JP6520008B2 - ジアシルグリセロールアシルトランスフェラーゼを用いた脂質の製造方法 - Google Patents
ジアシルグリセロールアシルトランスフェラーゼを用いた脂質の製造方法 Download PDFInfo
- Publication number
- JP6520008B2 JP6520008B2 JP2014159012A JP2014159012A JP6520008B2 JP 6520008 B2 JP6520008 B2 JP 6520008B2 JP 2014159012 A JP2014159012 A JP 2014159012A JP 2014159012 A JP2014159012 A JP 2014159012A JP 6520008 B2 JP6520008 B2 JP 6520008B2
- Authority
- JP
- Japan
- Prior art keywords
- gene
- acyl
- protein
- transformant
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images

Landscapes
- Enzymes And Modification Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
Description
生体内においてトリアシルグリセロールは、ジアシルグリセロールに脂肪酸由来のアシル−CoAからアシル基が転移して合成される。この転移反応にはジアシルグリセロールアシルトランスフェラーゼ(DGAT)が関与する。DGATは、哺乳類、植物、真菌などに存在し、DGAT1とDGAT2の2タイプが存在することが知られている。DGAT1とDGAT2は、共にジアシルグリセロールアシルトランスフェラーゼ活性を有するが、両者はアミノ酸配列の共通性に乏しい。
植物のDGAT1として、シロイヌナズナ(Genbank NP_179535.1)、Tung Tree(Genbank ABC94471.1)、ヤトロファ(Genbank ABB84383.1)、ヒマワリ(Genbank ADT91687.1)、ヒマ(Genbank XP_002514132.1)由来のものが知られている。また、植物のDGAT2として、シロイヌナズナ(Genbank NP_566952.1)、Tung Tree(Genbank ABC94473.1)、ヒマ(Genbank XP_002528531.1)、ヤトロファ(Genbank AEZ56254.1)、ヒマワリ(Genbank ADT91688.1)由来のものが知られている。
(1)宿主に下記(A)又は(B)のタンパク質をコードする遺伝子を導入して形質転換体を得る工程
(A) 配列番号1で表されるアミノ酸配列からなるタンパク質
(B) 配列番号1で表されるアミノ酸配列と80%以上の同一性を有するアミノ酸配列からなり、かつジアシルグリセロールアシルトランスフェラーゼ活性を有するタンパク質
(2)得られた形質転換体から脂質を採取する工程
本発明のジアシルグリセロールアシルトランスフェラーゼ、これをコードする遺伝子、形質転換体、及び製造方法は、脂質、特に中鎖脂肪酸及びこれを構成成分とする脂質の工業的生産に好適に用いることができる。
また本発明で、脂肪酸や脂肪酸を構成するアシル基の表記において、Cx:yとあるのは炭素原子数xで二重結合の数がyであることを表し、Cxは炭素原子数xの脂肪酸やアシル基であることを表す。
以下、ジアシルグリセロールアシルトランスフェラーゼ、これを用いた形質転換体、及び脂質の製造方法について順に説明する。
本発明のジアシルグリセロールアシルトランスフェラーゼは、配列番号1で表されるアミノ酸配列を有するタンパク質、及び当該タンパク質と機能的に均等なタンパク質である。具体的に、本発明のジアシルグリセロールアシルトランスフェラーゼには、以下の(A)又は(B)のタンパク質が包含される。
(A) 配列番号1で表されるアミノ酸配列からなるタンパク質
(B) 配列番号1で表されるアミノ酸配列と80%以上の同一性を有するアミノ酸配列からなり、かつジアシルグリセロールアシルトランスフェラーゼ活性を有するタンパク質
ジアシルグリセロールアシルトランスフェラーゼ(ジグリセリドアシルトランスフェラーゼ、DGAT)は、トリアシルグリセロール合成の最終段階に関与する酵素である。植物や藻類では、葉緑体にある脂肪酸合成系において各種アシル−ACPが合成され、このアシル−ACPから酵素アシル−ACPチオエステラーゼによって遊離脂肪酸が切り出される。遊離脂肪酸はアシル−CoAに変換されて、トリアシルグリセロール合成系に供される。トリアシルグリセロール合成系では、アシル−CoAのアシル基が、グリセロール骨格に導入される。最終段階では、ジアシルグリセロールにアシル−CoAからアシル基が転移してトリアシルグリセロールが合成される。この反応を触媒する酵素がDGATである。
このようにトリアシルグリセロールは、グリセロール骨格に3つのアシル基(脂肪酸残基)が導入されてなるが、導入されるアシル基の種類(炭素数及び不飽和度)は多岐にわたる。グリセロール骨格にどのような炭素数・不飽和度を有したアシル基を導入するかを決めているのは、各反応工程で働くアシルトランスフェラーゼであると考えられている。すなわち、トリアシルグリセロール合成の最終段階では、DGATの基質特異性によって、ジアシルグリセロールに結合する脂肪酸アシル−CoAの種類が変わると考えられている。
なお、タンパク質がジアシルグリセロールアシルトランスフェラーゼ活性を有することは、トリアシルグリセロール合成遺伝子欠損株を用いた系により確認することができる。トリアシルグリセロール合成遺伝子欠損株の一例としては、実施例で用いた酵母のΔdga1、Δlro1、Δare1、Δare2株が挙げられる。目的タンパク質をコードする遺伝子を当該欠損株において発現させ、トリアシルグリセロールの生成が確認されれば、当該タンパク質はジアシルグリセロールアシルトランスフェラ−ゼ活性を有すると判断できる。
本発明において「中鎖アシル−CoA特異的」なジアシルグリセロールアシルトランスフェラーゼとは、基質として中鎖アシル−CoAを、それ以外のアシル−CoAよりも優先的に利用する、すなわち、中鎖アシル−CoA由来のアシル基を優先的にジアシルグリセロールに転移する性質を有するジアシルグリセロールアシルトランスフェラーゼである。以下、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼを、中鎖特異的ジアシルグリセロールアシルトランスフェラーゼともいう。
また、本発明において、中鎖脂肪酸、中鎖アシル−CoA、中鎖アシル−ACPにおける中鎖とは、アシル基の炭素数が6以上14以下であることをいう。
ジアシルグリセロールアシルトランスフェラーゼの中鎖アシル−CoA特異性については、例えば、宿主細胞にジアシルグリセロールアシルトランスフェラーゼ遺伝子及びアシル−CoAを生産するアシル−ACPチオエステラーゼ遺伝子を導入し、これらの導入遺伝子が発現する条件で細胞を培養して、宿主細胞又は培養液中の脂肪酸組成の変化をガスクロマトグラフィー解析等の方法を用いて分析する方法、または精製したジアシルグリセロールアシルトランスフェラーゼタンパク質と、アシル−CoA及びジアシルグリセロールとを混合して反応させた溶液から薄層クロマトグラフィー法などによりトリアシルグリセロールを単離し、トリアシルグリセロール中の脂肪酸組成の変化をガスクロマトグラフィー解析等の方法を用いて分析する方法により確認することができる。
(B)において、ジアシルグリセロールアシルトランスフェラーゼ活性の点から、配列番号1で表されるアミノ酸配列との同一性は、85%以上であることが好ましく、90%以上であることがより好ましく、93%以上であることがさらに好ましく、95%以上であることがよりさらに好ましく、97%以上であることが特に好ましい。
本発明においてアミノ酸配列及び塩基配列の同一性はLipman-Pearson法(Science,227,1435,(1985))によって計算される。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(homology search)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
アミノ酸配列に欠失、置換、挿入、付加等の変異を導入する方法としては、例えば、アミノ酸配列をコードする塩基配列に変異を導入する方法が挙げられる。塩基配列に変異を導入する方法については、後述する。
本発明のジアシルグリセロールアシルトランスフェラーゼ遺伝子は、前記(A)又は(B)のタンパク質をコードする遺伝子である。
配列番号1に示すアミノ酸配列をコードする遺伝子の例として、配列番号2に示す塩基配列が挙げられる。配列番号2に示す塩基配列は、ココヤシ由来の野生型ジアシルグリセロールアシルトランスフェラーゼをコードする遺伝子の塩基配列の一例である。
(a) 配列番号2で表される塩基配列からなるDNA
(b) 配列番号2で表される塩基配列と70%以上の同一性を有する塩基配列からなり、かつジアシルグリセロールアシルトランスフェラーゼ活性を有するタンパク質をコードするDNA
後述の実施例に示すように、(a)のDNAからなる遺伝子は、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼをコードする遺伝子である。(b)のDNAからなる遺伝子も、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼをコードする遺伝子であることが好ましい。
本発明の形質転換体は、前記(A)又は(B)のタンパク質をコードする遺伝子と共に、脂質や脂肪酸の合成に関与する他の酵素をコードする遺伝子を共導入してなることが好ましい。脂質や脂肪酸の合成に関与する他の酵素としては、例えば、アセチル−CoAカルボキシラーゼ、β−ケトアシル−ACPシンターゼ、アシル−ACPチオエステラーゼが挙げられる。
アセチル−CoAカルボキシラーゼは脂肪酸合成の出発反応に関与し、アセチル−CoAとCO2との縮合反応により、マロニル−CoAを生成する。マロニル−CoAは、ACP(アシルキャリアプロテイン)に転移され、マロニル−ACPを生じる。
β−ケトアシル−ACPシンターゼは、脂肪酸合成経路においてアシル基の鎖長制御に関与する酵素である。植物の脂肪酸合成経路は葉緑体に局在し、マロニル−ACPを利用して炭素鎖の伸長反応を繰り返し、種々の鎖長のアシル−ACP(脂肪酸残基であるアシル基とACPとからなる複合体)が合成される。
アシル−ACPチオエステラーゼは、脂肪酸合成系で合成されたアシル−ACPのチオエステル結合を加水分解し、遊離の脂肪酸を生成する酵素である。アシル−ACPチオエステラーゼの作用によって、ACP上での脂肪酸合成が終了し、切り出された脂肪酸はトリアシルグリセロール等の合成に供される。
宿主にDGAT遺伝子とともにこれらの酵素をコードする遺伝子を導入することで、形質転換体の脂質や脂肪酸生産性を一層向上させることができる。
本発明で用いるアシル−ACPチオエステラーゼは、アシル−ACPチオエステラーゼ活性を有するタンパク質であればよい。本発明において、「アシル−ACPチオエステラーゼ活性を有する」とは、アシル−ACPのチオエステル結合を加水分解する活性を有することをいう。
上述のように、前記(A)のタンパク質は、中鎖特異的なジアシルグリセロールアシルトランスフェラーゼであるため、共導入するアシル−ACPチオエステラーゼも、中鎖アシル−ACPに特異的なチオエステラーゼ(以下、「中鎖特異的アシル−ACPチオエステラーゼ」という)が好ましい。本発明において「中鎖アシル−ACP特異的」なアシル−ACPチオエステラーゼとは、炭素数6〜14のアシル−ACPのチオエステル結合を選択的に加水分解する活性を有するアシル−ACPチオエステラーゼである。このようなアシル−ACPチオエステラーゼを用いることで、中鎖脂肪酸の生産性を一層向上させることができる。
具体的には、Umbellularia californicaアシル−ACPチオエステラーゼ(GenBank AAA34215.1);Cuphea calophylla subsp. mesostemonアシル−ACPチオエステラーゼ(GenBank ABB71581);Cocos nuciferaアシル−ACPチオエステラーゼ(CnFatB3:Jing et al. BMC Biochemistry 2011, 12:44参照、配列番号3、これをコードする遺伝子の塩基配列:配列番号4);Cinnamomum camphoraアシル−ACPチオエステラーゼ(GenBank AAC49151.1);Myristica fragransアシル−ACPチオエステラーゼ(GenBank AAB71729);Myristica fragransアシル−ACPチオエステラーゼ(GenBank AAB71730);Cuphea lanceolataアシル−ACPチオエステラーゼ(GenBank CAA54060);Cuphea hookerianaアシル−ACPチオエステラーゼ(GenBank Q39513);Ulumus americanaアシル−ACPチオエステラーゼ(GenBank AAB71731);Sorghum bicolorアシル−ACPチオエステラーゼ(GenBank EER87824);Sorghum bicolorアシル−ACPチオエステラーゼ(GenBank EER88593);Cocos nuciferaアシル−ACPチオエステラーゼ(CnFatB1:Jing et al. BMC Biochemistry 2011, 12:44参照);Cocos nuciferaアシル−ACPチオエステラーゼ(CnFatB2:Jing et al. BMC Biochemistry 2011, 12:44参照);Cuphea viscosissimaアシル−ACPチオエステラーゼ(CvFatB1:Jing et al. BMC Biochemistry 2011, 12:44参照);Cuphea viscosissimaアシル−ACPチオエステラーゼ(CvFatB2:Jing et al. BMC Biochemistry 2011, 12:44参照);Cuphea viscosissimaアシル−ACPチオエステラーゼ(CvFatB3:Jing et al. BMC Biochemistry 2011, 12:44参照);Elaeis guineensisアシル−ACPチオエステラーゼ(GenBank AAD42220);Desulfovibrio vulgarisアシル−ACPチオエステラーゼ(GenBank ACL08376);Bacteriodes fragilisアシル−ACPチオエステラーゼ(GenBank CAH09236);Parabacteriodes distasonisアシル−ACPチオエステラーゼ(GenBank ABR43801);Bacteroides thetaiotaomicronアシル−ACPチオエステラーゼ(GenBank AAO77182);Clostridium asparagiformeアシル−ACPチオエステラーゼ(GenBank EEG55387);Bryanthella formatexigensアシル−ACPチオエステラーゼ(GenBank EET61113);Geobacillus sp.アシル−ACPチオエステラーゼ(GenBank EDV77528);Streptococcus dysgalactiaeアシル−ACPチオエステラーゼ(GenBank BAH81730);Lactobacillus brevisアシル−ACPチオエステラーゼ(GenBank ABJ63754);Lactobacillus plantarumアシル−ACPチオエステラーゼ(GenBank CAD63310);Anaerococcus tetradiusアシル−ACPチオエステラーゼ(GenBank EEI82564);Bdellovibrio bacteriovorusアシル−ACPチオエステラーゼ(GenBank CAE80300);Clostridium thermocellumアシル−ACPチオエステラーゼ(GenBank ABN54268);Nannochloropsis oculataアシル−ACPチオエステラーゼ(配列番号5、これをコードする遺伝子の塩基配列:配列番号6);Nannochloropsis gaditanaアシル−ACPチオエステラーゼ(配列番号7、これをコードする遺伝子の塩基配列:配列番号8);Nannochloropsis granulataアシル−ACPチオエステラーゼ(配列番号9、これをコードする遺伝子の塩基配列:配列番号10);Symbiodinium microadriaticumアシル−ACPチオエステラーゼ(配列番号11、これをコードする遺伝子の塩基配列:配列番号12)、等が挙げられる。
また、これらと機能的に均等なタンパク質として、上述したいずれかのアシル−ACPチオエステラーゼのアミノ酸配列と50%以上(好ましくは70%以上、より好ましくは80%以上、さらに好ましくは90%以上、よりさらに好ましくは95%以上、96%以上、97%以上、98%以上、又は99%以上)の同一性を有するアミノ酸配列からなり、かつアシル−ACPチオエステラーゼ活性を有するタンパク質も用いることができる。
これらのアシル−ACPチオエステラーゼ及びそれらをコードする遺伝子の配列情報等は、例えば、国立生物工学情報センター(National Center for Biotechnology Information, NCBI)などから入手することができる。
また、大腸菌等の宿主細胞内で機能するプロモーターの下流にアシル−ACPチオエステラーゼ遺伝子を連結した融合遺伝子を宿主細胞へ導入し、導入したアシル−ACPチオエステラーゼ遺伝子が発現する条件で細胞を培養した後、細胞の破砕液に対し、Yuanらの方法(Yuan L, Voelker TA, Hawkins DJ. “Modification of the substrate specificity of an acyl-acyl carrier protein thioesterase by protein engineering” Proc Natl Acad Sci U S A. 1995 Nov 7;92(23), p.10639-10643)によって調製した各種アシル−ACPを基質とした反応を行うことにより、アシル−ACPチオエステラーゼ活性を測定することができる。
本発明の形質転換体は、宿主に前記(A)又は(B)のタンパク質をコードする遺伝子を導入して得られる。当該形質転換体では、宿主に比べ、中鎖脂肪酸又はそのエステル、及びこれらを構成成分とする脂質の生産能が有意に向上する。また、当該形質転換体では、宿主に比べ、中鎖脂肪酸含有量の高いトリアシルグリセロールを生産できる。また、当該形質転換体では、脂質中の脂肪酸組成が宿主に比べ改変される。
なお、宿主や形質転換体の脂肪酸及び脂質の生産能については、実施例で用いた方法により測定することができる。
前記微生物は原核生物、真核生物のいずれであってもよい。
原核生物として、エシェリキア(Escherichia)属に属する微生物やバシラス(Bacillus)属に属する微生物等を用いることができる。
真核微生物として、酵母や糸状菌等を用いることができ、サッカマイセス(Saccharomyces)属、クリベロマイセス(Kluyveromyces)属、シゾサッカロマイセス(Schizosaccharomyces)属、チゴサッカロマイセス(Zygosaccharomyces)属、ヤロウイア(Yarrowia)属、ハンゼヌラ(Hansenula)属、ピキア(Pichia)属、キャンディダ(Candida)属、又はモルチエラ(Mortierella)属に属する微生物を用いることが好ましい。なかでも、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、ヤロウイア・リポリティカ(Yarrowia lipolytica)、赤色酵母(Rhodosporidium toruloides)、またはモルチエラ・アルピナ(Mortierella alpina)が好ましい。
前記植物体としては、種子に脂質を高含有する観点から、シロイヌナズナ(Arabidopsis thaliana)、ナタネ、ココヤシ、パーム、クフェア、スナモモ、ダイズ、トウモロコシ、イネ、ヒマワリ、クスノキ、又はヤトロファが好ましく、シロイヌナズナがより好ましい。
具体的なベクターとしては、微生物を宿主とする場合には、例えば、pBluescript(pBS) II SK(-)(Stratagene社製)、pSTV系ベクター(タカラバイオ社製)、pUC系ベクター(宝酒造社製)、pET系ベクター(タカラバイオ社製)、pGEX系ベクター(GEヘルスケア社製)、pCold系ベクター(タカラバイオ社製)、pHY300PLK(タカラバイオ社製)、pUB110(Mckenzie,T. et al.,(1986),Plasmid 15(2);p.93-103)、pBR322(タカラバイオ社製)、pRS403(ストラタジーン社製)、又はpMW218/219(ニッポンジーン社製)が挙げられる。特に、宿主が大腸菌の場合は、pBluescript II SK(-)、又はpMW218/219が好ましく用いられる。
藻類を宿主とする場合には、例えば、pUC19(タカラバイオ社製)、P66(Chlamydomonas Center)、P-322(Chlamydomonas Center)、pPha-T1(Yangmin Gong, Xiaojing Guo, Xia Wan, Zhuo Liang, Mulan Jiang, “Characterization of a novel thioesterase (PtTE) from Phaeodactylum tricornutum”, Journal of Basic Microbiology, 2011 December, Volume 51, p.666-672.参照)、又はpJET1(コスモ・バイオ社製)が挙げられる。
植物細胞を宿主とする場合には、例えば、pRI系ベクター(タカラバイオ社製)、pBI系ベクター(クロンテック社製)、又はIN3系ベクター(インプランタイノベーションズ社製)が挙げられる。特に、宿主がシロイヌナズナの場合は、pRI系ベクター又はpBI系ベクターが好ましく用いられる。
具体的なプロモーターとしては、例えば、lacプロモーター、trpプロモーター、tacプロモーター、trcプロモーター、T7プロモーター、SpoVGプロモーター、カリフラワーモザイルウイルス35SRNAプロモーター、ハウスキーピング遺伝子プロモーター(例えば、チューブリンプロモーター、アクチンプロモーター、ユビキチンプロモーター等)、ナタネ由来Napin遺伝子プロモーター、植物由来Rubiscoプロモーター、又はナンノクロロプシス属由来のビオラキサンチン/クロロフィルa結合タンパク質遺伝子のプロモーターが挙げられる。
また、選択マーカーとしては、抗生物質耐性遺伝子(アンピシリン耐性遺伝子、クロラムフェニコール耐性遺伝子、エリスロマイシン耐性遺伝子、ネオマイシン耐性遺伝子、カナマイシン耐性遺伝子、スペクチノマイシン耐性遺伝子、テトラサイクリン耐性遺伝子、ブラストサイジンS耐性遺伝子、ビアラホス耐性遺伝子、ゼオシン耐性遺伝子、パロモマイシン耐性遺伝子、又はハイグロマイシン耐性遺伝子)等の薬剤耐性遺伝子が挙げられる。さらに、栄養要求性に関連する遺伝子の欠損等を選択マーカー遺伝子として使用することも可能である。
形質転換方法としては、宿主に目的遺伝子を導入しうる方法であれば特に限定されるものではない。例えば、カルシウムイオンを用いる方法、一般的なコンピテントセル形質転換方法(J.Bacterial.93,1925(1967))、プロトプラスト形質転換法(Mol.Gen.Genet.168,111(1979))、エレクトロポレーション法(FEMS Microbiol.Lett.55,135(1990))又はLP形質転換方法(T.Akamatsu及びJ.Sekiguchi,Archives of Microbiology,1987,146,p.353-357;T.Akamatsu及びH.Taguchi,Bioscience,Biotechnology,and Biochemistry,2001,65,4,p.823-829)等を用いることができる。宿主が植物の場合、アグロバクテリウムを用いた方法(C. R. Acad. Sci. Paris. Life Science 316, 1194(1993)等)、パーティクルガン法(BioRad社PDS-1000/He等)等を用いることができる。
次いで、上記で得られた形質転換体を用いて脂質を生産する。
本発明の製造方法は、前記(A)又は(B)のタンパク質をコードする遺伝子を導入した形質転換体から脂質を採取する工程を含む。当該工程は、脂質の生産性向上の観点から、前記(A)又は(B)のタンパク質をコードする遺伝子を導入した形質転換体を適切な条件にて培養して培養物を得る工程、及び得られた培養物から脂質を採取する工程を含むことが好ましい。なお、本発明において形質転換体を培養するとは、微生物、植物体、動物体、及びそれらの細胞や組織を培養、生育することをいい、植物体を土壌等で栽培することも含まれる。また、培養物には、培養液はもちろん、培養等した後の形質転換体そのものも含まれる。
また、脂質の生産効率の点から、培地中に、例えば脂肪酸生合成系に関与する前駆物質としてグリセロール、酢酸、又はマロン酸等を添加してもよい。
培地は天然海水又は人工海水をベースにしたものを使用してもよいし、市販の培養培地を使用してもよい。藻類の生育促進、中鎖脂肪酸の生産性向上のため、培地に、窒素源、リン源、金属塩、ビタミン類、微量金属等を適宜添加することができる。培地に接種する藻類の量は特に限定されないが、生育性の点から、培地当り1〜50%(vol/vol)が好ましい。培養温度は、藻類の増殖に悪影響を与えない範囲であれば特に制限されないが、通常、5〜40℃の範囲である。また、藻類の培養は、光合成ができるよう光照射下で行うことが好ましい。また、藻類の培養は、光合成ができるように二酸化炭素を含む気体の存在下、又は炭酸水素ナトリウムなどの炭酸塩を含む培地で行うことが好ましい。なお、培養は、通気攪拌培養、振とう培養又は静置培養のいずれでもよく、通気性の向上の観点から、振とう培養が好ましい。
本発明の製造方法において製造される脂質は、その利用性の点から、脂肪酸又はこれを構成成分とする脂質であることが好ましく、中鎖脂肪酸又はこれを構成成分とする脂質であることがより好ましい。
中鎖脂肪酸を構成成分とする脂質として好ましくは中鎖脂肪酸のエステルである。具体的には中鎖アシル基を有するトリアシルグリセロールが好ましい。
さらに界面活性剤等への利用性から、中鎖脂肪酸又はそのエステルは、炭素原子数12又は14の脂肪酸又はそのエステルがより好ましく、炭素数12の脂肪酸又はそのエステルがさらに好ましく、ラウリン酸又はそのエステルが特に好ましい。これらの高級脂肪酸を還元して得られる高級アルコールの誘導体は、界面活性剤として利用できる。
下記(1)及び(2)の工程を含む脂質の製造方法。
(1)宿主に下記(A)又は(B)のタンパク質をコードする遺伝子を導入して形質転換体を得る工程
(A) 配列番号1で表されるアミノ酸配列からなるタンパク質
(B) 配列番号1で表されるアミノ酸配列と80%以上の同一性を有するアミノ酸配列からなり、かつジアシルグリセロールアシルトランスフェラーゼ活性を有するタンパク質
(2)得られた形質転換体から脂質を採取する工程
前記(B)において、配列番号1で表されるアミノ酸配列との同一性が85%以上、好ましくは90%以上、より好ましくは93%以上、さらに好ましくは95%以上、よりさらに好ましくは97%以上である、<1>項記載の製造方法。
<3>
前記(A)又は(B)のタンパク質が、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼである、<1>又は<2>項記載の製造方法。
<4>
前記脂質が、中鎖脂肪酸又はそのエステルである、<1>〜<3>のいずれか1項記載の製造方法。
<5>
前記工程(1)において、宿主にアシル−ACPチオエステラーゼをコードする遺伝子を共導入する、<1>〜<4>のいずれか1項記載の製造方法。
<6>
前記アシル−ACPチオエステラーゼが、中鎖アシル−ACP特異的なアシル−ACPチオエステラーゼである、<5>項記載の製造方法。
<7>
前記宿主が微生物又は植物である、<1>〜<6>のいずれか1項記載の製造方法。
<8>
前記植物がシロイヌナズナである、<7>項記載の製造方法。
<9>
前記微生物が酵母である、<7>項記載の製造方法。
<10>
前記酵母がサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)である、<9>項記載の製造方法。
前記(A)又は(B)のタンパク質。
<12>
前記(B)において、配列番号1で表されるアミノ酸配列との同一性が85%以上、好ましくは90%以上、より好ましくは93%以上、さらに好ましくは95%以上、よりさらに好ましくは97%以上である、<11>項記載のタンパク質。
<13>
前記(B)のアミノ酸配列が、配列番号1のアミノ酸配列において1又は数個、好ましくは1〜20個、より好ましくは1〜10個、さらに好ましくは1〜5個、よりさらに好ましくは1〜3個、特に好ましくは1〜2個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列である、<11>又は<12>項記載のタンパク質。
<14>
前記(A)又は(B)のタンパク質が、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼである、<11>〜<13>のいずれか1項記載のタンパク質。
<11>〜<14>のいずれか1項記載のタンパク質をコードする遺伝子。
<16>
下記(a)又は(b)のDNAからなる遺伝子。
(a) 配列番号2で表される塩基配列からなるDNA
(b) 配列番号2で表される塩基配列と70%以上の同一性を有する塩基配列からなり、かつジアシルグリセロールアシルトランスフェラーゼ活性を有するタンパク質をコードするDNA
<17>
前記(b)において、配列番号2で表される塩基配列との同一性が80%以上、好ましくは90%以上、より好ましくは93%以上、さらに好ましくは95%以上、よりさらに好ましくは97%以上である、<16>項記載の遺伝子。
<18>
前記(b)の塩基配列が、配列番号2の塩基配列において1又は数個、好ましくは1〜20個、より好ましくは1〜10個、さらに好ましくは1〜5個、よりさらに好ましくは1〜3個、特に好ましくは1〜2個の塩基が欠失、置換、挿入、又は付加された塩基配列である、<16>又は<17>項記載の遺伝子。
<19>
前記(a)又は(b)の遺伝子が、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼをコードする遺伝子である、<16>〜<18>のいずれか1項記載の遺伝子。
宿主に、<15>〜<19>のいずれか1項記載の遺伝子を導入して得られる形質転換体。
<21>
宿主に<15>〜<19>のいずれか1項記載の遺伝子、及びアシル−ACPチオエステラーゼをコードする遺伝子を共導入して得られる、<20>項記載の形質転換体。
<22>
前記アシル−ACPチオエステラーゼが、中鎖アシル−ACP特異的なアシル−ACPチオエステラーゼである、<21>項記載の形質転換体。
<23>
前記宿主が微生物又は植物である、<20>〜<22>のいずれか1項記載の形質転換体。
<24>
前記植物がシロイヌナズナである、<23>項記載の形質転換体。
<25>
前記微生物が酵母である、<23>項記載の形質転換体。
<26>
前記酵母がサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)である、<25>項記載の形質転換体。
宿主に前記(A)又は(B)のタンパク質をコードする遺伝子を導入する工程を含む、脂質中の脂肪酸組成を改変する方法。
宿主に前記(A)又は(B)のタンパク質をコードする遺伝子を導入して形質転換体を得る工程を含む、脂質の生産性を向上させる方法。
<29>
前記脂質が中鎖脂肪酸又はそのエステルである、<28>項に記載の方法。
宿主に前記(a)又は(b)のタンパク質をコードする遺伝子を導入して形質転換する工程を含む、形質転換体の産生するトリアシルグリセロール中の中鎖脂肪酸含有量を向上させる方法。
脂質を製造するための、<20>〜<26>のいずれか1項に記載の形質転換体の使用。
<32>
前記脂質が中鎖脂肪酸又はそのエステルである、<31>項に記載の形質転換体の使用。
1.ココヤシのRNA抽出
ココヤシ固形胚乳を液体窒素で凍結した後、マルチビーズショッカー(安井器械製)を用いて破砕した。破砕した固形胚乳に対してフェノール/クロロホルム、50mM Tris-HCl (pH9)を添加しよく混ぜ、7500rpmで10分間遠心操作を行い、上清を回収した。回収した上清に再度同様のフェノール/クロロホルム処理を行った。上層を回収し、これに対してエタノール沈殿操作を行い、そこに含まれる核酸成分を精製した。RNA成分の精製のために、RNeasy Plant Mini Kit (Qiagen, Valencia, California製)を用いた。エタノール沈殿後の核酸ペレットに、1M DTTを1/100容量添加したRLT Bufferを加えてボルテックスしたものを、QIA shredder spin columnにアプライした。以降はキット添付のマニュアルに従って操作を行い、最終的に脱イオン水(dH2O)でココヤシ由来のtotal RNAを溶出した。得られたRNA溶液に対し、DNaseI(サーモサイエンティフィック製)をバッファーとともに添加し、1時間37℃にて処理を行った。その後、フェノール/クロロホルム処理・エタノール沈殿処理を行い、ココヤシ胚乳由来RNA溶液とした。
続いて、PrimeScript II 1st strand cDNA Synthesis Kit(タカラバイオ製)を用いて、得られたRNAからcDNAを調製した。
茨城県潮来市で採取した野生のアブラナ様植物から、Brassica rapa由来の Napin遺伝子プロモーターを、栃木県益子町で採取した野生のアブラナ様植物から、Napin遺伝子ターミネーターをそれぞれ取得した。PowerPlant DNA Isolation Kit(MO BIO Laboratories製, USA)を用いて、野生のアブラナ様植物のゲノムDNAを抽出した。得られたゲノムDNAをテンプレートとし、DNAポリメラーゼPrimeSTAR(タカラバイオ製)を用いたPCR反応により、上記プロモーターとターミネーター領域を増幅した。具体的には、プライマーNo.1とNo.2を用いてBrassica rapa由来のNapin遺伝子プロモーターを、プライマーNo.3とNo.4を用いてBrassica rapa由来のNapin遺伝子ターミネーターをそれぞれ増幅した。増幅が見られたPCR産物をテンプレートとし、Napin遺伝子プロモーターはプライマーNo.5とNo.6、Napin遺伝子ターミネーターはプライマーNo.3とNo.7を用いて、再度PCR反応を行った。Napin遺伝子プロモーター配列を配列番号14、Napin遺伝子ターミネーター配列を配列番号15に示す。これらの増幅断片をMighty TA-cloning Kit(タカラバイオ製)で処理した後、pMD20-Tベクター(タカラバイオ製)にライゲーション反応により挿入することで、Napin遺伝子プロモーターを含むプラスミドpPNapin1、およびNapin遺伝子ターミネーターを含むプラスミドpTNapin1をそれぞれ構築した。
次に、カリフォルニア・ベイ由来のアシル−ACPチオエステラーゼ(以下、BTEとも略記する)遺伝子の葉緑体移行シグナルペプチドをコードする遺伝子配列(配列番号16)を、Invitrogen社(Carlsbad, California)の提供する受託合成サービスを利用して取得した。取得した配列を含むプラスミドをテンプレートとし、PrimeSTAR、プライマーNo.12とNo.13を用いたPCR反応により、シグナルペプチドをコードする遺伝子断片を増幅した。Mighty TA-cloning Kit(タカラバイオ製)を用いて、増幅した遺伝子断片の両末端にデオキシアデニン(dA)を付加した後、pMD20-Tベクター(タカラバイオ製)にライゲーション反応により挿入し、プラスミドpSignalを構築した。
プラスミドpSignalを制限酵素Not Iで処理し、プラスミドp909PTnapinのNot Iサイトにライゲーション反応で連結し、プラスミドp909PTnapin-Sを得た。
下記の手順により、植物導入用ベクターpRI909が本来保持するカナマイシン耐性遺伝子を、Streptomyces hygroscopicus由来のビアラフォス耐性遺伝子(Bar遺伝子)に置換した。Bar遺伝子は、ホスフィノトリシンアセチル転移酵素をコードする。Streptomyces hygroscopicus由来のビアラフォス耐性遺伝子は、NCBIのGene Bankで開示された形質転換用ベクターpYW310(ACCESSION NO. DQ469641)の配列を参考に、Gene Script社の提供する受託合成サービスを利用して取得した(配列番号17)。人工合成遺伝子をテンプレートとし、PrimeSTAR、プライマーNo.18とNo.19を用いたPCR反応により、Bar遺伝子配列を増幅した。一方、pRI909をテンプレートとし、PrimeSTAR、プライマーNo.20とNo.21を用いたPCR反応により、pRI909ベクターからカナマイシン耐性遺伝子を除いた領域を増幅した。両増幅断片をNde IとSpe Iで処理し、ライゲーション反応で連結して、プラスミドpRI909 Barを構築した。
前記で構築したプラスミドp909CTEを、インプランタイノベーションズ(株)によるシロイヌナズナ形質転換受託サービスに供し、CTE遺伝子を導入したシロイヌナズナ(Arabidopsis thaliana, Colombia株)の形質転換体を得た。シロイヌナズナは、野生型及び形質転換体を、室温22℃、蛍光灯照明を用いて明期24時間(約4000ルクス)の条件で育成した。約2ヶ月の栽培の後、種子を収穫した。
プラスミドp909Poleosin-DGAT2096-TnapinをAgrobacterium tumefaciens GV3101株に導入し、これを用いてp909CTEを導入したシロイヌナズナを形質転換した。播種後1.5ヶ月程度育成したシロイヌナズナの花序を切除し、さらに6〜7日間育成したものに、プラスミドを導入したアグロバクテリウムを感染させた。得られた種子を、MS寒天培地(100μg/mlクラフォラン、7μg/mlビアラフォスを含む)に播種し、形質転換体を選抜した。得られた形質転換体を室温22℃、蛍光灯照明を用いて明期24時間の条件で育成し、約2ヶ月の栽培の後、種子を収穫した。
シロイヌナズナ種子を、マルチビーズショッカー(安井器械製)を用いて粉砕し、そこに20μlの7-ペンタデカノン(0.5mg/ml メタノール)(内部標準)と20μlの酢酸を添加したクロロホルム0.25ml、メタノール0.5mlを加え、十分に攪拌した後、15分間静置した。さらに、1.5%KCl 0.25mlとクロロホルム0.25mlを添加し、十分に攪拌した後、15分間静置した。室温、1500rpmで5分間遠心分離を行った後、下層部分を採取し、窒素ガスで乾燥した。乾燥したサンプルに、0.5N水酸化カリウム−メタノール溶液を100μl加え、70℃で30分間恒温することによりトリアシルグリセロールを加水分解した。3‐フッ化ホウ素メタノール錯体溶液を0.3ml添加して乾燥物を溶解し、80℃で10分間恒温することにより脂肪酸のメチルエステル化処理を行った。その後、飽和食塩水0.2mlとヘキサン0.3mlを添加し、十分に攪拌した後、30分間静置した。脂肪酸のメチルエステルが含まれるヘキサン層(上層部分)を採取し、ガスクロマトグラフィ(GC)分析に供した。
GCによりメチルエステル化した試料の分析を行った。使用したGCは、カラム:DB1-MS (J&W Scientific, Folsom, California)、分析装置:6890 (Agilent technology, Santa Clara, California)を用いて、[カラムオーブン温度:150℃保持0.5分→150〜320℃(20℃/分昇温)→320℃保持2分、注入口検出器温度:300℃、注入法:スプリットモード(スプリット比=75:1)、サンプル注入量5μl、カラム流速:0.3ml/min コンスタント、検出器:FID、キャリアガス:水素、メイクアップガス:ヘリウム]の条件で行った。
GC解析により得られた波形データのピーク面積より、各脂肪酸のメチルエステル量を定量した。なお、種子中の各脂質に対応するGCのピークは、各脂肪酸の標準品のメチルエステルの保持時間(Retention Time, RT)により同定した。また、各ピーク面積を内部標準である7-ペンタデカノンのピーク面積と比較することで試料間の補正を行い、解析に供した全種子中に含まれる脂肪酸量を算出した。
各形質転換シロイヌナズナ種子における、総脂肪酸中の各脂肪酸の割合を表2に示す。なお、表2は、独立した3ラインの平均値を示した。また、C18:nは、C18不飽和脂肪酸(C18:1〜C18:3)の総計を表す。
CTE遺伝子とCnDGAT2096遺伝子を共導入した形質転換体の種子では、CTE遺伝子のみを発現する形質転換体の種子に比べ、C12:0脂肪酸とC14:0脂肪酸の含有率が増加した。一方、C16:0脂肪酸の含有率は変化が見られなかった。これらの結果から、CnDGAT2096遺伝子がコードするタンパク質は、CTEにより増加したC12:0、C14:0及びC16:0脂肪酸を均等にトリアシルグリセロールへと導入しているのではなく、C12:0脂肪酸とC14:0脂肪酸を選択的にトリアシルグリセロールに導入していると考えられる。
1.酵母トリアシルグリセロール合成遺伝子欠損株の作製
酵母(Saccharomyces cerevisiae)のトリアシルグリセロール合成遺伝子を全て欠損させた株を下記の手順により取得した。トリアシルグリセロール合成遺伝子は、文献(Line Sandager, et al., J Biol Chem. Feb 22;277(8):6478-82, 2002)情報を参考に、DGA1、LRO1、ARE1、ARE2遺伝子を標的とした。遺伝子欠損はアミノ酸マーカー遺伝子をDNA断片として用いた相同組み換えによる一般的な手法で行った。
同様に、プライマーNo.40及びNo.41を用いたPCR反応により、LEU2遺伝子のORF上流700bp〜下流500bpの領域の遺伝子断片を、プライマーNo.42及びNo.43、又はプライマーNo.44及びNo.45を用いたPCR反応により、ARE2遺伝子上流500bp、又は下流500bpの領域の遺伝子断片をそれぞれ取得した。得られた遺伝子断片を、ΔARE1プラスミドと同様にpUC19ベクターに接続し、ΔARE2プラスミドを得た。
続いて、プライマーNo.46及びNo.47を用いたPCR反応により、MET15遺伝子のORF上流300bp〜下流300bpの領域の遺伝子断片を、プライマーNo.48及びNo.49、又はプライマーNo.50及びNo.51を用いたPCR反応により、LRO1遺伝子上流500bp、又は下流500bpの領域の遺伝子断片をそれぞれ取得した。得られた遺伝子断片を、ΔARE1プラスミドと同様にpUC19ベクターに接続し、ΔLRO1プラスミドを得た。
ΔARE1プラスミドを鋳型として、プライマーNo.34及びNo.37を用いたPCR反応により、遺伝子断片を取得した。これを用いて、酢酸リチウム法により、Saccharomyces cerevisiae BY4741ΔDGA1株(ΔDGA1::kanMX his3Δ1 leu2Δ0 met15Δ0 ura3Δ0)(Yeast Knock Out Strain Collection:Open Biosystems)を形質転換した。得られた株を、ΔARE2プラスミドを鋳型としプライマーNo.42及びNo.45を用いたPCR反応により取得した遺伝子断片を用いて、上記と同様の手法で形質転換した。得られた株をさらに、ΔLRO1プラスミドを鋳型としてプライマーNo.48及びNo.51を用いたPCR反応により取得した遺伝子断片を用いて、形質転換した。その結果、Saccharomyces cerevisiae Δdga1、Δlro1、Δare1、Δare2株を得た。
(1)DGAT遺伝子発現プラスミドの構築
Saccharomyces cerevisiaeのゲノムDNAを鋳型として、プライマーNo.52及びNo.53を用いたPCR反応により、URA3遺伝子のORF上流300bpからORF終止コドンまでの領域の遺伝子断片を、プライマーNo.54及びNo.55を用いたPCR反応により、CYC1遺伝子のORF下流500bpの領域の遺伝子断片を、プライマーNo.56及びNo.57を用いたPCR反応により、GAL1遺伝子のORF上流50bpの領域の遺伝子断片を、プライマーNo.58及びNo.59を用いたPCR反応により、URA3遺伝子のORF下流200bpの領域の遺伝子断片をそれぞれ取得した。
配列番号13に示すシロイヌナズナ由来のDGAT遺伝子(AtDGAT1遺伝子)を人工遺伝子の受託合成サービスを利用して取得し、プライマーNo.60及びNo.61を用いたPCR反応により、AtDGAT1遺伝子断片を取得した。得られた遺伝子断片をpUC19ベクターに接続し、AtDGAT1遺伝子発現プラスミドpUC-URAm-GAL1p-AtDGAT1を構築した。
また、実施例1で作製したココヤシcDNAをテンプレートとし、プライマーNo.62及びNo.63を用いたPCR反応により、ココヤシ由来のDGAT遺伝子(CnDGAT2096遺伝子、配列番号2)断片を取得した。得られた遺伝子断片をpUC19ベクターに接続し、CnDGAT2096遺伝子発現プラスミドpUC-URAm-GAL1p-CnDGATを構築した。
これらのプラスミドは、URA3プロモーター配列、URA3遺伝子断片、CYC1ターミネーター配列、GAL1プロモーター配列、各種DGAT遺伝子配列、及びURA3ターミネーター配列の順に連結したインサート配列と、pUC19ベクター配列とからなる。
なお、配列番号13に示すAtDGAT1遺伝子の塩基配列は、配列番号2に示すCnDGAT2096遺伝子の塩基配列と68%の同一性を示す。また、配列番号13の塩基配列がコードするアミノ酸配列は、配列番号1に示すCnDGAT2096のアミノ酸配列と66%の同一性を示す。
プラスミドpUC-URAm-GAL1p-AtDGAT1を鋳型とし、プライマーNo.52及びNo.59を用いてPCRを行った。得られた増幅DNA断片を用いて、酢酸リチウム法によりSaccharomyces cerevisiae Δdga1、Δlro1、Δare1、Δare2株を形質転換し、+AtDGAT1株を得た。
同様にpUC-URAm-GAL1p-CnDGATを鋳型とし、プライマーNo.52及びNo.59を用いてPCRを行うことでSaccharomyces cerevisiae Δdga1、Δlro1、Δare1、Δare2株を形質転換し、+CnDGAT株を得た。
各形質転換株を、Yeast Nitrogen base w/o Amino Acid 0.67%、Raffinose 2%からなる液体培地20mLに1白金耳植菌し、30℃で1日振とう培養した。培養液を遠心分離し、菌体を回収した。菌体をOD600=1となるよう、Yeast Nitrogen base w/o Amino Acid 0.67%、Galactose 2%からなる液体培地(Briji 58(シグマアルドリッチ社製)1%添加、及びラウリン酸またはミリスチン酸を2mM含有)20 mlに植菌した。24時間、30℃で培養した後、遠心分離によって菌体を集菌し、水洗後、凍結乾燥した。
凍結乾燥菌体を、8mLのクロロホルム・メタノール溶液(クロロホルム:メタノール=2:1)に懸濁し、薄層クロマトグラフィーを行い、脂質を分画した。薄層クロマトグラフィーは、シリカゲル60 プレート(メルク製)、展開溶媒ヘキサン:ジエチルエーテル:蟻酸=42:24:0.3の条件で行った。0.01%プリムリン(メタノール中に溶解)を噴霧し、UV照射して脂質を可視化した。
次いで、トリアシルグリセロール(TAG)画分のみをかきとり、かきとったシリカ粒子をKOHと3−フッ化ホウ素メタノールによってメチルエステルに誘導し、0.5N水酸化カリウム/メタノール溶液0.7mLを添加し、80℃で30分間恒温した。続いて1mLの14%三フッ化ホウ素溶液(SIGMA社製)を添加し、80℃にて10分間恒温した。その後、ヘキサン、飽和食塩水を各1mL添加し激しく撹拌し、室温にて30分間放置後、上層であるヘキサン層を回収して脂肪酸メチルエステルを得た。得られた脂肪酸メチルエステルをガスクロマトグラフィー(GC)分析に供した。
脂肪酸メチルエステル試料をGCにて分析した。使用したGCは、カラム:DB1-MS (J&W Scientific, Folsom, California)、分析装置:6890 (Agilent technology, Santa Clara, California)を用いて、[カラムオーブン温度:150℃保持0.5分→150〜320℃(20℃/分昇温)→320℃保持2分、注入口検出器温度:300℃、注入法:スプリットモード(スプリット比=75:1)、サンプル注入量5μl、カラム流速:0.3ml/min コンスタント、検出器:FID、キャリアガス:水素、メイクアップガス:ヘリウム]の条件で行った。
GC解析により得られた波形データのピーク面積より、実施例1と同様に、各脂肪酸のメチルエステル量を定量した。形質転換体から抽出したトリアシルグリセロール中の脂肪酸組成を、表3及び表4に示す。表3は各形質転換株をラウリン酸(C12:0)含有培地で培養した結果であり、表4は各形質転換株をミリスチン酸(C14:0)含有培地で培養した結果である。
また、表4から明らかなように、CnDGAT2096遺伝子を導入した株では、AtDGAT1遺伝子を導入した株と比べて、トリアシルグリセロール中のC14:0脂肪酸の含有量が増加していた。この結果から、CnDGAT2096遺伝子は、C14:0アシル基をジアシルグリセロールに選択的に導入してトリアシルグリセロールを合成していることがわかった。
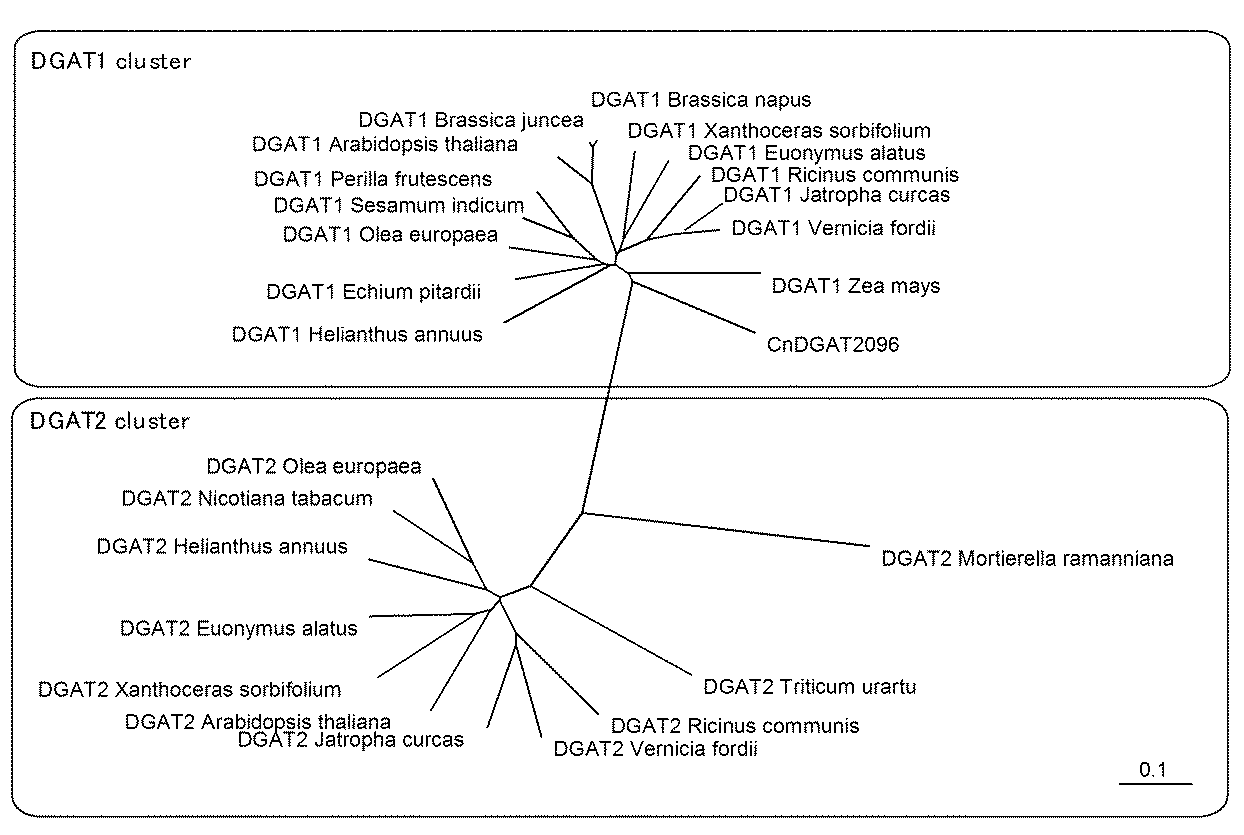
配列番号1に示すジアシルグリセロールアシルトランスフェラーゼの系統解析を行った。
系統解析には、配列番号1に示すアミノ酸配列とともに、表5に示すジアシルグリセロールアシルトランスフェラーゼのアミノ酸配列を用いた。表5に示すアミノ酸配列は、NCBI(National Center for Biotechnology Information)が提供するデータベース(http://www.ncbi.nlm.nih.gov/)、又は特許文献から取得した。
これらの配列をClustalX(http://www.clustal.org/)を用いたマルチプルアライメント解析に供し、近隣結合法(NJ法)にて系統樹を作製した。結果を図1に示す。
また、各配列について、配列番号1のアミノ酸配列との配列同一性を遺伝情報処理ソフトウェアGenetyx-Winを用いて調べ、表5に示した。なお、表5の「配列番号1のアミノ酸配列との同一性」において、*は全長アミノ酸配列ではなく、相同領域として選択された部分配列に対しての同一性を示す。例えば、DGAT2_Mortierella_ramannianaの全長配列は355アミノ酸であるが、この中から相同領域として77アミノ酸からなる部分配列が選択され、この部分配列と配列番号1の配列との同一性が18%であったことを示している。
Claims (16)
- 下記(1)及び(2)の工程を含む脂質の製造方法。
(1)宿主(但し、ヒトを除く)に下記(A)又は(B)のタンパク質をコードする遺伝子を導入して形質転換体を得る工程
(A) 配列番号1で表されるアミノ酸配列からなるタンパク質
(B) 配列番号1で表されるアミノ酸配列と90%以上の同一性を有するアミノ酸配列からなり、かつジアシルグリセロールアシルトランスフェラーゼ活性を有するタンパク質
(2)得られた形質転換体から脂質を採取する工程 - 前記(A)又は(B)のタンパク質が、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼである、請求項1記載の製造方法。
- 前記脂質が、中鎖脂肪酸又はそのエステルである、請求項1又は2記載の製造方法。
- 前記工程(1)において、宿主にアシル−ACPチオエステラーゼをコードする遺伝子を共導入する、請求項1〜3のいずれか1項記載の製造方法。
- 前記アシル−ACPチオエステラーゼが、中鎖アシル−ACP特異的なアシル−ACPチオエステラーゼである、請求項4記載の製造方法。
- 前記宿主が微生物又は植物である、請求項1〜5のいずれか1項記載の製造方法。
- 前記植物がシロイヌナズナである、請求項6記載の製造方法。
- 前記微生物が酵母である、請求項6記載の製造方法。
- 下記(A)又は(B)のタンパク質。
(A) 配列番号1で表されるアミノ酸配列からなるタンパク質
(B) 配列番号1で表されるアミノ酸配列と90%以上の同一性を有するアミノ酸配列からなり、かつジアシルグリセロールアシルトランスフェラーゼ活性を有するタンパク質 - 前記(A)又は(B)のタンパク質が、中鎖アシル−CoA特異的なジアシルグリセロールアシルトランスフェラーゼである、請求項9記載のタンパク質。
- 請求項9又は10記載のタンパク質をコードする遺伝子。
- 宿主(但し、ヒトを除く)に、請求項11記載の遺伝子を導入して得られる形質転換体。
- 宿主(但し、ヒトを除く)に請求項11記載の遺伝子、及びアシル−ACPチオエステラーゼをコードする遺伝子を共導入して得られる、請求項12記載の形質転換体。
- 前記宿主が微生物又は植物である、請求項12又は13記載の形質転換体。
- 前記植物がシロイヌナズナである、請求項14記載の形質転換体。
- 前記微生物が酵母である、請求項14記載の形質転換体。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2014159012A JP6520008B2 (ja) | 2014-08-04 | 2014-08-04 | ジアシルグリセロールアシルトランスフェラーゼを用いた脂質の製造方法 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2014159012A JP6520008B2 (ja) | 2014-08-04 | 2014-08-04 | ジアシルグリセロールアシルトランスフェラーゼを用いた脂質の製造方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2016034258A JP2016034258A (ja) | 2016-03-17 |
| JP6520008B2 true JP6520008B2 (ja) | 2019-05-29 |
Family
ID=55522574
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2014159012A Active JP6520008B2 (ja) | 2014-08-04 | 2014-08-04 | ジアシルグリセロールアシルトランスフェラーゼを用いた脂質の製造方法 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP6520008B2 (ja) |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP6985792B2 (ja) * | 2016-05-24 | 2021-12-22 | 花王株式会社 | 藻類の処理方法 |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| FI873252L (fi) * | 1986-07-31 | 1988-02-01 | Calgene Inc | Reglering av froespecifik transkription. |
| US6476294B1 (en) * | 1998-07-24 | 2002-11-05 | Calgene Llc | Plant phosphatidic acid phosphatases |
| JP5764339B2 (ja) * | 2010-05-06 | 2015-08-19 | 花王株式会社 | チオエステラーゼ及びそれを用いた脂肪酸又は脂質の製造方法 |
| JP6006553B2 (ja) * | 2012-07-10 | 2016-10-12 | 花王株式会社 | 組換えクラミドモナス属及びそれを用いたラウリン酸高含有油脂の製造方法 |
-
2014
- 2014-08-04 JP JP2014159012A patent/JP6520008B2/ja active Active
Also Published As
| Publication number | Publication date |
|---|---|
| JP2016034258A (ja) | 2016-03-17 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6310544B2 (ja) | β−ケトアシル−ACPシンターゼを用いた脂質の製造方法 | |
| JP6491881B2 (ja) | アシル−acpチオエステラーゼ | |
| JP6646580B2 (ja) | β−ケトアシル−ACPシンターゼを用いた中鎖脂肪酸の製造方法 | |
| JP6592434B2 (ja) | 脂質の製造方法 | |
| US9222101B2 (en) | Method of producing fatty acids or lipids containing fatty acids using thioesterase variants | |
| US8940514B2 (en) | Thioesterase and a method of producing fatty acids or lipids using the thioesterase | |
| JP6319889B2 (ja) | アシル−acpチオエステラーゼ改変体を用いた脂質の製造方法 | |
| JP6381139B2 (ja) | アシル−acpチオエステラーゼ | |
| JP6779664B2 (ja) | 脂質の製造方法 | |
| WO2017043419A1 (ja) | 脂質の製造方法 | |
| JP6520008B2 (ja) | ジアシルグリセロールアシルトランスフェラーゼを用いた脂質の製造方法 | |
| JP6969941B2 (ja) | 脂質の製造方法 | |
| JP6332855B2 (ja) | アシル−acpチオエステラーゼを用いた脂質の製造方法 | |
| WO2014045793A1 (ja) | チオエステラーゼ改変体を用いた脂質の製造方法 | |
| JP6226464B2 (ja) | チオエステラーゼ改変体を用いた脂質の製造方法 | |
| WO2016190239A1 (ja) | 脂質の製造方法 | |
| WO2017022587A1 (ja) | 脂質の製造方法 | |
| JP2016054703A (ja) | ラウリン酸の製造方法 | |
| JP2019047756A (ja) | 脂質の製造方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20170605 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20180605 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20180803 Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20180803 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20190115 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20190308 Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20190308 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20190326 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20190415 |
|
| R151 | Written notification of patent or utility model registration |
Ref document number: 6520008 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R151 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |