JP7680902B2 - タンパク質加水分解用アミノペプチダーゼ - Google Patents
タンパク質加水分解用アミノペプチダーゼ Download PDFInfo
- Publication number
- JP7680902B2 JP7680902B2 JP2021126487A JP2021126487A JP7680902B2 JP 7680902 B2 JP7680902 B2 JP 7680902B2 JP 2021126487 A JP2021126487 A JP 2021126487A JP 2021126487 A JP2021126487 A JP 2021126487A JP 7680902 B2 JP7680902 B2 JP 7680902B2
- Authority
- JP
- Japan
- Prior art keywords
- seq
- amino acid
- acid sequence
- polypeptide
- sequence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images


















Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/48—Hydrolases (3) acting on peptide bonds (3.4)
- C12N9/485—Exopeptidases (3.4.11-3.4.19)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12C—BEER; PREPARATION OF BEER BY FERMENTATION; PREPARATION OF MALT FOR MAKING BEER; PREPARATION OF HOPS FOR MAKING BEER
- C12C1/00—Preparation of malt
- C12C1/027—Germinating
- C12C1/047—Influencing the germination by chemical or physical means
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12C—BEER; PREPARATION OF BEER BY FERMENTATION; PREPARATION OF MALT FOR MAKING BEER; PREPARATION OF HOPS FOR MAKING BEER
- C12C11/00—Fermentation processes for beer
- C12C11/003—Fermentation of beerwort
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12C—BEER; PREPARATION OF BEER BY FERMENTATION; PREPARATION OF MALT FOR MAKING BEER; PREPARATION OF HOPS FOR MAKING BEER
- C12C5/00—Other raw materials for the preparation of beer
- C12C5/004—Enzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms, e.g. protozoa; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/14—Fungi; Culture media therefor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/80—Vectors or expression systems specially adapted for eukaryotic hosts for fungi
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/06—Preparation of peptides or proteins produced by the hydrolysis of a peptide bond, e.g. hydrolysate products
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y304/00—Hydrolases acting on peptide bonds, i.e. peptidases (3.4)
- C12Y304/11—Aminopeptidases (3.4.11)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/01—Fusion polypeptide containing a localisation/targetting motif
- C07K2319/02—Fusion polypeptide containing a localisation/targetting motif containing a signal sequence
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Food Science & Technology (AREA)
- Medicinal Chemistry (AREA)
- Mycology (AREA)
- Physiology (AREA)
- Physics & Mathematics (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Tropical Medicine & Parasitology (AREA)
- Botany (AREA)
- Virology (AREA)
- Enzymes And Modification Thereof (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Distillation Of Fermentation Liquor, Processing Of Alcohols, Vinegar And Beer (AREA)
- Seasonings (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Cereal-Derived Products (AREA)
Description
本願は、2015年6月26日に出願された米国仮特許出願第62/185503号お
よび2015年10月1日に出願された同第62/235937号の優先権および利益を
主張するものであり、各仮出願の発明の名称は「新規のタンパク質加水分解用アミノペプ
チダーゼ」である。
2016年6月10日に作成され、本明細書とともに提出された、192,505バイ
トのサイズを有する「20160610_NB40989-PCT sequence
listing prj_ST25.txt」という名のファイルに示された配列表は、
参照によりその全体が本明細書に組み込まれる。
従来、化学的加水分解により行われてきた。しかしながら、そのような化学的加水分解は
、加水分解中に得られるアミノ酸を大量に分解し、また、化学反応の過程で有害な副生物
を生成する。化学的加水分解により得られるタンパク質加水分解物を使用することの懸念
の増大が、酵素を使用した加水分解プロセスの開発に繋がっている。
している。アミノペプチダーゼ活性を有するポリペプチドは、ペプチド、ポリペプチドお
よびタンパク質のN末端からの1つ以上のアミノ酸残基の除去を触媒する。所望の特性お
よび高い加水分解度を有するタンパク質加水分解物を生成するには、一般に、ペプチダー
ゼ活性物を混合して使用する必要がある。単独で、または他の酵素と組み合わされて、食
品および餌料製品に使用されるタンパク質加水分解物の特性および加水分解度の改善に有
用な活性を有する単一成分のペプチダーゼ酵素を提供することが望まれる。
質および高い加水分解度を有するタンパク質加水分解物を得る方法を提供する。
明、およびその添付図面を参照することにより、より深く理解できるであろう。
のポリペプチドをコードする核酸配列、その核酸配列を含む核酸コンストラクト、ベクタ
ーおよび宿主細胞、ならびに、そのポリペプチドを生成する方法に関する。本開示はまた
、タンパク性基質から加水分解物を得る方法であって、タンパク性物質を、単独の、また
はプロテアーゼ、例えばエンドペプチダーゼと組み合わせたアミノペプチダーゼ活性を有
するポリペプチドに曝すことを含む方法に関する。本開示はまた、タンパク性基質から、
遊離グルタミン酸および/またはグルタミン酸残基を結合したペプチドに富む加水分解物
を得る方法であって、基質を、アミノペプチダーゼ活性を有するポリペプチドに曝すこと
を含む方法に関する。本開示はさらに、アミノペプチダーゼ活性を有するポリペプチドを
含む風味改善組成物に関する。組成物はさらに、追加の酵素活性を含んでもよい。
向上させるために使用され得る。あるいは、食品における風味改善は、本発明の方法によ
って得られる加水分解物の添加によって達成され得る。いくつかの実施形態において、本
明細書に記載のアミノペプチダーゼを使用して生成される加水分解物はまた、未処理の加
水分解物に比べて、苦味を低減することができる。
、産生宿主(例えば、トリコデルマ・リーゼイ(Trichoderma reesei
))におけるそれらの高収率での産生、ならびに、例えば苦味除去用、およびグルタミン
酸生成用のタンパク質加水分解物を生成するためのそれらの使用を提供する。実施例9に
示すように、本発明の2型アミノペプチダーゼは、1型アミノペプチダーゼと比べて、グ
ルタミン酸の放出が向上した。初期の結果は、驚くべきことに、これらのPepNが、他
の既知のアミノペプチダーゼと比べて、P1中のプロリンに対し、耐性を有することを示
唆している。したがって、本発明において使用するために教示されるプロリン耐性アミノ
ペプチダーゼは、広範なペプチドおよび/またはタンパク質基質に作用し得、そのような
広い基質特異性を有することに起因して、特定のアミノ酸(例えば、プロリンおよび/ま
たはリジンおよび/またはアルギニンおよび/またはグリシン)に富む基質の開裂を容易
には阻害されない。
優れた能力を有することを見出した。さらに、本発明者らは、驚いたことに、良好な能力
を有する一群の真菌2型アミノペプチダーゼを見出した。この一群の真菌2型アミノペプ
チダーゼは、それらの成熟アミノ酸配列中に長いN末端を有している。図9は、いくつか
の2型アミノペプチダーゼのアミノ酸配列のアライメントを示している。
プチドまたはタンパク質のN末端からのアミノ酸の切断を触媒するペプチダーゼ活性と定
義される。一般的な定義では、アミノペプチダーゼ活性は、ペプチド、オリゴペプチドま
たはタンパク質のN末端からアミノ酸X(ここで、Xは、Ala、Arg、Asn、As
p、Cys、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Ph
e、Pro、Ser、Thr、Trp、Tyr、およびValからなる群から選択される
アミノ酸残基、少なくともLeu、Glu、Gly、Ala、および/またはProの残
基であり得る)を開裂することができる。本発明のアミノペプチダーゼ活性を有するポリ
ペプチドが、ペプチド、ポリペプチド基質のN末端から開裂させるアミノ酸に関し非特異
的であり得ることは理解されよう。
位の保存残基I/VのN末端側に13を超える残基を含む予測成熟配列を有する、アミノ
ペプチダーゼ活性を有する単離ポリペプチド、またはそのフラグメントに関し、但し、そ
のフラグメントはアミノペプチダーゼ活性を有する。
くとも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より
好ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましく
は少なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミ
ノ酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプ
チド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリ
ペプチドは、配列番号1のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸
、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは
1個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明の
ポリペプチドは、配列番号1のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型
;およびそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する
。いくつかの実施形態において、本発明のポリペプチドは、配列番号1を含む。他の実施
形態において、本発明のポリペプチドは、配列番号1のアミノ酸配列、またはそのフラグ
メントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。配列番号1のフラ
グメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ末端から1個以上
のアミノ酸が削除されたポリペプチドである。いくつかの実施形態において、フラグメン
トは配列番号18の配列を含む。いくつかの実施形態において、ポリペプチドは配列番号
1のアミノ酸配列を有する。
とも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好
ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは
少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有するアミノ
酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプチ
ド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリペ
プチドは、配列番号2のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸、
より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは1
個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明のポ
リペプチドは、配列番号2のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型;
およびそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。
いくつかの実施形態において、本発明のポリペプチドは、配列番号2のアミノ酸配列を含
む。他の実施形態において、本発明のポリペプチドは、配列番号2のアミノ酸配列、また
はそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。配列
番号2のフラグメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ末端
から1個以上のアミノ酸が削除されたポリペプチドである。いくつかの実施形態において
、フラグメントは配列番号19の配列を含む。いくつかの実施形態において、ポリペプチ
ドは配列番号2のアミノ酸配列を有する。
とも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好
ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは
少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有するアミノ
酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプチ
ド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリペ
プチドは、配列番号3のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸、
より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは1
個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明のポ
リペプチドは、配列番号3のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型;
およびそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。
いくつかの実施形態において、本発明のポリペプチドは、配列番号3のアミノ酸配列を含
む。他の実施形態において、本発明のポリペプチドは、配列番号3のアミノ酸配列、また
はそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。配列
番号3のフラグメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ末端
から1個以上のアミノ酸が削除されたポリペプチドである。いくつかの実施形態において
、フラグメントは配列番号20の配列を含む。いくつかの実施形態において、ポリペプチ
ドは配列番号3のアミノ酸配列を有する。
くとも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より
好ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましく
は少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有するアミ
ノ酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプ
チド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリ
ペプチドは、配列番号4のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸
、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは
1個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明の
ポリペプチドは、配列番号4のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型
;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を有す
る。いくつかの実施形態において、本発明のポリペプチドは、配列番号4のアミノ酸配列
を含む。他の実施形態において、本発明のポリペプチドは、配列番号4のアミノ酸配列、
またはそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。
配列番号4のフラグメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ
末端から1個以上のアミノ酸が削除されたポリペプチドである。いくつかの実施形態にお
いて、フラグメントは配列番号21の配列を含む。いくつかの実施形態において、ポリペ
プチドは配列番号4のアミノ酸配列を有する。
とも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好
ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは
少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有するアミノ
酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプチ
ド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリペ
プチドは、配列番号5のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸、
より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは1
個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明のポ
リペプチドは、配列番号5のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型;
およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を有する
。いくつかの実施形態において、本発明のポリペプチドは、配列番号5のアミノ酸配列を
含む。他の実施形態において、本発明のポリペプチドは、配列番号5のアミノ酸配列、ま
たはそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。配
列番号5のフラグメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ末
端から1個以上のアミノ酸が削除されたポリペプチドである。いくつかの実施形態におい
て、フラグメントは配列番号22の配列を含む。いくつかの実施形態において、ポリペプ
チドは配列番号5のアミノ酸配列を有する。
とも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好
ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは
少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有するアミノ
酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプチ
ド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリペ
プチドは、配列番号6のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸、
より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは1
個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明のポ
リペプチドは、配列番号6のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型;
およびそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。
いくつかの実施形態において、本発明のポリペプチドは、配列番号6のアミノ酸配列を含
む。他の実施形態において、本発明のポリペプチドは、配列番号6のアミノ酸配列、また
はそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。配列
番号6のフラグメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ末端
から1個以上のアミノ酸が削除されたポリペプチドである。いくつかの実施形態において
、フラグメントは配列番号23の配列を含む。いくつかの実施形態において、ポリペプチ
ドは配列番号6のアミノ酸配列を有する。
とも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好
ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは
少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有するアミノ
酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプチ
ド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリペ
プチドは、配列番号7のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸、
より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは1
個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明のポ
リペプチドは、配列番号7のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型;
およびそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。
いくつかの実施形態において、本発明のポリペプチドは、配列番号7のアミノ酸配列を含
む。他の実施形態において、本発明のポリペプチドは、配列番号7のアミノ酸配列、また
はそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。配列
番号7のフラグメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ末端
から1個以上のアミノ酸が削除されたポリペプチドである。いくつかの実施形態において
、フラグメントは配列番号24の配列を含む。いくつかの実施形態において、ポリペプチ
ドは配列番号7のアミノ酸配列を有する。
とも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好
ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは
少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有するアミノ
酸配列を有する単離ポリペプチドであって、アミノペプチダーゼ活性を有するポリペプチ
ド(以下、「相同ポリペプチド」)に関する。いくつかの実施形態において、相同ポリペ
プチドは、配列番号8のアミノ酸配列と、5個のアミノ酸、好ましくは4個のアミノ酸、
より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ酸、最も好ましくは1
個のアミノ酸が異なるアミノ酸配列を有する。いくつかの実施形態において、本発明のポ
リペプチドは、配列番号8のアミノ酸配列、成熟アミノ酸配列または対立遺伝子変異型;
およびそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。
いくつかの実施形態において、本発明のポリペプチドは、配列番号8のアミノ酸配列を含
む。他の実施形態において、本発明のポリペプチドは、配列番号8のアミノ酸配列、また
はそのフラグメントを含み、そのフラグメントはアミノペプチダーゼ活性を有する。配列
番号8のフラグメントは、このアミノ酸配列のアミノ末端および/またはカルボキシ末端
から1個以上のアミノ酸が削除されたポリペプチドである。いくつかの実施形態において
、フラグメントは配列番号25の配列を含む。いくつかの実施形態において、ポリペプチ
ドは配列番号8のアミノ酸配列を有する。
少なくとも380個のアミノ酸残基、最も好ましくは、430個のアミノ酸残基を含む。
示す。対立遺伝子変異は、突然変異によって自然に生じ、集団内で表現型多型をもたらし
得る。
3、または配列番号4、または配列番号5、配列番号6、または配列番号7、または配列
番号8に示すポリペプチドのアミノ酸残基の全てを有するアミノペプチダーゼである。こ
の点に関し、例えば、親は 変異ポリペプチドの修飾(これは、変異型により、0、1も
しくはそれ以上であり得る)の全てを有するであろう。
は、中位のストリンジェンシーな条件下で、最も好ましくは、高ストリンジェンシーな条
件下で、オリゴヌクレオチドプローブとハイブリダイゼーションを行う核酸配列によって
コードされる、アミノペプチダーゼ活性を有する単離ポリペプチド、またはそのポリペプ
チドの対立遺伝子変異型およびフラグメント(このフラグメントはアミノペプチダーゼ活
性を有する)に関し、オリゴヌクレオチドプローブは、配列番号9、または配列番号10
、または配列番号11、または配列番号12、または配列番号13、配列番号14、また
は配列番号15、または配列番号16、またはその相補鎖の核酸配列のポリペチドコード
化部分と同じ条件下でハイブリダイゼーションを行う(J.Sambrook,E.F.
Fritsch,and T.Maniatus,1989,Molecular Cl
oning,A Laboratory Manual,2d edition,Col
d Spring Harbor,New York)。
配列番号11、または配列番号12、または配列番号13、配列番号14、または配列番
号15、または配列番号16に示す核酸配列のポリペプチドコード化部分に対応するオリ
ゴヌクレオチドプローブに、低~高ストリンジェンシーな条件下(すなわち、5X SS
PE、0.3%SDS、200Pg/ml断片処理済みの変性サケ精子DNA、ならびに
、それぞれ低、中および高ストリンジェンシーで25、35または50%のホルムアミド
中、42℃でプレハイブリダイゼーションおよびハイブリダイゼーション)で、標準サザ
ンブロット法にしたがいハイブリダイズすることを示す。
は配列番号5、配列番号6、もしくは配列番号7、もしくは配列番号8のアミノ酸配列、
またはその部分配列は、オリゴヌクレオチドプローブを設計するために使用され得、ある
いは例えば、配列番号9、もしくは配列番号10、もしくは配列番号11、もしくは配列
番号12、もしくは配列番号13、配列番号14、もしくは配列番号15、もしくは配列
番号16の核酸配列のポリペプチドコード化部分などの、本発明のポリペプチドをコード
する核酸配列、またはその部分配列は、当該技術分野でよく知られた方法にしたがい、異
なる属または種の菌株からアミノペプチダーゼ活性を有するポリペプチドをコードするD
NAを同定またはクローン化するために使用され得る特に、そのようなプローブは、その
中の対応する遺伝子を同定および単離するために、標準サザンブロット法にしたがって、
目的の属または種のゲノムまたはcDNAとハイブリダイゼーションするために使用する
ことができる。そのようなプローブは、全配列より相当に短い可能性があるが、長さが少
なくとも15個、好ましくは少なくとも25個、より好ましくは少なくとも40個のヌク
レオチドでなければならない。より長いプローブもまた使用することができる。DNAプ
ローブおよびRNAプローブの両方を使用できる。これらのプローブは、通常、対応する
遺伝子を検出するために(例えば、32P、3H、35S、ビオチンまたはアビジンを用
いて)標識される。
学ライブラリーによって、上記プローブとハイブリダイズし、かつアミノペプチダーゼ活
性を有するポリペプチドをコードするDNAをスクリーニングすることができる。そのよ
うな他の生物からのゲノムまたは他のDNAは、アガロースもしくはポリアクリルアミド
ゲル電気泳動法、または他の分離技術によって分離することができる。ライブラリー由来
のDNAまたは分離DNAは、ニトロセルロースまたは他の好適な担体材料に移して固定
化することができる。配列番号9、または配列番号10、または配列番号11、または配
列番号12、または配列番号13、配列番号14、または配列番号15、または配列番号
16の核酸配列のポリペプチドコード化部分と相同のクローンまたはDNAを同定するた
めに、サザンブロットにおいて担体材料が使用され、その担体材料は、30分間に、最終
的に3回、それぞれ、2×SSC、0.2%SDSを使用し、好ましくは少なくとも50
℃、より好ましくは少なくとも55℃、より好ましくは少なくとも60℃、より好ましく
は少なくとも65℃、より一層好ましくは少なくとも70℃、最も好ましくは少なくとも
75℃で洗浄される。オリゴヌクレオチドプローブがこれらの条件下でハイブリダイズす
る分子は、X線フィルムを使用して検出される。
の野生型アミノペプチダーゼポリペプチド配列と比較したときの置換を指す。比較は、変
異体アミノペプチダーゼポリペプチドと、配列番号1、または配列番号2、または配列番
号3、または配列番号4、または配列番号5、配列番号6、または配列番号7、または配
列番号8として示されているレファレンス配列を有する野生型アミノペプチダーゼの2つ
をアライニングすることによって行われる。
にあるアミノ酸に同じ修飾(通常、置換)を行うことを指す。
」という用語は、アミノペプチダーゼ配列が天然で、天然に会合し、天然において見出さ
れる少なくとも1つの他の構成成分を少なくとも実質的に含まないことを意味する。本発
明のアミノペプチダーゼ配列は、他の場合に物質が会合し得る1つ以上の汚染物質を実質
的に含まない形態で提供され得る。したがって、例えば、それは、1つ以上の潜在的に汚
染するポリペプチドおよび/または核酸分子を実質的に含まないことがある。
された」という用語は、所与の構成成分が高レベルで存在していることを意味する。構成
成分は、組成物中に存在する主要な構成成分であることが望ましい。アミノペプチダーゼ
は、少なくとも約90%、または少なくとも約95%、または少なくとも約98%のレベ
ルで存在していることが好ましく、前記レベルは、考慮される総組成物に対する乾燥重量
/乾燥重量基準で測定される。
に明記されない限り、複数形を含む。別途指示のない限り、核酸は5’から3’の方向に
左から右へと記載し、アミノ酸配列はアミノからカルボキシの方向に左から右へと記載す
る。別段の指示がない限り、本開示が本明細書に記載の特定の方法、プロトコルおよび試
薬に限定されないことに留意されたい。
意味を有するものとする。本明細書において他に特に定義しない限り、本明細書で使用す
る全ての技術用語および科学用語は、当業者が通常理解する意味と同一の意味を有する。
別途指示のない限り、本開示の実施には、分子生物学、タンパク質工学および微生物学に
おいて通常使用される従来技術が含まれる。本明細書に記載のものと類似の、または等価
な方法および材料が本開示の実施に使用されるが、いくつかの好適な方法および材料を本
明細書に記載する。この直下に定義した用語は、明細書全体を参照することにより、より
十分に定義される。
本発明の範囲は、本明細書中に定義した特定の特性を有するアミノペプチダーゼをコー
ドするヌクレオチド配列を包含する。
、例えばアスペルギルス・クラバタス(Aspergillus clavatus)か
ら得られるポリペプチドをコードする。いくつかの実施形態では、核酸配列は、ネオサル
トリア属(Neosartorya)、例えばアスペルギルス・フィッシェリ(Neos
artorya fischeri)から得られるポリペプチドをコードする。
位の保存残基I/VのN末端側に13を超える残基を含む予測成熟配列を有する、アミノ
ペプチダーゼ活性を有するポリペプチド、またはそのフラグメントをコードする核酸に関
し、但し、そのフラグメントはアミノペプチダーゼ活性を有する。
対し特定の核酸相同性を有するポリヌクレオチドである。いくつかの実施形態では、ポリ
ヌクレオチドは、配列番号9、または配列番号10、または配列番号11、または配列番
号12、または配列番号13、配列番号14、または配列番号15、または配列番号16
の核酸配列のポリペプチドコード化部分に対し、少なくとも50、60、65、70、7
5、80、85、90、91、92、93、94、95、96、97、98、99または
100%の同一性を有する核酸配列を含む。いくつかの実施形態では、ポリヌクレオチド
は、配列番号9、または配列番号10、または配列番号11、または配列番号12、また
は配列番号13、配列番号14、または配列番号15、または配列番号16からなる群か
ら選択される核酸配列を含む。他の実施形態では、本発明のポリヌクレオチドはまた、配
列番号9、または配列番号10、または配列番号11、または配列番号12、または配列
番号13、配列番号14、または配列番号15、または配列番号16からなる群から選択
される核酸配列に相補的な核酸配列を有し得る。いくつかの実施形態では、ポリヌクレオ
チドは、配列番号1、または配列番号2、または配列番号3、または配列番号4、または
配列番号5、配列番号6、または配列番号7、または配列番号8からなる群から選択され
るアミノ酸配列に対し少なくとも70、75、80、85、90、95、96、97、9
8、99または100%の同一性を有するアミノ酸配列を含む、組換えポリペプチドまた
はその活性フラグメントをコードする核酸配列を含む。いくつかの実施形態では、ポリヌ
クレオチドは、配列番号1、または配列番号2、または配列番号3、または配列番号4、
または配列番号5、配列番号6、または配列番号7、または配列番号8からなる群から選
択されるアミノ酸配列を含む、組換えポリペプチドまたはその活性フラグメントをコード
する核酸配列を含む。相同性は、アミノ酸配列アライメントにより、例えば、本明細書に
記載のBLAST、ALIGN、またはCLUSTALなどのプログラムを使用して決定
することができる。
番号3、または配列番号4、または配列番号5、配列番号6、または配列番号7、または
配列番号8と異なる、配列番号 1、または配列番号 2、または配列番号 3、または
配列番号 4、または配列番号 5、配列番号 6、または配列番号 7、または配列番
号 8のアミノ酸配列を有するポリペプチドをコードする核酸配列を包含する。本発明は
また、アミノペプチダーゼ活性を有する配列番号1、または配列番号2、または配列番号
3、または配列番号4、または配列番号5、配列番号6、または配列番号7、または配列
番号8のフラグメントをコードする、配列番号 9、または配列番号 10、または配列
番号 11、または配列番号 12、または配列番号 13、配列番号 14、または配
列番号15、または配列番号 16の部分配列に関する。配列番号9、または配列番号1
0、または配列番号11、または配列番号12、または配列番号13、配列番号14、ま
たは配列番号15、または配列番号16の部分配列は、5’末端および/または3’末端
から1個以上のヌクレオチドが欠失していることを除けば、配列番号9、または配列番号
10、または配列番号11、または配列番号12、または配列番号13、配列番号14、
または配列番号15、または配列番号16によって包含される核酸配列である。部分配列
は、好ましくは少なくとも990個、より好ましくは少なくとも1140個、最も好まし
くは少なくとも1290個のヌクレオチドを含有する。
配列またはポリヌクレオチド配列、ならびに、それらの変異体、相同体、フラグメントお
よび誘導体(例えば、それらの一部)を指す。ヌクレオチド配列は、ゲノムまたは合成ま
たは組換え起源のものであり得、それは、センス鎖を表すかアンチセンス鎖を表すかにか
かわらず二本鎖または一本鎖であり得る。
DNA、およびRNAが含まれる。好ましくは、それは本発明をコードするDNAを意味
し、より好ましくは、cDNA配列を意味する。
クレオチド配列は、その天然環境中に存在する場合、およびそれがその天然に会合する配
列(それもその天然環境中に存在する)に結合している場合、本発明による天然ヌクレオ
チド配列を含まない。参照の容易性のため、本出願人らは、この好ましい実施形態を「非
天然ヌクレオチド配列」と称する。これに関して、「天然ヌクレオチド配列」という用語
は、天然環境中に存在し、天然に会合するプロモーター(プロモーターもその天然環境中
に存在する)全体に作動可能に結合している場合、ヌクレオチド配列全体を意味する。し
かしながら、本発明の範囲に包含されるアミノ酸配列は、その天然生物中のヌクレオチド
配列の発現後に単離および/または精製することができる。しかしながら、好ましくは、
本発明の範囲に包含されるアミノ酸配列は、その天然生物中のヌクレオチド配列により発
現させることができるが、そのヌクレオチド配列は、それがその生物内で天然に会合する
プロモーターの制御下にない。
組換えDNA)を使用して調製される。しかしながら、本発明の代替的実施形態において
は、ヌクレオチド配列は、当該技術分野においてよく知られる化学的方法を使用して全部
または一部を合成することができる(Caruthers MH et al.,(19
80)Nuc Acids Res Symp Ser 215-23、およびHorn
T et al.,(1980)Nuc Acids Res Symp Ser 2
25-232を参照)。
号12、または配列番号13、配列番号14、または配列番号15、または配列番号16
の核酸配列に、少なくとも約50%、好ましくは約60%、好ましくは約70%、好まし
くは約80%、より好ましくは約90%、より一層好ましくは約95%、最も好ましくは
約97%の相同性を有する核酸配列に関する。本発明の目的のために、2つの核酸配列間
の相同性の程度は、CLUSTAL法(Higgins,1989,上記を参照)により
、同一テーブル、ギャップペナルティ10、およびギャップレングスペナルティ10を用
いて決定される。
似したポリペプチドの合成に必要であり得る。ポリペプチドに「実質的に類似した」とい
う用語は、非自然発生のポリペプチドの形態を指す。これらのポリペプチドは、何らかの
工学的な方法が加えられている点で、天然源から単離されたポリペプチドと相異し得る。
例えば、比活性、熱安定性、最適pHなどが、異なるポリペプチドの変異体を、例えば部
位特異的変異誘発法によって合成することは興味深いことであり得る。類似配列は、配列
番号9、または配列番号10、または配列番号11、または配列番号12、または配列番
号13、配列番号14、または配列番号15、または配列番号16の核酸配列のポリペプ
チドコード化部分、例えば、その部分配列として示された核酸配列に基づき、かつ/また
は核酸配列によってコードされるポリペプチドの他のアミノ酸配列を生じさせないが、酵
素の生成を目的とした宿主生物のコドンの使用に対応するヌクレオチド置換の導入によっ
て、もしくは、異なるアミノ酸配列を生じさせ得るヌクレオチド置換の導入によって構築
し得る。ヌクレオチド置換の概要については、例えば、Ford et al.,199
1,Protein Expression and Purification 2:
95-107を参照されたい。
もたらし得ることは、当業者には明らかであろう。本発明の単離核酸配列によってコード
され、したがって、好ましくは置換を受けることのないポリペプチドの活性に必須のアミ
ノ酸残基は、部位特異的変異誘発法またはアラニン置換誘発法などの当該技術分野で知ら
れる手順によって特定することができる(例えば、Cunningham and We
lls,1989,Science 244:1081-1085を参照)。後者の手法
では、分子中の正電荷を持つ全ての残基に変異を導入し、得られた分子のアミノペプチダ
ーゼ活性を試験して、分子の活性に不可欠なアミノ酸残基を特定する。基質と酵素との相
互作用部位もまた、核磁気共鳴分析法、結晶構造解析法または光親和性標識法のような手
法によって決定される三次元構造の解析によって決定される(例えば、de Vos e
t al.,1992,Science 255:306-312;Smith et
al.,1992,Journal of Molecular Biology 22
4:899-904;Wlodaver et al.,1992,FEBS Lett
ers 309:59-64を参照)。
一態様において、本発明はまた、本明細書に記載のアミノペプチダーゼ、ならびにアミ
ノ酸配列および/またはヌクレオチド配列を含む組成物に関する。
保存残基I/VのN末端側に13を超える残基を含む予測成熟配列を有する、アミノペプ
チダーゼ活性を有するポリペプチド、またはそのフラグメントを提供し、但し、そのフラ
グメントはアミノペプチダーゼ活性を有する。いくつかの実施形態では、組成物は、図9
の配列アライメントに示すように、67位の保存残基I/VのN末端側に13を超える残
基を含む予測成熟配列を有する、アミノペプチダーゼ活性を有するポリペプチドに対し、
少なくとも約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、
より好ましくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ま
しくは少なくとも約95%、さらに最も好ましくは少なくとも約97%の相同性を有する
ポリペプチドを含む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号1のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号1のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、但し、そのフラグメントはアミノペプチダーゼ
活性を有する。いくつかの実施形態では、組成物は配列番号1のアミノ酸配列を含む。他
の実施形態では、組成物は、配列番号1のアミノ酸配列またはそのフラグメントを有する
アミノペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有す
る。いくつかの実施形態では、組成物は、配列番号1のアミノ酸配列を有するポリペプチ
ドを含む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号2のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号2のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を
有する。いくつかの実施形態では、組成物は配列番号2のアミノ酸配列を含む。他の実施
形態では、組成物は、配列番号2のアミノ酸配列またはそのフラグメントを有するアミノ
ペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有する。い
くつかの実施形態では、組成物は、配列番号2のアミノ酸配列を有するポリペプチドを含
む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号3のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号3のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を
有する。いくつかの実施形態では、組成物は配列番号3のアミノ酸配列を含む。他の実施
形態では、組成物は、配列番号3のアミノ酸配列またはそのフラグメントを有するアミノ
ペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有する。い
くつかの実施形態では、組成物は、配列番号3のアミノ酸配列を有するポリペプチドを含
む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号4のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号4のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を
有する。いくつかの実施形態では、組成物は配列番号4のアミノ酸配列を含む。他の実施
形態では、組成物は、配列番号4のアミノ酸配列またはそのフラグメントを有するアミノ
ペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有する。い
くつかの実施形態では、組成物は、配列番号4のアミノ酸配列を有するポリペプチドを含
む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号5のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号5のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を
有する。いくつかの実施形態では、組成物は配列番号5のアミノ酸配列を含む。他の実施
形態では、組成物は、配列番号5のアミノ酸配列またはそのフラグメントを有するアミノ
ペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有する。い
くつかの実施形態では、組成物は、配列番号5のアミノ酸配列を有するポリペプチドを含
む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号6のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号6のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を
有する。いくつかの実施形態では、組成物は配列番号6のアミノ酸配列を含む。他の実施
形態では、組成物は、配列番号6のアミノ酸配列またはそのフラグメントを有するアミノ
ペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有する。い
くつかの実施形態では、組成物は、配列番号6のアミノ酸配列を有するポリペプチドを含
む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号7のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号7のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を
有する。いくつかの実施形態では、組成物は配列番号7のアミノ酸配列を含む。他の実施
形態では、組成物は、配列番号7のアミノ酸配列またはそのフラグメントを有するアミノ
ペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有する。い
くつかの実施形態では、組成物は、配列番号7のアミノ酸配列を有するポリペプチドを含
む。
も約50%、好ましくは少なくとも約60%、好ましくは少なくとも約70%、より好ま
しくは少なくとも約80%、より一層好ましくは少なくとも約90%、最も好ましくは少
なくとも約95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸
配列を有する、少なくとも1種のアミノペプチダーゼを含む一般組成物を提供する。いく
つかの実施形態では、組成物は、配列番号8のアミノ酸配列と、5個のアミノ酸、好まし
くは4個のアミノ酸、より好ましくは3個のアミノ酸、より一層好ましくは2個のアミノ
酸、最も好ましくは1個のアミノ酸が異なるアミノ酸配列を有する相同ポリペプチドを含
む。いくつかの実施形態では、組成物は、配列番号8のアミノ酸配列または対立遺伝子変
異型;およびそのフラグメントを含むが、そのフラグメントはアミノペプチダーゼ活性を
有する。いくつかの実施形態では、組成物は配列番号8のアミノ酸配列を含む。他の実施
形態では、組成物は、配列番号8のアミノ酸配列またはそのフラグメントを有するアミノ
ペプチダーゼを含むが、但し、そのフラグメントはアミノペプチダーゼ活性を有する。い
くつかの実施形態では、組成物は、配列番号8のアミノ酸配列を有するポリペプチドを含
む。
性を有するポリヌクレオチドを含む。いくつかの実施形態では、組成物は、配列番号9、
または配列番号10、または配列番号11、または配列番号12、または配列番号13、
配列番号14、または配列番号15、または配列番号16の核酸配列のポリペプチドコー
ド化部分の核酸配列に対し、少なくとも50、60、65、70、75、80、85、9
0、91、92、93、94、95、96、97、98、99または100%の同一性を
有する核酸配列を含むポリヌクレオチドを含む。いくつかの実施形態では、ポリヌクレオ
チドは、配列番号9、または配列番号10、または配列番号11、または配列番号12、
または配列番号13、配列番号14、または配列番号15、または配列番号16からなる
群から選択される核酸配列を含む。他の実施形態では、組成物は、配列番号9、または配
列番号10、または配列番号11、または配列番号12、または配列番号13、配列番号
14、または配列番号15、または配列番号16からなる群から選択される核酸配列に相
補的な核酸配列を有するポリヌクレオチドを含む。いくつかの実施形態では、組成物は、
配列番号1、または配列番号2、または配列番号3、または配列番号4、または配列番号
5、配列番号6、または配列番号7、または配列番号8からなる群から選択されるアミノ
酸配列に対し少なくとも70、75、80、85、90、95、96、97、98、99
または100%の同一性を有するアミノ酸配列を含む、組換えポリペプチドまたはその活
性フラグメントをコードする核酸配列を含むポリヌクレオチドを含む。いくつかの実施形
態では、組成物は、配列番号1、または配列番号2、または配列番号3、または配列番号
4、または配列番号5、配列番号6、または配列番号7、または配列番号8からなる群か
ら選択されるアミノ酸配列を含む、組換えポリペプチドまたはその活性フラグメントをコ
ードする核酸配列を含むポリヌクレオチドを含む。
ドラーゼ、カルボキシペプチダーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ
、シクロデキストリン・グリコシルトランスフェラーゼ、デオキシリボヌクレアーゼ、エ
ステラーゼ、アルファ-ガラクトシダーゼ、ベータ-ガラクトシダーゼ、グルコアミラー
ゼ、アルファ-グルコシダーゼ、ベータ-グルコシダーゼ、ハロペルオキダーゼ、インベ
ルターゼ、ラッカーゼ、リパーゼ、マンノシダーゼ、オキシダーゼ、ペクチン分解酵素、
ペプチドグルタミナーゼ、ペルオキシダーゼ、フィターゼ、ポリフェノールオキシダーゼ
、タンパク質分解酵素、リボヌクレアーゼ、トランスグルタミナーゼまたはキシラナーゼ
などの多数の酵素活性を含み得る。さらなる酵素は、アスペルギルス属(Aspergi
llus)、好ましくはアスペルギルス・アクレアタス(Aspergillus ac
uleatus)、アスペルギルス・アワモリ(Aspergillus awamor
i)、アスペルギルス・ニガー(Aspergillus niger)もしくはアスペ
ルギルス・オリゼ(Aspergillus oryzae)、またはトリコデルマ属(
Trichoderma)、またはフミコーラ属(Humicola)、好ましくはフミ
コーラ・インソレンス(Humicola insolens)、またはフザリウム属(
Fusarium)、好ましくはフザリウム・バクトリジオイデス(Fusarium
bactridioides)、フザリウム・セレアリス(Fusarium cere
alis)、フザリウム・クルークウェレンセ(Fusarium crookwell
ense)、フザリウム・クルモルム(Fusarium culmorum)、フザリ
ウム・グラミネアルム(Fusarium graminearum)、フザリウム・グ
ラミヌム(Fusarium graminum)、フザリウム・ヘテロスポルム(Fu
sarium heterosporum)、フザリウム・ネグンジ(Fusarium
negundi)、フザリウム・オキシスポルム(Fusarium oxyspor
um)、フザリウム・レチクラツム(Fusarium reticulatum)、フ
ザリウム・ロセウム(Fusarium roseum)、フザリウム・サンブシヌム(
Fusarium sambucinum)、フザリウム・サルコクロウム(Fusar
ium sarcochroum)、フザリウム・スポロトリキオイデス(Fusari
um sporotrichioides)、フザリウム・スルフレウム(Fusari
um sulphureum)、フザリウム・トルロスム(Fusarium toru
losum)、フザリウム・トリコテシオイデス(Fusarium trichoth
ecioides)もしくはフザリウム・ベネナツム(Fusarium venena
ium)に属する微生物によって産生され得る。
よび好適な担体を含む組成物に関する。本明細書中に記載のものを含む、当該技術分野で
知られる好適な担体が使用され得る。他の実施形態では、組成物は、さらにエンドペプチ
ダーゼを含む。いくつかの実施形態では、組成物はさらに、1種以上の非特異的に作用す
るエンドペプチダーゼおよび/またはエキソペプチダーゼ酵素を含む。いくつかの実施形
態では、組成物はさらに、1種以上の特異的に作用するエンドペプチダーゼおよび/また
はエキソペプチダーゼ酵素を含む。
ペプチダーゼ(EC 3.4.21.19);リシルエンドペプチダーゼ(EC 3.4
.21.50);ロイシルエンドペプチダーゼ(EC 3.4.21.57);グリシル
エンドペプチダーゼ(EC 3.4.22.25);プロリルエンドペプチダーゼ(EC
3.4.21.26);トリプシン(EC 3.4.21.4)もしくはトリプシン様
(リシン/アルギニン特異性)エンドペプチダーゼ、またはペプチジル-Aspメタロエ
ンドペプチダーゼ(EC 3.4.24.33)などのエンドペプチダーゼである。
ーゼ、ジペプチジルアミノペプチダーゼ、カルボキシペプチナーゼおよび他のアミノペプ
チダーゼからなる群から選択される。
ダーゼ酵素は、真菌性酸性エンドペプチダーゼ、中性メタロエンドペプチダーゼ、アルカ
リ性セリンエンドぺプチダーゼ、サブチリシン、ブロメライン、熱に安定な細菌性中性エ
ンドぺプチダーゼ、アルカリ性セリンエンドぺプチダーゼ(重複)からなる群から選択さ
れる。
はエキソペプチダーゼ酵素は、下記の1つ以上の市販品中の1種以上のプロテアーゼであ
ってよい。

素は、次の商業的に入手可能な製品の1つ以上に含まれるものであってよい:KANNA
SE(商標)、NOVOCARNETM Tender,およびNovozym 370
20、NOVO-PROTM D(いずれもNovozymesから入手可能);Bio
Sorb-ACDP(Noor Creations,India);ANGEL(登録
商標)Acid Protease(Angel Yeast Co、Ltd.,Chi
na)またはCOROLASE(登録商標)LAP(AB Enzymesから)。
とも1種を含む飼料添加物組成物および/または食品添加物組成物を提供する。
よび/または食品添加組成物が提供される。好適には、そのような食品添加組成物および
/または飼料添加組成物は、さらにアミノペプチダーゼを(任意選択的に、エンドプロテ
アーゼと組み合わされて)含み得る。
加剤の非限定的な例としては、塩(例えば、アルカリ塩、土類金属塩、付加的塩化物塩、
硫酸塩、硝酸塩、炭酸塩(ここで、対イオンの例は、カルシウム、カリウムおよびナトリ
ウムイオンである)、無機鉱物またはクレー(例えば、ゼオライト、カオリン、ベントナ
イト、タルクおよび/またはケイ酸塩)、炭水化物(例えば、スクロースおよび/または
デンプン)、着色顔料(例えば、二酸化チタン)、殺生物剤(例えば、RODALON(
登録商標)、PROXEL(登録商標))分散剤、消泡剤、還元剤、酸性剤、アルカリ性
剤、酵素安定剤(グリセロール、プロピレングリコール、ソルビトールなどのポリオール
、無機塩、糖類、糖もしくは糖アルコール、乳酸、ホウ酸、またはホウ酸誘導体、および
これらの組み合わせ)、酵素阻害剤、保存料(例えば、メチルパラベン、プロピルパラベ
ン、ベンゾエート、ソルベート、または他の食品に認可されている保存料)、ならびに、
これらの組み合わせが挙げられる。本調製物/組成物に使用し得る賦形剤としては、マル
トース、スクロース、グルコースシロップもしくは乾燥グルコースシロップなどのグルコ
ース、加工済みデンプン、ゼラチン化デンプン、L-乳酸、パルミチン酸アスコルビル、
トコフェロール、レシチン、クエン酸、クエン酸塩、リン酸、リン酸塩、アルギン酸ナト
リウム、カラゲナン、ローカストビーンガム、グアーガム、キサンタンガム、ペクチン、
カルボキシメチルセルロースナトリウム、モノグリセリドおよびジグリセリド、モノグリ
セリドおよびジグリセリドのクエン酸エステル、スクロースエステル、二酸化炭素、アル
ゴン、ヘリウム、窒素、亜酸化窒素、酸素、水素、ならびにオクテニルコハク酸デンプン
ナトリウムが挙げられる。本明細書中の実施例7に示すように、本発明によるアミノペプ
チダーゼは、塩化ナトリウムを含む組成物中で酵素活性を保持し得る。
本発明の生成物および/または組成物は、単独で使用されようと組成物中に含まれよう
と、任意の好適な形態で使用し得る。同様に、本発明のアミノペプチダーゼは、例えば、
エキソプロテアーゼおよび/またはエンドプロテアーゼと併用した場合、食品産業におい
て食品加工助剤または食品添加物(すなわち、食品成分、機能性食品成分または医薬品成
分などの成分)として使用するのに適した任意の形態で使用し得る。
たは制御放出用途のための錠剤、丸剤、カプセル剤、オビュール剤、液剤、または懸濁剤
のうちの1種以上が挙げられ、これらは香味料または着色剤を含有することができる。
分として使用する場合など)、その錠剤はまた、微結晶セルロース、ラクトース、クエン
酸ナトリウム、炭酸カルシウム、二塩基性リン酸カルシウム、およびグリシンなどの賦形
剤;デンプン(好ましくは、トウモロコシ、ジャガイモ、またはタピオカのデンプン)、
デンプングリコール酸ナトリウム、クロスカルメロースナトリウム、およびある種の複雑
ケイ酸塩などの崩壊剤;ポリビニルピロリドン、ヒドロキシプロピルメチルセルロース(
HPMC)、ヒドロキシプロピルセルロース(HPC)、スクロース、ゼラチン、および
アカシアゴムなどの造粒用結合剤;ステアリン酸マグネシウム、ステアリン酸、ベヘン酸
グリセリル、およびタルクなどの滑沢剤の1種以上を含有することができる。
、水、食塩水、アルコール、シリコーン、ワックス、ワセリン、植物油、ポリエチレング
リコール、プロピレングリコール、リポソーム、糖、ゼラチン、ラクトース、アミロース
、ステアリン酸マグネシウム、タルク、界面活性剤、ケイ酸、粘性パラフィン、香油、脂
肪酸モノグリセリドおよびジグリセリド、ペトロエスラル脂肪酸エステル、ヒドロキシメ
チルセルロース、ポリビニルピロリドンなどが挙げられる。
高分子量ポリエチレングリコールが挙げられる。
または組成物は、様々な甘味剤または香味剤、着色物質または染料と、また乳化剤および
/または懸濁化剤と、また水、エタノール、プロピレングリコール、およびグリセリンな
どの希釈剤と、ならびにこれらの組合せと併用することができる。
げられる。
いくつかの実施形態では、本発明のポリペプチドは、タンパク質加水分解物の生成にお
いて、例えば、加水分解度の増大のため、タンパク質加水分解物の一般的な苦味除去のた
め、および風味発生の増進のため、グルタミン酸塩の生成のため、ならびに/または、モ
ルト生成もしくは醸造中のFAN生成のようなその他の目的で使用することができる。
チドをプロテアーゼ(例えば、エンドペプチダーゼ)と併用して使用する方法に関する。
この方法は、タンパク質性基質を本ポリペプチドおよびエンドペプチダーゼで処理するこ
とを含む。基質は、それらの酵素で同時に処理してもよく連続的に処理してもよい。
タンパク質性基質に添加される。いくつかの実施形態では、本発明のポリペプチドは、タ
ンパク質性基質に、タンパク質100g当たり、約0.1~約100,000アミノペプ
チダーゼ単位の範囲、またはタンパク質100g当たり約1~約10,000アミノペプ
チダーゼ単位の範囲で添加される。本明細書で定義されるように、1アミノペプチダーゼ
単位(APU)は、特定の条件下、Ala-p-ニトロアニリド(Sigma Chem
ical Co.,St.Louis MO)から1分当たり1マイクロモルのp-ニト
ロアニリドを放出するのに必要な酵素量である。アミノペプチダーゼアッセイでは、ペプ
チド基質H-Ala-ニトロアニルドの加水分解をp-ニトロアニリド(pNA)の放出
によって測定する。pNAの波長405nmの吸光度をELISAリーダーにより測定す
る。20mMのCPBバッファー180μl、希釈酵素15μlおよび基質20μlによ
り、30℃で反応を進める。CPBバッファーは20mMクエン酸、20mMリン酸塩、
20mMホウ酸で構成され、pH9.0に調節される。基質は、1mlのDMSO(SI
GMA製のジメチルスルホキシド(カタログ番号 D2650))中のBACHEM製の
H-Ala-pNA(L-1070)20mgである。
ニホルミス(Bacillus licheniformis)もしくはバチルス・サブ
チリス(Bacillus subtilis)株、スタフィロコッカス(Staphy
lococcus)株、好ましくはスタフィロコッカス・アウレウス(Staphylo
coccus aureus)株、ストレプトミセス(Streptomyces)、好
ましくはストレプトミセス・サーモバラリス(Streptomyces thermo
vularis)もしくはストレプトミセス・グリセウス(Streptomyces
griseus)株、アスペルギルス(Aspergillus)株、好ましくはアスペ
ルギルス・アクレアタス(Aspergillus aculeatus)、アスペルギ
ルス・アワモリ(Aspergillus awamori)、アスペルギルス・フェチ
ヅス(Aspergillus foetidus)、アスペルギルス・ニドュランス(
Aspergillus nidulans)、アスペルギルス・ニガー(Asperg
illus niger)もしくはアスペルギルス・オリゼ(Aspergillus
oryzae)株、またはトリコデルマ(Trichoderma)株、好ましくはトリ
コデルマ・リーゼイ(Trichoderma reesei)株、フザリウム(Fus
arium)株、好ましくはフザリウム・ベネナツム(Fusarium venena
tum)株から得ることができる。いくつかの実施形態では、エンドペプチダーゼは、A
LPHALASE(登録商標)AFP、FOODPRO(登録商標)PAL、FOODP
RO(登録商標)PNL、FOODPRO(登録商標)AlkalineProteas
e、FOODPRO(登録商標)PXT、FOODPRO(登録商標)PBR、FOOD
PRO(登録商標)PHT、FOODPRO(登録商標)30L、およびFOODPRO
(登録商標)51FPからなる群から選択される。
効量で、好ましくは約0.05~約15AU/100gタンパク質の範囲で、より好まし
くは約0.1~約8AU/100gタンパク質の範囲で、タンパク質性基質に添加される
。1AU(アンソン単位)は、標準条件下(すなわち、25℃、pH7.5および反応時
間10分)、1分当たりフェノール試薬で1ミリグラム当量のチロシンと同じ色を示す量
のTCA可溶性生成物を遊離するような初速度でヘモグロビンを消化する酵素の量と定義
される。いくつかの実施形態では、エンドプロテアーゼは、タンパク質基質1kg当たり
約10~約3000mgの酵素、例えば、タンパク質基質1メートルトン(MT)当たり
0.01から3gの酵素の量で投与することができる。
剤が不活化しない都合の良い温度、好ましくは約20℃~約70℃の範囲の温度で行うこ
とができる。通常の手順に従えば、酵素製剤は、インキュベーション混合物の温度を、酵
素が不活化する温度、例えば、約70℃超にまで上昇させることによって、またはインキ
ュベーション混合物のpHを、酵素が不活化する程度、例えば、約4.0未満にまで低下
させることによって、適切に不活化することができる。
用されるとき、加水分解度(DH)は、タンパク質分解酵素によって加水分解されたタン
パク質中のアミノ結合の総数の割合である。一態様においては、本発明による酵素は、タ
ンパク質性基質のDHを少なくとも約5、7、10、12、15、17、20、22、2
5、27、30、35、40、45または50%上昇または増大させる。一例において、
この上昇は1型アミノペプチダーゼと比べてのものであるが、いかなる好適な比較も可能
である。好ましい実施形態では、タンパク質加水分解物は、Leu、Gly、Glu、S
er、Asp、Asn、Pro、Cys、Alaおよび/またはGlnの含有量が、例え
ば1.1倍に増大する。
つかの実施形態では、タンパク質加水分解物は、Leuの含有量が増大する。いくつかの
実施形態では、タンパク質加水分解物は、Glyの含有量が増大する。いくつかの実施形
態では、タンパク質加水分解物は、Serの含有量が増大する。いくつかの実施形態では
、タンパク質加水分解物は、Aspの含有量が増大する。いくつかの実施形態では、タン
パク質加水分解物は、Asnの含有量が増大する。いくつかの実施形態では、タンパク質
加水分解物は、Proの含有量が増大する。いくつかの実施形態では、タンパク質加水分
解物は、Cysの含有量が増大する。いくつかの実施形態では、タンパク質加水分解物は
、Alaの含有量が増大する。他のより好ましい実施形態では、タンパク質加水分解物は
、Glnの含有量が増大する。
明細書の実施例8に示すように、本発明による2種類のPepN 2酵素は、アスペルギ
ルス・オリゼ(Aspergillus oryzae)RIB40由来のロイシンアミ
ノペプチダーゼ2酵素(NCBIレファレンス配列:XP_001819545.1)(
配列番号17として以下に示す)より、低い生成物阻害を示した。

もいう―例えば、実施例を参照)のアミノペプチダーゼと比較して生成物阻害が低減した
、アミノペプチダーゼ活性を有する単離ポリペプチドを提供する。特に、本発明によるア
ミノペプチダーゼ活性を有する単離ポリペプチドは、2、3、4または5mM超の生成物
阻害定数(Ki)値を有し得る。好ましい実施形態では、アミノペプチダーゼ活性を有す
る単離ポリペプチドは、2型アミノペプチダーゼ酵素である。さらなる好ましい実施形態
では、2型アミノペプチダーゼ酵素は、本明細書で定義される配列番号1~8のいずれか
、特に配列番号1または5に示すとおりである。
プロリン残基を有するポリペプチドを加水分解することができ得る。特に、本発明による
アミノペプチダーゼ酵素は、N末端から番号を付与したときの2の位置にプロリン残基を
有するポリペプチドを、2時間のインキュベーションで出発濃度の半分超の濃度にまで加
水分解することができ得る加水分解度は当該技術分野で知られた任意の好適なプロトコル
、例えば、実施例10で示した方法で測定することができる。実施例10に示すように、
本発明による2種類のPepN2酵素(TRI032およびTRI035)は、ペプチド
TPAAARを徐々に加水分解して、2時間のインキュベーションで濃度を半分未満にま
で低下させることができるが、TRI063(アスペルギルス・オリゼ(A.oryza
e))およびCOROLASE(登録商標)LAPは、12時間以内にTPAAARの加
水分解を示すことはない。
に富む加水分解物を得る方法であって、基質を、アミノペプチダーゼ活性を有するポリペ
プチドの作用に曝すことを含む方法に関する。本発明はまた、遊離グルタミンもしくは遊
離グルタミン酸および/またはグルタミンもしくはグルタミン酸残基を結合したオリゴペ
プチドに富む加水分解物を得る方法であって、基質を、アミノペプチダーゼ活性を有する
ポリペプチドの作用に曝すことを含む方法に関する。
ペプチダーゼ活性を有するポリペプチドの作用に曝すことを含む方法に関する。
方法に関する。脱アミドプロセスは、基質を、アミノペプチダーゼ活性を有するポリペプ
チドの作用に曝すのと同時に、前にまたは後に行うことができる。
オリゴペプチドであれ、グルタミン酸(Glu)がタンパク質加水分解物の香味および食
味重要な役割を果たすため、香味が強化されたタンパク質加水分解物が生成される。いく
つかの実施形態では、本方法により、機能性が向上した、特に、溶解性、乳化性が向上し
、加水分解度が上昇し、かつ、起泡性が向上したタンパク質加水分解物もまた生成される
。
ン酸またはアスパラギン酸)への変換は、脱アミドとして知られている。脱アミドは、非
酵素的脱アミドプロセスとして、または酵素的脱アミドプロセスとして行い得る。
グルタミナーゼ、トランスグルタミナーゼおよび/またはペプチドグルタミナーゼに曝す
ことによって行われる。
Amano、日本)である。
0mgの量で投与することができる。いくつかの実施形態では、グルタミナーゼは、基質
タンパク質1g当たり約5mg~15mgの量で投与することができる。いくつかの実施
形態では、グルタミナーゼは、基質タンパク質1g当たり約10mgの量で投与すること
ができる。
プチダーゼ活性を有するポリペプチドの作用に曝すことを含む方法に関する。
P1050382およびJP5023182を参照)であっても、活性化第XIII因子
(例えば、国際公開第93/15234号パンフレットを参照)であっても、魚由来のも
の(例えば、EP555,649を参照)であっても、微生物から得られたもの(例えば
、EP379,606、国際公開第96/06931号パンフレット、国際公開第96/
22366号パンフレットを参照)であってもよい。いくつかの実施形態では、トランス
グルタミナーゼは、卵菌、例えば、フィトフトラ属(phytophthora)、好ま
しくはフィトフトラ・カクトルム(phytophthora cactorum)株、
または、ピシウム属(Pythium)、好ましくはピシウム・イレグラレ(Pythi
um irregulare)株、ピシウム・スピノサム(Pythium sp.)株
、ピシウム・インテルメディアム(Pythium intermedium)株、ピシ
ウム・ウルチマム(Pythium ultimum)株、またはピシウム・ペリイラム
(Pythium periilum)(もしくは、ピシウム・ペリクロカム(Pyth
ium periplocum))株から得られる。いくつかの実施形態では、トランス
グルタミナーゼは、細菌起源のものであり、バチルス属(Bacillus)、好ましく
はバチルス・サブチリス(Bacillus subtilis)株、ストレプトベルテ
ィシリウム属(Streptoverticillium)、好ましくは、ストレプトベ
ルティシリウム・モバラエンシス(Streptoverticillium moba
raensis)株、ストレプトベルティシリウム・グリセオバーティシラタム(Str
eptoverticillium griseoverticillatum)株、ま
たはストレプトベルティシリウム・シンナモネウム(Streptoverticill
ium cinnamoneum)株、およびストレプトミセス属(Streptomy
ces)、好ましくはストレプトミセス・リディカス(Streptomyces ly
dicus)株から得られる。
;EC.3.5.1.43)、またはペプチドグルタミナーゼII(タンパク質-グルタ
ミングルタミナーゼ;EC.3.5.1.44)であっても、あるいはこれらの混合物で
あってもよい。ペプチドグルタミナーゼは、アスペルギルス属(Aspergillus
)、好ましくはアスペルギルス・ヤポニカス(Aspergillus japonic
us)株、バチルス属(Bacillus)、好ましくはバチルス・サークランス(Ba
cillus circulans)株、クリプトコッカス属(Cryptococcu
s)、好ましくはクリプトコッカス・アルビダス(Cryptococcus albi
dus)、またはデバリオミセス属(Debaryomyces)、好ましくはデバリオ
ミセス・クロエケリ(Debaryomyces kloecheri)株から得ること
ができる。
る有効量で、好ましくは、基質量に対し酵素製剤約0.01~約5重量%の範囲で、より
好ましくは、約0.1~約1重量%の範囲で添加される。
る有効量で、好ましくは、基質100gに対し約0.01~約100,000ペプチドグ
ルタミナーゼ単位の範囲で、より好ましくは、約0.1~約10,000ペプチドグルタ
ミナーゼ単位の範囲で添加される。
nzymologia Vol.29 143頁)の手順にしたがって測定することがで
きる。この手順においては、1NのNaOHでpH6.5に調節した酵素試料0.5ml
を小さな容器に投入する。その後、この容器にpH10.8のホウ酸塩緩衝液1mlを加
える。放出されるアンモニアを5Nの硫酸で吸収させ、ネスラー試薬を用いて、混合物を
発色させ、420nmで測定する。1ペプチドグルタミナーゼ単位は、これらの条件下で
1分当たり1マイクロモルのアンモニアを生成することができる酵素量である。
る方法であって、モルト生成および/または醸造プロセス中に、基質を、アミノペプチダ
ーゼ活性を有するポリペプチドの作用に曝すことを含む方法に関する。
される。本発明のペプチドグルタミナーゼは、タンパク質性基質に、タンパク質の加水分
解プロセスで従来使用されている有効量で、好ましくは約0.001~約0.5AU/1
00g基質の範囲で、より好ましくは約0.01~約0.1AU/100g基質の範囲で
添加される。
結合したペプチドに富む加水分解物を生成する方法は、基質を、1種以上の非特異的に作
用するエンドペプチダーゼおよび/またはエキソペプチダーゼ酵素に曝すことを含む。こ
の工程は、タンパク質基質を本発明のポリペプチドに曝す工程と同時に行ってもよく、後
に行ってもよい。
キソペプチダーゼ酵素は、アスペルギルス(Aspergillus)株、またはバチル
ス(Bacillus)株から得られる。
ンパク質の加水分解プロセスで従来使用されている有効量で、好ましくは約 ~の範囲で
、基質に添加される。
タンパク質基質1kg当たり約50~約3000mgの酵素、例えば、タンパク質基質1
メートルトン(MT)当たり0.05から3gの酵素の量で投与することができる。
1MT当たり約4.0g未満の酵素の量で投与することができる。
ンパク質基質1MT当たり約0.5g~約5.0gの酵素で投与することができる。好適
には、エンドペプチダーゼおよび/またはエキソペプチダーゼは、タンパク質基質1MT
当たり約0.5g~約3.0gの酵素で投与することができる。より好適には、エンドプ
ロテアーゼは、タンパク質基質1MT当たり約1.0gから約2.0gの酵素において投
与することができる。
食品および/もしくは飼料添加物組成物1kg当たり約0.5mg~約2gの酵素量で投
与することができる。好適には、本発明のポリペプチドは、タンパク質基質ならびに/ま
たは食品および/もしくは飼料添加物組成物1kg当たり約1mg~約2gの酵素量で投
与することができる。より好適には、タンパク質基質ならびに/または食品および/もし
くは飼料添加物組成物1kg当たり約5mg~約1.5gの酵素量である。
0.5mg~約2gの酵素量で投与することができる。好適には、本発明のポリペプチド
は、タンパク質基質1kg当たり約1mg~約2gの酵素量で投与することができる。よ
り好適には、タンパク質基質1kg当たり約5mg~約1.5gの酵素量である。
~約500mgの酵素量で投与することができる。好適には、本発明のポリペプチドは、
タンパク質基質1kg当たり約50mg~約500mgの酵素量で投与することができる
。好適には、本発明のポリペプチドは、タンパク質基質1kg当たり約100mg~約4
50mgの酵素量で投与することができる。
℃の範囲の温度で行うことができる。酵素製剤は、その後、温度を、例えば約70℃を超
える温度にまで上昇させることによって、またはpHを、例えば約4.0未満にまで低下
させることによって不活化させることができる。
タンパク質(すなわち、ペプチド)、またはこれらの混合物から構成され得る。タンパク
質性基質は、野菜起源のものであっても動物起源のものであってもよい。いくつかの実施
形態では、タンパク質性基質は、野菜起源のもの、例えば、ダイズタンパク質、穀類タン
パク質、例えば小麦グルテン、トウモロコシグルテン、大麦、ライ麦、オート麦、コメ、
ゼイン、ハウチワマメ、綿実タンパク質、ラッカセイ、アルファルファタンパク質、エン
ドウ豆タンパク質、豆類タンパク質(fabaceous bean protein)
、ゴマタンパク質またはヒマワリ起源のものである。動物起源のタンパク質性基質は、ホ
エータンパク質、カゼイン、食肉タンパク質、魚タンパク質、赤血球、卵白、ゼラチン、
ラクトアルブミン、毛髪タンパク質または羽毛タンパク質であり得る。
本明細書に定義される特定の特性を有するタンパク質、または改変に好適なタンパク質
をコードするヌクレオチド配列は、前記タンパク質を産生する任意の細胞または生物から
同定および/または単離および/または精製することができる。当該技術分野においては
、ヌクレオチド配列の同定および/または単離および/または精製に関し、様々な方法が
よく知られている。
一例として、好適な配列が同定および/または単離および/または精製されたなら、よ
り多くの配列を調製するPCR増幅技術を使用することができる。
Aを使用して、ゲノムDNAおよび/またはcDNAライブラリーを構築することができ
る。酵素のアミノ酸配列がわかれば、標識オリゴヌクレオチドプローブを合成し、これを
用いて、生物から調製されたゲノムライブラリーから酵素をコードするクローンを同定す
ることができる。
あるいは、別の既知の酵素遺伝子と相同の配列を含有する標識オリゴヌクレオチドプロ
ーブを使用して、酵素をコードするクローンを同定することができる。後者の場合、より
低いストリンジェンシーのハイブリダイゼーションおよび洗浄条件が使用される。
Aの断片を挿入し、得られたゲノムDNAライブラリーにより酵素陰性細菌を形質転換し
、次いで、この形質転換された細菌を、酵素用基質(すなわち、マルトース)を含有する
寒天プレート上に形質転換された細菌をプレーティングし、これにより酵素を発現するク
ローンの同定を可能とすることにより同定することができる。
準的方法、例えば、Beucage S.L.et al.,(1981)Tetrah
edron Letters 22,p 1859-1869に記載されるホスホラミダ
イト法、またはMatthes et al.,(1984)EMBO J.3,p80
1-805に記載される方法による合成によって調製することができる。ホスホラミダイ
ト法においては、オリゴヌクレオチドを、例えば、自動DNA合成装置で合成し、精製し
、アニールし、ライゲートし、そして適切なベクターにクローニングする。
片をライゲートすることにより調製される混合ゲノムおよび合成起源、混合合成およびc
DNA起源、または混合ゲノムおよびcDNA起源のものであり得る。ライゲートされた
各断片は、ヌクレオチド配列全体の種々の部分に対応する。DNA配列はまた、例えば、
米国特許第4,683,202号明細書またはSaiki R K et al.,(S
cience(1988)239,pp487-491)に記載されるような、特異的プ
ライマーを使用するポリメラーゼ連鎖反応(PCR)により調製することができる。
本発明の範囲には、本明細書中に定義した特定の特性を有する酵素のアミノ酸配列もま
た包含される。
または用語「タンパク質」と同義である。一部の例では、用語「アミノ酸配列」、用語「
ペプチド」と同義である。一部の例では、用語「アミノ酸配列」は、用語「酵素」と同義
である。
することができ、または、組換えDNA技術の使用により調製することができる。
、トリペプチジルエキソペプチダーゼ、および他の形態のエンドプロテアーゼまたはエキ
ソプロテアーゼと共に使用することができる。したがって、本発明はまた、タンパク質の
組合せであって、本発明のアミノペプチダーゼ、および本発明の他のアミノペプチダーゼ
であり得る他の酵素を含む組み合わせをカバーする。この態様は、後述のセクションにお
いて考察する。
酵素ではない。これに関し、「天然酵素」という用語は、その天然環境中に存在し、その
天然ヌクレオチド配列により発現された酵素全体を意味する。
本発明はまた、本明細書に定義される特定の特性を有するポリペプチドのアミノ酸配列
とある程度の配列同一性または配列相同性を有する配列、またはそのようなポリペプチド
をコードする任意のヌクレオチド配列(以下、「相同配列」と称する)の使用を包含する
。ここで、「相同体」という用語は、対象アミノ酸配列および対象ヌクレオチド配列とあ
る相同性を有する実体を意味する。ここで、「相同性」という用語は、「同一性」と同じ
であると見なすことができる。
を保持し、かつ/またはアミノペプチダーゼの活性を向上させるポリペプチドを提供およ
び/またはコードするはずである。
好ましくは、少なくとも95または98%同一であり得るアミノ酸配列を含むと解釈され
る。通常、相同体は、対象アミノ酸配列と同一の活性部位などを含むであろう。相同性は
類似性(すなわち、類似の化学的特性/機能を有するアミノ酸残基)の観点から考慮する
こともできるが、本発明との関連では、配列同一性の観点から相同性を表現することが好
ましい。
(対象配列)と少なくとも75、80、85または90%同一、好ましくは、少なくとも
95または98%同一であり得るヌクレオチド配列を含むと解釈される。通常、相同体は
、対象配列と同じ、活性部位などをコードする配列を含むであろう。相同性は類似性(す
なわち、類似の化学的特性/機能を有するアミノ酸残基)の観点から考慮することもでき
るが、本発明との関連では、配列同一性の観点から相同性を表現することが好ましい。
グラムを使用して行うことができる。これらの市販のコンピュータプログラムは、2つ以
上の配列間の相同性率を計算することができる。
配列とアラインし、ある1つの配列中の各アミノ酸を他の配列中の対応するアミノ酸と1
回に1残基、直接比較する。これは「ギャップなし」アライメントと称される。通常、こ
のようなギャップなしアライメントは、比較的少数の残基にわたってのみ実施される。
る挿入または欠失が後続のアミノ酸残基をアライメントから外すことを考慮し得ず、した
がって、グローバルアライメントを実施した場合に相同性率を大きく低下させる可能性が
ある。このような理由から、ほとんどの配列比較法は、全相同性スコアを過度にペナルテ
ィ化せずに、あり得る挿入および欠失を考慮する最適なアライメントを作成するように設
計される。これは、配列アライメント中に「ギャップ」を挿入して局所的相同性の最大化
を試みることにより達成される。
少ないギャップを有する配列アライメント(2つの比較配列間のより高い関係性を反映す
る)が、多くのギャップを有するものより高いスコアを達成するように、そのアライメン
ト中で生じる各ギャップに対して「ギャップペナルティ」を割り当てる。通常、あるギャ
ップの存在に対して比較的高いコストを課し、そのギャップ中の後続の各残基に対してよ
り小さいペナルティを課す「アファインギャップコスト」が使用される。これは、最も一
般的に使用されるギャップスコアリングシステムである。高いギャップペナルティは、当
然ながら、より少数のギャップを有する最適化されたアライメントを生成するであろう。
大多数のアライメントプログラムは、ギャップペナルティが修飾されることを許容する。
しかし、配列比較のためにそのようなソフトウエアを使用する場合には、デフォルト値を
使用することが好ましい。
ライメントを作成することが要求される。このようなアライメントの実施に好適なコンピ
ュータプログラムは、Vector NTI(Invitrogen Corp.)であ
る。配列比較を実施し得るソフトウエアの例としては、例えば、BLASTパッケージ(
Ausubel et al.1999,Short Protocols in Mo
lecularBiology,4th Ed-Chapter 18を参照)、BLA
ST 2(FEMS Microbiol Lett 1999 174(2):247
-50;FEMS Microbiol Lett 1999 177(1):187-
8およびtachiana@ncbi.nlm.nih.govを参照)、FASTA(
Altschul et al,1990 J.Mol.Biol.403-410)、
およびAlignXが挙げられるが、これらに限定されない。少なくともBLAST、B
LAST 2およびFASTAは、オフラインおよびオンライン検索で入手可能である(
Ausubel et al.1999、上記、7-58~7-60頁を参照)。
体は、通常は、オールオアナッシングのペア比較に基づくものではない。代わりに、化学
的類似性または進化的距離に基づきそれぞれのペアワイズ比較にスコアを割り当てる数値
換算類似度(scaled similarity)スコア行列が一般に使用される。一
般に使用されるそのような行列の一例は、BLOSUM62行列(BLASTプログラム
スイートのためのデフォルト行列)である。Vector NTIプログラムは、一般に
、パブリックデフォルト値、または供給されている場合にはカスタムシンボル比較表を使
用する(さらなる詳細については、ユーザマニュアルを参照)。一部の用途については、
Vector NTIパッケージにためのデフォルト値を使用することが好ましい。
M(1988),Gene 73(1),237-244)と類似するアルゴリズムに基
づくVector NTI(Invitrogen Corp.)における多重アライメ
ント特徴を使用して算出することができる。
率を算出することが可能である。ソフトウエアは、通常、これを配列比較の一部として実
行し、数的結果を生成する。
ライメントに以下のパラメータを使用することが好ましい:
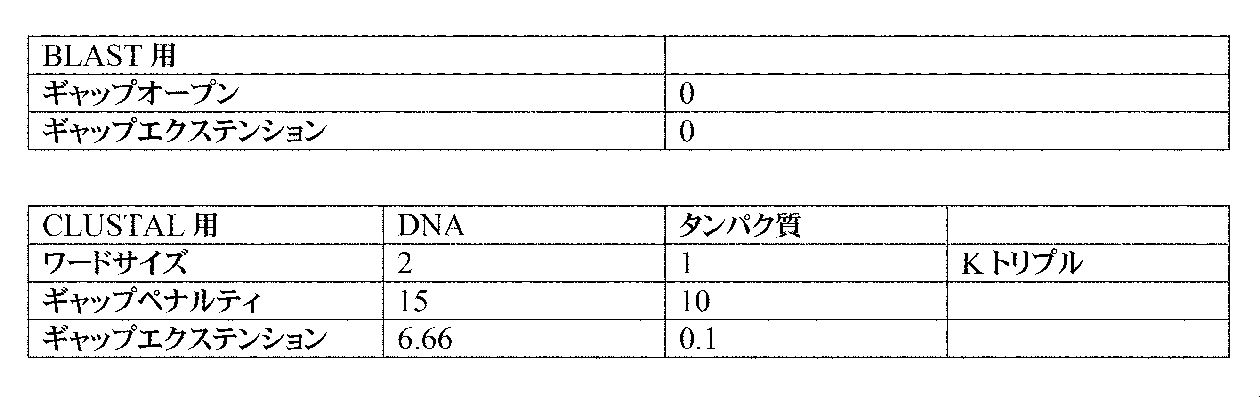
ティおよびギャップエクステンションを用いて使用することができる。
オチドにわたり、好ましくは、少なくとも30個の連続ヌクレオチドにわたり、好ましく
は、少なくとも40個の連続ヌクレオチドにわたり、好ましくは、少なくとも50個の連
続ヌクレオチドにわたり、好ましくは、少なくとも60個の連続ヌクレオチドにわたり、
好ましくは、少なくとも100個の連続ヌクレオチドにわたり決定される。
できる。
本発明はまた、タンパク質の任意のアミノ酸配列、またはそのようなパンパク質をコー
ドする任意のヌクレオチド配列の、変異体、相同体および誘導体の使用を包含する。
る相同性を有する実体を意味する。ここで、「相同性」という用語は、「同一性」と同じ
であると見なすことができる。
同一、好ましくは、少なくとも95、96、97、98または99%同一であり得るアミ
ノ酸配列を含むと解釈される。通常、相同体は、対象アミノ酸配列と同一の活性部位など
を含むであろう。相同性は類似性(すなわち、類似の化学的特性/機能を有するアミノ酸
残基)の観点から考慮することもできるが、本発明との関連では、配列同一性の観点から
相同性を表現することが好ましい。
列)と少なくとも75、80、85または90%同一、好ましくは、少なくとも95、9
6、97、98または99%同一であり得るヌクレオチド配列を含むと解釈される。通常
、相同体は、対象配列と同じ、活性部位などをコードする配列を含むであろう。相同性は
類似性(すなわち、類似の化学的特性/機能を有するアミノ酸残基)の観点から考慮する
こともできるが、本発明との関連では、配列同一性の観点から相同性を表現することが好
ましい。
グラムを使用して行うことができる。これらの市販のコンピュータプログラムは、2つ以
上の配列間の相同性率を計算することができる。相同性率は、連続配列にわたり計算する
ことができ、すなわち、ある1つの配列を他の配列とアラインし、ある1つの配列中の各
アミノ酸を他の配列中の対応するアミノ酸と1回に1残基、直接比較する。これは「ギャ
ップなし」アライメントと称される。通常、このようなギャップなしアライメントは、比
較的少数の残基にわたってのみ実施される。
る挿入または欠失が後続のアミノ酸残基をアライメントから外すことを考慮し得ず、した
がって、グローバルアライメントを実施した場合に相同性率を大きく低下させる可能性が
ある。このような理由から、ほとんどの配列比較法は、全相同性スコアを過度にペナルテ
ィ化せずに、あり得る挿入および欠失を考慮する最適なアライメントを作成するように設
計される。これは、配列アライメント中に「ギャップ」を挿入して局所的相同性の最大化
を試みることにより達成される。
少ないギャップを有する配列アライメント(2つの比較配列間のより高い関係性を反映す
る)が、多くのギャップを有するものより高いスコアを達成するように、そのアライメン
ト中で生じる各ギャップに対して「ギャップペナルティ」を割り当てる。通常、あるギャ
ップの存在に対して比較的高いコストを課し、そのギャップ中の後続の各残基に対してよ
り小さいペナルティを課す「アファインギャップコスト」が使用される。これは、最も一
般的に使用されるギャップスコアリングシステムである。高いギャップペナルティは、当
然ながら、より少数のギャップを有する最適化されたアライメントを生成するであろう。
大多数のアライメントプログラムは、ギャップペナルティが修飾されることを許容する。
しかし、配列比較のためにそのようなソフトウエアを使用する場合には、デフォルト値を
使用することが好ましい。例えば、GCG Wisconsin Bestfitパッケ
ージを使用する場合、アミノ酸配列のためのギャップペナルティのデフォルトは、ギャッ
プが-12および各エクステンションが-4である。
ライメントを作成することが要求される。このようなアライメントの実施に好適なコンピ
ュータプログラムは、GCG Wisconsin Bestfitパッケージ(Dev
ereux et al 1984 Nuc.Acids Research 12 p
387)である。配列比較を実施し得る他のソフトウエアの例としては、例えば、BLA
STパッケージ(Ausubel et al.,1999,上記を参照)、FASTA
(Altschul et al.,1990 J.Mol.Biol.403-410
)およびGENEWORKS比較ツールスイートが挙げられるが、これらに限定されない
。BLASTおよびFASTAの両者は、オフラインおよびオンライン検索で入手可能で
ある(Ausubel et al.,1999、上記を参照)。しかしながら、一部の
用途では、GCG Bestfitプログラムを使用することが好ましい。BLAST
2 Sequencesと称する新しいツールもまた、タンパク質およびヌクレオチド配
列の比較に利用可能である。
体は、通常は、オールオアナッシングのペア比較に基づくものではない。代わりに、化学
的類似性または進化的距離に基づきそれぞれのペアワイズ比較にスコアを割り当てる数値
換算類似度(scaled similarity)スコア行列が一般に使用される。一
般に使用されるそのような行列の一例は、BLOSUM62行列(BLASTプログラム
スイートのためのデフォルト行列)である。GCG Wisconsinプログラムは、
一般に、パブリックデフォルト値、または供給されている場合にはカスタムシンボル比較
表を使用する(さらなる詳細については、ユーザマニュアルを参照)。一部の用途では、
GCGパッケージのためのパブリックデフォルト値、または他のソフトウエアの場合には
、BLOSUM62などのデフォルト行列を使用することが好ましい。
M(1988)、上記を参照)と類似するアルゴリズムに基づくDNASIS(商標)(
Hitachi Software)における多重アライメント特徴を使用して算出する
ことができる。
率を算出することが可能である。ソフトウエアは、通常、これを配列比較の一部として実
行し、数的結果を生成する。
、挿入または置換も有し得る。意図的なアミノ酸置換は、その物質の二次的結合活性が保
持される限り、残基の極性、電荷、溶解度、疎水性、親水性および/または両親媒性の性
質の類似性に基づいて行うことができる。例えば、負荷電アミノ酸としては、アスパラギ
ン酸およびグルタミン酸が挙げられ;正荷電アミノ酸としては、リシンおよびアルギニン
が挙げられ;また、類似の親水性値を有する非荷電極性頭部基を持つアミノ酸としては、
ロイシン、イソロイシン、バリン、グリシン、アラニン、アスパラギン、グルタミン、セ
リン、トレオニン、フェニルアラニンおよびチロシンが挙げられる。
クの、好ましくは、第3列の同一行のアミノ酸を互いに置換することができる:
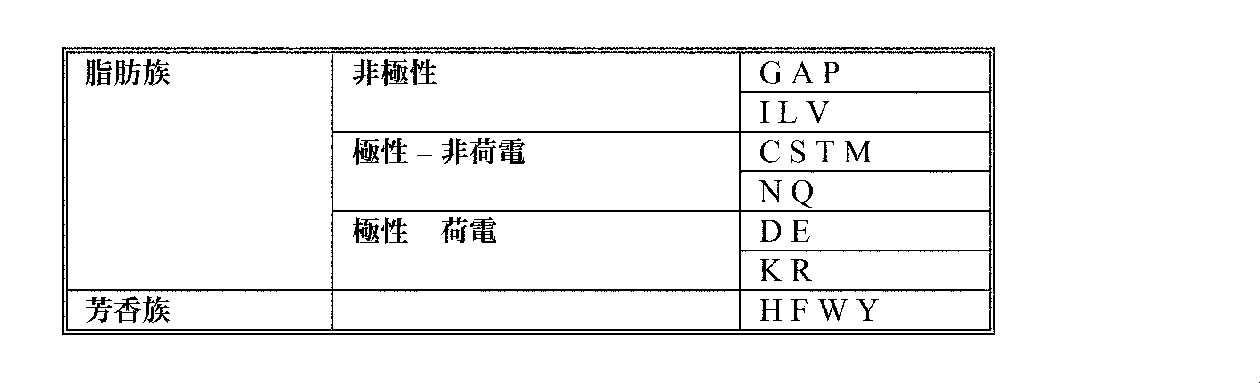
え(replacement))はいずれも、本明細書中では、既存のアミノ酸残基を代
替の残基と置き換えることを意味するのに使用される)、すなわち、塩基性のものと塩基
性のもの、酸性のものと酸性のもの、極性のものと極性のものなどの同種置換を包含する
。非相同置換、すなわち、ある分類の残基から他の分類の残基へ、または、オルニチン(
以下、Zと称する)、ジアミノ酪酸オルニチン(以下、Bと称する)、ノルロイシンオル
ニチン(以下、Oと称する)、ピリジルアラニン、チエニルアラニンおよびフェニルグリ
シンなどの非天然アミノ酸の混入を伴う置換もまた起こり得る。
、アルファ*およびアルファ二置換*アミノ酸、N-アルキルアミノ酸*、乳酸*、天然
アミノ酸のハロゲン化誘導体、例えば、トリフルオロチロシン*、p-Cl-フェニルア
ラニン*、p-Br-フェニルアラニン*、p-I-フェニルアラニン*、L-アリル-
グリシン*、β-アラニン*、L-α-アミノ酪酸*、L-γ-アミノ酪酸*、L-α-
アミノイソ酪酸*、L-ε-アミノカプロン酸#、7-アミノヘプタン酸*、L-メチオ
ニンスルホン*、L-ノルロイシン*、L-ノルバリン*、p-ニトロ-L-フェニルア
ラニン*、L-ヒドロキシプロリン#、L-チオプロリン*、フェニルアラニン(Phe
)のメチル誘導体、例えば、4-メチル-Phe*、ペンタメチル-Phe*、L-Ph
e(4-アミノ)#、L-Tyr(メチル)*、L-Phe(4-イソプロピル)*、L
-Tic(1,2,3,4-テトラヒドロイソキノリン-3-カルボン酸)*、L-ジア
ミノプロピオン酸およびL-Phe(4-ベンジル)*が挙げられる。記号*は、誘導体
の疎水性を示すために上記考察の目的(相同または非相同置換に関する)に利用されてお
り、一方、#は、誘導体の親水性を示すために利用されており、#*は、両親媒性を示す
。
なスペーサー基、例として、アルキル基、例えばメチル基、エチル基またはプロピル基を
、アミノ酸スペーサー、例えばグリシンまたはβ-アラニン残基に加えて含み得る。変異
のさらなる形態は、ペプトイド形態の1つ以上のアミノ酸残基の存在を含むが、当業者に
は十分に理解されよう。疑義を回避するため、「ペプトイド形態」は、α炭素置換基がα
炭素ではなく残基の窒素原子上に存在する変異アミノ酸残基を指すために使用される。ペ
プトイド形態のペプチドを調製する方法は当該技術分野、例えば、Simon RJ e
t al.,PNAS(1992)89(20),9367-9371、およびHorw
ell DC,Trends Biotechnol.(1995)13(4),132
-134において知られている。
クレオチドを含み得る。オリゴヌクレオチドに対する多数の異なるタイプの修飾が、当該
技術分野において知られている。これらとしては、メチルホスホネートおよびホスホロチ
オエート骨格、ならびに/または分子の3’および/もしくは5’末端におけるアクリジ
ンもしくはポリリジン鎖の付加が挙げられる。本発明の目的のため、本明細書に記載のヌ
クレオチド配列が当該技術分野において利用可能な任意の方法により修飾し得ることを理
解すべきである。このような修飾は、本発明のヌクレオチド配列のインビボ活性または寿
命を向上させるように実施することができる。
意の誘導体、断片もしくは誘導体の使用を包含する。配列がその断片に相補的である場合
、その配列をプローブとして使用して他の生物における類似のコード配列を同定するなど
のことが可能である。
数の手法で得ることができる。本明細書に記載の配列の他の変異体は、例えば、一連の個
体、例えば、異なる集団からの個体から作製されたDNAライブラリーをプロービングす
ることにより得ることができる。さらに、他の相同体を得ることができ、そのような相同
体およびその断片は、一般に、本明細書の配列表に示す配列に選択的にハイブリダイズし
得る。このような配列は、他の動物種から作製されたcDNAライブラリー、または他の
動物種からのゲノムDNAライブラリーをプロービングし、中~高ストリンジェンシーの
条件下で付属の配列表の配列のいずれか1つの全部または一部を含むプローブによりその
ようなライブラリーをプロービングすることにより得ることができる。類似の考察は、本
発明のポリペプチドまたはヌクレオチド配列の種相同体および対立遺伝子変異体を得るこ
とにも適用される。
異体および相同体内の配列を標的とするように設計されたプライマーを使用する縮重PC
Rを使用して得ることができる。保存配列は、例えば、いくつかの変異体/相同体からの
アミノ酸配列をアラインすることにより予測することができる。配列アライメントは、当
該技術分野で知られるコンピュータソフトウエアを使用して実施することができる。例え
ば、GCG Wisconsin PileUpプログラムが広く使用される。
列に対する単一配列プライマーを用いる配列のクローニングに使用されるものよりも低い
ストリンジェンシー条件において使用される。
誘発により得ることができる。これは、例えば、ポリヌクレオチド配列が発現する特定の
宿主細胞についてのコドン選好性を最適化するためにサイレントコドン配列変化が要求さ
れる場合に有用であり得る。制限酵素認識部位を導入するため、またはポリヌクレオチド
によりコードされるポリペプチドの特性もしくは機能を変更するために他の配列変化が望
まれることがある。
イマー、代替増幅反応のためのプライマー、プローブ、例えば、放射性もしくは非放射性
標識を使用する慣用の手段による顕示標識で標識されたものを産生するために使用するこ
とができ、またはポリヌクレオチドをベクター中にクローニングすることができる。この
ようなプライマー、プローブおよび他の断片は、少なくとも15、好ましくは、少なくと
も20、例えば、少なくとも25、30または40ヌクレオチド長であり、これらもまた
、本明細書において使用されるポリヌクレオチドという用語に包含される。
えにより、合成により、または当業者に利用可能な任意の手段により産生することができ
る。これらはまた、標準的手法によりクローニングすることもできる。
手段により産生される。自動化技術を使用してこれらを達成するための技術は、当該技術
分野において容易に利用可能である。
メラーゼ連鎖反応)クローニング技術を使用して産生される。プライマーは、増幅された
DNAを好適なクローニングベクター中にクローニングすることができるように、好適な
制限酵素認識部位を含有するように設計することができる。
本発明はまた、本発明の核酸配列に相補的な配列、または本発明の配列もしくは本発明
の配列に相補的な配列にハイブリダイズすることができる配列を包含する。
基対合を介して相補鎖と結合するプロセス」、およびポリメラーゼ連鎖反応(PCR)技
術において実施される増幅のプロセスを含む。
レオチド配列、またはその任意の誘導体、断片もしくは誘導体の使用を包含する。
得る配列に相補的な配列も包含する。
リンジェンシーな条件(例えば、50℃および0.2×SSC{1×SSC=0.15M
のNaCl、0.015Mのクエン酸Na3 pH7.0})下でハイブリダイズし得る
配列に相補的な配列を包含する。
高ストリンジェンシーな条件(例えば、65℃および0.1×SSC{1×SSC=0.
15MのNaCl、0.015Mのクエン酸Na3 pH7.0})下でハイブリダイズ
し得る配列に相補的な配列を包含する。
を含む)にハイブリダイズし得るヌクレオチド配列に関する。
を含む)にハイブリダイズし得る配列に相補的なヌクレオチド配列に関する。
イブリダイズし得るポリヌクレオチド配列もまた本発明の範囲内に含まれる。
び0.2×SSC)下で本発明のヌクレオチド配列にハイブリダイズすることができるヌ
クレオチド配列、またはその相補体をカバーする。
℃および0.1×SSC)下で本発明のヌクレオチド配列にハイブリダイズすることがで
きるヌクレオチド配列、またはその相補体をカバーする。
非限定的な例として、インビボまたはインビトロでヌクレオチド配列中に多数の部位特
異的またはランダムな変異を産生すること、および、続いて種々の手段によりコードされ
るポリペプチドの改善された機能性についてスクリーニングすることが可能である。
しくは天然変異体により組換えて、新たな変異体を産生することができる。このような新
たな変異体もまた、コードされるポリペプチドの改善された機能性についてスクリーニン
グされ得る。新たな好ましい変異体の産生は、当該技術分野において十分に確立された種
々の方法、例えば、エラー閾値変異誘発(Error Threshold Mutag
enesis)(国際公開第92/18645号パンフレット)、オリゴヌクレオチド媒
介ランダム変異誘発(米国特許第5,723,323号明細書)、DNAシャッフリング
(米国特許第5,605,793号明細書)、エキソ媒介遺伝子アセンブリ、国際公開第
00/58517号パンフレットにより達成することができる。これらおよび類似のラン
ダム指向分子進化法の適用により、タンパク質の構造または機能のいかなる予備知識もな
しで好ましい特徴を有する本発明の酵素の変異体の同定および選択が可能となり、予測可
能でないが、有益な変異または変異体の産生が可能となる。当該技術分野において、酵素
活性の最適化または改変のための分子進化の適用例が数多くあり、そのような例として、
限定はされないが、以下の1つ以上が挙げられる:
宿主細胞またはインビトロにおける発現および/または活性の最適化、
酵素活性の増大、基質および/または生成物特異性の改変、
酵素安定性または構造安定性の増大または低減、好ましい環境条件、例えば温度、pH、
基質における酵素の活性/特異性の改変。
タンパク質コードヌクレオチド配列を単離したなら、または推定タンパク質コードヌク
レオチド配列を同定したなら、本発明のタンパク質を調製するために配列を変異させるこ
とが望ましい場合がある。
クレオチドは、所望の変異部位をフランキングするヌクレオチド配列を含有する。
984)2,p646-649)に開示されている。酵素コードヌクレオチド配列中に突
然変異を導入する他の方法は、Nelson and Long(Analytical
Biochemistry(1989),180,p147-151)に記載されてい
る。
一態様において、本発明において使用される配列は、組換え配列、すなわち、組換えD
NA技術を使用して調製された配列である。
、例えば、J.Sambrook、E.F.Fritsch,and T.Maniat
is、1989,Molecular Cloning:A Laboratory M
anual, Second Edition,Books 1-3,Cold Spr
ing Harbor Laboratory Pressに説明されている。
一態様において、本発明において使用される配列は、合成配列、すなわち、インビトロ
化学または酵素的合成により調製された配列である。これとしては、限定されるものでは
ないが、宿主生物、例えば、メチロトローフ酵母のピキア属(Pichia)およびハン
セヌラ属(Hansenula)に最適なコドン使用頻度を用いて作製された配列が挙げ
られる。
酵素の発現
本発明において使用されるヌクレオチド配列は、組換え複製ベクター中に取り込むこと
ができる。ベクターは、ヌクレオチド配列を複製し、タンパク質/酵素形態で、適合性の
宿主細胞中で、および/または適合性の宿主細胞から発現させるために使用することがで
きる。
れる配列および/またはベクターに応じて、分泌、または、細胞内に含有させることがで
きる。コード配列は、特定の原核または真核細胞膜を介する物質コード配列の分泌を指示
するシグナル配列を用いて設計することができる。
「発現ベクター」という用語は、インビボまたはインビトロで発現し得るコンストラク
トを意味する。
れる」という用語は、好ましくは、ゲノム中への安定した取り込みをカバーする。
ド配列の発現を提供し得る調節配列に作動可能に結合しているベクター中に存在し得る。
下記の好適な宿主細胞中に形質転換することができる。
導入すべき宿主細胞に依存することが多い。
物質耐性、例えば、アンピシリン、カナマイシン、クロラムフェニコールまたはテトラサ
イクリン耐性を付与する遺伝子を含有し得る。あるいは、選択は、同時形質転換(国際公
開第91/17243号パンフレットに記載)により達成することができる。
を形質移入し、形質転換し、形質導入し、または感染させるために使用することができる
。
オチド配列を導入し、適合性の宿主細胞中にベクターを導入し、ベクターの複製を生じさ
せる条件下で宿主細胞を増殖させることにより、本発明のヌクレオチド配列を作製する方
法を提供する。
含み得る。このような配列の例は、プラスミドpUC19、pACYC177、pUB1
10、pE194、pAMB1およびpIJ702の複製起源である。
一部の適用では、本発明において使用されるヌクレオチド配列は、例えば、選択された
宿主細胞によるヌクレオチド配列の発現を提供し得る調節配列に作動可能に結合している
。例として、本発明は、そのような調節配列に作動可能に結合している本発明のヌクレオ
チド配列を含むベクターをカバーする。すなわち、ベクターは発現ベクターである。
でそれらを機能させる関係にある並置を指す。コード配列に「作動可能に結合している」
調節配列は、コード配列の発現が制御配列と適合する条件下で達成されるような手法でラ
イゲートされる。
シグナルを含む。
、RNAポリメラーゼ結合部位である。
ば、プロモーター、分泌リーダーおよびターミネーター領域の選択により達成することが
できる。
結合している。
。
例は、当該技術分野においてよく知られている。
らに含むことができる。例えば、特徴部は、プリブノーボックスまたはTATAボックス
などの保存領域であり得る。
「コンストラクト」という用語(「コンジュゲート」、「カセット」および「ハイブリ
ッド」などの用語と同義である)は、プロモーターに直接または間接的に付着している本
発明により使用されるヌクレオチド配列を含む。
配列、例えばSh1-イントロンまたはADHイントロンなどの、好適なスペーサー群の
提供である。同じことが、直接または間接的付着を含む、本発明に関連する「融合してい
る」という用語にも当てはまる。一部の場合において、これらの用語は、野生型遺伝子プ
ロモーターと通常会合するタンパク質をコードするヌクレオチド配列の天然の組合せを、
それらがいずれも天然環境に存在する場合には、カバーしない。
現し得る。
に作動可能に結合している本発明のヌクレオチド配列を含む。
「宿主細胞」という用語は、本発明との関連では、上記のヌクレオチド配列または発現
ベクターを含み、本明細書に定義される特定の特性を有するタンパク質の組換え産生に使
用される任意の細胞を含む。
ド配列により形質転換または形質移入された宿主細胞を提供する。細胞は、前記ベクター
と適合するように選択され、例えば、原核(例えば、細菌)、真菌、酵母または植物細胞
であり得る。
パク質のさらなるプロセシングについての所望度に応じて、酵母または他の真菌などの真
核宿主が好ましいことがある。一般に、酵母細胞は、操作がより容易なため、真菌細胞よ
りも好ましい。しかしながら、タンパク質のなかには、酵母細胞からの分泌が乏しかった
り、一部の場合、適切にプロセシングされなかったりする(例えば、酵母の超グリコシル
化)ものがある。これらの例においては、異なる真菌宿主生物を選択すべきである。
現産物に最適な生物学的活性を付与することが必要であり得る場合に、翻訳後修飾(例え
ば、ミリストイル化、グリコシル化、トランケーション、脂質化、およびチロシン、セリ
ンまたはトレオニンリン酸化)を提供し得る。
もよい。本発明はまた、親細胞の変異細胞を産生する方法であって、ポリペプチドをコー
ドする核酸配列、またはその制御配列を破壊または欠失させ、それにより変異細胞中のポ
リペプチドの産生量を親細胞より減少させることを含む方法に関する。
するポリペプチドの発現に必要な核酸配列を修飾または不活化することによって首尾よく
行うことができる。
「生物」という用語は、本発明との関連では、本発明によるポリペプチドをコードする
ヌクレオチド配列および/もしくはそれから得られた産物を含み得、かつ/またはプロモ
ーターが生物中に存在する場合に本発明によるヌクレオチド配列の発現を可能とし得る任
意の生物を含む。
プチドをコードするヌクレオチド配列および/もしくはそれから得られた産物を含み、か
つ/またはプロモーターが生物内の本発明によるヌクレオチド配列の発現を可能とし得る
任意の生物を含む。好ましくは、ヌクレオチド配列は、生物のゲノム中に取り込まれる。
ロモーター(これもその天然環境中に存在する)の制御下にある場合、天然環境中の天然
ヌクレオチドコード配列をカバーしない。
するヌクレオチド配列、本発明によるコンストラクト、本発明によるベクター、本発明に
よるプラスミド、本発明による細胞、本発明による組織、またはそれらの産物の1つ、ま
たは組合せを含む生物を含む。
プチドをコードするヌクレオチド配列を含み得る。
上記のとおり、宿主生物は、原核または真核生物であり得る。好適な原核宿主の例とし
ては、エシェリキア・コリ(E.coli)およびバチルス・サブティルス(Bacil
lus subtilis)が挙げられる。
ambrook et al(上記を参照)を参照されたい。原核宿主を使用する場合、
ヌクレオチド配列は、形質転換前に、イントロンの除去などにより、好適に改変する必要
があり得る。
プラスト形成およびプロトプラストの形質転換、そしてそれに続く細胞壁の再生を含む方
法により形質転換することができる。宿主微生物としてのアスペルギルス属(Asper
gillus)の使用が、欧州特許第0238023号明細書に記載されている。
Potrykus(Annu Rev Plant Physiol Plant Mo
l Biol[1991]42:205-225)およびChristou(Agro-
Food-Industry Hi-Tech March/April 1994 1
7-27)の論文に見出すことができる。植物形質転換に関するさらなる教示は、欧州特
許第0449375号明細書に見出すことができる。
。
宿主生物は、真菌、例えばカビであり得る。好適なそのような宿主の例としては、サー
モミセス属(Thermomyces)、アクレモニウム属(Acremonium)、
アスペルギルス属(Aspergillus)、ペニシリニウム属(Penicilli
um)、ムコル属(Mucor)、ニューロスポラ属(Neurospora)、トリコ
デルマ属(Trichoderma)などに属するメンバーが挙げられる。
65号明細書で考察されており、真菌の培養は、当該技術分野においてよく知られている
。アカパンカビ(N.crassa)に適用される技術の広範な概説は、例えば、Dav
is and de Serres,Methods Enzymol(1971)17
A:79-143に見出される。
7号明細書に概説されている。
ds Biotechnol 2002 May;20(5):200-6,Arche
r & Peberdy、Crit Rev Biotechnol(1997)17(
4):273-306に教示されている。
ク糸状菌の生産を包含する。
アスペルギルス・ニガー(Aspergillus niger)であり得る。
例えば、Turner,G.1994(Vectors for genetic ma
nipulation.In:Martinelli S.D.,Kinghorn J
.R.(Editors)Aspergillus:50 years on.Prog
ress in industrial microbiology vol 29.E
lsevier Amsterdam 1994.pp.641-666)の教示にした
がって調製することができる。
リコデルマ・リーゼイ(Trichoderma Reesei)であり得る。
他の実施形態において、トランスジェニック生物は、酵母であり得る。
(1995),49:341-54、およびCurr Opin Biotechnol
(1997)Oct;8(5):554-60で提供される。
es cerevisiae)またはピキア・パストリス(Pichia pastor
is)(FEMS Microbiol Rev(2000 24(1):45-66を
参照)の種は、異種遺伝子発現のためのビヒクルとして使用することができる。
中の異種遺伝子発現および遺伝子産物の分泌の原理の概説は、E Hinchcliff
e E Kenny(1993,“Yeast as a vehicle for t
he expression of heterologous genes”,Yea
sts,Vol 5,Anthony H Rose and J Stuart Ha
rrison,eds,2nd edition,Academic Press Lt
d.)に示されている。
発明によるトランスジェニックサッカロミセス(Saccharomyces)は、Hi
nnen et al.,(1978,PNAS USA 75,1929);Begg
s,J D(1978,Nature,275,104);およびIto,H et a
l(1983,J Bacteriology 153,163-168)の教示にした
がって調製することができる。
ナント抗生物質耐性マーカーを使用して選択することができる。
本発明のヌクレオチド配列により形質転換された宿主細胞は、コードされるポリペプチ
ドの産生をもたらし、細胞および/または培養培地からのポリペプチドの回収を容易にす
る条件下で培養することができる。
現の取得に好適な従来の任意の培地であってよい。
きる。
簡便に回収することができる。
タンパク質は、発現宿主から、タンパク質をより容易に回収することができる培養培地
中に分泌されることが望ましいことが多い。本発明によれば、分泌リーダー配列を所望の
発現宿主に基づき選択することができる。ハイブリッドシグナル配列も、本発明に関連し
て使用することができる。
laA-例えば、アスペルギルス属(Aspergillus)からの18アミノ酸型お
よび24アミノ酸型の両方)、a因子遺伝子(酵母、例えば、サッカロミセス属(Sac
charomyces)、クルイベロミセス属(Kluyveromyces)およびハ
ンセヌラ属(Hansenula))、またはα-アミラーゼ遺伝子(バチルス属(Ba
cillus))に由来するものである。
hods Enzymol(1990)182:132-43に概説されている。
アミノ酸配列の発現を検出および測定するための種々のプロトコルは、当該技術分野に
おいて知られている。例としては、酵素結合免疫吸着アッセイ(ELISA)、ラジオイ
ムノアッセイ(RIA)および蛍光活性化細胞ソーティング(FACS)が挙げられる。
よびアミノ酸アッセイにおいて使用することができる。
NJ)、Promega(Madison,WI)、およびUS Biochemica
l Corp(Cleveland,OH)が市販のキットおよびそれらの手順について
のプロトコルを供給している。
発光剤または発色剤、ならびに、基質、補因子、阻害剤、磁気粒子などが挙げられる。こ
のような標識の使用を教示する特許としては、米国特許第3,817,837号;米国特
許第850,752号;米国特許第3,939,350号;米国特許第3,996,34
5号;米国特許第4,277,437号;米国特許第4,275,149号および米国特
許第US4,366,241号が挙げられる。
うに産生することができる。
本発明において使用されるアミノ酸配列は、例えば、抽出および精製を補助する融合タ
ンパク質として産生することができる。融合タンパク質パートナーの例としては、グルタ
チオン-S-トランスフェラーゼ(GST)、6×His、GAL4(DNA結合および
/または転写活性化ドメイン)および(β-ガラクトシダーゼ)が挙げられる。融合タン
パク質パートナーおよび目的タンパク質配列間にタンパク質分解開裂部位を含めて融合タ
ンパク質配列の取り出しを可能とすることも好都合であり得る。
Biotechnol(1995)6(5):501-6に概説されている。
ンパク質をコードさせることができる。例えば、物質活性に影響し得る薬剤についてのペ
プチドライブラリーのスクリーニングのため、市販の抗体により認識される異種エピトー
プを発現するキメラ物質をコードさせることが有用であり得る。
本発明において使用される配列はまた、1種以上の追加の目的タンパク質(POI)ま
たは目的ヌクレオチド配列(NOI)と併用してもよい。
リコーゲンの代謝に関与するタンパク質または酵素、アセチルエステラーゼ、アミノペプ
チダーゼ、アミラーゼ、アラビナーゼ、アラビノフラノシダーゼ、カルボキシペプチダー
ゼ、カタラーゼ、セルラーゼ、キチナーゼ、キモシン、クチナーゼ、デオキシリボヌクレ
アーゼ、エピメラーゼ、エステラーゼ、α-ガラクトシダーゼ、β-ガラクトシダーゼ、
α-グルカナーゼ、グルカンリアーゼ、エンド-β-グルカナーゼ、グルコアミラーゼ、
グルコースオキシダーゼ、α-グルコシダーゼ、β-グルコシダーゼ、グルクロニダーゼ
、ヘミセルラーゼ、ヘキソースオキシダーゼ、ヒドロラーゼ、インベルターゼ、イソメラ
ーゼ、トリペプチジルエキソペプチダーゼ、ラッカーゼ、リパーゼ、リアーゼ、マンノシ
ダーゼ、オキシダーゼ、オキシドレダクターゼ、ペクテートリアーゼ、ペクチンアセチル
エステラーゼ、ペクチンデポリメラーゼ、ペクチンメチルエステラーゼ、ペクチン分解酵
素、ペルオキシダーゼ、フェノールオキシダーゼ、フィターゼ、ポリガラクツロナーゼ、
プロリンエンドプロテアーゼ、ラムノガラクツロナーゼ、リボヌクレアーゼ、タウマチン
、トランスフェラーゼ、エキソプロテアーゼ、トランスポートタンパク質、トランスグル
タミナーゼ、アミノペプチダーゼ、ヘキソースオキシダーゼ(D-ヘキソース:O2-オ
キシドレダクターゼ、EC 1.1.3.5)、またはこれらの組み合わせが挙げられる
。NOIは、それらの配列のいずれかのアンチセンス配列であってさえもよい。
のような配列は、例えば、英国特許出願第9821198.0号明細書に記載されるアス
ペルギルス・ニガー(Aspergillus niger)cypB遺伝子の産物とし
てのシャペロンタンパク質をコードすることができる。
発現を変える改変などの多くの理由から、その活性を改変するために操作することができ
る。さらなる例として、NOIはまた、特定の宿主細胞における発現を最適化するために
修飾され得る。他の配列の変化は、制限酵素認識部位を導入するために所望され得る。
ロチオエート骨格をその中に含み得る。
しては、限定はされないが、分子の5’および/または3’末端のフランキング配列の付
加、または分子骨格におけるホスホジエステラーゼ結合ではなくホスホロチオエートまた
は2’O-メチルの使用が挙げられる。
本発明は、特に記載のない限り、当業者の技能の範囲内である化学、分子生物学、微生
物学、組換えDNAおよび免疫学の慣用技術を用いる。このような技術は、文献に説明さ
れている。例えば、J.Sambrook,E.F.Fritsch,and T.Ma
niatis,1989,MolecularCloning:A Laborator
y Manual,Second Edition,Books1-3,Cold Sp
ring Harbor Laboratory Press;Ausubel,F.M
.etal.(1995 and periodic supplements;Cur
rent Protocols in Molecular Biology,ch.9
,13,and,16,John Wiley & Sons,New,York,N.
Y.);B.Roe,J.Crabtree,and,A.Kahn,1996,DNA
Isolation and Sequencing:Essential Tech
niques,John Wiley & Sons;M.J.Gait(Editor
),1984,Oligonucleotide Synthesis:A Pract
ical Approach, Irl Press;およびD.M.J.Lilley
,and J.E.Dahlberg,1992,Methods of Enzymo
logy:DNA Structure Part A:Synthesis and
Physical Analysis of DNA in Methods in E
nzymology,Academic Pressを参照されたい。これらのテキスト
全般はそれぞれ参照により本明細書に組み込まれる。
範囲を決して限定するものではない。添付の図面は、本開示の明細および説明に組込まれ
たそれらの一部と見なされるべきである。以下の実施例は、請求の範囲に記載の開示を説
明するために提示されたものであって、その開示を限定するものではない。
TRI036、TRI037、TRI038のクローニングおよび形質転換
トリコデルマ・リーゼイ(Trichoderma reesei)での発現のための
コドン最適化遺伝子として、真菌2型アミノペプチダーゼ(pepN_2、Merops
ファミリーM28.008.に属する;merops.sanger.ac.uk/)を
コードする合成遺伝子(TRI031、TRI032、TRI033、TRI034、T
RI035、TRI036、TRI037、TRI038)を、Geneart(Lif
e Technologies)から受注した。TRI031は、ネオサルトリア・フィ
ッシェリ(Neosartorya fischeri)NRRL181由来のNCBI
アクセッション番号:XP_001258675に対応し;TRI032は、ミセリオフ
トラ・テルモフィラ(Myceliophthora thermophila)ATC
C(登録商標)42464由来のNCBIアクセッション番号:XP_00366735
4に対応し;TRI033はフザリウム・オキシスポルム(Fusarium oxys
porum)Fo5176由来のNCBIアクセッション番号:EGU74500に対応
し;TRI034はフザリウム・オキシスポルム分化型クベンス(Fusarium o
xysporum f.sp.cubense)レース1由来のNCBIアクセッション
番号:ENH69875に対応し;TRI035はアスペルギルス・クラバタス(Asp
ergillus clavatus)NRRL1由来のNCBIアクセッション番号:
XP_001273779に対応する;TRI036はケトミウム・テルモフィルム(C
haetomium thermophilum)変種テルモフィルム(thermop
hilum)DSM1495からのNCBIアクセッション番号:EGS23402に対
応し;TRI037はアスペルギルス・テレウス(Aspergillus terre
us)NIH2624由来のNCBIアクセッション番号:XP_001217759に
対応し;かつTRI038はアスペルギルス・ニデュランス(Aspergillus
nidulans)FGSC A4由来のNCBIアクセッション番号:XP_6817
14に対応する。
B1およびattB2)(Life Technologies)を有するように発注し
、pDonr221 Gatewayベクター(Life Technologies)
中のプラスミドストックとして納品された。TRI031、TRI032、TRI033
、TRI034、TRI035、TRI036、TRI037、TRI038のアミノ酸
配列は、予測分泌シグナル配列(SignalP 4.0:膜貫通領域からシグナルペプ
チドを区別する。Thomas Nordahl Petersen, Soren B
runak, Gunnar von Heijne & Henrik Nielse
n.Nature Methods,8:785-786,2011)を有しており、こ
れらの内因性シグナル配列を、コザック配列を含有するリーダー配列、トリコデルマ・リ
ーゼイ(Trichoderma reesei)酸性真菌プロテアーゼ(AFP)から
の分泌シグナル配列およびトリコデルマ・リーゼイ(Trichoderma rees
ei)グルコアミラーゼ遺伝子(TrGA1)からのイントロンで置き換えた(図1を参
照)。
を使用してpDonr221ベクター中の合成遺伝子をdestination vec
tor pTrex8gMに組み換え、発現ベクターpTrex8gM_TRI031(
配列番号:9)、pTrex8gM_TRI032(配列番号10)、pTrex8gM
_TRI033(配列番号11)、pTrex8gM_TRI034(配列番号12)、
pTrex8gM_TRI035(配列番号13)、pTrex8gM_TRI036(
配列番号14)、pTrex8gM_TRI037(配列番号15)、およびpTrex
8gM_TRI038(配列番号16)を得た。
介プロトプラスト形質転換法を使用し、1.5-17μgの発現ベクターでトリコデルマ
・リーゼイ(Trichoderma reesei)株Cellulight(商標)
を個々に形質転換した。記載のように、形質転換には、僅かに変更したPEG-プロトプ
ラスト法を用いた。プロトプラストの調製には、10mMウリジンでpyr栄養要求性を
補完したトリコデルマ(Trichoderma)発芽培地により、胞子を26℃で約1
8時間成長させた。(トリコデルマ(Trichoderma))発芽培地:40mlの
50%グルコース、2g/Lのペプトン、15g/LのKH2PO4、5g/Lの(NH
4)2SO4、2.4mlの1M MgSO4、4.1mlの1M CaCl2、1ml
の400×トリコデルマ・リーゼイ(T.reesei)微量元素溶液{200g/Lの
FeSO4×7H2O、16g/LのZnSO4×7H2O、1.4g/LのMnSO4
×H2O、3.2g/LのCuSO4×5H2O、0.8g/LのH3BO3、175g
/Lのクエン酸})振盪速度200rpmで。発芽胞子を遠心分離により回収し、洗浄し
、45mg/mlの溶解酵素溶液(トリコデルマ・ハルジアヌム(Trichoderm
a harzianum)、Sigmaカタログ番号L1412)により処理して真菌細
胞壁を溶解させた。さらに、Penttilaeら(Gene 61(1987)155
-164)の記載のように、標準法によりプロトプラストを調製した。
トプラストを含有する形質転換混合物は、2mLの25%PEG溶液で処理し、2容量の
1.2Mのソルビトール/10mMのTris、pH7.5/10mMのCaCl2溶液
で希釈し、3%の選択的トップアガロース、1Mのソルビトール、10mMのNH4Cl
、1×MM溶液(2×MM溶液:30g/LのKH2PO4、20mLの1Mアセトアミ
ド、20mlの1M CsCl、6mlの20%MgSO4×7H2O、6mlの20%
CaCl2×2H2O、2mLのトリコデルマ・リーゼイ(T.reesei)微量元素
溶液(400×)、80mLの50%グルコース、全量を1リットルとし、pH4.5)
と混合し、10mMのNH4Clを含むMMプレート(MMプレート 2%寒天、1×M
M溶液)に注ぎ入れた。形質転換体は、MMプレート(ウリジン欠失)で原栄養成長する
ものを選択した。胞子が形成されるまで28℃で5~7日間、プレートをインキュベート
した。より良好な胞子形成のために、安定に見える形質転換体を10mMのNH4Clを
含む新しいMMプレートに移した。胞子が形成されたなら、0.85%NaCl、0.0
15%Tween(登録商標)80の溶液を使用して胞子を採取した。液体培養物を24
ウェルMTPフォーマット(スクリーニング用)、または振盪フラスコ(有効性確認試験
用)に植菌するために胞子懸濁液を使用した。前培養は3mLのYEG培地(5g/Lの
酵母エキス、22g/Lのグルコース、H2O)中。本培養は次の生産培地中(生産培地
:35g/Lの61%グルコース/ソホロース混合物、9g/Lのカザミノ酸、5g/L
の(NH4)2SO4、4.5g/LのKH2PO4、1g/LのCaCl2×2H2O
、1g/LのMgSO4×7H2O、33g/LのPIPPSバッファー、pH5.5、
2.5mL/Lの400×トリコデルマ・リーゼイ(T.reesei)微量元素(17
5g/Lのクエン酸、200g/LのFeSO4×7H2O、16g/LのZnSO4×
7H2O、3.2g/LのCuSO4×5H2O、1.4g/LのMnSO4×H2O、
0.8g/Lのホウ酸)pH5.5。変異体を生成するために、3mLの生産培地を24
ウェルMTPに加えた。振盪フラスコでは、容量をスケールアップした。
た。真空ろ過により培養上清を採取し、それらのパフォーマンスおよび発現レベルを試験
するために使用した。
れる分泌シグナルには、アンダーラインが付されている:

れる分泌シグナルには、アンダーラインが付されている。

れる分泌シグナルには、アンダーラインが付されている。

れる分泌シグナルには、アンダーラインが付されている。

れる分泌シグナルには、アンダーラインが付されている。

れる分泌シグナルには、アンダーラインが付されている。

れる分泌シグナルには、アンダーラインが付されている。

れる分泌シグナルには、アンダーラインが付されている。

。



列。



列。



列。



列。



列。



列。



列。


orya fischeri)由来のPepN 2(TRI031)の作動pHおよび温
度の最適値
グルタミン酸の遊離は、ダイズまたはグルテンの加水分解などの、野菜タンパク質の加
水分解における重要な品質パラメータである。遊離したグルタミン酸は、いわゆる「旨味
フレーバー」と捉えられ得る。グルタミン酸およびグルタミンの遊離の観点から、グルテ
ンの加水分解に最初に適用したpHの影響を調べるために、15~30分でグルテンを液
化し、その直後に1MのHClでグルテン加水分解の初期pHを調節した。グルテンをグ
ルタミナーゼ(Amano、日本)で処理した。さらに、グルテンを、FoodPro
Alkaline Protease、FoodPro PNLおよびPepN 2(ネ
オサルトリア・フィッシェリ(Neosartorya fischeri))で処理し
た。20時間後、限外ろ過により加水分解を停止させた(カットオフ10kDa、Sar
torius Stedium Biotech、Goettingen、German
y)。酵素によるグルタミン酸分析に透過性を使用した(酵素によるL-グルタミン酸分
析キット、Roche、Mannheim、Germany)。図2に示すように、Pe
pN 2はpH6.0と比較してpH範囲7.0~9.0でより高い効率を示した。
の影響を調べるために、グルテン懸濁液を15~30分で液化し、その直後に1MのHC
lでグルテン加水分解の初期pHをpH7.0に調節した。図3に示すように、40~6
0℃の温度範囲を使用した。グルテンをグルタミナーゼ(Amano、日本)と混合した
。さらに、グルテンをFoodPro(登録商標) Alkaline Proteas
e、FoodPro(登録商標) PNLおよびPepN 2(ネオサルトリア・フィッ
シェリ(Neosartorya fischeri))と混合した。加水分解を始めて
20時間後、限外ろ過により加水分解を停止させた(カットオフ10kDa、Sarto
rius Stedium、Goettingen、Germany)。酵素によるグル
タミン酸分析に透過性を使用した(酵素によるL-グルタミン酸分析キット、Roche
、Mannheim、Germany)。ネオサルトリア・フィッシェリ(Neosar
torya fischeri)由来のPepN 2は、60℃で最も高い効率を示した
。
rya fischeri)由来のPepN 2(TRI031)と、アスペルギルス・
クラバタス(Aspergillus clavatus)由来のPepN 2(TRI
035)との比較
加水分解度(DH)は、同量のタンパク質について、例えば、6MのHCl中、約12
0℃で約24時間行った酸加水分解と比較して、開裂したペプチド結合の相対量を表す。
したがって、PepN 1型またはPepN 2型などの一般的なPepNのアミノ酸遊
離効率の評価には、DHがより高いことが適用され得る。無細胞培養ブロスを濃縮し(カ
ットオフ10kDa、Sartorius、Goettingen、Germany)、
製造業者の説明通り使い捨て脱塩カラム(PD10、GE、Muenchen、Germ
any)を使用して脱塩した。基質として、予備加水分解した10%(w/w)Na-カ
ゼイン塩(DMK、Germany)、ホエータンパク質分離物(WPI;Arla、V
iby、Denmark)、ダイズタンパク質分離物(SPI;SUPRO(登録商標)
760、DuPont、Brabrand、Denmark)およびグルテン懸濁液を使
用した。予備加水分解は、Na-カゼイン塩、WPIおよびSPI懸濁液に対し、1%(
w/wタンパク質)のFOODPRO(登録商標)Alkaline Protease
、1%(w/wタンパク質)のFOODPRO(登録商標)PNLを用いて行った。予備
加水分解は全て55℃で18時間行い、その後、95℃で20分間不活化処理を行った。
使用した各PepN 2は、培養ブロスから最終加水分解へのアミノ酸のキャリーオーバ
ー除去の対照として、また、使用したエンドペプチダーゼを十分に不活化するために、9
5℃で15分間不活化した。加水分解は、96ウェルマイクロタイタープレートフォーマ
ット中で行い、下記表1に示すように、50μLのPepN 2原液と混合した150μ
Lの予備加水分解タンパク質懸濁液から構成された。加水分解は50℃で20時間行い、
その後、20μLの2M TCA(トリクロロ酢酸、Sigma-Aldrich、Sc
hnelldorf、Germany)の添加により加水分解を停止させた。ネオサルト
リア・フィッシェリ(Neosartorya fischeri)およびアスペルギル
ス・クラバタス(Aspergillus clavatus)由来のPepN 2型の
アミノ酸遊離効率を表1(B)および(A)に示す。特に、Na-カゼイン塩、WPIお
よびSPIの基質では、ネオサルトリア・フィッシェリ(N.fischeri)および
アスペルギルス・クラバタス(A.clavatus)由来のPepN 2型を使用する
と、非常に高いDHが達成された。さらに、PepN 2型原液濃度で達成されたDHは
、著しい増加であると考えられる。グルテンの加水分解では、0.02~0.3mg/m
L PepN 2のタンパク質範囲でDHが増加することがわかった。

33、TRI034、TRI035、TRI037およびTRI038と組み合わせて使
用したグルテンの加水分解
FOODPRO(登録商標)Alkaline Protease(FPAP)および
FOODPRO(登録商標)PNL(FPPNL)の添加によって予備加水分解したグル
テンの懸濁液を調製した。加水分解は55℃で行った。約18時間後、90℃に10分間
加熱することにより、加水分解を停止させた。50℃に冷却後、グルテンにグルタミナー
ゼを使用した。その後、150μLの予備加水分解したグルテンを96ウェルマイクロタ
イタープレートの各ウェルに移した。50μL(タンパク質濃度:0.5mg/mL)の
TRI032、TRI033、TRI034、TRI035、TRI037およびTRI
038を使用してグルタミン酸の遊離を行った。加水分解は18時間行い、20μLの2
M TCAの添加により停止させた。停止させた加水分解物をろ過(0.22mm)し、
酵素によるグルタミン酸分析キット(Roche、Mannheim、Germany)
によりさらに分析した。遊離したグルタミン酸の濃度を図4に示す。
ペプチドWHWLQLKPGQPMYをTRI031およびTRI035で加水分解し
た。20mMのCPB-バッファー(20mMクエン酸、20mMリン酸塩、20mMホ
ウ酸)中、55℃で、1μg/mLのアミノペプチダーゼと共に、ペプチド(1mg/m
L)をインキュベートした。示された時点で50μlの5%TFAでアリコット(50μ
L)を停止させ、LC-MS分析に供した。
Y(図5)をKPGQPMY(図6)へと、さらにQPMY(図7)へと加水分解できる
ことを示している。これはアスペルギルス・オリゼ(Aspergillus oryz
ae)のPepN 2について以前に記載されていること(A.M.Blinkovsk
y et al.,Biochimica et Biophysica Acta,1
480(2000)171-181、そこでは、アスペルギルス・オリゼ(Asperg
illus oryzae)のPepN 2は、ペプチドWHWLQLKPGQPMYの
ものを含め、調査したいかなるX-Pro結合も加水分解しないと主張された。)とは対
照的である。しかしながら、我々はまた、それらの酵素がペプチドをQPMYに開裂する
ことを見出している(図7)。
hermo Scientific、Bremen、Germany)と接続したAgi
lent 1100 LCシステム(Agilent Technologies)を使
用してキャピラリーLC-MS/MS分析を行った。試料を15cmのPhenomen
ix Jupiter 4μ Proteo 90A、C4分析用カラムに注入した。I
ONMAX(登録商標)イオン源(Thermo Scientific、San Jo
se)への流量16μL/minで、10分間の0~40%溶媒B H2O/CH3CN
/HCOOH(50/950/0.65、v/v/v)の勾配を使用して分離した。LT
Q Orbitrap Classic装置をデータ依存MS/MSモードで操作した。
ペプチド質量をOrbitrap(MSのスキャンは、m/z400において60000
の分解能で行った)により測定し、最大強度ペプチドのm/zを2つまで選択し、リニア
イオントラップ(LTQ)中でCIDを使用して断片化に供した。動的排除は、500質
量のリストサイズ、40秒の持続時間、およびリスト上の質量に対して±10ppmの排
除質量幅により有効とした。
yline 2.6.0.7176(MacLean,B.,et al.,「Skyl
ine:an open source document editor for c
reating and analyzing targeted proteomic
s experiments」,Bioinformatics,2010,26(7)
:p.966-8)を用いて生のファイルにアクセスした(Schilling,B.,
et al.,「Platform-independent and label-f
ree quantitation of proteomic data using
MS1 extracted ion chromatograms in skyl
ine:application to protein acetylation a
nd phosphorylation」,Mol Cell Proteomics,
2012.11(5):p.202-14)。プレカーサー同位体インポートフィルター
を分解能60,000の3つのカウント(M、M+1およびM+2)に設定し、最大強度
の電荷状態を使用した。基質および開裂生成物のペプチド配列を、Skylineに入力
し、それぞれの試料の強度を計算した。図はプログラムから直接コピーした(図8を参照
)。
加水分解度の、アスペルギルス・オリゼ(A.oryzae)およびラクトバチルス・ヘ
ルベティカス(L.helveticus)ATCC12046由来のPepNの加水分
解度との比較
Stressler,Eisele et al.(2013、下記)にしたがって、
PepN 1ラクトバチルス・ヘルベティカス(Lactobacillus helv
eticus)ATCC(登録商標)12046、PepN 1アスペルギルス・オリゼ
(Aspergillus oryzae)、およびアスペルギルス・クラバタス(As
pergillus clavatus)(TRI035)由来のPepN 2の酵素活
性を測定した。ラクトバチルス・ヘルベティカス(L.helveticus)からPe
pN 1を発現させ、Stressler、Eisele et al.(2013、下
記)に記載のように精製した。
ZYME(登録商標)500L(Sigma-Aldrich、Schnelldorf
、Germany)から、20mMのビス-トリス、pH6.5で平衡化したPD10カ
ラムで脱塩した後、精製した。アニオン交換クロマトグラフィーにより、Source
Q15、XK26/15(SQ15)カラム(GE-Lifesciences、USA
)を使用して試料を精製し、20mMのビス/トリス、pH6.5(バッファーA)で平
衡化させた。流量7mL/minで試料(30mL)をカラムに注入した。カラムをバッ
ファーAで洗浄し、結合したタンパク質を、0~0.5MのNaCl(20mM Bis
/Tris(pH6.5)中)の直線勾配により溶出させた(50分間)。全操作期間中
に約13mLの画分を回収し、氷上に保管した。アミノペプチダーゼ活性が最も高い画分
をまとめて、PD10(20mMのBis-Tris(pH6.0)中)で脱塩し、20
mMのBis/Tris、pH6.0(バッファーA)により平衡化したPoros Q
20 HR26/10、XK26/10カラム(GE-Lifesciences、US
A)による2回目の精製に供した。流量4mL/minでPepN1試料をカラムに注入
した。カラムをバッファーAで洗浄し、結合したタンパク質を、0~0.25MのNaC
l(20mM Bis/Tris(pH6.0)中)の直線勾配により溶出させた(30
分間)。全操作期間中に約8mLの画分を回収し、氷上に保管した。精製したアスペルギ
ルス・オリゼ(A.oryzae)由来のPepN 1は、SDS-PAGE電気泳動法
により約40kDaの分子量を有することがわかった。
-1の活性化範囲に標準化した。タンパク質の加水分解のために、1%(w/wWPI)
FOODPRO(登録商標)Alkaline Protease(DuPont、Br
abrand、Denmark)および1%(w/wWPI)FOODPRO(登録商標
)PNL(DuPont、Brabrand、Denmark)を、初期pH7.0でホ
エー乳漿タンパク質分離物懸濁液(WPI;LACPRODAN(登録商標)9224、
Arla Ingredients、Viby、Denmark)に加えた。エンドペプ
チダーゼの添加後、WPIを加え、200μLを96ウェルマイクロタイタープレートの
各ウェルに移した。その後、表2に示すように、5μLのPepN溶液を加えた。加水分
解を50℃、pH無制御で行い、18時間後に20μLの2M TCAを添加して停止さ
せた。DHの測定(Nielsen、Petersen et al.,2001、下記
)に先立って、全試料を0.22μmでろ過した。
ルギルス・オリゼ(A.oryzae)由来のPepN 1より、全ての用量で高いDH
を示し、ラクトバチルス・ヘルベティカス(L.helveticus)由来のPepN
1より、試験した4つの最大活性用量で高いDHを示すことがわかった。そして、2つ
の最大用量で、アスペルギルス・クラバタス(A.clavatus)由来のPepN
2は、PepN 1アミノペプチダーゼより、少なくとも25%高いDHを示した。

ナトリウムの影響
食品工業では、タンパク質の加水分解時の微生物による汚染のリスクを減じるために、
しばしば塩化ナトリウムを使用する。1%(w/wタンパク質)FOODPRO(登録商
標)Alkaline Protease(DuPont、Brabrand、Denm
ark)および1%(w/wタンパク質)FOODPRO(登録商標)PNL(DuPo
nt、Brabrand、Denmark)を添加することにより、予備加水分解した1
0%(w/w)ダイズ(SuPro(登録商標)760、DuPont、Brabran
d、Denmark)およびグルテン(Sigma-Aldrich、Schnelld
orf、Germany)を調製した。加水分解を、50℃、pH7.0(pH無制御)
で18時間行った(18時間後、熱による不活化;90℃;10分)。その後、加水分解
物を、塩化ナトリウム含有(185mM)加水分分解物と塩非含有加水分解物に分割した
。予備加水分解した各タンパク質懸濁液150μLを、表3に示すように、異なる方法で
標準化したアスペルギルス・クラバタス(Aspergillus clavatus)
PepN 2と混合した。50℃、pH無制御で加水分解を行い、18時間後に20μL
の2M TCAを添加して停止させた。DHの測定(Nielsen,Petersen
et al.2001、下記)に先立って、全試料を0.22μmでろ過した。表3に
示すように、185mMのNaClの添加は、ダイズおよびグルテンの加水分解における
加水分解度の測定値に全く影響しなかった。

od Science 66(5):642-646。
。
ス・オリゼ(A.oryzae))の生成物による阻害
この試験では、基質H-Ala-ニトロアニリド(pNA)の加水分解により酵素活性
を測定した。放出されたpNAの波長405nmにおける吸光度を、マイクロタイタープ
レートリーダーを使用して経時的に測定した。
エン酸塩、Na-リン酸塩およびNa-ホウ酸塩)pH9.0を使用した。酵素試料とし
てPepN 2 TRI031およびTRI035を使用した。20mgのH-Ala-
pNA基質(BACHEM、L-1070)を1mLのDMSO(ジメチルスルホキシド
;Sigmaカタログ番号D2650)中で可溶化した。96ウェルプレートのCost
ar assay plate 9017(Corning Inc)を使用し、これら
のプレートをMolecular DevicesのVERSAMAX(登録商標)リー
ダーで読み取った。阻害試験用に、各アミノ酸、すなわちリシン、ヒスチジン、ロイシン
、トリプトファン、プロリン、グリシン、セリン、アスパラギン、トレオニン、アスパラ
ギン酸、グルタミン酸の20mg/mL溶液10mLを調製した。トリプトファンおよび
プロリンはDMSOに溶解した。他のアミノ酸は全て、バッファーに溶解した。500μ
Lの各アミノ酸溶液を混合して、最終の阻害混合物を調製した。
、および20μLの阻害混合物をプレートのウェルに加え、混合した。各プレートには、
0.017、0.085、0.128、0.170、0.426、0.851、1.70
2、2.553mg/mLの阻害混合物濃度で、4種の酵素を全て含有させた。20μL
の基質を添加することにより、反応を開始させた。1つのプレートに、1つの基質濃度を
使用した。反応開始後、30℃に設定したマイクロプレートリーダーにできるだけ早くプ
レートを入れ、405nmにおける吸光度を30秒間隔で30分間測定した。各プレート
に1つの基質濃度(0.017、0.170、0.426、0.851、1.702、2
.553mg/mLのいずれか)とした6枚のプレートについて測定した。見かけの阻害
定数(Ki)を決定するために、データをGraphPad Prismにエクスポート
し、これにより阻害物質の濃度毎にミカエリス・メンテン曲線にフィッティングし、これ
に基づいて、Kiをmg/mlの単位で算出した。阻害物質の平均分子量133.9g/
molを使用して、単位をmMに変換した。異なる酵素についての、生成物による阻害の
みかけのKi値、およびそれぞれの標準偏差(SD)を表4に記す。

し、最大の見かけのKi値を有していることがわかる。それは、アスペルギルス・オリゼ
(A.oryzae)由来のTRI063と比較して、それらが生成物によって阻害され
にくいことを意味している。
1の活性
基質としてH-Glu-ニトロアニリド(pNA)およびH-Gln-ニトロアニリド
(pNA)を使用し、p-ニトロアニリド(pNA)の放出を405nmにおける吸光度
により測定した。バッファーとして20mMのCPB-バッファー(20mMのクエン酸
、20mMのNa-リン酸塩、20mMのホウ酸)pH9.0を使用した。10mgのH
-Glu-pNAおよびH-Gln-pNA(BACHEMまたはSchafer N;
Copenhagen、Denmarkから)を1mlのDMSO(SIGMA製のジメ
チルスルホキシド;カタログ番号D2650)に溶解した。
ing Inc.)中の、80μLのバッファー、10μLのH-Glu-pNAまたは
H-Gln-pNA基質、および10μLの適切に希釈したPepN 2を45℃でイン
キュベートし、SoftMaxPro5.4.1ソフトウエアで稼働するVERSAMA
X(登録商標)マイクロプレートリーダー(Molecular Devices)で、
30秒毎に30分間、405nmで測定した。10の時点から求めた曲線における直線部
分の最大傾斜として、活性を決定した。
epN 1ラクトバチルス・ヘルベティカス(L.helveticus)(Stres
sler、Eisele et al.、上記)、さらに、非精製PepN 1アスペル
ギルス・ソイエ(A.sojae)(COROLASE(登録商標)LAP、AB En
zymes)について行ったが、これらは、Ala-pNAまたはLeu-pNAについ
ては全て同等の活性を有した(実施例HのAla-pNAについての記載と同様ではある
が、阻害物非存在下で行った)。表5に示すように、2種のPepN 2アミノペプチダ
ーゼは、有意な活性を示さなかった2種のPepN 1酵素とは対照的に、Glu-pN
AおよびGln-pNAに対して活性を示した。これは、PepN 2アミノペプチダー
ゼが、GluおよびGlnの放出に関して、PepN 1アミノペプチダーゼよりはるか
に有効であることを示している。

ペプチドライブラリーXPAAAR(Xは全てシステイン以外のアミノ酸)を酵素試料
TRI032、TRI035、TRI063(アスペルギルス・オリゼ(A.oryza
e))およびCOROLASE(登録商標)LAPで加水分解した。3×20mMのCP
B-バッファー(20mMクエン酸、20mMリン酸塩、20mMホウ酸)中、55℃で
1μg/mLのアミノペプチダーゼと共に、ペプチドライブラリーXPAAAR(1mg
/mL)をインキュベートした。示された時点で50μlの5%TFAで50μLのアリ
コットを停止させ、LC-MS分析に供した。
hermo Scientific、Bremen、Germany)と接続したAgi
lent 1100 LCシステム(Agilent Technologies)を使
用してキャピラリーLC-MS/MS分析を行った。試料を15cmのPhenomen
ex Jupiter 4μ Proteo 90A、C4分析用カラムに注入した。I
ONMAX(登録商標)イオン源(Thermo Scientific、San Jo
se)への流量16μL/minで、10分間の0~40%溶媒B H2O/CH3CN
/HCOOH(50/950/0.65、v/v/v)の勾配を使用して分離した。LT
Q Orbitrap Classic装置をデータ依存MS/MSモードで操作した。
ペプチド質量をOrbitrap(MSのスキャンは、m/z400において60000
の分解能で行った)により測定し、最大強度ペプチドのm/zを2つまで選択し、リニア
イオントラップ(LTQ)中でCIDを使用して断片化に供した。動的排除は、500質
量のリストサイズ、40秒の持続時間、およびリスト上の質量に対して±10ppmの排
除質量幅により有効とした。
yline2.6.0.7176(MacLean,B.,et al.、上記)を用い
て生のファイルにアクセスした(Schilling,B.,et al.、上記を参照
)。プレカーサー同位体インポートフィルターを分解能60,000の3つのカウント(
M、M+1およびM+2)に設定し、最大強度の電荷状態を使用した。基質および開裂生
成物のペプチド配列を、Skylineに入力し、それぞれの試料の強度を計算した。3
種の同位体(M、M+1、およびM+2)のピーク面積の合計を基に図10を作成した。
基質標準曲線(Std3.125からStd50へ連続して2倍の濃度増加)を含めてお
り、これは基質濃度とピーク面積に直線関係があることを示している。
AAARを時間とともに加水分解し、2時間のインキュベーションで濃度を半分未満にま
で低下させることができるが、TRI063(アスペルギルス・オリゼ(A.oryza
e))およびCOROLASE(登録商標)LAPでは、12時間でTPAAARの加水
分解を引き起こさないことを示している。
そのような実施形態が例示のためにのみ提供されていることは明らかであろう。今や当業
者には、本発明から逸脱することなく、多くの変形、変更および置換が想起されるであろ
う。本発明を実施するにあたり、本明細書に記載の本発明の実施形態に対し、様々な代替
を使用し得ることは理解されるべきである。以下の請求項が本発明の範囲を定義し、また
、これらの請求項の範囲内の方法および構造、ならびにそれらの等価物が本発明の範囲に
よりカバーされることを意図している。
Claims (9)
- タンパク質性基質を、アミノペプチダーゼ活性を有する単離ポリペプチドに曝すことを含むタンパク質加水分解物の生成方法であって、
前記アミノペプチダーゼ活性を有する単離ポリペプチドは、配列番号1、配列番号2、または配列番号5のアミノ酸配列と少なくとも90%の同一性を有するアミノ酸配列を有し、N末端から番号を付与したときの2の位置にプロリン残基を有するポリペプチドを加水分解する、単離ポリペプチドである、方法。 - 前記タンパク質性基質をエンドペプチダーゼに曝すことをさらに含む請求項1に記載の方法。
- 前記加水分解物はLeu、Gly、Glu、Ser、Asp、Asn、Pro、Cys、Alaおよび/またはGlnに富んでいる請求項1または2に記載の方法。
- 前記加水分解物はGluおよび/またはGlnに富んでいる請求項1または2に記載の方法。
- 前記タンパク質基質はホエータンパク質分離物、カゼイン、ダイズタンパク質分離物、およびグルテンから選択される、請求項1に記載の方法。
- タンパク性基質から、遊離グルタミン酸および/またはグルタミン酸残基を結合したペプチドに富むタンパク質加水分解物を得る方法であって、前記基質を脱アミドプロセスに供し、かつアミノペプチダーゼ活性を有する単離ポリペプチドに曝すことを含む方法であって、
前記アミノペプチダーゼ活性を有する単離ポリペプチドは、配列番号1、配列番号2、または配列番号5のアミノ酸配列と少なくとも90%の同一性を有するアミノ酸配列を有し、N末端から番号を付与したときの2の位置にプロリン残基を有するポリペプチドを加水分解する、単離ポリペプチドである、方法 - 前記基質を1種以上の非特異的に作用するエンドペプチダーゼおよび/またはエキソペプチダーゼ酵素に曝すことをさらに含む請求項6に記載の方法。
- a)穀物を準備する工程、
b)モルト生成プロセス中に前記穀物に、アミノペプチダーゼ活性を有する単離ポリペプチドを有効量添加する工程、および
c)モルト化された穀物を得る工程
を含み、
前記アミノペプチダーゼ活性を有する単離ポリペプチドは、配列番号1、配列番号2、または配列番号5のアミノ酸配列と少なくとも90%の同一性を有するアミノ酸配列を有し、N末端から番号を付与したときの2の位置にプロリン残基を有するポリペプチドを加水分解する、単離ポリペプチドである、モルト化穀物の製造方法。 - 醸造中に遊離アミノ態窒素(FAN)を生成する方法であって、醸造プロセス中に、アミノペプチダーゼ活性を有する単離ポリペプチドを有効量添加する工程を含む方法であって、
前記アミノペプチダーゼ活性を有する単離ポリペプチドは、配列番号1、配列番号2、または配列番号5のアミノ酸配列と少なくとも90%の同一性を有するアミノ酸配列を有し、N末端から番号を付与したときの2の位置にプロリン残基を有するポリペプチドを加水分解する、単離ポリペプチドである、方法。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201562185503P | 2015-06-26 | 2015-06-26 | |
| US62/185,503 | 2015-06-26 | ||
| US201562235937P | 2015-10-01 | 2015-10-01 | |
| US62/235,937 | 2015-10-01 | ||
| JP2017567286A JP2018518976A (ja) | 2015-06-26 | 2016-06-27 | タンパク質加水分解用アミノペプチダーゼ |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2017567286A Division JP2018518976A (ja) | 2015-06-26 | 2016-06-27 | タンパク質加水分解用アミノペプチダーゼ |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2021184723A JP2021184723A (ja) | 2021-12-09 |
| JP7680902B2 true JP7680902B2 (ja) | 2025-05-21 |
Family
ID=56413854
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2017567286A Pending JP2018518976A (ja) | 2015-06-26 | 2016-06-27 | タンパク質加水分解用アミノペプチダーゼ |
| JP2021126487A Active JP7680902B2 (ja) | 2015-06-26 | 2021-08-02 | タンパク質加水分解用アミノペプチダーゼ |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2017567286A Pending JP2018518976A (ja) | 2015-06-26 | 2016-06-27 | タンパク質加水分解用アミノペプチダーゼ |
Country Status (8)
| Country | Link |
|---|---|
| US (1) | US11473073B2 (ja) |
| EP (1) | EP3314004A1 (ja) |
| JP (2) | JP2018518976A (ja) |
| CN (1) | CN107849595A (ja) |
| BR (1) | BR112017028035A2 (ja) |
| CA (1) | CA2990822A1 (ja) |
| MX (1) | MX2017016501A (ja) |
| WO (1) | WO2016210395A1 (ja) |
Families Citing this family (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| BR112019025950A2 (pt) | 2017-06-09 | 2020-09-15 | Novozymes A/S | polipeptídeo, uso e método para hidrolisar proteína |
| WO2021254943A1 (en) * | 2020-06-17 | 2021-12-23 | Dsm Ip Assets B.V. | Umami flavour composition |
| DK202400037Y4 (da) * | 2023-03-31 | 2025-02-12 | Dsm Ip Assets Bv | Hidtil ukendt enzymsammensætning og hidtil ukendt ølbrygningsfremgangsmåde |
| WO2025064561A1 (en) * | 2023-09-18 | 2025-03-27 | Manildra Milling Corporation | Wheat-based emulsifying agent |
| CN119955765B (zh) * | 2025-02-18 | 2025-09-16 | 山西启航合成生物科技有限公司 | 一种l-肌肽及其制备方法和氨肽酶 |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2000504571A (ja) | 1996-02-12 | 2000-04-18 | ギスト ブロカデス ベスローテン フェンノートシャップ | 発酵性麦汁の製造方法 |
| WO2014081700A1 (en) | 2012-11-20 | 2014-05-30 | Codexis, Inc. | Recombinant fungal polypeptides |
Family Cites Families (34)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| NL154598B (nl) | 1970-11-10 | 1977-09-15 | Organon Nv | Werkwijze voor het aantonen en bepalen van laagmoleculire verbindingen en van eiwitten die deze verbindingen specifiek kunnen binden, alsmede testverpakking. |
| US3817837A (en) | 1971-05-14 | 1974-06-18 | Syva Corp | Enzyme amplification assay |
| US3939350A (en) | 1974-04-29 | 1976-02-17 | Board Of Trustees Of The Leland Stanford Junior University | Fluorescent immunoassay employing total reflection for activation |
| US3996345A (en) | 1974-08-12 | 1976-12-07 | Syva Company | Fluorescence quenching with immunological pairs in immunoassays |
| US4275149A (en) | 1978-11-24 | 1981-06-23 | Syva Company | Macromolecular environment control in specific receptor assays |
| US4277437A (en) | 1978-04-05 | 1981-07-07 | Syva Company | Kit for carrying out chemically induced fluorescence immunoassay |
| US4366241A (en) | 1980-08-07 | 1982-12-28 | Syva Company | Concentrating zone method in heterogeneous immunoassays |
| JPS58149645A (ja) | 1982-03-01 | 1983-09-06 | Ajinomoto Co Inc | ゲル化物の製造法 |
| US4816567A (en) | 1983-04-08 | 1989-03-28 | Genentech, Inc. | Recombinant immunoglobin preparations |
| US4683202A (en) | 1985-03-28 | 1987-07-28 | Cetus Corporation | Process for amplifying nucleic acid sequences |
| EP0229046B1 (fr) | 1985-03-30 | 1994-05-04 | BALLIVET, Marc | Procede d'obtention d'adn, arn, peptides, polypeptides ou proteines, par une technique de recombinaison d'adn |
| JPS61279528A (ja) | 1985-06-05 | 1986-12-10 | Hokkai Can Co Ltd | 容器の製造方法 |
| DK122686D0 (da) | 1986-03-17 | 1986-03-17 | Novo Industri As | Fremstilling af proteiner |
| JPH0665280B2 (ja) | 1987-03-04 | 1994-08-24 | 味の素株式会社 | タンパクゲル化剤及びそれを用いるタンパクのゲル化方法 |
| KR100225087B1 (ko) | 1990-03-23 | 1999-10-15 | 한스 발터라벤 | 피타아제의 식물내 발현 |
| DK0531372T4 (da) | 1990-05-09 | 2004-08-09 | Novozymes As | Cellulasepræparat omfattende et endoglucanaseenzym |
| DE4112440C1 (ja) | 1991-04-16 | 1992-10-22 | Diagen Institut Fuer Molekularbiologische Diagnostik Gmbh, 4000 Duesseldorf, De | |
| DE69333718T2 (de) | 1992-01-14 | 2005-12-01 | Ajinomoto Co., Inc. | Gen, das für eine Fisch-Transglutaminase kodiert |
| BR9305768A (pt) | 1992-01-22 | 1997-01-28 | Novo Nordisk As | Fator XIII ativado composiçao e processo para a produçao do fator XIII ativado |
| CA2151154C (en) | 1992-12-10 | 1999-01-26 | William E. Hintz | Production of heterologous proteins in filamentous fungi |
| US5605793A (en) | 1994-02-17 | 1997-02-25 | Affymax Technologies N.V. | Methods for in vitro recombination |
| US5741665A (en) | 1994-05-10 | 1998-04-21 | University Of Hawaii | Light-regulated promoters for production of heterologous proteins in filamentous fungi |
| EP0777726B1 (en) | 1994-08-26 | 2006-06-07 | Novozymes A/S | Microbial transglutaminases, their production and use |
| CA2208730A1 (en) | 1995-01-19 | 1996-07-25 | Novo Nordisk A/S | Transglutaminases from oomycetes |
| CN1163607C (zh) * | 1997-05-16 | 2004-08-25 | 诺沃奇梅兹生物技术有限公司 | 具有氨肽酶活性的多肽及其编码核酸 |
| US6284219B1 (en) * | 1998-06-30 | 2001-09-04 | Phenome Sciences Inc. | In vivo determination of metabolic function for use in therapy management |
| US6881564B1 (en) * | 1999-09-10 | 2005-04-19 | The University Of Sydney | Dipeptidyl peptidases |
| JP2003219888A (ja) * | 2002-02-01 | 2003-08-05 | Kikkoman Corp | アミノペプチダーゼp、アミノペプチダーゼp遺伝子、組み換え体dna及びアミノペプチダーゼpの製造法 |
| US20050256057A1 (en) * | 2002-06-04 | 2005-11-17 | Luppo Edens | Protein hydrolysate rich in tripeptides |
| JP2007502991A (ja) * | 2003-08-20 | 2007-02-15 | バイオサイト インコーポレイテッド | 生物活性ナトリウム利尿ペプチドを測定するための、およびその治療可能性を向上させるための方法および組成物 |
| DK1673457T3 (da) * | 2003-08-25 | 2011-09-05 | Funzyme Biotechnologies Sa | Nye svampeproteiner og nukleinsyrer, der koder for disse |
| MX2008011665A (es) * | 2006-03-13 | 2008-10-08 | Becton Dickinson Co | Diagnóstico y pronóstico de los estados patológicos asociados a la dipeptidil-peptidasa. |
| WO2008150376A1 (en) * | 2007-05-21 | 2008-12-11 | Danisco Us, Inc., Genencor Division | Use of an aspartic protease (nsp24) signal sequence for heterologous protein expression |
| US8592194B2 (en) | 2007-10-09 | 2013-11-26 | Danisco Us Inc. | Glucoamylase variants with altered properties |
-
2016
- 2016-06-27 JP JP2017567286A patent/JP2018518976A/ja active Pending
- 2016-06-27 US US15/574,323 patent/US11473073B2/en active Active
- 2016-06-27 WO PCT/US2016/039494 patent/WO2016210395A1/en not_active Ceased
- 2016-06-27 EP EP16739307.3A patent/EP3314004A1/en active Pending
- 2016-06-27 CA CA2990822A patent/CA2990822A1/en active Pending
- 2016-06-27 MX MX2017016501A patent/MX2017016501A/es unknown
- 2016-06-27 CN CN201680037349.XA patent/CN107849595A/zh active Pending
- 2016-06-27 BR BR112017028035A patent/BR112017028035A2/pt not_active Application Discontinuation
-
2021
- 2021-08-02 JP JP2021126487A patent/JP7680902B2/ja active Active
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2000504571A (ja) | 1996-02-12 | 2000-04-18 | ギスト ブロカデス ベスローテン フェンノートシャップ | 発酵性麦汁の製造方法 |
| WO2014081700A1 (en) | 2012-11-20 | 2014-05-30 | Codexis, Inc. | Recombinant fungal polypeptides |
Non-Patent Citations (2)
| Title |
|---|
| Biochimica et Biophysica Acta, 2000年, Vol.1480, p.171-181 |
| Database NCBI Protein[online], Accession No. XP_003667354, <https://www.ncbi.nlm.nih.gov/protein/367036144?sat=46&satkey=73623056>04-JAN-2012uploaded, [retrieved 25 August 2022], Berka,R.M.,et al. Definition: peptidase [Thermothelomyces thermophila ATCC 42464] |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2016210395A1 (en) | 2016-12-29 |
| JP2021184723A (ja) | 2021-12-09 |
| CA2990822A1 (en) | 2016-12-29 |
| CN107849595A (zh) | 2018-03-27 |
| WO2016210395A8 (en) | 2017-12-07 |
| BR112017028035A2 (pt) | 2018-08-28 |
| MX2017016501A (es) | 2018-03-12 |
| US20180291360A1 (en) | 2018-10-11 |
| US11473073B2 (en) | 2022-10-18 |
| EP3314004A1 (en) | 2018-05-02 |
| JP2018518976A (ja) | 2018-07-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP7680902B2 (ja) | タンパク質加水分解用アミノペプチダーゼ | |
| JP6569662B2 (ja) | 新規タンパク質脱アミド酵素 | |
| Pasternack et al. | Bacterial pro‐transglutaminase from Streptoverticillium mobaraense: Purification, characterisation and sequence of the zymogen | |
| US7098001B2 (en) | Polypeptides having dipeptidyl aminopeptidase activity and nucleic acids encoding same | |
| AU2004266199A2 (en) | Novel fungal proteins and nucleic acids encoding same | |
| CN113646428B (zh) | 具有增加的n-末端氨基酸产率的蛋白质水解产物 | |
| CN108779154B (zh) | 增强的蛋白质产生及其方法 | |
| Ma et al. | Expression, purification and identification of a thermolysin-like protease, neutral protease I, from Aspergillus oryzae with the Pichia pastoris expression system | |
| CN119391735A (zh) | 不具有信号肽的天然分泌多肽的表达 | |
| US9382523B2 (en) | Method of producing a lipolytic enzyme | |
| Chen et al. | Alanine aminopeptidase from Bacillus licheniformis E7 expressed in Bacillus subtilis efficiently hydrolyzes soy protein to small peptides and free amino acids | |
| Li et al. | Functional expression of the keratinolytic serine protease gene sfp2 from Streptomyces fradiae var. k11 in Pichia pastoris | |
| Matsushita-Morita et al. | Overexpression and characterization of an extracellular leucine aminopeptidase from Aspergillus oryzae | |
| Ding et al. | Over-expression of a proline specific aminopeptidase from Aspergillus oryzae JN-412 and its application in collagen degradation | |
| WO2012127002A1 (en) | Sweet-tasting polypeptide from gram-positive bacteria | |
| CN111132556A (zh) | 用于对蛋白进行水解的多肽、用途和方法 | |
| US20110217414A1 (en) | Polypeptides having tyrosinase activity and polynucleotides encoding same | |
| US11447780B2 (en) | Preparation of wheat cysteine protease triticain-alpha produced in soluble form and method of producing same | |
| Wang et al. | Secretory expression, purification, characterization, and application of an Aspergillus oryzae prolyl aminopeptidase in Bacillus subtilis | |
| Song et al. | Functional expression and characterization of a novel aminopeptidase B from Aspergillus niger in Pichia pastoris | |
| Payie et al. | Construction, expression and characterization of a chimaeric mammalian-plant aspartic proteinase | |
| Tanaka et al. | High-level production and purification of clostripain expressed in a virulence-attenuated strain of Clostridium perfringens | |
| JP2023123807A (ja) | アルカリホスファターゼの製造方法及びそれを用いて得られるアルカリホスファターゼ、並びにその製造のためのベクター及び形質転換体 | |
| CN108103045B (zh) | 脂肪酶及其应用 | |
| Baltulionis | Altering the thermal lability properties of industrial proteases |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20210802 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20210802 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20220906 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20221206 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20230221 |
|
| RD03 | Notification of appointment of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7423 Effective date: 20230228 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20250305 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20250509 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7680902 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |