JP7075170B2 - 延長された単一ガイドrna及びその用途 - Google Patents
延長された単一ガイドrna及びその用途 Download PDFInfo
- Publication number
- JP7075170B2 JP7075170B2 JP2020561562A JP2020561562A JP7075170B2 JP 7075170 B2 JP7075170 B2 JP 7075170B2 JP 2020561562 A JP2020561562 A JP 2020561562A JP 2020561562 A JP2020561562 A JP 2020561562A JP 7075170 B2 JP7075170 B2 JP 7075170B2
- Authority
- JP
- Japan
- Prior art keywords
- base
- sequence
- sgrna
- target
- gene
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 108020005004 Guide RNA Proteins 0.000 title claims description 75
- 108090000623 proteins and genes Proteins 0.000 claims description 116
- 108091033409 CRISPR Proteins 0.000 claims description 47
- 239000002773 nucleotide Substances 0.000 claims description 44
- 125000003729 nucleotide group Chemical group 0.000 claims description 42
- 210000004027 cell Anatomy 0.000 claims description 40
- 239000000203 mixture Substances 0.000 claims description 40
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical class O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 claims description 34
- 230000000295 complement effect Effects 0.000 claims description 27
- 238000000034 method Methods 0.000 claims description 27
- 210000001161 mammalian embryo Anatomy 0.000 claims description 22
- 108010008532 Deoxyribonuclease I Proteins 0.000 claims description 13
- 102000007260 Deoxyribonuclease I Human genes 0.000 claims description 12
- 101710172430 Uracil-DNA glycosylase inhibitor Proteins 0.000 claims description 11
- 230000001915 proofreading effect Effects 0.000 claims description 6
- 101710199622 tRNA-specific adenosine deaminase Proteins 0.000 claims description 6
- 230000004913 activation Effects 0.000 claims description 3
- 230000002950 deficient Effects 0.000 claims description 3
- 210000005260 human cell Anatomy 0.000 claims description 3
- 102000007592 Apolipoproteins Human genes 0.000 claims description 2
- 108010071619 Apolipoproteins Proteins 0.000 claims description 2
- 210000004102 animal cell Anatomy 0.000 claims description 2
- 230000030648 nucleus localization Effects 0.000 claims description 2
- 101710163270 Nuclease Proteins 0.000 description 82
- 230000000694 effects Effects 0.000 description 69
- 230000035772 mutation Effects 0.000 description 66
- 108020004414 DNA Proteins 0.000 description 48
- 102000004169 proteins and genes Human genes 0.000 description 44
- 235000018102 proteins Nutrition 0.000 description 42
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 40
- 238000006467 substitution reaction Methods 0.000 description 35
- 108091079001 CRISPR RNA Proteins 0.000 description 23
- 229940104302 cytosine Drugs 0.000 description 20
- 150000007523 nucleic acids Chemical group 0.000 description 20
- 150000001413 amino acids Chemical class 0.000 description 19
- 108020004999 messenger RNA Proteins 0.000 description 19
- 235000001014 amino acid Nutrition 0.000 description 18
- 238000010457 gene scissor Methods 0.000 description 18
- 239000013598 vector Substances 0.000 description 18
- 241000196324 Embryophyta Species 0.000 description 17
- 210000003527 eukaryotic cell Anatomy 0.000 description 17
- 108091028113 Trans-activating crRNA Proteins 0.000 description 16
- 229940024606 amino acid Drugs 0.000 description 16
- 241001465754 Metazoa Species 0.000 description 15
- 102000053602 DNA Human genes 0.000 description 14
- 230000037429 base substitution Effects 0.000 description 13
- 102000004533 Endonucleases Human genes 0.000 description 12
- 108010042407 Endonucleases Proteins 0.000 description 12
- 108091028043 Nucleic acid sequence Proteins 0.000 description 12
- 241000124008 Mammalia Species 0.000 description 11
- 108020004511 Recombinant DNA Proteins 0.000 description 11
- 102000039446 nucleic acids Human genes 0.000 description 11
- 108020004707 nucleic acids Proteins 0.000 description 11
- 108020004705 Codon Proteins 0.000 description 10
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 10
- 238000012350 deep sequencing Methods 0.000 description 10
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 10
- 230000008859 change Effects 0.000 description 9
- 238000000338 in vitro Methods 0.000 description 9
- 108700028369 Alleles Proteins 0.000 description 8
- 238000006243 chemical reaction Methods 0.000 description 8
- 230000008685 targeting Effects 0.000 description 8
- 244000046052 Phaseolus vulgaris Species 0.000 description 7
- 235000010627 Phaseolus vulgaris Nutrition 0.000 description 7
- 108020004682 Single-Stranded DNA Proteins 0.000 description 7
- 241000193996 Streptococcus pyogenes Species 0.000 description 7
- CKLJMWTZIZZHCS-REOHCLBHSA-N aspartic acid group Chemical group N[C@@H](CC(=O)O)C(=O)O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 7
- 230000009615 deamination Effects 0.000 description 7
- 238000006481 deamination reaction Methods 0.000 description 7
- 238000004519 manufacturing process Methods 0.000 description 7
- 241000699666 Mus <mouse, genus> Species 0.000 description 6
- DRTQHJPVMGBUCF-XVFCMESISA-N Uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-XVFCMESISA-N 0.000 description 6
- 235000003704 aspartic acid Nutrition 0.000 description 6
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 6
- 210000002308 embryonic cell Anatomy 0.000 description 6
- 244000005700 microbiome Species 0.000 description 6
- 210000001938 protoplast Anatomy 0.000 description 6
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 5
- 108091027544 Subgenomic mRNA Proteins 0.000 description 5
- 229960005305 adenosine Drugs 0.000 description 5
- 229920002477 rna polymer Polymers 0.000 description 5
- 238000012360 testing method Methods 0.000 description 5
- 229940113082 thymine Drugs 0.000 description 5
- UHDGCWIWMRVCDJ-UHFFFAOYSA-N 1-beta-D-Xylofuranosyl-NH-Cytosine Natural products O=C1N=C(N)C=CN1C1C(O)C(O)C(CO)O1 UHDGCWIWMRVCDJ-UHFFFAOYSA-N 0.000 description 4
- UHDGCWIWMRVCDJ-PSQAKQOGSA-N Cytidine Natural products O=C1N=C(N)C=CN1[C@@H]1[C@@H](O)[C@@H](O)[C@H](CO)O1 UHDGCWIWMRVCDJ-PSQAKQOGSA-N 0.000 description 4
- 108010055325 EphB3 Receptor Proteins 0.000 description 4
- 102100031982 Ephrin type-B receptor 3 Human genes 0.000 description 4
- 241000699670 Mus sp. Species 0.000 description 4
- 108091034117 Oligonucleotide Proteins 0.000 description 4
- 238000004458 analytical method Methods 0.000 description 4
- 238000003776 cleavage reaction Methods 0.000 description 4
- UHDGCWIWMRVCDJ-ZAKLUEHWSA-N cytidine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O1 UHDGCWIWMRVCDJ-ZAKLUEHWSA-N 0.000 description 4
- 210000002257 embryonic structure Anatomy 0.000 description 4
- 230000002068 genetic effect Effects 0.000 description 4
- 238000001727 in vivo Methods 0.000 description 4
- 238000010428 oil painting Methods 0.000 description 4
- 210000003101 oviduct Anatomy 0.000 description 4
- 235000012736 patent blue V Nutrition 0.000 description 4
- 230000007017 scission Effects 0.000 description 4
- 239000004475 Arginine Substances 0.000 description 3
- 102000004190 Enzymes Human genes 0.000 description 3
- 108090000790 Enzymes Proteins 0.000 description 3
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- 241000605861 Prevotella Species 0.000 description 3
- 102000004389 Ribonucleoproteins Human genes 0.000 description 3
- 108010081734 Ribonucleoproteins Proteins 0.000 description 3
- 241000194017 Streptococcus Species 0.000 description 3
- 235000004279 alanine Nutrition 0.000 description 3
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 3
- DRTQHJPVMGBUCF-PSQAKQOGSA-N beta-L-uridine Natural products O[C@H]1[C@@H](O)[C@H](CO)O[C@@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-PSQAKQOGSA-N 0.000 description 3
- 210000002459 blastocyst Anatomy 0.000 description 3
- 230000003197 catalytic effect Effects 0.000 description 3
- 238000010586 diagram Methods 0.000 description 3
- 235000013922 glutamic acid Nutrition 0.000 description 3
- 239000004220 glutamic acid Substances 0.000 description 3
- 210000004962 mammalian cell Anatomy 0.000 description 3
- DRTQHJPVMGBUCF-UHFFFAOYSA-N uracil arabinoside Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-UHFFFAOYSA-N 0.000 description 3
- 229940045145 uridine Drugs 0.000 description 3
- 101710169336 5'-deoxyadenosine deaminase Proteins 0.000 description 2
- 101150001232 ALS gene Proteins 0.000 description 2
- 241000604451 Acidaminococcus Species 0.000 description 2
- 229930024421 Adenine Natural products 0.000 description 2
- GFFGJBXGBJISGV-UHFFFAOYSA-N Adenine Chemical compound NC1=NC=NC2=C1N=CN2 GFFGJBXGBJISGV-UHFFFAOYSA-N 0.000 description 2
- 102000055025 Adenosine deaminases Human genes 0.000 description 2
- 241000894006 Bacteria Species 0.000 description 2
- 108010051109 Cell-Penetrating Peptides Proteins 0.000 description 2
- 102000020313 Cell-Penetrating Peptides Human genes 0.000 description 2
- 102100026846 Cytidine deaminase Human genes 0.000 description 2
- 108010031325 Cytidine deaminase Proteins 0.000 description 2
- 102000000311 Cytosine Deaminase Human genes 0.000 description 2
- 108010080611 Cytosine Deaminase Proteins 0.000 description 2
- 241000589601 Francisella Species 0.000 description 2
- 108010070675 Glutathione transferase Proteins 0.000 description 2
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 2
- 102100029100 Hematopoietic prostaglandin D synthase Human genes 0.000 description 2
- 241000282412 Homo Species 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- 241000904817 Lachnospiraceae bacterium Species 0.000 description 2
- 101710175625 Maltose/maltodextrin-binding periplasmic protein Proteins 0.000 description 2
- 108091092724 Noncoding DNA Proteins 0.000 description 2
- 241000606860 Pasteurella Species 0.000 description 2
- 241000288906 Primates Species 0.000 description 2
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 2
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 2
- 241000283984 Rodentia Species 0.000 description 2
- 241000191967 Staphylococcus aureus Species 0.000 description 2
- 241000194022 Streptococcus sp. Species 0.000 description 2
- 108020005038 Terminator Codon Proteins 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 239000004473 Threonine Substances 0.000 description 2
- 108060008724 Tyrosinase Proteins 0.000 description 2
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 2
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 2
- 229960000643 adenine Drugs 0.000 description 2
- 125000003277 amino group Chemical group 0.000 description 2
- 238000012217 deletion Methods 0.000 description 2
- 230000037430 deletion Effects 0.000 description 2
- 230000005782 double-strand break Effects 0.000 description 2
- 238000002474 experimental method Methods 0.000 description 2
- 230000004927 fusion Effects 0.000 description 2
- 102000037865 fusion proteins Human genes 0.000 description 2
- 108020001507 fusion proteins Proteins 0.000 description 2
- 238000010362 genome editing Methods 0.000 description 2
- 210000004602 germ cell Anatomy 0.000 description 2
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 2
- FDGQSTZJBFJUBT-UHFFFAOYSA-N hypoxanthine Chemical compound O=C1NC=NC2=C1NC=N2 FDGQSTZJBFJUBT-UHFFFAOYSA-N 0.000 description 2
- 238000000520 microinjection Methods 0.000 description 2
- 238000010369 molecular cloning Methods 0.000 description 2
- 230000006780 non-homologous end joining Effects 0.000 description 2
- 239000013612 plasmid Substances 0.000 description 2
- BASFCYQUMIYNBI-UHFFFAOYSA-N platinum Chemical group [Pt] BASFCYQUMIYNBI-UHFFFAOYSA-N 0.000 description 2
- 108090000765 processed proteins & peptides Proteins 0.000 description 2
- 238000010188 recombinant method Methods 0.000 description 2
- 230000006798 recombination Effects 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 238000010189 synthetic method Methods 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 241001515965 unidentified phage Species 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- 235000014393 valine Nutrition 0.000 description 2
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 1
- 102000012758 APOBEC-1 Deaminase Human genes 0.000 description 1
- 108010079649 APOBEC-1 Deaminase Proteins 0.000 description 1
- 241000093740 Acidaminococcus sp. Species 0.000 description 1
- 206010001557 Albinism Diseases 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 240000002791 Brassica napus Species 0.000 description 1
- 235000011293 Brassica napus Nutrition 0.000 description 1
- 238000010356 CRISPR-Cas9 genome editing Methods 0.000 description 1
- 241001502303 Candidatus Methanoplasma Species 0.000 description 1
- 241001040999 Candidatus Methanoplasma termitum Species 0.000 description 1
- 241000223282 Candidatus Peregrinibacteria Species 0.000 description 1
- 108091026890 Coding region Proteins 0.000 description 1
- 241000195493 Cryptophyta Species 0.000 description 1
- 102220605874 Cytosolic arginine sensor for mTORC1 subunit 2_D10A_mutation Human genes 0.000 description 1
- 230000033616 DNA repair Effects 0.000 description 1
- 230000007018 DNA scission Effects 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 241000186394 Eubacterium Species 0.000 description 1
- 241000589599 Francisella tularensis subsp. novicida Species 0.000 description 1
- 108700007698 Genetic Terminator Regions Proteins 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 244000068988 Glycine max Species 0.000 description 1
- 235000010469 Glycine max Nutrition 0.000 description 1
- 102000006771 Gonadotropins Human genes 0.000 description 1
- 108010086677 Gonadotropins Proteins 0.000 description 1
- 206010020649 Hyperkeratosis Diseases 0.000 description 1
- UGQMRVRMYYASKQ-UHFFFAOYSA-N Hypoxanthine nucleoside Natural products OC1C(O)C(CO)OC1N1C(NC=NC2=O)=C2N=C1 UGQMRVRMYYASKQ-UHFFFAOYSA-N 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 1
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 1
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- 241001134638 Lachnospira Species 0.000 description 1
- 241001148627 Leptospira inadai Species 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 1
- 239000004472 Lysine Substances 0.000 description 1
- 241000542065 Moraxella bovoculi Species 0.000 description 1
- 108010077850 Nuclear Localization Signals Proteins 0.000 description 1
- 241000605894 Porphyromonas Species 0.000 description 1
- 241001135241 Porphyromonas macacae Species 0.000 description 1
- 241001135219 Prevotella disiens Species 0.000 description 1
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 1
- 101710149951 Protein Tat Proteins 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 102000003425 Tyrosinase Human genes 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 238000007792 addition Methods 0.000 description 1
- 150000003838 adenosines Chemical class 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 229940009098 aspartate Drugs 0.000 description 1
- CKLJMWTZIZZHCS-REOHCLBHSA-L aspartate group Chemical group N[C@@H](CC(=O)[O-])C(=O)[O-] CKLJMWTZIZZHCS-REOHCLBHSA-L 0.000 description 1
- 230000005540 biological transmission Effects 0.000 description 1
- 210000000170 cell membrane Anatomy 0.000 description 1
- 239000002299 complementary DNA Substances 0.000 description 1
- 238000012790 confirmation Methods 0.000 description 1
- 238000005520 cutting process Methods 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- -1 for example Proteins 0.000 description 1
- 239000012634 fragment Substances 0.000 description 1
- 125000000291 glutamic acid group Chemical group N[C@@H](CCC(O)=O)C(=O)* 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 210000002149 gonad Anatomy 0.000 description 1
- 239000002622 gonadotropin Substances 0.000 description 1
- 230000002363 herbicidal effect Effects 0.000 description 1
- 239000004009 herbicide Substances 0.000 description 1
- 125000000487 histidyl group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C([H])=N1 0.000 description 1
- 230000006801 homologous recombination Effects 0.000 description 1
- 238000002744 homologous recombination Methods 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 238000002347 injection Methods 0.000 description 1
- 239000007924 injection Substances 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 230000000813 microbial effect Effects 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 210000000633 nuclear envelope Anatomy 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 229910052697 platinum Inorganic materials 0.000 description 1
- 229920000724 poly(L-arginine) polymer Polymers 0.000 description 1
- 108010011110 polyarginine Proteins 0.000 description 1
- 230000023603 positive regulation of transcription initiation, DNA-dependent Effects 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 239000000047 product Substances 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 235000004252 protein component Nutrition 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- 230000001105 regulatory effect Effects 0.000 description 1
- 230000008439 repair process Effects 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 238000000926 separation method Methods 0.000 description 1
- 210000002966 serum Anatomy 0.000 description 1
- 230000005783 single-strand break Effects 0.000 description 1
- 125000006850 spacer group Chemical group 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 238000010361 transduction Methods 0.000 description 1
- 230000026683 transduction Effects 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 229940035893 uracil Drugs 0.000 description 1
- 108700026220 vif Genes Proteins 0.000 description 1
Images

















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/102—Mutagenizing nucleic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
- C12N15/902—Stable introduction of foreign DNA into chromosome using homologous recombination
- C12N15/907—Stable introduction of foreign DNA into chromosome using homologous recombination in mammalian cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
- C12N15/902—Stable introduction of foreign DNA into chromosome using homologous recombination
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Biomedical Technology (AREA)
- Zoology (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Molecular Biology (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Medicinal Chemistry (AREA)
- Cell Biology (AREA)
- Mycology (AREA)
- Crystallography & Structural Chemistry (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
Description
1)前記細胞をデアミナーゼコーディングDNA、RNA-ガイドヌクレアーゼコーディングDNA、及び延長されたガイドRNAコーディング遺伝子をそれぞれ含んだり、或いはこれらのうち2つ以上を含む組換えベクターで形質感染させたり、
2)前記細胞にデアミナーゼ、RNA-ガイドヌクレアーゼ、及び延長されたガイドRNA(例えば、デアミナーゼ、RNA-ガイドヌクレアーゼ、及び延長されたガイドRNAを含む混合物又は複合体形態のリボ核酸タンパク質)を直接注入したり、或いは
3)前記細胞にデアミナーゼコーディングmRNA、RNA-ガイドヌクレアーゼコーディングmRNA及びガイドRNAの混合物又はこれらのそれぞれを直接注入して行うことができる。
(1)D10、H840、又はD10+H840;
(2)D1135、R1335、T1337、又はD1135+R1335+T1337;又は
(3)(1)及び(2)残基の両方、でアミノ酸置換が起こったものであり得る。
(1)D10又はH840位置に突然変異(例えば、他のアミノ酸への置換)が導入されてエンドヌクレアーゼ活性を喪失しニッカーゼ活性を有する変形Cas9、又はストレプトコッカスピオゲネス(Streptococcus pyogenes)由来のCas9タンパク質に、D10及びH840位置にともに突然変異(例えば、他のアミノ酸への置換)が導入されてエンドヌクレアーゼ活性及びニッカーゼ活性をともに喪失した変形Cas9タンパク質;
(2)D1135、R1335及びT1337の一つ以上又はこれら全部に突然変異(例えば、他のアミノ酸への置換)が導入されて野生型と異なるPAM配列を認識する変形Cas9タンパク質;又は
(3)(1)及び(2)の突然変異がともに導入されてニッカーゼ活性を有し、野生型と異なるPAM配列を認識するか、エンドヌクレアーゼ活性及びニッカーゼ活性をともに喪失し、野生型と異なるPAM配列を認識する変形Cas9タンパク質であり得る。
標的配列とハイブリダイズ可能な部位(標的化配列)を含むCRISPR RNA(crRNA);
Casタンパク質、Cpf1などのようなヌクレアーゼと相互作用する部位を含むトランス活性化(trans-activating)crRNA(tracrRNA);及び
前記crRNA及びtracrRNAの主要部位(例えば、標的化配列を含むcrRNA部位及びヌクレアーゼと相互作用するtracrRNAの部位)が融合された形態の単一ガイドRNA(single guide RNA;sgRNA)からなる群から選ばれる1種以上であり得、
具体的にCRISPR RNA(crRNA)及びトランス活性化(trans-activating)crRNA(tracrRNA)を含む二重RNA(dual RNA)、又はcrRNA及びtracrRNAの主要部位を含む単一ガイドRNA(sgRNA)であり得る。
一例において、前記ガイドRNAは次の一般式1で表現され得る:
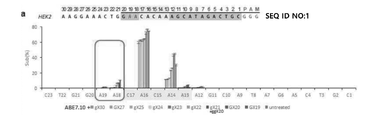
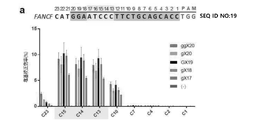
5’-(Ncas9)l-(GUUUUAGAGCUA)-(オリゴヌクレオチドリンカー)-(UAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC(配列番号60)-3’ (一般式1)
(Ncas9)lは、Ncas9が標的遺伝子(target gene)の標的部位(target site)上に結合(ハイブリダイズ)する標的化配列であり、その核酸配列は前記標的部位の配列によって決定され(すなわち、標的部位の配列とハイブリダイズ可能な配列である。)、lが前記標的化配列に含まれたヌクレオチド数を示すものであって、20であり得、5’-末端から最初の核酸は標的部位配列とマッチングであるグアニン(Gで表示;標的部位の対応位置がシトシン(C)である場合)又はミスマッチングであるグアニン(gで表示;標的部位の対応位置がシトシン(C)でない場合)であり;
前記オリゴヌクレオチドリンカーは、3~5個、例えば4個のヌクレオチドを含むものであり得、前記ヌクレオチドは互いに同一でも異なってもよく、A、U、C及びGからなる群からそれぞれ独立して選択され得る。
以下、実施例を用いて本発明をより詳細に説明する。これらの実施例は単に本発明をより具体的に説明するためのものであり、本発明の要旨によって本発明の範囲がこれらの実施例によって制限されないということは当業界における通常の知識を有する者にとって明らかであろう。
Claims (7)
- (i)デアミナーゼ又はこれをコードする遺伝子、
(ii)Cas9ニッカーゼ(nCas9)又は触媒活性欠乏Cas9(dCas9)又はこれをコードする遺伝子及び
(iii)標的配列とハイブリダイズ可能な延長されたガイドRNA又はこれをコードする遺伝子を含む塩基校正用組成物であって、
前記延長されたガイドRNAは、標的配列のPAM(Protospacer Adjacent Motif)から5’方向への20個のヌクレオチドの配列を有し、そして5’末端に標的配列と非相補的な2個のグアニン(G)をさらに含むことを特徴とする塩基校正用組成物。 - 前記デアミナーゼは、APOBEC1(apolipoprotein B editing complex 1)、AID(activation-induced deaminase)及びtadA(tRNA-specific adenosine deaminase)からなる群から選ばれることを特徴とする、請求項1に記載の組成物。
- ウラシルDNAグリコシラーゼ抑制剤(uracil DNA glycosylase inhibitor:UGI)又はこれをコードする遺伝子をさらに含むことを特徴とする、請求項1に記載の組成物。
- 核局在化配列(NLS)又はこれをコードする遺伝子をさらに含むことを特徴とする、請求項1に記載の組成物。
- 請求項1の塩基校正用組成物を非ヒト細胞に導入させる段階を含む塩基校正方法。
- 前記細胞はヒト細胞以外の動物細胞又は真核植物細胞であることを特徴とする、請求項5に記載の方法。
- 次の段階を含む突然変異が誘発されたヒト以外の哺乳動物又は真核植物の成体の作製方法;
(a)請求項1に記載の塩基校正用組成物を哺乳動物の胚又は真核植物の胚に導入させる段階;及び
(b)前記胚を成長させて成体を得る段階。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR20180008492 | 2018-01-23 | ||
| KR10-2018-0008492 | 2018-01-23 | ||
| PCT/KR2019/000962 WO2019147014A1 (ko) | 2018-01-23 | 2019-01-23 | 연장된 단일 가이드 rna 및 그 용도 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2021511824A JP2021511824A (ja) | 2021-05-13 |
| JP7075170B2 true JP7075170B2 (ja) | 2022-05-25 |
Family
ID=67394687
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020561562A Active JP7075170B2 (ja) | 2018-01-23 | 2019-01-23 | 延長された単一ガイドrna及びその用途 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20210032621A1 (ja) |
| EP (1) | EP3744844A4 (ja) |
| JP (1) | JP7075170B2 (ja) |
| KR (1) | KR20200103769A (ja) |
| CN (1) | CN111742051A (ja) |
| WO (1) | WO2019147014A1 (ja) |
Families Citing this family (18)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| IL294014B2 (en) | 2015-10-23 | 2024-07-01 | Harvard College | Nucleobase editors and their uses |
| IL308426A (en) | 2016-08-03 | 2024-01-01 | Harvard College | Adenosine nuclear base editors and their uses |
| US11661590B2 (en) | 2016-08-09 | 2023-05-30 | President And Fellows Of Harvard College | Programmable CAS9-recombinase fusion proteins and uses thereof |
| US11542509B2 (en) | 2016-08-24 | 2023-01-03 | President And Fellows Of Harvard College | Incorporation of unnatural amino acids into proteins using base editing |
| SG11201903089RA (en) | 2016-10-14 | 2019-05-30 | Harvard College | Aav delivery of nucleobase editors |
| WO2018119359A1 (en) | 2016-12-23 | 2018-06-28 | President And Fellows Of Harvard College | Editing of ccr5 receptor gene to protect against hiv infection |
| US11898179B2 (en) | 2017-03-09 | 2024-02-13 | President And Fellows Of Harvard College | Suppression of pain by gene editing |
| EP3592777A1 (en) | 2017-03-10 | 2020-01-15 | President and Fellows of Harvard College | Cytosine to guanine base editor |
| JP7191388B2 (ja) | 2017-03-23 | 2022-12-19 | プレジデント アンド フェローズ オブ ハーバード カレッジ | 核酸によってプログラム可能なdna結合蛋白質を含む核酸塩基編集因子 |
| US11560566B2 (en) | 2017-05-12 | 2023-01-24 | President And Fellows Of Harvard College | Aptazyme-embedded guide RNAs for use with CRISPR-Cas9 in genome editing and transcriptional activation |
| US11866726B2 (en) | 2017-07-14 | 2024-01-09 | Editas Medicine, Inc. | Systems and methods for targeted integration and genome editing and detection thereof using integrated priming sites |
| CN111801345A (zh) | 2017-07-28 | 2020-10-20 | 哈佛大学的校长及成员们 | 使用噬菌体辅助连续进化(pace)的进化碱基编辑器的方法和组合物 |
| US11319532B2 (en) | 2017-08-30 | 2022-05-03 | President And Fellows Of Harvard College | High efficiency base editors comprising Gam |
| CN111757937A (zh) | 2017-10-16 | 2020-10-09 | 布罗德研究所股份有限公司 | 腺苷碱基编辑器的用途 |
| WO2020191243A1 (en) | 2019-03-19 | 2020-09-24 | The Broad Institute, Inc. | Methods and compositions for editing nucleotide sequences |
| WO2021032155A1 (zh) * | 2019-08-20 | 2021-02-25 | 中国科学院遗传与发育生物学研究所 | 一种碱基编辑系统和其使用方法 |
| DE112021002672T5 (de) | 2020-05-08 | 2023-04-13 | President And Fellows Of Harvard College | Vefahren und zusammensetzungen zum gleichzeitigen editieren beider stränge einer doppelsträngigen nukleotid-zielsequenz |
| CN115678900A (zh) * | 2021-07-30 | 2023-02-03 | 中国科学院天津工业生物技术研究所 | 缩小碱基编辑器的编辑窗口的方法、碱基编辑器及用途 |
Citations (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2015133554A (ja) | 2014-01-10 | 2015-07-23 | 三菱電機株式会社 | 有線伝送装置及び終端抵抗の抵抗値の調整方法 |
| JP2016500003A (ja) | 2012-10-23 | 2016-01-07 | ツールゲン インコーポレイテッド | 標的dnaに特異的なガイドrnaおよびcasタンパク質コード核酸またはcasタンパク質を含む、標的dnaを切断するための組成物、ならびにその使用 |
| WO2016134081A1 (en) | 2015-02-18 | 2016-08-25 | Iowa State University Research Foundation, Inc. | Modification of transcriptional repressor binding site in nf-yc4 promoter for increased protein content and resistance to stress |
| JP2016536021A (ja) | 2013-11-07 | 2016-11-24 | エディタス・メディシン,インコーポレイテッド | CRISPR関連方法および支配gRNAのある組成物 |
| WO2017070632A2 (en) | 2015-10-23 | 2017-04-27 | President And Fellows Of Harvard College | Nucleobase editors and uses thereof |
| JP2017520243A (ja) | 2014-06-06 | 2017-07-27 | リジェネロン・ファーマシューティカルズ・インコーポレイテッドRegeneron Pharmaceuticals, Inc. | 標的遺伝子座を修飾するための方法及び組成物 |
| JP2017183724A (ja) | 2016-03-29 | 2017-10-05 | 住友化学株式会社 | 発光素子 |
Family Cites Families (11)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101611154A (zh) * | 2006-12-04 | 2009-12-23 | 艾博特公司 | 用于癌症治疗的成对诊断测定 |
| US20100076057A1 (en) * | 2008-09-23 | 2010-03-25 | Northwestern University | TARGET DNA INTERFERENCE WITH crRNA |
| US11053481B2 (en) * | 2013-12-12 | 2021-07-06 | President And Fellows Of Harvard College | Fusions of Cas9 domains and nucleic acid-editing domains |
| KR20160097327A (ko) * | 2013-12-12 | 2016-08-17 | 더 브로드 인스티튜트, 인코퍼레이티드 | 유전자 산물, 구조 정보 및 유도성 모듈형 cas 효소의 발현의 변경을 위한 crispr-cas 시스템 및 방법 |
| CN111471675A (zh) * | 2014-03-05 | 2020-07-31 | 国立大学法人神户大学 | 特异性转变靶向dna序列的核酸碱基的基因组序列的修饰方法、及其使用的分子复合体 |
| KR101785847B1 (ko) * | 2015-05-12 | 2017-10-17 | 연세대학교 산학협력단 | 선형 이중가닥 DNA를 활용한 CRISPR/Cas9 시스템을 이용한 표적 유전체 교정 |
| US9790490B2 (en) * | 2015-06-18 | 2017-10-17 | The Broad Institute Inc. | CRISPR enzymes and systems |
| WO2017099494A1 (ko) * | 2015-12-08 | 2017-06-15 | 기초과학연구원 | Cpf1을 포함하는 유전체 교정용 조성물 및 그 용도 |
| DK3447139T3 (da) * | 2016-04-21 | 2022-07-04 | Univ Kobe Nat Univ Corp | Fremgangsmåde til øgning af mutationsindføringseffektivitet i genomsekvensmodificeringsteknik, og molekylært kompleks til anvendelse dertil |
| US11192929B2 (en) * | 2016-12-08 | 2021-12-07 | Regents Of The University Of Minnesota | Site-specific DNA base editing using modified APOBEC enzymes |
| CN106834341B (zh) * | 2016-12-30 | 2020-06-16 | 中国农业大学 | 一种基因定点突变载体及其构建方法和应用 |
-
2019
- 2019-01-23 KR KR1020207021532A patent/KR20200103769A/ko not_active Application Discontinuation
- 2019-01-23 CN CN201980014548.2A patent/CN111742051A/zh active Pending
- 2019-01-23 JP JP2020561562A patent/JP7075170B2/ja active Active
- 2019-01-23 WO PCT/KR2019/000962 patent/WO2019147014A1/ko unknown
- 2019-01-23 EP EP19743145.5A patent/EP3744844A4/en active Pending
- 2019-01-23 US US16/964,277 patent/US20210032621A1/en active Pending
Patent Citations (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2016500003A (ja) | 2012-10-23 | 2016-01-07 | ツールゲン インコーポレイテッド | 標的dnaに特異的なガイドrnaおよびcasタンパク質コード核酸またはcasタンパク質を含む、標的dnaを切断するための組成物、ならびにその使用 |
| JP2016536021A (ja) | 2013-11-07 | 2016-11-24 | エディタス・メディシン,インコーポレイテッド | CRISPR関連方法および支配gRNAのある組成物 |
| JP2015133554A (ja) | 2014-01-10 | 2015-07-23 | 三菱電機株式会社 | 有線伝送装置及び終端抵抗の抵抗値の調整方法 |
| JP2017520243A (ja) | 2014-06-06 | 2017-07-27 | リジェネロン・ファーマシューティカルズ・インコーポレイテッドRegeneron Pharmaceuticals, Inc. | 標的遺伝子座を修飾するための方法及び組成物 |
| WO2016134081A1 (en) | 2015-02-18 | 2016-08-25 | Iowa State University Research Foundation, Inc. | Modification of transcriptional repressor binding site in nf-yc4 promoter for increased protein content and resistance to stress |
| WO2017070632A2 (en) | 2015-10-23 | 2017-04-27 | President And Fellows Of Harvard College | Nucleobase editors and uses thereof |
| JP2017183724A (ja) | 2016-03-29 | 2017-10-05 | 住友化学株式会社 | 発光素子 |
Non-Patent Citations (4)
| Title |
|---|
| BANNO S. et al.,Nature Microbiology,2018年02月05日,Vol.3,p.423-429, Supplementary Information |
| KATO-INUI T. et al.,Nucleic Acids Research,2018年04月17日,Vol.46 No.9,p.4677-4688 |
| RAN F. et al.,Nature Protocols,2013年,Vol.8 No.11,p.2281-2308 |
| RYU S. et al.,Nature Biotechnology,2018年04月27日,Vol.36 No.6,p.536-539 |
Also Published As
| Publication number | Publication date |
|---|---|
| US20210032621A1 (en) | 2021-02-04 |
| EP3744844A4 (en) | 2021-10-20 |
| JP2021511824A (ja) | 2021-05-13 |
| KR20200103769A (ko) | 2020-09-02 |
| EP3744844A1 (en) | 2020-12-02 |
| WO2019147014A1 (ko) | 2019-08-01 |
| CN111742051A (zh) | 2020-10-02 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7075170B2 (ja) | 延長された単一ガイドrna及びその用途 | |
| CN109517841B (zh) | 一种用于核苷酸序列修饰的组合物、方法与应用 | |
| US12024727B2 (en) | Enzymes with RuvC domains | |
| JP7502537B2 (ja) | Ruvcドメインを有する酵素 | |
| EP3152312B1 (en) | Methods and compositions for modifying a targeted locus | |
| KR102098915B1 (ko) | 키메라 게놈 조작 분자 및 방법 | |
| KR102151065B1 (ko) | 동물 배아의 염기 교정용 조성물 및 염기 교정 방법 | |
| AU2013355214B2 (en) | Crispr-based genome modification and regulation | |
| US20240209332A1 (en) | Enzymes with ruvc domains | |
| KR102116200B1 (ko) | 게놈 서열 변형 기법의 돌연변이 도입 효율을 증가시키는 방법 및 이에 사용되는 분자 복합체 | |
| US20220298494A1 (en) | Enzymes with ruvc domains | |
| CN117999351A (zh) | Ii类v型crispr系统 | |
| US20220220460A1 (en) | Enzymes with ruvc domains | |
| WO2021226369A1 (en) | Enzymes with ruvc domains | |
| KR102151064B1 (ko) | 매칭된 5' 뉴클레오타이드를 포함하는 가이드 rna를 포함하는 유전자 교정용 조성물 및 이를 이용한 유전자 교정 방법 | |
| KR102679001B1 (ko) | 신규의 개량된 염기 편집 또는 교정용 융합단백질 및 이의 용도 | |
| CA3225082A1 (en) | Enzymes with ruvc domains | |
| KR20180082981A (ko) | 중첩된 가이드핵산을 이용한 표적 핵산에 특정 핵산 서열을 삽입하기 위한 조성물 및 방법 | |
| CN115703842A (zh) | 高效率高精度的胞嘧啶c到鸟嘌呤g转变的碱基编辑器 | |
| GB2617659A (en) | Enzymes with RUVC domains |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20200828 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20200828 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20210928 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20211224 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20220412 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20220511 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7075170 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |