CN110914410A - γδT细胞扩增、组合物及其使用方法 - Google Patents
γδT细胞扩增、组合物及其使用方法 Download PDFInfo
- Publication number
- CN110914410A CN110914410A CN201880044728.0A CN201880044728A CN110914410A CN 110914410 A CN110914410 A CN 110914410A CN 201880044728 A CN201880044728 A CN 201880044728A CN 110914410 A CN110914410 A CN 110914410A
- Authority
- CN
- China
- Prior art keywords
- cells
- cell
- population
- isolated
- expanded
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images































































Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0636—T lymphocytes
- C12N5/0638—Cytotoxic T lymphocytes [CTL] or lymphokine activated killer cells [LAK]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K35/00—Medicinal preparations containing materials or reaction products thereof with undetermined constitution
- A61K35/12—Materials from mammals; Compositions comprising non-specified tissues or cells; Compositions comprising non-embryonic stem cells; Genetically modified cells
- A61K35/14—Blood; Artificial blood
- A61K35/17—Lymphocytes; B-cells; T-cells; Natural killer cells; Interferon-activated or cytokine-activated lymphocytes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/10—Cellular immunotherapy characterised by the cell type used
- A61K40/11—T-cells, e.g. tumour infiltrating lymphocytes [TIL] or regulatory T [Treg] cells; Lymphokine-activated killer [LAK] cells
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K40/00—Cellular immunotherapy
- A61K40/40—Cellular immunotherapy characterised by antigens that are targeted or presented by cells of the immune system
- A61K40/41—Vertebrate antigens
- A61K40/42—Cancer antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
- A61K45/06—Mixtures of active ingredients without chemical characterisation, e.g. antiphlogistics and cardiaca
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/06—Animal cells or tissues; Human cells or tissues
- C12N5/0602—Vertebrate cells
- C12N5/0634—Cells from the blood or the immune system
- C12N5/0636—T lymphocytes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2121/00—Preparations for use in therapy
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K2300/00—Mixtures or combinations of active ingredients, wherein at least one active ingredient is fully defined in groups A61K31/00 - A61K41/00
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/90—Serum-free medium, which may still contain naturally-sourced components
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2500/00—Specific components of cell culture medium
- C12N2500/99—Serum-free medium
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/21—Chemokines, e.g. MIP-1, MIP-2, RANTES, MCP, PF-4
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2301—Interleukin-1 (IL-1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2302—Interleukin-2 (IL-2)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2304—Interleukin-4 (IL-4)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2309—Interleukin-9 (IL-9)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2312—Interleukin-12 (IL-12)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2315—Interleukin-15 (IL-15)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2318—Interleukin-18 (IL-18)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2321—Interleukin-21 (IL-21)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/20—Cytokines; Chemokines
- C12N2501/23—Interleukins [IL]
- C12N2501/2333—Interleukin-33 (IL-33)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2501/00—Active agents used in cell culture processes, e.g. differentation
- C12N2501/50—Cell markers; Cell surface determinants
- C12N2501/599—Cell markers; Cell surface determinants with CD designations not provided for elsewhere
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Biomedical Technology (AREA)
- Immunology (AREA)
- Chemical & Material Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Biotechnology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Cell Biology (AREA)
- Hematology (AREA)
- Epidemiology (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- Pharmacology & Pharmacy (AREA)
- Medicinal Chemistry (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Developmental Biology & Embryology (AREA)
- Virology (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
本发明提供了从非造血组织来源扩增γδT细胞的方法。还提供了扩增的γδT细胞的组合物和使用扩增的γδT细胞的方法(例如,过继性T细胞疗法的一部分)。
Description
背景技术
对用于癌症的T细胞免疫疗法不断增长的兴趣集中在CD8+和CD4+αβT细胞亚型识别癌细胞并介导宿主保护性功能潜能的显著能力,特别是当通过PD-1、CTLA-4及其他受体所施加的抑制性途径的临床介导的拮抗作用脱抑制(de-repress)时。尽管如此,仍有许多问题。例如,似乎存在其中这些治疗效力似乎不良的许多主要临床情况。经常存在严重不良事件,预测效力或不良事件的能力受到极大限制,并且对于在常规、抗原特异性CD8+和CD4+αβT细胞应答激活之前,使得宿主能够感知肿瘤细胞(“免疫原性”)的相互作用的解释很少。
γδT细胞(γδT细胞)代表了在它们的表面上表达独特的,限定的γδT细胞受体(TCR)的T细胞亚型。这种TCR由一条γ和一条δ链组成。人γδT细胞可以广泛地分为一种或两种类型——外周血驻留的(resident)γδT细胞和非造血组织驻留的γδT细胞。大部分血液驻留的γδT细胞表达Vδ2 TCR,然而这在组织驻留的γδT细胞中不太常见,其更经常使用Vδ1和/或其他Vδ链。由于非造血组织驻留的γδT细胞不易于以较大数目获得,并且不存在常规分离或扩增规程,因此对它们尚未进行良好的鉴定和治疗应用研究。因此,在将非造血组织驻留的γδT细胞分离和扩增至足以研究和潜在用作疗法,例如,继承性T细胞疗法的量的方法领域中,仍未满足需要。
发明内容
本发明提供了使来自非造血组织来源(例如,非造血组织来源的γδT细胞,例如,非造血组织来源的Vδ1 T细胞)的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)扩增的方法。扩增方法包括在不存在明显TCR刺激和/或在存在IL-4、IL-15、IL-21和/或IL-2的情况下培养γδT细胞(例如,分离自非造血组织的基质细胞的γδT细胞)。还提供了扩增的γδT细胞(例如,非造血组织来源的γδT细胞,例如,非造血组织来源的Vδ1 T细胞)的组合物和使用扩增的γδT细胞的方法(例如,用于例如癌症治疗的过继性T细胞疗法的一部分)。
在一个方面,本发明的特征在于通过以下各项来扩增γδT细胞的方法:(i)提供获自非造血组织的γδT细胞群体;和(ii)在存在IL-2、IL-15和选自由以下各项组成的组的因子的情况下培养γδT细胞至少5天以产生扩增的γδT细胞群体:IL-4、IL-21、IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、人血小板裂解物(HPL)和基质细胞衍生因子-1(SDF-1)。例如,可以在存在IL-2、IL-15和IL-4;IL-2、IL-15和IL-21;或者IL-2、IL-15、IL-4和IL-21的情况下培养γδT细胞。
在另一个方面,本发明提供了通过以下各项来扩增γδT细胞的方法:(i)提供获自非造血组织的γδT细胞群体;和(ii)以对于产生γδT细胞的扩增群体有效的量,在存在IL-2、IL-4、IL-15和IL-21的情况下培养γδT细胞至少5天。
在上述方面中任一项的一些实施方式中,将γδT细胞同时暴露于IL-2、IL-4、IL-15和IL-21至少5天。在一些实施方式中,步骤(ii)包括在不存在外源TCR途径激动剂的情况下培养γδT细胞。在一些实施方式中,方法还包括,在步骤(i)之后,从非造血细胞分离γδT细胞以产生分离的γδT细胞群体,并且步骤(ii)包括:(a)在不存在明显基质细胞接触的情况下培养γδT细胞;(b)在不存在明显肿瘤细胞接触的情况下培养γδT细胞;和/或(c)在不存在明显饲养细胞接触的情况下培养γδT细胞。
在另一个方面,扩增γδT细胞的方法包括以下步骤:(i)提供非造血组织,组织包括非造血细胞和γδT细胞;(ii)从非造血细胞分离γδT细胞以获得分离的γδT细胞群体;和(iii)在存在IL-2、IL-15和选自IL-4、IL-21、IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、HPL和SDF-1组成的组的因子的情况下培养γδT细胞至少5天以产生扩增的γδT细胞群体。在一些实施方式中,步骤(ii)包括在存在IL-2、IL-15、IL-4和/或IL-21(例如,IL-2、IL-15和IL-4;IL-2、IL-15和IL-21;或者IL-2、IL-15、IL-4和IL-21)的情况下培养γδT细胞。在一些实施方式中,将γδT细胞同时暴露于IL-2、IL-15、IL-4和/或IL-21。在一些实施方式中,步骤(iii)包括在不存在与γδT细胞明显基质细胞接触的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在不存在外源TCR途径激动剂的情况下培养γδT细胞。
在另一个方面,本发明的特征在于扩增γδT细胞的方法,方法包括以下步骤:(i)提供获自非造血组织的γδT细胞群体;和(ii)以对于产生γδT细胞的扩增群体有效的量,在存在IL-2、IL-4、IL-15和IL-21的情况下培养γδT细胞至少5天。可以将γδT细胞同时暴露于IL-2、IL-4、IL-15和IL-21,例如,至少5天,或者可以在暴露于其他因子之前,将它们暴露于一种或多种因子。在一些实施方式中,步骤(ii)包括以对于产生γδT细胞的扩增群体有效的量在存在人重组IL-2、人重组IL-4、人重组IL-15和人重组IL-21的情况下培养γδT细胞至少5天。在一些实施方式中,步骤(ii)包括在不存在外源TCR途径激动剂(例如,抗CD3)的情况下,例如,在不存在明显TCR途径激活的情况下,培养γδT细胞。在一些实施方式中,步骤(ii)包括在存在1ng/mL至1,000ng/mL的浓度(例如,约10ng/mL或者约100ng/mL)的IL-21的情况下,培养γδT细胞。步骤(ii)还可以包括在存在选自IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、人血小板裂解物(HPL)和基质细胞衍生因子-1(SDF-1)组成的组的一种或多种因子的情况下培养γδT细胞。
在另一个方面,本文提供了通过以下步骤扩增γδT细胞的方法:(i)提供获自非造血组织的γδT细胞群体;和(ii)以对于产生γδT细胞的扩增群体有效的量,在存在IL-2和IL-15的情况下培养γδT细胞至少5天。在该方面的一些实施方式中,将γδT细胞同时暴露于IL-2和IL-15。步骤(ii)可以包括在不存在外源TCR途径激动剂或者在不存在明显的TCR途径激活的情况下培养γδT细胞。在一些实施方式中,步骤(ii)包括在存在选自IL-4、IL-21、IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、HPL和SDF-1组成的组的一种或多种因子的情况下培养γδT细胞。在一些实施方式中,在存在IL-4和/或IL-21的情况下培养γδT细胞。在一些实施方式中,步骤(ii)包括在存在IL-2、IL-4、IL-15和IL-21的情况下培养γδT细胞。IL-21可以处于1ng/mL至1,000ng/mL的浓度(例如,10ng/mL或100ng/mL)。
在另一个方面,本发明的特征在于通过以下步骤扩增γδT细胞的方法:(i)提供获自非造血组织的γδT细胞群体;和(ii)在不存在明显的TCR途径激活的情况下培养γδT细胞至少5天以产生γδT细胞的扩增群体。在一些实施方式中,步骤(ii)包括在存在IL-2和IL-15的情况下培养γδT细胞。在一些情况下,将γδT细胞同时暴露于IL-4和IL-15。在一些实施方式中,步骤(ii)包括在存在选自IL-4、IL-21、IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、HPL和SDF-1组成的组的一种或多种因子的情况下培养γδT细胞。例如,可以在存在IL-4、IL-21或两者的情况下培养γδT细胞。在一些实施方式中,在存在IL-2、IL-4、IL-15和IL-21的情况下培养γδT细胞。IL-21可以处于1ng/mL至1,000ng/mL的浓度(例如,10ng/mL或100ng/mL)。
在以上方面中的任一项的一些实施方式中,方法还包括,在步骤(i)之后,从非造血细胞分离γδT细胞以产生分离的γδT细胞群体,并且步骤(ii)包括在不存在明显基质细胞接触的情况下,在不存在明显肿瘤细胞接触的情况下和/或在不存在明显饲养细胞接触(例如,辐照的饲养细胞、B细胞或者抗原递呈细胞)的情况下培养γδT细胞。
在另一个方面,本文的特征在于通过以下步骤扩增γδT细胞的方法:(i)提供非造血组织,组织包括非造血细胞和γδT细胞;(ii)从非造血细胞分离γδT细胞以获得分离的γδT细胞群体;和(iii)在不存在与γδT细胞的明显基质细胞接触的情况下培养γδT细胞至少5天以产生γδT细胞的扩增群体。在一些实施方式中,步骤(iii)包括在不存在外源TCR途径激动剂和/或在不存在明显的TCR途径激活的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在存在IL-2和IL-15的情况下培养γδT细胞。例如,γδT细胞可以同时暴露于IL-2和IL-15。在一些实施方式中,步骤(iii)包括在存在选自IL-4、IL-21、IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、HPL和SDF-1组成的组的一种或多种因子的情况下培养γδT细胞。例如,在一些情况下,步骤(iii)包括在存在IL-4、IL-21或两者的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在存在IL-2、IL-4、IL-15和IL-21的情况下培养γδT细胞。IL-21可以处于1ng/mL至1,000ng/mL的浓度(例如,10ng/mL或100ng/mL)。
在另一个方面,本发明提供了通过以下步骤扩增γδT细胞的方法:(i)提供非造血组织,组织包括非造血细胞和γδT细胞;(ii)从非造血细胞分离γδT细胞以获得分离的γδT细胞群体;和(iii)在存在IL-2和IL-15的情况下培养γδT细胞至少5天以产生γδT细胞的扩增群体。在一些实施方式中,步骤(iii)包括在不存在与γδT细胞明显基质细胞接触的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在不存在外源TCR途径激动剂或在不存在明显的TCR途径激活的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在存在IL-2和IL-15的情况下培养γδT细胞。γδT细胞可以同时暴露于IL-2和IL-15。在一些情况下,步骤(iii)包括在存在选自IL-4、IL-21、IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、HPL和SDF-1组成的组的一种或多种因子的情况下培养γδT细胞。例如,可以在存在IL-4和/或IL-21的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在存在IL-2、IL-4、IL-15和IL-21的情况下培养γδT细胞。IL-21可以处于1ng/mL至1,000ng/mL的浓度(例如,10ng/mL或100ng/mL)。
在另一个方面,本文的特征在于通过以下步骤扩增γδT细胞的方法:(i)提供非造血组织,组织包括非造血细胞和γδT细胞;(ii)从非造血细胞分离γδT细胞以获得分离的γδT细胞群体;和(iii)在不存在明显的TCR途径激活的情况下培养γδT细胞至少5天以产生γδT细胞的扩增群体。在一些情况下,步骤(iii)包括在不存在外源TCR途径激动剂和/或在不存在与γδT细胞明显的基质细胞接触的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在存在IL-2和IL-15的情况下培养γδT细胞。例如,γδT细胞可以同时暴露于IL-2和IL-15。在一些实施方式中,步骤(iii)包括在存在选自IL-4、IL-21、IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、HPL和SDF-1组成的组的一种或多种因子的情况下培养γδT细胞。例如,可以在存在IL-2和/或IL-21的情况下培养γδT细胞。在一些实施方式中,步骤(iii)包括在存在IL-2、IL-4、IL-15和IL-21的情况下培养γδT细胞。IL-21可以处于1ng/mL至1,000ng/mL的浓度(例如,10ng/mL或100ng/mL)。
在以上方法中任一项的一些实施方式中,从非造血细胞分离γδT细胞的步骤包括在配置以有利于细胞从非造血组织流出(egress)的支架上培养γδT细胞和非造血细胞。在一些实施方式中,从非造血细胞分离γδT细胞的步骤包括在存在IL-2、IL-15或两者的情况下培养γδT细胞和非造血细胞。在一些实施方式中,分离的淋巴细胞群体包括分离的γδT细胞群体,并且分离的γδT细胞群体包括分离的Vδ1 T细胞和/或双阴性细胞(DN细胞)的群体。在一些情况下,在扩增步骤之前,1-10%的分离的淋巴细胞群体是γδT细胞。在一些实施方式中,在扩增步骤之前,1-10%的分离的淋巴细胞群体是Vδ1 T细胞。在扩增步骤之前,至少80%的分离的γδT细胞群体可以是Vδ1 T细胞,和/或小于10%的分离的γδT细胞群体可以是Vδ2 T细胞。在一些实施方式中,从分离的γδT细胞群体除去αβT细胞和/或NK细胞(例如,在扩增步骤之前)。
在一些实施方式中,在扩增步骤之前,分离的γδT细胞群体包括至少10%的CCR3+细胞,至少10%的CCR4+细胞,至少10%的CCR7+细胞,至少10%的CCR8+细胞或者至少10%的CD103+细胞。在一些实施方式中,在扩增步骤之前,相对于参比群体(例如,血液驻留的Vδ2T细胞的参比群体),分离的γδT细胞群体包括更高频率的CCR3+细胞、CCR4+细胞、CCR7+细胞和/或CCR8+细胞。在一些实施方式中,在扩增步骤之前,相对于参比群体(例如,血液驻留的Vδ2 T细胞的参比群体),分离的Vδ1 T细胞群体包括更高频率的NKG2D+细胞、CD56+细胞、CD69+细胞和/或TIM3+细胞。
在以上方面中任一项的一些实施方式中,在扩增步骤期间,在培养14天内,相对于扩增步骤之前的分离的γδT细胞群体,扩增的γδT细胞群体包括至少20倍的γδT细胞数目。另外或者作为另外一种选择,在扩增步骤期间,在培养21天内,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体可以包括至少50倍的γδT细胞数目。扩增的γδT细胞群体包括扩增的Vδ1 T细胞群体。在一些实施方式中,在扩增步骤期间,在培养14天内,相对于扩增之前的分离的Vδ1 T细胞群体,扩增的Vδ1 T细胞群体包括至少20倍的Vδ1 T细胞数目。另外或者作为另外一种选择,在扩增步骤期间,在培养21天内,相对于扩增之前的分离的Vδ1 T细胞群体,扩增的Vδ1 T细胞群体包括至少50倍的Vδ1 T细胞数目。
在以上方面中的任一项的一些实施方式中,扩增的γδT细胞群体表达CD27。例如,扩增的γδT细胞群体可以具有比分离的γδT细胞群体更大的中值CD27表面表达。在一些情况下,扩增的γδT细胞群体具有相对于分离的γδT细胞群体至少2倍的中值CD27表面表达。另外或者作为另外一种选择,扩增的γδT细胞群体可以具有相对于分离的γδT细胞群体更高频率的CD27+细胞。例如,扩增的γδT细胞群体可以具有相对于分离的γδT细胞群体至少5%更高频率的CD27+细胞。在一些实施方式中,扩增的Vδ1 T细胞群体表达CD27。在一些实施方式中,扩增的Vδ1 T细胞群体具有比分离的Vδ1 T细胞群体更大的中值CD27表面表达。例如,扩增的Vδ1 T细胞群体可以具有相对于分离的Vδ1 T细胞群体至少2倍的中值CD27表面表达。另外或者作为另外一种选择,扩增的Vδ1 T细胞群体可以具有相对于分离的Vδ1 T细胞群体更高频率的CD27+细胞。例如,扩增的Vδ1 T细胞群体可以具有相对于分离的Vδ1 T细胞群体至少5%更高频率的CD27+细胞。
在以上方面中的任一项的一些实施方式中,扩增的γδT细胞群体具有比分离的γδT细胞群体更低的中值TIGIT表面表达。例如,扩增的γδT细胞群体可以具有比分离的γδT细胞群体低至少50%的中值TIGIT表面表达。另外或者作为另外一种选择,扩增的γδT细胞群体可以具有比分离的γδT细胞群体更低频率的TIGIT+细胞。例如,扩增的γδT细胞群体可以具有比分离的γδT细胞群体至少20%更低频率的TIGIT+细胞。在一些实施方式中,扩增的Vδ1 T细胞群体具有比分离的Vδ1 T细胞群体更低的中值TIGIT表面表达。例如,扩增的Vδ1 T细胞群体可以具有比分离的Vδ1 T细胞群体低至少50%的中值TIGIT表面表达。另外或者作为另外一种选择,扩增的Vδ1 T细胞群体可以具有比分离的Vδ1 T细胞群体更低频率的TIGIT+细胞。例如,扩增的Vδ1 T细胞群体可以具有比分离的Vδ1 T细胞群体低至少20%更低频率的TIGIT+细胞。
在以上方面中的任一项的一些实施方式中,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的γδT细胞群体具有选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组的一种或多种标志物的更高的表面表达。另外或者作为另外一种选择,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的γδT细胞群体可以具有更高频率的表达选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组的一种或多种标志物的细胞。在一些实施方式中,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的γδT细胞群体具有选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组的一种或多种标志物的更低的表面表达。另外或者作为另外一种选择,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的γδT细胞群体可以具有更低频率的表达选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组的一种或多种标志物的细胞。
在一些实施方式中,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的Vδ1 T细胞群体具有选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组的一种或多种标志物的更高的表面表达。在一些实施方式中,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的γδT细胞群体具有更高频率的表达选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组的一种或多种标志物的细胞。在一些实施方式中,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的γδT细胞群体具有选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组的一种或多种标志物的更低的表面表达。在其他实施方式中,相对于参比群体(例如,相对于分离的γδT细胞群体,例如,相对于扩增步骤之前的分离的γδT细胞群体),扩增的γδT细胞群体具有更低频率的表达选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组的一种或多种标志物的细胞。
在以上方面中的任一项的一些实施方式中,步骤(iii)包括在不存在明显基质细胞接触的情况下,在不存在明显饲养细胞接触的情况下和/或在不存在明显肿瘤细胞接触的情况下培养γδT细胞。在一些实施方式中,非造血组织不是肿瘤组织。
在以上方面中的任一项的一些实施方式中,非造血组织是皮肤(例如,人皮肤,例如,通过钻孔活组织检查所获得的皮肤)。在其他实施方式中,非造血组织是肠组织。
在以上方面和实施方式中的任一项中,可以体外进行扩增γδT细胞的方法。
在以上方面和实施方式中的任一项中,提供非造血组织的步骤可以包括提供已获自受试者,例如,人或非人动物受试者的非造血组织。
在另一个方面,本发明的特征在于通过以上方面中任一项所述的方法所获得的扩增的γδT细胞。
在另一个方面,本发明提供了包括上述方面所述的扩增的γδT细胞的药物组合物。在一些实施方式中,所述药物组合物还包括其他治疗剂,其选自:免疫治疗剂、细胞毒性剂、生长抑制剂、放射疗法剂、抗血管生成剂或其两种或更多种试剂的组合。在一些情况下,其他治疗剂是免疫治疗剂(例如,IL-2,例如,低剂量IL-2,例如,0.3×106至3.0×106IU IL-2每天,例如,1.0×106IU IL-2每天)。
在另一个方面,本发明的特征在于以上方面所述的药物组合物用于在通过过继性T细胞疗法治疗受试者的方法中的使用。
在另一个方面,本发明的特征在于以上方面中的任一项所述的扩增的γδT细胞用于在通过过继性T细胞疗法治疗受试者的方法中的使用。
在另一个方面,本发明提供了以上方面中的任一项所述的扩增的γδT细胞或其药物组合物在生产用于受试者中癌症(例如,实体瘤)、感染(例如,巨细胞病毒(CMV)感染)或免疫病理学治疗的药物中的应用。
在另一个方面,本发明提供了以上方面中的任一项所述的扩增的γδT细胞或其药物组合物在用于受试者中癌症(例如,实体瘤)、感染(例如,巨细胞病毒(CMV)感染)或免疫病理学的治疗的方法中的使用。
在另一个方面,本发明的特征在于通过向对其有需要的受试者施用治疗有效量的通过根据任何上述实施方式所述的方法所获得的扩增的γδT细胞来通过过继性T细胞疗法治疗受试者的方法。在一些实施方式中,扩增的γδT细胞的治疗有效量为小于10×1012个细胞每剂量,或在治疗过程中小于10×1012个细胞。在一些实施方式中,方法还包括向对其有需要的受试者施用一种或多种其他治疗剂。其他治疗剂可以选自免疫治疗剂、细胞毒性剂、生长抑制剂、放射疗法剂、抗血管生成剂或其两种或更多种试剂的组合。可以与扩增的γδT细胞的施用同时,在其之前或之后施用其他治疗剂。在一些实施方式中,其他治疗剂是免疫治疗剂。在一个实施方式中,免疫治疗剂是IL-2(例如,低剂量IL-2,例如,0.3×106至3.0×106IU IL-2每天,例如,1.0×106IU IL-2每天)。这些实施方式适用于任何以上或以下与通过本文所公开的方法所获得的扩增的γδT细胞(包括这些γδT细胞的药物组合物)在通过过继性T细胞疗法治疗受试者的方法中的使用有关的方面。
在另一个方面,本发明的特征在于通过向对其有需要的受试者施用治疗有效量的任何上述方面所述的药物组合物来通过过继性T细胞疗法治疗受试者的方法。
在以上方面中的任一项的一些实施方式中,所述受试者是人(例如,人癌症患者(例如,正在治疗实体瘤的人癌症患者),或者正在治疗感染(例如,病毒,如CMV感染)的人癌症患者)。
在另一个方面,本发明的特征在于扩增γδT细胞的方法,其包括(i)提供获自非造血组织的γδT细胞群体;和(ii)以对于产生扩增的γδT细胞群体有效的量,在存在:(a)IL-2或IL-9;(b)IL-15;和(c)IL-21的情况下培养γδT细胞至少5天。在一些实施方式中,在步骤(ii)中,还在存在IL-4的情况下培养γδT细胞。
在另一个方面,本发明的特征在于通过(i)提供获自非造血组织的γδT细胞群体;和(ii)在存在IL-2、IL-15和选自IL-21、基质细胞来源的因子(SDF,例如,SDF-1)、IL-1β、IL-12、IL-18、和IL-33组成的组的因子的情况下培养γδT细胞至少5天以产生扩增的γδT细胞群体来扩增γδT细胞的方法。在一些实施方式中,步骤(ii)包括在不存在外源TCR途径激动剂的情况下培养γδT细胞。在一些实施方式中,步骤(ii)包括在无血清培养基中培养γδT细胞。在一些实施方式中,在步骤(i)之后,从非造血细胞分离γδT细胞以产生分离的γδT细胞群体。另外,步骤(ii)可以包括在不存在明显基质细胞接触的情况下培养γδT细胞;在不存在明显肿瘤细胞接触的情况下培养γδT细胞;和/或在不存在明显饲养细胞接触的情况下培养γδT细胞。
在另一个方面,本发明的特征在于通过以下步骤扩增γδT细胞的方法:(i)提供非造血组织,组织包括非造血细胞和γδT细胞;(ii)从非造血细胞分离γδT细胞以获得分离的γδT细胞群体;和(iii)在存在IL-2、IL-15和选自IL-21、SDF、IL-1β、IL-12、IL-18和IL-33的因子的情况下培养γδT细胞至少5天以产生扩增的γδT细胞群体。可以在存在IL-2、IL-15和IL-21的情况下培养γδT细胞。另外或者作为另外一种选择,可以在无血清培养基中培养γδT细胞。
在另一个方面,本发明的特征在于具有任何上述扩增的γδT细胞群体的表型的分离的γδT细胞群体。例如,在一些实施方式中,至少50%的分离的群体的γδT细胞表达CD27,但不显著表达TIGIT。在一些实施方式中,至少50%的分离的群体的γδT细胞表达Vδ1。
在另一个方面,本发明包括以上方面所述的分离的γδT细胞的药物组合物。
在另一个方面,本文提供了本文所描述的药物组合物的使用。
在另一个方面,本发明的特征在于通过向对其有需要的受试者施用治疗有效量的如上所述的扩增的γδT细胞,如上所述的分离的群体或者如上所述的药物组合物来通过过继性T细胞疗法治疗受试者的方法。
在本发明的每个方面内,本发明申请中所公开的任何实施方式可以与任何其他所公开的实施方式组合。
附图说明
图1A-图1D显示人的皮肤包含明显的驻留γδT细胞群体。图1A:使用Clark等人(Clark等人,Journal of Investigational Dermatology.2006.126(5):1059-70;“theClark protocol”)所发表的器官型细胞培养来分离皮肤驻留淋巴细胞。在CD45+细胞内,将抗CD3用于对T细胞染色,并且抗CD56抗体分别用于鉴别NK细胞、CD3-CD56+。在CD3+细胞内,将抗泛γδT(panγδT)细胞受体的抗体用于鉴别皮肤驻留的γδT细胞,并且将抗CD8α用于鉴别CD3+,泛γδTCR-门内的常规CD4和CD8阳性αβT细胞的比例。图1B显示了使用Clark规程的7-10个供体的这些实验的总结。使用该规程,人皮肤内的淋巴细胞仍与皮肤成纤维细胞接触,并且不添加细胞因子或者添加白介素-2(IL-2)、白介素-15(IL-15)或IL-2和IL-15,表明除当用IL-15或IL-2和IL-15添加培养时,γδT细胞群体稍微更大外,细胞因子的使用不会改变皮肤驻留的淋巴细胞组成,从而验证了Clark规程。如对于4个供体的总结中所表明的,显示了3周器官型皮肤培养后,使用细胞因子的淋巴细胞组成。图1C:皮肤驻留的γδ细胞主要包括表达Vδ1的γδT细胞(76.24%±17.3),较少的Vδ2 T细胞群体(3.06%±6.1)和对于Vδ1或Vδ2染色为阴性的泛γδTCR阳性细胞群体,其在本文中被称为双阴性(DN)γδT细胞(20.7%±13.97)。健康志愿者血液的对照染色显示出人γδT细胞的强区室化,因为在血液内,主要的γδT细胞群体表达Vδ2 TCR链。图1D:皮肤驻留的γδT细胞显示出先前与已长期激活的T细胞有关的标志物,尽管这些标志物是组织驻留性(residency)的特征指示,而不必需反应长期激活。柱状图显示了γδT细胞上所指明的标志物的染色(实心柱状图)相对于每种抗体的适当同种型对照(空心柱状图)。
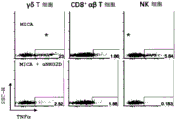
图2A至图2D显示通过Clark规程直接来源于人皮肤的皮肤驻留的γδT细胞显示出通过常规T细胞激活方式的激活的Th1-偏移应答,以及同样地,显示出通过单独的NKG2D配体激活时的Th1-偏移应答。图2A:皮肤驻留的γδT细胞显示出激活的和NK细胞相关的受体NKG2D(实心柱状图,相对于通过空心柱状图所表示的同种型)的强烈表达。通过作为已知的NKG2D受体的配体之一的板结合的重组MICA激活,皮肤γδT细胞在没有任何其他刺激并且独立于TCR连接的情况下应答,因为应答在存在NKG2D封闭抗体的情况下终止。在存在布雷菲德菌素A和100个单位IL-2/mL的情况下刺激细胞6小时,并且随后通过对CD107a染色分析脱颗粒。在表面染色和后续的胞内细胞因子染色之后,通过渗透性分析TNFα和INF-γ的产生。将佛波醇12-豆蔻酸酯13-乙酸酯(P)与离子霉素(I)结合用作T细胞激活的阳性对照。图2B:皮肤驻留的γδT细胞显示出TH1-偏移应答。使用Clark规程回收γδT细胞,并在存在布雷菲德菌素A的情况下用PMA和离子霉素刺激6h并对胞内细胞因子染色。通过刺激,新近分离自人皮肤的γδT细胞产生了TNFα和IFN-γ,但仅产生少量或不可检测的量的与Th2或Th-17细胞有关的细胞因子,例如,IL-4、IL-17A、IL-13、IL-22,然而常规CD4+αβT细胞显示出更多种类的细胞因子的生产。图2C:在直接来源于人皮肤的淋巴细胞中,γδT细胞、CD8a+常规αβT细胞和NK细胞表达了不同水平的NKG2D受体。在这些细胞中,NK细胞对对于单独的NKG2D配体的暴露应答,但是在T细胞内,在不存在任何TCR刺激的情况下,通过NKG2D配体刺激,仅γδT细胞群体显示出细胞因子应答(参见流式细胞术点图的上方的行)。可以使用可溶性抗NKG2D封闭抗体来阻断应答,这表示应答仅通过NKG2D受体介导。图2D:在皮肤驻留的γδT细胞中,仅Vδ1和DNγδT细胞显示出通过单独的重组MICA激活的先天样潜能(通过*表示)。皮肤内少量存在的表达Vδ2的T细胞未显示出这种应答。
图3A-图3D显示皮肤驻留的γδT细胞仅对具有强激活和增殖的皮肤基质的分离应答。图3A:使用Clark规程分离皮肤驻留的淋巴细胞。在3周器官型培养后,收获皮肤淋巴细胞并与任何残留的皮肤细胞,包括成纤维细胞分离,并以1百万个淋巴细胞/mL的密度将其置于补充有100U/mL IL-2的组织培养孔中。在另外3周后,使驻留的γδT细胞强烈扩增,并在皮肤淋巴细胞培养内富集。这种强烈的增殖现象是皮肤驻留的γδT细胞所独有的,并以3周内平均增殖127.18倍的大部分的Vδ1 T细胞为代表,然而常规的αβT细胞平均仅增殖5.21倍;相比差超过20倍。图3B:在14天内,皮肤驻留的Vδ1 T细胞通过标志物Ki-67(细胞周期的指示)的强烈上调来对组织损失应答(用边界为虚线的空心柱状图表示同种型对照;用空心柱状图表示第0天的Ki-67表达;用浅灰色柱状图表示第7天的Ki-67表达;用深灰色柱状图表示第14天的Ki-67表达)。此外,在与组织分离后,当与皮肤基质接触时,其中的大多数对IL-2受体α(CD25)为阴性的皮肤驻留的Vδ1 T细胞上调CD25(同种型对照:短划线柱状图,第0天染色:浅灰色柱状图,第7天染色:深灰色柱状图)。图3C:仅在以Vδ1 T细胞所表示的皮肤驻留的γδT细胞中观察到如通过Ki-67的中值荧光强度(MFI)所表示的高细胞周期速率,而在常规αβT细胞和NK细胞中均未观察到,其中MFI实际上在14天内降低。图3D:在3周培养之后,与基质细胞分离的皮肤淋巴细胞显示出强烈富集的驻留的γδT细胞群体。这种γδT细胞群体含有大部分的Vδ1阳性细胞(77.49%±17.04)和泛γδTCR阳性DN T细胞(21.46%±16.92)。在组织γδT细胞的3周扩增之后,在使用Clark规程新近收获的皮肤淋巴细胞中所观察到的初始的小Vδ2 T细胞群体减少并几乎丧失(0.6%±1.204)。
图4A和图4B显示皮肤驻留的γδT细胞对组织损失应答并且通过皮肤基质细胞,特别是成纤维细胞的接触依赖性机制而受到限制。图4A:在3周后,在如Clark规程中的器官型培养之后收获混合的皮肤淋巴细胞。然后,将混合的淋巴细胞接种到自体同源的皮肤成纤维细胞的汇合层的上方和转板中以控制通过成纤维细胞所产生的可溶性抑制剂的存在。14天后,对于γδT细胞和常规αβT细胞测量通过所存在的绝对细胞数所计算的倍数扩增。当与组织分离且在存在成纤维细胞时,但仅当不与自体同源的成纤维细胞直接细胞接触时,皮肤驻留的γδT细胞显示出强增殖应答。在任何所测试的条件下,常规αβT细胞不显示这种应答。图4B:将获自器官型培养的混合的淋巴细胞接种到补充有IL-2的自体同源的成纤维细胞单层上(浅灰色柱状图)或者接种到补充有IL-2的空孔中(深灰色柱状图)并培养7天。皮肤驻留的Vδ1 T细胞(左图)和泛γδTCR+DN T细胞(右图)在直接存在成纤维细胞的情况下保持休眠,但是当与皮肤器官型培养分离和不存在成纤维细胞时显示出强烈激活,如通过CD25、Th-1相关转录因子T-bet和细胞周期标志物Ki-67(短划线空心柱状图表示相应的同种型对照)的上调表达(MFI)所指示的。
图5A和图5B显示扩增的皮肤γδT细胞显示出脱抑制的迹象和强细胞毒性潜能的增加。图5A:在与器官型细胞培养分离后,使皮肤驻留的γδT细胞扩增14天。然后,使用流式细胞术,通过排除被全αβTCR单克隆抗体染色的所有常规T细胞,对γδT细胞进行阴性分选。然后,将150,000个分选的γδT细胞接种到96平底孔培养板上(重复两次)并在既不添加细胞因子,也不添加任何激活配体的情况下保持培养24小时。收获上清液并使用基于Affymetrix  的细胞因子阵列分析所产生的细胞因子。图5B:还将阴性分选的γδT细胞接种到1天前以10,000个细胞每孔的浓度接种的癌细胞系上。作为对照,使用了阴性分选的常规皮肤αβT细胞。以在存在100U/mL的IL-2,在存在和不存在NKG2D封闭抗体的情况下所指示的效应因子:靶标比接种T细胞。相对于常规αβT细胞,皮肤驻留的γδT细胞显示出优良的恶性细胞系杀死,如通过经ELISA所测量的半胱天冬氨酸酶切割的上皮细胞特异性细胞角蛋白18(CK18)的释放所显示的。细胞毒性至少部分通过NKG2D受体介导,如通过其在含有阻断NKG2D受体的抗体的培养中的减少所显示的。
的细胞因子阵列分析所产生的细胞因子。图5B:还将阴性分选的γδT细胞接种到1天前以10,000个细胞每孔的浓度接种的癌细胞系上。作为对照,使用了阴性分选的常规皮肤αβT细胞。以在存在100U/mL的IL-2,在存在和不存在NKG2D封闭抗体的情况下所指示的效应因子:靶标比接种T细胞。相对于常规αβT细胞,皮肤驻留的γδT细胞显示出优良的恶性细胞系杀死,如通过经ELISA所测量的半胱天冬氨酸酶切割的上皮细胞特异性细胞角蛋白18(CK18)的释放所显示的。细胞毒性至少部分通过NKG2D受体介导,如通过其在含有阻断NKG2D受体的抗体的培养中的减少所显示的。
图6A-图6D显示了人肠中的组织驻留的γδT细胞的分析。图6A:Clark规程的修改形式使得能够分离肠驻留的淋巴细胞。混合的肠淋巴细胞含有通常主要包括Vδ1 T细胞的大的组织驻留的γδT细胞群体,但是还含有Vδ2和双阴性γδT细胞。图6B:分离自肠器官型培养的γδT细胞显示出与皮肤来源的γδT细胞类似的应答,因为一旦将它们与肠基质分离,则它们随时间上调Ki-67。图6C:肠来源的γδT细胞通过产生IFN-γ和通过脱颗粒来对先天样刺激,如重组MICA应答,如通过CD107a上调所测量的。图6D:分离自肠器官型培养的γδT细胞显示出与皮肤来源的γδT细胞类似的应答,并且在细胞培养中随时间扩增,如通过缺少与肠基质接触的淋巴细胞培养中的整体富集所观察到的。
图7A和图7B显示了扩增的皮肤来源的γδT细胞的组织表型。图7A:对于皮肤归巢趋化因子受体CCR4和CCR8阳性的皮肤来源的γδT细胞染色。图7B:分别来源于皮肤或血液的扩增的γδT细胞上的表达水平是不同的。
图8显示在没有任何TCR刺激的情况下,皮肤来源的γδT细胞的脱抑制导致了自发的Th1细胞因子的产生,并且有趣地且与新鲜的TCR激活的γδT细胞相反,导致了特应性细胞因子IL-13的产生。与新近获得的γδT细胞一致,脱抑制且扩增的γδT细胞产生了可忽略的量的Th-2相关细胞因子,例如,IL-4和IL-5。使皮肤来源的γδT细胞扩增14天,并通过排除常规αβT细胞进行阴性分选。对于4个供体,在没有任何刺激或细胞因子添加的情况下,将150,000个混合的γδT细胞以1百万个细胞/毫升的密度在96孔平底板中孵育(重复两次)。24h后,收集上清液并使用Affymetrix的 -基细胞因子阵列进行分析。
-基细胞因子阵列进行分析。
图9显示扩增的和阴性分选的皮肤来源的γδT细胞对它们与之共培养的多种人肿瘤细胞系显示出强细胞毒性,如使用ELISA,通过靶细胞的半胱天冬氨酸酶-切割的细胞角蛋白18的释放所测量的。
图10A和10B显示新鲜的非扩增的皮肤来源的Vδ1 T细胞显示出在先T细胞激活的标志物。图10A:皮肤来源的Vδ1 T细胞表达高CD69和TIM3,以及低CD28。此外,它们显示出激活标志物NKG2D的高表达。在体外扩增期间,通过皮肤来源的Vδ1 T细胞维持这种表型。作为对比,来源于人血液的Vδ1 T细胞缺少这些激活的迹象,并且不表达CD69或TIM3。与皮肤来源的Vδ1 T细胞相比,血液来源的Vδ1 T细胞上的NKG2D表达要低得多,然而血液来源的Vδ1T细胞表达共刺激分子CD28。图10B:在不存在任何其他刺激,如T细胞受体的配体的情况下,仅皮肤来源的Vδ1T细胞对NKG2D配体,如重组MICA具有反应性。血液来源的Vδ1或Vδ2T细胞对先天样刺激未显示出这种反应性。如所指示的,将细胞与重组MICA或抗CD3抗体或两者一起接种到96孔板中。细胞在IL-2 100U/mL中,并且在最后4小时,在BFA中培养6h,然后对于IFN-γ进行表面抗原染色、渗透性和胞内染色。
图11显示了皮肤来源的Vδ1 T细胞表达较低水平的CD16,但是显示出显著的高亲合力IgG受体CD64的表面表达。因此,除直接细胞毒活性之外,组织来源的Vδ1 T细胞还可以用于提高单克隆抗体疗法,如CD20或Her2疗法的效力,因为它们将被所述抗体引导至恶性肿瘤和转移的位点,识别受调理的肿瘤细胞并通过抗体依赖性细胞介导的细胞毒性(ADCC)杀死它们。所显示的结果来自(4个中的)1个代表性供体。
图12显示了IL-2(左图)、IL-15(中图)和IL-2+IL-15(右图)中Vδ1T细胞的扩增。将新近分离的皮肤来源的淋巴细胞在96孔平底板中,在含有10%FCS和1%Pen/Strep的RPMI培养基中培养7天并分别添加IL-2、IL-15或IL-2+IL-15。IL-2和IL-15两者以及两种细胞因子的组合诱导Vδ1T细胞的增殖,如与在不存在任何基质细胞的情况下的同种型(真阴性)染色相比,通过Ki-67染色的迁移所指示的。Ki-67特异性地染色停留在细胞周期G0期的细胞并且通常与增殖相关。
图13显示了流式细胞术结果,其表明了在第21天,在扩增的Vδ1 T细胞表面上的CD9、CCR3和CD39的表达。扩增的皮肤来源的Vδ1 T细胞维持了高水平的细胞表面标志物CCR3、CD39和CD9,如通过(黑色柱状图)相对于等同的同种型染色(真阴性,空心柱状图)所指示的。
图14显示了皮肤来源的Vδ1 T细胞(深色柱)和血液来源的Vδ1 T细胞(浅色柱)中CCR3和CD9的mRNA表达。如本文所公开的,扩增皮肤来源的Vδ1 T细胞,并且使用板结合的VδT细胞受体的抗体(20μg/mL)扩增血液来源的Vδ1 T细胞。扩增后,使用荧光激活细胞分选术(FACS)分离Vδ1Τ细胞,并且对于两个组(血液=灰色vs.皮肤=黑色),从3个供体中分离RNA。对全mRNA测序,并且将所指示的mRNA的表达水平归一化并log2转化。以直接比较,并且以与GAPDH(在大部分人细胞中以高水平表达的共有看家基因)的比显示所有表达水平。
图15显示了皮肤来源的Vδ1 T细胞(深色柱)和血液来源的Vδ1 T细胞(浅色柱)中IL-13的mRNA表达。如本文所公开的,扩增皮肤来源的Vδ1 T细胞,并且使用板结合的高剂量的VδT细胞受体的抗体(20μg/mL)扩增血液来源的Vδ1 T细胞。扩增后,使用FACS分离Vδ1Τ细胞,并且对于两个组(血液=灰色vs.皮肤=黑色),从3个供体中分离RNA。对全mRNA测序,并且将IL-13的mRNA的表达水平归一化并log2转化。以直接比较,并且以与GAPDH的比显示表达水平。
图16A和图16B显示了在用PMA/离子霉素(图16A)或抗CD3(图16B)TCR刺激之后,皮肤来源的Vδ1 T细胞中细胞因子的产生。在分离和扩增之后,使用荧光激活细胞分选术(FACS)纯化皮肤来源的Vδ1 T细胞。对于三个供体,将150.000个Vδ1Τ细胞接种到96孔平底板中(重复两次),并且用板结合的抗CD3(5μg/mL)或者PMA/离子霉素刺激24小时。使用 平台分析上清液的所指示的细胞因子的绝对的量。
平台分析上清液的所指示的细胞因子的绝对的量。
图17A-图17H显示了扩增条件的结果。图17A和17B显示了分离的淋巴细胞(分离培养21天之后;图17A)和已在存在IL-2、IL-4、IL-15和IL-21中扩增20天的淋巴细胞(图17B)的代表性流式细胞术图以及设门方案。图17C显示了对由于IL-2(100U/mL)和IL-15(10ng/mL)的Vδ1 T细胞扩增所归一化的多种条件下的Vδ1 T细胞的倍数扩增。图17D显示了作为用IL-2+IL-15、IL2+IL-15+IL-4、IL-2+IL-15+IL-21和IL-2+IL-15+IL-4+IL-21处理的结果,相对于分离的群体的倍数扩增。图17E显示了第21天(扩增前)和第41天(扩增后)的绝对Vδ1+T细胞数目。如所指示的,在100U/mL IL-2、100U/mL IL-2+10ng/mL IL-15或者100U/mLIL-2+5ng/mL IL-4+10ng/mL IL-15+100ng/mL IL-21中进行扩增。(n=8-17)。图17F显示了在两个时间点,对于每种条件的平均(加SEM)Vδ1+T细胞数目。**p=0.001。学生氏非配对t检验。(n=8-17)。图17G显示了对单独使用100U/mL IL-2、5ng/mL IL-4和10ng/mL IL-15的扩增所归一化的使用不同浓度的IL-21的Vδ1+T细胞扩增(n=3)。图17H显示了对单独使用IL-2的扩增所归一化的使用100U/mL IL-2+5ng/mL IL-4+10ng/mL IL-15+10ng/mL IL-21的Vδ1+T细胞扩增。
图18A-图18D的特征在于通过扩增的Vδ1 T细胞的CD27的表达。图18A显示了对作为IL-2(100U/mL)和IL-15(10ng/mL)的结果的CD27表达所归一化的多种条件下的CD27的倍数表达(通过MFI)。图18B显示了由于用IL-2+IL-15、IL2+IL-15+IL-4、IL-2+IL-15+IL-21和IL-2+IL-15+IL-4+IL-21处理,相对于分离的群体的CD27 MFI。图18C显示了如通过流式细胞术所评价的,对单独使用100U/mL IL-2,5ng/mL IL-4和10ng/mL IL-15的扩增所归一化的使用不同浓度的IL-21的Vδ1+CD27表达(n=3)。图18D显示了如通过流式细胞术所评价的,对单独使用IL-2的扩增所归一化的使用100U/mL IL-2+5ng/mL IL-4+10ng/mL IL-15+10ng/mL IL-21的Vδ1+CD27表达。(n=4)。
图19的特征在于通过扩增的Vδ1 T细胞的TIGIT的表面表达。图19显示了对由于IL-2和IL-15的处理的TIGIT表达所归一化的在多种条件下的TIGIT(通过MFI)的倍数表达。
图20是显示作为CD27表达的函数的个体细胞的表面TIGIT表达的图。
图21是显示在存在或不存在血液来源的血清或血浆部份的情况下支持组织来源的γδT细胞的扩增和富集的细胞因子的使用的图。混合的淋巴细胞群体分离自含有在具有或不具有10%的人AB血清且含有IL-2、IL-4、IL-15和IL-21的培养基中扩增的2%的γδT细胞的组织样品。数据突出显示了相对于血清(295倍扩增,从2%富集至75%),在没有人血清的情况下的成功且等价的扩增(432倍)和富集(从2%富集至77%)。
图22A-图22D是显示来自分离的混合的淋巴细胞群体的组织来源的γδ细胞的示例性富集的图。细胞分离自单一人组织样品并复制,然后在含有10%血清(左列)或5%细胞疗法系统血清(CTSTM)(右列)的TexMACS培养基加IL-2、IL-4、IL-15和IL-21中扩增。如图所示,提供了分离和扩增的细胞培养物的谱图。图22A显示了含有相对低数目(<10%)的所期望的组织来源的γδT细胞的初始分离的培养。相反,图22B-图22D显示在扩增后,所得的细胞群体大量富集组织来源的γδT细胞。
图23是显示肠驻留Vδ1细胞上TIGIT的组成型表达的图。使用用于结肠上皮细胞内淋巴细胞释放的标准分离规程,使用分离自肠上皮细胞的Vδ1细胞产生数据。
图24A和图24B是显示如通过IFNγ(图24A)和TNFα(图24B)的表达所测量的,脊髓灰质炎病毒受体(PVR)特异性抑制TCR信号转导的图。将细胞与IL-2和IL-15一起孵育,并用抗CD3抗体激活。
图25A和图25B是显示如通过IFNγ(图25A)或TNFα(图25B)所测量的,PVR对TIGIT-阴性Vδ1+/Vδ3+细胞的抑制作用丧失的图。将细胞与IL-2、IL-15、IL-4和IL-21一起孵育,并用抗CD3抗体激活。
图26A和图26B是显示在皮肤来源的γδT细胞的扩增中,IL-9可以替代IL-2的功能的图。将来自三个供体(TS052、TS056和SK073)的皮肤组织置于9mm网格上并在补充有IL-2和IL-15的培养基中培养3周。然后,将分离的淋巴细胞在补充有IL-2、IL-4、IL-15和IL-21(左侧柱)或者IL-4、IL-9、IL-15和IL-21(右侧柱)的培养基中扩增。在扩增3周后,计算γδT细胞/网格(图26A)和Vδ1细胞/网格(图26B)的最终得率。在皮肤来源的γδT细胞的扩增中,用IL-9替代IL-2产生了等价的扩增效率。柱状图表示平均值+/-SEM。
图27A-图27C是显示如通过变化倍数(图27A)、γδTCR+T细胞的%(图27B)和Vδ1+T细胞的%(图27C)所测量的,在皮肤来源的γδT细胞的扩增中,IL-9可以替代IL-2的功能的图。皮肤组织来自6个供体(SK073、SK075、SK077、TS052、TS053和TS056)。2CK=2个细胞因子(IL-2+IL-15),4CK=4个细胞因子(IL-2+IL-15+IL-21+IL-4)。
具体实施方式
I.简介
本文提供了使来自非造血组织来源(例如,非造血组织来源的γδT细胞,例如,非造血组织来源的Vδ1 T细胞)的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或双阴性T细胞)扩增的方法。扩增方法包括在不存在明显的TCR刺激和/或在存在白介素-4(IL-4)、白介素-15(IL-15)、白介素-21(IL-21)和/或白介素-2(IL-2)的情况下,培养γδT细胞(例如,分离自非造血组织的基质细胞的γδT细胞)。还提供了扩增的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2T细胞,例如,Vδ1 T细胞和/或DN T细胞)的组合物和使用扩增的γδT细胞的方法(例如,作为(例如)用于癌症治疗的过继性T细胞疗法的一部分)。
II.定义
应理解本文所描述的本发明的方面和实施方式包括“包含”各方面和实施方式、“由各方面和实施方式组成”和“基本由各方面和实施方式组成”。除非另外说明,否则如本文所使用的,单数形成的“一个”、“一种”和“该”包括复数提及物。
如本文所使用的术语“约”是指该技术领域中的技术人员易于已知的各个值的惯例误差范围。在本文中提及“约”值或参数包括(并描述)了涉及该值或参数本身的实施方式。在一些情况下,“约”涵盖了对于给定值+20%,在一些情况下+10%,在一些情况下+5%,在一些情况下+1%,或者在一些情况下+0.1%的变化,因为这些变化对于所公开的方法的实施是适合的。
如本文所使用的,术语“基本的”和“基本地”是指显示所关心的特征或性质的总程度或等级或者接近总程度或等级的定性条件。生物领域的技术人员之一会理解生物和化学现象很少(如果发生过的话)是完全的和/或发生至完全或者实现或避免绝对结果。因此,术语“基本地”在本文中用于获取对多种生物和化学现象中所驻留的完全性的潜在缺少。当描述物理情况,如受体/配体相互作用或者细胞/细胞接触时,如果其功能结果是通过实施方法的人可用的常规方式可检测的,则所述情况为明显的。例如,“明显的TCR激活”是指细胞群体中可检测的TCR激活水平(例如,TCR激活的统计学显著程度)。在一些实施方式中,一旦暴露于多至0.1%、多至0.5%、多至1%、多至5%、多至10%、多至20%、多至30%或者多至40%的TCR途径激动剂(例如,抗体,例如,抗CD3,或凝集素)的EC50,则在各个细胞群体上显著激活TCR。同样地,“明显的细胞接触”(例如,明显的饲养细胞接触,明显的基质细胞接触或者明显的肿瘤细胞接触)是指能够在扩增细胞中引起可检测变化(例如,减少扩增)的细胞-至-细胞接触程度。在一些情况下,当污染细胞类型(例如,饲养细胞、基质细胞或肿瘤细胞)相对于扩增细胞群体以按数目计多至0.1%、多至0.5%、多至1%、多至5%、多至10%或多至20%存在于培养隔室中时,发生明显细胞接触。同样地,细胞“显著的数目”或者试剂“显著的量”是指如上定义,引起显著影响所需的数目或量。
如本文所使用的,“非造血细胞”包括基质细胞和上皮细胞。基质细胞是任何器官的非造血结缔组织细胞并且支持该器官实质细胞的功能。基质细胞的实例包括成纤维细胞、外膜细胞、间充质细胞、角化细胞、内皮细胞和非血液肿瘤细胞。上皮细胞是在整个身体的血管和器官的腔和表面上衬里的非造血细胞。它们的形状通常是鳞状、圆柱体或立方体的,并且可以作为单层细胞或者作为两个或多个细胞层布置。
如本文所使用的,“非造血组织驻留的γδT细胞”、“非造血组织来源的”和“非造血组织天然的γδT细胞”是指在组织移植时存在于非造血组织中的γδT细胞。非造血组织驻留的γδT细胞可以获自任何适合的人或非人动物非造血组织。非造血组织是血液或骨髓以外的组织。在一些实施方式中,γδT细胞未获自特定类型的生物流体样品,如血液或关节液。这些适合的人或非人动物非造血组织的实例包括皮肤或其部分(例如,真皮或表皮)、胃肠道(例如,胃肠上皮细胞、结肠、小肠、胃、阑尾、盲肠或直肠)、乳腺组织、肺(优选地其中组织不通过支气管肺泡灌洗获得)、前列腺、肝和胰脏。在一些实施方式中,非造血组织驻留的γδT细胞可以来源于淋巴组织,如胸腺、脾脏或扁桃腺。γδT细胞还可以存在于人癌症组织,例如,乳腺和前列腺。在一些实施方式中,γδT细胞不获自人癌症组织。可以通过标准技术,例如,通过移植(例如,活组织检查)获得非造血组织样品。非造血组织驻留的γδT细胞包括非Vδ2 T细胞,例如,Vδ1T细胞,双阴性(DN)T细胞、Vδ3 T细胞和Vδ5 T细胞。
如本文所使用的,“IL-2”是指用作一种或多种IL-2受体(IL-2R)亚基的激动剂的天然或重组IL-2或其变体(例如,其突变体、突变蛋白、类似物、亚基、受体复合物、片段、同种型和拟肽)。这些因子可以支持IL-2依赖性细胞系,CTLL-2(33;美国模式培养物保藏所 TIB 214)的增殖。成熟的人IL-2作为133氨基酸序列(缺少信号肽,由其他20个N末端氨基酸组成)存在,如Fujita等人,Cell 1986.46.3:401-407所述。IL-2突变蛋白是其中已对白细胞介素-2蛋白进行了特异性替换,同时保留了结合IL-2Rβ的能力的多肽,如US2014/0046026中所述。IL-2突变蛋白的特征可以在于天然IL-2多肽链中的一个或多个位点或其他残基处的氨基酸插入、缺失、替换和修饰。根据本发明公开,任何这些插入、缺失、替换和修饰产生了保留IL-2Rβ结合活性的IL-2突变蛋白。示例性突变蛋白可以包括1、2、3、4、5、6、7、8、9、10或更多个氨基酸取代。
TIB 214)的增殖。成熟的人IL-2作为133氨基酸序列(缺少信号肽,由其他20个N末端氨基酸组成)存在,如Fujita等人,Cell 1986.46.3:401-407所述。IL-2突变蛋白是其中已对白细胞介素-2蛋白进行了特异性替换,同时保留了结合IL-2Rβ的能力的多肽,如US2014/0046026中所述。IL-2突变蛋白的特征可以在于天然IL-2多肽链中的一个或多个位点或其他残基处的氨基酸插入、缺失、替换和修饰。根据本发明公开,任何这些插入、缺失、替换和修饰产生了保留IL-2Rβ结合活性的IL-2突变蛋白。示例性突变蛋白可以包括1、2、3、4、5、6、7、8、9、10或更多个氨基酸取代。
可以通过常规程序,如聚合酶链反应(PCR)获得编码人IL-2的核酸。人IL-2的氨基酸序列(Gene ID 3558)以登录位置号NP_000577.2GI:28178861存在于Genbank中。鼠科(小家鼠(Mus musculus))IL-2的氨基酸序列(Gene ID 16183)以登录位置号NP_032392.1GI:7110653存在于Genbank中。
IL-2还可以表示来源于多种哺乳动物种,包括(例如)人、猿、牛、猪、马和鼠的IL-2。变体可以保守地包括取代的序列,其表示给定氨基酸残基被具有类似生理化学特征的残基替代。保守替换的实例包括用一种脂肪族残基替换另一种,例如,用Ile、Val、Leu或Ala替换另一种,或者用一种极性残基替换另一种,如Lys和Arg;Glu和Asp;或者Gln和Asn之间的替换。其他这些保守替换,例如,具有类似疏水性特征的整个区域的替换是熟知的。天然存在的IL-2变体也涵盖在本发明中。这些变体的实例是由可变mRNA剪接事件所产生的或者由IL-2蛋白的蛋白水解切割所产生的蛋白,其中保留了IL-2的结合性。mRNA的可变剪接可以获得截短,但具有生物学活性的IL-2蛋白。可归因于蛋白水解的改变包括(例如)由于从IL-2蛋白的蛋白水解除去一个或多个末端氨基酸(通常1-10个氨基酸),在不同宿主细胞类型中表达时N-或C-末端中的差异。在一些实施方式中,可以修饰所述蛋白的末端或内部以改变其物理性质,例如,通过化学基团,如聚乙二醇修饰(Yang等人,Cancer 1995.76:687-694)。在一些实施方式中,可以用其他氨基酸修饰所述蛋白的末端或内部(Clark-Lewis等人,PNAS 1993.90:3574-3577)。
如本文所使用的,“IL-15”是指用作一种或多种IL-15受体(IL-15R)亚基的激动剂的天然或重组IL-15或其变体(例如,其突变体、突变蛋白、类似物、亚基、受体复合物、片段、同种型和拟肽)。如IL-2一样,IL-15是可以支持IL-2依赖性细胞系,CTLL-2增殖的已知的T细胞生长因子。Grabstein等人(Grabstein等人,Science 1994.264.5161:965-969)首先将IL-15报道为114-氨基酸成熟蛋白。如本文所使用的术语“IL-15”表示天然或重组IL-15及其突变蛋白、类似物、亚基或其复合物(例如,受体复合物,例如,sushi肽,如WO 2007/046006中所述),它们中的每一个可以刺激CTLL-2细胞的增殖。在CTLL-2增殖测定中,用IL-15的重组表达前体和成熟形式的框内融合转染的细胞上清液可以诱导CTLL-2的细胞增殖。
可以根据Grabstein等人(Grabstein等人,Science 1994.264.5161:965-969)所描述的程序或者通过常规程序,如聚合酶链反应(PCR)获得人IL-15。于1993年2月19日在 对人IL-15 cDNA进行了保藏,并且指定登录号为69245。
对人IL-15 cDNA进行了保藏,并且指定登录号为69245。
人IL-15的氨基酸序列(Gene ID 3600)以登录位置号NP000576.1GI:10835153(同种型1)和NP_751915.1 GI:26787986(同种型2)存在于Genbank中。鼠科(小家鼠(Musmusculus))的IL-15氨基酸序列(Gene ID 16168)以登录位置号NP_001241676.1GI:363000984存在于Genbank中。
IL-15还可以表示来源于多种哺乳动物种,包括(例如)人、猿、牛、猪、马和鼠的IL-15。如本文所提及的,IL-15“突变蛋白”或“变体”是与天然哺乳动物IL-15的序列显著同源,但是由于氨基酸缺失、插入或替换而具有不同于天然哺乳动物IL-15多肽的氨基酸序列的多肽。变体可以保守地包括取代的序列,其表示给定氨基酸残基被具有类似生理化学特征的残基替代。保守替换的实例包括用一种脂肪族残基替换另一种,例如,用Ile、Val、Leu或Ala替换另一种,或者用一种极性残基替换另一种,如Lys和Arg;Glu和Asp;或者Gln和Asn之间的替换。其他这些保守替换,例如,具有类似疏水性特征的整个区域的替换是熟知的。天然存在的IL-15变体也涵盖在本发明中。这些变体的实例是由可变mRNA剪接事件所产生的或者由IL-15蛋白的蛋白水解切割所产生的蛋白,其中保留了IL-15的结合性。mRNA的可变剪接可以获得截短,但具有生物学活性的IL-15蛋白。可归因于蛋白水解的改变包括(例如)由于从IL-15蛋白蛋白水解除去一个或多个末端氨基酸(通常1-10个氨基酸),在不同宿主细胞类型中表达时N-或C-末端中的差异。在一些实施方式中,可以修饰所述蛋白的末端以改变其物理性质,例如,通过化学基团,如聚乙二醇修饰(Yang等人,Cancer 1995.76:687-694)。在一些实施方式中,可以用其他氨基酸修饰所述蛋白的末端或内部(Clark-Lewis等人,PNAS 1993.90:3574-3577)。
如本文所使用的,“IL-4”是指用作一种或多种IL-4受体(IL-4R)亚基的激动剂的天然或重组IL-4或其变体(例如,其突变体、突变蛋白、类似物、亚基、受体复合物、片段、同种型和拟肽)。这些因子可以支持未处理过的辅助性T细胞(Th0细胞)分化为Th2细胞。成熟的人IL-4作为129氨基酸序列(缺少信号肽,由其他24个N末端氨基酸组成)存在。IL-4突变蛋白是其中已对白介素-4蛋白进行了特异性替换,同时保留了结合IL-4Rα的能力的多肽,如美国专利No.6,313,272中所描述的那些。IL-4突变蛋白的特征可以在于天然IL-4多肽链中的一个或多个位点或其他残基处的氨基酸插入、缺失、替换和修饰。根据本发明公开,任何这些插入、缺失、替换和修饰产生了保留IL-2Rα结合活性的IL-4突变蛋白。示例性突变蛋白可以包括1、2、3、4、5、6、7、8、9、10或更多个氨基酸取代。
可以通过常规程序,如聚合酶链反应(PCR)获得编码人IL-4的核酸。人IL-4的氨基酸序列(Gene ID 3565)以登录位置号NG_023252存在于Genbank中。鼠科(小家鼠(Musmusculus))IL-4的氨基酸序列(Gene ID 16189)以登录位置号NC_000077.6存在于Genbank中。
IL-4还可以表示来源于多种哺乳动物种,包括(例如)人、猿、牛、猪、马和鼠的IL-4。变体可以保守地包括取代的序列,其表示给定氨基酸残基被具有类似生理化学特征的残基替代。保守替换的实例包括用一种脂肪族残基替换另一种,例如,用Ile、Val、Leu或Ala替换另一种,或者用一种极性残基替换另一种,如Lys和Arg;Glu和Asp;或者Gln和Asn之间的替换。其他这些保守替换,例如,具有类似疏水性特征的整个区域的替换是熟知的。天然存在的IL-4变体也涵盖在本发明中。这些变体的实例是由可变mRNA剪接事件所产生的或者由IL-4蛋白的蛋白水解切割所产生的蛋白,其中保留了IL-4的结合性。mRNA的可变剪接可以获得截短,但具有生物学活性的IL-4蛋白。可归因于蛋白水解的改变包括(例如)由于从IL-4蛋白蛋白水解除去一个或多个末端氨基酸(通常1-10个氨基酸),在不同宿主细胞类型中表达时N-或C-末端中的差异。在一些实施方式中,可以修饰所述蛋白的末端以改变其物理性质,例如,通过化学基团,如聚乙二醇修饰(Yang等人,Cancer 1995.76:687-694)。在一些实施方式中,可以用其他氨基酸修饰所述蛋白的末端或内部(Clark-Lewis等人,PNAS1993.90:3574-3577)。
如本文所使用的,“IL-21”是指用作一种或多种IL-21受体(IL-21R)亚基的激动剂的天然或重组IL-21或其变体(例如,其突变体、突变蛋白、类似物、亚基、受体复合物、片段、同种型和拟肽)。这些因子可以支持自然杀伤(NK)和细胞毒性(CD8+)T细胞的增殖。成熟的人IL-21作为133氨基酸序列(缺少信号肽,由其他22个N末端氨基酸组成)存在。IL-21突变蛋白是其中已对白介素-21蛋白进行了特异性替换,同时保留了结合IL-21Rα的能力的多肽,如美国专利No.9,388,241中所描述的那些。IL-21突变蛋白的特征可以在于天然IL-21多肽链中的一个或多个位点或其他残基处的氨基酸插入、缺失、替换和修饰。根据本发明公开,任何这些插入、缺失、替换和修饰产生了保留IL-21R结合活性的IL-21突变蛋白。示例性突变蛋白可以包括1、2、3、4、5、6、7、8、9、10或更多个氨基酸取代。
可以通过常规程序,如聚合酶链反应(PCR)获得编码人IL-21的核酸。人IL-21的氨基酸序列(Gene ID 59067)以登录位置号NC_000004.12存在于Genbank中。鼠科(小家鼠(Mus musculus))IL-21的氨基酸序列(Gene ID 60505)以登录位置号NC_000069.6存在于Genbank中。
IL-21还可以表示来源于多种哺乳动物种,包括(例如)人、猿、牛、猪、马和鼠的IL-21。变体可以保守地包括取代的序列,其表示给定氨基酸残基被具有类似生理化学特征的残基替代。保守替换的实例包括用一种脂肪族残基替换另一种,例如,用Ile、Val、Leu或Ala替换另一种,或者用一种极性残基替换另一种,如Lys和Arg;Glu和Asp;或者Gln和Asn之间的替换。其他这些保守替换,例如,具有类似疏水性特征的整个区域的替换是熟知的。天然存在的IL-21变体也涵盖在本发明中。这些变体的实例是由可变mRNA剪接事件所产生的或者由IL-21蛋白的蛋白水解切割所产生的蛋白,其中保留了IL-21的结合性。mRNA的可变剪接可以获得截短,但具有生物学活性的IL-21蛋白。可归因于蛋白水解的改变包括(例如)由于从IL-21蛋白蛋白水解除去一个或多个末端氨基酸(通常1-10个氨基酸),在不同宿主细胞类型中表达时N-或C-末端中的差异。在一些实施方式中,可以修饰所述蛋白的末端以改变其物理性质,例如,通过化学基团,如聚乙二醇修饰(Yang等人,Cancer 1995.76:687-694)。在一些实施方式中,可以用其他氨基酸修饰所述蛋白的末端或内部(Clark-Lewis等人,PNAS 1993.90:3574-3577)。
如本文所使用的,“IL-9”是指用作一种或多种IL-9受体(IL-9R)亚基的激动剂的天然或重组IL-9或其变体(例如,其突变体、突变蛋白、类似物、亚基、受体复合物、片段、同种型和拟肽)。成熟的人IL-9作为144氨基酸序列存在。IL-9突变蛋白是其中已对白介素-9蛋白进行了特异性替换,同时保留了结合IL-9R的能力的多肽。IL-9突变蛋白的特征可以在于天然IL-9多肽链中的一个或多个位点或其他残基处的氨基酸插入、缺失、替换和修饰。根据本发明公开,任何这些插入、缺失、替换和修饰产生了保留IL-9R结合活性的IL-9突变蛋白。示例性突变蛋白可以包括1、2、3、4、5、6、7、8、9、10或更多个氨基酸取代。
可以通过常规程序,如聚合酶链反应(PCR)获得编码人IL-9的核酸。通过UniProtKB P15248提供了人IL-9的氨基酸序列。
IL-9还可以表示来源于多种哺乳动物种,包括(例如)人、猿、牛、猪、马和鼠的IL-9。变体可以保守地包括取代的序列,其表示给定氨基酸残基被具有类似生理化学特征的残基替代。保守替换的实例包括用一种脂肪族残基替换另一种,例如,用Ile、Val、Leu或Ala替换另一种,或者用一种极性残基替换另一种,如Lys和Arg;Glu和Asp;或者Gln和Asn之间的替换。其他这些保守替换,例如,具有类似疏水性特征的整个区域的替换是熟知的。天然存在的IL-9变体也涵盖在本发明中。这些变体的实例是由可变mRNA剪接事件所产生的或者由IL-9蛋白的蛋白水解切割所产生的蛋白,其中保留了IL-9的结合性。mRNA的可变剪接可以获得截短,但具有生物学活性的IL-9蛋白。可归因于蛋白水解的改变包括(例如)由于从IL-9蛋白蛋白水解除去一个或多个末端氨基酸(通常1-10个氨基酸),在不同宿主细胞类型中表达时N-或C-末端中的差异。在一些实施方式中,可以修饰所述蛋白的末端以改变其物理性质,例如,通过化学基团,如聚乙二醇修饰(Yang等人,Cancer 1995.76:687-694)。在一些实施方式中,可以用其他氨基酸修饰所述蛋白的末端或内部(Clark-Lewis等人,PNAS1993.90:3574-3577)。
以上因子中的任何一个或多个可以以对于产生扩增的γδT细胞群体有效的量包括在扩增规程中。如本文所使用的,短语“以对于……有效的量”是指引起可检测的结果的量(例如,相对于其起始群体,具有统计学显著提高的数目的细胞数目,例如,p<0.05)。在其中一次存在多个因子的情况下,有效量是指所有因子的综合影响(例如,IL-2和IL-15的综合影响,或者IL-2或IL-9、IL-4、IL-15和IL-21的综合影响)。
“T细胞受体(TCR)途径激动剂”或“激活TCR途径的试剂”是指通过TCR信号转导,引起T细胞,如αβT细胞和/或血液驻留的γδT细胞的增殖或其他激活后果的化合物。通过Src相关蛋白酪氨酸激酶(PTKs)、Lck和Fyn以及70kDA的ξ-链(TCR)相关蛋白激酶(ZAP70)顺序激活T细胞信号转导调节因子功能。这些PTK导致了包括T细胞接头激活剂(LAT)的多肽的磷酸化,其导致了通过胞外信号调节激酶(ERK)、c-Jun N末端激酶(JNK)和激活T细胞核因子(NFAT)的下游刺激。例如,通过CD28和CD45的共刺激可以提高磷酸化和提高TCR信号通路。因此,靶向TCR的一部分或共刺激途径的任何因子可以激活T细胞信号转导。TCR途径激动剂包括抗体(例如,单克隆抗体,例如,抗TCR Vδ1、抗TCRδTCS-1、抗TCR PANγδ和抗CD3)、凝集素(例如,植物凝集素,例如,伴刀豆凝集素A、来自菜豆(Phaseolus vulgaris)(PHA-P)、美洲商陆(Phytolacca Americana)、小麦(Triticum vulgaris)、小扁豆(Lens culinaris)、大豆(Glycine max)、朝鲜槐(Maackia amurensis)、豌豆(Pisum sativum)和欧洲接骨木(Sambucus nigra)的凝集素)、合成磷酸化抗原(例如,BrHPP(焦磷酸溴代醇)、2M3B1PP(2-甲基-3-丁烯基-1-焦磷酸盐)、HMBPP((E)-4-羟基-3-甲基-丁-2-烯焦磷酸酯)或者IPP(异戊烯焦磷酸酯))和N-二膦酸酯(例如,唑来膦酸)。TCR途径激动剂包括共受体激动剂,其包括抗体(例如,单克隆抗体,例如,抗CD2、抗CD6、抗CD9、抗CD28、抗CD43、抗CD94、抗CD160、抗SLAM、抗NKGD2、抗2B4、抗HLA-A、抗HLA-b、抗HLA-C和抗ICAM-3)和蛋白(例如,重组蛋白,例如,重组人蛋白,例如,CD7L、CD26、CD27L、CD30L、CD40L、OX40L、4-1BBL、ICAM-1、纤连蛋白、氢化可的松及其变体,例如,Fc-融合蛋白,例如,CD27L-Fc)。TCR途径激动剂可以是可溶性的或膜结合的,并且可以(例如)存在于细胞,如人工抗原递呈细胞(aAPC)上,如对于MHC或HLA复合物的情况。适合于激活T细胞信号转导的aAPC在本领域中是已知的。通过外源添加TCR途径激动剂激活T细胞的适合方法在本领域中是熟知的并且在Deniger等人的图1中得到总结(Deniger等人,Frontiers in Immunology.2014.5(636):1-10)。
“外源TCR途径激动剂”是指不来源于非造血组织或其供体的TCR途径激动剂(即它们不是外源添加的)。因此,会理解在本发明的一些实施方式中,TCR途径激动剂可以作为来自非造血组织的残余材料存在于培养物中(例如,可溶性纤连蛋白或细胞结合的ICAM-1)。在一些实施方式中,残余的TCR途径激动剂具有可忽略的浓度并且不显著激活T细胞。
如本文所使用的,“合成支架”、“支架”和“网格”是可互换使用的并且表示适合于支持细胞生长的非天然立体结构。移植物可以粘附至合成支架以有利于淋巴细胞从所述移植物流出到所述支架上。合成支架可以由天然和/或合成材料,如聚合物(例如,天然或合成聚合物,例如,聚乙烯吡咯烷酮、聚甲基丙烯酸甲酯、甲基纤维素、聚苯乙烯、聚丙烯、聚氨脂)、陶瓷(例如,磷酸三钙、铝酸钙、钙羟基磷灰石)或金属(钽、钛、铂以及与铂、铌、铪、钨相同元素族的金属及其合金的组合)构造成。根据本领域中已知的方法,可以将生物因子(例如,胶原蛋白(例如,胶原蛋白I或者胶原蛋白II)、纤连蛋白、层粘连蛋白、整合素、血管生成因子、抗炎因子、糖胺聚糖、vitrogens、抗体及其片段、细胞因子(例如,IL-2或IL-15及其组合)涂覆到支架表面上或包封在支架材料内以提高细胞粘附、迁移、存活或增殖。这种及其他方法可以用于从一些其他非造血组织类型,例如,肠、前列腺和乳腺分离淋巴细胞。考虑作为本发明的一部分使用的示例性合成支架包括在Clark规程中所使用的那种。
如本文所使用的,术语“分离”、“分离的”或“分离”是指破坏或阻止不同细胞群体之间的物理接触的操作(例如,造血细胞(例如,淋巴细胞)与非造血细胞的分离)。可以(例如)通过用移液器对混合细胞群体的强力吹打来破坏膜内结合,或者通过与(例如)趋化因子或细胞因子培养引起细胞群体从(例如)组织基质中“爬出”来进行分离,如通过Carrasco等人所述(Carrasco A等人,Journal of Immunological Methods 2013.389(1-2):29-37)。在培养期间,可以使用转板培养系统或者通过防止不同细胞群体之间物理接触的类似培养方法来维持分离。
如本文所使用的,“分离的γδ细胞群体”是指已与其来源的非造血组织分离,从而使其不与非造血细胞明显接触的造血细胞,包括γδ细胞群体(例如,根据本文所描述的任何分离规程)。同样地,“分离的Vδ1 T细胞群体”是指已与其来源的非造血组织分离,从而使其不与非造血细胞明显接触的造血细胞,包括Vδ1 T细胞群体(例如,根据本文所描述的任何分离规程)。因此,在这些情况下,分离是指造血细胞(例如,淋巴细胞)与非造血细胞(例如,基质细胞和/或上皮细胞)的分离。
术语“抗体”以其最广泛的含义使用并且具体地涵盖了单克隆抗体(包括全长单克隆抗体)、多克隆抗体、多重特异性抗体(例如,双重特异性抗体)和抗体片段,只要它们能够显示出所期望的生物活性。
如本文所使用的,“扩增步骤”是指在分离之后发生的培养期,其中通过细胞分裂使特定γδT细胞的数目增加。会理解可以在分离期期间发生细胞分裂,同时γδT细胞与基质细胞接触,但是扩增步骤直至分离完全后才开始。因此,当在“扩增步骤之前”的时间点鉴别分离的细胞群体时,它表示分离培养之后,扩增培养之前的时间点。
如本文所使用的,“扩增的γδ细胞群体”是指已在引起γδ细胞扩增,即γδ细胞数目增加的条件和持续时间培养的造血细胞,包括γδT细胞群体。同样地,如本文所使用的“扩增的Vδ1 T细胞群体”是指已在引起Vδ1 T细胞扩增,即Vδ1 T细胞数目增加的条件和持续时间培养的造血细胞,包括Vδ1 T细胞群体。
如本文所使用的,“饲养细胞”是指添加至培养中以向非造血组织来源的细胞提供细胞与细胞表面接触的任何外源细胞。饲养细胞可以是原代细胞(例如,来源于组织)或者来源于细胞系。饲养细胞可以是活细胞或辐照细胞,并且包括肿瘤细胞、成纤维细胞、B细胞及其他抗原递呈细胞。
术语“标志物”在本文中是指基于DNA、RNA、蛋白、碳水化合物、糖脂或细胞的分子标志物,并且可以通过标准方法(或者本文所公开的方法)检测其在患者样品中的表达或存在。
“表达”所关心的标志物的细胞或细胞群体是其中确定编码所述蛋白的mRNA,或者所述蛋白本身,包括其片段存在于所述细胞或所述群体中的细胞或细胞群体。可以通过多种方式检测标志物的表达。例如,在一些实施方式中,标志物的表达是指标志物在细胞上的表面密度。例如,如用作流式细胞术读数的平均荧光强度(MFI)代表了细胞群体上标志物的密度。本领域技术人员会理解MFI值取决于染色参数(例如,浓度、持续时间和温度)和荧光染料组成。然而,当在适当对照的背景中考虑时,MFI可以是定量的。例如,如果抗所述标志物的抗体的MFI显著高于在等同条件下染色的相同细胞群体上适当的同种型对照抗体的MFI,则可以将细胞群体称为表达所述标志物。另外或者作为另外一种选择,可以根据常规流式细胞术分析方法(例如,通过根据同种型或“荧光减一”(FMO)对照设置门),使用阳性和阴性门,基于逐个细胞,将细胞群体称为表达标志物。按照这个标准,如果对于所述标志物检测为阳性的细胞数目显著高于背景(例如,通过对同种型对照设门),则可以将所述群体称为“表达”所述标志物。
如本文所使用的,当作为阳性细胞百分比表示群体表达并且将百分比与参比群体的阳性细胞的相应百分比相比较时,百分比的差异是各个群体中每一个的亲代群体的百分比。例如,如果在10%的群体A的细胞上表达标志物并且在1%的群体B的细胞上表达相同标志物,则将群体A称为具有比群体B高9%频率的标志物阳性细胞(即,10%-1%,而不是10%÷1%)。当将亲代群体中的细胞数目乘以频率时,计算出细胞绝对数目的差异。在以上所提供的实例中,如果群体A中存在100个细胞,群体B中存在10个细胞,则群体A相对于群体B具有100倍数目的细胞,即(10%×100)÷(1%×10)。
标志物的表达水平可以是核酸表达水平(例如,DNA表达水平或RNA表达水平,例如,mRNA表达水平)。可以使用任何适合的确定核酸表达水平的方法。在一些实施方式中,使用qPCR、rtPCR、RNA-seq、多重qPCR或RT-qPCR、微阵列分析、基因表达系列分析(SAGE)、MassARRAY技术、原位杂交(例如,FISH)或其组合确定核酸表达水平。
如本文所使用的,细胞的“参比群体”是指对应于所关心的细胞的细胞群体,相对于其测量了所关心的细胞的表型。例如,可以将分离的非造血组织来源的γδ细胞群体上标志物的表达水平与造血组织来源的γδT细胞(例如,血液驻留的γδ细胞,例如,来源于相同供体或不同供体的血液驻留的γδ细胞)或者在不同条件(例如,在存在明显TCR激活的情况下,在存在外源TCR激活因子(例如,抗CD3)的情况下或者与基质细胞(例如,成纤维细胞)明显接触)下扩增的非造血组织来源的γδT细胞上的相同标志物的表达水平相比较。还可以将群体与较早状态下的自身相比较。例如,参比群体可以是其扩增之前的分离的细胞群体。在这种情况下,将扩增群体与扩增步骤之前其自身组成相比较,即在这种情况下,其过去的组成是参比群体。
“癌症”是指恶性癌细胞的异常增殖并且包括白血病,如急性髓细胞性白血病(AML)、慢性粒细胞性白血病(CML)、急性淋巴母细胞性白血病(ALL)和慢性淋巴细胞性白血病(CLL)、淋巴瘤,如霍奇金淋巴瘤、非霍奇金淋巴瘤和多发性骨髓瘤,以及实体癌,如肉瘤、皮肤癌、黑素瘤、膀胱癌、脑癌、乳腺癌、子宫癌、卵巢癌、前列腺癌、肺癌、结肠直肠癌、宫颈癌、肝癌、头颈癌、食道癌、胰脏癌、肾癌、肾上腺癌、胃癌、睾丸癌、胆囊和胆道癌、甲状腺癌、胸腺癌、骨癌和大脑癌。
癌症患者内的癌细胞可以在免疫学上不同于所述个体中的正常体细胞(例如,癌性肿瘤可以是免疫原性的)。例如,癌细胞可以能够在癌症患者中引起对所述癌细胞所表达的一种或多种抗原的全身性免疫应答。引起免疫应答的抗原可以是肿瘤抗原或者可以与正常细胞共有。患有癌症的患者可以显示出根据本领域中已知的临床标准,足以做出癌症诊断的至少一种可鉴别的病征、症状或实验室发现。这些临床标准的实例可见于医学教科书,如Harrison's Principles of Internal Medicine(Longo DL,Fauci AS,Kasper DL,Hauser SL,Jameson J,Loscalzo J主编.18e.New York,NY:McGraw-Hill;2012)。在一些情况下,个体中癌症的诊断可以包括获自所述个体的体液或组织样品中特定细胞类型(例如,癌细胞)的鉴别。
如本文所使用的,“实体瘤”是血液、骨髓或淋巴系统以外的身体组织的任何癌症。实体瘤还可以分为上皮细胞来源的那些和非上皮细胞来源的那些。上皮细胞实体瘤的实例包括胃肠道、结肠、乳腺、前列腺、肺、肾、肝、胰脏、卵巢、头和颈、口腔、胃、十二指肠、小肠、大肠、肛门、胆囊、唇、鼻咽、皮肤、子宫、男性生殖器、泌尿器、膀胱和皮肤的肿瘤。非上皮细胞来源的实体瘤包括肉瘤、脑肿瘤和骨肿瘤。
适合于如上所述的治疗的患者、受试者或个体可以是哺乳动物,如啮齿类(例如,豚鼠、仓鼠、大鼠、小鼠)、鼠科(例如,小鼠)、狗(例如,狗)、猫(例如,猫)、马(例如,马)、灵长类、猿(例如,猴或猿)、猴(例如,狨猴或狒狒)、猿(例如,大猩猩、黑猩猩、猩猩或长臂猿)或者人。
在一些实施方式中,患者、受试者或个体是人。在其他优选的实施方式中,可以使用非人哺乳动物,特别是通常用作显示人中治疗效力的模型的哺乳动物(例如,鼠、灵长类、猪、狗或兔)。
如本文所使用的,“治疗”(及其语法变化)是指临床干预,无论是人还是动物(例如,在兽医应用中),其中实现了一些所期望的治疗效果,例如,病况发展的抑制或延迟,并且包括在不存在治疗的情况下所预期的之外,发展速度的降低,发展速度的终止,病况的改善,病况的治愈或缓解(无论部分还是完全),病况的一种或多种症状和/或病征的预防、延迟、减轻或终止或者受试者或患者存活期的延长。
还包括作为预防措施的治疗(即预防)。例如,可以如本文所述治疗对癌症的发生或复发敏感或有此风险的患者、受试者或个体。这种治疗可以预防或延迟患者、受试者或个体中癌症的发生或复发。
具体地,治疗可以包括抑制癌症生长,包括癌症完全缓解和/或抑制癌症转移。癌症生长一般是指指示癌症内向更加发展的形式的变化的一些指标中的任一项。因此,用于测量癌症生长抑制的指标包括癌细胞存活的降低、肿瘤体积或形态的减少(例如,如使用计算机断层摄影(CT)、超声波扫描术或其他成象方法所确定的)、延迟的肿瘤生长、肿瘤脉管系统的破坏、迟发过敏性皮肤测试中改善的表现、溶胞T淋巴细胞活性的提高和肿瘤特异性抗原水平的降低。降低个体中癌性肿瘤中的免疫抑制可以改善所述个体抵抗癌症生长,具体地已存在于所述受试者中的癌症生长的能力,和/或降低所述个体中癌症生长的倾向。
在一些实施方式中,施用扩增的γδT细胞(例如,非造血组织来源的γδT细胞,例如,非造血组织来源的Vδ1 T细胞)以延迟疾病的发展或减缓疾病或病症的发展。
如本文所使用的,“施用”表示向患者提供疗法(例如,包含(例如)非造血组织来源的γδT细胞的过继性T细胞疗法)或者组合物(例如,药物组合物,例如,包含非造血组织来源的γδT细胞的药物组合物)剂量的方法。可以(例如)肌内、静脉内、皮内、经皮、动脉内、腹膜内、病灶内、颅内、关节内、前列腺内、胸膜内、气管内、鞘内、鼻内、阴道内、直肠内、局部、瘤内、腹膜、皮下、结膜下、血管内、粘膜、心包内、脐内、眼内、眶内、玻璃体内(例如,通过玻璃体内注射)、通过眼滴剂、口服、局部、经皮、通过吸入、通过注射、通过植入、通过输注、通过连续输注、通过直接浸洗靶细胞的局部灌注、通过导管、通过灌洗、在乳剂中或在脂质组合物中施用在本文方法中使用的组合物。还可以全身或局部施用在本文方法中使用的组合物。施用方法可以基于多种因素(例如,要施用的治疗剂或组合物和要治疗的病况、疾病或病症的严重程度)而不同。
“治疗有效量”是指治疗或预防哺乳动物中疾病或病症的治疗剂的量。就癌症而言,治疗有效量的所述治疗剂(例如,非造血组织来源的γδT)可以减少癌细胞的数目;降低原发肿瘤的尺寸;抑制(即在某种程度上减缓并且优选地终止)癌细胞向外周器官的浸润;抑制(即在某种程度上减缓并且优选地终止)肿瘤转移;在某种程度上抑制肿瘤生长;和/或在某种程度上减轻与所述病症有关的一种或多种症状。在药物可以防止生长和/或杀死已有癌细胞的程度,它可以是细胞抑制和/或细胞毒性的。对于癌症疗法,可以(例如)通过评价存活持续时间、疾病进展时间(TTP)、反应速率(例如,完全反应(CR)和部分反应(PR))、反应持续时间和/或生活质量来测量体内效力。
术语“同时”在本文中用于表示两种或更多种治疗剂的施用,其中施用中的至少一部分在时间上重叠。因此,同时施用包括当在一种或多种其他试剂的施用停止之后,继续一种或多种试剂的施用时的剂量施用方案。例如,在一些实施方式中,可以同时施用非造血组织来源的γδT细胞和IL-2。
术语“药物组合物”是指处于这种形式以允许包含其中的一种或多种活性成分的生物活性有效并且不包含对将施用所述制剂的患者具有不可接受的毒性的其他组分的制剂。
III.分离和使γδT细胞扩增的方法
本发明提供了用于分离和使来自可以从或已从患者移除的任何人或非人动物非造血组织的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2T细胞,例如,Vδ1 T细胞和/或DN T细胞)扩增的方法。在一些实施方式中,γδT细胞所来源并扩增的非造血组织是皮肤(例如,人皮肤),其可以通过本领域中已知的方法获得。在一些实施方式中,通过钻孔活组织检查获得皮肤。作为另外一种选择,可以将本文所提供的γδT细胞的分离和扩增方法应用于胃肠道(例如,结肠)、乳腺、肺、前列腺、肝、脾脏和胰脏。γδT细胞还可以存在于人癌症组织,例如,乳腺或前列腺肿瘤中。在一些实施方式中,γδT细胞可以来自人癌症组织(例如,实体瘤组织)。在其他实施方式中,γδT细胞可以来自人癌症组织以外的非造血组织(例如,无明显个数的肿瘤细胞的组织)。例如,γδT细胞可以来自与附近或相邻癌症组织分隔的皮肤(例如,健康皮肤)区域。
血液中主要的γδT细胞主要为Vδ2 T细胞,而非造血组织中主要的γδT细胞主要为Vδ1 T细胞,从而Vδ1 T细胞占非造血组织驻留的γδT细胞群体的约70-80%。然而,一些Vδ2 T细胞还存在于非造血组织中,例如,肠中,在此它们可以占约10-20%的γδT细胞(图6)。存在于非造血组织中的一些γδT细胞既不表达Vδ1,也不表达Vδ2 TCR,并且我们将它们命名为双阴性(DN)γδT细胞。这些DNγδT细胞可能大部分是Vδ3-表达T细胞,少数是Vδ5-表达T细胞。因此,通常存在于非造血组织中并且通过本发明所述的方法扩增的γδT细胞优选地为非Vδ2 T细胞,例如,Vδ1 T细胞,其中包括较少量的DNγδT细胞。
技术人员会理解某些非造血组织可以是高度血管化的,并且实际上,非造血组织样品易于被外周血驻留的细胞污染。为了避免或最大程度减少这种污染,根据本领域中已知的方法,可以小心以确保在分离和扩增培养中去掉外周血,如通过在适合的缓冲液中彻底清洗组织以除去血液驻留的细胞。例如,在一些实施方式中,不通过支气管肺泡灌洗获得分离自肺组织的γδT细胞群体。
从非造血组织分离非造血组织驻留的γδT细胞
在一些实施方式中,关键步骤是(例如)在培养数天或数周后,将非造血组织驻留的T细胞(例如,混合的淋巴细胞群体内的T细胞,其可以(例如)包括αβ细胞、自然杀伤(NK)细胞、B细胞以及γδ2和非γδ2 T细胞)与从中获得T细胞的组织的非造血细胞(例如,基质细胞,特别是成纤维细胞)的有意分离开。这允许在随后的数天和数周内非造血组织来源的Vδ1 T细胞和DNγδT细胞优先且快速扩增。
本发明提供了包括γδT细胞(例如,非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)与非造血组织(例如,皮肤,例如,通过钻孔活组织检查所获得的皮肤)分离的方法。在一个实施方式中,γδT细胞与非造血细胞的分离包括在配置以有利于细胞从非造血组织流出的合成支架上培养γδT细胞和非造血细胞。可以使用适合于淋巴细胞与实质组织分离的任何支架。合成支架可以由天然和/或合成材料,如聚合物(例如,天然或合成聚合物,例如,聚乙烯吡咯烷酮、聚甲基丙烯酸甲酯、甲基纤维素、聚苯乙烯、聚丙烯、聚氨脂)、陶瓷(例如,磷酸三钙、铝酸钙、钙羟基磷灰石)或金属(钽、钛、铂以及与铂、铌、铪、钨相同元素族的金属及其合金的组合)构造成。根据本领域中已知的方法,可以将生物因子(例如,胶原蛋白(例如,胶原蛋白I或者胶原蛋白II)、纤连蛋白、层粘连蛋白、整合素、血管生成因子、抗炎因子、糖胺聚糖、vitrogens、抗体及其片段、细胞因子(例如,IL-2或IL-15及其组合)、趋化因子和/或化学引诱剂涂覆到支架表面上或包封在支架材料内以提高细胞粘附、迁移、存活或增殖。在一些实施方式中,合成支架是如Clark规程中所描述的Cellfoam支架。作为另外一种选择,其他方法可以用于将淋巴细胞与一些其他非造血组织类型分离,例如,通过胞外基质组分的酶-基降解(例如,胶原酶)。
可以将分离培养实施从1小时(例如,就简单消化来说)至长达42天(就支架培养来说)的任何持续时间。在其中在支架上进行分离步骤的情况中,培养可以进行至少5天(例如,至少6天、至少7天、至少8天、至少9天、至少10天、至少12天、至少14天、至少16天、至少18天、至少20天、至少21天、至少24天、至少28天、至少30天、至少35天或者至少40天,例如,7至14天之间、14至21天之间或者21至35天之间,例如,约14天或约21天)。
在γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的分离期间,可以在存在生物因子的情况下培养非造血组织和其中来源的细胞以提高从组织的流出或促进一种或多种细胞亚群的存活力。在一些实施方式中,分离培养包括IL-2,例如,浓度至少10IU/mL(例如,10IU/mL至1,000IU/mL、20IU/mL至800IU/mL、25IU/mL至750IU/mL、30IU/mL至700IU/mL、40IU/mL至600IU/mL、50IU/mL至500IU/mL、75IU/mL至250IU/mL或者100IU/mL至200IU/mL、例如、10IU/mL至20IU/mL、20IU/mL至30IU/mL、30IU/mL至40IU/mL、40IU/mL至50IU/mL、50IU/mL至75IU/mL、75IU/mL至100IU/mL、100IU/mL至150IU/mL、150IU/mL至200IU/mL、200IU/mL至500IU/mL或者500IU/mL至1,000IU/mL)的IL-2。在一些实施方式中,分离培养包括约100IU/mL的浓度的IL-2。另外或者作为另外一种选择,分离培养可以包括IL-15,例如,至少0.1ng/mL的浓度(例如,0.1ng/mL至10,000ng/mL、1.0ng/mL至1,000ng/mL、5ng/mL至800ng/mL、10ng/mL至750ng/mL、20ng/mL至500ng/mL、50ng/mL至400ng/mL或100ng/mL至250ng/mL,例如,0.1ng/mL至1.0ng/mL、1.0ng/mL至5.0ng/mL、5.0ng/mL至10ng/mL、10ng/mL至20ng/mL、20ng/mL至50ng/mL、50ng/mL至100ng/mL、100ng/mL至200ng/mL、200ng/mL至500ng/mL或500ng/mL至1,000ng/mL)的IL-15。在一些实施方式中,分离培养包括约20ng/ml的浓度的IL-15。
在一些实施方式中,γδT细胞与非造血组织的分离包括在存在分别处于任何以上所列浓度的IL-2和IL-15的情况下的培养。在一些情况下,IL-2的浓度为约100IU/mL,IL-15的浓度为20ng/mL。
可以在不存在IL-6、IL-23和IL-1β的情况下或者在存在低浓度的这些细胞因子(例如,小于20ng/mL)的情况下培养γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞),因为这种细胞因子组合的添加可以起作用以降低非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的增殖。
一旦与非造血组织(例如,皮肤)分离,则γδT细胞通常将是含有(例如)αβT细胞、B细胞和自然杀伤(NK)细胞的较大的淋巴细胞群体的一部分。在一些实施方式中,在扩增之前,1%-10%的分离的淋巴细胞群体是γδT细胞(例如,在扩增之前,1%-10%的分离的皮肤来源的淋巴细胞群体是γδT细胞)。大多数情况下,γδT细胞群体(例如,皮肤来源的γδT细胞群体)将包括较大的Vδ1 T细胞群体。在一些实施方式中,在扩增之前,1-10%的分离的淋巴细胞(例如,皮肤来源的淋巴细胞)群体为Vδ1 T细胞(例如,在扩增之前,Vδ1 T细胞可以占分离的γδT细胞群体的50%以上,60%以上,70%以上,80%以上或90%以上)。在一些情况下,在扩增之前,小于10%的分离的γδT细胞群体为Vδ2 T细胞(例如,在扩增之前,小于10%的分离的皮肤来源的γδT细胞群体为Vδ2 T细胞)。
可以从分离的γδT细胞群体除去非Vδ1 T细胞或非DN T细胞,如Vδ2T细胞、αβT细胞、B细胞或者NK细胞(例如,在扩增之前、期间或之后)。
在扩增之前,分离的γδT细胞(例如,分离自皮肤的γδT细胞,例如,分离自皮肤的Vδ1 T细胞)具有不同于相应造血组织来源的细胞(例如,血液来源的γδT细胞,例如,血液来源的Vδ2 T细胞)的表型。例如,分离的γδT细胞群体可以表达比参比群体,例如,TCR激活的非造血组织驻留的γδT细胞群体或者相应造血组织来源的细胞(例如,血液来源的γδT细胞,例如,血液来源的Vδ2 T细胞)群体更高水平的CCR3、CCR4、CCR7、CCR8或CD103。在一些实施方式中,分离的γδT细胞群体包括至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的CCR3+细胞;至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的CCR4+细胞;至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的CCR7+细胞;至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的CCR8+细胞;和/或至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的CD103+细胞。分离的γδT细胞群体可以表达CCR3、CCR4、CCR7、CCR8或CD103中的一种或多种,两种或更多种,三种或更多种,四种或更多种,五种或更多种或者全部六种。
在一些实施方式中,分离的γδT细胞群体(例如,皮肤来源的γδT细胞,例如,皮肤来源的Vδ1 T细胞)表达比参比群体,例如,TCR激活的非造血组织驻留的γδT细胞群体或者相应造血组织来源的细胞(例如,血液来源的γδT细胞,例如,血液来源的Vδ2 T细胞)群体更高水平的NKGD2、CD56、CD69和/或TIM3。在一些实施方式中,分离的γδT细胞群体包括至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的NKGD2+细胞;至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的CD56+细胞;至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的CD69+细胞;和/或至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或以上的TIM3+细胞。分离的γδT细胞群体可以表达NKGD2、CD56、CD69和/或TIM3中的一种或多种,两种或更多种,三种或更多种,四种或更多种或者全部五种。
还可以通过功能鉴别分离的非造血组织来源的γδT细胞(例如,皮肤来源的γδT细胞,例如,皮肤来源的Vδ1 T细胞)群体。可以实施本领域中已知的和在实施例3中描述的功能测定来确定任何本发明所述的非造血组织来源的细胞(例如,分离的γδT细胞群体,例如,皮肤来源的Vδ1 T细胞,或者扩增的γδT细胞群体,例如,皮肤来源的Vδ1 T细胞)和参比细胞(例如,TCR激活的非造血组织驻留的γδT细胞群体或者相应造血组织来源的细胞,例如,血液来源的γδT细胞,例如,血液来源的Vδ2 T细胞群体)之间的功能差异。在一些实施方式中,分离的非造血组织来源的γδT细胞群体(例如,未与明显的TCR途径激活接触的分离的γδT细胞群体)分泌比参比群体(例如,TCR激活的非造血组织驻留的γδT细胞群体,例如,抗CD3激活的非造血组织驻留的γδT细胞群体)更高水平的IL-13。例如,相对于参比细胞群体(例如,TCR激活的非造血组织驻留的γδT细胞群体,例如,抗CD3激活的非造血组织驻留的γδT细胞群体),分离的非造血组织来源的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)可以分泌1.1倍、1.2倍、1.3倍、1.4倍、1.5倍、2倍、3倍、4倍、5倍、10倍、15倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍、100倍、200倍、300倍、400倍、500倍、1,000倍或更大浓度的IL-13。类似地,相对于参比细胞群体(例如,TCR激活的非造血组织驻留的γδT细胞群体,例如,抗CD3激活的非造血组织驻留的γδT细胞群体),分离群体中分泌IL-13的非造血组织来源的γδT细胞的个数或频率可以更大。例如,分离的γδT细胞群体内IL-13分泌细胞的频率(例如,分离的Vδ1 T细胞群体内IL-13分泌细胞的频率)可以比参比细胞群体(TCR激活的非造血组织驻留的γδT细胞群体,例如,抗CD3激活的非造血组织驻留的γδT细胞群体)更大。在一些实施方式中,分离的γδT细胞群体内IL-13分泌细胞的频率(例如,本发明所述的分离的DN T细胞或Vδ1 T细胞群体内IL-13分泌细胞的频率)比参比细胞群体(TCR激活的非造血组织驻留的γδT细胞群体,例如,抗CD3激活的非造血组织驻留的γδT细胞群体)大至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%、至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%。
非造血组织驻留的γδT细胞的扩增
本发明的特征在于扩增非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的方法。这些方法可以体外进行。在一些实施方式中,根据如上所述的方法,从已分离自非造血组织的γδT细胞群体扩增非造血组织驻留的γδT细胞。一般地,通过除去与基质细胞(例如,皮肤成纤维细胞)的物理接触,非造血组织驻留的γδT细胞能够自发扩增。因此,如上所述的基于支架的培养方法可以用于引起这种分离,从而导致γδT细胞去抑制以引发扩增。因此,在一些实施方式中,在扩增步骤期间无明显的TCR途径激活存在(例如,在培养中不包含外源TCR途径激活剂)。此外,本发明提供了扩增非造血组织驻留的γδT细胞的方法,其中方法不包括与饲养细胞、肿瘤细胞和/或抗原递呈细胞的接触。
本发明的发明人已开发了扩增规程,其包括在存在有效的生物因子混合物(cocktail)的情况下培养非造血组织驻留的γδT细胞以支持有效的γδT细胞扩增。在一个实施方式中,本发明提供了通过提供获自非造血组织的γδT细胞群体(例如,分离的非造血组织来源的γδT细胞群体,例如,根据本文所描述的方法分离的群体)和在存在IL-2、IL-4、IL-15和/或IL-21的情况下培养γδT细胞来扩增γδT细胞的方法。可以将这些细胞因子或其类似物以对于产生扩增的γδT细胞群体有效的量与所述细胞培养一段时间(例如,至少5天、至少6天、至少7天、至少8天、至少9天、至少10天、至少11天、至少12天、至少13天、至少14天、至少21天、至少28天或更长时间,例如,5天至40天、7天至35天、14天至28天或者约21天)。
在一些实施方式中,对于产生扩增的γδT细胞群体有效的IL-2的量为1IU/mL至2,000IU/mL(例如,5IU/mL至1,000IU/mL、10IU/mL至500IU/mL、20IU/mL至400IU/mL、50IU/mL至250IU/mL或约100IU/mL,例如,5IU/mL至10IU/mL、10IU/mL至20IU/mL、20IU/mL至30IU/mL、30IU/mL至40IU/mL、40IU/mL至50IU/mL、50IU/mL至60IU/mL、60IU/mL至70IU/mL、70IU/mL至80IU/mL、80IU/mL至90IU/mL、90IU/mL至100IU/mL、100IU/mL至120IU/mL、120IU/mL至140IU/mL、140IU/mL至150IU/mL、150IU/mL至175IU/mL、175IU/mL至200IU/mL、200IU/mL至300IU/mL、300IU/mL至400IU/mL、400IU/mL至500IU/mL、500IU/mL至1,000IU/mL、1,000IU/mL至1,500IU/mL、1,500IU/mL至2,000IU/mL或以上)。在一些实施方式中,对于产生扩增的γδT细胞群体有效的IL-2的量为约100IU/mL。
在一些实施方式中,对于产生扩增的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)群体有效的IL-4的量为至少0.1ng/mL(例如,0.1ng/mL至10,000ng/mL、1.0ng/mL至1,000ng/mL、5ng/mL至800ng/mL、10ng/mL至750ng/mL、20ng/mL至500ng/mL、50ng/mL至400ng/mL或100ng/mL至250ng/mL,例如,0.1ng/mL至1.0ng/mL、1.0ng/mL至5.0ng/mL、5.0ng/mL至10ng/mL、10ng/mL至20ng/mL、20ng/mL至50ng/mL、50ng/mL至100ng/mL、100ng/mL至200ng/mL、200ng/mL至500ng/mL或500ng/mL至1,000ng/mL)。在一些实施方式中,对于产生扩增的γδT细胞群体有效的IL-4的量为约5ng/mL。
在一些实施方式中,对于产生扩增的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)群体有效的IL-15的量为至少0.1ng/mL(例如,0.1ng/mL至10,000ng/mL、1.0ng/mL至1,000ng/mL、5ng/mL至800ng/mL、10ng/mL至750ng/mL、20ng/mL至500ng/mL、50ng/mL至400ng/mL或100ng/mL至250ng/mL、例如、0.1ng/mL至1.0ng/mL、1.0ng/mL至5.0ng/mL、5.0ng/mL至10ng/mL、10ng/mL至20ng/mL、20ng/mL至50ng/mL、50ng/mL至100ng/mL、100ng/mL至200ng/mL、200ng/mL至500ng/mL或500ng/mL至1,000ng/mL)。在一些实施方式中,对于产生扩增的γδT细胞群体有效的IL-15的量为约10ng/mL。
在一些实施方式中,对于产生扩增的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)群体有效的IL-21的量为至少0.1ng/mL(例如,0.1ng/mL至10,000ng/mL、1.0ng/mL至1,000ng/mL、5ng/mL至800ng/mL、10ng/mL至750ng/mL、20ng/mL至500ng/mL、50ng/mL至400ng/mL或100ng/mL至250ng/mL、例如、0.1ng/mL至1.0ng/mL、1.0ng/mL至5.0ng/mL、5.0ng/mL至10ng/mL、10ng/mL至20ng/mL、20ng/mL至50ng/mL、50ng/mL至100ng/mL、100ng/mL至200ng/mL、200ng/mL至500ng/mL或500ng/mL至1,000ng/mL)。在一些实施方式中,对于产生扩增的γδT细胞群体有效的IL-21的量为约10ng/mL。在其他实施方式中,对于产生扩增的γδT细胞群体有效的IL-21的量为约10ng/mL。
本文还提供了其他因子在非造血组织驻留的γδT细胞的扩增培养中的替代或添加。例如,在一些实施方式中,除了IL-2、IL-4、IL-15和IL-21中的任一项之外或者作为它们的替代,包括了选自IL-6、IL-7、IL-8、IL-9、IL-12、IL-18、IL-33、IGF-1、IL-1β、人血小板裂解物(HPL)和基质细胞衍生因子-1(SDF-1)组成的组中的任何一种或多种因子。在实施例3中的表2中提供了这些因子中每一个的适合的浓度。
会理解产生扩增的γδT细胞群体所需的以上细胞因子中的每一个的量将取决于一种或多种其他细胞因子的浓度。例如,如果IL-2的浓度升高或降低,则IL-15的浓度可以因此分别降低或提高。如上所述,对于产生扩增的群体有效的量在本文中表示所有因子对细胞扩增的综合影响。
在一些实施方式中,将γδT细胞同时暴露于每种因子(例如,将γδT细胞同时暴露于IL-2、IL-4、IL-15和IL-21,例如,至少5天)。在其他实例中,在与其他因子培养之前,将γδT细胞暴露于某些因子。例如,可以在扩增过程中逐渐向扩增培养提供其他因子,或者作为另外一种选择,可以将γδT细胞从一种因子或一组因子的培养转移至另一种。
在一些实施方式中,将γδT细胞在培养中扩增数小时(例如,约2、3、4、5、6、7、8、9、10、15、18或21小时)至约35天(例如,1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34或35天)的一段时期。在一个实施方式中,将γδT细胞扩增14至21天的一段时期。因此,包括分离培养期(例如,1至40天,例如,14至21天),在一些实施方式中,分离和扩增步骤可以持续28至56天,或者约41天。
扩增方法提供了数目比参比群体更大的扩增的γδT细胞群体。在一些实施方式中,扩增的γδT细胞群体的数目比扩增步骤之前的分离的γδT细胞群体的大。(例如,相对于扩增步骤之前的分离的γδT细胞群体,至少2倍的数目、至少3倍的数目、至少4倍的数目、至少5倍的数目、至少6倍的数目、至少7倍的数目、至少8倍的数目、至少9倍的数目、至少10倍的数目、至少15倍的数目、至少20倍的数目、至少25倍的数目、至少30倍的数目、至少35倍的数目、至少40倍的数目、至少50倍的数目、至少60倍的数目、至少70倍的数目、至少80倍的数目、至少90倍的数目、至少100倍的数目、至少200倍的数目、至少300倍的数目、至少400倍的数目、至少500倍的数目、至少600倍的数目、至少700倍的数目、至少800倍的数目、至少900倍的数目、至少1,000倍的数目、至少5,000倍的数目、至少10,000倍的数目或更多倍的数目)。
因此,本发明提供了以高速率(例如,通过除去基质细胞接触和/或TCR刺激,或者通过在存在有效量的因子的情况下培养)产生大的非造血组织来源的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)群体的方式。在一些实施方式中,本文所描述的扩增步骤以低群体倍增时间扩增γδT细胞,其通过下式表示:

考虑本文所提供的信息,例如,以下实施例3中所提供的信息,技术人员将认识到本发明提供了以小于5天(例如,小于4.5天、小于4.0天、小于3.9天、小于3.8天、小于3.7天、小于3.6天、小于3.5天、小于3.4天、小于3.3天、小于3.2天、小于3.1天、小于3.0天、小于2.9天、小于2.8天、小于2.7天、小于2.6天、小于2.5天、小于2.4天、小于2.3天、小于2.2天、小于2.1天、小于2.0天、小于46小时、小于42小时、小于38小时、小于35小时、小于32小时)的群体倍增时间扩增非造血组织来源的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的方法。
在一些实施方式中,在培养7天内,扩增的γδT细胞群体(例如,扩增的Vδ1 T细胞和/或DN T细胞群体)包括相对于扩增之前的分离的γδT细胞群体至少10倍数目的γδT细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少20倍、至少30倍、至少40倍、至少50倍、至少60倍、至少70倍、至少80倍、至少90倍、至少100倍、至少150倍、至少200倍、至少300倍、至少400倍、至少500倍、至少600倍、至少700倍、至少800倍、至少900倍、至少1,000倍、至少2,000倍、至少3,000倍、至少4,000倍、至少5,000倍、至少6,000倍、至少7,000倍或者至少8,000倍数目的γδT细胞)。在一些实施方式中,在培养14天内,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体(例如,扩增的Vδ1 T细胞和/或DN T细胞群体)包括至少20倍数目的γδT细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少30倍、至少40倍、至少50倍、至少60倍、至少70倍、至少80倍、至少90倍、至少100倍、至少150倍、至少200倍、至少300倍、至少400倍、至少500倍、至少600倍、至少700倍、至少800倍、至少900倍、至少1,000倍、至少2,000倍、至少3,000倍、至少4,000倍、至少5,000倍、至少6,000倍、至少7,000倍、至少8,000倍、至少9,000倍或者至少10,000倍数目的γδT细胞)。在一些实施方式中,在培养21天内,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体(例如,扩增的Vδ1 T细胞和/或DN T细胞群体)包括至少50倍数目的γδT细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少60倍、至少70倍、至少80倍、至少90倍、至少100倍、至少150倍、至少200倍、至少300倍、至少400倍、至少500倍、至少600倍、至少700倍、至少800倍、至少900倍、至少1,000倍、至少2,000倍、至少3,000倍、至少4,000倍、至少5,000倍、至少6,000倍、至少7,000倍、至少8,000倍、至少9,000倍或至少10,000倍数目的γδT细胞)。在一些实施方式中,在培养28天内,扩增的γδT细胞群体(例如,扩增的Vδ1 T细胞和/或DN T细胞群体)包括相对于扩增之前的分离的γδT细胞群体至少100倍数目的γδT细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少110倍、至少120倍、至少130倍、至少140倍、至少150倍、至少200倍、至少300倍、至少400倍、至少500倍、至少600倍、至少700倍、至少800倍、至少900倍、至少1,000倍、至少2,000倍、至少3,000倍、至少4,000倍、至少5,000倍、至少6,000倍、至少7,000倍、至少8,000倍、至少9,000倍、至少10,000倍、至少12,000倍或者至少15,000倍数目的γδT细胞)。
通过本文所提供的方法扩增的非造血组织来源的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)可以具有非常适合于抗肿瘤效力的表型。在一些实施方式中,扩增的γδT细胞群体(例如,皮肤来源的Vδ1 T细胞)具有比参比群体(例如,在扩增步骤之前分离的γδT细胞群体)更大的平均CD27表达。在一些实施方式中,扩增的γδT细胞群体具有相对于分离的γδT细胞群体至少2倍的平均CD27表达。(例如,相对于分离的γδT细胞群体,至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍、至少10倍、至少15倍、至少20倍、至少25倍、至少30倍、至少40倍、至少50倍、至少60倍、至少70倍、至少80倍、至少90倍、至少100倍、至少150倍、至少200倍、至少300倍、至少400倍、至少500倍、至少600倍、至少700倍、至少800倍、至少900倍、至少1,000倍、至少5,000倍、至少10,000倍、至少20,000倍或以上)。
扩增的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的单独的部分可以上调CD27,而另一个部分是CD27低或CD27-。在这种情况下,相对于分离的γδT细胞群体,扩增群体中CD27+细胞的频率可以更大。例如,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体可以具有至少5%更高频率的CD27+细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%更高频率的CD27+细胞)。在一些实施方式中,相对于分离的γδT细胞群体,扩增群体中CD27+细胞的数目可以升高。例如,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体可以具有至少2倍数目的CD27+细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%更高频率的CD27+细胞)。
在一些实施方式中,如本文所提供的扩增方法获得了相对于参比群体(例如,扩增步骤之前的分离的γδT细胞群体),具有低TIGIT表达的扩增的非造血组织来源的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)群体。在一些实施方式中,扩增的γδT细胞群体具有比参比群体(例如,在扩增步骤之前分离的γδT细胞群体)低的平均TIGIT表达。在一些实施方式中,扩增的γδT细胞群体具有比分离的γδT细胞群体低至少10%的平均TIGIT表达。(例如,比分离的γδT细胞群体低至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%)。
扩增的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的单独的部分可以表达TIGIT,例如,高水平的TIGIT,而另一个部分是TIGIT低或表达TIGIT-。在这种情况下,相对于分离的γδT细胞群体,扩增群体中TIGIT+细胞的频率可以更低。例如,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体可以具有至少5%更低频率的TIGIT+细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%更低频率的TIGIT+细胞)。在一些实施方式中,相对于扩增之前的分离的γδT细胞群体,扩增群体中TIGIT+细胞的数目可以降低。例如,相对于扩增之前的分离的γδT细胞群体中的TIGIT+细胞的数目,扩增的γδT细胞群体可以具有至少低10%的TIGIT+细胞(例如,相对于扩增之前的分离的γδT细胞群体中的TIGIT+细胞的数目,低至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%的TIGIT+细胞)。
在一些实施方式中,扩增的γδT细胞群体(例如,皮肤来源的γδT细胞或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)具有高数目或频率的CD27+细胞和低频率的TIGIT+细胞。在一些实施方式中,相对于参比群体(例如,相对于扩增之前的分离的γδT细胞群体),扩增的γδT细胞群体具有高频率的CD27+TIGIT-细胞。例如,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体可以具有至少5%更高频率的CD27+TIGIT-细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%更高频率的CD27+TIGIT-细胞)。在一些实施方式中,相对于分离的γδT细胞群体,扩增群体中CD27+TIGIT-细胞的数目可以升高。例如,相对于扩增之前的分离的γδT细胞群体,扩增的γδT细胞群体可以具有至少2倍数目的CD27+TIGIT-细胞(例如,相对于扩增之前的分离的γδT细胞群体,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%更高频率的CD27+TIGIT-细胞)。
在一些情况下,相对于参比群体,扩增的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)中CD27+γδT细胞群体上的平均TIGIT表达较低。在一些实施方式中,扩增的CD27+γδT细胞群体具有比参比群体(例如,在扩增步骤之前分离的CD27+γδT细胞群体)低的平均TIGIT表达。在一些实施方式中,扩增的CD27+γδT细胞群体具有比分离的CD27+γδT细胞群体低至少10%的平均TIGIT表达。(例如,比分离的CD27+γδT细胞群体低至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%)。
另外或者作为另外一种选择,相对于参比群体,扩增的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)中TIGIT-γδT细胞群体上的中值CD27表达较高。例如,扩增的TIGIT-γδT细胞群体可以具有相对于扩增之前的分离的TIGIT-γδT细胞群体至少5%更高频率的CD27+细胞(例如,相对于扩增之前的分离的TIGIT-γδT细胞群体,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%更高频率的CD27+细胞)。在一些实施方式中,相对于分离的TIGIT-γδT细胞群体,扩增群体中CD27+细胞的数目可以升高。例如,相对于扩增之前的分离的TIGIT-γδT细胞群体,扩增的TIGIT-γδT细胞群体可以具有至少2倍数目的CD27+细胞(例如,相对于扩增之前的分离的TIGIT-γδT细胞群体,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少60%、至少70%、至少80%、至少90%或多达100%更高频率的CD27+细胞)。
其他标志物表达的升高或降低可以另外或替代性地用于鉴别一种或多种扩增的非造血组织来源的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)群体,标志物包括CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30、CD2、NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64。在一些情况下,相对于分离的γδT细胞群体,例如,在扩增之前,扩增的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1T细胞和/或DN T细胞)具有更大的一种或多种标志物的平均表达,标志物选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组。另外或者作为另外一种选择,相对于分离的γδT细胞群体,扩增的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)可以具有更大的表达一种或多种标志物的细胞的频率,标志物选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组。在一些实施方式中,相对于分离的γδT细胞群体,扩增的γδT细胞群体(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)具有较低的一种或多种标志物的平均表达,标志物选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组。相对于分离的γδT细胞群体,扩增群体可以类似地具有较低频率的表达一种或多种标志物的细胞,标志物选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组。
因此,通过本发明所述的方法所产生的非造血组织驻留的γδT细胞可以具有以下性质中的一种或多种:(i)显示出表型CD69高、TIM3高和CD28低/缺乏;(ii)CCR3、CD39、CD11b和CD9中一种或多种的上调;(iii)在不存在TCR激动剂的情况下,对NKG2D配体响应而产生IFN-γ;(iv)在不存在TCR激动剂的情况下产生IL-13;(v)对TCR激活响应而产生IFN-γ、TNF-α和GM-CSF中的一种或多种;(vi)对TCR激活响应,不产生或不显著产生IL-17;(vii)在含有IL-2而无其他生长因子的培养基中生长;(viii)在不存在TCR激动剂的情况下显示出细胞毒性T细胞应答;和/或(ix)相对于正常细胞,对肿瘤细胞显示出选择性细胞毒性。
在一些情况下,通过本发明所述的方法所产生的非造血组织驻留的γδT细胞在不存在TCR激动剂的情况下产生IL-13,和/或在不存在TCR激动剂的情况下对NKG2D配体响应而产生IFN-γ。
适合于在γδT细胞增殖中使用的多种基础培养基,具体地,完全培养基,如AIM-V、Iscoves培养基和RPMI-1640(Life Technologies)是可用的。培养基可以补充有其他培养基因子,如血清、血清蛋白和选择性试剂,如抗生素。例如,在一些实施方式中,RPMI-1640培养基含有2mM谷氨酰胺、10%FBS、10mM HEPES,pH 7.2、1%青霉素-链霉素、丙酮酸钠(1mM;Life Technologies)、非必需氨基酸(例如,100μM Gly、Ala、Asn、Asp、Glu、Pro和Ser;1XMEM非必需氨基酸,Life Technologies)和10μl/Lβ-巯基乙醇。方便地,在37℃,在含有5%CO2的加湿的气氛中,在适合的培养基中培养细胞。
可以如本文所述,在任何适合的系统,包括搅拌罐发酵罐、气升式发酵罐、滚瓶、培养袋或皿及其他生物反应器,具体地中空纤维生物反应器中培养γδT细胞。这些系统的使用在本领域是熟知的。用于淋巴细胞培养的一般方法和技术在本领域中是熟知的。
本文所描述的方法可以包括不止一个选择步骤,例如,不止一个去除步骤。可以(例如)通过抗阴性选择细胞所独有的表面标志物的抗体组合来实现通过阴性选择的T细胞群体的富集。一种方法是通过使用抗阴性选择细胞上所存在的细胞表面标志物的单克隆抗体的混合物的阴性磁免疫粘附或流式细胞术的细胞分选和/或选择。
IV.药物组合物和治疗方法
通过本发明所述的方法所获得的γδT细胞可以用作药物,例如,用于过继性T细胞疗法的药物。这包括通过本发明所述的方法所获得的γδT细胞向患者的转移。疗法可以是自体同源的,即γδT细胞可以转移回到从中获得它们的相同患者,或者疗法可以是同种异体的,即可以将来自一个人的γδT细胞转移至不同的患者。在包括同种异体转移的情况下,γδT细胞可以明显不含αβT细胞。例如,可以(例如)在扩增之后,使用本领域中已知的任何合适的方法(例如,通过阴性选择,例如,使用磁珠),从γδT细胞群体除去αβT细胞。治疗方法可以包括;提供获自供体个体的非造血组织样品;培养来自如上所述的样品的γδT细胞以产生扩增群体;和将扩增的γδT细胞群体施用至受体个体。
要治疗的患者或受试者优选地是人癌症患者(例如,正在治疗实体瘤的人癌症患者),或者病毒感染患者(例如,CMV感染或HIV感染患者)。在一些情况下,患者已经和/或正在治疗实体瘤。
由于它们通常存在于非造血组织中,因此组织驻留的Vδ1 T和DNγδT细胞还比它们的全身血液驻留的对应物更可能归巢至或保留在肿瘤物质内,并且这些细胞的继承性转移可能对于靶向实体瘤和潜在地其他非造血组织相关免疫病理学更有效。
由于γδT细胞是非MHC限制的,因此它们不会将它们要转移至的宿主识别为外源的,这表示它们不易于导致移植物抗宿主病。这表示它们可以作为“成品”使用并转移至任何受体,例如,用于同种异体过继性T细胞疗法。
通过本发明方法所获得的非造血组织驻留的γδT细胞表达NKG2D并对与恶性肿瘤强烈相关的NKG2D配体(例如,MICA)应答。它们还在不存在任何激活作用的情况下表达细胞毒性谱,并因此可能对于杀死肿瘤细胞是有效的。例如,如本文所述,所获得的非造血组织驻留的γδT细胞可以在不存在任何激活作用的情况下表达IFN-γ、TNF-α、GM-CSF、CCL4、IL-13、颗粒溶素、颗粒酶A和B和穿孔素中的一种或多种,优选地,全部。可以不表达IL-17A。
因此,本文所报道的发现提供了通过本发明所述的方法所获得的非造血组织驻留的γδT细胞作为“成品”免疫疗法剂的临床应用的实践性和适合性的强有力的证据。与其他T细胞相比,这些细胞具有先天样杀死,不具有MHC限制并且显示出改善的向肿瘤的归巢和/或在肿瘤内的保留。
在一些实施方式中,患有非造血组织肿瘤的个体的治疗方法可以包括;提供获自供体个体的非造血组织的样品,培养来自如上所述的样品的γδT细胞以产生扩增群体,和;向所述患有肿瘤的个体施用扩增的γδT细胞群体。
药物组合物可以包括与一种或多种药物或生理学可用的载体、稀释剂或赋形剂结合的如本文所描述的扩增的非造血组织驻留的γδT细胞。这些组合物可以包括缓冲剂,如中性缓冲盐水、磷酸盐缓冲盐水等;碳水化合物,如葡萄糖、甘露糖、蔗糖或右旋糖酐、甘露糖醇;蛋白;多肽或氨基酸,如甘氨酸;抗氧化剂;螯合剂,如EDTA或谷胱甘肽;佐剂(例如,氢氧化铝);和防腐剂。可以在本发明所述的药物组合物中使用的冷冻保存溶液包括(例如)DMSO。可以将组合物配制(例如)用于静脉内施用。
在一个实施方式中,药物组合物明显不含,例如,不存在可检测水平的污染物,例如,内毒素或支原体。
在一些情况下,可以将通过任何上述方法所获得的治疗有效量的扩增的γδT细胞以治疗有效量施用于受试者(例如,用于癌症治疗,例如,用于实体瘤的治疗)。在一些情况下,扩增的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的治疗有效量小于10×1012个细胞每剂量(例如,小于9×1012个细胞每剂量,小于8×1012个细胞每剂量,小于7×1012个细胞每剂量,小于6×1012个细胞每剂量,小于5×1012个细胞每剂量,小于4×1012个细胞每剂量,小于3×1012个细胞每剂量,小于2×1012个细胞每剂量,小于1×1012个细胞每剂量,小于9×1011个细胞每剂量,小于8×1011个细胞每剂量,小于7×1011个细胞每剂量,小于6×1011个细胞每剂量,小于5×1011个细胞每剂量,小于4×1011个细胞每剂量,小于3×1011个细胞每剂量,小于2×1011个细胞每剂量,小于1×1011个细胞每剂量,小于9×1010个细胞每剂量,小于7.5×1010个细胞每剂量,小于5×1010个细胞每剂量,小于2.5×1010个细胞每剂量,小于1×1010个细胞每剂量,小于7.5×109个细胞每剂量,小于5×109个细胞每剂量,小于2.5×109个细胞每剂量,小于1×109个细胞每剂量,小于7.5×108个细胞每剂量,小于5×108个细胞每剂量,小于2.5×108个细胞每剂量,小于1×108个细胞每剂量,小于7.5×107个细胞每剂量,小于5×107个细胞每剂量,小于2.5×107个细胞每剂量,小于1×107个细胞每剂量,小于7.5×106个细胞每剂量,小于5×106个细胞每剂量,小于2.5×106个细胞每剂量,小于1×106个细胞每剂量,小于7.5×105个细胞每剂量,小于5×105个细胞每剂量,小于2.5×105个细胞每剂量或小于1×105个细胞每剂量)。
在一些实施方式中,在治疗过程中,扩增的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的治疗有效量小于10×1012个细胞(例如,在治疗过程中,小于9×1012个细胞,小于8×1012个细胞,小于7×1012个细胞,小于6×1012个细胞,小于5×1012个细胞,小于4×1012个细胞,小于3×1012个细胞,小于2×1012个细胞,小于1×1012个细胞,小于9×1011个细胞,小于8×1011个细胞,小于7×1011个细胞,小于6×1011个细胞,小于5×1011个细胞,小于4×1011个细胞,小于3×1011个细胞,小于2×1011个细胞,小于1×1011个细胞,小于9×1010个细胞,小于7.5×1010个细胞,小于5×1010个细胞,小于2.5×1010个细胞,小于1×1010个细胞,小于7.5×109个细胞,小于5×109个细胞,小于2.5×109个细胞,小于1×109个细胞,小于7.5×108个细胞,小于5×108个细胞,小于2.5×108个细胞,小于1×108个细胞,小于7.5×107个细胞,小于5×107个细胞,小于2.5×107个细胞,小于1×107个细胞,小于7.5×106个细胞,小于5×106个细胞,小于2.5×106个细胞,小于1×106个细胞,小于7.5×105个细胞,小于5×105个细胞,小于2.5×105个细胞或者小于1×105个细胞)。
在一些实施方式中,如本文所描述的扩增的非造血组织驻留的γδT细胞的剂量包括约1×106、1.1×106、2×106、3.6×106、5×106、1×107、1.8×107、2×107、5×107、1×108、2×108或5×108个细胞/kg。在一些实施方式中,扩增的非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2T细胞,例如,Vδ1 T细胞和/或DN T细胞)的剂量包括至少约1×106、1.1×106、2×106、3.6×106、5×106、1×107、1.8×107、2×107、5×107、1×108、2×108或5×108个细胞/kg。在一些实施方式中,扩增的非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的剂量包括多至约1×106、1.1×106、2×106、3.6×106、5×106、1×107、1.8×107、2×107、5×107、1×108、2×108或5×108个细胞/kg。在一些实施方式中,扩增的非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的剂量包括约1.1×106-1.8×107个细胞/kg。在一些实施方式中,扩增的非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的剂量包括约1×107、2×107、5×107、1×108、2×108、5×108、1×109、2×109或5×109个细胞。在一些实施方式中,扩增的非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的剂量包括至少约1×107、2×107、5×107、1×108、2×108、5×108、1×109、2×109或5×109个细胞。在一些实施方式中,扩增的非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)的剂量包括多至约1×107、2×107、5×107、1×108、2×108、5×108、1×109、2×109或5×109个细胞。
在一个实施方式中,向受试者施用104至106个扩增的非造血组织驻留的γδT细胞(例如,皮肤来源的γδT细胞和/或非Vδ2 T细胞,例如,Vδ1 T细胞和/或DN T细胞)每千克受试者体重。在一个实施方式中,所述受试者接受非造血组织驻留的γδT细胞群体的初始施用(例如,104至106个γδT细胞每kg受试者体重,例如,104至105个γδT细胞每kg受试者体重的初始施用)和一个或多个(例如,2、3、4或5个)扩增的非造血组织驻留的γδT细胞的后续施用(例如,一个或多个104至106个扩增的非造血组织驻留的γδT细胞每kg受试者体重,例如,104至105个扩增的非造血组织驻留的γδT细胞每kg受试者体重的后续施用)。在一个实施方式中,在先前施用之后小于15天,例如,14、13、12、11、10、9、8、7、6、5、4、3或2天,例如,在先前施用之后小于4、3或2天施用所述一个或多个后续施用。在一个实施方式中,所述受试者在至少三次γδT细胞群体施用过程中接受总计约106个γδT细胞每kg受试者体重,例如,所述受试者接受1×105个γδT细胞的初始剂量,3×105个γδT细胞的第二次施用,和6×105个γδT细胞的第三次施用,并且例如,在先前施用之后小于4、3或2天施用每个施用。
通过本发明所述的方法所获得的非造血组织驻留的γδT细胞还可以用于CAR-T疗法。这包括工程设计的T细胞受体(TCR)的产生以对T细胞重新编程以具有新的特异性,例如,单克隆抗体的特异性。工程设计的TCR可以使T细胞对恶性细胞特异,并因此对于癌症免疫疗法有用。例如,T细胞可以识别表达肿瘤抗原,如来自受试者组织的正常体细胞不表达的肿瘤相关抗原的癌细胞。因此,CAR-修饰的T细胞可以用于(例如)癌症患者的过继性T细胞疗法。
已描述了血液驻留的γδT细胞用于CAR的使用。然而,通过本发明所述的方法所获得的非造血组织驻留的γδT细胞可能是CAR-T法特别优良的载体,因为可以通过嵌合抗原特异性TCR转导它们,同时保留它们识别转化细胞的先天样能力,并且它们可能具有比血液驻留的γδT细胞或者常规的全身性αβT细胞更好的肿瘤渗透和保留能力。此外,它们对MHC依赖性抗原递呈的缺少降低了移植物抗宿主病的潜能并且使它们能够靶向表达低水平MHC的肿瘤。同样地,它们对常规共刺激的非依赖性(例如,通过CD28接合)提高了对表达低水平共刺激受体的配体的肿瘤的靶向。
在一些实施方式中,可以将一种或多种其他治疗剂施用于受试者。其他治疗剂可以选自免疫治疗剂、细胞毒性剂、生长抑制剂、放射疗法剂、抗血管生成剂或其两种或更多种试剂的组合。可以与扩增的γδT细胞的施用同时,在其之前或之后施用其他治疗剂。其他治疗剂可以是免疫治疗剂,其可以作用于受试者体内的靶标(例如,受试者自身的免疫系统)和/或作用于转移的γδT细胞。
组合物的施用可以以任何方便的形式进行。可以将本文所描述的组合物经动脉、皮下、皮内、瘤内、结内、髓内、肌内、通过静脉注射或腹膜内,例如,通过皮内或皮下注射施用于患者。可以将非造血组织驻留的γδT细胞的组合物直接注射到肿瘤、淋巴结或感染位点。
实施例
在大多数成年人中,Vδ2细胞仅以稳态包括小且剧烈变化的血液T细胞组分(0.01-5%),但是在通过多种因子,包括多种细菌和寄生生物的攻击后,细胞快速扩增,瞬间达到多达~25%的CD3+细胞。这种反应的主要基础在于Vδ2 TCR介导的低分子量“磷部分”的识别,包括羟基-甲基丁-2-烯基焦磷酸酯(HMBPP),它是用于通过(例如)香叶基化或法呢基化来修饰蛋白的胆固醇和其他脂质的关键微生物合成途径的中间产物。在灵长类中,这种合成是通过甲戊二羟酸途径发生的,其一个中间产物,异戊烯焦磷酸酯(IPP)以非常高的水平在病毒感染的和转化的细胞中表达,并且还是Vδ2 TCR介导的识别的靶标。
另外,一旦接合NKG2D配体,例如,MICA、MICB和ULBP,大部分Vδ2 T细胞表达高水平的可以激活或共刺激(与T细胞受体(TCR)一起)细胞溶胞潜能的NKG2D受体。那些配体是当细胞暴露于因子,如氧化或渗透应激或者紫外线时上调的主体蛋白。这些因子促进表皮生长因子受体(EGFR)途径的超高活性的信号转导,其通常在人实体瘤中也是失调的。
Vδ2 T细胞使用它们的TCR和/或NKG2D以及它们强大的溶胞能力来检测转化的细胞的能力以及向CD8+T细胞递呈抗原的外显潜能共同引起了以下观点:在临床上,可以利用Vδ2 T细胞来递送癌症免疫疗法。这可以通过细胞继承性转移来实现,在该方面,MHC对γδT细胞的限制的丧失显著且有利地限制了移植物抗宿主病(GvHD)的可能。为了实现这种情况,可以通过添加细胞因子,如白介素-2(IL-2)以及外源TCR激活剂,如磷酸化部分(例如,BrHPP),或者以及临床批准的二膦酸酯(例如,唑来膦酸)(其抑制甲戊二羟酸途径中的法呢基焦磷酸酯合酶,借此诱导TCR激活部分IPP的积累)来离体扩增血液驻留的Vγ9Vδ2γδT细胞。然而,通过因子,如BrHPP的Vγ9Vδ2细胞的长期激活可以逐渐导致细胞耗竭和减弱的细胞毒性潜能。
作为另外一种选择,可以使用HMBPP的药理学修饰形式或者临床批准的氨基二膦酸酯原位激活患者自身的γδT细胞。通过这些方法,已治疗了超过250位癌症患者,治疗效果看起来是安全的,但仅有非常少的完全缓解。有关细胞有限临床效力的一个主要问题在于通过长期抗原暴露,它们变得不可恢复地耗竭的倾向。第二个主要问题是它们表面上归巢至实体瘤和具有那些肿瘤的组织的低效率。
嵌合抗原受体T细胞(CAR-T)疗法在B细胞恶性肿瘤的临床过程中是有希望的。然而,对于实体瘤的治疗,到目前为止,CAR-T细胞的表现低于预期,其表现出较低的引起完全肿瘤反应的有效性和肿瘤外(off-tumor)细胞毒性的高发生率。对于外周血γδT细胞,CAR-T法对于实体瘤的成功的主要障碍在于全身性CAR-T细胞迁移至恶性肿瘤位点并以功能有效的形式存在于此的可能的低效率。另外,基于常规αβT细胞,CAR-T细胞必须克服肿瘤微环境中的免疫抑制信号,例如,通过PD1受体所传递的那些。
可能存在与γδT细胞用于CAR-T法有关的优势,因为可以用肿瘤反应性嵌合抗原特异性TCR转导它们,同时保留它们使用受体,如NKG2D识别转化的细胞的先天能力。因此,它们可以同时与肿瘤继承性(TCR)和先天性(NKG2D)介导的作用有关。然而,仍存在实质组织内人血液γδT细胞归巢至肿瘤并且在其中维持活性形式的表面低效率问题。这种考虑引起了对通常在非造血组织中驻留的γδT细胞的更详细的考虑。
这些T细胞作为它们发展的一部分迁移至非造血组织并因此不同于那些T细胞,例如,在全身性增强(致敏,priming)之后浸润组织的组织驻留的TCRαβ记忆T细胞(TRM细胞)。在小鼠中最好地研究了组织驻留的γδT细胞,其中已表明它们在皮肤、肠和生殖组织以及其他位点是普遍的。许多这些细胞已显示具有先天样功能潜能,其中它们可以对通过NKG2D受体激活的激发应答。本发明人最近获得了表明人的皮肤和肠同样有具有先天样活性的非造血组织驻留的γδT细胞的大区室的数据。在非造血组织内形成的恶性肿瘤、炎症、特异反应性、过敏症及其他病变的研究很大程度上不能考虑存在于其中发生病理性病变的组织内驻留的这些先天样人T细胞的潜在影响。
非造血组织内驻留的人γδT细胞的研究要差得多,因为它们的定位使得细胞更难以取样,并且因为尚未建立培养它们的方式。这种亚型包括具有非MHC-限制的溶胞潜能的多种细胞,由于它们不表达含有Vδ2-TCR,因此它们对低分子量的磷酸化部分完全不起反应。尽管对于这些细胞已知很少确切的TCR特异性,但是可用的数据表明所述细胞对自体抗原具有反应性,如内皮细胞蛋白C受体(EPCR),其通过巨细胞病毒(CMV)感染的细胞和通过多种实体瘤过表达。非造血组织相关的γδT细胞通常还表达NKG2D。考虑到这些性质以及细胞在非造血组织,如皮肤和肠内的细胞生理学驻留性,这些细胞向癌症患者的继承性转移对于靶向实体瘤和潜在地其他免疫病理学可能显著更有效。
将非Vδ2细胞用于免疫疗法需要原位扩增所述细胞或者在再输注前收获它们并离体扩增它们的方式。已采用了后一种方法,因为尚无已证明具有原位扩增大量非Vδ2 T细胞的能力的已知的TCR激活剂。为了克服非造血组织有限可用性的困难,一些研究人员已尝试扩增非常少的数目的来自血液的非Vδ2 T细胞,其中表达Vδ2的细胞是主要亚型,从而获得了以下假设:这些细胞与组织驻留的非Vδ2 T细胞是等价的。血液中存在的较少数目的非Vδ2 T细胞在活动性CMV感染期间显著扩增,与Vδ2 T细胞相比显示出对CMV优良的反应性,并且似乎能够在CMV子宫内感染的情况下保护人胎儿。另外,CMV反应性非Vδ2γδT细胞似乎在免疫抑制期间保护移植患者抵抗CMV的再次激活,并且通过对转化的细胞的交叉反应性,降低了继发性恶性肿瘤的风险。类似地,存在表明γδT细胞在控制HIV感染中起有益作用的数据,在这种情况下,相对于Vδ2 T细胞,非Vδ2γδT细胞在血液中扩增。
已通过添加直接激活TCR信号转导的外源因子,例如,通过使用如抗CD3抗体、泛γδ-TCR特异性抗体或植物凝集素(PHA)的试剂,或者通过将刺激的非Vδ2 T细胞与人工抗原递呈细胞(aAPC)共培养来离体扩增血液驻留的非Vδ2细胞,其中γδT细胞和aAPC之间的直接接触是非Vδ2 T细胞离体扩增所必需的。作为另外一种选择,已通过使用固定化的重组MICA(NKG2D配体),通过促进NKG2D受体的信号转导来扩增细胞,例如,如用于维持来自上皮癌浸润性淋巴细胞(TIL)的γδT细胞培养的离体增殖一样。总之,当前表达Vδ2的血液γδT细胞或非Vδ2血液γδT细胞的离体扩增方法需要添加试剂,从而不变地促进TCR和/或NKG2D受体以及补充细胞因子,如IL-2的激活。受体激活信号和细胞因子的这种组合反映了共同体所广泛采用的培养和扩增T细胞的标准方法。到目前为止,尚未描述显著扩增非造血组织中驻留的γδT细胞的方法。在本文中描述了这种方法。
作为人非造血组织驻留的γδT细胞(例如,皮肤γδT细胞)的表型和功能特征的一部分,本发明人已分离了通常在非造血组织内驻留的并且相对于αβT细胞和血液驻留的γδT细胞具有独特性质的不同且较大的γδT细胞群体。本发明人已发现细胞对NKG2D配体和细胞因子显示出强烈的不依赖于TCR的先天样反应。然而,对于原代αβT细胞扩增的工作通常使用与作为有益生长因子来源的其他支持细胞的共培养,本发明人已意外地显示皮肤及其他非造血组织中驻留的γδT细胞受与自体同源的皮肤成纤维细胞和潜在地其他基质组分,如角化细胞和内皮细胞接触时共培养这种细胞的严重且特异性地抑制。这些相互作用的减轻使得细胞能够以用于潜在临床应用的较大的量扩增。
此外,与目前扩增血液来源和肿瘤来源的γδT细胞的工作相反,本发明人已显示可以在不有意添加激活它们的TCR或NKG2D信号通路的任何外源试剂的情况下扩增这些非造血组织驻留的γδT细胞。
本文公开了有效且可重复地从人或非人动物的非造血组织,如皮肤和肠分离和扩增γδT细胞的新型方式。通过破坏非造血组织来源的非Vδ2 T细胞与自体同源的成纤维细胞和潜在地其他基质组分的接触来促进扩增,并且通过与IL-2、IL-15、IL-4和/或IL-21的培养来维持扩增。
描述了以下实施例以向本领域技术人员提供如何实施、制备和评价本文所主张的方法和化合物的完整公开和描述,并且以下实施例旨在是对本发明的纯粹示例,并且不意欲限制本发明人所认为的他们的发明的范围。
实施例1.分析方法
除非另有说明,否则使用以下方法来产生后续实施例的结果。
流式细胞术
使用以下抗体-荧光染料缀合物:Ki-67-BV421、CD3-BV510、Vδ1-PeVio770、TIM-3-PE、CD9-PE、CCR3-BV421和CD39-BV421进行流式细胞术。还使用eFluor770NIR对样品的存活力染色。商品化抗体购自Biolegend或Miltenyi。存活力染料(近IR)来自eBioscience。在使用Foxp3染色缓冲液组(eBioscience)固定和透性化的细胞上进行Ki-67染色。一旦完成每个实验,在PBS中清洗细胞群体并分成两份。用eFluor770NIR对细胞的存活力染色,并清洗,然后用TrueStain(Biolegend)染色以避免染色抗体的非特异性结合。将一半样品对所指示的表面标志物染色,并且将另一半仅对谱系标志物(CD3,Vδ1)染色,并且使用了所述表面标志物的等价的同种型对照。以相同浓度使用缀合至相同荧光染料的匹配的小鼠同种型抗体。同种型对照未结合至已知的人抗原并因此表示非特异性结合或假阳性。与其相应同种型对照或者如有指示,FMO相比较,显示了柱状图(图1D、2A、3B、4B、6B、7A和11-13)。数据总结表明对于所比较并因此处于高于同种型的水平的所指示的标志物染色为阳性的细胞百分比。在FlowJo(10.1版)上进行流式细胞术的数据分析。
RNA测序
将来自人皮肤和人血液Vδ1 T细胞的Vδ1 T细胞(在T细胞受体引起的扩增之后)分选(FACS)、离心并将细胞颗粒在RLT缓冲液中再悬浮。使用RNA-Micro-plus试剂盒(QIAGEN)制备RNA。使用具有RiboErase(HMR)的KAPA Stranded RNA-seq试剂盒(KAPA BIOSYSTEMS)来产生RNA文库。使用快速运行化学(rapid run chemistry)在HiSeq 2500(Illumina)上进行双端测序(读序长度:100bp)。使用RSEM(v1.2.11)以及Bowtie2对101个碱基对双端读序进行比对和定量。将读序与人转录组比对,并且已将计数值log2转化和分位数归一化。
细胞因子定量
用PMA和离子霉素或者板结合的抗CD3 mAb(OKT3,5μg/ml)刺激来自人皮肤的Vδ1T细胞24小时。然后,采集上清液并使用来自eBioscience的ProcartaPlex人细胞因子&趋化因子组1A(34plex)进行分析。使用Luminex FlexMap3D(Luminex Corp)分析测定。在Microsoft Excel中分析数据,并且显示3个供体(重复实验)的平均值。误差线表示标准偏差。
与成纤维细胞的共培养
对于每个分离培养,使用手术刀对两个培养皿(100×25mm,Corning)在几个位置进行刮划。将切碎的皮肤块置于刮划处。空气干燥5至10分钟后,皮肤块通常粘在培养皿上并添加10mL Skin-T培养基。每周更换一次培养基,并在生长3周之后,在用 (Life Technologies)处理之后,收获原代成纤维细胞。将成纤维细胞以1×104接种到48孔板中,或者就转板实验来说,以2×104接种到24孔板的下室中。2至3天后,成纤维细胞达到汇合并且使用添加2×105个混合的皮肤淋巴细胞(就48孔板来说)或者3×105个淋巴细胞(就24孔板、底孔板和转板来说)的RPMI和所指示的细胞因子起始共培养实验。
(Life Technologies)处理之后,收获原代成纤维细胞。将成纤维细胞以1×104接种到48孔板中,或者就转板实验来说,以2×104接种到24孔板的下室中。2至3天后,成纤维细胞达到汇合并且使用添加2×105个混合的皮肤淋巴细胞(就48孔板来说)或者3×105个淋巴细胞(就24孔板、底孔板和转板来说)的RPMI和所指示的细胞因子起始共培养实验。
血液来源的γδΤ细胞的扩增。
如果用TCR配体(就Vδ2来说,例如,IPP、HMBPP、二膦酸酯)或者交联TCR受体(mAb)或TCR相关激酶CD3的抗体添加刺激,则仅可以扩增PBMC内的血液来源的γδΤ细胞。还可以使用凝集素,如PHA实现相同的TCR交联效果。在不存在这些TCR刺激剂的添加的情况下,PBMC中的γδΤ细胞存活数日,但是不能扩增并且保持它们具有较少变化的初始T细胞亚型组成。
通过使全血在Ficoll上分层,然后以400g离心20分钟来分离红细胞、血浆和白色淋巴细胞/单核细胞,从而将来自健康志愿者的血液用于分离PBMC。通过移液器(stripett)小心收获白血球,并在冷PBS中清洗四次。将细胞以1×106个/mL的密度在具有10%热失活的胎牛血清(Life Technologies)、L-谷氨酰胺(292μg/ml;Life Technologies)、青霉素(100单位/ml;Life Technologies)、链霉素(100μg/ml;Life Technologies)、N-2-羟乙基哌嗪-N-2-乙烷磺酸(HEPES;0.01M;Life Technologies)、丙酮酸钠(1mM;LifeTechnologies)、最低必需培养基(MEM)非必需氨基酸(1X;Life Technologies)的RPMI-1640培养基(Life Technologies)中再悬浮并补充IL-2(100IU/mL)。在细胞转移前,将细胞转移至用泛γδTCR单克隆抗体(20μg/ml,B1克隆,Biolegend)涂覆的24孔板中90分钟。使细胞生长14天,每2-3天更换培养基并添加新鲜的细胞因子。一旦达到汇合,将细胞以1:1分开。在这些条件下,14天之后,通常通过它们的TCR(如果是CD69和CD25,如通过上调所指明的)高度激活初始较少的γδT细胞群体并且它们在很大程度上得到了富集,其主要包括Vδ2Τ细胞,但是还包括Vδ1 T细胞(多至所有γδT细胞的30%)。随后,可以使用FACS分离Vδ1 T细胞以用于功能或表型(例如,遗传)分析。
实施例2.从皮肤和肠分离非造血组织驻留的γδT细胞
使用Clark规程建立了立体皮肤移植规程。将尺寸为9mm×9mm×1.5mm的细胞泡沫基质(Cellfoam Matrices)(Cytomatrix Pty Ltd,Victoria,Australia)或等价物高压灭菌并在100mg/ml鼠尾胶原蛋白I(BD Biosciences)在PBS中的溶液中在室温下孵育30分钟,然后在PBS中清洗一次。在皮肤手术3-6小时内获得成年人皮肤样品。除去皮下脂肪并将剩余的皮肤组织切成测量为约1mm×1mm的碎片。将约5个皮肤碎片/外植体放置并下压到每个基质表面上。将每个基质置于含有2ml Skin-T培养基(具有10%热失活的胎牛血清(LifeTechnologies)、L-谷氨酰胺(292μg/ml;Life Technologies)、青霉素(100单位/ml;LifeTechnologies)、链霉素(100μg/ml;Life Technologies)、N-2-羟乙基哌嗪-N-2-乙烷磺酸(HEPES;0.01M;Life Technologies)、丙酮酸钠(1mM;Life Technologies)、最低必需培养基(MEM)非必需氨基酸(1X;Life Technologies)和3.5μl/L2-巯基乙醇(LifeTechnologies)的伊斯科夫氏改良达尔伯克氏培养基(IMDM;Life Technologies))的24孔板(Corning)的单独的孔中。对于培养的前7天,将两性霉素(2.5μg/ml;LifeTechnologies)加入至培养基。通过从每个孔中吸出上层1mL培养基并添加1ml新鲜的培养基,每周更新三次培养基。在培养开始时并且在每次培养基更新时,添加人重组IL-2(100IU/mL; Novartis Pharmaceutical UK Ltd)和人重组IL-15(20ng/mL;Biolegend)直至21-35天之后淋巴细胞分离为止,如表1所示。对于每个供体,在培养中设置了多达96个孔(4个24孔板)。
Novartis Pharmaceutical UK Ltd)和人重组IL-15(20ng/mL;Biolegend)直至21-35天之后淋巴细胞分离为止,如表1所示。对于每个供体,在培养中设置了多达96个孔(4个24孔板)。
为了分离淋巴细胞,将基质和培养基转移到含有10ml汉克平衡盐溶液(HBSS;LifeTechnologies)且具有0.01mM HEPES的50ml离心管(Corning)中(多至12个基质/管)。使用10ml移液管,用细胞悬液漂洗基质,并将所述细胞悬液通过70μm过滤器(BD Biosciences)进入到新的50ml离心管中。将基质漂洗另外重复2次。还将培养基从培养孔中吸出并通过70μm过滤器进入到新的50ml离心管中。用1ml 0.01mM HEPES/HBSS另外清洗孔两次并通过70μm过滤器。随后,通过离心(1600rpm,15分钟)分离细胞。将颗粒在Skin-T培养基中再悬浮。将最终的细胞颗粒在Skin-T培养基中再悬浮以用于后续的流式细胞术分析或功能研究。当需要细胞计数时,在此阶段通过(1)台盼蓝染色(0.4%)(Life Technologies)和血球计,或者(2) TT型细胞计数器和分析仪(Roche)对淋巴细胞计数。下表1显示了示例性研究的结果。
TT型细胞计数器和分析仪(Roche)对淋巴细胞计数。下表1显示了示例性研究的结果。
表1.每个供体的分离的淋巴细胞得率。

由于原代肠样品更易污染,因此在切碎并放置在支架上之前,将所获得的活组织检查样品首先在含有10%FCS、青霉素(500U/mL)、链霉素(500μg/mL)、庆大霉素(100μg/mL)、两性霉素B(12.5μg/mL)和甲硝唑(5μg/mL)的IMDM中清洗两次。将肠支架培养在Gut-T培养基(IMDM、10%FCS、青霉素100U/mL、链霉素100μg/mL、庆大霉素20μg/mL、甲硝唑1μg/mL)中生长。对于生长第1周,与皮肤类似,我们还使用了2.5μg/mL的两性霉素B。培养基含有IL-2(100IU/mL)和IL-15(20ng/mL)并且每周更换3次。由于肠结构比皮肤更疏松,因此在1周后收获淋巴细胞。
实施例3.非造血组织驻留的γδT细胞的表征
皮肤中富含人γδT细胞,其主要为Vδ2-,并且人γδT细胞参与人淋巴应激监视应答(lymphoid stress surveillance response)
图1A-图1D显示人的皮肤包括明显的驻留γδT细胞群体。使用Clark规程,我们使用补充有IL-2和IL-15的人残余皮肤样品以使得能够在3周的过程中长出组织驻留的淋巴细胞。平均得率为240,000个淋巴细胞/支架。与先前报道一致,表征了不同的皮肤驻留的淋巴细胞亚型,并且大部分细胞表达常规αβTCR,它们是组织驻留的“TRM”型中主要的。整体上,59.9%(±8.6%)的CD45+细胞是CD4+,和具有8.7%(±3.6%)的NK细胞部份的18.3%(±2.8%)的CD8+αβT细胞。另外,我们在我们的供体中发现了明显的γδT细胞群体(平均8.513%的CD45+细胞,±6.564%)(图1A和3D)。在器官型培养之后,这种淋巴细胞谱在约100个供体中是高度可重复的,并且与新水解的皮肤样品相当,仅在稍微提高的γδ群体方面有所不同,但是就实用性来说,提供了比标准组织消化规程大得多且更纯的淋巴细胞群体。根据与基于它们的TCRδ链的人γδT细胞的组织区室化有关的文献,大部分人皮肤γδT细胞表达与通过流式细胞术所表征的多种γ链配对的Vδ1 TCR链。与大部分外周血γδT细胞相反,这种链显示出与Vγ9连接的Vδ2链的单一特异性TCR杂二聚物并且它们在人皮肤样品中几乎是不存在的。然而,重要的是注意到既不表达Vδ1 TCR,也不表达Vδ2TCR的亚型,其在本文中被称为双阴性γδT细胞(DNγδT细胞;图1C)。
以这种方式生长的皮肤驻留的γδT细胞显示出缺少CD45RA的表达并且表达不同水平的共刺激性分子CCR7的非终末分化的记忆表型。相对于常规全身性T细胞,皮肤驻留的γδT细胞显示出高表面标志物CD69的表达以及程序性死亡受体1(PD-1)的表达,低至不存在的IL-2受体α(CD25)的水平,并且缺少共刺激性分子CD28,从而表明了上述或长期激活作用(图1D)。与它们的组织定位一致,Vδ1和DN细胞显示出皮肤和组织归巢标志物,如CCR4、CCR8和整合素αE(CD103;图7)的表达。这种组织-归巢标志物-组可以证明在免疫疗法环境中的有益性。另外,皮肤驻留的γδT细胞显示出对于激活受体NKG2D的高水平表达(图2A),其表示了这些细胞在淋巴应激监视应答中的可能作用。分别通过对DNA损伤、EGF-受体激活和氧胁迫响应的细胞来上调NKG2D配体,如MICA、MICB和ULBP,并因此可以使得表达NKG2D的T细胞能够表征并消除胁迫或转化的细胞,借此维持组织体内平衡。与该原理一致,我们发现通过暴露于NKG2D受体的重组配体(MICA、ULBP2)激活了通过本发明所述的方法扩展的皮肤驻留的γδT细胞,从而表明了如通过溶酶体相关膜蛋白CD107a的上调所测量的脱颗粒(图2A)。这种先天样特征是Vδ1+和DNγδT细胞所独有的,因为其他组织驻留的T细胞(图2C)和全身性γδT细胞缺少这种应答(图10B)。
整体上,当被PMA/离子霉素或者被NKG2D配体,例如,重组MICA蛋白(图2A和图2B)激活时,激活的皮肤驻留的Vδ1+和DNγδT细胞执行促炎性Th1偏移细胞因子程序(对IFN-γ、TNF-α和GM-CSF染色为阳性),借此认定了细胞的先天样应答。的确,通过抗体阻断NKG2D受体几乎完全终止了对MICA的应答(图2B和图2C)。
已知在某些疾病背景下,如银屑病和在一些肿瘤类型内,γδT细胞分泌IL-17。即使通过彻底激活,通过本发明所述的方法扩增的γδT细胞产生低水平的或者不产生IL-17(图2B和图8)。相反,通过TCR激活,组织驻留的表达CD4的αβT细胞的确产生IL-17(图2B)。一般地,与受限于与宿主保护有关的Th1偏移程序的Vδ1+和DNγδT细胞相比,αβT细胞显示出对PMA/离子霉素响应的更加不同的细胞因子谱(cytokine repertoire)。
与组织分离导致了人组织γδT细胞的激活和大量增殖
为了进一步研究人组织γδT细胞,将混合的皮肤淋巴细胞转移至细胞培养孔中并添加IL-2以随时间维持存活力。有趣地,通过与器官型培养中存在的基质和上皮细胞分离,Vδ1 T细胞独特地显示出在没有任何其他刺激的情况下的激活和增殖迹象。在7天内,Vδ1+和DNγδT细胞独特且大量上调核因子Ki-67并提高IL-2受体α的表面表达(CD25;图3B和4B)。显著地,在3周的过程中并且在仅存在IL-2的情况下,组织来源的Vδ1+和DNγδT细胞的生长超过了所有其他T细胞亚型,从而占所有皮肤淋巴细胞的多至65%,将数目平均提高了127.18倍,而αβT细胞仅增殖5.21倍(p=0.0124),如通过绝对细胞数目所测量的(图3A)。在Vδ1+和DNγδT细胞中,细胞周期相关的核因子KI-67的MFI在14天中从2,664.5(±1,876.1)提高至8,457.7(±4,574.2),而在相同时间过程中,在αβT细胞中,MFI从592.8(±390.5)降低至284.7(±140.1)(图3C)。还可以使用提高淋巴细胞的存活力和总数的其他重组IL-15来支持这种选择性的皮肤驻留的γδT细胞长出的现象。
成纤维细胞以接触依赖性的方式主动抑制皮肤γδT细胞
如上所述的Vδ1+和DNγδT细胞的显著扩增绝不会在其中存在彻底的成纤维细胞长出的器官型培养系统中发生。为了直接测试它们与Vδ1+和DNγδT细胞的共培养是否将抑制T细胞扩增,使自体同源的成纤维细胞长出。在3周支架培养后,将混合的皮肤淋巴细胞接种到空的或者含有先前建立的汇合的成纤维细胞单层的孔中,并且在每种情况下向培养基补充外源IL-2以维持T细胞生长。另外,使用转板以防止T细胞直接接触相同孔内的成纤维细胞,同时使它们受成纤维细胞分泌的可溶性因子影响。在14天的共培养中,Vδ1+和DNγδT细胞开始在其中没有成纤维细胞的孔中和其中阻止T细胞直接接触成纤维细胞的孔中增殖。如前所述,所有条件下的αβT细胞的增殖较低。当允许T细胞直接接触成纤维细胞时,Vδ1+和DNγδT细胞在2周内的生长速率显著降低,从无成纤维细胞接触的孔中的22.6(±8.07)降低至3.3(±0.17)倍(图4A)。通过与单独生长的淋巴细胞相比,在7天过程中,Vδ1 T细胞中的CD25、Ki-67和转录因子T-bet的上调的不存在进一步确认了这种接触介导的抑制(图4B)。一些形式的组织介导的免疫系统的控制似乎对于维持组织体内平衡将是很重要的,因为在没有它的情况下,将存在持续炎症的可能。通过基质成纤维细胞的Vδ1+和DNγδT细胞的抑制性调控将似乎是这种控制的实例。
总之,皮肤驻留的Vδ1+和DNγδT细胞的表型以及它们显著的功能潜能是对通常通过接触依赖性机制受它们相邻的皮肤成纤维细胞限制的激活前的T细胞的反映。通过将T细胞从与成纤维细胞的接触中释放,使所述机制失活,这将选择性地允许Vδ1+和DNγδT细胞扩增,然而皮肤中的其他T细胞是不受影响的。
接触介导的抑制的解除促进了通过皮肤Vδ1 T细胞的细胞毒性TH1偏移细胞因子应答
将混合的皮肤来源的淋巴细胞扩增14天,并且将荧光相关的细胞分选用于从γδT细胞中除去αβT细胞,从而使纯度达到高达90%。将那些高度富集的细胞以150,000个细胞/孔的浓度沉积在细胞培养孔中含有10%FCS的RPMI培养基中。24小时后收集上清液并使用 -基阵列评价广泛的效应因子细胞因子。完全出乎意料地,扩增的γδT细胞(仅通过它们与成纤维细胞的分离所引起)自发产生了大量TH1相关细胞因子,如IFN-γ(12,383.46±16,618.90pg/ml)、GM-CSF(4,316.73±4,534.96pg/ml)以及促炎趋化因子CCL4(14,877.34±10,935.64pg/ml)和CCL3(1,303.07±757.23pg/ml;图5A)。
-基阵列评价广泛的效应因子细胞因子。完全出乎意料地,扩增的γδT细胞(仅通过它们与成纤维细胞的分离所引起)自发产生了大量TH1相关细胞因子,如IFN-γ(12,383.46±16,618.90pg/ml)、GM-CSF(4,316.73±4,534.96pg/ml)以及促炎趋化因子CCL4(14,877.34±10,935.64pg/ml)和CCL3(1,303.07±757.23pg/ml;图5A)。
此外,在扩增期间并且与新分离的皮肤来源的TCR激活的γδT细胞相反,细胞产生了大量与特应性反应有关的自发IL-13。以低得多的水平或者完全没有产生其他细胞因子,如IL-17A(图8)。在用重组MICA(NKG2D配体)、抗CD3或PMA/离子霉素刺激后,可以进一步提高细胞的高效应因子潜能。为了评价扩增的γδT细胞对恶性靶细胞的细胞毒潜能,我们在24小时共培养实验中使用了已建立的转化细胞系。Vδ1+和DNγδT细胞以优于常规组织αβT细胞的剂量依赖性方式对HeLa细胞(宫颈)和CaCO2(结肠)显示出非常高的细胞毒活性(图5B)。此外,可以使用可溶性单克隆抗体,通过阻断NKG2D受体来强烈抑制γδ-细胞介导的细胞毒性,从而表明通过脱抑制的人皮肤来源的γδT细胞,该受体至少部分参与了肿瘤监视。使用其他靶标:HCT1954(乳腺癌)、MDAMB231(乳腺癌)和HCT116(结肠),进一步确认了这些细胞的细胞毒潜能(图9)。
通过(例如)TNFα、IFNγ和CD107a产量的增加,通过本发明所述的方法所产生的非造血组织驻留的γδT细胞还可以不同于其他血液来源的γδT细胞,其中在不存在任何T细胞受体刺激配体的情况下,它们对与恶性肿瘤强烈相关的NKG2D配体(MICA)应答(图2和图10)。它们还在不经历任何外源药理学或配体介导的T细胞受体激活的情况下进行细胞毒性T细胞应答,并因此在不存在刺激的情况下是细胞毒性的(图3和图5)。这表示与其他γδT细胞,与αβT细胞或者与NK细胞相比,通过本发明所述的方法所产生的非造血组织驻留的γδT细胞在它们的应答能力方面是独特的并且在不存在任何外源试剂激活的T细胞受体信号转导的情况下增殖(图3)。通过本发明所述的方法所产生的非造血组织驻留的γδT细胞还对CD69和PD-1染色阳性,其缺少CD28的表达并且仅表达低水平的CD25(参见图1D)。血液来源的γδT细胞不表达这种标志物的组合。此外,与血液来源的扩增的Vd2γδT细胞相比,它们显示出更高的组织归巢受体,如CCR4和CCR8的表达(图7B)。
人肠中组织驻留的γδT细胞
还表征了表达Vδ1 T细胞受体的人结肠来源的非造血组织驻留的γδT细胞群体(图6)。在3个供体中,使用与对于皮肤细胞所使用的相同的方法,在4至5周的时间过程中扩增这些细胞。在扩增期间,在它们与成纤维细胞富集的器官型细胞培养分离之后,结肠来源的Vδ1+和DNγδT细胞显示出类似的Ki-67上调模式。同样地,结肠驻留的Vδ1+和DNγδT细胞受NKG2D受体的配体的提供的强烈刺激。当与利妥昔单抗混合时,血液来源的γδT细胞是装备良好的以通过CD16表达执行抗体依赖性细胞介导的细胞毒性,从而证明了提高的靶向CD20阳性B系淋巴瘤的细胞毒性。类似地,当被单克隆抗体靶向时,可以更有效地杀死慢性淋巴细胞性白血病(CLL)和HER2阳性乳腺癌细胞。为了评价皮肤来源的Vδ1 T细胞靶向抗体调理的靶细胞的潜能,对三种IgG1相关Fc受体CD16、CD32和CD64的表达水平进行定量。皮肤来源的Vδ1 T细胞表达较低水平的Fc受体CD16,但是对于高亲合力IgG受体CD64显示出优良的表达水平(图11)。因此,通过抗体向恶性肿瘤和转移的位点的引导,识别受调理的肿瘤细胞并通过ADCC杀死靶标,组织来源的Vδ1 T细胞可以是装备良好的以用作单克隆抗体疗法,如CD20或Her2疗法的佐剂。
实施例4.非造血组织驻留的γδT细胞扩增条件的优化
皮肤来源的γδT细胞的扩增
在3至4周支架培养之后收获混合的淋巴细胞,将其在PBS中清洗,离心沉淀并以1×106个细胞/毫升的浓度在具有过滤的10%热失活的胎牛血清(Life Technologies)、L-谷氨酰胺(292μg/ml;Life Technologies)、青霉素(100单位/ml;Life Technologies)、链霉素(100μg/ml;Life Technologies)、N-2-羟乙基哌嗪-N-2-乙烷磺酸(HEPES;0.01M;LifeTechnologies)、丙酮酸钠(1mM;Life Technologies)、最低必需培养基(MEM)非必需氨基酸(1X;Life Technologies)和50μM 2-巯基乙醇(Life Technologies)的洛斯维帕克纪念研究所1640培养基(RPMI-1640;Life Technologies)中再悬浮。初始的Vδ1+细胞群体为1.12%的淋巴细胞。将细胞以2×105个细胞/孔接种到96孔平底板(Corning)中或者以2×106个细胞/孔接种到24孔板(Corning)中用于扩增,并以表2中所提供的浓度补充如所指示的因子。
表2.包含在扩增培养中的每个因子的浓度


每天通过显微镜监测细胞,并且每周提供三次新鲜的培养基和细胞因子。一旦完全汇合并且细胞聚集,则根据需要,将细胞1:1分开至另外的孔和板中。21天后,使用 (eBioscience)收获细胞,计数并使用流式细胞术分析。图17A和图17B显示了扩增前后的代表性流式细胞术图。除了每个处理组的扩增细胞的相对CD27和TIGIT表达,表3显示了对应于图17C的倍数-扩增值。使用扩增前后的Vδ1总数计算最终的Vδ1倍数扩增(%CD3+泛γδ+Vδ1+细胞÷100)×(总细胞数)。
(eBioscience)收获细胞,计数并使用流式细胞术分析。图17A和图17B显示了扩增前后的代表性流式细胞术图。除了每个处理组的扩增细胞的相对CD27和TIGIT表达,表3显示了对应于图17C的倍数-扩增值。使用扩增前后的Vδ1总数计算最终的Vδ1倍数扩增(%CD3+泛γδ+Vδ1+细胞÷100)×(总细胞数)。
表3.在存在多种因子的情况下,皮肤驻留的Vδ1 T细胞的扩增。

与单独的IL-2和IL-15中的扩增相比,其他因子的添加提高了Vδ1 T细胞的扩增,如图17C所示。考虑到Vδ1 T细胞对IL-2、IL-15、IL-4和IL-21响应的高得率,还研究了这些因子的组合,如图17D-17H所示。
使用平均荧光强度(MFI)确定了每个Vδ1+T细胞群体的初始和最终表型,包括CD27和TIGIT的表达,并且通过取中值MFI对每个组内的样品求平均值。表4显示了每个扩增条件下Vδ1+T细胞的CD27和TIGIT的表达。
表4.在存在多种因子的情况下扩增的皮肤驻留的Vδ1 T细胞的表型。
| 扩增因子 | CD27 MFI | TIGIT MFI |
| IL-2 | 113±5 | 7919±179 |
| IL-2、IL-15 | 226±33 | 4788±679 |
| IL-2、IL-15、IL-6 | 140±91 | 6663±767 |
| IL-2、IL-15、IL-7 | 128±115 | 6083±1813 |
| IL-1、IL-15、IL-4、IL-21 | 1284±1048 | 1721±308 |
| IL-2、IL-15、IL1β | -136±3.5 | 9453±390 |
| IL-2、IL-15、IL-18 | 121±75 | 4396±2782 |
| IL-2、IL-15、IL-9 | 260±21 | 5247±2333 |
| IL-2、IL-15、IGF-1 | 193±94 | 6584±1127 |
| IL-2、IL-15、IL-4、IL-21、IL-9 | 790±61 | 1389±803 |
| IL-2、IL-15、IL-4、IL-21、1%HPL | 990±258 | 1203±222 |
| IL-2、IL-15、IL-4、IL-21、5%HPL | 421±76 | 1045±798 |
相对于单独的IL-2和IL-15,其他因子的添加提高了CD27的表达,如通过平均荧光强度所测量的,如图18A所示。值得注意的,相对于单独的IL-2和IL-15,IL-4和IL-21的添加将CD27 MFI提高了约8倍。另外,相对于其其他组合,IL-2、IL-15、IL-4和IL-21的四因素组合获得了最高的CD27表达(图18B和图18D)。值得注意的,T细胞上CD27的低表达经常与具有较低的进一步长期增殖的潜能的耗尽的、终末分化的表型有关。
相对于扩增的Vδ1+T细胞的TIGIT表达,观察到了不同的趋势(图19)。具体地,结合IL-2和IL-15,TIGIT对IL-4和IL-21响应的表达降低。为了进一步研究该趋势,将TIGIT表达作为CD27表达的函数作图(图20)。在TIGIT和CD27之间观察到负相关。高TIGIT表达可以使T细胞对通过肿瘤微环境的抑制敏感,其中其配体脊髓灰质炎病毒受体(PVR;CD155)的表达可以较高。
实施例5. 4种细胞因子培养足以替代细胞培养血清
目前,当生产组织或肿瘤驻留或来源的T细胞时,复杂的血液来源的血清和血浆的添加是普通的。然而,由于这些成分驻留的批与批之间的差异、这些成分的高成本、考虑到先进治疗型医药产品(ATMP)行业的高需求所造成的有限供应以及来自这些成分的偶然因素交叉污染风险的提高,这些血清或血浆成分的使用可以是不希望的。因此,在成功表征支持如本文所描述的所期望的γδT细胞的扩增和富集的细胞因子之后,测试了这些细胞因子的使用是否可以消除对通常用于扩增组织来源的γδT细胞的复杂血清/血浆成分的任何需要。为了对其进行进一步测试,使免疫细胞从皮肤样品流出并测试是否仍需要血浆/血清来支持γδT细胞+/-血清的扩增和富集。因此,然后收获流出的细胞并在“第0天”将其接种到两种培养基上。第一培养基包括具有10%人来源的血清和细胞因子的不含动物来源的组分(ADCF)的培养基(TexMACS,Miltenyi)。第二培养基包含相同的ADCF培养基/细胞因子混合物,标准限定补充剂(CTSTM,含有纯化的人血清白蛋白、重组胰岛素和转铁蛋白),但无任何血清添加。图21显示了该研究的结果。意外地,在添加或不添加人血清的情况下,观察到了相当的富集和相当的倍数扩增。这表明有可能在不添加任何动物来源的成分或人血清的情况下扩增和富集组织来源的γδT细胞。
然后,对于来自从标准器官型培养收获的起始混合的淋巴细胞群体的γδT细胞的优先扩增评价了该方法。如果可实现,则这种优先扩增将是非常期望的,特别是如果所述规程产生了包含>50%的γδT细胞的最终扩增的淋巴细胞群体。的确,到目前为止,常规富集规程需要使用去除或富集技术,如磁免疫去除技术(例如,来自Miltenyi或Dynal)或者流式细胞分选技术(例如,来自BD Biosciences)以物理分离少数γδ细胞或物理去除大多数αβ细胞。相反,本研究评价了本发明方法是否可以富集存在于任何给定起始混合淋巴细胞群体中的γδ细胞而无需这种物理分离或去除规程。意外地,并且由于所述规程对于γδ细胞扩增的选择性高于也存在于起始群体中的其他细胞类型,使用这些扩增方法实现了这种富集。这导致了占存在于培养中的全部细胞的超过50%的更加富集、更纯的γδT细胞群体。在存在和/或不存在血清的情况下实现了这种富集,并且还在图21和图22A-图22D中进一步显示了这种富集,其中γδT细胞扩增了>100倍,从而在对于这种组织样品的所有情况下,将γδT细胞的纯度从<50%提高至>50%。
分离和扩增方法
使用Clark规程先前所述的等价方法并且在实施例2中,在支架培养3周后,从组织收获混合的淋巴细胞。所得的收获的细胞的谱图类似于实施例3中所描述的那些。将这些收获的细胞在HBSS+HEPES中清洗,离心沉淀并在含有(i)10%人AB血清(Life ScienceProduction)以及IL-2(100IU/L)、IL4(Biolegend,5ng/mL)、IL-15(Biolegend,20ng/mL)和IL-21(Biolegend,5ng/mL)或者(ii)5%CTSTM(Thermo Fisher Scientific)以及IL-2(100IU/L)、IL-4(Biolegend,5ng/mL)、IL-15(Biolegend,20ng/mL)和IL-21(Biolegend,5ng/mL)的TexMACS培养基(Miltenyi)中再悬浮。含有血清和不含血清的培养基两者还含有pen/strep抗生素(分别为100单位/ml,100ug/ml,Life Technologies)。然后,将细胞以2百万个细胞/孔接种到24孔板(Corning)中。一旦细胞变得汇合,则将它们通过以1/2至1/4之间的比例分开至含有相同培养基的新孔中来进行扩增/传代。在第21天,通过 细胞脱离溶液(Thermo-Fisher)收获细胞并通过流式细胞术对其进行分析以确定最终的细胞谱图,如图21和图22A-图22D中所提供的。
细胞脱离溶液(Thermo-Fisher)收获细胞并通过流式细胞术对其进行分析以确定最终的细胞谱图,如图21和图22A-图22D中所提供的。
实施例6.TIGIT表达的功能相关性
在肠驻留的Vδ1细胞上组成型表达TIGIT,如图23所示。使用通过常规组织消化分离自肠的Vδ1细胞产生数据。组织驻留的γδT细胞的组成型TIGIT表达不仅与皮肤来源的Vδ1细胞有关,而且TIGIT表达不是Clark规程(基于网格的分离程序)的假象。
此外,在仅与IL-2和IL-15孵育的细胞中,脊髓灰质炎病毒受体(PVR)特异性抑制TCR信号转导,如通过IFNγ表达(图24A)和TNFα(图24B)所测量的。在与IL-2、IL-15、IL-4和IL-21孵育的细胞中,PVR对TIGIT阴性Vδ1+/Vδ3+细胞的抑制作用丧失,如通过IFNγ(图25A)或TNFα(图25B)所测量的,从而表明从4种细胞因子混合物产生的TIGIT-阴性优先防止TIGIT介导的Vδ1+/Vδ3+T细胞激活的抑制。
实施例7.扩增培养中IL-9对IL-2的替代
将来自三个供体(TS052、TS056和SK073)的皮肤组织置于9mm网格上并在补充有IL-2和IL-15的培养基中培养3周。将分离的淋巴细胞在补充有IL-2、IL-4、IL-15和IL-21(图26A和26B,左侧柱)或者IL-4、IL-9、IL-15和IL-21(图26A和26B,右侧柱)的培养基中孵育。在扩增3周后,计算γδT细胞/网格(图26A)和Vδ1细胞/网格(图26B)的最终得率。
如图27A-27C所示,如通过变化倍数(图27A)、γδTCR+T细胞的%(图27B)和Vδ1+T细胞的%(图27C)所测量的,在皮肤γδT细胞的扩增中,IL-9足以替代IL-2的功能。皮肤组织来自6个供体(SK073、SK075、SK077、TS052、TS053和TS056)。2种细胞因子混合物包括IL-2和IL-15,而4种细胞因子混合物包括IL-2、IL-15、IL-21和IL-4。
如图28A和图28B所示,如通过Vδ1+T细胞上CD27表达的平均荧光强度(MFI)(图28A)和Vδ1+T细胞上CD27表达的归一化的MFI(图28B)所测量的,在皮肤γδT细胞的扩增中,IL-9足以替代IL-2的功能。与标准培养条件相比,在CD27表达中未观察到差异。皮肤组织来自4个供体(SK073、TS052、TS053和TS056)。2种细胞因子混合物包括IL-2和IL-15,而4种细胞因子混合物包括IL-2、IL-15、IL-21和IL-4。
皮肤来源的γδT细胞的扩增
在3周支架培养之后收获混合的淋巴细胞,将其在磷酸盐缓冲盐水(PBS)中清洗,离心沉淀并以1×106个细胞/毫升的浓度在具有过滤的10%热失活的FBS(LifeTechnologies)、L-谷氨酰胺(292μg/ml;Life Technologies)、青霉素(100单位/ml;LifeTechnologies)、链霉素(100μg/ml;Life Technologies)、N-2-羟乙基哌嗪-N-2-乙烷磺酸(HEPES;0.01M;Life Technologies)、丙酮酸钠(1mM;Life Technologies)、最低必需培养基(MEM)非必需氨基酸(1X;Life Technologies)和50μM 2-巯基乙醇(Life Technologies)的RPMI-1640中再悬浮。将细胞以2×106个细胞/孔接种到具有补充有如所指示的处于以下最终浓度的细胞因子:IL-2:100U/mL,IL-4:5ng/mL,IL-9:10ng/mL,IL-15:20ng/mL和IL-21:10ng/mL的培养基的24孔板(Corning)中以用于扩增。
通过显微镜每天监测细胞,并通过用1mL含有两倍浓度的细胞因子的新鲜培养基(即IL-2:200U/mL,IL-4:10ng/mL,IL-9:20ng/mL,IL-15:40ng/mL和IL-21:20ng/mL)更换1mL培养基每周提供三次新鲜的培养基和细胞因子。
一旦完全汇合并且细胞聚集,则根据需要,将细胞1:1分开至另外的孔和板中。21天后,使用 (eBioscience)收获细胞,计数并使用流式细胞术分析。使用扩增前后的数目计算最终的γδT细胞和Vδ1细胞数目,并且数据显示了扩增后最终的细胞得率/网格。
(eBioscience)收获细胞,计数并使用流式细胞术分析。使用扩增前后的数目计算最终的γδT细胞和Vδ1细胞数目,并且数据显示了扩增后最终的细胞得率/网格。
其他实施方式
在本说明书中提及的所有专利公开、专利和专利申请以每个独立的专利公开或专利申请具体且单独表明作为参考并入的相同程度作为参考并入本文。
尽管已结合其具体实施方式描述了本发明,但是会理解它能够进一步修饰,并且本发明申请旨在涵盖一般地,根据本发明的原理并且包括本发明所属领域内的已知或惯例实践内的与本发明公开的这些偏离所做的本发明的任何变化、使用或改变,并且可以适用于以上所述的基本特征,并处于权利要求的范围内。
其他实施方式在权利要求之内。
Claims (77)
1.一种扩增γδT细胞的方法,所述方法包括以下步骤:
(i)提供获自非造血组织的γδT细胞群体;和
(ii)在存在下列各项的情况下,以对于产生扩增的γδT细胞群体有效的量,培养所述γδT细胞至少5天:
(a)IL-2或IL-9;
(b)IL-15;和
(c)IL-21。
2.根据权利要求1所述的方法,其中步骤(ii)还包括在存在IL-4的情况下培养所述γδT细胞。
3.一种扩增γδT细胞的方法,所述方法包括以下步骤:
(i)提供获自非造血组织的γδT细胞群体;和
(ii)在存在IL-2、IL-15和选自IL-21、基质细胞来源的因子(SDF)、IL-1β、IL-12、IL-18和IL-33组成的组的因子的情况下培养所述γδT细胞至少5天以产生扩增的γδT细胞群体。
4.根据权利要求1-3中任一项所述的方法,其中步骤(ii)包括在不存在外源TCR途径激动剂的情况下培养所述γδT细胞。
5.根据权利要求1-4中任一项所述的方法,其中步骤(ii)包括在无血清培养基中培养所述γδT细胞。
6.根据权利要求1-5中任一项所述的方法,其中所述方法还包括,在步骤(i)之后,从非造血细胞分离所述γδT细胞以产生分离的γδT细胞群体,并且步骤(ii)包括:
(a)在不存在明显基质细胞接触的情况下培养所述γδT细胞;
(b)在不存在明显肿瘤细胞接触的情况下培养所述γδT细胞;和/或
(c)在不存在明显饲养细胞接触的情况下培养所述γδT细胞。
7.一种扩增γδT细胞的方法,所述方法包括以下步骤:
(i)提供非造血组织,组织包括非造血细胞和γδT细胞;
(ii)从非造血细胞分离γδT细胞以获得分离的γδT细胞群体;和
(iii)在存在IL-2、IL-15和选自IL-21、SDF、IL-1β、IL-12、IL-18和IL-33组成的组的因子的情况下培养所述γδT细胞至少5天以产生扩增的γδT细胞群体。
8.根据权利要求7所述的方法,其中所述非造血组织已获自人或非人动物受试者。
9.根据权利要求7或8所述的方法,其中步骤(ii)包括在存在IL-2、IL-15和IL-21的情况下培养所述γδT细胞。
10.根据权利要求7-9中任一项所述的方法,其中步骤(ii)包括在无血清培养基中培养所述γδT细胞。
11.根据权利要求7-10中任一项所述的方法,其中步骤(iii)包括在不存在与所述γδT细胞明显的基质细胞接触的情况下培养所述γδT细胞。
12.根据权利要求7-11中任一项所述的方法,其中步骤(iii)包括在不存在外源TCR途径激动剂的情况下培养所述γδT细胞。
13.根据权利要求6-12中任一项所述的方法,其中从非造血细胞分离所述γδT细胞的步骤包括在配置以有利于细胞从所述非造血组织流出的合成支架上培养所述γδT细胞和所述非造血细胞。
14.根据权利要求6-13中任一项所述的方法,其中从非造血细胞分离所述γδT细胞的步骤包括在存在IL-2和/或IL-15的情况下培养所述γδT细胞和所述非造血细胞。
15.根据权利要求6-14中任一项所述的方法,其中分离的淋巴细胞群体包括所述分离的γδT细胞群体,并且所述分离的γδT细胞群体包括分离的Vδ1T细胞群体。
16.根据权利要求15所述的方法,其中在扩增之前,1-10%的所述分离的淋巴细胞群体是γδT细胞。
17.根据权利要求15或16所述的方法,其中在扩增之前,1-10%的所述分离的淋巴细胞群体是Vδ1T细胞。
18.根据权利要求6-17中任一项所述的方法,其中在扩增之前,至少80%的所述分离的γδT细胞群体为Vδ1T细胞。
19.根据权利要求6-18中任一项所述的方法,其中在扩增之前,小于10%的所述分离的γδT细胞群体为Vδ2T细胞。
20.根据权利要求6-19中任一项所述的方法,其中从所述分离的γδT细胞群体除去αβT细胞和/或NK细胞。
21.根据权利要求6-20中任一项所述的方法,其中在扩增之前,所述分离的γδT细胞群体包括至少10%的CCR3+细胞,至少10%的CCR4+细胞,至少10%的CCR7+细胞,至少10%的CCR8+细胞或者至少10%的CD103+细胞。
22.根据权利要求6-21中任一项所述的方法,其中在扩增之前,相对于血液驻留的Vδ2T细胞的参比群体,所述分离的γδT细胞群体包括更高频率的CCR3+细胞、CCR4+细胞、CCR7+细胞和/或CCR8+细胞。
23.根据权利要求14-22中任一项所述的方法,其中相对于血液驻留的Vδ1T细胞的参比群体,所述分离的Vδ1T细胞群体包括更高频率的NKG2D+细胞、CD56+细胞、CD69+细胞和/或TIM3+细胞。
24.根据权利要求1-23中任一项所述的方法,其中在培养14天内,相对于扩增之前的所述分离的γδT细胞群体,所述扩增的γδT细胞群体包括至少20倍数目的γδT细胞。
25.根据权利要求1-24中任一项所述的方法,其中在培养21天内,相对于扩增之前的所述分离的γδT细胞群体,所述扩增的γδT细胞群体包括至少50倍数目的γδT细胞。
26.根据权利要求1-25中任一项所述的方法,其中所述扩增的γδT细胞群体包括扩增的Vδ1T细胞群体。
27.根据权利要求26所述的方法,其中在培养14天内,相对于扩增之前的所述分离的Vδ1T细胞群体,所述扩增的Vδ1T细胞群体包括至少20倍数目的Vδ1T细胞。
28.根据权利要求26或27所述的方法,其中在培养21天内,相对于扩增之前的所述分离的Vδ1T细胞群体,所述扩增的Vδ1T细胞群体包括至少50倍数目的Vδ1T细胞。
29.根据权利要求1-28中任一项所述的方法,其中所述扩增的γδT细胞群体表达CD27。
30.根据权利要求29所述的方法,其中所述扩增的γδT细胞群体具有比所述分离的γδT细胞群体更大的中值CD27表达。
31.根据权利要求30所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有至少2倍的中值CD27表达。
32.根据权利要求29所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有更高频率的CD27+细胞。
33.根据权利要求32所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有至少5%更高频率的CD27+细胞。
34.根据权利要求26-28中任一项所述的方法,其中所述扩增的Vδ1T细胞群体表达CD27。
35.根据权利要求34所述的方法,其中所述扩增的Vδ1T细胞群体具有比所述分离的Vδ1T细胞群体更大的平均CD27表达。
36.根据权利要求35所述的方法,其中相对于所述分离的Vδ1T细胞群体,所述扩增的Vδ1T细胞群体具有至少2倍的平均CD27表达。
37.根据权利要求34-36中任一项所述的方法,其中相对于所述分离的Vδ1T细胞群体,所述扩增的Vδ1T细胞群体具有更高频率的CD27+细胞。
38.根据权利要求37所述的方法,其中所述扩增的Vδ1T细胞群体具有相对于所述分离的Vδ1T细胞群体至少5%更高频率的CD27+细胞。
39.根据权利要求1-38中任一项所述的方法,其中所述扩增的γδT细胞群体具有比所述分离的γδT细胞群体更低的平均TIGIT表达。
40.根据权利要求39所述的方法,其中所述扩增的γδT细胞群体具有比所述分离的γδT细胞群体低至少50%的平均TIGIT表达。
41.根据权利要求39或40所述的方法,其中所述扩增的γδT细胞群体具有比所述分离的γδT细胞群体更低频率的TIGIT+细胞。
42.根据权利要求41所述的方法,其中所述扩增的γδT细胞群体具有比所述分离的γδT细胞群体至少20%更低频率的TIGIT+细胞。
43.根据权利要求26-28或者34-38中任一项所述的方法,其中所述扩增的Vδ1T细胞群体具有比所述分离的Vδ1T细胞群体更低的平均TIGIT表达。
44.根据权利要求43所述的方法,其中所述扩增的Vδ1T细胞群体具有比所述分离的Vδ1T细胞群体低至少50%的平均TIGIT表达。
45.根据权利要求26-28或者34-38中任一项所述的方法,其中所述扩增的Vδ1T细胞群体具有比所述分离的Vδ1T细胞群体更低频率的TIGIT+细胞。
46.根据权利要求45所述的方法,其中所述扩增的Vδ1T细胞群体具有比所述分离的Vδ1T细胞群体至少20%更低频率的TIGIT+细胞。
47.根据权利要求1-46中任一项所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有更高的选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组的一种或多种标志物的平均表达。
48.根据权利要求1-47中任一项所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有更高频率的表达一种或多种标志物的细胞,所述标志物选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组。
49.根据权利要求1-48中任一项所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有较低的选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组的一种或多种标志物的平均表达。
50.根据权利要求1-49中任一项所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有较低频率的表达一种或多种标志物的细胞,所述标志物选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组。
51.根据权利要求26-28、34-38、43或45中任一项所述的方法,其中相对于所述分离的Vδ1T细胞群体,所述扩增的Vδ1T细胞群体具有更高的选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组的一种或多种标志物的平均表达。
52.根据权利要求26-28、33-38、43、45或51中任一项所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有更高频率的表达一种或多种标志物的细胞,所述标志物选自CD124、CD215、CD360、CTLA4、CD1b、BTLA、CD39、CD45RA、Fas配体、CD25、ICAM-1、CD31、KLRG1、CD30和CD2组成的组。
53.根据权利要求26-28、34-38、43、45、51或52中任一项所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有较低的选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组的一种或多种标志物的平均表达。
54.根据权利要求26-28、34-38、43、45或51-53中任一项所述的方法,其中相对于所述分离的γδT细胞群体,所述扩增的γδT细胞群体具有较低频率的表达一种或多种标志物的细胞,所述标志物选自NKp44、NKp46、ICAM-2、CD70、CD28、CD103、NKp30、LAG3、CCR4、CD69、PD-1和CD64组成的组。
55.根据权利要求8-54中任一项所述的方法,其中步骤(iii)包括在不存在明显基质细胞接触的情况下培养所述γδT细胞。
56.根据权利要求8-55中任一项所述的方法,其中步骤(iii)包括在不存在明显饲养细胞接触的情况下培养所述γδT细胞。
57.根据权利要求8-56中任一项所述的方法,其中步骤(iii)包括在不存在明显肿瘤细胞接触的情况下培养所述γδT细胞。
58.根据权利要求1-57中任一项所述的方法,其中所述非造血组织不是肿瘤组织。
59.根据权利要求1-58中任一项所述的方法,其中所述非造血组织是皮肤。
60.一种通过根据权利要求1-59中任一项所述的方法获得的扩增的γδT细胞。
61.一种分离的γδT细胞群体,其中至少50%的所述分离群体的γδT细胞表达CD27,但不显著表达TIGIT。
62.根据权利要求61所述的分离的γδT细胞群体,其中至少50%的所述分离群体的γδT细胞表达Vδ1。
63.一种包含根据权利要求60所述的扩增的γδT细胞或者根据权利要求61或62所述的分离的γδT细胞群体的药物组合物。
64.根据权利要求63所述的药物组合物,用于在受试者中癌症或感染的治疗方法中使用。
65.根据权利要求63所述的药物组合物在生产用于治疗受试者中癌症或感染的药物中的应用。
66.一种通过过继性T细胞疗法治疗受试者的方法,其中所述方法包括向对其有需要的受试者施用治疗有效量的通过根据权利要求1-59中任一项所述的方法所获得的扩增的γδT细胞,根据权利要求60所述的扩增的γδT细胞,根据权利要求61或62所述的分离群体或者根据权利要求63所述的药物组合物。
67.根据权利要求66所述的方法,其中所述扩增的γδT细胞的治疗有效量为小于10×1012个细胞每剂量。
68.根据权利要求66或67所述的方法,还包括向对其有需要的受试者施用一种或多种其他治疗剂。
69.根据权利要求68所述的方法,其中所述一种或多种其他治疗剂选自免疫治疗剂、细胞毒性剂、生长抑制剂、放射疗法剂、抗血管生成剂及其组合组成的组。
70.根据权利要求68或69所述的方法,其中将所述一种或多种其他治疗剂与所述扩增的γδT细胞同时施用。
71.根据权利要求68或69所述的方法,其中在所述扩增的γδT细胞施用之后施用所述一种或多种其他治疗剂。
72.根据权利要求68-71中任一项所述的方法,其中所述其他治疗剂为免疫治疗剂。
73.一种通过过继性T细胞疗法治疗受试者的方法,其中所述方法包括向对其有需要的受试者施用治疗有效量的根据权利要求63所述的药物组合物。
74.根据权利要求66-73中任一项所述的方法,其中所述受试者为人。
75.根据权利要求74所述的方法,其中所述人是人癌症患者。
76.根据权利要求75所述的方法,其中所述人癌症患者正在治疗实体瘤。
77.根据权利要求76所述的方法,其中所述人是正在治疗病毒感染的人患者。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| GBGB1707048.3A GB201707048D0 (en) | 2017-05-03 | 2017-05-03 | Expansion of gamma delta cells, compositions, and methods of use thereof |
| GB1707048.3 | 2017-05-03 | ||
| PCT/EP2018/061413 WO2018202808A2 (en) | 2017-05-03 | 2018-05-03 | Expansion of gamma delta t cells, compositions, and methods of use thereof |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN110914410A true CN110914410A (zh) | 2020-03-24 |
Family
ID=59010925
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201880044728.0A Pending CN110914410A (zh) | 2017-05-03 | 2018-05-03 | γδT细胞扩增、组合物及其使用方法 |
Country Status (14)
| Country | Link |
|---|---|
| US (2) | US11655453B2 (zh) |
| EP (1) | EP3619298A2 (zh) |
| JP (2) | JP2020518289A (zh) |
| KR (2) | KR102812405B1 (zh) |
| CN (1) | CN110914410A (zh) |
| AU (1) | AU2018262698B2 (zh) |
| BR (1) | BR112019023076A2 (zh) |
| CA (1) | CA3062386A1 (zh) |
| EA (1) | EA201992610A1 (zh) |
| GB (1) | GB201707048D0 (zh) |
| IL (3) | IL312633B1 (zh) |
| MX (2) | MX2019013001A (zh) |
| PH (1) | PH12019502485A1 (zh) |
| WO (1) | WO2018202808A2 (zh) |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN109517793A (zh) * | 2018-11-30 | 2019-03-26 | 广州长峰生物技术有限公司 | 一种NK细胞和γδT细胞共培养的建立方法 |
| CN113234675A (zh) * | 2021-03-31 | 2021-08-10 | 广西医科大学 | 一种γδT细胞以及采用体外制备γδT细胞的方法 |
| CN115667501A (zh) * | 2020-05-12 | 2023-01-31 | 伽玛德治疗有限公司 | 用于分离γδT细胞的方法 |
Families Citing this family (18)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20060269973A1 (en) | 2004-11-24 | 2006-11-30 | Cassian Yee | Methods of using IL-21 for adoptive immunotherapy and identification of tumor antigens |
| GB201804701D0 (en) | 2018-03-23 | 2018-05-09 | Gammadelta Therapeutics Ltd | Lymphocytes expressing heterologous targeting constructs |
| WO2020095058A1 (en) * | 2018-11-08 | 2020-05-14 | GammaDelta Therapeutics Limited | Methods for isolating and expanding cells |
| GB201818243D0 (en) * | 2018-11-08 | 2018-12-26 | Gammadelta Therapeutics Ltd | Methods for isolating and expanding cells |
| GB201909144D0 (en) * | 2019-06-25 | 2019-08-07 | Autolus Ltd | Culture medium |
| MX2022005033A (es) | 2019-10-31 | 2022-09-09 | Morphosys Ag | Terapia combinada antitumoral que comprende un anticuerpo anti grupo de diferenciación 19 (cd19) y linfocitos t gamma delta. |
| JP2023504185A (ja) * | 2019-12-03 | 2023-02-01 | アディセット バイオ, インコーポレイテッド | γδT細胞集団を多価薬剤で増殖させるための方法及びその組成物 |
| US20210290674A1 (en) * | 2020-03-20 | 2021-09-23 | GammaDelta Therapeutics Ltd. | Methods for treating myeloid malignancies |
| MX2022011646A (es) | 2020-03-20 | 2022-10-13 | Gammadelta Therapeutics Ltd | Celulas t v delta1+ para el tratamiento de neoplasias mieloides. |
| GB202105113D0 (en) | 2021-04-09 | 2021-05-26 | Gammadelta Therapeutics Ltd | Novel method |
| US20240197876A1 (en) * | 2021-04-14 | 2024-06-20 | The Trustees Of The University Of Pennsylvania | Methods to determine treatment efficacy with gamma-delta t cells |
| IL307757A (en) * | 2021-04-16 | 2023-12-01 | Acepodia Biotechnologies Ltd | Novel compositions approved in gamma delta T cells, methods of preparation, and uses thereof |
| WO2023069322A1 (en) * | 2021-10-20 | 2023-04-27 | University Of Pittsburgh – Of The Commonwealth System Of Higher Education | Methods and materials for expanding tumor infiltrating gamma-delta t cells |
| KR20240130130A (ko) * | 2022-01-05 | 2024-08-28 | 인히브릭스 바이오사이언스, 인크. | 감마 델타 t 세포 결합 폴리펩티드 및 그의 용도 |
| WO2024193459A1 (en) * | 2023-03-17 | 2024-09-26 | Nanjing Legend Biotech Co., Ltd. | METHODS OF CULTURING Vδ1 T CELLS |
| WO2024209429A1 (en) | 2023-04-07 | 2024-10-10 | Takeda Pharmaceutical Company Limited | T cells for use in treating relapsed or refractory acute myeloid leukemia |
| WO2025017531A1 (en) | 2023-07-20 | 2025-01-23 | Takeda Pharmaceutical Company Limited | Grid-free methods for making gamma delta t cells |
| WO2025022361A1 (en) * | 2023-07-26 | 2025-01-30 | Janssen Biotech, Inc. | Materials and methods for cell growth |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20160175358A1 (en) * | 2014-11-17 | 2016-06-23 | Adicet Bio, Inc. | Engineered gamma delta t-cells |
| WO2016164705A1 (en) * | 2015-04-10 | 2016-10-13 | Omar Abdel-Rahman Ali | Immune cell trapping devices and methods for making and using the same |
| WO2016198480A1 (en) * | 2015-06-09 | 2016-12-15 | Lymphact - Lymphocyte Activation Technologies, S.A. | Methods for the production of tcr gamma delta+ t cells |
| CN106574244A (zh) * | 2014-06-11 | 2017-04-19 | 保利比奥斯博特有限公司 | 用细胞因子组合物扩增淋巴细胞用于主动细胞免疫治疗 |
Family Cites Families (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2000026347A1 (en) | 1998-11-04 | 2000-05-11 | Hemosol Inc. | Methods for the production of tcr gamma delta t cells |
| CN103635573B (zh) * | 2011-05-19 | 2016-05-11 | 分子医学研究所 | 包含γδT细胞的淋巴细胞系、其组合物及生产方法 |
| ES2539845B1 (es) | 2013-08-05 | 2016-04-13 | Universidad De Alicante | Procedimiento y síntesis de monolitos de carbón activo a partir de cascarilla de cacao |
| GB201506423D0 (en) | 2015-04-15 | 2015-05-27 | Tc Biopharm Ltd | Gamma delta T cells and uses thereof |
| US10829735B2 (en) | 2015-07-21 | 2020-11-10 | The Trustees Of The University Of Pennsylvania | Methods for improving the efficacy and expansion of immune cells |
| AU2016344779B2 (en) | 2015-10-30 | 2023-01-05 | Cancer Research Technology Limited | Expansion of non-haematopoietic tissue-resident gamma delta T cells and uses of these cells |
-
2017
- 2017-05-03 GB GBGB1707048.3A patent/GB201707048D0/en not_active Ceased
-
2018
- 2018-05-03 KR KR1020197035686A patent/KR102812405B1/ko active Active
- 2018-05-03 IL IL312633A patent/IL312633B1/en unknown
- 2018-05-03 IL IL319114A patent/IL319114A/en unknown
- 2018-05-03 MX MX2019013001A patent/MX2019013001A/es unknown
- 2018-05-03 WO PCT/EP2018/061413 patent/WO2018202808A2/en unknown
- 2018-05-03 JP JP2020512082A patent/JP2020518289A/ja active Pending
- 2018-05-03 AU AU2018262698A patent/AU2018262698B2/en active Active
- 2018-05-03 US US16/610,398 patent/US11655453B2/en active Active
- 2018-05-03 BR BR112019023076A patent/BR112019023076A2/pt unknown
- 2018-05-03 CA CA3062386A patent/CA3062386A1/en active Pending
- 2018-05-03 EA EA201992610A patent/EA201992610A1/ru unknown
- 2018-05-03 CN CN201880044728.0A patent/CN110914410A/zh active Pending
- 2018-05-03 EP EP18725128.5A patent/EP3619298A2/en active Pending
- 2018-05-03 IL IL270365A patent/IL270365B2/en unknown
- 2018-05-03 KR KR1020257016628A patent/KR20250080907A/ko active Pending
-
2019
- 2019-10-31 MX MX2024004897A patent/MX2024004897A/es unknown
- 2019-11-04 PH PH12019502485A patent/PH12019502485A1/en unknown
-
2023
- 2023-01-12 US US18/153,900 patent/US20230242878A1/en active Pending
- 2023-02-17 JP JP2023022974A patent/JP7698672B2/ja active Active
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN106574244A (zh) * | 2014-06-11 | 2017-04-19 | 保利比奥斯博特有限公司 | 用细胞因子组合物扩增淋巴细胞用于主动细胞免疫治疗 |
| US20160175358A1 (en) * | 2014-11-17 | 2016-06-23 | Adicet Bio, Inc. | Engineered gamma delta t-cells |
| WO2016164705A1 (en) * | 2015-04-10 | 2016-10-13 | Omar Abdel-Rahman Ali | Immune cell trapping devices and methods for making and using the same |
| WO2016198480A1 (en) * | 2015-06-09 | 2016-12-15 | Lymphact - Lymphocyte Activation Technologies, S.A. | Methods for the production of tcr gamma delta+ t cells |
Non-Patent Citations (1)
| Title |
|---|
| RACHAEL A CLARK 等: "A Novel Method for the Isolation of Skin Resident T Cells from Normal and Diseased Human Skin.", 《J INVEST DERMATOL.》 * |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN109517793A (zh) * | 2018-11-30 | 2019-03-26 | 广州长峰生物技术有限公司 | 一种NK细胞和γδT细胞共培养的建立方法 |
| CN115667501A (zh) * | 2020-05-12 | 2023-01-31 | 伽玛德治疗有限公司 | 用于分离γδT细胞的方法 |
| CN113234675A (zh) * | 2021-03-31 | 2021-08-10 | 广西医科大学 | 一种γδT细胞以及采用体外制备γδT细胞的方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| MX2024004897A (es) | 2024-05-07 |
| IL319114A (en) | 2025-04-01 |
| IL270365B1 (en) | 2024-06-01 |
| AU2018262698B2 (en) | 2025-07-03 |
| WO2018202808A9 (en) | 2019-03-07 |
| US11655453B2 (en) | 2023-05-23 |
| WO2018202808A3 (en) | 2019-01-03 |
| GB201707048D0 (en) | 2017-06-14 |
| PH12019502485A1 (en) | 2021-01-04 |
| KR102812405B1 (ko) | 2025-05-23 |
| KR20250080907A (ko) | 2025-06-05 |
| IL270365B2 (en) | 2024-10-01 |
| BR112019023076A2 (pt) | 2020-06-09 |
| MX2019013001A (es) | 2021-01-20 |
| IL312633A (en) | 2024-07-01 |
| JP7698672B2 (ja) | 2025-06-25 |
| JP2020518289A (ja) | 2020-06-25 |
| IL270365A (zh) | 2019-12-31 |
| CA3062386A1 (en) | 2018-11-08 |
| EA201992610A1 (ru) | 2020-04-23 |
| IL312633B1 (en) | 2025-04-01 |
| AU2018262698A1 (en) | 2019-12-12 |
| KR20200024770A (ko) | 2020-03-09 |
| EP3619298A2 (en) | 2020-03-11 |
| WO2018202808A2 (en) | 2018-11-08 |
| JP2023058687A (ja) | 2023-04-25 |
| US20210087528A1 (en) | 2021-03-25 |
| US20230242878A1 (en) | 2023-08-03 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7698672B2 (ja) | ガンマデルタt細胞の増幅、組成物およびその使用 | |
| JP7451468B2 (ja) | 非造血組織常在性γδ T細胞の増幅および該細胞の使用 | |
| RU2784566C2 (ru) | РАЗМНОЖЕНИЕ И ПРИМЕНЕНИЕ НЕГЕМОПОЭТИЧЕСКИХ ТКАНЕРЕЗИДЕНТНЫХ γδ Т-КЛЕТОК | |
| BR122025001918A2 (pt) | Métodos de expansão de células t gama delta, composições, seu uso e uso de células t gama delta expandidas | |
| EA044224B1 (ru) | ЭКСПАНСИЯ γδ-T-КЛЕТОК, КОМПОЗИЦИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | |
| EA049808B1 (ru) | ЭКСПАНСИЯ γΔ-Т-КЛЕТОК, КОМПОЗИЦИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | |
| HK1253954B (zh) | 非造血组织驻留γδ T细胞的扩增及这些细胞的用途 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |