KR20100080507A - 죽상경화증의 치료 - Google Patents
죽상경화증의 치료 Download PDFInfo
- Publication number
- KR20100080507A KR20100080507A KR1020107005287A KR20107005287A KR20100080507A KR 20100080507 A KR20100080507 A KR 20100080507A KR 1020107005287 A KR1020107005287 A KR 1020107005287A KR 20107005287 A KR20107005287 A KR 20107005287A KR 20100080507 A KR20100080507 A KR 20100080507A
- Authority
- KR
- South Korea
- Prior art keywords
- leu
- cetp
- ser
- prt
- artificial sequence
- Prior art date
Links
Images

































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/04—Peptides having up to 20 amino acids in a fully defined sequence; Derivatives thereof
- A61K38/08—Peptides having 5 to 11 amino acids
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/08—Linear peptides containing only normal peptide links having 12 to 20 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/04—Peptides having up to 20 amino acids in a fully defined sequence; Derivatives thereof
- A61K38/10—Peptides having 12 to 20 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
- A61K38/16—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- A61K38/17—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- A61K38/1703—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates
- A61K38/1709—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans from vertebrates from mammals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/0005—Vertebrate antigens
- A61K39/0012—Lipids; Lipoproteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/39—Medicinal preparations containing antigens or antibodies characterised by the immunostimulating additives, e.g. chemical adjuvants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
- A61K47/51—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent
- A61K47/62—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being a protein, peptide or polyamino acid
- A61K47/64—Drug-peptide, drug-protein or drug-polyamino acid conjugates, i.e. the modifying agent being a peptide, protein or polyamino acid which is covalently bonded or complexed to a therapeutically active agent
- A61K47/643—Albumins, e.g. HSA, BSA, ovalbumin or a Keyhole Limpet Hemocyanin [KHL]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K47/00—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient
- A61K47/50—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates
- A61K47/51—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent
- A61K47/62—Medicinal preparations characterised by the non-active ingredients used, e.g. carriers or inert additives; Targeting or modifying agents chemically bound to the active ingredient the non-active ingredient being chemically bound to the active ingredient, e.g. polymer-drug conjugates the non-active ingredient being a modifying agent the modifying agent being a protein, peptide or polyamino acid
- A61K47/64—Drug-peptide, drug-protein or drug-polyamino acid conjugates, i.e. the modifying agent being a peptide, protein or polyamino acid which is covalently bonded or complexed to a therapeutically active agent
- A61K47/646—Drug-peptide, drug-protein or drug-polyamino acid conjugates, i.e. the modifying agent being a peptide, protein or polyamino acid which is covalently bonded or complexed to a therapeutically active agent the entire peptide or protein drug conjugate elicits an immune response, e.g. conjugate vaccines
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K9/00—Medicinal preparations characterised by special physical form
- A61K9/0012—Galenical forms characterised by the site of application
- A61K9/0019—Injectable compositions; Intramuscular, intravenous, arterial, subcutaneous administration; Compositions to be administered through the skin in an invasive manner
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P3/00—Drugs for disorders of the metabolism
- A61P3/06—Antihyperlipidemics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P9/00—Drugs for disorders of the cardiovascular system
- A61P9/10—Drugs for disorders of the cardiovascular system for treating ischaemic or atherosclerotic diseases, e.g. antianginal drugs, coronary vasodilators, drugs for myocardial infarction, retinopathy, cerebrovascula insufficiency, renal arteriosclerosis
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/06—Linear peptides containing only normal peptide links having 5 to 11 amino acids
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55505—Inorganic adjuvants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55555—Liposomes; Vesicles, e.g. nanoparticles; Spheres, e.g. nanospheres; Polymers
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55566—Emulsions, e.g. Freund's adjuvant, MF59
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/60—Medicinal preparations containing antigens or antibodies characteristics by the carrier linked to the antigen
- A61K2039/6031—Proteins
- A61K2039/6081—Albumin; Keyhole limpet haemocyanin [KLH]
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Engineering & Computer Science (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Molecular Biology (AREA)
- Epidemiology (AREA)
- Immunology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Genetics & Genomics (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Gastroenterology & Hepatology (AREA)
- Microbiology (AREA)
- Heart & Thoracic Surgery (AREA)
- Cardiology (AREA)
- Mycology (AREA)
- Marine Sciences & Fisheries (AREA)
- Vascular Medicine (AREA)
- Urology & Nephrology (AREA)
- Zoology (AREA)
- Virology (AREA)
- Obesity (AREA)
- Hematology (AREA)
- Diabetes (AREA)
- Dermatology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Peptides Or Proteins (AREA)
- Plural Heterocyclic Compounds (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
Abstract
본 발명은 죽상경화증, 죽상경화증 위험 질환 및 죽상경화증 후유증을 예방하고/하거나 치료하기 위한 약제의 생성에서의 화합물의 용도에 관한 것이다.
Description
본 발명은 죽상경화증, 죽상경화증 위험 질환 및 죽상경화증 후유증의 예방 및 치료에 관한 것이다.
죽상경화증 후유증, 예를 들어, 말초 동맥 폐색 질환, 관상동맥 심질환 및 중풍성 뇌졸중은 여전히 미국, 유럽 및 아시아 대부분에서 주 사망 원인의 하나가 되고 있다. 죽상경화증의 발달은 성장 인자, 사이토카인 및 세포 상호작용의 복잡한 상호작용을 특징으로 하는, 동맥 혈관벽의 만성적인 진행성 염증이 존재하는 것으로 간주된다. "손상에 대한 반응(response-to-injury)"의 가설에 따르면, 내피의 손상이 질환의 초기 상태를 구성하고, 이로부터 세포 상호작용 연쇄 반응을 유발시켜 결국 죽상경화증 병변을 형성시키는 내피의 기능 이상이 야기된다. 이러한 "손상"을 촉진하는 위험 인자로서, 죽상경화증과 통계학적으로 유의하게 관련되는 외인성 및 내인성 영향이 언급된다. 이러한 내피 손상 인자의 가장 중요한 것들 중에서, 예를 들어 증가되거나 변형된 LDL, Lp(a), 동맥 고혈압, 당뇨병 및 과호모시스테인혈증이 언급된다. 내피는 단단하지만 오히려 매우 동적인 장벽을 구성하지 않기 때문에, 지질단백질의 증가된 투과성 이외에도 내피의 기능이상의 과정 중에 많은 분자 변형이 일어나며, 이러한 분자 변형은 단핵구, T-림프구 및 내피 세포의 상호작용에 결정적인 영향을 미친다. E, L 및 P 셀렉틴, 인테그린, ICMA-1, VCAM-1 및 혈소판 내피-세포 부착 분자-1 유형의 내피 부착 분자의 발현에 의해, 루멘 측부에서 단핵구 및 T-림프구의 부착이 일어난다. 내피 상으로의 후속하는 백혈구의 이동은 MCP-1, 인터루킨-8, PDGF, M-CSF 및 오스테오폰틴에 의해 매개된다. 소위 스캐빈저 수용체를 경유하여, 내막 내에 존재하는 대식세포 및 단핵구가 침투된 LDL 입자를 흡수할 수 있고, 이들을 세포질 내에 콜레스테롤 에스테르의 공포(vaculole)로서 침착시킨다. 이러한 방식으로 형성된 포말 세포는 혈관 내막 영역에 주로 덩어리져서 축적되어, 이미 유아기에 형성되는 "지방 선조(fatty streak)"를 형성한다. LDL은 저밀도의 지질단백질이며, 이는 트리글리세라이드가 풍부한 VLDL 입자로부터의 지질분해 효소의 대사 효과에 의해 형성된다. 중막의 평활근 세포 및 내피 세포에 대한 이들의 손상 특성 이외에도, LDL은 또한 단핵구에 대한 화학주성 효과를 가지며, 유전자 증폭을 통해 내피 세포의 MCP-1 및 MCSF의 발현을 증가시킬 수 있다. LDL과는 대조적으로, HDL은 소위 HDL 복합체의 형성 하에 아포지질단백질 E에 의해 매개된 로딩된 대식세포로부터 콜레스테롤 에스테르를 용해시킬 수 있다. SR-B1 수용체의 상호작용에 의해, 이들 콜레스테롤 에스테르-로딩된 입자는 간세포 또는 부신피질 세포에 결합하여, 각각 담즙산 및 스테로이드를 생성하기 위해 콜레스테롤을 전달할 수 있다. 이러한 메커니즘을 소위 콜레스테롤 역 전달(reverse cholesterol transport)이라고 하며, 이는 HDL의 보호 기능을 설명한다. 활성화된 대식세포는 HLA-DR을 통해 항원을 제시하여 CD4 및 CD8 림프구를 활성화시킬 수 있고, 이것에 의해 결과적으로 IFN-감마 및 TNF-알파와 같은 사이토카인의 분비가 자극되며, 또한 상기 활성화된 대식세포는 염증 반응을 증가시키는 데 기여한다. 질환의 추가 과정에서, 중막의 평활근 세포는 염증에 의해 변형된 내막 영역 내로 성장하기 시작한다. 이에 의해, 중간 병변이 이 단계에서 형성된다. 중간 병변으로부터 출발하여, 진행성의 복잡한 병변이 시간 경과에 따라 발달하며, 상기 병변은 루멘 측부 상에서 콜라겐이 풍부한 섬유소 캡, 세포 퇴적물 및 괴사성 코어에 의해 형태학적으로 특징지어진다. 리포이드 부분 및 세포수가 연속하여 증가하면, 내피가 인열될 것이고, 혈전 특성을 지닌 표면이 노출될 것이다. 이러한 인열시 혈소판의 부착 및 활성화로 인해, 사이토카인, 성장 인자 및 트롬빈을 함유하는 과립이 분비될 것이다. 대식세포의 단백질분해 효소는 섬유소 캡의 박막화를 담당하며, 상기 박막화에 의해 종국적으로 연속적인 혈전증 형성 및 혈관의 협착에 의해 플라크가 파괴되고 말초 혈관의 급성 허혈증이 야기될 것이다.
죽상경화증 병변의 형성을 담당하는 다양한 위험 인자가 있다. 과지질단백혈증, 동맥 고혈압 및 니코틴 남용이 이와 관련하여 특히 중요하다. 총 콜레스테롤 및 LDL 콜레스테롤에 있어서의 과다한 증가를 포함하는 질환은 가족 과콜레스테롤뇨혈증(FH)이다. 이는 가장 빈번하게 단세대로 유전되는 대사 질환에 속한다. 적절한 이종접합 형태는 1:500명의 빈도로 나타나고, 동종접합 형태는 더욱 드물게 1:일백만명의 빈도로 나타난다. 가족 과콜레스테롤뇨혈증은 염색체 19의 짧은 암(arm) 상의 LDL 수용체 유전자에서의 돌연변이로부터 기인한다. 이러한 돌연변이는 결실, 삽입 또는 점 돌연변이일 수 있다. 가족 과콜레스테롤뇨혈증에서 지질단백질의 특징적인 확인은 가장 일반적인 트리글리세라이드 및 VLDL 농도에서의 총 콜레스테롤 및 LDL 콜레스테롤의 증가이다. 종종 HDL이 저하되기도 한다. 표현형에서, 타입 IIAa-과지질단백혈증이 존재한다. 이종접합 형태에서는, 총 콜레스테롤이 정상 수준과 비교하여 2배 내지 3배 증가하고, 동종접합 형태에서는 5배 내지 6배 증가한다. 임상적으로 가족 과콜레스테롤뇨혈증은 초기 관상 경화증으로 자체적으로 드러난다. 일반적으로, 이종접합형의 인간에서, 관상동맥 심질환(CHD)의 최초 증상은 30 내지 40세 사이에 일어나며, 여성의 경우에는 이보다 평균 10년 뒤에 나타난다. 발병된 남자의 50%가 이들이 50세가 되기 전에 관상 동맥 경화증이 발병하여 사망한다. LDL 수준의 과다한 증가 이외에도, 저하된 HDL 농도가 죽상경화증의 급속한 진행의 원인이 된다. 죽상경화증 변화는 심장외 혈관, 예를 들어, 대동맥, 목동맥 및 말초 동맥에 대해서도 뚜렷하게 나타날 수 있다. 동종접합 형태의 질환에 대해서도, 관상 동맥 경화증이 초기 유아기에 이미 발달한다. 최초의 심근경색은 종종 10세 이전에 나타나며, 대부분의 경우 발병된 환자들은 이들이 20세가 되기 전에 사망한다. 황색종의 발달은 혈청 콜레스테롤의 수준 및 질환 지속기간의 함수이다. 20세 이상에서 발병된 이종접합형 개체의 약 75%가 힘줄 황색종을 나타낸다. 동종접합형의 개체들은 거의 100%가 피부 및 힘줄 황색종을 지니고 있다. 지질 침착물이 또한 눈꺼풀 상에 그리고 각막 내에 발생될 수 있다 (황색판종; 지질환(Arcus lipoides)). 그러나, 상기한 지질 침착물은 정상 콜레스테롤 수준에서도 확인되기 때문에, 과콜레스테롤뇨혈증의 특징적인 징후라고 보기는 어렵다. 또한, FH에 대해서도, 급성 관절염 및 건초염(tendosynovitides)이 빈번하게 발생한다. 개개의 지질단백질의 크기 및 밀도는 상이한데, 그 이유는 이들 지질단백질이 지질 및 단백질의 상이하게 큰 부분, 소위 아포단백질을 함유하기 때문이다. 단백질이 증가하고 지질 부분이 감소함에 따라 밀도가 증가한다. 지질단백질의 상이한 밀도 때문에, 이들을 초원심분리에 의해 다양한 분획으로 분리할 수 있다. 이는 지질단백질을, 하기한 이들의 주요 군으로 분류하는 데 대한 토대를 제공한다: 킬로마이크론(chylomicron), 매우 저밀도 지질단백질 (VLDL), 중간 밀도 지질단백질 (IDL), 저밀도 지질단백질 (LDL), 고밀도 지질단백질 (HDL), 지질단백질(a) (Lp(a)). 고도의 죽종형성 잠재력을 지닌 지질단백질 중에서, 주로 LDL, Lp(a) 및 VLDL이 있다. LDL은 대략 d = 1.006 내지 1.063 g/ml의 밀도를 갖고 있다. 코어는 에스테르화된 콜레스테롤 분자에 의해 형성된다. 이러한 고도의 소수성 코어는 인지질, 비에스테르화된 콜레스테롤 및 하나의 단일 Apo B100 분자의 외피에 의해 둘러싸여 있다. 게다가, 아포단백질 E는 LDL 입자의 표면 상에서 확인된다. LDL의 기능은 콜레스테롤을 말초 조직으로 전달하는 것이며, 여기서 아포단백질 B-100 분자에 의해 매개되어 콜레스테롤이 LDL 수용체를 통해 세포 내로 흡수된다. 포괄적인 역학 연구에서, 혈청 콜레스테롤 수준과 관상동맥 심질환의 발병율 사이의 양성 상관관계가 입증될 수 있었다. 160 mg/dl 초과의 LDL 콜레스테롤 수준은 높은 심장혈관 위험을 구성한다. LDL 콜레스테롤 수준 이외에도, 혈관 보호성 HDL 콜레스테롤 수준이 또한 심장혈관 질환에 대한 위험 프로파일을 예측함에 있어 중요한 역할을 한다. 35 mg/dl 미만의 수치가 증가된 위험과 관련되어 있다. VLDL은 저밀도 (d = 0.94 내지 1.006 g/ml) 및 높은 트리글리세라이드 부분을 지닌 지질단백질이다. 실질적으로, VLDL은 아포단백질 C, 및 작은 부분의 아포단백질 B-100 및 E를 함유한다. 킬로마이크론과는 다르게, VLDL은 식품 지질로 구성되는 것이 아니라, 내인적으로 형성된 트리글리세라이드로부터 간에서 합성되어 순환계로 분비된다. 킬로마이크론과 마찬가지로, 트리글리세라이드는 아포단백질 C-II-활성화된 지질단백질-리파아제에 의해 가수분해되고, 유리 지방산이 근육 및 지방 조직으로 공급된다. 남아있는 콜레스테롤 풍부 VLDL 잔여물을 소위 중간 밀도 지질단백질이라고 하는데, 그 이유는 이의 더욱 높은 밀도 때문이다. 지질단백질(a) (Lp(a))는 1.05 내지 1.12 g/ml의 밀도를 지니고 이의 조성도 LDL과 유사하다. 아포단백질 B-100 이외에도, 이의 단백질 부분은 Lp(a)의 특징인 아포단백질(a)로 구성된다. 현재까지, Lp(a)의 기능 및 생리에 대해서는 거의 알려진 바가 없다. 아포단백질(a) 분자가 플라스미노겐에 대해 고도의 서열 상동성을 갖고 있기 때문에, Lp(a)는 죽상경화판 상에서의 혈전의 형성을 촉진하고 또한 죽종형성 효과를 갖는 것으로 추정된다. Lp(a)는 아포단백질 B와 함께 죽상경화증 병변에서 확인된다. 후향 연구로부터 증가된 Lp(a)와 CHD 사이에서 상관관계가 입증되었다. 마찬가지로, 다수의 전향 연구의 메타분석으로부터 Lp(a)가 CHD의 발병에 대한 독립적인 위험 인자임이 밝혀졌다. 15 내지 35 mg/dl의 수준이 정상인 것으로 간주된다. 지금까지, Lp(a)는 식이 또는 약물 중 어느 것에 의해서도 영향받을 수 없었다. 따라서, 치료 수단이 추가의 위험 인자를 감소시키는 것으로 제한된다. 특히, LDL 콜레스테롤을 감소시키면 Lp(a)의 심장혈관 위험이 저하되는 것으로 보인다. 죽상경화증의 발병기전에서, 상당한 병태생리학적 중요성이 응고 인자로부터 비롯되고 있다. 역학 조사의 결과로부터, 혈장 내에서의 섬유소원 농도와 관상동맥 심질환, 주로 심근 경색의 발달 사이에서의 상관관계가 제안된다. 이러한 맥락에서, 증가된 섬유소원 수준 (> 300 mg/dl)이 심장혈관 질환에 대한 독립적인 지표 및 위험 인자임이 판명되었다. 또한, 고농도의 조직 플라스미노겐 활성인자 억제제 tPA-I이 CHD의 발병과 관련되어 있다. 과-트리글리세라이드혈증과 관상동맥 위험 사이의 관련성은 각 경우에 상이한데, 이는 혈액 지질의 상승 원인에 따라 달라진다. 트리글리세라이드가 독립적인 위험 인자로서 간주될 것인지 아닌지에 대한 논의에도 불구하고, 이들이 관상동맥 심질환의 발병기전에서 중요한 역할을 담당하고 있음이 논의되고 있다. 관상동맥 심질환의 발병율은 높은 LDL 콜레스테롤 및 높은 트리글리세라이드 수준을 나타내는 환자에서 가장 높다.
콜레스테롤 에스테르 전달 단백질 (CETP)은 지질단백질 사이에서 중성 지질과 인지질의 전달을 담당하며 HDL의 혈장 농도를 하향 조절하는 안정한 혈장 당단백질이다. CETP 지질 전달 활성의 억제는 이미 HDL 혈장 수준을 증가시키기 위한 치료법으로서 이미 제안되었다. 혈장에서 CETP 활성의 감소로부터 HDL 수준이 증가되는 것을 제안하는 많은 이유가 있다. 따라서, CETP는 HDL로부터 LDL 및 VLDL로의 콜레스테롤 에스테르의 전달에 의해 HDL 농도를 저하시킨다. 토끼 및 햄스터를 이용한 동물 실험에서, 항-CETP 모노클로날 항체, 안티센스 올리고누클레오티드 또는 CETP 억제제를 이용한 CETP의 일시적인 억제는 HDL 수준을 증가시켰다. CETP를 안티센스 올리고누클레오티드를 사용하여 계속하여 억제시켰더니 HDL 수준이 증가되었고, 이에 따라 죽상경화증에 대한 토끼 동물 모델에서 죽상경화증 병변이 감소되었다.
문헌에서, 여러 CETP 억제제가 기재되어 있으며, 이중 일부는 임상 시험중이다(예를 들어, 아나세트라피브(Anacetrapib)(Krishna R., Lancet 370 (9603) (2007): 1907-14) 및 토르세트라피브(Torcetrapib)(Sikorski, J.A., J.Med.Chem. 49 (1) (2006): 1-22)).
US 5,512,548호 및 WO 93/011782호에는, HDL로부터 VLDL 및 LDL로의 콜레스테롤 에스테르의 전달을 촉매하는 CETP를 억제할 수 있어, 환자에게 투여되는 경우에 항-죽상경화증 활성을 갖는 폴리펩티드 및 이의 유사체가 기재되어 있다. 상기 문헌에 따르면, 이러한 CETP 폴리펩티드 억제제는 다양한 공급원의 아포지질단백질 C 내지 I로부터 유래하며, 여기서 특히 아미노산 36까지의 N-말단 단편이 CETP 억제제로서 확인되었다.
또한 US 5,880,095 A호에는 개체에서 CETP의 활성을 억제할 수 있는 CETP-결합 펩티드가 기재되어 있다. CETP-억제 단백질은 돼지의 아포지질단백질 C-III의 N-말단 단편을 포함한다.
US 2006/0276400호 및 WO 96/034888호에는 CETP로부터 유래되고, T-세포 및/또는 B-세포 에피토프를 포함하는 펩티드가 기재되어 있다. 이러한 펩티드는 생체내에서 CETP 특이적 항체의 형성을 유도할 수 있다.
US 2004/0087481호 및 US 6,410,022 B1호에는 CETP-특이적인 면역 반응의 유도 때문에 심장혈관 질환, 예를 들어, 죽상경화증의 치료 및 예방에 사용될 수 있는 펩티드가 기재되어 있다. 이들 펩티드는 CETP로부터 유래하지 않는 T 헬퍼 세포 에피토프, 및 CETP로부터 발생하며 CETP로부터 직접 유래될 수 있는 하나 이상의 B-세포 에피토프를 포함한다. T 헬퍼 세포 에피토프는 유리하게는 파상풍 톡소이드로부터 유래하며, CETP의 적어도 하나의 B-세포 에피토프에 공유 결합한다. 유기체에 대해 이물질인 T 헬퍼 세포 에피토프를 사용함으로써, 개체의 체내에서 항체가 유도되고, 이로써 하나 이상의 CETP-B-세포 에피토프로 구성되는 펩티드 부분에 대해 항체가 유도되는 것이 가능하다.
문헌[Mao D et al (Vaccine 24(2006): 4942-4950)]에는 백신으로서 CETP의 B 세포 에피토프를 엔코딩하는 핵산 분자를 포함하는 플라스미드의 용도가 기재되어 있다.
WO 2006/029982호에는 죽상경화증의 치료 또는 예방을 위한 약제의 제조에 사용되는 CETP 미모토프(mimotope)가 기재되어 있다.
가장 최근에는, CETP에 관한 백신 접근법에 대한 제안이 이미 이루어졌다. 따라서, 예를 들어, 토끼가 항원으로서 콜레스테롤-에스테르 전달을 책임지는 CETP의 펩티드를 포함하는 백신으로 처리되었다. 면역화된 토끼는 감소된 CETP 활성, 및 증가된 HDL 및 감소된 LDL 값의 변경된 지질단백질 수준을 가졌다. 더욱이, 죽상경화증 모델의 처리된 시험 동물은 또한 대조 동물과 비교하여 감소된 죽상경화증 병변을 나타내었다.
Ⅱ기 임상 연구의 결과가 공개되었으며, 이러한 연구는 백신 CETi-1을 이용하여 미국 생명공학 회사 애번트(Avant)사에 의해 수행되었다(BioCentury Extra For Wednesday, October 22, 2003). 이러한 Ⅱ기 연구에서, 선행 Ⅰ기 연구에서와 같이, 임의의 의심스러운 부작용이 없이 매우 우수한 안전성 프로파일이 입증되었으며, 이는 기본적으로 항-CETP 예방접종 접근법으로부터 부작용이 예상되지 않는다는 결론이 도출되는 것을 가능케 한다. 그러나, 효능과 관련하여, 애번트사의 백신은 실망스러웠는데, 이는 상기 백신이 위약 치료에 의해 달성되는 것보다 현저하게 나은 증가된 HDL 수준을 야기시키지 않았기 때문이다.
CETi-1 백신의 문제점은 이러한 백신이 내인성 항원을 이용한다는 점이다. 인간 면역계는 내인성 구조와 관련하여 관용(tolerance)하는데, 이는 CETP가 아닌 대부분의 내인성 분자를 이용하여서는 자가항체가 형성되지 않는다는 점이 결정적이기 때문이다. 따라서, CETi-1 백신의 목표는 명백히 충분한 범위로 달성되지 않는 내인성 관용을 파괴하는 것이었다.
따라서, 면역계에 의해 외래인 것으로 간주되고, 이에 따라 자가 관용을 파괴하는 것이 필요하지 않도록 선택된 항-CETP 백신을 위한 항원을 제공하는 것이 본 발명의 목표이다. 이러한 항원은 죽상경화증, 죽상경화증 위험 질환 및 죽상경화증 후유증을 예방하고/하거나 치료하는데 사용될 수 있다.
따라서, 본 발명은, 콜레스테롤 에스테르 전달 단백질(CETP) 또는 CETP-에피토프의 4-머(mer) 내지 16-머 폴리펩티드 단편이 아니거나 이를 포함하지 않는 아미노산 서열 (Z1)nX1X2X3X4(Z2)m(여기서, Z1은 C가 아닌 아미노산 잔기이고, X1은 D, A, R, E, S, N, T 및 G로 구성된 군으로부터 선택된 아미노산 잔기이고, X2는 F, A, W, R, S, L, Q, V 및 M으로 구성된 군으로부터 선택된 아미노산 잔기이고, X3는 L, A, S, W, E, R, I 및 H로 구성된 군으로부터 선택된 아미노산 잔기이고, X4는 Q, A, H, D, K, R, S 및 E로 구성된 군으로부터 선택된 아미노산 잔기이고, Z2는 C가 아닌 아미노산 잔기이고, n은 0 내지 10, 바람직하게는 0 내지 9의 정수이고, m은 0 내지 3의 정수임)을 포함하는 화합물로서, 천연 CETP 당단백질에 특이적인 항체에 대한 결합 능력을 갖는 화합물, 또는 SYHATFL, TMAFPLN, HYHGAFL, EHHDIFL, TGLSVFL, WMPSLFY, SMPWWFF, TMPLLFW, DTWPGLE, SMPPIFY, MPLWWWD, SMPNLFY, RMPPIFY, NPFEVFL, TLPNWFW, SMPLTFY, SPHPHFL, NFMSIGL, SQFLASL, WSWPGLN, IAWPGLD, SKFMDTL, SMPMVFY, YEWVGLM, KGFLDHL, HQSDDKMPWWFF, YVWQDPSFTTFF, YVWQDPSFTTFF, LPQTHPLHLLED, GPVSIYADTDFL, DSNDTLTLAAFL, NGSPALSHMLFL, TDYDPMWVFFGY, IFPLDSQWQTFW, NESMPDLFYQPS, DWGDKYFSSFWN, VSAYNNV 및 WPLHLWQ로 구성된 군으로부터 선택된 아미노산 서열을 포함하는 화합물의 죽상경화증, 죽상경화증 위험 질환 및 죽상경화증 후유증을 예방하고/하거나 치료하기 위한 약제 생성에서의 용도에 관한 것이다.
본 발명은 상기 목적을 위해 CETP 미모토프를 제공한다. 이러한 미모토프는 CETP 효소 활성을 억제할 수 있는 항체를 유도할 수 있다. 본 발명에 따른 CETP 미모토프는 바람직하게는 아미노산 서열이 CETP 또는 CETP의 단편의 아미노산 서열로부터 변화된 항원성 폴리펩티드이다. 이러한 점에서, 본 발명의 미모토프는 하나 이상의 비천연 아미노산(즉, 20개의 "고전적인" 아미노산으로부터 유래되지 않음)을 포함할 수 있거나, 이들은 상기 비천연 아미노산으로 전부 어셈블리될 수 있다. 더욱이, 항-CETP 항체를 유도하는 본 발명의 항원은 D-아미노산 또는 L-아미노산 또는 DL-아미노산의 조합물로 어셈블리될 수 있고, 임의로 이들은 추가 개질, 고리 닫힘 또는 유도체화에 의해 변경될 수 있다. 적합한 항-CETP-항체-유도 항원이 시판되는 펩티드 라이브러리로부터 제공될 수 있다. 바람직하게는, 이러한 펩티드는 4개 이상의 아미노산 잔기, 특히 7개 이상의 아미노산 잔기 길이이며, 바람직한 길이는 16개 이하, 바람직하게는 14 또는 20개 이하의 아미노산(예를 들어, 5 내지 16개의 아미노산 잔기)일 수 있다. 그러나, 본 발명에 따르면, 보다 긴 펩티드가 또한 항-CETP-항체-유도 항원으로 매우 적절히 이용될 수 있다. 더욱이, 본 발명의 미모토프는 또한 폴리펩티드의 일부일 수 있고, 그 결과로 N-말단 및/또는 C-말단에 하나 이상의 추가 아미노산 잔기를 포함할 수 있다.
본 발명의 미모토프는 KLH 또는 다른 담체에 커플링된 C-FGFPEHLLVDFLQSLS(CETP 단백질의 16개의 C-말단 아미노산)의 포유동물로의 투여에 의해 수득될 수 있는 항체에 결합할 수 있다. 포유동물에 투여된 후, 미모토프는 해당 면역 반응을 유도할 수 있고, 상기 포유동물에서 CETP에 특이적이 항체가 생성된다.
본 발명의 CETP-미모토프(즉, 항-CETP-항체-유도 항원)는 파아지 라이브러리 또는 펩티드 라이브러리를 포함하는 다양한 방법에 의해 확인되고 제조될 수 있다. 이들은, 예를 들어, 대부분의 다양한 구조에 대한 조합 화학 또는 높은 처리량의 스크리닝 기술에 의해 생성되고 확인될 수 있다(Display: A Laboratory Manual by Carlos F. Barbas (Editor), et al.; Willats WG Phage display: practicalities and prospects. Plant Mol. Biol. 2002; 50(6):837-54).
더욱이, 본 발명에 따르면, 핵산에 기초한 항-CETP-항체-유도 항원("앱타머(aptamer)")가 또한 사용될 수 있고, 이들 역시 대부분의 다양한 (올리고누클레오티드) 라이브러리(예를 들어, 2-180개의 핵산 잔기)를 이용하여 발견될 수 있다(참조: Burgstaller et al., Curr. Opin. Drug Discov. Dev. 5(5) (2002), 690-700; Famulok et al., Acc. Chem. Res. 33 (2000), 591-599; Mayer et al., PNAS 98 (2001), 4961-4965, 등). 핵산에 기초한 항-CETP-항체-유도 항원에서, 핵산 백본이, 예를 들어, 천연 포스포디에스테르(phosphor-diester) 화합물, 또는 포스포로티오에이트 또는 조합물 또는 화학적 변이체(예를 들어, PNA로서)에 의해 제공될 수 있고, 여기서 본 발명에 따르면 염기로서 주로 U, T, A, C, G, H 및 mC가 사용될 수 있다. 본 발명에 따라 사용될 수 있는 누클레오티드의 2'-잔기는 바람직하게는 H, OH, F, Cl, NH2, O-메틸, O-에틸, O-프로필 또는 O-부틸이고, 여기서 핵산은 또한 다양하게 개질될 수 있으며, 즉 예를 들어 핵산은 보호기로 개질될 수 있는데, 이러한 보호기가 올리고누클레오티드 합성에 통상적으로 사용되기 때문이다. 따라서, 앱타머에 기초한 항-CETP-항체-유도 항원이 또한 본 발명의 범위 내에서 바람직한 항-CETP-항체-유도 항원이다.
본 발명에 따르면, 용어 "미모토프"는 이러한 미모토프가 미믹(mimic)으로 작용하는 에피토프와 동등한 위상을 갖는 형태를 갖는 분자를 의미한다. 미모토프는 요망되는 항원에 면역특이적으로 결합하는 항체의 동일한 항원 결합 영역에 결합한다. 미모토프는 이러한 미모토프가 미믹으로 작용하는 항원에 대해 반응성인 숙주에서 면역학적 반응을 유발할 것이다. 미모토프는 또한 에피토프 및 상기 에피토프에 결합하는 항체를 포함하는 시험관내 억제 검정(예를 들어, ELISA 억제 검정)에서 상기 미모토프가 미믹으로 작용하는 에피토프에 대한 경쟁자로 작용할 수 있다. 그러나, 본 발명의 미모토프는 포유동물에 투여시 특정 면역반응을 유도할 수 있으나, 시험관내 억제 검정에서 이러한 미모토프가 미믹으로 작용하는 에피토프의 결합을 반드시 방해하거나 이와 경쟁하지는 않는다.
본원에서 사용되는 용어 "에피토프"는 특정 항체 분자에 의해 인지되는 항원의 면역원성 영역을 의미한다. 일반적으로, 항원은 특정 에피토프를 인지하는 항체에 각각 결합할 수 있는 하나 이상의 에피토프를 가질 것이다.
본 발명에 개시된 아미노산 잔기에 대한 약어는 IUPAC 권고사항을 따른다:

본 발명의 미모토프는 당 분야에 널리 공지된 화학적 합성 방법에 의해 분리된 펩티드 또는 또 다른 펩티드 또는 폴리펩티드의 일부로서 합성적으로 생성될 수 있다. 대안적으로, 펩티드 미모토프는 펩티드 미모토프를 생성하는 미생물에서 생성될 수 있고, 이는 이후에 분리되고, 요망시 추가로 정제될 수 있다. 펩티드 미모토프는 미생물, 예를 들어, 박테리아, 효소 또는 진균, 진핵 세포, 예를 들어, 포유동물 또는 곤충 세포, 또는 재조합 바이러스 벡터, 예를 들어, 아데노바이러스, 폭스바이러스, 헤르페스바이러스, 셈리키 삼림열 바이러스(Simliki forest virus), 배큘로바이러스, 박테리오파아지, 신드비스 바이러스 또는 센다이 바이러스 벡터에서 생성될 수 있다. 펩티드 미모토프를 생성하기에 적합한 박테리아는 대장균(E.coli), B. 섭틸리스(B.subtilis) 또는 펩티드 미모토프와 같은 펩티드를 발현할 수 있는 임의의 다른 박테리아를 포함한다. 펩티드 미모토프를 발현하기에 적합한 효모 유형은 사카로마이세스 세레비지애(Saccharomyces cerevisiae), 스키조사카로마이세스 폼베(Schizosaccharomyces pombe), 캔디다(Candida), 피키아 파스토리스(Pichia pastoris) 또는 펩티드를 발현할 수 있는 임의의 다른 효모를 포함한다. 해당 방법은 당 분야에 널리 공지되어 있다. 재조합적으로 생성된 펩티드를 분리하고 정제하는 방법은 또한 당 분야에 널리 공지되어 있고, 이는, 예를 들어, 겔 여과, 친화성 크로마토그래피, 이온 교환 크로마토그래피 등을 포함한다.
펩티드 미모토프의 분리를 촉진하기 위해, 펩티드 미모토프가 친화성 크로마토그래피에 의한 분리를 가능하게 하는 이종성 폴리펩티드에 번역적으로 융합(공유 결합)된 융합 폴리펩티드가 제조될 수 있다. 통상적인 이종성 폴리펩티드는 His-Tag(예를 들어, His6; 6개의 히스티딘 잔기), GST-Tag(글루타티온-S-트랜스퍼라아제) 등이다. 융합 폴리펩티드는 미모토프의 정제를 촉진할 뿐만 아니라 정제 동안 미모토프 폴리펩티드가 분해되는 것을 방지할 수 있다. 정제 후에 이종성 폴리펩티드를 제거하는 것이 요망되는 경우, 융합 폴리펩티드는 펩티드 미모토프와 이종성 폴리펩티드 사이의 접합부에 절단 부위를 포함할 수 있다. 절단 부위는 상기 부위의 아미노산 서열에 특이적인 효소(예를 들어, 프로테아제)로 절단되는 아미노산 서열로 구성된다.
본 발명의 미모토프는 또한 N-말단 및/또는 C-말단에서 또는 이 부근에서 개질되며, 상기 위치에서 시스테인 잔기가 이에 결합된다. 한 바람직한 구체예에서, 말단에 위치된(펩티드의 N-말단 및 C-말단에 위치됨) 시스테인 잔기가 이황화결합을 통해 펩티드를 고리화시키는데 사용된다.
본 발명의 미모토프는 또한 다양한 검정 및 키트, 특히 면역학적 검정 및 키트에 사용될 수 있다. 따라서, 미모토프가 또 다른 펩티드 또는 폴리펩티드의 일부, 특히 면역학적 검정에서 리포터로 사용되는 효소일 수 있는 것이 특히 바람직하다. 이러한 리포터 효소는, 예를 들어, 알칼리성 포스파타아제 또는 호스라디쉬 퍼옥시다아제(horseradish peroxidase)를 포함한다.
용어 "죽상경화증 후유증" 또는 "죽상경화증의 후유증"은 죽상경화증의 결과인 질환을 의미한다. 이러한 질환은 특히 말초 동맥 폐색 질환, 관상동맥 심질환 및 중풍성 뇌졸중(apoplectic cerebral insultus)을 포함한다(예를 들어, 참조: Steinberg D. J. Lipid Res. (2005) 46: 179-190; Steinberg D et al. J. Lipid Res (2006) 47: 1339-1351).
본 발명의 또 다른 바람직한 구체예에 따르면, X1은 D이고, X4는 Q 또는 H, 바람직하게는 Q이다. 이러한 분자는 바람직하게는 이의 N-말단에, Xa가 P, Y, T 또는 K이고, Xb가 C가 아닌 아미노산 잔기이고, Xc가 H이고, Xd가 Y, L, H, V, T, I 또는 F이고, Xe가 Y, I, P, L, Q, S, R, T, F 또는 A이고, Xf가 A, W, V, Q, L, S, I, R 또는 T인, 서열 Xa Xb Xc Xd Xe Xf를 갖는 아미노산 잔기를 추가로 포함한다.
본 발명의 한 바람직한 구체예에 따르면, n은 7, 8 또는 9이고, Z1은 C가 아닌 아미노산 잔기이거나, F, G, F, A, P, W, Y, S, G, D, L, E, K, T, P, I 및 M으로 구성된 군, 바람직하게는 F, G, F, A, P, Y, T, S, G, K 및 D로 구성된 군으로부터 선택되고, Z2는 S, L, A, W, L, N, T, I, Y 및 H로 구성된 군으로부터 선택된다.
본 발명의 한 추가의 바람직한 구체예에 따르면, X1은 D, A, R, E 및 L로 구성된 군으로부터 선택되고, X2는 F, A, W, Q 및 R로 구성된 군으로부터 선택되고, X3는 L, A 및 S로 구성된 군으로부터 선택되고, X4는 Q, A 및 H로 구성된 군으로부터 선택된다.
본 발명의 한 바람직한 구체예에 따르면, X1은 D이고, X2는 F, Q 및 W로 구성된 군으로부터 선택되고, X3는 L 또는 S이고, X4는 Q 또는 H이다.
본 발명의 한 바람직한 구체예에 따르면, 화합물은, X8이 G, A, F, Y 및 K로 구성된 군으로부터 선택되고, X9이 E, Y, A, Q, K 및 S로 구성된 군으로부터 선택되고, X10이 H, V, L, F 및 I로 구성된 군으로부터 선택되고, X11이 L, W, S, I, F 및 Y로 구성된 군으로부터 선택되고, X12가 V, T, F 또는 I이고, X5가 S 또는 Y이고, X6가 L, A 또는 I이고, X7이 S, N 또는 T이고, o가 0 또는 1인, 아미노산 서열 FX8(F)oPX9HX10X11X12DX2X3X4X5X6X7을 포함한다.
본 발명의 화합물은 바람직하게는, X1이 D, S, N, T 및 G로 구성된 군으로부터 선택되고, X2가 F이고, X3가 L이고, X4가 Q, D, K, R, S 및 E로 구성된 군으로부터 선택되고, X5가 S 또는 T이고, X6가 L이고, X7이 C가 아닌 아미노산 잔기, 바람직하게는 S, T, A, M, F 및 W로 구성된 군으로부터 선택되는, 아미노산 서열 X1X2X3X4X5X6X7을 포함한다.
본 발명의 한 바람직한 구체예에 따르면, 아미노산 서열은 SSLELFL, SFLDTLT, NFLKTLS, DFLRTLT, AFLDTLV, TFLSSLA, GFLDSLM, SPHPHFL, SNFLKTL, TGFLATL, SDFLRAL, SANPRDFLETLF, RMFPESFLDTLW, TIYDSFLDSLAS, KPYLLKDFLEAL, AMGPYDALDLFL, TWNPIESFLESL, QYQTPLTFLEAL, RHISPATFLEAL, HTDSFLSTFYGD, ADSTFTSFLQTL, GPVSIYADTDFL, DSNDTLTLAAFL, TPTHYYADFSQL, LPGHLIWDSLHY, LPQTHPLHLLED, IPYHHLVDQLHH, YPYHVQVDVLQN, IPSHHLQDSLQL, EYAHHTSLDLRQ, EPLHFRSDRIQA, ATPSHLIIDRAQ, APKHLYADMSQA, FKPAHVSIDWLQ, MPAHLSRDLRQS, NPKHYSIDRHQA, SPQHLTTDRAQA, TPFHFAQDSWQW, TPTHYYADFSQLLS, TPTHYYADFSQSLS, GTPTHYYADFSQLL, GTPTHYYADFSQSL, FGTPTHYYADFSQSLS, FGFPTHYYADFSQSLS, LPGHLIWDSLHY, LPGHLIWDSLHYL, LPGHLIWDSLHYLS, LPGHLIWDSLHSL, LPGHLIWDSLHSLS, GLPGHLIWDSLHYL, GLPGHLIWDSLHSL, FGLPGHLIWDSLHSLS, FGFPGHLIWDSLHSLS, LPQTHPLHLLED, IPYHHLVDQLHH, IPYHHLVDQLHLS, IPYHHLVDQLHSLS, FGIPYHHLVDQLHHLS, FGFPYHHLVDQLHSLS, YPYHVQVDVLQN, YPYHVQVDVLQNLS, YPYHVQVDVLQSLS, FGYPYHVQVDVLQNLS, FGFPYHVQVDVLQSLS, IPSHHLQDSLQL, IPSHHLQDSLQLLS, IPSHHLQDSLQSLS, GIPSHHLQDSLQLL, FGIPSHHLQDSLQLLS, FGFPSHHLQDSLQSLS, EYAHHTSLDLRQ, EPLHFRSDRIQA, EPLHFRSDRIQALS, EPLHFRSDRIQSLS, GEPLHFRSDRIQAL, FGEPLHFRSDRIQALS, FGFPLHFRSDRIQSLS, APKHLYADMSQA, APKHLYADMSQALS, APKHLYADMSQSLS, GAPKHLYADMSQAL, FGFPKHLYADMSQSLS, MPAHLSRDLRQS, MPAHLSRDLRQSL, MPAHLSRDLRQSLS, GMPAHLSRDLRQSL, FGFPAHLSRDLRQSLS, NPKHYSIDRHQA, TPFHFAQDSWQW, TPFHFAQDSWQWLS, TPFHFAQDSWQSLS, GTPFHFAQDSWQWL, FGFPFHFAQDSWQSLS, ACSFAYLYRC, ACFMGDKWVC, ACVLYPKAIC, ACYMGQQFVC, ACLTAYLHWC, ACTLFPVAYC, ACWLFPYAHC, ACKSINMWLC, ACQTINRWLC, FGFPEHLLVDFLQSLS, FGFPEHLLVDFLQSLS, FPEHLLVDFLQSL, AGFPEHLLVDFLQSLS, FAFPEHLLVDFLQSLS, FGAPEHLLVDFLQSLS, FGFAEHLLVDFLQSLS, FGFPAHLLVDFLQSLS, FGFPEALLVDFLQSLS, FGFPEHALVDFLQSLS, FGFPEHLAVDFLQSLS, FGFPEHLLADFLQSLS, FGFPEHLLVAFLQSLS, FGFPEHLLVDALQSLS, FGFPEHLLVDFAQSLS, FGFPEHLLVDFLASLS, FGFPEHLLVDFLQALS, FGFPEHLLVDFLQSAS, FGFPEHLLVDFLQSLA, FAFPAHLLVDFLQALA, AAFPAHLLADFLQALA, SPQHLTTDRAQA, SPQHLTTDRAQALS, SPQHLTTDRAQSLS, GSPQHLTTDRAQAL, FGFPQHLTTDRAQSLS, FGFPQHLTTDWAQSLS, FGFPQHLTTDRLQSLS, FGFPQHLTTDWLQSLS, ATPSHLIIDRAQ, ATPSHLIIDRAQSLS, FGFPSHLIIDRAQSLS, FGFPSHLIIDWAQSLS, FGFPSHLIIDWLQSLS, FGFPSHLIIDWSQSLS, FATPSHLIIDWLQSLS, FKPAHVSIDWLQ, FKPAHVSIDWLQSLS, FGFPAHVSIDWLQSLS, AGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFAAHVSIDWLQSLS, FGFPAHVSADWLQSLS, FGFPAHVSIDWLQALS, FGFPAHVSIDWLQSLA, FAFPAHVSIDWLQALA, FGFAAHVSIDWLQSLS, FGFFAHVSIDWLQSLS, FGFPAHVSIRWLQSLS, FGFPAHVSIEWLQSLS, FGFPAHVSIDWLNSLS, FGFPAHVSIDWLHSLS, AGFPAHVSIDWLQSLS, PGFPAHVSIDWLQSLS, WGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FSFPAHVSIDWLQSLS, FYFPAHVSIDWLQSLS, FDFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFPAHVSIDWLQLLS, FGFPAHVSIDWLQWLS, FGFPAHVSIDWLQNLS, FGFPAHVSIDWLQTLS, FGFPAHVSIDWLQYLS, FGFPAHVSIDWLQSIS, FGFPAHVSIDWLQSLT, FGFPAHVSIDWLQSLY, FAFPAHVSIDWLQALA, FGFPAHVSIDRAQSLS, FGFPTHVSIDWLQSLS, FGFPFHVSIDWLQSLS, FGFPAHISIDWLQSLS, FGFPAHIIIDWLQSLS, FGFPAHLTTDWLQSLS, FGFPAHVFIDWLQSLS, FGFPAHVYIDWLQSLS, FGFPAHVSLDWLQSLS, FGFPAHVSADWLQSLS, TPTHYYADFSQSLS, FGFPAHVSIDWSQSLS, FGFPAHVSIDFSQSLS, FGFPSHIIIDWLQSLS, FGFPSHLIIEWLQSLS, AAFPAHLLADAAQALA, AAFPAHAAADFLQALA, AAFAAHLLADFLQAAA, AAAPAHLLVDAAQAAA, FAFPAHVFIDWLQSLS; FGFPAHVFIDWLQALS, FGFPAHVFIDWLQSLA, GFPAHVFIDWLQSLS, FPAHVFIDWLQSLS, PAHVFIDWLQSLS, FAFPAHVFIDWLQALA, FGFPEHLFVDFLQSLS, FGFPAHVHIDWLQSLS, FGFPAHVPIDWLQSLS, FGFPSHLFIDWAQSLS, PGFPAHVFIDWLQLIT, PAHVYIDWLQSLS, FGFPAHVYIDWLQ, FGFPAHVFIDWLQ, DFGFPSHLIIDWLQSLS, DFGFPAHVFIDWLQSLN, PSHLIIDWLQ, PAHVFIDWLQ, DFGFPAHVTIDWLQSLN, DFGFPAHVLIDWLQSLN, FGFPAHVYIDWLQSLS, FGFPAHVFIDWLQSLN 및 FGFPAHVFIDWLQSLA로 구성된 군으로부터 선택된다.
본 발명에 따라 사용되는 특히 바람직한 미모토프는 SANPRDFLETLF, RMFPESFLDTLW, SFLDTLT, NFLKTLS, DFLRTLT, TFLSSLA, GFLDSLM, FGFPYHVQVDVLQSLS, FGFPSHLIIDRAQSLS, FKPAHVSIDWLQSLS, FGFPAHVSIDWLQSLS, FGFPQHLTTDRAQSLS, FGFPTHYYADFSQSLS, FGFPGHLIWDSLHSLS, FGFPYHHLVDQLHSLS, FGFPSHHLQDSLQSLS, FGFPLHFRSDRIQSLS, FGFPKHLYADMSQSLS, FGFPAHLSRDLRQSLS 및 FGFPFHFAQDSWQSLS이다.
본 발명의 특히 바람직한 미모토프는 FGFPSHLIIDWLQSLS, FGFPAHVFIDWLQSLS 및 FGFPAHVYIDWLQSLS이다.
추가의 바람직한 미모토프는 FGFPAHVWIDWLQSLS, FGFPAHVFIDWLQSLN, FGFPAHFSIDWLQSLS, FGFPAHVSFDWLQSLS, FGFPEHVFIDWLQSLS, DFGFPAHVFIDWLQSLS, DFGFPSHLIIDWLQSLS, DFGFPAHVYIDWLQSLS, FGFPQHLFTDWLQSLS 및 FGFPKHLLVDFLQSLS이다.
본 발명의 한 바람직한 구체예에 따르면, 화합물은 약학적으로 허용되는 담체, 바람직하게는 키홀 림펫 헤모시아닌(Keyhole Limpet Hemocyanin, KLH), 파상풍 톡소이드, 알부민 결합 단백질, 우혈청 알부민, 덴드리머(dendrimer)(MAP; Biol. Chem. 358: 581), 펩티드 링커(또는 플랭킹(flanking) 영역) 뿐만 아니라 문헌[Singh et al., Nat. Biotech. 17 (1999), 1075-1081(특히, 이의 표 1에 기재된 것), 및 O'Hagan et al., Nature Reviews, Drug Discovery 2 (9) (2003), 727-735(특히, 이에 기재된 내인성 면역증강 화합물 및 전달 시스템)]에 기재된 애쥬번트 물질, 또는 이의 혼합물에 커플링된다. 이러한 상황에서 컨쥬게이션 화학(예를 들어, 이종이작용성 화합물, 예를 들어, GMBS, 및 또한 문헌["Bioconjugate Techniques", Greg T. Hermanson]에 기재된 다른 것들을 통함)이 당업자에게 공지된 반응으로부터 선택될 수 있다. 더욱이, 백신 조성물은 애쥬번트, 바람직하게는 저 가용성 알루미늄 조성물, 특히 수산화알루미늄과 함께 제형화될 수 있다. 물론, MF59 인산알루미늄, 인산칼슘, 사이토카인(예를 들어, IL-2, IL-12, GM-CSF), 사포닌(예를 들어, QS21), MDP 유도체, CpG 올리고(oligo), LPS, MPL, 폴리포스파젠, 에멀젼(예를 들어, 프로인트(Freund's), SAF), 리포솜, 바이로솜, iscoms, 코클리에이트(cochleate), PLG 마이크로입자, 폴록사머 입자, 바이러스-유사 입자, 이열성 장독소(LT), 콜레라 독소(CT), 돌연변이 독소(예를 들어, LTK63 및 LTR72), 마이크로입자 및/또는 중합화된 리포솜과 같은 애쥬번트가 또한 사용될 수 있다.
본 발명의 화합물은 바람직하게는 NHS-폴리 (에틸렌 옥시드)(PEO)(예를 들어, NHS-PEO4-말레이미드)로 구성된 군으로부터 선택되는 링커를 통해 담체 또는 애쥬번트에 결합된다.
본 발명의 화합물(미모토프) 및 약학적으로 허용되는 담체를 포함하는 백신은 임의의 적합한 적용 방법, 예를 들어, i.d., i.v., i.p., i.m., 비내, 경구, 피하 등, 및 임의의 적합한 전달 장치(O'Hagan et al., Nature Reviews, Drug Discovery 2 (9), (2003), 727-735)에 의해 투여될 수 있다. 본 발명의 화합물은 바람직하게는 정맥내, 피하, 피내 또는 근내 투여용으로 제형화된다(예를 들어, 참조:"Handbook of Pharmaceutical Manufacturing Formulations", Sarfaraz Niazi, CRC Press Inc, 2004).
통상적으로, 백신은 본 발명에 따른 화합물을 0.1 ng 내지 10 mg, 바람직하게는 10 ng 내지 1 mg, 특히 100 ng 내지 100 ㎍, 또는, 대안적으로, 예를 들어, 100 fmol 내지 10 μmol, 바람직하게는 10 pmol 내지 1 μmol, 특히 100 pmol 내지 100 nmol의 양으로 함유한다. 통상적으로, 백신은 또한 보조 물질, 예를 들어, 완충액, 안정화제 등을 함유할 수 있다.
본 발명의 또 다른 양태는 SYHATFL, TMAFPLN, HYHGAFL, EHHDIFL, SSLELFL, TGLSVFL, WMPSLFY, SMPWWFF, TMPLLFW, DTWPGLE, SMPPIFY, MPLWWWD, SMPNLFY, RMPPIFY, NPFEVFL, TLPNWFW, SMPLTFY, SFLDTLT, NFLKTLS, DFLRTLT, AFLDTLV, TFLSSLA, GFLDSLM, SPHPHFL, NFMSIGL, SQFLASL, SNFLKTL, TGFLATL, WSWPGLN, IAWPGLD, SKFMDTL, SDFLRAL, SMPMVFY, YEWVGLM, KGFLDHL, SANPRDFLETLF, RMFPESFLDTLW, TIYDSFLDSLAS, HQSDDKMPWWFF, KPYLLKDFLEAL, AMGPYDALDLFL, TWNPIESFLESL, YVWQDPSFTTFF, QYQTPLTFLEAL, RHISPATFLEAL, HTDSFLSTFYGD, YVWQDPSFTTFF, ADSTFTSFLQTL, GPVSIYADTDFL, DSNDTLTLAAFL, NGSPALSHMLFL, TDYDPMWVFFGY, IFPLDSQWQTFW, NESMPDLFYQPS, DWGDKYFSSFWN, VSAYNNV, WPLHLWQ, TPTHYYADFSQL, LPGHLIWDSLHY, LPQTHPLHLLED, IPYHHLVDQLHH, YPYHVQVDVLQN, IPSHHLQDSLQL, EYAHHTSLDLRQ, EPLHFRSDRIQA, ATPSHLIIDRAQ, APKHLYADMSQA, FKPAHVSIDWLQ, MPAHLSRDLRQS, NPKHYSIDRHQA, SPQHLTTDRAQA, TPFHFAQDSWQW, TPTHYYADFSQLLS, TPTHYYADFSQSLS, GTPTHYYADFSQLL, GTPTHYYADFSQSL, FGTPTHYYADFSQSLS, FGFPTHYYADFSQSLS, LPGHLIWDSLHY, LPGHLIWDSLHYL, LPGHLIWDSLHYLS, LPGHLIWDSLHSL, LPGHLIWDSLHSLS, GLPGHLIWDSLHYL, GLPGHLIWDSLHSL, FGLPGHLIWDSLHSLS, FGFPGHLIWDSLHSLS, LPQTHPLHLLED, IPYHHLVDQLHH, IPYHHLVDQLHLS, IPYHHLVDQLHSLS, FGIPYHHLVDQLHHLS, FGFPYHHLVDQLHSLS, YPYHVQVDVLQN, YPYHVQVDVLQNLS, YPYHVQVDVLQSLS, FGYPYHVQVDVLQNLS, FGFPYHVQVDVLQSLS, IPSHHLQDSLQL, IPSHHLQDSLQLLS, IPSHHLQDSLQSLS, GIPSHHLQDSLQLL, FGIPSHHLQDSLQLLS, FGFPSHHLQDSLQSLS, EYAHHTSLDLRQ, EPLHFRSDRIQA, EPLHFRSDRIQALS, EPLHFRSDRIQSLS, GEPLHFRSDRIQAL, FGEPLHFRSDRIQALS, FGFPLHFRSDRIQSLS, APKHLYADMSQA, APKHLYADMSQALS, APKHLYADMSQSLS, GAPKHLYADMSQAL, FGFPKHLYADMSQSLS, MPAHLSRDLRQS, MPAHLSRDLRQSL, MPAHLSRDLRQSLS, GMPAHLSRDLRQSL, FGFPAHLSRDLRQSLS, NPKHYSIDRHQA, TPFHFAQDSWQW, TPFHFAQDSWQWLS, TPFHFAQDSWQSLS, GTPFHFAQDSWQWL, FGFPFHFAQDSWQSLS, ACSFAYLYRC, ACFMGDKWVC, ACVLYPKAIC, ACYMGQQFVC, ACLTAYLHWC, ACTLFPVAYC, ACWLFPYAHC, ACKSINMWLC, ACQTINRWLC, FGFPEHLLVDFLQSLS, FGFPEHLLVDFLQSLS, FPEHLLVDFLQSL, AGFPEHLLVDFLQSLS, FAFPEHLLVDFLQSLS, FGAPEHLLVDFLQSLS, FGFAEHLLVDFLQSLS, FGFPAHLLVDFLQSLS, FGFPEALLVDFLQSLS, FGFPEHALVDFLQSLS, FGFPEHLAVDFLQSLS, FGFPEHLLADFLQSLS, FGFPEHLLVAFLQSLS, FGFPEHLLVDALQSLS, FGFPEHLLVDFAQSLS, FGFPEHLLVDFLASLS, FGFPEHLLVDFLQALS, FGFPEHLLVDFLQSAS, FGFPEHLLVDFLQSLA, FAFPAHLLVDFLQALA, AAFPAHLLADFLQALA, SPQHLTTDRAQA, SPQHLTTDRAQALS, SPQHLTTDRAQSLS, GSPQHLTTDRAQAL, FGFPQHLTTDRAQSLS, FGFPQHLTTDWAQSLS, FGFPQHLTTDRLQSLS, FGFPQHLTTDWLQSLS, ATPSHLIIDRAQ, ATPSHLIIDRAQSLS, FGFPSHLIIDRAQSLS, FGFPSHLIIDWAQSLS, FGFPSHLIIDWLQSLS, FGFPSHLIIDWSQSLS, FATPSHLIIDWLQSLS, FKPAHVSIDWLQ, FKPAHVSIDWLQSLS, FGFPAHVSIDWLQSLS, AGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFAAHVSIDWLQSLS, FGFPAHVSADWLQSLS, FGFPAHVSIDWLQALS, FGFPAHVSIDWLQSLA, FAFPAHVSIDWLQALA, FGFAAHVSIDWLQSLS, FGFFAHVSIDWLQSLS, FGFPAHVSIRWLQSLS, FGFPAHVSIEWLQSLS, FGFPAHVSIDWLNSLS, FGFPAHVSIDWLHSLS, AGFPAHVSIDWLQSLS, PGFPAHVSIDWLQSLS, WGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FSFPAHVSIDWLQSLS, FYFPAHVSIDWLQSLS, FDFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFPAHVSIDWLQLLS, FGFPAHVSIDWLQWLS, FGFPAHVSIDWLQNLS, FGFPAHVSIDWLQTLS, FGFPAHVSIDWLQYLS, FGFPAHVSIDWLQSIS, FGFPAHVSIDWLQSLT, FGFPAHVSIDWLQSLY, FAFPAHVSIDWLQALA, FGFPAHVSIDRAQSLS, FGFPTHVSIDWLQSLS, FGFPFHVSIDWLQSLS, FGFPAHISIDWLQSLS, FGFPAHIIIDWLQSLS, FGFPAHLTTDWLQSLS, FGFPAHVFIDWLQSLS, FGFPAHVYIDWLQSLS, FGFPAHVSLDWLQSLS, FGFPAHVSADWLQSLS, TPTHYYADFSQSLS, FGFPAHVWIDWLQSLS, FGFPAHVFIDWLQSLN, FGFPAHFSIDWLQSLS, FGFPAHVSFDWLQSLS, FGFPEHVFIDWLQSLS, DFGFPAHVFIDWLQSLS, DFGFPSHLIIDWLQSLS, DFGFPAHVYIDWLQSLS, FGFPQHLFTDWLQSLS, FGFPKHLLVDFLQSLS, FGFPAHVSIDWSQSLS, FGFPAHVSIDFSQSLS, FGFPSHIIIDWLQSLS, FGFPSHLIIEWLQSLS, AAFPAHLLADAAQALA, AAFPAHAAADFLQALA, AAFAAHLLADFLQAAA, AAAPAHLLVDAAQAAA, FAFPAHVFIDWLQSLS; FGFPAHVFIDWLQALS, FGFPAHVFIDWLQSLA, GFPAHVFIDWLQSLS, FPAHVFIDWLQSLS, PAHVFIDWLQSLS, FAFPAHVFIDWLQALA, FGFPEHLFVDFLQSLS, FGFPAHVHIDWLQSLS, FGFPAHVPIDWLQSLS, FGFPSHLFIDWAQSLS, PGFPAHVFIDWLQLIT, PAHVYIDWLQSLS, FGFPAHVYIDWLQ, FGFPAHVFIDWLQ, DFGFPSHLIIDWLQSLS, DFGFPAHVFIDWLQSLN, PSHLIIDWLQ, PAHVFIDWLQ, DFGFPAHVTIDWLQSLN, DFGFPAHVLIDWLQSLN, FGFPAHVYIDWLQSLS, FGFPAHVFIDWLQSLN 및 FGFPAHVFIDWLQSLA로 구성된 군으로부터 선택된 하나 이상의 아미노산 서열로 구성된 펩티드에 관한 것이다.
본 발명의 펩티드는 CETP에 대한 미모토프인 것으로 판명되었고, 따라서, 미모토프는 CETP 단편 C-FGFPEHLLVDFLQSLS(CETP 단백질의 16개의 C-말단 아미노산)에 결합하는 항체에 결합할 수 있었다.
또한, 본 발명의 또 다른 양태는 본 발명에 따른 하나 이상의 펩티드를 포함하는 약학적 제형에 관한 것이다.
본 발명의 펩티드는 개체에 투여될 수 있는 약학적 제형으로 제형화될 수 있다. 이러한 제형은, 예를 들어, 죽상경화증, 죽상경화증 위험 질환 및 죽상경화증 후유증을 예방하고/하거나 치료하는데 사용될 수 있다.
상기 제형 내의 펩티드는 본원에 개시된 펩티드의 풀(pool)로부터 조합될 수 있다. 또한, 본 발명의 펩티드 중 하나 이상을 포함하고, 이의 투여를 필요로 하는 개체에 개별적으로 또는 함께 투여될 수 있는 약학적 제형을 제공하는 것이 가능하다.
본 발명의 펩티드는 하나의 단일한 약학적 제형 또는 이의 2 또는 3개의 조합물로 혼합될 수 있다. 생성된 제형은 때에 맞춰 동일하거나 상이한 시기로 투여될 수 있다. 본 발명의 한 바람직한 구체예에 따르면, 제형에 존재하는 펩티드는 약학적으로 허용되는 담체, 바람직하게는 KLH(키홀 림펫 헤모시아닌)에 커플링된다.
본 발명은 하기 도면 및 실시예에 의해 추가로 예시되나, 이로 제한되는 것은 아니다.
도 1은 모노클로날 항체 "파울라(Paula)"를 이용한 파아지 디스플레이 라이브러리 Ph.D. 7의 스크리닝 후의 대표적 경쟁 ELISA의 결과를 도시한다.
도 2a 및 2b는 모노클로날 항체 "파울라"를 이용한 파아지 디스플레이 라이브러리 Ph.D. 12의 스크리닝 후의 2회의 통상적인 경쟁 ELISA의 결과를 도시한다.
도 3a 및 3b는 mAb 프리다(Frida)를 이용한 파아지 디스플레이 라이브러리 Ph.D. 7의 스크리닝 후의 2회의 대표적 경쟁 ELISA의 결과를 도시한다.
도 4a는 모노클로날 항체 "프리다"를 이용한 파아지 디스플레이 라이브러리 Ph.D. 12의 스크리닝 후의 대표적 경쟁 ELISA의 결과를 도시한다.
도 4b는 미모토프-BSA로 코팅된 ELISA 플레이트로의 모노클로날 항체 "프리다"의 결합을 도시한다.
도 5a 및 5b는 모노클로날 항체 "프리다"를 이용한 파아지 디스플레이 라이브러리 Ph.D. 12의 스크리닝 후의 대표적인 경쟁 ELISA의 결과를 도시한다.
도 6은 모노클로날 항체 "프리다"를 이용한 파아지 디스플레이 라이브러리 Ph.D. 12의 스크리닝 후의 2개의 미모토프의 경쟁 ELISA의 결과를 도시한다.
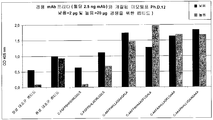
도 7a 내지 7d는 하기 미모토프-BSA 컨쥬게이트가 마우스에 주사되는 생체내 실험의 항체 역가(항 마우스 IgG)를 도시한다:

도 8a 및 8b는 모노클로날 항체 "프리다"를 이용한 파아지 디스플레이 라이브러리 Ph.D. 7C7의 스크리닝 후의 2회의 대표적 경쟁 ELISA의 결과를 도시한다.
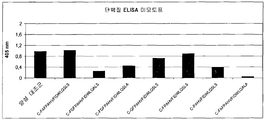
도 9는 "프리다"와 고리 미모토프 사이의 결합의 검출을 위한 시험관내 ELISA 시험을 도시한다.
도 10a 및 10b는 FGFPSHLIIDWLQSLS, FGFPAHVFIDWLQSLS 및 FGFPAHVYIDWLQSLS를 이용한 억제 ELISA 검정의 결과를 도시한다.
도 10a (코트 1 μM 펩티드, 검출 αIgG1)

도 10b (코트 1 μM 펩티드. 검출 αIgG1)

도 11은 마우스에 투여되는 본 발명의 미모토프에 의한 CETP에 특이적인 항체의 생체내 유도를 도시한다. Balb/c 마우스 / 30 ㎍ 펩티드, 2주 간격의 2회 주사. S3 = 3번째 주사 2주 후. 애쥬번트로서 명반(alum). 미모토프의 주사에 의해 유도된 본래의 에피토프(p4073)에 대한 역가. 웰 코팅: 50 ㎕의 1 μM p4073-BSA 또는 1 ㎍ / ml 불활성화된 KLH. 검출: αIgG:

도 12a 및 12b는 본 발명의 미모토프의 투여에 의한 CETP 특이적 항체의 생체내 유도를 도시한다. p4073에 대한 역가 및 이의 선택된 군의 CETP에 대한 역가에 대한 상관 관계(p4073에 대해 높은 역가를 나타냄): gr.4, gr.9, gr.10, gr.14, gr.16-20 / gr.1 (KLH), gr.2 (본래의 에피토프) - 대조군. 코팅: 각각 재조합 GST-CETP 또는 정제된 토끼 CETP:
도 12a

도 12b

도 13은 마우스에 투여되는 본 발명의 미모토프에 의한 CETP에 특이적인 항체의 생체내 유도를 도시한다.
각 군(각각 5마리의 Balb/c 마우스)의 혈청을 취합하고, 1:100으로 희석하고, 재조합 GST-CETP 또는 토끼 CETP로 각각 코팅된 ELISA 플레이트 상에서 시험하였다. 결합된 항체의 검출은 αlgG를 이용하였다.

도 14는 0.6 ㎕의 인간 혈청(내인성 CETP 활성을 가짐)이 KLH/명반(음성 대조군), p4703-KLH/명반(본래의 CETP 에피토프) 또는 p4361(또는 p4362 또는 p4325) 미모토프 각각으로 예방접종된 야생형 마우스(CETP 활성을 함유하지 않음)로부터의 혈청과 혼합되는 CETP 활성 검정을 도시한다. p4361-KLH/명반으로 예방접종된 마우스로부터의 1.2 ㎕ 및 0.6 ㎕ 혈청의 첨가가 CETP 활성을 완전히 억제하고, 0.2 ㎕ 혈청의 첨가가 KLH/명반-대조군 단독 또는 본래의 에피토프(p4073-KLH/명반)로 예방접종된 마우스로부터의 혈청의 첨가와 대조적으로 상기 활성을 현저히 감소시키는 것이 입증될 수 있었다.
도 15는 p4325-KLH/명반의 인간 혈청으로의 첨가가 CETP 활성을 현저히 억제하는 것을 도시한다.
도 16은 p4361-KLH/명반의 인간 혈청으로의 첨가가 CETP 활성을 현저히 억제하는 것을 도시한다.
도 17은 p4362-KLH/명반의 인간 혈청으로의 첨가가 CETP 활성을 현저히 억제하는 것을 도시한다.
도 18a는 미모토프를 이용한 억제 ELISA를 도시한다(코트(Coat.) 1μM 4073 펩티드, 검출 αIgG1).


도 18b는 미모토프를 이용한 억제 ELISA를 도시한다(코트 1 μM 4073 펩티드, 검출 αIgG1).


도 18c는 미모토프 특성규명을 위한 미모토프 스크린 PhD12 프리다 및 Ala-교환 / mAb 프리다를 이용한 억제 ELISA를 도시한다(코트 1 μM 4073. 검출 αIgG1).

도 19a는 펩티드 ELISA, C-DFGFPAHVYIDWLQSLS(p4628-KLH/명반)을 이용한 면역화, 본래의 에피토프에 대한 역가를 도시한다.
도 19b는 펩티드 ELISA, C-FGFPAHVFIDWLQSLN(p4474-KLH/명반)을 이용한 면역화, 본래의 에피토프에 대한 역가를 도시한다.
도 19c는 펩티드 ELISA, C-FGFPAHVFIDWLQSLN(p4474-KLH/명반)을 이용한 면역화, 주사된 미모토프에 대한 역가를 도시한다.
도 19d는 항-단백질 ELISA를 도시한다. 마우스에 애쥬번트로서 명반과 함께 KLH에 커플링된 30 ㎍의 지정된 미모토프를 3회 주사하였다. 각각의 군(5 마리의 마우스를 포함함)으로부터의 혈청을 풀링하고, 1:100으로 희석시키고, 정제된 토끼 CETP로 코팅된 ELISA 플레이트 상에서 시험하였다.
도 19e는 항-단백질 ELISA를 도시하며, 마우스에 애쥬번트로서 명반과 함께 KLH에 커플링된 30 ㎍의 지정된 미모토프를 3회 주사하였다. 마우스 혈청(1마리의 마우스)를 1:100으로 희석시키고, 정제된 토끼 CETP로 코팅된 ELISA 플레이트에서 시험하였다.
실시예:
고밀도 지질단백질(HDL)에서의 콜레스테롤의 혈장 농도와 관상동맥 심질환(CHD)의 발달 사이의 강한 역관계가 존재한다. 따라서, HDL이 감소하는 경우에 CHD에 대한 위험이 보다 높다. CHD를 갖는 환자의 33%는 낮은 혈장 수준의 HDL을 가지지만, HDL의 혈장 농도를 증가시키기 위한 효과적인 치료가 현재 존재하지 않는다. 식이 및 적당한 운동은 효과가 없고, 스타틴(statin)만이 HDL의 낮은 5 내지 7%의 증가를 달성하고, 니아신(niacin)은 부작용 및 이의 용도를 제한하는 순응도(compliance) 프로파일을 갖는다.
CETP 활성의 억제가 혈장 HDL 수준을 증가시키기 위한 치료 접근법으로 제안되어 왔다. CETP는 지질단백질 사이의 중성 지질 및 인지질의 전달을 촉진하고, 혈장 HDL의 농도를 조절하는 혈장 당단백질이다. CETP 활성의 억제는 여러 이유로 인해 혈장 HDL 농도를 증가시키는 것으로 예상된다. CETP는 HDL로부터 VLDL 및 LDL로 콜레스테릴 에스테르를 이동시킴으로써 HDL 농도를 낮춘다. 모노클로날 항체, 소분자(Sikorski, J.A., J.Med.Chem. 49 (1) (2006): 1-22) 또는 안티센스 올리고누클레오티드에 의한 토끼 및 햄스터에서의 CETP의 일시적 억제는 HDL을 증가시킨다. 안티센스 누클레오티드를 이용한 지속된 CETP 억제는 죽상경화증의 토끼 모델에서 혈장 HDL을 증가시키고, 죽상경화증 병변을 감소시켰다. CETP-트랜스제닉 마우스 및 래트는 감소된 혈장 HDL을 나타낸다. 감소된 CETP 활성을 갖는 인간은 상승된 혈장 HDL을 갖는다.
최근에, 백신 접근법이 제안되었다. 토끼를 중성 지질 전달 기능에 중요한 CETP의 영역을 함유하는 인간 CETP-유래 펩티드로 면역화시켰다. 예방접종된 토끼는 감소된 CETP 활성, 및 보다 낮은 LDL 및 보다 높은 HDL 농도의 변경된 지질단백질 프로파일을 가졌다. 더욱이, CETP 예방접종된 토끼는 대조 동물보다 작은 죽상경화증 병변을 갖는 것으로 나타났다.
상기 논의된 항-CETP 백신 접근법의 문제는 백신 제형이 자기 펩티드(self peptide)를 포함하고, 이에 따라 자기 항원에 대한 천연 관용(tolerance)을 파괴하여야 한다는 점이다. 본 발명은 예방접종에 사용될 수 있는 CETP 미모토프를 기재한다: 미모토프는 CETP에 대한 항체의 생성을 유도한다. CETP 미모토프는 자기 서열을 갖지 않고, 이에 따라 관용을 파괴할 필요가 없다. 따라서, 항-CETP 항체 반응의 유도가 크게 촉진된다. 미모토프는 모노클로날 항체(mAb) 및 (시판되는) 펩티드 라이브러리로 확인된다. CETP 활성을 중화시키는 항-CETP 모노클로날 항체가 사용된다. 이러한 mAb는 중성 지질 전달 활성에 필요한 CETP의 C-말단의 26개의 아미노산 내의 서열을 검출한다.
실시예 1: 파아지 디스플레이 라이브러리의 스크리닝에 사용되는 모노클로날 항체의 생성
A.) "융합체 F"로부터 유래된 2개의 항체:
Balb/c 마우스를 애쥬번트로서 KLH 및 명반에 커플링된 본래의 CETP 에피토프 C-FGFPEHLLVDFLQSLS(CETP 단백질의 16개의 C-말단 아미노산)로 면역화시켰다.
2개의 하이브리도마 클론(둘 모두 IgG1)을 정제시키고, 스크리닝에 사용하였다: F5AF9G4 ("파울라(Paula)") 및 F6F11D1("펠릭스(Pelix)").
이러한 2개의 모노클로날 항체는 ELISA에서 주사된 에피토프 뿐만 아니라 CETP 단백질을 인지한다. 이들은 또한 CETP 단백질(박테리아에서 발현된 재조합 단백질 뿐만 아니라 토끼 혈청으로부터 분리된 단백질)을 검출하기 위한 웨스턴 블롯(Western blot)에 사용될 수 있다. 둘 모두의 항체는 CETP 효소 활성을 억제하지 않는다(로아(Roar) CETP 활성 검정 키트로 시험됨, 예를 들어, US 5,585,235호; US 5,618,683호; US 5,770,355호 참조).
B.) "융합체 I"로부터 유래된 2개의 항체:
Balb/c 마우스를 애쥬번트로서 KLH 및 명반에 커플링된 본래의 CETP 에피토프 C-FGFPEHLLVDFLQSLS(CETP 단백질의 16개의 C-말단 아미노산)로 면역화시켰다.
2개의 하이브리도마 클론(둘 모두 IgG1)을 정제시키고, 스크리닝에 사용하였다: I2G6H5 ("프리다") 및 I2G6H7 ("제임스(James)").
이러한 2개의 모노클로날 항체는 ELISA에서 주사된 에피토프 뿐만 아니라 CETP 단백질을 인지한다. 이들은 또한 CETP 단백질(박테리아에서 발현된 재조합 단백질 뿐만 아니라 토끼 혈청으로부터 분리된 단백질)을 검출하기 위한 웨스턴 블롯에 사용될 수 있다. "융합체 F"로부터 유래된 항체(A 참조)와 대조적으로, 둘 모두의 항체 "프리다" 및 "제임스"는 CETP 효소 활성을 억제한다(로아 CETP 활성 검정 키트로 시험됨).
실시예 2: 미모토프의 파아지 디스플레이, 시험관내 억제 ELISA 및 생체내 시험
본 실시예에서 사용된 파아지 디스플레이 라이브러리는 다음과 같다:
Ph.D. 7: 뉴 잉글랜드 바이오랩스(New England BioLabs) E8102L (선형 7머(mer) 라이브러리)
Ph.D. C7C: 뉴 잉글랜드 바이오랩스 E8121L (7머 라이브러리, 고리화된 펩티드)
Ph.D. 12: 뉴 잉글랜드 바이오랩스 E8111L (선형 12머 라이브러리)
제조업체의 프로토콜(www.neb.com)에 따라 파아지 디스플레이를 수행하였다.
2 또는 3회의 이후의 라운드의 패닝(panning) 후, 단일 파아지 클론을 채집하고, 파아지 상층액을 패닝 절차에 사용되는 항체로 코팅된 플레이트 상의 ELISA에 적용시켰다. 상기 ELISA에서 양성(표적에 대한 강한 신호, 그러나 비특이적인 대조군에 대해서는 신호가 없음)인 파아지 클론을 서열분석하였다. DNA 서열로부터, 펩티드 서열을 추론하였다. 이러한 펩티드를 합성하고, 억제 ELISA에서 특성규명하였다.
1. 시험관내 억제 검정(ELISA)
다양한 양의 파아지 디스플레이로부터 유래된 펩티드(2 및 20 ㎍, 각각의 도면에 나타낸 바와 같음)를 스크리닝 절차에 사용되는 모노클로날 항체와 함께 인큐베이션하였다. ELISA 플레이트 상에 코팅된 본래의 CETP 에피토프(CETP 단백질의 C-말단의 16개의 아미노산)로의 항체의 후속 결합을 감소시키는 펩티드를 억제로 간주하였다(결과 참조: 도 19a 내지 19c)
2. 미모토프의 생체내 시험
억제 뿐만 아니라 몇몇 비-억제 펩티드를 KLH에 커플링시키고, 적절한 애쥬번트(마우스에 대해 수산화알루미늄 및 게르부(Gerbu) 100, 및 토끼에 대해 수산화알루미늄 또는 CFA/IFA)와 함께 마우스(야생형 또는 CETP-트랜스제닉 마우스; 옆구리로 피하 또는 귀로 피내 주사) 또는 토끼(옆구리로 피하 주사)에 주사하였다.
주사된 펩티드 뿐만 아니라 본래의 CETP 에피토프에 대한 역가를 결정하였다. 또한, 선택된 혈청에 대해 CETP 단백질에 대한 면역 반응을 또한 측정하였다(결과 참조: 도 7a 내지 7d 및 도 19a 내지 19e).
3. 결과
3.1. "융합체 F"로부터 유래된 2개의 항체 "파울라" 및 "펠릭스"를 이용한 스크리닝
3.1.1. 파아지 디스플레이 라이브러리 Ph.D. 7
3.1.1.1. 모노클로날 항체 "파울라"를 이용한 스크리닝
17개의 서열을 이러한 스크린에서 확인하였다:
P2_8 SYHATFL
P2_9 TMAFPLN
P2_11 HYHGAFL
P2_12 EHHDIFL
P2_15 SSLELFL
P2_16 TGLSVFL
P3_2 WMPSLFY
P3_6, 14, 28 SMPWWFF
P3_9 TMPLLFW
P3_13 DTWPGLE
P3_16 SMPPIFY
P3_17 MPLWWWD
P3_18 SMPNLFY
P3_19 RMPPIFY
P3_21 NPFEVFL
P3_25 TLPNWFW
P3_26 SMPLTFY
대표적 경쟁 ELISA의 결과가 도 1에 도시되어 있다.
3.1.1.2. 모노클로날 항체 "펠릭스"를 이용한 스크리닝
시험관내 경쟁 실험에서 모노클로날 항체 "펠릭스"의 결합을 억제하는 6개의 서열을 확인하였다:
F2-9 C SFLDTLT
F3-6 C NFLKTLS
F3-18 C DFLRTLT
F3-23 C AFLDTLV
F3-34 C TFLSSLA
F3-38 C GFLDSLM
시험관내 경쟁 실험에서 모노클로날 항체 "펠릭스"의 결합을 억제하지 않는 추가의 12개의 서열을 확인하였다:
F2-2+5 SPHPHFL
F2-6 NFMSIGL
F2-16/F3-30 SQFLASL
F2-29 SNFLKTL
F3-1-_ TGFLATL
F3-11-_ WSWPGLN
F3-17- IAWPGLD
F3-32- SKFMDTL
F3-41- SDFLRAL
F3-44-_ SMPMVFY
F3-49- YEWVGLM
F3-64- KGFLDHL
시험관내에서 모노클로날 항체 "펠릭스"의 결합을 억제하는 모든 미모토프를 KLH에 커플링시키고, 야생형 마우스(마우스는 CETP 단백질을 가지지 않음), CETP-tg 마우스 또는 토끼 각각에 피하(옆구리; s.c.) 또는 피내(i.d.) 주사하고, 시험되는 모든 애쥬번트(명반 및 CFA(완전 프로인트 애쥬번트); Gerbu)와 함께 주사된 펩티드에 대한 면역 반응을 유도시켰다.
상기 나열된 모든 시험관내 억제 미모토프에 대해, 본래의 CETP 에피토프에 반응하는 항체를 마우스 및 토끼에서 검출할 수 있었다.
6개의 미모토프 중 5개(하기 및 표 1 참조)에 대해, 정제된 인간 CETP 및 재조합적으로 발현된 인간 CETP와 반응하는 항체를 토끼 혈청으로부터의 ELISA에서 검출할 수 있었다:
F2-9 C SFLDTLT
F3-6 C NFLKTLS
F3-18 C DFLRTLT
F3-34 C TFLSSLA
F3-38 C GFLDSLM
옆구리에서의 피하 주사를 1주, 3주 및 7주에 마우스 당 30 ㎍의 펩티드-KLH를 이용하여 수행하였다. 귀에서의 피내 주사를 1주, 3주 및 6주에 마우스당 10 ㎍의 펩티드-KLH를 이용하여 수행하였다. 혈청을 3번째 주사 2주 후에 채취하였다. 명반을 가진 백신 제형(항상 마우스 당 1 mg): 250 ㎕ 이하, 한쪽 옆구리에 주사됨. 마우스당 1 ml를 갖는 명반 제형(각 옆구리로 500 ㎕)은 완충액으로서 1x PBS에 존재하였다.
게르부(Gerbu) 애쥬번트 100을 갖는 백신 제형(Gerbu Cat. Nr. #3100; 항상 마우스 당 50 ㎕ 애쥬번트): 200 ㎕, 완충액으로서 1x HEPES를 포함하여 100 ㎕가 각각의 옆구리로 주사됨.
표 1: 역가 결정의 결과



3.1.2. 파아지 디스플레이 라이브러리 Ph.D. 12
3.1.2.1. 모노클로날 항체 "파울라"를 이용한 스크리닝
이러한 스크린으로부터 유래된 20개의 아미노산 서열 중, 3개가 시험관내 억제 실험에서 억제하였다:
P12-19 SANPRDFLETLF
P12-21 RMFPESFLDTLW
P12-37 TIYDSFLDSLAS
억제하지 않는 펩티드는 다음과 같았다:
P12-5/44/46/49 HQSDDKMPWWFF
P12-9 KPYLLKDFLEAL
P12-24/43-_ AMGPYDALDLFL
P12-25 TWNPIESFLESL
P12-28+42 YVWQDPSFTTFF
P12-30 QYQTPLTFLEAL
P12-35- RHISPATFLEAL
P12-39- HTDSFLSTFYGD
P12-42- YVWQDPSFTTFF
P12-45- ADSTFTSFLQTL
P12-50-_ GPVSIYADTDFL
P12-51- _ DSNDTLTLAAFL
P12-52-_ NGSPALSHMLFL
P12-53- TDYDPMWVFFGY
P12-56- IFPLDSQWQTFW
P12-58- NESMPDLFYQPS
P12-61- DWGDKYFSSFWN
2개의 통상적인 경쟁 ELISA의 결과가 도 2A 및 2b에 도시되어 있다.
모든 3개의 미모토프를 KLH에 커플링시키고, 야생형 마우스(마우스는 CETP 단백질을 가지지 않음), CETP-tg 마우스 또는 토끼에 각각 주사하고, 시험되는 모든 애쥬번트(명반 및 CFA; 게르부(Gerbu))와 함께 주사된 펩티드에 대한 면역 반응을 유도시켰다.
미모토프 P12-19; C-SANPRDFLETLF 및 P12-21; C-RMFPESFLDTLW는 야생형 마우스 및 토끼에서 본래의 CETP 에피토프에 대한 면역 반응을 유도하였다.
이와 대조적으로, 미모토프 P12-37 C-TIYDSFLDSLAS는 본래의 에피토프에 대한 항체 반응을 유도하지 않았다.
3.2 "
융합체
I"로부터
유래된
2개의 항체 "
프리다
" 및 "제임스"를 이용한 스크리닝
3.2.1.
파아지
디스플레이 라이브러리
Ph
.D. 7
3.2.1.1.
모노클로날
항체 "
프리다
" 및 "제임스"를 이용한 스크리닝
상기 스크리닝에서 2개의 상이한 펩티드 서열을 확인하였고, 서열분석된 12개의 클론 중 11개가 동일한 서열을 가졌다. 이러한 펩티드는 시험관내 경쟁 실험에서 억제하지 않는다.
Fr7-2-2
Fr7-2B-65
Fr7-3-7
Fr7-3-13
Fr7-3-26
Fr7-3-32
Ja7-2-22
Ja7-3-28
Ja7-3-41
Ja7-3-52
Ja7-3-56 VSAYNNV
Ja7-3-89 WPLHLWQ
mAb "프리다"를 이용한 2개의 대표적 경쟁 ELISA의 결과가 도 3A 및 3b에 제시되어 있다. mAb "제임스"를 이용하여 동일한 패턴이 관찰되었다.
3.2.2.
파아지
디스플레이 라이브러리
Ph
.D. 12
3.2.2.1.
모노클로날
항체 "
프리다
"를 이용한 스크리닝
Fr12/2/6 TPTHYYADFSQL
Fr12/2/11 LPGHLIWDSLHY
Fr12/2/27 LPQTHPLHLLED
Fr12/3/1
Fr12/3/19
Fr12/3/88 IPYHHLVDQLHH
Fr12/3/26
Fr12/3/65 YPYHVQVDVLQN
Fr12/3/68 IPSHHLQDSLQL
Fr12/3/12 EYAHHTSLDLRQ
Fr12/3/83 EPLHFRSDRIQA
Fr12/3/55 ATPSHLIIDRAQ
Fr12/3/63 APKHLYADMSQA
Fr12/3/84 FKPAHVSIDWLQ
Fr12/3/47 MPAHLSRDLRQS
Fr12/3/80 NPKHYSIDRHQA
Fr12/3/40 SPQHLTTDRAQA
Fr12/3/35 TPFHFAQDSWQW
본 스크린에서 확인된 15개의 아미노산 서열 중 어느 것도 시험관내 경쟁 실험에서 억제하지 않았다. 그러나, 서열 분석은 다수의 미모토프에 대한 본래의 단백질 서열에 대해 다소 높은 상동성을 나타내었다. 한편, 몇몇 펩티드에 대해, 미모토프-BSA로 코팅된 ELISA 플레이트로의 모노클로날 항체 "프리다"의 결합이 관찰될 수 있었다(도 4a 및 4b 참조).
이는 고정된 미모토프로의 모노클로날 항체의 결합이 시험관내 경쟁 ELISA에서의 억제를 예측하는 것을 반드시 가능케 하는 것은 아님을 나타낸다.
본래의 서열 FGFPEHLLVDFLQSLS (CETP 단백질의 16개의 C-말단 AA)의 변이체를 이용한 시험관내 억제 실험은 N-말단으로부터 2개 이상의 아미노산 또는 C-말단으로부터 1개 이상의 아미노산을 제거하는 것이 억제를 중단시키는 것을 나타내었다(모노클로날 항체 "프리다" 및 "제임스"에 대해서임. "파울라" 및 "펠릭스"는 본래의 서열의 상이한 부분을 인지함).
또한, 동시에 N-말단으로부터 2개의 아미노산 및 C-말단으로부터 1개의 아미노산을 제거하는 것은 시험관내에서 더 이상 억제하지 않는 펩티드를 또한 발생시킨다.
C-FGFPEHLLVDFLQSLS "본래의" 서열(CETP로부터 유래된 펩티드)/ 시험관내에서 억제
C-GFPEHLLVDFLQSLS 서열 N-1 / 시험관내에서 억제
C-FPEHLLVDFLQSLS 서열 N-2 / 시험관내에서 억제
C-PEHLLVDFLQSLS 서열 N-3 / evtl. 시험관내에서 약간의 억제
C-FGFPEHLLVDFLQSL 서열 C-1 / 시험관내에서 억제
C-FGFPEHLLVDFLQS 서열 C-2 / 시험관내에서 억제하지 않음
C-FPEHLLVDFLQSL 서열 N-2 및 C-1 / 시험관내에서 억제하지 않음
"본래의" FGFPEHLLVDFLQSLS
Fr12/2/6 TPTHYYADFSQL
Fr12/2/11 LPGHLIWDSLHY
Fr12/2/27 LPQTHPLHLLED
Fr12/3/1 IPYHHLVDQLHH
Fr12/3/19 IPYHHLVDQLHH
Fr12/3/88 IPYHHLVDQLHH
Fr12/3/26 YPYHVQVDVLQN
Fr12/3/65 YPYHVQVDVLQN
Fr12/3/68 IPSHHLQDSLQL
Fr12/3/12 EYAHHTSLDLRQ
Fr12/3/83 EPLHFRSDRIQA
Fr12/3/55 ATPSHLIIDRAQ
Fr12/3/63 APKHLYADMSQA
Fr12/3/84 FKPAHVSIDWLQ
Fr12/3/47 MPAHLSRDLRQS
Fr12/3/80 NPKHYSIDRHQA
Fr12/3/40 SPQHLTTDRAQA
Fr12/3/35 TPFHFAQDSWQW
결과적으로, 주형으로 본래의 CETP 서열을 이용하여, 보다 긴 펩티드를 이용하여 시험관내 억제가 가능한지 확인하기 위해 본 파아지 디스플레이 과정에서 수득된 펩티드 서열을 N-말단 및/또는 C-말단에서 신장시켰다.
3.2.2.2.
미모토프
프리다
Ph
.D.12 및 이의
변이체
:
Fr12/2/6 TPTHYYADFSQL
Fr12/2/6 ext1 TPTHYYADFSQLLS
Fr12/2/6 ext2 TPTHYYADFSQSLS
Fr12/2/6 ext3 GTPTHYYADFSQLL
Fr12/2/6 ext4 GTPTHYYADFSQSL
Fr12/2/6 ext5 FGTPTHYYADFSQSLS
Fr12/2/6 ext6 FGFPTHYYADFSQSLS
Fr12/2/11 LPGHLIWDSLHY
Fr12/2/11 ext1 LPGHLIWDSLHYL
Fr12/2/11 ext2 LPGHLIWDSLHYLS
Fr12/2/11 ext3 LPGHLIWDSLHSL
Fr12/2/11 ext4 LPGHLIWDSLHSLS
Fr12/2/11 ext5 GLPGHLIWDSLHYL
Fr12/2/11 ext5 GLPGHLIWDSLHSL
Fr12/2/11 ext6 FGLPGHLIWDSLHSLS
Fr12/2/11 ext7 FGFPGHLIWDSLHSLS
Fr12/2/27 LPQTHPLHLLED
Fr12/3/1/19/88 ext1 IPYHHLVDQLHLS
Fr12/3/1/19/88 ext2 IPYHHLVDQLHSLS
Fr12/3/1/19/88 ext3 FGIPYHHLVDQLHHLS
Fr12/3/1/19/88 ext4 FGFPYHHLVDQLHSLS
Fr12/3/26/65ext1 YPYHVQVDVLQNLS
Fr12/3/26/65ext2 YPYHVQVDVLQSLS
Fr12/3/26/65ext3 FGYPYHVQVDVLQNLS
Fr12/3/26/65ext4 FGFPYHVQVDVLQSLS
Fr12/3/68 ext1 IPSHHLQDSLQLLS
Fr12/3/68 ext2 IPSHHLQDSLQSLS
Fr12/3/68 ext3 GIPSHHLQDSLQLL
Fr12/3/68 ext4 FGIPSHHLQDSLQLLS
Fr12/3/68 ext5 FGFPSHHLQDSLQSLS
Fr12/3/83 ext1 EPLHFRSDRIQALS
Fr12/3/83 ext2 EPLHFRSDRIQSLS
Fr12/3/83 ext3 GEPLHFRSDRIQAL
Fr12/3/83 ext4 FGEPLHFRSDRIQALS
Fr12/3/83 ext5 FGFPLHFRSDRIQSLS
Fr12/3/55 ext1 ATPSHLIIDRAQSLS
Fr12/3/55 ext2 FGFPSHLIIDRAQSLS
Fr12/3/55 ext2 R->W FGFPSHLIIDWAQSLS
Fr12/3/55 ext2 RA->WL FGFPSHLIIDWLQSLS
Fr12/3/63 ext1 APKHLYADMSQALS
Fr12/3/63 ext2 APKHLYADMSQSLS
Fr12/3/63 ext3 GAPKHLYADMSQAL
Fr12/3/63 ext4 FGFPKHLYADMSQSLS
Fr12/3/84 ext1 FKPAHVSIDWLQSLS
Fr12/3/84 ext2 FGFPAHVSIDWLQSLS
Fr12/3/47 ext1 MPAHLSRDLRQSL
Fr12/3/47 ext2 MPAHLSRDLRQSLS
Fr12/3/47 ext3 GMPAHLSRDLRQSL
Fr12/3/47 ext4 FGFPAHLSRDLRQSLS
Fr12/3/40 ext1 SPQHLTTDRAQALS
Fr12/3/40 ext2 SPQHLTTDRAQSLS
Fr12/3/40 ext3 GSPQHLTTDRAQAL
Fr12/3/40 ext4 FGFPQHLTTDRAQSLS
Fr12/3/35 ext1 TPFHFAQDSWQWLS
Fr12/3/35 ext2 TPFHFAQDSWQSLS
Fr12/3/35 ext3 GTPFHFAQDSWQWL
Fr12/3/35 ext4 FGFPFHFAQDSWQSLS
억제 ELISA의 대표적 예가 도 5A 및 5b에 도시되어 있다. 신장된 펩티드 Fr12/3/84 ext2 및 Fr12/3/55 ext3은 현저한 억제를 나타내었다:
C-FGFPSHLIIDRAQSLS Fr12/3/55 ext3
C-FGFPAHVSIDWLQSLS Fr12/3/84 ext2
3개의 추가 펩티드가 또한 본 검정에서 억제하였다:
C-FGFPYHVQVDVLQSLS Fr12/3/26/65ext4
C-FKPAHVSIDWLQSLS Fr12/3/84 ext1
C-FGFPQHLTTDRAQSLS Fr12/3/40 ext4
본래의 에피토프 및 파아지 디스플레이 스크린으로부터 유래된 모든 미모토프를 비교하는 서열 분석 후, 추가의 2개의 펩티드를 생성시켰다.
미모토프 Fr12/3/55 ext3 C-FGFPSHLIIDRAQSLS(ELISA에서 억제, 상기 참조)에 대해, 아미노산 교환을 억제 ELISA에서 시험하였다:
강한 억제:
C-FGFPAHVSIDWLQSLS Fr12/3/84 ext2
약간의 억제:
C-FGFPSHLIIDRAQSLS Fr12/3/55 ext3
변경된 서열을 갖는 펩티드(억제, 도 6 참조):
C-FGFPSHLIIDWAQSLS Fr12/3/55 ext2 R 대신 W
C-FGFPSHLIIDWLQSLS Fr12/3/55 ext2 RA 대신 WL
추가의 바람직한 미모토프를 하기 실시예 구성에 의해 특성규명하였다:


3.2.2.3.
미모토프의
생체내
시험
군당 5마리의 마우스의 암컷 Balb/c 마우스를 KLH에 커플링된 30 ㎍의 펩티드로 피하 면역화시켰다. 대조군에 KLH 또는 C-FGFPEHLLVDFLQSLS를 투여하였다. 애쥬번트로서 명반을 사용하였다. 투여된 펩티드는 모두 "프리다"에 결합할 수 있었고, CETP에 대한 면역 반응을 유도할 수 있었으나, 이러한 펩티드 중 일부는 시험관내(시험관내 억제 검정)에서 CETP의 "프리다"로의 결합을 억제하지 않았다. 항체 역가를 결정하기 위한 시험관내 ELISA 검정을 2주 간격의 2회의 예방접종(S2; 도 7a 내지 7d 참조) 후에 풀링된 혈청으로 수행하였다. ELISA 플레이트의 웰을 KLH(양성 대조군), 미모토프-BSA 컨쥬게이트, C-FGFPEHLLVDFLQSLS 및 관련이 없는 펩티드-BSA 컨쥬게이트(음성 대조군)로 코팅하였다. 항-마우스 IgG를 이용하여 검출을 수행하였다.
3.2.3.
파아지
디스플레이 라이브러리
Ph
.D. 7
C7
3.2.3.1.
모노클로날
항체 "
프리다
" 및 "제임스"를 이용한 스크리닝
Fr2-1 ACSFAYLYRC
Fr2-5
Fr2-6
Fr2-18
Fr2-19
Fr2-28
Ja2-5
Ja2-20
Ja2-23
Ja2-24
Ja2-30 ACFMGDKWVC
Fr2-7
Fr2-9 ACVLYPKAIC
Fr2-11
Ja2-19 ACYMGQQFVC
Fr2-16 ACLTAYLHWC
Fr2-20 ACTLFPVAYC
Fr2-25 ACWLFPYAHC
Fr2-26 ACKSINMWLC
Fr2-27 ACQTINRWLC
상기 미모토프-펩티드의 고리화 성질로 인해, 이들의 합성은 선형 펩티드의 합성보다 더욱 복잡하다. 9개의 고리 서열 중 7개를 억제 ELISA에서의 시험관내 분석을 위해 선택하였다(도 8a 및 8b 참조). 이러한 서열 중 어느 것도 본래의 CETP 에피토프에 대한 파아지 디스플레이 스크리닝에 사용되는 모노클로날 항체의 결합을 억제하지 않았다. 또한, 이러한 펩티드가 BSA에 커플링되고, ELISA 플레이트에 코팅되는 경우, 이들은 모노클로날 항체에 의해 검출되지 않았다(도 9 참조). 이는 Ph.D.7 또는 Ph.D.12 라이브러리로부터 유래된 미모토프를 이용한 데이터와 대조적이었으며, 이러한 펩티드가 BSA에 커플링되고, ELISA 플레이트 상에 코팅되는 경우 모노클로날 항체는 대부분의 확인된 미모토프에 결합하였다.
실시예
3:
CETP
활성 검정:
CETP 활성 검정을 시판(예를 들어, 로아(ROAR) CETP 활성 검정)되고, 예를 들어, US 5,585,235호, US 5,618,683호 및 US 5,770,355호에 기재된 검정을 이용하여 수행하였다. 검정을 제조업체의 권고에 따라 수행하였다.
SEQUENCE LISTING
<110> Affiris Forschungs- und Entwicklungs GmbH
<120> Treatment of atherosclerosis
<130> R 52450
<150> AT A 4258/2007
<151> 2007-08-10
<160> 238
<170> PatentIn version 3.5
<210> 1
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa is (Xaa)n and an amino acid residue other than C, wherein n
is an integer between 0 and 9
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa is an amino acid residue selected from the group consisting
of D, A, R, E, S, N, T and G
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> Xaa is an amino acid residue selected from the group consisting
of F, A, W, R, S, L, Q, V and M
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa is an amino acid residue selected from the group consisting
of L, A, S, W, E, R, I and H
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa is an amino acid residue selected from the group consisting
of Q, A, H, D, K, R, S and E
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa is (Xaa)m and an amino acid residue other than C, wherein m
is an integer between 0 and 9
<400> 1
Xaa Xaa Xaa Xaa Xaa Xaa
1 5
<210> 2
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 2
Ser Tyr His Ala Thr Phe Leu
1 5
<210> 3
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 3
Thr Met Ala Phe Pro Leu Asn
1 5
<210> 4
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 4
His Tyr His Gly Ala Phe Leu
1 5
<210> 5
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 5
Glu His His Asp Ile Phe Leu
1 5
<210> 6
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 6
Thr Gly Leu Ser Val Phe Leu
1 5
<210> 7
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 7
Trp Met Pro Ser Leu Phe Tyr
1 5
<210> 8
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 8
Ser Met Pro Trp Trp Phe Phe
1 5
<210> 9
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 9
Thr Met Pro Leu Leu Phe Trp
1 5
<210> 10
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 10
Asp Thr Trp Pro Gly Leu Glu
1 5
<210> 11
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 11
Ser Met Pro Pro Ile Phe Tyr
1 5
<210> 12
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 12
Met Pro Leu Trp Trp Trp Asp
1 5
<210> 13
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 13
Ser Met Pro Asn Leu Phe Tyr
1 5
<210> 14
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 14
Arg Met Pro Pro Ile Phe Tyr
1 5
<210> 15
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 15
Asn Pro Phe Glu Val Phe Leu
1 5
<210> 16
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 16
Thr Leu Pro Asn Trp Phe Trp
1 5
<210> 17
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 17
Ser Met Pro Leu Thr Phe Tyr
1 5
<210> 18
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 18
Ser Pro His Pro His Phe Leu
1 5
<210> 19
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 19
Asn Phe Met Ser Ile Gly Leu
1 5
<210> 20
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 20
Ser Gln Phe Leu Ala Ser Leu
1 5
<210> 21
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 21
Trp Ser Trp Pro Gly Leu Asn
1 5
<210> 22
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 22
Ile Ala Trp Pro Gly Leu Asp
1 5
<210> 23
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 23
Ser Lys Phe Met Asp Thr Leu
1 5
<210> 24
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 24
Ser Met Pro Met Val Phe Tyr
1 5
<210> 25
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 25
Tyr Glu Trp Val Gly Leu Met
1 5
<210> 26
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 26
Lys Gly Phe Leu Asp His Leu
1 5
<210> 27
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 27
His Gln Ser Asp Asp Lys Met Pro Trp Trp Phe Phe
1 5 10
<210> 28
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 28
Tyr Val Trp Gln Asp Pro Ser Phe Thr Thr Phe Phe
1 5 10
<210> 29
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 29
Tyr Val Trp Gln Asp Pro Ser Phe Thr Thr Phe Phe
1 5 10
<210> 30
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 30
Leu Pro Gln Thr His Pro Leu His Leu Leu Glu Asp
1 5 10
<210> 31
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 31
Gly Pro Val Ser Ile Tyr Ala Asp Thr Asp Phe Leu
1 5 10
<210> 32
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 32
Asp Ser Asn Asp Thr Leu Thr Leu Ala Ala Phe Leu
1 5 10
<210> 33
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 33
Asn Gly Ser Pro Ala Leu Ser His Met Leu Phe Leu
1 5 10
<210> 34
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 34
Thr Asp Tyr Asp Pro Met Trp Val Phe Phe Gly Tyr
1 5 10
<210> 35
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 35
Ile Phe Pro Leu Asp Ser Gln Trp Gln Thr Phe Trp
1 5 10
<210> 36
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 36
Asn Glu Ser Met Pro Asp Leu Phe Tyr Gln Pro Ser
1 5 10
<210> 37
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 37
Asp Trp Gly Asp Lys Tyr Phe Ser Ser Phe Trp Asn
1 5 10
<210> 38
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 38
Val Ser Ala Tyr Asn Asn Val
1 5
<210> 39
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 39
Trp Pro Leu His Leu Trp Gln
1 5
<210> 40
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 40
Cys Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu
1 5 10 15
Ser
<210> 41
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa is P, Y, T or K
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa is an amino acid residue other than C
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa is Y, L, H, V, T, I or F
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa is Y, I, P, L, Q, S, R, T, F or A
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa is A, W, V, Q, L, S, I, R or T
<400> 41
Xaa Xaa His Xaa Xaa Xaa
1 5
<210> 42
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa is selected from the group consisting of G, A, F, Y and K
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> F is (F)o, wherein o is 0 or 1
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa is selected from the group consisting of E, Y, A, Q, K and S
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa is selected from the group consisting of H, V, L, F and I
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> Xaa is selected from the group consisting of L, W, S, I, F and Y
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa is V, T, F or I
<220>
<221> MISC_FEATURE
<222> (11)..(11)
<223> Xaa is an amino acid residue selected from the group consisting
of F, A, W, R, S, L, Q, V and M
<220>
<221> MISC_FEATURE
<222> (12)..(12)
<223> Xaa is an amino acid residue selected from the group consisting
of L, A, S, W, E, R, I and H
<220>
<221> MISC_FEATURE
<222> (13)..(13)
<223> Xaa is an amino acid residue selected from the group consisting
of Q, A, H, D, K, R, S and E
<220>
<221> MISC_FEATURE
<222> (14)..(14)
<223> Xaa is S or Y
<220>
<221> MISC_FEATURE
<222> (15)..(15)
<223> Xaa is L, A or I
<220>
<221> MISC_FEATURE
<222> (16)..(16)
<223> Xaa is S, N or T
<400> 42
Phe Xaa Phe Pro Xaa His Xaa Xaa Xaa Asp Xaa Xaa Xaa Xaa Xaa Xaa
1 5 10 15
<210> 43
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa is selected from the group consisting of D, S, N, T and G
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa is selected from the group consisting of Q, D, K, R, S and E
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa is S or T
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> Xaa is an amino acid residue other than C, preferably selected
from the group consisting of S, T, A, M, F and W
<400> 43
Xaa Phe Leu Xaa Xaa Leu Xaa
1 5
<210> 44
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 44
Ser Ser Leu Glu Leu Phe Leu
1 5
<210> 45
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 45
Ser Phe Leu Asp Thr Leu Thr
1 5
<210> 46
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 46
Asn Phe Leu Lys Thr Leu Ser
1 5
<210> 47
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 47
Asp Phe Leu Arg Thr Leu Thr
1 5
<210> 48
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 48
Ala Phe Leu Asp Thr Leu Val
1 5
<210> 49
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 49
Thr Phe Leu Ser Ser Leu Ala
1 5
<210> 50
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 50
Gly Phe Leu Asp Ser Leu Met
1 5
<210> 51
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 51
Ser Pro His Pro His Phe Leu
1 5
<210> 52
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 52
Ser Asn Phe Leu Lys Thr Leu
1 5
<210> 53
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 53
Thr Gly Phe Leu Ala Thr Leu
1 5
<210> 54
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 54
Ser Asp Phe Leu Arg Ala Leu
1 5
<210> 55
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 55
Ser Ala Asn Pro Arg Asp Phe Leu Glu Thr Leu Phe
1 5 10
<210> 56
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 56
Arg Met Phe Pro Glu Ser Phe Leu Asp Thr Leu Trp
1 5 10
<210> 57
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 57
Thr Ile Tyr Asp Ser Phe Leu Asp Ser Leu Ala Ser
1 5 10
<210> 58
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 58
Lys Pro Tyr Leu Leu Lys Asp Phe Leu Glu Ala Leu
1 5 10
<210> 59
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 59
Ala Met Gly Pro Tyr Asp Ala Leu Asp Leu Phe Leu
1 5 10
<210> 60
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 60
Thr Trp Asn Pro Ile Glu Ser Phe Leu Glu Ser Leu
1 5 10
<210> 61
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 61
Gln Tyr Gln Thr Pro Leu Thr Phe Leu Glu Ala Leu
1 5 10
<210> 62
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 62
Arg His Ile Ser Pro Ala Thr Phe Leu Glu Ala Leu
1 5 10
<210> 63
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 63
His Thr Asp Ser Phe Leu Ser Thr Phe Tyr Gly Asp
1 5 10
<210> 64
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 64
Ala Asp Ser Thr Phe Thr Ser Phe Leu Gln Thr Leu
1 5 10
<210> 65
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 65
Gly Pro Val Ser Ile Tyr Ala Asp Thr Asp Phe Leu
1 5 10
<210> 66
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 66
Asp Ser Asn Asp Thr Leu Thr Leu Ala Ala Phe Leu
1 5 10
<210> 67
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 67
Thr Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Leu
1 5 10
<210> 68
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 68
Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Tyr
1 5 10
<210> 69
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 69
Leu Pro Gln Thr His Pro Leu His Leu Leu Glu Asp
1 5 10
<210> 70
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 70
Ile Pro Tyr His His Leu Val Asp Gln Leu His His
1 5 10
<210> 71
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 71
Tyr Pro Tyr His Val Gln Val Asp Val Leu Gln Asn
1 5 10
<210> 72
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 72
Ile Pro Ser His His Leu Gln Asp Ser Leu Gln Leu
1 5 10
<210> 73
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 73
Glu Tyr Ala His His Thr Ser Leu Asp Leu Arg Gln
1 5 10
<210> 74
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 74
Glu Pro Leu His Phe Arg Ser Asp Arg Ile Gln Ala
1 5 10
<210> 75
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 75
Ala Thr Pro Ser His Leu Ile Ile Asp Arg Ala Gln
1 5 10
<210> 76
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 76
Ala Pro Lys His Leu Tyr Ala Asp Met Ser Gln Ala
1 5 10
<210> 77
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 77
Phe Lys Pro Ala His Val Ser Ile Asp Trp Leu Gln
1 5 10
<210> 78
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 78
Met Pro Ala His Leu Ser Arg Asp Leu Arg Gln Ser
1 5 10
<210> 79
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 79
Asn Pro Lys His Tyr Ser Ile Asp Arg His Gln Ala
1 5 10
<210> 80
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 80
Ser Pro Gln His Leu Thr Thr Asp Arg Ala Gln Ala
1 5 10
<210> 81
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 81
Thr Pro Phe His Phe Ala Gln Asp Ser Trp Gln Trp
1 5 10
<210> 82
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 82
Thr Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Leu Leu Ser
1 5 10
<210> 83
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 83
Thr Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Ser Leu Ser
1 5 10
<210> 84
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 84
Gly Thr Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Leu Leu
1 5 10
<210> 85
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 85
Gly Thr Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Ser Leu
1 5 10
<210> 86
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 86
Phe Gly Thr Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Ser Leu Ser
1 5 10 15
<210> 87
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 87
Phe Gly Phe Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Ser Leu Ser
1 5 10 15
<210> 88
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 88
Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Tyr
1 5 10
<210> 89
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 89
Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Tyr Leu
1 5 10
<210> 90
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 90
Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Tyr Leu Ser
1 5 10
<210> 91
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 91
Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Ser Leu
1 5 10
<210> 92
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 92
Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Ser Leu Ser
1 5 10
<210> 93
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 93
Gly Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Tyr Leu
1 5 10
<210> 94
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 94
Gly Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Ser Leu
1 5 10
<210> 95
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 95
Phe Gly Leu Pro Gly His Leu Ile Trp Asp Ser Leu His Ser Leu Ser
1 5 10 15
<210> 96
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 96
Phe Gly Phe Pro Gly His Leu Ile Trp Asp Ser Leu His Ser Leu Ser
1 5 10 15
<210> 97
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 97
Leu Pro Gln Thr His Pro Leu His Leu Leu Glu Asp
1 5 10
<210> 98
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 98
Ile Pro Tyr His His Leu Val Asp Gln Leu His His
1 5 10
<210> 99
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 99
Ile Pro Tyr His His Leu Val Asp Gln Leu His Leu Ser
1 5 10
<210> 100
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 100
Ile Pro Tyr His His Leu Val Asp Gln Leu His Ser Leu Ser
1 5 10
<210> 101
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 101
Phe Gly Ile Pro Tyr His His Leu Val Asp Gln Leu His His Leu Ser
1 5 10 15
<210> 102
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 102
Phe Gly Phe Pro Tyr His His Leu Val Asp Gln Leu His Ser Leu Ser
1 5 10 15
<210> 103
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 103
Tyr Pro Tyr His Val Gln Val Asp Val Leu Gln Asn
1 5 10
<210> 104
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 104
Tyr Pro Tyr His Val Gln Val Asp Val Leu Gln Asn Leu Ser
1 5 10
<210> 105
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 105
Tyr Pro Tyr His Val Gln Val Asp Val Leu Gln Ser Leu Ser
1 5 10
<210> 106
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 106
Phe Gly Tyr Pro Tyr His Val Gln Val Asp Val Leu Gln Asn Leu Ser
1 5 10 15
<210> 107
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 107
Phe Gly Phe Pro Tyr His Val Gln Val Asp Val Leu Gln Ser Leu Ser
1 5 10 15
<210> 108
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 108
Ile Pro Ser His His Leu Gln Asp Ser Leu Gln Leu
1 5 10
<210> 109
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 109
Ile Pro Ser His His Leu Gln Asp Ser Leu Gln Leu Leu Ser
1 5 10
<210> 110
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 110
Ile Pro Ser His His Leu Gln Asp Ser Leu Gln Ser Leu Ser
1 5 10
<210> 111
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 111
Gly Ile Pro Ser His His Leu Gln Asp Ser Leu Gln Leu Leu
1 5 10
<210> 112
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 112
Phe Gly Ile Pro Ser His His Leu Gln Asp Ser Leu Gln Leu Leu Ser
1 5 10 15
<210> 113
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 113
Phe Gly Phe Pro Ser His His Leu Gln Asp Ser Leu Gln Ser Leu Ser
1 5 10 15
<210> 114
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 114
Glu Tyr Ala His His Thr Ser Leu Asp Leu Arg Gln
1 5 10
<210> 115
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 115
Glu Pro Leu His Phe Arg Ser Asp Arg Ile Gln Ala
1 5 10
<210> 116
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 116
Glu Pro Leu His Phe Arg Ser Asp Arg Ile Gln Ala Leu Ser
1 5 10
<210> 117
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 117
Glu Pro Leu His Phe Arg Ser Asp Arg Ile Gln Ser Leu Ser
1 5 10
<210> 118
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 118
Gly Glu Pro Leu His Phe Arg Ser Asp Arg Ile Gln Ala Leu
1 5 10
<210> 119
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 119
Phe Gly Glu Pro Leu His Phe Arg Ser Asp Arg Ile Gln Ala Leu Ser
1 5 10 15
<210> 120
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 120
Phe Gly Phe Pro Leu His Phe Arg Ser Asp Arg Ile Gln Ser Leu Ser
1 5 10 15
<210> 121
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 121
Ala Pro Lys His Leu Tyr Ala Asp Met Ser Gln Ala
1 5 10
<210> 122
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 122
Ala Pro Lys His Leu Tyr Ala Asp Met Ser Gln Ala Leu Ser
1 5 10
<210> 123
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 123
Ala Pro Lys His Leu Tyr Ala Asp Met Ser Gln Ser Leu Ser
1 5 10
<210> 124
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 124
Gly Ala Pro Lys His Leu Tyr Ala Asp Met Ser Gln Ala Leu
1 5 10
<210> 125
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 125
Phe Gly Phe Pro Lys His Leu Tyr Ala Asp Met Ser Gln Ser Leu Ser
1 5 10 15
<210> 126
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 126
Met Pro Ala His Leu Ser Arg Asp Leu Arg Gln Ser
1 5 10
<210> 127
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 127
Met Pro Ala His Leu Ser Arg Asp Leu Arg Gln Ser Leu
1 5 10
<210> 128
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 128
Met Pro Ala His Leu Ser Arg Asp Leu Arg Gln Ser Leu Ser
1 5 10
<210> 129
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 129
Gly Met Pro Ala His Leu Ser Arg Asp Leu Arg Gln Ser Leu
1 5 10
<210> 130
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 130
Phe Gly Phe Pro Ala His Leu Ser Arg Asp Leu Arg Gln Ser Leu Ser
1 5 10 15
<210> 131
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 131
Asn Pro Lys His Tyr Ser Ile Asp Arg His Gln Ala
1 5 10
<210> 132
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 132
Thr Pro Phe His Phe Ala Gln Asp Ser Trp Gln Trp
1 5 10
<210> 133
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 133
Thr Pro Phe His Phe Ala Gln Asp Ser Trp Gln Trp Leu Ser
1 5 10
<210> 134
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 134
Thr Pro Phe His Phe Ala Gln Asp Ser Trp Gln Ser Leu Ser
1 5 10
<210> 135
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 135
Gly Thr Pro Phe His Phe Ala Gln Asp Ser Trp Gln Trp Leu
1 5 10
<210> 136
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 136
Phe Gly Phe Pro Phe His Phe Ala Gln Asp Ser Trp Gln Ser Leu Ser
1 5 10 15
<210> 137
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 137
Ala Cys Ser Phe Ala Tyr Leu Tyr Arg Cys
1 5 10
<210> 138
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 138
Ala Cys Phe Met Gly Asp Lys Trp Val Cys
1 5 10
<210> 139
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 139
Ala Cys Val Leu Tyr Pro Lys Ala Ile Cys
1 5 10
<210> 140
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 140
Ala Cys Tyr Met Gly Gln Gln Phe Val Cys
1 5 10
<210> 141
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 141
Ala Cys Leu Thr Ala Tyr Leu His Trp Cys
1 5 10
<210> 142
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 142
Ala Cys Thr Leu Phe Pro Val Ala Tyr Cys
1 5 10
<210> 143
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 143
Ala Cys Trp Leu Phe Pro Tyr Ala His Cys
1 5 10
<210> 144
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 144
Ala Cys Lys Ser Ile Asn Met Trp Leu Cys
1 5 10
<210> 145
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 145
Ala Cys Gln Thr Ile Asn Arg Trp Leu Cys
1 5 10
<210> 146
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 146
Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 147
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 147
Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 148
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 148
Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu
1 5 10
<210> 149
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 149
Ala Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 150
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 150
Phe Ala Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 151
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 151
Phe Gly Ala Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 152
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 152
Phe Gly Phe Ala Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 153
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 153
Phe Gly Phe Pro Ala His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 154
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 154
Phe Gly Phe Pro Glu Ala Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 155
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 155
Phe Gly Phe Pro Glu His Ala Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 156
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 156
Phe Gly Phe Pro Glu His Leu Ala Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 157
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 157
Phe Gly Phe Pro Glu His Leu Leu Ala Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 158
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 158
Phe Gly Phe Pro Glu His Leu Leu Val Ala Phe Leu Gln Ser Leu Ser
1 5 10 15
<210> 159
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 159
Phe Gly Phe Pro Glu His Leu Leu Val Asp Ala Leu Gln Ser Leu Ser
1 5 10 15
<210> 160
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 160
Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Ala Gln Ser Leu Ser
1 5 10 15
<210> 161
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 161
Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Ala Ser Leu Ser
1 5 10 15
<210> 162
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 162
Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ala Leu Ser
1 5 10 15
<210> 163
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 163
Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Ala Ser
1 5 10 15
<210> 164
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 164
Phe Gly Phe Pro Glu His Leu Leu Val Asp Phe Leu Gln Ser Leu Ala
1 5 10 15
<210> 165
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 165
Phe Ala Phe Pro Ala His Leu Leu Val Asp Phe Leu Gln Ala Leu Ala
1 5 10 15
<210> 166
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 166
Ala Ala Phe Pro Ala His Leu Leu Ala Asp Phe Leu Gln Ala Leu Ala
1 5 10 15
<210> 167
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 167
Ser Pro Gln His Leu Thr Thr Asp Arg Ala Gln Ala
1 5 10
<210> 168
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 168
Ser Pro Gln His Leu Thr Thr Asp Arg Ala Gln Ala Leu Ser
1 5 10
<210> 169
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 169
Ser Pro Gln His Leu Thr Thr Asp Arg Ala Gln Ser Leu Ser
1 5 10
<210> 170
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 170
Gly Ser Pro Gln His Leu Thr Thr Asp Arg Ala Gln Ala Leu
1 5 10
<210> 171
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 171
Phe Gly Phe Pro Gln His Leu Thr Thr Asp Arg Ala Gln Ser Leu Ser
1 5 10 15
<210> 172
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 172
Phe Gly Phe Pro Gln His Leu Thr Thr Asp Trp Ala Gln Ser Leu Ser
1 5 10 15
<210> 173
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 173
Phe Gly Phe Pro Gln His Leu Thr Thr Asp Arg Leu Gln Ser Leu Ser
1 5 10 15
<210> 174
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 174
Phe Gly Phe Pro Gln His Leu Thr Thr Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 175
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 175
Ala Thr Pro Ser His Leu Ile Ile Asp Arg Ala Gln
1 5 10
<210> 176
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 176
Ala Thr Pro Ser His Leu Ile Ile Asp Arg Ala Gln Ser Leu Ser
1 5 10 15
<210> 177
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 177
Phe Gly Phe Pro Ser His Leu Ile Ile Asp Arg Ala Gln Ser Leu Ser
1 5 10 15
<210> 178
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 178
Phe Gly Phe Pro Ser His Leu Ile Ile Asp Trp Ala Gln Ser Leu Ser
1 5 10 15
<210> 179
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 179
Phe Gly Phe Pro Ser His Leu Ile Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 180
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 180
Phe Gly Phe Pro Ser His Leu Ile Ile Asp Trp Ser Gln Ser Leu Ser
1 5 10 15
<210> 181
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 181
Phe Ala Thr Pro Ser His Leu Ile Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 182
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 182
Phe Lys Pro Ala His Val Ser Ile Asp Trp Leu Gln
1 5 10
<210> 183
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 183
Phe Lys Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 184
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 184
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 185
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 185
Ala Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 186
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 186
Phe Ala Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 187
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 187
Phe Gly Ala Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 188
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 188
Phe Gly Phe Ala Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 189
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 189
Phe Gly Phe Pro Ala His Val Ser Ala Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 190
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 190
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ala Leu Ser
1 5 10 15
<210> 191
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 191
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ala
1 5 10 15
<210> 192
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 192
Phe Ala Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ala Leu Ala
1 5 10 15
<210> 193
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 193
Phe Gly Phe Ala Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 194
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 194
Phe Gly Phe Phe Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 195
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 195
Phe Gly Phe Pro Ala His Val Ser Ile Arg Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 196
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 196
Phe Gly Phe Pro Ala His Val Ser Ile Glu Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 197
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 197
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Asn Ser Leu Ser
1 5 10 15
<210> 198
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 198
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu His Ser Leu Ser
1 5 10 15
<210> 199
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 199
Ala Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 200
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 200
Pro Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 201
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 201
Trp Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 202
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 202
Phe Ala Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 203
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 203
Phe Ser Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 204
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 204
Phe Tyr Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 205
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 205
Phe Asp Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 206
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 206
Phe Gly Ala Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 207
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 207
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Leu Leu Ser
1 5 10 15
<210> 208
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 208
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Trp Leu Ser
1 5 10 15
<210> 209
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 209
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Asn Leu Ser
1 5 10 15
<210> 210
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 210
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Thr Leu Ser
1 5 10 15
<210> 211
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 211
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Tyr Leu Ser
1 5 10 15
<210> 212
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 212
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Ile Ser
1 5 10 15
<210> 213
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 213
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Thr
1 5 10 15
<210> 214
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 214
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ser Leu Tyr
1 5 10 15
<210> 215
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 215
Phe Ala Phe Pro Ala His Val Ser Ile Asp Trp Leu Gln Ala Leu Ala
1 5 10 15
<210> 216
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 216
Phe Gly Phe Pro Ala His Val Ser Ile Asp Arg Ala Gln Ser Leu Ser
1 5 10 15
<210> 217
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 217
Phe Gly Phe Pro Thr His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 218
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 218
Phe Gly Phe Pro Phe His Val Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 219
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 219
Phe Gly Phe Pro Ala His Ile Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 220
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 220
Phe Gly Phe Pro Ala His Ile Ile Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 221
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 221
Phe Gly Phe Pro Ala His Leu Thr Thr Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 222
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 222
Phe Gly Phe Pro Ala His Val Phe Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 223
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 223
Phe Gly Phe Pro Ala His Val Tyr Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 224
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 224
Phe Gly Phe Pro Ala His Val Ser Leu Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 225
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 225
Phe Gly Phe Pro Ala His Val Ser Ala Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 226
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 226
Thr Pro Thr His Tyr Tyr Ala Asp Phe Ser Gln Ser Leu Ser
1 5 10
<210> 227
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 227
Phe Gly Phe Pro Ala His Val Ser Ile Asp Trp Ser Gln Ser Leu Ser
1 5 10 15
<210> 228
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 228
Phe Gly Phe Pro Ala His Val Ser Ile Asp Phe Ser Gln Ser Leu Ser
1 5 10 15
<210> 229
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 229
Phe Gly Phe Pro Ala His Val Trp Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 230
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 230
Phe Gly Phe Pro Ala His Val Phe Ile Asp Trp Leu Gln Ser Leu Asn
1 5 10 15
<210> 231
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 231
Phe Gly Phe Pro Ala His Phe Ser Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 232
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 232
Phe Gly Phe Pro Ala His Val Ser Phe Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 233
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 233
Phe Gly Phe Pro Glu His Val Phe Ile Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 234
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 234
Asp Phe Gly Phe Pro Ala His Val Phe Ile Asp Trp Leu Gln Ser Leu
1 5 10 15
Ser
<210> 235
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 235
Asp Phe Gly Phe Pro Ser His Leu Ile Ile Asp Trp Leu Gln Ser Leu
1 5 10 15
Ser
<210> 236
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 236
Asp Phe Gly Phe Pro Ala His Val Tyr Ile Asp Trp Leu Gln Ser Leu
1 5 10 15
Ser
<210> 237
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 237
Phe Gly Phe Pro Gln His Leu Phe Thr Asp Trp Leu Gln Ser Leu Ser
1 5 10 15
<210> 238
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> CETP mimotope
<400> 238
Phe Gly Phe Pro Lys His Leu Leu Val Asp Phe Leu Gln Ser Leu Ser
1 5 10 15
Claims (15)
- 콜레스테롤 에스테르 전달 단백질(CETP) 또는 CETP-에피토프의 4-머(mer) 내지 16-머 폴리펩티드 단편이 아니거나 이를 포함하지 않는 아미노산 서열 (Z1)nX1X2X3X4(Z2)m(여기서, Z1은 C가 아닌 아미노산 잔기이고, X1은 D, A, R, E, S, N, T 및 G로 구성된 군으로부터 선택된 아미노산 잔기이고, X2는 F, A, W, R, S, L, Q, V 및 M으로 구성된 군으로부터 선택된 아미노산 잔기이고, X3는 L, A, S, W, E, R, I 및 H로 구성된 군으로부터 선택된 아미노산 잔기이고, X4는 Q, A, H, D, K, R, S 및 E로 구성된 군으로부터 선택된 아미노산 잔기이고, Z2는 C가 아닌 아미노산 잔기이고, n은 0 내지 10의 정수이고, m은 0 내지 3의 정수임)을 포함하는 화합물로서, 천연 CETP 당단백질에 특이적인 항체에 대한 결합 능력을 갖는 화합물, 또는 SYHATFL, TMAFPLN, HYHGAFL, EHHDIFL, TGLSVFL, WMPSLFY, SMPWWFF, TMPLLFW, DTWPGLE, SMPPIFY, MPLWWWD, SMPNLFY, RMPPIFY, NPFEVFL, TLPNWFW, SMPLTFY, SPHPHFL, NFMSIGL, SQFLASL, WSWPGLN, IAWPGLD, SKFMDTL, SMPMVFY, YEWVGLM, KGFLDHL, HQSDDKMPWWFF, YVWQDPSFTTFF, YVWQDPSFTTFF, LPQTHPLHLLED, GPVSIYADTDFL, DSNDTLTLAAFL, NGSPALSHMLFL, TDYDPMWVFFGY, IFPLDSQWQTFW, NESMPDLFYQPS, DWGDKYFSSFWN, VSAYNNV 및 WPLHLWQ로 구성된 군으로부터 선택된 아미노산 서열을 포함하는 화합물의 죽상경화증, 죽상경화증 위험 질환 및 죽상경화증 후유증을 예방하고/하거나 치료하기 위한 약제 생성에서의 용도.
- 제 1항에 있어서, 상기 화합물이 5 내지 16개의 아미노산 잔기를 포함하는 폴리펩티드임을 특징으로 하는 용도.
- 제 1항 또는 제 2항에 있어서, n이 7, 8 또는 9이고, Z1이 C가 아닌 아미노산 잔기이거나, F, G, A, W, Y, S, G, D, L, E, K, T, P, I, V 및 M으로 구성된 군, 바람직하게는 F, G, A, P, Y, T, S, G, K 및 D로 구성된 군으로부터 선택되고, Z2가 S, L, A, W, N, T, I, Y 및 H로 구성된 군으로부터 선택됨을 특징으로 하는 용도.
- 제 3항에 있어서, X1이 D, A, R, E 및 L로 구성된 군으로부터 선택되고, X2가 F, A, W, Q 및 R로 구성된 군으로부터 선택되고, X3가 L, A 및 S로 구성된 군으로부터 선택되고, X4가 Q, A 및 H로 구성된 군으로부터 선택됨을 특징으로 하는 용도.
- 제 4항에 있어서, X1이 D이고, X2가 F, Q 및 W로 구성된 군으로부터 선택되고, X3가 L 또는 S이고, X4가 Q 또는 H임을 특징으로 하는 용도.
- 제 5항에 있어서, 상기 화합물이, X8이 G, A, F, Y 및 K로 구성된 군으로부터 선택되고, X9이 E, Y, A, Q, K 및 S로 구성된 군으로부터 선택되고, X10이 H, V, L, F 및 I로 구성된 군으로부터 선택되고, X11이 L, W, S, I, F 및 Y로 구성된 군으로부터 선택되고, X12가 V, T, F 또는 I이고, X5가 S 또는 Y이고, X6가 L, A 또는 I이고, X7이 S, N 또는 T이고, o가 0 또는 1인, 아미노산 서열 FX8(F)oPX9HX10X11X12DX2X3X4X5X6X7을 포함함을 특징으로 하는 용도.
- 제 1항 또는 제 2항에 있어서, 상기 화합물이, X1이 D, S, N, T 및 G로 구성된 군으로부터 선택되고, X2가 F이고, X3가 L이고, X4가 Q, D, K, R, S 및 E로 구성된 군으로부터 선택되고, X5가 S 또는 T이고, X6가 L이고, X7이 C가 아닌 아미노산 잔기, 바람직하게는 S, T, A, M, F 및 W로 구성된 군으로부터 선택되는, 아미노산 서열 X1X2X3X4X5X6X7을 포함함을 특징으로 하는 용도.
- 제 1항 내지 제 7항 중 어느 한 항에 있어서, 상기 아미노산 서열이 SSLELFL, SFLDTLT, NFLKTLS, DFLRTLT, AFLDTLV, TFLSSLA, GFLDSLM, SPHPHFL, SNFLKTL, TGFLATL, SDFLRAL, SANPRDFLETLF, RMFPESFLDTLW, TIYDSFLDSLAS, KPYLLKDFLEAL, AMGPYDALDLFL, TWNPIESFLESL, QYQTPLTFLEAL, RHISPATFLEAL, HTDSFLSTFYGD, ADSTFTSFLQTL, GPVSIYADTDFL, DSNDTLTLAAFL, TPTHYYADFSQL, LPGHLIWDSLHY, LPQTHPLHLLED, IPYHHLVDQLHH, YPYHVQVDVLQN, IPSHHLQDSLQL, EYAHHTSLDLRQ, EPLHFRSDRIQA, ATPSHLIIDRAQ, APKHLYADMSQA, FKPAHVSIDWLQ, MPAHLSRDLRQS, NPKHYSIDRHQA, SPQHLTTDRAQA, TPFHFAQDSWQW, TPTHYYADFSQLLS, TPTHYYADFSQSLS, GTPTHYYADFSQLL, GTPTHYYADFSQSL, FGTPTHYYADFSQSLS, FGFPTHYYADFSQSLS, LPGHLIWDSLHY, LPGHLIWDSLHYL, LPGHLIWDSLHYLS, LPGHLIWDSLHSL, LPGHLIWDSLHSLS, GLPGHLIWDSLHYL, GLPGHLIWDSLHSL, FGLPGHLIWDSLHSLS, FGFPGHLIWDSLHSLS, LPQTHPLHLLED, IPYHHLVDQLHH, IPYHHLVDQLHLS, IPYHHLVDQLHSLS, FGIPYHHLVDQLHHLS, FGFPYHHLVDQLHSLS, YPYHVQVDVLQN, YPYHVQVDVLQNLS, YPYHVQVDVLQSLS, FGYPYHVQVDVLQNLS, FGFPYHVQVDVLQSLS, IPSHHLQDSLQL, IPSHHLQDSLQLLS, IPSHHLQDSLQSLS, GIPSHHLQDSLQLL, FGIPSHHLQDSLQLLS, FGFPSHHLQDSLQSLS, EYAHHTSLDLRQ, EPLHFRSDRIQA, EPLHFRSDRIQALS, EPLHFRSDRIQSLS, GEPLHFRSDRIQAL, FGEPLHFRSDRIQALS, FGFPLHFRSDRIQSLS, APKHLYADMSQA, APKHLYADMSQALS, APKHLYADMSQSLS, GAPKHLYADMSQAL, FGFPKHLYADMSQSLS, MPAHLSRDLRQS, MPAHLSRDLRQSL, MPAHLSRDLRQSLS, GMPAHLSRDLRQSL, FGFPAHLSRDLRQSLS, NPKHYSIDRHQA, TPFHFAQDSWQW, TPFHFAQDSWQWLS, TPFHFAQDSWQSLS, GTPFHFAQDSWQWL, FGFPFHFAQDSWQSLS, ACSFAYLYRC, ACFMGDKWVC, ACVLYPKAIC, ACYMGQQFVC, ACLTAYLHWC, ACTLFPVAYC, ACWLFPYAHC, ACKSINMWLC, ACQTINRWLC, FGFPEHLLVDFLQSLS, FGFPEHLLVDFLQSLS, FPEHLLVDFLQSL, AGFPEHLLVDFLQSLS, FAFPEHLLVDFLQSLS, FGAPEHLLVDFLQSLS, FGFAEHLLVDFLQSLS, FGFPAHLLVDFLQSLS, FGFPEALLVDFLQSLS, FGFPEHALVDFLQSLS, FGFPEHLAVDFLQSLS, FGFPEHLLADFLQSLS, FGFPEHLLVAFLQSLS, FGFPEHLLVDALQSLS, FGFPEHLLVDFAQSLS, FGFPEHLLVDFLASLS, FGFPEHLLVDFLQALS, FGFPEHLLVDFLQSAS, FGFPEHLLVDFLQSLA, FAFPAHLLVDFLQALA, AAFPAHLLADFLQALA, SPQHLTTDRAQA, SPQHLTTDRAQALS, SPQHLTTDRAQSLS, GSPQHLTTDRAQAL, FGFPQHLTTDRAQSLS, FGFPQHLTTDWAQSLS, FGFPQHLTTDRLQSLS, FGFPQHLTTDWLQSLS, ATPSHLIIDRAQ, ATPSHLIIDRAQSLS, FGFPSHLIIDRAQSLS, FGFPSHLIIDWAQSLS, FGFPSHLIIDWLQSLS, FGFPSHLIIDWSQSLS, FATPSHLIIDWLQSLS, FKPAHVSIDWLQ, FKPAHVSIDWLQSLS, FGFPAHVSIDWLQSLS, AGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFAAHVSIDWLQSLS, FGFPAHVSADWLQSLS, FGFPAHVSIDWLQALS, FGFPAHVSIDWLQSLA, FAFPAHVSIDWLQALA, FGFAAHVSIDWLQSLS, FGFFAHVSIDWLQSLS, FGFPAHVSIRWLQSLS, FGFPAHVSIEWLQSLS, FGFPAHVSIDWLNSLS, FGFPAHVSIDWLHSLS, AGFPAHVSIDWLQSLS, PGFPAHVSIDWLQSLS, WGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FSFPAHVSIDWLQSLS, FYFPAHVSIDWLQSLS, FDFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFPAHVSIDWLQLLS, FGFPAHVSIDWLQWLS, FGFPAHVSIDWLQNLS, FGFPAHVSIDWLQTLS, FGFPAHVSIDWLQYLS, FGFPAHVSIDWLQSIS, FGFPAHVSIDWLQSLT, FGFPAHVSIDWLQSLY, FAFPAHVSIDWLQALA, FGFPAHVSIDRAQSLS, FGFPTHVSIDWLQSLS, FGFPFHVSIDWLQSLS, FGFPAHISIDWLQSLS, FGFPAHIIIDWLQSLS, FGFPAHLTTDWLQSLS, FGFPAHVFIDWLQSLS, FGFPAHVYIDWLQSLS, FGFPAHVSLDWLQSLS, FGFPAHVSADWLQSLS, TPTHYYADFSQSLS, FGFPAHVWIDWLQSLS, FGFPAHVFIDWLQSLN, FGFPAHFSIDWLQSLS, FGFPAHVSFDWLQSLS, FGFPEHVFIDWLQSLS, DFGFPAHVFIDWLQSLS, DFGFPSHLIIDWLQSLS, DFGFPAHVYIDWLQSLS, FGFPQHLFTDWLQSLS, FGFPKHLLVDFLQSLS, FGFPAHVSIDWSQSLS, FGFPAHVSIDFSQSLS, FGFPSHIIIDWLQSLS, FGFPSHLIIEWLQSLS, AAFPAHLLADAAQALA, AAFPAHAAADFLQALA, AAFAAHLLADFLQAAA, AAAPAHLLVDAAQAAA, FAFPAHVFIDWLQSLS; FGFPAHVFIDWLQALS, FGFPAHVFIDWLQSLA, GFPAHVFIDWLQSLS, FPAHVFIDWLQSLS, PAHVFIDWLQSLS, FAFPAHVFIDWLQALA, FGFPEHLFVDFLQSLS, FGFPAHVHIDWLQSLS, FGFPAHVPIDWLQSLS, FGFPSHLFIDWAQSLS, PGFPAHVFIDWLQLIT, PAHVYIDWLQSLS, FGFPAHVYIDWLQ, FGFPAHVFIDWLQ, DFGFPSHLIIDWLQSLS, DFGFPAHVFIDWLQSLN, PSHLIIDWLQ, PAHVFIDWLQ, DFGFPAHVTIDWLQSLN, DFGFPAHVLIDWLQSLN, FGFPAHVYIDWLQSLS, FGFPAHVFIDWLQSLN 및 FGFPAHVFIDWLQSLA로 구성된 군으로부터 선택됨을 특징으로 하는 용도.
- 제 1항 내지 제 8항 중 어느 한 항에 있어서, 상기 화합물이 약학적으로 허용되는 담체, 바람직하게는 키홀 림펫 헤모시아닌(Keyhole Limpet Hemocyanin, KLH)에 커플링됨을 특징으로 하는 용도.
- 제 1항 내지 제 9항 중 어느 한 항에 있어서, 상기 화합물이 정맥내, 피하 또는 근내 투여용으로 제형화됨을 특징으로 하는 용도.
- 제 1항 내지 제 10항 중 어느 한 항에 있어서, 상기 화합물이 애쥬번트, 바람직하게는 수산화알루미늄과 함께 제형화됨을 특징으로 하는 용도.
- 제 1항 내지 제 11항 중 어느 한 항에 있어서, 상기 화합물이 0.1 ng 내지 10 mg, 바람직하게는 10 ng 내지 1 mg, 특히 100 ng 내지 10 ㎍의 양으로 포함됨을 특징으로 하는 용도.
- SYHATFL, TMAFPLN, HYHGAFL, EHHDIFL, SSLELFL, TGLSVFL, WMPSLFY, SMPWWFF, TMPLLFW, DTWPGLE, SMPPIFY, MPLWWWD, SMPNLFY, RMPPIFY, NPFEVFL, TLPNWFW, SMPLTFY, SFLDTLT, NFLKTLS, DFLRTLT, AFLDTLV, TFLSSLA, GFLDSLM, SPHPHFL, NFMSIGL, SQFLASL, SNFLKTL, TGFLATL, WSWPGLN, IAWPGLD, SKFMDTL, SDFLRAL, SMPMVFY, YEWVGLM, KGFLDHL, SANPRDFLETLF, RMFPESFLDTLW, TIYDSFLDSLAS, HQSDDKMPWWFF, KPYLLKDFLEAL, AMGPYDALDLFL, TWNPIESFLESL, YVWQDPSFTTFF, QYQTPLTFLEAL, RHISPATFLEAL, HTDSFLSTFYGD, YVWQDPSFTTFF, ADSTFTSFLQTL, GPVSIYADTDFL, DSNDTLTLAAFL, NGSPALSHMLFL, TDYDPMWVFFGY, IFPLDSQWQTFW, NESMPDLFYQPS, DWGDKYFSSFWN, VSAYNNV, WPLHLWQ, TPTHYYADFSQL, LPGHLIWDSLHY, LPQTHPLHLLED, IPYHHLVDQLHH, YPYHVQVDVLQN, IPSHHLQDSLQL, EYAHHTSLDLRQ, EPLHFRSDRIQA, ATPSHLIIDRAQ, APKHLYADMSQA, FKPAHVSIDWLQ, MPAHLSRDLRQS, NPKHYSIDRHQA, SPQHLTTDRAQA, TPFHFAQDSWQW, TPTHYYADFSQLLS, TPTHYYADFSQSLS, GTPTHYYADFSQLL, GTPTHYYADFSQSL, FGTPTHYYADFSQSLS, FGFPTHYYADFSQSLS, LPGHLIWDSLHY, LPGHLIWDSLHYL, LPGHLIWDSLHYLS, LPGHLIWDSLHSL, LPGHLIWDSLHSLS, GLPGHLIWDSLHYL, GLPGHLIWDSLHSL, FGLPGHLIWDSLHSLS, FGFPGHLIWDSLHSLS, LPQTHPLHLLED, IPYHHLVDQLHH, IPYHHLVDQLHLS, IPYHHLVDQLHSLS, FGIPYHHLVDQLHHLS, FGFPYHHLVDQLHSLS, YPYHVQVDVLQN, YPYHVQVDVLQNLS, YPYHVQVDVLQSLS, FGYPYHVQVDVLQNLS, FGFPYHVQVDVLQSLS, IPSHHLQDSLQL, IPSHHLQDSLQLLS, IPSHHLQDSLQSLS, GIPSHHLQDSLQLL, FGIPSHHLQDSLQLLS, FGFPSHHLQDSLQSLS, EYAHHTSLDLRQ, EPLHFRSDRIQA, EPLHFRSDRIQALS, EPLHFRSDRIQSLS, GEPLHFRSDRIQAL, FGEPLHFRSDRIQALS, FGFPLHFRSDRIQSLS, APKHLYADMSQA, APKHLYADMSQALS, APKHLYADMSQSLS, GAPKHLYADMSQAL, FGFPKHLYADMSQSLS, MPAHLSRDLRQS, MPAHLSRDLRQSL, MPAHLSRDLRQSLS, GMPAHLSRDLRQSL, FGFPAHLSRDLRQSLS, NPKHYSIDRHQA, TPFHFAQDSWQW, TPFHFAQDSWQWLS, TPFHFAQDSWQSLS, GTPFHFAQDSWQWL, FGFPFHFAQDSWQSLS, ACSFAYLYRC, ACFMGDKWVC, ACVLYPKAIC, ACYMGQQFVC, ACLTAYLHWC, ACTLFPVAYC, ACWLFPYAHC, ACKSINMWLC, ACQTINRWLC, FGFPEHLLVDFLQSLS, FGFPEHLLVDFLQSLS, FPEHLLVDFLQSL, AGFPEHLLVDFLQSLS, FAFPEHLLVDFLQSLS, FGAPEHLLVDFLQSLS, FGFAEHLLVDFLQSLS, FGFPAHLLVDFLQSLS, FGFPEALLVDFLQSLS, FGFPEHALVDFLQSLS, FGFPEHLAVDFLQSLS, FGFPEHLLADFLQSLS, FGFPEHLLVAFLQSLS, FGFPEHLLVDALQSLS, FGFPEHLLVDFAQSLS, FGFPEHLLVDFLASLS, FGFPEHLLVDFLQALS, FGFPEHLLVDFLQSAS, FGFPEHLLVDFLQSLA, FAFPAHLLVDFLQALA, AAFPAHLLADFLQALA, SPQHLTTDRAQA, SPQHLTTDRAQALS, SPQHLTTDRAQSLS, GSPQHLTTDRAQAL, FGFPQHLTTDRAQSLS, FGFPQHLTTDWAQSLS, FGFPQHLTTDRLQSLS, FGFPQHLTTDWLQSLS, ATPSHLIIDRAQ, ATPSHLIIDRAQSLS, FGFPSHLIIDRAQSLS, FGFPSHLIIDWAQSLS, FGFPSHLIIDWLQSLS, FGFPSHLIIDWSQSLS, FATPSHLIIDWLQSLS, FKPAHVSIDWLQ, FKPAHVSIDWLQSLS, FGFPAHVSIDWLQSLS, AGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFAAHVSIDWLQSLS, FGFPAHVSADWLQSLS, FGFPAHVSIDWLQALS, FGFPAHVSIDWLQSLA, FAFPAHVSIDWLQALA, FGFAAHVSIDWLQSLS, FGFFAHVSIDWLQSLS, FGFPAHVSIRWLQSLS, FGFPAHVSIEWLQSLS, FGFPAHVSIDWLNSLS, FGFPAHVSIDWLHSLS, AGFPAHVSIDWLQSLS, PGFPAHVSIDWLQSLS, WGFPAHVSIDWLQSLS, FAFPAHVSIDWLQSLS, FSFPAHVSIDWLQSLS, FYFPAHVSIDWLQSLS, FDFPAHVSIDWLQSLS, FGAPAHVSIDWLQSLS, FGFPAHVSIDWLQLLS, FGFPAHVSIDWLQWLS, FGFPAHVSIDWLQNLS, FGFPAHVSIDWLQTLS, FGFPAHVSIDWLQYLS, FGFPAHVSIDWLQSIS, FGFPAHVSIDWLQSLT, FGFPAHVSIDWLQSLY, FAFPAHVSIDWLQALA, FGFPAHVSIDRAQSLS, FGFPTHVSIDWLQSLS, FGFPFHVSIDWLQSLS, FGFPAHISIDWLQSLS, FGFPAHIIIDWLQSLS, FGFPAHLTTDWLQSLS, FGFPAHVFIDWLQSLS, FGFPAHVYIDWLQSLS, FGFPAHVSLDWLQSLS, FGFPAHVSADWLQSLS, TPTHYYADFSQSLS, FGFPAHVSIDWSQSLS, FGFPAHVSIDFSQSLS, FGFPSHIIIDWLQSLS, FGFPSHLIIEWLQSLS, AAFPAHLLADAAQALA, AAFPAHAAADFLQALA, AAFAAHLLADFLQAAA, AAAPAHLLVDAAQAAA, FAFPAHVFIDWLQSLS; FGFPAHVFIDWLQALS, FGFPAHVFIDWLQSLA, GFPAHVFIDWLQSLS, FPAHVFIDWLQSLS, PAHVFIDWLQSLS, FAFPAHVFIDWLQALA, FGFPEHLFVDFLQSLS, FGFPAHVHIDWLQSLS, FGFPAHVPIDWLQSLS, FGFPSHLFIDWAQSLS, PGFPAHVFIDWLQLIT, PAHVYIDWLQSLS, FGFPAHVYIDWLQ, FGFPAHVFIDWLQ, DFGFPSHLIIDWLQSLS, DFGFPAHVFIDWLQSLN, PSHLIIDWLQ, PAHVFIDWLQ, DFGFPAHVTIDWLQSLN, DFGFPAHVLIDWLQSLN, FGFPAHVYIDWLQSLS, FGFPAHVFIDWLQSLN 및 FGFPAHVFIDWLQSLA로 구성된 군으로부터 선택된 하나 이상의 아미노산 서열로 구성된 펩티드.
- 제 13항에 따른 하나 이상의 펩티드를 포함하는 약학적 제형.
- 제 14항에 있어서, 상기 펩티드가 약학적으로 허용되는 담체, 바람직하게는 KLH(키홀 림펫 헤모시아닌)에 커플링됨을 특징으로 하는 제형.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AT0125807A AT505574B1 (de) | 2007-08-10 | 2007-08-10 | Mimotope zur behandlung von atherosklerose |
| ATA1258/2007 | 2007-08-10 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20100080507A true KR20100080507A (ko) | 2010-07-08 |
Family
ID=39865273
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020107005287A KR20100080507A (ko) | 2007-08-10 | 2008-08-08 | 죽상경화증의 치료 |
Country Status (18)
| Country | Link |
|---|---|
| US (2) | US8618046B2 (ko) |
| EP (1) | EP2190451B1 (ko) |
| JP (2) | JP2010535818A (ko) |
| KR (1) | KR20100080507A (ko) |
| CN (1) | CN101835483B (ko) |
| AT (1) | AT505574B1 (ko) |
| AU (1) | AU2008286667B2 (ko) |
| CA (1) | CA2695988A1 (ko) |
| CY (1) | CY1114760T1 (ko) |
| DK (1) | DK2190451T3 (ko) |
| ES (1) | ES2439703T3 (ko) |
| HK (1) | HK1145634A1 (ko) |
| HR (1) | HRP20131229T1 (ko) |
| PL (1) | PL2190451T3 (ko) |
| PT (1) | PT2190451E (ko) |
| SI (1) | SI2190451T1 (ko) |
| TW (1) | TWI442933B (ko) |
| WO (1) | WO2009021254A1 (ko) |
Families Citing this family (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AT506535B1 (de) * | 2008-02-22 | 2010-04-15 | Affiris Forschungs & Entwicklungs Gmbh | Vaccine enthaltend alpha-synuclein-mimotope auf basis von peptiden |
| EP2532359A1 (en) | 2011-06-10 | 2012-12-12 | Affiris AG | CETP fragments |
| AU2014342528A1 (en) | 2013-10-28 | 2016-04-28 | Dots Technology Corp. | Allergen detection |
| CN105596811A (zh) * | 2015-11-10 | 2016-05-25 | 王冬梅 | 主治动脉粥样硬化的中药组合物及制法 |
| US11547746B2 (en) * | 2016-12-15 | 2023-01-10 | Talengen International Limited | Method for treating coronary atherosclerosis and complications thereof |
| WO2019020480A1 (en) * | 2017-07-24 | 2019-01-31 | INSERM (Institut National de la Santé et de la Recherche Médicale) | ANTIBODIES AND PEPTIDES FOR TREATING HCMV RELATED DISEASES |
| CN117244043B (zh) * | 2023-11-02 | 2024-09-20 | 徐州医科大学附属医院 | 破伤风类毒素在治疗血管内皮细胞损伤中的应用 |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6410022B1 (en) * | 1995-05-01 | 2002-06-25 | Avant Immunotherapeutics, Inc. | Modulation of cholesteryl ester transfer protein (CETP) activity |
| AT500835B1 (de) | 2004-09-13 | 2007-12-15 | Affiris Forschungs & Entwicklungs Gmbh | Cholinestertransport-protein-mimotop als atherosklerose-medikament |
| US20060276400A1 (en) | 2005-06-06 | 2006-12-07 | Hedy Adari | Modulation of cholesteryl ester transfer protein (CETP) activity |
| CN101384621A (zh) * | 2005-11-10 | 2009-03-11 | 受体生物公司 | 产生受体和配体同种型的方法 |
-
2007
- 2007-08-10 AT AT0125807A patent/AT505574B1/de not_active IP Right Cessation
-
2008
- 2008-08-08 DK DK08782809.1T patent/DK2190451T3/da active
- 2008-08-08 CN CN2008801113067A patent/CN101835483B/zh not_active Expired - Fee Related
- 2008-08-08 TW TW097130215A patent/TWI442933B/zh active
- 2008-08-08 PT PT87828091T patent/PT2190451E/pt unknown
- 2008-08-08 CA CA2695988A patent/CA2695988A1/en not_active Abandoned
- 2008-08-08 KR KR1020107005287A patent/KR20100080507A/ko not_active Application Discontinuation
- 2008-08-08 ES ES08782809.1T patent/ES2439703T3/es active Active
- 2008-08-08 JP JP2010520379A patent/JP2010535818A/ja active Pending
- 2008-08-08 WO PCT/AT2008/000281 patent/WO2009021254A1/en active Application Filing
- 2008-08-08 EP EP08782809.1A patent/EP2190451B1/en not_active Not-in-force
- 2008-08-08 SI SI200831094T patent/SI2190451T1/sl unknown
- 2008-08-08 PL PL08782809T patent/PL2190451T3/pl unknown
- 2008-08-08 AU AU2008286667A patent/AU2008286667B2/en not_active Ceased
- 2008-08-08 US US12/673,081 patent/US8618046B2/en not_active Expired - Fee Related
-
2010
- 2010-12-23 HK HK10112056.4A patent/HK1145634A1/xx not_active IP Right Cessation
-
2013
- 2013-09-25 JP JP2013198877A patent/JP2014040438A/ja active Pending
- 2013-11-21 US US14/086,159 patent/US20140179900A1/en not_active Abandoned
- 2013-12-27 HR HRP20131229AT patent/HRP20131229T1/hr unknown
-
2014
- 2014-01-16 CY CY20141100039T patent/CY1114760T1/el unknown
Also Published As
| Publication number | Publication date |
|---|---|
| CN101835483B (zh) | 2012-09-26 |
| AU2008286667A1 (en) | 2009-02-19 |
| ES2439703T3 (es) | 2014-01-24 |
| TW200911282A (en) | 2009-03-16 |
| AT505574A1 (de) | 2009-02-15 |
| JP2010535818A (ja) | 2010-11-25 |
| EP2190451A1 (en) | 2010-06-02 |
| CN101835483A (zh) | 2010-09-15 |
| DK2190451T3 (da) | 2013-11-04 |
| CA2695988A1 (en) | 2009-02-19 |
| CY1114760T1 (el) | 2016-12-14 |
| PT2190451E (pt) | 2013-12-23 |
| AU2008286667B2 (en) | 2013-04-04 |
| US20110275556A1 (en) | 2011-11-10 |
| SI2190451T1 (sl) | 2013-12-31 |
| TWI442933B (zh) | 2014-07-01 |
| US8618046B2 (en) | 2013-12-31 |
| US20140179900A1 (en) | 2014-06-26 |
| PL2190451T3 (pl) | 2014-03-31 |
| EP2190451B1 (en) | 2013-10-16 |
| AT505574B1 (de) | 2009-09-15 |
| HRP20131229T1 (hr) | 2014-01-31 |
| JP2014040438A (ja) | 2014-03-06 |
| HK1145634A1 (en) | 2011-04-29 |
| WO2009021254A1 (en) | 2009-02-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| EP1267908B1 (en) | Vaccine for the treatment of atherosclerosis | |
| KR101983685B1 (ko) | 백신 | |
| JP3839483B2 (ja) | コレステリルエステル転移タンパク質(cetp)活性の調節 | |
| US20140179900A1 (en) | Treatment of atherosclerosis with cholesterol ester transport protein mimotopes | |
| KR20070054206A (ko) | 죽상경화증의 치료 | |
| US20040052809A1 (en) | Vaccine | |
| JP2005508900A6 (ja) | ワクチン | |
| KR102101201B1 (ko) | Pcsk9 펩티드 백신 | |
| WO2004080375A2 (en) | Vaccine therapy of atherosclerosis | |
| RU2534883C2 (ru) | Вакцина для лечения и/или профилактики физического расстройства, связанного с ренин-активируемой ангиотензиновой системой и применение пептида для ее получения. | |
| WO2004081045A2 (en) | Vaccine related to modified apolipoprotein c-iii | |
| EP2717908B1 (en) | Cetp fragments | |
| TW201420112A (zh) | 胜肽、醫藥化合物及該醫藥化合物之使用 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| N231 | Notification of change of applicant | ||
| A201 | Request for examination | ||
| E902 | Notification of reason for refusal | ||
| E601 | Decision to refuse application |