JP2018522571A - Gitrl融合タンパク質およびその使用 - Google Patents
Gitrl融合タンパク質およびその使用 Download PDFInfo
- Publication number
- JP2018522571A JP2018522571A JP2018506513A JP2018506513A JP2018522571A JP 2018522571 A JP2018522571 A JP 2018522571A JP 2018506513 A JP2018506513 A JP 2018506513A JP 2018506513 A JP2018506513 A JP 2018506513A JP 2018522571 A JP2018522571 A JP 2018522571A
- Authority
- JP
- Japan
- Prior art keywords
- cells
- cell
- gitrl
- gitr
- domain
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 102000037865 fusion proteins Human genes 0.000 title claims description 381
- 108020001507 fusion proteins Proteins 0.000 title claims description 381
- 102100035283 Tumor necrosis factor ligand superfamily member 18 Human genes 0.000 title abstract description 49
- 101000597780 Mus musculus Tumor necrosis factor ligand superfamily member 18 Proteins 0.000 title abstract description 11
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 285
- 102000004196 processed proteins & peptides Human genes 0.000 claims abstract description 270
- 229920001184 polypeptide Polymers 0.000 claims abstract description 266
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 265
- 102000004169 proteins and genes Human genes 0.000 claims abstract description 242
- 238000000034 method Methods 0.000 claims abstract description 170
- 206010028980 Neoplasm Diseases 0.000 claims abstract description 161
- 230000027455 binding Effects 0.000 claims abstract description 155
- 241000282414 Homo sapiens Species 0.000 claims abstract description 86
- 238000005829 trimerization reaction Methods 0.000 claims abstract description 81
- 102000005962 receptors Human genes 0.000 claims abstract description 70
- 108020003175 receptors Proteins 0.000 claims abstract description 70
- 201000011510 cancer Diseases 0.000 claims abstract description 35
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 197
- 101000801234 Homo sapiens Tumor necrosis factor receptor superfamily member 18 Proteins 0.000 claims description 188
- 210000004027 cell Anatomy 0.000 claims description 164
- 102100033728 Tumor necrosis factor receptor superfamily member 18 Human genes 0.000 claims description 147
- 239000000427 antigen Substances 0.000 claims description 90
- 102000036639 antigens Human genes 0.000 claims description 90
- 108091007433 antigens Proteins 0.000 claims description 90
- 150000001413 amino acids Chemical group 0.000 claims description 76
- 238000011282 treatment Methods 0.000 claims description 73
- 238000003556 assay Methods 0.000 claims description 59
- 102000040430 polynucleotide Human genes 0.000 claims description 55
- 108091033319 polynucleotide Proteins 0.000 claims description 55
- 239000002157 polynucleotide Substances 0.000 claims description 55
- 230000000694 effects Effects 0.000 claims description 50
- 150000007523 nucleic acids Chemical class 0.000 claims description 50
- 210000000822 natural killer cell Anatomy 0.000 claims description 49
- 239000000203 mixture Substances 0.000 claims description 48
- 102100022153 Tumor necrosis factor receptor superfamily member 4 Human genes 0.000 claims description 42
- 239000003446 ligand Substances 0.000 claims description 41
- 102100027581 Forkhead box protein P3 Human genes 0.000 claims description 40
- 101000861452 Homo sapiens Forkhead box protein P3 Proteins 0.000 claims description 40
- 101710165473 Tumor necrosis factor receptor superfamily member 4 Proteins 0.000 claims description 40
- 102000039446 nucleic acids Human genes 0.000 claims description 39
- 108020004707 nucleic acids Proteins 0.000 claims description 39
- 239000013598 vector Substances 0.000 claims description 32
- 108050007222 Coronin Proteins 0.000 claims description 31
- 102000018123 coronin Human genes 0.000 claims description 31
- KRQUGYHEWIVMJV-UHFFFAOYSA-N coronin Natural products CCCCCCCCCCCCC=C/CCC1OC1CCC2OC2CCCCCCCCCCC3=CC(C)OC3=O KRQUGYHEWIVMJV-UHFFFAOYSA-N 0.000 claims description 31
- HSMPDPBYAYSOBC-UHFFFAOYSA-N khellin Chemical compound O1C(C)=CC(=O)C2=C1C(OC)=C1OC=CC1=C2OC HSMPDPBYAYSOBC-UHFFFAOYSA-N 0.000 claims description 31
- 230000001419 dependent effect Effects 0.000 claims description 30
- 230000004614 tumor growth Effects 0.000 claims description 30
- 239000013638 trimer Substances 0.000 claims description 26
- 230000035755 proliferation Effects 0.000 claims description 25
- 210000003289 regulatory T cell Anatomy 0.000 claims description 25
- 101000979342 Homo sapiens Nuclear factor NF-kappa-B p105 subunit Proteins 0.000 claims description 24
- 102100023050 Nuclear factor NF-kappa-B p105 subunit Human genes 0.000 claims description 24
- 102000047758 human TNFRSF18 Human genes 0.000 claims description 24
- 238000006467 substitution reaction Methods 0.000 claims description 24
- 102000004473 OX40 Ligand Human genes 0.000 claims description 23
- 239000000556 agonist Substances 0.000 claims description 23
- 108010042215 OX40 Ligand Proteins 0.000 claims description 22
- 239000003795 chemical substances by application Substances 0.000 claims description 20
- 230000035772 mutation Effects 0.000 claims description 20
- 239000005089 Luciferase Substances 0.000 claims description 19
- 230000004083 survival effect Effects 0.000 claims description 19
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 claims description 18
- 230000004913 activation Effects 0.000 claims description 18
- 230000004044 response Effects 0.000 claims description 18
- 102000004127 Cytokines Human genes 0.000 claims description 17
- 108090000695 Cytokines Proteins 0.000 claims description 17
- 108010076504 Protein Sorting Signals Proteins 0.000 claims description 15
- 230000006044 T cell activation Effects 0.000 claims description 15
- 210000002865 immune cell Anatomy 0.000 claims description 15
- 230000028993 immune response Effects 0.000 claims description 15
- 230000001404 mediated effect Effects 0.000 claims description 14
- 108060001084 Luciferase Proteins 0.000 claims description 13
- 230000003013 cytotoxicity Effects 0.000 claims description 13
- 231100000135 cytotoxicity Toxicity 0.000 claims description 13
- 238000012004 kinetic exclusion assay Methods 0.000 claims description 13
- 102100028681 C-type lectin domain family 4 member K Human genes 0.000 claims description 12
- 102000018658 Myotonin-Protein Kinase Human genes 0.000 claims description 12
- 239000003862 glucocorticoid Substances 0.000 claims description 12
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 claims description 12
- 230000000638 stimulation Effects 0.000 claims description 12
- 101710183165 C-type lectin domain family 4 member K Proteins 0.000 claims description 11
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 claims description 11
- 241000282567 Macaca fascicularis Species 0.000 claims description 11
- 108010072582 Matrilin Proteins Proteins 0.000 claims description 11
- 108010052185 Myotonin-Protein Kinase Proteins 0.000 claims description 11
- 108060008683 Tumor Necrosis Factor Receptor Proteins 0.000 claims description 11
- 102000003298 tumor necrosis factor receptor Human genes 0.000 claims description 11
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 10
- 230000009467 reduction Effects 0.000 claims description 10
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 claims description 9
- 108060008682 Tumor Necrosis Factor Proteins 0.000 claims description 9
- 102000000852 Tumor Necrosis Factor-alpha Human genes 0.000 claims description 9
- 125000000539 amino acid group Chemical group 0.000 claims description 9
- -1 aspartyl residue Chemical group 0.000 claims description 9
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 claims description 9
- 210000004899 c-terminal region Anatomy 0.000 claims description 9
- 238000010348 incorporation Methods 0.000 claims description 9
- 230000005764 inhibitory process Effects 0.000 claims description 9
- 230000003993 interaction Effects 0.000 claims description 9
- 230000037361 pathway Effects 0.000 claims description 9
- 230000001737 promoting effect Effects 0.000 claims description 9
- 229940104230 thymidine Drugs 0.000 claims description 9
- 241000282560 Macaca mulatta Species 0.000 claims description 8
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 8
- 230000037453 T cell priming Effects 0.000 claims description 8
- 231100000673 dose–response relationship Toxicity 0.000 claims description 8
- 206010009944 Colon cancer Diseases 0.000 claims description 7
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 claims description 7
- 206010035226 Plasma cell myeloma Diseases 0.000 claims description 7
- 230000019491 signal transduction Effects 0.000 claims description 7
- 101000738771 Homo sapiens Receptor-type tyrosine-protein phosphatase C Proteins 0.000 claims description 6
- 102000003814 Interleukin-10 Human genes 0.000 claims description 6
- 108090000174 Interleukin-10 Proteins 0.000 claims description 6
- 102100037422 Receptor-type tyrosine-protein phosphatase C Human genes 0.000 claims description 6
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 claims description 6
- 230000002708 enhancing effect Effects 0.000 claims description 6
- 201000005787 hematologic cancer Diseases 0.000 claims description 6
- 208000024200 hematopoietic and lymphoid system neoplasm Diseases 0.000 claims description 6
- 230000001939 inductive effect Effects 0.000 claims description 6
- 210000004881 tumor cell Anatomy 0.000 claims description 6
- 208000026310 Breast neoplasm Diseases 0.000 claims description 5
- 229940046168 CpG oligodeoxynucleotide Drugs 0.000 claims description 5
- 101000611183 Homo sapiens Tumor necrosis factor Proteins 0.000 claims description 5
- 108010002616 Interleukin-5 Proteins 0.000 claims description 5
- 208000034578 Multiple myelomas Diseases 0.000 claims description 5
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 claims description 5
- 102100040247 Tumor necrosis factor Human genes 0.000 claims description 5
- 210000001165 lymph node Anatomy 0.000 claims description 5
- 208000031261 Acute myeloid leukaemia Diseases 0.000 claims description 4
- 206010006187 Breast cancer Diseases 0.000 claims description 4
- 208000001333 Colorectal Neoplasms Diseases 0.000 claims description 4
- 208000017604 Hodgkin disease Diseases 0.000 claims description 4
- 208000021519 Hodgkin lymphoma Diseases 0.000 claims description 4
- 208000010747 Hodgkins lymphoma Diseases 0.000 claims description 4
- 108090000176 Interleukin-13 Proteins 0.000 claims description 4
- 108010002350 Interleukin-2 Proteins 0.000 claims description 4
- 108090000978 Interleukin-4 Proteins 0.000 claims description 4
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 claims description 4
- 206010068871 Myotonic dystrophy Diseases 0.000 claims description 4
- 208000015914 Non-Hodgkin lymphomas Diseases 0.000 claims description 4
- 206010033128 Ovarian cancer Diseases 0.000 claims description 4
- 238000006664 bond formation reaction Methods 0.000 claims description 4
- 210000002540 macrophage Anatomy 0.000 claims description 4
- 201000001441 melanoma Diseases 0.000 claims description 4
- 239000003607 modifier Substances 0.000 claims description 4
- 208000002154 non-small cell lung carcinoma Diseases 0.000 claims description 4
- 208000008443 pancreatic carcinoma Diseases 0.000 claims description 4
- 208000029729 tumor suppressor gene on chromosome 11 Diseases 0.000 claims description 4
- 208000024893 Acute lymphoblastic leukemia Diseases 0.000 claims description 3
- 208000014697 Acute lymphocytic leukaemia Diseases 0.000 claims description 3
- 208000010839 B-cell chronic lymphocytic leukemia Diseases 0.000 claims description 3
- 208000032791 BCR-ABL1 positive chronic myelogenous leukemia Diseases 0.000 claims description 3
- 206010005003 Bladder cancer Diseases 0.000 claims description 3
- 208000010833 Chronic myeloid leukaemia Diseases 0.000 claims description 3
- 208000000461 Esophageal Neoplasms Diseases 0.000 claims description 3
- 108010087819 Fc receptors Proteins 0.000 claims description 3
- 102000009109 Fc receptors Human genes 0.000 claims description 3
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 claims description 3
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 claims description 3
- 108090001007 Interleukin-8 Proteins 0.000 claims description 3
- 208000031422 Lymphocytic Chronic B-Cell Leukemia Diseases 0.000 claims description 3
- 208000033761 Myelogenous Chronic BCR-ABL Positive Leukemia Diseases 0.000 claims description 3
- 208000033776 Myeloid Acute Leukemia Diseases 0.000 claims description 3
- 230000006051 NK cell activation Effects 0.000 claims description 3
- 206010030155 Oesophageal carcinoma Diseases 0.000 claims description 3
- 206010061535 Ovarian neoplasm Diseases 0.000 claims description 3
- 206010061902 Pancreatic neoplasm Diseases 0.000 claims description 3
- 108091000080 Phosphotransferase Proteins 0.000 claims description 3
- 208000006664 Precursor Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 claims description 3
- 206010060862 Prostate cancer Diseases 0.000 claims description 3
- 208000000236 Prostatic Neoplasms Diseases 0.000 claims description 3
- 206010041067 Small cell lung cancer Diseases 0.000 claims description 3
- 208000024313 Testicular Neoplasms Diseases 0.000 claims description 3
- 206010057644 Testis cancer Diseases 0.000 claims description 3
- 208000024770 Thyroid neoplasm Diseases 0.000 claims description 3
- 208000032852 chronic lymphocytic leukemia Diseases 0.000 claims description 3
- 230000000295 complement effect Effects 0.000 claims description 3
- 238000012258 culturing Methods 0.000 claims description 3
- 201000004101 esophageal cancer Diseases 0.000 claims description 3
- 208000005017 glioblastoma Diseases 0.000 claims description 3
- 201000010536 head and neck cancer Diseases 0.000 claims description 3
- 208000014829 head and neck neoplasm Diseases 0.000 claims description 3
- 230000002489 hematologic effect Effects 0.000 claims description 3
- 206010073071 hepatocellular carcinoma Diseases 0.000 claims description 3
- 231100000844 hepatocellular carcinoma Toxicity 0.000 claims description 3
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 claims description 3
- 201000002528 pancreatic cancer Diseases 0.000 claims description 3
- 102000020233 phosphotransferase Human genes 0.000 claims description 3
- 208000000587 small cell lung carcinoma Diseases 0.000 claims description 3
- 239000003381 stabilizer Substances 0.000 claims description 3
- 201000003120 testicular cancer Diseases 0.000 claims description 3
- 201000002510 thyroid cancer Diseases 0.000 claims description 3
- 102000055008 Matrilin Proteins Human genes 0.000 claims description 2
- 206010057249 Phagocytosis Diseases 0.000 claims description 2
- 206010038389 Renal cancer Diseases 0.000 claims description 2
- 208000033878 Tertiary Lymphoid Structures Diseases 0.000 claims description 2
- 208000007097 Urinary Bladder Neoplasms Diseases 0.000 claims description 2
- 210000001772 blood platelet Anatomy 0.000 claims description 2
- 210000001072 colon Anatomy 0.000 claims description 2
- 238000004132 cross linking Methods 0.000 claims description 2
- 231100000433 cytotoxic Toxicity 0.000 claims description 2
- 230000001472 cytotoxic effect Effects 0.000 claims description 2
- 210000002889 endothelial cell Anatomy 0.000 claims description 2
- 210000003979 eosinophil Anatomy 0.000 claims description 2
- 210000000285 follicular dendritic cell Anatomy 0.000 claims description 2
- 210000003734 kidney Anatomy 0.000 claims description 2
- 210000001821 langerhans cell Anatomy 0.000 claims description 2
- 210000001616 monocyte Anatomy 0.000 claims description 2
- 210000000440 neutrophil Anatomy 0.000 claims description 2
- 230000008782 phagocytosis Effects 0.000 claims description 2
- 210000005134 plasmacytoid dendritic cell Anatomy 0.000 claims description 2
- 229940126625 tavolimab Drugs 0.000 claims description 2
- 201000005112 urinary bladder cancer Diseases 0.000 claims description 2
- 206010008342 Cervix carcinoma Diseases 0.000 claims 2
- 102000043136 MAP kinase family Human genes 0.000 claims 2
- 108091054455 MAP kinase family Proteins 0.000 claims 2
- 206010027406 Mesothelioma Diseases 0.000 claims 2
- 208000000102 Squamous Cell Carcinoma of Head and Neck Diseases 0.000 claims 2
- 208000005718 Stomach Neoplasms Diseases 0.000 claims 2
- 208000006105 Uterine Cervical Neoplasms Diseases 0.000 claims 2
- 201000005969 Uveal melanoma Diseases 0.000 claims 2
- 230000005888 antibody-dependent cellular phagocytosis Effects 0.000 claims 2
- 201000010881 cervical cancer Diseases 0.000 claims 2
- 230000007783 downstream signaling Effects 0.000 claims 2
- 206010017758 gastric cancer Diseases 0.000 claims 2
- 201000000459 head and neck squamous cell carcinoma Diseases 0.000 claims 2
- 201000011549 stomach cancer Diseases 0.000 claims 2
- 208000008839 Kidney Neoplasms Diseases 0.000 claims 1
- 208000008589 Obesity Diseases 0.000 claims 1
- 208000015634 Rectal Neoplasms Diseases 0.000 claims 1
- 210000002798 bone marrow cell Anatomy 0.000 claims 1
- 201000010982 kidney cancer Diseases 0.000 claims 1
- 235000020824 obesity Nutrition 0.000 claims 1
- 206010038038 rectal cancer Diseases 0.000 claims 1
- 201000001275 rectum cancer Diseases 0.000 claims 1
- 230000004936 stimulating effect Effects 0.000 claims 1
- 230000004927 fusion Effects 0.000 abstract description 75
- 235000018102 proteins Nutrition 0.000 description 184
- 241000699670 Mus sp. Species 0.000 description 99
- 235000001014 amino acid Nutrition 0.000 description 72
- 229940024606 amino acid Drugs 0.000 description 63
- 101000597779 Homo sapiens Tumor necrosis factor ligand superfamily member 18 Proteins 0.000 description 59
- 230000014509 gene expression Effects 0.000 description 44
- 229940027941 immunoglobulin g Drugs 0.000 description 40
- 125000003275 alpha amino acid group Chemical group 0.000 description 38
- 241000699666 Mus <mouse, genus> Species 0.000 description 36
- 210000000952 spleen Anatomy 0.000 description 26
- 239000012634 fragment Substances 0.000 description 25
- 208000036142 Viral infection Diseases 0.000 description 24
- 238000007792 addition Methods 0.000 description 24
- 230000009385 viral infection Effects 0.000 description 24
- 108020004414 DNA Proteins 0.000 description 20
- 230000004988 N-glycosylation Effects 0.000 description 20
- 238000002474 experimental method Methods 0.000 description 19
- 230000006870 function Effects 0.000 description 19
- 230000002829 reductive effect Effects 0.000 description 19
- 230000000259 anti-tumor effect Effects 0.000 description 18
- 238000000684 flow cytometry Methods 0.000 description 18
- 230000001965 increasing effect Effects 0.000 description 18
- 102200124876 rs28928896 Human genes 0.000 description 18
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 16
- 101710086766 FP protein Proteins 0.000 description 16
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 15
- 102000053830 human TNFSF18 Human genes 0.000 description 15
- 230000001976 improved effect Effects 0.000 description 15
- 230000003285 pharmacodynamic effect Effects 0.000 description 15
- 239000011780 sodium chloride Substances 0.000 description 15
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 description 14
- 108050005493 CD3 protein, epsilon/gamma/delta subunit Proteins 0.000 description 14
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 14
- 241001465754 Metazoa Species 0.000 description 14
- 238000000746 purification Methods 0.000 description 14
- 239000000126 substance Substances 0.000 description 14
- 238000012360 testing method Methods 0.000 description 14
- 238000004458 analytical method Methods 0.000 description 13
- 230000002601 intratumoral effect Effects 0.000 description 13
- 238000006206 glycosylation reaction Methods 0.000 description 12
- 239000000243 solution Substances 0.000 description 12
- 108091026890 Coding region Proteins 0.000 description 11
- 101000679851 Homo sapiens Tumor necrosis factor receptor superfamily member 4 Proteins 0.000 description 11
- 108010057466 NF-kappa B Proteins 0.000 description 11
- 230000008484 agonism Effects 0.000 description 11
- 230000013595 glycosylation Effects 0.000 description 11
- 238000002868 homogeneous time resolved fluorescence Methods 0.000 description 11
- 239000000178 monomer Substances 0.000 description 11
- 239000000047 product Substances 0.000 description 11
- 101100107610 Arabidopsis thaliana ABCF4 gene Proteins 0.000 description 10
- 102100033668 Cartilage matrix protein Human genes 0.000 description 10
- 108060003951 Immunoglobulin Proteins 0.000 description 10
- 101100068078 Saccharomyces cerevisiae (strain ATCC 204508 / S288c) GCN4 gene Proteins 0.000 description 10
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 10
- 102000018358 immunoglobulin Human genes 0.000 description 10
- 238000001727 in vivo Methods 0.000 description 10
- 238000013518 transcription Methods 0.000 description 10
- 230000035897 transcription Effects 0.000 description 10
- 238000002965 ELISA Methods 0.000 description 9
- 102000003945 NF-kappa B Human genes 0.000 description 9
- 241000700605 Viruses Species 0.000 description 9
- 238000004422 calculation algorithm Methods 0.000 description 9
- 238000012217 deletion Methods 0.000 description 9
- 230000037430 deletion Effects 0.000 description 9
- 210000003162 effector t lymphocyte Anatomy 0.000 description 9
- 102000050320 human TNFRSF4 Human genes 0.000 description 9
- 238000004020 luminiscence type Methods 0.000 description 9
- 238000002844 melting Methods 0.000 description 9
- 230000008018 melting Effects 0.000 description 9
- 108020004999 messenger RNA Proteins 0.000 description 9
- 230000007704 transition Effects 0.000 description 9
- 101100425749 Mus musculus Tnfrsf18 gene Proteins 0.000 description 8
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 8
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 8
- 239000013604 expression vector Substances 0.000 description 8
- 230000004048 modification Effects 0.000 description 8
- 238000012986 modification Methods 0.000 description 8
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 7
- 241000282412 Homo Species 0.000 description 7
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 7
- 101000764258 Mus musculus Tumor necrosis factor ligand superfamily member 4 Proteins 0.000 description 7
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 description 7
- 230000004071 biological effect Effects 0.000 description 7
- 230000008859 change Effects 0.000 description 7
- 239000003153 chemical reaction reagent Substances 0.000 description 7
- 238000000338 in vitro Methods 0.000 description 7
- 238000011534 incubation Methods 0.000 description 7
- 230000002401 inhibitory effect Effects 0.000 description 7
- 238000001990 intravenous administration Methods 0.000 description 7
- 239000013642 negative control Substances 0.000 description 7
- 239000002773 nucleotide Substances 0.000 description 7
- 125000003729 nucleotide group Chemical group 0.000 description 7
- 230000003612 virological effect Effects 0.000 description 7
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 6
- 101000860852 Homo sapiens Coronin-1A Proteins 0.000 description 6
- 230000006052 T cell proliferation Effects 0.000 description 6
- 210000000662 T-lymphocyte subset Anatomy 0.000 description 6
- 210000004369 blood Anatomy 0.000 description 6
- 239000008280 blood Substances 0.000 description 6
- 239000003636 conditioned culture medium Substances 0.000 description 6
- 230000003247 decreasing effect Effects 0.000 description 6
- 239000000539 dimer Substances 0.000 description 6
- 239000012636 effector Substances 0.000 description 6
- 238000010828 elution Methods 0.000 description 6
- 229960000310 isoleucine Drugs 0.000 description 6
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 6
- 210000004962 mammalian cell Anatomy 0.000 description 6
- 238000004519 manufacturing process Methods 0.000 description 6
- 239000003550 marker Substances 0.000 description 6
- 150000002482 oligosaccharides Polymers 0.000 description 6
- 239000008194 pharmaceutical composition Substances 0.000 description 6
- 210000004986 primary T-cell Anatomy 0.000 description 6
- RXWNCPJZOCPEPQ-NVWDDTSBSA-N puromycin Chemical compound C1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CO RXWNCPJZOCPEPQ-NVWDDTSBSA-N 0.000 description 6
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 6
- 230000011664 signaling Effects 0.000 description 6
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 6
- 230000003393 splenic effect Effects 0.000 description 6
- 208000024891 symptom Diseases 0.000 description 6
- 238000002054 transplantation Methods 0.000 description 6
- 102100028233 Coronin-1A Human genes 0.000 description 5
- 108010074328 Interferon-gamma Proteins 0.000 description 5
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 5
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 5
- 238000005119 centrifugation Methods 0.000 description 5
- 238000012512 characterization method Methods 0.000 description 5
- 230000009089 cytolysis Effects 0.000 description 5
- 201000010099 disease Diseases 0.000 description 5
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 5
- 239000002552 dosage form Substances 0.000 description 5
- GWQVMPWSEVRGPY-UHFFFAOYSA-N europium cryptate Chemical compound [Eu+3].N=1C2=CC=CC=1CN(CC=1N=C(C=CC=1)C=1N=C(C3)C=CC=1)CC(N=1)=CC(C(=O)NCCN)=CC=1C(N=1)=CC(C(=O)NCCN)=CC=1CN3CC1=CC=CC2=N1 GWQVMPWSEVRGPY-UHFFFAOYSA-N 0.000 description 5
- 238000009472 formulation Methods 0.000 description 5
- 230000005714 functional activity Effects 0.000 description 5
- 230000012010 growth Effects 0.000 description 5
- 208000015181 infectious disease Diseases 0.000 description 5
- 239000003112 inhibitor Substances 0.000 description 5
- 238000003780 insertion Methods 0.000 description 5
- 230000037431 insertion Effects 0.000 description 5
- 239000007928 intraperitoneal injection Substances 0.000 description 5
- 238000004949 mass spectrometry Methods 0.000 description 5
- 238000005259 measurement Methods 0.000 description 5
- 239000002609 medium Substances 0.000 description 5
- 125000002496 methyl group Chemical group [H]C([H])([H])* 0.000 description 5
- 238000012510 peptide mapping method Methods 0.000 description 5
- 239000013612 plasmid Substances 0.000 description 5
- 230000001105 regulatory effect Effects 0.000 description 5
- 230000003248 secreting effect Effects 0.000 description 5
- 241000894007 species Species 0.000 description 5
- 230000014616 translation Effects 0.000 description 5
- 108050003114 Coronin 1A Proteins 0.000 description 4
- 241000701022 Cytomegalovirus Species 0.000 description 4
- 239000004471 Glycine Substances 0.000 description 4
- 101001057504 Homo sapiens Interferon-stimulated gene 20 kDa protein Proteins 0.000 description 4
- 101001055144 Homo sapiens Interleukin-2 receptor subunit alpha Proteins 0.000 description 4
- 102100037850 Interferon gamma Human genes 0.000 description 4
- 102100027268 Interferon-stimulated gene 20 kDa protein Human genes 0.000 description 4
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 4
- 241000124008 Mammalia Species 0.000 description 4
- 108700026244 Open Reading Frames Proteins 0.000 description 4
- 238000000692 Student's t-test Methods 0.000 description 4
- 101710120037 Toxin CcdB Proteins 0.000 description 4
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 4
- 235000004279 alanine Nutrition 0.000 description 4
- 230000001580 bacterial effect Effects 0.000 description 4
- 239000012620 biological material Substances 0.000 description 4
- 230000015572 biosynthetic process Effects 0.000 description 4
- 229940098773 bovine serum albumin Drugs 0.000 description 4
- 239000000872 buffer Substances 0.000 description 4
- 238000004364 calculation method Methods 0.000 description 4
- 238000004113 cell culture Methods 0.000 description 4
- 238000004587 chromatography analysis Methods 0.000 description 4
- 230000001684 chronic effect Effects 0.000 description 4
- 230000008034 disappearance Effects 0.000 description 4
- 239000003814 drug Substances 0.000 description 4
- 239000003623 enhancer Substances 0.000 description 4
- 210000003527 eukaryotic cell Anatomy 0.000 description 4
- 238000003018 immunoassay Methods 0.000 description 4
- 239000007924 injection Substances 0.000 description 4
- 238000002347 injection Methods 0.000 description 4
- 238000001819 mass spectrum Methods 0.000 description 4
- 239000011159 matrix material Substances 0.000 description 4
- 230000003389 potentiating effect Effects 0.000 description 4
- 230000009696 proliferative response Effects 0.000 description 4
- 239000011347 resin Substances 0.000 description 4
- 229920005989 resin Polymers 0.000 description 4
- 230000028327 secretion Effects 0.000 description 4
- 238000001542 size-exclusion chromatography Methods 0.000 description 4
- 238000010186 staining Methods 0.000 description 4
- 239000000725 suspension Substances 0.000 description 4
- 230000001225 therapeutic effect Effects 0.000 description 4
- 210000001519 tissue Anatomy 0.000 description 4
- 238000013519 translation Methods 0.000 description 4
- 230000032258 transport Effects 0.000 description 4
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 3
- 108700028369 Alleles Proteins 0.000 description 3
- WVDDGKGOMKODPV-UHFFFAOYSA-N Benzyl alcohol Chemical compound OCC1=CC=CC=C1 WVDDGKGOMKODPV-UHFFFAOYSA-N 0.000 description 3
- 102100032912 CD44 antigen Human genes 0.000 description 3
- WQZGKKKJIJFFOK-QTVWNMPRSA-N D-mannopyranose Chemical compound OC[C@H]1OC(O)[C@@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-QTVWNMPRSA-N 0.000 description 3
- HVLSXIKZNLPZJJ-TXZCQADKSA-N HA peptide Chemical compound C([C@@H](C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@H]1N(CCC1)C(=O)[C@@H](N)CC=1C=CC(O)=CC=1)C1=CC=C(O)C=C1 HVLSXIKZNLPZJJ-TXZCQADKSA-N 0.000 description 3
- 101710154606 Hemagglutinin Proteins 0.000 description 3
- 241000238631 Hexapoda Species 0.000 description 3
- 101000868273 Homo sapiens CD44 antigen Proteins 0.000 description 3
- 101001018097 Homo sapiens L-selectin Proteins 0.000 description 3
- 102000003839 Human Proteins Human genes 0.000 description 3
- 108090000144 Human Proteins Proteins 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 3
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 3
- 102100033467 L-selectin Human genes 0.000 description 3
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 3
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 3
- 239000004472 Lysine Substances 0.000 description 3
- 239000012515 MabSelect SuRe Substances 0.000 description 3
- 101710093908 Outer capsid protein VP4 Proteins 0.000 description 3
- 101710135467 Outer capsid protein sigma-1 Proteins 0.000 description 3
- 101710176177 Protein A56 Proteins 0.000 description 3
- 239000012980 RPMI-1640 medium Substances 0.000 description 3
- 108700008625 Reporter Genes Proteins 0.000 description 3
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 3
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 3
- KZSNJWFQEVHDMF-UHFFFAOYSA-N Valine Natural products CC(C)C(N)C(O)=O KZSNJWFQEVHDMF-UHFFFAOYSA-N 0.000 description 3
- 238000013459 approach Methods 0.000 description 3
- 239000012911 assay medium Substances 0.000 description 3
- 238000002869 basic local alignment search tool Methods 0.000 description 3
- 229930189065 blasticidin Natural products 0.000 description 3
- 238000006243 chemical reaction Methods 0.000 description 3
- 238000007796 conventional method Methods 0.000 description 3
- 235000018417 cysteine Nutrition 0.000 description 3
- 238000001514 detection method Methods 0.000 description 3
- UQLDLKMNUJERMK-UHFFFAOYSA-L di(octadecanoyloxy)lead Chemical compound [Pb+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O UQLDLKMNUJERMK-UHFFFAOYSA-L 0.000 description 3
- 238000010586 diagram Methods 0.000 description 3
- 238000010494 dissociation reaction Methods 0.000 description 3
- 230000005593 dissociations Effects 0.000 description 3
- 229940079593 drug Drugs 0.000 description 3
- 238000005516 engineering process Methods 0.000 description 3
- 239000013613 expression plasmid Substances 0.000 description 3
- 238000001914 filtration Methods 0.000 description 3
- 239000007850 fluorescent dye Substances 0.000 description 3
- 150000004676 glycans Chemical class 0.000 description 3
- 239000003102 growth factor Substances 0.000 description 3
- 210000002443 helper t lymphocyte Anatomy 0.000 description 3
- 238000010211 hemagglutination inhibition (HI) assay Methods 0.000 description 3
- 239000000185 hemagglutinin Substances 0.000 description 3
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 3
- 230000005847 immunogenicity Effects 0.000 description 3
- 238000001802 infusion Methods 0.000 description 3
- 230000003834 intracellular effect Effects 0.000 description 3
- 238000007912 intraperitoneal administration Methods 0.000 description 3
- 150000002500 ions Chemical class 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 238000010369 molecular cloning Methods 0.000 description 3
- 238000000569 multi-angle light scattering Methods 0.000 description 3
- 229920001542 oligosaccharide Polymers 0.000 description 3
- 230000008520 organization Effects 0.000 description 3
- 239000002245 particle Substances 0.000 description 3
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 3
- 230000008488 polyadenylation Effects 0.000 description 3
- 239000002243 precursor Substances 0.000 description 3
- 230000008569 process Effects 0.000 description 3
- 230000000069 prophylactic effect Effects 0.000 description 3
- 229950010131 puromycin Drugs 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 238000010561 standard procedure Methods 0.000 description 3
- 238000007920 subcutaneous administration Methods 0.000 description 3
- 239000000758 substrate Substances 0.000 description 3
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 3
- 241000701161 unidentified adenovirus Species 0.000 description 3
- 125000002987 valine group Chemical group [H]N([H])C([H])(C(*)=O)C([H])(C([H])([H])[H])C([H])([H])[H] 0.000 description 3
- 230000003442 weekly effect Effects 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- 244000105975 Antidesma platyphyllum Species 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 108091035707 Consensus sequence Proteins 0.000 description 2
- SHZGCJCMOBCMKK-UHFFFAOYSA-N D-mannomethylose Natural products CC1OC(O)C(O)C(O)C1O SHZGCJCMOBCMKK-UHFFFAOYSA-N 0.000 description 2
- 102000053602 DNA Human genes 0.000 description 2
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- WSFSSNUMVMOOMR-UHFFFAOYSA-N Formaldehyde Chemical compound O=C WSFSSNUMVMOOMR-UHFFFAOYSA-N 0.000 description 2
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 2
- 102000003886 Glycoproteins Human genes 0.000 description 2
- 108090000288 Glycoproteins Proteins 0.000 description 2
- 108050008339 Heat Shock Transcription Factor Proteins 0.000 description 2
- 102000000039 Heat Shock Transcription Factor Human genes 0.000 description 2
- 101000766965 Homo sapiens C-type lectin domain family 4 member K Proteins 0.000 description 2
- 101001018382 Homo sapiens Cartilage matrix protein Proteins 0.000 description 2
- 101000901659 Homo sapiens Myotonin-protein kinase Proteins 0.000 description 2
- 101000713602 Homo sapiens T-box transcription factor TBX21 Proteins 0.000 description 2
- 101000914514 Homo sapiens T-cell-specific surface glycoprotein CD28 Proteins 0.000 description 2
- 101000611023 Homo sapiens Tumor necrosis factor receptor superfamily member 6 Proteins 0.000 description 2
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 2
- 241000725303 Human immunodeficiency virus Species 0.000 description 2
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 2
- SHZGCJCMOBCMKK-DHVFOXMCSA-N L-fucopyranose Chemical compound C[C@@H]1OC(O)[C@@H](O)[C@H](O)[C@@H]1O SHZGCJCMOBCMKK-DHVFOXMCSA-N 0.000 description 2
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 2
- 125000000174 L-prolyl group Chemical group [H]N1C([H])([H])C([H])([H])C([H])([H])[C@@]1([H])C(*)=O 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- KZSNJWFQEVHDMF-BYPYZUCNSA-N L-valine Chemical compound CC(C)[C@H](N)C(O)=O KZSNJWFQEVHDMF-BYPYZUCNSA-N 0.000 description 2
- 206010025323 Lymphomas Diseases 0.000 description 2
- 241000829100 Macaca mulatta polyomavirus 1 Species 0.000 description 2
- 206010027476 Metastases Diseases 0.000 description 2
- OVRNDRQMDRJTHS-UHFFFAOYSA-N N-acelyl-D-glucosamine Natural products CC(=O)NC1C(O)OC(CO)C(O)C1O OVRNDRQMDRJTHS-UHFFFAOYSA-N 0.000 description 2
- SQVRNKJHWKZAKO-PFQGKNLYSA-N N-acetyl-beta-neuraminic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)O[C@H]1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-PFQGKNLYSA-N 0.000 description 2
- MBLBDJOUHNCFQT-LXGUWJNJSA-N N-acetylglucosamine Natural products CC(=O)N[C@@H](C=O)[C@@H](O)[C@H](O)[C@H](O)CO MBLBDJOUHNCFQT-LXGUWJNJSA-N 0.000 description 2
- 241000283973 Oryctolagus cuniculus Species 0.000 description 2
- 108091093037 Peptide nucleic acid Proteins 0.000 description 2
- 108010047620 Phytohemagglutinins Proteins 0.000 description 2
- 239000002202 Polyethylene glycol Substances 0.000 description 2
- 239000004365 Protease Substances 0.000 description 2
- 108010026552 Proteome Proteins 0.000 description 2
- 241000700159 Rattus Species 0.000 description 2
- 208000006265 Renal cell carcinoma Diseases 0.000 description 2
- 206010039491 Sarcoma Diseases 0.000 description 2
- QAOWNCQODCNURD-UHFFFAOYSA-N Sulfuric acid Chemical compound OS(O)(=O)=O QAOWNCQODCNURD-UHFFFAOYSA-N 0.000 description 2
- 230000005867 T cell response Effects 0.000 description 2
- 102100036840 T-box transcription factor TBX21 Human genes 0.000 description 2
- 102100027213 T-cell-specific surface glycoprotein CD28 Human genes 0.000 description 2
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 2
- 239000004473 Threonine Substances 0.000 description 2
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 2
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 2
- 101710096997 Tumor necrosis factor ligand superfamily member 18 Proteins 0.000 description 2
- 102100040403 Tumor necrosis factor receptor superfamily member 6 Human genes 0.000 description 2
- MZVQCMJNVPIDEA-UHFFFAOYSA-N [CH2]CN(CC)CC Chemical group [CH2]CN(CC)CC MZVQCMJNVPIDEA-UHFFFAOYSA-N 0.000 description 2
- 230000021736 acetylation Effects 0.000 description 2
- 238000006640 acetylation reaction Methods 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 2
- 238000000540 analysis of variance Methods 0.000 description 2
- 239000012131 assay buffer Substances 0.000 description 2
- SQVRNKJHWKZAKO-UHFFFAOYSA-N beta-N-Acetyl-D-neuraminic acid Natural products CC(=O)NC1C(O)CC(O)(C(O)=O)OC1C(O)C(O)CO SQVRNKJHWKZAKO-UHFFFAOYSA-N 0.000 description 2
- 238000004166 bioassay Methods 0.000 description 2
- 230000000903 blocking effect Effects 0.000 description 2
- 230000037396 body weight Effects 0.000 description 2
- 239000012830 cancer therapeutic Substances 0.000 description 2
- 150000001720 carbohydrates Chemical group 0.000 description 2
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 2
- 230000004663 cell proliferation Effects 0.000 description 2
- 230000005859 cell recognition Effects 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 238000003776 cleavage reaction Methods 0.000 description 2
- 238000012875 competitive assay Methods 0.000 description 2
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 2
- 239000002299 complementary DNA Substances 0.000 description 2
- 239000012141 concentrate Substances 0.000 description 2
- 239000002872 contrast media Substances 0.000 description 2
- 230000001276 controlling effect Effects 0.000 description 2
- 239000013078 crystal Substances 0.000 description 2
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 2
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 2
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 2
- 229940088598 enzyme Drugs 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 239000012894 fetal calf serum Substances 0.000 description 2
- 235000009424 haa Nutrition 0.000 description 2
- 238000004128 high performance liquid chromatography Methods 0.000 description 2
- 210000004408 hybridoma Anatomy 0.000 description 2
- 230000002209 hydrophobic effect Effects 0.000 description 2
- 230000016784 immunoglobulin production Effects 0.000 description 2
- 238000002513 implantation Methods 0.000 description 2
- 238000000126 in silico method Methods 0.000 description 2
- 230000006698 induction Effects 0.000 description 2
- 230000004941 influx Effects 0.000 description 2
- 238000001361 intraarterial administration Methods 0.000 description 2
- 238000007918 intramuscular administration Methods 0.000 description 2
- 238000002372 labelling Methods 0.000 description 2
- 238000004811 liquid chromatography Methods 0.000 description 2
- 208000014018 liver neoplasm Diseases 0.000 description 2
- 230000004807 localization Effects 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- 238000001823 molecular biology technique Methods 0.000 description 2
- 238000010172 mouse model Methods 0.000 description 2
- 201000000050 myeloid neoplasm Diseases 0.000 description 2
- 238000001543 one-way ANOVA Methods 0.000 description 2
- 230000003287 optical effect Effects 0.000 description 2
- 210000001672 ovary Anatomy 0.000 description 2
- 230000036961 partial effect Effects 0.000 description 2
- 230000007170 pathology Effects 0.000 description 2
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 2
- 230000026731 phosphorylation Effects 0.000 description 2
- 238000006366 phosphorylation reaction Methods 0.000 description 2
- 230000001885 phytohemagglutinin Effects 0.000 description 2
- 239000000902 placebo Substances 0.000 description 2
- 229940068196 placebo Drugs 0.000 description 2
- 229920001223 polyethylene glycol Polymers 0.000 description 2
- 230000002062 proliferating effect Effects 0.000 description 2
- 108020001580 protein domains Proteins 0.000 description 2
- 238000001742 protein purification Methods 0.000 description 2
- 230000005855 radiation Effects 0.000 description 2
- 238000003127 radioimmunoassay Methods 0.000 description 2
- 208000015347 renal cell adenocarcinoma Diseases 0.000 description 2
- 238000004007 reversed phase HPLC Methods 0.000 description 2
- 230000002441 reversible effect Effects 0.000 description 2
- 230000007017 scission Effects 0.000 description 2
- 210000002966 serum Anatomy 0.000 description 2
- 239000002904 solvent Substances 0.000 description 2
- 206010041823 squamous cell carcinoma Diseases 0.000 description 2
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 2
- 239000006228 supernatant Substances 0.000 description 2
- 230000001629 suppression Effects 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 230000008685 targeting Effects 0.000 description 2
- 238000002560 therapeutic procedure Methods 0.000 description 2
- 125000003396 thiol group Chemical group [H]S* 0.000 description 2
- 229960000187 tissue plasminogen activator Drugs 0.000 description 2
- 230000005030 transcription termination Effects 0.000 description 2
- 238000010361 transduction Methods 0.000 description 2
- 230000026683 transduction Effects 0.000 description 2
- 238000012546 transfer Methods 0.000 description 2
- 230000009466 transformation Effects 0.000 description 2
- 230000009261 transgenic effect Effects 0.000 description 2
- 108020005087 unfolded proteins Proteins 0.000 description 2
- 239000004474 valine Substances 0.000 description 2
- 210000002845 virion Anatomy 0.000 description 2
- DQJCDTNMLBYVAY-ZXXIYAEKSA-N (2S,5R,10R,13R)-16-{[(2R,3S,4R,5R)-3-{[(2S,3R,4R,5S,6R)-3-acetamido-4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy}-5-(ethylamino)-6-hydroxy-2-(hydroxymethyl)oxan-4-yl]oxy}-5-(4-aminobutyl)-10-carbamoyl-2,13-dimethyl-4,7,12,15-tetraoxo-3,6,11,14-tetraazaheptadecan-1-oic acid Chemical compound NCCCC[C@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)CC[C@H](C(N)=O)NC(=O)[C@@H](C)NC(=O)C(C)O[C@@H]1[C@@H](NCC)C(O)O[C@H](CO)[C@H]1O[C@H]1[C@H](NC(C)=O)[C@@H](O)[C@H](O)[C@@H](CO)O1 DQJCDTNMLBYVAY-ZXXIYAEKSA-N 0.000 description 1
- UKAUYVFTDYCKQA-UHFFFAOYSA-N -2-Amino-4-hydroxybutanoic acid Natural products OC(=O)C(N)CCO UKAUYVFTDYCKQA-UHFFFAOYSA-N 0.000 description 1
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 1
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N 100676-05-9 Natural products OC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1 OWEGMIWEEQEYGQ-UHFFFAOYSA-N 0.000 description 1
- NHBKXEKEPDILRR-UHFFFAOYSA-N 2,3-bis(butanoylsulfanyl)propyl butanoate Chemical compound CCCC(=O)OCC(SC(=O)CCC)CSC(=O)CCC NHBKXEKEPDILRR-UHFFFAOYSA-N 0.000 description 1
- IEJPPSMHUUQABK-UHFFFAOYSA-N 2,4-diphenyl-4h-1,3-oxazol-5-one Chemical compound O=C1OC(C=2C=CC=CC=2)=NC1C1=CC=CC=C1 IEJPPSMHUUQABK-UHFFFAOYSA-N 0.000 description 1
- UAIUNKRWKOVEES-UHFFFAOYSA-N 3,3',5,5'-tetramethylbenzidine Chemical compound CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 UAIUNKRWKOVEES-UHFFFAOYSA-N 0.000 description 1
- BRMWTNUJHUMWMS-UHFFFAOYSA-N 3-Methylhistidine Natural products CN1C=NC(CC(N)C(O)=O)=C1 BRMWTNUJHUMWMS-UHFFFAOYSA-N 0.000 description 1
- YRNWIFYIFSBPAU-UHFFFAOYSA-N 4-[4-(dimethylamino)phenyl]-n,n-dimethylaniline Chemical compound C1=CC(N(C)C)=CC=C1C1=CC=C(N(C)C)C=C1 YRNWIFYIFSBPAU-UHFFFAOYSA-N 0.000 description 1
- XAUDJQYHKZQPEU-KVQBGUIXSA-N 5-aza-2'-deoxycytidine Chemical compound O=C1N=C(N)N=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 XAUDJQYHKZQPEU-KVQBGUIXSA-N 0.000 description 1
- 229940117976 5-hydroxylysine Drugs 0.000 description 1
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 1
- 108010085238 Actins Proteins 0.000 description 1
- 102000007469 Actins Human genes 0.000 description 1
- 102100034540 Adenomatous polyposis coli protein Human genes 0.000 description 1
- 229920000936 Agarose Polymers 0.000 description 1
- 241000239290 Araneae Species 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- DCXYFEDJOCDNAF-UHFFFAOYSA-N Asparagine Natural products OC(=O)C(N)CC(N)=O DCXYFEDJOCDNAF-UHFFFAOYSA-N 0.000 description 1
- 241000711404 Avian avulavirus 1 Species 0.000 description 1
- 241000304886 Bacilli Species 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 208000005440 Basal Cell Neoplasms Diseases 0.000 description 1
- 206010004146 Basal cell carcinoma Diseases 0.000 description 1
- 206010004593 Bile duct cancer Diseases 0.000 description 1
- 241000283690 Bos taurus Species 0.000 description 1
- 241000701822 Bovine papillomavirus Species 0.000 description 1
- 101100161935 Caenorhabditis elegans act-4 gene Proteins 0.000 description 1
- 101100180402 Caenorhabditis elegans jun-1 gene Proteins 0.000 description 1
- 241000282472 Canis lupus familiaris Species 0.000 description 1
- 101710132601 Capsid protein Proteins 0.000 description 1
- 241000700198 Cavia Species 0.000 description 1
- 241000282693 Cercopithecidae Species 0.000 description 1
- 241000631130 Chrysophyllum argenteum Species 0.000 description 1
- 101710094648 Coat protein Proteins 0.000 description 1
- 102000014196 Coronin 1A Human genes 0.000 description 1
- 241000699802 Cricetulus griseus Species 0.000 description 1
- 102100031096 Cubilin Human genes 0.000 description 1
- IGXWBGJHJZYPQS-SSDOTTSWSA-N D-Luciferin Chemical compound OC(=O)[C@H]1CSC(C=2SC3=CC=C(O)C=C3N=2)=N1 IGXWBGJHJZYPQS-SSDOTTSWSA-N 0.000 description 1
- 108010041986 DNA Vaccines Proteins 0.000 description 1
- 229940021995 DNA vaccine Drugs 0.000 description 1
- CYCGRDQQIOGCKX-UHFFFAOYSA-N Dehydro-luciferin Natural products OC(=O)C1=CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 CYCGRDQQIOGCKX-UHFFFAOYSA-N 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 108010016626 Dipeptides Proteins 0.000 description 1
- 206010061818 Disease progression Diseases 0.000 description 1
- 206010014733 Endometrial cancer Diseases 0.000 description 1
- 206010014759 Endometrial neoplasm Diseases 0.000 description 1
- 102100030751 Eomesodermin homolog Human genes 0.000 description 1
- 241000283086 Equidae Species 0.000 description 1
- 108050001049 Extracellular proteins Proteins 0.000 description 1
- 108091006020 Fc-tagged proteins Proteins 0.000 description 1
- 241000282326 Felis catus Species 0.000 description 1
- 108090000331 Firefly luciferases Proteins 0.000 description 1
- BJGNCJDXODQBOB-UHFFFAOYSA-N Fivefly Luciferin Natural products OC(=O)C1CSC(C=2SC3=CC(O)=CC=C3N=2)=N1 BJGNCJDXODQBOB-UHFFFAOYSA-N 0.000 description 1
- PNNNRSAQSRJVSB-SLPGGIOYSA-N Fucose Natural products C[C@H](O)[C@@H](O)[C@H](O)[C@H](O)C=O PNNNRSAQSRJVSB-SLPGGIOYSA-N 0.000 description 1
- 108010058643 Fungal Proteins Proteins 0.000 description 1
- 230000005526 G1 to G0 transition Effects 0.000 description 1
- 241000287828 Gallus gallus Species 0.000 description 1
- 101000581641 Gallus gallus Cartilage matrix protein Proteins 0.000 description 1
- 206010017993 Gastrointestinal neoplasms Diseases 0.000 description 1
- 108700039691 Genetic Promoter Regions Proteins 0.000 description 1
- 208000032612 Glial tumor Diseases 0.000 description 1
- 206010018338 Glioma Diseases 0.000 description 1
- 108010070675 Glutathione transferase Proteins 0.000 description 1
- 108010015899 Glycopeptides Proteins 0.000 description 1
- 102000002068 Glycopeptides Human genes 0.000 description 1
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 1
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 1
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 1
- 102100029100 Hematopoietic prostaglandin D synthase Human genes 0.000 description 1
- 102100021519 Hemoglobin subunit beta Human genes 0.000 description 1
- 108091005904 Hemoglobin subunit beta Proteins 0.000 description 1
- 241000711549 Hepacivirus C Species 0.000 description 1
- 206010019695 Hepatic neoplasm Diseases 0.000 description 1
- 241000700721 Hepatitis B virus Species 0.000 description 1
- 208000007514 Herpes zoster Diseases 0.000 description 1
- 108010093488 His-His-His-His-His-His Proteins 0.000 description 1
- 108010033040 Histones Proteins 0.000 description 1
- 101000924577 Homo sapiens Adenomatous polyposis coli protein Proteins 0.000 description 1
- 101100005713 Homo sapiens CD4 gene Proteins 0.000 description 1
- 101001064167 Homo sapiens Eomesodermin homolog Proteins 0.000 description 1
- 101000998953 Homo sapiens Immunoglobulin heavy variable 1-2 Proteins 0.000 description 1
- 101001008255 Homo sapiens Immunoglobulin kappa variable 1D-8 Proteins 0.000 description 1
- 101001047628 Homo sapiens Immunoglobulin kappa variable 2-29 Proteins 0.000 description 1
- 101001008321 Homo sapiens Immunoglobulin kappa variable 2D-26 Proteins 0.000 description 1
- 101001047619 Homo sapiens Immunoglobulin kappa variable 3-20 Proteins 0.000 description 1
- 101001008263 Homo sapiens Immunoglobulin kappa variable 3D-15 Proteins 0.000 description 1
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 1
- 101000699781 Homo sapiens Retrotransposon Gag-like protein 4 Proteins 0.000 description 1
- 101000764263 Homo sapiens Tumor necrosis factor ligand superfamily member 4 Proteins 0.000 description 1
- 101000851370 Homo sapiens Tumor necrosis factor receptor superfamily member 9 Proteins 0.000 description 1
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 1
- 108091006905 Human Serum Albumin Proteins 0.000 description 1
- 102000008100 Human Serum Albumin Human genes 0.000 description 1
- 241000701085 Human alphaherpesvirus 3 Species 0.000 description 1
- 241000701806 Human papillomavirus Species 0.000 description 1
- PMMYEEVYMWASQN-DMTCNVIQSA-N Hydroxyproline Chemical compound O[C@H]1CN[C@H](C(O)=O)C1 PMMYEEVYMWASQN-DMTCNVIQSA-N 0.000 description 1
- 102000009490 IgG Receptors Human genes 0.000 description 1
- 108010073807 IgG Receptors Proteins 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 102100036887 Immunoglobulin heavy variable 1-2 Human genes 0.000 description 1
- 102100022949 Immunoglobulin kappa variable 2-29 Human genes 0.000 description 1
- 102000008070 Interferon-gamma Human genes 0.000 description 1
- 108010050904 Interferons Proteins 0.000 description 1
- 102000014150 Interferons Human genes 0.000 description 1
- 108010063738 Interleukins Proteins 0.000 description 1
- 102000015696 Interleukins Human genes 0.000 description 1
- 108020004684 Internal Ribosome Entry Sites Proteins 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- AHLPHDHHMVZTML-BYPYZUCNSA-N L-Ornithine Chemical compound NCCC[C@H](N)C(O)=O AHLPHDHHMVZTML-BYPYZUCNSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- UKAUYVFTDYCKQA-VKHMYHEASA-N L-homoserine Chemical compound OC(=O)[C@@H](N)CCO UKAUYVFTDYCKQA-VKHMYHEASA-N 0.000 description 1
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 1
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 1
- 239000012097 Lipofectamine 2000 Substances 0.000 description 1
- 241000186781 Listeria Species 0.000 description 1
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 1
- DDWFXDSYGUXRAY-UHFFFAOYSA-N Luciferin Natural products CCc1c(C)c(CC2NC(=O)C(=C2C=C)C)[nH]c1Cc3[nH]c4C(=C5/NC(CC(=O)O)C(C)C5CC(=O)O)CC(=O)c4c3C DDWFXDSYGUXRAY-UHFFFAOYSA-N 0.000 description 1
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 1
- 102000043131 MHC class II family Human genes 0.000 description 1
- 108091054438 MHC class II family Proteins 0.000 description 1
- 101100005714 Macaca fascicularis CD4 gene Proteins 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- GUBGYTABKSRVRQ-PICCSMPSSA-N Maltose Natural products O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-PICCSMPSSA-N 0.000 description 1
- 201000005505 Measles Diseases 0.000 description 1
- 108090000143 Mouse Proteins Proteins 0.000 description 1
- 241001529936 Murinae Species 0.000 description 1
- 101000933447 Mus musculus Beta-glucuronidase Proteins 0.000 description 1
- 101000748848 Mus musculus Coronin-1A Proteins 0.000 description 1
- 101100445364 Mus musculus Eomes gene Proteins 0.000 description 1
- 101100153537 Mus musculus Tnfrsf4 gene Proteins 0.000 description 1
- JDHILDINMRGULE-LURJTMIESA-N N(pros)-methyl-L-histidine Chemical compound CN1C=NC=C1C[C@H](N)C(O)=O JDHILDINMRGULE-LURJTMIESA-N 0.000 description 1
- OVRNDRQMDRJTHS-RTRLPJTCSA-N N-acetyl-D-glucosamine Chemical compound CC(=O)N[C@H]1C(O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-RTRLPJTCSA-N 0.000 description 1
- OVRNDRQMDRJTHS-FMDGEEDCSA-N N-acetyl-beta-D-glucosamine Chemical compound CC(=O)N[C@H]1[C@H](O)O[C@H](CO)[C@@H](O)[C@@H]1O OVRNDRQMDRJTHS-FMDGEEDCSA-N 0.000 description 1
- 238000005481 NMR spectroscopy Methods 0.000 description 1
- 108091061960 Naked DNA Proteins 0.000 description 1
- 101710141454 Nucleoprotein Proteins 0.000 description 1
- 239000004677 Nylon Substances 0.000 description 1
- 108010038807 Oligopeptides Proteins 0.000 description 1
- 102000015636 Oligopeptides Human genes 0.000 description 1
- AHLPHDHHMVZTML-UHFFFAOYSA-N Orn-delta-NH2 Natural products NCCCC(N)C(O)=O AHLPHDHHMVZTML-UHFFFAOYSA-N 0.000 description 1
- UTJLXEIPEHZYQJ-UHFFFAOYSA-N Ornithine Natural products OC(=O)C(C)CCCN UTJLXEIPEHZYQJ-UHFFFAOYSA-N 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 229930182555 Penicillin Natural products 0.000 description 1
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- BELBBZDIHDAJOR-UHFFFAOYSA-N Phenolsulfonephthalein Chemical compound C1=CC(O)=CC=C1C1(C=2C=CC(O)=CC=2)C2=CC=CC=C2S(=O)(=O)O1 BELBBZDIHDAJOR-UHFFFAOYSA-N 0.000 description 1
- 241000709664 Picornaviridae Species 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 101710083689 Probable capsid protein Proteins 0.000 description 1
- 102000002067 Protein Subunits Human genes 0.000 description 1
- 108010001267 Protein Subunits Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 108091027981 Response element Proteins 0.000 description 1
- 102100029131 Retrotransposon Gag-like protein 4 Human genes 0.000 description 1
- 102100037486 Reverse transcriptase/ribonuclease H Human genes 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 241000714474 Rous sarcoma virus Species 0.000 description 1
- 208000004337 Salivary Gland Neoplasms Diseases 0.000 description 1
- 206010061934 Salivary gland cancer Diseases 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N Silicium dioxide Chemical compound O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- 229930006000 Sucrose Natural products 0.000 description 1
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 1
- 108090000925 TNF receptor-associated factor 2 Proteins 0.000 description 1
- 102100034779 TRAF family member-associated NF-kappa-B activator Human genes 0.000 description 1
- 108020005038 Terminator Codon Proteins 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- YZCKVEUIGOORGS-NJFSPNSNSA-N Tritium Chemical compound [3H] YZCKVEUIGOORGS-NJFSPNSNSA-N 0.000 description 1
- 102000004142 Trypsin Human genes 0.000 description 1
- 108090000631 Trypsin Proteins 0.000 description 1
- 102100036856 Tumor necrosis factor receptor superfamily member 9 Human genes 0.000 description 1
- 241000282458 Ursus sp. Species 0.000 description 1
- 206010046865 Vaccinia virus infection Diseases 0.000 description 1
- 206010047741 Vulval cancer Diseases 0.000 description 1
- 208000004354 Vulvar Neoplasms Diseases 0.000 description 1
- 101100445365 Xenopus laevis eomes gene Proteins 0.000 description 1
- 238000010521 absorption reaction Methods 0.000 description 1
- 230000001133 acceleration Effects 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 239000004480 active ingredient Substances 0.000 description 1
- 208000009956 adenocarcinoma Diseases 0.000 description 1
- 239000002671 adjuvant Substances 0.000 description 1
- 238000001042 affinity chromatography Methods 0.000 description 1
- 238000007818 agglutination assay Methods 0.000 description 1
- 125000001931 aliphatic group Chemical group 0.000 description 1
- 230000009435 amidation Effects 0.000 description 1
- 238000007112 amidation reaction Methods 0.000 description 1
- 150000003862 amino acid derivatives Chemical class 0.000 description 1
- 210000004102 animal cell Anatomy 0.000 description 1
- 239000003957 anion exchange resin Substances 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 230000005809 anti-tumor immunity Effects 0.000 description 1
- 229940088710 antibiotic agent Drugs 0.000 description 1
- 210000000612 antigen-presenting cell Anatomy 0.000 description 1
- 230000000890 antigenic effect Effects 0.000 description 1
- 230000007503 antigenic stimulation Effects 0.000 description 1
- 239000007864 aqueous solution Substances 0.000 description 1
- 239000007900 aqueous suspension Substances 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 235000009582 asparagine Nutrition 0.000 description 1
- 229960001230 asparagine Drugs 0.000 description 1
- 229940009098 aspartate Drugs 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 235000019445 benzyl alcohol Nutrition 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 230000002457 bidirectional effect Effects 0.000 description 1
- 208000026900 bile duct neoplasm Diseases 0.000 description 1
- 238000013357 binding ELISA Methods 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 201000001531 bladder carcinoma Diseases 0.000 description 1
- 201000000053 blastoma Diseases 0.000 description 1
- 108010006025 bovine growth hormone Proteins 0.000 description 1
- 210000000481 breast Anatomy 0.000 description 1
- 238000002619 cancer immunotherapy Methods 0.000 description 1
- 239000002775 capsule Substances 0.000 description 1
- 230000021523 carboxylation Effects 0.000 description 1
- 238000006473 carboxylation reaction Methods 0.000 description 1
- 125000002057 carboxymethyl group Chemical group [H]OC(=O)C([H])([H])[*] 0.000 description 1
- 239000012876 carrier material Substances 0.000 description 1
- 230000015556 catabolic process Effects 0.000 description 1
- 238000005341 cation exchange Methods 0.000 description 1
- 150000001768 cations Chemical class 0.000 description 1
- 230000003915 cell function Effects 0.000 description 1
- 230000010261 cell growth Effects 0.000 description 1
- 230000006037 cell lysis Effects 0.000 description 1
- 239000006285 cell suspension Substances 0.000 description 1
- 230000007969 cellular immunity Effects 0.000 description 1
- 230000005754 cellular signaling Effects 0.000 description 1
- 239000001913 cellulose Substances 0.000 description 1
- 229920002678 cellulose Polymers 0.000 description 1
- 210000003679 cervix uteri Anatomy 0.000 description 1
- 239000003638 chemical reducing agent Substances 0.000 description 1
- 239000005081 chemiluminescent agent Substances 0.000 description 1
- 208000006990 cholangiocarcinoma Diseases 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 238000012411 cloning technique Methods 0.000 description 1
- 208000029742 colonic neoplasm Diseases 0.000 description 1
- 238000004440 column chromatography Methods 0.000 description 1
- 230000009137 competitive binding Effects 0.000 description 1
- 230000002860 competitive effect Effects 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 238000002591 computed tomography Methods 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 230000004940 costimulation Effects 0.000 description 1
- 230000000139 costimulatory effect Effects 0.000 description 1
- 230000009260 cross reactivity Effects 0.000 description 1
- 230000016396 cytokine production Effects 0.000 description 1
- 230000001461 cytolytic effect Effects 0.000 description 1
- 238000006731 degradation reaction Methods 0.000 description 1
- YSMODUONRAFBET-UHFFFAOYSA-N delta-DL-hydroxylysine Natural products NCC(O)CCC(N)C(O)=O YSMODUONRAFBET-UHFFFAOYSA-N 0.000 description 1
- 239000005547 deoxyribonucleotide Substances 0.000 description 1
- 125000002637 deoxyribonucleotide group Chemical group 0.000 description 1
- 238000001212 derivatisation Methods 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 238000003745 diagnosis Methods 0.000 description 1
- 238000012631 diagnostic technique Methods 0.000 description 1
- 238000006471 dimerization reaction Methods 0.000 description 1
- 230000005750 disease progression Effects 0.000 description 1
- BFMYDTVEBKDAKJ-UHFFFAOYSA-L disodium;(2',7'-dibromo-3',6'-dioxido-3-oxospiro[2-benzofuran-1,9'-xanthene]-4'-yl)mercury;hydrate Chemical compound O.[Na+].[Na+].O1C(=O)C2=CC=CC=C2C21C1=CC(Br)=C([O-])C([Hg])=C1OC1=C2C=C(Br)C([O-])=C1 BFMYDTVEBKDAKJ-UHFFFAOYSA-L 0.000 description 1
- 239000002270 dispersing agent Substances 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- PMMYEEVYMWASQN-UHFFFAOYSA-N dl-hydroxyproline Natural products OC1C[NH2+]C(C([O-])=O)C1 PMMYEEVYMWASQN-UHFFFAOYSA-N 0.000 description 1
- 239000003937 drug carrier Substances 0.000 description 1
- 230000000857 drug effect Effects 0.000 description 1
- 230000000431 effect on proliferation Effects 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 201000008184 embryoma Diseases 0.000 description 1
- 239000006274 endogenous ligand Substances 0.000 description 1
- 230000009144 enzymatic modification Effects 0.000 description 1
- 230000001973 epigenetic effect Effects 0.000 description 1
- 238000011067 equilibration Methods 0.000 description 1
- YSMODUONRAFBET-UHNVWZDZSA-N erythro-5-hydroxy-L-lysine Chemical compound NC[C@H](O)CC[C@H](N)C(O)=O YSMODUONRAFBET-UHNVWZDZSA-N 0.000 description 1
- 210000003743 erythrocyte Anatomy 0.000 description 1
- 208000021045 exocrine pancreatic carcinoma Diseases 0.000 description 1
- 238000000605 extraction Methods 0.000 description 1
- 239000000835 fiber Substances 0.000 description 1
- 238000013467 fragmentation Methods 0.000 description 1
- 238000006062 fragmentation reaction Methods 0.000 description 1
- 238000010230 functional analysis Methods 0.000 description 1
- 230000005021 gait Effects 0.000 description 1
- 229930182830 galactose Natural products 0.000 description 1
- 239000000499 gel Substances 0.000 description 1
- 238000001502 gel electrophoresis Methods 0.000 description 1
- 238000012817 gel-diffusion technique Methods 0.000 description 1
- 238000002523 gelfiltration Methods 0.000 description 1
- 238000003197 gene knockdown Methods 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 235000013922 glutamic acid Nutrition 0.000 description 1
- 239000004220 glutamic acid Substances 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 244000144993 groups of animals Species 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 210000003630 histaminocyte Anatomy 0.000 description 1
- 229940121372 histone deacetylase inhibitor Drugs 0.000 description 1
- 239000003276 histone deacetylase inhibitor Substances 0.000 description 1
- 239000005556 hormone Substances 0.000 description 1
- 229940088597 hormone Drugs 0.000 description 1
- 102000045167 human CD207 Human genes 0.000 description 1
- 102000048595 human DMPK Human genes 0.000 description 1
- 102000051450 human TNFSF4 Human genes 0.000 description 1
- 229910052739 hydrogen Inorganic materials 0.000 description 1
- 239000001257 hydrogen Substances 0.000 description 1
- 125000001165 hydrophobic group Chemical group 0.000 description 1
- 229910052588 hydroxylapatite Inorganic materials 0.000 description 1
- 229960002591 hydroxyproline Drugs 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 210000001822 immobilized cell Anatomy 0.000 description 1
- 230000005931 immune cell recruitment Effects 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000000984 immunochemical effect Effects 0.000 description 1
- 230000002163 immunogen Effects 0.000 description 1
- 229940072221 immunoglobulins Drugs 0.000 description 1
- 238000001114 immunoprecipitation Methods 0.000 description 1
- 238000012405 in silico analysis Methods 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 206010022000 influenza Diseases 0.000 description 1
- 208000037797 influenza A Diseases 0.000 description 1
- 208000037798 influenza B Diseases 0.000 description 1
- 239000004615 ingredient Substances 0.000 description 1
- 229920000592 inorganic polymer Polymers 0.000 description 1
- 229960003130 interferon gamma Drugs 0.000 description 1
- 229940047124 interferons Drugs 0.000 description 1
- 229940047122 interleukins Drugs 0.000 description 1
- 238000007917 intracranial administration Methods 0.000 description 1
- 238000007919 intrasynovial administration Methods 0.000 description 1
- 238000007913 intrathecal administration Methods 0.000 description 1
- 108010003082 intrinsic factor-cobalamin receptor Proteins 0.000 description 1
- 229940065638 intron a Drugs 0.000 description 1
- 238000005342 ion exchange Methods 0.000 description 1
- 238000004255 ion exchange chromatography Methods 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 238000005304 joining Methods 0.000 description 1
- 208000022013 kidney Wilms tumor Diseases 0.000 description 1
- 210000003292 kidney cell Anatomy 0.000 description 1
- 238000012933 kinetic analysis Methods 0.000 description 1
- 238000011005 laboratory method Methods 0.000 description 1
- 208000032839 leukemia Diseases 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 210000004185 liver Anatomy 0.000 description 1
- 201000007270 liver cancer Diseases 0.000 description 1
- 239000012160 loading buffer Substances 0.000 description 1
- 238000011068 loading method Methods 0.000 description 1
- 238000001325 log-rank test Methods 0.000 description 1
- 210000004072 lung Anatomy 0.000 description 1
- 201000005296 lung carcinoma Diseases 0.000 description 1
- 239000012139 lysis buffer Substances 0.000 description 1
- 238000002595 magnetic resonance imaging Methods 0.000 description 1
- 238000002826 magnetic-activated cell sorting Methods 0.000 description 1
- 230000014759 maintenance of location Effects 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 210000001161 mammalian embryo Anatomy 0.000 description 1
- 108010082117 matrigel Proteins 0.000 description 1
- 230000009401 metastasis Effects 0.000 description 1
- 229930182817 methionine Natural products 0.000 description 1
- 230000011987 methylation Effects 0.000 description 1
- 238000007069 methylation reaction Methods 0.000 description 1
- 230000000813 microbial effect Effects 0.000 description 1
- 208000030427 mucinous ovarian cancer Diseases 0.000 description 1
- 210000003097 mucus Anatomy 0.000 description 1
- 238000002887 multiple sequence alignment Methods 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- 210000000066 myeloid cell Anatomy 0.000 description 1
- 229950006780 n-acetylglucosamine Drugs 0.000 description 1
- 239000007922 nasal spray Substances 0.000 description 1
- 238000011815 naïve C57Bl6 mouse Methods 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 230000036963 noncompetitive effect Effects 0.000 description 1
- 238000007899 nucleic acid hybridization Methods 0.000 description 1
- 238000011330 nucleic acid test Methods 0.000 description 1
- 235000015097 nutrients Nutrition 0.000 description 1
- 229920001778 nylon Polymers 0.000 description 1
- 238000006384 oligomerization reaction Methods 0.000 description 1
- 238000002515 oligonucleotide synthesis Methods 0.000 description 1
- 229920000620 organic polymer Polymers 0.000 description 1
- 229960003104 ornithine Drugs 0.000 description 1
- 238000004806 packaging method and process Methods 0.000 description 1
- 210000000496 pancreas Anatomy 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 201000010198 papillary carcinoma Diseases 0.000 description 1
- 239000008188 pellet Substances 0.000 description 1
- 229940049954 penicillin Drugs 0.000 description 1
- XYJRXVWERLGGKC-UHFFFAOYSA-D pentacalcium;hydroxide;triphosphate Chemical compound [OH-].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O XYJRXVWERLGGKC-UHFFFAOYSA-D 0.000 description 1
- 210000005259 peripheral blood Anatomy 0.000 description 1
- 239000011886 peripheral blood Substances 0.000 description 1
- 230000002093 peripheral effect Effects 0.000 description 1
- 239000008024 pharmaceutical diluent Substances 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- 229940124531 pharmaceutical excipient Drugs 0.000 description 1
- 229960003531 phenolsulfonphthalein Drugs 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000008363 phosphate buffer Substances 0.000 description 1
- 230000004962 physiological condition Effects 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- 229920002401 polyacrylamide Polymers 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 229920000136 polysorbate Polymers 0.000 description 1
- 229950008882 polysorbate Drugs 0.000 description 1
- 230000034190 positive regulation of NF-kappaB transcription factor activity Effects 0.000 description 1
- 230000004481 post-translational protein modification Effects 0.000 description 1
- 230000001124 posttranscriptional effect Effects 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 238000002360 preparation method Methods 0.000 description 1
- 239000003755 preservative agent Substances 0.000 description 1
- 230000002265 prevention Effects 0.000 description 1
- 238000004393 prognosis Methods 0.000 description 1
- 125000001500 prolyl group Chemical group [H]N1C([H])(C(=O)[*])C([H])([H])C([H])([H])C1([H])[H] 0.000 description 1
- 235000019419 proteases Nutrition 0.000 description 1
- 230000004952 protein activity Effects 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 230000009145 protein modification Effects 0.000 description 1
- 230000017854 proteolysis Effects 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 230000006337 proteolytic cleavage Effects 0.000 description 1
- 230000002285 radioactive effect Effects 0.000 description 1
- 108010027270 rat glucocorticoid-induced tumor necrosis factor receptor ligand Proteins 0.000 description 1
- 230000007420 reactivation Effects 0.000 description 1
- 238000003753 real-time PCR Methods 0.000 description 1
- 239000000018 receptor agonist Substances 0.000 description 1
- 229940044601 receptor agonist Drugs 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 201000010174 renal carcinoma Diseases 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 210000003935 rough endoplasmic reticulum Anatomy 0.000 description 1
- 238000005185 salting out Methods 0.000 description 1
- 150000003839 salts Chemical class 0.000 description 1
- 238000003118 sandwich ELISA Methods 0.000 description 1
- 238000010187 selection method Methods 0.000 description 1
- 238000001338 self-assembly Methods 0.000 description 1
- 238000002864 sequence alignment Methods 0.000 description 1
- SQVRNKJHWKZAKO-OQPLDHBCSA-N sialic acid Chemical compound CC(=O)N[C@@H]1[C@@H](O)C[C@@](O)(C(O)=O)OC1[C@H](O)[C@H](O)CO SQVRNKJHWKZAKO-OQPLDHBCSA-N 0.000 description 1
- 239000000741 silica gel Substances 0.000 description 1
- 229910002027 silica gel Inorganic materials 0.000 description 1
- 238000009097 single-agent therapy Methods 0.000 description 1
- 238000003998 size exclusion chromatography high performance liquid chromatography Methods 0.000 description 1
- 238000004513 sizing Methods 0.000 description 1
- 239000011734 sodium Substances 0.000 description 1
- 229910052708 sodium Inorganic materials 0.000 description 1
- 239000001488 sodium phosphate Substances 0.000 description 1
- 229910000162 sodium phosphate Inorganic materials 0.000 description 1
- 239000008279 sol Substances 0.000 description 1
- 230000003381 solubilizing effect Effects 0.000 description 1
- 238000000527 sonication Methods 0.000 description 1
- 125000006850 spacer group Chemical group 0.000 description 1
- 238000001228 spectrum Methods 0.000 description 1
- 210000004988 splenocyte Anatomy 0.000 description 1
- 239000007921 spray Substances 0.000 description 1
- 230000006641 stabilisation Effects 0.000 description 1
- 238000011105 stabilization Methods 0.000 description 1
- 230000010473 stable expression Effects 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 239000012089 stop solution Substances 0.000 description 1
- 229960005322 streptomycin Drugs 0.000 description 1
- 239000005720 sucrose Substances 0.000 description 1
- 125000004434 sulfur atom Chemical group 0.000 description 1
- 239000004094 surface-active agent Substances 0.000 description 1
- 230000002459 sustained effect Effects 0.000 description 1
- 239000003826 tablet Substances 0.000 description 1
- 229940124597 therapeutic agent Drugs 0.000 description 1
- 230000004797 therapeutic response Effects 0.000 description 1
- 125000000341 threoninyl group Chemical group [H]OC([H])(C([H])([H])[H])C([H])(N([H])[H])C(*)=O 0.000 description 1
- 238000004448 titration Methods 0.000 description 1
- 230000000699 topical effect Effects 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 108700012359 toxins Proteins 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 238000001890 transfection Methods 0.000 description 1
- 230000001052 transient effect Effects 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- 230000014621 translational initiation Effects 0.000 description 1
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 1
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 1
- 229910052722 tritium Inorganic materials 0.000 description 1
- 239000012588 trypsin Substances 0.000 description 1
- 238000013385 tryptic peptide mapping Methods 0.000 description 1
- 210000003171 tumor-infiltrating lymphocyte Anatomy 0.000 description 1
- 150000003668 tyrosines Chemical class 0.000 description 1
- 238000004704 ultra performance liquid chromatography Methods 0.000 description 1
- 238000000108 ultra-filtration Methods 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 238000011144 upstream manufacturing Methods 0.000 description 1
- 208000010570 urinary bladder carcinoma Diseases 0.000 description 1
- 229960005486 vaccine Drugs 0.000 description 1
- 208000007089 vaccinia Diseases 0.000 description 1
- 239000003981 vehicle Substances 0.000 description 1
- 230000035899 viability Effects 0.000 description 1
- 230000007419 viral reactivation Effects 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 238000012800 visualization Methods 0.000 description 1
- 239000011800 void material Substances 0.000 description 1
- 201000005102 vulva cancer Diseases 0.000 description 1
- 238000001262 western blot Methods 0.000 description 1
- 238000002424 x-ray crystallography Methods 0.000 description 1
- ONDPHDOFVYQSGI-UHFFFAOYSA-N zinc nitrate Chemical compound [Zn+2].[O-][N+]([O-])=O.[O-][N+]([O-])=O ONDPHDOFVYQSGI-UHFFFAOYSA-N 0.000 description 1
Images





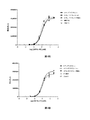















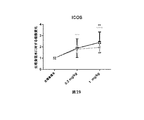
















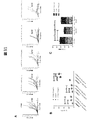


























Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/52—Cytokines; Lymphokines; Interferons
- C07K14/525—Tumour necrosis factor [TNF]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- A61K38/17—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- A61K38/19—Cytokines; Lymphokines; Interferons
- A61K38/191—Tumor necrosis factors [TNF], e.g. lymphotoxin [LT], i.e. TNF-beta
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
- A61K45/06—Mixtures of active ingredients without chemical characterisation, e.g. antiphlogistics and cardiaca
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
- A61K47/51—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent
- A61K47/68—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being an antibody, an immunoglobulin or a fragment thereof, e.g. an Fc-fragment
- A61K47/6801—Drug-antibody or immunoglobulin conjugates defined by the pharmacologically or therapeutically active agent
- A61K47/6803—Drugs conjugated to an antibody or immunoglobulin, e.g. cisplatin-antibody conjugates
- A61K47/6811—Drugs conjugated to an antibody or immunoglobulin, e.g. cisplatin-antibody conjugates the drug being a protein or peptide, e.g. transferrin or bleomycin
- A61K47/6813—Drugs conjugated to an antibody or immunoglobulin, e.g. cisplatin-antibody conjugates the drug being a protein or peptide, e.g. transferrin or bleomycin the drug being a peptidic cytokine, e.g. an interleukin or interferon
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
- A61P35/02—Antineoplastic agents specific for leukemia
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70575—NGF/TNF-superfamily, e.g. CD70, CD95L, CD153, CD154
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2878—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the NGF-receptor/TNF-receptor superfamily, e.g. CD27, CD30, CD40, CD95
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/62—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising only variable region components
- C07K2317/622—Single chain antibody (scFv)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/75—Agonist effect on antigen
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/30—Non-immunoglobulin-derived peptide or protein having an immunoglobulin constant or Fc region, or a fragment thereof, attached thereto
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/35—Fusion polypeptide containing a fusion for enhanced stability/folding during expression, e.g. fusions with chaperones or thioredoxin
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/70—Fusion polypeptide containing domain for protein-protein interaction
- C07K2319/73—Fusion polypeptide containing domain for protein-protein interaction containing coiled-coiled motif (leucine zippers)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/70—Fusion polypeptide containing domain for protein-protein interaction
- C07K2319/735—Fusion polypeptide containing domain for protein-protein interaction containing a domain for self-assembly, e.g. a viral coat protein (includes phage display)
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Immunology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Pharmacology & Pharmacy (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Gastroenterology & Hepatology (AREA)
- Epidemiology (AREA)
- Toxicology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Cell Biology (AREA)
- Oncology (AREA)
- Hematology (AREA)
- Peptides Or Proteins (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicinal Preparation (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
【選択図】図1
Description
本願とともに提出された、ASCIIテキストファイル(名称GITRLF−100P2_ST25.txt;サイズ:56,159バイト;および作成日:2016年6月15日)にて電子的に提出された配列表の内容は、その全体が参照により本明細書中に援用される。
用語「1つの(a)」または「1つの(an)」実体は、その実体の1つまたは複数を指し、例えば、「ポリペプチドサブユニット」は、1つまたは複数のポリペプチドサブユニットを表すと理解される。従って、用語「1つの(a)」(もしくは「1つの(an)」)、「1つまたは複数」および「少なくとも1つ」は、本明細書において互換的に使用することができる。

本開示は、限定はされないが、遺伝子導入された哺乳動物細胞培養液中で発現されるときの改善された収量;GITRに対する改善された結合親和性;様々な生物検定における改善された活性および/または精製または部分精製されるときのGITRLを含有する以前に開示されたポリペプチドと比べて改善された均一性を含む、改善された特性を有する多量体、例えば三量体または六量体タンパク質に組織化し得るGITRL融合ポリペプチドサブユニットに関する(例えば、Wyzgol et al.,J Immunol 2009;183:1851−1861を参照)。本明細書で提供されるGITRL融合ポリペプチドサブユニットは、IgG Fcドメイン、例えばヒトIgG Fcドメイン、三量体化ドメイン、およびヒトGITRLの受容体結合ドメインを含み得る。例示的な実施形態は、図7に概略的に図示される。典型的には、IgG Fcドメイン、三量体化ドメインおよびGITRL受容体結合ドメインは、N末端からC末端方向に配列される。例示的なGITRL IgG1融合ポリペプチドサブユニットは、配列番号6によって表される。
上記のようなGITRL融合ポリペプチドサブユニットは、六量体GITRL FPに自己組織化し得る。したがって、本開示は、上記のような6つのポリペプチドサブユニットを含む六量体タンパク質を提供する。実施例に記載の例示的なポリペプチドサブユニットは、本明細書中で「六量体GITRL FP」と称される六量体タンパク質に自己組織化する。六量体GITRL FPに自己組織化するGITRL融合ポリペプチドサブユニットのアミノ酸配列の非限定例が、配列番号6で提供される。にもかかわらず、当業者は、本開示に照らして、極めて多数の他の配列についても本明細書で示される六量体GITRL FPについての判定基準を満たすことを理解するであろう。配列番号6に対して少なくとも70%、80%、85%、90%、95%、96%、97%、98%、99%、99.5%、もしくは100%の配列同一性を有する6つのGITRL融合ポリペプチドサブユニットを含む六量体GITRL FPもまた、本明細書で提供される。
本開示は、本明細書で提供されるようなGITRL融合ポリペプチドサブユニットまたは六量体タンパク質、例えばGITRL FPをコードする核酸を含むポリヌクレオチドをさらに提供する。GITRL融合ポリペプチドサブユニットをコードする例示的なポリヌクレオチド配列は、配列番号5によって表される。特定の態様では、IgG Fcドメイン、三量体化ドメインおよびGITRL受容体結合ドメインをコードする核酸配列は、5’から3’の方向へ連結され、例えば5’から3’の方向へ隣接して連結される。他の態様では、提供されるポリヌクレオチドは、例えば分泌シグナルペプチドまたは膜局在化配列をコードするシグナル配列をさらに含み得る。ありとあらゆるGITRL融合ポリペプチドサブユニットまたは多量体、例えばそのサブユニットを含む六量体タンパク質をコードするポリヌクレオチドが、本開示によって提供される。
本明細書で提供されるような六量体タンパク質、例えば本明細書で提供されるようなGITRL FPを調製し、投与する方法は、それを必要とする被験者へは、例えばがん患者における免疫応答を増強する、例えば腫瘍成長を阻害もしくは低減するため、またはウイルス感染を有する患者へは、例えばウイルス負荷を低減するかまたはウイルス感染の再発を低減するため、当業者に周知であるかまたは当業者によって容易に決定され得る。本明細書に記載する六量体タンパク質、例えば、GITRL FPの投与経路は、例えば、経口、非経口、吸入または局所であってよい。本明細書で使用される非経口という用語は、例えば、静脈内、動脈内、腹腔内、筋肉内、皮下、直腸、または膣内投与を含む。これらの投与形態は全て、好適な形態として明らかに考慮されるが、別の投与形態の例として、特に、静脈内もしくは動脈内注射または滴注がある。通常、好適な医薬組成物は、限定されるものではないが、バッファー(例えば、酢酸、リン酸もしくはクエン酸バッファー)、界面活性剤(例えば、ポリソルベート)、安定剤(例えば、ヒトアルブミン)などを含み得る。本明細書の教示内容に適合可能な別の方法では、本明細書に記載する六量体タンパク質、例えば、本明細書に記載のGITRL FPは、有害な細胞集団の部位に直接送達され、これによって治療薬に対する患部組織の曝露を増大することができる。
本開示は、さらに、本明細書に記載する六量体タンパク質、例えば、GITRL FPを含み、本明細書に記載する方法を実施するために用いることができるキットも提供する。いくつかの態様では、キットは、少なくとも1種の本明細書に記載する精製六量体タンパク質、例えば、GITRL FPを1つまたは複数の容器内に含む。当業者は、本明細書で提供される開示した六量体タンパク質、例えば、GITRL FPが、当分野でよく知られている既定のキットフォーマットの1つに容易に組み込まれ得ることを容易に認識されよう。
本明細書に記載する六量体タンパク質、例えば、GITRL FPは、当分野で公知の任意の方法により、特異的および/または選択的結合についてアッセイすることができる。用いることができる免疫アッセイとして、いくつか挙げると、ウェスタンブロット、放射性免疫アッセイ、ELISA(酵素結合イムノソルベントアッセイ)、蛍光フォーカスアッセイ(FFA)、マイクロ中和アッセイ、血球凝集阻害アッセイ(HAI)、「サンドイッチ」免疫アッセイ、免疫沈降アッセイ、沈降素反応、ゲル拡散沈降反応、免疫核酸アッセイ、凝集アッセイ、補体結合アッセイ、免疫放射定量測定、蛍光免疫アッセイなどの技法を用いる競合または非競合アッセイシステムがあるが、これらに限定されるものではない。こうしたアッセイは、当分野ではよく知られている(例えば、Ausubel et al.,eds,(1994)Current Protocols in Molecular Biology(John Wiley & Sons,Inc.,NY)Vol.1を参照;これは、全体として参照により本明細書に組み込む)。FFA、マイクロ中和アッセイ、およびHAIは、以下の実施例に詳述する。
抗原活性化の間または後に抗原を経験したT細胞上、例えば抗原を経験したCD4+もしくはCD8+T細胞上のGITRに会合することによる被験者(例えば哺乳動物被験者、例えばヒト被験者)における抗原特異的免疫応答の増強は、多種多様な方法を用いて達成され得る。選択方法は主に、免疫応答を増強することが所望される場合の抗原のタイプに依存することになり、また利用可能な様々な方法が以下で考察される。いかなる方法が選択されても、本明細書で提供されるような六量体タンパク質、例えばGITRL FPは、被験者、例えばヒト患者に、被験者のT細胞に提示されるように、抗原によるT細胞の予備刺激の間または直後に投与され得る。OX40六量体タンパク質を用いて被験者、例えばヒト被験者における免疫応答を活性化する例示的な方法は、米国特許出願公開第2016/0024176号明細書(その全体が参照により本明細書中に援用される)中に提示されており、本明細書で提供されるGITRL FPとの併用に適応され得る。
T細胞プライミング剤と組み合わせて、六量体タンパク質または六量体タンパク質を含む組成物を有効量で被験者に投与するステップを含む、治療を必要とする被験者におけるがんを治療する方法もまた提供される。特定の態様では、T細胞プライミング剤は、DNAワクチン+アジュバントである。特定の態様では、この組み合わせは、例えば、E7合成ロングペプチド(SLP)とCpGオリゴデオキシヌクレオチドである。さらなる態様では、T細胞プライミング剤は、エピジェネティック修飾因子、例えば、5−アザ−2’−デオキシシチジンまたはヒストン修飾因子、例えばHDAC阻害剤である。さらなる態様では、T細胞プライミング剤は、ウイルス、例えば、ワクシニア(Vaccinia)、リステリア(Listeria)、またはニューキャッスル病ウイルス(Newcastle disease virus)である。
多量体ヒトGITRL−FP分子を作製するために用いることができる好適なヒト三量体化モチーフの同定
ヒト六量体GITRL FPを作製するための努力は、元々、GITRL三量体を安定化させ、六量体タンパク質を形成するため、GCN4 pII三量体化モチーフの使用に注目していた。しかし、このモチーフは、酵母タンパク質から誘導され、非ヒトモチーフを有するGITRL FPのヒトへの投与が免疫原性をもたらすことがある。したがって、ヒトタンパク質から誘導される三量体化モチーフを有する等価なGITRL FPを作製することが望ましいと考えられた。三量体コイルドコイルモチーフを形成する51タンパク質に由来するアミノ酸配列は、その三次元結晶構造またはオルソログの場合によって示される通り、Protein Data Bank(PDB;ワールドワイドウェブサイト「wwpdb.org」経由でインターネットによりアクセス可能)を介して同定し、経験による実験的試験におけるGITRL FP分子への組み込みとして、4つのコイルドコイルモチーフのサブセットを選択した。
コロニン1A(PDBコード:2akf)、マトリリン1(1aq5)、ランゲリン(3kqg)および筋強直性ジストロフィーキナーゼDMPK(PDBコード:1wt6)コイルドコイルモチーフの三次元結晶構造を、PyMol Visualisation Graphicsソフトウェアを用いて分析した。
51の候補配列を同定した三量体コイルドコイルタンパク質配列についてのProtein Data Bank(PDB)の検索後、多量体GITRL FPの作製に用いるため、4つの配列を選択した(表1−1)。これら4つの配列は、ヒトタンパク質のコロニン1a、マトリリン1、ランゲリンおよびDMPKに由来するコイルドコイル配列であった。マウスコロニン1a(PDBコード:2akf)由来のコイルドコイルモチーフの構造は、溶液中で安定な三量体であることが報告されている(Kammerer,R.A.et al.PNAS,vol:102,p13891−13896(2005))。ヒトコロニン1aのコイルドコイル配列は、マウスタンパク質に対して高い配列同一性(78.1%の配列同一性)を示すことから、三量体コイルドコイル構造を形成することが予測された。同様に、ニワトリマトリリン1(PDBコード:1aq5)由来のコイルドコイルモチーフの構造は、三量体であることが報告されている(Dames,S.A.et al.NAT.STRUCT.BIOL.5:687−691(1998);PDBコード:1aq5)。ヒトマトリリン1のコイルドコイル配列は、三量体コイルドコイル構造を、ニワトリオルソログに対するその高い配列同一性(60.0%の配列同一性)に基づいて形成することが予測された。ヒトランゲリン(PDBコード:3kqg)およびヒトDMPK(PDBコード:1wt6)のコイルドコイル配列もまた、三量体であることが報告されており(Feinberg,H.et al.,J.BIOL.CHEM.285:13285−13293(2010))、多量体ヒトGITRL融合タンパク質の作製に用いるために選択した。
より安定なGITRL三量体、ひいてはより安定な多量体GITRL FPを生成することになるモチーフのインシリコ予測は、表1−1中の配列をそれらのオリゴマー状態を予測するためのアルゴリズム(LOGICOIL)を用いて分析することによって行った。さらに、平行三量体の形成において得られたスコアを、2つのアルゴリズム(MARCOILおよびLOGICOIL)の組み合わせを介して、全ヒトプロテオームから同定されたコイルドコイルからのスコアと比べた。
ftpサイト「ftp.ncbi.nlm.nih.gov/refseq/H_sapiens/mRNA_Prot/human.protein.faa.gz.」からダウンロードしたヒトプロテオームに対して、ゲノムワイドな予測を実施した。このバージョンは、総数71861のタンパク質配列を含む。これらの配列をスキャンし、ワールドワイドウェブサイト「bcf.isb−sib.ch/Delorenzi/Marcoil/Marcoilcode.tar.gz」からダウンロードしたソフトウェアMARCOILを実行することにより、コイルドコイルモチーフを同定した。コイルドコイル配列のオリゴマー形成状態(平行もしくは抗平行二量体、三量体または四量体)を予測するため、特定の具体的な閾値(すなわち、0.01、0.10、0.50、0.90および0.99)を通るモチーフのすべてを、ワールドワイドウェブサイト「coiledcoils.chm.bris.ac.uk/LOGICOIL/LOGICOIL_Source.zip」経由でインターネットからダウンロードしたLOGICOILアルゴリズムを用いてさらに分析した。このパッケージをR言語(バージョン3.0.2−2013−09−25)において実行した。MARCOILからの出力をLOGICOILによって想定されるファイルフォーマットに変換するため、アドホックperlスクリプトを開発した。latticeパッケージ(バージョン0.20〜23)を用いて、ヒストグラムをRでプロットした。
Refseqからのすべてのヒト配列内のコイルドコイルモチーフを(MARCOILを用いて)予測した。異なる閾値に対するMARCOILアルゴリズムによって予測されるコイルドコイルモチーフの数を表1−2に示す。次に、0.99の閾値からのすべてのモチーフのオリゴマー状態を、表1−2での8つの選択した三量体コイルドコイルドメインとともに、LOGICOILを用いて予測した。
選択した三量体化モチーフの各々における配列は、GCN4 pII配列に加えて、適切な立体配置で、GITRL FPの他の要素(すなわち、短い可動性アミノ酸リンカー配列によって分離された、ヒトFcドメインとヒトGITRLの細胞外ドメインとの間にあるもの)をコードするDNAベクターにクローン化した。これらのベクターを、哺乳動物細胞を一過性に遺伝子導入するために用いて、組換え六量体GITRL FPタンパク質の分泌とその後の精製を可能にした。
懸濁CHO細胞に、PEIを用いて、異なる六量体GITRL FPをコードするDNAベクターを一過性に遺伝子導入し、80%の湿度下、140rpmで振盪しながら37℃で8日間成長させた。分泌タンパク質を含有する条件培地40mLを、1,600×gでの遠心分離および濾過により細胞および細胞片から分離した。タンパク質をMab SelectSure(商標)樹脂を用いて精製し、そのサイズおよび完全性を還元SDS−ポリアクリルアミドゲル電気泳動法により分析した。
組換え六量体GITRL FPタンパク質のすべてが類似レベルで発現したが、中でもコロニン1a wtモチーフを有する六量体GITRL−FPは、他のタンパク質よりも高収量をもたらすように思われた(表1−4:図1)。
GITRL FP変異体のGITRを発現する細胞への結合
六量体GITRL FP分子が細胞表面に発現されるGITRに結合し得るか否かを判定するため、六量体GITRL FP分子を、GITRを過剰発現する細胞とともにインキュベートし、それらの結合をそれらのFcドメインを介して検出した。
固定濃度のDyLight−649に複合した抗ヒトIgG抗体緩衝液、その後、セクション1.3中の上記の2倍希釈した精製六量体GITRL FPタンパク質およびアイソタイプ対照抗体(NIP228)を、2連ウェルで384ウェルの黒色壁付きの透明な底板に添加した。GITRを安定的に発現するCHO細胞をすべてのウェルに添加し、プレートを室温で4時間インキュベートした。CHO−GITR細胞への結合を、640nMのレーザーを用いるMirrorball(商標)プレート血球計数器を用いて測定し、蛍光を測定した。
結果を図2A〜Dに提示する。得られた結合特性は、六量体GITRL FPの濃度がDyLight−649に複合した検出抗体の濃度を超えるときに認められる「フック効果」を示す結果、六量体GITRL FPの濃度が高まるにつれて結合の低下が検出され、それ故、釣鐘状の結合特性がもたらされる。この現象にもかかわらず、異なる六量体GITRL FP分子は、酷似した結合特性をもたらす。
ヒトGITRLのヒトGITRへの結合に対する六量体GITRL FP分子の効果を、均一時間分解蛍光(HTRF)アッセイを用いて測定した。
六量体GITRL FP分子を、GITRL−HA(ヘマグルチニンタグ)のGITR−Fcへの結合が測定されるHTRFアッセイで滴定した。ヒトGITR Fcをユーロピウムクリプテートと複合させ、XL665と複合させた抗HA抗体を用いてGITRL−HAタンパク質を検出した。Prism 5.01ソフトウェア(GraphPad)を用いて分析データを4つのパラメータロジスティック方程式にカーブフィッティングすることにより、IC50値を判定した。
結果を図3A〜Dおよび表1−5に提示する。異なる三量体化モチーフを有する六量体GITRL FP分子はすべて、三量体GITRL−HAのGITR−Fcへの結合への強力な阻害剤である。GITRL FP(GCN4)よりも8倍低い効力を示すGITRL FP(ランゲリンwt)を例外として、大部分のGITRL FP変異体が、類似の阻害特性およびIC50値をもたらした。GITRL FP(コロニン1a)は、0.61nMで最低のIC50値を有した。
異なるGITRL FP分子(六量体GITRL IgG1 FP(配列番号6)および六量体GITRL IgG4 FP(配列番号40)の機能的活性を、GITRを安定的に発現するNFκBルシフェラーゼレポーター細胞を用いるアッセイにおいて測定した。GITRのアゴニズムおよびその後のNFκB経路の活性化によって駆動される発光を測定した。
6ポイントデータ曲線のため、GITRL FPタンパク質を4倍に連続希釈し、3通りに96ウェルプレートに添加した。次に、ヒトGITRを遺伝子導入したJurkat NF−κBルシフェラーゼレポーター細胞を、アッセイプレートのすべてのウェルに添加し、37℃で3時間インキュベートした。Steady−Glo試薬をアッセイプレートのすべてのウェルに添加することにより、ルシフェラーゼ発現を検出した。プレートを室温で5分間インキュベートし、次に発光を測定し、対数(アゴニスト)対応答のGraphPad Prism5.01(GraphPad)での可変勾配非線形曲線フィットを用いてEC50値を生成した。
結果を図4A〜Dおよび表1−6に示す。すべての六量体GITRL FPタンパク質がNF−κBシグナル伝達を誘導する。タンパク質は、GITRL FP(GCN4)に対して類似の効力特性およびEC50値を生成した。
GITRL FPタンパク質の融解温度を、放射特性が折り畳まれていないタンパク質の存在下で変化する蛍光色素(Sypro Orange)を用いて測定した。
多量体GITRL FP変異体の熱安定性を、融解温度(Tm)を計算するためのSypro Orangeに基づくアッセイを用いて評価した。異なるタンパク質を、96ウェルPCRプレートに懸濁する前、最初に2×PBSで0.5mg/mLまで希釈した。Sypro Orange(商標)をプレート上の各ウェルに添加し、次に密封した。プレートを、Chromo4(商標)連続蛍光検出器を用いてリアルタイム(商標)PCR装置上で読み取った。温度は、1℃ごとの読み取りと1秒の保持時間で20℃から90℃に高めるように設定した。アンフォールディング転移を、蛍光強度および蛍光誘導体を温度の関数としてプロットすることによって判定した。各多量体GITRL−FPタンパク質を二通りに分析した。
結果を図5A〜Dに示す。9つの多量体GITRL−FPタンパク質の各々における融解温度を下の表1−7にまとめる。すべての変異体は、他の大部分、すなわち低温(45〜50℃)で追加的な転移ピークを呈する(これは一部の構造的不安定性を示唆する)GITRL FP(ランゲリンwt)およびGITRL FP(ランゲリン変異体)の双方における62〜64℃と比べて59℃で単一の転移ピークを呈するGITRL FP(DMPK wt)以外では、類似の特性を有する。
8つのコイルドコイルドメインおよびそれら周囲の配列の予測される免疫原性を、ProPredアルゴリズムを用いて判定した。
ProPred[Singh&Raghava(2001)Bioinformatics 17(12)]を用いて、8つの選択したヒト三量体化モチーフ、および酵母GCN4 pIIのTスコアを評価した。任意のアミノ酸配列のTスコアにより、アミノ酸配列の全長に対して強力に結合するMHCクラスIIエピトープの数が定量される。ProPredが10,000の無作為に生成される9−mer配列セットを評価することによって得られる95パーセンタイル超のスコアに位置する結合スコアを戻す場合、9−merの部分配列は強力に結合するエピトープであると考えられる。Tスコアは、8つの最も一般的なヒト対立遺伝子に対して、モチーフに先行する2×[G4S]配列ならびにモチーフに後続する[G4]配列およびGITRLドメインの最初の4アミノ酸を含む各モチーフにつき計算した。次に、個別の対立遺伝子のTスコアの合計として、総Tスコアを形成した。
各三量体化モチーフおよびその周囲の配列における総Tスコアを表1−8に示す。最大スコアが8に等しく、それは配列内のすべての9−merが試験対象の8つの対立遺伝子すべてに対して強力に結合するエピトープである場合に対応することに注目されたし。興味深いことに、このグローバル分析によると、DMPK wtを除くほぼすべての候補がGCN4よりもやや強力な免疫原性特性を有することが予測される。マトリリンwtと変異体の双方は、最高の総Tスコアを有する。
方法
BioSep−Sec−S(商標)4000(ボイドボリューム、V0=5.7ml)でタンパク質を分析し、0.5ml/分の流速で30分間、すべてのランを実施した。UV280、屈折率および多角度レーザー光散乱検出器を用いて、溶出を監視した。試料を(1〜2mg/mlで)カラム上に負荷し、次にモル質量および粒子径を計算した。
多量体GITRL FP(マトリリン1 wt)
マトリリン−1三量体化モチーフを有するGITRL FPは、ゲル濾過カラム上に3つの異なるピークを生成したが、ピーク2は、グリコシル化六量体の予想されたモル質量に一致する312kDaの重量−平均モル質量を有し、総タンパク質質量の40.9%を占める一方、ピーク1は、六量体の二量体に対応する重量−平均モル質量(約610kDa)を有し、注射される総質量の41.3%を占める。ピーク3は、予想される質量(約215kDa)より低い質量を有するならば、それは断片化の結果または六量体からの二量体の欠損のいずれかであり得る。それは総タンパク質質量の17.7%を占める。したがって、このタンパク質製剤の組成物は異質性であるとともに、支配的なオリゴマー種は単一ではない。
UV280のオーバーレイ、屈折率および900レーザー散乱トレースの分析によると、このタンパク質の93.9%が300kDaの単分散種として16.4分で溶出することが示される。この観測質量は、六量体GITRL FPポリペプチドの予測された質量(274.95kDa)に一致する。計算質量と25kDaの観側質量との間の差異は、グリコシル化に起因する可能性が最も高い。14.7分後の小さいピークは、690kDaの重量−平均モル質量を有し、それは六量体の二量体に起因し得る。
同様に、GCN4 pII三量体化モチーフを有するGITRL FPのレーザー散乱および屈折率クロマトグラムの分析によると、このタンパク質の93.75%が、グリコシル化六量体に一致する、330kDaの単分散種として溶出することが示される。15分後のピーク溶出物は、総タンパク質のわずか6.25%を含有し、おそらくは六量体での二量体の存在を反映する約700kDaの重量−平均モル質量を有する。
図6は、3つの多量体GITRL FPタンパク質の各々に対する溶出ピークのモル質量組成を示す。3つすべてのタンパク質においては、約15分の保持時間を伴う初期ピークにわたるモル質量は、特に同一ピークの最初に溶出される分子の質量と終了時に溶出される分子の質量が100kDaより大きく異なるようなGITRL FP(マトリリン1 wt)の場合、非常に変化しやすい。約17分後の溶出ピークはそれらの分子組成においてあまり異質ではなく、GITRL FP(コロニン1a wt)における最大モル質量の変動は20kDa未満である。重量−平均モル質量におけるその変動は、同一ピーク内で溶出されている糖タンパク質のグリカン含量における差異に起因する可能性が最も高い。
多量体GITRL FP中の様々なコイルドコイル三量体化ドメインの性能を比較する表1−9に示すような収集データの分析において、コロニン1aコイルドコイルドメインは、改善された発現収量をもたらす。しかし、コロニン1aコイルドコイルモチーフおよび三量体コイルドコイルとして分類されるヒト配列からのモチーフの予測スコアの比較によると、コロニン1a wtが5のLOGICOILランキングを有したことが示される。
誘導および組成
配列番号6のアミノ酸配列を有する単量体サブユニットを含む六量体GITRL IgG1融合タンパク質を構築した。配列番号6の各GITRL IgG1 FP単量体サブユニットは、3つの異なるドメイン:1)ヒトIgG1 Fcドメイン;2)ヒトコロニン1Aタンパク質から誘導されるαヘリカルコイルドコイル三量体化ドメインおよび3)唯一の占有されるグリコシル化部位を排除するヒトGITRL ECDにおける単一の点突然変異(N161D)を有するヒトGITRL ECD、を含む。各ドメインは、可動性グリシンおよびセリンリッチもしくはグリシンリッチの例えば(Gly)4リンカーによって分離される(図8)。各単量体中のヒトGITRL ECDドメインは、溶液中に2つのさらなる単量体を伴う弱い非共有結合三量体を形成し、この会合は、ヒトコロニン1A三量体化ドメイン間の相互作用を通じて強化され、安定化された三量体構造をもたらす。各三量体中に存在するIgG1 Fcドメインの相互作用が、その後のGITRL三量体の二量体の形成につながり、最終的な六量体立体構造をもたらす。
ウエーブバッグバイオリアクター内、CD−CHO(商標)(Life Technologies Ltd,Paisley,UK)に類似した合成培地中の懸濁液中で成長するCHO細胞に、異なる六量体GITRL FPをコードするDNAベクターを遺伝子導入した。培養物を、CHO CD−Feed Efficient Feed A(商標)(Life Technologies Ltd,Paisley,UK)に類似した栄養素フィードを用いて、流加培養として10〜12日間維持し、次いでそれらを細胞除去用の濾過を用いて収集した。次に、馴化培地を精製前に冷蔵した。
配列番号6のGITRL IgG1 FPポリペプチドサブユニットのグリコシル化状態を、四重極飛行時間型(QTOF)質量分析(LC−QTOF MS)と共役した液体クロマトグラフィー、および質量分析を用いるペプチドマッピングを用いて判定した。結果を図9に示す。
リガンド−受容体結合に対する六量体GITRL IgG1 FPの効果
配列番号6の単量体GITRL IgG1 FPサブユニット配列を有する六量体GITRL IgG1 FPのヒトGITRLのヒトGITRへの結合に対する効果を、均一時間分解蛍光(HTRF)アッセイを用いて判定した。GITRL IgG1 FPを、GITRL−HA(ヘマグルチニンタグ)のGITR−Fcへの結合が測定されるHTRFアッセイで滴定した。ヒトGITR Fcをユーロピウムクリプテートと複合し、XL665と複合した抗HA抗体を用いて、GITRL−HAタンパク質を検出した。Prism 6.0xソフトウェア(GraphPad)を用いて、分析データを4つのパラメータロジスティック方程式にカーブフィッティングすることにより、IC50値を判定した。図10に示す代表的な結果は、GITRL IgG1 FPがGITRL−HAのGITRへの結合を0.562nMのIC50で阻害することを示す。アイソタイプ対照抗体NIP228では阻害が認められなかった。
この実験の目的は、GITRL融合タンパク質(FP)がGITR受容体を介してシグナルを送達する能力を判定することであった。
六量体GITRL FPを、hGITRおよび核内因子κB(NFκB)プロモーターに連結されたルシフェラーゼレポーター遺伝子を遺伝子導入したJurkat細胞に、溶液へ添加した。このアッセイでは、GITR受容体の活性化がNFκB経路を介したシグナル伝達をもたらし、次いで発光を介して測定可能なルシフェラーゼ活性における増強をもたらす。
六量体GITRL FPのアッセイ系への添加により、発光の増加がもたらされる。観察された効果は濃度依存性があり、EC50は約180pMであった。それに対し、アイソタイプ対照抗体の添加は、全く効果を有しなかった。結果を図11に示す。これらのデータは、六量体GITRL FPがGITR受容体の強力なアゴニストであることを示す。
この実験は、ヒトT細胞の増殖および機能に対する、六量体GITRL FPによって媒介されるGITRアゴニズムの影響を評価するために実行した。
GITR発現を上方制御するため、全ヒトT細胞を健常ヒト血液から単離し、プレートに結合した抗CD3の存在下での4日間の培養により、抗原を経験させた。抗原を経験した細胞を、六量体GITRL FPの存在下で亜最適濃度の抗CD3および抗CD28で再刺激する前、培地中での単独培養により2日間静止させた。細胞の増殖は、チミジンの細胞への取り込みを18時間かけて定量化するこによって評価した。インターフェロンγ(IFN−γ)の放出を、メソスケールディスカバリーを用いて定量化した。
抗CD3を抗CD28と一緒に添加した結果、最小レベルの増殖とIFN−γの最小放出がもたらされた。プレートに結合した六量体GITRL FPをプレートに結合した抗CD3および抗CD28と一緒に添加した結果、増殖のレベルおよびIFN−γの放出における濃度依存性増加がもたらされた。代表的なデータを図12および図13に示す。アイソタイプ対照抗体は、増殖またはサイトカインの放出に対する効果を全く有しなかった。
マウス六量体GITRL IgG1 FPは、FOXP3陽性制御性T細胞を含む腫瘍内CD4陽性T細胞を枯渇させ、腫瘍内でのCD8+T細胞対CD4+T細胞の比の増加をもたらすことがインビボで示されている。この実験は、六量体GITRL FPがADCCを介してヒトT細胞の枯渇を媒介する能力を判定し、生存細胞集団において生じるCD8:CD4比の変化を評価するために実行した。
GITR発現を上方制御するため、健常ヒト血液から単離した一次ヒトT細胞は、植物性血球凝集素(PHA)およびIL−2で抗原を経験させた。次にそれらを蛍光色素で標識し、指定濃度で、六量体GITRL FPとの指定される比で、一次NK細胞とともに24時間インキュベートした。フローサイトメトリーを用いて、アッセイの終了時に存在する生きたT細胞の百分率を定量化し、これを用いて、非処理ウェルに対する処理ウェルにおける溶解百分率を計算した。CD4およびCD8細胞の割合についても、フローサイトメトリーによって定量化した。
NK細胞は単独で一次T細胞の少量の溶解を媒介し、それはIgG1 Fcドメインを有する配列番号6で示されるGITRL融合ポリペプチドサブユニットを含む六量体GITRL FPを濃度依存的様式で添加することにより著しく増強された。NK細胞上のFcγ受容体に結合することができない、IgG4 Fcドメインを有する配列番号40のGITRL融合ポリペプチドサブユニットを含む六量体GITRL FPは、負の対照として用い、測定した溶解レベルに対する効果を全く有しなかった(図14)。
GITRは制御性T細胞(T−reg)上で増加したレベルで発現され、GITRを介するシグナル伝達がT−regの他のT細胞を抑制する能力に影響することが示唆されている。この実験は、T−reg機能に対する六量体GITRL FPの影響を評価するために実行した。
CD4+CD25−エフェクターT細胞およびCD4+CD25+T−regを健常ヒトドナーの末梢血から単離した。エフェクターT細胞は、指定される比での抗CD3抗体、抗CD28抗体、T−regおよび被験物質の存在下での培養前、CFSEで標識した。増殖するエフェクターT細胞の百分率をフローサイトメトリーによって分析した。
抗CD3および抗CD28の添加に応答して、エフェクターT細胞の増殖が認められた。増加する数のT−regの添加により、アッセイ中に分裂したエフェクターT細胞の百分率における減少がもたらされた。プレートに結合したアイソタイプ対照の添加により、分裂した細胞の百分率がさらに減少した一方、プレートに結合した六量体GITRL FPの添加により、T−regの不在下で認められる百分率まで分裂した細胞の百分率が回復した。結果を図16に示す。
アイソタイプ依存性の抗腫瘍活性
ヒトGITRLがマウスGITRと交差反応しないことから、ヒトGITRL FPはがんの免疫担当マウスモデルにおいて試験することができない。この試験を可能にするため、代理マウスのmGITRL IgG1 FPおよびmGITRL IgG2a FPを作製した。この試験は、がんのCT26モデルにおけるmGITRL FPの抗腫瘍活性を評価し、またこの活性の規模に対するFcアイソタイプの影響を判定するために実行した。
Balb/cマウスにCT26マウス結腸直腸がん細胞株を移植した。移植後6日目、動物にmGITRL FPを17日間で1回もしくは毎日のいずれかで腹腔内(i.p.)注射により投与した。mGITRL FPの2つの異なるバージョン(一方はmIgG2a Fcドメインを有し、もう一方はmIgG1 Fcドメインを有する)について試験した。5mg/kgおよび10mg/kgの2つの異なる用量レベルの各mGITRL FPを試験した。生理食塩水治療を負の対照として用いた。
生理食塩水治療群の中央値生存は22日であり、このグループ内のマウスのいずれも試験終了まで生存しなかった。10mg/kgのmGITRL FPでの治療により、中央値生存が32日まで延長され、試験終了時に50%生存率が得られた。5mg/kgのmGITRL FPで治療した群では、試験終了時に10匹中9匹のマウスが生存し、中央値生存時間を全く確定できなかった。5もしくは10mg/kgのmGITRL FP mIgG1での治療により、中央値生存が各々、28日および22.5日まで延長され、試験終了まで10匹中2匹のマウスの生存が得られた。結果を図17に示す。
この試験は、腫瘍保有マウスの脾臓および腫瘍におけるT細胞の活性化状態および割合に対するmGITRL FPの影響を判定するために実行した。
7〜9週齢のCT26腫瘍保有Balb/c雌マウスを、指定される通りの0.2mg/kgもしくは1mg/kgのいずれかのmGITRL FPの腹腔内注射により治療した。生理食塩水治療を負の対照として用いた。マウスを治療開始から7日後に屠殺し、フローサイトメトリーを用いて、脾臓および腫瘍における細胞の頻度および表現型を評価した。
mGITRL FPによる治療は、脾臓におけるすべてのT細胞サブセットでの増殖マーカーKi67の発現における増加をもたらし、これはこれらの細胞の増殖における増加を示唆する。図18を参照のこと。
細胞株および試薬
TC−1腫瘍株は、ATCC(カタログ番号CRL6475,Manassas,VA)から入手し、DMEM+10%FBS+1%ペニシリン/ストレプトマイシン中で維持した。CT26腫瘍株は、ATCC(Manassas,VA)から入手し、10%ウシ胎仔血清を添加したRPMI1640培地中で維持した。DTA−1およびアイソタイプ抗体は、Bio X Cell(West Lebanon,NH)から購入した。
TC−1実験では、Jackson Labs(Bar Harbor,ME)から入手した雌C57BL/6マウス(カタログ番号000664)マウスを用いた。CT26実験では、Envigo(Frederick,MD)から入手した雌Balb/Cマウスを用いた。マウスは、腫瘍移植時、6〜8週齢の間であった。すべての動物実験は、動物実験委員会(Institutional Animal Care and Use Committee)によって確立されたガイドラインに従って実行した。TC−1腫瘍移植においては、2×104個の生存可能なTC−1細胞を左後足蹠に皮下移植した。CT26腫瘍移植においては、5×105個の細胞を右側腹部に移植した。腫瘍成長は、ノギス1を用いた直接的測定により評価した。双方向的な測定結果を2〜4日ごとに収集し、腫瘍体積は体積=(長さ・幅2)/2を用いて計算した。腫瘍を6〜14日間発生させておき、次に腫瘍保有マウスを腫瘍体積により治療群に無作為化した。マウスは、IACUCプロトコルに従い、原発性腫瘍がTC−1/足蹠において1000mm3、CT26/側腹部において2000mm3を超えたとき、安楽死させた。PD試験においては、マウスを安楽死させ、腫瘍および脾臓を収集し、70uMのフィルター(Corning,Corning,NY)を通して粉砕し、単一の細胞懸濁液に処理した。
抗原特異的刺激のため、1〜2×106個の生細胞に、1ウェルあたり1ugのAH1ペプチド配列SPSYVYHQF(配列番号56)(Anaspec,Fremont,CA)を蒔いた。すべての株における染色の順序は、live/dead blue(Life Tech)、細胞外タンパク質、FOXP3 Fix/Permキット(Ebioscience)、それに続いて細胞内サイトカインであった。抗体は、GITR(クローンDTA−1)を含んだ。すべての試料は、LSR−IIまたはFortessa(BD San Jose,CA)のいずれかで走らせた。すべてのデータは、FlowJo(Treestar,Ashland,OR)を用いて分析した。
mGITRL−FPは極めて有効なGITRアゴニストであり、これはCT26腫瘍の拒絶をもたらす
GITRアゴニズムがBablcマウスにおけるCT26腫瘍の退縮および消失を引き起こす上で有効であることが以前に示されている。Balb/cマウスにCT26腫瘍細胞を移植し、6日後に腫瘍体積により無作為化し(群n=10)、mGITRL−FP IgG2a(Q2D×9)の単回投与または反復投与で治療した。用量範囲は、5mg/kg体重から0.04mg/kg体重に減少した。単回投与においては、mGITRL−FPの用量を1mg/kgまで減少させることで、30日目にかけてCT26マウス腫瘍の1つを残して全部が消失した(図40B)。最低用量として、0.2mg/kg群では3つの消失、0.1mg/kg群では2つの消失が得られた。mGITRL−FPのさらなる投与を追加することで有効性が高まり、0.04mg/kgの最低用量群を除く全部で100%の排出速度が認められた(図40C)。最低反復投与群では、4つの消失が認められた。
GITRアゴニストがTregを減少させるとともに、高結合活性のT細胞応答を増強することが以前に示されている。どの薬力学的効果をGITRL−FPが媒介する能力があるかを理解するため、CT26モデルを腫瘍退縮中に評価した。CT26を有するBalb/cマウスをmGITRL−FP IgG2aで治療し、投与開始から8日後、脾臓および腫瘍を収集した。試験の2つのアーム、すなわちTGI(腫瘍成長阻害)群(n=5)およびPD(薬力学)群(n=5)を含めた。mGITRL−FPで治療した単一マウス以外のすべてのCT26を治療した(図41A)。18日目、PD群を屠殺し、脾臓および腫瘍に存在する免疫細胞の両表現型を評価した。次に、CD8T細胞機能を、CT26免疫優勢エピトープAH1での再刺激によって評価した。mGITRL−FPが有意に増加することで、脾臓および腫瘍におけるTregの数が減少した(図41B)。mGITRL−FPは、CD8の百分率を変化させなかった一方、AH1に特異的な抗原であるCD8の数を有意に増加させた(図41C〜D)。脾臓では、全CD8の5〜10%が抗原特異的であり、IFNγおよびTNFαを産生する能力があった。腫瘍では、その数は5〜15%であった。これは腫瘍浸潤リンパ球と抹消貯蔵所(peripheral reservoir)の双方の有意な拡大である。mGITRL−FPがCD4 TregおよびCD8T細胞の双方に結合していたか否かを評価するため、DTA−1抗体を用いることを意図してGITRを染色した(図41E〜F)。Tregは、GITR MFI(平均蛍光強度)において75%を超える減少を示した。これと同じ効果がCD8T細胞上で認められ、これはmGITRL−FPがDTA−1のCD8T細胞上での結合を改変していることを示した。このデータに基づき、T細胞がKI−67増殖の増加を示すと仮定された。治療後8日目の評価後、CD4およびCD8T細胞の双方は、腫瘍と脾臓の双方において有意により多くのKI−67陽性細胞を示す(図41G〜H)。
次に、CT26を治療したマウスが同じ腫瘍の再負荷から保護されるか否かを検討した。これを評価するため、治療したマウスを、単回用量として1.0、0.5、もしくは0.2mg/kgのmGITRL−FP IgG2aで治療した群から再負荷した(図42A)。85日目での治療したマウスのみを除くすべてのマウスを、初期用量から減少させて再負荷した。CT26を最初に治療したすべてのマウスが腫瘍の再負荷から保護されたが、最低用量群では、小さい腫瘤が測定されたが迅速に排除された。この結果を精査するため、120日目にマウスを屠殺し、脾細胞を採取し、AH1ペプチドに対して再刺激した。AH1に特異的なT細胞の数において用量依存的増加が認められた(図42B〜C)。1mg/kgのmGITRL−FP群では、脾臓CD8T細胞の25%がCT26の単一エピトープに特異的である。これらのマウスは、再負荷中に腫瘍サイズにおける増加を全く示さなかった。最低用量0.2mg/kgは、6%の抗原特異的CD8を有し、120目までに除かれる再負荷時に少し成長しつつある腫瘤を示した。
組換えヒトGITRに対する配列番号6のGITRL IgG1 FP単量体サブユニットを含む六量体GITRL IgG1 FPの溶液のKD(解離定数)を、KinExA 3200装置(Sapidyne Instruments,Boise Idaho)を用いる動力学的排除アッセイ(KinExA)を用いて測定した。
組換えカニクイザルGITRに対する配列番号6のGITRL IgG1 FP単量体サブユニットを含む六量体GITRL IgG1 FPの溶液のKDを、ヒトGITRについてのセクション6.1に記載と同様の方法でKinExAを用いて測定した。六量体GITRL IgG1 FPのカニクイザルGITRへの結合におけるKDとして107nM得られた。
配列番号6のGITRL IgG1 FP単量体サブユニットを含む六量体GITRL IgG1 FPのカニクイザルGITRに対する交差反応性を、ELISAを用いて測定した。六量体GITRL IgG1 FPをビオチン化し、固定化されたカニクイザルGITR−Fへの結合を、ストレプトアビジン−HRPおよびTMB基質を用いて検出した。図21は、六量体GITRL IgG1 FPのカニクイザルGITRへの結合がヒトGITRに酷似し、かつ負の対照CD137−Fcタンパク質に対する結合が全く認められないことを示す。
カニクイザルにおける六量体GITRL IgG1融合タンパク質(FP)の薬力学的パラメータを決定するため、配列番号6のGITRL IgG1 FP単量体サブユニットを含む六量体GITRL IgG1 FPを、雄カニクイザルの群(5/群)に静脈内(IV)ボーラス注射によって投与した。
1日目、3日目および5日目、動物の別々の群に、1もしくは10mg/kgの六量体GITRL IgG1 FPを注射した。対照動物は、溶媒(20mMリン酸ナトリウム、230mMスクロース、0.02%P80、pH7.5)を受けた。薬力学的エンドポイントの測定のため、前投与期間中と1、3、5、9、11、15、18、22、および29日目に、血液試料を採取した。
細胞増殖を示す、%Ki67陽性T細胞亜集団、例えば(CD3+CD4+CD95highCD28+/dim/−全メモリーCD4+およびCD3+CD8+CD95highCD28+/dim/−全メモリーCD8+T細胞)における増加が認められた(図22)。この観察によると、11日目もしくは15日目の投与期間に最大に達し、次に試験終了に向けて減少した。
GITRL受容体結合ドメイン内に突然変異を有するhGITRL FP変異体の作製
hGITRL FP変異体を作製し、GITRに対して結合し、刺激するその能力について試験した。場合によっては、その熱安定性についても評価した。
遺伝子合成および標準のDNAクローニング技術を用いて、GITRL FP変異体をコードするDNAベクターを作製した。懸濁CHO細胞に、異なる六量体hGITRL融合タンパク質をコードするDNAベクターを、PEIを用いて一過性に遺伝子導入し、80%の湿度下、140rpmで振盪しながら37℃で8日間成長させた。分泌タンパク質を含有する40mLの条件培地を、1,600×gでの遠心分離および濾過により、細胞および細胞片から分離した。Mab SelectSure(商標)樹脂を用いてタンパク質を精製し、そのサイズおよび完全性を還元SDS−ポリアクリルアミドゲル電気泳動法によって分析した。
ヒトGITRLのヒトGITRへの結合に対するGITRL FP分子の効果を、均一時間分解蛍光(HTRF)アッセイを用いて判定した。
GITRL FP分子を、GITRL−HA(ヘマグルチニンタグ)のGITR−Fcへの結合が測定されるHTRFアッセイで滴定した。ヒトGITR Fcはユーロピウムクリプテートと複合させ、XL665と複合した抗HA抗体を用いて、GITRL−HAタンパク質を検出した。Prism5.01ソフトウェア(GraphPad)を用いて分析データを4つのパラメータロジスティック方程式にカーブフィッティングすることにより、IC50値を判定した。
突然変異した六量体GITRL FP分子(hGITRL−FP wt、N92D、N104DおよびN161D)はすべて、三量体GITRL−HAのGITR−Fcへの結合の強力な阻害剤であり、類似の阻害特性およびIC50値をもたらした(図23A、B)。
異なるhGITRL FP分子の機能的活性を、hGITRを安定的に発現するNFκB−ルシフェラーゼレポーター細胞を用いるアッセイにて測定した。hGITRのアゴニズムとその後のNFκB経路の活性化によって駆動される発光を測定した。
hGITRL FP分子を、hGITRL FPのhGITR−Fcへの結合が測定されるHTRFアッセイで滴定した。ヒトhGITR Fcはユーロピウムクリプテートと複合させ、XL665と複合した抗FLAG抗体を用いて、hGITRL FPタンパク質を検出した。Prism5.01ソフトウェア(GraphPad)を用いて分析データを1つの部位の飽和結合方程式にカーブフィッティングすることにより、KD値を判定した。
hGITRL FP wtおよびhGITRL FP N161Dにおける結合特性は酷似していた一方(各々、1.225nMおよび1.079nMのKD)、N129A突然変異hGITRL FPは、hGITRFcに対する結合低下を示した(KD=3.774nM)。N129A突然変異をN161D突然変異と組み合わせるとき、hGITRL FP分子の結合能力はさらなる負の影響を受けた(KD=11.85nM)(図24を参照)。
異なるhGITRL−FP分子の機能的活性を、hGITRを安定的に発現するNFκB−ルシフェラーゼレポーター細胞を用いるアッセイにて測定した。hGITRのアゴニズムとその後のNFκB経路の活性化によって駆動される発光を測定した。
6ポイントデータ曲線のため、hGITRL FPタンパク質を4倍に連続希釈し、3通りに96ウェルプレートに添加した。次に、ヒトGITRを遺伝子導入したJurkat NF−κBルシフェラーゼレポーター細胞を、アッセイプレートのすべてのウェルに添加し、37℃で3時間インキュベートした。Steady−Glo試薬をアッセイプレートのすべてのウェルに添加することにより、ルシフェラーゼ発現を検出した。プレートを室温で5分間インキュベートし、次に発光を測定し、対数(アゴニスト)対応答のGraphPad(商標)Prism5.01(GraphPad Software,Inc.La Jolla,CA USA)での可変勾配非線形曲線フィットを用いてEC50値を生成した。
すべてのGITRL FPタンパク質がNF−κBシグナル伝達を誘導する。N92DおよびN161D突然変異体hGITRL FPタンパク質は、GITRL FP(wt)と類似の効力特性およびEC50値をもたらした。N129AおよびN129A/N161D突然変異は、このアッセイにおけるhGITRL FPのGITRを刺激する能力に負の影響を与えた(図25A〜C)。
異なるhGITRL FP分子の機能的活性を、一次ヒトT細胞およびチミジン取り込みの読み出しを用いる同時刺激アッセイにて測定した。
ヒト末梢血単核球(PBMC)由来のCD3+T細胞を、マウス抗ヒトCD3抗体でコーティングしたTC処理プレート内でのインキュベーションを介して37℃で4日間刺激した。次に、それらを遠心分離によってペレット化し、アッセイ培地に再懸濁し、TC処理プレートに添加し、37℃で2日間インキュベートした(静止期)。hGITRL FP分子を、10ポイントにわたりアッセイ培地で2倍に連続希釈し、マウス抗ヒトCD3で予備コーティングしたTC処理プレートに3通りに添加した。2時間後、アッセイプレートを洗浄し、以前に調製したCD3+T細胞を各ウェルに添加し、37℃で4日間インキュベートした。4日後、アッセイ培地中のトリチウム標識チミジンを各ウェルに添加し、プレートを37℃でさらに18時間インキュベートした。インキュベーション後、チミジンの取り込みを、Topcount(商標)を用いて測定した。
試験したすべてのhGITRL FP突然変異体は、2つの独立実験において低下した活性を有するN92Dを例外として、hGITRL FP wtに等価な活性を示した(図26A〜C)。
GITRL FP wtおよびN92Dタンパク質の融解温度を蛍光色素(Sypro(商標)Orange)を用いて測定したが、その放射特性は折り畳まれていないタンパク質の存在下で変化する。
GITRL FP wtおよびN92D GITRL FP変異体の熱安定性を、融解温度(Tm)を計算するためのSypro Orangeに基づくアッセイを用いて評価した。タンパク質を、96ウェルPCRプレートに懸濁する前、最初に2×PBSで0.5mg/mLまで希釈した。Sypro(商標)Orangeをプレート上の各ウェルに添加し、次に密封した。プレートを、Chromo4(商標)連続蛍光検出器を用いてリアルタイムPCR装置上で読み取った。温度は、1℃ごとの読み取りと1秒の保持時間で20℃から90℃に高めるように設定した。アンフォールディング転移を、蛍光強度および蛍光誘導体を温度の関数としてプロットすることによって判定した。各GITRL FPタンパク質を二通りに分析した。
2つのGITRL FPタンパク質における融解温度を下の表4−1にまとめる。両変異体は67℃で転移ピークを有するが、hGITRL FPN92Dは、低温(54℃)で追加的な幅広い転移ピークを呈し、これは構造的不安定性を示唆する(図27)。
N104DGITRL FPタンパク質は、GITR/GITRL競合結合アッセイおよび一次T細胞再刺激アッセイにおいて野生型hGITRL FP対応物に等価な活性を示したが、これはN104がhGITRL FPの活性における主要な役割を果たさないことを示唆する。Aspに対するAsn161の突然変異は、N結合グリコシル化部位の除去を表し(実施例4におけるデータを参照)、この突然変異体(hGITRL FP N161D)はすべてのアッセイ(GITR/GITRL競合結合、直接結合、レポーターおよび一次T細胞再刺激アッセイ)において活性を保持したが、これはAsn161、およびこの部位のグリカンがGITRの結合およびアゴニズムなどの重要な機能に関与しないことを示唆する。
GITR結合ドメイン内部に2つの有望なN−グリコシル化コンセンサス部位(N161およびN129)が存在する。これらの部位、ならびにFcドメイン内部の基準のN−グリコシル化部位(IgGとの関連でのN297;成熟hGITRL FP配列内のN78)でのグリカンの存在および構造を質量分析によって測定した。
組換えタンパク質の発現および精製
組換えhGITRL FP wtおよびN161Dタンパク質を、親和性クロマトグラフィーとその後のサイズ排除クロマトグラフィーを用いて、関連タンパク質をコードするベクターを一過性に遺伝子導入したCHO細胞の条件培地から精製した。
hGITRL FP wtおよびhGITRL FP N161Dの試料を変性させ、還元し、還元したシステインをアルキル化した。次に、試料をトリプシンで消化した。37℃で4時間後、消化物を酸の添加によりクエンチした。ペプチドをUPLC上で逆相によって分離し、UV検出器および質量分析計を用いて測定した。得られたトリプシンペプチド配列を表5−1に提示する。
GITRLおよびFcドメインにおけるN−グリコシル化部位の占有、ならびに各部位での主要なオリゴ糖の構造を判定するため、ペプチドマッピング分析を用いた。hGITRL FP wtおよびhGITRL FP N161Dタンパク質の双方では、基準のFc N−グリコシル化部位(N78)がグリコシル化され、主要な炭水化物構造がCHO細胞内で発現されるIgG Fc領域に典型的な中性の二分岐複合型オリゴ糖であった。GITRL RBD内部のN129のN−グリコシル化コンセンサス部位は、hGITRL FP wtまたはhGITRL FP N161Dのいずれかにおける任意のオリゴ糖構造で占有されなかった。GITRL RBD内部のN161のN−グリコシル化コンセンサス部位はhGITRL FP wt中で占有され、主要な炭水化物構造は中性の帯電した二分岐複合型オリゴ糖であることが見出された。hGITRL FP N161Dタンパク質では、ペプチドマッピングにより、GITRL RBDからN161N−グリコシル化コンセンサス部位を除去する、N161Dアミノ酸置換が確認された。予想通り、N−グリコシル化がD161で全く検出されなかった。
GITRの標的化をより理解するため、マウスGITRL−FPの活性および薬力学的効果を、アゴニストのマウスOX40L FPを標的化するOX40の場合と比較した。
NF−κBレポーターアッセイ
Jurkat mGITRまたはヒトOX40 NFκB細胞を、指定の通り、50,000個の細胞(Jukat mGITR)または200,000個の細胞(JurkatヒトOX40)で、抗GITR抗体DTA−1(Biolegend)、NIP rIgG2bアイソタイプ対照、mGITRL−FP mIgG2aまたはプレートに固定化されたmOX40L−FP mIgG1もしくはアイソタイプ対照(タンパク質のOX40を介するシグナル伝達を不能にする(Y182A)位での単一アミノ酸突然変異を含む)と一緒に、96ウェルプレート内、37℃、5%CO2および85%の湿度下で培養した。mOX40L−FP mIgG1を有するアッセイにおいて、ルシフェリン基質を含有する定常Glo(登録商標)試薬(Promega)を5時間後または16時間後にプレートに添加し、プレートをプレートシェーカー上、暗所で30分間インキュベートした。Envisionプレートリーダー(Perkin Elmer)を用いて、アッセイシグナルを検出した。
5μgのタンパク質を、負荷緩衝液および還元剤と混合し、80℃で10分間変性させ、タンパク質分子量マーカー(Rainbow Marker、GE Healthcare)と一緒に、4〜20%トリス−グリシンSDS−ポリアクリルアミドゲル電気泳動ゲル(Thermo Fisher)上に負荷した。タンパク質を200Vで45分間電気泳動し、Instant Blueタンパク質染色(Sigma)を用いて染色した。
8〜10週齢BALB/cまたはC57BL/6雌マウスをCharles River UK Ltd.またはHarlan Laboratories Inc.から入手した。PBS中CT26またはB16F10−Luc2細胞の5×106個の細胞/mLまたは5×104個の細胞/mLの細胞密度での懸濁液100μLを、各動物の右側腹部に皮下注射した。B16F10−Luc2細胞株にルシフェラーゼレポーターをCAGプロモーターの制御下で組み込み、細胞を50%PBSおよび50%成長因子(還元)およびフェノールレッドフリーマトリゲル(Corning)に移植した。測定可能な腫瘍を腫瘍体積に基づいて無作為化した。各腫瘍の長さ(mm)および幅(mm)を電子ノギスで週3回測定した。腫瘍の体積(mm3)を、式(長さ[mm]×幅[mm]2)/2に基づいて計算した。腫瘍成長応答を、試験終了時に体積が200mm3未満であるように測定可能な腫瘍が存在しないかまたは持続的な腫瘍成長阻害が認められる場合の応答として分類した。各スパイダープロット上に示される退縮の数は、移植された腫瘍の総数に対する割合である。同系腫瘍の移植後6日目、またはそれらが200mm3の体積に達した時、マウスにmGITRL−FPまたはmOX40L−FPのいずれかを腹腔内投与した。B16F10−Luc2保有マウスにおける25mg/kgのmGITRL−FPの投与に限り、4用量に対して耐性を示した。
腫瘍、脾臓または腫瘍流入リンパ節を切断し、氷上のRPML−1640培地中に置いた。組織を、各々を40もしくは100μmのナイロンセルストレーナー(Falcon)を通過させることにより脱凝集させ、細胞を遠心分離によってペレット化し、赤血球溶解緩衝液(Sigma)に再懸濁した。室温で2分間のインキュベーション後、細胞を洗浄し、フローサイトメトリー緩衝液(Ebioscience)に再懸濁した。腫瘍組織試料を、穏やかなMACSディソシエーターおよび腫瘍解離キット(Miltenyi Biotec)を用いて製造業者の使用説明書に従って処理した。
Balb/cマウスに、5もしくは15mg/kgのmGITRL−FP mIgG2aを1回投与した。1群あたりマウス3匹を治療から5分後、0.5、1、2、6、24、72、144、および240時間後に屠殺した。血清試料を収集し、存在する循環mGITRL−FP mIgG2aのレベルを、捕獲と検出の双方として抗マウスGITRL mAb(R&D Systems)を用いてのサンドイッチELISAで評価した。
HEK293T−17細胞(ATCC,CRL−11268)を、DMEM(Invitrogen)+10%v/v熱不活化ウシ胎仔血清(HI FBS,HyClone)および1×非必須アミノ酸(NEAA,Invitrogen)で維持した。Jurkat細胞(ATCC,TIB−152)を、RPMI1640(Invitrogen)+10%v/vHI FBSで維持した。Jurkat mGITR NFκB細胞を、10%HI FBS、5μg/mLのブラストサイジン(Invitrogen)および5μg/mLのピューロマイシン(Invitrogen)を添加したRPMI1640で維持した。
Jurkat mGITR NFκB細胞株を、第3世代レンチウイルス系(Systems Biosciences)を用いるレンチウイルス形質導入によって作製した。マウスGITR遺伝子(NM_009400.2)をCAGプロモーターの制御下でのピューロマイシン耐性遺伝子を有する発現プラスミドにクローン化した。NFκBレポーター発現プラスミドを、ブラストサイジン耐性遺伝子と一緒に、最小プロモーター下でのホタルルシフェラーゼレポーター(luc2,Promega)の上流でNFκB応答要素の5つのコピーを発現するように設計した。各発現プラスミドを、パッケージングプラスミド(Systems Biosciences)と一緒に、リポフェクタミン2000(Invitrogen)を用いてHEK293T−17に同時導入した。ウイルス粒子を含有する上清を、遺伝子導入の48時間後に収集し、Jurkat細胞を形質導入するために用いた。ウイルス形質導入したJurkat細胞を、形質導入の48時間後から、選択抗生物質として5μg/mLのブラストサイジンおよび5μg/mLのピューロマイシンを添加した培地で培養した。JurkatヒトOX40 NFκB細胞株を、ヒトOX40を構成的に発現するように設計したレンチウイルスベクターを形質導入したことを除き、同様の手順を用いて作製した。マウスOX40LはヒトOX40に結合し、それ故、マウスOX40L融合タンパク質の活性を評価するため、ヒトOX40を発現するNFκBルシフェラーゼレポーターJurkat細胞株を用いることができる。
組換えマウスGITR−FcおよびOX40−Fc(R&D Systems)糖タンパク質を、PBS中1μg/mLで96ウェルプレート(Greiner)上に一晩コーティングした。プレートをPBST(PBS+0.01%ツイーン20)で洗浄し、室温、1%(w/v)BSAを含有するPBSで1時間ブロッキングし、PBSTで再び洗浄した。アッセイ緩衝液[PBS+1%ウシ血清アルブミン(BSA)]で希釈した1μg/mLのマウスGITRLまたはOX40L FPをウェルに25μL添加し、プレートを室温で2時間インキュベートした。3回洗浄後、アッセイ緩衝液で希釈した1μg/mLの西洋わさびペルオキシダーゼに結合した抗マウスFc抗体(Sigma)を各ウェルに25μL添加し、プレートを1時間インキュベートした。インキュベーション後、プレートをPBSTで3回洗浄し、テトラメチルベンジジン基質溶液(KPL)を各ウェルに25μL添加し、プレートを5分間インキュベートした。インキュベーション後、0.5Mの硫酸停止溶液をすべてのウェルに15μL添加した。450nMでの光学密度を、EnVisionプレートリーダー(PerkinElmer)を用いて測定した。
すべての統計学的分析は、Prism Statistical Softwareバージョン6を用いて実施した。
mGITRL−FPのインビトロ効力
インビボでの免疫細胞活性化および抗腫瘍活性に対するGITR受容体シグナル伝達の効果を検討するため、四量体mGITRL−FPを作製した。mGITRL−FPは、エフェクター細胞上のGITR受容体およびFcγRへの貪欲な結合を誘導するように設計した。その分子は、N末端からC末端にかけて、免疫グロブリンG(IgG)の結晶性断片(Fc)領域、イソロイシンジッパードメイン(ILZ)およびマウスGITRリガンドの細胞外(GITR結合)ドメイン(ECD)からなるものであった(図30A)。変性した精製mGITRL−FPをSDS−ポリアクリルアミドゲル電気泳動法によって可視化した時(図30B)、それは高度な均一性および予想される分子量48kDaよりもやや高い分子量を示し、それはおそらくはFcおよびmGITRLドメインのグリコシル化に起因する。mGITRL−FP mIgG2aおよび抗GITR抗体(DTA−1)の双方は、GITR依存性NFκBレポーター遺伝子細胞アッセイにおいてNF−κBシグナル伝達を誘導することができた一方、マウスOX40L FP mIgG1(mOX40L−FP mIgG1)、およびアイソタイプ対照では任意の検出可能なシグナルが欠如していた(図30C)。重要なことには、四量体mGITRL−FPは、このアッセイにおけるGITRアゴニズムに関連して0.05nMのEC50を示し、それは2.31nMのEC50を示したDTA−1よりも約50倍強力であった。
以前の試験では、FcγRを活性化することがDTA−1抗体の抗腫瘍活性にとって必要であり、かつこの抗体の抗腫瘍活性が、それがmIgG2a Fcを有するとき、rIgG2b FcまたはN297A突然変異体Fc(FcγRへの結合が欠如する)と比べて増強されることが示されている。mGITRL−FPの抗腫瘍活性に対するFcアイソタイプの影響を判定するため、CT26腫瘍保有マウスを、mIgG2aまたはmIgG1 Fcアイソタイプを有するmGITRL−FPで治療した。5もしくは10mg/kgのいずれかのアイソタイプでのマウスの治療により、腫瘍体積の減少によって明らかなように、生理食塩水で治療した対照と比べて著明な抗腫瘍活性が得られた(図31A)。しかし、抗腫瘍活性全体は、mGITRL−FP mIgG2aによる治療後、mGITRL−FP mIgG1の場合(7/20が全退縮、図31A)に対して増強した(11/20が全退縮)。
mGITRL−FP mIgG2aがmGITRL−FP mIgG1と比べて抗腫瘍活性の増強を誘導したことから、血液中のmGITRL−FP mIgG2aの曝露レベルおよび曝露時間がCT26腫瘍保有マウスにおける抗腫瘍応答にいかに関連するかを次に特徴づけた。第1に、1単回用量の生理食塩水対照または0.2、1、もしくは5mg/kgのmGITRL−FP mIgG2aでマウスを治療することによる、腫瘍成長に対する用量レベル増加の効果を測定した。0.2mg/kgのmGITRL−FP mIgG2aによる治療の結果、わずか1/10の完全退縮がもたらされたが、用量が0.2mg/kgから1mg/kgもしくは5mg/kgに上昇した(各々、6/10および9/10の退縮をもたらした)時、抗腫瘍免疫における用量依存的増加における証拠が存在した(図33A)。さらに、抗腫瘍活性はまた、0.2mg/kg投与の頻度を毎日1回(Q1D)または毎週1回(Q1W)に増加させることにより改善することができた(図33B)。これらの結果は、Q1Dスケジュールを用いるときに限って到達される、完全腫瘍退縮および最小残留腫瘍体積によって規定されるような最適な抗腫瘍活性を誘導するのに必要とされるmGITRL−FP mIgG2aの要求される曝露レベルが存在することを示した。この閾値を判定するため、mGITRL−FP mIgG2aの濃度を、2つの代替的な用量レベルのmGITRL−FPで治療したマウスの血液中で測定し、次にこれらの結果をPKモデルに組み込んだ。このモデルに基づき、Q1Dスケジュールによって持続される、また最適な抗腫瘍活性を維持するために必要と考えられるmGITRL−FP mIgG2aの血液濃度は1μg/mL以上であると計算された(図33C)。
OX40標的化に対して、GITRの関連機構を理解するため、mGITRL−FPの抗腫瘍活性を、類似のタンパク質構造を含み、GITR受容体と交差反応せず、OX40レポーター細胞アッセイにおいてNFκB発現を誘導することができるmOX40L−FPと直接的に比較した(図35AおよびB)。
mGITRL−FP mIgG2aでの治療後におけるCD4+T細胞上のOX40受容体の上方制御を仮定し(図34D)、T細胞活性化を最大化する可能性についてさらに検討した。mGITRL−FP mIgG2aおよびmOX40L−FP mIgG1を用いて、GITRおよびOX40経路を標的化する薬剤を組み合わせることの潜在的な利点を評価した。
材料および方法
細胞株および試薬
TC−1腫瘍株は、ATCC(カタログ番号CRL6475,Manassas,VA)から入手し、DMEM+10%FBS+1%ペニシリン/ストレプトマイシンで維持した。CT26腫瘍株は、ATCC(Manassas,VA)から入手し、10%ウシ胎仔血清を添加したRPMI1640培地で維持した。DTA−1およびアイソタイプ抗体は、Bio X Cell(West Lebanon,NH)から購入した。
45−merのHPV16−E7配列SSEEEDEIDGPAGQAEPDRAHYNIVTFCCKCDSTLRLCVQSTHVD(配列番号57)からなるE7合成ロングペプチド(SLP)を、New England Peptide(Gardner,MA)から合成した。E7 SLPを10μgもしくは3.3μgで投与し、全体積50μLで、Addavax(Life Technologies,Carlsbad、CA)およびPBS中、20μgのCpG ODN2395(TriLink,San Diego,CA)を用いて製剤化した。ワクチン接種を、尾の付け根の背側表面皮下に施した。
TC−1実験では、Jackson Labs(Bar Harbor,ME)から入手した雌C57BL/6マウス(カタログ番号000664)のマウスを用いた。CT26実験では、Envigo(Frederick、MD)から入手した雌Balb/Cマウスを用いた。マウスは、腫瘍移植の時点で6〜8週齢の間であった。すべての動物実験は、動物実験委員会(Institutional Animal Care and Use Committee)によって確立されたガイドラインに従って実行した。TC−1腫瘍移植においては、2×104個の生存可能TC−1細胞を左後足蹠に皮下移植した。CT26腫瘍移植においては、5×105個の細胞を右側腹部に移植した。腫瘍成長は、ノギス1を用いる直接的測定により評価した。双方向的な測定結果を2〜4日ごとに収集し、腫瘍体積は体積=(長さ・幅2)/2を用いて計算した。腫瘍を6〜14日間発生させておき、次に腫瘍保有マウスを腫瘍体積により治療群に無作為化した。マウスは、IACUCプロトコルに従い、原発性腫瘍がTC−1/足蹠において1000mm3、CT26/側腹部において2000mm3を超えたとき、安楽死させた。PD試験においては、マウスを安楽死させ、腫瘍および脾臓を収集し、70uMのフィルター(Corning(商標)、Corning,NY)を通して粉砕し、単一の細胞懸濁液に処理した。
抗原特異的刺激のため、1〜2×106個の生細胞に、1ウェルあたり1ugのAH1ペプチド配列SPSYVYHQF(配列番号56)(Anaspec,Fremont,CA)または10ugのE7ペプチド配列RAHYNIVTF(配列番号58)(Anaspec)およびタンパク質輸送阻害剤(Ebioscience,Santa Clara,CA)を蒔いた。5時間後、細胞を染色した。すべての染色における染色の順序は、live/dead blue(Life Tech)、細胞外タンパク質、FOXP3 Fix/Permキット(Ebioscience)、それに続いて細胞内サイトカインであった。抗体は、CD45(クローン30−F11)、CD4(クローンRM4−5)CD8(クローン53−6.7)、GITR(クローンDTA−1)、IFNγ(クローンXMG1.2)、TNFα(クローンMP6−XT22)、FOXP3(クローンFJK−16s)、KI−67(クローンSolA15)を含んだ。RAHYNIVTFを負荷したH2−Db E7デキストラマーをImmudex(Fairfax,VA)から購入し、我々は染色についてのそのプロトコルに従った。すべての試料は、LSR−IIまたはFortessa(BD San Jose,CA)のいずれかで走らせた。すべてのデータは、FlowJo(Treestar,Ashland,OR)を用いて分析した。
CT26は、高い腫瘍内CD45レベルおよび基礎CTLレベルを有する高免疫原性腫瘍モデルである。CT26は、低レベルのAH1特異的T細胞を自己刺激することが示されている。mGITRL−FPが抗原特異的応答の新規な生成をもたらし得るか否かを評価するため、E6/E7形質転換TC−1腫瘍モデルを用いた。このモデルは、公知のCD8T細胞エピトープを有し、そのE7特異的免疫応答は、E7を有する腫瘍からの保護をもたらし得る。TC−1腫瘍細胞をC57/b6マウスの足蹠に移植した。マウスは、14日目に測定し、腫瘍サイズに基づいて無作為化し、1mg/kg、5mg/kg、および20mg/kgのmGITRL−FP IgG2aで治療した(図43A)。腫瘍成長の遅延または阻害が任意の試験用量で認められた(図43B)。24日目、非治療および1mg/kg治療マウスのサブセットを屠殺し、脾臓および腫瘍を収集した。T細胞集団およびE7抗原特異的応答を評価した。CD4およびCD8T細胞上に認められたGITRレベルは、腫瘍内で有意に高めであった(図43C)。E7予備刺激を伴わない場合、mGITRL−FPは、TC−1腫瘍成長を遅延させるかまたは抗腫瘍免疫応答を生成することができなかった。
DNAワクチンの接種は、TC−1腫瘍モデルの腫瘍成長の阻止および阻害をもたらすE7特異的応答の生成において有効であることが示されている。この試験では、mGITRL−FPが抗腫瘍応答を駆動するのに予備刺激された抗原特異的T細胞を必要とし得ることが仮定された。この応答を生成するため、ナイーブC57/b6マウスの尾の付け根に、CpG(addavax)を有するE7合成ロングペプチド(SLP)を10ugでワクチン接種した。この期間中、マウスを3用量のmGITRL−FP IgG2aで治療した。7日後、脾細胞を採取し、T細胞のE7デキストラマーとの抗原特異性について評価した(図44A)。CD4およびCD8の百分率における差は認められなかったが、mGITRL−FPで治療した時、抗原特異的細胞における大幅な増加が認められた(図44B〜D)。加えて、ワクチン接種マウス単独に対するGITRレベルを評価し、抗原特異的Dex+細胞は、Dex−CD8T細胞よりも高いGITR MFIを有した(図44E〜F)。GITRレベルが腫瘍微小環境下で最高であるように思われたことから、この差が腫瘍を有するマウスにおいてさらに高めであることが仮定された。C57/b6マウスに、TC−1腫瘍を移植し、E7 SLP/CpG(Addavax)をワクチン接種した(図44G)。抗原特異的E7細胞の予備刺激が、双方の脾臓および腫瘍におけるデキストラマーによる測定として認められた(図44H〜I)。デキストラマーで陽性染色した細胞もまた、脾臓および腫瘍の双方におけるGITRについて高めであった(図44J〜K)。E7 SLPは、腫瘍の存在の有無にかかわらず、E7特異的応答を高度に予備刺激することができ、予備刺激した抗原特異的細胞は、他のCD8T細胞よりも高レベルのGITRを発現した。
mGITRL−FPがTC−1腫瘍細胞に対するE7特異的応答を拡大できたか否かを評価するため、14日目、TC−1腫瘍細胞を足蹠に移植し、マウスを腫瘍体積により無作為化した。尾の付け根に4用量でのmGITRL−FP IgG2aとともにCpG(Addavax)を加えた単回用量のE7 SLPの投与がワクチン接種日に開始した(図45A)。試験に、TGI(腫瘍成長阻害)群(n=10)およびPD(薬力学)群(n=5)の2つのアームを含めた。対照マウスの腫瘍は迅速に成長し、すべてのマウスが死亡し、中央値生存は27日であった。E7 SLP単独は、TC−1腫瘍を有するマウスに生存上の利点を提供し、85日目に10匹中1匹のマウスが生存し、腫瘍を含まず、中央値生存は46.5日であった(図45B)。E7 SLP+mGITRL−FPは、腫瘍成長を有意に遅延させ、85日目に3/10のマウスが腫瘍を含まず、5/10が生存し、中央値生存は80.5日であった(図45C〜D)。ワクチン接種単独は、腫瘍進行に遅延をもたらしたが、mGITRL−FPを最上部に添加することで進行がさらに遅延した。これは腫瘍特異的CD8の選択拡大が理由で生じると仮定された。21日目、PDマウス群を屠殺し、薬力学分析用に脾臓および腫瘍を採取した。足蹠からの腫瘍を、採取した組織量に応じてプールした(図45E)。単独またはmGITRL−FPとの組み合わせでのワクチン接種の結果、腫瘍内にCD45+細胞が有意に増加した(図45F)。抗原特異的機能を評価するため、腫瘍または脾臓の単一の細胞懸濁液を1ug/mLのE7ペプチドで再刺激した。ワクチン接種は抗原特異的細胞における基礎増加をもたらし、mGITRL−FPの添加により、これが脾臓および腫瘍の双方においてより高レベルにさらにブーストされた(図45G)。mGITRL−FPは、E7 SLPワクチンの存在時、脾臓におけるTregを枯渇させないように思われ、腫瘍におけるTregに限って枯渇させることができた(図45G)。
本願で開示されるヌクレオチドおよびタンパク質配列の注釈付きバージョンを表6−1に提示する。
Claims (136)
- IgG Fcドメイン;機能的多量体化ドメイン;およびグルココルチコイド誘導TNF受容体リガンド(GITRL)の受容体結合ドメインを含む単離された一本鎖ポリペプチドサブユニットであって、三量体または六量体タンパク質に自己組織化し得る、ポリペプチドサブユニット。
- 前記多量体化ドメインが三量体化ドメインである、請求項1に記載のポリペプチドサブユニット。
- アミノ末端からカルボキシ末端にかけて、前記IgG Fcドメイン、その後に前記多量体化または三量体化ドメイン、その後に前記GITRL受容体結合ドメインを含む、請求項1または2に記載のポリペプチドサブユニット。
- 前記IgG Fcドメインのカルボキシ末端が、第1のリンカー領域を介して前記多量体化または三量体化ドメインのアミノ末端に融合される、請求項1〜3のいずれか一項に記載のポリペプチドサブユニット。
- 前記多量体化または三量体化ドメインのカルボキシ末端が、第2のリンカー領域を介して前記GITRL受容体結合ドメインのアミノ末端に融合される、請求項1〜4のいずれか一項に記載のポリペプチドサブユニット。
- 前記IgG Fcドメインのカルボキシ末端が、前記多量体化または三量体化ドメインのアミノ末端に直接的に融合される、請求項1〜4のいずれか一項に記載のポリペプチドサブユニット。
- 前記IgG Fcドメインが、そのアミノ末端にIgGヒンジ領域を含む、請求項5または6に記載のポリペプチドサブユニット。
- 前記IgGヒンジ領域が、完全な重鎖間ジスルフィド結合形成を与える突然変異を含む、請求項5または6に記載のポリペプチドサブユニット。
- 前記IgGヒンジ領域が、IgG1ヒンジ領域、IgG4ヒンジ領域、またはその変異体を含む、請求項7または8に記載のポリペプチドサブユニット。
- 前記IgG4ヒンジ領域が、EU付番に従う228位にセリンからプロリンへの突然変異(S228P)を有する(IgG4P)、請求項9に記載のポリペプチドサブユニット。
- 前記IgG FcドメインがヒトIgG Fcドメインである、請求項1〜10のいずれか一項に記載のポリペプチドサブユニット。
- 前記多量体化または三量体化ドメインのカルボキシ末端が、前記GITRL受容体結合ドメインのアミノ末端に直接的に融合される、請求項1〜4または6〜10のいずれか一項に記載のポリペプチドサブユニット。
- 前記第1のリンカー領域、前記第2のリンカー領域、または前記第1および第2のリンカー領域が、存在する場合、(Gly4)nモチーフ、(Gly4Ser)nモチーフ(配列番号19)、Ser(Gly4Ser)nモチーフ(配列番号22)、GGGGSGGGGSGGGGSAL(配列番号23)、GGGGSGGGGSGGGGSA(配列番号24)、およびそれらの組み合わせ(式中、nは1、2、3、4、5、6、7、8、9および10からなる群から選択される正の整数である)を含有するリンカー領域からなる群から独立して選択される、請求項5〜12のいずれか一項に記載のポリペプチドサブユニット。
- 前記第1および第2のリンカー領域が、GGGGSGGGGSGGGGS(配列番号25)、およびGGGGSGGGGSGGGG(配列番号26)からなる群から独立して選択される、請求項13に記載のポリペプチドサブユニット。
- 前記第1のリンカー領域がGGGGSGGGGSGGGGSGGGGSGGGGS(配列番号20)であり、かつ前記第2のリンカー領域が(Gly4)モチーフである、請求項13または14に記載のポリペプチドサブユニット。
- 前記IgG Fcドメインが、IgG1、IgG2、IgG3、IgG4、IG4P Fcドメイン、またはその変異体を含む、請求項1〜15のいずれか一項に記載のポリペプチドサブユニット。
- 前記IgG Fcドメインが、252Y、254T、256E、およびそれらの組み合わせからなる群から選択される1つ以上のアミノ酸残基置換を有し、ここで前記残基はEU付番に従って付番される、請求項1〜16のいずれか一項に記載のポリペプチドサブユニット。
- 前記IgG FcドメインがCH2領域を含む、請求項1〜17のいずれか一項に記載のポリペプチドサブユニット。
- 前記IgG FcドメインがCH3領域をさらに含む、請求項18に記載のポリペプチドサブユニット。
- 前記IgG Fcドメインが、配列番号21に対して、少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するアミノ酸配列を含む、請求項19に記載のポリペプチドサブユニット。
- 前記IgG Fcドメインが配列番号21のアミノ酸配列を含む、請求項19に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、α−ヘリカルコイルドコイルドメイン、ロイシンジッパードメイン、またはそれらの組み合わせを含む、請求項1〜21のいずれか一項に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、マトリリン1、コロニン1a、筋強直性ジストロフィーキナーゼ(DMPK)、ランゲリン、またはそれらの組み合わせから誘導される、請求項22に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、マトリリン1またはコロニン1aから誘導される、請求項23に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、コロニン1aから誘導される、請求項24に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、配列番号11に対して少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するコロニン1a三量体化ドメインを含む、請求項25に記載のポリペプチドサブユニット。
- 前記コロニン1a三量体化ドメインが配列番号11のアミノ酸配列を含む、請求項26に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、配列番号10のコロニン1a三量体化ドメインを含む、請求項26に記載のポリペプチドサブユニット。
- 前記コロニン1a三量体化ドメインが、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、またはその任意の組み合わせもしくは変異体のアミノ酸配列を含む、請求項26に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、配列番号28に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するマトリリン1三量体化ドメインを含む、請求項24に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、配列番号30に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するDMPK三量体化ドメインを含む、請求項23に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、配列番号32に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するランゲリン三量体化ドメインを含む、請求項23に記載のポリペプチドサブユニット。
- 前記三量体化ドメインが、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、または配列番号33のアミノ酸配列を含む、請求項23に記載のポリペプチドサブユニット。
- 前記GITRL受容体結合ドメインが、配列番号34に対して、少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するアミノ酸配列を含む、請求項1〜33のいずれか一項に記載のポリペプチドサブユニット。
- 前記GITRL受容体結合ドメインが、配列番号37に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するアミノ酸配列を含み、ここで残基161はアスパラギル残基ではない、請求項1〜33のいずれか一項に記載のポリペプチドサブユニット。
- 前記GITRL受容体結合ドメインが配列番号35のアミノ酸配列を含む、請求項35に記載のポリペプチドサブユニット。
- 前記GITRL受容体結合ドメインが配列番号35のアミノ酸配列を含み、ここで残基161はアスパルチル残基である、請求項1〜33のいずれか一項に記載のポリペプチドサブユニット。
- 前記GITRL受容体結合ドメインが配列番号36のアミノ酸配列を含む、請求項37に記載のポリペプチドサブユニット。
- 前記GITRL受容体結合ドメインが配列番号37のアミノ酸配列を含む、請求項38に記載のポリペプチドサブユニット。
- 6つの前記ポリペプチドサブユニットから構築される六量体タンパク質が、ヒトGITRに特異的に結合し得る、請求項1〜39のいずれか一項に記載のポリペプチドサブユニット。
- 前記タンパク質がグリコシル化されない、請求項1〜40のいずれか一項に記載のポリペプチドサブユニット。
- 配列番号6に対して、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、もしくは少なくとも99%の配列同一性を有するアミノ酸配列を含む、請求項1〜41のいずれか一項に記載のポリペプチドサブユニット。
- 配列番号6のアミノ酸配列を含む、請求項42に記載のポリペプチドサブユニット。
- アゴニスト活性を有する、請求項1〜43のいずれか一項に記載のポリペプチドサブユニット。
- 関連の異種薬剤をさらに含む、請求項1〜44のいずれか一項に記載のポリペプチドサブユニット。
- 前記異種薬剤が異種ポリペプチドであり、かつ前記ポリペプチドサブユニットとペプチド結合を介して融合される、請求項45に記載のポリペプチドサブユニット。
- 前記異種ポリペプチドが前記IgG−FcドメインのN末端に融合されるか、GITRLの前記受容体結合ドメインのC末端に融合されるか、前記IgG−FcドメインのC末端および前記三量体化ドメインのN末端に融合されるか、またはGITRLの前記三量体化ドメインのC末端および前記受容体結合ドメインのN末端に融合される、請求項46に記載のポリペプチドサブユニット。
- 前記異種薬剤が、前記ポリペプチドサブユニットに化学的に結合される、請求項45に記載のポリペプチドサブユニット。
- 前記異種薬剤が、細胞傷害性分子、安定化剤、免疫応答修飾因子、または検出可能な薬剤を含む、請求項45〜48のいずれか一項に記載のポリペプチドサブユニット。
- 請求項1〜49のいずれか一項に記載の3つのポリペプチドサブユニットを含む三量体タンパク質。
- 請求項1〜49のいずれか一項に記載の6つのポリペプチドサブユニットを含む六量体タンパク質。
- ヒト、または非ヒト霊長類、任意選択的にはカニクイザル、アカゲザル、またはそれらの任意の組み合わせに由来するCD4+もしくはCD8+T細胞、B細胞、またはNK細胞で発現されるようなグルココルチコイド誘導TNF受容体(GITR)に特異的に結合し得、ここで前記CD4+もしくはCD8+T細胞またはB細胞が任意選択的に抗原を経験するか、または前記NK細胞が任意選択的に活性化される、請求項51に記載の六量体タンパク質。
- ヒト、または非ヒト霊長類、任意選択的にはカニクイザル、アカゲザル、またはそれらの任意の組み合わせに由来する一次CD4+もしくはCD8+T細胞で発現されるようなGITRに特異的に結合し得る、請求項52に記載の六量体タンパク質。
- 前記CD4+もしくはCD8+T細胞が抗原を経験する、請求項52または53のいずれか一項に記載の六量体タンパク質。
- ヒトGITRに対する結合親和性が、動力学的排除アッセイによって測定されるとき、約54nM〜約111nMである、請求項51または54に記載の六量体タンパク質。
- 前記結合親和性が、動力学的排除アッセイ(KinExA)において測定されるとき、約82nMである、請求項55に記載の六量体タンパク質。
- GITR陽性免疫細胞からの用量依存性増殖および用量依存性サイトカイン放出を誘導し得る、請求項51〜56のいずれか一項に記載の六量体タンパク質。
- 前記GITR陽性免疫細胞が、プレートに基づくアッセイにおけるT細胞である、請求項57に記載の六量体タンパク質。
- 前記細胞が、CD4+、CD8+、NK、またはB細胞である、請求項52〜58のいずれか一項に記載の六量体タンパク質。
- 前記GITR陽性免疫細胞、T細胞、CD4+、CD8+、またはB細胞が抗原を経験し、かつ/またはNK細胞が活性化される、請求項57〜59のいずれか一項に記載の六量体タンパク質。
- チミジン取り込みによって測定されるとき、約0.1〜約2.7nMのEC50で、一次抗原を経験したヒトT細胞の増殖を刺激し得る、請求項60に記載の六量体タンパク質。
- 前記EC50が約0.5nMである、請求項61に記載の六量体タンパク質。
- 前記サイトカインが、IFNγ、TNFα、IL−5、IL−10、IL−2、IL−4、IL−13、IL−8、IL−12p70、IL−1β、またはそれらの任意の組み合わせである、請求項57に記載の六量体タンパク質。
- GITR発現細胞における下流シグナル伝達経路を活性化し得、任意選択的には前記下流シグナル伝達経路はNFκB経路またはMAPK経路である、請求項51〜63のいずれか一項に記載の六量体タンパク質。
- 前記GITR発現細胞がT細胞である、請求項64に記載の六量体タンパク質。
- 前記GITR発現T細胞が、前記NFκBシグナル伝達経路の刺激に応答してルシフェラーゼを生成する、GITRを発現するJurkat NFκB−ルシフェラーゼレポーター細胞である、請求項64に記載の六量体タンパク質。
- がん治療を必要とする被験者への有効用量の投与が、前記被験者における腫瘍成長を阻害し得る、請求項51〜66のいずれか一項に記載の六量体タンパク質。
- 前記腫瘍成長阻害が、GITR陽性免疫細胞、任意選択的にはT細胞またはNK細胞の存在下で達成される、請求項67に記載の六量体タンパク質。
- 腫瘍成長が、アイソタイプに合致した対照の投与と比べて、少なくとも10%、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%もしくは少なくとも100%阻害される、請求項67または68に記載の六量体タンパク質。
- 前記ポリペプチドサブユニットが、IgG1、IgG2、IgG3、IgG4またはIgG4P Fcドメインを含み、かつGITRへの結合を通じてGITRを発現するCD4+もしくはCD8+T細胞の増殖およびサイトカイン活性化を誘導し得るが、CD4+もしくはCD8+T細胞に対する補体依存性または抗体依存性細胞傷害性を実質的に誘導しない、請求項51〜69のいずれか一項に記載の六量体タンパク質。
- 前記ポリペプチドサブユニットが、IgG1 Fcドメインを含み、かつ高レベルのGITRを発現する細胞に対してFc受容体依存性細胞傷害性を誘導する、請求項51〜69のいずれか一項に記載の六量体タンパク質。
- 前記ポリペプチドサブユニットが、IgG2、IgG3、IgG4、またはIG4P Fcドメインを含み、かつGITRへの結合を通じてGITR発現細胞の増殖を誘導し得る、請求項51〜69のいずれか一項に記載の六量体タンパク質。
- 前記Fc受容体依存性細胞傷害性が、抗体依存細胞傷害性または抗体依存性食作用である、請求項71に記載の六量体タンパク質。
- 高レベルのGITRを発現する前記細胞が腫瘍細胞である、請求項71に記載の六量体タンパク質。
- 高レベルのGITRを発現する前記細胞が、CD4+FOXP3+T細胞または抗原を経験したCD4+FOXP3−T細胞である、請求項71に記載の六量体タンパク質。
- 対照と比べて、CD4+FOXP3+制御性T細胞の頻度における減少をもたらす、請求項71に記載の六量体タンパク質。
- 対照と比べて、腫瘍内部のCD4+FOXP3+制御性T細胞の頻度における減少をもたらす、請求項71に記載の六量体タンパク質。
- 請求項51〜77のいずれか一項に記載の六量体タンパク質、および担体を含む組成物。
- 請求項1〜49のいずれか一項に記載のポリペプチドサブユニット、または請求項51〜78のいずれか一項に記載の六量体タンパク質をコードする核酸を含むポリヌクレオチド。
- 前記ポリペプチドサブユニットに作動可能に連結されるシグナルペプチドをコードする核酸配列をさらに含む、請求項79に記載のポリヌクレオチド。
- 前記シグナルペプチドをコードする前記核酸が、前記ポリペプチドサブユニットの前記IgG Fcドメインのアミノ末端をコードする核酸に作動可能に連結される、請求項80に記載のポリヌクレオチド。
- 配列番号5を含む、請求項79に記載のポリヌクレオチド。
- 請求項79〜82のいずれか一項に記載のポリヌクレオチドを含むベクター。
- 請求項79〜82のいずれか一項に記載のポリヌクレオチドまたは請求項83に記載のベクターを含む宿主細胞。
- 請求項1〜49のいずれか一項に記載のポリペプチドサブユニット、または請求項51〜77のいずれか一項に記載の六量体タンパク質を産生する方法であって、請求項84に記載の宿主細胞を、前記ポリヌクレオチドまたはベクターによってコードされる前記ポリペプチドサブユニットまたは六量体タンパク質が発現される条件下で培養するステップと、前記ポリペプチドサブユニットまたは六量体タンパク質を回収するステップと、を含む、方法。
- 抗原を経験したT細胞および/またはNK細胞の生存または増殖を促進するための方法であって、抗原を経験したT細胞および/またはNK細胞を、請求項51〜77のいずれか一項に記載の六量体タンパク質または請求項78に記載の組成物と接触させるステップを含み、ここで前記六量体タンパク質は前記T細胞および/またはNK細胞の表面上のGITRに特異的に結合し得る、方法。
- 活性化されたGITRを発現する免疫細胞からのサイトカイン放出を誘導する方法であって、これらの細胞を、請求項51〜77のいずれか一項に記載の六量体タンパク質または請求項78に記載の組成物と接触させるステップを含み、ここで前記六量体タンパク質はこれらの細胞の表面上のGITRに特異的に結合し得る、方法。
- 前記サイトカインが、IFNγ、TNFα、IL−10、GM−CSF、またはそれらの任意の組み合わせである、請求項87に記載の方法。
- 前記GITRを発現する免疫細胞が、抗原を経験したCD4+T細胞、抗原を経験したCD8+T細胞、活性化NK細胞またはそれらの組み合わせである、請求項87または88のいずれか一項に記載の方法。
- 前記活性化NK細胞、抗原を経験したCD4+もしくは抗原を経験したCD8+T細胞が、ヒトNK細胞、CD4+もしくはCD8+T細胞、カニクイザルNK細胞、CD4+もしくはCD8+T細胞、アカゲザルNK細胞、CD4+もしくはCD8+T細胞、またはそれらの組み合わせである、請求項86〜89のいずれか一項に記載の方法。
- T細胞またはNK細胞活性化を促進する方法であって、T細胞またはNK細胞を、請求項51〜77のいずれか一項に記載の六量体タンパク質または請求項78に記載の組成物と接触させるステップを含み、ここで前記六量体タンパク質は前記T細胞またはNK細胞の表面上のGITRに特異的に結合し得る、方法。
- 前記六量体タンパク質を、前記FcドメインとFcγRを発現する細胞との相互作用を通じて架橋するステップをさらに含む、請求項91に記載の方法。
- FcγRを発現する前記細胞が、B細胞、単球、マクロファージ、骨髄もしくは形質細胞様樹状細胞、濾胞樹状細胞、ランゲルハンス細胞、内皮細胞、NK細胞、好中球、好酸球、血小板、肥満細胞、一次ヒト腫瘍もしくは腫瘍灌流もしくは非灌流リンパ節に由来するCD45+細胞、他の二次もしくは三次リンパ様構造に由来するCD45+細胞、またはそれらの組み合わせである、請求項92に記載の方法。
- NK細胞またはT細胞活性化が、前記NFκBまたはMAPKシグナル伝達経路の刺激を通じて測定され得る、請求項91〜93のいずれか一項に記載の方法。
- 前記NKまたはT細胞が、活性化NK細胞、抗原を経験したCD4+T細胞、抗原を経験したCD8+T細胞、またはそれらの組み合わせである、請求項91〜94のいずれか一項に記載の方法。
- 前記NK細胞、CD4+もしくはCD8+T細胞が、一次ヒトNK細胞、CD4+もしくはCD8+T細胞、カニクイザルNK細胞、CD4+もしくはCD8+T細胞、アカゲザルNK細胞、CD4+もしくはCD8+T細胞、またはそれらの組み合わせである、請求項95に記載の方法。
- 前記接触させるステップが、前記六量体タンパク質または前記六量体タンパク質を含む組成物を有効量で被験者に投与するステップを含む、請求項86〜96のいずれか一項に記載の方法。
- 被験者におけるがんを治療する方法であって、請求項51〜77のいずれか一項に記載の六量体タンパク質、または請求項78に記載の組成物を有効量で治療を必要とする被験者に投与するステップを含む、方法。
- 前記被験者がヒト被験者またはイヌ被験体である、請求項98に記載の方法。
- 前記がんが固形腫瘍である、請求項98に記載の方法。
- 前記固形腫瘍が、結腸直腸がん、乳がん、肝細胞がん、非小細胞肺がん、小細胞肺がん、中皮腫、頭頚部がん、胃がん、膵がん、黒色腫、ぶどう膜黒色腫、腎臓がん、卵巣がん、子宮頸がん、神経膠芽腫、睾丸がん、甲状腺がん、前立腺がん、および食道がんからなる群から選択されるがんに関連する、請求項100に記載の方法。
- 前記がんが血液がんである、請求項98に記載の方法。
- 前記血液がんが、ホジキンリンパ腫、非ホジキンリンパ腫、多発性骨髄腫、急性リンパ性白血病、急性骨髄性白血病、慢性リンパ性白血病、および慢性骨髄性白血病からなる群から選択される、請求項102に記載の方法。
- 前記六量体タンパク質または組成物の投与により、腫瘍成長が阻害され得るか、腫瘍減少が促進され得るか、またはその双方である、請求項98〜103のいずれか一項に記載の方法。
- 腫瘍成長阻害が、NK細胞またはT細胞の存在下で達成される、請求項98〜104のいずれか一項に記載の方法。
- 腫瘍成長が、GITRL受容体結合ドメインが欠如したアイソタイプに合致した対照の投与と比べて、少なくとも10%、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%もしくは100%阻害される、請求項104または請求項105に記載の方法。
- 前記六量体タンパク質が、IgG1、IgG2、IgG3、IgG4またはIgG4P Fcドメインを含み、かつ抗原を経験したGITR発現免疫細胞の増殖をGITRへの結合を通じて誘導し得るが、前記活性化免疫細胞に対する補体依存性または抗体依存性細胞傷害性を実質的に誘導しない、請求項98〜106のいずれか一項に記載の方法。
- 前記GITR発現免疫細胞が、CD4+T細胞、NK細胞、またはCD8+T細胞である、請求項107に記載の方法。
- 前記六量体タンパク質が、IgG1 Fcドメインを含み、かつ高レベルのGITRを発現する細胞のNK細胞および/またはマクロファージ媒介性の抗体依存細胞傷害性(ADCC)または抗体依存性細胞食作用を誘導する、請求項98〜106のいずれか一項に記載の方法。
- 高レベルのGITRを発現する前記細胞が腫瘍細胞である、請求項109に記載の方法。
- 高レベルのGITRを発現する前記細胞がCD4+FOXP3+T細胞である、請求項109に記載の方法。
- 前記六量体タンパク質が、対照と比べて、CD4+FOXP3+制御性T細胞の頻度における減少をもたらす、請求項111に記載の方法。
- 前記六量体タンパク質が、対照と比べて、腫瘍内部のCD4+FOXP3+制御性T細胞の頻度における減少をもたらす、請求項111に記載の方法。
- 被験者における免疫応答を増強する方法であって、それを必要とする被験者に請求項51〜77のいずれか一項に記載の六量体タンパク質、または請求項78に記載の組成物を治療有効量で投与するステップを含む、方法。
- 前記被験者が、ヒト被験者またはイヌ被験体である、請求項114に記載の方法。
- それを必要とする前記被験者ががんを有する、請求項115に記載の方法。
- 前記がんが固形腫瘍である、請求項116に記載の方法。
- 前記被験者が、結腸直腸がん、乳がん、肝細胞がん、非小細胞肺がん(NSCLC)、小細胞肺がん、中皮腫、頭頚部がん、胃がん、膵がん、黒色腫、ぶどう膜黒色腫、腎臓がん、卵巣がん、子宮頸がん、神経膠芽腫、睾丸がん、甲状腺がん、前立腺がん、食道がん、膀胱がん、頭頸部扁平上皮がん(SCCHN)、結腸直腸がん(CRC)、血液がん、またはそれらの組み合わせを有する、請求項114〜117のいずれか一項に記載の方法。
- 前記がんが血液がんである、請求項118に記載の方法。
- 前記血液がんが、ホジキンリンパ腫、非ホジキンリンパ腫、多発性骨髄腫、急性リンパ性白血病、急性骨髄性白血病、慢性リンパ性白血病、および慢性骨髄性白血病からなる群から選択される、請求項119に記載の方法。
- 前記六量体タンパク質または組成物の投与により、腫瘍成長が阻害され得るか、腫瘍減少が促進され得るか、またはその双方である、請求項114〜120のいずれか一項に記載の方法。
- 腫瘍成長阻害が、NK細胞またはT細胞の存在下で達成される、請求項114〜121のいずれか一項に記載の方法。
- 腫瘍成長が、GITRL受容体結合ドメインが欠如したアイソタイプに合致した対照の投与と比べて、少なくとも10%、少なくとも20%、少なくとも30%、少なくとも40%、少なくとも50%、少なくとも60%、少なくとも70、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%もしくは100%阻害される、請求項121または122のいずれか一項に記載の方法。
- 前記六量体タンパク質が、IgG1、IgG2、IgG3、IgG4またはIgG4P Fcドメインを含み、かつ活性化されたGITR発現免疫細胞の増殖をGITRへの結合を通じて誘導し得るが、前記活性化免疫細胞に対する補体依存性または抗体依存性細胞傷害性を実質的に誘導しない、請求項114〜123のいずれか一項に記載の方法。
- 前記GITR発現免疫細胞が、CD4+T細胞、NK細胞、またはCD8+T細胞である、請求項124に記載の方法。
- 前記六量体タンパク質が、IgG1 Fcドメインを含み、かつ高レベルのGITRを発現する細胞のNK細胞および/またはマクロファージ媒介性の抗体依存細胞傷害性(ADCC)または抗体依存性細胞食作用を誘導する、請求項114〜123のいずれか一項に記載の方法。
- 高レベルのGITRを発現する前記細胞が腫瘍細胞である、請求項126に記載の方法。
- 高レベルのGITRを発現する前記細胞がCD4+FOXP3+T細胞である、請求項126に記載の方法。
- 前記六量体タンパク質が、対照と比べて、CD4+FOXP3+制御性T細胞の頻度における減少をもたらす、請求項126に記載の方法。
- 前記六量体タンパク質が、対照と比べて、腫瘍内部のCD4+FOXP3+制御性T細胞の頻度における減少をもたらす、請求項126に記載の方法。
- 被験者における固形腫瘍を治療する方法であって、請求項1に記載の単離された一本鎖ポリペプチドサブユニットおよびOX40アゴニストを前記被験者に投与するステップを含む、方法。
- 前記OX40アゴニストが、OX40リガンド融合タンパク質または抗OX40抗体の1つ以上である、請求項131に記載の方法。
- 前記OX40リガンド融合タンパク質がMEDI6383である、請求項132に記載の方法。
- 前記抗OX40抗体がMEDI0562である、請求項132に記載の方法。
- 被験者におけるがんを治療する方法であって、T細胞プライミング剤と組み合わせて、請求項51〜77のいずれか一項に記載の六量体タンパク質、または請求項78に記載の組成物を有効量で治療を必要とする被験者に投与するステップを含む、方法。
- 前記T細胞プライミング剤が、E7合成ロングペプチド(SLP)にCpGオリゴデオキシヌクレオチドを加えたものである、請求項136に記載の方法。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201562204212P | 2015-08-12 | 2015-08-12 | |
| US62/204,212 | 2015-08-12 | ||
| US201662350447P | 2016-06-15 | 2016-06-15 | |
| US62/350,447 | 2016-06-15 | ||
| PCT/EP2016/069175 WO2017025610A1 (en) | 2015-08-12 | 2016-08-11 | Gitrl fusion proteins and uses thereof |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2018522571A true JP2018522571A (ja) | 2018-08-16 |
| JP2018522571A5 JP2018522571A5 (ja) | 2019-09-12 |
Family
ID=56740217
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2018506513A Pending JP2018522571A (ja) | 2015-08-12 | 2016-08-11 | Gitrl融合タンパク質およびその使用 |
Country Status (17)
| Country | Link |
|---|---|
| US (2) | US10428131B2 (ja) |
| EP (1) | EP3334758A1 (ja) |
| JP (1) | JP2018522571A (ja) |
| KR (1) | KR20180033588A (ja) |
| CN (1) | CN108026158A (ja) |
| AU (1) | AU2016306625A1 (ja) |
| BR (1) | BR112018002039A2 (ja) |
| CA (1) | CA2994346A1 (ja) |
| CL (1) | CL2018000358A1 (ja) |
| CO (1) | CO2018002450A2 (ja) |
| HK (2) | HK1250044A1 (ja) |
| IL (1) | IL257113A (ja) |
| MX (1) | MX2018001787A (ja) |
| RU (1) | RU2018108236A (ja) |
| TW (1) | TW201718632A (ja) |
| WO (1) | WO2017025610A1 (ja) |
| ZA (1) | ZA201801640B (ja) |
Families Citing this family (48)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2934028A1 (en) * | 2013-12-17 | 2015-06-25 | Genentech, Inc. | Combination therapy comprising ox40 binding agonists and pd-1 axis binding antagonists |
| KR20160146747A (ko) | 2014-03-31 | 2016-12-21 | 제넨테크, 인크. | 항혈관신생제 및 ox40 결합 효능제를 포함하는 조합 요법 |
| MA41460A (fr) | 2015-02-03 | 2017-12-12 | Oncomed Pharm Inc | Agents de liaison à la tnfrsf et leurs utilisations |
| MY198560A (en) | 2015-10-02 | 2023-09-05 | Hoffmann La Roche | Bispecific antibodies specific for a costimulatory tnf receptor |
| WO2018027204A1 (en) | 2016-08-05 | 2018-02-08 | Genentech, Inc. | Multivalent and multiepitopic anitibodies having agonistic activity and methods of use |
| WO2018178074A1 (en) | 2017-03-29 | 2018-10-04 | F. Hoffmann-La Roche Ag | Trimeric antigen binding molecules specific for a costimulatory tnf receptor |
| WO2018185618A1 (en) | 2017-04-03 | 2018-10-11 | Novartis Ag | Anti-cdh6 antibody drug conjugates and anti-gitr antibody combinations and methods of treatment |
| AR111651A1 (es) | 2017-04-28 | 2019-08-07 | Novartis Ag | Conjugados de anticuerpos que comprenden agonistas del receptor de tipo toll y terapias de combinación |
| WO2018213297A1 (en) | 2017-05-16 | 2018-11-22 | Bristol-Myers Squibb Company | Treatment of cancer with anti-gitr agonist antibodies |
| WO2018215506A1 (en) * | 2017-05-23 | 2018-11-29 | Université De Genève | Beta-2-glycoprotein 1 derived peptide and use thereof for treating antiphospholipid syndrome |
| WO2018229715A1 (en) | 2017-06-16 | 2018-12-20 | Novartis Ag | Compositions comprising anti-cd32b antibodies and methods of use thereof |
| WO2018235056A1 (en) | 2017-06-22 | 2018-12-27 | Novartis Ag | IL-1BETA BINDING ANTIBODIES FOR USE IN THE TREATMENT OF CANCER |
| WO2018237173A1 (en) | 2017-06-22 | 2018-12-27 | Novartis Ag | ANTIBODY MOLECULES DIRECTED AGAINST CD73 AND CORRESPONDING USES |
| MX2019015885A (es) | 2017-06-22 | 2020-09-10 | Novartis Ag | Moleculas de anticuerpo que se unen a cd73 y usos de las mismas. |
| KR20200019865A (ko) | 2017-06-22 | 2020-02-25 | 노파르티스 아게 | 암 치료에 사용하기 위한 il-1베타 결합 항체 |
| CN111050791A (zh) | 2017-06-27 | 2020-04-21 | 诺华股份有限公司 | 用于抗tim-3抗体的给药方案及其用途 |
| AU2018302283A1 (en) | 2017-07-20 | 2020-02-06 | Novartis Ag | Dosage regimens of anti-LAG-3 antibodies and uses thereof |
| US20210040205A1 (en) | 2017-10-25 | 2021-02-11 | Novartis Ag | Antibodies targeting cd32b and methods of use thereof |
| RU2020119578A (ru) | 2017-11-16 | 2021-12-17 | Новартис Аг | Комбинированные терапии |
| US20210147547A1 (en) | 2018-04-13 | 2021-05-20 | Novartis Ag | Dosage Regimens For Anti-Pd-L1 Antibodies And Uses Thereof |
| TW202015726A (zh) | 2018-05-30 | 2020-05-01 | 瑞士商諾華公司 | Entpd2抗體、組合療法、及使用該等抗體和組合療法之方法 |
| WO2019232244A2 (en) | 2018-05-31 | 2019-12-05 | Novartis Ag | Antibody molecules to cd73 and uses thereof |
| BR112020026947B1 (pt) | 2018-07-10 | 2022-11-22 | Novartis Ag | Derivados de 3-(5-hidróxi-1-oxoisoindolin-2-il)piperidina-2,6-diona, e composição farmacêutica |
| AR116109A1 (es) | 2018-07-10 | 2021-03-31 | Novartis Ag | Derivados de 3-(5-amino-1-oxoisoindolin-2-il)piperidina-2,6-diona y usos de los mismos |
| WO2020021465A1 (en) | 2018-07-25 | 2020-01-30 | Advanced Accelerator Applications (Italy) S.R.L. | Method of treatment of neuroendocrine tumors |
| KR20210044243A (ko) * | 2018-08-13 | 2021-04-22 | 인히브릭스, 인크. | Ox40 결합 폴리펩티드 및 이의 용도 |
| EP3873532A1 (en) | 2018-10-31 | 2021-09-08 | Novartis AG | Dc-sign antibody drug conjugates |
| EP3897637A1 (en) | 2018-12-20 | 2021-10-27 | Novartis AG | Dosing regimen and pharmaceutical combination comprising 3-(1-oxoisoindolin-2-yl)piperidine-2,6-dione derivatives |
| WO2020128637A1 (en) | 2018-12-21 | 2020-06-25 | Novartis Ag | Use of il-1 binding antibodies in the treatment of a msi-h cancer |
| AU2019409139A1 (en) | 2018-12-21 | 2021-06-03 | Novartis Ag | Use of IL-1β binding antibodies |
| EP3897613A1 (en) | 2018-12-21 | 2021-10-27 | Novartis AG | Use of il-1beta binding antibodies |
| CA3119584A1 (en) | 2018-12-21 | 2020-06-25 | Novartis Ag | Use of il-1 beta antibodies in the treatment or prevention of myelodysplastic syndrome |
| EP3924054A1 (en) | 2019-02-15 | 2021-12-22 | Novartis AG | 3-(1-oxo-5-(piperidin-4-yl)isoindolin-2-yl)piperidine-2,6-dione derivatives and uses thereof |
| US20220144798A1 (en) | 2019-02-15 | 2022-05-12 | Novartis Ag | Substituted 3-(1-oxoisoindolin-2-yl)piperidine-2,6-dione derivatives and uses thereof |
| JP2022548881A (ja) | 2019-09-18 | 2022-11-22 | ノバルティス アーゲー | Entpd2抗体、組合せ療法並びに抗体及び組合せ療法を使用する方法 |
| KR20220087498A (ko) | 2019-10-21 | 2022-06-24 | 노파르티스 아게 | Tim-3 억제제 및 그의 용도 |
| KR20220103947A (ko) | 2019-10-21 | 2022-07-25 | 노파르티스 아게 | 베네토클락스 및 tim-3 억제제를 사용한 조합 요법 |
| KR20220116257A (ko) | 2019-12-20 | 2022-08-22 | 노파르티스 아게 | 골수섬유증 및 골수이형성 증후군을 치료하기 위한, 데시타빈 또는 항 pd-1 항체 스파르탈리주맙을 포함하거나 또는 포함하지 않는, 항 tim-3 항체 mbg453 및 항 tgf-베타 항체 nis793의 조합물 |
| WO2021144657A1 (en) | 2020-01-17 | 2021-07-22 | Novartis Ag | Combination comprising a tim-3 inhibitor and a hypomethylating agent for use in treating myelodysplastic syndrome or chronic myelomonocytic leukemia |
| EP4141120A4 (en) * | 2020-04-22 | 2024-05-22 | POSTECH Research and Business Development Foundation | CORONAVIRUS DISEASE 2019 (COVID-19) RECOMBINANT TRIMER-FORMING SPIKE PROTEIN, METHOD FOR MASS PRODUCTION OF RECOMBINANT SPIKE PROTEIN IN PLANTS, AND METHOD FOR PREPARING VACCINE COMPOSITION BASED ON SAME |
| KR20230024967A (ko) | 2020-06-11 | 2023-02-21 | 노파르티스 아게 | Zbtb32 억제제 및 이의 용도 |
| WO2021260528A1 (en) | 2020-06-23 | 2021-12-30 | Novartis Ag | Dosing regimen comprising 3-(1-oxoisoindolin-2-yl)piperidine-2,6-dione derivatives |
| CN116134027A (zh) | 2020-08-03 | 2023-05-16 | 诺华股份有限公司 | 杂芳基取代的3-(1-氧代异吲哚啉-2-基)哌啶-2,6-二酮衍生物及其用途 |
| WO2022043557A1 (en) | 2020-08-31 | 2022-03-03 | Advanced Accelerator Applications International Sa | Method of treating psma-expressing cancers |
| WO2022043558A1 (en) | 2020-08-31 | 2022-03-03 | Advanced Accelerator Applications International Sa | Method of treating psma-expressing cancers |
| TW202304979A (zh) | 2021-04-07 | 2023-02-01 | 瑞士商諾華公司 | 抗TGFβ抗體及其他治療劑用於治療增殖性疾病之用途 |
| AR125874A1 (es) | 2021-05-18 | 2023-08-23 | Novartis Ag | Terapias de combinación |
| CN113429488B (zh) * | 2021-07-14 | 2022-08-30 | 海正生物制药有限公司 | GITR/TGF-β双靶向融合蛋白及其用途 |
Family Cites Families (10)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5681718A (en) | 1986-03-14 | 1997-10-28 | Celltech Limited | Methods for enhanced production of tissue plasminogen activator in cell culture using alkanoic acids or salts thereof |
| DE19963859A1 (de) * | 1999-12-30 | 2001-07-12 | Apotech Res & Dev Ltd | Bi- oder Oligomer eines Di-, Tri-, Quattro- oder Pentamers von rekombinanten Fusionsproteinen |
| GB0216648D0 (en) | 2002-07-18 | 2002-08-28 | Lonza Biologics Plc | Method of expressing recombinant protein in CHO cells |
| CN1809589A (zh) * | 2003-05-23 | 2006-07-26 | Wyeth公司 | Gitr配体和gitr配体相关分子和抗体及其应用 |
| US20080187954A1 (en) | 2004-03-10 | 2008-08-07 | Lonza Ltd. | Method For Producing Antibodies |
| US20060280728A1 (en) * | 2005-05-06 | 2006-12-14 | Providence Health System | Trimeric OX40-immunoglobulin fusion protein and methods of use |
| JP5932217B2 (ja) * | 2007-07-12 | 2016-06-08 | ジーアイティーアール, インコーポレイテッド | Gitr結合分子を使用する併用療法 |
| TWI476001B (zh) * | 2011-12-26 | 2015-03-11 | Ind Tech Res Inst | 三倍體Fc融合蛋白及其用途 |
| WO2015116178A1 (en) | 2014-01-31 | 2015-08-06 | Thomas Jefferson University | Fusion proteins for modulating regulatory and effector t cells |
| CN106456734A (zh) | 2014-05-29 | 2017-02-22 | 免疫医疗有限责任公司 | Ox40l融合蛋白及其用途 |
-
2016
- 2016-08-11 CA CA2994346A patent/CA2994346A1/en not_active Abandoned
- 2016-08-11 TW TW105125874A patent/TW201718632A/zh unknown
- 2016-08-11 EP EP16753881.8A patent/EP3334758A1/en not_active Withdrawn
- 2016-08-11 MX MX2018001787A patent/MX2018001787A/es unknown
- 2016-08-11 CN CN201680045052.8A patent/CN108026158A/zh active Pending
- 2016-08-11 JP JP2018506513A patent/JP2018522571A/ja active Pending
- 2016-08-11 KR KR1020187006894A patent/KR20180033588A/ko unknown
- 2016-08-11 RU RU2018108236A patent/RU2018108236A/ru not_active Application Discontinuation
- 2016-08-11 BR BR112018002039A patent/BR112018002039A2/pt not_active Application Discontinuation
- 2016-08-11 WO PCT/EP2016/069175 patent/WO2017025610A1/en active Application Filing
- 2016-08-11 AU AU2016306625A patent/AU2016306625A1/en not_active Abandoned
- 2016-08-11 US US15/234,551 patent/US10428131B2/en not_active Expired - Fee Related
-
2018
- 2018-01-23 IL IL257113A patent/IL257113A/en unknown
- 2018-02-08 CL CL2018000358A patent/CL2018000358A1/es unknown
- 2018-03-05 CO CONC2018/0002450A patent/CO2018002450A2/es unknown
- 2018-03-09 ZA ZA2018/01640A patent/ZA201801640B/en unknown
- 2018-07-23 HK HK18109492.4A patent/HK1250044A1/zh unknown
- 2018-07-23 HK HK18109489.9A patent/HK1250043A1/zh unknown
-
2019
- 2019-09-03 US US16/559,152 patent/US20200140510A1/en not_active Abandoned
Also Published As
| Publication number | Publication date |
|---|---|
| ZA201801640B (en) | 2019-01-30 |
| HK1250043A1 (zh) | 2018-11-23 |
| HK1250044A1 (zh) | 2018-11-23 |
| CA2994346A1 (en) | 2017-02-16 |
| CL2018000358A1 (es) | 2018-07-06 |
| MX2018001787A (es) | 2018-06-06 |
| US20170073386A1 (en) | 2017-03-16 |
| KR20180033588A (ko) | 2018-04-03 |
| TW201718632A (zh) | 2017-06-01 |
| US20200140510A1 (en) | 2020-05-07 |
| EP3334758A1 (en) | 2018-06-20 |
| AU2016306625A1 (en) | 2018-03-29 |
| CO2018002450A2 (es) | 2018-11-22 |
| CN108026158A (zh) | 2018-05-11 |
| RU2018108236A (ru) | 2019-09-12 |
| US10428131B2 (en) | 2019-10-01 |
| BR112018002039A2 (pt) | 2018-09-18 |
| IL257113A (en) | 2018-03-29 |
| WO2017025610A1 (en) | 2017-02-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20200140510A1 (en) | Gitrl fusion proteins and uses thereof | |
| JP6666267B2 (ja) | Ox40l融合タンパク質およびその使用 | |
| CN108883173B (zh) | 抗体和其使用方法 | |
| JP6931329B2 (ja) | 免疫抑制機能を有する細胞に対するt細胞リダイレクト抗原結合分子を用いた併用療法 | |
| JP6212493B2 (ja) | 抗cd134(ox40)抗体およびその使用 | |
| AU2014238546B2 (en) | Humanized anti-CD134 (OX40) antibodies and uses thereof | |
| JP6976322B2 (ja) | 新規抗ctla4抗体 | |
| TW201713694A (zh) | 抗ox40抗體及其使用方法 | |
| JP2017532037A (ja) | ヒト化抗ox40抗体及びその使用 | |
| TW201735947A (zh) | 用於癌之治療的細胞傷害誘導治療劑 | |
| CN114507285A (zh) | Vista抗原结合分子 | |
| KR20200012907A (ko) | 변형된 IgG1 Fc 도메인 및 그와의 항-CD40 도메인 항체 융합체 | |
| EP3853247A2 (en) | Methods and uses of variant cd80 fusion proteins and related constructs | |
| WO2021198335A1 (en) | Bispecific antibodies comprising a modified c-terminal crossfab fragment | |
| TW202043284A (zh) | 多特異性結合蛋白 | |
| JP2023502776A (ja) | 癌細胞、免疫細胞および腫瘍微小環境へのシアリダーゼの免疫細胞送達 | |
| NZ718372B2 (en) | Anti-cd134 (ox40) antibodies and uses thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20190805 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20190805 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20200629 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20200728 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20210309 |