JP6971306B2 - 様々な疾患および障害の治療のためのmasp−3を阻害する組成物および方法 - Google Patents
様々な疾患および障害の治療のためのmasp−3を阻害する組成物および方法 Download PDFInfo
- Publication number
- JP6971306B2 JP6971306B2 JP2019505253A JP2019505253A JP6971306B2 JP 6971306 B2 JP6971306 B2 JP 6971306B2 JP 2019505253 A JP2019505253 A JP 2019505253A JP 2019505253 A JP2019505253 A JP 2019505253A JP 6971306 B2 JP6971306 B2 JP 6971306B2
- Authority
- JP
- Japan
- Prior art keywords
- masp
- seq
- antibody
- antigen
- binding fragment
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 101001055956 Homo sapiens Mannan-binding lectin serine protease 1 Proteins 0.000 title claims description 96
- 230000002401 inhibitory effect Effects 0.000 title claims description 83
- 239000000203 mixture Substances 0.000 title claims description 42
- 238000000034 method Methods 0.000 title description 73
- 238000011282 treatment Methods 0.000 title description 39
- 102100026061 Mannan-binding lectin serine protease 1 Human genes 0.000 title description 14
- 208000037765 diseases and disorders Diseases 0.000 title 1
- 230000037361 pathway Effects 0.000 claims description 306
- 230000027455 binding Effects 0.000 claims description 243
- 239000000427 antigen Substances 0.000 claims description 174
- 108091007433 antigens Proteins 0.000 claims description 173
- 102000036639 antigens Human genes 0.000 claims description 173
- 239000012634 fragment Substances 0.000 claims description 158
- 230000004913 activation Effects 0.000 claims description 121
- 230000000295 complement effect Effects 0.000 claims description 99
- 102000048783 human MASP1 Human genes 0.000 claims description 80
- 241000282414 Homo sapiens Species 0.000 claims description 76
- 102000012479 Serine Proteases Human genes 0.000 claims description 49
- 108010022999 Serine Proteases Proteins 0.000 claims description 49
- 125000000539 amino acid group Chemical group 0.000 claims description 34
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 claims description 28
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 25
- 102000039446 nucleic acids Human genes 0.000 claims description 14
- 108020004707 nucleic acids Proteins 0.000 claims description 14
- 150000007523 nucleic acids Chemical class 0.000 claims description 14
- 239000008194 pharmaceutical composition Substances 0.000 claims description 14
- 210000004369 blood Anatomy 0.000 claims description 12
- 239000008280 blood Substances 0.000 claims description 12
- 241001529936 Murinae Species 0.000 claims description 11
- 238000004519 manufacturing process Methods 0.000 claims description 9
- 241000124008 Mammalia Species 0.000 claims description 8
- 239000013604 expression vector Substances 0.000 claims description 6
- 239000000546 pharmaceutical excipient Substances 0.000 claims description 4
- 108010009817 Immunoglobulin Constant Regions Proteins 0.000 claims description 3
- 102000009786 Immunoglobulin Constant Regions Human genes 0.000 claims description 3
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 claims description 3
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 claims description 3
- 238000010367 cloning Methods 0.000 claims description 2
- 239000013599 cloning vector Substances 0.000 claims description 2
- 238000012258 culturing Methods 0.000 claims description 2
- 238000002955 isolation Methods 0.000 claims 1
- 102000004856 Lectins Human genes 0.000 description 190
- 108090001090 Lectins Proteins 0.000 description 190
- 239000002523 lectin Substances 0.000 description 190
- 102100026046 Mannan-binding lectin serine protease 2 Human genes 0.000 description 144
- 101710117460 Mannan-binding lectin serine protease 2 Proteins 0.000 description 139
- 210000002966 serum Anatomy 0.000 description 136
- 238000001994 activation Methods 0.000 description 122
- 230000024203 complement activation Effects 0.000 description 116
- 102100036050 Phosphatidylinositol N-acetylglucosaminyltransferase subunit A Human genes 0.000 description 103
- 201000003045 paroxysmal nocturnal hemoglobinuria Diseases 0.000 description 103
- 208000000733 Paroxysmal Hemoglobinuria Diseases 0.000 description 102
- 102100022133 Complement C3 Human genes 0.000 description 101
- 101000901154 Homo sapiens Complement C3 Proteins 0.000 description 101
- 241000699670 Mus sp. Species 0.000 description 98
- 239000003112 inhibitor Substances 0.000 description 96
- 230000002950 deficient Effects 0.000 description 85
- 206010018910 Haemolysis Diseases 0.000 description 78
- 230000001419 dependent effect Effects 0.000 description 70
- 210000003743 erythrocyte Anatomy 0.000 description 68
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 65
- 108090000623 proteins and genes Proteins 0.000 description 59
- 230000008021 deposition Effects 0.000 description 58
- 230000014207 opsonization Effects 0.000 description 55
- 206010064930 age-related macular degeneration Diseases 0.000 description 52
- 206010003246 arthritis Diseases 0.000 description 52
- 208000002780 macular degeneration Diseases 0.000 description 50
- 230000009089 cytolysis Effects 0.000 description 46
- 108090000056 Complement factor B Proteins 0.000 description 44
- 102000003712 Complement factor B Human genes 0.000 description 44
- 108090000581 Leukemia inhibitory factor Proteins 0.000 description 44
- 102100032352 Leukemia inhibitory factor Human genes 0.000 description 44
- 235000001014 amino acid Nutrition 0.000 description 41
- 230000001404 mediated effect Effects 0.000 description 41
- 230000000903 blocking effect Effects 0.000 description 40
- 230000008588 hemolysis Effects 0.000 description 40
- 229940024606 amino acid Drugs 0.000 description 39
- 238000003776 cleavage reaction Methods 0.000 description 39
- 230000000694 effects Effects 0.000 description 39
- 102000004169 proteins and genes Human genes 0.000 description 39
- 230000007017 scission Effects 0.000 description 39
- 102000016574 Complement C3-C5 Convertases Human genes 0.000 description 38
- 108010067641 Complement C3-C5 Convertases Proteins 0.000 description 38
- 150000001413 amino acids Chemical class 0.000 description 38
- 235000018102 proteins Nutrition 0.000 description 38
- 239000011575 calcium Substances 0.000 description 36
- 230000006870 function Effects 0.000 description 35
- 201000010099 disease Diseases 0.000 description 33
- 208000035475 disorder Diseases 0.000 description 32
- 108090000765 processed proteins & peptides Proteins 0.000 description 32
- 230000005764 inhibitory process Effects 0.000 description 31
- 241000699666 Mus <mouse, genus> Species 0.000 description 30
- 102000004196 processed proteins & peptides Human genes 0.000 description 30
- 241000588650 Neisseria meningitidis Species 0.000 description 27
- 230000006378 damage Effects 0.000 description 27
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 26
- 206010022822 Intravascular haemolysis Diseases 0.000 description 26
- 210000004027 cell Anatomy 0.000 description 26
- 229920001184 polypeptide Polymers 0.000 description 26
- 102000010911 Enzyme Precursors Human genes 0.000 description 23
- 108010062466 Enzyme Precursors Proteins 0.000 description 23
- 230000004154 complement system Effects 0.000 description 23
- 230000002829 reductive effect Effects 0.000 description 23
- 229960002224 eculizumab Drugs 0.000 description 22
- 238000006243 chemical reaction Methods 0.000 description 21
- 230000002757 inflammatory effect Effects 0.000 description 21
- 206010039073 rheumatoid arthritis Diseases 0.000 description 21
- 108010034753 Complement Membrane Attack Complex Proteins 0.000 description 20
- 241000283973 Oryctolagus cuniculus Species 0.000 description 20
- 208000007502 anemia Diseases 0.000 description 20
- 230000015572 biosynthetic process Effects 0.000 description 20
- 230000000977 initiatory effect Effects 0.000 description 20
- 102000035195 Peptidases Human genes 0.000 description 19
- 108091005804 Peptidases Proteins 0.000 description 19
- 206010063837 Reperfusion injury Diseases 0.000 description 19
- 239000012636 effector Substances 0.000 description 19
- 230000001225 therapeutic effect Effects 0.000 description 19
- 229920000392 Zymosan Polymers 0.000 description 18
- 208000015181 infectious disease Diseases 0.000 description 18
- 239000004365 Protease Substances 0.000 description 17
- DEFVIWRASFVYLL-UHFFFAOYSA-N ethylene glycol bis(2-aminoethyl)tetraacetic acid Chemical compound OC(=O)CN(CC(O)=O)CCOCCOCCN(CC(O)=O)CC(O)=O DEFVIWRASFVYLL-UHFFFAOYSA-N 0.000 description 17
- 244000052769 pathogen Species 0.000 description 17
- 239000000758 substrate Substances 0.000 description 17
- 239000003795 chemical substances by application Substances 0.000 description 16
- 238000001990 intravenous administration Methods 0.000 description 16
- 239000000047 product Substances 0.000 description 16
- 230000010410 reperfusion Effects 0.000 description 16
- 102000004190 Enzymes Human genes 0.000 description 15
- 108090000790 Enzymes Proteins 0.000 description 15
- 229940088598 enzyme Drugs 0.000 description 15
- 230000001965 increasing effect Effects 0.000 description 15
- 230000000302 ischemic effect Effects 0.000 description 15
- 230000008569 process Effects 0.000 description 15
- 235000000346 sugar Nutrition 0.000 description 15
- 230000008901 benefit Effects 0.000 description 14
- 238000002474 experimental method Methods 0.000 description 14
- 102000001554 Hemoglobins Human genes 0.000 description 13
- 108010054147 Hemoglobins Proteins 0.000 description 13
- 229920000057 Mannan Polymers 0.000 description 13
- 241000700159 Rattus Species 0.000 description 13
- 230000002949 hemolytic effect Effects 0.000 description 13
- 230000003993 interaction Effects 0.000 description 13
- 208000012947 ischemia reperfusion injury Diseases 0.000 description 13
- 230000007246 mechanism Effects 0.000 description 13
- 244000005700 microbiome Species 0.000 description 13
- 201000008482 osteoarthritis Diseases 0.000 description 13
- 108020004414 DNA Proteins 0.000 description 12
- 102000053602 DNA Human genes 0.000 description 12
- 206010015871 Extravascular haemolysis Diseases 0.000 description 12
- 230000035772 mutation Effects 0.000 description 12
- 102100022002 CD59 glycoprotein Human genes 0.000 description 11
- 102100031506 Complement C5 Human genes 0.000 description 11
- 102100025680 Complement decay-accelerating factor Human genes 0.000 description 11
- 241000282412 Homo Species 0.000 description 11
- 101000897400 Homo sapiens CD59 glycoprotein Proteins 0.000 description 11
- 101000941598 Homo sapiens Complement C5 Proteins 0.000 description 11
- 101000856022 Homo sapiens Complement decay-accelerating factor Proteins 0.000 description 11
- 241001465754 Metazoa Species 0.000 description 11
- 239000004074 complement inhibitor Substances 0.000 description 11
- 108090000062 ficolin Proteins 0.000 description 11
- 210000000056 organ Anatomy 0.000 description 11
- 230000004044 response Effects 0.000 description 11
- 101000909536 Homo sapiens Collectin-11 Proteins 0.000 description 10
- 206010061218 Inflammation Diseases 0.000 description 10
- 239000012190 activator Substances 0.000 description 10
- 238000004458 analytical method Methods 0.000 description 10
- 230000001580 bacterial effect Effects 0.000 description 10
- 238000000338 in vitro Methods 0.000 description 10
- 230000004054 inflammatory process Effects 0.000 description 10
- 208000014674 injury Diseases 0.000 description 10
- 239000007791 liquid phase Substances 0.000 description 10
- 238000010172 mouse model Methods 0.000 description 10
- 235000019419 proteases Nutrition 0.000 description 10
- 230000009885 systemic effect Effects 0.000 description 10
- 210000001519 tissue Anatomy 0.000 description 10
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 9
- 241000894006 Bacteria Species 0.000 description 9
- 229940124073 Complement inhibitor Drugs 0.000 description 9
- 101001056015 Homo sapiens Mannan-binding lectin serine protease 2 Proteins 0.000 description 9
- 108060003951 Immunoglobulin Proteins 0.000 description 9
- 241000282567 Macaca fascicularis Species 0.000 description 9
- 230000003321 amplification Effects 0.000 description 9
- 230000000844 anti-bacterial effect Effects 0.000 description 9
- 238000011260 co-administration Methods 0.000 description 9
- 230000036039 immunity Effects 0.000 description 9
- 102000018358 immunoglobulin Human genes 0.000 description 9
- 238000003199 nucleic acid amplification method Methods 0.000 description 9
- 230000036542 oxidative stress Effects 0.000 description 9
- 230000001717 pathogenic effect Effects 0.000 description 9
- 241000894007 species Species 0.000 description 9
- 239000000126 substance Substances 0.000 description 9
- 230000004083 survival effect Effects 0.000 description 9
- 208000011580 syndromic disease Diseases 0.000 description 9
- 102100034540 Adenomatous polyposis coli protein Human genes 0.000 description 8
- 102100024331 Collectin-11 Human genes 0.000 description 8
- 206010015866 Extravasation Diseases 0.000 description 8
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 8
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 8
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 8
- 208000027418 Wounds and injury Diseases 0.000 description 8
- 239000002299 complementary DNA Substances 0.000 description 8
- 239000003814 drug Substances 0.000 description 8
- 230000036251 extravasation Effects 0.000 description 8
- 230000014509 gene expression Effects 0.000 description 8
- 238000001361 intraarterial administration Methods 0.000 description 8
- 208000028867 ischemia Diseases 0.000 description 8
- 210000001503 joint Anatomy 0.000 description 8
- 238000011813 knockout mouse model Methods 0.000 description 8
- 210000004379 membrane Anatomy 0.000 description 8
- 239000012528 membrane Substances 0.000 description 8
- 239000002245 particle Substances 0.000 description 8
- 210000002381 plasma Anatomy 0.000 description 8
- 230000000451 tissue damage Effects 0.000 description 8
- 231100000827 tissue damage Toxicity 0.000 description 8
- 208000005590 Choroidal Neovascularization Diseases 0.000 description 7
- 206010060823 Choroidal neovascularisation Diseases 0.000 description 7
- 102000008186 Collagen Human genes 0.000 description 7
- 108010035532 Collagen Proteins 0.000 description 7
- 108700024394 Exon Proteins 0.000 description 7
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 7
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 7
- 241000191967 Staphylococcus aureus Species 0.000 description 7
- 101000978426 Sus scrofa Mannose-binding protein A Proteins 0.000 description 7
- 238000003556 assay Methods 0.000 description 7
- 150000001720 carbohydrates Chemical class 0.000 description 7
- 230000004087 circulation Effects 0.000 description 7
- 229920001436 collagen Polymers 0.000 description 7
- 230000007812 deficiency Effects 0.000 description 7
- 229940079593 drug Drugs 0.000 description 7
- 230000002147 killing effect Effects 0.000 description 7
- 230000035800 maturation Effects 0.000 description 7
- 230000000813 microbial effect Effects 0.000 description 7
- 230000004962 physiological condition Effects 0.000 description 7
- 230000009467 reduction Effects 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- 238000002560 therapeutic procedure Methods 0.000 description 7
- 238000001262 western blot Methods 0.000 description 7
- 102000000989 Complement System Proteins Human genes 0.000 description 6
- 108010069112 Complement System Proteins Proteins 0.000 description 6
- 102100024521 Ficolin-2 Human genes 0.000 description 6
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 6
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 6
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 6
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 6
- 208000006011 Stroke Diseases 0.000 description 6
- 208000000208 Wet Macular Degeneration Diseases 0.000 description 6
- 230000021917 activation of membrane attack complex Effects 0.000 description 6
- 235000014633 carbohydrates Nutrition 0.000 description 6
- 230000003247 decreasing effect Effects 0.000 description 6
- 230000007850 degeneration Effects 0.000 description 6
- 238000011161 development Methods 0.000 description 6
- 230000018109 developmental process Effects 0.000 description 6
- 230000002496 gastric effect Effects 0.000 description 6
- 229940027941 immunoglobulin g Drugs 0.000 description 6
- 230000002101 lytic effect Effects 0.000 description 6
- 230000002107 myocardial effect Effects 0.000 description 6
- 230000001681 protective effect Effects 0.000 description 6
- 239000000523 sample Substances 0.000 description 6
- 235000004400 serine Nutrition 0.000 description 6
- 230000002269 spontaneous effect Effects 0.000 description 6
- 238000006467 substitution reaction Methods 0.000 description 6
- 208000024891 symptom Diseases 0.000 description 6
- 108700029850 CR2-fH Proteins 0.000 description 5
- 101150018425 Cr1l gene Proteins 0.000 description 5
- 241000287828 Gallus gallus Species 0.000 description 5
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 5
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 5
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 5
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 5
- 101150104297 MASP1 gene Proteins 0.000 description 5
- 208000034762 Meningococcal Infections Diseases 0.000 description 5
- 108700022034 Opsonin Proteins Proteins 0.000 description 5
- 241000193998 Streptococcus pneumoniae Species 0.000 description 5
- 208000034841 Thrombotic Microangiopathies Diseases 0.000 description 5
- 230000001154 acute effect Effects 0.000 description 5
- 235000013330 chicken meat Nutrition 0.000 description 5
- 230000009137 competitive binding Effects 0.000 description 5
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 5
- 235000004554 glutamine Nutrition 0.000 description 5
- 210000002216 heart Anatomy 0.000 description 5
- 230000001771 impaired effect Effects 0.000 description 5
- 230000001976 improved effect Effects 0.000 description 5
- 238000002347 injection Methods 0.000 description 5
- 239000007924 injection Substances 0.000 description 5
- 238000007918 intramuscular administration Methods 0.000 description 5
- 108020004999 messenger RNA Proteins 0.000 description 5
- 238000007911 parenteral administration Methods 0.000 description 5
- 230000002263 peptidergic effect Effects 0.000 description 5
- 230000004224 protection Effects 0.000 description 5
- 210000003583 retinal pigment epithelium Anatomy 0.000 description 5
- 238000012552 review Methods 0.000 description 5
- 239000002904 solvent Substances 0.000 description 5
- 230000009870 specific binding Effects 0.000 description 5
- 229940031000 streptococcus pneumoniae Drugs 0.000 description 5
- 238000007920 subcutaneous administration Methods 0.000 description 5
- 238000001356 surgical procedure Methods 0.000 description 5
- 210000001179 synovial fluid Anatomy 0.000 description 5
- 150000007970 thio esters Chemical group 0.000 description 5
- 238000002054 transplantation Methods 0.000 description 5
- 208000009304 Acute Kidney Injury Diseases 0.000 description 4
- BPYKTIZUTYGOLE-IFADSCNNSA-N Bilirubin Chemical compound N1C(=O)C(C)=C(C=C)\C1=C\C1=C(C)C(CCC(O)=O)=C(CC2=C(C(C)=C(\C=C/3C(=C(C=C)C(=O)N\3)C)N2)CCC(O)=O)N1 BPYKTIZUTYGOLE-IFADSCNNSA-N 0.000 description 4
- 206010057248 Cell death Diseases 0.000 description 4
- 108090000909 Collectins Proteins 0.000 description 4
- 102000004405 Collectins Human genes 0.000 description 4
- 102100024508 Ficolin-1 Human genes 0.000 description 4
- 206010018364 Glomerulonephritis Diseases 0.000 description 4
- 102000003886 Glycoproteins Human genes 0.000 description 4
- 108090000288 Glycoproteins Proteins 0.000 description 4
- 208000032759 Hemolytic-Uremic Syndrome Diseases 0.000 description 4
- 208000003456 Juvenile Arthritis Diseases 0.000 description 4
- 206010059176 Juvenile idiopathic arthritis Diseases 0.000 description 4
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 4
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 4
- 102000003855 L-lactate dehydrogenase Human genes 0.000 description 4
- 108700023483 L-lactate dehydrogenases Proteins 0.000 description 4
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 4
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 4
- 108700041239 Mannose-Binding Protein Deficiency Proteins 0.000 description 4
- 102000004528 Mannose-Binding Protein-Associated Serine Proteases Human genes 0.000 description 4
- 108010042484 Mannose-Binding Protein-Associated Serine Proteases Proteins 0.000 description 4
- 102000007079 Peptide Fragments Human genes 0.000 description 4
- 108010033276 Peptide Fragments Proteins 0.000 description 4
- 206010035669 Pneumonia aspiration Diseases 0.000 description 4
- 241000288906 Primates Species 0.000 description 4
- 108010076504 Protein Sorting Signals Proteins 0.000 description 4
- 201000001263 Psoriatic Arthritis Diseases 0.000 description 4
- 208000036824 Psoriatic arthropathy Diseases 0.000 description 4
- 208000007536 Thrombosis Diseases 0.000 description 4
- 208000030886 Traumatic Brain injury Diseases 0.000 description 4
- 241000700605 Viruses Species 0.000 description 4
- 230000003213 activating effect Effects 0.000 description 4
- 239000003435 antirheumatic agent Substances 0.000 description 4
- 235000003704 aspartic acid Nutrition 0.000 description 4
- 201000009807 aspiration pneumonia Diseases 0.000 description 4
- 208000006673 asthma Diseases 0.000 description 4
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 4
- 208000019069 chronic childhood arthritis Diseases 0.000 description 4
- 230000007423 decrease Effects 0.000 description 4
- 230000007123 defense Effects 0.000 description 4
- 239000002988 disease modifying antirheumatic drug Substances 0.000 description 4
- 208000009190 disseminated intravascular coagulation Diseases 0.000 description 4
- 239000003937 drug carrier Substances 0.000 description 4
- 208000011325 dry age related macular degeneration Diseases 0.000 description 4
- 206010014801 endophthalmitis Diseases 0.000 description 4
- 210000002889 endothelial cell Anatomy 0.000 description 4
- 238000000684 flow cytometry Methods 0.000 description 4
- 230000005714 functional activity Effects 0.000 description 4
- 102000034356 gene-regulatory proteins Human genes 0.000 description 4
- 108091006104 gene-regulatory proteins Proteins 0.000 description 4
- 230000002068 genetic effect Effects 0.000 description 4
- 150000004676 glycans Chemical class 0.000 description 4
- 230000006698 induction Effects 0.000 description 4
- 230000000968 intestinal effect Effects 0.000 description 4
- 201000002215 juvenile rheumatoid arthritis Diseases 0.000 description 4
- 239000003446 ligand Substances 0.000 description 4
- 210000004185 liver Anatomy 0.000 description 4
- 108700008279 mouse MASP-3 Proteins 0.000 description 4
- 208000010125 myocardial infarction Diseases 0.000 description 4
- 230000002956 necrotizing effect Effects 0.000 description 4
- 229910052757 nitrogen Inorganic materials 0.000 description 4
- 230000001575 pathological effect Effects 0.000 description 4
- 210000001539 phagocyte Anatomy 0.000 description 4
- 230000001737 promoting effect Effects 0.000 description 4
- 102000005962 receptors Human genes 0.000 description 4
- 108020003175 receptors Proteins 0.000 description 4
- 238000011160 research Methods 0.000 description 4
- 230000009529 traumatic brain injury Effects 0.000 description 4
- 230000003442 weekly effect Effects 0.000 description 4
- 101150090724 3 gene Proteins 0.000 description 3
- 206010001052 Acute respiratory distress syndrome Diseases 0.000 description 3
- 208000024827 Alzheimer disease Diseases 0.000 description 3
- 239000004475 Arginine Substances 0.000 description 3
- 102000007350 Bone Morphogenetic Proteins Human genes 0.000 description 3
- 108010007726 Bone Morphogenetic Proteins Proteins 0.000 description 3
- 108010034358 Classical Pathway Complement C3 Convertase Proteins 0.000 description 3
- 102100023661 Coiled-coil domain-containing protein 115 Human genes 0.000 description 3
- 101710155594 Coiled-coil domain-containing protein 115 Proteins 0.000 description 3
- 102100025721 Cytosolic carboxypeptidase 2 Human genes 0.000 description 3
- -1 DAF Proteins 0.000 description 3
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- 208000009329 Graft vs Host Disease Diseases 0.000 description 3
- 101000932634 Homo sapiens Cytosolic carboxypeptidase 2 Proteins 0.000 description 3
- 101001041393 Homo sapiens Serine protease HTRA1 Proteins 0.000 description 3
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 3
- 239000004472 Lysine Substances 0.000 description 3
- 108010087870 Mannose-Binding Lectin Proteins 0.000 description 3
- 102000009112 Mannose-Binding Lectin Human genes 0.000 description 3
- 208000004451 Membranoproliferative Glomerulonephritis Diseases 0.000 description 3
- 101001033011 Mus musculus Granzyme C Proteins 0.000 description 3
- OVRNDRQMDRJTHS-FMDGEEDCSA-N N-acetyl-beta-D-glucosamine Chemical compound CC(=O)N[C@H]1[C@H](O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-FMDGEEDCSA-N 0.000 description 3
- 208000002193 Pain Diseases 0.000 description 3
- 102000005877 Peptide Initiation Factors Human genes 0.000 description 3
- 108010044843 Peptide Initiation Factors Proteins 0.000 description 3
- 241000283984 Rodentia Species 0.000 description 3
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 3
- 102100021119 Serine protease HTRA1 Human genes 0.000 description 3
- 206010052779 Transplant rejections Diseases 0.000 description 3
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Chemical compound CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 3
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 3
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 3
- 206010047115 Vasculitis Diseases 0.000 description 3
- 125000003277 amino group Chemical group 0.000 description 3
- 238000010171 animal model Methods 0.000 description 3
- 230000000890 antigenic effect Effects 0.000 description 3
- 208000007474 aortic aneurysm Diseases 0.000 description 3
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 3
- 229940112869 bone morphogenetic protein Drugs 0.000 description 3
- 239000000872 buffer Substances 0.000 description 3
- 230000002612 cardiopulmonary effect Effects 0.000 description 3
- 230000003197 catalytic effect Effects 0.000 description 3
- 230000006037 cell lysis Effects 0.000 description 3
- 210000000349 chromosome Anatomy 0.000 description 3
- 235000021310 complex sugar Nutrition 0.000 description 3
- 230000034994 death Effects 0.000 description 3
- 231100000517 death Toxicity 0.000 description 3
- 208000022401 dense deposit disease Diseases 0.000 description 3
- 238000001514 detection method Methods 0.000 description 3
- 230000003511 endothelial effect Effects 0.000 description 3
- 210000003414 extremity Anatomy 0.000 description 3
- 230000004438 eyesight Effects 0.000 description 3
- 102000037865 fusion proteins Human genes 0.000 description 3
- 108020001507 fusion proteins Proteins 0.000 description 3
- 239000000499 gel Substances 0.000 description 3
- 208000024908 graft versus host disease Diseases 0.000 description 3
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 3
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 3
- 210000004408 hybridoma Anatomy 0.000 description 3
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 3
- 238000001727 in vivo Methods 0.000 description 3
- 230000028709 inflammatory response Effects 0.000 description 3
- 229960000310 isoleucine Drugs 0.000 description 3
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 3
- 230000009456 molecular mechanism Effects 0.000 description 3
- 230000001338 necrotic effect Effects 0.000 description 3
- 239000013642 negative control Substances 0.000 description 3
- 210000000440 neutrophil Anatomy 0.000 description 3
- 244000045947 parasite Species 0.000 description 3
- 230000007170 pathology Effects 0.000 description 3
- 230000007310 pathophysiology Effects 0.000 description 3
- 239000002831 pharmacologic agent Substances 0.000 description 3
- 239000012071 phase Substances 0.000 description 3
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 3
- 230000002265 prevention Effects 0.000 description 3
- 230000017854 proteolysis Effects 0.000 description 3
- 238000003259 recombinant expression Methods 0.000 description 3
- 230000001105 regulatory effect Effects 0.000 description 3
- 230000008439 repair process Effects 0.000 description 3
- 210000001995 reticulocyte Anatomy 0.000 description 3
- 210000001525 retina Anatomy 0.000 description 3
- 229920002477 rna polymer Polymers 0.000 description 3
- 150000003384 small molecules Chemical class 0.000 description 3
- 229960002898 threonine Drugs 0.000 description 3
- 230000009261 transgenic effect Effects 0.000 description 3
- 230000007306 turnover Effects 0.000 description 3
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 3
- 230000002792 vascular Effects 0.000 description 3
- WQZGKKKJIJFFOK-SVZMEOIVSA-N (+)-Galactose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@H]1O WQZGKKKJIJFFOK-SVZMEOIVSA-N 0.000 description 2
- 108700028369 Alleles Proteins 0.000 description 2
- 108010003529 Alternative Pathway Complement C3 Convertase Proteins 0.000 description 2
- 208000003343 Antiphospholipid Syndrome Diseases 0.000 description 2
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 2
- 201000001320 Atherosclerosis Diseases 0.000 description 2
- 208000035913 Atypical hemolytic uremic syndrome Diseases 0.000 description 2
- 201000004569 Blindness Diseases 0.000 description 2
- 102000004506 Blood Proteins Human genes 0.000 description 2
- 108010017384 Blood Proteins Proteins 0.000 description 2
- 208000018240 Bone Marrow Failure disease Diseases 0.000 description 2
- 206010065553 Bone marrow failure Diseases 0.000 description 2
- 241000282693 Cercopithecidae Species 0.000 description 2
- 241001502567 Chikungunya virus Species 0.000 description 2
- 208000017667 Chronic Disease Diseases 0.000 description 2
- 208000006545 Chronic Obstructive Pulmonary Disease Diseases 0.000 description 2
- 108010078804 Classical Pathway Complement C5 Convertase Proteins 0.000 description 2
- 208000035473 Communicable disease Diseases 0.000 description 2
- 108010078015 Complement C3b Proteins 0.000 description 2
- 108010053085 Complement Factor H Proteins 0.000 description 2
- 102000016550 Complement Factor H Human genes 0.000 description 2
- 102000004127 Cytokines Human genes 0.000 description 2
- 108090000695 Cytokines Proteins 0.000 description 2
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 2
- 206010012689 Diabetic retinopathy Diseases 0.000 description 2
- 241000257465 Echinoidea Species 0.000 description 2
- 108010074864 Factor XI Proteins 0.000 description 2
- 108010080865 Factor XII Proteins 0.000 description 2
- 102000000429 Factor XII Human genes 0.000 description 2
- 241000192125 Firmicutes Species 0.000 description 2
- 206010017533 Fungal infection Diseases 0.000 description 2
- 208000008069 Geographic Atrophy Diseases 0.000 description 2
- 101000942967 Homo sapiens Leukemia inhibitory factor Proteins 0.000 description 2
- 101001056128 Homo sapiens Mannose-binding protein C Proteins 0.000 description 2
- PMMYEEVYMWASQN-DMTCNVIQSA-N Hydroxyproline Chemical compound O[C@H]1CN[C@H](C(O)=O)C1 PMMYEEVYMWASQN-DMTCNVIQSA-N 0.000 description 2
- 208000010159 IgA glomerulonephritis Diseases 0.000 description 2
- 206010021263 IgA nephropathy Diseases 0.000 description 2
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 2
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 2
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- 208000005777 Lupus Nephritis Diseases 0.000 description 2
- 108010068997 Mannose-Binding Lectins Proteins 0.000 description 2
- 102000001698 Mannose-Binding Lectins Human genes 0.000 description 2
- 201000009906 Meningitis Diseases 0.000 description 2
- 101001055959 Mus musculus Mannan-binding lectin serine protease 1 Proteins 0.000 description 2
- 206010028372 Muscular weakness Diseases 0.000 description 2
- OVRNDRQMDRJTHS-UHFFFAOYSA-N N-acelyl-D-glucosamine Natural products CC(=O)NC1C(O)OC(CO)C(O)C1O OVRNDRQMDRJTHS-UHFFFAOYSA-N 0.000 description 2
- 230000004988 N-glycosylation Effects 0.000 description 2
- 108091028043 Nucleic acid sequence Proteins 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 108010005642 Properdin Proteins 0.000 description 2
- 102100038567 Properdin Human genes 0.000 description 2
- 241000219061 Rheum Species 0.000 description 2
- 206010040047 Sepsis Diseases 0.000 description 2
- 241000130810 Streptococcus pneumoniae D39 Species 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 239000004473 Threonine Substances 0.000 description 2
- 206010043561 Thrombocytopenic purpura Diseases 0.000 description 2
- 201000007023 Thrombotic Thrombocytopenic Purpura Diseases 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 239000004480 active ingredient Substances 0.000 description 2
- 230000004721 adaptive immunity Effects 0.000 description 2
- 230000002411 adverse Effects 0.000 description 2
- 235000004279 alanine Nutrition 0.000 description 2
- 238000002266 amputation Methods 0.000 description 2
- 238000002399 angioplasty Methods 0.000 description 2
- 230000005875 antibody response Effects 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- 235000009582 asparagine Nutrition 0.000 description 2
- 229960001230 asparagine Drugs 0.000 description 2
- 210000000601 blood cell Anatomy 0.000 description 2
- 210000004556 brain Anatomy 0.000 description 2
- 210000001736 capillary Anatomy 0.000 description 2
- 239000003153 chemical reaction reagent Substances 0.000 description 2
- 210000000038 chest Anatomy 0.000 description 2
- 208000037976 chronic inflammation Diseases 0.000 description 2
- 208000037893 chronic inflammatory disorder Diseases 0.000 description 2
- 230000015271 coagulation Effects 0.000 description 2
- 238000005345 coagulation Methods 0.000 description 2
- 239000011248 coating agent Substances 0.000 description 2
- 238000000576 coating method Methods 0.000 description 2
- 102000006834 complement receptors Human genes 0.000 description 2
- 108010047295 complement receptors Proteins 0.000 description 2
- 235000018417 cysteine Nutrition 0.000 description 2
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 2
- 230000002354 daily effect Effects 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- 238000004090 dissolution Methods 0.000 description 2
- 239000006196 drop Substances 0.000 description 2
- 230000009977 dual effect Effects 0.000 description 2
- 230000004064 dysfunction Effects 0.000 description 2
- 230000008482 dysregulation Effects 0.000 description 2
- 239000002895 emetic Substances 0.000 description 2
- 239000002158 endotoxin Substances 0.000 description 2
- 230000008472 epithelial growth Effects 0.000 description 2
- 230000034841 erythrocyte clearance Effects 0.000 description 2
- 230000002349 favourable effect Effects 0.000 description 2
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 2
- 230000002538 fungal effect Effects 0.000 description 2
- BTCSSZJGUNDROE-UHFFFAOYSA-N gamma-aminobutyric acid Chemical compound NCCCC(O)=O BTCSSZJGUNDROE-UHFFFAOYSA-N 0.000 description 2
- 235000013922 glutamic acid Nutrition 0.000 description 2
- 239000004220 glutamic acid Substances 0.000 description 2
- 238000001631 haemodialysis Methods 0.000 description 2
- 208000014951 hematologic disease Diseases 0.000 description 2
- 230000000322 hemodialysis Effects 0.000 description 2
- 102000054960 human MASP2 Human genes 0.000 description 2
- 230000002209 hydrophobic effect Effects 0.000 description 2
- 208000013403 hyperactivity Diseases 0.000 description 2
- 230000008076 immune mechanism Effects 0.000 description 2
- 230000028993 immune response Effects 0.000 description 2
- 210000000987 immune system Anatomy 0.000 description 2
- 229940072221 immunoglobulins Drugs 0.000 description 2
- 238000010874 in vitro model Methods 0.000 description 2
- 238000011534 incubation Methods 0.000 description 2
- 230000001939 inductive effect Effects 0.000 description 2
- 239000003999 initiator Substances 0.000 description 2
- 230000015788 innate immune response Effects 0.000 description 2
- 238000007917 intracranial administration Methods 0.000 description 2
- 230000006623 intrinsic pathway Effects 0.000 description 2
- 238000003973 irrigation Methods 0.000 description 2
- 230000002262 irrigation Effects 0.000 description 2
- 210000003734 kidney Anatomy 0.000 description 2
- 230000003907 kidney function Effects 0.000 description 2
- 238000002647 laser therapy Methods 0.000 description 2
- 229920006008 lipopolysaccharide Polymers 0.000 description 2
- 231100000516 lung damage Toxicity 0.000 description 2
- 210000002540 macrophage Anatomy 0.000 description 2
- 230000004048 modification Effects 0.000 description 2
- 238000012986 modification Methods 0.000 description 2
- 210000001616 monocyte Anatomy 0.000 description 2
- 201000006417 multiple sclerosis Diseases 0.000 description 2
- 210000003205 muscle Anatomy 0.000 description 2
- 230000036473 myasthenia Effects 0.000 description 2
- 229950006780 n-acetylglucosamine Drugs 0.000 description 2
- 230000000269 nucleophilic effect Effects 0.000 description 2
- 239000002773 nucleotide Substances 0.000 description 2
- 125000003729 nucleotide group Chemical group 0.000 description 2
- 239000002674 ointment Substances 0.000 description 2
- 230000011164 ossification Effects 0.000 description 2
- 230000001590 oxidative effect Effects 0.000 description 2
- 230000000242 pagocytic effect Effects 0.000 description 2
- 230000003950 pathogenic mechanism Effects 0.000 description 2
- 230000010412 perfusion Effects 0.000 description 2
- 230000002093 peripheral effect Effects 0.000 description 2
- 108091008695 photoreceptors Proteins 0.000 description 2
- 230000035790 physiological processes and functions Effects 0.000 description 2
- 230000008488 polyadenylation Effects 0.000 description 2
- 102000054765 polymorphisms of proteins Human genes 0.000 description 2
- 102000040430 polynucleotide Human genes 0.000 description 2
- 108091033319 polynucleotide Proteins 0.000 description 2
- 239000002157 polynucleotide Substances 0.000 description 2
- 229920001282 polysaccharide Polymers 0.000 description 2
- 239000005017 polysaccharide Substances 0.000 description 2
- 230000004481 post-translational protein modification Effects 0.000 description 2
- 230000003389 potentiating effect Effects 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 230000000750 progressive effect Effects 0.000 description 2
- 230000000770 proinflammatory effect Effects 0.000 description 2
- 230000035755 proliferation Effects 0.000 description 2
- 235000019833 protease Nutrition 0.000 description 2
- 230000002797 proteolythic effect Effects 0.000 description 2
- 230000000284 resting effect Effects 0.000 description 2
- 208000013223 septicemia Diseases 0.000 description 2
- 238000002864 sequence alignment Methods 0.000 description 2
- 229940055944 soliris Drugs 0.000 description 2
- 238000005063 solubilization Methods 0.000 description 2
- 230000007928 solubilization Effects 0.000 description 2
- 238000010186 staining Methods 0.000 description 2
- 229910052717 sulfur Inorganic materials 0.000 description 2
- 230000008093 supporting effect Effects 0.000 description 2
- 230000008685 targeting Effects 0.000 description 2
- 238000012360 testing method Methods 0.000 description 2
- 235000008521 threonine Nutrition 0.000 description 2
- 230000008359 toxicosis Effects 0.000 description 2
- 238000013518 transcription Methods 0.000 description 2
- 230000035897 transcription Effects 0.000 description 2
- 238000011830 transgenic mouse model Methods 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- 238000012384 transportation and delivery Methods 0.000 description 2
- 230000008733 trauma Effects 0.000 description 2
- 210000004881 tumor cell Anatomy 0.000 description 2
- 229910052721 tungsten Inorganic materials 0.000 description 2
- 230000002485 urinary effect Effects 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- 238000007631 vascular surgery Methods 0.000 description 2
- 230000035899 viability Effects 0.000 description 2
- 230000004393 visual impairment Effects 0.000 description 2
- 239000002699 waste material Substances 0.000 description 2
- 210000005253 yeast cell Anatomy 0.000 description 2
- 229910052727 yttrium Inorganic materials 0.000 description 2
- HMLGSIZOMSVISS-ONJSNURVSA-N (7r)-7-[[(2z)-2-(2-amino-1,3-thiazol-4-yl)-2-(2,2-dimethylpropanoyloxymethoxyimino)acetyl]amino]-3-ethenyl-8-oxo-5-thia-1-azabicyclo[4.2.0]oct-2-ene-2-carboxylic acid Chemical compound N([C@@H]1C(N2C(=C(C=C)CSC21)C(O)=O)=O)C(=O)\C(=N/OCOC(=O)C(C)(C)C)C1=CSC(N)=N1 HMLGSIZOMSVISS-ONJSNURVSA-N 0.000 description 1
- 201000002884 3MC syndrome Diseases 0.000 description 1
- WLCZTRVUXYALDD-IBGZPJMESA-N 7-[[(2s)-2,6-bis(2-methoxyethoxycarbonylamino)hexanoyl]amino]heptoxy-methylphosphinic acid Chemical compound COCCOC(=O)NCCCC[C@H](NC(=O)OCCOC)C(=O)NCCCCCCCOP(C)(O)=O WLCZTRVUXYALDD-IBGZPJMESA-N 0.000 description 1
- 208000004476 Acute Coronary Syndrome Diseases 0.000 description 1
- 208000030090 Acute Disease Diseases 0.000 description 1
- 102000011767 Acute-Phase Proteins Human genes 0.000 description 1
- 108010062271 Acute-Phase Proteins Proteins 0.000 description 1
- QNAYBMKLOCPYGJ-UHFFFAOYSA-N Alanine Chemical compound CC([NH3+])C([O-])=O QNAYBMKLOCPYGJ-UHFFFAOYSA-N 0.000 description 1
- 108010089414 Anaphylatoxins Proteins 0.000 description 1
- 206010002556 Ankylosing Spondylitis Diseases 0.000 description 1
- 241000486679 Antitype Species 0.000 description 1
- 208000032467 Aplastic anaemia Diseases 0.000 description 1
- 101100235066 Arabidopsis thaliana LEA2 gene Proteins 0.000 description 1
- QYLJIYOGHRGUIH-CIUDSAMLSA-N Arg-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H](N)CCCNC(N)=N QYLJIYOGHRGUIH-CIUDSAMLSA-N 0.000 description 1
- 208000036487 Arthropathies Diseases 0.000 description 1
- 208000032116 Autoimmune Experimental Encephalomyelitis Diseases 0.000 description 1
- 241000193830 Bacillus <bacterium> Species 0.000 description 1
- 208000035143 Bacterial infection Diseases 0.000 description 1
- 208000009137 Behcet syndrome Diseases 0.000 description 1
- 208000019838 Blood disease Diseases 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 208000002381 Brain Hypoxia Diseases 0.000 description 1
- 201000006474 Brain Ischemia Diseases 0.000 description 1
- 235000012905 Brassica oleracea var viridis Nutrition 0.000 description 1
- 244000064816 Brassica oleracea var. acephala Species 0.000 description 1
- 108010074051 C-Reactive Protein Proteins 0.000 description 1
- 102100032752 C-reactive protein Human genes 0.000 description 1
- 208000016323 C3 glomerulonephritis Diseases 0.000 description 1
- 101150021919 CFB gene Proteins 0.000 description 1
- OYPRJOBELJOOCE-UHFFFAOYSA-N Calcium Chemical compound [Ca] OYPRJOBELJOOCE-UHFFFAOYSA-N 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 206010007556 Cardiac failure acute Diseases 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 206010008120 Cerebral ischaemia Diseases 0.000 description 1
- 206010009269 Cleft palate Diseases 0.000 description 1
- 102100030563 Coagulation factor XI Human genes 0.000 description 1
- 108091026890 Coding region Proteins 0.000 description 1
- 108010028778 Complement C4 Proteins 0.000 description 1
- 206010010356 Congenital anomaly Diseases 0.000 description 1
- 108091035707 Consensus sequence Proteins 0.000 description 1
- 206010011086 Coronary artery occlusion Diseases 0.000 description 1
- 241000557626 Corvus corax Species 0.000 description 1
- 208000025023 Cranial malformation Diseases 0.000 description 1
- PMATZTZNYRCHOR-CGLBZJNRSA-N Cyclosporin A Chemical compound CC[C@@H]1NC(=O)[C@H]([C@H](O)[C@H](C)C\C=C\C)N(C)C(=O)[C@H](C(C)C)N(C)C(=O)[C@H](CC(C)C)N(C)C(=O)[C@H](CC(C)C)N(C)C(=O)[C@@H](C)NC(=O)[C@H](C)NC(=O)[C@H](CC(C)C)N(C)C(=O)[C@H](C(C)C)NC(=O)[C@H](CC(C)C)N(C)C(=O)CN(C)C1=O PMATZTZNYRCHOR-CGLBZJNRSA-N 0.000 description 1
- 108010036949 Cyclosporine Proteins 0.000 description 1
- VVNCNSJFMMFHPL-VKHMYHEASA-N D-penicillamine Chemical compound CC(C)(S)[C@@H](N)C(O)=O VVNCNSJFMMFHPL-VKHMYHEASA-N 0.000 description 1
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 1
- 206010061818 Disease progression Diseases 0.000 description 1
- 238000002965 ELISA Methods 0.000 description 1
- 102000005593 Endopeptidases Human genes 0.000 description 1
- 108010059378 Endopeptidases Proteins 0.000 description 1
- 206010066919 Epidemic polyarthritis Diseases 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 108010008165 Etanercept Proteins 0.000 description 1
- 108010071241 Factor XIIa Proteins 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 108010073385 Fibrin Proteins 0.000 description 1
- 102000009123 Fibrin Human genes 0.000 description 1
- BWGVNKXGVNDBDI-UHFFFAOYSA-N Fibrin monomer Chemical compound CNC(=O)CNC(=O)CN BWGVNKXGVNDBDI-UHFFFAOYSA-N 0.000 description 1
- 108010049003 Fibrinogen Proteins 0.000 description 1
- 102000008946 Fibrinogen Human genes 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 206010059028 Gastrointestinal ischaemia Diseases 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 208000022461 Glomerular disease Diseases 0.000 description 1
- 229930186217 Glycolipid Natural products 0.000 description 1
- 229920002683 Glycosaminoglycan Polymers 0.000 description 1
- 102100030386 Granzyme A Human genes 0.000 description 1
- 208000010496 Heart Arrest Diseases 0.000 description 1
- 208000018565 Hemochromatosis Diseases 0.000 description 1
- HTTJABKRGRZYRN-UHFFFAOYSA-N Heparin Chemical compound OC1C(NC(=O)C)C(O)OC(COS(O)(=O)=O)C1OC1C(OS(O)(=O)=O)C(O)C(OC2C(C(OS(O)(=O)=O)C(OC3C(C(O)C(O)C(O3)C(O)=O)OS(O)(=O)=O)C(CO)O2)NS(O)(=O)=O)C(C(O)=O)O1 HTTJABKRGRZYRN-UHFFFAOYSA-N 0.000 description 1
- 101001009599 Homo sapiens Granzyme A Proteins 0.000 description 1
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 1
- 102100030357 Host cell factor 2 Human genes 0.000 description 1
- 108091012469 Host cell factor 2 Proteins 0.000 description 1
- 102000003839 Human Proteins Human genes 0.000 description 1
- 108090000144 Human Proteins Proteins 0.000 description 1
- 102000004157 Hydrolases Human genes 0.000 description 1
- 108090000604 Hydrolases Proteins 0.000 description 1
- 206010070511 Hypoxic-ischaemic encephalopathy Diseases 0.000 description 1
- 108010002162 IgK Proteins 0.000 description 1
- 206010061216 Infarction Diseases 0.000 description 1
- 102000051628 Interleukin-1 receptor antagonist Human genes 0.000 description 1
- 108700021006 Interleukin-1 receptor antagonist Proteins 0.000 description 1
- 108090001005 Interleukin-6 Proteins 0.000 description 1
- UETNIIAIRMUTSM-UHFFFAOYSA-N Jacareubin Natural products CC1(C)OC2=CC3Oc4c(O)c(O)ccc4C(=O)C3C(=C2C=C1)O UETNIIAIRMUTSM-UHFFFAOYSA-N 0.000 description 1
- 206010023203 Joint destruction Diseases 0.000 description 1
- 206010060820 Joint injury Diseases 0.000 description 1
- 108010076876 Keratins Proteins 0.000 description 1
- 102000011782 Keratins Human genes 0.000 description 1
- 125000000998 L-alanino group Chemical group [H]N([*])[C@](C([H])([H])[H])([H])C(=O)O[H] 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-N L-arginine Chemical compound OC(=O)[C@@H](N)CCCN=C(N)N ODKSFYDXXFIFQN-BYPYZUCNSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- 125000000393 L-methionino group Chemical group [H]OC(=O)[C@@]([H])(N([H])[*])C([H])([H])C(SC([H])([H])[H])([H])[H] 0.000 description 1
- FBOZXECLQNJBKD-ZDUSSCGKSA-N L-methotrexate Chemical compound C=1N=C2N=C(N)N=C(N)C2=NC=1CN(C)C1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 FBOZXECLQNJBKD-ZDUSSCGKSA-N 0.000 description 1
- 125000000174 L-prolyl group Chemical group [H]N1C([H])([H])C([H])([H])C([H])([H])[C@@]1([H])C(*)=O 0.000 description 1
- 125000000510 L-tryptophano group Chemical group [H]C1=C([H])C([H])=C2N([H])C([H])=C(C([H])([H])[C@@]([H])(C(O[H])=O)N([H])[*])C2=C1[H] 0.000 description 1
- 108090001030 Lipoproteins Proteins 0.000 description 1
- 102000004895 Lipoproteins Human genes 0.000 description 1
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 1
- 108700033009 MASP2 Deficiency Proteins 0.000 description 1
- 102000018721 Macroglobulins Human genes 0.000 description 1
- 108010091934 Macroglobulins Proteins 0.000 description 1
- 206010025421 Macule Diseases 0.000 description 1
- 101710085938 Matrix protein Proteins 0.000 description 1
- 102000012750 Membrane Glycoproteins Human genes 0.000 description 1
- 108010090054 Membrane Glycoproteins Proteins 0.000 description 1
- 101710127721 Membrane protein Proteins 0.000 description 1
- 208000036626 Mental retardation Diseases 0.000 description 1
- 108090000143 Mouse Proteins Proteins 0.000 description 1
- 208000034486 Multi-organ failure Diseases 0.000 description 1
- 208000010718 Multiple Organ Failure Diseases 0.000 description 1
- 241000699660 Mus musculus Species 0.000 description 1
- 101001056014 Mus musculus Mannan-binding lectin serine protease 2 Proteins 0.000 description 1
- 206010028289 Muscle atrophy Diseases 0.000 description 1
- 208000031888 Mycoses Diseases 0.000 description 1
- 102000005604 Myosin Heavy Chains Human genes 0.000 description 1
- 108010084498 Myosin Heavy Chains Proteins 0.000 description 1
- 201000002481 Myositis Diseases 0.000 description 1
- OVRNDRQMDRJTHS-CBQIKETKSA-N N-Acetyl-D-Galactosamine Chemical compound CC(=O)N[C@H]1[C@@H](O)O[C@H](CO)[C@H](O)[C@@H]1O OVRNDRQMDRJTHS-CBQIKETKSA-N 0.000 description 1
- MBLBDJOUHNCFQT-UHFFFAOYSA-N N-acetyl-D-galactosamine Natural products CC(=O)NC(C=O)C(O)C(O)C(O)CO MBLBDJOUHNCFQT-UHFFFAOYSA-N 0.000 description 1
- 241000921898 Neisseria meningitidis serogroup A Species 0.000 description 1
- 108020004485 Nonsense Codon Proteins 0.000 description 1
- 238000000636 Northern blotting Methods 0.000 description 1
- 208000008558 Osteophyte Diseases 0.000 description 1
- 241001494479 Pecora Species 0.000 description 1
- 208000018262 Peripheral vascular disease Diseases 0.000 description 1
- 206010036030 Polyarthritis Diseases 0.000 description 1
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 1
- 108010026552 Proteome Proteins 0.000 description 1
- 108010094028 Prothrombin Proteins 0.000 description 1
- 102100027378 Prothrombin Human genes 0.000 description 1
- 101001055958 Rattus norvegicus Mannan-binding lectin serine protease 1 Proteins 0.000 description 1
- 101001018257 Rattus norvegicus Mannan-binding lectin serine protease 2 Proteins 0.000 description 1
- 101000889463 Rattus norvegicus Trefoil factor 2 Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 208000033626 Renal failure acute Diseases 0.000 description 1
- 206010061481 Renal injury Diseases 0.000 description 1
- 208000013616 Respiratory Distress Syndrome Diseases 0.000 description 1
- 208000025747 Rheumatic disease Diseases 0.000 description 1
- 241000710942 Ross River virus Species 0.000 description 1
- 208000034189 Sclerosis Diseases 0.000 description 1
- 206010040070 Septic Shock Diseases 0.000 description 1
- 102000003667 Serine Endopeptidases Human genes 0.000 description 1
- 108090000083 Serine Endopeptidases Proteins 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- 230000006044 T cell activation Effects 0.000 description 1
- 108091008874 T cell receptors Proteins 0.000 description 1
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 1
- 108090000190 Thrombin Proteins 0.000 description 1
- 102000004357 Transferases Human genes 0.000 description 1
- 108090000992 Transferases Proteins 0.000 description 1
- 108090000631 Trypsin Proteins 0.000 description 1
- 102000004142 Trypsin Human genes 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 206010047571 Visual impairment Diseases 0.000 description 1
- 108700029631 X-Linked Genes Proteins 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 229960003697 abatacept Drugs 0.000 description 1
- 230000001594 aberrant effect Effects 0.000 description 1
- 230000002159 abnormal effect Effects 0.000 description 1
- 230000005856 abnormality Effects 0.000 description 1
- 238000009825 accumulation Methods 0.000 description 1
- 108091005646 acetylated proteins Proteins 0.000 description 1
- 206010051895 acute chest syndrome Diseases 0.000 description 1
- 201000011040 acute kidney failure Diseases 0.000 description 1
- 206010069351 acute lung injury Diseases 0.000 description 1
- 229960002964 adalimumab Drugs 0.000 description 1
- 230000033289 adaptive immune response Effects 0.000 description 1
- 238000009098 adjuvant therapy Methods 0.000 description 1
- 201000000028 adult respiratory distress syndrome Diseases 0.000 description 1
- 229960002833 aflibercept Drugs 0.000 description 1
- 108010081667 aflibercept Proteins 0.000 description 1
- 230000032683 aging Effects 0.000 description 1
- 125000001931 aliphatic group Chemical group 0.000 description 1
- 230000000735 allogeneic effect Effects 0.000 description 1
- 229960004238 anakinra Drugs 0.000 description 1
- 229940035676 analgesics Drugs 0.000 description 1
- 239000004037 angiogenesis inhibitor Substances 0.000 description 1
- 239000005557 antagonist Substances 0.000 description 1
- 239000000730 antalgic agent Substances 0.000 description 1
- 230000003302 anti-idiotype Effects 0.000 description 1
- 239000002260 anti-inflammatory agent Substances 0.000 description 1
- 229940121363 anti-inflammatory agent Drugs 0.000 description 1
- 230000001640 apoptogenic effect Effects 0.000 description 1
- YZXBAPSDXZZRGB-DOFZRALJSA-N arachidonic acid Chemical class CCCCC\C=C/C\C=C/C\C=C/C\C=C/CCCC(O)=O YZXBAPSDXZZRGB-DOFZRALJSA-N 0.000 description 1
- 210000001188 articular cartilage Anatomy 0.000 description 1
- 238000003149 assay kit Methods 0.000 description 1
- 206010003549 asthenia Diseases 0.000 description 1
- LMEKQMALGUDUQG-UHFFFAOYSA-N azathioprine Chemical compound CN1C=NC([N+]([O-])=O)=C1SC1=NC=NC2=C1NC=N2 LMEKQMALGUDUQG-UHFFFAOYSA-N 0.000 description 1
- 229960002170 azathioprine Drugs 0.000 description 1
- 208000022362 bacterial infectious disease Diseases 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 1
- 230000002146 bilateral effect Effects 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 238000012742 biochemical analysis Methods 0.000 description 1
- 239000003124 biologic agent Substances 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 230000005540 biological transmission Effects 0.000 description 1
- 229960000074 biopharmaceutical Drugs 0.000 description 1
- 230000017531 blood circulation Effects 0.000 description 1
- 230000023555 blood coagulation Effects 0.000 description 1
- 238000004820 blood count Methods 0.000 description 1
- 210000004204 blood vessel Anatomy 0.000 description 1
- 210000001185 bone marrow Anatomy 0.000 description 1
- 208000015322 bone marrow disease Diseases 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 229910052791 calcium Inorganic materials 0.000 description 1
- 125000001314 canonical amino-acid group Chemical group 0.000 description 1
- 102000023852 carbohydrate binding proteins Human genes 0.000 description 1
- 108091008400 carbohydrate binding proteins Proteins 0.000 description 1
- 125000002915 carbonyl group Chemical group [*:2]C([*:1])=O 0.000 description 1
- 230000000747 cardiac effect Effects 0.000 description 1
- 210000000845 cartilage Anatomy 0.000 description 1
- 239000003054 catalyst Substances 0.000 description 1
- 230000001364 causal effect Effects 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 230000005779 cell damage Effects 0.000 description 1
- 230000030833 cell death Effects 0.000 description 1
- 230000010261 cell growth Effects 0.000 description 1
- 210000000170 cell membrane Anatomy 0.000 description 1
- 210000002421 cell wall Anatomy 0.000 description 1
- 206010008118 cerebral infarction Diseases 0.000 description 1
- 230000000739 chaotic effect Effects 0.000 description 1
- 239000002820 chemotaxin Substances 0.000 description 1
- 230000035605 chemotaxis Effects 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 229960001265 ciclosporin Drugs 0.000 description 1
- 206010009259 cleft lip Diseases 0.000 description 1
- 230000001149 cognitive effect Effects 0.000 description 1
- 230000003920 cognitive function Effects 0.000 description 1
- 206010010121 compartment syndrome Diseases 0.000 description 1
- 230000001447 compensatory effect Effects 0.000 description 1
- 108010052926 complement C3d,g Proteins 0.000 description 1
- 201000008560 complement component 3 deficiency Diseases 0.000 description 1
- 230000008094 contradictory effect Effects 0.000 description 1
- 239000013078 crystal Substances 0.000 description 1
- 229930182912 cyclosporin Natural products 0.000 description 1
- 230000001461 cytolytic effect Effects 0.000 description 1
- 238000007822 cytometric assay Methods 0.000 description 1
- 230000007547 defect Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 230000001066 destructive effect Effects 0.000 description 1
- 230000009547 development abnormality Effects 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 230000003292 diminished effect Effects 0.000 description 1
- 230000008034 disappearance Effects 0.000 description 1
- 230000009266 disease activity Effects 0.000 description 1
- 230000005750 disease progression Effects 0.000 description 1
- 238000010494 dissociation reaction Methods 0.000 description 1
- 230000005593 dissociations Effects 0.000 description 1
- PMMYEEVYMWASQN-UHFFFAOYSA-N dl-hydroxyproline Natural products OC1C[NH2+]C(C([O-])=O)C1 PMMYEEVYMWASQN-UHFFFAOYSA-N 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 230000013020 embryo development Effects 0.000 description 1
- 201000002491 encephalomyelitis Diseases 0.000 description 1
- 229940066758 endopeptidases Drugs 0.000 description 1
- 210000002472 endoplasmic reticulum Anatomy 0.000 description 1
- 210000003989 endothelium vascular Anatomy 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- 230000003628 erosive effect Effects 0.000 description 1
- 229960000403 etanercept Drugs 0.000 description 1
- 230000003090 exacerbative effect Effects 0.000 description 1
- 201000010934 exostosis Diseases 0.000 description 1
- 208000012997 experimental autoimmune encephalomyelitis Diseases 0.000 description 1
- 238000013213 extrapolation Methods 0.000 description 1
- 230000001605 fetal effect Effects 0.000 description 1
- 229950003499 fibrin Drugs 0.000 description 1
- 229940012952 fibrinogen Drugs 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- 208000024386 fungal infectious disease Diseases 0.000 description 1
- 229930182830 galactose Natural products 0.000 description 1
- 238000012252 genetic analysis Methods 0.000 description 1
- 231100000852 glomerular disease Toxicity 0.000 description 1
- 239000003862 glucocorticoid Substances 0.000 description 1
- 229930004094 glycosylphosphatidylinositol Natural products 0.000 description 1
- 150000002343 gold Chemical class 0.000 description 1
- 229960001743 golimumab Drugs 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 208000018706 hematopoietic system disease Diseases 0.000 description 1
- 208000007475 hemolytic anemia Diseases 0.000 description 1
- 229960002897 heparin Drugs 0.000 description 1
- 229920000669 heparin Polymers 0.000 description 1
- 239000000710 homodimer Substances 0.000 description 1
- 230000007236 host immunity Effects 0.000 description 1
- 210000003917 human chromosome Anatomy 0.000 description 1
- 238000011577 humanized mouse model Methods 0.000 description 1
- 229910052739 hydrogen Inorganic materials 0.000 description 1
- 230000007062 hydrolysis Effects 0.000 description 1
- 238000006460 hydrolysis reaction Methods 0.000 description 1
- XXSMGPRMXLTPCZ-UHFFFAOYSA-N hydroxychloroquine Chemical compound ClC1=CC=C2C(NC(C)CCCN(CCO)CC)=CC=NC2=C1 XXSMGPRMXLTPCZ-UHFFFAOYSA-N 0.000 description 1
- 229960004171 hydroxychloroquine Drugs 0.000 description 1
- 229960002591 hydroxyproline Drugs 0.000 description 1
- 230000007124 immune defense Effects 0.000 description 1
- 230000001900 immune effect Effects 0.000 description 1
- 238000002991 immunohistochemical analysis Methods 0.000 description 1
- 230000002055 immunohistochemical effect Effects 0.000 description 1
- 238000011532 immunohistochemical staining Methods 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 230000002779 inactivation Effects 0.000 description 1
- 230000007574 infarction Effects 0.000 description 1
- 239000012678 infectious agent Substances 0.000 description 1
- 230000002458 infectious effect Effects 0.000 description 1
- 210000004969 inflammatory cell Anatomy 0.000 description 1
- 229960000598 infliximab Drugs 0.000 description 1
- 210000005007 innate immune system Anatomy 0.000 description 1
- 230000010354 integration Effects 0.000 description 1
- 208000037817 intestinal injury Diseases 0.000 description 1
- 210000000936 intestine Anatomy 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 230000009545 invasion Effects 0.000 description 1
- 238000011835 investigation Methods 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- 230000000622 irritating effect Effects 0.000 description 1
- 230000007654 ischemic lesion Effects 0.000 description 1
- 230000008407 joint function Effects 0.000 description 1
- 210000003127 knee Anatomy 0.000 description 1
- 230000002045 lasting effect Effects 0.000 description 1
- 201000010901 lateral sclerosis Diseases 0.000 description 1
- VHOGYURTWQBHIL-UHFFFAOYSA-N leflunomide Chemical compound O1N=CC(C(=O)NC=2C=CC(=CC=2)C(F)(F)F)=C1C VHOGYURTWQBHIL-UHFFFAOYSA-N 0.000 description 1
- 229960000681 leflunomide Drugs 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 210000005228 liver tissue Anatomy 0.000 description 1
- 230000033001 locomotion Effects 0.000 description 1
- 238000011866 long-term treatment Methods 0.000 description 1
- 230000007774 longterm Effects 0.000 description 1
- 210000004698 lymphocyte Anatomy 0.000 description 1
- 230000002934 lysing effect Effects 0.000 description 1
- 230000007257 malfunction Effects 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 238000007726 management method Methods 0.000 description 1
- 102000036209 mannose binding proteins Human genes 0.000 description 1
- 108020003928 mannose binding proteins Proteins 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 230000008774 maternal effect Effects 0.000 description 1
- 101150011109 mbl gene Proteins 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000010534 mechanism of action Effects 0.000 description 1
- 230000002503 metabolic effect Effects 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 229960000485 methotrexate Drugs 0.000 description 1
- 206010062198 microangiopathy Diseases 0.000 description 1
- 244000000010 microbial pathogen Species 0.000 description 1
- 210000000274 microglia Anatomy 0.000 description 1
- 150000002772 monosaccharides Chemical group 0.000 description 1
- 241000609532 mosquito-borne viruses Species 0.000 description 1
- 208000005264 motor neuron disease Diseases 0.000 description 1
- 208000029744 multiple organ dysfunction syndrome Diseases 0.000 description 1
- 230000020763 muscle atrophy Effects 0.000 description 1
- 230000003387 muscular Effects 0.000 description 1
- 201000000585 muscular atrophy Diseases 0.000 description 1
- 230000003680 myocardial damage Effects 0.000 description 1
- 208000037891 myocardial injury Diseases 0.000 description 1
- 208000031225 myocardial ischemia Diseases 0.000 description 1
- 230000016273 neuron death Effects 0.000 description 1
- 230000000422 nocturnal effect Effects 0.000 description 1
- 239000000041 non-steroidal anti-inflammatory agent Substances 0.000 description 1
- 229940021182 non-steroidal anti-inflammatory drug Drugs 0.000 description 1
- 230000037434 nonsense mutation Effects 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 150000003833 nucleoside derivatives Chemical class 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 230000003571 opsonizing effect Effects 0.000 description 1
- 230000008816 organ damage Effects 0.000 description 1
- 230000003204 osmotic effect Effects 0.000 description 1
- 230000001314 paroxysmal effect Effects 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 230000008807 pathological lesion Effects 0.000 description 1
- 229940005014 pegaptanib sodium Drugs 0.000 description 1
- 229960001639 penicillamine Drugs 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 210000005259 peripheral blood Anatomy 0.000 description 1
- 239000011886 peripheral blood Substances 0.000 description 1
- 230000002085 persistent effect Effects 0.000 description 1
- 230000000144 pharmacologic effect Effects 0.000 description 1
- 238000011458 pharmacological treatment Methods 0.000 description 1
- 230000000649 photocoagulation Effects 0.000 description 1
- 239000000049 pigment Substances 0.000 description 1
- 230000036470 plasma concentration Effects 0.000 description 1
- 208000030428 polyarticular arthritis Diseases 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 238000006116 polymerization reaction Methods 0.000 description 1
- 230000000379 polymerizing effect Effects 0.000 description 1
- 239000011148 porous material Substances 0.000 description 1
- 239000013641 positive control Substances 0.000 description 1
- 238000011533 pre-incubation Methods 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 238000004321 preservation Methods 0.000 description 1
- 238000012545 processing Methods 0.000 description 1
- 238000004393 prognosis Methods 0.000 description 1
- WJNRPILHGGKWCK-UHFFFAOYSA-N propazine Chemical compound CC(C)NC1=NC(Cl)=NC(NC(C)C)=N1 WJNRPILHGGKWCK-UHFFFAOYSA-N 0.000 description 1
- 230000009979 protective mechanism Effects 0.000 description 1
- 108020001580 protein domains Proteins 0.000 description 1
- 230000006337 proteolytic cleavage Effects 0.000 description 1
- 229940039716 prothrombin Drugs 0.000 description 1
- 150000003214 pyranose derivatives Chemical group 0.000 description 1
- 229960003876 ranibizumab Drugs 0.000 description 1
- 239000003642 reactive oxygen metabolite Substances 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 230000006798 recombination Effects 0.000 description 1
- 238000005215 recombination Methods 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000000306 recurrent effect Effects 0.000 description 1
- 238000002271 resection Methods 0.000 description 1
- 230000002207 retinal effect Effects 0.000 description 1
- 229960004641 rituximab Drugs 0.000 description 1
- 238000011808 rodent model Methods 0.000 description 1
- 229920006395 saturated elastomer Polymers 0.000 description 1
- 230000009291 secondary effect Effects 0.000 description 1
- 230000036303 septic shock Effects 0.000 description 1
- 230000001568 sexual effect Effects 0.000 description 1
- 230000035939 shock Effects 0.000 description 1
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 1
- 208000007056 sickle cell anemia Diseases 0.000 description 1
- 230000037432 silent mutation Effects 0.000 description 1
- 210000002460 smooth muscle Anatomy 0.000 description 1
- 239000007787 solid Substances 0.000 description 1
- 238000001228 spectrum Methods 0.000 description 1
- 230000006641 stabilisation Effects 0.000 description 1
- 238000011105 stabilization Methods 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 210000000130 stem cell Anatomy 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- NCEXYHBECQHGNR-QZQOTICOSA-N sulfasalazine Chemical compound C1=C(O)C(C(=O)O)=CC(\N=N\C=2C=CC(=CC=2)S(=O)(=O)NC=2N=CC=CC=2)=C1 NCEXYHBECQHGNR-QZQOTICOSA-N 0.000 description 1
- 229960001940 sulfasalazine Drugs 0.000 description 1
- NCEXYHBECQHGNR-UHFFFAOYSA-N sulfasalazine Natural products C1=C(O)C(C(=O)O)=CC(N=NC=2C=CC(=CC=2)S(=O)(=O)NC=2N=CC=CC=2)=C1 NCEXYHBECQHGNR-UHFFFAOYSA-N 0.000 description 1
- 230000000153 supplemental effect Effects 0.000 description 1
- 230000001629 suppression Effects 0.000 description 1
- 208000010648 susceptibility to HIV infection Diseases 0.000 description 1
- 230000002459 sustained effect Effects 0.000 description 1
- 230000008961 swelling Effects 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 210000005222 synovial tissue Anatomy 0.000 description 1
- 201000004595 synovitis Diseases 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 229940037128 systemic glucocorticoids Drugs 0.000 description 1
- 229940124597 therapeutic agent Drugs 0.000 description 1
- 125000003396 thiol group Chemical group [H]S* 0.000 description 1
- 229960004072 thrombin Drugs 0.000 description 1
- 230000002345 thrombinlike Effects 0.000 description 1
- 230000001732 thrombotic effect Effects 0.000 description 1
- 229960003989 tocilizumab Drugs 0.000 description 1
- 230000000699 topical effect Effects 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- FGMPLJWBKKVCDB-UHFFFAOYSA-N trans-L-hydroxy-proline Natural products ON1CCCC1C(O)=O FGMPLJWBKKVCDB-UHFFFAOYSA-N 0.000 description 1
- 238000011222 transcriptome analysis Methods 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 230000000472 traumatic effect Effects 0.000 description 1
- 238000011269 treatment regimen Methods 0.000 description 1
- 230000001960 triggered effect Effects 0.000 description 1
- 239000012588 trypsin Substances 0.000 description 1
- 229940046728 tumor necrosis factor alpha inhibitor Drugs 0.000 description 1
- 239000002451 tumor necrosis factor inhibitor Substances 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 description 1
- 230000008728 vascular permeability Effects 0.000 description 1
- 230000006442 vascular tone Effects 0.000 description 1
- 230000029812 viral genome replication Effects 0.000 description 1
- 208000029257 vision disease Diseases 0.000 description 1
- 238000002424 x-ray crystallography Methods 0.000 description 1
Images





































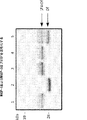























































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/02—Antithrombotic agents; Anticoagulants; Platelet aggregation inhibitors
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
- A61K39/39533—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals
- A61K39/3955—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum against materials from animals against proteinaceous materials, e.g. enzymes, hormones, lymphokines
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
- A61P11/06—Antiasthmatics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
- A61P11/16—Central respiratory analeptics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P13/00—Drugs for disorders of the urinary system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P13/00—Drugs for disorders of the urinary system
- A61P13/12—Drugs for disorders of the urinary system of the kidneys
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P19/00—Drugs for skeletal disorders
- A61P19/02—Drugs for skeletal disorders for joint disorders, e.g. arthritis, arthrosis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P21/00—Drugs for disorders of the muscular or neuromuscular system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P21/00—Drugs for disorders of the muscular or neuromuscular system
- A61P21/04—Drugs for disorders of the muscular or neuromuscular system for myasthenia gravis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/02—Drugs for disorders of the nervous system for peripheral neuropathies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
- A61P25/28—Drugs for disorders of the nervous system for treating neurodegenerative disorders of the central nervous system, e.g. nootropic agents, cognition enhancers, drugs for treating Alzheimer's disease or other forms of dementia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P27/00—Drugs for disorders of the senses
- A61P27/02—Ophthalmic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/06—Immunosuppressants, e.g. drugs for graft rejection
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P39/00—General protective or antinoxious agents
- A61P39/02—Antidotes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/06—Antianaemics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/08—Plasma substitutes; Perfusion solutions; Dialytics or haemodialytics; Drugs for electrolytic or acid-base disorders, e.g. hypovolemic shock
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/10—Drugs for disorders of the cardiovascular system for treating ischaemic or atherosclerotic diseases, e.g. antianginal drugs, coronary vasodilators, drugs for myocardial infarction, retinopathy, cerebrovascula insufficiency, renal arteriosclerosis
-
- B—PERFORMING OPERATIONS; TRANSPORTING
- B82—NANOTECHNOLOGY
- B82Y—SPECIFIC USES OR APPLICATIONS OF NANOSTRUCTURES; MEASUREMENT OR ANALYSIS OF NANOSTRUCTURES; MANUFACTURE OR TREATMENT OF NANOSTRUCTURES
- B82Y25/00—Nanomagnetism, e.g. magnetoimpedance, anisotropic magnetoresistance, giant magnetoresistance or tunneling magnetoresistance
-
- C—CHEMISTRY; METALLURGY
- C01—INORGANIC CHEMISTRY
- C01F—COMPOUNDS OF THE METALS BERYLLIUM, MAGNESIUM, ALUMINIUM, CALCIUM, STRONTIUM, BARIUM, RADIUM, THORIUM, OR OF THE RARE-EARTH METALS
- C01F17/00—Compounds of rare earth metals
- C01F17/20—Compounds containing only rare earth metals as the metal element
- C01F17/206—Compounds containing only rare earth metals as the metal element oxide or hydroxide being the only anion
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/40—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against enzymes
-
- C—CHEMISTRY; METALLURGY
- C09—DYES; PAINTS; POLISHES; NATURAL RESINS; ADHESIVES; COMPOSITIONS NOT OTHERWISE PROVIDED FOR; APPLICATIONS OF MATERIALS NOT OTHERWISE PROVIDED FOR
- C09K—MATERIALS FOR MISCELLANEOUS APPLICATIONS, NOT PROVIDED FOR ELSEWHERE
- C09K11/00—Luminescent, e.g. electroluminescent, chemiluminescent materials
- C09K11/08—Luminescent, e.g. electroluminescent, chemiluminescent materials containing inorganic luminescent materials
- C09K11/77—Luminescent, e.g. electroluminescent, chemiluminescent materials containing inorganic luminescent materials containing rare earth metals
-
- C—CHEMISTRY; METALLURGY
- C09—DYES; PAINTS; POLISHES; NATURAL RESINS; ADHESIVES; COMPOSITIONS NOT OTHERWISE PROVIDED FOR; APPLICATIONS OF MATERIALS NOT OTHERWISE PROVIDED FOR
- C09K—MATERIALS FOR MISCELLANEOUS APPLICATIONS, NOT PROVIDED FOR ELSEWHERE
- C09K11/00—Luminescent, e.g. electroluminescent, chemiluminescent materials
- C09K11/08—Luminescent, e.g. electroluminescent, chemiluminescent materials containing inorganic luminescent materials
- C09K11/77—Luminescent, e.g. electroluminescent, chemiluminescent materials containing inorganic luminescent materials containing rare earth metals
- C09K11/7728—Luminescent, e.g. electroluminescent, chemiluminescent materials containing inorganic luminescent materials containing rare earth metals containing europium
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/48—Hydrolases (3) acting on peptide bonds (3.4)
- C12N9/50—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25)
- C12N9/64—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25) derived from animal tissue
- C12N9/6421—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25) derived from animal tissue from mammals
- C12N9/6424—Serine endopeptidases (3.4.21)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y304/00—Hydrolases acting on peptide bonds, i.e. peptidases (3.4)
- C12Y304/21—Serine endopeptidases (3.4.21)
- C12Y304/21104—Mannan-binding lectin-associated serine protease-2 (3.4.21.104)
-
- H—ELECTRICITY
- H01—ELECTRIC ELEMENTS
- H01F—MAGNETS; INDUCTANCES; TRANSFORMERS; SELECTION OF MATERIALS FOR THEIR MAGNETIC PROPERTIES
- H01F1/00—Magnets or magnetic bodies characterised by the magnetic materials therefor; Selection of materials for their magnetic properties
- H01F1/0036—Magnets or magnetic bodies characterised by the magnetic materials therefor; Selection of materials for their magnetic properties showing low dimensional magnetism, i.e. spin rearrangements due to a restriction of dimensions, e.g. showing giant magnetoresistivity
- H01F1/0045—Zero dimensional, e.g. nanoparticles, soft nanoparticles for medical/biological use
- H01F1/0054—Coated nanoparticles, e.g. nanoparticles coated with organic surfactant
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/54—Medicinal preparations containing antigens or antibodies characterised by the route of administration
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/545—Medicinal preparations containing antigens or antibodies characterised by the dose, timing or administration schedule
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
-
- C—CHEMISTRY; METALLURGY
- C01—INORGANIC CHEMISTRY
- C01P—INDEXING SCHEME RELATING TO STRUCTURAL AND PHYSICAL ASPECTS OF SOLID INORGANIC COMPOUNDS
- C01P2002/00—Crystal-structural characteristics
- C01P2002/50—Solid solutions
- C01P2002/52—Solid solutions containing elements as dopants
- C01P2002/54—Solid solutions containing elements as dopants one element only
-
- C—CHEMISTRY; METALLURGY
- C01—INORGANIC CHEMISTRY
- C01P—INDEXING SCHEME RELATING TO STRUCTURAL AND PHYSICAL ASPECTS OF SOLID INORGANIC COMPOUNDS
- C01P2002/00—Crystal-structural characteristics
- C01P2002/80—Crystal-structural characteristics defined by measured data other than those specified in group C01P2002/70
- C01P2002/84—Crystal-structural characteristics defined by measured data other than those specified in group C01P2002/70 by UV- or VIS- data
-
- C—CHEMISTRY; METALLURGY
- C01—INORGANIC CHEMISTRY
- C01P—INDEXING SCHEME RELATING TO STRUCTURAL AND PHYSICAL ASPECTS OF SOLID INORGANIC COMPOUNDS
- C01P2004/00—Particle morphology
- C01P2004/60—Particles characterised by their size
- C01P2004/64—Nanometer sized, i.e. from 1-100 nanometer
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/24—Immunoglobulins specific features characterized by taxonomic origin containing regions, domains or residues from different species, e.g. chimeric, humanized or veneered
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/33—Crossreactivity, e.g. for species or epitope, or lack of said crossreactivity
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/34—Identification of a linear epitope shorter than 20 amino acid residues or of a conformational epitope defined by amino acid residues
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
- C07K2317/734—Complement-dependent cytotoxicity [CDC]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
Description
本願は、2016年8月1日に出願された米国特許仮出願第62/369,674号の恩典を主張し、また2016年11月8日に出願された米国特許仮出願第62/419,420号の恩典を主張し、さらに2017年3月29日に出願された米国特許仮出願第62/478,336号の恩典を主張するものであり、それらの3件全てはそれらの全体が参照により組み入れられる。
本願に関連する配列表はハードコピーの代わりにテキスト形式で提供され、本明細書に参照により組み入れられる。配列表を含むテキストファイルの名前は、MP_1_0254_US_Sequence_Listing_20170628_ST25であり、このファイルは191KBであり、2017年6月28日に作成されたものであり、また本明細書の出願と共にEFS-Webを介して提出されている。
補体系は、ヒトおよび他の脊椎動物において微生物感染および他の急性侵襲に対する免疫応答を開始、増幅、および組織化するための初期作用機構を提供する(M.K. Liszewski and J. P. Atkinson, 1993, in Fundamental Immunology, Third Edition, W.E, Paul編, Raven Press, Ltd., New York)。補体活性化は潜在的な病原体に対する有益な第一線の防御を提供するが、防御免疫応答を促進する補体の活性は宿主にとって潜在的な脅威となることもある(K.R, Kalli, et al., Springer Semin. Immunopathol. 15:417-431, 1994; B.P. Morgan, Eur. J. Clinical Investig. 24:219-228, 1994)。例えば、C3およびC5タンパク質分解産物は好中球を動員および活性化する。活性化好中球は宿主防御に不可欠であるが見境なく破壊酵素を放出し、臓器損傷を引き起こすことがある。さらに、補体活性化によって、近くの宿主細胞ならびに微生物標的の表面に溶解性補体成分が沈着し、その結果、宿主細胞が溶解することがある。
1つの局面において、本発明は、高い親和性(500pM未満のKDを有する)でヒトMASP-3のセリンプロテアーゼドメイン(SEQ ID NO:2のアミノ酸残基450〜728)に特異的に結合し、第二経路補体活性化を阻害する、単離されたモノクローナル抗体またはその抗原結合断片を提供する。いくつかの態様において、抗体または抗原結合断片は、以下の性質の少なくとも1つまたは複数を特徴とする:(a)プロD因子成熟を阻害すること;(b)ヒトMASP-1(SEQ ID NO:8)に結合しないこと;(c)哺乳動物対象において約1:1〜約2.5:1(MASP-3標的:mAb)のモル比で第二経路を阻害すること;(d)古典的経路を阻害しないこと;(e)溶血および/もしくはオプソニン化を阻害すること;(f)MASP-3セリンプロテアーゼ基質特異的切断を阻害すること;(g)溶血を減少させること、もしくはC3切断およびC3b表面沈着の減少;(h)活性化表面へのB因子および/もしくはBb因子沈着を減少させること;(i)プロD因子と比べて、静止レベル(循環中、かつ活性化表面の実験的添加なし)の活性D因子を減少させること;(j)活性化表面に応答して、プロD因子と比べて活性D因子のレベルを減少させること;(k)静止レベルおよび表面誘導レベルの液相Ba、Bb、C3b、もしくはC3aの生成を減少させること;ならびに/または(l)P因子沈着を減少させること。いくつかの態様において、ヒトMASP-3のセリンプロテアーゼドメイン中に位置するエピトープに特異的に結合し、該エピトープが、

(a)
(b)SEQ ID NO:125(DYYMN)に記載されるHC-CDR1、
(c)SEQ ID NO:137に記載されるHC-CDR1、SEQ ID NO:138に記載されるHC-CDR2、SEQ ID NO:140に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:206に記載されるLC-CDR1、SEQ ID NO:207に記載されるLC-CDR2、およびSEQ ID NO:208に記載されるLC-CDR3を含む、軽鎖可変領域;または
(d)SEQ ID NO:98に記載されるHC-CDR1、SEQ ID NO:99に記載されるHC-CDR2、SEQ ID NO:101に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:169に記載されるLC-CDR1、SEQ ID NO:171に記載されるLC-CDR2、およびSEQ ID NO:173に記載されるLC-CDR3を含む、軽鎖可変領域;または
(e)SEQ ID NO:103に記載されるHC-CDR1、SEQ ID NO:105に記載されるHC-CDR2、SEQ ID NO:107に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:176に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む、軽鎖可変領域;または
(f)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:116に記載されるHC-CDR2、SEQ ID NO:118に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:188に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:190に記載されるLC-CDR3を含む、軽鎖可変領域;または
(g)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:121に記載されるHC-CDR2、SEQ ID NO:123に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:191に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む、軽鎖可変領域
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片を提供する。
高い親和性(500pM未満のK D を有する)でヒトMASP-3のセリンプロテアーゼドメイン(SEQ ID NO:2のアミノ酸残基450〜728)に特異的に結合し、第二経路補体活性化を阻害する、単離されたモノクローナル抗体またはその抗原結合断片。
[本発明1002]
以下の性質の少なくとも1つまたは複数を特徴とする、本発明1001の単離された抗体またはその抗原結合断片:
(a)プロD因子成熟を阻害すること;
(b)ヒトMASP-1(SEQ ID NO:8)に結合しないこと;
(c)哺乳動物対象において約1:1〜約2.5:1(MASP-3標的:mAb)のモル比で第二経路を阻害すること;
(d)古典経路を阻害しないこと;
(e)溶血および/もしくはオプソニン化の阻害;
(f)MASP-3セリンプロテアーゼ基質特異的切断の阻害;
(g)溶血の減少、もしくはC3切断およびC3b表面沈着の減少;
(h)活性化表面へのB因子およびBb因子沈着の減少;
(i)プロD因子と比べた静止レベル(循環中、かつ活性化表面の実験的添加なし)の活性D因子の減少;
(j)活性化表面に応答した、プロD因子と比べた活性D因子のレベルの減少;
(k)静止レベルおよび表面誘導レベルの液相Ba、Bb、C3b、もしくはC3aの生成の減少;ならびに/または
(l)P因子沈着の減少。
[本発明1003]
前記ヒトMASP-3のセリンプロテアーゼドメイン中に位置するエピトープに特異的に結合し、該エピトープが、
[本発明1004]
SEQ ID NO:15の中のエピトープに結合する、本発明1003の抗体またはその抗原結合断片。
[本発明1005]
SEQ ID NO:10の中のエピトープに結合する、本発明1003の抗体またはその抗原結合断片。
[本発明1006]
SEQ ID NO:9の中のエピトープに結合する、本発明1003の抗体またはその抗原結合断片。
[本発明1007]
SEQ ID NO:12の中のエピトープにも結合する、本発明1005の抗体またはその抗原結合断片。
[本発明1008]
SEQ ID NO:11、SEQ ID NO:13、および/またはSEQ ID NO:14の少なくとも1つの中のエピトープにも結合する、本発明1006の抗体またはその抗原結合断片。
[本発明1009]
前記抗体が、
[本発明1010]
(a)SEQ ID NO:84(GKWIE)に記載されるHC-CDR1;
(b)SEQ ID NO:213(SYGXX、4位のXがMもしくはIでありかつ5位のXがSもしくはTである)に記載されるHC-CDR1;SEQ ID NO:74に記載されるHC-CDR2;およびSEQ ID NO:214(GGXAXDY、3位のXがEもしくはDでありかつ5位のXがMもしくはLである)に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:215(
(c)SEQ ID NO:209(XXDIN、1位のXがSもしくはTでありかつ2位のXがNもしくはDである)に記載されるHC-CDR1;SEQ ID NO:210(
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片。
[本発明1011]
前記LC-CDR1がSEQ ID NO:258を含む、本発明1010(a)の単離された抗体またはその抗原結合断片。
[本発明1012]
SEQ ID NO:30、SEQ ID NO:254、またはSEQ ID NO:255と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:45、SEQ ID NO:256、またはSEQ ID NO:280と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1010(a)の単離された抗体またはその抗原結合断片。
[本発明1013]
(a)の重鎖可変領域のHC-CDR1がSEQ ID NO:72(SYGMS)を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1014]
(a)の重鎖可変領域のHC-CDR1がSEQ ID NO:79(SYGIT)を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1015]
(a)の重鎖可変領域のHC-CDR3がSEQ ID NO:76(GGEAMDY)を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1016]
(a)の重鎖可変領域のHC-CDR3がSEQ ID NO:82(GGDALDY)を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1017]
(b)の軽鎖可変領域のLC-CDR1が、
[本発明1018]
前記軽鎖可変領域のLC-CDR1がSEQ ID NO:152を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1019]
(b)の軽鎖可変領域のLC-CDR3が、
[本発明1020]
(b)の軽鎖可変領域のLC-CDR3がSEQ ID NO:160(WQGTHFPYT)を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1021]
前記HC-CDR1がSEQ ID NO:72を含み、前記HC-CDR2がSEQ ID NO:74を含み、前記HC-CDR3がSEQ ID NO:76を含み、前記LC-CDR1がSEQ ID NO:153、SEQ ID NO:261、SEQ ID NO:262、またはSEQ ID NO:263を含み、前記LC-CDR2がSEQ ID NO:155を含み、かつ前記LC-CDR3がSEQ ID NO:157を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1022]
前記HC-CDR1がSEQ ID NO:79を含み、前記HC-CDR2がSEQ ID NO:74を含み、前記HC-CDR3がSEQ ID NO:82を含み、前記LC-CDR1がSEQ ID NO:159を含み、前記LC-CDR2がSEQ ID NO:155を含み、かつ前記LC-CDR3がSEQ ID NO:160を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1023]
SEQ ID NO:28、SEQ ID NO:251、またはSEQ ID NO:252と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:43、SEQ ID NO:253、またはSEQ ID NO:279と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1024]
SEQ ID NO:29と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:44と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1010(b)の単離された抗体またはその抗原結合断片。
[本発明1025]
(a)の重鎖可変領域のHC-CDR1がSEQ ID NO:56(TDDIN)を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1026]
(a)の重鎖可変領域のHC-CDR1がSEQ ID NO:62(SNDIN)を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1027]
(a)の重鎖可変領域のHC-CDR2が、
[本発明1028]
(a)の重鎖可変領域のHC-CDR2が、
[本発明1029]
(a)の重鎖可変領域のHC-CDR2が、
[本発明1030]
(a)の重鎖可変領域のHC-CDR2が、
[本発明1031]
(a)の重鎖可変領域のHC-CDR3がSEQ ID NO:60(LEDTY)を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1032]
(a)の重鎖可変領域のHC-CDR3がSEQ ID NO:65(VEDSY)を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1033]
(b)の軽鎖可変領域のLC-CDR1が、
[本発明1034]
(b)の軽鎖可変領域のLC-CDR1が、
[本発明1035]
前記HC-CDR1がSEQ ID NO:56を含み、前記HC-CDR2がSEQ ID NO:58を含み、前記HC-CDR3がSEQ ID NO:60を含み、かつ、前記LC-CDR1がSEQ ID NO:142、SEQ ID NO:257、SEQ ID NO:258、またはSEQ ID NO:259を含み、前記LC-CDR2がSEQ ID NO:144を含み、かつ前記LC-CDR3がSEQ ID NO:146を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1036]
前記HC-CDR1がSEQ ID NO:62を含み、前記HC-CDR2がSEQ ID NO:63、SEQ ID NO:67、またはSEQ ID NO:69を含み、かつ前記HC-CDR3がSEQ ID NO:65を含み、かつ、前記LC-CDR1がSEQ ID NO:149を含み、前記LC-CDR2がSEQ ID NO:144を含み、かつ前記LC-CDR3がSEQ ID NO:146を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1037]
SEQ ID NO:24、SEQ ID NO:248、またはSEQ ID NO:249と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:40、SEQ ID NO:250、またはSEQ ID NO:278と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1038]
SEQ ID NO:25と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:41と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1039]
SEQ ID NO:26と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:42と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1040]
SEQ ID NO:27と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:42と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1010(c)の単離された抗体またはその抗原結合断片。
[本発明1041]
(a)SEQ ID NO:91(GKWIE)に記載されるHC-CDR1、
(b)
(c)SEQ ID NO:125(DYYMN)に記載されるHC-CDR1、
(d)SEQ ID NO:132に記載されるHC-CDR1、SEQ ID NO:133に記載されるHC-CDR2、SEQ ID NO:135に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:203に記載されるLC-CDR1、SEQ ID NO:165に記載されるLC-CDR2、およびSEQ ID NO:204に記載されるLC-CDR3を含む、軽鎖可変領域;または
(e)SEQ ID NO:137に記載されるHC-CDR1、SEQ ID NO:138に記載されるHC-CDR2、SEQ ID NO:140に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:206に記載されるLC-CDR1、SEQ ID NO:207に記載されるLC-CDR2、およびSEQ ID NO:208に記載されるLC-CDR3を含む、軽鎖可変領域;または
(f)SEQ ID NO:98に記載されるHC-CDR1、SEQ ID NO:99に記載されるHC-CDR2、SEQ ID NO:101に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:169に記載されるLC-CDR1、SEQ ID NO:171に記載されるLC-CDR2、およびSEQ ID NO:173に記載されるLC-CDR3を含む、軽鎖可変領域;または
(g)SEQ ID NO:103に記載されるHC-CDR1、SEQ ID NO:105に記載されるHC-CDR2、SEQ ID NO:107に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:176に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む、軽鎖可変領域;または
(h)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:116に記載されるHC-CDR2、SEQ ID NO:118に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:188に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:190に記載されるLC-CDR3を含む、軽鎖可変領域;または
(i)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:121に記載されるHC-CDR2、SEQ ID NO:123に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:191に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む、軽鎖可変領域
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片。
[本発明1042]
SEQ ID NO:31と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:46と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(a)の単離された抗体またはその抗原結合断片。
[本発明1043]
SEQ ID NO:32と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:47と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(b)の抗体またはその抗原結合断片。
[本発明1044]
SEQ ID NO:33と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:48と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(c)の抗体またはその抗原結合断片。
[本発明1045]
SEQ ID NO:34と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:49と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(d)の抗体またはその抗原結合断片。
[本発明1046]
SEQ ID NO:35と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:50と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(e)の抗体またはその抗原結合断片。
[本発明1047]
SEQ ID NO:36と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:51と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(f)の抗体またはその抗原結合断片。
[本発明1048]
SEQ ID NO:37と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:52と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(g)の抗体またはその抗原結合断片。
[本発明1049]
SEQ ID NO:38と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:53と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(h)の抗体またはその抗原結合断片。
[本発明1050]
SEQ ID NO:39と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:54と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、本発明1041(i)の抗体またはその抗原結合断片。
[本発明1051]
ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、本発明1001、1010、または1041のいずれかの抗体またはその抗原結合断片。
[本発明1052]
単鎖抗体、scFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、本発明1001、1010、または1041のいずれかの抗体またはその抗原結合断片。
[本発明1053]
免疫グロブリン定常領域をさらに含む、本発明1001、1010、または1041のいずれかの抗体またはその抗原結合断片。
[本発明1054]
ヒト化されている、項目1〜23のいずれかの抗体またはその抗原結合断片。
[本発明1055]
前記抗体が、500pM未満の親和性で前記ヒトMASP-3のセリンプロテアーゼドメインに結合する、本発明1001、1010、または1041のいずれかの抗体またはその抗原結合断片。
[本発明1056]
前記抗体が哺乳動物血液中で第二経路活性化を阻害する、本発明1001、1010、または1041のいずれかの抗体またはその抗原結合断片。
[本発明1057]
本発明1001〜1056のいずれかの抗体またはその抗原結合断片の重鎖可変領域および/または軽鎖可変領域をコードする、単離されたDNA配列。
[本発明1058]
本発明1057のDNA配列の1つまたは複数を含む、クローニングベクターまたは発現ベクター。
[本発明1059]
本発明1058のクローニングベクターまたは発現ベクターの1つまたは複数を含む、宿主細胞。
[本発明1060]
本発明1059の宿主細胞を培養する工程、および前記抗体またはその抗原結合断片を単離する工程を含む、本発明1001〜1056のいずれかの抗体または抗原結合断片を産生するための方法。
[本発明1061]
本発明1001〜1056のいずれかの抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
[本発明1062]
それを必要とする哺乳動物対象に、該哺乳動物における第二経路補体活性化を阻害するのに十分な量の、高親和性MASP-3阻害抗体またはその抗原結合断片を含む本発明1061の組成物を投与する工程を含む、哺乳動物における第二経路補体活性化を阻害する方法。
[本発明1063]
前記抗体またはその抗原結合断片が、500pM未満の親和性でMASP-3に結合する、本発明1062の方法。
[本発明1064]
前記抗体が、約1:1〜約2.5:1(MASP-3標的:MASP-3阻害性mAb)のモル比で第二経路を阻害する、本発明1062または1063の方法。
[本発明1065]
前記抗体またはその抗原結合断片が、古典経路活性化に影響することなく、第二経路を選択的に阻害する、本発明1062の方法。
[本発明1066]
前記それを必要とする対象が、発作性夜間血色素尿症(PNH)、加齢黄斑変性症(AMD、ウェット型およびドライ型AMDを含む)、虚血再灌流障害、関節炎、播種性血管内凝固、血栓性微小血管症(溶血性尿毒症症候群(HUS)、非典型溶血性尿毒症症候群(aHUS)、血栓性血小板減少性紫斑病(TTP)、もしくは移植後TMAを含む)、喘息、デンスデポジット病、微量免疫型壊死性半月体形成性糸球体腎炎、外傷性脳損傷、誤嚥性肺炎、眼内炎、視神経脊髄炎、ベーチェット病、多発性硬化症、ギランバレー症候群、アルツハイマー病、筋萎縮性側索硬化症(ALS)、ループス腎炎、全身性エリテマトーデス(SLE)、糖尿病性網膜症、ブドウ膜炎、慢性閉塞性肺疾患(COPD)、C3糸球体症、移植片拒絶反応、移植片対宿主病(GVHD)、血液透析、敗血症、全身性炎症反応症候群(SIRS)、急性呼吸窮迫症候群(ARDS)、ANCA血管炎、抗リン脂質症候群、アテローム性動脈硬化症、IgA腎症、ならびに重症筋無力症からなる群より選択される第二経路疾患もしくは第二経路障害に罹患しているか、またはそれを発症する危険にある、本発明1062の方法。
本明細書に記載される発明のこれらおよび他の局面および態様は、以下の詳細な説明および図面を参照することによって明らかになるであろう。本明細書において参照される米国特許、米国特許出願公開公報、米国特許出願、外国特許、外国特許出願、および非特許刊行物はすべて、それぞれが個々に組み入れられるごとく、全体として参照により本明細書に組み入れられる。
SEQ ID NO:1 ヒトMASP-3 cDNA
SEQ ID NO:2 ヒトMASP-3タンパク質(リーダあり)
SEQ ID NO:3 マウスMASP-3タンパク質(リーダあり)
SEQ ID NO:4 ラットMASP-3タンパク質
SEQ ID NO:5 ニワトリMASP-3タンパク質
SEQ ID NO:6 ウサギMASP-3タンパク質
SEQ ID NO:7 カニクイザルMASP-3タンパク質
SEQ ID NO:8 ヒトMASP-1タンパク質(リーダあり)
ヒトMASP-3 SPドメインペプチド断片:
SEQ ID NO:9(ヒトMASP-3のアミノ酸498〜509、リーダあり)
SEQ ID NO:10(ヒトMASP-3のアミノ酸494〜508、リーダあり)
SEQ ID NO:11(ヒトMASP-3のアミノ酸544〜558、リーダあり)
SEQ ID NO:12(ヒトMASP-3のアミノ酸626〜638、リーダあり)
SEQ ID NO:13(ヒトMASP-3のアミノ酸639〜649、リーダあり)
SEQ ID NO:14(ヒトMASP-3のアミノ酸704〜713、リーダあり)
SEQ ID NO:15(ヒトMASP-3のアミノ酸498〜508、リーダあり)
SEQ ID NO:16(ヒトMASP-3のアミノ酸435〜447、リーダあり)
SEQ ID NO:17(ヒトMASP-3のアミノ酸454〜464、リーダあり)
SEQ ID NO:18(ヒトMASP-3のアミノ酸479〜493、リーダあり)
SEQ ID NO:19(ヒトMASP-3のアミノ酸514〜523、リーダあり)
SEQ ID NO:20(ヒトMASP-3のアミノ酸562〜571、リーダあり)
SEQ ID NO:21(ヒトMASP-3のアミノ酸583〜589、リーダあり)
SEQ ID NO:22(ヒトMASP-3のアミノ酸614〜623、リーダあり)
SEQ ID NO:23(ヒトMASP-3のアミノ酸667〜678、リーダあり)
SEQ ID NO:24〜39:重鎖可変領域 - マウス親
SEQ ID NO:24 4D5_VH
SEQ ID NO:25 1F3_VH
SEQ ID NO:26 4B6_VH
SEQ ID NO:27 1A10_VH
SEQ ID NO:28 10D12_VH
SEQ ID NO:29 35C1_VH
SEQ ID NO:30 13B1_VH
SEQ ID NO:31 1G4_VH
SEQ ID NO:32 1E7_VH
SEQ ID NO:33 2D7_VH
SEQ ID NO:34 49C11_VH
SEQ ID NO:35 15D9_VH
SEQ ID NO:36 2F5_VH
SEQ ID NO:37 1B11_VH
SEQ ID NO:38 2F2_VH
SEQ ID NO:39 11B6_VH
SEQ ID NO:40〜54:軽鎖可変領域 - マウス親
SEQ ID NO:40 4D5_VL
SEQ ID NO:41 1F3_VL
SEQ ID NO:42 4B6/1A10_VL
SEQ ID NO:43 10D12_VL
SEQ ID NO:44 35C1_VL
SEQ ID NO:45 13B1_VL
SEQ ID NO:46 1G4_VL
SEQ ID NO:47 1E7_VL
SEQ ID NO:48 2D7_VL
SEQ ID NO:49 49C11_VL
SEQ ID NO:50 15D9_VL
SEQ ID NO:51 2F5_VL
SEQ ID NO:52 1B11_VL
SEQ ID NO:53 2F2_VL
SEQ ID NO:54 11B6_VL
SEQ ID NO:55〜140:マウス親MASP-3 mAb由来の重鎖フレームワーク領域(FR)および相補性決定領域(CDR)
SEQ ID NO:141〜208:マウス親MASP-3 mAb由来の軽鎖FRおよびCDR
SEQ ID NO:209〜216:CDRコンセンサス配列
SEQ ID NO:217〜232:重鎖可変領域(マウス親)をコードするDNA
SEQ ID NO:233〜247:軽鎖可変領域(マウス親)をコードするDNA
SEQ ID NO:248:ヒト化4D5_VH-14(h4D5_VH-14)重鎖可変領域
SEQ ID NO:249:ヒト化4D5_VH-19(h4D5_VH-19)重鎖可変領域
SEQ ID NO:250:ヒト化4D5_VL-1(h4D5_VL-1)軽鎖可変領域
SEQ ID NO:251:ヒト化10D12_VH-45(h10D12_VH-45)重鎖可変領域
SEQ ID NO:252:ヒト化10D12_VH-49(h10D12_VH-49)重鎖可変領域
SEQ ID NO:253:ヒト化10D12_VL-21(h10D12-VL-21)軽鎖可変領域
SEQ ID NO:254:ヒト化13B1_VH-9(h13B1-VH-9)重鎖可変領域
SEQ ID NO:255:ヒト化13B1_VH-10(h13B1-VH-10)重鎖可変領域
SEQ ID NO:256:ヒト化13B1-VL-1(h13B1-VL-1)軽鎖可変領域
SEQ ID NO:257:4D5および13B1 LC-CDR1-NQ
SEQ ID NO:258:4D5および13B1 LC-CDR1-NA
SEQ ID NO:259:4D5および13B1 LC-CDR1-ST
SEQ ID NO:260:4D5、13B1親、およびバリアントのコンセンサスLC-CDR1
SEQ ID NO:261:10D12 LC-CDR1-DE
SEQ ID NO:262:10D12 LC-CDR1-DA
SEQ ID NO:263:10D12 LC-CDR1-GA
SEQ ID NO:264〜277:ヒト化4D5、10D12、および13B1のHC FRおよびCDR
SEQ ID NO:278:h4D5_VL-1-NA
SEQ ID NO:279:h10D12_VL-21-GA
SEQ ID NO:280:h13B1_VL-1-NA
SEQ ID NO:281〜287 ヒト化4D5、10D12、および13B1のLC FRおよびCDR
SEQ ID NO:288〜293:ヒト化4D5、10D12、13B1重鎖可変領域およびバリアントをコードするDNA
SEQ ID NO:294〜299:ヒト化4D5、10D12、13B1軽鎖可変領域およびバリアントをコードするDNA
SEQ ID NO:300:親DTLacO重鎖可変領域(VH)ポリペプチド
SEQ ID NO:301:MASP-3特異的クローンM3J5重鎖可変領域(VH)ポリペプチド
SEQ ID NO:302:MASP-3特異的クローンM3M1重鎖可変領域(VH)ポリペプチド
SEQ ID NO:303:親DTLacO軽鎖可変領域(VL)ポリペプチド
SEQ ID NO:304:MASP-3特異性クローンM3J5軽鎖可変領域(VL)ポリペプチド
SEQ ID NO:305:MASP-3特異性クローンM3M1軽鎖可変領域(VL)ポリペプチド
SEQ ID NO:306:MASP-3クローンD14重鎖可変領域(VH)ポリペプチド
SEQ ID NO:307:MASP-3クローンD14軽鎖可変領域(VL)ポリペプチド
SEQ ID NO:308:MASP-1クローン1E10重鎖可変領域(VH)ポリペプチド
SEQ ID NO:309:MASP-1クローン1E10軽鎖可変領域(VL)ポリペプチド
SEQ ID NO:310:ヒトIgG4定常領域
SEQ ID NO:311:ヒトIgG4定常領域、S228P変異あり
SEQ ID NO:312:ヒトIgG4定常領域、S228P変異_Xあり
SEQ ID NO:313:ヒトIgK定常領域
I. 定義
本明細書において特に定義されない限り、本明細書において使用される全ての用語は、本発明の当業者によって理解されるものと同じ意味を有する。本発明を説明するために明細書および特許請求の範囲において使用される用語を明確にするために、以下の定義を提供する。
i. 概略;レクチン経路は定義し直された
本明細書に記載されるように、本発明者らは、補体のレクチン経路が、いずれも糖質認識成分(MBL、CL-11、およびフィコリン)で形成されたレクチン経路活性化複合体によって駆動される、補体を活性化するための2つのエフェクターアーム:(i)「レクチン経路エフェクターアーム1」または「LEA-1」と呼ばれる、レクチン経路関連セリンプロテアーゼMASP-1およびMASP-3によって形成されたエフェクターアーム;ならびに(ii)本明細書では「レクチン経路エフェクターアーム2」または「LEA-2」と呼ばれるMASP-2駆動型活性化エフェクターアームを有するという驚くべき発見を達成した。LEA-1およびLEA-2はいずれも溶解および/またはオプソニン化を実施することができる。
レクチン経路の第一のエフェクターアーム、すなわちLEA-1は、レクチン経路関連セリンプロテアーゼMASP-1およびMASP-3によって形成される。本明細書に記載されるように、本発明者らはこれまでのところ、MASP-3の非存在かつMASP-1の存在において、第二経路が面構造上で実質的に活性化されないことを示した。これらの結果は、MASP-3が、第二経路を開始させる上で、これまで開示されたことがない役割を果たすことを実証し、これは、MASP-3のセリンプロテアーゼドメインを機能不全にする突然変異を有する珍しい3MC常染色体劣性障害の患者から採取されたMASP-3欠損3MC血清を使用して確認されている(Rooryck C, et al., Nat Genet. 43(3):197-203 (2011))。これらの新規な発見に基づき、従来から定義されるような第二経路を伴う補体活性化はMASP-3依存性であると予想される。事実、MASP-3、およびそのLEA-1活性化は、これまでわかりにくかった第二経路のイニシエーターということになり得る。
レクチン経路の第二のエフェクターアーム、すなわちLEA-2は、レクチン経路関連セリンプロテアーゼMASP-2によって形成される。MASP-2は、認識成分がそれぞれのパターンに結合した場合に活性化され、かつ、またMASP-1によっても活性化され得、その後、補体成分C4をC4aおよびC4bへと切断する。切断産物C4bが血漿C2に結合したのち、C4b結合C2は、C4b結合C2を酵素的に活性な複合体C4bC2aおよび小さなC2b切断断片へと転換する第二のMASP-2媒介性切断工程の基質になる。C4b2aは、豊富な血漿成分C3をC3aおよびC3bへと転換する、レクチン経路のC3転換C3コンバターゼである。C3bは、チオエステル結合を介して、近接する任意の面に結合する。いくつかのC3b断片がC3コンバターゼ複合体C4b2aに近接して結合する場合、このコンバターゼは、C5をC5bおよびC5aへと転換するようにその特異性を変化させて、C5コンバターゼ複合体C4b2a(C3b)nを形成する。このC5コンバターゼはMACの形成を開始することができるが、このプロセスは、それだけで溶解を促進するのには効果が不十分であると考えられる。むしろ、LEA-2によって産生される初期のC3bオプソニンが新たな第二経路C3コンバターゼおよびC5コンバターゼ部位の形成のための核を形成し、それが最終的に豊富なMAC形成および溶解を生じさせる。後者の事象は、LEA-2形成C3bと関連するB因子のD因子活性化によって媒介され、したがって、D因子の成熟におけるMASP-1の本質的役割のおかげでLEA-1に依存する。また、C4欠損マウスは虚血再灌流障害から保護されないが、一方、MASP-2欠損マウスは保護されることから(前記Schwaeble et al., PNAS, 2011)、虚血再灌流障害の病態生理学において重要な役割を果たす、C4の非存在においてC3を活性化するためのMASP-2依存性C4バイパス活性化経路がある。LEA-2はまた、プロトロンビンからトロンビンへの切断(共通経路)およびXII因子(ハーゲマン因子)をその酵素的に活性な形態XIIaへと転換するための切断を含む凝固経路に結び付いている。XIIa因子は他方でXI因子をXIaに切断する(内因性経路)。凝固カスケードの内因性経路活性化は、血栓形成にとってきわめて重要であるフィブリン形成を生じさせる。
(i)LEA-1:アクチベーター表面上のB因子の初期切断および活性化を通して第二経路コンバターゼC3bBbを生成することによって補体の活性化を開始し、かつ駆動し、次いで、それがC3b沈着および第二経路コンバターゼC3bBbの形成を触媒するMASP-3依存性活性化経路。MASP-3駆動型活性化経路は、微生物のオプソニン化および溶解において本質的な役割を果たし、細菌の表面上で第二経路を駆動して、膜侵襲複合体を生成するのに最適な活性化速度を生じさせる。
(ii)LEA-2:レクチン経路C3コンバターゼC4b2aの形成を生じさせ、C3切断産物C3bが蓄積すると、その後、C5コンバターゼC4b2a(C3b)nを生じさせるMASP-2依存性活性化経路。補体C4の非存在において、MASP-2は、C2および凝固因子XIを含む第二C3コンバターゼ複合体を形成することができる。
現在、3つのマンナン結合レクチン関連セリンプロテアーゼ(MASP-1、MASP-2、およびMASP-3)がヒト血清中でマンナン結合レクチン(MBL)と関連していることが公知である。マンナン結合レクチンはまた、最近の文献においては、「マンノース結合タンパク質」または「マンノース結合レクチン」とも呼ばれている。MBL-MASP複合体は、多様な微生物上に存在する糖質構造へのMBLの結合のおかげで、先天性免疫において重要な役割を果たす。MBLと糖質構造の特定のアレイとの相互作用がMASP酵素前駆体の活性化を生じさせ、それが他方で、補体成分C4およびC2を切断してC3コンバターゼC4b2bを形成することによって補体を活性化する(Kawasaki et al., J. Biochem 106:483-489 (1989); Matsushita & Fujita, J. Exp Med. 176:1497-1502 (1992); Ji et al., J. Immunol. 150:571-578 (1993))。
図2は、ヒトMASP-1ポリペプチド(SEQ ID NO:8)、ヒトMASP-3ポリペプチド(SEQ ID NO:2)、およびヒトMAp44ポリペプチドならびにそれらをコードするエキソンのドメイン構造を示す略図である。図2に示すように、セリンプロテアーゼMASP-1およびMASP-3は、C1rおよびC1sに見られるように配置された6つの別々ドメイン;すなわち、(I)N末端C1r/C1s/ウニVEGF/骨形成タンパク質(またはCUBI)ドメイン;(II)上皮成長因子(EGF)様ドメイン;(III)第二のCUBドメイン(CUBII);(IVおよびV)2つの補体制御タンパク質(CCP1およびCCP2)ドメイン;および(VI)セリンプロテアーゼ(SP)ドメインからなる。
図4は、ヒト(SEQ ID NO:2)、カニクイザル(SEQ ID NO:7)、ラット(SEQ ID NO:4)、ネズミ(SEQ ID NO:3)、ニワトリ(SEQ ID NO:5)およびウサギ(SEQ ID NO:6)からの完全長MASP-3タンパク質の比較を示すMASP-3の多種間アラインメントを提供する。図5は、ヒトのセリンプロテアーゼ(SP)ドメイン(SEQ ID NO:2のアミノ酸450〜728);ウサギのSPドメイン(SEQ ID NO:6のアミノ酸450〜728);ネズミのSPドメイン(SEQ ID NO:3のアミノ酸455〜733);ラットのSPドメイン(SEQ ID NO:4のアミノ酸455〜733)およびニワトリのSPドメイン(SEQ ID NO:5のアミノ酸448〜730)の多種間アラインメントを提供する。
補体のレクチン経路活性化ルートが、2つの独立したエフェクター機構:(i)LEA-2:補体駆動型オプソニン化、走化性(Schwaeble et al., PNAS 108:7523-7528 (2011))、および細胞溶解を媒介するMASP-2駆動型経路、ならびに(ii)LEA-1:アクチベーター表面上のB因子の切断および活性化によって第二経路コンバターゼC3bBbを生成することによって補体活性化を開始し、次いでそれがC3b沈着および第二経路コンバターゼC3bBbの形成を触媒し、それが結果として細胞溶解および微生物オプソニン化を生じさせることができる新規なMASP-3依存性活性化ルートを介して、補体を活性化するという結論を指摘する、いくつかの独立した線の強力な実験的証拠が本明細書に提供される。加えて、本明細書に記載されるように、MASP-1、MASP-3もしくはHTRA-1またはこれら3つのいずれかの組み合わせによるB因子および/またはD因子の別々のレクチン非依存性活性化が、第二経路を介する補体活性化を生じさせることもできる。
本明細書に記載されるように、高親和性MASP-3阻害抗体(たとえば、500pM未満の結合親和性を有する)は、MASP-3標的の濃度よりも低いモル濃度で(たとえば約1:1〜約2.5:1(MASP-3標的:mAb)のモル比で)哺乳動物対象、たとえばげっ歯類および非霊長類において第二経路を完全に阻害することが示されている(実施例11〜21を参照されたい)。実施例11に記載されるように、マウスへの高親和性MASP-3阻害抗体mAb 13B1の単回投与が、少なくとも14日間、全身性第二経路補体活性のほぼ完全な消失を生じさせた。さらに実施例12に記載されるように、PNHと関連する十分に確立された動物モデルにおいて実施された実験において、mAb 13B1がPNH様赤血球の生存率を有意に改善し、かつC5阻害よりも有意に良好にPNH様赤血球を保護するということが実証された。さらに、実施例13に記載されるように、関節炎のマウスモデルにおいて、mAb 13B1が疾患の発生率および重症度を下げることが実証された。この実施例における結果は、代表的な高親和性MASP-3阻害性mAb 13B1、10D12、および4D5が、霊長類において第二経路を遮断するのに非常に有効であることを実証する。カニクイザルへのmAb 13B1、10D12または4D5の単回投与は、およそ16日間持続する全身性第二経路活性の持続的消失を生じさせた。高親和性MASP-3阻害抗体で処置されたカニクイザルにおける第二経路消失の程度は、インビトロおよびインビボでD因子遮断によって達成される程度に匹敵し、このことはMASP-3阻害抗体によるD因子転換の完全な遮断を示した。したがって、高親和性MASP-3阻害性mAbは、第二経路活動亢進に関連する疾患に罹患している患者の治療において治療的有用性を有する。
PNHの概略
発作性夜間血色素尿症(PNH)は、時としてマルキアファーヴァ・ミケーリ症候群とも呼ばれ、後天的な、潜在的に命にかかわる血液疾患である。PNHは自然発症することがあり、これは「一次PNH」と呼ばれるか、または再生不良性貧血などの他の骨髄障害の状況では「二次PNH」と呼ばれる。症例の大半は一次PNHである。PNHは、補体誘導性の赤血球破壊(溶血)、少ない赤血球数(貧血)、血栓症、および骨髄機能不全を特徴とする。実験室におけるPNHの所見は、考えられる原因として自己反応性RBC結合抗体の非存在下で、血管内溶血性貧血:低ヘモグロビン、多量の乳酸デヒドロゲナーゼ、多数の網状赤血球数(破壊された細胞を交換するために未熟血球が骨髄によって放出される)、高ビリルビン(ヘモグロビンの破壊産物)と一致する変化を示す。
PNHにおける負の補体制御因子CD55およびCD59の不完全な表面発現の間の因果関係と、血管内溶血の阻止におけるエクリズマブの有効性の組み合わせから、PNHは、補体系によって媒介される状態であるとはっきりと定義される。このパラダイムは広く受け入れられているが、補体活性化を開始する事象、および関与する補体活性化経路がどういったものであるかは未解決のままである。CD55およびCD59は、全ての補体開始経路に共通する補体カスケード中の終末増幅段階を負に調節するので、補体活性化がレクチン経路によって開始されるか、古典経路によって開始されるか、第二経路の自発的代謝回転によって開始されるかに関係なく、これらの分子が欠損すると膜侵襲複合体の形成と膜組込みが悪化する。従って、PNH患者では、RBC表面上でのC3b沈着につながる、全ての補体活性化事象が、その後の増幅ならびに病理学的溶血(血管内溶血および/または血管外溶血)を誘発し、溶血発作を突然引き起こすことができる。PNH患者における溶血発作を誘発する分子事象のはっきりとした機構理解はまだなされていない。溶血発作を起こしているPNH患者における補体開始事象はまったく明らになってないので、PNHにおける補体活性化は低レベルの第二経路「チックオーバー」活性化により自然発生する可能性があり、その後に、CD55およびCD59の欠如による不適切な終末補体活性化制御によって増大するというのが主流となっている見解である。
このセクションは、PNHのインビトロモデルにおける溶血に対するLEA-2およびLEA-1遮断の阻害効果を記載する。この発見は、PNHの1つまたは複数の局面に罹患している患者を治療するためのLEA-2遮断物質(MASP-2に結合し、かつその機能を遮断する抗体を含むが、それに限定されない)およびLEA-1遮断物質(MASP-3、MASP-3または両方に結合し、かつそのMASP-1媒介性活性化の機能を遮断する抗体を含むが、それに限定されない)の有用性、ならびにエクリズマブのようなC5阻害因子による治療を受けるPNH患者においてC3断片媒介性血管外溶血の効果を緩和するためのLEA-2および/またはLEA-1および/またはMASP-3依存性レクチン非依存性補体活性化の阻害因子(MASP-2阻害因子、MASP-3阻害因子、およびMASP-2/MASP-3またはMASP-1/MASP-2二重または二重特異性阻害因子、ならびに、汎特異性MASP-1/MASP-2/MASP-3阻害因子を含む)の使用を裏付ける。
上に詳述したように、PNH患者は、循環からのRBCクリアランスの2つの別々の機構:膜侵襲複合体(MAC)の活性化による血管内溶血、ならびにC3bによるオプソニン化後の血管外溶血およびその後の細網内皮系による補体レセプター結合および取込み後のクリアランスにより、貧血になる。血管内溶血は、患者がエクリズマブで治療された場合に概ね予防される。エクリズマブは、補体開始活性化事象およびその後のオプソニン化の両方よりも下流で起こる終末溶解エフェクター機構を遮断するため、エクリズマブは血管外溶血を遮断しない(Risitano A.M. et. al., Blood 113:4094-100(2009))。その代わり、未治療PNH患者においては溶血を起こしたと考えられるRBCが、今や、活性化されたC3bタンパク質をその表面に蓄積できることができ、それが、細網内皮系による取込みを増強し、その血管外溶血を増強する。したがって、エクリズマブ治療は、実質的に、RBCの性質を血管内溶血から潜在的な血管外溶血に変える。結果として、エクリズマブで治療される一部のPNH患者は依然として貧血のままである。したがって、上流で補体活性化を遮断し、かつPNH RBCのオプソニン化を阻止する作用物質は、エクリズマブを用いてときおり見られる血管外溶血を遮断するのに特に適していることができるということになる。
PNHのインビトロモデルを使用して、本発明者らは、PNHにおける補体活性化および結果的な溶血が実際にLEA-2および/またはLEA-1活性化によって開始され、それが、第二経路の独立した機能ではないことを実証した。これらの研究は、Crry欠損マウスからのRBC(マウスにおける終末補体経路の重要な負の調節物質)およびPNH患者には存在しない同じ補体調節物質を欠くCD55/CD59欠損マウスからのRBCをはじめとする様々なマウス系統のマンナン感作RBCを使用した。マンナン感作Crry欠損RBCを補体充分なヒト血清に曝露すると、RBCは、3%の血清濃度で実質的に溶血したが(図19および20)、一方、補体欠損血清(HI:熱不活化)は溶血性ではなかった。驚いたことに、MASP-2抗体の添加によってLEA-2が遮断された補体充分な血清は溶血活性が低下しており、効果的な溶血のためには6%血清が必要であった。CD55/CD59欠損RBCを試験した場合にも同様な観察結果が得られた(図22)。MASP-2モノクローナル抗体で補充された補体充分なヒト血清(すなわち、LEA-2が抑制された血清)は、溶血の支持において未処理の血清よりも効果が約2倍の低さであった。さらに、未処理の血清に比べて未処理のWT RBCの効果的な溶血を促進するためには、より高濃度のLEA-2遮断血清(すなわち、抗MASP-2モノクローナル抗体で処理された)が必要であった(図21)。
本明細書に提示されるデータは、PNHにおける貧血に関して以下の病原性機構を示唆する。主としてであるがただし排他的でなくLEA-1によって開始される、終末補体成分の調節されない活性化およびMACの形成によるRBCの溶解による血管内溶血、ならびに、主としてLEA-2によって開始されると考えられる、C3bによるRBCのオプソニン化によって生じる血管外溶血。補体活性化を開始し、MAC形成および溶血を促進することにおけるLEA-2の認められる役割は明らかであるが、このプロセスは、溶血を生じさせるLEA-1開始補体活性化よりも効果が実質的に低いと考えられる。したがって、LEA-2遮断物質は、PNH患者における血管内溶血を有意に減少させると予想されるが、この治療活性は部分的でしかないと予想される。比較により、LEA-1遮断物質の場合に、PNH患者における血管内溶血のより実質的な減少が予想される。
本明細書に提示されるデータは、別々のクラスの治療剤によって別々に、または組み合わせて標的化することができる、PNHにおけるRBCクリアランスおよび貧血の以下の2つの病原性機構を明示する:主としてであるがただし排他的でなくLEA-1によって開始され、したがって、LEA-1遮断物質によって効果的に予防されると予想される血管内溶血;および、主としてLEA-2によって駆動され、したがってLEA-2遮断物質によって効果的に予防されるC3bオプソニン化による血管外溶血。
上に詳述したように、個々にLEA-1およびLEA-2を遮断し、ひいては組み合わさって、血管内溶血および血管外溶血を媒介するすべての補体活性化事象を遮断する薬理学的物質の組み合わせの使用は、PNH患者にとって最良の臨床転帰を提供すると予想される。この転帰は、例えば、LEA-1遮断活性を有する抗体と、LEA-2遮断活性を有する抗体との同時投与によって達成することができる。いくつかの態様において、LEA-1遮断活性およびLEA-2遮断活性が単一の分子実体に組み合わされ、LEA-1およびLEA-2複合遮断活性を有するそのような実体は、血管内溶血および血管外溶血を効果的に阻止し、PNHにおける貧血を予防する。そのような実体は、1つの抗原結合部位がMASP-1を特異的に認識し、LEA-1を遮断し、LEA-2を減少させ、第二の抗原結合部位がMASP-2を特異的に認識し、さらにLEA-2を遮断する二重特異性抗体を含み得るか、またはそれからなり得る。あるいは、そのような実体は二重特異性モノクローナル抗体からなり得、1つの抗原結合部位がMASP-3を特異的に認識し、ひいてはLEA-1を遮断し、第二の抗原結合部位がMASP-2を特異的に認識し、LEA-2を遮断する。そのような実体は最適には二重特異性モノクローナル抗体からなり得、1つの抗原結合部位がMASP-1およびMASP-3の両方を特異的に認識し、ひいてはLEA-1を遮断し、LEA-2を減少させ、その上、第二の抗原結合部位がMASP-2を特異的に認識し、さらにLEA-2を遮断する。また、タンパク質配列およびアーキテクチャ全体の類似性に基づき、機能的にMASP-1、MASP-2、およびMASP-3に特異的に結合し、ひいてはLEA-1およびLEA-2の機能的遮断を達成する、2つの同一の結合部位を有する従来の抗体を開発することができると考えることができる。汎MASP阻害活性を有するそのような抗体は、血管内溶血および血管外溶血の両方を阻止し、ひいてはPNH患者における貧血を効果的に治療すると予想される。
加齢黄斑変性症(AMD)は、高齢者における視力損傷および失明の主要な原因であり、先進国における失明症例の50%までを占める。成人におけるAMD罹患率は約3%であり、年齢とともに増加して、80歳超の人口のほぼ2/3が何らかの徴候を有する。米国においては175万超の個人が進行AMDを有し、人口高齢化とともに罹患率は高まり、2020年までにはほぼ300万に達すると予想されている(Friedman, D.S., et al., Arch. Ophthalmol. 122:564-572, 2004)。AMDは、表面を覆う中央網膜、すなわち黄斑の光受容体の変性および中心視力の損失を生じさせる網膜色素上皮(RPE)の異常である。AMDの早期および中期形態は、RPEに隣接する網膜下空間中のドルーゼン、すなわち脂質、タンパク質、リポタンパク質および壊死細胞片を含む黄色を帯びた物質の漸増的沈着ならびに網膜中の色素不規則性を特徴とする。進行AMDは、2つの臨床サブタイプ;非新生血管形成性地理的萎縮性(「ドライ型」)AMDおよび新生血管形成性滲出性(「ウェット型」)AMDからなる。ドライ型AMDが進行AMDの80〜90%を占めるが、突然かつ重篤な視覚損失の大部分はウェット型AMDの患者に起こる。2つのタイプのAMDが、類似した病態から生じる異なる表現型を表すのか、または2つの異なる状態を表すのかは不明である。現在、ドライ型AMDを治療するための治療法は米国薬品医薬品局(United States Food and Drug Administraion)(FDA)によって承認されていない。ウェット型AMDのためのFDA承認治療選択肢は、抗血管形成薬(ラニビズマブ、ペガプタニブナトリウム、アフリベルセプト)の硝子体内注射、レーザー療法、光力学的レーザー療法および埋め込み型望遠鏡を含む。
上述したヒトの遺伝的連鎖研究はAMD病原における補体系の重要な役割を示唆する。さらに、補体活性化産物は、ウェット型およびドライ型の両AMDにおける特徴的な病理学的病変であるドルーゼン中に豊富に存在する(Issa, P.C., et al., Graefes. Arch. Clin. Exp. Ophthalmol. 249:163-174, 2011)。しかし、補体活性化を開始する事象の性質および関与する補体活性化経路は不完全にしか理解されないままである。
組織虚血は広範囲の臨床障害の基礎である。血流の適時回復が虚血組織の保存に不可欠であるが、自然発生的に、または治療的介入によって起こることができる再灌流が、虚血再灌流(I/R)障害と呼ばれる現象である、さらなる組織損傷を招き得ることが長らく認識されている(Eltzschig, H. K. and Tobias, E., Nat. Med. 17:1391-1401, 2011)。I/R障害は、心臓(急性冠症候群)、腎臓(急性腎損傷)、腸(腸I/R)および脳(卒中)のように、単一の臓器を侵襲し得る。I/R障害はまた、大きな外傷および蘇生(多臓器不全)、循環停止(低酸素性脳障害、急性腎損傷)、末梢血管疾患および鎌状赤血球症(急性胸部症候群、急性腎損傷)ののちなど、複数の臓器を冒す場合もある。心臓手術(心肺バイパス後の急性心不全)、胸部手術(急性肺損傷)、末梢血管手術(コンパートメント症候群)、血管手術(急性腎損傷)および固形臓器移植(急性移植片不全)を含む大きな手術がI/R障害と関連する場合もある。現在、I/R障害を標的化する具体的な治療法は存在せず、虚血ゾーンにおける組織のサルベージを最大化し、これらの一般的状況における機能的転帰を改善するために有効な治療の必要性がある。
関節リウマチ(RA)は、全身的な症状発現も有し得る滑膜関節の慢性炎症性疾患である。RAは全世界人口の約1%を冒し、女性の方が2〜3倍かかりやすい。関節の炎症は、腫れ、痛みおよびこわばりとして現れる。疾患が進行するにつれ、関節の侵食および破壊が起こり得、その結果、可動域の障害および変形が生じる。RAの治療目標は、関節損傷の予防および抑制、関節機能損失および疾病進行の予防、症候の軽減および生活の質の改善ならびにドラッグフリー寛解の達成を含む。RAの薬理学的治療は、疾患修飾性抗リウマチ薬(DMARD)、鎮痛薬および抗炎症剤(糖質コルチコイドおよび非ステロイド系抗炎症薬)を含む。DMARDは、長期寛解を誘発し、不可逆性である関節破壊の進行を遅延または停止させることができるため、最も重要な治療である。旧来のDMARDの例は、小分子、例えばメトトレキサート、スルファサラジン、ヒドロキシクロロキン、金塩、レフルノミド、D-ペニシラミン、シクロスポリンおよびアザチオプリンを含む。旧来のDMARDが疾病を抑制するのに不十分である場合は、腫瘍壊死因子阻害因子(エタネルセプト、インフリキシマブ、アダリムマブ、セルトリズマブペゴルおよびゴリムマブ)、サイトカインアンタゴニスト(アナキンラおよびトシリズマブ)、リツキシマブおよびアバタセプトのような、炎症細胞または媒介物を標的化するいくつかの生物学的剤が利用可能な治療選択肢である。
播種性血管内凝固(DIC)は、出血および/または血栓症として臨床的に現れることができる凝固系の病理的過剰刺激の症候群である。DICは原発性状態としては起こらず、むしろ、組織損傷(外傷、火傷、熱中症、輸血反応、急性移植拒絶反応)、新生物、感染症、産科的状態(前置胎盤、羊水塞栓症、妊娠中毒症)ならびに雑多な状態、例えば心原性ショック、溺水、脂肪塞栓症、大動脈瘤を含む多様な疾患プロセスと関連して起こる。血小板減少症は、集中治療室中の患者において35%〜44%の発生率で頻繁に起こる異常であるが、DICはこれらの症例の約25%の病因である。すなわち、DICは重症患者の約10%において発症する(Levi, M. and Opal, S.M. Crit. Care 10:222-231, 2006)。DICの病態生理は、基礎にある疾患プロセスが生理学的凝固反応を開始させるということである。しかし、血栓症形成促進物質が正常な反対平衡機序を圧倒すると、微小循環中にフィブリンおよび血小板の不適切な沈着が起こり、臓器虚血、低フィブリノゲン血症および血小板減少を招くようになる。DICの診断は、検査値(プロトロンビン時間、部分トロンボプラスチン時間、フィブリン分解産物、Dダイマーまたは血小板数)の異常と共に、適切な基礎疾患またはプロセスにおける臨床所見に基づく。DICの一次治療は、誘因である基礎疾患に対処することである。臨床合併症を治療または予防するために、赤血球、血小板、新鮮な凍結血漿およびクリオプレシピテートの形態の血液製剤サポートが必要になる場合もある。
血栓性微小血管症(TMA)とは、血小板減少症、微小血管症性溶血性貧血および可変性臓器虚血を臨床的特徴とする障害の群をいう。TMAの特徴的な病理学的特徴は、血小板活性化ならびに小細動脈および細静脈中の微小血栓の形成である。古典的TMAは、溶血性尿毒症症候群(HUS)および血栓性血小板減少性紫斑病(TTP)である。HUSは、急性腎不全の存在によってTTPから区別される。HUSは、2つの形態:下痢関連(D+)または典型HUSおよび下痢を伴わない(D-)または非定型HUS(aHUS)で起こる。
D+HUSは、通常は大腸菌O157または別の志賀毒素産生性細菌株によって生じる前駆的下痢性疾患を伴い、子供におけるHUS症例の90%超を占め、子供における急性腎不全の最も一般的な原因である。大腸菌(Escherichia coli)O157によるヒト感染は相対的によくあるが、D+HUSまで進行する血性下痢の割合は、散発的症例においては3%〜7%の範囲であり、いくつかの大発生においては20%〜30%であった(Zheng, X.L. and Sadler, J. E., Annu. Rev. Pathol. 3:249-277, 2008)。HUSは通常、下痢発症後4〜6日で発症し、子供の約2/3は疾患の急性期において透析を必要とする。有効であることが示されている特定の治療法がないため、D+HUSの治療は支持的である。D+HUSの予後は良好であり、大多数の患者が腎機能を取り戻す。
非定型HUSは希少病であり、米国においては100万人あたり2人の推定発生率である(Loirat, C. and Fremeaux-Bacchi, V. Orphanet J. Rare Dis. 6:60-90, 2011)。非定型HUSは任意の年齢で発症することができるが、大多数の患者は小児期に発症を示す。非定型HUSは不均一である。いくつかの症例は家族性であり、いくつかは再発性であり、いくつかは感染症、一般的に上気道または胃腸炎によって誘発される。通常、aHUSの発症は突然であり、大部分の患者は入院時に透析を必要とする。さらなる腎症状発現が患者の約20%に見られ、中枢神経系、心筋梗塞、虚血性遠位壊疽または多臓器不全を含み得る。aHUSの治療は、臓器不全の場合の支持療法、血漿注入または血漿交換、および、エクリズマブ、最近米国および欧州連合において使用を承認された、C5を標的化するヒト化モノクローナル抗体、を含む。aHUSにおける予後はD+HUSにおける予後ほど良くなく、約25%が急性期に死亡し、大部分の生存者は末期腎疾患を発症する。
血栓性血小板減少性紫斑病(TTP)は、凝固カスケードまたは補体系を活性化する自己免疫または遺伝性機能不全によって生じる、生命を脅かす血液凝固系の障害である(George, JN, N Engl J Med; 354:1927-35, 2006)。これは体中の小さな血管中に多数の微細な血餅、すなわち血栓を生じさせ、これがTMAの特徴である。赤血球が剪断応力を受け、それが膜を損傷し、血管内溶血を招く。その結果として生じる血流の減少および内皮損傷が、脳、心臓および腎臓を含む臓器の損傷を生じさせる。TTPは、臨床的には血小板減少症、微小血管症性溶血性貧血、神経学的変化、腎不全および発熱を特徴とする。血漿交換の前の時代には、急性エピソード時の致死率は90%であった。血漿交換を実施してさえ、6ヶ月生存率は約80%である。
喘息は一般的な慢性炎症性気道疾患である。米国においては、18歳未満の子供700万人を含む約2500万人が喘息を有し、半分超が年に少なくとも一度は喘息発作を経験し、毎年170万を超える救急外来訪問数および450,000件の入院をもたらしている(ワールドワイドウェブ、gov/health/prof/lung/asthma/naci/asthma-info/index.htm.、2012年5月4日アクセス時点)。この疾患は不均一であり、複数の臨床表現型を有する。最も一般的な表現型はアレルギー性喘息である。他の表現型は、非アレルギー性喘息、アスピリン悪化呼吸器疾患、感染後喘息、職業性喘息、空気中の刺激物誘発性喘息および運動誘発性喘息を含む。アレルギー性喘息の主要な特徴は、多様な特定および不特定刺激への気道過敏性(AHR)、過度の気道粘液産生、肺好酸球増大および血清IgE濃度上昇を含む。喘息の症候は、咳、喘鳴、胸苦しさおよび息切れを含む。喘息治療の目標は、疾患を制御し、悪化、日常的な症候を最小限にし、患者が肉体的に活動的であることを可能にすることである。現在の治療指針は、喘息抑制が達成されるまでの段階的治療を奨励している。最初の治療ステップは、必要に応じて、速効性吸入β2アゴニストののち、長期管理薬、例えば吸入コルチコステロイド、長時間作用性吸入β2アゴニスト、ロイコトリエン修飾薬、テオフィリン、経口糖質コルチコステロイド、および抗IgEモノクローナル抗体である。
膜性増殖性糸球体腎炎(MPGN)は、メサンギウム細胞増殖と、メサンギウムの内皮下延長による糸球体毛細血管壁の肥厚とを形態学的特徴とする腎障害である。MPGNは原発性(特発性とも呼ばれる)または続発性に分類され、感染症、全身性免疫複合体疾患、新生物、慢性肝疾患などのような基礎疾患を有する。特発性MPGNは3つの形態学的タイプを含む。I型、すなわち古典的MPGNは、免疫複合体の内皮下沈着物および古典補体経路の活性化を特徴とする。II型、すなわちデンスデポジット病(DDD)は、さらなる膜内高密度沈着物を特徴とする。III型はさらなる上皮下沈着物を特徴とする。特発性MPGNは希であり、ネフローゼ症候群の原発性腎原因の約4〜7%しか占めない(Alchi, B. and Jayne, D. Pediatr. Nephrol. 25:1409-1418, 2010)。MPGNは、主として子供および若年成人を冒し、ネフローゼ症候群、急性腎炎症候群、無症候性タンパク尿および血尿または再発性肉眼的血尿として現れ得る。腎機能不全が大多数の患者において起こり、疾患は、ゆっくり進行する経過をたどり、患者の約40%が診断から10年以内に末期腎疾患を発症する(前記Alchi and Jayne, 2010)。現在の治療選択肢は、コルチコステロイド、免疫抑制剤、抗血小板レジメンおよび血漿交換を含む。
微量免疫型壊死性半月体形成性糸球体腎炎(NCGN)は、糸球体毛細血管壁が炎症の徴候を示すが、糸球体基底膜に対する微量の検出可能な免疫複合体沈着または抗体を有する急速進行性糸球体腎炎の一形態である。状態は腎機能の急速な低下を伴う。大部分のNCGN患者が、抗好中球細胞質自己抗体(ANCA)を有することがわかっており、したがって、ANCA関連血管炎と呼ばれる疾患の群に属する。血管炎は、血管壁の炎症およびフィブリノイド壊死を特徴とする血管の障害である。全身性血管炎は、血管サイズ:大、中、小に基づいて分類される。小血管血管炎のいくつかの形態は、ANCAの存在、すなわちヴェーゲナー肉芽腫症、顕微鏡的多発血管炎、チャーグ・ストラウス症候群および腎限定的血管炎(NCGN)を伴う。それらはまた、全身性エリテマトーデスのような基礎疾患の症状発現であることもできる。ANCAの標的抗原はプロテイナーゼ-3(PR3)およびミエロペルオキシダーゼ(MPO)を含む。微量免疫型NCGNは珍しく、英国ウェセックス(Wessex)において100万人あたり約4人の発生率が報告されている(Hedger, N. et al., Nephrol. Dial. Transplant. 15:1593-1599, 2000)。微量免疫型NCGN患者128名のウェセックス群においては、73%がANCA陽性であり、患者の59%が初期透析を必要とし、36%が長期透析を必要とした。微量免疫型NCGNの治療は、コルチコステロイドならびにシクロホスファミドおよびアザチオプリンのような免疫抑制剤を含む。ANCA関連血管炎のさらなる治療選択肢はリツキシマブおよび血漿交換を含む(Chen, M. and Kallenberg, C.G.M. Nat. Rev. Rheumatol. 6:653-664, 2010)。
外傷性脳損傷(TBI)は、毎年少なくとも1000万人の死亡または入院をもたらす主要な全世界的健康問題である(Langlois, J.A. et al., J. Head Trauma Rehabil. 21:375-378, 2006)。2003年、米国においては、120万件の救急外来訪問、29万件の入院および51,000件の死亡を含め、推定160万件のTBIがあった(Rutland-Brown, W. et al., J. Head Trauma Rehabil. 21:544-548, 2006)。米国におけるTBIの大多数は転倒および交通事故によって生じる。TBIは長期または一生涯の身体的、認知的、行動的および情動的後遺症をもたらす可能性がある。500万を超える米国人がTBI関連の長期または一生涯の身体障害とともに生きている(前記Langlois et al., 2006)。
多くの研究が、補体タンパク質と、アルツハイマー病、多発性硬化症、重症筋無力症、ギラン・バレー症候群、脳ループスおよび卒中を含む神経学的障害との関係を特定している(Wagner, E., et al., Nature Rev Drug Disc. 9:43-56, 2010において概説されている)。最近、シナプス除去におけるC1qおよびC3の役割が実証され、したがって、補体因子が正常なCNS機能および神経変性疾患の両方に関与する可能性が高い(Stevens, B. et al., Cell 131:1164-1178, 2007)。MASP-1およびMASP-3の遺伝子は脳および神経膠腫細胞株T98G中で広く発現し(Kuraya, M. et al., Int Immunol., 15:109-17, 2003)、CNSにおけるレクチン経路の役割と合致している。
誤嚥とは、口咽頭または胃いずれかの内容物の下気道への吸入と定義される。誤嚥は、誤嚥性(化学性)肺炎、原発性細菌性誤嚥性肺炎または化学性肺炎の続発性細菌感染の合併症を生じさせ得る。誤嚥の危険因子は、意識レベルの低下(例えば、頭部外傷、感覚中枢におけるアルコールまたは薬物誘発性の変化、卒中)、様々な胃腸および食道の異常ならびに神経筋疾患を含む。450万の市中肺炎症例の5〜15%が誤嚥性肺炎によるものであると推定される(Marik, P.E. New Engl. J. Med. 344:665-671, 2001)。化学性肺炎の治療は主として支持的であり、経験的抗生物質使用は議論の余地がある。細菌性誤嚥性肺炎の治療は、適切な抗生物質による治療であり、細菌性誤嚥性肺炎の治療は、誤嚥が市中で起こったのかまたは院内で起こったのかに基づき、というのはこれらの状況の間では考えられる原因生物が異なるからである。高リスク患者、例えば咽頭反射障害を有する介護施設中の高齢患者における誤嚥を防ぐための措置が講じられるべきである。有効な予防法であることが示されている措置は、摂食中にベッドの頭部を高くすること、歯科予防および良好な口腔衛生を含む。予防的抗生物質は有効性が示されておらず、耐性菌の出現を招き得ることから、推奨されない。
眼内炎は、眼窩洞の炎症状態であり、通常、感染によって生じる。眼内炎は、遠い感染源(例えば心内膜炎)からの生物の血行性伝播から生じる内因性眼内炎でもあり得るし、または眼科手術、異物および/もしくは鈍的もしくは穿通性眼部外傷の合併症として外部からの生物の直接接種から生じる外因性眼内炎でもあり得る。外因性眼内炎は内因性眼内炎よりもはるかに一般的であり、外因性眼内炎の大部分の症例は眼科手術ののち起こる。米国においては、白内障手術が眼内炎の主要な原因であり、この処置の0.1〜0.3%において発生し、過去10年間、発生率の明らかな増加が見られる(Taban, M. et al., Arch. Ophthalmol. 123:613-620, 2005)。術後眼内炎は、手術から2週間以内に急性に出る場合もあるし、または手術から数ヶ月後に遅発的に出る場合もある。急性眼内炎は一般に痛み、発赤、眼瞼の腫れおよび視力低下を呈する。遅発性眼内炎は急性型ほど一般的ではなく、患者は軽度の痛みおよびまぶしさしか訴えない場合もある。眼内炎の治療は、基礎にある原因に依存し、全身的および/または硝子体内抗生物質を含み得る。眼内炎は視力低下または失明を招く場合もある。
視神経脊髄炎(NMO)は、視神経および脊髄を標的化する自己免疫疾患である。これは、視神経炎として知られる視神経の炎症および脊髄炎として知られる脊髄の炎症をもたらす。NMOにおける脊髄病変は、脚または腕の衰弱または麻痺、失明、膀胱および腸の機能不全ならびに感覚機能不全を招き得る。
ベーチェット病またはベーチェット症候群は、しばしば粘膜潰瘍および眼の障害を呈する稀な免疫媒介性小血管全身性血管炎である。ベーチェット病(BD)は、再発性口腔潰瘍、性器潰瘍およびブドウ膜炎の三症候複合体を最初に記載したトルコ人皮膚科医Hulusi Behcetの名をとって1937年に命名された。BDは原因不明の全身性再発性炎症障害である。BDの炎症性血管周囲炎は、胃腸管系、肺系、筋骨格系、心血管系、および神経系を巻き込み得る。BDは、血管動脈瘤の破裂または重篤な神経学的合併症のせいで致命的になる可能性がある。視神経に血液を供給する血管の血管炎および閉塞から視神経症および萎縮が生じる場合もある。Al-Araji A, et al., Lancet Neurol., 8(2):192-204, 2009を参照されたい。
補体のレクチン経路が2つの主要な補体活性化アームLEA-1およびLEA-2で構成され、およびまた、レクチン非依存性補体活性化アームがあるという認識をもって、補体の免疫防御能力を完全に停止させることなく(すなわち、古典的経路を完全なままにしておく)、第二経路補体活性化と関連する病状、たとえば発作性夜間ヘモグロビン尿症(PNH)、加齢黄斑変性症(AMD、ウェット型およびドライ型AMDを含む)、虚血再灌流障害、関節炎、播種性血管内凝固、血栓性微小血管症(溶血性尿毒症症候群(HUS)、非典型溶血性尿毒症症候群(aHUS)、血栓性血小板減少性紫斑病(TTP)または移植後TMAを含む)、喘息、デンスデポジット病、微量免疫型壊死性半月体形成性糸球体腎炎、外傷性脳損傷、誤嚥性肺炎、眼内炎、視神経脊髄炎、ベーチェット病、多発性硬化症(MS)、ギランバレー症候群、アルツハイマー病、筋萎縮性側索硬化症(ALS)、ループス腎炎、全身性エリテマトーデス(SLE)、糖尿病性網膜症、ブドウ膜炎、慢性閉塞性肺疾患(COPD)、C3糸球体症、移植片拒絶反応、移植片対宿主病(GVHD)、血液透析、敗血症、全身性炎症反応症候群(SIRS)、急性呼吸窮迫症候群(ARDS)、ANCA血管炎、抗リン脂質症候群、アテローム性動脈硬化症、IgA腎症、ならびに重症筋無力症の、少なくとも1つを生じさせるこれらのエフェクタアームの1つまたは複数を特異的に阻害することが非常に望ましいということが理解されよう。これは、免疫複合体プロセシングを取り扱い、感染に対する宿主防御を支援するために、C1q依存性補体活性化系を完全なままにしておくと考えられる。
本明細書に記載されるように、本発明者らは、溶解を生じさせるLEA-1の活性化がMASP-3依存性であることを予想外に見いだした。さらに本明細書に記載されるように、生理学的条件下、MASP-3依存性LEA-1活性化はオプソニン化にも寄与し、それにより、LEA-2媒介補体活性化と共に付加的効果を提供する。本明細書において実証されるように、Ca++の存在においては、MASP-3がD因子-/-血清中でLEA-1の活性化を駆動することができるため、D因子は不要である。MASP-3、MASP-1およびHTRA-1はまた、プロD因子を活性D因子へと転換することができる。同様に、MASP-3(MASP-1およびMASP-2とは対照的に)は自己活性化酵素ではなく、MASP-1の支援なしにはその活性形態へと転換されることができないため、MASP-3活性化は、多くの場合、MASP-1に依存するように考えられる(Zundel, S. et al., J. Immunol. 172:4342-4350 (2004);Megyeri et al., J. Biol. Chem. 288:8922-8934 (2013)。MASP-3は自己活性化せず、多くの場合、MASP-1の活性がその酵素的に活性な形態へと転換されることを必要とするため、第二経路C3コンバターゼC3BbのMASP-3媒介活性化は、MASP-3チモーゲンまたはすでに活性化されたMASP-3を標的とすることによって、またはMASP-1媒介MASP-3活性化を標的とすることによって、またはその両方によって阻害することができる。理由は、多くの場合、MASP-1機能活性の非存在においては、MASP-3はそのチモーゲン形態にとどまり、第二経路C3コンバターゼ(C3bBb)の直接形成を通してLEA-1を駆動することができないからである。
本明細書における実施例11〜21に記載され、かつ以下の表2A、2Bおよび表3にまとめられるように、本発明者らは、ヒトMASP-3のセリンプロテアーゼドメイン中のエピトープに結合する驚くほど高親和性(すなわち≦500pM)のMASP-3阻害抗体を生成した。本明細書に記載されるように、本発明者らは、これらの高親和性MASP-3抗体が、ヒト血清、げっ歯類および非ヒト霊長類中で第二経路補体活性化を阻害することができることを実証した。これらの抗体の可変軽鎖および重鎖領域を配列決定し、単離し、Fabフォーマットおよび完全長IgGフォーマットの両方で分析した。実施例15に記載され、図50Aおよび50Bに示す樹状図に示されるように、抗体は、配列類似性にしたがって分類することができる。これらの抗体の重鎖可変領域および軽鎖可変領域の概要が図49Aおよび49Bに示され、かつ以下の表2Aおよび2Bに提供されている。代表的な高親和性MASP-3阻害抗体のヒト化バージョンが実施例19に記載されるように生成され、表3にまとめられている。
実施例18に記載され、図62に示され、かつ以下の表4にまとめられるように、本発明の高親和性MASP-3阻害抗体およびその抗原結合断片は、ヒトMASP-3のセリンプロテアーゼドメイン(SEQ ID NO:2のアミノ酸残基450〜728)内の1つまたは複数のエピトープを特異的に認識することがわかった。「特異的に認識する」とは、抗体が、任意の他の分子またはその一部に対するよりも有意に高い親和性で該エピトープに結合することをいう。
本発明の1つの局面において、抗体またはその機能的等価物は、CDRと指定される特定の超可変領域を含む。好ましくは、CDRは、Kabat CDR定義によるCDRである。CDRまたは超可変領域は、たとえば、他の抗体への配列アラインメントによって同定され得る。高親和性MASP-3阻害抗体のCDR領域が表18〜23に示されている。
1つの局面において、本発明は、(a)SEQ ID NO:209(XXDIN、1位のXはSまたはTでありかつ2位のXはNまたはDである)に記載されるHC-CDR1;SEQ ID NO:210(
別の局面において、本発明は、(a)SEQ ID NO:213(SYGXX、4位のXはMまたはIでありかつ5位のXはSまたはTである)に記載されるHC-CDR1;SEQ ID NO:74に記載されるHC-CDR2;およびSEQ ID NO:214(GGXAXDY、3位のXはEまたはDでありかつ5位のXはMまたはLである)に記載されるHC-CDR3を含む、重鎖可変領域;ならびに(b)SEQ ID NO:215(
1つの局面において、本発明は、(a)SEQ ID NO:84(GKWIE)に記載されるHC-CDR1;
1つの局面において、本発明は、(a)SEQ ID NO:91(GYWIE)に記載されるHC-CDR1;
別の局面において、本発明は、
(a)
(b)SEQ ID NO:125(DYYMN)に記載されるHC-CDR1、
(c)SEQ ID NO:137に記載されるHC-CDR1、SEQ ID NO:138に記載されるHC-CDR2、およびSEQ ID NO:140に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:206に記載されるLC-CDR1、SEQ ID NO:207に記載されるLC-CDR2、およびSEQ ID NO:208に記載されるLC-CDR3を含む、軽鎖可変領域;または
(d)SEQ ID NO:98に記載されるHC-CDR1、SEQ ID NO:99に記載されるHC-CDR2、およびSEQ ID NO:101に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:169に記載されるLC-CDR1、SEQ ID NO:171に記載されるLC-CDR2、およびSEQ ID NO:173に記載されるLC-CDR3を含む、軽鎖可変領域;または
(e)SEQ ID NO:103に記載されるHC-CDR1、SEQ ID NO:105に記載されるHC-CDR2、SEQ ID NO:107に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:176に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む軽鎖可変領域;または
(f)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:116に記載されるHC-CDR2、SEQ ID NO:118に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:188に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:190に記載されるLC-CDR3を含む、軽鎖可変領域;または
(g)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:121に記載されるHC-CDR2、およびSEQ ID NO:123に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:191に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む、軽鎖可変領域;または
(h)SEQ ID NO:132に記載されるHC-CDR1、SEQ ID NO:133に記載されるHC-CDR2、およびSEQ ID NO:135に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:203に記載されるLC-CDR1、SEQ ID NO:165に記載されるLC-CDR2、およびSEQ ID NO:204に記載されるLC-CDR3を含む、軽鎖可変領域
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片を提供する。
1つの態様において、本発明は、SEQ ID NO:24〜39、248〜249、251〜252、254〜255のいずれかと少なくとも80%、85%、90%、95%、98%、99%同一である配列を含むまたはそれからなる重鎖可変領域を含むか、または、SEQ ID NO:24、SEQ ID NO:25、SEQ ID NO:26、SEQ ID NO:27、SEQ ID NO:28、SEQ ID NO:29、SEQ ID NO:30、SEQ ID NO:31、SEQ ID NO:32、SEQ ID NO:33、SEQ ID NO:34、SEQ ID NO:35、SEQ ID NO:36、SEQ ID NO:37、SEQ ID NO:38、SEQ ID NO:39、SEQ ID NO:248、SEQ ID NO:249、SEQ ID NO:251、SEQ ID NO:252、SEQ ID NO:254、またはSEQ ID NO:255を含む重鎖可変領域を含む、高親和性MASP-3阻害抗体を提供する。
本明細書に記載されるように、本明細書に開示される高親和性MASP-3阻害抗体は、MASP-3のセリンプロテアーゼドメイン中の重複するエピトープを認識する。実施例18に記載され、図61A〜Eおよび62〜67に示され、表4および28にまとめられるように、交差競合分析およびpepscan結合分析が、高親和性MASP-3阻害抗体が交差競合し、MASP-3セリンプロテアーゼドメイン中に位置する共通のエピトープに結合することを示す。したがって、1つの態様において、本発明は、
SEQ ID NO:24に記載される重鎖可変領域およびSEQ ID NO:40に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:25に記載される重鎖可変領域およびSEQ ID NO:41に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:26に記載される重鎖可変領域およびSEQ ID NO:42に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:27に記載される重鎖可変領域およびSEQ ID NO:42に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:28に記載される重鎖可変領域およびSEQ ID NO:43に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:29に記載される重鎖可変領域およびSEQ ID NO:44に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:30に記載される重鎖可変領域およびSEQ ID NO:45に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:31に記載される重鎖可変領域およびSEQ ID NO:46に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:32に記載される重鎖可変領域およびSEQ ID NO:47に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:33に記載される重鎖可変領域およびSEQ ID NO:48に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:34に記載される重鎖可変領域およびSEQ ID NO:49に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:35に記載される重鎖可変領域およびSEQ ID NO:50に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:36に記載される重鎖可変領域およびSEQ ID NO:51に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:37に記載される重鎖可変領域およびSEQ ID NO:52に記載される軽鎖可変領域を含むモノクローナル抗体;
SEQ ID NO:38に記載される重鎖可変領域およびSEQ ID NO:53に記載される軽鎖可変領域を含むモノクローナル抗体;および
SEQ ID NO:39に記載される重鎖可変領域およびSEQ ID NO:54に記載される軽鎖可変領域を含むモノクローナル抗体
からなる群より選択される1つまたは複数によって認識されるヒトMASP-3のセリンプロテアーゼドメインの中のエピトープまたはその一部を特異的に認識する高親和性MASP-3阻害抗体を提供する。
・ 参照抗体によって認識されるエピトープを含むMASP-3またはその断片を提供する工程。
・ 試験抗体または参照抗体のいずれかが検出可能な標識で標識されている試験抗体および参照抗体を、該MASP-3に添加する工程。あるいは、両抗体は異なる検出可能な標識で標識されてもよい。
・ MASP-3における検出可能な標識の存在を検出する工程。
・ それにより、試験抗体が参照抗体を押し退け得るかどうかを検出する工程。
本発明のこの局面のいくつかの態様において、古典的補体経路の活性化から生じ得る炎症を減少させるために、本明細書に記載の高親和性MASP-3阻害抗体はエフェクター機能が低下している。IgG分子が古典的補体経路を誘発する能力は、この分子のFc部分の中にあることが示されている(Duncan, A.R.. et al., Nature 332:738-740 (1988))。この分子のFc部分が酵素切断によって除去されているIgG分子には、このエフェクター機能がない(Harlow, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, 1988を参照されたい)。従って、エフェクター機能を最小化する遺伝子操作Fc配列を有することによって、またはヒトIgG2もしくはIgG4アイソタイプにすることによって、この分子のFc部分を欠いた結果としてエフェクター機能が低下した抗体を作製することができる。
例えば以下の本明細書における実施例14に記載されるように、MASP-3抗体は、MASP-3ポリペプチド(例えば、完全長MASP-3)を用いてまたは抗原性MASP-3エピトープ含有ペプチド(例えば、MASP-3ポリペプチドの一部)を用いて作製することができる。免疫原性ペプチドは5アミノ酸残基と小さくてもよい。抗体を産生させるのに用いられるMASP-3のペプチドおよびポリペプチドは、天然ポリペプチドまたは組換えペプチドもしくは合成ペプチドおよび触媒的に不活性な組換えポリペプチドとして単離されてもよい。MASP-3抗体の作製において有用な抗原はまた、融合ポリペプチド、例えば、MASP-3ポリペプチドまたはその一部と免疫グロブリンポリペプチドまたはマルトース結合タンパク質との融合も含む。ポリペプチド免疫原は完全長分子またはその一部でもよい。ポリペプチド部分がハプテン様であれば、このような部分は、免疫のために、都合よく、巨大分子担体(例えば、キーホールリンペットヘモシアニン(KLH)、ウシ血清アルブミン(BSA)、または破傷風トキソイド)に接続または連結されてもよい。
本明細書で使用する「モノクローナル」という修飾語は、抗体が実質的に均一な抗体集団から得られているという特徴を示し、特定の方法による抗体の作製を必要とすると解釈してはならない。モノクローナル抗体は、連続培養細胞株による抗体分子の作製を提供する任意の技術、例えば、Kohler, G., et al., Nature 256:495 (1975)に記載のハイブリドーマ法を用いて得ることができる。または、モノクローナル抗体は、組換えDNA法(例えば、Cabillyに対する米国特許第4,816,567号を参照されたい)によって作られてもよい。モノクローナル抗体は、Clackson, T., et al., Nature 352:624-628 (1991)、およびMarks, J.D., et al., J. Mol Biol. 222:581-597 (1991)に記載の技術を用いてファージ抗体ライブラリーから単離することもできる。このような抗体は、IgG、IgM、IgE、IgA、IgDを含む任意の免疫グロブリンクラス、およびその任意のサブクラスの抗体でよい。
本発明の方法において有用なモノクローナル抗体には、重鎖および/または軽鎖の一部が、特定の種に由来する抗体または特定の抗体クラスもしくはサブクラスに属する抗体の対応する配列と同一または相同であるが、鎖の残りが、別の種に由来する抗体または別の抗体クラスもしくはサブクラスに属する抗体の対応する配列と同一または相同であるキメラ抗体、ならびにこのような抗体の断片が含まれる(Cabillyに対する米国特許第4,816,567号;およびMorrison, S.L., et al., Proc. Nat'l Acad. Sci. USA 81:6851-6855, (1984))。
MASP-3抗体は組換え法を用いて作ることもできる。例えば、ヒト抗体断片(VH、VL、Fv、D因子、Fab、またはF(ab') 2)を作製するようにヒト免疫グロブリン発現ライブラリー(例えば、Stratagene, Corp., La Jolla, CAから入手可能)を用いてヒト抗体を作ることができる。次いで、キメラ抗体の作製法に類似した技術を用いて、これらの断片を用いてヒト抗体全体を構築する。
本発明の方法において有用なMASP-3阻害物質は、インタクトな免疫グロブリン分子だけでなく、抗体断片から形成された、Fab、Fab'、F(ab) 2、F(ab') 2、およびFv断片、scFv断片、ダイアボディ、直鎖抗体、単鎖抗体分子、ならびに多重特異性抗体(例えば、二重特異性抗体および三重特異性抗体)を含む周知の断片も包含する。
または、重鎖Fv領域および軽鎖Fv領域が連結されている、MASP-3に特異的なペプチド単鎖結合分子を作製することができる。Fv断片は、単鎖抗原結合タンパク質(scFv)を形成するようにペプチドリンカーで連結されてもよい。これらの単鎖抗原結合タンパク質は、オリゴヌクレオチドで連結された、VHドメインをコードするDNAおよびVLドメインをコードするDNAを含む構造遺伝子を構築することによって調製される。構造遺伝子は発現ベクターに挿入され、その後に、大腸菌などの宿主細胞に導入される。組換え宿主細胞は、2つのVドメインを架橋するリンカーペプチドを有する1本のポリペプチド 鎖を合成する。scFvを作製するための方法は、例えば、Whitlow, et al.,「Methods:A Companion to Methods in Enzymology」2:97 (1991); Bird, et al., Science 242:423 (1988); Ladnerに対する米国特許第4,946,778号; Pack, P., et al., Bio/Technology 11:1271 (1993)に記載されている。
本発明の方法において有用な高親和性MASP-3阻害抗体は多重特異性(すなわち二重特異性および三重特性)抗体を包含する。二重特異性抗体は、少なくとも2つの異なる抗原への結合特異性を有するモノクロナールの好ましくはヒトまたはヒト化抗体である。1つの態様において、組成物および方法は、MASP-3のセリンプロテアーゼドメインへの結合特異性およびMASP-2への結合特異性(たとえば、MASP-2のCCP1-CCP2またはセリンプロテアーゼドメインの少なくとも1つに結合する)を含む二重特異性抗体の使用を含む。別の態様において、方法は、MASP-3のセリンプロテアーゼドメインへの結合特異性およびMASP-1への結合特異性(たとえば、MASP-1のセリンプロテアーゼドメインへの結合)を含む二重特異性抗体の使用を含む。別の態様において、方法は、MASP-3への結合特異性(たとえば、MASP-3のセリンプロテアーゼドメインに結合する)、MASP-2への結合特異性(たとえば、MASP-2のCCP1-CCP2またはセリンプロテアーゼドメインの少なくとも1つに結合する)およびMASP-1への結合特異性(たとえば、MASP-1のセリンプロテアーゼドメインに結合する)を含む三重特異性抗体の使用を含む。
投与
別の局面において、本発明は、それを必要とする対象、たとえば第二経路関連の疾患または状態、たとえばPNHのような溶血性疾患または加齢黄斑変性症(AMD)、虚血再灌流障害、関節炎、播種性血管内凝固、血栓性微小血管症(溶血性尿毒症症候群(HUS)、非典型溶血性尿毒症症候群(aHUS)または血栓性血小板減少性紫斑病(TTP)を含む)、喘息、デンスデポジット病、微量免疫型壊死性半月体形成性糸球体腎炎、外傷性脳損傷、誤嚥性肺炎、眼内炎、視神経脊髄炎、ベーチェット病、多発性硬化症(MS)、ギランバレー症候群、アルツハイマー病、筋萎縮性側索硬化症(ALS)、ループス腎炎、全身性エリテマトーデス(SLE)、糖尿病性網膜症、ブドウ膜炎、慢性閉塞性肺疾患(COPD)、C3糸球体症、移植片拒絶反応、移植片対宿主病(GVHD)、血液透析、敗血症、全身性炎症反応症候群(SIRS)、急性呼吸窮迫症候群(ARDS)、ANCA血管炎、抗リン脂質症候群、アテローム性動脈硬化症、IgA腎症、および重症筋無力症からなる群より選択される疾患または障害に罹患している対象において第二経路補体活性化の有害作用を阻害するための、高親和性MASP-3阻害抗体を含む組成物を提供する。
一般的に、MASP-3阻害抗体組成物、または、MASP-2阻害物質とMASP-3阻害物質の組み合わせを含む組成物は、他の任意の選択された治療剤と組み合わされてもよく、適宜、薬学的に許容される担体中に含まれる。担体は、無毒で、生体適合性があり、MASP-3阻害抗体またはMASP-2阻害物質(およびMASP-2阻害物質と組み合わされた他の任意の治療剤)の生物学的活性に悪影響を及ぼさないように選択される。ペプチド用の例示的な薬学的に許容される担体は、Yamadaに対する米国特許第5,211,657号に記載されている。本明細書に記載される、本発明において有用なMASP-3抗体は、経口投与、非経口投与、または外科的投与を可能にする、固体、半固体、ゲル、液体、または気体の形をした調製物、例えば、錠剤、カプセル、散剤、顆粒、軟膏、溶液、デポジトリ(depository)、吸入剤、および注射剤に製剤化されてもよい。本発明はまた、医療装置などをコーティングすることによる組成物の局所投与も意図する。
本明細書に記載される高親和性MASP-3阻害抗体に関してより具体的には、例示的な製剤を、水、油、食塩水、グリセロール、またはエタノールなどの滅菌液体でもよい薬学的担体と共に、生理学的に許容される希釈剤に溶解した化合物の注射投与量の溶液または懸濁液として非経口投与することができる。さらに、MASP-3抗体を含む組成物中には、補助物質、例えば、湿潤剤または乳化剤、界面活性剤、pH緩衝物質などが存在してもよい。薬学的組成物のさらなる成分には、石油(例えば、動物由来、野菜由来、または合成由来の石油)、例えば、ダイズ油および鉱油が含まれる。一般的に、グリコール、例えば、プロピレングリコールまたはポリエチレングリコールが注射液に好ましい液体担体である。
任意でMASP-2阻害物質と組み合わせたMASP-3阻害抗体を含む薬学的組成物は、局所投与方法または全身投与方法が、治療されている状態に最も適しているかどうかに応じて多くのやり方で投与することができる。さらに、本発明の組成物を移植可能な医療装置の表面に、または移植可能な医療装置の中にコーティングまたは組み込むことによって、本発明の組成物を送達することができる。
本明細書で使用する「全身送達」および「全身投与」という用語は、筋肉内(IM)投与経路、皮下投与経路、静脈内(IV)投与経路、動脈内投与経路、吸入投与経路、舌下投与経路、頬投与経路、局部投与経路、経皮投与経路、鼻投与経路、直腸投与経路、腟投与経路、および送達作用物質を1つまたは複数の目的の治療作用部位に効果的に分散させる他の投与経路を含む、経口経路および非経口経路を含むが、これらに限定されないことが意図される。本組成物の好ましい全身送達経路には、静脈内経路、筋肉内経路、皮下経路、動脈内経路、および吸入経路が含まれる。本発明の特定の組成物において用いられる選択された作用物質の正確な全身投与経路は、一つには、ある特定の投与経路に関連した代謝変換経路に対する作用物質感受性を説明するように決定されることが理解されると考えられる。例えば、ペプチド物質は、最も適切には、経口以外の経路によって投与することができる。
本明細書で使用する「局所」という用語は、目的の限局作用の部位の中に、または目的の限局作用の部位の周囲に薬物を適用することを包含し、例えば、皮膚または他の患部組織への局部送達、眼送達、くも膜下腔内(IT)、脳室内(ICV)、関節内、洞内、頭蓋内、もしくは小胞内の投与、留置、または灌注を含んでもよい。局所投与は、低用量の投与が全身副作用を回避するために、ならびに局所送達部位に活性物質を送達および濃縮するタイミングをより正確に制御するために好ましい場合がある。局所投与は、代謝、血流などの患者間のばらつきに関係なく標的部位において既知濃度を供給する。改善された投与量制御は直接的な送達方法によっても提供される。
予防的用途において、薬学的組成物は、第二経路関連の疾患または障害、たとえば発作性夜間ヘモグロビン尿症(PNH)、加齢黄斑変性症(AMD)、虚血再灌流障害、関節炎、播種性血管内凝固、血栓性微小血管症(溶血性尿毒症症候群(HUS)、非典型溶血性尿毒症症候群(aHUS)および血栓性血小板減少性紫斑病(TTP)を含む)、喘息、デンスデポジット病、微量免疫型壊死性半月体形成性糸球体腎炎、外傷性脳損傷、誤嚥性肺炎、眼内炎、視神経脊髄炎、ベーチェット病、多発性硬化症、ギランバレー症候群、アルツハイマー病、筋萎縮性側索硬化症(ALS)、ループス腎炎、全身性エリテマトーデス(SLE)、糖尿病性網膜症、ブドウ膜炎、慢性閉塞性肺疾患(COPD)、C3糸球体症、移植片拒絶反応、移植片対宿主病(GVHD)、血液透析、敗血症、全身性炎症反応症候群(SIRS)、急性呼吸窮迫症候群(ARDS)、ANCA血管炎、抗リン脂質症候群、アテローム性動脈硬化症、IgA腎症および重症筋無力症からなる群より選択される第二経路疾患または第二経路障害に罹患しやすい、またはそうでなければその危険のある対象に、状態の症状を発症するおそれを除く、または減らすのに十分な量で投与される。治療的用途において、薬学的組成物は、第二経路関連の疾患または障害、たとえば発作性夜間ヘモグロビン尿症(PNH)、加齢黄斑変性症(AMD)、虚血再灌流障害、関節炎、播種性血管内凝固、血栓性微小血管症(溶血性尿毒症症候群(HUS)、非典型溶血性尿毒症症候群(aHUS)、または血栓性血小板減少性紫斑病(TTP)を含む)、喘息、デンスデポジット病、微量免疫型壊死性半月体形成性糸球体腎炎、外傷性脳損傷、誤嚥性肺炎、眼内炎、視神経脊髄炎、ベーチェット病、多発性硬化症、ギランバレー症候群、アルツハイマー病、筋萎縮性側索硬化症(ALS)、ループス腎炎、全身性エリテマトーデス(SLE)、糖尿病性網膜症、ブドウ膜炎、慢性閉塞性肺疾患(COPD)、C3糸球体症、移植片拒絶反応、移植片対宿主病(GVHD)、血液透析、敗血症、全身性炎症反応症候群(SIRS)、急性呼吸窮迫症候群(ARDS)、ANCA血管炎、抗リン脂質症候群、アテローム性動脈硬化症、IgA腎症、および重症筋無力症からなる群より選択される第二経路疾患または第二経路障害の疑いがあるまたはすでにそれに罹患している対象に、状態の症状を軽減するのにまたは少なくとも部分的に減らすのに十分な治療的に有効な量で投与される。
以下の実施例は、本発明の実施について意図された最良の形態の単なる例示であり、本発明を限定すると解釈してはならない。本明細書中の全ての文献引用が明確に参照により組み入れられる。
本実施例は、MASP-2欠損マウスが、髄膜炎菌血清群Aまたは髄膜炎菌血清群Bのどちらかに感染した後に髄膜炎菌誘導死から保護されることを証明する。
MASP-2ノックアウトマウス(MASP-2 KOマウス)を、本明細書に参照として組み入れられるUS 7,919,094の実施例1に記載のように生成した。100μl体積で投与量2.6×107CFUの髄膜炎菌血清群A Z2491を腹腔内(i.p.)注射することによって、10週齢MASP-2 KOマウス(n=10)および野生型(WT)C57/BL6マウス(n=10)に接種した。感染用量を、最終濃度400mg/kgの鉄デキストランと一緒にマウスに投与した。72時間の期間にわたって感染後のマウスの生存をモニタリングした。
図6は、感染用量2.6×107cfuの髄膜炎菌血清群A Z2491を投与させた後のMASP-2 KOマウスおよびWTマウスの生存率(%)を図示したカプラン・マイヤープロットである。図6に示したように、感染後72時間の期間全体を通じて100%のMASP-2 KOマウスが生存した。対照的に、感染24時間後、WTマウスの80%しか生存しておらず(p=0.012)、感染72時間後、WTマウスの50%しか生存していなかった。これらの結果から、MASP-2欠損マウスは髄膜炎菌血清群A Z2491誘導死から保護されることが証明される。
本実施例は、髄膜炎菌感染後のMASP-2抗体の投与が髄膜炎菌感染マウスの生存率を増加させることを実証する。
参照により本明細書に組み入れられる米国特許第7,919,094号の実施例24に記載されているように、ラットMASP-2タンパク質を使用してFabファージディスプレイライブラリーをパンニングし、そこからFab2 #11を機能的に活性な抗体として同定した。ラットIgG2cおよびマウスIgG2aアイソタイプの完全長抗体をFab2 #11から生成した。マウスIgG2aアイソタイプの完全長MASP-2抗体を薬力学的パラメータに関して特徴決定した(米国特許第7,919,094号の実施例38に記載されているとおり)。
上記のように生成したFab2#11由来のマウスIgG2a完全長MASP-2抗体アイソタイプを、以下のように、髄膜炎菌感染マウスモデルにおいて試験した。
9週齢C57/BL6チャールズリバーマウスを、高用量(4×106cfu)の髄膜炎菌血清型B株MC58の腹腔内注射から3時間後、阻害性マウスMASP-2抗体(1.0mg/kg)(n=12)または対照アイソタイプ抗体(n=10)で処理した。
図10は、感染量4×106cfuの髄膜炎菌血清群B株MC58の投与ののち、阻害性MASP-2抗体(1.0mg/kg)または対照アイソタイプ抗体のいずれかを感染後3時間で投与したマウスの生存率%をグラフで示すカプラン・マイヤー(Kaplan-Meyer)プロットである。図10に示すように、MASP-2抗体で処理されたマウスは、90%が感染後72時間を通して生存した。対照的に、アイソタイプ抗体で処理されたマウスは、50%しか感染後72時間を通して生存しなかった。「*」という記号は、2つの生存曲線の比較によって決定されたp=0.0301を示す。
本実施例は、ヒト血清中の髄膜炎菌の補体依存性死滅がMASP-3依存性であることを実証する。
機能的MBLの血清レベルが低下した患者は、再発性の細菌および真菌感染への罹患性の増大を示す(Kilpatrick et al., Biochim Biophys Acta 1572:401-413 (2002))。髄膜炎菌がMBLによって認識されることは公知であり、MBL欠損血清が髄膜炎菌を溶解しないことが示されている。
1. 様々な補体欠損ヒト血清におけるおよびヒトMASP-2抗体で処理されたヒト血清における血清殺菌活性
この実験には、以下の補体欠損ヒト血清および対照ヒト血清を使用した。
図11は、表6に示すヒト血清試料中、様々な時点で回収された髄膜炎菌血清群B-MC58の生菌数のlog cfu/mLをグラフで示す。表7は、図11のスチューデントt検定の結果を提示する。
この実験には、以下の補体欠損ヒト血清および対照ヒト血清を使用した。
図12は、表8に示すヒト血清試料中、様々な時点で回収された髄膜炎菌血清群B-MC58の生菌数のlog cfu/mLをグラフで示す。図12に示すように、WT(NHS)血清が、髄膜炎菌に関して最高レベルの殺菌活性を有する。対照的に、MBL-/-およびMASP-3-/-(MASP-1充分である)ヒト血清は任意の殺菌活性を有しない。これらの結果は、ヒト20%(v/v)血清中の髄膜炎菌の補体依存性死滅がMASP-3およびMBL依存性であることを示す。表9は図12のスチューデントt検定の結果を提示する。
この実験には、以下の補体欠損マウス血清および対照マウス血清を使用した。
図13は、表10に示すマウス血清試料中、様々な時点で回収された髄膜炎菌血清群B-MC58の生菌数のlog cfu/mLをグラフで示す。図13に示すように、MASP-2-/-マウス血清は、WTマウス血清よりも髄膜炎菌に関して高いレベルの殺菌活性を有する。対照的に、MASP-1/3-/-マウス血清は任意の殺菌活性を有しない。「**」という記号はp=0.0058を示し、「***」という記号はp=0.001を示す。表11は、図13のスチューデントt検定の結果を提示する。
本実施例は、実施例1〜3に記載されるような、MASP-2 KOマウスにおいて認められた髄膜炎菌感染に対するMASP-3依存性抵抗の機構を決定するために実施された一連の実験を記載する。
以下のように、MASP-2 KOマウスにおいて認められた髄膜炎菌感染に対するMASP-3依存性抵抗の機構(上記実施例1〜3に記載)を決定するために一連の実験を実施した。
方法:
MASP-1/3欠損マウスがレクチン経路機能活性(「LEA-2」とも呼ばれる)を欠損しているかどうかを判定するために、参照により本明細書に組み入れられるSchwaeble W. et al., PNAS vol 108(18):7523-7528 (2011)に記載されているとおりに、レクチン活性化経路特異的アッセイ条件下(1%血漿)、試験される様々な補体欠損マウス系統からの血漿中のC3コンバターゼ活性の動態を測定するためのアッセイ法を実施した。
レクチン経路特異的条件下のC3活性化の動態(1%血清を有するマンナンコーティングされたプレート上のC3b沈着によって測定)を図14に示す。MASP-2-/-血漿中にはC3切断は見られなかった。B因子-/-(B因子-/-)血漿は、おそらくは増幅ループの損失のせいで、WT血漿の半分の速度でC3を切断した。C4-/-(T1/2=33分)およびMASP-1/3-/-欠損血漿(T1/2=49分)においてC3からC3bへのレクチン経路依存性転換の有意な遅延が見られた。MASP-1/3-/-血漿におけるC3活性化のこの遅延は、MASP-3依存性ではなくMASP-1依存性であることが示されている(Takahashi M. et al., J Immunol 180:6132-6138 (2008)を参照されたい)。これらの結果は、MASP-1/3-欠損マウスがレクチン経路機能活性(「LEA-2」とも呼ばれる)を欠損していないことを実証する。
原理:
MASP-3のセリンプロテアーゼをコードするエキソン中のフレームシフト突然変異によって生じる3MC症候群を有するMASP-3欠損患者の血清を試験することにより、第二経路活性化に対する遺伝性MASP-3欠損の影響を判定した。3MC症候群とは、特徴が重なるCarneavale症候群、Mingarelli症候群、Malpuech症候群およびMichels症候群の統一用語である。この珍しい常染色体劣性障害は、特徴的な顔面異形症、口唇裂および/または口蓋裂、頭蓋骨癒合症、学習障害、ならびに性器、四肢、および膀胱直腸異常を含む一連の発達的特徴を示す。Rooryck et al., Nature Genetics 43:197-203 (2011)は、3MC症候群の11家族を研究し、突然変異した2つの遺伝子COLEC11およびMASP-1を同定した。MASP-1遺伝子の突然変異は、MASP-3のセリンプロテアーゼドメインをコードするエキソンを機能不全にするが、MASP-1のセリンプロテアーゼドメインをコードするエキソンを機能不全にはしない。したがって、MASP-3のセリンプロテアーゼをコードするエキソン中に突然変異を有する3MC患者は、MASP-3を欠損しているが、MASP-1を充分に有する。
MASP-3欠損血清は、3MC患者、その3MC患者の母親および父親(両親とも、MASP-3セリンプロテアーゼドメインをコードするエキソンを機能不全にする突然変異を有するアレルに関してヘテロ接合性)ならびにC4欠損患者(両方のヒトC4遺伝子を欠損している)およびMBL欠損対象から得た。Bitter-Suermann et al., Eur. J. Immunol. 11:291-295 (1981))に記載されているような従来のAP特異的条件下(BBS/Mg++/EGTA、Ca++なし、ここで、BBS=スクロースを含有するバルビタール緩衝食塩水)、ザイモサンコーティングされたマイクロタイタープレート上、0.5〜25%の範囲の血清濃度で第二経路アッセイ法を実施し、時間の経過とともにC3b沈着を測定した。
図15は、ザイモサンコーティングされたマイクロタイタープレート上の第二経路駆動型C3b沈着のレベルを、MASP-3欠損、C4欠損およびMBL欠損対象から採取された血清試料中の血清濃度の関数としてグラフで示す。図15に示すように、MASP-3欠損患者の血清は、高い血清濃度(25%、12.5%、6.25%血清濃度)で残留第二経路(AP)活性を有するが、有意に高いAP50(すなわち、最大C3沈着の50%を達成するために必要な血清の9.8%)を有する。
方法:
MASP-2-/-、MASP-1/3-/-、およびWTマウスから採取された0%〜20%の範囲のマウス血清濃度を使用して、マンナン、ザイモサンおよび肺炎連鎖球菌D39でコーティングされたマイクロタイタープレート上のC3b沈着を測定した。「従来の」第二経路特異的条件下(すなわちCa++なしのBBS/EGTA/Mg++)またはレクチン経路および第二経路の両方が機能することを可能にする生理学的条件下(すなわちBBS/Mg++/Ca++)、C3b沈着アッセイ法を実施した。
図17Aは、従来の第二経路特異的条件下(すなわち、Ca++なしのBBS/EGTA/Mg++)またはレクチン経路および第二経路の両方が機能することを可能にする生理学的条件下(BBS/Mg++/Ca++)、マンナンコーティングされたマイクロタイタープレート上のC3b沈着のレベルを、WT、MASP-2欠損およびMASP-1/3欠損マウスから採取された血清試料中の血清濃度の関数としてグラフで示す。図17Bは、従来のAP特異的条件下(すなわち、Ca++なしのBBS/EGTA/Mg++)またはレクチン経路および第二経路の両方が機能することを可能にする生理学的条件下(BBS/Mg++/Ca++)、ザイモサンコーティングされたマイクロタイタープレート上のC3b沈着のレベルを、WT、MASP-2欠損およびMASP-1/3欠損マウスからの血清試料中の血清濃度の関数としてグラフで示す。図17Cは、従来のAP特異的条件下(すなわち、Ca++なしのBBS/EGTA/Mg++)またはレクチン経路および第二経路の両方が機能することを可能にする生理学的条件下(BBS/Mg++/Ca++)、肺炎連鎖球菌D39コーティングされたマイクロタイタープレート上のC3b沈着のレベルを、WT、MASP-2欠損およびMASP-1/3欠損マウスからの血清試料中の血清濃度の関数としてグラフで示す。
本実施例に記載された結果は、MASP-2阻害因子(またはMASP-2 KO)が、MASP-3駆動型第二経路活性化を促進することにより、髄膜炎菌感染からの有意な保護を提供することを実証する。マウス血清溶菌アッセイ法およびヒト血清溶菌アッセイ法の結果はさらに、髄膜炎菌に対する血清殺菌活性をモニターすることにより、髄膜炎菌に対する殺菌活性がMBL欠損(マウスMBL AおよびMBL C二重欠損血清およびヒトMBL欠損血清)中には存在しないことを示す。
本実施例は、発作性夜間血色素尿症(PNH)のマウスモデルから得られた血液試料に由来する赤血球の溶解に対するMASP-2欠損および/またはMASP-3欠損の阻害作用を証明する。
発作性夜間血色素尿症(PNH)はマルキアファーヴァ・ミケーリ症候群とも呼ばれ、補体誘導性の血管内溶血性貧血を特徴とする、後天的な、潜在的に命にかかわる血液疾患である。PNHの顕著な特徴は、補体制御因子CD55およびCD59がPNH赤血球上に存在しないために補体第二経路が無秩序に活性化した結果である慢性的な補体媒介性血管内溶血と、その後に起こるヘモグロビン尿症および貧血である。Lindorfer, M.A., et al., Blood 115(11)(2010)、Risitano, A.M, Mini-Reviews in Medicinal Chemistry, 11:528-535(2011)。PNHにおける貧血は血流中の赤血球の破壊が原因である。PNHの症状には、尿中のヘモグロビンの出現による赤色尿、背部痛、疲労、息切れ、および血栓症が含まれる。PNHは自然発症することがあり、これは「一次PNH」と呼ばれるか、または再生不良性貧血などの他の骨髄障害の状況では「二次PNH」と呼ばれる。PNHの治療には、貧血の場合は輸血、血栓症の場合は血液凝固阻止、および補体系を阻害することによって免疫破壊から血球を保護するモノクローナル抗体エクリズマブ(Soliris(登録商標))の使用(Hillmen P. et al., N. Engl. J. Med. 350(6) 552-9(2004))が含まれる。エクリズマブ(Soliris(登録商標))は、補体成分C5を標的とし、C5コンバターゼによるC5切断を遮断し、それによって、C5aの産生およびMACの集合を阻止するヒト化モノクローナル抗体である。エクリズマブによるPNH患者の治療は、乳酸デヒドロゲナーゼ(LDH)によって測定されるように血管内溶血を減少させ、そのため、患者の約半分におけるヘモグロビン安定化および輸血非依存性につながった(Hillmen P, et al., Mini-Reviews in Medicinal Chemistry, vol11(6)(2011))。エクリズマブ療法を受けているほぼ全員の患者においてLDHレベルが正常またはほぼ正常になったが(血管内溶血の管理のため)、患者の約1/3しかヘモグロビン値 約11gr/dLに達せず、エクリズマブを服用した残りの患者は中程度から重度の(すなわち、輸血依存性)貧血をほぼ同じ割合で示し続ける(Risitano A.M. et al., Blood 113:4094-100(2009))。Risitano et al., Mini-Reviews in Medicinal Chemistry 11:528-535(2011)に記載のように、エクリズマブを服用しているPNH患者は、PNH赤血球のかなりの部分に結合しているC3断片を含んだ(が、未治療患者は含んでいなかった)ことが証明された。このことから、膜に結合しているC3断片はPNH赤血球に対してオプソニンとして働き、その結果、特異的C3受容体を介して細網内皮細胞内に閉じ込められ、その後に、血管外溶血が起こるという結論が導かれた。従って、C3断片を介した血管外溶血を発症している患者は赤血球輸血を必要とし続けるので、これらの患者には、エクリズマブの使用の他に治療方針が必要とされる。
PNH動物モデル:
CrryおよびC3を欠損した(Crry/C3-/-)遺伝子標的化マウスおよびCD55/CD59欠損マウスから血液試料を採取した。これらのマウスは、赤血球上のそれぞれの表面補体制御因子を欠き、したがって、これらの赤血球はPNHヒト血球と同様に自発的に補体自己溶解を受けやすい。
1日目、ネズミRBC(±マンナンコーティング)の調製
材料は以下を含むものであった:新鮮なマウス血液、BBS/Mg++/Ca++(4.4mMバルビツール酸、1.8mMナトリウムバルビトン、145mM NaCl、pH7.4、5mM Mg++、5mM Ca++)、塩化クロム、CrCl3・6H2O(BBS/Mg++/Ca++中0.5mg/mL)、およびマンナン、BBS/Mg++/Ca++中100μg/mL。
材料は、BBS/ゼラチン/Mg++/Ca++(上記)、試験血清、96ウェル丸底および平底プレートならびに410〜414nmで96ウェルプレートを読み取る分光光度計を含むものであった。
上記プロトコールに詳述したように、CD55/CD59二重欠損マウスおよびCrry/C3二重欠損マウスから新鮮な血液を採取し、赤血球を調製した。赤血球を分割し、赤血球の半分をマンナンでコーティングし、他方の半分を未処理のままにし、最終濃度を108/mLに調節し、そのうち100μLを、上記のように実施した溶血アッセイ法に使用した。
最初の実験において、コーティングなしのWTマウス赤血球が任意のマウス血清中で溶解しないことがわかった。さらに、マンナンコーティングされたCrry-/-マウス赤血球は、WTマウス血清中ではゆっくりと溶解するが(37度で3時間超)、MBLヌル血清中では溶解しないことがわかった(データ示さず)。
マンナンコーティングされたCrry-/-マウス赤血球は、MBLを含む高希釈ヒト血清中では非常に良好に溶解するが、MBLを含まない高希釈ヒト血清中ではそうはならない。試験した各血清濃度での効率的な溶解は、第二経路がこの溶解に関与せず、または必要とされないことを暗示する。MBL欠損マウス血清およびヒト血清がマンナンコーティングされたCrry-/-マウス赤血球を溶解できないことは、古典経路もまた、観察された溶解とは関係がないことを示す。レクチン経路認識分子(すなわちMBL)が必要とされるため、この溶解はレクチン経路によって媒介される。
Crry/C3およびCD55/CD59二重欠損マウスから新鮮な血を採取し、マンナンコーティングされたCrry-/-マウス赤血球を、上記のような溶血アッセイ法において、以下のヒト血清:MASP-3-/-;MBLヌル;WT;ヒトMASP-2抗体で前処理されたNHS;および対照としての熱不活化NHSの存在において分析した。
マンナンコーティングされたCrry-/-マウス赤血球とともにNHSを、1/640まで(すなわち、1/40、1/80、1/160、1/320および1/640)希釈された希釈物、ヒトMBL-/-血清、ヒトMASP-3欠損血清(3MC患者からのもの)およびMASP-2 mAbで前処理されたNHSおよび対照として熱不活化NHSの中でインキュベートした。
Crry/C3およびCD55/CD59二重欠損マウスからの新鮮な血液から採取されたコーティングなしのCrry-/-マウス赤血球を、上記のような溶血アッセイ法において、以下の血清:MASP-3-/-;MBL-/-;WT;ヒトMASP-2抗体で前処理されたNHSおよび対照として熱不活化NHSの存在において分析した。
図21は、3MC(MASP-3-/-)患者、熱不活化(HI)NHS、MBL-/-、MASP-2抗体で前処理されたNHSおよびNHS対照からのヒト血清中、一定範囲の血清濃度にわたり、コーティングなしのネズミ赤血球の溶血(上清への溶解WTマウス赤血球のヘモグロビン放出を測光法によって測定)をグラフで示す。図21に示し、表13にまとめているように、MASP-3の阻害が非感作WTマウス赤血球の補体媒介性溶解を阻害することが実証される。
本実施例は、WTまたはMASP-1/3-/-マウス血清の存在における溶解に関してマンナンコーティングされたウサギ赤血球を試験する溶血アッセイ法を記載する。
1. マウスMASP-1/3欠損血清およびWT対照血清におけるウサギRBC(マンナンコーティングされた)の溶血アッセイ法
1日目、ウサギRBCの調製
材料は以下を含むものであった:新鮮なウサギ血液、BBS/Mg++/Ca++(4.4mMバルビツール酸、1.8mMナトリウムバルビトン、145mM NaCl、pH7.4、5mM Mg++、5mM Ca++)、0.1%ゼラチンを含むBBS/Mg++/Ca++、緩衝液に含まれた塩化クロム、すなわちCrCl3.6H2O(BBS/Mg++/Ca++中0.5mg/mL)およびマンナン、BBS/Mg++/Ca++中100μg/mL。
1. ウサギ全血(2mL)を、2つの1.5mLエッペンドルフ管に分割し、4℃の冷却エッペンドルフ遠心分離機中、8000rpm(約5.9rcf)で3分間遠心処理した。氷冷BBS/Mg++/Ca++中に再懸濁させたのち、RBCペレットを3回洗浄した。3回目の洗浄後、ペレットをBBS/Mg++/Ca++4mL中に再懸濁させた。このアリコート2mLを、コーティングなし対照として使用するために、15mLファルコンチューブに加えた。残り2mLのRBCアリコートをCrC13緩衝液2mL中に希釈し、マンナン溶液2mLを添加し、懸濁液を穏やかに混合しながら室温で5分間インキュベートした。BBS/0.1%ゼラチン/Mg++/Ca++7.5mLを混合物に加えることによって反応を停止させた。上記のように赤血球をペレット化し、RBCをBBS/0.1%ゼラチン/Mg++/Ca++で2回洗浄した。RBC懸濁液をBBS/0.1%ゼラチン/Mg++/Ca++中、4℃で貯蔵した。
2. 懸濁させたRBC100μLを水1.4mLで希釈し、8000rpm(約5.9rcf)で3分間スピンダウンし、上清のODを541nmで0.7に調節した(541nmで0.7のODは赤血球約109個/mLに相当)。
3. 再懸濁させたRBCをBBS/0.1%ゼラチン/Mg++/Ca++で108個/mLの濃度まで希釈した。
4. 試験血清の希釈物を氷冷BBS/ゼラチン/Mg++/Ca++中に調製し、各血清希釈物100μLを丸底プレートの対応するウェルにピペットで移した。適切に希釈したRBC100μL(108/mL)を各ウェルに加えた。完全な溶解のための対照として、精製水(100μL)を希釈RBC(100μL)と混合して100%溶解を生じさせ、一方、血清なしのBBS/0.1%ゼラチン/Mg++/Ca++(100μL)を陰性対照として使用した。次いで、プレートを37℃で1時間インキュベートした。
5. 丸底プレートを3250rpmで5分間遠心処理した。各ウェルからの上清(100μL)を平底プレートの対応するウェルに移し、ELISAリーダー中、415〜490nmでODを読み取った。結果を、490nmでのODに対する415nmでのODの比として報告する。
図23は、MASP-1/3-/-およびWT対照からの血清中の一定範囲の血清濃度にわたり、マウス血清によるマンナンコーティングされたウサギ赤血球の溶血(上清への溶解ウサギ赤血球のヘモグロビン放出を測光法によって測定)をグラフで示す。図23に示すように、MASP-3の阻害が、マンナンコーティングされたWTウサギ赤血球の補体媒介性溶解を阻止することが実証される。これらの結果はさらに、実施例5に記載されたようなPNHの1つまたは複数の局面の治療のためのMASP-3阻害因子の使用を裏付ける。
本実施例は、改変されたDT40細胞株DTLacOを使用するインビトロシステムを使用する、MASP-1およびMASP-3モノクローナル抗体の生成を記載する。
WO2009029315およびUS2010093033にさらに記載されているように、特定のポリペプチドの可逆性多様化誘発を可能にする改変されたDT40細胞株DTLacOを含むインビトロシステムを使用して、ヒトMASP-1およびMASP-3に対する抗体を生成した。DT40は、培養中にその重鎖および軽鎖免疫グロブリン(Ig)遺伝子を構成性突然変異させることが知られているニワトリB細胞株である。他のB細胞と同様に、この構成性突然変異誘発は、Ig遺伝子のV領域への突然変異、ひいては発現した抗体分子のCDRを標的化する。DT40細胞中の構成性突然変異誘発は、各機能的V領域よりも上流に位置する非機能的V遺伝子セグメント(疑似V遺伝子;ΨV)のアレイをドナー配列として使用する遺伝子変換によって起こる。ΨV領域の欠失は、以前、ヒトB細胞において一般に認められる機構である、多様化の機構における遺伝子転換から体細胞超変異への切替えを生じさせることが知られていた。DT40ニワトリB細胞リンパ腫システムが、エクスビボでの抗体進化のための有望な出発点であることが示されている(Cumbers, S. J. et al. Nat Biotechnol 20, 1129-1134 (2002); Seo, H. et al. Nat Biotechnol 23, 731-735 (2005))。DT40細胞は培養中、8〜10時間の倍加時間(ヒトB細胞システムの場合の20〜24時間に比べて)で強く増殖し、非常に効率的な相同遺伝子標的化を支援する(Buerstedde, J. M. et al. Embo J 9, 921-927 (1990))。DT40細胞は、多様化のための2つの別々の生理学的経路、それぞれ鋳型化突然変異および非鋳型化突然変異を創製する遺伝子転換および体細胞超変異にアクセスすることができることを条件に、非常に大きな潜在的V領域配列多様性を命令する(Maizels, N. Annu Rev Genet. 39, 23-46 (2005))。多様化した重鎖および軽鎖免疫グロブリン(Ig)は細胞表面表示IgMの形態で発現する。表面IgMは、構造的にIgG分子に似る二価形態を有する。特定の抗原への特異性をもってIgMを表示する細胞は、抗原の固定化可溶性バージョンまたは膜表示バージョンに結合させることによって単離することができる。しかし、抗体進化のためのDT40細胞の利用は実際には限られている。理由は、他の形質転換B細胞株と同様、多様化が生理学的速度の1%未満の速度でしか起こらないからである。
MASP-1およびMASP-3抗原結合についての選択
遺伝子標的化によって多様化させたDTLacO集団を、ヒトMASP-1(SEQ ID NO:8)およびMASP-3抗原(SEQ ID NO:2)と複合化したビーズに結合することによって最初の選択を実施し、その後、FACSにより、蛍光標識可溶性抗原を使用して選択した(Cumbers, S. J. et al. Nat Biotechnol 20, 1129-1134 (2002); Seo, H. et al. Nat Biotechnol 23, 731-735 (2005)。MASP-1とMASP-3との間で共有されるα鎖中の保存されたアミノ酸配列(図2に示す)および別々のβ鎖配列(図2に示す)のせいで、MASP-1およびMASP-3へのバインダのための別々の平行スクリーニングを実施して、MASP-1特異性mAb、MASP-3特異性mAbならびにMASP-1およびMASP-3の両方に結合することができる(二重特異性)mAbを同定した。2つの形態の抗原を使用して、バインダを選択し、スクリーニングした。まず、Fcドメインに融合した、完全長または断片のいずれかの組換えMASP-1またはMASP-3をDynal磁性プロテインGビーズに結合させるか、または、PECy5標識抗ヒトIgG(Fc)二次抗体を使用してFACSベースの選択に使用した。あるいは、MASP-1またはMASP-3タンパク質の組換えバージョンをDylight flourで直接標識し、選択およびスクリーニングに使用した。
PCR増幅V領域を293F細胞中のヒトIgG1の発現を支持するベクターにクローニングすることによって組換え抗体を生成した(Yabuki et al., PLoS ONE, 7(4):e36032 (2012))。MASP-1またはMASP-3を様々な濃度の蛍光標識可溶性抗原と結合させる抗体を発現するDTLacO細胞を染色することによって飽和結合反応速度を測定した。MASP-3依存性C3b沈着およびMASP-3依存性D因子切断を含むMASP-3特異性活性に関する機能アッセイ法を、それぞれ実施例8および9に記載するように実施した。MASP-1特異性活性、すなわちMASP-1依存性C3b沈着の阻害に関する機能アッセイ法を以下に記載するように実施した。
上記方法を使用して、数多くのMASP-1およびMASP-3結合抗体を生成した。FACS分析によって実証された結合を、MASP-3バインダのスクリーニングにおいて単離された代表的なクローンM3J5およびM3M1に関して記載する。
図26Aは、親DTLacO(SEQ ID NO:300)、MASP-3結合クローンM3J5(SEQ ID NO:301)およびM3M1(SEQ ID NO:302)ならびにMASP-1/MASP-3二重結合クローンD14(SEQ ID NO:306)および1E10(SEQ ID NO:308)に関する重鎖可変領域(VH)配列のアミノ酸アライメントを示す。
図26Bは、親DTLacO(SEQ ID NO:303)ならびにMASP-3結合クローンM3J5(SEQ ID NO:304)およびM3M1(SEQ ID NO:305)ならびにMASP-1/MASP-3二重結合クローンD14(SEQ ID NO:307)および1E10(SEQ ID NO:309)に関する軽鎖可変領域(VL)配列のアミノ酸アライメントを示す。
MASP-1は、MASP-2を活性化するその能力を介してLEA-2に寄与する(図1を参照されたい)。Wieslab(登録商標)補体システムスクリーニングMBLアッセイ法(Euro Diagnostica, Malmo, Sweden)は、LEA-2依存活性化(すなわち、従来のレクチン経路活性化)を単離する条件下、C5b-C9沈着を測定する。製造者の取り扱い指示に従って、代表的なクローン1E10を400nMの最終濃度で試験してアッセイ法を実施した。
上記の結果は、DTLacOプラットフォームが、LEA-1(以下、実施例8および9に示す)およびLEA-2(本実施例に示す)に対する阻害性を有するMASP-1およびMASP-3モノクローナル抗体の迅速なエクスビボ発見を可能にすることを示した。
黄色ブドウ球菌による3MC血清中の補体経路の分析
背景/原理:
MASP-3は、正常ヒト血清の存在または非存在において非固定化流体相マンナン、ザイモサンAまたはN-アセチルシステインへの曝露を経ても活性化されないことがわかった。しかし、組換えおよび天然のMASP-3は正常ヒト血清(NHS)または熱不活化ヒト血清(HIS)の存在および非存在において熱不活化黄色ブドウ球菌の表面で活性化されることがわかった(データ示さず)。また、正常ヒト血清の存在において黄色ブドウ球菌の表面でC3b沈着が起こり、フローサイトメーターを使用してその沈着をモニターすることができることがわかった。したがって、LEA-1に対するMASP-3の寄与を評価する手段として、本実施例に記載するように、黄色ブドウ球菌に対する第二経路(AP)応答を測定した。
組換えMASP-3:完全長組換えヒトMASP-3をコードするポリヌクレオチド配列、MASP-3の切断型セリンプロテアーゼ(SP)活性バージョン(CCP1-CCP2-SP)およびSP不活化形態のMASP-3(S679A)をpTriEx7哺乳動物発現ベクター(Invivogen)にクローニングした。得られた発現構築物は完全長MASP-3またはCCP1-CCP2-SP断片をアミノ末端Streptagおよびカルボキシ末端His6タグによってコードする。発現構築物を製造者によって提供されるプロトコールに従ってFreestyle 293-FまたはExpi293F細胞(Invitrogen)中にトランスフェクトした。5% CO2中37℃で3〜4日間の培養ののち、Streptactinアフィニティークロマトグラフィーを使用して組換えタンパク質を精製した。
最初の実験は、フローサイトメトリーアッセイ法がAP駆動型C3b沈着(AP-C3b)の存在または非存在を検出することができることを実証するために、以下のように実施した。以下の血清:正常ヒト血清、B因子(B因子)枯渇ヒト血清、D因子枯渇ヒト血清およびプロパージン枯渇ヒト血清(Complement Technology, Tyler, Texas., USAから入手)の5%を、Mg++/EGTA緩衝液またはEDTA中、試験抗体と4℃で一晩混合した。加熱殺菌黄色ブドウ球菌(108個/反応)を100μLの全量まで各混合物に加え、37℃で40分間回転流動させた。細菌を洗浄緩衝液中で洗浄し、細菌ペレットを洗浄緩衝液中に再懸濁させ、細菌表面のC3b沈着に関して各サンプルの80μLアリコートを分析し、それを、フローサイトメトリーを使用して、抗ヒトC3c(Dako, UK)で検出した。
1. 5%正常ヒト血清+EDTA
2. 5%正常ヒト血清+Mg/EGTA
3. 5%ヒト3MC(MASP-3-/-)血清+Mg++/EGTA
4. 5%ヒト3MC(MASP-3-/-)血清+Mg++/EGTAに加えて活性完全長rMASP-3
5. 5%ヒト3MC(MASP-3-/-)血清+Mg++/EGTAに加えて切断型活性rMASP-3(CCP1/CCP2/SP)
6. 5%ヒト3MC(MASP-3-/-)血清+Mg++/EGTAに加えて不活性rMASP-3(S679A)
7. 5%ヒト3MC(MASP-3-/-)血清+Mg++/EGTAに加えて活性完全長rMASP-1
B因子のMASP-3依存性活性化を分析するために、5%血清(正常ヒト血清または3MC患者血清のいずれか)と組換えタンパク質との様々な混合物を上記のようにアッセイした。各反応混合物から20μLを取り出し、タンパク質試料添加緩衝液に加えた。試料を70℃で10分間加熱し、SDS-PAGEゲルに添加した。B因子ポリクローナル抗体(R&D Systems)を使用してウェスタンブロット分析を実施した。B因子の活性化は、高めの分子量のプロB因子タンパク質に由来する低めの分子量の2つの切断産物(BbおよびBa)の形成によって明らかであった。
MASP-3依存性B因子活性化/切断のための最小要件を決定するために以下のアッセイ法を実施した。
図30は、B因子切断が分析されるクマシー染色SDS-PAGEゲルを示す。レーン1に示すように、B因子切断はC3、MASP-3およびプロD因子の存在において最適である。レーン2に示すように、C3は絶対に必要であるが、レーン4および5に示すように、C3が存在する限り、MASP-3またはプロD因子はいずれもB因子切断を媒介することができる。
本実施例に記載されるように、ヒト血清中の黄色ブドウ球菌におけるAP駆動型C3b沈着にはMASP-3が必要であることが実証された。したがって、実施例7に記載されるように同定された代表的なMASP-3 mAbがMASP-3の活性を阻害することができるかどうかを判定するために以下のアッセイ法を実施した。活性組換えMASP-3(CCP1-CCP2-SP)断片タンパク質(250ng)を3つの異なる濃度(0.5、2、および4μM)のアイソタイプ対照mAb、mAb1A5(MASP-3またはMASP-1に結合しないDTLacOプラットフォームから得られた対照)またはmAbD14(MASP-3に結合する)とともに氷上で1時間プレインキュベートした。酵素-mAb混合物を50μLの最終反応量で5% 3MC血清(MASP-3欠損)および5×107個の加熱殺菌黄色ブドウ球菌に曝露した。反応物を37℃で30分間インキュベートしたのち、C3b沈着の検出のために染色した。染色された細菌細胞をフローサイトメーターによって分析した。
要約すると、本実施例の結果は、MASP-3を欠損している血清中のAPの明らかな欠陥を実証する。したがって、B因子活性化およびC3b沈着を機能的終点として使用して、MASP-3がAPに対して非常に重要な寄与を成すことが実証された。さらに、MASP-3の触媒的に活性なC末端部分を含む機能的な組換えMASP-3の添加が、3MC患者からの血清中のB因子活性化およびC3b沈着における欠陥を補正する。逆に、本実施例においてさらに実証されるように、rMASP-3を有する3MC血清中のMASP-3抗体(例えばmAbD14)の添加はAP駆動型C3b沈着を阻害する。B因子活性化、ひいてはAPにおけるMASP-3の直接的な役割が、組換えMASP-3がC3とともに組換えB因子を活性化するのに十分であるという観察によって実証される。
本実施例は、MASP-1およびMASP-3がD因子を活性化することを実証する。
プロD因子の2つの異なる組換えバージョンを切断する能力に関して、組換えMASP-1およびMASP-3を試験した。第一のバージョン(プロD因子His)はN末端タグを欠くが、C末端Hisタグを有する。したがって、プロD因子のこのバージョンは、活性化中に切断によって除去される5アミノ酸プロペプチドを含む。第二のバージョン(STプロD因子His)はN末端上にStrep-TagII配列を有し、したがって、切断されるN末端断片を15アミノ酸まで増えている。STプロD因子はまた、His6タグをC末端に含む。STプロD因子Hisのプロペプチドの長さの増大が、プロD因子HIS形態で可能である分解に比較して、SDS-PAGEによる切断形態と非切断形態との分解を改善する。
図32はプロD因子基質切断のウェスタンブロット分析を示す。
実施例7に記載されたように同定された代表的なMASP-3およびMASP-1 mAbの、MASP-3依存性D因子切断に対する阻害効果を判定するために、以下のようにアッセイ法を実施した。活性組換えMASP-3タンパク質(80ng)を代表的なmAb D14、M3M1および対照抗体(MASP-1に特異的に結合するが、MASP-3には結合しない)1μgとともに室温で15分間プレインキュベートした。N末端Strepタグを有するプロD因子(ST-プロD因子-His、70ng)を加え、混合物を37℃で75分間インキュベートした。上記のように反応物を電気泳動させ、ブロッティングし、抗D因子で染色した。
本実施例は、MASP-3欠損がマンナンコーティングされたWTウサギ赤血球の補体媒介性溶解を防ぐことを実証する。
本明細書の実施例5および6に記載されるように、PNHのマウスモデルから採取された血液試料からの赤血球の溶解に対するMASP-2およびMASP-3欠損血清の効果が、PNHに罹患している対象を治療するためのMASP-2阻害および/またはMASP-3阻害の有効性を実証し、また、エクリズマブのようなC5阻害因子による治療を受けているPNH対象においてC3断片媒介性血管外溶血の効果を緩和するためのMASP-2の阻害因子および/またはMASP-3の阻害因子(二重または二重特異性MASP-2/MASP-3阻害因子を含む)の使用を裏付ける。
以下のように、3名の異なる3MC患者からMASP-3欠損血清を採取した。
3MC患者1は、MASP-3セリンプロテアーゼドメインをコードするエキソンを機能不全にする突然変異を有するアレルを含み、この3MC患者の母親および父親からも提供してもらった(両親とも、MASP-3セリンプロテアーゼドメインをコードするエキソンを機能不全にする突然変異を有するアレルに関してヘテロ接合性)。
3MC患者2は、MASP-1のエキソン12、すなわち、MASP-3のセリンプロテアーゼドメインをコードするエキソンにC1489T(H497Y)突然変異を有して、非機能的MASP-3を生じさせ、機能的MASP-1タンパク質を生じさせる。
3MC患者3は、MASP-1遺伝子中に確認された欠陥を有し、非機能的MASP-3および機能的MASP-1タンパク質を生じさせる。
Bitter-Suermann et al., Eur. J. Immunol. 11:291-295 (1981))に記載されているような従来のAP特異的条件下(BBS/Mg++/EGTA、Ca++なし、ここで、BBS=スクロースを含有するバルビタール緩衝食塩水)、ザイモサンコーティングされたマイクロタイタープレート上、0.5〜25%の範囲の血清濃度でAPアッセイ法を実施し、時間とともにC3b沈着を測定した。
図34は、ザイモサンコーティングされたマイクロタイタープレート上のAP駆動型C3b沈着のレベルを、MASP-3欠損対象(3MC)、C4欠損対象およびMBL欠損対象から採取された血清試料中の血清濃度の関数としてグラフで示す。図34に示し、以下の表15にまとめているように、患者2および患者3からのMASP-3欠損患者血清が、高い濃度(25%、12.5%、6.25%血清濃度)での残留AP活性を有し、かつ有意に高いAP50(すなわち、最大C3沈着の50%を達成するために必要な血清の8.2%および12.3%)を有する。
方法:3MC患者#2(MASP-3(-/-)、MASP-1(+/+))および3MC患者#3(MASP-3(-/-)、MASP-1(-/-))から血清を採取した。患者血清を、正常なドナーからの血清(W)とともに、SDS-ポリアクリルアミドゲルによって分離し、分解したタンパク質をポリフッ化ビニリデン膜上にブロッティングした。ヒトD因子特異的抗体によってヒトプロD因子(25,040Da)および/または成熟D因子(24,405Da)を検出した。
方法:3MC患者#2(MASP-3(-/-)、MASP-1(+/+))および3MC患者#3(MASP-3(-/-)、MASP-1(-/-))から採取した血清を、Wieslab補体システムスクリーン(Euro-Diagnostica, Malmo, Sweden)を製造者の取扱い指示に従って使用して、古典、レクチンおよび第二経路活性に関しても試験した。正常ヒト血清を対照として同時並行的に試験した。
方法:
Ca++の非存在における(すなわちEGTAを使用することによる)ウサギRBCの調製
ウサギ全血(2mL)を、2つの1.5mLエッペンドルフ管に分割し、4℃の冷却エッペンドルフ遠心分離機中、8000rpm(約5.9rcf)で3分間遠心処理した。氷冷BBS/Mg++/Ca++(4.4mMバルビツール酸、1.8mMナトリウムバルビトン、145mM NaCl、pH7.4、5mM Mg++、5mM Ca++)中に再懸濁させたのち、RBCペレットを3回洗浄した。3回目の洗浄後、ペレットをBBS/Mg++/Ca++4mL中に再懸濁させた。上記のように赤血球をペレット化し、RBCをBBS/0.1%ゼラチン/Mg++/Ca++で洗浄した。RBC懸濁液をBBS/0.1%ゼラチン/Mg++/Ca++中、4℃で貯蔵した。次いで、懸濁させたRBC100μLを水1.4mLで希釈し、8000rpm(約5.9rcf)で3分間スピンダウンし、上清のODを541nmで0.7に調節した(541nmで0.7のODは赤血球約109個/mlに相当)。その後、赤血球108個/mlの濃度を達成するために、OD 0.7で再懸濁させたRBC1mLをBBS/Mg++/EGTA 9mlに加えた。試験血清または血漿の希釈物を氷冷BBS、Mg++、EGTA中に調製し、各血清または血漿希釈物100μLを丸底プレートの対応するウェルにピペットで移した。適切に希釈したRBC 100μL(赤血球108個/ml)を各ウェルに加えた。ナノ水を使用して陽性対照(100%溶解)を製造し、血清または血漿なしのBBS/Mg++/EGTAによる希釈物を陰性対照として使用した。次いで、プレートを37℃で1時間インキュベートした。丸底プレートを3750rpmで5分間遠心処理した。次いで、各ウェルからの上清100μLを平底プレートの対応するウェルに移し、415〜490nmでODを読み取った。
図36は、Ca++の非存在下で測定した、正常な対象および2名の3MC患者(患者2および患者3)からの血清中、一定範囲の血清濃度にわたり、マンナンコーティングされたウサギ赤血球の溶血率(上清への溶解ウサギ赤血球のヘモグロビン放出を測光法によって測定)をグラフで示す。図36に示すように、MASP-3欠損が、正常ヒト血清に比べて、マンナンコーティングされた赤血球の補体媒介性溶解の割合を低下させることが実証される。正常ヒト血清からの2つの曲線と3MC患者からの2つの曲線との間の差は有意である(p=0.013、フリードマン試験)。
方法:
Bitter-Suermann et al., Eur. J. Immunol. 11:291-295 (1981))に記載されているような従来のAP特異的条件下(BBS/Mg++/EGTA、Ca++なし、ここで、BBS=スクロースを含有するバルビタール緩衝食塩水)、ザイモサンコーティングされたマイクロタイタープレート上、以下の血清試料中で、APアッセイ法を実施した:(1)完全長活性rMASP-3が0〜20μg/mLの範囲で加えられた3MC患者#2からの5%ヒト血清;(2)完全長活性rMASP-3が0〜20μg/mLの範囲で加えられた3MC患者#2からの10%ヒト血清;および(3)不活性rMASP-3A(S679A)が0〜20μg/mLの範囲で加えられた3MC患者#2からの5%ヒト血清。
図37は、ザイモサンコーティングされたマイクロタイタープレート上のAP駆動型C3b沈着のレベルを、ヒト3MC患者#2(MASP-3欠損)から採取された血清試料に加えられるrMASP-3タンパク質の濃度の関数としてグラフで示す。図37に示すように、活性組換えMASP-3タンパク質は、ザイモサンコーティングされたプレート上にAP駆動型C3b沈着を濃度依存的に再構成する。図37にさらに示すように、不活性rMASP-3(S679A)を含有する3MC血清中ではC3b沈着は認められなかった。
方法:
ウサギRBCを使用して、実験#2で上述された方法を使用して、以下の試験血清を、0〜12%の範囲で用いて溶血アッセイ法を実施した:(1)正常ヒト血清;(2)3MC患者血清;(3)3MC患者血清+活性完全長rMASP-3(20μg/ml);および(4)熱不活化ヒト血清。
図38は、Ca++の非存在において測定された、(1)正常ヒト血清;(2)3MC患者血清;(3)3MC患者血清+活性完全長rMASP-3(20μg/ml);および(4)熱不活化ヒト血清中、一定範囲の血清濃度にわたり、マンナンコーティングされたウサギ赤血球の溶血率(上清への溶解ウサギ赤血球のヘモグロビン放出を測光法によって測定)をグラフで示す。図38に示すように、rMASP-3を含む3MC血清中のウサギRBCの溶解率は、rMASP-3を含まない3MC血清中の溶解率に比べて有意に増大する(p=0.0006)。
方法:
3MC血清がLEA-2を欠損しているかどうかを調べるために、マンナンコーティングされたELISAプレートを使用してC3b沈着アッセイ法を実施した。クエン酸添加血漿を、BBS緩衝液中、連続希釈度(1:80から出発して1:160、1:320、1:640、1:1280、1:2560)に希釈し、マンナンコーティングされたプレート上に固定した。ニワトリ抗ヒトC3bアッセイ法を使用して、沈着したC3bを検出した。マンナンコーティングされたELISAプレート上のLEA-2駆動型C3b沈着(APおよびLEA-1が働くには血漿希釈度が高すぎる)を、正常ヒト対象(NHS)、2名の3MC患者(患者2および患者3)、患者3の両親およびMBL欠損対象からの血清中のヒト血清濃度の関数として評価した。
図40は、正常ヒト対象(NHS)、2名の3MC患者(患者2および患者3)、患者3の両親およびMBL欠損対象からの血清に関して、マンナンコーティングされたELISAプレート上のLEA-2駆動型(すなわち、MASP-2駆動型)C3b沈着のレベルを、BBS緩衝液中に希釈されたヒト血清の濃度の関数としてグラフで示す。これらのデータは患者2がMBL充分であることを示す。しかし、患者3および患者3の母親はMBL欠損であり、したがって、この人たちの血清はLEA-2を介してC3bをマンナン上に沈着させない。これらの血清中のMBLの置換は、患者3(MASP-3欠損を生じさせるSNPに関してホモ接合性である)およびその母親(突然変異MASP-3アレルに関してヘテロ接合性である)の血清中のLEA-2媒介性C3b沈着を回復させる(データ示さず)。この発見は、3MC血清がLEA-2を欠損しているのではなく、むしろ、機能的MASP-2を有すると考えられることを実証する。
これらの結果は、ザイモサンコーティングされたウェル上のC3b沈着の減少およびウサギ赤血球溶解の減少によって証明されるように、ヒト血清中のMASP-3欠損がAP活性の損失を生じさせることを実証する。機能的組換えヒトMASP-3で血清を補充することにより、両アッセイ法においてAPを回復させることができる。
本実施例は、キメラマウスV領域/ヒトIgG4定常領域抗ヒトMASP-3モノクローナル抗体(mAb M3-1、mAb 13B1とも呼ばれる)がMASP-3媒介第二経路補体(APC)活性化の強力な阻害物質であることを実証する。
キメラマウスV領域/ヒトIgG定常領域抗ヒトMASP-3モノクローナル抗体(mAb M3-1)の生成
MASP-1/3ノックアウトマウスをヒトMASP-3 CCP1-CCP2-SPドメイン(SEQ ID NO:2のアミノ酸301〜728)で免疫することにより、ネズミ抗ヒトMASP-3阻害抗体(mAb M3-1)を生成した(実施例14も参照されたい)。簡潔に記すと、免疫マウスからの脾細胞をP3/NS1/1-Ag4-1と融合し、得られたハイブリドーマクローンからの上清を、ヒトMASP-3に結合する抗体の産生に関しておよび補体プロD因子(プロCFD)からD因子(CFD)へのMASP-3媒介切断を遮断する能力に関してスクリーニングした。モノクローナル抗体(mAb)可変領域をRT-PCRによって単離し、配列決定し、ヒトIgG4発現ベクターへとクローニングした。キメラモノクローナル抗体を、一過的にトランスフェクトしたHEK293T細胞中で発現させ、精製し、マウスおよびヒトMASP-3への結合親和性に関してならびにプロCFDからCFDへのMASP-3媒介切断を阻害する能力に関して試験した。
以下にmAb M3-1の重鎖可変領域(VH)配列を提示する。SEQ ID NO:30のアミノ酸残基31〜35(H1)、50〜66(H2)および99〜102(H3)に対応するKabat CDR(31〜35(H1)、50〜65(H2)、および95〜102(H3)を下線で示す。
以下にmAb M3-1の軽鎖可変領域(VL)配列を提示する。SEQ ID NO:45のアミノ酸残基24〜40(L1)、56〜62(L2)および95〜102(L3)に対応するKabat CDR(24〜34(H1)、50〜56(H2)および89〜97(H3)を下線で示す。これらの領域は、Kabatシステムによって番号付けしてもChothiaシステムによって番号付けしても同じである。
ELISA実験において、M3-1の一価Fabバージョンを、組換え完全長ヒトおよびマウスMASP-3タンパク質への結合に関して試験した。複数の種由来のタンパク質に結合する抗MASP-3捕捉抗体で96ウェルプレートをコーティングすることにより、結合親和性測定を実施した。捕捉抗体は、MASP-1およびMASP-3のCCP1-CCP2領域に結合することが示されている。ヒトおよびマウスタンパク質の完全長バージョンを、捕捉抗体でコーティングされたELISAプレート上に固定化し、異なる濃度のM3-1 Fabを別々のウェル中で標的タンパク質に結合させた。HRPに結合した抗カッパ軽鎖抗体(Novus Biologicals NBP1-75064)を使用して結合M3-1を検出し、TMB基質試薬セット(BD Biosciences 555214)によって可視化した。
本開示に記載されるように、MASP-3は、少なくとも一部には、中心APC酵素であるCFDの活性化のその要求のために、APCの重要な調節物質であることがわかっている。同じく本開示に記載されるように、MASP-3は相対的に低い濃度で体内を循環し、遅い異化速度を有して、MASP-3抗体投与の静脈内、皮下および経口経路を通して炎症促進経路の長期的阻害を可能にする。ヒト血清中のMASP-3媒介CFD成熟を阻害し、APCを阻害するmAb M3-1の有効性を判定するために、以下の実験を実施した。正常なヒト血清は主に活性またはプロセシングされた(すなわち成熟した)CFDを含有するため、本発明者らは、CFD枯渇ヒト血清(Complement Technology A336)を組換え非プロセシング形態のCFD(プロCFD)によって再構成する実験を実施した。したがって、この実験システムにおいて、APC活性化はプロCFDから活性CFDへのプロセシングを必要とする。
インビボでAPCを阻害するmAb M3-1の有効性を判定するために、マウス群(n=4)に10mg/kg mAb M3-1の単回尾静脈内注射を施した。マウスから採取した血液を使用して血清を調製して、ザイモサン粒子上のC3(同じくC3bおよびiC3b)沈着のレベルを測定するエクスビボアッセイ法におけるAPC活性のフローサイトメトリー評価のためのマトリックスを提供した。投与前時点および複数の投与後時点(96時間、1週および2週)で採取した血液から調製した血清を7.5%に希釈し、ザイモサン粒子を添加してAPCを誘導した。抗体処置マウスを、単回静脈内用量の溶媒を投与された対照マウス群(n=4)と比較した。
本実施例は、キメラマウスV領域/ヒトIgG4定常領域抗ヒトMASP-3モノクローナル抗体(mAb M3-1、mAb 13B1とも呼ばれる)が、発作性夜間ヘモグロビン尿症(PNH)と関連するマウスモデルにおいて、Crryを欠く赤血球の生存率に明らかな利益を提供することを実証する。
実施例11および実施例14に記載したように、キメラマウスV領域/ヒトIgG4定常領域抗ヒトMASP-3モノクローナル抗体(mAb M3-1)を生成した。さらに実施例11に記載したように、mAb M3-1がマウスモデルにおいてインビボで強力なAPC阻害物質であることがわかった。本実施例は、PNHと関連するネズミモデルにおける有効性に関するmAb M3-1の分析を記載する。
PNHと関連するマウスモデルにおいて、マウスにおけるAPCの主要な細胞表面抑制物質を欠くCrry欠損マウスからの赤血球(RBC)をドナー細胞としての使用のために得た。野生型(WT)ドナーマウスから得たRBCを並行に実行した。これらのドナーRBCを、蛍光親油性色素(Sigma):WT(赤)およびCrry-(緑)で示差的に標識した。2つの異なる実験において、標識されたWTおよびCrry-ドナー細胞を1:1で混合し、野生型レシピエントマウスに静脈内注射し、20,000の生細胞イベントのフローサイトメトリー評価によってレシピエントマウスにおけるWTおよびCrry欠損RBC生存率(初期時点に対する)を測定した。最初の実験においては、mAb M3-1抗体の複数の投与前処置を与え、mAb M3-1の効果を、複数のマウス実験において有効性を示した(Wang et al., PNAS vol 92:8955-8959, 1995;Hugen et al., Kidney Int 71(7):646-54, 2007)C5阻害抗体である別の阻害補体抗体mAb BB5.1(Hycult Biotechから市販)の効果と比較した。C5阻害物質の投与は、PNHを有するヒト患者のための現在の治療標準である。第二の実験においては、mAb M3-1の処置前単回投与を評価した。
本実施例は、キメラMASP-3阻害性モノクローナル抗体(mAb M3-1、mAb 13B1とも呼ばれる)が、関節リウマチ(RA)のネズミモデルにおいて、コラーゲン抗体誘導関節炎(CAIA)における臨床スコアを減らすことを実証する実験を記載する。
CAIAは、十分に確立された関節炎の動物モデルである。RAへの洞察を提供することに加えて、CAIAモデルの病態はAPCとの確立された関係を有する。Bandaらは、B因子およびD因子のようなAPCの成分に欠損を有するマウスにおいて、CAIAモデルにおける結果の改善を実証した(Banda et al., J. Immunol vol 177:1904-1912, 2006およびBanda et al., Clinical & Exp Imunol vol 159:100-108, 2009)。APCマウスノックアウトは、WT対照と比べて、より低い関節炎(疾患)スコア、より低い発生率ならびに滑膜および周囲組織中でのより少ないC3およびH因子沈着を実証する。加えて、MASP1/3ノックアウトマウスにおいて、疾患活性スコア、関節中の補体C3組織沈着および組織病理学的損傷スコアが顕著に低下した(Banda et al., J Immunol vol 185:5598-5606, 2010)。したがって、CAIAにおける有効性に関してMASP-3阻害抗体mAb M3-1を分析した。
実施例11および実施例14に記載したように、キメラMASP-3モノクローナル抗体(mAb M3-1)を生成した。さらに実施例11に記載したように、mAb M3-1がマウスモデルにおいてインビボで強力なAPC阻害物質であることがわかった。
0=正常
1=1つの後および/または前足関節が冒されているか、またはごくわずかなびまん性紅斑および腫張がある
2=2つの後および/または前足関節が冒されているか、または軽度のびまん性紅斑および腫張がある
3=3つの後および/または前足関節が冒されているか、または中程度のびまん性紅斑および腫張がある
4=顕著なびまん性紅斑および腫張があるか、または4つの指関節が冒されている
5=重度のびまん性紅斑および足全体の重度の腫張があり、指を曲げられない
本実施例は高親和性抗ヒトMASP-3阻害抗体の生成を記載する。
MASP-3に対する限られた数の抗体が記載されている(Thiel et al., Mol. Immunol. 43:122, 2006;Moller-Kristensen et al., Int. Immunol. 19:141, 2006;Skjoedt et al., Immunobiol 215:921, 2010)。これらの抗体は、ウェスタンブロッティング、免疫沈降のような検出アッセイ法およびELISAアッセイ法における捕捉または検出試薬として有用であった。しかし、Thiel et al., 2006、Moller-Kristensen et al., 2006およびSkjoedt et al., 2010に記載された抗体は、MASP-3触媒活性を阻害するとは見いだされていない。
1. キメラマウスV領域/ヒトIgG定常領域抗ヒトMASP-3モノクローナル抗体の生成
Sigma Adjuvant System(Sigma-Aldrich, St Louis, MO)を使用して、7〜14週齢のC57BL/6、MASP-1/3ノックアウトマウスを、N末端にStrepTag IIエピトープタグを含むヒトMASP-3 CCP1/CCP2/SPポリペプチド(SEQ ID NO:2のアミノ酸残基299〜728);またはN末端にStrepTagIIを含むヒトMASP-3 SPドメイン(SEQ ID NO:2のアミノ酸残基450〜728)のいずれかで免疫した。マウス1匹あたり50μgの免疫原を腹腔内注射した。14日後、免疫マウスをアジュバント中のさらなる免疫原で追加免疫した。その後、数週間、マウスをPBS中の免疫原で14〜21日ごとに追加免疫した。マウスからの血清試料を尾部放血から定期的に調製し、抗原特異的抗体の存在に関してELISAによって試験した。脾臓融合の4日前に、有意な抗体価を有するマウスにPBS中の融合前免疫原追加免疫を施した。融合の3日前に、マウスの尾の付け根をPBS中50μgの抗CD40アゴニストmAb(R&D Systems, Minneapolis, MN)で皮下処置してB細胞数を増加させた(Rycyzyn et al., Hybridoma 27:25-30, 2008)。マウスを屠殺し、脾細胞を収穫し、50%ポリエチレングリコールまたは50%ポリエチレングリコール+10%DMSOを使用して、選択されたネズミ骨髄腫細胞株P3/NSI/1-AG4-1(NS-1)(ATCC No. TIB18)に融合した。融合物はハイブリドーマ細胞を生成し、それをHAT(ヒポキサンチン、アミノプテリンおよびチミジン)培地を含む96ウェル組織培養プレートに播種して、非融合細胞、骨髄腫ハイブリッドおよび脾ハイブリッドの増殖を阻害した。ハイブリドーマ選択後、MASP-3結合(ELISA)およびプロD因子活性化の阻害に関して培養上清をアッセイした。陽性ハイブリドーマを同定し、連続希釈法によってサブクローニングした。
表17に示すように、免疫されたMASP1/3 KOマウスから合計3328のハイブリドーマをスクリーニングし、そのうち>303がMASP-3に結合することがわかり、そのうち16がMASP-3に結合し、かつプロCFD活性化を阻害することがわかった。実施例11に記載されたmAb M3-1(13B1)は、表17に記載された16の機能的MASP-3阻害抗体の1つである。実施例15に記載されるように、16の機能的MASP-3阻害抗体はすべて異常に強い結合親和性(≦500pM)でヒトMASP-3に結合することがわかった。
本実施例は、MASP1/3ノックアウトマウスを免疫することによる、異常に強い結合親和性(すなわち、≦500pM〜20pMの範囲のナノモル未満の結合親和性)でヒトMASP-3を阻害する抗体の生成を記載する。本実施例に記載される抗体は、高い親和性(たとえば≦500pM)でヒトMASP-3に特異的に結合し、D因子成熟を阻害し、かつヒトMASP-1には結合しない。本明細書に記載されるように、ヒト、マウスおよびニワトリMASP-3のアミノ酸配列は、MASP-3のSPドメインが、特に活性部位において高度に保存されることを明らかにした(図4および5を参照されたい)。本実施例に記載されるように、MASP1/3 KOマウスにおいて異常に強い結合親和性を有するMASP-3阻害抗体を生成する能力は、一部には、野生型マウスにおいて非常に強力なMASP-3触媒部位特異的抗体の生成を妨げ得る免疫寛容の回避による可能性が高い。
本実施例は、高親和性抗ヒトMASP-3阻害性mAbのクローニングおよび配列分析を記載する。
組換え抗体のクローニングおよび精製:
実施例11および14に記載したハイブリドーマからRT-PCRを使用して重鎖および軽鎖可変領域をクローニングし、配列決定した。ヒトIgG4重鎖(SEQ ID NO:311)およびカッパ軽鎖(SEQ ID NO:313)定常領域に融合したマウスmAb可変領域からなるマウス−ヒトキメラmAbをExpi293F細胞中に組換えタンパク質として生成した。IgG4定常ヒンジ領域(SEQ ID NO:311)は安定化S228Pアミノ酸置換を含む。1つの態様において、キメラmAbを、S228Pアミノ酸置換を含み、かつ低いpHでFcRn相互作用を促進する変異も含むヒトIgG4定常ヒンジ領域(SEQ ID NO:312)に融合した。
SEQ ID NO:217:4D5重鎖可変領域(親)をコードするDNA
SEQ ID NO:233:4D5軽鎖可変領域(親)をコードするDNA
本実施例は、いくつかのインビトロアッセイ法における組換えの精製した高親和性MASP-3阻害抗体の機能的特性決定を記載する。
実施例11および14に記載したように生成した組換えMASP-3 mAbを、(i)ヒトMASP-3および他種MASP-3への結合;(ii)人工的基質の切断を阻害する能力;(iii)プロD因子からD因子への切断を阻害する能力;(iv)ヒト血清中の補体沈着の阻害および(v)ヒト血清中のウサギ赤血球溶解の阻害に関して以下のように特性決定した。
ELISAアッセイ法:
精製した組換えMASP-3 mAbを用いるMASP-3結合アッセイ法:
ヒトMASP-3:
ヒトMASP-3(CCP1-CCP2-SP断片)への16の精製した組換えMASP-3抗体の結合を測定するために、以下のようにサンドイッチELISAを実施した。ELISAプレートを、炭酸塩/重炭酸塩緩衝液中4℃で、4μg/mLの捕捉抗体αM3-259で一晩コーティングした。αM3-259は、ヒトMASP-3のCCP1-CCP2-SP領域で免疫したニワトリからの高結合活性組換えキメラニワトリ−ヒトMASP-3 mAbである。ドメインマッピング研究が、αM3-259が、ヒト、カニクイザル、マウス、ラット、およびイヌを含む複数の種由来のMASP-3のCCP1-CCP2領域に結合することを明らかにした。図51Cに示すように、αM3-259はMASP-1にも結合する。
図51Aおよび51Bは、ヒトMASP-3(CCP1-CCP2-SP)に対するMASP-3 mAb(精製した組換え体)の結合活性をグラフで示す。図51A、図51Bおよび表24に示すように、MASP-3 mAbは、ヒトMASP-3に関して、0.241nM〜0.023nMの範囲の高い結合活性を有する。これらの値は、以前に記載されたMASP-3 mAbに関して報告された値の10〜100倍の低さである(WO2013/192240に実施例15としても公表されている、本明細書における実施例7を参照されたい)。
MASP-3に対する高親和性MASP-3 mabの特異性を判定するために、結合実験を実施して、ヒトMASP-1およびヒトMASP-2への16の精製した組換えMASP-3抗体の結合を測定した。組換えMASP-1A(S646A、CCP1-CCP2-SP断片)およびMASP-2(CCP1-CCP2-SP断片)をプレート上に直接固定化したことを除き、MASP-3結合ELISAに関して記載したように結合を測定した。
図51Cは、代表的な精製した組換え高親和性抗ヒトMASP-3阻害抗体が、MASP-3への結合に選択的であり、ヒトMASP-1には結合しないことが示される、結合実験の結果をグラフで示す。
αM3-259を用いて組換え完全長マウスMASP-3(SEQ ID NO:3)をプレート上に捕捉したことを除き、ヒトMASP-3に関して先に記載したように、マウスMASP-3へのMASP-3 mAbの結合を測定した。両実験に使用された陰性対照mAbは、αM3-259と同じ免疫ニワトリから得られた組換えキメラニワトリ−ヒトmAb、mAb77であったが、mAb77はマウスMASP-3には結合しない。
図52は、マウス完全長MASP-3に対する代表的なMASP-3 mAb(精製した組換え体)の結合活性をグラフで示す。図52に示すように、試験したMASP-3 mAbの大部分はまた、マウスMASP-3に対して高い結合活性を有する。
背景/根拠:
その公知の天然基質(Iwaki et al., J. Immunol. 187:3751, 2011;Cortesio and Jiang, Arch. Biochem. Biophys. 449:164-170, 2006)に加えて、MASP-3は、様々なトリペプチド基質を加水分解することが示されている(Cortesio and Jiang, Ibid.)。非常に小さな基質として、これらの分子を使用してプロテアーゼの触媒部位をマッピングすることができる。トリペプチド切断の阻害は、抗体などの阻害物質が、触媒部位への小さな基質のアクセスを直接遮断するという、または同様にアクセスを拒否するSPドメインにおけるコンフォメーションシフトを生じさせるという指示である。そのようなものとして、抗体はまた、酵素の活性部位を妨害することによって大きな天然基質の触媒作用を遮断するものと予想することができる。機能的には、これは、MASP-3ヌルマウスまたは3MC患者(MASP-3を欠損)に非常に近いであろう。
組換えmAbの抗体価測定(666nMから0.91nMまで3倍希釈)をMASP-3 CCP1-CCP2-SP(197nM)とともに室温で15分間インキュベートした。トリペプチド基質BOC-V-P-R-AMC(t-ブチルオキシカルボニル-Val-Pro-Arg-7-アミノ-4-メチルクマリン)(R&D Systems, Cat. No. ES011)を最終濃度0.2mMで添加した。Arg-AMCアミド結合の加水分解が高蛍光基AMCを放出する。Spectramax M5e蛍光プレートリーダを使用して、励起380nm/発光460nm動力学値を37℃で5分ごと70分間記録した。
図53は、MASP-3モノクローナル抗体によるMASP-3依存性蛍光発生性トリペプチド切断の阻害を測定するアッセイ法の結果をグラフで示す。図53に示すように、試験したMASP-3 mAbは3つの異なる群に入る。
1. MASP-3によるペプチド切断の強力な阻害物質であるMASP-3 mAb:1A10(29.77nM)、1G4(29.64nM)、1F3(32.99nM)、4B6(26.03nM)、4D5(27.54nM)、10D12(30.94nM)、および13B1(30.13nM)。
2. MASP-3によるペプチド切断の弱いまたは非常に弱い阻害物質であるMASP-3 mAb:15D9、11B6、2F5、1E7、および2D7。
3. 中性であるか、またはMASP-3によるペプチド切断を刺激すると考えられる、MASP-3 mAb:1B11;2F2;77(対照mAb)。
方法:
活性組換えヒトMASP-3タンパク質(1反応あたり240ng)を、GVB++緩衝液中、代表的なMASP-3 mAbおよび対照mAb(これはMASP-1には結合するが、MASP-3には結合しない)とともに、総量9μLで室温にて15分間プレインキュベートした。次いで、プロD因子70ngをN末端Strep-tag IIエピトープタグ(ST−プロD因子−His)とともに各チューブに添加して、1反応あたりの最終量を10μLにした。反応物をサーモサイクラ中、37℃で6時間インキュベートした。次いで、各反応物の10分の1を12%Bis-Trisゲル上で電気泳動させて、プロD因子および活性D因子切断産物を分離した。分離したタンパク質をPVDF膜に移し、ビオチン化D因子抗体(R&D Systems)を用いる検出によるウェスタンブロットを使用して分析した。
図54は、インビトロアッセイ法において代表的なMASP-3 mAbがプロCFDからCFDへの組換えMASP-3媒介切断を遮断する能力を実証するウェスタンブロット分析を示す。図54に示すように、このアッセイ法において、代表的な高親和性MASP-3阻害性mAb 13B1、4B6、1G4、2D7、10D12、1A10、4D5、1E7、および1F3マウス−ヒトキメラmAbは、プロCFD切断の部分的ないし完全な阻害を示した。
方法:
様々な濃度のMASP-3 mAbを10%CFD枯渇ヒト血清(Complement Technology A336)およびGVB+Mg/EGTA(20nM)に添加し、氷上で30分間インキュベートしたのち、組換えSTプロD因子−His(2μg/mL最終)およびザイモサン(0.1mg/mL最終)を添加した。ザイモサン粒子は補体沈着のための活性化表面として機能する。混合物を37℃でインキュベートし、ザイモサン粒子の表面上の補体Bb因子(Quidel抗体A252)のフローサイトメトリー検出によってAPC活性を測定した。
図55Aは、D因子枯渇ヒト血清中、様々な濃度のMASP-3 mAb 1F3、1G4、2D7、および4B6の存在における、37℃で70分間のザイモサン粒子へのBb因子沈着のレベル(MFI単位で測定されフローサイトメトリー検出によって決定された)をグラフで示す。
方法:
別の実験状況においてAPCの阻害をモニターするために、本発明者らは、代表的なMASP-3 mAbがヒト血清中でウサギ赤血球の溶解を遮断する能力を評価した。様々な濃度のMASP-3 mAbを10%D因子枯渇ヒト血清およびGVB+Mg/EGTA(20nM)に添加し、氷上で30分間インキュベートしたのち、組換えST−プロB因子−His(2μg/mL最終)および赤血球(2.5×108個/mL最終)を添加した。混合物を37℃で70分間インキュベートし、反応物を希釈し、遊離ヘモグロビンのレベルを示す吸光度(A405)を測定することによって、APC媒介溶血を測定した。
図56Aは、CFD枯渇ヒト血清中、様々な濃度のMASP-3 mAb 1A10、1F3、4B6、4D5、1G4、および2F2の存在におけるウサギ赤血球溶解の阻害のレベルをグラフで示す。図56Bは、CFD枯渇ヒト血清中、様々な濃度のMASP-3 mAb 1B11、1E7、1G4、2D7、および2F5の存在におけるウサギ赤血球溶解の阻害のレベルをグラフで示す。結果を表27にまとめる。
方法:
代表的な組換えMASP-3 mAb(4D5)を、正常ヒト血清および3MC患者B(「患者B」)、血清中に検出可能なMASP-3を有さず、APCの欠損を示す個体由来の血清に由来するプロD因子の組換えMASP-3切断(および活性化)を遮断する能力に関して試験した。
図57は、3MC患者B血清中、活性rMASP-3、不活性rMASP-3、および活性rMASP-3+mAb 4D5の存在におけるプロD因子およびD因子のレベルを分析するウェスタンブロットを示す。図57に示すように、正常なヒト血清は主に成熟形態を含み、一方で、患者B血清は主にチモーゲン形態のD因子を含む。さらに図57に示すように、ザイモサンの存在における活性rMASP-3は、患者3血清中でプロD因子の切断を生じさせるが、一方で、不活性(チモーゲン)形態のMASP-3はそれを生じさせない。最後に、図57に示すように、MASP-3 mAb 4D5は、患者3血清中、活性rMASP-3の存在においてプロD因子の切断を遮断する。これらの結果はさらに、APCの活性化におけるプロD因子の切断におけるMASP-3の役割を実証し、MASP-3阻害性mAbがMASP-3媒介プロD因子切断を遮断し、それによってAPCを遮断することができることを実証する。
インビボでAPCを阻害する能力に関する代表的なMASP-3阻害性mAb 10D12および13B1の分析
1. インビボでのmAb M3-1(13B1)および10D12によるAPCの阻害:
方法:
インビボでAPCを阻害するMASP-3 mAb 13B1(M3-1)および10D12の有効性を判定するために、マウス群(n=4)に10mg/kg mAb 13B1の単回尾静脈内注射を施し、第二のマウス群(n=4)に10mg/kg mAb 10D12の単回尾静脈内注射を施した。マウスから採取した血液を使用して血清を調製して、ザイモサン粒子へのC3(同じくC3bおよびiC3b、Dako F020102-2)沈着のレベルを測定するエクスビボアッセイ法におけるAPC活性のフローサイトメトリー評価のためのマトリックスを提供した。投与前時点および複数の投与後時点(96時間、1週および2週)で収穫した血液から調製した血清を7.5%に希釈し、ザイモサン粒子(0.1mg/mL最終)を添加してAPCを誘導した。抗体処置マウスを、単回静脈内用量の溶媒を投与された対照マウス群(n=4)と比較した。
図58は、野生型マウスにおけるmAb M3-1(13B1)、mAb 10D12または溶媒の単回投与後の様々な時点におけるザイモサン粒子へのC3沈着のレベルをグラフで示す。図58に示すように、投与前時点で、3つの条件は匹敵しうるAPC活性レベルを示す。96時間および2つのより後の時点で、両mAb処置群は全身性APC活性のほぼ完全な消失を示すが、一方で、溶媒処置群のAPC活性は衰えないままである。
方法:
B因子チモーゲンから活性タンパク質分解酵素への転換中、B因子はD因子によってBa(〜30kDa)およびBb(〜60kDa)断片へと切断される。MASP-3 mAb 10D12で処置されたマウスから得られたマウス血清中のBa断片の状態を以下のように決定した。
APC活性化の測度として、図59は、mAb 10D12またはPBSで処置され、ザイモサンで刺激されたマウスから得られたマウス血清中のBa断片の状態を分析するウェスタンブロットを示す。図59の各レーンは異なるマウスを表し、レーンは、比較の目的で、代表的な溶媒マウスからの血清をMASP-3 mAb処置マウスからの血清の隣に示すよう交互に配置されている。溶媒またはmAb 10D12で処置されたマウスからの2つの対照条件が、血清試料中にザイモサンの非存在において存在するBaの基礎レベルを表すものとして、それぞれレーン1および2(ブロットの左側から開始して)に示されている。レーン3〜10はすべてザイモサンとのインキュベーション後に存在するBa断片のレベルを示す。すべての場合において、MASP-3 mAb処置マウスは溶媒処置マウスと比較してBa断片レベルの低下を実証する。
方法:
MASP-3阻害抗体によるAPC阻害の別の測度として、本発明者らは、代表的なMASP-3 mAb 10D12で処置されたマウスからの血清中、MASP-3抗体がウサギ赤血球の溶解を遮断する能力を、溶媒対照処置マウスからの血清中との比較において評価した。
図60は、MASP-3 mAb 10D12(10mg/kgまたは25mg/kg)で処置されたマウスまたは溶媒対照処置マウスからの20%血清による溶血の阻害のレベルをグラフで示す。図60に示すように、10mg/kgおよび25mg/kgのMASP-3 mAb 10D12で処置されたマウスからの血清は、溶媒処置マウスと比較して、1時間の試験期間中、より少ない全溶血を実証した。
本実施例に記載されるように、代表的な高親和性MASP-3阻害性mAb 13B1および10D12はインビボでAPCを阻害する。実施例12に記載したように、MASP-3モノクローナル抗体13B1(mAb M3-1とも呼ばれる)は、発作性夜間ヘモグロビン尿症(PNH)と関連するマウスモデルにおいて、Crryを欠く赤血球の生存率に対して明らかな利益を提供することがわかった。実施例13に記載したように、MASP-3 mAb M3-1が臨床関節炎スコアの発生率および重症度を用量依存的に低下させることがわかった。
本実施例は、高効力MASP-3阻害性mAbのエピトープ結合分析の結果を記載する。
方法:
96ウェルELISAアッセイプレートを、捕捉抗体αM3-259、MASP-1、およびMASP-3のCCP1-CCP2領域に結合することが示されているIgG4アイソタイプmAbでコーティングした。完全長ヒトMASP-3タンパク質を、捕捉抗体αM3-259を介してプレート上に固定化した。別個のコーティングされていないウェルにおいて、IgG4アイソタイプの1つの試験MASP-3 mAbの2倍希釈系を定濃度のIgG1アイソタイプの別の試験MASP-3抗体と混合した。コーティングされたウェルに混合物を添加し、捕捉されたMASP-3に結合させた。ヒトIgG1ヒンジ領域に対するHRP結合抗体(Southern Biotech 9052-05)およびTMB基質試薬セット(BD Biosciences 555214)を使用して、IgG1アイソフォームの検出によって2つの抗体の間の潜在的結合を測定した。
図61A〜61Eは競合結合分析の結果をグラフで示す。
方法:
16個のMASP-3 mAbのうち14個をPepscanによって評価して、それらが結合するMASP-3の領域を同定した。標的分子の線状および潜在的非連続性エピトープを再構築するために、SEQ ID NO:2(ヒトMASP-3)のアミノ酸残基299〜728に対応するペプチドのライブラリーを合成した。MASP-3のアミノ酸残基1〜298は免疫原中に存在せず、この分析には含まれなかった。
分析した各抗体に関するPepscanからのペプチド結合結果を以下に記載し、表4、表28、および図62〜67にまとめる。
中程度ストリンジェンシー条件下で試験した場合、抗体1F3、4B6、4D5、および1A10は非連続性エピトープ模倣体に結合し、また、単純な制約された線状模倣体にも結合した。データ分析は、抗体1F3、4B6、4D、5および1A10がすべてMASP-3のペプチドストレッチ
中程度ストリンジェンシー条件下で試験した場合、抗体10D12は、活性部位ヒスチジンH497に隣接する配列である、MASP-3のコア配列
中程度ストリンジェンシー条件下で試験した場合、抗体13B1は、MASP-3のペプチドストレッチ
低ストリンジェンシー条件下で試験した場合、抗体IG4は、MASP-3のペプチドストレッチ
高および低ストリンジェンシー条件それぞれの下で試験した場合、抗体1E7および2D7は、MASP-3のペプチドストレッチ
低ストリンジェンシー条件下で試験した場合、抗体2F5および15D9は、MASP-3のペプチドストレッチ
中程度ストリンジェンシー条件下で試験した場合、抗体1B11は、MASP-3のペプチドストレッチ
本実施例は、代表的なMASP-3 mAbのヒト化および潜在的な翻訳後修飾部位のエンジニアリングを記載する。
1. 代表的な高親和性MASP-3 mAbのヒト化
方法:
免疫原性リスクを減らすために、代表的な高親和性MASP-3阻害抗体4D5、10D12、および13B1をCDR移植法によってヒト化した。各MASP-3抗体のCDRをもっとも近いコンセンサスヒトフレームワーク配列中に移植した。バーニヤゾーン残基のいくつかをQuickchange部位特異的変異誘発(Agilent Technologies)によって改変した。得られたヒト化VHおよびVL領域をpcDNA3.1ベースのヒトIgG1またはIgG4およびIgK発現構築物に移入し、上記のように組換え抗体を発現させ、精製した。一価のFab断片を使用するELISAによってヒト化抗体の親和性を測定し、完全なIgG4フォーマットを使用するC3沈着アッセイ法によって効力を評価した。
mAb 4D5、10D12、および13B1の場合の重鎖可変領域および軽鎖可変領域の代表的なヒト化バージョンのアミノ酸配列を以下に提供する。CDR(Kabat)は下線で示されている。
代表的な高親和性MASP-3阻害性mAb 4D5、10D12、および13B1を翻訳後修飾に関して分析した。後続のグリシン、セリン、ヒスチジン、アラニン、またはアスパラギン(「NG」、「NS」、「NH」、「NA」、または「NN」モチーフ)を有するアスパラギン残基は、多くの場合、アスパラギン側鎖のアミド基の加水分解、すなわち「脱アミド化」を受けやすい。後続のグリシンまたはプロリン(「DG」または「DP」モチーフ)を有するアスパラギン酸残基は、多くの場合、相互転換、すなわち「異性化」を受けやすい。このような改変は電荷不均一性を生じさせ、結合界面で起こる場合には、抗体機能に影響を及ぼすおそれがある。また、このような改変は断片化、免疫原性および凝集の危険性を増大させるおそれもある。
多発性硬化症のマウスモデルにおける代表的なMASP-3阻害性mAb 13B1の分析
背景/根拠:後天性の炎症性かつ脱髄性自己免疫疾患である実験的自己免疫性脳脊髄炎(EAE)は、多発性硬化症(MS)の確立された動物モデルである。APCがEAEの発症/進行において重要な役割を果たすことを示唆する証拠が、B因子中和性抗体で処置されたマウスにおいて疾患が緩和されるという報告によって提供された(Hu et al., Mol. Immunol. 54:302, 2013)。本実施例は、EAEモデルにおける代表的な高親和性MASP-3阻害抗体13B1の分析を記載する。
EAE誘導:
この実験においては、Hooke Laboratories(Lawrence, MA)から購入したEAE誘導用キットを使用してEAEを誘導した。このキットは、完全フロイントアジュバント(CFA)中の神経抗原MOG35-55および百日咳毒素を含むものであった。
図68は、MASP-3阻害性mAb 13B1(10mg/kg)、B因子mAb 1379(40mg/kg)またはアイソタイプ対照mAb(10mg/kg)のいずれかで処置されたマウスにおけるEAEモデルの結果をグラフで示す。グラフ中、下向き矢印は抗B因子抗体の投与を示し、上向き矢印はmAb 13B1の最後の投与を示す。図68に示すように、MASP-3阻害性mAb 13B1およびB因子mAb 1379で処置されたマウスは、アイソタイプ対照と比較して、表35に示すパラメータにしたがってスコアリングされる臨床症状の改善を示した。
カニクイザルにおける代表的な高親和性MASP-3 mAbを用いる薬力学的実験
背景/根拠:げっ歯類実験において実証したように(図44)、高親和性MASP-3阻害抗体は、インビボで定常状態(静止)プロD因子成熟を阻害することができた。本実施例は、代表的な高親和性MASP-3阻害性mAbが非ヒト霊長類におけるAPC活性を阻害することができるかどうかを判定するためにカニクイザルにおいて実施された実験を記載する。
図69は、時間=0での高親和性MASP-3 mAb h13B1Xによる単回処置ののち経時的に3匹のカニクイザルの群から得られた血清試料中のAPC活性をグラフで示す。図は、APC阻害D因子mAbまたは中性アイソタイプ対照mAbのいずれかでスパイクされた5%血清中のザイモサン粒子の表面上の補体Bb因子を検出するフローサイトメトリーアッセイ法における平均MFIを示す。図69に示すように、カニクイザルは4時間という早さでAPC活性の減少を示す。MASP-3抗体処置がD因子阻害と同じくらい効果的にAPCを遮断する場合、2つのスパイクされた抗体条件は、投与後試料においては同一レベルのBb沈着阻害を示すが、投与前(または時間=0;図69)条件ではそれを示さない。図69に示すように、処置後72時間までに、APC活性は概ね、D因子mAbを血清試料に添加することによって達成される活性まで低下する。スパイクされたD因子抗体との比較により実験的に決定されるように、h13B1X処置によるほぼ完全な阻害は投与後336時間(14日間)まで持続する。したがって、これらの結果は、高親和性MASP-3阻害性mAbによる処置が非ヒト霊長類におけるAPCの完全な持続的阻害を提供することを実証する。
実施例11に記載したように、マウスへの高親和性MASP-3阻害抗体mAb 13B1の単回投与は、少なくとも14日間、全身性第二経路補体活性のほぼ完全な消失を生じさせた。さらに実施例12に記載したように、PNHと関連する十分に確立された動物モデルにおいて実施された実験において、mAb 13B1がPNH様赤血球の生存率を有意に改善し、C5阻害よりも有意に良好にPNH様赤血球を保護することが実証された。さらに、実施例13に記載したように、関節炎のマウスモデルにおいてmAb 13B1が疾患の発生率および重症度を下げることが実証された。本実施例における結果は、代表的な高親和性MASP-3阻害性mAb 13B1、10D12、および4D5が霊長類において第二経路を遮断するのに非常に有効であることを実証する。カニクイザルへのmAb 13B1、10D12または4D5の単回投与は、およそ16日間持続する全身性第二経路活性の持続的消失を生じさせた。高親和性MASP-3阻害抗体で処置されたカニクイザルにおける第二経路消失の程度は、インビトロでD因子遮断によって達成される程度に匹敵し、このことはMASP-3阻害抗体によるD因子転換の完全な遮断を示した。したがって、高親和性MASP-3阻害性mAbは、第二経路活動亢進に関連する疾患、たとえば発作性夜間ヘモグロビン尿症(PNH)、加齢黄斑変性症(AMD、ウェット型およびドライ型AMDを含む)、虚血再灌流障害、関節炎、播種性血管内凝固、血栓性微小血管症(溶血性尿毒症症候群(HUS)、非典型溶血性尿毒症症候群(aHUS)、血栓性血小板減少性紫斑病(TTP)または移植後TMAを含む)、喘息、デンスデポジット病、微量免疫型壊死性半月体形成性糸球体腎炎、外傷性脳損傷、誤嚥性肺炎、眼内炎、視神経脊髄炎、ベーチェット病、多発性硬化症、ギランバレー症候群、アルツハイマー病、筋萎縮性側索硬化症(ALS)、ループス腎炎、全身性エリテマトーデス(SLE)、糖尿病性網膜症、ブドウ膜炎、慢性閉塞性肺疾患(COPD)、C3糸球体症、移植片拒絶反応、移植片対宿主病(GVHD)、血液透析、敗血症、全身性炎症反応症候群(SIRS)、急性呼吸窮迫症候群(ARDS)、ANCA血管炎、抗リン脂質症候群、アテローム性動脈硬化症、IgA腎症、ならびに重症筋無力症に罹患している患者の治療において治療的有用性を有する。
本明細書の中で挙げられるすべての刊行物、特許出願、および特許は参照により本明細書に組み入れられる。
1A. 高い親和性(500pM未満のKDを有する)でヒトMASP-3のセリンプロテアーゼドメイン(SEQ ID NO:2のアミノ酸残基450〜728)に特異的に結合し、第二経路補体活性化を阻害する、単離されたモノクローナル抗体またはその抗原結合断片。
2A. 以下の性質の少なくとも1つまたは複数を特徴とする、項目1の単離された抗体もしくはその抗原結合断片:
(a)プロD因子成熟を阻害すること;
(b)ヒトMASP-1(SEQ ID NO:8)に結合しないこと;
(c)哺乳動物対象において約1:1〜約2.5:1(MASP-3標的:mAb)のモル比で第二経路を阻害すること;
(d)古典的経路を阻害しないこと;
(e)溶血および/もしくはオプソニン化の阻害;
(f)MASP-3セリンプロテアーゼ基質特異的切断の阻害;
(g)溶血の減少、もしくはC3切断およびC3b表面沈着の減少;
(h)活性化表面へのB因子およびBb因子沈着の減少;
(i)プロD因子と比べた静止レベル(循環中、かつ活性化表面の実験的添加なし)の活性D因子の減少;
(j)活性化表面に応答した、プロD因子と比べた活性D因子のレベルの減少;
(k)静止レベルおよび/もしくは表面誘導レベルの生成の液相Ba、Bb、C3b、もしくはC3aの減少;ならびに/または
(l)P因子沈着の減少。
3A. 前記ヒトMASP-3のセリンプロテアーゼドメイン中に位置するエピトープに特異的に結合し、該エピトープが、
4A. SEQ ID NO:15の中のエピトープに結合する、項目3の抗体またはその抗原結合断片。[すべてのI群abを含む]
5A. SEQ ID NO:9の中のエピトープに結合する、項目3の抗体または抗原結合断片。[10D12]
6A. SEQ ID NO:10の中のエピトープに結合する、項目3の抗体または抗原結合断片。[13B1]
7A. SEQ ID NO:12の中のエピトープにも結合する、項目6の抗体または抗原結合断片。[13B1]
8A. SEQ ID NO:10および/またはSEQ ID NO:12の中のエピトープにも結合する、項目3の抗体または抗原結合断片。[13B1]
9A. SEQ ID NO:9の中のエピトープに結合する、項目3の抗体または抗原結合断片。[1F3、4B6、4D5、1A10]
10A. SEQ ID NO:11、SEQ ID NO:13、および/またはSEQ ID NO:14の少なくとも1つの中のエピトープにも結合する、項目7の抗体または抗原結合断片。[1F3、4B6、4D5、1A10]
11A. SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:13、および/またはSEQ ID NO:14の少なくとも1つの中のエピトープにも結合する、項目7の抗体または抗原結合断片。[1F3、4B6、4D5、1A10]
12A. 前記抗体が
13A. SEQ ID NO:17の中のエピトープに結合する、項目12の抗体または抗原結合断片。[すべてのIIおよびIII群ab]
14A.
15A. SEQ ID NO:23の中のエピトープにも結合する、項目14の抗体または抗原結合断片。[1G4、1E7、2D7、15D9、2F5]
16A. SEQ ID NO:19および/またはSEQ ID NO:23の中のエピトープにも結合する、項目14の抗体または抗原結合断片。[Ig4、1E7、2D7]
17A. SEQ ID NO:18、SEQ ID NO:20、および/またはSEQ ID NO:23の中のエピトープにも結合する、項目14の抗体または抗原結合断片。[15D9、2F5]
18A. SEQ ID NO:18、SEQ ID NO:20、および/またはSEQ ID NO:23の少なくとも1つの中のエピトープにも結合する、項目14の抗体または抗原結合断片[15D9、2F5]。
19A. SEQ ID NO:16、SEQ ID NO:21、および/またはSEQ ID NO:22の少なくとも1つの中のエピトープにも結合する、項目14の抗体または抗原結合断片。[1B11]
20A. SEQ ID NO:16、SEQ ID NO:21、および/またはSEQ ID NO:22の少なくとも1つの中のエピトープにも結合する、項目14の抗体または抗原結合断片[1B11]。
21A. ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、項目1〜20のいずれかの抗体またはその抗原結合断片。
22A. 単鎖抗体、ScFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、項目1〜21のいずれかの抗体またはその抗原結合断片。
23A. 免疫グロブリン定常領域をさらに含む、項目1〜22のいずれかの抗体またはその抗原結合断片。
24A. ヒト化されている、項目1〜23のいずれかの抗体またはその抗原結合断片。
25A. 前記抗体が、500pM未満の親和性で前記ヒトMASP-3のセリンプロテアーゼドメインに結合する、項目1〜24のいずれかの抗体またはその抗原結合断片。
26A. 前記抗体が哺乳動物血液中で第二経路活性化を阻害する、項目1〜25のいずれかの抗体またはその抗原結合断片。
27A. 項目1A〜26Aのいずれかの抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
1B. (a)SEQ ID NO:209(XXDIN、1位のXがSまたはTでありかつ2位のXがNまたはDである)に記載されるHC-CDR1;SEQ ID NO:210(
(b)SEQ ID NO:212(
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片。
2B. (a)の重鎖可変領域のHC-CDR1がSEQ ID NO:56(TDDIN)を含む、項目1の単離された抗体またはその抗原結合断片。[4D5およびバリアント]
3B. (a)の重鎖可変領域のHC-CDR1がSEQ ID NO:62(SNDIN)を含む、項目1の単離された抗体またはその抗原結合断片。[1F3、4B6、および1A10]
4B. (a)の重鎖可変領域のHC-CDR2が
5B. (a)の重鎖可変領域のHC-CDR2が
6B. (a)の重鎖可変領域のHC-CDR2が
7B. (a)の重鎖可変領域のHC-CDR2が
8B. (a)の重鎖可変領域のHC-CDR3がSEQ ID NO:60(LEDTY)を含む、項目1の単離された抗体またはその抗原結合断片。[4D5およびバリアント]
9B. (a)の重鎖可変領域のHC-CDR3がSEQ ID NO:65(VEDSY)を含む、項目1の単離された抗体またはその抗原結合断片。[1F3、4B6、および1A10]
10B. (b)の軽鎖可変領域のLC-CDR1が、
11B. (b)の軽鎖可変領域のLC-CDR1が
12B. (b)の軽鎖可変領域のLC-CDR1が
13B. 前記HC-CDR1がSEQ ID NO:56を含み、前記HC-CDR2がSEQ ID NO:58を含み、前記HC-CDR3がSEQ ID NO:60を含み、かつ、前記LC-CDR1がSEQ ID NO:142、SEQ ID NO:257、SEQ ID NO:258、またはSEQ ID NO:259を含み、前記LC-CDR2がSEQ ID NO:144を含み、かつ前記LC-CDR3がSEQ ID NO:146を含む、項目1の単離された抗体またはその抗原結合断片。[LC-CDR1にバリアントを有する4D5の6つすべてのCDR]
14B. 前記HC-CDR1がSEQ ID NO:62を含み、前記HC-CDR2がSEQ ID NO:63、SEQ ID NO:67、またはSEQ ID NO:69を含み、前記HC-CDR3がSEQ ID NO:65を含み、かつ、前記LC-CDR1がSEQ ID NO:149を含み、前記LC-CDR2がSEQ ID NO:144を含み、かつ前記LC-CDR3がSEQ ID NO:146を含む、項目1の単離された抗体またはその抗原結合断片。[1F3、4B6、および1A10の6つすべてのCDR]
15B. ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、項目1〜14のいずれかの抗体またはその抗原結合断片。
16B. 単鎖抗体、ScFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、項目1〜15のいずれかの抗体またはその抗原結合断片。
17B. 免疫グロブリン定常領域をさらに含む、項目1〜16のいずれかの抗体またはその抗原結合断片。
18B. ヒト化されている、項目1〜17のいずれかの抗体またはその抗原結合断片。
19B. SEQ ID NO:24、SEQ ID NO:248、またはSEQ ID NO:249と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:40、SEQ ID NO:250、またはSEQ ID NO:278と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[4D5親、ヒト化および改変バージョン]。
20B. SEQ ID NO:25と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:41と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[1F3]。
21B. SEQ ID NO:26と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:42と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[4B6]。
22B. SEQ ID NO:27と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:42と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[1A10]。
23B. 前記抗体が500pM未満の親和性でヒトMASP-3に結合する、項目1〜22のいずれかの抗体またはその抗原結合断片。
24B. 前記抗体が哺乳動物血液中で第二経路活性化を阻害する、項目1〜23のいずれかの抗体またはその抗原結合断片。
25B. 項目1B〜24Bのいずれかの抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
1C. (a)SEQ ID NO:213(SYGXX、4位のXがMまたはIでありかつ5位のXがSまたはTである)に記載されるHC-CDR1;SEQ ID NO:74に記載されるHC-CDR2;およびSEQ ID NO:214(GGXAXDY、3位のXがEまたはDでありかつ5位のXがMまたはLである)に記載されるHC-CDR3を含む、重鎖可変領域;ならびに
(b)SEQ ID NO:215(
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片。
2C. (a)の重鎖可変領域のHC-CDR1がSEQ ID NO:72(SYGMS)を含む、項目1の単離された抗体またはその抗原結合断片。[10D12およびバリアント]
3C. (a)の重鎖可変領域のHC-CDR1がSEQ ID NO:79(SYGIT)を含む、項目1の単離された抗体またはその抗原結合断片。[35C1]
4C. (a)の重鎖可変領域のHC-CDR3がSEQ ID NO:76(GGEAMDY)を含む、項目1の単離された抗体またはその抗原結合断片。[10D12およびバリアント]
5C. (a)の重鎖可変領域のHC-CDR3がSEQ ID NO:82(GGDALDY)を含む、項目1の単離された抗体またはその抗原結合断片。[35C1]
6C. (b)の軽鎖可変領域のLC-CDR1が、
7C. 前記軽鎖可変領域のLC-CDR1が
8C. 前記軽鎖可変領域のLC-CDR1がSEQ ID NO:152を含む、項目1の単離された抗体またはその抗原結合断片。[35C1]
9C. (b)の軽鎖可変領域のLC-CDR3が
10C. (b)の軽鎖可変領域のLC-CDR3が
11C. 前記HC-CDR1がSEQ ID NO:72を含み、前記HC-CDR2がSEQ ID NO:74を含み、前記HC-CDR3がSEQ ID NO:76を含み、前記LC-CDR1がSEQ ID NO:153、SEQ ID NO:261、SEQ ID NO:262、またはSEQ ID NO:263を含み、前記LC-CDR2がSEQ ID NO:155を含み、かつ前記LC-CDR3がSEQ ID NO:157を含む、項目1の単離された抗体またはその抗原結合断片。[LC-CDR1にバリアントを有する10D12の6つすべてのCDR]
12C. 前記HC-CDR1がSEQ ID NO:79を含み、前記HC-CDR2がSEQ ID NO:74を含み、前記HC-CDR3がSEQ ID NO:82を含み、前記LC-CDR1がSEQ ID NO:159を含み、前記LC-CDR2がSEQ ID NO:155を含み、かつ前記LC-CDR3がSEQ ID NO:160を含む、項目1の単離された抗体またはその抗原結合断片。[35C1の6つすべてのCDR]
13C. ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、項目1〜12のいずれかの抗体またはその抗原結合断片。
14C. 単鎖抗体、ScFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、項目1〜13のいずれかの抗体またはその抗原結合断片。
15C. 免疫グロブリン定常領域をさらに含む、項目1〜14のいずれかの抗体またはその抗原結合断片。
16C. ヒト化されている、項目1〜15のいずれかの抗体またはその抗原結合断片。
17C. SEQ ID NO:28、SEQ ID NO:251、またはSEQ ID NO:252と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:43、SEQ ID NO:253またはSEQ ID NO:279と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[10D12親、ヒト化、およびバリアント]。
18C. SEQ ID NO:29と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:44と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[35C1]。
19C. 前記抗体が500pM未満の親和性でヒトMASP-3に結合する、項目1〜18のいずれかの抗体またはその抗原結合断片。
20C. 前記抗体が哺乳動物血液中で第二経路活性化を阻害する、項目1〜19のいずれかの抗体またはその抗原結合断片。
21C. 項目1C〜20Cのいずれかの抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
1D. (a)SEQ ID NO:84(GKWIE)に記載されるHC-CDR1;
(b)
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片。[13B1の6つすべてのCDR、およびLC-CDR1中のバリアント]
2D. 前記LC-CDR1がSEQ ID NO:258を含む、項目1の単離された抗体またはその抗原結合断片。[13B1 LC-CDR1 NAバリアント]
3D. ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、項目1〜2のいずれかの抗体またはその抗原結合断片。
4D. 単鎖抗体、ScFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、項目1〜3のいずれかの抗体またはその抗原結合断片。
5D. 免疫グロブリン定常領域をさらに含む、項目1〜4のいずれかの抗体またはその抗原結合断片。
6D. ヒト化されている、項目1〜5のいずれかの抗体またはその抗原結合断片。
7D. SEQ ID NO:30、SEQ ID NO:254、またはSEQ ID NO:255と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:45、SEQ ID NO:256、またはSEQ ID NO:280と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[13B1親、ヒト化、およびバリアント]。
8D. 前記抗体が500pM未満の親和性でヒトMASP-3に結合する、項目1〜7のいずれかの抗体またはその抗原結合断片。
9D. 前記抗体が哺乳動物血液中で第二経路活性化を阻害する、項目1〜8のいずれかの抗体またはその抗原結合断片。
10D. 項目1D〜9Dのいずれかの抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
1E. (a)SEQ ID NO:91(GYWIE)に記載されるHC-CDR1;
(b)SEQ ID NO:163(RSSQSLVQSNGNTYLH)に記載されるLC-CDR1、SEQ ID NO:165(KVSNRFS)に記載されるLC-CDR2、およびSEQ ID NO:167(SQSTHVPPT)に記載されるLC-CDR3を含む、軽鎖可変領域
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片。
2E. ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、項目1の抗体またはその抗原結合断片。
3E. 単鎖抗体、ScFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、項目1〜2のいずれかの抗体またはその抗原結合断片。
4E. 免疫グロブリン定常領域をさらに含む、項目1〜3のいずれかの抗体またはその抗原結合断片。
5E. ヒト化されている、項目1〜4のいずれか一つの抗体またはその抗原結合断片。
6E. SEQ ID NO:31と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:46と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の単離された抗体またはその抗原結合断片[1G4]。
7E. 前記抗体が500pM未満の親和性でヒトMASP-3に結合する、項目1〜6のいずれかの抗体またはその抗原結合断片。
8E. 前記抗体が哺乳動物血液中で第二経路活性化を阻害する、項目1〜7のいずれかの抗体またはその抗原結合断片。
9E. 項目1E〜8Eのいずれかの抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
1F. (a)
(b)SEQ ID NO:125(DYYMN)に記載されるHC-CDR1、
(c)SEQ ID NO:132に記載されるHC-CDR1、SEQ ID NO:133に記載されるHC-CDR2、SEQ ID NO:135に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:203に記載されるLC-CDR1、SEQ ID NO:165に記載されるLC-CDR2、およびSEQ ID NO:204に記載されるLC-CDR3を含む、軽鎖可変領域[49C11];または
(d)SEQ ID NO:137に記載されるHC-CDR1、SEQ ID NO:138に記載されるHC-CDR2、SEQ ID NO:140に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:206に記載されるLC-CDR1、SEQ ID NO:207に記載されるLC-CDR2、およびSEQ ID NO:208に記載されるLC-CDR3を含む、軽鎖可変領域[15D9];または
(e)SEQ ID NO:98に記載されるHC-CDR1、SEQ ID NO:99に記載されるHC-CDR2、SEQ ID NO:101に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:169に記載されるLC-CDR1、SEQ ID NO:171に記載されるLC-CDR2、およびSEQ ID NO:173に記載されるLC-CDR3を含む、軽鎖可変領域[2F5];または
(f)SEQ ID NO:103に記載されるHC-CDR1、SEQ ID NO:105に記載されるHC-CDR2、SEQ ID NO:107に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:176に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む、軽鎖可変領域[1B11];または
(g)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:116に記載されるHC-CDR2、SEQ ID NO:118に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:188に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:190に記載されるLC-CDR3を含む、軽鎖可変領域[2F2];または
(h)SEQ ID NO:114に記載されるHC-CDR1、SEQ ID NO:121に記載されるHC-CDR2、SEQ ID NO:123に記載されるHC-CDR3を含む、重鎖可変領域;ならびにSEQ ID NO:191に記載されるLC-CDR1、SEQ ID NO:178に記載されるLC-CDR2、およびSEQ ID NO:193に記載されるLC-CDR3を含む、軽鎖可変領域[11B6]
を含む、MASP-3に結合する単離された抗体またはその抗原結合断片。
2F. ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、項目1(a)〜(g)の抗体またはその抗原結合断片。
3F. 単鎖抗体、ScFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および全体抗体からなる群より選択される、項目1〜2のいずれかの抗体またはその抗原結合断片。
4F. 免疫グロブリン定常領域をさらに含む、項目1〜3のいずれかの抗体またはその抗原結合断片。
5F. ヒト化されている、項目1〜4のいずれかの抗体またはその抗原結合断片。
6F. SEQ ID NO:32と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:47と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[1E7]。
7F. SEQ ID NO:33と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:48と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[2D7]。
8F. SEQ ID NO:34と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:49と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[49C11]。
9F. SEQ ID NO:35と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:50と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[15D9]。
10F. SEQ ID NO:36と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:51と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[2F5]。
11F. SEQ ID NO:37と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:52と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[1B11]。
12F. SEQ ID NO:38と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:53と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[2F2]。
13F. SEQ ID NO:39と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である重鎖およびSEQ ID NO:54と少なくとも80%、85%、90%、95%、98%、99%、または100%同一である軽鎖を含む、項目1の抗体またはその抗原結合断片[11B6]。
14F. 前記抗体が、500pM未満の親和性でヒトMASP-3に結合する、項目1〜13のいずれかの抗体またはその抗原結合断片。
15F. 前記抗体が哺乳動物血液中で第二経路活性化を阻害する、項目1〜14のいずれかの抗体またはその抗原結合断片。
16F. 項目1F〜15Fのいずれかの抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
1. それを必要とする哺乳動物対象に、該哺乳動物における第二経路補体経路活性化を阻害するのに十分な量の、高親和性MASP-3阻害抗体またはその抗原結合断片を含む組成物を投与する工程を含む、哺乳動物における第二経路補体活性化を阻害する方法。
2. 前記抗体またはその抗原結合断片が、500pM未満の親和性でMASP-3に結合する、請求項1に記載の方法。
3. 前記抗体または前記抗原結合断片を含む前記組成物を投与する結果として、前記哺乳動物対象において以下の1つまたは複数が見られる、項目1の方法:
(a)D因子成熟の阻害;
(b)約1:1〜約2.5:1(MASP-3標的:mAb)のモル比で該対象に投与された場合の、第二経路の阻害;
(c)古典的経路が阻害されないこと;
(d)溶血および/もしくはオプソニン化の阻害;
(e)溶血の減少、もしくはC3切断およびC3b表面沈着の減少;
(f)活性化表面へのB因子およびBb因子沈着の減少;
(g)プロD因子と比べた静止レベル(循環中、かつ活性化表面の実験的添加なし)の活性D因子の減少;
(h)活性化表面に応答した、プロD因子と比べた活性D因子のレベルの減少;ならびに/または
(i)静止レベルおよび表面誘導レベルの液相Ba、Bb、C3b、もしくはC3aの生成の減少。
4. 前記抗体が約1:1〜約2.5:1(MASP-3標的:mAb)のモル比で第二経路を阻害する、項目1の方法。
5. 前記高親和性MASP-3抗体が請求項27A、25B、21C、10D、9E、または16Fのいずれかを特徴とする、項目1〜3のいずれかの方法。
6. 前記抗体またはその抗原結合断片が、古典的経路活性化に影響することなく、第二経路を選択的に阻害する、項目1〜4のいずれかの方法。
7. 前記哺乳動物対象が、発作性夜間ヘモグロビン尿症(PNH)、加齢黄斑変性症(AMD、ウェット型およびドライ型AMDを含む)、虚血再灌流障害、関節炎、播種性血管内凝固、血栓性微小血管症(溶血性尿毒症症候群(HUS)、非典型溶血性尿毒症症候群(aHUS)、血栓性血小板減少性紫斑病(TTP)もしくは移植後TMAを含む)、喘息、デンスデポジット病、微量免疫型壊死性半月体形成性糸球体腎炎、外傷性脳損傷、誤嚥性肺炎、眼内炎、視神経脊髄炎、ベーチェット病、多発性硬化症、ギランバレー症候群、アルツハイマー病、筋萎縮性側索硬化症(ALS)、ループス腎炎、全身性エリテマトーデス(SLE)、糖尿病性網膜症、ブドウ膜炎、慢性閉塞性肺疾患(COPD)、C3糸球体症、移植片拒絶反応、移植片対宿主病(GVHD)、血液透析、敗血症、全身性炎症反応症候群(SIRS)、急性呼吸窮迫症候群(ARDS)、ANCA血管炎、抗リン脂質症候群、アテローム性動脈硬化症、IgA腎症、ならびに重症筋無力症からなる群より選択される第二経路疾患もしくは第二経路障害に罹患しているか、またはそれを発症する危険にある、項目1〜6のいずれかの方法。
排他的所有権または特権を主張する本発明の態様は以下のとおりに定義される。
Claims (33)
- ヒトMASP-3のセリンプロテアーゼドメイン(SEQ ID NO:2のアミノ酸残基450〜728)に特異的に結合し、
SEQ ID NO:84(GKWIE)に記載されるHC-CDR1;
に記載されるHC-CDR2;およびSEQ ID NO:88(SEDV)に記載されるHC-CDR3を含む、重鎖可変領域;ならびに
に記載されるLC-CDR1;SEQ ID NO:144(WASTRES)に記載されるLC-CDR2;およびSEQ ID NO:161(KQSYNIPT)に記載されるLC-CDR3を含む、軽鎖可変領域
を含む、単離された抗体またはその抗原結合断片。 - ヒト抗体、ヒト化抗体、キメラ抗体、ネズミ抗体、およびこれらのいずれかの抗原結合断片からなる群より選択される、請求項1に記載の抗体またはその抗原結合断片。
- 単鎖抗体、scFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、請求項1に記載の抗体またはその抗原結合断片。
- 免疫グロブリン定常領域をさらに含む、請求項1に記載の抗体またはその抗原結合断片。
- ヒト化されている、請求項1に記載の抗体またはその抗原結合断片。
- 前記抗体が、500pM未満の親和性で前記ヒトMASP-3のセリンプロテアーゼドメインに結合する、請求項1に記載の抗体またはその抗原結合断片。
- 前記抗体が哺乳動物血液中で第二経路活性化を阻害する、請求項1に記載の抗体またはその抗原結合断片。
- 請求項1に記載の抗体またはその抗原結合断片の重鎖可変領域および軽鎖可変領域をコードする、単離された核酸、または
該重鎖可変領域をコードする核酸と該軽鎖可変領域をコードする核酸とを組み合わせた核酸。 - 請求項8に記載の核酸の1つまたは複数を含む、クローニングベクターまたは発現ベクター。
- 請求項9に記載のクローニングベクターまたは発現ベクターの1つまたは複数を含む、宿主細胞。
- 請求項10に記載の宿主細胞を培養する工程、および前記抗体またはその抗原結合断片を単離する工程を含む、請求項1に記載の抗体または抗原結合断片を産生するための方法。
- 請求項1に記載の抗体または抗原結合断片と、薬学的に許容される賦形剤とを含む、組成物。
- 請求項12に記載の組成物を含む、哺乳動物における第二経路補体活性化を阻害するための薬学的組成物。
- 前記LC-CDR1がSEQ ID NO:258を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記HC-CDR2がSEQ ID NO:86を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記HC-CDR2がSEQ ID NO:275を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記LC-CDR1がSEQ ID NO:142を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記LC-CDR1がSEQ ID NO:257を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記LC-CDR1がSEQ ID NO:259を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記HC-CDR1がSEQ ID NO:84を含み、前記HC-CDR2がSEQ ID NO:86を含み、前記HC-CDR3がSEQ ID NO:88を含み、前記LC-CDR1がSEQ ID NO:258を含み、前記LC-CDR2がSEQ ID NO:144を含み、かつ前記LC-CDR3がSEQ ID NO:161を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記HC-CDR1がSEQ ID NO:84を含み、前記HC-CDR2がSEQ ID NO:275を含み、前記HC-CDR3がSEQ ID NO:88を含み;前記LC-CDR1がSEQ ID NO:258を含み、前記LC-CDR2がSEQ ID NO:144を含み、かつ前記LC-CDR3がSEQ ID NO:161を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記重鎖可変領域が、SEQ ID NO:30に記載されるアミノ酸配列を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記軽鎖可変領域が、SEQ ID NO:45に記載されるアミノ酸配列を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記重鎖可変領域が、SEQ ID NO:30に記載されるアミノ酸配列を含み、かつ前記軽鎖可変領域が、SEQ ID NO:45に記載されるアミノ酸配列を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記重鎖可変領域が、SEQ ID NO:254またはSEQ ID NO:255に記載されるアミノ酸配列を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- 前記軽鎖可変領域が、SEQ ID NO:256またはSEQ ID NO:280に記載されるアミノ酸配列を含む、請求項1記載の単離された抗体またはその抗原結合断片。
- SEQ ID NO:254またはSEQ ID NO:255を含む重鎖可変領域、およびSEQ ID NO:256またはSEQ ID NO:280を含む軽鎖可変領域を含み、ヒトMASP-3に結合する単離された抗体またはその抗原結合断片。
- 前記重鎖可変領域がSEQ ID NO:254を含み、かつ前記軽鎖可変領域がSEQ ID NO:256を含む、請求項27記載の単離された抗体またはその抗原結合断片。
- 前記重鎖可変領域がSEQ ID NO:255を含み、かつ前記軽鎖可変領域がSEQ ID NO:256を含む、請求項27記載の単離された抗体またはその抗原結合断片。
- 前記重鎖可変領域がSEQ ID NO:254を含み、かつ前記軽鎖可変領域がSEQ ID NO: 280を含む、請求項27記載の単離された抗体またはその抗原結合断片。
- 前記重鎖可変領域がSEQ ID NO:255を含み、かつ前記軽鎖可変領域がSEQ ID NO: 280を含む、請求項27記載の単離された抗体またはその抗原結合断片。
- 前記抗体が、単鎖抗体、scFv、Fab断片、Fab'断片、F(ab')2断片、ヒンジ領域を欠く一価抗体、および抗体全体からなる群より選択される、請求項27記載の単離された抗体。
- 免疫グロブリン定常領域をさらに含む、請求項27記載の単離された抗体。
Priority Applications (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2021178522A JP7350822B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
| JP2021178521A JP7350821B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201662369674P | 2016-08-01 | 2016-08-01 | |
| US62/369,674 | 2016-08-01 | ||
| US201662419420P | 2016-11-08 | 2016-11-08 | |
| US62/419,420 | 2016-11-08 | ||
| US201762478336P | 2017-03-29 | 2017-03-29 | |
| US62/478,336 | 2017-03-29 | ||
| PCT/US2017/044714 WO2018026722A1 (en) | 2016-08-01 | 2017-07-31 | Compositions and methods of inhibiting masp-3 for the treatment of various diseases and disorders |
Related Child Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021178521A Division JP7350821B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
| JP2021178522A Division JP7350822B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2019533982A JP2019533982A (ja) | 2019-11-28 |
| JP6971306B2 true JP6971306B2 (ja) | 2021-11-24 |
Family
ID=61073545
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019505253A Active JP6971306B2 (ja) | 2016-08-01 | 2017-07-31 | 様々な疾患および障害の治療のためのmasp−3を阻害する組成物および方法 |
| JP2021178522A Active JP7350822B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
| JP2021178521A Active JP7350821B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
Family Applications After (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021178522A Active JP7350822B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
| JP2021178521A Active JP7350821B2 (ja) | 2016-08-01 | 2021-11-01 | 様々な疾患および障害の治療のためのmasp-3を阻害する組成物および方法 |
Country Status (25)
| Country | Link |
|---|---|
| US (4) | US10639369B2 (ja) |
| EP (1) | EP3490603A4 (ja) |
| JP (3) | JP6971306B2 (ja) |
| KR (3) | KR102553777B1 (ja) |
| CN (4) | CN116333147A (ja) |
| AU (4) | AU2017306069B2 (ja) |
| BR (1) | BR112019001247A2 (ja) |
| CA (3) | CA3173979A1 (ja) |
| CL (1) | CL2019000230A1 (ja) |
| CO (1) | CO2019001212A2 (ja) |
| CR (1) | CR20190105A (ja) |
| CU (1) | CU24568B1 (ja) |
| EC (1) | ECSP19015192A (ja) |
| GE (1) | GEP20247593B (ja) |
| IL (3) | IL298689B2 (ja) |
| JO (6) | JOP20170154B1 (ja) |
| MA (1) | MA45798A (ja) |
| MX (3) | MX2019000965A (ja) |
| PE (1) | PE20190389A1 (ja) |
| PH (1) | PH12019500204A1 (ja) |
| SA (1) | SA519400950B1 (ja) |
| SG (3) | SG10202010279YA (ja) |
| TW (4) | TW202214689A (ja) |
| UY (1) | UY37349A (ja) |
| WO (1) | WO2018026722A1 (ja) |
Families Citing this family (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2017035405A1 (en) | 2015-08-26 | 2017-03-02 | Achillion Pharmaceuticals, Inc. | Amino compounds for treatment of immune and inflammatory disorders |
| WO2017035401A1 (en) | 2015-08-26 | 2017-03-02 | Achillion Pharmaceuticals, Inc. | Amide compounds for treatment of immune and inflammatory disorders |
| WO2018160891A1 (en) | 2017-03-01 | 2018-09-07 | Achillion Pharmaceutical, Inc. | Pharmaceutical compounds for treatment of medical disorders |
| CA3053818A1 (en) | 2017-03-01 | 2018-09-07 | Achillion Pharmaceuticals, Inc. | Aryl, heteroaryl, and heterocyclic pharmaceutical compounds for treatment of medical disorders |
| EP3814374A4 (en) * | 2018-05-25 | 2022-03-09 | Achillion Pharmaceuticals, Inc. | BIOMARKERS OF NEPHROPATHY ASSOCIATED WITH THE ALTERNATIVE COMPLEMENT PATHWAY |
| KR20210057086A (ko) | 2018-09-06 | 2021-05-20 | 아칠리온 파르마세우티칼스 인코포레이티드 | 다니코판의 형태체 형태 |
| WO2020051532A2 (en) | 2018-09-06 | 2020-03-12 | Achillion Pharmaceuticals, Inc. | Macrocyclic compounds for the treatment of medical disorders |
| BR112021005506A2 (pt) | 2018-09-25 | 2021-06-15 | Achillion Pharmaceuticals, Inc. | formas mórficas de inibidores de fator complementar d |
| CN110244053B (zh) * | 2019-05-09 | 2022-03-11 | 北京大学第三医院(北京大学第三临床医学院) | 用于诊断狼疮肾炎并肺动脉高压疾病的分子标志物及其用途 |
| CN112812182B (zh) * | 2019-11-15 | 2023-01-06 | 深圳宾德生物技术有限公司 | 一种靶向fgfr4的单链抗体、嵌合抗原受体、嵌合抗原受体t细胞及其制备方法和应用 |
| CA3161701A1 (en) * | 2020-01-21 | 2021-07-29 | Zhaoli LI | Semg2 antibody and use thereof |
| CA3189666A1 (en) * | 2020-08-18 | 2022-02-24 | William Jason Cummings | Monoclonal antibodies, compositions and methods for detecting complement factor d |
| CN112950324B (zh) * | 2021-03-15 | 2022-06-03 | 重庆邮电大学 | 一种知识图谱辅助的成对排序个性化电商推荐方法及系统 |
| CN115304670A (zh) * | 2021-05-07 | 2022-11-08 | 华南农业大学 | 一种猫冠状病毒核衣壳蛋白的鼠源单克隆抗体及其编码基因和应用 |
| WO2023173036A2 (en) * | 2022-03-10 | 2023-09-14 | Omeros Corporation | Masp-2 and masp-3 inhibitors, and related compositions and methods, for treatment of sickle cell disease |
Family Cites Families (39)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US4331647A (en) | 1980-03-03 | 1982-05-25 | Goldenberg Milton David | Tumor localization and therapy with labeled antibody fragments specific to tumor-associated markers |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| US4946778A (en) | 1987-09-21 | 1990-08-07 | Genex Corporation | Single polypeptide chain binding molecules |
| US5223409A (en) | 1988-09-02 | 1993-06-29 | Protein Engineering Corp. | Directed evolution of novel binding proteins |
| US5211657A (en) | 1988-11-07 | 1993-05-18 | The United States Government As Represented By The Secretary Of The Department Of Health And Human Services | Laminin a chain deduced amino acid sequence, expression vectors and active synthetic peptides |
| US5530101A (en) | 1988-12-28 | 1996-06-25 | Protein Design Labs, Inc. | Humanized immunoglobulins |
| US5549910A (en) | 1989-03-31 | 1996-08-27 | The Regents Of The University Of California | Preparation of liposome and lipid complex compositions |
| JP3218637B2 (ja) | 1990-07-26 | 2001-10-15 | 大正製薬株式会社 | 安定なリポソーム水懸濁液 |
| JP2958076B2 (ja) | 1990-08-27 | 1999-10-06 | 株式会社ビタミン研究所 | 遺伝子導入用多重膜リポソーム及び遺伝子捕捉多重膜リポソーム製剤並びにその製法 |
| EP0672142B1 (en) | 1992-12-04 | 2001-02-28 | Medical Research Council | Multivalent and multispecific binding proteins, their manufacture and use |
| US5856121A (en) | 1994-02-24 | 1999-01-05 | Case Western Reserve University | Growth arrest homebox gene |
| US6074642A (en) * | 1994-05-02 | 2000-06-13 | Alexion Pharmaceuticals, Inc. | Use of antibodies specific to human complement component C5 for the treatment of glomerulonephritis |
| US5741516A (en) | 1994-06-20 | 1998-04-21 | Inex Pharmaceuticals Corporation | Sphingosomes for enhanced drug delivery |
| US5795587A (en) | 1995-01-23 | 1998-08-18 | University Of Pittsburgh | Stable lipid-comprising drug delivery complexes and methods for their production |
| US5731168A (en) | 1995-03-01 | 1998-03-24 | Genentech, Inc. | Method for making heteromultimeric polypeptides |
| US5738868A (en) | 1995-07-18 | 1998-04-14 | Lipogenics Ltd. | Liposome compositions and kits therefor |
| US7273925B1 (en) | 1998-12-15 | 2007-09-25 | Brigham And Women's Hospital, Inc. | Methods and products for regulating lectin complement pathway associated complement activation |
| US20040031072A1 (en) * | 1999-05-06 | 2004-02-12 | La Rosa Thomas J. | Soy nucleic acid molecules and other molecules associated with transcription plants and uses thereof for plant improvement |
| CA2391402A1 (en) | 1999-12-02 | 2001-06-07 | Jens Christian Jensenius | Masp-3, a complement-fixing enzyme, and uses for it |
| SG98393A1 (en) | 2000-05-19 | 2003-09-19 | Inst Materials Research & Eng | Injectable drug delivery systems with cyclodextrin-polymer based hydrogels |
| ES2601143T3 (es) | 2002-07-19 | 2017-02-14 | Omeros Corporation | Copolímeros tribloque biodegradables, métodos de síntesis de los mismos, e hidrogeles y biomateriales preparados a partir de los mismos |
| US20040115194A1 (en) * | 2002-09-06 | 2004-06-17 | Yi Wang | Method of treatment of asthma using antibodies to complement component C5 |
| AU2004216176B2 (en) | 2003-02-21 | 2008-04-03 | Genentech, Inc. | Methods for preventing and treating tissue damage associated with ischemia-reperfusion injury |
| DE102004017370A1 (de) | 2004-04-08 | 2005-10-27 | Nutrinova Nutrition Specialties & Food Ingredients Gmbh | PUFA-PKS Gene aus Ulkenia |
| US7919094B2 (en) | 2004-06-10 | 2011-04-05 | Omeros Corporation | Methods for treating conditions associated with MASP-2 dependent complement activation |
| US7612181B2 (en) | 2005-08-19 | 2009-11-03 | Abbott Laboratories | Dual variable domain immunoglobulin and uses thereof |
| WO2007024715A2 (en) | 2005-08-19 | 2007-03-01 | Abbott Laboratories | Dual variable domain immunoglobin and uses thereof |
| CA2685714C (en) | 2007-05-31 | 2016-04-12 | University Of Washington | Inducible mutagenesis of target genes |
| AU2010272483B2 (en) | 2009-07-17 | 2016-07-21 | Omeros Corporation | MASP isoforms as inhibitors of complement activation |
| EP2462161B1 (en) * | 2009-08-06 | 2017-03-08 | Immunas Pharma, Inc. | Antibodies that specifically bind to a beta oligomers and use thereof |
| US8163283B2 (en) * | 2009-09-03 | 2012-04-24 | Vancouver Biotech Ltd. | Monoclonal antibodies against gonadotropin-releasing hormone receptor |
| MX336682B (es) * | 2010-03-05 | 2016-01-27 | Hoffmann La Roche | Anticuerpos contra csf-1r humanos y usos de los mismos. |
| US20120122107A1 (en) | 2010-11-16 | 2012-05-17 | Aarhus Universitet | Homeostatic multidomain protein, and uses for it |
| CN107011443B (zh) * | 2011-05-04 | 2021-04-30 | 奥默罗斯公司 | 用于抑制masp-2依赖的补体活化的组合物 |
| SI2833907T1 (en) * | 2012-04-06 | 2018-07-31 | Omeros Corporation | COMPOSITION AND METHODS OF INJECTION OF MASP-1 AND / OR MASP-3 FOR TREATMENT OF PAROXYSIMAL NIGHT CHEMOGLOBINURIA |
| BR112014031522A2 (pt) * | 2012-06-18 | 2017-08-01 | Omeros Corp | métodos para inibir a ativação de complemento dependente de masp-3, para inibir a ativação de complemento dependente de masp-2 e para fabricar um medicamento |
| ES2760023T3 (es) * | 2013-02-20 | 2020-05-12 | Univ Pennsylvania | Tratamiento del cáncer utilizando receptor de antígeno quimérico anti-EGFRvIII humanizado |
| AU2014248515B2 (en) * | 2013-03-13 | 2019-03-07 | Prothena Biosciences Limited | Tau immunotherapy |
| KR102448454B1 (ko) * | 2014-01-29 | 2022-09-28 | 다나-파버 캔서 인스티튜트 인크. | Muc1-c/세포외 도메인 (muc1-c/ecd)에 대한 항체 |
-
2017
- 2017-07-03 JO JOP/2017/0154A patent/JOP20170154B1/ar active
- 2017-07-30 TW TW110149485A patent/TW202214689A/zh unknown
- 2017-07-30 TW TW110149487A patent/TWI820555B/zh active
- 2017-07-30 TW TW106125649A patent/TWI756248B/zh active
- 2017-07-30 TW TW112116731A patent/TW202334243A/zh unknown
- 2017-07-31 GE GEAP201715019A patent/GEP20247593B/en unknown
- 2017-07-31 MA MA045798A patent/MA45798A/fr unknown
- 2017-07-31 WO PCT/US2017/044714 patent/WO2018026722A1/en active Application Filing
- 2017-07-31 SG SG10202010279YA patent/SG10202010279YA/en unknown
- 2017-07-31 BR BR112019001247A patent/BR112019001247A2/pt active Search and Examination
- 2017-07-31 CA CA3173979A patent/CA3173979A1/en active Pending
- 2017-07-31 JP JP2019505253A patent/JP6971306B2/ja active Active
- 2017-07-31 CN CN202210980626.8A patent/CN116333147A/zh active Pending
- 2017-07-31 MX MX2019000965A patent/MX2019000965A/es unknown
- 2017-07-31 CN CN202210979607.3A patent/CN116327918A/zh active Pending
- 2017-07-31 CN CN201780047995.9A patent/CN109715209B/zh active Active
- 2017-07-31 CN CN202210979606.9A patent/CN116333146A/zh active Pending
- 2017-07-31 EP EP17837477.3A patent/EP3490603A4/en active Pending
- 2017-07-31 IL IL298689A patent/IL298689B2/en unknown
- 2017-07-31 KR KR1020227010749A patent/KR102553777B1/ko active IP Right Grant
- 2017-07-31 SG SG11201900606VA patent/SG11201900606VA/en unknown
- 2017-07-31 CA CA3031980A patent/CA3031980C/en active Active
- 2017-07-31 KR KR1020227010743A patent/KR102624948B1/ko active IP Right Grant
- 2017-07-31 PE PE2019000261A patent/PE20190389A1/es unknown
- 2017-07-31 IL IL264592A patent/IL264592B2/en unknown
- 2017-07-31 CR CR20190105A patent/CR20190105A/es unknown
- 2017-07-31 US US15/665,030 patent/US10639369B2/en active Active
- 2017-07-31 AU AU2017306069A patent/AU2017306069B2/en active Active
- 2017-07-31 KR KR1020197005706A patent/KR102382804B1/ko active IP Right Grant
- 2017-07-31 SG SG10202010277SA patent/SG10202010277SA/en unknown
- 2017-07-31 CU CU2019000006A patent/CU24568B1/es unknown
- 2017-07-31 IL IL298690A patent/IL298690B2/en unknown
- 2017-07-31 CA CA3122348A patent/CA3122348A1/en active Pending
- 2017-08-01 UY UY0001037349A patent/UY37349A/es active IP Right Grant
-
2019
- 2019-01-22 MX MX2023000482A patent/MX2023000482A/es unknown
- 2019-01-22 MX MX2023000525A patent/MX2023000525A/es unknown
- 2019-01-23 SA SA519400950A patent/SA519400950B1/ar unknown
- 2019-01-28 PH PH12019500204A patent/PH12019500204A1/en unknown
- 2019-01-29 CL CL2019000230A patent/CL2019000230A1/es unknown
- 2019-02-12 CO CONC2019/0001212A patent/CO2019001212A2/es unknown
- 2019-02-28 EC ECSENADI201915192A patent/ECSP19015192A/es unknown
-
2020
- 2020-04-01 US US16/837,600 patent/US10745274B1/en active Active
- 2020-06-23 US US16/909,578 patent/US11027015B2/en active Active
- 2020-07-08 AU AU2020204551A patent/AU2020204551B2/en active Active
- 2020-10-06 AU AU2020250188A patent/AU2020250188B2/en active Active
-
2021
- 2021-04-27 US US17/241,628 patent/US11883493B2/en active Active
- 2021-11-01 JP JP2021178522A patent/JP7350822B2/ja active Active
- 2021-11-01 JP JP2021178521A patent/JP7350821B2/ja active Active
-
2022
- 2022-10-03 JO JOP/2022/0245A patent/JOP20220245A1/ar unknown
- 2022-10-03 JO JOP/2022/0242A patent/JOP20220242A1/ar unknown
- 2022-10-03 JO JOP/2022/0241A patent/JOP20220241A1/ar unknown
- 2022-10-03 JO JOP/2022/0244A patent/JOP20220244A1/ar unknown
- 2022-10-03 JO JOP/2022/0243A patent/JOP20220243A1/ar unknown
- 2022-10-17 AU AU2022256090A patent/AU2022256090A1/en active Pending
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6771531B2 (ja) | 様々な疾患および障害の治療のためにmasp−1および/またはmasp−2および/またはmasp−3を阻害する組成物および方法 | |
| JP6971306B2 (ja) | 様々な疾患および障害の治療のためのmasp−3を阻害する組成物および方法 | |
| NZ751019B2 (en) | Compositions and methods of inhibiting masp-3 for the treatment of various diseases and disorders |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20190322 Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20190213 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20190322 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20200316 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20200612 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20200914 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20210203 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20210319 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20210430 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210721 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20211004 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20211101 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 6971306 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |