CN115279902A - 用于调控基因表达的系统 - Google Patents
用于调控基因表达的系统 Download PDFInfo
- Publication number
- CN115279902A CN115279902A CN202080072368.2A CN202080072368A CN115279902A CN 115279902 A CN115279902 A CN 115279902A CN 202080072368 A CN202080072368 A CN 202080072368A CN 115279902 A CN115279902 A CN 115279902A
- Authority
- CN
- China
- Prior art keywords
- polya
- aptamer
- nucleic acid
- ligand
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images

















































































































































































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/115—Aptamers, i.e. nucleic acids binding a target molecule specifically and with high affinity without hybridising therewith ; Nucleic acids binding to non-nucleic acids, e.g. aptamers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/16—Aptamers
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/50—Vector systems having a special element relevant for transcription regulating RNA stability, not being an intron, e.g. poly A signal
Landscapes
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Wood Science & Technology (AREA)
- Organic Chemistry (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Zoology (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Microbiology (AREA)
- Plant Pathology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Physics & Mathematics (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
描述了与基因表达调控有关的组合物和方法。在一些实施方案中,本公开提供了用于使用核酸构建体调控基因表达的组合物和方法。在一些实施方案中,本公开认识到替代剪接在用核酸构建体调控核酸构建体中的效用。在一些实施方案中,本公开认识到利用配体结合适体调控基因表达的效用。
Description
相关申请的交叉引用
本申请要求于2019年8月30日提交的美国临时申请号62/894,611、2019年9月23日提交的美国临时申请号62/904,635和2020年6月24日提交的美国临时申请号63/043,504的优先权,所述申请各自的内容以引用方式整体并入本文。
政府许可权
本发明是在由美国国家卫生研究院(National Institutes of Health)颁发的EB013584在政府支持下完成的。政府对本发明具有一定的权利。
背景技术
可通过提高灵敏度和减少泄漏来改进用于调节基因表达的基于核酸的构建体。
发明内容
本公开认识到与可调控基因产物表达相关的核酸构建体的发现。在一些实施方案中,本公开提供了用于使用核酸构建体调控基因表达的组合物和方法。在一些实施方案中,本公开认识到替代剪接在调控核酸构建体中的基因表达中的效用。在一些实施方案中,本公开认识到利用配体结合适体调控基因表达的效用。
在一些实施方案中,本公开提供了一种用于调节基因表达的系统,所述系统包含polyA适体多核苷酸,所述polyA适体多核苷酸在5'至3'方向上包含:5'剪接供体位点;工程内含子;第一3'剪接受体位点;polyA开关,其包含具有一个或多个配体结合口袋的两个或更多个配体结合适体,和其中的至少一个polyA切割信号;第二3'剪接受体位点;以及编码可表达多肽的核酸序列。
在一些实施方案中,本公开的polyA适体多核苷酸包含两个配体结合适体。在一些实施方案中,polyA适体多核苷酸包含三个配体结合适体。在一些实施方案中,polyA适体多核苷酸包含polyA开关,所述polyA开关包含三向接合部(three way junction)。在一些实施方案中,三向接合部包括一个或多个RNA双链茎的接合部。在一些实施方案中,三向接合部的部分是单链的。在一些实施方案中,RNA双链茎包含配体结合适体。在一些实施方案中,编码可表达多肽的核酸序列包含5’UTR。
在一些实施方案中,本公开提供了一种用于调节细胞中基因产物表达的方法。方法包括以下步骤:向细胞中引入系统,所述系统在5'至3'方向上包含:5'剪接供体位点;工程内含子;第一3'剪接受体位点;polyA开关,其包含具有一个或多个配体结合口袋的两个或更多个配体结合适体,和其中的至少一个polyA切割信号;第二3'剪接受体位点。在一些实施方案中,通过本文所述的方法表达的基因产物对于细胞是外源性的。在一些实施方案中,通过本文所述的方法表达的基因产物对于细胞是内源性的。在一些实施方案中,本公开提供的方法发生在个体的一个或多个细胞中,配体是葡萄糖,个体患有糖尿病、糖尿病前期或糖尿病并发症,且/或可表达的多核苷酸是胰岛素。在一些实施方案中,本公开提供的方法发生在个体的一个或多个细胞中,可表达的多核苷酸是治疗性基因产物,诸如人生长激素、凝血因子X或抗肌萎缩蛋白。在一些实施方案中,本公开提供的方法发生在个体的一个或多个细胞中,配体是癌症生物标志物的基因产物,并且可表达的多核苷酸是自杀基因。在一些实施方案中,本公开提供的方法发生在个体中,可表达的多核苷酸是报告基因,并且报告基因的表达位置和/或强度提供关于配体在个体的一个或多个细胞中的空间分布、时间波动或两者的信息。在一些实施方案中,本公开提供的方法发生在个体、组织或细胞中,其中可表达的多核苷酸编码可检测的基因产物,并且其中对相应的个体、组织或细胞进行成像。
附图说明
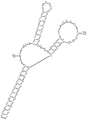
图1A至图1C提供了本文所述的polyA适体多核苷酸方面的示意图。图1A描绘了基于配体诱导的替代剪接和polyA信号切割的“混合”开关机制。图1B描绘了Y形polyA开关的配置。对Y形结构的不同部分的名称进行标记。图C示出了具有代表性的Y形polyA开关Y196CAA的配置。
图2A至图2C展示了另外的Y形结构的结果,所述Y形结构不同地配置并且polyA切割信号不同地定位。polyA信号由红线指示。三向接合部由方框指示。图2A和图2B示出了替代的Y形配置,其中三个适体(适体A、B和C)在三向接合部周围不同地排列。图2C示出了三个相互堆叠的适体而没有三向接合部。
图3A至图3C展示了本文所述的polyA适体多核苷酸中polyA切割信号数量的修饰结果。图3A示出了位于两个不同茎上的2个polyA信号(红色方框)。图3B仅示出了部分埋在臂1-2中的一个polyA信号。图3C示出了臂1-2中嵌入2个polyA信号(红色方框)。
图4A至图4L展示了本文所述的polyA适体多核苷酸的三向接合部的修饰结果。图4L示出了最佳的三向接合部序列。
图5展示了相对于本文所述的polyA适体多核苷酸的三向接合部的位置的polyA信号的修饰结果。
图6A至图6B展示了本文所述的polyA适体多核苷酸的第三双链茎(在图1B中称为臂3-1和3-2)的修饰结果。图6A展示了臂3-1的修饰结果。图6B展示了臂3-2的修饰结果。
图7A至图7B展示了本文所述的polyA适体多核苷酸的第二双链茎(在图1B中称为臂2-1和2-2)的修饰结果。图7A展示了臂2-2的修饰结果。图7B展示了臂2-1的修饰结果。
图8展示了本文所述的polyA适体多核苷酸的第一双链茎(在图1B中称为臂1-2)上部的修饰结果。
图9展示了本文所述的polyA适体多核苷酸的第一双链茎(在图1B中称为臂1-1)下部的修饰结果。
图10A至图10B展示了本文所述的polyA适体多核苷酸的适体方向的修饰结果。图10A示出了在适体B的方向反转的情况下的结果。图10B示出了在适体A的方向反转的情况下的结果。
图11A至图11B展示了本文所述的polyA适体多核苷酸中每个适体的贡献。图11A示出了通过A到C点突变(由箭头指示)使每个适体失活的作用。图11B示出了删除适体A对诱导作用的影响。
图12A至图12D展示了在本文所述的polyA适体多核苷酸之后修饰可表达多核苷酸的5'UTR的结果。图12A示出了使用不同亲本构建体在可表达多核苷酸的5'UTR中插入CAA重复序列(下划线)的结果。图12B示出了使用S56作为亲本构建体测试具有强3'剪接位点的新5'UTR序列的结果。图12C示出了将非结构化间隔区序列插入Y305和Y300的5'UTR的结果。图12D示出了在5'UTR中的3'剪接位点之前插入CAA重复序列。
图13A至图13B示出了本文所述的polyA适体多核苷酸的G quad序列的重要性。图13A示出了G-quad序列对使用Y196CAA作为亲本构建体的诱导作用的影响。图13B示出了使用S56作为亲本构建体测试不同G-quad序列以替换4MAZ G-quad的结果。
图14展示了本文所述的polyA适体多核苷酸的四环素诱导的替代剪接的确认。在不存在Tc的情况下,IVS2剪接的RNA通过polyA切割降解(泳道1和3)。Tc的存在诱导Y196CAA-2MAZ和Y196CAA-4MAZ(泳道2和4)中的替代剪接。配体诱导的替代剪接在4MAZ的存在下更加明显。
图15A至图15G展示了本文所述的polyA适体多核苷酸的第一3'剪接受体位点的修饰结果。图15A示出了将IVS2 3'剪接位点移动到Y196CAA-4MAZ的臂1-1中的结果。图15B示出了当完全嵌入到适体A附近的臂1-1中(红色箭头)时,第一3'剪接位点受到强烈抑制,从而导致非常低的诱导作用。通过删除适体A的部分序列来降低适体A的钳制效应可恢复诱导作用。图15C示出了沿S9m的臂1移动IVS 3'剪接位点(蓝色方框)的结果,并且图15D示出了将IVS 3'剪接位点放置在臂1-2的凸出中的结果。图15E示出了通过突变IVS2 3'剪接位点之后的碱基来改变预测的剪接强度的结果。图15F示出了将mini-IVS2 3'剪接位点进一步移动进入或远离臂1-1中的适体A的结果。图15G示出了第一3'剪接位点(CAGNNN)之后的三个碱基的随机化。
图16A至图16C示出了本文所述的polyA适体多核苷酸的第二3'剪接受体位点的修饰结果。图16A示出了修饰5'UTR以改变替代的3'剪接位点的强度的结果。图12B示出了在5'UTR(TAGNNN)中'TAG'之后的三个碱基随机化以调节替代的3'剪接位点的强度以便改进诱导作用的结果。图12C示出了将选自随机化的最佳TAGNNN序列合并到Y329 5'UTR中的结果。
图17A和图17B展示了本文所述的polyA适体多核苷酸的工程内含子大小的修饰结果。图17A示出了改变IVS2内含子的大小和剪接元件的结果。图17B示出了从具有较短工程内含子的构建体(S159、S164和S169)中去除CAA重复序列的结果。
图18A至图18C展示了将上游开放阅读框(μORF)包含在本文所述的polyA适体多核苷酸中的结果。图18A示出了将上游开放阅读框包含在polyA适体中的示意图。插入的上游ATG起始密码子是加框的。图18B示出了用上游开放阅读框微调构建体的5'UTR序列的结果。图18C示出了包含上游开放阅读框的一种代表性混合开关(hybrid switch)。
图19A至图19E展示了本文所述的polyA适体多核苷酸在存在配体的情况下控制可表达多肽的基因表达的能力。图19A示出了代表性S系列构建体与Y196CAA-4MAZ的性能。图19B示出了通过显微镜观察到的代表性S系列构建体与Y196CAA-4MAZ的剂量反应。图19C示出了Y300和Y301的性能。图19D示出了通过荧光素酶报告基因测定确定的Y362和Y367的剂量反应。图19E示出了Y362和Y367对1ug/ml四环素的反应,如通过使用eGFP报告基因信号的荧光激活细胞分选(FACS)确定的。所有结果中的“诱导倍数”计算为在存在与不存在四环素的情况下转基因表达的比率。
图20展示了本文所述的polyA适体多核苷酸作为内源开关发挥作用以控制基因组中内源基因的表达的能力。
图21描绘了组合三个位置处的单碱基变化的Y形polyA开关的配置。此处显示的Y387构建体含有所有三个变化。
图22表明,三个单碱基变化的组合在低药物浓度下显著增加可表达多肽的诱导表达。使用四种不同的亲本构建体(Y359、Y360、Y361、Y362C)证明单碱基变化对诱导作用的影响。在所有四种不同的亲本构建体中,这些单碱基变化对诱导作用的影响都是相似的。上图示出了具有标准变化的诱导倍数。下图绘制了每个构建体的诱导倍数。
图23A和图23B展示了从构建体Y362和Y386诱导表达的剂量反应分析,构建体Y362和Y386包含组合三个位置处的单碱基变化的Y形polyA开关。图23A示出了使用最大诱导倍数作为EC100参考,四环素的诱导作用在低至0.5μg/ml至1μg/ml Tc时达到最大水平的50%(EC50)。图23B示出了使用亲本构建体(HDM-Luc,其具有类似的序列但没有Y形结构)的最大表达水平作为EC100参考的类似计算。在这种情况下,通过低至0.5μg/ml至1.2μg/ml的四环素达到EC50。
具体实施方式
在一些实施方案中,本公开提供了用于可调控基因产物表达的组合物和方法。在一些实施方案中,用于可调控基因产物表达的组合物和方法包含polyA适体多核苷酸。在一些实施方案中,polyA适体多核苷酸包含一个或多个剪接供体位点、一个或多个剪接受体位点、工程内含子;polyA开关;和编码可表达多肽的核酸序列等。在一些实施方案中,polyA开关包含至少一个配体结合适体。在一些实施方案中,polyA开关包含至少一种polyA切割信号。在一些实施方案中,polyA适体多核苷酸包含RNA双链茎。
适体
适体是像受体一样折叠并与特定配体结合的短RNA序列。用于产生对特定配体具有高亲和力的适体的有效体外进化方法已得到很好的建立。适体的结合亲和力通常可达到纳摩尔范围,与抗体的结合亲和力相当。在这方面,适体可被视为由RNA制成的抗体。适体与抗体的区别在于它的小尺寸(通常小于50个碱基)和它的模块化性质。这些特征使适体能够在不失去其结合功能的情况下与其他RNA结构整合并控制其他RNA结构。已经证明,适体可将自切割RNA核酶转化为以配体依赖性方式运行,并在试管和细胞中像分子开关一样发挥作用。
在一些实施方案中,polyA适体多核苷酸包含一个或多个RNA双链茎。在一些实施方案中,RNA双链茎是由包含在单个polyA适体多核苷酸内的互补核酸的分子内碱基配对形成的核酸结构。在一些实施方案中,RNA双链茎也可称为臂。在一些实施方案中,polyA适体多核苷酸包含一个或多个RNA双链茎。在一些实施方案中,polyA适体多核苷酸包含两个RNA双链茎。在一些实施方案中,polyA适体多核苷酸包含三个RNA双链茎。在一些实施方案中,RNA双链茎包含配体结合适体。在一些实施方案中,polyA适体多核苷酸包含两个配体结合适体。在一些实施方案中,polyA适体多核苷酸包含三个配体结合适体。
在一些实施方案中,至少两个RNA双链茎连接以形成接合部。在一些实施方案中,RNA双链茎的接合部包含单链区。在一些实施方案中,三个RNA茎相交以形成三向接合部。在一些实施方案中,三向接合部包含至少一个单链区。在一些实施方案中,三向接合部包含一个、两个或三个单链区。
在一些实施方案中,双链RNA茎的序列选自以下中的一个:
| SEQ ID NO.: | 序列(5'到3') |
| 2 | GGGUGUUUGUGGC |
| 3 | CACGAGAUCUGG |
| 4 | GCGUUUUAUACUU |
| 5 | CUCUGCAGAUGUU |
在一些实施方案中,由RNA双链茎的接合部形成的单链区包含至少一种核酸。在一些实施方案中,由RNA双链茎的接合部形成的单链区包含一种、两种、三种、四种、五种、六种、七种、八种、九种、十种、十一种、十二种或更多种核酸。在一些实施方案中,三向接合部包含第一单链区、第二单链区和第三单链区。在一些实施方案中,第一单链区包含至少一种选自C和A的碱基。在一些实施方案中,第二单链区包含至少一种选自C和A的碱基。
在一些实施方案中,RNA双链茎的长度为30、20、10或5个碱基对。在一些实施方案中,RNA双链茎的长度为5至30、10至30、20至30、5至10、5至20、5至30或10至20个碱基对。在一些实施方案中,RNA双链茎的长度为最多30个碱基对。在一些实施方案中,RNA双链茎的长度为小于30、20或10个碱基对。
在一些实施方案中,polyA适体多核苷酸包含一个或多个适体。在一些实施方案中,polyA适体多核苷酸包含两个适体。在一些实施方案中,polyA适体多核苷酸包含三个适体。
在一些实施方案中,包含在本文所述的polyA适体多核苷酸中的适体包含至少一个单链区和至少一个适体RNA双链茎。在一些实施方案中,适体RNA双链茎包含单链区。在一些实施方案中,适体RNA具有序列为AATAAGATTACCGAAAGGCAATCTTATT的RNA双链茎(例如,臂2-2)。在一些实施方案中,适体RNA具有序列为CCAGATCGAATTCGATCTGG的RNA双链茎(例如,臂3-2)。在一些实施方案中,适体RNA具有长度在6-10;7-11;8-12;9-13;10-14个碱基对的长度范围内的RNA双链茎。
PolyA切割信号
根据各种实施方案,可使用多种polyA信号中的任何一种(例如,由polyA信号序列编码)。通过非限制性实例的方式,哺乳动物细胞中使用的polyA信号序列包括:AAUAAA、AUUAAA、AGUAAA、ACUAAA、UAUAAA、CAUAAA、GAUAAA、AAUAUA、AAUACA和AAUAGA。在一些实施方案中,polyA开关可包括两个或更多个polyA信号序列(例如,3、4、5、6、7、8、9、10或更多个)。
多腺苷酸化是存在于所有哺乳动物细胞中的基础mRNA加工机制。通常,哺乳动物polyA信号存在于3'非翻译区(UTR)。相比之下,本公开提供了包含存在于表达构建体中可表达多核苷酸诸如基因的3'非翻译区(UTR)以外的位置处的polyA切割信号的组合物和方法。当polyA信号在5’UTR中人工形成(细胞中通常不存在)时,polyA信号的有效切割会导致在所述位点添加polyA尾。这导致与转基因序列相关的mRNA的后半部分被去除和降解,并因此导致基因表达缺失。在一些实施方案中,polyA信号存在于编码可表达多核苷酸(mRNA)的核酸序列的翻译起始位点的上游,所述可表达多核苷酸编码表达多肽。在一些实施方案中,polyA信号位于mRNA的5'UTR中。在一些实施方案中,三向接合部的单链区包含全部或部分polyA切割信号。在一些实施方案中,三向接合部的第三单链区包含全部或部分polyA切割信号。在一些实施方案中,RNA双链茎包含全部或部分polyA切割信号。在一些实施方案中,第三RNA双链茎包含全部或部分polyA切割信号。在一些实施方案中,如本文所用的polyA切割信号的一部分包括一个、两个、三个或四个核苷酸。在一些实施方案中,polyA切割信号具有AAUAAA的序列。在一些实施方案中,polyA切割信号具有AUUAAA、AGUAAA、ACUAAA、UAUAAA、CAUAAA、GAUAAA、AAUAUA、AAUACA、AAUAGA、AAAAAG或ACUAAA的序列。在构建体中使用两个或更多个polyA信号的实施方案中,polyA信号可以相同或可以不同。在特定实施方案中,可表达的多核苷酸能够由RNA聚合酶II转录。
在一些实施方案中,5'UTR中polyA切割信号的存在靶向polyA信号之后的mRNA的后半部分以进行降解,并且在本公开的各种组合物和方法中利用了这种能力。在一些实施方案中,5'UTR中polyA切割信号的存在导致由polyA适体多核苷酸编码的前mRNA/mRNA的切割。在一些实施方案中,由polyA适体多核苷酸编码的前mRNA/mRNA的切割导致前mRNA/mRNA的后半部分降解。在一些实施方案中,由polyA适体多核苷酸编码的前mRNA/mRNA的切割导致多肽不表达。
在特定实施方案中,polyA切割信号在包含一个或多个配体可结合的至少一个配体结合适体的polyA适体多核苷酸内。在一些实施方案中,配体与配体结合适体的结合确定替代剪接之后polyA切割信号是否存在于前mRNA/mRNA中。在一些实施方案中,配体与配体结合适体的结合确定替代剪接之后前mRNA/mRNA是否被切割。在一些实施方案中,配体与配体结合适体的结合确定替代剪接之后可表达多肽是否表达。
工程内含子
在一些实施方案中,polyA适体多核苷酸包含工程内含子。在一些实施方案中,工程内含子包含一个或多个剪接位点。在一些实施方案中,剪接位点是或包含剪接供体位点(例如,包含GU序列)。在一些实施方案中,剪接位点是或包含剪接受体位点(例如,包含AG序列)。在一些实施方案中,工程内含子中的剪接位点起作用(例如,彼此结合和/或与一个或多个内源剪接位点结合),以从polyA适体多核苷酸切除工程内含子。
在一些实施方案中,工程内含子的前面是5'剪接供体位点。在一些实施方案中,polyA适体多核苷酸包含工程内含子的5'的区域中的5'剪接供体位点。在一些实施方案中,polyA适体多核苷酸包含工程内含子的3'的第一3'剪接受体位点。在一些实施方案中,本文所述的polyA适体多核苷酸的工程内含子包含5'剪接供体位点和第一3'剪接受体位点。在一些实施方案中,polyA适体多核苷酸包含编码可表达多肽的核酸序列。在一些实施方案中,polyA适体多核苷酸包含紧邻编码可表达多肽的核酸序列的5'的第二3'剪接受体位点。
在一些实施方案中,polyA适体多核苷酸包含剪接供体位点的5'的启动子。示例性启动子包括例如CMV、E1F、VAV、TCRvβ、MCSV、SV40启动子、RSV启动子和PGK启动子。
在一些实施方案中,在不存在与配体结合适体结合的配体的情况下,由本文所述的polyA适体多核苷酸编码的前mRNA的剪接发生在5'剪接供体位点与第一3'剪接受体位点之间。在一些实施方案中,由本文所述的polyA适体多核苷酸编码的前mRNA的5'剪接供体位点与第一3'剪接受体位点之间的剪接导致包含编码可表达多肽的核酸序列的5'UTR之前的polyA切割信号的mRNA。在一些实施方案中,在编码可表达多肽的核酸序列的5'UTR之前存在polyA切割信号导致polyA切割位点处的切割以及编码可表达多肽的序列的降解。
在一些实施方案中,在存在与配体结合适体结合的配体的情况下,由本文所述的polyA适体多核苷酸编码的前mRNA的剪接发生在5'剪接供体位点与第二3'剪接受体位点之间。在一些实施方案中,在5'剪接供体位点与第二3'剪接受体位点之间的由本文所述的polyA适体多核苷酸编码的前mRNA的剪接导致包含编码可表达多肽的核酸序列的mRNA。在一些实施方案中,在5'剪接供体位点与第二3'剪接受体位点之间的由所描述的polyA适体多核苷酸编码的前mRNA的剪接导致通过剪接去除polyA切割信号。在一些实施方案中,由本文所述的polyA适体多核苷酸编码的前mRNA的5'剪接供体位点与第二3'剪接受体位点之间的剪接导致可表达多肽的表达。
在一些实施方案中,polyA适体多核苷酸包含两个或更多个配体结合适体。在一些实施方案中,两个或更多个配体结合适体中的每一个结合不同的配体。在一些实施方案中,polyA适体多核苷酸包含两个或更多个单独的polyA开关。在一些实施方案中,第一polyA开关包含结合第一配体的第一适体,并且第二polyA开关包含结合第二配体的第二适体。在一些实施方案中,第一适体和第二适体不相同,并且第一配体和第二配体不相同。在一些实施方案中,第一适体和第二适体不相同,并且第一配体和第二配体相同。
在一些实施方案中,工程内含子是任何序列。在一些实施方案中,工程内含子的长度为大约100、200、300、400或500个核苷酸。在一些实施方案中,工程内含子的长度在100-200;110-200;120-200;130-200;140-200;150-200;160-200;170-200;或180-200个碱基的范围内。在一些实施方案中,工程内含子的长度为最多100、110、120、130、140、150、160、170、180、190、200、210、220个碱基。在一些实施方案中,工程内含子具有以下序列:
GTGAGTCTTAAGCCAGCTACCATTCTGCTTTTATTTTATCGTTGGGATAAGGCTGGATTATTCTGAGTCCAAGCTAGGCCCTTTTGCTAATCATCTTCATACCTCTTATCTTCCTCTGCAG(SEQ ID NO.:1)
在一些实施方案中,工程内含子具有以下序列:
GTGAGTCTATGGGACCCTTGATGTTTTCTTTCCCCTTCTTTTCTATGGTTAAGTTCATGTCATAGGAAGGGGAGAAGTAACAGGGTACACATATTGACCAAATCAGGGTAATTTTGCATTTGTAATTTTAAAAAATGCTTTCTTCTTTTAATATACTTTTTTGTTTATCTTATTTCTAATACTTTCCCTAATCTCTTTCTTTCAGGGCAATAATGATACAATGTATCATGCCTCTTTGCACCATTCTAAAGAATAACAGTGATAATTTCTGGGTTAAGGCAATAGCAATATTTCTGCATATAAATATTTCTGCATATAAATTGTAACTGATGTAAGAGGTTTCATATTGCTAATAGCAGCTACAATCCAGCTACCATTCTGCTTTTATTTTATGGTTGGGATAAGGCTGGATTATTCTGAGTCCAAGCTAGGCCCTTTTGCTAATCATGTTCATACCTCTTATCTTCCTCCCACAG(SEQ ID NO.:49)
如本文所用,内含子可指DNA序列或其对应的RNA序列。
在一些实施方案中,polyA适体多核苷酸包含另外的序列以促进、调控或协助polyA适体多核苷酸内的polyA信号切割。在一些实施方案中,polyA适体多核苷酸包含在编码可表达多肽的核酸序列的5’和polyA切割信号的3'的G-U丰富区。在一些实施方案中,polyA适体多核苷酸包含另外的序列以促进、调控或协助polyA适体多核苷酸内的剪接。在一些实施方案中,polyA适体多核苷酸包含能够调节替代剪接强度的核酸三联体序列。在一些实施方案中,核酸三联体序列在相对于5'UTR中的第二3'受体位点的3'。在一些实施方案中,核酸三联体序列在工程内含子的3'。在一些实施方案中,核酸三联体序列的序列包含任何三个核苷酸。在一些实施方案中,核酸三联体序列的序列包括TAG、TCT、TTC、TTG、TGA、TGC、TCC、ACA、AAC、ACC、AGC、AGG、CCT、CCC、TTT、TGA、TCT、TAC、CAC或CAT。
在一些实施方案中,polyA适体多核苷酸包含在编码可表达多肽的核酸序列的5’和polyA切割信号的3'的G-U丰富区。在一些实施方案中,polyA适体多核苷酸包含在编码可表达多肽的核酸序列的5’和G-U丰富区的3'的G丰富区。在一些实施方案中,G丰富区在本领域中应理解为MAZ序列。在一些实施方案中,polyA适体多核苷酸包含一个或多个G丰富区。在一些实施方案中,polyA适体多核苷酸包含一个或多个连续G丰富区。在一些实施方案中,polyA适体多核苷酸包含一个或多个MAZ序列。在一些实施方案中,polyA适体多核苷酸包含一个或多个连续MAZ序列。在一些实施方案中,polyA适体多核苷酸包含一个、两个、三个、四个、五个、六个MAZ序列。连续的MAZ可被一个或多个间隔区序列分开。在一些实施方案中,G富含区的序列是AACGGGGGAGGGGGAGGAAAGGGGGAGGGGGAGGAAAGGGGGAGGGGGAGGAAAGGGGGAGGGGGA(SEQ ID NO.:47)。
在一些实施方案中,polyA适体多核苷酸包含一个或多个起始密码子。在一些实施方案中,polyA适体多核苷酸包含一个或多个框外起始密码子。在一些实施方案中,框外起始密码子相对于编码可表达多肽的核酸序列的编码序列是框外的。在一些实施方案中,polyA适体多核苷酸包含至少一个框外起始密码子。在一些实施方案中,polyA适体多核苷酸包含工程内含子的3'的第一3'剪接受体位点的3'的至少一个框外起始密码子。
可表达多肽
在一些实施方案中,polyA适体多核苷酸包含编码可表达多肽的核酸序列。在一些实施方案中,编码可表达多肽的核酸序列包含5’UTR。在一些实施方案中,编码可表达多肽的核酸序列的5’UTR包含3’剪接受体位点。在一些实施方案中,编码可表达多肽的核酸序列的5’UTR包含分支点和3’剪接受体位点。分支点在本领域中应理解为包含参与对5'供体剪接位点发起亲核攻击的一个或多个核苷酸。在一些实施方案中,编码可表达多肽的核酸序列的5'UTR不包含分支点。在一些实施方案中,编码可表达多肽的核酸序列的5’UTR包含间隔区序列。在一些实施方案中,间隔区序列包含至少一个CAA重复。在一些实施方案中,编码可表达多肽的核酸序列的5'UTR具有GCGGCCGCCTTAATTAACAGTGTTCACTAGAGCCAACAACAACAACAACAACAACAACAACAACGACACC(SEQ ID NO.:48)的序列。
在一些实施方案中,本公开中设想的编码可表达多肽的核酸序列可以是编码任何多肽的任何核酸序列或任何基因。在一些实施方案中,编码非编码RNA的核酸序列。在一些实施方案中,本公开中设想的编码可表达多肽的核酸序列可以是外源核酸。在一些实施方案中,本公开中设想的编码可表达多肽的核酸序列可以是对于已引入polyA适体多核苷酸的受试者是内源性的基因。在一些实施方案中,将本公开的polyA适体多核苷酸引入到个体基因组的调控目的基因表达的区域中。因此,在一些实施方案中,本公开的polyA适体多核苷酸可用于调控对于个体是内源性的基因的表达。在一些实施方案中,本公开的polyA适体多核苷酸的编码可表达多肽的核酸序列是内源核酸序列。
在一些实施方案中,可表达多肽是胰岛素。在一些实施方案中,可表达多肽是人生长激素。在一些实施方案中,可表达多肽是凝血因子X。在一些实施方案中,可表达多肽是抗肌萎缩蛋白。在一些实施方案中,可表达多肽是自杀蛋白。在一些实施方案中,自杀蛋白是诱导细胞死亡的蛋白。示例性自杀蛋白包括混合谱系激酶结构域样假激酶(MLKL)、受体相互作用的丝氨酸/苏氨酸-蛋白激酶3(RIPK3)、受体相互作用的丝氨酸/苏氨酸-蛋白激酶1(RIPK1)、具有死亡结构域的Fas相关蛋白(FADD)或gasdermin D(GSDMD)、半胱氨酸-天冬氨酸蛋白酶、半胱氨酸天冬氨酸酶或半胱氨酸依赖性天冬氨酸定向蛋白酶(CASPASE-1或CASP-1)、CASPASE-4、CASPASE-5、CASPASE-12、PYCARD/ASC(含有PYD和CARD结构域的/具有死亡结构域的Fas相关蛋白)或其变体。
在一些实施方案中,可表达多肽是可检测的基因产物。在一些实施方案中,可检测的基因产物是报告基因。在一些实施方案中,报告基因是能够提供可检测信号和/或包括产生可检测信号的能力(例如通过催化将化合物转化为可检测产物的反应)的蛋白质。可检测信号可包括例如荧光或发光。可检测信号、检测它们的方法以及将它们并入试剂(例如包含报告蛋白的多肽)中的方法是本领域众所周知的。在任何方面的一些实施方案中,可检测信号可包括可通过光谱、光化学、生物化学、免疫化学、电磁、放射化学或化学手段检测的信号(诸如荧光、化学荧光或化学发光)或通过任何其他适当的手段检测的信号。在任何方面的一些实施方案中,报告蛋白选自由以下组成的组:荧光素酶、纳米荧光素酶、β-内酰胺酶、β-半乳糖苷酶、辣根过氧化物酶、碱性磷酸酶、过氧化氢酶、碳酸酐酶、绿色荧光蛋白、红色荧光蛋白、青色荧光蛋白、黄色荧光蛋白、胰蛋白酶、蛋白酶、补充并激活截短报告蛋白的肽、激酶。
在一些实施方案中,本公开的polyA适体多核苷酸的活性或功能通过可表达多肽的表达来测量。在一些实施方案中,本公开的polyA适体多核苷酸的活性或功能通过倍数诱导来测量。在一些实施方案中,倍数诱导计算为存在配体的情况下的可表达多肽与不存在配体的情况下的可表达多肽的比率。在一些实施方案中,倍数诱导计算为存在适体的情况下的可表达多肽与存在不同适体的情况下的可表达多肽的比率。在一些实施方案中,倍数诱导计算为存在包含至少一个剪接受体位点和一个剪接供体位点的适体的情况下的可表达多肽与存在不具有剪接位点的不同适体的情况下的可表达多肽的比率。在一些实施方案中,倍数诱导计算为在引入polyA适体多核苷酸之前内源基因的表达与在引入调控同一内源基因表达的polyA适体多核苷酸之后内源基因的表达的比率。
配体
根据各种实施方案,可选择配体以促进所提供系统的所需最终目的。因此,配体可以是或包含多肽、核酸、小分子、药物、代谢物或其组合。在一些实施方案中,配体可以是或包含细胞代谢物、异常细胞蛋白质或由病原生物体(例如,病毒、细菌或真菌)表达的蛋白质。例如,在一些实施方案中,配体可以是外源施用的小分子,使得可以在特定治疗环境中根据需要容易地调节系统的剂量和功能。例如,在一些实施方案中,配体是四环素或其衍生物。在一些实施方案中,可选择配体使得可表达多肽的表达响应于特定的生物条件(例如,感染、肿瘤发生、高或低葡萄糖)而发生,例如,作为可检测细胞、组织或受试者中的一种或多种细胞内“特征”的生物传感器系统。因此,在一些实施方案中,配体对于受试者是内源性的(例如,内源蛋白)。在一些实施方案中,配体是新霉素或其衍生物。在一些实施方案中,配体是茶碱或其衍生物。在一些实施方案中,配体是葡萄糖。在一些实施方案中,配体是癌症生物标志物。
载体
在一些实施方案中,本公开的polyA适体多核苷酸可通过载体引入。在一些实施方案中,载体可为病毒载体。合适的病毒载体包括但不限于慢病毒载体、逆转录病毒载体、α病毒、picornal(例如,脊髓灰质炎)疫苗、腺病毒、腺相关病毒、疱疹病毒和禽痘病毒载体。
包括治疗的示例性用途
根据本公开,polyA适体多核苷酸和/或包括一种或多种polyA适体多核苷酸的系统可用于多种应用中的任一种。例如,在一些实施方案中,本公开的polyA适体多核苷酸用于治疗患有疾病的个体,例如,通过提供由可表达多核苷酸编码的治疗性蛋白质的可控表达。在一些实施方案中,疾病是由遗传病症引起的某种或某些蛋白质的缺乏。在一些实施方案中,疾病是糖尿病、糖尿病前期或糖尿病并发症。在一些实施方案中,疾病是癌症。在一些实施方案中,疾病是肌营养不良。在一些实施方案中,疾病是遗传性因子X缺乏。在一些实施方案中,本公开的polyA适体多核苷酸与疾病的其他治疗组合提供。在一些实施方案中,本公开的polyA适体多核苷酸用于诱导将细胞重编程成多能干细胞(诱导多能干细胞或iPSC)。在一些实施方案中,在疾病的其他治疗之前、期间或之后引入或施用本公开的polyA适体多核苷酸。在一些实施方案中,治疗性蛋白质可以是或包含胰岛素、生长激素、抗肌萎缩蛋白、白蛋白、因子IX、Oct4、Sox2、Klf4、cMyc及其任何组合。
在一些实施方案中,包含polyA适体多核苷酸的系统可用于提供关于疗法在特定受试者中是否有效的信息。在需要确定一种或多种疗法在受试者中是否有效的一些实施方案中,在提供疗法之前可在受试者中使用系统,以诸如检测用于疗法的特定指示化合物的存在或不存在,然后在提供一次或多次疗法之后,可在受试者中使用所述系统来检测特定指示化合物的存在或不存在。在其他实施方案中,直到在向受试者提供一次或多次疗法之后,系统才用于监测疗法,以诸如鉴定指示疗法功效的特定化合物的存在或不存在。
在一些实施方案中,polyA适体多核苷酸和/或包括一种或多种polyA适体多核苷酸的系统可用作生物传感器。根据各种实施方案,所提供的系统可提供关于特定环境(例如,细胞内、细胞外和/或环境的环境)的空间和/或时间信息。例如,在一些实施方案中,包含至少一种polyA适体多核苷酸的系统可用于检测受试者中的一种或多种特定分子特征以及允许产生所需的可表达多肽以便响应于一种或多种分子特征的存在而实现所需生物学状态。在一些实施方案中,分子特征可以是或包括:特定内源基因产物(例如,疾病相关基因产物/蛋白质)的存在、毒素的存在、外源基因产物的存在、代谢物(例如,来自环境污染物的代谢物)的存在及其任何组合。
在一些实施方案中,polyA适体多核苷酸可包含一个或多个报告基因部分(例如,报告基因产物,例如,成像报告基因)。在一些实施方案中,包含在polyA适体多核苷酸中的可表达多核苷酸编码报告基因产物(例如,蛋白质)。在一些实施方案中,报告基因产物可以是或包含荧光素酶、绿色荧光蛋白、红色荧光蛋白、β-半乳糖苷酶、红外荧光蛋白、近红外荧光蛋白、视蛋白及其任何组合。
在一些实施方案中,包含polyA适体多核苷酸的系统可编码报告基因产物和治疗基因产物。在一些此类实施方案中,报告基因产物和治疗基因产物的表达可由同一适体控制。在一些实施方案中,报告基因产物和治疗基因产物的表达可由不同的适体控制。
范例
本实施例描述了高度响应的基因调控机制,所述机制利用药物诱导的替代剪接的力量来控制polyA切割。图1提供了本公开的一些实施方案的表示。如图1A所示,当在转基因的5'UTR处人工创建工程短内含子(mini-IVS2)和新的polyA信号(红色)时,内含子的有效剪接和polyA信号的切割导致mRNA的后半部分被破坏,并因此导致基因表达丧失。特定配体与作为Y形开关(绿色)的一部分设计的适体的结合有效地诱导了替代剪接。配体诱导的替代剪接导致Y形结构和人工5'UTR polyA信号的去除。这反过来又导致完整mRNA的保存,并因此导致诱导的基因表达。应注意到,第二3'剪接位点(3'ss)构建在5'UTR序列中。此3'剪接位点仅在配体(例如,四环素,“Tc”)与适体结合后被激活。Y结构旁边的4MAZ序列是为了加强配体结合时的替代剪接。
图1B提供了如本文所述的包含三个适体的polyA开关的展示。每个适体位于Y形RNA结构的一个臂上。这种Y形设计具有几个重要的优点:它包含3个适体来控制战略性地放置在中央三向接合部处的polyA信号(pA)。通过这样做,它利用了由三种不同适体产生的四环素结合效应的联合力量;Y形结构紧凑,并且需要整体较短的序列来包含3个适体;Y形结构被设计来在RNA生物合成期间进行固有折叠。三个适体以正向-正反方向排列,以最小化适体之间替代折叠的机会。此外,已知长于35bp的双链RNA茎在细胞中引起先天免疫反应。因此,Y结构中的所有茎都被制成显著短于35pb,以消除先天免疫反应。
图1C提供了如本文所述的polyA开关的核酸序列的实例(Y196CAA)。设计并测试了多于370个结构,以广泛探测Y形结构的每个部件的影响。这些包括(1)每个臂的长度、(2)每个臂的序列、(3)每个臂的环、以及(4)放置polyA信号的中央三向接合部的序列和大小。在这些非限制性实施例中进一步描述了这些部件的修饰效果。
实施例1:PolyA切割信号的调节
位置
制备构建体以测试另外的Y形结构,所述Y形结构不同地配置并且polyA切割信号不同地定位。制备四种不同的构建体:B1-B4,其中polyA信号(红色)放置在适体C附近并被三向接合部钳制(图2A;示出了B1构建体)。这些不显示或显示出极小的诱导作用。制备在三向接合部附近具有polyA信号的另外四种构建体:T1-T4(图2B)。这些也显示出极小或中度的诱导作用。图2C例证了一个polyA开关,其中3个适体彼此堆叠而没有三向接合部。对于这种配置,观察到极小的诱导作用。图1B中示出的特定Y形配置(其中polyA信号放置在三向接合部附近)用于另外的测试。在这种配置中,三向接合部以不同的方向弯曲,以提供钳制polyA信号的独特几何形状。每个臂的稳定性由两个因素决定:碱基对的数量和碱基对的组成(例如,G-C比A-U或G-U对更稳定)。
PolyA切割信号的数量
进行了测试以评估Y形结构中一个或多个polyA信号的最佳数量。图3A展示了对Y系列中的三种结构的测试,其中2个polyA信号由红色方框指示。Y1示出约12倍的诱导作用,在这三个构建体中最高。在所述组中,臂3-1的大部分是A-U或G-U对,因此需要较长的茎才能达到一定的稳定性。如图所示,本文例证的构建体的臂包含双链核酸茎。较短的臂3-1提供较低的诱导作用。图3B进一步展示了臂长的影响。Y5至Y9仅具有一个polyA信号(红色方框),其中臂3-1(蓝色方框)和臂2-1(绿色方框)的长度可变。臂3-1和臂2-1的长度从Y5至Y9逐步缩短1bp。这一polyA配置导致更好的诱导作用。图3C表明,当臂1-2中的一行中存在2个polyA信号(Y6mut)时,诱导作用降低大约一半。Y6mut:与Y6相同,除了臂1-2中嵌入2个polyA信号(红色方框)。基于这些结果,确定polyA信号的最佳数量和位置:单个polyA信号部分嵌入臂1-2和三向接合部中。所述配置用作进一步优化的基础。
实施例2:三向接合部的优化
修改三向接合部的环境直接影响polyA信号的钳制。因此,Y形开关的性能对三向接合部的任何变化都非常敏感。围绕三向接合部进行了广泛的突变/插入/缺失研究,以鉴定最佳序列。图4A显示出Y22中的U到G突变使诱导作用加倍,可能是因为所述突变在臂3-1上产生了加强了polyA信号的钳制的新的G-U碱基对。图4B提供了显示不同三向接合部序列对诱导作用的影响的实例。图4C比较了三向接合部的方框1中具有3个碱基的构建体与具有1个碱基的构建体。Y107至Y110是在方框1中具有3个碱基的Y79的衍生物。Y107至Y110在方框1中仅具有一个碱基。Y107的性能与Y79相似,表明方框1中的一个未配对碱基就足够了。图4D示出了将一个碱基插入到三向接合部的方框2中的结果,这会导致三向接合部中折叠的细微变化。结果表明,最佳配置是方框2中的一个未配对碱基。对于图4E中的构建体,方框1和方框2中的单碱基是随机的。对16个组合进行了测试,并且结果显示出与同一天测试的亲本Y79相比,Y127、Y130和Y134是其中最好的。图4F示出了使用Y130作为基础的构建体的进一步优化。经测试的修饰都没有导致显著的改进。图4G示出了导致诱导作用几乎没有变化的相对于Y143进行的另外修饰。图4H示出了相对于Y147进行的另外修饰。与Y147相比,Y163略微提高了诱导作用,而Y162略微降低了诱导作用。图4I示出了相对于Y163进行的另外修饰。与Y163相比,Y177略微提高了诱导作用,而Y178略微降低了诱导作用。图4J示出了相对于Y152进行的修饰。与Y152相比,这些修饰导致显著改进。特别地,Y166几乎使诱导作用加倍。Y166充当进一步优化的新基础。图4K示出了相对于Y166进行的另外修饰。与Y166相比,这些修饰导致显著改进。它们还充当优化的新基础。
Y174、Y175、Y176和Y177(参见图4L)是最佳三向接合部序列。所有这些构建体在方框1和方框2中都具有单碱基C或A。在这些构建体中,polyA信号AAUAAA(红色方框)的前3个碱基在三向接合部的口袋中是打开的。polyA信号的最后2个碱基嵌入臂1-2中。
改变polyA信号相对于三向接合部口袋的位置可改变诱导能力(图5)。在Y135至Y140中,相对于Y101进行了改变,三向接合部的口袋沿polyA信号移动。结果,polyA信号在臂1-2中嵌入更深。这些修饰导致较低的诱导作用。Y101mut是Y101的衍生物,在臂2-1中含有去除潜在的3'剪接位点的翻转的C-G对(由红色箭头指示)。构建体Y141至Y159基于Y101mut。三向接合部口袋沿polyA信号移动。沿polyA信号移动三向接合部口袋的诱导结果在图5的最后部分中示出。
实施例3:双链茎
如本文所述的PolyA适体多核苷酸构建体包含核酸(例如,RNA)双链茎。此类双链区在本公开中也称为臂。长度、稳定性和核苷酸组成的修饰可影响polyA适体多核苷酸的强度和有效性。
早期的结果(使用构建体Y1至Y9,图3)指示臂3-1的稳定性需要在一定范围内。臂3是非常敏感的区域,因为它非常接近polyA信号。臂3稳定性的微小变化会导致polyA信号钳制的显著变化,从而导致诱导作用。以Y35为基础,我们进行了许多修饰以优化臂3。图6A至图6B展示了基于臂3变化的诱导作用变化。在这些图中,亲代构建体位于右侧,并且修饰结果显示在左侧。图6A示出了臂3-1的修饰结果。臂3-2强度降低的构建体Y43至Y45基于Y35;臂3-2强度降低的构建体Y188C和Y189C基于Y175;臂3-2强度降低的构建体Y188D和Y189D基于Y176。通过在不同位置将G-C对更改为G-U对而具有较弱臂3-2强度的构建体Y219A至Y224A基于Y197。图6B示出了臂3-2的修饰结果。构建体Y201至Y203基于Y175。具有较弱臂3-2的构建体Y216B至Y217B基于Y208。结果表明,增加臂3-2的长度和改变环序列大大降低诱导作用。
这些修饰中的大多数显著降低诱导作用,并且没有一个超过Y35。因此,Y35的臂3代表了所测试的构建体的Y形结构的最佳臂3序列。用于臂3修饰的一些其他亲本构建体(诸如Y175、Y197和Y210)都共享相同的Y35的臂3序列。
对是臂2(即臂2-1和臂2-2)的双链茎的修饰会改变臂2的稳定性。所述修饰包括长度、序列的变化以及在茎中产生错配的点突变(图7)。
图7A示出了臂2-2的修饰结果。构建体Y48至Y53基于Y35。图7B示出了臂2-1修饰的结果。这些修饰的结果指示,与臂3相比,诱导作用对臂2稳定性的变化不太敏感。这大概是因为臂2没有直接连接到polyA信号。尽管如此,臂2需要一定程度的稳定性才能实现良好的诱导作用。不稳定的臂2导致非常低的诱导作用。这些结果中显示的臂2的序列是凭经验确定的。一些臂2序列已经在最佳稳定性范围内,并且代表导致非常有效的诱导作用的接近最佳序列。稳定性的进一步增加会提高或降低诱导作用。
图8示出了各种修饰臂1-2的结果。图9示出了臂1-1的各种修饰结果。
实施例4:适体的方向
每个适体相对于其他适体的方向可能对polyA适体多核苷酸的功能产生影响。图10A示出了基于Y35的构建体Y54至Y57的结果,其中适体B方向反转。反转适体B的方向在很大程度上消除诱导作用。图10B示出了基于Y196CAA的构建体Y240至Y252的结果,其中适体A方向反转。反转适体A的方向完全消除诱导作用,无论臂1-2的长度如何。
实施例5:每个适体对诱导作用的贡献
图11A展示了Y形结构的每个适体对诱导作用的贡献。Y形结构的每个适体可通过结合口袋中的A到C突变(箭头)禁用,这消除了与其配体四环素的结合。NA:适体A被禁用;NB:适体B被禁用;NC:适体C被禁用;NAB:适体A和B被禁用;NBC:适体B和C被禁用;NAC:适体A和C被禁用。这些结果指示适体C对最终诱导作用的贡献最重要。之后是适体B,然后是适体A。
图11B展示了从Y形结构中去除适体A的影响。方框指示为每个构建体去除的序列。尽管与亲本Y196CAA相比水平显著降低,但去除适体A保留适度诱导作用。
实施例6:5'UTR的修饰
图12A表明,在5'UTR中插入CAA重复序列(带下划线)可改变诱导作用水平。在这里,在Y196、Y208、Y209和Y211中插入CAA重复序列都导致更高的诱导作用。将含有CAA重复序列的间隔区序列插入Y301的5'UTR中会导致对诱导作用产生不同的影响。这些间隔区序列彼此之间仅略有不同,但导致诱导作用差异很大,这表明所述区域对变化非常敏感。图12B示出了使用S56作为亲本构建体测试具有强3'剪接位点的新5'UTR序列的一些实例。图12C示出了在不使用CAA重复序列的情况下,将固有非结构化的RNA序列添加到翻译起始ATG附近的5'UTR的结果。这些构建体基于Y300和Y305。在基于Y300的构建体中,Y329是最好的。虽然它没有超过Y305的性能,但它具有不使用CAA重复序列的优点。图12D示出了CAA重复序列的插入位置也显著影响诱导作用。
实施例7:G Quad序列的重要性
我们测试了G-quad序列对诱导作用的影响。图13A显示出3MAZ或CD44 G-quad与使用Y196CAA作为亲本的2MAZ相比达到相似的诱导作用水平。然而,4MAZ由于其有效诱导替代剪接的能力使诱导作用显著加倍。图13B示出了当使用S56构建体作为亲本测试不同的G-quad序列以替换4MAZ G-quad时的诱导作用结果。在这些构建体中,4MAZ被以下替换:一个CD44 G-quad‘TGGTGGTGGAATGGT’(S177)、两个CD44 G-quad‘TGGTGGTGGAATGGTAAATGGTGGTGGAATGGT’(S178)或四个CD44 G-quad‘TGGTGGTGGAATGGTAAATGGTGGTGGAATGGTAAATGGTGGTGGAATGGTAAATGG TGGTGGAATGGT’(S179)。结果表明4MAZ的作用是独特的,并且不能被其他G-quad序列替换。4MAZ序列具有独特的特性,并且是混合开关的关键元件,需要有效的polyA信号切割和Tc诱导的替代剪接。图14进一步展示了4MAZ序列的重要性。RT-PCR揭示了药物诱导的替代剪接的机制。在不存在Tc的情况下,IVS2剪接的RNA通过polyA切割降解(泳道1和3)。Tc的存在诱导Y196CAA-2MAZ和Y196CAA-4MAZ(泳道2和4)中的替代剪接。Sanger测序证实了Tc诱导的带(下带)含有预期的替代剪接RNA接合。与Y196CAA-2MAZ相比,Tc诱导的替代剪接在Y196CAA-4MAZ中更为明显(泳道4与2)。在这种诱导的替代剪接下,在存在Tc的情况下,polyA信号和Y形结构都被去除,并且对蛋白质表达的诱导作用显著增加。
实施例8:调节第一3'剪接受体位点
为了进一步优化Tc诱导的替代剪接机制,我们广泛探讨了IVS2 3'剪接位点位置和周围序列/结构的影响。修饰包括:将IVS23'剪接位点嵌入臂1;将IVS2 3'剪接位点移动接近或远离适体结合位点;将IVS2 3'剪接位点放在臂1的松散凸出中;改变携带IVS23'剪接位点的臂1的长度或稳定性;改变IVS2 3'剪接位点的剪接强度。图15A示出了将IVS2 3'剪接位点逐渐移动到Y196CAA-4MAZ的臂1-1中的结果(S1-S4)。它还表明,当IVS2 3'剪接位点从CAG突变为CCC时(S5),诱导作用几乎被消除。图15B表明,当IVS 3'剪接位点完全嵌入适体A的Tc结合口袋附近的臂1-1中时(红色箭头;S9),所述剪接位点被强烈抑制,从而导致非常低的诱导作用。这表明适体对IVS2 3'剪接位点的钳制不能太强。此外,通过删除适体A的部分序列(S9m)来降低适体A的钳制效应可恢复诱导作用。沿着S9m的臂1移动IVS 3'剪接位点会导致S19与亲本S9m相比更短且具有相似的诱导作用水平(图15C)。图15D展示了当IVS2 3'剪接位点CAG放置在臂1-2的凸出中时对诱导作用的影响。S47至S50基于S19。在1ug/mL Tc时,S47至S50中的大多数产生较低的诱导作用。在5ug/mL Tc时,除了S50之外,S47至S50与S19相比具有相似或更高的诱导作用。图15E示出了通过突变IVS2 3'剪接位点之后的碱基来改变预测的剪接强度的结果。在基于S9m和基于Y196CAA-4MAZ的配置中,改变IVS2 3'剪接位点的强度不会显著改变诱导作用。图15F示出了将mini-IVS2 3'剪接位点进一步移动进入或远离茎的结果,这都导致较低的诱导作用。图15G示出了在mini-IVS2的3'剪接位点的cag之后的三个碱基随机化以选择具有最高性能的序列的效果。这组构建体(特别是Y362、Y366和Y367)表现出卓越的开关效率,超过了Y300和Y301的性能。通过测试鉴定最佳NNN序列:基于Y344的:Y359(CAT)、Y360(TTT)、Y361(TGA)、Y362(TCT);基于Y358的:Y363(CAT)、Y366(TAC)、Y367(TTT)
实施例9:调节5'UTR中的第二3'剪接受体位点
进行测定以测试调节5'UTR中的第二3'剪接受体位点强度的效果。位于4MAZ之后和起始密码子ATG之前的Y196CAA-4MAZ的5'UTR序列具有以下序列:gcggccgccaacaacaacaacaacaacaacaacaacaacaacaacaacataacagtgttcactagcaacctcaaacagacaccATG。添加另外的分支点(S10)、ppt(S11)或将CAG突变为CCC(S12)或AAG(S13)都会导致诱导作用降低(图16A)。为了在不存在Tc的情况下激活正确的3'剪接位点(IVS2 3'剪接位点),以及在存在Tc的情况下激活正确的3'剪接位点(第二替代3'剪接位点),我们使用具有短内含子的构建体作为起点,并且使用随机化方法来选择5'UTR(TAGNNN)中的TAG之后的最佳三个碱基以改进诱导作用(图16B)。我们还将这三个最佳碱基(NNN)插入Y329的5'UTR以评估性能(图16C)。在这些中,Y344的性能最好。
实施例10:内含子大小
我们测试了通过减小IVS2内含子的大小来缩短混合开关整体大小的效果。图17A示出了示例性内含子序列。构建体S164至S168与S159至S163相似,但在IVS2 3'剪接位点之前的相同位置插入了分支点TACTAAC。以S164的内含子序列为例:Gtgagtctatgccagctacca ttctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatCttc aTACTAACctcttatcttcctctgCAG。构建体S169至S173与S159至S163相似,但在IVS23'剪接位点之前的相同位置插入了分支点TACTAAC和再一个3'剪接位点CAG。以S169的内含子序列为例:GTgagtctatgccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagcTACTAACttttcctgtgcttcttcagacctcttatcttcctctgCAG。将IVS2内含子大小从476个碱基减少到120-200个碱基显著降低了诱导作用(图16B)。添加了不同的剪接元件以加强IVS2内含子剪接的Y164到Y173的结果与没有添加这些元件的结果相比导致更低的诱导作用。这表明在存在Tc的情况下,缩短IVS2内含子的元件或向IVS2内含子中添加元件会改变3'剪接位点激活的选择。先前我们已经证明CAA重复序列会改变5'UTR中3'剪接位点的剪接强度。在这里,将从S159、S164和S169中去除CAA重复序列(红色)。与S56相比,S192(具有120个碱基内含子)在1ug/mL Tc下的诱导作用更好,并且在5ug/mL Tc下的诱导作用相似。由于更短的内含子而更紧凑的S192用作进一步修饰的新基础。
实施例11:上游框外AUG(μORF)的添加
引入上游框外AUG来构建S192,以测试IVS2剪接转录物对报告基因翻译的影响。修饰包括:(1)紧接IVS2 3'剪接位点之后将TAC更改为ATG以创建新的起始密码子(红色方框),(2)更改臂1另一侧的相应碱基以保持茎中的碱基配对,以及(3)将臂2-1中的框内终止密码子tga突变为aga(红色箭头),因此从这个新ATG的翻译可产生相当长的蛋白质。参见图18A。
显示了IVS2 3'剪接位点CAG之后的序列。新μORF加了下划线:ctgCAGATGttcctcg agatctggggaggtgaagaatacgaccacctaataagattaccgaaaggcaatcttattaaaacataccagatctt gagagggtgtttgtggcaaaacataccagatcgaattcgatctggggaggtgaagaatacgaccacctgctacaag tacctaataaaCATtagCGGaGaaacataccactgtgtgttggttttttgtgtgttaacgggggagggggaggaaa gggggagggggaggaaagggggagggggaggaaagggggagggggagcggccgccataacagtgttcactagcaaccTcaaacagacaccATG。这种方法显著降低了IVS2剪接转录物的泄漏表达,因此显著增加了S206的结果所示的诱导作用。
通过基于S206(图18B)对5'UTR序列进行微调来进一步优化此构建体。所有这些构建体都表现出非常好的诱导作用。由于较短的内含子和部分缺失的适体A,这些构建体更紧凑。它们在低至1ug/mL的Tc浓度下表现非常好,并且在5ug/mL下达到高达约700倍的诱导作用。
综上所述,在优化Tc对IVS2 3'剪接位点与替代3'剪接位点之间剪接选择的影响的过程中,我们发现放置IVS2 3'剪接位点的最佳位置是将其嵌入Y结构的臂1中。为了将IVS2 3'剪接位点放置在所述位置,从Y结构中删除适体A。创建消除IVS2剪接转录物中的报告基因翻译的上游框外AUG(μORF)可降低泄漏表达。与Y196CAA-4MAZ相比,S222(图17C)在较低的药物浓度、较高的基因表达水平下显示现出较高的诱导倍数,并且也许更重要的是,S222对Tc高度敏感,并且在低Tc浓度下表现良好。
构建体性能
图19A展示了代表性S系列构建体相对于Y196CAA-4MAZ的性能比较。图18B示出了通过显微镜观察到的来自混合开关构建体的表达的剂量反应。
为了避免上游开放阅读框(μORF)的蛋白质翻译产生的潜在免疫原性,我们构建了旨在超越S222的性能的另一个不含μORF的混合开关。为了构建这个新的混合开关,我们回到了Y196CAA-4MAZ设计,因为它有3个适体,而S222中有2个适体。为了进一步改进Y196CAA-4MAZ,我们(1)使用具有120个碱基的mini-IVS2内含子,(2)优化mini-IVS2序列的3'剪接位点,(3)优化含有下游替代3'剪接位点的5'UTR序列。这些努力导致产生在性能上超过S222的一组构建体。四环素的诱导作用非常有效,在低至0.5μg/ml至1μg/ml的药物浓度下,它们诱导基因表达达到最大水平的50%(EC50)。这种四环素浓度可使用FDA批准的剂量在人血清中常规实现,并且比先前使用任何基于RNA的基因调控技术实现的浓度低一个数量级。图19C展示了这些新构建体的性能与S222的性能的比较。Y300的5'UTR序列:gcggccgcCataacagtgttcactagcaTccCcaaacagacaccATG。Y301:基于Y300,具有修饰的5’UTR gcggccTTaATtaacagtgttcactaggacaccATG。
图19D展示了通过荧光素酶测定确定的Y362和Y367的性能。图19E示出了Y362和Y367对1ug/ml四环素的反应,如通过使用eGFP报告基因信号的荧光激活细胞分选(FACS)确定的。所有结果中的“诱导倍数”计算为在存在与不存在四环素的情况下转基因表达的比率。
实施例12:核糖开关在内源性位置的插入
Y形polyA开关在与CRISPR结合时创建强大的技术平台来控制哺乳动物基因组中任何内源基因的表达。图20提供了使用干细胞膜蛋白CD133来展示原理的示意图。内源性CD133的条件基因表达是通过使用CRISPR-Cas9和修复矩阵在CD133的5'UTR处插入Y196核糖开关来实现的。图20A顶部:三个gRNA(g1、g2和g3)用于指定CD133的翻译起始附近的CRISPR-Cas9切割的位置。图20A底部:两侧为CD133的上游和下游同源序列的含有mini-CMV启动子、IVS2内含子和Y196核糖开关的修复矩阵用于修复。图20B提供了实验程序的示意图。Y196核糖开关首先通过CRISPR-Cas9插入亲本CD133-细胞。然后,成功工程化的细胞以剂量依赖性的方式对Tc作出反应,以开启CD133表达。针对CD133蛋白的FITC缀合的抗体用于标记和分离对Tc作出反应的细胞。图19C显示出内源性CD133的条件表达受Tc调控。工程细胞克隆(在这种情况下为293T细胞)中的CD133表达显示出几乎没有或没有背景泄漏。CD133表达是由Tc特异性诱导的,但不是其类似物Doxy。ND:无药物治疗,Tc:四环素,Doxy:强力霉素。在使用或不使用药物的情况下处理细胞克隆2天,然后收获用于流动分析。X轴显示出单个细胞的抗体染色强度。图20D如预期的那样显示出,由Tc诱导的CD133蛋白(如FITC-抗CD133抗体所揭示的)与正常内源性CD133蛋白一样定位于细胞膜。在使用或不使用2μg/ml药物的情况下处理稳定的细胞克隆2天,然后收获用于图像流动分析(Amnis)。同样,诱导作用显然是特定于Tc而不是Doxy。
所描述的数据代表了高度响应的基因调控机制,所述机制利用药物诱导的替代剪接的力量来控制polyA切割。工程化的组合创建了敏感的基于RNA的开关,所述开关可由小分子药物控制,并且能够严格调控哺乳动物细胞中的基因表达。与其他报道的方法相比,本文所述的这种混合开关技术表现出非常低的泄漏表达,并且有效地开启了人细胞中接近700倍的转基因表达。此外,四环素的诱导作用非常有效,在低至0.5μg/ml至1μg/ml的药物浓度下,它诱导基因表达达到最大水平的50%(EC50)。这种四环素浓度可使用FDA批准的剂量在人血清中常规实现,并且比先前使用其他基于RNA的基因调控技术实现的浓度低一个数量级。
因此,这种混合开关技术有利地安全地用于人患者以控制治疗基因或转基因的表达。因此,本公开满足了本领域长期以来的需求,即提供高效且非免疫原性的技术,以对人类食用安全的药物浓度调控细胞中的目的基因。
实施例13:三个位置的单碱基变化的组合
测试了对Y形结构序列的三个碱基变化的组合,以确定对poly A适体的诱导作用性能的累积影响。如图21所示,这三种突变由臂1-1中的“A”缺失;关闭臂2-2中的未配对中断的“A”到“G”变化;和在polyA信号之前的三向接合部中的“A”插入组成。这些突变是使用四种不同的亲本构建体实现的,所述亲本构建体在mini-IVS2内含子后面具有不同的碱基。如表1所示,总共设计了12个构建体来探测累积效应。

图22表明,三个单碱基变化的组合在较低药物浓度下显著增加诱导作用。另外,图23A和图23B展示了构建体Y362和Y387的剂量反应分析。仅使用1ug/ml四环素,Y362和Y387在293T细胞中有效地开启高达650至700倍的转基因表达。对于两种构建体,使用最大诱导倍数作为EC100参考,四环素的诱导作用在低至0.5μg/ml至1μg/ml Tc时达到最大水平的50%(EC50)(图23A)。使用亲本构建体(HDM-Luc,其具有类似的序列但没有Y形结构)的最大表达水平作为EC100参考的计算也显示出低至0.5μg/ml至1.2μg/ml的类似EC50值(图23B)。Y387是特别有效的设计,因为它的EC50值为0.5μg/ml,无论所使用的EC100参考是多少。
实施例14:方法
在此提交的附图中描述的测定如下进行:
荧光素酶测定
将细胞以25000-30000个细胞/孔的密度接种在96孔板中。孵育24小时后,用50ngDNA载体转染每个孔,并与不含四环素或含有不同浓度的四环素的培养基再孵育18小时。使用Polarstar Omega读板器(BMG Labtech,USA)以相对光单位(RLU)测量荧光素酶活性。为了制备36mL的测定缓冲液,将144μL 1M DTT、108μL 0.1M ATP、252μL 0.1M荧光素和360μl0.05M辅酶A添加到35mL基本缓冲液(25mM Tricine、0.5mM EDTA-Na2、0.54mM三磷酸钠、16.3mM MgSO4.7H2O和0.8%Triton X-100)。移除细胞培养基之后,向每个孔中添加40μL测定缓冲液,并且用Polarstar Omega读板器读取荧光素酶活性两次。诱导倍数计算为存在与不存在四环素时转基因表达的比率。
RT-PCR
在不存在或存在四环素的情况下,将用相应构建体转染的细胞在培养基中于37℃生长18小时。根据RiboPureTM RNA纯化试剂盒(Ambion,Austin,TX)提供的方案分离总RNA。对于RT-PCR,根据制造商的方案使用SuperScript III(invitrogen,Carlsbad,CA)进行RT,并且使用靶向转录物和报告基因开头的引物进行PCR。
荧光显微镜
将细胞以1.2×105个细胞/孔的密度接种在12孔板中。孵育24小时后,用500ngDNA载体转染每个孔,并与不含四环素或含有不同浓度的四环素的培养基再孵育18小时。图像是在荧光显微镜(Zeiss Axiovert 40CFL)上以200倍的放大倍数拍摄的。
实施例15:示例性构建体序列
以下序列是本文所述的系统的部件的实施方案的另外实例。所述序列作为DNA序列提供,当转录时形成RNA适体的部件:
+1:转录起始
黑色:5'前导RNA序列
下划线:IVS2内含子或mini-IVS2内含子
粗体:Y形polyA开关(4MAZ带下划线)
斜体:5'UTR
ATG:翻译起始以粗体显示
Y196CAA-4MAZ


Y208

Y209


Y211

Y226


Y227


Y300

Y329

Y305


Y305D1

Y305D2

Y305D3


Y305D4

Y305D5


Y305D6

Y305D7

Y301


Y305D9

Y305D10

Y305D11


Y305D12

Y305D13


Y344

Y359

Y360


Y361

Y362

Y358


Y363

Y366


Y367

Y375

Y376


S206

S210

S211


S212

S213

S214

S215

S222

S223

S272

Y387


Y392

Y393

Y394


Y395

Y396

Y397

等效方案
本领域技术人员仅仅使用常规试验将认识到或者能够确定本文所描述的发明的具体实施方案的很多等效方案。本发明的范围不旨在限于上述说明,而是如以下权利要求所述。
Claims (61)
1.一种用于调节基因表达的系统,所述系统包含polyA适体多核苷酸,
所述polyA适体多核苷酸在5'至3'方向上包含:
a)5'剪接供体位点;
b)工程内含子;
c)第一3'剪接受体位点;
d)polyA开关,其包含具有一个或多个配体结合口袋的两个或更多个配体结合适体,和其中的至少一个polyA切割信号;
e)第二3'剪接受体位点;以及
f)编码可表达多肽的核酸序列。
2.如权利要求1所述的系统,其中所述polyA开关包含两个配体结合适体。
3.如权利要求1所述的系统,其中所述polyA开关包含三个配体结合适体。
4.如权利要求1所述的系统,其中所述polyA开关包含三向接合部。
5.如权利要求4所述的系统,其中所述三向接合部包含第一双链RNA茎、第二双链RNA茎和第三双链RNA茎的接合部。
6.如权利要求5所述的系统,其中所述第一双链RNA茎不包含配体结合适体。
7.如权利要求5所述的系统,其中所述第一双链RNA茎、第二双链RNA茎和第三双链RNA茎中的每一个都包含配体结合适体。
8.如权利要求5所述的系统,其中所述三向接合部包含至少一个单链区。
9.如权利要求8所述的系统,其中所述三向接合部包含第一单链区、第二单链区和第三单链区。
10.如权利要求9所述的系统,其中所述第一单链区位于所述第一双链RNA茎与所述第二双链RNA茎之间。
11.如权利要求9所述的系统,其中所述第二单链区位于所述第二双链RNA茎与所述第三双链RNA茎之间。
12.如权利要求9所述的系统,其中所述第三单链区位于所述第一适体的所述第三双链RNA茎与所述第一双链RNA茎之间。
13.如前述权利要求中任一项所述的系统,其中所述第一适体和所述第二适体在5'至3'方向上处于相同方向。
14.如前述权利要求中任一项所述的系统,其中所述第三适体在5'至3'方向上相对于所述第一适体和所述第二适体处于相反方向。
15.如权利要求1所述的系统,其中所述polyA切割信号的一个或多个核苷酸在所述三向接合部、所述第三双链RNA茎、所述第三单链区或所述第一双链RNA茎内。
16.如权利要求15所述的系统,其中所述第三单链区包含所述polyA切割信号的前四个碱基。
17.如权利要求15所述的系统,其中所述第一双链RNA茎包含所述polyA切割信号的最后两个碱基。
18.如权利要求15所述的系统,其中所述第一双链RNA茎包含所述polyA切割信号的全部。
19.如权利要求3所述的系统,其中所述第三适体的结合口袋与所述三向接合部之间的双链RNA茎的长度在10与15个碱基对之间。
20.如权利要求10所述的系统,其中所述第一单链区包含至少一个选自C和A的碱基。
21.如权利要求11所述的系统,其中所述第二单链区包含至少一个选自C和A的碱基。
22.如权利要求5所述的系统,其中所述第二双链RNA茎的序列是SEQ ID NO.:3。
23.如权利要求5所述的系统,其中所述第三双链RNA茎的序列是SEQ ID NO.:2。
24.如权利要求5所述的系统,其中所述第一双链RNA茎的序列是SEQ ID NO.:4。
25.如权利要求5所述的系统,其中所述第一双链RNA茎的序列是SEQ ID NO.:5。
26.如权利要求1所述的系统,其中所述编码可表达多肽的核酸序列还包含5'UTR。
27.如权利要求26所述的系统,其中所述5'UTR还包含CAA重复序列。
28.如权利要求26所述的系统,其中所述5'UTR还包含一个或多个3'剪接受体位点。
29.如权利要求26所述的系统,其中所述工程化5'UTR具有序列SEQ ID NO.:48。
30.如权利要求1所述的系统,其还包含所述编码可表达多肽的核酸序列的5'和所述polyA切割信号的3'的G-U丰富区。
31.如权利要求29所述的系统,其中所述3'受体位点之后是调节替代剪接的强度的核酸三联体序列。
32.如权利要求31所述的系统,其中所述核酸三联体在相对于所述5'UTR中的所述第二3'受体位点的3'并且具有选自以下的序列:TAG、TCT、TTC、TTG、TGA、TGC、TCC、ACA、AAC、ACC、AGC、AGG、CCT和CCC。
33.如权利要求1所述的系统,其还包含所述编码可表达多肽的核酸序列的5'和所述G-U丰富区的3'的G丰富区。
34.如权利要求33所述的系统,其中所述G丰富区包含4MAZ序列。
35.如权利要求1所述的系统,其中所述工程内含子具有长度为100与200个碱基之间的序列。
36.如权利要求1所述的系统,其中所述工程内含子具有序列SEQ ID NO 1。
37.如权利要求1所述的系统,其中所述工程内含子之后是调节内含子剪接的强度的核酸三联体序列。
38.如权利要求37所述的系统,其中所述核酸三联体序列是选自以下的序列:TTT、TGA、TCT、TAC、CAC和CAT。
39.如权利要求1所述的系统,其中所述系统包含选自下组的序列:SEQ ID NO.:6至SEQID NO.:56。
40.如权利要求39所述的系统,其中所述系统包含选自下组的序列:SEQ ID NO.:6;SEQID NO.:13;SEQ ID NO.:14;SEQ ID NO.:28;SEQ ID NO.:32;SEQ ID NO.:33;SEQIDNO.:36;SEQ ID NO.:38;SEQ ID NO.:44;SEQ ID NO.:46;SEQ ID NO.:50;
NO.:51;NO.:52;NO.:53;NO.:54;NO.:55;NO.:56。
41.一种用于递送如权利要求1所述的系统的载体。
42.如权利要求41所述的载体,其中所述载体是病毒载体。
43.如权利要求42所述的载体,其中所述载体选自腺病毒载体、慢病毒载体;腺相关病毒载体、脊髓灰质炎病毒载体和逆转录病毒载体。
44.一种用于调节细胞中基因产物表达的方法,所述方法包括以下步骤:
向所述细胞中引入系统,所述系统在5'至3'方向上包含:
a)5'剪接供体位点
b)工程内含子
c)第一3'剪接受体位点
d)polyA开关,其包含具有一个或多个配体结合口袋的两个或更多个配体结合适体,和其中的至少一个polyA切割信号;以及
e)第二3'剪接受体位点。
45.如权利要求44所述的方法,其中所述基因产物对于所述细胞是外源性的。
46.如权利要求45所述的方法,其中所述系统还包含紧邻e)的剪接位点的3'的编码所述基因产物的核酸序列。
47.如权利要求44所述的方法,其中所述基因产物对于所述细胞是内源性的。
48.如权利要求47所述的方法,其中所述方法不包括施用所述配体以抑制所述内源基因产物的表达。
49.如权利要求44所述的方法,其中所述系统还包含a)的剪接位点的5'的启动子。
50.如权利要求49所述的方法,其中所述启动子是CMV启动子。
51.如前述权利要求中任一项所述的方法,其中所述方法发生在个体的一个或多个细胞中,所述配体是葡萄糖,所述个体患有糖尿病、糖尿病前期或糖尿病并发症,且/或所述可表达的多核苷酸是胰岛素。
52.如前述权利要求中任一项所述的方法,其中所述方法发生在个体的一个或多个细胞中,所述配体是癌症生物标志物的基因产物,并且所述可表达的多核苷酸是自杀基因。
53.如前述权利要求中任一项所述的方法,其中所述方法发生在个体中,所述可表达的多核苷酸是报告基因,并且所述报告基因的表达位置和/或强度提供关于配体在所述个体的一个或多个细胞中的空间分布、时间波动或两者的信息。
54.如前述权利要求中任一项所述的方法,其中所述方法发生在个体、组织或细胞中,其中所述可表达的多核苷酸编码可检测的基因产物,并且其中对相应的所述个体、组织或细胞进行成像。
55.如权利要求50所述的方法,其中a)的载体和/或b)的细胞在疗法之前、疗法期间和/或疗法之后提供给所述个体。
56.一种编码poly A适体多核苷酸的核酸分子,所述poly A适体多核苷酸在5'至3'方向上包含:
a)5'剪接供体位点;
b)工程内含子;
c)第一3'剪接受体位点;
d)polyA开关,其包含具有一个或多个配体结合口袋的两个或更多个配体结合适体,和其中的至少一个polyA切割信号;
e)第二3'剪接受体位点;以及
f)编码可表达多肽的核酸序列。
57.如权利要求56所述的核酸分子,其中所述核酸是DNA。
58.如权利要求56所述的核酸分子,其中所述核酸是RNA。
59.一种用于递送如权利要求56所述的核酸的载体。
60.如权利要求59所述的载体,其中所述载体是病毒载体。
61.如权利要求59所述的载体,其中所述载体选自腺病毒载体、慢病毒载体;腺相关病毒载体、脊髓灰质炎病毒载体和逆转录病毒载体。
Applications Claiming Priority (7)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962894611P | 2019-08-30 | 2019-08-30 | |
| US62/894,611 | 2019-08-30 | ||
| US201962904635P | 2019-09-23 | 2019-09-23 | |
| US62/904,635 | 2019-09-23 | ||
| US202063043504P | 2020-06-24 | 2020-06-24 | |
| US63/043,504 | 2020-06-24 | ||
| PCT/US2020/048561 WO2021041924A2 (en) | 2019-08-30 | 2020-08-28 | System for regulating gene expression |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN115279902A true CN115279902A (zh) | 2022-11-01 |
Family
ID=74683442
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN202080072368.2A Pending CN115279902A (zh) | 2019-08-30 | 2020-08-28 | 用于调控基因表达的系统 |
Country Status (11)
| Country | Link |
|---|---|
| US (1) | US20220290147A1 (zh) |
| EP (1) | EP4022065A4 (zh) |
| JP (1) | JP2022546408A (zh) |
| KR (1) | KR20220049619A (zh) |
| CN (1) | CN115279902A (zh) |
| AU (1) | AU2020335909A1 (zh) |
| BR (1) | BR112022003512A2 (zh) |
| CA (1) | CA3152513A1 (zh) |
| IL (1) | IL290944A (zh) |
| MX (1) | MX2022002390A (zh) |
| WO (1) | WO2021041924A2 (zh) |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA3227613A1 (en) | 2021-08-11 | 2023-02-16 | William Dowdle | Inducible systems for altering gene expression in hypoimmunogenic cells |
Family Cites Families (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2007529223A (ja) * | 2004-03-15 | 2007-10-25 | バイオジェン・アイデック・エムエイ・インコーポレイテッド | 選択的スプライシングを用いて真核細胞においてポリペプチドマルチマーを発現するための方法および構築物 |
| EP1752536A4 (en) * | 2004-05-11 | 2008-04-16 | Alphagen Co Ltd | POLYNUCLEOTIDE CAUSING RNA INTERFERENCE AND METHOD OF REGULATING GENE EXPRESSION WITH THE USE OF THE SAME |
| US8178503B2 (en) * | 2006-03-03 | 2012-05-15 | International Business Machines Corporation | Ribonucleic acid interference molecules and binding sites derived by analyzing intergenic and intronic regions of genomes |
| US20150284738A1 (en) * | 2006-12-29 | 2015-10-08 | Rodina Holding S.A. | Artificial dna sequence with optimized leader function in 5' (5'-utr) for the improved expression of heterologous proteins in plants |
| US9347073B2 (en) * | 2012-02-10 | 2016-05-24 | The Board Of Trustees Of The Leland Stanford Junior University | Mini-intronic plasmid vectors |
| US20150141320A1 (en) * | 2012-05-16 | 2015-05-21 | Rana Therapeutics, Inc. | Compositions and methods for modulating gene expression |
| US9567581B2 (en) * | 2012-08-07 | 2017-02-14 | The General Hospital Corporation | Selective reactivation of genes on the inactive X chromosome |
| EP3719134B1 (en) * | 2015-03-11 | 2023-08-23 | The United States of America, as represented by The Secretary, Department of Health and Human Services | Rp2 vectors for treating x-linked retinitis pigmentosa |
| CA3005224A1 (en) * | 2015-11-12 | 2017-05-18 | Baylor College Of Medicine | Exogenous control of mammalian gene expression through aptamer-mediated modulation of polyadenylation |
-
2020
- 2020-08-28 AU AU2020335909A patent/AU2020335909A1/en active Pending
- 2020-08-28 MX MX2022002390A patent/MX2022002390A/es unknown
- 2020-08-28 CN CN202080072368.2A patent/CN115279902A/zh active Pending
- 2020-08-28 US US17/638,619 patent/US20220290147A1/en active Pending
- 2020-08-28 KR KR1020227010164A patent/KR20220049619A/ko unknown
- 2020-08-28 CA CA3152513A patent/CA3152513A1/en active Pending
- 2020-08-28 JP JP2022513155A patent/JP2022546408A/ja active Pending
- 2020-08-28 BR BR112022003512A patent/BR112022003512A2/pt active Search and Examination
- 2020-08-28 WO PCT/US2020/048561 patent/WO2021041924A2/en active Application Filing
- 2020-08-28 EP EP20856728.9A patent/EP4022065A4/en active Pending
-
2022
- 2022-02-27 IL IL290944A patent/IL290944A/en unknown
Also Published As
| Publication number | Publication date |
|---|---|
| BR112022003512A2 (pt) | 2022-05-17 |
| JP2022546408A (ja) | 2022-11-04 |
| WO2021041924A2 (en) | 2021-03-04 |
| CA3152513A1 (en) | 2021-03-04 |
| US20220290147A1 (en) | 2022-09-15 |
| EP4022065A4 (en) | 2024-02-14 |
| AU2020335909A1 (en) | 2022-04-14 |
| IL290944A (en) | 2022-04-01 |
| EP4022065A2 (en) | 2022-07-06 |
| MX2022002390A (es) | 2022-05-13 |
| WO2021041924A3 (en) | 2021-04-08 |
| KR20220049619A (ko) | 2022-04-21 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR102455623B1 (ko) | CRISPR/Cas12f1 시스템 효율화를 위한 엔지니어링 된 가이드 RNA 및 그 용도 | |
| RU2577971C2 (ru) | Система для увеличения экспрессии генов и вектор, содержащий указанную систему | |
| BR112020024292A2 (pt) | rna circular para translação em células eucarióticas | |
| Guo et al. | Multiplexed genome regulation in vivo with hyper-efficient Cas12a | |
| US9879254B2 (en) | Targeting RNAs to microvesicles | |
| KR20220066225A (ko) | 선택적 유전자 조절을 위한 조성물 및 방법 | |
| JP2002525066A (ja) | 核酸の特異的な細胞の局在化のための転写法 | |
| KR20230076820A (ko) | 진핵생물 유전체 공학을 위한 합성 미니어처 crispr-cas(casmini) 시스템 | |
| EP2914721B1 (en) | A rna trans-splicing molecule (rtm) for use in the treatment of cancer | |
| CN115279902A (zh) | 用于调控基因表达的系统 | |
| US20230383275A1 (en) | Sgrna targeting aqp1 rna, and vector and use thereof | |
| CN116783295A (zh) | 指导rna的新型设计及其用途 | |
| KR20200011135A (ko) | 융합 펩타이드 나노 어셈블리와 칼슘 이온을 이용한 핵산 세포 형질도입법과 그의 응용 | |
| JP5738862B2 (ja) | 細胞への核酸導入方法および核酸複合体 | |
| KR20220119084A (ko) | 폴리뉴클레오티드를 엑소솜에 전달하기 위한 핵산 구축물 | |
| JP4709971B2 (ja) | 細胞内へ核酸を導入する為の新規な分子並びに細胞内へ導入する核酸および細胞内へ核酸を導入する為の新規な方法 | |
| Reza et al. | Triplex-mediated genome targeting and editing | |
| CN112771165A (zh) | 通过血脑屏障的核酸适配体及其应用 | |
| CN117230043B (zh) | Cas13蛋白及其应用 | |
| US20240115739A1 (en) | Synthetic cas12a for enhanced multiplex gene control and editing | |
| CN116949092A (zh) | 一种转座子系统及其应用 | |
| Dietz et al. | Muscle Specific Promotors for Gene Therapy–A Comparative Study in Proliferating and Differentiated Cells | |
| WO2022170007A1 (en) | Fusion proteins for crispr-based transcriptional repression | |
| WO2024040202A1 (en) | Fusion proteins and uses thereof for precision editing | |
| WO2023164482A2 (en) | Treatment for nucleotide repeat expansion disease |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |