KR20220009977A - 개선된 프라이머 인식을 갖는 phi29 dna 폴리머라아제 돌연변이체 - Google Patents
개선된 프라이머 인식을 갖는 phi29 dna 폴리머라아제 돌연변이체 Download PDFInfo
- Publication number
- KR20220009977A KR20220009977A KR1020217040028A KR20217040028A KR20220009977A KR 20220009977 A KR20220009977 A KR 20220009977A KR 1020217040028 A KR1020217040028 A KR 1020217040028A KR 20217040028 A KR20217040028 A KR 20217040028A KR 20220009977 A KR20220009977 A KR 20220009977A
- Authority
- KR
- South Korea
- Prior art keywords
- lys
- phi29
- dna polymerase
- ile
- leu
- Prior art date
Links
Images
















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1241—Nucleotidyltransferases (2.7.7)
- C12N9/1252—DNA-directed DNA polymerase (2.7.7.7), i.e. DNA replicase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/26—Preparation of nitrogen-containing carbohydrates
- C12P19/28—N-glycosides
- C12P19/30—Nucleotides
- C12P19/34—Polynucleotides, e.g. nucleic acids, oligoribonucleotides
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6844—Nucleic acid amplification reactions
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6869—Methods for sequencing
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y207/00—Transferases transferring phosphorus-containing groups (2.7)
- C12Y207/07—Nucleotidyltransferases (2.7.7)
- C12Y207/07007—DNA-directed DNA polymerase (2.7.7.7), i.e. DNA replicase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2521/00—Reaction characterised by the enzymatic activity
- C12Q2521/10—Nucleotidyl transfering
- C12Q2521/101—DNA polymerase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2533/00—Reactions characterised by the enzymatic reaction principle used
- C12Q2533/10—Reactions characterised by the enzymatic reaction principle used the purpose being to increase the length of an oligonucleotide strand
- C12Q2533/101—Primer extension
Abstract
야생형 효소와 비교하여 개선된 프라이머 인식을 갖는 박테리오파지 Phi29 DNA 폴리머라아제가 본원에 개시된다. 특정 돌연변이체는 돌연변이 K64R 또는 M97K 중 하나 또는 둘 모두를 포함한다. 제공된 돌연변이체는 야생형 Phi29 DNA 폴리머라아제보다 더 짧고 더 긴 무작위 합성 DNA 프라이머를 보다 효율적으로 사용하여, 다중 변위 증폭 (MDA) 반응에서 더 많은 증폭 산물을 생성할 수 있다. 본 발명의 돌연변이체는 야생형 Phi29 DNA 폴리머라아제로 실행한 반응과 비교하여 더 적은 편향과 더 양호한 커버리지로 인간 게놈 DNA 를 증폭시킨다.
Description
관련
출원에 대한 교차 참조
본 출원은 2019 년 5 월 17 일에 출원한 미국 가출원 번호 62/849,252 의 우선일의 이득을 주장하며, 이의 내용은 본원에 그 전체가 포함된다.
서열 목록
본 출원은 본원에 참조로 포함되는 컴퓨터 판독가능한 형태의 서열 목록을 포함한다.
Phi29 DNA 폴리머라아제 (Phi29 DNApol) 는 선형 dsDNA 분자의 양 말단에서 단백질-프라이밍된 개시 및 각 DNA 가닥의 완전 신장 모두를 촉진시켜 박테리오파지 게놈 (19285 bp) 을 복제하는 것을 담당하는 단량체성 효소 (66 kDa) 이다 (Blanco and Salas, 1984; 1985). Phi29 DNApol 은 DNA 폴리머라아제의 패밀리 B 에 속하며 (Bernad et al, 1987), 손바닥 (palm), 엄지 (thumb) 및 손가락 (finger) 서브도메인 뿐만 아니라 TPR1 과 TPR2 로 지칭되는 2 개의 추가적인 도메인을 함유하는 일반적인 오른손 방향 접힘을 나타낸다 (Rodriguez et al, 2005; Kamtekar et al, 2006; Berman et al, 2007). Phi29 DNApol 은 수많은 DNA 증폭 및 DNA 서열분석 기술 및 플랫폼에서 그의 응용을 가능하게 하는 고유한 특성: 가공성 인자의 부재 하에 DNA 결합 이벤트 당 70000 개 초과의 뉴클레오티드를 혼입할 수 있게 하는 고가공성 DNA 합성 (Blanco et al, 1989); 헬리카아제-유형 효소의 분재 하에 이중-가닥 DNA 의 풀림과 커플링된 중합을 허용하는 예외적인 가닥-변위 (Blanco et al, 1989); 매우 낮은 오류 삽입률 (10-4 내지 10-6) 로의 합성의 높은 충실도 및 충실도를 혼입된 106 내지 108 개 뉴클레오티드에서 1 개 오류까지 집합적으로 향상시키는 삽입된 오류의 효율적 교정 (Esteban et al, 1993 and 1994) 을 나타낸다.
이러한 특성은 Phi29 DNApol 이 등온 다중 변이 증폭 (multiple displacement amplification) (MDA) (Dean et al, 2002) 및 롤링 서클 증폭 (rolling circle amplification) (RCA) (Lizardi et al, 1998) 을 위한 최상의 선택이 되게 한다. 이들 DNA 증폭 기술은 Phi29 DNApol 과 무작위 합성 프라이머 (RP), 주로 헥사뉴클레오티드 또는 6량체, 또는 반응 동안 제자리 (in situ) 에서 DNA 프라이머를 합성할 수 있는 DNA 프리마아제의 조합을 기반으로 한다 (Picher et al, 2016).
특정 샘플 (예를 들어, 단일 세포) 로부터 이용가능한 DNA 의 양이 서열분석 과정에 충분하지 않기 때문에, DNA 의 증폭이 현재 서열분석 기술에 종종 요구된다. 불행하게도, DNA 증폭은 오차가 도입되고, 비대칭 (편향) 이 발생하고, 심지어 미세한 수준의 오염 DNA 의 동시 증폭을 촉진시킬 위험이 있다. 따라서, 증폭의 품질을 결정하는 주요 매개변수는 반응 생성물 내의 오염물 및 인공물의 부재, 커버리지 폭 및 균일성, 낮은 뉴클레오티드 오류율, 및 단일 뉴클레오티드 변이체 (SNV), 카피 수 변이체 (CNV) 및 구조적 변이체를 회수하는 능력이다.
무작위 6량체를 기반으로 하는 현재의 MDA 방법에서 잠재적인 증폭 편향에 대한 공급원은 올리고뉴클레오티드의 상이한 서열-의존적 하이브리드화 동역학에서 발생하는 프라이밍 불균등이다. 보다 더 중요한 것은, 자가-페어링 6량체의 지수적 증폭에 의해 야기된 프라이머-유래 투입-독립적 DNA 증폭 인공물을 생성하는 경향이다.
40℃ 의 반응 온도를 갖는 6량체 대신 더 긴 프라이머를 사용하는 것은 DNA 증폭 인공물을 상당히 감소시키는 것으로 나타났다 (Alsmadi et al, 2009). 이러한 거동 배후의 가장 가능성 있는 이유는 더 높은 온도가 프라이머의 안정한 자가-페어링 및 그에 따른 이들의 후속적 증폭의 가능성을 감소시킨다는 것이다. 그러나, 40℃ (Phi29 DNApol 의 최적 온도보다 10℃ 높음) 만큼 높은 온도에서 증폭 반응을 실행하기 위해서는 열안정성 또는 내열성 Phi29 DNApol 변이체가 요구된다. 이와 관련하여, 일부 돌연변이된 Phi29 DNApol 이 개선된 열안정성을 나타내는 것으로 기재되었다 (Povilaitis et al, 2016).
변형된 DNA 중합효소는 DNA 서열분석, DNA 증폭, 라이브러리 제조, DNA 유전자형 분석 등과 같은 다수의 적용에 유용할 수 있다. 본 발명은 개선된 특성을 부여하는 돌연변이를 포함하는 재조합 Phi29 DNA 폴리머라아제를 제공하며, 이들 또는 다른 적용에 특히 바람직하다. 이러한 아미노산 서열 변화는 더 짧은 무작위 합성 프라이머를 사용함으로써 다중 변위 DNA 증폭 (MDA) 에서의 성능을 개선시킬 수 있고, 이는 감소된 증폭 인공물, 더 양호한 서열-의존적 하이브리드화 동역학을 초래하고, 따라서 개선된 커버리지 폭 및 균일성을 초래한다. "Phi29" 는 때때로 "Φ29" 로 표기된다.
재조합 Phi29 DNA 폴리머라제는 K64R 및 M97K 로 이루어지는 군으로부터 1 또는 2 개의 돌연변이를 포함한다.
본 명세서에 포함되고 본 명세서의 일부를 형성하는 첨부 도면은 예시적인 구현예를 도시하고, 상세한 설명과 함께, 또한 관련 기술 분야의 통상의 기술자가 이들 구현예 및 당업자에게 명백할 다른 구현예를 제조 및 이용할 수 있게 하는 역할을 한다. 본 발명은 하기 도면과 관련하여 보다 구체적으로 설명될 것이다:
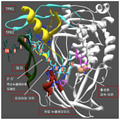
도 1. DNA 및 dNTP 와 복합된 Phi29 DNApol 의 3D-구조 (PDB id: 2PYL). 대부분의 단백질은 엄지 (THUMB) (진녹색), TPR2 (시안) 및 TPR1 (황색) 서브도메인을 제외하고는 백색으로 표시된다. N-말단 3'-5' 엑소뉴클레아제 도메인은 완전히 도시되지 않았다 (2 개 세그먼트만이 표시되어 있으며 (황색), 하나는 Arg96 을 함유하고 다른 하나는 Lys64 를 함유함). 프라이머 가닥 (시안) 은 각각의 뉴클레오티드 위치에 상응하는 수를 나타낸다. 주형 가닥 (연녹색), 유입 뉴클레오티드 (마젠타) 및 2 개의 활성화 금속 이온 (베이지) 가 또한 표시된다.
도 2. A ) 결정 구조에서 유래한 바와 같은, 프라이머 가닥의 첫 번째 10 개 뉴클레오티드와의 상호작용에 관여하는 야생형 (WT) Phi29 DNApol 아미노산 잔기의 개략도 (PDB id: 2PYL). 1 로 번호 매겨진 뉴클레오티드는 가장 3'-말단이며, 흔히 "프라이머 말단" 으로 기재되고, 효소 활성 위치에 가장 가까운 것이다. B ) 개략도는 표시된 상이한 돌연변이 (마젠타) 에 의해 기원하는, 프라이머 가닥과의 신규한 상호작용의 이득을 나타낸다. 유색 화살표는 상호작용이 포스포디에스테르 (적색), 당 (오렌지색) 또는 염기 (녹색) 을 포함하는지를 나타낸다. 돌연변이체 T499K 및 T499R 은 상보성/주형 가닥의 동일한 위치와 상호작용하는 것으로 예측된다 (점선 화살표로 표시함).
도 3. 반응에서 투입물로서 1 ng 의 인간 게놈 DNA 를 사용하는, TthPrimPol 또는 상이한 길이의 무작위 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol 또는 설계된 돌연변이체의 증폭 효율.
도 4. 제공된 데옥시뉴클레오티드 (dNTP) 농도의 함수로서의 WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 엑소뉴클레아제 및 폴리머라아제 활성의 균형.
도 5. 반응에서 투입물로서 1 ng 의 인간 게놈 DNA 를 사용하는, 상이한 길이의 무작위 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 증폭 효율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 6. 낮은 이온 강도 조건 하에 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 또는 이중 돌연변이체 K64R / M97K 를 갖는 투입 DNA 의 부재 하에 관찰되는 증폭 수율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 7. 높은 이온 강도 조건 하에 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 또는 이중 돌연변이체 K64R / M97K 를 갖는 투입 DNA 의 부재 하에 관찰되는 증폭 수율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 8. 높은 이온 강도 조건 하에 반응에서 투입물로서 1 ng 의 인간 게놈 DNA 를 사용하는, 상이한 길이의 무작위 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 증폭 효율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 9. 낮은 및 높은 이온 강도 조건 하에 반응에서 상이한 인간 게놈 DNA 투입물 (1, 10, 100 pg 및 1 ng) 을 사용하는, 상이한 길이의 무작위 프라이머 (4량체 (4N), 5량체 (5N), 6량체 (6N)) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 증폭 효율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 10. Phi29 DNApol 변이체 및 TthPrimPol 을 조합하는 다중 변위 증폭 (MDA) 에 의한 1, 10, 100 pg 및 1 ng 의 인간 게놈 DNA 의 증폭. 각각의 DNA 양에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 11. 커버리지 차이가 관찰될 수 있는 조건에 도달하기 위해 각각의 경우 상이한 인간 게놈 DNA 투입 및 Phi29 DNApol 변이체와의 조합으로의, 6량체 (6N), 5량체 (5N) 및 4량체 (4N) 를 사용하는 증폭 반응의 CovCheck 분석으로부터 수득한 추정 커버리지 값.
도 12. Phi29 DNApol 변이체와 조합으로 TthPrimPol 에 의해 실행된 증폭 반응의 CovCheck 분석으로부터 수득한 추정 커버리지 값.
도 1. DNA 및 dNTP 와 복합된 Phi29 DNApol 의 3D-구조 (PDB id: 2PYL). 대부분의 단백질은 엄지 (THUMB) (진녹색), TPR2 (시안) 및 TPR1 (황색) 서브도메인을 제외하고는 백색으로 표시된다. N-말단 3'-5' 엑소뉴클레아제 도메인은 완전히 도시되지 않았다 (2 개 세그먼트만이 표시되어 있으며 (황색), 하나는 Arg96 을 함유하고 다른 하나는 Lys64 를 함유함). 프라이머 가닥 (시안) 은 각각의 뉴클레오티드 위치에 상응하는 수를 나타낸다. 주형 가닥 (연녹색), 유입 뉴클레오티드 (마젠타) 및 2 개의 활성화 금속 이온 (베이지) 가 또한 표시된다.
도 2. A ) 결정 구조에서 유래한 바와 같은, 프라이머 가닥의 첫 번째 10 개 뉴클레오티드와의 상호작용에 관여하는 야생형 (WT) Phi29 DNApol 아미노산 잔기의 개략도 (PDB id: 2PYL). 1 로 번호 매겨진 뉴클레오티드는 가장 3'-말단이며, 흔히 "프라이머 말단" 으로 기재되고, 효소 활성 위치에 가장 가까운 것이다. B ) 개략도는 표시된 상이한 돌연변이 (마젠타) 에 의해 기원하는, 프라이머 가닥과의 신규한 상호작용의 이득을 나타낸다. 유색 화살표는 상호작용이 포스포디에스테르 (적색), 당 (오렌지색) 또는 염기 (녹색) 을 포함하는지를 나타낸다. 돌연변이체 T499K 및 T499R 은 상보성/주형 가닥의 동일한 위치와 상호작용하는 것으로 예측된다 (점선 화살표로 표시함).
도 3. 반응에서 투입물로서 1 ng 의 인간 게놈 DNA 를 사용하는, TthPrimPol 또는 상이한 길이의 무작위 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol 또는 설계된 돌연변이체의 증폭 효율.
도 4. 제공된 데옥시뉴클레오티드 (dNTP) 농도의 함수로서의 WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 엑소뉴클레아제 및 폴리머라아제 활성의 균형.
도 5. 반응에서 투입물로서 1 ng 의 인간 게놈 DNA 를 사용하는, 상이한 길이의 무작위 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 증폭 효율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 6. 낮은 이온 강도 조건 하에 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 또는 이중 돌연변이체 K64R / M97K 를 갖는 투입 DNA 의 부재 하에 관찰되는 증폭 수율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 7. 높은 이온 강도 조건 하에 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 또는 이중 돌연변이체 K64R / M97K 를 갖는 투입 DNA 의 부재 하에 관찰되는 증폭 수율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 8. 높은 이온 강도 조건 하에 반응에서 투입물로서 1 ng 의 인간 게놈 DNA 를 사용하는, 상이한 길이의 무작위 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 증폭 효율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 9. 낮은 및 높은 이온 강도 조건 하에 반응에서 상이한 인간 게놈 DNA 투입물 (1, 10, 100 pg 및 1 ng) 을 사용하는, 상이한 길이의 무작위 프라이머 (4량체 (4N), 5량체 (5N), 6량체 (6N)) 와 조합으로의, WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K 및 이중 돌연변이체 K64R / M97K 의 증폭 효율. 각각의 프라이머 길이 N 에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 10. Phi29 DNApol 변이체 및 TthPrimPol 을 조합하는 다중 변위 증폭 (MDA) 에 의한 1, 10, 100 pg 및 1 ng 의 인간 게놈 DNA 의 증폭. 각각의 DNA 양에 대해 컬럼은 좌측에서 우측으로 하기 순서로 나타낸다: WT Phi29 DNApol, 돌연변이체 K64R, 돌연변이체 M97K, 이중 돌연변이체 K64R / M97K.
도 11. 커버리지 차이가 관찰될 수 있는 조건에 도달하기 위해 각각의 경우 상이한 인간 게놈 DNA 투입 및 Phi29 DNApol 변이체와의 조합으로의, 6량체 (6N), 5량체 (5N) 및 4량체 (4N) 를 사용하는 증폭 반응의 CovCheck 분석으로부터 수득한 추정 커버리지 값.
도 12. Phi29 DNApol 변이체와 조합으로 TthPrimPol 에 의해 실행된 증폭 반응의 CovCheck 분석으로부터 수득한 추정 커버리지 값.
발명의 상세한 설명
I.
정의
"단리된" 은 분자가 존재하는 우세한 종류, 즉 몰 기준으로, 조성물 중의 임의의 다른 개별 거대분자 종류보다 더 풍부한 종류라는 것을 의미한다. 일반적으로, 단리된 분자는 정제된 관심 대상 종류인 조성물 중에 존재하는 거대분자 종류의 80% 초과, 90% 초과, 95% 초과, 98% 초과 또는 99% 초과를 포함할 수 있다. 용매 종류, 소분자 (<500 달톤), 안정화제 (예를 들어, BSA) 및 원소 이온 종류는 이 정의의 목적을 위한 거대분자 종류로 간주되지 않는다.
본원에서 사용되는 용어 "재조합 핵산" 은 자연에서 서로 정상적으로 부착되지 않는 둘 이상의 부착된 뉴클레오티드 서열을 포함하는 핵산 분자를 지칭한다.
본원에서 사용되는 용어 "재조합 세포" 는 재조합 핵산을 포함하는 세포, 예를 들어, 동물, 식물, 진균 또는 미생물 (예를 들어, 박테리아) 세포를 지칭한다.
둘 이상의 뉴클레오티드 서열 또는 아미노산 서열 사이의 서열 관계를 기술하기 위해 사용된 용어는 "참조 서열", "~에서 선택된", "비교 윈도우", "동일한", "서열 동일성 백분율", "실질적으로 동일한", "상보적인" 및 "실질적으로 상보적인” 을 포함한다.
"참조 서열" 은 서열 비교의 기초로 사용되는 정의된 서열이며, 더 큰 서열, 예를 들어 완전한 cDNA, 단백질 또는 유전자 서열의 서브세트일 수 있다.
2 개의 핵산 또는 폴리펩티드는 각각 (1) 2 개의 핵산 간에 유사한 서열 (즉, 완전한 핵산 또는 폴리펩티드 서열의 오직 일부) 또는 (2) 2 개의 핵산 사이에서 분기되는 서열을 포함할 수 있기 때문에, 2 개 (또는 이상) 의 핵산 또는 폴리펩티드 간의 서열 비교는 전형적으로 "비교 윈도우 (comparison window)" 에 대해 2 개의 핵산의 서열을 비교하여 서열 유사성의 국소 영역을 확인하고 비교함으로써 수행된다.
"비교 윈도우" 는 참조 서열과 비교되는 전형적으로 적어도 12 개의 연속 뉴클레오티드 또는 4 개의 연속 아미노산 잔기의 개념적 세그먼트를 지칭한다. 비교 윈도우는 종종 적어도 15 개 또는 적어도 25 개의 뉴클레오티드 또는 적어도 5 개 또는 적어도 8 개의 아미노산의 길이를 갖는다. 비교 윈도우는 두 서열의 최적 정렬을 위해 참조 서열 (부가 또는 결실을 포함하지 않음) 과 비교하여 약 20% 이하의 부가 또는 결실 (즉, 갭) 을 포함할 수 있다. 비교 윈도우를 정렬하기 위한 최적의 서열 정렬은 알고리즘의 컴퓨터화된 구현 (GAP, BESTFIT, FASTA, 및 TFASTA - Wisconsin Genetics Software Package Release 7.0, Genetics Computer Group, 575 Science Dr., Madison, WI) 에 의해 또는 검사에 의해 수행될 수 있고, 다양한 방법 중 하나에 의해 생성된 최상의 정렬 (즉, 비교 윈도우에 대해 가장 높은 상동성 백분율을 초래함) 이 선택된다.
대상 뉴클레오티드 서열 또는 아미노산 서열은 뉴클레오티드 또는 아미노산 서열의 길이에 걸쳐 최대 상응성을 위해 정렬될 때 두 서열이 동일한 경우 참조 서열과 "동일" 하다.
두 서열 사이의 "서열 동일성 백분율" 은 비교 윈도우에 걸쳐 2 개의 최적으로 정렬된 서열을 비교하고, 두 서열에서 동일한 뉴클레오티드 또는 아미노산이 발생하는 위치의 수를 결정하여 매칭되는 위치의 수를 산출하고, 비교 윈도우 내의 총 위치 수 (즉, 윈도우 크기) 에 의해 매칭된 위치 수를 나누고, 결과에 100 을 곱하여 서열 동일성의 백분율을 산출함으로써 계산된다.
달리 지정하지 않는 한, 두 서열을 비교하는데 사용되는 비교 윈도우는 더 짧은 서열의 길이이다.
방법은 Natl. Acad. Sci. USA 85:2444; Higgins & Sharp (1988) Gene 73:237-244; Higgins & Sharp, CABIOS 5:151-153 (1989); Corpet et al. (1988) Nucleic Acids Research 16:10881-90; Huang et al. (1992) Computer Applications in the Biosciences 8:155-65; 및 Pearson et al. (1994) Methods in Molecular Biology 24:307-31 에서 추가로 기재되어 있다. 정렬은 또한 종종 검사 및 수동 정렬에 의해 수행된다.
대상 아미노산 서열 또는 뉴클레오티드 서열이 비교 윈도우에 걸쳐 적어도 80% 서열 동일성을 갖는 경우, 대상 뉴클레오티드 서열 또는 아미노산 서열은 참조 서열과 "실질적으로 동일" 하다. 따라서, 참조 서열과 적어도 85% 서열 동일성, 적어도 90% 서열 동일성, 적어도 95% 서열 동일성, 적어도 98% 서열 동일성 또는 적어도 99% 서열 동일성을 갖는 서열이 또한 "실질적으로 동일" 하다. 서로 동일한 2 개의 서열은 물론, 또한 "실질적으로 동일" 하다.
본원에서 사용되는 용어 "전사 조절 서열" 은 작동가능하게 연결되는 제 2 뉴클레오티드 서열의 전사를 조절하는 제 1 뉴클레오티드 서열을 지칭한다.
본원에서 사용되는 바와 같이, 뉴클레오티드 서열은 전사 조절 서열이 세포에서 뉴클레오티드 서열의 전사를 조절하도록 기능할 때 전사 조절 서열과 "작동가능하게 연결" 된다. 이것은 폴리머라제와 프로모터 사이의 상호작용을 통해 뉴클레오티드 서열의 전사를 촉진하는 것을 포함한다.
"프로모터" 는 DNA 에서 뉴클레오티드 서열의 RNA 전사체로의 전사를 촉진시키기에 적어도 충분한 전사 조절 서열이다. 프로모터로부터 전사된 전사체는 전형적으로 전사 시작 위치의 프로모터 다운스트림으로부터의 서열 뿐만 아니라 mRNA 의 경우 아미노산 서열을 인코딩하는 다운스트림 서열을 포함한다. 프로모터는 전사 시작 위치의 바로 업스트림에서 예측가능한 위치로 인해 가장 특성화된 전사 조절 서열이다. 프로모터는 RNA 폴리머라제의 인식, 결합 및 전사 개시 활성을 조절하는 서열을 포함한다. 이들 서열은 시스 작용할 수 있거나 트랜스 작용 인자에 반응할 수 있다. 조절의 특성에 따라, 프로모터는 구성적이거나 조절될 수 있다. 이들은 종종 2 개의 분리된 세그먼트: 코어 및 연장된 프로모터 부위를 갖는 것으로 기술된다.
코어 프로모터는 RNA 폴리머라제 인식, 결합 및 전사 개시에 충분한 서열을 포함한다. 코어 프로모터는 전사 시작 위치, RNA 폴리머라제 결합 위치 및 다른 일반적인 전사 결합 위치를 포함하고, 사전-개시 복합체 형태 및 일반적인 전사 기작이 어셈블리되는 곳이다. 사전-개시 복합체는 일반적으로 전사 시작 위치 (TSS) 의 50 개 뉴클레오티드 (nt) 내에 있다.
코어 프로모터는 또한 mRNA 를 폴리펩티드로 번역하는데 필요한 리보솜 결합 위치에 대한 서열을 포함한다.
연장된 프로모터 부위는 소위 근접 프로모터를 포함하며, 이는 전사 시작 위치의 약 250 개 뉴클레오티드 업스트림으로 (즉, -250 nt) 연장된다. 많은 유전자가 추가 업스트림에 위치한 전사 조절 요소를 갖는 것으로 밝혀졌다. 특히, 유전자의 대부분의 전사 조절 요소를 포함하는 단편은 전사 시작 위치의 700 nt 까지 또는 그 이상의 업스트림까지 연장될 수 있다. 특정 유전자에서, 전사 조절 서열은 전사 시작 위치의 수천 개의 뉴클레오티드 업스트림에서 발견되었다.
본원에서 사용되는 바와 같이, 제 1 뉴클레오티드 서열이 사실상 제 2 뉴클레오티드 서열에 부착되지 않은, 예를 들어 이와 작동가능하게 연결되지 않은 경우, 제 1 뉴클레오티드 서열은 제 2 뉴클레오티드 서열에 대해 "이종" 이다. 연장하여, 폴리펩티드는 전사 조절 서열과 이종인 뉴클레오티드 서열에 의해 인코딩되는 경우 전사 조절 서열에 대해 "이종" 이다.
본원에서 사용되는 용어 "대립유전자 변이체" 는 유전자의 자연 발생 변이를 지칭한다.
본원에서 사용되는 용어 "인공 변이체" 는 자연 발생 유전자 또는 단백질에 대한 하나 이상의 유전자 변형을 포함하는 유전자 또는 단백질을 지칭한다.
본원에서 사용되는 용어 "돌연변이" 는 일반적으로, 야생형과 비교하여 뉴클레오티드 서열에서의 변경, 변이 또는 다형성을 지칭한다. 이러한 변경, 변이 또는 다형성은 참조 게놈, 예를 들어 게놈 데이터베이스에 관련하여 존재할 수 있다. 돌연변이는 단일 뉴클레오티드 변이 (SNV), 치환, 삽입 또는 결실 (집합적으로 "삽입-결실" (indel) 로도 지칭함), 및 반복물을 포함하지만 이에 제한되지는 않는다.
II.
개요
서열-의존적 하이브리드화 동역학에서 유래한 증폭 인공물 및 증폭 편향을 감소시키기 위한 신규한 전략은 현재의 금 본위 6량체보다 더 짧은 DNA 프라이머의 사용을 이용할 수 있다. 더 짧은 DNA 프라이머를 인식하고, 안정적으로 결합하고 효율적으로 사용할 수 있는 Phi29 DNApol 변이체를 수득하는 것을 필요로 하는 이러한 전략은, 현재의 DNA 증폭 기술을 상당히 개선시킬 것이다.
DNA 및 유입 뉴클레오티드와 복합체화된 Phi29 DNApol 의 3D-구조의 이용가능성 (Berman et al, 2007) 은 프라이머 가닥과의 상호작용에 직접적으로 관여하는 아미노산 잔기의 상세한 조사를 수행하게 하였다 (도 1). 프라이머 가닥의 이들 리간드 (도 2A 에서의 개략도 참조) 는 하기와 같다:
ㆍ R96 (프라이머의 뉴클레오티드 7 과 8 사이에서 포스포디에스테르와 상호작용함).
ㆍ R306 (프라이머의 뉴클레오티드 8 과 9 사이에서 포스포디에스테르와 상호작용함).
ㆍ R308 (프라이머의 뉴클레오티드 9 와 10 사이에서 포스포디에스테르와 상호작용함).
ㆍ K498 (프라이머의 제 1 3' 뉴클레오티드의 당과 상호작용함).
ㆍ Y500 (프라이머의 뉴클레오티드 1 과 2 사이에서 포스포디에스테르와 상호작용함).
ㆍ K529 (프라이머의 뉴클레오티드 1 과 2 사이에서 포스포디에스테르와 상호작용함).
이들 소수의 접촉을 기반으로 하여, Phi29 DNApol 은 프라이머 가닥의 첫 번째 10 개 염기에 걸쳐 있는 직접적 상호작용을 확립하는데, 이는 이러한 크기가 프라이머에 최대 결합 안정성을 부여할 것임을 시사한다. 뉴클레오티드 3 과 6 사이의 간격에서 접촉이 부족하다는 것은 상당히 놀라운 일이다. 명백하게, Phi29 DNApol 를 사용하는 현재의 MDA 절차는 무작위 6량체의 제공을 기반으로 하며, 이는 2 개의 제 1 뉴클레오티드 사이의 포스포디에스테르 결합 및 3'-말단 뉴클레오티드의 염기와의 접촉에 의해서만 불량하게 안정화될 것이다. 따라서, 6량체는 Phi29 DNApol 에 의해 결합 및 연장되는 초기 프라이머로서 사용하기에 최적의 크기를 갖지 않는다. 매우 가능성 있게, 프라이머-이량체로 공지된 자가-하이브리드화 인공물을 최소화하면서, 효율적이고 심지어 증폭을 가능하게 하는 충분히 짧은 간격으로 임의의 DNA 샘플에서 상보체를 갖도록 이들 차선의 프라이머를 선택하였다.
한편, 대안적인 TruePrime DNA 증폭 기술 (Picher et al, 2016) 은 요구에 따라서 DNA 프라이머를 합성하기 위해 DNA 프리마아제 (TthPrimPol) 를 이용하지만, TthPrimPol 에 의해 Phi29 DNApol 로 전달되는 최적의 프라이머 크기가 무엇인지, 그리고 Phi29 DNApol 에 의한 최적의 신장을 위해 요구되는 최소 크기보다 더 짧게 유지되는 이러한 프라이머의 운명이 무엇인지는 확립되지 않았다.
이러한 정보 및 통고를 기반으로 하여, 본 발명자들은 이상적으로는 4 내지 6 개 뉴클레오티드의 한계에서 짧은 프라이머에 대한 개선된 친화성을 갖는 Phi29 DNApol 돌연변이체 (본 발명의 변이체) 를 생성할 가능성을 조사하였다. 이러한 목적을 위해, 본 발명자들은 2 가지 상이한 접근방식을 따랐다: 1) 일부 기존 상호작용을 보강하는 것, 2) 프라이머 부위에서 새로운 (존재하지 않는) 효소:DNA 리간드를 생성하는 것.
이러한 개선된 변이체는 RPs-기반 MDA 절차에서 유용하며, 아마도 프라이머-이량체 인공물 및 증폭 키메라의 형성을 감소시킬 것으로 예상된다. 또한, TruePrime DNA 증폭 기술과 관련하여, TthPrimPol 에 의해 생성될 수 있는 짧은 프라이머의 사용은 증폭 효율을 증가시킬 수 있고/있거나 개선된 커버리지를 유도할 수 있다.
다시금, Phi29 DNApol 의 3D-구조의 상세한 분석 (Berman et al, 2007) 은 "기능 획득 (gain of function)" 돌연변이에 대한 후보로서 5 개 아미노산 잔기의 선택을 가능하게 하였다. 이들 잔기는: Lys64 (ExoII 모티프에 위치함), Met97 (Arg96, WT Phi29 DNApol 의 프라이머 리간드에 이웃함), Thr499 (Lys498 및 Tyr500, WT Phi29 DNApol 의 2 개 프라이머 리간드에 이웃함), Thr534 및 Lys538 (Lys529, WT Phi29 DNApol 의 프라이머 리간드에 근접함) 이다. 이들 잔기에서 선택된 돌연변이 (도 2B 에서 요약됨) 는 하기와 같다:
ㆍ K64R, 프라이머 가닥의 잔기 4 와 5 사이에서 포스포디에스테르와의 상호작용 획득을 생성함.
ㆍ K64KG; K64KK; L63LG; L63LH, Lys64 에 측면위치하는 +1 삽입 돌연변이로서, B 패밀리 DNA 폴리머라아제의 상이한 ExoII 모티프에서 관찰된 이종성과 일치하게 설계됨. 이들 변화는 프라이머 가닥의 잔기 4 및 5 와의 상호작용을 획득할 것으로 예상됨.
ㆍ R96K, 프라이머의 잔기 7 과 8 사이에서 포스포디에스테르와의 상호작용을 약화시키는 것으로 예상됨.
ㆍ M97K, 프라이머 가닥의 뉴클레오티드 5 의 질소 염기와의 상호작용 획득을 생성함.
ㆍ M97R, 프라이머 가닥의 아미노산 잔기 4 및 5 의 염기와의 상호작용 획득을 생성함.
ㆍ T499K, 주형 가닥의 아미노산 잔기 5 의 당과의 상호작용 획득을 생성함.
ㆍ T499R, 주형 가닥의 아미노산 잔기 4 및 5 의 당과의 상호작용 획득을 생성함.
ㆍ K529R, 프라이머 가닥의 잔기 1 과 3 사이에서 포스포디에스테르 결합과의 이중 상호작용 획득을 생성함.
ㆍ T534K, 프라이머 가닥의 아미노산 잔기 4 의 당과의 상호작용 획득을 생성함.
ㆍ T534R, 프라이머 가닥의 잔기 3 과 4 사이에서 포스포디에스테르와의 상호작용 획득을 생성함.
ㆍ K538R, 프라이머 가닥의 잔기 2 와 3 사이에서 포스포디에스테르와의 상호작용 획득을 생성함.
짧은 프라이머에 대한 Phi29 DNApol 의 친화성을 증가시키도록 설계된, 표시한 돌연변이를 하기 표준 프로토콜에 따라 발현시키고 정제하여 WT Phi29 DNApol 을 수득하였다. 도입된 돌연변이에 의해 유래하는 프라이머 가닥과의 상호작용의 임의의 특정한 획득이 전좌, 가공성 또는 적절한 (TthPrimPol) 및 무작위-프라이밍된 DNA 증폭 기술로서 Phi29 DNApol 특징에 대해 불리한 효과를 가질지 예상할 수 없다.
III.
핵산, 발현 구축물, 재조합 세포 및 돌연변이체
폴리머라아제
폴리펩티드
A.
핵산
개선된 프라이머 인식을 갖는 돌연변이체 Phi29 폴리머라아제를 인코딩하는 뉴클레오티드 서열을 갖는 핵산이 본원에 제공된다. 야생형 Phi29 폴리머라아제에 대한 뉴클레오티드 서열은 SEQ ID NO.: 1 에 제공된다. 돌연변이체 Phi29 폴리머라아제 서열을 인코딩하는 핵산은 돌연변이 K64R 및 M97K 중 하나 또는 돌 모두를 갖는다. 일부 구현예에서 이들 돌연변이 중 하나 또는 둘 모두를 인코딩하는 뉴클레오티드 서열은 SEQ ID NO.: 1 의 서열과 실질적으로 동일하다.
B.
발현 구축물
본원에 기재된 바와 같은 돌연변이체 Phi29 폴리머라아제를 인코딩하는 뉴클레오티드 서열에 작동가능하게 연결된 전사 조절 서열을 포함하는 발현 구축물이 또한 본원에 제공된다. 발현 구축물은 플라스미드의 형태 또는 관심 세포에서의 발현에 적절한 임의의 다른 형태를 취할 수 있다.
C.
재조합 세포
본원에 기재된 바와 같은 발현 구축물을 포함하는 재조합 세포가 또한 본원에 제공된다. 특정 구현예에서 세포는 박테리아 세포이다. 이러한 재조합 세포는 본 개시물의 핵산 분자를 재생성하고 본 개시물의 돌연변이체 Phi29 폴리머라제를 생성하는데 유용하다. 돌연변이체 Phi29 폴리머라제는 발현 구축물을 포함하는 재조합 세포를 배양함으로써 생성될 수 있다. 사용된 전사 조절 서열은 구성적 프로모터를 포함할 수 있다.
D.
돌연변이체
Phi29
폴리머라아제
개선된 프라이머 인식을 갖는 돌연변이체 Phi29 폴리머라아제가 또한 본원에 제공된다. 본 개시물의 돌연변이체 Phi29 폴리머라아제는 SEQ ID NO.: 1 (UniProtKB - P03680 으로서 수탁됨) 의 아미노산 서열과 실질적으로 동일하며 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두를 포함하는 아미노산 서열을 갖는다.
실질적으로 동일한 아미노산 서열을 갖는 폴리머라아제는, 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두를 포함하는 경우, 대립유전자 변이체와 같은 자연 발생 서열을 기반으로 할 수 있다. 이러한 변이체는 야생형 서열 SEQ ID NO.: 1 과 비교하여 최대 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 또는 1 개 또는 그 이하의 아미노산 치환, 부가 또는 결실을 가질 수 있는데, 단, 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두가 존재한다.
바람직하게는, 본 발명의 DNA 폴리머라아제의 아미노산 서열은 SEQ ID NO: 2, SEQ ID NO: 3 또는 SEQ ID NO: 4 와 적어도 80% 의 동일성을 갖는다. 보다 바람직하게는, 본 발명의 폴리머라아제의 아미노산 서열은 SEQ ID NO: 2, SEQ ID NO: 3 또는 SEQ ID NO: 4 와 적어도 90% 의 동일성을 갖는다. 보다 더 바람직하게는, 본 발명의 폴리머라아제의 아미노산 서열은 SEQ ID NO: 2, SEQ ID NO: 3 또는 SEQ ID NO: 4 이다.
IV.
사용 방법
본원에 기재된 돌연변이체 Phi29 폴리머라아제를 사용하여 프라이머 연장 및/또는 핵산 중합을 수행하는 방법이 본원에 제공된다. 프라이머 연장 방법은 핵산 복제, 증폭 및 서열분석에 유용하다.
프라이머 연장은 핵산 분자 주형에 대한 프라이머의 하이브리드화에 이어서 프라이머의 3' 말단에 뉴클레오티드를 부가하는 폴리머라아제에 의해 촉매화되는 중합 반응을 포함한다. 프라이머는 반응 혼합물에 외인적으로 첨가될 수 있거나, 프리마아제/폴리머라아제에 의해 생성될 수 있다. 프리마아제는 핵산 주형에 상보적인 프라이머로 지칭되는 올리고뉴클레오티드의 합성을 촉매화하는 효소이다. 하나의 이러한 프리마아제는 예컨대 TthPrimPol 이다.
합성 프라이머는 전형적으로 핵산 증폭에 사용된다. 이러한 프라이머는 전형적으로 약 6 내지 약 25 개 뉴클레오티드 길이이다. 특정 서열이 증폭되는 경우, 프라이머는 표적 서열에 상보적인 서열을 가질 수 있다. 전체 게놈 증폭 또는 다른 비방향성 증폭 방법론을 위해, 무작위 프라이머가 사용될 수 있다. 무작위 프라이머는 전형적으로 각각의 염기가 세트 내의 하나 이상의 프라이머에서의 올리고뉴클레오티드 내의 각각의 위치에 존재하는 올리고뉴클레오티드의 집합 또는 세트를 포함한다. 특정 상황에서, 하나 이상의 위치 (예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9 또는 10) 는 고정된 염기 또는 2 또는 3 개의 염기의 조합에 의해 채워질 수 있다.
A.
증폭
예를 들어 Mullis (US 5,656,493) 에 의해 도입된 바와 같은 폴리머라아제 연쇄 반응 (PCR) 에 의한 핵산의 증폭은 의학 및 생물학적 연구에서 사용되는 필수 기법이다. 이는 핵산의 클로닝, 조작 또는 서열분석, 유전자의 DNA-기반 기능적 및 계통학적 분석, 질환의 검출 및 진단과 같은 다양한 응용 뿐만 아니라 법의학 및 친자 검사에서 성공적으로 사용되어 왔다.
B.
롤링 서클 증폭
롤링 서클 증폭은 단일 가닥, 공유 폐쇄 DNA 분자와 같은 공유 폐쇄 DNA 분자를 증폭하는 방법이다. 주형 DNA 분자는 프라이머, 예를 들어 프리마아제/폴리머라아제에 의해 제공된 프라이머로 프라이밍된다. DNA 폴리머라아제는 폐쇄 DNA 분자 주변의 프라이머에 대해 프라이머 연장을 수행한다. 폴리머라아제는 하이브리드화된 카피를 대체하고 주형 주변의 폴리뉴클레오티드 연장을 지속하여, 연접 증폭 산물을 생성한다.
C.
다중 변위 증폭 (
MDA
)
다중 변위 증폭 (MDA) 은 주형으로부터의 프라이밍 및 연장이 ssDNA 사슬을 생성하여 이것이 가닥-변위 합성에 의해 연속적으로 재프라이밍 및 카피되어 다중 분지형 DNA 구조를 생성할 수 있는, 등온, 비-PCR 기반 DNA 증폭 방법이다. 이중-가닥 DNA 샘플의 초기 변성 후, 다중 가닥 변위 증폭 (MDA) 은 DNA 합성이 증폭된 분자에서의 많은 위치로부터 연속적으로 프라이밍 및 연장될 수 있으므로, 추가 라운드의 변성 필요 없이, 다중-분지형 구조를 생성한다. 분지들은 새로운 프라이머가 하나의 DNA 분자 주형으로부터 분지된 영역으로 연장됨에 따라 서로 변위된다. MDA 는 예를 들어, 2011 년 4 월 21 일에 공개된 WO2011/047307A1 ("다중 변위 증폭") 에서 추가로 기재되어 있다. MDA 는 자가-생성된 ssDNA 주형 상의 다수의 프라이밍 위치에서의 프라이머를 연장시키는 등온 중합으로서 간략히 기재될 수 있다.
특정 구현예에서 MDA 는 초기 주형 및 이의 증폭된 카피 상의 다수 위치에서 증폭을 프라이밍하기 위한 프라이머로서 무작위 3량체, 4량체, 5량체, 6량체, 7량체 또는 8량체를 이용한다. 개시된 방법의 특정 구현예에서, 프라이밍은 DNA 프리마아제/폴리머라아제, 예컨대 TthPrimPol 로 달성된다.
특정 구현예에서, 이중-가닥, 선형 폴리뉴클레오티드의 증폭은: 1) 무작위 합성 프라이머 및/또는 DNA-유도 프리마아제/폴리머라아제, 예컨대 TthPrimPol; 2) 가닥-변위 활성을 갖는 변형된 DNA 폴리머라아제, 예컨대 Phi29 DNApol; 3) dNTP 를 사용하는 것을 포함한다. 특정 구현예에서, dNTP 기질은 변형되지 않는다. 다른 구현예에서, dNTP 는 표지된 기, 예를 들어 형광 분자의 부착에 의해 변형될 수 있다. 본원에서 사용되는 용어 "표지" 는 핵산 분자와 같은 분자에 부착된 화학적 모이어티를 지칭한다. 검출가능한 표지는 예를 들어 형광 표지, 발광 표지, 효소 표지, 비색 표지, 예컨대 콜로이드성 금 또는 착색된 유리 또는 플라스틱 비드 및 방사성 표지를 포함한다. 조합하여, 이들 3 개 시약은 무작위 합성 프라이머에 의해 또는 프리마아제/폴리머라아제에 의해 다중 프라이밍되고 DNA 폴리머라아제에 의해 연장된 주어진 DNA 의 다중 변위 증폭 (MDA) 을 촉진시킨다. 또한, 무작위 합성 프라이머 및/또는 프리마아제/폴리머라아제 및 DNA 폴리머라아제의 조합은 프리마아제/폴리머라아제 및/또는 무작위 올리고뉴클레오티드 프라이머로의 증폭된 분자의 프라이밍 및 DNA 폴리머라아제에 의한 프라이머 연장을 통해 다중 가닥 변위 증폭에 영향을 줄 수 있다.
1.
가닥 변위 활성을 갖는 DNA
폴리머라아제
MDA 로서의 증폭 방법은 가닥 변위 활성을 갖는 DNA 폴리머라아제, 예를 들어,단일-가닥 DNA 예를 들어 우선적으로 이중-가닥 DNA 에 강한 결합을 갖는 폴리머라아제를 이용할 수 있다. 가닥 변위 활성은 프라이머 위치를 연장하면서 DNA 분자의 하이브리드화된 가닥을 변위하는데 유용할 수 있다.
본원에 개시된 방법에서 유용한 가닥 변위 활성을 갖는 DNA 폴리머라아제는 예를 들어 Phi29 DNApol 을 포함한다. Phi29 DNApol 은 예를 들어 New England Biolabs (Ipswich, MA, USA), ThermoFisher Scientific (Waltham, MA, USA and Expedeon (Cambridge, UK) 에서 상업적으로 입수할 수 있다. Phi29 DNApol 은 단일 효소 : DNA 결합 이벤트로부터 70 kb 보다 긴 DNA 단편을 생성할 수 있는, DNA 중합에 결합된 고유한 높은 진행성 및 가닥-배위 능력 모두를 갖는다 (Blanco et al., 1989). 이러한 잠재성은 Phi29 DNApol 이 헤어핀 루프와 같은 2 차 구조를 함유하는 DNA 주형을 복제할 수 있게 한다. 효소는 또한 3' → 5' 엑소뉴클레아제 교정 활성을 가지며 (Blanco and Salas, 1985; Garmendia et al., 1992), Taq DNA 폴리머라아제-기반 방법과 비교하여 최대 1000 배 더 높은 충실도를 제공한다.
2.
데옥시리보뉴클레오시드
트리포스페이트
프라이머 생성 및 프라이머 연장은 단지 데옥시리보뉴클레오티드 기질 예를 들어 dNTP 를 제공하여, DNA 프라이머를 합성할 수 있는 TthPrimPol 로서의 특화된 DNA 프리마아제/폴리머라아제 (Picher et al, 2016) 및 Phi29 DNApol 로서의 신장 DNA 폴리머라아제의 조합에 의해 달성될 수 있다. 전형적으로, 이들은 4 개의 표준 염기, A, T, G 및 C 를 포함한다. 그러나, 특정 구현예에서 이노신과 같은 비-자연적 뉴클레오티드가 포함될 수 있다. 특정 구현예에서 뉴클레오티드는 이들이 혼입되는 폴리뉴클레오티드의 검출 또는 포획을 위한 표지를 가질 수 있다.
D.
DNA 서열분석
현재, 흔히 "1 세대 서열분석", "2 세대 서열분석" (종종 "차세대 서열분석" 또는 NGS 로 불림), 및 단일 분자 서열분석 (SMS) 로도 공지되는 "3 세대 서열분석" 으로 분류되는 다수의 상이한 서열분석 기법이 존재한다. 1 세대 서열분석은 Maxam 과 Gilbert (Maxam and Gilbert, 1977) 또는 Sanger (Sanger et al, 1977; Sanger and Coulson, 1978) 의 방법을 주로 지칭하며, 이들 중 후자만이 오늘날 사용된다.
2 세대 또는 차세대 서열분석은 염기 위치의 진보된 기술적 (광학적) 검출 방법을 사용하여 동시에 많은 서열을 생성하는 기법을 지칭한다. 기존 방법에 대한 개요는 (Metzker, 2010) 에서 제공된다.
3 세대 또는 단일 분자 서열분석 (SMS) 기법은 사전 증폭을 필요로 하지 않으며, DNA 의 클론 또는 앙상블 (ensemble) 이 아니라 그 서열이 폴리머라아제의 활성의 결과로서 종종 "실시간" 으로 카피/판독되고 온라인 기록되는 단일 분자이다 (Sam et al, 2011; Thompson and Milos, 2011).
본원에서 사용되는 용어 "고처리량 서열분석" 은 수천 개의 핵산 분자의 동시 또는 거의 동시 서열분석을 지칭한다. 고처리량 서열분석을 위한 플랫폼은 제한없이, 대규모 병렬형 시그너처 서열분석 (massively parallel signature sequencing) (MPSS), 폴로니 (Polony) 서열분석, 454 파이로시퀀싱 (pyrosequencing), Illumina (Solexa) 서열분석, SOLiD 서열분석, 이온 토렌트 반도체 서열분석 (Ion Torrent semiconductor sequencing), DNA 나노볼 (nanoball) 서열분석 (Complete Genomics/BGI Shenzhen), 헬리스토프 (Heliscope) 단일 분자 서열분석, 단일 분자 실시간 (single molecule real time) (SMRT) 서열분석 (PacBio), 및 나노포어 DNA 서열분석 (예를 들어, Oxford Nanopore) 을 포함한다.
본원에 기재된 방법은 제한없이, 전체 게놈 서열분석, 엑솜 (exome) 서열분석 및 앰플리콘 서열분석에 사용될 수 있다. 그러나, 증폭된 분자 자체는 특정 앰플리콘의 증폭을 거칠 수 있다. 게놈에서의 유전자 서열에 대한 베이트 (bait) 를 사용하는 서열 포획을 사용하여 엑솜을 나타내는 증폭된 분자를 단리할 수 있다. mRNA 를 이중 가닥 cDNA 로 역전사함으로써, 증폭된 전사체를 서열분석을 위해 생성할 수 있다.
V.
키트
본원에 개시된 방법을 수행하는데 사용하기 위한 키트가 또한 본원에 제공된다. 본원에 사용되는 용어 "키트" 는 함께 사용하는 것으로 의도된 품목의 집합을 지칭한다.
본원에 개시된 특정 키트는: (1) PrimPol 효소 (예를 들어, TthPrimPol); (2) DNA 폴리머라아제 (예를 들어, Phi29 DNApol); (3) 무작위 3량체; (4) 무작위 4량체; (5) 무작위 5량체; (6) 무작위 7량체; (7) 무작위 8량체; (8) 무작위 프라이머; (9) dNTP; (10) 반응 완충제; (11) 임의의 상술한 요소와 함께 사용하기 위한 완충제에서 선택되는 2, 3, 4, 5, 6, 7 개 요소를 포함한다. 키트는 또한 시약을 보유하기 위한 용기를 포함할 수 있다. 용기 자체는 운송 용기 내에 위치할 수 있다. 용기는 수 배송에 의해 또는 통상의 수송사, 예컨대 국가 우편 시스템 또는 배송 서비스 예컨대 FedEx 에 의해 전송될 수 있다. 키트는 또한 상자 또는 가방과 같은 채혈된 혈액을 중앙 시설에 운송하기 위한 용기를 포함할 수 있다. 키트는 또한 전형적으로, 데이터 분석 및 해석을 위한 소프트웨어 뿐만 아니라 사용을 위한 지시사항을 포함한다.
예시적
구현예
1. 돌연변이 K64R 또는 M97K 중 하나 또는 둘 모두를 포함하는 Phi29 유형 DNA 폴리머라아제.
2. SEQ ID NO: 2; SEQ ID NO: 3 또는 SEQ ID NO: 4 와 적어도 80% 의 동일성을 갖는 아미노산 서열을 갖는 Phi29 유형 DNA 폴리머라아제.
3. 주형 DNA 를 복제, 증폭 또는 서열분석하기 위한 방법으로서, 상기 DNA 를 적어도:
a) 구현예 1 내지 2 중 어느 하나에 따른 DNA 폴리머라아제, b) 완충제, c) 마그네슘 클로라이드, d) 프라이머 및 e) 뉴클레오시드 트리포스페이트
를 포함하는 반응 혼합물과 접촉시키는 것을 포함하는 방법.
4. 하기를 포함하는, 구현예 3 에 따른 방법을 실행하기 위한 키트:
a) 구현예 1 내지 2 중 어느 하나에 따른 DNA 폴리머라아제, b) 완충제 및 c) 마그네슘 클로라이드.
5. 구현예 1 내지 2 중 어느 하나에 따른 DNA 폴리머라아제, 및 하기 중 하나 이상을 포함하는, 구현예 3 에 따른 방법을 실행하기 위한 키트:
(a) PrimPol 효소 (예를 들어, TthPrimPol); (b) 무작위 3량체; (c) 무작위 4량체; (d) 무작위 5량체; (e) 무작위 7량체; (f) 무작위 8량체; (g) dNTP; (h) 반응 완충제; (i) 임의의 상술한 요소와 함께 사용하기 위한 완충제.
6. Phi29 유형 DNA 폴리머라아제로서, Phi29 유형 DNA 폴리머라아제가 SEQ ID NO:1 과 적어도 80%, 85%, 90%, 95%, 98% 또는 99%, 99.5% 서열 동일성을 갖는 아미노산 서열을 갖고, Phi29 유형 DNA 폴리머라아제가 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두를 포함하는 것인 Phi29 유형 DNA 폴리머라아제.
7. 구현예 6 에 있어서, SEQ ID NO:2, SEQ ID NO:3 또는 SEQ ID NO:4 의 서열을 갖는 Phi29 유형 DNA 폴리머라아제.
8. 구현예 6 에 있어서, 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두에 추가로 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 또는 1 개 이하의 아미노산 치환, 부가 또는 결실을 갖는 Phi29 유형 DNA 폴리머라아제.
9. Phi29 유형 DNA 폴리머라아제를 인코딩하는 뉴클레오티드 서열을 포함하는 단리된 핵산 분자로서, Phi29 유형 DNA 폴리머라아제가 SEQ ID NO:1 과 적어도 80%, 85%, 90%, 95%, 98% 또는 99%, 99.5% 서열 동일성을 갖는 아미노산 서열을 갖고, Phi29 유형 DNA 폴리머라아제가 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두를 포함하는, 단리된 핵산 분자.
10. 구현예 9 에 있어서, Phi29 유형 DNA 폴리머라아제가 SEQ ID NO:2, SEQ ID NO:3 또는 SEQ ID NO:4 의 서열을 갖는, 단리된 핵산 분자.
11. 구현예 9 에 있어서, Phi29 유형 DNA 폴리머라아제가 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두에 추가로 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 또는 1 개 이하의 아미노산 치환, 부가 또는 결실을 갖는, 단리된 핵산 분자.
12. 구현예 9-11 중 어느 하나의 Phi29 유형 DNA 폴리머라아제와 작동가능하게 연결되는 전사 조절 서열을 포함하는 재조합 핵산.
13. 구현예 12 에 있어서, 전사 조절 서열이 박테리아 또는 포유동물 프로모터를 포함하는 재조합 핵산.
14. 구현예 12 에 있어서, 플라스미드 벡터, 바이러스 벡터, 코스미드 및 트랜스포존에서 선택되는 벡터에 함유되는 재조합 핵산.
15. 구현예 14 에 있어서, Phi29 유형 DNA 폴리머라아제를 인코딩하는 뉴클레오티드 서열에 관련된 위치에 있는 클로닝 위치를 포함하여 클로닝 위치 내로 삽입된 전사 조절 서열이 Phi29 유형 DNA 폴리머라아제를 인코딩하는 뉴클레오티드 서열과 작동가능하게 연결되게 되는 재조합 핵산.
16. 구현예 12-15 중 어느 하나의 재조합 핵산을 포함하는 재조합 세포.
17. 하기 단계를 포함하는 방법:
a) 핵산 주형 분자를 구현예 1, 2, 6-8 중 어느 하나의 Phi29 유형 DNA 폴리머라아제, 및 프라이머 연장에 충분한 시약과 접촉시키는 단계; 및
b) 핵산 주형을 사용하여 폴리머라아제로 프라이머 연장을 수행하는 단계.
18. 구현예 17 에 있어서, 프라이머 연장에 충분한 시약이 올리고뉴클레오티드 프라이머를 포함하는 방법.
19. 구현예 18 에 있어서, 올리고뉴클레오티드 프라이머가 3량체, 4량체, 5량체, 6량체, 6량체, 8량체, 9량체 또는 10량체 중 하나 이상을 포함하는 방법.
20. 구현예 19 에 있어서, 프라이머가 무작위 프라이머인 방법.
21. 구현예 18 에 있어서, 올리고뉴클레오티드 프라이머가 5 내지 25 개 뉴클레오티드의 링크 (link) 를 갖는 방법.
22. 구현예 17 에 있어서, 프라이머 연장에 충분한 시약은 프리마아제/폴리머라아제 (예를 들어, TthPrimPol) 를 포함하는 방법.
23. 구현예 17 에 있어서, 프라이머 연장이 대략 31℃, 32℃, 33℃, 34℃, 35℃, 36℃, 37℃, 38℃, 39℃, 40℃, 41℃ 또는 42℃ 중 임의의 온도에서 또는 그 이상의 온도에서 수행되는 방법.
24. 구현예 17 에 있어서, 주형 핵산 분자가 1 ng, 100 pg, 10 pg 또는 1 pg 이하의 양으로 존재하는 방법.
25. 구현예 17 에 있어서, 프라이머 연장이 (i) 다중 변위 증폭 ("MDA") 또는 (2) 롤링 서클 증폭을 포함하는 방법.
26. 구현예 17 에 있어서, 프라이머 연장이 다중 어닐링 및 루핑 기반 증폭 사이클 (multiple annealing and looping-based amplification cycle) (MALBAC) 을 포함하는 방법.
27. 구현예 17-26 중 어느 하나에 있어서, Phi29 유형 DNA 폴리머라아제가 치환 K64R 및 M97K 둘 모두를 포함하는 방법.
실시예
실시예
1: 다중 변위 증폭 반응에서 WT
Phi29
DNA
pol
보다 더
짧은 무작위 합성
프라이머를
사용할 수 있는 돌연변이체를 검출하기 위한 스크리닝
도 3 에서는 Phi29 DNApol 변이체 및 TthPrimPol 또는 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 를 조합한 다중 변위 증폭 (MDA) 에 의한 1 ng 의 인간 게놈 DNA 의 증폭을 나타낸다.
도 3 에서 관찰되는 바와 같이, WT Phi29 DNApol 은 인간 게놈 DNA 를 증폭시키기 위해 5량체, 6량체, 7량체 및 8량체 뿐만 아니라 TthPrimPol 을 효율적으로 사용하였다. 3량체 및 4량체는 증폭을 실행하기에 적합하지 않았다.
생성된 Phi29 DNApol 변이체의 군으로부터, 이들 중 6 개 (K538R, T534K, T534R, L63LH, K64KG 및 K64KK) 는 프라이머 크기 또는 TthPrimPol 의 대안적 사용과는 독립적으로 MDA 에서 완전히 불활성이었다. 또 다른 세트의 돌연변이체 (K529R, M97R, R96K, L63LG 및 T499K) 는 WT Phi29 DNApol 보다 더 나쁜 증폭 성능을 나타내었는데, 이는 특정 프라이머 크기를 사용하는데 더 낮은 증폭 수율 및/또는 한계를 보여준다. 예를 들어, 돌연변이체 M97R 은 5량체 및 6량체를 효율적으로 사용할 수 있었던 반면, 7량체 및 8량체는 증폭을 촉발시키지 않았다. 유사하게, 돌연변이체 R96K 는 무작위 합성 프라이머의 세트로부터 6량체만을 사용할 수 있었다. 놀랍게도, 삽입 돌연변이체 L63LG 는 5량체, 6량체, 7량체 및 8량체로 DNA 를 증폭할 수 있었으나, TthPrimPol 과의 조합은 어떠한 증폭된 물질도 생성하지 않았다. 반대로, 돌연변이체 T499K 는 TthPrimPol 의 존재 하에 DNA 를 단지 약간 증폭시킬 수 있었던 반면, 무작위 합성 프라이머 중 아무 것도 MDA 를 촉진하지 않았다.
돌연변이체 T499R 은 WT Phi29 DNApol 과 대략 유사한 거동을 나타내었다.
마지막으로, 돌연변이체 K64R 및 M97K 는 WT Phi29 DNApol 에 대해 상당한 개선을 나타내었다. 이들 모두는 4량체를 사용할 수 있는 유일한 것들인 한편, WT Phi29 DNApol 및 나머지 돌연변이체는 어떠한 증폭 수율도 나타내지 않았다.
2 개의 "기능 획득" 돌연변이를 동일한 폴리펩티드에 도입하여 이중 돌연변이체 K64R / M97K 를 생성하였는데, 이는 하기 실시예에서 나타낸 바와 같이 WT Phi29 DNApol 및 단일 돌연변이체 K64R 및 M97K 와 비교하여 깊이 특징화되었다.
실시예
2:
Phi29
DNApol
돌연변이체
M97K
및 이중 돌연변이체
K64R
/
M97K 에서
엑소뉴클레아제
활성에 비해
폴리머라아제
활성이 선호됨
도 4 에서는 WT Phi29 DNApol 과 관련된 가장 관련있는 본 발명의 돌연변이체 (K64R, M97K 및 돌연변이체 K64R / M97K) 의 3'-5' 엑소뉴클레아제와 5'-3' 중합 활성 사이에 동적 평형의 분석을 나타낸다. 주형 (5' AGAAGTGTATCTGGTACTCACTGTGATC, SEQ ID NO: 6) 에 하이브리드화된 5'-표지된 프라이머 (5' GATCACAGTGAGTAC, SEQ ID NO: 5) 에 의해 형성된 DNA 이중체를 사용하여, dNTP 농도 (0, 10, 25, 50, 100 및 500 nM) 의 함수로서 DNA 합성과 DNA 분해 사이의 커플링을 분석하였다. dNTP 의 부재시에는 프라이머-말단의 엑소뉴클레오리틱 분해가 관찰된다. 분해 패턴은 WT Phi29 DNApol 에 대한 본 발명의 변이체의 엑소뉴클레아제 활성 수준을 반영한다. dNTP 의 농도가 증가함에 따라, 활성은 5'-3' 중합에 의해 점진적으로 극복되며, 순 dNMP 혼입은 표지된 프라이머의 크기 증가로서 관찰될 수 있어, 각각의 돌연변이체에 대한 프라이머의 효율적인 신장을 수득하기 위해 필요한 dNTP 의 농도를 규정한다. 도 4 에서 관찰되는 바와 같이, 돌연변이체 K64R 은 WT 효소에 의해 표시되는 바와 대략 유사한 Pol/Exo 평형을 나타내어, 25 nM dNTP 에서 28 량체 위치에 도달하였다. 한편, 돌연변이체 M97K 및 돌연변이체 K64R / M97K 는 시험한 최저 dNTP 농도 (10 nM) 로 동일한 위치 (28-량체) 에 도달하였는데, 이는 이들 돌연변이체의 폴리머라아제 활성이 엑소뉴클레아제에 비해 선호된다는 것을 나타낸다.
실시예
3: 본 발명의 돌연변이체 (
K64R
,
M97K
및
K64R
/
M97K
) 는 다중 변위 증폭 반응에서 WT
Phi29
DNApol
보다 더
짧은 무작위 합성
프라이머를
사용할 수 있음
도 5 에서는 Phi29 DNApol 선택 변이체 (K64R, M97K 및 이중 돌연변이체 K64R / M97K) 및 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 를 조합한 다중 변위 증폭에 의한 1 ng 의 인간 게놈 DNA 의 증폭을 나타낸다. 나타낸 증폭 수율은 각각의 조건 당 3 반복을 포함한 2 개의 독립적 실험의 평균이다. 2 개 실험으로부터의 표준 편차를 나타낸다.
도 5 에서 관찰된 바와 같이, 시험한 효소 중 아무 것도 무작위 합성 3량체를 사용하여 게놈 DNA 를 효율적으로 증폭시킬 수 없었다. 이중 돌연변이체 K64R / M97K 만이 1 μg 에 가까운 수율을 나타내었다.
3 개의 본 발명의 변이체는 4량체를 사용하여 증폭을 촉발할 수 있는 한편, WT Phi29 DNApol 로는 증폭이 관찰되지 않았다. 변이체 K64R 은 최저 증폭 수율 (2.7 μg) 을 나타내었고, 돌연변이체 M97K 는 약간 더 높은 수율 (3.8 μg) 을 나타내었고, 이중 돌연변이체 K64R / M97K 는 훨씬 더 높은 수율 (12.9 μg) 을 나타내었다. 이중 돌연변이체에서 관찰된 최고 수율은 동일한 폴리펩티드에서 둘 모두의 돌연변이의 상승 효과를 나타낸다.
WT Phi29 DNApol 및 3 개의 본 발명의 변이체는 증폭을 시작하기 위해 무작위 5량체를 효율적으로 사용할 수 있었다. 또한, 이중 돌연변이체 K64R / M97K 는 단일 변이체 및 WT 효소의 성능을 명백히 극복하면서, 20 μg 초과의 증폭된 DNA 인 최고 수율을 생성하였다.
무작위 6량체의 경우, 모든 경우에 증폭 수율이 더 높지만, 유사한 비교 패턴이 관찰된다.
무작위 7량체를 사용하여, WT Phi29 DNApol 은 6량체를 사용하여 수득한 결과에 대해 동일한 수율을 유지하는 한편, 3 개의 본 발명의 변이체는 증폭 효율을 감소시켜 무작위 5량체로 수득한 것들과 유사한 DNA 수준을 생성하는 경향이 있었다.
8량체의 경우, K64R 및 M97K 단일 돌연변이체 둘 모두는 WT Phi29 DNApol 보다 더 낮은 증폭 수율을 나타내었다. 반면, 이중 돌연변이체 K64R / M97K 는 시험한 모든 조건에서 발생한 바와 같이 WT Phi29 DNApol 을 명백히 극복하였으며, 증폭을 개시하는데 사용한 무작위 합성 프라이머의 길이에 관계없이 강건하고 효율적인 증폭 값이 확인된다.
실시예
4: 비-주형 대조군 (NTC) 에서 투입
DNA 의
부재 하에 관찰된 배경 증폭에서의 이온 강도의 효과
도 6 에서는 WT Phi29 DNApol 또는 선택된 본 발명의 변이체 (K64R, M97K 및 K64R / M97K) 및 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 를 조합할 때 투입 DNA 의 부재 하에 관찰된 증폭 수율을 나타낸다.
시험한 낮은 이온 강도 조건 (20 mM KCl; 57 mM NaCl) 하에, M97K 단일 돌연변이체 및 K64R / M97K 이중 돌연변이체 둘 모두는 5량체 및 6량체 (그러나 이중 돌연변이체의 경우에는 4량체) 를 사용하는 경우 투입 DNA 의 부재 하에 상당한 증폭 수율을 나타내었다 (도 6 참조). 그러나, 증폭 수율은 DNA (1 ng) 가 동일한 조건에서 투입물로서 사용되는 경우 수득된 것보다 상당히 더 낮은데 (도 5 참조), 이는 상이한 증폭 메커니즘이 수반되었음을 나타낸다. 투입 DNA 의 부재 하 Phi29 DNApol 의 프라이머-이량체 증폭 능력은 당업계에 잘 알려져 있으며 (Alsmadi et al, 2009), 프라이머-이량체의 안정성은 시험한 조건 하에 M97K 단일 돌연변이체 및 K64R / M97K 이중 돌연변이체에서 향상될 수 있다.
도 7 에서는 투입 DNA 의 부재 하에, 그러나 암모늄 술페이트 [(NH4)2SO4] 의 첨가에 의해 이온 강도 조건을 증가시키면서 관찰된 증폭 수율을 나타낸다. 암모늄 술페이트 (45 mM) 의 존재시, 투입 DNA 의 부재 하 관찰된 증폭 수준은 모든 변이체 및 모든 프라이머 크기에서 완전히 제거된다.
실시예
5: 높은 이온 강도 조건은 상이한 크기의 무작위
프라이머를
사용하여
DNA 의
증폭에 대한 이중 돌연변이체
K64R
/
M97K 의
강건성
및 효율을 강화함
도 8 에서는 높은 이온 강도 조건 (20 mM KCl; 57 mM NaCl; 45 mM (NH4)2SO4) 하 WT Phi29 DNApol 또는 선택된 본 발명의 변이체 (K64R, M97K 및 K64R / M97K) 및 상이한 크기의 무작위 합성 프라이머: 3량체 (3N), 4량체 (4N), 5량체 (5N), 6량체 (6N), 7량체 (7N) 또는 8량체 (8N) 를 조합한 다중 변위 증폭 (MDA) 에 의한 1 ng 의 인간 게놈 DNA 의 증폭을 나타낸다. 나타낸 증폭 수율은 각각의 조건 당 3 반복을 포함한 2 개의 독립적 실험의 평균이다. 2 개 실험으로부터의 표준 편차를 나타낸다.
도 8 에서 관찰된 바와 같이, 시험한 효소 중 아무 것도 무작위 합성 3량체를 사용하여 게놈 DNA 를 효율적으로 증폭시킬 수 없었다. 이중 돌연변이체 K64R / M97K 만이 600 ng 에 가까운 수율을 나타내었다.
이전 조건 (도 5 참조) 에서 관찰된 것과 대조적으로, 4량체는 M97K 단일 돌연변이체 및 K64R / M97K 이중 돌연변이체에 의해서만 효율적으로 사용된 한편, 단일 돌연변이체 K64R 은 1 μg 에 가까운 작은 수율만을 생성하였다. 현저하게, M97K 단일 돌연변이체 및 K64R / M97K 이중 돌연변이체에 대해 관찰된 증폭 수율은 암모늄 술페이트의 부재 하에 수득된 것과 비교하여 증가하였다 (각각 4 내지 12, 및 13 내지 16 μg).
5량체 및 6량체의 경우, M97K 단일 돌연변이체 및 K64R / M97K 이중 돌연변이체는 유사한 결과를 나타내어, WT 효소 또는 K64R 변이체로 수득한 증폭 수율을 명백히 넘어섰다. 암모늄 술페이트의 부재 하에 나타난 바와 같이, WT Phi29 DNApol 은 K64R 변이체보다 더 높은 수율을 나타내었다.
7량체의 경우, 이중 돌연변이체 K64R / M97K 만이 더 짧은 무작위 합성 프라이머로 및/또는 암모늄 술페이트의 부재 하에 수득한 증폭 수율을 유지하였다. WT Phi29 DNApol 및 K64R 변이체 둘 모두는 수율을 상당히 감소시켰으며, 이들 조건에서 동일한 값을 나타내었다. M97K 돌연변이체로 수득한 수율은 또한 이전 조건과 비교하여 감소하였다.
마지막으로, 8량체는 이중 돌연변이체 K64R / M97K 에 의해서만 효율적으로 사용된 한편, 다른 3 개의 효소는 매우 낮은 증폭 수율을 나타내었다.
Phi29 DNApol 이중 돌연변이체 K64R / M97K 는 낮은 및 높은 이온 강도 조건 모두에서 온전한 증폭 성능을 유지하는데, 이는 효소와 프라이머 뉴클레오티드 5 로부터의 질소 염기의 추가적인 접촉 및 뉴클레오티드 4 와 5 사이의 포스포디에스테르 결합에 의해 얻어지는 기능 획득의 결과일 수 있다 (도 2 참조). 이러한 추가적인 접촉은 효소가 상이한 이온 강도 조건 하에 상이한 크기의 프라이머를 능숙하게 안정화시킬 수 있게 한다.
실시예
6: 이중 돌연변이
K64R
/
M97K 는
시험한
프라이머
크기에 관계없이 미량의 DNA 를 매우 민감하게 증폭시킴
도 9 에서는 낮은 (20 mM KCl; 57 mM NaCl) 또는 높은 (20 mM KCl; 57 mM NaCl; 45 mM (NH4)2SO4) 이온 강도 조건 하 WT Phi29 DNApol 또는 선택된 본 발명의 변이체 (K64R, M97K 및 K64R / M97K) 및 상이한 크기의 무작위 합성 프라이머: 4량체 (4N), 5량체 (5N) 또는 6량체 (6N) 를 조합한 다중 변위 증폭 (MDA) 에 의한 상이한 양의 인간 게놈 DNA (1, 10, 100 pg 및 1 ng) 의 증폭을 나타낸다.
낮은 이온 강도 조건 (도 9, 상단 패널) 하에, 이중 돌연변이체 K64R / M97K 는 시험한 모든 조건에서 가장 일관적이며 가장 높은 증폭 수율을 생성한다.
무작위 합성 4량체의 경우, 이전에 나타낸 바와 같이 (도 5 참조), WT Phi29 DNApol 은 시험한 DNA 투입물 중 어느 것도 증폭시킬 수 없었다. 변이체 K64R 은 1 ng 의 DNA 투입만으로도 검출가능한 수율을 생성하였고, 더 낮은 양의 DNA 를 증폭시키는 민감도가 결여되었다. 반대로, M97K 및 M97K / K64R 돌연변이체 둘 모두는 시험한 DNA 투입물을 효율적으로 증폭시켰고, 이중 돌연변이체는 모든 경우에 더 높은 수율을 생성하였다.
무작위 합성 5량체의 경우, 모든 효소는 이들을 사용하여 증폭을 시작할 수 있었으나, 상이한 수준의 민감도 및 효율을 나타내었다. WT Phi29 DNApol 은 DNA 투입량이 감소하는 경우 증폭에서 상당한 감소를 보인 한편, 3 개의 본 발명의 변이체는 시험한 모든 조건에서 합리적인 효율을 유지하였다. 이중 돌연변이체 M97K / K64R 은 DNA 투입량과 관계없이 3 개의 본 발명의 변이체 중에서 가장 높은 증폭 효율을 나타내었으며, 따라서 최상의 민감도를 나타내었다.
무작위 합성 6량체의 경우, 모든 효소는 시험한 각각의 DNA 투입물의 증폭을 개시하기 위해 이들을 효율적으로 사용할 수 있었으며, 모든 경우에 주목할만한 증폭 수율을 나타내었다. 3 개의 본 발명의 변이체는 WT Phi29 DNApol 을 넘어섰으며, 낮은 DNA 투입물이 분석된 경우 더 높은 증폭 수율을 나타내었다. 5량체로 발생하는 바와 같이, 이중 돌연변이체 M97K / K64R 은 DNA 투입량과 관계없이 3 개의 본 발명의 변이체 중에서 가장 높은 증폭 효율을 나타내었다.
이전에 나타낸 바와 같이 (도 6 참조), M97K 및 M97K / K64R 변이체 둘 모두는 5량체 및 6량체를 사용하는 경우, 투입 DNA (비-주형 대조군, NTC) 의 부재 하에 상당한 증폭 수율을 나타내었다. 이러한 이유로, 높은 이온 강도 조건 (20 mM KCl; 57 mM NaCl; 45 mM (NH4)2SO4) 하에 동일한 민감도 분석을 실행하여, 프라이머-이량체 증폭에서 유래한 이러한 인공적 효과를 방지하였다.
높은 이온 강도 조건 (도 9, 하단 패널 참조) 하에, 무작위 합성 4량체는 낮은 이온 강도 조건과 비교하여 시험한 변이체 중에서 유사한 사용 패턴을 나타내었으나, 증폭 수율은 대부분의 경우 증가하였다. 이러한 규칙에 대한 예외는, 1 pg 및 10 pg DNA 투입으로 더 낮은 수율을 나타낸 변이체 M97K 였다.
높은 이온 강도 조건 하 무작위 합성 5량체의 경우, M97K 및 M97K / K64R 변이체는 민감도 및 효율 면에서 최상의 성능을 나타내었으며, 동일한 DNA 투입량으로의 낮은 이온 강도 조건과 비교하여 더 높은 증폭 수율을 나타내었다. 제한된 양의 DNA 를 시험한 경우 (1 및 10 pg) 반응의 이온 강도 증가는 변이체 K64R 의 증폭 효율 감소를 생성한 한편, 효율은 다른 두 투입물과 유사 (100 pg) 하거나 더 높았다 (1 ng). 놀랍게도, WT Phi29 DNApol 은 모든 경우에 이러한 조건 하에서 K64R 변이체를 넘어섰다.
높은 이온 강도 조건 하 무작위 합성 6량체의 경우, 이중 돌연변이체 K64R / M97K 는 낮은 이온 강도 조건 하에 수득한 결과와 비교하여 모든 DNA 투입으로 관찰된 수율을 증가시킨 유일한 변이체였다. 단일 돌연변이체 M97K 는 가장 낮은 투입 (1 및 10 pg) 으로 더 낮은 수율을 나타낸 한편, 100 pg 및 1 ng DNA 투입으로 수율을 증가시켰는데, 이는 민감도 감소를 나타낸다. 변이체 K64R 및 WT Phi29 DNApol 은 유사한 거동을 나타내었다. 5량체로 발생하는 바와 같이, WT Phi29 DNApol 은 모든 경우에 이러한 조건 하에 K64R 변이체보다 더 높은 증폭 수율을 생성하였다.
요약하면, 이중 돌연변이체 K64R / M97K 는 시험한 모든 DNA 프라이머를 사용한 증폭 반응 동안 낮은 이온 강도 조건 및 높은 이온 강도 조건 둘 모두 하에서 증폭 효율 및 민감도 면에서 최상을 성능을 나타내었다.
실시예
7:
프라이머가
Tth
PrimPol
에 의해 생성될 때 본 발명의
변이체
사용에 의해 증폭 효율 및 민감도는 변형되지 않음
도 10 에서는 WT Phi29 DNApol 또는 선택된 본 발명의 변이체 (K64R, M97K 및 K64R / M97K) 및 TthPrimPol, Phi29 DNApol 에 대한 반응 과정에서 프라이머를 합성할 수 있는 DNA 프리마아제를 조합한 다중 변위 증폭 (MDA) 에 의한 1, 10, 100 pg 및 1 ng 의 인간 게놈 DNA 의 증폭을 나타낸다 (Picher et al, 2016).
도 10 에서 관찰되는 바와 같이, WT Phi29 DNA pol 과 시험한 본 발명의 변이체 사이에 상당한 수율 차이는 없으며, 이는 이러한 셋업 하에 유사한 민감도 및 효율 수준을 초래한다.
실시예
8: 선택된 본 발명의
변이체
(
K64R
,
M97K
및
K64R
/
M97K
) 는 CovCheck 기술에 의해 측정된 증폭 커버리지를 개선시킴
CovCheck 기술은 각각의 인간 염색체로부터의 작은 부분을 증폭시키는 24 개의 상이한 프라이머 쌍을 포함하는 PCR 패널을 사용하여 전체 게놈 증폭의 커버리지 분석을 가능하게 한다. CovCheck 기술은 CovCheck 커버리지 값을 로우-패스 (low-pass) 전체 게놈 서열분석을 통해 수득한 실제 커버리지와 비교하고, 우수한 상관 값을 수득함으로써 검증되었다 (https://www.expedeon.com/products/genomics/dna-rna-products/covcheck-pcr-kits/).
각각의 변이체로 수득한 증폭 커버리지를 분석하기 위해, 제한된 양의 투입 물질을 선택하였다: 30 pg 의 인간 게놈 DNA. 이러한 DNA 양은 5 개 인간 이배체 게놈과 동등하며, 이는 각각의 염색체의 충분한 카피가 증폭에 이용가능할 것임을 보장하는 가장 낮은 양일 수 있다. 이 수준 아래에서는, 특정 부위 또는 완전한 염색체는 정제된 DNA 샘플에서의 분자의 무작위 분포로 인해 증폭을 위한 투입물에 부재할 수 있어, 증폭 실패로 인한 것이 아니라 주형의 부재로 인해 증폭된 물질에서 커버되지 않는 부위를 초래한다.
도 11 에서는 WT Phi29 DNApol 또는 선택된 본 발명의 변이체 및 30 pg 의 인간 게놈 DNA 투입과 조합으로 6량체, 5량체 및 4량체를 사용한 증폭 반응의 CovCheck 분석으로부터 수득한 추정 커버리지 값을 나타낸다. 커버리지 값은 조건 당 6 개의 독립적 반응의 평균이다.
무작위 합성 6량체의 경우, WT Phi29 DNApol 에 의해 수득한 값과 비교하여, 3 개의 본 발명의 변이체를 사용하는 경우에 증폭 커버리지가 개선된다.
무작위 합성 5량체의 경우, 모든 효소가 이들 조건에서 90% 를 넘는 커버리지 값을 나타내었다. 따라서, 유의한 차이를 관찰할 수 없었다. 그러나, M97K 변이체는 시험한 6 개 복제물에서 완벽한 커버리지로 두드러졌다.
무작위 합성 4량체의 경우, M97K 및 M97K / K64R 변이체만이 증폭된 DNA 를 생성하였으며, 이는 4량체와 조합시 WT Phi29 DNApol 및 K64R 변이체에 의해 나타난 증폭 민감도와 일치한다 (도 9). CovCheck 분석으로, 두 경우 모두에서 우수한 증폭 커버리지가 드러났으며 (99%), 이는 증폭 커버리지 및 균일성을 최대화하기 위해 가능한 최단 프라이머와 조합으로 본 발명의 변이체를 사용하고, 증폭 편향 및 서열의 손실을 방지하는 이점을 지적한다.
효소 (TthPrimPol) 를 사용하여 Phi29 DNApol 에 대한 DNA 프라이머를 생성하는 경우, 도 12 는 증폭 반응에서 투입물로서 30 pg (5 게놈 당량) 의 인간 게놈 DNA 를 사용하여 TthPrimPol 와 WT Phi29 DNApol 또는 본 발명의 변이체의 조합에 의해 실행된 증폭 반응의 CovCheck 분석으로부터 수득한 추정 커버리지 값을 나타낸다. 커버리지 값은 조건 당 12 개의 독립적 반응의 평균이다. CovCheck 분석으로, 또한 본 발명의 변이체 사용시 증폭 커버리지의 개선이 드러났으며, 이는 본래 DNA 투입물에 대해 증폭된 물질의 균일성을 향상시키는 것들의 이점을 지지한다.
참고문헌 (본원에 참조로 포함됨)



달리 명시되지 않는 한, 본원에 사용된 바와 같이, 하기의 의미가 적용된다. "~일 수 있다" 라는 단어는 의무적 의미 (즉, 필수적이라는 의미) 보다는 허용적 의미 (즉, 가능성을 갖는 의미) 로 사용된다. "포함하다", "포함하는" 및 "포함되다" 등의 단어는 포함하는 것을 의미하지만 이에 제한되지는 않는다. 단수형은 복수형의 대상을 포함한다. 따라서, 예를 들어, "하나의 요소" 에 대한 언급은 하나 이상의 요소, 예컨대 "하나 이상" 에 대한 다른 용어 및 구절의 사용에도 불구하고 2 개 이상의 요소의 조합을 포함한다. 달리 언급되지 않는 한, 용어 "또는" 은 비-배타적, 즉 "및" 과 "또는" 을 모두 포함한다. 수식어와 시퀀스 사이의 용어 "~중 어느 것" 은 수식어가 시퀀스의 각 멤버를 수식한다는 것을 의미한다. 따라서 예를 들어, 어구 "적어도 1, 2 또는 3 중 어느 것" 은 "적어도 1, 적어도 2 또는 적어도 3" 을 의미한다. 특정 구현예에서, 다양한 요소들을 "포함하는" 발명들은 또한 이러한 요소들로 "본질적으로 이루어질 수 있다". 용어 "~로 본질적으로 이루어지는" 은 청구된 조합의 기본적이고 신규한 특징에 실질적으로 영향을 주지 않는 언급된 요소 및 추가 요소의 포함을 의미한다.
상세한 설명 및 도면이 본 발명을 개시된 특정 형태로 제한하는 것을 의도하지 않지만, 반면, 청구범위에 의해 정의된 바와 같은 본 발명의 취지 및 범주 내에 있는 모든 변형, 등가물 및 대안물을 커버함이 이해되어야 한다. 본 발명의 다양한 양태의 추가 변형 및 대안적 구현예는 이러한 설명을 고려하여 당업자에게 명백할 것이다. 따라서, 상세한 설명 및 도면은 단지 예시적인 것으로 이해되어야 하며, 당업자에게 본 발명을 실행하는 일반적인 방식을 교시하기 위한 것이다. 본원에 나타내고 기재한 본 발명의 형태가 구현예의 예로서 취해질 것임이 이해되어야 한다. 요소 및 물질은 본원에 예시되고 기재된 것들을 대체할 수 있고, 부분 및 과정은 역전되거나 생략될 수 있으며, 본 발명의 특정한 특징은 독립적으로 활용될 수 있어, 이들 모두는 본 발명의 상세한 설명의 이득을 가진 후 당업자에게 명백할 것이다. 뒤이어지는 청구범위에서 기재한 바와 같이, 본 발명의 취지 및 범주에서 벗어나지 않으면서 본원에 기재된 요소에 변화가 행해질 수 있다. 본원에서 사용된 제목들은 단지 조직상의 목적을 위한 것이며 상세한 설명의 범주를 제한하는 것으로 사용되는 것을 의미하지 않는다.
본 명세서에 언급된 모든 공개물, 특허 및 특허 출원은 각각의 개별적 공개물, 특허 또는 특허 출원이 본원에 참조로 포함되는 것으로 구체적이고 개별적으로 표시되는 것과 동일한 정도로 본원에 참조로 포함된다.
서열 목록
이탤릭체의 아미노산은 일부
구현예에서
표시되지 않는다.
SEQ
ID NO 1: 야생형
Phi29
DNA
폴리머라아제
(
UniProtKB
- P03680)



SEQ
ID NO 2:
K64R
Phi29DNApol
돌연변이체



SEQ
ID NO 3:
M97K
Phi29DNApol
돌연변이체



SEQ
ID NO 4:
K64R
/
M97K
Phi29DNApol
이중 돌연변이체



SEQUENCE LISTING
<110> 4BASEBIO SL
<120> PHI29 DNA POLYMERASE MUTANTS WITH IMPROVED PRIMER RECOGNITION
<130> 4BA17083PCT
<140> US62/849,252
<141> 2019-05-17
<160> 6
<170> PatentIn version 3.5
<210> 1
<211> 575
<212> PRT
<213> Bacillus subtilis
<220>
<223> Phi29DNApol wild type
<400> 1
Met Lys His Met Pro Arg Lys Met Tyr Ser Cys Asp Phe Glu Thr Thr
1 5 10 15
Thr Lys Val Glu Asp Cys Arg Val Trp Ala Tyr Gly Tyr Met Asn Ile
20 25 30
Glu Asp His Ser Glu Tyr Lys Ile Gly Asn Ser Leu Asp Glu Phe Met
35 40 45
Ala Trp Val Leu Lys Val Gln Ala Asp Leu Tyr Phe His Asn Leu Lys
50 55 60
Phe Asp Gly Ala Phe Ile Ile Asn Trp Leu Glu Arg Asn Gly Phe Lys
65 70 75 80
Trp Ser Ala Asp Gly Leu Pro Asn Thr Tyr Asn Thr Ile Ile Ser Arg
85 90 95
Met Gly Gln Trp Tyr Met Ile Asp Ile Cys Leu Gly Tyr Lys Gly Lys
100 105 110
Arg Lys Ile His Thr Val Ile Tyr Asp Ser Leu Lys Lys Leu Pro Phe
115 120 125
Pro Val Lys Lys Ile Ala Lys Asp Phe Lys Leu Thr Val Leu Lys Gly
130 135 140
Asp Ile Asp Tyr His Lys Glu Arg Pro Val Gly Tyr Lys Ile Thr Pro
145 150 155 160
Glu Glu Tyr Ala Tyr Ile Lys Asn Asp Ile Gln Ile Ile Ala Glu Ala
165 170 175
Leu Leu Ile Gln Phe Lys Gln Gly Leu Asp Arg Met Thr Ala Gly Ser
180 185 190
Asp Ser Leu Lys Gly Phe Lys Asp Ile Ile Thr Thr Lys Lys Phe Lys
195 200 205
Lys Val Phe Pro Thr Leu Ser Leu Gly Leu Asp Lys Glu Val Arg Tyr
210 215 220
Ala Tyr Arg Gly Gly Phe Thr Trp Leu Asn Asp Arg Phe Lys Glu Lys
225 230 235 240
Glu Ile Gly Glu Gly Met Val Phe Asp Val Asn Ser Leu Tyr Pro Ala
245 250 255
Gln Met Tyr Ser Arg Leu Leu Pro Tyr Gly Glu Pro Ile Val Phe Glu
260 265 270
Gly Lys Tyr Val Trp Asp Glu Asp Tyr Pro Leu His Ile Gln His Ile
275 280 285
Arg Cys Glu Phe Glu Leu Lys Glu Gly Tyr Ile Pro Thr Ile Gln Ile
290 295 300
Lys Arg Ser Arg Phe Tyr Lys Gly Asn Glu Tyr Leu Lys Ser Ser Gly
305 310 315 320
Gly Glu Ile Ala Asp Leu Trp Leu Ser Asn Val Asp Leu Glu Leu Met
325 330 335
Lys Glu His Tyr Asp Leu Tyr Asn Val Glu Tyr Ile Ser Gly Leu Lys
340 345 350
Phe Lys Ala Thr Thr Gly Leu Phe Lys Asp Phe Ile Asp Lys Trp Thr
355 360 365
Tyr Ile Lys Thr Thr Ser Glu Gly Ala Ile Lys Gln Leu Ala Lys Leu
370 375 380
Met Leu Asn Ser Leu Tyr Gly Lys Phe Ala Ser Asn Pro Asp Val Thr
385 390 395 400
Gly Lys Val Pro Tyr Leu Lys Glu Asn Gly Ala Leu Gly Phe Arg Leu
405 410 415
Gly Glu Glu Glu Thr Lys Asp Pro Val Tyr Thr Pro Met Gly Val Phe
420 425 430
Ile Thr Ala Trp Ala Arg Tyr Thr Thr Ile Thr Ala Ala Gln Ala Cys
435 440 445
Tyr Asp Arg Ile Ile Tyr Cys Asp Thr Asp Ser Ile His Leu Thr Gly
450 455 460
Thr Glu Ile Pro Asp Val Ile Lys Asp Ile Val Asp Pro Lys Lys Leu
465 470 475 480
Gly Tyr Trp Ala His Glu Ser Thr Phe Lys Arg Ala Lys Tyr Leu Arg
485 490 495
Gln Lys Thr Tyr Ile Gln Asp Ile Tyr Met Lys Glu Val Asp Gly Lys
500 505 510
Leu Val Glu Gly Ser Pro Asp Asp Tyr Thr Asp Ile Lys Phe Ser Val
515 520 525
Lys Cys Ala Gly Met Thr Asp Lys Ile Lys Lys Glu Val Thr Phe Glu
530 535 540
Asn Phe Lys Val Gly Phe Ser Arg Lys Met Lys Pro Lys Pro Val Gln
545 550 555 560
Val Pro Gly Gly Val Val Leu Val Asp Asp Thr Phe Thr Ile Lys
565 570 575
<210> 2
<211> 575
<212> PRT
<213> Artificial Sequence
<220>
<223> K64R Phi29DNApol mutant
<400> 2
Met Lys His Met Pro Arg Lys Met Tyr Ser Cys Asp Phe Glu Thr Thr
1 5 10 15
Thr Lys Val Glu Asp Cys Arg Val Trp Ala Tyr Gly Tyr Met Asn Ile
20 25 30
Glu Asp His Ser Glu Tyr Lys Ile Gly Asn Ser Leu Asp Glu Phe Met
35 40 45
Ala Trp Val Leu Lys Val Gln Ala Asp Leu Tyr Phe His Asn Leu Arg
50 55 60
Phe Asp Gly Ala Phe Ile Ile Asn Trp Leu Glu Arg Asn Gly Phe Lys
65 70 75 80
Trp Ser Ala Asp Gly Leu Pro Asn Thr Tyr Asn Thr Ile Ile Ser Arg
85 90 95
Met Gly Gln Trp Tyr Met Ile Asp Ile Cys Leu Gly Tyr Lys Gly Lys
100 105 110
Arg Lys Ile His Thr Val Ile Tyr Asp Ser Leu Lys Lys Leu Pro Phe
115 120 125
Pro Val Lys Lys Ile Ala Lys Asp Phe Lys Leu Thr Val Leu Lys Gly
130 135 140
Asp Ile Asp Tyr His Lys Glu Arg Pro Val Gly Tyr Lys Ile Thr Pro
145 150 155 160
Glu Glu Tyr Ala Tyr Ile Lys Asn Asp Ile Gln Ile Ile Ala Glu Ala
165 170 175
Leu Leu Ile Gln Phe Lys Gln Gly Leu Asp Arg Met Thr Ala Gly Ser
180 185 190
Asp Ser Leu Lys Gly Phe Lys Asp Ile Ile Thr Thr Lys Lys Phe Lys
195 200 205
Lys Val Phe Pro Thr Leu Ser Leu Gly Leu Asp Lys Glu Val Arg Tyr
210 215 220
Ala Tyr Arg Gly Gly Phe Thr Trp Leu Asn Asp Arg Phe Lys Glu Lys
225 230 235 240
Glu Ile Gly Glu Gly Met Val Phe Asp Val Asn Ser Leu Tyr Pro Ala
245 250 255
Gln Met Tyr Ser Arg Leu Leu Pro Tyr Gly Glu Pro Ile Val Phe Glu
260 265 270
Gly Lys Tyr Val Trp Asp Glu Asp Tyr Pro Leu His Ile Gln His Ile
275 280 285
Arg Cys Glu Phe Glu Leu Lys Glu Gly Tyr Ile Pro Thr Ile Gln Ile
290 295 300
Lys Arg Ser Arg Phe Tyr Lys Gly Asn Glu Tyr Leu Lys Ser Ser Gly
305 310 315 320
Gly Glu Ile Ala Asp Leu Trp Leu Ser Asn Val Asp Leu Glu Leu Met
325 330 335
Lys Glu His Tyr Asp Leu Tyr Asn Val Glu Tyr Ile Ser Gly Leu Lys
340 345 350
Phe Lys Ala Thr Thr Gly Leu Phe Lys Asp Phe Ile Asp Lys Trp Thr
355 360 365
Tyr Ile Lys Thr Thr Ser Glu Gly Ala Ile Lys Gln Leu Ala Lys Leu
370 375 380
Met Leu Asn Ser Leu Tyr Gly Lys Phe Ala Ser Asn Pro Asp Val Thr
385 390 395 400
Gly Lys Val Pro Tyr Leu Lys Glu Asn Gly Ala Leu Gly Phe Arg Leu
405 410 415
Gly Glu Glu Glu Thr Lys Asp Pro Val Tyr Thr Pro Met Gly Val Phe
420 425 430
Ile Thr Ala Trp Ala Arg Tyr Thr Thr Ile Thr Ala Ala Gln Ala Cys
435 440 445
Tyr Asp Arg Ile Ile Tyr Cys Asp Thr Asp Ser Ile His Leu Thr Gly
450 455 460
Thr Glu Ile Pro Asp Val Ile Lys Asp Ile Val Asp Pro Lys Lys Leu
465 470 475 480
Gly Tyr Trp Ala His Glu Ser Thr Phe Lys Arg Ala Lys Tyr Leu Arg
485 490 495
Gln Lys Thr Tyr Ile Gln Asp Ile Tyr Met Lys Glu Val Asp Gly Lys
500 505 510
Leu Val Glu Gly Ser Pro Asp Asp Tyr Thr Asp Ile Lys Phe Ser Val
515 520 525
Lys Cys Ala Gly Met Thr Asp Lys Ile Lys Lys Glu Val Thr Phe Glu
530 535 540
Asn Phe Lys Val Gly Phe Ser Arg Lys Met Lys Pro Lys Pro Val Gln
545 550 555 560
Val Pro Gly Gly Val Val Leu Val Asp Asp Thr Phe Thr Ile Lys
565 570 575
<210> 3
<211> 575
<212> PRT
<213> Artificial Sequence
<220>
<223> M97K Phi29DNApol mutant
<400> 3
Met Lys His Met Pro Arg Lys Met Tyr Ser Cys Asp Phe Glu Thr Thr
1 5 10 15
Thr Lys Val Glu Asp Cys Arg Val Trp Ala Tyr Gly Tyr Met Asn Ile
20 25 30
Glu Asp His Ser Glu Tyr Lys Ile Gly Asn Ser Leu Asp Glu Phe Met
35 40 45
Ala Trp Val Leu Lys Val Gln Ala Asp Leu Tyr Phe His Asn Leu Lys
50 55 60
Phe Asp Gly Ala Phe Ile Ile Asn Trp Leu Glu Arg Asn Gly Phe Lys
65 70 75 80
Trp Ser Ala Asp Gly Leu Pro Asn Thr Tyr Asn Thr Ile Ile Ser Arg
85 90 95
Lys Gly Gln Trp Tyr Met Ile Asp Ile Cys Leu Gly Tyr Lys Gly Lys
100 105 110
Arg Lys Ile His Thr Val Ile Tyr Asp Ser Leu Lys Lys Leu Pro Phe
115 120 125
Pro Val Lys Lys Ile Ala Lys Asp Phe Lys Leu Thr Val Leu Lys Gly
130 135 140
Asp Ile Asp Tyr His Lys Glu Arg Pro Val Gly Tyr Lys Ile Thr Pro
145 150 155 160
Glu Glu Tyr Ala Tyr Ile Lys Asn Asp Ile Gln Ile Ile Ala Glu Ala
165 170 175
Leu Leu Ile Gln Phe Lys Gln Gly Leu Asp Arg Met Thr Ala Gly Ser
180 185 190
Asp Ser Leu Lys Gly Phe Lys Asp Ile Ile Thr Thr Lys Lys Phe Lys
195 200 205
Lys Val Phe Pro Thr Leu Ser Leu Gly Leu Asp Lys Glu Val Arg Tyr
210 215 220
Ala Tyr Arg Gly Gly Phe Thr Trp Leu Asn Asp Arg Phe Lys Glu Lys
225 230 235 240
Glu Ile Gly Glu Gly Met Val Phe Asp Val Asn Ser Leu Tyr Pro Ala
245 250 255
Gln Met Tyr Ser Arg Leu Leu Pro Tyr Gly Glu Pro Ile Val Phe Glu
260 265 270
Gly Lys Tyr Val Trp Asp Glu Asp Tyr Pro Leu His Ile Gln His Ile
275 280 285
Arg Cys Glu Phe Glu Leu Lys Glu Gly Tyr Ile Pro Thr Ile Gln Ile
290 295 300
Lys Arg Ser Arg Phe Tyr Lys Gly Asn Glu Tyr Leu Lys Ser Ser Gly
305 310 315 320
Gly Glu Ile Ala Asp Leu Trp Leu Ser Asn Val Asp Leu Glu Leu Met
325 330 335
Lys Glu His Tyr Asp Leu Tyr Asn Val Glu Tyr Ile Ser Gly Leu Lys
340 345 350
Phe Lys Ala Thr Thr Gly Leu Phe Lys Asp Phe Ile Asp Lys Trp Thr
355 360 365
Tyr Ile Lys Thr Thr Ser Glu Gly Ala Ile Lys Gln Leu Ala Lys Leu
370 375 380
Met Leu Asn Ser Leu Tyr Gly Lys Phe Ala Ser Asn Pro Asp Val Thr
385 390 395 400
Gly Lys Val Pro Tyr Leu Lys Glu Asn Gly Ala Leu Gly Phe Arg Leu
405 410 415
Gly Glu Glu Glu Thr Lys Asp Pro Val Tyr Thr Pro Met Gly Val Phe
420 425 430
Ile Thr Ala Trp Ala Arg Tyr Thr Thr Ile Thr Ala Ala Gln Ala Cys
435 440 445
Tyr Asp Arg Ile Ile Tyr Cys Asp Thr Asp Ser Ile His Leu Thr Gly
450 455 460
Thr Glu Ile Pro Asp Val Ile Lys Asp Ile Val Asp Pro Lys Lys Leu
465 470 475 480
Gly Tyr Trp Ala His Glu Ser Thr Phe Lys Arg Ala Lys Tyr Leu Arg
485 490 495
Gln Lys Thr Tyr Ile Gln Asp Ile Tyr Met Lys Glu Val Asp Gly Lys
500 505 510
Leu Val Glu Gly Ser Pro Asp Asp Tyr Thr Asp Ile Lys Phe Ser Val
515 520 525
Lys Cys Ala Gly Met Thr Asp Lys Ile Lys Lys Glu Val Thr Phe Glu
530 535 540
Asn Phe Lys Val Gly Phe Ser Arg Lys Met Lys Pro Lys Pro Val Gln
545 550 555 560
Val Pro Gly Gly Val Val Leu Val Asp Asp Thr Phe Thr Ile Lys
565 570 575
<210> 4
<211> 575
<212> PRT
<213> Artificial Sequence
<220>
<223> K64R / M97K Phi29DNApol double mutant
<400> 4
Met Lys His Met Pro Arg Lys Met Tyr Ser Cys Asp Phe Glu Thr Thr
1 5 10 15
Thr Lys Val Glu Asp Cys Arg Val Trp Ala Tyr Gly Tyr Met Asn Ile
20 25 30
Glu Asp His Ser Glu Tyr Lys Ile Gly Asn Ser Leu Asp Glu Phe Met
35 40 45
Ala Trp Val Leu Lys Val Gln Ala Asp Leu Tyr Phe His Asn Leu Arg
50 55 60
Phe Asp Gly Ala Phe Ile Ile Asn Trp Leu Glu Arg Asn Gly Phe Lys
65 70 75 80
Trp Ser Ala Asp Gly Leu Pro Asn Thr Tyr Asn Thr Ile Ile Ser Arg
85 90 95
Lys Gly Gln Trp Tyr Met Ile Asp Ile Cys Leu Gly Tyr Lys Gly Lys
100 105 110
Arg Lys Ile His Thr Val Ile Tyr Asp Ser Leu Lys Lys Leu Pro Phe
115 120 125
Pro Val Lys Lys Ile Ala Lys Asp Phe Lys Leu Thr Val Leu Lys Gly
130 135 140
Asp Ile Asp Tyr His Lys Glu Arg Pro Val Gly Tyr Lys Ile Thr Pro
145 150 155 160
Glu Glu Tyr Ala Tyr Ile Lys Asn Asp Ile Gln Ile Ile Ala Glu Ala
165 170 175
Leu Leu Ile Gln Phe Lys Gln Gly Leu Asp Arg Met Thr Ala Gly Ser
180 185 190
Asp Ser Leu Lys Gly Phe Lys Asp Ile Ile Thr Thr Lys Lys Phe Lys
195 200 205
Lys Val Phe Pro Thr Leu Ser Leu Gly Leu Asp Lys Glu Val Arg Tyr
210 215 220
Ala Tyr Arg Gly Gly Phe Thr Trp Leu Asn Asp Arg Phe Lys Glu Lys
225 230 235 240
Glu Ile Gly Glu Gly Met Val Phe Asp Val Asn Ser Leu Tyr Pro Ala
245 250 255
Gln Met Tyr Ser Arg Leu Leu Pro Tyr Gly Glu Pro Ile Val Phe Glu
260 265 270
Gly Lys Tyr Val Trp Asp Glu Asp Tyr Pro Leu His Ile Gln His Ile
275 280 285
Arg Cys Glu Phe Glu Leu Lys Glu Gly Tyr Ile Pro Thr Ile Gln Ile
290 295 300
Lys Arg Ser Arg Phe Tyr Lys Gly Asn Glu Tyr Leu Lys Ser Ser Gly
305 310 315 320
Gly Glu Ile Ala Asp Leu Trp Leu Ser Asn Val Asp Leu Glu Leu Met
325 330 335
Lys Glu His Tyr Asp Leu Tyr Asn Val Glu Tyr Ile Ser Gly Leu Lys
340 345 350
Phe Lys Ala Thr Thr Gly Leu Phe Lys Asp Phe Ile Asp Lys Trp Thr
355 360 365
Tyr Ile Lys Thr Thr Ser Glu Gly Ala Ile Lys Gln Leu Ala Lys Leu
370 375 380
Met Leu Asn Ser Leu Tyr Gly Lys Phe Ala Ser Asn Pro Asp Val Thr
385 390 395 400
Gly Lys Val Pro Tyr Leu Lys Glu Asn Gly Ala Leu Gly Phe Arg Leu
405 410 415
Gly Glu Glu Glu Thr Lys Asp Pro Val Tyr Thr Pro Met Gly Val Phe
420 425 430
Ile Thr Ala Trp Ala Arg Tyr Thr Thr Ile Thr Ala Ala Gln Ala Cys
435 440 445
Tyr Asp Arg Ile Ile Tyr Cys Asp Thr Asp Ser Ile His Leu Thr Gly
450 455 460
Thr Glu Ile Pro Asp Val Ile Lys Asp Ile Val Asp Pro Lys Lys Leu
465 470 475 480
Gly Tyr Trp Ala His Glu Ser Thr Phe Lys Arg Ala Lys Tyr Leu Arg
485 490 495
Gln Lys Thr Tyr Ile Gln Asp Ile Tyr Met Lys Glu Val Asp Gly Lys
500 505 510
Leu Val Glu Gly Ser Pro Asp Asp Tyr Thr Asp Ile Lys Phe Ser Val
515 520 525
Lys Cys Ala Gly Met Thr Asp Lys Ile Lys Lys Glu Val Thr Phe Glu
530 535 540
Asn Phe Lys Val Gly Phe Ser Arg Lys Met Lys Pro Lys Pro Val Gln
545 550 555 560
Val Pro Gly Gly Val Val Leu Val Asp Asp Thr Phe Thr Ile Lys
565 570 575
<210> 5
<211> 15
<212> DNA
<213> Artificial Sequence
<220>
<223> 5'-labelled primer of DNA duplex
<400> 5
gatcacagtg agtac 15
<210> 6
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> template of DNA duplex
<400> 6
agaagtgtat ctggtactca ctgtgatc 28
Claims (27)
- 돌연변이 K64R 또는 M97K 중 하나 또는 둘 모두를 포함하는 Phi29 유형 DNA 폴리머라아제.
- SEQ ID NO: 2; SEQ ID NO: 3 또는 SEQ ID NO: 4 와 적어도 80% 의 동일성을 갖는 아미노산 서열을 갖는 Phi29 유형 DNA 폴리머라아제.
- 주형 DNA 를 복제, 증폭 또는 서열분석하기 위한 방법으로서, 상기 DNA 를 적어도:
a) 제 1 항 또는 제 2 항에 따른 DNA 폴리머라아제,
b) 완충제,
c) 마그네슘 클로라이드,
d) 프라이머 및
e) 뉴클레오시드 트리포스페이트
를 포함하는 반응 혼합물과 접촉시키는 것을 포함하는 방법. - 하기를 포함하는, 제 3 항에 따른 방법을 실행하기 위한 키트:
a) 제 1 항 또는 제 2 항에 따른 DNA 폴리머라아제, b) 완충제 및 c) 마그네슘 클로라이드. - 제 1 항 또는 제 2 항에 따른 DNA 폴리머라아제, 및 하기 중 하나 이상을 포함하는, 제 3 항에 따른 방법을 실행하기 위한 키트:
(a) PrimPol 효소 (예를 들어, TthPrimPol);
(b) 무작위 3량체;
(c) 무작위 4량체;
(d) 무작위 5량체;
(e) 무작위 7량체;
(f) 무작위 8량체;
(g) dNTP;
(h) 반응 완충제;
(i) 임의의 상술한 요소와 함께 사용하기 위한 완충제. - Phi29 유형 DNA 폴리머라아제로서, Phi29 유형 DNA 폴리머라아제가 SEQ ID NO:1 과 적어도 80%, 85%, 90%, 95%, 98% 또는 99%, 99.5% 서열 동일성을 갖는 아미노산 서열을 갖고, Phi29 유형 DNA 폴리머라아제가 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두를 포함하는 것인 Phi29 유형 DNA 폴리머라아제.
- 제 6 항에 있어서, SEQ ID NO:2, SEQ ID NO:3 또는 SEQ ID NO:4 의 서열을 갖는 Phi29 유형 DNA 폴리머라아제.
- 제 6 항에 있어서, 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두에 추가로 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 또는 1 개 이하의 아미노산 치환, 부가 또는 결실을 갖는 Phi29 유형 DNA 폴리머라아제.
- Phi29 유형 DNA 폴리머라아제를 인코딩하는 뉴클레오티드 서열을 포함하는 단리된 핵산 분자로서, Phi29 유형 DNA 폴리머라아제가 SEQ ID NO:1 과 적어도 80%, 85%, 90%, 95%, 98% 또는 99%, 99.5% 서열 동일성을 갖는 아미노산 서열을 갖고, Phi29 유형 DNA 폴리머라아제가 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두를 포함하는 것인 단리된 핵산 분자.
- 제 9 항에 있어서, Phi29 유형 DNA 폴리머라아제가 SEQ ID NO:2, SEQ ID NO:3 또는 SEQ ID NO:4 의 서열을 갖는 것인 단리된 핵산 분자.
- 제 9 항에 있어서, Phi29 유형 DNA 폴리머라아제가 아미노산 치환 K64R 및 M97K 중 하나 또는 둘 모두에 추가로 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2 또는 1 개 이하의 아미노산 치환, 부가 또는 결실을 갖는 것인 단리된 핵산 분자.
- 제 9 항 내지 제 11 항 중 어느 한 항의 Phi29 유형 DNA 폴리머라아제와 작동가능하게 연결되는 전사 조절 서열을 포함하는 재조합 핵산.
- 제 12 항에 있어서, 전사 조절 서열이 박테리아 또는 포유동물 프로모터를 포함하는 것인 재조합 핵산.
- 제 12 항에 있어서, 플라스미드 벡터, 바이러스 벡터, 코스미드 및 트랜스포존에서 선택되는 벡터에 함유되는 재조합 핵산.
- 제 14 항에 있어서, Phi29 유형 DNA 폴리머라아제를 인코딩하는 뉴클레오티드 서열에 관련된 위치에 있는 클로닝 위치를 포함하여 클로닝 위치 내로 삽입된 전사 조절 서열이 Phi29 유형 DNA 폴리머라아제를 인코딩하는 뉴클레오티드 서열과 작동가능하게 연결되게 되는 재조합 핵산.
- 제 12 항 내지 제 15 항 중 어느 한 항의 재조합 핵산을 포함하는 재조합 세포.
- 하기 단계를 포함하는 방법:
a) 핵산 주형 분자를 제 1 항, 제 2 항, 제 6 항 내지 제 8 항 중 어느 한 항의 Phi29 유형 DNA 폴리머라아제, 및 프라이머 연장에 충분한 시약과 접촉시키는 단계; 및
b) 핵산 주형을 사용하여 폴리머라아제로 프라이머 연장을 수행하는 단계. - 제 17 항에 있어서, 프라이머 연장에 충분한 시약이 올리고뉴클레오티드 프라이머를 포함하는 것인 방법.
- 제 18 항에 있어서, 올리고뉴클레오티드 프라이머가 3량체, 4량체, 5량체, 6량체, 6량체, 8량체, 9량체 또는 10량체 중 하나 이상을 포함하는 것인 방법.
- 제 19 항에 있어서, 프라이머가 무작위 프라이머인 방법.
- 제 18 항에 있어서, 올리고뉴클레오티드 프라이머가 5 내지 25 개 뉴클레오티드의 링크 (link) 를 갖는 것인 방법.
- 제 17 항에 있어서, 프라이머 연장에 충분한 시약이 프리마아제/폴리머라아제 (예를 들어, TthPrimPol) 를 포함하는 것인 방법.
- 제 17 항에 있어서, 프라이머 연장이 대략 31℃, 32℃, 33℃, 34℃, 35℃, 36℃, 37℃, 38℃, 39℃, 40℃, 41℃ 또는 42℃ 중 임의의 온도에서 또는 그 이상의 온도에서 수행되는 방법.
- 제 17 항에 있어서, 주형 핵산 분자가 1 ng, 100 pg, 10 pg 또는 1 pg 이하의 양으로 존재하는 것인 방법.
- 제 17 항에 있어서, 프라이머 연장이 (i) 다중 변위 증폭 ("MDA") 또는 (2) 롤링 서클 증폭 (rolling circle amplification) 을 포함하는 방법.
- 제 17 항에 있어서, 프라이머 연장이 다중 어닐링 및 루핑 기반 증폭 사이클 (multiple annealing and looping-based amplification cycle) (MALBAC) 을 포함하는 방법.
- 제 17 항 내지 제 26 항 중 어느 한 항에 있어서, Phi29 유형 DNA 폴리머라아제가 치환 K64R 및 M97K 둘 모두를 포함하는 것인 방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962849252P | 2019-05-17 | 2019-05-17 | |
| US62/849,252 | 2019-05-17 | ||
| PCT/EP2020/063740 WO2020234200A1 (en) | 2019-05-17 | 2020-05-15 | Phi29 dna polymerase mutants with improved primer recognition |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20220009977A true KR20220009977A (ko) | 2022-01-25 |
Family
ID=71016482
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020217040028A KR20220009977A (ko) | 2019-05-17 | 2020-05-15 | 개선된 프라이머 인식을 갖는 phi29 dna 폴리머라아제 돌연변이체 |
Country Status (9)
| Country | Link |
|---|---|
| US (1) | US20220235337A1 (ko) |
| EP (1) | EP3969575A1 (ko) |
| JP (1) | JP2022533366A (ko) |
| KR (1) | KR20220009977A (ko) |
| CN (1) | CN114174502A (ko) |
| AU (1) | AU2020277641A1 (ko) |
| CA (1) | CA3140244A1 (ko) |
| SG (1) | SG11202112620TA (ko) |
| WO (1) | WO2020234200A1 (ko) |
Families Citing this family (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN113122517B (zh) * | 2021-03-24 | 2023-02-14 | 深圳清华大学研究院 | 聚合酶突变体及其应用 |
Family Cites Families (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5656493A (en) | 1985-03-28 | 1997-08-12 | The Perkin-Elmer Corporation | System for automated performance of the polymerase chain reaction |
| ES2628739T3 (es) | 2009-10-15 | 2017-08-03 | Ibis Biosciences, Inc. | Amplificación por desplazamiento múltiple |
| US9422535B2 (en) * | 2013-04-25 | 2016-08-23 | Thermo Fisher Scientific Baltics Uab | phi29 DNA polymerase mutants having increased thermostability and processivity |
| US10280411B2 (en) * | 2015-10-27 | 2019-05-07 | Pacific Biosciences of California, In.c | Methods, systems, and reagents for direct RNA sequencing |
| KR20230052995A (ko) * | 2016-12-19 | 2023-04-20 | 퀀텀-에스아이 인코포레이티드 | 시퀀싱 반응을 위한 중합 효소 |
| CN110719955B (zh) * | 2017-07-28 | 2023-05-02 | 深圳华大智造科技股份有限公司 | 提高了热稳定性的Phi29 DNA聚合酶突变体及其应用 |
| JP7256280B2 (ja) * | 2018-10-11 | 2023-04-11 | 深▲せん▼華大生命科学研究院 | 熱安定性が向上したPhi29 DNAポリメラーゼ突然変異体及びそのシーケンシングにおける応用 |
-
2020
- 2020-05-15 WO PCT/EP2020/063740 patent/WO2020234200A1/en unknown
- 2020-05-15 KR KR1020217040028A patent/KR20220009977A/ko unknown
- 2020-05-15 EP EP20730981.6A patent/EP3969575A1/en active Pending
- 2020-05-15 US US17/611,861 patent/US20220235337A1/en active Pending
- 2020-05-15 CA CA3140244A patent/CA3140244A1/en active Pending
- 2020-05-15 SG SG11202112620TA patent/SG11202112620TA/en unknown
- 2020-05-15 CN CN202080036838.XA patent/CN114174502A/zh active Pending
- 2020-05-15 JP JP2021568525A patent/JP2022533366A/ja active Pending
- 2020-05-15 AU AU2020277641A patent/AU2020277641A1/en active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| EP3969575A1 (en) | 2022-03-23 |
| CA3140244A1 (en) | 2020-11-26 |
| SG11202112620TA (en) | 2021-12-30 |
| AU2020277641A1 (en) | 2021-12-09 |
| WO2020234200A1 (en) | 2020-11-26 |
| US20220235337A1 (en) | 2022-07-28 |
| JP2022533366A (ja) | 2022-07-22 |
| CN114174502A (zh) | 2022-03-11 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US10704091B2 (en) | Genotyping by next-generation sequencing | |
| JP6998404B2 (ja) | 標的ヌクレオチド配列の富化及び決定方法 | |
| JP6690005B2 (ja) | ナノポアシーケンシングのためのポリメラーゼ−鋳型複合体 | |
| JP6902052B2 (ja) | 複数のリガーゼ組成物、システム、および方法 | |
| EP2668294B1 (en) | Paired end bead amplification and high throughput sequencing | |
| US20230357733A1 (en) | Reverse Transcriptase and Methods of Use | |
| US20210261944A1 (en) | Compositions and methods for ordered and continuous complementary DNA (cDNA) synthesis across non-continuous templates | |
| EP3485034B1 (en) | System and method for transposase-mediated amplicon sequencing | |
| KR20220009977A (ko) | 개선된 프라이머 인식을 갖는 phi29 dna 폴리머라아제 돌연변이체 | |
| EP3350326B1 (en) | Compositions and methods for polynucleotide assembly | |
| CN110446791B (zh) | 多核苷酸衔接子及其使用方法 | |
| KR20230124636A (ko) | 멀티플렉스 반응에서 표적 서열의 고 감응성 검출을위한 조성물 및 방법 | |
| US20120135472A1 (en) | Hot-start pcr based on the protein trans-splicing of nanoarchaeum equitans dna polymerase | |
| WO2002090538A1 (fr) | Procede de synthese d'acide nucleique | |
| JP6416885B2 (ja) | 高温で活性なhivタイプ1グループo逆転写酵素 | |
| NL2031471B1 (en) | Methods and kits for enzymatic synthesis of g4-prone polynucleotides | |
| CN114901820B (zh) | 构建基因突变文库的方法 | |
| EP3673084B1 (en) | Method for introducing mutations | |
| US20210355518A1 (en) | Generating nucleic acids with modified bases using recombinant terminal deoxynucleotidyl transferase | |
| WO2024059719A2 (en) | Compositions for preventing repetitive addition of switching oligonucleotides and nonspecific primer extension during cdna synthesis and methods of use thereof | |
| WO2003099990A2 (en) | Compositions and methods for cloning nucleic acids |