JP5604657B2 - 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 - Google Patents
植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 Download PDFInfo
- Publication number
- JP5604657B2 JP5604657B2 JP2009060154A JP2009060154A JP5604657B2 JP 5604657 B2 JP5604657 B2 JP 5604657B2 JP 2009060154 A JP2009060154 A JP 2009060154A JP 2009060154 A JP2009060154 A JP 2009060154A JP 5604657 B2 JP5604657 B2 JP 5604657B2
- Authority
- JP
- Japan
- Prior art keywords
- amino acid
- seq
- gene
- protein
- acid sequence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images













Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/90—Isomerases (5.)
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A40/00—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production
- Y02A40/10—Adaptation technologies in agriculture, forestry, livestock or agroalimentary production in agriculture
- Y02A40/146—Genetically Modified [GMO] plants, e.g. transgenic plants
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Molecular Biology (AREA)
- Biomedical Technology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Cell Biology (AREA)
- Plant Pathology (AREA)
- Medicinal Chemistry (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Enzymes And Modification Thereof (AREA)
Description
(b)配列番号5に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、プロテインホスファターゼ2C活性を有するタンパク質
(c)配列番号4に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされプロテインホスファターゼ2C活性を有するタンパク質
(b)配列番号32に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、グルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質
(c)配列番号31に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ、グルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質
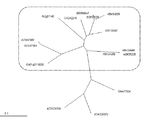
植物体内において過剰発現させるプロテインホスファターゼ2C遺伝子は、配列番号1〜3に示すアミノ酸配列からなる3つの共通配列をN末端側からこの順で有するプロテインホスファターゼ2Cをコードする。なお、参考文献(TRENDS in Plant Science Vol.9 No.5 May 2004 Pages 236-243)の第237頁に挙げられたFigure1.topographic cladogramにおいてグループEとして分類された遺伝子群は、配列番号1〜3に示すアミノ酸配列からなる3つの共通配列をN末端側からこの順で有するプロテインホスファターゼ2Cをコードする遺伝子である。なお、参考文献によれば、シロイヌナズナにおいては、76個のプロテインホスファターゼ2C遺伝子が予測されており、これら遺伝子についてT-Coffeeソフトウェア(参考文献;Notredame, C. et al. 2000 T-Coffee: a novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 302, 205-247)を用いて系統樹を作成した結果が参考文献のFigure1として開示されている。この系統樹において、グループEに分類されたプロテインホスファターゼ2C遺伝子は、配列番号1〜3に示すアミノ酸配列からなる3つの共通配列をN末端側からこの順で有するプロテインホスファターゼ2Cをコードしている。配列番号1〜3に示すアミノ酸配列からなる3つの共通配列は、上述した分類におけるグループEに特徴的な配列であって、他のグループとの明確な区別基準となる配列である。
このグループは、上記参考文献(1)で示された中性非極性アミノ酸のうち、脂肪属性の疎水性側鎖をもつアミノ酸のグループであり、V(Val、バリン)、L(Leu、ロイシン)、I(Ile、イソロイシン)及びM(Met、メチオニン)から構成される。参考文献(1)による中性非極性アミノ酸と分類されるもののうちFGACWPは以下理由で、この「脂肪族疎水性アミノ酸グループ」には含めない。G(Gly、グリシン)やA(Ala、アラニン)はメチル基以下の大きさで非極性の効果が弱いからである。C(Cys、システイン)はS-S結合に重要な役目を担う場合があり、また、酸素原子や窒素原子と水素結合を形成する特性があるからである。F(Phe、フェニルアラニン)やW(Trp、トリプトファン)は側鎖がとりわけ大きな分子量をもち、かつ、芳香族の効果が強いからである。P(Pro、プロリン)はイミノ酸効果が強く、ポリペプチドの主鎖の角度を固定してしまうからである。
このグループは、中性極性アミノ酸のうちヒドロキシメチレン基を側鎖に持つアミノ酸のグループであり、S(Ser、セリン)とT(Thr、スレオニン)から構成される。SとTの側鎖に存在する水酸基は、糖の結合部位であるため、あるポリペプチド(タンパク質)が特定の活性を持つために重要な部位である場合が多い。
このグループは、酸性であるカルボキシル基を側鎖に持つアミノ酸のグループであり、D(Asp、アスパラギン酸)とE(Glu、グルタミン酸)から構成される。
このグループは、塩基性アミノ酸のグループであり、K(Lys、リジン)とR(Arg、アルギニン)から構成される。これらKとRは、pHの広い範囲で正に帯電し塩基性の性質をもつ。一方、塩基性アミノ酸に分類されるH(His、ヒスチジン)はpH7においてほとんどイオン化されないので、このグループには分類されない。
このグループは、全てα位の炭素元素に側鎖としてメチレン基が結合しその先に極性基を有すると言う特徴を持つ。非極性基であるメチレン基の物理的大きさが酷似している特徴を持ち、N(Asn、アスパラギン、極性基はアミド基)、D(Asp、アスパラギン酸、極性基はカルボキシル基)とH(His、ヒスチジン、極性基はイミダゾール基)から成る。
このグループは、全てα位の炭素元素に側鎖としてジメチレン基以上の直鎖炭化水素が結合しその先に極性基を有すると言う特徴を持つ。非極性基であるジメチレン基の物理的大きさが酷似している特徴を持つ。E(Glu、グルタミン酸、極性基はカルボキシル基)、K(Lys、リジン、極性基はアミノ基)、Q(Gln、グルタミン、極性基はアミド基)、R(Arg、アルギニン、極性基はイミノ基とアミノ基)から成る。
このグループには、側鎖にベンゼン核を持つ芳香族アミノ酸であり、芳香族特有の化学的性質を特徴とする。F(Phe、フェニルアラニン)、Y(Tyr、チロシン)、W(Trp、トリプトファン)から成る。
このグループには、側鎖に環状構造を持つと同時に極性も持つアミノ酸で、H(H、ヒスチジン、環状構造と極性基は共にイミダゾール基)、Y(Tyr、チロシン、環状構造はベンゼン核で極性基は水酸基)から成る。
植物体内において過剰発現させるグルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼ遺伝子は、グルタチオンに結合することで活性が制御され、葉緑体等のプラスチド内においてフルクトース1,6-ビスリン酸をジヒドロキシアセトンリン酸とグリセルアルデヒド3-リン酸とに変換する反応(可逆反応)を触媒する活性を有する酵素をコードする遺伝子である。グルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼ遺伝子は、上記活性を有する酵素をコードする遺伝子であれば、如何なる植物由来の遺伝子であっても良いし、植物から単離した遺伝子に突然変異を導入した変異型遺伝子であっても良い。
発現ベクターは、過剰発現を可能とするプロモーターと、上述したプロテインホスファターゼ2C遺伝子及びグルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼ遺伝子を含むように構築する。なお、プロテインホスファターゼ2C遺伝子を含む発現ベクターと、グルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼ遺伝子を含む発現ベクターとを別個に準備しても良い。
上述した発現ベクターは、一般的な形質転換方法によって対象の植物内に導入される。発現ベクターを植物細胞に導入する方法(形質転換方法)は特に限定されるものではなく、植物細胞に応じた適切な従来公知の方法を用いることができる。具体的には、例えば、アグロバクテリウムを用いる方法や直接植物細胞に導入する方法を用いることができる。アグロバクテリウムを用いる方法としては、例えば、Bechtold, E., Ellis, J. and Pelletier, G. (1993) In Planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis plants. C.R. Acad. Sci. Paris Sci. Vie, 316, 1194-1199. あるいは、Zyprian E, Kado Cl, Agrobacterium-mediated plant transformation by novel mini-T vectors in conjunction with a high-copy vir region helper plasmid. Plant Molecular Biology, 1990, 15(2), 245-256.に記載された方法を用いることができる。
ナス科:タバコ(Nicotiana tabacum)、ナス(Solanum melongena)、ジャガイモ(Solaneum tuberosum)、トマト(Lycopersicon lycopersicum)、トウガラシ(Capsicum annuum)、ペチュニア(Petunia)など。
マメ科:ダイズ(Glycine max)、エンドウ(Pisum sativum)、ソラマメ(Vicia faba)、フジ(Wisteria floribunda)、ラッカセイ(Arachis hypogaea)、ミヤコグサ(Lotus corniculatus var. japonicus)、インゲンマメ(Phaseolus vulgaris)、アズキ(Vigna angularis)、アカシア(Acacia)など。
キク科:キク(Chrysanthemum morifolium)、ヒマワリ(Helianthus annuus)など。
ヤシ科:アブラヤシ(Elaeis guineensis、Elaeis oleifera)、ココヤシ(Cocos nucifera)、ナツメヤシ(Phoenix dactylifera)、ロウヤシ(Copernicia)など。
ウルシ科:ハゼノキ(Rhus succedanea)、カシューナットノキ(Anacardium occidentale)、ウルシ(Toxicodendron vernicifluum)、マンゴー(Mangifera indica)、ピスタチオ(Pistacia vera)など。
ウリ科:カボチャ(Cucurbita maxima、Cucurbita moschata、Cucurbita pepo)、キュウリ(Cucumis sativus)、カラスウリ(Trichosanthes cucumeroides)、ヒョウタン(Lagenaria siceraria var. gourda)など。
バラ科:アーモンド(Amygdalus communis)、バラ(Rosa)、イチゴ(Fragaria)、サクラ(Prunus)、リンゴ(Malus pumila var. domestica)など。
ナデシコ科:カーネーション(Dianthus caryophyllus)など。
ヤナギ科:ポプラ(Populus trichocarpa、Populus nigra、Populus tremula) など。
イネ科:トウモロコシ(Zea mays)、イネ(Oryza sativa)、オオムギ(Hordeum vulgare)、コムギ(Triticum aestivum)、タケ(Phyllostachys)、サトウキビ(Saccharum officinarum)、ネピアグラス(Pennisetum pupureum)、エリアンサス(Erianthus ravenae)、ミスキャンタス(ススキ)(Miscanthus virgatum)、ソルガム(Sorghum)スイッチグラス(Panicum)など。
ユリ科:チューリップ(Tulipa)、ユリ(Lilium)など。
上述した形質転換処理後、植物体のなかから適切な形質転換体を選抜する選抜工程を、従来公知の方法で行うことができる。選抜の方法は特に限定されるものではなく、例えば、ハイグロマイシン耐性等の薬剤耐性を基準として選抜してもよいし、形質転換体を育成した後に、植物体そのもの、または任意の器官や組織の重量を測定して野生型と比較して有意に増産しているものを選抜してもよい。
1.材料および方法
1−1.実験材料
実験材料にはシロイヌナズナ変異体(Activation-tag T-DNA lines: Weigel T-DNS lines、計20072系統)の種子を用いた。なお、種子はNottingham Arabidopsis Stock Centre(NASC) より購入した。実験材料として使用した種子についてはWeigel, D. et al., 2000, Plant Physiol. 122, 1003-1013を参考とすることができる。
1−2−1.塩耐性変異体の選抜
Weigel T-DNA linesの種子を、NaCl 125 mMあるいは150 mMを含む改変MS寒天(1%)培地〔B5培地のビタミン、ショ糖10 g/l、寒天(細菌培地用;和光純薬工業社製)8 g/L〕に無菌播種し、22℃、30〜100μmol/m2/secの照明下(16時間明期/8時間暗期のサイクル)で培養した。播種後2〜4週間後、塩耐性変異体候補を選抜した。なお、MS培地はMurashige, T. et al., 1962, Physiol. Plant. 15, 473-497を参照する。また、B5培地についてはGamborg, O.L. et al. , 1968, Experimental Cell Research 50, 151-158を参照する。
選抜した耐塩性シロイヌナズナ系統のゲノムへのT-DNA挿入部位を、TAIL-PCR法により決定した。まず、栽培したシロイヌナズナから幼葉を採取し、液体窒素凍結下で粉砕した。QIAGEN社製DNA調製キット( DNeasy Plant Mini Kit)を用いてキット添付の標準プロトコルに従ってDNAを調製した。
Weigel T-DNA linesで用いられているアクティベーションタギング用ベクター(pSKI015 : GenBank accession No.AF187951)の左のT-DNA配列(T-DNA left border)付近に3種類の特異的プライマーTL1、TL2及びTL3を設定し、任意プライマーP1を用いて、以下のPCR反応液及び反応条件下でTAIL-PCR(島本功、佐々木卓治監修, 新版, 植物のPCR実験プロトコール, 2000, 83-89pp, 秀潤社, 東京 ; Liu, Y.G. and Whttier, R.F., 1995, Genomics 25, 674-681 ; Liu, Y.G. et al., Plant J., 8, 457-463, 1995)を行い、T-DNAに隣接するゲノムDNAを増幅した。
TL2: 5'-CGC TGC GGA CAT CTA CAT TTT TG-3'(配列番号25)
TL3: 5'-TCC CGG ACA TGA AGC CAT TTA C-3'(配列番号26)
P1: 5'-NGT CGA SWG ANA WGA A-3'(配列番号27)
1回目のPCR反応液組成及び反応条件をそれぞれ表1及び表2に示す。

アクティベートされている遺伝子を、1−2−3.により判明したT-DNA挿入部位(At3g05630)近傍10Kb以内に存在する推定Open reading frame(ORF)遺伝子の配列から予測した。
1−2−4.でアクティベート(活性化)されていると予測したPP2C(protein phosphatase 2C)遺伝子(At3g05640)のORF領域を含む断片を増幅するために、TAIR(http://www.arabidopsis.org/home.html)で公開されている配列情報を基にPCR用プライマー5640PF1及び5640PR1を設計・合成した。なおこれらプライマーの末端には、発現ベクターへ導入するときに必要となる制限酵素サイト(Bsr G IまたはSal I)を付加するように設計した。
5'-ACG CGT CGA CAT GGG ACA TTT CTC TTC CAT GTT CAA CGG-3'
5640PR1(配列番号30):
5'-TGT ACA TGT ACA CTA TAG AGA TGG CGA CGA CGA TGA AGA ATG G-3'
タバコモザイクウイルス由来のomega配列を含む植物発現用ベクターpBI121に、PP2C(protein phosphatase 2C)遺伝子(At3g05640)のORFを含む断片を挿入したコンストラクトの作製を行った。
1−2−6.で作製した植物発現用ベクターをエレクトロポレーション法(Plant Molecular Biology Mannal, Second Edition , B. G. Stanton and A. S. Robbert, Kluwer Acdemic Publishers 1994)により、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)C58C1株に導入した。次いで植物発現用ベクターが導入されたアグロバクテリウム・ツメファシエンスを、Cloughらにより記載された浸潤法(Steven J. Clough and Andrew F. Bent, 1998, The Plant Journal 16, 735-743)により、野生型シロイヌナズナ エコタイプCol-0に導入した。
1−2−7.で作製したT2種子を無菌播種し、バーミキュライト混合土を入れた直径50mmのポットに移植した。比較対象として非組換えシロイヌナズナを移植した。これを22℃、16時間明期8時間暗期、光強度約30〜45μmol m-2 s-1で栽培し移植後合計11週間栽培した。栽培後、地上部の植物体を紙袋に入れ、22℃、湿度60%の条件で2週間乾燥させた後に、全バイオマス量及び種子量を電子天秤で秤量した。
上記1−2−8.の結果として、野生型及びPP2C(protein phosphatase 2C)遺伝子(At3g05640)のORFを含む断片を導入した形質転換植物体の地上部を撮影した写真を図5に示す。また、結果として、地上部の全バイオマス量及び種子量を測定した結果を図6及び図7に示す。
本実施例1では、PP2C(protein phosphatase 2C)遺伝子(At3g05640)を導入した形質転換植物に対して、更にグルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼ遺伝子(以下、FBA1遺伝子)を導入した形質転換植物を作製した。
2−1.実験材料
実験材料には、1−2−7.で作製したPP2C(protein phosphatase 2C)遺伝子(At3g05640)のORFを含む断片を導入した形質転換植物体のT3世代以降の種子を用いた。野生型シロイヌナズナは、エコタイプCol-0を用いた。
2−2−1.FBA1遺伝子(At2g01140)の取得
4週齢のシロイヌナズナの野生型Columbia(Col-0)からトータルRNAを単離し、Prost arfirststrand RT-PCRキット(Stratagene社製)を用いてRT−PCR(テンプレートRNA量5.0μg)を行い、cDNAを作製した。
1F−1:5’-GGATCCTATGGCGTCTGCTAG-3’(配列番号44)
1R−1:5’-ATCTGCAACGGTCTCGGGAGA-3’(配列番号45)
1F−2:5’-GTGTGGTCCGAGGTGTTCTTCT-3’(配列番号46)
1R−2:5’-GAGCTCGAGTAGGTGTAACCCTTG-3’(配列番号47)
植物発現用ベクターpMAT137-HM(Matsuoka K. and Nakamura K., 1991, Proc. Natl. Acad. Sci. U S A 88, 834-838)に、2−2−1.で取得したFBA1遺伝子(At2g01140)を含む断片を挿入したコンストラクトの作製を行った。
2−2−2.で作製した植物発現用ベクターpMAT137-Hmをエレクトロポレーション法(Plant Molecular Biology Mannal, Second Edition , B. G. Stanton and A. S. Robbert, Kluwer Acdemic Publishers 1994)により、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)C58C1株に導入した。次いで、植物発現用ベクターが導入されたアグロバクテリウム・ツメファシエンスを、Cloughらにより記載された浸潤法(Steven J. Clough and Andrew F. Bent, 1998, The Plant Journal 16, 735-743)により、野生型シロイヌナズナエコタイプCol-0に導入した。
2−2−3.で作製したT2種子を播種し、正方形のプラスチックポット(6.5×6.5×5cm)の中に底からバーミキュライト(旭工業社製)、クレハ培養土(クレハ園芸培土、呉羽化学社製)、バーミキュライトを2:2:1の割合で3層に入れた土壌に播種し、生育温度22℃、長日(16−h明/8−h暗)条件で生育させた。対象には参考例1における1−2−7.で作製したPP2C遺伝子を導入した形質転換植物体を用いた。これを22℃、16時間明期8時間暗期、光強度約70〜100μmol m-2 s-1で栽培し移植後合計11週間栽培した。栽培後、地上部の植物体を紙袋に入れ、22℃、湿度60%の条件で2週間乾燥させた後に、全バイオマス量を電子天秤で秤量した。
上記2−2−4.の結果として、野生型、PP2C(protein phosphatase 2C)遺伝子(At3g05640)のORFを含む断片を導入した形質転換植物体及びPP2C(protein phosphatase 2C)遺伝子(At3g05640)のORFを含む断片を導入した形質転換植物体にFBA1遺伝子(At2g01140)を導入した植物体の地上部を撮影した写真を図8に示す。また、結果として、地上部の全バイオマス量を測定した結果を図9に示す。なお、図9に示したバイオマス量は、3個体を含むポット毎のバイオマス量を6個のポットについて測定し、その平均値を示している。
Claims (6)
- 以下の(a)〜(c)のいずれかのタンパク質からなるプロテインホスファターゼ2Cをコードする遺伝子及びグルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼをコードする遺伝子を過剰発現させた植物体。
(a)配列番号5及び53〜62からなる群から選ばれる1つのアミノ酸配列を含むタンパク質
(b)配列番号5及び53〜62からなる群から選ばれる1つのアミノ酸配列において1〜20個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、プロテインホスファターゼ2C活性を有するタンパク質
(c)配列番号4に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされプロテインホスファターゼ2C活性を有するタンパク質 - 上記グルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼは、以下の(a)〜(c)のいずれかのタンパク質からなることを特徴とする請求項1記載の植物体。
(a)配列番号32〜43からなる群から選ばれる1つのアミノ酸配列を含むタンパク質
(b)配列番号32〜43からなる群から選ばれる1つのアミノ酸配列において1〜20個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、グルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質
(c)配列番号31に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされグルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質 - 以下の(a)〜(c)のいずれかのタンパク質からなるプロテインホスファターゼ2Cをコードする遺伝子及びグルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼをコードする遺伝子を過剰発現させる、バイオマス量及び/又は種子量を増産させる方法。
(a)配列番号5及び53〜62からなる群から選ばれる1つのアミノ酸配列を含むタンパク質
(b)配列番号5及び53〜62からなる群から選ばれる1つのアミノ酸配列において1〜20個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、プロテインホスファターゼ2C活性を有するタンパク質
(c)配列番号4に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされプロテインホスファターゼ2C活性を有するタンパク質 - 上記グルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼは、以下の(a)〜(c)のいずれかのタンパク質からなることを特徴とする請求項3記載の方法。
(a)配列番号32〜43からなる群から選ばれる1つのアミノ酸配列を含むタンパク質
(b)配列番号32〜43からなる群から選ばれる1つのアミノ酸配列において1〜20個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、グルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質
(c)配列番号31に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされグルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質 - 以下の(a)〜(c)のいずれかのタンパク質からなるプロテインホスファターゼ2Cをコードする遺伝子及びグルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼをコードする遺伝子を過剰発現させた形質転換植物を準備する工程と、
前記形質転換植物の後代植物のバイオマス量及び/又は種子量を測定し、当該バイオマス量及び/又は種子量が有意に向上した系統を選抜する工程とを含む、植物体の製造方法。
(a)配列番号5及び53〜62からなる群から選ばれる1つのアミノ酸配列を含むタンパク質
(b)配列番号5及び53〜62からなる群から選ばれる1つのアミノ酸配列において1〜20個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、プロテインホスファターゼ2C活性を有するタンパク質
(c)配列番号4に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされプロテインホスファターゼ2C活性を有するタンパク質 - 上記グルタチオン結合性プラスチド型フルクトース1,6-ビスリン酸アルドラーゼは、以下の(a)〜(c)のいずれかのタンパク質からなることを特徴とする請求項5記載の製造方法。
(a)配列番号32〜43からなる群から選ばれる1つのアミノ酸配列を含むタンパク質
(b)配列番号32〜43からなる群から選ばれる1つのアミノ酸配列において1〜20個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、グルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質
(c)配列番号31に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされグルタチオンが結合することによりフルクトース1,6-ビスリン酸アルドラーゼ活性を示すタンパク質
Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2009060154A JP5604657B2 (ja) | 2009-03-12 | 2009-03-12 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
| US13/256,190 US9546376B2 (en) | 2009-03-12 | 2010-03-10 | Gene for increasing the production of plant biomass and/or seeds and method for use thereof |
| PCT/JP2010/053939 WO2010104092A1 (ja) | 2009-03-12 | 2010-03-10 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
| BRPI1009784-8A BRPI1009784A2 (pt) | 2009-03-12 | 2010-03-10 | Gene para aumentar a produção de biomassa vegetal e/ou sementes e método para aplicação das mesmas. |
| CN2010800113921A CN102395675A (zh) | 2009-03-12 | 2010-03-10 | 使植物的生物量和/或种子量增产的基因及其利用方法 |
| AU2010222106A AU2010222106B2 (en) | 2009-03-12 | 2010-03-10 | Gene for increasing the production of plant biomass and/or seeds and method for use thereof |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2009060154A JP5604657B2 (ja) | 2009-03-12 | 2009-03-12 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2010207199A JP2010207199A (ja) | 2010-09-24 |
| JP2010207199A5 JP2010207199A5 (ja) | 2012-01-12 |
| JP5604657B2 true JP5604657B2 (ja) | 2014-10-08 |
Family
ID=42728381
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2009060154A Expired - Fee Related JP5604657B2 (ja) | 2009-03-12 | 2009-03-12 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US9546376B2 (ja) |
| JP (1) | JP5604657B2 (ja) |
| CN (1) | CN102395675A (ja) |
| AU (1) | AU2010222106B2 (ja) |
| BR (1) | BRPI1009784A2 (ja) |
| WO (1) | WO2010104092A1 (ja) |
Families Citing this family (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2009113684A1 (ja) | 2008-03-14 | 2009-09-17 | トヨタ自動車株式会社 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
| JP5604657B2 (ja) | 2009-03-12 | 2014-10-08 | トヨタ自動車株式会社 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
| JP5787413B2 (ja) * | 2010-09-22 | 2015-09-30 | 国立大学法人 奈良先端科学技術大学院大学 | 塊茎生産能または匍匐枝形成能が野生株に比して向上している匍匐枝形成植物の作製方法、当該方法によって作製された匍匐枝形成植物 |
| US9238819B2 (en) * | 2011-05-04 | 2016-01-19 | Versitech Limited | Method for speeding up plant growth and improving yield by altering expression levels of kinases and phosphatases |
| JP6397610B2 (ja) | 2013-09-10 | 2018-09-26 | 株式会社豊田中央研究所 | 師部組織増産に適した植物体及びその利用 |
| CN115028696B (zh) * | 2021-02-20 | 2023-05-02 | 中国农业大学 | 一种与果实品质相关的蛋白及其编码基因的应用 |
| CN118703533B (zh) * | 2024-08-01 | 2025-11-21 | 湖南农业大学 | OsGAT3基因在调控水稻Cd、Cu或Mn含量及株高中的应用 |
Family Cites Families (24)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP1591522A3 (en) | 1993-10-06 | 2005-11-09 | New York University | Transgenic plants that exhibit enhanced nitrogen assimilation |
| WO1998010082A1 (en) | 1994-09-01 | 1998-03-12 | University Of Florida Research Foundation, Inc. | Materials and methods for increasing corn seed weight |
| US6252139B1 (en) | 1996-07-18 | 2001-06-26 | The Salk Institute For Biological Studies | Method of increasing growth and yield in plants |
| JP2001505410A (ja) | 1996-09-05 | 2001-04-24 | ユニバーシティー オブ フロリダ リサーチ ファウンデーション インク. | トウモロコシの種子重量を増加させるための材料および方法 |
| HUP0001660A3 (en) | 1997-03-26 | 2002-03-28 | Univ Cambridge Tech | Plants with modified growth |
| US7598361B2 (en) * | 1997-11-24 | 2009-10-06 | Monsanto Technology Llc | Nucleic acid molecules and other molecules associated with the sucrose pathway |
| US20100293663A2 (en) | 1998-06-16 | 2010-11-18 | Thomas La Rosa | Nucleic Acid Molecules and Other Molecules Associated with Plants and Uses Thereof for Plant Improvement |
| US20020040490A1 (en) | 2000-01-27 | 2002-04-04 | Jorn Gorlach | Expressed sequences of arabidopsis thaliana |
| AU8681101A (en) | 2000-08-24 | 2002-03-04 | Scripps Research Inst | Stress-regulated genes of plants, transgenic plants containing same, and methodsof use |
| AU2001289843A1 (en) * | 2001-08-28 | 2002-02-13 | Bayer Cropscience Ag | Polypeptides for identifying herbicidally active compounds |
| JP2005185101A (ja) | 2002-05-30 | 2005-07-14 | National Institute Of Agrobiological Sciences | 植物の全長cDNAおよびその利用 |
| US20070136839A1 (en) | 2003-10-14 | 2007-06-14 | Ceres, Inc. | Promoter, promoter control elements, and combinations, and uses thereof |
| JP2005130770A (ja) | 2003-10-30 | 2005-05-26 | Ajinomoto Co Inc | 植物体あたり得られるデンプン量が増加したバレイショおよびその作出法 |
| EP1732378B1 (en) | 2004-04-02 | 2013-01-16 | CropDesign N.V. | Plants overexpressing a NAP1-like protein with increased seed yield a method for making the same |
| EP2489726A3 (en) * | 2005-01-12 | 2012-11-28 | Monsanto Technology LLC | Genes and uses for plant improvement |
| ES2373408T3 (es) * | 2006-02-09 | 2012-02-03 | Japan Science And Technology Agency | Planta que tiene una mejor capacidad de crecimiento y de resistencia a las enfermedades y método para la producción de la misma. |
| CA2644273A1 (en) * | 2006-04-05 | 2008-03-27 | Metanomics Gmbh | Process for the production of a fine chemical |
| ES2513143T3 (es) | 2006-12-11 | 2014-10-24 | Japan Science And Technology Agency | Regulador del crecimiento vegetal y sus usos |
| US7951789B2 (en) | 2006-12-28 | 2011-05-31 | Idenix Pharmaceuticals, Inc. | Compounds and pharmaceutical compositions for the treatment of viral infections |
| MX2009007556A (es) | 2007-01-16 | 2009-08-12 | Japan Science & Tech Agency | Planta con rendimiento incrementado de semillas. |
| WO2009113684A1 (ja) * | 2008-03-14 | 2009-09-17 | トヨタ自動車株式会社 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
| JP5604657B2 (ja) | 2009-03-12 | 2014-10-08 | トヨタ自動車株式会社 | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 |
| JP5519192B2 (ja) | 2009-06-04 | 2014-06-11 | トヨタ自動車株式会社 | 種子のタンパク質含量を増産させる遺伝子及びその利用方法 |
| JP5250807B2 (ja) | 2009-09-11 | 2013-07-31 | トヨタ自動車株式会社 | 植物のバイオマス量及び/又は種子量を増産させる方法、バイオマス量及び/又は種子量を増産できる植物の製造方法 |
-
2009
- 2009-03-12 JP JP2009060154A patent/JP5604657B2/ja not_active Expired - Fee Related
-
2010
- 2010-03-10 WO PCT/JP2010/053939 patent/WO2010104092A1/ja not_active Ceased
- 2010-03-10 US US13/256,190 patent/US9546376B2/en not_active Expired - Fee Related
- 2010-03-10 CN CN2010800113921A patent/CN102395675A/zh active Pending
- 2010-03-10 AU AU2010222106A patent/AU2010222106B2/en not_active Ceased
- 2010-03-10 BR BRPI1009784-8A patent/BRPI1009784A2/pt not_active Application Discontinuation
Also Published As
| Publication number | Publication date |
|---|---|
| WO2010104092A1 (ja) | 2010-09-16 |
| CN102395675A (zh) | 2012-03-28 |
| AU2010222106B2 (en) | 2013-08-29 |
| AU2010222106A1 (en) | 2011-11-03 |
| US9546376B2 (en) | 2017-01-17 |
| US20120005787A1 (en) | 2012-01-05 |
| JP2010207199A (ja) | 2010-09-24 |
| BRPI1009784A2 (pt) | 2015-08-25 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5403628B2 (ja) | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 | |
| JP5250807B2 (ja) | 植物のバイオマス量及び/又は種子量を増産させる方法、バイオマス量及び/又は種子量を増産できる植物の製造方法 | |
| JP5454086B2 (ja) | 植物に環境ストレス耐性を付与する遺伝子及びその利用方法 | |
| JP5604657B2 (ja) | 植物のバイオマス量及び/又は種子量を増産させる遺伝子及びその利用方法 | |
| JP5672004B2 (ja) | 植物のバイオマス量を増産させる遺伝子及びその利用方法 | |
| JP5212955B2 (ja) | 植物のバイオマス量を増産させる遺伝子及びその利用方法 | |
| JP5686977B2 (ja) | 植物の油脂生産性を増大させる遺伝子及びその利用方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20111117 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20111117 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20131029 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20131227 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20140708 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20140729 |
|
| R151 | Written notification of patent or utility model registration |
Ref document number: 5604657 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R151 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| LAPS | Cancellation because of no payment of annual fees |