JP2022526297A - タウ凝集に関連する遺伝的脆弱性を明らかにするためのcrispr/casドロップアウトスクリーニングプラットフォーム - Google Patents
タウ凝集に関連する遺伝的脆弱性を明らかにするためのcrispr/casドロップアウトスクリーニングプラットフォーム Download PDFInfo
- Publication number
- JP2022526297A JP2022526297A JP2021556467A JP2021556467A JP2022526297A JP 2022526297 A JP2022526297 A JP 2022526297A JP 2021556467 A JP2021556467 A JP 2021556467A JP 2021556467 A JP2021556467 A JP 2021556467A JP 2022526297 A JP2022526297 A JP 2022526297A
- Authority
- JP
- Japan
- Prior art keywords
- protein
- cell population
- tau
- repeat domain
- reporter
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
- 108091033409 CRISPR Proteins 0.000 title claims abstract description 158
- 230000002776 aggregation Effects 0.000 title claims abstract description 112
- 238000004220 aggregation Methods 0.000 title claims abstract description 111
- 230000002068 genetic effect Effects 0.000 title claims abstract description 40
- 238000012216 screening Methods 0.000 title claims description 70
- 238000010354 CRISPR gene editing Methods 0.000 title abstract description 44
- 238000000034 method Methods 0.000 claims abstract description 215
- 230000004913 activation Effects 0.000 claims abstract description 30
- 210000004027 cell Anatomy 0.000 claims description 696
- 108090000623 proteins and genes Proteins 0.000 claims description 678
- 108020005004 Guide RNA Proteins 0.000 claims description 419
- 102000004169 proteins and genes Human genes 0.000 claims description 357
- 102000013498 tau Proteins Human genes 0.000 claims description 352
- 108010026424 tau Proteins Proteins 0.000 claims description 352
- 230000008685 targeting Effects 0.000 claims description 105
- 150000007523 nucleic acids Chemical class 0.000 claims description 104
- 102000039446 nucleic acids Human genes 0.000 claims description 98
- 108020004707 nucleic acids Proteins 0.000 claims description 98
- 102000035181 adaptor proteins Human genes 0.000 claims description 93
- 108091005764 adaptor proteins Proteins 0.000 claims description 93
- 230000029279 positive regulation of transcription, DNA-dependent Effects 0.000 claims description 86
- 230000035772 mutation Effects 0.000 claims description 71
- 231100000225 lethality Toxicity 0.000 claims description 57
- 230000027455 binding Effects 0.000 claims description 54
- 238000002866 fluorescence resonance energy transfer Methods 0.000 claims description 48
- 108091032973 (ribonucleotides)n+m Proteins 0.000 claims description 42
- 238000002474 experimental method Methods 0.000 claims description 42
- 108010082025 cyan fluorescent protein Proteins 0.000 claims description 40
- 108091005957 yellow fluorescent proteins Proteins 0.000 claims description 40
- 230000014509 gene expression Effects 0.000 claims description 35
- 108091079001 CRISPR RNA Proteins 0.000 claims description 32
- 108091026890 Coding region Proteins 0.000 claims description 27
- 239000003550 marker Substances 0.000 claims description 26
- 230000035897 transcription Effects 0.000 claims description 26
- 238000013518 transcription Methods 0.000 claims description 26
- 241000282414 Homo sapiens Species 0.000 claims description 25
- 238000010361 transduction Methods 0.000 claims description 20
- 230000026683 transduction Effects 0.000 claims description 20
- 210000005260 human cell Anatomy 0.000 claims description 19
- 102100032606 Heat shock factor protein 1 Human genes 0.000 claims description 17
- 101000867525 Homo sapiens Heat shock factor protein 1 Proteins 0.000 claims description 17
- 108091006047 fluorescent proteins Proteins 0.000 claims description 17
- 102000034287 fluorescent proteins Human genes 0.000 claims description 17
- RXWNCPJZOCPEPQ-NVWDDTSBSA-N puromycin Chemical compound C1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CO RXWNCPJZOCPEPQ-NVWDDTSBSA-N 0.000 claims description 16
- 238000000338 in vitro Methods 0.000 claims description 15
- 108700009124 Transcription Initiation Site Proteins 0.000 claims description 14
- 210000004962 mammalian cell Anatomy 0.000 claims description 14
- 238000011144 upstream manufacturing Methods 0.000 claims description 14
- 241000193996 Streptococcus pyogenes Species 0.000 claims description 13
- 108010084455 Zeocin Proteins 0.000 claims description 12
- CWCMIVBLVUHDHK-ZSNHEYEWSA-N phleomycin D1 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC[C@@H](N=1)C=1SC=C(N=1)C(=O)NCCCCNC(N)=N)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C CWCMIVBLVUHDHK-ZSNHEYEWSA-N 0.000 claims description 12
- 108010077850 Nuclear Localization Signals Proteins 0.000 claims description 11
- 208000015181 infectious disease Diseases 0.000 claims description 11
- 210000003527 eukaryotic cell Anatomy 0.000 claims description 10
- 230000003612 virological effect Effects 0.000 claims description 9
- 101001091269 Escherichia coli Hygromycin-B 4-O-kinase Proteins 0.000 claims description 8
- 108010025815 Kanamycin Kinase Proteins 0.000 claims description 8
- 101001091268 Streptomyces hygroscopicus Hygromycin-B 7''-O-kinase Proteins 0.000 claims description 8
- 229940079593 drug Drugs 0.000 claims description 8
- 239000003814 drug Substances 0.000 claims description 8
- 229950010131 puromycin Drugs 0.000 claims description 8
- 108010045647 puromycin N-acetyltransferase Proteins 0.000 claims description 8
- 239000013603 viral vector Substances 0.000 claims description 8
- 108010045123 Blasticidin-S deaminase Proteins 0.000 claims description 7
- 108091006106 transcriptional activators Proteins 0.000 claims description 7
- 210000004899 c-terminal region Anatomy 0.000 claims description 6
- 230000001965 increasing effect Effects 0.000 claims description 5
- 102000007999 Nuclear Proteins Human genes 0.000 claims description 4
- 108010089610 Nuclear Proteins Proteins 0.000 claims description 4
- 239000001963 growth medium Substances 0.000 claims description 4
- 230000030648 nucleus localization Effects 0.000 claims description 4
- 230000015271 coagulation Effects 0.000 claims description 3
- 238000005345 coagulation Methods 0.000 claims description 3
- 238000010586 diagram Methods 0.000 abstract description 9
- 230000002195 synergetic effect Effects 0.000 abstract description 5
- 101710196377 Protein enabled Proteins 0.000 abstract description 2
- 235000018102 proteins Nutrition 0.000 description 317
- 125000003729 nucleotide group Chemical group 0.000 description 78
- 239000002773 nucleotide Substances 0.000 description 77
- 108091027544 Subgenomic mRNA Proteins 0.000 description 68
- 230000000295 complement effect Effects 0.000 description 50
- 238000003776 cleavage reaction Methods 0.000 description 30
- 230000004048 modification Effects 0.000 description 30
- 238000012986 modification Methods 0.000 description 30
- 230000007017 scission Effects 0.000 description 30
- 238000006467 substitution reaction Methods 0.000 description 30
- 235000001014 amino acid Nutrition 0.000 description 23
- 108090000765 processed proteins & peptides Proteins 0.000 description 23
- 101710163270 Nuclease Proteins 0.000 description 21
- 150000001413 amino acids Chemical class 0.000 description 21
- 229940024606 amino acid Drugs 0.000 description 20
- 201000010099 disease Diseases 0.000 description 19
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 19
- 230000001665 lethal effect Effects 0.000 description 18
- 229920001184 polypeptide Polymers 0.000 description 18
- 102000004196 processed proteins & peptides Human genes 0.000 description 18
- 239000013598 vector Substances 0.000 description 18
- 238000013459 approach Methods 0.000 description 17
- 238000012360 testing method Methods 0.000 description 17
- 108091028043 Nucleic acid sequence Proteins 0.000 description 15
- 230000030833 cell death Effects 0.000 description 15
- 231100000518 lethal Toxicity 0.000 description 15
- 108700024394 Exon Proteins 0.000 description 14
- 108091005461 Nucleic proteins Proteins 0.000 description 14
- 108020004999 messenger RNA Proteins 0.000 description 14
- 101710124574 Synaptotagmin-1 Proteins 0.000 description 13
- 102100035100 Transcription factor p65 Human genes 0.000 description 13
- 239000012634 fragment Substances 0.000 description 13
- 102000040430 polynucleotide Human genes 0.000 description 13
- 108091033319 polynucleotide Proteins 0.000 description 13
- 239000002157 polynucleotide Substances 0.000 description 13
- 230000005782 double-strand break Effects 0.000 description 12
- 230000000694 effects Effects 0.000 description 12
- 108020001507 fusion proteins Proteins 0.000 description 12
- 102000037865 fusion proteins Human genes 0.000 description 12
- 230000001404 mediated effect Effects 0.000 description 12
- 238000007481 next generation sequencing Methods 0.000 description 12
- 210000001519 tissue Anatomy 0.000 description 12
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 11
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 11
- 108700039887 Essential Genes Proteins 0.000 description 11
- 108700008625 Reporter Genes Proteins 0.000 description 11
- 101000910035 Streptococcus pyogenes serotype M1 CRISPR-associated endonuclease Cas9/Csn1 Proteins 0.000 description 11
- 125000003275 alpha amino acid group Chemical group 0.000 description 11
- 238000004458 analytical method Methods 0.000 description 11
- 230000003197 catalytic effect Effects 0.000 description 11
- 238000001514 detection method Methods 0.000 description 11
- 230000037361 pathway Effects 0.000 description 11
- 230000008569 process Effects 0.000 description 11
- 241000699666 Mus <mouse, genus> Species 0.000 description 10
- 241000700159 Rattus Species 0.000 description 10
- 230000008859 change Effects 0.000 description 10
- 230000006870 function Effects 0.000 description 10
- 230000001717 pathogenic effect Effects 0.000 description 10
- 238000012546 transfer Methods 0.000 description 10
- 102000053602 DNA Human genes 0.000 description 9
- 101710115937 Microtubule-associated protein tau Proteins 0.000 description 9
- 102100040243 Microtubule-associated protein tau Human genes 0.000 description 9
- 239000000306 component Substances 0.000 description 9
- -1 eBFP2 Proteins 0.000 description 9
- 239000000203 mixture Substances 0.000 description 9
- 230000001105 regulatory effect Effects 0.000 description 9
- 108091023037 Aptamer Proteins 0.000 description 8
- 108020004414 DNA Proteins 0.000 description 8
- 241000713666 Lentivirus Species 0.000 description 8
- 230000003176 fibrotic effect Effects 0.000 description 8
- 230000000415 inactivating effect Effects 0.000 description 8
- 230000003834 intracellular effect Effects 0.000 description 8
- 230000004770 neurodegeneration Effects 0.000 description 8
- 208000015122 neurodegenerative disease Diseases 0.000 description 8
- 230000006780 non-homologous end joining Effects 0.000 description 8
- 239000002245 particle Substances 0.000 description 8
- 239000000047 product Substances 0.000 description 8
- 108700028369 Alleles Proteins 0.000 description 7
- 108010043121 Green Fluorescent Proteins Proteins 0.000 description 7
- 102000004144 Green Fluorescent Proteins Human genes 0.000 description 7
- 241000283984 Rodentia Species 0.000 description 7
- 230000004849 abnormal protein aggregation Effects 0.000 description 7
- 238000001727 in vivo Methods 0.000 description 7
- 210000004940 nucleus Anatomy 0.000 description 7
- 108010054624 red fluorescent protein Proteins 0.000 description 7
- 230000008439 repair process Effects 0.000 description 7
- 238000001890 transfection Methods 0.000 description 7
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 6
- 208000024827 Alzheimer disease Diseases 0.000 description 6
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 6
- 101000891579 Homo sapiens Microtubule-associated protein tau Proteins 0.000 description 6
- 108700005090 Lethal Genes Proteins 0.000 description 6
- 108060001084 Luciferase Proteins 0.000 description 6
- 108700019146 Transgenes Proteins 0.000 description 6
- 235000004279 alanine Nutrition 0.000 description 6
- 230000015572 biosynthetic process Effects 0.000 description 6
- 230000003833 cell viability Effects 0.000 description 6
- 238000000684 flow cytometry Methods 0.000 description 6
- 108010021843 fluorescent protein 583 Proteins 0.000 description 6
- 238000002744 homologous recombination Methods 0.000 description 6
- 230000006801 homologous recombination Effects 0.000 description 6
- 102000057063 human MAPT Human genes 0.000 description 6
- 230000001939 inductive effect Effects 0.000 description 6
- 238000003780 insertion Methods 0.000 description 6
- 230000037431 insertion Effects 0.000 description 6
- 230000010354 integration Effects 0.000 description 6
- 230000007246 mechanism Effects 0.000 description 6
- 108010006654 Bleomycin Proteins 0.000 description 5
- 102220605874 Cytosolic arginine sensor for mTORC1 subunit 2_D10A_mutation Human genes 0.000 description 5
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 5
- 206010029350 Neurotoxicity Diseases 0.000 description 5
- 108091034117 Oligonucleotide Proteins 0.000 description 5
- 206010044221 Toxic encephalopathy Diseases 0.000 description 5
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical class O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 5
- 229960001561 bleomycin Drugs 0.000 description 5
- OYVAGSVQBOHSSS-UAPAGMARSA-O bleomycin A2 Chemical compound N([C@H](C(=O)N[C@H](C)[C@@H](O)[C@H](C)C(=O)N[C@@H]([C@H](O)C)C(=O)NCCC=1SC=C(N=1)C=1SC=C(N=1)C(=O)NCCC[S+](C)C)[C@@H](O[C@H]1[C@H]([C@@H](O)[C@H](O)[C@H](CO)O1)O[C@@H]1[C@H]([C@@H](OC(N)=O)[C@H](O)[C@@H](CO)O1)O)C=1N=CNC=1)C(=O)C1=NC([C@H](CC(N)=O)NC[C@H](N)C(N)=O)=NC(N)=C1C OYVAGSVQBOHSSS-UAPAGMARSA-O 0.000 description 5
- 108091005948 blue fluorescent proteins Proteins 0.000 description 5
- 210000000349 chromosome Anatomy 0.000 description 5
- 210000000805 cytoplasm Anatomy 0.000 description 5
- 239000005090 green fluorescent protein Substances 0.000 description 5
- 230000006872 improvement Effects 0.000 description 5
- 238000000520 microinjection Methods 0.000 description 5
- 239000003607 modifier Substances 0.000 description 5
- 210000002569 neuron Anatomy 0.000 description 5
- 230000007135 neurotoxicity Effects 0.000 description 5
- 231100000228 neurotoxicity Toxicity 0.000 description 5
- 238000010606 normalization Methods 0.000 description 5
- HMFHBZSHGGEWLO-UHFFFAOYSA-N pentofuranose Chemical group OCC1OC(O)C(O)C1O HMFHBZSHGGEWLO-UHFFFAOYSA-N 0.000 description 5
- 229920002401 polyacrylamide Polymers 0.000 description 5
- 230000002829 reductive effect Effects 0.000 description 5
- 230000005783 single-strand break Effects 0.000 description 5
- 241000894007 species Species 0.000 description 5
- KDCGOANMDULRCW-UHFFFAOYSA-N 7H-purine Chemical compound N1=CNC2=NC=NC2=C1 KDCGOANMDULRCW-UHFFFAOYSA-N 0.000 description 4
- 208000004051 Chronic Traumatic Encephalopathy Diseases 0.000 description 4
- 108020004705 Codon Proteins 0.000 description 4
- 230000004568 DNA-binding Effects 0.000 description 4
- 108010053770 Deoxyribonucleases Proteins 0.000 description 4
- 102000016911 Deoxyribonucleases Human genes 0.000 description 4
- 108091005941 EBFP Proteins 0.000 description 4
- 102000004190 Enzymes Human genes 0.000 description 4
- 108090000790 Enzymes Proteins 0.000 description 4
- 206010016654 Fibrosis Diseases 0.000 description 4
- 101000860092 Francisella tularensis subsp. novicida (strain U112) CRISPR-associated endonuclease Cas12a Proteins 0.000 description 4
- UYTPUPDQBNUYGX-UHFFFAOYSA-N Guanine Natural products O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 4
- 239000005089 Luciferase Substances 0.000 description 4
- 101710125418 Major capsid protein Proteins 0.000 description 4
- 102000029749 Microtubule Human genes 0.000 description 4
- 108091022875 Microtubule Proteins 0.000 description 4
- 208000018737 Parkinson disease Diseases 0.000 description 4
- 241000194017 Streptococcus Species 0.000 description 4
- 208000034799 Tauopathies Diseases 0.000 description 4
- DRTQHJPVMGBUCF-XVFCMESISA-N Uridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-XVFCMESISA-N 0.000 description 4
- 125000000539 amino acid group Chemical group 0.000 description 4
- 206010002026 amyotrophic lateral sclerosis Diseases 0.000 description 4
- 230000005540 biological transmission Effects 0.000 description 4
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 4
- 238000006243 chemical reaction Methods 0.000 description 4
- 238000012217 deletion Methods 0.000 description 4
- 230000037430 deletion Effects 0.000 description 4
- 208000017004 dementia pugilistica Diseases 0.000 description 4
- 230000034431 double-strand break repair via homologous recombination Effects 0.000 description 4
- 238000004520 electroporation Methods 0.000 description 4
- 210000002257 embryonic structure Anatomy 0.000 description 4
- 108010045262 enhanced cyan fluorescent protein Proteins 0.000 description 4
- 230000005284 excitation Effects 0.000 description 4
- 230000004761 fibrosis Effects 0.000 description 4
- 230000037433 frameshift Effects 0.000 description 4
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 4
- 150000002632 lipids Chemical class 0.000 description 4
- 238000001638 lipofection Methods 0.000 description 4
- 210000001161 mammalian embryo Anatomy 0.000 description 4
- 210000004688 microtubule Anatomy 0.000 description 4
- 210000003470 mitochondria Anatomy 0.000 description 4
- 230000009437 off-target effect Effects 0.000 description 4
- 230000036961 partial effect Effects 0.000 description 4
- 229920000642 polymer Polymers 0.000 description 4
- 230000002062 proliferating effect Effects 0.000 description 4
- 230000010076 replication Effects 0.000 description 4
- 238000013207 serial dilution Methods 0.000 description 4
- 210000001082 somatic cell Anatomy 0.000 description 4
- 230000000638 stimulation Effects 0.000 description 4
- 239000000126 substance Substances 0.000 description 4
- 238000010200 validation analysis Methods 0.000 description 4
- 238000012795 verification Methods 0.000 description 4
- YMHOBZXQZVXHBM-UHFFFAOYSA-N 2,5-dimethoxy-4-bromophenethylamine Chemical compound COC1=CC(CCN)=C(OC)C=C1Br YMHOBZXQZVXHBM-UHFFFAOYSA-N 0.000 description 3
- 101000860090 Acidaminococcus sp. (strain BV3L6) CRISPR-associated endonuclease Cas12a Proteins 0.000 description 3
- 239000004475 Arginine Substances 0.000 description 3
- 101150018129 CSF2 gene Proteins 0.000 description 3
- 101710132601 Capsid protein Proteins 0.000 description 3
- 101710094648 Coat protein Proteins 0.000 description 3
- MYMOFIZGZYHOMD-UHFFFAOYSA-N Dioxygen Chemical compound O=O MYMOFIZGZYHOMD-UHFFFAOYSA-N 0.000 description 3
- 201000011240 Frontotemporal dementia Diseases 0.000 description 3
- 239000004471 Glycine Substances 0.000 description 3
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 3
- 241000282412 Homo Species 0.000 description 3
- 101000834898 Homo sapiens Alpha-synuclein Proteins 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 239000004472 Lysine Substances 0.000 description 3
- 241001465754 Metazoa Species 0.000 description 3
- 101710141454 Nucleoprotein Proteins 0.000 description 3
- 229910019142 PO4 Inorganic materials 0.000 description 3
- 241001494479 Pecora Species 0.000 description 3
- 101710083689 Probable capsid protein Proteins 0.000 description 3
- 230000004570 RNA-binding Effects 0.000 description 3
- 241000203644 Streptoalloteichus hindustanus Species 0.000 description 3
- 101000910045 Streptococcus thermophilus (strain ATCC BAA-491 / LMD-9) CRISPR-associated endonuclease Cas9 2 Proteins 0.000 description 3
- 102000040945 Transcription factor Human genes 0.000 description 3
- 108091023040 Transcription factor Proteins 0.000 description 3
- 241000545067 Venus Species 0.000 description 3
- 239000012190 activator Substances 0.000 description 3
- 230000004520 agglutination Effects 0.000 description 3
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 3
- 101150038738 ble gene Proteins 0.000 description 3
- 108010083912 bleomycin N-acetyltransferase Proteins 0.000 description 3
- 238000007385 chemical modification Methods 0.000 description 3
- 239000003153 chemical reaction reagent Substances 0.000 description 3
- 102000021178 chitin binding proteins Human genes 0.000 description 3
- 108091011157 chitin binding proteins Proteins 0.000 description 3
- 238000012258 culturing Methods 0.000 description 3
- 238000009826 distribution Methods 0.000 description 3
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 3
- 231100000221 frame shift mutation induction Toxicity 0.000 description 3
- 238000009396 hybridization Methods 0.000 description 3
- 230000002209 hydrophobic effect Effects 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 239000002502 liposome Substances 0.000 description 3
- 235000018977 lysine Nutrition 0.000 description 3
- 239000011159 matrix material Substances 0.000 description 3
- 239000000178 monomer Substances 0.000 description 3
- 230000016273 neuron death Effects 0.000 description 3
- 238000010899 nucleation Methods 0.000 description 3
- 210000000056 organ Anatomy 0.000 description 3
- 229910052760 oxygen Inorganic materials 0.000 description 3
- 239000001301 oxygen Substances 0.000 description 3
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 3
- 239000010452 phosphate Substances 0.000 description 3
- 239000002243 precursor Substances 0.000 description 3
- 230000006798 recombination Effects 0.000 description 3
- 238000005215 recombination Methods 0.000 description 3
- 239000000758 substrate Substances 0.000 description 3
- 238000003786 synthesis reaction Methods 0.000 description 3
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical group [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 3
- 230000002103 transcriptional effect Effects 0.000 description 3
- 108091006107 transcriptional repressors Proteins 0.000 description 3
- 241001515965 unidentified phage Species 0.000 description 3
- 229940035893 uracil Drugs 0.000 description 3
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 2
- LRFVTYWOQMYALW-UHFFFAOYSA-N 9H-xanthine Chemical compound O=C1NC(=O)NC2=C1NC=N2 LRFVTYWOQMYALW-UHFFFAOYSA-N 0.000 description 2
- 241000093740 Acidaminococcus sp. Species 0.000 description 2
- 102100026882 Alpha-synuclein Human genes 0.000 description 2
- 241000272517 Anseriformes Species 0.000 description 2
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 2
- 108091005950 Azurite Proteins 0.000 description 2
- 102100026189 Beta-galactosidase Human genes 0.000 description 2
- 101710201279 Biotin carboxyl carrier protein Proteins 0.000 description 2
- 241000283690 Bos taurus Species 0.000 description 2
- 241000589875 Campylobacter jejuni Species 0.000 description 2
- 241000700198 Cavia Species 0.000 description 2
- 241000579895 Chlorostilbon Species 0.000 description 2
- 108091005960 Citrine Proteins 0.000 description 2
- 241000699800 Cricetinae Species 0.000 description 2
- 101150074775 Csf1 gene Proteins 0.000 description 2
- 102000005636 Cyclic AMP Response Element-Binding Protein Human genes 0.000 description 2
- 108010045171 Cyclic AMP Response Element-Binding Protein Proteins 0.000 description 2
- 238000007399 DNA isolation Methods 0.000 description 2
- 230000033616 DNA repair Effects 0.000 description 2
- 206010061818 Disease progression Diseases 0.000 description 2
- 241000282326 Felis catus Species 0.000 description 2
- 241000589599 Francisella tularensis subsp. novicida Species 0.000 description 2
- 241000588088 Francisella tularensis subsp. novicida U112 Species 0.000 description 2
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 2
- 102000053187 Glucuronidase Human genes 0.000 description 2
- 108010060309 Glucuronidase Proteins 0.000 description 2
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 2
- 108010070675 Glutathione transferase Proteins 0.000 description 2
- 102100029100 Hematopoietic prostaglandin D synthase Human genes 0.000 description 2
- 108010068250 Herpes Simplex Virus Protein Vmw65 Proteins 0.000 description 2
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 2
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 2
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 2
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 2
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- 241000689670 Lachnospiraceae bacterium ND2006 Species 0.000 description 2
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 2
- 101150070547 MAPT gene Proteins 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 102100025169 Max-binding protein MNT Human genes 0.000 description 2
- 241001193016 Moraxella bovoculi 237 Species 0.000 description 2
- 102000007474 Multiprotein Complexes Human genes 0.000 description 2
- 108010085220 Multiprotein Complexes Proteins 0.000 description 2
- 241000699670 Mus sp. Species 0.000 description 2
- 241000588650 Neisseria meningitidis Species 0.000 description 2
- 208000009869 Neu-Laxova syndrome Diseases 0.000 description 2
- WSDRAZIPGVLSNP-UHFFFAOYSA-N O.P(=O)(O)(O)O.O.O.P(=O)(O)(O)O Chemical compound O.P(=O)(O)(O)O.O.O.P(=O)(O)(O)O WSDRAZIPGVLSNP-UHFFFAOYSA-N 0.000 description 2
- 238000012408 PCR amplification Methods 0.000 description 2
- 108010029485 Protein Isoforms Proteins 0.000 description 2
- 102000001708 Protein Isoforms Human genes 0.000 description 2
- 108091034057 RNA (poly(A)) Proteins 0.000 description 2
- 238000003559 RNA-seq method Methods 0.000 description 2
- PYMYPHUHKUWMLA-LMVFSUKVSA-N Ribose Natural products OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PYMYPHUHKUWMLA-LMVFSUKVSA-N 0.000 description 2
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 2
- 241000700584 Simplexvirus Species 0.000 description 2
- 101100166144 Staphylococcus aureus cas9 gene Proteins 0.000 description 2
- 241000194020 Streptococcus thermophilus Species 0.000 description 2
- 241000187191 Streptomyces viridochromogenes Species 0.000 description 2
- 241000203587 Streptosporangium roseum Species 0.000 description 2
- 241000282898 Sus scrofa Species 0.000 description 2
- 102000002933 Thioredoxin Human genes 0.000 description 2
- 108091028113 Trans-activating crRNA Proteins 0.000 description 2
- 241000186064 Trueperella pyogenes Species 0.000 description 2
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 2
- 230000002159 abnormal effect Effects 0.000 description 2
- 230000003213 activating effect Effects 0.000 description 2
- 238000007792 addition Methods 0.000 description 2
- 210000004504 adult stem cell Anatomy 0.000 description 2
- 230000004075 alteration Effects 0.000 description 2
- 235000009582 asparagine Nutrition 0.000 description 2
- 229960001230 asparagine Drugs 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 108010005774 beta-Galactosidase Proteins 0.000 description 2
- DRTQHJPVMGBUCF-PSQAKQOGSA-N beta-L-uridine Natural products O[C@H]1[C@@H](O)[C@H](CO)O[C@@H]1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-PSQAKQOGSA-N 0.000 description 2
- 210000004556 brain Anatomy 0.000 description 2
- 230000030570 cellular localization Effects 0.000 description 2
- 210000003169 central nervous system Anatomy 0.000 description 2
- 238000012512 characterization method Methods 0.000 description 2
- 229930002875 chlorophyll Natural products 0.000 description 2
- 235000019804 chlorophyll Nutrition 0.000 description 2
- 239000011035 citrine Substances 0.000 description 2
- 210000000078 claw Anatomy 0.000 description 2
- 230000003013 cytotoxicity Effects 0.000 description 2
- 231100000135 cytotoxicity Toxicity 0.000 description 2
- 238000007405 data analysis Methods 0.000 description 2
- 230000000779 depleting effect Effects 0.000 description 2
- 230000001627 detrimental effect Effects 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- 238000007847 digital PCR Methods 0.000 description 2
- 230000005750 disease progression Effects 0.000 description 2
- 239000010976 emerald Substances 0.000 description 2
- 229910052876 emerald Inorganic materials 0.000 description 2
- 239000003623 enhancer Substances 0.000 description 2
- 230000007613 environmental effect Effects 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 230000001605 fetal effect Effects 0.000 description 2
- 210000002950 fibroblast Anatomy 0.000 description 2
- 238000001917 fluorescence detection Methods 0.000 description 2
- 238000010448 genetic screening Methods 0.000 description 2
- 235000013922 glutamic acid Nutrition 0.000 description 2
- 239000004220 glutamic acid Substances 0.000 description 2
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 2
- 235000004554 glutamine Nutrition 0.000 description 2
- 230000005847 immunogenicity Effects 0.000 description 2
- 230000001976 improved effect Effects 0.000 description 2
- 238000002347 injection Methods 0.000 description 2
- 239000007924 injection Substances 0.000 description 2
- 230000003993 interaction Effects 0.000 description 2
- 229960000310 isoleucine Drugs 0.000 description 2
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 2
- 238000012417 linear regression Methods 0.000 description 2
- 244000144972 livestock Species 0.000 description 2
- 238000004519 manufacturing process Methods 0.000 description 2
- 239000000693 micelle Substances 0.000 description 2
- 239000004005 microsphere Substances 0.000 description 2
- 239000002105 nanoparticle Substances 0.000 description 2
- 210000002682 neurofibrillary tangle Anatomy 0.000 description 2
- 230000007170 pathology Effects 0.000 description 2
- 230000002085 persistent effect Effects 0.000 description 2
- 125000002467 phosphate group Chemical group [H]OP(=O)(O[H])O[*] 0.000 description 2
- 239000013612 plasmid Substances 0.000 description 2
- 229920000747 poly(lactic acid) Polymers 0.000 description 2
- 230000008488 polyadenylation Effects 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 230000035755 proliferation Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 230000003252 repetitive effect Effects 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 229910052594 sapphire Inorganic materials 0.000 description 2
- 239000010980 sapphire Substances 0.000 description 2
- 230000019491 signal transduction Effects 0.000 description 2
- 230000009870 specific binding Effects 0.000 description 2
- 230000009897 systematic effect Effects 0.000 description 2
- 238000010381 tandem affinity purification Methods 0.000 description 2
- 230000001225 therapeutic effect Effects 0.000 description 2
- 108060008226 thioredoxin Proteins 0.000 description 2
- 229940094937 thioredoxin Drugs 0.000 description 2
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 2
- 238000003151 transfection method Methods 0.000 description 2
- 230000001052 transient effect Effects 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- GWBUNZLLLLDXMD-UHFFFAOYSA-H tricopper;dicarbonate;dihydroxide Chemical compound [OH-].[OH-].[Cu+2].[Cu+2].[Cu+2].[O-]C([O-])=O.[O-]C([O-])=O GWBUNZLLLLDXMD-UHFFFAOYSA-H 0.000 description 2
- 241000701161 unidentified adenovirus Species 0.000 description 2
- 241001430294 unidentified retrovirus Species 0.000 description 2
- DRTQHJPVMGBUCF-UHFFFAOYSA-N uracil arabinoside Natural products OC1C(O)C(CO)OC1N1C(=O)NC(=O)C=C1 DRTQHJPVMGBUCF-UHFFFAOYSA-N 0.000 description 2
- 229940045145 uridine Drugs 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- UVBYMVOUBXYSFV-XUTVFYLZSA-N 1-methylpseudouridine Chemical compound O=C1NC(=O)N(C)C=C1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 UVBYMVOUBXYSFV-XUTVFYLZSA-N 0.000 description 1
- UVBYMVOUBXYSFV-UHFFFAOYSA-N 1-methylpseudouridine Natural products O=C1NC(=O)N(C)C=C1C1C(O)C(O)C(CO)O1 UVBYMVOUBXYSFV-UHFFFAOYSA-N 0.000 description 1
- 102100027831 14-3-3 protein theta Human genes 0.000 description 1
- KQPKMEYBZUPZGK-UHFFFAOYSA-N 4-[(4-azido-2-nitroanilino)methyl]-5-(hydroxymethyl)-2-methylpyridin-3-ol Chemical compound CC1=NC=C(CO)C(CNC=2C(=CC(=CC=2)N=[N+]=[N-])[N+]([O-])=O)=C1O KQPKMEYBZUPZGK-UHFFFAOYSA-N 0.000 description 1
- ZAYHVCMSTBRABG-UHFFFAOYSA-N 5-Methylcytidine Natural products O=C1N=C(N)C(C)=CN1C1C(O)C(O)C(CO)O1 ZAYHVCMSTBRABG-UHFFFAOYSA-N 0.000 description 1
- ZAYHVCMSTBRABG-JXOAFFINSA-N 5-methylcytidine Chemical compound O=C1N=C(N)C(C)=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 ZAYHVCMSTBRABG-JXOAFFINSA-N 0.000 description 1
- 241000007910 Acaryochloris marina Species 0.000 description 1
- 241001135192 Acetohalobium arabaticum Species 0.000 description 1
- 241001464929 Acidithiobacillus caldus Species 0.000 description 1
- 241000605222 Acidithiobacillus ferrooxidans Species 0.000 description 1
- 101710159080 Aconitate hydratase A Proteins 0.000 description 1
- 101710159078 Aconitate hydratase B Proteins 0.000 description 1
- 241000251468 Actinopterygii Species 0.000 description 1
- 241000059559 Agriotes sordidus Species 0.000 description 1
- 241000640374 Alicyclobacillus acidocaldarius Species 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 241000190857 Allochromatium vinosum Species 0.000 description 1
- 241000147155 Ammonifex degensii Species 0.000 description 1
- 241000192542 Anabaena Species 0.000 description 1
- 241000620196 Arthrospira maxima Species 0.000 description 1
- 240000002900 Arthrospira platensis Species 0.000 description 1
- 235000016425 Arthrospira platensis Nutrition 0.000 description 1
- 241001495183 Arthrospira sp. Species 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 241000906059 Bacillus pseudomycoides Species 0.000 description 1
- 241000157302 Bison bison athabascae Species 0.000 description 1
- 241000823281 Burkholderiales bacterium Species 0.000 description 1
- 101150069031 CSN2 gene Proteins 0.000 description 1
- 102000000584 Calmodulin Human genes 0.000 description 1
- 108010041952 Calmodulin Proteins 0.000 description 1
- 241001496650 Candidatus Desulforudis Species 0.000 description 1
- 241001040999 Candidatus Methanoplasma termitum Species 0.000 description 1
- 241000223282 Candidatus Peregrinibacteria Species 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 241000283707 Capra Species 0.000 description 1
- 102100028892 Cardiotrophin-1 Human genes 0.000 description 1
- 102100025064 Cellular tumor antigen p53 Human genes 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 108091005944 Cerulean Proteins 0.000 description 1
- 241000282994 Cervidae Species 0.000 description 1
- 108010035563 Chloramphenicol O-acetyltransferase Proteins 0.000 description 1
- 108020004638 Circular DNA Proteins 0.000 description 1
- 241000193163 Clostridioides difficile Species 0.000 description 1
- 241000193155 Clostridium botulinum Species 0.000 description 1
- 241000699802 Cricetulus griseus Species 0.000 description 1
- 241000065719 Crocosphaera Species 0.000 description 1
- 108091005943 CyPet Proteins 0.000 description 1
- XFXPMWWXUTWYJX-UHFFFAOYSA-N Cyanide Chemical compound N#[C-] XFXPMWWXUTWYJX-UHFFFAOYSA-N 0.000 description 1
- 241000159506 Cyanothece Species 0.000 description 1
- 206010011732 Cyst Diseases 0.000 description 1
- 102220605872 Cytosolic arginine sensor for mTORC1 subunit 2_D16A_mutation Human genes 0.000 description 1
- 102220605836 Cytosolic arginine sensor for mTORC1 subunit 2_E1369R_mutation Human genes 0.000 description 1
- 102220605919 Cytosolic arginine sensor for mTORC1 subunit 2_E1449H_mutation Human genes 0.000 description 1
- 102220605899 Cytosolic arginine sensor for mTORC1 subunit 2_R1556A_mutation Human genes 0.000 description 1
- HMFHBZSHGGEWLO-SOOFDHNKSA-N D-ribofuranose Chemical compound OC[C@H]1OC(O)[C@H](O)[C@@H]1O HMFHBZSHGGEWLO-SOOFDHNKSA-N 0.000 description 1
- 101710155335 DELLA protein SLR1 Proteins 0.000 description 1
- 108010009540 DNA (Cytosine-5-)-Methyltransferase 1 Proteins 0.000 description 1
- 102100036279 DNA (cytosine-5)-methyltransferase 1 Human genes 0.000 description 1
- 102100024810 DNA (cytosine-5)-methyltransferase 3B Human genes 0.000 description 1
- 101710123222 DNA (cytosine-5)-methyltransferase 3B Proteins 0.000 description 1
- 230000007018 DNA scission Effects 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 206010012289 Dementia Diseases 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- 101150068427 EP300 gene Proteins 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 241000326311 Exiguobacterium sibiricum Species 0.000 description 1
- 108060002716 Exonuclease Proteins 0.000 description 1
- 241000192016 Finegoldia magna Species 0.000 description 1
- 108090000331 Firefly luciferases Proteins 0.000 description 1
- 241000589601 Francisella Species 0.000 description 1
- 108700036482 Francisella novicida Cas9 Proteins 0.000 description 1
- 241000589602 Francisella tularensis Species 0.000 description 1
- 101150106478 GPS1 gene Proteins 0.000 description 1
- 241000287828 Gallus gallus Species 0.000 description 1
- KOSRFJWDECSPRO-WDSKDSINSA-N Glu-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(O)=O KOSRFJWDECSPRO-WDSKDSINSA-N 0.000 description 1
- 108060003760 HNH nuclease Proteins 0.000 description 1
- 102000029812 HNH nuclease Human genes 0.000 description 1
- 241000700721 Hepatitis B virus Species 0.000 description 1
- 108091027305 Heteroduplex Proteins 0.000 description 1
- 101710121996 Hexon protein p72 Proteins 0.000 description 1
- 102000008157 Histone Demethylases Human genes 0.000 description 1
- 108010074870 Histone Demethylases Proteins 0.000 description 1
- 102100022846 Histone acetyltransferase KAT2B Human genes 0.000 description 1
- 102100038885 Histone acetyltransferase p300 Human genes 0.000 description 1
- 241001272567 Hominoidea Species 0.000 description 1
- 101000916283 Homo sapiens Cardiotrophin-1 Proteins 0.000 description 1
- 101000721661 Homo sapiens Cellular tumor antigen p53 Proteins 0.000 description 1
- 101000744174 Homo sapiens DNA-3-methyladenine glycosylase Proteins 0.000 description 1
- 101001047006 Homo sapiens Histone acetyltransferase KAT2B Proteins 0.000 description 1
- 101000615488 Homo sapiens Methyl-CpG-binding domain protein 2 Proteins 0.000 description 1
- 101000615495 Homo sapiens Methyl-CpG-binding domain protein 3 Proteins 0.000 description 1
- 101001111984 Homo sapiens N-acylneuraminate-9-phosphatase Proteins 0.000 description 1
- 101000602926 Homo sapiens Nuclear receptor coactivator 1 Proteins 0.000 description 1
- 101000651467 Homo sapiens Proto-oncogene tyrosine-protein kinase Src Proteins 0.000 description 1
- 101000631695 Homo sapiens Succinate dehydrogenase assembly factor 3, mitochondrial Proteins 0.000 description 1
- 101000801209 Homo sapiens Transducin-like enhancer protein 4 Proteins 0.000 description 1
- 101000802101 Homo sapiens mRNA decay activator protein ZFP36L2 Proteins 0.000 description 1
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 1
- 208000026350 Inborn Genetic disease Diseases 0.000 description 1
- 108020004684 Internal Ribosome Entry Sites Proteins 0.000 description 1
- 241001430080 Ktedonobacter racemifer Species 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- 241001112693 Lachnospiraceae Species 0.000 description 1
- 241000448224 Lachnospiraceae bacterium MA2020 Species 0.000 description 1
- 241000186673 Lactobacillus delbrueckii Species 0.000 description 1
- 241000186869 Lactobacillus salivarius Species 0.000 description 1
- 241000254158 Lampyridae Species 0.000 description 1
- 108091026898 Leader sequence (mRNA) Proteins 0.000 description 1
- 241001148627 Leptospira inadai Species 0.000 description 1
- 241001134698 Lyngbya Species 0.000 description 1
- 101710175625 Maltose/maltodextrin-binding periplasmic protein Proteins 0.000 description 1
- 241000501784 Marinobacter sp. Species 0.000 description 1
- 102000006890 Methyl-CpG-Binding Protein 2 Human genes 0.000 description 1
- 108010072388 Methyl-CpG-Binding Protein 2 Proteins 0.000 description 1
- 102100021299 Methyl-CpG-binding domain protein 2 Human genes 0.000 description 1
- 102100021291 Methyl-CpG-binding domain protein 3 Human genes 0.000 description 1
- 108060004795 Methyltransferase Proteins 0.000 description 1
- 241000192710 Microcystis aeruginosa Species 0.000 description 1
- 241000190928 Microscilla marina Species 0.000 description 1
- 102000009664 Microtubule-Associated Proteins Human genes 0.000 description 1
- 108010020004 Microtubule-Associated Proteins Proteins 0.000 description 1
- 101100219625 Mus musculus Casd1 gene Proteins 0.000 description 1
- 102100023906 N-acylneuraminate-9-phosphatase Human genes 0.000 description 1
- 241000167285 Natranaerobius thermophilus Species 0.000 description 1
- 101100385413 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) csm-3 gene Proteins 0.000 description 1
- 241000192147 Nitrosococcus Species 0.000 description 1
- 241000919925 Nitrosococcus halophilus Species 0.000 description 1
- 241000203619 Nocardiopsis dassonvillei Species 0.000 description 1
- 108091007494 Nucleic acid- binding domains Proteins 0.000 description 1
- 108700026244 Open Reading Frames Proteins 0.000 description 1
- 241000192520 Oscillatoria sp. Species 0.000 description 1
- 102100035593 POU domain, class 2, transcription factor 1 Human genes 0.000 description 1
- 101710084414 POU domain, class 2, transcription factor 1 Proteins 0.000 description 1
- 241000142651 Pelotomaculum thermopropionicum Species 0.000 description 1
- 108010088535 Pep-1 peptide Proteins 0.000 description 1
- 101100440941 Petroselinum crispum CPRF1 gene Proteins 0.000 description 1
- 241001135648 Petrotoga Species 0.000 description 1
- 241000286209 Phasianidae Species 0.000 description 1
- 241001599925 Polaromonas naphthalenivorans Species 0.000 description 1
- 241001472610 Polaromonas sp. Species 0.000 description 1
- 229920002873 Polyethylenimine Polymers 0.000 description 1
- 241000878522 Porphyromonas crevioricanis Species 0.000 description 1
- 241000605861 Prevotella Species 0.000 description 1
- 241001302521 Prevotella albensis Species 0.000 description 1
- 241001135219 Prevotella disiens Species 0.000 description 1
- 102000055027 Protein Methyltransferases Human genes 0.000 description 1
- 108700040121 Protein Methyltransferases Proteins 0.000 description 1
- 101710149951 Protein Tat Proteins 0.000 description 1
- 102100027384 Proto-oncogene tyrosine-protein kinase Src Human genes 0.000 description 1
- 241000590028 Pseudoalteromonas haloplanktis Species 0.000 description 1
- 229930185560 Pseudouridine Natural products 0.000 description 1
- PTJWIQPHWPFNBW-UHFFFAOYSA-N Pseudouridine C Natural products OC1C(O)C(CO)OC1C1=CNC(=O)NC1=O PTJWIQPHWPFNBW-UHFFFAOYSA-N 0.000 description 1
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 1
- 102000009572 RNA Polymerase II Human genes 0.000 description 1
- 108010009460 RNA Polymerase II Proteins 0.000 description 1
- 102000014450 RNA Polymerase III Human genes 0.000 description 1
- 108010078067 RNA Polymerase III Proteins 0.000 description 1
- 230000026279 RNA modification Effects 0.000 description 1
- 230000006819 RNA synthesis Effects 0.000 description 1
- 102000044126 RNA-Binding Proteins Human genes 0.000 description 1
- 101710105008 RNA-binding protein Proteins 0.000 description 1
- 101150065817 ROM2 gene Proteins 0.000 description 1
- 238000011529 RT qPCR Methods 0.000 description 1
- 101100047461 Rattus norvegicus Trpm8 gene Proteins 0.000 description 1
- 241000242739 Renilla Species 0.000 description 1
- 235000001537 Ribes X gardonianum Nutrition 0.000 description 1
- 235000001535 Ribes X utile Nutrition 0.000 description 1
- 235000016919 Ribes petraeum Nutrition 0.000 description 1
- 244000281247 Ribes rubrum Species 0.000 description 1
- 235000002355 Ribes spicatum Nutrition 0.000 description 1
- 102000006382 Ribonucleases Human genes 0.000 description 1
- 108010083644 Ribonucleases Proteins 0.000 description 1
- 108091028664 Ribonucleotide Proteins 0.000 description 1
- 108020004422 Riboswitch Proteins 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- 108091027967 Small hairpin RNA Proteins 0.000 description 1
- 241000191967 Staphylococcus aureus Species 0.000 description 1
- 241000194022 Streptococcus sp. Species 0.000 description 1
- 241001633172 Streptococcus thermophilus LMD-9 Species 0.000 description 1
- 241001518258 Streptomyces pristinaespiralis Species 0.000 description 1
- 241000271567 Struthioniformes Species 0.000 description 1
- 102100028996 Succinate dehydrogenase assembly factor 3, mitochondrial Human genes 0.000 description 1
- 241000282887 Suidae Species 0.000 description 1
- 241000192560 Synechococcus sp. Species 0.000 description 1
- 210000001744 T-lymphocyte Anatomy 0.000 description 1
- 101710137500 T7 RNA polymerase Proteins 0.000 description 1
- 108700026226 TATA Box Proteins 0.000 description 1
- 101710192266 Tegument protein VP22 Proteins 0.000 description 1
- 241000206213 Thermosipho africanus Species 0.000 description 1
- 108091036066 Three prime untranslated region Proteins 0.000 description 1
- 102000006601 Thymidine Kinase Human genes 0.000 description 1
- 108020004440 Thymidine kinase Proteins 0.000 description 1
- 102000005747 Transcription Factor RelA Human genes 0.000 description 1
- 108010031154 Transcription Factor RelA Proteins 0.000 description 1
- 102100022972 Transcription factor AP-2-alpha Human genes 0.000 description 1
- 101710189834 Transcription factor AP-2-alpha Proteins 0.000 description 1
- 102100038313 Transcription factor E2-alpha Human genes 0.000 description 1
- 102100033763 Transducin-like enhancer protein 4 Human genes 0.000 description 1
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 1
- 102000004887 Transforming Growth Factor beta Human genes 0.000 description 1
- 108091023045 Untranslated Region Proteins 0.000 description 1
- 208000036142 Viral infection Diseases 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- 208000027418 Wounds and injury Diseases 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 241001673106 [Bacillus] selenitireducens Species 0.000 description 1
- 241001531273 [Eubacterium] eligens Species 0.000 description 1
- 238000010521 absorption reaction Methods 0.000 description 1
- 108020002494 acetyltransferase Proteins 0.000 description 1
- 102000005421 acetyltransferase Human genes 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 210000001789 adipocyte Anatomy 0.000 description 1
- 238000005054 agglomeration Methods 0.000 description 1
- 230000004931 aggregating effect Effects 0.000 description 1
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 1
- PPQRONHOSHZGFQ-LMVFSUKVSA-N aldehydo-D-ribose 5-phosphate Chemical compound OP(=O)(O)OC[C@@H](O)[C@@H](O)[C@@H](O)C=O PPQRONHOSHZGFQ-LMVFSUKVSA-N 0.000 description 1
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 1
- 125000003277 amino group Chemical group 0.000 description 1
- 210000004102 animal cell Anatomy 0.000 description 1
- 238000000137 annealing Methods 0.000 description 1
- 229940011019 arthrospira platensis Drugs 0.000 description 1
- 229940009098 aspartate Drugs 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 238000003556 assay Methods 0.000 description 1
- 230000003376 axonal effect Effects 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- WGDUUQDYDIIBKT-UHFFFAOYSA-N beta-Pseudouridine Natural products OC1OC(CN2C=CC(=O)NC2=O)C(O)C1O WGDUUQDYDIIBKT-UHFFFAOYSA-N 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 230000002457 bidirectional effect Effects 0.000 description 1
- 238000005415 bioluminescence Methods 0.000 description 1
- 230000029918 bioluminescence Effects 0.000 description 1
- 244000309464 bull Species 0.000 description 1
- 210000004900 c-terminal fragment Anatomy 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 239000000969 carrier Substances 0.000 description 1
- 101150055766 cat gene Proteins 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 108020001778 catalytic domains Proteins 0.000 description 1
- 229920006317 cationic polymer Polymers 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 210000004671 cell-free system Anatomy 0.000 description 1
- 230000033077 cellular process Effects 0.000 description 1
- 230000010094 cellular senescence Effects 0.000 description 1
- 239000003795 chemical substances by application Substances 0.000 description 1
- 235000013330 chicken meat Nutrition 0.000 description 1
- ATNHDLDRLWWWCB-AENOIHSZSA-M chlorophyll a Chemical compound C1([C@@H](C(=O)OC)C(=O)C2=C3C)=C2N2C3=CC(C(CC)=C3C)=[N+]4C3=CC3=C(C=C)C(C)=C5N3[Mg-2]42[N+]2=C1[C@@H](CCC(=O)OC\C=C(/C)CCC[C@H](C)CCC[C@H](C)CCCC(C)C)[C@H](C)C2=C5 ATNHDLDRLWWWCB-AENOIHSZSA-M 0.000 description 1
- 239000001752 chlorophylls and chlorophyllins Substances 0.000 description 1
- 239000002299 complementary DNA Substances 0.000 description 1
- 230000000536 complexating effect Effects 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 210000002808 connective tissue Anatomy 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 101150055601 cops2 gene Proteins 0.000 description 1
- 239000008358 core component Substances 0.000 description 1
- 238000012937 correction Methods 0.000 description 1
- 208000031513 cyst Diseases 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- 238000004163 cytometry Methods 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 239000007857 degradation product Substances 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 239000000412 dendrimer Substances 0.000 description 1
- 229920000736 dendritic polymer Polymers 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 238000006471 dimerization reaction Methods 0.000 description 1
- 230000009977 dual effect Effects 0.000 description 1
- 210000003981 ectoderm Anatomy 0.000 description 1
- 210000001671 embryonic stem cell Anatomy 0.000 description 1
- 210000001900 endoderm Anatomy 0.000 description 1
- 210000002889 endothelial cell Anatomy 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 230000004049 epigenetic modification Effects 0.000 description 1
- 210000002919 epithelial cell Anatomy 0.000 description 1
- 210000000981 epithelium Anatomy 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 102000013165 exonuclease Human genes 0.000 description 1
- 210000001808 exosome Anatomy 0.000 description 1
- 238000000799 fluorescence microscopy Methods 0.000 description 1
- 239000007850 fluorescent dye Substances 0.000 description 1
- 238000001215 fluorescent labelling Methods 0.000 description 1
- 229940118764 francisella tularensis Drugs 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 230000004927 fusion Effects 0.000 description 1
- 230000004545 gene duplication Effects 0.000 description 1
- 208000016361 genetic disease Diseases 0.000 description 1
- 210000004602 germ cell Anatomy 0.000 description 1
- 210000001654 germ layer Anatomy 0.000 description 1
- 108010055341 glutamyl-glutamic acid Proteins 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 210000003494 hepatocyte Anatomy 0.000 description 1
- 125000002887 hydroxy group Chemical group [H]O* 0.000 description 1
- 230000028993 immune response Effects 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 208000014674 injury Diseases 0.000 description 1
- 230000015788 innate immune response Effects 0.000 description 1
- 210000002510 keratinocyte Anatomy 0.000 description 1
- 210000003292 kidney cell Anatomy 0.000 description 1
- 101150066555 lacZ gene Proteins 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 238000011551 log transformation method Methods 0.000 description 1
- 238000004020 luminiscence type Methods 0.000 description 1
- 102100034703 mRNA decay activator protein ZFP36L2 Human genes 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 239000002609 medium Substances 0.000 description 1
- 210000003716 mesoderm Anatomy 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 108091070501 miRNA Proteins 0.000 description 1
- 239000002679 microRNA Substances 0.000 description 1
- 230000025608 mitochondrion localization Effects 0.000 description 1
- 108091005601 modified peptides Proteins 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 210000005087 mononuclear cell Anatomy 0.000 description 1
- 210000003205 muscle Anatomy 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- 231100000219 mutagenic Toxicity 0.000 description 1
- 230000003505 mutagenic effect Effects 0.000 description 1
- 210000004898 n-terminal fragment Anatomy 0.000 description 1
- 210000000653 nervous system Anatomy 0.000 description 1
- 230000001537 neural effect Effects 0.000 description 1
- 230000000626 neurodegenerative effect Effects 0.000 description 1
- 210000004498 neuroglial cell Anatomy 0.000 description 1
- 108091027963 non-coding RNA Proteins 0.000 description 1
- 102000042567 non-coding RNA Human genes 0.000 description 1
- 210000000633 nuclear envelope Anatomy 0.000 description 1
- 239000002777 nucleoside Substances 0.000 description 1
- 125000003835 nucleoside group Chemical group 0.000 description 1
- 210000003463 organelle Anatomy 0.000 description 1
- 210000001672 ovary Anatomy 0.000 description 1
- 238000004806 packaging method and process Methods 0.000 description 1
- 230000001575 pathological effect Effects 0.000 description 1
- 150000004713 phosphodiesters Chemical class 0.000 description 1
- 150000003013 phosphoric acid derivatives Chemical class 0.000 description 1
- 230000004962 physiological condition Effects 0.000 description 1
- 108010011110 polyarginine Proteins 0.000 description 1
- 230000003389 potentiating effect Effects 0.000 description 1
- 230000002028 premature Effects 0.000 description 1
- 230000001737 promoting effect Effects 0.000 description 1
- 230000004952 protein activity Effects 0.000 description 1
- 230000004845 protein aggregation Effects 0.000 description 1
- 230000004850 protein–protein interaction Effects 0.000 description 1
- PTJWIQPHWPFNBW-GBNDHIKLSA-N pseudouridine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1C1=CNC(=O)NC1=O PTJWIQPHWPFNBW-GBNDHIKLSA-N 0.000 description 1
- 238000003908 quality control method Methods 0.000 description 1
- 230000004044 response Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 230000001177 retroviral effect Effects 0.000 description 1
- 239000002336 ribonucleotide Substances 0.000 description 1
- 125000002652 ribonucleotide group Chemical group 0.000 description 1
- 150000003290 ribose derivatives Chemical class 0.000 description 1
- 230000003248 secreting effect Effects 0.000 description 1
- 230000001235 sensitizing effect Effects 0.000 description 1
- 230000001568 sexual effect Effects 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 210000004683 skeletal myoblast Anatomy 0.000 description 1
- 239000004055 small Interfering RNA Substances 0.000 description 1
- 210000000329 smooth muscle myocyte Anatomy 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 238000003153 stable transfection Methods 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 210000000130 stem cell Anatomy 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 230000002123 temporal effect Effects 0.000 description 1
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 description 1
- 229940113082 thymine Drugs 0.000 description 1
- 230000005026 transcription initiation Effects 0.000 description 1
- 230000002463 transducing effect Effects 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 238000003146 transient transfection Methods 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 230000005945 translocation Effects 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 241001529453 unidentified herpesvirus Species 0.000 description 1
- 230000009385 viral infection Effects 0.000 description 1
- 238000011179 visual inspection Methods 0.000 description 1
- 229940075420 xanthine Drugs 0.000 description 1
- 210000005253 yeast cell Anatomy 0.000 description 1
Images

































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0684—Cells of the urinary tract or kidneys
- C12N5/0686—Kidney cells
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1079—Screening libraries by altering the phenotype or phenotypic trait of the host
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1082—Preparation or screening gene libraries by chromosomal integration of polynucleotide sequences, HR-, site-specific-recombination, transposons, viral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1086—Preparation or screening of expression libraries, e.g. reporter assays
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/68—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving proteins, peptides or amino acids
- G01N33/6893—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving proteins, peptides or amino acids related to diseases not provided for elsewhere
- G01N33/6896—Neurological disorders, e.g. Alzheimer's disease
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2510/00—Genetically modified cells
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N21/00—Investigating or analysing materials by the use of optical means, i.e. using sub-millimetre waves, infrared, visible or ultraviolet light
- G01N21/62—Systems in which the material investigated is excited whereby it emits light or causes a change in wavelength of the incident light
- G01N21/63—Systems in which the material investigated is excited whereby it emits light or causes a change in wavelength of the incident light optically excited
- G01N21/64—Fluorescence; Phosphorescence
- G01N21/6428—Measuring fluorescence of fluorescent products of reactions or of fluorochrome labelled reactive substances, e.g. measuring quenching effects, using measuring "optrodes"
- G01N2021/6439—Measuring fluorescence of fluorescent products of reactions or of fluorochrome labelled reactive substances, e.g. measuring quenching effects, using measuring "optrodes" with indicators, stains, dyes, tags, labels, marks
- G01N2021/6441—Measuring fluorescence of fluorescent products of reactions or of fluorochrome labelled reactive substances, e.g. measuring quenching effects, using measuring "optrodes" with indicators, stains, dyes, tags, labels, marks with two or more labels
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N21/00—Investigating or analysing materials by the use of optical means, i.e. using sub-millimetre waves, infrared, visible or ultraviolet light
- G01N21/62—Systems in which the material investigated is excited whereby it emits light or causes a change in wavelength of the incident light
- G01N21/63—Systems in which the material investigated is excited whereby it emits light or causes a change in wavelength of the incident light optically excited
- G01N21/64—Fluorescence; Phosphorescence
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Biomedical Technology (AREA)
- Zoology (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Urology & Nephrology (AREA)
- Bioinformatics & Computational Biology (AREA)
- Crystallography & Structural Chemistry (AREA)
- Medicinal Chemistry (AREA)
- Immunology (AREA)
- Cell Biology (AREA)
- Hematology (AREA)
- Pathology (AREA)
- General Physics & Mathematics (AREA)
- Analytical Chemistry (AREA)
- Virology (AREA)
- Food Science & Technology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Neurosurgery (AREA)
- Neurology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
Description
この出願は、2019年3月18日に出願された米国出願第62/820,101号の利益を主張し、これはすべての目的のためにその全体が参照により本明細書に組み込まれる。
ファイル544640SEQLIST.txtに記述された配列表は、75.7キロバイトであり、2020年3月16日に作成され、参照により本明細書に組み込まれる。
本明細書で互換的に使用される、「タンパク質」、「ポリペプチド」、および「ペプチド」という用語は、コード化および非コード化アミノ酸ならびに化学的または生化学的に修飾または誘導体化したアミノ酸を含む、任意の長さのアミノ酸のポリマー形態を含む。これらの用語には、修飾されたペプチド骨格を有するポリペプチドなどの修飾されたポリマーも含まれる。「ドメイン」という用語は、特定の機能または構造を有するタンパク質またはポリペプチドの任意の部分を指す。
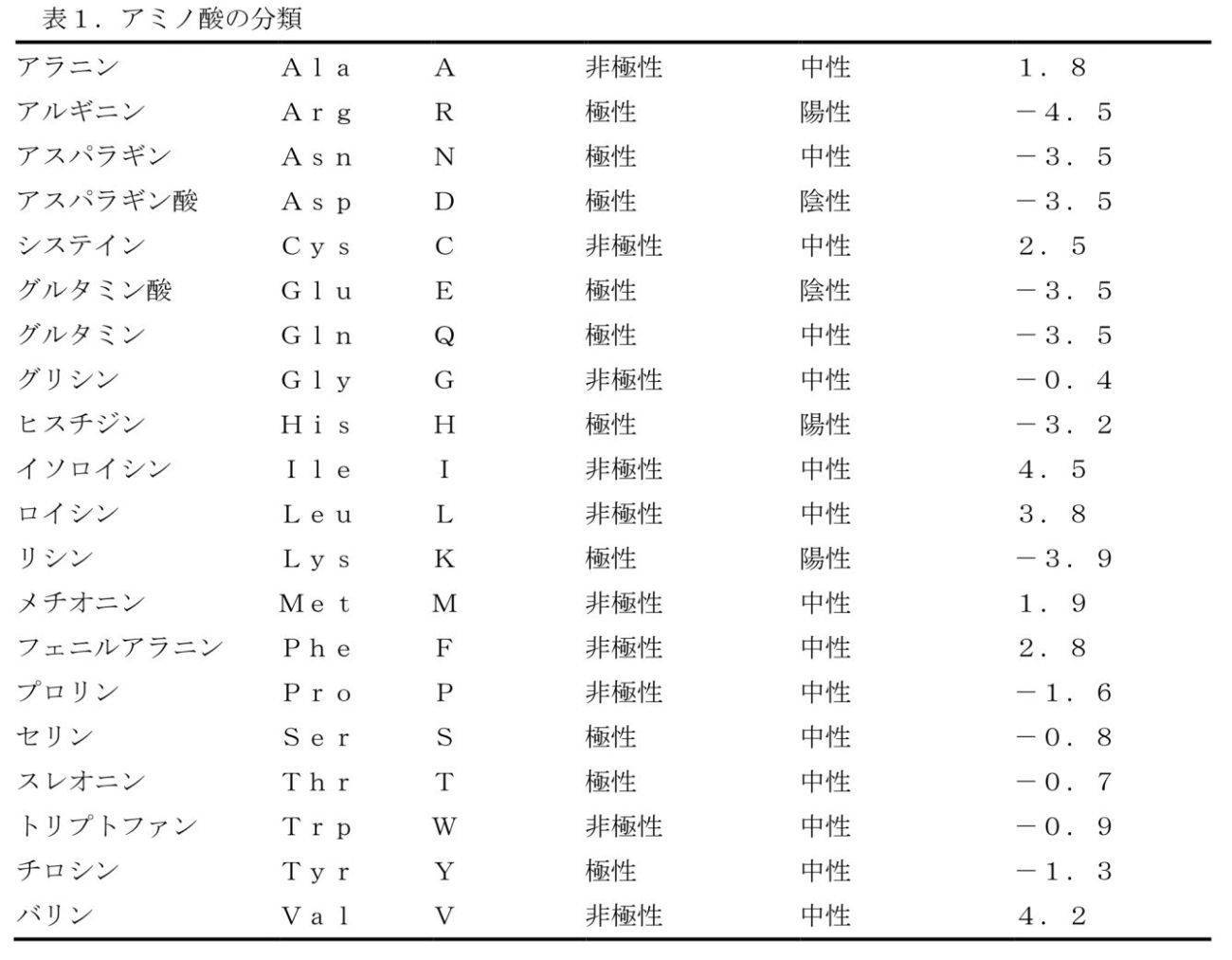
[発明を実施するための形態]
Casタンパク質対応タウバイオセンサー細胞、ならびにタウ凝集に関連する遺伝的脆弱性をスクリーニングするためにそのような細胞を作製および使用する方法が提供される。CRISPR/Cas相乗的活性化メディエーター(SAM)対応タウバイオセンサー細胞、ならびにタウ凝集に関連する遺伝的脆弱性をスクリーニングするためにそのような細胞を作製および使用する方法もまた提供される。
A.Cas/Tauバイオセンサー細胞およびSAM/タウバイオセンサー細胞
第1のレポーターに連結された第1のタウリピートドメイン(例えば、タウ微小管結合ドメイン(MBD)を含む)および第2のレポーターに連結された第2のタウリピートドメインを発現するだけでなく、Cas9などのCasタンパク質も発現する細胞が本明細書に開示される。第1のレポーターに連結された第1のタウリピートドメイン(例えば、タウ微小管結合ドメイン(MBD)を含む)および第2のレポーターに連結された第2のタウリピートドメインを発現するだけでなく、1つ以上の転写活性化ドメインに融合したヌクレアーゼ不活性Casタンパク質を含むキメラCasタンパク質、および1つ以上の転写活性化ドメインに融合したアダプタータンパク質を含むキメラアダプタータンパク質も発現する細胞もまた本明細書に開示される。第1のレポーターに連結された第1のタウリピートドメインは、安定して発現され得、第2のレポーターに連結された第2のタウリピートドメインは、安定して発現され得る。例えば、第1のレポーターに連結された第1のタウリピートドメインをコードするDNAは、ゲノム的に組み込むことができ、第2のレポーターに連結された第2のタウリピートドメインをコードするDNAは、ゲノム的に組み込むことができる。同様に、Casタンパク質は、Cas/タウバイオセンサー細胞において安定して発現され得る。例えば、Casタンパク質をコードするDNAは、ゲノム的に組み込むことができる。同様に、キメラCasタンパク質および/またはキメラアダプタータンパク質は、SAM/タウバイオセンサー細胞において安定して発現され得る。例えば、キメラCasタンパク質をコードするDNAは、ゲノム的に組み込むことができ、かつ/またはキメラアダプタータンパク質をコードするDNAは、ゲノム的に組み込むことができる。
微小管関連タンパク質タウは、微小管の集合および安定性を促進するタンパク質であり、主にニューロンにおいて発現される。タウは、ニューロンの微小管を安定させ、したがって軸索伸長を促進する役割を果たす。アルツハイマー病(AD)およびタウオパチーと呼ばれる関連する神経変性疾患のファミリーでは、タウタンパク質は、異常に過剰リン酸化され、神経原線維変化として現れるフィラメントの束(対らせん状フィラメント)に凝集する。タウオパチーは、脳内の異常なタウ沈着を特徴とする不均一な神経変性状態のグループである。
本明細書に開示されるCas/タウバイオセンサー細胞はまた、Casタンパク質をコードする核酸(DNAまたはRNA)を含む。任意に、Casタンパク質は、安定して発現される。任意に、細胞は、ゲノム的に組み込まれたCasコード配列を含む。同様に、本明細書に開示されるSAM/タウバイオセンサー細胞はまた、1つ以上の転写活性化ドメイン(例えば、VP64)に融合したヌクレアーゼ不活性Casタンパク質を含むキメラCasタンパク質をコードする核酸(DNAまたはRNA)も含む。任意に、キメラCasタンパク質は、安定して発現される。任意に、細胞は、ゲノム的に組み込まれたキメラCasコード配列を含む。
本明細書に開示されるSAM/タウバイオセンサー細胞は、1つ以上の転写活性化ドメイン(例えば、VP64)に融合したヌクレアーゼ不活性Casタンパク質を含むキメラCasタンパク質をコードする核酸(DNAまたはRNA)だけでなく、任意に、1つ以上の転写活性化ドメインに融合された(例えば、p65およびHSF1に融合された)アダプタータンパク質(例えば、MS2コートタンパク質(MCP))を含むキメラアダプタータンパク質をコードする核酸(DNAまたはRNA)も含み得る。任意に、キメラCasタンパク質および/またはキメラアダプタータンパク質は、安定して発現される。任意に、細胞は、ゲノム的に組み込まれたキメラCasタンパク質コード配列および/またはゲノム的に組み込まれたキメラアダプタータンパク質コード配列を含む。
本明細書に開示されるCas/タウバイオセンサー細胞は、任意の細胞型であり得、インビトロ、エクスビボ、またはインビボであり得る。Cas/タウバイオセンサー細胞株または細胞集団は、モノクローナル細胞株または細胞集団であり得る。同様に、本明細書に開示されるSAM/タウバイオセンサー細胞は、任意の細胞型であり得、インビトロ、エクスビボ、またはインビボであり得る。SAM/タウバイオセンサー細胞株または細胞集団は、モノクローナル細胞株または細胞集団であり得る。細胞は、任意の供給源からのものであり得る。例えば、細胞は、真核細胞、動物細胞、植物細胞、または真菌(例えば、酵母)細胞であり得る。そのような細胞は、魚細胞もしくは鳥細胞であり得るか、またはそのような細胞は、ヒト細胞、非ヒト哺乳動物細胞、げっ歯類細胞、マウス細胞、もしくはラット細胞などの哺乳動物細胞であり得る。哺乳動物には、例えば、ヒト、非ヒト霊長類、サル、類人猿、ネコ、イヌ、ウマ、雄牛、シカ、バイソン、ヒツジ、げっ歯類(例えば、マウス、ラット、ハムスター、ギニアブタ)、家畜(例えば、雌牛および去勢雄牛などのウシ種、ヒツジおよびヤギなどのヒツジ種、ブタおよびイノシシなどのブタ種)が含まれる。鳥には、例えば、鶏、七面鳥、ダチョウ、ガチョウ、およびアヒルが含まれる。家畜および農業動物も含まれる。「非ヒト動物」という用語は、ヒトを除外する。特定の例では、Cas/タウバイオセンサー細胞は、ヒト細胞(例えば、HEK293T細胞)である。同様に、特定の例では、SAM/タウバイオセンサー細胞は、ヒト細胞(例えば、HEK293T細胞)である。
本明細書に開示されるCas/タウバイオセンサー細胞は、任意の既知の手段によって生成され得る。第1のレポーターに連結された第1のタウリピートドメイン、第2のレポーターに連結された第2のタウリピートドメイン、およびCasタンパク質は、任意の既知の手段によって任意の形態(例えば、DNA、RNA、またはタンパク質)で細胞に導入され得る。同様に、本明細書に開示されるSAM/タウバイオセンサー細胞は、任意の既知の手段によって生成され得る。第1のレポーターに連結された第1のタウリピートドメイン、第2のレポーターに連結された第2のタウリピートドメイン、キメラCasタンパク質、およびキメラアダプタータンパク質は、任意の既知の手段によって任意の形態(例えば、DNA、RNA、またはタンパク質)で細胞に導入され得る。「導入すること」は、配列が細胞の内部にアクセスできるように、核酸またはタンパク質を細胞に提示することを含む。本明細書で提供される方法は、核酸またはタンパク質を細胞に導入するための特定の方法に依存せず、核酸またはタンパク質が少なくとも1つの細胞の内部にアクセスできることのみに依存する。核酸およびタンパク質を様々な細胞型に導入するための方法は既知であり、例えば、安定したトランスフェクション法、一過性トランスフェクション法、およびウイルス媒介法が含まれる。任意に、標的化ベクターを使用することができる。
本明細書に開示されるCas/タウバイオセンサー細胞およびそれらの細胞を培養するための培地を含むインビトロ培養物または組成物もまた、本明細書に開示される。本明細書に開示されるSAM/タウバイオセンサー細胞およびそれらの細胞を培養するための培地を含むインビトロ培養物または組成物もまた、本明細書に開示される。細胞は、Agg[-]細胞またはAgg[+]細胞であり得る。
本明細書に開示されるCRISPRnスクリーニング方法は、ゲノムワイドgRNAノックアウトライブラリなどのCRISPRガイドRNA(gRNA)ノックアウトライブラリを利用する。Cas9などのCasヌクレアーゼは、特定の標的配列を標的とするように設計されたgRNAを介して、特定のゲノム遺伝子座で二本鎖切断を誘導するようにプログラムされ得る。Casタンパク質の標的化特異性は短いgRNAによって付与されるため、プールされたゲノムスケールの機能的スクリーニングが可能である。そのようなライブラリは、mRNAを標的化することによりタンパク質発現を低減するshRNAライブラリなどのライブラリに比べていくつかの利点を有する。対照的に、gRNAライブラリは、遺伝子のゲノムコード領域に導入されたフレームシフト変異を介してノックアウトを達成する。
「ガイドRNA」または「gRNA」は、Casタンパク質(例えば、Cas9タンパク質)に結合し、かつCasタンパク質を標的DNA内の特定の位置に標的化するRNA分子である。ガイドRNAは、「DNA標的化セグメント」および「タンパク質結合セグメント」の2つのセグメントを含み得る。「セグメント」は、RNA中のヌクレオチドの連続したストレッチなどの、分子のセクションまたは領域を含む。Cas9のものなど、いくつかのgRNAは、「アクチベーターRNA」(例えば、tracrRNA)および「ターゲッターRNA」(例えば、CRISPR RNAまたはcrRNA)の2つの別個のRNA分子を含み得る。他のgRNAは、単一RNA分子(単一RNAポリヌクレオチド)であり、これはまた「単一分子gRNA」、「単一ガイドRNA」、または「sgRNA」と呼ばれ得る。例えば、WO2013/176772、WO2014/065596、WO2014/089290、WO2014/093622、WO2014/099750、WO2013/142578、およびWO2014/131833を参照されたく、それらの各々は、すべての目的のためにその全体が参照により本明細書に組み込まれる。例えば、Cas9の場合、単一ガイドRNAは、tracrRNAに融合したcrRNAを含み得る(例えば、リンカーを介して)。例えば、Cpf1の場合、標的配列への結合を達成するためにcrRNAのみが必要とされる。「ガイドRNA」および「gRNA」という用語は、二重分子(すなわち、モジュラー)gRNAおよび単一分子gRNAの両方を含む。
ガイドRNAの標的DNAには、結合に十分な条件が存在する場合に、gRNAのDNA標的セグメントが結合するDNAに存在する核酸配列が含まれる。好適なDNA/RNA結合条件には、細胞に通常存在する生理学的条件が含まれる。他の好適なDNA/RNA結合条件(例えば、無細胞系における条件)は、当該技術分野で知られている(例えば、Molecular Cloning:A Laboratory Manual,3rd Ed.(Sambrook et al.,Harbor Laboratory Press 2001)を参照されたく、すべての目的のためにその全体が参照により本明細書に組み込まれる)。gRNAに相補的であり、かつそれとハイブリダイズする標的DNAの鎖は、「相補的鎖」と呼ばれることができ、「相補的鎖」に相補的である(したがって、Casタンパク質またはgRNAに相補的ではない)標的DNAの鎖は、「非相補鎖」または「テンプレート鎖」と呼ばれることができる。
本明細書に開示されるCas/タウバイオセンサー細胞株は、タウ凝集に関連する遺伝的脆弱性を明らかにするためのドロップアウトスクリーニングの方法で使用することができる。そのような方法は、本明細書の他の場所に開示されるように、Cas/タウバイオセンサー細胞の凝集陽性および凝集陰性集団を提供することと、複数の固有のガイドRNAを含むライブラリを各集団に導入することと、各細胞集団における時間経過にわたる複数の時点での複数の固有のガイドRNAの各々の存在量(読み取りカウント)を決定することとを含み得る。
添付の配列表に列記されているヌクレオチドおよびアミノ酸配列は、ヌクレオチド塩基のための標準的な文字略語、およびアミノ酸のための三文字表記を使用して示されている。ヌクレオチド配列は、配列の5’末端から始まって先へ(すなわち、各行で左から右へ)進み3’末端に至る標準規則に従っている。各ヌクレオチド配列の1本の鎖のみが示されているが、相補鎖は、示されている鎖を任意に参照することによって含まれると理解される。アミノ酸配列をコードするヌクレオチド配列が提供される場合、同じアミノ酸配列をコードするそのコドン縮重バリアントもまた提供されることが理解される。アミノ酸配列は、配列のアミノ末端から始まって先へ(すなわち、各行で左から右へ)進みカルボキシ末端に至る標準規則に従っている。


タンパク質の異常な凝集または線維化は、特にアルツハイマー病(AD)、パーキンソン病(PD)、前頭側頭型認知症(FTD)、筋萎縮性側索硬化症(ALS)、慢性外傷性脳症(CTE)、クロイツフェルト・ヤコブ病(CJD)などのいくつかの神経変性疾患を含む、多くの疾患の決定的な特徴である。これらの疾患の多くでは、ある特定のタンパク質の不溶性凝集体への線維化は、疾患の特徴であるだけでなく、神経毒性の原因因子としても関係している。さらに、これらの疾患は、ステレオタイプのパターンに従って中枢神経系を介する凝集の病状の伝播によって特徴付けられ、このプロセスは疾患の進行と相関している。したがって、異常なタンパク質凝集のプロセス、または凝集体の細胞間増殖を変更する遺伝子および遺伝子経路の同定は、神経変性疾患の病因をよりよく理解する上で、ならびに治療的介入のための戦略を考案する上で大いに価値がある。

実施例1で同定された脆弱性を調査するために、プールされたゲノムワイドCRISPRヌクレアーゼ(CRISPRn)スクリーニングを行い、タウ凝集に関連する遺伝的脆弱性を明らかにした。具体的には、凝集体あり(Agg[+])または凝集体なし(Agg[-])のいずれかのCas9発現タウ-CFP/タウ-YFPバイオセンサー細胞に、sgRNAあたり500X細胞の掲示で各標的遺伝子にノックアウト変異を導入するためのレンチウイルス送達アプローチを使用して、2つのヒトゲノムワイドCRISPR sgRNAライブラリで形質導入した。図4および9を参照されたい。CRISPR sgRNAライブラリは、機能的ノックアウトのために5’構成的エクソンを標的とし、遺伝子あたり平均3~4個のsgRNAをカバーする。sgRNAは、オフターゲットゲノム配列とのミスマッチが2つ以下のsgRNAを回避することにより、オフターゲット効果を回避するように設計された。ライブラリは、1000個の非標的化対照sgRNAを有する19,050個のヒト遺伝子および1864個のmiRNAをカバーする。ライブラリを、感染多重度(MOI)<0.3で、sgRNAあたり>500細胞のカバレッジで形質導入した。タウバイオセンサー細胞を、ピューロマイシンの選択の下で増殖させ、細胞ごとに固有のsgRNAの組み込みおよび発現を有する細胞を選択した。ピューロマイシンの選択は、1μg/mLでの形質導入の24時間後に開始した。


転写活性化されたときに疾患関連タンパク質凝集体との合成致死性を呈する遺伝子を同定するために、転写活性化(hSAM)CRISPRa sgRNAライブラリを用いてゲノムワイドスクリーニングを行うためのプラットフォームが開発された。プラットフォームは、転写活性化されたときに、特に異常なタンパク質凝集の状況で細胞死を引き起こす遺伝子を同定する。そのような遺伝子の同定は、神経変性疾患の状況において凝集体関連神経毒性のメカニズム、およびニューロンの死を促進する遺伝的経路を解明し得る。
プールされたゲノムワイド転写活性化(hSAM)CRISPRaスクリーニングを行い、実施例3で開発されたクローン細胞株を使用して細胞生存率に影響するタウの遺伝的修飾因子を明らかにした。具体的には、凝集体あり(Agg[+])または凝集体なし(Agg[-])のいずれかのSAM発現(すなわち、dCas9-VP64/MS2-P65-HSF1発現)タウ-CFP/タウ-YFPバイオセンサー細胞に、sgRNAあたり500X細胞の掲示で各標的遺伝子を転写活性化するためのレンチウイルス送達アプローチを使用して、ヒトゲノムワイドCRISPR hSAM sgRNAライブラリで形質導入した。CRISPR hSAM sgRNAライブラリは、転写開始部位の200bp上流内の部位を標的とし、遺伝子あたりの平均カバレッジは3~4のsgRNAである。sgRNAは、オフターゲットゲノム配列とのミスマッチが2つ以下のsgRNAを回避することにより、オフターゲット効果を回避するように設計された。ライブラリは18,946のヒト遺伝子をカバーする。ライブラリを、感染多重度(MOI)<0.3で、sgRNAあたり>500細胞のカバレッジで形質導入した。タウバイオセンサー細胞を、ゼオシンの選択の下で増殖させ、細胞ごとに固有のsgRNAの組み込みおよび発現を含む細胞を選択した。
Claims (97)
- タウ凝集に関連する遺伝的脆弱性をスクリーニングする方法であって、
(a)凝集陽性細胞集団および凝集陰性細胞集団を提供することであって、各細胞集団が、Casタンパク質、第1のレポーターに連結された第1のタウリピートドメイン、および第2のレポーターに連結された第2のタウリピートドメインを含み、
前記凝集陽性細胞集団において、前記第1のレポーターに連結された前記第1のタウリピートドメインおよび前記第2のレポーターに連結された前記第2のタウリピートドメインが、凝集状態で安定して存在し、
前記凝集陰性細胞集団において、前記第1のレポーターに連結された前記第1のタウリピートドメインおよび前記第2のレポーターに連結された前記第2のタウリピートドメインが、凝集状態で安定して存在しない、提供することと、
(b)複数の遺伝子を標的とする複数の固有のガイドRNAを含むライブラリを各細胞集団に導入することであって、前記複数の固有のガイドRNAが、前記Casタンパク質と複合体を形成し、前記Casタンパク質が、前記複数の遺伝子を切断して遺伝子機能のノックアウトをもたらす、導入することと、
(c)各細胞集団における時間経過にわたる複数の時点での前記複数の固有のガイドRNAの各々の存在量を決定することと、を含み、
前記凝集陰性細胞集団ではないが前記凝集陽性細胞集団におけるガイドRNAの枯渇、または前記凝集陰性細胞集団に対して、前記凝集陽性細胞集団における時間経過にわたるガイドRNAのより劇的な枯渇パターンが、前記ガイドRNAによって標的とされた前記遺伝子が、タウタンパク質凝集体との合成致死性を呈し、タウ凝集に関連する遺伝的脆弱性であることを示す、方法。 - 前記Casタンパク質が、Cas9タンパク質である、請求項1に記載の方法。
- 前記Casタンパク質が、Streptococcus pyogenes Cas9である、請求項2に記載の方法。
- 前記Casタンパク質が、配列番号21を含み、任意に、前記Casタンパク質が、配列番号22に示される配列を含むコード配列によってコードされる、請求項3に記載の方法。
- 前記Casタンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインが、前記細胞集団において安定して発現される、先行請求項のいずれかに記載の方法。
- 前記Casタンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインをコードする核酸が、前記細胞集団においてゲノム的に組み込まれる、先行請求項のいずれかに記載の方法。
- 各ガイドRNAが、構成的エクソンを標的とする、先行請求項のいずれかに記載の方法。
- 各ガイドRNAが、5’構成的エクソンを標的とする、請求項7に記載の方法。
- 各ガイドRNAが、第1のエクソン、第2のエクソン、または第3のエクソンを標的とする、先行請求項のいずれかに記載の方法。
- タウ凝集に関連する遺伝的脆弱性をスクリーニングする方法であって、
(a)凝集陽性細胞集団および凝集陰性細胞集団を提供することであって、各細胞集団が、1つ以上の転写活性化ドメインに融合したヌクレアーゼ不活性Casタンパク質を含むキメラCasタンパク質と、1つ以上の転写活性化ドメインに融合したアダプタータンパク質を含むキメラアダプタータンパク質と、第1のレポーターに連結された第1のタウリピートドメインと、第2のレポーターに連結された第2のタウリピートドメインと、を含み、
前記凝集陽性細胞集団において、前記第1のレポーターに連結された前記第1のタウリピートドメインおよび前記第2のレポーターに連結された前記第2のタウリピートドメインが、凝集状態で安定して存在し、
前記凝集陰性細胞集団において、前記第1のレポーターに連結された前記第1のタウリピートドメインおよび前記第2のレポーターに連結された前記第2のタウリピートドメインが、凝集状態で安定して存在しない、提供することと、
(b)複数の遺伝子を標的とする複数の固有のガイドRNAを含むライブラリを各細胞集団に導入することであって、前記複数の固有のガイドRNAが、前記キメラCasタンパク質および前記キメラアダプタータンパク質と複合体を形成し、前記複合体が、前記複数の遺伝子の転写を活性化して遺伝子発現の増加をもたらす、導入することと、
(c)各細胞集団における時間経過にわたる複数の時点での前記複数の固有のガイドRNAの各々の存在量を決定することと、を含み、
前記凝集陰性細胞集団ではないが前記凝集陽性細胞集団におけるガイドRNAの枯渇、または前記凝集陰性細胞集団に対して、前記凝集陽性細胞集団における時間経過にわたるガイドRNAのより劇的な枯渇パターンが、前記ガイドRNAによって標的とされた前記遺伝子の活性化が、タウタンパク質凝集体との合成致死性を呈し、タウ凝集に関連する遺伝的脆弱性であることを示す、方法。 - 前記Casタンパク質が、Cas9タンパク質である、請求項10に記載の方法。
- 前記Casタンパク質が、Streptococcus pyogenes Cas9である、請求項11に記載の方法。
- 前記キメラCasタンパク質が、VP64転写活性化ドメインに融合した前記ヌクレアーゼ不活性Casタンパク質を含み、任意に、前記キメラCasタンパク質が、N末端からC末端に、前記ヌクレアーゼ不活性Casタンパク質、核局在化シグナル、および前記VP64転写活性化因子ドメインを含む、請求項10~12のいずれか一項に記載の方法。
- 前記アダプタータンパク質が、MS2コートタンパク質であり、前記キメラアダプタータンパク質中の前記1つ以上の転写活性化ドメインが、p65転写活性化ドメインおよびHSF1転写活性化ドメインを含み、任意に、前記キメラアダプタータンパク質が、N末端からC末端に、前記MS2コートタンパク質、核局在化シグナル、前記p65転写活性化ドメイン、および前記HSF1転写活性化ドメインを含む、請求項10~13のいずれか一項に記載の方法。
- 前記キメラCasタンパク質が、配列番号36を含み、任意に、前記キメラCasタンパク質が、配列番号38に示される配列を含むコード配列によってコードされる、請求項10~14のいずれか一項に記載の方法。
- 前記キメラアダプタータンパク質が、配列番号37を含み、任意に、前記キメラアダプタータンパク質が、配列番号39に示される配列を含むコード配列によってコードされる、請求項10~15のいずれか一項に記載の方法。
- 前記キメラCasタンパク質、前記キメラアダプタータンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインが、前記細胞集団において安定して発現される、請求項10~16のいずれか一項に記載の方法。
- 前記キメラCasタンパク質、前記キメラアダプタータンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインをコードする核酸が、前記細胞集団においてゲノム的に組み込まれる、請求項10~17のいずれか一項に記載の方法。
- 各ガイドRNAが、転写開始部位の200bp上流内のガイドRNA標的配列を標的とする、請求項10~18のいずれか一項に記載の方法。
- 各ガイドRNAが、前記キメラアダプタータンパク質が特異的に結合することができる1つ以上のアダプター結合エレメントを含み、任意に、各ガイドRNAが、前記キメラアダプタータンパク質が特異的に結合することができる2つのアダプター結合エレメントを含み、任意に、第1のアダプター結合エレメントが、前記1つ以上のガイドRNAの各々の第1のループ内にあり、第2のアダプター結合エレメントが、前記1つ以上のガイドRNAの各々の第2のループ内にあり、任意に、前記アダプター結合エレメントが、配列番号33に示される配列を含み、
任意に、1つ以上のガイドRNAの各々が、トランス活性化CRISPR RNA(tracrRNA)部分に融合したCRISPR RNA(crRNA)部分を含む単一ガイドRNAであり、前記第1のループが、配列番号17の残基13~16に対応するテトラループであり、前記第2のループが、配列番号17の残基53~56に対応するステムループ2である、請求項10~19のいずれか一項に記載の方法。 - 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、ヒトタウリピートドメインである、先行請求項のいずれかに記載の方法。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、凝集促進変異を含む、先行請求項のいずれかに記載の方法。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、タウP301S変異を含む、請求項22に記載の方法。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、タウ4リピートドメインを含む、先行請求項のいずれかに記載の方法。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、配列番号11を含む、先行請求項のいずれかに記載の方法。
- 前記第1のタウリピートドメインおよび前記第2のタウリピートドメインが、同じである、先行請求項のいずれかに記載の方法。
- 前記第1のタウリピートドメインおよび前記第2のタウリピートドメインが、同じであり、各々が、タウP301S変異を含むタウ4リピートドメインを含む、先行請求項のいずれかに記載の方法。
- 前記第1のレポーターおよび前記第2のレポーターが、蛍光タンパク質である、先行請求項のいずれかに記載の方法。
- 前記第1のレポーターおよび前記第2のレポーターが、蛍光共鳴エネルギー移動(FRET)対である、請求項28に記載の方法。
- 前記第1のレポーターが、シアン蛍光タンパク質(CFP)であり、前記第2のレポーターが、黄色蛍光タンパク質(YFP)である、請求項29に記載の方法。
- 前記細胞が、真核細胞である、先行請求項のいずれかに記載の方法。
- 前記細胞が、哺乳動物細胞である、請求項31に記載の方法。
- 前記細胞が、ヒト細胞である、請求項32に記載の方法。
- 前記細胞が、HEK293T細胞である、請求項33に記載の方法。
- 前記複数の固有のガイドRNAが、前記細胞の大部分が前記固有のガイドRNAのうちの1つのみを受け取るように選択された濃度で導入される、先行請求項のいずれかに記載の方法。
- 前記複数の固有のガイドRNAが、100個以上の遺伝子、1000個以上の遺伝子、または10000個以上の遺伝子を標的とする、先行請求項のいずれかに記載の方法。
- 前記ライブラリが、ゲノムワイドライブラリである、先行請求項のいずれかに記載の方法。
- 複数の標的配列が、前記標的化された複数の遺伝子の各々において平均して標的化される、先行請求項のいずれかに記載の方法。
- 少なくとも3個の標的配列が、前記標的化された複数の遺伝子の各々において平均して標的化される、請求項38に記載の方法。
- 約3~約6個の標的配列が、前記標的化された複数の遺伝子の各々において平均して標的化される、請求項39に記載の方法。
- 前記複数の固有のガイドRNAが、ウイルス形質導入によって前記細胞集団に導入される、先行請求項のいずれかに記載の方法。
- 前記複数の固有のガイドRNAの各々が、別個のウイルスベクター内にある、請求項41に記載の方法。
- 前記複数の固有のガイドRNAが、レンチウイルス形質導入によって前記細胞集団に導入される、請求項41または42に記載の方法。
- 前記細胞集団が、約0.3未満の感染多重度で感染する、請求項41~43のいずれか一項に記載の方法。
- 前記複数の固有のガイドRNAが、選択マーカーとともに前記細胞集団に導入され、ステップ(b)が、前記選択マーカーを含む細胞を選択することをさらに含む、先行請求項のいずれかに記載の方法。
- 前記選択マーカーが、薬物に対する耐性を付与し、任意に、前記選択マーカーが、ピューロマイシンまたはゼオシンに対する耐性を付与する、請求項45に記載の方法。
- 前記選択マーカーが、ネオマイシンホスホトランスフェラーゼ、ハイグロマイシンBホスホトランスフェラーゼ、ピューロマイシン-N-アセチルトランスフェラーゼ、およびブラストサイジンSデアミナーゼから選択される、請求項46に記載の方法。
- ステップ(b)において前記複数の固有のガイドRNAが導入される前記細胞集団が各々、固有のガイドRNAあたり約500個を超える細胞を含む、先行請求項のいずれかに記載の方法。
- ステップ(c)の前記時間経過が、約1週間超である、先行請求項のいずれかに記載の方法。
- ステップ(c)の前記時間経過が、約2週間超である、請求項49に記載の方法。
- ステップ(c)の前記時間経過が、約10~約15の細胞倍加を含む、先行請求項のいずれかに記載の方法。
- ステップ(c)の前記複数の時点が、少なくとも3つの時点を含む、先行請求項のいずれかに記載の方法。
- ステップ(c)の前記複数の時点が、約4つの時点または約6つの時点を含む、請求項52に記載の方法。
- ステップ(c)の各時点の間が約1日超である、先行請求項のいずれかに記載の方法。
- ステップ(c)の各時点の間が約2日超である、請求項54に記載の方法。
- ステップ(c)の各時点の間が約3~約4日である、請求項55に記載の方法。
- 前記遺伝子を標的とするガイドRNAが、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない場合、遺伝子がステップ(c)でタウタンパク質凝集体との合成致死性を呈するとみなされる、先行請求項のいずれかに記載の方法。
- 前記遺伝子を標的とするガイドRNAが、前記凝集陰性細胞集団に対して、前記凝集陽性細胞集団において前記時間経過にわたってより劇的な枯渇パターンを有する場合、遺伝子がステップ(c)でタウタンパク質凝集体との合成致死性を呈するとみなされる、請求項57に記載の方法。
- 各時点での前記ガイドRNAの存在量が以前の時点での前記ガイドRNAの存在量以下である場合、ガイドRNAがステップ(c)で枯渇しているとみなされる、先行請求項のいずれかに記載の方法。
- 前記第2の時点の後の各時点での前記ガイドRNAの存在量が2つの時点前の時点の存在量以下である場合、ガイドRNAがステップ(c)で枯渇しているとみなされる、先行請求項のいずれかに記載の方法。
- 前記遺伝子を標的とする前記ライブラリ内の前記ガイドRNAの約30%超が、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない場合、遺伝子がステップ(c)でタウタンパク質凝集体との合成致死性を呈するとみなされる、先行請求項のいずれかに記載の方法。
- 遺伝子が、以下の状況:
(1)前記ライブラリ内に前記遺伝子を標的とする1つのガイドRNAが存在し、前記1つのガイドRNAが、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない、
(2)前記ライブラリ内に前記遺伝子を標的とする2つのガイドRNAが存在し、前記2つのガイドRNAの少なくとも1つが、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない、
(3)前記ライブラリ内に前記遺伝子を標的とする3つのガイドRNAが存在し、前記3つのガイドRNAの少なくとも1つが、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない、
(4)前記ライブラリ内に前記遺伝子を標的とする4つのガイドRNAが存在し、前記4つのガイドRNAの少なくとも2つが、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない、
(5)前記ライブラリ内に前記遺伝子を標的とする5つのガイドRNAが存在し、前記5つのガイドRNAの少なくとも2つが、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない、および
(6)前記ライブラリ内に前記遺伝子を標的とする6つのガイドRNAが存在し、前記6つのガイドRNAの少なくとも3つが、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない、のうちのいずれか1つにおいて、ステップ(c)でタウタンパク質凝集体との合成致死性を呈するとみなされる、請求項61に記載の方法。 - 前記方法が、少なくとも3つの異なる実験で少なくとも3回繰り返され、遺伝子が、前記少なくとも3つの異なる実験の約50%超でタウタンパク質凝集体との合成致死性を呈するとみなされる場合、タウタンパク質凝集体との合成致死性を呈するとみなされる、先行請求項のいずれかに記載の方法。
- ステップ(c)の前記時間経過が、約2週間超であり、
ステップ(c)の前記複数の時点が、約6つの時点を含み、
ステップ(c)の各時点の間が約3~約4日であり、
前記第2の時点の後の各時点での前記ガイドRNAの存在量が2つの時点前の時点の存在量以下である場合、ガイドRNAがステップ(c)で枯渇しているとみなされ、
前記遺伝子を標的とする前記ライブラリ内の前記ガイドRNAの約30%超が、前記凝集陽性細胞集団において枯渇しているが、前記凝集陰性細胞集団においては枯渇していない場合、遺伝子がステップ(c)でタウタンパク質凝集体との合成致死性を呈するとみなされる、先行請求項のいずれかに記載の方法。 - Casタンパク質、第1のレポーターに連結された第1のタウリピートドメイン、および第2のレポーターに連結された第2のタウリピートドメインを含む、1つ以上の細胞集団。
- 前記Casタンパク質が、Cas9タンパク質である、請求項65に記載の細胞集団。
- 前記Casタンパク質が、Streptococcus pyogenes Cas9である、請求項66に記載の細胞集団。
- 前記Casタンパク質が、配列番号21を含み、任意に、前記Casタンパク質が、配列番号22に示される配列を含むコード配列によってコードされる、請求項67に記載の細胞集団。
- 前記Casタンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインが、前記細胞において安定して発現される、請求項65~68のいずれか一項に記載の細胞集団。
- 前記Casタンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインをコードする核酸が、前記細胞においてゲノム的に組み込まれる、請求項65~69のいずれか一項に記載の細胞集団。
- 1つ以上の転写活性化ドメインに融合したヌクレアーゼ不活性Casタンパク質を含むキメラCasタンパク質と、1つ以上の転写活性化ドメインに融合したアダプタータンパク質を含むキメラアダプタータンパク質と、第1のレポーターに連結された第1のタウリピートドメインと、第2のレポーターに連結された第2のタウリピートドメインと、を含む、1つ以上の細胞集団。
- 前記Casタンパク質が、Cas9タンパク質である、請求項71に記載の細胞集団。
- 前記Casタンパク質が、Streptococcus pyogenes Cas9である、請求項72に記載の細胞集団。
- 前記キメラCasタンパク質が、VP64転写活性化ドメインに融合した前記ヌクレアーゼ不活性Casタンパク質を含み、任意に、前記キメラCasタンパク質が、N末端からC末端に、前記ヌクレアーゼ不活性Casタンパク質、核局在化シグナル、および前記VP64転写活性化因子ドメインを含む、請求項71~73のいずれか一項に記載の細胞集団。
- 前記アダプタータンパク質が、MS2コートタンパク質であり、前記キメラアダプタータンパク質中の前記1つ以上の転写活性化ドメインが、p65転写活性化ドメインおよびHSF1転写活性化ドメインを含み、任意に、前記キメラアダプタータンパク質が、N末端からC末端に、前記MS2コートタンパク質、核局在化シグナル、前記p65転写活性化ドメイン、および前記HSF1転写活性化ドメインを含む、請求項71~74のいずれか一項に記載の細胞集団。
- 前記キメラCasタンパク質が、配列番号36を含み、任意に、前記キメラCasタンパク質が、配列番号38に示される配列を含むコード配列によってコードされる、請求項71~75のいずれか一項に記載の細胞集団。
- 前記キメラアダプタータンパク質が、配列番号37を含み、任意に、前記キメラアダプタータンパク質が、配列番号39に示される配列を含むコード配列によってコードされる、請求項71~76のいずれか一項に記載の細胞集団。
- 前記キメラCasタンパク質、前記キメラアダプタータンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインが、前記細胞において安定して発現される、請求項71~77のいずれか一項に記載の細胞集団。
- 前記キメラCasタンパク質、前記キメラアダプタータンパク質、前記第1のレポーターに連結された前記第1のタウリピートドメイン、および前記第2のレポーターに連結された前記第2のタウリピートドメインをコードする核酸が、前記細胞においてゲノム的に組み込まれる、請求項71~78のいずれか一項に記載の細胞集団。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、ヒトタウリピートドメインである、請求項65~79のいずれか一項に記載の細胞集団。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、凝集促進変異を含む、請求項65~80のいずれか一項に記載の細胞集団。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、タウP301S変異を含む、請求項81に記載の細胞集団。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、タウ4リピートドメインを含む、請求項65~82のいずれか一項に記載の細胞集団。
- 前記第1のタウリピートドメインおよび/または前記第2のタウリピートドメインが、配列番号11を含む、請求項65~83のいずれか一項に記載の細胞集団。
- 前記第1のタウリピートドメインおよび前記第2のタウリピートドメインが、同じである、請求項65~84のいずれか一項に記載の細胞集団。
- 前記第1のタウリピートドメインおよび前記第2のタウリピートドメインが、同じであり、各々が、タウP301S変異を含むタウ4リピートドメインを含む、請求項65~85のいずれか一項に記載の細胞集団。
- 前記第1のレポーターおよび前記第2のレポーターが、蛍光タンパク質である、請求項65~86のいずれか一項に記載の細胞集団。
- 前記第1のレポーターおよび前記第2のレポーターが、蛍光共鳴エネルギー移動(FRET)対である、請求項87に記載の細胞集団。
- 前記第1のレポーターが、シアン蛍光タンパク質(CFP)であり、前記第2のレポーターが、黄色蛍光タンパク質(YFP)である、請求項88に記載の細胞集団。
- 前記細胞が、真核細胞である、請求項65~89のいずれか一項に記載の細胞集団。
- 前記細胞が、哺乳動物細胞である、請求項90に記載の細胞集団。
- 前記細胞が、ヒト細胞である、請求項91に記載の細胞集団。
- 前記細胞が、HEK293T細胞である、請求項92に記載の細胞集団。
- 前記細胞が、インビトロである、請求項65~93のいずれか一項に記載の細胞集団。
- 前記第1のレポーターに連結された前記第1のタウリピートドメインおよび前記第2のレポーターに連結された前記第2のタウリピートドメインが、凝集状態で安定して存在していない、請求項65~94のいずれか一項に記載の細胞集団。
- 前記第1のレポーターに連結された前記第1のタウリピートドメインおよび前記第2のレポーターに連結された前記第2のタウリピートドメインが、凝集状態で安定して存在する、請求項65~95のいずれか一項に記載の細胞集団。
- 請求項65~96のいずれか一項に記載の細胞集団および培養培地を含む、インビトロ培養物。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962820101P | 2019-03-18 | 2019-03-18 | |
| US62/820,101 | 2019-03-18 | ||
| PCT/US2020/023121 WO2020190927A1 (en) | 2019-03-18 | 2020-03-17 | Crispr/cas dropout screening platform to reveal genetic vulnerabilities associated with tau aggregation |
Publications (3)
| Publication Number | Publication Date |
|---|---|
| JP2022526297A true JP2022526297A (ja) | 2022-05-24 |
| JPWO2020190927A5 JPWO2020190927A5 (ja) | 2023-09-27 |
| JP7389135B2 JP7389135B2 (ja) | 2023-11-29 |
Family
ID=70285886
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2021556467A Active JP7389135B2 (ja) | 2019-03-18 | 2020-03-17 | タウ凝集に関連する遺伝的脆弱性を明らかにするためのcrispr/casドロップアウトスクリーニングプラットフォーム |
Country Status (11)
| Country | Link |
|---|---|
| US (2) | US11781131B2 (ja) |
| EP (1) | EP3769090B1 (ja) |
| JP (1) | JP7389135B2 (ja) |
| KR (1) | KR20210141536A (ja) |
| CN (1) | CN113711046B (ja) |
| AU (1) | AU2020241856A1 (ja) |
| CA (1) | CA3127814C (ja) |
| ES (1) | ES2969026T3 (ja) |
| IL (1) | IL286359B1 (ja) |
| SG (1) | SG11202108090XA (ja) |
| WO (1) | WO2020190927A1 (ja) |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA3127813A1 (en) | 2019-03-18 | 2020-09-24 | Regeneron Pharmaceuticals, Inc. | Crispr/cas screening platform to identify genetic modifiers of tau seeding or aggregation |
| SG11202108090XA (en) | 2019-03-18 | 2021-08-30 | Regeneron Pharma | Crispr/cas dropout screening platform to reveal genetic vulnerabilities associated with tau aggregation |
| KR20220024053A (ko) | 2019-06-14 | 2022-03-03 | 리제너론 파마슈티칼스 인코포레이티드 | 타우병증의 모델 |
| CN114426982B (zh) * | 2022-02-22 | 2024-03-22 | 上海交通大学 | 一种细胞代谢重编程的方法及其应用 |
Family Cites Families (46)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2001271553A1 (en) * | 2000-06-23 | 2002-01-08 | Genetastix Corporation | High throughput generation and screening of fully human antibody repertoire in yeast |
| CN101333555B (zh) * | 2007-12-25 | 2012-03-21 | 浙江大学 | 新型的g蛋白偶联受体细胞水平筛选系统构建与应用 |
| JP2011517838A (ja) | 2008-04-11 | 2011-06-16 | ユーティーシー パワー コーポレイション | マニホルド・サンプを備えたバイポーラプレートおよび燃料電池 |
| CN103025344B (zh) | 2010-05-17 | 2016-06-29 | 桑格摩生物科学股份有限公司 | 新型dna-结合蛋白及其用途 |
| WO2013141680A1 (en) | 2012-03-20 | 2013-09-26 | Vilnius University | RNA-DIRECTED DNA CLEAVAGE BY THE Cas9-crRNA COMPLEX |
| US9637739B2 (en) | 2012-03-20 | 2017-05-02 | Vilnius University | RNA-directed DNA cleavage by the Cas9-crRNA complex |
| ES2960803T3 (es) | 2012-05-25 | 2024-03-06 | Univ California | Métodos y composiciones para la modificación de ADN diana dirigida por RNA y para la modulación de la transcripción dirigida por RNA |
| RU2018132044A (ru) | 2012-07-03 | 2018-10-19 | Вашингтон Юниверсити | Антитела против тау |
| WO2014033644A2 (en) | 2012-08-28 | 2014-03-06 | Novartis Ag | Methods of nuclease-based genetic engineering |
| ES2926021T3 (es) | 2012-10-23 | 2022-10-21 | Toolgen Inc | Composición para escindir un ADN objetivo que comprende un ARN guía específico para el ADN objetivo y ácido nucleico codificador de proteína Cas o proteína Cas, y uso de la misma |
| US9910048B2 (en) | 2012-12-03 | 2018-03-06 | Washington University | Method for detection of aggregates in biological samples |
| EP3617309A3 (en) | 2012-12-06 | 2020-05-06 | Sigma Aldrich Co. LLC | Crispr-based genome modification and regulation |
| EP3144390B1 (en) | 2012-12-12 | 2020-03-18 | The Broad Institute, Inc. | Engineering of systems, methods and optimized guide compositions for sequence manipulation |
| CN114634950A (zh) | 2012-12-12 | 2022-06-17 | 布罗德研究所有限公司 | 用于序列操纵的crispr-cas组分系统、方法以及组合物 |
| SG10201912328UA (en) | 2012-12-12 | 2020-02-27 | Broad Inst Inc | Delivery, Engineering and Optimization of Systems, Methods and Compositions for Sequence Manipulation and Therapeutic Applications |
| EP2931899A1 (en) | 2012-12-12 | 2015-10-21 | The Broad Institute, Inc. | Functional genomics using crispr-cas systems, compositions, methods, knock out libraries and applications thereof |
| US8697359B1 (en) | 2012-12-12 | 2014-04-15 | The Broad Institute, Inc. | CRISPR-Cas systems and methods for altering expression of gene products |
| IL308158A (en) | 2012-12-17 | 2023-12-01 | Harvard College | RNA-guided human genome engineering |
| EP2922393B2 (en) | 2013-02-27 | 2022-12-28 | Helmholtz Zentrum München - Deutsches Forschungszentrum für Gesundheit und Umwelt (GmbH) | Gene editing in the oocyte by cas9 nucleases |
| US10119133B2 (en) | 2013-03-15 | 2018-11-06 | The General Hospital Corporation | Using truncated guide RNAs (tru-gRNAs) to increase specificity for RNA-guided genome editing |
| EP2981617B1 (en) | 2013-04-04 | 2023-07-05 | President and Fellows of Harvard College | Therapeutic uses of genome editing with crispr/cas systems |
| US9267135B2 (en) | 2013-06-04 | 2016-02-23 | President And Fellows Of Harvard College | RNA-guided transcriptional regulation |
| EP3725885A1 (en) | 2013-06-17 | 2020-10-21 | The Broad Institute, Inc. | Functional genomics using crispr-cas systems, compositions methods, screens and applications thereof |
| US20160237455A1 (en) | 2013-09-27 | 2016-08-18 | Editas Medicine, Inc. | Crispr-related methods and compositions |
| WO2015065964A1 (en) | 2013-10-28 | 2015-05-07 | The Broad Institute Inc. | Functional genomics using crispr-cas systems, compositions, methods, screens and applications thereof |
| WO2015122922A1 (en) | 2014-02-14 | 2015-08-20 | Ipierian, Inc. | Tau peptides, anti-tau antibodies, and methods of use thereof |
| US20150376586A1 (en) | 2014-06-25 | 2015-12-31 | Caribou Biosciences, Inc. | RNA Modification to Engineer Cas9 Activity |
| WO2016011080A2 (en) | 2014-07-14 | 2016-01-21 | The Regents Of The University Of California | Crispr/cas transcriptional modulation |
| WO2016049258A2 (en) | 2014-09-25 | 2016-03-31 | The Broad Institute Inc. | Functional screening with optimized functional crispr-cas systems |
| WO2016106236A1 (en) | 2014-12-23 | 2016-06-30 | The Broad Institute Inc. | Rna-targeting system |
| US9790490B2 (en) | 2015-06-18 | 2017-10-17 | The Broad Institute Inc. | CRISPR enzymes and systems |
| US10676735B2 (en) | 2015-07-22 | 2020-06-09 | Duke University | High-throughput screening of regulatory element function with epigenome editing technologies |
| CA3006781A1 (en) | 2015-12-07 | 2017-06-15 | Arc Bio, Llc | Methods and compositions for the making and using of guide nucleic acids |
| KR102617874B1 (ko) | 2016-03-30 | 2023-12-22 | 인텔리아 테라퓨틱스, 인크. | Crispr/cas 성분을 위한 지질 나노입자 제제 |
| WO2017172764A1 (en) | 2016-04-01 | 2017-10-05 | The Regents Of The University Of California | Modified cell line and method of determining tauopathies |
| CN109072191B (zh) * | 2016-04-04 | 2024-03-22 | 苏黎世联邦理工学院 | 用于蛋白生产和文库产生的哺乳动物细胞系 |
| EP3565836A1 (en) | 2017-01-04 | 2019-11-13 | H. Lundbeck A/S | Antibodies specific for hyperphosphorylated tau for the treatment of ocular diseases |
| GB201708998D0 (en) | 2017-06-06 | 2017-07-19 | Univ Leuven Kath | Methods for identifying activators and suppressors |
| WO2019010384A1 (en) | 2017-07-07 | 2019-01-10 | The Broad Institute, Inc. | METHODS FOR DESIGNING GUIDE SEQUENCES FOR GUIDED NUCLEASES |
| SG11201912236YA (en) * | 2017-07-31 | 2020-01-30 | Regeneron Pharma | Crispr reporter non-human animals and uses thereof |
| MX2020001178A (es) | 2017-07-31 | 2020-09-25 | Regeneron Pharma | Celulas madre embrionarias de raton transgenico con cas y ratones y usos de los mismos. |
| KR20240038811A (ko) | 2018-03-19 | 2024-03-25 | 리제너론 파마슈티칼스 인코포레이티드 | CRISPR/Cas 시스템을 사용한 동물에서의 전사 조절 |
| CA3127813A1 (en) | 2019-03-18 | 2020-09-24 | Regeneron Pharmaceuticals, Inc. | Crispr/cas screening platform to identify genetic modifiers of tau seeding or aggregation |
| SG11202108090XA (en) | 2019-03-18 | 2021-08-30 | Regeneron Pharma | Crispr/cas dropout screening platform to reveal genetic vulnerabilities associated with tau aggregation |
| EP3942558A1 (en) | 2019-03-18 | 2022-01-26 | Regeneron Pharmaceuticals, Inc. | Methods and systems for crispr selection |
| KR20220024053A (ko) | 2019-06-14 | 2022-03-03 | 리제너론 파마슈티칼스 인코포레이티드 | 타우병증의 모델 |
-
2020
- 2020-03-17 SG SG11202108090XA patent/SG11202108090XA/en unknown
- 2020-03-17 KR KR1020217032083A patent/KR20210141536A/ko not_active Application Discontinuation
- 2020-03-17 CN CN202080028975.9A patent/CN113711046B/zh active Active
- 2020-03-17 WO PCT/US2020/023121 patent/WO2020190927A1/en unknown
- 2020-03-17 JP JP2021556467A patent/JP7389135B2/ja active Active
- 2020-03-17 CA CA3127814A patent/CA3127814C/en active Active
- 2020-03-17 US US16/821,384 patent/US11781131B2/en active Active
- 2020-03-17 IL IL286359A patent/IL286359B1/en unknown
- 2020-03-17 EP EP20718947.3A patent/EP3769090B1/en active Active
- 2020-03-17 ES ES20718947T patent/ES2969026T3/es active Active
- 2020-03-17 AU AU2020241856A patent/AU2020241856A1/en active Pending
-
2023
- 2023-08-31 US US18/459,091 patent/US20230416728A1/en active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| CN113711046A (zh) | 2021-11-26 |
| CA3127814A1 (en) | 2020-09-24 |
| WO2020190927A1 (en) | 2020-09-24 |
| US11781131B2 (en) | 2023-10-10 |
| IL286359A (en) | 2021-10-31 |
| CN113711046B (zh) | 2023-08-29 |
| ES2969026T3 (es) | 2024-05-16 |
| AU2020241856A1 (en) | 2021-08-12 |
| IL286359B1 (en) | 2024-02-01 |
| EP3769090A1 (en) | 2021-01-27 |
| KR20210141536A (ko) | 2021-11-23 |
| US20200299681A1 (en) | 2020-09-24 |
| CA3127814C (en) | 2024-05-14 |
| EP3769090C0 (en) | 2023-11-15 |
| EP3769090B1 (en) | 2023-11-15 |
| US20230416728A1 (en) | 2023-12-28 |
| SG11202108090XA (en) | 2021-08-30 |
| JP7389135B2 (ja) | 2023-11-29 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN116590257B (zh) | VI-E型和VI-F型CRISPR-Cas系统及其用途 | |
| JP7389135B2 (ja) | タウ凝集に関連する遺伝的脆弱性を明らかにするためのcrispr/casドロップアウトスクリーニングプラットフォーム | |
| JP7461368B2 (ja) | タウの播種または凝集の遺伝的修飾因子を同定するためのcrispr/casスクリーニングプラットフォーム | |
| US20230212542A1 (en) | Novel crispr dna targeting enzymes and systems | |
| US20030143597A1 (en) | Methods for making polynucleotide libraries, polynucleotide arrays, and cell libraries for high-throughput genomics analysis | |
| US20240076613A1 (en) | Models of tauopathy | |
| WO2023274226A1 (en) | Crispr/cas system and uses thereof | |
| RU2808829C2 (ru) | Платформа crispr/cas для исключающего скрининга для выявления генетических уязвимостей, связанных с агрегацией тау-белка | |
| RU2819350C2 (ru) | Скрининговая платформа crispr/cas для идентификации генетических модификаторов засева или агрегации тау-белка | |
| JP2024071489A (ja) | タウの播種または凝集の遺伝的修飾因子を同定するためのcrispr/casスクリーニングプラットフォーム | |
| JPWO2020122195A1 (ja) | ゲノム編集された細胞の製造方法 | |
| US20040137490A1 (en) | Methods for making polynucleotide libraries, polynucleotide arrays, and cell libraries for high-throughput genomics analysis | |
| WO2023164482A2 (en) | Treatment for nucleotide repeat expansion disease |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20230306 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20230306 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20230919 |
|
| A871 | Explanation of circumstances concerning accelerated examination |
Free format text: JAPANESE INTERMEDIATE CODE: A871 Effective date: 20230919 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20231025 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20231116 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7389135 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |