CN101646772A - 抗伤寒沙门氏菌和其它肠细菌病原体的基于番木瓜花叶病毒的疫苗 - Google Patents
抗伤寒沙门氏菌和其它肠细菌病原体的基于番木瓜花叶病毒的疫苗 Download PDFInfo
- Publication number
- CN101646772A CN101646772A CN200880010037A CN200880010037A CN101646772A CN 101646772 A CN101646772 A CN 101646772A CN 200880010037 A CN200880010037 A CN 200880010037A CN 200880010037 A CN200880010037 A CN 200880010037A CN 101646772 A CN101646772 A CN 101646772A
- Authority
- CN
- China
- Prior art keywords
- papmv
- vlp
- antigen
- ompc
- coat protein
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 241000709995 Papaya mosaic virus Species 0.000 title claims abstract description 396
- 229960005486 vaccine Drugs 0.000 title claims abstract description 123
- 241000305071 Enterobacterales Species 0.000 title claims abstract description 65
- 241000293871 Salmonella enterica subsp. enterica serovar Typhi Species 0.000 title claims description 104
- 244000052769 pathogen Species 0.000 title description 11
- 108091007433 antigens Proteins 0.000 claims abstract description 273
- 102000036639 antigens Human genes 0.000 claims abstract description 272
- 239000000427 antigen Substances 0.000 claims abstract description 270
- 101710132601 Capsid protein Proteins 0.000 claims abstract description 210
- 101710094648 Coat protein Proteins 0.000 claims abstract description 209
- 101710125418 Major capsid protein Proteins 0.000 claims abstract description 209
- 101710141454 Nucleoprotein Proteins 0.000 claims abstract description 209
- 101710083689 Probable capsid protein Proteins 0.000 claims abstract description 209
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 claims abstract description 203
- 230000004927 fusion Effects 0.000 claims abstract description 18
- 108700028353 OmpC Proteins 0.000 claims description 209
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 197
- 108700006385 OmpF Proteins 0.000 claims description 124
- 238000000034 method Methods 0.000 claims description 103
- 238000002360 preparation method Methods 0.000 claims description 77
- 239000000203 mixture Substances 0.000 claims description 68
- 108700030444 papaya mosiac virus coat Proteins 0.000 claims description 62
- 241001465754 Metazoa Species 0.000 claims description 61
- 108010013381 Porins Proteins 0.000 claims description 60
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 48
- 239000012634 fragment Substances 0.000 claims description 45
- 230000004044 response Effects 0.000 claims description 43
- 230000000890 antigenic effect Effects 0.000 claims description 41
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 39
- PXFBZOLANLWPMH-UHFFFAOYSA-N 16-Epiaffinine Natural products C1C(C2=CC=CC=C2N2)=C2C(=O)CC2C(=CC)CN(C)C1C2CO PXFBZOLANLWPMH-UHFFFAOYSA-N 0.000 claims description 28
- 241001467018 Typhis Species 0.000 claims description 18
- 230000001939 inductive effect Effects 0.000 claims description 14
- 229920001184 polypeptide Polymers 0.000 claims description 14
- 230000004048 modification Effects 0.000 claims description 13
- 238000012986 modification Methods 0.000 claims description 13
- 101100518271 Salmonella typhi ompC gene Proteins 0.000 claims description 12
- 101100350286 Salmonella typhi ompF gene Proteins 0.000 claims description 8
- 108091033319 polynucleotide Proteins 0.000 claims description 5
- 102000040430 polynucleotide Human genes 0.000 claims description 5
- 239000002157 polynucleotide Substances 0.000 claims description 5
- 241000894007 species Species 0.000 claims description 5
- 102000007739 porin activity proteins Human genes 0.000 claims 2
- 101150010487 are gene Proteins 0.000 claims 1
- 241000700605 Viruses Species 0.000 abstract description 54
- 201000010099 disease Diseases 0.000 abstract description 29
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 abstract description 29
- 201000008297 typhoid fever Diseases 0.000 abstract description 16
- 208000037386 Typhoid Diseases 0.000 abstract description 15
- 239000002245 particle Substances 0.000 abstract description 14
- 230000027455 binding Effects 0.000 abstract description 11
- 230000028993 immune response Effects 0.000 abstract description 8
- 230000002068 genetic effect Effects 0.000 abstract description 3
- 230000021615 conjugation Effects 0.000 abstract 1
- 230000003389 potentiating effect Effects 0.000 abstract 1
- 210000004027 cell Anatomy 0.000 description 99
- 235000001014 amino acid Nutrition 0.000 description 93
- 229940024606 amino acid Drugs 0.000 description 92
- 150000001413 amino acids Chemical class 0.000 description 89
- 241000699666 Mus <mouse, genus> Species 0.000 description 78
- 241000894006 Bacteria Species 0.000 description 75
- 230000036039 immunity Effects 0.000 description 64
- 108090000623 proteins and genes Proteins 0.000 description 63
- 238000012360 testing method Methods 0.000 description 62
- 102000017033 Porins Human genes 0.000 description 58
- 125000003295 alanine group Chemical group N[C@@H](C)C(=O)* 0.000 description 56
- 102000004169 proteins and genes Human genes 0.000 description 55
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 50
- 241000607142 Salmonella Species 0.000 description 50
- 210000001744 T-lymphocyte Anatomy 0.000 description 50
- 235000018102 proteins Nutrition 0.000 description 46
- 208000015181 infectious disease Diseases 0.000 description 44
- 239000002671 adjuvant Substances 0.000 description 42
- 230000000968 intestinal effect Effects 0.000 description 40
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 35
- 206010022000 influenza Diseases 0.000 description 35
- 230000008521 reorganization Effects 0.000 description 35
- 239000002953 phosphate buffered saline Substances 0.000 description 33
- 239000000523 sample Substances 0.000 description 33
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 32
- 239000002158 endotoxin Substances 0.000 description 29
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 28
- 239000002773 nucleotide Substances 0.000 description 28
- 125000003729 nucleotide group Chemical group 0.000 description 28
- ROHFNLRQFUQHCH-YFKPBYRVSA-N L-leucine Chemical compound CC(C)C[C@H](N)C(O)=O ROHFNLRQFUQHCH-YFKPBYRVSA-N 0.000 description 27
- COLNVLDHVKWLRT-QMMMGPOBSA-N L-phenylalanine Chemical compound OC(=O)[C@@H](N)CC1=CC=CC=C1 COLNVLDHVKWLRT-QMMMGPOBSA-N 0.000 description 26
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 25
- AGPKZVBTJJNPAG-WHFBIAKZSA-N L-isoleucine Chemical compound CC[C@H](C)[C@H](N)C(O)=O AGPKZVBTJJNPAG-WHFBIAKZSA-N 0.000 description 25
- 230000001681 protective effect Effects 0.000 description 25
- 241000588921 Enterobacteriaceae Species 0.000 description 24
- 239000003795 chemical substances by application Substances 0.000 description 24
- 229920006008 lipopolysaccharide Polymers 0.000 description 24
- 150000007523 nucleic acids Chemical class 0.000 description 24
- 238000002965 ELISA Methods 0.000 description 23
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 22
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 22
- 102220539644 Piwi-like protein 1_K97V_mutation Human genes 0.000 description 22
- DCXYFEDJOCDNAF-REOHCLBHSA-N L-asparagine Chemical compound OC(=O)[C@@H](N)CC(N)=O DCXYFEDJOCDNAF-REOHCLBHSA-N 0.000 description 21
- 230000001580 bacterial effect Effects 0.000 description 21
- 238000002983 circular dichroism Methods 0.000 description 20
- 210000004443 dendritic cell Anatomy 0.000 description 20
- 238000007912 intraperitoneal administration Methods 0.000 description 20
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 20
- 239000000243 solution Substances 0.000 description 20
- 101150013553 CD40 gene Proteins 0.000 description 19
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 19
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 19
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 19
- 230000001717 pathogenic effect Effects 0.000 description 19
- 210000002966 serum Anatomy 0.000 description 19
- 239000006228 supernatant Substances 0.000 description 19
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 18
- 241000699670 Mus sp. Species 0.000 description 18
- 230000008878 coupling Effects 0.000 description 18
- 238000010168 coupling process Methods 0.000 description 18
- 238000005859 coupling reaction Methods 0.000 description 18
- 230000003053 immunization Effects 0.000 description 18
- 238000002649 immunization Methods 0.000 description 18
- 239000003550 marker Substances 0.000 description 18
- CKLJMWTZIZZHCS-REOHCLBHSA-N L-aspartic acid Chemical compound OC(=O)[C@@H](N)CC(O)=O CKLJMWTZIZZHCS-REOHCLBHSA-N 0.000 description 17
- 230000003321 amplification Effects 0.000 description 17
- 230000008859 change Effects 0.000 description 17
- 238000013016 damping Methods 0.000 description 17
- 238000003199 nucleic acid amplification method Methods 0.000 description 17
- 108020004707 nucleic acids Proteins 0.000 description 17
- 102000039446 nucleic acids Human genes 0.000 description 17
- 108010058846 Ovalbumin Proteins 0.000 description 16
- 238000004458 analytical method Methods 0.000 description 16
- 239000012530 fluid Substances 0.000 description 16
- 229940092253 ovalbumin Drugs 0.000 description 16
- 238000001556 precipitation Methods 0.000 description 16
- 230000009182 swimming Effects 0.000 description 16
- 241000588747 Klebsiella pneumoniae Species 0.000 description 15
- 230000000694 effects Effects 0.000 description 15
- 239000000463 material Substances 0.000 description 15
- 239000002609 medium Substances 0.000 description 15
- 229940031348 multivalent vaccine Drugs 0.000 description 15
- 238000011282 treatment Methods 0.000 description 15
- 108010088729 HLA-A*02:01 antigen Proteins 0.000 description 14
- 241000607768 Shigella Species 0.000 description 14
- 239000002253 acid Substances 0.000 description 14
- -1 albumen Proteins 0.000 description 14
- 210000003719 b-lymphocyte Anatomy 0.000 description 14
- 238000006243 chemical reaction Methods 0.000 description 14
- 238000010561 standard procedure Methods 0.000 description 14
- 241000712461 unidentified influenza virus Species 0.000 description 14
- 108091028043 Nucleic acid sequence Proteins 0.000 description 13
- 230000005875 antibody response Effects 0.000 description 13
- 238000010790 dilution Methods 0.000 description 13
- 239000012895 dilution Substances 0.000 description 13
- 238000005516 engineering process Methods 0.000 description 13
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 13
- 210000005105 peripheral blood lymphocyte Anatomy 0.000 description 13
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 12
- 210000001185 bone marrow Anatomy 0.000 description 12
- 230000008569 process Effects 0.000 description 12
- 238000011160 research Methods 0.000 description 12
- 239000000725 suspension Substances 0.000 description 12
- 238000005199 ultracentrifugation Methods 0.000 description 12
- 230000003612 virological effect Effects 0.000 description 12
- 108010075704 HLA-A Antigens Proteins 0.000 description 11
- 102000011786 HLA-A Antigens Human genes 0.000 description 11
- 108091034117 Oligonucleotide Proteins 0.000 description 11
- 241000283973 Oryctolagus cuniculus Species 0.000 description 11
- 238000004140 cleaning Methods 0.000 description 11
- 108010021083 hen egg lysozyme Proteins 0.000 description 11
- 230000001965 increasing effect Effects 0.000 description 11
- 238000002347 injection Methods 0.000 description 11
- 239000007924 injection Substances 0.000 description 11
- 238000011081 inoculation Methods 0.000 description 11
- 239000002243 precursor Substances 0.000 description 11
- 238000000746 purification Methods 0.000 description 11
- 238000011725 BALB/c mouse Methods 0.000 description 10
- 241000588914 Enterobacter Species 0.000 description 10
- 241000588748 Klebsiella Species 0.000 description 10
- 108700026244 Open Reading Frames Proteins 0.000 description 10
- 241000607734 Yersinia <bacteria> Species 0.000 description 10
- 230000000240 adjuvant effect Effects 0.000 description 10
- 239000003153 chemical reaction reagent Substances 0.000 description 10
- 230000008034 disappearance Effects 0.000 description 10
- 239000013604 expression vector Substances 0.000 description 10
- 150000002460 imidazoles Chemical class 0.000 description 10
- 230000001976 improved effect Effects 0.000 description 10
- 238000001338 self-assembly Methods 0.000 description 10
- 230000009466 transformation Effects 0.000 description 10
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 9
- 108020004705 Codon Proteins 0.000 description 9
- 241000196324 Embryophyta Species 0.000 description 9
- 241000293869 Salmonella enterica subsp. enterica serovar Typhimurium Species 0.000 description 9
- 210000004369 blood Anatomy 0.000 description 9
- 239000008280 blood Substances 0.000 description 9
- 238000012239 gene modification Methods 0.000 description 9
- 230000005017 genetic modification Effects 0.000 description 9
- 235000013617 genetically modified food Nutrition 0.000 description 9
- 210000004698 lymphocyte Anatomy 0.000 description 9
- 238000001228 spectrum Methods 0.000 description 9
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 8
- 241000588724 Escherichia coli Species 0.000 description 8
- 102100037850 Interferon gamma Human genes 0.000 description 8
- 108010074328 Interferon-gamma Proteins 0.000 description 8
- 241000588769 Proteus <enterobacteria> Species 0.000 description 8
- 206010040047 Sepsis Diseases 0.000 description 8
- 241000282898 Sus scrofa Species 0.000 description 8
- 238000011156 evaluation Methods 0.000 description 8
- 239000000499 gel Substances 0.000 description 8
- 230000005847 immunogenicity Effects 0.000 description 8
- 238000007789 sealing Methods 0.000 description 8
- 230000028327 secretion Effects 0.000 description 8
- 239000000126 substance Substances 0.000 description 8
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 8
- 206010012735 Diarrhoea Diseases 0.000 description 7
- 102100025137 Early activation antigen CD69 Human genes 0.000 description 7
- 101000934374 Homo sapiens Early activation antigen CD69 Proteins 0.000 description 7
- 241000607762 Shigella flexneri Species 0.000 description 7
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 7
- 239000000284 extract Substances 0.000 description 7
- 230000001900 immune effect Effects 0.000 description 7
- 210000000987 immune system Anatomy 0.000 description 7
- 230000002688 persistence Effects 0.000 description 7
- 239000000047 product Substances 0.000 description 7
- 230000002441 reversible effect Effects 0.000 description 7
- 102220023256 rs387907547 Human genes 0.000 description 7
- 210000000952 spleen Anatomy 0.000 description 7
- HNDVDQJCIGZPNO-YFKPBYRVSA-N L-histidine Chemical compound OC(=O)[C@@H](N)CC1=CN=CN1 HNDVDQJCIGZPNO-YFKPBYRVSA-N 0.000 description 6
- 241000712899 Lymphocytic choriomeningitis mammarenavirus Species 0.000 description 6
- 101710116435 Outer membrane protein Proteins 0.000 description 6
- 241000009328 Perro Species 0.000 description 6
- 229920001213 Polysorbate 20 Polymers 0.000 description 6
- 241000288906 Primates Species 0.000 description 6
- 241000607132 Salmonella enterica subsp. enterica serovar Gallinarum Species 0.000 description 6
- 241000607479 Yersinia pestis Species 0.000 description 6
- 238000003556 assay Methods 0.000 description 6
- 230000001419 dependent effect Effects 0.000 description 6
- 238000006073 displacement reaction Methods 0.000 description 6
- 239000003814 drug Substances 0.000 description 6
- 150000002148 esters Chemical class 0.000 description 6
- 238000000684 flow cytometry Methods 0.000 description 6
- 235000011194 food seasoning agent Nutrition 0.000 description 6
- 102000037865 fusion proteins Human genes 0.000 description 6
- 108020001507 fusion proteins Proteins 0.000 description 6
- HNDVDQJCIGZPNO-UHFFFAOYSA-N histidine Natural products OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 6
- 230000002163 immunogen Effects 0.000 description 6
- 239000007788 liquid Substances 0.000 description 6
- 239000006166 lysate Substances 0.000 description 6
- 238000004519 manufacturing process Methods 0.000 description 6
- 239000003921 oil Substances 0.000 description 6
- 235000019198 oils Nutrition 0.000 description 6
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 6
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 6
- 238000012545 processing Methods 0.000 description 6
- 239000002516 radical scavenger Substances 0.000 description 6
- 230000001105 regulatory effect Effects 0.000 description 6
- 230000003248 secreting effect Effects 0.000 description 6
- 238000000926 separation method Methods 0.000 description 6
- 208000013223 septicemia Diseases 0.000 description 6
- 238000006467 substitution reaction Methods 0.000 description 6
- 230000004083 survival effect Effects 0.000 description 6
- 241000193830 Bacillus <bacterium> Species 0.000 description 5
- 101150083464 CP gene Proteins 0.000 description 5
- 241000283707 Capra Species 0.000 description 5
- 241000148131 Colibacter Species 0.000 description 5
- 108020004414 DNA Proteins 0.000 description 5
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 5
- 239000004278 EU approved seasoning Substances 0.000 description 5
- 108010067770 Endopeptidase K Proteins 0.000 description 5
- 241000588722 Escherichia Species 0.000 description 5
- 241000282326 Felis catus Species 0.000 description 5
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 5
- 241001494479 Pecora Species 0.000 description 5
- 241000392514 Salmonella enterica subsp. enterica serovar Dublin Species 0.000 description 5
- 229930006000 Sucrose Natural products 0.000 description 5
- CZMRCDWAGMRECN-UGDNZRGBSA-N Sucrose Chemical compound O[C@H]1[C@H](O)[C@@H](CO)O[C@@]1(CO)O[C@@H]1[C@H](O)[C@@H](O)[C@H](O)[C@@H](CO)O1 CZMRCDWAGMRECN-UGDNZRGBSA-N 0.000 description 5
- 239000007983 Tris buffer Substances 0.000 description 5
- 241000607447 Yersinia enterocolitica Species 0.000 description 5
- 125000000539 amino acid group Chemical group 0.000 description 5
- 230000002238 attenuated effect Effects 0.000 description 5
- 230000015572 biosynthetic process Effects 0.000 description 5
- 229910052799 carbon Inorganic materials 0.000 description 5
- 230000015556 catabolic process Effects 0.000 description 5
- 150000001875 compounds Chemical class 0.000 description 5
- 239000007859 condensation product Substances 0.000 description 5
- 238000006731 degradation reaction Methods 0.000 description 5
- 239000003085 diluting agent Substances 0.000 description 5
- 238000004043 dyeing Methods 0.000 description 5
- 238000010828 elution Methods 0.000 description 5
- 235000013305 food Nutrition 0.000 description 5
- 108010037896 heparin-binding hemagglutinin Proteins 0.000 description 5
- 230000003308 immunostimulating effect Effects 0.000 description 5
- 238000001727 in vivo Methods 0.000 description 5
- 230000008595 infiltration Effects 0.000 description 5
- 238000001764 infiltration Methods 0.000 description 5
- 150000002632 lipids Chemical class 0.000 description 5
- 201000001441 melanoma Diseases 0.000 description 5
- 238000012544 monitoring process Methods 0.000 description 5
- 230000007935 neutral effect Effects 0.000 description 5
- 210000000056 organ Anatomy 0.000 description 5
- 102000013415 peroxidase activity proteins Human genes 0.000 description 5
- 108040007629 peroxidase activity proteins Proteins 0.000 description 5
- 235000021110 pickles Nutrition 0.000 description 5
- 239000013612 plasmid Substances 0.000 description 5
- 239000013641 positive control Substances 0.000 description 5
- 239000000843 powder Substances 0.000 description 5
- 238000010010 raising Methods 0.000 description 5
- 230000009257 reactivity Effects 0.000 description 5
- UCSJYZPVAKXKNQ-HZYVHMACSA-N streptomycin Chemical compound CN[C@H]1[C@H](O)[C@@H](O)[C@H](CO)O[C@H]1O[C@@H]1[C@](C=O)(O)[C@H](C)O[C@H]1O[C@@H]1[C@@H](NC(N)=N)[C@H](O)[C@@H](NC(N)=N)[C@H](O)[C@H]1O UCSJYZPVAKXKNQ-HZYVHMACSA-N 0.000 description 5
- 238000010254 subcutaneous injection Methods 0.000 description 5
- 239000007929 subcutaneous injection Substances 0.000 description 5
- 239000005720 sucrose Substances 0.000 description 5
- 210000001519 tissue Anatomy 0.000 description 5
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 5
- 238000002211 ultraviolet spectrum Methods 0.000 description 5
- 241001515965 unidentified phage Species 0.000 description 5
- 229940098232 yersinia enterocolitica Drugs 0.000 description 5
- 239000000120 Artificial Saliva Substances 0.000 description 4
- 241000283724 Bison bonasus Species 0.000 description 4
- 241000283690 Bos taurus Species 0.000 description 4
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 4
- 235000009467 Carica papaya Nutrition 0.000 description 4
- HEDRZPFGACZZDS-UHFFFAOYSA-N Chloroform Chemical compound ClC(Cl)Cl HEDRZPFGACZZDS-UHFFFAOYSA-N 0.000 description 4
- 102000004127 Cytokines Human genes 0.000 description 4
- 108090000695 Cytokines Proteins 0.000 description 4
- FBPFZTCFMRRESA-JGWLITMVSA-N D-glucitol Chemical compound OC[C@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-JGWLITMVSA-N 0.000 description 4
- 208000004232 Enteritis Diseases 0.000 description 4
- 102000004190 Enzymes Human genes 0.000 description 4
- 108090000790 Enzymes Proteins 0.000 description 4
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 4
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 4
- 241000700721 Hepatitis B virus Species 0.000 description 4
- 241000238631 Hexapoda Species 0.000 description 4
- 241000725303 Human immunodeficiency virus Species 0.000 description 4
- 241000701806 Human papillomavirus Species 0.000 description 4
- 206010028980 Neoplasm Diseases 0.000 description 4
- 241001503436 Plasmodiophora brassicae Species 0.000 description 4
- 206010035664 Pneumonia Diseases 0.000 description 4
- 241000709992 Potato virus X Species 0.000 description 4
- 102000004245 Proteasome Endopeptidase Complex Human genes 0.000 description 4
- 108090000708 Proteasome Endopeptidase Complex Proteins 0.000 description 4
- 241001112090 Pseudovirus Species 0.000 description 4
- 241001138501 Salmonella enterica Species 0.000 description 4
- 230000004913 activation Effects 0.000 description 4
- 238000013459 approach Methods 0.000 description 4
- 239000000872 buffer Substances 0.000 description 4
- 241001233037 catfish Species 0.000 description 4
- 238000004113 cell culture Methods 0.000 description 4
- 238000001142 circular dichroism spectrum Methods 0.000 description 4
- 238000003501 co-culture Methods 0.000 description 4
- 238000000576 coating method Methods 0.000 description 4
- 238000012258 culturing Methods 0.000 description 4
- 230000004069 differentiation Effects 0.000 description 4
- 239000006185 dispersion Substances 0.000 description 4
- 231100000284 endotoxic Toxicity 0.000 description 4
- 230000002346 endotoxic effect Effects 0.000 description 4
- 229940088598 enzyme Drugs 0.000 description 4
- 239000008103 glucose Substances 0.000 description 4
- 239000008187 granular material Substances 0.000 description 4
- 229960001438 immunostimulant agent Drugs 0.000 description 4
- 239000003022 immunostimulating agent Substances 0.000 description 4
- 238000000338 in vitro Methods 0.000 description 4
- 238000003780 insertion Methods 0.000 description 4
- 230000037431 insertion Effects 0.000 description 4
- 238000005259 measurement Methods 0.000 description 4
- 230000001404 mediated effect Effects 0.000 description 4
- 229930182817 methionine Natural products 0.000 description 4
- 235000006109 methionine Nutrition 0.000 description 4
- 235000013336 milk Nutrition 0.000 description 4
- 239000008267 milk Substances 0.000 description 4
- 210000004080 milk Anatomy 0.000 description 4
- 239000013642 negative control Substances 0.000 description 4
- 230000036961 partial effect Effects 0.000 description 4
- 239000008194 pharmaceutical composition Substances 0.000 description 4
- 229920000642 polymer Polymers 0.000 description 4
- 238000010926 purge Methods 0.000 description 4
- 108010028349 saliva Orthana Proteins 0.000 description 4
- 239000012266 salt solution Substances 0.000 description 4
- 238000012216 screening Methods 0.000 description 4
- 239000002904 solvent Substances 0.000 description 4
- 125000006850 spacer group Chemical group 0.000 description 4
- 238000007920 subcutaneous administration Methods 0.000 description 4
- 239000000375 suspending agent Substances 0.000 description 4
- 208000024891 symptom Diseases 0.000 description 4
- 238000013518 transcription Methods 0.000 description 4
- 230000035897 transcription Effects 0.000 description 4
- 235000013311 vegetables Nutrition 0.000 description 4
- 239000003981 vehicle Substances 0.000 description 4
- 210000002845 virion Anatomy 0.000 description 4
- 239000000080 wetting agent Substances 0.000 description 4
- 244000215068 Acacia senegal Species 0.000 description 3
- QTBSBXVTEAMEQO-UHFFFAOYSA-M Acetate Chemical compound CC([O-])=O QTBSBXVTEAMEQO-UHFFFAOYSA-M 0.000 description 3
- 241000251468 Actinopterygii Species 0.000 description 3
- 240000001436 Antirrhinum majus Species 0.000 description 3
- 241000219173 Carica Species 0.000 description 3
- 241000588919 Citrobacter freundii Species 0.000 description 3
- FBPFZTCFMRRESA-FSIIMWSLSA-N D-Glucitol Natural products OC[C@H](O)[C@H](O)[C@@H](O)[C@H](O)CO FBPFZTCFMRRESA-FSIIMWSLSA-N 0.000 description 3
- 238000001712 DNA sequencing Methods 0.000 description 3
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 3
- 239000006144 Dulbecco’s modified Eagle's medium Substances 0.000 description 3
- 241000283073 Equus caballus Species 0.000 description 3
- LYCAIKOWRPUZTN-UHFFFAOYSA-N Ethylene glycol Chemical compound OCCO LYCAIKOWRPUZTN-UHFFFAOYSA-N 0.000 description 3
- 241000287828 Gallus gallus Species 0.000 description 3
- 102000006354 HLA-DR Antigens Human genes 0.000 description 3
- 108010058597 HLA-DR Antigens Proteins 0.000 description 3
- 101000914484 Homo sapiens T-lymphocyte activation antigen CD80 Proteins 0.000 description 3
- 102100022297 Integrin alpha-X Human genes 0.000 description 3
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 3
- FFEARJCKVFRZRR-BYPYZUCNSA-N L-methionine Chemical compound CSCC[C@H](N)C(O)=O FFEARJCKVFRZRR-BYPYZUCNSA-N 0.000 description 3
- 241000239218 Limulus Species 0.000 description 3
- 108090001074 Nucleocapsid Proteins Proteins 0.000 description 3
- 235000019483 Peanut oil Nutrition 0.000 description 3
- 229930182555 Penicillin Natural products 0.000 description 3
- JGSARLDLIJGVTE-MBNYWOFBSA-N Penicillin G Chemical compound N([C@H]1[C@H]2SC([C@@H](N2C1=O)C(O)=O)(C)C)C(=O)CC1=CC=CC=C1 JGSARLDLIJGVTE-MBNYWOFBSA-N 0.000 description 3
- DNIAPMSPPWPWGF-UHFFFAOYSA-N Propylene glycol Chemical compound CC(O)CO DNIAPMSPPWPWGF-UHFFFAOYSA-N 0.000 description 3
- 241001354013 Salmonella enterica subsp. enterica serovar Enteritidis Species 0.000 description 3
- 241001546666 Salmonella enterica subsp. enterica serovar Newport Species 0.000 description 3
- 102100027222 T-lymphocyte activation antigen CD80 Human genes 0.000 description 3
- 229920004890 Triton X-100 Polymers 0.000 description 3
- 206010046793 Uterine inflammation Diseases 0.000 description 3
- 108010015780 Viral Core Proteins Proteins 0.000 description 3
- 108020000999 Viral RNA Proteins 0.000 description 3
- 241000607477 Yersinia pseudotuberculosis Species 0.000 description 3
- 241001148129 Yersinia ruckeri Species 0.000 description 3
- 238000010521 absorption reaction Methods 0.000 description 3
- WQZGKKKJIJFFOK-VFUOTHLCSA-N beta-D-glucose Chemical compound OC[C@H]1O[C@@H](O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-VFUOTHLCSA-N 0.000 description 3
- 230000003115 biocidal effect Effects 0.000 description 3
- 210000001124 body fluid Anatomy 0.000 description 3
- 239000010839 body fluid Substances 0.000 description 3
- 244000309466 calf Species 0.000 description 3
- 201000011510 cancer Diseases 0.000 description 3
- 229940041514 candida albicans extract Drugs 0.000 description 3
- KRKNYBCHXYNGOX-UHFFFAOYSA-N citric acid Chemical compound OC(=O)CC(O)(C(O)=O)CC(O)=O KRKNYBCHXYNGOX-UHFFFAOYSA-N 0.000 description 3
- 239000011248 coating agent Substances 0.000 description 3
- 239000002299 complementary DNA Substances 0.000 description 3
- 235000009508 confectionery Nutrition 0.000 description 3
- 239000013256 coordination polymer Substances 0.000 description 3
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 3
- 230000009849 deactivation Effects 0.000 description 3
- 239000008367 deionised water Substances 0.000 description 3
- 230000029087 digestion Effects 0.000 description 3
- 239000000975 dye Substances 0.000 description 3
- 208000001848 dysentery Diseases 0.000 description 3
- 239000000839 emulsion Substances 0.000 description 3
- 108010030074 endodeoxyribonuclease MluI Proteins 0.000 description 3
- 208000010227 enterocolitis Diseases 0.000 description 3
- 238000003114 enzyme-linked immunosorbent spot assay Methods 0.000 description 3
- 239000000706 filtrate Substances 0.000 description 3
- 235000019688 fish Nutrition 0.000 description 3
- 210000003495 flagella Anatomy 0.000 description 3
- 210000001035 gastrointestinal tract Anatomy 0.000 description 3
- 150000004676 glycans Chemical class 0.000 description 3
- PCHJSUWPFVWCPO-UHFFFAOYSA-N gold Chemical compound [Au] PCHJSUWPFVWCPO-UHFFFAOYSA-N 0.000 description 3
- 239000010931 gold Substances 0.000 description 3
- 229910052737 gold Inorganic materials 0.000 description 3
- 230000036541 health Effects 0.000 description 3
- 238000003119 immunoblot Methods 0.000 description 3
- 238000003126 immunogold labeling Methods 0.000 description 3
- 230000003993 interaction Effects 0.000 description 3
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 3
- 230000002045 lasting effect Effects 0.000 description 3
- 239000007937 lozenge Substances 0.000 description 3
- 210000004962 mammalian cell Anatomy 0.000 description 3
- 239000011159 matrix material Substances 0.000 description 3
- 230000007246 mechanism Effects 0.000 description 3
- 229910052751 metal Inorganic materials 0.000 description 3
- 239000002184 metal Substances 0.000 description 3
- 238000002156 mixing Methods 0.000 description 3
- 231100000252 nontoxic Toxicity 0.000 description 3
- 230000003000 nontoxic effect Effects 0.000 description 3
- 239000000312 peanut oil Substances 0.000 description 3
- 229940049954 penicillin Drugs 0.000 description 3
- 238000002823 phage display Methods 0.000 description 3
- 229920001282 polysaccharide Polymers 0.000 description 3
- 239000005017 polysaccharide Substances 0.000 description 3
- 239000011734 sodium Substances 0.000 description 3
- 239000000600 sorbitol Substances 0.000 description 3
- 230000000638 stimulation Effects 0.000 description 3
- 229960005322 streptomycin Drugs 0.000 description 3
- 239000003826 tablet Substances 0.000 description 3
- 239000003053 toxin Substances 0.000 description 3
- 231100000765 toxin Toxicity 0.000 description 3
- 239000000304 virulence factor Substances 0.000 description 3
- 230000007923 virulence factor Effects 0.000 description 3
- 239000012138 yeast extract Substances 0.000 description 3
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 3
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 2
- PUPZLCDOIYMWBV-UHFFFAOYSA-N (+/-)-1,3-Butanediol Chemical compound CC(O)CCO PUPZLCDOIYMWBV-UHFFFAOYSA-N 0.000 description 2
- WHTVZRBIWZFKQO-AWEZNQCLSA-N (S)-chloroquine Chemical compound ClC1=CC=C2C(N[C@@H](C)CCCN(CC)CC)=CC=NC2=C1 WHTVZRBIWZFKQO-AWEZNQCLSA-N 0.000 description 2
- GVJHHUAWPYXKBD-UHFFFAOYSA-N (±)-α-Tocopherol Chemical compound OC1=C(C)C(C)=C2OC(CCCC(C)CCCC(C)CCCC(C)C)(C)CCC2=C1C GVJHHUAWPYXKBD-UHFFFAOYSA-N 0.000 description 2
- VBICKXHEKHSIBG-UHFFFAOYSA-N 1-monostearoylglycerol Chemical compound CCCCCCCCCCCCCCCCCC(=O)OCC(O)CO VBICKXHEKHSIBG-UHFFFAOYSA-N 0.000 description 2
- 206010000234 Abortion spontaneous Diseases 0.000 description 2
- 241000724328 Alfalfa mosaic virus Species 0.000 description 2
- NLXLAEXVIDQMFP-UHFFFAOYSA-N Ammonia chloride Chemical compound [NH4+].[Cl-] NLXLAEXVIDQMFP-UHFFFAOYSA-N 0.000 description 2
- 241000272525 Anas platyrhynchos Species 0.000 description 2
- 241001056488 Anatis Species 0.000 description 2
- 208000004429 Bacillary Dysentery Diseases 0.000 description 2
- 239000002028 Biomass Substances 0.000 description 2
- 241000723655 Cowpea mosaic virus Species 0.000 description 2
- 241000699800 Cricetinae Species 0.000 description 2
- 206010011409 Cross infection Diseases 0.000 description 2
- 241000607473 Edwardsiella <enterobacteria> Species 0.000 description 2
- 241000588697 Enterobacter cloacae Species 0.000 description 2
- 241000283074 Equus asinus Species 0.000 description 2
- 208000005577 Gastroenteritis Diseases 0.000 description 2
- 206010017915 Gastroenteritis shigella Diseases 0.000 description 2
- 229930182566 Gentamicin Natural products 0.000 description 2
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 2
- 239000004471 Glycine Substances 0.000 description 2
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 2
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 2
- 229920000084 Gum arabic Polymers 0.000 description 2
- 102000002812 Heat-Shock Proteins Human genes 0.000 description 2
- 108010004889 Heat-Shock Proteins Proteins 0.000 description 2
- 208000032759 Hemolytic-Uremic Syndrome Diseases 0.000 description 2
- 101000725916 Homo sapiens Putative tumor antigen NA88-A Proteins 0.000 description 2
- MHAJPDPJQMAIIY-UHFFFAOYSA-N Hydrogen peroxide Chemical compound OO MHAJPDPJQMAIIY-UHFFFAOYSA-N 0.000 description 2
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 2
- 102000001706 Immunoglobulin Fab Fragments Human genes 0.000 description 2
- 108010054477 Immunoglobulin Fab Fragments Proteins 0.000 description 2
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 2
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 2
- 108010050904 Interferons Proteins 0.000 description 2
- 102000014150 Interferons Human genes 0.000 description 2
- 108010002350 Interleukin-2 Proteins 0.000 description 2
- 102000004388 Interleukin-4 Human genes 0.000 description 2
- 108090000978 Interleukin-4 Proteins 0.000 description 2
- XEEYBQQBJWHFJM-UHFFFAOYSA-N Iron Chemical compound [Fe] XEEYBQQBJWHFJM-UHFFFAOYSA-N 0.000 description 2
- 241001429318 Johnsongrass mosaic virus Species 0.000 description 2
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 2
- DAQAKHDKYAWHCG-UHFFFAOYSA-N Lactacystin Natural products CC(=O)NC(C(O)=O)CSC(=O)C1(C(O)C(C)C)NC(=O)C(C)C1O DAQAKHDKYAWHCG-UHFFFAOYSA-N 0.000 description 2
- FYYHWMGAXLPEAU-UHFFFAOYSA-N Magnesium Chemical compound [Mg] FYYHWMGAXLPEAU-UHFFFAOYSA-N 0.000 description 2
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 2
- 241000124008 Mammalia Species 0.000 description 2
- 201000009906 Meningitis Diseases 0.000 description 2
- 241000282341 Mustela putorius furo Species 0.000 description 2
- 206010029803 Nosocomial infection Diseases 0.000 description 2
- 206010030113 Oedema Diseases 0.000 description 2
- 101710203379 Outer membrane porin C Proteins 0.000 description 2
- 241001479489 Peponocephala electra Species 0.000 description 2
- 108010004729 Phycoerythrin Proteins 0.000 description 2
- 241000235648 Pichia Species 0.000 description 2
- 206010035148 Plague Diseases 0.000 description 2
- 229920001214 Polysorbate 60 Polymers 0.000 description 2
- 239000004793 Polystyrene Substances 0.000 description 2
- 241000125945 Protoparvovirus Species 0.000 description 2
- 102100027596 Putative tumor antigen NA88-A Human genes 0.000 description 2
- 241000700159 Rattus Species 0.000 description 2
- 101000833955 Salmonella typhi Glutamine synthetase adenylyl transferase Proteins 0.000 description 2
- 241000607720 Serratia Species 0.000 description 2
- 241000607764 Shigella dysenteriae Species 0.000 description 2
- CDBYLPFSWZWCQE-UHFFFAOYSA-L Sodium Carbonate Chemical compound [Na+].[Na+].[O-]C([O-])=O CDBYLPFSWZWCQE-UHFFFAOYSA-L 0.000 description 2
- 229920002472 Starch Polymers 0.000 description 2
- 108091081024 Start codon Proteins 0.000 description 2
- 239000012505 Superdex™ Substances 0.000 description 2
- 241000723873 Tobacco mosaic virus Species 0.000 description 2
- 229920001615 Tragacanth Polymers 0.000 description 2
- 239000013504 Triton X-100 Substances 0.000 description 2
- 108090000631 Trypsin Proteins 0.000 description 2
- 102000004142 Trypsin Human genes 0.000 description 2
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 2
- 239000006035 Tryptophane Substances 0.000 description 2
- COQLPRJCUIATTQ-UHFFFAOYSA-N Uranyl acetate Chemical compound O.O.O=[U]=O.CC(O)=O.CC(O)=O COQLPRJCUIATTQ-UHFFFAOYSA-N 0.000 description 2
- 206010000210 abortion Diseases 0.000 description 2
- 231100000176 abortion Toxicity 0.000 description 2
- 235000010489 acacia gum Nutrition 0.000 description 2
- 239000000205 acacia gum Substances 0.000 description 2
- 150000007513 acids Chemical class 0.000 description 2
- 230000033289 adaptive immune response Effects 0.000 description 2
- 210000005006 adaptive immune system Anatomy 0.000 description 2
- 108091005764 adaptor proteins Proteins 0.000 description 2
- 102000035181 adaptor proteins Human genes 0.000 description 2
- 230000001464 adherent effect Effects 0.000 description 2
- 238000001042 affinity chromatography Methods 0.000 description 2
- 230000002776 aggregation Effects 0.000 description 2
- 238000004220 aggregation Methods 0.000 description 2
- 239000000556 agonist Substances 0.000 description 2
- 239000003513 alkali Substances 0.000 description 2
- 238000010171 animal model Methods 0.000 description 2
- 238000000137 annealing Methods 0.000 description 2
- 239000003963 antioxidant agent Substances 0.000 description 2
- 230000003078 antioxidant effect Effects 0.000 description 2
- 235000006708 antioxidants Nutrition 0.000 description 2
- 239000000074 antisense oligonucleotide Substances 0.000 description 2
- 238000012230 antisense oligonucleotides Methods 0.000 description 2
- 239000007864 aqueous solution Substances 0.000 description 2
- 125000003118 aryl group Chemical group 0.000 description 2
- 238000003149 assay kit Methods 0.000 description 2
- 239000000305 astragalus gummifer gum Substances 0.000 description 2
- 239000002585 base Substances 0.000 description 2
- 230000009286 beneficial effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 230000005540 biological transmission Effects 0.000 description 2
- 230000000903 blocking effect Effects 0.000 description 2
- 239000007853 buffer solution Substances 0.000 description 2
- 102220369445 c.668T>C Human genes 0.000 description 2
- 239000001506 calcium phosphate Substances 0.000 description 2
- 229910000389 calcium phosphate Inorganic materials 0.000 description 2
- 235000011010 calcium phosphates Nutrition 0.000 description 2
- 150000001720 carbohydrates Chemical class 0.000 description 2
- 235000014633 carbohydrates Nutrition 0.000 description 2
- 239000011545 carbonate/bicarbonate buffer Substances 0.000 description 2
- 239000000969 carrier Substances 0.000 description 2
- 230000007969 cellular immunity Effects 0.000 description 2
- 229960003677 chloroquine Drugs 0.000 description 2
- WHTVZRBIWZFKQO-UHFFFAOYSA-N chloroquine Natural products ClC1=CC=C2C(NC(C)CCCN(CC)CC)=CC=NC2=C1 WHTVZRBIWZFKQO-UHFFFAOYSA-N 0.000 description 2
- 238000011097 chromatography purification Methods 0.000 description 2
- 230000000052 comparative effect Effects 0.000 description 2
- 230000000295 complement effect Effects 0.000 description 2
- 238000005336 cracking Methods 0.000 description 2
- 230000006378 damage Effects 0.000 description 2
- 230000034994 death Effects 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- 238000000502 dialysis Methods 0.000 description 2
- 238000001035 drying Methods 0.000 description 2
- 238000002337 electrophoretic mobility shift assay Methods 0.000 description 2
- 239000002095 exotoxin Substances 0.000 description 2
- 231100000776 exotoxin Toxicity 0.000 description 2
- MHMNJMPURVTYEJ-UHFFFAOYSA-N fluorescein-5-isothiocyanate Chemical compound O1C(=O)C2=CC(N=C=S)=CC=C2C21C1=CC=C(O)C=C1OC1=CC(O)=CC=C21 MHMNJMPURVTYEJ-UHFFFAOYSA-N 0.000 description 2
- 230000006870 function Effects 0.000 description 2
- 230000000799 fusogenic effect Effects 0.000 description 2
- 239000007903 gelatin capsule Substances 0.000 description 2
- 238000002523 gelfiltration Methods 0.000 description 2
- 229960002518 gentamicin Drugs 0.000 description 2
- 239000011521 glass Substances 0.000 description 2
- 235000011187 glycerol Nutrition 0.000 description 2
- 230000012010 growth Effects 0.000 description 2
- 229920000591 gum Polymers 0.000 description 2
- 238000010438 heat treatment Methods 0.000 description 2
- BXWNKGSJHAJOGX-UHFFFAOYSA-N hexadecan-1-ol Chemical class CCCCCCCCCCCCCCCCO BXWNKGSJHAJOGX-UHFFFAOYSA-N 0.000 description 2
- FBPFZTCFMRRESA-UHFFFAOYSA-N hexane-1,2,3,4,5,6-hexol Chemical compound OCC(O)C(O)C(O)C(O)CO FBPFZTCFMRRESA-UHFFFAOYSA-N 0.000 description 2
- 238000000703 high-speed centrifugation Methods 0.000 description 2
- 229910052739 hydrogen Inorganic materials 0.000 description 2
- 239000001257 hydrogen Substances 0.000 description 2
- 230000002209 hydrophobic effect Effects 0.000 description 2
- 230000002458 infectious effect Effects 0.000 description 2
- 108010061181 influenza matrix peptide (58-66) Proteins 0.000 description 2
- 239000003112 inhibitor Substances 0.000 description 2
- 229940079322 interferon Drugs 0.000 description 2
- 238000007913 intrathecal administration Methods 0.000 description 2
- 238000010253 intravenous injection Methods 0.000 description 2
- DAQAKHDKYAWHCG-RWTHQLGUSA-N lactacystin Chemical compound CC(=O)N[C@H](C(O)=O)CSC(=O)[C@]1([C@@H](O)C(C)C)NC(=O)[C@H](C)[C@@H]1O DAQAKHDKYAWHCG-RWTHQLGUSA-N 0.000 description 2
- 235000012204 lemonade/lime carbonate Nutrition 0.000 description 2
- 230000003902 lesion Effects 0.000 description 2
- 230000000527 lymphocytic effect Effects 0.000 description 2
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 2
- 239000011777 magnesium Substances 0.000 description 2
- 229910052749 magnesium Inorganic materials 0.000 description 2
- HQKMJHAJHXVSDF-UHFFFAOYSA-L magnesium stearate Chemical compound [Mg+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O HQKMJHAJHXVSDF-UHFFFAOYSA-L 0.000 description 2
- 238000012423 maintenance Methods 0.000 description 2
- 239000002480 mineral oil Substances 0.000 description 2
- 235000010446 mineral oil Nutrition 0.000 description 2
- 208000015994 miscarriage Diseases 0.000 description 2
- 229940031346 monovalent vaccine Drugs 0.000 description 2
- 210000003928 nasal cavity Anatomy 0.000 description 2
- 230000009871 nonspecific binding Effects 0.000 description 2
- 239000000346 nonvolatile oil Substances 0.000 description 2
- 206010034674 peritonitis Diseases 0.000 description 2
- 230000002085 persistent effect Effects 0.000 description 2
- 239000000546 pharmaceutical excipient Substances 0.000 description 2
- COLNVLDHVKWLRT-UHFFFAOYSA-N phenylalanine Natural products OC(=O)C(N)CC1=CC=CC=C1 COLNVLDHVKWLRT-UHFFFAOYSA-N 0.000 description 2
- 229920002223 polystyrene Polymers 0.000 description 2
- 238000002203 pretreatment Methods 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 230000003449 preventive effect Effects 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 238000007639 printing Methods 0.000 description 2
- 239000012460 protein solution Substances 0.000 description 2
- 239000010453 quartz Substances 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 230000002829 reductive effect Effects 0.000 description 2
- 208000023504 respiratory system disease Diseases 0.000 description 2
- 230000000717 retained effect Effects 0.000 description 2
- 238000003757 reverse transcription PCR Methods 0.000 description 2
- 229940007046 shigella dysenteriae Drugs 0.000 description 2
- 201000005113 shigellosis Diseases 0.000 description 2
- VYPSYNLAJGMNEJ-UHFFFAOYSA-N silicon dioxide Inorganic materials O=[Si]=O VYPSYNLAJGMNEJ-UHFFFAOYSA-N 0.000 description 2
- 235000020183 skimmed milk Nutrition 0.000 description 2
- 229910052708 sodium Inorganic materials 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 239000007787 solid Substances 0.000 description 2
- 210000004988 splenocyte Anatomy 0.000 description 2
- 208000000995 spontaneous abortion Diseases 0.000 description 2
- 239000007921 spray Substances 0.000 description 2
- 239000008107 starch Substances 0.000 description 2
- 235000019698 starch Nutrition 0.000 description 2
- 238000005728 strengthening Methods 0.000 description 2
- 239000000758 substrate Substances 0.000 description 2
- 239000006188 syrup Substances 0.000 description 2
- 235000020357 syrup Nutrition 0.000 description 2
- 230000009885 systemic effect Effects 0.000 description 2
- 230000000699 topical effect Effects 0.000 description 2
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 2
- 230000001960 triggered effect Effects 0.000 description 2
- XFNJVJPLKCPIBV-UHFFFAOYSA-N trimethylenediamine Chemical compound NCCCN XFNJVJPLKCPIBV-UHFFFAOYSA-N 0.000 description 2
- 239000012588 trypsin Substances 0.000 description 2
- 229960004799 tryptophan Drugs 0.000 description 2
- 210000004881 tumor cell Anatomy 0.000 description 2
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 2
- 208000019206 urinary tract infection Diseases 0.000 description 2
- 230000000007 visual effect Effects 0.000 description 2
- 210000002268 wool Anatomy 0.000 description 2
- WRIDQFICGBMAFQ-UHFFFAOYSA-N (E)-8-Octadecenoic acid Natural products CCCCCCCCCC=CCCCCCCC(O)=O WRIDQFICGBMAFQ-UHFFFAOYSA-N 0.000 description 1
- DNIAPMSPPWPWGF-GSVOUGTGSA-N (R)-(-)-Propylene glycol Chemical compound C[C@@H](O)CO DNIAPMSPPWPWGF-GSVOUGTGSA-N 0.000 description 1
- GEYOCULIXLDCMW-UHFFFAOYSA-N 1,2-phenylenediamine Chemical compound NC1=CC=CC=C1N GEYOCULIXLDCMW-UHFFFAOYSA-N 0.000 description 1
- 229940058015 1,3-butylene glycol Drugs 0.000 description 1
- IXPNQXFRVYWDDI-UHFFFAOYSA-N 1-methyl-2,4-dioxo-1,3-diazinane-5-carboximidamide Chemical compound CN1CC(C(N)=N)C(=O)NC1=O IXPNQXFRVYWDDI-UHFFFAOYSA-N 0.000 description 1
- OTLLEIBWKHEHGU-UHFFFAOYSA-N 2-[5-[[5-(6-aminopurin-9-yl)-3,4-dihydroxyoxolan-2-yl]methoxy]-3,4-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-3,5-dihydroxy-4-phosphonooxyhexanedioic acid Chemical compound C1=NC=2C(N)=NC=NC=2N1C(C(C1O)O)OC1COC1C(CO)OC(OC(C(O)C(OP(O)(O)=O)C(O)C(O)=O)C(O)=O)C(O)C1O OTLLEIBWKHEHGU-UHFFFAOYSA-N 0.000 description 1
- IVLXQGJVBGMLRR-UHFFFAOYSA-N 2-aminoacetic acid;hydron;chloride Chemical compound Cl.NCC(O)=O IVLXQGJVBGMLRR-UHFFFAOYSA-N 0.000 description 1
- LQJBNNIYVWPHFW-UHFFFAOYSA-N 20:1omega9c fatty acid Natural products CCCCCCCCCCC=CCCCCCCCC(O)=O LQJBNNIYVWPHFW-UHFFFAOYSA-N 0.000 description 1
- XMTQQYYKAHVGBJ-UHFFFAOYSA-N 3-(3,4-DICHLOROPHENYL)-1,1-DIMETHYLUREA Chemical compound CN(C)C(=O)NC1=CC=C(Cl)C(Cl)=C1 XMTQQYYKAHVGBJ-UHFFFAOYSA-N 0.000 description 1
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 1
- 229940090248 4-hydroxybenzoic acid Drugs 0.000 description 1
- NWAGXLBTAPTCPR-UHFFFAOYSA-N 5-(2,5-dioxopyrrolidin-1-yl)oxy-5-oxopentanoic acid Chemical class OC(=O)CCCC(=O)ON1C(=O)CCC1=O NWAGXLBTAPTCPR-UHFFFAOYSA-N 0.000 description 1
- NVWKYNBWYYJZTK-UHFFFAOYSA-N 8-(2,5-dioxopyrrolidin-1-yl)oxy-8-oxooctanoic acid Chemical class OC(=O)CCCCCCC(=O)ON1C(=O)CCC1=O NVWKYNBWYYJZTK-UHFFFAOYSA-N 0.000 description 1
- QSBYPNXLFMSGKH-UHFFFAOYSA-N 9-Heptadecensaeure Natural products CCCCCCCC=CCCCCCCCC(O)=O QSBYPNXLFMSGKH-UHFFFAOYSA-N 0.000 description 1
- 208000004998 Abdominal Pain Diseases 0.000 description 1
- 206010000087 Abdominal pain upper Diseases 0.000 description 1
- 206010000228 Abortion infected Diseases 0.000 description 1
- 235000006491 Acacia senegal Nutrition 0.000 description 1
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 1
- 241000607525 Aeromonas salmonicida Species 0.000 description 1
- 101710153593 Albumin A Proteins 0.000 description 1
- 108010088751 Albumins Proteins 0.000 description 1
- 102000009027 Albumins Human genes 0.000 description 1
- GUBGYTABKSRVRQ-XLOQQCSPSA-N Alpha-Lactose Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@H](O)[C@H](O)[C@H]1O GUBGYTABKSRVRQ-XLOQQCSPSA-N 0.000 description 1
- 239000005995 Aluminium silicate Substances 0.000 description 1
- 241000252082 Anguilla anguilla Species 0.000 description 1
- 102100025511 Anti-Muellerian hormone type-2 receptor Human genes 0.000 description 1
- 101710089052 Anti-Muellerian hormone type-2 receptor Proteins 0.000 description 1
- 239000004475 Arginine Substances 0.000 description 1
- CIWBSHSKHKDKBQ-JLAZNSOCSA-N Ascorbic acid Chemical compound OC[C@H](O)[C@H]1OC(=O)C(O)=C1O CIWBSHSKHKDKBQ-JLAZNSOCSA-N 0.000 description 1
- 241000972773 Aulopiformes Species 0.000 description 1
- 241000271566 Aves Species 0.000 description 1
- 241000711404 Avian avulavirus 1 Species 0.000 description 1
- 108050001427 Avidin/streptavidin Proteins 0.000 description 1
- 244000063299 Bacillus subtilis Species 0.000 description 1
- 241000710073 Bean yellow mosaic virus Species 0.000 description 1
- 208000031462 Bovine Mastitis Diseases 0.000 description 1
- 206010006187 Breast cancer Diseases 0.000 description 1
- 208000026310 Breast neoplasm Diseases 0.000 description 1
- 210000003771 C cell Anatomy 0.000 description 1
- PNLAUONNUXHSMX-UHFFFAOYSA-N C.CP Chemical compound C.CP PNLAUONNUXHSMX-UHFFFAOYSA-N 0.000 description 1
- 108010029697 CD40 Ligand Proteins 0.000 description 1
- 102100032937 CD40 ligand Human genes 0.000 description 1
- 108090000565 Capsid Proteins Proteins 0.000 description 1
- 241000252229 Carassius auratus Species 0.000 description 1
- 240000006432 Carica papaya Species 0.000 description 1
- 206010008342 Cervix carcinoma Diseases 0.000 description 1
- 108091006146 Channels Proteins 0.000 description 1
- 241001227713 Chiron Species 0.000 description 1
- 241000588923 Citrobacter Species 0.000 description 1
- 108020004635 Complementary DNA Proteins 0.000 description 1
- 108020004394 Complementary RNA Proteins 0.000 description 1
- 206010010774 Constipation Diseases 0.000 description 1
- FBPFZTCFMRRESA-KVTDHHQDSA-N D-Mannitol Chemical compound OC[C@@H](O)[C@@H](O)[C@H](O)[C@H](O)CO FBPFZTCFMRRESA-KVTDHHQDSA-N 0.000 description 1
- 241000238557 Decapoda Species 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 238000009007 Diagnostic Kit Methods 0.000 description 1
- ZNZYKNKBJPZETN-WELNAUFTSA-N Dialdehyde 11678 Chemical compound N1C2=CC=CC=C2C2=C1[C@H](C[C@H](/C(=C/O)C(=O)OC)[C@@H](C=C)C=O)NCC2 ZNZYKNKBJPZETN-WELNAUFTSA-N 0.000 description 1
- 206010012741 Diarrhoea haemorrhagic Diseases 0.000 description 1
- 241000199914 Dinophyceae Species 0.000 description 1
- 101100347633 Drosophila melanogaster Mhc gene Proteins 0.000 description 1
- 241000949274 Edwardsiella ictaluri Species 0.000 description 1
- 241000607471 Edwardsiella tarda Species 0.000 description 1
- 108010000912 Egg Proteins Proteins 0.000 description 1
- 102000002322 Egg Proteins Human genes 0.000 description 1
- 206010014896 Enterocolitis haemorrhagic Diseases 0.000 description 1
- 239000004593 Epoxy Substances 0.000 description 1
- 241000010756 Escherichia coli O157:H7 str. EDL933 Species 0.000 description 1
- VGGSQFUCUMXWEO-UHFFFAOYSA-N Ethene Chemical compound C=C VGGSQFUCUMXWEO-UHFFFAOYSA-N 0.000 description 1
- 239000005977 Ethylene Substances 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 108010009066 Gastric Mucins Proteins 0.000 description 1
- 102000009338 Gastric Mucins Human genes 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- SXRSQZLOMIGNAQ-UHFFFAOYSA-N Glutaraldehyde Chemical compound O=CCCCC=O SXRSQZLOMIGNAQ-UHFFFAOYSA-N 0.000 description 1
- 244000068988 Glycine max Species 0.000 description 1
- 235000010469 Glycine max Nutrition 0.000 description 1
- 108010074032 HLA-A2 Antigen Proteins 0.000 description 1
- 102000025850 HLA-A2 Antigen Human genes 0.000 description 1
- 102000004447 HSP40 Heat-Shock Proteins Human genes 0.000 description 1
- 108010042283 HSP40 Heat-Shock Proteins Proteins 0.000 description 1
- 206010019233 Headaches Diseases 0.000 description 1
- 239000005057 Hexamethylene diisocyanate Substances 0.000 description 1
- 108010033040 Histones Proteins 0.000 description 1
- 101000911390 Homo sapiens Coagulation factor VIII Proteins 0.000 description 1
- 101001002709 Homo sapiens Interleukin-4 Proteins 0.000 description 1
- 101000878605 Homo sapiens Low affinity immunoglobulin epsilon Fc receptor Proteins 0.000 description 1
- 101000917826 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-a Proteins 0.000 description 1
- 101000917824 Homo sapiens Low affinity immunoglobulin gamma Fc region receptor II-b Proteins 0.000 description 1
- 102000017727 Immunoglobulin Variable Region Human genes 0.000 description 1
- 108010067060 Immunoglobulin Variable Region Proteins 0.000 description 1
- 244000283207 Indigofera tinctoria Species 0.000 description 1
- 206010061218 Inflammation Diseases 0.000 description 1
- 108020005350 Initiator Codon Proteins 0.000 description 1
- 108010063738 Interleukins Proteins 0.000 description 1
- 102000015696 Interleukins Human genes 0.000 description 1
- 101710198693 Invasin Proteins 0.000 description 1
- 239000007760 Iscove's Modified Dulbecco's Medium Substances 0.000 description 1
- ONIBWKKTOPOVIA-BYPYZUCNSA-N L-Proline Chemical compound OC(=O)[C@@H]1CCCN1 ONIBWKKTOPOVIA-BYPYZUCNSA-N 0.000 description 1
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 1
- GUBGYTABKSRVRQ-QKKXKWKRSA-N Lactose Natural products OC[C@H]1O[C@@H](O[C@H]2[C@H](O)[C@@H](O)C(O)O[C@@H]2CO)[C@H](O)[C@@H](O)[C@H]1O GUBGYTABKSRVRQ-QKKXKWKRSA-N 0.000 description 1
- 235000019687 Lamb Nutrition 0.000 description 1
- 241001071864 Lethrinus laticaudis Species 0.000 description 1
- ROHFNLRQFUQHCH-UHFFFAOYSA-N Leucine Natural products CC(C)CC(N)C(O)=O ROHFNLRQFUQHCH-UHFFFAOYSA-N 0.000 description 1
- 108090001030 Lipoproteins Proteins 0.000 description 1
- 102000004895 Lipoproteins Human genes 0.000 description 1
- 102100038007 Low affinity immunoglobulin epsilon Fc receptor Human genes 0.000 description 1
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 1
- 102000004317 Lyases Human genes 0.000 description 1
- 108090000856 Lyases Proteins 0.000 description 1
- OFOBLEOULBTSOW-UHFFFAOYSA-N Malonic acid Chemical compound OC(=O)CC(O)=O OFOBLEOULBTSOW-UHFFFAOYSA-N 0.000 description 1
- 229930195725 Mannitol Natural products 0.000 description 1
- PQMWYJDJHJQZDE-UHFFFAOYSA-M Methantheline bromide Chemical compound [Br-].C1=CC=C2C(C(=O)OCC[N+](C)(CC)CC)C3=CC=CC=C3OC2=C1 PQMWYJDJHJQZDE-UHFFFAOYSA-M 0.000 description 1
- YXOLAZRVSSWPPT-UHFFFAOYSA-N Morin Chemical compound OC1=CC(O)=CC=C1C1=C(O)C(=O)C2=C(O)C=C(O)C=C2O1 YXOLAZRVSSWPPT-UHFFFAOYSA-N 0.000 description 1
- 241001529936 Murinae Species 0.000 description 1
- 208000000112 Myalgia Diseases 0.000 description 1
- 108010064696 N,O-diacetylmuramidase Proteins 0.000 description 1
- XUYPXLNMDZIRQH-LURJTMIESA-N N-acetyl-L-methionine Chemical compound CSCC[C@@H](C(O)=O)NC(C)=O XUYPXLNMDZIRQH-LURJTMIESA-N 0.000 description 1
- 101150028431 N5 gene Proteins 0.000 description 1
- 241001077878 Neurolaena lobata Species 0.000 description 1
- 244000061176 Nicotiana tabacum Species 0.000 description 1
- 235000002637 Nicotiana tabacum Nutrition 0.000 description 1
- 108091092724 Noncoding DNA Proteins 0.000 description 1
- 239000005642 Oleic acid Substances 0.000 description 1
- ZQPPMHVWECSIRJ-UHFFFAOYSA-N Oleic acid Natural products CCCCCCCCC=CCCCCCCCC(O)=O ZQPPMHVWECSIRJ-UHFFFAOYSA-N 0.000 description 1
- 206010031009 Oral pain Diseases 0.000 description 1
- 229930040373 Paraformaldehyde Natural products 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- ISWSIDIOOBJBQZ-UHFFFAOYSA-N Phenol Natural products OC1=CC=CC=C1 ISWSIDIOOBJBQZ-UHFFFAOYSA-N 0.000 description 1
- 206010035717 Pneumonia klebsiella Diseases 0.000 description 1
- 229920003171 Poly (ethylene oxide) Polymers 0.000 description 1
- 108091036414 Polyinosinic:polycytidylic acid Proteins 0.000 description 1
- 108010021757 Polynucleotide 5'-Hydroxyl-Kinase Proteins 0.000 description 1
- 102000008422 Polynucleotide 5'-hydroxyl-kinase Human genes 0.000 description 1
- 101000714429 Potato virus X Coat protein Proteins 0.000 description 1
- 101900267314 Potato virus Y Capsid protein Proteins 0.000 description 1
- ONIBWKKTOPOVIA-UHFFFAOYSA-N Proline Natural products OC(=O)C1CCCN1 ONIBWKKTOPOVIA-UHFFFAOYSA-N 0.000 description 1
- 239000004365 Protease Substances 0.000 description 1
- 229940079156 Proteasome inhibitor Drugs 0.000 description 1
- 241000589516 Pseudomonas Species 0.000 description 1
- 239000012980 RPMI-1640 medium Substances 0.000 description 1
- 241001068263 Replication competent viruses Species 0.000 description 1
- 101710141795 Ribonuclease inhibitor Proteins 0.000 description 1
- 229940122208 Ribonuclease inhibitor Drugs 0.000 description 1
- 102100037968 Ribonuclease inhibitor Human genes 0.000 description 1
- 241000283984 Rodentia Species 0.000 description 1
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 1
- 241000235070 Saccharomyces Species 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 241000533331 Salmonella bongori Species 0.000 description 1
- 241000522522 Salmonella enterica subsp. enterica serovar Abortusovis Species 0.000 description 1
- 241000596091 Salmonella enterica subsp. enterica serovar Anatum Species 0.000 description 1
- 241000607683 Salmonella enterica subsp. enterica serovar Pullorum Species 0.000 description 1
- 241000132553 Salmonella enterica subsp. enterica serovar Typhisuis Species 0.000 description 1
- 241000277331 Salmonidae Species 0.000 description 1
- 239000012506 Sephacryl® Substances 0.000 description 1
- 208000002359 Septic Abortion Diseases 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 108010017898 Shiga Toxins Proteins 0.000 description 1
- 241000147000 Shigella flexneri 2a Species 0.000 description 1
- 244000061456 Solanum tuberosum Species 0.000 description 1
- 235000002595 Solanum tuberosum Nutrition 0.000 description 1
- 235000021355 Stearic acid Nutrition 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- 230000006044 T cell activation Effects 0.000 description 1
- 230000024932 T cell mediated immunity Effects 0.000 description 1
- 229940124929 TYPHIM Vi Drugs 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 241000656145 Thyrsites atun Species 0.000 description 1
- 108020005202 Viral DNA Proteins 0.000 description 1
- 108010067390 Viral Proteins Proteins 0.000 description 1
- 229930003427 Vitamin E Natural products 0.000 description 1
- YVNQAIFQFWTPLQ-UHFFFAOYSA-O [4-[[4-(4-ethoxyanilino)phenyl]-[4-[ethyl-[(3-sulfophenyl)methyl]amino]-2-methylphenyl]methylidene]-3-methylcyclohexa-2,5-dien-1-ylidene]-ethyl-[(3-sulfophenyl)methyl]azanium Chemical compound C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C(=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S(O)(=O)=O)C)C=2C(=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S(O)(=O)=O)C)C=C1 YVNQAIFQFWTPLQ-UHFFFAOYSA-O 0.000 description 1
- 230000003187 abdominal effect Effects 0.000 description 1
- 150000008065 acid anhydrides Chemical class 0.000 description 1
- 238000010306 acid treatment Methods 0.000 description 1
- 230000002378 acidificating effect Effects 0.000 description 1
- 210000002659 acromion Anatomy 0.000 description 1
- 230000003213 activating effect Effects 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 238000001261 affinity purification Methods 0.000 description 1
- 150000001335 aliphatic alkanes Chemical class 0.000 description 1
- 229940037003 alum Drugs 0.000 description 1
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 1
- 229910021502 aluminium hydroxide Inorganic materials 0.000 description 1
- 235000012211 aluminium silicate Nutrition 0.000 description 1
- 235000019270 ammonium chloride Nutrition 0.000 description 1
- KLOHDWPABZXLGI-YWUHCJSESA-M ampicillin sodium Chemical compound [Na+].C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C([O-])=O)(C)C)=CC=CC=C1 KLOHDWPABZXLGI-YWUHCJSESA-M 0.000 description 1
- 244000037640 animal pathogen Species 0.000 description 1
- 230000000636 anti-proteolytic effect Effects 0.000 description 1
- 238000002617 apheresis Methods 0.000 description 1
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 1
- 235000003704 aspartic acid Nutrition 0.000 description 1
- 238000000211 autoradiogram Methods 0.000 description 1
- 239000000688 bacterial toxin Substances 0.000 description 1
- 238000002869 basic local alignment search tool Methods 0.000 description 1
- 235000013871 bee wax Nutrition 0.000 description 1
- 239000012166 beeswax Substances 0.000 description 1
- OQFSQFPPLPISGP-UHFFFAOYSA-N beta-carboxyaspartic acid Natural products OC(=O)C(N)C(C(O)=O)C(O)=O OQFSQFPPLPISGP-UHFFFAOYSA-N 0.000 description 1
- 239000003139 biocide Substances 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 238000010241 blood sampling Methods 0.000 description 1
- 208000037815 bloodstream infection Diseases 0.000 description 1
- 238000009835 boiling Methods 0.000 description 1
- 235000019437 butane-1,3-diol Nutrition 0.000 description 1
- PMSGJXMYHUSZEI-UHFFFAOYSA-N butanedioic acid;pyrrolidine-2,5-dione Chemical compound O=C1CCC(=O)N1.OC(=O)CCC(O)=O PMSGJXMYHUSZEI-UHFFFAOYSA-N 0.000 description 1
- 210000004899 c-terminal region Anatomy 0.000 description 1
- 238000010804 cDNA synthesis Methods 0.000 description 1
- BPKIGYQJPYCAOW-FFJTTWKXSA-I calcium;potassium;disodium;(2s)-2-hydroxypropanoate;dichloride;dihydroxide;hydrate Chemical compound O.[OH-].[OH-].[Na+].[Na+].[Cl-].[Cl-].[K+].[Ca+2].C[C@H](O)C([O-])=O BPKIGYQJPYCAOW-FFJTTWKXSA-I 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 238000012754 cardiac puncture Methods 0.000 description 1
- 239000005018 casein Substances 0.000 description 1
- BECPQYXYKAMYBN-UHFFFAOYSA-N casein, tech. Chemical compound NCCCCC(C(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(CC(C)C)N=C(O)C(CCC(O)=O)N=C(O)C(CC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(C(C)O)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=N)N=C(O)C(CCC(O)=O)N=C(O)C(CCC(O)=O)N=C(O)C(COP(O)(O)=O)N=C(O)C(CCC(O)=N)N=C(O)C(N)CC1=CC=CC=C1 BECPQYXYKAMYBN-UHFFFAOYSA-N 0.000 description 1
- 235000021240 caseins Nutrition 0.000 description 1
- 239000003054 catalyst Substances 0.000 description 1
- 201000010881 cervical cancer Diseases 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 239000007979 citrate buffer Substances 0.000 description 1
- 210000001072 colon Anatomy 0.000 description 1
- 230000024203 complement activation Effects 0.000 description 1
- 239000003184 complementary RNA Substances 0.000 description 1
- 230000003750 conditioning effect Effects 0.000 description 1
- 210000002808 connective tissue Anatomy 0.000 description 1
- 238000010276 construction Methods 0.000 description 1
- 239000000356 contaminant Substances 0.000 description 1
- 229940028617 conventional vaccine Drugs 0.000 description 1
- NKLPQNGYXWVELD-UHFFFAOYSA-M coomassie brilliant blue Chemical compound [Na+].C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=2C=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=C1 NKLPQNGYXWVELD-UHFFFAOYSA-M 0.000 description 1
- 230000000139 costimulatory effect Effects 0.000 description 1
- 239000003431 cross linking reagent Substances 0.000 description 1
- 230000009260 cross reactivity Effects 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 210000004748 cultured cell Anatomy 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 238000007405 data analysis Methods 0.000 description 1
- 238000000354 decomposition reaction Methods 0.000 description 1
- 230000018044 dehydration Effects 0.000 description 1
- 238000006297 dehydration reaction Methods 0.000 description 1
- 230000030609 dephosphorylation Effects 0.000 description 1
- 238000006209 dephosphorylation reaction Methods 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 238000001514 detection method Methods 0.000 description 1
- 238000010586 diagram Methods 0.000 description 1
- 150000004985 diamines Chemical class 0.000 description 1
- 238000001085 differential centrifugation Methods 0.000 description 1
- KZNICNPSHKQLFF-UHFFFAOYSA-N dihydromaleimide Natural products O=C1CCC(=O)N1 KZNICNPSHKQLFF-UHFFFAOYSA-N 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- FPAFDBFIGPHWGO-UHFFFAOYSA-N dioxosilane;oxomagnesium;hydrate Chemical compound O.[Mg]=O.[Mg]=O.[Mg]=O.O=[Si]=O.O=[Si]=O.O=[Si]=O.O=[Si]=O FPAFDBFIGPHWGO-UHFFFAOYSA-N 0.000 description 1
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 1
- 238000007598 dipping method Methods 0.000 description 1
- 239000006196 drop Substances 0.000 description 1
- 229940079593 drug Drugs 0.000 description 1
- 238000012377 drug delivery Methods 0.000 description 1
- 238000001962 electrophoresis Methods 0.000 description 1
- 238000004520 electroporation Methods 0.000 description 1
- 239000012149 elution buffer Substances 0.000 description 1
- 239000003974 emollient agent Substances 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 238000005538 encapsulation Methods 0.000 description 1
- 206010014599 encephalitis Diseases 0.000 description 1
- 210000001163 endosome Anatomy 0.000 description 1
- 230000002708 enhancing effect Effects 0.000 description 1
- 238000001976 enzyme digestion Methods 0.000 description 1
- 210000003743 erythrocyte Anatomy 0.000 description 1
- 210000000416 exudates and transudate Anatomy 0.000 description 1
- 238000009313 farming Methods 0.000 description 1
- 239000000194 fatty acid Substances 0.000 description 1
- 230000002550 fecal effect Effects 0.000 description 1
- 238000000855 fermentation Methods 0.000 description 1
- 230000004151 fermentation Effects 0.000 description 1
- 238000001914 filtration Methods 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 125000000524 functional group Chemical group 0.000 description 1
- WIGCFUFOHFEKBI-UHFFFAOYSA-N gamma-tocopherol Natural products CC(C)CCCC(C)CCCC(C)CCCC1CCC2C(C)C(O)C(C)C(C)C2O1 WIGCFUFOHFEKBI-UHFFFAOYSA-N 0.000 description 1
- 239000007789 gas Substances 0.000 description 1
- 230000002496 gastric effect Effects 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 238000010353 genetic engineering Methods 0.000 description 1
- 229960002989 glutamic acid Drugs 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 125000005456 glyceride group Chemical group 0.000 description 1
- 125000005908 glyceryl ester group Chemical group 0.000 description 1
- 229940075507 glyceryl monostearate Drugs 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- 239000003979 granulating agent Substances 0.000 description 1
- 239000001963 growth medium Substances 0.000 description 1
- 239000007902 hard capsule Substances 0.000 description 1
- 231100000869 headache Toxicity 0.000 description 1
- 208000027909 hemorrhagic diarrhea Diseases 0.000 description 1
- RRAMGCGOFNQTLD-UHFFFAOYSA-N hexamethylene diisocyanate Chemical compound O=C=NCCCCCCN=C=O RRAMGCGOFNQTLD-UHFFFAOYSA-N 0.000 description 1
- NAQMVNRVTILPCV-UHFFFAOYSA-N hexane-1,6-diamine Chemical compound NCCCCCCN NAQMVNRVTILPCV-UHFFFAOYSA-N 0.000 description 1
- 102000057593 human F8 Human genes 0.000 description 1
- 102000055229 human IL4 Human genes 0.000 description 1
- 230000004727 humoral immunity Effects 0.000 description 1
- 230000008348 humoral response Effects 0.000 description 1
- 150000002433 hydrophilic molecules Chemical class 0.000 description 1
- 125000001165 hydrophobic group Chemical group 0.000 description 1
- 238000007654 immersion Methods 0.000 description 1
- 230000000984 immunochemical effect Effects 0.000 description 1
- 230000003116 impacting effect Effects 0.000 description 1
- 230000006872 improvement Effects 0.000 description 1
- 230000002779 inactivation Effects 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- 239000003701 inert diluent Substances 0.000 description 1
- 230000002757 inflammatory effect Effects 0.000 description 1
- 230000004054 inflammatory process Effects 0.000 description 1
- 238000001802 infusion Methods 0.000 description 1
- 229940102223 injectable solution Drugs 0.000 description 1
- 210000000936 intestine Anatomy 0.000 description 1
- 150000002500 ions Chemical class 0.000 description 1
- 229910052742 iron Inorganic materials 0.000 description 1
- 230000001788 irregular Effects 0.000 description 1
- AGPKZVBTJJNPAG-UHFFFAOYSA-N isoleucine Natural products CCC(C)C(N)C(O)=O AGPKZVBTJJNPAG-UHFFFAOYSA-N 0.000 description 1
- 229960000310 isoleucine Drugs 0.000 description 1
- FZWBNHMXJMCXLU-BLAUPYHCSA-N isomaltotriose Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1OC[C@@H]1[C@@H](O)[C@H](O)[C@@H](O)[C@@H](OC[C@@H](O)[C@@H](O)[C@H](O)[C@@H](O)C=O)O1 FZWBNHMXJMCXLU-BLAUPYHCSA-N 0.000 description 1
- QXJSBBXBKPUZAA-UHFFFAOYSA-N isooleic acid Natural products CCCCCCCC=CCCCCCCCCC(O)=O QXJSBBXBKPUZAA-UHFFFAOYSA-N 0.000 description 1
- 210000004731 jugular vein Anatomy 0.000 description 1
- NLYAJNPCOHFWQQ-UHFFFAOYSA-N kaolin Chemical compound O.O.O=[Al]O[Si](=O)O[Si](=O)O[Al]=O NLYAJNPCOHFWQQ-UHFFFAOYSA-N 0.000 description 1
- 230000002147 killing effect Effects 0.000 description 1
- 239000008101 lactose Substances 0.000 description 1
- 231100000225 lethality Toxicity 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 238000004811 liquid chromatography Methods 0.000 description 1
- 229940057995 liquid paraffin Drugs 0.000 description 1
- 239000012160 loading buffer Substances 0.000 description 1
- 239000000314 lubricant Substances 0.000 description 1
- 201000003265 lymphadenitis Diseases 0.000 description 1
- 239000012139 lysis buffer Substances 0.000 description 1
- 235000019359 magnesium stearate Nutrition 0.000 description 1
- 239000000594 mannitol Substances 0.000 description 1
- 235000010355 mannitol Nutrition 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 210000001806 memory b lymphocyte Anatomy 0.000 description 1
- 150000002742 methionines Chemical class 0.000 description 1
- 229920000609 methyl cellulose Polymers 0.000 description 1
- 239000001923 methylcellulose Substances 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- ZAHQPTJLOCWVPG-UHFFFAOYSA-N mitoxantrone dihydrochloride Chemical compound Cl.Cl.O=C1C2=C(O)C=CC(O)=C2C(=O)C2=C1C(NCCNCCO)=CC=C2NCCNCCO ZAHQPTJLOCWVPG-UHFFFAOYSA-N 0.000 description 1
- 239000003068 molecular probe Substances 0.000 description 1
- 239000001788 mono and diglycerides of fatty acids Substances 0.000 description 1
- 229940045641 monobasic sodium phosphate Drugs 0.000 description 1
- 210000001616 monocyte Anatomy 0.000 description 1
- 210000000865 mononuclear phagocyte system Anatomy 0.000 description 1
- UXOUKMQIEVGVLY-UHFFFAOYSA-N morin Natural products OC1=CC(O)=CC(C2=C(C(=O)C3=C(O)C=C(O)C=C3O2)O)=C1 UXOUKMQIEVGVLY-UHFFFAOYSA-N 0.000 description 1
- 235000007708 morin Nutrition 0.000 description 1
- 208000030194 mouth disease Diseases 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- 239000007923 nasal drop Substances 0.000 description 1
- 239000007922 nasal spray Substances 0.000 description 1
- 229920001206 natural gum Polymers 0.000 description 1
- 210000000822 natural killer cell Anatomy 0.000 description 1
- 239000007764 o/w emulsion Substances 0.000 description 1
- QIQXTHQIDYTFRH-UHFFFAOYSA-N octadecanoic acid Chemical compound CCCCCCCCCCCCCCCCCC(O)=O QIQXTHQIDYTFRH-UHFFFAOYSA-N 0.000 description 1
- OQCDKBAXFALNLD-UHFFFAOYSA-N octadecanoic acid Natural products CCCCCCCC(C)CCCCCCCCC(O)=O OQCDKBAXFALNLD-UHFFFAOYSA-N 0.000 description 1
- ZQPPMHVWECSIRJ-KTKRTIGZSA-N oleic acid Chemical compound CCCCCCCC\C=C/CCCCCCCC(O)=O ZQPPMHVWECSIRJ-KTKRTIGZSA-N 0.000 description 1
- 230000003287 optical effect Effects 0.000 description 1
- 229940126578 oral vaccine Drugs 0.000 description 1
- 210000004279 orbit Anatomy 0.000 description 1
- 210000004681 ovum Anatomy 0.000 description 1
- 238000004091 panning Methods 0.000 description 1
- FJKROLUGYXJWQN-UHFFFAOYSA-N papa-hydroxy-benzoic acid Natural products OC(=O)C1=CC=C(O)C=C1 FJKROLUGYXJWQN-UHFFFAOYSA-N 0.000 description 1
- 229920002866 paraformaldehyde Polymers 0.000 description 1
- 244000045947 parasite Species 0.000 description 1
- 238000003909 pattern recognition Methods 0.000 description 1
- 230000008447 perception Effects 0.000 description 1
- 238000009521 phase II clinical trial Methods 0.000 description 1
- 229960003742 phenol Drugs 0.000 description 1
- YBYRMVIVWMBXKQ-UHFFFAOYSA-N phenylmethanesulfonyl fluoride Chemical class FS(=O)(=O)CC1=CC=CC=C1 YBYRMVIVWMBXKQ-UHFFFAOYSA-N 0.000 description 1
- 150000003016 phosphoric acids Chemical class 0.000 description 1
- 229920002401 polyacrylamide Polymers 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 108010094020 polyglycine Proteins 0.000 description 1
- 229920000232 polyglycine polymer Polymers 0.000 description 1
- 229940115272 polyinosinic:polycytidylic acid Drugs 0.000 description 1
- 229940031937 polysaccharide vaccine Drugs 0.000 description 1
- 239000001267 polyvinylpyrrolidone Substances 0.000 description 1
- 235000013855 polyvinylpyrrolidone Nutrition 0.000 description 1
- 229920000036 polyvinylpyrrolidone Polymers 0.000 description 1
- 244000144977 poultry Species 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 235000019419 proteases Nutrition 0.000 description 1
- 239000003207 proteasome inhibitor Substances 0.000 description 1
- 229960000856 protein c Drugs 0.000 description 1
- 238000002818 protein evolution Methods 0.000 description 1
- 230000002797 proteolythic effect Effects 0.000 description 1
- 238000000197 pyrolysis Methods 0.000 description 1
- 239000001397 quillaja saponaria molina bark Substances 0.000 description 1
- 229940047431 recombinate Drugs 0.000 description 1
- 238000011084 recovery Methods 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 230000003362 replicative effect Effects 0.000 description 1
- 108091008146 restriction endonucleases Proteins 0.000 description 1
- 239000003161 ribonuclease inhibitor Substances 0.000 description 1
- 230000000630 rising effect Effects 0.000 description 1
- DPCQJLQPDJPRCM-UHFFFAOYSA-N s-acetyl ethanethioate Chemical group CC(=O)SC(C)=O DPCQJLQPDJPRCM-UHFFFAOYSA-N 0.000 description 1
- 150000003873 salicylate salts Chemical class 0.000 description 1
- 235000019515 salmon Nutrition 0.000 description 1
- 229940098714 salmonella enterica subsp. enterica serovar typhi Drugs 0.000 description 1
- 229930182490 saponin Natural products 0.000 description 1
- 150000007949 saponins Chemical class 0.000 description 1
- 238000012163 sequencing technique Methods 0.000 description 1
- 239000003352 sequestering agent Substances 0.000 description 1
- 239000008159 sesame oil Substances 0.000 description 1
- 235000011803 sesame oil Nutrition 0.000 description 1
- 230000001568 sexual effect Effects 0.000 description 1
- 238000012868 site-directed mutagenesis technique Methods 0.000 description 1
- 235000010413 sodium alginate Nutrition 0.000 description 1
- 239000000661 sodium alginate Substances 0.000 description 1
- 229940005550 sodium alginate Drugs 0.000 description 1
- 229910001467 sodium calcium phosphate Inorganic materials 0.000 description 1
- 235000017550 sodium carbonate Nutrition 0.000 description 1
- 229910000029 sodium carbonate Inorganic materials 0.000 description 1
- 239000001509 sodium citrate Substances 0.000 description 1
- AJPJDKMHJJGVTQ-UHFFFAOYSA-M sodium dihydrogen phosphate Chemical compound [Na+].OP(O)([O-])=O AJPJDKMHJJGVTQ-UHFFFAOYSA-M 0.000 description 1
- 239000001488 sodium phosphate Substances 0.000 description 1
- 239000012064 sodium phosphate buffer Substances 0.000 description 1
- 239000007901 soft capsule Substances 0.000 description 1
- 238000002798 spectrophotometry method Methods 0.000 description 1
- 238000005507 spraying Methods 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 239000008117 stearic acid Substances 0.000 description 1
- 230000001954 sterilising effect Effects 0.000 description 1
- 238000004659 sterilization and disinfection Methods 0.000 description 1
- 210000001562 sternum Anatomy 0.000 description 1
- 208000002254 stillbirth Diseases 0.000 description 1
- 231100000537 stillbirth Toxicity 0.000 description 1
- 230000004936 stimulating effect Effects 0.000 description 1
- 238000003756 stirring Methods 0.000 description 1
- 238000003860 storage Methods 0.000 description 1
- 229940031626 subunit vaccine Drugs 0.000 description 1
- 229960002317 succinimide Drugs 0.000 description 1
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 1
- 238000001356 surgical procedure Methods 0.000 description 1
- 238000002560 therapeutic procedure Methods 0.000 description 1
- 230000008719 thickening Effects 0.000 description 1
- 210000001694 thigh bone Anatomy 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 238000001890 transfection Methods 0.000 description 1
- 238000003146 transient transfection Methods 0.000 description 1
- 230000014616 translation Effects 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- ODLHGICHYURWBS-LKONHMLTSA-N trappsol cyclo Chemical compound CC(O)COC[C@H]([C@H]([C@@H]([C@H]1O)O)O[C@H]2O[C@@H]([C@@H](O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O[C@H]3O[C@H](COCC(C)O)[C@H]([C@@H]([C@H]3O)O)O3)[C@H](O)[C@H]2O)COCC(O)C)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O)[C@@H]3O[C@@H]1COCC(C)O ODLHGICHYURWBS-LKONHMLTSA-N 0.000 description 1
- HRXKRNGNAMMEHJ-UHFFFAOYSA-K trisodium citrate Chemical compound [Na+].[Na+].[Na+].[O-]C(=O)CC(O)(CC([O-])=O)C([O-])=O HRXKRNGNAMMEHJ-UHFFFAOYSA-K 0.000 description 1
- 229940038773 trisodium citrate Drugs 0.000 description 1
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 1
- 125000000430 tryptophan group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C2=C([H])C([H])=C([H])C([H])=C12 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 239000013598 vector Substances 0.000 description 1
- 210000003462 vein Anatomy 0.000 description 1
- 238000012795 verification Methods 0.000 description 1
- 230000029812 viral genome replication Effects 0.000 description 1
- 239000000277 virosome Substances 0.000 description 1
- 235000019165 vitamin E Nutrition 0.000 description 1
- 229940046009 vitamin E Drugs 0.000 description 1
- 239000011709 vitamin E Substances 0.000 description 1
- 239000003643 water by type Substances 0.000 description 1
- 238000009736 wetting Methods 0.000 description 1
Images

















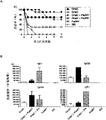











Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/025—Enterobacteriales, e.g. Enterobacter
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/02—Bacterial antigens
- A61K39/025—Enterobacteriales, e.g. Enterobacter
- A61K39/0275—Salmonella
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/145—Orthomyxoviridae, e.g. influenza virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/385—Haptens or antigens, bound to carriers
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/39—Medicinal preparations containing antigens or antibodies characterised by the immunostimulating additives, e.g. chemical adjuvants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P1/00—Drugs for disorders of the alimentary tract or the digestive system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/46—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- C07K14/47—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
- C07K14/4701—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals not used
- C07K14/4748—Tumour specific antigens; Tumour rejection antigen precursors [TRAP], e.g. MAGE
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5256—Virus expressing foreign proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5258—Virus-like particles
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55516—Proteins; Peptides
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/60—Medicinal preparations containing antigens or antibodies characteristics by the carrier linked to the antigen
- A61K2039/6031—Proteins
- A61K2039/6075—Viral proteins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/77—Internalization into the cell
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/70—Fusion polypeptide containing domain for protein-protein interaction
- C07K2319/735—Fusion polypeptide containing domain for protein-protein interaction containing a domain for self-assembly, e.g. a viral coat protein (includes phage display)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16134—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/26011—Flexiviridae
- C12N2770/26022—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/26011—Flexiviridae
- C12N2770/26023—Virus like particles [VLP]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/26011—Flexiviridae
- C12N2770/26034—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Immunology (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Organic Chemistry (AREA)
- Virology (AREA)
- Epidemiology (AREA)
- Mycology (AREA)
- Microbiology (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biochemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Zoology (AREA)
- Communicable Diseases (AREA)
- Oncology (AREA)
- Pulmonology (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Peptides Or Proteins (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Abstract
本发明提供一种抗原提呈系统(APS),该抗原提呈系统包含与番木瓜花叶病毒(PapMV)或源自番木瓜花叶病毒的病毒样颗粒(VLP)组合的一种或多种肠细菌抗原。APS中包含的PapMV或VLP能够增强针对所述一种或多种肠细菌抗原的免疫应答。所述APS可以用作(例如)针对肠细菌疾病(例如伤寒)的疫苗。APS中包含的一种或多种抗原能够与PapMV或PapMV VLP的外壳蛋白接合,或者它们可以是非接合的(即,所述抗原与PapMV或PapMV VLP分离),或者所述APS既包含接合的抗原也包含非接合的抗原。所述接合可以(例如)通过与外壳蛋白的基因融合来实现,或者通过共价、非共价或亲和方式结合来实现。
Description
技术领域
本发明涉及疫苗制剂和佐剂的领域,具体而言,本发明涉及基于植物病毒颗粒的疫苗。
背景技术
肠杆菌科细菌包括一大类革兰氏阴性菌,并且包括众多对人类医学和兽医学具有重要临床意义的病原体。肠细菌病原体是肠道感染和全身性感染的致病因子,并且其包括肠道沙门氏菌(Salmonellaenterica)(伤寒和食源性胃肠炎)、致病性大肠杆菌(pathogenicEscherichia coli)(各种食源性肠道感染、尿路感染、脑膜炎、和败血症)、志贺菌属种(Shigella spp.) (细菌性痢疾)、耶尔森菌属种(Yersinia spp.)(鼠疫和小肠结肠炎)以及克雷白氏杆菌(Klebsiella)和肠杆菌属种(Enterobacter spp.)(肺炎和血液感染)。这些肠细菌品种还与医源性感染和革兰氏阴性菌败血症相关。此外,在治疗肠细菌引起的疾病时,对抗生素的多重耐药性已经成为一个主要问题(Paterson,2006,AJIC 34:20-28)。
人体的至少5种胃肠疾病是由大肠杆菌引起的。大肠杆菌菌株的致病性是由于其内存在一种或多种毒力因子而产生。这些毒力因子包括:侵染因子(侵染素)、不耐热肠毒素和耐热肠毒素、细菌外毒素(verotoxin)以及定殖因子或粘附素。最近发现的大肠杆菌疾病是由血清型01 57:H7的菌株引起的出血性结肠炎。这些菌株产生细菌外毒素,并且该疾病以痛苦的腹部绞痛和出血性腹泻为特征。该相同菌株还与人体的溶血性尿毒性综合征(HUS)有关。
小肠结肠炎耶尔森菌(Yersinia enterocolitica)亦引起人体中的腹泻。此外,腹泻还与迟缓爱德华菌(Edwardsiella tarda)和柠檬酸杆菌(Citrobacter)的菌株引起的感染有关。肺炎克雷白氏杆菌(Klebsiella pneumoniae)和阴沟肠杆菌(Enterobacter cloacae)的菌株已经从患有热带口炎性腹泻的病人中分离出来。
伤寒(一种严重的全身性感染)是通过肠细菌伤寒沙门氏菌(S.typhi)对网状内皮系统的急性感染而引起的。每年至少发生1600万例的伤寒,其中导致60万例死亡。这种感染通过粪-口途径传播并且通常通过摄入污染的食物或水而进入人体。在这种细菌穿过肠壁之后,其在24至72小时内繁殖并进入血液内,从而导致伤寒和菌血症。在10至14天的时期内,伤寒的早期症状包括:头疼、发热、便秘、胃疼、厌食和肌痛。
肠杆菌科细菌还代表非人类动物中的一大类病原体。例如,肺炎克雷白氏杆菌是灵长类动物中呼吸系统疾病的常见病因,并且假结核耶尔森菌(Yersinia pseudotuberculosis)与小肠结肠炎和腹膜炎有关。灵长类动物中最经常诊断出的腹泻病是由志贺氏杆菌、大肠杆菌和沙门氏菌引起的。猫狗等家庭宠物易患由大肠杆菌引起的诸如膀胱炎和其它泌尿生殖道感染之类的疾病。变形杆菌(proteus)类引起猫狗中的其它疾病。
牛犊等家畜易患全身性大肠杆菌病和新生牛犊腹泻(犊牛白痢),这些疾病是由含有K99菌毛粘附素的耐热肠毒素性大肠杆菌血清型引起的。牛的乳腺炎最通常由大肠杆菌和沙雷氏菌(较少见的是克雷白氏杆菌和弗氏柠檬酸杆菌(Citrobacter freundii))的感染而引起的。都柏林沙门氏菌(Salmonella dublin)和鼠沙门氏菌(S.typhimurium)也是奶牛中的重要病原体。大肠杆菌的感染在小猪中引起腹泻,或在轻度腹泻之后发生水肿。猪大肠杆菌菌株通过存在K88菌毛粘附素来表征,所述K88菌毛粘附素的抗原性不同于上述的K99抗原。猪的乳腺炎和子宫炎由肺炎克雷白氏杆菌(K.pneumoniae)引起,并且其肠炎和淋巴腺炎由小肠结肠炎耶尔森菌(Yersinia enterocolitica)引起。猪霍乱沙门氏菌(S.choleraesuis)和猪伤寒沙门氏菌(S.typhisuis)亦引起猪的疾病。
沙门氏菌(例如鼠沙门氏菌、新港沙门氏菌(S.newport)和鸭沙门氏菌(S.anatum))引起马的肠炎,这会导致高的致死率和脓毒性流产。马驹和母马会遭受肺炎克雷白氏杆菌的感染,这可导致子宫炎(母马)和肺炎(马驹)。绵羊亦会患肠杆菌科细菌引起的各种疾病。例如,产生耐热肠毒素的大肠杆菌菌株引起羊羔的幼羊腹泻。这些菌株的大多数含有K-99菌毛粘附素。沙门氏菌性流产通常由流产沙门氏菌(Salmonella abortusovis)、鼠沙门氏菌或都柏林沙门氏菌引起,这些细菌还导致死产和羊毛损伤。
大肠杆菌引起的败血症是导致小鸡死亡的重要原因。肠道沙门氏菌血清型以及鸡白痢沙门氏菌(Salmonella pullorum)和鸡沙门氏菌(Salmonella gallinarum)是家禽的主要病原体。
渔业的巨大损失亦可归因于各种肠细菌的感染。例如,鲑鱼和鳟鱼孵化场的红嘴病(redmouth disease)的爆发是由鲁氏耶尔森菌(Yersinia ruckeri)引起的。迟缓爱德华菌是鳗鲡、鲶鱼和金鱼的病原体,并且鲶鱼爱德华菌(Edwardsiella ictaluri)对鲶鱼具有致病性。
尽管抗生素治疗肠杆菌感染是可行的,但是肠杆菌科细菌中耐药性的出现和蔓延使得传染病的治疗复杂化,并有产生对目前所有可用的药剂都具有耐药性的菌种的危险(Paterson,2006,AJIC34:20-28)。因此,人们已经研究了用于预防和治疗细菌感染的疫苗。例如,美国专利申请No.10/394,517(公开号为2003/0215464)描述了缺失功能性脂蛋白(LPS)的减毒肠杆菌科细菌。这些减毒细菌不能够引起免疫应答,因而只是潜在的候选疫苗。
人们还开发了抗伤寒的疫口服的减毒活galE突变体Ty21a( 疫苗,由位于瑞士的Berne市的Berna Biotech有限公司生产)在流行病区有效,但是它没有被许可用于小于六岁龄的儿童。此外,需要3至4次的给药以实现部分保护的免疫应答(Levine等,1999,Vaccine 1:S22-S27)。Vi荚膜多糖疫苗(Vi capsularpolysaccharide vaccine)(ViCPS)(Typhim ViTM;Aventis Pasteur)被许可用于超过2岁龄的儿童;Vi的一次注射提供与Ty21a疫苗类似的保护作用,但是仅仅短期地达到该效果。缺少持久性免疫是该疫苗的主要缺点(Lin等,2001,N Engl J Med.344:1263-9;Sabitha等,2004,Indian J.Med.Sci.58:141-149)。此外,这些疫苗通常在50%至80%的接受者中实现最佳的保护作用(Crump等,2004,Bulletin ofthe World Health Organisation,82;346-353;Levine等,1999,Vaccine1:S22-S27;Lin等,2001,N Engl J Med.344:1263-9;Sabitha等,2004,Indian J.Med.Sci.58:141-149)。基于伤寒沙门氏菌(Ty-800)的减毒菌株的新候选疫苗已经由Emergent BioSolutions公司(位于美国马里兰州的Gaithersburg市)开发。该候选疫苗目前在越南(流行病区)处于二期临床试验阶段(Crump等,2004,ibid.)。关于Ty21a疫苗,该疫苗通过口服途径给药,并且可能不适于儿童服用。
疫苗,由位于瑞士的Berne市的Berna Biotech有限公司生产)在流行病区有效,但是它没有被许可用于小于六岁龄的儿童。此外,需要3至4次的给药以实现部分保护的免疫应答(Levine等,1999,Vaccine 1:S22-S27)。Vi荚膜多糖疫苗(Vi capsularpolysaccharide vaccine)(ViCPS)(Typhim ViTM;Aventis Pasteur)被许可用于超过2岁龄的儿童;Vi的一次注射提供与Ty21a疫苗类似的保护作用,但是仅仅短期地达到该效果。缺少持久性免疫是该疫苗的主要缺点(Lin等,2001,N Engl J Med.344:1263-9;Sabitha等,2004,Indian J.Med.Sci.58:141-149)。此外,这些疫苗通常在50%至80%的接受者中实现最佳的保护作用(Crump等,2004,Bulletin ofthe World Health Organisation,82;346-353;Levine等,1999,Vaccine1:S22-S27;Lin等,2001,N Engl J Med.344:1263-9;Sabitha等,2004,Indian J.Med.Sci.58:141-149)。基于伤寒沙门氏菌(Ty-800)的减毒菌株的新候选疫苗已经由Emergent BioSolutions公司(位于美国马里兰州的Gaithersburg市)开发。该候选疫苗目前在越南(流行病区)处于二期临床试验阶段(Crump等,2004,ibid.)。关于Ty21a疫苗,该疫苗通过口服途径给药,并且可能不适于儿童服用。
尽管减毒细菌作为用于预防和治疗传染病的疫苗或疫苗载体的潜在广泛应用比其它疫苗具有显著优势,但是关于使用减毒细菌的安全问题并不能忽视。因此,据认为,开发用于传染病的可替代治疗方案(包括结合有特异性抗原的新的疫苗平台)是未来5-10年中改善全球卫生状况的重点(Nikaido,2003,Microbiol Mol Biol Rev.67:593-656)。在这发面,已经有人体对表面细菌抗原的免疫应答的研究报导。例如,Vi(荚膜多糖)、H(鞭毛成分)以及O(脂多糖[LPS])细菌抗原引起针对伤寒沙门氏菌的抗体应答的能力提供了使用特异性抗原开发可替代方案值得进行研究的证据(在文献Calderon等,1986,Infection and Immunity,52:209-212;Ortiz等,1989,J Clin.Microbiol.27:1640-1645中有所讨论)。最近,研究人员发现,孔蛋白也是引起对由多种革兰氏阴性菌引起的感染的特异性保护性免疫应答的重要抗原(Humphries等,2006,Vaccine 24:36-44;Kim等,1999,J Immunol.162:6855-6866)。孔蛋白是革兰氏阴性菌的三聚体的、暴露的外膜蛋白(OMP),其起着相对非特异的通道的作用,从而容许小的亲水性分子通过外膜(Nikaido,2003,Microbiol Mol Biol Rev.67:593-656)。
对孔蛋白的免疫应答似乎既涉及体液介导的免疫途径,又涉及细胞介导的免疫途径。例如,伤寒的严重或恢复期的患者表现出高水平的孔蛋白特异性抗体(Calderon等,1986,Infect.Immun.52:209-212;Ortiz等,1989,J Clin.Microbiol.27:1640-1645)。此外,伤寒患者以及用Ty21a口服疫苗免疫的志愿者已经显示出孔蛋白特异性细胞免疫应答(Salerno-Goncalves等,2002,J Immunol.169:2196-203)。伤寒沙门氏菌的两种主要孔蛋白OmpC和OmpF已经显示出能够提高鼠的持久性抗体应答(Secundino等,2006,Immunology 117:59)。因此,综上所述,这些数据表明,孔蛋白是针对肠细菌感染的细胞免疫应答和体液免疫应答的重要靶点。
在各种疫苗开发的新方法中,作为有前景的策略,已经出现由病毒核壳体(viral nucleocapsid)制成的病毒样颗粒(VLP)。迄今为止,两种VLP疫苗(乙型肝炎病毒(HBV)疫苗和人乳头瘤病毒(HPV)疫苗)已经显示出在人体中起有效作用(Fagan等,1987,J.Med.Virol,21:49-56;Harper等,2004,Lancet,364:1757-1765)。例如,由人乳头瘤病毒(HPV)的主核壳体蛋白L1制成的VLP在抗妇女子宫颈癌的发展时显示出提供100%的保护作用(Ault,K.A.,2006,Obstet.Gynecol.Surv.61:S26-S31;Harper等,2004,Lancet 364:1757-1765,另外请参见国际专利申请PCT/US01/18701(WO 02/04007))。诸如噬菌体Qβ(Maurer等,2005,Eur.J.Immunol.35:2031-2040)、由病毒核心蛋白制成的乙型肝炎病毒VLP(Mihailova等,2006,Vaccine24:4369-4377;Pumpens等,2002,Intervirology 45:24-32)和细小病毒VLP(Antonis等,2006,Vaccine 24:5481-5490;Ogasawara等,2006,InVivo 20:319-324)等平台也已经显示出携带抗原表位以及引起强的抗体应答的能力。类似地,美国专利No.6,627,202描述了用作抗原表位输送系统的HBV核心蛋白,其含有通过HBV壳体结合肽交联的抗原(包括靶向于或源自各种病毒和细菌的抗原)。
已经描述有使用来自植物病毒的VLP作为抗原表位提呈系统(epitope presentation system)。植物病毒主要由高免疫原性的蛋白组成,并具有复杂的、重复的和结晶的组织。此外,它们在种系起源学上明显不同于动物免疫系统,这使得它们成为开发疫苗的良好的候选物。例如,豇豆花叶病毒(CPMV)、约翰逊草花叶病毒(JGMV)、烟草花叶病毒X(TMVX)和苜蓿花叶病毒(AIMV)已经被修饰用于目标抗原表位的提呈(Canizares,M.C.等,2005,Immunol.Cell.Biol.83:263-270;Brennan等,2001,Molec.Biol.17:15-26;Saini和Vrati,2003,J.Virol.77:3487-3494)。国际专利申请PCT/GB97/010659(WO 97/39134)描述了包含外壳蛋白和非病毒蛋白的嵌合的病毒样颗粒,其可以用于(例如)肽抗原表位的提呈。国际专利申请PCT/US01/07355(WO 01/66778)描述了一种植物病毒外壳蛋白,特别是在N-端处通过连接子与目标多肽融合的烟草花叶病毒外壳蛋白,其可以包含病原性微生物的抗原表位。国际专利申请PCT/US01/20272(WO 02/00169)描述了含有马铃薯病毒Y外壳蛋白或与外源肽融合的截断的菜豆黄花叶病毒外壳蛋白的疫苗,特别是新城疫病毒(Newcastle Disease Virus)的抗原表位或人类免疫缺陷病毒(HIV)的抗原表位。此外,美国专利No.6,042,832描述了将融合苜蓿花叶病毒或烟草线条病毒壳体蛋白的多肽(例如病原体抗原表位)施用给动物以提高免疫应答的方法。
已经描述有源自番木瓜花叶病毒(PapMV)的外壳蛋白的VLP及其作为免疫增强剂的用途(国际专利申请No.PCT/CA03/00985(WO 2004/004761))。PapMV在大肠杆菌中的表达导致由数百个以重复的和结晶的方式组织的CP亚单元构成的VLP的自组装(Tremblay等,2006,FEBS J 273:14)。对PapMV CP缺失构建体的表达和纯化的研究进一步表明,CP亚单元的自组装(或多聚化)对于功能是重要的(Lecours等,2006,Protein Expression andPurification,47:273-280)。已经表明,PapMV VLP(其含有源自gp100或流感病毒M1蛋白的抗原表位)能够引起抗原表位的MHC I类交叉提呈,从而导致特异性人体T细胞的扩增(Leclerc,D.等,J.Virol,2007,81(3):1319-26;在2006年11月22日印刷之前电子出版)。
已经描述有源自携带各种抗原决定簇(其来自HIV、HCV、EBV或流感病毒)的马铃薯病毒X(PVX)的VLP(欧洲专利申请No.1167 530)。携带HIV抗原表位的PVX VLP能够在鼠体内通过体液介导的途径和细胞介导的途径来诱导抗体的产生。额外的佐剂可以与PVX VLP联用,以增强这种效果。
已经报导,乙型肝炎病毒核心蛋白或细小病毒VLP即使不携带遗传信息时,也会诱导CTL应答(Ruedl等,2002,Eur.J.Immunol.32;818-825;Martinez等,2003,Virology,305;428-435),并且它们在其嵌入的细胞内不能主动复制。还描述有携带有来自淋巴细胞性脉络丛脑膜炎病毒(lymphocytic choriomeningitis virus)(LCMV)或卵清白蛋白的抗原表位的VLP通过树突状细胞在体内的交叉提呈(Ruedl等,2002,ibid.;Moron等.,2003,J.Immunol.171:2242-2250)。还描述有携带有来自LCMV的抗原表位的乙型肝炎病毒核心蛋白VLP启动CTL应答的能力,但是,当单独施用时该VLP不能够诱导CTL应答并且不能介导对病毒攻击的有效保护。只有当该VLP与抗CD40抗体或CpG寡核苷酸联用时才能诱导有效的CTL应答(Storni等.,2002,J.Immunol.168:2880-2886)。较早的报导表明,携带有来自LCMV的肽的猪细小病毒样颗粒(PPMV)能够保护小鼠抵抗致命的LCMV攻击(Sedlik等,2000,J.Virol.74:5769-5775)。
人们还描述了融合有亲和肽的番木瓜花叶病毒VLP的用途(Morin等,2007,J.Biotechnology,128:423-434),所述的VLP能够结合根肿菌(Plasmodiophora brassicae)的休眠孢子。该VLP显示出能够以高亲和力结合根肿菌孢子,并且被提出作为用于检测根肿菌的抗体的替代物。
提供上述背景信息的目的是使人们了解申请人认为可能与本发明有关的信息。提供上述信息既不是意图承认,也不应该解释为其构成了针对本发明的现有技术。
发明概述
本发明的一个目的是提供抗伤寒沙门氏菌和其它肠细菌病原体的基于番木瓜花叶病毒的疫苗。根据本发明的一个方面,提供一种抗原提呈系统,其包含与番木瓜花叶病毒(PapMV)或者源自PapMV外壳蛋白的VLP组合的一种或多种肠细菌抗原,其中,所述PapMV或VLP能够增强针对所述一种或多种肠细菌抗原的免疫应答。
根据本发明的另一个方面,提供一种疫苗组合物,其包含本发明的一种或多种抗原提呈系统和可药用的载体。
根据本发明的另一个方面,提供一种多肽,其包含与一种或多种亲和肽融合的番木瓜花叶病毒外壳蛋白,所述一种或多种亲和肽能够结合肠细菌抗原。
根据本发明的另一个方面,提供一种多肽,其包含与一种或多种肠细菌抗原融合的番木瓜花叶病毒外壳蛋白。
根据另一个方面,提供一种编码本发明多肽的多核苷酸。
根据另一个方面,提供本发明的抗原提呈系统在需要其的动物中用于诱导针对肠细菌的免疫应答中的用途。
根据另一个方面,提供本发明的疫苗在需要其的动物中用于诱导针对肠细菌的免疫应答中的用途。
根据另一个方面,提供一种本发明的抗原提呈系统在制备疫苗中的用途。
根据本发明的另一个方面,提供一种用于免疫动物以抗伤寒沙门氏菌感染的疫苗,所述疫苗包含抗原提呈系统,所述抗原提呈系统包含与番木瓜花叶病毒(PapMV)或源自PapMV外壳蛋白的VLP组合的抗原,所述抗原源自伤寒沙门氏菌OmpC或OmpF,其中所述PapMV或VLP能够在所述动物中增强针对所述抗原的免疫应答
根据本发明的另一个方面,提供一种在动物中诱导针对肠细菌的免疫应答的方法,所述方法包括给所述动物施用有效量的本发明的抗原提呈系统。
根据本发明的另一个方面,提供一种在动物中诱导针对肠细菌的免疫应答的方法,所述方法包括给所述动物施用有效量的本发明的疫苗。
附图简要说明
在下面结合附图的详细描述中,本发明的这些特征以及其它特征将变得显而易见。
图1示出(A)番木瓜花叶病毒外壳(或壳体)蛋白的氨基酸序列(GenBank登录号NP_044334.1;SEQ ID NO:1)、(B)编码番木瓜花叶病毒外壳蛋白的核苷酸序列(GenBank登录号NC 001748(核苷酸5889-6536);SEQ ID NO:2)、以及(C)修饰的PapMV的外壳蛋白CPΔN5的氨基酸序列(SEQ ID NO:3)。
图2示出:(A)WT PapMV病毒、(B)CPΔN5PapMV病毒样颗粒(VLP)、(C)E128A VLP、(D)K97A纯化蛋白、(E)免疫金标记的重组CP ΔN5VLP(其含有具有6XH标记物的抗体)的电子显微照片,其中横杠表示200nm;以及(F)PapMV病毒和重组CPΔN5VLP以及E128A VLP的平均长度(n=150)。
图3示出野生型(WT)PapMV和包含重组突变体PapMV外壳蛋白CP ΔN5、E128A或K97A的PapMV病毒样颗粒(VLP)的圆二色(CD)谱;(A)显示WT病毒(黑线)、CPΔN5(虚线)和E128A VLP(灰线)的远紫外光谱,(B)显示通过乙酸法从WT病毒分离的盘状物(黑线)和CPΔN5蛋白(盘状物)的高速上清液(虚线)的远紫外光谱,(C)显示CPΔN5蛋白(盘状物)(虚线)和K97A蛋白(灰线)的远紫外光谱,(D)显示CPΔN5和E128A VLP在250nm和350nm之间的远紫外光谱,以及(E)显示CPΔN5和K97A盘状物在250nm和350nm之间的远紫外光谱。
图4示出重组突变体PapMV外壳蛋白、CP ΔN5和K97A的凝胶过滤分析的结果;(A)示出CPΔN5纯化蛋白的蛋白洗脱曲线(黑线;灰线:分子量标记物),以及(B)示出CPΔN5纯化蛋白(黑线)和K97A纯化蛋白(灰线)的蛋白洗脱曲线。
图5示出重组突变体PapMV外壳蛋白和野生型(WT)PapMV外壳蛋白的圆二色(CD)谱温度诱导变性曲线;(A)显示CPΔN5和E128A突变体VLP以及WT病毒的曲线,以及(B)显示CPΔN5和WT盘状物的曲线。光谱以平均残基椭圆率的单位进行显示。通过记录在222nm处的[Q](其为温度的函数)来监测解折叠(unfolding)。所有显示的CD谱都是采用在pH 7.2的10mM NaP缓冲液中浓度为1mg/毫升的蛋白产生的。
图6示出:(A)PapMV外壳蛋白(PapMV)和修饰的PapMV构建体的C-端序列的比对,其中所述修饰的PapMV构建体含有来自流感病毒M1蛋白的HLA-A*0201限制抗原表位(57至65位,标记为PapMV Flu)或来自gp100的HLA-A*0201限制抗原表位(在210位处具有M的209-217位;标记为PapMV gp100);以及(B)各种重组的自组装PapMV制剂的电子显微照片。
图7示出重组的PapMV外壳蛋白(PapMV)和外壳蛋白融合体(PapMV-gp100和PapMV-FIu)在合成2个月和7个月之后的稳定性的SDS-PAGE分析结果。其还显示将7个月的原蛋白制剂在23℃或37℃下再培养7天的结果,以及在加入加样缓冲液和在沸水中培养之前将其用蛋白酶K处理5分钟的结果。
图8示出当对两种不同来源的APC进行冲击时,来自两种抗原的MHC I类抗原表位能够被交叉提呈。从HLA-A*0201供体制备CD40活化的B淋巴细胞和DC,并用所示的制剂在不同浓度下冲击18小时。清洗冲击后的细胞并加入gp100-特异性T细胞(A)或FLU-特异性T细胞(B),以另外冲击18小时。通过用ELISA分析测量的IFN-γ分泌量来显示T细胞活化的证据。
图9示出得自(A)和(B)的结果,其中(A)为:将从HLA-A*02阳性或阴性的供体制备的经PapMV Flu或PapMV gp100冲击的CD40活化的B淋巴细胞与FLU-特异性T细胞(左栏)或gp100-特异性T细胞(右栏)共培养,和(B)为:将HLA-A*0201+和gp100+黑素瘤细胞系、或经PapMV Flu冲击的HLA-A*02+CD40活化的B细胞与封闭MHC I类、II类或HLA-DR提呈的抗体培养。然后添加FLU-特异性T细胞或gp 100-特异性T细胞。结果以基于IFN-γ分泌测定法的识别百分比表示,100%对应于阳性对照的分泌量。
图10显示由PapMV介导的MHC I类交叉提呈是蛋白酶体非依赖性的。将HLA-A*0201+和gp100+黑素瘤细胞系、或经PapMV Flu冲击的HLA-A*02+CD40活化的B细胞与封闭MHC I类、II类或HLA-DR提呈的抗体培养。然后添加FLU-特异性T细胞或gp100-特异性T细胞。结果以基于IFN-γ分泌测定法的识别百分比表示,100%对应于阳性对照的分泌量。
图11示出用PapMV Flu冲击的APC使对源自流感病毒M1蛋白的HLA-A*0201抗原表位特异性的T淋巴细胞扩增的分析。(A)将来自HLA-A2+正常供体的PBMC与自体性CD40活化的B细胞(模拟物)或者用PapMV、PapMV Flu或PapMV gp100冲击的CD40活化的B细胞共培养。将培养的细胞在第7天再次刺激并且在不同时间添加IL-2。在第15天通过将其与T2细胞(其经过源自Flu或gp100的HLA-A*0201肽冲击)共培养来评价扩增细胞的特异性。通过ELISPOT测定分泌IFN-γ的细胞的频率。(B)将扩增培养的T细胞与受冲击的CD40活化的B淋巴细胞共培养。
图12示出按照图11中的方式处理的扩增的T细胞的IFN-γ分泌的分析结果。
图13示出在CD-1小鼠中隆起的背部气囊中,PapMV外壳蛋白和LPS的促炎反应的比较。数据以5只小鼠的平均值±SEM表示。结果代表两个相同但独立的试验。
图14示出在分别皮下注射多聚的PapMV-E2、单体的对应物(PapMVCP27-215-E2)、单独的HCV E2肽或PBS的C3H/HeJ小鼠中,对PapMV外壳蛋白特异性的IgG抗体的应答(ELISA板涂上PapMV-CP或PapMV27-215)。数据代表了4只小鼠的抗体效价的平均值。这些结果代表两个相同但独立的试验。图表上的黑箭头表示第15天的加强注射。
图15示出在分别用抗原、抗原加PapMV、抗原加弗氏完全佐剂(FCA)或抗原加源自大肠杆菌O111:B4的LPS进行免疫的BALB/c小鼠(每组三只)中,PapMV在第0天增强抗体对模型抗原(A)鸡卵溶菌酶(HEL)和(B)卵清蛋白(OVA)的应答的能力。图中示出了2个试验的代表性结果。对于(C)IgG1、(D)IgG2a和(E)IgG2b而言,通过ELISA用模型抗原(HEL或OVA)将从免疫的动物采集的血清中的抗体同型化。
图16示出:(A)野生型PapMV外壳蛋白和重组构建体的C-端的氨基酸序列,所述重组构建体在PapMV外壳蛋白的C-端包含融合体,所述融合体为亲和肽与OmpC的融合体或亲和肽与OmpF的融合体(分别为构建体PapMV OmpC和PapMV OmpF);(B)显示纯化的蛋白PapMV、PapMV OmpC、PapMV OmpF、OmpC和OmpF的SDS-PAGE图[第一泳道:分子量标记物,第二泳道:PapMV VLP,第三泳道:PapMV OmpC VLP,第四泳道:PapMV OmpF VLP,第五泳道:纯化的OmpC,第六泳道:纯化的OmpF];和(C)重组PapMVOmpC VLP和PapMV OmpF VLP的高速粒状沉淀的电子显微照片。
图17示出PapMV VLP与它们各自的抗原高亲和力结合;(A)为显不PapMV OmpC VLP与OmpC抗原高亲和力结合的ELISA;(B)为显示PapMV OmpF VLP与OmpF抗原高亲和力结合的ELISA。
图18示出小鼠中抗伤寒沙门氏菌攻击的保护试验的结果。(A)示出单用OmpC免疫的小鼠和用含有OmpC+PapMV OmpC VLP的制剂免疫的小鼠中,抗100LD50的伤寒沙门氏菌的保护能力;(B)示出单用OmpF免疫的小鼠和用含有OmpF+PapMV OmpF VLP的制剂免疫的小鼠中,抗100LD50的伤寒沙门氏菌的保护能力;(C)示出单用OmpC免疫的小鼠和用含有OmpC+PapMV OmpC VLP的制剂免疫的小鼠中,抗500LD50的伤寒沙门氏菌的保护能力;并且(D)示出单用OmpF免疫的小鼠和用含有OmpF+PapMV OmpF VLP的制剂免疫的小鼠中,抗500LD50的伤寒沙门氏菌的保护能力。
图19示出:针对OmpC的抗体应答的评价(A-D);在用OmpC或包含OmpC+PapMV OmpC VLP的疫苗免疫的小鼠之间测量针对同种型的IgG1(A)、IgG2a(B)、IgG2b(C)和IgG3(D)的OmpC的IgG应答,以及(E)与单用OmpC或PapMV OmpC免疫的情况相比,用OmpC和PapMV OmpC共免疫之后再用伤寒沙门氏菌攻击有利于小鼠对伤寒沙门氏菌感染的持久性主动保护(如存活率%所示)。
图20示出PapMV病毒增加了小鼠中的OmpC孔蛋白的保护能力;(A)对雌性BALB/c小鼠(每组10只)单用10μg OmpC或与30μg PapMV联用进行腹腔内免疫。在第15天单用OmpC给予加强免疫。将对照小鼠注射盐水(ISS)或PapMV。在第21天,用100 LD50(实心符号)或500(空心符号)LD50的伤寒沙门氏菌对试验组进行攻击,并记录攻击10天之后的存活率。对照组用20 LD50的伤寒沙门氏菌攻击。图中示出了3次试验的代表性结果。(B)将雌性BALB/c小鼠(每组5只)在第0天单用10μg OmpC或者与30μg PapMV或弗氏不完全佐剂(IFA)(1∶1,v/v)联用进行腹腔内免疫。在第15天仅用OmpC对全部小鼠给予加强免疫。对照小鼠仅注射盐水(等渗盐水(ISS))。在初次免疫后的第21天通过酶联免疫吸附试验(ELISA)测量抗体效价。
图21示出(A)源自肠道沙门氏菌亚种(Salmonella enterica subsp.enterica serovar Typhi)Ty2的OmpC前体的氨基酸序列(SEQ IDNO:39)(GenBank登录号P0A264);和(B)源自肠道沙门氏菌亚种CT18的OmpF前体蛋白的氨基酸序列(SEQ ID NO:40)(GenBank登录号CAD05399)。
图22示出(A)和(B)的氨基酸序列,其中(A)为:包含用于结合OmpC的亲和肽的PapMV外壳蛋白[SEQ ID NO:42],和(B)包含用于结合OmpF的亲和肽的PapMV外壳蛋白[SEQ ID NO:43]。克隆型序列和野生型序列的不同以黑体和下划线标记出;所述亲和肽序列加有下划线,并且组氨酸标记物显示为斜体。
图23示出:(A)野生型PapMV外壳蛋白和重组构建体的C-端的氨基酸序列,所述重组构建体包含在PapMV外壳蛋白和位于该蛋白的C-端的6XHis标记物(6His)之间融合的伤寒沙门氏菌孔蛋白的环6肽(以黑体和下划线示出);(B)显示重组蛋白的纯化结果的SDS-PAGE凝胶图。1.分子量标记物,2.在诱导蛋白PapMV-环6表达之前的细菌的总细菌裂解液,3.在用IPTG诱导蛋白PapMV-环6表达之后的细菌的总细菌裂解液,4.含有纯化的PapMV-环6蛋白的VLP的高速粒状沉淀,以及(C)显示PapMV病毒、没有任何融合的PapMV重组VLP以及含有环6肽的PapMV重组VLP(PapMV-环6)的电子显微照片。
图24示出:(A)源自包含伤寒沙门氏菌孔蛋白的环6肽的PapMV外壳蛋白(PapMV-环6)的外壳蛋白的氨基酸序列[SEQ IDNO:46],其中环6肽序列加下划线,并且组氨酸标记物用斜体示出,以及(B)编码PapMV-环6蛋白的核苷酸序列[SEQ ID NO:47]。
图25为显示对PapMV-环6构建体的抗体应答的结果。将5只Balb/c小鼠用100μg的PapMV-环6每隔两周进行皮下免疫。在第28天进行最后的采血,并使用以下的条件进行ELISA,所述条件为:(A)合成的和纯化的环6肽,(B)PapMV平台和(C)源自伤寒沙门氏菌的纯化的OmpC。
图26示出在小鼠中抗伤寒沙门氏菌攻击的保护试验的结果。(A)示出在小鼠中分别用OmpF和含有OmpF+PapMV OmpF VLP或OmpF+PapMV的制剂抗77LD50的伤寒沙门氏菌攻击的保护能力,和(B)示出在小鼠中用OmpF和含有OmpF+PapMV OmpF VLP或OmpF+PapMV的制剂抗378LD50的伤寒沙门氏菌攻击的保护能力。
图27示出:(A)PapMV SM外壳蛋白和重组构建体的C-端的氨基酸序列,所述重组构建体包含亲和肽与OmpC在PapMV SM外壳蛋白的C-端处的融合体;(B)显示纯化的蛋白PapMV SM OmpC的SDS-PAGE图[第一泳道:分子量标记物,第二泳道:用IPTG诱导前的细菌裂解液,第三泳道:表达高量的PapMV SM CP的细菌的裂解液,第四泳道:纯化的PapMV SM OmpC蛋白PapMV OmpF];以及(C)重组PapMV SM OmpC的高速粒状沉淀的放大100,000倍的电子显微照片。
图28示出:(A)包含亲和肽的PapMV SM外壳蛋白(PapMVSM OmpC)的氨基酸序列[SEQ ID NO:48],所述亲和肽用于结合OmpC,以及(B)编码PapMV SM OmpC蛋白的核苷酸序列[SEQ IDNO:49]。
图29示出在小鼠中抗伤寒沙门氏菌攻击的保护试验的结果,(A)示出在小鼠中用OmpC和用含有OmpC+PapMV SM OmpCVLP或OmpC+PapMV VLP的制剂抗105LD50的伤寒沙门氏菌的保护能力,以及(B)示出在小鼠中用OmpC和含有OmpC+PapMV SMOmpC VLP或OmpC+PapMV VLP的制剂抗520LD50的伤寒沙门氏菌的保护能力。
图30示出伤寒沙门氏菌孔蛋白的稳定性的评价结果。(A)示出纯化的孔蛋白(OmpC+OmpF)(5μg)的SDS-PAGE图,所述孔蛋白为1999年11月9日制备的批次ISIPOR(Salazar-Gonzales等,2004,Immunology Letters 93),(B)示出2008年1月的相同的ISIPOR批次(0.2μg;泳道2)的SDS-PAGE图,以及(C)示出纯化的OmpC蛋白、2007年4月30日纯化的批次-1239的SDS-PAGE图[左手泳道:2007年4月30日进行操作的凝胶;泳道1:分子量标记物,泳道2:OmpC(10μg);右手泳道:2008年1月18日进行操作的凝胶,泳道1:OmpC(10μg)]。
图31示出PapMV在体内引起了树突状细胞(DC)、巨噬细胞和B细胞中的共刺激分子(co-stimulatory molecule)的上调。将BALB/c小鼠用30μg经盐水稀释的PapMV(灰线)或50μg的聚肌胞苷酸(黑线)腹腔内(i.p.)免疫,并且将对照小鼠注射盐水(实心柱状图)。在免疫后24小时,采集淋巴结和脾脏的细胞并染色。进行流式细胞术以分析细胞群,所述细胞群定义为:CD11c+CD11b-(DC)、CD11c-CD11b+(巨噬细胞)和CD11c-B220+(B细胞)。图中示出两个试验的代表性结果。FITC,异硫氰酸荧光素;MHC II,主要组织相容性复合体II类。
发明详述
本发明提供一种抗原提呈系统(APS),该抗原提呈系统包含与番木瓜花叶病毒(PapMV)或源自番木瓜花叶病毒的VLP组合的一种或多种来自肠杆菌科细菌的抗原。根据本发明的一个实施方案,该APS能够在动物内诱导体液免疫应答和/或细胞免疫应答。因此该APS适合用作抗细菌性疾病或病症的疫苗,其可能需要免疫系统的这两种分支之一或二者的主动参与。
在本发明的一个实施方案中,该APS包含一种或多种伤寒沙门氏菌(S.typhi)抗原,并且适用作治疗和/或预防伤寒的疫苗。
在本发明的另一个实施方案中,该APS包含一种或多种外膜蛋白(Omp)抗原。在一个具体的实施方案中,该APS包含一种或多种OmpC和/或OmpF抗原。如本领域中已知的,在肠杆菌科的成员中,这两种蛋白的区域是保守的。因此,在另一个实施方案中,本发明提供包含一种或多种OmpC和/或OmpF抗原的APS,该APS适用作多价疫苗,以对由多种肠细菌品种引起的感染提供保护作用。在另外的实施方案中,本发明提供包含一种或多种OmpC和/或OmpF抗原的APS,该APS适用作单价疫苗。
定义
除非另有定义,否则本文使用的所有的技术术语和科学术语具有与本发明所属领域的普通技术人员的一般理解相同的含义。
本文使用的术语“约”是指由给定值大致偏差+/-10%。应当理解,无论是否特指,这样的偏差总是包括在本文提供的任何给定值中。
本文使用的术语“佐剂”是指扩大、激发、促进、增强和/或调节动物内的免疫应答的试剂。佐剂本身可能或不可能对免疫应答产生作用。
本文使用的“嵌合蛋白”是在在正常编码两种单独的蛋白或蛋白片段的两种或多种基因天然地或由于人工干预而发生重组,以提供编码这两种蛋白中的每一种的全部或部分的组合的蛋白(“嵌合蛋白”)的多核苷酸时而产生的蛋白。在本发明的上下文中,“融合蛋白”被视为“嵌合蛋白”。
本文使用的表述“融合外壳蛋白”是指这样的融合蛋白,其中融合体中的一种蛋白为PapMV外壳蛋白。
本文使用的术语“免疫应答”是指动物的免疫系统在响应抗原或抗原物质时发生的反应性变化,并且可能涉及抗体的产生、细胞介导免疫的诱导、补体活化、产生免疫耐受性或者它们的组合。
本文使用的术语“有效的免疫保护应答”和“免疫保护”是指这样的免疫应答,其针对一种或多种抗原,从而使接种的动物免受疾病和/或病原体感染。对于本发明的目的而言,免受疾病和/或病原体感染不仅包括绝对预防疾病或感染,还包括与未接种的感染或患病的动物相比,接种的动物在疾病或感染的程度或速率上任何可检测的降低,或者接种的动物在疾病或由病原体感染所导致的任何症状或病症的严重程度方面任何可检测的降低。可以在动物中诱导有效的免疫保护应答,其中该动物之前未患有疾病、之前未感染病原体和/或在接种时没有疾病或感染。还可以在接种时已经患病或感染病原体的动物中诱导有效的免疫保护应答。免疫保护可以是一种或多种机制(包括体液免疫和/或细胞免疫)的结果。
本文中可交替使用的术语“免疫刺激”和“免疫性刺激”是指与动物病原体或疾病无关的分子(例如PapMV或PapMV VLP)通过刺激免疫系统和/或提高免疫系统应答感染或疾病的能力而对病原体感染或疾病提供保护的能力。免疫性刺激可以具有预防效果、治疗效果或它们的组合。
“重组病毒”是这样的病毒,其中天然存在的病毒的遗传物质与其它遗传物质组合。
本文使用的应用于对象的术语“天然存在”是指这样的事实:该对象能够在自然界中找到。例如,能够从天然来源中分离出来、并且未经人工在实验室中有意修饰的有机体(包括病毒)或有机体中存在的多肽或多核苷酸序列就是天然存在的。
本文使用的术语“多肽”或“肽”意指其中具有通过肽键连接的至少4个氨基酸的分子。
本文使用的表达方式“病毒核酸”可以是病毒的基因组(或其主要部分)、或碱基序列与该基因组互补的核酸分子。与病毒RNA互补的DNA分子也被视为病毒核酸,碱基序列与病毒DNA互补的RNA分子也是如此。
本文使用的术语“病毒样颗粒”(VLP)是指具有与病毒颗粒类似的物理外观的自组装颗粒。该VLP可以含有或不含有病毒核酸。VLP通常不能复制。
本文使用的术语“假病毒”是指包含核酸序列(例如DNA或RNA)的VLP,包括质粒形式的核酸。假病毒通常不能复制。
本文使用的术语“疫苗”是指能够产生免疫应答的物质。
本文使用的术语“免疫原”和“抗原”是指能够单独或与佐剂组合在对象中诱导免疫应答的一个分子、多个分子、分子的一部分或多部分、或分子的组合、直到并包括整个细胞和组织。该免疫原/抗原可以包含单个抗原表位,或者可以包含多个抗原表位。因此,该术语涵盖肽、碳水化合物、蛋白、核酸、各种微生物(包括病毒、细菌和寄生物)的整体或部分。半抗原亦被视为包含在本文使用的术语“免疫原”和“抗原”中。
本文中可交替使用的术语“免疫”和“接种”是指为提高免疫应答的目的而将疫苗施用给对象,并且其可具有预防效果、治疗效果或二者的组合。根据待治疗的对象,可以使用各种方法实现免疫,所述方法包括但不限于:腹腔注射(i.p.)、静脉注射(i.v.)、肌内注射(i.m.)、口服、鼻腔内施用、喷雾施用以及浸渍施用。
本文使用的术语“初次(prime)”及其语法变体是指在用抗原进行加强接种之前,在动物内刺激和/或激活针对抗原的免疫应答。
本文针对疾病或病原体使用的术语“治疗”、“经治疗的”或“治疗的”是指增强对象对疾病或病原体感染的抵抗力的治疗(即降低了对象染病或感染病原体的可能性)、以及在对象已经染病或感染之后为了对抗疾病或感染而进行的治疗(例如,使疾病或感染减轻、消除、改善或稳定)。
本文使用的术语“受试者”或“患者”是指需要治疗的动物。
本文使用的术语“动物”是指人以及非人类动物,其包括但不限于:哺乳动物、禽类和鱼类,并且涵盖家养、农场、动物园、实验室以及野生的动物,例如,奶牛、猪、马、山羊、绵羊和其它有蹄类动物、狗、猫、鸡、鸭、非人类的灵长类动物、豚鼠、兔子、雪貂、大鼠、仓鼠和小鼠。
本文涉及核酸或氨基酸序列而使用的术语“基本上相同”表示,当使用(例如)下文所述的方法进行最佳比对时,该核酸或氨基酸序列与确定的第二核酸或氨基酸序列(或“参考序列”)具有至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%的序列同源性。“基本上同源”可用于指各种类型和长度的序列,例如全长序列、功能域、编码序列和/或调节序列、启动子以及基因组序列。例如,可以通过本领域技术人员的技能范围内的各种方法、采用公开可得的计算机软件来确定两种氨基酸或核酸序列的同源百分比,所述软件例如为:Smith Waterman Alignment(Smith,T.F.and M.S.Waterman(1981)JMoI Biol 147:195-7);并入到GeneMatcher PlusTM(Schwarz和Dayhof(1979)Atlas of Protein Sequence and Structure,Dayhof,M.O.,Ed pp353-358)的“BestFit”(Smith和Waterman,Advances in AppliedMathematics,482-489(1981));BLAST程序(Basic Local AlignmentSearch Tool(Altschul,S.F.,W.Gish等.(1990)J Mol Biol 215:403-10),以及它们的各种变体,包括:BLAST-2、BLAST-P、BLAST-N、BLAST-X、WU-BLAST-2、ALIGN、ALIGN-2、CLUSTAL,以及Megalign(DNASTAR)软件。此外,本领域技术人员能够确定用于测量比对的合适的参数,其包括为了在比较的序列长度内实现最大比对而需要的算法。通常,对于氨基酸序列而言,比较序列的长度至少为10个氨基酸。本领域技术人员会理解,实际长度取决于所比较的序列的全部长度,并且可以是至少20、至少30、至少40、至少50、至少60、至少70、至少80、至少90、至少100、至少110、至少120、至少130、至少140、至少150或至少200个氨基酸,或其可以为氨基酸序列的全长。对于核酸而言,比较序列的长度通常为至少25个核苷酸,但是其可以是至少50、至少100、至少125、至少150、至少200、至少250、至少300、至少350、至少400、至少450、至少500、至少550、或至少600个核苷酸,或其可以为核酸序列的全长。
术语“对应于”或“相当于”表示核酸序列与参考核酸序列的全部或部分相同。与此截然不同的是,本文使用的术语“互补”表示核酸序列与参考核酸序列的互补链的全部或部分相同。为了解释,核酸序列“TATAC”对应于参考序列“TATAC”并且与参考序列“GTATA”互补。
抗原提呈系统(APS)
本发明的抗原提呈系统(APS)包含与番木瓜花叶病毒(PapMV)或源自PapMV外壳蛋白的VLP组合的一种或多种肠细菌抗原。“源自”的含义是指,VLP包含这样的外壳蛋白,如下面更详细的描述的那样,该外壳蛋白具有与野生型外壳蛋白的序列基本上相同的氨基酸序列,并且可以任选地包含一种或多种与外壳蛋白连接的抗原。PapMV外壳蛋白可以是野生型外壳蛋白或其修饰形式(其能够多聚化并且自组装而形成VLP)。如下面详细描述的那样,在本发明的一个实施方案中,APS包含源自肠细菌伤寒沙门氏菌(S.typhi)的一种或多种抗原。在另一个实施方案中,APS包含源自肠杆菌科的不同细菌的一种或多种抗原。
APS所包含的一种或多种抗原可以与PapMV或PapMV VLP的外壳蛋白接合,或者它们可以是非接合的(即抗原与PapMV或PapMV VLP分离),或APS可以既包含接合抗原,也包含不接合的抗原。在后一种情况下,不接合的抗原被称为额外的分离抗原(AIA)。AIA可以与接合抗原相同或不同。接合可以是(例如)通过与外壳蛋白基因融合,或通过共价、非共价或亲和方式结合而实现的。
因此,包含在APS中的PapMV或PapMV VLP起到能够增强动物内的免疫应答的免疫增强剂的作用,并且可任选地被用作APS所包含的一种或多种抗原的载体。根据本发明的一个实施方案,APS能够诱导动物内的体液免疫应答和/或细胞免疫应答,因此适用作(例如)抗肠细菌感染的疫苗。
番木瓜花叶病毒(PapMV)和PapMV VLP
本发明的APS包含PapMV或PapMV VLP。PapMV VLP由重组PapMV外壳蛋白形成,该重组PapMV外壳蛋白已经多聚化并自组装而形成VLP。当组装后,各VLP包含外壳蛋白亚单元的长的螺旋状阵列。野生型病毒包含超过1200个外壳蛋白亚单元,并且长约500nm。但是,短于或者长于野生型病毒的PapMV VLP仍然是有效的。在本发明的一个实施方案中,VLP包含至少40个外壳蛋白亚单元。在另一个实施方案中,VLP包含约40个到约1600个的外壳蛋白亚单元。在可供选用的实施方案中,VLP的长度为至少40nm。在另一个实施方案中,VLP的长度在约40nm和约600nm之间。
本发明的VLP可以由多个具有相同的氨基酸序列的重组外壳蛋白制备,使得在组装后,最终的VLP包含相同的外壳蛋白亚单元,或者VLP可以由多个具有不同的氨基酸序列的重组外壳蛋白制备,使得在组装后,最终的VLP包含不同的外壳蛋白亚单元。
用于形成VLP的外壳蛋白可以是整个的PapMV外壳蛋白或其部分,或可以是PapMV外壳蛋白的基因修饰形式,该基因修饰形式(例如)包含一个或多个氨基酸的缺失、插入、替换等,条件是该外壳蛋白保持多聚化并组装成VLP的能力。野生型PapMV外壳(或壳体)蛋白的氨基酸序列是本领域中已知的(参见,Sit等,1989,J.Gen.Virol.,70:2325-2331,以及GenBank登录号NP_044334.1),并且在本文中表示为SEQ ID NO:1(参见图1A)。PapMV外壳蛋白的核苷酸序列也是本领域中已知的(参见,Sit等,ibid.,以及GenBank登录号NC_001748(核苷酸5889-6536)),并且在本文中表示为SEQID NO:2(参见图1B)。
如上所述,VLP所包含的重组PapMV外壳蛋白的氨基酸序列不必与亲本序列精确对应,即,其可以是修饰的或“变体序列”。例如,重组蛋白可以通过置换、插入或缺失一个或多个氨基酸残基来突变,使得该位置的残基不对应于亲本(参考)序列。但是,本领域技术人员将理解,这样的突变不是广泛的并且不会显著影响重组外壳蛋白多聚化并组装成VLP的能力。可以(例如)通过使用标准技术的电子显微镜法(例如本文中的实施例给出的示例性方法)来评价PapMV外壳蛋白的变体形式组装成多聚体和VLP的能力。
因此,本发明还涵盖为保持了多聚化并组装成VLP的能力的野生型蛋白的片段(即功能性片段)的重组外壳蛋白。例如,片段可以包含在蛋白的N-端、C-端或内部缺失一个或多个氨基酸、或它们的组合。一般而言,功能性片段的长度为至少100个氨基酸。在本发明对一个实施方案中,功能性片段的长度为至少150个氨基酸、至少160个氨基酸、至少170个氨基酸、至少180个氨基酸、至少190个氨基酸。为了保持蛋白多聚化的能力,在蛋白的N-端所进行的缺失应该通常缺失少于25个氨基酸。
根据本发明,当重组外壳蛋白包含变体序列时,该变体序列与参考序列至少约70%相同。在一个实施方案中,该变体序列与参考序列至少约75%相同。在其它的实施方案中,该变体序列与参考序列至少约80%、至少约85%、至少约90%、至少约95%、至少约97%相同。在一个具体的实施方案中,该参考氨基酸序列为SEQ ID NO:1。
在本发明对一个实施方案中,VLP包含PapMV外壳蛋白的基因修饰的形式(即变体)。在另一个实施方案中,PapMV外壳蛋白被基因修饰以在蛋白的N-端或C-端缺失氨基酸,和/或包括一个或多个氨基酸置换。在另外的实施方案中,PapMV外壳蛋白被基因修饰以在蛋白的N-端或C-端缺失约1个至约10个氨基酸。
在具体的实施方案中,该PapMV外壳蛋白被基因修饰以去除位于靠近蛋白的N-端(即在SEQ ID NO:1的1和6位)并且能够启动转录的两个蛋氨酸密码子中的一个。去除一个转录启动密码子容许制备同质的蛋白组。可以通过(例如)以下方法去除选择的蛋氨酸密码子,所述方法为:置换组成该密码子的一个或多个核苷酸,使得该密码子编码不同于蛋氨酸的氨基酸,或变成无意义的密码子。可供选用的方式是,可以删除密码子的全部或部分,或编码包含所选密码子的蛋白的核酸的5’区域。在本发明的具体的实施方案中,该PapMV外壳蛋白被基因修饰以从蛋白的N-端缺失1个到5个氨基酸。在另外的实施方案中,经基因修饰的PapMV外壳蛋白具有与SEQID NO:3基本上相同的氨基酸序列。
当重组蛋白包含含有一个或多个氨基酸置换的变体序列时,这些置换可以是“保守”置换或“非保守”置换。保守置换涉及到用具有类似侧链性质的另一种残基来替换一种氨基酸残基。如本领域中所已知,20种天然存在的氨基酸可以根据其侧链的理化性质分组。合适的分组包括:丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、蛋氨酸、苯丙氨酸和色氨酸(疏水性侧链);甘氨酸、丝氨酸、苏氨酸、半胱氨酸、酪氨酸、天冬酰胺和谷氨酰胺(极性,无电荷侧链);天门冬氨酸和谷氨酸(酸性侧链)和赖氨酸、精氨酸和组氨酸(碱性侧链)。另一组氨基酸是苯丙氨酸、色氨酸和酪氨酸(芳香侧链)。保守置换涉及到用相同组的另一种氨基酸来置换氨基酸。非保守置换涉及到用具有不同的侧链性质的另一种残基来替换一种氨基酸残基,例如,用中性或碱性的残基来替换酸性残基,用酸性或碱性残基来替换中性残基,用亲水性残基来替换疏水性残基等等。
在本发明的一个实施方案中,变体序列包含一个或多个非保守置换。用另一种不同性质的氨基酸来替换一种氨基酸可以改善外壳蛋白的性质。例如,如本文所述,外壳蛋白的128位残基的突变会促进蛋白组装为VLP。因此,在本发明的一个实施方中,外壳蛋白包含外壳蛋白的128位残基的突变,其中该位点的谷氨酸残基被中性残基置换。在另一个实施方案中,在128位的谷氨酸残基被丙氨酸残基置换。
同样,编码重组外壳蛋白的核酸序列不必与亲本参考序列精确对应,而是可以通过遗传码的简并性而变化,和/或使得其编码上述的变体氨基酸序列。因此,在本发明的一个实施方案中,编码重组外壳蛋白的核酸序列与参考序列至少约70%相同。在另一个实施方案中,编码重组外壳蛋白的核酸序列与参考序列至少约75%相同。在其它的实施方案中,编码重组外壳蛋白的核酸序列与参考序列至少约80%、至少约85%或至少约90%相同。在具体的实施方案中,该参考核酸序列是SEQ ID NO:2。
如下面更详细地描述的那样,PapMV VLP外壳蛋白可以任选地与一种或多种抗原、亲和肽或其它短肽序列发生基因融合,以促进与一种或多种抗原的连接。
抗原
本发明的APS包含源自肠杆菌科的革兰氏阴性菌的一种或多种抗原。合适的肠杆菌科细菌的例子包括(但不限于):大肠杆菌属(Escherichia)、沙门氏菌属(Salmonella)、耶尔森菌属(Yersinia)、志贺菌属(Shigella)、变形杆菌属(Proteus)、克雷白氏杆菌属(Klebsiella)以及肠杆菌属(Enterobacter)的品种。该抗原可以是源自肠细菌的免疫原、抗原表位、模拟抗原表位或抗原决定簇,并且可以是(例如)肽、蛋白、核酸、或细菌表面抗原成分(SBAC)(例如脂多糖(LPS)、荚膜抗原(本质上是蛋白质或多糖)、或鞭毛成分),或其可以是细菌的灭活或减毒形式。
可以基于所需要的APS的最终用途来选择候选抗原,例如,其针对何种细菌,是打算用作多价疫苗还是单价疫苗,和/或所施用的动物。合适的候选抗原可以被本领域技术人员容易地选择。多种抗原性肠细菌成分是本领域已知的,并且是包含在本发明的APS中的合适的候选抗原。本领域技术人员也会理解,可以基于使用本领域已知的免疫学技术测试抗原诱导动物内的免疫应答的能力,来选择用于包含在本发明的APS中的适合候选抗原。
在本发明的一个实施方案中,候选抗原选自大肠杆菌属(Escherichia)、沙门氏菌属(Salmonella)、耶尔森菌属(Yersinia)、志贺菌属(Shigella)、变形杆菌属(Proteus)、克雷白氏杆菌属(Klebsiella)或肠杆菌属(Enterobacter)中的品种。在本发明的另一个实施方案中,候选抗原选自大肠杆菌属(Escherichia)、沙门氏菌属(Salmonella)、耶尔森菌属(Yersinia)、志贺菌属(Shigella)、克雷白氏杆菌属(Klebsiella)或肠杆菌属(Enterobacter)中的品种。在另一个实施方案中,候选抗原选自大肠杆菌属(Escherichia)或沙门氏菌属(Salmonella)中的品种。在另一个实施方案中,候选抗原选自沙门氏菌属(Salmonella)中的品种。在具体的实施方案中,候选抗原是伤寒沙门氏菌(S.typhi)抗原。
合适的候选细菌表面抗原成分(SBAC)的例子包括:各种肠杆菌H(鞭毛成分)抗原和O(LPS)抗原、伤寒沙门氏菌Vi(荚膜多糖)抗原、大肠杆菌K和CFA(荚膜成分)抗原、以及大肠杆菌菌毛粘附素抗原(K88和K99)。合适的候选抗原蛋白的例子包括:外膜蛋白(Omp),其也被称为孔蛋白(Secundino等,2006,Immunology117:59);相关的孔蛋白,如伤寒沙门氏菌铁调节的外膜蛋白(IROMP,Sood等,2005,Mol Cell Biochem 273:69-78)和热休克蛋白(HSP),所述热休克蛋白包括(但不限于)伤寒沙门氏菌HSP40(Sagi等,2006,Vaccine 24:7135-7141)。合适的孔蛋白的非限定性的例子包括OmpC和OmpF,其在多种沙门氏菌属和大肠杆菌属的品种中有发现。在其它肠杆菌科中也发现有OmpC和OmpF的直系同源物(Orthologue),并且该直系同源物是用于本发明的合适的候选抗原蛋白。此外,基于肠杆菌科的孔蛋白中发现的序列的保守区域,Omp1B(弗氏志贺菌(Shigella flexneri))、OmpC2(鼠疫耶尔森菌(Yersinia pestis))、OmpD(肠道沙门氏菌(S.enterica))、OmpK36(肺炎克雷白氏杆菌(Klebsiella pneumonie))、OmpN((大肠杆菌))和OmpS(肠道沙门氏菌(S.enterica))是合适的候选物(Diaz-Quinonez等,2004,Infect.and Immunity 72:3059-3062)。
还适合作为候选抗原的是金属转运蛋白,例如:大肠杆菌SitABCD、MntH和FeoB金属转运蛋白(Sabri等,2008,Infect.Immun.76:601-611;在2007年11月9日印刷前电子出版)。
源自各种肠杆菌科的抗原蛋白的序列是本领域中已知的,并且易于从国家生物技术信息中心(NCBI)维护的Genbank数据库中获取。例如:GenBank登录号P0A264(SEQ ID NO:39;亦见图21A所示)和GenBank登录号NP_804453:OmpC(肠道沙门氏菌亚种Ty2);GenBank登录号CAD05399(SEQ ID NO:40;亦见图21B所示):OmpF前体蛋白(肠道沙门氏菌亚种CT18);GenBank登录号16761195:OmpC(鼠伤寒沙门氏菌);GenBank登录号47797:OmpC(伤寒沙门氏菌);GenBank登录号8953564:OmpC(明尼苏达伤寒沙门氏菌);GenBank登录号19743624:OmpC(都柏林伤寒沙门氏菌);GenBank登录号19743622:OmpC(鸡伤寒沙门氏菌);GenBank登录号26248604:OmpC(大肠杆菌);GenBank登录号24113600:Omp1B(福氏志贺菌);GenBank登录号16764875:OmpC2(鼠疫杆菌);GenBank登录号16764916:OmpD(鼠伤寒沙门氏菌);GenBank登录151149831:OmpK36(肺炎克雷白氏杆菌);GenBank登录号3273514:OmpN(大肠杆菌);以及GenBank登录号16760442:OmpS(伤寒沙门氏菌)。
在本发明的一个实施方案中,包含在APS中的抗原为蛋白抗原。该蛋白抗原可以是全长蛋白、基本上全长的蛋白(例如,N-端和/或C-端缺失约25个或更少的氨基酸的蛋白)、蛋白的抗原片段、或者它们的组合。在应用时,全长蛋白可以是蛋白的前体形式或蛋白的成熟(加工)形式。抗原片段可以包含一种或多种抗原表位,因此其长度的大小可以为数个氨基酸(例如,至少4个氨基酸)的肽至数百个氨基酸的多肽。在本发明的一个实施方案中,抗原片段的长度在约4个氨基酸至约250个氨基酸之间。在另一个实施方案中,抗原片段的长度在约5个氨基酸至约200个氨基酸之间。在其它的实施方案中,抗原片段的长度在约5个氨基酸至约150个氨基酸之间,在约5个氨基酸至约100个氨基酸之间,在约5个氨基酸至约75个氨基酸之间,在约5个氨基酸至约70个氨基酸之间,在约5个氨基酸至约60个氨基酸之间,以及在约5个氨基酸至约50个氨基酸之间。
在本发明的一个实施方案中,包含在APS中的抗原是全长或基本上全长的蛋白。在另一个实施方案中,包含在APS中的抗原是全长或基本上全长的孔蛋白。在另外的实施方案中,包含在APS中的抗原是全长或基本上全长的OmpC和/或OmpF蛋白,或全长或基本上全长的OmpC和/或OmpF的直系同源物。
如表1所示,伤寒沙门氏菌OmpC蛋白和源自其它肠杆菌科的OmpC直系同源物的序列高度保守。该序列在Salmonella spp.之间甚至更加高度保守(参见表1)。同样,伤寒沙门氏菌OmpF蛋白和源自其它的沙门氏菌品种的OmpF直系同源物的序列高度保守(参见表2)。因此,在本发明的一个实施方案中,包含在APS中的至少一个抗原是全长或基本上全长的OmpC或OmpC直系同源物,其具有与图21A所示的伤寒沙门氏菌OmpC蛋白序列[SEQ ID NO:39]至少约75%相同的氨基酸序列。在另一个实施方案中,包含在APS中的至少一个抗原是全长或基本上全长的OmpC或OmpC直系同源物,其具有与图21A所示的伤寒沙门氏菌OmpC蛋白序列[SEQ ID NO:39]至少约78%相同的氨基酸序列。在本发明的另一个实施方案中,包含在APS中的至少一个抗原是全长或基本上全长的OmpC或OmpC直系同源物,其具有与图21A所示的伤寒沙门氏菌OmpC蛋白序列[SEQ ID NO:39]至少约80%相同的氨基酸序列。
在本发明的另一个实施方案中,包含在APS中的至少一个抗原是全长或基本上全长的OmpF或OmpF直系同源物,其具有与图21B所示的伤寒沙门氏菌OmpF蛋白序列[SEQ ID NO:40]至少约75%相同的氨基酸序列。在本发明的另一个实施方案中,包含在APS中的至少一个抗原是全长或基本上全长的OmpF或OmpF直系同源物,其具有与图21B所示的伤寒沙门氏菌蛋白OmpF序列[SEQ ID NO:40]至少约78%相同的氨基酸序列。在本发明的另一个实施方案中,包含在APS中的至少一个抗原是全长或基本上全长的OmpF或OmpF直系同源物,其具有与图21B所示的伤寒沙门氏菌OmpF蛋白序列[SEQ ID NO:40]至少约80%相同的氨基酸序列。
在本发明的具体的实施方案中,包含在APS中的抗原是全长或基本上全长的伤寒沙门氏菌OmpC蛋白和/或OmpF蛋白。
表1:OmpC和源自多种肠杆菌科的OmpC直系同源物的序列同一性
| 有机体 | 蛋白 | 参考 | 同一性%1 |
| 鼠沙门氏菌(Salmonella typhimurium)LT2 | OmpC | P0A263 | 100 |
| 邦戈尔沙门氏菌(Salmonella bongori) | ORF_2828(推定OmpC) | coliBase2:GL0268093 | 99 |
| 肠炎沙门氏菌(Salmonella enteritidis)PT4 | ORF_1402(推定OmpC) | coliBase2:GL0633863 | 98 |
| 鸡沙门氏菌(Salmonella gallinarum)287/91 | ORF_222(推定OmpC) | coliBase2:GL0641663 | 98 |
| 大肠杆菌O157:H7 EDL933 | OmpC | Q8XE41 | 80 |
| 痢疾志贺菌(Shigella dysenteriae) | ORF 14 | coliBase2: | 78 |
| M131649(M131) | (推定OmpC) | GL0181393 | |
| 弗氏志贺菌(Shigella flexneri)2a 2457T | Omp1b | Q83QU7 | 78 |
1同一性%相对于伤寒沙门氏菌OmpC蛋白(GenBank登录号P0A264)而言,并且使用BLASTP 2.2.3[2002年4月24日]程序来测定(Altschul,S.F等,(1997),Nucleic Acids Res.25:3389-3402)。
2Nucleic Acids Research,2004,Vol.32,Database issueD296-D299。
32007年1月23日的GL号。
表2:OmpF和源自多种肠杆菌科的OmpF直系同源物的序列同一性
| 有机体 | 蛋白 | 参考 | 同一性%1 |
| 肠炎沙门氏菌PT4 | ORF 34 | coliBase2:GL0607313 | 100 |
| 鸡沙门氏菌287/91 | ORF 21 | coliBase2:GL0692163 | 99 |
| 鼠沙门氏菌DT104 | ORF 287 | coliBase2:GL00443623 | 99 |
| 邦戈尔沙门氏菌 | ORF 1160 | coliBase2:GL0253983 | 98 |
| 大肠杆菌DH10B | ORF 2 | coliBBase2:GL0376943 | 58 |
| 弗氏志贺菌2a 2457T | OmpF | Q83RY7 | 58 |
1同一性%相对于伤寒沙门氏菌OmpF蛋白(GenBank登录号CAD05399)而言,并且使用BLASTP 2.2.3[2002年4月24日]程序来测定(Altschul,S.F.等,(1997),Nucleic Acids Res.25:3389-3402)。
2Nucleic Acids Research,2004,Vol.32,Database issueD296-D299。
32007年1月23日的GL号。
在可供选用的实施方案中,包含在APS中的抗原是蛋白的抗原片段。上述蛋白的多个抗原区域已经被鉴定并且是用于本发明的APS的合适的候选抗原。例如,外膜蛋白(Omp)的表面外露环构成了用于包含在本发明的APS中的合适的候选抗原片段。从伤寒沙门氏菌分离的OmpC的环6(L6)肽和环7(L7)肽是候选抗原的非限定性例子。已经有伤寒沙门氏菌OmpC的L6肽和L7肽的序列的报导并分别鉴定为:GTSNGSNPST(SEQ ID NO:4)和KDISNGYGASYGDQ(SEQ ID NO:5)(Paniagua-Solis等,1996,FEMS 141:31-36)。包含如SEQ ID NO:4和SEQ ID NO:5所列出的序列的全部或部分的抗原片段也是合适的候选物。例如,具有序列GTSNGSNPSTSYGFAN[SEQID NO:50]的孔蛋白片段(其包含L6肽的关键氨基酸)和具有序列QSKGKDISNGYGASYGDQD[SEQ ID NO:54]的孔蛋白片段(其包含L7肽的关键氨基酸)。
其它的抗原性抗原表位也是已知的。例如,除了L6肽和L7肽以外的OmpC的特异性片段可用作包含在本发明的APS中的候选抗原,其包括:OmpC-73肽[SEQ ID NO:6]和OmpC-132肽[SEQ IDNO:7](参见表3)(Diaz-Quinonez等,2004,Infect.and Immunity72:3059-3062)。这些肽的序列在不同的肠杆菌科细菌之间是高度保守的,从而使这些肽适用作包含在APS(希望使其用于多价疫苗)中的候选抗原。其它抗原表位序列(如SEQ ID NO:8、9和10)的例子如表3所示。作为伤寒沙门氏菌特有的潜在B细胞抗原表位,伤寒沙门氏菌OmpC蛋白的可变区也已经被鉴定(Arockiasamy和Krishnaswamy,1995,J.Biosci,20:235-243)(参见表3,SEQ IDNO:11、12、13、14、15、16、17和18)。包含这些区域的抗原片段也是包含在本发明的APS中的候选抗原。
表3:源自OmpC和其它肠杆菌科的孔蛋白的候选抗原表位的序列
| 蛋白 | 菌株 | 抗原表位序列 | SEQ ID NO |
| OmpC | 鼠伤寒沙门氏菌;伤寒沙门氏菌;明尼苏达伤寒沙门氏菌;都柏林伤寒沙门氏菌; | TRVAFAGL | 6 |
| 鸡伤寒沙门氏菌和大肠杆菌 | |||
| Omp1b | 弗氏志贺菌 | TRVAFAGL | 6 |
| OmpS | 伤寒沙门氏菌 | TRLAFAGL | 9 |
| OmpN | 大肠杆菌 | TRLAFAGL | 9 |
| OmpD | 鼠伤寒沙门氏菌 | TRLAFAGL | 9 |
| OmpC2 | 鼠疫耶尔森菌 | TRLGFAGL | 10 |
| OmpK36 | 肺炎克雷白氏杆菌 | TRLAFAGL | 9 |
| OmpC | 鼠伤寒沙门氏菌;伤寒沙门氏菌;明尼苏达伤寒沙门氏菌;都柏林伤寒沙门氏菌;鸡伤寒沙门氏菌和大肠杆菌 | RNTDFFGL | 7 |
| Omp1b | 弗氏志贺菌 | RNSDFFGL | 8 |
| OmpS | 伤寒沙门氏菌 | RNTDFFGL | 7 |
| OmpN | 大肠杆菌 | RNTDFFGL | 7 |
| OmpD | 鼠伤寒沙门氏菌 | RNTDFFGL | 7 |
| OmpC2 | 鼠疫耶尔森菌 | RNTDFFGL | 7 |
| OmpK36 | 肺炎克雷白氏杆菌 | RN SDFFGL | 8 |
| OmpC(氨基酸45-51) | 伤寒沙门氏菌 | SDDKGSD | 11 |
| OmpC(氨基酸84-92) | 伤寒沙门氏菌 | NQTEGSNDS | 12 |
| OmpC(氨基酸114-147) | 伤寒沙门氏菌 | NYGVTYDVTSWTDVLPEFGGDTYGADNFMQQRGN | 13 |
| OmpC(氨基酸175-190) | 伤寒沙门氏菌 | GSVSGENTNGRSLLNQ | 14 |
| OmpC(氨基酸218-236) | 伤寒沙门氏菌 | RTADQNNTANARLYGNGDR | 15 |
| OmpC(氨基酸262-282) | 伤寒沙门氏菌 | NATRFGTSNGSNPSTSYGFAN | 16 |
| OmpC(氨基酸312-328) | 伤寒沙门氏菌 | DISNGYGASYGDQDIVK | 17 |
| OmpC(氨基酸351-368) | 伤寒沙门氏菌 | NLLDKNDETRDAGINTDD | 18 |
在本发明的一个实施方案中,APS包含全长的Omp或含有数个上述的抗原表位的片段。在另一个实施方案中,抗原包含在变体之间保守的序列,例如,氨基酸TRXXFAGL(SEQ ID NO:19,其中X可以是V、L或G),或氨基酸RNXDFFGL(SEQ ID NO:20,其中X可以是T或S)所限定的保守区域。
在本发明的一个实施方案中,所选的包含在APS中的至少一个抗原是孔蛋白抗原。根据另一个实施方案,至少一个抗原是整个孔蛋白。根据又一个实施方案,包含在APS中的候选抗原为OmpC和/或OmpF抗原。在另一个实施方案中,包含在APS中的候选抗原是OmpC肽或OmpF肽,例如:环6肽、环7肽、OmpC-73肽或OmpC-132肽,或它们的变体。在另一个实施方案中,候选抗原包含与下面所列出的序列基本上相同的氨基酸序列或者它们的片段:SEQ IDNO:4、SEQ ID NO:5、SEQ ID NO:6、SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ ID NO:10、SEQ ID NO:11、SEQ ID NO:12、SEQID NO:13、SEQ ID NO:14、SEQ ID NO:15、SEQ ID NO:16、SEQ IDNO:17、SEQ ID NO:18、SEQ ID NO:19或SEQ ID NO:20。在另一个实施方案中,候选抗原包含与SEQ ID NO:4或SEQ ID NO:5所列出的序列基本上相同的氨基酸序列或者它们的片段。
如上所述,所选抗原的大小可以变化并且所选抗原的大小对本发明的实施并非关键的。抗原可以是对于T细胞、B细胞、NK细胞和巨噬细胞的表面结构、或者与细胞表面结构有关的I类或II类APC而言特异性的,或者被它们所识别。在一个实施方案中,本发明特别适用于小的免疫原性弱的抗原。所选的抗原应该不干扰PapMV或PapMV VLP的组装,或者不干扰PapMV或PapMV VLP免疫增强免疫应答的能力。本领域技术人员使用标准技术(例如下面的部分以及实施例中所述的那些)能够容易地从上述候选抗原中选出合适的抗原。
本发明的APS可以包含一种或多种抗原,各抗原均具有能够触发特异性免疫应答的单个抗原表位,或者该APS可以包含一种或多种抗原,其中各抗原包含多个抗原表位。APS所包含的抗原可以是相同的,或者它们可以是不同的,并且当存在多个蛋白抗原时,它们可以源自单一一种蛋白或源自多种蛋白,包括源自多于一种类型的蛋白的抗原(例如,源自Omp和HSP的抗原)。还涵盖包含代表肠细菌抗原蛋白的组合(例如,源自多于一种肠细菌的抗原)的抗原的APS,也涵盖包含源自蛋白的相同的保守区域、但是代表了不同的肠细菌中所见的蛋白的变体的抗原的APS。
在本发明的一个具体的实施方案中,APS包含在肠杆菌科的不同成员或者肠杆菌科的亚群之间保守的一种或多种抗原。例如,OmpC或其片段,和/或OmpF或其片段。这样的抗原适合包含在打算用作多价疫苗的APS中。序列比较表明OmpC/OmpF和它们的肠细菌直系同源物之间高度保守(包括成段的相同的氨基酸)。事实上,在所有的沙门氏菌属中,OmpC和OmpF显示了98%-100%的同一性(参见表1)。这些蛋白与源自其它肠细菌(包括大肠杆菌、志贺菌属、肺炎克雷白氏杆菌和产气肠杆菌)的直系同源物的比较表明与源自伤寒沙门氏菌的蛋白序列具有75%-85%的同一性(参见表2)。
本发明还涵盖了APS可以包含这样的抗原:该抗原为与一种或多种细菌表面抗原成分(SBAC)(例如,O抗原)组合的整个蛋白或其片段。因此,根据一个实施方案,该APS包含一种或多种SBAC与一种或多种抗原蛋白、肽、或其片段的组合。
如上所述,本发明的APS可以包含一种或多种接合抗原或一种或多种额外的分离抗原(AIA)。所述AIA可以与APS的接合抗原相同或不同。本发明涵盖接合抗原和AIA的各种组合,并且包括下面的非限定性的例子:源自第一肠细菌品种的一种或多种接合抗原和源自相同的肠细菌品种的一种或多种AIA;源自第一肠细菌品种的一种或多种接合抗原和源自另一种肠细菌品种的一种或多种AIA;一种或多种接合蛋白或蛋白片段抗原和一种或多种整个蛋白或蛋白片段的AIA;一种或多种接合SBAC抗原和一种或多种SBAC AIA;一种或多种接合蛋白或蛋白片段抗原和一种或多种SBAC AIA(或反之亦然);一种或多种接合的整个蛋白抗原和一种或多种整个蛋白AIA;一种或多种接合的整个蛋白抗原和作为所述整个蛋白的肽的一种或多种AIA。一种或多种接合抗原和AIA可以属于同一蛋白族(例如,Omp)或者它们可以属于不同的族(例如,Omp和HSP)。
在本发明的一个实施方案中,APS包含一种或多种接合抗原和一种或多种作为整个蛋白的AIA。根据另一个实施方案,至少一种AIA是整个的孔蛋白。根据另一个实施方案,至少一种AIA是OmpC或者OmpF。在另一个实施方案中,至少一种AIA是OmpC的肽(例如,环6肽、环7肽、OMPC-73肽、或OMPC-132肽、或者它们的变体)或OmpF的肽。在另一个实施方案中,至少一种AIA包含与下列序列基本上相同的氨基酸序列:SEQ ID NO:4、SEQ ID NO:5、SEQ ID NO:6、SEQ ID NO:7、SEQ ID NO:8、SEQ ID NO:9、SEQ IDNO:10、SEQ ID NO:11、SEQ ID NO:12、SEQ ID NO:13、SEQ IDNO:14、SEQ ID NO:15、SEQ ID NO:16、SEQ ID NO:17、SEQ IDNO:18、SEQ ID NO:19或者SEQ ID NO:20。在另一个实施方案中,由APS所包含的接合抗原和AIA是相同的。
抗原-PapMV组合和抗原PapMV-VLP组合
如上所述,由APS所包含的一种或多种抗原可以与PapMV或PapMV VLP的外壳蛋白接合,或者它们可以以非接合的形式存在于APS中(即抗原与PapMV或PapMV VLP仅简单地组合),或者它们可以同时以接合和非接合的形式存在。接合可以通过(例如)抗原与外壳蛋白发生基因融合来实现,或者通过共价、非共价或亲和方式结合来实现。但是,抗原与PapMV或VLP的组合不应影响宿主免疫系统对该抗原的识别或者不应影响PapMV或VLP增强免疫应答的能力。
根据本发明的一个实施方案,一种或多种抗原与PapMV VLP的外壳蛋白接合。由于VLP包含自组装外壳蛋白的多种拷贝,因此抗原与外壳蛋白的连接容许抗原以有组织的方式在VLP的表面上提呈。根据该实施方案,所述VLP包含这样的外壳蛋白,该外壳蛋白包括作为重组PapMV外壳蛋白的第一部分,所述第一部分与包含一种或多种抗原的第二部分接合。由于接合抗原的外壳蛋白保持了与其它融合外壳蛋白或野生型外壳蛋白组装而形成免疫原-载体VLP的能力,因此选择不会干扰上述能力的抗原用于接合。
为了容许抗原在VLP的表面上提呈并提高抗原的免疫识别性,优选使抗原与外壳蛋白的设置在VLP的外表面上的区域连接。因此可以将抗原插至邻近外壳蛋白的氨基-(N-)端或羧基-(C)端处或者与其连接,或者可以将其插至外壳蛋白的设置在VLP的外表面上的内环中或与其连接。这样的环的例子如SEQ ID NO:1所列的PapMV外壳蛋白的包含第49至52个氨基酸的区域。在本发明的一个实施方案中,抗原存在于PapMV外壳蛋白的C-端。
根据本发明的一个实施方案,所述APS包含与一种或多种抗原基因融合的PapMV外壳蛋白。为了避免抗原可能干扰PapMV外壳蛋白-抗原融合而自组装成VLP能力,所选的用于与PapMV外壳蛋白基因融合的抗原长度通常小于约50个氨基酸。在本发明的一个实施方案中,所选的用于与PapMV外壳蛋白基因融合的抗原长度通常小于约45个氨基酸。
如下文更详细描述的那样,较大的抗原可以通过与PapMV或VLP的简单组合、或者通过化学交联或亲和连接而容易地包含在APS内。后一种连接方法能够促进抗原与PapMV的结合并且在施用给动物时产生针对抗原的改善的免疫应答。
在另一个实施方案中,通过共价或非共价的(例如,离子、疏水、氢键等)连接方式使抗原与外壳蛋白化学交联。如本领域已知的那样,可以修饰抗原和/或外壳蛋白以促进这样的交联,例如,通过将官能团或化学部分添加至蛋白和/或抗原(例如在C-端、或N端、或内部位置)中来实现。示例性的修饰包括:添加诸如S-乙酰基巯基琥珀酸酐(SAMSA)或S-乙酰基硫代乙酸酯(SATA)等官能团,或添加一种或多种半胱氨酸残基。其它的交联剂是本领域中已知的并且很多是市售可得的(例如,参见Pierce Chemical公司和Sigma-Aldrich公司的商品目录)。其例子包括(但不限于):二胺,例如1,6-己二胺、1,3-丙二胺和1,3-二氨基乙烷;二醛,例如戊二醛;琥珀酰亚胺酯,例如乙二醇-双(琥珀酸N-羟基琥珀酰亚胺酯)、二琥珀酰亚胺基戊二酸酯、二琥珀酰亚胺基辛二酸酯、N-(g-马来酰亚胺基丁酰氧基)磺基琥珀酰亚胺酯和乙二醇双(琥珀酰亚胺琥珀酸酯);二异氰酸酯,例如六亚甲基二异氰酸酯;双环氧乙烷,1,4-丁二醇二缩水甘油醚;二羧酸,例如琥珀酰二水杨酸酯;3-马来酰亚胺基丙酸N-羟基琥珀酰亚胺酯等等。上述交联剂中许多都含有将抗原与VLP隔开的间隔物。本发明亦涵盖使用其它间隔物。各种间隔物在本领域中是已知的,并且包括(但不限于):6-氨基己酸;1,3-丙二胺;1,3-二氨基乙烷;以及短的氨基酸序列,例如1至5个氨基酸的聚甘氨酸序列。
为了促进一种或多种抗原与VLP的外壳蛋白的共价连接,外壳蛋白可以与暴露于VLP表面的短肽或氨基酸连接子(amino acidlinker)基因融合并提供用于抗原化学连接的合适位点。例如,包含半胱氨酸残基的短肽,或具有能够形成共价键的侧链的其他氨基酸残基(例如,酸性或碱性的残基)或者可以容易修饰而形成共价键的氨基酸残基在本领域内是已知的。所述氨基酸连接子或肽的长度可以(例如)在1个和约20个氨基酸之间。在一个实施方案中,所述外壳蛋白与含有一个或多个赖氨酸残基的短肽融合,所述短肽可通过使用上述的合适交联剂与(例如)抗原中的半胱氨酸残基共价结合。在一个具体的实施方案中,外壳蛋白与甘氨酸和赖氨酸残基的短肽序列融合。在另一个实施方案中,所述肽包含序列:GGKGG。
在本发明的另一个实施方案中,抗原通过存在于外壳蛋白表面上的亲和部分连接。根据该实施方案,PapMV VLP包含亲和部分(例如肽),在自组装后所述亲和部分暴露于VLP表面并能够特异地结合抗原。所述亲和部分可以与PapMV或VLP基因融合(在肽或蛋白片段的情况下),或者与PapMV或VLP共价或非共价连接。抗原与亲和部分的结合不应干扰宿主免疫系统对抗原的识别。根据本发明的一个实施方案,所述PapMV VLP包含能够结合整个蛋白的亲和部分。根据本发明的另一个实施方案,所述PapMV VLP包含能够结合蛋白片段或肽的亲和部分。根据本发明的另一个实施方案,所述PapMV VLP包含能够结合细菌表面抗原成分(SBAC)的亲和部分。
在本发明的一个具体的实施方案中,提供包含PapMV VLP的APS,所述PapMV VLP包含与孔蛋白结合的亲和肽。在本发明的另一个实施方案中,提供包含PapMV VLP的APS,所述PapMV VLP包含与OmpC或OmpF结合的亲和肽。
合适的亲和部分的例子包括(但不限于):抗体和抗体片段(例如Fab片段、Fab’片段、Fab’-SH片段、F(ab’)2片段、Fv片段、双抗体、以及单链Fv(scFv)分子)、链霉抗生物素蛋白(以结合生物素标记的抗原)、特异地结合抗原的亲和肽或蛋白片段。
可以通过本领域已知的技术(例如噬菌体或酵母展示技术)来选择用作亲和部分的合适的肽或抗体(包括抗体片段)。所述肽可以是天然存在的、重组的、合成的或者这些的组合。例如,所述肽可以是天然存在的蛋白或多肽的片段。术语“肽”还包括肽的类似物、肽的衍生物以及类肽化合物。这样的化合物是本领域中众所周知的,并且较天然存在的肽,其可以更有优势,包括(例如):更好的化学稳定性、增强的耐蛋白降解性、提高的药理学性能(例如半衰期、吸收性、效价和效能)和/或降低的抗原性。
合适的肽的长度可以在约3个氨基酸至约50个氨基酸的范围内。根据本发明的一个实施方案,亲和结合肽的长度至少为5个氨基酸。根据本发明的另一个实施方案,亲和结合肽的长度至少为7个氨基酸。根据本发明的另一个实施方案,所述亲和结合肽的长度在约5至约50个氨基酸之间。根据本发明的另一个实施方案,所述亲和结合肽的长度在约7至约50个氨基酸之间。在本发明的其它的实施方案中,所述亲和结合肽的长度在约5至约45个氨基酸之间,在约5至约40个氨基酸之间,在约5至约35个氨基酸之间,在约5至约30个氨基酸之间。根据本发明的具体的实施方案,所述亲和结合肽的长度为3、4、5、6、7、8、9、10、11、12、13、14或15个氨基酸。如本领域的技术人员将会理解的那样,所选的用于使抗原与亲和部分结合的肽的长度不应干扰PapMV VLP自组装的能力,或者一旦结合,其不应干扰宿主免疫系统识别抗原的能力。
PapMV或VLP所包含的亲和部分可以是单个肽或者可以是肽的串联或多重排列。为了促进大的抗原的结合,如果需要,可以在亲和部分和外壳蛋白之间引入间隔物。合适的间隔物包括短的中性氨基酸(例如甘氨酸)片段。例如,约3个至约10个中性氨基酸的片段。
使用标准技术(例如,参见Current Protocols in Immunology,ed.Coligan等,J.Wiley&Sons,New York,NY)和/或市售可得的噬菌体展示试剂盒(例如,可得自New England Biolabs的Ph.D.系列试剂盒,和可得自Novagen公司的T7- 试剂盒),可以将噬菌体展示技术用于选择与目标抗原蛋白结合的特定肽。下面的实施例VII还提供了通过噬菌体展示技术来选择肽的例子。
试剂盒),可以将噬菌体展示技术用于选择与目标抗原蛋白结合的特定肽。下面的实施例VII还提供了通过噬菌体展示技术来选择肽的例子。
表4示出了通过噬菌体展示技术鉴定的结合孔蛋白的代表性肽。本领域技术人员将理解,这些肽仅仅是例子,并且其它对目标孔蛋白具有亲和性的肽也可以容易地使用本领域已知的技术来鉴别。还涵盖表4中所列出的序列的保持结合孔蛋白能力的截断形式,其包含(例如)至少4个连续的氨基酸。根据本发明的具体的实施方案,提供包含PapMV VLP的APS,所述PapMV VLP包含一种或多种亲和肽,所述亲和肽包含下面所列出的序列的全部或部分:SEQ IDNO:21、SEQ ID NO:22、SEQ ID NO:23、SEQ ID NO:24、SEQ ID NO:25或SEQ ID NO:26。
表4:通过噬菌体展示技术和亲和肽的频率来选择的亲和肽
| 靶 | 选择的亲和肽 | SEQ ID NO |
| OmpC | SLSLIQT | 21 |
| OmpC | EAKGLIR | 22 |
| OmpC | TATYLLD | 23 |
| OmpF | FHENWPS | 24 |
| OmpF | FHEFWPT | 25 |
| OmpF | FHEXWPT,其中X是N或F | 26 |
APS的制备
本发明提供APS,所述APS包含PapMV或源自重组的PapMV外壳蛋白的PapMV VLP。本发明还提供重组的PapMV VLP,所述重组的PapMV VLP包含与外壳蛋白基因融合的一种或多种抗原或亲和部分。这些重组的外壳蛋白能够多聚化并组装成VLP。下文和实施例中描述了使抗原或用于连接抗原的亲和肽和外壳蛋白基因融合的方法。将各种分子与蛋白化学交联的方法是本领域中众所周知的并可以使用。
番木瓜花叶病毒
PapMV是本领域中已知的,并且可以以ATCC No.PV-204TM得自(例如)美国典型培养物保藏中心(ATCC)。该病毒可以依照标准程序(例如,参见Erickson,J.W.&Bancroft,J.B.,1978,Virology90:36-46)保存在宿主植物(例如番木瓜(Carica papaya)和金鱼草(Antirrhinum majus))中并从其中纯化。
PapMV VLP
本领域的技术人员可以采用野生型蛋白的序列,通过标准的基因工程技术容易地制得用于制备本发明的VLP的重组外壳蛋白。蛋白基因工程的方法是本领域中已知的(例如,参见Ausubel等.(1994&updates)Current Protocols in Molecular Biology,John Wiley&Sons,New York),并且野生型PapMV外壳蛋白的序列也是已知的(参见SEQ ID NO:1和2)。
可以使用标准技术(参见,例如,Ausubel等,ibid.)实现编码野生型蛋白的核酸序列的分离和克隆。例如,可以通过以下的方法从PapMV中直接获得核酸序列,所述方法为:通过标准方法提取RNA,然后从RNA模板合成cDNA(例如,通过RT-PCR实现)。PapMV可以通过标准技术从感染的植物叶子(其表现出花斑症状)提纯(例如,参见下文提供的实施例I)。
然后将编码外壳蛋白的核酸序列直接插入到合适的表达载体或经过一个或多个亚克隆步骤后插入到合适的表达载体。本领域技术人员将会理解,所用的精确的载体并非本发明的关键。合适的载体的例子包括(但不限于):质粒、噬粒(phagemid)、柯斯质粒(cosmid)、噬菌体、杆状病毒、逆转录病毒或DNA病毒。然后可以按照下面更详细描述的方法将该外壳蛋白表达和纯化。
可供选用的另外一种方式是,通过本领域中已知的标准体外定点突变技术,可以进一步改造编码外壳蛋白的核酸序列以引入一个或多个突变(例如前面描述的那些)。可以通过一个或多个组成编码序列的合适核苷酸的缺失、插入、置换、逆转或其组合来引入突变。例如,这可以通过基于PCR的技术实现上述过程,为此,应设计包含错配、插入或缺失一个或多个核苷酸的引物。突变的存在可以通过多种标准技术来验证,例如通过限制性分析法或通过DNA测序法来验证。
如上所述,还可以改造外壳蛋白以产生包含一种或多种抗原或亲和肽(其与外壳蛋白融合)的融合蛋白。制备融合蛋白的方法是本领域技术人员所熟知的。可以将编码融合蛋白的DNA序列插入如上所述的合适的表达载体。
本领域普通技术人员将会理解,可以以诸多方式改变编码外壳蛋白或融合蛋白的DNA而不影响编码的蛋白的活性。例如,DNA序列的变化可以优化用于表达蛋白的宿主细胞内的优先密码子,或可以含有其它的有利于表达的序列改变。
本领域技术人员将会理解,表达载体还可以包含有效转录编码外壳蛋白或融合蛋白的DNA序列所需要的调节元件,例如转录元件。可以包含于载体中的调节元件的例子包括(但不限于):启动子、增强子、终止子和多聚腺苷酸化信号。因此,本发明提供包含调节元件的载体,所述调节元件与编码经基因改造的外壳蛋白的核酸序列操作地连接。本领域技术人员将会理解,合适的调节元件的选择取决于用于表达基因改造的外壳蛋白所选的宿主细胞,并且上述调节元件可以来自多种来源,其包括细菌、真菌、病毒、哺乳动物或昆虫的基因。
在本发明的上下文中,所述表达载体可以额外地含有有利于表达的蛋白纯化的异源核酸序列。这种异源核酸序列的例子包括(但不限于):亲和标记物(例如金属亲和标记物、组氨酸标记物)、编码抗生物素蛋白/链霉抗生物素蛋白的序列、编码谷胱甘肽-S-转移酶(GST)的序列和编码生物素的序列。在使用前,可以根据本领域中已知的技术,将所述异源核酸序列编码的氨基酸从表达的外壳蛋白中除去。可供选用的另外一种方式是,如果对应于异源核酸序列表达的氨基酸不干扰随后组装成VLP,可以将其保留在外壳蛋白。
在本发明的一个实施方案中,外壳蛋白被表达为组氨酸标记的蛋白。所述组氨酸标记物可以位于外壳蛋白的羧基端或氨基端。
可以通过本领域中已知的各种方法之一将表达载体引入合适的宿主细胞或组织。这样的方法通常可以在Ausubel等(ibid.)中具有描述,并且包括(例如):稳定或瞬时转染法、脂质转染法、电穿孔法和重组病毒载体感染法。本领域技术人员将会理解,用于表达外壳蛋白的合适的宿主细胞的选择取决于所选的载体。宿主细胞的例子包括(但不限于):细菌、酵母、昆虫、植物和哺乳动物细胞。精确的宿主细胞并非本发明的关键。所述外壳蛋白可以在原核宿主(例如,大肠杆菌、杀鲑气单胞菌或枯草杆菌)或在真核宿主(例如,拟酵母菌(Saccharomyce)或毕赤酵母属(Pichia);哺乳动物细胞,如COS、NIH3T3、CHO、BHK、293或HeLa细胞;昆虫细胞或植物细胞)中制备。
如果需要,可以通过本领域中已知的标准技术从宿主细胞中纯化外壳蛋白(例如,参见Current Protocols in Protein Science,ed.Coligan,J.E.等,Wiley&Sons,New York,NY),并使用完整的蛋白或其蛋白水解片段通过标准的肽测序技术进行测序以证实蛋白的同一性。
本发明的重组外壳蛋白能够多聚化并组装成VLP。一般而言,组装在表达外壳蛋白的宿主细胞中发生。可以通过标准技术(例如下文的实施例部分提供的那些)从宿主细胞中分离VLP。可以通过标准技术(例如层析技术)进一步纯化VLP,以去除污染的宿主细胞蛋白或其它化合物(例如LPS)。在本发明的一个实施方案中,所述VLP被纯化以去除LPS。
在本发明的一个实施方案中,所述外壳蛋白在宿主细胞中组装以提供重组病毒并且能够用于制备包含核酸和融合蛋白的感染性病毒颗粒。这可以使感染性病毒颗粒能够感染邻近的细胞并在其中表达融合蛋白。在该实施方案中,用于复制病毒的宿主细胞可以是容许病毒复制的植物细胞、昆虫细胞、哺乳动物细胞或细菌细胞。该细胞可以是用于病毒(病毒样颗粒源于该病毒)的天然的宿主细胞,但这并不是必需的。宿主细胞可以用颗粒形式的病毒(即,包含核酸和蛋白的组装的棒)或可供选用的核酸形式的病毒(即,RNA,如病毒RNA;cDNA或由cDNA制备的失控转录物)初始感染,条件是用于初始感染的病毒核酸能够复制并产生具有嵌合蛋白的整个病毒颗粒。
重组和修饰的外壳蛋白的特征
可以通过标准技术,例如,通过电子显微镜观察纯化的蛋白(例如,参见实施例I),来分析重组外壳蛋白和共价连接有抗原或亲和肽的外壳蛋白的多聚化并自组装成VLP的能力。此外,可以使用超速离心将VLP分离为粒状沉淀,而将较小的聚集物(20聚体和更小的聚集物)留在上清液中,并且可以使用圆二色(CD)光谱来比较重组或修饰的蛋白与WT病毒的二级结构(例如,参见实施例I)。
如果需要,可以通过本领域中已知的技术测定VLP和PapMV的稳定性,所述技术例如为SDS-PAGE法和蛋白酶K降解分析法(参见实施例II)。根据本发明的一个实施方案,本发明的PapMV和PapMV VLP在高温下是稳定的,并且能够在室温下容易地贮藏。
PapMV或VLP贮备物的制备
可以通过标准技术制备重组PapMV或VLP的贮备物。例如,含有重组外壳蛋白的PapMV或假病毒可以在合适的宿主(例如番木瓜或金鱼草)中增殖,使得可以收获足够的PapMV或假病毒。
可以从合适的宿主细胞(例如大肠杆菌)中制备PapMV VLP贮备物,所述合适的宿主细胞被表达载体或编码组成VLP的重组外壳蛋白的载体转化或转染。然后按照本领域中已知的那样,将所述宿主细胞在适于编码的蛋白表达的条件下培养。所表达的外壳蛋白在宿主细胞中多聚化并组装成VLP,并可以通过标准技术从细胞中分离(例如,通过将细胞裂解并使细胞裂解液进行一个或多个层析纯化步骤)。
如下文提供的实施例所示的那样,PapMV VLP是稳定的结构,因此VLP贮备物可以容易地在室温下或冰箱中贮藏。
效能评价
为了评价本发明的APS作为疫苗的效能,可以进行攻击研究。该研究涉及通过标准技术将几组测试动物(例如小鼠)接种本发明的APS。平行地制备包含未接种动物和/或接种市售可得的疫苗的对照组、或其它阳性对照。在接种后的合适的一段时间,用合适的肠细菌攻击动物。采集接种之前和之后以及攻击之后的动物血样,然后分析针对病毒的抗体应答。抗体应答的合适的测试方式包括(但不限于):免疫印迹分析法和酶联免疫吸附测定法(ELISA)。
还可以通过本领域中已知的技术(包括下面提供的实施例中描述的那些)来评价细胞免疫应答。例如,在体外和体内通过树突状细胞将在PapMV VLP上表达的抗原表位处理并交叉提呈至特异性T淋巴细胞。用于评价细胞(T淋巴细胞)免疫的诱导的其它有用的技术包括:(例如)通过ELISA监测T淋巴细胞的增殖和IFN-γ分泌物的释放,以监测细胞因子的诱导(参见实施例III)。
疫苗组合物
本发明提供适于用作治疗或预防肠细菌感染的疫苗的组合物,所述组合物包含一种或多种本发明的APS以及一种或多种无毒的可药用的载体、稀释剂和/或赋形剂。如果需要,可以在该组合物中包含其它的活性成分、佐剂和/或免疫增强剂。
该组合物可以被配制为适合于各种途径施用。例如,该组合物可以被配制为用于口服、局部施用、鼻腔施用、直肠施用、或胃肠外施用,包括吸入或喷雾施用。本文使用的术语“胃肠外”包括:皮下注射、静脉注射、肌内注射、鞘内注射(intrathecal injection)、胸骨内注射(intrasternal injection)或输注技术。在本发明的一个实施方案中,该组合物被配制为用于局部施用、直肠施用或胃肠外施用、或通过吸入或喷雾而施用。在本发明的另一个实施方案中,该组合物被配制为胃肠外施用。
所述组合物优选包含有效量的一种或多种本发明的APS。本文所用的术语“有效量”是指表现出可检测的免疫应答所需要的APS的量。对于给定的指标,可以用(例如)细胞培养测定或者动物模型(通常为啮齿动物、兔、犬、猪或灵长类)来最初估测APS的有效量。所述动物模型还可以用于确定施用的合适浓度和途径。然后这样的信息可以用于确定对要治疗的动物(包括人)施用的有用剂量和施用途径。在本发明的一个实施方案中,单位剂量包含约10μg至约10mg的外壳蛋白。在另一个实施方案中,单位剂量包含约10μg至约5mg的外壳蛋白。在另外的实施方案中,单位剂量包含约40μg至约2mg的外壳蛋白。可以使用一种或多种剂量免疫动物,并且这些剂量可以在同一天施用或在数天或数周的期间内施用。
如上所述,本发明的APS可以包含多个抗原(接合和/或非接合的形式),并因此单一一种APS可以提供多价疫苗制剂。还可以通过使用包含在肠杆菌科的不同品种之间保守的抗原(接合和/或非接合的形式)的APS来提供多价疫苗。还涵盖包含多种APS的多价疫苗组合物,其中各APS包含不同的抗原。多价疫苗可用于(例如)提供针对肠杆菌科的多于一个品种的防护。除了具有更高价的疫苗以外,多价疫苗制剂还包括二价制剂和三价制剂。本发明的一个实施方案提供一种多价疫苗。本发明的另一个实施方案提供一种包含在多个肠杆菌科的品种之间保守的抗原的多价疫苗。另一个实施方案提供这样一种多价疫苗,该多价疫苗包含多个(即2个或更多个)APS,各APS包含不同的抗原。在本发明的另一个实施方案中,提供这样一种多价疫苗,该多价疫苗包含含有接合的OmpC或OmpF抗原和非接合的OmpC或OmpF抗原的APS。
包含多个(即2个或更多个)APS(各APS包含不同的抗原)的疫苗制剂由于在制剂中具有更高数目的抗原表位,因此可以提供增强的保护作用。因此本发明的一个实施方案提供包含2个或更多个APS的疫苗制剂,各APS包含不同的抗原。在另一个实施方案中,提供这样一种疫苗制剂,该疫苗制剂包含含有OmpC抗原的APS和含有OmpF抗原的APS。在一个具体的实施方案中,疫苗制剂所包含的2个或更多个APS既包含接合抗原,又包含非接合抗原。
用于口服使用的组合物可以被配制为(例如)片剂、锭剂(troche)、糖锭(lozenge)、水性或油性悬浮液、分散性粉末剂或颗粒剂、乳剂、硬胶囊剂或软胶囊剂、或糖浆剂、或酏剂。所述组合物可以根据本领域中已知的用于制备药物组合物的标准方法来制备,并且可以含有选自由甜味剂、调味剂、着色剂以及防腐剂所组成的组中的一种或多种试剂,以提供药学上美观和可口的制剂。片剂含有与合适的无毒的可药用的赋形剂混合的APS,所述赋形剂包括(例如):惰性稀释剂,例如碳酸钙、碳酸钠、乳糖、磷酸钙或磷酸钠;成粒剂和崩解剂,例如玉米淀粉或海藻酸;粘合剂,例如淀粉、明胶或阿拉伯胶;以及润滑剂,例如硬脂酸镁、硬脂酸或滑石粉。所述片剂可以是无包衣的,或者它们可以通过已知的技术包衣以延缓在胃肠道中的分解和吸收,从而在延长的时间内提供持续的作用。例如,可以使用诸如单硬脂酸甘油酯或二硬脂酸甘油酯等延时材料。
用于口服使用的组合物还可以表现为硬明胶胶囊的形式,其中所述APS与惰性固态稀释剂(例如,碳酸钙、磷酸钙或高岭土)混合;或表现为软明胶胶囊的形式,其中活性成分与水介质或油介质(例如,花生油、液态石蜡或橄榄油)混合。
用于鼻腔施用的组合物可以包括(例如):鼻腔喷雾剂、滴鼻剂、悬浮液、溶液、凝胶剂、膏剂、乳膏和粉剂。该组合物被配制为用于通过合适的市售可得的鼻腔喷射装置(例如,AccusprayTM(得自Becton Dickinson公司))施用。其它的鼻腔施用方法是本领域中已知的。
配制为水性悬浮液的组合物含有与一种或多种合适的赋形剂混合的APS,所述赋形剂例如为:悬浮剂,例如羧甲基纤维素钠、甲基纤维素、羟丙基甲基纤维素、海藻酸钠、聚乙烯吡咯烷酮、羟丙基-β-环糊精、黄蓍胶以及阿拉伯胶;分散剂或润湿剂,例如天然存在的磷脂(例如卵磷脂),或者环氧烷烃与脂肪酸的缩合产物(例如,聚氧乙烯硬脂酸酯),或者环氧乙烷与长链脂肪醇的缩合产物(例如十七碳乙氧基十六醇(hepta-decaethyleneoxycetanol)),或者环氧乙烷与源自脂肪酸和己糖醇的偏酯的缩合产物(例如聚氧乙烯山梨糖醇单油酸酯),或者环氧乙烷与源自脂肪酸和己糖醇酐的偏酯的缩合产物(例如聚氧乙烯脱水山梨糖醇单油酸酯)。所述水性悬浮液还可以含有一种或多种防腐剂(例如对羟基苯乙酯、或对羟苯甲酸正丙酯)、一种或多种着色剂、一种或多种调味剂或者一种或多种甜味剂(例如蔗糖或糖精)。
可以通过将APS悬浮于植物油(如花生油、橄榄油、芝麻油或椰子油)或矿物油(如液体石蜡)中而将组合物配制成油性悬浮液。油性悬浮液可含有增稠剂,例如蜂蜡、硬石蜡或十六醇。可以任选地添加将上述的甜味剂和/或调味剂以提供可口的口服制剂。可通过添加抗氧化剂(例如抗坏血酸)来使这些组合物保存。
所述组合物可以配制为分散性粉末剂或颗粒剂,所述粉末剂或颗粒剂随后可通过加水而制备水性悬浮液。这些分散性粉末剂或颗粒剂提供了与一种或多种分散剂或湿润剂、悬浮剂和/或防腐剂混合的APS。合适的分散剂或润湿剂和悬浮剂示例为上文所提到的那些。在这些组合物中,还可以包含其它赋形剂,例如甜味剂、调味剂以及着色剂。
本发明的组合物还可以配制成水包油型乳剂。油相可以是植物油(如橄榄油或花生油)、或矿物油(如液体石蜡)、或可以是这些油的混合物。这些组合物中所包含的合适乳化剂包括:天然存在的树胶,如阿拉伯树胶或黄蓍胶;天然存在的磷脂,如大豆、卵磷脂;或衍生自脂肪酸和己糖醇、酐的酯或偏酯,例如脱水山梨糖醇单油酸酯,以及所述偏酯与环氧乙烷的缩合产物,例如聚氧乙烯脱水山梨糖醇单油酸酯。所述乳剂还可任选地含有甜味剂和调味剂。
可通过将APS与一种或多种甜味剂(例如甘油、丙二醇、山梨糖醇或蔗糖)组合而将所述组合物配制成糖浆剂或酏剂。这些制剂还可任选地含有一种或多种缓和剂(demulcent)、防腐剂、调味剂和/或着色剂。
可根据本领域已知的方法并使用上述合适的一种或多种分散剂或润湿剂和/或悬浮剂将所述组合物配制为无菌注射水性悬浮液或油性悬浮液。无菌注射制剂可以是在无毒的胃肠外可接受的稀释剂或溶剂(例如1,3-丁二醇)中的无菌注射溶液或悬浮液。所用的可接受载体和溶剂包括(但不限于):水、林格氏溶液(Ringer’s solution)、乳酸盐林格氏溶液和等渗氯化钠溶液。其它例子包括常规用作溶剂或悬浮介质的无菌非挥发性油,以及多种刺激性小的非挥发性油(包括(例如)合成的单甘油酯或二甘油酯)。诸如油酸等脂肪酸也可以用于注射剂的制备。
本发明的组合物可以任选地含有:防腐剂,如抗微生物剂、抗氧化剂、螯合剂和惰性气体;和/或稳定剂,如碳水化合物(例如山梨糖醇、甘露醇、淀粉、蔗糖、葡萄糖或葡聚糖)、蛋白(例如白蛋白或酪蛋白)或含有蛋白的试剂(例如胎牛血清或脱脂牛奶);以及合适的缓冲液(例如磷酸盐缓冲液)。可以根据熟知的参数来调节该组合物的pH和各种组分的具体浓度。
此外,可以任选地将具有佐剂活性的一种或多种化合物添加至疫苗组合物。合适的佐剂包括(例如):明矾佐剂(例如氢氧化铝、磷酸盐或氧化物);油-乳液(例如 或
或 );皂角苷、或维生素E溶解物(solubilisate)。已知病毒颗粒也具有佐剂的性质(Adjuvant and Antigen Delivery Properties of Virosomes,Glück,R等,2005,Current Drug Delivery,2:395-400),并且可以与本发明的APS联用。
);皂角苷、或维生素E溶解物(solubilisate)。已知病毒颗粒也具有佐剂的性质(Adjuvant and Antigen Delivery Properties of Virosomes,Glück,R等,2005,Current Drug Delivery,2:395-400),并且可以与本发明的APS联用。
如本文中以上所述和指出的,PapMV和PapMV VLP具有佐剂性质。因此,在本发明的一个实施方案中,所述疫苗组合物包含额外的PapMV或PapMV VLP作为佐剂。在一些实施方案中,使用PapMV或PapMV VLP可以提供超过市售可得的佐剂的优点,因为已经观察到,当通过注射施用时,PapMV或PapMV VLP不会引起明显的局部毒性。
本发明还包括经调理的疫苗组合物,例如,该疫苗组合物包含由以前免疫过疫苗、抗原或PapMV VLP的动物或人体中分离的抗体。基于由以前免疫过疫苗、抗原或PapMV VLP的动物或人体中分离的抗体的重组抗体也可以用于调理该疫苗组合物。
本发明还包括含有本发明的APS的疫苗组合物和市售可得的肠细菌疫苗的组合。这种疫苗包括Vi和Ty21a伤寒疫苗。
其它的药物组合物和制备药物组合物的方法是本领域中已知的并(例如)在文献“Remington:The Science and Practice of Pharmacy”(以前为“Remingtons Pharmaceutical Sciences”);Gennaro,A.,Lippincott,Williams&Wilkins,Philadelphia,PA(2000)中有所描述。
疫苗稳定性
如前文和本文提供的实施例所述,PapMV VLP是稳定的结构,因此提供稳定的疫苗组合物。源自伤寒沙门氏菌的孔蛋白在冰箱或室温下亦表现出长期的稳定性(例如,参见下面的实施例XIII和图30)。因此,本发明的一个实施方案提供一种疫苗组合物,所述疫苗组合物含有PapMV、或PapMV VLP以及一种或多种伤寒沙门氏菌孔蛋白,其中,在不存在常规的防腐剂或存在少量的合适去污剂的情况下,所述疫苗组合物在冰箱中(例如,约1℃至约10℃)、在较长的时间(例如六个月或更长)内是稳定的。在另一个实施方案中,本发明提供一种疫苗组合物,所述疫苗组合物含有PapMV、或PapMV VLP以及一种或多种伤寒沙门氏菌孔蛋白,所述疫苗组合物在冰箱中、在至少6个月至约36个月内是稳定的。在另一个实施方案中,本发明提供一种疫苗组合物,所述疫苗组合物含有PapMV、或PapMV VLP以及一种或多种伤寒沙门氏菌孔蛋白,所述疫苗组合物在冰箱中在约5年内是稳定的。在另一个实施方案中,本发明提供一种疫苗组合物,所述疫苗组合物含有PapMV、或PapMV VLP以及一种或多种伤寒沙门氏菌孔蛋白,其中在不存在常规的防腐剂或存在少量的合适去污剂的情况下,所述疫苗组合物在常温下在至少1个月至6个月内是稳定的。
应用和用途
本发明提供了本文描述的APS的多种应用和用途。非限定性的例子包括将APS作为针对一种或多种肠细菌感染的疫苗的用途,以及将APS用于筛选针对肠细菌的抗体的用途。因此本发明还提供在动物内诱导针对一种或多种肠细菌抗原的免疫应答的方法。此外,本发明的APS用于制备药物(例如佐剂、疫苗和/或药物组合物)的用途也在本发明的范围内。
本发明的APS适合用于人类和非人类动物,包括家畜和农畜。APS的施用方案不必与任何其它通常可接受的接种程序不同。可以以足以产生有效的免疫应答的量而单次施用APS,或者可供选用的另外一种方式是,可以使用其他的方案,其中在初始施用APS后,只用抗原或用APS进行加强。类似地,在初始施用后,如果抗体效价落在可接受的水平以下,可以用APS或抗原进行加强。
当APS包含非接合的抗原时,根据需要免疫应答的受试者的需要,PapMV或VLP组分可以与抗原同时施用,或者其可以在施用抗原之前或之后施用。
本发明的一个实施方案提供包含APS的疫苗和常规的肠细菌疫苗(例如Vi或Ty21a伤寒疫苗)的联用。根据该实施方案,所述APS疫苗可以与常规的疫苗同时施用(例如,通过联用两种组合物来实现),或者其可以在施用常规的疫苗之前或之后施用。
本发明的一个实施方案提供所述APS作为人用肠细菌疫苗的用途。本发明的另一个实施方案提供所述APS作为针对下列菌属的品种的人用肠细菌疫苗的用途:埃希氏菌属、沙门氏菌属、耶尔森菌属、志贺菌属、变形杆菌属、克雷白氏杆菌属、或肠杆菌属。本发明的又一个实施方案提供所述APS作为针对下列属种的人用肠细菌疫苗的用途:沙门氏菌属种、大肠杆菌、耶尔森菌属种、志贺菌属种、克雷白氏杆菌属种、或肠杆菌属种。另一个实施方案提供所述APS作为针对下列属种的人用肠细菌疫苗的用途:沙门氏菌属种、大肠杆菌、或志贺菌属种。用于所述用途的APS包含选自孔蛋白抗原、HSP抗原和SBAC抗原中的一种或多种抗原。
根据所述APS中包含的抗原,所述疫苗用于治疗和/预防肠细菌引起的疾病和病症,例如伤寒、食源性胃肠炎、食源性肠道感染、尿路感染、脑膜炎、败血症、细菌性痢疾、瘟疫、小肠结肠炎、肺炎、血流感染、医源性感染、和革兰氏阴性菌败血症。
在本发明的一个实施方案中,所述APS包含一种或多种OmpC抗原、OmpF抗原、源自OmpC直系同源物的抗原、或源自OmpF直系同源物的抗原。本发明的一个实施方案提供APS作为针对各种沙门氏菌属种、大肠杆菌和志贺菌属种的肠细菌疫苗的用途,所述APS包含一种或多种OmpC和/或OmpF抗原、或其直系同源物。在本发明的一个具体的实施方案中,提供包含OmpC或其直系同源物的APS单独地、或与包含OmpF或其直系同源物的APS联用作为多价疫苗的用途。
本发明的另一个实施方案提供APS作为针对伤寒沙门氏菌和用于治疗或预防人伤寒的疫苗的用途。在一个实施方案中,所述疫苗既适用于成人,也适用于儿童。在本发明的一个具体的实施方案中,提供一种用作人用伤寒疫苗的APS,所述APS包含至少一种源自伤寒沙门氏菌的孔蛋白的抗原。如上所述,所述APS可以任选地包含一种或多种AIA(所述AIA为孔蛋白抗原)和/或源自不同的伤寒沙门氏菌的细菌成分(即源自其它蛋白,如HSP;和/或源自SBAC)的抗原。在一个具体的实施方案中,提供包含OmpC或其直系同源物的ASP单独地或与包含OmpF或其直系同源物的ASP联用作为伤寒疫苗的用途。在本发明的另一个实施方案中,提供一种APS作为人伤寒疫苗的用途,所述APS包含至少一种氨基酸序列基本等同于下述序列中的任意一种的抗原:SEQ ID NO:4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20、或者它们的变体。在另一个实施方案中,提供一种APS作为人伤寒疫苗的用途,所述APS包含至少一种氨基酸序列基本等同于SEQ ID NO:4、或5、或者它们的变体的抗原。
本发明的可供选用的另外一种实施方案提供所述APS作为非人类动物(例如奶牛、猪、马、山羊、绵羊、犬、猫、鸡、鸭、火鸡、非人类的灵长类、豚鼠、兔、雪貂、大鼠、仓鼠、小鼠、鱼或禽)的肠细菌疫苗的用途。该疫苗可用于治疗和/或预防肠细菌疾病或病症,例如,呼吸系统疾病、肠炎、腹膜炎、膀胱炎和其它泌尿生殖道感染、全身性大肠杆菌病、牛腹泻、水肿、乳腺炎、淋巴结炎、肠炎、败血性流产、子宫炎、肺炎、腹泻、死胎、羊毛损伤、和败血症。
对于在非人类动物中的用途,所述APS可以包含上文鉴定的人用的一种或多种抗原,其包括(但不限于):OmpC抗原、OmpF抗原、HSP抗原和SBAC抗原。在本发明的另一个实施方案中,提供APS在非人类动物中作为肠细菌疫苗的用途,其中所述肠细菌疫苗针对医学相关的肠杆菌科中的一种或多种菌种。上述菌种的例子包括(例如):在灵长类动物中的肺炎克雷白氏杆菌、假结核耶尔森菌、志贺菌属种、大肠杆菌、沙门氏菌属种;在猫和犬中的大肠杆菌和变形杆菌属种;在马中的伤寒沙门氏菌、新港沙门氏菌、鸭沙门氏菌和肺炎克雷白氏杆菌;在禽中的大肠杆菌、肠道沙门氏菌、鸡白痢沙门氏菌和鸡沙门氏菌;在绵羊中的大肠杆菌、流产沙门氏菌、伤寒沙门氏菌和都柏林沙门氏菌;在奶牛中的大肠杆菌、沙雷氏菌属种、克雷白氏杆菌属种、弗氏柠檬酸杆菌、都柏林沙门氏菌和伤寒沙门氏菌;在猪中的大肠杆菌、肺炎克雷白氏杆菌、小肠结肠炎耶尔森菌、猪霍乱沙门氏菌、猪伤寒沙门氏菌、鼠沙门氏菌和德尔卑沙门氏菌;以及在鱼中的鲁氏耶尔森菌、迟缓爱德华菌、和鲶鱼爱德华菌。
如本文所指出,PapMV VLP能够增强针对抗原的体液和/或CTL应答。因此,在本发明的一个实施方案中,提供一种包含APS的疫苗,所述APS包含能够产生体液应答和/或CTL应答的伤寒沙门氏菌抗原。
本发明还提供所述APS作为筛选试剂的用途,例如,筛选肠细菌(例如伤寒沙门氏菌)的抗体。所述APS可以容易地适用于常规的免疫技术(例如酶联免疫吸附测定(ELISA)或免疫印迹分析),因此可用于诊断和研究的领域中。
试剂盒
本发明还提供包含一种或多种用作肠细菌疫苗的APS的试剂盒。在一个实施方案中,本发明提供包含一种或多种用作针对伤寒的疫苗的APS的试剂盒,所述APS包含伤寒沙门氏菌抗原。所述试剂盒的单独的组分被封装在单独的容器中,并且这种容器可附带监管药品或生物制品的生产、使用或销售的政府机构规定形式的通知书,该通知书反映了政府机构对生产、使用或销售的许可。该试剂盒可以可任选地包含列出疫苗的使用方法或施用方案的说明或指导。
当试剂盒的一种或多种组分以溶液(例如水溶液或无菌水溶液)形式提供时,容器构件本身可以是吸入器、注射器、吸量管、眼滴管或其他类似的装置,所述溶液可以从这些装置中施用给受试者或用于与试剂盒内的其它组分混合。
所述试剂盒的组分还可以以干燥或冻干形式提供,并且所述试剂盒还可另外含有用于冻干组分的还原的合适溶剂。不管容器的数量或类型如何,本发明的试剂盒还可含有辅助组合物施用给受试者的工具。这种工具可以是吸入器、注射器、吸量管、镊子、测量勺、眼滴管或类似的医学上批准的递送载体。
本发明还提供了包含用于抗体检测的本发明的一种或多种APS的筛选试剂盒。该试剂盒可以是诊断试剂盒或用于研究目的的试剂盒。所述试剂盒的单独的组分封装在单独的容器中,并且这种容器可附带监管生物制品的生产、使用或销售的政府机构规定形式的通知书,该通知书反映了政府机构对生物制品的生产、使用或销售的许可。该试剂盒可以可任选地包含列出疫苗的使用方法或施用方案的说明或指导。
为了更好地理解本文所述的发明而给出了下面的实施例。应当理解,这些实施例旨在描述本发明的示例性实施方案,并且无意于以任何方式限制本发明的范围。
实施例
实施例I:突变对RNA结合和PapMV外壳蛋白的自组装的影响
按照文献Tremblay,M-H.等,2006,FEBS J.,273:14-25所列的方法,使用PapMV外壳蛋白(CP ΔN5;如SEQ ID NO:3所示)制备一系列突变体,所述PapMV外壳蛋白具有在2位处插入的丙氨酸,并且其N-端的5个氨基酸被从WT序列中删除。根据PapMV CP的氨基酸90至169之间的19个马铃薯X病毒CP的氨基酸序列的比对(其显示了在大多数马铃薯X病毒中,对应于PapMV CP的128位的氨基酸是A,但是在PapMV中是E)以及带电荷的残基R104、K133、K137和R161可能涉及与基因组RNA的相互作用并且在病毒基因组的组装和封装中起重要作用的报导(Abouhaidar,&Lai,1989,J.Gen.Virol.70,1871-5),制备如下的突变体:E128A;K97A;R104K105R108/A;R118D120K121/A;K133K137/A;D142D145/A;E148A;R161A或E166E168/A。
电子显微镜检测和免疫金标记
将VLP或病毒在10mM Tris-HCl(pH 8)中稀释,并在涂覆碳的formvar栅格上吸附3分钟。将栅格用8mL的BSA(10mg/毫升)封闭30秒并用PBS清洗。在室温下将栅格与在PBS中1∶10稀释的兔抗6XH标记物抗体(得自位于美国宾夕法尼亚州Pittsburgh市的Amersham公司)培养30分钟。然后将栅格用PBS清洗3次,并在室温下与在PBS中1∶20稀释的接合有6nm金颗粒的驴抗兔抗体(Jackson Immuno Research,West Baltimore Pike,West Grove,PA,USA)培养30分钟。然后如上所述用去离子水清洗栅格并染色。
圆二色谱
在20℃下用Olis RSM 1000(Olis,Conway DriveSuites A&B,Bogart,GA,USA)快速扫描单色仪记录CD谱。对于远紫外CD(260nm-190nm),使用通路长度为0.1cm的恒温石英皿。使用公式[θ]MRW=[θ]*MRW/(10×c×1)来计算平均残基椭圆率值([θ]MRW,单位为deg×cm2×dmol),其中[θ]是椭圆率(单位为度),MRW是蛋白中的残基的平均分子量(本研究中使用108),c是单位为g/毫升的蛋白浓度,并且l是单位为cm的通路长度(Johnson,W.C.(1996)Circular Dichroism Instrumentation in Circular Dichroism andthe Conformational Analysis of Biomolecules(Fasman,G.D.,ed)pp.635-652,Kluwer Academic/Plenum Publishers)。
在室温下用Jasco Model J-710仪器(Jasco,Commerce Dr.Easton,MD,USA)记录近紫外CD谱(250nm-350nm)。使用石英比色杯进行记录(通路长度为0.1cm)。以速率为100nm/分钟、步长为0.2nm的10次扫描将光谱平均化。棒状和盘状样品的浓度分别为1.5mg/毫升和5.5mg/毫升。
RNA转录物和电泳迁移率变动分析(EMSA)
使用RiboMAXTM Large Scale RNA Production System-T7试剂盒(Promega P1300,Madison,WI,USA)和T7启动子之前的PapMV的5’端的80nt克隆通过体外转录来制备RNA探针。所述克隆在体外转录之前用EcoRI线性化。在用于DNA/RNA纯化的G50Quick SpinColumn(Roche 1273965)上将所述RNA转录物纯化。使用相同的方法制备用于体外组装分析的PapMV的5’端的1800个核苷酸的转录物。使用虾碱性磷酸酶(Fermentas,Hanover,MD,USA,EFO511)使该RNA探针脱磷酸化,并用T4多核苷酸激酶(NEB,Ipswich,MA,USA,M0201S)使其进行γ32P-ATP标记。然后如前所述使用G-50Quick Spin Column将该探针纯化。将标记的RNA与重组蛋白在室温下培养60分钟。每一个反应均使用165fmol的RNA和各种量的在体外的组装缓冲液中的纯化重组蛋白,所述组装缓冲液含有7.5U的核糖核酸酶抑制剂(Amersham Biosciences 27-0816O1)。反应的终体积是10μL;在上样至5%的非变性聚丙烯酰胺凝胶之前,向样品中加入2μL的上样染料。在0.5X的Tris-硼酸盐-EDTA缓冲液中在10mA的条件下进行90分钟电泳。将凝胶干燥并在Kodak BioMaxMS膜(Amersham Biosciences V8326886)上进行16小时的自动射线照相并显影。
PapMV的纯化和盘状物的分离
通过差速离心从显示出花斑症状的感染的番木瓜叶子中纯化PapMV。将感染的叶子(100g)在盛有100ml的50mM Tris-HCl(pH 8.0)(含有10mM EDTA)的市售混合器中研磨。将研磨的叶子通过纱布过滤,向滤液中加入1%的Triton X-100,并将滤液轻轻搅拌10min。逐滴加入氯仿至其体积相当于滤液体积的1/4。在4℃下将溶液再搅拌30min,并在10000g下离心20min以去除沉淀物。将上清液进行高速(100000g)离心120min。使病毒粒状沉淀悬浮并通过蔗糖垫(30%蔗糖)在100000g下进行另一次高速离心达3.5h。将最终的病毒粒状沉淀悬浮于10mL 50nM的Tris(pH 8.0)中。如果存留有颜色,则使用氯仿进行额外的净化。通过超速离心收集纯化的病毒。按照前面所述的方法(Erickson等,1976,Virology.72,514-7),通过乙酸降解法进行盘状物的分离。
胰蛋白酶消化
在37℃下,将10μg蛋白以50μL的体积在pH 8.5的100mMTris HCl缓冲液中培养120分钟,所述缓冲液含有0.2μg胰蛋白酶(Roche,Indianapolis,IN,USA,1418475)。通过加入10μL的上样染料来终止反应,所述上样染料含有5%的SDS、5mM DTT和40%的甘油。在上样至SDS-PAGE凝胶之前将样品煮沸5分钟。通过考马斯亮蓝(Coomassie blue)染色使所述蛋白可视化。
结果
该PapMV CP在CP的开放阅读框(ORF)的1位和6位具有两个M残基。尚不清楚是否这两个起始密码子在病毒的复制期间都被使用。但是,已经显示,纯化病毒的CP的一大部分在N-端都缺失了数个氨基酸(Zhang等,1993,J.MoI.Biol.234,885-7)。为了确保在大肠杆菌中只产生一种开放阅读框,将N-端的5个氨基酸去除,使得M6起到起始密码子的作用。在NcoI位点引入起始密码子引入了在所有的构建体中均发现的额外的A。在蛋白的C-端引入6XH标记物以有利于纯化过程。所述重组蛋白CP ΔN5在大肠杆菌BL21(pLysS)中表达,并显示比从纯化病毒中提取的WT CP稍大的分子量(MW)。所观察到的两种蛋白之间的差异可能是由融合在C-端的6XH标记物引起的。用Ni2+柱将所述重组蛋白亲和纯化,并用1M咪唑洗脱。据估测,纯化重组蛋白的收率为40mg/升至50mg/升。使用针对WT PapMV CP而出现的抗体的免疫印迹分析证明该纯化的蛋白确为PapMV CP。
制备具有一个、二个或三个A置换的9个不同的突变体。五个突变体(R118-D120-K121/A、K133-K137/A、D142-D145/A、R161A和E166-E167-R168/A)产生不稳定的蛋白并且检测不到或以非常低的水平表达。这可能是在该保守区域的诱变影响了该CP的自然折叠。突变体K97A、R104K105R108/A、E128A和E148A能够以与CPΔN5类似的水平表达,并且如CPΔN5所示易于使用6XH标记物纯化。但是,在透析期间咪唑的去除使得突变体R104K105R108/A和E148A聚集并沉淀,并且这种突变可能影响蛋白的折叠。
如纯化重组蛋白的电子显微照片所示(图2B),构建体CPΔN5在大肠杆菌中自组装成VLP。该VLP与天然的病毒颗粒的形状和直径相似(比较图2A和2B)。为了分析和定量以VLP形式存在的纯化蛋白的比例,通过在100,000g下超速离心2小时来分离VLP和较小的聚集体。发现大多数CPΔN5蛋白(80%)在上清液中。发现VLP在粒状沉淀中,并占总的纯化重组蛋白的20%。但是,纯化的蛋白K97A在超速离心后留在上清液中。与之相反,发现重组蛋白E128A在超速离心后全部在粒状沉淀中。
电子显微镜检测显示,从高速粒状沉淀中分离的E128A VLP与WT病毒类似(图2C)。分别测量CPΔN5和E128A的150个VLP的长度并确定平均长度(图2F)。如图所示,CPΔN5VLP的长度(50nm)比天然病毒(长度为500nm)短10倍。但是,E128A VLP约比CPΔN5VLP长3倍(图2C和2F),这表明该突变体能够更有效地支持组装的起始和延伸。最后,纯化K97A蛋白的电子显微照片显示了直径为15nm至50nm的无组织的聚集体(图2D)。该聚集体的轮廓是不规则的,从而表明该蛋白不能够自组织成VLP。
使用Ni2+柱对VLP的纯化是有效的,从而表明6XH标记物位于表面并能够与亲和柱相互作用。为了证实这个假设,对CPΔN5VLP进行免疫金标记试验,其方法为:使用抗6XHis标记物兔抗血清,之后使用金颗粒标记的二级驴抗兔抗体。正如预料的那样,该VLP饰有金颗粒(图2E),从而表明肽(6XH)与C-端的融合是允许的并且暴露于VLP的表面。该C-端融合的肽的表面暴露表明了C-端作为连接免疫原的合适位点是适宜的。
圆二色(CD)谱用于比较重组蛋白和WT病毒的二级结构。据估测,CP ΔN5的二级结构为49%的α-螺旋和15%的无规卷曲。CPΔN5VLP和WT病毒的CD谱显示出略微不同的曲线(图3A)。WT病毒在208nm处测得的CD信号比CPΔN5VLP更明显(图3A)。有趣的是,E128A VLP在208nm处测得的CD信号与WT病毒重叠(图3A)。
分离的CPΔN5盘状物(纯化的蛋白的高速上清液)的CD信号与使用乙酸方法从纯化的病毒中分离的盘状物的CD信号一致(图3B)。有趣的是注意到在208nm处测得的盘状物的CD信号通常不如VLP明显。该结果表明当盘状物组装成VLP时,α-螺旋的含量增加。最后,将纯化的K97A蛋白的折叠与CPΔN5的高速上清液相比较。两种蛋白显示了一致的CD曲线(图3E)。
还获得了250nm和350nm之间的光谱以测量蛋白中芳香族残基和色氨酸残基的吸收。这些残基的环境的变化影响记录的信号并显示了三级结构的变化。CPΔN5和E128A的VLP看起来是类似的(图3D),表明这两种VLP的三级结构相同。CPΔN5盘状物和K97A的光谱非常相似,这表明两种蛋白具有相似的三级结构(图3E)。样品之间的曲线强度的微小差异可能是由于蛋白浓度的微小变化所致。
在超速离心之后,发现80%的纯化重组蛋白CPΔN5和全部的K97A蛋白以多聚体(盘状物)的形式存在于上清液中。为了测量这些蛋白的多聚化的水平,使用SuperdexTM 200分析CPΔN5和纯化K97A蛋白的高速上清液。对于CPΔN5,大多数蛋白以450kDa的高分子量复合物(相当于约20个亚单元(该蛋白亚单元的分子量为23kDa)的多聚体)形式洗脱。收集在81.27ml处洗脱的第二个峰,并上样至SuperdexTM 75柱以提高分离度。洗脱的蛋白为39kDa的蛋白。将从得自该峰的样品进行SDS-PAGE,并显示出独特的条带;该样品小于CPΔN5,因而对应于CPΔN5的降解产物。可能的情况是,该降解的蛋白不能够形成高分子量的复合物并以二聚体留在溶液中。
K97A的洗脱曲线可以划分成3个主要的峰(图4B)。第二个峰在50ml处洗脱并与CPΔN5盘状物重叠(图4A),这可能对应于盘状物结构。第一个峰在41ml和43ml之间洗脱,并对应于尺寸大于700kDa的聚集的物质。该洗脱曲线较宽,并显示出肩峰,这表明该物质并不是均匀的,并且可能对应于K97A盘状物通过非特异性相互作用粘合在一起的聚集体。第三个峰没有进一步分析,并且可能对应于CPΔN5构建体所显示的截断的蛋白。这些结果证实,K97A能够与其它蛋白亚单元形成盘状物,但是不能够在大肠杆菌中组装成VLP。
通过亲和色谱将CPΔN5盘状物从纯化蛋白的高速上清液中分离并用于体外的组装测试。通过凝胶过滤分离直径为17nm的盘状物。在室温下将50ml浓度为1mg/毫升的CPΔN5盘状物与0.05mg的RNA培养30分钟。电子显微镜检测显示该RNA和该蛋白组装成规则长度(150nm)的VLP,该长度相当于在体外的组装测试中使用的RNA(PapMV的5’端1800nt)的长度。该结果清楚地显示,在体外,约20个亚单元的盘状物是VLP的组成部件(building block)。纯化的K97A重组蛋白不能在这些条件下与RNA组装成VLP。
K97A和E128A突变体显示了对PapMV CP完全相反的效果。为了评价CPΔN5和E128A VLP是否含有RNA,以分光光度法测量不同的VLP的280/260比率,并与纯化的病毒的蛋白相比较(表5)。正如预料的那样,由于该VLP具有较低水平的RNA,因此显示出比盘状物小的280/260比率。E128A VLP的280/260比率与纯化的病毒相当。有趣的是,CPΔN5VLP的该比率高出了50%。CPΔN5和K97A的分离的盘状物的280/260比率与用乙酸从纯化的病毒中提取的盘状物相当。
表5:PapMV VLP的OD 280/260

为了评价形成VLP的能力与CP对RNA的亲和性是否直接相关,从CPΔN5和E128A突变体中分离盘状物并与K97A相比较。将CPΔN5的高速上清液用于分离450kDa的多聚体(盘状物)。由于E128A突变体在大肠杆菌中只形成VLP,因此通过乙酸处理使这些VLP分裂并分离E128A盘状物(Abouhaidar&Bancroft,1978,Virology.90,54-9)。将不同量的盘状物在含有165fmol的32P标记的RNA探针的10μL的体积中培养,所述RNA探针由PapMV的5’端非编码区的80个核苷酸的转录物制得。通过电泳迁移率变动分析法(EMSA)来分离蛋白-RNA复合物。CPΔN5的盘状物以协作的方式与该探针相互作用,并且引起500ng(22pmol)的蛋白的变动。该结果显示,只有当摩尔比(盘状物/RNA)达到1,000(这对应于与RNA亲和性较弱)时,分离后不含RNA的CPΔN5盘状物才能够在体外与RNA相互作用。当以E128A突变体的分离盘状物和相同的探针进行类似的试验时,该E128A盘状物比CPΔN5更有效地结合RNA。仅仅50ng的蛋白就足以生成蛋白RNA的复合物。即使在高浓度(多达1500ng)下,纯化的K97A蛋白在同样条件的下也不能引起蛋白RNA的复合物的形成。用从CPΔN5 VLP中提取的RNA(按照前述标记)重复EMSA,并且用该不含PapMV封装信号的RNA得到了同样的结果。
为了评价VLP的稳定性,并测量盘状物组装成棒状结构是否提高了该复合物的稳定性,通过CD分光光度法监测它们的耐热性。在表现出任何的疲劳(fatigue)迹象之前,CPΔN5 VLP和病毒都耐高温(超过60℃)(图5A)。纯化的PapMV是所测试的最稳定的结构并且能够耐接近100℃的温度。报导的PapMV的失活温度是70℃。CPΔN5 VLP比WT病毒更敏感,这可是由于存在位于C-端得6XHis标记物所致。E128A VLP似乎对热更为敏感并在42℃下表现出疲劳的迹象(图5A)。盘状物在40℃时迅速变性(图5B)。该结果显示,盘状物封装为棒状结构显著地提高了稳定性。
据估测,用胰蛋白酶处理PapMV会导致C-端的氨基酸198处发生断裂。在这些条件下,余下的蛋白耐蛋白酶。对纯化的病毒颗粒和重组VLP以及盘状物进行类似的分析,并且显示PapMV似乎不受胰蛋白酶的影响。电子显微镜检测证实处理的病毒与未处理的病毒外观一致,然而,从CPΔN5和CPΔN5 VLP分离的盘状物都对胰蛋白酶非常敏感,并产生对应于降解片段的数个较低分子量的条带,这表明VLP的表面暴露有数个带正电荷的残基并且是可利用的。E128A显示了与WT病毒类似的耐胰蛋白酶性。
实施例II:PapMV gp100和PapMV Flu VLP的制备和改造
PapMV CP基因的克隆
将CPΔN5PapMV外壳蛋白(CP)基因用于下面的实施例,并且按照文献Tremblay,M-H.等,.2006,FEBS J.,273:14-25中所述的方法进行制备。简言之,使用下面的寡核苷酸引物,通过RT-PCR由分离的病毒RNA扩增CP基因。
正向CPΔN5引物:
5′AGTCCCATGGATCCAACGTCCAATCTTCTG-3′[SEQ IDNO:27]
反向CPΔN5引物:
5′-ATGCGGATCCTTACTAATGGTGATGGTGATGGTGTTCGGGGGGTGGAAG-3′[SEQ ID NO:28]
将PCR产物用NcoI和BamHI消化,并插入载体pET-3d内以制备CPΔN5PapMV VLP克隆,其中N-端的5个氨基酸从WT序列中缺失。PapMV VLP还在重组蛋白的2位具有插入的丙氨酸。CPΔN5PapMV VLP克隆的氨基酸序列如图1C[SEQ ID NO:3]所示。
PapMV gp100和PapMV Flu构建体的克隆和改造
选择源自确切的肿瘤抗原gp100的9聚体(9-mer)HLA-A*0201抗原表位(IMDQVPFSV;SEQ ID NO:51)和源自流感病毒M1蛋白的9聚体HLA-A*0201抗原表位(GILGFVFTL;SEQ ID NO:52)。HLA-A*0201抗原表位的N-端和C-端侧的侧面连接来自各自的天然序列的5个残基,以有利于通过蛋白酶体进行天然加工(参见图6A)。
为了制备PapMV gp100构建体,使用下面的寡核苷酸:
gp100正义寡核苷酸
5′-CTAGTTCTTCTGCGTTCACCATCATGGACCAGGTTCCGTTCTCTGTTTCTGTTTCTCAGCTGA-3′[SEQ ID NO:29],和
gp100反义寡核苷酸
5′-CTAGTCAGCTGAGAAACAGAAACAGAGAACGGAACCTGGTCCATGATGGTGAACGCAGAAGAA-3′[SEQ IDNO:30]。
将这两个寡核苷酸退火,并克隆至CPΔN5PapMV CP克隆的SepI和MluI位点,其中所述CPΔN5PapMV CP克隆已经用相同的酶线性化。
为了制备PapMV FLU构建体,使用下面的寡核苷酸:
FLU正义寡核苷酸
5′-CTAGTTCTCCGCTGACCAAAGGTATCCTGGGTTTCGTTTTCACCCTGACCGTTCCGTCTGAAA-3′[SEQ ID NO:31],和
FLU反义寡核苷酸
5′-CTAGTTTCAGACGGAACGGTCAGGGTGAAAACGAAACCCAGGATACCTTTGGTCAGCGGAGAA-3′[SEQ IDNO:32]。
如针对gp100构建体所述的那样,将这两个寡核苷酸退火并在CPΔN5PapMV CP的C-端处克隆。
得到的PapMV gp100和PapMV FLU构建体包含PapMV CP基因,其各自的肽在C-端融合并具有6XH标记物以容易进行纯化过程。通过DNA测序证实PapMV克隆的序列。
PapMV、PapMV FLU和PapMV gp100在大肠杆菌中的表达
大肠杆菌表达株BL21(DE3)RIL(Stratagene,La Jolla,CA)被含有上述构建体之一的质粒pET-3d转化,并保持在含有氨苄青霉素(50μg/毫升)的2xYT培养基中。细菌细胞在37℃下生长至在600nm下的光密度为0.6,并用1mm异丙基-b-d-硫代半乳糖苷(IPTG)诱导蛋白表达。诱导在25℃下进行16小时。通过以6000r.p.m离心15分钟来收集细菌。将粒状沉淀在用冰冷却的裂解缓冲液(50mmNaH2PO4 pH 8.0、300mm NaCl,10mm咪唑、20μm苯甲磺酰氟、1mg/毫升裂解酶)中重新悬浮,并以一次通过弗氏细胞压碎器(FrenchPress)使细菌裂解。将裂解液在13000r.p.m.下离心2次,每次30min,以清除细胞碎片。在4℃和温和搅拌的条件下,将上清液与1mLNi-NTA(Qiagen,Valencia,CA)培养4小时。将裂解液上样至柱子上,并以3×15mL洗脱缓冲液(50mm NaH2PO4 pH 8.0,300mmNaCl)清洗珠子,所述缓冲液含有浓度增加的咪唑(10mm,20mm和50mm)。然后用15mL工作缓冲液(10mm Tris/HCl pH 8或10mm磷酸钠缓冲液pH 7.2)清洗珠子。进行2次清洗步骤以去除LPS污染物,一次用pH 8的10mM Tris-HCl、50mM咪唑、0.5%TritonX100,另一次用pH 8的10mM Tris-HCl、50mM咪唑、1%两性清洗剂。蛋白以含1M咪唑的工作缓冲液洗脱。将洗脱的蛋白在Beckman 50.2TI转子中进行高速的超速离心(100000g)120min。将VLP粒状沉淀在无内毒素的PBS(Sigma公司)中重新悬浮。最后,使用0.45μm的过滤器过滤蛋白溶液。通过BCA蛋白试剂盒(Pierce公司)评价蛋白浓度。按照制造商的说明书(Cambrex),用鲎试验评价纯化的蛋白中的LPS的水平,并且评价结果为低于0.005内毒素单位(EU)/μg蛋白。该过程得到大于20mg的纯化VLP/升细菌培养物。
电子显微镜检测和SDS-PAGE
将蛋白用PBS稀释,并在涂覆碳的formvar栅格上吸附3分钟。将栅格用去离子水清洗2次并在室温下用0.1%的醋酸双氧铀染色10分钟。然后在Jeol JEM220FS透射电子显微镜上观察该栅格。使用Adobe Photoshop软件评价100个VLP的平均长度。
使用得自Bio-Rad公司(Hercules,CA)的mini-protean系统,按照本领域中描述的方法(Lepage和Lapointe,2006,Cancer Res.66:2423-2432)进行SDS-PAGE分析。通过考马斯亮蓝染色(Bio-Rad)显示蛋白。在一些试验中,添加最终浓度为13μg/毫升的蛋白酶K(Invitrogen)。
结果
在大肠杆菌中制备的不同PapMV VLP的电子显微镜检测分析(图6B)显示出长度在80nm至200nm范围内的典型的长棒状结构,并且PapMV VLP的直径为15nm;而改造的PapMV gp100和Flu VLP的直径为16nm。所述PapMV CP能够在大肠杆菌中自发地组装成尺寸和形状与PapMV VLP类似的VLP。
将新鲜的和7个月的VLP制剂在4℃的PBS中进行SDS-PAGE分析(图7)。没有在凝胶上检测到降解的证据。此外,将制剂在室温下或在37℃继续培养7天,而没有任何明显的降解。最后,将PapMV制剂与蛋白酶K培养(作为降解的阳性对照),其导致了改造的PapMV VLP的快速降解。没有融合肽的PapMV VLP对蛋白酶K更具耐性,这表明C-端处的融合可能局部地使该区域不稳定,并且增加了对该酶的易感性。
实施例III:在PapMV VLP上表达的gp100和流感病毒M1抗原表位的体外加工和交叉提呈
肽
Gp100肽和Flu肽由GLBiochem上海有限公司合成,并在DMSO(Sigma)中重新悬浮。
培养基和细胞培养物
按照本领域中描述的方法(Lapointe等,2003,Can.Res.63:653-662),在完全培养基(Iscove′s Modified Dulbecco′s Medium;Invitrogen;Carlsbad,CA;和Wisent;St-Bruno,Québec,Canada)中培养T淋巴细胞、树突状细胞(DC)和CD40刺激的B淋巴细胞(CD40-B),所述完全培养基补加7.5%的人血清(热灭活,从正常的供体制备)、2mM的L-谷氨酰胺、100U/毫升的青霉素/链霉素和10μg/毫升的庆大霉素(后面的三者得自Invitrogen和Wisent公司)。
按照本领域中描述的方法(Lapointe等,2003,Can.Res.63:653-662;Lepage and Lapointe,2006,Cancer Res.66:2423-2432),通过加入500ng/毫升的可溶的CD40L三聚体(Immunex Corporation;Seattle,WA)和500U/毫升的重组人IL-4(Peprotech;Rocky Hill,NJ),将CD40活化的B细胞由外周血单核细胞(PBMC)扩增和培养。
通过改变由文献“Sallusto等,1994,J.Exp.Med.179:1109-1118”所述的原始方案,由PBMC制备DC,所述PBMC是由源自正常供体的血液净化制剂(apheresis preparation)中收集的(Lapointe等,2000,Eur.J.Immunol.30:3291-3298)。简言之,通过离心淋巴细胞分离培养基(Wisent)从血液中富集PBMC。在37℃下,通过在组织培养瓶或培养板中进行2小时贴壁(adherence)来富集单核细胞(在T-25中3×107个细胞,在6孔平底培养板中1.5×107个细胞/孔、或在24孔平底培养板中5×106个细胞/孔,其全部得自Costar,Corning,NY)。将贴壁的细胞用PBS(Wisent)清洗一次然后在完全培养基中培养,所述完全培养基补加100ng/毫升的GM-CSF(1,000U/毫升)和500ng/毫升的IL-4(1,000U/毫升)(其均得自Peprotech,Rocky Hill,NJ)。在第3和第5天再次添加GM-CSF和IL-4。在第6天添加PapMVVLP(按照实施例II描述的方法制备)并在第7天收集以用于识别和扩增试验。
黑色素瘤细胞系1088mel在Surgery Branch(NCI/NIH)得到。SK23、T2和乳腺肿瘤细胞系MDA231从美国典型培养物保藏中心(ATCC;Manassas,VA)获得。将所有的肿瘤细胞系培养于RPMI1640培养基,所述培养基补加10%热灭活的FBS、2mM L-谷氨酰胺、100U/毫升的青霉素/链霉素和10μg/毫升的庆大霉素。
PapMV冲击的APC的交叉提呈
通过gp100-特异性T细胞或流感病毒M1-特异性T细胞分析不同的靶细胞,以用于确定抗原表位的MHC I类提呈。Gp 100-特异性CD8+T细胞克隆由National Cancer Institute;NIH,Bethesda,MD友善提供,并且其对209-217位处的天然HLA-A*0201-限制性抗原表位(ITD QVP FSV;SEQ ID NO:53)和在210位处用M修饰的形式(IMDQVP FSV;SEQ ID NO:51)均具有特异性,这提高了肽/MHC复合物的稳定性。使用在文献“Dudley等,1999,J.Immunother,22:288-298”中所描述的快速扩增方案扩增gp100-特异性T细胞。
按照以下方法制备对流感病毒M1衍生的HLA-A*0201-限制性抗原表位(59-67;GIL GFV FTL;SEQ ID NO:52)具有特异性的T细胞系。以特异性的抗体(OneLambda,Canoga Park,CA),通过流式细胞仪(FACScalibur;BD Biosciences,Mississauga,ON)鉴定源自HLA-A*02+正常供体的PBMC。在48孔培养板的多个孔中,将以上制备的PBMC用1μM合成FLU肽在上述培养基内进行刺激。7天后将培养物用肽冲击(以1μM的浓度进行3小时,之后用PBS清洗3次)的自体PBMC或CD40刺激的B淋巴细胞培养物再次刺激。然后每2-3天以150IU/毫升的量添加白细胞介素(IL)-2(Chiron,Emeryville,CA),并将培养物保持在0.5×106个细胞/毫升和2×106个细胞/毫升之间。在按照本领域描述的方法(Lapointe等,2001,J.Immunol.167:4758-4764)将各培养物与肽冲击的T2细胞共培养之后,使用偶联抗体(Endogen,Woburn,MA)通过ELISA进行干扰素(IFN)-γ分泌测定来评价其特异性。
为了评价由PapMV CP介导的交叉提呈,用各种形式的修饰PapMV CP以10μg/毫升-50μg/毫升的量将CD40活化的B细胞或DC冲击20小时。收集细胞,用PBS清洗2次,并接种至96孔培养板中的完全培养基(4×104-10×104个细胞/孔)内。将gp100-特异性T细胞或流感病毒M1-特异性T细胞以2×104-10×104个细胞/孔的量添加到完全培养基内,并保持20小时。收集培养物上清液并按照本领域中描述的方法(Lepag和Lapointe,2006,Cancer Res.66:2423-2432)通过ELISA评价干扰素(IFN)-γ。在一些试验中,将APC用50μM-70μM的氯喹(Sigma)或20μg/毫升-25μg/毫升的乳胞素或1.3μM-3.3μM的MG-132(后两种得自Calbiochem公司,SanDiego,CA)预处理1小时。用PBS清洗细胞并在含有1/20初始抑制剂浓度的培养基中重新悬浮。将处理的细胞用不同的PapMV变体冲击并且按照上述的方法进行MHC I类介导的提呈的分析。
T细胞扩增
用各种形式的修饰PapMV CP(如前面的实施例II所描述)将CD40活化的B细胞或DC冲击20小时。收集细胞,用PBS清洗2次。在48孔培养板的单个孔中,将冲击的APC(2×105-5×105)与自体的PBMC(2×106个细胞)在500μL完全培养基中共培养。当培养基变黄时,去除200μL培养基,并加入400μL新鲜的完全培养基。在第7至10天向单独的培养物中加入新鲜的经PapMV冲击的APC(5×105;冲击20小时并用PBS清洗2次)。然后每3天以100IU/毫升的量添加IL-2。
然后在第15至20天测量T细胞的特异性。简言之,按照本领域中描述的方法(Lapointe等,2001,J.Immunol.167:4758-4764)将扩增的细胞与经肽冲击的T2细胞、或经PapMV冲击的APC共培养。然后按照本领域中描述的方法(Lepage和Lapointe,2006,ibid.)使用偶联抗体(Endogen;Woburn,MA)通过ELISA评价IFN-γ的分泌。可供选用的另外一种方式是,按照制造商的说明,使用偶联抗体(MABTECH)通过ELISPOT分析法来评价抗原-特异性T细胞的频率。使用自动计数器(得自CTL Technologies公司,Cleveland,OH)计算斑点。
结果
DC在本领域中被定义为最佳的APC,并且在当前的试验以及本领域中已经证明在CD40刺激之后扩增的B淋巴细胞是有效的APC。将这些APC用PapMV VLP、PapMV gp100和PapMV-Flu冲击,并与对源自gp100和流感病毒M1蛋白的MHC I类抗原表位具有特异性的确定的T细胞共培养。如图8A所呈现,用PapMV gp100冲击的APC被gp100-特异性T细胞克隆所识别。相反地,只有用PapMV Flu冲击的APC被流感病毒M1特异性T细胞所识别(图8B)。各T细胞培养物的特异性通过用对应于各抗原表位的合成肽冲击的CD40活化B细胞来证实。此外,向特异性T细胞中添加PapMV-Flu不能刺激它们分泌IFN-γ,表明APC对于肽识别是必需的。
为了证实HLA-A*02是参与肽提呈的限制性要素,用由2个额外的HLA-A*02供体以及对该等位基因阴性的其他2个供体制备的APC进行类似的试验。如图9A所示,对于PapMV-Flu冲击的HLA-A*02+供体而言,FLU-特异性T细胞最具反应性(左栏)。其与供体#3具有弱的反应性可以通过这样的事实来解释:该FLU T-细胞系是异源的,并且对可能由另一种MHC I类等位基因提呈的肽具有特异性的T-细胞可能存在于培养物中。此外,gp100特异性T-细胞克隆只与PapMV gp100冲击的HLA-A*02+APC具有反应性(图9A;右栏),这进一步证实了表达相关限制性要素的APC对抗原提呈是必需的。最后,根据本领域中所实施的方法(Lapointe等,2003,Can.Res.63:2836-2843;Lapointe等,2001,J.Immunol.167:4758-4764),使用封闭MHC I类、或MHC II类或HLA-DR提呈的抗体来控制MHC I类的提呈。作为对照,将表达HLA-A*0201和gp100的黑素瘤细胞系与gp100-特异性的T-细胞克隆共培养,并且正如预料的那样,只有封闭MHC I类提呈的抗体丧失了识别(图9B;右部)。如本领域中所证明的那样(Dudley等,1999,J.Immunother.22:288-298;Lapointe等,2001,ibid.),将gp100-特异性T-细胞克隆与细胞系HLA-A*02-/gp100+、或+/-、或-/-共培养均不能激发IFN-γ的分泌。当将一组封闭性抗体施加至PapMV Flu冲击的APC时,只有抗-MHC I类会降低特异性T细胞系的识别(图9B;左部)。后面的这些数据显示,源自PapMV-Flu的FLU肽通过MHC I类(特别是HLA-A*02)提呈。
为了确定PapMV VLP的加工是否由蛋白酶体介导,使用两种不同的蛋白酶体抑制剂,即,乳胞素和MG-132,并且通过封闭来自gp100(黑素瘤细胞)的HLA-A*0201抗原表位的典型MHC I类提呈来控制其活性(图10,右部)。当使用PapMV Flu冲击的APC时,M1肽的提呈不受两种抑制剂的影响(图10,左部)。正如预料的那样,氯喹预处理(其中和了核内体的pH)对FLU抗原表位的交叉提呈有微弱的负面影响,而类似的处理没有改变黑素瘤细胞的典型的MHC I类提呈。总而言之,这些数据表明由PapMV VLP介导的MHCI类的交叉提呈是蛋白酶体非依赖性的。
还评价了用PapMV VLP冲击的APC是否具有扩增源自异源的T细胞群(源自PBMC)的抗原特异性T淋巴细胞的能力。将DC和CD40活化的B淋巴细胞用PapMV VLP冲击,并根据上述方案将冲击的APC与自体PBMC共培养。首先通过ELISPOT分析法来评价特异性扩增的T细胞的频率。如图11A所示,当将PapMV Flu冲击的APC与PBMC一起使用时,产生对HLA-A*0201流感病毒M1肽特异性的细胞。正如预料的那样,未冲击的APC或用PapMV或PapMV gp100冲击的APC不能产生Flu特异性T细胞。正如预料的那样,对于没有黑素瘤的健康的正常供体,没有产生gp100-特异性T细胞。然后评价扩增的T细胞对各种冲击的APC的反应性,并且当使用通过经FLU肽或PapMV Flu冲击的APC扩增的T细胞来扩增T细胞时,IFN-γ的分泌似乎一样高(>5000;图11B)。用PapMV、PapMV gp100或PapMV Flu冲击的APC不能产生对PapMV CP特异性的T细胞,这表明针对PapMV CP的细胞超前免疫(cellularpre-immunity)是微弱的。最后,将扩增的T细胞与用不同量的HLA-A*0201流感病毒M1肽冲击的T2细胞共培养(图12)。在两个独立的试验中,由于T细胞具有识别用0.1nM和0.01nM的肽冲击的T2细胞的能力,因此产生了具有高亲和力的高反应性T细胞。分泌量大多>100000pg/毫升(这是很高的),并且用流感病毒M1肽冲击的APC培养的T细胞通常亲和力较低。从这些试验中可得到结论:经PapMV冲击的APC具有扩增高亲和力的特异性T细胞的能力。有趣的是,没有先前存在针对PapMV CP的细胞超前免疫的证据。
实施例IV:PapMVCP-E2和PapMVCP27-215-E2的制备和改造
该实施例证明,PapMV外壳蛋白的多聚化对PapMV VLP的免疫原性是关键的。
PapMV外壳蛋白的克隆和改造
按照实施例IV描述的方法克隆PapMV CP基因。为了制备
PapMVCP-E2构建体,使用下面的寡核苷酸:
5′-GATCACTAGTGTGGTGGTGGGTACCACCGATCGTAGCGGTGCGCCGACCTACAGCTGGGGTGCGAACGATACGCGTCATG-3′[SEQ ID NO:33],和
5′-CATGACGCGTATCGTTCGCACCCCAGCTGTAGGTCGGCGCACCGCTACGATCGGTGGTACCCACCACCACACTAGTGATC-3′[SEQ ID NO:34]。
在连接至经SpeI/MluI线性化的PapMVCP克隆之前,将这两个寡核苷酸一起退火并用SpeI和MluI消化。
通过以下方法由PapMVCP-E2质粒构建用于截断的外壳蛋白E2融合体(PapMVCP27-215-E2)的表达载体,所述方法为:首先制备下面的用于PCR的两个寡核苷酸(包含NocI限制性位点),所述寡核苷酸被设计为缺失PapMV CP的前26个氨基酸:
正向引物:
5′-AGTCCCATGGCCGATCCAACGTCCAATCTTCTG-3′[SEQID NO:35],和
反向引物:
5′-ACGTCCATGGTATATCTCCTTCTTAAAG-3′[SEQ IDNO:36]。
然后将PCR产物自连接。按照与用于构建PapMVCP27-215-E2克隆相同的程序由PapMVCP质粒获得用于PapMVCP27-215的表达载体。通过DNA测序验证所有的PapMV克隆的序列。
PapMVCP-E2和PapMVCP27-215-E2的表达和纯化
按照实施例IV中描述的方法进行PapMVCP构建体的表达和纯化。E2肽由GLBiochem上海有限公司合成,并在无内毒素的PBS(Sigma公司)中重新悬浮。在使用之前,用0.45μm的过滤器过滤蛋白溶液。使用BCA蛋白试剂盒(Pierce公司)评价蛋白的量。按照制造商的说明(Cambrex公司),用鲎试验评价纯化的蛋白中的LPS的水平,并且结果为低于0.005内毒素单位(EU)/μg蛋白。
SDS-PAGE、电印迹和电子显微镜检测
按照前面的实施例中描述的方法进行SDS-PAGE和电印迹。将蛋白以PBS稀释并在涂覆碳的formvar栅格上吸附3分钟。将栅格用去离子水清洗2次并在室温下用0.1%的醋酸双氧铀染色10分钟。然后在Jeol JEM220FS透射电子显微镜上观察该栅格。使用AdobePhotoshop软件来评价100个VLP的平均VLP长度。
免疫
将5只4至8周龄的C3H/HeJ小鼠(得自Charles RiversLaboratories)皮下注射25μg的PapMVCP-E2、PapMVCP27-215-E2或相同量的E2肽(2μg)或无内毒素的PBS(Sigma公司)。初次免疫2周后给予一次加强剂量。在不同的时间点获得血样并贮存于-20℃直至分析。所有的试验方案经Laval大学的动物保护委员会批准。
ELISA定量
将在pH9.6的0.1M NaHCO3缓冲液中被稀释至浓度为1μg/毫升的P3、P3E2、PapMVCP、PapMVCP27-215或PapMVCP-E2以100μl/孔-200μl/孔的量,涂覆在96孔Costar高结合性培养板(得自Corning公司,NY,USA)上过夜。在37℃下将该培养板用PBS/0.1%Tween-20/2%BSA(150μl/孔)封闭1小时。在用PBS/0.1%Tween-20清洗3次之后,添加从1∶50开始以2倍次序稀释的血清并在37℃下培养1小时。培养之后,将培养板清洗3次,并与在PBS/0.1%Tween-20/2%BSA中以1/10,000稀释的100μl的偶联过氧化酶的山羊抗小鼠IgG、IgG1、IgG2a、IgG2b(其均得自Jackson Immunoresarch公司)、IgG3(得自Rockland公司)在37℃下培养1小时。清洗3次之后,根据制造商的说明用100μl的TMB-S检测IgG的存在;通过添加100μL 0.18mM的H2SO4终止反应并且在450nm处读取OD。该结果表示为抗体的终点效价,并且在OD值为用PBS小鼠的血清的1∶50稀释液得到的背景值的3倍时测定。
关于确定人血清中的抗体水平,应用相同的条件,不同之处在于使用以1/80000稀释的偶联过氧化酶的山羊抗人IgG作为二级抗体。源自感染HCV患者的血清由B.Willems(Hopital Saint Luc,CHUM)提供:该结果表示为抗体的终点效价,并且在OD值为15个未感染的患者的血清池(pool of sera)的血清的1∶25稀释液得到的背景值的3倍时测定。
脾细胞的再激发
在第65天将CD-1小鼠(22周龄)杀死之前,在第0、15、30、45天将其免疫PapMVCP-E2(25μg)。取出脾脏并悬浮于DMEM(2×105个细胞/孔)。用高渗的氯化铵溶液去除红血球。清洗脾细胞并重新悬浮于200μl的DMEM培养基(补加10%FBS[HyClone]、100U/毫升青霉素、100μg/毫升链霉素、2mM L-谷氨酰胺、1mM丙酮酸钠和50μM β-巯基丙醇的DMEM)中,直到在96孔平底培养小板(Costar)中的浓度为2.5×105个细胞/毫升。将样品与25μg/毫升的PapMVCP或PapMVCP-E2VLP培养4天。使用Concavalin A(ConA-10μg/毫升)和PBS作为阳性和阴性对照。收集细胞培养物的上清液,并使用liquid mouse 10细胞因子试剂盒(得自Qiagen公司)测量细胞因子。
小鼠中的气囊
在10至12周龄的CD-1小鼠(Charles River Laboratories)中凸起气囊。通过在第0、3天皮下注射3ml无菌空气在背部凸起气囊。在第7天,将1ml的重组PapMVCP(1μg/毫升至10μg/毫升)、LPS(10μg/毫升)或PBS注射进气囊。处理6小时后,通过CO2窒息杀死小鼠。用1mL PBS-5mM EDTA清洗该气囊1次,然后用2mLPBS-5mM EDTA清洗该气囊2次,并将渗出液在室温下以500×g离心5min。在醋酸蓝(acetic blue)染色之后用血细胞计数器对细胞计数。
骨髓细胞提取和在APC中的分化
从BALN/c小鼠的大腿骨中获得骨髓祖细胞(progenitors cell),并在树突状细胞分化骨髓培养基(其含有95%的RPMI和1%青霉素-链霉素并补加5%的X63-GM-CSF培养基培养物的上清液;X63-GM247CSF细胞系由B.Ludewig,Research Department,Kantonal Hospital St.Gallen,Switzerland提供)中培养6天。在第2和4天部分置换培养基。在第6天,用下列培养基来置换上述培养基:没有LX63调节的培养基。在第7天,使用FITC抗-CD11c和PE-Cy5.5抗-CD11b表面标记物(BD Biosciences)通过流式细胞术来验证APC的富集。该制剂含有25%的CD11c+Cd11b+细胞和超过80%的Cd11b+细胞。该制剂在本文中被称为APC。
流式细胞术
为了评价PapMVCP-E2或PapMVCP27-215-E2在APC中的内化,将1百万个源自骨髓的APC在37℃下与25μg的PapMVCP-E2或PapMVCP27-215-E2培养2小时。简言之,在4℃下将细胞用PBS10%、FBS和抗-CD 16/CD32(1μg/百万个细胞)封闭15min。在用PBS清洗2次之后,在室温下将细胞用PBS/2%多聚甲醛固定10min。在用渗透缓冲液(PBS 10%;FBS 0.2%;Triton X-100)清洗2次之后,在4℃下将细胞与在渗透缓冲液中经1/200稀释的兔多克隆抗体培养45min。在用渗透缓冲液清洗2次之后,在4℃将细胞与渗透缓冲液中经1/5000稀释的二级抗体抗兔IgG alexa 488(MolecularProbes)培养45min。在用PBS清洗之后,立即用EPICS-XL细胞荧光计分析细胞。使用WINMDI2.8进行数据分析。用于检测的兔多克隆Ab以我们自己的设备制备:将兔免疫前的血清用作阴性对照。
共聚焦显微镜
按照与前面描述相同的分化方案,使APC在底部包括无菌载玻片的12孔培养板(Cornin,NY,USA)中生长(200,000个细胞/孔)。对于抗原内化的研究,使用5μg抗原/200000个细胞。固定、渗透、初级和二级抗体培养的步骤如流式细胞术中所描述的那样。立即通过Fluoview Fv300共聚焦显微镜用X60的油浸物镜来分析载玻片。通过x-z切片依次地获得荧光图像,以避免非特异性的波道干扰。然后将图像用Image J软件数字化处理。
统计分析
将非参数的Krustal-Wallis和Dunn′s多重比较测试用于统计分析。P<0.05的值被认为具有统计学显著意义。使用程序PRISM 3.03进行统计分析。
结果
纯化的重组蛋白表现出预期的23kDa的分子量(PapMVCP27-215-E2),对于PapMVCP和PapMVCP-E2而言分子量是26kDa,并且内毒素水平总是在0.005EU/μg蛋白以下。电子显微镜(EM)观察证实,在PapMVCP的C-端处E2肽并没有影响蛋白自组装成与重组的PapMVCP VLP类似的VLP的能力。正如所预料的那样,PapMVCP27-215-E2不能形成VLP,并保持为以前所显示的那样的单体形式(Leclerc等,1998,J Biol Chem,273:29015-21)。VLP的长度是可变的,尺寸在201±80nm的范围内。201nm长的蛋白代表560个CP拷贝,所述CP以重复和结晶的形式呈现E2肽。
为了测试PapMVCP VLP的促炎性能,使用小鼠气囊模型。注射LP含量极低(<0.005EU/μg)的10μg PapMVCP VLP不能在处理6小时之后引起白细胞汇集至CD 1小鼠的气囊内。与此形成对比的是,以1,000和1EU的剂量注射LPS非常有效地引起白细胞的汇集(图13)。该结果表明PapMVCP VLP在6小时后并不促炎,并且在PapMVCP蛋白样品中极低水平的LPS不会在随后的试验中发挥任何显著的免疫原性效果。
测试单体形式(PapMVCP27-215-E2)和多聚形式(PapMVCP-E2)形式在源自骨髓的APC中内化的能力,所述APC富集在源自骨髓的树突状细胞(BMDDC)中。流式细胞术显示多聚形式和单体形式均能有效地对APC进行免疫标记(对于PapMVCP-E2VLP而言为95.3%,并且对于PapMVCP27-215-E2VLP而言为92.6%)。为了使重组蛋白和APC之间的相互作用可视化,通过共聚焦显微镜观察经处理的APC。在两种情况下,免疫标记的PapMVCP信号清楚地为泡状(胞质内和细胞核周围)。两种重组蛋白都在APC中有效地内化。
为了检验PapMVCP VLP诱导免疫应答的能力,将C3H/HeJ小鼠皮下注射25μg的重组VLP(PapMVCP-E2)或25μg的单体形式(PapMVCP27-215-E2)。各剂量中存在的E2肽的量估计为2μg。初次免疫后第15天给予加强剂量。分析小鼠血清的抗PapMVCP、PapMVCP27-215和抗E2肽的抗体。在第12天,在用PapMVCP-E2免疫的小鼠中清晰地检测到抗-CP IgG,而在用PapMVCP27-215-E2接种的小鼠的血清中,即使在第15天给予加强剂量之后,也仅仅检测到微弱的抗-CP水平(图14)。
实施例V:PapMV的佐剂效果
按照实施例I描述的方法纯化PapMV。
抗原
不含LPS的OVA Grade VI购自Sigma-Aldrich Chemical公司(StLouis,Mo)。鸡卵白溶菌酶(HEL)购自Research Organics公司(Cleveland,OH)。源自大肠杆菌O111:B4的LPS购自Sigma-Aldrich公司(St Louis,MO)。
免疫
将6-8周龄的BALB/c小鼠饲养并保持在墨西哥国立自治大学(UNAM)医学院实验医学系的动物实验室中,并遵照良好实验室规范指南进行饲养。为了研究佐剂的效果,在第0天单用2mg的OVA或HEL或者与30mg的PapMV、CFA 1∶1(v/v)或5mg的来自大肠杆菌O111:B4的LPS(Sigma-Aldrich)联合对各组小鼠进行腹腔内免疫。对照小鼠仅注射盐水溶液。如图15所示,在不同的时间从眼眶后静脉窦(retro-orbital sinus)采集血样。将各种血清样品贮存于-20℃下直至分析。每次试验使用3只小鼠。
通过ELISA测定抗体效价
用在0.1M碳酸盐-碳酸氢盐缓冲液(pH9.5)中的1mg/毫升的PapMV、100mg/毫升的HEL或150mg/毫升的OVA涂覆96孔高结合性聚苯乙烯培养板(New York,NY)。将培养板在37℃下培养1小时,然后在4℃下过夜。在次日使用之前,将培养板用含有0.05%Tween-20的PBS(pH7.2)(PBS-T)(Sigma-Aldrich)清洗3次。在37℃下用5%的稀释于PBS中的脱脂奶粉(PBS-M)封闭非特异性结合1小时。清洗之后,将小鼠血清以1∶40在PBS-M中稀释,并将2倍次序稀释液加到孔内。将培养板在37℃下培养1小时,然后用PBS-T清洗4次。添加偶联过氧化酶的兔抗小鼠IgM(优选以1∶1000稀释)IgG、IgG1、IgG2a、IgG2b抗体(Zymed公司,San Francisco,CA)或Ig3(优选以1∶3000稀释)(Rockland公司,Gilbertsville,PA),然后将培养板在37℃下培养1小时并用PBS-T清洗3次。使用在0.1M柠檬酸盐缓冲液(含有30%的过氧化氢,pH5.6)中的邻苯二胺(0.5mg/毫升;Sigma-Aldrich公司)作为酶底物。用2.5N的H2SO4终止反应,并使用装有BIOLINX 2.22软件的自动ELISA培养板读取器(Dynex Technologies MRII公司,Chantilly,VA,USA)测定在490nm处的吸光度。抗体效价以-Log2稀释度×40计算。阳性效价被定义为在阴性对照的平均值以上3SD。
结果
当将佐剂与免疫原性差的疫苗同时施用时,观察到固有免疫应答向抗体应答的转译(translation)。佐剂是能够增强或提高抗体或细胞针对抗原的免疫应答的物质。为了确定PapMV是否是能够促进对其它抗原的持久性抗体应答的佐剂,单用OVA或HEL或与下列佐剂联用免疫BALB/c小鼠:PapMV、CFA或LPS。在图15A和15B所表示的时间点,通过ELISA测量对OVA或HEL特异性的IgG抗体效价。在免疫动物内,根据对OVA和HEL的总IgG应答观察到PapMV的佐剂效果。在免疫后的第30天,观察到PapMV对HEL引起佐剂效果,此时与单用HEL引起的抗体效价相比,抗体效价增加了8倍。该抗体效价的差异维持至第120天但是在第400天则消失(图15A)。尽管LPS仅仅在免疫后的前30天引起了佐剂效果,但是CFA在免疫后的第8天至第400天试验的终结显示了最强的佐剂效果(图15A)。对于用OVA免疫,仅仅直至初次免疫后的第120天才观察到对总IgG抗体效价的佐剂效果,之后抗体效价随时间下降,并且与第400天的OVA相比,其抗体效价高4倍(图15B)。在第20天进行进一步的分析以鉴别OVA和与佐剂共免疫的OVA(其中PapMV并不表现出总IgG应答的佐剂效果)诱导何种IgG亚类。PapMV、LPS和CFA产生了OVA-特异性的IgG2a和IgG2b抗体效价,而单独的OVA只产生了IgG1-特异性抗体效价(图15C-E)。当将OVA与所用的任何一种佐剂共免疫时,没有观察到对IgG1的佐剂效果。这些结果显示,PapMV、LPS、和CFA对IgG亚类针对OVA应答的具有佐剂效果。此外,PapMV显示了引起对模型抗原的特异性抗体效价持久性增加的佐剂性能。总而言之,这些数据表明PapMV具有内在的佐剂性能,其可能介导了所观察到的固有应答向持久的抗原特异性抗体应答的转译。
实施例VI:伤寒沙门氏菌孔蛋白的纯化
下列纯化步骤用于纯化OmpC和OmpF。该纯化步骤基于文献Secundino等.(2006),Immunology 117:59描述的方法。
将两种蛋白从伤寒沙门氏菌中共纯化。使用敲除突变体的伤寒沙门氏菌实现对OmpC和OmpF各自的纯化,其中OmpC[STYC171(OmpC-)]或者OmpF[STYF302(OmpF-)]的开放阅读框被中断。就共纯化而言,用于从敲除突变体形式的细菌中纯化各蛋白的步骤如下所示。该步骤概括如下:
在37℃、200rpm的条件下使菌株伤寒沙门氏菌9、12、Vi:d(ATCC 9993)在补加酵母提取物、镁和葡萄糖的基本培养基A(Minimal medium A)中生长。10L补加酵母提取物、镁和葡萄糖的基本培养基A的配方是:5.0g的脱水柠檬酸钠(NaC6H5O7:2H2O)、31.0g的磷酸二氢钠(NaH2PO4)、70.0g的磷酸一氢钠(Na2HPO4)、10.0g的(NH4)2SO4、200mL 5%的酵母提取物溶液(在300mL中15.0g)。每个4L的锥形烧瓶分配1.434L培养基。在121℃、15磅压力/平方英寸的条件下进行15min灭菌。然后各烧瓶中加入以下物质:6.0mL 25%的无菌MgSO4溶液和60.0mL 12.5%的葡萄糖溶液。将烧瓶与伤寒沙门氏菌培养物培养过夜并且当OD540达到1.0时,停止培养,并将培养物在4℃下以7,500rpm离心15min。最后将粒状沉淀重新悬浮于100mL的pH7.7的Tris-HCl(6g,Tris碱/L),并将生物质(biomass)在冰中超声90min然后在4℃下以7,500rpm离心20min。向每10ml上清液中加入以下物质:2.77mL MgCl21M、25mL RNaseA(10,000U/毫升)、25mL DNaseA(10,000U/毫升)。然后将该混合物在37℃下以120rpm培养30min。
按照以下方法从该混合物中提取孔蛋白:
1.在4℃下以45,000rpm进行超速离心45min,保留粒状沉淀。
2.将粒状沉淀重新悬浮于10mL 2%的Tris-HCl-SDS中,之后均质化。
3.在32℃、120rpm的条件下进行30min的培养步骤。
4.在20℃下以40,000rpm进行超速离心30min,保留粒状沉淀。
5.将粒状沉淀重新悬浮于5mL 2%的Tris-HCl-SDS中,之后均质化。
6.在32℃、120rpm的条件下进行30min的培养步骤。
7.在20℃下以40,000rpm进行超速离心30min,保留粒状沉淀。
8.将粒状沉淀重新悬浮于20mL 1%的Nikaido缓冲液-SDS中,之后均质化。[对于1L的Nikaido缓冲液:将6.0g Tris碱、10.0g SDS、23.4g NaCl、1.9g EDTA溶解于水中,并将pH调至pH 7.7。然后加人0.5mL β-巯基乙醇溶液]
9.在37℃、120rpm的条件下培养混合物120min。
10.在20℃下以40,000rpm进行超速离心45min。回收含有孔蛋白提取物的上清液。
使用快速蛋白液相色谱法(FPLC)从该上清液中纯化孔蛋白。在纯化过程中使用不含β-巯基乙醇的0.5X Nikaido缓冲液(参见上文)。使用Sephacryl S-200(FPLC WATERS 650E)以10ml/分钟的流速分离孔蛋白。使用22mL上清液上柱。在260nm和280nm处监测洗脱组分。保留含有纯化的孔蛋白的主峰并在4℃下贮存。纯化的孔蛋白在长时间内(超过一年)稳定。
结果:图16B示出通过上述步骤纯化的孔蛋白OmpC和OmpF的SDS-PAGE图。
实施例VII:包含亲和肽的PapMV VLP的制备和改造
亲和肽的选择
使用Ph.D-7噬菌体展示肽实验室试剂盒(New England Biolabs公司)来选择针对纯化的OmpC和OmpF的特异性肽。所采用的方案是被称为“淘洗”(panning)的体外选择过程,其按照制造商的说明书进行。简言之,将2×1011的噬菌体加到结合到ELISA板的孔的底部的10μg纯化的OmpC或OmpF中,并将孔的内容物在室温下轻轻混合1小时。通过在室温下培养10min,用1mL 200mM的甘氨酸-HCl(pH2.2)洗脱没有结合的噬菌体。为了中和上清液并避免杀死噬菌体,加入150μL的1M的Tris-HCl(pH9.1)。然后扩增洗脱的噬菌体,并使其通过额外的结合/扩增循环,以富集有利于结合序列的噬菌体群。用于第一轮淘洗的清洗缓冲液含有0.1%的Tween20,并将其浓度在随后几轮中增加至0.5%。在每一轮的淘洗之间,将选择的噬菌体在大肠杆菌ER2738中扩增。该循环重复3次以选择对各孔蛋白具有最高亲和性的那些蛋白。这样鉴定的肽如表6所示。
表6:OmpC和OmpF亲和肽的序列和出现频率
| 靶蛋白 | 肽的序列 | 频率 | SEQ ID NO |
| OmpC | SLSLIQT | 1/8 | 21 |
| OmpC | EAKGLIR | 6/8 | 22 |
| OmpC | TATYLLD | 1/8 | 23 |
| OmpF | FHENWPS | 3/5 | 24 |
| OmpF | FHEFWPT | 2/5 | 25 |
与选择的亲和肽融合的高亲和力PapMV VLP的改造、表达和纯化
从以上淘洗过程中鉴定的那些肽中选择出一种亲和肽以用于各孔蛋白(OmpC和OmpF)。在PapMV外壳蛋白(CP)的C-端处克隆相应的DNA序列。使用PapMv CP CPΔN5(Tremblay,M-H.等,2006,FEBS J.,273:14-25;参见实施例I)作为模板。使用PCR将编码所选的各肽的序列引入并克隆至pET-3D表达载体(Stratagene公司,La Jolla,CA)。简言之,在PCR反应中,使用正向引物(SEQID NO:37;如下)和引物PapOmpC(SEQ ID NO:38;如下),并且使用PapMV CP基因PapMV CP CPΔN5作为模板。
正向引物:
5′-ATCGCCATGGCATCCACACCCAACATAGCCTTCCCCGCCATCACC-3′[SEQ ID NO:37]
PapOmpC(反向引物):
3′-GGTTAAGGAAGGTGGGGGGCTTCTCCGCTTCCCCAACTAAGCATGGTAGTGGTAGTGGTAATCATTCCTAGGTGAC-5′[SEQID NO:38]
得到的PCR片段含有在PapMV CP的C-端融合的肽EAKGLIR。以相同的方法,使用正向引物(SEQ ID NO:37)和引物PapOmpF(SEQID NO:41;如下)通过PCR在PapMV CP的C-端处引入融合的肽FHENWPS。
PapOmpF(反向引物):
3′-GGTTAAGGAAGGTGGGGGGCTTAAAGTACTCTTAACCGGAAGCGTGGTAGTGGTAGTGGTAATCATTCCTAGGTGAC-5′[SEQ ID NO:41]
将两个PCR片段分别用限制性酶NcoI和BamHI消化并克隆至以相同的酶消化的pET3-D载体。对克隆进行测序以证实该肽在PapMV CP的框架内。
按照前述的方法(Tremblay,M-H等,2006,FEBS J.,273:14-25;Secundino等,2006,Immunology 117:59),将包含亲和肽的改造的PapMV CP在大肠杆菌BL21RIL中表达。简言之,通过弗氏细胞压碎器使细菌裂解,并上Ni2+柱上,用pH 8的10mM Tris-HCl、50mM咪唑、0.5%Triton X100清洗,然后用pH 8的10mM Tris-HCl、50mM咪唑、1%两性清洗剂清洗,以除去内毒素污染物。将洗脱的蛋白在Beckman 50.2TI转子中进行高速离心(100000g)120min。将VLP粒状沉淀在无内毒素的PBS(Sigma)中重新悬浮。在使用前将蛋白用0.45μm的过滤器过滤。通过SDS-PAGE测定蛋白的纯度。使用BCA蛋白试剂盒(Pierce)评价蛋白的量。按照制造商的说明(Cambrex),用鲎试验评价纯化的蛋白中的LPS的水平,并且结果为低于0.005内毒素单位(EU)/μg蛋白。
该两种PapMV外壳蛋白的序列如图22所示(SEQ ID NO:42-包含OmpC亲和肽的PapMV外壳蛋白(图22A),和SEQ ID NO:43-包含OmpF亲和肽的PapMV外壳蛋白(图22B))。与野生型相比,在包含OmpC亲和肽的PapMV外壳蛋白的外壳蛋白序列中观察到两个氨基酸差异(在图22A中以黑体和下划线标出),这可能是在PRC反应期间引入的。
ELISA
对于各验,使用10μg的各个靶蛋白(OmpC或OmpF)涂覆ELISA板。将增加量的PapMV VLP用于结合分析。使用针对PapMVCP的多克隆鼠抗体和与偶联过氧化酶的二级抗小鼠抗体来揭示VLP对它们的靶蛋白的亲和件。
结果
亲和肽的选择
使用噬菌体展示技术来选择与OmpC或OmpF结合的特异性肽。对结合至OmpC的8个噬菌体和结合至OmpF的5个噬菌体进行测序。这些肽的序列和出现频率如表6所示。肽EAKGLIR[SEQ IDNO:22]显示了最高的频率,因此被选为OmpC的亲和肽。肽FHENWPS[SEQ ID NO:23]在OmpF筛选中频率最高,因此被选为OmpF的亲和肽。有趣的是,OmpF的两种亲和肽是同源的,这是因为7个氨基酸中有5个是相同的并在亲和肽的相同的位点出现。
高亲和力PapMV VLP的合成
将肽序列EAKGLIR[SEQ ID NO:22]和FHENWPS[SEQ IDNO:23]融合至PapMV外壳蛋白的C-端(图16A)。所述融合肽连接有6XH标记物以利于纯化过程(Tremblay,M-H.等,2006,ibid.)。将重组构建体在大肠杆菌中表达并通过使用Ni2+柱的亲和色谱来纯化。将该蛋白用500mM的咪唑洗脱、透析并以100,000g超速离心,以使VLP形成粒状沉淀(图16B)。电子显微镜(EM)观察证实,在PapMV CP的C-端引入所述肽并不影响该蛋白自组装成VLP的能力(图16C)。VLP的长度是可变的,尺寸在201±80nm范围内。201nm长度的蛋白代表560个CP拷贝,所述CP以重复的形式呈现该肽。
通过ELISA型结合试验来显示各PapMV VLP与它们各自的抗原的高亲和力。对于两种VLP,已经清楚地证明其与它们各自的抗原的结合,并且所述的结合随着在分析中使用的VLP的量而增加(图17A-B)。因此,推断当在溶液中以1∶1的比率混合时,PapMV VLP将结合同类的抗原以形成复合体。
实施例VIII:具有高亲和力PapMV VLP对伤寒沙门氏菌的免疫
小鼠
使用6-8周龄的雌性BALB/c小鼠(Harlan,Mexico或CharlesRiver,Canada),并饲养于墨西哥国家自立大学(UNAM)的医学院实验医学系的动物设施或者拉瓦尔大学附属医院(Centre Hospitalier de1’UniversitéLaval)的动物设施内。
攻击试验
在不存在外部佐剂的情况下,用10μg OmpC、10μg OmpC+10μg PapOmpC、10μg OmpF、10μg OmpF+10μg PapOmpF、10μgPapOmpC、10μg PapOmpF或盐水(ISS)对小鼠(每组10只)进行腹腔内(i.p)免疫(第0天)。在第15天,分别用无佐剂的10μg OmpC或10μg OmpF使小鼠接受腹腔内加强免疫。在第25或140天,用100或500LD50的伤寒沙门氏菌(ATCC 9993)对小鼠进行腹腔内攻击,所述伤寒沙门氏菌重新悬浮于500μL含有5%胃膜素(Sigma)的TE缓冲液(50mM Tris,pH 7.2、5mM EDTA)内。将保护率定义为攻击10天之后的存活百分比。1LD50在90000CFU的条件下确定。
免疫
在不存在外部佐剂的情况下,用10μg OmpC、10μg OmpC+10μg PapOmpC、10μg PapOmpC或等渗盐水(ISS)对小鼠(每组5只)进行腹腔内(i.p)免疫(第0天)。在第15天,用无佐剂的10μg OmpC使小鼠接受腹腔内加强免疫。在图18和19所示的不同的时间从颈静脉采集血样。将各种血清样品在-20℃下贮存直至分析。
ELISA
用在0.1M的碳酸盐-碳酸氢盐缓冲液(pH9.5)中的10μg/毫升的OmpC涂覆96孔高结合性聚苯乙烯培养板(Nunc)。将培养板在37℃下培养1小时,然后在4℃下过夜。将培养板用蒸馏水-0.1%Tween-20清洗4次。在37℃下用封闭缓冲液(PBS pH 7.4-2%BSA(Sigma))封闭非特异性结合1小时。清洗之后,将混合的小鼠血清以1∶40在封闭缓冲液中稀释,并将2倍次序稀释液加到孔内。将培养板在37℃下培养1.5小时,然后清洗4次。添加HRP偶联的山羊抗小鼠IgG1、IgG2a、IgG2b(Jackson Immunochemicals)或IgG3(Rockland)(1∶10000),并在37℃下培养1小时,然后清洗4次。作为检测体系,使用TMB过氧化酶底物(Fitzgerald)。在37℃下于黑暗中培养10分钟之后,用2.5N的H2SO4终止反应,并使用自动ELISA培养板读取器测定在450nm处的吸光度。抗体效价以-Log2稀释度×40计算。将阳性效价定义为在阴性对照的平均值以上的3SD。
被动免疫和攻击
在不存在外部佐剂的情况下,用10μg OmpC、10μg OmpC+10μg PapMV OmpC、10μg PapMV OmpC或等渗盐水(ISS)对小鼠(每组5只)进行腹腔内免疫(第0天)。在第15天,用无佐剂的10μg OmpC使小鼠接受腹腔内加强免疫。在第23天进行心脏穿刺,并收集各组的血清样品,然后在-20℃下贮存。使未试验过的小鼠(每组5只)腹腔内接受200μL收集的补体失活的免疫血清。在转移3小时后,按照上述的方法将小鼠用重新悬浮在粘蛋白中的100LD50的伤寒沙门氏菌攻击。将保护率定义为攻击10天后的存活百分比。
结果
PapMV VLP提高了孔蛋白的保护能力
之前已表明纯化的蛋白OmpC和OmpF在小鼠中提供对伤寒沙门氏菌攻击的保护,其中单用OmpC提供对100LD50的60%保护率(Secundino等,2006,Immunology 117:59)。为了提高每一种孔蛋白(OmpC和OmpF)各自的免疫原性,制备包含PapMV VLP的疫苗制剂。测试两种不同的制剂PapMV OmpC VLP+OmpC和PapMVOmpF+OmpF在小鼠中对100和500LD50的伤寒沙门氏菌的保护能力,并将结果与单用OmpC或OmpF免疫小鼠获得的结果相比较。所述PapMV VLP与其孔蛋白的比率保持在1∶1。
在用100LD50的伤寒沙门氏菌攻击的情况下,向OmpC添加PapMV OmpC VLP将OmpC的保护能力从70%提高为100%(图18A)。当用500LD50攻击小鼠时,保护效力的提高甚至更大,其中当OmpC与PapMV OmpC VLP联用时,观察到保护率从30%增加至90%(图18C)。类似地,使用100LD50的伤寒沙门氏菌攻击时,PapMVOmpF VLP将OmpF的保护能力从60%提高至90%(图18B)。但是,当用500LD50的伤寒沙门氏菌进行攻击时,只观察到较小的差异(图18D)。结果表明对于针对伤寒沙门氏菌攻击的保护,OmpC是比OmpF更好的抗原。在两种情况下,PapMV VLP显著地提高了孔蛋白的保护能力。
为了确定PapMV VLP是否提高了对OmpC的抗体效价,测量使用10μg OmpC或使用含有10μg OmpC和10μg PapMV OmpC VLP的接合疫苗接种的小鼠的IgG效价。在任何一种处理情况下都没有发现不同的IgG同种型IgG1、IgG2a、IgG2b、IgG3的效价的显著差异(图19A-D),这表明在使用PapMV VLP的情况下,所观察到的保护的提高可能与CTL应答和/或抗体在抗伤寒沙门氏菌感染时的结合效力的提高有关,而非增加了抗体本身的产率。
接种PapMV VLP提高了对孔蛋白的记忆应答
为了评价包含PapMV VLP以及OmpC的疫苗制剂的记忆应答,单用OmpC或用包含PapMV OmpC VLP和OmpC的疫苗制剂每隔两周免疫小鼠两次,接着在第15天单用OmpC加强免疫。在第140天,将小鼠用100LD50的伤寒沙门氏菌攻击。结果清楚地显示,用包含PapMV OmpC VLP和OmpC的疫苗制剂初次显著地提高(3倍提高)了接种小鼠的保护能力(图19E)。因而,该试验证明,PapMVVLP不仅提高了小鼠对伤寒沙门氏菌攻击的保护,还提供了更好的记忆应答。
实施例IX:PapMV和OmpC的组合针对伤寒沙门氏菌的保护能力
按照实施例I描述的方法纯化PapMV。
保护试验
在第0天用10μg OmpC或者之前已经与30μg PapMV在4℃下培养1小时的10μg OmpC腹腔内接种BALB/c小鼠(每组10只)。在第15天单用10μg OmpC进行加强接种。对照小鼠仅注射盐水。在第21天,将小鼠用悬浮于5%的粘蛋白(如上所述)中的100和500LD50的伤寒沙门氏菌(STYC302DompF菌株)攻击,并按照之前描述的方法(Isibasi等,1992,Vaccine 10:811-813;Isibasi等,1988,Infect.lmmun.56:2953-2959),监测攻击10天之后的存活率。
结果
为了测试从受感染的植物中分离的PapMV病毒在增强由OmpC提供的保护方面的佐剂能力,将用OmpC免疫和用混合有PapMV纯化病毒的OmpC免疫、随后都用伤寒沙门氏菌攻击的小鼠进行比较。与单用OmpC的情况相比,当使用混合有PapMV纯化病毒的OmpC时,在用100LD50或500LD50的伤寒沙门氏菌攻击之后,观察到存活率高出30%(图20A)。在第0天和第15天仅用PapMV免疫并在第21天攻击的小鼠中没有观察到保护(图21A),在用PapMV免疫并在24小时后攻击的小鼠中也没有观察到保护。这些数据排除了观察到的保护是由于PapMV诱导炎症增强所致的想法。为了测试增加的保护是否与对OmpC特异性的抗体效价的增加相关,测试了PapMV对OmpC-特异性抗体效价的佐剂效果。PapMV与OmpC共免疫引起了抗OmpC的IgG1、IgG2a、IgG2b和IgG3的效价增加(图21B)。这些结果进一步证实,PapMV的佐剂性能增强了由OmpC引发的固有免疫应答以及适应性免疫应答,从而实现对伤寒沙门氏菌攻击的保护。因此,可以将PapMV和伤寒沙门氏菌OmpC孔蛋白的同时施用视为增强对伤寒沙门氏菌攻击的保护能力。
实施例V-IX概括的试验结果表明,PapMV具有内在的佐剂性能,该性能能够引起抗原特异性免疫球蛋白的转换,提供对模型抗原的持久性抗体应答,并且增强了单用OmpC或OmpF的保护能力。这些数据表明,PapMV和PapMV VLP增强了由OmpC孔蛋白引发的固有免疫应答和适应性免疫应答向对伤寒沙门氏菌攻击的保护的转译。
实施例X:含有伤寒沙门氏菌孔蛋白抗原表位的PapMV VLP的免疫原效果
已经表明,暴露于细菌表面上的伤寒沙门氏菌孔蛋白的环6和环7对应的两个小的抗原表位参与通过孔蛋白免疫引发的保护机制(Panigua-Solis等,1995,Immunol Infect.Dis.5:244-249)。这些区域仅存在于伤寒沙门氏菌的孔蛋白中,因此没有发现其与其它革兰氏阴性菌的孔蛋白具有交叉反应性。因此,包含这些抗原表位的孔蛋白片段代表了用于开发利用PapMV VLP作为佐剂-载体的重组亚单位疫苗的优异候选抗原。
重组PapMV VLP的制备
将PapMV外壳蛋白进行改造,以在其C-端处引入氨基酸序列GTSNGSNPSTSYGFAN[SEQ ID NO:50],该序列包含源自伤寒沙门氏菌孔蛋白的免疫显性的环6抗原表位(参见图23A)。使用前面的实施例所述的方法将该重组蛋白在大肠杆菌中表达(参见图23B)。简言之,在PCR反应中,以PapMV CP CPΔN5基因为模板,使用正向引物(SEQ ID NO:44;如下)和反向引物(SEQ ID NO:45;如下)。
正向引物:
5′-CTAGTGGTACTTCTAACGGTTCTAACCCGTCTACTTCTTACGGTTTCGCGAACA-3′[SEQ ID NO:44]
反向引物:
5′-CTAGTGTTCGCGAAACCGTAAGAAGTAGACGGGTTAGAACCGTTAGAAGTACCA-3′[SEQ ID NO:45]
如前面的实施例所述,通过Ni2+亲和性标记物层析从大肠杆菌中纯化重组蛋白,并且其表现出自组装成与从受感染的植物叶子中纯化的天然病毒颗粒类似的VLP(参见图23C)。
包含环6肽的PapMV外壳蛋白的氨基酸序列(PapMV-环6)如图24A(SEQ ID NO:46)所示,并且编码PapMV-环6蛋白的核苷酸序列如图24B(SEQ ID NO:47)所示。
Balb/c小鼠中对PapMV-环6的免疫应答
使用6-8周龄的雌性BALB/c小鼠(Harlan,Mexico或CharlesRiver,Canada),并将其饲养在拉瓦尔大学附属医院的动物设施内。在不存在外部佐剂的情况下,用100μg PapMV-环6对小鼠通过皮下途径(s.c.)进行免疫(第0天)。在第15天,用100μg PapMV-环6使小鼠接受皮下加强免疫。在第28天采集最终的血样并使用合成的环6肽和纯化的OmpC蛋白通过标准的ELISA测定免疫应答。
结果证实,PapMV平台具有高度的免疫原性(图25A),因为其在5只小鼠的4只中触发了环6特异性抗体的产生。尽管PapMV-环6VLP在5只处理的小鼠中触发了针对PapMV平台的大量抗体(图25B),但是同样的血清并未表现出对纯化的OmpC蛋白的反应(图25C)。该结果表明在PapMV CP融合体中的环6肽的结构可能采用了与该肽在OmpC蛋白的固有环境中略微不同的构型。
为了容许环6肽序列在融合至PapMV CP时采用其固有构型,将上文的序列(SEQ ID NO:50)克隆至PapMV CP的残基49-52之间的内环中。使用生物信息学技术鉴定用于此目的的环。
实施例XI:与伤寒沙门氏菌OmpF组合的PapMV VLP的保护能力
本试验使用实施例VII、VIII和IX中所描述的下列蛋白:
·PapMV CPΔN5VLP(“PapMV”)(参见图1C)
·包含针对伤寒沙门氏菌OmpF的亲和肽的PapMV OmpF VLP(参见图22B)
·OmpF(参见图21B)
在第0天,将Balb/c小鼠(每组10只)按照如下方法进行腹腔内(i.p.)免疫:
组1-10μg OmpF
组2-10μg OmpF+10μg PapMV
组3-10μg OmpF+10μg PapMV OmpF
组4-10μg PapMV
组5-10μg PapMV OmpF
在第15天,对组1、2和3使用10μg OmpF进行第二免疫(加强免疫)。组4用10μg PapMV加强免疫,组5用10μg PapMV OmpF加强免疫。在第21天用伤寒沙门氏菌进行攻击。通过试验确立,粘蛋白中的90000CFU的伤寒沙门氏菌对应于1LD50。在第31天杀死所有的小鼠。
结果
在77LD50(图26A)的条件下,所有含有OmpF的制剂提供100%的保护率。正如预料的那样,用不含OmpF的制剂接种的小鼠在攻击(剂量低至14LD50)数天内全部死亡。但是,当用更高剂量的伤寒沙门氏菌(378LD50)攻击小鼠时,在含有OmpF的制剂之间观察到差异。如图26B所示,在该剂量下最有效的制剂是用PapMV OmpF+OmpF的组合(组3)第一免疫和单用OmpF加强免疫。与单用PapMV作为OmpF的佐剂相比,可能是PapMV OmpF VLP对其靶OmpF具有高的亲和力,从而提高了保护能力。
该数据与实施例VIII中所示的情况相一致,并且证实了PapMVOmpF提高了OmpF对伤寒沙门氏菌攻击的保护能力。此外,该数据显示,PapMV OmpF是比没有亲和肽的PapMV VLP更好的佐剂。如上所述,可能是PapMV OmpF VLP对OmpF较强的结合力促进了VLP-OmpF复合体的形成,这又使VLP分子的佐剂能力最大化。
实施例XII:与伤寒沙门氏菌OmpC组合的PapMV VLP的保护能力
本试验使用实施例VII、VIII和IX中所描述的下列蛋白:
·PapMV CPΔN5VLP(“PapMV”)(参见图1C)
·OmpC(参见图21A)
构建包含OmpC亲和肽EAKGLIR[SEQ ID NO:22]的另一个PapMV CP融合体,所述融合体包含额外的4个氨基酸,在亲和肽的每一侧各有2个(如图27A所示,在N-端一侧是TR并且在C-端一侧是TS)。这些氨基酸是限制性位点SpeI-MluI存在的结果,其中所述限制性位点被用于与PapMV CP融合的亲和肽的克隆。将上述构建体命名为“PapMV SM OmpC”。图28A提供了PapMV SM OmpC蛋白的完整氨基酸序列(SEQ ID NO:48),并且图28B提供了编码PapMV SM OmpC蛋白的核苷酸序列(SEQ ID NO:49)。
按照上述实施例描述的方法纯化PapMV SM OmpC蛋白(参见图27B)。正如预料的那样,通过电子显微镜,纯化的该蛋白显示了VLP的形成(图27C)。
免疫
在第0天,将Balb/c小鼠(每组10只)按照如下方法进行腹腔内(i.p.)免疫:
组1-10μg OmpC
组2-10μg OmpC+10μg PapMV
组3-10μg OmpC+10μg PapMV OmpC
组4-10μg PapMV
组5-10μg PapMV OmpC
在第15天,在组1、2和3中使用10μg OmpC进行第二免疫(加强免疫)。组4用10μg PapMV加强免疫,组5用10μg PapMV OmpC加强免疫。在第21天用伤寒沙门氏菌进行攻击。通过试验确立,粘蛋白中90000CFU的伤寒沙门氏菌对应于1LD50。在第31天杀死所有的小鼠。
结果
在105LD50(图29A)的条件下,所有含有OmpC的制剂提供100%的保护率。正如预料的那样,用不含OmpC的制剂接种的小鼠在攻击(剂量低至20LD50)的数天内全部死亡。但是,当用更高剂量的伤寒沙门氏菌(520LD50)攻击小鼠时,在含有OmpC的制剂之间观察到差异。如图29B所示,在该剂量下最有效的制剂是用PapMV+OmpC的组合(组2)第一免疫和单用OmpC加强免疫。包含与OmpC结合的PapMV SM OmpC VLP的制剂(组3)也表现良好,但是似乎效果略逊于PapMV+OmpC的组合。但是,在本试验中,不能确定效力的差异是否足以具有统计学显著意义。
总而言之,如实施例VII、XI和XII所示,具有或没有亲和肽的重组PapMV VLP能够用作分别提高对伤寒沙门氏菌的两种不同抗原OmpC和OmpF的保护能力的强的佐剂。
实施例XIII:伤寒沙门氏菌孔蛋白的稳定性
通过标准的SDS-PAGE评价伤寒沙门氏菌的纯化孔蛋白的稳定性。使用2种孔蛋白样品。第一种样品包含OmpC以及OmpF纯化蛋白(批次ISIPOR)并在1999年11月9日制备(该制剂与文献Salazar-Gonzales等,2004,Immunology Letters 93的图1(b)中所示的制剂相一致)。第二种样品仅包含纯化的OmpC蛋白(批次1239)并在2007年4月30日制备。
将两种样品在4±3℃下避光贮存于PBS溶液(pH 7.4±0.1)中。通过下列条件进行SDS-PAGE:
·12%的丙烯酰胺。
·在上样至凝胶上之前,通过加入β-巯基乙醇并在95℃加热5min将所有的样品变性。
该结果如图30所示。从该图可以看出,两种孔蛋白制剂都十分稳定。由于没有通过SDS-PAGE看到降解迹象(参照图30A和30B),因此ISIPOR似乎在超过9年(从1999至2008)的时间内都非常稳定。纯化的OmpC也表现出高的稳定性,其中蛋白在贮存9个月之后保持完整(参照图30C的左栏和右栏)。
实施例XIV:PapMV在体内引起了DC、巨噬细胞和B细胞的活化标志物的上调
按照实施例I中描述的方法纯化PapMV。
DC、巨噬细胞和B淋巴细胞的体内活化试验
用分别混悬在500μl盐水中的30μg PapMV和50μg聚肌胞苷酸(Poly I:C)腹腔内免疫BALB/c小鼠(7周龄)。使用盐水(500μl)作为对照,并在免疫24小时后获取淋巴结和脾脏。从这些器官制备细胞悬浮液,并使用藻红蛋白(PE)-CD 11c、别藻蓝蛋白-CD11b、B220-多甲藻叶绿素蛋白(PerCP)、FITC-CD80、FITC-CD86、FITC-CD40、FITC-CD69和FITC-主要组织相容性复合体II类(MHCII类)抗体(全部购自BD Pharmingen公司)进行流式细胞术染色。
结果
据信PapMV作为病原相关分子模式(PAMP)而被免疫系统识别,因此,如同其它的模式识别受体(PRR)激动剂那样,它也应该促进不成熟的DC成熟化或者活化APC。为了表征PapMV在体内刺激APC的能力,分离用PapMV免疫的小鼠的淋巴结和脾脏,并分析共刺激和活化分子(CD80、CD86、CD40、CD69以及MHC II类)的上调。如图31所示,PapMV引起了淋巴结(LN)DC(CD11c+CD11b-)中的CD80和CD69以及脾脏DC中的CD40和CD69的上调。对于巨噬细胞(CD11c-CD11b+),在LN中观察到CD80和CD69的上调,而在脾脏中观察到CD40和CD69的上调。在LN B淋巴细胞(CD11c-B220+)中CD69上调,而在脾脏B细胞中CD40和CD69上调。TLR-3激动剂(聚肌胞苷酸)引起所测试的标志物的上调,但是所观察到的曲线与由PapMV所引起的情况显著不同(图31)。
APC针对T细胞的抗原提呈已经显示出固有免疫系统对活化适应性免疫系统的重要性。观察到载有PapMV的APC有效地引起抗体应答。这些数据以及PapMV对APC引起的效果(图31)表明,PapMV刺激的APC提供了促进有效的T细胞依赖性抗体应答所需要的抗原和细胞因子环境,并且这可能是由PapMV传递的内在佐剂信号转译成已观察到的持久性抗体应答所涉及的机制。PapMV可能作为PAMP和作为抗原(Pamptigen)被免疫系统所感知,同时活化固有免疫系统和适应性免疫系统,这将有利于诱导记忆间隔(memorycompartment)。Pamptigen还可以结合特异性B细胞上的BCR和PRR(例如TLR),从而导致抗体分泌性细胞的分化或记忆B细胞的产生。Pamptigen可以降低活化免疫应答所需要的抗原阈值。因此,少量的持久性抗原可以保持高的抗体效价。
PapMV的强的免疫原性和内在佐剂性能可以转译成针对PapMV和共免疫的模型抗原的持久的特异性抗体应答(参见实施例IX)。由于PapMV可以用经济的过程制备并且在室温下稳定,因此其非常适合于开发诱导持久性免疫的新的佐剂和疫苗平台。
尽管已经结合某些的具体实施方案描述了本发明,但是在不背离本发明的精神和范围的情况下,其各种修改对本领域的技术人员是显而易见的。对本领域技术人员显而易见的所有这些修改都意图包含在所附的权利要求的范围内。
序列表
<110>弗利亚生物技术公司
康斯坦丁诺·三世·罗伯托·洛佩斯-马西亚斯
<120>抗伤寒沙门氏菌和其它肠细菌病原体的基于番木瓜花叶病毒的疫苗
<130>VI-090902-59
<140>PCT/CA2008/000154
<141>2008-01-28
<150>60/886,873
<151>2007-01-26
<160>54
<170>FastSEQ forWindows Version 4.0
<210>1
<211>215
<212>PRT
<213>番木瓜花叶病毒
<400>1
Met Ser Lys Ser Ser Met Ser Thr Pro Asn Ile Ala Phe Pro Ala Ile
1 5 10 15
Thr Gln Glu Gln Met Ser Ser Ile Lys Val Asp Pro Thr Ser Asn Leu
20 25 30
Leu Pro Ser Gln Glu Gln Leu Lys Ser Val Ser Thr Leu Met Val Ala
35 40 45
Ala Lys Val Pro Ala Ala Ser Val Thr Thr Val Ala Leu Glu Leu Val
50 55 60
Asn Phe Cys Tyr Asp Asn Gly Ser Ser Ala Tyr Thr Thr Val Thr Gly
65 70 75 80
Pro Ser Ser Ile Pro Glu Ile Ser Leu Ala Gln Leu Ala Ser Ile Val
85 90 95
Lys Ala Ser Gly Thr Ser Leu Arg Lys Phe Cys Arg Tyr Phe Ala Pro
100 105 110
Ile Ile Trp Asn Leu Arg Thr Asp Lys Met Ala Pro Ala Asn Trp Glu
115 120 125
Ala Ser Gly Tyr Lys Pro Ser Ala Lys Phe Ala Ala Phe Asp Phe Phe
130 135 140
Asp Gly Val Glu Asn Pro Ala Ala Met Gln Pro Pro Ser Gly Leu Ile
145 150 155 160
Arg Ser Pro Thr Gln Glu Glu Arg Ile Ala Asn Ala Thr Asn Lys Gln
165 170 175
Val His Leu Phe Gln Ala Ala Ala Gln Asp Asn Asn Phe Thr Ser Asn
180 185 190
Ser Ala Phe Ile Thr Lys Gly Gln Ile Ser Gly Ser Thr Pro Thr Ile
195 200 205
Gln Phe Leu Pro Pro Pro Glu
210 215
<210>2
<211>648
<212>DNA
<213>番木瓜花叶病毒
<400>2
atgtctaagt caagtatgtc cacacccaac atagccttcc ccgccatcac ccaggaacag 60
atgagctcga ttaaggtcga tccaacgtcc aatcttctgc cctcccaaga gcagttaaag 120
tcagtgtcca ccctcatggt agctgctaag gttccagcag ccagtgttac aactgtggca 180
ttggagttgg tcaacttctg ctatgacaat gggtccagcg cgtacaccac agtgactggc 240
ccatcatcaa taccggagat atcactggca caattggcta gtattgtcaa agcttccggc 300
acttccctta gaaaattctg ccggtacttc gcgccaataa tctggaatct gaggacggac 360
aaaatggctc ctgccaattg ggaggcttca ggatacaagc caagcgccaa atttgccgcg 420
ttcgacttct tcgacggggt ggagaatccg gcggccatgc aacccccttc gggactaatc 480
aggtcgccga cccaggaaga gcggattgcc aatgctacca acaaacaggt gcatctcttc 540
caagccgcgg cacaggacaa caactttacc agcaactccg ccttcatcac caaaggccaa 600
atttctgggt caaccccaac catccaattc cttccacccc ccgaataa 648
<210>3
<211>211
<212>PRT
<213>人工序列
<220>
<223>修饰的PapMV外壳蛋白CPdeltaN5
<400>3
Met Ala Ser Thr Pro Asn Ile Ala Phe Pro Ala Ile Thr Gln Glu Gln
1 5 10 15
Met Ser Ser Ile Lys Val Asp Pro Thr Ser Asn Leu Leu Pro Ser Gln
20 25 30
Glu Gln Leu Lys Ser Val Ser Thr Leu Met Val Ala Ala Lys Val Pro
35 40 45
Ala Ala Ser Val Thr Thr Val Ala Leu Glu Leu Val Asn Phe Cys Tyr
50 55 60
Asp Asn Gly Ser Ser Ala Tyr Thr Thr Val Thr Gly Pro Ser Ser Ile
65 70 75 80
Pro Glu Ile Ser Leu Ala Gln Leu Ala Ser Ile Val Lys Ala Ser Gly
85 90 95
Thr Ser Leu Arg Lys Phe Cys Arg Tyr Phe Ala Pro Ile Ile Trp Asn
100 105 110
Leu Arg Thr Asp Lys Met Ala Pro Ala Asn Trp Glu Ala Ser Gly Tyr
115 120 125
Lys Pro Ser Ala Lys Phe Ala Ala Phe Asp Phe Phe Asp Gly Val Glu
130 135 140
Asn Pro Ala Ala Met Gln Pro Pro Ser Gly Leu Thr Arg Ser Pro Thr
145 150 155 160
Gln Glu Glu Arg Ile Ala Asn Ala Thr Asn Lys Gln Val His Leu Phe
165 170 175
Gln Ala Ala Ala Gln Asp Asn Asn Phe Ala Ser Asn Ser Ala Phe Ile
180 185 190
Thr Lys Gly Gln Ile Ser Gly Ser Thr Pro Thr Ile Gln Phe Leu Pro
195 200 205
Pro Pro Glu
210
<210>4
<211>10
<212>PRT
<213>人工序列
<220>
<223>源自伤寒沙门氏菌孔蛋白的环6肽
<400>4
Gly Thr Ser Asn Gly Ser Asn Pro Ser Thr
1 5 10
<210>5
<211>14
<212>PRT
<213>人工序列
<220>
<223>源自伤寒沙门氏菌孔蛋白的环7肽
<400>5
Lys Asp Ile Ser Asn Gly Tyr Gly Ala Ser Tyr Gly Asp Gln
1 5 10
<210>6
<211>8
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>6
Thr Arg Val Ala Phe Ala Gly Leu
1 5
<210>7
<211>8
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>7
Arg Asn Thr Asp Phe Phe Gly Leu
1 5
<210>8
<211>8
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>8
Arg Asn Ser Asp Phe Phe Gly Leu
1 5
<210>9
<211>8
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>9
Thr Arg Leu Ala Phe Ala Gly Leu
1 5
<210>10
<211>8
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>10
Thr Arg Leu Gly Phe Ala Gly Leu
1 5
<210>11
<211>7
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>11
Ser Asp Asp Lys Gly Ser Asp
1 5
<210>12
<211>9
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>12
Asn Gln Thr Glu Gly Ser Asn Asp Ser
1 5
<210>13
<211>34
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>13
Asn Tyr Gly Val Thr Tyr Asp Val Thr Ser Trp Thr Asp Val Leu Pro
1 5 10 15
Glu Phe Gly Gly Asp Thr Tyr Gly Ala Asp Asn Phe Met Gln Gln Arg
20 25 30
Gly Asn
<210>14
<211>16
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>14
Gly Ser Val Ser Gly Glu Asn Thr Asn Gly Arg Ser Leu Leu Asn Gln
1 5 10 15
<210>15
<211>19
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>15
Arg Thr Ala Asp Gln Asn Asn Thr Ala Asn Ala Arg Leu Tyr Gly Asn
1 5 10 15
Gly Asp Arg
<210>16
<211>21
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>16
Asn Ala Thr Arg Phe Gly Thr SerAsn Gly Ser Asn Pro Ser Thr Ser
1 5 10 15
Tyr Gly Phe Ala Asn
20
<210>17
<211>17
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>17
Asp Ile Ser Asn Gly Tyr Gly Ala Ser Tyr Gly Asp Gln Asp Ile Val
1 5 10 15
Lys
<210>18
<211>18
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<400>18
Asn Leu Leu Asp Lys Asn Asp Glu Thr Arg Asp Ala Gly Ile Asn Thr
1 5 10 15
Asp Asp
<210>19
<211>8
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<223>Xaa可以是VAL、LEU或GLY
<400>19
Thr Arg Xaa Xaa Phe Ala Gly Leu
1 5
<210>20
<211>8
<212>PRT
<213>人工序列
<220>
<223>肠细菌孔蛋白抗原表位序列
<223>Xaa可以是THR或SER
<400>20
Arg Asn Xaa Asp Phe Phe Gly Leu
1 5
<210>21
<211>7
<212>PRT
<213>人工序列
<220>
<223>亲和肽
<400>21
Ser Leu Ser Leu Ile Gln Thr
1 5
<210>22
<211>7
<212>PRT
<213>人工序列
<220>
<223>亲和肽
<400>22
Glu Ala Lys Gly Leu Ile Arg
1 5
<210>23
<211>7
<212>PRT
<213>人工序列
<220>
<223>亲和肽
<400>23
Thr Ala Thr Tyr Leu Leu Asp
1 5
<210>24
<211>7
<212>PRT
<213>人工序列
<220>
<223>亲和肽
<400>24
Phe His Glu Asn Trp Pro Ser
1 5
<210>25
<211>7
<212>PRT
<213>人工序列
<220>
<223>亲和肽
<400>25
Phe His Glu Phe Trp Pro Thr
1 5
<210>26
<211>7
<212>PRT
<213>人工序列
<220>
<223>亲和肽
<223>Xaa可以是ASN或PHE
<400>26
Phe His Glu Xaa Trp Pro Thr
1 5
<210>27
<211>30
<212>DNA
<213>人工序列
<220>
<223>引物
<400>27
agtcccatgg atccaacgtc caatcttctg 30
<210>28
<211>49
<212>DNA
<213>人工序列
<220>
<223>引物
<400>28
atgcggatcc ttactaatgg tgatggtgat ggtgttcggg gggtggaag 49
<210>29
<211>63
<212>DNA
<213>人工序列
<220>
<223>寡核苷酸
<400>29
ctagttcttc tgcgttcacc atcatggacc aggttccgtt ctctgtttct gtttctcagc 60
tga 63
<210>30
<211>63
<212>DNA
<213>人工序列
<220>
<223>寡核苷酸
<400>30
ctagtcagct gagaaacaga aacagagaac ggaacctggt ccatgatggt gaacgcagaa 60
gaa 63
<210>31
<211>63
<212>DNA
<213>人工序列
<220>
<223>寡核苷酸
<400>31
ctagttctcc gctgaccaaa ggtatcctgg gtttcgtttt caccctgacc gttccgtctg 60
aaa 63
<210>32
<211>63
<212>DNA
<213>人工序列
<220>
<223>寡核苷酸
<400>32
ctagtttcag acggaacggt cagggtgaaa acgaaaccca ggataccttt ggtcagcgga 60
gaa 63
<210>33
<211>80
<212>DNA
<213>人工序列
<220>
<223>寡核苷酸
<400>33
gatcactagt gtggtggtgg gtaccaccga tcgtagcggt gcgccgacct acagctgggg 60
tgcgaacgat acgcgtcatg 80
<210>34
<211>80
<212>DNA
<213>人工序列
<220>
<223>寡核苷酸
<400>34
catgacgcgt atcgttcgca ccccagctgt aggtcggcgc accgctacga tcggtggtac 60
ccaccaccac actagtgatc 80
<210>35
<211>33
<212>DNA
<213>人工序列
<220>
<223>引物
<400>35
agtcccatgg ccgatccaac gtccaatctt ctg 33
<210>36
<211>28
<212>DNA
<213>人工序列
<220>
<223>引物
<400>36
acgtccatgg tatatctcct tcttaaag 28
<210>37
<211>45
<212>DNA
<213>人工序列
<220>
<223>引物
<400>37
atcgccatgg catccacacc caacatagcc ttccccgccatcacc 45
<210>38
<211>76
<212>DNA
<213>人工序列
<220>
<223>引物
<400>38
ggttaaggaa ggtggggggc ttctccgctt ccccaactaa gcatggtagt ggtagtggta 60
atcattccta ggtgac 76
<210>39
<211>378
<212>PRT
<213>伤寒沙门氏菌Ty2
<400>39
Met Lys Val Lys Val Leu Ser Leu Leu Val Pro Ala Leu Leu Val Ala
1 5 10 15
Gly Ala Ala Asn Ala Ala Glu Ile Tyr Asn Lys Asp Gly Asn Lys Leu
20 25 30
Asp Leu Phe Gly Lys Val Asp Gly Leu His Tyr Phe Ser Asp Asp Lys
35 40 45
Gly Ser Asp Gly Asp Gln Thr Tyr Met Arg Ile Gly Phe Lys Gly Glu
50 55 60
Thr Gln Val Asn Asp Gln Leu Thr Gly Tyr Gly Gln Trp Glu Tyr Gln
65 70 75 80
Ile Gln Gly Asn Gln Thr Glu Gly Ser Asn Asp Ser Trp Thr Arg Val
85 90 95
Ala Phe Ala Gly Leu Lys Phe Ala Asp Ala Gly Ser Phe Asp Tyr Gly
100 105 110
Arg Asn Tyr Gly Val Thr Tyr Asp Val Thr Ser Trp Thr Asp Val Leu
115 120 125
Pro Glu Phe Gly Gly Asp Thr Tyr Gly Ala Asp Asn Phe Met Gln Gln
130 135 140
Arg Gly Asn Gly Tyr Ala Thr Tyr Arg ASn Thr Asp Phe Phe Gly Leu
145 150 155 160
Val Asp Gly Leu Asp Phe Ala Leu Gln Tyr Gln Gly Lys Asn Gly Ser
165 170 175
Val Ser Gly Glu Asn Thr Asn Gly Arg Ser Leu Leu Asn Gln Asn Gly
180 185 190
Asp Gly Tyr Gly Gly Ser Leu Thr Tyr Ala Ile Gly Glu Gly Phe Ser
195 200 205
Val Gly Gly Ala Ile Thr Thr Ser Lys Arg Thr Ala Asp Gln Asn Asn
210 215 220
Thr Ala Asn Ala Arg Leu Tyr Gly Asn Gly Asp Arg Ala Thr Val Tyr
225 230 235 240
Thr Gly Gly Leu Lys Tyr Asp Ala Asn Asn Ile Tyr Leu Ala Ala Gln
245 250 255
Tyr Ser Gln Thr Tyr Asn Ala Thr Arg Phe Gly Thr Ser Asn Gly Ser
260 265 270
Asn Pro Ser Thr Ser Tyr Gly Phe Ala Asn Lys Ala Gln Asn Phe Glu
275 280 285
Val Val Ala Gln Tyr Gln Phe Asp Phe Gly Leu Arg Pro Ser Val Ala
290 295 300
Tyr Leu Gln Ser Lys Gly Lys Asp Ile Ser Asn Gly Tyr Gly Ala Ser
305 310 315 320
Tyr Gly Asp Gln Asp Ile Val Lys Tyr Val Asp Val Gly Ala Thr Tyr
325 330 335
Tyr Phe Asn Lys Asn Met Ser Thr Tyr Val Asp Tyr Lys Ile Asn Leu
340 345 350
Leu Asp Lys Asn Asp Phe Thr Arg Asp Ala Gly Ile Asn Thr Asp Asp
355 360 365
Ile Val Ala Leu Gly Leu Val Tyr Gln Phe
370 375
<210>40
<211>363
<212>PRT
<213>伤寒沙门氏菌CT18
<400>40
Met Met Lys Arg Lys Ile Leu Ala Ala Val Ile Pro Ala Leu Leu Ala
1 5 10 15
Ala Ala Thr Ala Asn Ala Ala Glu Ile Tyr Asn Lys Asp Gly Asn Lys
20 25 30
Leu Asp Leu Tyr Gly Lys Ala Val Gly Arg His Val Trp Thr Thr Thr
35 40 45
Gly Asp Ser Lys Asn Ala Asp Gln Thr Tyr Ala Gln Ile Gly Phe Lys
50 55 60
Gly Glu Thr Gln Ile Asn Thr Asp Leu Thr Gly Phe Gly Gln Trp Glu
65 70 75 80
Tyr Arg Thr Lys Ala Asp Arg Ala Glu Gly Glu Gln Gln Asn Ser Asn
85 90 95
Leu Val Arg Leu Ala Phe Ala Gly Leu Lys Tyr Ala Glu Val Gly Ser
100 105 110
Ile Asp Tyr Gly Arg Asn Tyr Gly Ile Val Tyr Asp Val Glu Ser Tyr
115 120 125
Thr Asp Met Ala Pro Tyr Phe Ser Gly Glu Thr Trp Gly Gly Ala Tyr
130 135 140
Thr Asp Asn Tyr Met Thr Ser Arg Ala Gly Gly Leu Leu Thr Tyr Arg
145 150 155 160
Asn Ser Asp Phe Phe Gly Leu Val Asp Gly Leu SerPhe Gly Ile Gln
165 170 175
Tyr Gln Gly Lys Asn Gln Asp Asn His Ser Ile Asn Ser Gln Asn Gly
180 185 190
Asp Gly Val Gly Tyr Thr Met Ala Tyr Glu Phe Asp Gly Phe Gly Val
195 200 205
Thr Ala Ala Tyr Ser Asn Ser Lys Arg Thr Asn Asp Gln Gln Asp Arg
210 215 220
Asp Gly Asn Gly Asp Arg Ala Glu Ser Trp Ala Val Gly Ala Lys Tyr
225 230 235 240
Asp Ala Asn Asn Val Tyr Leu Ala Ala Val Tyr Ala Glu Thr Arg Asn
245 250 255
Met Ser Ile Val Glu Asn Thr Val Thr Asp Thr Val Glu Met Ala Asn
260 265 270
Lys Thr Gln Asn Leu Glu Val Val Ala Gln Tyr Gln Phe Asp Phe Gly
275 280 285
Leu Arg Pro Ala Ile Ser Tyr Val Gln Ser Lys Gly Lys Gln Leu Asn
290 295 300
Gly Ala Asp Gly Ser Ala Asp Leu Ala Lys Tyr Ile Gln Ala Gly Ala
305 310 315 320
Thr Tyr Tyr Phe Asn Lys Asn Met Asn Val Trp Val Asp Tyr Arg Phe
325 330 335
Asn Leu Leu Asp Glu Asn Asp Tyr Ser Ser Ser Tyr Val Gly Thr Asp
340 345 350
Asp Gln Ala Ala Val Gly Ile Thr Tyr Gln Phe
355 360
<210>41
<211>77
<212>DNA
<213>人工序列
<220>
<223>引物
<400>41
ggttaaggaa ggtggggggc ttaaagtact cttaaccgga agcgtggtag tggtagtggt 60
aatcattcct aggtgac 77
<210>42
<211>224
<212>PRT
<213>人工序列
<220>
<223>包含用于与OmpC结合的亲和肽的PapMV外壳蛋白
<400>42
Met Ala Ser Thr Pro Asn Ile Ala Phe Pro Ala Ile Thr Gln Glu Gln
1 5 10 15
Met Ser Ser Ile Glu Val Asp Pro Thr Ser Asn Leu Leu Pro Ser Gln
20 25 30
Glu Gln Leu Lys Ser Val Ser Thr Leu Met Val Ala Ala Lys Val Pro
35 40 45
Ala Ala Ser Val Thr Thr Val Ala Leu Glu Leu Val Asn Phe Cys Tyr
50 55 60
Asp Asn Gly Ser Ser Ala Tyr Thr Thr Val Thr Gly Pro Ser Ser Ile
65 70 75 80
Pro Glu Ile Ser Leu Ala Gln Leu Ala Ser Ile Val Lys Ala Ser Gly
85 90 95
Thr Ser Leu Arg Lys Phe Cys Arg Tyr Phe Ala Pro Ile Ile Trp Asn
100 105 110
Leu Arg Thr Asp Lys Met Ala Pro Ala Asn Trp Glu Ala Ser Gly Tyr
115 120 125
Lys Pro Ser Ala Lys Phe Ala Ala Phe Asp Phe Phe Asp Gly Val Glu
130 135 140
Asn Pro Ala Ala Met Gln Pro Pro Ser Gly Leu Thr Arg Ser Pro Thr
145 150 155 160
Gln Glu Glu Arg Ile Ala Asn Ala Thr Asn Lys Gln Val His Leu Phe
165 170 175
Gln Ala Ala Ala Gln Gly Asn Asn Phe Ala Ser Asn Ser Ala Phe Ile
180 185 190
Thr Lys Gly Gln Ile Ser Gly Ser Thr Pro Thr Ile Gln Phe Leu Pro
195 200 205
Pro Pro Glu Glu Ala Lys Gly Leu Ile Arg His His His His His His
210 215 220
<210>43
<211>224
<212>PRT
<213>人工序列
<220>
<223>包含用于与OmpF结合的亲和肽的PapMV外壳蛋白
<400>43
Met Ala Ser Thr Pro Asn Ile Ala Phe Pro Ala Ile Thr Gln Glu Gln
1 5 10 15
Met Ser Ser Ile Lys Val Asp Pro Thr Ser Asn Leu Leu Pro Ser Gln
20 25 30
Glu Gln Leu Lys Ser Val Ser Thr Leu Met Val Ala Ala Lys Val Pro
35 40 45
Ala Ala Ser Val Thr Thr Val Ala Leu Glu Leu Val Asn Phe Cys Tyr
50 55 60
Asp Asn Gly Ser Ser Ala Tyr Thr Thr Val Thr Gly Pro Ser Ser Ile
65 70 75 80
Pro Glu Ile Ser Leu Ala Gln Leu Ala Ser Ile Val Lys Ala Ser Gly
85 90 95
Thr Ser Leu Arg Lys Phe Cys Arg Tyr Phe Ala Pro Ile Ile Trp Asn
100 105 110
Leu Arg Thr Asp Lys Met Ala Pro Ala Asn Trp Glu Ala Ser Gly Tyr
115 120 125
Lys Pro Ser Ala Lys Phe Ala Ala Phe Asp Phe Phe Asp Gly Val Glu
130 135 140
Asn Pro Ala Ala Met Gln Pro Pro Ser Gly Leu Thr Arg Ser Pro Thr
145 150 155 160
Gln Glu Glu Arg Ile Ala Asn Ala Thr Asn Lys Gln Val His Leu Phe
165 170 175
Gln Ala Ala Ala Gln Asp Asn Asn Phe Ala Ser Asn Ser Ala Phe Ile
180 185 190
Thr Lys Gly Gln Ile Ser Gly Ser Thr Pro Thr Ile Gln Phe Leu Pro
195 200 205
Pro Pro Glu Phe His Glu Asn Trp Pro Ser His His His His His His
210 215 220
<210>44
<211>54
<212>DNA
<213>人工序列
<220>
<223>引物
<400>44
ctagtggtac ttctaacggt tctaacccgt ctacttctta cggtttcgcg aaca 54
<210>45
<211>54
<212>DNA
<213>人工序列
<220>
<223>引物
<400>45
ctagtgttcg cgaaaccgta agaagtagac gggttagaac cgttagaagt acca 54
<210>46
<211>237
<212>PRT
<213>人工序列
<220>
<223>包含伤寒沙门氏菌孔蛋白环6肽的PapMV外壳蛋白
<400>46
Met Ala Ser Thr Pro Asn Ile Ala Phe Pro Ala Ile Thr Gln Glu Gln
1 5 10 15
Met Ser Ser Ile Lys Val Asp Pro Thr Ser Asn Leu Leu Pro Ser Gln
20 25 30
Glu Gln Leu Lys Ser Val Ser Thr Leu Met Val Ala Ala Lys Val Pro
35 40 45
Ala Ala Ser Val Thr Thr Val Ala Leu Glu Leu Val Asn Phe Cys Tyr
50 55 60
Asp Asn Gly Ser Ser Ala Tyr Thr Thr Val Thr Gly Pro Ser Ser Ile
65 70 75 80
Pro Glu Ile Ser Leu Ala Gln Leu Ala Ser Ile Val Lys Ala Ser Gly
85 90 95
ThrSer Leu Arg Lys Phe Cys Arg Tyr Phe Ala Pro Ile Ile Trp Asn
100 105 110
Leu Arg Thr Asp Lys Met Ala Pro Ala Asn Trp Glu Ala Ser Gly Tyr
115 120 125
Lys Pro Ser Ala Lys Phe Ala Ala Phe Asp Phe Phe Asp Gly Val Glu
130 135 140
Asn Pro Ala Ala Met Gln Pro Pro Ser Gly Leu Thr Arg Ser Pro Thr
145 150 155 160
Gln Glu Glu Arg Ile Ala Asn Ala Thr Asn Lys Gln Val His Leu Phe
165 170 175
Gln Ala Ala Ala Gln Asp Asn Asn Phe Ala Ser Asn Ser Ala Phe Ile
180 185 190
Thr Lys Gly Gln Ile Ser Gly Ser Thr Pro Thr Ile Gln Phe Leu Pro
195 200 205
Pro Pro Glu Thr Ser Gly Thr Ser Asn Gly Ser Asn Pro Ser Thr Ser
210 215 220
Tyr Gly Phe Ala Asn Thr Arg His His His His His His
225 230 235
<210>47
<211>714
<212>DNA
<213>人工序列
<220>
<223>编码包含伤寒沙门氏菌孔蛋白环6肽的PapMV外壳蛋白的核苷酸序列
<400>47
atggcatcca cacccaacat agccttcccc gccatcaccc aggaacaaat gagctcgatt 60
aaggtcgatc caacgtccaa tcttctgccc tcccaagagc agttaaagtc agtgtccacc 120
ctcatggtag ctgctaaggt tccagcagcc agtgttacaa ctgtggcatt ggagttggtt 180
aacttctgct atgacaatgg gtccagcgcg tacaccacag tgactggccc atcatcaata 240
ccggagatat cactggcaca attggccagc attgtcaaag cttccggcac ttcccttagg 300
aaattctgcc ggtacttcgc gccaataatc tggaatctga ggacggacaa aatggctcct 360
gccaattggg aggcctcagg atacaagcca agcgccaaat ttgccgcgtt cgacttcttc 420
gacggggtgg agaatccggc ggccatgcaa cccccttcgg gactaaccag gtcgccgacc 480
caggaagagc ggattgccaa tgccaccaac aaacaggtgc atctcttcca agccgcggca 540
caggacaaca actttgccag caactccgcc ttcatcacca aaggccaaat ttctgggtca 600
accccaacca tccaattcct tccacccccc gaaactagtg gtacttctaa cggttctaac 660
ccgtctactt cttacggttt cgcgaacacg cgtcaccatc accatcacca ttag 714
<210>48
<211>228
<212>PRT
<213>人工序列
<220>
<223>包含用于与OmpC结合的亲和肽的PapMV SM外壳蛋白
<400>48
Met Ala Ser Thr Pro Asn Ile Ala Phe Pro Ala Ile Thr Gln Glu Gln
1 5 10 15
Met Ser Ser Ile Lys Val Asp Pro Thr Ser Asn Leu Leu Pro Ser Gln
20 25 30
Glu Gln Leu Lys Ser Val Ser Thr Leu Met Val Ala Ala Lys Val Pro
35 40 45
Ala Ala Ser Val Thr Thr Val Ala Leu Glu Leu Val Asn Phe Cys Tyr
50 55 60
Asp Asn Gly Ser Ser Ala Tyr Thr Thr Val Thr Gly Pro Ser Ser Ile
65 70 75 80
Pro Glu Ile Ser Leu Ala Gln Leu Ala Ser Ile Val Lys Ala Ser Gly
85 90 95
Thr Ser Leu Arg Lys Phe Cys Arg Tyr Phe Ala Pro Ile Ile Trp Asn
100 105 110
Leu Arg Thr Asp Lys Met Ala Pro Ala Asn Trp Glu Ala Ser Gly Tyr
115 120 125
Lys Pro Ser Ala Lys Phe Ala Ala Phe Asp Phe Phe Asp Gly Val Glu
130 135 140
Asn Pro Ala Ala Met Gln Pro Pro Ser Gly Leu Thr Arg Ser Pro Thr
145 150 155 160
Gln Glu Glu Arg Ile Ala Asn Ala Thr Asn Lys Gln Val His Leu Phe
165 170 175
Gln Ala Ala Ala Gln Asp Asn Asn Phe Ala Ser Asn Ser Ala Phe Ile
180 185 190
Thr Lys Gly Gln Ile Ser Gly Ser Thr Pro Thr Ile Gln Phe Leu Pro
195 200 205
Pro Pro Glu Thr Ser Glu Ala Lys Gly Leu Ile Arg Thr Arg His His
210 215 220
His His His His
225
<210>49
<211>687
<212>DNA
<213>人工序列
<220>
<223>编码包含用于与OmpC结合的亲和肽的PapMV SM外壳蛋白的核苷酸序列
<400>49
atggcatcca cacccaacat agccttcccc gccatcaccc aggaacaaat gagctcgatt 60
aaggtcgatc caacgtccaa tcttctgccc tcccaagagc agttaaagtc agtgtccacc 120
ctcatggtag ctgctaaggt tccagcagcc agtgttacaa ctgtggcatt ggagttggtt 180
aacttctgct atgacaatgg gtccagcgcg tacaccacag tgactggccc atcatcaata 240
ccggagatat cactggcaca attggccagc attgtcaaag cttccggcac ttcccttagg 300
aaattctgcc ggtacttcgc gccaataatc tggaatctga ggacggacaa aatggctcct 360
gccaattggg aggcctcagg atacaagcca agcgccaaat ttgccgcgtt cgacttcttc 420
gacggggtgg agaatccggc ggccatgcaa cccccttcgg gactaaccag gtcgccgacc 480
caggaagagc ggattgccaa tgccaccaac aaacaggtgc atctcttcca agccgcggca 540
caggacaaca actttgccag caactccgcc ttcatcacca aaggccaaat ttctgggtca 600
accccaacca tccaattcct tccacccccc gaaactagtg aggcgaaggg gttgattcgt 660
acgcgtcacc atcaccatca ccattag 687
<210>50
<211>16
<212>PRT
<213>人工序列
<220>
<223>包含源自伤寒沙门氏菌孔蛋白的环6抗原表位的肽
<400>50
Gly Thr Ser Asn Gly Ser Asn Pro Ser Thr Ser Tyr Gly Phe Ala Asn
1 5 10 15
<210>51
<211>9
<212>PRT
<213>人工序列
<220>
<223>源自gp100肿瘤抗原的抗原表位
<400>51
Ile Met Asp Gln Val Pro Phe Ser Val
1 5
<210>52
<211>9
<212>PRT
<213>人工序列
<220>
<223>源自流感病毒M1蛋白的抗原表位
<400>52
Gly Ile Leu Gly Phe Val Phe Thr Leu
1 5
<210>53
<211>9
<212>PRT
<213>人工序列
<220>
<223>源自gp100肿瘤抗原的抗原表位
<400>53
Ile Thr Asp Gln Val Pro Phe Ser Val
1 5
<210>54
<211>19
<212>PRT
<213>人工序列
<220>
<223>源自伤寒沙门氏菌孔蛋白的环7肽
<400>54
Gln Ser Lys Gly Lys Asp Ile Ser Asn Gly Tyr Gly Ala Ser Tyr Gly
1 5 10 15
Asp Gln Asp
Claims (48)
1.一种抗原提呈系统,其包含与番木瓜花叶病毒(PapMV)或源自PapMV外壳蛋白的VLP组合的一种或多种肠细菌抗原,其中所述PapMV或VLP能够增强针对所述一种或多种肠细菌抗原的免疫应答。
2.根据权利要求1所述的抗原提呈系统,其中所述VLP源自修饰的PapMV外壳蛋白,所述修饰的PapMV外壳蛋白能够多聚化以形成所述VLP。
3.根据权利要求2所述的抗原提呈系统,其中所述抗原提呈系统包含这样的VLP,所述VLP包含与SEQ ID NO:3所列出的序列基本上相同的氨基酸序列。
4.根据权利要求1、2或3中任一项所述的抗原提呈系统,其中所述一种或多种抗原与所述PapMV或VLP的外壳蛋白是接合的。
5.根据权利要求1、2或3中任一项所述的抗原提呈系统,其中所述一种或多种抗原与所述PapMV或VLP是非接合的。
6.根据权利要求4所述的抗原提呈系统,其中所述一种或多种抗原与所述外壳蛋白基因融合。
7.根据权利要求4所述的抗原提呈系统,其中所述一种或多种抗原是通过亲和结合与所述外壳蛋白连接的。
8.根据权利要求7所述的抗原提呈系统,其中所述亲和结合是通过与所述外壳蛋白基因融合的亲和肽实现的,所述亲和肽能够结合所述一种或多种抗原。
9.根据权利要求1、2、3、4、5、6、7或8中任一项所述的抗原提呈系统,其中所述一种或多种抗原是孔蛋白或其抗原片段。
10.根据权利要求1、2、3、4、5、6、7、8或9中任一项所述的抗原提呈系统,其中所述一种或多种抗原选自由OmpC、OmpC的片段、OmpF和OmpF的片段所组成的组。
11.根据权利要求1、2、3、4、5、6、7、8、9或10中任一项所述的抗原提呈系统,其中所述一种或多种抗原是伤寒沙门氏菌抗原。
12.根据权利要求1所述的抗原提呈系统,其中所述抗原提呈系统包含这样的VLP,所述VLP包含与SEQ ID NO:42、SEQ IDNO:43或SEQ ID NO:48所列出的序列基本上相同的氨基酸序列。
13.一种疫苗组合物,其包含:
权利要求1、2、3、4、5、6、7、8、9、10、11或12中任一项所述的抗原提呈系统;以及
可药用的载体。
14.一种多肽,其包含与一种或多种亲和肽融合的番木瓜花叶病毒外壳蛋白,所述一种或多种亲和肽能够结合肠细菌抗原。
15.根据权利要求14所述的多肽,其中所述抗原是伤寒沙门氏菌抗原。
16.根据权利要求14或15所述的多肽,其中所述一种或多种亲和肽包含这样的序列,所述序列选自SEQ ID NO:21、SEQ IDNO:22、SEQ ID NO:23、SEQ ID NO:24和SEQ ID NO:25所列出的序列。
17.根据权利要求14或15所述的多肽,其中所述多肽包含与SEQ ID NO:42、SEQ ID NO:43或SEQ ID NO:48所列出的序列基本上相同的氨基酸序列。
18.一种多肽,其包含与一种或多种肠细菌抗原融合的番木瓜花叶病毒外壳蛋白。
19.根据权利要求18所述的多肽,其中所述一种或多种抗原是伤寒沙门氏菌抗原。
20.根据权利要求18或19所述的多肽,其中所述一种或多种抗原在所述PapMV外壳蛋白的C-端处或内环中融合。
21.一种多核苷酸,所述多核苷酸编码权利要求13、14、15、16、17、18、19或20中任一项所述的多肽。
22.根据权利要求1、2、3、4、5、6、7、8、9、10、11或12中任一项所述的抗原提呈系统在需要其的动物中用于诱导针对肠细菌的免疫应答中的用途。
23.根据权利要求13所述的疫苗在需要其的动物中用于诱导针对肠细菌的免疫应答中的用途。
24.根据权利要求22或23所述的用途,其中所述肠细菌是伤寒沙门氏菌。
25.根据权利要求22、23或24中任一项所述的用途,其中所述动物是人类。
26.根据权利要求22、23或24中任一项所述的用途,其中所述动物是非人类动物。
27.根据权利要求1、2、3、4、5、6、7、8、9、10、11或12中任一项所述的抗原提呈系统在制备疫苗中的用途。
28.根据权利要求27所述的用途,其中所述疫苗在需要其的动物中用于诱导针对肠细菌的免疫应答。
29.根据权利要求27或28所述的用途,其中所述肠细菌是伤寒沙门氏菌。
30.根据权利要求27、28或29中任一项所述的用途,其中所述动物是人类。
31.根据权利要求27、28或29中任一项所述的用途,其中所述动物是非人类动物。
32.一种用于免疫动物以抗伤寒沙门氏菌感染的疫苗,所述疫苗包含抗原提呈系统,所述抗原提呈系统包含与番木瓜花叶病毒(PapMV)或源自PapMV外壳蛋白的VLP组合的抗原,所述抗原源自伤寒沙门氏菌OmpC或OmpF,其中所述PapMV或VLP能够在所述动物中增强针对所述抗原的免疫应答。
33.根据权利要求32所述的疫苗,其中所述抗原提呈系统包含这样的VLP,所述VLP包含与SEQ ID NO:42、SEQ ID NO:43或SEQID NO:48所列出的序列基本上相同的氨基酸序列。
34.一种在动物中诱导针对肠细菌的免疫应答的方法,所述方法包括给所述动物施用有效量的根据权利要求1所述的抗原提呈系统。
35.根据权利要求34所述的方法,其中所述VLP源自修饰的PapMV外壳蛋白,所述修饰的PapMV外壳蛋白能够多聚化以形成所述VLP。
36.根据权利要求35所述的方法,其中所述抗原提呈系统包含这样的VLP,所述VLP包含与SEQ ID NO:3所列出的序列基本上相同的氨基酸序列。
37.根据权利要求34所述的方法,其中所述一种或多种抗原与所述PapMV或VLP的外壳蛋白是接合的。
38.根据权利要求34所述的方法,其中所述一种或多种抗原与所述PapMV或VLP是非接合的。
39.根据权利要求37所述的方法,其中所述一种或多种抗原与所述外壳蛋白是基因融合的。
40.根据权利要求37所述的方法,其中所述一种或多种抗原通过亲和结合与所述外壳蛋白连接。
41.根据权利要求40所述的方法,其中所述亲和结合是通过与所述外壳蛋白基因融合的亲和肽实现的,所述亲和肽能够结合所述一种或多种抗原。
42.根据权利要求34所述的方法,其中所述一种或多种抗原是孔蛋白或其抗原片段。
43.根据权利要求34所述的方法,其中所述一种或多种抗原选自由OmpC、OmpC的片段、OmpF和OmpF的片段所组成的组。
44.根据权利要求34所述的方法,其中所述一种或多种抗原是伤寒沙门氏菌抗原。
45.根据权利要求34所述的方法,其中所述抗原提呈系统包含这样的VLP,所述VLP包含与SEQ ID NO:42、SEQ ID NO:43或SEQID NO:48所列出的序列基本上相同的氨基酸序列。
46.根据权利要求34所述的方法,其中所述肠细菌是伤寒沙门氏菌。
47.根据权利要求34所述的方法,其中所述动物是人类。
48.根据权利要求34所述的方法,其中所述动物是非人类动物。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US88687307P | 2007-01-26 | 2007-01-26 | |
| US60/886,873 | 2007-01-26 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN101646772A true CN101646772A (zh) | 2010-02-10 |
Family
ID=39644061
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN200880010037A Pending CN101646772A (zh) | 2007-01-26 | 2008-01-28 | 抗伤寒沙门氏菌和其它肠细菌病原体的基于番木瓜花叶病毒的疫苗 |
Country Status (7)
| Country | Link |
|---|---|
| US (1) | US20100316665A1 (zh) |
| EP (1) | EP2082042A4 (zh) |
| JP (1) | JP2010516713A (zh) |
| CN (1) | CN101646772A (zh) |
| CA (1) | CA2676481A1 (zh) |
| MX (1) | MX2009007942A (zh) |
| WO (1) | WO2008089569A1 (zh) |
Cited By (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104459126A (zh) * | 2014-12-16 | 2015-03-25 | 四川大学 | 一种基于免疫磁珠富集沙门氏菌的方法 |
| CN112566494A (zh) * | 2018-06-12 | 2021-03-26 | 肯塔基生物处理股份有限公司 | 病毒和抗原纯化和偶联 |
| US11529413B2 (en) | 2018-06-12 | 2022-12-20 | Kbio Holdings Limited | Virus and antigen purification and conjugation |
| US11690907B2 (en) | 2018-06-12 | 2023-07-04 | Kbio Holdings Limited | Vaccines formed by virus and antigen conjugation |
| US11696948B2 (en) | 2018-06-12 | 2023-07-11 | Kbio Holdings Limited | Vaccines formed by virus and antigen conjugation |
Families Citing this family (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US8101189B2 (en) | 2002-07-05 | 2012-01-24 | Folia Biotech Inc. | Vaccines and immunopotentiating compositions and methods for making and using them |
| ES2562774T3 (es) * | 2006-11-15 | 2016-03-08 | Folia Biotech Inc. | Vacunas para la gripe basadas en el virus del mosaico de la papaya |
| CA2728374A1 (en) * | 2008-06-17 | 2010-01-14 | Universite Laval | Compositions comprising salmonella porins and uses thereof as adjuvants and vaccines |
| US20110206727A1 (en) * | 2008-07-30 | 2011-08-25 | Denis Leclerc | Multivalent Vaccines Based on Papaya Mosaic Virus and Uses Thereof |
| CA2774640C (en) * | 2009-09-18 | 2019-11-26 | Fraunhofer Usa Inc. | Virus like particles comprising target proteins fused to plant viral coat proteins |
| GB2498323B8 (en) * | 2009-11-05 | 2014-08-06 | Hal Padgett | Generation of antigenic virus-like particles through protein-protein linkages |
| AU2012255595A1 (en) * | 2011-05-13 | 2013-12-12 | Folia Biotech Inc. | Virus-like particles and process for preparing same |
| EP3257868A4 (en) * | 2015-02-13 | 2018-07-11 | National University Corporation Kyoto Institute of Technology | Peptides having affinity for polydimethylsiloxane, and uses thereof |
| CN115851771B (zh) * | 2022-08-03 | 2023-06-20 | 扬州大学 | 一种不表达Peg菌毛的鸡伤寒沙门菌弱毒分离株及其应用 |
Family Cites Families (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU750702B2 (en) * | 1997-05-01 | 2002-07-25 | Chiron Corporation | Use of virus-like particles as adjuvants |
| CZ20011907A3 (cs) * | 1998-12-04 | 2002-01-16 | Biogen, Inc. | Částice jádra HBV s imunogenními sloľkami navázanými prostřednictvím peptidových ligandů |
| EP2272525A3 (en) * | 2002-07-05 | 2011-11-16 | Folia Biotech Inc. | Adjuvant viral particle |
| US8101189B2 (en) * | 2002-07-05 | 2012-01-24 | Folia Biotech Inc. | Vaccines and immunopotentiating compositions and methods for making and using them |
| US7285264B2 (en) * | 2003-09-08 | 2007-10-23 | E.I. Du Pont De Nemours And Company | Peptide-based body surface coloring reagents |
| US8071552B2 (en) * | 2004-04-05 | 2011-12-06 | Universite Bordeaux 2 | Peptides and peptidomimetics binding to CD23 |
| ES2562774T3 (es) * | 2006-11-15 | 2016-03-08 | Folia Biotech Inc. | Vacunas para la gripe basadas en el virus del mosaico de la papaya |
| EP2078086A4 (en) * | 2006-11-15 | 2010-06-30 | Folia Biotech Inc | PAPAYAMOSAIC VIRUS-BASED IMMUNOGENE AFFINITY-CONJUGATED ANTIGENSYSTEMS AND THEIR USES |
-
2008
- 2008-01-28 MX MX2009007942A patent/MX2009007942A/es not_active Application Discontinuation
- 2008-01-28 CN CN200880010037A patent/CN101646772A/zh active Pending
- 2008-01-28 US US12/449,140 patent/US20100316665A1/en not_active Abandoned
- 2008-01-28 WO PCT/CA2008/000154 patent/WO2008089569A1/en active Application Filing
- 2008-01-28 EP EP08706301A patent/EP2082042A4/en not_active Withdrawn
- 2008-01-28 JP JP2009546624A patent/JP2010516713A/ja active Pending
- 2008-01-28 CA CA002676481A patent/CA2676481A1/en not_active Abandoned
Cited By (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104459126A (zh) * | 2014-12-16 | 2015-03-25 | 四川大学 | 一种基于免疫磁珠富集沙门氏菌的方法 |
| CN112566494A (zh) * | 2018-06-12 | 2021-03-26 | 肯塔基生物处理股份有限公司 | 病毒和抗原纯化和偶联 |
| US11485956B2 (en) | 2018-06-12 | 2022-11-01 | Kbio Holdings Limited | Virus and antigen purification and conjugation |
| US11529413B2 (en) | 2018-06-12 | 2022-12-20 | Kbio Holdings Limited | Virus and antigen purification and conjugation |
| US11655461B2 (en) | 2018-06-12 | 2023-05-23 | Kbio Holdings Limited | Antigen purification |
| US11690907B2 (en) | 2018-06-12 | 2023-07-04 | Kbio Holdings Limited | Vaccines formed by virus and antigen conjugation |
| US11696948B2 (en) | 2018-06-12 | 2023-07-11 | Kbio Holdings Limited | Vaccines formed by virus and antigen conjugation |
Also Published As
| Publication number | Publication date |
|---|---|
| US20100316665A1 (en) | 2010-12-16 |
| AU2008209284A8 (en) | 2009-09-24 |
| AU2008209284A2 (en) | 2009-09-24 |
| WO2008089569A9 (en) | 2008-09-12 |
| AU2008209284A9 (en) | 2009-10-08 |
| WO2008089569A1 (en) | 2008-07-31 |
| MX2009007942A (es) | 2010-06-01 |
| EP2082042A1 (en) | 2009-07-29 |
| EP2082042A4 (en) | 2010-08-18 |
| JP2010516713A (ja) | 2010-05-20 |
| CA2676481A1 (en) | 2008-07-31 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN101646772A (zh) | 抗伤寒沙门氏菌和其它肠细菌病原体的基于番木瓜花叶病毒的疫苗 | |
| US8101189B2 (en) | Vaccines and immunopotentiating compositions and methods for making and using them | |
| EP2069503B1 (en) | Papaya mosaic virus-based vaccines for influenza | |
| US20110206727A1 (en) | Multivalent Vaccines Based on Papaya Mosaic Virus and Uses Thereof | |
| US20100047264A1 (en) | Immunogenic Affinity-Conjugated Antigen Systems Based on Papaya Mosaic Virus and Uses Thereof | |
| BR112012005703A2 (pt) | particuça do tipo virus, composição imunogênia e medicamento | |
| Smith et al. | Display of peptides on the surface of tobacco mosaic virus particles | |
| JP2010538619A (ja) | Malvaモザイクウイルスおよびウイルス様粒子およびこれらの使用 | |
| US20130280298A1 (en) | Immunogenic Affinity-Conjugated Antigen Systems Based on Papaya Mosaic Virus and Uses Thereof | |
| CN102127554A (zh) | 一种日本脑炎颗粒疫苗及其制备方法和应用 | |
| CN111607605A (zh) | 一种多价表位和亚单位疫苗的构建方法 | |
| KR102451412B1 (ko) | 세포성, 체액성 면역 반응의 동시 유도 및 광범위한 방어능을 갖는 신규 면역 증강용 단백질 및 이를 포함하는 구제역 백신 조성물 | |
| CN107970444A (zh) | 复合佐剂及含有该复合佐剂的疫苗 | |
| EP3994152A1 (en) | Arthrospira platensis non-parenteral therapeutic delivery platform | |
| KR101991577B1 (ko) | 다수의 에피토프로 구성된 재조합 항원 단백질 및 이의 제조방법 | |
| KR102320271B1 (ko) | 부유세포 배양에 적응된 A/ASIA/Sea-97 유전형 구제역 바이러스 백신주 및 이를 포함하는 구제역 바이러스 백신 조성물 | |
| KR100829380B1 (ko) | 파스튜렐라 멀토시다 균주의 재조합 외막단백질 h를이용한 가금 콜레라 백신 | |
| US20150056231A1 (en) | Recombinant papaya mosaic virus coat proteins and uses thereof in influenza vaccines | |
| CN104593386A (zh) | 甲型副伤寒沙门菌ompN基因原核表达系统及其重组表达蛋白的应用 | |
| EP4208472A1 (en) | Ompa mutations enhance omv production in bordetella pertussis | |
| CN103524615A (zh) | 一种抗兔病毒性出血症病毒rhdv卵黄抗体的制备方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| C02 | Deemed withdrawal of patent application after publication (patent law 2001) | ||
| WD01 | Invention patent application deemed withdrawn after publication |
Open date: 20100210 |