JP4059404B2 - 甲状腺機能を刺激する活性を持つ抗体 - Google Patents
甲状腺機能を刺激する活性を持つ抗体 Download PDFInfo
- Publication number
- JP4059404B2 JP4059404B2 JP2006257126A JP2006257126A JP4059404B2 JP 4059404 B2 JP4059404 B2 JP 4059404B2 JP 2006257126 A JP2006257126 A JP 2006257126A JP 2006257126 A JP2006257126 A JP 2006257126A JP 4059404 B2 JP4059404 B2 JP 4059404B2
- Authority
- JP
- Japan
- Prior art keywords
- seq
- variable region
- amino acid
- binds
- acid sequence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
Images


Landscapes
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Peptides Or Proteins (AREA)
- Investigating Or Analysing Biological Materials (AREA)
Description
アルギニン R or Arg
アスパラギン N or Asn
アスパラギン酸 D or Asp
システイン C or Cys
グルタミン Q or Gln
グルタミン酸 E or Glu
グリシン G or Gly
ヒスチジン H or His
イソロイシン I or Ile
ロイシン L or Leu
リジン K or Lys
メチオニン M or Met
フェニルアラニン F or Phe
プロリン P or Pro
セリン S or Ser
トレオニン T or Thr
トリプトファン W or Trp
チロシン Y or Tyr
バリン V or Val
CDR1;31−35番アミノ酸/148−162番塩基
CDR2;50−65番アミノ酸/205−252番塩基
CDR3;98−112番アミノ酸/349−393番塩基
rB6B7軽鎖(アミノ酸配列:配列18、塩基配列:配列2)
CDR1;24−34番アミノ酸/130−162番塩基
CDR2;50−56番アミノ酸/208−228番塩基
CDR3;89−96番アミノ酸/325−345番塩基
CDR1;31−35番アミノ酸/148−162番塩基
CDR2;50−66番アミノ酸/205−255番塩基
CDR3;99−106番アミノ酸/352−375番塩基
r101−2軽鎖(アミノ酸配列:配列20、塩基配列:配列4)
CDR1;24−35番アミノ酸/130−165番塩基
CDR2;51−57番アミノ酸/201−231番塩基
CDR3;90−98番アミノ酸/328−354番塩基
本発明のイムノグロブリン可変領域は、実際の患者組織に由来するものであるので病原性に深く関与する抗体に近い反応性を備えているものと推測できる。このような抗体は、競合法やインヒビションアッセイによる免疫分析に有用である。臨床的に分析意義の大きいと思われる病原性に関連する抗体に反応性が近いので、これらの抗体を高い精度で分析することが可能となるためである。
本発明のイムノグロブリン可変領域は、遺伝子操作によって高度に安定な品質を実現するので、標準物質として有用である。利用分野1に説明した免疫分析において、正確な定量を行うためには安定した標準物質が不可欠である。本発明はこのような要求を満足する。
本発明のTSABは、甲状腺機能低下症状を改善するために用いることができる。ヒトのリンパ球に由来する本発明のイムノグロブリン可変領域は、ヒトに投与した場合でも異物として認識されにくいため、免疫システムで排除されることなく確実にTRへ到達できる。
本発明のTSABは、TRに対してTSHと同じ甲状腺刺激活性を持っている。したがってTRにおけるシグナル伝達のモデル系の構成に有用である。TSH分子と、本発明によるTSABの分子構造を比較する事により、シグナル伝達に必要な構造の解析が期待できる。
本発明のTSABは、患者の体内に存在する病原性の自己抗体と同様の反応性を持っているものと推定される。したがって、生体外(in vitro)におけるTSAB阻害剤のスクリーニングに有用である。TR発現細胞を利用してTSAB活性を決定する技術は知られているので、スクリーニングすべき候補化合物と本発明のTSABの存在下でTR発現細胞を培養し、TSABのみの存在下で培養したものと比較すれば、TSAB阻害活性を指標とするスクリーニングを容易に行う事が可能である。
本発明によって得られるTSABの可変領域は、抗イディオタイプ抗体を得るための免疫原として有用である。抗体の可変領域を認識する抗体は、免疫原として作用させた抗体のリガンドの立体構造を模倣していると言われている。こうして得られた抗体に対する抗体は、抗イディオタイプ抗体と呼ばれている。本発明においては抗イディオタイプ抗体の可変領域はTSABのリガンドであるTRの構造を模倣するものと考えられるから、様々な利用用途が考えられる。まずTSABの検出のための抗原として有用である。このような用途に用いるときには、抗イディオタイプ抗体を生体外でしか利用しないため、その由来はヒトに限定されない。
あるいは抗イディオタイプ抗体のCDRのみをヒト型の抗体のCDRと置換した、いわゆるヒト化抗体を用いれば、更に安全性の高い治療薬とすることができる。生体内に導入されたTSABの抗イディオタイプ抗体は、血液中のTSABを中和するばかりでなくTSABを産生する抗体産生細胞の表面に露出したTSABの可変領域に結合することによって細胞障害作用を発揮し、結果として病原性のTSAB産生を抑制する。
バセドウ病患者の末梢血リンパ球から樹立したTSAB産生B6B7株および101−2株から、PCR法によって抗体の可変領域をコードする遺伝子ライブラリーを調製した。B6B7はIgGタイプの抗体を、101−2はIgMタイプの抗体を産生するクローンで、いずれの抗体もTSAB活性を備えている。
Cγ:5'- GAGAGAGAGAGAGAGAGCGCCTGAATTCCACGACACCGTCACCG-3'
Cμ:5'-AACGGCCACGCTGCTCGTATC-3'
Cκ:5'- GAGAGAGAGAGAGAGAGAATTCTGTAGGTGCTGTCCTTGCTGTCCTG-3'
アダプター:5'-CGATAAGCTTGGATCCTCGAG-3'
アダプター:5'-CTCGAGGATCCAAG-3'
Jκプライマー:5'-GGCGAATTCTTACGTTTGATCTCCACCTTGGTCCC-3'
3’配列決定用プライマー:5'-TCAGAATGGAATGTGCAG-3'
再構築したイムノグロブリン全分子をコードするDNAを含むベクターpSRIG−neo(20μg)でマウス・ミエローマP3X63−Ag8.653を形質転換し、10%のウシ胎児血清および1mg/mLのG418(GIBCO−BRL製)を含むRPMI1640培地で培養した。G418耐性細胞をサブクローニングしIgGの産生を指標としてスクリーニングして、高い濃度でIgGを分泌するものについてTRAB活性を測定した。
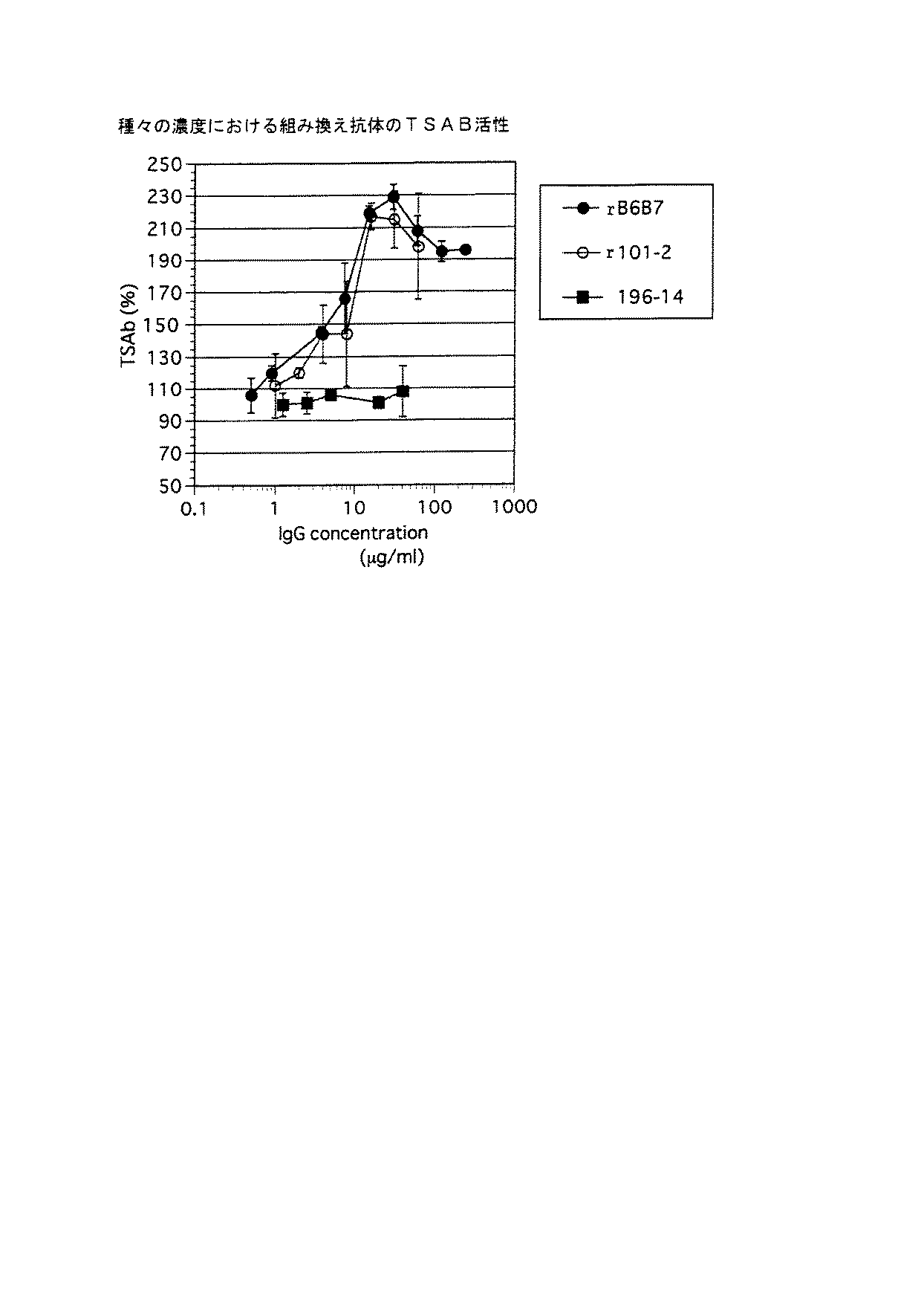
2.で得たTRに結合する抗体TRAB(rB6B7およびr101−2)を精製し、更にその甲状腺刺激活性を分析(TSAB検定)した。イムノグロブリン発現ベクターで形質転換した細胞を無血清培地S−CloneSF−B(三光純薬工業製、商品名)で培養し上清を集めた。150mLの培養上清を2mLのプロテインAセファロース4Fast Flow(ファルマシア製)で精製し、ポリエチレングリコールで濃縮した。これを改変ハンクス培地に対して透析し、得られた精製抗体についてTSAB活性、TBII活性を分析した。
Claims (6)
- 次の工程1)-4)で構成されるサイロトロピン受容体に結合する物質を免疫学的に検出する方法であって、サイロトロピン受容体と結合する標識抗体として甲状腺機能を刺激する活性を持つ可変領域が配列番号17に示すアミノ酸配列の重鎖および配列番号18に示すアミノ酸配列の軽鎖の両方を具備するrB6B7、若しくは可変領域が配列番号19に示すアミノ酸配列の重鎖および配列番号20に示すアミノ酸配列の軽鎖の両方を具備するr101−2を用いる検出方法
1)サイロトロピン受容体抗原を用意する工程
2)サイロトロピン受容体と結合する物質を検出すべき試料を1)の抗原と接触させる工程
3)更にサイロトロピン受容体と結合する標識抗体を接触させる工程
4)抗原と結合した標識抗体、または結合しなかった標識抗体に由来する標識を測定し、サイロトロピン受容体と結合する物質を検出する工程 - 次の工程1)-3)で構成されるサイロトロピン受容体に結合する物質を免疫学的に検出する方法であって、サイロトロピン受容体と結合する標識抗体として甲状腺機能を刺激する活性を持つ可変領域が配列番号17に示すアミノ酸配列の重鎖および配列番号18に示すアミノ酸配列の軽鎖の両方を具備するrB6B7、若しくは可変領域が配列番号19に示すアミノ酸配列の重鎖および配列番号20に示すアミノ酸配列の軽鎖の両方を具備するr101−2を用いる検出方法
1)サイロトロピン受容体抗原を用意する工程
2) サイロトロピン受容体に結合する物質を検出すべき試料と、サイロトロピン受容体と結合する標識抗体を同時に1)の抗原と接触させる工程
3)抗原と結合した標識抗体、または結合しなかった標識抗体に由来する標識を測定し、サイロトロピン受容体に結合する物質を検出する工程 - 前記サイロトロピン受容体と結合する物質成分が、サイロトロピン受容体に結合して甲状腺機能を刺激する活性を持つ自己抗体またはサイロトロピン受容体を刺激する自己抗体の拮抗物質である、請求項1または2記載の方法
- 標識抗体として、配列番号1に示す塩基配列によってコードされるアミノ酸配列からなるポリペプチドを含むイムノグロブリン重鎖可変領域および配列番号2に示す塩基配列によってコードされるアミノ酸配列からなるポリペプチドを含むイムノグロブリン軽鎖可変領域の両方を具備するrB6B7、若しくは配列番号3に示す塩基配列によってコードされるアミノ酸配列からなるポリペプチドを含むイムノグロブリン重鎖可変領域および配列番号4に示す塩基配列によってコードされるアミノ酸配列からなるポリペプチドを含むイムノグロブリン軽鎖可変領域の両方を具備するr101−2を用いる請求項1〜3記載の検出方法
- 重鎖および軽鎖の両方を具備する標識抗体のイムノグロブリン重鎖可変領域の相補性決定領域(CDR1、CDR2、CDR3)および軽鎖可変領域の相補性決定領域(CDR1、CDR2、CDR3)が、
CDR1(配列番号5)、CDR2(配列番号6)、CDR3(配列番号7)からなる配列番号17に示すrB6B7重鎖可変領域
および
CDR1(配列番号8)、CDR2(配列番号9)、CDR3(配列番号10)からなる配列番号18に示すrB6B7軽鎖可変領域
若しくは
CDR1(配列番号11)、CDR2(配列番号12)、CDR3(配列番号13)からなる配列番号19に示すr101−2重鎖可変領域
および
CDR1(配列番号14)、CDR2(配列番号15)、CDR3(配列番号16)からなる配列番号20に示すr101−2軽鎖可変領域
である請求項1〜4記載の検出方法 - 請求項1〜5記載のサイロトロピン受容体に結合する物質を免疫学的に検出する方法の標準物質であって、サイロトロピン受容体と結合する標識抗体として甲状腺機能を刺激する活性を持つ、配列番号1および配列番号2、若しくは配列番号3および配列番号4に示す塩基配列を発現させることによって得られる重鎖および軽鎖の両方を具備する組み換え体を含む標準物質
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2006257126A JP4059404B2 (ja) | 2006-09-22 | 2006-09-22 | 甲状腺機能を刺激する活性を持つ抗体 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2006257126A JP4059404B2 (ja) | 2006-09-22 | 2006-09-22 | 甲状腺機能を刺激する活性を持つ抗体 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP7328235A Division JPH09140386A (ja) | 1995-11-22 | 1995-11-22 | 甲状腺機能を刺激する活性を持つ抗体 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2007075115A JP2007075115A (ja) | 2007-03-29 |
| JP4059404B2 true JP4059404B2 (ja) | 2008-03-12 |
Family
ID=37936031
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2006257126A Expired - Lifetime JP4059404B2 (ja) | 2006-09-22 | 2006-09-22 | 甲状腺機能を刺激する活性を持つ抗体 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP4059404B2 (ja) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US10131709B2 (en) | 2011-12-28 | 2018-11-20 | Immunoqure Ag | Nucleic acid molecules encoding monoclonal antibodies specific for IL-22 |
| WO2013098419A1 (en) * | 2011-12-28 | 2013-07-04 | Immunoqure Ag | Method of providing monoclonal auto-antibodies with desired specificity |
| CN111647624A (zh) * | 2020-06-10 | 2020-09-11 | 成都和同易创生物科技有限公司 | Tsab人源单克隆抗体重组载体、重组抗体及其制备方法 |
-
2006
- 2006-09-22 JP JP2006257126A patent/JP4059404B2/ja not_active Expired - Lifetime
Also Published As
| Publication number | Publication date |
|---|---|
| JP2007075115A (ja) | 2007-03-29 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| RU2765306C2 (ru) | Антитело против в7-н3, его антигенсвязывающий фрагмент и их медицинское применение | |
| DK2279412T3 (en) | PRESENT UNKNOWN COMPOSITIONS AND PROCEDURES FOR TREATING IMMUNRATED DISEASES | |
| KR101720394B1 (ko) | 친화성 복합체에 대한 항체 | |
| TWI801862B (zh) | 抗tigit的抗體、其製備方法和應用 | |
| CN114478753A (zh) | 非免疫原性单结构域抗体 | |
| CN112020511A (zh) | 用于兽医用途的白介素-31单克隆抗体 | |
| JP2009278998A (ja) | チロトロフィン(tsh)受容体の抗原決定領域、その使用およびその抗体 | |
| JP2009278998A5 (ja) | ||
| CN110914304A (zh) | Cd96抗体、其抗原结合片段及医药用途 | |
| EP1414861A2 (en) | Humanized antibodies against icam-1, their production and uses | |
| AU2002363027A1 (en) | Humanized antibodies against ICAM-1, their production and uses | |
| WO1997002290A1 (fr) | ANTICORPS DE LIGAND ANTIFas ET PROCEDE D'EPREUVE LE FAISANT INTERVENIR | |
| JP4374316B2 (ja) | β−アミロイドまたはその誘導体に対する抗体およびその用途 | |
| JP4984160B2 (ja) | 抗体の作製方法 | |
| JP4059404B2 (ja) | 甲状腺機能を刺激する活性を持つ抗体 | |
| CN115298216A (zh) | 抗体或其抗原结合片段、其制备方法及医药用途 | |
| EP2727937A1 (en) | Soluble integrin 4 mutant | |
| KR20210108972A (ko) | 항페리오스틴 항체 및 이의 용도 | |
| CN113912716B (zh) | 针对α-突触核蛋白抗原的抗体及其应用 | |
| CA2291798A1 (en) | Monoclonal antibody against human telomerase catalytic subunit | |
| CN115947854A (zh) | 抗人cd40蛋白单克隆抗体、制备方法及其应用 | |
| WO2020239014A1 (zh) | 抗cgrp抗体及其应用 | |
| CN111892657A (zh) | 用于检测米田堡血型抗原的抗体及其片段、试剂盒及方法 | |
| JPH09140386A (ja) | 甲状腺機能を刺激する活性を持つ抗体 | |
| JPH0216997A (ja) | γ−ANPを認識するモノクローナル抗体 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20070305 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070426 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20070615 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070719 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20070816 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20070910 |
|
| A911 | Transfer of reconsideration by examiner before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20071023 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20071114 |
|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20071120 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20071212 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20071213 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| S531 | Written request for registration of change of domicile |
Free format text: JAPANESE INTERMEDIATE CODE: R313531 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| R360 | Written notification for declining of transfer of rights |
Free format text: JAPANESE INTERMEDIATE CODE: R360 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| R370 | Written measure of declining of transfer procedure |
Free format text: JAPANESE INTERMEDIATE CODE: R370 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| S531 | Written request for registration of change of domicile |
Free format text: JAPANESE INTERMEDIATE CODE: R313531 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101228 Year of fee payment: 3 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20111228 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121228 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121228 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121228 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121228 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131228 Year of fee payment: 6 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |