JP2005296009A - クエン酸シンターゼ活性を有する新規耐熱性タンパク質 - Google Patents
クエン酸シンターゼ活性を有する新規耐熱性タンパク質 Download PDFInfo
- Publication number
- JP2005296009A JP2005296009A JP2005080333A JP2005080333A JP2005296009A JP 2005296009 A JP2005296009 A JP 2005296009A JP 2005080333 A JP2005080333 A JP 2005080333A JP 2005080333 A JP2005080333 A JP 2005080333A JP 2005296009 A JP2005296009 A JP 2005296009A
- Authority
- JP
- Japan
- Prior art keywords
- protein
- citrate synthase
- gene
- present
- amino acid
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Withdrawn
Links
Images




Classifications
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02P—CLIMATE CHANGE MITIGATION TECHNOLOGIES IN THE PRODUCTION OR PROCESSING OF GOODS
- Y02P20/00—Technologies relating to chemical industry
- Y02P20/50—Improvements relating to the production of bulk chemicals
- Y02P20/52—Improvements relating to the production of bulk chemicals using catalysts, e.g. selective catalysts
Landscapes
- Enzymes And Modification Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
【課題】 クエン酸シンターゼ活性を有する新規耐熱性タンパク質を提供する。
【解決手段】 配列番号1のアミノ酸配列からなるタンパク質は、クエン酸シンターゼ活性を有する。このタンパク質は、超好熱性古細菌であって、好気性thermoacidophilic crenarchaeonの1種であるスルホロブス・トコダイイ(Sulfolobus tokodaii)種7(JCM10545)の遺伝子配列から、クエン酸シンターゼ活性を有するタンパク質をコードすると推定される遺伝子(配列番号2)をクローニングし、これを大腸菌を用いて発現させることにより得たものである。図1は、前記タンパク質のSDS−PAGE写真である。
【選択図】 図1
【解決手段】 配列番号1のアミノ酸配列からなるタンパク質は、クエン酸シンターゼ活性を有する。このタンパク質は、超好熱性古細菌であって、好気性thermoacidophilic crenarchaeonの1種であるスルホロブス・トコダイイ(Sulfolobus tokodaii)種7(JCM10545)の遺伝子配列から、クエン酸シンターゼ活性を有するタンパク質をコードすると推定される遺伝子(配列番号2)をクローニングし、これを大腸菌を用いて発現させることにより得たものである。図1は、前記タンパク質のSDS−PAGE写真である。
【選択図】 図1
Description
本発明は、クエン酸シンターゼ活性を有する新規耐熱性タンパク質に関する。なお、本出願は、国の委託の成果に係る出願である。
クエン酸シンターゼは、水およびアセチル補酵素Aの存在下、オキサロ酢酸に作用し、クエン酸と補酵素Aとを生成する反応を触媒する機能を有する酵素であり、クエン酸回路におけるクエン酸生成の最終反応に関与する酵素である。また、クエン酸シンターゼの工業的利用も提案されている(例えば、特許文献1参照)。クエン酸シンターゼとしては、様々の生物由来のものが知られているが、その中で、耐熱性菌由来のものも知られている。前記耐熱性菌としては、超好熱性古細菌(hyperthermophlic archaeum)に属する菌である、サーモプロテウス・テナックス(Thermoproteus tenax)と、スルホロブス・ソルファタリカス(Sulfolobus solfataricus)とがあり、これらの菌由来のクエン酸シンターゼが知られている(例えば、非特許文献1および非特許文献2参照)。このような耐熱性菌由来のクエン酸シンターゼは、工業的用途を広げるものと期待されているが、前記2種類の菌由来のものだけでは、未だ不十分である。
他方、前記の菌以外の超好熱性古細菌(例えば、非特許文献3参照)についての研究があり、スルホロブス属細菌の1種であるスルホロブス・トコダイイ(Sulfolobus tokodaii)(JCM10545)(例えば、非特許文献4参照)は、その遺伝子が既に解析されている(例えば、非特許文献5参照)。したがって、この超好熱古細菌が、クエン酸シンターゼを産生するとすれば、それは優れた耐熱性を有すると予想され、これによって工業的用途さらに広がる可能性がある。
特表2002−508519号公報
Journal of Molecular Biology, 304(4), 657-668, 2000
Biochemistry, 39(9), 2283-2296, 2000
Advances in Protein Chemistry, Volume 48, Enzymes and Proteins from Hyperthermophilic Microorganisms (M.Adams ed.), Academic Press (1996)
Suzuki, T. et al., Extremophiles, 2002 Feb;6(1):39-44
Kawarabayashi,Y. et al., "Complete genome sequence of an aerobic thermoacidophilic crenarchaeon, Sulfolobus tokodaii strain7", DNA Res. 8(4), 123-140 (2001)
本発明は、このような事情に鑑みなされたものであり、クエン酸シンターゼ活性を持つ新規耐熱性タンパク質の提供を、その目的とする。
前記目的を達成するために、超好熱性古細菌であるスルホロブス・トコダイイ(Sulfolobus tokodaii)(JCM10545)のゲノム情報について調べたところ、この細菌が、クエン酸シンターゼを産生する可能性があることを突き止めた。この知見に基づき、さらに研究を重ねたところ、この細菌の遺伝子から、クエン酸シンターゼ活性を持つ新規耐熱性タンパク質を発現させることに成功し、本発明に到達した。
すなわち、本発明のタンパク質は、下記の(a)または(b)のタンパク質である。
(a) 配列番号1のアミノ酸配列からなる耐熱性タンパク質。
(b) 配列番号1のアミノ酸配列において、1つ若しくは数個のアミノ酸残基が、欠失、置換、付加若しくは挿入されたアミノ酸配列からなり、クエン酸シンターゼ活性を有する耐熱性タンパク質。
(a) 配列番号1のアミノ酸配列からなる耐熱性タンパク質。
(b) 配列番号1のアミノ酸配列において、1つ若しくは数個のアミノ酸残基が、欠失、置換、付加若しくは挿入されたアミノ酸配列からなり、クエン酸シンターゼ活性を有する耐熱性タンパク質。
なお、スルホロブス・トコダイイ(Sulfolobus tokodaii)(JCM10545)は、理化学研究所生物基盤研究部微生物系統保存施設に保存されており、第三者の要求により分譲可能である。
スルホロブス・トコダイイ(Sulfolobus tokodaii)(JCM10545)の生育温度は80℃であり、生育限界温度が87℃であるから、本発明のタンパク質は、80〜87℃の高温であっても活性がある。
本発明のタンパク質の酵素反応の温度条件は、好ましくは、55〜95℃の範囲であり、より好ましくは、65〜85℃の範囲である。
前記クエン酸シンターゼ活性は、水およびアセチル補酵素Aの存在下、オキサロ酢酸に作用し、クエン酸と補酵素Aとを生成する機能である。
前述のように、本発明の新規耐熱性タンパク質は、超好熱性古細菌由来であり、具体的には、スルホロブス・トコダイイ(Sulfolobus tokodaii)(JCM10545)由来である。但し、本発明のタンパク質は、この菌が産生するものに限定されず、遺伝子工学的手法により、他の生物が産生するものであってもよい。
つぎに、本発明の発現ベクターは、前記本発明のタンパク質をコードするDNAまたは配列番号2に記載のDNAを含むベクターである。
つぎに、本発明の形質転換体は、前記本発明のベクターにより形質転換された形質転換体である。なお、宿主は特に制限されず、例えば、大腸菌等がある。
つぎに、本発明のタンパク質の製造方法は、前記本発明の形質転換体を培養する工程と、前記培養工程において発現した前記タンパク質を回収する工程とを含む製造方法である。
つぎに、本発明の製造方法は、酵素反応により、オキサロ酢酸、アセチル補酵素Aおよび水からクエン酸および補酵素Aを製造する方法であって、前記酵素として前記本発明のタンパク質を用い、温度55〜95℃の条件で前記酵素反応を行う製造方法である。なお、前記温度条件は、65〜85℃の範囲であることがより好ましい。このように、前記本発明のタンパク質を用いれば、温度55〜95℃の高温領域で酵素反応を実施でき、この結果、工業的な用途が広がる。なお、この製造方法において、前記酵素反応のpHは、pH7以上の範囲が好ましい。
以下、本発明について、さらに詳細に説明する。
本発明者らは、海洋底から採取された超好熱性古細菌であって、好気性thermoacidophilic crenarchaeonの1種であるスルホロブス・トコダイイ(Sulfolobus tokodaii)種7(JCM10545)の遺伝子配列からクエン酸シンターゼ活性を示すと推定される遺伝子(配列番号2)をクローニングし、これを大腸菌を用いて発現させることにより、本発明の新規耐熱性タンパク質を得るに至った。遺伝子のクローニング方法は、後記した実施例1に記載した通り実施した。クローニングされた遺伝子の塩基配列は配列番号2に示す通りとおりであり、また、その推定アミノ酸配列は配列番号1に示す通りである。なお、本発明の耐熱性タンパク質は、クエン酸シンターゼ活性を有していれば、配列番号1のアミノ酸配列において、一つ以上若しくは数個のアミノ酸残基が、欠質、置換、付加若しくは挿入されていてもよい。このアミノ酸配列における「アミノ酸の欠失、置換、付加若しくは挿入」は、当業者に公知の方法(例えば、突然変異誘発法)に従って実施することができる。
本発明のタンパク質は、前述の本発明のタンパク質の製造方法により製造可能であるが、これに限定されず、他の製造方法で製造されてもよい。例えば配列番号1に示すように、そのアミノ酸配列が決定されているタンパク質については、その配列を元に当業者に公知の手法、例えば、個々のアミノ酸を化学的に重合してタンパク質を合成する方法に従って調製することができる。
本発明のタンパク質をコードする遺伝子の一例としては、配列番号2に示す遺伝子がある。前記遺伝子は、例えば、後記する実施例2に示すように超好熱性古細菌スルホロブス・トコダイイ(Sulfolobus tokodaii)(JCM10545)のゲノムから、例えば配列番号2で示される塩基配列の一部をプライマーとして用いるPCR法あるいは該DNA断片をプローブとして用いるハイブリダイゼーション法により調製することができる。また、その塩基配列をもとに、当業者に公知である核酸化学合成法等に従って前記遺伝子を得ることもできるが、これらに限定されない。
本発明の発現ベクターは、前記遺伝子もしくは配列番号2のDNAを適当なベクターに挿入することによって得ることができる。本発明の遺伝子を挿入するためのベクターは、宿主中で複製可能なものであれば、特に制限されるものではなく、例えば、プラスミドDNA、ファージDNA、AcMNPVなどのバキュロウイルスなどが挙げられる。プラスミドDNAは、大腸菌やアグロバクテリウムからアルカリ抽出法またはその変法などにより調製することができる。また、市販プラスミドとして、例えばpET-11a(Novagen社製)あるいはバチルス属の宿主を用いた分泌型のプラスミドなどを用いてもよい。これらのプラスミドは、アンピシリン耐性遺伝子、カナマイシン耐性遺伝子、クロラムフェニコール耐性遺伝子などが含まれていてもよい。
ベクターへの遺伝子等の挿入は、例えば、精製された遺伝子の塩基配列を適当な制限酵素で切断し、適当なベクターDNAの制限酵素部位またはマルチクローニングサイトに挿入してベクターに連結する方法などを用いることができるが、これらに限定されない。また、本発明の遺伝子の機能が発揮されるように、本発明の発現ベクターには本発明の遺伝子のほか、プロモーター、ターミネーター、リボソーム結合配列などを組み込んでいてもよい。さらに、本発明の遺伝子も他のタンパク質のコードする配列を融合したものを挿入してもよい。
前記発現ベクターで宿主生物を形質転換すれば、本発明の形質転換体が得られる。宿主生物としては、本発明の遺伝子を発現できるものであれば、特に制限されるものではなく、例えば、大腸菌などの原核細胞が挙げられるが、これらに限定されない。形質転換法としては、既に公知である塩化カルシウム法などを使用することができるが、これらの方法に限定されない。
本発明のタンパク質の製造方法は、前記形質転換体を培養する工程と、前記培養工程において発現した前記タンパク質を回収する工程とを含む製造方法である。前記培養する方法は、宿主細胞の培養に用いられる通常の方法に従って行われる。大腸菌等の微生物を宿主とした形質転換体を培養する培地としては、微生物が資化し得る炭素源、窒素源、無機塩類などを含有し、形質転換体の培養を効率的に行えるものであれば、天然培地、合成培地などのいずれを用いてもよい。本発明のタンパク質の回収は、特に制限されない。前記タンパク質が菌体内または細胞内に生産される場合には、菌体または細胞を破砕することによって前記タンパク質を回収する。また、本発明の前記タンパク質が菌体外または細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離などにより菌体または細胞を除去した後、タンパク質の単離精製に用いられる一般的な生化学的方法、例えば、硫酸アンモニウム沈殿、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィーなどを単独でまたは適宜組み合わせて用いることにより、培養物中から本発明のタンパク質を単離精製することができる。なお、培養液をそのまま使用する場合、熱処理をすることにより、他のタンパク質が失活するので、実質上、本発明のタンパク質のみの酵素液として使用できる。
以下に実施例により本発明をさらに詳細に説明するが、本発明はこれらにより限定されない。
染色体DNAの調製
スルホロブス・トコダイイ(Sulfolobus tokodaii(JCM10545))をL培地中で37℃にて一晩培養して集菌したものに、SSC溶液(0.15M NaCl、0.015M クエン酸ナトリウム)10mL、0.5M EDTA0.2mL、100mg/mL ニワトリ卵白リゾチーム0.1mLおよび10%非イオン性界面活性剤Brij-58を0.5mL加え、0℃で30分間放置した後、プロテイナーゼK(Merck社製)5mgを10%SDS0.2mLに溶かした溶液を加え、37℃で2、3日間放置した。この溶液に水飽和フェノール、クロロホルム、イソアミルアルコールの混合溶液を加えて、37℃で1時間放置した後、水層を分取し、そこへエタノールを加えてDNAを沈殿濃縮した。このDNAの沈殿をTE溶液(10mM Tris−HCl(pH7.5)、1mM EDTA(pH8.0))10mLに溶解し、リボヌクレアーゼ0.25mL(最終濃度0.25mg/mL)を加えて、37℃で一晩放置した後、エタノールで沈殿させた。次いで、DNAをTE溶液5mLに溶解した後、260nmの吸光度より、DNA濃度を決定した(Clarke,L. & Carbon,J. (1979) Methods Enzymol. 68, 396-408)。
スルホロブス・トコダイイ(Sulfolobus tokodaii(JCM10545))をL培地中で37℃にて一晩培養して集菌したものに、SSC溶液(0.15M NaCl、0.015M クエン酸ナトリウム)10mL、0.5M EDTA0.2mL、100mg/mL ニワトリ卵白リゾチーム0.1mLおよび10%非イオン性界面活性剤Brij-58を0.5mL加え、0℃で30分間放置した後、プロテイナーゼK(Merck社製)5mgを10%SDS0.2mLに溶かした溶液を加え、37℃で2、3日間放置した。この溶液に水飽和フェノール、クロロホルム、イソアミルアルコールの混合溶液を加えて、37℃で1時間放置した後、水層を分取し、そこへエタノールを加えてDNAを沈殿濃縮した。このDNAの沈殿をTE溶液(10mM Tris−HCl(pH7.5)、1mM EDTA(pH8.0))10mLに溶解し、リボヌクレアーゼ0.25mL(最終濃度0.25mg/mL)を加えて、37℃で一晩放置した後、エタノールで沈殿させた。次いで、DNAをTE溶液5mLに溶解した後、260nmの吸光度より、DNA濃度を決定した(Clarke,L. & Carbon,J. (1979) Methods Enzymol. 68, 396-408)。
発現プラスミドの構築と遺伝子発現
1.発現プラスミドの構築
耐熱性クエン酸シンターゼ遺伝子の翻訳領域の前後に制限酵素NdeIおよびBamHI、NotIサイトを含むDNAを構築する目的で下記のDNAプライマーを合成し、このプライマーを用いたPCRで耐熱性クエン酸シンターゼ遺伝子の翻訳領域の前後に制限酵素サイトを導入した。用いたDNAポリメラーゼはKOD Dash(東洋紡社製)であった。
Forward primer(配列番号3):5'-ATATCATATGGAAATCAAAAAAGGGCTTGAAGACGTTTAT-3'
Reverse primer(配列番号4):5'-ATATGGATCCGCGGCCGCTTATTACCTACTATCAATTGGAAT-3'
1.発現プラスミドの構築
耐熱性クエン酸シンターゼ遺伝子の翻訳領域の前後に制限酵素NdeIおよびBamHI、NotIサイトを含むDNAを構築する目的で下記のDNAプライマーを合成し、このプライマーを用いたPCRで耐熱性クエン酸シンターゼ遺伝子の翻訳領域の前後に制限酵素サイトを導入した。用いたDNAポリメラーゼはKOD Dash(東洋紡社製)であった。
Forward primer(配列番号3):5'-ATATCATATGGAAATCAAAAAAGGGCTTGAAGACGTTTAT-3'
Reverse primer(配列番号4):5'-ATATGGATCCGCGGCCGCTTATTACCTACTATCAATTGGAAT-3'
PCR反応後、Ex Taq(宝酒造社製)を用いて増幅断片の3’末端側にデオキシアデノシンを付加した後、pGEM-T Easy Vector(Promega社製)と、T4リガーゼで15℃、30分間反応させ連結した。連結したDNAを大腸菌DH5αのコンピテントセルに導入し、形質転換体のコロニーを得た。得られた形質転換体をアンピシリンを含むLB培地(18mL)で24時間培養し、その培養液からプラスミドを改変アルカリSDS法で精製した。プラスミド中に期待される大きさのインサートが存在することを、アガロース電気泳動で確認した。精製プラスミドのインサートの塩基配列は、BigDye Terminator kit(登録商標:Applied Biosystems社製)とABI PRISM 3700 DNA Analyzer(登録商標:Applied Biosystems社製)を用いて決定し、インサートの塩基配列が、耐熱性クエン酸シンターゼ遺伝子の正しい配列であることを確認した。正しい配列を有するプラスミドの一部を制限酵素NdeIとBamHIで完全分解(37℃で2時間)した後、アガロース電気泳動により、耐熱性クエン酸シンターゼ遺伝子を精製した。pET-11a(Novagen社製)を制限酵素NdeIとBamHIで切断・精製した後、上記の構造遺伝子とT4リガーゼで反応させ連結した。連結したDNAの一部を大腸菌DH5αのコンピテントセルに導入し、アンピシリンを含むLB寒天プレートに適量まき、37℃で一晩培養し、形質転換体のコロニーを得た。得られた形質転換体をアンピシリンを含むLB培地(18mL)で24時間培養し、その培養液から発現プラスミドを改変アルカリSDS法で精製した。
2.組換え遺伝子の発現
大腸菌Rosetta-gami(DE3)(Novagen社製)のコンピテントセルを融解して、ファルコンチューブに0.1mL移した。その中に上記1.の精製発現プラスミドの溶液0.002mLを加え氷中に20分間放置した後、42℃でヒートショックを90秒間行い、氷中に1分間放置した後、クロラムフェニコールとアンピシリンを含むLB寒天プレートに適量まき、37℃で一晩培養し、形質転換体を得た。得られた形質転換体をアンピシリンを含むLB培地(5mL)で18時間培養し、耐熱性クエン酸シンターゼ遺伝子を発現した。培養後、遠心分離(13,000G、10分)で集菌した。集菌した菌体に、破砕液(20mM Tris−HCl、100mM KCl、pH7.5)を0.2mL加え、超音波発生器で細胞を破砕し、その懸濁液を0.1mLずつ2本のサンプルチューブに分けた。一方のサンプルチューブは遠心分離(13,000G、10分)して上清と沈殿に分け、沈殿は破砕液0.1mLで懸濁した。もう一方のサンプルチューブは、熱処理(70℃、10分)を施した後、遠心分離(13,000G、10分)して上清と沈殿に分け、沈殿は破砕液0.1mLで懸濁した。これらの試料の一部をSDS−ポリアクリルアミドゲル電気泳動(PAGE)で分析し、発現を確認できた。この結果を、図1のSDS−PAGE写真に示す。
耐熱性クエン酸シンターゼの発現が見られた試料についてSDSポリアクリルアミドゲル電気泳動を行った後、エレクトロブロッティングによりPVDF膜に転写し、染色によって可視化された目的組換えタンパク質である耐熱性クエン酸シンターゼのバンドを切り出し、プロテインシーケンサーModel492Procise(Applied Biosystems社製)を用いて、アミノ末端配列を解析した結果、配列番号5に示すように7残基のアミノ末端配列が決定できた。この配列により、耐熱性クエン酸シンターゼであることを確認できた。このタンパク質は、373アミノ酸残基より構成されており、その推定分子量は43.1kDであり、図1の結果とほぼ一致した。
大腸菌Rosetta-gami(DE3)(Novagen社製)のコンピテントセルを融解して、ファルコンチューブに0.1mL移した。その中に上記1.の精製発現プラスミドの溶液0.002mLを加え氷中に20分間放置した後、42℃でヒートショックを90秒間行い、氷中に1分間放置した後、クロラムフェニコールとアンピシリンを含むLB寒天プレートに適量まき、37℃で一晩培養し、形質転換体を得た。得られた形質転換体をアンピシリンを含むLB培地(5mL)で18時間培養し、耐熱性クエン酸シンターゼ遺伝子を発現した。培養後、遠心分離(13,000G、10分)で集菌した。集菌した菌体に、破砕液(20mM Tris−HCl、100mM KCl、pH7.5)を0.2mL加え、超音波発生器で細胞を破砕し、その懸濁液を0.1mLずつ2本のサンプルチューブに分けた。一方のサンプルチューブは遠心分離(13,000G、10分)して上清と沈殿に分け、沈殿は破砕液0.1mLで懸濁した。もう一方のサンプルチューブは、熱処理(70℃、10分)を施した後、遠心分離(13,000G、10分)して上清と沈殿に分け、沈殿は破砕液0.1mLで懸濁した。これらの試料の一部をSDS−ポリアクリルアミドゲル電気泳動(PAGE)で分析し、発現を確認できた。この結果を、図1のSDS−PAGE写真に示す。
耐熱性クエン酸シンターゼの発現が見られた試料についてSDSポリアクリルアミドゲル電気泳動を行った後、エレクトロブロッティングによりPVDF膜に転写し、染色によって可視化された目的組換えタンパク質である耐熱性クエン酸シンターゼのバンドを切り出し、プロテインシーケンサーModel492Procise(Applied Biosystems社製)を用いて、アミノ末端配列を解析した結果、配列番号5に示すように7残基のアミノ末端配列が決定できた。この配列により、耐熱性クエン酸シンターゼであることを確認できた。このタンパク質は、373アミノ酸残基より構成されており、その推定分子量は43.1kDであり、図1の結果とほぼ一致した。
組換え大腸菌の大量培養
実施例2と同様にして調製したプラスミドDNA(pET−11aベクター)を用いて、大腸菌DH5α株を常法に従い形質転換した。形質転換された大腸菌DH5αからアルカリSDS法を用いてプラスミドDNAを抽出した。このプラスミドDNAを用いて、大腸菌ロゼッタ・ガミ(Rosetta−Gami)(DE3)株を形質転換した。プレート上に生えてきたコロニーを3白金耳量とり、5mLのLBL培地(1%ペプトン、0.5%酵母抽出液、0.5%NaCl、0.1%ラクトース、50μg/mLアンピシリン、40μg/mLクロラムフェニコール)に植菌して、培養開始直前まで約6時間、37℃で前培養した。この前培養液を全量、3Lの4×LBL培地(4%ペプトン、2%酵母抽出液、 2%NaCl、50μg/mLアンピシリン)に加え、高密度培養槽(ABLE社製)にて37℃、pH7.2、圧力0.02Paでコンピュータプログラム制御し、培養した。pHは、オートクレーブ済みの2M HCl(和光純薬社製)および2M NaOH(和光純薬社製)で調整した。集菌約19時間前にオートクレーブ済みの300mL発現誘導液(10%ラクトース、20%グリセロール)を加えた。集菌約19時間前に培養温度を30℃とし、以降は30℃とした。大腸菌の生育度が定常期に入ったところ(培養開始43時間後)で大型遠心分離機(Beckman社製、商品名AvantiHP−30I)を用い集菌した。回収した菌体は、−30℃で保存した。この時、菌体を少量、別に取り、150mM NaCl、20mM Tris−HCl(pH8)、5mM β−メルカプトエタノール(和光純薬社製)に溶解・懸濁し、超音波破砕装置(TOMY社製、商品名UD−201)で破砕した。この溶液を2等分し、一方を9100G、4℃で10分間、遠心分離し、上清と沈殿に分け、他方を75℃に設定した恒温槽(TAITEC社製、商品名DryThermoUnit DTU−1C)で10分間、加熱した後、9100G、4℃で10分間、遠心分離し、上清と沈殿に分けた。これら4種の上清および沈殿(沈殿は、菌体破砕液にて再懸濁)に変性剤(62.5mM Tris−HCl(pH6.8)、10%グリセロール、2%SDS、2.4%β−メルカプトエタノール、0.005%ブロムフェノールブルー(和光純薬社製))を加え、95℃で5分間加熱し、変性させた。これらの変性させたタンパク質溶液を12.5%または15%(発現させるタンパク質の分子量により異なる)ポリアクリルアミドゲルに加え、SDS−PAGEにて電気泳動を行った。染色液(和光純薬社製、商品名Quick−CBB)を用い、電気泳動後のゲルを染色・脱色し、目的タンパク質の発現を確認した。
実施例2と同様にして調製したプラスミドDNA(pET−11aベクター)を用いて、大腸菌DH5α株を常法に従い形質転換した。形質転換された大腸菌DH5αからアルカリSDS法を用いてプラスミドDNAを抽出した。このプラスミドDNAを用いて、大腸菌ロゼッタ・ガミ(Rosetta−Gami)(DE3)株を形質転換した。プレート上に生えてきたコロニーを3白金耳量とり、5mLのLBL培地(1%ペプトン、0.5%酵母抽出液、0.5%NaCl、0.1%ラクトース、50μg/mLアンピシリン、40μg/mLクロラムフェニコール)に植菌して、培養開始直前まで約6時間、37℃で前培養した。この前培養液を全量、3Lの4×LBL培地(4%ペプトン、2%酵母抽出液、 2%NaCl、50μg/mLアンピシリン)に加え、高密度培養槽(ABLE社製)にて37℃、pH7.2、圧力0.02Paでコンピュータプログラム制御し、培養した。pHは、オートクレーブ済みの2M HCl(和光純薬社製)および2M NaOH(和光純薬社製)で調整した。集菌約19時間前にオートクレーブ済みの300mL発現誘導液(10%ラクトース、20%グリセロール)を加えた。集菌約19時間前に培養温度を30℃とし、以降は30℃とした。大腸菌の生育度が定常期に入ったところ(培養開始43時間後)で大型遠心分離機(Beckman社製、商品名AvantiHP−30I)を用い集菌した。回収した菌体は、−30℃で保存した。この時、菌体を少量、別に取り、150mM NaCl、20mM Tris−HCl(pH8)、5mM β−メルカプトエタノール(和光純薬社製)に溶解・懸濁し、超音波破砕装置(TOMY社製、商品名UD−201)で破砕した。この溶液を2等分し、一方を9100G、4℃で10分間、遠心分離し、上清と沈殿に分け、他方を75℃に設定した恒温槽(TAITEC社製、商品名DryThermoUnit DTU−1C)で10分間、加熱した後、9100G、4℃で10分間、遠心分離し、上清と沈殿に分けた。これら4種の上清および沈殿(沈殿は、菌体破砕液にて再懸濁)に変性剤(62.5mM Tris−HCl(pH6.8)、10%グリセロール、2%SDS、2.4%β−メルカプトエタノール、0.005%ブロムフェノールブルー(和光純薬社製))を加え、95℃で5分間加熱し、変性させた。これらの変性させたタンパク質溶液を12.5%または15%(発現させるタンパク質の分子量により異なる)ポリアクリルアミドゲルに加え、SDS−PAGEにて電気泳動を行った。染色液(和光純薬社製、商品名Quick−CBB)を用い、電気泳動後のゲルを染色・脱色し、目的タンパク質の発現を確認した。
組換えタンパク質の精製
実施例3において、−30℃で保存してあった菌体を、150mM NaCl、20mM Tris−HCl(pH8)、5mM β−メルカプトエタノールに溶解・懸濁し、超音波破砕装置(TOMY社製、商品名UD−201)で破砕し、75℃に設定した恒温槽(TAITEC社製、商品名DryThermoUnit DTU−1C)で、10分間、加熱した後、すばやく冷却した。次に、この破砕菌体液を大型遠心分離機(Beckman社製、商品名Avanti HP−30I)を用いて、100,000Gで1時間、遠心分離し、上清を回収した。
つぎに、この上清をタンパク質精製装置(Amersham Biosciences社製、AKTATM explorer)を用いて、1.2M 硫安、20mM Tris−HCl(pH8.0)、5mM β−メルカプトエタノールの緩衝溶液に置換した後、疎水交換カラム(Amersham Biosciences社製、RESOURCETMPhe 6mL)に通した。1.2M→0の硫安濃度勾配で溶出させ、各画分をSDS−PAGEにて確認し、目的タンパク質の画分を回収した。
つぎに、回収した画分を、20mM MES(2−morpholinoethanesulfonic acid)(pH6.0)、5mM β−メルカプトエタノールの緩衝溶液に置換した後、陽イオン交換カラム(Amersham Biosciences社製、RESOURCETM S 6mL)に通した。塩化ナトリウムで溶出を行い、溶出してきた各画分をSDS−PAGEにて確認し、目的タンパク質の画分を回収した。
つぎに、回収した画分を、10mMのリン酸緩衝溶液に置換した後、ヒドロキシアパタイトカラム(Bioscale社製、商品名Ceramic Hydroxyapatite、Type I Column、CHT−101)に通した。高濃度リン酸緩衝溶液で溶出を行い、溶出してきた各画分をSDS−PAGE電気泳動にて確認し、目的タンパクの画分を回収した。
つぎに、回収した画分を、遠心濃縮チューブ(ミリポア社製、VIVASPINTM 10000)を用いて遠心分離して濃縮し、タンパク質精製装置(GILSON社製)を用いて、20mM Tris−HCl(pH9.0)、5mM β−メルカプトエタノール、150mM NaClで平衡化したゲルろ過カラム(Amersham Biosciences社製、HiLoadTM 16/60、SuperdexTM 200)に通した。溶出してきた各画分をSDS−PAGE電気泳動にて確認し、目的タンパク質の画分を回収した。
つぎに、回収した画分を遠心濃縮チューブ(ミリポア社製、VIVASPINTM 10000)を用いて遠心分離して濃縮し、活性測定に供した。
実施例3において、−30℃で保存してあった菌体を、150mM NaCl、20mM Tris−HCl(pH8)、5mM β−メルカプトエタノールに溶解・懸濁し、超音波破砕装置(TOMY社製、商品名UD−201)で破砕し、75℃に設定した恒温槽(TAITEC社製、商品名DryThermoUnit DTU−1C)で、10分間、加熱した後、すばやく冷却した。次に、この破砕菌体液を大型遠心分離機(Beckman社製、商品名Avanti HP−30I)を用いて、100,000Gで1時間、遠心分離し、上清を回収した。
つぎに、この上清をタンパク質精製装置(Amersham Biosciences社製、AKTATM explorer)を用いて、1.2M 硫安、20mM Tris−HCl(pH8.0)、5mM β−メルカプトエタノールの緩衝溶液に置換した後、疎水交換カラム(Amersham Biosciences社製、RESOURCETMPhe 6mL)に通した。1.2M→0の硫安濃度勾配で溶出させ、各画分をSDS−PAGEにて確認し、目的タンパク質の画分を回収した。
つぎに、回収した画分を、20mM MES(2−morpholinoethanesulfonic acid)(pH6.0)、5mM β−メルカプトエタノールの緩衝溶液に置換した後、陽イオン交換カラム(Amersham Biosciences社製、RESOURCETM S 6mL)に通した。塩化ナトリウムで溶出を行い、溶出してきた各画分をSDS−PAGEにて確認し、目的タンパク質の画分を回収した。
つぎに、回収した画分を、10mMのリン酸緩衝溶液に置換した後、ヒドロキシアパタイトカラム(Bioscale社製、商品名Ceramic Hydroxyapatite、Type I Column、CHT−101)に通した。高濃度リン酸緩衝溶液で溶出を行い、溶出してきた各画分をSDS−PAGE電気泳動にて確認し、目的タンパクの画分を回収した。
つぎに、回収した画分を、遠心濃縮チューブ(ミリポア社製、VIVASPINTM 10000)を用いて遠心分離して濃縮し、タンパク質精製装置(GILSON社製)を用いて、20mM Tris−HCl(pH9.0)、5mM β−メルカプトエタノール、150mM NaClで平衡化したゲルろ過カラム(Amersham Biosciences社製、HiLoadTM 16/60、SuperdexTM 200)に通した。溶出してきた各画分をSDS−PAGE電気泳動にて確認し、目的タンパク質の画分を回収した。
つぎに、回収した画分を遠心濃縮チューブ(ミリポア社製、VIVASPINTM 10000)を用いて遠心分離して濃縮し、活性測定に供した。
活性測定
1.測定方法
実施例4において精製したタンパク質は、クエン酸シンターゼであり、クエン酸シンターゼは、生体内で、オキサロ酢酸+アセチル補酵素A+H2O→クエン酸+補酵素A+H+の反応を触媒する酵素である。前記実施例4において精製したタンパク質について、下記の種々条件下で、その活性を測定した。この活性測定は、つぎの通りである。すなわち、100mMのKCl(和光純薬社製)と、2mMのEDTA(pH8.0、ナカライテスク社製)と、0.2mMのオキサロ酢酸(ナカライテスク社製)と、0.2mMのアセチル補酵素A(SIGMA社製)と、9.32nMの前記実施例4において精製したタンパク質とを含む50mMの緩衝溶液(後述のとおり、各測定によって種類が異なる)を3分間反応させた。この反応系に0.1mMのDTNB(5,5'−ジチオビス−2−ニトロ安息香酸(和光純薬社製))を共存させておき、これと反応生成物である補酵素Aとが反応してできる化合物の412nmでの吸光度(モル吸光係数13,600M-1cm-1)を定量することにより、反応の進行度(前記実施例4において精製したタンパク質の活性に対応する)を観測した。進行度の指標として見かけの速度定数kappを用いた。各測定値は、前記実施例4において精製したタンパク質を加えず、緩衝溶液に50mM Tris−HCl(25℃でのpH9.0)、150mM NaClを用いて前記測定を行ったコントロール実験結果を差し引いた値である。なお、分光光度計には、HITACHI社製、商品名U−3300を、インキュベータには、NESLAB社製、商品名RTE−111を用いた。
1.測定方法
実施例4において精製したタンパク質は、クエン酸シンターゼであり、クエン酸シンターゼは、生体内で、オキサロ酢酸+アセチル補酵素A+H2O→クエン酸+補酵素A+H+の反応を触媒する酵素である。前記実施例4において精製したタンパク質について、下記の種々条件下で、その活性を測定した。この活性測定は、つぎの通りである。すなわち、100mMのKCl(和光純薬社製)と、2mMのEDTA(pH8.0、ナカライテスク社製)と、0.2mMのオキサロ酢酸(ナカライテスク社製)と、0.2mMのアセチル補酵素A(SIGMA社製)と、9.32nMの前記実施例4において精製したタンパク質とを含む50mMの緩衝溶液(後述のとおり、各測定によって種類が異なる)を3分間反応させた。この反応系に0.1mMのDTNB(5,5'−ジチオビス−2−ニトロ安息香酸(和光純薬社製))を共存させておき、これと反応生成物である補酵素Aとが反応してできる化合物の412nmでの吸光度(モル吸光係数13,600M-1cm-1)を定量することにより、反応の進行度(前記実施例4において精製したタンパク質の活性に対応する)を観測した。進行度の指標として見かけの速度定数kappを用いた。各測定値は、前記実施例4において精製したタンパク質を加えず、緩衝溶液に50mM Tris−HCl(25℃でのpH9.0)、150mM NaClを用いて前記測定を行ったコントロール実験結果を差し引いた値である。なお、分光光度計には、HITACHI社製、商品名U−3300を、インキュベータには、NESLAB社製、商品名RTE−111を用いた。
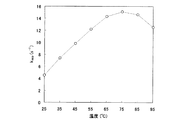
2.至適温度
25、35、45、55、65、75、85、95℃の各温度において、前記測定を行い、kappを求めた。緩衝溶液には、各温度でpHが7.5になるように調製したTris−HClを用いた。測定結果を、下記表1および図2のグラフに示す。図示のとおり、65〜85℃において特に高い活性を示した。
25、35、45、55、65、75、85、95℃の各温度において、前記測定を行い、kappを求めた。緩衝溶液には、各温度でpHが7.5になるように調製したTris−HClを用いた。測定結果を、下記表1および図2のグラフに示す。図示のとおり、65〜85℃において特に高い活性を示した。
(表1)
温度(℃) k app (s -1 )
25 4.54
35 7.37
45 9.79
55 12.2
65 14.3
75 15.1
85 14.6
95 12.5
温度(℃) k app (s -1 )
25 4.54
35 7.37
45 9.79
55 12.2
65 14.3
75 15.1
85 14.6
95 12.5
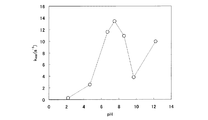
3.至適pH
pH2.2、4.7、6.7、7.5、8.6、9.7、12.2の各条件で、前記測定を行い、kappを求めた。反応温度は、55℃とした。測定に用いた緩衝溶液と目的のpHとの関係を下記表2に示す。
pH2.2、4.7、6.7、7.5、8.6、9.7、12.2の各条件で、前記測定を行い、kappを求めた。反応温度は、55℃とした。測定に用いた緩衝溶液と目的のpHとの関係を下記表2に示す。
(表2)
pH 緩衝溶液
2.2 Gly−HCl
4.7 MES−NaOH
6.7 MES−NaOH
7.5 Tris−HCl
8.6 Tris−HCl
9.7 Gly−NaOH
12.2 KCl−NaOH
pH 緩衝溶液
2.2 Gly−HCl
4.7 MES−NaOH
6.7 MES−NaOH
7.5 Tris−HCl
8.6 Tris−HCl
9.7 Gly−NaOH
12.2 KCl−NaOH
前記kappの測定結果を、下記表3および図3のグラフに示す。図示のとおり、pH9.7付近で活性の低下が見られるが、これは基質の自然分解が原因と考えられた。この点を除外して考えると、高いpH領域(pH7以上)において強い活性を示していることが分かる。
(表3)
pH k app (s -1 )
2.2 0.263
4.7 2.54
6.7 11.6
7.5 13.4
8.6 10.9
9.7 3.80
12.2 9.95
pH k app (s -1 )
2.2 0.263
4.7 2.54
6.7 11.6
7.5 13.4
8.6 10.9
9.7 3.80
12.2 9.95
4.熱安定性
前記測定に先立ち、前記実施例4において精製したタンパク質を、95℃で、3時間および6時間前処理した。前記前処理物を用いて、前記測定を行い、kappを求めた。反応温度は、55℃、pH7.5とした。なお、前処理時間0時間は、前処理を行っていないものである。測定結果を、下記表4に示す。
前記測定に先立ち、前記実施例4において精製したタンパク質を、95℃で、3時間および6時間前処理した。前記前処理物を用いて、前記測定を行い、kappを求めた。反応温度は、55℃、pH7.5とした。なお、前処理時間0時間は、前処理を行っていないものである。測定結果を、下記表4に示す。
(表4)
前処理時間(時間) k app (s -1 )
0 13.4
3 2.37
6 1.64
前処理時間(時間) k app (s -1 )
0 13.4
3 2.37
6 1.64
前処理時間0時間のkappを基準(100%)として各前処理時間での残存活性を求めた。測定結果を、図4のグラフに示す。図示のとおり、前処理を行うことで、活性の低下は見られるものの、完全に活性を失うことはないことが分かった。
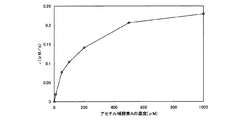
5.アセチル補酵素Aに対するk cat 、K M 、k cat /K M の算出
前記測定において、オキサロ酢酸の濃度は、0.2mMのまま固定して、アセチル補酵素Aの濃度を変えることで、アセチル補酵素Aの濃度と反応速度vとの関係を求めた。反応温度は、75℃、pH7.5とした。測定結果を、下記表5および図5のグラフに示す。
前記測定において、オキサロ酢酸の濃度は、0.2mMのまま固定して、アセチル補酵素Aの濃度を変えることで、アセチル補酵素Aの濃度と反応速度vとの関係を求めた。反応温度は、75℃、pH7.5とした。測定結果を、下記表5および図5のグラフに示す。
(表5)
アセチル補酵素Aの濃度(μM) v(μM/s)
0 0
10 0.0118
50 0.0772
100 0.104
200 0.141
500 0.206
1000 0.229
アセチル補酵素Aの濃度(μM) v(μM/s)
0 0
10 0.0118
50 0.0772
100 0.104
200 0.141
500 0.206
1000 0.229
上記測定結果より、IGOR Pro(ヒューリンクス社製)を用いて、アセチル補酵素Aに対するkcat、KM、kcat/KMを算出した。算出結果を、下記表6に示す。
(表6)
k cat (s -1 ) K M (μM) k cat /K M (s -1 /M)
28.0(58.4) 146 1.92×105(4.00×105)
k cat (s -1 ) K M (μM) k cat /K M (s -1 /M)
28.0(58.4) 146 1.92×105(4.00×105)
上記表6において、括弧内の数値は、オキサロ酢酸に対する動力学定数(kcat)を用いたときのアセチル補酵素Aに対する動力学定数である。
6.オキサロ酢酸に対するk cat 、K M 、k cat /K M の算出
前記測定において、アセチル補酵素Aの濃度を、1.5mMに固定して、オキサロ酢酸の濃度を変えることで、オキサロ酢酸の濃度と反応速度vとの関係を求めた。反応温度は、75℃、pH7.5とした。この測定においては、前記コントロール実験におけるオキサロ酢酸塩濃度を0μMとした。測定結果を、下記表7および図6のグラフを示す。
前記測定において、アセチル補酵素Aの濃度を、1.5mMに固定して、オキサロ酢酸の濃度を変えることで、オキサロ酢酸の濃度と反応速度vとの関係を求めた。反応温度は、75℃、pH7.5とした。この測定においては、前記コントロール実験におけるオキサロ酢酸塩濃度を0μMとした。測定結果を、下記表7および図6のグラフを示す。
(表7)
オキサロ酢酸の濃度(μM) v(μM/s)
0 0
10 0.00917
50 0.0583
100 0.240
200 0.342
500 0.424
1000 0.459
オキサロ酢酸の濃度(μM) v(μM/s)
0 0
10 0.00917
50 0.0583
100 0.240
200 0.342
500 0.424
1000 0.459
上記測定結果より、IGOR Pro(ヒューリンクス社製)を用いて、オキサロ酢酸に対するkcat、KM、kcat/KMを算出した。算出結果を、下記表8に示す。
(表8)
k cat (s -1 ) K M (μM) k cat /K M (s -1 /M)
58.4 315 1.85×105
k cat (s -1 ) K M (μM) k cat /K M (s -1 /M)
58.4 315 1.85×105
7.二次構造の熱安定性
この測定では、前記測定方法に代えて、5μMの前記実施例4において精製したタンパク質、50mMのリン酸ナトリウム(25℃でのpH7.5)、100mMのKClからなる反応溶液を用意し、下記の種々条件下、前記タンパク質溶液のCD(円偏光二色性)の測定を行った。測定には、円偏光二色性分散計(日本分光社製、商品名J−720W)および光路長0.1cmのセルを用いた。
この測定では、前記測定方法に代えて、5μMの前記実施例4において精製したタンパク質、50mMのリン酸ナトリウム(25℃でのpH7.5)、100mMのKClからなる反応溶液を用意し、下記の種々条件下、前記タンパク質溶液のCD(円偏光二色性)の測定を行った。測定には、円偏光二色性分散計(日本分光社製、商品名J−720W)および光路長0.1cmのセルを用いた。
7−1.温度変化測定
前記反応溶液を、pH7.0で25℃から95℃まで1℃/minで温度変化させたときの222nmの円偏光二色性スペクトルの変化を測定した。なお、ここで用いた222nmは、α−へリックス由来の吸収波長である。測定結果を、図7のグラフに示す。図示のとおり、25℃から95℃までの連続温度変化を見ても、前記実施例4において精製したタンパク質の二次構造がほぼ保持されていることが分かった。
前記反応溶液を、pH7.0で25℃から95℃まで1℃/minで温度変化させたときの222nmの円偏光二色性スペクトルの変化を測定した。なお、ここで用いた222nmは、α−へリックス由来の吸収波長である。測定結果を、図7のグラフに示す。図示のとおり、25℃から95℃までの連続温度変化を見ても、前記実施例4において精製したタンパク質の二次構造がほぼ保持されていることが分かった。
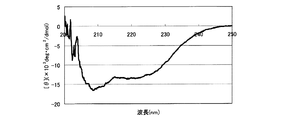
7−2.スペクトル測定
前記反応溶液の95℃、pH7.0での遠紫外部(200〜250nm)のスペクトルの測定を行った。測定結果を、図8のグラフに示す。なお、図8における残基モル楕円率[θ](deg・cm2/dmol)は、[θ]=100θ/(l・c)により求めた。ここで、θは、実測値(mdeg)、lは、光路長(cm)、cは、残基モル濃度(M)である。図示のとおり、209nm、220nmで負の極大値を示したことから、前記実施例4において精製したタンパク質の二次構造は、95℃でも保持されていることが分かった。
前記反応溶液の95℃、pH7.0での遠紫外部(200〜250nm)のスペクトルの測定を行った。測定結果を、図8のグラフに示す。なお、図8における残基モル楕円率[θ](deg・cm2/dmol)は、[θ]=100θ/(l・c)により求めた。ここで、θは、実測値(mdeg)、lは、光路長(cm)、cは、残基モル濃度(M)である。図示のとおり、209nm、220nmで負の極大値を示したことから、前記実施例4において精製したタンパク質の二次構造は、95℃でも保持されていることが分かった。
本発明により、クエン酸シンターゼ活性を有する新規耐熱性タンパク質が提供できる。本発明のタンパク質は、高温下で使用することが可能であり、工業的用途が広がると共に、基質濃度の増加、反応効率の向上、混入微生物の除去、保存期間および耐用期間の延長などの多くの利点がもたらされる。
配列番号1:耐熱性クエン酸シンターゼのアミノ酸配列
配列番号2:耐熱性クエン酸シンターゼのアミノ酸配列をコードする塩基配列
配列番号3:耐熱性クエン酸シンターゼの構造遺伝子の末端に制限酵素部位NdeIおよびBamHI、NotIを導入するための順方向プライマー
配列番号4:耐熱性クエン酸シンターゼの構造遺伝子の末端に制限酵素部位NdeIおよびBamHI、NotIを導入するための逆方向プライマー
配列番号5:N末端アミノ酸配列
配列番号2:耐熱性クエン酸シンターゼのアミノ酸配列をコードする塩基配列
配列番号3:耐熱性クエン酸シンターゼの構造遺伝子の末端に制限酵素部位NdeIおよびBamHI、NotIを導入するための順方向プライマー
配列番号4:耐熱性クエン酸シンターゼの構造遺伝子の末端に制限酵素部位NdeIおよびBamHI、NotIを導入するための逆方向プライマー
配列番号5:N末端アミノ酸配列
Claims (9)
- 下記の(a)または(b)の耐熱性タンパク質
(a) 配列番号1のアミノ酸配列からなる耐熱性タンパク質。
(b) 配列番号1のアミノ酸配列において、1つ若しくは数個のアミノ酸残基が、欠失、置換、付加若しくは挿入されたアミノ酸配列からなり、クエン酸シンターゼ活性を有する耐熱性タンパク質。 - 前記クエン酸シンターゼ活性が、アセチル補酵素A、オキサロ酢酸および水に作用して、クエン酸および補酵素Aを生成する機能である請求項1記載のタンパク質。
- 超好熱性古細菌由来である請求項1または2記載のタンパク質。
- 超好熱性古細菌が、スルホロブス・トコダイイ(Sulfolobus tokodaii)(JCM10545)である請求項3記載のタンパク質。
- 請求項1から4のいずれかに記載のタンパク質をコードするDNAまたは配列番号2に記載のDNAを含むベクター。
- 請求項5記載のベクターにより形質転換された形質転換体。
- 請求項1から4のいずれかに記載のタンパク質の製造方法であって、請求項6記載の形質転換体を培養する工程と、前記培養工程において発現した前記タンパク質を回収する工程とを含む製造方法。
- 酵素反応により、オキサロ酢酸、アセチル補酵素Aおよび水からクエン酸および補酵素Aを製造する方法であって、前記酵素として請求項1から4のいずれかに記載のタンパク質を用い、温度55〜95℃の条件で前記酵素反応を行う製造方法。
- 前記酵素反応のpHが、pH7以上の範囲である請求項8記載の製造方法。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2005080333A JP2005296009A (ja) | 2004-03-19 | 2005-03-18 | クエン酸シンターゼ活性を有する新規耐熱性タンパク質 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2004080862 | 2004-03-19 | ||
| JP2005080333A JP2005296009A (ja) | 2004-03-19 | 2005-03-18 | クエン酸シンターゼ活性を有する新規耐熱性タンパク質 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2005296009A true JP2005296009A (ja) | 2005-10-27 |
Family
ID=35328300
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2005080333A Withdrawn JP2005296009A (ja) | 2004-03-19 | 2005-03-18 | クエン酸シンターゼ活性を有する新規耐熱性タンパク質 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP2005296009A (ja) |
-
2005
- 2005-03-18 JP JP2005080333A patent/JP2005296009A/ja not_active Withdrawn
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JPWO2007142210A1 (ja) | 光学活性アルコールの製造方法 | |
| EP3922718B1 (en) | Heat-resistant dna polymerase mutant having high amplification activity | |
| JP2002247991A (ja) | タンパク質の耐熱性を向上させる方法、該方法により耐熱性の向上したタンパク質、および該タンパク質をコードする核酸 | |
| EP1487974A4 (en) | NAD PHOSPHITE OXYDOREDUCTASE, NEW CATALYST FROM BACTERIA USEFUL FOR REGENERATING NAD (P) H | |
| JP2005296010A (ja) | 2−イソプロピルリンゴ酸シンターゼ活性を有する新規耐熱性タンパク質 | |
| JP4280827B2 (ja) | アセチルグルタミン酸キナーゼ活性を有する新規耐熱性タンパク質 | |
| JP2004298185A (ja) | ホスホグリセリン酸デヒドロゲナーぜ活性を有する新規耐熱性タンパク質 | |
| JP2005296009A (ja) | クエン酸シンターゼ活性を有する新規耐熱性タンパク質 | |
| JP4250762B2 (ja) | 6−n−ヒドロキシアミノプリンの代謝活性を有する新規耐熱性タンパク質 | |
| JP4292294B2 (ja) | ピロリン−5−カルボン酸レダクターゼ活性を有する新規耐熱性タンパク質 | |
| JP4280826B2 (ja) | シスタチオニン−γ−シンターゼ活性を有する新規耐熱性タンパク質 | |
| CN114480345A (zh) | MazF突变体、重组载体、重组工程菌及其应用 | |
| JP2005296008A (ja) | 水銀レダクターゼ活性を有する新規耐熱性タンパク質 | |
| JP2005296012A (ja) | ホスホエノールピルビン酸シンターゼ活性を有する新規耐熱性タンパク質 | |
| JP2005296011A (ja) | アデニロコハク酸リアーゼ活性を有する新規耐熱性タンパク質 | |
| JP2000069971A (ja) | 新規な耐熱性蟻酸脱水素酵素 | |
| JP2006288390A (ja) | Gdp−マンノースピロホスホリラーゼ活性を有する新規耐熱性タンパク質 | |
| JP2006254869A (ja) | アミロ−α−1,6−グリコシダーゼ活性を有する新規耐熱性タンパク質 | |
| JP2004298186A (ja) | Dna相同的組換え修復活性を有する新規耐熱性タンパク質 | |
| JP2006288391A (ja) | エノラーゼ活性を有する新規耐熱性タンパク質 | |
| JP2006288389A (ja) | Gmpシンターゼ活性を有する新規耐熱性タンパク質 | |
| JP5935382B2 (ja) | RrhJ1IIヌクレアーゼおよびその遺伝子 | |
| JP2004298189A (ja) | フマル酸ヒドラターゼ活性を有する新規耐熱性タンパク質 | |
| JP2004298184A (ja) | ホスホグリセリン酸キナーゼ活性を有する新規耐熱性タンパク質 | |
| JP2005261348A (ja) | アコニット酸ヒドラターゼ活性を有する新規耐熱性タンパク質 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A300 | Withdrawal of application because of no request for examination |
Free format text: JAPANESE INTERMEDIATE CODE: A300 Effective date: 20080603 |