JP2009079062A - 抗cd40モノクローナル抗体 - Google Patents
抗cd40モノクローナル抗体 Download PDFInfo
- Publication number
- JP2009079062A JP2009079062A JP2008275853A JP2008275853A JP2009079062A JP 2009079062 A JP2009079062 A JP 2009079062A JP 2008275853 A JP2008275853 A JP 2008275853A JP 2008275853 A JP2008275853 A JP 2008275853A JP 2009079062 A JP2009079062 A JP 2009079062A
- Authority
- JP
- Japan
- Prior art keywords
- antibody
- variable region
- chain variable
- seq
- hybridoma
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images

















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2878—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the NGF-receptor/TNF-receptor superfamily, e.g. CD27, CD30, CD40, CD95
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P19/00—Drugs for skeletal disorders
- A61P19/02—Drugs for skeletal disorders for joint disorders, e.g. arthritis, arthrosis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P25/00—Drugs for disorders of the nervous system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P29/00—Non-central analgesic, antipyretic or antiinflammatory agents, e.g. antirheumatic agents; Non-steroidal antiinflammatory drugs [NSAID]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/06—Immunosuppressants, e.g. drugs for graft rejection
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/08—Antiallergic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P7/00—Drugs for disorders of the blood or the extracellular fluid
- A61P7/04—Antihaemorrhagics; Procoagulants; Haemostatic agents; Antifibrinolytic agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/10—Drugs for disorders of the cardiovascular system for treating ischaemic or atherosclerotic diseases, e.g. antianginal drugs, coronary vasodilators, drugs for myocardial infarction, retinopathy, cerebrovascula insufficiency, renal arteriosclerosis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/20—Immunoglobulins specific features characterized by taxonomic origin
- C07K2317/21—Immunoglobulins specific features characterized by taxonomic origin from primates, e.g. man
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/74—Inducing cell proliferation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/30—Non-immunoglobulin-derived peptide or protein having an immunoglobulin constant or Fc region, or a fragment thereof, attached thereto
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Immunology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Engineering & Computer Science (AREA)
- Biophysics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Biochemistry (AREA)
- Rheumatology (AREA)
- Diabetes (AREA)
- Hematology (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
- Neurosurgery (AREA)
- Urology & Nephrology (AREA)
- Virology (AREA)
- Biomedical Technology (AREA)
- Neurology (AREA)
- Transplantation (AREA)
- Heart & Thoracic Surgery (AREA)
- Cardiology (AREA)
- Vascular Medicine (AREA)
- Pain & Pain Management (AREA)
- Pulmonology (AREA)
- Dermatology (AREA)
- Orthopedic Medicine & Surgery (AREA)
- Physical Education & Sports Medicine (AREA)
Abstract
【課題】従来の抗ヒトCD40抗体に比べてより治療効果の高いことが期待されるアゴニスティックな抗ヒトCD40抗体又はその機能的断片の提供。
【解決手段】CD40に対し、アゴニスティックに作用する抗体又はその機能的断片。
【選択図】なし
【解決手段】CD40に対し、アゴニスティックに作用する抗体又はその機能的断片。
【選択図】なし
Description
本発明は、ヒトのB細胞、樹状細胞(DC)等の表面に存在するヒトCD40抗原を認識する抗体又はその機能的断片に関する。具体的には、本発明は樹状細胞(DC)表面のヒトCD40抗原に対して実質的にアンタゴニスティックである抗ヒトCD40抗体又はその機能的断片、および従来の抗ヒトCD40抗体に比べてより治療効果の高いことが期待されるアゴニスティックな抗ヒトCD40抗体又はその機能的断片に関する。
1. CD40
CD40は分子量50kDaの細胞膜表面に存在する抗原であり、B細胞、樹状細胞(DC)、ある種の癌細胞、そして胸腺上皮細胞に発現している。CD40はB細胞やDCの増殖、分化に重要な働きをしていることが知られている。CD40は、ヒトB細胞表面に発現する抗原として同定され(非特許文献1及び2を参照)、アミノ酸配列の相同性から、CD40は、低親和性NGFレセプターやTNFレセプター、CD27、OX40、CD30などが属しているTNFレセプターファミリーの1つのメンバーとして考えられている。ヒトおよびマウスのCD40に対するリガンド(CD40L)は、近年遺伝子クローニングされ、II型膜蛋白質であること、及び活性化したCD4+T細胞に発現していることが分かった。CD40Lは、強力な活性化シグナルをヒトおよびマウスのB細胞に導入することも分かっている。
CD40は分子量50kDaの細胞膜表面に存在する抗原であり、B細胞、樹状細胞(DC)、ある種の癌細胞、そして胸腺上皮細胞に発現している。CD40はB細胞やDCの増殖、分化に重要な働きをしていることが知られている。CD40は、ヒトB細胞表面に発現する抗原として同定され(非特許文献1及び2を参照)、アミノ酸配列の相同性から、CD40は、低親和性NGFレセプターやTNFレセプター、CD27、OX40、CD30などが属しているTNFレセプターファミリーの1つのメンバーとして考えられている。ヒトおよびマウスのCD40に対するリガンド(CD40L)は、近年遺伝子クローニングされ、II型膜蛋白質であること、及び活性化したCD4+T細胞に発現していることが分かった。CD40Lは、強力な活性化シグナルをヒトおよびマウスのB細胞に導入することも分かっている。
また、樹状細胞にはB細胞よりも多くのCD40発現が確認されており、重要な役割を担っていることが明らかとなってきた。CD40がCD40Lと結合すると、抗原提示細胞(APC)の活性化、すなわちCD80(B7-1)やCD86(B7-2)などの補助刺激分子の発現、あるいはIL-12の産生が増強される(非特許文献3及び4を参照)。樹状細胞は強い抗原提示能を有し、強力なヘルパーT(Th)細胞活性化能を持っている。また、ナイーブ(naive) Th細胞のTh1又はTh2細胞への分化を樹状細胞が制御していると考えられている。ミエロイド系樹状細胞である末梢血単球をGM-CSF及びIL-4とともに培養してCD40Lにより成熟させた樹状細胞(DC1)はin vitroにおいて、IL-12産生能を有し、異系naive Th細胞を刺激活性化し、IFNγ産生T細胞を誘導する(すなわちTh1への分化を促す)。この作用は抗IL-12抗体により阻害されることから、IL-12を介した反応と考えられる。一方リンパ組織T領域や、末梢血に存在するplasmacytoid T 細胞をIL-3、CD40リガンドとともに培養したリンパ球系樹状細胞(DC2)は、IL-12産生能は有さず、そして異系naive Th細胞を刺激活性化し、IL-4産生T細胞を誘導し、Th2への分化を促進することが示されている。Th1細胞は細胞性免疫の活性化にかかわり、Th2細胞は液性免疫能を高めると同時に細胞性免疫能の抑制に関与すると考えられている。Th1細胞のヘルプで活性化された細胞傷害性T細胞(CTL)は、細胞質内で増殖する病原体(多くのウィルス、リステリア菌、結核菌、およびトキソプラズマ原虫など)や腫瘍細胞を除去することができる。
膜表面に発現したCD40を認識する抗CD40モノクローナル抗体が、B細胞に対していろいろな生物活性を示すことは示されてきた。抗CD40モノクローナル抗体は、CD40とCD40Lとの相互作用に対してアゴニスティックなものと、アンタゴニスティックなものとに大別される。
2. アゴニスティック抗体
アゴニスティック抗体の作用として、B細胞の活性化が知られている。たとえば、抗CD40抗体が細胞接着を誘導する(非特許文献5及び6を参照)、細胞の大きさを増進する(非特許文献6及び7を参照)、抗IgM抗体、抗CD20抗体またはphorbol esterのみで活性化されたB細胞の分裂を誘導する(非特許文献8〜10を参照)、IL4存在下でB細胞の分裂を誘導する(非特許文献7及び11を参照)、IL-4刺激、T細胞除去培養細胞のIgE(非特許文献12及び13を参照)、IgG、IgM(非特許文献13を参照)の発現を誘導する、IL-4によるB細胞からの可溶性CD23/FceRIIの分泌と(非特許文献14及び15を参照)細胞上の発現増強(非特許文献16を参照)をする、IL-6の生産を促進する(非特許文献17を参照)ことが報告されている。さらには、CDw32+接着細胞存在下で、IL-4及び抗CD40抗体を添加することにより、ヒト初代培養B細胞から、B細胞クローンを樹立することや(非特許文献18を参照)、胚中心の中心細胞のアポトーシスが、抗原レセプターの働きにかかわらず、CD40を介して阻害されること(非特許文献19を参照)が報告されている。以上のようにCD40は、ヒトB細胞表面に発現する抗原として同定されたため、単離された抗体の多くは、主にヒトB細胞に対する増殖分化誘導機能、癌細胞における細胞死誘導活性を指標に評価されてきた(非特許文献20〜22を参照)。
アゴニスティック抗体の作用として、B細胞の活性化が知られている。たとえば、抗CD40抗体が細胞接着を誘導する(非特許文献5及び6を参照)、細胞の大きさを増進する(非特許文献6及び7を参照)、抗IgM抗体、抗CD20抗体またはphorbol esterのみで活性化されたB細胞の分裂を誘導する(非特許文献8〜10を参照)、IL4存在下でB細胞の分裂を誘導する(非特許文献7及び11を参照)、IL-4刺激、T細胞除去培養細胞のIgE(非特許文献12及び13を参照)、IgG、IgM(非特許文献13を参照)の発現を誘導する、IL-4によるB細胞からの可溶性CD23/FceRIIの分泌と(非特許文献14及び15を参照)細胞上の発現増強(非特許文献16を参照)をする、IL-6の生産を促進する(非特許文献17を参照)ことが報告されている。さらには、CDw32+接着細胞存在下で、IL-4及び抗CD40抗体を添加することにより、ヒト初代培養B細胞から、B細胞クローンを樹立することや(非特許文献18を参照)、胚中心の中心細胞のアポトーシスが、抗原レセプターの働きにかかわらず、CD40を介して阻害されること(非特許文献19を参照)が報告されている。以上のようにCD40は、ヒトB細胞表面に発現する抗原として同定されたため、単離された抗体の多くは、主にヒトB細胞に対する増殖分化誘導機能、癌細胞における細胞死誘導活性を指標に評価されてきた(非特許文献20〜22を参照)。
抗CD40抗体がDCを成熟させることが示された(非特許文献23を参照)。さらに、抗原特異的CD8T細胞プライミングにおけるCD4T細胞の役割は、CD40-CD40Lシグナリングを介したDCの活性化にあることが報告され、抗CD40モノクローナル抗体(mAb)により、樹状細胞(DC)の活性化におけるCD4ヘルパーT細胞の役割を代替できることが示された(非特許文献24を参照)。また、マウスにおいて抗CD40抗体の投与によりCD40を発現する腫瘍細胞のみならず非発現腫瘍細胞からも生体を防御可能であることが示された(非特許文献25を参照)。
今まで報告されてきた抗体はDC細胞に対する効果を指標に単離されているものは少ない。しかしDC細胞の機能修飾という点からは、B細胞に作用する抗体を選別したのでは、治療薬としては不充分である可能性が高い。マウスCD40に対する抗体のモノクローナル抗体では、抗体の認識するエピトープによって、DCには反応するが血管内皮細胞には反応しないクローンと、逆にDCには反応しないが血管内皮細胞には反応するクローンとが存在すると報告されている(非特許文献26を参照)。ヒトCD40抗体に関してもエピトープによってDCへの結合や作用の異なることが推測される。
抗CD40抗体又はCD40リガンドによりCD40を発現するリンパ腫細胞株の増殖を抑制し、細胞死を誘導できることが分かっている(非特許文献23及び27〜29を参照)。アゴニスティック抗体において興味深いことは、抗体の働きがCD40Lの機能と必ずしも一致しないことである。B細胞に対する活性化作用とB細胞腫瘍に対する増殖抑制作用も、必ずしも一致しない。DC活性化能と腫瘍細胞増殖抑制作用を併せ持つ抗体の開発が望ましい。また、アゴニスティック抗体の中には、CD40LのCD40への結合を、阻害するものも、しないものも存在する(非特許文献30を参照)。例えば、G28-5(ATCC No.HB-9110)の生産する抗体はCD40Lと競合するため、CD40Lとの併用効果はない。抗体によってCD40発現細胞の活性化の程度にも差がある。単独で弱いアゴニスティック活性を示す抗体でも、CD40リガンドと併用することによって、CD40リガンド単独による活性よりも、抗体存在下で活性が著しく促進する場合がある。逆に、単独でアゴニスティック活性を示す抗体でも、CD40リガンドを阻害することによって、CD40リガンド単独による活性よりも、抗体存在下での活性を低下させてしまう場合がある(非特許文献31を参照)。CD40リガンドと競合しない抗体では、それ自身による腫瘍細胞の増殖抑制作用は弱くとも、CD40リガンド存在下ではより強い増殖抑制ができることが示されている(非特許文献29を参照)。このことから、CD40に結合し、それ自身で細胞増殖を抑制する抗体でありながら、CD40リガンドのCD40に対する結合は阻害しないような抗体を開発することが望ましい。このような特徴を生かすことにより、可溶性CD40Lよりも有効な治療薬を開発できる可能性がある。たとえば、可溶性CD40LはCD40と結合し活性化するが、同時に生体内に存在しているCD40Lの働きを抑制する。CD40Lと競合しない抗体ならば、このようなことは起こらず、相乗効果により、より良い治療効果が期待できる。
3. アンタゴニスティック抗体
一方、上記のように、CD40が免疫反応において重要な役割を担っていることから、CD40とそのリガンドの結合を阻害することで、臓器移植時の免疫抑制や自己免疫疾患の治療薬が開発できると期待される。Sawada-Haseらは、クローン病患者の末梢血中の単球ではCD40を強く発現する細胞の割合が上昇していることを報告している。しかしながら、CD40とそのリガンドの結合を阻害する抗体については、まだ良く分かっていない。そのような阻害性抗体は、たとえば、CD40の機能解析や、CD40の活性化が必要な疾患の治療に有効である可能性がある。また、CD40リガンドに対する阻害抗体も、CD40とCD40リガンドとの結合が関与する疾患薬として有効である可能性が示されている。しかしながら、CD40Lは活性化した血小板に発現するという報告(非特許文献32を参照)があるため、抗CD40L抗体を治療薬として使った場合、血栓を引き起こす危険性が存在することが報告されている(非特許文献33を参照)。このような観点から、CD40とそのリガンドの結合を阻害する抗体治療薬としては、抗CD40L抗体よりも、むしろCD40に対する抗体の方が安全性に優れると期待できる。抗CD40抗体としてはCD40LのCD40への結合を抑制し、なおかつ、抗体自身がCD40を活性化しないことが必要とされる。
一方、上記のように、CD40が免疫反応において重要な役割を担っていることから、CD40とそのリガンドの結合を阻害することで、臓器移植時の免疫抑制や自己免疫疾患の治療薬が開発できると期待される。Sawada-Haseらは、クローン病患者の末梢血中の単球ではCD40を強く発現する細胞の割合が上昇していることを報告している。しかしながら、CD40とそのリガンドの結合を阻害する抗体については、まだ良く分かっていない。そのような阻害性抗体は、たとえば、CD40の機能解析や、CD40の活性化が必要な疾患の治療に有効である可能性がある。また、CD40リガンドに対する阻害抗体も、CD40とCD40リガンドとの結合が関与する疾患薬として有効である可能性が示されている。しかしながら、CD40Lは活性化した血小板に発現するという報告(非特許文献32を参照)があるため、抗CD40L抗体を治療薬として使った場合、血栓を引き起こす危険性が存在することが報告されている(非特許文献33を参照)。このような観点から、CD40とそのリガンドの結合を阻害する抗体治療薬としては、抗CD40L抗体よりも、むしろCD40に対する抗体の方が安全性に優れると期待できる。抗CD40抗体としてはCD40LのCD40への結合を抑制し、なおかつ、抗体自身がCD40を活性化しないことが必要とされる。
ヒトCD40に特異的に結合し、CD40LのCD40への結合を抑制し、なおかつ、CD40を活性化しない抗体についての報告は、過去に膨大な量の研究がなされてきたにもかかわらず、5D12と命名されたマウス抗ヒトCD40抗体のわずか1例のみである(非特許文献34を参照)。まして、B細胞に対して中和活性を示す抗体が、DC細胞に対しても同様に、CD40リガンドの作用を中和できるかどうかは知られていなかった。さらに、ビオチン化した抗マウスCD40抗体の作用が、アビジンでの架橋によって増強されることが報告されている(非特許文献35を参照)。本発明者は可溶性リガンドに予め遺伝子工学的に付与された、タグ(FLAG)に対する抗体(M2)を用いて、可溶性CD40リガンドのB細胞株(Ramos細胞)に対する作用を増強し、その中和活性を測定したところ、5D12(ATCC No.HB-11339)がわずかにしか中和活性を示さないことを確認した。
本発明者は新たに、アンタゴニスト抗体である5D12が架橋によって、CD40Lの非存在下であっても、抗体単独でアゴニスト活性を持つことを見出した。従来、マウスCD40抗体の作用がビオチンとアビジン架橋によって増強されることが報告されている(非特許文献35を参照)。また、プレートに固相化した抗免疫グロブリン抗体を用いて、CD40抗体を固相化することで、腫瘍細胞の増殖抑制活性が増加することが知られていたが、これは固相化による効果と考えられていた。しかし、培養液中に抗免疫グロブリン抗体を添加し、抗CD40抗体を架橋することによって、アンタゴニスティック抗体であってもアゴニスティック活性を示す可能性があることは知られていなかった。治療に用いる抗体に抗原性があれば、ヒトの体内でCD40抗体に結合する抗体が生じ、CD40抗体が架橋されることによって、あたかもCD40リガンドのような活性を生じる、全く逆の作用が懸念される。したがって治療薬の安全性の点から、抗体の抗原性を低く押さえることが非常に重要であると考えられる。マウス抗体の可変領域の配列をもとに、ヒト化技術により、治療薬として開発する場合、ヒト化抗体は免疫原性があることが知られているため、投与後に抗ヒト化抗CD40抗体ができる可能性があり、アゴニスティック抗体となる危険がある。また抗原性が低くとも、抗体レセプター(FcR)によって抗CD40抗体が架橋される可能性もある。これらの点から、アンタゴニスティック抗体は、CD40に特異的に結合し、CD40Lの結合を抑制し、かつ、架橋によってもCD40を活性化しないヒト抗体であって、FcRへの結合が弱いことが望ましいと考えられる。
E. A. Clark et. al., Proc. Natl.Acad. Sci. USA 83: 4494, 1986
I. Stamenkovic et. al., EMBO J. 8:1403, 1989
Caux, C., et al., J.Exp.Med., 180:1263,1994
Shu, U.,et al., Eur.J.Immunol.,25:1125,1995
Barrett et al., J. Immunol. 146: 1722, 1991
Gordon et al., J. Immunol. 140: 1425, 1988
Valle et al., Eur. J. Immunol. 19: 1463, 1989
Clark and Ledbetter, Proc. Natl. Acad. Sci. USA 83: 4494, 1986
Gordon et al., LEUCOCYTE TYPING III. A. J. McMicheal ed. Oxford University Press. Oxford, p.426
Paulie et al., J. Immunol.142: 590,1989
Gordon et al., Eur. J. Immunol. 17: 1535, 1987
Jabara et al., J. Exp. Med. 172: 1861, 1990
Gascan et al., J Immunol. 147:8,1991
Gordon and Guy, Immunol. Today 8:339, 1987
Cairns et al., Eur. J. Immunol. 18: 349, 1988
Challa A, Allergy, 54: 576, 1999
Clark and Shu, J. Immunol. 145: 1400, 1990
Bancherauet al., Science 241:70,1991
Liu et al., Nature 342: 929,1989
Katira, A. et. al., LEUKOCYTE TYPING V. S. F. Schlossossman, et. al. eds.p.547. Oxford University Press. Oxford
W. C. Flansow et. al., LEUKOCYTE TYPING V. S. F. Schlossossman, et. al. eds.p.555. Oxford University Press. Oxford
J. D. Pound et. al., International Immunology, 11: 11, 1999
Z. H. Zhou et. al., Hybridoma, 18:471 1999
Shoenberger, S.P., et.al., Nature, 480,1998
French,R.R., et.al., Nature Medicine, 5,1999
Van Den Berg, TK, et.al., Immunology, 88:294, 1996
Funakoshi S et al., Blood, 83: 2782, 1994
Funakoshi S et al., Journal of Immunotherapy, 19,93,1996
Joseph A et al., Cancer Research, 60: 3225, 2000
Challa A et al., Allergy, 54: 576, 1999
Pound et al.,International Immunology, 11:11,1999
V. Henn et. al., Nature 391: 591, 1998
T. Kawai et. al., Nat. Medi. 6: 114, 2000
J. Kwekkeboom et. Al., Immunology 79: 439, 1993
Johnson et al., Eur J Immunol, 24,: 1835, 1994
上述のように、近年DC細胞の機能解析が進み、CD40はDC細胞の働きを制御する、重要な遺伝子として認識され始めた。このような背景から、本発明は、DC細胞を用いた評価系を採用することにより、樹状細胞(DC)表面のヒトCD40抗原に対しても実質的にアンタゴニスティックである抗ヒトCD40抗体又はその機能的断片、および従来の抗ヒトCD40抗体に比べてより治療効果の高いことが期待されるアゴニスティックな抗ヒトCD40抗体又はその機能的断片を提供することを目的とする。
本発明者らは、ヒトCD40に対する抗体の作製に関して鋭意研究した結果、従来知られている抗CD40抗体に比して、より疾患への治療効果が高いと考えられる新規なアゴニスティック抗体、及びアンタゴニスティック抗体を作製することに成功し、本発明を完成した。すなわち、本発明は以下の通りである。
(1)以下の(a)〜(f)から選ばれる少なくとも1つの性質を有する、ヒトCD40に対する抗体又はその機能的断片。
(a) 樹状細胞に作用し、LPS及びIFNγの存在下でIL-12を生産させる
(b) 樹状細胞に作用し、該樹状細胞を成熟させる活性がG28-5抗体よりも高い
(c) B細胞樹立細胞株に対して、CD95の発現を促進する活性がG28-5抗体よりも高い
(d) B細胞樹立細胞株に対して増殖を抑制する活性がG28-5抗体よりも高い
(e) B細胞樹立細胞株に対して細胞死を誘導する
(f) CD40リガンドのCD40に対する結合を阻害しない
(a) 樹状細胞に作用し、LPS及びIFNγの存在下でIL-12を生産させる
(b) 樹状細胞に作用し、該樹状細胞を成熟させる活性がG28-5抗体よりも高い
(c) B細胞樹立細胞株に対して、CD95の発現を促進する活性がG28-5抗体よりも高い
(d) B細胞樹立細胞株に対して増殖を抑制する活性がG28-5抗体よりも高い
(e) B細胞樹立細胞株に対して細胞死を誘導する
(f) CD40リガンドのCD40に対する結合を阻害しない
(2)上記本発明の抗体又はその機能的断片は、樹状細胞の成熟が、20μg/ml以下の濃度で行われるものである。また、B細胞樹立細胞株に対してCD95の発現の促進は、20μg/ml以下の抗体濃度で行われる。B細胞樹立細胞株としては、例えばRamos又はHS-Sultonが挙げられる。
(3)また、上記本発明の抗体又はその機能的断片は、1x106個/mlの濃度の樹状細胞に、0.1μg/ml以上の濃度の抗体が添加された場合は、100pg/ml以上のIL-12の生産をもたらすものであり、1μg/ml以上の濃度の抗体が添加された場合は、1000pg/ml以上、好ましくは10000pg/ml以上のIL-12の生産をもたらすものである。
(4)さらに、上記本発明の抗体又はその機能的断片は、B細胞樹立細胞株(Ramos細胞)に対するCD95の発現促進が、抗体濃度0.01μg/mlから10μg/mlの範囲において、対照であるG28-5抗体の約2倍から3倍以上となるものである。例えば、抗体濃度0.01μg/mlでは、対照であるG28-5抗体の約2倍から6倍以上であり、抗体濃度0.1μg/mlでは、対照であるG28-5抗体の約2倍から7倍以上であり、抗体濃度1μg/mlでは、対照であるG28-5抗体の約2倍から7倍以上であり、そして抗体濃度10μg/mlでは、対照であるG28-5抗体の約2倍から6倍以上である。
(5)ハイブリドーマKM302-1(受託番号FERM BP-7578)、KM341-1-19(受託番号FERM BP-7759)、2105(受託番号FERM BP-8024)またはF1-102(受託番号ATCC PTA-3337)により生産される抗体の重鎖可変領域および軽鎖可変領域のアミノ酸配列を有する抗体又はその機能的断片。

(6)受託番号ATCC PTA-3302およびATCC PTA-3303であるプラスミドDNAにそれぞれコードされる、ハイブリドーマF2-103の産生する抗体の重鎖可変領域および軽鎖可変領域、受託番号ATCC PTA-3304およびATCC PTA-3305であるプラスミドDNAにそれぞれコードされる、ハイブリドーマF5-77の産生する抗体の重鎖可変領域および軽鎖可変領域、または受託番号ATCC PTA-3306およびATCC PTA-3307であるプラスミドDNAにそれぞれコードされる、ハイブリドーマF5-157の産生する抗体の重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有する抗体又はその機能的断片。
(7)配列番号28および30にそれぞれ示される、ハイブリドーマKM341-1-19の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号32および34にそれぞれ示される、ハイブリドーマ2105の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号36および38にそれぞれ示される、ハイブリドーマ110の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号40および42にそれぞれ示される、ハイブリドーマ115の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号52および54にそれぞれ示される、ハイブリドーマKM643-4-11の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号60および62にそれぞれ示される、ハイブリドーマF2-103の産生する抗体の重鎖可変領域および軽鎖可変領域、または配列番号64および66にそれぞれ示される、ハイブリドーマF5−77の産生する抗体の重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有する抗体又はその機能的断片。
(8)配列番号27および29にそれぞれ示される、ハイブリドーマKM341-1-19から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号31および33にそれぞれ示される、ハイブリドーマ2105から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号35および37にそれぞれ示される、ハイブリドーマ110から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号39および41にそれぞれ示される、ハイブリドーマ115から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号51および53にそれぞれ示される、ハイブリドーマKM643-4-11から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号59および61にそれぞれ示される、ハイブリドーマF2-103から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、または配列番号63および65にそれぞれ示される、ハイブリドーマF5−77から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有する抗体又はその機能的断片。
(9)以下の(g)〜(j)から選ばれる少なくとも1つの性質を有する、ヒトCD40に対する抗体又はその機能的断片。
(g) CD40に対するリガンドの作用を中和する
(h) B細胞樹立細胞株上のCD40に対するリガンドがCD40発現細胞に与える影響の1つ以上を中和又は緩和し、かつ、抗免疫グロブリン抗体による架橋によって前記B細胞樹立細胞株上のCD40に対するアゴニスティックな作用が5D12より弱い
(i) B細胞樹立細胞株に対して、CD40リガンドが、CD95の発現を増加させる作用を緩和又は中和する
(j) 樹状細胞に発現するCD40に対してアンタゴニスティックな作用を有する
(g) CD40に対するリガンドの作用を中和する
(h) B細胞樹立細胞株上のCD40に対するリガンドがCD40発現細胞に与える影響の1つ以上を中和又は緩和し、かつ、抗免疫グロブリン抗体による架橋によって前記B細胞樹立細胞株上のCD40に対するアゴニスティックな作用が5D12より弱い
(i) B細胞樹立細胞株に対して、CD40リガンドが、CD95の発現を増加させる作用を緩和又は中和する
(j) 樹状細胞に発現するCD40に対してアンタゴニスティックな作用を有する
(10) 上記(9)の抗体又はその機能的断片は、飽和量のCD40L発現細胞を添加した1x106個/mlの濃度のRamos細胞に、0.1 μg/mlの濃度の抗体が添加された場合は、Ramos細胞のCD95の発現を対照の約10%以下に抑制しうるものであり、 1 μg/mlの濃度の抗体が添加された場合は、Ramos細胞のCD95の発現が陰性コントロールと同レベルまで抑制されるものであり、10μg/mlの濃度の抗体が添加された場合は、Ramos細胞のCD95の発現が陰性コントロールと同レベルまで抑制されるものである。
(11)上記(9)の抗体又はその機能的断片は、インビトロにおいて、可溶性CD40L(1μg/ml)を添加した1x105個の扁桃腺B細胞に、0.001μg/mlから10μg/mlの濃度の抗体が添加された場合に、扁桃腺B細胞の増殖が約80から95%以上抑制されるものである。例えば、0.01μg/mlから10μg/mlの濃度の抗体が添加された場合は、扁桃腺B細胞の増殖が約95%以上抑制されるものであり、特に、0.001μg/mlの濃度の抗体が添加された場合は、扁桃腺B細胞の増殖が約80%以上抑制されるものである。
(12)ハイブリドーマKM281-1-10(受託番号FERM BP-7579)、4D11(受託番号FERM BP-7758)またはF4-465(受託番号ATCC PTA-3338)により生産される抗体の重鎖可変領域および軽鎖可変領域のアミノ酸配列を有する抗体又はその機能的断片。
(13)配列番号44および46にそれぞれ示される、ハイブリドーマKM281-1-10の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号48および50にそれぞれ示される、ハイブリドーマ4D11の産生する抗体の重鎖可変領域および軽鎖可変領域、または配列番号56および58にそれぞれ示される、ハイブリドーマF4-465の産生する抗体の重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有する抗体又はその機能的断片。
(14)配列番号43および45にそれぞれ示される、ハイブリドーマKM281-1-10から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号47および49にそれぞれ示される、ハイブリドーマ4D11の産生する抗体の重鎖可変領域および軽鎖可変領域、または配列番号55および57にそれぞれ示される、ハイブリドーマF4-465から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有する抗体又はその機能的断片。
(15)上記(1)〜(14)の抗体又はその機能的断片としては、ヒト抗体のものが挙げられる。
(16)上記(1)〜(15)のいずれかの抗体又はその機能的断片を有効成分として含む医薬組成物。
(17)上記(1)〜(8)のいずれかに記載の抗体又はその機能的断片を有効成分として含む、免疫賦活化剤、抗腫瘍剤又は自己免疫疾患治療剤。
(18)上記(9)〜(14)のいずれかに記載の抗体又はその機能的断片を有効成分として含む、免疫抑制剤、自己免疫疾患治療剤、アレルギー治療剤又は血液凝固第VIII因子阻害症候群治療剤。
(16)上記(1)〜(15)のいずれかの抗体又はその機能的断片を有効成分として含む医薬組成物。
(17)上記(1)〜(8)のいずれかに記載の抗体又はその機能的断片を有効成分として含む、免疫賦活化剤、抗腫瘍剤又は自己免疫疾患治療剤。
(18)上記(9)〜(14)のいずれかに記載の抗体又はその機能的断片を有効成分として含む、免疫抑制剤、自己免疫疾患治療剤、アレルギー治療剤又は血液凝固第VIII因子阻害症候群治療剤。
(19)ここで、本発明のモノクローナル抗体の認識するヒトCD40のエピトープは、ヒトCD40の一次アミノ酸配列から得られたオーバーラップする合成オリゴペプチドへの結合を調べるなどの周知の方法によって決定することができる(例えばEd Harlow and David Lane(eds.), Antibodies: A Laboratory Manual, 1988 Cold Spring Harbor Laboratory Press; US Patent No. 4708871)。ファージディスプレイによるペプチドライブラリーキット(New England Biolabs)をエピトープマッピングに用いることもできる。本発明は、上述した個々のハイブリドーマによって産生される抗体又はその機能的断片の認識するヒトCD40の新規なエピトープを認識する抗体又はその機能的断片をも包含する。
(20)本発明はさらに、上述した個々のハイブリドーマから単離された、抗体の重鎖および/または軽鎖の少なくとも可変領域をコードする核酸(RNAまたはcDNA)、該核酸を含むベクター、該核酸を保持する宿主細胞を提供する。
本発明により、CD40に対する抗体が提供される。本発明の抗体は、CD40に対してアゴニスティックに作用するもの及びアンタゴニスティックに作用するものの両者が含まれるため、それぞれ、免疫賦活剤、免疫抑制剤等として有用である。
以下、本発明を詳細に説明する。本明細書は、本願の優先権の基礎であるPCT出願PCT/US01/13672(2001年4月27日出願)、日本国特許出願2001-142482号(2001年5月11日出願)、日本国特許出願2001-310535号(2001年10月5日出願)及び米国特許出願USSN 10/040,244(2001年10月26日出願)の明細書及び/又は図面に記載される内容を包含する。
発明者らは、後述するように、B細胞上のCD40に対してアンタゴニスティックである公知のモノクローナル抗体5D12(ATCC No.HB-11339)が、DC細胞上のCD40に対してはアンタゴニスティックではなかったという知見を得た。さらに発明者は、CD40Lの作用をブロックするアンタゴニスティック抗体であっても、多くのモノクローナル抗体が抗免疫グロブリン抗体による架橋によって、それ自身がアゴニスティック活性を示してしまうことを見出した。
1.定義
本明細書で使用する用語の定義は以下のとおりである。
本発明でいう「ヒトCD40」とは、クラークら(E. A. Clark et. al., Proc. Natl. Acad. Sci. USA 83: 4494, 1986)又はスタメンコビックら(I. Stamenkovic et. al., EMBO J. 8:1403, 1989)により示されているアミノ酸配列を有するポリペプチドを意味し、特にB細胞、DC、マクロファージ、内皮細胞、上皮細胞、あるいはそれらの腫瘍細胞表面に発現する抗原ポリペプチドである。
「抗CD40モノクローナル抗体」とは、細胞発現CD40、全長CD40又は部分長CD40に対するモノクローナル抗体のいずれをも意味するが、より好ましくはCD40の細胞外部分に結合し、CD40を発現している細胞にアゴニスティックまたはアンタゴニスティックな作用をもたらすモノクローナル抗体を意味する。
本明細書で使用する用語の定義は以下のとおりである。
本発明でいう「ヒトCD40」とは、クラークら(E. A. Clark et. al., Proc. Natl. Acad. Sci. USA 83: 4494, 1986)又はスタメンコビックら(I. Stamenkovic et. al., EMBO J. 8:1403, 1989)により示されているアミノ酸配列を有するポリペプチドを意味し、特にB細胞、DC、マクロファージ、内皮細胞、上皮細胞、あるいはそれらの腫瘍細胞表面に発現する抗原ポリペプチドである。
「抗CD40モノクローナル抗体」とは、細胞発現CD40、全長CD40又は部分長CD40に対するモノクローナル抗体のいずれをも意味するが、より好ましくはCD40の細胞外部分に結合し、CD40を発現している細胞にアゴニスティックまたはアンタゴニスティックな作用をもたらすモノクローナル抗体を意味する。
さらに、本発明で「抗体」とは、イムノグロブリンを構成する重鎖可変領域及び重鎖定常領域並びに軽鎖の可変領域及び軽鎖の定常領域をコードする遺伝子(「抗体遺伝子」と総称する)に由来するものである。本発明の抗体には、いずれのイムノグロブリンクラス及びアイソタイプを有する抗体をも包含する。本発明における抗体の「機能的断片」とは、前記で定義した抗体の一部分(部分断片)であって、抗体の抗原への作用を1つ以上保持するものを意味し、具体的にはF(ab')2、Fab'、Fab、Fv、ジスルフィド結合FV、一本鎖FV(scFV)、およびこれらの重合体等が挙げられる(D.J.King., Applications and Engineering of Monoclonal Antibodies., 1998 T.J.International Ltd)。
本発明で「ヒト抗体」とは、ヒト由来の抗体遺伝子の発現産物である抗体を意味する。
「アゴニスティック」とは、B細胞、腫瘍細胞又は樹状細胞などの細胞表面上に発現するCD40に、そのリガンドが結合することを促進する作用、あるいは、CD40リガンドがCD40発現細胞に与える影響の1つ以上を、CD40を発現する細胞に与える作用を意味し、「アゴニスティック抗体」とは、そのようなアゴニスティック作用を有する抗体を意味する。
「アゴニスティック」とは、B細胞、腫瘍細胞又は樹状細胞などの細胞表面上に発現するCD40に、そのリガンドが結合することを促進する作用、あるいは、CD40リガンドがCD40発現細胞に与える影響の1つ以上を、CD40を発現する細胞に与える作用を意味し、「アゴニスティック抗体」とは、そのようなアゴニスティック作用を有する抗体を意味する。
「アンタゴニスティック」とは、B細胞、腫瘍細胞又は樹状細胞などの細胞表面上に発現するCD40にそのリガンドが結合することを阻害する作用、あるいは、CD40リガンドがCD40発現細胞に与える影響の1つ以上を中和する作用を意味し、「アンタゴニスティック抗体」とはそのような作用を有する抗体を意味する。
本発明で言う「樹状細胞(DC)」とは、樹状白血球とも呼ばれ、強力な抗原提示機能を有する細胞の一群を指す。用いる樹状細胞は、例えば骨髄、さい帯血又は末梢血中に含まれるCD34陽性前駆細胞を培養することによって誘導される。あるいは、末梢血中のCD14陽性単球をGM-CSF及びIL-4の存在下で培養することにより得られる。
「未熟なDC」(Immmature DC)とは、CD14陰性、CD1a強陽性、CD83,CD86陽性、MHCクラスII陽性であるDCを意味する。
「成熟DC」とは、CD14陰性、CD1a陽性でCD83、CD86、MHCクラスII強陽性となったものを意味する。
「成熟DC」とは、CD14陰性、CD1a陽性でCD83、CD86、MHCクラスII強陽性となったものを意味する。
本発明で「DCを活性化する」とは、CD40による刺激に応答してDCが起こす変化を意味し、例えば 未成熟DCを成熟化させ、CD80、CD86、HLA-ClassIIを高発現させる、さらにはIL-12の産生を増強する、あるいはT細胞が共存する場合には、T細胞に刺激を与え、増殖を促すことを意味する。
本発明でB細胞、B細胞株を活性化するとは、CD40による刺激に応答して細胞が起こす変化を意味し、例えばDNA合成を生じ、チミジン取り込みを促進し、CD95の発現量を増加させることをいう。
2.抗体の取得
本件発明の抗体を取得するためには、抗原として組換え体で生産・精製した可溶性ヒトCD40、又はヒトCD40を発現する遺伝子組換えマウス細胞株をマウスに免疫することが好適である。免疫に使用するマウスはヒト抗体を産生するマウス(Tomizuka. et al., Proc Natl Acad Sci USA., 2000 Vol97:722)であることが望ましい。組換え体で生産・精製した可溶性ヒトCD40に結合するモノクローナル抗体を選別することで、B細胞特異的に反応するクローンを選抜するよりも、B細胞以外の細胞に発現するCD40にも反応する抗体を得やすいと考えられる。免疫したマウスのリンパ節の細胞や脾細胞を使用し、モノクローナル抗体の製造において慣用されているケーラー及びミルシュタインらの方法(Nature., 1975 Vol.256:495)を用いてハイブリドーマの作製を行うことができる。
本件発明の抗体を取得するためには、抗原として組換え体で生産・精製した可溶性ヒトCD40、又はヒトCD40を発現する遺伝子組換えマウス細胞株をマウスに免疫することが好適である。免疫に使用するマウスはヒト抗体を産生するマウス(Tomizuka. et al., Proc Natl Acad Sci USA., 2000 Vol97:722)であることが望ましい。組換え体で生産・精製した可溶性ヒトCD40に結合するモノクローナル抗体を選別することで、B細胞特異的に反応するクローンを選抜するよりも、B細胞以外の細胞に発現するCD40にも反応する抗体を得やすいと考えられる。免疫したマウスのリンパ節の細胞や脾細胞を使用し、モノクローナル抗体の製造において慣用されているケーラー及びミルシュタインらの方法(Nature., 1975 Vol.256:495)を用いてハイブリドーマの作製を行うことができる。
また、BIAcore 2000(ビアコア社)などの表面プラズモン共鳴測定装置を用いて可溶性CD40LのCD40への結合を解析し、CD40Lと競合しない抗体を選別する。またBリンパ腫の細胞増殖抑制を抗体単独で抑制する抗体を選別した。さらに抗体の選別をDCに作用するか否かを指標として行うことにより、CD40Lと競合せず、樹状細胞又はB細胞に働き、かつCD40発現癌細胞の増殖を抑制するという利点を備えた抗体を作製・選別しうる。
本発明の抗体は、取得されたハイブリドーマを培養することによって得られる。また、B細胞やハイブリドーマ等の抗体産生細胞からヒトモノクローナル抗体又はその可変領域をコードする遺伝子をクローニングし、適当なベクターに組み込んで、これを宿主(例えば哺乳類細胞株、大腸菌、酵母細胞、昆虫細胞、植物細胞など)に導入し、遺伝子組換え技術を用いて産生させた組換型抗体を調製することができる(P.J.Delves., ANTIBODY PRODUCTION ESSENTIAL TECHNIQUES., 1997 WILEY、P.Shepherd and C.Dean., Monoclonal Antibodies., 2000 OXFORD UNIVERSITY PRESS, J.W.Goding., Monoclonal Antibodies: principles and practice., 1993 ACADEMIC PRESS)。さらに、トランスジェニック動物作製技術を用いて目的抗体の遺伝子が内在性遺伝子に組み込まれたトランスジェニックなウシ、ヤギ、ヒツジまたはブタを作製し、そのトランスジェニック動物のミルク中からその抗体遺伝子に由来するモノクローナル抗体を大量に取得することも可能である。ハイブリドーマをインビトロで培養する場合には、培養する細胞種の特性、試験研究の目的及び培養方法等の種々条件に合わせて、ハイブリドーマを増殖、維持及び保存させ、培養上清中にモノクローナル抗体を産生させるために用いられるような既知栄養培地又は既知の基本培地から誘導調製されるあらゆる栄養培地を用いて実施することが可能である。
3.スクリーニング
アゴニスティック抗体の選抜には、ヒトBリンフォーマを用いた解析によって行い、CD95発現を促進する抗体の選別が可能である。さらに純化したDCの培養液に抗体を添加し、成熟させる抗体を選ぶ。あるいは、未熟なDC細胞を用いた混合リンパ球反応でT細胞増殖活性を示す抗体を選ぶ。さらに成熟したDCに添加しIL-12生産促進作用のある抗体を選ぶ。また、CD40を発現する腫瘍細胞の増殖を抑制もしくは細胞死誘導活性を持つ抗体を選ぶ。CD40Lとの競合はBIACoreのような表面プラズモン共鳴測定装置を用いて、可溶性CD40と可溶性CD40リガンドの結合を抗体が阻害するかどうかで判別できる。あるいは、B細胞株に対するCD40リガンドの作用を増強するかどうかで判別する。
アゴニスティック抗体の選抜には、ヒトBリンフォーマを用いた解析によって行い、CD95発現を促進する抗体の選別が可能である。さらに純化したDCの培養液に抗体を添加し、成熟させる抗体を選ぶ。あるいは、未熟なDC細胞を用いた混合リンパ球反応でT細胞増殖活性を示す抗体を選ぶ。さらに成熟したDCに添加しIL-12生産促進作用のある抗体を選ぶ。また、CD40を発現する腫瘍細胞の増殖を抑制もしくは細胞死誘導活性を持つ抗体を選ぶ。CD40Lとの競合はBIACoreのような表面プラズモン共鳴測定装置を用いて、可溶性CD40と可溶性CD40リガンドの結合を抗体が阻害するかどうかで判別できる。あるいは、B細胞株に対するCD40リガンドの作用を増強するかどうかで判別する。
アンタゴニスティック抗体の選抜には、ヒトBリンフォーマを用いた解析によって行い、さらにFLAGをタグとして有する可溶性CD40Lを抗FLAG抗体存在下で添加することで、可溶性CD40Lの、ヒトBリンフォーマ細胞上のCD40への結合をより強力に阻害する抗体の選別が可能である。可溶性CD40Lの代りに、CD40Lをコードする遺伝子の導入により、細胞表面に多くのCD40Lを発現している組換え細胞を用いることも可能である。次いでヒト抗体を抗ヒトIgG抗体で架橋することで、架橋によってBリンパ腫を活性化してしまうクローンを除外する。さらに純化し成熟させたDC細胞を用いた混合リンパ球反応でT細胞増殖抑制活性を示す抗体や、あるいは成熟したDCにCD40リガンドを添加した場合にIL-12生産抑制作用のある抗体を選ぶ。
以上のようにして得られた抗体は、少なくとも以下のいずれかの治療効果上有用と考えられる性質を有するものである。
(1) アゴニスティック抗体の場合
(a) 樹状細胞に作用し、LPS(リポポリサッカライド)及びIFNγの存在化でIL-12を生産させる。この場合のLPS濃度は10pg/mlから10ng/mlであり、IFNγの濃度は10-4Mから10-2Mである。1μg/ml以上の抗体濃度、好ましくは0.1μg/ml以上の抗体濃度において、公知のアゴニスティック抗CD40抗体であるG28-5抗体を対照とした試験よりもIL-12の生産量が多い。1x106個/mlの濃度の樹状細胞に抗体濃度0.1μg/ml以上が添加された場合に100pg/ml以上のIL-12が生産され、あるいは1μg/ml以上が添加された場合に1000pg/ml以上の、好ましくは10000pg/ml以上のIL-12が生産される(実施例9、実施例13を参照のこと)。
(1) アゴニスティック抗体の場合
(a) 樹状細胞に作用し、LPS(リポポリサッカライド)及びIFNγの存在化でIL-12を生産させる。この場合のLPS濃度は10pg/mlから10ng/mlであり、IFNγの濃度は10-4Mから10-2Mである。1μg/ml以上の抗体濃度、好ましくは0.1μg/ml以上の抗体濃度において、公知のアゴニスティック抗CD40抗体であるG28-5抗体を対照とした試験よりもIL-12の生産量が多い。1x106個/mlの濃度の樹状細胞に抗体濃度0.1μg/ml以上が添加された場合に100pg/ml以上のIL-12が生産され、あるいは1μg/ml以上が添加された場合に1000pg/ml以上の、好ましくは10000pg/ml以上のIL-12が生産される(実施例9、実施例13を参照のこと)。
(b) 樹状細胞に結合し、樹状細胞を成熟させる作用を有する。しかも、20μg/ml以下の濃度、好ましくは0.1〜15μg/ml、さらに好ましくは5〜15μg/mlの濃度で抗体を樹状細胞とともに培養すると、その成熟させる活性はG28-5抗体よりも高い結果が得られる(実施例9を参照のこと)。
(c) B細胞樹立細胞株に対して、CD95の発現を促進する活性がG28-5抗体よりも高い。この場合、10μg/ml以上、好ましくは1μg/ml以上、さらに好ましくは0.1μg/ml以上、より好ましくは0.01μg/ml以上、最も好ましくは0.001μg/mlを超える抗体濃度において、G28-5抗体を対照とした試験よりもCD95の発現を促進する活性がG28-5抗体よりも高い。CD95の発現を促進する活性について、抗体濃度10μg/ml、1μg/ml、0.1μg/ml、0.01μg/mlにおいてG28-5抗体を対照とした試験の倍率は以下の通りである(表1)。
CD95の発現が促進するということは、抗体がB細胞樹立細胞株を活性化したことを意味する。ここで、B細胞樹立細胞株としては、Ramos細胞、HS-Sulton細胞が挙げられる。なお、Ramos細胞はバーキットリンパ腫であり、ヒトの胚中心のB細胞のモデル細胞である。HS-Sulton細胞はバーキットリンパ腫である(実施例6、実施例12を参照のこと)。
(d) B細胞樹立細胞株(Ramos細胞又はHS-Sulton細胞)に対してDNA合成、チミジン取り込み、増殖を抑制する活性がG28-5抗体よりも高い。この場合の抗体濃度は、少なくとも0.05μg/ml、好ましくは0.1〜15μg/mlである(実施例8を参照のこと)。
(e) B細胞樹立細胞株に対して細胞死を誘導する(実施例16を参照のこと)。
(f) CD40リガンドのCD40に対する結合を阻害しない。「阻害しない」とは、CD40に予め抗体が結合した状態であっても(つまり抗体が存在する場合であっても)、該抗体が存在しない場合と同程度に、CD40LがCD40に結合できることを意味する。CD40リガンドとCD40はいずれかあるいは双方が膜発現型あるいは可溶性タンパクであっても良い(実施例11を参照のこと)。
上記性質を有する抗体は、例えば、ハイブリドーマKM302-1(FERM BP-7578)、ハイブリドーマKM341-1-19(FERM BP-7759)により産生される。
ハイブリドーマKM341-1-19により産生されるモノクローナル抗体の重鎖(H鎖)および軽鎖(L鎖)可変領域の塩基配列およびアミノ酸配列が決定された(実施例17)。本発明はハイブリドーマKM341-1-19の生産するモノクローナル抗体の少なくとも重鎖可変領域、もしくは重鎖全長をコードするDNA、ならびに軽鎖可変領域をコードするDNAを提供する。当該DNAは実施例17において記載されたものの他、コドン縮重により同じアミノ酸配列をコードする他のDNAをも包含する。また、本発明は、実施例17において開示された少なくとも重鎖可変領域のアミノ酸配列、もしくは重鎖全長のアミノ酸配列と、軽鎖可変領域のアミノ酸配列によって規定されるモノクローナル抗体もしくはその機能的断片を提供する。
(2) アンタゴニスティック抗体の場合
(g) CD40に対するリガンドの作用を中和する。ここで、「リガンドの作用」とは、T細胞又は他の細胞上に発現するリガンドの作用、及びCD40に対する遊離のリガンドの作用のいずれをも意味する(実施例7、実施例14を参照のこと)。
(g) CD40に対するリガンドの作用を中和する。ここで、「リガンドの作用」とは、T細胞又は他の細胞上に発現するリガンドの作用、及びCD40に対する遊離のリガンドの作用のいずれをも意味する(実施例7、実施例14を参照のこと)。
(h) B細胞樹立細胞株上のCD40に対するリガンドがCD40発現細胞に与える影響の1つ以上を中和し、かつ、抗免疫グロブリン抗体による架橋によって前記B細胞樹立細胞株上のCD40に対するアゴニスティックな作用を示さない。その作用は5D12よりも弱い。「CD40発現細胞に与える影響」とは、発現細胞の活性化を意味し、B細胞においてはチミジン取り込み、増殖を、B細胞樹立細胞株においてはCD95の発現増強などを意味する。また、DCにおいては成熟化、CD86、HLA-DR発現増強、共存するT細胞のチミジン取り込み、増殖の促進、IL-12やIL-10の生産などを意味する。抗免疫グロブリン抗体による架橋は、培養液中に0.1μg/ml以上の抗免疫グロブリン抗体を存在させることにより行われる(実施例7を参照のこと)。
(i) B細胞樹立細胞株に対して、架橋したCD40Lあるいは、細胞に発現させたCD40LがCD95の発現を増加させる活性を緩和又は中和する。タグに対する抗体などで架橋し、作用が増強されたCD40Lに対しても、活性を緩和あるいは中和する。CD40に対するリガンド(遊離のリガンド、特定の細胞に発現するリガンドの両者を含む)が、CD40発現細胞に結合すると、細胞内でシグナル伝達を起こして、最終的に細胞表面上にCD95(Fas)を発現する。従って、本発明のアンタゴニスティック抗体は、CD40に結合することにより、上記シグナル伝達を阻害してCD95の発現を中和する。この場合の抗体濃度は、1μg/ml以上、好ましくは0.1μg/ml以上である(実施例7、実施例14を参照のこと)。
(j) DC上のCD40に対してアンタゴニスティックである。すなわち、CD40LがDCを活性化する活性を緩和又は中和する。DCが共存するT細胞上のリガンドによって刺激されると、T細胞を活性化し、チミジン取り込みなどを促進させる。異なった個体由来のDCとT細胞とを共存させる混合リンパ球反応では、DCとT細胞が相互作用を起こすことで、T細胞の活性化が起こる。本発明のアンタゴニスティック抗体は、CD40に結合することにより、上記相互作用を阻害し、結果としてチミジンの取り込みが抑制される。この場合の抗体濃度は、少なくとも0.001μg/mlであり、好ましくは0.1〜10μg/mlである(実施例10を参照のこと)。
上記アンタゴニスティック抗体は、例えば、ハイブリドーマKM281-1-10(FERM BP-7579)、KM281-2-10-1-2(FERM BP-7580)(2001年5月9日付、独立行政法人産業技術総合研究所 特許生物寄託センター(茨城県つくば市東1丁目1番地1中央第6))、4D11(FERM BP-7758)により産生される。
(3) 本発明の抗体は、当業者に周知である遺伝子工学的改変、すなわち抗体重鎖のサブクラスを規定する領域を他のサブクラスを規定する領域に置換することにより、異なるサブクラスのものに変換しうる(例えば、EP 314161公報を参照のこと)。重鎖(heavy chain)の可変領域を、そのまま他のサブクラスの定常領域(constant region)につなげることができる。例えば、本件発明の抗体のサブクラスをIgG2あるいはIgG4に変換することにより、Fcレセプターに対する結合度を低下させることが可能である。具体的には、Kabatら(Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institute of Health, Bethesda, Md.(1991))による、ヒト抗体重鎖、EU index 118(Ala),119(Ser)部位にNheI site(GCTAGC)を導入し、この制限酵素を使って消化することにより、アミノ酸を変えずに、他のサブクラスのIgGとつなぎかえることができる。さらに、定常領域のアミノ酸配列を人為的に改変すること、あるいはそのような配列を有する定常領域配列と本発明の抗体の可変領域とを結合することにより、Fcレセプターに対する結合度を低下させるこや(Lund J., et al., J. Immunol. 1991 vol147:2657-2662 )、CDC活性を増加または減少することも可能である(Tao M., et. Al., J. Exp. Med. 1991 vol 1025-1028, Idusogie E E., et. Al., J. Immunol. 2001 vol166:2571-5) 。また、ADCCやCDCなどの作用を避けるために、予めサブクラスがIgG2やIgG4の抗体のみを選別することも可能である。また、本件発明の抗体に、放射性核種、細菌毒素、化学療法剤、プロドラッグなどを結合させることにより癌などの疾患の治療効果をさらに増強することも可能である。
4.医薬組成物
また、本発明の抗体の精製された製剤を含有する医薬組成物もまた、本発明の範囲内に含まれる。このような医薬組成物は、好ましくは、抗体に加えて、生理学的に許容され得る希釈剤またはキャリアを含んでおり、他の抗体または抗生物質のような他の薬剤との混合物であってもよい。適切なキャリアには、生理的食塩水、リン酸緩衝生理食塩水、リン酸緩衝生理食塩水グルコース液、および緩衝生理食塩水が含まれるが、これらに限定されるものではない。或いは、抗体は凍結乾燥(フリーズドライ)し、必要とされるときに上記のような緩衝水溶液を添加することにより再構成して使用してもよい。投与経路は、経口ルート、並びに静脈内、筋肉内、皮下および腹腔内の注射または配薬を含む非経腸的ルートである。
また、本発明の抗体の精製された製剤を含有する医薬組成物もまた、本発明の範囲内に含まれる。このような医薬組成物は、好ましくは、抗体に加えて、生理学的に許容され得る希釈剤またはキャリアを含んでおり、他の抗体または抗生物質のような他の薬剤との混合物であってもよい。適切なキャリアには、生理的食塩水、リン酸緩衝生理食塩水、リン酸緩衝生理食塩水グルコース液、および緩衝生理食塩水が含まれるが、これらに限定されるものではない。或いは、抗体は凍結乾燥(フリーズドライ)し、必要とされるときに上記のような緩衝水溶液を添加することにより再構成して使用してもよい。投与経路は、経口ルート、並びに静脈内、筋肉内、皮下および腹腔内の注射または配薬を含む非経腸的ルートである。
この場合、本発明の抗体の有効量と適切な希釈剤及び薬理学的に使用し得るキャリアとの組合せとして投与される有効量は、1回につき体重1kgあたり0.1mg〜100mgであり、2日から8週間間隔で投与される。
本発明の抗体を含む医薬組成物を使用する場合は、アゴニスティック抗体については、免疫賦活化剤(抗ウィルス剤、抗感染症剤)、抗腫瘍剤、自己免疫疾患治療剤であり、これらの疾患が複数併発してもよい。あるいは、ガン特異的ペプチドなどのワクチンとアジュバントとして併用することもできる。また、アンタゴニスティック抗体については、臓器移植時における免疫抑制剤(膵島移植や腎臓などの移植時における拒絶反応、GVHDの予防又は治療剤)として、あるいは自己免疫疾患(例えば、リウマチ、動脈硬化治療薬、多発性硬化症、全身性エリトマトーデス、特発性血小板減少症、クローン病など)治療剤、喘息などアレルギー治療剤、血液凝固第VIII因子阻害症候群の治療薬として有用であり、これらの疾患が複数併発してもよい。
CD40が関与する疾患の治療手段として、抗CD40抗体を用いる場合、DC細胞の機能を指標に、抗体を選択することにより、より良い治療効果をもたらす抗体が得られると期待できる。
アゴニスティック抗体では、よりDCの活性化ができる抗体を選抜することにより、免疫増強の作用の強い抗体が得られることが期待される。さらに成熟DCのIL-12の生産促進を指標とすることで、CTL誘導作用の強い抗体が得られる。CTL誘導により、ウィルス感染細胞や腫瘍細胞を除去する効果の高い抗体が得られる。また、相乗効果が期待できることからCD40に結合する抗体でありながら、CD40リガンドのCD40に対する結合は阻害しないような抗体が望ましい。癌治療を考えた場合、CD40を発現する癌細胞に直接細胞死を誘導あるいは増殖を抑制し、かつDC細胞を効率的に活性化する抗体があれば、相乗的な効果が期待され、CD40を発現しない腫瘍にも使用可能な治療薬となると考えられる。この抗体はウィルス感染症の治療薬や抗腫瘍薬として有用と考えられる。
一方、CD40に特異的に結合し、CD40Lの結合を抑制し、なおかつ、CD40を活性化しない抗体についても、B細胞に対するリガンドの作用を抑制するのみならず、DC細胞への作用を抑制できることが望まれる。しかし、今まで得られた、抗体はB細胞に対する効果を指標にとられてきたことから、樹状細胞にも強い抑制作用を持つ抗体を取得し、医薬品として開発することには大きな意義がある。さらに上述のように抗CD40抗体は架橋によって、全く逆の作用が懸念されるため、架橋によってもCD40を活性化しない抗体が必要とされる。これまでに報告されたヒトCD40に対する、マウス等の非ヒト哺乳動物由来のモノクローナル抗体、およびマウスモノクローナル抗体の可変領域とヒト免疫グロブリンの定常領域からなるキメラ抗体やCDRグラフティングによるヒト型化抗体は、抗原性を持つことが懸念されるため、CD40リガンドとの結合を阻害する抗体としてはヒト抗体が望ましい。
以下、実施例により本発明をさらに具体的に説明する。但し、本発明はこれら実施例に技術的範囲が限定されるものではない。
〔実施例1〕 抗原の作製
(1)細胞
EL-4細胞は、マウス由来のT細胞樹立株であり、容易に入手することができる(ATCC No.:TIB-39)。Ramos B 細胞(ATCC No.:CRL-1596)並びにマウス抗CD40抗体産生ハイブリドーマ G28-5(HB-9110)及び5D12(HB-11339)は、ATCCから購入した。
(1)細胞
EL-4細胞は、マウス由来のT細胞樹立株であり、容易に入手することができる(ATCC No.:TIB-39)。Ramos B 細胞(ATCC No.:CRL-1596)並びにマウス抗CD40抗体産生ハイブリドーマ G28-5(HB-9110)及び5D12(HB-11339)は、ATCCから購入した。
(2) 抗原の発現と精製
ヒトCD40cDNA(Genbank Accession Number: NM_001250)を鋳型として、以下のプライマーを用いて、PCRにより(95℃ 5秒、55℃ 30秒、72℃ 30秒)×20サイクルの条件で細胞外領域を増幅した。
プライマー1:5'-CCCAGATCTGTCCATCCAGAACCACCCACTGCATGCAGAG-3'(配列番号1)
プライマー2:5'-ACAAGATCTGGGCTCTACGTATCTCAGCCGATCCTGGGGAC-3'(配列番号2)
ヒトCD40cDNA(Genbank Accession Number: NM_001250)を鋳型として、以下のプライマーを用いて、PCRにより(95℃ 5秒、55℃ 30秒、72℃ 30秒)×20サイクルの条件で細胞外領域を増幅した。
プライマー1:5'-CCCAGATCTGTCCATCCAGAACCACCCACTGCATGCAGAG-3'(配列番号1)
プライマー2:5'-ACAAGATCTGGGCTCTACGTATCTCAGCCGATCCTGGGGAC-3'(配列番号2)
増幅したcDNAは、pFastBacベクター(Gibco BRL)の、メリチンシグナル配列の後、ヒトIgG1由来FC又はマウスIgG2a由来FC領域の前に挿入した。CD40生産のため、組換えバキュロウィルスは、使用説明書にしたがって作成した。Tn5細胞を組替えウィルスで感染させた後、4日間培養した。上清は、0.22nmフィルターで処理した後、ProteinG sepharose(Amersham Pharmacia)を添加し、4℃で静かに振盪した。一晩後、sepharoseをカラムに移し、20倍容量のPBSで洗浄した。ヒトCD40 FC蛋白質は、20mMグリシン緩衝液(pH3.0)により溶出した。CD40を細胞表面に発現させるためのベクターは、Randolph J. Noelleから入手した(Inui, S et al.,EJI, 20,1747-1753,1990)。全長cDNAはXbaIにより酵素切断し、pCDNA3(INVITROGEN)に挿入した。ベクターをEL-4細胞に導入し、0.5mg/mlのG418(Gibco BRL)存在下で培養することにより安定発現株を得た。CD40の発現は、FITC-結合抗ヒトCD40抗体(Pharmingen)を使い、FACS解析により確認した。
〔実施例2〕 免疫用マウスの作製
免疫に用いたマウスは、内因性Ig重鎖及びκ軽鎖破壊の両者についてホモ接合体の遺伝的背景を有しており、かつ、ヒトIg重鎖遺伝子座を含む14番染色体断片(SC20)及びヒトIgκ鎖トランスジーン(KCo5)を同時に保持する。このマウスはヒトIg重鎖遺伝子座を持つ系統Aのマウスと、ヒトIgκ鎖トランスジーンを持つ系統Bのマウスとの交配により作製した。系統Aは、内因性Ig重鎖及びκ軽鎖破壊の両者についてホモ接合体であり、子孫伝達可能な14番染色体断片 (SC20)を保持するマウス系統であり、例えば富塚らの報告(Tomizuka. et al., Proc Natl Acad Sci USA., 2000 Vol97:722)に記載されている。A系統マウスを免疫することによって、以下に示すハイブリドーマ、F2-103, F5-77を取得した。また、系統Bは内因性Ig重鎖及びκ軽鎖破壊の両者についてホモ接合体であり、ヒトIgκ鎖トランスジーン(KCo5)を保持するマウス系統(トランスジェニックマウス)であり、例えばFishwildらの報告(Nat Biotechnol., 1996 Vol14:845)に記載されている。
免疫に用いたマウスは、内因性Ig重鎖及びκ軽鎖破壊の両者についてホモ接合体の遺伝的背景を有しており、かつ、ヒトIg重鎖遺伝子座を含む14番染色体断片(SC20)及びヒトIgκ鎖トランスジーン(KCo5)を同時に保持する。このマウスはヒトIg重鎖遺伝子座を持つ系統Aのマウスと、ヒトIgκ鎖トランスジーンを持つ系統Bのマウスとの交配により作製した。系統Aは、内因性Ig重鎖及びκ軽鎖破壊の両者についてホモ接合体であり、子孫伝達可能な14番染色体断片 (SC20)を保持するマウス系統であり、例えば富塚らの報告(Tomizuka. et al., Proc Natl Acad Sci USA., 2000 Vol97:722)に記載されている。A系統マウスを免疫することによって、以下に示すハイブリドーマ、F2-103, F5-77を取得した。また、系統Bは内因性Ig重鎖及びκ軽鎖破壊の両者についてホモ接合体であり、ヒトIgκ鎖トランスジーン(KCo5)を保持するマウス系統(トランスジェニックマウス)であり、例えばFishwildらの報告(Nat Biotechnol., 1996 Vol14:845)に記載されている。
系統Aの雄マウスと系統Bの雌マウス、あるいは系統Aの雌マウスと系統Bの雄マウスの交配により得られた、血清中にヒトIg重鎖及びκ軽鎖が同時に検出される個体(Ishida&Lonberg, IBC's 11th Antibody Engineering, Abstract 2000)を、以下の免疫実験に用いた。なお、前記ヒト抗体産生マウスは、契約を結ぶことによって、麒麟麦酒株式会社より入手可能である。前記マウスを免疫することによって、以下に示すハイブリドーマ、KM302-1、KM341-1-19、KM643-4-11、2053、2105、3821、3822、285、110、115、 KM281-1-10、KM281-2-10-1-2、KM283-5、KM292-1-24、KM225-2-56、KM341-6-9、4D11、5H10、11E1、5G3、3811、3411、3417を取得した。また、Kuroiwaらが報告しているヒト抗体Lambda鎖を保持するキメラマウス(Kuroiwa et. Al., Nat Biotechnol., 2000 vol 18:1086)も以下の免疫実験に用いた。このマウスからはハイブリドーマF4-465を取得した。
〔実施例3〕 ヒトCD40に対するヒトモノクローナル抗体の調製
本実施例におけるモノクローナル抗体の作製は、単クローン抗体実験操作入門(安東民衛ら著作、講談社発行 1991)に記載される一般的方法に従って調製した。免疫原としてのヒトCD40は、実施例1で調製したヒトCD40ヒトFCとCD40発現EL-4細胞を用いた。被免疫動物は、実施例2で作製したヒト免疫グロブリンを産生するヒト抗体産生マウスを用いた。
本実施例におけるモノクローナル抗体の作製は、単クローン抗体実験操作入門(安東民衛ら著作、講談社発行 1991)に記載される一般的方法に従って調製した。免疫原としてのヒトCD40は、実施例1で調製したヒトCD40ヒトFCとCD40発現EL-4細胞を用いた。被免疫動物は、実施例2で作製したヒト免疫グロブリンを産生するヒト抗体産生マウスを用いた。
ヒト抗体産生マウスに、CD40:hFcを1個体当たり2から100μg/回で免疫した。抗原溶液は初回を除いて等量の不完全フロイントアジュバント(シグマ)と混合し、皮下に数カ所に分けて注入した。約10日から3週間ごとに3-4回免疫を行った。初回は不完全フロイントアジュバント(シグマ)を用いた。マウスの尾から採血し、血清中のCD40に対するヒト抗体γおよびκを、ELISAを用いて測定した。脾臓を摘出する3-4日前に尾静脈より、PBSに溶解したCD40:Fcを20μg注射し最終免疫した。
ヒト抗体産生マウスに、ヒトCD40を発現するマウスEL-4細胞を免疫した。EL-4細胞108個/mlでPBSに懸濁し、予めPBSでエマルジョン化した等量のRIBIアジュバントと緩やかに混合した。細胞は約10日から3週間ごとに3-5回免疫を行った。アジュバントを用いない場合は、8000radでX線照射し用いた。
免疫されたマウスから脾臓を外科的に取得し、回収した脾臓細胞をマウスミエローマSP2/0(ATCC No.: CRL1581)と5:1で混合し、融合剤としてポリエチレングリコール1500(Boehringer Mannheim社製)を用いて細胞融合させることにより多数のハイブリドーマを作製した。ハイブリドーマの選択は、10%のウシ胎児血清(Fetal Calf Serum、FCS)とヒポキサンチン(H)、アミノプテリン(A)、チミジン(T)を含有するHAT含有DMEM培地(Gibco BRL社製)中で培養することにより行った。さらに、HT含有DMEM培地を用いて限界希釈法によりシングルクローンにした。培養は、96ウェルマイクロタイタープレート(ベクトンディッキンソン社製)中で行った。抗ヒトCD40ヒトモノクローナル抗体を産生するハイブリドーマクローンの選択(スクリーニング)は、実施例4で後述する酵素標識免疫吸着アッセイ(ELISA)および蛍光活性化セルソーター(FACS)により測定することにより行った。
ヒトモノクローナル抗体産生ハイブリドーマのELISAによるスクリーニングは、以下に述べる3種類のELISAおよびFACS解析により、ヒト免疫グロブリンγ鎖(hIgγ)及びヒト免疫グロブリン軽鎖κを有し、かつヒトCD40に特異的な反応性を有するヒトモノクローナル抗体を産生する多数のハイブリドーマを得た。なお、本実施例を含め以下のいずれの実施例中、並びに実施例における試験結果として示した表または図中においては、各々の本発明のヒト抗ヒトCD40モノクローナル抗体を産生するハイブリドーマクローンは記号を用いて命名した。また、当該記号の次に「抗体」を付したものは、それぞれのハイブリドーマにより産生される抗体、または当該ハイブリドーマから単離された抗体遺伝子(全長あるいは可変領域)を保持する宿主細胞により生産された組換え抗体を意味する。また文脈上明らかな範囲において、ハイブリドーマクローンの名称が抗体の名称をあらわす場合がある。
以下のハイブリドーマクローンはシングルクローンを表わす。
アゴニスティック抗体:
KM302-1、KM341-1-19、KM643-4-11、2053、2105、3821、3822、285、110、115、F1-102、F2-103、F5-77、F5-157
アンタゴニスティック抗体:
KM281-1-10、KM281-2-10-1-2、KM283-5、KM292-1-24、KM225-2-56、KM341-6-9、4D11、5H10、11E1、5G3、3811、3411、3417、F4-465
アゴニスティック抗体:
KM302-1、KM341-1-19、KM643-4-11、2053、2105、3821、3822、285、110、115、F1-102、F2-103、F5-77、F5-157
アンタゴニスティック抗体:
KM281-1-10、KM281-2-10-1-2、KM283-5、KM292-1-24、KM225-2-56、KM341-6-9、4D11、5H10、11E1、5G3、3811、3411、3417、F4-465
それらの内3つのハイブリドーマクローンKM 302-1、KM 281-1-10およびKM 281-2-10-1-2を平成13年(2001年)5月9日付で、クローンKM341-1-19および4D11を平成13年(2001年)9月27日付で、クローン2105を平成14年(2002年)4月17日付で独立行政法人産業技術総合研究所 特許生物寄託センター(茨城県つくば市東1丁目1番地1中央第6)にブダペスト条約に基づき国際寄託した。F2-103、F5-77およびF5-157の重鎖および軽鎖の可変領域を有するプラスミドを2001年4月19日付で, ハイブリドーマクローンF1-102およびF4-465を2001年4月24日付でATCC (American Type Culture Collection, 10801 University Blvd., Manassas, Virginia,USA)(アメリカ国立菌培養収集所、アメリカ合衆国 ヴァージニア州 20110-2209 マナサス 10801 ユニバーシティブルバード)にブダペスト条約に基づき国際寄託した(表2)。
〔実施例4〕 ハイブリドーマのスクリーニング
ヒト免疫グロブリンγ鎖を有するモノクローナル抗体の検出
実施例1で作製したヒトCD40マウスFC(1μg/ml) 50μl/ウェルを、ELISA用96穴マイクロプレート(Maxisorp、Nunc社製)の各ウェルに加え、4℃でインキュベートし、ヒトCD40マウスFCをマイクロプレートに吸着させた。次いで、上清を捨て、各ウェルにブロッキング試薬(ブロックエース、 大日本製薬)を加え室温でインキュベートしブロックした。各ウェルに、各々のハイブリドーマの培養上清(50μl)を加え、反応させた後、各ウェルを、0.1%Tween20含有リン酸緩衝液(PBS-T)で洗浄した。次いで、過酸化酵素で標識されたヤギ抗ヒトIgG(γ)抗体(シグマ、A0170)を1%FBS含有PBS-Tで5,000倍に希釈した溶液(50μl/ウェル)を、各ウェルに加え、インキュベートした。マイクロプレートを、PBS-Tで3回洗浄後、発色基質液(TMB、50μl/ウェル、住友ベークライト社製)を各ウェルに加え、室温下で30分間インキュベートした。各ウェルに、停止液(50μl/ウェル)を加え、反応を止めた。波長450nmでの吸光度をマイクロプレートリ−ダ−で測定した。陽性wellの培養上清をFACSにて解析し,Ramos細胞を染色するwellを選び、その細胞を限界希釈法でクローニングし、1wellについて1クローンの細胞を得た。ヒトCD40マウスFCを用いたELISAでhκ陽性を確認した。その結果、20個体のマウスから173クローンの抗ヒトCD40抗体が取得できた。その一部を表3(アゴニスティック抗体)及び表4(アンタゴニスティック抗体)に示す。アゴニスティック抗体のうち、少なくともKM341-1-19と2105は、CD40Lが発現している細胞、CD40が発現している細胞および抗体を用いた競合試験でリガンドとの顕著な競合を示さなかった。
ヒト免疫グロブリンγ鎖を有するモノクローナル抗体の検出
実施例1で作製したヒトCD40マウスFC(1μg/ml) 50μl/ウェルを、ELISA用96穴マイクロプレート(Maxisorp、Nunc社製)の各ウェルに加え、4℃でインキュベートし、ヒトCD40マウスFCをマイクロプレートに吸着させた。次いで、上清を捨て、各ウェルにブロッキング試薬(ブロックエース、 大日本製薬)を加え室温でインキュベートしブロックした。各ウェルに、各々のハイブリドーマの培養上清(50μl)を加え、反応させた後、各ウェルを、0.1%Tween20含有リン酸緩衝液(PBS-T)で洗浄した。次いで、過酸化酵素で標識されたヤギ抗ヒトIgG(γ)抗体(シグマ、A0170)を1%FBS含有PBS-Tで5,000倍に希釈した溶液(50μl/ウェル)を、各ウェルに加え、インキュベートした。マイクロプレートを、PBS-Tで3回洗浄後、発色基質液(TMB、50μl/ウェル、住友ベークライト社製)を各ウェルに加え、室温下で30分間インキュベートした。各ウェルに、停止液(50μl/ウェル)を加え、反応を止めた。波長450nmでの吸光度をマイクロプレートリ−ダ−で測定した。陽性wellの培養上清をFACSにて解析し,Ramos細胞を染色するwellを選び、その細胞を限界希釈法でクローニングし、1wellについて1クローンの細胞を得た。ヒトCD40マウスFCを用いたELISAでhκ陽性を確認した。その結果、20個体のマウスから173クローンの抗ヒトCD40抗体が取得できた。その一部を表3(アゴニスティック抗体)及び表4(アンタゴニスティック抗体)に示す。アゴニスティック抗体のうち、少なくともKM341-1-19と2105は、CD40Lが発現している細胞、CD40が発現している細胞および抗体を用いた競合試験でリガンドとの顕著な競合を示さなかった。
ヒト免疫グロブリン軽鎖κ(Igκ)を有するモノクローナル抗体の検出は、過酸化酵素で標識したヤギ抗ヒトIgκ抗体(1,000倍希釈、50μl/ウェル、Southern Biotechnology社製)を用いた他は、上述したヒト免疫グロブリンγ鎖のELISAと同様に行った。
各モノクローナル抗体のサブクラス同定は、それぞれ過酸化酵素で標識したヒツジ抗ヒトIgG1抗体、ヒツジ抗ヒトIgG2抗体、ヒツジ抗ヒトIgG3抗体又はヒツジ抗ヒトIgG4抗体(各2,000倍希釈、50μl/ウェル、The Binding Site社製)を用いた他は、上述したヒト免疫グロブリンγ鎖のELISAと同様に行った。
ヒトCD40発現細胞に対する各モノクローナル抗体の反応試験
CD40が発現していると報告されているRamos細胞株に対する各モノクローナル抗体の反応性の検討をFACS解析で行った。
CD40が発現していると報告されているRamos細胞株に対する各モノクローナル抗体の反応性の検討をFACS解析で行った。
2x106/mlの濃度でRamos細胞株を0.1%NaN3、2%FCS含有PBSの染色バッファー(SB)に浮遊させた。細胞浮遊液(100μl/ウェル)を96-well 丸底プレート(ベクトンディッキンソン社製)に分注した。各々のハイブリドーマの培養上清(50μl)を加え、氷温下30分間インキュベートした。陰性コントロールとしてヒト血清アルブミンに対するヒトIgG1抗体を用い、ハイブリドーマ培養培地で2μg/mlの濃度に調製し、50μl添加後氷温下15分間インキュベートした。SBで洗浄した後、250倍希釈したR-PE蛍光標識抗ヒト抗体(Southern Biotechnology社製)50μl を加え、氷温下15分間インキュベートした。SBで2回洗浄した後、300-500μlのFACS緩衝液に懸濁し、FACS(FACSort、FACScan、ベクトンディッキンソン社製)で各細胞の蛍光強度を測定した。その結果、Ramos細胞株に結合活性を有する抗体を選別した。
〔実施例5〕 各抗体の調製
モノクローナル抗体を含む培養上清の調製は以下の方法にて行った。
G28-5抗体生産ハイブリドーマはATCCより入手した(ATCC No.HB-9110)。抗CD40抗体産生ハイブリドーマをウシインシュリン(5μg/ml、Gibco BRL社製)、ヒトトランスフェリン(5μg/ml、Gibco BRL社製)、エタノールアミン(0.01mM、シグマ社製)、亜セレン酸ナトリウム(2.5x10-5 nM、シグマ社製)含有eRDF培地(極東製薬社製)に馴化した。スピナ−フラスコにて培養し、ハイブリドーマの生細胞率が90%になった時点で培養上清を回収した。回収した上清は、10μm と0.2μmのフィルター(ゲルマンサイエンス社製)に供し、ハイブリドーマ等の雑排物を除去した。
モノクローナル抗体を含む培養上清の調製は以下の方法にて行った。
G28-5抗体生産ハイブリドーマはATCCより入手した(ATCC No.HB-9110)。抗CD40抗体産生ハイブリドーマをウシインシュリン(5μg/ml、Gibco BRL社製)、ヒトトランスフェリン(5μg/ml、Gibco BRL社製)、エタノールアミン(0.01mM、シグマ社製)、亜セレン酸ナトリウム(2.5x10-5 nM、シグマ社製)含有eRDF培地(極東製薬社製)に馴化した。スピナ−フラスコにて培養し、ハイブリドーマの生細胞率が90%になった時点で培養上清を回収した。回収した上清は、10μm と0.2μmのフィルター(ゲルマンサイエンス社製)に供し、ハイブリドーマ等の雑排物を除去した。
上記培養上清からの抗CD40抗体の精製は以下の方法で行った。抗CD40抗体を含む培養上清をHyper D Protein A カラム(日本ガイシ製)あるいはマウスIgG1の精製にはProtein Gカラム(アマシャムファルマシアバイオテク)を用い、付属の説明書に従い吸着緩衝液としてPBS(-)、溶出緩衝液として0.1 M クエン酸ナトリウム緩衝液(pH 3)を用いてアフィニティー精製した。溶出画分は1 MTris-HCl (pH 8.0) あるいはNa2HPO4溶液を添加してpH7.2 付近に調整した。調製された抗体溶液は、透析膜(10000カット、Spectrum Laboratories社製)あるいはSPカラム(アマシャムファルマシアバイオテク)を用いてPBS(-)に置換し、孔径0.22μm のメンブランフィルターMILLEX-GV(MILLIPORE 製)でろ過滅菌した。精製抗体の濃度は280 nmの吸光度を測定し、1 mg/ml を1.45 OD として算出した。
〔実施例6〕 Ramos細胞における抗CD40アゴニスティック抗体によるCD95発現促進
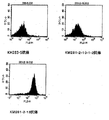
5.0x105個/mlのRamos細胞懸濁液を96ウエルプレートに100μl/wellで播種した(1ウェルあたり5x104個)。ハイブリドーマ培養上清又は精製抗体を20μg/mlに培地で希釈し、96ウエルプレートに100μl/wellの濃度で添加した。一晩培養後、細胞を集めR-PE標識抗CD95抗体(Pharmingen NJ)を用い、FACSCanあるいはFACSsort(ベクトンデッキンソン)を使って解析した。図1に結果を示す。図中横軸はCD95の発現強度を示す。抗体添加を太線、未添加を細線により示した。KM302-1抗体は、公知の抗体であるG28-5抗体に比して、CD95発現をより促進することが示された。すなわち、より効果の高いアゴニスティックであることが示された。
5.0x105個/mlのRamos細胞懸濁液を96ウエルプレートに100μl/wellで播種した(1ウェルあたり5x104個)。ハイブリドーマ培養上清又は精製抗体を20μg/mlに培地で希釈し、96ウエルプレートに100μl/wellの濃度で添加した。一晩培養後、細胞を集めR-PE標識抗CD95抗体(Pharmingen NJ)を用い、FACSCanあるいはFACSsort(ベクトンデッキンソン)を使って解析した。図1に結果を示す。図中横軸はCD95の発現強度を示す。抗体添加を太線、未添加を細線により示した。KM302-1抗体は、公知の抗体であるG28-5抗体に比して、CD95発現をより促進することが示された。すなわち、より効果の高いアゴニスティックであることが示された。
〔実施例7〕 Ramos細胞における抗CD40アンタゴニスティック抗体によるCD95発現抑制
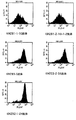
1.0x106個/mlのRamos細胞懸濁液を96ウエルプレートに50μl/wellで播種した。ハイブリドーマ培養上清又は精製抗体を2μg/mlに培地で調整し、96ウエルプレートに100μl/well添加した。可溶性CD40リガンド(ALEXIS CORPORATION)を4μg/mlと抗FLAG抗体(M2、シグマ)4μg/mlとを培地に添加し、96ウエルプレートに50μl/well添加した。一晩培養後、細胞を集めR-PE標識抗CD95抗体(Pharmingen NJ)を用い、FACSを使って解析した。図2A、図2B、図3に結果を示す。図中横軸はCD95の発現強度を示す。KM281-1-10、KM281-2-10-1-2、KM283-5、KM292-1-24、KM225-2-56の各ハイブリドーマが生産した抗体では陰性コントロールと同程度までCD95の発現を抑制した。
1.0x106個/mlのRamos細胞懸濁液を96ウエルプレートに50μl/wellで播種した。ハイブリドーマ培養上清又は精製抗体を2μg/mlに培地で調整し、96ウエルプレートに100μl/well添加した。可溶性CD40リガンド(ALEXIS CORPORATION)を4μg/mlと抗FLAG抗体(M2、シグマ)4μg/mlとを培地に添加し、96ウエルプレートに50μl/well添加した。一晩培養後、細胞を集めR-PE標識抗CD95抗体(Pharmingen NJ)を用い、FACSを使って解析した。図2A、図2B、図3に結果を示す。図中横軸はCD95の発現強度を示す。KM281-1-10、KM281-2-10-1-2、KM283-5、KM292-1-24、KM225-2-56の各ハイブリドーマが生産した抗体では陰性コントロールと同程度までCD95の発現を抑制した。
図3において、KM281-1-10抗体(下パネル)は、公知の抗体である5D12抗体(中央パネル)がわずかにしかCD95発現を抑制しなかったのに比して、CD95発現をより抑制することが示された。すなわち、より効果の高いアンタゴニスティックであることが示された。これによってヒトモノクローナル抗体がアンタゴニスティック抗体であることが示された。
抗免疫グロブリン抗体による架橋の影響
1.0x106個/mlのRamos細胞懸濁液を96ウエルプレートに50μl/wellで播種した。ハイブリドーマ培養上清又は精製抗体を2μg/mlに培地で調整し、96ウエルプレートに100μl/well添加した。抗ヒトIgG抗体(シグマ、I3382)あるいは抗マウスIgG抗体(Biosource、AMI3401)を4μg/mlとなるよう培地に添加し、96ウエルプレートに50μl/well添加した。一晩培養後、細胞を集めR-PE標識抗CD95抗体(Pharmingen NJ)を用い、FACSを使って解析した。図4、図5に結果を示す。図中横軸はCD95の発現強度を示す。KM281-1-10、KM281-2-10-1-2の各ハイブリドーマが生産した抗体ではCD95の発現を抑制した。しかし、5D12、KM283-5、KM292-1-24、KM225-2-56の各ハイブリドーマが生産した抗体ではCD95の発現を逆に増強した。
1.0x106個/mlのRamos細胞懸濁液を96ウエルプレートに50μl/wellで播種した。ハイブリドーマ培養上清又は精製抗体を2μg/mlに培地で調整し、96ウエルプレートに100μl/well添加した。抗ヒトIgG抗体(シグマ、I3382)あるいは抗マウスIgG抗体(Biosource、AMI3401)を4μg/mlとなるよう培地に添加し、96ウエルプレートに50μl/well添加した。一晩培養後、細胞を集めR-PE標識抗CD95抗体(Pharmingen NJ)を用い、FACSを使って解析した。図4、図5に結果を示す。図中横軸はCD95の発現強度を示す。KM281-1-10、KM281-2-10-1-2の各ハイブリドーマが生産した抗体ではCD95の発現を抑制した。しかし、5D12、KM283-5、KM292-1-24、KM225-2-56の各ハイブリドーマが生産した抗体ではCD95の発現を逆に増強した。
〔実施例8〕 Ramos細胞における抗CD40アゴニスティック抗体による増殖抑制
1.0x105個/mlのRamos細胞、HS-Sulton懸濁液を96ウエルプレートに100μl/wellで播種した。精製抗体あるいは可溶性CD40リガンドと抗FLAG抗体(M2)との等量混合物を培地に添加し、2日培養後、100μCi/mlの3H-Thymidine (Amersham Pharmacia)を10μl添加した。18時間後、Macro96 Harvester(SKATRON)を用いて、Printed Filtermat A(Wallac)にハーベストし、乾燥後Betap;Scint(Wallac)に良く浸し、パッケージング後、1205 BETAPLATE液体シンチレーションカウンターで活性測定した。図6に結果を示す。図中、縦軸は細胞による3Hチミジンの取り込み量を、横軸は培養液中の抗体あるいはCD40L濃度を示す。Ramos細胞、HS-Sulton細胞ともに、KM302-1抗体を添加した場合に、従来のG28-5抗体やCD40Lと比較して、チミジンの取り込みが低く、KM302-1抗体が腫瘍細胞の増殖を効果的に抑えられるアゴニスティック抗体であることが示された。
1.0x105個/mlのRamos細胞、HS-Sulton懸濁液を96ウエルプレートに100μl/wellで播種した。精製抗体あるいは可溶性CD40リガンドと抗FLAG抗体(M2)との等量混合物を培地に添加し、2日培養後、100μCi/mlの3H-Thymidine (Amersham Pharmacia)を10μl添加した。18時間後、Macro96 Harvester(SKATRON)を用いて、Printed Filtermat A(Wallac)にハーベストし、乾燥後Betap;Scint(Wallac)に良く浸し、パッケージング後、1205 BETAPLATE液体シンチレーションカウンターで活性測定した。図6に結果を示す。図中、縦軸は細胞による3Hチミジンの取り込み量を、横軸は培養液中の抗体あるいはCD40L濃度を示す。Ramos細胞、HS-Sulton細胞ともに、KM302-1抗体を添加した場合に、従来のG28-5抗体やCD40Lと比較して、チミジンの取り込みが低く、KM302-1抗体が腫瘍細胞の増殖を効果的に抑えられるアゴニスティック抗体であることが示された。
〔実施例9〕 CD40アゴニスティック抗体による樹状細胞の活性化
(1) 材料及び方法
組換え型ヒトIL-4はGenzyme techne社より購入した。抗ヒトCD14 MACS beadsはMiltenyi Biotech GmbH社より購入した。LymphoprepはNycomed Pharma ASより購入した。培養に用いた培地は、DC誘導時にはRPMI1640(Gibco BRL)に10% heat inactivated FCS(Cell Culture Technologies), 10mM HEPES(Sigma), 55μM 2-mercaptoethanol (Gibco BRL), 硫酸ストレプトマイシン(明治製菓)を加えて用いた。染色時の細胞洗浄には2% FCS (Cell Culture Technologies), 0.02% Azaid 添加PBS (Sigma)を用いた。細胞凍結時には日本全薬工業社製のセルバンカーを用いた。
(1) 材料及び方法
組換え型ヒトIL-4はGenzyme techne社より購入した。抗ヒトCD14 MACS beadsはMiltenyi Biotech GmbH社より購入した。LymphoprepはNycomed Pharma ASより購入した。培養に用いた培地は、DC誘導時にはRPMI1640(Gibco BRL)に10% heat inactivated FCS(Cell Culture Technologies), 10mM HEPES(Sigma), 55μM 2-mercaptoethanol (Gibco BRL), 硫酸ストレプトマイシン(明治製菓)を加えて用いた。染色時の細胞洗浄には2% FCS (Cell Culture Technologies), 0.02% Azaid 添加PBS (Sigma)を用いた。細胞凍結時には日本全薬工業社製のセルバンカーを用いた。
(2) 単球由来DCの誘導
末梢血よりLymphoprepを用いた密度勾配遠心により単核球を調製した(PBMC)。これを抗ヒトCD14 MACS beadsでポジティブセレクションすることによりCD14陽性画分と陰性画分に分離した。陽性画分に組換え型ヒトGM-CSF (50ng/ml)と組換え型ヒトIL-4 (100ng/ml)を添加して、10%FCSを添加したRPMI1640培地にて6well プレートで培養した。培養開始時の細胞濃度は1×106/mlで1 wellに3mlずつで行った。培養中は2日に1度の培地交換を行った。培地交換は培養液の1割を遠心チューブにとり遠心し、上清を除去後、とった培養液の2倍容量の新しい培養液(サイトカインなどを上記濃度で含む)で懸濁し、各wellに戻した。培養6日目には細胞を回収し、細胞数を計算後、上記培地に1×106/mlの濃度で懸濁し、抗CD40抗体又はそのアイソタイプ対照を添加して24 wellプレートでさらに4日間の培養を行った。この間培地交換は行わなかった(1ウェルあたりの細胞数1x106個、細胞濃度1x106個/ml)。
末梢血よりLymphoprepを用いた密度勾配遠心により単核球を調製した(PBMC)。これを抗ヒトCD14 MACS beadsでポジティブセレクションすることによりCD14陽性画分と陰性画分に分離した。陽性画分に組換え型ヒトGM-CSF (50ng/ml)と組換え型ヒトIL-4 (100ng/ml)を添加して、10%FCSを添加したRPMI1640培地にて6well プレートで培養した。培養開始時の細胞濃度は1×106/mlで1 wellに3mlずつで行った。培養中は2日に1度の培地交換を行った。培地交換は培養液の1割を遠心チューブにとり遠心し、上清を除去後、とった培養液の2倍容量の新しい培養液(サイトカインなどを上記濃度で含む)で懸濁し、各wellに戻した。培養6日目には細胞を回収し、細胞数を計算後、上記培地に1×106/mlの濃度で懸濁し、抗CD40抗体又はそのアイソタイプ対照を添加して24 wellプレートでさらに4日間の培養を行った。この間培地交換は行わなかった(1ウェルあたりの細胞数1x106個、細胞濃度1x106個/ml)。
(3) 細胞染色及びフローサイトメーターによる解析
染色には抗HLA-DR抗体(アイソタイプ対照: rat IgG2a)、抗CD86 抗体(アイソタイプ対照:ラットIgG1)、抗CD83抗体(アイソタイプ対照:ラットIgG2b)を用いた。まず、抗体を加え30分、4℃でインキュベートした後、3回の洗浄し、Beckton Dickinson社製FACS Caliburを用いて解析を行った。
染色には抗HLA-DR抗体(アイソタイプ対照: rat IgG2a)、抗CD86 抗体(アイソタイプ対照:ラットIgG1)、抗CD83抗体(アイソタイプ対照:ラットIgG2b)を用いた。まず、抗体を加え30分、4℃でインキュベートした後、3回の洗浄し、Beckton Dickinson社製FACS Caliburを用いて解析を行った。
(4) Mature DCに対するIL-12 secretion能の亢進
上記のように未成熟 DCを得た後、LPS (400pg/ml)とIFNγ (10-3M)を加えさらに2日間の培養後成熟 DCを得た。これに抗CD40抗体またはアイソタイプ対照を10μg/ml加え、24時間後の上清についてIL-12の産生をELISA (Pharmingen社製)法にて測定した。
上記のように未成熟 DCを得た後、LPS (400pg/ml)とIFNγ (10-3M)を加えさらに2日間の培養後成熟 DCを得た。これに抗CD40抗体またはアイソタイプ対照を10μg/ml加え、24時間後の上清についてIL-12の産生をELISA (Pharmingen社製)法にて測定した。
(5) 結果および考察
アゴニスティック抗体であるKM302-1抗体が、DCの成熟化に及ぼす抗体の影響を図7に、成熟DCのIL-12生産に及ぼす影響を図8に示す。G28-5抗体を対照として成熟の度合いを比べた。CD86、HLA-DRの発現を見た結果、G28-5抗体と比較して、KM302-1抗体ではさらに発現が上昇し、すなわち成熟度が亢進させることが示された。また、IL-12の分泌については成熟DCへのKM302-1抗体の処理によって亢進することが示された。以上よりKM302-1抗体が、DCに対するアゴニスティック抗体として作用することが示された。
アゴニスティック抗体であるKM302-1抗体が、DCの成熟化に及ぼす抗体の影響を図7に、成熟DCのIL-12生産に及ぼす影響を図8に示す。G28-5抗体を対照として成熟の度合いを比べた。CD86、HLA-DRの発現を見た結果、G28-5抗体と比較して、KM302-1抗体ではさらに発現が上昇し、すなわち成熟度が亢進させることが示された。また、IL-12の分泌については成熟DCへのKM302-1抗体の処理によって亢進することが示された。以上よりKM302-1抗体が、DCに対するアゴニスティック抗体として作用することが示された。
〔実施例10〕 DC-MLR
正常ヒトから採取した血液(末梢血)を、2000rpm 10分遠心し、血清を吸い取った。血球分画をPBSで再懸濁し、Ficoll(Amersham Pharmacia)の上に静かに載せた。2000rpm 30分遠心し、中間層のPBMC部分を回収する。PBSで2回洗浄した後、MACSを用い特定の細胞分離に使用した。
正常ヒトから採取した血液(末梢血)を、2000rpm 10分遠心し、血清を吸い取った。血球分画をPBSで再懸濁し、Ficoll(Amersham Pharmacia)の上に静かに載せた。2000rpm 30分遠心し、中間層のPBMC部分を回収する。PBSで2回洗浄した後、MACSを用い特定の細胞分離に使用した。
DC培養のための単球の分離は、MACS(Miltenyi Biotec GmbH)を用い添付の説明書にしたがって行った。簡単に説明すると、PBMC 1x108個に対して、MACS Buffer 800μl,MACS CD14 (Miltenyi Biotec GmbH、502-01)200μlを添加し、4℃で15分処理した。MACS LS カラムに細胞を吸着させ、洗浄した。カラムに吸着した細胞を単球として回収した。カラムに吸着しなかった細胞に、MACS HLA-DR(Miltenyi Biotec GmbH、461-01)を添加し、BSカラムでHLR−DR陽性細胞を除去し、T細胞画分とした。FACSによりCD3陽性細胞の比率を計測し、T細胞画分全体の細胞数より実質的なT細胞数を算出した。得られた単球は、6ウエル培養皿に、1x106細胞/mlの濃度で100ng/mlのIL−4(R&D system), 50ng/ml のG-CSF(KIRIN), 10%のFCS(SIGMA)を含むR0培地(PPMI培地に、β-メルカプトエタノール(Gibco),HEPES(SIGMA)を添加したもの)で培養した。培養5日後に、10ng/mlのLPS(DIFCO)を添加し、成熟DCに分化させた。
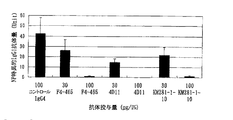
MLRは、異なるヒトから分離した、T細胞と成熟DCを混合することにより行った。T細胞/DCの細胞比をそれぞれ、1:80とし、T細胞数は、2x105細胞/ウエルとする。まず最初に、DCに抗体を加え30分間反応させる。その後、T細胞を加え、4日培養した後に、100μCi/mlの3H-Thymidine (Amersham Pharmacia)を10μl添加した。14時間後、Macro96 Harvester(SKATRON)を用いて、Printed Filtermat A (Wallac)にハーベストし、乾燥後Betap;Scint (Wallac)に良く浸し、パッケージング後、1205 BETAPLATE液体シンチレーションカウンターで活性測定した。また未成熟DCを用いたMLRは、異なるヒトから分離した、T細胞と成熟DCを混合することにより行った。T細胞/DCの細胞比をそれぞれ、1:40として同様に行った。図9、図10に結果を示す。KM281-1-10抗体の添加によりチミジンの取り込みが低く、MLRが抑えられることが示された。また、図10においてKM283-5、5D12抗体はDC-MLRを抑制できず、すなわちKM281-1-10抗体のみがDCに対するCD40リガンドの働きを中和するアンタゴニスティック抗体であることが示された。さらに、アゴニスティック抗体であるKM302-1抗体が、未成熟DCを用いたMLRに及ぼす抗体の影響を調べた結果を図11に示す。DCが活性化することにより、T細胞との相互作用が促進され、チミジンの取り込みが増加している。これによりKM302-1は未成熟DCに作用するアゴニスティック抗体であることが示された。
〔実施例11〕 CD40抗体によるCD40LのCD40への結合に及ぼす影響
BIAcore 2000(ビアコア)を用いて、抗CD40抗体を、固定化されたCD40ヒトFCに結合させた後、可溶性CD40LのCD40への結合量の変化を測定した。装置に付属した説明書に従い、可溶性CD40ヒトFCをCMチップ(CM5,ビアコア)に固定化した。次いで25(μg/ml)の抗CD40抗体を添加し、CD40に結合させた。さらに10(μg/ml)の可溶性CD40Lを添加し結合させた。CD40L添加前後の結合量の差を測定した。コントロ−ルIgGを添加した場合、CD40Lの結合量は100RUとなった。KM302-1抗体添加後では、CD40Lの結合量は110RUとなり、KM283-5抗体添加後では、18RUとなった。これにより、KM302-1抗体はCD40LのCD40への結合を阻害しないことが示された。
BIAcore 2000(ビアコア)を用いて、抗CD40抗体を、固定化されたCD40ヒトFCに結合させた後、可溶性CD40LのCD40への結合量の変化を測定した。装置に付属した説明書に従い、可溶性CD40ヒトFCをCMチップ(CM5,ビアコア)に固定化した。次いで25(μg/ml)の抗CD40抗体を添加し、CD40に結合させた。さらに10(μg/ml)の可溶性CD40Lを添加し結合させた。CD40L添加前後の結合量の差を測定した。コントロ−ルIgGを添加した場合、CD40Lの結合量は100RUとなった。KM302-1抗体添加後では、CD40Lの結合量は110RUとなり、KM283-5抗体添加後では、18RUとなった。これにより、KM302-1抗体はCD40LのCD40への結合を阻害しないことが示された。
〔実施例12〕Ramos細胞における抗CD40アゴニスティック抗体によるCD95発現促進
実施例4で得られたハイブリドーマの精製抗体を実施例6の方法に従って解析し、アゴニスト抗体を生産するクローンを選別した(1ウェルあたりの細胞数5x104個、細胞濃度2.5x105個/ml)。図12に結果を示す。図中横軸は培養液中の抗体濃度を、縦軸は平均蛍光強度、すなわちCD95発現強度を示す。KM341-1-19,2105抗体は、0.01μg/ml以上の濃度で公知のマウス抗体であるG28-5抗体に比して、Ramos細胞に対してCD95発現をより促進することが示された。すなわち、より効果の高いアゴニスティックであることが示された。また、0.01μg/mlでKM341-1-19, 2105抗体のRamos細胞にCD95発現を増進するアゴニスト活性は10μg/mlのG28-5抗体の活性より高かった(図12)。それぞれの抗体濃度において、CD95の発現がG28-5抗体を添加した場合の何倍に相当するかを表5にまとめた。
実施例4で得られたハイブリドーマの精製抗体を実施例6の方法に従って解析し、アゴニスト抗体を生産するクローンを選別した(1ウェルあたりの細胞数5x104個、細胞濃度2.5x105個/ml)。図12に結果を示す。図中横軸は培養液中の抗体濃度を、縦軸は平均蛍光強度、すなわちCD95発現強度を示す。KM341-1-19,2105抗体は、0.01μg/ml以上の濃度で公知のマウス抗体であるG28-5抗体に比して、Ramos細胞に対してCD95発現をより促進することが示された。すなわち、より効果の高いアゴニスティックであることが示された。また、0.01μg/mlでKM341-1-19, 2105抗体のRamos細胞にCD95発現を増進するアゴニスト活性は10μg/mlのG28-5抗体の活性より高かった(図12)。それぞれの抗体濃度において、CD95の発現がG28-5抗体を添加した場合の何倍に相当するかを表5にまとめた。
〔実施例13〕CD40アゴニスティック抗体による樹状細胞の活性化
実施例9の方法に従って、CD40アゴニスティック抗体による成熟DCのIL-12およびIL-10生産に及ぼす影響を調べた。IL-10はELISA (Pharmingen社製)法にて測定した。図13および図14に結果を示す。KM341-1-19抗体の処理によってIL-12の分泌が亢進することが示された。一方、5000radでX線照射した、CD40リガンド発現組換えL細胞を2x105個/ml共存させた場合でも、IL-12とIL-10の培養液中濃度は254、51pg/mlであり、KM341-1-19抗体を1μg/ml添加した場合より少なかった。
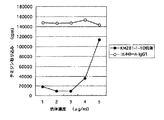
実施例9の方法に従って、CD40アゴニスティック抗体による成熟DCのIL-12およびIL-10生産に及ぼす影響を調べた。IL-10はELISA (Pharmingen社製)法にて測定した。図13および図14に結果を示す。KM341-1-19抗体の処理によってIL-12の分泌が亢進することが示された。一方、5000radでX線照射した、CD40リガンド発現組換えL細胞を2x105個/ml共存させた場合でも、IL-12とIL-10の培養液中濃度は254、51pg/mlであり、KM341-1-19抗体を1μg/ml添加した場合より少なかった。
以上よりKM341-1-19抗体が、DCに対して効果的なアゴニスティック抗体として作用することが示された。0.1μg/mlでKM341-1-19抗体の成熟DCにIL-12を分泌させるアゴニスト活性は100μg/mlのG28-5抗体の活性より高く、1μg/mlでKM341-1-19抗体の成熟DCにIL-12を分泌させるアゴニスト活性は100μg/mlのG28-5抗体の活性より100倍以上高かった(図13)。また、1μg/mlでKM341-1-19抗体の成熟DCにIL-10を分泌させるアゴニスト活性は100μg/mlのG28-5抗体の活性より10倍以上高かった(図14)。また、KM341-1-19抗体は、サブクラスがIgG2であり、IgG1やIgG3よりFcレセプターへの結合性が低く、NK細胞のキラー活性の感作や補体系の活性化も弱い。このことから抗体によって、CD40発現細胞の機能あるいは細胞そのものが減少してしまうという危険性が低いと考えられる。またFcレセプターによる架橋を受けにくいことから、体内でのアゴニスト活性が、架橋によって大きく変動することがなく、薬効を制御し易いと期待される。
〔実施例14〕Ramos細胞における抗CD40アンタゴニスティック抗体によるCD95発現抑制
1.0x106個/mlのRamos細胞懸濁液を平底96ウェルプレートに50μl/wellで播種した(1ウェルあたり細胞数5x104個)。培地で希釈した精製抗体を96ウェルプレートに100μl/well添加した。ヒトCD40リガンドを発現する組換えマウスL細胞(Spriggs, M.K. et. al., J.Exp.Med., 176: 1543,1992; Garrone, P. et. al., J.Exp.Med., 182: 1265,1995などを参照のこと)を1.0x105個/mlに調製し、50μl/wellで添加した(1ウェルあたりRamos細胞の細胞数5x104個、細胞濃度2.5x105個/ml、1ウェルあたりマウスL細胞の細胞数5x103個、細胞濃度2.5x104個/ml)。一晩培養後、細胞を集めR-PE標識抗CD95抗体を用い、FACSを用いて解析した。図15に結果を示す。図中縦軸は平均蛍光強度、すなわちCD95の発現強度を示す。公知の抗体である5D12抗体がわずかにしか抑制しなかったのに比較して、4D11抗体では0.1μg/mlの濃度でもCD40L発現細胞を添加しない陰性コントロールと同程度までCD95の発現を抑制した。また、1μg/mlの濃度では,4D11,F4-465,KM281-1-10がCD40L発現細胞を添加しない陰性コントロールと同程度までCD95の発現を抑制した。これによって4D11、F4-465,KM281-1-10抗体が、より効果的なアンタゴニスティック抗体であることが示された。それぞれの抗体濃度において、アンタゴニスティック抗体無添加の対照を100とした平均蛍光強度の相対値を表6に示す。
1.0x106個/mlのRamos細胞懸濁液を平底96ウェルプレートに50μl/wellで播種した(1ウェルあたり細胞数5x104個)。培地で希釈した精製抗体を96ウェルプレートに100μl/well添加した。ヒトCD40リガンドを発現する組換えマウスL細胞(Spriggs, M.K. et. al., J.Exp.Med., 176: 1543,1992; Garrone, P. et. al., J.Exp.Med., 182: 1265,1995などを参照のこと)を1.0x105個/mlに調製し、50μl/wellで添加した(1ウェルあたりRamos細胞の細胞数5x104個、細胞濃度2.5x105個/ml、1ウェルあたりマウスL細胞の細胞数5x103個、細胞濃度2.5x104個/ml)。一晩培養後、細胞を集めR-PE標識抗CD95抗体を用い、FACSを用いて解析した。図15に結果を示す。図中縦軸は平均蛍光強度、すなわちCD95の発現強度を示す。公知の抗体である5D12抗体がわずかにしか抑制しなかったのに比較して、4D11抗体では0.1μg/mlの濃度でもCD40L発現細胞を添加しない陰性コントロールと同程度までCD95の発現を抑制した。また、1μg/mlの濃度では,4D11,F4-465,KM281-1-10がCD40L発現細胞を添加しない陰性コントロールと同程度までCD95の発現を抑制した。これによって4D11、F4-465,KM281-1-10抗体が、より効果的なアンタゴニスティック抗体であることが示された。それぞれの抗体濃度において、アンタゴニスティック抗体無添加の対照を100とした平均蛍光強度の相対値を表6に示す。
〔実施例15〕抗CD40アゴニスティック抗体によるRamos細胞移植モデルにおける抗腫瘍効果
生後5週齢のC.B.17/Icr-scidJc1マウス(日本クレア)に抗アシアロGM1抗体を静脈注射した。1日後に腫瘍細胞としてRamos細胞をマウス1匹当たり5x106個静脈注射した。1日後にKM302-1抗体あるいは陰性対照として抗ヒトアルブミンヒトIgG抗体を静脈注射した。投与量はKM302-1抗体をマウス1匹当たり1、10、100μg、陰性対照抗体100μgをそれぞれ5個体のマウスに1回投与した。結果を図16に示す。陰性対照投与群は移植34日後にすべて死亡したのに対して、KM302-1抗体10μg、100μgを投与された群では5匹すべて生存しており、KM302-1抗体の抗腫瘍効果が確認された。KM302-1抗体はIgG4サブクラスであり、Fcレセプターを介した抗体依存性細胞傷害機構(ADCC)や補体系の活性化が弱いにもかかわらず、10μg単回投与で担ガンマウスの生存期間の延長が見られた。
生後5週齢のC.B.17/Icr-scidJc1マウス(日本クレア)に抗アシアロGM1抗体を静脈注射した。1日後に腫瘍細胞としてRamos細胞をマウス1匹当たり5x106個静脈注射した。1日後にKM302-1抗体あるいは陰性対照として抗ヒトアルブミンヒトIgG抗体を静脈注射した。投与量はKM302-1抗体をマウス1匹当たり1、10、100μg、陰性対照抗体100μgをそれぞれ5個体のマウスに1回投与した。結果を図16に示す。陰性対照投与群は移植34日後にすべて死亡したのに対して、KM302-1抗体10μg、100μgを投与された群では5匹すべて生存しており、KM302-1抗体の抗腫瘍効果が確認された。KM302-1抗体はIgG4サブクラスであり、Fcレセプターを介した抗体依存性細胞傷害機構(ADCC)や補体系の活性化が弱いにもかかわらず、10μg単回投与で担ガンマウスの生存期間の延長が見られた。
〔実施例16〕抗CD40アゴニスティック抗体によるRamos細胞の増殖抑制
10%FBS添加RPMI1640培地で1x104個/mlに調製したRamos細胞懸濁液100μlを96ウェルプレートに分注し、20μg/mlに培地を用いて調製したKM341-1-19抗体あるいは可溶性リガンド溶液を添加した。可溶性リガンドにはリガンドと同濃度(反応溶液中の濃度が10μg/ml)の抗FLAG抗体(M2)を共存させ活性を強めた。5日間培養後、各ウェルに20μlのMTS試薬(Promega)を加え2から3時間反応させた。波長490nmで細胞や抗体を含まない培地のみのウェルとの吸光度の差を測定し、生存している細胞数を測定した。また、同様に96ウェルU底プレートを用いてG28-5抗体と増殖抑制作用を比較した。2μg/mlに培地を用いて調製したKM341-1-19抗体あるいはG28-5抗体を添加した。結果を図17に示す。KM341-1-19抗体を添加したウェルでは死細胞が観察され、細胞数はG28-5抗体やリガンド添加のウェルに比較して顕著に少なく、吸光度も低くなり、腫瘍細胞の増殖が抑制され、細胞死が誘導されることが示された。
10%FBS添加RPMI1640培地で1x104個/mlに調製したRamos細胞懸濁液100μlを96ウェルプレートに分注し、20μg/mlに培地を用いて調製したKM341-1-19抗体あるいは可溶性リガンド溶液を添加した。可溶性リガンドにはリガンドと同濃度(反応溶液中の濃度が10μg/ml)の抗FLAG抗体(M2)を共存させ活性を強めた。5日間培養後、各ウェルに20μlのMTS試薬(Promega)を加え2から3時間反応させた。波長490nmで細胞や抗体を含まない培地のみのウェルとの吸光度の差を測定し、生存している細胞数を測定した。また、同様に96ウェルU底プレートを用いてG28-5抗体と増殖抑制作用を比較した。2μg/mlに培地を用いて調製したKM341-1-19抗体あるいはG28-5抗体を添加した。結果を図17に示す。KM341-1-19抗体を添加したウェルでは死細胞が観察され、細胞数はG28-5抗体やリガンド添加のウェルに比較して顕著に少なく、吸光度も低くなり、腫瘍細胞の増殖が抑制され、細胞死が誘導されることが示された。
〔実施例17〕抗体遺伝子のcDNAクローニング
KM341-1-19, 2105, 110, 115, KM281-1-10 ,4D11, KM643-4-11, F4-465、F2-103, F5-77抗体を生産するハイブリドーマを培養し、細胞を遠心によって集めた。これにTRIZOL(Gibco BRL)を添加し、取扱説明書にしたがってTotalRNAを抽出した。抗体cDNAの可変領域のクローニングは、CLONTECH社のSMART RACE cDNA amplification Kit を用い、添付の説明書にしたがって行った。5μgのtotal RNAを鋳型として1st Strand cDNAを作製した。KM341-1-19, 2105, 110, 115, KM281-1-10 ,4D11, KM643-4-11, F2-103, F5-77 重鎖(H鎖)の増幅には、Takara社のZ−Taqを用い,UMPとhh6プライマーで、98℃1秒、68℃30秒のサイクルを30回繰り返した。さらに、この反応液1μlを鋳型とし、NUMPとhh3プライマーを用いて、98℃1秒、68℃30秒のサイクルを20回繰り返した。F4-465重鎖の増幅には、UMPとhh2プライマーを用い、Advantage 2 PCR kit (Clonthech, cat#1910)を使って、94℃5秒、72℃3分 5サイクル、94℃5秒、70℃0秒、72℃3分、5サイクル、94℃5秒、68℃10秒、72℃3分、25サイクルで行った。
KM341-1-19, 2105, 110, 115, KM281-1-10 ,4D11, KM643-4-11, F4-465、F2-103, F5-77抗体を生産するハイブリドーマを培養し、細胞を遠心によって集めた。これにTRIZOL(Gibco BRL)を添加し、取扱説明書にしたがってTotalRNAを抽出した。抗体cDNAの可変領域のクローニングは、CLONTECH社のSMART RACE cDNA amplification Kit を用い、添付の説明書にしたがって行った。5μgのtotal RNAを鋳型として1st Strand cDNAを作製した。KM341-1-19, 2105, 110, 115, KM281-1-10 ,4D11, KM643-4-11, F2-103, F5-77 重鎖(H鎖)の増幅には、Takara社のZ−Taqを用い,UMPとhh6プライマーで、98℃1秒、68℃30秒のサイクルを30回繰り返した。さらに、この反応液1μlを鋳型とし、NUMPとhh3プライマーを用いて、98℃1秒、68℃30秒のサイクルを20回繰り返した。F4-465重鎖の増幅には、UMPとhh2プライマーを用い、Advantage 2 PCR kit (Clonthech, cat#1910)を使って、94℃5秒、72℃3分 5サイクル、94℃5秒、70℃0秒、72℃3分、5サイクル、94℃5秒、68℃10秒、72℃3分、25サイクルで行った。
hh6プライマー:5'-GGT CCG GGA GAT CAT GAG GGT GTC CTT-3'(配列番号3)
hh3プライマー:5'-GTG CAC GCC GCT GGT CAG GGC GCC TG-3'(配列番号4)
hh2プライマー:5'-GCT GGA GGG CAC GGT CAC CAC GCT G-3' (配列番号5)
hh3プライマー:5'-GTG CAC GCC GCT GGT CAG GGC GCC TG-3'(配列番号4)
hh2プライマー:5'-GCT GGA GGG CAC GGT CAC CAC GCT G-3' (配列番号5)
この後、増幅したPCR産物をPCR purification kit(QIAGEN)により精製し、hh4をプライマーとして塩基配列の決定を行った。あるいは、PCRScript(Stratagene, Lajolla, CA)又はPCR−Blunt(Invitrogene, Carlsbad, CA)にサブクローニングし配列を決定した。配列情報を基に、抗体重鎖特異的プライマーを合成した。KM341-1-19の場合341Hプライマー、2105の場合は2105Hsalプライマー, 110,115の場合には110Hsalプライマー, KM281-1-10の場合には281Hsalプライマー,4D11の場合は4D11Salプライマー、 KM643-4-11の場合には643Hsalプライマー ,F4-465の場合はH11-9 5'プライマー、F2-103の場合はF2-103Hプライマー、F5-77の場合はF5-77Hプライマーを合成した。抗体重鎖特異的プライマーとhh4を用いて、1stStrandcDNAからcDNAを増幅し、これを鋳型として抗体特異的プライマーを使って逆方向からの配列を決定した。
hh4プライマー:5'-GGTGCCAGGGGGAAGACCGATGG-3’(配列番号6)
341Hプライマー:5'-atatgtcgacGCTGAATTCTGGCTGACCAGGGCAG-3’(配列番号7)
2105Hsal:atatgtcgacTCCCAGGTGTTTCCATTCAGTGATCAG (配列番号8)
110Hsal:atatgtcgacTTCCATTCGGTGATCAGCACTGAACAC (配列番号9)
281Hsal: atatgtcgacTTTGAGAGTCCTGGACCTCCTGTG (配列番号10)
4D11Sal: atatgtcgacGAGTCATGGATCTCATGTGCAAG (配列番号11)
643Hsal: atatgtcgacCCAGGGCAGTCACCAGAGCTCCAGAC (配列番号12)
H11-9 5':-ACC GTG TCG ACT ACG CGG GAG TGA CT (配列番号13)
F2-103H: accgtgtcgacgctgatcaggactgcaca(配列番号14)
F5-77H: accgtgtcgacggtgatcaggactgaacag(配列番号15)
341Hプライマー:5'-atatgtcgacGCTGAATTCTGGCTGACCAGGGCAG-3’(配列番号7)
2105Hsal:atatgtcgacTCCCAGGTGTTTCCATTCAGTGATCAG (配列番号8)
110Hsal:atatgtcgacTTCCATTCGGTGATCAGCACTGAACAC (配列番号9)
281Hsal: atatgtcgacTTTGAGAGTCCTGGACCTCCTGTG (配列番号10)
4D11Sal: atatgtcgacGAGTCATGGATCTCATGTGCAAG (配列番号11)
643Hsal: atatgtcgacCCAGGGCAGTCACCAGAGCTCCAGAC (配列番号12)
H11-9 5':-ACC GTG TCG ACT ACG CGG GAG TGA CT (配列番号13)
F2-103H: accgtgtcgacgctgatcaggactgcaca(配列番号14)
F5-77H: accgtgtcgacggtgatcaggactgaacag(配列番号15)
KM341-1-19、2105、110、115、KM281-1-10、4D11、KM643-4-11、F2-103、F5-77の軽鎖(L鎖)は、UMPとhk2プライマーを使って、98℃1秒、68℃30秒のサイクルを30回繰り返して増幅した。F4-465の軽鎖は、UMPとhL2プライマーを使って、98℃1秒、68℃30秒のサイクルを30回繰り返して増幅した。増幅したPCR産物を、PCR purification kitにより精製し、hk6もしくはhL2プライマーを用いて塩基配列を決定した。この配列を基に、軽鎖特異的プライマーを合成した。KM341-1-19の場合341Kプライマー、2105の場合は2053KBglプライマー, 110,115の場合には110KBglプライマー, KM281-1-10の場合には281KBglプライマー,4D11の場合は4D11KBgl、 KM643-4-11の場合には643KBglプライマー、 F4-465の場合はLamda 5'プライマー,F-103,F5-77の場合はF2-103Kプライマーを合成した。
341−1−19、110、115、KM643-4-11、KM281-1-10、4D11、2105の場合は、軽鎖特異的プライマーとhk6プライマーを使って1stStrand cDNAからcDNAを増幅し、これを鋳型として両方向からの配列を決定した。F4-465、F2-103,F5-77については、PCRScript(Stratagene, Lajolla, CA)又はPCR−Blunt(Invitrogene, Carlsbad, CA)にサブクローニングし配列を決定した。
hk2プライマー:5'-GTT GAA GCT CTT TGT GAC GGG CGA GC-3'(配列番号16)
hL2プライマー:5'- TCT TCT CCA CGG TGC TCC CTT CAT-3'(配列番号17)
341Kプライマー:5'-atatagatctGAACTGCTCAGTTAGGACCCAGAGG-3'(配列番号18)
2053KBgl: atatagatctCGCGGGGAAGGAGACTGCTCAGTT (配列番号19)
110KBgl: atatagatctAGTCAGACCCAGTCAGGACACAGC (配列番号20)
281KBgl: atatagatctGAGCTGCTCAGTTAGGACCCAGAGGG (配列番号21)
4D11KBgl: atatagatctTAAGCAAGTGTAACAACTCAGAGTAC (配列番号22)
643KBgl: atatagatctGAGGAACTGCTCAGTTAGGACCCAGAGG (配列番号23)
Lamda 5':-AACTCCAGATCTGCCTCAGGAAGCAGCATC (配列番号24)
F2-103K: aactccagatctagggcaagcagtggtaac(配列番号25)
hk6プライマー:5'-TGGCGGGAAGATGAAGACAGATGGTG-3'(配列番号26)
hL2プライマー:5'- TCT TCT CCA CGG TGC TCC CTT CAT-3'(配列番号17)
341Kプライマー:5'-atatagatctGAACTGCTCAGTTAGGACCCAGAGG-3'(配列番号18)
2053KBgl: atatagatctCGCGGGGAAGGAGACTGCTCAGTT (配列番号19)
110KBgl: atatagatctAGTCAGACCCAGTCAGGACACAGC (配列番号20)
281KBgl: atatagatctGAGCTGCTCAGTTAGGACCCAGAGGG (配列番号21)
4D11KBgl: atatagatctTAAGCAAGTGTAACAACTCAGAGTAC (配列番号22)
643KBgl: atatagatctGAGGAACTGCTCAGTTAGGACCCAGAGG (配列番号23)
Lamda 5':-AACTCCAGATCTGCCTCAGGAAGCAGCATC (配列番号24)
F2-103K: aactccagatctagggcaagcagtggtaac(配列番号25)
hk6プライマー:5'-TGGCGGGAAGATGAAGACAGATGGTG-3'(配列番号26)
341−1−19 のH鎖全長、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号27の5'末端から50番目のアデニン(A)からはじまるATGコドンであり、終止コドンは1472番目のチミン(T)からはじまるTGAである。抗体可変領域と定常領域の境界は5'末端から493番目のアデニン(A)と494番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号28のN末端から148番目のセリン(S)残基までであり、149番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号28のN末端より20番目のセリン(S)までと予測された。成熟体のN末端は配列番号28の21番目のグルタミン(Q)であるものと考えられる。
H鎖DNAの翻訳開始点は、配列番号27の5'末端から50番目のアデニン(A)からはじまるATGコドンであり、終止コドンは1472番目のチミン(T)からはじまるTGAである。抗体可変領域と定常領域の境界は5'末端から493番目のアデニン(A)と494番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号28のN末端から148番目のセリン(S)残基までであり、149番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号28のN末端より20番目のセリン(S)までと予測された。成熟体のN末端は配列番号28の21番目のグルタミン(Q)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号29の5'末端から29番目のAからはじまるATGコドンであり、可変領域は5'末端から400番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号30のN末端から124番目のリジン(K)までである。精製されたL鎖蛋白質のN末端分析により、L鎖のシグナル配列は配列番号30のN末端より20番目のグリシン(G)までであり、成熟体のN末端は配列番号30の21番目のグルタミン酸(E)であることが明らかとなった。
341−1−19 H鎖(配列番号27):
GTCGACGCTGAATTCTGGCTGACCAGGGCAGCCACCAGAGCTCCAGACAATGTCTGTCTCCTTCCTCATCTTCCTGCCCGTGCTGGGCCTCCCATGGGGTGTCCTGTCACAGGTCCAACTGCAGCAGTCAGGTCCAGGACTGGTGAAGCCCTCGCAGACCCTCTCACTCACCTGTGCCATCTCCGGGGACAGTGTCTCTAGCAACAGTGCTACTTGGAACTGGATCAGGCAGTCCCCATCGAGAGACCTTGAGTGGCTGGGAAGGACATACTACAGGTCCAAGTGGTATCGTGATTATGTAGGATCTGTGAAAAGTCGAATAATCATCAACCCAGACACATCCAACAACCAGTTCTCCCTGCAGCTGAACTCTGTGACTCCCGAGGACACGGCTATATATTACTGTACAAGAGCACAGTGGCTGGGAGGGGATTACCCCTACTACTACAGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCTTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCGCCCTGCTCCAGGAGCACCTCCGAGAGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCTCTGACCAGCGGCGTGCACACCTTCCCAGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAACTTCGGCACCCAGACCTACACCTGCAACGTAGATCACAAGCCCAGCAACACCAAGGTGGACAAGACAGTTGAGCGCAAATGTTGTGTCGAGTGCCCACCGTGCCCAGCACCACCTGTGGCAGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCACGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCACGGGAGGAGCAGTTCAACAGCACGTTCCGTGTGGTCAGCGTCCTCACCGTTGTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAACCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACACCTCCCATGCTGGACTCAGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGAGGATCC
GTCGACGCTGAATTCTGGCTGACCAGGGCAGCCACCAGAGCTCCAGACAATGTCTGTCTCCTTCCTCATCTTCCTGCCCGTGCTGGGCCTCCCATGGGGTGTCCTGTCACAGGTCCAACTGCAGCAGTCAGGTCCAGGACTGGTGAAGCCCTCGCAGACCCTCTCACTCACCTGTGCCATCTCCGGGGACAGTGTCTCTAGCAACAGTGCTACTTGGAACTGGATCAGGCAGTCCCCATCGAGAGACCTTGAGTGGCTGGGAAGGACATACTACAGGTCCAAGTGGTATCGTGATTATGTAGGATCTGTGAAAAGTCGAATAATCATCAACCCAGACACATCCAACAACCAGTTCTCCCTGCAGCTGAACTCTGTGACTCCCGAGGACACGGCTATATATTACTGTACAAGAGCACAGTGGCTGGGAGGGGATTACCCCTACTACTACAGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCTTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCGCCCTGCTCCAGGAGCACCTCCGAGAGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCTCTGACCAGCGGCGTGCACACCTTCCCAGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAACTTCGGCACCCAGACCTACACCTGCAACGTAGATCACAAGCCCAGCAACACCAAGGTGGACAAGACAGTTGAGCGCAAATGTTGTGTCGAGTGCCCACCGTGCCCAGCACCACCTGTGGCAGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACGTGCGTGGTGGTGGACGTGAGCCACGAAGACCCCGAGGTCCAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCACGGGAGGAGCAGTTCAACAGCACGTTCCGTGTGGTCAGCGTCCTCACCGTTGTGCACCAGGACTGGCTGAACGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGGCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAACCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACACCTCCCATGCTGGACTCAGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGAGGATCC
341−1−19 H鎖アミノ酸配列(配列番号28)
MSVSFLIFLPVLGLPWGVLSQVQLQQSGPGLVKPSQTLSLTCAISGDSVSSNSATWNWIRQSPSRDLEWLGRTYYRSKWYRDYVGSVKSRIIINPDTSNNQFSLQLNSVTPEDTAIYYCTRAQWLGGDYPYYYSMDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSNFGTQTYTCNVDHKPSNTKVDKTVERKCCVECPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
MSVSFLIFLPVLGLPWGVLSQVQLQQSGPGLVKPSQTLSLTCAISGDSVSSNSATWNWIRQSPSRDLEWLGRTYYRSKWYRDYVGSVKSRIIINPDTSNNQFSLQLNSVTPEDTAIYYCTRAQWLGGDYPYYYSMDVWGQGTTVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSNFGTQTYTCNVDHKPSNTKVDKTVERKCCVECPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
341−1−19 L鎖(配列番号29):
ACTGCTCAGTTAGGACCCAGAGGGAACCATGGAAGCCCCAGCTCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAACACTTTCGGCCCTGGGACCAAAGTGGATATCAAACGTACG
ACTGCTCAGTTAGGACCCAGAGGGAACCATGGAAGCCCCAGCTCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAACACTTTCGGCCCTGGGACCAAAGTGGATATCAAACGTACG
341−1−19 L鎖アミノ酸配列(配列番号30)
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNTFGPGTKVDIKRT
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNTFGPGTKVDIKRT
2105のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号31の5'末端から70番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から495番目のアデニン(A)と496番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号32のN末端から142番目のセリン(S)残基までであり、149番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号32のN末端より19番目のcystein(C)までと予測された。成熟体のN末端は配列番号32の20番目のGlutamic acid(E)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号33の5'末端から28番目のAからはじまるATGコドンであり、可変領域は5'末端から405番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号34のN末端から126番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号34のN末端より20番目のGlycine(G)までと予測された。成熟体のN末端は配列番号34の21番目のGlutamic acid(E)であるものと考えられる。
2105 H鎖(配列番号:31)
CTGAACACAGACCCGTCGACTCCCAGGTGTTTCCATTCAGTGATCAGCACTGAACACAGAGGACTCACCATGGAGTTGGGACTGAGCTGGATTTTCCTTTTGGCTATTTTAAAAGGTGTCCAGTGTGAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGATTATGCCATGCACTGGGTCCGGCAAGCTCCAGGGAAGGGCCTGGAGTGGGTCTCAGGTATTAGTTGGAATAGTGGTAGCTTGGTGCATGCGGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTGTATCTGCAAATGAACAGTCTGAGAGCTGAGGACACGGCCTTGTATTACTGTGCAAGAGATAGGCTATTTCGGGGAGTTAGGTACTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGCACCAAGG
CTGAACACAGACCCGTCGACTCCCAGGTGTTTCCATTCAGTGATCAGCACTGAACACAGAGGACTCACCATGGAGTTGGGACTGAGCTGGATTTTCCTTTTGGCTATTTTAAAAGGTGTCCAGTGTGAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGCAGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTGATGATTATGCCATGCACTGGGTCCGGCAAGCTCCAGGGAAGGGCCTGGAGTGGGTCTCAGGTATTAGTTGGAATAGTGGTAGCTTGGTGCATGCGGACTCTGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACTCCCTGTATCTGCAAATGAACAGTCTGAGAGCTGAGGACACGGCCTTGTATTACTGTGCAAGAGATAGGCTATTTCGGGGAGTTAGGTACTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGCACCAAGG
2105 H鎖アミノ酸配列(配列番号:32)
MELGLSWIFLLAILKGVQCEVQLVESGGGLVQPGRSLRLSCAASGFTFDDYAMHWVRQAPGKGLEWVSGISWNSGSLVHADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCARDRLFRGVRYYGMDVWGQGTTVTVSSASTK
MELGLSWIFLLAILKGVQCEVQLVESGGGLVQPGRSLRLSCAASGFTFDDYAMHWVRQAPGKGLEWVSGISWNSGSLVHADSVKGRFTISRDNAKNSLYLQMNSLRAEDTALYYCARDRLFRGVRYYGMDVWGQGTTVTVSSASTK
2105 L鎖(配列番号:33)
CTGCTCAGTTAGGACCCAGAGGGAACCATGGAAGCCCCAGCTCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCCACTGGCTCACTTTCGGCGGGGGGACCAAGGTGGAGATCAAACGTACGGTG
CTGCTCAGTTAGGACCCAGAGGGAACCATGGAAGCCCCAGCTCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCCACTGGCTCACTTTCGGCGGGGGGACCAAGGTGGAGATCAAACGTACGGTG
2105 L鎖アミノ酸配列(配列番号:34)
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSHWLTFGGGTKVEIKRTV
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSHWLTFGGGTKVEIKRTV
110のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号35の5'末端から60番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から479番目のアデニン(A)と480番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号36のN末端から140番目のセリン(S)残基までであり、141番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号36のN末端より19番目のCystein(C)までと予測された。成熟体のN末端は配列番号36の20番目のグルタミン(Q)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号37の5'末端から35番目のAからはじまるATGコドンであり、可変領域は5'末端から421番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号38のN末端から129番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号38のN末端より22番目のCystein(C)までと予測された。成熟体のN末端は配列番号38の23番目のvaline(V)であるものと考えられる。
110 H鎖(配列番号:35)
CTGAACACAGACCCGTCGACTTCCATTCGGTGATCAGCACTGAACACAGAGGACTCACCATGGAGTTTGGGCTGAGCTGGGTTTTCCTCGTTGCTCTTTTAAGAGGTGTCCAGTGTCAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCGTCTGGATTCACCTTCAGTAGCTATGGCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATGGTATGATGGAAGTATTAAATACTATGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCGAGAGAGGGCTACAATATTTTGACTGGTTATTTTGGCTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGGG
CTGAACACAGACCCGTCGACTTCCATTCGGTGATCAGCACTGAACACAGAGGACTCACCATGGAGTTTGGGCTGAGCTGGGTTTTCCTCGTTGCTCTTTTAAGAGGTGTCCAGTGTCAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCGTCTGGATTCACCTTCAGTAGCTATGGCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATGGTATGATGGAAGTATTAAATACTATGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCGAGAGAGGGCTACAATATTTTGACTGGTTATTTTGGCTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGGG
110 H鎖アミノ酸配列(配列番号:36)
MEFGLSWVFLVALLRGVQCQVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVIWYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAREGYNILTGYFGYWGQGTLVTVSSASTK
MEFGLSWVFLVALLRGVQCQVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVIWYDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAREGYNILTGYFGYWGQGTLVTVSSASTK
110 L鎖(配列番号:37)
TCACAGATCTAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGCTCCCAGGTGCCAGATGTGTCATCTGGATGACCCAGTCTCCATCCTTACTCTCTGCATCTACAGGAGACAGAGTCACCATCAGTTGTCGGATGAGTCAGGGCATTAGCAGTGATTTAGCCTGGTATCAGCAAAAACCAGGGAAAGCCCCTGAGCTCCTGATCTCTGCTGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCTGCCTGCAGTCTGAAGATTTTGCAACTTATTACTGTCAACAGTATTATAGTTTTCCGTGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACG
TCACAGATCTAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGCTCCCAGGTGCCAGATGTGTCATCTGGATGACCCAGTCTCCATCCTTACTCTCTGCATCTACAGGAGACAGAGTCACCATCAGTTGTCGGATGAGTCAGGGCATTAGCAGTGATTTAGCCTGGTATCAGCAAAAACCAGGGAAAGCCCCTGAGCTCCTGATCTCTGCTGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCTGCCTGCAGTCTGAAGATTTTGCAACTTATTACTGTCAACAGTATTATAGTTTTCCGTGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACG
110 L鎖アミノ酸配列(配列番号:38)
MDMRVPAQLLGLLLLWLPGARCVIWMTQSPSLLSASTGDRVTISCRMSQGISSDLAWYQQKPGKAPELLISAASTLQSGVPSRFSGSGSGTDFTLTISCLQSEDFATYYCQQYYSFPWTFGQGTKVEIKRT
MDMRVPAQLLGLLLLWLPGARCVIWMTQSPSLLSASTGDRVTISCRMSQGISSDLAWYQQKPGKAPELLISAASTLQSGVPSRFSGSGSGTDFTLTISCLQSEDFATYYCQQYYSFPWTFGQGTKVEIKRT
115のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号39の5'末端から60番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から479番目のアデニン(A)と480番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号40のN末端から140番目のセリン(S)残基までであり、141番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号40のN末端より19番目のCystein(C)までと予測された。成熟体のN末端は配列番号40の20番目のグルタミン(Q)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号41の5'末端から35番目のAからはじまるATGコドンであり、可変領域は5'末端から421番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号42のN末端から129番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号42のN末端より22番目のCystein(C)までと予測された。成熟体のN末端は配列番号42の23番目のvaline(V)であるものと考えられる。
115 H鎖(配列番号:39)
CTGAACACAGACCCGTCGACTTCCATTCGGTGATCAGCACTGAACACAGAGGACTCACCATGGAGTTTGGGCTGAGCTGGGTTTTCCTCGTTGCTCTTTTAAGAGGTGTCCAGTGTCAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCGTCTGGATTCACCTTCAGTAGCTATGGCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATGGAATGATGGAAGTATTAAATACTATGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCGAGAGAGGGCTACAATATTTTGACTGGTTATTTTGGCTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGG
CTGAACACAGACCCGTCGACTTCCATTCGGTGATCAGCACTGAACACAGAGGACTCACCATGGAGTTTGGGCTGAGCTGGGTTTTCCTCGTTGCTCTTTTAAGAGGTGTCCAGTGTCAGGTGCAGCTGGTGGAGTCTGGGGGAGGCGTGGTCCAGCCTGGGAGGTCCCTGAGACTCTCCTGTGCAGCGTCTGGATTCACCTTCAGTAGCTATGGCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATGGAATGATGGAAGTATTAAATACTATGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCGAGAGAGGGCTACAATATTTTGACTGGTTATTTTGGCTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGG
115 H鎖アミノ酸配列(配列番号:40)
MEFGLSWVFLVALLRGVQCQVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVIWNDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAREGYNILTGYFGYWGQGTLVTVSSASTK
MEFGLSWVFLVALLRGVQCQVQLVESGGGVVQPGRSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAVIWNDGSIKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAREGYNILTGYFGYWGQGTLVTVSSASTK
115 L鎖(配列番号:41)
TCACAGATCTAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGCTCCCAGGTGCCAGATGTGTCATCTGGATGACCCAGTCTCCATCCTTACTCTCTGCATCTACAGGAGACAGAGTCACCATCAGTTGTCGGATGAGTCAGGGCATTAGCAGTGATTTAGCCTGGTATCAGCAAAAACCAGGGAAAGCCCCTGAGCTCCTGATCTCTGCTGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCTGCCTGCAGTCTGAAGATTTTGCAACTTATTACTGTCAACAGTATTATAGTTTTCCGTGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACG
TCACAGATCTAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGCTCCCAGGTGCCAGATGTGTCATCTGGATGACCCAGTCTCCATCCTTACTCTCTGCATCTACAGGAGACAGAGTCACCATCAGTTGTCGGATGAGTCAGGGCATTAGCAGTGATTTAGCCTGGTATCAGCAAAAACCAGGGAAAGCCCCTGAGCTCCTGATCTCTGCTGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGTGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCTGCCTGCAGTCTGAAGATTTTGCAACTTATTACTGTCAACAGTATTATAGTTTTCCGTGGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACG
115 L鎖アミノ酸配列(配列番号:42)
MDMRVPAQLLGLLLLWLPGARCVIWMTQSPSLLSASTGDRVTISCRMSQGISSDLAWYQQKPGKAPELLISAASTLQSGVPSRFSGSGSGTDFTLTISCLQSEDFATYYCQQYYSFPWTFGQGTKVEIKRT
MDMRVPAQLLGLLLLWLPGARCVIWMTQSPSLLSASTGDRVTISCRMSQGISSDLAWYQQKPGKAPELLISAASTLQSGVPSRFSGSGSGTDFTLTISCLQSEDFATYYCQQYYSFPWTFGQGTKVEIKRT
281−1−10のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号43の5'末端から52番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から468番目のアデニン(A)と469番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号44のN末端から139番目のセリン(S)残基までであり、140番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号44のN末端より19番目のセリン(S)までと予測された。成熟体のN末端は配列番号44の20番目のグルタミン(Q)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号45の5'末端から41番目のAからはじまるATGコドンであり、可変領域は5'末端から424番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号46のN末端から128番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号46のN末端より20番目のGlycine(G)までと予測された。成熟体のN末端は配列番号46の21番目のGlutamic acid(E)であるものと考えられる。
281−1−10 H鎖(配列番号:43)
CTGAACACAGACCCGTCGACTTTGAGAGTCCTGGACCTCCTGTGCAAGAACATGAAACATCTGTGGTTCTTCCTTCTCCTGGTGGCAGCTCCCAGATGGGTCCTGTCCCAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGTGGCTCCATCAGTGGTTACTACTGGAGCTGGATCCGGCAGCCCCCAGGGAAGGGACTGGAGTGGATTGGGTATATCTATTACAGTGGGAGCACCAACTACAATCCCTCCCTCAAGAGTCGAGTCACCATATCAGTAGACACGTCCAAGAACCAGTTCTCCCTGAAGCTGAATTCTGTGACCGCTGCGGACACGGCCGTGTATTACTGTGCGAGAGCCCCCTTGCACGGTGACTACAAATGGTTCCACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGG
CTGAACACAGACCCGTCGACTTTGAGAGTCCTGGACCTCCTGTGCAAGAACATGAAACATCTGTGGTTCTTCCTTCTCCTGGTGGCAGCTCCCAGATGGGTCCTGTCCCAGGTGCAGCTGCAGGAGTCGGGCCCAGGACTGGTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGTGGCTCCATCAGTGGTTACTACTGGAGCTGGATCCGGCAGCCCCCAGGGAAGGGACTGGAGTGGATTGGGTATATCTATTACAGTGGGAGCACCAACTACAATCCCTCCCTCAAGAGTCGAGTCACCATATCAGTAGACACGTCCAAGAACCAGTTCTCCCTGAAGCTGAATTCTGTGACCGCTGCGGACACGGCCGTGTATTACTGTGCGAGAGCCCCCTTGCACGGTGACTACAAATGGTTCCACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGG
281−1−10 H鎖アミノ酸配列(配列番号:44)
MKHLWFFLLLVAAPRWVLSQVQLQESGPGLVKPSETLSLTCTVSGGSISGYYWSWIRQPPGKGLEWIGYIYYSGSTNYNPSLKSRVTISVDTSKNQFSLKLNSVTAADTAVYYCARAPLHGDYKWFHPWGQGTLVTVSSASTK
MKHLWFFLLLVAAPRWVLSQVQLQESGPGLVKPSETLSLTCTVSGGSISGYYWSWIRQPPGKGLEWIGYIYYSGSTNYNPSLKSRVTISVDTSKNQFSLKLNSVTAADTAVYYCARAPLHGDYKWFHPWGQGTLVTVSSASTK
281−1−10 L鎖(配列番号:45)
TCACAGATCTGAGCTGCTCAGTTAGGACCCAGAGGGAACCATGGAAACCCCAGCGCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACGCAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCAGCTACTTAGCCTGGTACCAGCAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGGTGCATCCAGCAGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAGCCTGAAGATTTTGCAGTGTATTACTGTCAGCAGTATGGTAGCTCACCGATCACCTTCGGCCAAGGGACACGACTGGAGATCAAACGTACG
TCACAGATCTGAGCTGCTCAGTTAGGACCCAGAGGGAACCATGGAAACCCCAGCGCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACGCAGTCTCCAGGCACCCTGTCTTTGTCTCCAGGGGAAAGAGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCAGCTACTTAGCCTGGTACCAGCAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGGTGCATCCAGCAGGGCCACTGGCATCCCAGACAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGACTGGAGCCTGAAGATTTTGCAGTGTATTACTGTCAGCAGTATGGTAGCTCACCGATCACCTTCGGCCAAGGGACACGACTGGAGATCAAACGTACG
281−1−10 L鎖アミノ酸配列(配列番号:46)
METPAQLLFLLLLWLPDTTGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPITFGQGTRLEIKRT
METPAQLLFLLLLWLPDTTGEIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPITFGQGTRLEIKRT
4D11のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号47の5'末端から16番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から456番目のアデニン(A)と457番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号48のN末端から147番目のセリン(S)残基までであり、148番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号48のN末端より26番目のセリン(S)までと予測された。成熟体のN末端は配列番号48の27番目のグルタミン(Q)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号49の5'末端から59番目のAからはじまるATGコドンであり、可変領域は5'末端から442番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号50のN末端から128番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号50のN末端より22番目のCystein(C)までと予測された。成熟体のN末端は配列番号50の23番目のAlanine(A)であるものと考えられる。
4D11 H鎖(配列番号:47)
ATATGTCGACGAGTCATGGATCTCATGTGCAAGAAAATGAAGCACCTGTGGTTCTTCCTCCTGCTGGTGGCGGCTCCCAGATGGGTCCTGTCCCAGCTGCAGCTGCAGGAGTCGGGCCCAGGACTACTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGCGGCTCCATCAGCAGTCCTGGTTACTACGGGGGCTGGATCCGCCAGCCCCCAGGGAAGGGGCTGGAGTGGATTGGGAGTATCTATAAAAGTGGGAGCACCTACCACAACCCGTCCCTCAAGAGTCGAGTCACCATATCCGTAGACACGTCCAAGAACCAGTTCTCCCTGAAGCTGAGCTCTGTGACCGCCGCAGACACGGCTGTGTATTACTGTACGAGACCTGTAGTACGATATTTTGGGTGGTTCGACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGC
ATATGTCGACGAGTCATGGATCTCATGTGCAAGAAAATGAAGCACCTGTGGTTCTTCCTCCTGCTGGTGGCGGCTCCCAGATGGGTCCTGTCCCAGCTGCAGCTGCAGGAGTCGGGCCCAGGACTACTGAAGCCTTCGGAGACCCTGTCCCTCACCTGCACTGTCTCTGGCGGCTCCATCAGCAGTCCTGGTTACTACGGGGGCTGGATCCGCCAGCCCCCAGGGAAGGGGCTGGAGTGGATTGGGAGTATCTATAAAAGTGGGAGCACCTACCACAACCCGTCCCTCAAGAGTCGAGTCACCATATCCGTAGACACGTCCAAGAACCAGTTCTCCCTGAAGCTGAGCTCTGTGACCGCCGCAGACACGGCTGTGTATTACTGTACGAGACCTGTAGTACGATATTTTGGGTGGTTCGACCCCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGC
4D11 H鎖アミノ酸配列(配列番号:48)
MDLMCKKMKHLWFFLLLVAAPRWVLSQLQLQESGPGLLKPSETLSLTCTVSGGSISSPGYYGGWIRQPPGKGLEWIGSIYKSGSTYHNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCTRPVVRYFGWFDPWGQGTLVTVSSAS
MDLMCKKMKHLWFFLLLVAAPRWVLSQLQLQESGPGLLKPSETLSLTCTVSGGSISSPGYYGGWIRQPPGKGLEWIGSIYKSGSTYHNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCTRPVVRYFGWFDPWGQGTLVTVSSAS
4D11 L鎖(配列番号:49)
AGATCTTAAGCAAGTGTAACAACTCAGAGTACGCGGGGAGACCCACTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTTCTGCTGCTCTGGCTCCCAGGTGCCAGATGTGCCATCCAGTTGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGGGCATTAGCAGTGCTTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCTCCTAAGCTCCTGATCTATGATGCCTCCAATTTGGAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTATTACTGTCAACAGTTTAATAGTTACCCGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACG
AGATCTTAAGCAAGTGTAACAACTCAGAGTACGCGGGGAGACCCACTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTTCTGCTGCTCTGGCTCCCAGGTGCCAGATGTGCCATCCAGTTGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGGGCATTAGCAGTGCTTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCTCCTAAGCTCCTGATCTATGATGCCTCCAATTTGGAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTATTACTGTCAACAGTTTAATAGTTACCCGACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACG
4D11 L鎖アミノ酸配列(配列番号:50)
MDMRVPAQLLGLLLLWLPGARCAIQLTQSPSSLSASVGDRVTITCRASQGISSALAWYQQKPGKAPKLLIYDASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFNSYPTFGQGTKVEIKRT
MDMRVPAQLLGLLLLWLPGARCAIQLTQSPSSLSASVGDRVTITCRASQGISSALAWYQQKPGKAPKLLIYDASNLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFNSYPTFGQGTKVEIKRT
KM643-4-11のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号51の5'末端から1番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から447番目のアデニン(A)と448番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号52のN末端から149番目のセリン(S)残基までであり、150番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号52のN末端より20番目のセリン(S)までと予測された。成熟体のN末端は配列番号52の21番目のグルタミン(Q)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号53の5'末端から38番目のAからはじまるATGコドンであり、可変領域は5'末端から409番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号54のN末端から124番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号54のN末端より20番目のGlycine(G)までと予測された。成熟体のN末端は配列番号54の21番目のGlutamic acid(E)であるものと考えられる。
KM643-4-11 H鎖(配列番号:51)
ATGTCTGTCTCCTTCCTCATCTTCCTGCCCGTGCTGGGCCTCCCATGGGGTGTCCTGTCACAGGTACAGCTGCAGCAGTCAGGTCCAGGACTGGTGAAGCCCTCGCAGACCCTCTCATTCACCTGTGCCATCTCCGGGGACAGTGTCTCTAGCAACAGTGCTGCTTGGAACTGGATCAGGCAGTCCCCATCGAGAGGCCTTGAGTGGCTGGGAAGGACATACTACAGGTCCAAGTGGTATAAAGATTATGCAGTATCTGTGAAAAGTCGAATAACCATCAACCCAGACACATCCAAGAACCAGTTCTCCCTGCAGCTGAACTCTGTGACCCCCGAGGACACGGCTGTGTATTACTGTGCAAGAGGGTATTACTATGGTTCGGGGAGCTATCCCTACTACTACCAAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGC
ATGTCTGTCTCCTTCCTCATCTTCCTGCCCGTGCTGGGCCTCCCATGGGGTGTCCTGTCACAGGTACAGCTGCAGCAGTCAGGTCCAGGACTGGTGAAGCCCTCGCAGACCCTCTCATTCACCTGTGCCATCTCCGGGGACAGTGTCTCTAGCAACAGTGCTGCTTGGAACTGGATCAGGCAGTCCCCATCGAGAGGCCTTGAGTGGCTGGGAAGGACATACTACAGGTCCAAGTGGTATAAAGATTATGCAGTATCTGTGAAAAGTCGAATAACCATCAACCCAGACACATCCAAGAACCAGTTCTCCCTGCAGCTGAACTCTGTGACCCCCGAGGACACGGCTGTGTATTACTGTGCAAGAGGGTATTACTATGGTTCGGGGAGCTATCCCTACTACTACCAAATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGC
KM643-4-11 H鎖アミノ酸配列(配列番号:52)
MSVSFLIFLPVLGLPWGVLSQVQLQQSGPGLVKPSQTLSFTCAISGDSVSSNSAAWNWIRQSPSRGLEWLGRTYYRSKWYKDYAVSVKSRITINPDTSKNQFSLQLNSVTPEDTAVYYCARGYYYGSGSYPYYYQMDVWGQGTTVTVSSAS
MSVSFLIFLPVLGLPWGVLSQVQLQQSGPGLVKPSQTLSFTCAISGDSVSSNSAAWNWIRQSPSRGLEWLGRTYYRSKWYKDYAVSVKSRITINPDTSKNQFSLQLNSVTPEDTAVYYCARGYYYGSGSYPYYYQMDVWGQGTTVTVSSAS
KM643-4-11 L鎖(配列番号:53)
AATTGAGGAACTGCTCAGTTAGGACCCAGAGGGAACCATGGAAGCCCCAGCTCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGTGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAACACTTTCGGCGGAGGGACCAAGGTGGAGATCAAACGAAC
AATTGAGGAACTGCTCAGTTAGGACCCAGAGGGAACCATGGAAGCCCCAGCTCAGCTTCTCTTCCTCCTGCTACTCTGGCTCCCAGATACCACCGGAGAAATTGTGTTGACACAGTCTCCAGCCACCCTGTCTTTGTCTCCAGGGGAAAGTGCCACCCTCTCCTGCAGGGCCAGTCAGAGTGTTAGCAGCTACTTAGCCTGGTACCAACAGAAACCTGGCCAGGCTCCCAGGCTCCTCATCTATGATGCATCCAACAGGGCCACTGGCATCCCAGCCAGGTTCAGTGGCAGTGGGTCTGGGACAGACTTCACTCTCACCATCAGCAGCCTAGAGCCTGAAGATTTTGCAGTTTATTACTGTCAGCAGCGTAGCAACACTTTCGGCGGAGGGACCAAGGTGGAGATCAAACGAAC
KM643-4-11 L鎖アミノ酸配列(配列番号:54)
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGESATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNTFGGGTKVEIKR
MEAPAQLLFLLLLWLPDTTGEIVLTQSPATLSLSPGESATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNTFGGGTKVEIKR
F4-465のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号55の5'末端から47番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から484番目のアデニン(A)と445番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号56のN末端から146番目のセリン(S)残基までであり、147番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号56のN末端より19番目のセリン(S)までと予測された。成熟体のN末端は配列番号56の20番目のグルタミン(Q)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号57の5'末端から81番目のAからはじまるATGコドンであり、可変領域は5'末端から440番目の(C)までである。アミノ酸配列において、可変領域は配列番号58のN末端から120番目のThreonine(T)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号58のN末端より19番目のAlanine(G)までと予測された。成熟体のN末端は配列番号58の20番目のSerine(S)であるものと考えられる。
F4-465 H鎖(配列番号:55)
CTGAACACAGACCCGTCGACTACGCGGGAGACCACAGCTCCACACCATGGACTGGACCTGGAGGATCCTATTCTTGGTGGCAGCAGCAACAGGTGCCCACTCCCAGGTGCAGCTGGTGCAATCTGGGTCTGAGTTGAAGAAGCCTGGGGCCTCAGTGAAGGTCCCCTGCAAGGCTTCTGGATACACCTTCACTAGCTATGCTATGAATTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGATGGGATGGATCAACACCAACACTGGGAACCCAACGTATGCCCAGGGCTTCACAGGACGGTTTGTCTTCTCCTTGGACACCTCTGTCAGCACGGCATATCTGCAGATCAGCAGCCTAAAGGCTGAGGACACTGCCGTGTATTACTGTGCGAGAGAGGTAGTACCAGTTGCTATGAGGGTAACTCACTACTACTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGCACCAA
CTGAACACAGACCCGTCGACTACGCGGGAGACCACAGCTCCACACCATGGACTGGACCTGGAGGATCCTATTCTTGGTGGCAGCAGCAACAGGTGCCCACTCCCAGGTGCAGCTGGTGCAATCTGGGTCTGAGTTGAAGAAGCCTGGGGCCTCAGTGAAGGTCCCCTGCAAGGCTTCTGGATACACCTTCACTAGCTATGCTATGAATTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGATGGGATGGATCAACACCAACACTGGGAACCCAACGTATGCCCAGGGCTTCACAGGACGGTTTGTCTTCTCCTTGGACACCTCTGTCAGCACGGCATATCTGCAGATCAGCAGCCTAAAGGCTGAGGACACTGCCGTGTATTACTGTGCGAGAGAGGTAGTACCAGTTGCTATGAGGGTAACTCACTACTACTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGCACCAA
F4-465 H鎖アミノ酸配列(配列番号:56)
MDWTWRILFLVAAATGAHSQVQLVQSGSELKKPGASVKVPCKASGYTFTSYAMNWVRQAPGQGLEWMGWINTNTGNPTYAQGFTGRFVFSLDTSVSTAYLQISSLKAEDTAVYYCAREVVPVAMRVTHYYYGMDVWGQGTTVTVSSAST
MDWTWRILFLVAAATGAHSQVQLVQSGSELKKPGASVKVPCKASGYTFTSYAMNWVRQAPGQGLEWMGWINTNTGNPTYAQGFTGRFVFSLDTSVSTAYLQISSLKAEDTAVYYCAREVVPVAMRVTHYYYGMDVWGQGTTVTVSSAST
F4-465 L鎖(配列番号:57)
CTGGGTACGGTAACCGTCAGATCGCCTGGAGACGCCATCACAGATCTGCCTCAGGAAGCAGCATCGGAGGTGCCTCAGCCATGGCATGGATCCCTCTCTTCCTCGGCGTCCTTGTTTACTGCACAGGATCCGTGGCCTCCTATGAGCTGACTCAGCCACCCTCAGTGTCCGTGGCCCCAGGACAGACAGCCAGCATCACCTGTTCTGGAGATAAATTGGGGGATAATTTTACTTGCTGGTATCAGCAGAAGCCAGGCCAGTCCCCTGTGCTGGTCATCTTTCAGGATTGGAAGCGGCGCCCAGGGATCCCTGCGCGATTCTCTGGCTCCAAGTCTGGGAACACAGCCACTCTGACCATCAGCGGGACCCAGGCTATGGATGAGGCTGACTATTACTGTCAGGCGTGGGACATCAGCACTGTGGTATTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCCTGGAAGGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTACCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATGAATTCAGATCCGTTAACGGTTACCAACTACCTAGACTGGATTCGTGACCAACATA
CTGGGTACGGTAACCGTCAGATCGCCTGGAGACGCCATCACAGATCTGCCTCAGGAAGCAGCATCGGAGGTGCCTCAGCCATGGCATGGATCCCTCTCTTCCTCGGCGTCCTTGTTTACTGCACAGGATCCGTGGCCTCCTATGAGCTGACTCAGCCACCCTCAGTGTCCGTGGCCCCAGGACAGACAGCCAGCATCACCTGTTCTGGAGATAAATTGGGGGATAATTTTACTTGCTGGTATCAGCAGAAGCCAGGCCAGTCCCCTGTGCTGGTCATCTTTCAGGATTGGAAGCGGCGCCCAGGGATCCCTGCGCGATTCTCTGGCTCCAAGTCTGGGAACACAGCCACTCTGACCATCAGCGGGACCCAGGCTATGGATGAGGCTGACTATTACTGTCAGGCGTGGGACATCAGCACTGTGGTATTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAGCCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCCTGGAAGGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTACCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATGAATTCAGATCCGTTAACGGTTACCAACTACCTAGACTGGATTCGTGACCAACATA
F4-465 L鎖アミノ酸配列(配列番号:58)
MAWIPLFLGVLVYCTGSVASYELTQPPSVSVAPGQTASITCSGDKLGDNFTCWYQQKPGQSPVLVIFQDWKRRPGIPARFSGSKSGNTATLTISGTQAMDEADYYCQAWDISTVVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
MAWIPLFLGVLVYCTGSVASYELTQPPSVSVAPGQTASITCSGDKLGDNFTCWYQQKPGQSPVLVIFQDWKRRPGIPARFSGSKSGNTATLTISGTQAMDEADYYCQAWDISTVVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
F2-103のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号59の5'末端から32番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から463番目のアデニン(A)と464番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号60のN末端から144番目のセリン(S)残基までであり、145番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号60のN末端より19番目のCystein(C)までと予測された。成熟体のN末端は配列番号60の20番目のGlutamic acid(E)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号61の5'末端から29番目のAからはじまるATGコドンであり、可変領域は5'末端から415番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号62のN末端から129番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号62のN末端より22番目のCystein(C)までと予測された。成熟体のN末端は配列番号62の23番目のAsp(D)であるものと考えられる。
F2-103 H鎖(配列番号:59)
GCTGATCAGGACTGCACACAGAGAACTCACCATGGAGTTTGGGCTGAGCTGGGTTTTCCTTGTTGCTATTTTAAAAGGTGTCCAGTGTGAGGTGCAGCTGGTGGAGTCCGGGGGAGGCTTAGTTCAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGTCTCTGGATTCACCTTCAGTACCTACTGGATGCACTGGGTCCGCCAAGCTCCAGGGAAGGGGCTGGTGTGGGTCTCACGTATTAATAGTGATGGGAGTAGCACAACCTACGCGGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACACGCTGTATCTGCAAATGAACAGTCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCAAGAGATAGAGTACTATGGATCGGGGAGTTATCCTACTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCT
GCTGATCAGGACTGCACACAGAGAACTCACCATGGAGTTTGGGCTGAGCTGGGTTTTCCTTGTTGCTATTTTAAAAGGTGTCCAGTGTGAGGTGCAGCTGGTGGAGTCCGGGGGAGGCTTAGTTCAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGTCTCTGGATTCACCTTCAGTACCTACTGGATGCACTGGGTCCGCCAAGCTCCAGGGAAGGGGCTGGTGTGGGTCTCACGTATTAATAGTGATGGGAGTAGCACAACCTACGCGGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAACGCCAAGAACACGCTGTATCTGCAAATGAACAGTCTGAGAGCCGAGGACACGGCTGTGTATTACTGTGCAAGAGATAGAGTACTATGGATCGGGGAGTTATCCTACTACGGTATGGACGTCTGGGGCCAAGGGACCACGGTCACCGTCTCCTCAGCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCT
F2-103 H鎖アミノ酸配列(配列番号:60)
MEFGLSWVFLVAILKGVQCEVQLVESGGGLVQPGGSLRLSCAVSGFTFSTYWMHWVRQAPGKGLVWVSRINSDGSSTTYADSVKGRFTISRDNAKNTLYLQMNSLRAEDTAVYYCARDRVLWIGELSYYGMDVWGQGTTVTVSSASTKGPSVFPLAPSSKSTS
MEFGLSWVFLVAILKGVQCEVQLVESGGGLVQPGGSLRLSCAVSGFTFSTYWMHWVRQAPGKGLVWVSRINSDGSSTTYADSVKGRFTISRDNAKNTLYLQMNSLRAEDTAVYYCARDRVLWIGELSYYGMDVWGQGTTVTVSSASTKGPSVFPLAPSSKSTS
F2-103 L鎖(配列番号:61)
GGGGAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGCTCCCAGGTGCCAAATGTGACATCCAGATGACCCAGTCTCCTTCCACCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAACTGGTTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGCTCTATAAGGCATCTGGTTTAGAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAACAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAACAGTCTAATAGTTATTCGTGGACGTTCGGCCACGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGA
GGGGAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGCTCCCAGGTGCCAAATGTGACATCCAGATGACCCAGTCTCCTTCCACCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCCAGTCAGAGTATTAGTAACTGGTTGGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGCTCTATAAGGCATCTGGTTTAGAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGAATTCACTCTCACCATCAACAGCCTGCAGCCTGATGATTTTGCAACTTATTACTGCCAACAGTCTAATAGTTATTCGTGGACGTTCGGCCACGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGA
F2-103 L鎖アミノ酸配列(配列番号:62)
MDMRVPAQLLGLLLLWLPGAKCDIQMTQSPSTLSASVGDRVTITCRASQSISNWLAWYQQKPGKAPKLLLYKASGLESGVPSRFSGSGSGTEFTLTINSLQPDDFATYYCQQSNSYSWTFGHGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGL
MDMRVPAQLLGLLLLWLPGAKCDIQMTQSPSTLSASVGDRVTITCRASQSISNWLAWYQQKPGKAPKLLLYKASGLESGVPSRFSGSGSGTEFTLTINSLQPDDFATYYCQQSNSYSWTFGHGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGL
F5-77のH鎖可変領域、及びL鎖可変領域をコードするDNA並びにH鎖及びL鎖のアミノ酸配列をそれぞれ以下に示す。
H鎖DNAの翻訳開始点は、配列番号63の5'末端から100番目のアデニン(A)からはじまるATGコドンであり、抗体可変領域と定常領域の境界は5'末端から528番目のアデニン(A)と529番目のグアニン(G)間に位置する。アミノ酸配列において、H鎖可変領域は配列番号64のN末端から143番目のセリン(S)残基までであり、144番目のアラニン(A)以降が定常領域である。遺伝子配列予測ソフトウェア(Signal P ver.2)により、H鎖のシグナル配列は配列番号64のN末端より19番目のCystein(C)までと予測された。成熟体のN末端は配列番号64の20番目のGlutamic acid(E)であるものと考えられる。
L鎖DNAの翻訳開始点は、配列番号65の5'末端から59番目のAからはじまるATGコドンであり、可変領域は5'末端から445番目のアデニン(A)までである。アミノ酸配列において、可変領域は配列番号66のN末端から129番目のリジン(K)までである。遺伝子配列予測ソフトウェア(Signal P ver.2)により、L鎖のシグナル配列は配列番号66のN末端より22番目のCystein(C)までと予測された。成熟体のN末端は配列番号66の23番目のAsp(D)であるものと考えられる。
F5-77 H鎖(配列番号:63)
GGTCTATATAAGCAGAGCTGGGTACGTCCTCACATTCAGTGATCAGCACTGAACACAGACCCGTCGACGGTGATCAGGACTGAACAGAGAGAACTCACCATGGAGTTTGGGCTGAGCTGGCTTTTTCTTGTGGCTATTTTAAAAGGTGTCCAGTGTGAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGCACATACTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGATGGGGGGTACTATGGTTCGGGGAGTTATGGGTACTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCC
GGTCTATATAAGCAGAGCTGGGTACGTCCTCACATTCAGTGATCAGCACTGAACACAGACCCGTCGACGGTGATCAGGACTGAACAGAGAGAACTCACCATGGAGTTTGGGCTGAGCTGGCTTTTTCTTGTGGCTATTTTAAAAGGTGTCCAGTGTGAGGTGCAGCTGTTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACCTTTAGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAGGGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGCACATACTACGCAGACTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGATGGGGGGTACTATGGTTCGGGGAGTTATGGGTACTTTGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCC
F5-77 H鎖アミノ酸配列(配列番号:64)
MEFGLSWLFLVAILKGVQCEVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDGGYYGSGSYGYFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFP
MEFGLSWLFLVAILKGVQCEVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDGGYYGSGSYGYFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFP
F5-77 L鎖(配列番号:65)
CAAGCAGTGGTAACAACGCAGAGTACGCGGGGGGAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGTTCCCAGGTTCCAGATGCGACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGGATCTGTAGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGGTATTAGCAGCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGGATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATTTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAACAGGCTAGCAGTTTCCCTCGGACATTCGGCCAAGGGACCAAGGTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGA
CAAGCAGTGGTAACAACGCAGAGTACGCGGGGGGAGTCAGACCCAGTCAGGACACAGCATGGACATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGTTCCCAGGTTCCAGATGCGACATCCAGATGACCCAGTCTCCATCTTCCGTGTCTGGATCTGTAGGAGACAGAGTCACCATCACTTGTCGGGCGAGTCAGGGTATTAGCAGCTGGTTAGCCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGGATCCAGTTTGCAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATTTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCAACAGGCTAGCAGTTTCCCTCGGACATTCGGCCAAGGGACCAAGGTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGA
F5-77 L鎖アミノ酸配列(配列番号:66)
MDMRVPAQLLGLLLLWFPGSRCDIQMTQSPSSVSGSVGDRVTITCRASQGISSWLAWYQQKPGKAPKLLIYAGSSLQSGVPSRFSGSGFGTDFTLTISSLQPEDFATYYCQQASSFPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGL
MDMRVPAQLLGLLLLWFPGSRCDIQMTQSPSSVSGSVGDRVTITCRASQGISSWLAWYQQKPGKAPKLLIYAGSSLQSGVPSRFSGSGFGTDFTLTISSLQPEDFATYYCQQASSFPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGL
[実施例18]抗体たんぱく質の動物細胞における発現
前記で取得した抗体の可変領域を含むDNA断片をN5KG1(IDEC Pharmaceuticals, US patent6001358)などの適当なベクターに組み込むことにより、抗体発現ベクターを作製する。発現のための宿主細胞には、例えばCHO-Ras(Katakura Y., et al., Cytotechnology, 31: 103-109, 1999)が好適に用いられる。宿主細胞へのベクターの導入は例えばエレクトロポレーションにより実施できる。抗体発現ベクター約2μgを制限酵素で線状化し、Bio-Rad electrophoreterをもちいて350V、500μFの条件で、4x107個のCHO−Ras細胞に遺伝子を導入し、96well culture plateに播種する。発現ベクターの選択マーカーに対応した薬剤を添加して培養を継続する。コロニーが観察されたら、実施例4に示した方法によって、抗体発現株を選別する。選別した細胞からの抗体精製は、実施例5にしたがって行うことができる。
前記で取得した抗体の可変領域を含むDNA断片をN5KG1(IDEC Pharmaceuticals, US patent6001358)などの適当なベクターに組み込むことにより、抗体発現ベクターを作製する。発現のための宿主細胞には、例えばCHO-Ras(Katakura Y., et al., Cytotechnology, 31: 103-109, 1999)が好適に用いられる。宿主細胞へのベクターの導入は例えばエレクトロポレーションにより実施できる。抗体発現ベクター約2μgを制限酵素で線状化し、Bio-Rad electrophoreterをもちいて350V、500μFの条件で、4x107個のCHO−Ras細胞に遺伝子を導入し、96well culture plateに播種する。発現ベクターの選択マーカーに対応した薬剤を添加して培養を継続する。コロニーが観察されたら、実施例4に示した方法によって、抗体発現株を選別する。選別した細胞からの抗体精製は、実施例5にしたがって行うことができる。
〔実施例19〕CD40アンタゴニスティック抗体による抗原特異的抗体産生抑制作用
マウス内因性CD40破壊についてホモ接合体の遺伝子背景を有し、かつ、ヒトCD40遺伝子のトランスジーンを有しているマウス(Yasui. et al. Int. Immunol. 2002 Vol14 : 319)に4-hydroxy-3-nitrophenylacetyl-chiken γ-globulin conjugates (NP-CGG:大阪大学微生物病研究所 菊谷 仁教授より分与)とアラム(ARAM: Antigen recognition activation motif)の複合体を100μg(NP-CGG量として)腹腔内注射することにより感作した。各モノクローナル抗体は抗原感作直前に30または100μgの量を尾静脈内投与した。陰性対照として抗ヒトアルブミンヒトIgG4抗体100μgを投与した。感作7日後、眼窩静脈叢より採血し、血清中のNP特異的IgG1およびIgM抗体量をELISA法により測定した。ELISA法はNPを結合した牛血清アルブミン(NP-BSA:2.5μg/ml)50μl/ウェルを、ELISA用96穴マイクロプレート(Maxisorp、Nunc社製)の各ウェルに加え、4℃でインキュベートし、NP-BSAを吸着させた。次いで、上清を捨て、各ウェルにブロッキング試薬(スーパーブロック、Pierce社製)を加え室温でインキュベートしブロックした後、各ウェルを0.1%Tween20含有リン酸緩衝液(PBS-T)で3回洗浄した。次いで、各ウェルに10%ブロックエース含有PBS-Tで希釈した各血清(50μl/ウェル)を加え、37℃で2時間インキュベートし反応させた。マイクロプレートをPBS-Tで3回洗浄後、アルカリフォスファターゼで標識されたヤギ抗マウスIgG1またはIgM抗体(コスモバイオ、1070-04または1020-04)を10%ブロックエース含有PBS-Tで1,000倍に希釈した溶液(50μl/ウェル)を、各ウェルに加え、37℃で2時間インキュベートした。次いで、マイクロプレートをPBS-Tで3回洗浄後、発色基質液(50μl/ウェル、Sigma104、phosphatase substrate)を各ウェルに加え、波長405nmでの吸光度をマイクロプレートリーダーで測定した。その結果を図18、図19に示す。図中縦軸はNP-CGGをC57BL/6マウスに2回注射した後採血してプールした血清について、IgG1抗体の場合は10,000倍希釈したものを、IgM抗体の場合は100倍したものをそれぞれ1ユニットとして換算した値を示す。F4-465、4D11およびKM281-1-10抗体は100μgの投与によりNP特異的なIgG1およびIgM抗体産生を強く抑制した。
マウス内因性CD40破壊についてホモ接合体の遺伝子背景を有し、かつ、ヒトCD40遺伝子のトランスジーンを有しているマウス(Yasui. et al. Int. Immunol. 2002 Vol14 : 319)に4-hydroxy-3-nitrophenylacetyl-chiken γ-globulin conjugates (NP-CGG:大阪大学微生物病研究所 菊谷 仁教授より分与)とアラム(ARAM: Antigen recognition activation motif)の複合体を100μg(NP-CGG量として)腹腔内注射することにより感作した。各モノクローナル抗体は抗原感作直前に30または100μgの量を尾静脈内投与した。陰性対照として抗ヒトアルブミンヒトIgG4抗体100μgを投与した。感作7日後、眼窩静脈叢より採血し、血清中のNP特異的IgG1およびIgM抗体量をELISA法により測定した。ELISA法はNPを結合した牛血清アルブミン(NP-BSA:2.5μg/ml)50μl/ウェルを、ELISA用96穴マイクロプレート(Maxisorp、Nunc社製)の各ウェルに加え、4℃でインキュベートし、NP-BSAを吸着させた。次いで、上清を捨て、各ウェルにブロッキング試薬(スーパーブロック、Pierce社製)を加え室温でインキュベートしブロックした後、各ウェルを0.1%Tween20含有リン酸緩衝液(PBS-T)で3回洗浄した。次いで、各ウェルに10%ブロックエース含有PBS-Tで希釈した各血清(50μl/ウェル)を加え、37℃で2時間インキュベートし反応させた。マイクロプレートをPBS-Tで3回洗浄後、アルカリフォスファターゼで標識されたヤギ抗マウスIgG1またはIgM抗体(コスモバイオ、1070-04または1020-04)を10%ブロックエース含有PBS-Tで1,000倍に希釈した溶液(50μl/ウェル)を、各ウェルに加え、37℃で2時間インキュベートした。次いで、マイクロプレートをPBS-Tで3回洗浄後、発色基質液(50μl/ウェル、Sigma104、phosphatase substrate)を各ウェルに加え、波長405nmでの吸光度をマイクロプレートリーダーで測定した。その結果を図18、図19に示す。図中縦軸はNP-CGGをC57BL/6マウスに2回注射した後採血してプールした血清について、IgG1抗体の場合は10,000倍希釈したものを、IgM抗体の場合は100倍したものをそれぞれ1ユニットとして換算した値を示す。F4-465、4D11およびKM281-1-10抗体は100μgの投与によりNP特異的なIgG1およびIgM抗体産生を強く抑制した。
〔実施例20〕CD40アンタゴニスティック抗体による扁桃腺B細胞の増殖抑制
ヒト扁桃腺はチルドレンズ・ホスピタル(サン ディエゴ、米国カリフォルニア州)より入手した。扁桃腺を小片に裁断し、細かく刻み、70マイクロメーターのナイロンメッシュ細胞ストレーナーを通すことにより、細胞懸濁液を調製した。PBSにて数回洗浄の後、細胞数を計数し、90%ヒト血清(ICN)、10%DMSOで凍結保存した。細胞は融解の後、10%ヒト血清、2.5マイクログラム/mlのアンフォテリシン(ファンギゾン、ギブコ/BRL)を添加した標準的RPMI培地に再懸濁して使用した。
ヒト扁桃腺はチルドレンズ・ホスピタル(サン ディエゴ、米国カリフォルニア州)より入手した。扁桃腺を小片に裁断し、細かく刻み、70マイクロメーターのナイロンメッシュ細胞ストレーナーを通すことにより、細胞懸濁液を調製した。PBSにて数回洗浄の後、細胞数を計数し、90%ヒト血清(ICN)、10%DMSOで凍結保存した。細胞は融解の後、10%ヒト血清、2.5マイクログラム/mlのアンフォテリシン(ファンギゾン、ギブコ/BRL)を添加した標準的RPMI培地に再懸濁して使用した。
1x105個の細胞を96穴ウェルに加え、抗ヒトCD40抗体を0.01、0.1、1.0および10マイクログラム/mlの濃度で添加した。試験は各濃度につき三連で実施した。flag標識したCD40L(Alexis)1マイクログラム/ml、CD40Lエンハンサー抗体(Alexis)1マイクログラム/mlを各ウェルに添加して3日間培養の後、1マイクロCiの[3H]チミジンを各ウェルに添加した。12〜15時間後に細胞を回収、液体シンチレーションカウンターにより扁桃腺B細胞の増殖を計測した。B細胞がCD40Lの刺激により増殖し、抗体を添加されなかった場合のカウントを100、CD40Lが添加されず、B細胞が刺激されなかった場合のカウントを0とした。例えば相対的なカウントの値として30が計測された場合、本実験では、70%の増殖阻害が起こったと表現することとした。
公知のアンタゴニスティック抗体である5D12は、抗体濃度100マイクログラム/mlでも50%を超える増殖抑制を示さなかった。F4-465はわずか0.01マイクログラム/mlの濃度で約80%の増殖抑制を示し、0.1〜10マイクログラム/mlの濃度で約95%の増殖抑制を示した(図20)。
本明細書で引用した全ての刊行物、特許及び特許出願は、そのまま参考として本明細書に取り入れるものとする。
本明細書で引用した全ての刊行物、特許及び特許出願は、そのまま参考として本明細書に取り入れるものとする。
配列番号1〜26:合成DNA
Claims (15)
- ハイブリドーマ2105(受託番号FERM BP-8024)またはF1-102(受託番号ATCC PTA-3337)により生産される抗体の重鎖可変領域および軽鎖可変領域のアミノ酸配列を有するヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片。
- 受託番号ATCC PTA-3302およびATCC PTA-3303であるプラスミドDNAにそれぞれコードされる、ハイブリドーマF2-103の産生する抗体の重鎖可変領域および軽鎖可変領域、受託番号ATCC PTA-3304およびATCC PTA-3305であるプラスミドDNAにそれぞれコードされる、ハイブリドーマF5-77の産生する抗体の重鎖可変領域および軽鎖可変領域、または受託番号ATCC PTA-3306およびATCC PTA-3307であるプラスミドDNAにそれぞれコードされる、ハイブリドーマF5-157の産生する抗体の重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有するヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片。
- 配列番号32および34にそれぞれ示される、ハイブリドーマ2105の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号36および38にそれぞれ示される、ハイブリドーマ110の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号40および42にそれぞれ示される、ハイブリドーマ115の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号52および54にそれぞれ示される、ハイブリドーマKM643-4-11の産生する抗体の重鎖可変領域および軽鎖可変領域、配列番号60および62にそれぞれ示される、ハイブリドーマF2-103の産生する抗体の重鎖可変領域および軽鎖可変領域、または配列番号64および66にそれぞれ示される、ハイブリドーマF5−77の産生する抗体の重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有するヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片。
- 配列番号31および33にそれぞれ示される、ハイブリドーマ2105から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号35および37にそれぞれ示される、ハイブリドーマ110から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号39および41にそれぞれ示される、ハイブリドーマ115から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号51および53にそれぞれ示される、ハイブリドーマKM643-4-11から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、配列番号59および61にそれぞれ示される、ハイブリドーマF2-103から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、または配列番号63および65にそれぞれ示される、ハイブリドーマF5−77から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有するヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片。
- ハイブリドーマ2053または285により産生される抗体の重鎖可変領域および軽鎖可変領域のアミノ酸配列を有するヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片。
- ハイブリドーマKM281-1-10(受託番号FERM BP-7579)または4D11(受託番号FERM BP-7758)により生産される抗体の重鎖可変領域および軽鎖可変領域のアミノ酸配列を有するヒトCD40に対する抗体又はアンタゴニスティックな作用を有するその機能的断片。
- 配列番号44および46にそれぞれ示される、ハイブリドーマKM281-1-10の産生する抗体の重鎖可変領域および軽鎖可変領域、または配列番号48および50にそれぞれ示される、ハイブリドーマ4D11の産生する抗体の重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有するヒトCD40に対する抗体又はアンタゴニスティックな作用を有するその機能的断片。
- 配列番号43および45にそれぞれ示される、ハイブリドーマKM281-1-10から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域、または配列番号47および49にそれぞれ示される、ハイブリドーマ4D11の産生する抗体の重鎖可変領域および軽鎖可変領域から単離された核酸配列にコードされる重鎖可変領域および軽鎖可変領域の、マチュア部分のアミノ酸配列を有するヒトCD40に対する抗体又はアンタゴニスティックな作用を有するその機能的断片。
- ハイブリドーマKM281-2-10-1-2、KM283-5、KM292-1-24、KM225-2-56、5H10、11E1または5G3により産生される抗体の重鎖可変領域および軽鎖可変領域のアミノ酸配列を有するヒトCD40に対する抗体又はアンタゴニスティックな作用を有するその機能的断片。
- ヒト抗体である、請求項1〜5のいずれかに記載のヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片。
- ヒト抗体である、請求項6〜9のいずれかに記載のヒトCD40に対する抗体又はアンタゴニスティックな作用を有するその機能的断片。
- 請求項1〜5および10のいずれかに記載のヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片を有効成分として含む医薬組成物。
- 請求項6〜9および11のいずれかに記載のヒトCD40に対する抗体又はアンタゴニスティックな作用を有するその機能的断片を有効成分として含む医薬組成物。
- 請求項1〜5および10のいずれかに記載のヒトCD40に対する抗体又はアゴニスティックな作用を有するその機能的断片を有効成分として含む、免疫賦活化剤、抗腫瘍剤又は自己免疫疾患治療剤。
- 請求項6〜9および11のいずれかに記載のヒトCD40に対する抗体又はアンタゴニスティックな作用を有するその機能的断片を有効成分として含む、免疫抑制剤、自己免疫疾患治療剤、アレルギー治療剤又は血液凝固第VIII因子阻害症候群治療剤。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2008275853A JP2009079062A (ja) | 2001-04-27 | 2008-10-27 | 抗cd40モノクローナル抗体 |
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| PCT/US2001/013672 WO2001083755A2 (en) | 2000-04-28 | 2001-04-27 | Human anti-cd40 antibodies and methods of making and using same |
| JP2001142482 | 2001-05-11 | ||
| JP2001310535 | 2001-10-05 | ||
| US10/040,244 US20030059427A1 (en) | 2000-04-28 | 2001-10-26 | Isolation and characterization of highly active anti-CD40 antibody |
| JP2008275853A JP2009079062A (ja) | 2001-04-27 | 2008-10-27 | 抗cd40モノクローナル抗体 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2007193819A Division JP4242437B2 (ja) | 2001-04-27 | 2007-07-25 | 抗cd40モノクローナル抗体 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2009242792A Division JP2010013486A (ja) | 2001-04-27 | 2009-10-21 | 抗cd40モノクローナル抗体 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2009079062A true JP2009079062A (ja) | 2009-04-16 |
Family
ID=30773322
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2008275853A Pending JP2009079062A (ja) | 2001-04-27 | 2008-10-27 | 抗cd40モノクローナル抗体 |
| JP2009242792A Pending JP2010013486A (ja) | 2001-04-27 | 2009-10-21 | 抗cd40モノクローナル抗体 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2009242792A Pending JP2010013486A (ja) | 2001-04-27 | 2009-10-21 | 抗cd40モノクローナル抗体 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US7063845B2 (ja) |
| JP (2) | JP2009079062A (ja) |
| KR (1) | KR100918540B1 (ja) |
| CA (1) | CA2445349C (ja) |
| ES (1) | ES2491265T3 (ja) |
| TW (3) | TW200911837A (ja) |
Families Citing this family (41)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20030059427A1 (en) * | 2000-04-28 | 2003-03-27 | Force Walker R. | Isolation and characterization of highly active anti-CD40 antibody |
| AU2001296507A1 (en) * | 2000-10-02 | 2002-04-15 | Chiron Corporation | Human anti-cd40 antibodies |
| US20020197256A1 (en) * | 2001-04-02 | 2002-12-26 | Genentech, Inc. | Combination therapy |
| EP2011802A3 (en) * | 2001-04-27 | 2009-04-15 | Kyowa Hakko Kirin Co., Ltd. | Anti-CD40 monoclonal antibody |
| AR039067A1 (es) * | 2001-11-09 | 2005-02-09 | Pfizer Prod Inc | Anticuerpos para cd40 |
| US7378492B2 (en) * | 2002-02-20 | 2008-05-27 | Incyte Corporation | CD40-related receptor that binds CD40L |
| US8277810B2 (en) | 2003-11-04 | 2012-10-02 | Novartis Vaccines & Diagnostics, Inc. | Antagonist anti-CD40 antibodies |
| US20050136055A1 (en) * | 2003-12-22 | 2005-06-23 | Pfizer Inc | CD40 antibody formulation and methods |
| US8513008B2 (en) | 2004-10-07 | 2013-08-20 | Argos Therapeutics, Inc. | Mature dendritic cell compositions and methods for culturing same |
| CN101080487B (zh) * | 2004-10-07 | 2012-11-14 | 阿戈斯治疗公司 | 成熟树突细胞组合物及其培养方法 |
| DK1889065T3 (da) * | 2005-05-18 | 2013-09-02 | Xoma Technology Ltd | Metoder til diagnostisering og behandling af sygdomme med en autoimmun- og/eller inflammationskomponent |
| PT3248613T (pt) | 2005-07-18 | 2022-03-16 | Seagen Inc | Conjugados de ligante de fármaco e beta-glucuronida |
| CN101351478A (zh) * | 2005-11-01 | 2009-01-21 | 诺华有限公司 | 抗cd40抗体的应用 |
| WO2007053767A1 (en) * | 2005-11-01 | 2007-05-10 | Novartis Ag | Uses of anti-cd40 antibodies |
| CN101426817B (zh) | 2006-04-21 | 2013-07-10 | 诺华有限公司 | 拮抗性抗-cd40抗体药物组合物 |
| CN101479375B (zh) * | 2006-05-03 | 2016-03-30 | 科罗拉多州立大学董事会 | Cd40激动剂抗体/1型干扰素协同佐剂组合、包含前述的结合物及其作为增强细胞免疫的治疗剂的用途 |
| US20090074711A1 (en) * | 2006-09-07 | 2009-03-19 | University Of Southhampton | Human therapies using chimeric agonistic anti-human cd40 antibody |
| AR083847A1 (es) | 2010-11-15 | 2013-03-27 | Novartis Ag | Variantes de fc (fragmento constante) silenciosas de los anticuerpos anti-cd40 |
| WO2012075111A1 (en) | 2010-11-30 | 2012-06-07 | Novartis Ag | Uses of anti-cd40 antibodies in combination therapy for b cell-related cancers |
| AU2012229236B2 (en) | 2011-03-11 | 2017-05-18 | Beth Israel Deaconess Medical Center, Inc. | Anti-CD40 antibodies and uses thereof |
| EP3508500A1 (en) | 2011-04-29 | 2019-07-10 | Apexigen, Inc. | Anti-cd40 antibodies and methods of use |
| SG11201401518TA (en) | 2011-10-28 | 2014-05-29 | Teva Pharmaceuticals Australia Pty Ltd | Polypeptide constructs and uses thereof |
| KR102270618B1 (ko) | 2012-10-30 | 2021-06-30 | 아펙시젠, 인코포레이티드 | 항-cd40 항체 및 사용 방법 |
| PT2992013T (pt) | 2013-04-29 | 2020-03-05 | Teva Pharmaceuticals Australia Pty Ltd | Anticorpos anti-cd38 e fusões a interferão alfa-2b atenuado |
| US11117975B2 (en) | 2013-04-29 | 2021-09-14 | Teva Pharmaceuticals Australia Pty Ltd | Anti-CD38 antibodies and fusions to attenuated interferon alpha-2B |
| GB201316644D0 (en) * | 2013-09-19 | 2013-11-06 | Kymab Ltd | Expression vector production & High-Throughput cell screening |
| UA119352C2 (uk) | 2014-05-01 | 2019-06-10 | Тева Фармасьютикалз Острейліа Пті Лтд | Комбінація леналідоміду або помалідоміду і конструкції анти-cd38 антитіло-атенуйований інтерферон альфа-2b та спосіб лікування суб'єкта, який має cd38-експресуючу пухлину |
| CA2965414C (en) | 2014-10-29 | 2024-01-09 | Teva Pharmaceuticals Australia Pty Ltd | Interferon .alpha.2.beta. variants |
| EP3831847A1 (en) | 2014-10-29 | 2021-06-09 | Seagen Inc. | Dosage and administration of non-fucosylated anti-cd40 antibodies |
| AU2016315873B2 (en) | 2015-09-04 | 2022-08-18 | Primatope Therapeutics Inc. | Humanized anti-CD40 antibodies and uses thereof |
| MA45488A (fr) | 2015-10-22 | 2018-08-29 | Juno Therapeutics Gmbh | Procédés, kits et appareil de culture de cellules |
| WO2017156349A1 (en) | 2016-03-10 | 2017-09-14 | Cold Genesys, Inc. | Methods of treating solid or lymphatic tumors by combination therapy |
| EA201892040A1 (ru) | 2016-03-25 | 2019-04-30 | Сиэтл Дженетикс, Инк. | Способ получения пегилированных соединений лекарственный препарат - линкер и их промежуточных соединений |
| EP3445783A2 (en) * | 2016-04-18 | 2019-02-27 | Celldex Therapeutics, Inc. | Agonistic antibodies that bind human cd40 and uses thereof |
| CN117717604A (zh) | 2016-07-19 | 2024-03-19 | 梯瓦制药澳大利亚股份有限公司 | 抗cd47联合治疗 |
| CA3056134A1 (en) | 2017-03-24 | 2018-09-27 | Seattle Genetics, Inc. | Process for the preparation of glucuronide drug-linkers and intermediates thereof |
| EP3604384B1 (en) | 2017-03-30 | 2021-09-08 | NOF Corporation | Hydrophilic polymer derivative having self-immolative acetal linker and composite using same |
| JP6990355B2 (ja) | 2017-03-30 | 2022-01-12 | 日油株式会社 | ヘテロ二官能性単分散ポリエチレングリコール及びそれを用いた複合体 |
| TW201842335A (zh) | 2017-04-27 | 2018-12-01 | 德商朱諾醫療公司 | 寡聚粒子試劑及其使用方法 |
| WO2019176875A1 (ja) | 2018-03-13 | 2019-09-19 | 日油株式会社 | 主鎖および側鎖に単分散ポリエチレングリコールを有するヘテロ二官能性化合物 |
| JPWO2021060439A1 (ja) | 2019-09-26 | 2021-04-01 |
Family Cites Families (18)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5247069A (en) | 1986-06-13 | 1993-09-21 | Oncogen | Ligands and methods for augmenting B-cell proliferation |
| EP0434879A1 (en) | 1989-12-14 | 1991-07-03 | Schering-Plough | Method of making factor dependent human B cell lines |
| US6150584A (en) * | 1990-01-12 | 2000-11-21 | Abgenix, Inc. | Human antibodies derived from immunized xenomice |
| US5874082A (en) * | 1992-07-09 | 1999-02-23 | Chiron Corporation | Humanized anti-CD40 monoclonal antibodies and fragments capable of blocking B cell proliferation |
| US5397703A (en) | 1992-07-09 | 1995-03-14 | Cetus Oncology Corporation | Method for generation of antibodies to cell surface molecules |
| DK0945465T3 (da) | 1992-07-09 | 2007-01-15 | Novartis Vaccines & Diagnostic | Antagonistiske monoklonale antistoffer mod humant CD40 |
| DK0724456T3 (da) | 1993-10-01 | 2004-04-13 | Immunex Corp | CD40-Antistoffer |
| DE69637481T2 (de) | 1995-04-27 | 2009-04-09 | Amgen Fremont Inc. | Aus immunisierten Xenomäusen stammende menschliche Antikörper gegen IL-8 |
| AU2466895A (en) | 1995-04-28 | 1996-11-18 | Abgenix, Inc. | Human antibodies derived from immunized xenomice |
| ATE455179T1 (de) | 1997-02-28 | 2010-01-15 | Kyowa Hakko Kirin Co Ltd | Humanes chromosomfragment und ein vektor, der dies enthält |
| US6051228A (en) | 1998-02-19 | 2000-04-18 | Bristol-Myers Squibb Co. | Antibodies against human CD40 |
| IL139601A0 (en) | 1998-05-26 | 2002-02-10 | Genitrix Llc | Compositions and methods of modulating an immune response to an antigen |
| AU4954899A (en) | 1998-06-26 | 2000-01-17 | Trustees Of Dartmouth College | Methods and compositions for modulating antigen-specific immunological (humoral)responses by targeting such antigen to apcs in conjunction with anti-cd40 ligan d administration |
| US6946129B1 (en) | 1999-06-08 | 2005-09-20 | Seattle Genetics, Inc. | Recombinant anti-CD40 antibody and uses thereof |
| DE60035057T2 (de) | 1999-10-04 | 2008-01-31 | Novartis Vaccines and Diagnostics, Inc., Emeryville | CD40 Antagonist zur Behandlung von Psoriasis |
| AU3662101A (en) | 2000-02-01 | 2001-08-14 | Tanox Inc | Cd40-binding apc-activating molecules |
| AU2001259215A1 (en) | 2000-04-28 | 2001-11-12 | La Jolla Institute For Allergy And Immunology | Human anti-cd40 antibodies and methods of making and using same |
| AU2001296507A1 (en) | 2000-10-02 | 2002-04-15 | Chiron Corporation | Human anti-cd40 antibodies |
-
2001
- 2001-04-27 US US09/844,684 patent/US7063845B2/en not_active Expired - Lifetime
-
2002
- 2002-04-26 TW TW097138439A patent/TW200911837A/zh unknown
- 2002-04-26 ES ES08017499.8T patent/ES2491265T3/es not_active Expired - Lifetime
- 2002-04-26 TW TW091108894A patent/TWI327574B/zh not_active IP Right Cessation
- 2002-04-26 TW TW095131602A patent/TWI332510B/zh not_active IP Right Cessation
- 2002-04-26 CA CA2445349A patent/CA2445349C/en not_active Expired - Lifetime
-
2003
- 2003-10-27 KR KR1020037014090A patent/KR100918540B1/ko active IP Right Grant
-
2008
- 2008-10-27 JP JP2008275853A patent/JP2009079062A/ja active Pending
-
2009
- 2009-10-21 JP JP2009242792A patent/JP2010013486A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| US7063845B2 (en) | 2006-06-20 |
| CA2445349A1 (en) | 2002-11-07 |
| TWI332510B (en) | 2010-11-01 |
| US20020142358A1 (en) | 2002-10-03 |
| KR20030096337A (ko) | 2003-12-24 |
| CA2445349C (en) | 2013-08-20 |
| KR100918540B1 (ko) | 2009-09-21 |
| JP2010013486A (ja) | 2010-01-21 |
| TW200716675A (en) | 2007-05-01 |
| ES2491265T3 (es) | 2014-09-05 |
| TWI327574B (ja) | 2010-07-21 |
| TW200911837A (en) | 2009-03-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP3847715B2 (ja) | 抗cd40モノクローナル抗体 | |
| EP2009027B1 (en) | Anti-CD40 monoclonal antibody | |
| JP2009079062A (ja) | 抗cd40モノクローナル抗体 | |
| JP4242437B2 (ja) | 抗cd40モノクローナル抗体 | |
| JP4242388B2 (ja) | 抗cd40抗体の変異体 | |
| CA2447602C (en) | Anti-trail-r antibodies | |
| AU2006203620B2 (en) | Anti-CD40 monoclonal anitbody | |
| KR100937068B1 (ko) | 항-cd40 모노클로날 항체 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Written amendment |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20090213 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20090324 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20090721 |