WO2018003339A1 - 細胞特異的にヌクレアーゼを制御する方法 - Google Patents
細胞特異的にヌクレアーゼを制御する方法 Download PDFInfo
- Publication number
- WO2018003339A1 WO2018003339A1 PCT/JP2017/018742 JP2017018742W WO2018003339A1 WO 2018003339 A1 WO2018003339 A1 WO 2018003339A1 JP 2017018742 W JP2017018742 W JP 2017018742W WO 2018003339 A1 WO2018003339 A1 WO 2018003339A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- mirna
- nuclease
- protein
- nucleic acid
- acid sequence
- Prior art date
Links
Images






























Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/11—Antisense
- C12N2310/113—Antisense targeting other non-coding nucleic acids, e.g. antagomirs
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/14—Type of nucleic acid interfering N.A.
- C12N2310/141—MicroRNAs, miRNAs
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2330/00—Production
- C12N2330/10—Production naturally occurring
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/10—Cells modified by introduction of foreign genetic material
Definitions
- the present invention relates to a method for controlling a nuclease in a cell-specific manner and a miRNA-responsive RNA used in the method.
- CRISPR / Cas9 System cleaves a target gene with a nuclease Cas9 protein (Cas9) and a single guide RNA (sgRNA) incorporating 20 bases complementary to the target sequence (for example, Non-Patent Document 1). reference).
- Cas9 nuclease Cas9 protein
- sgRNA single guide RNA
- CRISPR / Cas9 System for cell therapy. For example, by placing a Cas9 gene or sgRNA under the control of a cancer cell-specific promoter, CRISPR / Cas9 System is activated specifically for cancer cells to induce cell death (for example, see Non-Patent Document 2). .
- the present inventors have determined that a nucleic acid sequence specifically recognized by miRNA that has been developed by the present inventors is linked with a nucleic acid sequence encoding a nuclease. It has been found that by using this method, the above-mentioned problems can be solved and the nuclease activity can be controlled in response to cell-specific miRNA.
- a method for controlling a nuclease in a cell-specific manner including a step of introducing a miRNA-responsive mRNA encoding a nuclease into a cell, wherein the miRNA-responsive mRNA is: a method comprising: (i) a nucleic acid sequence specifically recognized by miRNA; and (ii) a nucleic acid sequence corresponding to the coding region of said nuclease.
- the nucleic acid sequence specifically recognized by the miRNA is either a miR-302a target sequence or a miR-21 target sequence.
- the nuclease is a Cas9 protein or a variant thereof, The method according to [1] or [2], further comprising a step of introducing sgRNA having a guide sequence specifically recognized by the nuclease target gene into the cell group.
- the method according to [4], wherein the undifferentiated cells are cells that express miR-302a.
- the miRNA-responsive mRNA according to any one of [1] to [5], wherein the miRNA-responsive mRNA comprises a nucleic acid sequence in which (i) and (ii) are linked in the 5 ′ to 3 ′ direction.
- An miRNA-responsive mRNA comprising (i) a nucleic acid sequence specifically recognized by miRNA, and (ii) a nucleic acid sequence corresponding to the coding region of nuclease.
- the miRNA-responsive mRNA according to [7] wherein the nucleic acid sequence specifically recognized by the miRNA is either a miR-302a target sequence or a miR-21 target sequence.
- the miRNA-responsive mRNA according to [9], A kit for controlling a nuclease in a cell-specific manner, comprising a sgRNA having a guide sequence specifically recognized by a target gene of the nuclease.
- a) Trigger protein-responsive mRNA encoding nuclease b) a method for controlling a nuclease in a cell-specific manner, comprising the step of introducing into the cell a miRNA-responsive mRNA encoding the trigger protein,
- the a) protein-responsive mRNA encoding nuclease is: (ia) a nucleic acid sequence that specifically binds to the protein; (iia) a miRNA-responsive mRNA comprising a nucleic acid sequence corresponding to the coding region of the nuclease and encoding the trigger protein; (ib) a nucleic acid sequence specifically recognized by miRNA; (iib) A method comprising a nucleic acid sequence corresponding
- the trigger protein includes an L7Ae protein or a derivative thereof
- the nucleic acid sequence that specifically binds to the protein includes a K-turn sequence or a derivative thereof.
- the nucleic acid sequence specifically recognized by the miRNA is either a miR-302a target sequence or a miR-21 target sequence.
- the nuclease is a Cas9 protein or a variant thereof, The method according to any one of [11] to [13], further comprising the step of introducing sgRNA comprising a guide sequence specifically recognized by the nuclease target gene into the cell.
- the a) protein-responsive mRNA encoding the trigger protein comprises a nucleic acid sequence in which (ia) and (iia) are linked in the 5 ′ to 3 ′ direction; Any one of [11] to [16], wherein the b) miRNA-responsive mRNA encoding the nuclease comprises a nucleic acid sequence in which (ib) and (iib) are linked in the 5 ′ to 3 ′ direction.
- a) Trigger protein-responsive mRNA encoding a nuclease comprising: (ia) a nucleic acid sequence that specifically binds to the protein; and (iia) a nucleic acid sequence corresponding to the coding region of the nuclease; b) miRNA-responsive mRNA encoding a trigger protein including: a nuclease control agent comprising: (ib) a nucleic acid sequence specifically recognized by miRNA; and (iib) a nucleic acid sequence corresponding to the coding region of the trigger protein.
- nuclease controlling agent according to [18], wherein the trigger protein includes an L7Ae protein or a derivative thereof, and the nucleic acid sequence that specifically binds to the protein includes a K-turn sequence or a derivative thereof.
- the nucleic acid sequence specifically recognized by the miRNA is either the miR-302a target sequence or the miR-21 target sequence [18] or [ [19] The nuclease controlling agent according to [19].
- nuclease controlling agent according to any one of [18] to [21], A kit for controlling a nuclease in a cell-specific manner, comprising a sgRNA having a guide sequence specifically recognized by a target gene of the nuclease.
- nuclease control method of the present invention it has become possible to control nuclease activity in response to the expression of miRNA indicating the state of cells.
- nuclease can be introduced into cells with mRNA, and the nuclease can be controlled in a cell-specific manner. Therefore, clinical application is expected from the viewpoint of safety and specificity.
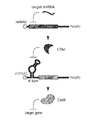
- FIG. 1A is a conceptual diagram showing a construct of miRNA-responsive mRNA encoding a nuclease used in the first embodiment of the present invention.
- FIG. 1 (b) is a conceptual diagram showing the reaction mechanism of miRNA-responsive cas9 mRNA in microRNA (miRNA) -responsive CRISPR / Cas9 System.
- FIG. 2 (a) is a photograph showing a fluorescent gene knockout by miRNA-responsive CRISPR / Cas9 mRNA introduced into HeLa cells as a fluorescence image.
- FIG. 2 (b) is a diagram showing the knockout of the fluorescent gene by miRNA-responsive CRISPR / Cas9 mRNA introduced into HeLa cells as a histogram.
- FIG. 1A is a conceptual diagram showing a construct of miRNA-responsive mRNA encoding a nuclease used in the first embodiment of the present invention.
- FIG. 1 (b) is a conceptual diagram showing the reaction mechanism of miRNA-responsive cas9 m
- FIG. 2C is a graph in which GFP negative population of the histogram shown in FIG. 2B is defined as Cas9 activity.
- FIG. 3 is a graph showing that miR-21-responsive Cas9 mRNA introduced into HeLa cells is translationally suppressed by miR-21 endogenous to HeLa cells and the expression of Cas9 is controlled.
- FIG. 4 (a) is a graph showing Cas9 activity calculated from EGFP activity assay that miR-302a-responsive Cas9 mRNA introduced into iPS cells was translationally suppressed by endogenous miR-302a of iPS cells. is there.
- FIG. 4 (b) is a gel image showing the results of T7E1 assay when miR-302a-responsive Cas9 mRNA is introduced into iPS cells. The gel image and its quantitative value are correlated with Cas9 activity and GFP negative population. It shows that there is.
- FIG. 4C is a graph showing indelsind (Cas9 activity) calculated from the gel image shown in FIG.
- Fig. 5 (a) is a schematic diagram of Cell killing system
- Fig. 5 (b) shows the rate of cell death induction when sgRNA targeting miR-21-responsive Cas9 RNA and AluI is introduced into HeLa cells. It is a graph to show.
- FIG. 4C is a graph showing indelsind (Cas9 activity) calculated from the gel image shown in FIG.
- Fig. 5 (a) is a schematic diagram of Cell killing system
- Fig. 5 (b) shows the rate of cell death induction when sgRNA targeting miR-21-responsive Cas9
- FIG. 6A is a graph showing RelativeelCas9 activity calculated from EGFP activity assay when miR-302a-responsive Cas9 mRNA is introduced into mDA cells.
- FIG. 6 (b) is a gel image of T7E1 assay when miR-302a-responsive Cas9 mRNA is introduced into mDA cells.
- FIG. 6C is a graph showing Relative Cas9 activity based on T7E1 assay.
- FIG. 7 (a) shows the result of evaluating the co-culture of iPS_GFP and HeLa_GFP by the change in fluorescence intensity by GFP knockout.
- FIG. 7B is a graph showing Cas9 activity calculated based on the result of FIG. FIG.
- FIG. 8A is a conceptual diagram showing two constructs, miRNA-responsive mRNA encoding a trigger protein and trigger protein-responsive mRNA encoding a nuclease, used in the second embodiment of the present invention.
- FIG. 8 (b) is a graph showing that Cas9 activity is increased by introducing the mRNA set of the second embodiment by endogenous miR-21 in HeLa cells.
- FIG. 8 (c) shows the intracellular expression when two constructs of miRNA-responsive mRNA encoding the trigger protein schematically shown in FIG. 8 (a) and trigger protein-responsive mRNA encoding nuclease are used. It is a graph which shows having increased Cas9 activity in response to miRNA of.
- FIG. 8 (b) is a graph showing that Cas9 activity is increased by introducing the mRNA set of the second embodiment by endogenous miR-21 in HeLa cells.
- FIG. 8 (c) shows the intracellular expression when two constructs of miRNA-responsive mRNA encoding the
- FIG. 9A shows the results of measuring the expression level of miR-21 with ⁇ HeLa-EGFP, iPS-EGFP, and mDA-EGFP, respectively.
- FIG. 9B shows the results of measuring the expression level of miR-302a with HeLa-EGFP, iPS-EGFP, and mDA-EGFP.
- Figure 10 shows whether Control Cas9 mRNA, miR-21-responsive Cas9 RNA mRNA and miR-302a-responsive Cas9 mRNA are introduced into HeLa cells (sgRNA is also introduced), and whether the Cas9 protein expression is affected is simple Western ( Wes) shows the results of the study.
- FIG. 11 (a) is a PCR product sequencing scheme showing the sequence around the target sequence.
- FIG. 11 (a) is a PCR product sequencing scheme showing the sequence around the target sequence.
- FIG. 11 (b) shows that the PCR product contains a mutation in the target region of the EGFP gene.
- FIG. 12 (a) shows the results of verifying the expression variation of a gene whose function is known to be controlled by miR-21 by qPCR using HeLa-EGFP cells.
- FIG. 12 (b) shows the result of verifying the expression variation of a gene whose function is known to be controlled by miR-302a by qPCR using iPS-EGFP cells.
- FIG. 13 (a) shows the transfection efficiency of HeLa-EGFP by BFPBmRNA and sgRNA, respectively.
- FIG. 13 (b) shows the transfection efficiency of iPS-EGFP by each of BFP mRNA and sgRNA.
- FIG. 14 (a) shows Cas9 activity when “GG” is removed from the 5 ′ end of sgRNA targeting EGFP.
- FIG. 14 (b) shows the results when miR-302 responsive Cas9 mRNA was modified to 4x miR-302 responsive Cas9 mRNA in order to reduce the leakage of Cas9 activity.
- FIG. 15 (a) shows a photograph of cells into which 4x miR-302 responsive Cas9 mRNA was introduced.
- FIG. 15 (b) shows a histogram of Cas9 Activity in cells into which 4x miR-302 responsive Cas9 mRNA was introduced.
- FIG. 15 (c) shows the quantification result of Cas9 Activity in cells into which 4x miR-302 responsive Cas9 mRNA was introduced.
- the present invention provides a method for controlling a nuclease in a cell-specific manner.
- a method for controlling a nuclease in a cell-specific manner is at least by using a nucleic acid sequence specifically recognized by miRNA in conjunction with a nucleic acid sequence encoding a nuclease.
- a nucleic acid sequence specifically recognized by miRNA and a nucleic acid sequence encoding nuclease can be included in one mRNA to control the nuclease in a cell-specific manner.
- mRNA containing a nucleic acid sequence specifically recognized by miRNA and mRNA containing a nucleic acid sequence encoding a nuclease can be separately contained, and a combination thereof can control a nuclease in a cell-specific manner.
- description will be divided into embodiments.
- the present invention is a method for controlling a nuclease in a cell-specific manner, comprising the step of introducing a miRNA-responsive mRNA encoding a nuclease into a cell.
- controlling a nuclease in a cell-specific manner means controlling the activity of a nuclease based on the expression state of a cellular endogenous miRNA.
- the cell in the present invention is not particularly limited and may be any cell.
- it may be a cell collected from a multicellular species or a cell obtained by culturing an isolated cell.
- the cells are particularly cells collected from mammals (eg, humans, mice, monkeys, pigs, rats, etc.) or cells obtained by culturing cells isolated from mammals or mammalian cell lines. is there.
- somatic cells include keratinized epithelial cells (eg, keratinized epidermal cells), mucosal epithelial cells (eg, epithelial cells of the tongue surface), exocrine glandular epithelial cells (eg, mammary cells), hormone-secreting cells (eg, , Adrenal medullary cells), metabolic and storage cells (eg, hepatocytes), luminal epithelial cells that make up the interface (eg, type I alveolar cells), luminal epithelial cells of the inner chain (eg, blood vessels Endothelial cells), ciliated cells with transport ability (eg, airway epithelial cells), cells for extracellular matrix secretion (eg, fibroblasts), contractile cells (eg, smooth muscle cells), blood and immune system Cells (eg, T lymphocytes), sensory cells (eg, sputum cells), autonomic nervous system neurons (eg, cholinergic neurons), sensory organs and peripheral neuron support cells (e
- undifferentiated progenitor cells include tissue stem cells (somatic stem cells) such as neural stem cells, hematopoietic stem cells, mesenchymal stem cells, and dental pulp stem cells.
- the cells in the present invention may be cells that have been subjected to artificial manipulation after collecting somatic cells, such as a cell group comprising iPS cells prepared from the somatic cells, or ES cells or iPS cells.
- the cell group which can contain the cell differentiated other than the desired cell may be sufficient.
- the cells in the present invention are particularly preferably in a viable state.
- the cell being in a viable state means a cell in a state where metabolic capacity is maintained.
- miRNA-responsive mRNA is introduced into a cell, it is a cell that can be used in subsequent applications without losing its natural characteristics, and in a viable state, particularly while maintaining its ability to divide.
- nuclease activity can be controlled in response to miRNA expressed in cells by using miRNA-responsive mRNA encoding nuclease, which will be described in detail below. Controlling the activity of nuclease refers to decreasing or increasing the expression level of nuclease, thereby decreasing or increasing the activity of nuclease.
- miRNA-responsive mRNA encoding a nuclease is also referred to as miRNA-responsive mRNA or miRNA switch, and refers to mRNA comprising the following nucleic acid sequences (i) and (ii): (i) specific by miRNA (Ii) a nucleic acid sequence corresponding to the coding region of a nuclease.
- the nucleic acid sequence specifically recognized by the (i) miRNA and the nucleic acid sequence corresponding to the coding region of (ii) nuclease are operably linked.
- Fig.1 (a) is a figure which shows typically an example of miRNA-responsive mRNA which can be used in the method which concerns on this invention.
- a sequence that responds to miRNA is incorporated into 5 ′ UTR, and a gene encoding nuclease is inserted into the protein coding region.
- the “miRNA” in the present invention is a short non-coding RNA (20-25 bases) existing in cells involved in regulation of gene expression through inhibition of translation from mRNA to protein and degradation of mRNA. .
- This miRNA is transcribed as a single-difference pri-miRNA that can take a hairpin loop structure containing miRNA and its complementary strand, and a part of it is cut into a pre-miRNA by an enzyme called Drosha in the nucleus. After being transported outside the nucleus, it is further cleaved by Dicer and functions.
- the miRNA of (i) is specifically expressed or specifically expressed in a specific cell for the purpose of controlling the nuclease activity, particularly suppressing or activating nuclease expression.
- MiRNA that has not been selected can be selected as appropriate.
- MiRNAs that are specifically expressed are 10% or more, 20% or more, 30% or more, 40% or more, 50% or more, 60% or more, 70% in certain cells compared to other cells.
- miRNAs that are highly expressed at a rate of 80% or more, 90% or more, or more are exemplified.
- Such miRNAs are listed in database information (for example, http://www.mirbase.org/ or http://www.microrna.org/) and / or described in the database. Can be selected appropriately from miRNAs described in literature information
- the nucleic acid sequence specifically recognized by miRNA is preferably, for example, a sequence that is completely complementary to the miRNA.
- it may have a mismatch (mismatch) with a completely complementary sequence.
- the mismatch from the sequence that is completely complementary to the miRNA may be any mismatch that can be normally recognized by the miRNA in the desired cell, and the mismatch of about 40 to 50% in the original function in the cell in vivo. There may be.
- mismatch is not particularly limited, but 1 base, 2 bases, 3 bases, 4 bases, 5 bases, 6 bases, 7 bases, 8 bases, 9 bases, or 10 bases or 1% of the total recognition sequence, 5% %, 10%, 20%, 30%, or 40% discrepancy.
- the miRNA target sequence on the mRNA ⁇ ⁇ ⁇ provided by the cell in particular, on the part other than the seed region, that is, on the ⁇ ⁇ 5 'side of the target sequence corresponding to about 3' side 16 base of the miRNA
- a region may contain a number of mismatches, and portions of the seed region may contain no mismatches, or may contain 1 base, 2 bases, or 3 bases mismatches.
- Such a sequence may be any base length including the number of bases to which RISC specifically binds, and the length is not particularly limited, but is preferably a sequence of 18 bases or more and less than 24 bases, more preferably 20 It is a sequence of not less than 22 bases.
- the nucleic acid sequence specifically recognized by miRNA introduces miRNA-responsive RNA having the sequence into the desired cell and other cells, and the expression of the corresponding marker gene is suppressed only in the desired cell. By confirming this, it can be appropriately determined and used.
- nucleic acid sequence corresponding to the coding region of nuclease used in the present invention is a gene that encodes a protein that functions as an enzyme that is translated in a cell and degrades a nucleic acid.
- nucleic acid sequences corresponding to the nuclease coding region include a gene encoding ClusteredClusterRegularly Interspaced Short Palindromic Repeats-AssociatedProteins 9 (Cas9), a gene encoding a transcriptional activator-like effector nuclease (TALEN), and a homing endonuclease And a gene encoding a zinc finger nuclease, but are not limited thereto.
- a mutant or derivative having the same function can be used.
- a gene encoding a Cas9 mutant or a gene encoding a Cas9 mutant fusion protein also corresponds to the coding region of the nuclease. Included in the nucleic acid sequence.
- these nucleases can be designed specifically for target nucleic acids that are degraded by nucleases.
- sgRNA is specifically designed as a target nucleic acid that is degraded by the nuclease. Details will be described later.
- a nucleic acid sequence specifically recognized by miRNA and a nucleic acid sequence corresponding to the coding region of nuclease is operably linked means that an open reading frame encoding nuclease (including an initiation codon).
- the miRNA-responsive mRNA preferably comprises a Cap structure (7-methyl guanosine 5 ′ phosphate), an open reading frame encoding a nuclease, and a poly A tail in the 5 ′ to 3 ′ direction from the 5 ′ end, It comprises at least one miRNA target sequence in the 5′UTR, in the 3′UTR, and / or in the open reading frame.
- the location of the miRNA target sequence in the mRNA may be 5'UTR, 3'UTR, or within the open reading frame (3 'to the start codon), and all of these include miRNA
- the target sequence may be provided.
- the number of miRNA target sequences may be one, two, three, four, five, six, seven, eight or more.
- the nucleic acid sequences (i) and (ii) are linked in this order in the 5 ′ to 3 ′ direction.
- the number of bases and the kind of base between the cap structure and the target sequence of miRNA may be arbitrary as long as they do not constitute a stem structure or a three-dimensional structure.
- the number of bases between the cap structure and the miRNA target sequence can be designed to be 0 to 50 bases, preferably 10 to 30 bases.
- the number of bases and the type of base between the miRNA target sequence and the start codon may be arbitrary as long as they do not constitute a stem structure or a three-dimensional structure, and the number of bases between the miRNA target sequence and the start codon is 0. It can be designed with an arrangement of -50 bases, preferably 10-30 bases.
- an AUG serving as an initiation codon does not exist in the miRNA target sequence in the miRNA-responsive mRNA.
- the target sequence of miRNA exists in 5'UTR and AUG is included in the target sequence, it is designed to be in-frame in relation to the marker gene linked to the 3 'side. It is preferable.
- AUG in the target sequence can be converted to GUG and used.
- the location of the target sequence in the 5 ′ UTR can be changed as appropriate.
- the number of bases between the cap structure and the AUG sequence in the target sequence is 0 to 60 bases, for example, 0 to 15 bases, 10 to 20 bases, 20 to 30 bases, 30 to 40 bases, 40 to 50 bases , Can be designed in an arrangement of 50 to 60 bases.
- miRNA-responsive mRNA to be introduced into a cell may be one type, or two, three, four, five, six, seven, or more than eight types. Good.
- miRNA-responsive mRNAs for example, mRNAs that have different miRNA target site sequences and respond to different miRNAs can be used.
- miRNA-responsive mRNA can be introduced in the form of mRNA as a method for introducing it into cells.
- RNA may be introduced into somatic cells by methods such as lipofection and microinjection, and RNA containing 5-methylcytidine and pseudoruidine (TriLink Biotechnologies) may be used to suppress degradation (Warren L, ( 2010) Cell Stem Cell. 7: 618-630).
- the positions of the modified bases can be all or part of the uridine and cytidine independently, and if they are part of the base, they can be random positions at an arbitrary ratio.
- miRNA-responsive mRNA can be introduced in the form of DNA such as a vector, and the same introduction method as described above can be used.
- multiple mRNAs should be added to the cell group.
- Co-introduction is preferred. This is because the ratio of two or more co-introduced mRNAs in cells is maintained in individual cells, and the activity ratio of proteins expressed from these mRNAs is constant in the cell population.
- the introduction amount at this time varies depending on the cell group to be introduced, the mRNA to be introduced, the introduction method and the kind of the introduction reagent, and those skilled in the art can appropriately select these to obtain a desired translation amount of nuclease.
- control mRNA refers to mRNA that does not have a miRNA target site. That is, the control mRNA is mRNA that is introduced into the cell and translated without being affected by the expression level of the endogenous miRNA. In a preferred embodiment, the control mRNA is introduced into a cell group together with the miRNA-responsive RNA, and can function as a control for confirming and identifying the cell into which the miRNA-responsive RNA is introduced. The amount of control mRNA introduced can also be appropriately selected by those skilled in the art to obtain a desired translation amount.
- the miRNA-responsive RNA introduced into the cell contains miRNA that specifically recognizes the target sequence in the cell, translation is suppressed and the expression level of the nuclease decreases, thereby reducing the nuclease activity. Therefore, the nuclease activity can be reduced specifically for cells expressing a specific miRNA. As a result, the cleavage activity of the target gene that is the target of the nuclease is reduced, and the influence on the function of the target gene is reduced. On the other hand, if miRNA that specifically recognizes the target sequence does not exist in the cell, miRNA-responsive mRNA is not suppressed and nuclease is expressed in the cell.
- disconnection of the target gene used as the target of a nuclease advances,
- the fate of the cell relevant to the function of the said gene can be controlled by impairing the function of the said gene. Therefore, in one example, cells in which miRNA that specifically recognizes the target sequence does not exist can lead to cell death.
- FIG. 1 (b) is a diagram schematically showing the action in cells when miRNA-responsive Cas9 mRNA encoding Cas9 protein and sgRNA are introduced into the cells.
- Target miRNA When there is no target miRNA that binds to miRNA-responsive Cas9 mRNA (Target miRNA (-)), Cas9 protein is translated from miRNA-responsive mRNA. Therefore, a guide strand (sgRNA) co-introduced with miRNA-responsive mRNA and Cas9 protein form a complex, and genome editing, specifically cleaving target DNA, is performed.
- the target miRNA is present (Target miRNA (+)
- the miRNA-responsive Cas9 mRNA binds to the 5'UTR of miRNA-responsive ⁇ Cas9 mRNA, resulting in translational suppression and degradation of mRNAmswitch, and miRNA-responsive-Cas9 mRNA Cas9 Protein translation is suppressed. Therefore, since no Cas9 / sgRNA complex is formed, genome editing, specifically, cleavage of the target DNA is not performed.
- miRNA-responsive RNA encoding Cas9 protein can be designed as described above.
- miRNA-responsive mRNA9 encoding Cas9 protein is also indicated as miRNA-responsive Cas9 mRNA.
- miRNA include miR-302a and miR-21.
- miR-302a is known as a miRNA endogenous to pluripotent stem cells such as iPS cells and ES cells. Therefore, miR-302a-responsive mRNA encoding Cas9 protein is introduced into a group of cells including pluripotent stem cells such as iPS cells and differentiated cells, thereby expressing Cas9 protein specifically in pluripotent stem cells.
- the target gene can be cleaved by causing the Cas9 protein to function in differentiated cells. That is, it is possible to control the activity of the Cas9 protein according to the degree of cell differentiation or the cell initialization (reprogramming) state.
- miR-21 is known to be specifically expressed in Hela cells. These sequences are shown in Table 1 below. Table 2 shows examples of miR target sequences specifically recognized by miR-302a and miR-21.
- miR-302a target sequence miR-302a-responsive Cas 9 mRNA that suppresses the expression of Cas9 protein in response to cell endogenous miR-302a
- miR-21 target sequence miR-21 Specific examples of miR-21-responsive Ca 9 mRNA that suppresses the expression of Cas9 protein in response to -21 include the sequences shown in Table 3 below, but are not limited thereto. In the sequences of Table 3, the underlined portion and the double underlined portion indicate the miRNA target sequence, AUG indicates the start codon, and the dotted line indicates 3′UTR.
- SgRNA can be designed to incorporate a base sequence of about 20, for example, about 18-22, in the vicinity of the 5 ′ end, which specifically recognizes the target gene of Cas9.
- the base sequence that specifically recognizes the Cas9 target gene is preferably a sequence that is completely complementary to the target gene.
- it may have a mismatch (mismatch) with a completely complementary sequence.
- the mismatch from the sequence that is completely complementary to the target gene may be the same as defined for miRNA and its target sequence.
- a sequence of about 1 to 5 bases may be present on the 5 'side of the base sequence that specifically recognizes the target gene of Cas9.
- the 3 ′ sequence of the base sequence that specifically recognizes the target gene is not limited to the specific sequence shown in Table 4 below, and is known to function in the CRISPR / Cas9sSystem.
- the sequence of sgRNA may be used, and examples are known in which Tetraloop, stem loop2, and 3 'ends are modified, but are not limited to specific ones as long as the target gene can be cleaved together with Cas9.
- the following sgRNA shown in Table 4 can be used, but the sgRNA that can be used in the method of the present invention is not limited to these.
- the underlined portion indicates a sequence complementary to the Cas9 target gene.
- SEQ ID NO: 8 is sgRNA targeting DMD (Duchenne muscular dystrophy) gene
- SEQ ID NO: 9 is sgRNA targeting Alu1 repeat sequence
- SEQ ID NOs: 10, 11 are sgRNA targeting EGFP gene. is there.
- target sequences that can be used to cause cell death in a cell-specific manner using Cas9 include the following Alu1 repeat sequences, as well as repeat sequences such as Telomere and (AC) n. It is not limited to.
- the step of introducing miRNA-responsive Cas9 mRNA and sgRNA designed according to the above into cells can be performed by the method described above.
- the introduction amount at that time can be determined as appropriate so that those skilled in the art can achieve the desired function of Cas9, and is not limited.
- a kit can be obtained by combining desired miRNA-responsive Cas9 mRNA and sgRNA specifically recognizing a Cas9 target gene sequence.
- the activity of Cas9 can be suppressed specifically for cells expressing a specific miRNA, and the activity of Cas9 can be maintained only in cells that do not express a specific miRNA.
- Cas9 activity can be suppressed specifically in undifferentiated cells expressing miR-302a, and Cas9 activity can be maintained in differentiated cells that hardly express miR-302a.
- genome editing can be performed specifically for cells that express specific miRNAs. .
- the present invention provides a cell-specific method for controlling a nuclease, comprising a) a trigger protein-responsive mRNA encoding the nuclease, and b) an miRNA-responsive mRNA encoding the trigger protein. And the step of introducing into a cell.
- controlling a nuclease in a cell-specific manner means controlling the activity of a nuclease based on the expression state of a cellular endogenous miRNA.
- miRNA expressed in a cell by using a) a trigger protein-responsive mRNA encoding a nuclease and b) an miRNA-responsive mRNA encoding the trigger protein, which will be described in detail below.
- controlling the activity of nuclease means increasing the expression level of nuclease and thereby increasing the activity of nuclease. .
- the trigger protein-responsive mRNA encoding a nuclease means an mRNA comprising the following nucleic acid sequences (ia) and (iia): (ia) a nucleic acid sequence that specifically binds to the trigger protein; And (iia) a nucleic acid sequence corresponding to the coding region of the nuclease.
- the nucleic acid sequence that specifically binds to the (ia) trigger protein and the nucleic acid sequence that corresponds to the coding region of (iia) nuclease are operably linked.
- FIG. 8A is a diagram schematically showing an example of a trigger protein-responsive mRNA that can be used in the method according to the present invention.
- a sequence that responds to the trigger protein is incorporated into 5 ′ UTR, and a gene encoding a nuclease is inserted into the protein coding region.
- the nucleic acid sequence that specifically binds to the trigger protein of (ia) is an RNA containing a sequence that forms an RNA-protein binding motif.
- the RNA containing the sequence forming the RNA-protein binding motif is the RNA side included in the binding motif between RNA and protein in a natural or known RNA-protein complex, or in vitro evolution method (in vitro selection method)
- a sequence that forms a natural RNA-protein binding motif is usually composed of about 5 to 30 bases, and is specific to a protein having a specific amino acid sequence noncovalently, that is, by hydrogen bonding. It is known to form bonds.
- the sequences that form such natural RNA-protein binding motifs are listed in Tables 5 and 6 below and the database available on the website: http: // gibk26. bse. kyutech. ac. jp / jouhou / image / dna-protein / RNA / RNA. From html, a motif that causes a desired structural change can be appropriately selected and obtained.
- RNA-protein binding motif preferably used in this embodiment can be estimated from the three-dimensional structure of a homologous protein that has already undergone an X-ray crystal structure analysis or NMR structure analysis or a structural analysis. Motif. Furthermore, the protein is preferably a motif that specifically recognizes the secondary structure and base sequence of RNA.
- the RNA containing a sequence that forms an artificial RNA-protein binding motif is the RNA side in the binding motif between RNA and protein in an artificially designed RNA-protein complex.
- the base sequence of such RNA is usually composed of about 10 to 80 bases, and forms a specific bond with a specific amino acid sequence of a specific protein non-covalently, that is, by hydrogen bonding.
- An RNA aptamer that specifically binds to a specific protein is exemplified as the RNA containing a sequence that forms such an artificial RNA-protein binding motif.
- An RNA aptamer that specifically binds to a desired target protein can be obtained by, for example, an evolutionary engineering method known as in vitro selection method or SELEX method.
- the trigger protein at this time is a protein to which the RNA aptamer binds.
- the RNA sequences listed in Table 7 below are known, and these can also be used as sequences forming the RNA-protein binding motif of the present invention.
- the sequence forming the RNA-protein binding motif preferably has a dissociation constant Kd with the corresponding trigger protein of about 0.1 nM to about 1 ⁇ M.
- variants of such sequences are also encompassed by the sequences according to the present invention.
- the term “variant” as used herein refers to a variant having a dissociation constant Kd of 10%, 20%, 30%, 40% or 50% or more higher than a protein that specifically binds to a sequence that forms an RNA-protein binding motif. Or 10%, 20%, 30%, 40% or 50% or less of the mutants. Such mutants can be appropriately selected and used as long as an RNA-protein complex can be formed.
- the base sequence of such a variant can hybridize under stringent conditions with a nucleic acid (complementary strand) having a sequence complementary to the sequence (positive strand) forming the RNA-protein binding motif.
- a base sequence of a degree may be used.
- the stringent conditions here are the melting temperature (Tm) of the nucleic acid to be bound as taught by Berger and Kimmel (1987, GuideGto Molecular Cloning Techniques Methods in Enzymology, Vol.152, Academic Press, San Diego CA). Can be determined based on For example, as washing conditions after hybridization, the conditions of about “1 ⁇ SSC, 0.1% SDS, 37 ° C.” can be mentioned.
- the complementary strand is preferably one that maintains a hybridized state with the target positive strand even when washed under such conditions.
- washing is performed under more severe hybridization conditions such as “0.5 ⁇ SSC, 0.1% SDS, 42 ° C.”, more strictly “0.1 ⁇ SSC, 0.1% SDS, 65 ° C.”
- the conditions for maintaining the hybridized state between the positive strand and the complementary strand can be mentioned.
- it comprises a base sequence having at least 90%, preferably at least 95%, 96%, 97%, 98% or 99% sequence identity with the RNA sequence contained in the RNA-protein binding motif described above. .
- Such mutants can maintain a constant bond with a protein that specifically binds to a sequence that forms an RNA-protein binding motif and contribute to the formation of an RNA-protein complex.
- RNA-protein binding motif examples include L7Ae (Moore T et al., Structure Vol. 12, pp. 807-818 (2004)) shown in Table 8 below.
- binding sequences include boxC motif (5′-GGCGUGAUGAGGC-3 ′) (SEQ ID NO: 40), kink-loop (SEQ ID NO: 41), and kink-loop 2 (SEQ ID NO: 42).
- MS2 stem loop motif 22: Keryer-Bibens C, Barreau C, Osborne HB (2008) Tethering of proteins to RNAs by bacteriophage proteins. Biol Cell 100: 125-138
- Fr15 24: Batey RT, Williamson JR (1996) Interaction of the Bacillus stearothermophilus ribosomal protein S15 with 16 S rRNA: I. Defining the minimal RNA . J Mol Biol 261: 536-549).
- a further specific example is an enzyme that performs aminoacylation, which is known to have Threonyl-tRNA synthetase (Cell (Cambridge, Mass.) V97, which binds to its own mRNA and has feedback inhibition that inhibits translation.
- Cell Cell (Cambridge, Mass.) V97, which binds to its own mRNA and has feedback inhibition that inhibits translation.
- pp.371-381 (1999) which is a sequence to which 5′-GGCGGUAUGUGAUCUUGUCGUUGGGUCACCACUGCGCC-3 ′ (SEQ ID NO: 43), and variants thereof are present.
- R9-2 5′-GGGGUGUCGAGCUAGGGAAGAAAGCCGGGGGCUGCAGAUAAUGUUAUAGC-3 ′ (SEQ ID NO: 44), which is a base sequence forming an RNA-protein binding motif derived from Bcl-2 family CED-9, which is a cancer cell-specific endogenous protein , And variants thereof, nucleotide sequences derived from aptamers of RNA sequences that bind to NF-kappaB, and variants thereof.
- nucleic acid sequence corresponding to the coding region of nuclease is a gene that encodes a protein that functions as an enzyme that is translated in a cell and degrades the nucleic acid, and has been described in the first embodiment. The description is omitted here.
- the nucleic acid sequence that specifically binds to the trigger protein and the nucleic acid sequence corresponding to the coding region of the nuclease are operably linked to each other, that is, an open reading frame that encodes a nuclease (including an initiation codon).
- an open reading frame that encodes a nuclease including an initiation codon.
- the trigger protein responsive mRNA encoding the nuclease is preferably in the 5 ′ to 3 ′ orientation from the 5 ′ end, in the Cap structure (7 methylguanosine 5 ′ phosphate), the open reading frame encoding the nuclease, and A nucleic acid sequence comprising a poly A tail and specifically binding to at least one trigger protein within the 5′UTR, within the 3′UTR, and / or within the open reading frame.
- the position of the nucleic acid sequence that specifically binds to the trigger protein in the mRNA may be 5'UTR, 3'UTR, or within the open reading frame (3 'side of the start codon) All of these may have a nucleic acid sequence that specifically binds to the trigger protein.
- the number of nucleic acid sequences that specifically bind to the trigger protein may be one, two, three, four, five, six, seven, eight or more.
- the nucleic acid sequences (ia) and (iia) are linked in this order in the 5 ′ to 3 ′ direction.
- the number of bases and the type of base between the cap structure and the nucleic acid sequence that specifically binds to the trigger protein may be arbitrary as long as they do not constitute a stem structure or a three-dimensional structure.
- the number of bases between the cap structure and the nucleic acid sequence that specifically binds to the trigger protein can be designed to be 0 to 50 bases, preferably 10 to 30 bases.
- the number of bases and the type of base between the nucleic acid sequence that specifically binds to the trigger protein and the start codon may be arbitrary as long as they do not constitute a stem structure or a three-dimensional structure, and specifically bind to the trigger protein.
- the number of bases between the nucleic acid sequence to be started and the start codon can be designed to be 0 to 50 bases, preferably 10 to 30 bases.
- an AUG serving as a start codon does not exist in the nucleic acid sequence that specifically binds to the trigger protein in the trigger protein-responsive mRNA.
- the nucleic acid sequence that specifically binds to the trigger protein is present in the 5 ′ UTR and AUG is included in the nucleic acid sequence, it is in-frame in relation to the marker gene linked to the 3 ′ side. It is preferable to be designed so that Alternatively, when AUG is included in the nucleic acid sequence that specifically binds to the trigger protein, it is also possible to convert the AUG in the nucleic acid sequence that specifically binds to the trigger protein into GUG for use.
- the location of the nucleic acid sequence that specifically binds to the trigger protein in the 5 ′ UTR can be changed as appropriate.
- the number of bases between the cap structure and the AUG sequence in the nucleic acid sequence that specifically binds to the trigger protein is 0 to 60 bases, such as 0 to 15 bases, 10 to 20 bases, 20 to 30 bases, 30 It can be designed in an arrangement of ⁇ 40 bases, 40 ⁇ 50 bases, 50 ⁇ 60 bases.
- miRNA-responsive mRNA encoding a trigger protein is also referred to as miRNA-responsive mRNA or miRNA switch, and means mRNA comprising the following nucleic acid sequences (ib) and (iib): (ib) miRNA And (iib) a nucleic acid sequence corresponding to the coding region of the trigger protein.
- the nucleic acid sequence specifically recognized by the (ib) miRNA and the nucleic acid sequence corresponding to the coding region of the trigger protein (iib) are operably linked.
- MRNA2 of Fig.8 (a) is a figure which shows typically an example of miRNA-responsive mRNA which can be used in the method which concerns on this embodiment.
- miRNA-responsive mRNA a sequence that responds to miRNA is incorporated into 5 ′ UTR, and a gene encoding a trigger protein is inserted into the protein coding region.
- RNA target sequence (B) The definition of “miRNA” in the miRNA-responsive mRNA encoding the trigger protein and the definition of the nucleic acid sequence specifically recognized by the miRNA are the same as those described in the first embodiment, and the description is omitted. To do. Also in this embodiment, (ib) a nucleic acid sequence specifically recognized by miRNA is also referred to as a miRNA target sequence.
- the nucleic acid sequence corresponding to the coding region of the trigger protein is the coding sequence of the trigger protein.
- the trigger protein is determined by the relationship between (ia) the nucleic acid sequence that specifically binds to the trigger protein, and is designed by selecting a combination that specifically binds the RNA sequence of (ia) and (iib) the trigger protein. can do.
- the nucleic acid sequence that specifically binds to (i) trigger protein is boxC motif (SEQ ID NO: 40), kink-loop (SEQ ID NO: 41), kink-loop2 (SEQ ID NO: 42)
- the trigger protein is L7Ae (Moore T et al., Structure Vol. 12, pp. 807-818 (2004)).
- the corresponding trigger protein can be used for the nucleic acid sequence that specifically binds to the (i) sputum trigger protein exemplified above.
- a nucleic acid sequence specifically recognized by miRNA (miRNA target sequence) and (iib) a nucleic acid sequence corresponding to the coding region of the trigger protein is functionally linked to the trigger protein.
- miRNA target sequence miRNA target sequence
- a nucleic acid sequence corresponding to the coding region of the trigger protein is functionally linked to the trigger protein.
- the miRNA-responsive mRNA encoding the trigger protein is preferably a Cap structure (7 methylguanosine 5 ′ phosphate), an open reading frame encoding the trigger protein, in the 5 ′ to 3 ′ direction from the 5 ′ end, and It comprises a poly A tail and comprises at least one miRNA target sequence in the 5′UTR, in the 3′UTR, and / or in the open reading frame.
- the location of the miRNA target sequence in the mRNA may be 5'UTR, 3'UTR, or within the open reading frame (3 'to the start codon), and all of these include miRNA
- the target sequence may be provided.
- the number of miRNA target sequences may be one, two, three, four, five, six, seven, eight or more.
- the nucleic acid sequences (ib) and (iib) are linked in this order in the 5 ′ to 3 ′ direction.
- the design of the nucleic acid sequence of (ib) and (iib), the number of bases between the Cap structure, the number and arrangement of the sequence of (ib), and the same sequence as the start codon is the same as in the first embodiment or a ) It may be similar to the design in the trigger protein responsive mRNA encoding nuclease.
- the trigger protein-responsive mRNA encoding a nuclease and b) the miRNA-responsive mRNA encoding the trigger protein can be designed as described above, they can be prepared by a normal genetic engineering technique. Two of these two mRNAs function as a set. Two or more sets of mRNAs each having different miRNA target sequences, trigger proteins, and nucleases can be designed for cell introduction.
- the mRNA set of a) and b) is preferably co-introduced when introduced into cells. At this time, as in the first embodiment, a control mRNA can also be co-introduced.
- nuclease control by such a set of a) and b) mRNA will be described.
- the miRNA-responsive mRNA introduced into the cell contains miRNA that specifically recognizes the target sequence in the cell, translation is suppressed and the expression level of the trigger protein decreases.
- the trigger protein When the trigger protein is present, it binds to the trigger protein-responsive mRNA and is inhibited from translation.
- the expression level of the trigger protein decreases, the translation amount of the trigger protein-responsive mRNA increases and promotes nuclease translation.
- Nuclease activity can be improved.
- the cleavage activity of the target gene that is the target of the nuclease is also improved, and the target gene is cleaved to cause loss of its function.
- the miRNA-responsive mRNA is not suppressed in translation and the trigger protein is expressed in the cell. Then, the trigger protein suppresses the translation of the trigger protein-responsive mRNA, and the expression level of the nuclease decreases. As a result, the probability that the nuclease will cleave the target gene is reduced and the function of the target gene is maintained.
- FIG. 11 (a) is a diagram schematically showing the action when L7Ae-responsive mRNA encoding Cas9 protein and miRNA-responsive mRNA encoding L7Ae are introduced into cells.
- PCR amplification was performed in the following cycle (after 94 ° C for 2 min, 13 cycles of 98 ° C for 10 sec and 68 ° C for 10 sec, stored at 4 ° C).
- the gene encoding Cas9 protein can be obtained at the following cycle (94 ° C) using the corresponding primer and KOD-Plus-Neo (KOD-401, TOYOBO) from the template plasmid (pHL-EF1a-SphcCas9-iC-A). After 2 min, 98 cycles of 98 ° C for 10 sec and 68 ° C for 140 sec were performed and stored at 4 ° C) to perform PCR amplification.
- Cas9mRNA The template for Cas9mRNA responding to miRNA under the conditions of Control (Cas9mRNA) is 94 ° C (2mmin, followed by 20 cycles of 98 ° C (10msec) and 68 ° C (140msec) at 4 ° C). (After 94 ° C for 2 min, 98 ° C for 10 sec, 60 ° C for 30 sec, and 68 ° C for 140 sec for 20 cycles and stored at 4 ° C).
- the template of sgRNA is 20 cycles of 2 primers (98 ° C 30 sec, 98 ° C 10 sec, 57 ° C 30 sec, 68 ° C, 6 ⁇ sec), reacted at 72 ° C 10 min, then 4 ° (Stored in C).
- PCR products were purified using MiniElute PCR purification Kit (QIAGEN). However, those using a plasmid in the PCR reaction were treated with the restriction enzyme Dpn I before purification. Corresponding primers and oligonucleotide sequences are shown in Tables 9A and B.
- Cas9 mRNA and sgRNA Cas9 mRNA was produced by MegaScript kit (Ambion). At this time, modified bases pseudoridine-5′-triphosphate and 5-methylcytidine-5′-triphosphate (TriLink Bio Technologies) were added in place of UTP and CTP, respectively, to suppress the immune reaction. GTP was diluted 5-fold with Anti Reverse Cap Analog (TriLink Bio Technologies). The sgRNA was prepared by MEGAshortscript kit (Ambion) using natural bases (ATP, GTP, CTP, UTP). The reaction mixture was incubated at 37 ° C for 6 hours, TURBO DNase (Amibion) was added, and then incubated at 37 ° C for an additional 30 minutes.
- modified bases pseudoridine-5′-triphosphate and 5-methylcytidine-5′-triphosphate TriLink Bio Technologies
- the obtained mRNA was purified by FavorPrep Blood / Cultured Cells total RNA extraction column (Favorgen Biotech) and incubated at 37 ° C. for 30 minutes using Antarctic phosphatase (New England Biolabs). Then, it further refine
- the coding region of Cas9 mRNA and the sequences of 5′UTR and 3′UTR are shown in Tables 10A and 10B below.
- iPS_GFP (AAVS1-CAG :: GFP iPS cells) were provided by Woltjen Lab (CiRA, Kyoto University, Japan). iPS_GFP was cultured on a laminin-coated ⁇ laminin-511 E8 (iMatrix-511, nippi) ⁇ plate using StemFit (Ajinomoto) medium. HeLa_GFP was cultured in a medium in which FMEM (JBS, final concentration 10%) and Hygromycin B (50 mg / mL) were added to DMEM High Glucose (nakarai tesque). Normal HeLa cells were cultured in a medium in which FMEM (JBS, final concentration 10%) was added to DMEM High Glucose (nakarai tesque). All cells were cultured under conditions of 37 ° C and 5% CO 2 .
- FMEM JBS, final concentration 10%
- Hygromycin B 50 mg / mL
- Differentiation induction Differentiation induction from iPS_GFP to mDA_GFP
- iPS_GFP cells were replated (5x10 6 cells / well) in 6-well plates coated with laminin using differentiation induction medium, and Morizana et al., Neural Development: Methods and Protocols, Methods in Molecular Biology, vol. 1018, DOI 10.1007 / 978-1-62703-444-9_2 was cultured according to a modified protocol. Thereafter, the medium was changed every day. It was used for each experiment after 14 days.
- the composition of the differentiation induction medium is shown in Table 11 below.
- Transfected normal HeLa and HeLa_GFP were seeded in 24-well plates.
- iPS_GFP and mDA_GFP were seeded in a laminin-coated 24-well plate (cell number: 5 ⁇ 10 4 cells / well).
- Transfection was performed using a Stemfect RNA transfection kit (Stemgent) according to the protocol (refer to each experimental section for the amount of transgene). The medium was changed 4 hours after transfection (except mDA_GFP).
- Cell killing was analyzed 48 hours after transfection, T7E1 assay, EGFP activity assay, and co-culture were analyzed 72 hours after transfection. Before entering each analysis, the cells were photographed with an IX81 microscope (Olympus) (FIG. 2).
- T7E1 assay 100 ng Cas9 mRNA and 300 ng (iPS_GFP) or 100 ng (mDA_GFP) sgRNA were used.
- the transfected cells were washed with PBS and then treated with 200 ⁇ L Accumax (Funakoshi) for 10 minutes at 37 ° C. and 5% CO 2 .
- Cells were collected in a 1.5 mL tube and centrifuged (room temperature, 1000 rpm, 5 minutes). After discarding the supernatant, the precipitated cells were washed with PBS and centrifuged under the same conditions as before.
- Proteinase K in Lysis buffer (1 M Tris-HCl (pH.7.6) [final 0.05 M], 0.5 M EDTA [final 0.02 M], 5 M NaCl [0.1 M], 10% SDS [final 1%], D2W) 500 mL of (x100; final 1x) was added and treated at 55 degrees for 3 hours or more. Thereafter, genomic DNA was extracted using PCI. The target sequence was amplified from the extracted genomic DNA by Nested PCR.
- Second PCR was performed under the conditions of (94 ° C 2 min, 98 ° C 10 sec, 60 ° C 30 sec, 68 ° C 15 sec, 35 cycles, 72 ° C 3 min reaction, and stored at 4 ° C) went.
- the PCR product was purified using the MiniElute PCR purification Kit (QIAGEN).
- Indels 100 ⁇ (1-sqrt (1-(b + c) / (a + b + c))) a: Band intensity of PCR product not cleaved with restriction enzyme b, c: Band intensity of PCR product cleaved with restriction enzyme
- Cas9 activity assay 100 ng Cas9 mRNA and 300 ng (iPS_GFP, HeLa_GFP) or 100 ng (mDA_GFP) sgRNA and 5 pmol miRNA inhibitor (mirVana) or negative control (miRNA inhibitor and negative control are optional) were used. Cells were washed with PBS. Thereafter, HeLa_GFP was treated with 100 ⁇ L of 0.25% trypsin-EDTA at 37 ° C. and 5% CO 2 for 5 minutes, and then 100 ⁇ L of medium was added. iPS_GFP and mDA_GFP were treated with 200 ⁇ L Accumax (Funakoshi) at 37 ° C. and 5% CO 2 for 10 minutes.
- iPS_GFP and mDA_GFP were treated with 200 ⁇ L Accumax (Funakoshi) at 37 ° C. and 5% CO 2 for 10 minutes.
- CYTOX Red dead-cell stain ThermoFisher 15 minutes, protected from light and left at room temperature. It measured by Aria-II (BD), Accuri (BD), and LSR (BD). EGFP negative cells (%) were defined as Cas9 activity (%).
- Cell killing system 10 ng Cas9 mRNA and 300 ng sgRNA were used. Prior to washing with PBS, the medium was collected in a 1.5 mL tube. After washing with PBS, normal HeLa was treated with 100 ⁇ L of 0.25% trypsin-EDTA at 37 ° C. and 5% CO 2 for 5 minutes. Thereafter, 100 ⁇ L of medium was added, and the cells were collected into the 1.5 mL tube and centrifuged (room temperature, 1000 rpm, 5 minutes). After discarding the supernatant, the precipitated cells were washed with PBS and centrifuged under the same conditions as before.
- Co-culture 50 ng Cas9 mRNA and 150 ng sgRNA were used.
- the cells were collected in a 1.5 mL tube by the same method as Cas9 activity assay in which iPS_GFP cells and HeLa_GFP cells were seeded at a ratio of 3 to 2, and then centrifuged (room temperature, 1000 rpm, 5 minutes). After removing the supernatant, Alexa Fluor® 647 Mouse anti-Human TRA-1-60 Antigen was stained according to the protocol (using twice the amount of antibody as the protocol) and measured by LSR.
- FIG. 2 (a) is a fluorescence microscope image of HeLa cells into which miRNA-responsive mRNA has been introduced.
- the fluorescence of GFP is weaker than the untreated or negative control for the target sequence.
- a target targeting the DMD gene was used.
- FIG. 2A one of the three experiments was shown as a representative.
- FIG. 2 (c) is a graph in which the GFP negative population in FIG. 2 (b) is defined as Cas9 activity.
- the following Cas9 activity shall be calculated by this method unless otherwise specified. From the above, when various miRNA-responsive mRNAs and control mRNAs were introduced into HeLa cells, it was found that only Cas9 mRNA that responds to miR-21 had low Cas9 activity, and Cas9 mRNA that responded to miR-21 was miR-21. The possibility of being controlled by was shown.
- FIG. 4 (a) shows the results evaluated by Cas9 activity assay
- FIG. 4 (b) shows the gel images evaluated by T7E1 assay. The gel image is shown representatively from one of the three experiments.
- FIG. 4 (c) shows the result of calculating Indels (defined as Cas9 activity) from T7E1 assay. The calculation formula is as described in detail in the item of T7E1 ⁇ assay.
- FIG. 5 is a diagram schematically showing a scheme for causing cell death using the miRNA-responsive CRISPR / Cas9 system of the present invention.
- FIG. 5 (a) schematically shows that Cas9 fragments the genome by making the sgRNA sequence a repeat sequence on the genome. When the genome is fragmented, cells cause cell death.
- miR-21-responsive Cas9 mRNA was used and Alu1 was targeted and cell death control was verified. The experiment verified whether cell death could be controlled by using Cas9 mRNA in response to miR-21 in HeLa cells according to the method described in Cell Killing Sysetem. The results are shown in FIG.
- mi-21-responsive Cas9 mRNA is designed so that the translation of Cas9 ⁇ ⁇ mRNA is suppressed and Cas9 activity is reduced in cells where miR-21 is specifically expressed.
- FIG. 5 (b) shows that cell death is induced in Control Cas9 mRNACa (Cas9 activity is high).
- Cas9 mRNA responding to miR-21 (Cas9 activity is low) had the same dead cell rate as Untreated. This result shows that cell death was not induced by miR-21-responsive Cas9 mRNA.
- FIG. 6 shows the results of an experiment in which miR-302a-responsive Cas9 mRNA was introduced into mDA cells.
- miRNA responsive mRNA control, 302a-responsive, 4x miR-302a-responsive
- sgRNA one targeting GFP or one targeting DMD as negative control
- 4x RmiR-302a-responsive Cas9 5mRNA is a 5'UTR with 4 miR-302a-5p target sequences inserted.
- mDA cells midbrain dopaminergic neurons
- FIG. 6B shows a representative gel image
- the miRNA-responsive CRISPR / Cas9 system regulates Cas9 activity in response to the state of cells, that is, miRNA.
- Differentiating iPS cells reduces the activity of endogenous miR-302a-5p, so the activity of Cas9 mRNA in response to miR-302a is low in iPS but high in mDA (Control Cas9 activity similar to mRNA) is predicted.
- Relative Cas9 activity was calculated, the activity recovered with Cas9 mRNA responding to miR-302a, and the activity was close to that of Control Cas9 mRNA.
- FIG. 7 shows the result of co-culture.
- iPS_GFP and HeLa_GFP were seeded on a 24 well plate at 5x10 5 cells / well 17-24 hours before transfection.
- 50 ng of mRNA and 150 ng of sgRNA were transfected using a stemfect transfection reagent according to the protocol.
- As the mRNA control Cas9 mRNA and 4x miR-302a-responsive Cas9 mRNA were used.
- As sgRNA one targeting the GFP gene and one targeting the DMD gene as negative control were used.
- FIG.7 (b) is a graph which shows Cas9 activity computed by the following formula
- the results in FIG. 7 suggested the possibility of genome editing only for target cell populations from different cell populations.
- this experiment by utilizing the high expression (activity) of miR-302a in HeLa cells and iPS cells, while preventing iGFP cells from causing GFP knockout, In cells, we succeeded in causing GFP knockout.
- Transfected normal HeLa and HeLa_GFP were seeded in 24-well plates.
- iPS_GFP and mDA_GFP were seeded in a laminin-coated 24-well plate (cell number: 5 ⁇ 10 4 cells / well).
- Transfection was performed using a Stemfect RNA transfection kit (Stemgent) according to the protocol (refer to each experimental section for the amount of transgene). The medium was changed 4 hours after transfection (except mDA_GFP). Evaluation of gene expression level, Evaluation of Cas9 protein expression level, Evaluation of transfection efficiency is 24 hours after transfection, Cell killing is 48 hours after transfection, T7E1 assay, EGFP activity assay, co-culture, On-system are transfected Analysis was performed after 72 hours. Before entering each analysis, the cells were photographed with an IX81 microscope (Olympus).
- On-system MRNA was designed as outlined in FIG. 11A.
- a target site of miRNA a sequence complementary to miR-21 was used.
- L7Ae protein was used, and as a sequence that specifically binds to L7Ae protein, kinkturn motif was used, and mRNA was prepared in the same manner as the previous method.
- the sequence of the prepared mRNA (Kt-Cas9 mRNA) is as shown in Tables 12A and B below, in which the L7Ae binding motif is double-underlined and the Cas9 coding region is italicized.
- sequences of mRNA are as shown in Table 13 below.
- the target sequence of miR-21-5p is double-underlined, L7Ae or Tag
- the BFP code region is shown in italics.
- HeLa_GFP was seeded at 5x10 5 cells / well in a 24-well plate 17-24 hours before transfection.
- OFF condition Control L7Ae mRNA 15 ng + kt-Cas9 mRNA (having kt motif in 5'UTR) 5 ng + sgRNA 150 ng
- ON condition miR-21-responsive L7Ae mRNA 15 ng + kt-Cas9 mRNA 5 ng + sgRNA 150 ng
- Transfection was performed using stemfect transfection reagent according to the protocol. Three days after transfection, measurement was performed by Accuri.
- RNA expression level Three types of cells were used: HeLa-EGFP, iPS-EGFP, and mDA-EGFP.
- TaqMan registered trademark
- Measurements were performed using MicroRNA Cells-to-C T TM Kit (Ambion). The cell lysate was subjected to reverse transcription using has-miR-21-5p (Assay ID: 000397), 302a-5p (Assay ID: 002381) and RNU6B (Assay ID: 001093) Taqman probes (Applied Biosystems). I did it.
- qPCR was performed using Taqman probe and using StepOne Plus Real-Time PCR System (Applied Biosystems). Target miRNA was normalized with RNU6B. Furthermore, mDA-EGFP was normalized so as to be 1.
- Cas9 protein expression level 100 ng Cas9 mRNA and 300 ng sgRNA were used. 24 hours after transfection, PBS was washed, and the cells were lysed and collected with 50 ⁇ L of M-PER cocktail (mixed with M-PER Mammalian Protein Extraction Reagent [ThermoFisher], protease inhibitor, PMSF). After shaking for 5 minutes, the solution was collected in a 1.5 mL tube. After centrifugation (12400 rpm, 4 ° C, 5 min), the supernatant was recovered in a new 1.5 mL tube, and the protein concentration was measured by the BCA method.
- M-PER cocktail mixed with M-PER Mammalian Protein Extraction Reagent [ThermoFisher], protease inhibitor, PMSF.
- GAPDH was used as a loading control.
- Primary antibody Cas9 antibody (50-fold dilution, Active Motif), GAPDH antibody (100-fold dilution, Santa Cruz).
- Secondary antibody anti-mouse, anti-rabbit (ProteinSimple)
- the second PCR product (iPS-EGFP) obtained in the T7E1 assay was inserted into the pUC19 vector, and the sequence was read with a sequencer.
- the primer used in the second PCR was phosphorylated with T4PNK, and the second PCR product was subjected to PCR again. Thereafter, it was inserted into the pUC19 vector, and the sequence was determined using M13 Fwd New primer, T7E1 Fwd primer, and T7E1 Rev primer.
- the sequencer used was an Applies Biosystems 3500xL Genetic analyzer.
- HeLa-EGFP and iPS-EGFP were used. 100 ng BFP mRNA and 300 ng Cy5 labeled sgRNA were used. 24 hours after transfection, cells were washed 3 times with PBS. Thereafter, the cells were collected in a 1.5 mL tube (see the Cas9 activity assay for how to remove the cells) and measured by LSR.
- Cas9 activity assay (modified sgRNA with iPS-EGFP: 1, 2) It carried out like Cas9 activity assay.
- FIGS. ON-system is a system in which Cas9 activity is increased by endogenous miRNA.
- FIG. 8A is a conceptual diagram showing an outline of ON-system. MRNA1 expressing Cas9 (sequence binding to L7Ae, an RNA-binding protein derived from archaea: K-turn is encoded in 5'UTR) is mRNA2 expressing L7Ae (expression is controlled by miRNA) The translation is controlled by.
- FIG. 8 (b) shows that Cas9 activity was increased by endogenous miR-21 in HeLa cells. That is, Cas9 activity was increased by the target miRNA (here miR-21).
- FIG. 9A shows the results of measuring the expression level of miR-21
- FIG. 9B shows the results of measuring the expression level of miR-302a with HeLa-EGFP, iPS-EGFP, and mDA-EGFP.
- miR-21 iPS-EGFP cells show high expression of miR-302a.
- Control Cas9 mRNA and miR-21-responsive Cas9 mRNA and miR-302a-responsive Cas9 mRNA were introduced into HeLa cells (sgRNA was also introduced), and the effect on Cas9 protein expression was examined by Simple Western (Wes) did.
- the results are shown in FIG.
- Cas9 protein was detected in Control and miR-302a-responsive Cas9 mRNA, but not in miR-21-responsive Cas9 mRNA.
- miRNA-21-responsive Cas9 mRNA in HeLa cells is inhibited from miR-21 translation, indicating that Cas9 protein expression is almost absent.
- GAPDH was used as a loading control.
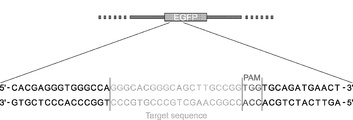
- FIG. 11A is a scheme showing a sequence around the target sequence.
- FIG. 11 (b) shows that there is a mutation in the target region of the EGFP gene. That is, the decrease in the fluorescence intensity of EGFP indicates that the EGFP gene is knocked out by the CRISPR / Cas9 system. “ ⁇ ” in the figure indicates base deletion and “+” indicates base insertion. The number on the rightmost side indicates the number of colonies / total number of colonies in the obtained sequence.
- the green letters are “target sequence”, the blue letters (TGG and ACC in FIG.
- FIG. 12 (a) and FIG. 12 (b) show the results of verifying the expression variation of genes whose functions are known to be controlled by miR-21 and miR-302a by qPCR. Comparing the expression levels of Control Cas9 mRNA and miRNA-responsive Cas9 mRNA into cells, the difference in the expression levels was not seen. That is, it is shown that the miRNA-responsive CRISPR / Cas9 system hardly affects the functions of endogenous miR-21 and miR-302a.
- FIG. 13 (a) shows the transfection efficiency of HeLa-EGFP and FIG. 13 (b) shows the transfection efficiency of iPS-EGFP. Transfection efficiency was measured for each of BFP mRNA and sgRNA (labelled with Cy5). The transfection efficiency was about 90% or more.
- FIG. 14 (a) shows the Cas9 activity when “GG” is removed from the 5 ′ end of sgRNA targeting EGFP.
- the Cas9 activity which was about 30%, has increased to nearly 70%, which indicates that the Cas9 activity is improved by modifying the guide strand.
- Cas9 activity is close to 30% even in the negative control of the inhibitor.
- FIG. 14 (b) modifies miR-302 responsive Cas9 mRNA to reduce the leak of Cas9 activity seen in FIG. 14 (a), resulting in 4x miR-302 responsive Cas9 mRNA. It was found that 50 ng of Cas9 mRNA and 300 ng of sgRNA maintained high activity while further reducing leakage.
- Cas9 activity assay (modified sgRNA with iPS-EGFP: 2) 100 ng Cas9 mRNA and 300 ng sgRNA were used.
- EGFP activity assay iPS-EGFP: 1
- miR-302 responsive Cas9 mRNA was modified and 4x miR-302 responsive Cas9 mRNA ( This is the result when the sequence that responds to miR-302 is inserted in 5'UTR in tandem.
- FIG. 15A is a photograph of a cell
- FIG. 15B is a histogram at that time
- FIG. 15C is a quantitative result. It was found that the leakage of Cas9 activity was suppressed by increasing the sequence that responded to miRNA.
Abstract
miRNAによって特異的に認識される核酸配列と、ヌクレアーゼをコードする核酸配列とを連動させることにより、細胞特異的なmiRNAに応答してヌクレアーゼの活性を制御する方法を提供する。
Description
本発明は細胞特異的にヌクレアーゼを制御する方法、及び当該方法に用いるmiRNA-responsive mRNAに関する。
CRISPR/Cas9 Systemは、ヌクレアーゼであるCas9蛋白質(Cas9)と標的配列に相補的な20塩基を組み込んだシングルガイドRNA(sgRNA)により、目的遺伝子を切断するものである(例えば、非特許文献1を参照)。近年、このCRISPR/Cas9 Systemを細胞治療に用いようとする動向がある。例えば、ガン細胞特異的プロモーターの支配下にCas9遺伝子やsgRNAを置くことでガン細胞特異的にCRISPR/Cas9 Systemを起動させ細胞死を誘発させるというものである(例えば、非特許文献2を参照)。
Le Cong et al., Science 339, 819 (2013)
Yuchen Liu et al. Yuchen Liu et al. 1466996882191_0 5, 2014
しかし、CRISPR/Cas9 Systemを用いたCas9とsgRNAの細胞内導入には、プラスミドが用いられてきたため、それらがゲノムに挿入され変異が生じる危険性があった。また、プロモーターを用いる方法には、(1)プロモーターの特異性が低い(ガン特異的に高発現する一方で、正常細胞での多少発現してしまうことがある)、(2)ガン細胞特異的プロモーターは一般的に遺伝子発現が弱い、という問題点がある。上記技術は、これらを解決するために遺伝子回路(AND gate)を用いる方法を試みたが、(1)適切なプロモーター同士の組み合わせを探す必要がある、(2)プロモーターを変えるために、その都度、プラスミドを作製する必要がある、という問題が挙げられる。
本発明者らは上記の課題を解決すべく鋭意検討を行った結果、本発明者らにより開発されてきたmiRNAによって特異的に認識される核酸配列を、ヌクレアーゼをコードする核酸配列と連動して用いることにより、上記課題を解決し、細胞特異的なmiRNAに応答してヌクレアーゼの活性を制御する方法とすることを見出した。
すなわち、本発明は次に記載の事項を提供するものである。
[1] ヌクレアーゼをコードするmiRNA応答性mRNAを、細胞に導入する工程を含む、細胞特異的にヌクレアーゼを制御する方法であって、当該miRNA応答性mRNAが下記:
(i) miRNAによって特異的に認識される核酸配列、および
(ii) 前記ヌクレアーゼのコード領域に対応する核酸配列を含む、方法。
[2] 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[1]に記載の方法。
[3] 前記ヌクレアーゼが、Cas9タンパク質またはその変異体であり、
さらに、前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAを前記細胞群に導入する工程を含む、[1]または[2]に記載の方法。
[4] 未分化細胞特異的にヌクレアーゼを制御する、[3]に記載の方法。
[5] 前記未分化細胞が、miR-302aを発現する細胞である、[4]に記載の方法。
[6] 前記miRNA応答性mRNAが、(i)および(ii)が5'から3'の方向に連結されている核酸配列を含む、[1]から[5]のいずれか1項に記載の方法。
[7] (i) miRNAによって特異的に認識される核酸配列、および
(ii) ヌクレアーゼのコード領域に対応する核酸配列を含む、miRNA応答性mRNA。
[8] 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[7]に記載のmiRNA応答性mRNA。
[9] 前記ヌクレアーゼが、Cas9タンパク質またはその変異体である、[7]または[8]に記載のmiRNA応答性mRNA。
[10] [9]に記載のmiRNA応答性mRNAと、
前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAと
を含む、細胞特異的にヌクレアーゼを制御するためのキット。
[11] a)ヌクレアーゼをコードするトリガータンパク質応答性mRNAと、
b)前記トリガータンパク質をコードするmiRNA応答性mRNAと
を、細胞に導入する工程を含む、細胞特異的にヌクレアーゼを制御する方法であって、
前記a)ヌクレアーゼをコードするタンパク質応答性mRNAが下記:
(ia) 前記タンパク質に特異的に結合する核酸配列と、
(iia) 前記ヌクレアーゼのコード領域に対応する核酸配列と
を含み
前記b)トリガータンパク質をコードするmiRNA応答性mRNAが下記:
(ib) miRNAによって特異的に認識される核酸配列と、
(iib) 前記トリガータンパク質のコード領域に対応する核酸配列を含む、方法。
[12] 前記トリガータンパク質が、L7Aeタンパク質もしくはその誘導体を含み、前記タンパク質に特異的に結合する核酸配列が、K-turn配列もしくはその誘導体を含む、[11]に記載の方法。
[13] 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[11]または[12]に記載の方法。
[14] 前記ヌクレアーゼが、Cas9タンパク質またはその変異体であり、
さらに、前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAを前記細胞に導入する工程を含む、[11]~[13]のいずれか1項に記載の方法。
[15] 未分化細胞特異的にヌクレアーゼを制御する、[14]に記載の方法。
[16] 前記未分化細胞が、miR-302aを発現する細胞である、[15]に記載の方法。
[17] 前記a)トリガータンパク質をコードするタンパク質応答性mRNAが、(ia) および(iia) が5'から3'の方向に連結されている核酸配列を含み、
前記b)ヌクレアーゼをコードするmiRNA応答性mRNAが、(ib)および(iib)が5'から3'の方向に連結されている核酸配列を含む、[11]~[16]のいずれか1項に記載の方法。
[18] a)下記を含むヌクレアーゼをコードするトリガータンパク質応答性mRNA:
(ia) 前記タンパク質に特異的に結合する核酸配列、及び
(iia) 前記ヌクレアーゼのコード領域に対応する核酸配列と、
b)下記を含むトリガータンパク質をコードするmiRNA応答性mRNA:
(ib) miRNAによって特異的に認識される核酸配列、及び
(iib) 前記トリガータンパク質のコード領域に対応する核酸配列と
を含むヌクレアーゼ制御剤。
[19] 前記トリガータンパク質が、L7Aeタンパク質もしくはその誘導体を含み、前記タンパク質に特異的に結合する核酸配列が、K-turn配列もしくはその誘導体を含む、[18]に記載のヌクレアーゼ制御剤。
[20] 前記トリガータンパク質をコードするmiRNA応答性mRNAにおいて、前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[18]または[19]に記載のヌクレアーゼ制御剤。
[21] 前記トリガータンパク質応答性mRNAにおいて、前記ヌクレアーゼが、Cas9タンパク質またはその変異体である、[18]~[20]のいずれか1項に記載のヌクレアーゼ制御剤。
[22] [18]~[21]のいずれか1項に記載のヌクレアーゼ制御剤と、
前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAと
を含む、細胞特異的にヌクレアーゼを制御するためのキット。
[1] ヌクレアーゼをコードするmiRNA応答性mRNAを、細胞に導入する工程を含む、細胞特異的にヌクレアーゼを制御する方法であって、当該miRNA応答性mRNAが下記:
(i) miRNAによって特異的に認識される核酸配列、および
(ii) 前記ヌクレアーゼのコード領域に対応する核酸配列を含む、方法。
[2] 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[1]に記載の方法。
[3] 前記ヌクレアーゼが、Cas9タンパク質またはその変異体であり、
さらに、前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAを前記細胞群に導入する工程を含む、[1]または[2]に記載の方法。
[4] 未分化細胞特異的にヌクレアーゼを制御する、[3]に記載の方法。
[5] 前記未分化細胞が、miR-302aを発現する細胞である、[4]に記載の方法。
[6] 前記miRNA応答性mRNAが、(i)および(ii)が5'から3'の方向に連結されている核酸配列を含む、[1]から[5]のいずれか1項に記載の方法。
[7] (i) miRNAによって特異的に認識される核酸配列、および
(ii) ヌクレアーゼのコード領域に対応する核酸配列を含む、miRNA応答性mRNA。
[8] 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[7]に記載のmiRNA応答性mRNA。
[9] 前記ヌクレアーゼが、Cas9タンパク質またはその変異体である、[7]または[8]に記載のmiRNA応答性mRNA。
[10] [9]に記載のmiRNA応答性mRNAと、
前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAと
を含む、細胞特異的にヌクレアーゼを制御するためのキット。
[11] a)ヌクレアーゼをコードするトリガータンパク質応答性mRNAと、
b)前記トリガータンパク質をコードするmiRNA応答性mRNAと
を、細胞に導入する工程を含む、細胞特異的にヌクレアーゼを制御する方法であって、
前記a)ヌクレアーゼをコードするタンパク質応答性mRNAが下記:
(ia) 前記タンパク質に特異的に結合する核酸配列と、
(iia) 前記ヌクレアーゼのコード領域に対応する核酸配列と
を含み
前記b)トリガータンパク質をコードするmiRNA応答性mRNAが下記:
(ib) miRNAによって特異的に認識される核酸配列と、
(iib) 前記トリガータンパク質のコード領域に対応する核酸配列を含む、方法。
[12] 前記トリガータンパク質が、L7Aeタンパク質もしくはその誘導体を含み、前記タンパク質に特異的に結合する核酸配列が、K-turn配列もしくはその誘導体を含む、[11]に記載の方法。
[13] 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[11]または[12]に記載の方法。
[14] 前記ヌクレアーゼが、Cas9タンパク質またはその変異体であり、
さらに、前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAを前記細胞に導入する工程を含む、[11]~[13]のいずれか1項に記載の方法。
[15] 未分化細胞特異的にヌクレアーゼを制御する、[14]に記載の方法。
[16] 前記未分化細胞が、miR-302aを発現する細胞である、[15]に記載の方法。
[17] 前記a)トリガータンパク質をコードするタンパク質応答性mRNAが、(ia) および(iia) が5'から3'の方向に連結されている核酸配列を含み、
前記b)ヌクレアーゼをコードするmiRNA応答性mRNAが、(ib)および(iib)が5'から3'の方向に連結されている核酸配列を含む、[11]~[16]のいずれか1項に記載の方法。
[18] a)下記を含むヌクレアーゼをコードするトリガータンパク質応答性mRNA:
(ia) 前記タンパク質に特異的に結合する核酸配列、及び
(iia) 前記ヌクレアーゼのコード領域に対応する核酸配列と、
b)下記を含むトリガータンパク質をコードするmiRNA応答性mRNA:
(ib) miRNAによって特異的に認識される核酸配列、及び
(iib) 前記トリガータンパク質のコード領域に対応する核酸配列と
を含むヌクレアーゼ制御剤。
[19] 前記トリガータンパク質が、L7Aeタンパク質もしくはその誘導体を含み、前記タンパク質に特異的に結合する核酸配列が、K-turn配列もしくはその誘導体を含む、[18]に記載のヌクレアーゼ制御剤。
[20] 前記トリガータンパク質をコードするmiRNA応答性mRNAにおいて、前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、[18]または[19]に記載のヌクレアーゼ制御剤。
[21] 前記トリガータンパク質応答性mRNAにおいて、前記ヌクレアーゼが、Cas9タンパク質またはその変異体である、[18]~[20]のいずれか1項に記載のヌクレアーゼ制御剤。
[22] [18]~[21]のいずれか1項に記載のヌクレアーゼ制御剤と、
前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAと
を含む、細胞特異的にヌクレアーゼを制御するためのキット。
本発明に係るヌクレアーゼの制御方法によれば、細胞の状態を示すmiRNAの発現に応答して、ヌクレアーゼ活性を制御することが可能になった。本発明の方法においては、細胞内へのヌクレアーゼの導入をmRNAにより行い、かつ、細胞特異的にヌクレアーゼを制御することができる。ゆえに、安全性や特異性の観点から、臨床への応用が期待される。
以下に本発明を、実施態様を示して詳細に説明する。しかしながら、本発明は以下の実施態様に限定されるものではない。
本発明は、細胞特異的にヌクレアーゼを制御する方法を提供するものである。細胞特異的にヌクレアーゼを制御する方法は、少なくとも、miRNAによって特異的に認識される核酸配列を、ヌクレアーゼをコードする核酸配列と連動して用いることによる。例えば、miRNAによって特異的に認識される核酸配列と、ヌクレアーゼをコードする核酸配列とを1つのmRNAに含めて細胞特異的にヌクレアーゼを制御することができる。あるいは、miRNAによって特異的に認識される核酸配列を含むmRNAと、ヌクレアーゼをコードする核酸配列を含むmRNAとを別個に含み、これらの組み合わせにより細胞特異的にヌクレアーゼを制御することができる。以下に、実施の形態に分けて記載する。
[第1実施形態:OFF switchヌクレアーゼ制御]
本発明は、一実施形態によれば、細胞特異的にヌクレアーゼを制御する方法であって、ヌクレアーゼをコードするmiRNA応答性mRNAを細胞に導入する工程を含む。本発明において、細胞特異的にヌクレアーゼを制御するとは、細胞内在性のmiRNAの発現状態に基づいて、ヌクレアーゼの活性を制御することをいう。
本発明は、一実施形態によれば、細胞特異的にヌクレアーゼを制御する方法であって、ヌクレアーゼをコードするmiRNA応答性mRNAを細胞に導入する工程を含む。本発明において、細胞特異的にヌクレアーゼを制御するとは、細胞内在性のmiRNAの発現状態に基づいて、ヌクレアーゼの活性を制御することをいう。
本発明における、細胞とは、特に限定されるものではなく任意の細胞であってよい。例えば、多細胞生物種から採取した細胞であってもよく、単離された細胞を培養することによって得られる細胞であってもよい。当該細胞は、特には、哺乳動物(例えば、ヒト、マウス、サル、ブタ、ラット等)採取した細胞、若しくは哺乳動物より単離された細胞又は哺乳動物細胞株を培養することによって得られる細胞である。体細胞としては、例えば、角質化する上皮細胞(例、角質化表皮細胞)、粘膜上皮細胞(例、舌表層の上皮細胞)、外分泌腺上皮細胞(例、乳腺細胞)、ホルモン分泌細胞(例、副腎髄質細胞)、代謝・貯蔵用の細胞(例、肝細胞)、境界面を構成する内腔上皮細胞(例、I型肺胞細胞)、内鎖管の内腔上皮細胞(例、血管内皮細胞)、運搬能をもつ繊毛のある細胞(例、気道上皮細胞)、細胞外マトリックス分泌用細胞(例、線維芽細胞)、収縮性細胞(例、平滑筋細胞)、血液と免疫系の細胞(例、Tリンパ球)、感覚に関する細胞(例、桿細胞)、自律神経系ニューロン(例、コリン作動性ニューロン)、感覚器と末梢ニューロンの支持細胞(例、随伴細胞)、中枢神経系の神経細胞とグリア細胞(例、星状グリア細胞)、色素細胞(例、網膜色素上皮細胞)、およびそれらの前駆細胞(組織前駆細胞)等が挙げられる。
細胞の分化の程度や細胞を採取する動物の齢などに特に制限はなく、未分化な前駆細胞(体性幹細胞も含む)であっても、最終分化した成熟細胞であっても、同様に本発明における細胞として用いることができる。ここで未分化な前駆細胞としては、たとえば神経幹細胞、造血幹細胞、間葉系幹細胞、歯髄幹細胞等の組織幹細胞(体性幹細胞)が挙げられる。
細胞の分化の程度や細胞を採取する動物の齢などに特に制限はなく、未分化な前駆細胞(体性幹細胞も含む)であっても、最終分化した成熟細胞であっても、同様に本発明における細胞として用いることができる。ここで未分化な前駆細胞としては、たとえば神経幹細胞、造血幹細胞、間葉系幹細胞、歯髄幹細胞等の組織幹細胞(体性幹細胞)が挙げられる。
本発明における細胞は、前期体細胞を採取後に人為的な操作を加えた細胞であってよく、例えば、前記体細胞から調製したiPS細胞を含んでなる細胞群であり、あるいはES 細胞やiPS細胞などによって例示される多能性幹細胞を分化させた後に得られる細胞群であって、所望する細胞以外に分化された細胞を含み得る細胞群であってもよい。本発明における細胞は、特には生存状態にあることが好ましい。本発明において、細胞が生存状態にあるとは、代謝能を維持した状態の細胞を意味する。本発明においては、細胞にmiRNA応答性mRNAを導入した後にも、その生来の特性を失うことなく、生存状態のまま、特に分裂能を維持したまま、続く用途に用いることができる細胞である。
上記のような細胞において、細胞内で発現されるmiRNAの種類や量は、細胞種に特異的であることが知られている。本発明においては、以下に詳述する、ヌクレアーゼをコードするmiRNA応答性mRNAを用いることにより、細胞内で発現されるmiRNAに応答して、ヌクレアーゼの活性を制御することができる。ヌクレアーゼの活性を制御するとは、ヌクレアーゼの発現量を低下させ、あるいは増加させて、これにより、ヌクレアーゼの活性を低下させ、あるいは上昇させることをいう。
miRNA-応答性mRNA
本発明において、ヌクレアーゼをコードするmiRNA応答性mRNAは、miRNA-responsive mRNA、あるいはmiRNAスイッチとも言い、以下の(i)および(ii)の核酸配列を含むmRNAを意味する:(i) miRNAによって特異的に認識される核酸配列、および(ii) ヌクレアーゼのコード領域に対応する核酸配列。
当該(i)miRNAによって特異的に認識される核酸配列と(ii) ヌクレアーゼのコード領域に対応する核酸配列は、機能的に連結されている。図1(a)は、本発明に係る方法において使用しうるmiRNA-responsive mRNAの一例を模式的に示す図である。当該miRNA-responsive mRNAにおいては、5'UTRにmiRNAに応答する配列が組みこまれ、蛋白質コーディングリージョンにヌクレアーゼをコードする遺伝子が挿入されている。
本発明において、ヌクレアーゼをコードするmiRNA応答性mRNAは、miRNA-responsive mRNA、あるいはmiRNAスイッチとも言い、以下の(i)および(ii)の核酸配列を含むmRNAを意味する:(i) miRNAによって特異的に認識される核酸配列、および(ii) ヌクレアーゼのコード領域に対応する核酸配列。
当該(i)miRNAによって特異的に認識される核酸配列と(ii) ヌクレアーゼのコード領域に対応する核酸配列は、機能的に連結されている。図1(a)は、本発明に係る方法において使用しうるmiRNA-responsive mRNAの一例を模式的に示す図である。当該miRNA-responsive mRNAにおいては、5'UTRにmiRNAに応答する配列が組みこまれ、蛋白質コーディングリージョンにヌクレアーゼをコードする遺伝子が挿入されている。
本発明における「miRNA」とは、mRNAからタンパク質への翻訳の阻害やmRNAの分解を通して、遺伝子の発現調節に関与する、細胞内に存在する短鎖(20-25塩基)のノンコーディングRNAである。このmiRNAは、miRNAとその相補鎖を含むヘアピンループ構造を取ることが可能な一本差のpri-miRNAとして転写され、核内にあるDroshaと呼ばれる酵素により一部が切断されpre-miRNAとなって核外に輸送された後、さらにDicerによって切断されて機能する。
(i)のmiRNAとしては、ある特定の細胞においてヌクレアーゼ活性を制御、特にはヌクレアーゼの発現を抑制または活性化することを目的として、その細胞で特異的に発現している、あるいは特異的に発現していないmiRNAを適宜選択することができる。特異的に発現しているmiRNAとは、他の細胞と比較してある特定の細胞において、10%以上、20%以上、30%以上、40%以上、50%以上、60%以上、70%以上、80%以上、90%以上あるいはそれ以上の割合で高く発現しているmiRNAが例示される。このようなmiRNAは、データベースの情報(例えば、http://www.mirbase.org/又はhttp://www.microrna.org/)に登録されたmiRNA、及び/または当該データベースに記載されている文献情報に記載されたmiRNAより適宜選択することができる
本発明において、miRNAによって特異的に認識される核酸配列は、例えば、当該miRNAに完全に相補的な配列であることが好ましい。あるいは、当該miRNAにおいて認識され得る限り、完全に相補的な配列との不一致(ミスマッチ)を有していても良い。当該miRNAに完全に相補的な配列からの不一致は、所望の細胞において、通常にmiRNAが認識し得る不一致であれば良く、生体内における細胞内の本来の機能では、40~50% 程度の不一致があっても良い。このような不一致は、特に限定されないが、1塩基、2塩基、3塩基、4塩基、5塩基、6塩基、7塩基、8塩基、9塩基、若しくは10塩基又は全認識配列の1%、5%、10%、20%、30%、若しくは40%の不一致が例示される。また、特には、細胞が備えているmRNA 上のmiRNA標的配列のように、特に、シード領域以外の部分に、すなわち miRNA の 3' 側 16 塩基程度に対応する、標的配列内の 5' 側の領域に、多数の不一致を含んでもよく、シード領域の部分は、不一致を含まないか、1塩基、2塩基、若しくは3塩基の不一致を含んでもよい。このような配列は、RISCが特異的に結合する塩基数を含む塩基長であればよく、長さは別段限定されないが、好ましくは、18塩基以上、24塩基未満の配列、より好ましくは、20塩基以上、22塩基未満の配列である。本発明において、miRNAによって特異的に認識される核酸配列は、所望の細胞および他の細胞へ当該配列を有するmiRNA-responsive mRNAを導入し、所望の細胞においてのみ対応するマーカー遺伝子の発現が抑制されることを確認することによって、適宜決定して用いることができる。
本発明において使用される前記(ii)の「ヌクレアーゼのコード領域に対応する核酸配列」とは、細胞内で翻訳されて核酸を分解する酵素として機能するタンパク質をコードする遺伝子である。ヌクレアーゼのコード領域に対応する核酸配列としては、一例としては、Clustered Regularly Interspaced Short Palindromic Repeats-AssociatedProteins 9(Cas9)をコードする遺伝子、転写活性化因子様エフェクターヌクレアーゼ(TALEN)コードする遺伝子、ホーミングエンドヌクレアーゼをコードする遺伝子、ジンクフィンガーヌクレアーゼをコードする遺伝子を含むが、これらに限定されない。なお、いずれのヌクレアーゼについても、同様の機能を備える変異体や誘導体を用いることもでき、例えば、Cas9変異体をコードする遺伝子またはCas9変異体融合タンパク質をコードする遺伝子も、ヌクレアーゼのコード領域に対応する核酸配列に含まれる。また、これらのヌクレアーゼは、ヌクレアーゼにより分解する標的核酸に特異的に設計することができる。なお、ヌクレアーゼがCas9の場合には、sgRNAを、ヌクレアーゼにより分解する標的核酸に特異的に設計する。詳細は後述する。
本発明において、miRNAによって特異的に認識される核酸配列とヌクレアーゼのコード領域に対応する核酸配列が機能的に連結されるとは、ヌクレアーゼをコードするオープンリーディングフレーム(ただし、開始コドンを含む。)の5'UTR内、3'UTR内、及び/または当該オープンリーディングフレーム内に、少なくとも1つのmiRNAの標的配列を備えることを意味する。miRNA-responsive mRNAは、好ましくは、5'末端から、5'から3'の向きに、Cap構造(7メチルグアノシン5'リン酸)、ヌクレアーゼをコードするオープンリーディングフレーム並びに、ポリAテイルを備え、5'UTR内、3'UTR内、及び/またはオープンリーディングフレーム内に少なくとも1つのmiRNAの標的配列を備える。mRNAにおけるmiRNAの標的配列の位置は、5'UTRであっても、3'UTRであってもよく、オープンリーディングフレーム内(開始コドンの3'側)であってもよく、これらのすべてにmiRNAの標的配列を備えていてもよい。したがって、miRNA標的配列の数は、1つ、2つ、3つ、4つ、5つ、6つ、7つ、8つあるいはそれ以上であっても良い。
好ましくは、miRNA-responsive mRNAは、(i)および(ii)の核酸配列が、5'から3'の方向にこの順序で連結されている。このとき、cap構造とmiRNAの標的配列との間の塩基数及び塩基の種類は、ステム構造や立体構造を構成しない限り、任意であってよい。例えば、cap構造とmiRNA標的配列との間の塩基数は、0~50塩基、好ましくは、10~30塩基となるように設計することができる。また、miRNA標的配列と開始コドンとの間の塩基数及び塩基の種類は、ステム構造や立体構造を構成しない限り、任意であってよく、miRNA標的配列と開始コドンと間の塩基数は、0~50塩基、好ましくは、10~30塩基となるような配置にて設計することができる。
本発明において、miRNA-responsive mRNA 中のmiRNA標的配列内には、開始コドンとなるAUGが存在しないことが好ましい。例えば、miRNAの標的配列が5'UTRに存在し、かつ、当該標的配列内にAUGを含む場合には、3'側に連結されるマーカー遺伝子との関係上でインフレームとなるように設計されることが好ましい。あるいは、標的配列内にAUGを含む場合、標的配列内のAUGをGUGに変換して使用することも可能である。また、標的配列内のAUGの影響を最小限に留めるために、5'UTR内における標的配列の配置場所を適宜変更することができる。例えば、cap構造と標的配列内のAUG配列との間の塩基数が、0~60塩基、例えば、0~15塩基、10~20塩基、20~30塩基、30~40塩基、40~50塩基、50~60塩基となるような配置にて設計され得る。
本発明の方法において、細胞に導入するmiRNA-responsive mRNAは、1種類であってもよく、2種、3種、4種、5種、6種、7種、あるいは8種類以上であってもよい。2種以上のmiRNA-responsive mRNAを導入する場合には、例えば、miRNA標的部位の配列が異なり、異なるmiRNAに応答するmRNAを用いることができる。異なる複数のmiRNA-responsive mRNAを用いることにより、例えば、miRNA-responsive mRNAが導入される細胞群に含まれうる複数種の細胞に対して、ヌクレアーゼの活性を制御することが可能になる。
miRNA-responsive mRNAの導入
本発明において、miRNA-responsive mRNAを細胞に導入する方法としてはmRNAの形態で導入することができる。例えばリポフェクション、マイクロインジェクションなどの手法によって体細胞内に導入しても良く、分解を抑制するため、5-メチルシチジンおよびpseudouridine (TriLink Biotechnologies)を取り込ませたRNAを用いても良い(Warren L, (2010) Cell Stem Cell.7:618-630)。修飾塩基の位置は、ウリジン、シチジンいずれの場合も、独立に、全てあるいは一部とすることができ、一部である場合には、任意の割合でランダムな位置とすることができる。あるいは、miRNA-responsive mRNAは、ベクターなどのDNAの形態で導入することもでき、導入法は、上記と同様のものを用いることができる。
本発明において、miRNA-responsive mRNAを細胞に導入する方法としてはmRNAの形態で導入することができる。例えばリポフェクション、マイクロインジェクションなどの手法によって体細胞内に導入しても良く、分解を抑制するため、5-メチルシチジンおよびpseudouridine (TriLink Biotechnologies)を取り込ませたRNAを用いても良い(Warren L, (2010) Cell Stem Cell.7:618-630)。修飾塩基の位置は、ウリジン、シチジンいずれの場合も、独立に、全てあるいは一部とすることができ、一部である場合には、任意の割合でランダムな位置とすることができる。あるいは、miRNA-responsive mRNAは、ベクターなどのDNAの形態で導入することもでき、導入法は、上記と同様のものを用いることができる。
異なる2種以上のmiRNA-responsive mRNAを導入する場合、あるいはmiRNA-responsive mRNAと、後述するコントロールとなるmRNA(以下、コントロールmRNAとも指称する)とを用いる場合には、複数のmRNAを細胞群に共導入することが好ましい。共導入した2種以上のmRNAの細胞内での割合は個々の細胞で維持されるため、これらのmRNAから発現するタンパク質の活性比は、細胞集団内において一定となるためである。この時の導入量は、導入される細胞群、導入するmRNA、導入方法および導入試薬の種類により異なり、所望の翻訳量のヌクレアーゼを得るために当業者は適宜これらを選択することができる。本発明において、コントロールmRNAとは、miRNAの標的部位を有さないmRNAをいう。すなわち、コントロールmRNAは、細胞内在性のmiRNAの発現量の影響を受けることなく、細胞に導入されて翻訳されるmRNAである。コントロールmRNAは、好ましい態様においては、miRNA-responsive mRNAとともに細胞群に導入されて、miRNA-responsive mRNAが導入された細胞を確認し、識別するためのコントロールとして機能し得る。コントロールmRNAの導入量もまた、所望の翻訳量を得るために当業者は適宜これらを選択することができる。
細胞内に導入された当該miRNA-responsive mRNAは、細胞内に、標的配列を特異的に認識するmiRNAが存在すると、翻訳抑制されてヌクレアーゼの発現量が低下し、これによりヌクレアーゼ活性も低下する。したがって、特定のmiRNAを発現する細胞特異的に、ヌクレアーゼ活性を低下させることができる。これにより、ヌクレアーゼの標的となる標的遺伝子の切断活性は低下した状態となり、標的遺伝子の機能への影響が小さい状態となる。一方、細胞内に、標的配列を特異的に認識するmiRNAが存在しないと、miRNA-responsive mRNAは翻訳抑制されることがなく、細胞内でヌクレアーゼが発現する。これにより、当該細胞において、ヌクレアーゼの標的となる標的遺伝子の切断が進行し、当該遺伝子の機能を損なわせることにより、当該遺伝子の機能に関連する細胞の運命を制御することができる。したがって、一例においては、標的配列を特異的に認識するmiRNAが存在しない細胞を、細胞死に至らせることができる。
次に、より具体的な例として、ヌクレアーゼとして、Cas9タンパク質を用いる場合の本発明の方法を説明する。本発明は、一実施態様によれば、Cas9タンパク質をコードするmiRNA-responsive mRNAを細胞に導入する工程と、前記Cas9タンパク質の標的配列によって特異的に認識されるガイド配列を備えるsgRNAを細胞に導入する工程とを含む。図1(b)は、Cas9タンパク質をコードするmiRNA-responsive Cas9 mRNAと、sgRNAとを細胞に導入した場合の、細胞内での作用を模式的に示す図である。miRNA-responsive Cas9 mRNAに結合する標的miRNAが存在しないとき(Target miRNA(-))は、miRNA-responsive mRNAからCas9蛋白質が翻訳される。よって、miRNA-responsive mRNAと共導入したガイド鎖(sgRNA)とCas9蛋白質が複合体を形成し、ゲノム編集、具体的には標的DNAの切断が行われる。標的miRNAが存在するとき(Target miRNA(+))は、標的miRNAがmiRNA-responsive Cas9 mRNA の5'UTRに結合することにより翻訳抑制やmRNA switchの分解が起こり、miRNA-responsive Cas9 mRNAからのCas9蛋白質の翻訳が抑制される。よって、Cas9/sgRNA複合体が形成されないため、ゲノム編集、具体的には標的DNAの切断は行われない。
ここで、Cas9タンパク質をコードするmiRNA-responsive mRNAは、先に述べた通りに設計することができる。なお、Cas9タンパク質をコードするmiRNA-responsive mRNA を以下、miRNA-responsive Cas9 mRNA とも表示する。miRNAの一例としては、miR-302a、miR-21が挙げられる。miR-302aは、iPS細胞、ES細胞などの多能性幹細胞内在性のmiRNAとして知られている。したがって、Cas9タンパク質をコードするmiR-302a-responsive mRNAを、iPS細胞などの多能性幹細胞と、分化細胞とを含む細胞群に導入することにより、多能性幹細胞特異的にCas9タンパク質の発現を抑制し、sgRNAの標的遺伝子機能を損なわずに維持することができ、分化細胞においてはCas9タンパク質を機能させて、標的遺伝子を切断することができる。すなわち、細胞の分化の度合い、あるいは細胞の初期化(リプログラミング)状態に応じて、Cas9タンパク質の活性を制御することが可能である。一方、miR-21は、Hela細胞に特異的に発現していることが知られている。これらの配列を下記の表1に示す。また、miR-302a 及びmiR-21により特異的に認識されるmiR標的配列の一例を表2に示す。


miR-302a標的配列を備え、細胞内在性のmiR-302aに応答してCas9タンパク質の発現が抑制されるmiR-302a-responsive Cas 9 mRNA、及びmiR-21標的配列を備え、細胞内在性のmiR-21に応答してCas9タンパク質の発現が抑制されるmiR-21-responsive Cas 9 mRNAとしては、具体的には、以下の表3に示す配列が挙げられるが、これらには限定されない。表3の配列中、下線部及び二重下線部は、miRNA標的配列を、AUGは開始コドンを、点下線部は3'UTRをそれぞれ示す。





sgRNAは、Cas9の標的となる遺伝子を特異的に認識する、20程度、例えば、18~22の程度の塩基配列を5'末端近傍に組み込むように設計することができる。Cas9の標的となる遺伝子を特異的に認識する塩基配列は、標的となる遺伝子に完全に相補的な配列であることが好ましい。あるいは、当該標的となる遺伝子において認識され得る限り、完全に相補的な配列との不一致(ミスマッチ)を有していても良い。当該標的となる遺伝子に完全に相補的な配列からの不一致については、miRNAとその標的配列について定義したのと同様であってよい。また、このCas9の標的となる遺伝子を特異的に認識する塩基配列の5’側に、1~5塩基程度の配列があってもよい。さらに、標的となる遺伝子を特異的に認識する塩基配列の3'側の配列は、以下の表4に示す具体的な配列に限定されず、CRISPR/Cas9 Systemにおいて機能することが知られているsgRNAの配列であってよく、Tetraloop,stem loop2, 3’末端を改変した例が知られているが、Cas9とともに、標的遺伝子の切断が可能なものであれば、特定のものには限定されない。
例えば、本発明においては、以下の、表4に示すsgRNAを用いることができるが、本発明の方法に使用しうるsgRNAは、これらには限定されない。表4の配列中、下線部は、Cas9の標的遺伝子に相補的な配列を示す。表中、配列番号8は、DMD (Duchenne muscular dystrophy)遺伝子を標的とするsgRNA、配列番号9は、Alu1反復配列を標的とするsgRNA、配列番号10、11は、EGFP遺伝子を標的とするsgRNAである。

Cas9により、例えば、細胞特異的に細胞死を起こさせるために用いることができる標的配列としては、下記のAlu1反復配列の他、Telomereや(AC)nのような反復配列が挙げられるが、これらには限定されない。
上記に従って設計されたmiRNA-responsive Cas9 mRNAとsgRNAとを細胞に導入する工程は、先に説明した方法により実施することができる。また、その際の導入量は、当業者が所望のCas9による機能を発揮するために、適宜決定することができ、限定されるものではない。
所望のmiRNA-responsive Cas9 mRNAと、Cas9の標的となる遺伝子配列を特異的に認識するsgRNAとを組み合わせて、キットとすることができる。これにより、例えば、特定のmiRNAを発現する細胞特異的に、Cas9の活性を抑制することができ、特定のmiRNAを発現しない細胞においてのみ、Cas9の活性を維持することができる。一例としては、miR-302aを発現する未分化細胞特異的にCas9の活性を抑制し、miR-302aをほとんど発現しない分化細胞においてはCas9の活性を維持することができる。また、既存のONスイッチとくみあわせることで、特定のmiRNAを発現する細胞特異的に、ゲノム編集することも可能である。
[第2実施形態:ON switchヌクレアーゼ制御]
本発明は、第2実施形態によれば、細胞特異的にヌクレアーゼを制御する方法であって、a)ヌクレアーゼをコードするトリガータンパク質応答性mRNAと、b)前記トリガータンパク質をコードするmiRNA応答性mRNAとを細胞に導入する工程を含む。本発明において、細胞特異的にヌクレアーゼを制御するとは、細胞内在性のmiRNAの発現状態に基づいて、ヌクレアーゼの活性を制御することをいう。
本発明は、第2実施形態によれば、細胞特異的にヌクレアーゼを制御する方法であって、a)ヌクレアーゼをコードするトリガータンパク質応答性mRNAと、b)前記トリガータンパク質をコードするmiRNA応答性mRNAとを細胞に導入する工程を含む。本発明において、細胞特異的にヌクレアーゼを制御するとは、細胞内在性のmiRNAの発現状態に基づいて、ヌクレアーゼの活性を制御することをいう。
本実施形態においても、細胞の定義は、先の第1実施形態と同様であり、説明を省略する。本実施形態においては、以下に詳述する、a)ヌクレアーゼをコードするトリガータンパク質応答性mRNAと、b)前記トリガータンパク質をコードするmiRNA応答性mRNAとを用いることにより、細胞内で発現されるmiRNAに応答して、ヌクレアーゼの活性を制御することができる。本実施形態において、ヌクレアーゼの活性を制御するとは、ヌクレアーゼの発現量を増加させて、これにより、ヌクレアーゼの活性を上昇させることをいう。
a)ヌクレアーゼをコードするトリガータンパク質応答性mRNA
本実施形態において、ヌクレアーゼをコードする、トリガータンパク質応答性mRNAは、以下の(ia)および(iia)の核酸配列を含むmRNAを意味する:(ia) トリガータンパク質に特異的に結合する核酸配列、および(iia)ヌクレアーゼのコード領域に対応する核酸配列。
当該(ia) トリガータンパク質に特異的に結合する核酸配列と(iia)ヌクレアーゼのコード領域に対応する核酸配列は、機能的に連結されている。図8(a)のmRNA1は、本発明に係る方法において使用しうるトリガータンパク質応答性mRNAの一例を模式的に示す図である。当該トリガータンパク質応答性mRNAにおいては、5'UTRにトリガータンパク質に応答する配列が組みこまれ、蛋白質コーディングリージョンにヌクレアーゼをコードする遺伝子が挿入されている。
本実施形態において、ヌクレアーゼをコードする、トリガータンパク質応答性mRNAは、以下の(ia)および(iia)の核酸配列を含むmRNAを意味する:(ia) トリガータンパク質に特異的に結合する核酸配列、および(iia)ヌクレアーゼのコード領域に対応する核酸配列。
当該(ia) トリガータンパク質に特異的に結合する核酸配列と(iia)ヌクレアーゼのコード領域に対応する核酸配列は、機能的に連結されている。図8(a)のmRNA1は、本発明に係る方法において使用しうるトリガータンパク質応答性mRNAの一例を模式的に示す図である。当該トリガータンパク質応答性mRNAにおいては、5'UTRにトリガータンパク質に応答する配列が組みこまれ、蛋白質コーディングリージョンにヌクレアーゼをコードする遺伝子が挿入されている。
(ia)のトリガータンパク質に特異的に結合する核酸配列は、RNA-タンパク質結合モチーフを形成する配列を含むRNAである。RNA-タンパク質結合モチーフを形成する配列を含むRNAとは、天然または既知のRNA-タンパク質複合体における、RNAとタンパク質との結合モチーフに含まれるRNA側、または試験管内進化法(in vitro selection法)により得られた人工的なRNA-タンパク質結合モチーフに含まれるRNA側である。したがって、トリガータンパク質は、RNA-タンパク質結合モチーフを形成する配列を含むRNAとは、天然または既知のRNA-タンパク質複合体における、RNAとタンパク質との結合モチーフに含まれるタンパク質側を含む。
天然のRNA-タンパク質結合モチーフを形成する配列は、通常、約5~30塩基で構成されており、特定のアミノ酸配列を保有するタンパク質と、非共有結合的に、すなわち水素結合により、特異的な結合を形成することが知られている。このような天然のRNA-タンパク質結合モチーフを形成する配列は、以下の表5及び表6、及びウェブサイト上で利用できるデータベース:http://gibk26.bse.kyutech.ac.jp/jouhou/image/dna-protein/RNA/RNA.htmlから、所望の構造変化を生ずるモチーフを適宜選択して入手することができる。本実施形態において好ましく用いられるRNA-タンパク質結合モチーフは、X線結晶構造解析またはNMRによる構造解析が既に行われているモチーフ、あるいは構造解析がなされている相同タンパク質の立体構造から立体構造を推定可能なモチーフである。さらに、タンパク質がRNAの二次構造及び塩基配列を特異的に認識するモチーフであることが好ましい。


人工のRNA-タンパク質結合モチーフを形成する配列を含むRNAとは、人工的に設計したRNA-タンパク質複合体における、RNAとタンパク質との結合モチーフ中のRNA側である。このようなRNAの塩基配列は、通常、約10~80塩基で構成されており、特定のタンパク質の特定のアミノ酸配列と、非共有結合的に、すなわち水素結合により、特異的な結合を形成するよう設計する。このような人工的なRNA-タンパク質結合モチーフを形成する配列を含むRNAとしては、特定のタンパク質に特異的に結合するRNAアプタマーが例示される。標的となる所望のタンパク質に対し特異的に結合するRNAアプタマーは、例えば、in vitro selection法またはSELEX法として知られている進化工学的手法より得ることができる。このときのトリガータンパク質は、当該RNAアプタマーが結合するタンパク質となる。例えば、以下の表7に挙げるRNA配列が知られており、これらもまた本発明のRNA-タンパク質結合モチーフを形成する配列として用いることができる。

本実施形態において、RNA-タンパク質結合モチーフを形成する配列は、対応するトリガータンパク質との解離定数Kdが、約0.1nM~約1μM程度であるものが好ましい。
また、これらのRNA-タンパク質結合モチーフを形成する配列自体に加え、このような配列の変異体も本発明による当該配列に包含される。ここでいう変異体とは、RNA-タンパク質結合モチーフを形成する配列に特異的に結合するタンパク質との間の解離定数Kdが10%、20%、30%、40%または50%以上高い変異体もしくは10%、20%、30%、40%または50%以下の変異体である。このような変異体は、RNA-タンパク質複合体が形成できる限り、適宜選択して用いることができる。また、このような変異体の塩基配列は、当該RNA-タンパク質結合モチーフを形成する配列(正鎖)に対する相補的な配列を有する核酸(相補鎖)とストリンジェントな条件でハイブリダイズすることができる程度の塩基配列でもよい。ここでストリンジェントな条件は、Berger and Kimmel(1987,Guide to Molecular Cloning Techniques Methods in Enzymology,Vol.152,Academic Press,San Diego CA)に教示されるように、結合する核酸の融解温度(Tm)に基づいて決定することができる。例えばハイブリダイズ後の洗浄条件として、通常「1×SSC、0.1%SDS、37℃」程度の条件を挙げることができる。相補鎖は、かかる条件で洗浄しても対象とする正鎖とハイブリダイズ状態を維持するものであることが好ましい。特に制限されないが、より厳しいハイブリダイズ条件として「0.5×SSC、0.1%SDS、42℃」程度の洗浄条件、さらに厳しくは「0.1×SSC、0.1%SDS、65℃」程度の洗浄条件で洗浄しても正鎖と相補鎖とがハイブリダイズ状態を維持する条件を挙げることができる。具体的には、上述のRNA-タンパク質結合モチーフに含まれるRNAの配列と少なくとも90%、好ましくは少なくとも95%、96%、97%、98%または99%の配列同一性を有する塩基配列からなる。かかる変異体は、RNA-タンパク質結合モチーフを形成する配列に特異的に結合するタンパク質との間で、一定の結合を保持し、RNA-タンパク質複合体の形成に寄与することができる。
本実施形態によるRNA-タンパク質結合モチーフを形成する配列の具体的な例としては、下記表8に示す、L7Ae(Moore T et al., Structure Vol. 12, pp. 807-818 (2004))が結合する配列である、boxC motif(5’-GGCGUGAUGAGC-3’)(配列番号40)、kink-loop(配列番号41)、kink-loop2(配列番号42)が挙げられる。

別の具体例としては、MS2コートタンパク質が特異的に結合する配列であるMS2ステムループモチーフ(22:Keryer-Bibens C, Barreau C, Osborne HB (2008) Tethering of proteins to RNAs by bacteriophage proteins. Biol Cell 100:125-138)、バチルスのリボソームタンパク質S15が結合する配列であるFr15(24:Batey RT, Williamson JR (1996) Interaction of the Bacillus stearothermophilus ribosomal protein S15 with 16 S rRNA: I. Defining the minimal RNA site. J Mol Biol 261:536-549)が挙げられる。
さらなる具体例には、アミノアシル化を行う酵素であって、自身のmRNAに結合し、翻訳を阻害するフィードバック阻害を持つことが知られているThreonyl-tRNA synthetase(Cell (Cambridge, Mass.) v97, pp.371-381 (1999))が結合する配列である、5’-GGCGUAUGUGAUCUUUCGUGUGGGUCACCACUGCGCC-3’(配列番号43)、およびその変異体がある。また、癌細胞特異的な内在性タンパク質であるBcl-2ファミリーCED-9由来のRNA-タンパク質結合モチーフを形成する塩基配列である、R9-2;5’-GGGUGCUUCGAGCGUAGGAAGAAAGCCGGGGGCUGCAGAUAAUGUAUAGC-3’ (配列番号44)、およびその変異体、NF-kappaBに結合するRNA配列のアプタマー由来の塩基配列およびその変異体が挙げられる。
次に、(iia)の「ヌクレアーゼのコード領域に対応する核酸配列」とは、細胞内で翻訳されて核酸を分解する酵素として機能するタンパク質をコードする遺伝子であり、第1実施形態において説明したのと同様であって、ここでは説明を省略する。
本発明において、トリガータンパク質に特異的に結合する核酸配列とヌクレアーゼのコード領域に対応する核酸配列が機能的に連結されるとは、ヌクレアーゼをコードするオープンリーディングフレーム(ただし、開始コドンを含む。)の5'UTR内、3'UTR内、及び/または当該オープンリーディングフレーム内に、少なくとも1つのトリガータンパク質に特異的に結合する核酸配列を備えることを意味する。ヌクレアーゼをコードする、トリガータンパク質応答性mRNAは、好ましくは、5'末端から、5'から3'の向きに、Cap構造(7メチルグアノシン5'リン酸)、ヌクレアーゼをコードするオープンリーディングフレーム並びに、ポリAテイルを備え、5'UTR内、3'UTR内、及び/またはオープンリーディングフレーム内に少なくとも1つのトリガータンパク質に特異的に結合する核酸配列を備える。mRNAにおけるトリガータンパク質に特異的に結合する核酸配列の位置は、5'UTRであっても、3'UTRであってもよく、オープンリーディングフレーム内(開始コドンの3'側)であってもよく、これらのすべてにトリガータンパク質に特異的に結合する核酸配列を備えていてもよい。したがって、トリガータンパク質に特異的に結合する核酸配列の数は、1つ、2つ、3つ、4つ、5つ、6つ、7つ、8つあるいはそれ以上であっても良い。
好ましくは、ヌクレアーゼをコードする、トリガータンパク質応答性mRNAは、(ia)および(iia)の核酸配列が、5'から3'の方向にこの順序で連結されている。このとき、cap構造とトリガータンパク質に特異的に結合する核酸配列との間の塩基数及び塩基の種類は、ステム構造や立体構造を構成しない限り、任意であってよい。例えば、cap構造とトリガータンパク質に特異的に結合する核酸配列との間の塩基数は、0~50塩基、好ましくは、10~30塩基となるように設計することができる。また、トリガータンパク質に特異的に結合する核酸配列と開始コドンとの間の塩基数及び塩基の種類は、ステム構造や立体構造を構成しない限り、任意であってよく、トリガータンパク質に特異的に結合する核酸配列と開始コドンと間の塩基数は、0~50塩基、好ましくは、10~30塩基となるような配置にて設計することができる。
本発明において、トリガータンパク質応答性mRNA 中のトリガータンパク質に特異的に結合する核酸配列内には、開始コドンとなるAUGが存在しないことが好ましい。例えば、トリガータンパク質に特異的に結合する核酸配列が5'UTRに存在し、かつ、当該核酸配列内にAUGを含む場合には、3'側に連結されるマーカー遺伝子との関係上でインフレームとなるように設計されることが好ましい。あるいは、トリガータンパク質に特異的に結合する核酸配列内にAUGを含む場合、トリガータンパク質に特異的に結合する核酸配列内のAUGをGUGに変換して使用することも可能である。また、トリガータンパク質に特異的に結合する核酸配列内のAUGの影響を最小限に留めるために、5'UTR内におけるトリガータンパク質に特異的に結合する核酸配列の配置場所を適宜変更することができる。例えば、cap構造とトリガータンパク質に特異的に結合する核酸配列内のAUG配列との間の塩基数が、0~60塩基、例えば、0~15塩基、10~20塩基、20~30塩基、30~40塩基、40~50塩基、50~60塩基となるような配置にて設計され得る。
b)トリガータンパク質をコードするmiRNA応答性mRNA
本実施形態において、トリガータンパク質をコードするmiRNA応答性mRNAは、miRNA-responsive mRNA、あるいはmiRNAスイッチとも言い、以下の(ib)および(iib)の核酸配列を含むmRNAを意味する:(ib) miRNAによって特異的に認識される核酸配列、及び(iib) 前記トリガータンパク質のコード領域に対応する核酸配列を含む。
当該(ib)miRNAによって特異的に認識される核酸配列と(iib) トリガータンパク質のコード領域に対応する核酸配列は、機能的に連結されている。図8(a)のmRNA2は、本実施形態に係る方法において使用しうるmiRNA-responsive mRNAの一例を模式的に示す図である。当該miRNA-responsive mRNAにおいては、5'UTRにmiRNAに応答する配列が組みこまれ、蛋白質コーディングリージョンにトリガータンパク質をコードする遺伝子が挿入されている。
本実施形態において、トリガータンパク質をコードするmiRNA応答性mRNAは、miRNA-responsive mRNA、あるいはmiRNAスイッチとも言い、以下の(ib)および(iib)の核酸配列を含むmRNAを意味する:(ib) miRNAによって特異的に認識される核酸配列、及び(iib) 前記トリガータンパク質のコード領域に対応する核酸配列を含む。
当該(ib)miRNAによって特異的に認識される核酸配列と(iib) トリガータンパク質のコード領域に対応する核酸配列は、機能的に連結されている。図8(a)のmRNA2は、本実施形態に係る方法において使用しうるmiRNA-responsive mRNAの一例を模式的に示す図である。当該miRNA-responsive mRNAにおいては、5'UTRにmiRNAに応答する配列が組みこまれ、蛋白質コーディングリージョンにトリガータンパク質をコードする遺伝子が挿入されている。
(b)トリガータンパク質をコードするmiRNA応答性mRNAにおける「miRNA」の定義、miRNAによって特異的に認識される核酸配列の定義については、第1実施形態において説明したのと同様であり、説明を省略する。また、本実施形態においても、(ib) miRNAによって特異的に認識される核酸配列を、miRNAの標的配列、とも指称する。
(iib)トリガータンパク質のコード領域に対応する核酸配列は、トリガータンパク質のコーディング配列である。トリガータンパク質は、(ia) トリガータンパク質に特異的に結合する核酸配列との関係で決定され、(ia)のRNA配列と、(iib)トリガータンパク質とが特異的に結合する組み合わせを選択して設計することができる。例えば、(ia) トリガータンパク質に特異的に結合する核酸配列が、boxC motif(配列番号40)、kink-loop(配列番号41)、kink-loop2(配列番号42)の場合、トリガー蛋白質は、L7Ae(Moore T et al., Structure Vol. 12, pp. 807-818 (2004))である。他にも、先に例示した(ia) トリガータンパク質に特異的に結合する核酸配列に対し、対応するトリガータンパク質を用いることができる。
本発明において、(ib) miRNAによって特異的に認識される核酸配列(miRNAの標的配列)、及び(iib)トリガータンパク質のコード領域に対応する核酸配列が機能的に連結されるとは、トリガータンパク質をコードするオープンリーディングフレーム(ただし、開始コドンを含む。)の5'UTR内、3'UTR内、及び/または当該オープンリーディングフレーム内に、少なくとも1つのmiRNAの標的配列を備えることを意味する。トリガータンパク質をコードするmiRNA応答性mRNAは、好ましくは、5'末端から、5'から3'の向きに、Cap構造(7メチルグアノシン5'リン酸)、トリガータンパク質をコードするオープンリーディングフレーム並びに、ポリAテイルを備え、5'UTR内、3'UTR内、及び/またはオープンリーディングフレーム内に少なくとも1つのmiRNAの標的配列を備える。mRNAにおけるmiRNAの標的配列の位置は、5'UTRであっても、3'UTRであってもよく、オープンリーディングフレーム内(開始コドンの3'側)であってもよく、これらのすべてにmiRNAの標的配列を備えていてもよい。したがって、miRNAの標的配列の数は、1つ、2つ、3つ、4つ、5つ、6つ、7つ、8つあるいはそれ以上であっても良い。
好ましくは、トリガータンパク質をコードするmiRNA応答性mRNAは、(ib)および(iib)の核酸配列が、5'から3'の方向にこの順序で連結されている。(ib)および(iib)の核酸配列や、Cap構造との間の塩基数、(ib)の配列の数や配置について、及び開始コドンと同じ配列についての設計は、第1実施形態、あるいはa)ヌクレアーゼをコードするトリガータンパク質応答性mRNAにおける設計と同様であってよい。
a)ヌクレアーゼをコードするトリガータンパク質応答性mRNA、及びb)トリガータンパク質をコードするmiRNA応答性mRNAは、上記のように設計ができれば、通常の遺伝子工学的手法により作製することができる。これらの2種のmRNAは2つで1セットとして機能する。細胞導入のために、miRNAの標的配列、トリガータンパク質、及びヌクレアーゼがそれぞれ異なっている2セット以上のmRNAを設計することもできる。a)、b)のmRNAのセットは、細胞内に導入する際に、共導入することが好ましい。また、この際、第1実施形態と同様に、コントロールmRNAも共導入することができる。
次に、このようなa)、b)のmRNAのセットによる、ヌクレアーゼの制御について説明する。細胞内に導入された当該miRNA-responsive mRNAは、細胞内に、標的配列を特異的に認識するmiRNAが存在すると、翻訳抑制されてトリガータンパク質の発現量が低下する。トリガータンパク質が存在すると、トリガータンパク質応答性mRNAに結合して翻訳抑制されるところ、トリガータンパク質の発現量が低下すると逆にトリガータンパク質応答性mRNAの翻訳量が増加し、ヌクレアーゼの翻訳を促進して、ヌクレアーゼ活性を向上させることができる。これにより、ヌクレアーゼの標的となる標的遺伝子の切断活性も向上し、標的遺伝子が切断されその機能消失を引き起こす。一方、細胞内に、標的配列を特異的に認識するmiRNAが存在しないと、miRNA-responsive mRNAは翻訳抑制されることがなく、細胞内でトリガータンパク質が発現する。すると、トリガータンパク質がトリガータンパク質応答性mRNAの翻訳を抑制して、ヌクレアーゼの発現量が低下する。その結果、ヌクレアーゼが標的遺伝子を切断する確率は減少し、標的遺伝子の機能は維持される。
次に、このようなa)、b)のmRNAのセットによる、ヌクレアーゼの制御について説明する。細胞内に導入された当該miRNA-responsive mRNAは、細胞内に、標的配列を特異的に認識するmiRNAが存在すると、翻訳抑制されてトリガータンパク質の発現量が低下する。トリガータンパク質が存在すると、トリガータンパク質応答性mRNAに結合して翻訳抑制されるところ、トリガータンパク質の発現量が低下すると逆にトリガータンパク質応答性mRNAの翻訳量が増加し、ヌクレアーゼの翻訳を促進して、ヌクレアーゼ活性を向上させることができる。これにより、ヌクレアーゼの標的となる標的遺伝子の切断活性も向上し、標的遺伝子が切断されその機能消失を引き起こす。一方、細胞内に、標的配列を特異的に認識するmiRNAが存在しないと、miRNA-responsive mRNAは翻訳抑制されることがなく、細胞内でトリガータンパク質が発現する。すると、トリガータンパク質がトリガータンパク質応答性mRNAの翻訳を抑制して、ヌクレアーゼの発現量が低下する。その結果、ヌクレアーゼが標的遺伝子を切断する確率は減少し、標的遺伝子の機能は維持される。
次に、より具体的な例として、ヌクレアーゼとして、Cas9タンパク質を用いる場合の本発明の方法を説明する。本発明は、一実施態様によれば、Cas9タンパク質をコードするトリガー蛋白質応答性 mRNAと、トリガータンパク質をコードするmiRNA応答性mRNA、加えて前記Cas9タンパク質の標的配列によって特異的に認識されるガイド配列を備えるsgRNAを細胞に共導入する工程を含む。図11(a)は、Cas9タンパク質をコードするL7Ae-responsive mRNAと、L7AeをコードするmiRNA応答性mRNAとを細胞に導入する場合の作用を模式的に示す図である。所定のmiRNAが存在すると、前段落に示したメカニズムにてヌクレアーゼ活性が向上し、sgRNAの標的遺伝子のゲノム編集、具体的にはsgRNAの標的DNAの切断活性を向上させることができる。
以下、本発明を、実施例を用いてより詳細に説明する。しかしながら、下記の実施例は、本発明を限定するものではない。
IVT (in vitro transcription) に用いる鋳型DNAの作製
5'UTRの鋳型(miRNA標的配列なし)と3'UTRの鋳型は対応するプライマーとKOD-Plus-Neo(KOD-401,TOYOBO)を用いて、以下のサイクルで(94°C 2min後、98°C 10 sec、68°C 10 secを13サイクル行い、4°Cで保存)PCR増幅を行った。Cas9蛋白をコードする遺伝子は、鋳型プラスミド(pHL-EF1a-SphcCas9-iC-A) から対応するプライマーとKOD-Plus-Neo(KOD-401,TOYOBO)を用いて、以下のサイクルで(94°C 2 min後、98°C 10 sec、68°C 140 secを20サイクル行い、4°Cで保存)PCR増幅を行った。
5'UTRの鋳型(miRNA標的配列なし)と3'UTRの鋳型は対応するプライマーとKOD-Plus-Neo(KOD-401,TOYOBO)を用いて、以下のサイクルで(94°C 2min後、98°C 10 sec、68°C 10 secを13サイクル行い、4°Cで保存)PCR増幅を行った。Cas9蛋白をコードする遺伝子は、鋳型プラスミド(pHL-EF1a-SphcCas9-iC-A) から対応するプライマーとKOD-Plus-Neo(KOD-401,TOYOBO)を用いて、以下のサイクルで(94°C 2 min後、98°C 10 sec、68°C 140 secを20サイクル行い、4°Cで保存)PCR増幅を行った。
上記で作製したPCR産物とそれぞれ対応するプライマー(miRNA標的配列を挿入するときは、5'UTRの代わりにオリゴDNAを使用)を用いてIVTの鋳型となるFull DNA TemplateをPCRにより作製した。Control Cas9 mRNAの鋳型は、(94°C 2 min後、98°C 10 sec, 68°C 140 secを20サイクル行い、4°Cで保存)の条件で、miRNAに応答するCas9 mRNAの鋳型は、(94°C 2 min後、98°C 10 sec, 60°C 30 sec, 68°C 140 secを20サイクル行い、4°Cで保存)の条件で行った。 sgRNAの鋳型は、2つのprimerを(98°C 30 sec, 98°C 10 sec, 57°C 30 sec,68°C, 6 secを20サイクル行い、72°C 10 minで反応後、4°Cで保存)の条件でPCRにより作製した。
全てのPCR産物は、MiniElute PCR purification Kit (QIAGEN)により精製した。但し、PCR反応でプラスミドを用いたものに関しては、精製前に制限酵素Dpn Iによる処理を施した。対応するプライマー及びオリゴヌクレオチド配列を表9A、Bに示す。


Cas9 mRNAとsgRNAの作製と精製
Cas9 mRNAはMegaScript kit (Ambion)により作製した。この時、免疫反応を抑えるため修飾塩基pseudouridine-5'-triphosphateと5-methylcytidine-5'-triphosphate (TriLink Bio Technologies)をUTPとCTPの代わりにそれぞれ加えた。またGTPは、Anti Reverse Cap Analog (TriLink Bio Technologies)で5倍希釈した。sgRNAは天然の塩基(ATP, GTP, CTP, UTP)を用いMEGAshortscript kit (Ambion)により作製した。反応混合液を37°Cで6時間インキュベートして、TURBO DNase (Amibion)を加えた後、37°Cでさらに30分インキュベートした。得られたmRNAは、FavorPrep Blood / Cultured Cells total RNA extraction column (Favorgen Biotech)で精製し、Antarctic phosphatase (New England Biolabs)を用いて37C°で30分インキュベートした。その後、RNeasy Mini Elute Cleanup Kit (QIAGEN)により、さらに精製した。sgRNAは精製後さらにウレアページ(10%)による切り出し精製を行った。Cas9 mRNAのcoding region、並びに5'UTR、3'UTRの配列を下記表10A、10Bに示す。
Cas9 mRNAはMegaScript kit (Ambion)により作製した。この時、免疫反応を抑えるため修飾塩基pseudouridine-5'-triphosphateと5-methylcytidine-5'-triphosphate (TriLink Bio Technologies)をUTPとCTPの代わりにそれぞれ加えた。またGTPは、Anti Reverse Cap Analog (TriLink Bio Technologies)で5倍希釈した。sgRNAは天然の塩基(ATP, GTP, CTP, UTP)を用いMEGAshortscript kit (Ambion)により作製した。反応混合液を37°Cで6時間インキュベートして、TURBO DNase (Amibion)を加えた後、37°Cでさらに30分インキュベートした。得られたmRNAは、FavorPrep Blood / Cultured Cells total RNA extraction column (Favorgen Biotech)で精製し、Antarctic phosphatase (New England Biolabs)を用いて37C°で30分インキュベートした。その後、RNeasy Mini Elute Cleanup Kit (QIAGEN)により、さらに精製した。sgRNAは精製後さらにウレアページ(10%)による切り出し精製を行った。Cas9 mRNAのcoding region、並びに5'UTR、3'UTRの配列を下記表10A、10Bに示す。


培養細胞
iPS_GFP (AAVS1-CAG::GFP iPS cells) はWoltjen Lab (CiRA, Kyoto University, Japan)より供与されたものである。iPS_GFPは、StemFit (Ajinomoto)培地を用いてラミニンコート{laminin-511 E8 (iMatrix-511, nippi)}したプレートで培養した。HeLa_GFPはDMEM High Glucose (nakarai tesque)にFBS (JBS、終濃度10%)とHygromycin B (50 mg / mL)を加えた培地で培養した。Normal HeLa cellsはDMEM High Glucose (nakarai tesque)にFBS (JBS、終濃度10%)を加えた培地で培養した。全ての細胞は37°C、5% CO2の条件で培養した。
分化誘導(iPS_GFPからmDA_GFPへの分化誘導)
Day0にiPS_GFP細胞を、分化誘導培地を用いてラミニンコートした6-wellプレートに再播種(5x106 cells/well)し、Morizana et al., Neural Development: Methods and Protocols, Methods in Molecular Biology, vol.1018,DOI 10.1007/978-1-62703-444-9_2 を改変したプロトコールにより培養した。以後、毎日培地交換を行った。14日後にそれぞれの実験に用いた。分化誘導培地の組成を下記の表11に示した。

Day0にiPS_GFP細胞を、分化誘導培地を用いてラミニンコートした6-wellプレートに再播種(5x106 cells/well)し、Morizana et al., Neural Development: Methods and Protocols, Methods in Molecular Biology, vol.1018,DOI 10.1007/978-1-62703-444-9_2 を改変したプロトコールにより培養した。以後、毎日培地交換を行った。14日後にそれぞれの実験に用いた。分化誘導培地の組成を下記の表11に示した。
トランスフェクション
normal HeLaとHeLa_GFP は24-wellプレートに播種した。iPS_GFPとmDA_GFPはラミニンコートした24-wellプレートに播種した(細胞数:5x104cells/well)。 トランスフェクションはStemfect RNA transfection kit (Stemgent)を用いプロトコールに従い行った(導入遺伝子量はそれぞれの実験項を参照)。トランスフェクション後4時間で培地を交換した(但し、mDA_GFPを除く)。Cell killingはトランスフェクション48時間後に、T7E1 assay、EGFP activity assay、co-cultureはトランスフェクション72時間後に解析を行った。それぞれの解析に入る前に、IX81 microscope (Olympus)で細胞を撮影した(図2)。
normal HeLaとHeLa_GFP は24-wellプレートに播種した。iPS_GFPとmDA_GFPはラミニンコートした24-wellプレートに播種した(細胞数:5x104cells/well)。 トランスフェクションはStemfect RNA transfection kit (Stemgent)を用いプロトコールに従い行った(導入遺伝子量はそれぞれの実験項を参照)。トランスフェクション後4時間で培地を交換した(但し、mDA_GFPを除く)。Cell killingはトランスフェクション48時間後に、T7E1 assay、EGFP activity assay、co-cultureはトランスフェクション72時間後に解析を行った。それぞれの解析に入る前に、IX81 microscope (Olympus)で細胞を撮影した(図2)。
T7E1 assay
100 ngのCas9 mRNAと300 ng (iPS_GFP)もしくは100 ng (mDA_GFP)のsgRNAを使用した。トランスフェクション後の細胞をPBSで洗浄後、200 μL Accumax (Funakoshi) で37°C、5% CO2 の条件で10分間処理した。1.5 mL Tubeに細胞を回収し遠心した(室温、1000 rpmで5分)。上精を廃棄後、沈殿した細胞をPBSで洗浄し、先ほどと同じ条件で遠心した。Lysis buffer (1 M Tris-HCl (pH.7.6)[final 0.05 M], 0.5 M EDTA [final 0.02 M], 5 M NaCl [0.1 M], 10% SDS[final 1%], D2W)にproteinase K (x100; final 1x)を500mL加えて55度で3時間以上処理した。その後、PCIを用いてゲノムDNAの抽出を行った。Nested PCRにより、抽出したゲノムDNAから標的配列を増幅した。First PCRは、(94°C 2 min後、98°C 10 sec, 60°C 30 sec, 68°C 30 secを20サイクル行い、72°C 3 min反応後、4°C で保存)、Second PCRは、(94°C 2 min後、98°C 10 sec, 60°C 30 sec,68°C 15 secを35サイクル行い、72°C 3 min反応後、4°C で保存)の条件で行った。PCR産物は、MiniElute PCR purification Kit (QIAGEN)により精製した。精製後、PCR産物の変性と再会合を、(95°C 5 min後、 95°C から85°Cまで毎秒2度で冷却し、85°Cから25°Cまでは毎秒0.1°Cで冷却し、その後4°Cで保存)の条件で行った。反応後、制限酵素T7 Endonuclease Iによる処理を施した(37℃で 15 min)。15分後、0.5 M EDTAを加え反応を停止させ、5%ポリアクリルアミドゲル電気泳動を行い、SYBER GREEN mixture (I+II =1:1)で染色し撮影した。PCRで使用したプライマーを表9に示した。
Indels (Cas9 activity)を以下の公式により算出した。
Indels = 100 × (1 - sqrt(1 - (b + c)/(a + b + c)))
a : 制限酵素で切断されなかったPCR産物のバンド強度
b,c : 制限酵素で切断されたPCR産物のバンド強度
100 ngのCas9 mRNAと300 ng (iPS_GFP)もしくは100 ng (mDA_GFP)のsgRNAを使用した。トランスフェクション後の細胞をPBSで洗浄後、200 μL Accumax (Funakoshi) で37°C、5% CO2 の条件で10分間処理した。1.5 mL Tubeに細胞を回収し遠心した(室温、1000 rpmで5分)。上精を廃棄後、沈殿した細胞をPBSで洗浄し、先ほどと同じ条件で遠心した。Lysis buffer (1 M Tris-HCl (pH.7.6)[final 0.05 M], 0.5 M EDTA [final 0.02 M], 5 M NaCl [0.1 M], 10% SDS[final 1%], D2W)にproteinase K (x100; final 1x)を500mL加えて55度で3時間以上処理した。その後、PCIを用いてゲノムDNAの抽出を行った。Nested PCRにより、抽出したゲノムDNAから標的配列を増幅した。First PCRは、(94°C 2 min後、98°C 10 sec, 60°C 30 sec, 68°C 30 secを20サイクル行い、72°C 3 min反応後、4°C で保存)、Second PCRは、(94°C 2 min後、98°C 10 sec, 60°C 30 sec,68°C 15 secを35サイクル行い、72°C 3 min反応後、4°C で保存)の条件で行った。PCR産物は、MiniElute PCR purification Kit (QIAGEN)により精製した。精製後、PCR産物の変性と再会合を、(95°C 5 min後、 95°C から85°Cまで毎秒2度で冷却し、85°Cから25°Cまでは毎秒0.1°Cで冷却し、その後4°Cで保存)の条件で行った。反応後、制限酵素T7 Endonuclease Iによる処理を施した(37℃で 15 min)。15分後、0.5 M EDTAを加え反応を停止させ、5%ポリアクリルアミドゲル電気泳動を行い、SYBER GREEN mixture (I+II =1:1)で染色し撮影した。PCRで使用したプライマーを表9に示した。
Indels (Cas9 activity)を以下の公式により算出した。
Indels = 100 × (1 - sqrt(1 - (b + c)/(a + b + c)))
a : 制限酵素で切断されなかったPCR産物のバンド強度
b,c : 制限酵素で切断されたPCR産物のバンド強度
Cas9 activity assay
100 ngのCas9 mRNAと300 ng (iPS_GFP, HeLa_GFP)もしくは100 ng (mDA_GFP)のsgRNAと5 pmolの miRNA inhibitor(mirVana)もしくはnegative control (miRNA inhibitorとnegative controlは任意)を使用した。細胞をPBSで洗浄した。その後、HeLa_GFPは100 μL 0.25% trypsin-EDTAで37°C、5% CO2 の条件で5分間処理した後、100 μLの培地を加えた。iPS_GFPとmDA_GFPは200 μL Accumax (Funakoshi) で37°C、5% CO2 の条件で10分間処理した。1.5 mLチューブに細胞を回収し、CYTOX Red dead-cell stain (ThermoFisher)で処理した(15分間、遮光して室温放置)。Aria-II (BD)、Accuri(BD)、LSR(BD)により測定した。 EGFP negative cells (%)をCas9 activity (%)と定義した。
100 ngのCas9 mRNAと300 ng (iPS_GFP, HeLa_GFP)もしくは100 ng (mDA_GFP)のsgRNAと5 pmolの miRNA inhibitor(mirVana)もしくはnegative control (miRNA inhibitorとnegative controlは任意)を使用した。細胞をPBSで洗浄した。その後、HeLa_GFPは100 μL 0.25% trypsin-EDTAで37°C、5% CO2 の条件で5分間処理した後、100 μLの培地を加えた。iPS_GFPとmDA_GFPは200 μL Accumax (Funakoshi) で37°C、5% CO2 の条件で10分間処理した。1.5 mLチューブに細胞を回収し、CYTOX Red dead-cell stain (ThermoFisher)で処理した(15分間、遮光して室温放置)。Aria-II (BD)、Accuri(BD)、LSR(BD)により測定した。 EGFP negative cells (%)をCas9 activity (%)と定義した。
Cell killing system
10 ngのCas9 mRNAと300 ngのsgRNAを使用した。PBSで洗浄する前に、培地を1.5 mLチューブに回収した。PBSでの洗浄後、normal HeLaを100 μL 0.25% trypsin-EDTAで37°C、5% CO2 の条件で5分間処理した。その後、100 μLの培地を加えて細胞を先ほどの1.5 mLチューブに回収し遠心した(室温、1000 rpmで5分)。上精を廃棄後、沈殿した細胞をPBSで洗浄し、先ほどと同じ条件で遠心した。上精を廃棄後、53 μLの染色試薬の混合物{annexin V, Alexa Fluor 488 conjugate (Life Technologies)、 Annexin-binding buffer 5x (Life Technologies)、 SYTOX Red dead-cell stain}で染色した。 Accuri (BD)により測定した。
10 ngのCas9 mRNAと300 ngのsgRNAを使用した。PBSで洗浄する前に、培地を1.5 mLチューブに回収した。PBSでの洗浄後、normal HeLaを100 μL 0.25% trypsin-EDTAで37°C、5% CO2 の条件で5分間処理した。その後、100 μLの培地を加えて細胞を先ほどの1.5 mLチューブに回収し遠心した(室温、1000 rpmで5分)。上精を廃棄後、沈殿した細胞をPBSで洗浄し、先ほどと同じ条件で遠心した。上精を廃棄後、53 μLの染色試薬の混合物{annexin V, Alexa Fluor 488 conjugate (Life Technologies)、 Annexin-binding buffer 5x (Life Technologies)、 SYTOX Red dead-cell stain}で染色した。 Accuri (BD)により測定した。
Co-culture
50 ngのCas9 mRNAと150 ngのsgRNAを使用した。iPS_GFP細胞とHeLa_GFP細胞を3対2の比率で播種したCas9 activity assayと同様の方法で細胞を1.5 mLチューブに回収したのち、遠心を行った(室温、1000 rpmで5分)。上精を除去したのち、Alexa FluorR 647 Mouse anti-Human TRA-1-60 Antigen をプロトコールに従い染色(プロトコールの2倍量の抗体を使用)し、LSRで測定した。
50 ngのCas9 mRNAと150 ngのsgRNAを使用した。iPS_GFP細胞とHeLa_GFP細胞を3対2の比率で播種したCas9 activity assayと同様の方法で細胞を1.5 mLチューブに回収したのち、遠心を行った(室温、1000 rpmで5分)。上精を除去したのち、Alexa FluorR 647 Mouse anti-Human TRA-1-60 Antigen をプロトコールに従い染色(プロトコールの2倍量の抗体を使用)し、LSRで測定した。
結果
HeLa細胞におけるmiRNA-responsive CRISPR/Cas9 Systemの挙動を図2に示す。HeLa細胞では、内在性miR-21の活性が高いことが知られている。図2(a)は、miRNA-responsive mRNAを導入したHeLa細胞の蛍光顕微鏡画像である。miRNA に応答しないControlと、miRNA-302aに応答するCas9 mRNAでは、GFPの蛍光がUntreatedや、標的配列に対するネガティブコントロールに比べ弱くなっている。なお、ネガティブコントロールとしては、DMD遺伝子を標的としたものを用いた。一方、miRNA-21に応答するCas9 mRNAでは蛍光に変化が見られなかった。なお、図2(a)は、3回行った実験のうちの1回を代表として記載した。また、図2(b)から、上記蛍光画像と、Aria-II解析による蛍光強度の定量結果が一致していることが明らかになった。2(b)も、3回行った実験のうちの1回を代表として記載した。図2(c)は、上記図2(b)のGFP negative populationをCas9 activityと定義しグラフ化した図である。なお、以降のCas9 activityは特別な記述がない限り、この方法で算出したものとする。以上より、HeLa細胞に各種miRNA-responsive mRNA及びコントロールmRNAを導入した場合に、miR-21に応答するCas9 mRNAのみがCas9 activityが低いことがわかり、miR-21に応答するCas9 mRNA がmiR-21によって制御されている可能性が示された。
HeLa細胞におけるmiRNA-responsive CRISPR/Cas9 Systemの挙動を図2に示す。HeLa細胞では、内在性miR-21の活性が高いことが知られている。図2(a)は、miRNA-responsive mRNAを導入したHeLa細胞の蛍光顕微鏡画像である。miRNA に応答しないControlと、miRNA-302aに応答するCas9 mRNAでは、GFPの蛍光がUntreatedや、標的配列に対するネガティブコントロールに比べ弱くなっている。なお、ネガティブコントロールとしては、DMD遺伝子を標的としたものを用いた。一方、miRNA-21に応答するCas9 mRNAでは蛍光に変化が見られなかった。なお、図2(a)は、3回行った実験のうちの1回を代表として記載した。また、図2(b)から、上記蛍光画像と、Aria-II解析による蛍光強度の定量結果が一致していることが明らかになった。2(b)も、3回行った実験のうちの1回を代表として記載した。図2(c)は、上記図2(b)のGFP negative populationをCas9 activityと定義しグラフ化した図である。なお、以降のCas9 activityは特別な記述がない限り、この方法で算出したものとする。以上より、HeLa細胞に各種miRNA-responsive mRNA及びコントロールmRNAを導入した場合に、miR-21に応答するCas9 mRNAのみがCas9 activityが低いことがわかり、miR-21に応答するCas9 mRNA がmiR-21によって制御されている可能性が示された。
miRNA-21に対するインヒビターを用いることで、miRNA-21に応答するCas9 mRNAが内在性のmiR-21により制御されているのかをHeLa細胞を用いて検証した。結果を図3に示す。miRNAに対するインヒビターのネガティブコントロール(以下Negative controlと図では表記)では、Cas9 activityが低いのに対してmiR-21 inhibitorを用いるとCas9 activityが上昇(rescue)したことが明らかとなった。つまり、miRNA-21に応答するCas9 mRNAは、内在性のmiR-21により制御されていることが明らかになった。
miRNA-302aに対するインヒビターを用いることで、miRNA-302aに応答するCas9 mRNAが内在性のmiR-302aにより制御されているのかをiPS細胞を用いて検証した。iPS細胞は、miR-302aの活性が高いことが知られている。結果を図4に示す。Negative controlではCas9 activityが低いのに対して、miR-302a inhibitorを用いるとCas9 activityがrescueされた。つまり、miRNA-302aに応答するCas9 mRNAは内在性のmiR-302aにより制御されていることが判明した。図4(a)は、Cas9 activity assayにより評価した結果を、図4(b)は、T7E1 assayにより評価したゲル画像を示す。ゲル画像は、3回行った実験のうちの1回を代表として図示した。図4(c)は、T7E1 assayよりIndels (Cas9 activityと定義)を算出した結果を示す。算出の公式については、T7E1 assayの項目において詳述したとおりである。
図5は、本発明のmiRNA-responsive CRISPR/Cas9 Systemを用いて、細胞死を引き起こすスキームを模式的に示す図である。図5(a)は、sgRNAの配列をゲノム上のリピート配列にすることでCas9がゲノムを断片化することを模式的に示す。ゲノムが断片化されることにより細胞は細胞死を起こす。本実験では、miR-21-responsive Cas9 mRNAを用い、Alu1を標的として、細胞死の制御を検証した。実験は、Cell Killing Sysetemに記載の方法に従い、HeLa細胞においてmiR-21に応答するCas9 mRNAを用いることにより細胞死が制御できるのかを検証した。結果を、図5(b)に示す。mi-21-responsive Cas9 mRNA は、miR-21が特異的に発現している細胞において、Cas9 mRNAの翻訳が抑制され、Cas9活性が低下するように設計されている。図5(b)から、ControlのCas9 mRNA (Cas9 activityは高い)では細胞死が誘導されていることがわかる。一方、miR-21に応答するCas9 mRNA(Cas9 activityは低い)ではUntreatedと同じくらいの死細胞率だった。この結果より、miR-21-responsive Cas9 mRNAにより、細胞死が誘導されなかったことがわかる。これより、miRNA-responsive CRISPR/Cas9 Systemを用いることで細胞死が制御可能であることが実証された。
図6は、miR-302a-responsive Cas9 mRNAをmDA細胞へ導入した実験の結果を示す。miRNA responsive mRNA (control, 302a-responsive, 4x miR-302a-responsive)と、sgRNA (GFP を標的としているもの、もしくは negative control として DMD を標的としたもの)を調製した。4x miR-302a-responsive Cas9 mRNAは、5'UTR に miR-302a-5p 標的配列を 4 個挿入したものである。mDA細胞(midbrain dopaminergic neurons)は、先の方法に従ってiPS細胞から分化誘導したものを用いた。miRNA responsive mRNA、sgRNAのそれぞれ100 ngをmDA細胞に共導入し、Cas9 activity assayに従って、GFP knockout による蛍光強度の変化を評価した。結果を図6に示す。図6(a)は、Relative Cas9 activity を示し、Cas9 activity を Cas9 mRNA = 1 としてグラフ化したものである。各回に1つのサンプルで実験を行い、独立してその実験を3回行った(n=3)。エラーバーは、平均±標準偏差を示す。
図6(a)と同様にして調製した、miRNA responsive mRNA、sgRNAを、それぞれ100 ngをmDA細胞に共導入し、T7E1 assay により評価した。図6(b)は、代表的なゲル画像、図6(c)は、T7E1 assay に基づくCas9 activity を Cas9 mRNA = 1 として表したRelative Cas9 activity を示す。各回に1つのサンプルで実験を行い、独立してその実験を3回行った(n=3)。エラーバーは、平均±標準偏差を示す。
図6の結果から、本発明に係るmiRNA-responsive CRISPR/Cas9 Systemが、細胞の状態、つまり、miRNAに呼応してCas9活性を制御していることが明らかになった。iPS細胞を分化させる(今回はmDA)ことで、内在性のmiR-302a-5pの活性が低下するため、miR-302aに応答するCas9 mRNAの活性が iPSでは低いがmDAでは高いこと(Control Cas9 mRNAと同様の活性)が予測される。実際に、Relative Cas9 activityを算出したところ、miR-302aに応答するCas9 mRNAで活性が回復し、Control Cas9 mRNAに近い活性を示した。
図7は、co-cultureの結果を示す。この実験では、iPS_GFP と HeLa_GFP をトランスフェクション 17-24 時間前に 24 well plate に 5x105 cells/well で播種した。この時、 iPS : HeLa = 3 : 2 の比率(細胞数)で行った。mRNA 50ng, sgRNA 150ng を、stemfect transfection reagent をプロトコールに従い使用しトランスフェクションを行った。mRNA としては、control Cas9 mRNA, 4x miR-302a-responsive Cas9 mRNA を用いた。sgRNAとしては、GFP遺伝子を標的とするもの、及びnegative control としてDMD遺伝子を標的としたものを用いた。トランスフェクションから 3 日後、細胞を Alexa FluorR 647 Mouse anti-Human TRA-1-60 Antigen で染色後、LSR により測定した。GFP knockout による蛍光強度の変化により評価した結果を図7(a)に示す。各回に1つのサンプルで実験を行い、独立してその実験を3回行い(n=3)、代表的なドットプロットを図示した。各パネルにおいて、縦軸は GFPの蛍光強度、横軸は TRA-1-60蛍光強度を示す。右下に、TRA-1 により分離できる細胞集団の分布を示した。
図7(b)は、図7(a)の結果に基づいて、以下の式により算出されるCas9 activity を示すグラフである。エラーバーは、平均±標準偏差を示す。
Cas9 activity (%) = Q4/(Q1+Q4) x 100: HeLa
Cas9 activity (%) = Q3/(Q2+Q3) x 100: iPS
Cas9 activity (%) = Q4/(Q1+Q4) x 100: HeLa
Cas9 activity (%) = Q3/(Q2+Q3) x 100: iPS
図7の結果から、異なる細胞集団から標的とする細胞集団に対してのみゲノム編集を行える可能性が示唆された。つまり、本実験では、HeLa細胞とiPS細胞において、iPS細胞で、miR-302aの発現(活性)が高いことを利用しすることにより、iPS細胞ではGFP knockoutを起こらないようにする一方で、HeLa細胞ではGFP knockoutを引き起こすことに成功した。
トランスフェクション
normal HeLaとHeLa_GFP は24-wellプレートに播種した。iPS_GFPとmDA_GFPはラミニンコートした24-wellプレートに播種した(細胞数:5x104 cells/well)。 トランスフェクションはStemfect RNA transfection kit (Stemgent)を用いプロトコールに従い行った(導入遺伝子量はそれぞれの実験項を参照)。トランスフェクション後4時間で培地を交換した(但し、mDA_GFPを除く)。Evaluation of gene expression level, Evaluation of Cas9 protein expression level, Evaluation of transfection efficiencyはトランスフェクション24時間後に、Cell killingはトランスフェクション48時間後に、T7E1 assay、 EGFP activity assay、co-culture、On-systemはトランスフェクション72時間後に解析を行った。それぞれの解析に入る前に、IX81 microscope (Olympus)で細胞を撮影した。
normal HeLaとHeLa_GFP は24-wellプレートに播種した。iPS_GFPとmDA_GFPはラミニンコートした24-wellプレートに播種した(細胞数:5x104 cells/well)。 トランスフェクションはStemfect RNA transfection kit (Stemgent)を用いプロトコールに従い行った(導入遺伝子量はそれぞれの実験項を参照)。トランスフェクション後4時間で培地を交換した(但し、mDA_GFPを除く)。Evaluation of gene expression level, Evaluation of Cas9 protein expression level, Evaluation of transfection efficiencyはトランスフェクション24時間後に、Cell killingはトランスフェクション48時間後に、T7E1 assay、 EGFP activity assay、co-culture、On-systemはトランスフェクション72時間後に解析を行った。それぞれの解析に入る前に、IX81 microscope (Olympus)で細胞を撮影した。
On-system
図11Aに概要を示すようにmRNAを設計した。miRNAの標的部位としては、miR-21に相補的な配列を用いた。miRNAに応答して発現抑制されるトリガータンパク質としては、L7Aeタンパク質を用い、L7Aeタンパク質に特異的に結合する配列としては、キンクターンモチーフを用い、先の方法と同様にして、mRNAを作製した。作製したmRNA(Kt-Cas9 mRNA)の配列は以下の表12A、Bの通りであり、表中、L7Ae 結合モチーフは二重下線で、Cas9コード領域はイタリック体で示す。また、mRNA(L7Ae mRNA、miR-21-responsive L7Ae、Tag BFP mRNA)の配列は以下の表13の通りであり、表中、miR-21-5pの標的配列は二重下線で、L7AeまたはTag BFPのコード領域はイタリック体で示す。
図11Aに概要を示すようにmRNAを設計した。miRNAの標的部位としては、miR-21に相補的な配列を用いた。miRNAに応答して発現抑制されるトリガータンパク質としては、L7Aeタンパク質を用い、L7Aeタンパク質に特異的に結合する配列としては、キンクターンモチーフを用い、先の方法と同様にして、mRNAを作製した。作製したmRNA(Kt-Cas9 mRNA)の配列は以下の表12A、Bの通りであり、表中、L7Ae 結合モチーフは二重下線で、Cas9コード領域はイタリック体で示す。また、mRNA(L7Ae mRNA、miR-21-responsive L7Ae、Tag BFP mRNA)の配列は以下の表13の通りであり、表中、miR-21-5pの標的配列は二重下線で、L7AeまたはTag BFPのコード領域はイタリック体で示す。



HeLa_GFP をトランスフェクション 17-24 時間前に 24 well plate に 5x105 cells/well で播種した。
OFF 状態の条件:Control L7Ae mRNA 15 ng + kt-Cas9 mRNA (kt motifを5’UTRに持つ)5 ng + sgRNA 150 ng
ON 状態の条件:miR-21-responsive L7Ae mRNA 15 ng + kt-Cas9 mRNA 5 ng + sgRNA 150 ng
stemfect transfection reagent をプロトコールに従い使用しトランスフェクションを行った。トランスフェクションから 3 日後、Accuri により測定した。
OFF 状態の条件:Control L7Ae mRNA 15 ng + kt-Cas9 mRNA (kt motifを5’UTRに持つ)5 ng + sgRNA 150 ng
ON 状態の条件:miR-21-responsive L7Ae mRNA 15 ng + kt-Cas9 mRNA 5 ng + sgRNA 150 ng
stemfect transfection reagent をプロトコールに従い使用しトランスフェクションを行った。トランスフェクションから 3 日後、Accuri により測定した。
Evaluation of miRNA expression level
HeLa-EGFP, iPS-EGFP, mDA-EGFPの3種類の細胞を用いた。TaqMan(登録商標) MicroRNA Cells-to-CT(商標) Kit (Ambion)を用いて測定を行なった。細胞溶解液を has-miR-21-5p (Assay ID: 000397), 302a-5p (Assay ID: 002381) そして RNU6B (Assay ID: 001093) Taqman probes (Applied Biosystems)のそれぞれを用いて逆転写反応を行なった。qPCRはTaqman probeを用いて行い、StepOne Plus Real-Time PCR System (Applied Biosystems)を用いて行なった。標的miRNAをRNU6Bによりノーマライズした。さらに、mDA-EGFPを1となるようにノーマライズした。
HeLa-EGFP, iPS-EGFP, mDA-EGFPの3種類の細胞を用いた。TaqMan(登録商標) MicroRNA Cells-to-CT(商標) Kit (Ambion)を用いて測定を行なった。細胞溶解液を has-miR-21-5p (Assay ID: 000397), 302a-5p (Assay ID: 002381) そして RNU6B (Assay ID: 001093) Taqman probes (Applied Biosystems)のそれぞれを用いて逆転写反応を行なった。qPCRはTaqman probeを用いて行い、StepOne Plus Real-Time PCR System (Applied Biosystems)を用いて行なった。標的miRNAをRNU6Bによりノーマライズした。さらに、mDA-EGFPを1となるようにノーマライズした。
Evaluation of Cas9 protein expression level
100 ngのCas9 mRNAと300 ngの sgRNAを使用した。トランスフェクションから24時間後、PBS washを行い、50 μLの M-PER cocktail (M-PER Mammalian Protein Extraction Reagent [ThermoFisher], protease inhibitor, PMSFを混合したもの) により細胞を溶解し回収した。5分間振盪した後、1.5 mLチューブに溶液を回収した。遠心(12400 rpm, 4°C, 5 min)後、上清を新しい1.5 mL チューブに回収してタンパク質濃度をBCA法により測定した。0.5 mg/mLにタンパク質溶液を希釈後、Wes (ProteinSimple) のプロトコールに従いタンパク質の検出を行なった。GAPDHはローディングコントロールとして使用した。一次抗体:Cas9 antibody (50倍希釈、Active Motif), GAPDH antibody (100倍希釈、Santa Cruz)。二次抗体:anti-mouse, anti-rabbit (ProteinSimple)
100 ngのCas9 mRNAと300 ngの sgRNAを使用した。トランスフェクションから24時間後、PBS washを行い、50 μLの M-PER cocktail (M-PER Mammalian Protein Extraction Reagent [ThermoFisher], protease inhibitor, PMSFを混合したもの) により細胞を溶解し回収した。5分間振盪した後、1.5 mLチューブに溶液を回収した。遠心(12400 rpm, 4°C, 5 min)後、上清を新しい1.5 mL チューブに回収してタンパク質濃度をBCA法により測定した。0.5 mg/mLにタンパク質溶液を希釈後、Wes (ProteinSimple) のプロトコールに従いタンパク質の検出を行なった。GAPDHはローディングコントロールとして使用した。一次抗体:Cas9 antibody (50倍希釈、Active Motif), GAPDH antibody (100倍希釈、Santa Cruz)。二次抗体:anti-mouse, anti-rabbit (ProteinSimple)
Sequencing
T7E1 assayで得られた 2回目のPCR産物(iPS-EGFP)をpUC19ベクターに挿入後、シーケンサーにより配列を読んだ。まず2回目のPCRで用いたプライマーをT4PNKによりリン酸化をし、2回目のPCR産物に対して再度、PCRをかけた。その後pUC19ベクターに挿入し、M13 FwdNew primer, T7E1 Fwd primer, T7E1 Rev primerを用いて配列を決定した。シーケンサーはApplies Biosystems 3500xL Genetic analyzerを使用した。
T7E1 assayで得られた 2回目のPCR産物(iPS-EGFP)をpUC19ベクターに挿入後、シーケンサーにより配列を読んだ。まず2回目のPCRで用いたプライマーをT4PNKによりリン酸化をし、2回目のPCR産物に対して再度、PCRをかけた。その後pUC19ベクターに挿入し、M13 FwdNew primer, T7E1 Fwd primer, T7E1 Rev primerを用いて配列を決定した。シーケンサーはApplies Biosystems 3500xL Genetic analyzerを使用した。
Evaluation of gene expression level
HeLa-EGFPもしくはiPS-EGFPを用いた。また、100 ng Cas9 mRNA, 300 ng sgRNAを用いた。Trizol (ThermoFisher)をプロトコールに従い用いることで、total RNAを細胞より抽出した。また、TURBO DNase Inactivation Kit (Ambion)を用い、ゲノムDNAを除去した。ReverTra Acr(登録商標)qPCR RT Master Mix (TOYOBO)を用いて、上記処理を施したサンプル (250もしくは300 ng) を逆転写した。qPCRをTHUNDERBIRD(登録商標) SYBR(登録商標) qPCR Mix (TOYOBO)を用いて行なった。反応は、StepOne Plus Real-Time PCR System (Applied Biosystems)を使用した。 標的mRNAはGAPDHでノーマライズした。さらに、Control Cas9 mRNAでの遺伝子発現が1となるようにノーマライズした。
HeLa-EGFPもしくはiPS-EGFPを用いた。また、100 ng Cas9 mRNA, 300 ng sgRNAを用いた。Trizol (ThermoFisher)をプロトコールに従い用いることで、total RNAを細胞より抽出した。また、TURBO DNase Inactivation Kit (Ambion)を用い、ゲノムDNAを除去した。ReverTra Acr(登録商標)qPCR RT Master Mix (TOYOBO)を用いて、上記処理を施したサンプル (250もしくは300 ng) を逆転写した。qPCRをTHUNDERBIRD(登録商標) SYBR(登録商標) qPCR Mix (TOYOBO)を用いて行なった。反応は、StepOne Plus Real-Time PCR System (Applied Biosystems)を使用した。 標的mRNAはGAPDHでノーマライズした。さらに、Control Cas9 mRNAでの遺伝子発現が1となるようにノーマライズした。
Evaluation of transfection efficiency
HeLa-EGFPとiPS-EGFPを用いた。100 ngのBFP mRNAと300 ngのCy5でラベル化したsgRNAを用いた。トランスフェクションから24時間後、細胞をPBSで3回ウォッシュした。その後、細胞を1.5mLチューブに回収(細胞の剥がし方は、Cas9 activity assayの項参照)して、LSRにより測定した。
HeLa-EGFPとiPS-EGFPを用いた。100 ngのBFP mRNAと300 ngのCy5でラベル化したsgRNAを用いた。トランスフェクションから24時間後、細胞をPBSで3回ウォッシュした。その後、細胞を1.5mLチューブに回収(細胞の剥がし方は、Cas9 activity assayの項参照)して、LSRにより測定した。
Cas9 activity assay (iPS-EGFPでsgRNAを改変: 1, 2)
Cas9 activity assayと同様にして実施した。
Cas9 activity assayと同様にして実施した。
結果を図8~15に示す。ON-systemは内在性のmiRNAによってCas9の活性が上昇するシステムである。図8(a)はON-systemの概要を示す概念図である。Cas9を発現するmRNA1(古細菌由来のRNA結合タンパク質であるL7Aeが結合する配列: K-turnを5’UTRにコードしている)はL7Aeを発現するmRNA2(miRNAによって発現が制御されている)によって翻訳が制御されている。図8(b)はHeLa細胞の内在性のmiR-21により、Cas9活性が上昇したことを示している。つまり標的miRNA(ここではmiR-21)により、Cas9活性が上昇したことを示している。
miRNA expression level
図9(a)はmiR-21、図9(b)はmiR-302aの発現レベルを HeLa-EGFP, iPS-EGFP, mDA-EGFPでそれぞれ測定した結果である。HeLa-EGFP細胞ではmiR-21のiPS-EGFP細胞ではmiR-302aの発現が高いことを示している。
図9(a)はmiR-21、図9(b)はmiR-302aの発現レベルを HeLa-EGFP, iPS-EGFP, mDA-EGFPでそれぞれ測定した結果である。HeLa-EGFP細胞ではmiR-21のiPS-EGFP細胞ではmiR-302aの発現が高いことを示している。
Evaluation of Cas9 protein expression level
Control Cas9 mRNAとmiR-21-responsive Cas9 mRNAとmiR-302a-responsive Cas9 mRNAをHeLa細胞に導入(sgRNAも一緒に導入)し、Cas9タンパク質の発現に影響があるのかをSimple Western (Wes) により検討した。結果を図10に示す。ControlやmiR-302a-responsive Cas9 mRNAではCas9タンパク質が検出されたが、miR-21-responsive Cas9 mRNAでは検出されなかった。つまり、HeLa細胞においてmiRNA-21-responsive Cas9 mRNAではmiR-21による翻訳抑制が起こるためCas9タンパク質の発現がほぼみられなくなることを示している。また、GAPDHをローディングコントロールとして用いた。
Control Cas9 mRNAとmiR-21-responsive Cas9 mRNAとmiR-302a-responsive Cas9 mRNAをHeLa細胞に導入(sgRNAも一緒に導入)し、Cas9タンパク質の発現に影響があるのかをSimple Western (Wes) により検討した。結果を図10に示す。ControlやmiR-302a-responsive Cas9 mRNAではCas9タンパク質が検出されたが、miR-21-responsive Cas9 mRNAでは検出されなかった。つまり、HeLa細胞においてmiRNA-21-responsive Cas9 mRNAではmiR-21による翻訳抑制が起こるためCas9タンパク質の発現がほぼみられなくなることを示している。また、GAPDHをローディングコントロールとして用いた。
Sequencing
図4の(b)、(c)に示すT7E1より得られたPCR産物の配列を読んだ。図11(a)はスキームズであり、標的配列周辺の配列を示している。図11(b)は、EGFP遺伝子の標的領域に変異が入っていることを示している。つまり、EGFPの蛍光強度の減少はCRISPR/Cas9 systemによるEGFP遺伝子のノックアウトであることを示している。
図中にある「Δ」は塩基の欠失を「+」は塩基の挿入を示している。また、一番右側にある数字は、 得られた配列のコロニー数/総コロニー数、を示している。
緑文字は「標的配列」、青文字(図11(a)のTGG、ACC)は「PAM (Protospacer Adjascent motif)」、紫文字(図11(b)のCCA)は「PAMに対して相補な配列」、赤字(Control Cas9 mRNA 2行目の、5’側から9、10塩基、12~18塩基の-、3行目の、5’側から19塩基から41塩基、4行目の、5’側から19塩基から35塩基、miR-302a-responsive Cas9 mRNA+miR-302inhibitor 2行目の5’側から20塩基、3行目の5’側から17塩基の右側の-)は「変異」を示している。また、図中の塩基または「-」表示の間の縦線は、青字、赤字などの字色の変わる境界を示している。
図4の(b)、(c)に示すT7E1より得られたPCR産物の配列を読んだ。図11(a)はスキームズであり、標的配列周辺の配列を示している。図11(b)は、EGFP遺伝子の標的領域に変異が入っていることを示している。つまり、EGFPの蛍光強度の減少はCRISPR/Cas9 systemによるEGFP遺伝子のノックアウトであることを示している。
図中にある「Δ」は塩基の欠失を「+」は塩基の挿入を示している。また、一番右側にある数字は、 得られた配列のコロニー数/総コロニー数、を示している。
緑文字は「標的配列」、青文字(図11(a)のTGG、ACC)は「PAM (Protospacer Adjascent motif)」、紫文字(図11(b)のCCA)は「PAMに対して相補な配列」、赤字(Control Cas9 mRNA 2行目の、5’側から9、10塩基、12~18塩基の-、3行目の、5’側から19塩基から41塩基、4行目の、5’側から19塩基から35塩基、miR-302a-responsive Cas9 mRNA+miR-302inhibitor 2行目の5’側から20塩基、3行目の5’側から17塩基の右側の-)は「変異」を示している。また、図中の塩基または「-」表示の間の縦線は、青字、赤字などの字色の変わる境界を示している。
Evaluation of gene expression level
miR-21、miR-302aのそれぞれにより機能が制御されていることが知られている遺伝子の発現変動をqPCRにより検証した結果を図12(a)、図12(b)に示す。Control Cas9 mRNAとmiRNA-responsive Cas9 mRNAを細胞に導入した時の遺伝子発現レベルをそれぞれ比べると、その発現レベルの差はほぼ見られなかった。つまり、miRNA-responsive CRISPR/Cas9 systemが内在性のmiR-21, miR-302aの機能にほとんど影響を与えていないことを示している。
miR-21、miR-302aのそれぞれにより機能が制御されていることが知られている遺伝子の発現変動をqPCRにより検証した結果を図12(a)、図12(b)に示す。Control Cas9 mRNAとmiRNA-responsive Cas9 mRNAを細胞に導入した時の遺伝子発現レベルをそれぞれ比べると、その発現レベルの差はほぼ見られなかった。つまり、miRNA-responsive CRISPR/Cas9 systemが内在性のmiR-21, miR-302aの機能にほとんど影響を与えていないことを示している。
Evaluation of transfection efficiency
図13(a)はHeLa-EGFPの図13(b)はiPS-EGFPのトランスフェクション効率を示す。BFP mRNAとsgRNA(Cy5でラベリング)のそれぞれでトランスフェクション効率を測定してある。トランスフェクション効率は約90%以上であった。
図13(a)はHeLa-EGFPの図13(b)はiPS-EGFPのトランスフェクション効率を示す。BFP mRNAとsgRNA(Cy5でラベリング)のそれぞれでトランスフェクション効率を測定してある。トランスフェクション効率は約90%以上であった。
EGFP activity assay (iPS-EGFPでsgRNAを改変: 1)
図14(a)はEGFPを標的としたsgRNAの5’末端から「GG」を除いた時のCas9活性を示している。図4(a)では約30%であったCas9活性が70%近くまで上昇していることより、ガイド鎖を改変することによりCas9活性が改善しているのがわかる。しかしながら、インヒビターのネガティブコントロールでもCas9活性が30%近くある。図14(b)は図14(a)で見られたCas9活性のリークを減らすために、miR-302 responsive Cas9 mRNAを改変し、4x miR-302 responsive Cas9 mRNAにしてある。Cas9 mRNAを50 ng、sgRNAを300 ngにすることで、リークをさらに減らしつつ、高い活性を保っていることがわかった。
図14(a)はEGFPを標的としたsgRNAの5’末端から「GG」を除いた時のCas9活性を示している。図4(a)では約30%であったCas9活性が70%近くまで上昇していることより、ガイド鎖を改変することによりCas9活性が改善しているのがわかる。しかしながら、インヒビターのネガティブコントロールでもCas9活性が30%近くある。図14(b)は図14(a)で見られたCas9活性のリークを減らすために、miR-302 responsive Cas9 mRNAを改変し、4x miR-302 responsive Cas9 mRNAにしてある。Cas9 mRNAを50 ng、sgRNAを300 ngにすることで、リークをさらに減らしつつ、高い活性を保っていることがわかった。
Cas9 activity assay (iPS-EGFPでsgRNAを改変: 2)
100 ngのCas9 mRNAと300 ngのsgRNAを用いた。EGFP activity assay (iPS-EGFPでsgRNAを改変: 1)の図14(a)で見られたCas9活性のリークをなくすため、miR-302 responsive Cas9 mRNAを改変し、4x miR-302 responsive Cas9 mRNA(miR-302に応答する配列を5’UTRに4個タンデムに挿入してある)にした時の結果である。図15(a)は細胞の写真、図15(b)はその時のヒストグラム、図15(c)は定量結果である。miRNAに応答する配列を増やすことで、Cas9活性のリークを抑えていることがわかった。
100 ngのCas9 mRNAと300 ngのsgRNAを用いた。EGFP activity assay (iPS-EGFPでsgRNAを改変: 1)の図14(a)で見られたCas9活性のリークをなくすため、miR-302 responsive Cas9 mRNAを改変し、4x miR-302 responsive Cas9 mRNA(miR-302に応答する配列を5’UTRに4個タンデムに挿入してある)にした時の結果である。図15(a)は細胞の写真、図15(b)はその時のヒストグラム、図15(c)は定量結果である。miRNAに応答する配列を増やすことで、Cas9活性のリークを抑えていることがわかった。
Claims (22)
- ヌクレアーゼをコードするmiRNA応答性mRNAを、細胞に導入する工程を含む、細胞特異的にヌクレアーゼを制御する方法であって、当該miRNA応答性mRNAが下記:
(i) miRNAによって特異的に認識される核酸配列、および
(ii) 前記ヌクレアーゼのコード領域に対応する核酸配列を含む、方法。 - 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、請求項1に記載の方法。
- 前記ヌクレアーゼが、Cas9タンパク質またはその変異体であり、
さらに、前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAを前記細胞に導入する工程を含む、請求項1または2に記載の方法。 - 未分化細胞特異的にヌクレアーゼを制御する、請求項3に記載の方法。
- 前記未分化細胞が、miR-302aを発現する細胞である、請求項4に記載の方法。
- 前記miRNA応答性mRNAが、(i)および(ii)が5'から3'の方向に連結されている核酸配列を含む、請求項1から5のいずれか1項に記載の方法。
- (i) miRNAによって特異的に認識される核酸配列、および
(ii) ヌクレアーゼのコード領域に対応する核酸配列を含む、miRNA応答性mRNA。 - 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、請求項7に記載のmiRNA応答性mRNA。
- 前記ヌクレアーゼが、Cas9タンパク質またはその変異体である、請求項7または8に記載のmiRNA応答性mRNA。
- 請求項9に記載のmiRNA応答性mRNAと、
前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAと
を含む、細胞特異的にヌクレアーゼを制御するためのキット。 - a)ヌクレアーゼをコードするトリガータンパク質応答性mRNAと、
b)前記トリガータンパク質をコードするmiRNA応答性mRNAと
を、細胞に導入する工程を含む、細胞特異的にヌクレアーゼを制御する方法であって、
前記a)ヌクレアーゼをコードするタンパク質応答性mRNAが下記:
(ia) 前記タンパク質に特異的に結合する核酸配列と、
(iia) 前記ヌクレアーゼのコード領域に対応する核酸配列と
を含み
前記b)トリガータンパク質をコードするmiRNA応答性mRNAが下記:
(ib) miRNAによって特異的に認識される核酸配列と、
(iib) 前記トリガータンパク質のコード領域に対応する核酸配列を含む、方法。 - 前記トリガータンパク質が、L7Aeタンパク質もしくはその誘導体を含み、前記タンパク質に特異的に結合する核酸配列が、K-turn配列もしくはその誘導体を含む、請求項11に記載の方法。
- 前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、請求項11または12に記載の方法。
- 前記ヌクレアーゼが、Cas9タンパク質またはその変異体であり、
さらに、前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAを前記細胞に導入する工程を含む、請求項11~13のいずれか1項に記載の方法。 - 未分化細胞特異的にヌクレアーゼを制御する、請求項14に記載の方法。
- 前記未分化細胞が、miR-302aを発現する細胞である、請求項15に記載の方法。
- 前記a)トリガータンパク質をコードするタンパク質応答性mRNAが、(ia) および(iia) が5'から3'の方向に連結されている核酸配列を含み、
前記b)ヌクレアーゼをコードするmiRNA応答性mRNAが、(ib)および(iib)が5'から3'の方向に連結されている核酸配列を含む、請求項11~16のいずれか1項に記載の方法。 - a)下記を含むヌクレアーゼをコードするトリガータンパク質応答性mRNA:
(ia) 前記タンパク質に特異的に結合する核酸配列、及び
(iia) 前記ヌクレアーゼのコード領域に対応する核酸配列と、
b)下記を含むトリガータンパク質をコードするmiRNA応答性mRNA:
(ib) miRNAによって特異的に認識される核酸配列、及び
(iib) 前記トリガータンパク質のコード領域に対応する核酸配列と
を含むヌクレアーゼ制御剤。 - 前記トリガータンパク質が、L7Aeタンパク質もしくはその誘導体を含み、前記タンパク質に特異的に結合する核酸配列が、K-turn配列もしくはその誘導体を含む、請求項18に記載のヌクレアーゼ制御剤。
- 前記トリガータンパク質をコードするmiRNA応答性mRNAにおいて、前記miRNAによって特異的に認識される核酸配列が、miR-302a標的配列またはmiR-21標的配列のいずれかである、請求項18または19に記載のヌクレアーゼ制御剤。
- 前記トリガータンパク質応答性mRNAにおいて、前記ヌクレアーゼが、Cas9タンパク質またはその変異体である、請求項18~20のいずれか1項に記載のヌクレアーゼ制御剤。
- 請求項18~21のいずれか1項に記載のヌクレアーゼ制御剤と、
前記ヌクレアーゼの標的遺伝子によって特異的に認識されるガイド配列を備えるsgRNAと
を含む、細胞特異的にヌクレアーゼを制御するためのキット。
Priority Applications (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US16/313,328 US20190256830A1 (en) | 2016-06-27 | 2017-05-18 | Method for cell-specifically controlling nuclease |
| JP2018524954A JP7362097B2 (ja) | 2016-06-27 | 2017-05-18 | 細胞特異的にヌクレアーゼを制御する方法 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2016-126944 | 2016-06-27 | ||
| JP2016126944 | 2016-06-27 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2018003339A1 true WO2018003339A1 (ja) | 2018-01-04 |
Family
ID=60786037
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2017/018742 WO2018003339A1 (ja) | 2016-06-27 | 2017-05-18 | 細胞特異的にヌクレアーゼを制御する方法 |
Country Status (3)
| Country | Link |
|---|---|
| US (1) | US20190256830A1 (ja) |
| JP (1) | JP7362097B2 (ja) |
| WO (1) | WO2018003339A1 (ja) |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2015006747A2 (en) * | 2013-07-11 | 2015-01-15 | Moderna Therapeutics, Inc. | Compositions comprising synthetic polynucleotides encoding crispr related proteins and synthetic sgrnas and methods of use. |
| WO2015168547A2 (en) * | 2014-05-01 | 2015-11-05 | Andre Lieber | In vivo gene engineering with adenoviral vectors |
| WO2016010119A1 (ja) * | 2014-07-16 | 2016-01-21 | 国立大学法人京都大学 | 分化細胞の抽出方法 |
| WO2016040395A1 (en) * | 2014-09-08 | 2016-03-17 | Massachusetts Institute Of Technology | Rna-based logic circuits with rna binding proteins, aptamers and small molecules |
-
2017
- 2017-05-18 US US16/313,328 patent/US20190256830A1/en not_active Abandoned
- 2017-05-18 WO PCT/JP2017/018742 patent/WO2018003339A1/ja active Application Filing
- 2017-05-18 JP JP2018524954A patent/JP7362097B2/ja active Active
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2015006747A2 (en) * | 2013-07-11 | 2015-01-15 | Moderna Therapeutics, Inc. | Compositions comprising synthetic polynucleotides encoding crispr related proteins and synthetic sgrnas and methods of use. |
| WO2015168547A2 (en) * | 2014-05-01 | 2015-11-05 | Andre Lieber | In vivo gene engineering with adenoviral vectors |
| WO2016010119A1 (ja) * | 2014-07-16 | 2016-01-21 | 国立大学法人京都大学 | 分化細胞の抽出方法 |
| WO2016040395A1 (en) * | 2014-09-08 | 2016-03-17 | Massachusetts Institute Of Technology | Rna-based logic circuits with rna binding proteins, aptamers and small molecules |
Non-Patent Citations (3)
| Title |
|---|
| HIROSAWA, MOE ET AL.: "Cell-type-specific genome editing with a microRNA-responsive CRISPR-Cas9 switch", NUCLEIC ACIDS RESEARCH, vol. 45, no. 13, 19 May 2017 (2017-05-19), pages e118, XP055444821, ISSN: 1362-4962, DOI: 10.1093/nar/gkx309 * |
| HIROSAWA, MOE ET AL.: "Jinki RNA o Kiban to shity CRISPER/CAS9 System no Kochiku", THE RNA SOCIETY OF JAPAN NENKAI YOSHISHU, vol. 17, 2015, pages 243 * |
| KARAGIANNIS, PETER ET AL.: "RNA-based gene circuits for cell regulation", PROC. JPN. ACAD. SER. B, vol. 92, no. 9, November 2016 (2016-11-01), pages 412 - 422, XP055601170, ISSN: 1349-2896, DOI: 10.2183/pjab.92.412 * |
Also Published As
| Publication number | Publication date |
|---|---|
| JP7362097B2 (ja) | 2023-10-17 |
| US20190256830A1 (en) | 2019-08-22 |
| JPWO2018003339A1 (ja) | 2019-11-07 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR102382772B1 (ko) | 화학적으로 변형된 가이드 rna를 사용하는 고 특이성 게놈 편집 | |
| US10760081B2 (en) | Compositions and methods for enhancing CRISPR activity by POLQ inhibition | |
| US9783801B2 (en) | Methods for creating and identifying functional RNA interference elements | |
| EP2925866B1 (en) | Circular rna for inhibition of microrna | |
| EP3222723A1 (en) | Development of universal cancer drugs and vaccines | |
| JP4517061B2 (ja) | ダンベル型dnaの効率的な製造方法 | |
| TW201842923A (zh) | 包含mir-302前驅體的組合物在製造用於肺癌治療之藥物上的用途 | |
| US20200165607A1 (en) | Composition and method of using mir-302 precursors as anti-cancer drugs for treating human lung cancer | |
| EP3384026A1 (en) | Use of microrna precursors as drugs for inducing cd34-positive adult stem cell expansion | |
| US20170342418A1 (en) | Use of microrna precursors as drugs for inducing cd34-positive adult stem cell expansion | |
| WO2018003339A1 (ja) | 細胞特異的にヌクレアーゼを制御する方法 | |
| EP2882859A2 (en) | Production and utilization of a novel anti-cancer drug in therapy | |
| JP6779513B2 (ja) | インビボクローニング可能な細胞株をスクリーニングするための方法、インビボクローニング可能な細胞株の製造方法、細胞株、インビボクローニング方法、及びインビボクローニングを行うためのキット | |
| WO2020241748A1 (ja) | 標的遺伝子を編集する蛋白質を細胞特異的に制御する方法 | |
| KR102193864B1 (ko) | 마이크로rna의 비정규 표적을 억제하는 rna 간섭 유도 핵산 및 그 용도 | |
| KR102193873B1 (ko) | 마이크로rna의 비정규 표적을 억제하는 rna 간섭 유도 핵산 및 그 용도 | |
| KR102240833B1 (ko) | let-7 억제제가 도입된 엑소좀을 이용한 골분화 억제방법 | |
| Hwang et al. | Precise editing of pathogenic nucleotide repeat expansions in iPSCs using paired prime editor | |
| EP3653710A1 (en) | HIGH EXPRESSION mRNA SWITCH | |
| JP2017528151A (ja) | 干渉性分子のスクリーニング方法 | |
| Schneider | The long non-coding RNA" upstream to Slitrk3"(RUS) affects chromatin organization by binding to Brd2 and Smarca5 | |
| Chaudhry | Evaluation of CRISPR-Cas9 approaches to investigate microRNA targeting in human chondrocytes | |
| Yiu | Investigating the role of non-coding RNAs in doxorubicin-induced cardiotoxicity | |
| WO2018223843A1 (zh) | 用于dna编辑的系统及其应用 | |
| Peach | Analysis and kinase-mediated decay of RNAs with 5'-hydroxyl termini |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 17819706 Country of ref document: EP Kind code of ref document: A1 |
|
| ENP | Entry into the national phase |
Ref document number: 2018524954 Country of ref document: JP Kind code of ref document: A |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| 122 | Ep: pct application non-entry in european phase |
Ref document number: 17819706 Country of ref document: EP Kind code of ref document: A1 |