JP2021524451A - 最適化済みgp41結合分子及びその使用 - Google Patents
最適化済みgp41結合分子及びその使用 Download PDFInfo
- Publication number
- JP2021524451A JP2021524451A JP2020564553A JP2020564553A JP2021524451A JP 2021524451 A JP2021524451 A JP 2021524451A JP 2020564553 A JP2020564553 A JP 2020564553A JP 2020564553 A JP2020564553 A JP 2020564553A JP 2021524451 A JP2021524451 A JP 2021524451A
- Authority
- JP
- Japan
- Prior art keywords
- binding
- domain
- epitope
- molecule
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 230000027455 binding Effects 0.000 title claims abstract description 627
- 101800001690 Transmembrane protein gp41 Proteins 0.000 title claims abstract description 300
- 208000031886 HIV Infections Diseases 0.000 claims abstract description 82
- 239000012636 effector Substances 0.000 claims abstract description 69
- 238000000034 method Methods 0.000 claims abstract description 52
- 238000011282 treatment Methods 0.000 claims abstract description 27
- 239000008194 pharmaceutical composition Substances 0.000 claims abstract description 23
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 384
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 371
- 229920001184 polypeptide Polymers 0.000 claims description 367
- 210000004027 cell Anatomy 0.000 claims description 162
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 157
- 238000006467 substitution reaction Methods 0.000 claims description 53
- 102100029185 Low affinity immunoglobulin gamma Fc region receptor III-B Human genes 0.000 claims description 40
- 101000917839 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-B Proteins 0.000 claims description 33
- 101000917858 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 claims description 30
- 239000000203 mixture Substances 0.000 claims description 30
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 claims description 25
- 230000004048 modification Effects 0.000 claims description 24
- 238000012986 modification Methods 0.000 claims description 24
- 150000001413 amino acids Chemical class 0.000 claims description 21
- 101001109501 Homo sapiens NKG2-D type II integral membrane protein Proteins 0.000 claims description 12
- 102100022680 NKG2-D type II integral membrane protein Human genes 0.000 claims description 12
- 210000002966 serum Anatomy 0.000 claims description 12
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 claims description 11
- 108010088751 Albumins Proteins 0.000 claims description 10
- 102000009027 Albumins Human genes 0.000 claims description 10
- 108010004217 Natural Cytotoxicity Triggering Receptor 1 Proteins 0.000 claims description 9
- 102100032870 Natural cytotoxicity triggering receptor 1 Human genes 0.000 claims description 8
- 239000012190 activator Substances 0.000 claims description 6
- AUZONCFQVSMFAP-UHFFFAOYSA-N disulfiram Chemical compound CCN(CC)C(=S)SSC(=S)N(CC)CC AUZONCFQVSMFAP-UHFFFAOYSA-N 0.000 claims description 6
- 239000003937 drug carrier Substances 0.000 claims description 6
- 208000032420 Latent Infection Diseases 0.000 claims description 5
- 208000037771 disease arising from reactivation of latent virus Diseases 0.000 claims description 5
- 230000003053 immunization Effects 0.000 claims description 5
- 230000002265 prevention Effects 0.000 claims description 4
- PHEDXBVPIONUQT-UHFFFAOYSA-N Cocarcinogen A1 Natural products CCCCCCCCCCCCCC(=O)OC1C(C)C2(O)C3C=C(C)C(=O)C3(O)CC(CO)=CC2C2C1(OC(C)=O)C2(C)C PHEDXBVPIONUQT-UHFFFAOYSA-N 0.000 claims description 3
- 108700010908 HIV-1 proteins Proteins 0.000 claims description 3
- DNVXATUJJDPFDM-KRWDZBQOSA-N JQ1 Chemical compound N([C@@H](CC(=O)OC(C)(C)C)C1=NN=C(N1C=1SC(C)=C(C)C=11)C)=C1C1=CC=C(Cl)C=C1 DNVXATUJJDPFDM-KRWDZBQOSA-N 0.000 claims description 3
- 229960005549 JQ1 Drugs 0.000 claims description 3
- 229960005520 bryostatin Drugs 0.000 claims description 3
- MJQUEDHRCUIRLF-TVIXENOKSA-N bryostatin 1 Chemical compound C([C@@H]1CC(/[C@@H]([C@@](C(C)(C)/C=C/2)(O)O1)OC(=O)/C=C/C=C/CCC)=C\C(=O)OC)[C@H]([C@@H](C)O)OC(=O)C[C@H](O)C[C@@H](O1)C[C@H](OC(C)=O)C(C)(C)[C@]1(O)C[C@@H]1C\C(=C\C(=O)OC)C[C@H]\2O1 MJQUEDHRCUIRLF-TVIXENOKSA-N 0.000 claims description 3
- MUIWQCKLQMOUAT-AKUNNTHJSA-N bryostatin 20 Natural products COC(=O)C=C1C[C@@]2(C)C[C@]3(O)O[C@](C)(C[C@@H](O)CC(=O)O[C@](C)(C[C@@]4(C)O[C@](O)(CC5=CC(=O)O[C@]45C)C(C)(C)C=C[C@@](C)(C1)O2)[C@@H](C)O)C[C@H](OC(=O)C(C)(C)C)C3(C)C MUIWQCKLQMOUAT-AKUNNTHJSA-N 0.000 claims description 3
- 229960002563 disulfiram Drugs 0.000 claims description 3
- PGHMRUGBZOYCAA-ADZNBVRBSA-N ionomycin Chemical compound O1[C@H](C[C@H](O)[C@H](C)[C@H](O)[C@H](C)/C=C/C[C@@H](C)C[C@@H](C)C(/O)=C/C(=O)[C@@H](C)C[C@@H](C)C[C@@H](CCC(O)=O)C)CC[C@@]1(C)[C@@H]1O[C@](C)([C@@H](C)O)CC1 PGHMRUGBZOYCAA-ADZNBVRBSA-N 0.000 claims description 3
- PGHMRUGBZOYCAA-UHFFFAOYSA-N ionomycin Natural products O1C(CC(O)C(C)C(O)C(C)C=CCC(C)CC(C)C(O)=CC(=O)C(C)CC(C)CC(CCC(O)=O)C)CCC1(C)C1OC(C)(C(C)O)CC1 PGHMRUGBZOYCAA-UHFFFAOYSA-N 0.000 claims description 3
- 229960005184 panobinostat Drugs 0.000 claims description 3
- FWZRWHZDXBDTFK-ZHACJKMWSA-N panobinostat Chemical compound CC1=NC2=CC=C[CH]C2=C1CCNCC1=CC=C(\C=C\C(=O)NO)C=C1 FWZRWHZDXBDTFK-ZHACJKMWSA-N 0.000 claims description 3
- PHEDXBVPIONUQT-RGYGYFBISA-N phorbol 13-acetate 12-myristate Chemical compound C([C@]1(O)C(=O)C(C)=C[C@H]1[C@@]1(O)[C@H](C)[C@H]2OC(=O)CCCCCCCCCCCCC)C(CO)=C[C@H]1[C@H]1[C@]2(OC(C)=O)C1(C)C PHEDXBVPIONUQT-RGYGYFBISA-N 0.000 claims description 3
- OHRURASPPZQGQM-GCCNXGTGSA-N romidepsin Chemical compound O1C(=O)[C@H](C(C)C)NC(=O)C(=C/C)/NC(=O)[C@H]2CSSCC\C=C\[C@@H]1CC(=O)N[C@H](C(C)C)C(=O)N2 OHRURASPPZQGQM-GCCNXGTGSA-N 0.000 claims description 3
- 229960003452 romidepsin Drugs 0.000 claims description 3
- OHRURASPPZQGQM-UHFFFAOYSA-N romidepsin Natural products O1C(=O)C(C(C)C)NC(=O)C(=CC)NC(=O)C2CSSCCC=CC1CC(=O)NC(C(C)C)C(=O)N2 OHRURASPPZQGQM-UHFFFAOYSA-N 0.000 claims description 3
- 108010091666 romidepsin Proteins 0.000 claims description 3
- WAEXFXRVDQXREF-UHFFFAOYSA-N vorinostat Chemical group ONC(=O)CCCCCCC(=O)NC1=CC=CC=C1 WAEXFXRVDQXREF-UHFFFAOYSA-N 0.000 claims description 3
- 229960000237 vorinostat Drugs 0.000 claims description 3
- 101710114816 Gene 41 protein Proteins 0.000 claims description 2
- 101000935040 Homo sapiens Integrin beta-2 Proteins 0.000 claims description 2
- 102100025390 Integrin beta-2 Human genes 0.000 claims description 2
- 241000713772 Human immunodeficiency virus 1 Species 0.000 abstract description 68
- 238000012063 dual-affinity re-targeting Methods 0.000 abstract description 15
- 230000005847 immunogenicity Effects 0.000 abstract description 12
- 230000002829 reductive effect Effects 0.000 abstract description 12
- 238000000375 direct analysis in real time Methods 0.000 abstract description 11
- 102100024952 Protein CBFA2T1 Human genes 0.000 description 101
- 241000282414 Homo sapiens Species 0.000 description 98
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 76
- 108050005493 CD3 protein, epsilon/gamma/delta subunit Proteins 0.000 description 76
- 239000000427 antigen Substances 0.000 description 44
- 108091007433 antigens Proteins 0.000 description 44
- 102000036639 antigens Human genes 0.000 description 44
- 210000001744 T-lymphocyte Anatomy 0.000 description 43
- 235000018102 proteins Nutrition 0.000 description 40
- 108090000623 proteins and genes Proteins 0.000 description 40
- 102000004169 proteins and genes Human genes 0.000 description 40
- 125000001495 ethyl group Chemical group [H]C([H])([H])C([H])([H])* 0.000 description 33
- 235000001014 amino acid Nutrition 0.000 description 29
- 206010028980 Neoplasm Diseases 0.000 description 28
- 230000006870 function Effects 0.000 description 27
- 108091008874 T cell receptors Proteins 0.000 description 26
- 102000016266 T-Cell Antigen Receptors Human genes 0.000 description 25
- 239000003814 drug Substances 0.000 description 25
- 101000946843 Homo sapiens T-cell surface glycoprotein CD8 alpha chain Proteins 0.000 description 23
- 241000725303 Human immunodeficiency virus Species 0.000 description 23
- 102100034922 T-cell surface glycoprotein CD8 alpha chain Human genes 0.000 description 23
- 102100034428 Dual specificity protein phosphatase 1 Human genes 0.000 description 22
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 22
- 230000001225 therapeutic effect Effects 0.000 description 21
- 239000004472 Lysine Substances 0.000 description 20
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 20
- 229940024606 amino acid Drugs 0.000 description 20
- 239000000833 heterodimer Substances 0.000 description 19
- 102000005962 receptors Human genes 0.000 description 19
- 108020003175 receptors Proteins 0.000 description 19
- 101800001707 Spacer peptide Proteins 0.000 description 18
- 230000000694 effects Effects 0.000 description 18
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 17
- 235000018417 cysteine Nutrition 0.000 description 16
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 16
- 201000010099 disease Diseases 0.000 description 16
- 102100027088 Dual specificity protein phosphatase 5 Human genes 0.000 description 15
- 201000011510 cancer Diseases 0.000 description 15
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 15
- 210000000822 natural killer cell Anatomy 0.000 description 15
- 101000934346 Homo sapiens T-cell surface antigen CD2 Proteins 0.000 description 14
- 108060003951 Immunoglobulin Proteins 0.000 description 14
- 102100029205 Low affinity immunoglobulin gamma Fc region receptor II-b Human genes 0.000 description 14
- 102100025237 T-cell surface antigen CD2 Human genes 0.000 description 14
- 210000004899 c-terminal region Anatomy 0.000 description 14
- 102000018358 immunoglobulin Human genes 0.000 description 14
- 241000894007 species Species 0.000 description 14
- 239000003795 chemical substances by application Substances 0.000 description 13
- 230000002147 killing effect Effects 0.000 description 13
- 238000004519 manufacturing process Methods 0.000 description 13
- 230000035772 mutation Effects 0.000 description 13
- 230000008685 targeting Effects 0.000 description 13
- 229940079593 drug Drugs 0.000 description 12
- 230000003993 interaction Effects 0.000 description 12
- 229940124597 therapeutic agent Drugs 0.000 description 12
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 11
- 125000000539 amino acid group Chemical group 0.000 description 11
- 230000015572 biosynthetic process Effects 0.000 description 11
- 230000022534 cell killing Effects 0.000 description 11
- 108091033319 polynucleotide Proteins 0.000 description 11
- 102000040430 polynucleotide Human genes 0.000 description 11
- 239000002157 polynucleotide Substances 0.000 description 11
- 230000003612 virological effect Effects 0.000 description 11
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 10
- 241001465754 Metazoa Species 0.000 description 10
- 241000699666 Mus <mouse, genus> Species 0.000 description 10
- 101000917824 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-b Proteins 0.000 description 9
- 241000283984 Rodentia Species 0.000 description 9
- 102100036011 T-cell surface glycoprotein CD4 Human genes 0.000 description 9
- 238000001727 in vivo Methods 0.000 description 9
- 230000001965 increasing effect Effects 0.000 description 9
- 208000015181 infectious disease Diseases 0.000 description 9
- 230000001404 mediated effect Effects 0.000 description 9
- 102220005411 rs35873730 Human genes 0.000 description 9
- 239000000126 substance Substances 0.000 description 9
- 208000024891 symptom Diseases 0.000 description 9
- 238000002560 therapeutic procedure Methods 0.000 description 9
- 241000282412 Homo Species 0.000 description 8
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 8
- 238000003556 assay Methods 0.000 description 8
- 210000003719 b-lymphocyte Anatomy 0.000 description 8
- 238000006243 chemical reaction Methods 0.000 description 8
- 238000010276 construction Methods 0.000 description 8
- 108020001507 fusion proteins Proteins 0.000 description 8
- 210000004602 germ cell Anatomy 0.000 description 8
- 238000005734 heterodimerization reaction Methods 0.000 description 8
- 210000000987 immune system Anatomy 0.000 description 8
- 230000031998 transcytosis Effects 0.000 description 8
- 101100334515 Homo sapiens FCGR3A gene Proteins 0.000 description 7
- 102000037865 fusion proteins Human genes 0.000 description 7
- 101150026046 iga gene Proteins 0.000 description 7
- 230000003472 neutralizing effect Effects 0.000 description 7
- 230000001737 promoting effect Effects 0.000 description 7
- 230000009467 reduction Effects 0.000 description 7
- 108010021472 Fc gamma receptor IIB Proteins 0.000 description 6
- 102100022102 Histone-lysine N-methyltransferase 2B Human genes 0.000 description 6
- 101001045848 Homo sapiens Histone-lysine N-methyltransferase 2B Proteins 0.000 description 6
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 6
- 102100034349 Integrase Human genes 0.000 description 6
- 241000288906 Primates Species 0.000 description 6
- 230000003213 activating effect Effects 0.000 description 6
- 238000007792 addition Methods 0.000 description 6
- 239000002671 adjuvant Substances 0.000 description 6
- 238000013459 approach Methods 0.000 description 6
- 230000000295 complement effect Effects 0.000 description 6
- 208000035475 disorder Diseases 0.000 description 6
- 230000004927 fusion Effects 0.000 description 6
- 210000004379 membrane Anatomy 0.000 description 6
- 239000012528 membrane Substances 0.000 description 6
- 210000004877 mucosa Anatomy 0.000 description 6
- 230000003389 potentiating effect Effects 0.000 description 6
- 241000282693 Cercopithecidae Species 0.000 description 5
- 208000035473 Communicable disease Diseases 0.000 description 5
- 108010021468 Fc gamma receptor IIA Proteins 0.000 description 5
- 108010087819 Fc receptors Proteins 0.000 description 5
- 102000009109 Fc receptors Human genes 0.000 description 5
- 102100026122 High affinity immunoglobulin gamma Fc receptor I Human genes 0.000 description 5
- 101100334524 Homo sapiens FCGR3B gene Proteins 0.000 description 5
- 101000913074 Homo sapiens High affinity immunoglobulin gamma Fc receptor I Proteins 0.000 description 5
- 101000917826 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-a Proteins 0.000 description 5
- 108010073807 IgG Receptors Proteins 0.000 description 5
- 241000700605 Viruses Species 0.000 description 5
- 230000004913 activation Effects 0.000 description 5
- 210000004369 blood Anatomy 0.000 description 5
- 239000008280 blood Substances 0.000 description 5
- 238000012512 characterization method Methods 0.000 description 5
- 230000003013 cytotoxicity Effects 0.000 description 5
- 231100000135 cytotoxicity Toxicity 0.000 description 5
- 239000012642 immune effector Substances 0.000 description 5
- 230000002163 immunogen Effects 0.000 description 5
- 229940121354 immunomodulator Drugs 0.000 description 5
- 238000002360 preparation method Methods 0.000 description 5
- 210000001519 tissue Anatomy 0.000 description 5
- 239000013598 vector Substances 0.000 description 5
- 125000001433 C-terminal amino-acid group Chemical group 0.000 description 4
- 241000196324 Embryophyta Species 0.000 description 4
- 108090000288 Glycoproteins Proteins 0.000 description 4
- 102000003886 Glycoproteins Human genes 0.000 description 4
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 4
- 102100034353 Integrase Human genes 0.000 description 4
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 4
- 241000124008 Mammalia Species 0.000 description 4
- 241001529936 Murinae Species 0.000 description 4
- 108010001657 NK Cell Lectin-Like Receptor Subfamily K Proteins 0.000 description 4
- 102000000812 NK Cell Lectin-Like Receptor Subfamily K Human genes 0.000 description 4
- 230000002494 anti-cea effect Effects 0.000 description 4
- 230000036436 anti-hiv Effects 0.000 description 4
- 230000000890 antigenic effect Effects 0.000 description 4
- 230000004071 biological effect Effects 0.000 description 4
- 238000010367 cloning Methods 0.000 description 4
- 150000001875 compounds Chemical class 0.000 description 4
- 238000006471 dimerization reaction Methods 0.000 description 4
- 230000008030 elimination Effects 0.000 description 4
- 238000003379 elimination reaction Methods 0.000 description 4
- 108010078428 env Gene Products Proteins 0.000 description 4
- 210000000981 epithelium Anatomy 0.000 description 4
- 239000000710 homodimer Substances 0.000 description 4
- 230000028993 immune response Effects 0.000 description 4
- 230000036039 immunity Effects 0.000 description 4
- 229940072221 immunoglobulins Drugs 0.000 description 4
- 230000001976 improved effect Effects 0.000 description 4
- 238000001802 infusion Methods 0.000 description 4
- 108091008042 inhibitory receptors Proteins 0.000 description 4
- 239000000463 material Substances 0.000 description 4
- 238000006386 neutralization reaction Methods 0.000 description 4
- 230000036961 partial effect Effects 0.000 description 4
- 239000000546 pharmaceutical excipient Substances 0.000 description 4
- 230000002285 radioactive effect Effects 0.000 description 4
- 238000003786 synthesis reaction Methods 0.000 description 4
- 239000013638 trimer Substances 0.000 description 4
- 241000283690 Bos taurus Species 0.000 description 3
- 241000282465 Canis Species 0.000 description 3
- 102000001301 EGF receptor Human genes 0.000 description 3
- 108060006698 EGF receptor Proteins 0.000 description 3
- 101710121417 Envelope glycoprotein Proteins 0.000 description 3
- 101710091045 Envelope protein Proteins 0.000 description 3
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Natural products NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 3
- 101800000194 Growth hormone-binding protein Proteins 0.000 description 3
- 102400001066 Growth hormone-binding protein Human genes 0.000 description 3
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 3
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- 102000043129 MHC class I family Human genes 0.000 description 3
- 108091054437 MHC class I family Proteins 0.000 description 3
- 241000282567 Macaca fascicularis Species 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- 101710188315 Protein X Proteins 0.000 description 3
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 3
- 102000004584 Somatomedin Receptors Human genes 0.000 description 3
- 108010017622 Somatomedin Receptors Proteins 0.000 description 3
- 230000006044 T cell activation Effects 0.000 description 3
- 239000004480 active ingredient Substances 0.000 description 3
- 230000001154 acute effect Effects 0.000 description 3
- 235000004279 alanine Nutrition 0.000 description 3
- 230000000259 anti-tumor effect Effects 0.000 description 3
- 210000000612 antigen-presenting cell Anatomy 0.000 description 3
- 230000000112 colonic effect Effects 0.000 description 3
- 230000021615 conjugation Effects 0.000 description 3
- 231100000433 cytotoxic Toxicity 0.000 description 3
- 229940127089 cytotoxic agent Drugs 0.000 description 3
- 231100000599 cytotoxic agent Toxicity 0.000 description 3
- 230000001472 cytotoxic effect Effects 0.000 description 3
- 238000013461 design Methods 0.000 description 3
- 229910003460 diamond Inorganic materials 0.000 description 3
- 239000010432 diamond Substances 0.000 description 3
- 239000000539 dimer Substances 0.000 description 3
- 230000009977 dual effect Effects 0.000 description 3
- 238000005516 engineering process Methods 0.000 description 3
- 210000003979 eosinophil Anatomy 0.000 description 3
- 230000005714 functional activity Effects 0.000 description 3
- 210000003630 histaminocyte Anatomy 0.000 description 3
- 210000004408 hybridoma Anatomy 0.000 description 3
- 210000002865 immune cell Anatomy 0.000 description 3
- 238000000338 in vitro Methods 0.000 description 3
- 239000004615 ingredient Substances 0.000 description 3
- 230000000977 initiatory effect Effects 0.000 description 3
- 238000002347 injection Methods 0.000 description 3
- 239000007924 injection Substances 0.000 description 3
- 230000003834 intracellular effect Effects 0.000 description 3
- 239000003446 ligand Substances 0.000 description 3
- 239000008176 lyophilized powder Substances 0.000 description 3
- 210000002540 macrophage Anatomy 0.000 description 3
- 235000013336 milk Nutrition 0.000 description 3
- 239000008267 milk Substances 0.000 description 3
- 210000004080 milk Anatomy 0.000 description 3
- 210000005087 mononuclear cell Anatomy 0.000 description 3
- 239000013642 negative control Substances 0.000 description 3
- 108010068617 neonatal Fc receptor Proteins 0.000 description 3
- 210000000440 neutrophil Anatomy 0.000 description 3
- 239000002773 nucleotide Substances 0.000 description 3
- 125000003729 nucleotide group Chemical group 0.000 description 3
- 238000011275 oncology therapy Methods 0.000 description 3
- 238000002823 phage display Methods 0.000 description 3
- 238000003259 recombinant expression Methods 0.000 description 3
- 238000005215 recombination Methods 0.000 description 3
- 230000006798 recombination Effects 0.000 description 3
- 230000003248 secreting effect Effects 0.000 description 3
- 238000013207 serial dilution Methods 0.000 description 3
- 230000011664 signaling Effects 0.000 description 3
- 238000010532 solid phase synthesis reaction Methods 0.000 description 3
- 230000004083 survival effect Effects 0.000 description 3
- 239000003053 toxin Substances 0.000 description 3
- 231100000765 toxin Toxicity 0.000 description 3
- 108700012359 toxins Proteins 0.000 description 3
- 210000004881 tumor cell Anatomy 0.000 description 3
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- 101100208114 Arabidopsis thaliana TRX8 gene Proteins 0.000 description 2
- 208000003950 B-cell lymphoma Diseases 0.000 description 2
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 2
- 241000282472 Canis lupus familiaris Species 0.000 description 2
- 108010022366 Carcinoembryonic Antigen Proteins 0.000 description 2
- 102100025475 Carcinoembryonic antigen-related cell adhesion molecule 5 Human genes 0.000 description 2
- 102000000844 Cell Surface Receptors Human genes 0.000 description 2
- 108010001857 Cell Surface Receptors Proteins 0.000 description 2
- 206010009944 Colon cancer Diseases 0.000 description 2
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 2
- 108091035707 Consensus sequence Proteins 0.000 description 2
- 108020004414 DNA Proteins 0.000 description 2
- 206010061818 Disease progression Diseases 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- 241001482630 Epinnula magistralis Species 0.000 description 2
- 241000283073 Equus caballus Species 0.000 description 2
- 241000282326 Felis catus Species 0.000 description 2
- 102100021260 Galactosylgalactosylxylosylprotein 3-beta-glucuronosyltransferase 1 Human genes 0.000 description 2
- 239000004471 Glycine Substances 0.000 description 2
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 2
- 101000894906 Homo sapiens Galactosylgalactosylxylosylprotein 3-beta-glucuronosyltransferase 1 Proteins 0.000 description 2
- 101000604674 Homo sapiens Immunoglobulin kappa variable 4-1 Proteins 0.000 description 2
- 101000946860 Homo sapiens T-cell surface glycoprotein CD3 epsilon chain Proteins 0.000 description 2
- 102000009490 IgG Receptors Human genes 0.000 description 2
- 102100038198 Immunoglobulin kappa variable 4-1 Human genes 0.000 description 2
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 2
- 102000003855 L-lactate dehydrogenase Human genes 0.000 description 2
- 108700023483 L-lactate dehydrogenases Proteins 0.000 description 2
- 206010025323 Lymphomas Diseases 0.000 description 2
- 102000043131 MHC class II family Human genes 0.000 description 2
- 108091054438 MHC class II family Proteins 0.000 description 2
- 102220482767 Mitochondrial coenzyme A diphosphatase NUDT8_D79A_mutation Human genes 0.000 description 2
- 108091028043 Nucleic acid sequence Proteins 0.000 description 2
- 241000283973 Oryctolagus cuniculus Species 0.000 description 2
- 208000002193 Pain Diseases 0.000 description 2
- 206010037660 Pyrexia Diseases 0.000 description 2
- 206010040047 Sepsis Diseases 0.000 description 2
- 108700042075 T-Cell Receptor Genes Proteins 0.000 description 2
- 102100035794 T-cell surface glycoprotein CD3 epsilon chain Human genes 0.000 description 2
- 206010058874 Viraemia Diseases 0.000 description 2
- 230000009471 action Effects 0.000 description 2
- 239000003708 ampul Substances 0.000 description 2
- 238000004458 analytical method Methods 0.000 description 2
- 230000001130 anti-lysozyme effect Effects 0.000 description 2
- 102000025171 antigen binding proteins Human genes 0.000 description 2
- 108091000831 antigen binding proteins Proteins 0.000 description 2
- 239000002246 antineoplastic agent Substances 0.000 description 2
- 235000003704 aspartic acid Nutrition 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 2
- 230000033228 biological regulation Effects 0.000 description 2
- 230000005540 biological transmission Effects 0.000 description 2
- 238000002619 cancer immunotherapy Methods 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 2
- 238000003776 cleavage reaction Methods 0.000 description 2
- 239000012141 concentrate Substances 0.000 description 2
- 239000013256 coordination polymer Substances 0.000 description 2
- 238000004132 cross linking Methods 0.000 description 2
- 230000009089 cytolysis Effects 0.000 description 2
- 239000002619 cytotoxin Substances 0.000 description 2
- 238000012217 deletion Methods 0.000 description 2
- 230000037430 deletion Effects 0.000 description 2
- 230000001419 dependent effect Effects 0.000 description 2
- 238000010586 diagram Methods 0.000 description 2
- 239000003085 diluting agent Substances 0.000 description 2
- 230000005750 disease progression Effects 0.000 description 2
- 239000002552 dosage form Substances 0.000 description 2
- 231100000673 dose–response relationship Toxicity 0.000 description 2
- 230000002708 enhancing effect Effects 0.000 description 2
- 210000002919 epithelial cell Anatomy 0.000 description 2
- 238000000684 flow cytometry Methods 0.000 description 2
- 239000012634 fragment Substances 0.000 description 2
- 238000010353 genetic engineering Methods 0.000 description 2
- 102000054766 genetic haplotypes Human genes 0.000 description 2
- 230000012010 growth Effects 0.000 description 2
- 230000002519 immonomodulatory effect Effects 0.000 description 2
- 230000005931 immune cell recruitment Effects 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 238000002649 immunization Methods 0.000 description 2
- 229940127121 immunoconjugate Drugs 0.000 description 2
- 230000002637 immunotoxin Effects 0.000 description 2
- 239000002596 immunotoxin Substances 0.000 description 2
- 229940051026 immunotoxin Drugs 0.000 description 2
- 231100000608 immunotoxin Toxicity 0.000 description 2
- 239000007943 implant Substances 0.000 description 2
- 230000006872 improvement Effects 0.000 description 2
- 238000010348 incorporation Methods 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 239000002502 liposome Substances 0.000 description 2
- 239000007788 liquid Substances 0.000 description 2
- 238000004020 luminiscence type Methods 0.000 description 2
- 210000004698 lymphocyte Anatomy 0.000 description 2
- 230000007246 mechanism Effects 0.000 description 2
- 230000010534 mechanism of action Effects 0.000 description 2
- 229910021645 metal ion Inorganic materials 0.000 description 2
- 239000000178 monomer Substances 0.000 description 2
- 239000002071 nanotube Substances 0.000 description 2
- 102000039446 nucleic acids Human genes 0.000 description 2
- 108020004707 nucleic acids Proteins 0.000 description 2
- 150000007523 nucleic acids Chemical class 0.000 description 2
- 238000005457 optimization Methods 0.000 description 2
- 230000001717 pathogenic effect Effects 0.000 description 2
- 239000002243 precursor Substances 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 238000012545 processing Methods 0.000 description 2
- 230000000069 prophylactic effect Effects 0.000 description 2
- 230000017854 proteolysis Effects 0.000 description 2
- 238000000163 radioactive labelling Methods 0.000 description 2
- 238000010188 recombinant method Methods 0.000 description 2
- 230000001105 regulatory effect Effects 0.000 description 2
- 230000004044 response Effects 0.000 description 2
- FGDZQCVHDSGLHJ-UHFFFAOYSA-M rubidium chloride Chemical compound [Cl-].[Rb+] FGDZQCVHDSGLHJ-UHFFFAOYSA-M 0.000 description 2
- 230000007017 scission Effects 0.000 description 2
- 210000003491 skin Anatomy 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 125000006850 spacer group Chemical group 0.000 description 2
- 230000000087 stabilizing effect Effects 0.000 description 2
- 230000000699 topical effect Effects 0.000 description 2
- 238000001890 transfection Methods 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 230000009261 transgenic effect Effects 0.000 description 2
- 230000001960 triggered effect Effects 0.000 description 2
- 210000000605 viral structure Anatomy 0.000 description 2
- 210000002845 virion Anatomy 0.000 description 2
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- 208000030507 AIDS Diseases 0.000 description 1
- 108010066676 Abrin Proteins 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- 208000023275 Autoimmune disease Diseases 0.000 description 1
- 108090001008 Avidin Proteins 0.000 description 1
- 208000028564 B-cell non-Hodgkin lymphoma Diseases 0.000 description 1
- 102100038080 B-cell receptor CD22 Human genes 0.000 description 1
- 230000003844 B-cell-activation Effects 0.000 description 1
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 description 1
- 102100026189 Beta-galactosidase Human genes 0.000 description 1
- 108030001720 Bontoxilysin Proteins 0.000 description 1
- 208000011691 Burkitt lymphomas Diseases 0.000 description 1
- 102100032529 C-type lectin domain family 1 member B Human genes 0.000 description 1
- 108010041397 CD4 Antigens Proteins 0.000 description 1
- 101150013553 CD40 gene Proteins 0.000 description 1
- 102100027217 CD82 antigen Human genes 0.000 description 1
- 102000000905 Cadherin Human genes 0.000 description 1
- 108050007957 Cadherin Proteins 0.000 description 1
- UXVMQQNJUSDDNG-UHFFFAOYSA-L Calcium chloride Chemical compound [Cl-].[Cl-].[Ca+2] UXVMQQNJUSDDNG-UHFFFAOYSA-L 0.000 description 1
- OKTJSMMVPCPJKN-NJFSPNSNSA-N Carbon-14 Chemical compound [14C] OKTJSMMVPCPJKN-NJFSPNSNSA-N 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010067225 Cell Adhesion Molecules Proteins 0.000 description 1
- 102000016289 Cell Adhesion Molecules Human genes 0.000 description 1
- 206010057248 Cell death Diseases 0.000 description 1
- 102000004127 Cytokines Human genes 0.000 description 1
- 108090000695 Cytokines Proteins 0.000 description 1
- 101710112752 Cytotoxin Proteins 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 1
- 102100025137 Early activation antigen CD69 Human genes 0.000 description 1
- 102100029722 Ectonucleoside triphosphate diphosphohydrolase 1 Human genes 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 241000282324 Felis Species 0.000 description 1
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 1
- 102100021186 Granulysin Human genes 0.000 description 1
- 101710168479 Granulysin Proteins 0.000 description 1
- 102000001398 Granzyme Human genes 0.000 description 1
- 108060005986 Granzyme Proteins 0.000 description 1
- 208000037357 HIV infectious disease Diseases 0.000 description 1
- 102100030595 HLA class II histocompatibility antigen gamma chain Human genes 0.000 description 1
- 208000002250 Hematologic Neoplasms Diseases 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 108010088652 Histocompatibility Antigens Class I Proteins 0.000 description 1
- 102000008949 Histocompatibility Antigens Class I Human genes 0.000 description 1
- 108010027412 Histocompatibility Antigens Class II Proteins 0.000 description 1
- 102000018713 Histocompatibility Antigens Class II Human genes 0.000 description 1
- 101000884305 Homo sapiens B-cell receptor CD22 Proteins 0.000 description 1
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 description 1
- 101001095043 Homo sapiens Bone marrow proteoglycan Proteins 0.000 description 1
- 101000942284 Homo sapiens C-type lectin domain family 1 member B Proteins 0.000 description 1
- 101000914469 Homo sapiens CD82 antigen Proteins 0.000 description 1
- 101000934374 Homo sapiens Early activation antigen CD69 Proteins 0.000 description 1
- 101001012447 Homo sapiens Ectonucleoside triphosphate diphosphohydrolase 1 Proteins 0.000 description 1
- 101001082627 Homo sapiens HLA class II histocompatibility antigen gamma chain Proteins 0.000 description 1
- 101001027081 Homo sapiens Killer cell immunoglobulin-like receptor 2DL1 Proteins 0.000 description 1
- 101000945371 Homo sapiens Killer cell immunoglobulin-like receptor 2DL2 Proteins 0.000 description 1
- 101000945331 Homo sapiens Killer cell immunoglobulin-like receptor 2DL4 Proteins 0.000 description 1
- 101000777628 Homo sapiens Leukocyte antigen CD37 Proteins 0.000 description 1
- 101001137987 Homo sapiens Lymphocyte activation gene 3 protein Proteins 0.000 description 1
- 101001131990 Homo sapiens Peroxidasin homolog Proteins 0.000 description 1
- 101000582986 Homo sapiens Phospholipid phosphatase-related protein type 3 Proteins 0.000 description 1
- 101000633786 Homo sapiens SLAM family member 6 Proteins 0.000 description 1
- 101000851376 Homo sapiens Tumor necrosis factor receptor superfamily member 8 Proteins 0.000 description 1
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 1
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 1
- 101900082162 Human immunodeficiency virus type 1 group M subtype B Surface protein gp120 Proteins 0.000 description 1
- 102000009786 Immunoglobulin Constant Regions Human genes 0.000 description 1
- 108010009817 Immunoglobulin Constant Regions Proteins 0.000 description 1
- 108700005091 Immunoglobulin Genes Proteins 0.000 description 1
- 206010062016 Immunosuppression Diseases 0.000 description 1
- 102000003814 Interleukin-10 Human genes 0.000 description 1
- 108090000174 Interleukin-10 Proteins 0.000 description 1
- 102220478985 Interleukin-4 receptor subunit alpha_L64A_mutation Human genes 0.000 description 1
- 102000004889 Interleukin-6 Human genes 0.000 description 1
- 108090001005 Interleukin-6 Proteins 0.000 description 1
- 102100037363 Killer cell immunoglobulin-like receptor 2DL1 Human genes 0.000 description 1
- 102100033599 Killer cell immunoglobulin-like receptor 2DL2 Human genes 0.000 description 1
- 102100033633 Killer cell immunoglobulin-like receptor 2DL4 Human genes 0.000 description 1
- 241000235058 Komagataella pastoris Species 0.000 description 1
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 1
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 1
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 1
- 108090001090 Lectins Proteins 0.000 description 1
- 102000004856 Lectins Human genes 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 102100031586 Leukocyte antigen CD37 Human genes 0.000 description 1
- 108060001084 Luciferase Proteins 0.000 description 1
- 239000005089 Luciferase Substances 0.000 description 1
- 102100020862 Lymphocyte activation gene 3 protein Human genes 0.000 description 1
- 208000030289 Lymphoproliferative disease Diseases 0.000 description 1
- 241000282560 Macaca mulatta Species 0.000 description 1
- 206010065764 Mucosal infection Diseases 0.000 description 1
- 241000699660 Mus musculus Species 0.000 description 1
- 230000006051 NK cell activation Effects 0.000 description 1
- BPUNWVGCTMEMKQ-UHFFFAOYSA-N Neopulchellin Natural products CC1CC2OC(=O)C(=C)C2CC2(C)C(O)CC(O)C12 BPUNWVGCTMEMKQ-UHFFFAOYSA-N 0.000 description 1
- 244000061176 Nicotiana tabacum Species 0.000 description 1
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 1
- 208000015914 Non-Hodgkin lymphomas Diseases 0.000 description 1
- 102220473319 PCNA-associated factor_I65A_mutation Human genes 0.000 description 1
- 102000004503 Perforin Human genes 0.000 description 1
- 108010056995 Perforin Proteins 0.000 description 1
- KHGNFPUMBJSZSM-UHFFFAOYSA-N Perforine Natural products COC1=C2CCC(O)C(CCC(C)(C)O)(OC)C2=NC2=C1C=CO2 KHGNFPUMBJSZSM-UHFFFAOYSA-N 0.000 description 1
- 102100034601 Peroxidasin homolog Human genes 0.000 description 1
- 206010057249 Phagocytosis Diseases 0.000 description 1
- 108010004729 Phycoerythrin Proteins 0.000 description 1
- 241000276498 Pollachius virens Species 0.000 description 1
- 241000589516 Pseudomonas Species 0.000 description 1
- 241000508269 Psidium Species 0.000 description 1
- 241000700159 Rattus Species 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 241000219061 Rheum Species 0.000 description 1
- 102100029197 SLAM family member 6 Human genes 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 108010084592 Saporins Proteins 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 108010029180 Sialic Acid Binding Ig-like Lectin 3 Proteins 0.000 description 1
- 102000001555 Sialic Acid Binding Ig-like Lectin 3 Human genes 0.000 description 1
- 241000580858 Simian-Human immunodeficiency virus Species 0.000 description 1
- 108700011201 Streptococcus IgG Fc-binding Proteins 0.000 description 1
- 230000024932 T cell mediated immunity Effects 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 101710120037 Toxin CcdB Proteins 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 1
- 102100036857 Tumor necrosis factor receptor superfamily member 8 Human genes 0.000 description 1
- 206010067584 Type 1 diabetes mellitus Diseases 0.000 description 1
- 241000700618 Vaccinia virus Species 0.000 description 1
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 1
- HCHKCACWOHOZIP-IGMARMGPSA-N Zinc-65 Chemical compound [65Zn] HCHKCACWOHOZIP-IGMARMGPSA-N 0.000 description 1
- KJNGJIPPQOFCSK-WQEMXFENSA-N [85SrH2] Chemical compound [85SrH2] KJNGJIPPQOFCSK-WQEMXFENSA-N 0.000 description 1
- 238000010521 absorption reaction Methods 0.000 description 1
- 230000021736 acetylation Effects 0.000 description 1
- 238000006640 acetylation reaction Methods 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 230000003044 adaptive effect Effects 0.000 description 1
- 230000001464 adherent effect Effects 0.000 description 1
- 230000009824 affinity maturation Effects 0.000 description 1
- 230000001270 agonistic effect Effects 0.000 description 1
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 1
- 239000002168 alkylating agent Substances 0.000 description 1
- 229940100198 alkylating agent Drugs 0.000 description 1
- 230000000735 allogeneic effect Effects 0.000 description 1
- 239000004037 angiogenesis inhibitor Substances 0.000 description 1
- 229940121369 angiogenesis inhibitor Drugs 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 230000003042 antagnostic effect Effects 0.000 description 1
- 230000000781 anti-lymphocytic effect Effects 0.000 description 1
- 230000000840 anti-viral effect Effects 0.000 description 1
- 230000009830 antibody antigen interaction Effects 0.000 description 1
- 239000003080 antimitotic agent Substances 0.000 description 1
- 238000011225 antiretroviral therapy Methods 0.000 description 1
- 229940124522 antiretrovirals Drugs 0.000 description 1
- 239000003903 antiretrovirus agent Substances 0.000 description 1
- 230000006907 apoptotic process Effects 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 206010003246 arthritis Diseases 0.000 description 1
- 230000005784 autoimmunity Effects 0.000 description 1
- 230000001580 bacterial effect Effects 0.000 description 1
- 210000004082 barrier epithelial cell Anatomy 0.000 description 1
- 210000003651 basophil Anatomy 0.000 description 1
- 235000013527 bean curd Nutrition 0.000 description 1
- 108010005774 beta-Galactosidase Proteins 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 230000008512 biological response Effects 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 230000037396 body weight Effects 0.000 description 1
- 239000000872 buffer Substances 0.000 description 1
- 239000001110 calcium chloride Substances 0.000 description 1
- 229910001628 calcium chloride Inorganic materials 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- PASHVRUKOFIRIK-UHFFFAOYSA-L calcium sulfate dihydrate Chemical compound O.O.[Ca+2].[O-]S([O-])(=O)=O PASHVRUKOFIRIK-UHFFFAOYSA-L 0.000 description 1
- 208000035269 cancer or benign tumor Diseases 0.000 description 1
- 150000001720 carbohydrates Chemical class 0.000 description 1
- 235000014633 carbohydrates Nutrition 0.000 description 1
- 229910002091 carbon monoxide Inorganic materials 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 230000020411 cell activation Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 238000002659 cell therapy Methods 0.000 description 1
- 230000010307 cell transformation Effects 0.000 description 1
- 230000036755 cellular response Effects 0.000 description 1
- 230000005754 cellular signaling Effects 0.000 description 1
- 230000008859 change Effects 0.000 description 1
- 230000009920 chelation Effects 0.000 description 1
- 230000002759 chromosomal effect Effects 0.000 description 1
- 230000001684 chronic effect Effects 0.000 description 1
- 208000032852 chronic lymphocytic leukemia Diseases 0.000 description 1
- 208000029742 colonic neoplasm Diseases 0.000 description 1
- 230000009918 complex formation Effects 0.000 description 1
- 230000000536 complexating effect Effects 0.000 description 1
- 230000004940 costimulation Effects 0.000 description 1
- 230000000139 costimulatory effect Effects 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 150000001945 cysteines Chemical class 0.000 description 1
- 238000004163 cytometry Methods 0.000 description 1
- 239000002254 cytotoxic agent Substances 0.000 description 1
- 238000002784 cytotoxicity assay Methods 0.000 description 1
- 231100000263 cytotoxicity test Toxicity 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- 230000003111 delayed effect Effects 0.000 description 1
- 210000004443 dendritic cell Anatomy 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- 238000003745 diagnosis Methods 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 230000003292 diminished effect Effects 0.000 description 1
- 206010013023 diphtheria Diseases 0.000 description 1
- 230000006806 disease prevention Effects 0.000 description 1
- 238000010494 dissociation reaction Methods 0.000 description 1
- 230000005593 dissociations Effects 0.000 description 1
- 229960003722 doxycycline Drugs 0.000 description 1
- XQTWDDCIUJNLTR-CVHRZJFOSA-N doxycycline monohydrate Chemical compound O.O=C1C2=C(O)C=CC=C2[C@H](C)[C@@H]2C1=C(O)[C@]1(O)C(=O)C(C(N)=O)=C(O)[C@@H](N(C)C)[C@@H]1[C@H]2O XQTWDDCIUJNLTR-CVHRZJFOSA-N 0.000 description 1
- 238000012377 drug delivery Methods 0.000 description 1
- 238000009510 drug design Methods 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 239000008393 encapsulating agent Substances 0.000 description 1
- 210000001163 endosome Anatomy 0.000 description 1
- 239000003623 enhancer Substances 0.000 description 1
- 108700004025 env Genes Proteins 0.000 description 1
- 101150030339 env gene Proteins 0.000 description 1
- 230000002255 enzymatic effect Effects 0.000 description 1
- 230000004890 epithelial barrier function Effects 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 231100000776 exotoxin Toxicity 0.000 description 1
- 239000002095 exotoxin Substances 0.000 description 1
- 239000000835 fiber Substances 0.000 description 1
- GNBHRKFJIUUOQI-UHFFFAOYSA-N fluorescein Chemical compound O1C(=O)C2=CC=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 GNBHRKFJIUUOQI-UHFFFAOYSA-N 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 238000010230 functional analysis Methods 0.000 description 1
- 125000000524 functional group Chemical group 0.000 description 1
- 230000000855 fungicidal effect Effects 0.000 description 1
- 239000000417 fungicide Substances 0.000 description 1
- 210000004392 genitalia Anatomy 0.000 description 1
- 230000035784 germination Effects 0.000 description 1
- WHUUTDBJXJRKMK-VKHMYHEASA-L glutamate group Chemical group N[C@@H](CCC(=O)[O-])C(=O)[O-] WHUUTDBJXJRKMK-VKHMYHEASA-L 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 210000002443 helper t lymphocyte Anatomy 0.000 description 1
- 210000003958 hematopoietic stem cell Anatomy 0.000 description 1
- 208000002672 hepatitis B Diseases 0.000 description 1
- 125000005842 heteroatom Chemical group 0.000 description 1
- 230000000423 heterosexual effect Effects 0.000 description 1
- 239000003276 histone deacetylase inhibitor Substances 0.000 description 1
- 230000000652 homosexual effect Effects 0.000 description 1
- 230000003054 hormonal effect Effects 0.000 description 1
- 102000053350 human FCGR3B Human genes 0.000 description 1
- 208000033519 human immunodeficiency virus infectious disease Diseases 0.000 description 1
- 238000011577 humanized mouse model Methods 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 230000036737 immune function Effects 0.000 description 1
- 238000011502 immune monitoring Methods 0.000 description 1
- 238000003018 immunoassay Methods 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 230000001506 immunosuppresive effect Effects 0.000 description 1
- 239000012678 infectious agent Substances 0.000 description 1
- 230000002458 infectious effect Effects 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- 238000003780 insertion Methods 0.000 description 1
- 230000037431 insertion Effects 0.000 description 1
- 102000006495 integrins Human genes 0.000 description 1
- 108010044426 integrins Proteins 0.000 description 1
- 229940076144 interleukin-10 Drugs 0.000 description 1
- 229940100601 interleukin-6 Drugs 0.000 description 1
- 230000000968 intestinal effect Effects 0.000 description 1
- 210000004347 intestinal mucosa Anatomy 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- YWXYYJSYQOXTPL-SLPGGIOYSA-N isosorbide mononitrate Chemical compound [O-][N+](=O)O[C@@H]1CO[C@@H]2[C@@H](O)CO[C@@H]21 YWXYYJSYQOXTPL-SLPGGIOYSA-N 0.000 description 1
- 239000002523 lectin Substances 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- 230000029226 lipidation Effects 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 238000011068 loading method Methods 0.000 description 1
- 238000003670 luciferase enzyme activity assay Methods 0.000 description 1
- HWYHZTIRURJOHG-UHFFFAOYSA-N luminol Chemical compound O=C1NNC(=O)C2=C1C(N)=CC=C2 HWYHZTIRURJOHG-UHFFFAOYSA-N 0.000 description 1
- 230000036210 malignancy Effects 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 238000007726 management method Methods 0.000 description 1
- PWHULOQIROXLJO-BJUDXGSMSA-N manganese-54 Chemical compound [54Mn] PWHULOQIROXLJO-BJUDXGSMSA-N 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 210000003622 mature neutrocyte Anatomy 0.000 description 1
- 230000034217 membrane fusion Effects 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- 150000002739 metals Chemical class 0.000 description 1
- 239000010445 mica Substances 0.000 description 1
- 229910052618 mica group Inorganic materials 0.000 description 1
- 238000002715 modification method Methods 0.000 description 1
- 238000007479 molecular analysis Methods 0.000 description 1
- 230000009149 molecular binding Effects 0.000 description 1
- 230000004001 molecular interaction Effects 0.000 description 1
- 238000012544 monitoring process Methods 0.000 description 1
- 238000002625 monoclonal antibody therapy Methods 0.000 description 1
- 238000010172 mouse model Methods 0.000 description 1
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 1
- 210000002200 mouth mucosa Anatomy 0.000 description 1
- 229960003816 muromonab-cd3 Drugs 0.000 description 1
- 208000025113 myeloid leukemia Diseases 0.000 description 1
- DDOVBCWVTOHGCU-QMXMISKISA-N n-[(e,2s,3r)-3-hydroxy-1-[(2r,3r,4s,5r,6r)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxynonadec-4-en-2-yl]octadecanamide Chemical compound CCCCCCCCCCCCCCCCCC(=O)N[C@H]([C@H](O)\C=C\CCCCCCCCCCCCCC)CO[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O DDOVBCWVTOHGCU-QMXMISKISA-N 0.000 description 1
- 230000000174 oncolytic effect Effects 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- 230000003204 osmotic effect Effects 0.000 description 1
- 230000001590 oxidative effect Effects 0.000 description 1
- 230000005298 paramagnetic effect Effects 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 244000052769 pathogen Species 0.000 description 1
- 231100000255 pathogenic effect Toxicity 0.000 description 1
- 230000035515 penetration Effects 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 229930192851 perforin Natural products 0.000 description 1
- 230000000737 periodic effect Effects 0.000 description 1
- 210000005105 peripheral blood lymphocyte Anatomy 0.000 description 1
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 1
- 230000002085 persistent effect Effects 0.000 description 1
- 230000008782 phagocytosis Effects 0.000 description 1
- 239000002831 pharmacologic agent Substances 0.000 description 1
- 238000001050 pharmacotherapy Methods 0.000 description 1
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 1
- 230000026731 phosphorylation Effects 0.000 description 1
- 238000006366 phosphorylation reaction Methods 0.000 description 1
- 239000013612 plasmid Substances 0.000 description 1
- 210000004180 plasmocyte Anatomy 0.000 description 1
- 238000003752 polymerase chain reaction Methods 0.000 description 1
- 102000054765 polymorphisms of proteins Human genes 0.000 description 1
- 230000002064 post-exposure prophylaxis Effects 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 238000011533 pre-incubation Methods 0.000 description 1
- 238000004393 prognosis Methods 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000002062 proliferating effect Effects 0.000 description 1
- 230000035755 proliferation Effects 0.000 description 1
- 230000020978 protein processing Effects 0.000 description 1
- 238000001742 protein purification Methods 0.000 description 1
- 230000005180 public health Effects 0.000 description 1
- 239000012857 radioactive material Substances 0.000 description 1
- 238000003127 radioimmunoassay Methods 0.000 description 1
- 230000007115 recruitment Effects 0.000 description 1
- 210000000664 rectum Anatomy 0.000 description 1
- 238000007634 remodeling Methods 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 208000030925 respiratory syncytial virus infectious disease Diseases 0.000 description 1
- 230000000284 resting effect Effects 0.000 description 1
- 230000000717 retained effect Effects 0.000 description 1
- 238000012552 review Methods 0.000 description 1
- 102220012488 rs397516129 Human genes 0.000 description 1
- 102220078865 rs776745618 Human genes 0.000 description 1
- 229940102127 rubidium chloride Drugs 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 230000028327 secretion Effects 0.000 description 1
- 210000004739 secretory vesicle Anatomy 0.000 description 1
- 208000013223 septicemia Diseases 0.000 description 1
- 239000007790 solid phase Substances 0.000 description 1
- 239000000243 solution Substances 0.000 description 1
- 230000009870 specific binding Effects 0.000 description 1
- 239000003381 stabilizer Substances 0.000 description 1
- 238000012289 standard assay Methods 0.000 description 1
- 239000008227 sterile water for injection Substances 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 238000002626 targeted therapy Methods 0.000 description 1
- 238000012360 testing method Methods 0.000 description 1
- 230000009258 tissue cross reactivity Effects 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 238000011830 transgenic mouse model Methods 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 230000005909 tumor killing Effects 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 229960005486 vaccine Drugs 0.000 description 1
- 239000004474 valine Substances 0.000 description 1
- 239000003981 vehicle Substances 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- 238000005406 washing Methods 0.000 description 1
- 238000009736 wetting Methods 0.000 description 1
- 239000000080 wetting agent Substances 0.000 description 1
Images




















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1036—Retroviridae, e.g. leukemia viruses
- C07K16/1045—Lentiviridae, e.g. HIV, FIV, SIV
- C07K16/1063—Lentiviridae, e.g. HIV, FIV, SIV env, e.g. gp41, gp110/120, gp160, V3, PND, CD4 binding site
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/395—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum
- A61K39/42—Antibodies; Immunoglobulins; Immune serum, e.g. antilymphocytic serum viral
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
- A61K45/06—Mixtures of active ingredients without chemical characterisation, e.g. antiphlogistics and cardiaca
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2809—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against the T-cell receptor (TcR)-CD3 complex
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2815—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against CD8
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/283—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against Fc-receptors, e.g. CD16, CD32, CD64
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/46—Hybrid immunoglobulins
- C07K16/468—Immunoglobulins having two or more different antigen binding sites, e.g. multifunctional antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/24—Immunoglobulins specific features characterized by taxonomic origin containing regions, domains or residues from different species, e.g. chimeric, humanized or veneered
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/31—Immunoglobulins specific features characterized by aspects of specificity or valency multispecific
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/35—Valency
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/567—Framework region [FR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/62—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising only variable region components
- C07K2317/626—Diabody or triabody
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
Abstract
本発明は、免疫原性が低減された最適化済みHIV‐1 gp41結合分子を対象とする。より具体的には、本発明は、gp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインであって、レシピエント被験者への投与時に1つ以上の該ドメインの免疫原性を低減するように最適化されているgp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインを含む、最適化済みgp41結合分子に関する。本発明は特に、((DART(登録商標)ダイアボディを含む)二重特異性ダイアボディ、BiTE(登録商標)、二重特異性抗体、(TRIDENT(商標)分子を含む)3価結合分子等を含む)多重特異性gp41結合分子である、gp41結合分子であって:(i)1つ以上の上述のような最適化済みgp41結合可変ドメイン;及び(ii)エフェクタ細胞の表面上に存在する分子のエピトープに結合できるドメインを含む、gp41結合分子に関する。本発明はまた、上述のようなgp41結合分子のいずれを含む医薬組成物、及びHIV‐1感染の治療における上述のようなgp41結合分子のいずれの使用を伴う方法も対象とする。
Description
[関連出願の相互参照]
本出願は、米国特許出願第62/673,462号(2019年5月18日出願;係属中)に対する優先権を主張するものであり、上記出願は参照によりその全体が本出願に援用される。
本出願は、米国特許出願第62/673,462号(2019年5月18日出願;係属中)に対する優先権を主張するものであり、上記出願は参照によりその全体が本出願に援用される。
[配列表の参照]
本出願は、連邦規則法典第37巻第1.821節以下による1つ以上の配列表を含み、これらの配列表は、コンピュータ可読媒体(ファイル名:1301_0155PCT_配列_Listing_ST25.txt、2019年5月11日作成、サイズ:158,835バイト)において開示されており、上記ファイルはその全体が本出願に援用される。
本出願は、連邦規則法典第37巻第1.821節以下による1つ以上の配列表を含み、これらの配列表は、コンピュータ可読媒体(ファイル名:1301_0155PCT_配列_Listing_ST25.txt、2019年5月11日作成、サイズ:158,835バイト)において開示されており、上記ファイルはその全体が本出願に援用される。
本発明は、免疫原性が低減された最適化済みHIV‐1 gp41結合分子を対象とする。より具体的には、本発明は、gp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインであって、レシピエント被験者への投与時に1つ以上の該ドメインの免疫原性を低減するように最適化されているgp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインを含む、最適化済みgp41結合分子に関する。本発明は特に、((DART(登録商標)ダイアボディを含む)二重特異性ダイアボディ、BiTE(登録商標)、二重特異性抗体、(TRIDENT(商標)分子を含む)3価結合分子等を含む)多重特異性gp41結合分子である、gp41結合分子であって:(i)1つ以上の上述のような最適化済みgp41結合可変ドメイン;及び(ii)エフェクタ細胞の表面上に存在する分子のエピトープに結合できるドメインを含む、gp41結合分子に関する。本発明はまた、上述のようなgp41結合分子のいずれを含む医薬組成物、及びHIV‐1感染の治療における上述のようなgp41結合分子のいずれの使用を伴う方法も対象とする。
高活性抗レトロウイルス療法(Highly Active Antiretroviral Therapy:HAART)は、感染した個体におけるウイルス負荷の低減及びHIV 1型(HIV‐1)感染の影響の改善に有効であった。しかしながら、HAARTはHIV‐1感染を根絶することはできず、感染細胞の除去を加速しない。感染した個体において、HIV‐1はHAARTとってアクセス不可能である静止記憶CD4+T細胞内に統合されたプロウイルスとして、潜伏状態で生存する。よって、このような療法の投与にもかかわらず、HIV‐1ウイルスは、治療を回避したHIV‐1感染細胞の潜伏リザーバ内において、個体内で生存する。CD8+T細胞は、このような潜伏状態で感染している細胞内のHIV‐1の排除に関して、限定された能力しか有しない。
よって、HIV‐1感染個体の治療のための治療剤、特にウイルス感染細胞を標的とし、HIV‐1感染細胞の潜伏リザーバを減少させる可能性があり、反復投与に好適である作用剤に対する需要が存在する。
HIV‐1ウイルスのエンベロープ糖タンパク質(envelope glycoprotein:Env)は、(CD4受容体に結合するウイルスの能力による)ウイルスの感染性にとって、及び膜融合の促進のために、重要である。HIV‐1 Env遺伝子産物は、2つのサブユニットgp120及びgp41の三量体複合体からなる。Envタンパク質は、グリコシル化gp160前駆体タンパク質として合成され、これは三量体へと折り畳まれ、タンパク質分解的に切断されて、成熟したgp120及びgp41タンパク質がもたらされる。切断されたEnvは、細胞表面からのビリオンの発芽のために、他のウイルス成分と共に集合し、感染細胞の表面上に存在する(非特許文献1)。よって、HIV Env及び特にそのgp41サブユニットは、抗体仲介型細胞殺滅による持続性HIV感染リザーバの治療的排除のための、極めて特異的なウイルス標的である(非特許文献2)。
Miranda、L., et al. , 2002 "Cell Surface Expression Of The HIV-1 Envelope Glycoproteins Is Directed From Intracellular CTLA-4-Containing Regulated Secretory Granules." Proc Natl Acad Sci U S A. 99(12): 8031-8036
Sloane, D. , et al. , 2015, "Targeting HIV Reservoir in Infected CD4 T Cells by Dual-Affinity Re-targeting Molecules (DARTs) that Bind HIV Envelope and Recruit Cytotoxic T Cells" PLOS Pathogens | DOI:10.1371/journal.ppat.1005233
しかしながら、これまでの全ての進歩にも関わらず、抗HIV活性が強化された、及び/又は免疫原性が低減された、最適化済みHIV gp41結合分子に対する需要は依然として残っている。本発明はこの需要、並びにHIV‐1の治療及び予防のための改良された治療法に対する需要に対処する。
本発明は、免疫原性が低減された最適化済みHIV‐1 gp41結合分子を対象とする。より具体的には、本発明は、gp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインであって、レシピエント被験者への投与時に該ドメインの免疫原性を低減するように最適化されているgp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインを含む、最適化済みgp41結合分子に関する。本発明は特に、((DART(登録商標)ダイアボディを含む)二重特異性ダイアボディ、BiTE(登録商標)、二重特異性抗体、(TRIDENT(商標)分子を含む)3価結合分子等を含む)多重特異性gp41結合分子である、gp41結合分子であって:(i)1つ以上の上述のような最適化済みgp41結合可変ドメイン;及び(ii)エフェクタ細胞の表面上に存在する分子のエピトープに結合できるドメインを含む、gp41結合分子に関する。本発明はまた、上述のようなgp41結合分子のいずれを含む医薬組成物、及びHIV‐1感染の治療における上述のようなgp41結合分子のいずれの使用を伴う方法も対象とする。
詳細には、本発明は、可変軽鎖(VL)ドメイン及び可変重鎖(VH)ドメインを含むgp41結合分子を提供し、上記VLドメインは配列番号57のアミノ酸配列を含み、及び/又は上記VHドメインは配列番号58のアミノ酸配列を含む。
本発明は更に、上記gp41結合分子が:
(a)配列番号57のアミノ酸配列を含む上記VLドメイン;及び
(b)配列番号58のアミノ酸配列を含む上記VHドメイン
を含む、上述のgp41結合分子の実施形態に関する。
(a)配列番号57のアミノ酸配列を含む上記VLドメイン;及び
(b)配列番号58のアミノ酸配列を含む上記VHドメイン
を含む、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が、抗体であるか、上記抗体のgp41エピトープ結合部分を含む、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が:
(a)二重特異性抗体;又は
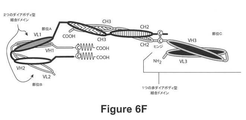
(b)2つ、3つ、4つ若しくは5つのポリペプチド鎖を含む共有結合複合体である、ダイアボディ;又は
(c)3つ、4つ、5つ、若しくは6つ以上のポリペプチド鎖を含む共有結合複合体である、3価結合性分子
である、上述のgp41結合分子の実施形態に関する。
(a)二重特異性抗体;又は
(b)2つ、3つ、4つ若しくは5つのポリペプチド鎖を含む共有結合複合体である、ダイアボディ;又は
(c)3つ、4つ、5つ、若しくは6つ以上のポリペプチド鎖を含む共有結合複合体である、3価結合性分子
である、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が上記ダイアボディであり、アルブミン結合ドメイン(Albumin‐Binding Domain:ABD)を含む、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子がFc領域を含む、上述のgp41結合分子の実施形態に関する。本発明は更に、上記Fc領域が、変異型Fc領域であって:
(a)FcγRに対する上記変異型Fc領域の親和性を低減する1つ以上のアミノ酸修飾;及び/又は
(b)上記変異型Fc領域の血清半減期を向上させる1つ以上のアミノ酸修飾
を含む、変異型Fc領域である、上述のgp41結合分子の実施形態に関する。
(a)FcγRに対する上記変異型Fc領域の親和性を低減する1つ以上のアミノ酸修飾;及び/又は
(b)上記変異型Fc領域の血清半減期を向上させる1つ以上のアミノ酸修飾
を含む、変異型Fc領域である、上述のgp41結合分子の実施形態に関する。
本発明は更に、FcγRに対する上記変異型Fc領域の親和性を低減する上記修飾が、L234A;L235A;又はL234A及びL235Aの置換を含み、上記番号付与はKabat中のEUインデックスの番号付与である、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記変異型Fc領域の血清半減期を向上させる上記修飾が、M252Y;M252Y及びS254T;M252Y及びT256E;M252Y、S254T及びT256E;又はK288D及びH435Kの置換を含み、上記番号付与はKabat中のEUインデックスの番号付与である、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が二重特異性であり、gp41のエピトープに免疫特異的に結合できる1つのエピトープ結合部位と、エフェクタ細胞の表面上に存在する分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位とを含む、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が二重特異性であり、gp41のエピトープに免疫特異的に結合できる2つのエピトープ結合部位と、エフェクタ細胞の表面上に存在する分子のエピトープに免疫特異的に結合できる2つのエピトープ結合部位とを含む、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が三重特異性であり:
(a)gp41のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;
(b)エフェクタ細胞の表面上に存在する第1の分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;及び
(c)エフェクタ細胞の表面上に存在する第2の分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位
を含む、上述のgp41結合分子の実施形態に関する。
(a)gp41のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;
(b)エフェクタ細胞の表面上に存在する第1の分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;及び
(c)エフェクタ細胞の表面上に存在する第2の分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位
を含む、上述のgp41結合分子の実施形態に関する。
本発明は更に、エフェクタ細胞の表面上に存在する上記第1の分子がCD3であり、エフェクタ細胞の表面上に存在する上第2の分子がCD8である、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が三重特異性であり:
(a)gp41の第1のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;
(b)gp41の第2のエピトープ、又は異なるHIV‐1タンパク質のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;及び
(c)エフェクタ細胞の表面上に存在する分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位
を含み、gp41の上記第1のエピトープと上記第2のエピトープとは異なるものである、上述のgp41結合分子の実施形態に関する。
(a)gp41の第1のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;
(b)gp41の第2のエピトープ、又は異なるHIV‐1タンパク質のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;及び
(c)エフェクタ細胞の表面上に存在する分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位
を含み、gp41の上記第1のエピトープと上記第2のエピトープとは異なるものである、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が、gp41とエフェクタ細胞の表面上に存在する分子とに同時に結合できる、上述のgp41結合分子の実施形態に関する。本発明は更に、エフェクタ細胞の表面上に存在する上記分子が、CD2、CD3、CD8、CD16、TCR、NKp46、又はNKG2Dである、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記エフェクタ細胞が、細胞傷害性T細胞又はナチュラルキラー(NK)細胞である、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が、gp41を発現する細胞と細胞傷害性T細胞との協調的結合を仲介する、上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が、第1のポリペプチド鎖、第2のポリペプチド鎖、及び第3のポリペプチド鎖を含み:
I)(a)上記第1のポリペプチド鎖は配列番号113を含み;
(b)上記第2のポリペプチド鎖は配列番号114を含み;及び
(c)上記第3のポリペプチド鎖は配列番号112を含むか;
又は
II)(a)上記第1のポリペプチド鎖は配列番号115を含み;
(b)上記第2のポリペプチド鎖は配列番号116を含み;及び
(c)上記第3のポリペプチド鎖は配列番号112を含む、
上述のgp41結合分子の実施形態に関する。
I)(a)上記第1のポリペプチド鎖は配列番号113を含み;
(b)上記第2のポリペプチド鎖は配列番号114を含み;及び
(c)上記第3のポリペプチド鎖は配列番号112を含むか;
又は
II)(a)上記第1のポリペプチド鎖は配列番号115を含み;
(b)上記第2のポリペプチド鎖は配列番号116を含み;及び
(c)上記第3のポリペプチド鎖は配列番号112を含む、
上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が、第1のポリペプチド鎖、第2のポリペプチド鎖、第3のポリペプチド鎖、及び第4のポリペプチド鎖を含み:
(a)上記第1のポリペプチド鎖は配列番号113を含み;
(b)上記第2のポリペプチド鎖は配列番号114を含み;
(c)上記第3のポリペプチド鎖は配列番号117を含み;及び
(d)上記第4のポリペプチド鎖は配列番号118を含む、
上述のgp41結合分子の実施形態に関する。
(a)上記第1のポリペプチド鎖は配列番号113を含み;
(b)上記第2のポリペプチド鎖は配列番号114を含み;
(c)上記第3のポリペプチド鎖は配列番号117を含み;及び
(d)上記第4のポリペプチド鎖は配列番号118を含む、
上述のgp41結合分子の実施形態に関する。
本発明は更に、上記分子が、第1のポリペプチド鎖、第2のポリペプチド鎖、第3のポリペプチド鎖、及び第4のポリペプチド鎖を含み:
I)(a)上記第1のポリペプチド鎖は配列番号113を含み;
(b)上記第2のポリペプチド鎖は配列番号114を含み;
(c)上記第3のポリペプチド鎖は配列番号119を含み;及び
(d)上記第4のポリペプチド鎖は配列番号120を含むか;
又は
II)(a)上記第1のポリペプチド鎖は配列番号115を含み;
(b)上記第2のポリペプチド鎖は配列番号116を含み;
(c)上記第3のポリペプチド鎖は配列番号119を含み;及び
(d)上記第4のポリペプチド鎖は配列番号120を含む、
上述のgp41結合分子の実施形態に関する。
I)(a)上記第1のポリペプチド鎖は配列番号113を含み;
(b)上記第2のポリペプチド鎖は配列番号114を含み;
(c)上記第3のポリペプチド鎖は配列番号119を含み;及び
(d)上記第4のポリペプチド鎖は配列番号120を含むか;
又は
II)(a)上記第1のポリペプチド鎖は配列番号115を含み;
(b)上記第2のポリペプチド鎖は配列番号116を含み;
(c)上記第3のポリペプチド鎖は配列番号119を含み;及び
(d)上記第4のポリペプチド鎖は配列番号120を含む、
上述のgp41結合分子の実施形態に関する。
本発明は更に、有効量の請求項1〜22のいずれか1項に記載のgp41結合分子と、薬学的に許容可能なキャリアとを含む、医薬組成物に関する。
本発明は更に、それを必要とする被験者においてHIV‐1感染を治療又は予防するための方法であって、上述のgp41結合分子のうちのいずれか1つ又は上述の医薬組成物を治療的有効量で含む組成物を上記被験者に投与するステップを含む、方法に関する。
本発明は更に、潜伏感染活性化剤(ボリノスタット、ロミデプシン、パノビノスタット、ジスルフィラム、JQ1、ブリオスタチン、PMA、イオノマイシン、又はこれらのいずれの組み合わせ等)を投与するステップを更に含む、HIV‐1感染を治療又は予防するための上記方法に関する。
本発明は、免疫原性が低減された最適化済みHIV‐1 gp41結合分子を対象とする。より具体的には、本発明は、gp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインであって、レシピエント被験者への投与時に該ドメインの免疫原性を低減するように最適化されているgp41結合可変軽鎖(VL)ドメイン及び/又はgp41結合可変重鎖(VH)ドメインを含む、最適化済みgp41結合分子に関する。本発明は特に、((DART(登録商標)ダイアボディを含む)二重特異性ダイアボディ、BiTE(登録商標)、二重特異性抗体、(TRIDENT(商標)分子を含む)3価結合分子等を含む)多重特異性gp41結合分子である、gp41結合分子であって:(i)1つ以上の上述のような最適化済みgp41結合可変ドメイン;及び(ii)エフェクタ細胞の表面上に存在する分子のエピトープに結合できるドメインを含む、gp41結合分子に関する。本発明はまた、上述のようなgp41結合分子のいずれを含む医薬組成物、及びHIV‐1感染の治療における上述のようなgp41結合分子のいずれの使用を伴う方法も対象とする。
I.抗体及び他の結合分子
A.抗体
本発明のgp41結合分子は、抗体であってよく、又は(例えば抗体ポリペプチドの断片化、切断等によって)gp41結合抗体から誘導可能であってよく、若しくは抗体分子のポリペプチド鎖のうちの1つ以上のアミノ酸配列の使用によって得られるものであってよく、若しくはこのようなポリペプチドをコードするポリヌクレオチドによって発現されるものであってよく、若しくはこのようなポリヌクレオチドのヌクレオチド配列から得られるものであってよい。
抗体(antibody)は、炭水化物、ポリヌクレオチド、脂質、ポリペプチド等の分子の特定のドメイン又は部分又は立体配座(「エピトープ(epitope)」に特異的に結合できる、免疫グロブリン分子である。エピトープ含有分子は、免疫原性活性を有してよく、従ってエピトープ含有分子は動物の体内で抗体産生応答を誘発する。このような分子は「抗原(antigen)」と呼ばれる。本明細書において使用される場合、用語「抗体(antibody及びantibodies)」は、モノクローナル抗体、多重特異性抗体、ヒト抗体、ヒト化抗体、合成抗体、キメラ抗体、ポリクローナル抗体、ラクダ化抗体、単鎖Fv(scFv)、単鎖抗体、Fab部分、F(ab’)部分、ジスルフィド結合二重特異性Fv(sdFv)、細胞内抗体、及び以上のうちのいずれかのエピトープ結合ドメインを包含する。このような免疫グロブリン分子は、いずれのタイプ(例えばIgG、IgE、IgM、IgD、IgA及びIgY)、クラス(例えばIgG1、IgG2、IgG3、IgG4、IgA1及びIgA2)若しくはサブクラス、又は種(ウシ、ウマ、ネコ、イヌ、げっ歯類、(例えばカニクイザル等のサル、ヒトを含む)霊長類等)のものとすることができる。
A.抗体
本発明のgp41結合分子は、抗体であってよく、又は(例えば抗体ポリペプチドの断片化、切断等によって)gp41結合抗体から誘導可能であってよく、若しくは抗体分子のポリペプチド鎖のうちの1つ以上のアミノ酸配列の使用によって得られるものであってよく、若しくはこのようなポリペプチドをコードするポリヌクレオチドによって発現されるものであってよく、若しくはこのようなポリヌクレオチドのヌクレオチド配列から得られるものであってよい。
抗体(antibody)は、炭水化物、ポリヌクレオチド、脂質、ポリペプチド等の分子の特定のドメイン又は部分又は立体配座(「エピトープ(epitope)」に特異的に結合できる、免疫グロブリン分子である。エピトープ含有分子は、免疫原性活性を有してよく、従ってエピトープ含有分子は動物の体内で抗体産生応答を誘発する。このような分子は「抗原(antigen)」と呼ばれる。本明細書において使用される場合、用語「抗体(antibody及びantibodies)」は、モノクローナル抗体、多重特異性抗体、ヒト抗体、ヒト化抗体、合成抗体、キメラ抗体、ポリクローナル抗体、ラクダ化抗体、単鎖Fv(scFv)、単鎖抗体、Fab部分、F(ab’)部分、ジスルフィド結合二重特異性Fv(sdFv)、細胞内抗体、及び以上のうちのいずれかのエピトープ結合ドメインを包含する。このような免疫グロブリン分子は、いずれのタイプ(例えばIgG、IgE、IgM、IgD、IgA及びIgY)、クラス(例えばIgG1、IgG2、IgG3、IgG4、IgA1及びIgA2)若しくはサブクラス、又は種(ウシ、ウマ、ネコ、イヌ、げっ歯類、(例えばカニクイザル等のサル、ヒトを含む)霊長類等)のものとすることができる。
用語「モノクローナル抗体(monoclonal antibody)」は均質な抗体の集団を指し、上記モノクローナル抗体は、抗原の選択的結合に関わる(自然に発生する又は自然に発生しない)アミノ酸から構成される。モノクローナル抗体は特異性が高く、単一のエピトープ(又は抗原部位)に対して指向性を有する。用語「モノクローナル抗体」は、完全なモノクローナル抗体及び全長モノクローナル抗体だけでなく、これらの部分(Fab、Fab’、F(ab’)2、Fv等)、単鎖(scFv)、その突然変異体、抗体部分を含む融合タンパク質、ヒト化モノクローナル抗体、キメラモノクローナル抗体、並びに必要な特異性及び抗原への結合能力を有する抗原認識部位を含む免疫グロブリン分子の他のいずれの修飾構成を包含する。抗原の源又は抗原を作製する方法(例えば、ハイブリドーマ、ファージ選択、組み換え発現、遺伝子導入動物等による)に関して、限定は意図されていない。この用語は、免疫グロブリン全体、及び「抗体」の定義において上述した部分等を含む。
モノクローナル抗体の作製方法は当該技術分野において公知である。採用してよい1つの方法は、Kohler, G. et al. (1975) “Continuous Cultures Of Fused Cells Secreting Antibody Of Predefined Specificity,” Nature 256:495-497の方法又はその修正例である。典型的には、モノクローナル抗体はマウス、ラット又はウサギにおいて発現する。上記抗体は、動物を、所望のエピトープを含有する免疫原性量の細胞、細胞抽出物又はタンパク製剤で免疫化することによって産生される。免疫原は、一次細胞、培養された細胞株、癌細胞、タンパク質、ペプチド、核酸又は組織とすることができるが、これらに限定されない。免疫化に使用してよい細胞は、これらを免疫原として使用するよりもある期間(例えば少なくとも24時間)だけ前に、培養してよい。細胞は単独で、又はRibi等の非変性アジュバントと組み合わせて、免疫原として使用してよい(Jennings, V.M. (1995) “Review of Selected Adjuvants Used in Antibody Production,” ILAR J. 37(3):119-125を参照)。一般に細胞は、免疫原として使用される際、完全な状態、及び好ましくは生存できる状態に維持しなければならない。完全な細胞は、破裂した細胞よりも、免疫性を与えられた動物が抗原をより良好に検出できるようにすることができる。変性又は強いアジュバント、例えばフロイントアジュバントの使用は、細胞を破裂させる場合があり、従って推奨されない。免疫原は、2週間に1回若しくは1週間に1回等、周期的な間隔で複数回投与してよく、又は動物中(例えば組織組み換え中)に生存能力を維持できるように投与してよい。あるいは、所望の病原性エピトープに対する免疫特異性を有する既存のモノクローナル抗体及び他の同等の抗体は、当該技術分野で公知のいずれの手段によって、組み換え配列及び産生できる。一実施形態では、このような抗体を配列し、続いてポリヌクレオチド配列を発現又は繁殖のためのベクターにクローン化する。関心対象の抗体をエンコードする配列は、宿主細胞中のベクター中に保持され、続いて上記宿主細胞を、将来使用するために膨張させて冷凍できる。このような抗体のポリヌクレオチド配列は、以下で詳述するように、本発明の単一特異性又は多重特異性(例えば二重特異性、三重特異性及び四重特異性)分子、並びに親和性最適化済み、キメラ抗体、ヒト化抗体及び/又はイヌ化抗体を生成することによって、抗体の親和性又は他の特徴を改善するための、遺伝子操作のために使用してよい。
本発明の抗体及び結合分子は、その結合ドメインを介して、「免疫特異的な(immunospecific)」様式でエピトープに結合する。本明細書中で使用される場合、分子は、別の分子のあるエピトープに、代替的なエピトープに対してよりも頻繁に、迅速に、長期間、及び/又は高い親和性で結合又は連結する場合に、該エピトープに免疫特異的な様式で(又は「免疫特異的に(immunospecifically)」)結合すると表現される。例えば、ウイルスエピトープに免疫特異的に結合する抗体は、該抗体が他のウイルスエピトープ又は非ウイルスエピトープに結合するよりも、高い親和性で、高い結合活性で、より迅速に、及び/又はより長期間結合する抗体である。この定義を読むと、例えば第1の標的に免疫特異的に結合する抗体(又は部分若しくはエピトープ)は、第2の標的に特異的又は優先的に結合するものであっても、そうでなくてもよいことも理解される。従って「免疫特異的結合(immunospecific binding)」は、必ずしも排他的な結合を要求しない(ただしこれを含み得る)。一般には、結合に関する言及は「免疫特異的な」結合を意味するが、必ずしもそうではない。天然抗体は1つのエピトープ種にしか結合できない(即ちこれらは「単一特異性(monospecific)」である)が、天然抗体は、該種の複数の複製に免疫特異的に結合できる(即ち「2価性(bivalency)」又は「多価性(multivalency)」を示す)。2つの分子は、これらの結合が、それぞれのリガンドに受容体が結合する際の特異性を示す場合に、「生理特異的に(physiospecific)」様式で互いに結合できると表現される。
この数十年、抗体の治療能力に対する関心が再燃しており、抗体は、バイオテクノロジー由来薬物の主要な分類の1つとなっている(Chan, C.E. et al. (2009) “The Use Of Antibodies In The Treatment Of Infectious Diseases,” Singapore Med. J. 50(7):663-666)。200を超える抗体ベースの薬物について、その使用が承認されているか、又は開発中である。
1.抗体の一般的な構造的属性
自然発生した免疫グロブリン(例えばIgG)の基本的な構造単位は、2つの長い「重鎖」と複合体化した2つの短い「軽鎖」からなる四量体であり、通常約150,000Daの糖タンパク質として表される。各鎖は、「可変ドメイン」を含むアミノ末端(「N末端」)部分と、少なくとも1つの「定常ドメイン」を含むカルボキシ末端(「C末端」)部分とからなる。IgG軽鎖は、単一の「軽鎖可変ドメイン」(「VL」)と、単一の「軽鎖定常ドメイン」(「CL」)とからなる。よって、IgG分子の軽鎖の構造は、n‐VL‐CL‐cである(ここでn及びcはそれぞれ、ポリペプチドのN末端及びC末端を表す)。IgG重鎖は、単一の「重鎖可変ドメイン」(「VH」)、3つの「重鎖定常ドメイン」(「CH1」、「CH2」及び「CH3」)、並びにCH1ドメインとCH2ドメインとの間に位置する「ヒンジ」領域(「H」)からなる。よって、IgG重鎖の構造は、n‐VH‐CH1‐H‐CH2‐CH3‐cである(ここでn及びcはそれぞれ、ポリペプチドのN末端及びC末端を表す)。無傷の修飾されていない抗体(例えばIgG抗体)の、抗原のエピトープに結合する能力は、可変ドメインの存在及びその配列に左右される。特段の記載がない限り、本明細書に記載のタンパク質分子のドメインの順序は、「N末端からC末端への(N‐terminal to C‐terminal)」方向である。
自然発生した免疫グロブリン(例えばIgG)の基本的な構造単位は、2つの長い「重鎖」と複合体化した2つの短い「軽鎖」からなる四量体であり、通常約150,000Daの糖タンパク質として表される。各鎖は、「可変ドメイン」を含むアミノ末端(「N末端」)部分と、少なくとも1つの「定常ドメイン」を含むカルボキシ末端(「C末端」)部分とからなる。IgG軽鎖は、単一の「軽鎖可変ドメイン」(「VL」)と、単一の「軽鎖定常ドメイン」(「CL」)とからなる。よって、IgG分子の軽鎖の構造は、n‐VL‐CL‐cである(ここでn及びcはそれぞれ、ポリペプチドのN末端及びC末端を表す)。IgG重鎖は、単一の「重鎖可変ドメイン」(「VH」)、3つの「重鎖定常ドメイン」(「CH1」、「CH2」及び「CH3」)、並びにCH1ドメインとCH2ドメインとの間に位置する「ヒンジ」領域(「H」)からなる。よって、IgG重鎖の構造は、n‐VH‐CH1‐H‐CH2‐CH3‐cである(ここでn及びcはそれぞれ、ポリペプチドのN末端及びC末端を表す)。無傷の修飾されていない抗体(例えばIgG抗体)の、抗原のエピトープに結合する能力は、可変ドメインの存在及びその配列に左右される。特段の記載がない限り、本明細書に記載のタンパク質分子のドメインの順序は、「N末端からC末端への(N‐terminal to C‐terminal)」方向である。
(a)定常ドメイン
(i)軽鎖定常ドメイン
好ましいCLドメインは、ヒトIgG CLκドメインである。例示的なヒトCLκドメインのアミノ酸配列は、(配列番号1):
RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ WKVDNALQSG
NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT HQGLSSPVTK
SFNRGEC
である。
(i)軽鎖定常ドメイン
好ましいCLドメインは、ヒトIgG CLκドメインである。例示的なヒトCLκドメインのアミノ酸配列は、(配列番号1):
RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ WKVDNALQSG
NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT HQGLSSPVTK
SFNRGEC
である。
あるいは、例示的なCLドメインは、ヒトIgG CLλドメインである。例示的なヒトCLλドメインのアミノ酸配列は、(配列番号2):
QPKAAPSVTL FPPSSEELQA NKATLVCLIS DFYPGAVTVA WKADSSPVKA
GVETTPSKQS NNKYAASSYL SLTPEQWKSH RSYSCQVTHE GSTVEKTVAP
TECS
である。
QPKAAPSVTL FPPSSEELQA NKATLVCLIS DFYPGAVTVA WKADSSPVKA
GVETTPSKQS NNKYAASSYL SLTPEQWKSH RSYSCQVTHE GSTVEKTVAP
TECS
である。
(ii)重鎖CH1ドメイン
抗体の2つの重鎖のCH1ドメインは、抗体の軽鎖の「CL」定常領域と複合体化し、介在ヒンジドメインを介して重鎖CH2ドメインに付着する。
抗体の2つの重鎖のCH1ドメインは、抗体の軽鎖の「CL」定常領域と複合体化し、介在ヒンジドメインを介して重鎖CH2ドメインに付着する。
例示的なCH1ドメインは、ヒトIgG1 CH1ドメインである。例示的なヒトIgG1 CH1ドメインのアミノ酸配列は、(配列番号3):
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKRV
である。
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKRV
である。
例示的なCH1ドメインは、ヒトIgG2 CH1ドメインである。例示的なヒトIgG2 CH1ドメインのアミノ酸配列は、(配列番号4):
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSNFGTQT YTCNVDHKPS NTKVDKTV
である。
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSNFGTQT YTCNVDHKPS NTKVDKTV
である。
例示的なCH1ドメインは、ヒトIgG3 CH1ドメインである。例示的なヒトIgG3 CH1ドメインのアミノ酸配列は、(配列番号5):
ASTKGPSVFP LAPCSRSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YTCNVNHKPS NTKVDKRV
である。
ASTKGPSVFP LAPCSRSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YTCNVNHKPS NTKVDKRV
である。
例示的なCH1ドメインは、ヒトIgG4 CH1ドメインである。例示的なヒトIgG4 CH1ドメインのアミノ酸配列は、(配列番号6):
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSSLGTKT YTCNVDHKPS NTKVDKRV
である。
ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV
HTFPAVLQSS GLYSLSSVVT VPSSSLGTKT YTCNVDHKPS NTKVDKRV
である。
(b)重鎖ヒンジ領域
1つの例示的なヒンジドメインは、ヒトIgG1ヒンジドメインである。例示的なヒトIgG1ヒンジドメインのアミノ酸配列は、(配列番号7):EPKSCDKTHTCPPCPである。
1つの例示的なヒンジドメインは、ヒトIgG1ヒンジドメインである。例示的なヒトIgG1ヒンジドメインのアミノ酸配列は、(配列番号7):EPKSCDKTHTCPPCPである。
別の例示的なヒンジドメインは、ヒトIgG2ヒンジドメインである。例示的なヒトIgG2ヒンジドメインのアミノ酸配列は、(配列番号8):ERKCCVECPPCPである。
別の例示的なヒンジドメインは、ヒトIgG3ヒンジドメインである。例示的なヒトIgG3ヒンジドメインのアミノ酸配列は、(配列番号9):
ELKTPLGDTT HTCPRCPEPK SCDTPPPCPR CPEPKSCDTP PPCPRCPEPK
SCDTPPPCPR CP
である。
ELKTPLGDTT HTCPRCPEPK SCDTPPPCPR CPEPKSCDTP PPCPRCPEPK
SCDTPPPCPR CP
である。
別の例示的なヒンジドメインは、ヒトIgG4ヒンジドメインである。例示的なヒトIgG4ヒンジドメインのアミノ酸配列は、(配列番号10):ESKYGPPCPSCPである。本明細書中で記載されるように、IgG4ヒンジドメインは、S228P置換等の安定化突然変異を含んでよい。例示的なS228P‐安定化ヒトIgG4ヒンジドメインのアミノ酸配列は、(配列番号11):ESKYGPPCPPCPである。
(c)重鎖CH2及びCH3ドメイン
2つの重鎖のCH2及びCH3ドメインは、相互作用してIgG抗体の「Fcドメイン(Fc Domain)」を形成し、これは、Fcγ受容体(FcγR)を含むがこれに受容されない細胞Fc受容体によって認識されるドメインである。本明細書中で使用される場合、用語「Fc領域」は、IgG重鎖のC末端領域を定義するために使用される。Fc領域の一部分(Fc領域全体を包含する一部分を含む)を、本明細書では「Fcドメイン」と呼ぶ。Fc領域は、そのアミノ酸配列が、他のIgGアイソタイプに対してよりも、あるIgGアイソタイプに対して最も相同性が高い場合に、この特定のIgGアイソタイプ、クラス又はサブクラスのものであると表現される。抗体は、診断学におけるその公知の使用法に加えて、治療剤として有用であることが示されている。
2つの重鎖のCH2及びCH3ドメインは、相互作用してIgG抗体の「Fcドメイン(Fc Domain)」を形成し、これは、Fcγ受容体(FcγR)を含むがこれに受容されない細胞Fc受容体によって認識されるドメインである。本明細書中で使用される場合、用語「Fc領域」は、IgG重鎖のC末端領域を定義するために使用される。Fc領域の一部分(Fc領域全体を包含する一部分を含む)を、本明細書では「Fcドメイン」と呼ぶ。Fc領域は、そのアミノ酸配列が、他のIgGアイソタイプに対してよりも、あるIgGアイソタイプに対して最も相同性が高い場合に、この特定のIgGアイソタイプ、クラス又はサブクラスのものであると表現される。抗体は、診断学におけるその公知の使用法に加えて、治療剤として有用であることが示されている。
例示的なヒトIgG1のCH2‐CH3ドメインのアミノ酸配列は、(配列番号12):
231 240 250 260 270 280
APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
290 300 310 320 330
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
340 350 360 370 380
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
390 400 410 420 430
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
440 447
ALHNHYTQKS LSLSPGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
231 240 250 260 270 280
APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
290 300 310 320 330
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
340 350 360 370 380
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
390 400 410 420 430
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
440 447
ALHNHYTQKS LSLSPGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
例示的なヒトIgG2のCH2‐CH3ドメインのアミノ酸配列は、(配列番号13):
231 240 250 260 270 280
APPVA-GPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVQFNWYVD
290 300 310 320 330
GVEVHNAKTK PREEQFNSTF RVVSVLTVVH QDWLNGKEYK CKVSNKGLPA
340 350 360 370 380
PIEKTISKTK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDISVE
390 400 410 420 430
WESNGQPENN YKTTPPMLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
440 447
ALHNHYTQKS LSLSPGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
231 240 250 260 270 280
APPVA-GPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVQFNWYVD
290 300 310 320 330
GVEVHNAKTK PREEQFNSTF RVVSVLTVVH QDWLNGKEYK CKVSNKGLPA
340 350 360 370 380
PIEKTISKTK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDISVE
390 400 410 420 430
WESNGQPENN YKTTPPMLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
440 447
ALHNHYTQKS LSLSPGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
例示的なヒトIgG3のCH2‐CH3ドメインのアミノ酸配列は、(配列番号14):
231 240 250 260 270 280
APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVQFKWYVD
290 300 310 320 330
GVEVHNAKTK PREEQYNSTF RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
340 350 360 370 380
PIEKTISKTK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
390 400 410 420 430
WESSGQPENN YNTTPPMLDS DGSFFLYSKL TVDKSRWQQG NIFSCSVMHE
440 447
ALHNRFTQKS LSLSPGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
231 240 250 260 270 280
APELLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVQFKWYVD
290 300 310 320 330
GVEVHNAKTK PREEQYNSTF RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
340 350 360 370 380
PIEKTISKTK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
390 400 410 420 430
WESSGQPENN YNTTPPMLDS DGSFFLYSKL TVDKSRWQQG NIFSCSVMHE
440 447
ALHNRFTQKS LSLSPGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
例示的なヒトIgG4のCH2‐CH3ドメインのアミノ酸配列は、(配列番号15):
231 240 250 260 270 280
APEFLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSQED PEVQFNWYVD
290 300 310 320 330
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
340 350 360 370 380
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLTCLVK GFYPSDIAVE
390 400 410 420 430
WESNGQPENN YKTTPPVLDS DGSFFLYSRL TVDKSRWQEG NVFSCSVMHE
440 447
ALHNHYTQKS LSLSLGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
231 240 250 260 270 280
APEFLGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSQED PEVQFNWYVD
290 300 310 320 330
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
340 350 360 370 380
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLTCLVK GFYPSDIAVE
390 400 410 420 430
WESNGQPENN YKTTPPVLDS DGSFFLYSRL TVDKSRWQEG NVFSCSVMHE
440 447
ALHNHYTQKS LSLSLGX
であり、これはKabatに記載のEUインデックスによって番号付与されており、Xはリシン(K)であるか、又は不在である。
本明細書全体を通して、IgG重鎖の定常領域の残基の番号付与は、Kabat et al.、Sequences of Proteins of Immunological Interest、5th Ed. Public Health Service、NH1、MD (1991)(「Kabat」、これは参照により明示的に本明細書に援用される)によるEUインデックスの番号付与である。用語「Kabatに記載のEUインデックス(EU index as in Kabat)」は、ヒトIgG1 EU抗体の定常ドメインの番号付与を指す。多型は、抗体定常領域内の多数の異なる位置(例えばKabatに記載のEUインデックスによる番号付与で270位、272位、312位、315位、356位及び358位を含むがこれらに限定されないFc位置)において観察されており、従ってここで提示される配列と従来技術の配列との間にはわずかな差異が存在し得る。ヒト免疫グロブリンの多型形態は、十分に特性決定されている。現在、18個のGmアロタイプが公知である:G1m(1,2,3,17)又はG1m(a,x,f,z)、G2m(23)又はG2m(n)、G3m(5,6,10,11,13,14,15,16,21,24,26,27,28)又はG3m(b1,c3,b3,b0,b3,b4,s,t,g1,c5,u,v,g5)(Lefranc, et al., The human IgG subclasses: molecular analysis of structure, function and regulation. Pergamon, Oxford, pp. 43-78 (1990); Lefranc, G. et al., 1979, Hum. Genet.: 50, 199-211)。特に、本発明の抗体が、いずれの免疫グロブリン遺伝子のいずれのアロタイプ、アイソアロタイプ又はハプロタイプを組み込むことができ、本明細書中で提示される配列のアロタイプ、アイソアロタイプ又はハプロタイプに限定されないと考えられる。更に、発現系によっては、CH3ドメインのC末端アミノ酸残基(上記太字)は、翻訳後に除去できる。従ってCH3ドメインのC末端残基は、本発明のgp41結合分子の任意のアミノ酸残基である。本発明によって具体的に包含されるのは、CH3ドメインのC末端残基が欠けたgp41結合分子である。また本発明によって具体的に包含されるのは、CH3ドメインのC末端リシン残基を含む構造である。
(d)可変ドメイン
IgG分子の可変ドメインは、エピトープと接触することになる抗体のアミノ酸残基を含有する3つの相補性決定領域(「CDR」)、及びフレームワーク領域(「FR」)と呼ばれる介在性非CDRセグメントからなり、上記フレームワークセグメントは、一般にCDRループの構造を維持してCDRの位置を決定することにより、このような接触を可能とする(ただし特定のフレームワーク残基もエピトープに接触し得る)。従って、VL及びVHドメインは、構造n‐FR1‐CDR1‐FR2‐CDR2‐FR3‐CDR3‐FR4‐cを有する。CDRのアミノ酸配列は、ある抗体がある特定のエピトープに結合できるかどうかを決定する。抗体軽鎖と抗体重鎖との相互作用、特に、それぞれ別個のポリペプチド上に存在するこれらのVLドメインとVHドメインとの相互作用は、抗体のエピトープ結合部位を形成する。
IgG分子の可変ドメインは、エピトープと接触することになる抗体のアミノ酸残基を含有する3つの相補性決定領域(「CDR」)、及びフレームワーク領域(「FR」)と呼ばれる介在性非CDRセグメントからなり、上記フレームワークセグメントは、一般にCDRループの構造を維持してCDRの位置を決定することにより、このような接触を可能とする(ただし特定のフレームワーク残基もエピトープに接触し得る)。従って、VL及びVHドメインは、構造n‐FR1‐CDR1‐FR2‐CDR2‐FR3‐CDR3‐FR4‐cを有する。CDRのアミノ酸配列は、ある抗体がある特定のエピトープに結合できるかどうかを決定する。抗体軽鎖と抗体重鎖との相互作用、特に、それぞれ別個のポリペプチド上に存在するこれらのVLドメインとVHドメインとの相互作用は、抗体のエピトープ結合部位を形成する。
免疫グロブリンの成熟した重鎖及び軽鎖の可変ドメインからのアミノ酸は、鎖内でのアミノ酸の位置によって指定される。Kabatは、抗体に関する多数のアミノ酸配列を記載し、各サブグループに関するアミノ酸コンセンサス配列を同定し、残基番号を各アミノ酸に割り当てており、またCDRは、Kabatによって定義されるように同定される(Chothia, C. & Lesk, A. M.(1987) “Canonical Structures For The Hypervariable Regions Of Immunoglobulins,” J. Mol. Biol. 196:901-917)によって定義されるCDRH1は、残基5個分前から始まることが理解されるだろう)。Kabatの番号付与スキームは、その大要に含まれていない抗体に対して、保存されたアミノ酸を参照して上記抗体をKabatのコンセンサス配列のうちの1つと整列させることにより、拡張可能である。残基番号を割り当てるこの方法は、当該技術分野において標準的なものとなっており、キメラ変異型又はヒト化変異型を含む異なる抗体の同一の位置のアミノ酸が容易に同定される。例えば、ヒト抗体軽鎖の50位のアミノ酸は、マウス抗体軽鎖の50位のアミノ酸と同一位置を占める。
抗体の軽鎖の第1、第2及び第3のCDRである(又は第1、第2及び第3のCDRとして機能し得る)ポリペプチドは、本明細書ではそれぞれCDRL1ドメイン、CDRL2ドメイン及びCDRL3ドメインと呼ばれる。同様に、抗体の重鎖の第1、第2及び第3のCDRである(又は第1、第2及び第3のCDRとして機能し得る)ポリペプチドは、本明細書ではそれぞれCDRH1ドメイン、CDRH2ドメイン及びCDRH3ドメインと呼ばれる。よって、CDRL1ドメイン、CDRL2ドメイン、CDRL3ドメイン、CDRH1ドメイン、CDRH2ドメイン、及びCDRH3ドメインという用語は、あるタンパク質に組み込まれた場合に、当該タンパク質が、軽鎖及び重鎖若しくはダイアボディ若しくは単鎖結合分子(例えばscFv、BiTe(登録商標)等)を有する抗体であるか、又は別のタイプのタンパク質であるかにかかわらず、当該タンパク質を、特定のエピトープに結合できるようにする、ポリペプチドを対象としている。
用語「エピトープ結合ドメイン(Epitope Binding Domain)」(例えばgp41結合ドメイン)は、結合分子の、あるエピトープ(例えばgp41のエピトープ)に免疫特異的に結合できる能力に寄与する、該結合分子の部分(又はこのような部分のアミノ酸配列を有するポリペプチド)を指す。エピトープ結合ドメインは、抗体のVL若しくはVHドメイン、若しくは抗体のCDRドメインのうちの1、2、3、4若しくは5個を含有してよく、又は抗体のCDRドメインのうちの6個全てを含有してよく、このようなエピトープに免疫特異的に結合できるものの、このような抗体のエピトープとは異なるエピトープに対する免疫特異性、親和性又は選択性を呈してもよい。エピトープ結合ドメインは、CDRの一部のみ、即ちCDR残基の、結合に必要なサブセットのみを含有してよく、これはSDRと呼ばれる(Kim, J.H. et al. (2012) “Humanization By CDR Grafting And Specificity-Determining Residue Grafting,” Methods Mol. Biol. 907:237-245; Kim, K.S. et al. (2010) “Construction Of A Humanized Antibody To Hepatitis B Surface Antigen By Specificity-Determining Residues (SDR)-Grafting And De-Immunization,” Biochem. Biophys. Res. Commun. 396(2):231-237; Kashmiri, S.V. et al. (2005) “SDR Grafting - A New Approach To Antibody Humanization,” Methods 36(1):25-34; Gonzales, N.R. et al. (2004) “SDR Grafting Of A Murine Antibody Using Multiple Human Germline Templates To Minimize Its Immunogenicity,” Mol. Immunol. 41:863-872)。しかしながら好ましくは、エピトープ結合ドメインは、このような抗体のCDRドメインのうちの6個全てを含有することになる。あるエピトープ結合ドメインは、単一のポリペプチド鎖(例えばscFv)であってよく、又はそれぞれがアミノ末端及びカルボキシ末端を有する、ジスルフィド結合によって互いに共有結合できる2つ以上のポリペプチド鎖(例えばダイアボディ、Fab部分、Fab2部分等)を含んでよい。
2.抗体のヒト化
本発明はまた特に、ある抗体のVL又はVHドメイン、好ましくはある抗体のVL及びVHドメインの両方を含む結合分子を包含する。好ましくは、このような抗体はヒト化抗体である。モノクローナル抗体は典型的には、マウス又はウサギ等の非ヒト種の体内で調製される。このような抗体の変異型及び/又は定常ドメインは、免疫原として認識され得るため、これらに対する免疫応答が引き起こされる。しかしながら、このような分子は、1つ以上のアミノ酸置換を導入して、上記抗体をヒトによって生成される抗体により近い抗体とすることにより「ヒト化(humanize)」でき、これによって上記抗体の免疫原性を低減又は排除できる。用語「ヒト化(humanized)」抗体は、キメラ分子であって、一般に組み換え技術を用いて調製され、非ヒト種からの免疫グロブリンのエピトープ結合ドメインと、残りの、ヒト免疫グロブリンの構造及び/又は配列に基づく分子の免疫グロブリン構造とを有する、キメラ分子を指す。このような抗体の可変ドメインのポリヌクレオチド配列は、上記誘導体を生成するため及び上記抗体の親和性又は他の特徴を改善するための遺伝子操作のために使用できる。様々な抗体に対するこのアプローチの適用は、LoBuglio, A.F. et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224; Sato, K. et al. (1993) Cancer Res 53:851-856. Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; Kettleborough, C. A. et al. (1991) “Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation,” Protein Engineering 4:773-3783; Maeda, H. et al. (1991) “Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity,” Human Antibodies Hybridoma 2:124-134; Gorman, S. D. et al. (1991) “Reshaping A Therapeutic CD4 Antibody,” Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) “Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo,” Bio/Technology 9:266-271; Co, M. S. et al. (1991) “Humanized Antibodies For Antiviral Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) “Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; 及びCo, M.S. et al. (1992) “Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen,” J. Immunol. 148:1149-1154によって報告されている。いくつかの実施形態では、ヒト化抗体は全てのCDR配列を保存する(例えば、マウス抗体からの6つのCDR全てを含有するヒト化マウス抗体)。他の実施形態では、ヒト化抗体は、オリジナルの抗体に対して配列が異なる改変された1つ以上のCDR(1つ、2つ、3つ、4つ、5つ及び/又は6つ)を有する。
本発明はまた特に、ある抗体のVL又はVHドメイン、好ましくはある抗体のVL及びVHドメインの両方を含む結合分子を包含する。好ましくは、このような抗体はヒト化抗体である。モノクローナル抗体は典型的には、マウス又はウサギ等の非ヒト種の体内で調製される。このような抗体の変異型及び/又は定常ドメインは、免疫原として認識され得るため、これらに対する免疫応答が引き起こされる。しかしながら、このような分子は、1つ以上のアミノ酸置換を導入して、上記抗体をヒトによって生成される抗体により近い抗体とすることにより「ヒト化(humanize)」でき、これによって上記抗体の免疫原性を低減又は排除できる。用語「ヒト化(humanized)」抗体は、キメラ分子であって、一般に組み換え技術を用いて調製され、非ヒト種からの免疫グロブリンのエピトープ結合ドメインと、残りの、ヒト免疫グロブリンの構造及び/又は配列に基づく分子の免疫グロブリン構造とを有する、キメラ分子を指す。このような抗体の可変ドメインのポリヌクレオチド配列は、上記誘導体を生成するため及び上記抗体の親和性又は他の特徴を改善するための遺伝子操作のために使用できる。様々な抗体に対するこのアプローチの適用は、LoBuglio, A.F. et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224; Sato, K. et al. (1993) Cancer Res 53:851-856. Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; Kettleborough, C. A. et al. (1991) “Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation,” Protein Engineering 4:773-3783; Maeda, H. et al. (1991) “Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity,” Human Antibodies Hybridoma 2:124-134; Gorman, S. D. et al. (1991) “Reshaping A Therapeutic CD4 Antibody,” Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) “Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo,” Bio/Technology 9:266-271; Co, M. S. et al. (1991) “Humanized Antibodies For Antiviral Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) “Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; 及びCo, M.S. et al. (1992) “Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen,” J. Immunol. 148:1149-1154によって報告されている。いくつかの実施形態では、ヒト化抗体は全てのCDR配列を保存する(例えば、マウス抗体からの6つのCDR全てを含有するヒト化マウス抗体)。他の実施形態では、ヒト化抗体は、オリジナルの抗体に対して配列が異なる改変された1つ以上のCDR(1つ、2つ、3つ、4つ、5つ及び/又は6つ)を有する。
抗体をヒト化する際の一般原理は、抗体のエピトープ結合ドメインの塩基配列を保持しながら、抗体の非ヒト残部をヒト抗体配列と交換するステップを伴う。モノクローナル抗体をヒト化するためには、4つの一般的なステップが存在する。上記ステップは以下の通りである:(1)開始抗体の軽鎖及び重鎖可変ドメインのヌクレオチド及び予測されるアミノ酸配列を決定するステップ;(2)ヒト化抗体又はイヌ化抗体を設計するステップ、即ちヒト化又はイヌ化プロセス中に使用する抗体フレームワーク領域を決定するステップ;(3)実際のヒト化又はイヌ化法/技術;並びに(4)ヒト化抗体のトランスフェクション及び発現。例えば米国特許第4,816,567号;米国特許第5,807,715号;米国特許第5,866,692号;及び米国特許第6,331,415号を参照。
非ヒト免疫グロブリン由来の抗原結合ドメインを含む多数のヒト化抗体分子が記述されており、これらは、げっ歯類又は修飾げっ歯類可変ドメインと、ヒト定常ドメインに融合した、これらに関連する相補性決定領域(CDR)とを有するキメラ抗体を含む(例えばWinter et al. (1991) “Man‐made Antibodies,” Nature 349:293-299; Lobuglio et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224 (1989); Shaw et al. (1987) “Characterization Of A Mouse/Human Chimeric Monoclonal Antibody (17-1A) To A Colon Cancer Tumor-Associated Antigen,” J. Immunol. 138:4534-4538;及びBrown et al. (1987) “Tumor-Specific Genetically Engineered Murine/Human Chimeric Monoclonal Antibody,” Cancer Res. 47:3577-3583を参照)。他の参照文献は、適切なヒト抗体定常ドメインと融合する前にヒト支持性フレームワーク領域(FR)にグラフと重合される、げっ歯類CDRについて説明している(例えば、Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536;及びJones et al. (1986) “Replacing The Complementarity-Determining Regions In A Human Antibody With Those From A Mouse,”Nature 321:522-525を参照)。別の参照文献は、組み換えによってベニアリングされたげっ歯類フレームワーク領域によって支持されたげっ歯類CDRについて説明している。例えば欧州公開特許第519,596号を参照。これらの「ヒト化」分子は、ヒトレシピエントのこれらの部分の治療的応用の期間及び効果を制限する、げっ歯類抗ヒト抗体分子に対する望ましくない免疫学的応答を最小化するよう設計される。抗体をヒト化するために利用してよい他の方法は、Daugherty et al. (1991) “Polymerase Chain Reaction Facilitates The Cloning, CDR-Grafting, And Rapid Expression Of A Murine Monoclonal Antibody Directed Against The CD18 Component Of Leukocyte Integrins,” Nucl. Acids Res. 19:2471-2476並びに米国特許第6,180,377号;米国特許第6,054,297号;米国特許第5,997,867号;及び米国特許第5,866,692号によって開示されている。本発明は特に、「ヒト化」抗体のVL及び/又はVHドメインを含む(抗体及びダイアボディを含む)結合分子を包含する。
このような成功にもかかわらず、治療用途に最適化された、安定した官能性ヘテロ二量体非単一特異性ダイアボディの生産は、ポリペプチド鎖で採用するドメインを注意深く考察し配置することによって、更に改善できる。よって本発明は、gp41と、免疫エフェクタ細胞の表面上に存在する分子とに同時に結合できる、安定した治療に有用なヘテロ二量体ダイアボディ及びヘテロ二量体Fcダイアボディを、共有結合によって形成するよう特に設計された、特定のポリペプチドの提供を対象とする。
B.二重特異性抗体
上述のように、天然抗体は1つのエピトープ種にしか結合できない(即ちこれらは単一特異性である)が、上記種の複数の複製に結合できる(即ち2価性又は多価性を示す)。抗体の、抗原のエピトープに結合する能力は、該抗体のVL及びVHドメインの存在及びそのアミノ酸配列に依存する。抗体の軽鎖と重鎖との相互作用、特にそのVLドメインとVHドメインとの相互作用は、IgG等の天然抗体の2つのエピトープ結合ドメインのうちの1つを形成する。
上述のように、天然抗体は1つのエピトープ種にしか結合できない(即ちこれらは単一特異性である)が、上記種の複数の複製に結合できる(即ち2価性又は多価性を示す)。抗体の、抗原のエピトープに結合する能力は、該抗体のVL及びVHドメインの存在及びそのアミノ酸配列に依存する。抗体の軽鎖と重鎖との相互作用、特にそのVLドメインとVHドメインとの相互作用は、IgG等の天然抗体の2つのエピトープ結合ドメインのうちの1つを形成する。
抗体の官能性は、2つの別個の異なる抗原(若しくは同一の抗原の異なるエピトープ)に同時に結合できる多重特異性抗体系分子を生成することによって、並びに/又は同一のエピトープ及び/若しくは抗原に関して更に高い価数(即ち3つ以上の結合ドメイン)を有する抗体系分子を生成することによって、増強できる。
天然抗体より高い能力を有する分子を提供するために、広範な組み換え二重特異性抗体フォーマットが開発されてきた(例えば国際公開第2008/003116号、国際公開第2009/132876号、国際公開第2008/003103号、国際公開第2007/146968号、国際公開第2009/018386号、国際公開第2012/009544号、国際公開第2013/070565号を参照)。このようなアプローチの大半は、更なる結合ドメイン(例えばscFv、VL、VH等)を抗体コア(IgA、IgD、IgE、IgG若しくはIgM)へ若しくは上記抗体コア内に融合させるため、又は複数の抗原結合部分(例えば2つのFab部分若しくはscFv)を互いに融合させるために、リンカーペプチドを使用する。代替的なフォーマットは、結合タンパク質(例えばscFv、VL、VH等)を、CH2‐CH3ドメイン又は代替となるポリペプチド等の二量体化ドメインに融合させるために、リンカーペプチドを使用する(国際公開第2005/070966号、国際公開第2006/107786A号、国際公開第2006/107617A号、国際公開第2007/046893号)。典型的には、このようなアプローチは妥協及びトレードオフを伴う。例えば国際公開第2013/174873号、国際公開第2011/133886号、及び国際公開第2010/136172号は、リンカーの使用によって、治療的設定に問題が生じる場合があることを開示しており、2つ以上の抗原に結合できるように、CL及びCH1ドメインがそれぞれの自然位置から切り替わり、かつVL及びVHドメインが多様化されている、三重特異性抗体を教示している(国際公開第2008/027236号;国際公開第2010/108127号)。従って、これらの文書に開示されている分子は、結合特異性を、追加の抗原種に結合できる能力と交換している。国際公開第2013/163427号及び国際公開第2013/119903号は、CH2ドメインを修飾して、結合ドメインを含む融合タンパク付加物を含有させるステップを開示している。この文書は、CH2が、エフェクタ機能の仲介において最低限の役割しか果たさないらしいことを記載している。国際公開第2010/028797号、国際公開第2010028796号、及び国際公開第2010/028795号は、Fc領域が追加のVL及びVHドメインで置換されて、3価結合分子を形成している、組み換え抗体を開示している。国際公開第2003/025018号及び国際公開第2003012069号は、個々の鎖がscFvドメインを含有する組み換えダイアボディを開示している。国際公開第2013/006544号は、単一のポリペプチド鎖として合成された後、タンパク質分解に供されることによってヘテロ二量体構造が得られる、多価fab分子を開示している。従って、これらの文書に開示されている分子は、エフェクタ機能を仲介する能力の全て又はある程度を、追加の抗原種に結合できる能力と交換している。国際公開第2014/022540号、国際公開第2013/003652号、国際公開第2012/162583号、国際公開第2012/156430号、国際公開第2011/086091号、国際公開第2008/024188号、国際公開第2007/024715号、国際公開第2007/075270号、国際公開第1998/002463号、国際公開第1992/022583号、及び国際公開第1991/003493号は、追加の結合ドメイン又は官能基を抗体又は抗体部分に付加するステップ(例えば抗体の軽鎖にダイアボディを付加するステップ、又は抗体の軽鎖及び重鎖に追加のVL及びVHドメインを付加するステップ、又は異種融合タンパク質を互いに対して付加するステップ若しくは複数のFabドメインを互いに対して連鎖させるステップ)を開示している。従って、これらの文書に開示されている分子は、ナイーブ抗体構造を、追加の抗原種に結合できる能力と交換している。
II.二重特異性ダイアボディ
本技術分野は更に、2つ以上の異なるエピトープ種と結合できる(即ち2価性又は多価性に加えて二重特異性又は多重特異性を呈することができる)という点でこのような天然抗体とは異なるダイアボディを産生できる可能性に注目している(例えばHolliger et al. (1993) “’Diabodies’: Small Bivalent And Bispecific Antibody Fragments,” Proc. Natl. Acad. Sci. (U.S.A.) 90:6444-6448; 米国公開特許第2004/0058400号(Hollinger et al.);米国公開特許第2004/0220388号(Mertens et al.);Alt et al. (1999) FEBS Lett. 454(1-2):90-94;Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672;国際公開第02/02781号(Mertens et al.);Olafsen, T. et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Protein Eng Des Sel. 17(1):21-27;Wu, A. et al. (2001) “Multimerization Of A Chimeric Anti-CD20 Single Chain Fv-Fv Fusion Protein Is Mediated Through Variable Domain Exchange,” Protein Engineering 14(2):1025-1033;Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992;Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588;Baeuerle, P.A. et al. (2009) “Bispecific T cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944を参照)。
本技術分野は更に、2つ以上の異なるエピトープ種と結合できる(即ち2価性又は多価性に加えて二重特異性又は多重特異性を呈することができる)という点でこのような天然抗体とは異なるダイアボディを産生できる可能性に注目している(例えばHolliger et al. (1993) “’Diabodies’: Small Bivalent And Bispecific Antibody Fragments,” Proc. Natl. Acad. Sci. (U.S.A.) 90:6444-6448; 米国公開特許第2004/0058400号(Hollinger et al.);米国公開特許第2004/0220388号(Mertens et al.);Alt et al. (1999) FEBS Lett. 454(1-2):90-94;Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672;国際公開第02/02781号(Mertens et al.);Olafsen, T. et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Protein Eng Des Sel. 17(1):21-27;Wu, A. et al. (2001) “Multimerization Of A Chimeric Anti-CD20 Single Chain Fv-Fv Fusion Protein Is Mediated Through Variable Domain Exchange,” Protein Engineering 14(2):1025-1033;Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992;Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588;Baeuerle, P.A. et al. (2009) “Bispecific T cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944を参照)。
ダイアボディの設計は、軽鎖及び/又は重鎖可変ドメインが短い連鎖ペプチドを用いて連結されている、単鎖可変ドメイン部分(scFv)の構造に基づく。Bird et al. (1988) (“Single-Chain Antigen-Binding Proteins,” Science 242:423-426)は、一方の可変ドメインのカルボキシ末端と他方の可変ドメインのアミノ末端との間の約3.5nmを埋める連結ペプチドの例を記載している。他の配列のリンカーも設計及び使用されている(Bird et al. (1988) “Single-Chain Antigen-Binding Proteins,” Science 242:423-426)。リンカーは、薬剤の付着又は剛性支持体の付着といった追加の機能のために修飾できる。単鎖変異体は、組み換えによって又は合成によって産生できる。scFvの合成産生に関しては、自動化されたシンセサイザを使用できる。scFvの組み換え産生に関しては、scFvをエンコードするポリヌクレオチドを含有する好適なプラスミドを、酵母、植物、昆虫若しくは哺乳類細胞等の真核細胞又は大腸菌等の原核細胞である好適な宿主細胞に導入できる。関心対象のscFvをエンコードするポリヌクレオチドは、ポリヌクレオチドのライゲーション等の従来の操作によって作製できる。得られたscFvは、当該技術分野で公知の標準的なタンパク質精製技術を用いて単離できる。
非単一特異性「ダイアボディ(diabody)」の提供は、異なるエピトープを発現する細胞を共連結(co‐ligate)して共存させることができる能力という、抗体を上回る有意な利点を提供する。従って二重特異性ダイアボディは、療法及び免疫診断を含む広範な用途を有する。二重特異性は、様々な用途におけるダイアボディの設計及び操作の大幅な柔軟性を可能とし、これにより、多量体抗原の結合活性の上昇、異なる複数の抗原の架橋、及び両標的抗原の存在に基づく特定の細胞タイプに対する指向性標的化を提供する。当該技術分野において公知のダイアボディ分子は、(〜50kDa以下の小さいサイズのダイアボディに関して)その2価性、低い解離率及び循環からの迅速な排除により、腫瘍撮像の分野での特定の使用も示している(Fitzgerald et al. (1997) “Improved Tumour Targeting By Disulphide Stabilized Diabodies Expressed In Pichia pastoris,” Protein Eng. 10:1221-1225)。
二重特異性ダイアボディを産生する能力により、異なる細胞上に存在する受容体の共連結(例えば細胞傷害性T細胞と、疾患抗原を発現する癌細胞又は病原体感染細胞等の標的細胞との架橋)によって、2つの細胞を共連結するために、上記ダイアボディを(「トランス(trans)」として)使用できるようになる(Staerz et al. (1985) “Hybrid Antibodies Can Target Sites For Attack By T Cells,” Nature 314:628-631, and Holliger et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody,” Protein Eng. 9:299-305; Marvin et al. (2005) “Recombinant Approaches To IgG-Like Bispecific Antibodies,” Acta Pharmacol. Sin. 26:649-658; 非特許文献2)。あるいは又は更に、二重特異性(又は多重特異性)ダイアボディを(「シス」として)用いて、同一の細胞の表面上に存在する受容体等の分子を共連結できる。異なる細胞及び/又は受容体の共連結は、エフェクタ機能及び/又は免疫細胞シグナリングを変調するために有用である。エピトープ結合ドメインを含む多重特異性分子(例えば二重特異性ダイアボディ)は、Tリンパ球、ナチュラルキラー(NK)細胞、抗原提示細胞又は他の単核細胞上で発現される、CD2、CD3、CD8、CD16、TCR、NKG2D等といったいずれの免疫細胞の表面決定因子、又はCD19、CD20、CD22、CD30、CD37、CD40及びCD74といったB細胞の表面決定因子を対象としてよい(Moore, P.A. et al. (2011) “Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T cell Killing Of B-Cell Lymphoma,” Blood 117(17):4542-4551; Cheson, B.D. et al. (2008) “Monoclonal Antibody Therapy For B-Cell Non-Hodgkin’s Lymphoma,” N. Engl. J. Med. 359(6):613-626; Castillo, J. et al. (2008) “Newer Monoclonal Antibodies For Hematological Malignancies,” Exp. Hematol. 36(7):755-768)。特に、免疫エフェクタ細胞上に存在する細胞表面受容体を対象とするエピトープ結合ドメインは、標的転換細胞殺滅を仲介できる多重特異性結合分子の生成に有用である。
多くの研究において、エフェクタ細胞決定因子、例えばFcγ受容体(FcγR)に結合するダイアボディは、エフェクタ細胞を活性化させることも分かった(Holliger et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T cells Mediated By A Bispecific Diabody,” Protein Eng. 9:299-305; Holliger et al. (1999) “Carcinoembryonic Antigen (CEA)-Specific T cell Activation In Colon Carcinoma Induced By Anti-CD3 x Anti-CEA Bispecific Diabodies And B7 x Anti-CEA Bispecific Fusion Proteins,” Cancer Res. 59:2909-2916;国際公開第2006/113665号;国際公開第2008/157379号;国際公開第2010/080538号;国際公開第2012/018687号;国際公開第2012/162068号)。通常、エフェクタ細胞の活性化は、Fcドメイン‐FcγR相互作用によるエフェクタ細胞への抗原結合抗体の結合によってトリガされる。従ってこれに関して、ダイアボディ分子は、Fcドメインを含むかどうかとは無関係に、(例えば当該技術分野において公知の、又は本明細書に例示されているいずれのエフェクタ機能アッセイ(例えばADCCアッセイ)においてアッセイされるように)Ig様官能性を示し得る。腫瘍とエフェクタ細胞とを架橋させることによって、ダイアボディは腫瘍細胞近傍にエフェクタ細胞を移動させるだけではなく、効果的な腫瘍殺滅をもたらす(例えばCao et al. (2003) "Bispecific Antibody Conjugates In Therapeutics, "Adv. Drug. Deliv. Rev. 55:171-197を参照)。
しかしながら、上述の二重特異性ダイアボディの利点は顕著なコストにつながる。このような非単一特異性ダイアボディの形成は、2つ以上の別個の異なるポリペプチドの良好な集合を必要とする(即ち上記形成は、ダイアボディが、異なるポリペプチド鎖種のヘテロ二量体形成によって形成されることを必要とする)。この事実は、同一のポリペプチド鎖のホモ二量体形成によって形成される単一特異性ダイアボディとは対照的である。非単一特異性ダイアボディを形成するために少なくとも2つの異なるポリペプチド(即ち2つのポリペプチド種)を提供しなければならないため、及びこのようなポリペプチドのホモ二量体形成は不活性分子をもたらす(Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588)ため、このようなポリペプチドの産生は、同一種のポリペプチド間での共有結合を防止できる(即ちこれらのホモ二量体化を防止できる)ような方法で達成しなければならない(Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588)。従って本技術分野は、このようなポリペプチドの非共有結合的連結を教示している(例えばOlafsen et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Prot. Engr. Des. Sel. 17:21-27; Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588; Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672を参照)。
しかしながら、本技術分野は、非共有結合的に連結したポリペプチドで構成される二重特異性ダイアボディが不安定であり、非官能性単一ポリペプチド鎖モノマーへと容易に分解することを認識している(例えばLu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672を参照)。
この課題をものともせず、本技術分野は、DART(登録商標)ダイアボディと呼ばれる、安定した共有結合ヘテロ二量体性非単一特異性ダイアボディの開発に成功した。例えば、Liu. L et al. (2017) “MGD011, A CD19 x CD3 Dual-Affinity Retargeting Bi-specific Molecule Incorporating Extended Circulating Half-life for the Treatment of B-Cell Malignancies,” Clin Cancer Res. 23(6):1506-1518; Tsai, P. et al. (2016) “CD19xCD3 DART Protein Mediates Human B-Cell Depletion In Vivo In Humanized BLT Mice,” Mol. Ther. Oncolytics 3:15024. doi: 10.1038/mto.2015.24; Chen, X. et al. (2016) “Mechanistic Projection of First-in-Human Dose for Bispecific Immunomodulatory P-Cadherin LP-DART: An Integrated PK/PD Modeling Approach,” Clin. Pharmacol. Ther. 100(3):232-241;非特許文献2; Al Hussaini, M. et al. (2015) “Targeting CD123 In AML Using A T-Cell Directed Dual-Affinity Re-Targeting (DART(R)) Platform,” Blood pii: blood-2014-05-575704; Chichili, G.R. et al. (2015) “A CD3xCD123 Bispecific DART For Redirecting Host T Cells To Myelogenous Leukemia: Preclinical Activity And Safety In Nonhuman Primates,” Sci. Transl. Med. 7(289):289ra82; Johnson, S. et al. (2010) “Effector Cell Recruitment With Novel Fv-Based Dual-Affinity Re-Targeting Protein Leads To Potent Tumor Cytolysis And In Vivo B-Cell Depletion,” J. Molec. Biol. 399(3):436-449; Veri, M.C. et al. (2010) “Therapeutic Control Of B Cell Activation Via Recruitment Of Fcgamma Receptor IIB (CD32B) Inhibitory Function With A Novel Bispecific Antibody Scaffold,” Arthritis Rheum. 62(7):1933-1943; Moore, P.A. et al. (2011) “Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T cell Killing Of B-Cell Lymphoma,” Blood 117(17):4542-4551;米国特許第9,932,400号;米国特許第9,908,938号;米国特許第9,889,197号;米国特許第9,884,921号;米国特許第9,822,181号;米国特許第9,296,816号;米国特許第9,284,375号;米国特許第8,795,667号;米国特許第8,187,593号;米国特許第8,669,349号;米国特許第8,784,808号;米国特許第8,795,667号;米国特許第8,802,091号;米国特許第8,802,093号;米国特許第8,946,387号;及び米国特許第8,968,730号、並びに米国公開特許第2009/0060910号;米国公開特許第2010/0174053号;米国公開特許第2011/0081347号;米国公開特許第2011/0097323号;米国公開特許第2011/0117089号;米国公開特許第2012/0034221号;米国公開特許第2012/0294796号;米国公開特許第2013/0149236号;米国公開特許第2013/0295121号;米国公開特許第2014/0017237号;米国公開特許第2014/0099318号;米国公開特許第2014/0255407号;米国公開特許第2015/0175697号;米国公開特許第2016/0017038号;米国公開特許第2016/0159908号;米国公開特許第2016/0159908号;米国公開特許第2016/0194396号;米国公開特許第2016/0194396号;米国公開特許第2016/0200827号;米国公開特許第2016/0200827号;米国公開特許第2016/0222105号;米国公開特許第2016/0222105号;米国公開特許第2016/0333111号;米国公開特許第2016/0355586号;米国公開特許第2016/0355586号;米国公開特許第2017/0157251号;米国公開特許第2017/0157251号;米国公開特許第2017/0198037号;米国公開特許第2017/0198037号;米国公開特許第2017/0198045号;米国公開特許第2017/0204176号;米国公開特許第2017/0204176号;米国公開特許第2017/0247452号;及び米国公開特許第2018/0094072号;欧州特許第1868650号;欧州特許第2158221号;欧州特許第2247304号;欧州特許第2252631号;欧州特許第2282770号;欧州特許第2328934号;欧州特許第2376109号;欧州特許第2542256号;欧州特許第2601216号;欧州特許第2714079号;欧州特許第2714733号;欧州特許第2786762号;欧州特許第2839842号;欧州特許第2840091号;並びに国際公開第2006/113665号;国際公開第2008/157379号;国際公開第2010/027797号;国際公開第2010/033279号;国際公開第2010/080538号;国際公開第2011/109400号;国際公開第2012/018687号;国際公開第2012/162067号;国際公開第2012/162068号;国際公開第2014/159940号;国際公開第2015/021089号;国際公開第2015/026892号;及び国際公開第2015/026894号を参照。このようなダイアボディは、2つ以上の共有結合的に複合体化したポリペプチドを含み、1つ以上のシステイン残基を、採用したポリペプチド種(これらはジスルフィド結合を形成でき、これによってこのようなポリペプチド鎖の1つ以上のペアを互いに共有結合させることができる)それぞれの中へと組み込むステップを伴う。例えば、このような構造のC末端へのシステイン残基の追加は、関与するポリペプチド鎖間のジスルフィド結合を可能とすることが分かっており、これは、ダイアボディの結合特性に干渉することなく、得られるダイアボディを安定化させる。
最も単純なDART(登録商標)は、2つのポリペプチド鎖を含み、これらはそれぞれ3つのドメインを含む(図1A〜1B)。第1のポリペプチド鎖は:(i)第1の免疫グロブリンの軽鎖可変ドメイン(VL1)の結合領域を含むドメイン;(ii)第2の免疫グロブリンの重鎖可変ドメイン(VH2)の結合領域を含む、第2のドメイン;並びに(iii)上記第2のポリペプチド鎖とのヘテロ二量体化を促進する役割、及びダイアボディの第2のポリペプチド鎖に対する第1のポリペプチド鎖の共有結合を促進する役割を果たす、第3のドメイン(「ヘテロ二量体促進ドメイン(Heterodimer‐Promoting Domain)を含む。第2のポリペプチド鎖は:相補的な第1のドメイン(VL2ドメイン);相補的な第2のドメイン(VH1ドメイン);並びに第1のポリペプチド鎖の第3のドメインと複合体化して、第1のポリペプチド鎖とのヘテロ二量体化及び共有結合を促進する、第3のドメイン(「ヘテロ二量体促進ドメイン」)を含有する。このような分子は、安定かつ強力であり、2つ以上の抗原を同時に結合する能力を有する。一実施形態では、第1及び第2のポリペプチド鎖の第3のドメインはそれぞれ、システイン(「Cマーク(Cを丸で囲んだマーク)」)残基を含有し、このシステイン残基は、ジスルフィド結合を介してポリペプチドを一体に結合させる役割を果たす。
上記ポリペプチド鎖のうちの一方又は両方の第3のドメインは更に、CH2‐CH3ドメインの配列を有してよく、これにより、ダイアボディのポリペプチドの複合体化によって、細胞(Bリンパ球、樹状細胞、ナチュラルキラー細胞、マクロファージ、好中球、好酸球、好塩基球及び肥満細胞等)のFc受容体に結合できるFcドメインが形成される)。このような分子の多数の変形例が記載されており(例えば米国公開特許第2013‐0295121号;米国公開特許第2010‐0174053号;米国公開特許第2007‐0004909号;米国公開特許第2009‐0060910号;欧州公開特許第2714079号;欧州公開特許第2601216号;欧州公開特許第2376109号;欧州公開特許第2158221号;及び国際公開第2012/162068号;国際公開第2012/018687号;国際公開第2010/080538号;国際公開第2006/113665号を参照)、また本明細書中で提供される。このような分子の多数の変異型が記載されており(例えば、米国公開特許第2015/0175697号;米国公開特許第2014/0255407号;米国公開特許第2014/0099318号;米国公開特許第2013/0295121号;米国公開特許第2010/0174053号;米国公開特許第2009/0060910号;米国公開特許第2007‐0004909号;欧州公開特許第2714079号;欧州公開特許第2601216号;欧州公開特許第2376109号;欧州公開特許第2158221号;欧州公開特許第1868650号;及び国際公開第2012/162068号;国際公開第2012/018687号;国際公開第2010/080538号;国際公開第2006/113665号参照)、また本明細書中で提供される。
二重特異性又は4価分子が望ましいもののFcは必要ない場合の用途のために、代替的な構成が当該技術分野において公知であり、これは、「BiTE(登録商標)抗体」とも呼ばれる、二重特異性T細胞エンゲージャ分子(例えば国際公開第1993/11161号;及び国際公開第2004/106381号参照)並びに「TandAb(登録商標)」とも呼ばれる4価タンデム抗体(例えば米国公開特許第2011‐0206672号;欧州公開特許第2371866号;国際公開第1999/057150号;国際公開第2003/025018号;及び国際公開第2013/013700号参照)を含むが、これに限定されない。BiTE(登録商標)は、タンデム結合されたscFvを含む単一のポリペプチド鎖から形成され、またTandAb(登録商標)は、VH1、VL2、VH2、及びVL2ドメインをそれぞれ有する2つの同一の鎖のホモ二量体化によって形成される。
III.本発明の実施形態
本発明は、「エピトープ結合分子(Epitope‐Binding Molecule)」を対象とし、これは、上記分子がエピトープに結合することを可能にする「エピトープ結合部位(Epitope‐Binding Site)」を形成するために十分な、CDRL1ドメイン、CDRL2ドメイン、CDRL3ドメイン、CDRH1ドメイン、CDRH2ドメイン、若しくはCDRH3ドメイン、又はこれらのいずれの組み合わせ若しくは部分的組み合わせといった「エピトープ結合ドメイン(Epitope‐Binding Domain)」を含む。例えば、「gp41エピトープ結合分子(gp41‐Epitope‐Binding Molecule)」は、上記分子がgp41のエピトープに結合することを可能にする「gp41エピトープ結合部位(gp41‐Epitope‐Binding Site)」を形成するために十分な、CDRL1ドメイン、CDRL2ドメイン、CDRL3ドメイン、CDRH1ドメイン、CDRH2ドメイン、若しくはCDRH3ドメイン、又はこれらのいずれの組み合わせ若しくは部分的組み合わせといった「エピトープ結合ドメイン(Epitope‐Binding Domain)」を含む。しかしながら好ましくは、このようなgp41結合分子は、gp41のエピトープに免疫特異的に結合する「抗gp41抗体(anti‐gp41 antibody)」のCDRL1ドメイン、CDRL2ドメイン、CDRL3ドメイン、CDRH1ドメイン、CDRH2ドメイン、及びCDRH3ドメインを有する。いくつかの実施形態では、このような分子は、このような抗gp41抗体のgp41‐VL結合ドメインのみ、又はこのような抗gp41抗体のgp41‐VH結合ドメインのみを含んでよく、これらのドメインは他の結合ドメインと相互作用して、gp41のエピトープへの免疫特異的結合を仲介できるか、又はこれらのドメインだけで、このような結合を仲介するのに十分である場合もある。いくつかの実施形態では、このような分子は、抗gp41抗体のgp41‐VL結合ドメイン及び抗gp41抗体のgp41‐VH結合ドメインの両方を含んでよく、これらは同一の抗体由来であっても、異なる抗体由来であってもよい。
本発明は、「エピトープ結合分子(Epitope‐Binding Molecule)」を対象とし、これは、上記分子がエピトープに結合することを可能にする「エピトープ結合部位(Epitope‐Binding Site)」を形成するために十分な、CDRL1ドメイン、CDRL2ドメイン、CDRL3ドメイン、CDRH1ドメイン、CDRH2ドメイン、若しくはCDRH3ドメイン、又はこれらのいずれの組み合わせ若しくは部分的組み合わせといった「エピトープ結合ドメイン(Epitope‐Binding Domain)」を含む。例えば、「gp41エピトープ結合分子(gp41‐Epitope‐Binding Molecule)」は、上記分子がgp41のエピトープに結合することを可能にする「gp41エピトープ結合部位(gp41‐Epitope‐Binding Site)」を形成するために十分な、CDRL1ドメイン、CDRL2ドメイン、CDRL3ドメイン、CDRH1ドメイン、CDRH2ドメイン、若しくはCDRH3ドメイン、又はこれらのいずれの組み合わせ若しくは部分的組み合わせといった「エピトープ結合ドメイン(Epitope‐Binding Domain)」を含む。しかしながら好ましくは、このようなgp41結合分子は、gp41のエピトープに免疫特異的に結合する「抗gp41抗体(anti‐gp41 antibody)」のCDRL1ドメイン、CDRL2ドメイン、CDRL3ドメイン、CDRH1ドメイン、CDRH2ドメイン、及びCDRH3ドメインを有する。いくつかの実施形態では、このような分子は、このような抗gp41抗体のgp41‐VL結合ドメインのみ、又はこのような抗gp41抗体のgp41‐VH結合ドメインのみを含んでよく、これらのドメインは他の結合ドメインと相互作用して、gp41のエピトープへの免疫特異的結合を仲介できるか、又はこれらのドメインだけで、このような結合を仲介するのに十分である場合もある。いくつかの実施形態では、このような分子は、抗gp41抗体のgp41‐VL結合ドメイン及び抗gp41抗体のgp41‐VH結合ドメインの両方を含んでよく、これらは同一の抗体由来であっても、異なる抗体由来であってもよい。
本発明の一実施形態は、「多重特異性(multispecific)」gp41結合分子に関し、これらは二重特異性であり、「第1のエピトープ」及び「第2のエピトープ」に結合でき、これらのエピトープは互いに同一ではない。エピトープを「第1のエピトープ」、「第2のエピトープ」等として表記することは、これらのエピトープの説明に先行する基礎を提供することのみを目的としており、実質的な区別を意味するものではない。このような多重特異性分子は、第1のエピトープに結合できる「VL1」/「VH1」ドメイン、及び第2のエピトープに結合できる「VL2」/「VH2」ドメインを含む。表記法「VL1」及び「VH1」はそれぞれ、このような二重特異性分子の第1のエピトープに結合する可変軽鎖ドメイン及び可変重鎖ドメインを指す。同様に、表記法「VL2」及び「VH2」はそれぞれ、このような二重特異性分子の第2のエピトープに結合する軽鎖可変ドメイン及び重鎖可変ドメインを指す。ある特定のエピトープが第1のエピトープ、第2のエピトープ等のいずれとして指定されるかは無関係であり、このような表記法は、本発明の結合分子のポリペプチド鎖のドメインの存在及び配向のみに関連する。一実施形態では、このようなエピトープのうちの一方はヒトgp41のエピトープであり、もう一方はgp41の異なるエピトープであるか、又はgp41ではない分子のエピトープである。特定の実施形態では、このようなエピトープのうちの一方はヒトgp41のエピトープであり、もう一方は、Tリンパ球、ナチュラルキラー(NK)細胞、又は他の単核細胞といったエフェクタ細胞の表面上に存在する分子(例えばCD2、CD3、CD8、CD16、T細胞受容体(TCR)、NKG2D等)のエピトープである。特定の実施形態では、多重特異性分子は3つ以上のエピトープ結合ドメインを含む。特定の実施形態では、本発明の多重特異性分子は三重特異性であり、第1のエピトープ、第2のエピトープ、及び「第3のエピトープ」に結合できる。このような三重特異性分子は、第1のエピトープに結合できるVL1/VH1ドメイン、第2のエピトープに結合できるVL2/VH2ドメイン、並びに第3のエピトープに結合できる「VL3」及び「VH3」ドメインを含む。他の実施形態では、本発明の多重特異性分子は四重特異性であり、第1のエピトープ、第2のエピトープ、第3のエピトープ、及び「第4のエピトープ」に結合できる。このような四重特異性分子は、第1のエピトープに結合できるVL1/VH1ドメイン、第2のエピトープに結合できるVL2/VH2ドメイン、第3のエピトープに結合できるVL3/VH3ドメイン、並びに第4のエピトープに結合できる「VL4」及び「VH4」ドメインを含む。このような多重特異性分子は、gp41の少なくとも1つのエピトープ、及びgp41ではない分子の少なくとも1つのエピトープに結合し、また更に、gp41の更なるエピトープ及び/又はgp41ではない分子の更なるエピトープに結合してよい。本発明は特に、本明細書中で提供される方法のうちのいずれかを用いて生成される、二重特異性ダイアボディ、BiTE(登録商標)、抗体、TandAb(登録商標)、及び3価分子を包含する。
多重特異性gp41結合分子
一実施形態では、本発明のgp41結合分子は、2つ以上の異なるエピトープに結合できる多重特異性gp41結合分子となり:
(1)gp41のあるエピトープに免疫特異的に結合する第1のエピトープ結合ドメイン(即ちgp41結合ドメイン;好ましくは本発明の最適化済みgp41結合VLドメイン及び/又は本発明の最適化済みgp41結合VHドメインを含む);並びに
(2)第2のエピトープに免疫特異的に結合する第2のエピトープ結合ドメインであって、上記第2のエピトープは、(i)gp41の異なるエピトープ、又は(ii)gp41ではない分子のエピトープである、第2のエピトープ結合ドメイン;並びに
(3)任意に、第3の更なるエピトープ、第3の更なるエピトープ及び第4の更なるエピトープ等に免疫特異的に結合する、1つ以上の追加のエピトープ結合ドメインであって、このような追加のエピトープ結合ドメインは、第1のエピトープ又は第2のエピトープと同一であっても異なっていてもよい、追加のエピトープ結合ドメイン
を含む。
一実施形態では、本発明のgp41結合分子は、2つ以上の異なるエピトープに結合できる多重特異性gp41結合分子となり:
(1)gp41のあるエピトープに免疫特異的に結合する第1のエピトープ結合ドメイン(即ちgp41結合ドメイン;好ましくは本発明の最適化済みgp41結合VLドメイン及び/又は本発明の最適化済みgp41結合VHドメインを含む);並びに
(2)第2のエピトープに免疫特異的に結合する第2のエピトープ結合ドメインであって、上記第2のエピトープは、(i)gp41の異なるエピトープ、又は(ii)gp41ではない分子のエピトープである、第2のエピトープ結合ドメイン;並びに
(3)任意に、第3の更なるエピトープ、第3の更なるエピトープ及び第4の更なるエピトープ等に免疫特異的に結合する、1つ以上の追加のエピトープ結合ドメインであって、このような追加のエピトープ結合ドメインは、第1のエピトープ又は第2のエピトープと同一であっても異なっていてもよい、追加のエピトープ結合ドメイン
を含む。
このようなgp41結合分子は好ましくは、標的細胞又は免疫エフェクタ細胞に固有の抗原のセットを認識するエピトープ結合ドメインの組み合わせを含む。特に本発明は、gp41のエピトープと、エフェクタ細胞の表面上に存在する分子(即ちエフェクタ細胞分子(Effector Cell Molecule:ECM))、特にTリンパ球、ナチュラルキラー(NK)細胞、又は他の単核細胞の表面上に存在する分子のエピトープとに結合できる、多重特異性gp41結合分子に関する。例えば本発明のこのようなgp41結合分子は、gp41のあるエピトープに免疫特異的に結合する第1のエピトープ結合ドメインと、ECM、特にCD2、CD3、CD8、CD16、T細胞受容体(TCR)、又はNKG2Dのエピトープに免疫特異的に結合する第2のエピトープ結合ドメインとを含むように構築できる。本発明の他の実施形態では、このような結合分子は更に、このような分子が、ECMの追加のエピトープ、及び/又はHIV‐1感染細胞の表面上に存在する分子の追加のエピトープ(例えばgp41の第2のエピトープ、若しくはHIV‐1エンベロープタンパク質の他のエピトープ)に結合できるようにするために十分な、エピトープ結合ドメインを含有する。本発明はまた、1つ以上のこのような分子を含む医薬組成物も対象とする。
一実施形態では、このようなgp41結合分子は二重特異性であるものの1価であり、従って、gp41の単一のエピトープのみ、及びエフェクタ細胞の表面上に存在する分子(ECM)の単一のエピトープのみに結合する能力を有する。あるいは、このような分子は多重特異性かつ多価であってよく、即ち合計1つ、2つ、3つ、又は4つのエピトープに結合でき、これは:gp41の1つ、2つ、又は3つのエピトープ(2つ又は3つのgp41エピトープは同一であっても異なっていてもよい)、及び1つ以上のECMの3つ、2つ、又は1つのエピトープ;あるいはgp41の1つ又は2つのエピトープ(2つのgp41エピトープは同一であっても異なっていてもよい)、及び1つ以上のECMの2つ又は1つのエピトープ、及び特に1つ以上のこのような分子がHIV‐1感染細胞の表面上で発現される場合には任意に1つ以上の異なるHIV‐1分子の1つ又は2つのエピトープに結合するように、いずれの方法で割り当てることができる。
よって、このような分子が単一のECMのみに免疫特異的に結合できる場合、これらの分子は:
(1)1つのみのgp41エピトープ、並びにECMの1つ、2つ、又は3つのエピトープ(2つのエピトープは同一であっても異なっていてもよく、3つのエピトープは同一であっても、異なっていても、同一である2つのエピトープ及び異なる1つのエピトープであってもよい);あるいは
(2)2つのみのgp41エピトープ(2つのエピトープは同一であっても異なっていてもよい)、及びECMの1つ又は2つのエピトープ(2つのエピトープは同一であっても異なっていてもよい);あるいは
(3)3つのgp41エピトープ(3つのエピトープは同一であっても、異なっていても、同一である2つのエピトープ及び異なる1つのエピトープであってもよい)、並びにECMの1つのエピトープ
に免疫特異的に結合できるものとなり得、あるいは任意に、これらの分子は、1つ又は2つのgp41エピトープ(これらのエピトープは同一であっても異なっていてもよい)、及び異なるHIV‐1分子の2つ又は1つのエピトープ(2つのエピトープは同一であっても異なっていてもよい)に結合できるものとなり得る。
(1)1つのみのgp41エピトープ、並びにECMの1つ、2つ、又は3つのエピトープ(2つのエピトープは同一であっても異なっていてもよく、3つのエピトープは同一であっても、異なっていても、同一である2つのエピトープ及び異なる1つのエピトープであってもよい);あるいは
(2)2つのみのgp41エピトープ(2つのエピトープは同一であっても異なっていてもよい)、及びECMの1つ又は2つのエピトープ(2つのエピトープは同一であっても異なっていてもよい);あるいは
(3)3つのgp41エピトープ(3つのエピトープは同一であっても、異なっていても、同一である2つのエピトープ及び異なる1つのエピトープであってもよい)、並びにECMの1つのエピトープ
に免疫特異的に結合できるものとなり得、あるいは任意に、これらの分子は、1つ又は2つのgp41エピトープ(これらのエピトープは同一であっても異なっていてもよい)、及び異なるHIV‐1分子の2つ又は1つのエピトープ(2つのエピトープは同一であっても異なっていてもよい)に結合できるものとなり得る。
同様に、このような分子が2つの異なるECM(例えば第1のECM及び第2のECM)に免疫特異的に結合できる場合、これらの分子は:
(1)1つのみのgp41エピトープ、及び第1のECMの1つ又は2つのエピトープ(2つの第1のECMエピトープは同一であっても異なっていてもよい)、及び第2のECMの2つ又は1つのエピトープ(2つの第2のECMエピトープは同一であっても異なっていてもよい);あるいは
(2)2つのみのgp41エピトープ(2つのエピトープは同一であっても異なっていてもよい)、及び第1のECMの1つのエピトープ、及び第2のECMの1つのエピトープ
に免疫特異的に結合できるものとなり得、あるいは任意に、これらの分子は、1つのgp41エピトープ、及び異なるHIV‐1分子の1つのエピトープに結合できるものとなり得る。
(1)1つのみのgp41エピトープ、及び第1のECMの1つ又は2つのエピトープ(2つの第1のECMエピトープは同一であっても異なっていてもよい)、及び第2のECMの2つ又は1つのエピトープ(2つの第2のECMエピトープは同一であっても異なっていてもよい);あるいは
(2)2つのみのgp41エピトープ(2つのエピトープは同一であっても異なっていてもよい)、及び第1のECMの1つのエピトープ、及び第2のECMの1つのエピトープ
に免疫特異的に結合できるものとなり得、あるいは任意に、これらの分子は、1つのgp41エピトープ、及び異なるHIV‐1分子の1つのエピトープに結合できるものとなり得る。
2つのエピトープに結合できるこのような多重特異性分子の非限定的な例は以下に記載されており:
(1)gp41の1つのエピトープ、及びECMの1つのエピトープであるCD3に結合する、DART‐A;並びに
(2)gp41の1つのエピトープ、及びECMの1つのエピトープであるCD16に結合する、DART‐B
を含む。
(1)gp41の1つのエピトープ、及びECMの1つのエピトープであるCD3に結合する、DART‐A;並びに
(2)gp41の1つのエピトープ、及びECMの1つのエピトープであるCD16に結合する、DART‐B
を含む。
3つのエピトープに結合できるこのような多重特異性分子の非限定的な例は以下に記載されており:
(1)gp41の1つのエピトープ、並びに2つの異なるECMのエピトープ(CD3及びCD8)に結合する、TRIDENT‐A;
(2)gp41の1つのエピトープ、ECMの1つのエピトープ(CD3)、及び異なるHIV‐1分子の1つのエピトープ(gp120)に結合する、TRIDENT‐B;
(3)gp41の1つのエピトープ、ECMの1つのエピトープ(CD16)、及び異なるHIV‐1分子の1つのエピトープ(gp120)に結合する、TRIDENT‐C;及び
(4) gp41の2つのエピトープ、並びにECMの1つのエピトープ(CD3)に結合する、TRIDENT‐D
を含む。
(1)gp41の1つのエピトープ、並びに2つの異なるECMのエピトープ(CD3及びCD8)に結合する、TRIDENT‐A;
(2)gp41の1つのエピトープ、ECMの1つのエピトープ(CD3)、及び異なるHIV‐1分子の1つのエピトープ(gp120)に結合する、TRIDENT‐B;
(3)gp41の1つのエピトープ、ECMの1つのエピトープ(CD16)、及び異なるHIV‐1分子の1つのエピトープ(gp120)に結合する、TRIDENT‐C;及び
(4) gp41の2つのエピトープ、並びにECMの1つのエピトープ(CD3)に結合する、TRIDENT‐D
を含む。
表1は更に、本発明の例示的な分子の結合特異性の可能な組み合わせを示す。

より複雑な分子を形成することにより、1つ以上のECMと、任意に5つ以上のエピトープ結合ドメインを有する1つ以上の異なるHIV‐1分子とに結合できる、gp41結合分子を得ることができる。よって、本発明の分子が結合できるエピトープ又は追加のエピトープの性質に課される制限は、このような追加の結合能力によって、上記分子又はgp41のエピトープに結合できる上記分子の結合ドメインの、このような結合が阻害されず、また上記分子又はECMのエピトープに結合できる上記分子の結合ドメインの、このような結合が阻害されないこと以外には存在しない。よって、本発明のgp41結合分子は、代替的な又は追加のエピトープ結合ドメインを有してよい。一例として、本発明は、gp41のエピトープに免疫特異的に結合できる第1のエピトープ結合ドメイン、及びECMのエピトープに免疫特異的に結合できる第2のエピトープ結合ドメイン、及び異なるECM又は任意に異なるHIV‐1分子に免疫特異的に結合できる第3のエピトープ結合ドメインを含む、gp41結合分子を考える。
1.Fcドメインを含まない多重特異性gp41結合ダイアボディ
一実施形態では、本発明の多重特異性gp41結合分子は二重特異性ダイアボディであり、第1のエピトープ及び第2のエピトープ両方に結合できるドメインを含むものの、Fcドメインを含まず、従ってFc‐FcγR相互作用によるFcγR分子への結合が不可能である。しかしながら、このような分子は、そのgp41結合ドメインのSDR又はCDRを介してgp41に結合できる。よって、Fcドメインの不在は、このような分子が、阻害性受容体CD32B等のFcγRに結合するのを防止する役割を果たす。
一実施形態では、本発明の多重特異性gp41結合分子は二重特異性ダイアボディであり、第1のエピトープ及び第2のエピトープ両方に結合できるドメインを含むものの、Fcドメインを含まず、従ってFc‐FcγR相互作用によるFcγR分子への結合が不可能である。しかしながら、このような分子は、そのgp41結合ドメインのSDR又はCDRを介してgp41に結合できる。よって、Fcドメインの不在は、このような分子が、阻害性受容体CD32B等のFcγRに結合するのを防止する役割を果たす。
二重特異性ダイアボディのこのような実施形態の第1のポリペプチド鎖は、好ましくは、N末端からC末端への方向に:N末端;第1又は第2のエピトープに結合できるモノクローナル抗体のVLドメイン(即ちVLgp41又はVL第2のエヒ゜トーフ゜);第1の介在スペーサペプチド(リンカー1);(上記第1のポリペプチド鎖がVL gp41を含有する場合は)第2のエピトープに結合できるモノクローナル抗体のVHドメイン、又は(上記第1のポリペプチド鎖がVL第2のエヒ゜トーフ゜を含有する場合は)gp41のエピトープに結合できるモノクローナル抗体のVHドメイン;任意にシステイン残基を含有する第2の介在スペーサペプチド(リンカー2);ヘテロ二量体促進ドメイン;及びC末端を含む(図1A〜1B)。
二重特異性ダイアボディのこの実施形態の第2のポリペプチド鎖は、N末端からC末端への方向に:N末端;第1又は第2のエピトープに結合できるモノクローナル抗体のVLドメイン(即ちVLgp41又はVL第2のエヒ゜トーフ゜;及び上記VLドメインは上記ダイアボディの上記第1のポリペプチド鎖に含まれないよう選択される);介在スペーサペプチド(リンカー1);第1又は第2のエピトープに結合できるモノクローナル抗体のVHドメイン(即ちVHgp41又はVH第2のエヒ゜トーフ゜;及び上記VHドメインは上記ダイアボディの上記第1のポリペプチド鎖に含まれないよう選択される);任意にシステイン残基を含有する第2の介在スペーサペプチド(リンカー2);ヘテロ二量体促進ドメイン;及びC末端を含む(図1A〜1B)。ある特定のエピトープに対して特異的な、採用されるVL及びVHドメインは、好ましくは同一のモノクローナル抗体から得られる、又は同一のモノクローナル抗体に由来する。しかしながらこれらのドメインは、これらのドメインが連結して、上記エピトープに免疫特異的に結合できる官能性結合ドメインを形成する場合、異なるモノクローナル抗体に由来してもよい。このような異なる抗体は、本明細書では「対応する(corresponding)」抗体と呼ばれる。
このようなダイアボディの第1のポリペプチド鎖のVLドメインは、上記ダイアボディの第2のポリペプチド鎖のVHドメインと相互作用して、上記エピトープのうちの1つ(例えば第1のエピトープ又は第2のエピトープ)に対して特異的な第1の官能性エピトープ結合ドメインを形成する。同様に、第2のポリペプチド鎖のVLドメインは、第1のポリペプチド鎖のVHドメインと相互作用して、他方のエピトープ(即ち第2のエピトープ又は第1のエピトープ)に対して特異的な第2の官能性エピトープ結合ドメインを形成する。よって、第1及び第2のポリペプチド鎖のVL及びVHドメインの選択は、ダイアボディのこれら2つのポリペプチド鎖が合わせて、第1のエピトープ及び第2のエピトープの両方に結合できるVL及びVHドメインを含む(即ちこれらが合わせて、VL第1のエヒ゜トーフ゜/VH第1のエヒ゜トーフ゜及びVL第2のエヒ゜トーフ゜/VH第2のエヒ゜トーフ゜、例えばVLgp41/VHgp41及びVL第2のエヒ゜トーフ゜/VH第2のエヒ゜トーフを含む)ように、「調整(coordinate)」される。
最も好ましくは、介在スペーサペプチド(即ち上述のVLドメインとVHドメインとを隔てる「リンカー1」)の長さは、上記ポリペプチド鎖の上記VL及びVHドメインが互いに対して結合する(例えば0、1、2、3、4、5、6、7、8又は9個の介在リンカーアミノ酸残基からなる)のを実質的に又は完全に防止するよう選択される。従って第1のポリペプチド鎖のVL及びVHドメインは、互いに対して結合することが実質的に又は全くできない。同様に、第2のポリペプチド鎖のVL及びVHドメインは、互いに対して結合することが実質的に又は全くできない。好ましい介在スペーサペプチド(リンカー1)は、配列(配列番号16):GGGSGGGGを有する。
上記第2の介在スペーサペプチド(リンカー2)の長さ及び組成は、上述のような二量体化を促進する1つ以上のポリペプチドドメイン(即ち「ヘテロ二量体促進ドメイン」)の選択に基づいて選択される。典型的には、上記第2の介在スペーサペプチド(「リンカー2」)の長さは、3〜20個のアミノ酸残基である。特に、採用した1つ以上のヘテロ二量体促進ドメインがシステイン残基を含まない場合、システイン含有第2の介在スペーサペプチド(リンカー2)が利用される。システイン含有第2の介在スペーサペプチド(リンカー2)は、1つ、2つ、3つ又は4つ以上のシステインを含有する。好ましいシステイン含有スペーサペプチド(リンカー2)は、配列GGCGGG(配列番号17)を有する。あるいは、リンカー2はシステインを含まず(例えば、GGG、GGGS(配列番号18)、LGGGSG(配列番号19)、GGGSGGGSGGG(配列番号20)、ASTKG(配列番号21)、LEPKSS(配列番号22)、APSSS(配列番号23)等)、以下で説明されるシステイン含有ヘテロ二量体促進ドメインが使用される。任意に、システイン含有リンカー2及びシステイン含有ヘテロ二量体促進ドメインの両方が使用される。
ヘテロ二量体促進ドメインは、一方のポリペプチド鎖上にGVEPKSC(配列番号24)又はVEPKSC(配列番号25)又はAEPKSC(配列番号26)、及び他方のポリペプチド鎖上にGFNRGEC(配列番号27)又はFNRGEC(配列番号28)であってもよい(米国特許第2007/0004909号)。
ある好ましい実施形態では、ヘテロ二量体促進ドメインは、対向する電荷のタンデム反復コイルドメイン、例えば、グルタミン酸残基がpH7において負の電荷を形成する「Eコイル」ヘテロ二量体促進ドメイン(配列番号29:EVAALEK-EVAALEK-EVAALEK-EVAALEK)、又はリシン残基がpH7において正の電荷形成する「Kコイル」ヘテロ二量体促進ドメイン(配列番号30:KVAALKE-KVAALKE-KVAALKE-KVAALKE)を含む。このような荷電ドメインの存在により、第1のポリペプチドと第2のポリペプチドとの間の連結が促進され、従ってヘテロ二量体形成が促進される。上述のEコイル及びKコイルの配列の、1つ以上のシステイン残基を含むような修飾を含む、ヘテロ二量体促進ドメインを利用してよい。このようなシステイン残基の存在により、一方のポリペプチド鎖に存在するコイルが、他方のポリペプチド鎖に存在する相補的なコイルと共有結合し、これにより上記ポリペプチド鎖を互いに共有結合させて、ダイアボディの安定性を向上できる。特に好ましい例は、アミノ酸配列EVAACEK-EVAALEK-EVAALEK-EVAALEK(配列番号31)を有する修飾されたEコイルと、アミノ酸配列KVAACKE-KVAALKE-KVAALKE-KVAALKE(配列番号32)を有する修飾されたKコイルとを含む、ヘテロ二量体促進ドメインである。
国際公開第2012/018687号に開示されているように、ダイアボディのインビボ薬物動態特性を改善するために、ダイアボディを、ダイアボディの末端のうちの1つ又は複数に血清結合タンパク質のポリペプチド部分を含有するように修飾してよい。最も好ましくは、このような血清結合タンパク質のポリペプチド部分は、ダイアボディのポリペプチド鎖のC末端に配置されることになる。アルブミンは、血漿中に最も豊富なタンパク質であり、ヒトにおいて19日の半減期を有する。アルブミンは複数の小さな分子結合ドメインを有し、これによりアルブミンは他のタンパク質に非共有結合して血清半減期を延長できる。連鎖球菌株G148のタンパク質Gのアルブミン結合ドメイン3(ABD3)は、安定した3重螺旋束を形成する46個のアミノ酸残基からなり、広範なアルブミン結合特異性を有する(Johansson, M.U. et al. (2002) “Structure, Specificity, And Mode Of Interaction For Bacterial Albumin-Binding Modules,” J. Biol. Chem. 277(10):8114-8120)。従って、ダイアボディのインビボ薬物動態特性を改善するための、血清結合タンパク質の特に好ましいポリペプチド部分は、連鎖球菌タンパク質Gからのアルブミン結合ドメイン(ABD)、及びより好ましくは、連鎖球菌株G148のタンパク質Gのアルブミン結合ドメイン3(ABD3)(配列番号33):LAEAKVLANR ELDKYGVSDY YKNLIDNAKS AEGVKALIDE ILAALPである。
国際公開第2012/162068号(参照により本出願に援用される)に開示されているように、配列番号33の「脱免疫化(deimmunized)」変異型は、MHCクラスII結合を減衰させる又は排除する能力を有する。組み合わさった突然変異の結果に基づいて、以下の置換の組み合わせが、このような脱免疫化ABDを形成するための好ましい置換であると考えられる:66D/70S+71A;66S/70S+71A;66S/70S+79A;64A/65A/71A;64A/65A/71A+66S;64A/65A/71A+66D;64A/65A/71A+66E;64A/65A/79A+66S;64A/65A/79A+66D;64A/65A/79A+66E。修飾L64A、I65A及びD79A、又は修飾N66S、T70S及びD79Aを有する変異型ABD。アミノ酸配列:
LAEAKVLANR ELDKYGVSDY YKNLID 66NAKS 70 A 71EGVKALIDE ILAALP(配列番号34)
又はアミノ酸配列:
LAEAKVLANR ELDKYGVSDY YKNA 64 A 65NNAKT VEGVKALIA 79E ILAALP(配列番号35)
又はアミノ酸配列:
LAEAKVLANR ELDKYGVSDY YKNLIS 66NAKS 70VEGVKALIA 79E ILAALP(配列番号36)
を有する、変異型脱免疫化ABDが特に好ましい。というのは、このような脱免疫化ABDは、MHCクラスII結合の減衰を提供しながら、略野生型の結合を呈するためである。従って、ABDを有するこのようなダイアボディの上記第1のポリペプチド鎖は、好ましくはこのようなポリペプチド鎖のEコイル(又はKコイル)ドメインに対してC末端に位置決めされることによって、上記Eコイル(又はKコイル)ドメインとABD(これは好ましくは脱免疫化ABDである)との間に介在する、第3のリンカー(リンカー3)を含有する。このようなリンカー3の好ましい配列は、配列番号18:GGGSである。
LAEAKVLANR ELDKYGVSDY YKNLID 66NAKS 70 A 71EGVKALIDE ILAALP(配列番号34)
又はアミノ酸配列:
LAEAKVLANR ELDKYGVSDY YKNA 64 A 65NNAKT VEGVKALIA 79E ILAALP(配列番号35)
又はアミノ酸配列:
LAEAKVLANR ELDKYGVSDY YKNLIS 66NAKS 70VEGVKALIA 79E ILAALP(配列番号36)
を有する、変異型脱免疫化ABDが特に好ましい。というのは、このような脱免疫化ABDは、MHCクラスII結合の減衰を提供しながら、略野生型の結合を呈するためである。従って、ABDを有するこのようなダイアボディの上記第1のポリペプチド鎖は、好ましくはこのようなポリペプチド鎖のEコイル(又はKコイル)ドメインに対してC末端に位置決めされることによって、上記Eコイル(又はKコイル)ドメインとABD(これは好ましくは脱免疫化ABDである)との間に介在する、第3のリンカー(リンカー3)を含有する。このようなリンカー3の好ましい配列は、配列番号18:GGGSである。
2.Fcドメインを含む多重特異性gp41結合ダイアボディ
本発明の一実施形態は、Fcドメインを含み、またgp41のエピトープと第2のエピトープ(例えば異なるHIV‐1エンベロープタンパク質のエピトープ、又はエフェクタ細胞の表面上で発現されるエピトープ)とに同時に結合できる、多重特異性ダイアボディ(例えば二重特異性、三重特異性、四重特異性等)に関する。このような分子のFcドメインは、いずれのアイソタイプ(例えばIgG1、IgG2、IgG3、又はIgG4)のものであってよい。上記分子は更に、CH1ドメイン及び/又はヒンジドメインを含んでよい。CH1ドメイン及び/又はヒンジドメインが存在する場合、これらはいずれのアイソタイプ(例えばIgG1、IgG2、IgG3、又はIgG4)のものであってよく、好ましくは所望のFcドメインと同一のアイソタイプのものである。
本発明の一実施形態は、Fcドメインを含み、またgp41のエピトープと第2のエピトープ(例えば異なるHIV‐1エンベロープタンパク質のエピトープ、又はエフェクタ細胞の表面上で発現されるエピトープ)とに同時に結合できる、多重特異性ダイアボディ(例えば二重特異性、三重特異性、四重特異性等)に関する。このような分子のFcドメインは、いずれのアイソタイプ(例えばIgG1、IgG2、IgG3、又はIgG4)のものであってよい。上記分子は更に、CH1ドメイン及び/又はヒンジドメインを含んでよい。CH1ドメイン及び/又はヒンジドメインが存在する場合、これらはいずれのアイソタイプ(例えばIgG1、IgG2、IgG3、又はIgG4)のものであってよく、好ましくは所望のFcドメインと同一のアイソタイプのものである。
上記ダイアボディポリペプチド鎖のうちの一方又は両方にIgG CH2‐CH3ドメインを追加して、上記ダイアボディ鎖の複合体化によってFcドメインが形成されるようにすると、上記ダイアボディの生物学的半減期が増大し、及び/又は価数が変化する。このようなダイアボディは、ポリペプチド鎖を互いに共有結合させて、gp41及び第2のエピトープに同時に結合できる共有結合ダイアボディを形成できる配列を有する、2つ以上のポリペプチド鎖を含む。上記ダイアボディポリペプチドの両方にIgG CH2‐CH3ドメインを組み込むと、2鎖二重特異性Fcドメイン含有ダイアボディの形成が可能となる(図2)。
あるいは、上記ダイアボディポリペプチドのうちの一方にIgG CH2‐CH3ドメインを組み込むと、より複雑な4鎖二重特異性Fcドメイン含有ダイアボディの形成が可能となる(図3A〜3C)。図3Cは、定常軽鎖(CL)ドメイン及び定常重鎖CH1ドメインを有する代表的な4鎖ダイアボディを示すが、このようなドメインの部分及び他のポリペプチドを代わりに採用してもよい(例えば図3A及び3B、米国公開特許第2013‐0295121号;米国公開特許第2010‐0174053号及び米国公開特許第2009‐0060910号;欧州公開特許第2714079号;欧州公開特許第2601216号;欧州公開特許第2376109号;欧州公開特許第2158221号、及び国際公開第2012/162068号;国際公開第2012/018687号;国際公開第2010/080538号を参照)。従って例えば、CH1ドメインの代わりに、ヒトIgGのヒンジドメイン由来のアミノ酸配列GVEPKSC(配列番号24)、VEPKSC(配列番号25)又はAEPKSC(配列番号26)を有するペプチドを採用してよく、またCLドメインの代わりに、ヒトκ軽鎖のC末端6アミノ酸、GFNRGEC(配列番号27)又はFNRGEC(配列番号28)を採用してよい。4鎖ダイアボディを含有する代表的なペプチドを図3Aに示す。あるいは、又は更に、対向する電荷のタンデムコイルドメイン、例えば「Eコイル」螺旋ドメイン(配列番号29:EVAALEK-EVAALEK-EVAALEK-EVAALEK又は配列番号30:EVAACEK-EVAALEK-EVAALEK-EVAALEK);及び「Kコイル」ドメイン(配列番号31:KVAALKE-KVAALKE-KVAALKE-KVAALKE又は配列番号32:KVAACKE-KVAALKE-KVAALKE-KVAALKE)を含むペプチドを採用してよい。4鎖ダイアボディを含有する代表的なコイルドメインを図3Bに示す。
本発明のFcドメイン含有ダイアボディ分子は、更なる介在スペーサペプチド(リンカー)を含んでよく、このようなリンカーは一般に、ヘテロ二量体促進ドメイン(例えばEコイル若しくはKコイル)とCH2‐CH3ドメインとの間、及び/又はCH2‐CH3ドメインと可変ドメイン(即ちVH若しくはVL)との間に組み込まれる。典型的には、この追加のリンカーは3〜20個のアミノ酸残基を含み、任意にIgGヒンジドメインの全体又は一部分(好ましくは、1個、2個、3個、若しくは4個以上のシステイン残基を有するIgGヒンジドメインのシステイン含有部分)を含有してよい。本発明の二重特異性Fcドメイン含有ダイアボディ分子において採用できるリンカーとしては:GGGS(配列番号18)、LGGGSG(配列番号19)、GGGSGGGSGGG(配列番号20)、ASTKG(配列番号21)、LEPKSS(配列番号22)、APSSS(配列番号23)、APSSSPME(配列番号37)、VEPKSADKTHTCPPCP(配列番号38)、LEPKSADKTHTCPPCP(配列番号39)、DKTHTCPPCP(配列番号40)、scFvリンカー:GGGGSGGGGSGGGGS(配列番号41);「長い(long)」リンカー:GGGGSGGGSGGG(配列番号42)、GGC、及びGGGが挙げられる。クローニングを容易にするために、GGG又はGGCの代わりにLEPKSS(配列番号22)を使用してよい。更に、ペプチドGGG又はLEPKSS(配列番号22)の直後にDKTHTCPPCP(配列番号40)を続けることによって、代替リンカー:GGGDKTHTCPPCP(配列番号43);及びLEPKSSDKTHTCPPCP(配列番号44)を形成してよい。本発明の二重特異性Fcドメイン含有分子は、リンカーに加えて又はリンカーに代えて、IgGヒンジドメインを組み込んでよい。例示的なヒンジドメインとしては:IgG1からのEPKSCDKTHTCPPCP(配列番号7);IgG2からのERKCCVECPPCP(配列番号8);IgG3からのELKTPLGDTTHTCPRCPEPKSCDTPPPCPRCPEPKSCDTPP PCPRCPEPKSCDTPPPCPRCP(配列番号9);IgG4からのESKYGPPCPSCP(配列番号10);及び鎖交換を低減するための安定化S228P置換(下線を付して示されている)を含むIgG4ヒンジ変異型からのESKYGPPCPPCP(配列番号11)が挙げられる(Kabatに記載のEUインデックスに従って番号付与)。
図3A〜3Cに提示されているように、本発明のFcドメイン含有ダイアボディは4つの異なる鎖を含んでよい。このようなダイアボディの第1及び第3のポリペプチド鎖は、4つのドメイン:(i)VL1含有ドメイン;(ii)VH2含有ドメイン;(iii)ヘテロ二量体促進ドメイン;及び(iv)CH2‐CH3配列を含有するドメインを含有する。第2及び第4のポリペプチド鎖は:(i)VL2含有ドメイン;(ii)VH1含有ドメイン;及び(iii)ヘテロ二量体促進ドメインを含有し、上記ヘテロ二量体促進ドメインは、第1/第3のポリペプチド鎖と第2/第4のポリペプチド鎖との二量体化を促進する。第3及び第4のポリペプチド鎖のVL及び/又はVHドメイン、並びに第1及び第2のポリペプチド鎖のVL及び/又はVHドメインは、同一であっても異なっていてもよく、これにより、単一特異性、二重特異性又は四重特異性の4価結合が可能となる。表記法「VL3」及び「VH3」はそれぞれ、このようなダイアボディの「第3のエピトープ」に結合する軽鎖可変ドメイン及び重鎖可変ドメインを指す。同様に、表記法「VL4」及び「VH4」はそれぞれ、このようなダイアボディの「第4のエピトープ」に結合する軽鎖可変ドメイン及び重鎖可変ドメインを指す。本発明の代表的な4鎖二重特異性Fcドメイン含有ダイアボディのポリペプチド鎖の一般構造を表2に提示する。
ある具体的実施形態では、本発明のダイアボディは、合計4つのポリペプチド鎖からなる、二重特異性、4価(即ち4つのエピトープ結合ドメインを有する)、Fc含有ダイアボディである(図3A〜3C)。本発明の二重特異性、4価、Fc含有ダイアボディは、2つの第1のエピトープ結合ドメイン及び2つの第2のエピトープ結合ドメインを含む。
更なる実施形態では、本発明のFcドメイン含有ダイアボディは、3つのポリペプチド鎖を含んでよい。このようなダイアボディの第1のポリペプチドは、3つのドメイン:(i)VL1含有ドメイン;(ii)VH2含有ドメイン;及び(iii)CH2‐CH3配列を含有するドメインを含有する。このようなダイアボディの第2のポリペプチドは:(i)VL2含有ドメイン;(ii)VH1含有ドメイン;並びに(iii)上記ダイアボディの第1のポリペプチド鎖とのヘテロ二量体化及び共有結合を促進するドメインを含有する。このようなダイアボディの第3のポリペプチドは、CH2‐CH3配列を含む。従ってこのようなダイアボディの上記第1及び第2のポリペプチド鎖は、一体に連結して、上記第1又は第2のエピトープに結合できるVL1/VH1エピトープ結合ドメイン、及び上記エピトープ農地のもう一方に結合できるVL2/VH2エピトープ結合ドメインを形成する。上記第1及び第2のポリペプチドは、それぞれの第3のドメインのシステイン残基が関わるジスルフィド結合によって、互いに結合する。特に上記第1及び第3のポリペプチド鎖は、互いに複合体化して、ジスルフィド結合によって安定化されたFcドメインを形成する。このような二重特異性ダイアボディは、強度が増強されている。図4A及び4Bは、このようなダイアボディの構造を示す。このようなFcドメイン含有ダイアボディは、2つの配向(表3)のうちのいずれを有してよい。
ある具体的実施形態では、本発明のダイアボディは、合計3つのポリペプチド鎖からなる、二重特異性、2価(即ち2つのエピトープ結合ドメインを有する)、Fc含有ダイアボディである(図4A〜4B)。本発明の二重特異性、2価Fc含有ダイアボディは、第1又は第2のエピトープに対して免疫特異的な1つのエピトープ結合ドメイン、及び上記エピトープのうちのもう一方に対して特異的に結合できるVL2/VH2エピトープ結合ドメインを含む。
更なる実施形態では、上記Fcドメイン含有ダイアボディは、合計5つのポリペプチド鎖を含んでよい。ある特定の実施形態では、上記5つのポリペプチド鎖のうちの2つは、同一のアミノ酸配列を有する。このようなダイアボディの第1のポリペプチド鎖は:(i)VH1含有ドメイン;(ii)CH1含有ドメイン;及び(iii)CH2‐CH3配列を含有するドメインを含有する。上記第1のポリペプチド鎖は、VH1及び重鎖定常領域を含有する抗体の重鎖であってよい。このようなダイアボディの第2及び第5のポリペプチド鎖は:(i)VL1含有ドメイン;及び(ii)CL含有ドメインを含有する。このようなダイアボディの上記第2及び/又は第5のポリペプチド鎖は、上記第1/第3のポリペプチド鎖のVH1に対して相補的なVL1を含有する抗体の軽鎖であってよい。上記第1、第2及び/又は第5のポリペプチド鎖は、自然に発生する抗体から単離できる。あるいはこれらは、組み換えによって構成できる。このようなダイアボディの第3のポリペプチド鎖は:(i)VH1含有ドメイン;(ii)CH1含有ドメイン;(iii)CH2‐CH3配列を含有するドメイン;(iv)VL2含有ドメイン;(v)VH3含有ドメイン;及び(vi)ヘテロ二量体促進ドメインを含有し、上記ヘテロ二量体促進ドメインは、上記第3の鎖と上記第4の鎖との二量体化を促進する。このようなダイアボディの第4のポリペプチドは:(i)VL3含有ドメイン;(ii)VH2含有ドメイン;並びに(iii)上記ダイアボディの第3のポリペプチド鎖とのヘテロ二量体化及び共有結合を促進するドメインを含有する。
従って、このようなダイアボディの上記第1及び第2のポリペプチド鎖と上記第3及び第5のポリペプチド鎖とは、互いに連結して、第1のエピトープに結合できる2つのVL1/VH1エピトープ結合ドメインを形成する。このようなダイアボディの上記第3及び第4のポリペプチド鎖は、互いに連結して、第2のエピトープに結合できるVL2/VH2エピトープ結合ドメイン、及び第3のエピトープに結合できるVL3/VH3結合ドメインを形成する。上記第1及び第3のポリペプチドは、それぞれの定常領域のシステイン残基が関わるジスルフィド結合によって、互いに結合する。特に上記第1及び第3のポリペプチド鎖は、互いに複合体化して、Fcドメインを形成する。このような多重特異性ダイアボディは、強度が増強されている。図5は、このようなダイアボディの構造を示す。VL1/VH1、VL2/VH2及びVL3/VH3ドメインは、同一であっても異なっていてもよく、これにより、単一特異性、二重特異性又は三重特異性である結合が可能となることが理解されるだろう。
上記ポリペプチド鎖のVL及びVHドメインは、所望のエピトープに特異的なVL/VH結合ドメインを形成するよう選択される。上記ポリペプチド鎖の連結によって形成されるVL/VH結合ドメインは、同一であっても異なっていてもよく、これにより、単一特異性、二重特異性、三重特異性又は四重特異性である4価結合が可能となる。特に上記VL及びVHドメインは、多価ダイアボディが、第1のエピトープに関する2つの結合ドメイン及び第2のエピトープに関する2つの結合ドメイン、又は第1のエピトープに関する3つの結合ドメイン及び第2のエピトープに関する1つの結合ドメイン、又は(図5に示すように)第1のエピトープに関する2つの結合ドメイン、第2のエピトープに関する1つの結合ドメイン、及び第3のエピトープに関する1つの結合ドメインを含むように選択してよい。本発明の代表的な5鎖Fcドメイン含有ダイアボディのポリペプチド鎖の一般構造を表4に示す。
ある具体的実施形態では、本発明のダイアボディは、第1のエピトープに免疫特異的な2つのエピトープ結合ドメイン、及び第2のエピトープに特異的な2つのエピトープ結合ドメインを有する、合計5つのポリペプチド鎖からなる、二重特異性、4価(即ち4つのエピトープ結合ドメインを有する)、Fc含有ダイアボディである。別の実施形態では、本発明の二重特異性、4価、Fc含有ダイアボディは、第1のエピトープに免疫特異的な3つのエピトープ結合ドメイン、及び第2のエピトープに特異的な1つのエピトープ結合ドメインを含む。上述のように、VL及びVHドメインは、三重特異性結合が可能となるように選択してよい。従って本発明は、三重特異性4価Fc含有ダイアボディも包含する。本発明の三重特異性4価Fc含有ダイアボディは、第1のエピトープに対して免疫特異的な2つのエピトープ結合ドメイン、第2の分子に対して免疫特異的な1つのエピトープ結合ドメイン、及び第3のエピトープに対して免疫特異的な1つのエピトープ結合ドメインを含む。
従来の免疫機能では、抗体‐抗原複合体と免疫系の細胞との相互作用は、抗体依存性細胞傷害性、肥満細胞脱顆粒及び食作用等のエフェクタ機能から、リンパ球増殖及び抗体分泌を制御するもの等の免疫調節性シグナルにまで及ぶ、広範な応答をもたらす。これらの相互作用は全て、抗体又は免疫複合体のFcドメインの、造血細胞上の特殊化された細胞表面受容体への結合によって開始される。上述のように、抗体及び免疫複合体によってトリガされる細胞応答の多様性は、3つのFc受容体:FcγRI(CD64)、FcγRII(CD32)及びFcγRIII(CD16)の構造不均質性によって得られる。FcγRI(CD64)、FcγRIIA(CD32A)及びFcγRIII(CD16)は活性化(即ち免疫系増強)受容体であり;FcγRIIB(CD32B)は阻害性(即ち免疫系減衰)受容体である。更に、新生児Fc受容体(FcRn)との相互作用は、エンドソームから細胞表面へのIgG分子の再循環及び血中への放出を仲介する。例示的な野生型IgG1(配列番号12)、IgG2(配列番号13)、IgG3(配列番号14)及びIgG4(配列番号15)のアミノ酸配列は、既に提示されている。
Fcドメインの修飾は、表現型の変化、例えば血清半減期の変化、安定性の変化、細胞酵素に対する感受性の変化、又はエフェクタ機能の変化につながり得る。従って、本発明のFcドメイン含有結合分子をエフェクタ機能に関して修飾して、例えば癌の治療におけるこのような分子の有効性を増強することが望ましい場合がある。例えば、作用機序が、標的抗原を担持する細胞の殺滅ではなく遮断又は拮抗作用を伴う抗体の場合といった、特定の場合においては、Fcドメイン仲介型エフェクタ機能の低減又は削減が望ましい。エフェクタ機能の上昇は、FcγRが低いレベルで発現する腫瘍及び外来細胞、例えばFcγRIIBのレベルが低い腫瘍特異性B細胞(例えば非ホジキンリンパ腫、CLL及びバーキットリンパ腫)といった、望ましくない細胞を対象とする場合に一般に望ましい。エフェクタ機能活性が与えられた又は変更された本発明のこのような分子は、エフェクタ機能活性の効率の増強が望ましい疾患、障害又は感染の治療及び/又は予防のために有用である。
従って特定の実施形態では、本発明のgp41結合Fcドメイン含有分子のFcドメインは、操作された可変Fc領域であってよい。本発明の二重特異性Fcドメイン含有分子のFcドメインは、1つ以上のFc受容体(例えば1つ以上のFcγR)に結合できる能力を有してよいが、より好ましくは、上記変異型Fcドメインは、(野生型Fcドメインが呈する結合に対して)FcγRIA(CD64)、FcγRIIA(CD32A)、FcγRIIB(CD32B)、FcγRIIIA(CD16A)若しくはFcγRIIIB(CD16B)への結合が変化しており、例えばある活性化受容体への結合が増強されており、及び/又は1つ以上の阻害性受容体に結合する能力が低下しているか若しくは上記能力を有しない。従って、本発明の二重特異性Fcドメイン含有分子のFcドメインは、完全FcドメインのCH2ドメインのうちのある程度若しくは全体及び/若しくはCH3ドメインのうちのある程度若しくは全体を含んでよく、又は(例えば完全FcドメインのCH2若しくはCH3ドメインに対する1つ以上の挿入及び/若しくは1つ以上の欠失を含んでよい)変異型CH2及び/若しくは変異型CH3配列を含んでよい。このようなFcドメインは、非Fcポリペプチド部分を含んでよく、又は自然に発生しない完全Fcドメインの部分を含んでよく、又はCH2及び/若しくはCH3ドメインの自然に発生しない配向を含んでよい(例えば2つのCH2ドメイン若しくは2つのCH3ドメイン、若しくはN末端からC末端への方向において、CH3ドメインと、これが連結したCH2ドメイン、等)。
変化したエフェクタ機能として表されるFcドメイン修飾は当該技術分野において公知であり、活性化型受容体に対する結合を増大させる修飾(例えばFcγRIIA(CD16A))、及び阻害型受容体に対する結合を低下させる修飾(例えばFcγRIIB(CD32B))が含まれる(例えばStavenhagen, J.B. et al. (2007) “Fc Optimization Of Therapeutic Antibodies Enhances Their Ability To Kill Tumor Cells In Vitro And Controls Tumor Expansion In Vivo Via Low-Affinity Activating Fcgamma Receptors,” Cancer Res. 57(18):8882-8890を参照)。表5は、活性化受容体への結合を増大させる、及び/又は阻害性受容体への結合を低下させる例示的な修飾の、単一、二重、三重、四重及び五重置換のリスト((EUインデックスに従った)番号付与及び置換は、上で提示した配列番号12のアミノ酸配列に対するものである)を示す。
CD32Bへの結合が低下した、及び/又はCD16Aへの結合が増大した、ヒトIgG1 Fcドメインの例示的な変異型は、F243L、R292P、Y300L、V305I又はP396L置換を含有し、ここでの番号付与は、Kabat中のEUインデックスの番号付与である。これらのアミノ酸置換は、いずれの組み合わせで、ヒトIgG1 Fcドメイン内に存在してよい。一実施形態では、上記変異型ヒトIgG1 Fcドメインは、F243L、R292P及びY300L置換を含有する。別の実施形態では、上記変異型ヒトIgG1 Fcドメインは、F243L、R292P、Y300L、V305I及びP396L置換を含有する。
特定の実施形態では、本発明のFcドメイン含有結合分子のFcドメインに関して、(野生型IgG1 Fcドメイン(配列番号12)が呈する結合に対して)FcγRIA(CD64)、FcγRIIA(CD32A)、FcγRIIB(CD32B)、FcγRIIIA(CD16A)又はFcγRIIIB(CD16B)への結合が低下している(又はこれらに略結合しない)ことが好ましい。ある具体的実施形態では、本発明のFcドメイン含有結合分子は、抗体依存性細胞仲介型細胞傷害性(ADCC)エフェクタ機能が低下したIgG Fcドメインを含む。ある好ましい実施形態では、このような結合分子のCH2‐CH3ドメインは、以下の置換:L234A、L235A、D265A、N297Q、及びN297Gのうちのいずれの1つ、2つ、3つ又は4つを含み、ここでの番号付与は、Kabat中のEUインデックスの番号付与である。別の実施形態では、CH2‐CH3ドメインは、N297Q置換、N297G置換、L234A及びL235A置換又はD265A置換を含有するが、それはこれらの突然変異がFcR結合を消失させるためである。あるいは、(野生型IgG1 Fcドメイン(配列番号12)が呈する結合及びエフェクタ機能に対して)FcγRIIIA(CD16A)に対する結合が元来低い(又は略結合しない)、及び/又はエフェクタ機能が元来低い、天然のFcドメインのCH2‐CH3ドメインを利用する。ある具体的実施形態では、本発明のFcドメイン含有結合分子は、IgG2 Fcドメイン(配列番号13)、IgG3 Fcドメイン(配列番号14)、又はIgG4 Fcドメイン(配列番号15)を含む。IgG4 Fcドメインを利用する場合、本発明は、上述のヒンジ領域S228P置換(例えば配列番号11参照)等の安定化突然変異の導入も包含する。N297G、N297Q、L234A、L235A及びD265A置換はエフェクタ機能を消失させるため、エフェクタ機能が望まれる状況下では、これらの置換は採用しないことが好ましい。
エフェクタ機能が低下したか又は消失した、本発明のFcドメイン含有分子のCH2及びCH3ドメインに関する好ましいIgG1配列は、L234A/L235Aの置換を含む(配列番号45):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
Fcドメインを含むタンパク質の血清半減期は、FcRnに関するFcドメインの結合親和性を増大させることによって増大させることができる。本明細書において使用される場合、用語「半減期(half‐life)」は、投与後の分子の平均生存時間の尺度となる、分子の薬物動態特性を意味する。半減期は、血清中で(即ち循環半減期)又は他の組織中で測定した場合に、分子の既知の量の50パーセント(50%)が被験者の身体(例えばヒト患者若しくは他の哺乳類)又はその特定の体腔から排除されるために必要な時間として表すことができる。一般に、半減期の増大は、投与される分子の循環における平均滞留時間(MRT)の増大につながる。
いくつかの実施形態では、本発明のFcドメイン含有結合分子は、野生型Fc領域に対して少なくとも1つのアミノ酸修飾を含み、従って(野生型Fcドメインを含む場合に対して)増大した半減期を有する、変異型Fcドメインを含む。いくつかの実施形態では、本発明のFcドメイン含有結合分子は、238、250、252、254、256、257、256、265、272、286、288、303、305、307、308、309、311、312、317、340、356、360、362、376、378、380、382、413、424、428、433、434、435及び436からなる群から選択される1つ以上の位置に半減期延長アミノ酸置換を含む、変異型IgG Fcドメインを含み、ここでの番号付与は、Kabat中のEUインデックスの番号付与である。Fcドメイン含有分子の半減期を増大させることができる多数の突然変異が当該技術分野において公知であり、例えばM252Y、S254T、T256E及びこれらの組み合わせが挙げられる。例えば:米国特許第6,277,375号;米国特許第7,083,784号;米国特許第7,217,797号;米国特許第8,088,376号;米国公開特許第2002/0147311号;米国公開特許第2007/0148164号;並びに国際公開第98/23289号;国際公開第2009/058492号;及び国際公開第2010/033279号(これらは参照によりその全体が本出願に援用される)に記載されている突然変異を参照。
いくつかの実施形態では、半減期が増強された本発明のFcドメイン含有結合分子は、Fcドメイン残基250、252、254、256、257、288、307、308、309、311、378、428、433、434、435及び436のうちの2つ以上における置換、特にT250Q、M252Y、S254T、T256E、K288D、T307Q、V308P、A378V、M428L、N434A、H435K及びY436Iから選択される2つ以上の置換を含む変異型Fcドメインを有し、ここでの番号付与は、Kabat中のEUインデックスの番号付与である。ある具体的実施形態では、上記分子は:
(A)M252Y、S254T及びT256E;
(B)M252Y及びS254T;
(C)M252Y及びT256E;
(D)T250Q及びM428L;
(E)T307Q及びN434A;
(F)A378V及びN434A;
(G)N434A及びY436I;
(H)V308P及びN434A;又は
(I)K288D及びH435K
の置換を含む変異型IgG Fc領域を有してよい。
(A)M252Y、S254T及びT256E;
(B)M252Y及びS254T;
(C)M252Y及びT256E;
(D)T250Q及びM428L;
(E)T307Q及びN434A;
(F)A378V及びN434A;
(G)N434A及びY436I;
(H)V308P及びN434A;又は
(I)K288D及びH435K
の置換を含む変異型IgG Fc領域を有してよい。
ある好ましい実施形態では、本発明のgp41結合Fcドメイン含有分子は、置換:M252Y、S254T及びT256Eのうちのいずれの1つ、2つ又は3つを含む変異型IgG Fc領域を有する。本発明は更に:
(A)エフェクタ機能及び/又はFcγRを変化させる、1つ以上の突然変異;並びに
(B)血清半減期を延長させる、1つ以上の突然変異
を含む変異型Fc領域を有する、gp41結合分子を包含する。
(A)エフェクタ機能及び/又はFcγRを変化させる、1つ以上の突然変異;並びに
(B)血清半減期を延長させる、1つ以上の突然変異
を含む変異型Fc領域を有する、gp41結合分子を包含する。
半減期の増大を提供する(及びカニクイザルFcRn及びヒトFcRnの両方への結合が10倍増大した本発明のFcドメイン含有分子のCH2及びCH3ドメインに関するIgG1配列(Dall’Acqua, W.F. et al. (2006) “Properties of Human IgG1s Engineered for Enhanced Binding to the Neonatal Fc Receptor (FcRn),” J. Biol. Chem. 281(33):23514-23524)は、置換M252Y/S254T/T256Eを含む(配列番号46):
APELLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
APELLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
置換L234A/L235Aによって提供されるエフェクタ機能の低下又は消失と、置換M252Y/S254T/T256Eによって提供される血清半減期の増大とが組み合わさった、本発明のFcドメイン含有分子のCH2及びCH3ドメインに関する別のIgG1配列は、配列番号47:
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
によって提供され、ここでXはリシン(K)であるか、又は不在である。
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
によって提供され、ここでXはリシン(K)であるか、又は不在である。
アミノ酸配列が異なる複数のFcドメイン含有ポリペプチド鎖を有することが望まれる(例えばFcドメイン含有ポリペプチド鎖が同一ではないことが望まれる)特定の抗体、ダイアボディ及び3価結合分子に関して、同一の鎖(例えば2つの第1のポリペプチド鎖、又は2つの第3のポリペプチド鎖)のCH2‐CH3ドメイン間でのホモ二量体化の発生を低減又は防止することが望ましい。このようなポリペプチド鎖のCH2及び/又はCH3ドメインは配列において同一である必要はなく、有利には2つのポリペプチド鎖間のヘテロ二量体複合体形成を促進するために修飾される。例えば、CH2又はCH3ドメインにアミノ酸置換(好ましくは「ノブ(knob)」を形成する嵩高な側鎖基、例えばトリプトファンを含むアミノ酸による置換)を導入して、立体障害により、同様に変異させたドメインとの相互作用を防止し、上記変化させたドメインを、相補的な又は適応した変異(例えばグリシンによる置換)を施されたドメイン、即ち「ホール(hole)」と対合させて、ヘテロ二量体化を促進できる。このような一連の突然変異は、Fcドメインを形成するCH2‐CH3ドメインを含むポリペプチドのいずれのペアに施すことができる。ホモ二量体化を抑えてヘテロ二量体化を促進するためのタンパク質加工の方法は、特に免疫グロブリン様分子の加工に関して当該技術分野で公知であり、本明細書に包含される(例えばRidgway et al. (1996) “‘Knobs-Into-Holes’ Engineering Of Antibody CH3 Domains For Heavy Chain Heterodimerization,” Protein Engr. 9:617-621; Atwell et al. (1997) “Stable Heterodimers From Remodeling The Domain Interface Of A Homodimer Using A Phage Display Library,” J. Mol. Biol. 270: 26-35;及びXie et al. (2005) “A New Format Of Bispecific Antibody: Highly Efficient Heterodimerization, Expression And Tumor Cell Lysis,” J. Immunol. Methods 296:95-101を参照(これらはそれぞれ参照によりその全体が本明細書に援用される))。
好ましいノブは、IgG Fcドメインを修飾して修飾基T366Wを含有させることによって生成される。好ましいホールは、IgG Fcドメインを修飾して修飾基T366S、L368A及びY407Vを含有させることによって生成される。ホール担持ポリペプチド鎖ホモ二量体を、二重特異性ヘテロ二量体Fcドメイン含有分子から精製するのを補助するために、好ましくは、ポリペプチド鎖のホール担持CH2及びCH3ドメインのタンパク質A結合ドメインを、位置435(H435R)におけるアミノ酸置換によって変異させる。このようにして、ホール担持ポリペプチド鎖ホモ二量体はタンパク質Aに結合せず、その一方で二重特異性ヘテロ二量体は、タンパク質A結合ドメインを介してタンパク質Aに結合する能力を有したままとなる。代替実施形態では、ホール担持ポリペプチド鎖は、434位及び435位にアミノ酸置換を組み込んでよい(N434A/N435K)。
本発明のFcドメイン含有分子のあるFcドメイン含有ポリペプチド鎖のCH2及びCH3ドメインに関する好ましいIgG1アミノ酸配列は、「ノブ担持」配列(配列番号48)を有する:
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
M252Y/S254T/T256E置換及び「ノブ担持」配列を有する、本発明のFcドメイン含有分子のあるFcドメイン含有ポリペプチド鎖のCH2及びCH3ドメインに関する別のIgG1アミノ酸配列は、配列番号49:
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
であり、ここでXはリシン(K)であるか、又は不在である。
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE
ALHNHYTQKS LSLSPGX
であり、ここでXはリシン(K)であるか、又は不在である。
2つのポリペプチド鎖を有する本発明のFcドメイン含有分子の他方のFcドメイン含有ポリペプチド鎖(又は3つ、4つ、若しくは5つのポリペプチド鎖を有するFcドメイン含有分子の第3のポリペプチド鎖)のCH2及びCH3ドメインに関する好ましいIgG1アミノ酸配列は、「ホール担持」配列を有する(配列番号50):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE
ALHNRYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE
ALHNRYTQKS LSLSPGX
ここでXはリシン(K)であるか、又は不在である。
M252Y/S254T/T256E置換及び「ホール担持」配列を有する、本発明のFcドメイン含有分子の他方のFcドメイン含有ポリペプチド鎖のCH2及びCH3ドメインに関する別のIgG1アミノ酸配列は、配列番号51:
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE
ALHNRYTQKS LSLSPGX
であり、ここでXはリシン(K)であるか、又は不在である。
APEAAGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSHED PEVKFNWYVD
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA
PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE
ALHNRYTQKS LSLSPGX
であり、ここでXはリシン(K)であるか、又は不在である。
本発明のFcドメイン含有分子の上記1つのFcドメイン含有ポリペプチドのCH2及びCH3ドメインに関するIgG4アミノ酸配列は、Y252/T254/E256を有することにより、(IgG1 CH2及びCH3ドメインに比べて)血清半減期が増大する(配列番号52):
APEFLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSQED PEVQFNWYVD
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSRL TVDKSRWQEG NVFSCSVMHE
ALHNHYTQKS LSLSLGX
ここでXはリシン(K)であるか、又は不在である。
APEFLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSQED PEVQFNWYVD
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLTCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSRL TVDKSRWQEG NVFSCSVMHE
ALHNHYTQKS LSLSLGX
ここでXはリシン(K)であるか、又は不在である。
このようなIgG4 CH2‐CH3アミノ酸配列の「ノブ担持」変異型は、配列番号53:
APEFLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSQED PEVQFNWYVD
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLWCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSRL TVDKSRWQEG NVFSCSVMHE
ALHNHYTQKS LSLSLGX
ここでXはリシン(K)であるか、又は不在である。
APEFLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSQED PEVQFNWYVD
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLWCLVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLYSRL TVDKSRWQEG NVFSCSVMHE
ALHNHYTQKS LSLSLGX
ここでXはリシン(K)であるか、又は不在である。
このようなIgG4 CH2‐CH3アミノ酸配列の「ホール担持」変異型は、配列番号54:
APEFLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSQED PEVQFNWYVD
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLSCAVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLVSRL TVDKSRWQEG NVFSCSVMHE
ALHNRYTQKS LSLSLGX
ここでXはリシン(K)であるか、又は不在である。
APEFLGGPSV FLFPPKPKDT LYITREPEVT CVVVDVSQED PEVQFNWYVD
GVEVHNAKTK PREEQFNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKGLPS
SIEKTISKAK GQPREPQVYT LPPSQEEMTK NQVSLSCAVK GFYPSDIAVE
WESNGQPENN YKTTPPVLDS DGSFFLVSRL TVDKSRWQEG NVFSCSVMHE
ALHNRYTQKS LSLSLGX
ここでXはリシン(K)であるか、又は不在である。
後に記載されるように、配列番号51、配列番号52、配列番号53、及び配列番号54のCH2‐CH3ドメインは、アラニンによる234位の置換及びアラニンによる235位の置換を含み、従って、(野生型Fcドメイン(配列番号12)が示す結合と比べて)FcγRIA(CD64)、FcγRIIA(CD32A)、FcγRIIB(CD32B)、FcγRIIIA(CD16A)又はFcγRIIIB(CD16B)への結合が低下した(又は結合を実質的に示さない)Fcドメインを形成する。本発明はまた、野生型アラニン残基、Fcドメインのエフェクタ機能及び/又はFγR結合活性を修正する代替的な及び/又は更なる置換を含む、このようなCH2‐CH3ドメインも包含する。本発明はまた、1つ以上の半減期延長アミノ酸置換を更に含む、このようなCH2‐CH3ドメインも包含する。特に本発明は、M252Y/S254T/T256Eを更に含む、このようなホール担持及びノブ担持CH2‐CH3ドメインを包含する。
第1のポリペプチド鎖が、配列番号48又は配列番号49のもの等の「ノブ担持」CH2‐CH3配列を有することが好ましい。しかしながら、理解されるように、第1のポリペプチド鎖中に「ホール担持」CH2‐CH3ドメイン(例えば配列番号50又は配列番号51)を採用でき、この場合「ノブ担持」CH2‐CH3ドメイン(例えば配列番号48又は配列番号49)は、2つのポリペプチド鎖を有する本発明のFcドメイン含有分子の第2のポリペプチド鎖中(又は3つ、4つ若しくは5つのポリペプチド鎖を有するFcドメイン含有分子の第3のポリペプチド鎖中)に採用される。
他の実施形態では、本発明は、国際公開第2007/110205号;国際公開第2011/143545号;国際公開第2012/058768号;国際公開第2013/06867号;国際公開第2014/081955号;国際公開第2016/086189号(これらは全て、参照によりその全体が本出願に援用される)において開示されているもの等の当該技術分野で公知の突然変異を用いて、ホモ二量体化よりもヘテロ二量体化を指向するよう操作されたCH2及び/又はCH3ドメインを含む、Fcドメイン含有結合分子を包含する。
3.Fcドメインを含有する3価結合分子
本発明の更なる実施形態は、Fcドメインを含み、また第1のエピトープ、第2のエピトープ及び第3のエピトープ(ここで上記エピトープのうちの少なくとも1つは、別のエピトープと同一ではない)に同時に結合できる、三重特異性3価結合性分子に関する。このような三重特異性3価結合性分子は3つのエピトープ結合ドメインを含み、そのうちの2つは、結合ドメインA及び結合ドメインBを提供するダイアボディ型結合ドメインであり、1つは、結合ドメインCを提供するFab型結合ドメイン(又はscFv型結合ドメイン)である(例えば図6A〜6F、並びに国際公開第2015/184207号及び国際公開第2015/184203号を参照)。このような3価結合分子は従って、第1のエピトープに結合できる「VL1」/「VH1」ドメイン、並びに第2のエピトープに結合できる「VL2」/「VH2」ドメイン、並びに上記3価結合分子の第3のエピトープに結合できる「VL3」及び「VH3」ドメインを含む。「ダイアボディ型結合ドメイン」は、上述のように、ダイアボディ中に存在するエピトープ結合ドメインのタイプであり、単一のポリペプチド鎖は、VLドメイン及びVHドメインの両方を含むが、これらのドメインは相互作用してエピトープ結合部位を形成することはない。「Fab型結合ドメイン」及び「scFv型結合ドメイン」はそれぞれ、上述のように、抗体中に存在するエピトープ結合ドメインのタイプであり、(1)単一のポリペプチド鎖はVLドメイン若しくはVHドメインを含み、かつ第2のポリペプチド鎖は対応するVHドメイン若しくはVLドメインを含み、これによってエピトープ結合部位が形成されるか;又は(2)単一のポリペプチド鎖はVLドメイン及びVHドメインの両方を含み、これらのドメインが相互作用してエピトープ結合部位を形成する。Fab型結合ドメインもまた、Fab型結合ドメインを形成する2つのポリペプチド鎖が単一のエピトープ結合ドメインしか含まないが、ダイアボディ型結合ドメインを形成する2つのポリペプチド鎖は少なくとも2つのエピトープ結合部位を含むという点で、ダイアボディ型結合ドメインとは異なる。同様に、scFv型結合ドメインも、これらが単一のエピトープ結合ドメインしか含まないという点で、ダイアボディ型結合ドメインとは異なる。よって本明細書において使用される場合、Fab型及びscFv型結合ドメインは、ダイアボディ型結合ドメインとは別個である。
本発明の更なる実施形態は、Fcドメインを含み、また第1のエピトープ、第2のエピトープ及び第3のエピトープ(ここで上記エピトープのうちの少なくとも1つは、別のエピトープと同一ではない)に同時に結合できる、三重特異性3価結合性分子に関する。このような三重特異性3価結合性分子は3つのエピトープ結合ドメインを含み、そのうちの2つは、結合ドメインA及び結合ドメインBを提供するダイアボディ型結合ドメインであり、1つは、結合ドメインCを提供するFab型結合ドメイン(又はscFv型結合ドメイン)である(例えば図6A〜6F、並びに国際公開第2015/184207号及び国際公開第2015/184203号を参照)。このような3価結合分子は従って、第1のエピトープに結合できる「VL1」/「VH1」ドメイン、並びに第2のエピトープに結合できる「VL2」/「VH2」ドメイン、並びに上記3価結合分子の第3のエピトープに結合できる「VL3」及び「VH3」ドメインを含む。「ダイアボディ型結合ドメイン」は、上述のように、ダイアボディ中に存在するエピトープ結合ドメインのタイプであり、単一のポリペプチド鎖は、VLドメイン及びVHドメインの両方を含むが、これらのドメインは相互作用してエピトープ結合部位を形成することはない。「Fab型結合ドメイン」及び「scFv型結合ドメイン」はそれぞれ、上述のように、抗体中に存在するエピトープ結合ドメインのタイプであり、(1)単一のポリペプチド鎖はVLドメイン若しくはVHドメインを含み、かつ第2のポリペプチド鎖は対応するVHドメイン若しくはVLドメインを含み、これによってエピトープ結合部位が形成されるか;又は(2)単一のポリペプチド鎖はVLドメイン及びVHドメインの両方を含み、これらのドメインが相互作用してエピトープ結合部位を形成する。Fab型結合ドメインもまた、Fab型結合ドメインを形成する2つのポリペプチド鎖が単一のエピトープ結合ドメインしか含まないが、ダイアボディ型結合ドメインを形成する2つのポリペプチド鎖は少なくとも2つのエピトープ結合部位を含むという点で、ダイアボディ型結合ドメインとは異なる。同様に、scFv型結合ドメインも、これらが単一のエピトープ結合ドメインしか含まないという点で、ダイアボディ型結合ドメインとは異なる。よって本明細書において使用される場合、Fab型及びscFv型結合ドメインは、ダイアボディ型結合ドメインとは別個である。
典型的には、本発明の3価gp41結合分子は、4つの異なるポリペプチド鎖を含む(図6A〜6B参照)が、上記分子は、例えばこれらのポリペプチド鎖を(例えばペプチド結合によって)互いに融合させることによって、又はこれらのポリペプチドを「分割(dividing)」して追加のポリペプチド鎖を形成することによって、又はより少数若しくは追加のポリペプチド鎖をジスルフィド結合によって連結することによって、より少数又はより多数のポリペプチド鎖を含むことができる。図6C〜6Fは、3つのポリペプチド鎖を有する分子を概略的に示すことによって、本発明のこの態様を図示している。図6A〜6Fに提示されているように、本発明の3価結合分子は、上記ダイアボディ型結合ドメインがFcドメインに対するN末端(図6A、6C、及び6D)又はC末端(図6B、6E、及び6F)となる交互の配向を有してよい。3価結合分子の生成に有用なCH2及びCH3ドメインは上述されており、ノブ担持及びホール担持ドメインを含む。
特定の実施形態では、本発明のこのような3価gp41結合分子の第1のポリペプチド鎖は:(i)VL1含有ドメイン;(ii)VH2含有ドメイン;(iii)ヘテロ二量体促進ドメイン;及び(iv)CH2‐CH3配列を含有するドメインを含有する。上記VL1及びVL2ドメインは、表5(図6A及び6Bも参照)に提示されるように、上記CH2‐CH3含有ドメインに対してN末端又はC末端に位置する。このような実施形態の第2のポリペプチド鎖は:(i)VL2含有ドメイン;(ii)VH1含有ドメイン;及び(iii)ヘテロ二量体促進ドメインを含有する。このような実施形態の第3のポリペプチド鎖は:(i)VH3含有ドメイン;(ii)CH1含有ドメイン;及び(iii)CH2‐CH3配列を含有するドメインを含有する。上記第3のポリペプチド鎖は、VH3及び重鎖定常領域を含有する抗体、又は上記ドメインを含有するポリペプチドの、重鎖であってよい。このような実施形態の第4のポリペプチドは:(i)VL3含有ドメイン;及び(ii)CL含有ドメインを含有する。上記第4のポリペプチド鎖は、上記第3のポリペプチド鎖のVH3に対して相補的なVL3を含有する抗体、又は上記ドメインを含有するポリペプチドの、軽鎖であってよい。上記第3又は第4のポリペプチド鎖は、自然に発生する抗体から単離できる。あるいはこれらは、組み換えによって、合成によって、又は他の手段によって構成できる。
上記第1及び第2のポリペプチド鎖の軽鎖可変ドメインは、介在スペーサペプチドによって、このようなポリペプチド鎖の重鎖可変ドメインから隔てられ、上記介在スペーサリンカーは、これらのVL1/VH2(又はこれらのVL2/VH1)ドメインを一体に連結して、第1又は第2のエピトープに結合できるエピトープ結合ドメインを形成することを可能にするには短すぎる長さを有する。この目的のために好ましい介在スペーサペプチド(リンカー1)は、配列(配列番号16):GGGSGGGGを有する。上記3価結合分子の他のドメインは、任意にシステイン残基を含む、1つ以上の介在スペーサペプチド(リンカー)によって隔てられていてよい。特に上述のように、このようなリンカーは典型的には可変ドメイン(即ちVH又はVL)とペプチドヘテロ二量体促進ドメイン(例えばEコイル又はKコイル)との間、及び上記ペプチドヘテロ二量体促進ドメイン(例えばEコイル又はKコイル)とCH2‐CH3ドメインとの間に組み込まれる。3価結合分子の生成に有用な例示的なリンカーは上で提示されており、また国際公開第2015/184207号;及び国際公開第2015/184203号にも提示されている。従ってこのような3価結合分子の第1及び第2のポリペプチド鎖は、一体に連結して、第1のエピトープに結合できるVL1/VH1結合ドメイン、及び第2のエピトープに結合できるVL2/VH2結合ドメインを形成する。このような3価結合分子の第3及び第4のポリペプチド鎖は、一体に連結して、第3のエピトープに結合できるVL3/VH3結合ドメインを形成する。
上述のように、本発明の3価gp41結合分子は、3つのポリペプチドを含んでよい。3つのポリペプチド鎖を含む3価結合分子は、(例えば介在スペーサペプチド(リンカー4)を用いて)第4のポリペプチドN末端のドメインを第3のポリペプチドのVH3含有ドメインに連結することによって得ることができる。あるいは、以下の3つのドメイン:(i)VL3含有ドメイン;(ii)VH3含有ドメイン;及び(iii)CH2‐CH3配列を含有するドメインを含有する本発明の3価結合分子の第3のポリペプチド鎖が利用され、ここでVL3及びVH3は、これらのドメインが連結してエピトープ結合ドメインを形成できるようにするために十分な長さ(少なくとも9以上のアミノ酸残基)を有する介在スペーサペプチドによって、互いから隔てられる。この目的に関して好ましい1つの介在スペーサペプチドは、配列:GGGGSGGGGSGGGGS(配列番号41)を有する。
このような3価結合性分子のVL1/VH1、VL2/VH2及びVL3/VH3ドメインは異なっていてよく、それによって単一特異性、二重特異性又は三重特異性の結合が可能となることが理解されるだろう。特に、VL及びVHドメインは、3価結合分子が第1のエピトープに対する2つの結合ドメイン及び第2のエピトープに対する1つの結合ドメイン、又は第1のエピトープに対する1つの結合ドメイン及び第2のエピトープに対する2つの結合ドメイン、又は第1のエピトープに対する1つの結合ドメイン、第2のエピトープに対する1つの結合ドメイン、及び第3のエピトープに対する1つの結合ドメイン、を含むように選択される。
本発明の代表的な3価結合性分子のポリペプチド鎖の一般構造を、図6A〜6F及び表6で提供する。
上述のように、このような3価結合分子は、3つ、4つ、5つ又は6つ以上のポリペプチド鎖を含んでよい。
IV.本発明の例示的なgp41結合分子
本発明は、7B2抗体の結合特異性を有する最適化済みgp41結合ドメインを有することによって、gp41 HIV‐1エンベロープ(Env)タンパク質に結合できる、gp41結合分子(例えば抗体、ダイアボディ、scFv、抗体、TandAb(登録商標)、trident(商標)等)を対象とする。
本発明は、7B2抗体の結合特異性を有する最適化済みgp41結合ドメインを有することによって、gp41 HIV‐1エンベロープ(Env)タンパク質に結合できる、gp41結合分子(例えば抗体、ダイアボディ、scFv、抗体、TandAb(登録商標)、trident(商標)等)を対象とする。
A.HIVのエンベロープ(Env)糖タンパク質
HIVのエンベロープ(Env)糖タンパク質は、増殖性感染細胞の表面上で発現される。HIV 1型(HIV‐1)は、トランスサイトーシスとして知られるプロセスでの全ての伝達において、粘膜を通って宿主に入る(Shen, R. et al. (2010) “GP41-Specific Antibody Blocks Cell-Free HIV Type 1 Transcytosis Through Human Rectal Mucosa And Model Colonic Epithelium,” J. Immunol. 184(7):3648-3655)。腸及び生殖器上皮を通したHIV‐1トランスサイトーシスは、gp41 envタンパク質(Bomsel, M. et al. (1998) “Intracellular Neutralization Of HIV Transcytosis Across Tight Epithelial Barriers By Anti-HIV Envelope Protein dIgA Or IgM,” Immunity 9:277-287; Alfsen, A. et al. (2002) “HIV-1 Gp41 Envelope Residues 650-685 Exposed On Native Virus Act As A Lectin To Bind Epithelial Cell Galactosyl Ceramide,” J. Biol. Chem. 277:25649-25659; Alfsen, A. et al. (2001) “Secretory IgA Specific For A Conserved Epitope On gp41 Envelope Glycoprotein Inhibits Epithelial Transcytosis Of HIV-1,” J. Immunol. 166:6257-6265; Tudor, D. et al. (2009) “HIV-1 gp41-Specific Monoclonal Mucosal IgAs Derived from Highly Exposed But IgG-Seronegative Individuals Block HIV-1 Epithelial Transcytosis And Neutralize CD4+ Cell Infection: An IgA Gene And Functional Analysis,” Mucosal Immunol. 2:412-426)、gp120、及びgp160を含むウイルス成分と、グリコスフィンゴ脂質ガラクトシルセラミド、共受容体CCR5、ヘパリン硫酸塩プロテオグリカン付着受容体、シンデカン及びアグリンを含む宿主上皮細胞受容体及び付着分子(Shen, R. et al. (2010) “GP41-Specific Antibody Blocks Cell-Free HIV Type 1 Transcytosis Through Human Rectal Mucosa and Model Colonic Epithelium,” J. Immunol. 184(7):3648-3655)とを伴うことが報告されている。
HIVのエンベロープ(Env)糖タンパク質は、増殖性感染細胞の表面上で発現される。HIV 1型(HIV‐1)は、トランスサイトーシスとして知られるプロセスでの全ての伝達において、粘膜を通って宿主に入る(Shen, R. et al. (2010) “GP41-Specific Antibody Blocks Cell-Free HIV Type 1 Transcytosis Through Human Rectal Mucosa And Model Colonic Epithelium,” J. Immunol. 184(7):3648-3655)。腸及び生殖器上皮を通したHIV‐1トランスサイトーシスは、gp41 envタンパク質(Bomsel, M. et al. (1998) “Intracellular Neutralization Of HIV Transcytosis Across Tight Epithelial Barriers By Anti-HIV Envelope Protein dIgA Or IgM,” Immunity 9:277-287; Alfsen, A. et al. (2002) “HIV-1 Gp41 Envelope Residues 650-685 Exposed On Native Virus Act As A Lectin To Bind Epithelial Cell Galactosyl Ceramide,” J. Biol. Chem. 277:25649-25659; Alfsen, A. et al. (2001) “Secretory IgA Specific For A Conserved Epitope On gp41 Envelope Glycoprotein Inhibits Epithelial Transcytosis Of HIV-1,” J. Immunol. 166:6257-6265; Tudor, D. et al. (2009) “HIV-1 gp41-Specific Monoclonal Mucosal IgAs Derived from Highly Exposed But IgG-Seronegative Individuals Block HIV-1 Epithelial Transcytosis And Neutralize CD4+ Cell Infection: An IgA Gene And Functional Analysis,” Mucosal Immunol. 2:412-426)、gp120、及びgp160を含むウイルス成分と、グリコスフィンゴ脂質ガラクトシルセラミド、共受容体CCR5、ヘパリン硫酸塩プロテオグリカン付着受容体、シンデカン及びアグリンを含む宿主上皮細胞受容体及び付着分子(Shen, R. et al. (2010) “GP41-Specific Antibody Blocks Cell-Free HIV Type 1 Transcytosis Through Human Rectal Mucosa and Model Colonic Epithelium,” J. Immunol. 184(7):3648-3655)とを伴うことが報告されている。
よって、gp41は、細胞傷害性免疫複合体(immunoconjugate:IC)のための標的として使用でき、ここでは、毒素、薬物、又は放射性核種を含む細胞殺滅部分が、gp41結合分子に化学的又は遺伝子的に連結される(例えばPincus, S.H. et al. (2017) “Design and In Vivo Characterization of Immunoconjugates Targeting HIV gp160,” J. Virol. 91(3). pii: e01360-16. doi: 10.1128/JVI.01360-16を参照)。
B.抗体7B2
抗体7B2(Genbank受託番号JX188438及びJX188439)は、この分子の免疫優性ヘリックス‐ループ‐ヘリックス領域の598〜604において、HIV gp41に結合する、抗HIV env ヒトIgG1抗体である(Sadraeian, M. et al. (2017) “Selective Cytotoxicity Of A Novel Immunotoxin Based On Pulchellin A Chain For Cells Expressing HIV Envelope,” Sci. Rep. 7(1):7579 doi: 10.1038/s41598-017-08037-3)。上記抗体は、エプスタイン・バー(EB)ウイルスB細胞の形質転換及びヘテロハイブリドーマの産生を用いて、HIV‐1慢性感染被験者から単離された(Pincus, S.H. et al. (2003) “In Vivo Efficacy Of Anti-Glycoprotein 41, But Not Anti-Glycoprotein 120, Immunotoxins In A Mouse Model Of HIV Infection,” J. Immunol. 170(4):2236-2241)。抗体7B2は、ウイルス粒子及び感染細胞の両方を認識できることが分かっている(Santra, S. et al. (2015) “Human Non-neutralizing HIV-1 Envelope Monoclonal Antibodies Limit the Number of Founder Viruses during SHIV Mucosal Infection in Rhesus Macaques,” PLoS Pathog. 11(8):e1005042. doi: 10.1371/journal.ppat.1005042; Tay, M.Z. et al. (2016) “Antibody-Mediated Internalization of Infectious HIV-1 Virions Differs among Antibody Isotypes and Subclasses,” PLoS Pathog. 12(8):e1005817. doi: 10.1371/journal.ppat.1005817)。
抗体7B2(Genbank受託番号JX188438及びJX188439)は、この分子の免疫優性ヘリックス‐ループ‐ヘリックス領域の598〜604において、HIV gp41に結合する、抗HIV env ヒトIgG1抗体である(Sadraeian, M. et al. (2017) “Selective Cytotoxicity Of A Novel Immunotoxin Based On Pulchellin A Chain For Cells Expressing HIV Envelope,” Sci. Rep. 7(1):7579 doi: 10.1038/s41598-017-08037-3)。上記抗体は、エプスタイン・バー(EB)ウイルスB細胞の形質転換及びヘテロハイブリドーマの産生を用いて、HIV‐1慢性感染被験者から単離された(Pincus, S.H. et al. (2003) “In Vivo Efficacy Of Anti-Glycoprotein 41, But Not Anti-Glycoprotein 120, Immunotoxins In A Mouse Model Of HIV Infection,” J. Immunol. 170(4):2236-2241)。抗体7B2は、ウイルス粒子及び感染細胞の両方を認識できることが分かっている(Santra, S. et al. (2015) “Human Non-neutralizing HIV-1 Envelope Monoclonal Antibodies Limit the Number of Founder Viruses during SHIV Mucosal Infection in Rhesus Macaques,” PLoS Pathog. 11(8):e1005042. doi: 10.1371/journal.ppat.1005042; Tay, M.Z. et al. (2016) “Antibody-Mediated Internalization of Infectious HIV-1 Virions Differs among Antibody Isotypes and Subclasses,” PLoS Pathog. 12(8):e1005817. doi: 10.1371/journal.ppat.1005817)。
7B2抗体のVLドメインのアミノ酸配列(配列番号55)を以下及び図7Aに示す(CDRL残基は下線を付して示されている):
DIVMTQSPDS LAVSPGERAT IHCKSSQTLL YSSNNRHSIA WYQQRPGQPP
KLLLYWASMR LSGVPDRFSG SGSGTDFTLT INNLQAEDVA IYYCHQYSSH
PPTFGHGTRV EIK
DIVMTQSPDS LAVSPGERAT IHCKSSQTLL YSSNNRHSIA WYQQRPGQPP
KLLLYWASMR LSGVPDRFSG SGSGTDFTLT INNLQAEDVA IYYCHQYSSH
PPTFGHGTRV EIK
7B2抗体のVHドメインのアミノ酸配列(配列番号56)を以下及び図7Bに示す(CDRH残基は下線を付して示されている):
QVQLVQSGGG VFKPGGSLRL SCEASGFTFT EYYMTWVRQA PGKGLEWLAY
ISKNGEYSKY SPSSNGRFTI SRDNAKNSVF LQLDRLSADD TAVYYCARAD
GLTYFSELLQ YIFDLWGQGA RVTVSS
QVQLVQSGGG VFKPGGSLRL SCEASGFTFT EYYMTWVRQA PGKGLEWLAY
ISKNGEYSKY SPSSNGRFTI SRDNAKNSVF LQLDRLSADD TAVYYCARAD
GLTYFSELLQ YIFDLWGQGA RVTVSS
C.最適化済み7B2GL
最適化済み7B2GL抗体のVLドメインのアミノ酸配列(配列番号57)を以下及び図7Aに示す(CDRL残基は下線を付して示されている):
DIVMTQSPDS LAVSLGERAT INCKSSQTLL YSSNNRHSIA WYQQKPGQPP
KLLIYWASMR LSGVPDRFSG SGSGTDFTLT ISSLQAEDVA VYYCHQYSSH
PPTFGQGTKV EIK
最適化済み7B2GL抗体のVLドメインのアミノ酸配列(配列番号57)を以下及び図7Aに示す(CDRL残基は下線を付して示されている):
DIVMTQSPDS LAVSLGERAT INCKSSQTLL YSSNNRHSIA WYQQKPGQPP
KLLIYWASMR LSGVPDRFSG SGSGTDFTLT ISSLQAEDVA VYYCHQYSSH
PPTFGQGTKV EIK
最適化済み7B2GL抗体のVHドメインのアミノ酸配列(配列番号58)を以下及び図7Bに示す(CDRH残基は下線を付して示されている):
QVQLVESGGG LVKPGGSLRL SCAASGFTFS EYYMTWIRQA PGKGLEWVSY
ISKNGEYSKY SPSSNGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCARAD
GLTYFSELLQ YIFDLWGQGT LVTVSS
QVQLVESGGG LVKPGGSLRL SCAASGFTFS EYYMTWIRQA PGKGLEWVSY
ISKNGEYSKY SPSSNGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCARAD
GLTYFSELLQ YIFDLWGQGT LVTVSS
V.例示的なエフェクタ細胞結合能力
本明細書に記載されているように、本発明は、gp41のエピトープ及びエフェクタ細胞表面分子(「ECM])のエピトープに結合できる多重特異性gp41結合分子に関する。本明細書中で使用される場合、用語「エフェクタ細胞(effector cell)」は、標的細胞(例えば外来細胞、感染細胞又は癌細胞)の殺滅を直接的又は間接的に仲介する細胞を指す。エフェクタ細胞の例としては、ヘルパーT細胞、細胞傷害性T細胞、ナチュラルキラー(NK)細胞、形質細胞(抗体分泌B細胞)、マクロファージ及び顆粒球が挙げられる。好ましいエフェクタ細胞表面分子(「ECM」)としては、CD2、CD3、CD8、CD16、TCR及びNKG2D受容体が挙げられる。従って、上記分子のエピトープに、又は他のエフェクタ細胞表面分子に免疫特異的に結合できる分子を、本発明の原理に従って使用してよい。標的細胞の標的転換殺滅を仲介できる分子の構築にVH及び/若しくはVLドメイン、並びに/又はVL領域のCDRLのうちの1つ、2つ、若しくは3つ全て、並びに/又はVHドメインのCDRHのうちの1つ、2つ、若しくは3つ全てを使用してよい例示的な抗体を、以下に提供する。
本明細書に記載されているように、本発明は、gp41のエピトープ及びエフェクタ細胞表面分子(「ECM])のエピトープに結合できる多重特異性gp41結合分子に関する。本明細書中で使用される場合、用語「エフェクタ細胞(effector cell)」は、標的細胞(例えば外来細胞、感染細胞又は癌細胞)の殺滅を直接的又は間接的に仲介する細胞を指す。エフェクタ細胞の例としては、ヘルパーT細胞、細胞傷害性T細胞、ナチュラルキラー(NK)細胞、形質細胞(抗体分泌B細胞)、マクロファージ及び顆粒球が挙げられる。好ましいエフェクタ細胞表面分子(「ECM」)としては、CD2、CD3、CD8、CD16、TCR及びNKG2D受容体が挙げられる。従って、上記分子のエピトープに、又は他のエフェクタ細胞表面分子に免疫特異的に結合できる分子を、本発明の原理に従って使用してよい。標的細胞の標的転換殺滅を仲介できる分子の構築にVH及び/若しくはVLドメイン、並びに/又はVL領域のCDRLのうちの1つ、2つ、若しくは3つ全て、並びに/又はVHドメインのCDRHのうちの1つ、2つ、若しくは3つ全てを使用してよい例示的な抗体を、以下に提供する。
A.例示的なCD2結合能力
一実施形態では、本発明の分子は、上述のようなエフェクタ細胞の表面上に存在するCD2のエピトープに結合できる。CD2は、T細胞及びナチュラルキラー(NK)細胞上に見られる細胞付着分子である。CD2は、恐らくNK細胞ナノチューブ形成の促進因子として、NK細胞の細胞傷害性を強化する(Mace, E.M. et al. (2014) “Cell Biological Steps and Checkpoints in Accessing NK Cell Cytotoxicity,” Immunol. Cell. Biol. 92(3):245-255; Comerci, C.J. et al. (2012) “CD2 Promotes Human Natural Killer Cell Membrane Nanotube Formation,” PLoS One 7(10):e47664:1-12)。CD2に特異的に結合する分子としては、抗CD2抗体「CD2 mAb Lo‐CD2a」が挙げられる。
一実施形態では、本発明の分子は、上述のようなエフェクタ細胞の表面上に存在するCD2のエピトープに結合できる。CD2は、T細胞及びナチュラルキラー(NK)細胞上に見られる細胞付着分子である。CD2は、恐らくNK細胞ナノチューブ形成の促進因子として、NK細胞の細胞傷害性を強化する(Mace, E.M. et al. (2014) “Cell Biological Steps and Checkpoints in Accessing NK Cell Cytotoxicity,” Immunol. Cell. Biol. 92(3):245-255; Comerci, C.J. et al. (2012) “CD2 Promotes Human Natural Killer Cell Membrane Nanotube Formation,” PLoS One 7(10):e47664:1-12)。CD2に特異的に結合する分子としては、抗CD2抗体「CD2 mAb Lo‐CD2a」が挙げられる。
CD2 mAb Lo‐CD2aのVHドメインのアミノ酸配列(ATCC受託番号:HB‐11423;配列番号59)を以下に示す(CDRH残基は下線を付して示されている):
EVQLQQSGPE LQRPGASVKL SCKASGYIFT EYYMYWVKQR PKQGLELVGR
IDPEDGSIDY VEKFKKKATL TADTSSNTAY MQLSSLTSED TATYFCARGK
FNYRFAYWGQ GTLVTVSS
EVQLQQSGPE LQRPGASVKL SCKASGYIFT EYYMYWVKQR PKQGLELVGR
IDPEDGSIDY VEKFKKKATL TADTSSNTAY MQLSSLTSED TATYFCARGK
FNYRFAYWGQ GTLVTVSS
CD2 mAb Lo‐CD2aのVLドメインのアミノ酸配列(ATCC受託番号:11423;配列番号60)を以下に示す(CDRL残基は下線を付して示されている):
DVVLTQTPPT LLATIGQSVS ISCRSSQSLL HSSGNTYLNW LLQRTGQSPQ
PLIYLVSKLE SGVPNRFSGS GSGTDFTLKI SGVEAEDLGV YYCMQFTHYP
YTFGAGTKLE LK
DVVLTQTPPT LLATIGQSVS ISCRSSQSLL HSSGNTYLNW LLQRTGQSPQ
PLIYLVSKLE SGVPNRFSGS GSGTDFTLKI SGVEAEDLGV YYCMQFTHYP
YTFGAGTKLE LK
B.例示的なCD3結合能力
一実施形態では、本発明の分子はCD3のエピトープに結合できる。CD3は、4つの別個の鎖で構成されたT細胞共受容体である(Wucherpfennig, K.W. et al. (2010) “Structural Biology Of The T-Cell Receptor: Insights Into Receptor Assembly, Ligand Recognition, And Initiation Of Signaling,” Cold Spring Harb. Perspect. Biol. 2(4):a005140; pages 1-14)。哺乳類では、この複合体は、1つのCD3γ鎖、1つのCD3δ鎖、及び2つのCD3ε鎖を含有する。これらの鎖は、T細胞受容体(TCR)として知られる分子と連結して、Tリンパ球の活性化シグナルを生成する。CD3の不在下では、TCRは適切に集合せず、崩壊する(Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170-177)。CD3は、全ての成熟T細胞の膜に結合し、また他の細胞タイプには事実上結合しないことが分かっている(Janeway, C.A. et al. (2005) In: Immunobiology: The Immune System In Health And Disease,” 6th ed. Garland Science Publishing, NY, pp. 214- 216; Sun, Z. J. et al. (2001) “Mechanisms Contributing To T Cell Receptor Signaling And Assembly Revealed By The Solution Structure Of An Ectodomain Fragment Of The CD3ε:γ Heterodimer,” Cell 105(7):913-923; Kuhns, M.S. et al. (2006) “Deconstructing The Form And Function Of The TCR/CD3 Complex,” Immunity. 2006 Feb;24(2):133-139を参照)。CD3に特異的に結合する分子としては、抗CD3抗体「CD3 mAb 1」及び「OKT3」が挙げられる。抗CD3抗体CD3 mAb 1は、非ヒト霊長類(例えばカニクイザル)に結合できる。
一実施形態では、本発明の分子はCD3のエピトープに結合できる。CD3は、4つの別個の鎖で構成されたT細胞共受容体である(Wucherpfennig, K.W. et al. (2010) “Structural Biology Of The T-Cell Receptor: Insights Into Receptor Assembly, Ligand Recognition, And Initiation Of Signaling,” Cold Spring Harb. Perspect. Biol. 2(4):a005140; pages 1-14)。哺乳類では、この複合体は、1つのCD3γ鎖、1つのCD3δ鎖、及び2つのCD3ε鎖を含有する。これらの鎖は、T細胞受容体(TCR)として知られる分子と連結して、Tリンパ球の活性化シグナルを生成する。CD3の不在下では、TCRは適切に集合せず、崩壊する(Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170-177)。CD3は、全ての成熟T細胞の膜に結合し、また他の細胞タイプには事実上結合しないことが分かっている(Janeway, C.A. et al. (2005) In: Immunobiology: The Immune System In Health And Disease,” 6th ed. Garland Science Publishing, NY, pp. 214- 216; Sun, Z. J. et al. (2001) “Mechanisms Contributing To T Cell Receptor Signaling And Assembly Revealed By The Solution Structure Of An Ectodomain Fragment Of The CD3ε:γ Heterodimer,” Cell 105(7):913-923; Kuhns, M.S. et al. (2006) “Deconstructing The Form And Function Of The TCR/CD3 Complex,” Immunity. 2006 Feb;24(2):133-139を参照)。CD3に特異的に結合する分子としては、抗CD3抗体「CD3 mAb 1」及び「OKT3」が挙げられる。抗CD3抗体CD3 mAb 1は、非ヒト霊長類(例えばカニクイザル)に結合できる。
ヒト化CD3 mAb 1のVHドメインのアミノ酸配列(配列番号61)を以下に示す(CDRH残基は下線を付して示されている):
ヒト化CD3 mAb 1のVLドメインのアミノ酸配列(配列番号62)を以下に示す(CDRL残基は下線を付して示されている):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI
GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF
GGGTKLTVLG
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI
GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF
GGGTKLTVLG
ヒト化変異型「CD3 mAb 1(D65G)」を組み込むこともできる。CD3 mAb 1(D65G)は、CD3 mAb 1のVLドメイン(配列番号62)と、D65G置換(Kabat第65位、配列番号63の残基68に対応)を有するVH CD3 mAb 1ドメインとを含む。
CD3 mAb 1(D65G)のVHドメインのアミノ酸配列(配列番号63)を以下に示す(CDRH残基は下線を付して示されており、置換された位置(D65G)は二重下線を付して示されている):
あるいは、CD3 mAb 1のヒト化親和性変異型を組み込むこともできる。変異型としては、「CD3 mAb 1 Low」と呼ばれる低親和性変異型、及び「CD3 mAb 1 Fast」と呼ばれる、より高いオフレートを有する変異型が挙げられる。CD mAb1のVLドメイン(配列番号62)はCD3 mAb 1 Low及びCD3 mAb1 Fastと共通であり、上で提供されている。CD3 mAb 1 Low及びCD3 mAb1 FastそれぞれのVHドメインのアミノ酸配列を以下で提供する。
抗ヒトCD3 mAb 1 Lowの可変重鎖ドメインのアミノ酸配列(配列番号64)を以下に示す(CDRH残基は下線を付して示されている):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR
IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR
HGNFGNSYVT WFAYWGQGTL VTVSS
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR
IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR
HGNFGNSYVT WFAYWGQGTL VTVSS
抗ヒトCD3 mAb 1 Fastの可変重鎖ドメインのアミノ酸配列(配列番号65)を以下に示す(CDRH残基は下線を付して示されている):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR
IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR
HKNFGNSYVT WFAYWGQGTL VTVSS
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR
IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR
HKNFGNSYVT WFAYWGQGTL VTVSS
利用可能な別の抗CD3抗体は、抗体ムロモナブ‐CD3「OKT3」である(Xu et al. (2000) “In Vitro Characterization Of Five Humanized OKT3 Effector Function Variant Antibodies,” Cell. Immunol. 200:16-26); Norman, D.J. (1995) “Mechanisms Of Action And Overview Of OKT3,” Ther. Drug Monit. 17(6):615-620; Canafax, D.M. et al. (1987) “Monoclonal Antilymphocyte Antibody (OKT3) Treatment Of Acute Renal Allograft Rejection,” Pharmacotherapy 7(4):121-124; Swinnen, L.J. et al. (1993) “OKT3 Monoclonal Antibodies Induce Interleukin-6 And Interleukin-10: A Possible Cause Of Lymphoproliferative Disorders Associated With Transplantation,” Curr. Opin. Nephrol. Hypertens. 2(4):670-678)。
OKT3のVHドメインのアミノ酸配列(配列番号66)を以下に示す(CDRH残基は下線を付して示されている):
QVQLQQSGAE LARPGASVKM SCKASGYTFT RYTMHWVKQR PGQGLEWIGY
INPSRGYTNY NQKFKDKATL TTDKSSSTAY MQLSSLTSED SAVYYCARYY
DDHYCLDYWG QGTTLTVSS
QVQLQQSGAE LARPGASVKM SCKASGYTFT RYTMHWVKQR PGQGLEWIGY
INPSRGYTNY NQKFKDKATL TTDKSSSTAY MQLSSLTSED SAVYYCARYY
DDHYCLDYWG QGTTLTVSS
OKT3のVLドメインのアミノ酸配列(配列番号67)を以下に示す(CDRL残基は下線を付して示されている):
QIVLTQSPAI MSASPGEKVT MTCSASSSVS YMNWYQQKSG TSPKRWIYDT
SKLASGVPAH FRGSGSGTSY SLTISGMEAE DAATYYCQQW SSNPFTFGSG
TKLEINR
QIVLTQSPAI MSASPGEKVT MTCSASSSVS YMNWYQQKSG TSPKRWIYDT
SKLASGVPAH FRGSGSGTSY SLTISGMEAE DAATYYCQQW SSNPFTFGSG
TKLEINR
利用可能な更なる抗CD3抗体としては、限定するものではないが、国際公開第2008/119566号;及び国際公開第2005/118635号に記載されているものが挙げられる。
C.例示的なCD8結合能力
一実施形態では、本発明の分子は、上述のようなエフェクタ細胞の表面上に存在するCD8のエピトープに結合できる。CD8は、細胞傷害性T細胞上で発現される、2つの別個の鎖で構成されたT細胞共受容体である(Leahy, D.J., (1995) “A Structural View of CD4 and CD8,” FASEB J., 9:17-25)。CD8+ T細胞の活性化は、標的細胞の表面上に配列された抗原:主要組織適合遺伝子複合体クラスI(major histocompability class I:MHC I)と、CD8+ T細胞の表面上に配列されたCD8とT細胞受容体との複合体との間の、共刺激相互作用によって仲介されることが分かっている(Gao, G., and Jakobsen, B., (2000). "Molecular interactions of coreceptor CD8 and MHC class I: the molecular basis for functional coordination with the T-Cell Receptor". Immunol Today 21: 630-636)。特定の免疫系細胞によってしか発現されないMHC II分子とは異なり、MHC I分子は極めて広範に発現される。よって細胞傷害性T細胞は、多様な細胞タイプに結合できる。活性化された細胞傷害性T細胞は、細胞毒素パーフォリン、グランザイム、及びグラニュライシンの放出によって細胞殺滅を仲介する。CD8に特異的に結合する分子としては、抗CD8抗体「OKT8」及び「TRX2」が挙げられる。
一実施形態では、本発明の分子は、上述のようなエフェクタ細胞の表面上に存在するCD8のエピトープに結合できる。CD8は、細胞傷害性T細胞上で発現される、2つの別個の鎖で構成されたT細胞共受容体である(Leahy, D.J., (1995) “A Structural View of CD4 and CD8,” FASEB J., 9:17-25)。CD8+ T細胞の活性化は、標的細胞の表面上に配列された抗原:主要組織適合遺伝子複合体クラスI(major histocompability class I:MHC I)と、CD8+ T細胞の表面上に配列されたCD8とT細胞受容体との複合体との間の、共刺激相互作用によって仲介されることが分かっている(Gao, G., and Jakobsen, B., (2000). "Molecular interactions of coreceptor CD8 and MHC class I: the molecular basis for functional coordination with the T-Cell Receptor". Immunol Today 21: 630-636)。特定の免疫系細胞によってしか発現されないMHC II分子とは異なり、MHC I分子は極めて広範に発現される。よって細胞傷害性T細胞は、多様な細胞タイプに結合できる。活性化された細胞傷害性T細胞は、細胞毒素パーフォリン、グランザイム、及びグラニュライシンの放出によって細胞殺滅を仲介する。CD8に特異的に結合する分子としては、抗CD8抗体「OKT8」及び「TRX2」が挙げられる。
OKT8のVHドメインのアミノ酸配列(配列番号68)を以下に示す(CDRH残基は下線を付して示されている):
QVQLLESGPE LLKPGASVKM SCKASGYTFT DYNMHWVKQS HGKSLEWIGY
IYPYTGGTGY NQKFKNKATL TVDSSSSTAY MELRSLTSED SAVYYCARNF
RYTYWYFDVW GQGTTVTVSS
QVQLLESGPE LLKPGASVKM SCKASGYTFT DYNMHWVKQS HGKSLEWIGY
IYPYTGGTGY NQKFKNKATL TVDSSSSTAY MELRSLTSED SAVYYCARNF
RYTYWYFDVW GQGTTVTVSS
OKT8のVLドメインのアミノ酸配列(配列番号69)を以下に示す(CDRL残基は下線を付して示されている):
DIVMTQSPAS LAVSLGQRAT ISCRASESVD SYDNSLMHWY QQKPGQPPKV
LIYLASNLES GVPARFSGSG SRTDFTLTID PVEADDAATY YCQQNNEDPY
TFGGGTKLEI KR
DIVMTQSPAS LAVSLGQRAT ISCRASESVD SYDNSLMHWY QQKPGQPPKV
LIYLASNLES GVPARFSGSG SRTDFTLTID PVEADDAATY YCQQNNEDPY
TFGGGTKLEI KR
TRX2のVHドメインのアミノ酸配列(配列番号70)を以下に示す(CDRH残基は下線を付して示されている):
QVQLVESGGG VVQPGRSLRL SCAASGFTFS DFGMNWVRQA PGKGLEWVAL
IYYDGSNKFY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKPH
YDGYYHFFDS WGQGTLVTVS S
QVQLVESGGG VVQPGRSLRL SCAASGFTFS DFGMNWVRQA PGKGLEWVAL
IYYDGSNKFY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKPH
YDGYYHFFDS WGQGTLVTVS S
TRX2のVLドメインのアミノ酸配列(配列番号71)を以下に示す(CDRL残基は下線を付して示されている):
DIQMTQSPSS LSASVGDRVT ITCKGSQDIN NYLAWYQQKP GKAPKLLIYN
TDILHTGVPS RFSGSGSGTD FTFTISSLQP EDIATYYCYQ YNNGYTFGQG
TKVEIK
DIQMTQSPSS LSASVGDRVT ITCKGSQDIN NYLAWYQQKP GKAPKLLIYN
TDILHTGVPS RFSGSGSGTD FTFTISSLQP EDIATYYCYQ YNNGYTFGQG
TKVEIK
D.例示的なCD16結合能力
一実施形態では、本発明の分子はCD16のエピトープに結合できる。CD16は、好中球、好酸球、ナチュラルキラー(NK)細胞、及び凝集しているものの単量体ではないヒトIgGに結合する組織マクロファージによって発現される(Peltz, G.A. et al. (1989) “Human Fc Gamma RIII: Cloning, Expression, And Identification Of The Chromosomal Locus Of Two Fc Receptors For IgG,” Proc. Natl. Acad. Sci. (U.S.A.) 86(3):1013-1017; Bachanova, V. et al. (2014) “NK Cells In Therapy Of Cancer,” Crit. Rev. Oncog. 19(1-2):133-141; Miller, J.S. (2013) “Therapeutic Applications: Natural Killer Cells In The Clinic,” Hematology Am. Soc. Hematol. Educ. Program. 2013:247-253; Youinou, P. et al. (2002) “Pathogenic Effects Of Anti-Fc Gamma Receptor IIIB (CD16) On Polymorphonuclear Neutrophils In Non-Organ-Specific Autoimmune Diseases,” Autoimmun Rev. 1(1-2):13-19; Peipp, M. et al. (2002) “Bispecific Antibodies Targeting Cancer Cells,” Biochem. Soc. Trans. 30(4):507-511)。CD16に特異的に結合する分子としては、抗CD16抗体「3G8」及び「A9」が挙げられる。ヒト化3G8(「h3G8」)抗体は、国際公開第03/101485号に記載されている。
一実施形態では、本発明の分子はCD16のエピトープに結合できる。CD16は、好中球、好酸球、ナチュラルキラー(NK)細胞、及び凝集しているものの単量体ではないヒトIgGに結合する組織マクロファージによって発現される(Peltz, G.A. et al. (1989) “Human Fc Gamma RIII: Cloning, Expression, And Identification Of The Chromosomal Locus Of Two Fc Receptors For IgG,” Proc. Natl. Acad. Sci. (U.S.A.) 86(3):1013-1017; Bachanova, V. et al. (2014) “NK Cells In Therapy Of Cancer,” Crit. Rev. Oncog. 19(1-2):133-141; Miller, J.S. (2013) “Therapeutic Applications: Natural Killer Cells In The Clinic,” Hematology Am. Soc. Hematol. Educ. Program. 2013:247-253; Youinou, P. et al. (2002) “Pathogenic Effects Of Anti-Fc Gamma Receptor IIIB (CD16) On Polymorphonuclear Neutrophils In Non-Organ-Specific Autoimmune Diseases,” Autoimmun Rev. 1(1-2):13-19; Peipp, M. et al. (2002) “Bispecific Antibodies Targeting Cancer Cells,” Biochem. Soc. Trans. 30(4):507-511)。CD16に特異的に結合する分子としては、抗CD16抗体「3G8」及び「A9」が挙げられる。ヒト化3G8(「h3G8」)抗体は、国際公開第03/101485号に記載されている。
マウス3G8のVHドメインのアミノ酸配列(配列番号72)を以下に示す(CDRH残基は下線を付して示されている):
QVTLKESGPG ILQPSQTLSL TCSFSGFSLR TSGMGVGWIR QPSGKGLEWL
AHIWWDDDKR YNPALKSRLT ISKDTSSNQV FLKIASVDTA DTATYYCAQI
NPAWFAYWGQ GTLVTVSA
QVTLKESGPG ILQPSQTLSL TCSFSGFSLR TSGMGVGWIR QPSGKGLEWL
AHIWWDDDKR YNPALKSRLT ISKDTSSNQV FLKIASVDTA DTATYYCAQI
NPAWFAYWGQ GTLVTVSA
マウス3G8のVLドメインのアミノ酸配列(配列番号73)を以下に示す(CDRL残基は下線を付して示されている):
DTVLTQSPAS LAVSLGQRAT ISCKASQSVD FDGDSFMNWY QQKPGQPPKL
LIYTTSNLES GIPARFSASG SGTDFTLNIH PVEEEDTATY YCQQSNEDPY
TFGGGTKLEI K
DTVLTQSPAS LAVSLGQRAT ISCKASQSVD FDGDSFMNWY QQKPGQPPKL
LIYTTSNLES GIPARFSASG SGTDFTLNIH PVEEEDTATY YCQQSNEDPY
TFGGGTKLEI K
h3G8のVHドメインのアミノ酸配列(配列番号74)を以下に示す(CDRH残基は下線を付して示されている):
QVTLRESGPA LVKPTQTLTL TCTFSGFSLS TSGMGVGWIR QPPGKALEWL
AHIWWDDDKR YNPALKSRLT ISKDTSKNQV VLTMTNMDPV DTATYYCAQI
NPAWFAYWGQ GTLVTVSS
QVTLRESGPA LVKPTQTLTL TCTFSGFSLS TSGMGVGWIR QPPGKALEWL
AHIWWDDDKR YNPALKSRLT ISKDTSKNQV VLTMTNMDPV DTATYYCAQI
NPAWFAYWGQ GTLVTVSS
マウスh3G8のVLドメインのアミノ酸配列(配列番号75)を以下に示す(CDRL残基は下線を付して示されている):
DIVMTQSPDS LAVSLGERAT INCKASQSVD FDGDSFMNWY QQKPGQPPKL
LIYTTSNLES GVPDRFSGSG SGTDFTLTIS SLQAEDVAVY YCQQSNEDPY
TFGQGTKLEI K
DIVMTQSPDS LAVSLGERAT INCKASQSVD FDGDSFMNWY QQKPGQPPKL
LIYTTSNLES GVPDRFSGSG SGTDFTLTIS SLQAEDVAVY YCQQSNEDPY
TFGQGTKLEI K
A9のVHドメインのアミノ酸配列(配列番号76)を以下に示す(CDRH残基は下線を付して示されている):
QVQLQQSGAE LVRPGTSVKI SCKASGYTFT NYWLGWVKQR PGHGLEWIGD
IYPGGGYTNY NEKFKGKATV TADTSSRTAY VQVRSLTSED SAVYFCARSA
SWYFDVWGAR TTVTVSS
QVQLQQSGAE LVRPGTSVKI SCKASGYTFT NYWLGWVKQR PGHGLEWIGD
IYPGGGYTNY NEKFKGKATV TADTSSRTAY VQVRSLTSED SAVYFCARSA
SWYFDVWGAR TTVTVSS
A9のVLドメインのアミノ酸配列(配列番号77)を以下に示す(CDRL残基は下線を付して示されている):
DIQAVVTQES ALTTSPGETV TLTCRSNTGT VTTSNYANWV QEKPDHLFTG
LIGHTNNRAP GVPARFSGSL IGDKAALTIT GAQTEDEAIY FCALWYNNHW
VFGGGTKLTVL
DIQAVVTQES ALTTSPGETV TLTCRSNTGT VTTSNYANWV QEKPDHLFTG
LIGHTNNRAP GVPARFSGSL IGDKAALTIT GAQTEDEAIY FCALWYNNHW
VFGGGTKLTVL
米国特許第9,035,026号は、CD16Aに結合できるもののCD16Bには結合できない抗CD16抗体を記載している。このような抗体は、疾患関連細胞を対象とした二重又は多重特異性分子の成分として特に有用である。というのは、上記抗体は主にNK細胞を動員し、循環する可溶性CD16Bによって結合されることがなく、又は好中球若しくは活性化された好酸球への結合によってNK細胞の結合から逸らされることがないためである。
米国特許第9,035,026号の抗CD16A抗体のVHドメインのアミノ酸配列(配列番号78)を以下に示す(CDRH残基は下線を付して示されている):
EVQLVQSGAE VKKPGESLKV SCKASGYTFT SYYMHWVRQA PGQGLEWMGI
INPSGGSTSY AQKFQGRVTM TRDTSTSTVY MELSSLRSED TAVYYCARGS
AYYYDFADYW GQGTLVTVSS
EVQLVQSGAE VKKPGESLKV SCKASGYTFT SYYMHWVRQA PGQGLEWMGI
INPSGGSTSY AQKFQGRVTM TRDTSTSTVY MELSSLRSED TAVYYCARGS
AYYYDFADYW GQGTLVTVSS
米国特許第9,035,026号の上述の抗CD16A抗体のに関する7個の好適なVLドメインのアミノ酸配列を、以下に配列番号79〜85として示す:
SYELMQPPSV SVSSGQTASI PCSGDKLEEK YVSWYQQRPG QSPVLVIYQD
NKRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号79)
SYELTQPLSE SVAQGQTARI TCGGNNIESR NVHWYQQKPG QAPVLVIYRD
NNRPSGIPER FSGSNSGNMA TLTISRAQAG DAADYYCQVW DNYTVLFGGG
TKLTVL(配列番号80)
SYELTQPPSV AVAPGKTARI TCGGNNIGSK NVHWYQQKPG QAPVLVIYRD
SNRPSGIPER FSGSNSGNTA TLTISRAQAG DEADFYCQVW DNYIVLFGGG
TKLTVL(配列番号81)
QAVLTQPPSV SVAPGQTARI PCEGNNIGSK NVHWYRQKPG QVPVLVMYDD
SDRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号82)
QPVLTQPLSV SVAPGQTARI TCGGNNIGSK NVHWYQQKPG QAPVLVIYRD
SSRPSGIPER LSGSNSGDTA TLTISRAQAG DEADYYCQVW DDYIVVFGGG
TKLTVL(配列番号83)
SYELTQPPSV SVTPGQTATI TCGANDIGKR NVHWYQQRPG QSPVLVIYQD
NKRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号84)
QPVLTQPSSV SVAPGQTATI SCGGHNIGSK NVHWYQQRPG QSPVLVIYQD
NKRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号85)
SYELMQPPSV SVSSGQTASI PCSGDKLEEK YVSWYQQRPG QSPVLVIYQD
NKRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号79)
SYELTQPLSE SVAQGQTARI TCGGNNIESR NVHWYQQKPG QAPVLVIYRD
NNRPSGIPER FSGSNSGNMA TLTISRAQAG DAADYYCQVW DNYTVLFGGG
TKLTVL(配列番号80)
SYELTQPPSV AVAPGKTARI TCGGNNIGSK NVHWYQQKPG QAPVLVIYRD
SNRPSGIPER FSGSNSGNTA TLTISRAQAG DEADFYCQVW DNYIVLFGGG
TKLTVL(配列番号81)
QAVLTQPPSV SVAPGQTARI PCEGNNIGSK NVHWYRQKPG QVPVLVMYDD
SDRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号82)
QPVLTQPLSV SVAPGQTARI TCGGNNIGSK NVHWYQQKPG QAPVLVIYRD
SSRPSGIPER LSGSNSGDTA TLTISRAQAG DEADYYCQVW DDYIVVFGGG
TKLTVL(配列番号83)
SYELTQPPSV SVTPGQTATI TCGANDIGKR NVHWYQQRPG QSPVLVIYQD
NKRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号84)
QPVLTQPSSV SVAPGQTATI SCGGHNIGSK NVHWYQQRPG QSPVLVIYQD
NKRPSGIPER FSGSNSGNTA TLTISGTQAM DEADYYCQVW DNYSVLFGGG
TKLTVL(配列番号85)
更なる実施形態では、ヒト化抗CD16抗体hCD16‐M1又はヒト化抗CD16抗体hCD16‐M2のCD16結合ドメインを、gp41結合ドメインと協調して使用して、CD16及びgp41に結合できる多重特異性結合分子を生成してよい。hCD16‐M1のVHドメインのアミノ酸配列(配列番号127)を以下に示す(CDRH残基は下線を付して示されている):
EVQLVESGGG LVKPGGSLRL SCAASGFTFS NYGMSWVRQA PGKGLEWVAT
ISGGGSYTFY PDSVKGRFTI SRDNAKNSLY LQMNSLRTED TALYYCVRQS
ARAPEPYWGQ GTLVTVSS
EVQLVESGGG LVKPGGSLRL SCAASGFTFS NYGMSWVRQA PGKGLEWVAT
ISGGGSYTFY PDSVKGRFTI SRDNAKNSLY LQMNSLRTED TALYYCVRQS
ARAPEPYWGQ GTLVTVSS
hCD16‐M1のVLドメインのアミノ酸配列(配列番号128)を以下に示す(CDRL残基は下線を付して示されている):
DIQMTQSPSF LSASVGDRVT ITCRASQNVG THVAWYQQKP GKAPKSLLYS
ASYRYSGVPS RFSGSGSGTD FTLTISSLQS EDIATYYCQQ YKSYPLTFGQ
GTKLEIK
DIQMTQSPSF LSASVGDRVT ITCRASQNVG THVAWYQQKP GKAPKSLLYS
ASYRYSGVPS RFSGSGSGTD FTLTISSLQS EDIATYYCQQ YKSYPLTFGQ
GTKLEIK
hCD16‐M2は、マウス抗ヒトCD16モノクローナル抗体CD16‐M2のヒト化誘導体である。ヒト化は、2つの好適なVHドメイン(hCD16‐M2 VH1及びhCD16‐M2 VH2)をもたらした。これらはいずれも、得られるヒト化VLドメイン(hCD16‐M2 VL1)と共に使用してよい。
hCD16‐M2のVHドメインVH1のアミノ酸配列(配列番号129)を以下に示す(CDRH残基は下線を付して示されている):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT SSAMHWVRQA PGQGLEWMGY
INHYNDGIKY NERFKGRVTI TADKSTSTAY MELSSLRSED TAVYYCATGY
RYASWFASWG QGTLVTVSS
QVQLVQSGAE VKKPGASVKV SCKASGYTFT SSAMHWVRQA PGQGLEWMGY
INHYNDGIKY NERFKGRVTI TADKSTSTAY MELSSLRSED TAVYYCATGY
RYASWFASWG QGTLVTVSS
hCD16‐M2のVHドメインVH2のアミノ酸配列(配列番号130)を以下に示す(CDRH残基は下線を付して示されている):
認識されるように、hCD16‐M2 VH1のアミノ酸配列(配列番号129)は、CDRH3の直前の残基にT98R置換(上では四角で囲んで示されている)有するという点で、hCD16‐M2 VH2のアミノ酸配列(配列番号130)とは異なる。
VLドメインのアミノ酸配列hCD16‐M2 VL1(配列番号131)を以下に示す(CDRL残基は下線を付して示されている):
EIVLTQSPAT LSVSPGERAT LSCRASQNIG TSIHWYQQKP DQSPKLLIKS
VSESISGVPS RFSGSGSGTD FTLTINSLEA EDFATYYCQQ SNSWPLTFGQ
GTKLEIK
EIVLTQSPAT LSVSPGERAT LSCRASQNIG TSIHWYQQKP DQSPKLLIKS
VSESISGVPS RFSGSGSGTD FTLTINSLEA EDFATYYCQQ SNSWPLTFGQ
GTKLEIK
利用可能な更なる抗CD16抗体としては、限定するものではないが、市販の抗体DJ130c(Tamm, A. et al. (1996) “The Binding Epitopes Of Human CD16 (Fc Gamma RIII) Monoclonal Antibodies. Implications For Ligand Binding,” J. Immunol. 157(4):1576-1581)、eBioCB16(ThermoFisher)、若しくは1D3(Abcam)、又は国際公開第03/101485号及び国際公開第2006/125668号に記載されているものが挙げられる。
E.例示的なTCR結合能力
一実施形態では、本発明の分子は、T細胞受容体(TCR)のエピトープに結合できる。T細胞受容体は、CD4+又はCD8+T細胞によってネイティブに発現され、これらの細胞が、抗原提示細胞のMHCクラスI若しくはクラスIIタンパク質によって結合及び提示される抗原ペプチドを認識できるようにする。TCRによるpMHC(ペプチド‐MHC)複合体の認識は、細胞免疫応答の伝播を開始させ、これは、サイトカインの産生及び抗原提示細胞の溶解につながる(例えばArmstrong, K.M. et al. (2008) “Conformational Changes And Flexibility In T-Cell Receptor Recognition Of Peptide-MHC Complexes,” Biochem. J. 415(Pt 2):183-196; Willemsen, R. (2008) “Selection Of Human Antibody Fragments Directed Against Tumor T-Cell Epitopes For Adoptive T-Cell Therapy,” Cytometry A. 73(11):1093-1099; Beier, K.C. et al. (2007) “Master Switches Of T-Cell Activation And Differentiation,” Eur. Respir. J. 29:804-812; Mallone, R. et al. (2005) “Targeting T Lymphocytes For Immune Monitoring And Intervention In Autoimmune Diabetes,” Am. J. Ther. 12(6):534-550を参照)。CD3は、TCRに結合する受容体である(Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170-177; Guy, C.S. et al. (2009) “Organization Of Proximal Signal Initiation At The TCR:CD3 Complex,” Immunol. Rev. 232(1):7-21; St. Clair, E.W. (Epub 2009 Oct 12) “Novel Targeted Therapies For Autoimmunity,” Curr. Opin. Immunol. 21(6):648-657; Baeuerle, P.A. et al. (Epub 2009 Jun 9) “Bispecific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944; Smith-Garvin, J.E. et al. (2009) “T Cell Activation,” Annu. Rev. Immunol. 27:591-619; Renders, L. et al. (2003) “Engineered CD3 Antibodies For Immunosuppression,” Clin. Exp. Immunol. 133(3):307-309)。
一実施形態では、本発明の分子は、T細胞受容体(TCR)のエピトープに結合できる。T細胞受容体は、CD4+又はCD8+T細胞によってネイティブに発現され、これらの細胞が、抗原提示細胞のMHCクラスI若しくはクラスIIタンパク質によって結合及び提示される抗原ペプチドを認識できるようにする。TCRによるpMHC(ペプチド‐MHC)複合体の認識は、細胞免疫応答の伝播を開始させ、これは、サイトカインの産生及び抗原提示細胞の溶解につながる(例えばArmstrong, K.M. et al. (2008) “Conformational Changes And Flexibility In T-Cell Receptor Recognition Of Peptide-MHC Complexes,” Biochem. J. 415(Pt 2):183-196; Willemsen, R. (2008) “Selection Of Human Antibody Fragments Directed Against Tumor T-Cell Epitopes For Adoptive T-Cell Therapy,” Cytometry A. 73(11):1093-1099; Beier, K.C. et al. (2007) “Master Switches Of T-Cell Activation And Differentiation,” Eur. Respir. J. 29:804-812; Mallone, R. et al. (2005) “Targeting T Lymphocytes For Immune Monitoring And Intervention In Autoimmune Diabetes,” Am. J. Ther. 12(6):534-550を参照)。CD3は、TCRに結合する受容体である(Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170-177; Guy, C.S. et al. (2009) “Organization Of Proximal Signal Initiation At The TCR:CD3 Complex,” Immunol. Rev. 232(1):7-21; St. Clair, E.W. (Epub 2009 Oct 12) “Novel Targeted Therapies For Autoimmunity,” Curr. Opin. Immunol. 21(6):648-657; Baeuerle, P.A. et al. (Epub 2009 Jun 9) “Bispecific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944; Smith-Garvin, J.E. et al. (2009) “T Cell Activation,” Annu. Rev. Immunol. 27:591-619; Renders, L. et al. (2003) “Engineered CD3 Antibodies For Immunosuppression,” Clin. Exp. Immunol. 133(3):307-309)。
T細胞受容体に特異的に結合する分子は、抗TCR抗体「BMA031」を含む(欧州特許第0403156号;Kurrle, R. et al. (1989) “BMA 031 - A TCR-Specific Monoclonal Antibody For Clinical Application,” Transplant Proc. 21(1 Pt 1):1017-1019; Nashan, B. et al. (1987) “Fine Specificity Of A Panel Of Antibodies Against The TCR/CD3 Complex,” Transplant Proc. 19(5):4270-4272; Shearman, C.W. et al. (1991) “Construction, Expression, And Biologic Activity Of Murine/Human Chimeric Antibodies With Specificity For The Human α/β T Cell,” J. Immunol. 146(3):928-935; Shearman, C.W. et al. (1991) “Construction, Expression And Characterization of Humanized Antibodies Directed Against The Human α/β T Cell Receptor,” J. Immunol. 147(12):4366-4373)。
BMA031のVHドメインのアミノ酸配列(配列番号86)を以下に示す(CDRH残基は下線を付して示されている):
QVQLVQSGAE VKKPGASVKV SCKASGYKFT SYVMHWVRQA PGQGLEWIGY
INPYNDVTKY NEKFKGRVTI TADKSTSTAY LQMNSLRSED TAVHYCARGS
YYDYDGFVYW GQGTLVTVSS
QVQLVQSGAE VKKPGASVKV SCKASGYKFT SYVMHWVRQA PGQGLEWIGY
INPYNDVTKY NEKFKGRVTI TADKSTSTAY LQMNSLRSED TAVHYCARGS
YYDYDGFVYW GQGTLVTVSS
BMA031のVLドメインのアミノ酸配列(配列番号87)を以下に示す(CDRL残基は下線を付して示されている):
EIVLTQSPAT LSLSPGERAT LSCSATSSVS YMHWYQQKPG KAPKRWIYDT
SKLASGVPSR FSGSGSGTEF TLTISSLQPE DFATYYCQQW SSNPLTFGQG
TKLEIK
EIVLTQSPAT LSLSPGERAT LSCSATSSVS YMHWYQQKPG KAPKRWIYDT
SKLASGVPSR FSGSGSGTEF TLTISSLQPE DFATYYCQQW SSNPLTFGQG
TKLEIK
F.例示的なNKG2D結合能力
一実施形態では、本発明の分子はNKG2D受容体のエピトープに結合できる。NKG2D受容体は、全てのヒト(及び他の哺乳類)のナチュラルキラー細胞上(Bauer, S. et al. (1999) “Activation Of NK Cells And T Cells By NKG2D, A Receptor For Stress-Inducible MICA,” Science 285(5428):727-729; Jamieson, A.M. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29)並びに全てのCD8+T細胞上(Groh, V. et al. (2001) “Costimulation Of CD8αβ T Cells By NKG2D Via Engagement By MIC Induced On Virus-Infected Cells,” Nat. Immunol. 2(3):255-260; Jamieson, A.M. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29)で発現する。NKG2D受容体に特異的に結合する分子としては、抗NKG2D抗体「KYK‐1.0」及び「KYK‐2.0」が挙げられる(Kwong, KY et al. (2008) “Generation, Affinity Maturation, And Characterization Of A Human Anti-Human NKG2D Monoclonal Antibody With Dual Antagonistic And Agonistic Activity,” J. Mol. Biol. 384:1143-1156)。
一実施形態では、本発明の分子はNKG2D受容体のエピトープに結合できる。NKG2D受容体は、全てのヒト(及び他の哺乳類)のナチュラルキラー細胞上(Bauer, S. et al. (1999) “Activation Of NK Cells And T Cells By NKG2D, A Receptor For Stress-Inducible MICA,” Science 285(5428):727-729; Jamieson, A.M. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29)並びに全てのCD8+T細胞上(Groh, V. et al. (2001) “Costimulation Of CD8αβ T Cells By NKG2D Via Engagement By MIC Induced On Virus-Infected Cells,” Nat. Immunol. 2(3):255-260; Jamieson, A.M. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29)で発現する。NKG2D受容体に特異的に結合する分子としては、抗NKG2D抗体「KYK‐1.0」及び「KYK‐2.0」が挙げられる(Kwong, KY et al. (2008) “Generation, Affinity Maturation, And Characterization Of A Human Anti-Human NKG2D Monoclonal Antibody With Dual Antagonistic And Agonistic Activity,” J. Mol. Biol. 384:1143-1156)。
KYK‐1.0のVHドメインのアミノ酸配列(配列番号88)を以下に示す(CDRH残基は下線を付して示されている):
EVQLVESGGG VVQPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF
IRYDGSNKYY ADSVKGRFTI SRDNSKNTKY LQMNSLRAED TAVYYCAKDR
FGYYLDYWGQ GTLVTVSS
EVQLVESGGG VVQPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF
IRYDGSNKYY ADSVKGRFTI SRDNSKNTKY LQMNSLRAED TAVYYCAKDR
FGYYLDYWGQ GTLVTVSS
KYK‐1.0のVLドメインのアミノ酸配列(配列番号89)を以下に示す(CDRL残基は下線を付して示されている):
QPVLTQPSSV SVAPGETARI PCGGDDIETK SVHWYQQKPG QAPVLVIYDD
DDRPSGIPER FFGSNSGNTA TLSISRVEAG DEADYYCQVW DDNNDEWVFG
GGTQLTVL
QPVLTQPSSV SVAPGETARI PCGGDDIETK SVHWYQQKPG QAPVLVIYDD
DDRPSGIPER FFGSNSGNTA TLSISRVEAG DEADYYCQVW DDNNDEWVFG
GGTQLTVL
KYK‐2.0のVHドメインのアミノ酸配列(配列番号90)を以下に示す(CDRH残基は下線を付して示されている):
QVQLVESGGG LVKPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF
IRYDGSNKYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKDR
GLGDGTYFDY WGQGTTVTVS S
QVQLVESGGG LVKPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF
IRYDGSNKYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKDR
GLGDGTYFDY WGQGTTVTVS S
KYK‐2.0のVLドメインのアミノ酸配列(配列番号91)を以下に示す(CDRL残基は下線を付して示されている):
QSALTQPASV SGSPGQSITI SCSGSSSNIG NNAVNWYQQL PGKAPKLLIY
YDDLLPSGVS DRFSGSKSGT SAFLAISGLQ SEDEADYYCA AWDDSLNGPV
FGGGTKLTVL
QSALTQPASV SGSPGQSITI SCSGSSSNIG NNAVNWYQQL PGKAPKLLIY
YDDLLPSGVS DRFSGSKSGT SAFLAISGLQ SEDEADYYCA AWDDSLNGPV
FGGGTKLTVL
G.例示的なNKp46結合能力
一実施形態では、本発明の分子は、NKp46(CD335)のエピトープに結合できる。NKp46は、標的細胞の排除に関わる主要なNK細胞活性化受容体である(Sivori S, et al. (1997). "p46, a Novel Natural Killer Cell-specific Surface Molecule That Mediates Cell Activation," J. Exp. Med. 186 (7): 1129-36.)。NKp46は、全てのNK細胞サブセット上のみで発現する(Narni-Mancinelli E, et al. (2011). “Fate mapping analysis of lymphoid cells expressing the NKp46 cell surface receptor,” Proc Natl Acad Sci U S A 108:18324-9)。NKp46に特異的に結合する分子としては、抗NKp46抗体「BAB281」及び「NKp46‐3」が挙げられる(国際公開第2015/197593号)。
一実施形態では、本発明の分子は、NKp46(CD335)のエピトープに結合できる。NKp46は、標的細胞の排除に関わる主要なNK細胞活性化受容体である(Sivori S, et al. (1997). "p46, a Novel Natural Killer Cell-specific Surface Molecule That Mediates Cell Activation," J. Exp. Med. 186 (7): 1129-36.)。NKp46は、全てのNK細胞サブセット上のみで発現する(Narni-Mancinelli E, et al. (2011). “Fate mapping analysis of lymphoid cells expressing the NKp46 cell surface receptor,” Proc Natl Acad Sci U S A 108:18324-9)。NKp46に特異的に結合する分子としては、抗NKp46抗体「BAB281」及び「NKp46‐3」が挙げられる(国際公開第2015/197593号)。
BAB281のVHドメインのアミノ酸配列(配列番号92)を以下に示す(CDRH残基は下線を付して示されている):
QIQLVQSGPE LQKPGETVKI SCKASGYTFT NYGMNWVKQA PGKGLKWMGW
INTNTGEPTY AEEFKGRFAF SLETSASTAY LQINNLKNED TATYFCARDY
LYYFDYWGQG TTLTVSS
QIQLVQSGPE LQKPGETVKI SCKASGYTFT NYGMNWVKQA PGKGLKWMGW
INTNTGEPTY AEEFKGRFAF SLETSASTAY LQINNLKNED TATYFCARDY
LYYFDYWGQG TTLTVSS
BAB281のVLドメインのアミノ酸配列(配列番号93)を以下に示す(CDRL残基は下線を付して示されている):
DNIVMTQSPK SMSMSVGERV TLTCKASENV VTYVSWYQQK PEQSPKLLIY
GASNRYTGVP DRFTGSGSAT DFTLTISSVQ AEDLADYHCG QGYSYPYTFG
GGTKLEIK
DNIVMTQSPK SMSMSVGERV TLTCKASENV VTYVSWYQQK PEQSPKLLIY
GASNRYTGVP DRFTGSGSAT DFTLTISSVQ AEDLADYHCG QGYSYPYTFG
GGTKLEIK
NKp46‐3のVHドメインのアミノ酸配列(配列番号94)を以下に示す(CDRH残基は下線を付して示されている):
EVQLQQSGPE LVKPGASVKI SCKTSGYTFT EYTMHWVKQS HGKSLEWIGG
ISPNIGGTSY NQKFKGKATL TVDKSSSTAY MELRSLTSED SAVYYCARRG
GSFDYWGQGT TLTVSS
EVQLQQSGPE LVKPGASVKI SCKTSGYTFT EYTMHWVKQS HGKSLEWIGG
ISPNIGGTSY NQKFKGKATL TVDKSSSTAY MELRSLTSED SAVYYCARRG
GSFDYWGQGT TLTVSS
NKp46‐3のVLドメインのアミノ酸配列(配列番号95)を以下に示す(CDRL残基は下線を付して示されている):
DIVMTQSPAT LSVTPGDRVS LSCRASQSIS DYLHWYQQKS HESPRLLIKY
ASQSISGIPS RFSGSGSGSD FTLSINSVEP EDVGVYYCQN GHSFPLTFGA
GTKLELK
DIVMTQSPAT LSVTPGDRVS LSCRASQSIS DYLHWYQQKS HESPRLLIKY
ASQSISGIPS RFSGSGSGSD FTLSINSVEP EDVGVYYCQN GHSFPLTFGA
GTKLELK
NKp46に結合する更なる抗体は、国際公開第2015/197593号及び国際公開第2017/016805号に記載されている。ナチュラルキラー細胞の細胞表面に結合する他の例示的な抗体としては、抗体:A1、AC2、EPR3678(2)、EPR20461、EPR20627及びIMG17B5F11(CD39に結合);TB01、HNK‐1/Leu‐7及びNK1(CD57に結合);FN50(CD69に結合);5B5、B‐L2、TS82b及びC33(CD82に結合);3B3、B199.2及びEP7169(CD161に結合);17D9(CLEC1Bに結合);2F9(KIR2DL1に結合);EPR8825(KIR2DL2に結合);mAb 33(KIR2DL4に結合);11E3、17B4、EPR4392(2)、EPR20261及びEPR20627(リンパ球活性化遺伝子3に結合);A10、C7、CX5、1D11及びMM0489‐10R27(NKG2Dに結合);BMK13(PRG2に結合);EPR9916(SLAMF6に結合);等が挙げられる。このようなNK細胞表面分子それぞれに結合可能な抗体は、Abcam plc及び他の供給元から市販されており、本発明の目的に容易に適合させることができる。
VI.例示的な代替HIV‐1結合分子
成熟ヒト免疫不全ウイルス1型(HIV‐1)エンベロープ(Env)糖タンパク質三量体は、ウイルスgp160前駆体タンパク質の切断によって生じる、非共有結合したgp120/gp41ヘテロ二量体の3つのコピーで構成される。一実施形態では、本発明の分子は、HIV‐1 Envタンパク質gp120、gp160及び/又はgp41のエピトープに結合でき、これは7B2のエピトープとは異なる。
成熟ヒト免疫不全ウイルス1型(HIV‐1)エンベロープ(Env)糖タンパク質三量体は、ウイルスgp160前駆体タンパク質の切断によって生じる、非共有結合したgp120/gp41ヘテロ二量体の3つのコピーで構成される。一実施形態では、本発明の分子は、HIV‐1 Envタンパク質gp120、gp160及び/又はgp41のエピトープに結合でき、これは7B2のエピトープとは異なる。
モノクローナル抗体A32は、HIV‐1 Env gp120のC1領域中の立体配座エピトープを認識し(Wyatt et al. (1995) “Involvement Of The V1/V2 Variable Loop Structure In The Exposure Of Human Immunodeficiency Virus Type 1 gp120 Epitopes Induced By Receptor Binding,” J. Virol. 69:5723-5733)、強力なADCC活性を仲介して、HIV‐1感染個体において検出可能なADCC仲介型Ab活性の大部分をブロックできる(Ferrari, G. et al. (2011) “An HIV-1 gp120 Envelope Human Monoclonal Antibody That Recognizes a C1 Conformational Epitope Mediates Potent Antibody-Dependent Cellular Cytotoxicity (ADCC) Activity and Defines a Common ADCC Epitope in Human HIV-1 Serum,” J. Virol. 85(14):7029-7036)。
フレームワーク領域1及び/又は4に若干の変化を有する抗体A32の複数のVHドメインが、当該技術分野において報告されている(例えばタンパク質データベース受託番号PDB:4YBL_H、米国公開特許第2015/0239961号、及び国際公開第2006/044410号を参照)。これらの変異型抗体A32 VHドメインのいずれを、本発明に従って使用してよい。A32の例示的なVHドメインのアミノ酸配列(配列番号96)を以下に示す(CDRH残基は下線を付して示されている):
QVQLQESGPG LVKPSQTLSL SCTVSGGSSS SGAHYWSWIR QYPGKGLEWI
GYIHYSGNTY YNPSLKSRIT ISQHTSENQF SLKLNSVTVA DTAVYYCARG
TRLRTLRNAF DIWGQGTLVT VSS
QVQLQESGPG LVKPSQTLSL SCTVSGGSSS SGAHYWSWIR QYPGKGLEWI
GYIHYSGNTY YNPSLKSRIT ISQHTSENQF SLKLNSVTVA DTAVYYCARG
TRLRTLRNAF DIWGQGTLVT VSS
A32のVLドメインのアミノ酸配列(配列番号97)を以下に示す(CDRL残基は下線を付して示されている):
QSALTQPPSA SGSPGQSVTI SCTGTSSDVG GYNYVSWYQH HPGKAPKLII
SEVNNRPSGV PDRFSGSKSG NTASLTVSGL QAEDEAEYYC SSYTDIHNFV
FGGGTKLTVL
QSALTQPPSA SGSPGQSVTI SCTGTSSDVG GYNYVSWYQH HPGKAPKLII
SEVNNRPSGV PDRFSGSKSG NTASLTVSGL QAEDEAEYYC SSYTDIHNFV
FGGGTKLTVL
A32のVLドメインのアミノ酸配列(配列番号97)を、A32の例示的なVHドメイン(配列番号96)と共に、又は変異型抗体A32 VHドメイン(例えばタンパク質データベース受託番号PDB:4YBL_H、米国公開特許第2015/0239961号、及び国際公開第2006/044410号を参照)のいずれと共に使用して、抗HIV‐1 Env gp120エピトープ結合部位を形成できる。
モノクローナル抗体10‐1074は、HIV‐1 Env gp120のV3ループのベースを標的とし(例えば国際公開第2014/063059号を参照)、単離されている最も強力な抗体HIV‐1中和抗体の1つであり、また初期段階の臨床試験においてある程度の生体内活性を示した(Caskey, M., et al., (2017) “Antibody 10-1074 Suppresses Viremia In HIV-1-infected individuals.” Nat Med. 23:185-191)。
10‐1074のVHドメインのアミノ酸配列(配列番号98)を以下に示す(CDRH残基は下線を付して示されている):
QVQLQESGPG LVKPSETLSV TCSVSGDSMN NYYWTWIRQS PGKGLEWIGY
ISDRESATYN PSLNSRVVIS RDTSKNQLSL KLNSVTPADT AVYYCATARR
GQRIYGVVSF GEFFYYYSMD VWGKGTTVTV SS
QVQLQESGPG LVKPSETLSV TCSVSGDSMN NYYWTWIRQS PGKGLEWIGY
ISDRESATYN PSLNSRVVIS RDTSKNQLSL KLNSVTPADT AVYYCATARR
GQRIYGVVSF GEFFYYYSMD VWGKGTTVTV SS
10‐1074のVLドメインのアミノ酸配列(配列番号99)を以下に示す(CDRL残基は下線を付して示されている):
SYVRPLSVAL GETARISCGR QALGSRAVQW YQHRPGQAPI LLIYNNQDRP
SGIPERFSGT PDINFGTRAT LTISGVEAGD EADYYCHMWD SRSGFSWSFG
GATRLTVL
SYVRPLSVAL GETARISCGR QALGSRAVQW YQHRPGQAPI LLIYNNQDRP
SGIPERFSGT PDINFGTRAT LTISGVEAGD EADYYCHMWD SRSGFSWSFG
GATRLTVL
モノクローナル抗体3BNC117は、gp120のCD4結合部位を標的とし、幅広い中和性を有する抗HIV‐1抗体であり(例えば国際公開2013/016468号を参照)、これは初期段階の臨床試験においてある程度の生体内活性を示した(Caskey, M., et al., (2015) “Viraemia Suppressed In HIV-1-Infected Humans By Broadly Neutralizing Antibody 3BNC117,” Nature 522:487-491)。
3BNC117のVHドメインのアミノ酸配列(配列番号100)を以下に示す(CDRH残基は下線を付して示されている):
QVQLLQSGAA VTKPGASVRV SCEASGYNIR DYFIHWWRQA PGQGLQWVGW
INPKTGQPNN PRQFQGRVSL TRHASWDFDT FSFYMDLKAL RSDDTAVYFC
ARQRSDYWDF DVWGSGTQVT VSS
QVQLLQSGAA VTKPGASVRV SCEASGYNIR DYFIHWWRQA PGQGLQWVGW
INPKTGQPNN PRQFQGRVSL TRHASWDFDT FSFYMDLKAL RSDDTAVYFC
ARQRSDYWDF DVWGSGTQVT VSS
3BNC117のVLドメインのアミノ酸配列(配列番号101)を以下に示す(CDRL残基は下線を付して示されている):
DIQMTQSPSS LSASVGDTVT ITCQANGYLN WYQQRRGKAP KLLIYDGSKL
ERGVPSRFSG RRWGQEYNLT INNLQPEDIA TYFCQVYEFV VPGTRLDLK
DIQMTQSPSS LSASVGDTVT ITCQANGYLN WYQQRRGKAP KLLIYDGSKL
ERGVPSRFSG RRWGQEYNLT INNLQPEDIA TYFCQVYEFV VPGTRLDLK
モノクローナル抗体PGT121及びPGT145は、Env認識に関してgp120グリカンに強く依存する、幅広い中和性を有するHIV‐1抗体である(Mouquet H, et al., (2012) “Complex-type N-glycan recognition by potent broadly neutralizing HIV antibodies.” Proc Natl Acad Sci U S A 109: E3268-3277; Yasmeen, A., et al. (2014) “Differential Binding Of Neutralizing And Non-Neutralizing Antibodies To Native-Like Soluble HIV-1 Env Trimers, Uncleaved Env Proteins, And Monomeric Subunits.” Retrovirology 11:41; 国際公開第2012/030904号)。
PGT121のVHドメインのアミノ酸配列(配列番号102)を以下に示す(CDRH残基は下線を付して示されている):
QMQLQESGPG LVKPSETLSL TCSVSGASIS DSYWSWIRRS PGKGLEWIGY
VHKSGDTNYS PSLKSRVNLS LDTSKNQVSL SLVAATAADS GKYYCARTLH
GRRIYGIVAF NEWFTYFYMD VWGNGTQVTVSS
QMQLQESGPG LVKPSETLSL TCSVSGASIS DSYWSWIRRS PGKGLEWIGY
VHKSGDTNYS PSLKSRVNLS LDTSKNQVSL SLVAATAADS GKYYCARTLH
GRRIYGIVAF NEWFTYFYMD VWGNGTQVTVSS
PGT121のVLドメインのアミノ酸配列(配列番号103)を以下に示す(CDRL残基は下線を付して示されている):
SDISVAPGET ARISCGEKSL GSRAVQWYQH RAGQAPSLII YNNQDRPSGI
PERFSGSPDS PFGTTATLTI TSVEAGDEAD YYCHIWDSRV PTKWVFGGGT
TLTVL
SDISVAPGET ARISCGEKSL GSRAVQWYQH RAGQAPSLII YNNQDRPSGI
PERFSGSPDS PFGTTATLTI TSVEAGDEAD YYCHIWDSRV PTKWVFGGGT
TLTVL
PGT145のVHドメインのアミノ酸配列(配列番号104)を以下に示す(CDRH残基は下線を付して示されている):
QVQLVQSGAE VKKPGSSVKV SCKASGNSFS NHDVHWVRQA TGQGLEWMGW
MSHEGDKTGL AQKFQGRVTI TRDSGASTVY MELRGLTADD TAIYYCLTGS
KHRLRDYFLY NEYGPNYEEW GDYLATLDVW GHGTAVTVSS
QVQLVQSGAE VKKPGSSVKV SCKASGNSFS NHDVHWVRQA TGQGLEWMGW
MSHEGDKTGL AQKFQGRVTI TRDSGASTVY MELRGLTADD TAIYYCLTGS
KHRLRDYFLY NEYGPNYEEW GDYLATLDVW GHGTAVTVSS
PGT145のVLドメインのアミノ酸配列(配列番号105)を以下に示す(CDRL残基は下線を付して示されている):
EVVITQSPLF LPVTPGEAAS LSCKCSHSLQ HSTGANYLAW YLQRPGQTPR
LLIHLATHRA SGVPDRFSGS GSGTDFTLKI SRVESDDVGT YYCMQGLHSP
WTFGQGTKVE IK
EVVITQSPLF LPVTPGEAAS LSCKCSHSLQ HSTGANYLAW YLQRPGQTPR
LLIHLATHRA SGVPDRFSGS GSGTDFTLKI SRVESDDVGT YYCMQGLHSP
WTFGQGTKVE IK
モノクローナル抗体VRC01は、gp120のCD4結合部位を対象とする、幅広い中和性を有する抗HIV‐1抗体である(Wu, X. et al. (2010) “Rational Design Of Envelope Identifies Broadly Neutralizing Human Monoclonal Antibodies To HIV-1,” Science 329:856-861 and Zhou, T. et al. (2010) “Structural Basis For Broad And Potent Neutralization Of HIV-1 By Antibody VRC01,” Science 329:811-817; 国際公開第2013/163427号)。
VRC01のVHドメインのアミノ酸配列(配列番号106)を以下に示す(CDRH残基は下線を付して示されている):
QVQLVQSGGQ MKKPGESMRI SCRASGYEFI DCTLNWIRLA PGKRPEWMGW
LKPRGGAVNY ARPLQGRVTM TRDVYSDTAF LELRSLTVDD TAVYFCTRGK
NCDYNWDFEH WGRGTPVIVS S
QVQLVQSGGQ MKKPGESMRI SCRASGYEFI DCTLNWIRLA PGKRPEWMGW
LKPRGGAVNY ARPLQGRVTM TRDVYSDTAF LELRSLTVDD TAVYFCTRGK
NCDYNWDFEH WGRGTPVIVS S
VRC01のVLドメインのアミノ酸配列(配列番号107)を以下に示す(CDRL残基は下線を付して示されている):
EIVLTQSPGT LSLSPGETAI ISCRTSQYGS LAWYQQRPGQ APRLVIYSGS
TRAAGIPDRF SGSRWGPDYN LTISNLESGD FGVYYCQQYE FFGQGTKVQV
DIKR
EIVLTQSPGT LSLSPGETAI ISCRTSQYGS LAWYQQRPGQ APRLVIYSGS
TRAAGIPDRF SGSRWGPDYN LTISNLESGD FGVYYCQQYE FFGQGTKVQV
DIKR
モノクローナル抗体10E8は、幅広い中和性を有するHIV‐1 gp41膜近位外部領域(membrane‐proximal external region:MPER)特異性抗体である(Huang,J.,et al. 2012. “Broad And Potent Neutralization Of HIV-1 By A gp41-Specific Human Antibody.” Nature. 491:406-12).
10E8のVHドメインのアミノ酸配列(配列番号108)を以下に示す(CDRH残基は下線を付して示されている):
EVQLVESGGG LVKPGGSLRL SCSASGFDFD NAWMTWVRQP PGKGLEWVGR
ITGPGEGWSV DYAAPVEGRF TISRLNSINF LYLEMNNLRM EDSGLYFCAR
TGKYYDFWSG YPPGEEYFQD WGRGTLVTVS S
EVQLVESGGG LVKPGGSLRL SCSASGFDFD NAWMTWVRQP PGKGLEWVGR
ITGPGEGWSV DYAAPVEGRF TISRLNSINF LYLEMNNLRM EDSGLYFCAR
TGKYYDFWSG YPPGEEYFQD WGRGTLVTVS S
10E8のVLドメインのアミノ酸配列(配列番号109)を以下に示す(CDRL残基は下線を付して示されている):
SYELTQETGV SVALGRTVTI TCRGDSLRSH YASWYQKKPG QAPILLFYGK
NNRPSGVPDR FSGSASGNRA SLTISGAQAE DDAEYYCSSR DKSGSRLSVF
GGGTKLTVL
SYELTQETGV SVALGRTVTI TCRGDSLRSH YASWYQKKPG QAPILLFYGK
NNRPSGVPDR FSGSASGNRA SLTISGAQAE DDAEYYCSSR DKSGSRLSVF
GGGTKLTVL
本発明は具体的には、上述の抗HIV‐1モノクローナル抗体の、VL及び/若しくはVHドメイン、並びに/又はVL領域のCDRLのうちの1つ、2つ、若しくは3つ、及び/若しくはVHドメインのCDRHのうちの1つ、2つ、若しくは3つを含む、多重特異性gp41結合分子を含み、またこれを包含する。
VII.例示的な多重特異性gp41結合分子
ここで提供されるのは、gp41のエピトープに対して免疫特異的である第1のエピトープ結合ドメイン(即ちgp41結合ドメイン)と、ECMのエピトープに対して免疫特異的である第2のエピトープ結合ドメインとが組み込まれた、一連の例示的なgp41結合分子である。任意に、このような分子には、gp41の異なるエピトープ、並びに/又は異なるHIV‐1分子のエピトープ、並びに/又はECMの異なるエピトープ及び/若しくは異なるECMのエピトープに対して免疫特異的である、第3のエピトープ結合ドメイン(又は第3のエピトープ結合ドメイン及び第4のエピトープ結合ドメイン)が組み込まれる。特に好ましいのは、7B2GLの最適化済みgp41結合ドメインを含むこのような分子である。このような例示的なgp41結合分子の共有ダイアボディ構造及び配列を、表7にまとめ、以下で詳細に説明する。認識されるように、類似の分子は、(これらの例示的なgp41結合分子中に使用されているVL及びVHドメインの代わりに所望の抗体のVL及びVHドメインを使用することによって)同様に構築できる。
ここで提供されるのは、gp41のエピトープに対して免疫特異的である第1のエピトープ結合ドメイン(即ちgp41結合ドメイン)と、ECMのエピトープに対して免疫特異的である第2のエピトープ結合ドメインとが組み込まれた、一連の例示的なgp41結合分子である。任意に、このような分子には、gp41の異なるエピトープ、並びに/又は異なるHIV‐1分子のエピトープ、並びに/又はECMの異なるエピトープ及び/若しくは異なるECMのエピトープに対して免疫特異的である、第3のエピトープ結合ドメイン(又は第3のエピトープ結合ドメイン及び第4のエピトープ結合ドメイン)が組み込まれる。特に好ましいのは、7B2GLの最適化済みgp41結合ドメインを含むこのような分子である。このような例示的なgp41結合分子の共有ダイアボディ構造及び配列を、表7にまとめ、以下で詳細に説明する。認識されるように、類似の分子は、(これらの例示的なgp41結合分子中に使用されているVL及びVHドメインの代わりに所望の抗体のVL及びVHドメインを使用することによって)同様に構築できる。
A.gp41×CD3結合分子、DART‐1
「DART‐1」と呼称されるgp41×CD3結合分子が、第1の例示的な二重特異性gp41結合分子である。DART‐1は、gp41及びCD3抗原に結合できるFcドメイン含有二重特異性ダイアボディである。DART‐1は3つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2のVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、CD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。上記3つのポリペプチド鎖は、連結して、gp41のエピトープ及びCD3抗原のエピトープに免疫特異的に結合できる共有結合ダイアボディを形成する(例えば図4Aを参照)。
「DART‐1」と呼称されるgp41×CD3結合分子が、第1の例示的な二重特異性gp41結合分子である。DART‐1は、gp41及びCD3抗原に結合できるFcドメイン含有二重特異性ダイアボディである。DART‐1は3つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2のVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、CD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。上記3つのポリペプチド鎖は、連結して、gp41のエピトープ及びCD3抗原のエピトープに免疫特異的に結合できる共有結合ダイアボディを形成する(例えば図4Aを参照)。
DART‐1の第1のポリペプチド鎖は、配列番号110:
のアミノ酸配列を有する。
DART‐1の第1のポリペプチド鎖(配列番号110)の残基1〜113は、7B2のVLドメイン(配列番号55)に対応する。第1のポリペプチド鎖の残基114〜121(二重下線部)は、リンカー1(GGGSGGGG;配列番号16)に対応する。第1のポリペプチド鎖の残基122〜246は、CD3 mAb 1(D65G)のVHドメイン(配列番号63)に対応する。残基247〜251は、リンカー(配列番号21、下線部)に対応する。第1のポリペプチド鎖の残基252〜279は、システイン含有Eコイル(配列番号31)に対応する。第1のポリペプチド鎖の残基280〜292は、リンカー(配列番号40)に対応する。第1のポリペプチド鎖の残基293〜509は、「ノブ担持」ドメイン(配列番号48)に対応し、その最後の残基はリシンである。
DART‐1の第2のポリペプチド鎖は、配列番号111:
のアミノ酸配列を有する。
DART‐1の第2のポリペプチド鎖(配列番号111)の残基1〜110は、CD3 mAb 1のVLドメイン(配列番号62)に対応する。第2のポリペプチド鎖の残基111〜118(二重下線部)は、リンカー1(GGGSGGGG;配列番号16)に対応する。第2のポリペプチド鎖の残基119〜244は、7B2のVHドメイン(配列番号56)に対応する。第2のポリペプチド鎖の残基245〜249は、リンカー(配列番号21、下線部)に対応する。第2のポリペプチド鎖の残基250〜277は、システイン含有Kコイル(配列番号32)に対応する。
DART‐1の第3のポリペプチド鎖は、配列番号112:
DKTHTCPPCP APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED
PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK
CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK
GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG
NVFSCSVMHE ALHNRYTQKS LSLSPGK
のアミノ酸配列を有する。
DKTHTCPPCP APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED
PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK
CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK
GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG
NVFSCSVMHE ALHNRYTQKS LSLSPGK
のアミノ酸配列を有する。
DART‐1の第3のポリペプチド鎖(配列番号112)の残基1〜10は、リンカー(配列番号40)に対応する。第3のポリペプチド鎖の残基11〜227は、「ホール担持」IgG1 CH2‐CH3ドメイン(配列番号50)に対応し、H435R置換(下線を付して示されている)を含有し、その最後の残基はリシンである。上述のように、H435R置換は、上記分子の、タンパク質Aに結合する能力を排除する。
認識されるように、DART‐1の第3のポリペプチド鎖は、いずれのエピトープ結合部位を含有せず、よって、図4A〜4Bで提供されている一般構造を有する様々なgp41結合分子中で使用できる。従ってDART‐1の第3のポリペプチド鎖は、本明細書では「共通のダイアボディポリペプチド鎖(Common Diabody Polypeptide Chain)」と呼ばれる。
B.gp41×CD3結合分子、DART‐A
「DART‐A」と呼称される第2の例示的なgp41×CD3結合分子は、上述のDART‐1と同様であるが、親7B2 VL及びVHドメインの代わりに抗ヒトgp41抗体7B2GLのVL及びVHドメインを含有する。
「DART‐A」と呼称される第2の例示的なgp41×CD3結合分子は、上述のDART‐1と同様であるが、親7B2 VL及びVHドメインの代わりに抗ヒトgp41抗体7B2GLのVL及びVHドメインを含有する。
DART‐Aの第1のポリペプチド鎖は、配列番号113:
のアミノ酸配列を有する。
DART‐Aの第1のポリペプチド鎖(配列番号113)の残基1〜113は、7B2GLのVLドメイン(配列番号57)に対応する。第1のポリペプチド鎖の残基114〜121(二重下線部)は、リンカー1(GGGSGGGG;配列番号16)に対応する。第1のポリペプチド鎖の残基122〜246は、CD3 mAb 1(D65G)のVHドメイン(配列番号63)に対応する。残基247〜251は、リンカー(配列番号21、下線部)に対応する。第1のポリペプチド鎖の残基252〜279は、システイン含有Eコイル(配列番号31)に対応する。第1のポリペプチド鎖の残基280〜292は、リンカー(配列番号40)に対応する。第1のポリペプチド鎖の残基293〜509は、「ノブ担持」ドメイン(配列番号48)に対応し、その最後の残基はリシンである。
DART‐Aの第2のポリペプチド鎖は、配列番号114:
のアミノ酸配列を有する。
DART‐Aの第2のポリペプチド鎖(配列番号114)の残基1〜110は、CD3 mAb 1のVLドメイン(配列番号62)に対応する。第2のポリペプチド鎖の残基111〜118(二重下線部)は、リンカー1(GGGSGGGG;配列番号16)に対応する。第2のポリペプチド鎖の残基119〜244は、7B2GLのVHドメイン(配列番号58)に対応する。第2のポリペプチド鎖の残基245〜249は、リンカー(配列番号21、下線部)に対応する。第2のポリペプチド鎖の残基250〜277は、システイン含有Kコイル(配列番号32)に対応する。
DART‐Aの第3のポリペプチド鎖のアミノ酸配列は、DART‐1の第3のポリペプチド鎖(即ち配列番号112)のアミノ酸配列と同一である。
C.gp41×CD16結合分子、DART‐B
「DART‐B」と呼称されるgp41×CD16結合分子が、第3の例示的な二重特異性gp41結合分子である。DART‐Bは、上述のDART‐Aと同様であるが、DART‐AのCD3結合ドメインの代わりにCD16結合ドメインを含む。よってDART‐Bは3つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、抗ヒトCD16抗体3G8のVL及びVHドメインを含む(従ってCD16抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
「DART‐B」と呼称されるgp41×CD16結合分子が、第3の例示的な二重特異性gp41結合分子である。DART‐Bは、上述のDART‐Aと同様であるが、DART‐AのCD3結合ドメインの代わりにCD16結合ドメインを含む。よってDART‐Bは3つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、抗ヒトCD16抗体3G8のVL及びVHドメインを含む(従ってCD16抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
DART‐Bの第1のポリペプチド鎖は、配列番号115:
のアミノ酸配列を有する。
DART‐Bの第1のポリペプチド鎖(配列番号115)の残基1〜113は、7B2GLのVLドメイン(配列番号57)に対応する。第1のポリペプチド鎖の残基114〜121(二重下線部)は、リンカー1(GGGSGGGG;配列番号16)に対応する。第1のポリペプチド鎖の残基122〜239は、h3G8のVHドメイン(配列番号74)に対応する。残基240〜244は、リンカー(配列番号21、下線部)に対応する。第1のポリペプチド鎖の残基245〜272は、システイン含有Eコイル(配列番号31)に対応する。第1のポリペプチド鎖の残基273〜285は、リンカー(配列番号40)に対応する。第1のポリペプチド鎖の残基286〜502は、「ノブ担持」ドメイン(配列番号48)に対応し、その最後の残基はリシンである。
DART‐Bの第2のポリペプチド鎖は、配列番号116:
のアミノ酸配列を有する。
DART‐Bの第2のポリペプチド鎖(配列番号116)の残基1〜111はh3G8のVLドメイン(配列番号75)に対応する。第2のポリペプチド鎖の残基112〜119(二重下線部)は、リンカー1(GGGSGGGG;配列番号16)に対応する。第2のポリペプチド鎖の残基120〜245は、7B2GLのVHドメイン(配列番号58)に対応する。第2のポリペプチド鎖の残基246〜250は、リンカー(配列番号21、下線部)に対応する。第2のポリペプチド鎖の残基251〜278は、システイン含有Kコイル(配列番号32)に対応する。
DART‐Bの第3のポリペプチド鎖のアミノ酸配列は、DART‐1の第3のポリペプチド鎖(即ち配列番号112)のアミノ酸配列と同一である。
D.gp41×CD3×CD8結合分子、TRIDENT‐A
「TRIDENT‐A」と呼称されるgp41×CD3×CD8結合分子が、第1の例示的な3価gp41結合分子である。TRIDENT‐Aは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、CD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインと、TRX8のVL及びVHドメインを含む(従ってCD8抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。これら4つのポリペプチド鎖は連結して、gp41のエピトープ、CD3抗原のエピトープ、及びCD8抗原のエピトープに免疫特異的に結合できる共有結合3化分子を形成する(例えば図6Aを参照)。
「TRIDENT‐A」と呼称されるgp41×CD3×CD8結合分子が、第1の例示的な3価gp41結合分子である。TRIDENT‐Aは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、CD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインと、TRX8のVL及びVHドメインを含む(従ってCD8抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。これら4つのポリペプチド鎖は連結して、gp41のエピトープ、CD3抗原のエピトープ、及びCD8抗原のエピトープに免疫特異的に結合できる共有結合3化分子を形成する(例えば図6Aを参照)。
TRIDENT‐Aの第1のポリペプチド鎖のアミノ酸配列は、上述のDART‐Aダイアボディの第1のポリペプチド鎖のアミノ酸配列(配列番号113)と同一である。同様に、TRIDENT‐Aの第2のポリペプチド鎖のアミノ酸配列は、上述のDART‐Aダイアボディの第2のポリペプチド鎖のアミノ酸配列(配列番号114)と同一である。
TRIDENT‐Aの第3のポリペプチド鎖は、配列番号117:
QVQLVESGGG VVQPGRSLRL SCAASGFTFS DFGMNWVRQA PGKGLEWVAL
IYYDGSNKFY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKPH
YDGYYHFFDS WGQGTLVTVS SASTKGPSVF PLAPSSKSTS GGTAALGCLV
KDYFPEPVTV SWNSGALTSG VHTFPAVLQS SGLYSLSSVV TVPSSSLGTQ
TYICNVNHKP SNTKVDKRVE PKSCDKTHTC PPCPAPEAAG GPSVFLFPPK
PKDTLMISRT PEVTCVVVDV SHEDPEVKFN WYVDGVEVHN AKTKPREEQY
NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK ALPAPIEKTI SKAKGQPREP
QVYTLPPSRE EMTKNQVSLS CAVKGFYPSD IAVEWESNGQ PENNYKTTPP
VLDSDGSFFL VSKLTVDKSR WQQGNVFSCS VMHEALHNRY TQKSLSLSPG
K
のアミノ酸配列を有する。
QVQLVESGGG VVQPGRSLRL SCAASGFTFS DFGMNWVRQA PGKGLEWVAL
IYYDGSNKFY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKPH
YDGYYHFFDS WGQGTLVTVS SASTKGPSVF PLAPSSKSTS GGTAALGCLV
KDYFPEPVTV SWNSGALTSG VHTFPAVLQS SGLYSLSSVV TVPSSSLGTQ
TYICNVNHKP SNTKVDKRVE PKSCDKTHTC PPCPAPEAAG GPSVFLFPPK
PKDTLMISRT PEVTCVVVDV SHEDPEVKFN WYVDGVEVHN AKTKPREEQY
NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK ALPAPIEKTI SKAKGQPREP
QVYTLPPSRE EMTKNQVSLS CAVKGFYPSD IAVEWESNGQ PENNYKTTPP
VLDSDGSFFL VSKLTVDKSR WQQGNVFSCS VMHEALHNRY TQKSLSLSPG
K
のアミノ酸配列を有する。
TRIDENT‐Aの第3のポリペプチド鎖の残基1〜121は、抗CD8抗体TRX2のVHドメイン(配列番号70)に対応する。残基121〜219は、IgG1 CH1ドメイン(配列番号3)に対応する。残基220〜234は、IgG1ヒンジドメイン(配列番号7)に対応する。残基235〜451は、IgG1「ホール担持」CH2‐CH3ドメイン(配列番号50)に対応する。
TRIDENT‐Aの第4のポリペプチド鎖は、配列番号118:
DIQMTQSPSS LSASVGDRVT ITCKGSQDIN NYLAWYQQKP GKAPKLLIYN
TDILHTGVPS RFSGSGSGTD FTFTISSLQP EDIATYYCYQ YNNGYTFGQG
TKVEIKRTVA APSVFIFPPS DEQLKSGTAS VVCLLNNFYP REAKVQWKVD
NALQSGNSQE SVTEQDSKDS TYSLSSTLTL SKADYEKHKV YACEVTHQGL
SSPVTKSFNR GEC
のアミノ酸配列を有する。
DIQMTQSPSS LSASVGDRVT ITCKGSQDIN NYLAWYQQKP GKAPKLLIYN
TDILHTGVPS RFSGSGSGTD FTFTISSLQP EDIATYYCYQ YNNGYTFGQG
TKVEIKRTVA APSVFIFPPS DEQLKSGTAS VVCLLNNFYP REAKVQWKVD
NALQSGNSQE SVTEQDSKDS TYSLSSTLTL SKADYEKHKV YACEVTHQGL
SSPVTKSFNR GEC
のアミノ酸配列を有する。
TRIDENT‐Aの第4のポリペプチド鎖の残基1〜106は、抗CD8抗体TRX2のVLドメイン(配列番号71)に対応する。残基107〜213は、CLκドメイン(配列番号1)に対応する。
E.gp41×CD3×gp120結合分子、TRIDENT‐B
「TRIDENT‐B」と呼称される第2の例示的な3価gp41×CD3×gp120結合分子は、上述のTRIDENT‐Aと同様であるが、TRIDENT‐AのTRX8 VL及びVHドメインの代わりに、抗ヒトgp120抗体A32のVL及びVHドメインを含有する。従ってTRIDENT‐Bは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、CD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインと、A32のVL及びVHドメインを含む(従ってgp120抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
「TRIDENT‐B」と呼称される第2の例示的な3価gp41×CD3×gp120結合分子は、上述のTRIDENT‐Aと同様であるが、TRIDENT‐AのTRX8 VL及びVHドメインの代わりに、抗ヒトgp120抗体A32のVL及びVHドメインを含有する。従ってTRIDENT‐Bは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、CD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインと、A32のVL及びVHドメインを含む(従ってgp120抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
TRIDENT‐Bの第1のポリペプチド鎖のアミノ酸配列は、上述のDART‐Aダイアボディの第1のポリペプチド鎖のアミノ酸配列(配列番号113)と同一である。同様に、TRIDENT‐Bの第2のポリペプチド鎖のアミノ酸配列は、上述のDART‐Aダイアボディの第2のポリペプチド鎖のアミノ酸配列(配列番号114)と同一である。
TRIDENT‐Bの第3のポリペプチド鎖は、配列番号119:
QVQLQESGPG LVKPSQTLSL SCTVSGGSSS SGAHYWSWIR QYPGKGLEWI
GYIHYSGNTY YNPSLKSRIT ISQHTSENQF SLKLNSVTVA DTAVYYCARG
TRLRTLRNAF DIWGQGTLVT VSSASTKGPS VFPLAPSSKS TSGGTAALGC
LVKDYFPEPV TVSWNSGALT SGVHTFPAVL QSSGLYSLSS VVTVPSSSLG
TQTYICNVNH KPSNTKVDKR VEPKSCDKTH TCPPCPAPEA AGGPSVFLFP
PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE
QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR
EPQVYTLPPS REEMTKNQVS LSCAVKGFYP SDIAVEWESN GQPENNYKTT
PPVLDSDGSF FLVSKLTVDK SRWQQGNVFS CSVMHEALHN RYTQKSLSLS
PGK
のアミノ酸配列を有する。
QVQLQESGPG LVKPSQTLSL SCTVSGGSSS SGAHYWSWIR QYPGKGLEWI
GYIHYSGNTY YNPSLKSRIT ISQHTSENQF SLKLNSVTVA DTAVYYCARG
TRLRTLRNAF DIWGQGTLVT VSSASTKGPS VFPLAPSSKS TSGGTAALGC
LVKDYFPEPV TVSWNSGALT SGVHTFPAVL QSSGLYSLSS VVTVPSSSLG
TQTYICNVNH KPSNTKVDKR VEPKSCDKTH TCPPCPAPEA AGGPSVFLFP
PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE
QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR
EPQVYTLPPS REEMTKNQVS LSCAVKGFYP SDIAVEWESN GQPENNYKTT
PPVLDSDGSF FLVSKLTVDK SRWQQGNVFS CSVMHEALHN RYTQKSLSLS
PGK
のアミノ酸配列を有する。
TRIDENT‐Bの第3のポリペプチド鎖の残基1〜123は、抗gp120抗体A32のVHドメイン(配列番号96)に対応する。残基124〜221は、IgG1 CH1ドメイン(配列番号3)に対応する。残基222〜236は、IgG1ヒンジドメイン(配列番号7)に対応する。残基237〜453は、IgG1「ホール担持」CH2‐CH3ドメイン(配列番号50)に対応する。
TRIDENT‐Bの第4のポリペプチド鎖は、配列番号120:
QSALTQPPSA SGSPGQSVTI SCTGTSSDVG GYNYVSWYQH HPGKAPKLII
SEVNNRPSGV PDRFSGSKSG NTASLTVSGL QAEDEAEYYC SSYTDIHNFV
FGGGTKLTVL RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ
WKVDNALQSG NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT
HQGLSSPVTK SFNRGEC
のアミノ酸配列を有する。
QSALTQPPSA SGSPGQSVTI SCTGTSSDVG GYNYVSWYQH HPGKAPKLII
SEVNNRPSGV PDRFSGSKSG NTASLTVSGL QAEDEAEYYC SSYTDIHNFV
FGGGTKLTVL RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ
WKVDNALQSG NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT
HQGLSSPVTK SFNRGEC
のアミノ酸配列を有する。
TRIDENT‐Bの第4のポリペプチド鎖の残基1〜110は、抗gp120抗体A32のVLドメイン(配列番号97)に対応する。残基111〜217は、CLκドメイン(配列番号1)に対応する。
F.gp41×CD16×gp120結合分子、TRIDENT‐C
「TRIDENT‐C」と呼称される第3の例示的な3価gp41×CD16×gp120結合分子は、上述のTRIDENT‐Bと同様であるが、TRIDENT‐BのCD3 mAb 1(D65G) VL及びVHドメインの代わりに、抗ヒトCD16抗体h3G8のVL及びVHドメインを含有する。従ってTRIDENT‐Cは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、h3G8のVL及びVHドメインを含む(従ってCD16抗原のエピトープに対して免疫特異的である)1つの結合ドメインと、A32のVL及びVHドメインを含む(従ってgp120抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
「TRIDENT‐C」と呼称される第3の例示的な3価gp41×CD16×gp120結合分子は、上述のTRIDENT‐Bと同様であるが、TRIDENT‐BのCD3 mAb 1(D65G) VL及びVHドメインの代わりに、抗ヒトCD16抗体h3G8のVL及びVHドメインを含有する。従ってTRIDENT‐Cは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)1つの結合ドメインと、h3G8のVL及びVHドメインを含む(従ってCD16抗原のエピトープに対して免疫特異的である)1つの結合ドメインと、A32のVL及びVHドメインを含む(従ってgp120抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
TRIDENT‐Cの第1のポリペプチド鎖のアミノ酸配列は、上述のDART‐Bダイアボディの第1のポリペプチド鎖のアミノ酸配列(配列番号115)と同一である。同様に、TRIDENT‐Cの第2のポリペプチド鎖のアミノ酸配列は、上述のDART‐Bダイアボディの第2のポリペプチド鎖のアミノ酸配列(配列番号116)と同一である。
TRIDENT‐Cの第3及び第4のポリペプチドのアミノ酸配列は、上述のTRIDENT‐Bの第3及び第4のポリペプチドのアミノ酸配列(配列番号119及び配列番号120)と同一である。
G.gp41×CD3×gp41結合分子、TRIDENT‐D
「TRIDENT‐D」と呼称される第4の例示的な3価gp41×CD3×gp41結合分子は、上述のTRIDENT‐Aと同様であるが、TRIDENT‐AのTRX2抗CD8 VL及びVHドメインの代わりに、抗ヒトgp41抗体7B2GLのVL及びVHドメインを含有する。従ってTRIDENT‐Dは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)2つの結合ドメインと、hCD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
「TRIDENT‐D」と呼称される第4の例示的な3価gp41×CD3×gp41結合分子は、上述のTRIDENT‐Aと同様であるが、TRIDENT‐AのTRX2抗CD8 VL及びVHドメインの代わりに、抗ヒトgp41抗体7B2GLのVL及びVHドメインを含有する。従ってTRIDENT‐Dは4つのポリペプチド鎖からなり、また抗ヒトgp41抗体7B2GLのVL及びVHドメインを含む(従ってgp41のエピトープに対して免疫特異的である)2つの結合ドメインと、hCD3 mAb 1のVL及びVHドメインを含む(従ってCD3抗原のエピトープに対して免疫特異的である)1つの結合ドメインとを有する。
TRIDENT‐Dの第1のポリペプチド鎖のアミノ酸配列は、上述のDART‐Aダイアボディの第1のポリペプチド鎖のアミノ酸配列(配列番号113)と同一である。同様に、TRIDENT‐Dの第2のポリペプチド鎖のアミノ酸配列は、上述のDART‐Aダイアボディの第2のポリペプチド鎖のアミノ酸配列(配列番号114)と同一である。
TRIDENT‐Dの第3のポリペプチド鎖は、配列番号121:
QVQLVESGGG LVKPGGSLRL SCAASGFTFS EYYMTWIRQA PGKGLEWVSY
ISKNGEYSKY SPSSNGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCARAD
GLTYFSELLQ YIFDLWGQGT LVTVSSASTK GPSVFPLAPS SKSTSGGTAA
LGCLVKDYFP EPVTVSWNSG ALTSGVHTFP AVLQSSGLYS LSSVVTVPSS
SLGTQTYICN VNHKPSNTKV DKRVEPKSCD KTHTCPPCPA PEAAGGPSVF
LFPPKPKDTL MISRTPEVTC VVVDVSHEDP EVKFNWYVDG VEVHNAKTKP
REEQYNSTYR VVSVLTVLHQ DWLNGKEYKC KVSNKALPAP IEKTISKAKG
QPREPQVYTL PPSREEMTKN QVSLSCAVKG FYPSDIAVEW ESNGQPENNY
KTTPPVLDSD GSFFLVSKLT VDKSRWQQGN VFSCSVMHEA LHNRYTQKSL
SLSPGK
のアミノ酸配列を有する。
QVQLVESGGG LVKPGGSLRL SCAASGFTFS EYYMTWIRQA PGKGLEWVSY
ISKNGEYSKY SPSSNGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCARAD
GLTYFSELLQ YIFDLWGQGT LVTVSSASTK GPSVFPLAPS SKSTSGGTAA
LGCLVKDYFP EPVTVSWNSG ALTSGVHTFP AVLQSSGLYS LSSVVTVPSS
SLGTQTYICN VNHKPSNTKV DKRVEPKSCD KTHTCPPCPA PEAAGGPSVF
LFPPKPKDTL MISRTPEVTC VVVDVSHEDP EVKFNWYVDG VEVHNAKTKP
REEQYNSTYR VVSVLTVLHQ DWLNGKEYKC KVSNKALPAP IEKTISKAKG
QPREPQVYTL PPSREEMTKN QVSLSCAVKG FYPSDIAVEW ESNGQPENNY
KTTPPVLDSD GSFFLVSKLT VDKSRWQQGN VFSCSVMHEA LHNRYTQKSL
SLSPGK
のアミノ酸配列を有する。
TRIDENT‐Dの第3のポリペプチド鎖の残基1〜126は、最適化済み抗gp41抗体7B2GLのVHドメイン(配列番号58)に対応する。残基127〜224は、IgG1 CH1ドメイン(配列番号3)に対応する。残基225〜239は、IgG1ヒンジドメイン(配列番号7)に対応する。残基240〜456は、IgG1「ホール担持」CH2‐CH3ドメイン(配列番号50)に対応する。
TRIDENT‐Dの第4のポリペプチド鎖は、配列番号122:
DIVMTQSPDS LAVSLGERAT INCKSSQTLL YSSNNRHSIA WYQQKPGQPP
KLLIYWASMR LSGVPDRFSG SGSGTDFTLT ISSLQAEDVA VYYCHQYSSH
PPTFGQGTKV EIKRTVAAPS VFIFPPSDEQ LKSGTASVVC LLNNFYPREA
KVQWKVDNAL QSGNSQESVT EQDSKDSTYS LSSTLTLSKA DYEKHKVYAC
EVTHQGLSSP VTKSFNRGEC
のアミノ酸配列を有する。
DIVMTQSPDS LAVSLGERAT INCKSSQTLL YSSNNRHSIA WYQQKPGQPP
KLLIYWASMR LSGVPDRFSG SGSGTDFTLT ISSLQAEDVA VYYCHQYSSH
PPTFGQGTKV EIKRTVAAPS VFIFPPSDEQ LKSGTASVVC LLNNFYPREA
KVQWKVDNAL QSGNSQESVT EQDSKDSTYS LSSTLTLSKA DYEKHKVYAC
EVTHQGLSSP VTKSFNRGEC
のアミノ酸配列を有する。
TRIDENT‐Dの第4のポリペプチド鎖の残基1〜113は、抗gp41抗体7B2GLのVLドメイン(配列番号57)に対応する。残基114〜220は、CLκドメイン(配列番号1)に対応する。
H.代替的な多重特異性gp41結合分子
本開示に鑑みて認識されるように、上述の分子中に存在するVL及びVHドメインの代わりに、代替的なECM及び/又はHIV‐1抗原のVL及びVHドメインを採用することによって、7B2GLのVL及びVHドメインを含み、かつ代替的なECM及び/又はHIV‐1抗原のための結合部位を含む、上述の例示的な分子のうちのいずれの一般構造を有する更なる多重特異性gp41結合分子を構築できる。数多くのECM及びHIV‐1結合部位に由来するVL及びVHドメインが本明細書中に提供されており、更なるVL及びVHドメインが当該技術分野において公知である。同様に、本明細書中で提供されているように、特に本明細書中で提供されている代替的なリンカー及び/又はヘテロ二量体促進ドメインを組み込んだ、代替的な多重特異性gp41結合分子も、同様に構築できる。
本開示に鑑みて認識されるように、上述の分子中に存在するVL及びVHドメインの代わりに、代替的なECM及び/又はHIV‐1抗原のVL及びVHドメインを採用することによって、7B2GLのVL及びVHドメインを含み、かつ代替的なECM及び/又はHIV‐1抗原のための結合部位を含む、上述の例示的な分子のうちのいずれの一般構造を有する更なる多重特異性gp41結合分子を構築できる。数多くのECM及びHIV‐1結合部位に由来するVL及びVHドメインが本明細書中に提供されており、更なるVL及びVHドメインが当該技術分野において公知である。同様に、本明細書中で提供されているように、特に本明細書中で提供されている代替的なリンカー及び/又はヘテロ二量体促進ドメインを組み込んだ、代替的な多重特異性gp41結合分子も、同様に構築できる。
VIII.産生方法
最も好ましくは、本発明の分子は、当該技術分野において公知であるように、上記ポリペプチドをエンコードする核酸分子の組み換え発現によって産生される。
最も好ましくは、本発明の分子は、当該技術分野において公知であるように、上記ポリペプチドをエンコードする核酸分子の組み換え発現によって産生される。
本発明のポリペプチドは、固相ペプチド合成を用いて便利に調製できる(Merrifield, B. (1986) “Solid Phase Synthesis,” Science 232(4748):341-347; Houghten, R.A. (1985) “General Method For The Rapid Solid-Phase Synthesis Of Large Numbers Of Peptides: Specificity Of Antigen-Antibody Interaction At The Level Of Individual Amino Acids,” Proc. Natl. Acad. Sci. (U.S.A.) 82(15): 5131-5135; Ganesan, A. (2006) “Solid-Phase Synthesis In The Twenty-First Century,” Mini Rev. Med. Chem. 6(1):3-10)。
当該技術分野において公知の方法を用いて、抗体を組み換えによって作製し、発現させてよい。抗体は、まず宿主動物から作製された抗体を単離し、そのDNA配列を取得し、上記DNA配列を使用して宿主細胞(例えばCHO細胞)内で抗体を組み換え発現させることによって作製できる。採用できる別の方法は、植物(例えばタバコ)又は遺伝子組み換え動物のミルク中で抗体配列を発現させることである。植物又はミルク中で抗体を組み換え発現させるための好適な方法は、開示されている(例えばPeeters et al. (2001) "Production Of Antibodies And Antibody Fragments In Plants," Vaccine 19:2756; Lonberg, N. et al. (1995) "Human Antibodies From Transgenic Mice," Int. Rev. Immunol 13:65-93; and Pollock et a/.(1999) "Transgenic Milk As A Method For The Production Of Recombinant Antibodies," J. Immunol Methods 231 : 147-157を参照)。例えばヒト化、単鎖等の抗体の誘導体を作製するための好適な方法は、当該技術分野において公知であり、また上述されている。別の代替案では、抗体はファージディスプレイ技術によって、組み換えによって作製できる(例えば米国特許第5,565,332号;米国特許第5,580,717号;米国特許第5,733,743号;米国特許第6,265,150号;及びWinter, G. et al. (1994) "Making Antibodies By Phage Display Technology," Annu. Rev. Immunol. 12.433-455を参照)。
関心対象のポリヌクレオチドを含有するベクター(例えば本発明の結合分子のポリペプチドをエンコードするポリヌクレオチド)は:電気穿孔;塩化カルシウム、塩化ルビジウム、リン酸カルシウム、DEAE‐デキストラン又は他の物質を使用したトランスフェクション;微粒子銃;リポフェクション;及び感染(例えばベクターがワクシニアウイルス等の感染性因子である場合)を含む多数の適切な手段のうちのいずれによって、宿主細胞に導入できる。ベクター又はポリヌクレオチドの導入の選択は、宿主細胞の特徴に左右される場合が多い。
ポリペプチド、又はタンパク質を発現する目的で、異種DNAを過剰発現できるいずれの宿主細胞を使用できる。好適な哺乳類宿主細胞の非限定的な例としては、COS、HeLa及びCHO細胞が挙げられるが、これらに限定されない。
本発明は、本発明の結合分子のアミノ酸配列を含むポリペプチドを含む。本発明のポリペプチドは、当該技術分野において公知の手順で産生できる。上記ポリペプチドは、抗体のタンパク質分解若しくは他の分解によって、上述のような組み換え法(即ち単一若しくは融合ポリペプチド)によって、又は化学的合成によって産生できる。上記抗体のポリペプチド、特にアミノ酸最高約50個分の比較的短いポリペプチドは、化学的合成によって便利に作製される。化学的合成方法は当該技術分野において公知であり、市販されている。
本発明は、本開示の結合分子(上記分子の特性に有意な影響を及ぼさない、機能的に同等のポリペプチド、及び活性が増強した又は低下した変異型を含む)を含んでよい。ポリペプチドの修飾は、当該技術分野において慣用的に実践されており、本明細書で詳細に説明する必要はない。修飾されたポリペプチドの例としては、アミノ酸残基の保存的置換を有するポリペプチド、機能的活性を大幅に又は劣化するように変化させない1つ以上の欠失若しくは追加、又は化学的類似体の使用が挙げられる。互いを保存的に置換できるアミノ酸残基としては:グリシン/アラニン;セリン/トレオニン;バリン/イソロイシン/ロイシン;アスパラギン/グルタミン;アスパラギン酸/グルタミン酸;リジン/アルギニン;及びフェニルアラニン/チロシンが挙げられるが、これらに限定されない。これらのポリペプチドとしては、グリコシル化及び非グリコシル化ポリペプチド、並びに例えば異なる複数の糖によるグリコシル化、アセチル化及びリン酸化といった、その他の変換後修飾を有するポリペプチドも挙げられる。好ましくは、アミノ酸置換は保存的であり、即ち置換されたアミノ酸は、オリジナルのアミノ酸と同様の、電荷、サイズ等の化学的特性を有する。このような保存的置換は当該技術分野において公知であり、その例は上述されている。アミノ酸修飾は、1つ以上のアミノ酸を変化させるか又は修飾することから、可変ドメイン等の領域の完全な再設計にまで及んでよい。可変ドメインの変化は、結合親和性及び/又は特異性を変化させる。他の修飾方法としては、酵素的手段、酸化的置換及びキレート化を含むがこれらに限定されない、当該技術分野において公知の連結技術の使用が挙げられる。修飾は例えば、イムノアッセイのための標識の付与、例えばラジオイムノアッセイのための放射性部分の付与等のために使用できる。修飾されたポリペプチドは、当該技術分野において確立された手順を用いて作製され、また当該技術分野において公知の標準的なアッセイを用いてスクリーニングできる。
一実施形態では、軽鎖、重鎖又は軽鎖及び重鎖の両方を含む融合ポリペプチドが提供される。別の実施形態では、融合ポリペプチドは、異種免疫グロブリン定常領域を含有する。別の実施形態では、融合ポリペプチドは、公的に寄託されたハイブリドーマから産生された抗体のVH及びVLドメインを含有する。本発明の目的のために、抗体融合タンパク質は、該タンパク質をgp41及びECM(例えばCD3、CD16等)の両方に免疫特異的に結合できるようにし、かつ天然分子においては上記抗体融合タンパク質が結合しない別のアミノ酸配列、例えば異種配列又は別の領域(例えば脱免疫化アルブミン結合ドメイン、タンパク質A認識配列、ペプチドタグ等)由来の同種配列を含有する、ポリペプチドドメインを含有する。
本発明は特に、診断又は治療用部分にコンジュゲートする上記結合分子(例えば抗体、ダイアボディ、3価結合分子等)を包含する。診断を目的として、本発明の結合分子を、検出可能な物質と連結させてよい。このような結合分子は、臨床試験手順の一部としての疾患の発症又は進行の監視及び/又は予後判定、例えば特定の療法の有効性の決定に有用である。検出可能な物質の例としては、様々な酵素(例えば西洋ワサビペルオキシダーゼ、βガラクトシダーゼ等)、補欠分子族(例:アビジン/ビオチン)、蛍光物質(例えばアンベリフェロン、フルオレセイン又はフィコエリスリン)、発光材料(例えばルミノール)、生物発光材料(例えばルシフェラーゼ又はエクオリン)、放射性材料(例えば炭素14、マンガン54、ストロンチウム85又は亜鉛65)、陽電子放出金属、及び非放射性常磁性金属イオンが挙げられる。上記検出可能な物質は、直接的に、又は当該技術分野において公知の技法を用いて、介在物(例えばリンカー)を介して間接的に、結合分子と連結又はコンジュゲートさせてよい。
治療を目的として、本発明の結合分子を、細胞毒素等の治療成分(例えば細胞増殖抑制剤若しくは細胞破壊剤)、治療剤又はαエミッタ等の放射性金属イオンとコンジュゲートさせてよい。細胞毒素又は細胞傷害剤としては、シュードモナス外毒素、ジフテリア毒素、ボツリヌス毒素A〜F、リシンアブリン、サポリン、及びこれらの作用剤の細胞傷害性断片といった、細胞に有害ないずれの作用剤が挙げられる。治療剤としては、障害を予防的又は治療的に処置するための治療効果を有するいずれの作用剤が挙げられる。このような治療剤は、化学治療剤、タンパク質又はポリペプチド治療剤であってよく、所望の生物活性を有する、及び/又は所与の生物応答を修正する治療剤を含んでよい。治療剤の例としては、アルキル化剤、血管新生阻害剤、抗有糸分裂剤、ホルモン療法剤、及び細胞増殖性疾患の治療に有用な抗体が挙げられる。治療用部分は、直接的に、又は当該技術分野において公知の技法を用いて、介在物(例えばリンカー)を介して間接的に、結合分子と連結又はコンジュゲートさせてよい。
IX.本発明の結合分子の使用
上述のように、gp41及びECMに結合できる分子は、細胞表面上にこのようなgp41を発現する標的細胞(即ち病原体感染細胞)の標的転換殺滅を仲介できる。このような分子は、治療目的のために、例えば感染症、特に潜伏HIV‐1感染を有する被験者において使用できる。よって本発明の結合分子は、gp41の発現に関連する又は上記発現を特徴とする疾患又は状態、特に潜伏HIV‐1感染を治療する能力を有する。よって、限定するものではないが、本発明の結合分子は、HIV‐1感染、特に潜伏HIV‐1感染の治療に採用できる。
上述のように、gp41及びECMに結合できる分子は、細胞表面上にこのようなgp41を発現する標的細胞(即ち病原体感染細胞)の標的転換殺滅を仲介できる。このような分子は、治療目的のために、例えば感染症、特に潜伏HIV‐1感染を有する被験者において使用できる。よって本発明の結合分子は、gp41の発現に関連する又は上記発現を特徴とする疾患又は状態、特に潜伏HIV‐1感染を治療する能力を有する。よって、限定するものではないが、本発明の結合分子は、HIV‐1感染、特に潜伏HIV‐1感染の治療に採用できる。
特に本発明は、gp41に結合できる分子が、gp41に結合できる抗体のエピトープ結合ドメインを含み、かつ(標的転換細胞殺滅(例えば標的転換T細胞殺滅)を仲介することによって)gp41発現性標的細胞の標的転換殺滅を仲介するために、免疫エフェクタ細胞の表面上のECM(特にCD3及び/又はCD16)に結合できるエピトープ結合ドメインを含む、上記方法を包含する。
特定の態様では、本発明は、本発明のgp41結合分子、特に限定するものではないが二重特異性及び三重特異性分子(例えば二重特異性抗体、二重特異性ダイアボディ、3価結合分子等)といった多重特異性gp41結合分子の、ある個体のHIV‐1感染を治療及び予防する方法における使用を提供し、これは、上記個体に、治療有効量の本発明のgp41結合分子を含む組成物を、薬学的に許容可能な形態で投与するステップを含む。特定の実施形態では、上記gp41結合分子は、異なる複数のHIV‐1エピトープ、好ましくはHIV‐1エンベロープ上に存在する異なる複数のエピトープに結合する。
本明細書に記載の様々なgp41結合分子は、以下を含むがこれらに限定されない状況において、有用性を有する:
i)HIV‐1感染への予想される既知の曝露の状況において、本発明のgp41結合分子を殺菌剤として予防的に(例えばIV、局所、又は経鼻)投与できる;
ii)レイプ被害者若しくは商業的セックスワーカーの状況、又はコンドームによる保護のないいずれの同性愛者若しくは異性愛者間での伝染の状況といった、既知の又は疑わしい曝露の状況において、本発明のgp41結合分子を曝露後予防法として例えばIV又は局所投与できる;
iii)急性HIV‐1感染(AHI)の状況において、本発明のgp41結合分子を、初期ウイルス負荷の制御のため、又はウイルス感染CD4 T細胞の排除のために、AHIの治療として投与できる。
i)HIV‐1感染への予想される既知の曝露の状況において、本発明のgp41結合分子を殺菌剤として予防的に(例えばIV、局所、又は経鼻)投与できる;
ii)レイプ被害者若しくは商業的セックスワーカーの状況、又はコンドームによる保護のないいずれの同性愛者若しくは異性愛者間での伝染の状況といった、既知の又は疑わしい曝露の状況において、本発明のgp41結合分子を曝露後予防法として例えばIV又は局所投与できる;
iii)急性HIV‐1感染(AHI)の状況において、本発明のgp41結合分子を、初期ウイルス負荷の制御のため、又はウイルス感染CD4 T細胞の排除のために、AHIの治療として投与できる。
本発明によると、本発明のgp41結合分子は、被験者若しくは被験者の免疫系/細胞がHIV‐1に接触する前、又はこのような接触の約48時間以内に投与できる。この時間枠内での投与により、被験者の脆弱な細胞がHIVに感染するのを最大限阻害できる。
更に、様々な形態の本発明のgp41結合分子を、慢性又は急性HIV‐1感染被験者に投与でき、これを用いて残存するウイルス感染細胞を殺滅できる。これは、これらのgp41結合細胞がウイルス感染細胞の表面に結合して、上記感染細胞の標的転換殺滅を仲介できることによる。
特定の実施形態では、本発明のgp41結合分子は、潜伏感染活性化剤と組み合わせて投与でき、これにより、被験者の体内に存在し得るHIV感染細胞の潜伏リザーバを活性化できる。休止細胞中の潜伏プロウイルスHIV‐1 DNAを活性化させることによって、以前は不活性であった細胞が新たなウイルスの産生を開始し、これが、本発明のgp41結合分子によって増強された免疫系によって認識されて排除されることが期待される。潜伏感染活性化剤の非限定的な例は、HDAC阻害剤、例えばボリノスタット、ロミデプシン、パノビノスタット、ジスルフィラム、JQ1、ブリオスタチン、PMA、イオノマイシン、又はこれらのいずれの組み合わせである。Bullen et al. Nature Medicine 20、425-429 (2014)を参照。
特定の実施形態では、本発明のgp41結合分子は、抗レトロウイルス剤と組み合わせて投与できる。
ある特定の実施形態では、gp41及びECMに結合できる分子は、二重特異性抗体、BiTe(登録商標)、又はTandAb(登録商標)である。
ある特定の実施形態では、gp41及びECMに結合できる分子は、二重特異性ダイアボディである。
ある特定の実施形態では、gp41及びECMに結合できる分子は、3価結合分子である。
本明細書中で使用される場合、用語「療法を提供する(providing a therapy)」、及び「治療する(treating)」は:疾患に由来する症状の低減;感染症の症状(例えばウイルス量、発熱、疼痛、敗血症等)の軽減;疾患に由来する症状の低減;レシピエント被験者のQOLの向上;被験者の疾患の治療のために提供される他の薬物の用量の低減;標的化及び/若しくは内在化等による、別の薬物の効果の増強;疾患の進行の遅延:並びに/又はレシピエント被験者の生存期間の延長といったいずれの臨床的結果を含むがこれに限定されない、有益な又は所望の結果のいずれの兆候に関連する、組成物のいずれの投与を指す。
治療の被験者としては、動物、最も好ましくは非霊長類(例えばウシ、ウマ、ネコ、イヌ、げっ歯類)又は霊長類(例えばカニクイザル等のサル、ヒト等)といった哺乳類種が挙げられる。ある好ましい実施形態では、被験者はヒトである。
本発明の様々な実施形態によって治療できる例示的な障害としては、限定するものではないが、HIV‐1感染(特に標的転換細胞殺滅を仲介できる分子によって結合されるgp41の発現に関連する潜伏HIV‐1感染)が挙げられる。様々な実施形態では、本発明は、被験者の疾患又は障害の治療、予防又は管理のための方法及び組成物を包含し、これは、治療的有効量の本発明の結合分子を被験者に投与するステップを含む。このような分子は、HIV‐1負荷の低減、又はHIV感染細胞の排除のために特に有用である。特定の作用機序による束縛を意図したものではないが、上記分子は、標的細胞に対するエフェクタ機能を仲介してよく、標的細胞、架橋細胞表面抗原及び/又は標的細胞上の受容体に対する免疫系の活性化を促進してよく、またアポトーシス又は負の成長調節シグナリングを増強してよく、又はこれらの組み合わせを実現してよく、これにより、標的細胞の除去及び/又はその個数の低減をもたらす。
X.医薬組成物
本発明は、本発明のgp41結合分子を含む、組成物を包含する。本発明の組成物は、医薬組成物の製造に使用できるバルク薬剤組成物(例えば不純又は非滅菌組成物)、及び単位剤形の調製に使用できる医薬組成物(即ち被験者又は患者への投与に好適な組成物)を含む。これらの組成物は、予防的又は治療的有効量のgp41に結合でき、かつECMに結合できることにより、標的細胞(例えばHIV‐1感染細胞等)の標的転換殺滅を仲介できる分子(即ちgp41×ECM結合分子)、又はこれらの薬剤の組み合わせと、薬学的に許容可能なキャリアとを含む。好ましくは、本発明の組成物は、予防的又は治療的有効量の本発明の結合分子のうちの1つ又は複数と、薬学的に許容可能なキャリアとを含む。ある好ましい態様では、上記組成物は略精製済みである(即ち上記組成物の効果を制限する、又は望ましくない副作用を生成する物質を略含まない)。
本発明は、本発明のgp41結合分子を含む、組成物を包含する。本発明の組成物は、医薬組成物の製造に使用できるバルク薬剤組成物(例えば不純又は非滅菌組成物)、及び単位剤形の調製に使用できる医薬組成物(即ち被験者又は患者への投与に好適な組成物)を含む。これらの組成物は、予防的又は治療的有効量のgp41に結合でき、かつECMに結合できることにより、標的細胞(例えばHIV‐1感染細胞等)の標的転換殺滅を仲介できる分子(即ちgp41×ECM結合分子)、又はこれらの薬剤の組み合わせと、薬学的に許容可能なキャリアとを含む。好ましくは、本発明の組成物は、予防的又は治療的有効量の本発明の結合分子のうちの1つ又は複数と、薬学的に許容可能なキャリアとを含む。ある好ましい態様では、上記組成物は略精製済みである(即ち上記組成物の効果を制限する、又は望ましくない副作用を生成する物質を略含まない)。
このような組成物の様々な処方を、投与に用いてよい。1つ以上の薬理学的有効成分に加えて、本発明の組成物は、好適な、薬学的に許容可能なキャリアを含有してよく、上記キャリアは、当該技術分野において公知の賦形剤及び助剤を含み、また薬理学的に有効な物質の投与を促進する、又は上記活性化合物の、作用部位への送達のために薬学的に使用できる調製物への加工を促進する、比較的不活性の物質である。例えば賦形剤は、形状若しくは粘度を与えることができ、又は希釈剤として作用できる。好適な賦形剤としては、安定化剤、湿潤及び乳化剤、浸透圧を変化させるための塩、カプセル化剤、緩衝液、並びに皮膚浸透促進剤が挙げられるが、これらに限定されない。
ある具体的実施形態では、用語「薬学的に許容可能な(pharmaceutically acceptable)」は、動物、より詳細にはヒトにおける使用に関して、連邦規制当局若しくは州政府によって承認されている、又は米国薬局方若しくは他の一般に認められた薬局方に記載されていることを意味する。用語「キャリア(carrier)」は、希釈剤、アジュバント(例えばフロイントアジュバント(完全及び不完全))、賦形剤、又は治療薬の投与に用いられるビヒクルを指す。一般に、本発明の組成物の成分は、例えば活性作用剤の量を示すアンプル又はサシェ等の気密性コンテナ中の凍結乾燥粉末又は無水濃縮物として、別個に、又は単位剤形にまとめて混合された状態で供給される。組成物を点滴によって投与する場合、組成物は、滅菌された薬学的グレードの水又は食塩水を内包する点滴ボトルを用いて吐出できる。組成物を注射によって投与する場合、投与前に成分を混合できるように、注射用無菌水又は食塩水のアンプルを提供できる。
本発明はまた、単独の又は上述のような薬学的に許容可能なキャリアと組み合わされた本発明のgp41結合分子で充填された1つ以上のコンテナを含む、医薬パック又はキットも提供する。更に、疾患の治療に有用な1つ以上の他の予防剤又は治療剤も、上記医薬パック又はキットに含めることができる。本発明はまた、本発明の医薬組成物の成分のうちの1つ又は複数で充填された1つ以上のコンテナを含む、医薬パック又はキットも提供する。任意に、このような1つ以上のコンテナは、医薬製品又は生物学的製品の製造、使用又は販売を規制する政府機関によって規定された形式の通知と関連するものとすることができ、上記通知は、ヒトへの投与のための製造、使用又は販売の機関による承認を反映したものである。
本発明は、上述の方法において使用できるキットを提供する。キットは、本発明の結合分子のいずれを含むことができる。このキットは更に、癌の治療に有用な1つ以上の他の予防剤及び/又は治療剤を、1つ以上のコンテナ内に含むことができる
XI.投与方法
本発明の組成物は、有効量の本発明のgp41結合分子を含む医薬組成物を、被験者に投与することによる、疾患、障害又は感染症に関連する1つ以上の症状の治療、予防及び改善のために提供できる。ある好ましい態様では、上記組成物は実質的に精製される(即ち上記組成物の効果を制限する、又は望ましくない副作用を生成する物質を実質的に含まない)。ある具体的実施形態では、被験者は動物、好ましくは非霊長類(例えばウシ属、ウマ科、ネコ科、イヌ科、げっ歯類等)又は霊長類(例えばカニクイザル等のサル、ヒト等)といった哺乳類である。ある好ましい実施形態では、被験者はヒトである。
本発明の組成物は、有効量の本発明のgp41結合分子を含む医薬組成物を、被験者に投与することによる、疾患、障害又は感染症に関連する1つ以上の症状の治療、予防及び改善のために提供できる。ある好ましい態様では、上記組成物は実質的に精製される(即ち上記組成物の効果を制限する、又は望ましくない副作用を生成する物質を実質的に含まない)。ある具体的実施形態では、被験者は動物、好ましくは非霊長類(例えばウシ属、ウマ科、ネコ科、イヌ科、げっ歯類等)又は霊長類(例えばカニクイザル等のサル、ヒト等)といった哺乳類である。ある好ましい実施形態では、被験者はヒトである。
本発明のgp41結合分子又は上記gp41結合分子を含む組成物を投与する方法としては:非経口投与(例えば皮内、筋肉内、腹腔内、静脈内及び皮下);硬膜外;並びに粘膜(例えば鼻腔内及び経口経路)が挙げられるがこれらに限定されない。ある具体的実施形態では、本発明の結合分子は、筋肉内、静脈内又は皮下投与される。組成物は、いずれの便利な経路によって、例えば点滴又はボーラス注射によって、上皮又は粘膜皮膚層(例えば口腔粘膜、直腸及び腸粘膜等)を通した吸収によって投与してよく、他の生物学的活性作用剤と共に投与してよい。投与は全身性又は局所性とすることができる。
本発明はまた、本発明のgp41結合分子の調製物を、この分子の量を示すアンプル又はサシェ等の気密性コンテナ内に包装することも提供する。一実施形態では、上記gp41結合分子は、気密性コンテナ内の乾燥滅菌凍結乾燥粉末又は無水濃縮物として供給され、例えば水又は食塩水を用いて、被験者への投与に適切な濃度へと再構成できる。好ましくは、本発明のgp41結合分子は、気密性コンテナ内の乾燥滅菌凍結乾燥粉末として供給される。
凍結乾燥された本発明のgp41結合分子の調製物は、その元々のコンテナ内において2℃〜8℃で保管するべきであり、またこの分子は、再構成後12時間以内、好ましくは6時間以内、5時間以内、3時間以内又は1時間以内に投与するべきである。ある代替実施形態では、このような分子は、この分子、融合タンパク質又はコンジュゲート分子の量及び濃度を示す気密性コンテナ内に、液体形態で供給される。好ましくは、このような結合分子は、液体形態で提供される場合、気密性コンテナ内に供給される。
障害に関連する1つ以上の症状の治療、予防及び改善に効果的な本発明の上記調製物の量は、標準的な臨床技術によって決定できる。処方において採用するべき正確な用量は、投与経路及び状態の重篤度にも左右され、施術者の判断及び各レシピエント被験者の状況に従って決定するべきである。効果的な用量は、インビトロ又は動物モデル試験系から得られた用量‐応答曲線から外挿できる。
本明細書において使用される場合、医薬組成物の「有効量(effective amount)」は:疾患によってもたらされる症状の低減;感染の症状(例えばウイルス負荷、発熱、疼痛、敗血症等)を減弱させる疾患に起因する症状の減弱;これによる、疾患に罹患したヒトのQOLの上昇;疾患を治療するために必要な他の投薬量の低減;標的化及び/若しくは内在化等による別の投薬の効果の増強;疾患の進行の遅延;並びに/又は個体の生存期間の延長を含むがこれらに限定されない、有益な又は所望の結果を得るために十分な量である。単独で投与される個々の有効成分に対して適用される場合、この用語は、上記成分のみを指す。組み合わせに対して適用される場合、この用語は、治療効果をもたらす複数の有効成分の合計量を指し、上記有効成分が組み合わせた状態で投与されるか、順次投与されるか、又は同時に投与されるかにかかわらない。
有効量は、1回又は複数回の投与で投与できる。本発明の目的のために、薬剤、化合物又は医薬組成物の有効量は、直接的又は間接的な:HIV‐1感染細胞の殺滅及び/若しくは増殖の低減;HIV‐1ウイルスの増殖(又はその影響)の低減;並びにHIV仲介型疾患の進展の低減及び/又は遅延に十分な量である。いくつかの実施形態では、薬剤、化合物又は医薬組成物の有効量は、別の薬剤、化合物又は医薬組成物と組み合わせて達成してもしなくてもよい。従って「有効量」は、1つ以上の化学療法剤を投与する文脈で考えることができ、また単一の作用剤が、1つ以上の他の作用剤と組み合わせて所望の結果を達成できる又は所望の結果を達成する場合に、有効量で与えられているものと考えることができる。個別の必要量は異なるが、各成分の有効量の最適な範囲の決定は当業者の能力の範囲内である。
本発明に包含される結合分子に関して、レシピエント被験者に投与される投薬量は好ましくは、レシピエント被験者の体重(kg)に基づいて決定される。本発明に包含される結合分子に関して、レシピエント被験者に投与される投薬量は典型的には、被験者の体重に対して約0.01μg/kg〜約300mg/kg以上である。
本発明の結合分子の投与の用量及び頻度は、例えば脂質化等の改質によって分子の取り込み及び組織貫通を増強することによって低減又は変更できる。
レシピエント被験者に投与される本発明の結合分子の投薬量は、単一作用剤による治療としての使用のために計算してよい。あるいは、上記分子は他の治療用組成物と併用され、レシピエント被験者に投与される投薬量は、上記分子を単一作用剤による治療として使用する場合よりも小さくなる。
本発明の医薬組成物は、治療が必要な領域に局所的に投与してよく、これは、以下に限定されるものではないが、例えば局所的点滴によって、注射によって、又はインプラントを用いて達成してよく、上記インプラントは多孔性、非多孔性又はゼラチン材料性であり、SIALASTIC(登録商標)膜等の膜又は繊維を含む。好ましくは、本発明の分子を投与する際、この分子が吸収されない材料を使用するよう注意しなければならない。
本発明の組成物は、小嚢、特にリポソーム中で送達できる(Langer (1990) “New Methods Of Drug Delivery,” Science 249:1527-1533); Treat et al., in Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein and Fidler (eds.), Liss, New York, pp.353-365(1989); Lopez-Berestein,同上、pp.317-327参照)。
治療的又は予防的有効量の本発明の結合分子を用いた被験者の治療は、単回治療を含むことができ、又は好ましくは一連の複数回の治療を含むことができる。ある好ましい例では、被験者は、本発明の医薬組成物を用いて、約1〜10週間、好ましくは2〜8週間、より好ましくは約3〜7週間、更に好ましくは約4、5又は6週間にわたって処置される。本発明の医薬組成物は、1日1回投与でき、この投与は1週間に1回、1週間に2回、2週間に1回、1ヶ月に1回、6週間に1回、2ヶ月に1回、1年に2回又は1年に1回等のスケジュールで行われる。あるいは本発明の医薬組成物は、1日2回投与でき、この投与は1週間に1回、1週間に2回、2週間に1回、1ヶ月に1回、6週間に1回、2ヶ月に1回、1年に2回又は1年に1回等のスケジュールで行われる。あるいは、本発明の医薬組成物は、1日3回投与でき、この投与は1週間に1回、1週間に2回、2週間に1回、1ヶ月に1回、6週間に1回、2ヶ月に1回、1年に2回又は1年に1回等のスケジュールで行われる。治療に使用される上記分子の有効な用量設定は、特定の治療の期間に亘って増減させてよいことも理解されるだろう。
これまで本発明を概説してきたが、本発明は、以下の実施例を参照することによって更に容易に理解されるだろう。これらの実施例は例示として提供されており、特段の記載がない限り、本発明を限定することを意図したものではない。
実施例1
最適化
「7B2 IgG」と呼称される抗HIV‐1 Env(gp41タンパク質)抗体(GenBank受託番号AFQ31503;Buchacher, A. et al. (1994) “Generation Of Human Monoclonal Antibodies Against HIV-1 Proteins; Electrofusion And Epstein-Barr Virus Transformation For Peripheral Blood Lymphocyte Immortalization,” AIDS Res. Hum. Retroviruses 10(4):359-369; Shen, R. (2010) “GP41-Specific Antibody Blocks Cell-Free HIV-1 Type 1 Transcytosis Through Human Rectal Mucosa And Model Colonic Epithelium,” J. Immunol. 184(7):3648-3655)は、ヒトIgG生殖細胞系列レパートリーでは珍しいフレームワーク領域中に多数のアミノ酸残基を含むことが分かっている。このような珍しい非生殖細胞系列の残基の存在は、免疫原性となり得、レシピエント被験者への反復投与時に抗薬物抗体の生成をもたらす可能性がある。これらのドメインの免疫原性の可能性を低減する、更には排除するために、7B2可変領域を、そのフレームワーク領域に突然変異を導入することで、上述のような珍しいアミノ酸残基を生殖細胞系列アミノ酸残基に置換することにより、最適化した。
最適化
「7B2 IgG」と呼称される抗HIV‐1 Env(gp41タンパク質)抗体(GenBank受託番号AFQ31503;Buchacher, A. et al. (1994) “Generation Of Human Monoclonal Antibodies Against HIV-1 Proteins; Electrofusion And Epstein-Barr Virus Transformation For Peripheral Blood Lymphocyte Immortalization,” AIDS Res. Hum. Retroviruses 10(4):359-369; Shen, R. (2010) “GP41-Specific Antibody Blocks Cell-Free HIV-1 Type 1 Transcytosis Through Human Rectal Mucosa And Model Colonic Epithelium,” J. Immunol. 184(7):3648-3655)は、ヒトIgG生殖細胞系列レパートリーでは珍しいフレームワーク領域中に多数のアミノ酸残基を含むことが分かっている。このような珍しい非生殖細胞系列の残基の存在は、免疫原性となり得、レシピエント被験者への反復投与時に抗薬物抗体の生成をもたらす可能性がある。これらのドメインの免疫原性の可能性を低減する、更には排除するために、7B2可変領域を、そのフレームワーク領域に突然変異を導入することで、上述のような珍しいアミノ酸残基を生殖細胞系列アミノ酸残基に置換することにより、最適化した。
図7A〜7Bに示すように、9個の突然変異をVLフレームワーク領域に導入し(図7A)、17個のアミノ酸置換をVHフレームワーク領域に導入する(図7B)ことによって、それぞれ「7B2GL VL」及び「7B2GL VH」と呼称される、完全に生殖細胞系列化されたVL及びVHドメインを生成した。得られたVL/VHドメインは、「7B2GL」と総称される。
抗体7B2及び抗体7B2GLのVLドメインのアミノ酸配列は、上で提示されている(それぞれ配列番号55及び配列番号57)。フレームワーク1〜3供与体のVLドメインのアミノ酸配列(IGKV4‐1)及びフレームワーク4供与体のフレームワーク4領域のアミノ酸配列(IGKJ1‐01)は:
IGKV4‐1(配列番号123):
DIVMTQSPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP
KLLIYWASTR ESGVPDRFSG SGSGTDFTLT ISSLQAEDVA VYYCQQYYST
P
及びIGKJ1‐01(配列番号124):
FGQGTKVEIK
である。
IGKV4‐1(配列番号123):
DIVMTQSPDS LAVSLGERAT INCKSSQSVL YSSNNKNYLA WYQQKPGQPP
KLLIYWASTR ESGVPDRFSG SGSGTDFTLT ISSLQAEDVA VYYCQQYYST
P
及びIGKJ1‐01(配列番号124):
FGQGTKVEIK
である。
抗体7B2及び抗体7B2GLのVHドメインのアミノ酸配列は、上で提示されている(それぞれ配列番号56及び配列番号58)。フレームワーク1〜3供与体のVHドメインのアミノ酸配列(IGVH3‐11)及びフレームワーク4供与体のフレームワーク4領域のアミノ酸配列(IGHJ4‐01)は:
IGVH3‐11(配列番号125):
QVQLVESGGG LVKPGGSLRL SCAASGFTFS DYYMSWIRQA PGKGLEWVSY
ISSSSSYTNY ADSVKGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCAR
及びIGHJ4‐01(配列番号126):
WGQGTLVTVSS
である。
IGVH3‐11(配列番号125):
QVQLVESGGG LVKPGGSLRL SCAASGFTFS DYYMSWIRQA PGKGLEWVSY
ISSSSSYTNY ADSVKGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCAR
及びIGHJ4‐01(配列番号126):
WGQGTLVTVSS
である。
完全に生殖細胞系列化された可変領域を含むIgG(「7B2GL IgG」と呼称される)の結合を、Attana Cell A200 QCMによって調査した。簡潔に述べると、25、50、及び100nMのHIV‐1 JRFL gp140組み換えタンパク質(膜透過及びC末端アミノ酸残基が除かれているが、7B2によって認識されるgp41エピトープを保持している、HIV‐1 Envタンパク質)に、抗ヒトIgG Fc表面によってコーティングされたAttanaセンサチップ上に個別に捕捉された7B2 IgG(図8A)又は7B2GL IgG(図8B)上を通過させる。動態パラメータ(ka、kd、KD)の分析のために、当てはめモデルとして1:1結合を選択した。というのは、これが数学的に最小限しか操作されないモデルであるためである。しかしながら、JRFL gp140は単量体(20〜30%)、二量体(40%)及び三量体(30%)の混合物であるため、このモデルは相互作用を完全に記述しない。従って、表8で提供されている動態パラメータ(1:1結合の当てはめを用いたka、kd、KD)は主に、7B2及び7B2GL構成物の互いに対するAg結合活性を直接比較する目的において有用である。抗原結合にとって重要であることが知られており(Foote and Winter (1992) “Antibody Framework Residues Affecting The Conformation Of The Hypervariable Loops,” J. Mol. Biol. 224:487-499)、かつ典型的には変化しない(例えばChromikova et al. (2015) “Introduction Of Germline Residues Improves The Stability Of Anti-HIV mAb 2G12-IgM,” Biochim. Biophys. Acta 1854:1536-1544を参照)Vernierゾーン(例えばVHドメインのKabat30位、48位、49位、及び79位、並びにVLドメインのKabat48位)に、多数の変化が位置しているものの、これらの研究は、26個の生殖細胞の突然変異全てが十分な耐容性を有していたこと、及び完全に生殖細胞系列化された抗体が、野生型抗体7B2と略同一のエピトープ結合動態を示したことを示している。
実施例2
HIV×CD3結合分子
HIV Env gp41タンパク質及び例示的な免疫エフェクタ細胞標的CD3に結合できる、2つのHIV×CD3結合分子を生成した。特に、7B2又は7B2GL可変ドメインを含み、かつ3つのポリペプチド鎖を有する、(「DART‐1」及び「DART‐A」と呼称される)2つの二重特異性ダイアボディを生成した。これらはそれぞれCD3 mAb 1(D65G)可変ドメインを含む。DART‐1及びDART‐Aは、HIV及びCD3に結合できる、図4Aに示されている一般構造を有するFcドメイン含有二重特異性ダイアボディである。これらの例示的な分子のドメイン及びアミノ酸配列については、上で詳述されている。
HIV×CD3結合分子
HIV Env gp41タンパク質及び例示的な免疫エフェクタ細胞標的CD3に結合できる、2つのHIV×CD3結合分子を生成した。特に、7B2又は7B2GL可変ドメインを含み、かつ3つのポリペプチド鎖を有する、(「DART‐1」及び「DART‐A」と呼称される)2つの二重特異性ダイアボディを生成した。これらはそれぞれCD3 mAb 1(D65G)可変ドメインを含む。DART‐1及びDART‐Aは、HIV及びCD3に結合できる、図4Aに示されている一般構造を有するFcドメイン含有二重特異性ダイアボディである。これらの例示的な分子のドメイン及びアミノ酸配列については、上で詳述されている。
DART‐1及びDART‐Aの生物学的活性を、多数のアッセイで調査した。DART‐1及びDART‐Aの、HEK/D371細胞の表面上で発現されるgp41に結合する能力を、フローサイトメトリーで評価した。簡潔に述べると、gp140発現性HEK293/D371細胞(Dr. John Kappes(アラバマ大学バーミンガム校)によって得られた、1μg/mLのドキシサイクリンを用いた24時間の予備インキュベーションの後の、HIV‐1 CM244(サブタイプAE)gp140エンベロープのドキシサイクリン誘導性発現)を、DART‐1、DART‐A、又はRSV×CD3陰性対照の連続希釈物(10μg/mL〜0.0024μg/mL)を用いてインキュベートした。洗浄後、細胞を、Alexa 488‐抗ヒトIgG1 Fc二次抗体を用いてインキュベートし、(例えばGuava EasyCyte 8HT(Millipore)、及びGraphPad Prism 7でプロットされたMFIを用いて)フローサイトメトリーで分析した。図9に示すように、DART‐1及びDART‐Aは、同様の用量依存的様式で、gp140発現性HEK293/D371細胞に結合する。
DART‐1及びDART‐Aの、T細胞を活性化させる能力を、Jurkat T細胞レポータアッセイ(Promega)を用いて評価した。簡潔に述べると、Jurkat IL2 Luc2Pレポータ細胞を、DART‐1、DART‐A、又はRSV×CD3陰性対照の連続希釈物(10μg/mL〜0.0024μg/mL)の存在下で、gp140発現性HEK293/D371細胞(20:1)と共培養し、ONE‐GLO(商標)ルシフェラーゼアッセイシステム(Promega)を用いて発光を測定した。図10に示すように、DART‐1及びDART‐Aは、同等のT細胞活性化活性を示す。
DART‐1及びDART‐Aの、T細胞標的転換標的細胞殺滅を仲介する能力を、CTLアッセイを用いて評価した。簡潔に述べると、(例えばDynabeads(登録商標)Untouched(商標)ヒトT細胞キット(Invitrogen)を用いて)Pan T細胞を健康なヒトPBMCから単離した。gp140発現性HEK293/D371(1〜4×105細胞/mL)を用いて評価した。簡潔に述べると、Jurkat IL2 Luc2Pレポータ細胞を、DART‐1、DART‐A、又はRSV×CD3陰性対照の連続希釈物と、ヒトT細胞とで、エフェクタ:標的(E:T)比=10:1で、又は指示されているような様々なE:T比で処理し、37℃、5%CO2で24時間インキュベートした。細胞傷害性は、(例えばCytoTox 96(登録商標)非放射性細胞傷害性アッセイ(Promega)を用いて)乳酸デヒドロゲナーゼ(LDH)放出によって測定される。図11に示すように、DART‐1及びDART‐Aは、同等のT細胞標的転換標的細胞殺滅活性を示す。
これらの研究の結果は、最適化されかつ完全に生殖細胞系列化された7B2GLドメインを生成するために7B2のVL及びVHドメインに導入される多数の置換が、gp140結合、又は上述のアッセイにおける生物学的活性に影響を及ぼさないことを実証している。
本明細書において言及されている全ての公刊物及び特許は、個々の公刊物又は特許出願それぞれの全体が参照により本明細書に援用されていることが具体的かつ独立に指示されている場合と同程度に、参照により本明細書に援用されている。本発明をその具体的実施形態に関して説明したが、更なる修正形態が可能であり、本出願は、本発明が属する分野の公知の方法又は慣例の範囲内であるような、及びこれまでに挙げた必須の特徴に適用できるような、本開示からの逸脱を含む、本発明の原理に概ね従う本発明のいずれの変形、使用又は改変を包含することを意図していることを理解されたい。
Claims (26)
- 可変軽鎖(VL)ドメイン及び可変重鎖(VH)ドメインを含む、gp41結合分子であって、前記VLドメインは配列番号57のアミノ酸配列を含み、及び/又は前記VHドメインは配列番号58のアミノ酸配列を含む、gp41結合分子。
- 前記gp41結合分子は:
(a)配列番号57のアミノ酸配列を含む前記VLドメイン;及び
(b)配列番号58のアミノ酸配列を含む前記VHドメイン
を含む、請求項1に記載のgp41結合分子。 - 前記分子は、抗体であるか、前記抗体のgp41エピトープ結合部分を含む、請求項1又は2に記載のgp41結合分子。
- 前記分子は:
(a)二重特異性抗体;又は
(b)2つ、3つ、4つ若しくは5つのポリペプチド鎖を含む共有結合複合体である、ダイアボディ;又は
(c)3つ、4つ、5つ、若しくは6つ以上のポリペプチド鎖を含む共有結合複合体である、3価結合性分子
である、請求項1〜3のいずれか1項に記載のgp41結合分子。 - 前記分子は前記ダイアボディであり、アルブミン結合ドメイン(ABD)を含む、請求項4に記載のgp41結合分子。
- 前記分子はFc領域を含む、請求項1〜4のいずれか1項に記載のgp41結合分子。
- 前記Fc領域は、変異型Fc領域であって:
(a)FcγRに対する前記変異型Fc領域の親和性を低減する1つ以上のアミノ酸修飾;及び/又は
(b)前記変異型Fc領域の血清半減期を向上させる1つ以上のアミノ酸修飾
を含む、変異型Fc領域である、請求項5に記載のgp41結合分子。 - FcγRに対する前記変異型Fc領域の親和性を低減する前記修飾は、L234A;L235A;又はL234A及びL235Aの置換を含み、前記番号付与はKabat中のEUインデックスの番号付与である、請求項7に記載のgp41結合分子。
- 前記変異型Fc領域の血清半減期を向上させる前記修飾は、M252Y;M252Y及びS254T;M252Y及びT256E;M252Y、S254T及びT256E;又はK288D及びH435Kの置換を含み、前記番号付与はKabat中のEUインデックスの番号付与である、請求項7又は8に記載のgp41結合分子。
- 前記分子は二重特異性であり、gp41のエピトープに免疫特異的に結合できる1つのエピトープ結合部位と、エフェクタ細胞の表面上に存在する分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位とを含む、請求項1〜9のいずれか1項に記載のgp41結合分子。
- 前記分子は二重特異性であり、gp41のエピトープに免疫特異的に結合できる2つのエピトープ結合部位と、エフェクタ細胞の表面上に存在する分子のエピトープに免疫特異的に結合できる2つのエピトープ結合部位とを含む、請求項1〜9のいずれか1項に記載のgp41結合分子。
- 前記分子は三重特異性であり:
(a)gp41のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;
(b)エフェクタ細胞の表面上に存在する第1の分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;及び
(c)エフェクタ細胞の表面上に存在する第2の分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位
を含む、請求項1〜9のいずれか1項に記載のgp41結合分子。 - 前記分子は三重特異性であり:
(a)gp41の第1のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;
(b)gp41の第2のエピトープ、又は異なるHIV‐1タンパク質のエピトープに免疫特異的に結合できる1つのエピトープ結合部位;及び
(c)エフェクタ細胞の表面上に存在する分子のエピトープに免疫特異的に結合できる1つのエピトープ結合部位
を含み、gp41の前記第1のエピトープと前記第2のエピトープとは異なるものである、請求項1〜9のいずれか1項に記載のgp41結合分子。 - 前記分子は、gp41とエフェクタ細胞の表面上に存在する分子とに同時に結合できる、請求項10〜13のいずれか1項に記載のgp41結合分子。
- エフェクタ細胞の表面上に存在する前記分子は、CD2、CD3、CD8、CD16、TCR、NKp46、又はNKG2Dである、請求項10〜14のいずれか1項に記載のgp41結合分子。
- 前記エフェクタ細胞は、細胞傷害性T細胞又はナチュラルキラー(NK)細胞である、請求項10〜15のいずれか1項に記載のgp41結合分子。
- エフェクタ細胞の表面上に存在する前記分子はCD3である、請求項15に記載のgp41結合分子。
- エフェクタ細胞の表面上に存在する前記第1の分子はCD3であり、エフェクタ細胞の表面上に存在する前記第2の分子はCD18である、請求項12に記載のgp41結合分子。
- 前記分子は、gp41を発現する細胞と細胞傷害性T細胞との協調的結合を仲介する、請求項10〜18のいずれか1項に記載のgp41結合分子。
- 前記分子は、第1のポリペプチド鎖、第2のポリペプチド鎖、及び第3のポリペプチド鎖を含み:
I)(a)前記第1のポリペプチド鎖は配列番号113を含み;
(b)前記第2のポリペプチド鎖は配列番号114を含み;及び
(c)前記第3のポリペプチド鎖は配列番号112を含むか;
又は
II)(a)前記第1のポリペプチド鎖は配列番号115を含み;
(b)前記第2のポリペプチド鎖は配列番号116を含み;及び
(c)前記第3のポリペプチド鎖は配列番号112を含む、
請求項1〜4及び6〜9のいずれか1項に記載のgp41結合分子。 - 前記分子は、第1のポリペプチド鎖、第2のポリペプチド鎖、第3のポリペプチド鎖、及び第4のポリペプチド鎖を含み:
(a)前記第1のポリペプチド鎖は配列番号113を含み;
(b)前記第2のポリペプチド鎖は配列番号114を含み;
(c)前記第3のポリペプチド鎖は配列番号117を含み;及び
(d)前記第4のポリペプチド鎖は配列番号118を含む、
請求項1〜4及び6〜9のいずれか1項に記載のgp41結合分子。 - 前記分子は、第1のポリペプチド鎖、第2のポリペプチド鎖、第3のポリペプチド鎖、及び第4のポリペプチド鎖を含み:
I)(a)前記第1のポリペプチド鎖は配列番号113を含み;
(b)前記第2のポリペプチド鎖は配列番号114を含み;
(c)前記第3のポリペプチド鎖は配列番号119を含み;及び
(d)前記第4のポリペプチド鎖は配列番号120を含むか;
又は
II)(a)前記第1のポリペプチド鎖は配列番号115を含み;
(b)前記第2のポリペプチド鎖は配列番号116を含み;
(c)前記第3のポリペプチド鎖は配列番号119を含み;及び
(d)前記第4のポリペプチド鎖は配列番号120を含む、
請求項1〜4及び6〜9のいずれか1項に記載のgp41結合分子。 - 有効量の請求項1〜22のいずれか1項に記載のgp41結合分子と、薬学的に許容可能なキャリアとを含む、医薬組成物。
- HIV‐1感染の治療又は予防を必要とする被験者においてHIV‐1感染を治療又は予防するための方法であって、請求項1〜22のいずれか1項に記載のgp41結合分子又は請求項23に記載の医薬組成物を治療的有効量で含む組成物を前記被験者に投与するステップを含む、方法。
- 前記方法は更に、潜伏感染活性化剤を投与するステップを含む、請求項24に記載の方法。
- 前記潜伏感染活性化剤は、ボリノスタット、ロミデプシン、パノビノスタット、ジスルフィラム、JQ1、ブリオスタチン、PMA、イオノマイシン、又はこれらのいずれの組み合わせである、請求項25に記載の方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201862673462P | 2018-05-18 | 2018-05-18 | |
| US62/673,462 | 2018-05-18 | ||
| PCT/US2019/032030 WO2019222104A1 (en) | 2018-05-18 | 2019-05-13 | Optimized gp41-binding molecules and uses thereof |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2021524451A true JP2021524451A (ja) | 2021-09-13 |
| JPWO2019222104A5 JPWO2019222104A5 (ja) | 2022-05-16 |
Family
ID=68540936
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2020564553A Pending JP2021524451A (ja) | 2018-05-18 | 2019-05-13 | 最適化済みgp41結合分子及びその使用 |
Country Status (12)
| Country | Link |
|---|---|
| US (1) | US20210246194A1 (ja) |
| EP (1) | EP3794027A4 (ja) |
| JP (1) | JP2021524451A (ja) |
| CN (1) | CN112533945A (ja) |
| AU (1) | AU2019269383A1 (ja) |
| BR (1) | BR112020023432A2 (ja) |
| CA (1) | CA3100398A1 (ja) |
| IL (1) | IL278832A (ja) |
| MX (1) | MX2020012309A (ja) |
| SG (1) | SG11202011355QA (ja) |
| WO (1) | WO2019222104A1 (ja) |
| ZA (1) | ZA202007056B (ja) |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP3148580B1 (en) * | 2014-05-29 | 2021-01-20 | MacroGenics, Inc. | Tri-specific binding molecules that specifically bind to multiple cancer antigens and methods of use thereof |
| AP2017009823A0 (en) * | 2014-09-26 | 2017-03-31 | Macrogenics Inc | Bi-specific monovalent diabodies that are capable of binding cd19 and cd3, and uses thereof |
| AU2018385409A1 (en) | 2017-12-12 | 2020-07-02 | Macrogenics Inc. | Bispecific CD 16-binding molecules and their use in the treatment of disease |
| CN111647078B (zh) * | 2020-06-22 | 2022-04-26 | 中国医学科学院医学生物学研究所 | 一种抗hiv的单克隆抗体及其制备方法和应用 |
Family Cites Families (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| BRPI0816785A2 (pt) * | 2007-09-14 | 2017-05-02 | Adimab Inc | bibliotecas de anticorpos sintéticos racionalmente desenhadas, e, usos para as mesmas |
| WO2014159940A1 (en) * | 2013-03-14 | 2014-10-02 | Macrogenics, Inc. | Bispecific molecules that are immunoreactive with immune effector cells that express an activating receptor |
| US9988438B2 (en) * | 2013-07-25 | 2018-06-05 | Board of Supervisor of Louisiana State University and Agricultural and Mechanical College | Anti-HIV dual specificity antibodies and methods of HIV treatment |
| UA116479C2 (uk) * | 2013-08-09 | 2018-03-26 | Макродженікс, Інк. | БІСПЕЦИФІЧНЕ МОНОВАЛЕНТНЕ Fc-ДІАТІЛО, ЯКЕ ОДНОЧАСНО ЗВ'ЯЗУЄ CD32B I CD79b, ТА ЙОГО ЗАСТОСУВАННЯ |
| EP3148580B1 (en) * | 2014-05-29 | 2021-01-20 | MacroGenics, Inc. | Tri-specific binding molecules that specifically bind to multiple cancer antigens and methods of use thereof |
| EP3201227A4 (en) * | 2014-09-29 | 2018-04-18 | Duke University | Bispecific molecules comprising an hiv-1 envelope targeting arm |
| WO2016168758A1 (en) * | 2015-04-17 | 2016-10-20 | Igm Biosciences, Inc. | Multi-valent human immunodeficiency virus antigen binding molecules and uses thereof |
| KR20180068999A (ko) * | 2015-10-06 | 2018-06-22 | 알렉터 엘엘씨 | 항-trem2 항체 및 그의 사용방법 |
| SG11201810883TA (en) * | 2016-06-07 | 2019-01-30 | Macrogenics Inc | Combination therapy |
-
2019
- 2019-05-13 JP JP2020564553A patent/JP2021524451A/ja active Pending
- 2019-05-13 SG SG11202011355QA patent/SG11202011355QA/en unknown
- 2019-05-13 WO PCT/US2019/032030 patent/WO2019222104A1/en unknown
- 2019-05-13 CN CN201980048041.9A patent/CN112533945A/zh active Pending
- 2019-05-13 MX MX2020012309A patent/MX2020012309A/es unknown
- 2019-05-13 BR BR112020023432-0A patent/BR112020023432A2/pt unknown
- 2019-05-13 CA CA3100398A patent/CA3100398A1/en active Pending
- 2019-05-13 AU AU2019269383A patent/AU2019269383A1/en not_active Abandoned
- 2019-05-13 EP EP19802868.0A patent/EP3794027A4/en active Pending
- 2019-05-13 US US17/055,805 patent/US20210246194A1/en active Pending
-
2020
- 2020-11-12 ZA ZA2020/07056A patent/ZA202007056B/en unknown
- 2020-11-18 IL IL278832A patent/IL278832A/en unknown
Non-Patent Citations (7)
| Title |
|---|
| GUIDO FERRARI ET AL., NATURE REVIES DRUG DISCOVERY, vol. 15, JPN6023014359, December 2016 (2016-12-01), pages 823 - 834, ISSN: 0005032635 * |
| JAMES J. STEINHARDT ET AL., NATURE COMMUNICATIONS, vol. 9, JPN6023014360, 28 February 2018 (2018-02-28), pages 877, ISSN: 0005032636 * |
| MARK L CHIU AND GARY L GILILAND, CURRENT OPINION IN STRUCTURAL BIOLOGY, vol. 38, JPN6023014358, June 2016 (2016-06-01), pages 163 - 173, ISSN: 0005032633 * |
| RUIZHONG SHEN ET AL., THE JOURNAL OF IMMUNOLOGY, vol. 184, no. 7, JPN6023014362, 1 April 2010 (2010-04-01), pages 3648 - 3655, ISSN: 0005032638 * |
| SAMPA SANTRA ET AL,, PLOS PATHOGENS, JPN7023001438, 3 August 2015 (2015-08-03), pages 1 - 38, ISSN: 0005032632 * |
| ULRICH BRINKMANN AND ROLAND E. KONTERMANN, MABS, vol. 9, no. 2, JPN7023001439, 2017, pages 182 - 212, ISSN: 0005032634 * |
| YAOXING HUANG ET AL., CELL, vol. 165, no. 7, JPN6023014361, 16 June 2016 (2016-06-16), pages 1621 - 1631, ISSN: 0005032637 * |
Also Published As
| Publication number | Publication date |
|---|---|
| MX2020012309A (es) | 2021-06-08 |
| ZA202007056B (en) | 2021-10-27 |
| WO2019222104A1 (en) | 2019-11-21 |
| EP3794027A1 (en) | 2021-03-24 |
| CN112533945A (zh) | 2021-03-19 |
| SG11202011355QA (en) | 2020-12-30 |
| US20210246194A1 (en) | 2021-08-12 |
| EP3794027A4 (en) | 2022-02-09 |
| IL278832A (en) | 2021-01-31 |
| AU2019269383A1 (en) | 2020-12-10 |
| BR112020023432A2 (pt) | 2021-02-23 |
| CA3100398A1 (en) | 2019-11-21 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| TWI788327B (zh) | 能夠結合cd137和腫瘤抗原的雙特異性結合分子及其用途 | |
| TWI773646B (zh) | 結合lag-3的分子和其使用方法 | |
| JP6647222B2 (ja) | 三重特異性結合分子及びその使用方法 | |
| JP2019517539A (ja) | 併用療法 | |
| JP2019506444A (ja) | Pd‐1及びctla‐4との免疫応答性を有する二重特異性並びにその使用方法 | |
| TW201730212A (zh) | Ror1-結合分子及其使用方法 | |
| JP7337079B2 (ja) | 変異型cd3結合ドメイン、及び疾患の治療のための併用療法におけるその使用 | |
| JP2021524451A (ja) | 最適化済みgp41結合分子及びその使用 | |
| TWI814758B (zh) | 雙特異性cd16-結合分子及其在疾病治療中的用途 | |
| KR102630036B1 (ko) | Adam9-결합 분자, 및 이것의 사용 방법 | |
| JP7378567B2 (ja) | Lag‐3結合分子及びその使用方法 | |
| AU2021224787A1 (en) | CD137 binding molecules and uses thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20220506 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20220506 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20230410 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20231110 |