JP5190263B2 - 超高スループットの光学−ナノ細孔dna読み取りプラットフォーム - Google Patents
超高スループットの光学−ナノ細孔dna読み取りプラットフォーム Download PDFInfo
- Publication number
- JP5190263B2 JP5190263B2 JP2007525792A JP2007525792A JP5190263B2 JP 5190263 B2 JP5190263 B2 JP 5190263B2 JP 2007525792 A JP2007525792 A JP 2007525792A JP 2007525792 A JP2007525792 A JP 2007525792A JP 5190263 B2 JP5190263 B2 JP 5190263B2
- Authority
- JP
- Japan
- Prior art keywords
- designed
- dissociation
- polymer
- molecule
- design
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
Images














Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6813—Hybridisation assays
- C12Q1/6827—Hybridisation assays for detection of mutation or polymorphism
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6869—Methods for sequencing
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/483—Physical analysis of biological material
- G01N33/487—Physical analysis of biological material of liquid biological material
- G01N33/48707—Physical analysis of biological material of liquid biological material by electrical means
- G01N33/48721—Investigating individual macromolecules, e.g. by translocation through nanopores
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y10—TECHNICAL SUBJECTS COVERED BY FORMER USPC
- Y10T—TECHNICAL SUBJECTS COVERED BY FORMER US CLASSIFICATION
- Y10T436/00—Chemistry: analytical and immunological testing
- Y10T436/14—Heterocyclic carbon compound [i.e., O, S, N, Se, Te, as only ring hetero atom]
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y10—TECHNICAL SUBJECTS COVERED BY FORMER USPC
- Y10T—TECHNICAL SUBJECTS COVERED BY FORMER US CLASSIFICATION
- Y10T436/00—Chemistry: analytical and immunological testing
- Y10T436/14—Heterocyclic carbon compound [i.e., O, S, N, Se, Te, as only ring hetero atom]
- Y10T436/142222—Hetero-O [e.g., ascorbic acid, etc.]
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y10—TECHNICAL SUBJECTS COVERED BY FORMER USPC
- Y10T—TECHNICAL SUBJECTS COVERED BY FORMER US CLASSIFICATION
- Y10T436/00—Chemistry: analytical and immunological testing
- Y10T436/14—Heterocyclic carbon compound [i.e., O, S, N, Se, Te, as only ring hetero atom]
- Y10T436/142222—Hetero-O [e.g., ascorbic acid, etc.]
- Y10T436/143333—Saccharide [e.g., DNA, etc.]
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Physics & Mathematics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- General Health & Medical Sciences (AREA)
- Immunology (AREA)
- Biomedical Technology (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Analytical Chemistry (AREA)
- Biochemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Genetics & Genomics (AREA)
- Microbiology (AREA)
- Hematology (AREA)
- Nanotechnology (AREA)
- Spectroscopy & Molecular Physics (AREA)
- Urology & Nephrology (AREA)
- Food Science & Technology (AREA)
- Medicinal Chemistry (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Description
本発明は、ポリマー分子を分析するためのシステムを提供する。詳しくは、本発明は、一本鎖のポリヌクレオチド分子の配列決定に関する。
WatsonおよびCrickが1953年にDNA分子の構造を解明して以来、遺伝学研究者は、個々のDNA分子を配列決定する、迅速かつ効率的な方法を見出そうとしてきた。Sanger/BarrellおよびMaxam/Gilbertは、1975と1977との間に、配列決定技術において、主要なブレークスルーを代表する2つの新規のDNA配列決定法を開発した。現在広範に使用される方法は全て、Sanger/Barrell法に基づき、最近23年間におけるDNA配列決定の開発は、大体、この方法の改変であった。
Technology Review、1999 Mar/Apr 102(2):64〜68
本発明は、ポリマー分子を分析するため方法に関する。これらの方法は、単一の分子の感度でのDNA分子およびRNA分子の高スループットな読み取りに使用される。本発明の方法は、以下を包含する:(1)DNA(またはRNA)二本鎖の電気的に制御された解離(unzipping)、(2)1つ以上の分子シグナル検出を使用する、分子の正体(identity)(またはコード)の読み取り、および(3)核酸とハイブリダイズしたプローブとの間の分子の相互作用(例えば、強力に相互作用するプローブは、移行速度にさらなる低下を生じる)による、ナノ細孔通過プロセス(nano pore threading process)の制御された減速。
本発明は、ポリマー分子を分析する方法に関する。これらの方法は、単一の分子の感度での、DNA分子およびRNA分子の高スループットな読み取りに使用される。本発明の方法は、以下を包含する:(1)DNA(またはRNA)二本鎖の電気的に制御された解離、(2)1つ以上の分子シグナル検出を使用する分子の正体(またはコード)の読み取り、および(3)核酸とハイブリダイズしたプローブとの間の分子の相互作用(例えば、強力に相互作用するプローブは、移行速度にさらなる低下を生じる)による、ナノ細孔通過プロセスの制御された減速。
時間依存的な電場によるナノ細孔測定(例えば、配列決定法)を可能にするために、解離法を開発した。本解離法は、解離電圧を可変にしたまま、細孔への侵入速度を最適化する高い電圧を使用する。したがって、数百の個々の分子を、短い間の時間(数秒間)で調べることができる。本方法は、ポリヌクレオチドの通過の間に、熟練者が、ナノ細孔を横切って印加される電場を迅速に変化させることを可能にする。
PAGEで精製されたssDNAおよびDNAヘアピン(Eurogentec,San Diego,CA)を、10mM Tris、1mM EDTA、pH 8.5溶液中で緩衝化させ、測定の前に、10分間、75℃まで加熱し、その後、4℃まで急冷した。ヘアピン分子は、3’の一本鎖の突出(50マーのポリ−dA)、以下の配列(自己相補的な部分には下線が施してある)を有し、介在する6塩基ループを含む10bpのヘリックス部分から構成されている:

(ssDNAのトランスロケーション時間の分布)
各々個々のDNAのトランスロケーション時間(tT)は、α−HLの細孔のチャネル部分内へ最初の数塩基のDNA分子の侵入から、チャネルの他方の側から分子が抜け出るまでの時間間隔として規定される。これらの事象は、開口細孔電流の約10%までの、細孔を通って流れるイオン電流の急激な低下により明確に観察される。ヘアピンの場合に、DNAは、その一本鎖の3’突出によりチャネルに侵入することしかできない。なぜなら、その他方の末端のループは、細孔に侵入するには大きすぎるからである。この場合において、tTは、上記の3つの連続的なプロセスの合計である(すなわち、tT=ts1+tunzip+ts2、ここで、ts1は、一本鎖の突出(ポリ(dA)50)のスライド時間であり、ts2は、解離したヘアピン+ループ(26塩基)のスライド時間であり、そしてtunzipは解離時間である)。
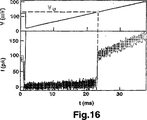
定常電圧(または、力)でのDNAヘアピンの解離を研究した。電圧の急激な変化をナノ細孔を横切って印加し、そして解離動態を測定した。この実験で使用した代表的な電圧および電流のトレースを図13に示す。DNAは、時間t=0でチャネルに侵入する。図10に説明されるように、分子は、短時間の間、細孔内に引き込まれ、保持される。t=0.8msにおいて、解離電圧(90mV)を印加するが、解離がt=tU=70ms(電流の急激な上昇により示される)で生じるまで、電流はブロックされる(より低いレベル)。解離電圧VUを印加すると、電流は、その適切な閉鎖細孔レベルまでわずかに上昇し(細孔電流の拡大図については、図11を参照のこと)、t〜70ms(解離電圧の印加から測定される)まで、この閉鎖レベルに留まった。この時点において、電流は、この電圧に対応する開口細孔電流レベルまで急激に上昇した。解離したヘアピンのスライド時間は、図13の時間スケールで決定するには短すぎるので、この移行は、ヘアピンの解離の瞬間を示す。1分間あたり約100回の個別の解離事象を累積する自動化手順を使用して、解離プロセスを反復した。
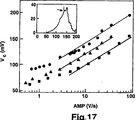
能動的な制御法によって、任意の時間依存的な電圧V(t)を印加して、結合の切断の動態を測定することができる。特に、力分光法の測定を、代表的に、一定の負荷速度または
Claims (40)
- 標的ポリヌクレオチドを分析する方法であって、該方法は:
(a)複数の設計モノマーを提供する工程であって、ここで、1つ以上の設計モノマーは、所定のヌクレオチド塩基と対応し、設計モノマーの各々は、1つ以上のシグナル分子と関連付けられる、工程;
(b)リガーゼ酵素を使用して、標的ポリヌクレオチドのヌクレオチド配列に対応する配列内に複数の設計モノマーを連結することにより、設計ポリマーを形成する工程;
(c)該設計モノマーと関連付けられたシグナル分子を、該設計ポリマーにアニーリングさせて、設計ポリマー−シグナル複合体を形成する工程;および
(d)ナノ細孔を使用して該複合体の一方の末端から該シグナル分子を順次除去することによる、該複合体を分析に供する工程であって、該複合体からの該シグナル分子の除去は、該標的ポリヌクレオチド内のヌクレオチド塩基配列を同定するために用いられるシグナルのパターンを生成する、工程、
を包含する、方法。 - 前記標的ポリヌクレオチドが、DNA、RNAおよびPNAからなる群より選択される、請求項1に記載の方法。
- 前記標的ポリヌクレオチドがDNAである、請求項2に記載の方法。
- 前記DNAが、cDNAまたはgDNAのいずれかである、請求項3に記載の方法。
- 前記設計モノマーが、5ヌクレオチド塩基〜10ヌクレオチド塩基に及ぶヌクレオチド塩基の範囲を有する、請求項1に記載の方法。
- 前記設計モノマーが、DNA、RNAまたはPNAを含む、請求項1に記載の方法。
- 前記標的ポリヌクレオチド内のヌクレオチドの各々が、二進コードにより表される、請求項1に記載の方法。
- 前記ヌクレオチドが、アデニン、シトシン、ウラシル(uricil)、グアニンおよびチミンからなる群より選択される、請求項7に記載の方法。
- 前記二進コードが2つの分子ビーコンから構成され、ここで、該分子ビーコンの特定の配列が、前記ヌクレオチドに対応する、請求項7に記載の方法。
- 前記シグナル分子が前記設計モノマーにハイブリダイズされる、請求項1に記載の方法。
- 前記シグナル分子と前記設計モノマーとの間のハイブリダイゼーションのパーセントが、90%以上である、請求項10に記載の方法。
- 前記シグナル分子と前記設計モノマーとの間のハイブリダイゼーションのパーセントが、80%〜90%の間である、請求項10に記載の方法。
- 前記シグナル分子と前記設計モノマーとの間のハイブリダイゼーションのパーセントが、70%〜80%の間である、請求項10に記載の方法。
- 前記シグナル分子と前記設計モノマーとの間のハイブリダイゼーションのパーセントが、60%〜70%の間である、請求項10に記載の方法。
- 前記シグナル分子が分子ビーコンである、請求項1に記載の方法。
- 前記分子ビーコンが、1つ以上のフルオロフォアと1つ以上のクエンチャーとを含む、請求項15に記載の方法。
- 前記分子ビーコンの5’末端上のヌクレオチド配列が、該分子ビーコンの3’末端上のヌクレオチド配列と相補的である、請求項16に記載の方法。
- 2つの分子ビーコンが存在し、各々の分子ビーコンが特定の設計モノマーに対応する、請求項15に記載の方法。
- 第一の分子ビーコンが第一の設計モノマーに対応し、第二の分子ビーコンが第二の設計モノマーに対応する、請求項18に記載の方法。
- ナノ細孔システムへ前記設計ポリマー−シグナル分子複合体を導入する工程をさらに包含する、請求項1に記載の方法。
- 前記ナノ細孔システムが、複数のナノ細孔と1つ以上の検出システムとを備える、請求項20に記載の方法。
- 前記ナノ細孔システムが、1つ以上のチャネルと脂質二重層とを含む、請求項21に記載の方法。
- 前記ナノ細孔が、α溶血素、バクテリオファージλに対するレセプター、グラミシジン、バリノマイシン、OmpF、OmpC、PhoE、Tsx、F線毛およびミトコンドリアポーリン(VDAC)からなる群より選択される、請求項1に記載の方法。
- 前記ナノ細孔がα溶血素である、請求項23に記載の方法。
- 前記α溶血素が前庭とチャネルとを含む、請求項24に記載の方法。
- 前記前庭が、2.5nmの直径を有する、請求項25に記載の方法。
- 前記チャネルが、1.5nmの直径を有する、請求項25に記載の方法。
- 前記チャネルが、0.5nm〜2.0nmの範囲に及ぶ細孔サイズを有する、請求項22に記載の方法。
- 電圧を使用して前記ナノ細孔システム内のナノ細孔を通して前記設計ポリマーをトランスロケーションさせることによる前記設計ポリマー−シグナル分子複合体の解離をさらに包含し、ここで、該設計ポリマーの一本鎖部分が、前記ナノ細孔システムのチャネルに侵入し、それを通過する、請求項20に記載の方法。
- 前記解離が、前記ナノ細孔システムを横切って確立された電場によりもたらされる、請求項29に記載の方法。
- 前記チャネルへ、かつ該チャネルを通っての前記一本鎖の設計ポリマーの移動が、前記ナノ細孔システムを横切る電場により促進される、請求項29に記載の方法。
- 前記解離プロセスが1つ以上のシグナル分子の遊離をもたらす、請求項29に記載の方法。
- 前記遊離されたシグナル分子が、検出可能なエネルギーを放出する分子とクエンチャー分子とを含み、該遊離されたシグナル分子が自己ハイブリダイズし、それにより該放出する分子をクエンチする、請求項32に記載の方法。
- 前記放出する分子がフルオロフォアである、請求項33に記載の方法。
- 前記フルオロフォアが、TMR、Cy5およびCPMからなる群より選択される、請求項34に記載の方法。
- 前記クエンチング分子が、ダブシル(Dabcyl)、ダブシル(Dabsyl)、および、メチルレッドからなる群より選択される、請求項34に記載の方法。
- 工程(d)の分析が、前記シグナル分子からのエネルギーの検出を包含する、請求項1に記載の方法。
- 前記検出が蛍光の検出を包含する、請求項37に記載の方法。
- 前記検出システムが光学的な検出システムである、請求項21に記載の方法。
- 前記光学的な検出システムが蛍光検出器である、請求項39に記載の方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US60126404P | 2004-08-13 | 2004-08-13 | |
| US60/601,264 | 2004-08-13 | ||
| PCT/US2005/028566 WO2006020775A2 (en) | 2004-08-13 | 2005-08-12 | An ultra high-throughput opti-nanopore dna readout platform |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2011186120A Division JP2011254837A (ja) | 2004-08-13 | 2011-08-29 | 超高スループットの光学−ナノ細孔dna読み取りプラットフォーム |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2008509671A JP2008509671A (ja) | 2008-04-03 |
| JP5190263B2 true JP5190263B2 (ja) | 2013-04-24 |
Family
ID=35908145
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2007525792A Expired - Fee Related JP5190263B2 (ja) | 2004-08-13 | 2005-08-12 | 超高スループットの光学−ナノ細孔dna読み取りプラットフォーム |
| JP2011186120A Pending JP2011254837A (ja) | 2004-08-13 | 2011-08-29 | 超高スループットの光学−ナノ細孔dna読み取りプラットフォーム |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2011186120A Pending JP2011254837A (ja) | 2004-08-13 | 2011-08-29 | 超高スループットの光学−ナノ細孔dna読み取りプラットフォーム |
Country Status (8)
| Country | Link |
|---|---|
| US (2) | US7972858B2 (ja) |
| EP (1) | EP1784754A4 (ja) |
| JP (2) | JP5190263B2 (ja) |
| CN (2) | CN101103357B (ja) |
| AU (1) | AU2005272823B2 (ja) |
| CA (1) | CA2577079C (ja) |
| IL (2) | IL181262A (ja) |
| WO (1) | WO2006020775A2 (ja) |
Families Citing this family (146)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN101103357B (zh) | 2004-08-13 | 2012-10-03 | 哈佛学院院长等 | 超高处理量光学-纳米孔dna读出平台 |
| GB0523282D0 (en) | 2005-11-15 | 2005-12-21 | Isis Innovation | Methods using pores |
| CN101495656B (zh) * | 2006-06-07 | 2017-02-08 | 纽约哥伦比亚大学理事会 | 采用带修饰的核苷酸通过纳米通道进行dna序列测定 |
| CN101680873B (zh) | 2007-04-04 | 2015-11-25 | 加利福尼亚大学董事会 | 使用纳米孔的组合物、设备、系统和方法 |
| US9121843B2 (en) | 2007-05-08 | 2015-09-01 | Trustees Of Boston University | Chemical functionalization of solid-state nanopores and nanopore arrays and applications thereof |
| US20110105366A1 (en) * | 2007-06-18 | 2011-05-05 | Illumina, Inc. | Microfabrication methods for the optimal patterning of substrates |
| WO2009046094A1 (en) | 2007-10-01 | 2009-04-09 | Nabsys, Inc. | Biopolymer sequencing by hybridization of probes to form ternary complexes and variable range alignment |
| GB2453377A (en) | 2007-10-05 | 2009-04-08 | Isis Innovation | Transmembrane protein pores and molecular adapters therefore. |
| WO2009092035A2 (en) | 2008-01-17 | 2009-07-23 | Sequenom, Inc. | Methods and compositions for the analysis of biological molecules |
| US20110229877A1 (en) | 2008-07-07 | 2011-09-22 | Oxford Nanopore Technologies Limited | Enzyme-pore constructs |
| AU2009269792A1 (en) * | 2008-07-07 | 2010-01-14 | Oxford Nanopore Technologies Limited | Base-detecting pore |
| US9650668B2 (en) | 2008-09-03 | 2017-05-16 | Nabsys 2.0 Llc | Use of longitudinally displaced nanoscale electrodes for voltage sensing of biomolecules and other analytes in fluidic channels |
| CN102186989B (zh) | 2008-09-03 | 2021-06-29 | 纳伯塞斯2.0有限责任公司 | 用于流体通道中生物分子和其它分析物的电压感测的纵向移位纳米级电极的使用 |
| US8262879B2 (en) | 2008-09-03 | 2012-09-11 | Nabsys, Inc. | Devices and methods for determining the length of biopolymers and distances between probes bound thereto |
| GB0820927D0 (en) | 2008-11-14 | 2008-12-24 | Isis Innovation | Method |
| KR101797773B1 (ko) * | 2009-01-30 | 2017-11-15 | 옥스포드 나노포어 테크놀로지즈 리미티드 | 막횡단 시퀀싱에서 핵산 구축물용 어댑터 |
| US8731843B2 (en) * | 2009-02-03 | 2014-05-20 | Complete Genomics, Inc. | Oligomer sequences mapping |
| WO2010091023A2 (en) * | 2009-02-03 | 2010-08-12 | Complete Genomics, Inc. | Indexing a reference sequence for oligomer sequence mapping |
| WO2010091021A2 (en) * | 2009-02-03 | 2010-08-12 | Complete Genomics, Inc. | Oligomer sequences mapping |
| GB0905140D0 (en) | 2009-03-25 | 2009-05-06 | Isis Innovation | Method |
| US9017937B1 (en) | 2009-04-10 | 2015-04-28 | Pacific Biosciences Of California, Inc. | Nanopore sequencing using ratiometric impedance |
| US8986928B2 (en) | 2009-04-10 | 2015-03-24 | Pacific Biosciences Of California, Inc. | Nanopore sequencing devices and methods |
| WO2010127045A2 (en) | 2009-04-29 | 2010-11-04 | Complete Genomics, Inc. | Method and system for calling variations in a sample polynucleotide sequence with respect to a reference polynucleotide sequence |
| US8926904B2 (en) | 2009-05-12 | 2015-01-06 | Daniel Wai-Cheong So | Method and apparatus for the analysis and identification of molecules |
| KR20110018763A (ko) * | 2009-08-18 | 2011-02-24 | 삼성전자주식회사 | 표적 물질을 기판에 고정하는 방법 및 장치 |
| CA2808576A1 (en) | 2009-09-30 | 2011-04-07 | Quantapore, Inc. | Ultrafast sequencing of biological polymers using a labeled nanopore |
| KR101814056B1 (ko) | 2009-12-01 | 2018-01-02 | 옥스포드 나노포어 테크놀로지즈 리미티드 | 생화학적 분석 기구 |
| US20110192723A1 (en) * | 2010-02-08 | 2011-08-11 | Genia Technologies, Inc. | Systems and methods for manipulating a molecule in a nanopore |
| US8324914B2 (en) | 2010-02-08 | 2012-12-04 | Genia Technologies, Inc. | Systems and methods for characterizing a molecule |
| US20110287414A1 (en) * | 2010-02-08 | 2011-11-24 | Genia Technologies, Inc. | Systems and methods for identifying a portion of a molecule |
| US9678055B2 (en) | 2010-02-08 | 2017-06-13 | Genia Technologies, Inc. | Methods for forming a nanopore in a lipid bilayer |
| CA2995178C (en) * | 2010-02-08 | 2023-09-12 | Genia Technologies, Inc. | Method and system for forming a nanopore in a lipid bilayer |
| US9605307B2 (en) | 2010-02-08 | 2017-03-28 | Genia Technologies, Inc. | Systems and methods for forming a nanopore in a lipid bilayer |
| EP4268944A3 (en) * | 2010-02-23 | 2024-03-20 | University of Washington | Analyte sequencing with nanopores |
| EP2366993A1 (en) * | 2010-03-08 | 2011-09-21 | Nxp B.V. | A sensor and a method of assembling a sensor |
| AU2011238582A1 (en) * | 2010-03-30 | 2012-10-25 | Trustees Of Boston University | Tools and method for nanopores unzipping-dependent nucleic acid sequencing |
| JP5764296B2 (ja) * | 2010-03-31 | 2015-08-19 | 株式会社日立ハイテクノロジーズ | 生体ポリマーの特性解析法 |
| US8652779B2 (en) | 2010-04-09 | 2014-02-18 | Pacific Biosciences Of California, Inc. | Nanopore sequencing using charge blockade labels |
| GB2492729A (en) * | 2010-05-11 | 2013-01-09 | Univ Boston | Use of Nanopore arrays for multiplex sequencing of nucleic acids |
| WO2012009578A2 (en) | 2010-07-14 | 2012-01-19 | The Curators Of The University Of Missouri | Nanopore-facilitated single molecule detection of nucleic acids |
| US20120077691A1 (en) | 2010-09-24 | 2012-03-29 | Full Spectrum Genetics, Inc. | Method of analyzing binding interactions |
| US8715933B2 (en) | 2010-09-27 | 2014-05-06 | Nabsys, Inc. | Assay methods using nicking endonucleases |
| EP2622343B1 (en) | 2010-10-01 | 2016-01-20 | Oxford Nanopore Technologies Limited | Biochemical analysis apparatus using nanopores |
| US8725422B2 (en) | 2010-10-13 | 2014-05-13 | Complete Genomics, Inc. | Methods for estimating genome-wide copy number variations |
| WO2012067911A1 (en) | 2010-11-16 | 2012-05-24 | Nabsys, Inc. | Methods for sequencing a biomolecule by detecting relative positions of hybridized probes |
| US10443096B2 (en) | 2010-12-17 | 2019-10-15 | The Trustees Of Columbia University In The City Of New York | DNA sequencing by synthesis using modified nucleotides and nanopore detection |
| GB2500360B (en) | 2010-12-22 | 2019-10-23 | Genia Tech Inc | Nanopore-based single DNA molecule characterization, identification and isolation using speed bumps |
| US8962242B2 (en) | 2011-01-24 | 2015-02-24 | Genia Technologies, Inc. | System for detecting electrical properties of a molecular complex |
| US9110478B2 (en) | 2011-01-27 | 2015-08-18 | Genia Technologies, Inc. | Temperature regulation of measurement arrays |
| WO2012109574A2 (en) | 2011-02-11 | 2012-08-16 | Nabsys, Inc. | Assay methods using dna binding proteins |
| CA2826374C (en) | 2011-02-11 | 2024-01-23 | Oxford Nanopore Technologies Limited | Mutant pores |
| WO2012121756A1 (en) * | 2011-03-04 | 2012-09-13 | Quantapore, Inc. | Apparatus and methods for performing optical nanopore detection or sequencing |
| WO2012177792A2 (en) | 2011-06-24 | 2012-12-27 | Sequenom, Inc. | Methods and processes for non-invasive assessment of a genetic variation |
| AU2012288629B2 (en) | 2011-07-25 | 2017-02-02 | Oxford Nanopore Technologies Limited | Hairpin loop method for double strand polynucleotide sequencing using transmembrane pores |
| US9367663B2 (en) | 2011-10-06 | 2016-06-14 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| WO2013052907A2 (en) | 2011-10-06 | 2013-04-11 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US10424394B2 (en) | 2011-10-06 | 2019-09-24 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US10196681B2 (en) | 2011-10-06 | 2019-02-05 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US9984198B2 (en) | 2011-10-06 | 2018-05-29 | Sequenom, Inc. | Reducing sequence read count error in assessment of complex genetic variations |
| US8688388B2 (en) | 2011-10-11 | 2014-04-01 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US20130244340A1 (en) * | 2012-01-20 | 2013-09-19 | Genia Technologies, Inc. | Nanopore Based Molecular Detection and Sequencing |
| LT2805280T (lt) | 2012-01-20 | 2022-12-27 | Sequenom, Inc. | Diagnostikos būdai, kurie atsižvelgia į eksperimentines sąlygas |
| US8986629B2 (en) | 2012-02-27 | 2015-03-24 | Genia Technologies, Inc. | Sensor circuit for controlling, detecting, and measuring a molecular complex |
| EP2820155B1 (en) | 2012-02-28 | 2017-07-26 | Population Genetics Technologies Ltd. | Method for attaching a counter sequence to a nucleic acid sample |
| WO2013154999A2 (en) | 2012-04-09 | 2013-10-17 | The Trustees Of Columbia University In The City Of New York | Method of preparation of nanopore and uses thereof |
| AU2013246702B2 (en) | 2012-04-10 | 2017-06-15 | Oxford Nanopore Technologies Limited | Mutant lysenin pores |
| CN104428425A (zh) | 2012-05-04 | 2015-03-18 | 考利达基因组股份有限公司 | 测定复杂肿瘤全基因组绝对拷贝数变异的方法 |
| US10504613B2 (en) | 2012-12-20 | 2019-12-10 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US9920361B2 (en) | 2012-05-21 | 2018-03-20 | Sequenom, Inc. | Methods and compositions for analyzing nucleic acid |
| US9012022B2 (en) * | 2012-06-08 | 2015-04-21 | Illumina, Inc. | Polymer coatings |
| WO2013185137A1 (en) | 2012-06-08 | 2013-12-12 | Pacific Biosciences Of California, Inc. | Modified base detection with nanopore sequencing |
| EP2861768A4 (en) | 2012-06-15 | 2016-03-02 | Genia Technologies Inc | CHIP SETUP AND HIGH ACCURACY NUCLEIC ACID SEQUENCING |
| CN103509852B (zh) * | 2012-06-18 | 2014-11-19 | 北京大学 | 一种基于纳米孔器件对生物分子探针标定dna的特异位点进行检测的方法 |
| US20150119259A1 (en) | 2012-06-20 | 2015-04-30 | Jingyue Ju | Nucleic acid sequencing by nanopore detection of tag molecules |
| US10497461B2 (en) | 2012-06-22 | 2019-12-03 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US11155860B2 (en) | 2012-07-19 | 2021-10-26 | Oxford Nanopore Technologies Ltd. | SSB method |
| US10482994B2 (en) | 2012-10-04 | 2019-11-19 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US9365896B2 (en) | 2012-10-19 | 2016-06-14 | Agilent Technologies, Inc. | Addition of an adaptor by invasive cleavage |
| US9651539B2 (en) | 2012-10-28 | 2017-05-16 | Quantapore, Inc. | Reducing background fluorescence in MEMS materials by low energy ion beam treatment |
| US9605309B2 (en) | 2012-11-09 | 2017-03-28 | Genia Technologies, Inc. | Nucleic acid sequencing using tags |
| US9914966B1 (en) | 2012-12-20 | 2018-03-13 | Nabsys 2.0 Llc | Apparatus and methods for analysis of biomolecules using high frequency alternating current excitation |
| US10294516B2 (en) | 2013-01-18 | 2019-05-21 | Nabsys 2.0 Llc | Enhanced probe binding |
| US20130309666A1 (en) | 2013-01-25 | 2013-11-21 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| US9759711B2 (en) | 2013-02-05 | 2017-09-12 | Genia Technologies, Inc. | Nanopore arrays |
| EP2964779B1 (en) | 2013-03-08 | 2018-08-29 | Oxford Nanopore Technologies Limited | Use of spacer elements in a nucleic acid to control movement of a helicase |
| GB201314695D0 (en) | 2013-08-16 | 2013-10-02 | Oxford Nanopore Tech Ltd | Method |
| WO2014186036A1 (en) | 2013-03-14 | 2014-11-20 | Allegro Diagnostics Corp. | Methods for evaluating copd status |
| US10732183B2 (en) | 2013-03-15 | 2020-08-04 | The Trustees Of Columbia University In The City Of New York | Method for detecting multiple predetermined compounds in a sample |
| US11976329B2 (en) | 2013-03-15 | 2024-05-07 | Veracyte, Inc. | Methods and systems for detecting usual interstitial pneumonia |
| WO2014144564A2 (en) | 2013-03-15 | 2014-09-18 | Veracyte, Inc. | Biomarkers for diagnosis of lung diseases and methods of use thereof |
| AU2014227754B2 (en) | 2013-03-15 | 2018-03-01 | The Curators Of The University Of Missouri | Encoded nanopore sensor for multiplex nucleic acids detection |
| GB201313477D0 (en) | 2013-07-29 | 2013-09-11 | Univ Leuven Kath | Nanopore biosensors for detection of proteins and nucleic acids |
| LT2981921T (lt) | 2013-04-03 | 2023-02-27 | Sequenom, Inc. | Neinvazinio genetinių variacijų vertinimo būdai ir procesai |
| CN105358714B (zh) | 2013-05-04 | 2020-03-24 | 斯坦福大学托管董事会 | 从含有少量靶标dna的样品富集dna测序文库 |
| US9862997B2 (en) | 2013-05-24 | 2018-01-09 | Quantapore, Inc. | Nanopore-based nucleic acid analysis with mixed FRET detection |
| IL309903A (en) | 2013-05-24 | 2024-03-01 | Sequenom Inc | Methods and processes for non-invasive evaluation of genetic variations |
| KR102299305B1 (ko) | 2013-06-21 | 2021-09-06 | 시쿼넘, 인코포레이티드 | 유전적 변이의 비침습 평가를 위한 방법 및 프로세스 |
| US20150073724A1 (en) | 2013-07-29 | 2015-03-12 | Agilent Technologies, Inc | Method for finding variants from targeted sequencing panels |
| US10726942B2 (en) | 2013-08-23 | 2020-07-28 | Complete Genomics, Inc. | Long fragment de novo assembly using short reads |
| BR112016007401B1 (pt) | 2013-10-04 | 2023-04-11 | Sequenom, Inc. | Método para determinar a presença ou ausência de uma aneuploidia cromossômica em uma amostra |
| CA2925111C (en) | 2013-10-07 | 2024-01-16 | Sequenom, Inc. | Methods and processes for non-invasive assessment of chromosome alterations |
| US9551697B2 (en) | 2013-10-17 | 2017-01-24 | Genia Technologies, Inc. | Non-faradaic, capacitively coupled measurement in a nanopore cell array |
| EP3640349A3 (en) | 2013-10-23 | 2020-07-29 | Roche Sequencing Solutions, Inc. | High speed molecular sensing with nanopores |
| US9567630B2 (en) | 2013-10-23 | 2017-02-14 | Genia Technologies, Inc. | Methods for forming lipid bilayers on biochips |
| US11479766B2 (en) | 2013-12-05 | 2022-10-25 | New England Biolabs, Inc. | Methods for labeling a population of RNA molecules |
| GB201403096D0 (en) | 2014-02-21 | 2014-04-09 | Oxford Nanopore Tech Ltd | Sample preparation method |
| CN106715453B (zh) | 2014-03-24 | 2021-04-30 | 哥伦比亚大学董事会 | 用于生产带标签的核苷酸的化学方法 |
| US10934581B2 (en) | 2014-04-30 | 2021-03-02 | International Business Machines Corporation | Bow tie DNA compositions and methods |
| US10167503B2 (en) | 2014-05-02 | 2019-01-01 | Oxford Nanopore Technologies Ltd. | Mutant pores |
| US10457979B2 (en) * | 2014-05-14 | 2019-10-29 | Stratos Genomics, Inc. | Translocation control for sensing by a nanopore |
| EP3760739A1 (en) | 2014-07-30 | 2021-01-06 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| CN114605507A (zh) | 2014-09-01 | 2022-06-10 | 弗拉芒区生物技术研究所 | 突变csgg孔 |
| AU2015319825A1 (en) * | 2014-09-26 | 2017-04-27 | Nooma Bio, Inc. | Target sequence detection by nanopore sensing of synthetic probes |
| WO2016055778A1 (en) | 2014-10-07 | 2016-04-14 | Oxford Nanopore Technologies Limited | Mutant pores |
| ES2789000T3 (es) | 2014-10-10 | 2020-10-23 | Quantapore Inc | Análisis de polinucleótidos basado en nanoporos con marcadores fluorescentes que se inactivan mutuamente |
| GB201418159D0 (en) | 2014-10-14 | 2014-11-26 | Oxford Nanopore Tech Ltd | Method |
| JP6757316B2 (ja) | 2014-10-24 | 2020-09-16 | クアンタポール, インコーポレイテッド | ナノ構造のアレイを使用するポリマーの効率的光学分析 |
| CN107206043A (zh) | 2014-11-05 | 2017-09-26 | 维拉赛特股份有限公司 | 使用机器学习和高维转录数据在经支气管活检上诊断特发性肺纤维化的系统和方法 |
| US20160125130A1 (en) | 2014-11-05 | 2016-05-05 | Agilent Technologies, Inc. | Method for assigning target-enriched sequence reads to a genomic location |
| JP2015126744A (ja) * | 2015-03-23 | 2015-07-09 | 株式会社日立ハイテクノロジーズ | 生体ポリマーの特性解析装置及び生体ポリマーの特性解析チップ |
| US20190078145A1 (en) | 2015-12-08 | 2019-03-14 | Quantapore, Inc. | Method of translocating nucleic acids through nanopores |
| WO2017123647A1 (en) | 2016-01-15 | 2017-07-20 | Quantapore, Inc. | Optically-based nanopore analysis with reduced background |
| EP3417065A4 (en) * | 2016-02-18 | 2019-07-17 | President and Fellows of Harvard College | METHOD AND SYSTEMS FOR MOLECULAR RECORDING BY CRISPR-CAS SYSTEM |
| US11486873B2 (en) | 2016-03-31 | 2022-11-01 | Ontera Inc. | Multipore determination of fractional abundance of polynucleotide sequences in a sample |
| EP4180531A3 (en) | 2016-05-12 | 2023-08-23 | Trustees of Boston University | Nasal epithelium gene expression signature and classifier for the prediction of lung cancer |
| GB201609220D0 (en) | 2016-05-25 | 2016-07-06 | Oxford Nanopore Tech Ltd | Method |
| EP3464612A4 (en) | 2016-05-31 | 2019-11-27 | Quantapore Inc. | BICOLOR SEQUENCING BY NANOPORES |
| EP3482196B1 (en) | 2016-07-05 | 2022-02-23 | Quantapore, Inc. | Optically based nanopore sequencing |
| EP3491560A1 (en) | 2016-07-27 | 2019-06-05 | Sequenom, Inc. | Genetic copy number alteration classifications |
| CA3031812A1 (en) | 2016-08-19 | 2018-02-22 | Quantapore, Inc. | Optically-based nanopore sequencing using quenching agents |
| AU2017324949B2 (en) | 2016-09-07 | 2024-05-23 | Veracyte, Inc. | Methods and systems for detecting usual interstitial pneumonia |
| WO2018067457A1 (en) | 2016-10-03 | 2018-04-12 | So Daniel Wai Cheong | Method and apparatus for the analysis and identification of molecules |
| EP3565533A4 (en) | 2017-01-05 | 2021-02-17 | The University of Chicago | INHIBITION OF ENTERIC INFECTION BY MODULATION OF MICROBIOTA |
| US11694768B2 (en) | 2017-01-24 | 2023-07-04 | Sequenom, Inc. | Methods and processes for assessment of genetic variations |
| EP4276197A3 (en) | 2017-11-16 | 2024-01-17 | New England Biolabs, Inc. | Mapping the location, type and strand of damaged nucleotides in double-stranded dna |
| CN107937485A (zh) * | 2017-12-29 | 2018-04-20 | 澳门大学 | 一种检测和分析dna的方法 |
| GB201807793D0 (en) | 2018-05-14 | 2018-06-27 | Oxford Nanopore Tech Ltd | Method |
| CN108932401B (zh) * | 2018-06-07 | 2021-09-24 | 江西海普洛斯生物科技有限公司 | 一种测序样本的标识方法及其应用 |
| EP3863636A1 (en) | 2018-10-08 | 2021-08-18 | Revolution Medicines, Inc. | Shp2 inhibitor compositions for use in treating cancer |
| US11493499B1 (en) | 2018-10-30 | 2022-11-08 | Seagate Technology Llc | Event timing detection for DNA sequencing |
| US20220002798A1 (en) * | 2018-12-07 | 2022-01-06 | Bgi Shenzhen | Nanopore sequencing method |
| IL294484A (en) | 2020-01-07 | 2022-09-01 | Revolution Medicines Inc | Dosage for shp2 inhibitor and methods for treating cancer |
| US11674947B2 (en) | 2020-06-13 | 2023-06-13 | International Business Machines Corporation | Nanopore structures |
| US20230314432A1 (en) | 2020-07-10 | 2023-10-05 | Covid Diagnostics Ltd. | Compositions, methods, and systems for detecting immune response |
| US20220364991A1 (en) * | 2021-03-24 | 2022-11-17 | Northeastern University | Method and System for Decoding Information Stored on a Polymer Sequence |
| WO2023164462A2 (en) * | 2022-02-22 | 2023-08-31 | Naio, Inc. | Compositions, systems, and methods for data storage by modifying and reading clusters of convertible monomers in polymers |
Family Cites Families (16)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6087186A (en) * | 1993-07-16 | 2000-07-11 | Irori | Methods and apparatus for synthesizing labeled combinatorial chemistry libraries |
| US5795782A (en) | 1995-03-17 | 1998-08-18 | President & Fellows Of Harvard College | Characterization of individual polymer molecules based on monomer-interface interactions |
| WO1998035012A2 (en) * | 1997-02-12 | 1998-08-13 | Chan Eugene Y | Methods and products for analyzing polymers |
| US6130046A (en) * | 1998-05-04 | 2000-10-10 | Affymetrix, Inc. | Techniques for synthesis integrity evaluation utilizing cycle fidelity probes |
| NO986133D0 (no) | 1998-12-23 | 1998-12-23 | Preben Lexow | FremgangsmÕte for DNA-sekvensering |
| EP2383776B1 (en) * | 1999-06-22 | 2015-02-25 | President and Fellows of Harvard College | Solid state nanopore device for evaluating biopolymers |
| WO2001018247A2 (en) | 1999-09-03 | 2001-03-15 | Lifebeam Technologies, Inc. | Optical system for rapid polymer analysis |
| EP1285252A1 (en) | 2000-04-24 | 2003-02-26 | Eagle Research & Development, LLC | An ultra-fast nucleic acid sequencing device and a method for making and using the same |
| US6413792B1 (en) | 2000-04-24 | 2002-07-02 | Eagle Research Development, Llc | Ultra-fast nucleic acid sequencing device and a method for making and using the same |
| WO2002042496A2 (en) * | 2000-11-27 | 2002-05-30 | The Regents Of The University Of California | Methods and devices for characterizing duplex nucleic acid molecules |
| US20030087253A1 (en) * | 2001-03-30 | 2003-05-08 | Chao Zhang | Polynucleotide markers for ovarian cancer |
| EP1409735A4 (en) * | 2001-06-25 | 2005-10-05 | Georgia Tech Res Inst | DOUBLE-RESONANCE ENERGY TRANSFER NUCLEIC PROBES |
| GB2378245A (en) * | 2001-08-03 | 2003-02-05 | Mats Nilsson | Nucleic acid amplification method |
| US20040110179A1 (en) * | 2002-03-15 | 2004-06-10 | Shuber Anthony P. | Method for alteration detection |
| US7005264B2 (en) | 2002-05-20 | 2006-02-28 | Intel Corporation | Method and apparatus for nucleic acid sequencing and identification |
| CN101103357B (zh) | 2004-08-13 | 2012-10-03 | 哈佛学院院长等 | 超高处理量光学-纳米孔dna读出平台 |
-
2005
- 2005-08-12 CN CN200580032954XA patent/CN101103357B/zh not_active Expired - Fee Related
- 2005-08-12 CA CA2577079A patent/CA2577079C/en not_active Expired - Fee Related
- 2005-08-12 AU AU2005272823A patent/AU2005272823B2/en not_active Ceased
- 2005-08-12 WO PCT/US2005/028566 patent/WO2006020775A2/en active Application Filing
- 2005-08-12 CN CN2012102981578A patent/CN102925549A/zh active Pending
- 2005-08-12 EP EP05785616A patent/EP1784754A4/en not_active Withdrawn
- 2005-08-12 JP JP2007525792A patent/JP5190263B2/ja not_active Expired - Fee Related
- 2005-08-12 US US11/573,627 patent/US7972858B2/en not_active Expired - Fee Related
-
2007
- 2007-02-11 IL IL181262A patent/IL181262A/en not_active IP Right Cessation
-
2011
- 2011-01-12 IL IL210586A patent/IL210586A0/en unknown
- 2011-06-30 US US13/173,247 patent/US8802838B2/en not_active Expired - Fee Related
- 2011-08-29 JP JP2011186120A patent/JP2011254837A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| AU2005272823B2 (en) | 2012-04-12 |
| CA2577079C (en) | 2014-05-20 |
| CN101103357B (zh) | 2012-10-03 |
| EP1784754A4 (en) | 2009-05-27 |
| IL181262A0 (en) | 2008-03-20 |
| CN102925549A (zh) | 2013-02-13 |
| IL181262A (en) | 2012-06-28 |
| US7972858B2 (en) | 2011-07-05 |
| JP2011254837A (ja) | 2011-12-22 |
| WO2006020775A2 (en) | 2006-02-23 |
| JP2008509671A (ja) | 2008-04-03 |
| US20110257043A1 (en) | 2011-10-20 |
| AU2005272823A1 (en) | 2006-02-23 |
| IL210586A0 (en) | 2011-07-31 |
| WO2006020775A3 (en) | 2006-06-15 |
| US20090029477A1 (en) | 2009-01-29 |
| US8802838B2 (en) | 2014-08-12 |
| CN101103357A (zh) | 2008-01-09 |
| CA2577079A1 (en) | 2006-02-23 |
| EP1784754A2 (en) | 2007-05-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP5190263B2 (ja) | 超高スループットの光学−ナノ細孔dna読み取りプラットフォーム | |
| McNally et al. | Optical recognition of converted DNA nucleotides for single-molecule DNA sequencing using nanopore arrays | |
| US20190360035A1 (en) | Nanopore-based nucleic acid analysis with mixed fret detection | |
| JP5372570B2 (ja) | ナノポアを用いたバイオポリマー決定方法、システム、及びキット | |
| US20140087474A1 (en) | Apparatus and methods for performing optical nanopore detection or sequencing | |
| AU2010301128B2 (en) | Ultrafast sequencing of biological polymers using a labeled nanopore | |
| JP2021503969A (ja) | 出現(emergence)による核酸のシーケンシング | |
| JP6740222B2 (ja) | 互いに消光する蛍光標識とのナノポアを基礎にしたポリマー分析 | |
| EP3387108B1 (en) | Method of translocating nucleic acids through nanopores | |
| CN114207149A (zh) | 根据突现的测序 | |
| US20190033286A1 (en) | Redundant polymer analysis by translocation reversals | |
| Lee et al. | Rapid DNA sequencing by direct nanoscale reading of nucleotide bases on individual DNA chains | |
| AU2012201675A1 (en) | An ultra high-throughput opti-nanopore DNA readout platform | |
| Linh et al. | Genotyping single nucleotide polymorphisms using different molecular beacon multiplexed within a suspended core optical fiber | |
| ZIXIN | Large Scale Genome Mapping on An Array of Nanochannels |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20080725 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20091029 |
|
| A711 | Notification of change in applicant |
Free format text: JAPANESE INTERMEDIATE CODE: A711 Effective date: 20091030 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20091030 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20110428 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20110727 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20110803 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20110829 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A821 Effective date: 20110829 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20111213 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20120313 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20120501 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20120801 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20121228 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20130128 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20160201 Year of fee payment: 3 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| LAPS | Cancellation because of no payment of annual fees |