JP5974397B2 - イヌ科動物に感染することができるインフルエンザウイルス、その使用 - Google Patents
イヌ科動物に感染することができるインフルエンザウイルス、その使用 Download PDFInfo
- Publication number
- JP5974397B2 JP5974397B2 JP2012149223A JP2012149223A JP5974397B2 JP 5974397 B2 JP5974397 B2 JP 5974397B2 JP 2012149223 A JP2012149223 A JP 2012149223A JP 2012149223 A JP2012149223 A JP 2012149223A JP 5974397 B2 JP5974397 B2 JP 5974397B2
- Authority
- JP
- Japan
- Prior art keywords
- virus
- seq
- dogs
- canine
- influenza
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images











Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/145—Orthomyxoviridae, e.g. influenza virus
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
- A61P31/16—Antivirals for RNA viruses for influenza or rhinoviruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/02—Immunomodulators
- A61P37/04—Immunostimulants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
- C07K14/08—RNA viruses
- C07K14/11—Orthomyxoviridae, e.g. influenza virus
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N7/00—Viruses; Bacteriophages; Compositions thereof; Preparation or purification thereof
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/569—Immunoassay; Biospecific binding assay; Materials therefor for microorganisms, e.g. protozoa, bacteria, viruses
- G01N33/56983—Viruses
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5252—Virus inactivated (killed)
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5254—Virus avirulent or attenuated
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/54—Medicinal preparations containing antigens or antibodies characterised by the route of administration
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/54—Medicinal preparations containing antigens or antibodies characterised by the route of administration
- A61K2039/541—Mucosal route
- A61K2039/543—Mucosal route intranasal
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/55—Medicinal preparations containing antigens or antibodies characterised by the host/recipient, e.g. newborn with maternal antibodies
- A61K2039/552—Veterinary vaccine
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55555—Liposomes; Vesicles, e.g. nanoparticles; Spheres, e.g. nanospheres; Polymers
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/70—Multivalent vaccine
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16121—Viruses as such, e.g. new isolates, mutants or their genomic sequences
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16134—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2760/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses negative-sense
- C12N2760/00011—Details
- C12N2760/16011—Orthomyxoviridae
- C12N2760/16111—Influenzavirus A, i.e. influenza A virus
- C12N2760/16141—Use of virus, viral particle or viral elements as a vector
- C12N2760/16143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/005—Assays involving biological materials from specific organisms or of a specific nature from viruses
- G01N2333/08—RNA viruses
- G01N2333/11—Orthomyxoviridae, e.g. influenza virus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2500/00—Screening for compounds of potential therapeutic value
- G01N2500/10—Screening for compounds of potential therapeutic value involving cells
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2800/00—Detection or diagnosis of diseases
- G01N2800/12—Pulmonary diseases
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Virology (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Immunology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Microbiology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- Pulmonology (AREA)
- General Engineering & Computer Science (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Epidemiology (AREA)
- Mycology (AREA)
- Biophysics (AREA)
- Communicable Diseases (AREA)
- Hematology (AREA)
- Urology & Nephrology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Oncology (AREA)
- Physics & Mathematics (AREA)
- Gastroenterology & Hepatology (AREA)
- Pathology (AREA)
- Food Science & Technology (AREA)
- Cell Biology (AREA)
Description
本出願は2006年4月21日に出願された米国特許出願第11/409,416号の一部継続出願である;また本出願は2005年10月19日に出願された米国特許出願第60/728,449号、2005年12月29日に出願された60/754,881、2006年1月14日に出願された同60/759,162号、2006年1月23日に出願された同60/761,451号、および2006年3月3日に出願された同60/779,080号に対する優先権を主張し、各出願の開示は簡単な概要、発明の詳細な説明、実施例、特許請求の範囲、要約、図、表、核酸配列、アミノ酸配列および図を含めて、参照によりその全体が本明細書に組み入れられる。
「ケンネルコフ」、つまり感染性気管気管支炎(ITB)は、主として咳を特徴とするイヌの急性伝染性呼吸器感染症である(Fordら、1998年)。イヌITBは世界中で最も蔓延しているイヌの感染性呼吸器疾患の一つと考えられており、イヌが犬舎のような過密集団の環境下で飼育されると大流行が流行割合に達することがある。大半の大流行は、イヌ-イヌ間の直接的な接触または呼吸器分泌物のエアロゾル化に起因する(Fordら、1998年)。臨床徴候は、上下気道の上皮にコロニーを形成する細菌性およびウイルス性物質の一つまたは組み合わせに感染することによって引き起こされる。イヌパラインフルエンザウイルス(CPiV)および気管支敗血症菌(ボルデテラ・ブロンキセプチカ(Bordetella bronchiseptica))は感染したイヌから分離される最も一般的な生物であるが、イヌジステンパーウイルス(CDV)ならびに1型および2型イヌアデノウイルス(CAV-1、CAV-2)などの複数のその他のウイルス、ならびに連鎖球菌(Streptococcus sp.)、パスツレラ・ムルトシダ(Pasteurella multocoda)および大腸菌(Escherichia coli)などの細菌が臨床上の経過および帰結に影響を及ぼし得る(Fordら、1998年)。過密集団では罹患率の高い大流行が極めて効率的および急速に発生するが、合併症である呼吸器感染症および死亡は稀である。生命の危険性のある二次的細菌性肺炎が発症する可能性があるが、ITB症例の大部分は自己限定的であり、何ら治療することなく回復する(Fordら、1998年)。
本発明は、イヌ科に感染してイヌ科に呼吸器疾患を引き起こすことのできる分離されたインフルエンザウイルスに関する。本発明は、本発明のインフルエンザウイルスに対する免疫応答を誘発するための組成物および方法にも関する。本発明は、さらに、本発明のウイルスを同定して、本発明のウイルスの動物への感染を診断するための組成物および方法にも関する。
ワクチンをイヌに投与するための装置、
ワクチンのイヌへの投与を支援する薬学的に許容される賦形剤、
ワクチンに対するイヌの免疫応答を促進する薬学的に許容される賦形剤、
イヌによってワクチンと同時に摂取される食物、および/または
イヌによってワクチンと同時に摂取されるごちそう(treat)。
本発明は、イヌ科に感染して呼吸器疾患を引き起こすことのできる分離されたインフルエンザウイルスに関する。一つの態様において、本発明のインフルエンザウイルスは配列番号:2、4、6、8、10、12、14、16、18、20、22、24、26、28、30、32、33、34、48、50、52、54、56、58、60、62、64、66、68、70、72、74、76もしくは78、またはその機能性および/もしくは免疫原性断片もしくは変異型のいずれかに示されるアミノ酸配列を持つタンパク質をコードするポリヌクレオチドを含む。具体的な態様において、ポリヌクレオチドは配列番号:1、3、5、7、9、11、13、15、17、19、21、23、25、27、29、31、47、49、51、53、55、57、59、61、63、65、67、69、71、73、75もしくは77、またはその断片もしくは変異型のいずれかに示されるヌクレオチド配列を含む。本発明のインフルエンザウイルスは、H1、H2、H3、H4、H5、H6、H7、H8、およびH9、H10、H11、H12、H13、H14、H15もしくはH16のHA亜型、またはN1、N2、N3、N4、N5、N6、N7、N8もしくはN9のNA亜型を含むことができる。具体的な態様において、本発明のインフルエンザウイルスはH3亜型である。ウイルスは、本明細書に開示される方法に従って、感染したイヌから分離して細胞または卵で培養することができる。一つの例示的態様において、インフルエンザウイルスはインフルエンザAウイルスである。
Tm=81.5 C+16.6 Log[Na+]+0.41(%G+C)-0.61(%ホルムアミド)-600/塩基対の二本鎖の長さ。
(1)1×SSPE、0.1%SDSにおいて室温で15分ずつ2回(低ストリンジェントな洗浄)
(2)0.2×SSPE、0.1%SDSにおいてTm-20Cで15分間1回(中ストリンジェントな洗浄)。
グレイハウンドからの血液および鼻腔スワブの採取
呼吸器疾患の大流行が発生しているレース用犬舎の臨床的に疾患または正常なグレイハウンドから、頚静脈穿刺により急性期および回復期の血液試料を採取した。回復期試料は急性試料の4〜12週後に採取した。血清を回収し、-80℃で保存した。鼻腔スワブを採取して、細菌分離のための提出まで活性炭を加えたAmies輸送培地(Becton Dickinson Biosciences)中に置いた。
2004年1月のフロリダトラックでの大流行において死亡した8頭中5頭のグレイハウンドについて、University of Florida College of Veterinary Medicine(UF CVM)のAnatomic Pathology Serviceにて徹底した死亡後検査を実施した。もう1頭のイヌの死亡後検査は個人の動物病院で実施されて、組織を病理組織学的診断のためにUF CVMに提出した。組織は10%中性緩衝ホルマリン中で固定して、パラフィンに包埋し、5μmの切片を病理組織学的診断のためのヘマトキシリン・エオジンで染色するか、または以下に記載する通り免疫組織化学的検査に備えて加工した。未固定の組織は細菌培養に提出して、同じく-80℃で保存した。
急性期および回復期の対の血清試料は、イヌジステンパーウイルス、2型アデノウイルスおよびパラインフルエンザウイルスに対する血清中和試験のため、Cornell University College of Veterinary MedicineのAnimal Health Diagnostic Laboratory(AHDL)に送付した。抗体価は、細胞培養物のウイルス感染を阻害する血清の最終希釈として表される。急性期および回復期試料間の抗体価の≧4倍の増加として定義される血清変換はウイルス感染を示した。これらのウイルス性病原体に対する血清変換は検出されなかった。
対の鼻腔スワブおよび死亡後組織は、細菌の分離および同定のためにUF CVMのDiagnostic Clinical Microbiology/Parasitology/Serology Serviceに送付した。試料は、非選択培地ならびにボルデテラ属(Regan-Lowe; Remel)およびマイコプラズマ属(Remel)の選択培地で培養した。すべての培養物は、増殖なしと報告する前に21日間維持した。数頭のグレイハウンドから得られた鼻腔スワブは、細菌培養のためにKansas State University College of Veterinary MedicineのDepartment of Diagnostic Medicine/Pathobiologyにも送付した。臨床的に疾患が認められて試験が行われた70頭のイヌの内、気管支敗血症菌(Bordetella bronchiseptica)が1頭のイヌの鼻腔から分離されて、マイコプラズマ属は33頭のイヌの鼻腔から回収された。パスツレラ・ムルトシダは、膿性鼻分泌物の見られるイヌの鼻腔から広く回収された。2004年1月の大流行で死亡したイヌの内の2頭では死亡後に肺に若干の大腸菌の増殖が認められ、1頭では大腸菌およびストレプトコッカス・カニス(Streptococcus canis)の若干の増殖が見られ、またもう1頭では緑膿菌(Pseudomonas aeruginosa)および酵母の若干の増殖が認められた。死亡したイヌの気管または肺からは、気管支敗血症菌およびマイコプラズマは分離されなかった。
凍結組織を融解して、0.5%ウシ血清アルブミン(BSA)および抗生物質を強化した10倍量の最小必須培地(MEM)を加えてホモジナイズした。遠心分離により固体の細片を除去して、上清を培養細胞または10日齢の孵化鶏卵に接種した。死亡したグレイハウンド由来の組織ホモジネートを、広範囲のウイルス性病原体の複製を支持する種々の細胞培養物に接種した。細胞培養物には、Vero(アフリカミドリザル腎上皮細胞、ATCC No. CCL-81)、A-72(イヌ腫瘍線維芽細胞、CRL-1542)、HRT-18(ヒト直腸上皮細胞、CRL-11663)、MDCK(イヌ腎上皮細胞、CCL-34)、イヌ一次腎上皮細胞(AHDL, Cornell University)、イヌ一次肺上皮細胞(AHDL)、およびウシ一次精巣細胞(AHDL)が含まれた。MDCKおよびHRT細胞は2.5μg/mL TPCK処理トリプシン(Sigma)を強化したMEMで培養し、残りの細胞株は10%ウシ胎児血清および抗生物質を強化したMEMで培養した。細胞は、25cm2のフラスコを用いて5%CO2含有加湿大気中、37℃で培養した。対照培養には強化MEMを接種した。培養物は形態学的変化を1日1回観察して、接種後5日に回収した。回収した液体および細胞は遠心分離により清澄化して、最初の接種において記載した通り、新鮮な細胞に接種し;2回の盲目継代を行った。清澄化した上清の血球凝集活性は、記載される通り(Burlesonら、1992年;Kendalら、1982年)、ニワトリまたはシチメンチョウの赤血球を用いて測定した。鶏胚を用いたウイルス分離では、組織ホモジネートの0.1mLを尿膜嚢に接種して、35℃にて48時間、インキュベートした。2回の盲目継代後、尿膜腔液の血球凝集活性を、記載される通り(Burlesonら、1992年;Kendalら、1982年)、測定した。
製造業者の説明書に従ってRNeasyキット(Qiagen, Valencia, CA)を用いて、組織培養上清または尿膜腔液から全RNAを抽出した。全RNA(10ng)は、一段階RT-PCRキット(Qiagen, Valencia, CA)を製造業者の説明書に従って使用して、cDNAに逆転写した。cDNA内の8つのインフルエンザウイルス遺伝子のコード領域のPCR増幅は、汎用遺伝子特異的プライマーセットを用いて既報(Klimovら、1992a)の通り実施した。得られたDNA単位複製配列は、サイクルシーケンシングダイターミネーターケミストリー(ABI)を用いたApplied Biosystems 3100自動DNAシーケンサーでの自動シーケンシングのための鋳型として用いられた。ヌクレオチド配列は、GCG Package(著作権)、バージョン10.0(Accelyrs)(Womble, 2000)を用いて解析した。ヌクレオチド配列からの系統発生の推定およびブーツストラップの算出にはPhylogeny Inference Package(著作権)バージョン3.5を用いた(Felsenstein、1989年)。系統樹は、MEGA(著作権)プログラム(Kumarら、2004年)において実行されたTamura-Neiガンマモデルを用いて近隣結合法により得られた系統樹と比較して、PAUP(著作権)4.0ベータプログラム(Sinauer Associates)により確認した。
6カ月齢の4頭の特定病原体除去ビーグル犬[(雌雄各2頭(Liberty Research)]を使用した。理学的検査、ならびに詳細な血球計測/型別計測を含むベースラインの血液検査、血清化学的検査パネル、および尿検査より、動物が健常であることが明らかとなった。動物は、実験動物管理認定協会より認定されたBSL 2-強化施設内で群飼いした。ベースラインの直腸温度は1日2回、7日間、記録した。イヌは、気管内チューブの挿管に備えて、プロポフォール(ディプリバン(登録商標)、Zeneca Pharmaceuticals、0.4mg/kg体重〜効果)の静脈内注入により麻酔した。各イヌにA/イヌ/フロリダ/43/2004(イヌ/FL/04)(H3N8)ウイルスを106.6の半数組織培養物感染用量(TCID50)の総用量で接種して、半量は気管内チューブを介して気管末端に投与して、残りの半量はカテーテルを介して深部鼻気道に投与した。理学的検査および直腸温の記録は、1日2回、接種後(p.i.)14日間実施した。p.i.0、3、5、7、10および14日に頚静脈穿刺により血液試料(4mL)を採取した。鼻腔および口咽頭の標本は、各イヌからp.i.0〜5、7、10および14日にポリエステルスワブ(Fisher Scientific)を用いて採取した。スワブはウイルス輸送培地(Remel)に回収して、-80℃で保存した。2頭のイヌ(雌雄各1頭)はp.i.5日にBeuthanasia-D(登録商標)液(1mL/5kg体重;Schering-Plough Animal Health Corp)の静脈内接種により安楽死させて、残りの2頭は14日目に死亡後検査のために安楽死させた。組織学的検査のための組織は既述の通り加工した。ウイルス培養用の組織は-80℃で保存した。この試験は、University of Florida Institutional Animal Care and Use Committeeにより承認された。
肺ホモジネート、および遠心分離によるスワブ輸送培地の清澄化により調製したスワブ抽出液の段階希釈は、0.5%BSAおよび抗生物質を強化したMEMで調製した。プラークアッセイ法は、6穴組織培養プレート中の単層のMDCK細胞を用いて既報(Burlesonら、1992年)の通り実施した。接種した単層細胞層に0.8%アガロースおよびTPCK-トリプシン 1.5μg/mLを含む強化MEMを重層した。細胞は、固定およびクリスタルバイオレットでの染色前に、5%CO2を含む加湿大気中で37℃にて72時間、培養した。ウイルス濃度は、組織1グラム当たりまたはスワブ当たりのプラーク形成単位(PFU)として表した。
グレイハウンドおよびビーグル犬由来の脱パラフィンおよび再水和した5μm肺組織切片をBond-Rite(商標)スライドガラス(Richard-Allan Scientific, Kalamazoo, MI)にセットして、プロテイナーゼK(DakoCytomation, Carpenteria, CA)、ペルオキシダーゼ遮断試薬(Dako(登録商標)En Vision(商標)Peroxidase Kit, Dako Corp.)で順次処理した。切片を、イヌジステンパーウイルス(VMRD, Inc.)、2型イヌアデノウイルス(VMRD, Inc.)、イヌパラインフルエンザウイルス(VMRD, Inc.)、またはインフルエンザA H3(Chemicon International, Inc.)に対するモノクローナル抗体の1:500希釈と共に室温で2時間インキュベートした。対照には、同一の切片とマウスIgG(1mg/mL、Serotec, Inc.)のインキュベート、およびモノクローナル抗体と通常のイヌ肺切片のインキュベートが含まれた。一次抗体で処理した後、切片は製造業者の説明書に従って二次イムノペルオキシダーゼおよびペルオキシダーゼ基質試薬(Dako(登録商標)EnVision(商標)Peroxidase Kit, Dako Corp.)と共にインキュベートした。切片をヘマトキシリンで対比染色して、清澄化剤#2およびBluing試薬(Richard-Allan Scientific, Kalamazoo, MI)で処理して脱水し、Permount(ProSciTech)を用いてカバーガラスをかけた。
血清試料は、56℃での60分間の熱失活前に、レセプター破壊酵素(RDE, Denka)(血清1容積:RDE 3容積)と共にインキュベートした。インフルエンザA/イヌ/FL/04(H3N8)ウイルスはMDCK細胞を用いて37℃にて36〜48時間、増殖させた。ウイルス培養の上清を回収して、遠心分離により清澄化して-80℃で保存した。HIアッセイ法は既報(Kendalら、1982年)の通り実施した。つまり、25μl中4血球凝集単位のウイルスをマイクロタイターウェルの段階希釈した等量の血清に加えて、室温で30分間、インキュベートした。等量の0.5% v/vシチメンチョウ赤血球を加えて、30分後に血球凝集力価を目視により求めた。エンドポイントのHI力価は、血球凝集を完全に阻害する血清の最終希釈として定義された。血清変換は、急性期および回復期の対の試料間におけるHI力価の≧4倍の増大として定義された。単一試料の血清陽性は≧1:32のHI抗体価として定義された。
A/イヌ/FL/04(H3N8)に対する血清抗体反応の中和は、既報のMNアッセイ法(Roweら、1999年)により検出された。但し、イヌ血清はアッセイ前に上記のようにRDE処理した。エンドポイントの力価は、100 TCID50のウイルスを50%中和する血清の最高希釈として定義された。血清変換は、急性期および回復期の対の試料間におけるMN力価の≧4倍の増大として定義された。単一試料の血清陽性は≧1:80のMN力価として定義された。
2004年1月に、フロリダのトラックの2箇所の犬舎およびこれらの犬舎にイヌを供給する地元飼育場で飼育されている22頭のレース用グレイハウンドにおいて、呼吸器疾患の大流行が発生した。各犬舎建物には約60頭、飼育場には約300頭のイヌがいた。大流行は6日間起こって、その後、新しい症例は特定されなかった。22頭中14頭のイヌが39.5〜41.5℃の発熱を示し、10〜14日間にわたって咳をして、最終的に回復した。残りの8頭の内、見かけ上健康であった6頭のイヌは予想に反して死亡して、口および鼻から出血が見られた。その他の2頭のイヌは、急激な悪化のために、口および鼻からの出血発生の24時間以内に安楽死させた。これらの双方のイヌは41℃の熱があった。8頭中4頭の死亡が犬舎施設で発生して、4頭は飼育場で発生した。死亡の50%が大流行の3日目に発生した。22頭のイヌは17カ月齢から4歳であったが、73%は17〜33カ月齢であった。
グレイハウンドの臨床的および病理学的観察所見におけるイヌ/FL/04ウイルスの役割について検討するため、本発明者らはインフルエンザA H3に対するモノクローナル抗体を用いて肺組織の免疫組織化学的染色(IHC)を実施した。ウイルス性H3抗原は気管支および細気管支の上皮細胞、気管支腺上皮細胞の細胞質に一貫して検出され、またマクロファージは気道腔および肺胞腔に検出された(図2A)。これらのデータは、多くのイヌにおけるH3亜型のインフルエンザウイルスの肺感染の診断を支持する。
呼吸器疾患大流行の病因論におけるイヌ/FL/04様ウイルスの関与について調べるため、本発明者らは血球凝集阻害(HI)およびマイクロ中和(MN)により病気のイヌ11頭および無症状の接触犬16頭から採取した急性期および回復期の対の血清を分析した。急性期から回復期までのイヌ/FL/04に対する抗体価の≧4倍の増加として定義される血清変換が、両アッセイ法において、11頭中8頭(73%)の病犬で発生した(表1)。血清変換はHIアッセイ法では16頭中6頭(38%)の無症状接触犬で発生し、MNアッセイ法では16頭中8頭(50%)が血清変換を示した(表1)。血清変換のデータは、イヌのイヌ/FL/04様ウイルスの感染が大半の動物において呼吸器疾患の発症と時間的に一致することを実証した。
イヌ/FL/04ウイルスのイヌへの感染能力についてより理解するために、4頭の6カ月齢の純系ビーグル犬に106.6半数組織培養物感染用量(TCID50)を気管内または鼻腔内経路によりそれぞれ接種した。すべてのイヌが接種後(p.i.)最初の2日間に発熱(直腸温 ≧39℃)を発症したが、14日間の観察期間を通して咳または鼻分泌物などの呼吸器症状を示したものはいなかった。ウイルス排出は、鼻および口咽頭のスワブにおけるウイルスの定量によって調べた。4頭中2頭のイヌのみが検出可能量のウイルスを排出した。1頭のイヌはp.i.1日および2日にウイルスを排出し(スワブ当たり1.0〜2.5 log10 PFU)、その他のイヌは接種後連続4日間にわたってウイルスを排出した(スワブ当たり1.4〜4.5 log10 PFU)。p.i. 5日における2頭のイヌの死亡後検査より、グレイハウンドにおける自然発生性疾患に見られる所見とほぼ等しい壊死性増殖性気管炎、気管支炎および細気管支炎が認められたが、肺出血または気管支肺炎は認められなかった。IHCにより、ウイルスH3抗原が気管支、細気管支および気管支腺の上皮細胞の細胞質に検出された(図2B)。感染性ウイルスが1頭のイヌの肺組織から回収された。p.i.14日における残りの2頭の死亡後検査より、呼吸組織における最小限の組織学的変化、IHCによるウイルスH3抗原の不在、および肺ホモジネートからのウイルスの無回収が確認された。これらの後者の2頭のイヌにおける血清変換は、MNアッセイ法においてp.i.7日に検出され、14日までに抗体価はさらに2〜3倍に増加した。これらの結果より、熱性反応、肺実質におけるウイルス抗原および感染性ウイルスの存在、インフルエンザに典型的な病理組織学的所見、ならびに血清変換に示されるように、イヌ/FL/04感染に対するイヌの感受性が確立された。実験的に接種したビーグル犬において重度の疾患および死亡を再現できなかったことは、自然感染したグレイハウンドの大集団が無症候性であったことを考えると、驚くべきことではない。
2004年1月の大流行以前にイヌ/FL/04様インフルエンザウイルスがフロリダのグレイハウンド集団で広まっていたかどうかを調べるため、65頭のレース用グレイハウンドに由来する保存血清についてHIおよびMNアッセイ法を用いてイヌ/FL/04に対する抗体の存在を調べた。1996年〜1999年に試料採取した33頭のイヌでは、検出可能な抗体はなかった。2000年〜2003年に試料採取した32頭の内、9頭は双方のアッセイ法において血清陽性であった−2000年に1頭、2002年に2頭、および2003年に6頭(表5)。血清陽性のイヌは1999年から2003年まで病因不明の呼吸器疾患の大流行に関与したフロリダのトラックで飼育されており、イヌ/FL/04様ウイルスがこれらの大流行の原因物質であった可能性があることを示唆する。この可能性をさらに調べるために、本発明者らは2003年3月に出血性気管支肺炎のために死亡したグレイハウンドから採取された保存組織を調べた。1頭のイヌに由来する肺ホモジネートをMDCK細胞および鶏胚に接種したところ、H3N8インフルエンザウイルスが回収されて、A/イヌ/フロリダ/242/2003(イヌ/FL/03)と命名された。イヌ/FL/03の完全ゲノムの配列解析はイヌ/FL/04に対して>99%同一性を示し(表4)、イヌ/FL/04様ウイルスが2004年以前にグレイハウンドに感染していたことを示す。
2004年6月から8月にフロリダ、テキサス、アラバマ、アーカンソー、ウェストバージニアおよびカンザスの14箇所のトラックで何千頭ものレース用グレイハウンドに呼吸器疾患の大流行が発生した。
b 臨床的疾病犬と接触して飼育されている無症候性のイヌの数。
c A/イヌ/フロリダ/43/2004ウイルスを用いた血球凝集阻害(HI)アッセイ法。
d A/イヌ/フロリダ/43/2004ウイルスを用いたマイクロ中和(MN)アッセイ法
e 急性期および回復期の対の血清の抗体価が少なくとも4倍に増加したイヌのパーセント。
f 回復期血清において陽性抗体価(HI抗体価 ≧32:MN抗体価 ≧80)を伴うイヌのパーセント。
g 回復期血清の抗体価幾何学平均。
† 共通のウマH3 HAからの変化なしを示す。
b ポリクローナル抗血清はフェレットにおいて作出されたが、その他のすべての抗血清はヒツジまたはヤギにおいて作出された。
b 該当なし:N8ノイラミニダーゼはヒトまたはブタのウイルスではこれまで報告されなかった。
b 6頭の2003年血清陽性犬の範囲を含む、血清陽性犬におけるマイクロ中和試験抗体価。
b 急性期および回復期血清の抗体価における≧4倍の増加を示すイヌのパーセント。
c 回復期血清において陽性の抗体価(HI抗体価>16)を示したイヌのパーセント。
d 回復期血清における抗体価幾何学平均
イヌの組織
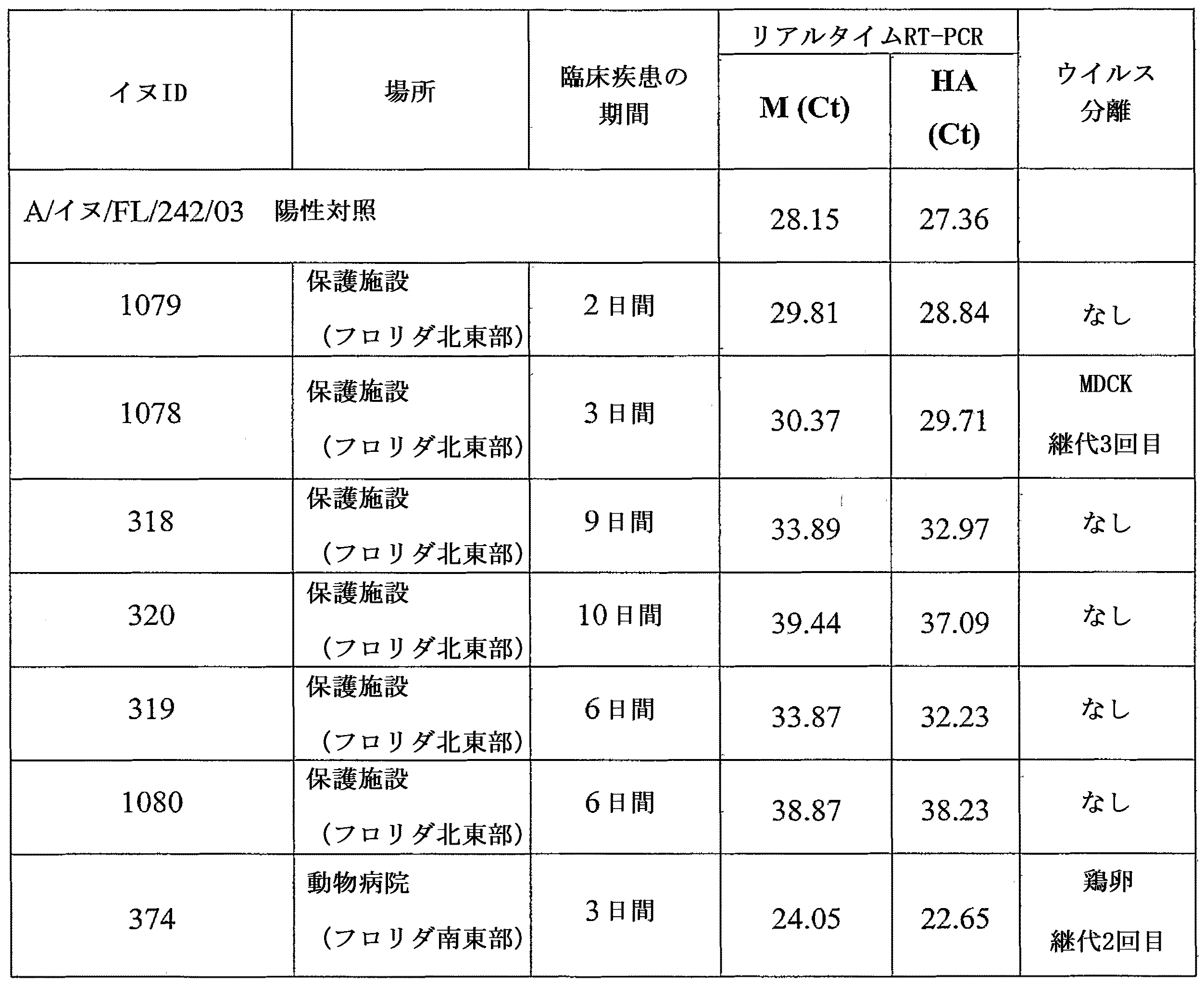
フロリダ北東部の保護施設におけるインフルエンザ大流行で2005年4月/5月に死亡した6頭の雑種犬およびフロリダ南東部の動物病院でのインフルエンザ大流行において2005年5月に死亡した1頭のペットのヨークシャーテリアについて、University of Florida College of Veterinary MedicineのAnatomic Pathology Serviceにて死亡後検査を実施した。組織は10%中性緩衝ホルマリン中で固定して、パラフィンに包埋し、病理組織学的診断のための5μmの切片をヘマトキシリン・エオジンで染色した。未固定の組織は、ウイルス学的分析まで-80℃で保存した。
7頭の各イヌに由来する凍結肺組織を融解して、使い捨ての組織グラインダー(Kendall, Lifeline Medical Inc., Danbury, CT)を用いて、0.5%ウシ血清アルブミン(BSA)および抗生物質(ゲンタマイシンおよびシプロフロキサシン)を強化した最小必須培地(MEM)を加えてホモジナイズした。市販のキット(RNeasy(登録商標)Mini Kit, QIAGEN Inc., Valencia, CA)を製造業者の説明書に従って用いて全RNAを抽出し、最終液量 60μLの緩衝液で溶出した。呼吸器疾患を伴わないイヌから採取した肺組織からも、全RNAを抽出した。
ROXをパッシブレファレンス色素として含むQuantiTect(登録商標)プローブRT-PCRキット(QIAGEN Inc., Valencia, CA)を用いて、イヌ組織試料から抽出された全RNAに対して一段階定量的リアルタイムRT-PCRを実施した。つまり、2つのプライマー-プローブセットを各試料中のインフルエンザA配列の検出に用いた(表7)。1つのプライマー-プローブセットはイヌヘマグルチニン(H3)遺伝子配列に選択的であった。他方のプライマー-プローブセットはA型インフルエンザウイルスのマトリックス(M)遺伝子の高度に保存的領域をターゲットとした。各リアルタイムRT-PCR反応において、最終液量 25μL中、2X QuantiTech(登録商標) Probe RT-PCR Master Mix 12.5μL、QuantiTech(登録商標)RT Mix 0.25μL、フォワードおよびリバースプライマー(それぞれ、0.4μMの最終濃度)、プローブ(0.1μMの最終濃度)、ならびにRNaseフリーの水を含む反応混合液に抽出した全RNA 5μLを加えた。イヌ組織試料から抽出されたRNAの存在に関する内因性の内部対照である18S rRNAの検出には、TaqMan(登録商標)リボソームRNA対照試薬(Applied Biosystems, Foster City, CA)を製造業者の説明書に従って使用した。
7頭の各イヌに由来する凍結肺組織を融解して、0.5%(BSA)および抗生物質(ゲンタマイシンおよびシプロフロキサシン)を強化した10倍量のDulbecco修正イーグル培地(DMEM)を加えてホモジナイズした。遠心分離により固体の細片を除去して、上清を1μg/mL TPCK処理したトリプシン(Sigma-Aldrich Corp., St. Louis, MO)および抗生物質(ゲンタマイシンおよびシプロフロキサシン)を加えたDMEMで培養したMadin-Darbyイヌ腎(MDCK)細胞に接種した。細胞は、25cm2のフラスコを用いて5%CO2含有加湿大気中、37℃にて培養した。培養物は形態学的変化を1日1回観察して、接種後5日に回収した。回収した培養物を遠心分離により清澄化して、上清を初回接種において記載した通り新鮮なMDCK細胞に接種し;血球凝集またはRT-PCRによりインフルエンザウイルスの証拠を示さなかった試料についてはさらに2回の継代を行った。清澄化した上清の血球凝集活性は、既報の通り(Burleson, F.ら、1992年;Kendal, P.ら、1982年)、0.5%シチメンチョウ赤血球を用いて測定した。RT-PCRは、下記の通り、実施した。
ホモジネートは、MDCK細胞の接種に関して上記の通り、凍結肺組織から調製した。ホモジネート(0.2mL)を10日齢の孵化鶏卵の尿嚢に接種した。35℃にて48時間インキュベート後、尿膜腔液の回収前に一晩、卵を4℃で冷却した。清澄化した上清の血球凝集活性は、既報の通り(Burleson, F.ら、1992年;Kendal, P.ら、1982年)、0.5%シチメンチョウ赤血球を用いて測定した。RT-PCRは、下記の通り、実施した。初回接種後にインフルエンザウイルスの証拠を示さなかった試料については、孵化鶏卵を用いたさらに2回の継代を行った。
製造業者の説明書に従ってQIAamp(登録商標)ウイルスRNAミニキット(QIAGEN Inc., Valencia, CA)を用いて、MDCK上清または尿膜腔液からウイルスRNAを抽出した。ウイルスRNAは、QIAGEN(登録商標)一段階RT-PCRキット(QIAGEN Inc., Valencia, CA)を製造業者の説明書に従って使用して、cDNAに逆転写した。cDNA内の8つのインフルエンザウイルス遺伝子のコード領域のPCR増幅は、汎用遺伝子特異的プライマーセット(プライマー配列は要求に応じて与えられる)を用いて既報(Klimov, A.ら、1992b)の通り実施した。得られたDNA単位複製配列は、サイクルシーケンシングダイターミネーターケミストリー(Applied Biosystems, Foster City, CA)を用いたABI PRISM(登録商標)3100自動DNAシーケンサーでの自動シーケンシングのための鋳型として用いられた。ヌクレオチド配列はLasergene 6 Package(登録商標)(DNASTAR, Inc., Madison, WI)を用いて解析した。ヌクレオチド配列からの系統発生の推定およびブーツストラップの算出にはPHYLIPバージョン3.5(著作権)ソフトウェアプログラムを用いた(Felsenstein, J.、1989年)。系統樹は、MEGA(著作権)プログラム(Kumar, S.ら、2004年)において実行されたTamura-Neiモデルを用いて近隣結合法により得られた系統樹と比較して、PAUP(著作権)4.0ベータプログラム(Sinauer Associates, Inc., Sunderland, MA)により確認した。
血清試料は、56℃での30分間の熱失活前に、レセプター破壊酵素(RDE、デンカ生検株式会社、東京、日本)(血清1容積:RDE 3容積)と共にインキュベートした。インフルエンザA/イヌ/ジャクソンビル/05(H3N8)ウイルスはMDCK細胞を用いて5%CO2中、37℃にて72時間、増殖させた。ウイルス培養の上清を回収して、遠心分離により清澄化して-80℃で保存した。HIアッセイ法で用いられるその他のウイルスはすべて、10日齢の孵化鶏卵で培養し、尿膜腔液を回収して-80℃で保存した。HIアッセイ法は既報(Kendal, P.ら、1982年)の通り実施した。つまり、25μl中4血球凝集単位のウイルスを96穴プラスチックプレートにて段階希釈した等量の血清に加えて、室温で30分間、インキュベートした。等量の0.5%シチメンチョウ赤血球を加えて、30分後に血球凝集力価を目視により求めた。エンドポイントのHI力価は、血球凝集を完全に阻害する血清の最終希釈として定義された。
2005年4月および5月に、既報(Crawford, P.C.ら、2005年)の呼吸器疾患大流行がフロリダ北東部の保護施設に収容されていたイヌで発生した。この大流行には3カ月齢から9歳までの齢期範囲の少なくとも58頭が関与し、純系のイヌおよび雑種犬が含まれた。最も一般的な臨床徴候は、7〜21日間にわたる膿性鼻分泌物および咳であった。≧7日にわたって臨床疾患が認められた43頭の内、41頭はイヌ/FL/04(H3N8)に対して32から>1024のHI抗体価を示した。少なくとも10頭の犬が肺炎に進行し、この内、6頭は安楽死させた。これらの6頭の雑種犬は、4カ月齢から3歳までの雌雄各3頭であった。臨床徴候の期間は、安楽死の時点で2〜10日であった。死亡後検査において、これらのイヌは肺のうっ血および浮腫を示した。呼吸器の組織学的検査では、鼻炎、気管炎、気管支炎、細気管支炎および化膿性気管支肺炎が認められた。気管、気管支、細気管支および気管支腺に、上皮細胞の壊死およびびらんが認められた。呼吸組織に好中球およびマクロファージの浸潤が見られた。
7頭のイヌから得られた肺組織を、A型インフルエンザのM遺伝子およびイヌH3N8インフルエンザAウイルスのH3遺伝子を検出する定量的リアルタイムRT-PCRアッセイ法で解析した。7頭すべてのイヌの肺がインフルエンザAのM遺伝子およびイヌインフルエンザH3遺伝子の双方に関して陽性であった(表8)。MDCK細胞で3回継代後、3日間の肺炎の後に死亡した保護施設のイヌの肺からインフルエンザAのH3N8亜型のウイルスが分離された。このウイルスはA/イヌ/ジャクソンビル/05(H3N8)(イヌ/Jax/05)と命名された。孵化鶏卵で2回継代後、同じく3日間の肺炎の後に死亡したペットのイヌの肺からインフルエンザAのH3N8亜型のウイルスが分離された。このウイルスはA/イヌ/マイアミ/05(H3N8)(イヌ/マイアミ/05)と命名された。
イヌ/Jax/05およびイヌ/マイアミ/05の配列解析の結果、それらのヘマグルチニン(HA)遺伝子は2004年および2005年のトラックでのインフルエンザ大流行時に肺炎で死亡したレース用グレイハウンドの肺から回収されたイヌ/FL/04、イヌ/TX/04およびイヌ/アイオワ/05分離物と98%一致することが明らかとなった(Crawford, P.C, ら、2005年;Yoon K-Yら、2005年)。さらに、イヌ/Jax/05およびイヌ/マイアミ/05のHA遺伝子は、2000年以降に分離された現代のウマインフルエンザウイルスと98%一致した。HA遺伝子の系統学的比較では、イヌ/Jax/05およびイヌ/マイアミ/05ウイルスはイヌ/FL/04、イヌ/TX/04、およびイヌ/アイオワ/05のグレイハウンド分離物、ならびに現代のウマ分離物とクラスターを形成し、1990年代初期に分離された古いウマウイルスとは異なる群を形成することが示された(図4)。さらに、イヌ/Jax/05、イヌ/マイアミ/05およびイヌ/アイオワ/05分離物は、イヌ/FL/04またはイヌ/FL/03よりもイヌ/Tx/04により密接に関連した。2005年の分離物は先の2003年および2004年イヌウイルスから枝分れしたと思われる亜型を形成し、約10箇所の節減的に情報価値のある部位が異なる。これらの相違は、外部の感染源から定期的に再導入されるのとは対照的に、イヌからイヌへと水平伝播するという仮説を支持する。2003年から2005年までの突然変異の蓄積は、イヌインフルエンザウイルスに生じたと予想されるように、ウイルスが新しい宿主に伝達された後に受けなければならない適応の進行中の過程を例示するものである。
6検体のすべてのイヌ分離物には、それらを現代のウマインフルエンザと区別する保存的アミノ酸置換があった(表9)。これらの保存的置換はI15M、N83S、W222L、I328TおよびN483Tであった。成熟HAタンパク質を系統学的に比較した結果、イヌ/Jax/05、イヌ/マイアミ/05およびイヌ/アイオワ/05ウイルスはイヌ/TX/04分離物と共に亜型を形成することが示された(図4)。この亜型とその他のイヌウイルスを区別する3つのアミノ酸変化(L118V、K261NおよびG479E)があった(表9)。2005年の分離物をそれらのルーツであるイヌ/TX/04と区別する2つのアミノ酸変化(F79LおよびG218E)があった。さらに、グレイハウンド以外のイヌからの2005年分離物であるイヌ/Jax/05およびイヌ/マイアミ/05は1個のアミノ酸変化であるR492Kがイヌ/アイオワ/05グレイハウンド分離物と異なった。最後に、イヌ/Jax/05は一つのアミノ酸S107Pにおいてイヌ/マイアミ/05とは異なった。その他のすべてのH3N8ウマおよびイヌウイルスにおいて、Tを持つA/ウマ/チーリン/1/89(Guo Y.ら、1992年)を除いてSは107の位置に保存される。
血球凝集阻害(HI)試験は、以前および現代のウマインフルエンザウイルスおよびイヌインフルエンザウイルス、ならびにインフルエンザウイルスに感染したウマおよびイヌから2005年に採取された血清を用いて実施された(表10)。イヌ/FL/04に対して免疫したフェレットの血清も分析に含めた。ウマ血清のHI抗体価は現代のウマウイルスを以前の分離物と比較して調べた結果、8〜16倍高かったが、イヌウイルスを用いて調べると少なくとも1/4低下した。イヌ血清は以前のウマウイルスとは非反応性であったが、現代のウマ分離物およびイヌ分離物を用いて調べると抗体価は4倍に増加した。これは、イヌインフルエンザウイルスに対して免疫されたフェレットの血清についても観察された。これらの血清反応パターンは、イヌインフルエンザウイルスおよび現代のウマインフルエンザウイルスの間の抗原類似性を実証するものであり、系統学的解析と一致した。イヌ/マイアミ/05分離物に対するウマ、イヌおよびフェレット血清の抗体価は、2003年および2004年のイヌ分離物における抗体価とほぼ等しかった。しかし、イヌ/Jax/05分離物の抗体価は1/2〜1/4であった。このことは、イヌ/Jax/05が抗原的にはその他のイヌ分離物とは異なり、成熟HAの107位における単一のアミノ酸の変化に一部関連し得ることを示唆する。
b 大文字は固定された核酸残基を示す。
b イヌ/FL/04ウイルスで免疫したフェレットの血清
イヌインフルエンザウイルス接種物
ウイルス接種物は、Madin-Darbyイヌ腎(MDCK)上皮細胞に既報(Crawfordら、2005年)のオリジナル分離物の3回継代物であるA/イヌ/FL/43/04(H3N8)のストックを接種して調製した。1μg/mL TPCK処理トリプシン(Sigma-Aldrich Corp., St. Louis, MO)および抗生物質(ゲンタマイシンおよびシプロフロキサシン)を強化したDulbecco最小必須培地(DMEM)中の接種されたMDCK細胞は、250cm2のフラスコに入れて5%CO2を含む加湿大気中、37℃にて生育させた。培養物は形態学的変化を1日1回観察して、接種後5日に回収した。回収した培養物は遠心分離によって清澄化して、上清をイヌの接種まで-80℃で保存した。ReedおよびMuench法によるウイルス力価の測定には、上清の画分を使用した。力価は、1mL当たり107半数組織培養物感染用量(TCID50)のA/イヌ/フロリダ/43/04(イヌ/FL/2004)であった。
コロニーで繁殖した4カ月齢の8頭の雑種犬(Marshall BioResources, North Rose, NY)(雌雄各4頭)をUniversity of Florida Institutional Animal Care and Use Committeeにより承認された実験的接種試験に使用した。イヌの体重は13〜17kgであった。イヌは、理学的検査、ベースラインの血液検査、および接種前2週間の体温記録に基づいて、健常であった。すべてのイヌは、施設への到着時および2週間後に採取した対の血清試料について実施した血清学的検査に基づいて、イヌインフルエンザウイルスへの曝露歴はなかった。イヌは、気管内チューブの挿管に備えて、プロポフォール(ディプリバン(登録商標)、Zeneca Pharmaceuticals、行うために0.4mg/kg体重)の静脈内注入により麻酔した。6頭(雌雄各3頭)のイヌに、気管内チューブに挿入した直径の小さなゴム製カテーテルを介して5mL無菌生理食塩液中107 TCID50のイヌ/FL/04ウイルスを遠位気管に投与して、それぞれ接種した。2頭のイヌ(雌雄各1頭)には等量の無菌生理食塩液を偽接種した。偽接種した対照のイヌは、ウイルス接種したイヌとは別の部屋に収容して、別の担当者がケアした。理学的検査および直腸温の記録は、1日2回、接種後(p.i.)6日間実施した。
ウイルス排出をモニターするために、p.i.0〜6日にポリエステルスワブ(Fisher Scientific International Inc., Pittsburgh, PA)を用いて口咽頭標本を各イヌから1日2回、採取した。スワブは、0.5%ウシ血清アルブミン(BSA)を含む無菌リン酸緩衝生理食塩液(PBS)1mL中に置いた。直腸スワブは0日から6日まで1日1回、各イヌから採取した。スワブ抽出液は、スワブ輸送培地を遠心分離によって清澄化して調製した。スワブ抽出液の画分は、Directigen(商標)の市販の免疫アッセイキット(BD, Franklin Lakes, NJ)を製造業者の説明書に従って用いて、インフルエンザAウイルス核タンパク質について直ちに検査した。残りの抽出液は、その他のウイルス学的アッセイ法まで-80℃にて保存した。
p.i. 1日に、偽接種した1頭およびウイルス接種した1頭のイヌをBeuthanasia-D(登録商標)液(1mL/5kg体重;Schering-Plough Animal Health Corp)の静脈内注入により安楽死させた。p.i. 2日から5日まで毎日、ウイルス接種した1頭のイヌを同様に安楽死させた。p.i. 6日に偽接種およびウイルス接種した残りのイヌを安楽死させた。詳細な死亡後検査は1名の試験担当者(WLC)により実施された。組織は10%中性緩衝ホルマリン中で固定して、パラフィンに包埋し、5μmの切片を病理組織学的診断のためにヘマトキシリン・エオジンで染色するか、または以下に記載する通り免疫組織化学的検査に備えて加工した。未固定の肺組織は、細菌の分離および同定のため、University of Florida College of Veterinary MedicineのDiagnostic Clinical Microbiology/Parasitology/Serology Serviceに送付した。試料は、非選択培地およびボルデテラ属(Regan-Lowe; Remel, Lenexa, KS)およびマイコプラズマ属(Remel)の選択培地で培養した。すべての培養物は、増殖なしと報告する前に21日間維持した。未固定の組織は、同じくウイルス学的分析まで-80℃で保存した。
脱パラフィンおよび再水和した5μmの気管および肺組織切片をBond-Rite(商標)スライドガラス(Richard-Allan Scientific, Kalamazoo, MI)にセットして、プロテイナーゼK(DAKOCytomation, Carpenteria, CA)ペルオキシダーゼ遮断試薬(DAKO(登録商標)EnVision(商標)ペルオキシダーゼキット、DAKO Corp., Carpenteria, CA)で順次処理した。切片は、インフルエンザA H3に対するモノクローナル抗体の1:500希釈(Chemicon International, Inc., Ternecula, CA)と共に室温で2時間、インキュベートした。対照には、同一の切片とマウスIgG(1mg/mL、Serotec, Inc. Raleigh, NC)のインキュベート、およびモノクローナル抗体と正常なイヌ肺切片のインキュベートが含まれた。一次抗体で処理した後、切片は製造業者の説明書に従って二次イムノペルオキシダーゼおよびペルオキシダーゼ基質試薬(Dako(登録商標)EnVision(商標)ペルオキシダーゼキット、Dako Corp.)と共にインキュベートした。ヘマトキシリンで切片を対比染色して、清澄化剤#2およびBluing試薬(Richard-Allan Scientific, Kalamazoo, MI)で処理して脱水し、Permount(ProSciTech, Queensland, Australia)を用いてカバーガラスをかけた。
各イヌに由来する肺および気管の組織を融解して、使い捨ての組織グラインダー(Kendall, Lifeline Medical Inc., Danbury, CT)を用いて、0.5%ウシ血清アルブミン(BSA)および抗生物質(ゲンタマイシンおよびシプロフロキサシン)を強化した最小必須培地(MEM)を加えてホモジナイズした。市販のキット(RNeasy(登録商標)ミニキット、QIAGEN Inc., Valencia, CA)を製造業者の説明書に従って用いて、組織ホモジネートならびに口咽頭および直腸スワブ抽出液から全RNAを抽出して、最終液量 60μLの緩衝液で溶出した。
ROXをパッシブレファレンス色素として含むQuantiTect(登録商標)プローブRT-PCRキット(QIAGEN Inc., Valencia, CA)およびA型インフルエンザウイルスのマトリックス(M)遺伝子の高度保存領域をターゲットとするプライマー-プローブセット(Payungporn S.ら、2006年a;Payungporn S.ら、2006年b)を用いて、全RNAに対して一段階定量的リアルタイムRT-PCRを実施した。各リアルタイムRT-PCR反応において、最終液量 25μL中、2X QuantiTech(登録商標) Probe RT-PCR Master Mix 12.5μL、QuantiTech(登録商標)RT Mix 0.25μL、フォワードおよびリバースプライマー(それぞれ、0.4μMの最終濃度)、プローブ(0.1μMの最終濃度)、ならびにRNaseフリーの水を含む反応混合液に抽出した全RNA 5μLを加えた。スワブおよび組織試料から抽出されたRNAの存在に関する内因性の内部対照として、また標準化対照としてのGAPDHの検出には、TaqMan(登録商標)GAPDH対照試薬(Applied Biosystems, Foster City, CA)を製造業者の説明書に従って使用した。
ウイルス接種したイヌから得られた肺および気管の凍結組織を融解して、0.5%BSAおよび抗生物質を強化した10倍量のDMEM中でホモジナイズした。遠心分離により固体の細片を除去して、上清を上記の通り1μg/mLのTPCK処理したトリプシン(Sigma-Aldrich Corp., St. Louis, MO)および抗生物質を強化したDMEMで培養したMDCK細胞に接種した。細胞は、25cm2のフラスコを用いて5%CO2含有加湿大気中、37℃にて培養した。培養物は形態学的変化を1日1回観察して、接種後5日に回収した。回収した培養物を遠心分離により清澄化して、上清を初回接種において記載した通り新鮮なMDCK細胞に接種し;血球凝集またはRT-PCRによりインフルエンザウイルスの証拠を示さなかった試料についてはさらに2回の継代を行った。清澄化した上清の血球凝集活性は、上記の通り0.5%シチメンチョウ赤血球(Crawfordら、2005年)を用いて測定した。RT-PCRは、下記の通り、実施した。
製造業者の説明書に従ってQIAamp(登録商標)ウイルスRNAミニキット(QIAGEN Inc., Valencia, CA)を用いて、MDCK上清からウイルスRNAを抽出した。ウイルスRNAは、QIAGEN(登録商標)一段階RT-PCRキット(QIAGEN Inc., Valencia, CA)を製造業者の説明書に従って使用して、cDNAに逆転写した。cDNA内の8つのインフルエンザウイルス遺伝子のコード領域のPCR増幅は、汎用遺伝子特異的プライマーセット(プライマー配列は要求に応じて与えられる)を用いて既報(Crawfordら、2005年)の通り実施した。得られたDNA単位複製配列は、サイクルシーケンシングダイターミネーターケミストリー(Applied Biosystems, Foster City, CA)を用いたABI PRISM(登録商標)3100自動DNAシーケンサーでの自動シーケンシングのための鋳型として用いられた。ヌクレオチド配列はLasergene 6 Package(登録商標)(DNASTAR, Inc., Madison, WI)を用いて解析した。呼吸器での複製中に何らかの変化が生じたか否かを調べるために、感染犬から回収されたウイルスのヌクレオチド配列を接種物中のウイルスの配列と比較した。
ウイルス接種された6頭のすべてのイヌが最初のp.i. 2日間に発熱(直腸温 ≧39℃)を発症したが、6日間の観察期間を通して咳または鼻分泌物などの呼吸器症状を示したものはいなかった。偽接種したイヌは臨床的に健常な状態を維持した。
インフルエンザAの核タンパク質がp.i. 24時間にウイルス接種したイヌの1頭から採取された口咽頭スワブで検出された。p.i. 72、84および120時間に1頭のイヌから、またp.i. 108、120および132時間にはもう1頭のイヌから採取された口咽頭スワブが定量的リアルタイムRT-PCRによりウイルスに関して陽性であった(表11)。スワブ抽出液1μL当たりのインフルエンザM遺伝子コピーの絶対数は、p.i.3日から6日まで経時的に増加した。直腸スワブには、ウイルスは検出されなかった。
特定病原体除去ビーグルを用いた過去の実験的感染(Crawfordら、2005年)とは対照的に、ウイルス接種した雑種犬は、p.i.1日から6日までの肺の肉眼的および組織学的分析により示される通り、肺炎を発症した。肺炎に加えて、イヌは、自然感染したイヌにおける報告(Crawfordら、2005年)と同様に鼻炎、気管炎、気管支炎および細気管支炎を呈した。気道の裏打ち細胞および気管支腺に上皮の壊死およびびらんが認められ、粘膜下組織には好中球およびマクロファージの浸潤が見られた(図5、上段)。免疫組織化学的検査により、気管支、細気管支および気管支腺の上皮細胞にウイルス性H3抗原が検出された(図5、下段)。細菌の重複感染はなかった。2頭の偽接種犬の呼吸組織は正常であった。
気管および肺は、p.i. 1日から6日まですべてのイヌにおいて定量的リアルタイムRT-PCRによりウイルスに関して陽性であった(表12)。気管ホモジネート1μL当たりのインフルエンザM遺伝子コピーの絶対数はp.i. 1日から5日まで増加して、その後、6日には減少した。肺ホモジネート1μL当たりのM遺伝子コピーの絶対数はp.i. 1日から6日まで減少した。一般に、気管には6日間の各p.i.日に肺よりも≧1log10多いウイルスが含まれた。
b 標準化比は、各スワブ抽出液についてM(Ct)をGAPDH(Ct)で割って算出した。
c スワブ抽出液1μL当たりのマトリックス遺伝子コピーの絶対数。
b 標準化比は、各組織ホモジネートについてM(Ct)をGAPDH(Ct)で割って算出した。
c 組織ホモジネート1μL当たりのマトリックス遺伝子コピーの絶対数。
ウイルス株
イヌインフルエンザウイルス株ならびにトリ、ウマおよびヒト由来のウイルス株(表15に列記)を孵化鶏卵またはMDCK細胞で増殖させて、それらの感染性をニワトリ胚におけるエンドポイント希釈またはプラーク法により定量した。シチメンチョウ赤血球(red blood cell)の赤血球(erythrocytes)を用いた血球凝集試験により、迅速なウイルスの定量が実施された。
2005年にウイルス性呼吸器疾患が疑われる症例から採取された合計60頭のイヌの肺組織について、イヌインフルエンザウイルスの存在を調べた。
20〜30mgの重量の肺組織のブロックを使い捨ての組織グラインダー(Kendal)に入れてホモジナイズした。製造業者の説明書に従って市販のキット(RNeasyミニキット、Qiagen, Valencia, CA)を用いて全RNAを抽出し、最終液量 60μLの液量で溶出した。
CLUSTAL Xプログラム(バージョン1.8)を用いて、様々な亜型および多様な動物種に由来するH3およびM遺伝子のマルチプル配列アラインメントを実施した。マトリックス(M)プライマーおよびプローブはインフルエンザAウイルスの異なる亜型に対応する公知の配列を通じての保存領域より選択され、H3ヘマグルチニン遺伝子特異的なプライマーおよびプローブはウマおよびイヌインフルエンザAウイルス遺伝子に特異的にマッチして、相同のトリおよびヒト遺伝子にはミスマッチするように選択した(表13)。プライマー設計ソフトウェア(OLIGOS バージョン9.1)およびEXIQON(http://lnatools.com)より提供されるウェブ上の解析ツールをTmの算出ならびに二次構造および自己ハイブリダイゼーションの予測に使用した。18S rRNAの保存領域を、イヌ組織試料から抽出されたRNAの存在に関する内因性内部対照として使用した。Pre-Developed TaqMan(登録商標)Assay Reagents for Eukaryotic 18S rRNA(VIC/TAMRA) (Applied Biosystems)を組織試料における18S rRNAのリアルタイム検出に使用した。
一段階定量的リアルタイムRT-PCRは、ROXをパッシブレファレンス色素として含むQuantitectプローブRT-PCRキット(Qiagen, Valencia, CA)を用いて実施した 各リアルタイムRT-PCR反応において、最終液量 20μL中、2X QuantiTechプローブRT-PCRマスターミックス 10μL、QuantiTech RT Mix 0.2μL、プライマー(H3遺伝子については最終濃度0.4μM、M遺伝子については最終濃度0.6μM)、プローブ(H3遺伝子については最終濃度0.1μM、M遺伝子については最終濃度0.2μM)、RNaseフリーの水を含む反応混合液と化合するための鋳型としてRNA試料の5μLを使用した。一段階リアルタイムRT-PCRはMx3005PリアルタイムQPCRシステム(Stratagene)を用いて実施した。サイクリング条件は、50℃、30分間の逆転写過程を含んだ。HotStarTaq DNAポリメラーゼを活性化するための95℃、15分間の1回目変性段階の後、増幅は変性(94℃、15秒間)およびアニーリング/伸長(60℃、30秒間)を含む40サイクルの期間中に実施された。FAM(H3およびM検出のため発光波長516nm)およびVIC(18S rRNA検出のため発光波長555nm)の蛍光シグナルはサイクル当たり1回ずつ、伸長段階の終了時に得た。リアルタイムPCRアッセイ法のデータの取得および解析はMx3005Pソフトウェア バージョン2.02(Stratagene)を用いて実施した。
各プライマー/プローブセットの特異性について試験するために、インフルエンザAウイルスの複数の公知の亜型から抽出されたRNAをリアルタイムRT-PCRアッセイ法における鋳型として用いた(表15)。
イヌインフルエンザAウイルス(A/イヌ/フロリダ/242/2003(H3N8))の遺伝子は、T7プロモーターと結合したプライマーを用いることによってH3(nt 1〜487)およびM(nt 1〜276)のPCR単位複製配列を作出するために使用した(表13)。続いて、リボプローブインビトロ転写システム-T7(Promega)を製造業者の説明書に従って用いることによって、H3およびM遺伝子の精製PCR単位複製配列をインビトロにおける転写のための鋳型として用いた。転写されたRNAの濃度は、260nmでの吸光度を測定して算出した。次に、感受性試験を実施するためにRNAを108〜10コピー/μLの範囲で10倍段階希釈した。さらに、リアルタイムRT-PCRの全体の性能を調べるために、初回RNA鋳型濃度(コピー/μL)の対数を各希釈より得られた閾値サイクル(Ct)に対してプロットして標準曲線を作成した。
106.67EID50/mL(HA=64)のA/ワイオミング/3/2003 (H3N2)および107.17EID50/mL(HA=16)のA/イヌ/フロリダ/242/2003(H3N8)を含む2つのウイルス株のストックウイルスを検出閾値アッセイ法に使用した。高速インフルエンザA抗原検出キットであるDirectigen Flu A(Becton, Dickinson and Company)を製造業者の説明書に従って使用して、標本のリン酸緩衝生理食塩液(PBS)(125μL)の対数希釈を用いた。それぞれのDirectigen Flu A検査装置には膜の中央部に紫色のドットとして現れるH1N1インフルエンザ抗原スポットがあり、核タンパク質(NP)に対するモノクローナル抗体に基づいて検査の完全性が示される。ドットを取り巻く紫色の三角形の出現は、試験標本におけるインフルエンザNPの存在を示す。三角形からの紫色のシグナルの強度は、+(三角形の輪郭)、++(明るく着色した三角形)、+++(暗紫色の三角形)および++++(極めて暗紫色の三角形)としてスコア化される。ウイルスRNAはQIAampウイルスRNAミニキット(Qiagen, Valencia, CA)を用いて各ウイルス希釈液の125μL画分からウイルスRNAを抽出し、最終液量50μLに溶出した。抽出したウイルスRNAの5μL容量を、Directigen Flu Aキットを用いた比較感受性試験においてリアルタイムRT-PCRにより試験した。
イヌインフルエンザに関するリアルタイムRT-PCRアッセイ法は、宿主細胞由来の18S rRNAならびにインフルエンザAウイルスゲノム由来のMおよびH3をターゲットとする3つの分子プローブからの情報に依拠する(表14)。宿主遺伝子の増幅は標本の品質および完全性に関するレポーターである。イヌインフルエンザ(H3N8)ウイルスを含む臨床的、剖検または実験的試料は3つのプローブを用いて増幅シグナルが得られることが期待された。Mおよび18S rRNAプローブを用いた場合は増幅シグナルを与えるがH3については陰性である標本は、ヒト、ブタもしくはトリ由来、またはH3以外の亜型に由来するインフルエンザウイルスH3亜型の指標である。これらの稀なケースは、シーケンシングによって分析することができる単位複製配列cDNAを作出するためのHA汎用プライマーを用いたRT-PCRにより解決することができる。適切に採取および取り扱われたインフルエンザAウイルスを含まない標本は、18S rRNA単位複製配列シグナルのみを与える。18S rRNAプローブおよびH3プローブのみが増幅シグナルを与える状況は、そうでないことが証明されない限り、技術が不完全であることの指標であり;Mプローブでの偽陰性またはH3での偽陽性のいずれかが実証されなければならない。最後に、3つのプローブを用いて増幅シグナルを与えられない標本は、不完全な試料の採取、変性、RNA抽出の不備、またはPCRで用いられたポリメラーゼの阻害物質の存在が示唆される。
フロリダで大流行したインフルエンザから分離されたイヌインフルエンザ(canine influenza)(イヌインフルエンザ(canine flu))ウイルスはH3N8型インフルエンザウイルスであることが観察されていて、ウマインフルエンザウイルス株であるA/ウマ/オハイオ/03と密接に関連した(Crawford et al., SCIENCE Vol. 309, September 2005、参照により全体が本特許に組み入れられる)。この試験では、イヌにおいてインフルエンザ様疾病を誘発するためにウマインフルエンザウイルス株であるA/ウマ/オハイオ/03を使用することの可能性について検討された。
雌雄の10頭の13週齢のビーグル犬を供給業者から入手して、BSL-2施設内において個別のケージに入れて飼育した。イヌは各群5頭の2群に無作為に振り分けた。表26に示す通り、一方の群は気管内刺激に供して、他方の群は口鼻腔内刺激に供した。14週齢の時点でイヌを刺激した。
この実験の結果を表27にまとめる。インフルエンザ関連性の臨床徴候が刺激後数頭のイヌにおいて観察された。これらの徴候には、発熱(>103○F;>39.4℃)および咳嗽が含まれた。2群の5頭中1頭(即ち、20%)に比して、1群では5頭中2頭(即ち、40%)が発熱(>103○F;>39.4℃)を示した。刺激後に口鼻腔刺激群の1頭がくしゃみを示して、もう1頭が咳嗽を示した。1群では、10〜80のHI力価範囲、幾何学平均力価(GMT)20が観察された。2群では40〜160の力価範囲、GMT 86が観察された。各群の1頭のイヌがインフルエンザと適合性するまたはインフルエンザに疾患特異的な病理組織学的病変を示した。
** 直腸温?≧103○F;≧39.4℃
フロリダで大流行したインフルエンザから分離されたイヌインフルエンザ(イヌインフルエンザ)ウイルスはH3N8インフルエンザウイルスであることが観察されていて、配列類似性に基づいて、ウマインフルエンザウイルスのA/ウマ/オハイオ/03と密接に関連した。以下の試験は、実験的に不活化したウマインフルエンザウイルスワクチンの有効性を調べるために実施された。
雌雄の9頭の7週齢のビーグル犬を供給業者から入手して、BSL-2施設内において個別のケージに入れて飼育した。これらのイヌは、表28に示す通り、2群に無作為に振り分けた。
この実験の結果を表29および30にまとめる。ワクチン接種した全てのイヌがワクチン接種後に血清転換を示した。40〜640のHI力価範囲および129のGMTがウマインフルエンザウイルスA/ウマ/オハイオ/03でのワクチン接種後期間に観察されて、イヌインフルエンザ分離物であるA/イヌ/フロリダ/242/03では160〜320のHI力価および211の幾何平均力価が観察された。6頭中2頭のワクチン接種犬が>103○F(>39.4℃)の発熱を1日示して、刺激後、いずれのイヌにもその他の臨床徴候は観察されなかった。
ワクチン接種した全てのイヌが不活化してCARBIGEN(商標)でアジュバント化したウマインフルエンザウイルスに反応した。イヌインフルエンザウイルス分離物を用いたHI力価の結果は、不活化したウマインフルエンザワクチンがイヌインフルエンザウイルスに対して検出可能な量の交差反応抗体を誘導したことを示唆する。この実験で用いられた刺激ウイルスがビーグル犬に注目に値するいかなる臨床的な疾病も誘発しなかったにも関わらず、イヌインフルエンザウイルス分離物を用いたHI力価に基づいて、不活化したウマワクチンは、イヌにおいて、H3N8型イヌインフルエンザウイルスによって引き起こされる「イヌインフルエンザ」疾病から潜在的にイヌを保護することができる交差反応抗体を誘導するために用いることができると結論付けられた。
** ワクチン接種にはCARBIGEN(商標)でアジュバント化した不活化ウマインフルエンザウイルスOhio 03ワクチンを使用した。
*** 刺激後7日に安楽死させた。
** ワクチン接種にはCARBIGEN(商標)でアジュバント化した不活化ウマインフルエンザウイルスOhio 03ワクチンを使用した。
フロリダでのインフルエンザ大流行から分離されたイヌインフルエンザウイルスは、多くのH3N8型ウマインフルエンザウイルス分離物と密接に関連すると特徴付けられた。DNAおよびアミノ酸配列類似性の分析から、イヌインフルエンザウイルスはウマインフルエンザウイルスのA/ウマ/オハイオ/03と極めて類似することが示された。市販のウマインフルエンザワクチンのイヌにおける有効性を調べるために、イヌにおいて以下の試験が実施された。
雌雄の約16カ月齢の雑種20頭およびビーグル犬20頭を試験に使用した。イヌは各群6〜7頭の6群(表31)に無作為に振り分けた。1および4群のイヌには、16および17カ月齢の時点で、市販の不活化してアジュバント化したウマインフルエンザワクチン(EQUICINE(商標), Intervet Inc., Millsboro, DE)を皮下(SQ)経路によりワクチン接種した。2および5群のイヌに修飾ウマ/ケンタッキー/91生インフルエンザワクチンを1mlの液量で鼻腔内投与(単一の鼻孔)によりワクチン接種した。標準的プロトコル(SAM 124, CVB, USDA, Ames, IA)を用いてH3N8ウマインフルエンザウイルスおよびイヌインフルエンザウイルスを用いたHI力価の測定に備えて、ワクチン接種日、1回目ワクチン接種後(1、2、4および5群)および2回目ワクチン接種後(1および4群)7および14日に血液試料を採取した。
** EQUICINE II(商標)はIntervet Inc.より液体ワクチンとして販売されている。EQUICINE II(商標)は、不活化されたA/ペンシルバニア/63インフルエンザ(つまり、「A/Pa/63」)ウイルスおよびA/ウマ/ケンタッキー/93インフルエンザ(つまり、「A/KY/93」)ウイルスをカルボポール(即ち、HAVLOGEN(登録商標)(Intervet Inc.))と共に含む。より具体的には、EQUICINE II(商標)の1回用量は、不活化したA/Pa/63?106.0 EID50、不活化したA/KY/93?106.7 EID50、体積に基づく0.25%カルボポール、および1mlの総液量を作成するために十分なPBSを含む。
*** A/KY/91は水で再溶解した凍結乾燥ワクチンである。このような再溶解は、ワクチン投与量を総液量1mlとするために十分なワクチン級の水を用いて実施された。ワクチンはウマ/ケンタッキー/91インフルエンザ(つまり「A/KY/91」)ウイルスを含んだ。このワクチンについては、例えば、参照によりそれらの全体が本特許に組み入れられる米国特許第6,436,408号、同6,398,774号および同6,177,082号において考察される。再溶解した場合、ワクチンの1回用量はA/KY/91 1ml当たり107.2 TCID50、N-Z AMINE AS(商標) 1ml当たり0.015g、ゼラチン 1ml当たり0.0025g、およびDラクトース 1ml当たり0.04gを含んだ。N-Z AMINE AS(商標)は、カゼインの酵素加水分解によって産生されるアミノ酸およびペプチドの精製したソースである。N-Z AMINE AS(商標)はKerry Bio-Science(Norwich, NY,USA)より販売されている。
ワクチン接種した全てのイヌがワクチン接種後に血清転換を示して、HI力価はウマインフルエンザウイルス(H3N8型)を用いたA/KY/91ワクチン群のイヌにおける10〜40に比して、EQUICINE II(商標)ワクチン群では10〜80であった。
血清学により、市販のウマインフルエンザワクチンを用いたイヌのワクチン接種は中等度のインフルエンザ抗体反応を刺激することが示された。H3N8型インフルエンザウイルスでの刺激後のイヌにおけるインフルエンザ関連性の臨床徴候の発現には、若干の品種差がある可能性がある。弱毒化生ウマインフルエンザワクチン(A/KY/91)は、雑種犬において直腸温の点で臨床的疾病の発症からの有意(P<0.05)な保護を示した。同様に、弱毒化生ウイルスワクチンは鼻分泌物中のインフルエンザウイルスの排出を予防した。
試験のためにイヌにおいて疾病を誘発することが成功しなかった報告書を考慮して、イヌにおけるイヌインフルエンザ刺激モデルを開発するためにイヌインフルエンザウイルスであるH3N8を用いることの可能性を以下の試験で検討した。
10頭の雌雄の雑種犬を、市販業者から入手して、BSL-2施設にてケージで飼育した。イヌは各群5頭の2群に無作為に振り分けた。表33に示す通り、一方の群は気管内/鼻腔内刺激に供して、他方の群は供された。
1および2群の全てのイヌが24〜48時間以内にイヌインフルエンザの臨床徴候を発症した。各イヌが以下の臨床徴候の2つまたはそれよりも多くを示した:発熱(>103.0○F;>39.4℃)、咳嗽、漿液性または粘液膿性の眼分泌物、漿液性または粘液膿性の鼻分泌物、嘔吐、下痢、抑鬱、体重低下、空嘔吐、喀血および聴取可能なラ音。1群の5頭中5頭および2群の5頭中4頭のイヌの肺組織が以下の1つまたは複数を含む病理組織学的病変を示した:びまん性膿性気管支肺炎、腔内の好中球浸出物のプラグならびに粘膜および細気管支周辺組織における顕著な単核細胞凝集を伴う気管支炎/細気管支炎、多数の泡沫性マクロファージを伴う肺胞内の混合浸出物、リンパ細胞性および形質細胞性ならびに顆粒球性の細胞浸潤、ならびにインフルエンザウイルス感染と適合性またはインフルエンザウイルス感染に疾病特異的なII型肺細胞の増殖を伴う肺胞隔壁の肥厚。気管組織の試料は正常であった。
本試験で用いられたもののようなH3N8イヌインフルエンザ分離物は、本試験で記載された方法または同様の方法の一つを用いてイヌにおいてイヌインフルエンザ疾病を誘発するために使用することができる。
イヌにおけるイヌインフルエンザ刺激モデルを開発するためのイヌインフルエンザウイルスH3N8を使用する可能性について、以下の試験でさらに詳しく検討した。
15頭の17〜18週齢の雑種犬および5頭の15週齢のビーグル犬を市販業者から入手して、BSL-2施設においてケージで飼育した。雑種犬は、各群5頭の3群(1群〜3群)に無作為に振り分けた。ビーグル犬は全て、表34に示す通り1つの群(4群)に振り分けた。
1および4群のイヌの80%(5頭中4頭)、2および3群のイヌの100%が48時間以内にイヌインフルエンザの臨床徴候を発症した。各イヌは以下の臨床徴候の1つまたは複数を示した:発熱(>103.0○F;>39.4℃)、咳嗽、漿液性または粘液膿性の眼分泌物、漿液性または粘液膿性の鼻分泌物、嘔吐、下痢、抑鬱、体重低下、空嘔吐およびラ音。ビーグル犬に観察された臨床徴候は一般に雑種犬に比してより軽度かつ短期間であった。
本試験で用いられたもののようなH3N8イヌインフルエンザ分離物は、104.8〜106.8 TCID50の刺激用量範囲で、本試験で記載された方法または同様の方法を用いてイヌにおいてイヌインフルエンザ様またはケンネルコフ様の疾病を誘発するために用いられ得る。雑種犬およびビーグル犬に認められた臨床徴候にはいくつかの相違があった。一般に、ビーグル犬は雑種犬に比してより軽度のインフルエンザ関連性の臨床徴候を示す傾向がある。
以下の試験は、H3N8ウマインフルエンザワクチンのイヌにおけるイヌインフルエンザウイルスに対する有効性を調べるために実施された。
17頭の14週齢の雑種犬および10頭の8週齢のビーグル犬を市販業者から入手した。イヌは表35に示す通り5試験群に無作為に振り分けて、研究施設内で飼育した。
2回ワクチン接種群(1および4群)の全てのイヌがイヌインフルエンザウイルス分離物に対してHI抗体価反応を発現した(表37)。刺激後、全ての群において刺激後14日の力価が約4倍に増大したことは、全てのイヌが刺激ウイルスに曝露されたことを間接的に示した。全てのイヌがイヌインフルエンザの以下の徴候の1つまたは複数を発症した:発熱(>103.0○F;>39.4℃)、咳嗽、漿液性または粘液膿性の眼分泌物、漿液性または粘液膿性の鼻分泌物、嘔吐、下痢、抑鬱、体重低下、および呼吸困難。ワクチン接種群は、齢期の対応する対照群に比して、程度の軽い臨床徴候を示した(表38)。8週齢(P=0.040)および14週齢(P=0.003)のイヌ(それぞれ、4群および1群)の双方において、2回用量のワクチン接種に起因して臨床徴候の有意な軽減が見られた。この実験では、1回用量のワクチン接種は臨床徴候の有意な軽減を示さなかった(P=0.294)(2群)。
この試験の結果は以下を実証する。(1)不活化したH3N8ウマインフルエンザウイルスはワクチン接種したイヌにおいてイヌインフルエンザウイルス交差反応性のHI抗体反応を誘発することができる、(2)H3N8ウマインフルエンザウイルスワクチンの使用はイヌにおいてイヌインフルエンザウイルス疾病の程度を軽減することができる、および(3)H3N8ウマインフルエンザウイルスワクチンの使用は鼻および/または口分泌物中のウイルス排出を抑制することができる。
** 2回目ワクチン接種−1群および4群;1回目ワクチン接種−2群
*** 刺激日
「+/−」 軽度の病変(不明確)
「−」 正常
イヌにおけるイヌインフルエンザウイルスに対する多価H3N8ウマインフルエンザワクチンの有効性を調べるために、以下の試験が実施された。
17頭の15週齢のビーグル犬を市販業者から入手した。イヌは表41に示す通り3群に無作為に振り分けて、研究施設にて飼育した。
ワクチン接種した全てのイヌがイヌインフルエンザウイルス分離物に対してHI抗体力価反応を発現した(表43)。刺激後に、全ての群において刺激前のHI力価に比して刺激後14日のHI力価が約4倍に増大したことは全てのイヌが刺激ウイルスに曝露されたことを間接的に示す。全てのイヌがイヌインフルエンザ疾病の徴候を発症して、各イヌは以下の臨床徴候の1つまたは複数を示した:発熱(>103.0○F;>39.4℃)、咳嗽、漿液性または粘液膿性の眼分泌物、漿液性または粘液膿性の鼻分泌物、嘔吐、下痢、抑鬱、体重低下および呼吸困難。ワクチン接種群は、齢期の対応する対照群に比して、程度の軽い臨床徴候を示した(表44)。イヌにおける2回用量のワクチン接種に起因して、臨床徴候の有意(P=0.028)な軽減が見られた(1群)。1回用量のワクチン接種は臨床徴候の有意な軽減を示さなかった(P=0.068)(2群)。
この試験の結果は、1)不活化したH3N8ウマインフルエンザウイルスがワクチン接種したイヌにおいてイヌインフルエンザウイルス交差反応性HI抗体反応を誘発することができる、および2)H3N8ウマインフルエンザウイルスワクチンの使用がイヌにおいてイヌインフルエンザウイルス疾病の程度を軽減することができることを実証する。
** 2回目ワクチン接種−1群;1回目ワクチン接種−2群
*** 刺激日
「+/−」 軽度の病変(不明確)
「−」 正常
(1)イヌにおけるイヌインフルエンザウイルスに対する一価対多価H3N8ウマインフルエンザワクチンの有効性、および(2)ワクチン有効性に対する投与経路の影響を調べるために、以下の試験が実施された。
30頭の10週齢のビーグル犬を市販業者から入手した。イヌは表46に示す通り6群に無作為に振り分けて、研究施設にて飼育した。
ワクチンの種類に関わらず、SQ経路を介してワクチン接種された全てのイヌがイヌインフルエンザウイルス分離物に対してHI抗体価反応を発現した(表48)。ワクチンの種類に関わらず、INワクチン接種群(即ち、1、3および5群)のイヌには、ワクチン接種後期間中、イヌインフルエンザウイルス分離物に対してHI抗体価反応を発現したイヌはいなかった。しかし、全てのイヌにおいて刺激後14日までに抗体価が4倍に増加し、間接的に、全てのイヌが刺激ウイルスに曝露されたことが示された(表47)。
この試験の結果は以下を実証する。(1)不活化したH3N8ウマインフルエンザウイルスはSQ経路を介してワクチン接種したイヌにおいてイヌインフルエンザウイルス交差反応性HI抗体反応を誘発することができる、(2)一価(VAX-1およびVAX-2)または多価ワクチン(VAX-3)のいずれの鼻腔内投与もイヌでは有効でなかった、および(3)多価ワクチン(VAX-3)の皮下投与はイヌにおいてイヌインフルエンザウイルス疾病の程度を有意(P=0.016)に軽減させた。
** 2回目ワクチン接種
*** 刺激日
「+/−」 軽度の病変(不明確)
「−」 正常
イヌインフルエンザ疾病はH3N8インフルエンザウイルス(CIV)によって引き起こされる。CIVはウマH3N8ウイルスに非常に密接に関連して(Crawfordら、2005年)、曝露された全てのイヌに感染する。曝露されたイヌの約80%が臨床徴候を発症した。以下の試験では、不活化したH3N8ウマインフルエンザウイルスワクチンおよびイヌインフルエンザウイルスワクチンの有効性について調べた。
35頭のビーグル犬および5頭の雑種犬がこの試験で使用された。ビーグル犬は3群に無作為に振り分けられた(表51)。雑種犬は全て、対照群(3群)に振り分けられた。全てのイヌが標準的なグロースダイエットで飼育されて、飲水は自由に与えられた。
ワクチン接種した全てのイヌ(1および2群)がイヌインフルエンザウイルス分離物に対してHI抗体価反応を発現した(表53)。全てのイヌがイヌインフルエンザの以下の徴候の1つまたは複数を発症した:発熱(>103.0○F;>39.4℃)、咳嗽、漿液性または粘液膿性の眼分泌物、漿液性または粘液膿性の鼻分泌物、嘔吐、下痢、抑鬱、および食欲不振。ワクチン接種群は、齢期の対応する対照群に比して、程度の軽い臨床徴候を示した(表54)。VAX-1(1群)またはVAX-2(2群)のいずれかをワクチン接種したイヌにおいて、臨床徴候の有意(P<0.001)な軽減が見られた。
この試験の結果は以下を実証する。(1)不活化したH3N8ウマインフルエンザウイルスおよびイヌインフルエンザウイルスのワクチンはワクチン接種したイヌにおいてイヌインフルエンザウイルス反応性HI抗体反応を誘発することができる、(2)H3N8ウマインフルエンザウイルスまたはイヌインフルエンザウイルスワクチンの使用はイヌにおいてイヌインフルエンザウイルス疾病の程度を軽減することができる、ならびに(3)H3N8ウマインフルエンザウイルスまたはイヌインフルエンザウイルスワクチンの使用は鼻および/または口分泌物中のウイルス排出を抑制することができる。
** 2回目ワクチン接種
*** 刺激日
P−鼻もしくは口または鼻腔および口腔のスワブからウイルスが分離された。
米国特許第5,106,739号
米国特許第5,034,322号
米国特許第6,455,760号
米国特許第6,696,623号
米国特許第4,683,202号
米国特許第4,683,195号
米国特許第4,800,159号
米国特許第4,965,188号
米国特許第5,994,056号
米国特許第6,814,934号
米国特許第6,436,408号
米国特許第6,398,774号
米国特許第6,177,082号
公開された米国特許出願第20040078841号
公開された米国特許出願第20040067506号
公開された米国特許出願第20040019934号
公開された米国特許出願第20030177536号
公開された米国特許出願第20030084486号
公開された米国特許出願第20040123349号
配列番号:1は、本発明に従って使用することができるPB2タンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:2は、配列番号:1によってコードされるアミノ酸配列である。
配列番号:3は、本発明に従って使用することができるPB1タンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:4は、配列番号:3によってコードされるアミノ酸配列である。
配列番号:5は、本発明に従って使用することができるPAタンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:6は、配列番号:5によってコードされるアミノ酸配列である。
配列番号:7は、本発明に従って使用することができるNSタンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:8は、配列番号:7によってコードされるアミノ酸配列である。
配列番号:9は、本発明に従って使用することができるNPタンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:10は、配列番号:9によってコードされるアミノ酸配列である。
配列番号:11は、本発明に従って使用することができるNAタンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:12は、配列番号:11によってコードされるアミノ酸配列である。
配列番号:13は、本発明に従って使用することができるMAタンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:14は、配列番号:13によってコードされるアミノ酸配列である。
配列番号:15は、本発明に従って使用することができるHAタンパク質をコードするイヌインフルエンザウイルス(フロリダ/43/04)のヌクレオチド配列である。
配列番号:16は、配列番号:15によってコードされるアミノ酸配列である。
配列番号:17は、本発明に従って使用することができるPB2タンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:18は、配列番号:17によってコードされるアミノ酸配列である。
配列番号:19は、本発明に従って使用することができるPB1タンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:20は、配列番号:19によってコードされるアミノ酸配列である。
配列番号:21は、本発明に従って使用することができるPAタンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:22は、配列番号:21によってコードされるアミノ酸配列である。
配列番号:23は、本発明に従って使用することができるNSタンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:24は、配列番号:23によってコードされるアミノ酸配列である。
配列番号:25は、本発明に従って使用することができるNPタンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:26は、配列番号:25によってコードされるアミノ酸配列である。
配列番号:27は、本発明に従って使用することができるNAタンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:28は、配列番号:27によってコードされるアミノ酸配列である。
配列番号:29は、本発明に従って使用することができるMAタンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:30は、配列番号:29によってコードされるアミノ酸配列である。
配列番号:31は、本発明に従って使用することができるHAタンパク質をコードするイヌインフルエンザウイルス(FL/242/03)のヌクレオチド配列である。
配列番号:32は、配列番号:31によってコードされるアミノ酸配列である。
配列番号:33は、N末端の16のアミノ酸シグナル配列が除去されている配列番号:16に示すHAタンパク質の成熟型である。
配列番号:34は、N末端の16のアミノ酸シグナル配列が除去されている配列番号:32に示すHAタンパク質の成熟型である。
配列番号:35は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:36は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:37は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:38は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:39は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:41は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:42は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:43は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:44は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:45は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:46は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:47は、本発明に従って使用することができるPB2タンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:48は、配列番号:47によってコードされるアミノ酸配列である。
配列番号:49は、本発明に従って使用することができるPB1タンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:50は、配列番号:49によってコードされるアミノ酸配列である。
配列番号:51は、本発明に従って使用することができるPAタンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:52は、配列番号:51によってコードされるアミノ酸配列である。
配列番号:53は、本発明に従って使用することができるNSタンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:54は、配列番号:53によってコードされるアミノ酸配列である。
配列番号:55は、本発明に従って使用することができるNPタンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:56は、配列番号:55によってコードされるアミノ酸配列である。
配列番号:57は、本発明に従って使用することができるNAタンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:58は、配列番号:57によってコードされるアミノ酸配列である。
配列番号:59は、本発明に従って使用することができるMAタンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:60は、配列番号:59によってコードされるアミノ酸配列である。
配列番号:61は、本発明に従って使用することができるHAタンパク質をコードするイヌインフルエンザウイルス(マイアミ/2005)のヌクレオチド配列である。
配列番号:62は、配列番号:61によってコードされるアミノ酸配列である。
配列番号:63は、本発明に従って使用することができるPB2タンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:64は、配列番号:63によってコードされるアミノ酸配列である。
配列番号:65は、本発明に従って使用することができるPB1タンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:66は、配列番号:65によってコードされるアミノ酸配列である。
配列番号:67は、本発明に従って使用することができるPAタンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:68は、配列番号:67によってコードされるアミノ酸配列である。
配列番号:69は、本発明に従って使用することができるNSタンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:70は、配列番号:69によってコードされるアミノ酸配列である。
配列番号:71は、本発明に従って使用することができるNPタンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:72は、配列番号:71によってコードされるアミノ酸配列である。
配列番号:73は、本発明に従って使用することができるNAタンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:74は、配列番号:73によってコードされるアミノ酸配列である。
配列番号:75は、本発明に従って使用することができるMAタンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:76は、配列番号:75によってコードされるアミノ酸配列である。
配列番号:77は、本発明に従って使用することができるHAタンパク質をコードするイヌインフルエンザウイルス(ジャクソンビル/2005)のヌクレオチド配列である。
配列番号:78は、配列番号:77によってコードされるアミノ酸配列である。
配列番号:79は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:80は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:81は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:82は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:83は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:84は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:85は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:86は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:87は、本発明に従って用いることができるオリゴヌクレオチドである。
配列番号:88は、本発明に従って用いることができるオリゴヌクレオチドである。
Claims (25)
- 配列番号:78を有する、イヌ/Jax/05と命名されるウイルス分離物のヘマグルチニン(HA)ポリペプチドとまたは、配列番号:62を有する、イヌ/マイアミ/05と命名されるウイルス分離物のHAポリペプチドと99%もしくはそれよりも高いアミノ酸配列同一性を持つHAポリペプチドをコードするポリヌクレオチドを含む、または
配列番号:78を有する、イヌ/Jax/05と命名されるウイルス分離物のヘマグルチニン(HA)ポリペプチドとまたは、配列番号:62を有する、イヌ/マイアミ/05と命名されるウイルス分離物のHAポリペプチドと99%もしくはそれよりも高いアミノ酸配列同一性を持つHAポリペプチドを含む、
イヌ科動物に感染することができる、分離されたインフルエンザウイルスであって、不活化または弱毒化された、前記インフルエンザウイルス。 - H3のHA血清型を持つ、請求項1記載のインフルエンザウイルス。
- H3N8の血清型を持つ、請求項1記載のインフルエンザウイルス。
- 配列番号:62もしくは78に示されるアミノ酸配列またはそれらの機能性および/もしくは免疫原性断片を持つポリペプチドをコードするポリヌクレオチドを含む、または
該ポリヌクレオチドが、配列番号:62もしくは78に示されるアミノ酸配列と99%もしくはそれよりも高い配列同一性を持つポリペプチドをコードする、
請求項1記載のインフルエンザウイルス。 - 配列番号:61もしくは77のいずれかに示されるヌクレオチド配列を持つポリヌクレオチドを含む、または
該ポリヌクレオチドが配列番号:61もしくは77のいずれかに示されるヌクレオチド配列と98%もしくはそれよりも高い配列同一性を持つ、
請求項1記載のインフルエンザウイルス。 - イヌ/Jax/05と命名されるウイルス分離物またはイヌ/マイアミ/05と命名されるウイルス分離物である、請求項1記載のインフルエンザウイルス。
- 免疫原がイヌ科動物に感染することができるインフルエンザウイルスに対する免疫応答を誘発することができる、請求項1記載のインフルエンザウイルスの免疫原を含む組成物。
- 免疫原が無細胞のウイルス全体もしくはその一部;ウイルスポリヌクレオチド;ウイルスタンパク質;ウイルスポリペプチドもしくはペプチド;ウイルス感染細胞;組換え型ウイルスベクターをベースとする構築物;リアソータントウイルス;または該ウイルスの裸の核酸を含む、請求項7記載の組成物。
- 前記ウイルスタンパク質、ポリペプチドまたはペプチドが、配列番号:62もしくは78のいずれかに示されるアミノ酸配列またはその機能性および/もしくは免疫原性断片を含む、または
前記ポリヌクレオチドが配列番号:62もしくは78のいずれかに示されるアミノ酸配列と99%もしくはそれよりも高い配列同一性を持つポリペプチドをコードする、
請求項8記載の組成物。 - 前記ウイルスポリヌクレオチドが、配列番号:62もしくは78のいずれかに示されるアミノ酸配列、またはその機能性および/もしくは免疫原性断片を含むポリペプチドをコードする、請求項8記載の組成物。
- ウイルスが、配列番号:78を有する、イヌ/Jax/05と命名されるウイルス分離物のヘマグルチニン(HA)ポリペプチドとまたは、配列番号:62を有する、イヌ/マイアミ/05と命名されるウイルス分離物のHAポリペプチドと99%もしくはそれよりも高いアミノ酸配列同一性を持つHAポリペプチドをコードするポリヌクレオチドを含む、または
ウイルスが、配列番号:78を有する、イヌ/Jax/05と命名されるウイルス分離物のヘマグルチニン(HA)ポリペプチドとまたは、配列番号:62を有する、イヌ/マイアミ/05と命名されるウイルス分離物のHAポリペプチドと99%もしくはそれよりも高いアミノ酸配列同一性を持つHAポリペプチドを含む、
イヌインフルエンザワクチンであって、
以下を含む、イヌインフルエンザワクチン:
少なくとも1つのH3インフルエンザウイルス抗原および/または少なくとも1つのH7インフルエンザウイルス抗原の治療上有効量、ならびに
少なくとも1つの薬学的に許容される賦形剤。 - 以下のイヌインフルエンザウイルス分離物の1つまたは複数に由来する1つまたは複数の抗原を含む、請求項11記載のワクチン:イヌ/Jax/05、およびイヌ/マイアミ/05。
- ウイルス抗原が不活化ウイルスを含む、請求項11記載のワクチン。
- 以下のイヌインフルエンザウイルス分離物の1つまたは複数に由来する1つまたは複数の抗原を含む、請求項13記載のワクチン:イヌ/Jax/05、およびイヌ/マイアミ/05。
- ウイルス抗原が弱毒化生ウイルスを含む、請求項11記載のワクチン。
- 非ヒト動物においてイヌ科動物に感染することができるインフルエンザウイルスに対する免疫応答を誘発するための方法であって、請求項7記載の組成物の有効量を該動物に投与する工程を含む、方法。
- 組成物が配列番号:62もしくは78のいずれかに示されるアミノ酸配列、またはその機能性および/もしくは免疫原性断片を含むウイルスタンパク質、ポリペプチドまたはペプチドを含む、請求項16記載の方法。
- 組成物が、イヌ科動物に感染することができるインフルエンザウイルスまたは該ウイルスの一部を含み、
インフルエンザウイルスが、イヌ/Jax/05と命名されるウイルス分離物、もしくはイヌ/マイアミ/05と命名されるウイルス分離物、または該ウイルス分離物のいずれかの一部からなる群より選択される、
請求項16記載の方法。 - 免疫応答がインフルエンザウイルスによる動物の感染を予防または阻害する防御免疫応答である、請求項16記載の方法。
- インフルエンザウイルスがH3より選択されるHA血清型を持つ、請求項16記載の方法。
- 組成物がアジュバントをさらに含む、請求項16記載の方法。
- 動物がイヌ科である、請求項16記載の方法。
- イヌ科動物が飼い慣らされたイヌである、請求項22記載の方法。
- 組成物が非経口的、皮下、腹腔内、筋肉内、鼻腔内または経口的に投与される、請求項16記載の方法。
- インフルエンザウイルスが不活化または弱毒化された、請求項18記載の方法。
Applications Claiming Priority (12)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US72844905P | 2005-10-19 | 2005-10-19 | |
| US60/728,449 | 2005-10-19 | ||
| US75488105P | 2005-12-29 | 2005-12-29 | |
| US60/754,881 | 2005-12-29 | ||
| US75916206P | 2006-01-14 | 2006-01-14 | |
| US60/759,162 | 2006-01-14 | ||
| US76145106P | 2006-01-23 | 2006-01-23 | |
| US60/761,451 | 2006-01-23 | ||
| US77908006P | 2006-03-03 | 2006-03-03 | |
| US60/779,080 | 2006-03-03 | ||
| US40941606A | 2006-04-21 | 2006-04-21 | |
| US11/409,416 | 2006-04-21 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2008536829A Division JP2009512449A (ja) | 2005-10-19 | 2006-10-19 | イヌにおける呼吸器疾患管理のための材料および方法 |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015075626A Division JP6220361B2 (ja) | 2005-10-19 | 2015-04-02 | イヌ科動物に感染することができるインフルエンザウイルス、その使用 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2012254080A JP2012254080A (ja) | 2012-12-27 |
| JP5974397B2 true JP5974397B2 (ja) | 2016-08-23 |
Family
ID=37885846
Family Applications (3)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2008536829A Withdrawn JP2009512449A (ja) | 2005-10-19 | 2006-10-19 | イヌにおける呼吸器疾患管理のための材料および方法 |
| JP2012149223A Active JP5974397B2 (ja) | 2005-10-19 | 2012-07-03 | イヌ科動物に感染することができるインフルエンザウイルス、その使用 |
| JP2015075626A Active JP6220361B2 (ja) | 2005-10-19 | 2015-04-02 | イヌ科動物に感染することができるインフルエンザウイルス、その使用 |
Family Applications Before (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2008536829A Withdrawn JP2009512449A (ja) | 2005-10-19 | 2006-10-19 | イヌにおける呼吸器疾患管理のための材料および方法 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015075626A Active JP6220361B2 (ja) | 2005-10-19 | 2015-04-02 | イヌ科動物に感染することができるインフルエンザウイルス、その使用 |
Country Status (14)
| Country | Link |
|---|---|
| EP (2) | EP1945659B9 (ja) |
| JP (3) | JP2009512449A (ja) |
| KR (2) | KR101548436B1 (ja) |
| CN (3) | CN101563361B (ja) |
| AU (1) | AU2006304747B2 (ja) |
| BR (1) | BRPI0617735B1 (ja) |
| CA (2) | CA2626489C (ja) |
| ES (2) | ES2393406T3 (ja) |
| IL (2) | IL190906A (ja) |
| MX (3) | MX395177B (ja) |
| NO (2) | NO346351B1 (ja) |
| NZ (4) | NZ735684A (ja) |
| RU (2) | RU2520081C2 (ja) |
| WO (1) | WO2007047938A2 (ja) |
Families Citing this family (22)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| MX358750B (es) * | 2005-04-21 | 2018-08-31 | Univ Florida | Materiales y metodos para el control de enfermedades respiratorias en caninos. |
| US20080241184A1 (en) * | 2005-08-25 | 2008-10-02 | Jules Maarten Minke | Canine influenza vaccines |
| US7682619B2 (en) | 2006-04-06 | 2010-03-23 | Cornell Research Foundation, Inc. | Canine influenza virus |
| US7674469B2 (en) | 2006-10-25 | 2010-03-09 | Internet International B.V. | Feline influenza vaccine and method of use |
| BR112012022680B1 (pt) * | 2010-03-10 | 2021-03-02 | Intervet International B.V | uso de uma vacina inativada contra vírus da influenza canina |
| CN101838708B (zh) * | 2010-03-30 | 2012-07-18 | 华南农业大学 | 一种检测犬流感的方法 |
| CN102220293B (zh) * | 2011-05-26 | 2012-12-19 | 中国农业科学院上海兽医研究所 | 犬流感重组病毒及其制备方法和应用 |
| CN103242433B (zh) * | 2012-02-14 | 2016-12-14 | 中国医学科学院病原生物学研究所 | 一种腺病毒非结构蛋白免疫原、其抗体及应用 |
| US10080792B2 (en) | 2013-09-23 | 2018-09-25 | Engen Bio, Inc. | Influenza vaccine and therapy |
| CN104436157A (zh) | 2013-09-23 | 2015-03-25 | 恩金生物有限公司 | 流感疫苗和治疗 |
| CA2967532A1 (en) | 2014-11-24 | 2016-06-02 | Intervet International B.V. | Inactivated equine influenza virus vaccines |
| JP2015120709A (ja) * | 2015-01-09 | 2015-07-02 | ラボラトリオ アヴィメキシコ エスエー ディーイー シーヴィーLaboratorio Avi−Mex,S.A. De C.V. | 遺伝子組み換え不活性化ウィルスベクターワクチン |
| CN104911150B (zh) * | 2015-05-27 | 2018-05-01 | 华南农业大学 | 一种h3n2犬流感病毒的单克隆抗体杂交瘤细胞株的建立及其单克隆抗体的制备与应用 |
| CN106636475B (zh) * | 2017-03-01 | 2020-05-22 | 中国农业大学 | 一种检测北美h3n8亚型犬流感病毒的引物组及其应用 |
| EP3930749A1 (en) * | 2019-02-27 | 2022-01-05 | University of Rochester | Multivalent live-attenuated influenza vaccine for prevention and control of equine influenza virus (eiv) in horses |
| WO2021236488A1 (en) * | 2020-05-18 | 2021-11-25 | Thomas Jefferson University | Viral sample collection |
| CA3194830A1 (en) * | 2020-10-08 | 2022-04-14 | Paula WALKER | Methods and apparatuses for detecting respiratory infections |
| CN112410467A (zh) * | 2020-11-23 | 2021-02-26 | 深圳市赛格诺生物科技有限公司 | 一种检测甲型和乙型流感病毒的冻干型荧光rt-pcr试剂及方法 |
| WO2024196133A1 (ko) * | 2023-03-21 | 2024-09-26 | 고려대학교 산학협력단 | 교차 면역원성을 갖는 h3 아형 인플루엔자 바이러스 및 이를 포함하는 백신 |
| KR102746574B1 (ko) | 2023-11-27 | 2024-12-24 | 김태윤 | 반려동물의 호흡기 건강 개선용 조성물 |
| KR20250108327A (ko) | 2024-01-08 | 2025-07-15 | 건국대학교 산학협력단 | 반려동물 치료를 위한 코삽입관 고정장치 |
| US12233100B1 (en) | 2024-10-31 | 2025-02-25 | Tae Yoon Kim | Composition for improving respiratory health of companion animals |
Family Cites Families (40)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5034322A (en) | 1983-01-17 | 1991-07-23 | Monsanto Company | Chimeric genes suitable for expression in plant cells |
| US4683202A (en) | 1985-03-28 | 1987-07-28 | Cetus Corporation | Process for amplifying nucleic acid sequences |
| US4965188A (en) | 1986-08-22 | 1990-10-23 | Cetus Corporation | Process for amplifying, detecting, and/or cloning nucleic acid sequences using a thermostable enzyme |
| US4683195A (en) | 1986-01-30 | 1987-07-28 | Cetus Corporation | Process for amplifying, detecting, and/or-cloning nucleic acid sequences |
| US4800159A (en) | 1986-02-07 | 1989-01-24 | Cetus Corporation | Process for amplifying, detecting, and/or cloning nucleic acid sequences |
| US5106739A (en) | 1989-04-18 | 1992-04-21 | Calgene, Inc. | CaMv 355 enhanced mannopine synthase promoter and method for using same |
| US5994056A (en) | 1991-05-02 | 1999-11-30 | Roche Molecular Systems, Inc. | Homogeneous methods for nucleic acid amplification and detection |
| US5484719A (en) | 1991-08-26 | 1996-01-16 | Edible Vaccines, Inc. | Vaccines produced and administered through edible plants |
| US5612487A (en) | 1991-08-26 | 1997-03-18 | Edible Vaccines, Inc. | Anti-viral vaccines expressed in plants |
| UA48104C2 (uk) | 1991-10-04 | 2002-08-15 | Новартіс Аг | Фрагмент днк, який містить послідовність,що кодує інсектицидний протеїн, оптимізовану для кукурудзи,фрагмент днк, який забезпечує направлену бажану для серцевини стебла експресію зв'язаного з нею структурного гена в рослині, фрагмент днк, який забезпечує специфічну для пилку експресію зв`язаного з нею структурного гена в рослині, рекомбінантна молекула днк, спосіб одержання оптимізованої для кукурудзи кодуючої послідовності інсектицидного протеїну, спосіб захисту рослин кукурудзи щонайменше від однієї комахи-шкідника |
| RU2193065C2 (ru) * | 1994-03-14 | 2002-11-20 | Мерк энд Ко. Инк. | Конструкция днк (варианты), днк-вектор, иммуногенная композиция против вируса гриппа, способ индукции иммунного ответа, вакцина и способ вакцинации |
| GB9524395D0 (en) | 1995-11-29 | 1996-01-31 | Nickerson Biocem Ltd | Promoters |
| WO1997041236A1 (en) * | 1996-04-29 | 1997-11-06 | Andreas Zurbriggen | Polynucleotide vaccine against canine distemper |
| FR2750865B1 (fr) * | 1996-06-27 | 1998-12-04 | Rhone Merieux | Vaccin vivant recombinant a base d'herpesvirus canin, notamment contre la maladie de carre, la rage ou le virus parainfluenza de type 2 |
| US6252139B1 (en) | 1996-07-18 | 2001-06-26 | The Salk Institute For Biological Studies | Method of increasing growth and yield in plants |
| US6063385A (en) | 1997-11-07 | 2000-05-16 | Wisconsin Alumni Research Foundation | DNA vaccine for parvovirus |
| US6472375B1 (en) | 1998-04-16 | 2002-10-29 | John Wayne Cancer Institute | DNA vaccine and methods for its use |
| GB9809666D0 (en) * | 1998-05-06 | 1998-07-01 | Isis Innovation | Modified viruses |
| US6177082B1 (en) * | 1998-08-13 | 2001-01-23 | The University Of Pittsburgh-Of The Commonwealth System Of Higher Education | Cold-adapted equine influenza viruses |
| CA2348665C (en) | 1998-11-24 | 2003-04-15 | Pioneer Hi-Bred International, Inc. | Root-preferred promoters and their use |
| EP1035209A1 (en) * | 1999-03-06 | 2000-09-13 | ARTEMIS Pharmaceuticals GmbH | Stable recombinant influenza viruses free of helper viruses |
| GB9916794D0 (en) * | 1999-07-16 | 1999-09-22 | Isis Innovation | In vitro virus reconstitution |
| JP2003508086A (ja) * | 1999-07-30 | 2003-03-04 | アイシス イノヴェイション リミテッド | ワクチンとして有用な弱毒化インフルエンザウイルス |
| US6398774B1 (en) | 1999-09-29 | 2002-06-04 | Heska Corporation | Intranasal delivery system |
| DE19960843A1 (de) | 1999-12-16 | 2001-06-28 | Florian Grundler | Wurzelspezifischer Promotor |
| AU4930001A (en) * | 2000-04-03 | 2001-10-15 | Univ Pittsburgh | Cold-adapted equine influenza viruses |
| US20050089985A1 (en) * | 2000-06-23 | 2005-04-28 | American Cyanamid Company | Rescue of canine distemper virus from cdna |
| US6455760B1 (en) | 2000-11-16 | 2002-09-24 | The Salk Institute Of Biological Studies | Expression of flavin-containing monoxygenases in plants |
| US20040067506A1 (en) | 2000-12-04 | 2004-04-08 | Ben Scheres | Novel root specific promoter driving the expression of a novel lrr receptor-like kinase |
| EP2283858A3 (en) * | 2001-07-27 | 2011-03-23 | Wyeth LLC | West Nile Vaccine |
| EP3009507B1 (en) * | 2002-02-13 | 2020-06-24 | Wisconsin Alumini Research Foundation | Signal for packaging of influenza virus vectors |
| US7285656B2 (en) | 2002-04-26 | 2007-10-23 | Her Majesty The Queen In Right Of Canada, As Represented By The Minister Of Natural Resources Canada, Canadian Forest Service | Root-specific conifer gene promoter and its use |
| CN1678345A (zh) * | 2002-04-26 | 2005-10-05 | 米迪缪尼疫苗股份有限公司 | 制备流感病毒的多质粒系统 |
| AU2003294533A1 (en) * | 2002-12-19 | 2004-07-14 | Universite Laval | Molecular methods and compositions for detecting and quantifying respiratory viruses |
| US20040123349A1 (en) | 2002-12-20 | 2004-06-24 | Qi Xie | SINAT5, an Arabidopsis thaliana gene promotes ubiquitin related degradation |
| JP2004285019A (ja) * | 2003-03-25 | 2004-10-14 | Kyoto Prefecture | 生体外異物の不活化方法 |
| US8592197B2 (en) | 2003-07-11 | 2013-11-26 | Novavax, Inc. | Functional influenza virus-like particles (VLPs) |
| ATE469972T1 (de) * | 2003-12-23 | 2010-06-15 | Medimmune Llc | Multiplasmid-system zur erzeugung des grippevirus |
| WO2006073436A2 (en) * | 2004-04-29 | 2006-07-13 | The Trustees Of Columbia University In The City Of New York | Mass tag pcr for multiplex diagnostics |
| MX358750B (es) * | 2005-04-21 | 2018-08-31 | Univ Florida | Materiales y metodos para el control de enfermedades respiratorias en caninos. |
-
2006
- 2006-10-19 NZ NZ735684A patent/NZ735684A/en unknown
- 2006-10-19 CA CA2626489A patent/CA2626489C/en active Active
- 2006-10-19 ES ES06826359T patent/ES2393406T3/es active Active
- 2006-10-19 ES ES11003016.0T patent/ES2496315T3/es active Active
- 2006-10-19 MX MX2015003583A patent/MX395177B/es unknown
- 2006-10-19 EP EP06826359.9A patent/EP1945659B9/en active Active
- 2006-10-19 EP EP11003016.0A patent/EP2407480B1/en not_active Not-in-force
- 2006-10-19 NZ NZ567809A patent/NZ567809A/en unknown
- 2006-10-19 RU RU2008119461/10A patent/RU2520081C2/ru not_active Application Discontinuation
- 2006-10-19 AU AU2006304747A patent/AU2006304747B2/en not_active Ceased
- 2006-10-19 KR KR1020137010121A patent/KR101548436B1/ko active Active
- 2006-10-19 CN CN200680041958.9A patent/CN101563361B/zh active Active
- 2006-10-19 WO PCT/US2006/041061 patent/WO2007047938A2/en not_active Ceased
- 2006-10-19 KR KR1020087011965A patent/KR101597534B1/ko not_active Expired - Fee Related
- 2006-10-19 NZ NZ71775106A patent/NZ717751A/en unknown
- 2006-10-19 JP JP2008536829A patent/JP2009512449A/ja not_active Withdrawn
- 2006-10-19 BR BRPI0617735-2A patent/BRPI0617735B1/pt not_active IP Right Cessation
- 2006-10-19 CA CA3090231A patent/CA3090231A1/en active Pending
- 2006-10-19 MX MX2008005234A patent/MX2008005234A/es active IP Right Grant
- 2006-10-19 NO NO20082257A patent/NO346351B1/no unknown
- 2006-10-19 CN CN201310741621.0A patent/CN104017775B/zh active Active
- 2006-10-19 NZ NZ627888A patent/NZ627888A/en unknown
- 2006-10-19 CN CN202310548441.4A patent/CN117587038A/zh active Pending
-
2008
- 2008-04-16 IL IL190906A patent/IL190906A/en active IP Right Grant
- 2008-04-21 MX MX2011001779A patent/MX341842B/es unknown
-
2012
- 2012-07-03 JP JP2012149223A patent/JP5974397B2/ja active Active
-
2013
- 2013-05-09 IL IL226253A patent/IL226253A/en active IP Right Grant
-
2014
- 2014-01-17 RU RU2014101481A patent/RU2711807C2/ru active
-
2015
- 2015-04-02 JP JP2015075626A patent/JP6220361B2/ja active Active
-
2022
- 2022-06-09 NO NO2022022C patent/NO2022022I1/no unknown
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6220361B2 (ja) | イヌ科動物に感染することができるインフルエンザウイルス、その使用 | |
| US11160859B2 (en) | Materials and methods for respiratory disease control in canines | |
| JP6301855B2 (ja) | イヌにおける呼吸器疾患管理のための材料および方法 | |
| AU2020264347B2 (en) | Influenza viruses able to infect canids, uses thereof | |
| US11865172B2 (en) | Materials and methods for respiratory disease control in canines | |
| RU2802222C2 (ru) | Вирус гриппа, способный инфицировать собачьих, и его применение |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20131127 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140221 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140226 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20140522 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20141203 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20150402 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20150521 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20150522 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20150805 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20150827 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20151030 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20160609 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20160627 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20160629 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20160627 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 5974397 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |