであるヒスチジンは、残基(-R)にイミダゾール環(pKa3 6.04)を有するので、エンドソームとリソソームにおいて緩衝能力(buffering capacity)を増加させる効果があり、リポソームをはじめとする非ウイルス性遺伝子伝達体(non-viral gene carrier)においてエンドソーム脱出効率を高めるためにヒスチジン修飾を利用できるという点が知られている(Novel histidine-conjugated galactosylated cationic liposomes for efficient hepatocyte selective gene transfer in human hepatoma HepG2 cells.J.Controlled Release 118,pp262-270)。
前記アミン基又はポリヒスチジン(polyhistidine)基は、一つ以上のリンカーを通じて親水性物質又は親水性物質ブロックと連結され得る。
本発明の構造式(1)による二重鎖オリゴヌクレオチド構造体の親水性物質にアミン基又はポリヒスチジン(polyhistidine)基が導入される場合には、構造式(9)のような構造を有することができる。
前記構造式(9)において、A、B、R、Xは、Y構造式(1)における定義と同一であり、
Pは、アミン基又はポリヒスチジン基を意味し、J
1とJ
2はリンカーであり、J
1及びJ
2は独立して、単純共有結合、PO
3-、SO
3、CO
2、C
2-12アルキル、アルケニル、アルキニルから選択可能であるが、これに限定されず、用いられる親水性物質に応じて、本発明の目的に符合するJ
1とJ
2はいかなるリンカーでも用いられ得ることは、通常の技術者には明らかであろう。
好ましくは、アミン基が導入された場合には、J2は単純共有結合又はPO3-、J1はC6アルキルであることが好ましいが、これに限定されない。
また、ポリヒスチジン(polyhistidine)基が導入された場合には、構造式(9)では、J
2は単純共有結合又はPO
3-、J
1は化合物(4)が好ましいが、これに限定されない。
また、構造式(9)による二重鎖オリゴヌクレオチド構造体の親水性物質が構造式(5)又は構造式(6)による親水性物質ブロックであり、これにアミン基又はポリヒスチジン(polyhistidine)基が導入される場合には、構造式(10)又は構造式(11)のような構造を有することができる。
前記構造式(10)及び構造式(11)において、X、R、Y、B、A’、J、m及びnは、構造式(5)又は構造式(6)における定義と同一であり、P、J
1及びJ
2は構造式(9)における定義と同一である。
特に、前記構造式(10)及び構造式(11)において、親水性物質は、アンフィレグリン特異的二重鎖オリゴヌクレオチドセンス鎖の3’末端に結合した形態であることが好ましく、この場合、前記構造式(9)~構造式(11)は、次の構造式(12)~構造式(14)の形態を有することができる。
前記構造式(12)~構造式(14)において、X、R、Y、B、A、A’、J、m、n、P、J
1及びJ
2は、前記構造式(9)~構造式(11)における定義と同一であり、5’及び3’は、アンフィレグリン特異的二重鎖オリゴヌクレオチドセンス鎖の5’末端及び3’末端を意味する。
本発明で導入可能なアミン基には1次~3次アミン基を用いることができ、1次アミン基が用いられることが特に好ましい。前記導入されたアミン基はアミン塩として存在してもよいが、例えば、1次アミン基の塩は、NH3
+の形態で存在できる。
また、本発明で導入可能なポリヒスチジン基は、3~10個のヒスチジンを含むことが好ましく、特に好ましくは、5~8個、最も好ましくは6個のヒスチジンを含むことができる。ヒスチジンの他に一つ以上のシステインがさらに含まれてもよい。
一方、本発明に係るアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体及び該構造体から形成されたナノ粒子にターゲッティングモイエティが備えられると、効率的にターゲット細胞への伝達を促進し、比較的低い濃度の投与量でもターゲット細胞に伝達され、高いターゲット遺伝子発現調節機能を示すことができ、他の臓器及び細胞への非特異的なアンフィレグリン特異的二重鎖オリゴヌクレオチドの伝達を防止することができる。
これによって、本発明は、前記構造式(1)~構造式(4)、構造式(7)及び構造式(8)による構造体にリガンド(L)、特に、受容体媒介内包作用(receptor-mediated endocytosis,RME)を通じてターゲット細胞内在化(internalization)を増進させる受容体と特異的に結合する特性を持つリガンド(ligand)がさらに結合した二重鎖オリゴRNA、構造体を提供し、例えば、構造式(1)による二重鎖オリゴRNA構造体にリガンドが結合した形態は、下記構造式(15)のような構造を有する。
前記構造式(15)において、A、B、X及びYは、前記構造式(1)における定義と同一であり、Lは、受容体媒介内包作用(receptor-mediated endocytosis,RME)を通じてターゲット細胞内在化(internalization)を増進させる受容体と特異的に結合する特性を持つリガンドを意味し、iは、1~5の整数、好ましくは1~3の整数である。
前記構造式(15)におけるリガンドは、好ましくはターゲット細胞特異的に細胞内在化(internalization)を増進させるRME特性を持つターゲット受容体特異的抗体、アプタマー又はペプチド;又は葉酸(Folate、一般に、folateとfolic acidは同じ意味で使われており、本発明における葉酸は、自然状態又は人体で活性化状態であるfolateを意味する。)、N-アセチルガラクトサミン(N-acetyl Galactosamine,NAG)などのヘキソアミン(hexoamine)、ブドウ糖(glucose)、マンノース(mannose)をはじめとする糖や炭水化物(carbohydrate)などの化学物質などから選択可能であるが、これに限定されるものではない。
また、前記構造式(15)における親水性物質Aは、構造式(5)及び構造式(6)による親水性物質ブロックの形態で用いることができる。
本発明のさらに他の態様として、本発明は、アンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体を製造する方法を提供する。
本発明に係るアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体を製造する過程は、例えば、
(1)固形支持体(solid support)に基づいて親水性物質を結合させる段階;
(2)前記親水性物質が結合した固形支持体に基づいてオリゴヌクレオチド単一鎖を合成する段階;
(3)前記オリゴヌクレオチド単一鎖の5’末端に疎水性物質を共有結合させる段階;
(4)前記オリゴヌクレオチド単一鎖の配列と相補的な配列のオリゴヌクレオチド単一鎖を合成する段階;
(5)合成が完了すると、固形支持体からオリゴヌクレオチド-高分子構造体及びオリゴヌクレオチド単一鎖を分離精製する段階;
(6)製造されたオリゴヌクレオチド-高分子構造体と相補的な配列のオリゴヌクレオチド単一鎖のアニールによって二重鎖オリゴヌクレオチド構造体を製造する段階;を含んでなり得る。
本発明における固形支持体(solid support)は、CPG(Controlled Pore Glass)が好ましいが、これに限定されず、ポリスチレン(PS)、ポリメチルメタクリレート(PMMA)シリカゲル、セルロースペーパーなどが用いられ得る。CPGの場合、直径は40~180μmが好ましく、500~3000Åの孔隙サイズを有することが好ましい。前記段階(5)の後、製造が完了すると、精製されたRNA-高分子構造体及びオリゴヌクレオチド単一鎖は、MALDI-TOF質量分析機で分子量を測定し、目的とするオリゴヌクレオチド-高分子構造体及びオリゴヌクレオチド単一鎖が製造されたかを確認することができる。前記製造方法において、(2)段階で合成されたオリゴヌクレオチド単一鎖の配列と相補的な配列のオリゴヌクレオチド単一鎖を合成する段階(4)は、(1)段階の前、又は(1)段階~(5)段階のいずれか一過程中に行われても構わない。
また、(2)段階で合成されたオリゴヌクレオチド単一鎖と相補的配列を含むオリゴヌクレオチド単一鎖は、5’末端にリン酸基が結合した形態で用いられたことを特徴とする製造方法も可能である。
一方、本発明のアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体にさらにリガンドが結合した二重鎖オリゴヌクレオチド構造体の製造方法を提供する。
リガンドが結合したアンフィレグリン特異的二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体を製造する方法は、例えば、
(1)官能基が結合している固形支持体に親水性物質を結合させる段階;
(2)官能基-親水性物質が結合している固形支持体に基づいてオリゴヌクレオチド単一鎖を合成する段階;
(3)前記オリゴヌクレオチド単一鎖の5’末端に疎水性物質を共有結合させる過程で合成する段階;
(4)前記オリゴヌクレオチド単一鎖の配列と相補的な配列のオリゴヌクレオチド単一鎖を合成する段階;
(5)合成が完了すると、固形支持体から官能基-オリゴヌクレオチド-高分子構造体及び相補的な配列のオリゴヌクレオチド単一鎖を分離する段階;
(6)前記官能基を用いて親水性物質の末端にリガンドを結合させてリガンド-オリゴヌクレオチド-高分子構造体単一鎖を製造する段階;
(7)製造されたリガンド-オリゴヌクレオチド-高分子構造体と相補的な配列のオリゴヌクレオチド単一鎖のアニールを通じてリガンド-二重鎖オリゴヌクレオチド構造体を製造する段階;
を含んでなり得る。
前記(6)段階後に、製造が完了すると、リガンド-オリゴヌクレオチド-高分子構造体及び相補的な配列のオリゴヌクレオチド単一鎖を分離精製した後、MALDI-TOF質量分析機で分子量を測定し、目的するリガンド-オリゴヌクレオチド-高分子構造体及び相補的なオリゴヌクレオチドが製造されたかを確認することができる。製造されたリガンド-オリゴヌクレオチド-高分子構造体と相補的な配列のオリゴヌクレオチド単一鎖のアニールを通じてリガンド-二重鎖オリゴヌクレオチド構造体を製造することができる。前記製造方法において、(3)段階で合成されたオリゴヌクレオチド単一鎖の配列と相補的な配列のオリゴヌクレオチド単一鎖を合成する段階(4)は、独立した合成過程として(1)段階の前、又は(1)段階~(6)段階のいずれか一過程中に行われても構わない。
本発明は、さらに他の観点において、本発明に係る二重鎖オリゴヌクレオチド構造体を含むナノ粒子に関する。本発明に係る二重鎖オリゴヌクレオチド構造体の場合、疎水性物質の疎水性相互作用によって自己組立ナノ粒子を形成するが(大韓民国登録特許公報第1224828号)、このようなナノ粒子は、体内への伝達効率及び体内における安定性に極めて優れるだけでなく、粒子サイズの均一性にも優れているので、QC(Quality Control)が容易であり、薬物としての製造工程が簡単である。
本発明において、前記ナノ粒子は、互いに異なる配列を含む二重鎖オリゴヌクレオチドを含む二重鎖オリゴヌクレオチド構造体が混合されてなることを特徴とし、例えば、前記ナノ粒子は、配列番号10~12から選択されるいずれか一つの配列を含むセンス鎖とこれに相補的な配列を含むアンチセンス鎖を含む同種のアンフィレグリン特異的二重鎖オリゴヌクレオチドを含むことができるが、さらに他の態様として、配列番号10~12から選択されるいずれか一つの配列を含むセンス鎖とこれに相補的な配列を含むアンチセンス鎖を含む異種のアンフィレグリン特異的二重鎖オリゴヌクレオチドを共に含んでもよく、本発明で開示していないアンフィレグリン特異的二重鎖オリゴヌクレオチドを共に含んでもよいだろう。
本発明は、他の観点において、本発明に係る二重鎖オリゴヌクレオチド、二重鎖オリゴヌクレオチド構造体又は二重鎖オリゴヌクレオチド構造体を含むナノ粒子を有効成分として含有する線維症又は呼吸器疾患の予防又は治療用医薬組成物に関する。
本発明に係る線維症又は呼吸器疾患の予防又は治療用医薬組成物は、結合組織再形成(tissue remodeling)、特に、肺動脈再形成(pulmonary artery remodeling)及び気道再形成(airway remodeling)を抑制し、線維症又は呼吸器疾患の予防又は治療に効果を奏する。
本発明において、呼吸器疾患は、慢性閉塞性疾患(COPD)、喘息、急慢性気管支炎、アレルギー鼻炎、鎮咳去痰、気管支炎、細気管支炎、咽喉炎、扁桃炎又は喉頭炎であることを特徴とし、線維症は、特発性肺線維化症(IPF)、肝線維症(liver fibrosis)、肝硬変(cirrhosis)、骨髄線維症(myelofibrosis)、心筋線維症(myocardial fibrosis)、腎臓線維症(renal fibrosis)、肺線維症(pulmonary fibrosis)、心臓線維症(cardiac fibrosis)及び放射線誘発線維症(radiation-induced fibrosis)からなる群から選ばれることを特徴とし得るが、これに限定されるものではない。本発明において、前記放射線誘発線維症は、癌、腫瘍などの治療のためにしばしば用いられる放射線療法によって頻繁に誘発される副作用であり、放射線線維化症候群(radiation fibrosis syndrom,RFS)と同じ意味で使用可能である。
本発明の組成物には、投与のために、前記記載された有効成分に加えて、薬剤学的に許容可能な担体を1種以上さらに含んで製造することができる。薬剤学的に許容可能な担体は、本発明の有効成分と両立可能でなければならず、食塩水、滅菌水、リンゲル液、緩衝食塩水、デキストロース溶液、マルトデキストリン溶液、グリセロール、エタノール及びこれら成分のいずれか1つの成分又は2つ以上の成分を混合して使用することができ、必要によって、抗酸化剤、緩衝液、静菌剤などの他の通常の添加剤を添加できる。また、希釈剤、分散剤、界面活性剤、結合剤及び潤滑剤をさらに添加して、水溶液、懸濁液、乳濁液などの注射用剤形に製剤化できる。特に、凍結乾燥(lyophilized)した形態の剤形で製剤化して提供することが好ましい。凍結乾燥剤形の製造のために、本発明の属する技術の分野に通常知られている方法を用いることができ、凍結乾燥のための安定化剤が追加されてもよい。当分野における適切な方法で又はレミングトンの薬科学(Remington’s pharmaceutical Science,Mack Publishing company,Easton PA)に開示されている方法を用いて、各疾病に応じて又は成分に応じて好ましく製剤化できる。
本発明の組成物は、通常の患者の症候と疾病の深刻度に基づいて本技術分野における通常の専門家が決定できる。また、散剤、錠剤、カプセル剤、液剤、注射剤、軟膏剤、シロップ剤などの様々な形態で製剤化でき、単位投与量又は多回投与量の容器、例えば、密封したアンプル及び瓶などとして提供され得る。
本発明の組成物は、経口又は非経口投与が可能である。本発明に係る組成物の投与経路は、これらに限定されるものではないが、例えば、口腔、吸入用、静脈内、筋肉内、動脈内、骨髄内、硬膜内、心臓内、経皮、皮下、腹腔内、臓管、舌下又は局所投与が可能である。特に、呼吸器疾患治療のために気管支内点滴注入を通じた肺への投与も可能である。本発明に係る組成物の投与量は、患者の体重、年齢、性別、健康状態、食餌、投与時間、方法、排泄率又は疾病の重症度などによってその範囲が様々であり、本技術分野における通常の専門家が容易に決定できる。また、臨床投与のために公知の技術を用いて本発明の組成物を適切な剤形に製剤化できる。
本発明は、他の観点において、本発明に係る医薬組成物を含む凍結乾燥した形態の剤形に関する。
本発明は、他の観点において、本発明に係る線維症又は呼吸器疾患の予防又は治療用医薬組成物を、線維症又は呼吸器疾患の予防又は治療を必要とする個体に投与する段階を含む線維症又は呼吸器疾患の予防又は治療方法に関する。
本発明において、呼吸器疾患は、慢性閉塞性疾患(COPD)、喘息、急慢性気管支炎、アレルギー鼻炎、鎮咳去痰、気管支炎、細気管支炎、咽喉炎、扁桃炎又は喉頭炎であることを特徴とし、線維症は、特発性肺線維化症(IPF)、肝硬変(cirrhosis)、骨髄線維症(myelofibrosis)、心筋線維症(myocardial fibrosis)、腎臓線維症(renal fibrosis)、肺線維症(pulmonary fibrosis)、心臓線維症(cardiac fibrosis)、肝線維症(liver fibrosis)、又は放射線誘発線維症(radiation-induced fibrosis)であることを特徴とし得るが、これに限定されるものではない。
本発明は、さらに他の観点において、線維症又は呼吸器疾患の予防又は治療に用いるための前記二重鎖オリゴヌクレオチド、これを含む二重鎖オリゴヌクレオチド構造体及び前記二重鎖オリゴヌクレオチド構造体を含むナノ粒子に関する。
本発明は、さらに他の観点において、線維症又は呼吸器疾患の予防又は治療に用いるための前記薬学的組成物に関する。
本発明は、さらに他の観点において、線維症又は呼吸器疾患の予防又は治療のための薬物の製造のための前記二重鎖オリゴヌクレオチド、これを含む二重鎖オリゴヌクレオチド構造体及び前記二重鎖オリゴヌクレオチド構造体を含むナノ粒子の用途に関する。
以下、実施例を用いて本発明をより詳細に説明する。これらの実施例は、単に本発明をより具体的に説明するためのものであり、本発明の範囲がこれらの実施例によって制限されないということは、当該技術分野における通常の知識を有する者にとって明らかであろう。したがって、本発明の実質的な範囲は、添付する請求項とそれらの等価物によって定義されるといえよう。
本発明では、アンフィレグリンの発現を阻害できる3個の特異的な配列を同定し、それらがアンフィレグリンを暗号化するmRNAと相補的に結合してアンフィレグリンの発現を効果的に抑制することによって、線維症と呼吸器疾患関連疾患を効果的に治療できることを確認した。
実施例1.アンフィレグリンをターゲットとするSAMiRNAスクリーニングのためのアルゴリズムと候補配列選定
SAMiRNA基礎治療剤高速大量スクリーニング(SAMiRNA based drug high-throughput screening)は、全mRNAに対して1-塩基(1-base)又は2-塩基(2-base)スライディングウィンドウアルゴリズム(sliding window algorithm)を適用して可能な全ての候補配列を生成し、相同性フィルタリング(homology filtering)を行って不要な候補配列を除去し、最終的に選別された全SAMiRNAに対して当該遺伝子発現阻害の程度を確認する方法である。
まず、アンフィレグリンに対するSAMiRNA候補配列に対するデザイン過程は、ヒトアンフィレグリンmRNAであるNM_001657.3(1290bp)に対して1-塩基又は2-塩基スライディングウィンドウアルゴリズムを適用して、19個の塩基で構成された1,257個のSAMiRNA候補配列を最終選別し、アンフィレグリン阻害程度実験を行った。
実施例2.二重鎖オリゴRNA構造体の合成
本発明で製造された二重鎖オリゴRNA構造体(SAMiRNA)は、下記構造式のような構造を有する。
モノSAMiRNA(n=4)二重鎖オリゴ構造体のセンス鎖は、3,4,6-トリアセチル-1-ヘキサ(エチレングリコール)-N-アセチルガラクトサミン-CPG(1-Hexa(Ethylene Glycol)-N-Acetyl Galactosamine-CPG)を支持体とし、親水性物質単量体であるDMTヘキサエチレングリコールホスホロアミダート(Demethoxytrityl hexaethylene glycol phosphoramidate)3個を、前記反応によって連続して結合した後、RNA又はDNA合成を行った後、さらに5’末端部位に疎水性物質である二硫化結合が含まれているC
24(C
6-S-S-C
18)を結合させることで、3’末端にNAG-ヘキサエチレングリコール-(-PO
3-ヘキサエチレングリコール)3が結合しており、5’末端にC
24(C
6-S-S-C
18)が結合しているモノSAMiRNA(n=4)のセンス鎖を作った。
合成が完了すると、60℃の温湯器(water bath)で28%(v/v)アンモニア(ammonia)を処理し、合成されたRNA単一鎖及びオリゴ(DNA又はRNA)-高分子構造体をCPGから分離した後、脱保護(deprotection)反応によって保護残基を除去した。保護残基が除去されたRNA単一鎖及びオリゴ(DNA又はRNA)-高分子構造体は、70℃のオーブンでN-メチルピロリドン(N-methylpyrolidon)、トリエチルアミン(triethylamine)及びトリエチルアミントリヒドロフルオリド(triethylaminetrihydrofluoride)を体積比10:3:4で処理し、2’TBDMS(tert-butyldimethylsilyl)を除去した。これらの反応物から、高速液体クロマトグラフィー(High Performance Liquid Chromatography,HPLC)でRNA単一鎖、オリゴ(DNA又はRNA)-高分子構造体及びリガンドが結合したオリゴ(DNA又はRNA)-高分子構造体を分離し、これをMALDI-TOF質量分析機(MALDI TOF-MS、SHIMADZU、日本)で分子量を測定し、合成しようとする塩基配列及びオリゴ-高分子構造体と符合するか確認した。その後、それぞれの二重鎖オリゴ構造体を製造するためにセンス鎖とアンチセンス鎖を同量混合して1Xアニーリングバッファ(30mM HEPES、100mM酢酸カリウム(Potassium acetate)、2mM酢酸マグネシウム(Magnesium acetate)、pH7.0~7.5)に入れ、90℃恒温水槽で3分反応させた後、さらに37℃で反応させ、目的とするSAMiRNAを製造した。製造された二重鎖オリゴRNA構造体は、電気泳動によってアニールを確認した。
実施例3.ヒト(Human)アンフィレグリンをターゲットとしてRNAiを誘導するSAMiRNAナノ粒子の高速大量スクリーニング(HTS)
3.1 SAMiRNAナノ粒子の製造
実施例2で合成されたアンフィレグリン配列をターゲットとする1,257種のSAMiRNAを1X DPBS(Dulbe cco’s Phosphate Buffered Saline)(WELGENE,KR)に溶かして凍結乾燥器(LGJ-100F,CN)で5日間凍結乾燥させた。凍結乾燥したナノ粒子パウダーを脱塩蒸留水(deionized distilled water(Bioneer,KR))1.429mlに溶かして均質化し、本発明のための実験に使用した。
3.2 SAMiRNAナノ粒子の細胞内処理方法
アンフィレグリンの発現を抑制するSAMiRNAの発掘のために、ヒト由来肺癌細胞株であるA549を用い、A549細胞株は、10%ウシ胎児血清(Hyclone,US)及び1%ペニシリン-ストレプトマイシン(Hyclone,US)が含まれたGibcoTMHam’s F-12K(Kaighn’s)培地(Thermo,US)を用いて37℃、5% CO2条件で培養した。上と同じ培地を用いてA549細胞株を96ウェルプレート(Costar,US)に2×104cells/wellの条件で分注し、翌日、前記実施例3.1で脱塩蒸留水(deionized distilled water)で均質化したSAMiRNAを1X DPBSで希釈し、細胞に500nM又は1000nMとなるように処理した。SAMiRNAの処理は、12時間ごとに1回ずつ処理する条件で総4回処理し、37℃、5% CO2条件で培養した。
3.3 ヒト(Human)アンフィレグリンmRNA発現抑制効能分析を用いたSAMiRNAスクリーニング
実施例3-2でSAMiRNA処理された細胞株から全RNAを抽出してcDNAを製造した後、実時間PCR(real-time PCR)を用いてアンフィレグリン遺伝子のmRNA発現量を定量した。
アンフィレグリン遺伝子のmRNA発現量分析のために、AccuPower(R)Dual-HotStart RT-qPCRキット(Bioneer,Korea)の各ウェルにAREG順方向プライマー300nM、AREG逆方向プライマー300nM、AREGプローブ300nM、RPL13A順方向プライマー300nM、RPL13A逆方向プライマー300nM、RPL13Aプローブ300nM、TBP順方向プライマー400nM、TBP逆方向プライマー400nM、TBPプローブ300nMが追加されて乾燥製造された(表2、高速大量スクリーニング(HTS)実験に用いられたプライマー及び加水分解プローブ配列)。製造されたキットの性能は、A549細胞全RNAを用いて検量曲線を作成し、PCR増幅効率(表3)で判定した。RT-qPCRは、95℃5分、(95℃5秒、58℃15秒)×45サイクルで反応をし、各サイクルごとに蛍光値が検出(detection)されるプロトコルを用いた。
SAMiRNA処理された96ウェルプレート(Costar,US)は全RNA抽出からRT-qPCRまで自動化装備であるExiStation HTTM KoreaにHT DNA/RNA抽出キット(Bioneer,Korea)とアンフィレグリン分析のためのプライマー及びプローブを含んで別途に製造されたAccuPower(R)Dual-HotStart RT-qPCRキット(Bioneer,Korea)を、自動化プログラムにしたがって全RNA抽出及びワンステップRT-qPCRが行われた。
qPCRアレイ後、導出される2遺伝子のCt値を2(-Delta Delta C(T))法[Livak KJ,Schmittgen TD.2001.Analys is of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T))Method.Methods.Dec;25(4):4 02-8]を用いて、相対定量分析により、対照群に対して実験群であるアンフィレグリンmRNAの相対的な量(%)を計算した。
高効率のSAMiRNAを選別するために、最終濃度500nM又は1000nMの濃度でアンフィレグリンmRNA発現量が、対照群と対比して発現量の減少効率が最も良い配列、すなわち、配列番号1~14の配列をそれぞれセンス鎖として有するSAMiRNA 14種を選別した。
図1に示すように、アンフィレグリンを標的にする1,257種のSAMiRNAから、アンフィレグリン遺伝子発現を最も効果的に抑制するSAMiRNA 14種を最終選別した。当該SAMiRNAの配列情報は、下記表4の通りである。
実施例4.ヒト(Human)アンフィレグリンをターゲットとしてRNAiを誘導するSAMiRNAナノ粒子スクリーニング
実施例3で選別された配列番号1~14の配列をそれぞれセンス鎖として有するSAMiRNAを用いて肺癌細胞株であるA549を処理し、前記細胞株においてアンフィレグリンmRNAの発現様相を分析した。
4.1 SAMiRNAナノ粒子の細胞内処理方法
アンフィレグリンの発現を抑制するSAMiRNA発掘のために、ヒト由来肺癌細胞株であるA549を用いたが、A549細胞株は、10%ウシ胎児血清(Hyclone,US)及び1%ペニシリン-ストレプトマイシン(Hyclone,US)が含まれたGibcoTMHam’s F-12K(Kaighn’s)培地(Thermo,US)を用いて37℃、5% CO2条件で培養した。同上の培地を用いてA549細胞株を12ウェルプレート(Costar,US)に8×104cells/wellの条件で分注し、翌日、前記実施例3.1で脱塩蒸留水(deionized distilled water)で均質化したSAMiRNAを1X DPBSで希釈し、細胞に200nM又は600nMとなるように処理した。SAMiRNAの処理は、12時間ごとに1回ずつ処理する条件で総4回処理し、37℃、5% CO2条件で培養した。
4.2 ヒト(Human)アンフィレグリンmRNA発現抑制効能分析を用いたSAMiRNAスクリーニング
実施例4-1でSAMiRNA処理された細胞株から全RNAを抽出してcDNAを製造した後、実時間PCR(real-time PCR)を用いてアンフィレグリン遺伝子のmRNA発現量を定量した。
4-2-1 SAMiRNA処理された細胞からRNA分離及びcDNA製造
RNA抽出キット(AccuPrep Cell total RNA extraction kit,BIONEER、韓国)を用いて、前記実施例4-1でSAMiRNA処理された細胞株から全RNAを抽出し、抽出されたRNAはRNA逆転写酵素(AccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20,Bioneer、韓国)を用いて、下記のような方法でcDNAを製造した。具体的に、0.25mlエッペンドルフチューブに含まれたAccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20(Bioneer、韓国)に1チューブ当たりに抽出された1μgずつのRNAを入れ、総体積が20μlとなるように、DEPC(diethyl pyrocarbonate)処理された蒸留水を添加した。これを遺伝子増幅器(MyGenieTM96Gradient Thermal Block,BIONEER、韓国)で37℃で30秒間RNAとプライマーを混成化し、48℃で4分間cDNAを製造する2つの過程を12回反復した後、95℃で5分間酵素を不活性化させて増幅反応を終了した。
4-2-2 ヒト(Human)アンフィレグリンmRNAの相対定量分析
実施例4-2-1で製造されたcDNAを鋳型とし、SYBRグリーン方式の実時間qPCRを用いて、SAMiRNA対照試料(control sample)に対するアンフィレグリンmRNAの相対的発現率を下記のような方法で分析した。96ウェルプレートの各ウェルに前記実施例4-2-1で製造されたcDNAを蒸留水で5倍希釈し、アンフィレグリンmRNA発現量分析のために希釈されたcDNA 3μlとAccuPower(R) 2X GreenStarTM qPCR MasterMix(BIONEER、韓国)を25μl、蒸留水19μl、アンフィレグリンqPCRプライマー(配列番号17及び18(表5);10pmole/μlそれぞれ、BIONEER、韓国)を3μl入れて混合液を作った。一方、アンフィレグリンmRNAの発現量を正規化するために、ハウスキーピング遺伝子(housekeeping gene、以下、HK遺伝子)であるGAPDH(Glyceraldehyde-3-Phosphate Dehydrogenase)を標準遺伝子とした。前記混合液が入っている96ウェルプレートをExicyclerTM Real-Time Quantitative Thermal Block(BIONEER、韓国)を用いて下記のような反応を行った:95℃で15分間反応して酵素の活性化及びcDNAの二次構造をなくした後、94℃で30秒変性(denaturing)、58℃で30秒アニーリング(annealing)、72℃で30秒延長(extension)、SYBRグリーンスキャン(SYBR green scan)の4つの過程を42回繰り返し行い、72℃で3分間最終延長を行った後、55℃で1分間温度を維持し、55℃から95℃までメルティングカーブ(melting curve)を分析した。
PCRが終了した後、それぞれ得たターゲット遺伝子のCt(threshold cycle)値は、GAPDH遺伝子によって補正されたターゲット遺伝子のCt値を求めた後、遺伝子発現阻害を起こさないコントロール配列であるSAMiRNA(SAMiCONT)が処理された実験群を対照群にしてΔCt値の差を求めた。前記ΔCt値と計算式2(-ΔCt)×100を用いて、アンフィレグリン特異的SAMiRNAが処理された細胞のターゲット遺伝子の発現量を相対定量した。
高効率のSAMiRNAを選別するために、最終濃度200nM又は600nMの濃度でアンフィレグリンmRNA発現量が対照群に比して発現量の減少効率が最も良い配列、すなわち、配列番号10、11及び12の配列をそれぞれセンス鎖として有するSAMiRNA 3種を選別した。
図3に示すように、アンフィレグリンを標的とする14種のSAMiRNAから、アンフィレグリン遺伝子発現を最も効果的に抑制するSAMiRNA 3種を最終選別した。当該SAMiRNAの配列情報は、下記表6の通りである。
実施例5.肺癌細胞株(A549)から選別されたSAMiRNAを用いたヒト(Human)アンフィレグリンの発現抑制
実施例4で選別された配列番号10、11及び12の配列をそれぞれセンス鎖として有するSAMiRNAを用いて肺癌細胞株であるA549を処理し、前記細胞株においてアンフィレグリンmRNAの発現様相を分析してSAMiRNAのIC50値を確認した。
5.1 SAMiRNAナノ粒子の製造及び粒度分析
実施例2で合成されたアンフィレグリン配列をターゲットとする3種のSAMiRNAを1X DPBS(WELGENE,KR)に溶かして凍結乾燥器(LGJ-100F,CN)で5日間凍結乾燥させた。凍結乾燥したナノ粒子パウダーを脱塩蒸留水(deionized distilled water(Bioneer,KR))2mlに溶かして均質化し、本発明のための実験に使用した。製造されたSAMiRNAナノ粒子の粒度分析のために、Zetasizer Nano ZS(Malvern,UK)を用いてSAMiRNAのサイズ及び多分散指数(polydispersity index)を測定した結果、選別されたSAMiRNAに対するナノ粒子のサイズ及び多分散指数は下記表7の通りであり、グラフは、図2のように測定された。
5.2 SAMiRNAナノ粒子の細胞内処理方法
アンフィレグリンの発現を抑制する選別されたSAMiRNA効果を確認するために、ヒト由来肺癌細胞株であるA549を用いたが、A549細胞株は、10%ウシ胎児血清(Hyclone,US)及び1%ペニシリン-ストレプトマイシン(Hyclone,US)が含まれたGibcoTMHam’s F-12K(Kaighn ’s)培地(Thermo,US)を用いて37℃、5% CO2条件で培養した。同上の培地を用いてA549細胞株を12ウェルプレート(Costar,US)に8×104cells/wellの条件で分注し、翌日、前記実施例5.1で脱塩蒸留水(deionized distilled water)で均質化したSAMiRNAを1X DPBSで希釈し、細胞に12.5nM、25nM、50nM、100nM、200nM、600nM又は1200nMとなるように処理した。SAMiRNAの処理は、12時間ごとに1回ずつ処理する条件で総4回処理し、37℃、5% CO2条件で培養した。
5.3 ヒト(Human)アンフィレグリンmRNA発現抑制効能分析を用いたSAMiRNA IC50確認
実施例5-2でSAMiRNA処理された細胞株から全RNAを抽出してcDNAを製造した後、実時間PCR(real-time PCR)を用いてアンフィレグリン遺伝子のmRNA発現量を相対定量した。
5-3-1 SAMiRNA処理された細胞からRNA分離及びcDNA製造
RNA抽出キット(AccuPrep Cell total RNA extraction kit,BIONEER、韓国)を用いて、前記実施例5-2でSAMiRNA処理された細胞株から全RNAを抽出し、抽出されたRNAは、RNA逆転写酵素(AccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20,Bioneer、韓国)を用いて、下記のような方法でcDNAを製造した。具体的に、0.25mlエッペンドルフチューブに含まれたAccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20(Bioneer、韓国)に1チューブ当たりに抽出された1μgずつのRNAを入れ、総体積が20μlとなるようにDEPC(diethyl pyrocarbonate)処理された蒸留水を添加した。これを遺伝子増幅器(MyGenieTM96Gradient Thermal Block,BIONEER、韓国)で37℃で30秒間RNAとプライマーを混成化し、48℃で4分間cDNAを製造する2つの過程を12回反復した後、95℃で5分間酵素を不活性化させて増幅反応を終了した。
5-3-2 ヒト(Human)アンフィレグリンmRNAの相対定量分析
実施例5-3-1で製造されたcDNAを鋳型とし、SYBRグリーン方式の実時間qPCRを用いて、SAMiRNA対照試料に対比するアンフィレグリンmRNAの相対的発現率を下記のような方法で分析した。96ウェルプレートの各ウェルに前記実施例5-3-1で製造されたcDNAを蒸留水で5倍希釈し、アンフィレグリンmRNA発現量分析のために希釈されたcDNA 3μlとAccuPower(R) 2XGreenStarTM qPCR MasterMix(BIONEER、韓国)を25μl、蒸留水19μl、アンフィレグリンqPCRプライマー(配列番号17及び18(表5);10pmole/μlそれぞれ、BIONEER、韓国)を3μl入れて混合液を作った。一方、アンフィレグリンmRNAの発現量を正規化するために、ハウスキーピング遺伝子(housekeeping gene、以下、HK遺伝子)であるGAP DH(Glyceraldehyde-3-Phosphate Dehydrogenase)を標準遺伝子とした。前記混合液が入っている96ウェルプレートをExicyclerTMReal-Time Quantitative Thermal Block(BIONEER、韓国)を用いて下記のような反応を行った:95℃で15分間反応して酵素の活性化及びcDNAの二次構造をなくした後、94℃で30秒変性(denaturing)、58℃で30秒アニーリング(annealing)、72℃で30秒延長(extension)、SYBRグリーンスキャン(SYBR green scan)の4つの過程を42回繰り返し行い、72℃で3分間最終延長を行った後、55℃で1分間温度を維持し、55℃から95℃までメルティングカーブ(melting curve)を分析した。
PCRが終了した後、それぞれ得たターゲット遺伝子のCt(threshold cycle)値は、GAPDH遺伝子によって補正されたターゲット遺伝子のCt値を求めた後、遺伝子発現阻害を起こさないコントロール配列であるSAMiRNA(SAMiCONT)が処理された実験群を対照群にしてΔCt値の差を求めた。前記ΔCt値と計算式2(-ΔCt)×100を用いて、アンフィレグリン特異的SAMiRNAが処理された細胞のターゲット遺伝子の発現量を相対定量した。
その結果、配列番号10、11及び12の配列をそれぞれセンス鎖として有するアンフィレグリン特異的SAMiRNAはいずれも100nMの低濃度においても50%以上のアンフィレグリンmRNA発現量の減少を示し、非常に高効率でアンフィレグリン発現を阻害する効果を示すことを確認した。それぞれのIC50は、配列番号10の配列をセンス鎖として有するアンフィレグリン特異的SAMiRNAの場合、図4に見られるように28.75nM、配列番号11の配列をセンス鎖として有するアンフィレグリン特異的SAMiRNAの場合、図5に見られるように26.04nM、配列番号12の配列をセンス鎖として有するアンフィレグリン特異的SAMiRNAの場合、図6に見られるように12.07nMと確認された。特に、配列番号12の配列をセンス鎖として有するアンフィレグリン特異的SAMiRNAの場合、図6に見られるように、25nMの低濃度においても50%以上のアンフィレグリンmRNA発現量の減少を示し、選別された3種の配列のうち最も効果的にアンフィレグリン遺伝子発現を阻害する効果を示すことを確認した。
実施例6.ヒト末梢血液単核細胞(peripheral blood mononuclear cell,PBMC)を用いたin vitro細胞毒性評価
SAMi-hAREGによる内在免疫関連サイトカインのmRNA増加量を確認するために、10% FBS(fetal bivine serum;HycloneTM)を含むRPMI1640(HycloneTM)培地にePBMC(R)Cryopreserved Human PBMC(ヒト末梢血液単核細胞、Cellular Technology Limited,USA)を12ウェルプレート(Costar USA)にウェル当たり5×105cellとなるように分注した。細胞が安定化するように1時間37℃、5% CO2培養器に培養した後、分注されたPBMCにSAMi-CON(DNA/RNA)、SAMi-hAREG#10(DNA/RNA)、SAMi-hAREG#11(DNA/RNA)、SAMi-hAREG#12(DNA/RNA)、SAMi-CON(RNA/RNA)、SAMi-hAREG#10(RNA/RNA)、SAMi-hAREG#11(RNA/RNA)、SAMi-hAREG#12(RNA/RNA)を2.5μMとなるようにそれぞれ処理し、6時間37℃5% CO2培養器に培養した。陽性対照群として20μg/mlのコンカナバリン(Concanavalin)A(Sigma Aldrich,USA)を使用した。
その後、全ての細胞を回収し、RNA抽出キット(RNeasy Mini Kit,Qiagen,Germany)とRNase-Free DNaseセット(Qiagen,Germany)でメーカー提供プロトコルにしたがって全RNAを抽出した。
抽出されたRNA200ngと脱イオン殺菌DW(Deionized sterile DW)(Bioneer,Korea)、そしてRNA逆転写酵素(AccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20,Bioneer、韓国)を混合し、これを遺伝子増幅器(MyGenieTM96 Gradient Thermal Block,BIONEER、韓国)を用いて(37℃30秒、48℃4分、55℃30秒)×12サイクル、95℃、5分のような条件で反応して総20μl cDNAを合成した。
合成されたcDNAは、RPL13A、I L1B、IL6、IFNG、TNF、IL12B遺伝子に対するqPCRプライマーと混合した後、ExicyclerTM96 Real-Time Quantitative Thermal Block(Bioneer,Korea)を用いて95℃5分、(95℃5秒、58℃15秒)×45サイクル条件で増幅した。
qPCRアレイ後、導出される2つの遺伝子のCt値を、2(-Delta Delta C(T))法[Livak KJ,Schmittgen TD.2001.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T))Method.Methods.Dec;25(4):402-8]を用いて相対定量分析により、対照群対比実験群のmRNAの相対的な量(%)を計算した。
その結果、図7に見られるように、ヒト末梢血液単核細胞(peripheral blood mononuclear cell,human PBMC)においてアンフィレグリン特異的SAMiRNA #10、SAMiRNA #11及びSAMiRNA #12による内在免疫関連サイトカインが発現しないことが確認された。
実施例7.選別された配列番号10、11及び12の配列をセンス鎖として含むDNA/RNAハイブリッド及びRNA/RNAハイブリッドSAMiRNAによるヒト(Human)アンフィレグリンの発現抑制比較分析
実施例4で選別された配列番号10、11及び12の配列をそれぞれセンス鎖として有するアンフィレグリン特異的SAMiRNAを含む二重鎖オリゴDNA/RNAハイブリッド及びRNA/RNAハイブリッドを用いて肺癌細胞株であるA549を処理し、前記細胞株においてアンフィレグリンmRNAの相対的な発現量(%)を比較分析した。
7.1 SAMiRNAナノ粒子の細胞内処理方法
アンフィレグリンの発現を抑制するSAMiRNA発掘のために、ヒト由来肺癌細胞株であるA549を用いたが、A549細胞株は10%ウシ胎児血清(Hyclone,US)及び1%ペニシリン-ストレプトマイシン(Hyclone,US)が含まれたGibcoTMHam’s F-12K(Kaighn’s)培地(Thermo,US)を用いて37℃、5% CO2条件で培養した。同上の培地を用いてA549細胞株を12ウェルプレート(Costar,US)に8×104cells/wellの条件で分注し、翌日、前記実施例3.1で脱塩蒸留水(deionized distilled water)で均質化したSAMiRNAを1X DPBSで希釈し、細胞に200nM、600nM又は1200nMとなるように処理した。SAMiRNAの処理は、12時間ごとに1回ずつ処理する条件で総4回処理し、37℃、5% CO2条件で培養した。
7.2 ヒト(Human)アンフィレグリンmRNA発現抑制効能分析を用いたSAMiRNAスクリーニング
実施例7-1でSAMiRNA処理された細胞株から全RNAを抽出してcDNAを製造した後、実時間PCR(real-time PCR)を用いてアンフィレグリン遺伝子のmRNA発現量を定量した。
7-2-1 SAMiRNA処理された細胞からRNA分離及びcDNA製造
RNA抽出キット(AccuPrep Cell total RNA extraction kit,BIONEER、韓国)を用いて、前記実施例7-1でSAMiRNA処理された細胞株から全RNAを抽出し、抽出されたRNAはRNA逆転写酵素(AccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20,Bioneer、韓国)を用いて、下記のような方法でcDNAを製造した。具体的に、0.25mlエッペンドルフチューブに含まれたAccuPower¢ c RocketScriptTMCycle RT Premix with oligo(dT)20(Bioneer、韓国)に1チューブ当たりに抽出された1μgずつのRNAを入れ、総体積が20μlとなるように、DEPC(diethyl pyrocarbonate)処理された蒸留水を添加した。これを遺伝子増幅器(MyGenieTM96 Gradient Thermal Block,BIONEER、韓国)で37℃で30秒間RNAとプライマーを混成化し、48℃で4分間cDNAを製造する2つの過程を12回反復した後、95℃で5分間酵素を不活性化させて増幅反応を終了した。
7-2-2 ヒト(Human)アンフィレグリンmRNAの相対定量分析
実施例7-2-1で製造されたcDNAを鋳型とし、SYBRグリーン方式の実時間qPCRを用いて、SAMiRNA対照試料に対比するアンフィレグリンmRNAの相対的発現率を、下記のような方法で分析した。96ウェルプレートの各ウェルに前記実施例7-2-1で製造されたcDNAを蒸留水で5倍希釈し、アンフィレグリンmRNA発現量分析のために希釈されたcDNA 3μlとAccuPower(R) 2X GreenStarTM qPCR MasterMix(BIONEER、韓国)を25μl、蒸留水19μl、アンフィレグリンqPCRプライマー(配列番号17及び18(表5);10pmole/μlそれぞれ、BIONEER、韓国)を3μl入れて混合液を作った。一方、アンフィレグリンmRNAの発現量を正規化するために、ハウスキーピング遺伝子(housekeeping gene、以下、HK遺伝子)であるGAPDH(Glyceraldehyde-3-Phosphate Dehydrogenase)を標準遺伝子とした。前記混合液が入っている96ウェルプレートをExicyclerTMReal-Time Quantitative Thermal Block(BIONEER、韓国)を用いて下記のような反応を行った:95℃で15分間反応して酵素の活性化及びcDNAの二次構造をなくした後、94℃で30秒変性(denaturing)、58℃で30秒アニーリング(annealing)、72℃で30秒延長(extension)、SYBRグリーンスキャン(SYBR green scan)の4つの過程を42回繰り返し行い、72℃で3分間最終延長を行った後、55℃で1分間温度を維持し、55℃から95℃までメルティングカーブ(melting curve)を分析した。
PCRが終了した後、それぞれ得たターゲット遺伝子のCt(threshold cycle)値は、GAPDH遺伝子によって補正されたターゲット遺伝子のCt値を求めた後、遺伝子発現阻害を起こさないコントロール配列であるSAMiRNA(SAMiCONT)が処理された実験群を対照群としてΔCt値の差を求めた。前記ΔCt値と計算式2(-ΔCt)×100を用いて、アンフィレグリン特異的SAMiRNAが処理された細胞のターゲット遺伝子の発現量を定量した。
二重鎖オリゴDNA/RNAハイブリッド及びRNA/RNAハイブリッドの中から高効率のSAMiRNAを選別するために、最終濃度200nM、600nM、1200nMの濃度でアンフィレグリンmRNA発現量が対照群に比して発現量の減少効率が最も良い配列、すなわち、配列番号12の配列をセンス鎖として有するDNA/RNAハイブリッドであるSAMiRNA(90%以上の遺伝子発現を抑制)を最終選別した。
図8に示すように、選別された3種のアンフィレグリン特異的SAMiRNAを含む二重鎖オリゴDNA/RNAハイブリッド及びRNA/RNAハイブリッドから、アンフィレグリン遺伝子発現を最も効果的に抑制するDNA/RNAハイブリッドSAMiRNA 12種を最終選別した。
実施例8.マウス(Mouse)アンフィレグリンをターゲットとしてRNAiを誘導するSAMiRNAナノ粒子の高速大量スクリーニング(HTS)
siRNA治療剤の場合、異なる種(Strain)に共通適用可能な最適配列発掘が容易でない。この場合、治療効果を分析する動物モデル(in vivo efficacy test確認)特異的なsiRNA配列(surrogate sequ ence;mouse gene-specific siRNA)をデザインし、当該遺伝子の発現阻害による薬剤学的効能(pharmacologically active)及び当該遺伝子発現阻害による毒性部分を検証するように、米FDAガイドラインが存在する(presentation by Robert T.Dorsam Ph.D.Pharmacology/Toxicology Reviewer,FDA/CDER)。
既存アルゴリズム基盤siRNAデザインプログラム(自社保有のTurbo-si-designer)を用いて既存に発掘したスクリーニングを補完し、SAMiRNA基盤配列発掘高速大量スクリーニング(SAMiRNA based siRNA sequence high-throughput screening)を行った。ターゲット遺伝子全体を対象に19mer長さのsiRNAを1塩基スライディングウィンドウスキャニング(1 base sliding window scanning)して(前記ヒトアンフィレグリンターゲットスクリーニングと同じ方法)、可能なマウスアンフィレグリン遺伝子(NM_009704.4)完全転写(full transcript)配列を対象に総1190個の候補siRNA配列を生成し、相同性フィルタリング(blast sequence homology filtering)を行って、不要な他の遺伝子発現に影響を与える候補配列を除去し、最終的に選別された237種のSAMiRNAを合成後、マウス由来NIH3T3細胞株に10% FBSが存在する細胞培養条件で1uM濃度で処理し、in vitro発現阻害効果を表8(qPCR用プライマー配列情報)のプライマーを用いて1次スクリーニングした。(図9)
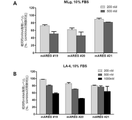
その後、追加スクリーニングによって前記NIH3T3細胞株から選別された2種の配列(配列番号19、20)及び先行マイルストーン研究によって発掘した配列番号21のマウスSAMiRNA-アンフィレグリンに対して、マウス肺線維芽細胞の細胞株であるMLg細胞株において、10% FBSが存在する細胞培養条件で200nM及び500nM処理濃度で処理して追加スクリーニングを行った結果、配列番号20が最も優れた発現阻害効果を示すことを確認した。(図10a)
さらに、マウス肺上皮細胞株(Lung epithelial cell line)であるLA-4細胞株に、選別された2種配列(配列番号19、20)及び先行マイルストーン研究によって発掘した配列番号21のマウスSAMiRNA-アンフィレグリンを、10% FBSが存在する細胞培養条件で200nM、500nM及び1000nM処理濃度で処理して追加発現阻害効果を確認した結果、配列番号20から最も優れた発現阻害効果を再確認した。(図10のb)
図10に示すように、マウスアンフィレグリンを標的とする237種のSAMiRNAから、アンフィレグリン遺伝子発現を最も効果的に抑制するSAMiRNA 2種を最終選別した。当該SA MiRNAの配列情報は、下記表9の通りである。
実施例9.シリカ(Silica)誘発肺線維症マウスモデルで微静脈投与法によるSAMiRNA-mAREGの効能検証
シリカ(Silicon dioxide,SIGMA、韓国)で誘発した肺線維化症動物モデルにおけるSAMi-mAREGに効能分析のために実施した。実験をするために、7週齢のマウスを入手して1週間純化させた。モデルを誘発するためにシリカ(3mg)を溶かし、マウスに器官内注射方法を用いた。誘発3日後、異常症状を示さないマウスを選別して正常群、実験群に生理食塩液(PBS)を投与した群、実験群にSAMiRNA-Controlを投与した群、実験群(SAMi-mAREG#20)に1mg/kgと5mg/kgを各2日間隔で3回投与した。そして、モデル誘発後14日目にマウスを犠牲した。
9-1.シリカ(Silica)で誘導された肺線維化症動物モデルにおけるSAMiRNAに対する遺伝子発現分析
犠牲させたマウスの肺組織を得てホモゲナイザーを用いて組織を粉砕した。RNA抽出キット(AccuPrep Cell total RNA extraction kit,BIONEER、韓国)を用いて、前記実施例7-1でSAMiRNA処理された細胞株から全RNAを抽出し、抽出されたRNAは、RNA逆転写酵素(AccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20,Bioneer、韓国)を用いて、下記のような方法でcDNAを製造した。製造されたcDNAを鋳型とし、SYBRグリーン方式の実時間qPCRを用いて各群の全mRNAの相対的発現率を下記のような方法で分析した。96ウェルプレートの各ウェルに製造されたcDNAを蒸留水で5倍希釈し、アンフィレグリンmRNA発現量分析のために、希釈されたcDNA 3μlとAccuPower(R) 2X GreenStarTM qPCR MasterMix(BIONEER、韓国)を25μl、蒸留水19μl、アンフィレグリンqPCRプライマー(配列番号24及び25(表8);10pmole/μlそれぞれ、BIONEER、韓国)を3μl入れて混合液を作った。一方、アンフィレグリン、フィブロネクチン及びコラーゲン3α1 mRNAの発現量を正規化するために、ハウスキーピング遺伝子(housekeeping gene、以下、HK遺伝子)であるRPL13Aを標準遺伝子とした。
PCRが終了した後、それぞれ得たターゲット遺伝子のCt(thre shold サイクル)値は、RPL13A遺伝子によって補正されたターゲット遺伝子のCt値を求めた後、各群を比較してΔCt値の差を求めた。前記ΔCt値と計算式2(-ΔCt)×100を用いて、アンフィレグリン、フィブロネクチン及びコラーゲン3α1遺伝子の発現量を相対定量した。
その結果、アンフィレグリンの場合、シリカモデルに生理食塩液を処理した群とシリカモデル群にSAMiRNA-Control投与した群に比して、SAM iRNA-AREG 1mg/kg及びSAMiRNA-AREG 5mg/kgを処理した群において減少効果を確認した。また、フィブロネクチンとコラーゲン3α1の場合、シリカモデルに生理食塩液を処理した群とシリカモデル群にSAMiRNA-Control投与した群に比して、SAMiRNA-AREG 1mg/kg及びSAMiRNA-AREG 5mg/kgを投与した群から濃度依存的に減少効果を確認した。
9-2.シリカで誘発した肺線維化症動物モデルにおけるSAMiRNAに対する組織病理学的分析
シリカモデルに対するSAMiRNA-AREGが細胞の基質成分の発現に影響を与えるかどうかを検証するために免疫組織化学染色(immunohistochemical staining)を行った。動物モデルを各群別に犠牲させ、組織固定、水洗、脱水、透明、浸透、包埋及び薄切過程を通じてパラフィン切片を作製した。パラフィン切片を薄切機で薄く切って組織をスライドに接着させる。肺組織を病理学的に観察するために、ヘマトキシリン及びエオシン(H&E)染色を行い、コラーゲン1の発現程度を確認するために、マッソントリクローム(Masson’s trichrome)染色を行った。そして、アンフィレグリン発現程度を分析するために免疫組織化学染色を行った。
ヘマトキシリン及びエオシン染色から、正常モデルの肺組織に比べて、シリカで誘発した肺組織に損傷が起きたことを分かった。しかし、SAMiRNA-AREGを投与したマウスの肺組織では、正常モデルの肺組織のように肺組織の損傷が相対的に少なく起きたことが分かった。そして、線維化が発生した程度を確認するために、マッソントリクローム染色を行った。シリカモデル群に生理食塩液を投与した群、SAMiRNA-Controlを投与した群に比して、SAMiRNA-AREGを投与した群では、肺組織間の間質(interstitium)で線維化の発現程度が減少したことを確認した。また、アンフィレグリンに対する免疫組織化学染色から、シリカで誘発した動物群では細胞間の間質において多く発現していることが分かった。しかし、SAMiRNA-AREGを投与した群では、肺組織間の間質にAREGの発現が減少したことが確認できた。
結論的に、シリカ誘発肺線維症マウスモデルに、マウスSAMiRNA-AREGを、微静脈投与によって3回(10日、12日、14日)、1mg/kg及び5mg/kg投与濃度で反復投与し、肺組織からターゲット遺伝子及び線維化マーカー遺伝子発現阻害効能及びH&E染色及びマッソントリクローム染色を行った。ターゲット遺伝子であるアンフィレグリン及び線維化マーカー遺伝子であるコラーゲン及びフィブロネクチンの発現を確認した結果、シリカ誘発において発現が増加し、SAMiRNA処理によって発現抑制効能が濃度依存的に確認された。また、組織染色の結果、シリカ誘発マウスにPBS又は対照群であるSAMiRNA-Controlを処理した群では、肺組織への細胞の浸潤(infiltration)及びコラーゲンが増加したが、実験群であるSAMiRNA-AREG処理によって、濃度依存的に細胞浸潤(Cellular infiltration)及びコラーゲンが、対照群であるDPBSを処理した群と比較される程度に著しく減少することを確認した。(図11)
そして、シリカ誘発肺線維化症モデルにおけるAREG免疫組織化学染色(Immunohistochemistry staining)を行った。図11で、シリカ誘発肺線維症マウスモデルにSAMiRNA-AREG 1mg/kg、5mg/kgを投与した組織においてAREG発現程度をIHC染色を用いて確認した。シリカ+PBS、シリカ+SAMi-Contを投与したモデルにおいて、正常肺組織に比べてAREGの発現が間質位置(interstitial site)に多く増加したことを確認した。一方、SAMiRNA-AREGを1mg/kgと5mg/kg投与した組織におけるAREG発現を確認した結果、シリカ誘発した肺組織に比べてAREG発現が顕著に減少したことが確認できた(図11)。
実施例10.ブレオマイシン(Bleomycin)誘発肺線維症マウスモデルで微静脈投与法によるSAMiRNA-mAREGの効能検証
ブレオマイシン誘発肺線維症マウスモデルに、マウスSAMiRNA-AREG(S AMi-mAREG#20)を、微静脈投与によって3回(8日、1日、12日)、5mg/kg濃度で反復投与してSircol assay分析を行った結果、対照群であるSAM iRNA-Contに比して、実験群であるSAMiRNA-AREG投与した群においてコラーゲンタンパク質の量が>40%減少することを確認した。また、同一実験群の肺組織からRNAを抽出し、ターゲット遺伝子であるアンフィレグリン及び線維化マーカー遺伝子であるコラーゲンの発現阻害効能を確認した結果、対照群に比して発現阻害効能が確認された。試験物質の場合、既存配列に比べて新規発掘した配列番号20に対する効能が同等以上と確認された。肺組織からH&E染色及びコラーゲン特異的染色のためのマッソントリクローム染色を行った。組織染色の結果、ブレオマイシン誘発マウスにPBS又は対照群であるSAMiRNA-Controlを処理した群では、肺組織への細胞の浸潤(infiltration)が増加し、コラーゲン蓄積による染色が増加することを確認した。実験群であるSAMiRNA-AREG処理によって細胞浸潤及びコラーゲン蓄積が顕著に減少することを確認した(図12)。組織染色方法及びターゲット遺伝子発現確認方法は、実施例8と同一に行った。
実施例11.SAMiRNA-AREGを投与したマウスに発生したUUO(Unilateral ureteral obstruction)によって誘導された腎臓線維症(renal fibrosis)に対する効果確認
UUO手術で誘発した腎臓線維症動物モデルにおけるSAMiRNA-AREG(SAMi-mAREG#20)に効能分析をするために行った。まず、腎臓線維症動物モデルを作製するために、アイフラン液(ハナ製薬、韓国)でマウスを呼吸麻酔させた。皮膚と腹膜を切開して4-0シルクで左側腎臓の尿管を2カ所で縛り、尿路感染を防ぐために2カ所の中間部位を切った。そして、右側部位は、同じ方法で手術したが、尿管は縛らなかった。同様に、正常群のモデル群も開腹して左側腎臓の尿管を確認したが、縛らなかった。そして、感染を防ぐために、腹膜と皮膚に縫合を施した。モデル誘発6時間後にSAMiRNA-AREG 1mg/kg、5mg/kgを、一番目の投与を行った。24時間後に2番目の投与を行った。最終2回投与を行い、最後投与後24時間後に動物を犠牲させた。動物モデルの群は、正常群、UUOモデル群に生理食塩液を投与した群、UUOモデル群にSAMiRNA-AREG 1mg/kg、5mg/kgを投与した群の合計4群である。
11-1. UUO(Unilateral ureteral obstruction)によって誘導された腎臓線維症(renal fibrosis)におけるSAMiRNAに対する遺伝子発現分析
犠牲させたマウスの肺組織を得てホモゲナイザーを用いて組織を粉砕した。RNA抽出キット(AccuPrep Cell total RNA extraction kit,BIONEER、韓国)を用いて、前記実施例7-1でSAMiRNA処理された細胞株から全RNAを抽出し、抽出されたRNAはRNA逆転写酵素(AccuPower(R)RocketScriptTMCycle RT Premix with oligo(dT)20,Bioneer、韓国)を用いて、下記のような方法でcDNAを製造した。製造されたcDNAを鋳型とし、SYBRグリーン方式の実時間qPCRを用いて各群の全mRNAの相対的発現率を、下記のような方法で分析した。96ウェルプレートの各ウェルに製造されたcDNAを蒸留水で5倍希釈し、アンフィレグリンmRNA及び線維化標識子の発現量分析のために、希釈されたcDNA 3μlとAccuPower(R) 2X GreenStarTM qPCR MasterMix(BIONEER、韓国)を25μl、蒸留水19μl、アンフィレグリン及び線維化標識子のqPCRプライマー(配列番号24及び25(表8);10pmole/μlそれぞれ、BIONEER、韓国)を3μl入れて混合液を作った。一方、形質転換生長因子1、アンフィレグリン、フィブロネクチン、コラーゲン1、平滑筋アクチン及びコラーゲン3α1 mRNAの発現量を正規化するために、ハウスキーピング遺伝子(housekeeping gene、以下、HK遺伝子)であるRPL13Aを標準遺伝子とした。また、炎症性因子の効能を検証するためにCCR2遺伝子も共に確認した。
PCRが終了した後、それぞれ得たターゲット遺伝子のCt(threshold cycle)値は、RPL13A遺伝子を用いて補正されたターゲット遺伝子のCt値を求めた後、各群を比較してΔCt値の差を求めた。前記ΔCt値と計算式2(-ΔCt)×100を用いて形質転換生長因子1、アンフィレグリン、フィブロネクチン、コラーゲン1、コラーゲン3α1、及びCCR2遺伝子の発現量を定量した。
その結果、アンフィレグリンの場合、正常群に比してUUOモデル群に生理食塩液を投与した群では、60倍以上増加した。しかし、SAMiRNA-AREG 1mg/kg及びSAMiRNA-AREG 5mg/kgを処理した群ではアンフィレグリンの遺伝子発現が4倍減少したことを確認した。また、フィブロネクチンと形質転換生長因子1の場合、UUOモデル群に比べてSAMiRNA-AREG 1mg/kg及びSAMiRNA-AREG 5mg/kgを投与した群において濃度依存的に減少したことを確認した。また、コラーゲン3α1、コラーゲン1、及び平滑筋アクチンの場合、SAMiRNA-AREG 1mg/kgを投与した群ではUUOモデルに比して減少する傾向はあった。一方、SAMiRNA-AREG 5mg/kgを投与した群では、SAMiRNA-AREG 1mg/kg投与した群に比べて優れた減少効果を確認した。また、CCR2は、UUOモデルでは約6倍以上増加したが、SAMiRNA-AREGを投与した群では減少効果を確認した。
以上、本発明内容における特定の部分を詳細に記述したが、当業界における通常の知識を有する者にとって、このような具体的技術は単に好ましい実施態様であるだけで、これによって本発明の範囲が制限されるものでない点は明らかであろう。したがって、本発明の実質的な範囲は、添付する請求項とそれらの等価物によって定義されるといえよう。