KR20200123458A - 돌연변이를 도입하는 방법 - Google Patents
돌연변이를 도입하는 방법 Download PDFInfo
- Publication number
- KR20200123458A KR20200123458A KR1020207027106A KR20207027106A KR20200123458A KR 20200123458 A KR20200123458 A KR 20200123458A KR 1020207027106 A KR1020207027106 A KR 1020207027106A KR 20207027106 A KR20207027106 A KR 20207027106A KR 20200123458 A KR20200123458 A KR 20200123458A
- Authority
- KR
- South Korea
- Prior art keywords
- nucleic acid
- acid molecule
- target nucleic
- mutation
- sequence
- Prior art date
Links
Images
















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6844—Nucleic acid amplification reactions
- C12Q1/686—Polymerase chain reaction [PCR]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/66—General methods for inserting a gene into a vector to form a recombinant vector using cleavage and ligation; Use of non-functional linkers or adaptors, e.g. linkers containing the sequence for a restriction endonuclease
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/10—Processes for the isolation, preparation or purification of DNA or RNA
- C12N15/1034—Isolating an individual clone by screening libraries
- C12N15/1065—Preparation or screening of tagged libraries, e.g. tagged microorganisms by STM-mutagenesis, tagged polynucleotides, gene tags
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1241—Nucleotidyltransferases (2.7.7)
- C12N9/1252—DNA-directed DNA polymerase (2.7.7.7), i.e. DNA replicase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6844—Nucleic acid amplification reactions
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2521/00—Reaction characterised by the enzymatic activity
- C12Q2521/10—Nucleotidyl transfering
- C12Q2521/101—DNA polymerase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2525/00—Reactions involving modified oligonucleotides, nucleic acids, or nucleotides
- C12Q2525/10—Modifications characterised by
- C12Q2525/101—Modifications characterised by incorporating non-naturally occurring nucleotides, e.g. inosine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2525/00—Reactions involving modified oligonucleotides, nucleic acids, or nucleotides
- C12Q2525/10—Modifications characterised by
- C12Q2525/179—Modifications characterised by incorporating arbitrary or random nucleotide sequences
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2525/00—Reactions involving modified oligonucleotides, nucleic acids, or nucleotides
- C12Q2525/10—Modifications characterised by
- C12Q2525/191—Modifications characterised by incorporating an adaptor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2525/00—Reactions involving modified oligonucleotides, nucleic acids, or nucleotides
- C12Q2525/10—Modifications characterised by
- C12Q2525/197—Modifications characterised by incorporating a spacer/coupling moiety
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2527/00—Reactions demanding special reaction conditions
- C12Q2527/137—Concentration of a component of medium
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2531/00—Reactions of nucleic acids characterised by
- C12Q2531/10—Reactions of nucleic acids characterised by the purpose being amplify/increase the copy number of target nucleic acid
- C12Q2531/113—PCR
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Genetics & Genomics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Molecular Biology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biophysics (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Biomedical Technology (AREA)
- Immunology (AREA)
- Analytical Chemistry (AREA)
- Medicinal Chemistry (AREA)
- Plant Pathology (AREA)
- Bioinformatics & Computational Biology (AREA)
- Gastroenterology & Hepatology (AREA)
- Crystallography & Structural Chemistry (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
- Enzymes And Modification Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
본 발명은 (a) 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및 (b) 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 포함하는 적어도 하나의 타겟 핵산 분자로 돌연변이를 도입하는 방법에 관한 것이다. 본 발명은 또한 하나 이상의 핵산 분자(들)에 돌연변이를 도입하는 방법에서 저편향 DNA 중합효소의 용도, 샘플 태그 (sample tag) 그룹, 샘플 태그 그룹을 설계하는 방법, 컴퓨터 판독 가능 매체, 및 타겟 핵산 분자를 우선적으로 (preferentially) 증폭하는 방법에 관한 것이다.
Description
본 발명은 하나 이상의 핵산 분자(들)에 돌연변이를 도입하는 방법, 하나 이상의 핵산 분자(들)에 돌연변이를 도입하는 방법에서 저편향 DNA 중합효소(low bias DNA polymerase)의 용도, 샘플 태그 (sample tag) 그룹, 샘플 태그 그룹을 설계하는 방법, 컴퓨터 판독 가능 매체 및 타겟 핵산 분자를 우선적으로 (preferentially) 증폭하는 방법에 관한 것이다.
DNA 중합효소 (DNA polymerases)를 사용하여 핵산 서열에 돌연변이를 도입할 수 있다. 이것은 여러 응용 분야에서 유용할 수 있다. 예를 들어 돌연변이 유발 (mutagenesis) 기술은 돌연변이 유발 (SAM) 기술에 의해 보조되는 시퀀싱을 포함한 응용 분야에서 유용할 수 있으며 단백질의 활성에 영향을 미치는 돌연변이를 찾기 위해 단백질 서열에 돌연변이를 도입하는데 유용할 수 있다.
돌연변이는 정확도 (fidelity)가 낮은 DNA 중합효소를 사용하여 도입될 수 있다. 저정확도 DNA 중합효소 (Low fidelity DNA polymerases)는 복제 중에 실수를 하여 돌연변이 도입을 유발한다. 그러나, 많은 저정확도 DNA 중합효소는 돌연변이 반응 (복제 라운드) 당 2% 미만의 비율로만 돌연변이를 도입하며, 일부 응용 분야의 경우 더 높은 돌연변이 발생률이 유용하다. 또한, 저정확도 DNA 중합효소는 편향된 (biased) 방식으로 돌연변이를 도입할 수 있다. 이러한 DNA 중합효소는 고편향 DNA 중합 효소 (high bias DNA polymerases)라고 불린다.
돌연변이는 dPTP와 같은 뉴클레오티드 유사체의 존재 하에, DNA 중합효소를 사용하여, 서열을 복제함으로써 도입될 수 있다. DNA 중합효소는 천연 뉴클레오티드 대신에 뉴클레오티드 유사체를 통합시킬 (incorporate) 수 있다. 그런 다음, 뒤에 이어지는 복제주기에서, 뉴클레오티드 유사체는 원래 서열에 존재하지 않았던 천연 뉴클레오티드와 쌍을 이루어 돌연변이를 도입할 수 있다. 뉴클레오티드 유사체의 존재 하에 서열을 복제하여 돌연변이를 도입하면 더 높은 돌연변이 비율을 달성할 수 있다.
일반적으로 사용되는 DNA 중합효소 (예를 들어, Taq 중합효소)는 천연 뉴클레오티드 대신 뉴클레오티드 유사체를 통합시키는데 사용될 수 있다. 그러나, 이러한 중합효소는 고편향 중합효소이다. 고편향 DNA 중합효소는 돌연변이 편향 (mutation bias) 및 주형 증폭 편향 (template amplication bias)의 두 가지의 가능한 편향을 나타낼 수 있다.
일부 고편향 중합효소는 4개의 모든 천연 뉴클레오티드 (아데닌, 시토신, 구아닌 및 티민)를 균등하게 무작위로 (uniformly at random) 돌연변이시키지 않기 때문에, 높은 돌연변이 편향을 가진다. 예를 들어, 고편향 DNA 중합효소는 일부 뉴클레오티드를 다른 것보다 더 큰 빈도로 돌연변이시킬 수 있다. 아데닌/티민 쌍은 두 개의 수소 결합으로 연결되는 반면, 구아닌/시토신 쌍은 세 개의 수소 결합으로 연결된다. 따라서, 고편향 DNA 중합효소가 구아닌/사이토신 쌍보다 아데닌/티민 쌍에 돌연변이를 도입할 가능성이 더 높을 수 있다.
높은 돌연변이 편향을 갖는 고편향 중합효소는 뉴클레오티드 유사체를 무작위로 통합시키지 못할 수 있다. 예를 들어, 고편향 중합효소는 특정 염기를 뉴클레오티드 유사체로 대체하는 것을 선호할 수 있다. DPTP는 두 가지의 서로 다른 토토머 (tautomeric) 형태, 이미노 형태 (imino form)와 아미노 형태 (amino form) 사이에서 상호전환 될 수 있다. 이미노 토토머는 아데닌과 왓슨-크릭 염기쌍을 형성할 수 있는 반면, 아미노 형태는 구아닌과 왓슨-크릭 염기쌍을 형성할 수 있다 (Kong Thoo Lin P, Brown DM (1989)."Synthesis and duplex stability of oligonucleotides containing cytosine-thymine analogues". Nucleic Acids Research. 17:10373-10383; Stone MJ et al. (1991). "Molecular basis for methoxyamine-initiated mutagenesis: 1H nuclear magnetic resonance studies of base-modified oligodeoxynucleotides." Journal of Molecular Biology. 222:711 -723; Nedderman ANR et al. (1993). "Molecular basis for methoxyamine initiated mutagenesis: 1H nuclear magnetic resonance studies of oligonucleotide duplexes containing base-modified cytosine residues ". Journal of Molecular Biology. 230:1068-1076; Moore MH et al. (1995). "Direct observation of two base-pairing modes of a cytosine-thymine analogue with guanine in a DNAZ-form duplex. Significance for base analogue mutagenesis". Journal of Molecular Biology. 251:665-673). 이는 실질적으로 dPTP의 존재 하에서의 복제는 뉴클레오티드 서열에서 아데닌, 시토신, 구아닌 또는 티민 대신에 치환 (substitutions)을 도입하는데 사용될 수 있음을 의미한다. 그러나, 수용액에서 dPTP의 이미노 형태의 아미노 형태에 대한 비율은 약 10:1인 것으로 나타났다 (Harris VH et al. (2003)."The effect of tautomeric constant on the specificity of nucleotide incorporation during DNA replication: support for the rare tautomer hypothesis of substitution mutagenesis". Journal of Molecular Biology. 326:1389-1401). 따라서, Taq 중합효소와 같은 중합효소를 사용하여 dPTP를 사용하여 돌연변이를 도입하면, 구아닌과 시토신의 치환보다 훨씬 더 자주 아데닌과 티민의 치환을 도입한다 (Zaccolo M et al. (1996). "An approach to random mutagenesis of DNA using mixtures of triphosphate derivatives of nucleoside analogues". Journal of Molecular Biology. 255:589-603; Harris VH et al. (2003). “The effect of tautomeric constant on the specificity of nucleotide incorporation during DNA replication: support for the rare tautomer hypothesis of substitution mutagenesis". Journal of Molecular Biology. 326:1389-1401).
두번째로, 고편향 중합효소는 주형 증폭 편향을 보여줄 수 있다. 즉, 다른 것보다 PCR 주기 당 더 높은 성공률로 일부 주형 핵산 분자를 복제할 수 있다. PCR의 여러 주기에서 이러한 편향은 주형 간의 카피 수 (copy number)에 극심한 차이를 만들 수 있다. 주형 핵산 분자의 영역은 이차 구조를 형성할 수 있거나 다른 것보다 일부 뉴클레오티드 (예를 들어, 구아닌 또는 시토신 뉴클레오티드)를 더 높은 비율로 포함할 수 있다. 고편향 중합효소는, 예를 들어, 아데닌 및 티민이 풍부한 주형 핵산 분자와 비교하여 구아닌 및 시토신이 풍부한 주형 핵산 분자를 증폭하는데 더 효과적일 수 있거나 이차 구조를 형성하지 않는 주형 핵산 분자를 증폭하는데 더 효과적일 수 있다.
돌연변이 유발의 많은 응용은 돌연변이 유발이 낮은 편향 (돌연변이 편향 및 주형 증폭 편향 둘 다)으로 수행될 수 있다면 더 효과적이다.
많은 2세대 시퀀싱 플랫폼이 짧은 핵산 단편만 시퀀싱 할 수 있고, 시퀀싱 단계를 위한 충분한 핵산 분자를 제공하기 위해 시퀀싱 과정 중에 증폭될 타겟 핵산 분자를 필요로 하여 게놈 서열의 정확한 조립이 어렵다는 것이 입증되었다. 사용자가 더 큰 핵산 서열을 시퀀싱하고자 하는 경우, 이는 타겟 핵산 분자의 영역을 시퀀싱함으로써 달성할 수 있다. 그런 다음 사용자는 상기 영역의 서열 (sequences of the regions)로부터 전체 핵산 서열의 서열을 계산적으로 (computationally) 조립하여야만 한다.
영역의 서열을 사용한 핵산 서열의 조립은 어려울 수 있다. 특히, 서열의 긴 영역이 서로 매우 유사한 경우 두 영역의 서열이 모두 동일한 원래 주형 핵산 분자의 복제물 (replicates)인지 또는 두 가지 다른 원래 주형 핵산 분자의 서열에 해당하는지 여부를 결정하는 것이 어려울 수 있다. 비슷하게, 두 영역의 서열이 주형 핵산 분자의 동일한 부분의 복제 서열에 해당하는지, 또는 실제로 주형 핵산 분자 내에서 두 개의 다른 반복 (repeats)에 해당하는지 여부를 결정하는 것이 어려울 수 있다. 이러한 어려움은 증폭 전에 타겟 핵산 분자에 돌연변이를 도입하여 회피할 수 있다. 그렇게 되면 사용자는 동일한 돌연변이 패턴을 갖는 단편들이 동일한 원래의 주형 핵산 분자의 동일한 부분에서 유래했을 가능성이 있음을 확인할 수 있다. 이러한 유형의 시퀀싱 방법은 때때로 돌연변이 유발을 이용한 시퀀싱 (sequencing aided by mutagenesis: SAM)이라고도 불린다.
전술한 시퀀싱 방법은 타겟 핵산 분자에 도입되는 돌연변이가 균등하게 무작위일 때 더 효과적이다. 돌연변이가 균등하게 무작위이면, 예를 들어, 주형 핵산 분자의 어떤 부분이 고유한 돌연변이 패턴을 가질 확률이 더 높다. 따라서, 균등하게 무작위로 돌연변이를 도입할 수 있는 (낮은 돌연변이 편향을 가짐) DNA 중합효소의 규명이 필요하다.
또한, 주형 증폭 편향 (template amplification bias)이 높은 DNA 중합효소를 이용한 시퀀싱 방법은 제한적일 수 있다. 높은 주형 증폭 편향을 갖는 DNA 중합효소는 일부 타겟 핵산 분자를 다른 것보다 더 잘 복제 및/또는 돌연변이시킬 것이므로, 이러한 고편향 DNA 중합효소 (high bias DNA polymerase)를 사용하는 시퀀싱 방법은 일부 타겟 핵산 분자를 잘 시퀀싱하지 못할 수 있다.
본 발명자들은 저편향 중합효소 (낮은 주형 증폭 편향 및 낮은 돌연변이 편향을 모두 가짐)인 중합효소를 규명하였으며, 따라서 이는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법에 특히 유용하다.
사용자는 한번에 하나 이상의 샘플에서 본 발명의 방법을 사용하기를 바랄 수 있다. 그러한 경우, 어떤 타겟 핵산 분자가 어떤 원본 샘플에서 유래되었는지 확인할 수 있는 것이 사용자에게 유리하다. 이러한 확인은 샘플 태그로 타겟 핵산 분자를 표지 (labelling)하여 달성할 수 있다. 그러나, 샘플 태그는 그 자체가 그 방법 중에 돌연변이될 수 있으므로, 본 발명자들은 돌연변이가 되더라도 서로 구별될 수 있는 샘플 태그를 설계하는 방법을 밝혀냈다.
사용자는 또한 본 발명의 방법이 확실히, 짧은 핵산 분자와 비교하여 우선적으로 긴 타겟 핵산 분자를 돌연변이 및 증폭시키는데 사용되기를 바랄 수 있다. 본 발명자들은 타겟 핵산 분자의 각 말단에 특별한 프라이머 결합 부위를 도입함으로써 이것이 달성될 수 있음을 발견하였다.
따라서, 본 발명의 일 양상에서, 하기를 포함하는 적어도 하나의 타겟 핵산 분자로 돌연변이를 도입하는 방법을 제공한다:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 저편향 DNA 중합효소 (low bias DNA polymerase)를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계.
본 발명의 제2 양상에서, 적어도 하나의 타겟 핵산 분자로 돌연변이를 도입하는 방법에서 저편향 DNA 중합효소의 용도를 제공한다.
본 발명의 제3 양상에서, 본 발명의 돌연변이를 도입하는 방법을 포함하는 적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법을 제공한다.
본 발명의 제4 양상에서, 본 발명의 돌연변이를 도입하는 방법을 포함하는 단백질을 엔지니어링 (engineering)하는 방법을 제공한다.
본 발명의 제5 양상에서, 샘플 태그 그룹 (a group of sample tags)으로서, 각 샘플 태그는 적어도 1개의 낮은 확률 돌연변이 차이 또는 적어도 3개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 (substantially) 다른 것인 샘플 태그 그룹을 제공한다.
본 발명의 제6 양상에서, 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법에 사용하기에 적합한 샘플 태그 그룹을 설계하는 방법으로서, 하기를 포함하는 방법을 제공한다:
a. 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 분석하여, 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 낮은 확률 돌연변이의 평균 개수를 결정하는 단계; 및
b. 샘플 태그 그룹의 서열을 결정하는 단계로서, 각 샘플 태그는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 낮은 확률 돌연변이의 평균 개수보다 더 많은 낮은 확률 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 단계
본 발명의 제7 양상에서, 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법으로서, 하기를 포함하는 방법을 제공한다:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하기 위해, DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하여 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 단계로서,
상기 단계 b. 는 불균등 농도의 dNTP를 사용하여 수행하는 것인 단계.
본 발명의 제8 양상에서, 본 발명의 샘플 태그 그룹을 설계하는 방법으로 얻을 수 있는 샘플 태그 그룹을 제공한다.
본 발명의 제9 양상에서, 본 발명의 샘플 태그 그룹을 설계하는 방법을 수행하도록 구성된 컴퓨터 판독가능 매체를 제공한다.
본 발명의 제10 양상에서, 길이가 1 kbp보다 큰 타겟 핵산 분자를 우선적으로 (preferentially) 증폭하는 방법으로서, 하기를 포함하는 방법을 제공한다:
a. 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계;
b. 타겟 핵산 분자의 3' 말단에 제1 어댑터 및 타겟 핵산 분자의 5' 말단에 제2 어댑터를 도입하는 단계; 및
c. 상기 제1 어댑터의 일부에 상보적인 프라이머들을 사용하여 상기 타겟 핵산 분자를 증폭하는 단계로서,
상기 제1 어댑터 및 제2 어댑터는 서로에게 어닐링(annealing)하는 것인 단계.
도 1은 dPTP의 존재 또는 부재 하에 3 가지 서로 다른 중합효소로 달성된 돌연변이 수준을 나타낸다. 패널 A는 Taq (Jena Biosciences)를 사용하여 얻은 데이터를 나타내고, 패널 B는 LongAmp (New England Biolabs)를 사용하여 얻은 데이터를 나타내고, 패널 C는 Primestar GXL (Takara)을 사용하여 얻은 데이터를 나타낸다. 짙은 회색 막대는 dPTP 부재 하에 얻은 결과를 나타내고 옅은 회색 막대는 0.5mM dPTP 존재 하에 얻은 결과를 나타낸다.
도 2는 다양한 G+C 함량을 가진 주형에서 Thermococcus 중합효소 (Primestar GXL; Takara)를 사용하여 dPTP 돌연변이 유발에 의해 얻은 돌연변이 비율을 나타낸다. 관찰된 돌연변이 비율의 중앙값은 S. aureus (33% GC)의 낮은 GC 주형의 경우 ~7% 인 반면, 다른 주형의 중앙값은 약 8% 였다.
도 3은 시퀀스 목록이다.
도 4는 서로 어닐링하는 제1 프라이머 결합 부위 및 제2 프라이머 결합 부위가 사용될 때의 자가 어닐링 (self-annealing) 핵산 분자를 묘사한 것이다.
도 6은 서로 어닐링하는 어댑터 (우측 선) 또는 표준 어댑터 (좌측 선)를 사용하여 증폭된 타겟 핵산 분자의 크기를 묘사한 것이다.
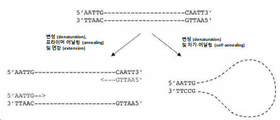
도 7은 뉴클레오티드 유사체 dPTP (도 7에서 "P"로 지칭됨)를 사용한 돌연변이를 그림으로 나타낸 것이다.
도 2는 다양한 G+C 함량을 가진 주형에서 Thermococcus 중합효소 (Primestar GXL; Takara)를 사용하여 dPTP 돌연변이 유발에 의해 얻은 돌연변이 비율을 나타낸다. 관찰된 돌연변이 비율의 중앙값은 S. aureus (33% GC)의 낮은 GC 주형의 경우 ~7% 인 반면, 다른 주형의 중앙값은 약 8% 였다.
도 3은 시퀀스 목록이다.
도 4는 서로 어닐링하는 제1 프라이머 결합 부위 및 제2 프라이머 결합 부위가 사용될 때의 자가 어닐링 (self-annealing) 핵산 분자를 묘사한 것이다.
도 6은 서로 어닐링하는 어댑터 (우측 선) 또는 표준 어댑터 (좌측 선)를 사용하여 증폭된 타겟 핵산 분자의 크기를 묘사한 것이다.
도 7은 뉴클레오티드 유사체 dPTP (도 7에서 "P"로 지칭됨)를 사용한 돌연변이를 그림으로 나타낸 것이다.
일반적인 정의
달리 정의되지 않는 한, 본 명세서에서 사용된 기술 및 과학 용어는 본 발명이 속하는 기술 분야의 통상의 기술자에 의해 일반적으로 이해되는 것과 동일한 의미를 갖는다.
일반적으로, 용어 "포함하는 (comprising)"은 포함하지만 이에 제한되지 않음을 의미하는 것으로 의도된다. 예를 들어, 소정의 단계를 "포함하는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법"이라는 문구는 상기 방법이 언급된 단계를 포함하지만, 추가 단계가 수행될 수 있음을 의미하는 것으로 해석되어야 한다.
본 발명의 일부 구현예에서, "포함하는"이라는 단어는 "이루어진 (consisting of)"이라는 문구로 대체된다. 용어 "이루어진"은 제한하려는 의도이다. 예를 들어, 소정의 단계로 "이루어진 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법"이라는 문구는 상기 방법이 언급된 단계를 포함하고 추가 단계가 수행되지 않음을 의미하는 것으로 이해되어야 한다.
본 발명의 목적상, 2개 서열 (예를 들어 2개의 폴리뉴클레오티드 서열)의 동일성 백분율 (percent identity)을 결정하기 위해, 상기 서열들은 최적의 비교 목적을 위해 정렬된다 (예를 들어, 제2 서열과의 최적 정렬을 위해 제 1 서열에 갭이 도입될 수 있음). 그런 다음 각 위치에서의 뉴클레오티드 또는 아미노산 잔기를 비교한다. 제1 서열의 위치가 제2 서열의 해당 위치와 동일한 잔기에 의해 점유되면, 상기 잔기는 해당 위치에서 동일하다. 두 서열 사이의 동일성 백분율은 서열들이 공유하는 동일한 위치 수의 함수이다 (즉, 동일성 % = 동일한 위치 수/총 위치 수 x 100). 일반적으로 서열 비교는 참조 서열 (reference sequence)의 길이에 걸쳐 수행된다. 예를 들어, 테스트 서열이 서열번호 2 (참조 서열)와 적어도 95% 동일한지 여부를 평가하기 위해, 당업자는 서열번호 2의 길이에 걸쳐 정렬을 수행하고 테스트 서열에서 얼마나 많은 위치가 서열번호 2의 위치와 동일한지 확인할 것이다. 적어도 80%의 위치가 동일하다면, 상기 테스트 서열은 서열번호 2와 적어도 80% 동일하다. 만약 상기 서열이 서열번호 2보다 짧다면, 갭 (gap)들은 동일하지 않은 위치로 간주되어야 한다.
당업자는 2개 서열 사이의 상동성 (homology) 또는 동일성 (identity)을 결정하기 위해 이용가능한 여러 컴퓨터 프로그램을 알고 있다. 예를 들어, 2개 서열 간의 서열 비교 및 동일성 백분율 결정은 수학적 알고리즘을 사용하여 수행할 수 있다. 일 구체예에서, 두 아미노산 또는 핵산 서열 간의 동일성 백분율은 Blosum 62 matrix 또는 PAM250 matrix, 및 갭 가중치 (gap weight) 16, 14, 12, 10, 8, 6, 또는 4 및 길이 가중치 (length weight) 1, 2, 3, 4, 5, 또는 6을 이용한, Accelrys GCG 소프트웨어 패키지 (http://www.accelrys.com/products/gcg/에서 이용 가능)의 GAP 프로그램에 통합된 Needleman and Wunsch (1970) 알고리즘을 사용하여 결정된다.
적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법
일 양상에서, 본 발명은 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 제공한다. 다른 양상에서, 본 발명은 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법에서 저편향 DNA 중합효소의 용도를 제공한다.
돌연변이는 치환 (substitution) 돌연변이, 삽입 (insertion) 돌연변이 또는 결실 (deletion) 돌연변이 일 수 있다. 본 발명의 목적상, 용어 "치환 돌연변이"는 뉴클레오티드가 다른 뉴클레오티드로 대체되는 것을 의미하는 것으로 해석되어야 한다. 예를 들어, 서열 ATCC에서 서열 AGCC로의 변환은 치환 돌연변이이다. 본 발명의 목적상, 용어 "삽입 돌연변이"는 적어도 하나의 뉴클레오티드가 서열에 추가되는 것을 의미하는 것으로 해석되어야 한다. 예를 들어, 서열 ATCC의 서열 ATTCC로의 전환은 삽입 돌연변이의 예이다 (추가적인 T 뉴클레오티드가 삽입됨). 본 발명의 목적상, 용어 "결실 돌연변이"는 적어도 하나의 뉴클레오티드가 서열로부터 제거되는 것을 의미하는 것으로 해석되어야 한다. 예를 들어, 서열 ATTCC에서 ATCC로의 변환은 결실 돌연변이의 예이다 (T 뉴클레오티드가 제거됨). 상기 돌연변이는 바람직하게는 치환 돌연변이이다.
본 발명의 목적상, "핵산 분자 (nucleic acid molecule)"는 임의의 길이의 뉴클레오티드의 중합체 형태 (polymeric form)를 지칭한다. 상기 뉴클레오티드는 데옥시리보뉴클레오티드 (deoxyribonucleotides), 리보뉴클레오티드 (ribonucleotides) 또는 이의 유사체 (analogs)일 수 있다. 바람직하게는, 상기 타겟 핵산 분자는 데옥시리보뉴클레오티드 또는 리보뉴클레오티드로 구성된다. 보다 더 바람직하게는, 상기 타겟 핵산 분자는 데옥시리보뉴클레오티드로 구성되며, 즉 타겟 핵산 분자는 DNA 분자이다.
상기 적어도 하나의 "타겟 핵산 분자 (target nucleic acid molecule)"는 상기 방법의 사용자가 돌연변이를 도입하고자 하는 임의의 핵산 분자일 수 있다. 타겟 핵산 분자는 염색체와 같은 더 큰 핵산 분자의 일부를 형성할 수 있다. 타겟 핵산 분자는 유전자, 다수의 유전자 (multiple genes) 또는 유전자 단편 (fragment)을 포함할 수 있다. 타겟 핵산 분자는 크기가 1 kbp 초과, 1.5 kbp 초과, 2 kbp 초과, 4 kbp 초과, 5 kbp 초과, 7 kbp 초과, 8 kbp 초과, 1 kbp 내지 50 kbp, 또는 1kbps 내지 20kbps 일 수 있다.
용어 "적어도 하나의 타겟 핵산 분자"는 용어 "적어도 하나의 타겟 핵산 분자들"과 상호교환 가능한 것으로 간주된다.
"적어도 하나의 타겟 핵산 분자"는 단일 가닥일 수 있거나, 또는 이중 가닥 복합체의 일부일 수 있다. 예를 들어, 만약 적어도 하나의 타겟 핵산 분자가 데옥시리보뉴클레오티드로 구성되는 경우, 이는 이중 가닥 DNA 복합체의 일부를 형성할 수 있다. 이 경우, 하나의 가닥 (예를 들어 코딩 (coding) 가닥)은 적어도 하나의 타겟 핵산 분자로 간주될 것이고, 다른 가닥은 적어도 하나의 타겟 핵산 분자에 상보적인 핵산 분자이다.
적어도 하나의 타겟 핵산 분자로 돌연변이를 도입하는 방법은 하기를 포함할 수 있다:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 저편향 DNA 중합효소 (low bias DNA polymerase)를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계.
적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계
적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법은 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계를 포함 할 수 있다.
적어도 하나의 샘플은 적어도 하나의 타겟 핵산 분자를 포함하는 임의의 샘플을 포함할 수 있다. 적어도 하나의 샘플은 임의의 출처로부터 얻을 수 있다. 예를 들어, 상기 적어도 하나의 샘플은 인간으로부터 유래된 핵산 샘플, 예를 들어 인간 환자의 피부 면봉에서 추출된 샘플을 포함할 수 있다. 대안적으로는, 상기 적어도 하나의 샘플은 물 공급원의 샘플과 같은 다른 출처에서 유래될 수 있다. 이러한 샘플에는 수십억 개의 주형 핵산 분자가 포함될 수 있다. 본 발명의 방법을 사용하여 이들 수십억 개의 타겟 핵산 분자 각각을 동시에 돌연변이시키는 것이 가능할 것이며, 따라서 본 발명의 방법에 사용될 수 있는 타겟 핵산 분자의 수에는 상한 (upper limit)이 없다.
일 구체예에서, 단계 a. 하나 이상의 샘플을 제공하는 것을 포함한다. 예를 들어, 단계 a.는 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 15, 20, 25, 50, 75 또는 100개의 샘플을 제공하는 것을 포함할 수 있다. 선택적으로, 단계 a.는 2000개 미만, 1000개 미만, 750개 미만 또는 500개 미만의 샘플을 제공하는 것을 포함한다. 다른 구현예에서, 단계 a.는 2 내지 100개, 2 내지 75개, 2 내지 50개, 2 내지 25개, 5 내지 15개, 또는 7 내지 15개 샘플을 제공하는 것을 포함한다.
저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계
본 발명의 방법은 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 포함할 수 있다.
적어도 하나의 타겟 핵산 분자를 증폭하는 것은
적어도 하나의 타겟 핵산 분자 및/또는 상기 적어도 하나의 타겟 핵산 분자의 복제물 (replicates)에 상보적인 적어도 하나의 핵산 분자를 제공하기 위해 적어도 하나의 타겟 핵산 분자를 복제하는 것을 지칭한다. 저편향 DNA 중합효소를 사용한 적어도 하나의 타겟 핵산 분자의 증폭은, 적어도 하나의 타겟 핵산 분자의 복제 수를 증가시키고, 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입한다. 돌연변이가 도입되기 때문에, 상기 복제물은 원래의 적어도 하나의 타겟 핵산 분자와 반드시 동일하지는 않다. 상기 원래의 적어도 하나의 타겟 핵산 분자 및 적어도 하나의 타겟 핵산 분자의 복제물은 "적어도 하나의 돌연변이된 타겟 핵산 분자"로 통틀어 지칭될 수 있다.
예를 들어, 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 것은 적어도 하나의 타겟 핵산 분자의 복제물 생성 (generation)을 촉매하기 위해 저편향 DNA 중합효소에 적합한 조건 하에서 적어도 하나의 타겟 핵산 분자를 포함하는 샘플을 저편향 DNA 중합효소 및 적합한 프라이머들과 함께 배양하는 것을 포함할 수 있다.
적합한 프라이머들은 적어도 하나의 타겟 핵산 분자에 인접한 (flanking) 영역 또는 적어도 하나의 타겟 핵산 분자에 상보적인 핵산 분자에 인접한 영역에 상보적인 짧은 핵산 분자를 포함한다. 예를 들어, 상기 타겟 핵산 분자가 염색체의 일부인 경우, 상기 프라이머는 타겟 핵산 분자 3'말단의 바로 3' 위치의 염색체 영역 및 타겟 핵산 분자 5' 말단의 바로 5'위치인 영역에 상보적인 핵산 분자에 상보적일 수 있고, 또는 상기 프라이머는 타겟 핵산 분자에 상보적인 핵산 분자 3'말단의 바로 3' 위치의 염색체 영역 및 타겟 핵산 분자에 상보적인 핵산 분자 5' 말단의 바로 5'위치인 영역에 상보적인 핵산 분자에 상보적일 수 있다. 대안적으로는, 사용자는 적어도 하나의 타겟 핵산 분자에 인접한 영역에 프라이머 결합 부위 (짧은 핵산 서열)를 도입할 수 있다. 이는 "바코드, 샘플 및 어댑터" 섹션에서 보다 자세히 기재하였다.
적합한 조건은 저편향 DNA 중합효소가 적어도 하나의 타겟 핵산 분자의 복제물 생성을 촉매할 수 있는 온도를 포함한다. 예를 들어, 40℃ 내지 90℃, 50℃ 내지 80℃, 60℃ 내지 70℃, 또는 약 68℃의 온도가 사용될 수 있다.
적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 다수 회의 복제 라운드 (round)를 포함할 수 있다. 예를 들어, 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 바람직하게는 하기를 포함한다:
i) 적어도 하나의 타겟 핵산 분자에 상보적인 적어도 하나의 핵산 분자를 제공하기 위해 상기 적어도 하나의 타겟 핵산 분자를 복제하는 라운드; 및
ii) 상기 적어도 하나의 타겟 핵산 분자의 복제물 (replicates)을 제공하기 위해 상기 적어도 하나의 타겟 핵산 분자를 복제하는 라운드.
선택적으로, 상기 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 적어도 하나의 타겟 핵산 분자를 복제하는 라운드를 적어도 2, 적어도 4, 적어도 6, 적어도 8, 또는 적어도 10회 포함한다. 상기 적어도 하나의 타겟 핵산 분자를 복제하는 이러한 라운드 중 일부는 60℃ 내지 80℃의 온도에서 복제 라운드를 적어도 1, 적어도 2, 적어도 3, 적어도 4, 적어도 5, 또는 적어도 6회 포함한다.
선택적으로, 상기 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 중합효소 연쇄반응 (polymerase chain reaction: PCR)을 사용하여 수행된다. PCR은 핵산 분자를 복제하기 위해 하기의 다수 회의 라운드를 포함하는 과정이다:
a) 용융 (melting);
b) 어닐링 (annealing);
c) 연장 (extension); 및
d) 신장 (elongation).
상기 핵산 분자 (예를 들어, 상기 적어도 하나의 타겟 핵산 분자)는 적합한 프라이머들 및 본 발명의 저편향 DNA 중합효소와 같은 중합효소와 혼합된다. 용융 단계에서, 상기 핵산 분자는 90℃ 이상의 온도로 가열되어 이중-가닥 핵산 분자가 변성 (denature)된다 (두 가닥으로 분리됨). 어닐링 단계에서, 상기 핵산 분자는 프라이머들이 핵산 분자에 어닐링할 수 있도록 75℃ 미만, 예를 들어 55℃ 내지 70℃, 약 55℃, 또는 약 68℃의 온도로 냉각된다. 연장 단계에서, 상기 핵산 분자는 60℃초과의 온도로 가열되어 DNA 중합효소가 주형 가닥에 상보적인 뉴클레오티드를 추가하는 프라이머 연장을 촉매할 수 있도록 한다. 신장 단계에서, 상기 핵산 분자는 DNA 중합효소가 높은 활성을 갖는 온도, 예를 들어 60℃ 내지 70℃로 가열되어 새로운 핵산 가닥을 완성하기 위해 추가적인 상보적 핵산의 추가를 촉매할 수 있도록 한다.
선택적으로, 본 발명의 방법은 저편향 DNA 중합효소를 사용하는 다수 회의 라운드를 포함한다.
저편향 DNA 중합효소
본 발명의 방법은 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 포함할 수 있다.
본 발명에 따르면, "저편향 DNA 중합효소"는 (a) 낮은 돌연변이 편향, 및/또는 (b) 낮은 주형 증폭 편향을 나타내는 DNA 중합효소이다.
낮은 돌연변이 편향
낮은 돌연변이 편향을 나타내는 저편향 DNA 중합효소는 아데닌과 티민, 아데닌과 구아닌, 아데닌과 시토신, 티민과 구아닌, 티민과 시토신 또는 구아닌과 시토신을 비슷한 비율로 돌연변이시킬 수 있는 DNA 중합효소이다. 일 구체예에서, 상기 저편향 DNA 중합효소는 유사한 비율로 아데닌, 티민, 구아닌 및 시토신을 돌연변이시킬 수 있다.
선택적으로, 상기 저편향 DNA 중합효소는 아데닌과 티민, 아데닌과 구아닌, 아데닌과 시토신, 티민과 구아닌, 티민과 시토신, 또는 구아닌과 시토신을 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비 (rate ratio)로 돌연변이시킬 수 있다. 바람직하게는, 상기 저편향 DNA 중합효소는 구아닌과 아데닌을 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비로 돌연변이시킬 수 있다. 바람직하게는, 상기 저편향 DNA 중합효소는 티민과 시토신을 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비로 돌연변이시킬 수 있다.
이러한 구체예에서, 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계에서, 상기 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 아데닌과 티민, 아데닌과 구아닌, 아데닌과 시토신, 티민과 구아닌, 티민과 시토신, 또는 구아닌과 시토신 뉴클레오티드를 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비로 돌연변이시킨다. 바람직하게는, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 구아닌과 아데닌 뉴클레오티드를 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비로 돌연변이시킨다. 바람직하게는, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 티민과 시토신 뉴클레오티드를 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비로 돌연변이 시킨다.
선택적으로, 저편향 DNA 중합효소는 아데닌, 티민, 구아닌, 및 시토신을 각각 .5-1.5:0.5-1.5:0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4:0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2:0.8-1.2:0.8-1.2, 또는 약 1:1:1:1 의 비율비로 돌연변이시킬 수 있다. 바람직하게는, 상기 저편향 DNA 중합효소는 아데닌, 티민, 구아닌 및 시토신을 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3의 비율비로 돌연변이시킬 수 있다.
이러한 구체예에서, 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계에서, 상기 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 아데닌, 티민, 구아닌, 및 시토신 뉴클레오티드를 각각 0.5-1.5:0.5-1.5:0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4:0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2:0.8-1.2:0.8-1.2, 또는 약 1:1:1:1의 비율비로 돌연변이시킬 수 있다. 바람직하게는, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 아데닌, 티민, 구아닌, 및 시토신 뉴클레오티드를 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3의 비율비로 돌연변이시킬 수 있다.
상기 아데닌, 티민, 시토신, 및/또는 구아닌은 또 다른 뉴클레오티드로 치환될 수 있다. 예를 들어, 만약 저편향 DNA 중합효소가 아데닌을 돌연변이시킬 수 있는 경우, 저편향 DNA 중합효소의 존재 하에서 상기 적어도 하나의 타겟 핵산 분자의 증폭은 상기 핵산 분자의 적어도 하나의 아데닌 뉴클레오티드를 티민, 구아닌, 또는 시토신으로 치환할 수 있다. 유사하게, 만약 저편향 DNA 중합효소가 티민을 돌연변이시킬 수 있는 경우, 저편향 DNA 중합효소의 존재 하에서 상기 적어도 하나의 타겟 핵산 분자의 증폭은 상기 핵산 분자의 적어도 하나의 티민 뉴클레오티드를 아데닌, 구아닌, 또는 시토신으로 치환할 수 있다. 만약 저편향 DNA 중합효소가 구아닌을 돌연변이시킬 수 있는 경우, 저편향 DNA 중합효소의 존재 하에서 상기 적어도 하나의 타겟 핵산 분자의 증폭은 상기 핵산 분자의 적어도 하나의 구아닌 뉴클레오티드를 티민, 아데닌, 또는 시토신으로 치환할 수 있다. 만약 저편향 DNA 중합효소가 시토신을 돌연변이시킬 수 있는 경우, 저편향 DNA 중합효소의 존재 하에서 상기 적어도 하나의 타겟 핵산 분자의 증폭은 상기 핵산 분자의 적어도 하나의 시토신 뉴클레오티드를 티민, 구아닌, 또는 아데닌으로 치환할 수 있다.
저편향 DNA 중합효소는 뉴클레오티드를 직접 치환할 수는 없지만, 상보적인 가닥의 상응하는 뉴클레오티드를 대체함으로써 해당 뉴클레오티드를 돌연변이시킬 수 있다. 예를 들어, 타겟 핵산 분자가 티민을 포함하는 경우, 적어도 하나의 핵산 분자에 상보적인 적어도 하나의 핵산 분자의 상응하는 위치에 존재하는 아데닌 뉴클레오티드가 있을 것이다. 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에 상보적인 적어도 하나의 핵산 분자의 아데닌 뉴클레오티드를 구아닌으로 대체할 수 있으므로, 상기 적어도 하나의 핵산 분자에 상보적인 적어도 하나의 핵산 분자가 복제되었을 때, 이는 원래 티민이 있었던 상응하는 복제된 적어도 하나의 타겟 핵산 분자에 시토신이 존재하게 한다 (티민에서 시토신으로의 치환).
일 구체예에서, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산에서 1% 내지 15%, 2% 내지 10%, 또는 약 8%의 뉴클레오티드를 돌연변이시킨다. 이러한 구체예에서, 저편향 DNA 중합효소를 사용하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 적어도 하나의 타겟 핵산 분자에서 1% 내지 15%, 2% 내지 10%, 또는 약 8%의 뉴클레오티드가 돌연변이로 되도록 수행된다. 예를 들어, 만약 사용자가 타겟 핵산 분자에서 8%의 뉴클레오티드를 돌연변이시키고, 저편향 DNA 중합효소가 복제 라운드 당 1%의 뉴클레오티드를 돌연변이시키기를 바라는 경우, 저편향 DNA 중합효소를 사용하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 복제 라운드를 8회 포함할 수 있다.
일 구체예에서, 상기 저편향 DNA 중합효소는 복제 라운드 당 적어도 하나의 타겟 핵산 분자 중의 0% 내지 3%, 0% 내지 2%, 0.1% 내지 5%, 0.2% 내지 3%, 또는 약 1.5%의 뉴클레오티드를 돌연변이시킬 수 있다. 일 구체예에서, 상기 저편향 DNA 중합효소는 복제 라운드 당 적어도 하나의 타겟 핵산 분자 중의 0% 내지 3%, 0% 내지 2%, 0.1% 내지 5%, 0.2% 내지 3%, 또는 약 1.5%의 뉴클레오티드를 돌연변이시킨다. 각 라운드 별로 발생하는 돌연변이의 실제 양은 다를 수 있으나, 평균이 0% 내지 3%, 0% 내지 2%, 0.1% 내지 5%, 0.2% 내지 3%, 또는 약 1.5%일 수 있다.
DNA 중합효소가 뉴클레오티드를 돌연변이시킬 수 있는지 여부 및, 만약 그렇다면, 어떤 비율로 돌연변이 시키는지
저편향 DNA 중합효소가 복제 라운드 당 적어도 하나의 타겟 핵산 분자에서 소정 비율의 뉴클레오티드를 돌연변이시킬 수 있는지 여부는 저편향 DNA 중합효소의 존재 하에 알려진 서열의 핵산 분자를 정해진 횟수의 복제 라운드만큼 증폭함으로써 결정될 수 있다. 그 결과로 생성된 증폭된 핵산 분자를 시퀀싱하고, 복제 라운드 당 돌연변이된 뉴클레오티드의 백분율을 계산할 수 있다. 예를 들어, 알려진 서열의 핵산 분자는 저편향 DNA 중합효소의 존재 하에 PCR을 10회 라운드로 사용하여 증폭될 수 있다. 생성된 핵산 분자는 그 다음 시퀀싱 될 수 있다. 생성된 핵산 분자가 원래 알려진 서열의 상응하는 뉴클레오티드와 다른 10%의 뉴클레오티드를 포함하는 경우, 사용자는 저편향 DNA 중합효소가 적어도 하나의 타겟 핵산에서 복제 라운드 당 평균적으로 뉴클레오티드의 1%를 돌연변이시킬 수 있음을 이해할 것이다. 유사하게, 저편향 DNA 중합효소가 주어진 방법에서 적어도 하나의 타겟 핵산 분자에 있는 뉴클레오티드의 소정 비율을 돌연변이시키는지 여부를 확인하기 위해, 사용자는 알려진 서열의 핵산 분자에 대해 상기 방법을 수행하고 상기 방법이 완료되면 돌연변이되는 뉴클레오티드의 백분율을 결정하기 위해 시퀀싱을 사용할 수 있다.
저편향 DNA 중합효소는, 핵산 분자를 증폭시키는데 사용될 때, 그것이 일부 경우의 뉴클레오티드가 치환 또는 결실된 핵산 분자를 제공할 때, 핵산 분자, 예를 들어 아데닌을 돌연변이시킬 수 있다. 바람직하게는, 용어 "돌연변이시키다(mutate)"는 치환 돌연변이의 도입을 지칭하고, 일부 구체예에서 "돌연변이시키다"라는 용어는 "~의 치환을 도입하다 (introduces substitutions of)"로 대체될 수 있다.
저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 수행할 때, 그 단계가 일부 경우의 뉴클레오티드가 돌연변이된 적어도 하나의 타겟 핵산 분자를 유발할 경우, 저편향 DNA 중합효소는 본 발명의 방법에서 적어도 하나의 타겟 핵산 분자에서 아데닌과 같은 뉴클레오티드를 돌연변이시킨다. 저편향. 예를 들어, 만약 저편향 DNA 중합효소가 적어도 하나의 타겟 핵산 분자에서 아데닌을 돌연변이시키면, 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 수행할 때, 이 단계는 적어도 하나의 아데닌이 치환되거나 결실된 적어도 하나의 타겟 핵산 분자를 야기한다.
DNA 중합효소가 소정 돌연변이를 도입할 수 있는지 여부를 결정하기 위해, 당업자는 알려진 서열의 핵산 분자를 사용하여 DNA 중합효소를 테스트하기 만하면 된다. 알려진 서열의 적합한 핵산 분자는 E. coli MG1655와 같은 알려진 서열의 박테리아 게놈으로부터의 단편이다. 당업자는 저편향 DNA 중합효소의 존재 하에서 PCR을 사용하여 알려진 서열의 핵산 분자를 증폭할 수 있다. 그런 다음 당업자는 증폭된 핵산 분자를 시퀀싱하고 그 서열이 원래의 알려진 서열과 동일한지 여부를 결정할 수 있다. 아니면, 당업자는 돌연변이의 성격 (nature)을 결정할 수 있다. 예를 들어, 만약 당업자가 DNA 중합효소가 뉴클레오티드 유사체를 사용하여 아데닌을 돌연변이시킬 수 있는지 여부를 결정하기를 바라는 경우, 당업자는 뉴클레오티드 유사체의 존재 하에 PCR을 사용하여 알려진 서열의 핵산 분자를 증폭하고, 그 결과 생성된 증폭된 핵산 분자를 시퀀싱할 수 있을 것이다. 만약 증폭된 DNA가 알려진 서열에서 아데닌 뉴클레오티드에 상응하는 위치에 돌연변이를 갖는 경우, 당업자는 상기 DNA 중합효소가 뉴클레오티드 유사체를 사용하여 아데닌을 돌연변이시킬 수 있음을 알 것이다.
비율비는 유사한 방식으로 계산할 수 있다. 예를 들어, 당업자가 구아닌 및 사이토신 뉴클레오티드가 돌연변이되는 비율비를 결정하기를 바라는 경우, 당업자는 저편향 DNA 중합효소의 존재 하에 PCR을 사용하여 알려진 서열을 갖는 핵산 분자를 증폭할 수 있다. 그런 다음 당업자는 증폭된 핵산 분자를 시퀀싱하고 얼마나 많은 구아닌 뉴클레오티드가 치환 또는 결실되었는지 및 얼마나 많은 시토신 뉴클레오티드가 치환 또는 결실되었는지 확인할 수 있다. 상기 비율비는 치환 또는 결실된 구아닌 뉴클레오티드 수에 대한 치환 또는 결실된 시토신 뉴클레오티드 수의 비율이다. 예를 들어, 만약 16개의 구아닌 뉴클레오티드가 대체 또는 결실되고 8개의 시토신 뉴클레오티드가 대체 또는 결실 된 경우, 구아닌 및 시토신 뉴클레오티드는 각각 16:8 또는 2:1의 비율로 돌연변이된 것이다.
뉴클레오티드 유사체의 사용
저편향 DNA 중합효소는 뉴클레오티드를 다른 뉴클레오티드로 직접 대체할 수는 없지만 (적어도 높은 빈도로는 아님), 상기 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 여전히 핵산 분자를 돌연변이시킬 수 있다. 저편향 DNA 중합효소는 뉴클레오티드를 다른 천연 뉴클레오티드 (즉, 시토신, 구아닌, 아데닌 또는 티민) 또는 뉴클레오티드 유사체로 대체할 수 있다.
예를 들어, 저편향 DNA 중합효소는 고정확도 DNA 중합효소 (high fidelity DNA polymerase)일 수 있다. 고정확도 DNA 중합효소는 매우 정확하기 때문에, 일반적으로 돌연변이를 거의 도입하지 않는 경향이 있다. 그러나, 본 발명자들은 일부 고정확도 DNA 중합효소가 타겟 핵산 분자에 뉴클레오티드 유사체를 도입할 수 있기 때문에, 여전히 타겟 핵산 분자를 돌연변이시킬 수 있음을 발견하였다.
일 구체예에서, 뉴클레오티드 유사체의 부재하에, 고정확도 DNA 중합효소는 복제 라운드 당 0.01% 미만, 0.0015% 미만, 0.001% 미만, 0% 내지 0.0015%, 또는 0% 내지 0.001%의 돌연변이를 도입한다.
일 구체예에서, 저편향 DNA 중합효소는 뉴클레오티드 유사체를 적어도 하나의 타겟 핵산 분자에 통합시킬 (incorporate) 수 있다. 일 구체예에서, 저편향 DNA 중합효소는 뉴클레오티드 유사체를 적어도 하나의 타겟 핵산 분자에 통합시킨다. 일 구체예에서, 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 아데닌, 티민, 구아닌 및/또는 시토신을 돌연변이시킬 수 있다. 일 구체예에서, 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 적어도 하나의 타겟 핵산 분자에서 아데닌, 티민, 구아닌 및/또는 시토신을 돌연변이시킨다. 일 구체예에서, DNA 중합효소는 구아닌, 시토신, 아데닌 및/또는 티민을 뉴클레오티드 유사체로 대체한다. 일 구체예에서, DNA 중합효소는 구아닌, 시토신, 아데닌 및/또는 티민을 뉴클레오티드 유사체로 대체할 수 있다.
뉴클레오티드 유사체를 적어도 하나의 타겟 핵산 분자에 통합시키는 것은 뉴클레오티드를 돌연변이시키는데 사용될 수 있는데, 이는 이들이 기존 뉴클레오티드를 대신하여 통합될 수 있고 반대 가닥의 뉴클레오티드와 쌍을 이룰 수 있기 때문이다. 예를 들어, dPTP는 피리미딘 뉴클레오티드 대신 핵산 분자에 포함될 수 있다 (티민 또는 시토신을 대체할 수 있음); 도 7 참조. 핵산 가닥에 들어가면, 이미노 토토머 (imino tautomeric) 형태일 때 아데닌과 쌍을 이룰 수 있다. 따라서, 상보적 가닥이 형성될 때, 상기 상보적 가닥은 dPTP에 상보적인 위치에 존재하는 아데닌을 가질 수 있다. 유사하게, 핵산 가닥에 들어가면, 아미노 토토머 (amino tautomeric) 형태일 때 구아닌과 쌍을 이룰 수 있다. 따라서, 상보적 가닥이 형성 될 때, 상기 상보적 가닥은 dPTP에 상보적인 위치에 존재하는 구아닌을 가질 수 있다.
예를 들어, dPTP가 본 발명의 적어도 하나의 타겟 핵산 분자에 도입되는 경우, 적어도 하나의 타겟 핵산 분자에 상보적인 적어도 하나의 핵산 분자가 형성될 때, 상기 적어도 하나의 타겟 핵산 분자에 상보적인 적어도 하나의 타겟 핵산 분자는 적어도 하나의 타겟 핵산 분자에서 dPTP에 상보적인 위치에 (dPTP가 그것의 아미노 또는 이미노 형태인지에 따라) 아데닌 또는 구아닌을 포함할 것이다. 적어도 하나의 타겟 핵산 분자에 상보적인 적어도 하나의 핵산 분자가 복제될 때, 적어도 하나의 타겟 핵산 분자에 대하여 생성된 복제물은 적어도 하나의 타겟 핵산 분자에서 dPTP에 상응하는 위치에 티민 또는 사이토신을 포함할 것이다. 따라서, 티민 또는 시토신에 대한 돌연변이는 돌연변이된 적어도 하나의 타겟 핵산 분자에 도입될 수 있다.
대안적으로는, dPTP가 적어도 하나의 타겟 핵산 분자에 상보적인 적어도 하나의 핵산 분자에 도입되는 경우, 적어도 하나의 타겟 핵산 분자의 복제물이 형성될 때, 적어도 하나의 타겟 핵산 분자의 복제물은 적어도 하나의 타겟 핵산 분자에 상보적인 적어도 하나의 핵산 분자에서 dPTP에 상보적인 위치에 (dPTP의 토토머 형태에 따라) 아데닌 또는 구아닌을 포함할 것이다. 따라서, 아데닌 또는 구아닌으로의 돌연변이는 돌연변이된 적어도 하나의 타겟 핵산 분자에 도입될 수 있다.
일 구체예에서, 저편향 DNA 중합효소는 시토신 또는 티민을 뉴클레오티드 유사체로 대체할 수 있다. 다른 구체예에서, 상기 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 구아닌 또는 아데닌 뉴클레오티드를 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비로 도입할 수 있다. 상기 구아닌 또는 아데닌 뉴클레오티드는, 그들을 dPTP와 같은 뉴클레오티드 유사체와 대향하여 짝짓는 저편향 DNA 중합효소에 의해 도입될 수 있다. 다른 구체예에서, 상기 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 구아닌 또는 아데닌 뉴클레오티드를 각각 0.7-1.3:0.7-1.3의 비율비로 도입할 수 있다.
당업자는 통상적인 방법을 사용하여, 저편향 DNA 중합효소가 뉴클레오티드 유사체를 적어도 하나의 타겟 핵산 분자에 통합시킬 수 있는지 또는 적어도 하나의 타겟 핵산에서 통상적인 방법으로 뉴클레오티드 유사체를 사용하여 아데닌, 티민, 구아닌 및/또는 시토신을 돌연변이시킬 수 있는지 여부를 결정할 수 있다.
예를 들어, 저편향 DNA 중합효소가 뉴클레오티드 유사체를 적어도 하나의 타겟 핵산 분자에 통합시킬 수 있는지 여부를 결정하기 위해, 통상의 기술자는 저편향 DNA 중합효소를 사용하여 복제 라운드 2회로 핵산 분자를 증폭할 수 있다. 제1 복제 라운드는 뉴클레오티드 유사체의 존재 하에 일어나야 하고, 제2 라운드는 뉴클레오티드 유사체의 부재 하에 일어나야 한다. 그 결과 증폭된 핵산 분자를 시퀀싱하여 돌연변이가 도입되었는지 여부를 확인할 수 있고, 만약 도입되었다면 얼마나 많은 돌연변이가 도입되었는지 확인할 수 있다. 사용자는 뉴클레오티드 유사체 없이 실험을 반복하고 뉴클레오티드 유사체가 있는 경우와 없는 경우 도입된 돌연변이의 수를 비교하여야 한다. 뉴클레오티드 유사체로 도입된 돌연변이의 수가 뉴클레오티드 유사체 없이 도입된 돌연변이의 수보다 현저히 많으면, 사용자는 저편향 DNA 중합효소가 뉴클레오티드 유사체를 통합시킬 수 있다는 결론을 내릴 수 있다. 유사하게, 당업자는 DNA 중합효소가 뉴클레오티드 유사체를 통합시킬 수 있는지 또는 뉴클레오티드 유사체를 사용하여 아데닌, 티민, 구아닌 및/또는 시토신을 돌연변이시키는지를 결정할 수 있다. 당업자는 뉴클레오티드 유사체의 존재 하에서 방법을 수행하고, 상기 방법이 원래 아데닌, 티민, 구아닌 및/또는 시토신이 차지하는 위치에서 돌연변이를 유발하는지 확인하기만 하면 된다.
사용자가 뉴클레오티드 유사체를 사용하여 적어도 하나의 타겟 핵산 분자를 돌연변이시키기를 바라는 경우, 상기 방법은 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 포함할 수 있으며, 여기서 저편향 DNA 중합효소를 사용하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 뉴클레오티드 유사체의 존재 하에 수행되고, 상기 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 제공한다.
적합한 뉴클레오티드 유사체는 dPTP (2'deoxy-P-nucleoside-5'-triphosphate), 8-Oxo-dGTP (7,8-dihydro-8-oxoguanine), 5Br-dUTP (5-bromo-2'-deoxy-uridine-5'-triphosphate), 2OH-dATP (2-hydroxy-2'-deoxyadenosine-5'-triphosphate), dKTP (9-(2-Deoxy-β-D-ribofuranosyl)-N6-methoxy-2,6,-diaminopurine-5'-triphosphate) 및 dITP (2'-deoxyinosine 5'-trisphosphate)를 포함한다. 뉴클레오티드 유사체는 dPTP일 수 있다. 뉴클레오티드 유사체는 표 1에 기재된 치환 돌연변이를 도입하는데 사용될 수 있다.
| 뉴클레오티드 | 치환 |
| 8-oxo-dGTP | A:T에서 C:G로 및 T:A에서 G:C로 |
| dPTP | A:T에서 G:C로 및 G:C에서 A:T로 |
| 5Br-dUTP | A:T에서 G:C로 및 T:A에서 C:G로 |
| 2OH-dATP | A:T에서 C:G로, G:C에서 T:A 및 A:T에서 G:C로 |
| dITP | A:T에서 G:C로 및 G:C에서 A:T로 |
| dKTP | A:T에서 G:C로 및 G:C에서 A:T로 |
상이한 뉴클레오티드 유사체는 적어도 하나의 타겟 핵산 분자에 상이한 돌연변이를 도입하기 위해 단독으로 또는 조합하여 사용될 수 있다. 따라서, 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 구아닌에서 아데닌으로의 치환 돌연변이, 사이토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 도입할 수 있다. 상기 저편향 DNA 중합효소는, 선택적으로 뉴클레오티드 유사체를 사용하여, 구아닌에서 아데닌으로의 치환 돌연변이, 사이토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 도입할 수 있다.
상기 저편향 DNA 중합효소는 구아닌에서 아데닌으로의 치환 돌연변이, 사이토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 각각 0.5-1.5:0.5-1.5:0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4:0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2:0.8-1.2:0.8-1.2, 또는 약 1:1:1:1의 비율비로 도입할 수 있다. 바람직하게는, 상기 저편향 DNA 중합효소는 구아닌에서 아데닌으로의 치환 돌연변이, 사이토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 각각 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3의 비율비로 도입할 수 있다. 상기 저편향 DNA 중합효소가 치환 돌연변이를 도입할 수 있는지 여부와 그 비율비가 어떠한지를 결정하는 적합한 방법은 "DNA 중합효소가 뉴클레오티드를 돌연변이시킬 수 있는지 여부 및, 만약 그렇다면, 어떤 비율로 돌연변이 시키는지"라는 표제 하에 기재하였다.
일부 방법에서, 상기 저편향 DNA 중합효소가 구아닌에서 아데닌으로의 치환 돌연변이, 사이토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 각각 0.5-1.5:0.5-1.5:0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4:0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2:0.8-1.2:0.8-1.2, 또는 약 1:1:1:1의 비율비로 도입할 수 있다. 바람직하게는, 상기 저편향 DNA 중합효소는 구아닌에서 아데닌으로의 치환 돌연변이, 사이토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 각각 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3의 비율비로 도입한다. 상기 저편향 DNA 중합효소가 치환 돌연변이를 도입할 수 있는지 여부와 그 비율비가 어떠한지를 결정하는 적합한 방법은 "DNA 중합효소가 뉴클레오티드를 돌연변이시킬 수 있는지 여부 및, 만약 그렇다면, 어떤 비율로 돌연변이 시키는지"라는 표제 하에 기재하였다.
일반적으로, 저편향 DNA 중합효소가 뉴클레오티드 유사체를 사용하여 돌연변이를 도입하는 경우, 복제 라운드 1회 이상이 필요하다. 제1 복제 라운드에서 저편향 DNA 중합효소는 뉴클레오티드 대신 뉴클레오티드 유사체를 도입하고, 제2 라운드에서는 그 뉴클레오티드 유사체가 천연 뉴클레오티드와 쌍을 이루어 상보적 가닥에 치환 돌연변이를 도입한다. 제2 복제 라운드는 뉴클레오티드 유사체의 존재 하에 수행될 수 있다. 그러나, 상기 방법은 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 추가로 포함할 수 있다. 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 저편향 DNA 중합효소를 사용하여 수행 될 수 있다.
선택적으로, 상기 방법은 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하고, 상기 방법은 저편향 DNA 중합효소를 사용하여 상기 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 더 포함한다.
낮은 주형 증폭 편향
저편향 DNA 중합효소는 낮은 주형 증폭 편향을 가질 수 있다. 저편향 DNA 중합효소는 사이클 당 성공률이 비슷한 다른 타겟 핵산 분자를 증폭할 수 있는 경우 낮은 주형 증폭 편향을 갖는다. 고편향 DNA 중합효소는 G:C 함량이 높거나 이차 구조를 많이 포함하는 주형 핵산 분자를 증폭하는 데 어려움을 겪을 수 있다. 일 구체예에서, 본 발명의 저편향 DNA 중합효소는 길이가 25,000개 미만, 10,000개 미만, 1 내지 15,000개, 또는 1 내지 10,000개 뉴클레오티드인 주형 핵산 분자에 대한 낮은 주형 증폭 편향을 갖는다.
일 구체예에서, DNA 중합효소가 낮은 주형 증폭 편향을 갖는지 여부를 결정하기 위해, 당업자는 DNA 중합효소를 사용하여 다양한 서열을 증폭할 수 있고, 결과적으로 증폭된 DNA를 시퀀싱함으로써 상이한 서열이 상이한 수준에서 증폭되는지를 확인할 수 있다. 예를 들어, 당업자는 GC 함량이 높은 핵산 분자, GC 함량이 낮은 핵산 분자, 이차 구조를 많이 포함하는 핵산 분자 및 이차 구조를 적게 포함하는 핵산 분자를 포함하는 서로 다른 특성을 갖는 다양한 짧은 핵산 분자 (아마도 50개 뉴클레오티드)를 선택할 수 있다. 그런 다음 사용자는 DNA 중합효소를 사용하여 이러한 서열을 증폭하고 각 핵산 분자가 증폭되는 수준을 정량화 할 수 있다. 일 구체예에서, 수준이 서로의 25%, 20%, 10% 또는 5% 이내이면, 상기 DNA 중합 효소는 낮은 주형 증폭 편향을 갖는다.
대안적으로는, 일 구체예에서, DNA 중합효소는 0.1 미만, 0.09 미만 또는 0.08 미만의 Kolmolgorov-Smirnov D 값으로 7-10kbp 단편을 증폭할 수 있는 경우 낮은 주형 증폭 편향을 갖는다. 특정 저편향 DNA 중합효소가 7-10kbp 단편을 증폭할 수 있는 Kolmolgorov-Smirnov D 값은 실시예 4에 제공된 분석을 사용하여 결정될 수 있다.
저편향 DNA 중합효소는 고정확도 DNA 중합효소일 수 있다. 고정확도 DNA 중합효소는 오류 발생 가능성이 높지 않은 DNA 중합효소이므로, 뉴클레오티드 유사체의 부재 하에서 타겟 핵산 분자의 증폭에 사용시 일반적으로 많은 수의 돌연변이를 도입하지 않는다. 오류가 발생하기 쉬운 (error-prone) DNA 중합효소가 더 효과적이라고 간주되기 때문에, 고정확도 DNA 중합효소는 돌연변이를 도입하는 방법에 일반적으로 사용되지 않는다. 그러나, 본 출원은 특정 고정확도 중합효소가 뉴클레오티드 유사체를 사용하여 돌연변이를 도입할 수 있고, 이러한 돌연변이는 Taq 중합효소와 같은 오류가 발생하기 쉬운 DNA 중합 효소에 비해 낮은 편향으로 도입될 수 있음을 입증하였다.
고정확도 DNA 중합효소에는 추가적인 이점이 있다. 고정확도 DNA 중합효소는 뉴클레오티드 유사체와 함께 사용할 때 돌연변이를 도입하는 데 사용될 수 있지만, 뉴클레오티드 유사체의 부재 하에서 타겟 핵산 분자를 매우 정확하게 복제할 수 있다. 이는 사용자가 적어도 하나의 타겟 핵산 분자를 높은 효과로 돌연변이시키고 동일한 DNA 중합효소를 사용하여 높은 정확도로 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭할 수 있음을 의미한다. 저정확도 DNA 중합효소를 사용하여 타겟 핵산 분자를 돌연변이시키는 경우, 이는 타겟 핵산 분자가 증폭되기 전에 반응 혼합물에서 제거되어야 할 수 있다.
고정확도 DNA 중합효소는 교정 (proof-reading) 활성을 가질 수 있다. 교정 확성은 DNA 중합효소가 타겟 핵산 분자를 높은 정확도로 증폭하는 데 도움이 될 수 있다. 예를 들어, 저편향 DNA 중합효소는 교정 도메인을 포함할 수 있다. 교정 도메인은 상기 중합효소에 의해 추가된 뉴클레오티드가 올바른지 (상보적 가닥의 상응하는 핵산과 올바르게 짝을 이루는지 확인) 확인하고, 그렇지 않은 경우 이를 핵산 분자에서 제거할 수 있다. 본 발명자들은 놀랍게도 일부 DNA 중합효소에서 교정 도메인이 천연 뉴클레오티드와 뉴클레오티드 유사체의 쌍을 받아들임을 발견하였다. 적합한 교정 도메인의 구조 및 순서는 당업자에게 알려져 있다. 교정 도메인을 포함하는 DNA 중합효소에는 Pfu 중합효소 (Pyrococcus furiosus에서 유래됨), T4 중합효소 (박테리오파지 T4에서 유래됨) 및 하기에서 더 자세히 설명된 Thermococcal 중합효소와 같은 DNA 중합효소 패밀리 Ⅰ, Ⅱ 및 Ⅲ의 멤버들이 포함된다.
일 구체예에서, 뉴클레오티드 유사체의 부재 하에, 상기 고정확도 DNA 중합효소는 복제 라운드 당 0.01% 미만, 0.0015% 미만, 0.001% 미만, 0% 내지 0.0015%, 또는 0% 내지 0.001%의 돌연변이를 도입한다.
또한, 상기 저편향 DNA 돌연변이는 진행도 증진 (processivity enhancing) 도메인을 포함할 수 있다. 진행도 증진 도메인은 DNA 중합효소가 타겟 핵산 분자를 더 빠르게 증폭할 수 있도록 한다. 이는 본 발명의 방법이 더 빨리 수행될 수 있도록 하므로 이점이 있다.
써모코커스 중합효소
일 구체예에서, 저편향 DNA 중합효소는 서열번호 2, 서열번호 4, 서열번호 6 또는 서열번호 7을 포함하는 폴리펩티드의 단편 (fragment) 또는 변이체 (variant)이다. 상기 서열번호 2, 4, 6 및 7의 폴리펩티드는 써모코커스 중합효소이다. 상기 서열번호 2, 서열번호 4, 서열번호 6 또는 서열번호 7의 중합효소는 고정확도를 갖는 저편향 DNA 중합효소이고, 이들은 dPTP와 같은 뉴클레오티드 유사체를 포함시킴으로써 타겟 핵산 분자를 돌연변이시킬 수 있다. 상기 서열번호 2, 서열번호 4, 서열번호 6 또는 서열번호 7의 중합효소는 돌연변이 편향이 낮고 주형 증폭 편향이 낮기 때문에 특히 유리하다. 이들은 또한 매우 진행성 (processive)이고 교정 도메인을 포함하는 고정확도 중합효소로서, 이는 뉴클레오티드 유사체의 부재 하에서 이들이 돌연변이된 타겟 핵산 분자를 빠르고 정확하게 증폭할 수 있음을 의미한다.
저편향 DNA 중합효소는 적어도 400, 적어도 500, 적어도 600, 적어도 700, 또는 적어도 750개의 연속하는, 하기 아미노산의 단편을 포함한다:
a. 서열번호 2의 서열;
b. 서열번호 2와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
c. 서열번호 4의 서열;
d. 서열번호 4와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
e. 서열번호 6의 서열;
f. 서열번호 6과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
g. 서열번호 7의 서열; 또는
h. 서열번호 7과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열.
바람직하게는, 상기 저편향 DNA 중합효소는 적어도 700개의 연속하는, 하기 아미노산의 단편을 포함한다:
a. 서열번호 2의 서열;
b. 서열번호 2와 적어도 98%, 또는 적어도 99% 동일한 서열;
c. 서열번호 4의 서열;
d. 서열번호 4와 적어도 98%, 또는 적어도 99% 동일한 서열;
e. 서열번호 6의 서열;
f. 서열번호 6과 적어도 98%, 또는 적어도 99% 동일한 서열;
g. 서열번호 7의 서열; 또는
h. 서열번호 7과 적어도 98%, 또는 적어도 99% 동일한 서열.
상기 저편향 DNA 중합효소는 하기를 포함할 수 있다:
a. 서열번호 2의 서열;
b. 서열번호 2와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
c. 서열번호 4의 서열;
d. 서열번호 4와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
e. 서열번호 6의 서열;
f. 서열번호 6과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
g. 서열번호 7의 서열; 또는
h. 서열번호 7과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열.
바람직하게는, 상기 저편향 DNA 중합효소는 하기를 포함할 수 있다:
a. 서열번호 2의 서열;
b. 서열번호 2와 적어도 98%, 또는 적어도 99% 동일한 서열;
c. 서열번호 4의 서열;
d. 서열번호 4와 적어도 98%, 또는 적어도 99% 동일한 서열;
e. 서열번호 6의 서열;
f. 서열번호 6과 적어도 98%, 또는 적어도 99% 동일한 서열;
g. 서열번호 7의 서열; 또는
h. 서열번호 7과 적어도 98%, 또는 적어도 99% 동일한 서열.
상기 저편향 DNA 중합효소는 써모코커스 중합효소, 또는 이의 유도체 (derivative)일 수 있다. 서열번호 2, 4, 6 및 7의 DNA 중합효소는 써모코커스 중합효소이다. 써모코커스 중합효소는 이들이 일반적으로 고정확도 중합효소로서 낮은 돌연변이 편향 및 주형 증폭 편향으로 뉴클레오티드 유사체를 사용하여 돌연변이를 도입하는데 사용될 수 있다는 점에서 유리하다.
써모코커스 중합효소는 써모코커스 속 (genus)의 균주로부터 분리된 중합효소의 폴리펩티드 서열을 갖는다. 써모코커스 중합효소의 유도체는 써모코커스 중합효소의 적어도 400, 적어도 500, 적어도 600, 적어도 700, 적어도 750개의 연속하는 아미노산의 단편일 수 있고, 또는 써모코커스 중합효소의 적어도 400, 적어도 500, 적어도 600, 적어도 700, 적어도 750개의 연속하는 아미노산의 단편과 적어도 95%, 적어도 98%, 적어도 99%, 또는 100% 동일할 수 있다. 상기 써모코커스 중합효소의 유도체는 써모코커스 중합효소와 적어도 95%, 적어도 98%, 적어도 99%, 또는 100% 동일할 수 있다. 상기 써모코커스 중합효소의 유도체는 써모코커스 중합효소와 적어도 98% 동일할 수 있다.
임의의 균주로부터의 써모코커스 중합효소는 본 발명의 맥락에서 효과적일 수 있다. 일 구체예에서, 상기 써모코커스 중합효소는 T.kodakarensis, T.siculi, T.celer 및 T.sp KS-1로 이루어진 군으로부터 선택된 써모코커스 균주로부터 유래된 것이다. 이들 균주로부터 유래된 써모코커스 중합효소를 서열번호 2, 서열번호 4, 서열번호 6 및 서열번호 7에 나타내었다.
선택적으로, 상기 저편향 DNA 중합효소는 50℃ 내지 90℃, 60℃ 내지 80℃, 또는 약 68℃의 온도에서 높은 촉매 활성을 갖는 중합효소이다.
바코드, 샘플 태그 및 어댑터
상기 방법은 타겟 핵산 분자에 바코드를 도입하는 단계를 더 포함할 수 있다. 본 발명의 목적상, 바코드는 뉴클레오티드의 축퇴 (degenerate) 또는 무작위로 생성된 서열이다. 용어 "바코드 (barcode)"는 "고유 분자 표지"(unique molecular identifiers: UMIs) 또는 "고유 분자 태그"(unique molecular tags: UMTs)라는 용어와 동의어이다. 상기 방법은 타겟 핵산 분자에 1개, 2개 또는 그 이상의 바코드를 도입하는 단계를 포함할 수 있다. 바람직한 구체예에서, 상기 방법은 다양한 바코드를 타겟 핵산 분자에 도입하는 단계를 포함하여, 바코드가 도입된 후, 대부분의 원래 타겟 핵산 분자가 다른 원래 타겟 핵산 분자에 비해 고유한 바코드를 포함하도록 한다.
바코드를 타겟 핵산 분자에 도입하는 것은 본 발명의 돌연변이를 도입하는 방법이 서열을 결정하는 방법의 일부로서 사용되는 경우 유용할 수 있다. 바코드의 사용은 사용자가 적어도 하나의 타겟 핵산 분자 (또는 증폭 또는 단편화된 적어도 하나의 타겟 핵산 분자)의 각 서열이 유래된 원래의 적어도 하나의 타겟 핵산 분자를 확인하는 데 도움이 될 수 있다. 각각의 원래 타겟 핵산 분자에 사용된 바코드가 다른 경우, 사용자는 바코드 또는 타겟 핵산 분자를 시퀀싱할 수 있으며, 동일한 바코드를 포함하는 타겟 핵산 분자의 서열이 동일한 원래 타겟 핵산 분자에서 유래한 타겟 핵산 분자의 서열일 가능성이 높다.
적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법은 샘플 태그를 타겟 핵산 분자에 도입하는 단계를 포함할 수 있다. 샘플 태그는 알려진 (특정된) 서열의 짧은 일련의 핵산이다. 예를 들어, 본 발명의 방법은 상이한 샘플에서 채취한 다수 개의 타겟 핵산 분자에 대해 수행될 수 있다. 이러한 샘플은 풀링될 (pooled) 수 있지만, 풀링 전에, 샘플 태그가 샘플의 타겟 핵산 분자에 도입될 수 있다 (타겟 핵산 분자가 샘플 태그로 표지됨). 다른 샘플로부터의 타겟 핵산 분자는 다른 샘플 태그로 표지될 수 있다. 선택적으로, 동일한 샘플의 타겟 핵산 분자는 동일한 샘플 태그 또는 샘플 태그의 동일한 하위 그룹 (sup-group)의 샘플 태그로 표지된다. 예를 들어, 사용자가 2개의 샘플을 사용하기로 결정한 경우, 제1 샘플의 타겟 핵산 분자는 특정된 서열을 갖는 제1 샘플 태그로 표지될 수 있고 제2 샘플의 타겟 핵산 분자는 두번째로 특정된 서열을 갖는 제2 샘플 태그로 표지될 수 있다. 유사하게, 사용자가 2개의 샘플을 사용하기로 결정한 경우, 제1 샘플의 타겟 핵산 분자는 제1 샘플 태그 하위 그룹의 샘플 태그로 표지될 수 있으며 제2 샘플의 타겟 핵산 분자는 샘플 태그의 두 번째 하위 그룹의 샘플 태그로 표지될 수 있다. 사용자는 제1 샘플 태그 또는 샘플 태그의 첫번째 하위 그룹의 샘플 태그를 포함하는 임의의 타겟 핵산 분자가 제1 샘플에서 유래하고, 제2 샘플 태그 또는 샘플 태그의 두번째 하위 그룹의 샘플 태그를 포함하는 임의의 타겟 핵산 분자가 제2 샘플에서 유래할 것을 이해할 수 있다. 타겟 핵산 분자를 시퀀싱함으로써 타겟 핵산 분자를 표지하는데 어떤 태그가 사용되었는지 결정할 수 있다. 적합한 시퀀싱 방법은 하기에서 더 자세히 설명하였다.
일 구체예에서, 상기 샘플 태그는 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계 전에 도입된다 (타겟 핵산 분자는 샘플 태그로 표지됨). 이는 샘플이 상기 방법의 초기 단계에서 풀링될 수 있음을 의미하여, 취급 시간 (handling time), 필요한 시약의 수 및 시료 취급 실수의 발생가능성을 감소시키므 유리하다. 그러나, 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계 전에 샘플 태그가 도입되면, 상기 샘플 태그가 저편향 DNA 중합효소에 의해 돌연변이될 가능성이 있다. 본 발명자들은 돌연변이된 경우에도 서로 구별될 수 있도록 설계된 샘플 태그 그룹을 설계하였다.
일 구체예에서, 샘플 태그 그룹이 사용되고 다른 샘플로부터의 타겟 핵산 분자는 상기 그룹의 서로 다른 샘플 태그로 표지된다. 동일한 샘플로부터의 타겟 핵산 분자는 그룹의 동일한 샘플 태그 또는 그룹의 동일한 샘플 태그 하위 그룹의 샘플 태그로 표지될 수 있다. 예를 들어, 샘플 태그 그룹이 A, B, C 및 D라는 샘플 태그를 포함하는 경우, 제1 샘플의 모든 타겟 핵산 분자는 A 또는 A/B를 사용하여 표지될 수 있으며, 제2 샘플의 모든 타겟 핵산 분자는 샘플은 C 또는 C/D를 사용하여 표지될 수 있다. 샘플 태그 그룹의 각 샘플 태그는 적어도 1개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 (substantially) 다를 수 있다. 샘플 태그 그룹의 각 샘플 태그는 적어도 1개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 다를 수 있다.
일 양상에서, 본 발명은 샘플 태그 그룹을 제공하며, 여기서 그룹의 각 샘플 태그는 적어도 1개의 낮은 확률 돌연변이 차이에 의해 그룹의 모든 다른 샘플 태그와 실질적으로 다르다. 각 샘플 태그는 적어도 1개의 낮은 확률 돌연변이 차이로 그룹의 모든 다른 샘플 태그와 다를 수 있다.
"적어도 1개의 낮은 확률 돌연변이 차이에 의해 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른"이라는 용어는 샘플 태그가 적어도 1개의 낮은 확률 돌연변이에 의해 돌연변이되는 경우에도 태그가 여전히 거의 서로 다를 수 있도록 (실질적으로 모든 태그 또는 모든 다른 태그) 각 태그를 설계하였음을 의미한다. 일 구체예에서, 용어 "실질적으로 모든 다른 샘플 태그"는 다른 샘플 태그의 적어도 90%, 적어도 95%, 또는 적어도 98%를 지칭한다. 낮은 확률 돌연변이는 본 발명의 돌연변이를 도입하는 방법에서 드물게 발생하는 돌연변이이다. 예를 들어, 낮은 확률 돌연변이는 전이 돌연변이 또는 인델 돌연변이일 수 있다. 본 발명의 돌연변이를 도입하는 방법이 뉴클레오티드 유사체로서 dPTP를 사용하여 수행될 때 전이 돌연변이 및 인델 돌연변이가 드물게 발생한다. 전이 돌연변이는 퓨린 뉴클레오티드를 피리미딘 뉴클레오티드 (아데닌에서 시토신, 아데닌에서 티민, 구아닌에서 시토신 또는 구아닌에서 티민) 또는 피리미딘 뉴클레오티드를 퓨린 뉴클레오티드 (사이토신에서 아데닌, 시토신에서 구아닌, 티민에서 아데닌, 또는 티민에서 구아닌)로 대체하는 것이다. 인델 돌연변이는 결실 돌연변이 또는 삽입 돌연변이이다. 적합한 태그는 통계적 방법을 사용하여 계산적으로 설계될 수 있다. 예를 들어, 당업자는 본 발명의 돌연변이를 도입하는 방법을 수행하고, 핵산 분자 산물을 시퀀싱하여 도입된 돌연변이의 유형이 무엇인지를 결정할 수 있을 것이다. 가장 빈번하게 발생하는 돌연변이는 높은 확률 돌연변이이고, 가장 적게 발생하는 돌연변이는 낮은 확률 돌연변이이다.
사용자는 본 발명의 샘플 태그 그룹을 설계하기 위한 방법을 사용하여 적합한 샘플 태그를 생산할 수 있다.
선택적으로, 각 샘플 태그는 적어도 2개, 적어도 3개, 적어도 4개, 적어도 5개, 3 내지 50개, 3 내지 25개, 또는 3 내지 10개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다르다. 선택적으로, 각 샘플 태그는 적어도 2개, 적어도 3개, 적어도 4개, 적어도 5개, 3 내지 50개, 3 내지 25개, 또는 3 내지 10개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 다르다.
각 샘플 태그는 적어도 2개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다를 수 있다. 높은 확률 돌연변이 차이는 본 발명의 돌연변이를 도입하는 방법에서 자주 발생하는 돌연변이이다. 예를 들어, 높은 확률 돌연변이 차이는 전이 돌연변이일 수 있다. 전이 돌연변이는 퓨린 뉴클레오티드를 다른 퓨린 뉴클레오티드 (아데닌에서 구아닌 또는 구아닌에서 아데닌)로 대체하거나 피리미딘 뉴클레오티드를 다른 피리미딘 뉴클레오티드로 (사이토신에서 티민 또는 티민에서 시토신) 대체하는 것이다.
각 샘플 태그는 적어도 2개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 다를 수 있다, 예를 들어, 각 샘플 태그는 적어도 2개의 높은 확률 돌연변이에 의해 돌연변이되는 경우에도, 태그가 여전히 거의 서로 다를 수 있도록 설계된 것이다.
선택적으로, 각 샘플 태그는 적어도 3개, 2 내지 50개, 3 내지 25개, 또는 3 내지 10개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다를 수 있다. 선택적으로, 각 샘플 태그는 적어도 3개, 2 내지 50개, 3 내지 25개, 또는 3 내지 10개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 다를 수 있다.
일 구체예에서, 각 샘플 태그는 적어도 8개 뉴클레오티드, 적어도 10개 뉴클레오티드, 적어도 12개 뉴클레오티드, 8 내지 50개 뉴클레오티드, 10 내지 50개 뉴클레오티드, 또는 10 내지 50개 뉴클레오티드 길이이다.
적합한 샘플 태그는 서열 번호 8 내지 136의 태그이다.
상기 방법은 타겟 핵산 분자 각각에 어댑터 (adapters)를 도입하는 단계를 더 포함할 수 있다. 상기 어댑터는 프라이머 결합 부위를 포함할 수 있다. 본 발명의 목적상, 프라이머 결합 부위는 프라이머들이 특이적으로 혼성화하기에 충분히 긴 뉴클레오티드의 공지된 서열이다. 선택적으로, 프라이머 결합 부위는 적어도 8개, 적어도 10개, 적어도 12개, 8 내지 50개, 또는 10 내지 25개 길이의 뉴클레오티드이다.
상기 방법은 적어도 하나의 타겟 핵산 분자의 3' 말단에 제1 어댑터 및 적어도 하나의 타겟 핵산 분자의 5' 말단에 제2 어댑터를 도입하는 단계를 포함할 수 있고, 상기 제1 어댑터 및 제2 어댑터는 서로에게 어닐링 (anneal)할 수 있다.
일 양상에서, 본 발명은 길이가 1 kbp보다 큰 타겟 핵산 분자를 우선적으로 (preferentially) 증폭하는 방법으로서, 하기를 포함하는 방법을 제공한다:
a. 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계;
b. 타겟 핵산 분자의 3' 말단에 제1 어댑터 및 타겟 핵산 분자의 5' 말단에 제2 어댑터를 도입하는 단계; 및
c. 상기 제1 어댑터의 일부에 상보적인 프라이머들을 사용하여 상기 타겟 핵산 분자를 증폭하는 단계로서,
상기 제1 어댑터 및 제2 어댑터는 서로에게 어닐링 (anneal)하는 것인 단계.
제2 어댑터는 제1 프라이머 결합 부위에 상보적인 부분을 포함할 수 있고, 상기 제1 어댑터는 제1 프라이머 결합 부위를 포함할 수 있다.
본 발명자들은 적어도 하나의 타겟 핵산 분자에서 서로 어닐링 할 수 있는 제1 어댑터 및 제2 어댑터를 도입함으로써, 이들이 본 발명의 방법이 긴 타겟 핵산을 우선적으로 증폭 및/또는 돌연변이시키는 것을 보장할 수 있음을 발견하였다. 제1 어댑터가 제2 어댑터에 어닐링 할 수 있다면, 본 발명의 방법에서 이들은 그렇게 하여 적어도 하나의 타겟 핵산 분자에서 자가 어닐링하게 (self-annealed) 된다 (도 5에 나타냄). 자가 어닐링한 타겟 핵산 분자들은 복제되지 않으므로 본 발명의 방법에 의해 증폭 및/또는 돌연변이되지 않는다. 본 발명의 방법 동안 제1 어댑터 및 제2 어댑터가 서로 어닐링하게 될 가능성은 더 긴 타겟 핵산 분자보다 짧은 타겟 핵산 분자에서 더 높을 것이다. 이러한 이유로, 본 발명의 적어도 하나의 타겟 핵산 분자에 대한 제1 어댑터 및 제2 어댑터의 첨가는 더 큰 적어도 하나의 타겟 핵산 분자를 우선적으로 증폭하는데 사용될 수 있다.
핵산 분자를 우선적으로 증폭하는 방법은 1.5 kbp보다 긴 타겟 핵산 분자를 우선적으로 증폭하는 방법일 수 있다. 상기 방법은 타겟 핵산 분자를 시퀀싱하는 단계를 더 포함할 수 있다. 가능한 시퀀싱 방법의 예는 맥삼 길버트 (Maxam Gilbert) 시퀀싱, 생어 (Sanger) 시퀀싱, 나노포어 (nanopore) 시퀀싱 또는 브리지 (bridge) PCR을 포함하는 시퀀싱을 포함한다. 전형적인 구체예에서, 시퀀싱 단계는 브릿지 PCR을 수반한다. 선택적으로, 브리지 PCR 단계는 5초 초과, 10초 초과, 15초 초과 또는 20초 초과의 연장 (extension) 시간을 사용하여 수행된다. 브리지 PCR 사용의 예는 Illumina Genome Analyzer Sequencer에 있다.
사용자는 제1 어댑터와 제2 어댑터가 서로 어닐링 할 수 있는지 여부를 결정할 수 있다. 일 구체예에서, 사용자는 제1 어댑터를 포함하는 핵산 분자를 제공하고, 제2 어댑터를 포함하는 프라이머가 PCR 조건 하에서 핵산 분자의 복제를 개시할 수 있는지 여부를 확인함으로써 제1 어댑터 및 제2 어댑터가 서로 어닐링 할 수 있는지 여부를 확인할 수 있다.
대안적으로는, 일 구체예에서, 제1 어댑터 및 제2 어댑터는 이들이 다음 조건 하에서 혼성화하는 경우 서로 어닐링 할 수 있는 것으로 간주될 수 있다: 등몰 (equimolar) 농도의 두 프라이머가 혼합된 다음 (예를 들어, 50μM), 95℃와 같은 고온에서 5분 동안 배양하여 상기 프라이머들이 단일 가닥인지 확인한다. 그런 다음, 용액을 약 45분에 걸쳐 천천히 실온 (25℃)으로 냉각한다.
상기 방법은 서로 동일하거나, 또는 실질적으로 서로 동일한 프라이머를 사용하여 타겟 핵산 분자를 증폭하는 단계를 포함할 수 있다. 상기 프라이머는 제1 어댑터의 일부에 상보적일 수 있다. 2개의 프라이머는 동일한 서열을 갖거나, 또는 1개, 2개 또는 3개의 뉴클레오티드가 다른 서열을 갖는 경우 서로 "실질적으로 동일"하다. 바람직한 구체예에서, 본 발명의 방법은 서열이 동일하거나 단일 뉴클레오티드 차이로 서로 다른 프라이머들을 사용하여 타겟 핵산 분자를 증폭하는 단계를 포함한다.
일 구체예에서, 제1 어댑터 및 제2 어댑터는 서로 상보적이거나 실질적으로 서로 상보적인 서열을 포함한다. 제1 어댑터가 제2 어댑터와 적어도 80%, 적어도 90%, 적어도 95%, 또는 적어도 99% 동일한 핵산 분자에 상보적인 경우, 제1 어댑터는 제2 어댑터에 실질적으로 상보적일 수 있다.
사용자는 프라이머 결합 부위를 포함하는 프라이머들을 사용할 수 있으며, 이러한 프라이머는 마지막 복제 라운드에서 생성된 적어도 하나의 타겟 핵산 분자의 복제물을 우선적으로 증폭하는데 사용될 수 있다. 예를 들어, 제3 프라이머 결합 부위를 포함하는 제1 프라이머 세트가 복제 라운드에 사용될 수 있다. 추가 복제 라운드에서 제3 프라이머 결합 부위에 결합하는 제2 프라이머 세트가 사용될 수 있다. 제2 프라이머 세트는 제1 프라이머 세트를 사용하여, 이전 복제 라운드에서 생성된 적어도 하나의 타겟 핵산 분자의 복제물만을 복제할 것이다.
제3 및 추가적인 프라이머 세트가 사용될 수 있다. 이전 복제물을 우선적으로 복제하는 것은 각각의 증폭된 타겟 핵산 분자가 높은 수준의 돌연변이를 포함하도록 보장할 수 있기 때문에 유리하다 (저편향 DNA 중합효소에 의해 적어도 1회의 복제 라운드에 노출된 적어도 하나의 타겟 핵산 분자만이 복제되기 때문).
따라서, 본 발명의 상기 방법은 하기를 포함할 수 있다:
(a) 적어도 하나의 타겟 핵산 분자 또는 타겟 핵산 분자들의 3' 말단에 제1 프라이머 결합 부위를 포함하는 제1 어댑터 및 적어도 하나의 타겟 핵산 분자 또는 타겟 핵산 분자들의 5' 말단에 상기 제1 프라이머 결합 부위에 상보적인 부분을 포함하는 제2 어댑터를 도입하는 단계로서, 상기 제1 어댑터 및 제2 어댑터는 서로 어닐링 할 수 있는 것인 단계;
(b) 제1 프라이머 결합 부위에 상보적이고 제2 프라이머 결합 부위를 포함하는 제1 프라이머 세트를 사용하여 타겟 핵산 분자를 증폭하는 단계로서, 선택적으로 저편향 DNA 중합효소를 사용하는 것인 단계; 및
(c) 상기 제2 프라이머 결합 부위에 상보적인 제2 프라이머 세트를 사용하여 타겟 핵산 분자를 증폭하는 단계로서, 선택적으로 저편향 DNA 중합효소를 사용하는 것인 단계.
제2 프라이머 세트는 제3 프라이머 결합 부위를 포함할 수 있고, 추가적인 증폭 단계는 제3 또는 추가적인 프라이머 결합 부위에 상보적인 제3 또는 추가적인 프라이머 세트를 사용하여 수행될 수 있다.
바코드, 샘플 태그 및/또는 어댑터는 후속적인 어댑터 라이게이션 (ligation) (선택적으로 스티키-엔드 (sticky-end) 라이게이션)과 함께 타겟 핵산의 PCR, 태그맨테이션 (tagmentation) 및 물리적 쉬어링 (shearing) 또는 제한 분해 (restriction digestion)를 포함하는 임의의 적합한 방법을 사용하여 도입될 수 있다. 예를 들어, PCR은 적어도 하나의 타겟 핵산 분자에 혼성화 할 수 있는 제1 프라이머 세트를 사용하여 적어도 하나의 타겟 주형 핵산 분자에서 수행될 수 있다. 바코드, 샘플 태그 및 어댑터는 바코드, 샘플 태그 및/또는 어댑터를 포함하는 부분 (5'말단 부분)을 포함하는 프라이머, 및 상기 적어도 하나의 타겟 핵산 분자에 혼성화할 수 있는 (선택적으로 상보적인) 서열을 갖는 부분 (3' 말단 부분)을 사용하여 PCR에 의해, 적어도 하나의 타겟 핵산 분자 각각에 도입될 수 있다. 이러한 프라이머는 타겟 핵산 분자에 혼성화하고, PCR 프라이머 연장 (extension)은 바코드, 샘플 태그 및/또는 어댑터를 포함하는 핵산 분자를 제공한다. 이들 프라이머를 사용한 PCR의 추가 사이클은 바코드, 샘플 태그 및/또는 어댑터를 적어도 하나의 타겟 핵산 분자의 다른 말단에 추가하는데 사용될 수 있다. 프라이머들은 축퇴 (degenerate)될 수 있다, 즉, 상기 프라이머의 3' 말단 부분은 유사할 수 있지만 서로 동일하지는 않다.
바코드, 샘플 태그 및/또는 어댑터는 태그맨테이션을 사용하여 도입될 수 있다. 바코드, 샘플 태그 및/또는 어댑터는 직접 태그맨테이션을 사용하거나, 또는 태그맨테이션으로 정의된 서열을 도입하고 뒤이어 상기 정의된 서열에 혼성화할 수 있는 부분을 포함하는 프라이머들, 및 바코드, 샘플 태그 및/또는 어댑터를 포함하는 부분을 사용하는 2회 사이클의 PCR로 도입될 수 있다. 상기 바코드, 샘플 태그 및/또는 어댑터는 원래의 적어도 하나의 타겟 핵산 분자의 제한 분해에 뒤이어 바코드, 샘플 태그 및/또는 어댑터를 포함하는 핵산의 라이게이션에 의해 도입될 수 있다. 원래의 적어도 하나의 핵산 분자의 제한 분해는 상기 분해가 시퀀싱 될 영역을 포함하는 핵산 분자 (적어도 하나의 타겟 주형 핵산 분자)를 생성하도록 수행되어야 한다. 바코드, 샘플 태그 및/또는 어댑터는 적어도 하나의 타겟 핵산 분자의 쉬어링, 뒤이은 말단 복구 (end repair), A-tailing 및 그 후 바코드, 샘플 태그 및/또는 어댑터를 포함하는 핵산 서열의 라이게이션에 의해 도입될 수 있다.
적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법
본 발명의 일 양상은 본 발명의 돌연변이를 도입하는 방법을 포함하는 적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법에 관한 것이다.
전술한 바와 같이, 본 발명의 돌연변이를 도입하는 방법은, 돌연변이는 당업자가 서열을 조립할 수 있게 하기 때문에, 적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법의 일부로서 유용할 수 있다.
배경기술 섹션에서 기재한 바와 같이, 시퀀싱 방법은 시퀀싱될 하나 이상의 타겟 핵산 분자에 돌연변이를 도입하는 단계를 통합함으로써 개선될 수 있다. 사용자는 종종 시퀀싱하기 전에 적어도 하나의 타겟 핵산 분자를 증폭 및/또는 단편화한다. 그 다음, 사용자는 증폭되거나 단편화된 적어도 하나의 타겟 핵산 분자의 영역의 서열로부터 타겟 핵산 분자 중 적어도 하나에 대한 컨센서스 (consensus) 서열을 조립할 것이다. 증폭 또는 단편화 이전에 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 것은 사용자가 증폭되거나 단편화된 적어도 하나의 타겟 핵산 분자의 영역의 각 서열이 유래된 원래의 적어도 하나의 주형 핵산 분자가 어떤 것인지 확인하는데 도움을 주고, 이로부터 컨센서스 서열의 정확성을 개선한다.
도입되는 돌연변이가 더 무작위적일수록, 증폭되거나 단편화된 적어도 하나의 타겟 핵산 분자의 각 서열이 유래된 원래의 적어도 하나의 타겟 핵산 분자가 어떤 것인지를 확인하기가 더 쉬워진다. 저편향 DNA 중합효소를 사용하는 본 발명의 돌연변이를 도입하는 방법은, 실질적으로 무작위 방식으로 돌연변이를 도입하는 데 사용될 수 있으므로, 적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법에 포함될 대상으로서 이상적이다.
상기 적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법은 하기 단계를 포함할 수 있다:
a. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해, 본 발명의 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 수행하는 단계;
b. 돌연변이된 서열 리드(reads)를 제공하기 위해, 상기 적어도 하나의 돌연변이된 타겟 핵산 분자의 영역을 시퀀싱하는 단계; 및
c. 상기 돌연변이된 서열 리드를 사용하여 적어도 하나의 타겟 핵산 분자의 적어도 한 부분의 서열을 조립하는 단계
일반적으로, 시퀀싱 단계는 임의의 시퀀싱 방법을 사용하여 수행될 수 있다. 가능한 시퀀싱 방법의 예는 맥삼 길버트 시퀀싱, 생어 시퀀싱, 나노포어 시퀀싱 또는 브리지 PCR을 포함하는 시퀀싱을 포함한다. 전형적인 구체예에서, 시퀀싱 단계는 브릿지 PCR을 수반한다. 선택적으로, 브리지 PCR 단계는 5초 초과, 10초 초과, 15초 초과 또는 20초 초과의 연장 (extension) 시간을 사용하여 수행된다. 브리지 PCR 사용의 예는 Illumina Genome Analyzer Sequencer에 있다.
상기 방법은 돌연변이된 서열 리드를 제공하기 위해 적어도 하나의 돌연변이된 타겟 핵산 분자의 시퀀싱 영역을 포함할 수 있다. 상기 영역은 적어도 하나의 돌연변이된 타겟 핵산 분자의 실질적인 부분을 포함할 수 있는 단편에 상응할 수 있다. 전체 적어도 하나의 돌연변이된 타겟 핵산 분자는 어떤 이유로 서열화 될 수 없을 수도 있지만, 사용자는 여전히 적어도 하나의 돌연변이된 타겟 핵산 분자의 일부의 서열이 유용하다는 것을 알 수 있다. 적어도 하나의 돌연변이된 타겟 핵산 분자의 영역은 적어도 하나의 돌연변이된 타겟 핵산 분자의 전체 길이를 포함할 수 있다.
상기 방법은 돌연변이된 서열 리드로부터 적어도 하나의 타겟 핵산 분자의 적어도 일부에 대한 서열을 조립하는 단계를 포함할 수 있다. 상기 서열은 돌연변이된 서열 리드를 정렬하고 동일한 돌연변이 패턴을 공유하는 리드를 함께 그룹화함으로써 조립될 수 있다. 서열은 동일한 그룹의 돌연변이된 서열 리드로부터 조립될 것이다. 조립은 Clustal W2, IDBA-UD 또는 SOAPdenovo와 같은 소프트웨어를 사용하여 수행될 수 있다.
적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법은 하기 단계를 포함할 수 있다:
a. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해 본 발명에 따른 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 수행하는 단계;
b. 적어도 하나의 단편화된 (fragmented) 및/또는 증폭된 돌연변이된 타겟 핵산 분자를 제공하기 위해, 상기 적어도 하나의 돌연변이된 타겟 핵산 분자를 단편화 및/또는 증폭하는 단계;
c. 돌연변이된 서열 리드를 제공하기 위해, 상기 적어도 하나의 단편화된 및/또는 증폭된 돌연변이된 타겟 핵산 분자의 영역을 시퀀싱하는 단계; 및
d. 상기 돌연변이된 서열 리드를 사용하여 적어도 하나의 핵산 분자의 적어도 한 부분의 서열을 조립하는 단계.
적어도 하나의 돌연변이된 타겟 핵산 분자를 증폭하는 단계는 PCR과 같은 임의의 적합한 증폭 기술을 통해 수행될 수 있다. 적합하게는, 상기 PCR은 "저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계"라는 표제 하에 기재된 조건 하에서 저편향 DNA 중합효소를 사용하여 수행될 수 있다.
적어도 하나의 돌연변이된 타겟 핵산 분자를 단편화하는 단계는 임의의 적절한 방법을 사용하여 수행될 수 있다. 예를 들어, 단편화는 제한 분해를 사용하거나 적어도 하나의 돌연변이된 타겟 핵산 분자의 적어도 하나의 내부 영역에 상보적인 프라이머들과 함께 PCR을 사용하여 수행될 수 있다. 바람직하게는, 단편화는 임의의 단편을 생성하는 기술을 사용하여 수행된다. 용어 "임의적 단편 (arbitrary fragment)"은 무작위로 생성된 단편, 예를 들어 태그맨테이션에 의해 생성된 단편을 의미한다. 제한효소를 사용하여 생성된 단편은 제한 분해가 사용되는 제한 효소로 정의되는 특정 DNA 서열에서 발생하기 때문에 "임의적"이지 않다. 보다 더 바람직하게는, 단편화는 태그맨테이션에 의해 수행된다. 단편화가 태그맨테이션에 의해 수행되는 경우, 태그맨테이션 반응은 어댑터 영역을 적어도 하나의 돌연변이된 타겟 핵산 분자로 선택적으로 도입한다. 이 어댑터 영역은 예를 들어, 적어도 하나의 돌연변이된 타겟 핵산 분자가 Illumina 기술을 사용하여 시퀀싱될 수 있도록 어댑터를 코딩할 수 있는 짧은 DNA 서열이다.
상기 태그맨테이션 단계는 적어도 하나의 돌연변이된 단편화된 타겟 핵산 분자를 농축 (enriching)하는 추가적인 단계를 포함할 수 있다. 상기 적어도 하나의 돌연변이된 단편화된 타겟 핵산 분자를 농축하는 단계는 PCR에 의해 수행될 수 있다. 적합하게는, 상기 PCR은 "저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계"라는 표제 하에 기재된 조건 하에서 저편향 DNA 중합효소를 사용하여 수행될 수 있다.
단백질을 엔지니어링하는 방법
본 발명의 돌연변이를 도입하는 방법은 단백질을 엔지니어링하는 방법의 일부로서 유용할 수 있다. 예를 들어, 단백질 엔지니어링은 단백질의 활성을 증가 또는 감소시키거나 구조를 변경하는 돌연변이를 찾는 것을 포함할 수 있다. 단백질 엔지니어링의 일환으로, 사용자는 단백질을 무작위로 돌연변이시키고 돌연변이가 단백질의 활성 또는 구조에 어떤 영향을 미치는지 확인하기를 바랄 수 있다. 본 방법은 고도로 무작위적인 돌연변이 유발을 야기하는 방법이며, 따라서 단백질을 엔지니어링하는 방법의 일부로서 유리하게 사용될 수 있다.
따라서, 본 발명의 일 양상에서 본 발명의 돌연변이를 도입하는 방법을 포함하는 단백질을 엔지니어링하는 방법을 제공한다.
상기 방법은 하기 단계를 포함할 수 있다:
a. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해 본 발명의 돌연변이를 도입하는 방법을 수행하는 단계;
b. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자를 벡터에 삽입하는 단계; 및
c. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질을 발현하는 단계.
상기 방법은 하기 단계를 포함할 수 있다:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 제공하기 위해, 상기 뉴클레오티드 유사체 존재 하에 저편향 DNA 중합효소를 사용하여 상기 적어도 하나의 타겟 핵산 분자를 증폭하는 단계;
c. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해 뉴클레오티드 유사체 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계;
d. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자를 벡터에 삽입하는 단계; 및
e. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질을 발현하는 단계.
임의의 적합한 벡터가 사용될 수 있다. 선택적으로 벡터는 플라스미드, 바이러스, 코스미드 또는 인공 염색체이다. 전형적으로, 벡터는 삽입된 서열에 작동 가능하게 연결된 제어 서열 (control sequence)을 추가로 포함하여, 폴리펩티드의 발현을 허용한다. 바람직하게는, 본 발명의 벡터는 폴리펩티드의 발현을 허용하기 위해 필요하고 올바른 방향성 (orientation)으로 위치하는 적절한 개시제 (initiators), 프로모터 (promoters), 인핸서 (enhances) 및 기타 요소들을 더 포함한다.
선택적으로, 적어도 하나의 돌연변이된 타겟 핵산 분자를 발현하는 단계는 상기 벡터로 박테리아 세포를 형질전환 (transforming)하거나, 진핵세포를 형질감염 (transfecting)하거나 또는 진핵세포를 형질도입 (transducing)함으로써 달성된다. 선택적으로, 박테리아 세포는 Escherichia coli (E.coli) 세포이다.
예를 들어, 적어도 하나의 돌연변이된 타겟 핵산 분자를 발현하는 단계는 적어도 하나의 돌연변이된 타겟 핵산 분자를 플라스미드 벡터에 삽입하고 플라스미드로 E.coli 를 형질전환하는 것을 포함할 수 있다. 상기 플라스미드는 lac 또는 T7 프로모터 (Dubendorff JW, Studier FW (1991))와 같은 E.coli에서 발현하기에 적합한 제어 요소를 포함할 수 있다. "Controlling basal expression in an inducible T7 expression system by blocking the target T7 promoter with lac repressor" Journal of Molecular Biology. 219 (1): 45-59.)). 적합한 발현 기술은 Sambrook, J. et al., (1989) Molecular Cloning : A Laboratory Manual Second Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York에 기재되어 있다.
대안적으로는, 적어도 하나의 돌연변이된 타겟 핵산 분자를 발현하는 단계는 인 비트로 방법을 사용하여 타겟 핵산 분자를 증폭하는 단계로부터 직접 생성된 단편을 발현하는 것을 포함할 수 있다.
상기 방법은 적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질의 활성을 테스트하거나 구조를 평가하는 단계를 더 포함할 수 있다.
적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질의 활성을 테스트하거나 구조를 평가하는 단계는 임의의 여러 주지의 기술을 사용하여 수행될 수 있다. 예를 들어, 당업자는 핵 자기 공명 (NMR) 기술, 초-저온 전자 현미경과 같은 현미경 기술, 소각 X선 산란 기술, 또는 X선 결정학을 포함하는 단백질의 구조를 평가하는데 적합한 기술을 알고 있을 것이다.
유사하게, 당업자는 단백질의 활성을 평가하기 위해 사용될 수 있는 기술을 알고있을 것이다. 사용되는 방법은 적어도 하나의 돌연변이된 타겟 핵산 분자에 의해 코딩되는 단백질에 따라 달라질 것이다. 예를 들어, 적어도 하나의 돌연변이된 타겟 핵산 분자에 의해 코딩되는 단백질이 혈액 응고 인자인 경우, 당업자는 예를 들어 발색성 응고 분석 (chromogenic clotting assay)을 사용하여 단백질의 응고 활성을 테스트 할 것이다. 대안적으로는, 적어도 하나의 돌연변이된 타겟 핵산 분자에 의해 코딩된 단백질이 효소인 경우, 통상의 기술자는 반응을 촉매하는 속도를 측정, 예를 들어 효소에 의해 촉매된 반응의 출발 생성물의 농도 감소 또는 최종 생성물의 농도 증가를 측정하여, 효소의 활성을 테스트 할 수 있다.
샘플 태그 그룹을 설계하는 방법
일 양상에서, 본 발명은 하기를 포함하는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법에 사용하기에 적합한 샘플 태그 그룹을 설계하는 방법을 더 제공한다:
a. 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 분석하고 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 낮은 확률 돌연변이의 평균 개수를 결정하는 단계; 및
b. 샘플 태그 그룹의 서열을 결정하는 단계로서 각 샘플 태그는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 낮은 확률 돌연변이의 평균 개수보다 큰 낮은 확률 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 단계.
예를 들어, 사용자는 무작위 서열을 생성하기 위해 컴퓨터 프로그램을 사용하여 제1 추정 (putative) 샘플 태그를 생성할 수 있다. 제1 추정 샘플 태그를 샘플 태그 그룹에 추가한다. 그런 다음 사용자는 동일한 방식으로 제2 추정 샘플 태그를 생성하고, 제2 추정 샘플 태그의 서열을 제1 추정 샘플 태그와 비교하여 제2 샘플 태그가 제1 샘플 태그와 다른지 여부를 확인하여서 상당 수의 낮은 확률 돌연변이가 제2 추정 샘플 태그에 도입되더라도 이는 여전히 제1 추정 샘플 태그와 다를 수 있다. 그렇다면, 제2 추정 샘플 태그를 샘플 태그 그룹에 추가한다. 그렇지 않다면, 제2 추정 샘플 태그는 폐기한다. 이를 제3 및 추가적인 추정 샘플 태그에 대해 반복한다.
전술한 바와 같이, 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법에서 샘플 태그를 적어도 하나의 타겟 핵산 분자에 추가하는 것이 유리하다. 그러나, 돌연변이가 도입되기 전에 샘플 태그가 추가되면, 이는 샘플 태그가 돌연변이되어 동일하거나 다른 샘플에서 유래한 타겟 핵산 분자를 구별하는데 사용할 수 없음을 의미할 수 있다. 이는 샘플 태그가 돌연변이로 된 경우에도 사용자가 구분할 수 있도록 샘플 태그를 서로 충분히 다르게 설계하여 방지할 수 있다.
상기 방법은 하기 단계를 더 포함할 수 있다:
a. (i) 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 분석하고 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 높은 확률 돌연변이의 평균 개수를 결정하는 단계; 및
(ii) 샘플 태그 그룹의 서열을 결정하는 단계로서 각 샘플 태그는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 높은 확률 돌연변이의 평균 개수보다 큰 높은 확률 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 단계.
낮은 확률 돌연변이는 전환 돌연변이 또는 인델 돌연변이일 수 있다. 높은 확률 돌연변이는 전이 돌연변이일 수 있다.
상기 방법은 컴퓨터에서 구현되는 (computer implemented) 방법일 수 있다.
본 발명의 다른 양상에서, 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법에 사용하기에 적합한 샘플 태그 그룹을 설계하는 방법을 수행하도록 구성된 컴퓨터 판독가능 매체 (computer-readable medium)이 제공된다.
본 발명의 다른 양상에서, 본 발명의 샘플 태그를 설계하는 단계를 통해 얻을 수 있는 샘플 태그 그룹이 제공된다. 선택적으로, 샘플 태그 그룹은 본 발명의 샘플 태그를 설계하는 방법을 통해 얻을 수 있다.
불균등 농도의 dNTP의 사용
저편향 DNA 중합효소를 사용한 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 불균등 농도의 dNTP를 사용하여 수행될 수 있다.
본 발명의 일 양상에서, 하기 단계를 포함하는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법이 제공된다:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하기 위해 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하여 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 단계로서,
상기 단계 b. 는 불균등 농도의 dNTP를 사용하여 수행되는 것인 단계.
DNA 중합효소 (예를 들어, 저편향 DNA 중합효소)를 사용하여 적어도 하나의 타겟 핵산을 증폭할 수 있도록, 상기 타겟 핵산은 DNA 복제가 발생하기에 적합한 조건 하, 예를 들어 PCR 기계에서 DNA 중합효소 및 dNTP에 노출될 수 있다. 적어도 하나의 타겟 핵산을 증폭하는 단계가 불균등 농도 (unequal concentrations)의 dNTP를 사용하여 수행되는 경우, 타겟 핵산은 DNA 중합효소 (예를 들어, 저편향 DNA 중합효소) 및 dNTP에 노출되며, 여기서 dNTP의 농도는 서로에 대하여 상대적으로 다르다.
용어 dNTP는 데옥시뉴클레오티드를 의미한다. 구체적으로, 본 출원의 맥락에서 용어 "dNTPs"는 dTTP (데옥시티미딘 트리포스페이트) 또는 dUTP (데옥시우리딘), dGTP (데옥시구아니딘 트리포스페이트), dCTP (데옥시시티딘 트리포스페이트), 및 dATP (데옥시아데노신 트리포스페이트)를 포함하는 용액을 의미하는 것으로 의도된다. 선택적으로, "dNTPs"는 dTTP (데옥시티미딘 트리포스페이트), dGTP (데옥시구아니딘 트리포스페이트), dCTP (데옥시시티딘 트리포스페이트), 및 dATP (데옥시아데노신 트리포스페이트)를 포함하는 용액을 지칭한다.
"불균등 농도의 dNTPs"라는 문구는 4개의 dNTP가 서로에 대해 서로 다른 농도로 용액에 존재함을 의미한다. 예를 들어, 하나의 dNTP가 다른 3개의 dNTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, 2개의 dNTP가 다른 2개의 dNTPS에 비해 (보다) 더 높은 농도로 존재할 수 있고, 3개의 dNTP가 다른 하나의 dNTP에 비해 (보다) 더 높은 농도로 존재할 수 있다.
DGTP는 dCTP, dTTP 및 dATP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dGTP는 dTTP 및 dATP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dGTP는 dATP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dGTP는 dTTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dCTP는 dGTP, dTTP 및 dATP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dCTP는 dTTP 및 dATP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dCTP는 dATP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dCTP는 dTTP에 비해 (보다) 더 높은 농도로 존재할 수 있으며, dTTP는 dGTP, dCTP 및 dATP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dTTP는 dGTP 및 dCTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dTTP는 dCTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dTTP는 dGTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dATP는 dGTP, dTTP 및 dCTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dATP는 dGTP 및 dCTP에 비해 (보다) 더 높은 농도로 존재할 수 있으며, dATP는 dGTP에 비해 (보다) 더 높은 농도로 존재할 수 있으며, dATP는 dGTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dCTP 및 dATP는 dGTP 및 dCTP에 비해 (보다) 더 높은 농도로 존재할 수 있고, dGTP 및 dCTP는 dATP 및 dTTP에 비해 (보다) 더 높은 농도로 존재할 수 있다.
사용자는 편리한 방법으로 불균등 농도의 dNTP 용액을 준비할 수 있다. DATP, dTTP, dGTP 및 dTTP 용액은 상업적으로 쉽게 이용가능하며 사용자는 적절한 비율로 이들을 혼합하기만 하면 된다.
선택적으로, 상기 방법은:
(i) 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 추가적인 단계를 포함하고 상기 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 추가적인 단계는 불균등 농도의 dNTP를 사용하여 수행되는 것인 단계; 또는
(ii) 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하고, 저편향 DNA 중합효소를 사용하여 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 추가적인 단계를 포함하며, 상기 저편향 DNA 중합효소를 사용하여 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 추가적인 단계는 불균등 농도의 dNTP를 사용하여 수행되는 것인 단계.
선택적으로, 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하기 위해 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하여 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 것은 뉴클레오티드 유사체의 존재 하에 수행된다. 선택적으로, 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하기 위해 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하여 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 것은 뉴클레오티드 유사체의 부재 하에 수행되고, 선택적으로 이 단계는 불균등 농도의 dNTP를 사용하여 수행된다.
적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하기 위해 뉴클레오티드 유사체가 사용되는 경우, 이는 일반적으로 2개의 증폭 단계를 포함할 것이다. 첫번째 증폭 단계에서, 뉴클레오티드 유사체는 타겟 핵산 분자에 통합된다 (a mutation step, 돌연변이 단계). 두번째 증폭 단계에서, 뉴클레오티드 유사체는 천연 뉴클레오티드와 쌍을 이루어, 타겟 핵산 분자의 한 가닥에 돌연변이를 도입한다 (a recovery step, 회복 단계). 타겟 핵산 분자가 추가로 증폭될 때, 이 돌연변이는 타겟 핵산 분자의 두 가닥 모두로 전달된다. 선택적으로, 첫번째 (mutation) 증폭 단계 및 두번째 (recovery) 증폭 단계는 모두 불균등 농도의 dNTP를 사용하여 수행될 수 있다. 선택적으로 불균등 농도의 dNTP는 첫번째 (mutation) 증폭 단계와 두번째 (recovery) 증폭 단계에서 다르다. 예를 들어, 불균등 농도의 dNTP는 첫번째 (mutation) 증폭 단계에서 다른 dNTP보다 낮은 농도의 dTTP를 포함할 수 있고, 불균등 농도의 dNTP는 두번째 (recovery) 증폭 단계에서 다른 dNTP보다 낮은 농도의 dATP를 포함할 수 있다. 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계 또는 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하는 단계는 하나 이상의 "mutation steps"에 해당할 수 있다. 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 추가 단계 또는 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 추가 단계는 하나 이상의 "recovery steps"에 해당할 수 있다.
선택적으로, 상기 뉴클레오티드 유사체는 dPTP이다.
일 구체예에서, 불균등 농도의 dNTP는 도입된 돌연변이의 프로필 (profile)을 변경하기 위해 사용된다. 불균등 농도의 dNTP는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 것을 포함하는 방법에 사용된다. 따라서, 상기 방법은 돌연변이를 포함하는 타겟 핵산 분자 (예를 들어, 본원에 기재된 돌연변이된 타겟 핵산 분자)를 생성한다. 상기 방법에 의해 주어진 타겟 핵산 분자에 도입되는 돌연변이의 수, 돌연변이의 유형, 및 각 돌연변이의 위치는 도입되는 "돌연변이의 프로필 (profile of mutations)"로 지칭될 수 있다. 용어 "돌연변이의 유형"은 돌연변이의 특성, 즉 치환 돌연변이, 삽입 돌연변이 또는 결실 돌연변이이고, 치환 돌연변이인 경우 시작 뉴클레오티드가 무엇이고 상기 시작 뉴클레오티드가 무엇으로 돌연변이 되었는지 (예를 들어, A에서 G로의 돌연변이는 G로 돌연변이된 A 시작 뉴클레오티드를 가짐)? 를 지칭하는 것으로 의도된다.
사용자는 테스트 타겟 핵산 분자를 복제한 다음, 복제물의 일부를 본 발명의 돌연변이를 도입하는 방법을 포함하는 방법에 적용하고, 복제물 일부는 남겨둠 (돌연변이시키지 않음)으로써 주어진 방법에 의해 도입되는 "돌연변이의 프로필"을 결정할 수 있다. 그 다음 사용자는 본 발명의 돌연변이를 도입하는 방법을 포함하는 방법에 적용된 복제물 및 남겨둔 복제물을 시퀀싱 할 수 있다. 마지막으로, 사용자는 본 발명의 돌연변이를 도입하는 방법을 포함하는 방법에 적용된 복제물의 서열, 및 돌연변이 수. 돌연변이 유형 및 각 돌연변이의 위치를 결정하기 위해 남겨둔 복제물을 정렬할 수 있다. 대안적으로는, 사용자는 알려진 서열의 테스트 타겟 핵산 분자를 사용할 수 있다. 사용자는 본 발명의 돌연변이를 도입하는 방법을 포함하는 방법에 테스트 타겟 핵산 분자를 적용한 다음, 어떤 돌연변이 프로필이 도입되었는지 확인하기 위해 결과적인 돌연변이된 타겟 핵산 분자를 시퀀싱하기만 하면 된다.
사용자는 여러 가지 방법으로 돌연변이 프로필을 변경하기를 바랄 수 있다. 예를 들어, 전술한 바와 같이, 돌연변이 편향을 감소시킬 수 있는 것이 유리하다. 따라서, 일 구체예에서, 도입되는 돌연변이의 프로필에서 편향을 감소시키기 위해 불균등 농도의 dNTP를 사용하였다. 다른 구체예에서, 상기 방법은 낮은 편향 돌연변이 프로필로 돌연변이를 도입하는 방법이다.
본 출원은 불균등 농도의 dNTP를 사용하여 도입되는 돌연변이의 프로필에서 편향을 감소시키는 데 사용될 수 있음을 보여준다. 예를 들어, DNA 중합효소 (예를 들어, 전술한 저편향 DNA 중합효소)를 사용하여 타겟 핵산 분자를 돌연변이시키고, 다른 돌연변이에 비해 더 많은 수의 G에서 A로의 돌연변이를 도입하면, 사용자는 다른 dNTP와 비교하여 dATP의 농도를 감소시킬 수 있고, 이는 A 뉴클레오티드가 dGTP 대신 통합되는 빈도를 감소시키고 또한 G에서 A로의 돌연변이 수를 감소시킬 수 있다.
유사하게, 타겟 핵산 분자에 돌연변이를 도입할 때 뉴클레오티드 유사체가 사용되는 경우, dNTP의 상대적인 농도를 변경하여 돌연변이 프로필을 변경할 수 있다. 예를 들어, dPTP는 G에서 A로, C에서 T로, A에서 G로, T에서 C 로의 돌연변이를 도입하는 데 사용할 수 있다. 위에서 자세히 기재한 바와 같이, dPTP는 T 뉴클레오티드 또는 C 뉴클레오티를 대신할 수 있으며, dPTP가 아미노 또는 이미노 형태인지 여부에 따라, 후속적으로 A 뉴클레오티드 또는 G 뉴클레오티드와 쌍을 이룰 수 있다. 이는 두 가지 시나리오로 이어진다. 첫번째 시나리오에서, dPTP는 (예를 들어) 센스 가닥에서 T를 대체한 다음 (mutation step), 안티센스 가닥에서 A (돌연변이 없음) 또는 G (A에서 G 돌연변이)와 쌍을 이룰 수 있다. dPTP가 안티센스 가닥에서 T를 대체하고 G와 쌍을 이루는 경우, 돌연변이체 G는 C와 쌍을 이루어 센스 가닥의 복제물에서 T에서 C 돌연변이를 도입한다 (recovery step). 반대로, dPTP는 안티센스 가닥에서 T를 대체할 수 있으며, 이는 센스 가닥에서 A에서 G로의 돌연변이 및 안티센스 가닥의 복제물에서 T에서 C로의 돌연변이로 이어질 수 있다. 두번째 시나리오에서 dPTP는 (예를 들어) 센스 가닥에서 C를 대체한 다음, 안티센스 가닥에서 A (G에서 A 돌연변이) 또는 G (돌연변이 없음)와 쌍을 이룰 수 있다 (mutation step). dPTP가 안티센스 가닥에서 C를 대체하고 A와 쌍을 이루는 경우, 돌연변이체 A는 T와 쌍을 이루어 센스 가닥의 복제물에서 C에서 T 돌연변이를 도입한다 (recovery step). 반대로, dPTP는 안티센스 가닥에서 C를 대체할 수 있으며, 이는 센스 가닥에서 G에서 A로의 돌연변이 및 안티센스 가닥의 복제물에서 C에서 T로의 돌연변이로 이어질 수 있다.
본 출원은 G에서 A로 및 C에서 T로의 돌연변이 비율이 A에서 G로 및 T에서 C로의 돌연변이의 비율보다 높으면 다른 dNTP에 비해 (바람직하게는 dCTP의 농도와 비교하여) dTTP의 농도를 감소시키는 것은 dTTP 대신 dPTP가 통합되는 것을 장려하여, 두번째 시나리오에 비해 상술한 첫번째 시나리오의 경우를 증가시킴을 입증하였고, 이는 첫번째 시나리오에서 A에서 G로 및 T에서 C로의 돌연변이의 도입이 증가함을 의미한다. 유사하게, 본 출원은 회복 단계 (recovery step) 동안 dATP의 수준이 감소하면, G에서 A로, C에서 T로의 돌연변이 수준이 증가함을 입증하였다. 이는 상기 두번째 시나리오에서 dATP가 다른 dNTP에 비해 (바람직하게는 dGTP의 농도와 비교하여) 더 낮은 농도로 존재하는 경우 C 뉴클레오티드 대신 통합된 dPTP가 G와 더 자주 쌍을 이룰 것이고, G에서 A로 및 C에서 T로의 돌연변이 도입이 감소할 것임을 의미한다. 상기 두가지 시나리오를 도 7에 나타내었다.
본원에 개시된 저편향 DNA 중합효소조차도 타겟 핵산 분자에 돌연변이를 작은 편향으로 도입한다. 본 출원은 저편향 DNA 중합효소로 불균등 농도의 dNTP를 사용하면 사실상 모든 돌연변이 편향을 제거할 수 있음을 증명하였다.
본 출원에 제공된 정보에 기초하여, 다양한 dNTP의 농도를 변경하는 것이 뉴클레오티드 유사체가 사용되는지 여부에 따라 돌연변이 프로필에 어떻게 영향을 미칠 것인지를 결정하는 것은 당업자의 능력 내에 있다. 따라서, 일부 구체예에서, 불균등 농도의 dNTP를 사용하는 방법은 도입되는 돌연변이의 프로필에서 편향을 감소시키기 위해 수준이 증가 또는 감소되어야하는 dNTP를 확인하는 단계를 포함한다.
선택적으로, 불균등 농도의 dNTP는 다른 dNTP에 비해 낮은 농도의 dTTP를 포함한다. 전술한 바와 같이, 이는 dPTP가 뉴클레오티드 유사체로 사용될 때 도입되는 T에서 C로 및 A에서 G로의 돌연변이의 비율을 증가시킬 수 있다. 선택적으로, 상기 불균등 농도의 dNTP는 dTTP를 dATP, dCTP 또는 dGTP 농도의 75% 미만, 70% 미만, 60% 미만, 55% 미만, 25% 내지 75%, 25% 내지 70, 25% 내지 60%, 또는 약 50%의 농도로 포함한다. 선택적으로, 상기 불균등 농도의 dNTP는 dTTP를 dCTP 농도의 60% 미만의 농도로 포함한다. 선택적으로, 상기 불균등 농도의 dNTP는 dTTP를 dCTP 농도의 25% 내지 60%의 농도로 포함한다.
선택적으로, 불균등 농도의 dNTP는 다른 dNTP에 비해 낮은 농도의 dATP를 포함한다. 전술한 바와 같이, 이는 dPTP가 뉴클레오티드 유사체로 사용될 때 도입되는 G에서 A로 및 C에서 T로의 돌연변이의 비율을 감소시킬 수 있다. 선택적으로, 상기 불균등 농도의 dNTP는 dATP를 dTTP, dCTP 또는 dGTP 농도의 75% 미만, 70% 미만, 60% 미만, 55% 미만, 25% 내지 75%, 25% 내지 70, 25% 내지 60%, 또는 약 50%의 농도로 포함한다. 선택적으로, 상기 불균등 농도의 dNTP는 dATP를 dGTP 농도의 75% 미만, 70% 미만, 60% 미만, 55% 미만, 25% 내지 75%, 25% 내지 70, 25% 내지 60%, 또는 약 50%의 농도로 포함한다. 선택적으로, 상기 불균등 농도의 dNTP는 dATP를 dGTP 농도의 60% 미만의 농도로 포함한다. 선택적으로, 상기 불균등 농도의 dNTP는 dATP를 dGTP 농도의 25% 내지 60%의 농도로 포함한다.
위의 두 시나리오에 기재된 바와 같이, dPTP를 뉴클레오티드 유사체로 사용할 때 dTTP를 감소시키면 타겟 핵산 분자의 T 뉴클레오티드를 dPTP로 대체하도록 장려하여 T에서 C로 및 A에서 G로의 돌연변이가 증가한다. 따라서, 다른 dNTP보다 더 낮은 농도의 dTTP를 포함하는 불균등 농도의 dNTP는 바람직하게는 돌연변이 유발 단계 (예를 들어 dPTP의 존재 하의 PCR 단계)에서 사용된다. 유사하게, dPTP를 뉴클레오티드 유사체로 사용할 때 dATP를 감소시키면 C 뉴클레오티드를 대체하고 dATP와 쌍을 이루는 dPTP의 수가 감소하여 G에서 A로, C에서 T로의 돌연변이가 증가한다. dATP 및 dPTP의 짝지음 (pairing)은 복구 단계 (recovery step)에서 발생하는 경향이 있으므로, 복구 단계에서 dATP를 감소시키면 G에서 A로, C에서 T로의 돌연변이 수가 증가한다. 따라서, 선택적으로, 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하거나 뉴클레오티드 유사체의 부재 하에 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 불균등 농도의 dNTP를 사용하여 수행된다. 불균등 농도의 dNTP는 다른 dNTP에 비해 낮은 농도의 dATP를 포함한다.
실시예
실시예 1 - 다른 중합효소의 PrimeStar GXL을 사용한 핵산 분자 돌연변이화
DNA 분자를 적절한 크기 (예를 들어, 10kb)로 단편화 (fragmented)하고 태그맨테이션(tagmentation)을 사용하여 각 말단에 정의된 서열 프라이밍 부위 (어댑터)를 부착하였다.
첫번째 단계는 DNA를 단편화하기 위한 태그맨테이션이다. 4μl 이하 부피의 하나 이상의 박테리아 균주에 포함된 50ng의 고 분자량 게놈 DNA를 다음 조건 하에서 태그맨테이션하였다. 50ng DNA를 4μl Nextera Transposase (1:50으로 희석), 및 8μl 2X 태그맨테이션 버퍼 (20mM Tris [pH7.6], 20mM MgCl, 20% (v/v) 디메틸포름아미드)와 총 부피 16μl로 혼합하였다. 반응물 (reaction)을 55℃에서 5분 동안 배양하고, 4μl의 NT 버퍼 (또는 0.2% SDS)를 반응물에 첨가하고 상기 반응물을 실온에서 5분 동안 배양하였다.
상기 태그맨테이션 반응물을 0.6 부피의 비드를 사용하는 left side size selection에 대한 제조업체의 지침에 따라 SPRIselect 비드 (Beckman Coulter)를 사용하여 세척하고, DNA를 분자생물학용수(molecular grade water) 중에 용출하였다.
그 다음 제한된 6회 사이클 동안 표준 dNTP 및 dPTP의 조합과 함께 PCR을 수행하였다. Primestar GXL를 사용하여, 12.5ng의 태그맨테이션 및 정제된 DNA를 1 x GXL 버퍼, dATP, dTTP, dGTP 및 dCTP 각각 200μM, 0.5mM dPTP 및 0.4μM 맞춤형 프라이머들을 포함하는 부피 25μl의 총 반응물에 추가하였다 (표 2).
| i7 맞춤형 인덱스 프라이머 | CAAGCAGAAGACGGCATACGAGAT | NNNNNN | XXXXXX | GTCTCGTGGGCTCGG |
| i5 맞춤형 인덱스 프라이머 | AATGATACGGCGACCACCGAGATCTACAC | XXXXXX | NNNNNN | TCGTCGGCAGCGTC |
표 2. 10kbp 주형에서 mutagenesis PCR에 사용되는 맞춤형 프라이머. XXXXXX는 정의되는, 샘플-특이적 6-8nt의 바코드 서열이다. NNNNNN은 무작위 뉴클레오티드의 6nt 영역이다.
상기 반응물은 Primestar GXL의 존재 하에 다음과 같은 열 순환 (thermal cycling)이 수행된다. 68℃에서 3분 동안 초기 갭 연장 (extention) 후, 98℃에서 10초, 55℃에서 15초 및 68℃에서 10분으로 6회 사이클을 수행하였다.
다음 단계는 주형에서 dPTP를 제거하고 이를 전이 (transition) 돌연변이로 대체하기 위한 ("recovery PCR") dPTP 없는 PCR이다. PCR 반응물을 SPRIselect 비드로 세척하여 과량의 dPTP 및 프라이머들을 제거한 다음, dPTP 포함 사이클 (dPTP incorporation cycles) 동안 도입된 단편 말단에 어닐링하는 프라이머들을 사용하여 추가적으로 10회 (최소 1회, 최대 20회)의 증폭을 수행하였다 (표 3).
| i7 flow cell 프라이머 | CAAGCAGAAGACGGCATACGA |
| i5 flow cell 프라이머 | AATGATACGGCGACCACCGA |
이어서, 예를 들어 7-10 kb의 원하는 크기 범위에서 증폭되고 돌연변이된 단편의 크기를 선택하기 위한 겔 추출 단계를 수행하였다. 겔 추출은 수동 또는 BluePippin과 같은 자동화 시스템을 통해 수행할 수 있다. 이어서 16-20 사이클의 추가 PCR ("enrichment PCR") 단계를 수행하였다.
정의된 개수의 긴 돌연변이 된 주형을 증폭한 후, 주형의 무작위 단편화 (random fragmentation)를 수행하여 시퀀싱을 위한 겹치는 (overlapping) 더 짧은 단편 그룹을 생성하였다. 단편화는 태그맨테이션에 의해 수행되었다.
PCR 증폭을 위해 상기 반응물을 3개의 풀 (pool)로 나눈 것을 제외하고는, 이전 단계의 긴 DNA 단편에 대하여 표준 태그맨테이션 반응 (예를 들어, Nextera XT 또는 Nextera Flex)을 수행하였다. 이는 원래 주형 (샘플 바코드 포함)의 각 말단에서 유래된 단편뿐만 아니라 양 말단에서 새롭게 태그맨테이션된 긴 주형의 내부 (internal) 단편을 선택적으로 증폭할 수 있게 한다. 이를 통해 Illumina 기기 (예를 들어, MiSeq 또는 HiSeq)에서의 시퀀싱을 위한 3개의 풀을 효과적으로 생성하였다.
상기 방법은 표준 Taq (Jena Biosciences)와 Taq 및 LongAmp (New England Biolabs)라는 교정 중합효소 (DeepVent)의 혼합물을 사용하여 반복하였다.
이 실험에서 얻은 데이터를 도 1에 나타내었다. 대조군에서는 dPTP가 사용되지 않았다. 리드들 (Reads)은 E. coli 게놈에 대해 맵핑되었으며, 중간값 약 8%의 돌연변이율이 달성되었다.
실시예 2 - 서로 다른 DNA 중합효소의 돌연변이 빈도 비교
다양한 DNA 중합효소로 돌연변이를 유발시켰다 (표 4). 실시예 1의 방법에 기재된 바와 같이 E. coli 균주 MG1655의 게놈 DNA를 태그맨테이션하여 긴 단편을 생성하고 비드로 세척하였다. 이어서, 0.5 mM dPTP, SPRIselect 비드 정제 및 비드 정제의 존재 하에 6 사이클의 "mutagenesis PCR" 및 dPTP 부재 하에 추가적인 14-16 사이클의 "recovery PCR"을 수행하였다. 생성된 긴 돌연변이된 주형에 대하여 표준 태그맨테이션 반응을 수행하고 (실시예 1 참조) Illumina MiSeq 기기에서 "내부" 단편을 증폭 및 시퀀싱하였다.
돌연변이율은 표 4에 나타내었으며, 알려진 참조 게놈 (reference genome)에서 DNA의 Illumina 시퀀싱을 사용하여 측정된 dPTP 돌연변이 유발 반응을 통한 염기 치환 빈도를 표준화하였다. Taq 중합효소의 경우, 써모코커스 (Thermococcus) 중합효소에 최적화된 버퍼에서 사용하는 경우에도, 주형 G+C 부위에서 ~12%의 돌연변이만이 발생하였다. 써모코커스-유사 (Thermococcus-like) 중합효소는 주형 G+C 부위에서 58~69%의 돌연변이를 유발하는 반면, Pyrococcus에서 유래된 중합효소는 주형 G+C 부위에서 88%의 돌연변이를 발생시킨다.
효소들은 Jena Biosciences (Taq), Takara (Primestar 변이체), Merck Millipore (KOD DNA 중합효소) 및 New England Biolabs (Phusion)에서 제공받았다.
이 실험에서 Taq는 제공된 버퍼, 및 Primestar GXL 버퍼 (Takara)로 테스트하였다. 다른 모든 반응은 각 중합효소에 대해 표준 공급된 버퍼를 사용하여 수행하였다.
| 돌연변이 비율 (관찰된 총 돌연변이에 대한 %) | ||||||
| 중합효소 1 | 기원 | A → G | T → C | G → A | C → T | 그 외 (전환) |
| Taq (표준 버퍼) | Thermus aquaticus | 43.1 | 41.7 | 6.3 | 6.1 | 2.7 |
| Taq (써모코커스 버퍼2) |
Thermus aquaticus | 48.9 | 47.5 | 2.9 | 0.7 | 0.0 |
| Primestar GXL | Thermococcus | 21.5 | 20.1 | 29.5 | 28.9 | 0.0 |
| Primestar HS | Thermococcus | 16.3 | 15.2 | 30.1 | 38.4 | 0.0 |
| Primestar Max | Thermococcus | 16.5 | 14.6 | 33.2 | 35.7 | 0.0 |
| KOD DNA polymerase | Thermococcus | 20.5 | 16.1 | 31.8 | 31.5 | 0.0 |
| Phusion | Pyrococcus | 5.4 | 6.4 | 44.1 | 44.1 | 0.0 |
실시예 3 - dPTP 돌연변이 유발 비율의 결정
단일 세트의 반응 조건 하에서 써모코커스 (Thermococcus) 중합효소 (Primestar GXL; Takara)를 사용하여 G+C 함량 수준이 다른 (33-66%) 다양한 게놈 DNA 샘플에서 dPTP 돌연변이 유발을 수행하였다. "recovery PCR"의 10회 사이클을 수행한 것을 제외하고는, 실시예 3의 방법에 기재된 바와 같이 돌연변이 유발 및 시퀀싱을 수행하였다. 예측한 바와 같이, 돌연변이 비율은 G+C 함량의 다양성에도 불구하고 샘플 간에 거의 비슷하였다 (비율 중간값 7-8%) (도 2).
실시예 4 - 주형 증폭 편향의 측정
주형 증폭 편향 (Template amplification bias)은 두 가지 중합효소: Illumina 시퀀싱 프로토콜에서 일반적으로 사용되는 교정 중합효소 인 Kapa HiFi, 및 긴 단편을 증폭하는 능력으로 알려진 KOD 패밀리 중합효소 인 PrimeStar GXL에 대해 측정하였다. 첫번째 실험에서 Kapa HiFi를 크기가 약 2kbp인 제한된 개수의 E. coli 게놈 DNA 주형 증폭에 사용하였었다. 그 다음, 이들 증폭된 단편의 말단을 시퀀싱하였다. 유사한 실험을 E. coli로부터의 약 7-10kbp의 단편에 대해 PrimeStar GXL로 수행하였다. 각 말단 서열 리드의 위치를 E. coli 참조 게놈에 대해 맵핑하여 결정하였다. 인접한 단편 말단 사이의 거리를 측정하였다. 이들 거리를 균일 분포에서 무작위로 샘플링된 거리 세트와 비교하였다. 비교는 비모수 (nonparametric) Kolmolgorov-Smirnov 검정, D를 통해 수행하였다. 두 샘플이 동일한 분포에서 유래하는 경우, D값은 0에 가까워진다. 저편향 PrimeStar 중합효소의 경우, 50,000개 게놈 위치의 균일한 무작위 샘플과 비교할 때, 50,000개의 단편 말단에서 측정 시 D=0.07임을 확인하였다. Kapa HiFi 중합효소의 경우 50,000개의 단편 말단에서 D=0.14임을 확인하였다.
실시예 5 - 더 긴 주형의 우선적인 증폭을 위한 2개의 동일한 프라이머 결합 부위 및 단일 프라이머 서열의 사용
전술한 바와 같이, 태그맨테이션은 DNA 분자를 단편화하고 단편의 말단에 프라이머 결합 부위 (어댑터)를 동시에 도입하기 위해 사용할 수 있다. Nextera 태그맨테이션 시스템 (Illumina)은 2개의 고유한 어댑터 (여기에서 X 및 Y라고 함) 중 하나가 로딩된 트랜스포사제 (transposase) 효소를 사용한다. 이는 일부는 동일한 말단 서열 (X-X, Y-Y)이 있고 일부는 고유한 말단 (X-Y)이 있는 산물 (products)의 무작위 혼합물을 생성한다. 표준 Nextera 프로토콜은 각 말단에 서로 다른 어댑터를 포함하는 "X-Y" 산물을 선택적으로 증폭하기 위해 2개의 구별되는 프라이머 서열을 사용한다 (Illumina 기술을 사용한 시퀀싱에 필요). 그러나, 단일 프라이머 서열을 사용하여 동일한 말단 어댑터로 "X-X" 또는 "Y-Y" 단편을 증폭할 수도 있다.
동일한 말단 어댑터를 포함하는 긴 돌연변이된 주형을 생성하기 위해, 50ng의 고 분자량 게놈 DNA (E. coli 균주 MG1655)를 먼저 태그맨테이션한 다음 실시예 1에 기재된 바와 같이 SPRIselect 비드로 세척하였다. 이어서, 표준 dNTP와 dPTP의 조합으로 "mutagenesis PCR" 5회 사이클을 수행하였으며, 이는 단일 프라이머 서열을 사용한 것을 제외하고는 실시예 1에 자세히 설명된 대로 수행하였다 (표 5).
PCR 반응물을 SPRIselect 비드로 세척하여 과잉 dPTP 및 프라이머들을 제거한 다음, dPTP 부재 하에 추가적으로 10회 사이클의 "recovery PCR"을 수행하여 주형의 예?媤? 전이 돌연변이로 대체하였다. Recovery PCR은 dPTP 포함 사이클 동안 도입된 단편 말단에 어닐링하는 단일 프라이머를 사용하여 수행하였고, 그에 따라 이전 PCR 단계에서 생성된 돌연변이된 주형의 선택적인 증폭을 가능하게 하였다.
| 프라이머 이름 | 단계 | 서열 | |||
| single_mut | mutagenesis | TCGGTCTGCGCCTCTAGC | NNN | XXXXXXXXXXXXX | GTCTCGTGGGCTCGGAG |
| single_rec | recovery | CAAGCAGAAGACGGCATACGAGAT | TCGGTCTGCGCCTCTAGC | ||
표 5. 양 말단에서 동일한 기본 어댑터 구조가 있는 돌연변이된 주형을 생성하기 위해 사용된 프라이머. 프라이머 "single_mut"은 Nextera 태그맨테이션에 의해 생성된 DNA 단편에 대한 mutagenesis PCR에 사용되었다. 상기 프라이머는 단편 말단에 추가적인 프라이머 결합 부위를 도입하는 5' 부분을 포함한다. 프라이머 "single_rec"은 이 부위에 어닐링 (annealing)할 수 있고, recovery PCR 동안 single_mut 프라이머로 생성된 돌연변이된 주형을 선택적으로 증폭하는데 사용되었다. XXXXXXXXXXXXX는 정의된, 샘플-특이적 13nt의 바코드 서열이다. NNN은 무작위 뉴클레오티드의 3nt 영역이다.
대조군으로서, mutagenesis PCR (표 2에 나타냄) 및 recovery PCR (표 3) 둘 다 동안에 2개의 별개의 프라이머 서열을 사용한 것을 제외하고는, 각 말단에 서로 다른 어댑터를 가진 돌연변이된 주형을 전술한 것과 동일한 프로토콜을 사용하여 생성하였다. 최종 PCR 산물은 SPRIselect 비드로 세척하고 2100 Bioanalzyer System (Agilent)을 사용하여 고감도 DNA 칩에서 분석하였다. 도 xxx에 나타낸 바와 같이, 동일한 말단 어댑터로 생성된 주형은 이중 어댑터 (dual adapters)를 포함하는 대조군 샘플보다 평균적으로 현저히 길었다. 대조군 주형은 최소 ~800bp 크기까지 감지될 수 있었던 반면, 단일 어댑터 샘플의 경우 2000bp 미만의 주형이 관찰되지 않았다.
동일한 말단 어댑터 (파란색)가 있는 돌연변이된 주형과 이중 어댑터가 있는 대조군 주형을 Agilent 2100 Bioanalyzer (고감도 DNA 키트)에 러닝하여 크기 프로필 (profile)을 비교하였다. 동일한 말단 어댑터의 사용은 < 2kbp의 주형의 증폭을 억제하였다. 상기 데이터를 도 6에 나타내었다.
실시예 8 - PCR 동안 자연적인 dNTP 수준을 변경에 의한,
써모코커스
중합효소의 돌연변이 편향의 추가적인 감소
써모코커스 중합효소는 다른 DNA 중합효소에 비해 훨씬 더 균형 잡힌 돌연변이 프로필을 생성하지만, G 및 C 부위에서는 돌연변이에 대한 적은 양의 편향을 보인다 (표 4 참조). 이러한 남아 있는 편향을 제거하기 위해, mutagenesis 및 recovery PCR 단계 동안 자연적인 dNTP의 농도를 변경하여 다른 뉴클레오티드의 상대적 포함 비율 (relative incorporation rates)에 영향을 미치는 효과를 테스트하였다.
먼저, PCR 반응에서의 개별 뉴클레오티드 농도가 다양하다는 점을 제외하고는, 실시예 5에 약술된 접근법을 사용하여 박테리아 게놈 DNA (E. coli 균주 MG1655)로부터 긴 돌연변이된 주형을 제조하였다. 이는 4개의 천연 뉴클레오티드 (New England Biolabs에서 구입)의 개별 용액을 표준 최종 농도 200μM 또는 160μM (표준 대비 80%) 또는 100μM (50%)의 더 낮은 농도로 PCR 혼합물에 각각 추가하여 수행하였다. 반응 당 하나의 뉴클레오티드만을 변경하였다. 대조군으로서, Primestar GXL 중합 효소 (Takara)가 제공된 등몰 (equimolar) dNTP 혼합물을 사용하여, 모든 천연 뉴클레오티드를 동일한 최종 농도인 200μM로 첨가하였다. 표 5에 나타낸 프라이머들을 사용하여 5회 사이클의 mutagenesis PCR 및 12회 사이클의 recovery PCR을 수행하였다. 다음으로, 생성된 긴 돌연변이된 주형에 대하여 표준 태그맨테이션 (실시예 1 참조)을 수행하고 Illumina MiSeq 기기에서 "내부" 단편을 증폭 및 시퀀싱하였다. 돌연변이 빈도는 알려진 참조 서열과 비교하여 결정하였다.
표 6에 나타낸 바와 같이 mutagenesis 및/또는 recovery PCR 동안 개별 dNTP의 농도 변화는 관찰된 돌연변이 프로필을 변경시켰다. 중요한 것은, 돌연변이 유발 동안 dTTP의 양을 50%로 제한하면 각 뉴클레오티드에 대해 사실상 동일한 돌연변이 빈도를 생성함을 발견하였다(표 3). 이는, 써모코커스 중합효소에 남아 있는 돌연변이 편향이 dNTP 수준의 변화를 통해 제거될 수 있음을 확인시켜준다.
|
돌연변이 빈도
(관찰된 총 돌연변이에 대한 %) |
||||
| 처리 (Treatment ) | A → G | T → C | G →A | C → T |
| 등몰 (Equimolar) dNTP 대조군 |
17.4 | 16.8 | 32.1 | 33.7 |
| 80% dTTP (mutagenesis) | 13.9 | 13.8 | 36.1 | 36.2 |
| 50% dTTP (mutagenesis) | 23.7 | 24.9 | 25.3 | 26.2 |
| 80% dATP (recovery) | 13.4 | 12.5 | 36.7 | 37.3 |
| 50% dATP (recovery) | 18.9 | 19.1 | 31.4 | 30.6 |
| 80% dTTP (mutagenesis) 및 80% dATP (recovery) |
17.8 | 15.0 | 34.0 | 33.2 |
| 50% dTTP (mutagenesis) 및50% dATP (recovery) | 34.4 | 34.7 | 15.4 | 15.5 |
<110> LONGAS TECHNOLOGIES PTY LTD
<120> METHOD FOR INTRODUCING MUTATIONS
<130> PM037854
<140> TBC
<141> 2019-02-19
<150> GB 1802744.1
<151> 2018-02-20
<160> 136
<170> PatentIn version 3.5
<210> 1
<211> 2325
<212> DNA
<213> Artificial Sequence
<220>
<223> DNA polymerase from Thermococcus sp. KS-1
<400> 1
atgatcctcg acactgacta cataactgag aatggaaaac ccgtcataag gattttcaag 60
aaggagaacg gcgagtttaa gattgagtac gataggactt ttgaacccta catttacgcc 120
ctcctgaagg acgattctgc cattgaggag gtcaagaaga taaccgccga gaggcacgga 180
acggttgtaa cggttaagcg ggctgaaaag gttcagaaga agttcctcgg gagaccagtt 240
gaggtctgga aactctactt tactcaccct caggacgtcc cagcgataag ggacaagata 300
cgagagcatc cagcagttat tgacatctac gagtacgaca tacccttcgc caagcgctac 360
ctcatagaca agggattagt gccaatggaa ggcgacgagg agctgaaaat gcttgccttt 420
gatatcgaga cgctctacca tgagggcgag gagttcgccg aggggccaat ccttatgata 480
agctacgccg acgaggaagg ggccagggtg ataacgtgga agaacgcgga tctgccctac 540
gttgacgtcg tctcgacgga gagggagatg ataaagcgct tcctaaaggt ggtcaaagag 600
aaagatcctg acgtcctaat aacctacaac ggcgacaact tcgacttcgc ctacctaaaa 660
aaacgctgtg aaaagcttgg aataaacttc acgctcggaa gggacggaag cgagccgaag 720
attcagagga tgggcgacag gtttgccgtc gaagtgaagg gacggataca cttcgatctc 780
tatcctgtga taagacggac gataaacctg cccacataca cgcttgaggc cgtttatgaa 840
gccgtcttcg gtcagccgaa ggagaaggtc tacgctgagg agatagctac agcttgggag 900
agcggtgaag gccttgagag agtagccaga tactcgatgg aagatgcgaa ggtcacatac 960
gagcttggga aggagttttt ccctatggag gcccagcttt ctcgcttaat cggccagtcc 1020
ctctgggacg tctcccgctc cagcactggc aacctcgttg agtggttcct cctcaggaag 1080
gcctacgaga ggaatgagct ggccccgaac aagcccgatg aaaaggagct ggccagaaga 1140
cgacagagct atgaaggagg ctatgtaaaa gagcccgaga gagggttgtg ggagaacata 1200
gtgtacctag attttagatc tctgtacccc tcaatcatca tcacccacaa cgtctcgccg 1260
gatactctca acagggaagg atgcaaggaa tatgacgttg ccccccaggt cggtcaccgc 1320
ttctgcaagg acttcccagg atttatcccg agcctgcttg gagacctcct agaggagagg 1380
cagaagataa agaagaagat gaaggccacg attgacccga tcgagaggaa gctcctcgat 1440
tacaggcaga gggccatcaa gatcctggcc aacagctact acggttacta cggctatgca 1500
agggcgcgct ggtactgcaa ggagtgtgca gagagcgtaa cggcctgggg aagggagtac 1560
ataacgatga ccatcagaga gatagaggaa aagtacggct ttaaggtaat ctacagcgac 1620
accgacggat tttttgccac aatacctgga gccgatgctg aaaccgtcaa aaagaaggcg 1680
atggagttcc tcaagtatat caacgccaaa ctcccgggcg cgcttgagct cgagtacgag 1740
ggcttctaca aacgcggctt cttcgtcacg aagaagaagt acgcggtgat agacgaggaa 1800
ggcaagataa caacgcgcgg acttgagatt gtgaggcgcg actggagcga gatagcgaaa 1860
gagacgcagg cgagggttct tgaagctttg ctaaaggacg gtgacgtcga gaaggccgtg 1920
aggatagtca aagaagttac cgaaaagctg agcaagtacg aggttccgcc ggagaagctg 1980
gtgatccacg agcagataac gagggattta aaggactaca aggcaaccgg tccccacgtt 2040
gccgttgcca agaggttggc cgcgagagga gtcaaaatac gccctggaac ggtgataagc 2100
tacatcgtgc tcaagggctc tgggaggata ggcgacaggg cgataccgtt cgacgagttc 2160
gacccgacga agcacaagta cgacgccgag tactacattg agaaccaggt tctcccagcc 2220
gttgagagaa ttctgagagc cttcggttac cgcaaggaag acctgcgcta ccagaagacg 2280
agacaggttg gtctgggagc ctggctgaag ccgaagggaa cttga 2325
<210> 2
<211> 774
<212> PRT
<213> Artificial Sequence
<220>
<223> DNA polymerase from Thermococcus sp. KS-1
<400> 2
Met Ile Leu Asp Thr Asp Tyr Ile Thr Glu Asn Gly Lys Pro Val Ile
1 5 10 15
Arg Ile Phe Lys Lys Glu Asn Gly Glu Phe Lys Ile Glu Tyr Asp Arg
20 25 30
Thr Phe Glu Pro Tyr Ile Tyr Ala Leu Leu Lys Asp Asp Ser Ala Ile
35 40 45
Glu Glu Val Lys Lys Ile Thr Ala Glu Arg His Gly Thr Val Val Thr
50 55 60
Val Lys Arg Ala Glu Lys Val Gln Lys Lys Phe Leu Gly Arg Pro Val
65 70 75 80
Glu Val Trp Lys Leu Tyr Phe Thr His Pro Gln Asp Val Pro Ala Ile
85 90 95
Arg Asp Lys Ile Arg Glu His Pro Ala Val Ile Asp Ile Tyr Glu Tyr
100 105 110
Asp Ile Pro Phe Ala Lys Arg Tyr Leu Ile Asp Lys Gly Leu Val Pro
115 120 125
Met Glu Gly Asp Glu Glu Leu Lys Met Leu Ala Phe Asp Ile Glu Thr
130 135 140
Leu Tyr His Glu Gly Glu Glu Phe Ala Glu Gly Pro Ile Leu Met Ile
145 150 155 160
Ser Tyr Ala Asp Glu Glu Gly Ala Arg Val Ile Thr Trp Lys Asn Ala
165 170 175
Asp Leu Pro Tyr Val Asp Val Val Ser Thr Glu Arg Glu Met Ile Lys
180 185 190
Arg Phe Leu Lys Val Val Lys Glu Lys Asp Pro Asp Val Leu Ile Thr
195 200 205
Tyr Asn Gly Asp Asn Phe Asp Phe Ala Tyr Leu Lys Lys Arg Cys Glu
210 215 220
Lys Leu Gly Ile Asn Phe Thr Leu Gly Arg Asp Gly Ser Glu Pro Lys
225 230 235 240
Ile Gln Arg Met Gly Asp Arg Phe Ala Val Glu Val Lys Gly Arg Ile
245 250 255
His Phe Asp Leu Tyr Pro Val Ile Arg Arg Thr Ile Asn Leu Pro Thr
260 265 270
Tyr Thr Leu Glu Ala Val Tyr Glu Ala Val Phe Gly Gln Pro Lys Glu
275 280 285
Lys Val Tyr Ala Glu Glu Ile Ala Thr Ala Trp Glu Ser Gly Glu Gly
290 295 300
Leu Glu Arg Val Ala Arg Tyr Ser Met Glu Asp Ala Lys Val Thr Tyr
305 310 315 320
Glu Leu Gly Lys Glu Phe Phe Pro Met Glu Ala Gln Leu Ser Arg Leu
325 330 335
Ile Gly Gln Ser Leu Trp Asp Val Ser Arg Ser Ser Thr Gly Asn Leu
340 345 350
Val Glu Trp Phe Leu Leu Arg Lys Ala Tyr Glu Arg Asn Glu Leu Ala
355 360 365
Pro Asn Lys Pro Asp Glu Lys Glu Leu Ala Arg Arg Arg Gln Ser Tyr
370 375 380
Glu Gly Gly Tyr Val Lys Glu Pro Glu Arg Gly Leu Trp Glu Asn Ile
385 390 395 400
Val Tyr Leu Asp Phe Arg Ser Leu Tyr Pro Ser Ile Ile Ile Thr His
405 410 415
Asn Val Ser Pro Asp Thr Leu Asn Arg Glu Gly Cys Lys Glu Tyr Asp
420 425 430
Val Ala Pro Gln Val Gly His Arg Phe Cys Lys Asp Phe Pro Gly Phe
435 440 445
Ile Pro Ser Leu Leu Gly Asp Leu Leu Glu Glu Arg Gln Lys Ile Lys
450 455 460
Lys Lys Met Lys Ala Thr Ile Asp Pro Ile Glu Arg Lys Leu Leu Asp
465 470 475 480
Tyr Arg Gln Arg Ala Ile Lys Ile Leu Ala Asn Ser Tyr Tyr Gly Tyr
485 490 495
Tyr Gly Tyr Ala Arg Ala Arg Trp Tyr Cys Lys Glu Cys Ala Glu Ser
500 505 510
Val Thr Ala Trp Gly Arg Glu Tyr Ile Thr Met Thr Ile Arg Glu Ile
515 520 525
Glu Glu Lys Tyr Gly Phe Lys Val Ile Tyr Ser Asp Thr Asp Gly Phe
530 535 540
Phe Ala Thr Ile Pro Gly Ala Asp Ala Glu Thr Val Lys Lys Lys Ala
545 550 555 560
Met Glu Phe Leu Lys Tyr Ile Asn Ala Lys Leu Pro Gly Ala Leu Glu
565 570 575
Leu Glu Tyr Glu Gly Phe Tyr Lys Arg Gly Phe Phe Val Thr Lys Lys
580 585 590
Lys Tyr Ala Val Ile Asp Glu Glu Gly Lys Ile Thr Thr Arg Gly Leu
595 600 605
Glu Ile Val Arg Arg Asp Trp Ser Glu Ile Ala Lys Glu Thr Gln Ala
610 615 620
Arg Val Leu Glu Ala Leu Leu Lys Asp Gly Asp Val Glu Lys Ala Val
625 630 635 640
Arg Ile Val Lys Glu Val Thr Glu Lys Leu Ser Lys Tyr Glu Val Pro
645 650 655
Pro Glu Lys Leu Val Ile His Glu Gln Ile Thr Arg Asp Leu Lys Asp
660 665 670
Tyr Lys Ala Thr Gly Pro His Val Ala Val Ala Lys Arg Leu Ala Ala
675 680 685
Arg Gly Val Lys Ile Arg Pro Gly Thr Val Ile Ser Tyr Ile Val Leu
690 695 700
Lys Gly Ser Gly Arg Ile Gly Asp Arg Ala Ile Pro Phe Asp Glu Phe
705 710 715 720
Asp Pro Thr Lys His Lys Tyr Asp Ala Glu Tyr Tyr Ile Glu Asn Gln
725 730 735
Val Leu Pro Ala Val Glu Arg Ile Leu Arg Ala Phe Gly Tyr Arg Lys
740 745 750
Glu Asp Leu Arg Tyr Gln Lys Thr Arg Gln Val Gly Leu Gly Ala Trp
755 760 765
Leu Lys Pro Lys Gly Thr
770
<210> 3
<211> 2325
<212> DNA
<213> Artificial Sequence
<220>
<223> DNA polymerase from Thermococcus celer
<400> 3
atgatcctcg acgctgacta catcaccgaa gatgggaagc ccgtcgtgag gatattcagg 60
aaggagaagg gcgagttcag aatcgactac gacagggact tcgagcccta catctacgcc 120
ctcctgaagg acgattcggc catcgaggag gtgaagagga taaccgttga gcgccacggg 180
aaggccgtca gggttaagcg ggtggagaag gtcgaaaaga agttcctcaa caggccgata 240
gaggtctgga agctctactt caatcacccg caggacgttc cggcgataag ggacgagata 300
aggaagcatc cggccgtcgt tgatatctac gagtacgaca tccccttcgc caagcgctac 360
ctcatcgata aggggctcgt cccgatggag ggggaggagg agctcaaact gatggccttc 420
gacatcgaga ccctctacca cgagggagac gagttcgggg aggggccgat cctgatgata 480
agctacgccg acggggacgg ggcgagggtc ataacctgga agaagatcga cctcccctac 540
gtcgacgtcg tctcgaccga gaaggagatg ataaagcgct tcctccaggt ggtgaaggag 600
aaggacccgg acgtgctcgt aacttacaac ggcgacaact tcgacttcgc ctacctgaag 660
agacgctccg aggagcttgg attgaagttc atcctcggga gggacgggag cgagcccaag 720
atccagcgca tgggcgaccg cttcgccgtc gaggtgaagg ggaggataca cttcgacctc 780
tacccggtga taaggcgcac cgtgaacctg ccgacctaca cgctcgaggc ggtctacgag 840
gccatcttcg ggaggccaaa ggagaaggtc tacgccgggg agatagtgga ggcctgggaa 900
accggcgagg gtcttgagag ggttgcccgc tactccatgg aggacgcaaa ggttaccttc 960
gagctcggga gggagttctt cccgatggag gcccagctct cgaggctcat cggccagggt 1020
ctctgggacg tctcccgctc gagcaccggc aacctggtcg agtggttcct cctgaggaag 1080
gcctacgaga ggaacgaact ggccccgaac aagccgagcg gccgggaagt ggagatcagg 1140
aggcgtggct acgccggtgg ttacgttaag gagccggaga ggggtttatg ggagaacatc 1200
gtgtacctcg actttcgctc tctttacccc tccatcatca taacccacaa cgtctcgccc 1260
gataccctaa acagggaggg ctgtgagaac tacgacgtcg ccccccaggt ggggcataag 1320
ttctgcaaag attttccggg cttcatcccg agcctgctcg gaggcctgct tgaggagagg 1380
cagaagataa agcggaggat gaaggcctct gtggatcccg ttgagcggaa gctcctcgat 1440
tacaggcaga gggccatcaa gatactggcc aacagcttct acggatacta cggctacgcg 1500
agggcgaggt ggtactgcag ggagtgcgcg gagagcgtta ccgcctgggg cagggagtac 1560
atcgataggg tcatcaggga gctcgaggag aagttcggct tcaaggtgct ctacgcggac 1620
acggacggac tgcacgccac gatccccggg gcggacgccg ggaccgtcaa ggagagggcg 1680
agggggttcc tgagatacat caaccccaag ctccccggcc tcctggagct cgagtacgag 1740
gggttctacc tgaggggttt cttcgtgacg aagaagaagt acgcggtcat agacgaggag 1800
ggcaagataa ccacgcgcgg cctcgagata gtcaggcggg actggagcga ggtggccaag 1860
gagacgcagg cgagggtcct ggaggcgata ctgaggcacg gtgacgtcga ggaggccgtt 1920
agaatcgtca gggaggtaac cgaaaagctg agcaagtacg aggttccgcc ggagaaactg 1980
gtgatccacg agcagataac gagggatttg agggactaca aagccacggg accgcacgtg 2040
gcggtggcga agcgcctggc cgggaggggg gtaaggatac gccccgggac ggtgataagc 2100
tacatcgtcc tcaagggctc cggaaggata ggggacaggg cgattccctt cgacgagttc 2160
gacccgacta agcacaggta cgacgccgac tactacatcg agaaccaggt tctgccagcc 2220
gtcgagagga tcctgaaggc cttcggctac cgcaaggagg acctgaaata ccagaagacg 2280
aggcaggtgg gcctgggtgc gtggctcaac gcggggaagg ggtga 2325
<210> 4
<211> 774
<212> PRT
<213> Artificial Sequence
<220>
<223> DNA polymerase from Thermococcus celer
<400> 4
Met Ile Leu Asp Ala Asp Tyr Ile Thr Glu Asp Gly Lys Pro Val Val
1 5 10 15
Arg Ile Phe Arg Lys Glu Lys Gly Glu Phe Arg Ile Asp Tyr Asp Arg
20 25 30
Asp Phe Glu Pro Tyr Ile Tyr Ala Leu Leu Lys Asp Asp Ser Ala Ile
35 40 45
Glu Glu Val Lys Arg Ile Thr Val Glu Arg His Gly Lys Ala Val Arg
50 55 60
Val Lys Arg Val Glu Lys Val Glu Lys Lys Phe Leu Asn Arg Pro Ile
65 70 75 80
Glu Val Trp Lys Leu Tyr Phe Asn His Pro Gln Asp Val Pro Ala Ile
85 90 95
Arg Asp Glu Ile Arg Lys His Pro Ala Val Val Asp Ile Tyr Glu Tyr
100 105 110
Asp Ile Pro Phe Ala Lys Arg Tyr Leu Ile Asp Lys Gly Leu Val Pro
115 120 125
Met Glu Gly Glu Glu Glu Leu Lys Leu Met Ala Phe Asp Ile Glu Thr
130 135 140
Leu Tyr His Glu Gly Asp Glu Phe Gly Glu Gly Pro Ile Leu Met Ile
145 150 155 160
Ser Tyr Ala Asp Gly Asp Gly Ala Arg Val Ile Thr Trp Lys Lys Ile
165 170 175
Asp Leu Pro Tyr Val Asp Val Val Ser Thr Glu Lys Glu Met Ile Lys
180 185 190
Arg Phe Leu Gln Val Val Lys Glu Lys Asp Pro Asp Val Leu Val Thr
195 200 205
Tyr Asn Gly Asp Asn Phe Asp Phe Ala Tyr Leu Lys Arg Arg Ser Glu
210 215 220
Glu Leu Gly Leu Lys Phe Ile Leu Gly Arg Asp Gly Ser Glu Pro Lys
225 230 235 240
Ile Gln Arg Met Gly Asp Arg Phe Ala Val Glu Val Lys Gly Arg Ile
245 250 255
His Phe Asp Leu Tyr Pro Val Ile Arg Arg Thr Val Asn Leu Pro Thr
260 265 270
Tyr Thr Leu Glu Ala Val Tyr Glu Ala Ile Phe Gly Arg Pro Lys Glu
275 280 285
Lys Val Tyr Ala Gly Glu Ile Val Glu Ala Trp Glu Thr Gly Glu Gly
290 295 300
Leu Glu Arg Val Ala Arg Tyr Ser Met Glu Asp Ala Lys Val Thr Phe
305 310 315 320
Glu Leu Gly Arg Glu Phe Phe Pro Met Glu Ala Gln Leu Ser Arg Leu
325 330 335
Ile Gly Gln Gly Leu Trp Asp Val Ser Arg Ser Ser Thr Gly Asn Leu
340 345 350
Val Glu Trp Phe Leu Leu Arg Lys Ala Tyr Glu Arg Asn Glu Leu Ala
355 360 365
Pro Asn Lys Pro Ser Gly Arg Glu Val Glu Ile Arg Arg Arg Gly Tyr
370 375 380
Ala Gly Gly Tyr Val Lys Glu Pro Glu Arg Gly Leu Trp Glu Asn Ile
385 390 395 400
Val Tyr Leu Asp Phe Arg Ser Leu Tyr Pro Ser Ile Ile Ile Thr His
405 410 415
Asn Val Ser Pro Asp Thr Leu Asn Arg Glu Gly Cys Glu Asn Tyr Asp
420 425 430
Val Ala Pro Gln Val Gly His Lys Phe Cys Lys Asp Phe Pro Gly Phe
435 440 445
Ile Pro Ser Leu Leu Gly Gly Leu Leu Glu Glu Arg Gln Lys Ile Lys
450 455 460
Arg Arg Met Lys Ala Ser Val Asp Pro Val Glu Arg Lys Leu Leu Asp
465 470 475 480
Tyr Arg Gln Arg Ala Ile Lys Ile Leu Ala Asn Ser Phe Tyr Gly Tyr
485 490 495
Tyr Gly Tyr Ala Arg Ala Arg Trp Tyr Cys Arg Glu Cys Ala Glu Ser
500 505 510
Val Thr Ala Trp Gly Arg Glu Tyr Ile Asp Arg Val Ile Arg Glu Leu
515 520 525
Glu Glu Lys Phe Gly Phe Lys Val Leu Tyr Ala Asp Thr Asp Gly Leu
530 535 540
His Ala Thr Ile Pro Gly Ala Asp Ala Gly Thr Val Lys Glu Arg Ala
545 550 555 560
Arg Gly Phe Leu Arg Tyr Ile Asn Pro Lys Leu Pro Gly Leu Leu Glu
565 570 575
Leu Glu Tyr Glu Gly Phe Tyr Leu Arg Gly Phe Phe Val Thr Lys Lys
580 585 590
Lys Tyr Ala Val Ile Asp Glu Glu Gly Lys Ile Thr Thr Arg Gly Leu
595 600 605
Glu Ile Val Arg Arg Asp Trp Ser Glu Val Ala Lys Glu Thr Gln Ala
610 615 620
Arg Val Leu Glu Ala Ile Leu Arg His Gly Asp Val Glu Glu Ala Val
625 630 635 640
Arg Ile Val Arg Glu Val Thr Glu Lys Leu Ser Lys Tyr Glu Val Pro
645 650 655
Pro Glu Lys Leu Val Ile His Glu Gln Ile Thr Arg Asp Leu Arg Asp
660 665 670
Tyr Lys Ala Thr Gly Pro His Val Ala Val Ala Lys Arg Leu Ala Gly
675 680 685
Arg Gly Val Arg Ile Arg Pro Gly Thr Val Ile Ser Tyr Ile Val Leu
690 695 700
Lys Gly Ser Gly Arg Ile Gly Asp Arg Ala Ile Pro Phe Asp Glu Phe
705 710 715 720
Asp Pro Thr Lys His Arg Tyr Asp Ala Asp Tyr Tyr Ile Glu Asn Gln
725 730 735
Val Leu Pro Ala Val Glu Arg Ile Leu Lys Ala Phe Gly Tyr Arg Lys
740 745 750
Glu Asp Leu Lys Tyr Gln Lys Thr Arg Gln Val Gly Leu Gly Ala Trp
755 760 765
Leu Asn Ala Gly Lys Gly
770
<210> 5
<211> 2328
<212> DNA
<213> Artificial Sequence
<220>
<223> DNA polymerase from Thermococcus siculi
<400> 5
atgatcctcg acacggacta catcacggaa gatgggaaac ccgtcataag gatattcaag 60
aaagagaacg gcgagttcaa gatcgagtac gacaggactt ttgaacccta catctacgcc 120
ctcctgaagg acgactccgc gattgaggat gttaaaaaga taaccgccga gaggcacgga 180
acggtggtga aggtcaagcg cgccgaaaag gtgcagaaga agttcctagg caggccggtt 240
gaagtctgga agctctactt cacccacccc caagatgtcc cggcgataag ggacaagatt 300
aggaagcatc cagctgtaat tgacatctac gagtacgaca taccattcgc caagcgctac 360
ctcatcgaca agggcctgat tccgatggag ggtgaagaag agcttaagat gctcgccttc 420
gacattgaga cgctctacca tgagggtgag gagttcgccg aggggcctat tctgatgata 480
agctacgccg acgagagcga ggcacgcgtc atcacctgga agaaaatcga cctcccctac 540
gttgacgtcg tctcaacgga gaaggagatg ataaagcgct tcctccgcgt tgtgaaggag 600
aaagatcccg atgtcctcat aacctacaac ggcgacaact tcgacttcgc ctacctgaag 660
aagcgctgtg aaaagcttgg aataaacttc ctccttggaa gggacgggag cgagccgaag 720
atccagagaa tgggtgaccg cttcgccgtt gaggtgaagg ggaggataca cttcgacctc 780
tatcctgtaa taaggcgcac gataaacctg ccgacctaca tgcttgaggc agtctacgag 840
gccatctttg ggaagccaaa ggagaaggtt tacgccgagg agatagccac cgcttgggaa 900
accggagagg gccttgagag ggtggctcgc tactctatgg aggacgcgaa ggtcacgttt 960
gagcttggaa aggagttctt cccgatggag gcccaacttt cgaggttggt cggccagagc 1020
ttctgggatg tcgcgcgctc aagcacgggc aatctggtcg agtggttcct cctcaggaag 1080
gcctacgaga ggaacgagct ggctccaaac aagccctctg gaagggaata tgacgagagg 1140
cgcggtggat acgccggcgg ctacgtcaag gaaccggaaa agggcctgtg ggagaacata 1200
gtctacctcg actataaatc tctctacccc tcaatcatca tcacccacaa cgtctcgccc 1260
gataccctca accgcgaggg ctgtaaggag tatgacgtag ctccacaggt cggccaccgc 1320
ttctgcaagg actttccagg cttcatcccg agcctgctcg gggatctcct ggaggagagg 1380
cagaagataa agaggaagat gaaggcaaca attgacccga tcgagagaaa gctccttgat 1440
tacaggcaac gggccatcaa gatccttcta aatagttttt acggctacta cggctacgca 1500
agggctcgct ggtactgcaa ggagtgtgcc gagagcgtta cggcatgggg aagggaatat 1560
atcaccatga caatcaggga aatagaagag aagtatggct ttaaagtact ttatgcggac 1620
actgacggct tcttcgcgac gattcccggg gaagatgccg agaccatcaa aaagagggcg 1680
atggagttcc tcaagtacat aaacgccaaa ctccccggtg cgctcgaact tgagtacgag 1740
gacttctaca ggcgcggctt cttcgtcacc aagaagaaat acgcggttat cgacgaggag 1800
ggcaagataa caacgcgcgg gctggagatc gtcaggcgcg actggagcga gatagccaag 1860
gagacgcagg cgcgggttct ggaggccctt ctgaaggacg gtgacgtcga agaggccgtg 1920
agcatagtca aagaagtgac cgagaagctg agcaagtacg aggttccgcc ggagaagctc 1980
gttatccacg agcagataac gcgcgagctg aaggactaca aggcaacggg accacacgtg 2040
gcgatagcga agaggttagc cgcgagaggc gtcaaaatcc gccccgggac agtcatcagc 2100
tacatcgtgc tcaagggctc cgggaggata ggcgacaggg cgattccctt cgacgagttc 2160
gaccccacga agcacaagta cgatgcagag tactacatcg agaaccaggt tctacctgcc 2220
gtcgagagga ttctgaaggc cttcggctat cgcggtgagg agctcagata ccagaagacg 2280
aggcaggttg gacttggggc gtggctgaag ccgaagggga aggggtga 2328
<210> 6
<211> 775
<212> PRT
<213> Artificial Sequence
<220>
<223> DNA polymerase from Thermococcus siculi
<400> 6
Met Ile Leu Asp Thr Asp Tyr Ile Thr Glu Asp Gly Lys Pro Val Ile
1 5 10 15
Arg Ile Phe Lys Lys Glu Asn Gly Glu Phe Lys Ile Glu Tyr Asp Arg
20 25 30
Thr Phe Glu Pro Tyr Ile Tyr Ala Leu Leu Lys Asp Asp Ser Ala Ile
35 40 45
Glu Asp Val Lys Lys Ile Thr Ala Glu Arg His Gly Thr Val Val Lys
50 55 60
Val Lys Arg Ala Glu Lys Val Gln Lys Lys Phe Leu Gly Arg Pro Val
65 70 75 80
Glu Val Trp Lys Leu Tyr Phe Thr His Pro Gln Asp Val Pro Ala Ile
85 90 95
Arg Asp Lys Ile Arg Lys His Pro Ala Val Ile Asp Ile Tyr Glu Tyr
100 105 110
Asp Ile Pro Phe Ala Lys Arg Tyr Leu Ile Asp Lys Gly Leu Ile Pro
115 120 125
Met Glu Gly Glu Glu Glu Leu Lys Met Leu Ala Phe Asp Ile Glu Thr
130 135 140
Leu Tyr His Glu Gly Glu Glu Phe Ala Glu Gly Pro Ile Leu Met Ile
145 150 155 160
Ser Tyr Ala Asp Glu Ser Glu Ala Arg Val Ile Thr Trp Lys Lys Ile
165 170 175
Asp Leu Pro Tyr Val Asp Val Val Ser Thr Glu Lys Glu Met Ile Lys
180 185 190
Arg Phe Leu Arg Val Val Lys Glu Lys Asp Pro Asp Val Leu Ile Thr
195 200 205
Tyr Asn Gly Asp Asn Phe Asp Phe Ala Tyr Leu Lys Lys Arg Cys Glu
210 215 220
Lys Leu Gly Ile Asn Phe Leu Leu Gly Arg Asp Gly Ser Glu Pro Lys
225 230 235 240
Ile Gln Arg Met Gly Asp Arg Phe Ala Val Glu Val Lys Gly Arg Ile
245 250 255
His Phe Asp Leu Tyr Pro Val Ile Arg Arg Thr Ile Asn Leu Pro Thr
260 265 270
Tyr Met Leu Glu Ala Val Tyr Glu Ala Ile Phe Gly Lys Pro Lys Glu
275 280 285
Lys Val Tyr Ala Glu Glu Ile Ala Thr Ala Trp Glu Thr Gly Glu Gly
290 295 300
Leu Glu Arg Val Ala Arg Tyr Ser Met Glu Asp Ala Lys Val Thr Phe
305 310 315 320
Glu Leu Gly Lys Glu Phe Phe Pro Met Glu Ala Gln Leu Ser Arg Leu
325 330 335
Val Gly Gln Ser Phe Trp Asp Val Ala Arg Ser Ser Thr Gly Asn Leu
340 345 350
Val Glu Trp Phe Leu Leu Arg Lys Ala Tyr Glu Arg Asn Glu Leu Ala
355 360 365
Pro Asn Lys Pro Ser Gly Arg Glu Tyr Asp Glu Arg Arg Gly Gly Tyr
370 375 380
Ala Gly Gly Tyr Val Lys Glu Pro Glu Lys Gly Leu Trp Glu Asn Ile
385 390 395 400
Val Tyr Leu Asp Tyr Lys Ser Leu Tyr Pro Ser Ile Ile Ile Thr His
405 410 415
Asn Val Ser Pro Asp Thr Leu Asn Arg Glu Gly Cys Lys Glu Tyr Asp
420 425 430
Val Ala Pro Gln Val Gly His Arg Phe Cys Lys Asp Phe Pro Gly Phe
435 440 445
Ile Pro Ser Leu Leu Gly Asp Leu Leu Glu Glu Arg Gln Lys Ile Lys
450 455 460
Arg Lys Met Lys Ala Thr Ile Asp Pro Ile Glu Arg Lys Leu Leu Asp
465 470 475 480
Tyr Arg Gln Arg Ala Ile Lys Ile Leu Leu Asn Ser Phe Tyr Gly Tyr
485 490 495
Tyr Gly Tyr Ala Arg Ala Arg Trp Tyr Cys Lys Glu Cys Ala Glu Ser
500 505 510
Val Thr Ala Trp Gly Arg Glu Tyr Ile Thr Met Thr Ile Arg Glu Ile
515 520 525
Glu Glu Lys Tyr Gly Phe Lys Val Leu Tyr Ala Asp Thr Asp Gly Phe
530 535 540
Phe Ala Thr Ile Pro Gly Glu Asp Ala Glu Thr Ile Lys Lys Arg Ala
545 550 555 560
Met Glu Phe Leu Lys Tyr Ile Asn Ala Lys Leu Pro Gly Ala Leu Glu
565 570 575
Leu Glu Tyr Glu Asp Phe Tyr Arg Arg Gly Phe Phe Val Thr Lys Lys
580 585 590
Lys Tyr Ala Val Ile Asp Glu Glu Gly Lys Ile Thr Thr Arg Gly Leu
595 600 605
Glu Ile Val Arg Arg Asp Trp Ser Glu Ile Ala Lys Glu Thr Gln Ala
610 615 620
Arg Val Leu Glu Ala Leu Leu Lys Asp Gly Asp Val Glu Glu Ala Val
625 630 635 640
Ser Ile Val Lys Glu Val Thr Glu Lys Leu Ser Lys Tyr Glu Val Pro
645 650 655
Pro Glu Lys Leu Val Ile His Glu Gln Ile Thr Arg Glu Leu Lys Asp
660 665 670
Tyr Lys Ala Thr Gly Pro His Val Ala Ile Ala Lys Arg Leu Ala Ala
675 680 685
Arg Gly Val Lys Ile Arg Pro Gly Thr Val Ile Ser Tyr Ile Val Leu
690 695 700
Lys Gly Ser Gly Arg Ile Gly Asp Arg Ala Ile Pro Phe Asp Glu Phe
705 710 715 720
Asp Pro Thr Lys His Lys Tyr Asp Ala Glu Tyr Tyr Ile Glu Asn Gln
725 730 735
Val Leu Pro Ala Val Glu Arg Ile Leu Lys Ala Phe Gly Tyr Arg Gly
740 745 750
Glu Glu Leu Arg Tyr Gln Lys Thr Arg Gln Val Gly Leu Gly Ala Trp
755 760 765
Leu Lys Pro Lys Gly Lys Gly
770 775
<210> 7
<211> 774
<212> PRT
<213> Artificial Sequence
<220>
<223> DNA polymerase from Thermococcus kodakarensis
<400> 7
Met Ile Leu Asp Thr Asp Tyr Ile Thr Glu Asp Gly Lys Pro Val Ile
1 5 10 15
Arg Ile Phe Lys Lys Glu Asn Gly Glu Phe Lys Ile Glu Tyr Asp Arg
20 25 30
Thr Phe Glu Pro Tyr Phe Tyr Ala Leu Leu Lys Asp Asp Ser Ala Ile
35 40 45
Glu Glu Val Lys Lys Ile Thr Ala Glu Arg His Gly Thr Val Val Thr
50 55 60
Val Lys Arg Val Glu Lys Val Gln Lys Lys Phe Leu Gly Arg Pro Val
65 70 75 80
Glu Val Trp Lys Leu Tyr Phe Thr His Pro Gln Asp Val Pro Ala Ile
85 90 95
Arg Asp Lys Ile Arg Glu His Pro Ala Val Ile Asp Ile Tyr Glu Tyr
100 105 110
Asp Ile Pro Phe Ala Lys Arg Tyr Leu Ile Asp Lys Gly Leu Val Pro
115 120 125
Met Glu Gly Asp Glu Glu Leu Lys Met Leu Ala Phe Asp Ile Glu Thr
130 135 140
Leu Tyr Glu Glu Gly Glu Glu Phe Ala Glu Gly Pro Ile Leu Met Ile
145 150 155 160
Ser Tyr Ala Asp Glu Glu Gly Ala Arg Val Ile Thr Trp Lys Asn Val
165 170 175
Asp Leu Pro Tyr Val Asp Val Val Ser Thr Glu Arg Glu Met Ile Lys
180 185 190
Arg Phe Leu Arg Val Val Lys Glu Lys Asp Pro Asp Val Leu Ile Thr
195 200 205
Tyr Asn Gly Asp Asn Phe Asp Phe Ala Tyr Leu Lys Lys Arg Cys Glu
210 215 220
Lys Leu Gly Ile Asn Phe Ala Leu Gly Arg Asp Gly Ser Glu Pro Lys
225 230 235 240
Ile Gln Arg Met Gly Asp Arg Phe Ala Val Glu Val Lys Gly Arg Ile
245 250 255
His Phe Asp Leu Tyr Pro Val Ile Arg Arg Thr Ile Asn Leu Pro Thr
260 265 270
Tyr Thr Leu Glu Ala Val Tyr Glu Ala Val Phe Gly Gln Pro Lys Glu
275 280 285
Lys Val Tyr Ala Glu Glu Ile Thr Thr Ala Trp Glu Thr Gly Glu Asn
290 295 300
Leu Glu Arg Val Ala Arg Tyr Ser Met Glu Asp Ala Lys Val Thr Tyr
305 310 315 320
Glu Leu Gly Lys Glu Phe Leu Pro Met Glu Ala Gln Leu Ser Arg Leu
325 330 335
Ile Gly Gln Ser Leu Trp Asp Val Ser Arg Ser Ser Thr Gly Asn Leu
340 345 350
Val Glu Trp Phe Leu Leu Arg Lys Ala Tyr Glu Arg Asn Glu Leu Ala
355 360 365
Pro Asn Lys Pro Asp Glu Lys Glu Leu Ala Arg Arg Arg Gln Ser Tyr
370 375 380
Glu Gly Gly Tyr Val Lys Glu Pro Glu Arg Gly Leu Trp Glu Asn Ile
385 390 395 400
Val Tyr Leu Asp Phe Arg Ser Leu Tyr Pro Ser Ile Ile Ile Thr His
405 410 415
Asn Val Ser Pro Asp Thr Leu Asn Arg Glu Gly Cys Lys Glu Tyr Asp
420 425 430
Val Ala Pro Gln Val Gly His Arg Phe Cys Lys Asp Phe Pro Gly Phe
435 440 445
Ile Pro Ser Leu Leu Gly Asp Leu Leu Glu Glu Arg Gln Lys Ile Lys
450 455 460
Lys Lys Met Lys Ala Thr Ile Asp Pro Ile Glu Arg Lys Leu Leu Asp
465 470 475 480
Tyr Arg Gln Arg Ala Ile Lys Ile Leu Ala Asn Ser Tyr Tyr Gly Tyr
485 490 495
Tyr Gly Tyr Ala Arg Ala Arg Trp Tyr Cys Lys Glu Cys Ala Glu Ser
500 505 510
Val Thr Ala Trp Gly Arg Glu Tyr Ile Thr Met Thr Ile Lys Glu Ile
515 520 525
Glu Glu Lys Tyr Gly Phe Lys Val Ile Tyr Ser Asp Thr Asp Gly Phe
530 535 540
Phe Ala Thr Ile Pro Gly Ala Asp Ala Glu Thr Val Lys Lys Lys Ala
545 550 555 560
Met Glu Phe Leu Lys Tyr Ile Asn Ala Lys Leu Pro Gly Ala Leu Glu
565 570 575
Leu Glu Tyr Glu Gly Phe Tyr Glu Arg Gly Phe Phe Val Thr Lys Lys
580 585 590
Lys Tyr Ala Val Ile Asp Glu Glu Gly Lys Ile Thr Thr Arg Gly Leu
595 600 605
Glu Ile Val Arg Arg Asp Trp Ser Glu Ile Ala Lys Glu Thr Gln Ala
610 615 620
Arg Val Leu Glu Ala Leu Leu Lys Asp Gly Asp Val Glu Lys Ala Val
625 630 635 640
Arg Ile Val Lys Glu Val Thr Glu Lys Leu Ser Lys Tyr Glu Val Pro
645 650 655
Pro Glu Lys Leu Val Ile His Glu Gln Ile Thr Arg Asp Leu Lys Asp
660 665 670
Tyr Lys Ala Thr Gly Pro His Val Ala Val Ala Lys Arg Leu Ala Ala
675 680 685
Arg Gly Val Lys Ile Arg Pro Gly Thr Val Ile Ser Tyr Ile Val Leu
690 695 700
Lys Gly Ser Gly Arg Ile Gly Asp Arg Ala Ile Pro Phe Asp Glu Phe
705 710 715 720
Asp Pro Thr Lys His Lys Tyr Asp Ala Glu Tyr Tyr Ile Glu Asn Gln
725 730 735
Val Leu Pro Ala Val Glu Arg Ile Leu Arg Ala Phe Gly Tyr Arg Lys
740 745 750
Glu Asp Leu Arg Tyr Gln Lys Thr Arg Gln Val Gly Leu Ser Ala Trp
755 760 765
Leu Lys Pro Lys Gly Thr
770
<210> 8
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 8
tagaattgaa gaa 13
<210> 9
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 9
tggccatagc tac 13
<210> 10
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 10
gtcatctgcg acc 13
<210> 11
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 11
ttcgcgcttg gac 13
<210> 12
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 12
cgcgaaccgt tag 13
<210> 13
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 13
ttgcagcctc taa 13
<210> 14
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 14
tctactagta cga 13
<210> 15
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 15
gtaggttcta ctg 13
<210> 16
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 16
gccaatatca agt 13
<210> 17
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 17
ctatcttgct ggt 13
<210> 18
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 18
gttctcatag gta 13
<210> 19
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 19
gtctatgaac caa 13
<210> 20
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 20
cggagcgctt att 13
<210> 21
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 21
tatgccatga gga 13
<210> 22
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 22
atacgactcg gag 13
<210> 23
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 23
gatggaactc agc 13
<210> 24
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 24
ggacctgcat gaa 13
<210> 25
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 25
tagactggaa ctt 13
<210> 26
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 26
gaattacctc gtt 13
<210> 27
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 27
aggatcaggc tac 13
<210> 28
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 28
acgcgtagaa gag 13
<210> 29
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 29
cttcgagact tac 13
<210> 30
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 30
gacggctaac tcc 13
<210> 31
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 31
ttagcattct ctt 13
<210> 32
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 32
gcaaggcata gta 13
<210> 33
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 33
acctagatat gga 13
<210> 34
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 34
acgccaaggc gta 13
<210> 35
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 35
tatgacggat ccg 13
<210> 36
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 36
cctccattag aga 13
<210> 37
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 37
attgaatact ctg 13
<210> 38
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 38
gagatgagaa gaa 13
<210> 39
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 39
tctgagtagc cgg 13
<210> 40
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 40
aataggtagt acg 13
<210> 41
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 41
gtcgaagaag tcc 13
<210> 42
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 42
tactgcatct cgt 13
<210> 43
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 43
gacgtattag agc 13
<210> 44
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 44
cctgcattat tcg 13
<210> 45
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 45
acgaatgatg ctc 13
<210> 46
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 46
tactagcaga gat 13
<210> 47
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 47
ctcctcatct tcc 13
<210> 48
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 48
tcctctgcgc tgc 13
<210> 49
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 49
ccttctcagt ccg 13
<210> 50
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 50
cagcttcata gcg 13
<210> 51
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 51
ttgactctcg cgc 13
<210> 52
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 52
tatcctgagc gat 13
<210> 53
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 53
aacgcctagc cga 13
<210> 54
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 54
ccgaagacgt cat 13
<210> 55
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 55
gagttctcca gat 13
<210> 56
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 56
tgcatccgcg ctt 13
<210> 57
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 57
cctgaactca agt 13
<210> 58
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> sample tag sequence
<400> 58
ggtcgtatgc gta 13
<210> 59
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 59
aggcctctct acc 13
<210> 60
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 60
gtactccatc caa 13
<210> 61
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 61
cagcggacgc gct 13
<210> 62
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 62
atctctctta gca 13
<210> 63
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 63
aagcaataat aat 13
<210> 64
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 64
aaggcgactc cga 13
<210> 65
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 65
acgtctctag gag 13
<210> 66
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 66
ccatcagacc tct 13
<210> 67
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 67
acttaatcgt act 13
<210> 68
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 68
tggaattctc caa 13
<210> 69
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 69
ccatacgatc agg 13
<210> 70
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 70
ttatggagca ata 13
<210> 71
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 71
gctcggcgtt cga 13
<210> 72
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 72
ttggccagtc gct 13
<210> 73
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 73
cagatacgta gag 13
<210> 74
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 74
aatgctatta tcc 13
<210> 75
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 75
gcagcatgcc gat 13
<210> 76
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 76
ggagagttac ctc 13
<210> 77
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 77
gagagtccat gat 13
<210> 78
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 78
caatctattc tga 13
<210> 79
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 79
gctcttagta tcc 13
<210> 80
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 80
ccatagttat ggt 13
<210> 81
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 81
tgcgagatcg aag 13
<210> 82
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 82
agagaagtcg agt 13
<210> 83
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 83
ggtaactcca tat 13
<210> 84
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 84
tgctattcca ggc 13
<210> 85
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 85
aaccgcgagg ctc 13
<210> 86
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 86
ttctagagat acc 13
<210> 87
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 87
ttcgctcaag tat 13
<210> 88
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 88
cagagaaggc gca 13
<210> 89
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 89
tagaattggc ctc 13
<210> 90
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 90
ggccattctc cag 13
<210> 91
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 91
tccaacgcgc gtt 13
<210> 92
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 92
gccgcagatt acg 13
<210> 93
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 93
gcagttcgaa cgc 13
<210> 94
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 94
ttctctctgc agg 13
<210> 95
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 95
taagctacca gcg 13
<210> 96
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 96
ctgcatgagg ttg 13
<210> 97
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 97
ttgcctagcg agg 13
<210> 98
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 98
caactgaatt agg 13
<210> 99
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 99
aagcggtcct ctt 13
<210> 100
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 100
aatggaagga ccg 13
<210> 101
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 101
gagttagtaa gtt 13
<210> 102
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 102
ttcctaattc caa 13
<210> 103
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 103
gttctggttc gct 13
<210> 104
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 104
gttcatctct tcc 13
<210> 105
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 105
attccgagga aga 13
<210> 106
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 106
cttagccgag aga 13
<210> 107
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 107
gtctgctacg ctt 13
<210> 108
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 108
atggcgccgc gca 13
<210> 109
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 109
taattggtta tct 13
<210> 110
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 110
tcggttataa gtc 13
<210> 111
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 111
tgcctgagaa cgt 13
<210> 112
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 112
agatgcggtt aac 13
<210> 113
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 113
atggaatagg cga 13
<210> 114
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 114
agagatgcga tcg 13
<210> 115
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 115
ctccaactaa cgt 13
<210> 116
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 116
gccttgctac tgg 13
<210> 117
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 117
cttcgtctct acg 13
<210> 118
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 118
acgctcatag cct 13
<210> 119
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 119
gtcgaagata agg 13
<210> 120
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 120
gccggagtcc tcg 13
<210> 121
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 121
tatacggcga cct 13
<210> 122
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 122
aggtagatat tcg 13
<210> 123
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 123
ttaaggtact gct 13
<210> 124
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 124
cggatctggt ata 13
<210> 125
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 125
gaggtctcgg agg 13
<210> 126
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 126
ggcatcgatg gac 13
<210> 127
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 127
gatctccgat ata 13
<210> 128
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 128
gattcggaat act 13
<210> 129
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 129
ctgcgatccg gcc 13
<210> 130
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 130
gatccggttg caa 13
<210> 131
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 131
cgtcaggctt gac 13
<210> 132
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 132
tcggcaaggc gag 13
<210> 133
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 133
gaacggcgaa cgc 13
<210> 134
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 134
cctcaagcgg act 13
<210> 135
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 135
gaagccagat ggt 13
<210> 136
<211> 13
<212> DNA
<213> Artificial Sequence
<220>
<223> Sample tag sequence
<400> 136
tgctcatacc aat 13
Claims (105)
- 하기를 포함하는 적어도 하나의 타겟 핵산 분자로 돌연변이를 도입하는 방법:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 저편향 DNA 중합효소 (low bias DNA polymerase)를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계. - 적어도 하나의 타겟 핵산 분자로 돌연변이를 도입하는 방법에서의 저편향 DNA 중합효소의 용도.
- 청구항 2에 있어서, 상기 적어도 하나의 타겟 핵산 분자로 돌연변이를 도입하는 방법은 하기를 포함하는 것인 용도:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계. - 청구항 1 내지 3 중 어느 한 항에 있어서, 상기 돌연변이는 치환 돌연변이인 방법 또는 용도.
- 청구항 1 내지 4 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 아데닌, 티민, 구아닌, 및 시토신 뉴클레오티드를 각각 0.5-1.5:0.5-1.5:0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4:0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2:0.8-1.2:0.8-1.2, 또는 약 1:1:1:1의 비율비 (rate ratio)로 돌연변이시키는 것인 방법 또는 용도.
- 청구항 1 내지 5 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 아데닌, 티민, 구아닌, 및 시토신 뉴클레오티드를 각각 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3의 비율비로 돌연변이시키는 것인 방법 또는 용도.
- 청구항 1 내지 6 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에서 1% 내지 15%, 2% 내지 10%, 또는 약 8%의 뉴클레오티드를 돌연변이시키는 것인 방법 또는 용도.
- 청구항 1 내지 7 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 복제 라운드 당 적어도 하나의 타겟 핵산 분자 중의 0% 내지 3%, 또는 0% 내지 2%의 뉴클레오티드를 돌연변이시키는 것인 방법 또는 용도.
- 청구항 1 내지 8 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 적어도 하나의 타겟 핵산 분자에 뉴클레오티드 유사체를 통합시키는 (incorporates) 것인 방법 또는 용도.
- 청구항 1 내지 9 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 적어도 하나의 타겟 핵산 분자에서 아데닌, 티민, 구아닌, 및/또는 시토신을 돌연변이시키는 것인 방법 또는 용도.
- 청구항 1 내지 10 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 구아닌, 시토신, 아데닌, 및/또는 티민을 뉴클레오티드 유사체로 대체하는 것인 방법 또는 용도.
- 청구항 1 내지 11 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 구아닌 또는 아데닌 뉴클레오티드를 각각 0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2, 또는 약 1:1의 비율비로 도입하는 것인 방법 또는 용도.
- 청구항 1 내지 12 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 뉴클레오티드 유사체를 사용하여 구아닌 또는 아데닌 뉴클레오티드를 각각 0.7-1.3:0.7-1.3의 비율비로 도입하는 것인 방법 또는 용도.
- 청구항 9 내지 13 중 어느 한 항에 있어서, 상기 방법은 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 포함하며, 상기 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 뉴클레오티드 유사체의 존재 하에 수행되는 것이고, 상기 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 제공하는 것인 방법 또는 용도.
- 청구항 9 내지 14 중 어느 한 항에 있어서, 상기 뉴클레오티드 유사체는 dPTP 것인 방법 또는 용도.
- 청구항 15에 있어서, 상기 저편향 DNA 중합효소는 구아닌에서 아데닌으로의 치환 돌연변이, 시토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 도입하는 것인 방법 또는 용도.
- 청구항 16에 있어서, 상기 저편향 DNA 중합효소는 구아닌에서 아데닌으로의 치환 돌연변이, 시토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 각각 0.5-1.5:0.5-1.5:0.5-1.5:0.5-1.5, 0.6-1.4:0.6-1.4:0.6-1.4:0.6-1.4, 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3, 0.8-1.2:0.8-1.2:0.8-1.2:0.8-1.2, 또는 약 1:1:1:1의 비율비로 도입하는 것인 방법 또는 용도.
- 청구항 16 또는 17에 있어서, 상기 저편향 DNA 중합효소는 구아닌에서 아데닌으로의 치환 돌연변이, 시토신에서 티민으로의 치환 돌연변이, 아데닌에서 구아닌으로의 치환 돌연변이, 및 티민에서 시토신으로의 치환 돌연변이를 각각 0.7-1.3:0.7-1.3:0.7-1.3:0.7-1.3의 비율비로 도입하는 것인 방법 또는 용도.
- 청구항 1 내지 18 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 고정확도 DNA 중합효소인 것인 방법 또는 용도.
- 청구항 19에 있어서, 상기 고정확도 DNA 중합효소는, 뉴클레오티드 유사체의 부재 하에, 복제 라운드 당 0.01% 미만, 0.0015% 미만, 0.001% 미만, 0% 내지 0.0015%, 또는 0% 내지 0.001%의 돌연변이를 도입하는 것인 방법 또는 용도.
- 청구항 14 또는 15에 있어서, 상기 방법은 뉴클레오티드 유사체의 부재 하에 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 더 포함하는 것인 방법 또는 용도.
- 청구항 21에 있어서, 상기 뉴클레오티드 유사체의 부재 하에 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 저편향 DNA 중합효소를 사용하여 수행되는 것인 방법 또는 용도.
- 청구항 1 내지 22 중 어느 한 항에 있어서, 상기 방법은 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하고, 상기 방법은 저편향 DNA 중합효소를 사용하여 상기 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 단계를 더 포함하는 것인 방법 또는 용도.
- 청구항 1 내지 23 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 낮은 주형 증폭 편향 (template amplification bias)을 갖는 것인 방법 또는 용도.
- 청구항 1 내지 24 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 교정(proof-reading) 도메인 및/또는 진행도 증진(processivity enhancing) 도메인을 포함하는 것인 방법 또는 용도.
- 청구항 1 내지 25 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 적어도 400, 적어도 500, 적어도 600, 적어도 700, 또는 적어도 750개의 연속하는, 하기 아미노산의 단편을 포함하는 것인 방법 또는 용도:
a. 서열번호 2의 서열;
b. 서열번호 2와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
c. 서열번호 4의 서열;
d. 서열번호 4와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
e. 서열번호 6의 서열;
f. 서열번호 6과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
g. 서열번호 7의 서열; 또는
h. 서열번호 7과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열. - 청구항 26에 있어서, 상기 저편향 DNA 중합효소는 하기를 포함하는 것인 방법 또는 용도:
a. 서열번호 2의 서열;
b. 서열번호 2와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
c. 서열번호 4의 서열;
d. 서열번호 4와 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
e. 서열번호 6의 서열;
f. 서열번호 6과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열;
g. 서열번호 7의 서열; 또는
h. 서열번호 7과 적어도 95%, 적어도 98%, 또는 적어도 99% 동일한 서열. - 청구항 27에 있어서, 상기 저편향 DNA 중합효소는 서열번호 2와 적어도 98% 동일한 서열을 포함하는 것인 방법 또는 용도.
- 청구항 27에 있어서, 상기 저편향 DNA 중합효소는 서열번호 4와 적어도 98% 동일한 서열을 포함하는 것인 방법 또는 용도.
- 청구항 27에 있어서, 상기 저편향 DNA 중합효소는 서열번호 6과 적어도 98% 동일한 서열을 포함하는 것인 방법 또는 용도.
- 청구항 27에 있어서, 상기 저편향 DNA 중합효소는 서열번호 7과 적어도 98% 동일한 서열을 포함하는 것인 방법 또는 용도.
- 청구항 1 내지 31 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소는 써모코커스 (thermococcal) 중합효소, 또는 그의 유도체 (derivative)인 것인 방법 또는 용도.
- 청구항 32에 있어서, 상기 저편향 DNA 중합효소는 써모코커스 중합효소인 것인 방법 또는 용도.
- 청구항 32 또는 33에 있어서, 상기 써모코커스 중합효소는 T.kodakarensis, T.siculi, T.celer 및 T.sp KS-1로 이루어진 군으로부터 선택된 써모코커스 균주로부터 유래된 것인 방법 또는 용도.
- 청구항 1 내지 34 중 어느 한 항에 있어서, 적어도 하나의 타겟 핵산 분자에 바코드 (barcodes)를 도입하는 단계를 더 포함하는 방법 또는 용도.
- 청구항 1 내지 35 중 어느 한 항에 있어서, 적어도 하나의 타겟 핵산 분자에 샘플 태그 (sample tags)를 도입하는 단계를 더 포함하는 방법 또는 용도.
- 청구항 36에 있어서, 샘플 태그 그룹이 사용되고, 서로 다른 샘플로부터의 타겟 핵산 분자가 상기 그룹으로부터의 서로 다른 샘플 태그로 표지되는 것인 방법 또는 용도.
- 청구항 37에 있어서, 각 샘플 태그는 적어도 1개의 낮은 확률 돌연변이 차이 또는 적어도 3개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 (substantially) 다른 것인 방법 또는 용도.
- 청구항 38에 있어서, 각 샘플 태그는 적어도 3개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 방법 또는 용도.
- 청구항 38 또는 39에 있어서, 각 샘플 태그는 3 내지 25개, 또는 3 내지 10개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 방법 또는 용도.
- 청구항 38 내지 40 중 어느 한 항에 있어서, 상기 낮은 확률 돌연변이는 전환 (transversion) 돌연변이 또는 인델 (indel) 돌연변이인 것인 방법 또는 용도.
- 청구항 37 내지 41 중 어느 한 항에 있어서, 각 샘플 태그는 적어도 5개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 방법 또는 용도.
- 청구항 42에 있어서, 각 샘플 태그는 5 내지 25개, 또는 5 내지 10개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 방법 또는 용도.
- 청구항 38 내지 43 중 어느 한 항에 있어서, 상기 높은 확률 돌연변이는 전이 (transition) 돌연변이인 것인 방법 또는 용도.
- 청구항 37 내지 44 중 어느 한 항에 있어서, 상기 샘플 태그 그룹은 청구항 71 내지 75 중 어느 한 항의 방법으로 얻을 수 있는 것인 방법 또는 용도.
- 청구항 1 내지 45 중 어느 한 항에 있어서, 적어도 하나의 타겟 핵산 분자 각각에 어댑터 (adapters)를 도입하는 단계를 더 포함하는 것인 방법 또는 용도.
- 청구항 46에 있어서, 적어도 하나의 타겟 핵산 분자의 3' 말단에 제1 어댑터 및 적어도 하나의 타겟 핵산 분자의 5' 말단에 제2 어댑터를 도입하는 단계를 포함하고, 상기 제1 어댑터 및 제2 어댑터는 서로에게 어닐링 (annealing)하는 것인 방법 또는 용도.
- 청구항 47에 있어서, 상기 적어도 하나의 타겟 핵산 분자는, 서로 동일하고 상기 제1 어댑터의 일부에 상보적인(complementary) 프라이머들을 사용하여 증폭되는 것인 방법 또는 용도.
- 청구항 47 또는 48에 있어서, 상기 제1 어댑터는 제2 어댑터에 적어도 80%, 적어도 90%, 적어도 95%, 적어도 99%, 또는 100% 동일한 핵산 분자에 상보적인 것인 방법 또는 용도.
- 청구항 48 또는 49에 있어서, 상기 프라이머는 제2 프라이머 결합 부위를 포함하고, 상기 방법은 상기 프라이머를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계, 상기 프라이머를 제거하는 단계, 및 상기 제2 프라이머 결합 부위에 어닐링하는 제2 프라이머 세트를 사용하여 적어도 하나의 타겟 핵산 분자를 추가로 증폭하는 단계를 더 포함하는 것인 방법 또는 용도.
- 청구항 1 내지 50 중 어느 한 항에 있어서, 상기 방법은 상기 타겟 핵산 분자 각각에 바코드, 샘플 태그 및 어댑터를 도입하는 단계를 더 포함하는 것인 방법 또는 용도.
- 청구항 1 내지 51 중 어느 한 항에 있어서, 상기 바코드, 샘플 태그 및/또는 어댑터는 태그멘테이션(tagmentation)에 의해, 또는 쉬어링(shearing) 및 라이게이션 (ligation)에 의해 도입되는 것인 방법 또는 용도.
- 청구항 1 내지 52 중 어느 한 항에 있어서, 상기 적어도 하나의 타겟 핵산 분자는 1 kbp 초과, 1.5 kbp 초과, 2 kbp 초과, 4 kbp 초과, 5kbp 초과, 7 kbp 초과, 또는 8 kbp를 초과하는 것인 방법 또는 용도.
- 청구항 1 또는 3 내지 53 중 어느 한 항의 돌연변이를 도입하는 방법을 포함하는 적어도 하나의 타겟 핵산 분자의 서열을 결정하는 방법.
- 청구항 54에 있어서, 하기 단계를 포함하는 방법:
a. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해, 청구항 1 또는 3 내지 53 중 어느 한 항의 방법을 수행하는 단계;
b. 돌연변이된 서열 리드(reads)를 제공하기 위해, 상기 적어도 하나의 돌연변이된 타겟 핵산 분자의 영역을 시퀀싱하는 단계; 및
c. 상기 돌연변이된 서열 리드를 사용하여 적어도 하나의 타겟 핵산 분자의 적어도 일부의 서열을 조립하는 단계. - 청구항 54에 있어서, 하기 단계를 포함하는 방법:
a. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해, 청구항 1 또는 3 내지 53 중 어느 한 항의 방법을 수행하는 단계;
b. 적어도 하나의 단편화된(fragmented) 및/또는 증폭된 돌연변이된 타겟 핵산 분자를 제공하기 위해, 상기 적어도 하나의 돌연변이된 타겟 핵산 분자를 단편화 및/또는 증폭하는 단계;
c. 돌연변이된 서열 리드를 제공하기 위해, 상기 적어도 하나의 단편화된 및/또는 증폭된 돌연변이된 타겟 핵산 분자의 영역을 시퀀싱하는 단계; 및
d. 상기 돌연변이된 서열 리드를 사용하여 적어도 하나의 핵산 분자의 적어도 한 부분의 서열을 조립하는 단계. - 청구항 1 또는 3 내지 53 중 어느 한 항의 돌연변이를 도입하는 방법을 포함하는 단백질을 엔지니어링 (engineering)하는 방법.
- 청구항 57에 있어서, 하기 단계를 포함하는 방법:
a. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해, 청구항 1 또는 3 내지 53의 방법을 수행하는 단계;
b. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자를 벡터에 삽입하는 단계; 및
c. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질을 발현하는 단계. - 청구항 58에 있어서, 하기 단계를 포함하는 방법:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 제공하기 위해, 상기 뉴클레오티드 유사체 존재 하에 저편향 DNA 중합효소를 사용하여 상기 적어도 하나의 타겟 핵산 분자를 증폭하는 단계;
c. 적어도 하나의 돌연변이된 타겟 핵산 분자를 제공하기 위해, 뉴클레오티드 유사체 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계;
d. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자를 벡터에 삽입하는 단계; 및
e. 상기 적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질을 발현하는 단계. - 청구항 58 또는 59에 있어서, 상기 방법은 상기 적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질의 활성을 테스트하거나 구조를 평가하는 단계를 더 포함하는 것인 방법.
- 청구항 58 내지 60 중 어느 한 항에 있어서, 상기 벡터는 플라스미드, 바이러스, 코스미드 (cosmid), 또는 인공 염색체 (artificial chromosome)인 것인 방법.
- 청구항 58 내지 61 중 어느 한 항에 있어서, 상기 적어도 하나의 돌연변이된 타겟 핵산 분자가 코딩하는 단백질을 발현하는 단계는 상기 벡터로 박테리아 세포를 형질전환 (transforming)하거나, 진핵세포를 형질감염 (transfecting)하거나 또는 진핵세포를 형질도입 (transducing)함으로써 이루어지는 것인 방법.
- 샘플 태그 그룹으로서, 상기 각 샘플 태그는 적어도 1개의 낮은 확률 돌연변이 차이 또는 적어도 3개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 샘플 태그 그룹.
- 청구항 63에 있어서, 상기 각 샘플 태그는 적어도 3개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 샘플 태그 그룹.
- 청구항 63 또는 64에 있어서, 상기 각 샘플 태그는 3 내지 25개, 또는 3 내지 10개의 낮은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 샘플 태그 그룹.
- 청구항 63 내지 65 중 어느 한 항에 있어서, 상기 낮은 확률 돌연변이는 전환 돌연변이 또는 인델 돌연변이인 것인 샘플 태그 그룹.
- 청구항 63 내지 66 중 어느 한 항에 있어서, 상기 각 샘플 태그는 적어도 5개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 샘플 태그 그룹.
- 청구항 63 내지 67 중 어느 한 항에 있어서, 상기 각 샘플 태그는 5 내지 25개, 또는 5 내지 10개의 높은 확률 돌연변이 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 샘플 태그 그룹.
- 청구항 63 내지 68에 있어서, 상기 높은 확률 돌연변이는 전이 돌연변이인 것인 샘플 태그 그룹.
- 청구항 63 내지 69 중 어느 한 항에 있어서, 상기 각 샘플 태그는 적어도 8개 뉴클레오티드, 적어도 10개 뉴클레오티드, 적어도 12개 뉴클레오티드, 8 내지 50개의 뉴클레오티드, 10 내지 50개의 뉴클레오티드, 또는 10 내지 50개의 뉴클레오티드 길이인 것인 샘플 태그 그룹.
- 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법에 사용하기에 적합한 샘플 태그 그룹을 설계하는 방법으로서, 하기 단계를 포함하는 방법:
a. 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 분석하고 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 낮은 확률 돌연변이의 평균 개수를 결정하는 단계; 및
b. 샘플 태그 그룹의 서열을 결정하는 단계로서, 상기 각 샘플 태그는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 낮은 확률 돌연변이의 평균 개수보다 더 많은 낮은 확률 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 단계. - 청구항 71에 있어서, 다음을 더 포함하는 방법:
a. (i) 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법을 분석하고 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 높은 확률 돌연변이의 평균 개수를 결정하는 단계; 및
(ii) 샘플 태그 그룹의 서열을 결정하는 단계로서, 상기 각 샘플 태그는 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법 중에 발생하는 높은 확률 돌연변이의 평균 개수보다 더 많은 높은 확률 차이로 그룹 내의 모든 다른 샘플 태그와 실질적으로 다른 것인 단계. - 청구항 71 또는 72에 있어서, 상기 낮은 확률 돌연변이는 전환 돌연변이 또는 인델 돌연변이인 것인 방법.
- 청구항 72 내지 73 중 어느 한 항에 있어서, 상기 높은 확률 돌연변이는 전이 돌연변이인 것인 방법.
- 청구항 71 내지 74 중 어느 한 항에 있어서, 컴퓨터에서 구현되는 (computer implemented) 것인 방법.
- 청구항 1 내지 75 중 어느 한 항에 있어서, 상기 저편향 DNA 중합효소를 사용하여 적어도 하나의 타겟 핵산 분자를 증폭하는 단계는 불균등 농도의 dNTP를 사용하여 수행되는 것인 방법 또는 용도.
- 청구항 1 내지 76 중 어느 한 항에 있어서,
(i) 상기 방법은 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 추가 단계를 포함하고, 상기 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 추가 단계는 불균등 농도의 dNTP를 사용하여 수행되는 것이고; 또는
(ii) 상기 방법은 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하고, 상기 방법은 저편향 DNA 중합효소를 사용하여 상기 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 추가 단계를 포함하고, 상기 저편향 DNA 중합효소를 사용하여 상기 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 추가 단계는 불균등 농도의 dNTP를 사용하여 수행되는 것인 방법. - 하기 단계를 포함하는, 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 방법,:
a. 적어도 하나의 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계; 및
b. 돌연변이된 적어도 하나의 타겟 핵산 분자를 제공하기 위해, DNA 중합효소를 사용하여 상기 적어도 하나의 타겟 핵산 분자를 증폭하여 적어도 하나의 타겟 핵산 분자에 돌연변이를 도입하는 단계로서,
상기 단계 b. 는 불균등 농도의 dNTP를 사용하여 수행되는 것인 단계. - 청구항 78에 있어서, 상기 단계 b. 는 뉴클레오티드 유사체의 존재 하에 수행되는 것인 방법.
- 청구항 79에 있어서, 상기 뉴클레오티드 유사체는 dPTP인 것인 방법.
- 청구항 79 또는 80에 있어서, 상기 방법은 뉴클레오티드 유사체의 부재 하에 상기 돌연변이된 적어도 하나의 타겟 핵산 분자를 증폭하는 추가 단계 c. 를 포함하는 것인 방법.
- 청구항 81에 있어서, 상기 단계 c. 는 불균등 농도의 dNTP를 사용하여 수행되는 것인 방법.
- 청구항 76 내지 82 중 어느 한 항에 있어서, 불균등 농도의 dNTP는 도입된 돌연변이의 프로필 (profile)을 변경하기 위해 사용되는 것인 방법.
- 청구항 83에 있어서, 불균등 농도의 dNTP는 도입된 돌연변이의 프로필에서 편향 (bias)을 줄이기 위해 사용되는 것인 방법.
- 청구항 1 내지 84 중 어느 한 항에 있어서, 상기 방법은 저편향 돌연변이 프로필로 돌연변이를 도입하는 방법인 것인 방법.
- 청구항 76 내지 85 중 어느 한 항에 있어서, 상기 불균등 농도의 dNTP는 dATP, dCTP, dTTP 및 dGTP를 포함하고 dATP, dCTP, dTTP 또는 dGTP 중 1개 또는 2개는 다른 dNTP보다 낮은 농도인 것인 방법.
- 청구항 76 내지 86 중 어느 한 항에 있어서, 불균등 농도의 dNTP의 사용은 도입된 돌연변이의 프로필에서 편향을 줄이기 위해 그 수준이 증가 또는 감소되어야 하는 dNTP를 확인하는 단계를 포함하는 것인 방법.
- 청구항 76 내지 87 중 어느 한 항에 있어서, 불균등 농도의 dNTP의 사용은 다른 dNTP보다 낮은 농도의 dTTP를 포함하는 것인 방법.
- 청구항 88에 있어서, 상기 불균등 농도의 dNTP는 dTTP를 dATP, dCTP 또는 dGTP 농도의 75% 미만, 70% 미만, 60% 미만, 55% 미만, 25% 내지 75%, 25% 내지 70, 25% 내지 60%, 또는 약 50%의 농도로 포함하는 것인 방법.
- 청구항 89에 있어서, 상기 불균등 농도의 dNTP는 dTTP를 dCTP 농도의 75% 미만, 70% 미만, 60% 미만, 55% 미만, 25% 내지 75%, 25% 내지 70, 25% 내지 60%, 또는 약 50%의 농도로 포함하는 것인 방법.
- 청구항 90에 있어서, 상기 불균등 농도의 dNTP는 dTTP를 dCTP 농도의 60% 미만의 농도로 포함하는 것인 방법.
- 청구항 87에 있어서, 상기 불균등 농도의 dNTP는 dTTP를 dCTP 농도의 25% 내지 60%의 농도로 포함하는 것인 방법.
- 청구항 77, 또는 81 내지 92 중 어느 한 항에 있어서, 상기 뉴클레오티드 유사체의 부재 하에 상기 뉴클레오티드 유사체를 포함하는 적어도 하나의 타겟 핵산 분자를 증폭하는 단계 또는 상기 돌연변이된 적어도 하나의 타겟 핵산 분자를 뉴클레오티드 유사체의 부재 하에 증폭하는 단계는 불균등 농도의 dNTP를 사용하여 수행되는 것인 방법.
- 청구항 93에 있어서, 상기 불균등 농도의 dNTP는 dATP를 다른 dNTP보다 낮은 농도로 포함하는 것인 방법.
- 청구항 94에 있어서, 상기 불균등 농도의 dNTP는 dATP를 dTTP, dCTP 또는 dGTP 농도의 75% 미만, 70% 미만, 60% 미만, 55% 미만, 25% 내지 75%, 25% 내지 70, 25% 내지 60%, 또는 약 50%의 농도로 포함하는 것인 방법.
- 청구항 95에 있어서, 상기 불균등 농도의 dNTP는 dATP를 dGTP 농도의 75% 미만, 70% 미만, 60% 미만, 55% 미만, 25% 내지 75%, 25% 내지 70, 25% 내지 60%, 또는 약 50%의 농도로 포함하는 것인 방법.
- 청구항 96에 있어서, 상기 불균등 농도의 dNTP는 dATP를 dGTP 농도의 60% 미만의 농도로 포함하는 것인 방법.
- 청구항 96 또는 97에 있어서, 상기 불균등 농도의 dNTP는 dATP를 dGTP 농도의 25% 내지 60%의 농도로 포함하는 것인 방법.
- 청구항 71 내지 74 중 어느 한 항의 방법으로 얻을 수 있는 샘플 태그 그룹.
- 청구항 71 내지 74 중 어느 한 항의 방법을 수행하도록 구성된 컴퓨터 판독가능 매체.
- a. 타겟 핵산 분자를 포함하는 적어도 하나의 샘플을 제공하는 단계;
b. 타겟 핵산 분자의 3' 말단에 제1 어댑터 및 타겟 핵산 분자의 5' 말단에 제2 어댑터를 도입하는 단계; 및
c. 상기 제1 어댑터의 일부에 상보적인 프라이머들을 사용하여 상기 타겟 핵산 분자를 증폭하는 단계를 포함하고,
상기 제1 어댑터 및 제2 어댑터는 서로에게 어닐링(anneal)하는 것인,
길이가 1 kbp보다 큰 타겟 핵산 분자를 우선적으로 (preferentially) 증폭하는 방법. - 청구항 101에 있어서, 상기 프라이머들은 서로 동일한 것인 방법.
- 청구항 101 또는 102에 있어서, 상기 제1 어댑터는 제2 어댑터에 적어도 80%, 적어도 90%, 적어도 95%, 적어도 99%, 또는 100% 동일한 핵산 분자에 상보적인 것인 방법.
- 청구항 101 내지 103 중 어느 한 항에 있어서, 상기 방법은 길이가 1.5 kbp보다 큰 타겟 핵산 분자를 우선적으로 증폭하는 방법인 것인 방법.
- 청구항 101 내지 104 중 어느 한 항에 있어서, 상기 타겟 핵산 분자를 시퀀싱하는 단계를 더 포함하는 것인 방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| GB1802744.1 | 2018-02-20 | ||
| GBGB1802744.1A GB201802744D0 (en) | 2018-02-20 | 2018-02-20 | Method |
| PCT/GB2019/050443 WO2019162657A1 (en) | 2018-02-20 | 2019-02-19 | Method for introducing mutations |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20200123458A true KR20200123458A (ko) | 2020-10-29 |
Family
ID=61783821
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020207027106A KR20200123458A (ko) | 2018-02-20 | 2019-02-19 | 돌연변이를 도입하는 방법 |
Country Status (12)
| Country | Link |
|---|---|
| US (2) | US11421238B2 (ko) |
| EP (2) | EP4008792A1 (ko) |
| JP (2) | JP7413283B2 (ko) |
| KR (1) | KR20200123458A (ko) |
| CN (1) | CN112088219A (ko) |
| AU (1) | AU2019223315A1 (ko) |
| CA (1) | CA3091770A1 (ko) |
| DK (1) | DK3673084T3 (ko) |
| ES (1) | ES2889548T3 (ko) |
| GB (1) | GB201802744D0 (ko) |
| SG (1) | SG11202007924TA (ko) |
| WO (1) | WO2019162657A1 (ko) |
Family Cites Families (15)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5780231A (en) * | 1995-11-17 | 1998-07-14 | Lynx Therapeutics, Inc. | DNA extension and analysis with rolling primers |
| WO2002079502A1 (en) | 2001-03-28 | 2002-10-10 | The University Of Queensland | A method for nucleic acid sequence analysis |
| EP2292767B1 (en) | 2004-06-04 | 2013-11-20 | Takara Bio, Inc. | Polypepdides having DNA polymerase activity |
| US9481912B2 (en) * | 2006-09-12 | 2016-11-01 | Longhorn Vaccines And Diagnostics, Llc | Compositions and methods for detecting and identifying nucleic acid sequences in biological samples |
| JP5279339B2 (ja) | 2008-05-16 | 2013-09-04 | タカラバイオ株式会社 | 逆転写反応用組成物 |
| WO2011106368A2 (en) | 2010-02-23 | 2011-09-01 | Illumina, Inc. | Amplification methods to minimise sequence specific bias |
| AU2011253984A1 (en) * | 2010-12-07 | 2012-06-28 | Biobalance Llc | Method For Identifying E. Coli M-17 |
| US9977861B2 (en) | 2012-07-18 | 2018-05-22 | Illumina Cambridge Limited | Methods and systems for determining haplotypes and phasing of haplotypes |
| HUE058149T2 (hu) * | 2014-01-31 | 2022-07-28 | Swift Biosciences Inc | Javított eljárások DNS-szubsztrátok feldolgozására |
| CN104232761A (zh) | 2014-08-27 | 2014-12-24 | 武汉凯吉盈科技有限公司 | 基因全片段快速测序方法 |
| EP3957742A1 (en) | 2014-10-10 | 2022-02-23 | Cold Spring Harbor Laboratory | Random nucleotide mutation for nucleotide template counting and assembly |
| WO2017059582A1 (zh) | 2015-10-09 | 2017-04-13 | 云南省烟草农业科学研究院 | 一种抗烟草花叶病毒的N'au基因及其克隆方法和应用 |
| CN107760771A (zh) | 2017-11-16 | 2018-03-06 | 杭州迪安医学检验中心有限公司 | 一种乳腺癌易感基因brca1和brca2全基因捕获引物、试剂盒及其方法 |
| CN107955806B (zh) | 2017-12-15 | 2020-03-31 | 中国科学院深海科学与工程研究所 | 一种深渊海参来源的超氧化物歧化酶Cu,Zn SOD的制备方法及其应用 |
| EP4343770A2 (en) | 2018-10-26 | 2024-03-27 | The Board of Trustees of the Leland Stanford Junior University | Methods and uses of introducing mutations into genetic material for genome assembly |
-
2018
- 2018-02-20 GB GBGB1802744.1A patent/GB201802744D0/en not_active Ceased
-
2019
- 2019-02-19 JP JP2020566338A patent/JP7413283B2/ja active Active
- 2019-02-19 ES ES19707096T patent/ES2889548T3/es active Active
- 2019-02-19 US US16/971,293 patent/US11421238B2/en active Active
- 2019-02-19 EP EP21190654.0A patent/EP4008792A1/en active Pending
- 2019-02-19 CA CA3091770A patent/CA3091770A1/en active Pending
- 2019-02-19 SG SG11202007924TA patent/SG11202007924TA/en unknown
- 2019-02-19 DK DK19707096.4T patent/DK3673084T3/da active
- 2019-02-19 KR KR1020207027106A patent/KR20200123458A/ko unknown
- 2019-02-19 CN CN201980027144.7A patent/CN112088219A/zh active Pending
- 2019-02-19 WO PCT/GB2019/050443 patent/WO2019162657A1/en unknown
- 2019-02-19 EP EP19707096.4A patent/EP3673084B1/en active Active
- 2019-02-19 AU AU2019223315A patent/AU2019223315A1/en active Pending
-
2022
- 2022-06-20 US US17/807,835 patent/US20220348940A1/en active Pending
-
2023
- 2023-12-27 JP JP2023221021A patent/JP2024038164A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| US20220348940A1 (en) | 2022-11-03 |
| JP2021514205A (ja) | 2021-06-10 |
| ES2889548T3 (es) | 2022-01-12 |
| DK3673084T3 (da) | 2021-11-08 |
| US11421238B2 (en) | 2022-08-23 |
| CN112088219A (zh) | 2020-12-15 |
| WO2019162657A1 (en) | 2019-08-29 |
| EP3673084A1 (en) | 2020-07-01 |
| GB201802744D0 (en) | 2018-04-04 |
| EP3673084B1 (en) | 2021-08-11 |
| EP4008792A1 (en) | 2022-06-08 |
| CA3091770A1 (en) | 2019-08-29 |
| SG11202007924TA (en) | 2020-09-29 |
| US20210010008A1 (en) | 2021-01-14 |
| AU2019223315A1 (en) | 2020-10-01 |
| JP7413283B2 (ja) | 2024-01-15 |
| JP2024038164A (ja) | 2024-03-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20230099446A1 (en) | Fast pcr for str genotyping | |
| RU2698125C2 (ru) | Библиотеки для секвенирования нового поколения | |
| EP2971080B1 (en) | Methods for amplification and sequencing using thermostable tthprimpol | |
| EP2173872B1 (en) | Copy dna and sense rna | |
| JP2003526378A (ja) | 高忠実度逆転写酵素およびその使用 | |
| CA2406939A1 (en) | Thermostable reverse transcriptases and uses thereof | |
| US8114653B2 (en) | Thermostable Y-family polymerases and chimeras | |
| EP1546355A2 (en) | Methods of use for thermostable rna ligases | |
| CN111684064A (zh) | 突变dna聚合酶 | |
| KR20220113522A (ko) | 주형-비의존성 핵산을 합성하는 방법 및 키트 | |
| WO2022144003A1 (zh) | 一种用于高通量靶向测序的多重pcr文库构建方法 | |
| EP3673084B1 (en) | Method for introducing mutations | |
| US20230257803A1 (en) | Strand displacing amplification enzymes | |
| CN114686565A (zh) | 一种用于高通量靶向测序的单管嵌套多重pcr扩增方法 | |
| CN112074612A (zh) | 一种具有更高特异性的核酸扩增方法 | |
| JPWO2019162657A5 (ko) | ||
| ZA200610533B (en) | Thermostable Y-family polymerases and chimeras |