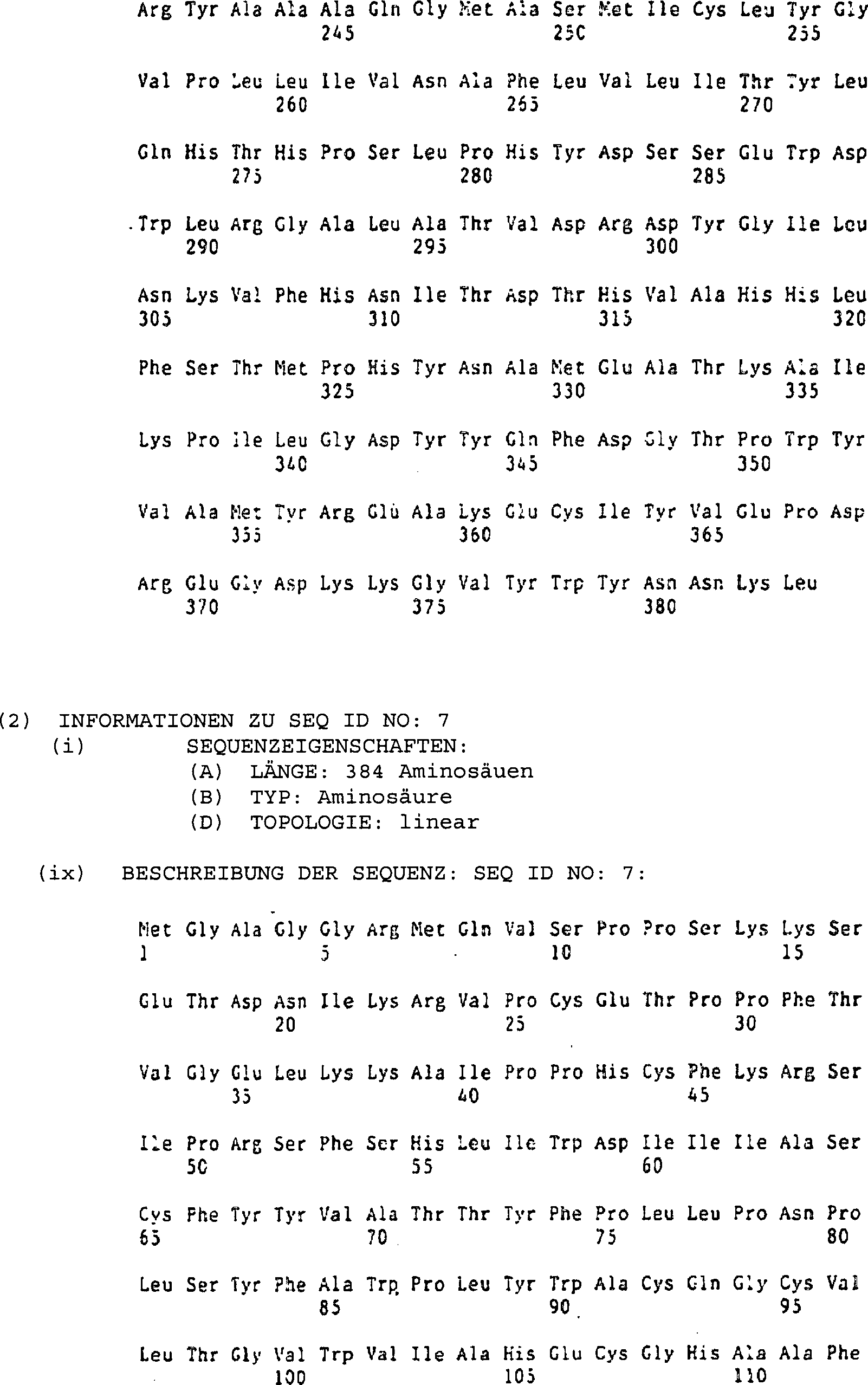-
TECHNISCHES
GEBIET
-
Die
vorliegende Erfindung betrifft die Identifizierung von Nukleinsäuresequenzen
und Nukleinsäure-Konstrukten
und damit in Verbindung stehende Verfahren, sowie die Verwendung
dieser Sequenzen und Konstrukte, um genetisch modifizierte Pflanzen
zum Zwecke der Veränderung
der Fettsäurezusammensetzung
von pflanzlichen Ölen,
Wachsen und verwandten Verbindungen herzustellen.
-
DEFINITIONEN
-
Gegenstand
dieser Erfindung ist eine Klasse von Enzymen, die eine Hydroxylgruppe
in mehrere verschiedene Fettsäuren
einbringen, was zur Herstellung von mehreren verschiedenen Arten
von hydroxylierten Fettsäuren
führt.
Diese Enzyme katalysieren insbesondere die Hydroxylierung von Ölsäure zu 12-Hydroxyölsäure sowie
die Hydroxylierung von Icosensäure
zu 14-Hydroxyicosensäure.
Andere Fettsäuren,
wie Palmitoleinsäure
oder Erucasäure
können
ebenfalls Substrate sein. Da es nicht möglich ist, sich auf dieses
Enzym durch Hinweis auf ein einzelnes Substrat oder Produkt zu beziehen,
wird im Rahmen dieser Anmeldung auf dieses Enzym als kappa-Hydroxylasen
Bezug genommen, um klarzustellen, dass das Enzym das Hydroxyl drei
Kohlenstoffatome distal (d.h. entfernt vom Carboxylkohlenstoff der
Acylkette) von einer Doppelbindung einbringt, die in der Nähe des Zentrums
der Acylkette lokalisiert ist.
-
Die
folgenden Fettsäuren
sind ebenfalls Gegenstand der Erfindung: Rizinolsäure, 12-Hydroxy-cis-9-octadecensäure (12OH-18:1cisΔ9);
Lesquerolsäure,
14-Hydroxy-cis-11-icosensäure
(14OH-20:1cisΔ11); Densipolsäure, 12-Hydroxy-cis-9,15-octadecadiensäure (12OH-18:2cisΔ9,15);
Auricolsäure,
14-Hydroxy-cis-11,17-icosadiensäure, (14OH-20:2cisΔ11,17);
Hydroxyerucasäure,
16-Hydroxy-cis-13-docosensäure (16OH-22:1cisΔ13);
Hydroxypalmi toleinsäure,
12-Hydroxy-cis-9-hexadecensäure
(12OH-16:1cisΔ9);
Icosensäure (20:1cisΔ11).
Es wird angemerkt, dass Icosensäure
in einigen Ländern
Eicosensäure
genannt wird.
-
HINTERGRUND
-
Ausführliche
Analysen der Fettsäurezusammensetzung
von Samenölen
von verschiedenen Spezies höherer
Pflanzen haben dazu geführt,
dass mindestens 33 strukturell verschiedene monohydroxylierte Pflanzenfettsäuren und
12 verschiedene polyhydroxylierte Pflanzenfettsäuren identifiziert wurden,
die von einer oder von mehreren Pflanzen-Spezies akkumuliert werden
(Überblick
in van de Loo et al., 1993). Rizinolsäure, der hauptsächliche
Bestandteil des Samenöls
der Rizinuspflanze Rizinus communis (L.), ist von kommerzieller Bedeutung.
Es wurde eine Gen dieser Spezies kloniert, das für eine Fettsäure-Hydroxylase
kodiert; dieses Gen wurde dazu verwendet, Rizinolsäure in transgenen
Pflanzen einer anderen Spezies zu produzieren. Der wissenschaftliche
Beweis, der die Behauptung stützt,
wurde 1995 veröffentlicht.
Gegenstand dieser Erfindung ist die Verwendung des Rizinus-Hydroxylase-Gens,
um damit auch andere hydroxylierte Fettsäuren wie Lesquerolsäure, Densipolsäure, Hydroxypalmitoleinsäure, Hydroxyerucasäure und
Auricolsäure
in transgenen Pflanzen zu produzieren. Darüber hinaus ist die Identifizierung
eines Gens, das für
eine homologe Hydroxylase aus Lesquerella fendleri kodiert, sowie
die Verwendung dieses Gens zur Herstellung dieser hydroxylierten Fettsäuren in
transgenen Pflanzen ebenfalls Gegenstand dieser Erfindung.
-
Rizinus
ist eine kleinere Ölsamenpflanze.
Ungefähr
50 % des Samengewichtes besteht aus Öl (Triacylglycerol), in dem
85 bis 90 % der gesamten Fettsäuren
aus der hydroxylierten Fettsäure
Rizinolsäure
bestehen. Öl,
das aus Rizinus-Samen gepresst oder extrahiert wurde, hat zahlreiche
industrielle Anwendungen, die auf den Eigenschaften basieren, welche
von der hydroxylierten Fettsäure
herrühren.
Die wichtigsten Verwendungen sind die Herstellung von Farben und
Lacken, Nylon-artigen synthetischen Polymeren, Harzen, Schmiermitteln
und Kosmetika (Atsmon, 1989). Neben dem Öl enthält der Rizinus-Samen das extrem
toxische Protein Rizin, allergene Proteine sowie das Alkaloid Rizinin.
Diese Bestandteile schließen
die Verwendung von unbehandeltem Samen-Mehl (nach Ölextraktion)
als Viehfutter aus, was normalerweise eine wichtigen ökonomischen
Aspekt der Ölsamenverwendung
darstellt. Darüber
hinaus hat Rizinus wegen der wechselhaften Beschaffenheit der Rizinuspflanzen
und den fehlenden Aufwendungen bei der Züchtung nur wenige agronomisch vorteilhafte
Eigenschaften. Aus der Gesamtschau dieser Gründe wird Rizinus in den Vereinigten
Staaten nicht mehr angebaut; die Entwicklung einer alternativen,
heimischen Bezugsquelle für
hydroxylierte Fettsäuren
wäre daher
attraktiv. Die Herstellung von Rizinolsäure, dem wichtigen Bestandteil
des Rizinusöls,
in einer etablierten Ölsamenpflanze
durch genetische Veränderung
würde ein
besonders wirksames Mittel zur Erzeugung einer heimischen Bezugsquelle
darstellen.
-
Da
es keine praktische Bezugsquelle für Lesquerolsäure, Densipolsäure und
Auricolsäure
aus Pflanzen gibt, die an moderne landwirtschaftliche Verfahren
angepasst sind, werden diese Fettsäuren derzeit von der Industrie
nicht im großen
Maßstab
verwendet. Die Fettsäuren
würden
jedoch ähnlich
wie Rizinolsäure
verwendet werden, wenn sie in größeren Mengen
hergestellt werden könnten
und zu Kosten, die denjenigen der anderen aus Pflanzen erhaltenen
Fettsäuren ähneln (Smith,
1985). Pflanzen-Spezies, wie bestimmte Spezies der Gattung Lesquerella,
die einen hohen Anteil dieser Fettsäuren akkumulieren, sind nicht
domestiziert worden und werden gegenwärtig nicht als praktische Bezugsquelle
für Fettsäuren in
Betracht gezogen (Hirsinger, 1989). Diese Erfindung stellt einen
nützlichen
Schritt zur eventuellen Herstellung dieser und anderer hy droxylierter
Fettsäuren
in transgenen Pflanzen von landwirtschaftlicher Bedeutung dar.
-
Die
taxonomischen Beziehungen zwischen Pflanzen mit ähnlichen oder identischen Arten
von ungewöhnlichen
Fettsäuren
sind untersucht worden (van de Loo et al., 1993). In einigen Fällen treten
bestimmte Fettsäuren
vorwiegend oder ausschließlich
in verwandten Taxa auf. In anderen Fällen scheint es keine direkte Verbindung
zwischen taxonomischer Verwandtschaft und dem Auftreten von ungewöhnlichen
Fettsäuren
zu geben. In diesem Zusammenhang wurde kürzlich Rizinolsäure in 12
Gattungen aus 10 Familien identifiziert (zusammengefasst in van
de Loo et al., 1993). Es scheint daher, dass die Fähigkeit
zur Synthese hydroxylierter Fettsäuren während der Radiation der Angiospermen
mehrere Male unabhängig
aufgetreten ist. Dies deutete für
uns darauf hin, dass die Enzyme, welche Hydroxyl-Gruppen in Fettsäuren einbringen,
durch geringfügige Modifikationen
eines verwandten Enzyms entstanden sind. Wie vorliegend tatsächlich gezeigt
wird, ist die Sequenzähnlichkeit
zwischen Δ12-Fettsäure-Desaturasen
und der kappa-Hydroxylase aus Rizinus so hoch, dass es auf Basis
der Hinweise in der schriftlichen Literatur nicht möglich ist,
eindeutig zu bestimmen, ob ein bestimmtes Enzym eine Desaturase
oder eine Hydroxylase ist. Dementsprechend zeigt die Patentanmeldung (WO
94/11516), die behauptet, die Isolierung und Verwendung von Δ12-Fettsäure-Desaturasen zu lehren, nicht,
wie man zwischen einer Hydroxylase und einer Desaturase unterscheiden
kann. Da es für
die genetische Manipulation von Pflanzenölen wichtig ist, diese Aktivitäten unterscheiden
zu können,
ist die Brauchbarkeit dieser Anmeldung auf diejenigen Beispiele
beschränkt,
in denen ein direkter experimenteller Beweis (z.B. eine veränderte Fettsäurezusammensetzung
in transgenen Pflanzen) geliefert wird, der die Zuweisung zu einer Funktion
stützt.
Ein Verfahren zur Unterscheidung zwischen Fettsäure-Desaturasen und Fettsäure-Hydroxylasen
auf Basis der Aminosäuresequenz
des Enzyms ist ebenfalls Gegenstand dieser Erfindung.
-
Ein
Merkmal hydroxylierter oder anderer ungewöhnlicher Fettsäuren ist,
dass sie im Allgemeinen auf Samen-Triacylglycerole beschränkt sind,
die weitgehend von den polaren Lipiden durch unbekannte Mechanismen
ausgeschlossen werden (Battey und Ohlrogge, 1989; Prasad et al.,
1987). Dies ist sehr verblüffend, da
Diacylglycerol sowohl ein Vorläufer
für Triacylglycerol
als auch für
polare Lipide ist. Von Rizinus-Mikrosomen wurden einige Hinweise
darauf erhalten, dass der Pool der Rizinoleoylenthaltenden polaren
Lipiden durch eine Präferenz
der Diacylglycerol-Acyltransferase für Rizinoleat-enthaltende Diacylglycerole
minimiert ist (Bafor et al., 1991). Analysen von vegetativen Geweben
haben zu einigen Berichten über
ungewöhnliche
Fettsäuren
geführt,
die sich von denen unterscheiden, die in der Cuticula (cuticle)
auftreten. Die Cuticula enthält
verschiedene hydroxylierte Fettsäuren,
die miteinander verestert sind, wobei ein Polyester mit hohem Molekulargewicht
gebildet wird, der strukturellen Aufgaben dient. Es gibt eine kleine
Anzahl von anderen Ausnahmen, in denen ungewöhnliche Fettsäuren in
nicht-Samengeweben vorgefunden werden.
-
Die
Biosynthese von Rizinolsäure
aus Ölsäure im sich
entwickelnden Endosperm von Rizinus (Ricinus communis) wurde durch
eine Vielzahl von Verfahren untersucht. Morris, (1967) zeigte in
Doppel-Markierungsstudien, dass die Hydroxylierung unmittelbar durch
Hydroxylsubstitution und nicht über
einen ungesättigtes
Intermediat, ein Keto-Intermediat oder ein Epoxy-Intermediat verläuft. Die Hydroxylierung unter
Verwendung von Oleyl-CoA als Vorläufer kann in ungereinigten
Präparationen
oder Mikrosomen gezeigt werden, wobei jedoch die Aktivität in Mikrosomen
nicht stabil und veränderlich
ist, und die Isolierung von Mikrosomen einen beträchtlichen
und zuweilen sogar vollständigen
Verlust der Aktivität
mit sich bringt (Galliard und Stumpf, 1966; Moreau und Stumpf, 1981). Ölsäure kann
Oleyl-CoA als Vorläufer
ersetzen, jedoch nur in Gegenwart von CoA, Mg2+ und
ATP (Galliard und Stumpf, 1966), was darauf verweist, dass die Aktivierung
zu einem Acyl-CoA notwendig ist. Es konnte jedoch keine Radioaktivität in Rizinoleoyl-CoA
(Moreau und Stumpf, 1981) nachgewiesen werden. Diese und andere
kürzlich
gemachten Beobachtungen (Bafor et al., 1991) sind als Beleg interpretiert
worden, dass das Substrat für
die Rizinus-Oleat-Hydroxylase Ölsäure ist,
die mit Phosphatidylcholin oder einem anderen Phospholipid verestert
ist.
-
Die
Hydroxylase ist empfindlich gegenüber Cyanid und Azid, und eine
Dialyse gegen Metall-Chelatoren verringert die Aktivität, welche
durch Zugabe von FeSO4 wieder hergestellt
werden kann, was darauf verweist, dass Eisen an der Enzymaktivität beteiligt
ist (Galliard und Stumpf, 1966). Die Synthese von Rizinolsäure erfordert
molekularen Sauerstoff (Galliard und Stumpf, 1966; Moreau und Stumpf
1981) und NAD(P)H, um Cytochrom b5 zu reduzieren, von dem man annimmt,
dass es sich um den intermediären
Elektronendonor der Hydroxylasereaktion handelt (Smith et al., 1992).
Kohlenstoffmonoxid inhibiert die Hydroxylierung nicht, was darauf
verweist, dass Cytochrom P450 nicht beteiligt ist (Galliard und
Stumpf, 1966; Moreau und Stumpf, 1981). Daten einer Untersuchung
zur Substratspezifität
der Hydroxylase zeigen, dass alle Substratparameter (d.h. Kettenlänge und
Doppelbindungsposition in Bezug auf die beiden Enden) wichtig sind;
Abweichungen hinsichtlich dieser Parametern verursachen eine relativ
zur Ölsäure verringerte
Aktivität
(Howling et al., 1972). Die Position, an der das Hydroxyl eingebracht
wurde, wurde jedoch durch den Teil mit der Doppelbindung bestimmt,
und liegt immer 3 Kohlenstoffatome distal. Das Acyl-Hydroxylase-Enzym
aus Rizinus kann somit abhängig
von der Verfügbarkeit
von Substanzen eine Familie von verschiedenen hydroxylierten Fettsäuren produzieren.
Das Enzym wird daher der Einfachheit halber im Rahmen der vorliegenden
Anmel dung als kappa-Hydroxylase bezeichnet (und nicht als Oleat-Hydroxylase), um
auf die breite Substratspezifität
zu verweisen.
-
Die
kappa-Hydroxylase aus Rizinus weist zahlreiche oberflächliche Ähnlichkeiten
mit den mikrosomalen Fettsäure-Desaturasen
auf (Browse und Somerville, 1991). Pflanzen verfügen über eine mikrosomale Oleat-Desaturase,
die an der Position Δ12
aktiv ist. Das Substrat dieses Enzyms (Schmidt et al., 1993) und
das Substrat der Hydroxylase (Bafor et al., 1991) scheint eine Fettsäure zu sein,
die mit der sn-2-Position von Phosphatidylcholin verestert ist.
Wenn Oleat das Substrat ist, tritt die Modifikation an derselben
Position (Δ12) in
der Kohlenstoffkette auf und erfordert dieselben Cofaktoren, nämlich Elektronen
von NADH über
Cytochrom b5 und molekularen Sauerstoff.
Keines der Enzyme wird durch Kohlenstoffmonoxid, dem charakteristischen
Inhibitor der Cytochrom P450-Enzyme, inhibiert (Moreau und Stumpf,
1981).
-
Es
scheint keine veröffentlichten
biochemischen Studien über
die Eigenschaften des Hydroxylase-Enzyms (der Hydroxylase-Enzyme) in Lesquerella
zu geben.
-
WO-A-93/11245
und WO-A-94/11516 offenbaren eine Reihe von Nukleinsäurefragmenten,
die für Fettsäure-Desaturasen
oder für
mit Fettsäure-Desaturasen
verwandte Enzyme kodieren, zur Herstellung von transgenen Pflanzen,
die in der Lage sind, Öle
mit veränderten
Mengen an ungesättigten
Fettsäuren
zu produzieren.
-
Konzeptionelle
Basis der Erfindung
-
Ein
cDNA-Klon aus Rizinus kann für
die Herstellung von Rizinolsäure
in transgenen Pflanzen verwendet werden. Wie oben erwähnt, haben
biologische Untersuchungen anderer vorgeschlagen, dass die Rizinus-Hydroxylase
keine strikte Spezifität
für Ölsäure aufweisen
könnte,
sondern auch die Hydroxylierung anderer Fettsäuren, wie Icosensäure (20:1cisΔ11)
katalysieren könnte
(Howling et al., 1972). Basierend auf diesen Studien würde man
erwarten, dass die Expression der Rizinus-Hydroxylase in transgenen
Pflanzen von Spezies wie Brassica napus und Arabidopsis thaliana,
die Fettsäuren
wie Icosensäure
(20:1cisΔ11)
und Erucasäure (13-Docosensäure; 22:1cisΔ13)
akkumulieren, aufgrund der Aktivität der Hydroxylase gegenüber diesen
Fettsäuren
zur Akkumulierung einiger der hydroxylierten Derivate dieser Fettsäuren führt. Wir
haben nunmehr einen weiteren direkten Beleg für eine solche Behauptung erhalten,
wobei der Beleg auf der Produktion von Rizinol-, Lesquerol-, Densipol-,
und Auricol-Fettsäure in transgenen
Arabidopsis-Pflanzen basiert und vorliegend als Beispiel 1 eingefügt wurde.
-
Der
Rizinus-Hydroxylase-Klon und die Rizinus-Hydroxylase-Sequenzen, die von
diesem erhalten werden, könnten
dazu verwendet werden, andere Hydroxylase-Klone von Pflanzen-Spezies
wie Lesquerella fendleri zu erhalten, von denen bekannt ist, dass
sie hydroxylierte Fettsäuren
in Samenölen
akkumulieren. Wir haben ein Beispiel für eine solche Verwendung zur
Isolierung eines neuen Hydroxylase-Gens von Lesquerella fendleri
bereitgestellt.
-
Im
Hinblick auf den hohen Grad an Sequenzähnlichkeit zwischen Δ12-Fettsäure-Denaturasen
und der Rizinus-Hydroxylase (van de Loo et al., 1995) muss die Gültigkeit
von Ansprüchen
für die
Verwendung von Desaturase- oder Hydroxylase-Genen oder davon abgeleiteten
Sequenzen zur Identifizierung von Genen mit identischer Funktion
aus anderen Arten mit Skepsis betrachtet werden. In dieser Anmeldung
wird ein Verfahren gelehrt, durch das Hydroxylase-Gene von Desaturasen
unterschieden werden können,
und es werden Verfahren beschrieben, durch die Δ12-Denaturasen durch Modifikation
des Gens, welches für
die Desaturasen ko diert, zu Hydroxylasen umgewandelt werden können. Eine
mechanistische Basis für
die ähnlichen
Reaktionsmechanismen von Desaturasen und Hydroxylasen wurde in einer
früheren
Patentanmeldung dargestellt. Die vorliegenden Belege verweisen darauf,
dass die Fettsäure-Desaturasen
einen ähnlichen
Reaktionsmechanismus wie das bakterielle Enzym Methan-Monooxygenase
aufweisen, das eine Reaktion katalysiert, bei der es zu einem Transfer
eines Sauerstoffatoms (CH4 → CH3OH) kommt (van de Loo et al., 1993). Der
Cofaktor des Hydroxylase-Bestandteils der Methan-Monooxygenase wird
als μ-Oxo-verbrücktes Di-Eisencluster (FeOFe)
bezeichnet. Die zwei Eisenatome des FeOFe-Clusters sind durch Stickstoff-
oder Sauerstoffatome gebunden (liganded), die von Proteinen stammen,
und sie sind über
das kovalent-verbrückende
Sauerstoffatom streng Redoxgekoppelt. Das FeOFe-Cluster akzeptiert
zwei Elektronen, die es zum Di-Eisen (diferrous)-Zustand reduzieren,
bevor die Bindung von Sauerstoff erfolgt. Bei der Bindung von Sauerstoff
ist es wahrscheinlich, dass auch eine heterolytische Spaltung erfolgt,
die zu einer hochvalenten Oxo-Eisen-reaktiven Spezies führt, welche
durch Resonanz-Umlagerung stabilisiert wird, die in dem streng gekoppelten
FeOFe-Cluster möglich
sind. Der stabilisierte hochvalente Oxo-Eisen-Zustand der Methan-Monooxygenase ist
in der Lage, Protonen von Methan zu extrahieren, gefolgt von einem
Sauerstofftransfer, was zu Methanol führt. Es wurde gezeigt, dass
der FeOFe-Cofaktor unmittelbar für
die Fettsäuremodifikation
bei Pflanzen relevant ist, indem gezeigt wurde, dass die Stearoyl-ACP-Desaturase
aus Rizinus diese Art von Cofaktor enthält (Fox et al., 1993).
-
Auf
Basis der obigen Überlegungen
haben wir angenommen, dass die Rizinus-Oleat-Hydroxylase eine strukturell
modifizierte Fettsäure-Desaturase
ist, wobei dies auf drei Argumenten basiert. Das erste Argument betrifft
die taxonomische Verteilung von Pflanzen, die Rizinolsäure enthalten.
Rizinolsäure
wird in 12 Gattungen von 10 Familien höherer Pflanzen gefunden (zusam mengefasst
in van de Loo et al., 1993). Daher werden Pflanzen, in denen Rizinolsäure auftritt,
im gesamten Reich der Pflanzen gefunden, wobei jedoch nahe Verwandte
dieser Pflanzen die ungewöhnliche
Fettsäure
nicht enthalten. Dieses Muster verweist darauf, dass die Fähigkeit
Rizinolsäure
zu synthetisieren, mehrere Male unabhängig entstanden ist (und verloren
wurde) und somit eine recht kürzlich
stattgefundene Verzweigung darstellt. Mit anderen Worten ist die
Fähigkeit
Rizinolsäure
zu synthetisieren schnell entstanden, was darauf hindeutet, dass
eine verhältnismäßig geringfügige genetische Änderung
in der Struktur des Ausgangsenzyms notwendig war, um dies zu erreichen.
-
Das
zweite Argument ist, dass zahlreiche biochemische Eigenschaften
der kappa-Hydroxylase aus Rizinus denen von mikrosomalen Desaturasen ähneln, wie
oben dargestellt wird (z.B. wirken beide bevorzugt auf Fettsäuren, die
mit der sn-2-Position von Phosphatidylcholin verestert sind, beide
verwenden Cytochrom b5 als intermediären Elektronendonor, beide
werden durch Cyanid gehemmt, beide benötigen molekularen Sauerstoff
als Substrat, von beiden wird angenommen, dass sie im endoplasmatischen
Retikulum lokalisiert sind).
-
Das
dritte Argument stammt aus der obigen Diskussion bezüglich der
Oxygenase-Cofaktoren, in der vorgeschlagen wird, dass die Membran-gebundenen
Fettsäure-Desaturasen
aus Pflanzen einen μ-Oxo-verbrückten Cofaktor
vom Di-Eisen-Cluster-Typ aufweisen, und dass solche Cofaktoren in
der Lage sind, sowohl die Fettsäure-Desaturierungen
als auch die Fettsäure-Hydroxylierungen
abhängig
von den Elektronen- und Struktur-Eigenschaften des Aktivitätszentrums
des Proteins zu katalysieren.
-
In
der Zusammenschau dieser drei Argumente wurde angenommen, dass die
kappa-Hydroxylase aus dem Endosperm von Rizinus homolog zu der mikrosomalen
Oleat-Δ12-Desaturase
ist, die in allen Pflanzen gefunden wird. Der Beleg, der diese Hypothese
unterstützt,
ist offenbart worden.
-
Mehrere
Gene, die für
mikrosomale Δ12-Desaturasen
aus verschiedenen Spezies kodieren, sind kürzlich kloniert worden (Okuley
at al., 1994), und wesentliche Informationen bezüglich der Struktur dieser Enzyme sind
nunmehr bekannt. Somit wird in der folgenden Erfindung gezeigt,
wie die strukturellen Informationen bezüglich der Fettsäure-Desaturasen
zu verwenden sind, um kappa-Hydroxylase-Gene gemäß der Erfindung zu isolieren.
Dieses Beispiel zeigt das Verfahren, durch das jedes Gen für eine Kohlenstoffmonoxid-insensitive pflanzliche
Fettsäure-Hydroxylase durch
den Fachmann identifiziert werden kann.
-
KURZE BESCHREIBUNG
DER ERFINDUNG
-
Zusammenfassend
gesagt stellt die vorliegende Erfindung ein isoliertes Nukleinsäurefragment
bereit, das eine Nukleinsäuresequenz
umfasst, welche für
eine Fettsäure-Hydroxylase
mit einer Aminosäure-Identität von 60
% oder mehr in Bezug auf die in SEQ ID NO: 4 dargestellte Aminosäuresequenz
kodiert.
-
Die
Aminosäuresequenz-Identität ist vorzugsweise
90 % oder mehr in Bezug auf die SEQ ID NO: 4 dargestellte Aminosäuresequenz.
-
Eine
solche Aminosäure-Identität kann 100
% betragen.
-
Die
Erfindung umfasst ein isoliertes Nukleinsäurefragment, das für ein Polypeptid
kodiert, welches Fettsäure-Hydroxylase-Aktivität aufweist
und eine Nukleinsäuresequenz-Identität von 90
% oder mehr (wahlweise 100 %) in Bezug auf die in SEQ ID NO: 1,
2 oder 3 dargestellte Nukleotidsequenz aufweist.
-
Bei
dem isolierten Nukleinsäurefragment
kann es sich um ein solches handeln, das aus einer Öl-produzierenden
Pflanzen-Spezies isoliert ist.
-
Die
Erfindung umfasst ein chimäres
Gen, das in der Lage ist, veränderte
Mengen Rizinolsäure
in einer transformierten Pflanzenzelle zu bewirken, wobei das chimäre Gen ein
wie oben beschriebenes erfindungsgemäßes Nukleinsäurefragment
umfasst, wobei das Fragment in funktionsfähiger Weise mit geeigneten
regulatorischen Sequenzen verknüpft
ist.
-
Die
Erfindung umfasst ein chimäres
Gen, das in der Lage ist, veränderte
Mengen Lesquerolsäure
in einer transformierten Pflanzenzelle zu bewirken, wobei das chimäre Gen ein
wie oben beschriebenes erfindungsgemäßes Nukleinsäurefragment
umfasst, wobei das Fragment in funktionsfähiger Weise mit geeigneten regulatorischen
Sequenzen verknüpft
ist.
-
Die
Erfindung umfasst ein chimäres
Gen, das in der Lage ist, veränderte
Mengen an Fettsäuren
in einer transformierten Pflanzenzelle zu bewirken, wobei das chimäre Gen ein
wie oben beschriebenes erfindungsgemäßes Nukleinsäurefragment
umfasst, wobei das Fragment in funktionsfähiger Weise mit geeigneten regulatorischen
Sequenzen verknüpft
ist.
-
Die
Erfindung umfasst Pflanzen, die irgendeines dieser chimären Gene
umfassen, z.B. Raps, Crambe, Brassica juncea, Brassica nigra, Sumpfblume,
Flachs, Sonnenblume, Distel, Baumwolle, Cuphea, Sojabohne, Erdnuss,
Kokosnuss, Ölpalme
oder Mais.
-
Bei
dem erfindungsgemäßen isolierten
Nukleinsäurefragment
kann es sich um ein solches handeln, das von Ricinus communis (L.)
(Rizinus) erhältlich
ist.
-
Die
Erfindung umfasst ein Verfahren zur Herstellung von Samenöl, das veränderte Mengen
an hydroxylierten Fettsäuren
enthält,
bei dem man:
- (a) eine Pflanzenzelle einer Öl-produzierenden
Spezies mit einem chimären
Gen transformiert, das eine wie oben beschriebene erfindungsgemäße isolierte
Nukleinsäure
enthält;
- (b) ausgehend von den transformierten Pflanzenzellen von Schritt
(a) fertile Pflanzen anzüchtet;
- (c) Nachkommen-Samen von den fertilen Pflanzen von Schritt (b)
auf die gewünschten
Mengen hydroxylierter Fettsäuren
untersucht; und
- (d) die Nachkommen-Samen von Schritt (c) weiterverarbeitet,
um Samenöl
zu erhalten, das veränderte Mengen
ungesättigter
Fettsäuren
enthält.
-
Die
Nutzpflanze kann ausgewählt
sein aus der Gruppe bestehend aus Raps, Crambe, Brassica juncea,
Canola, Flachs, Sonnenblume, Distel, Baumwolle, Cuphea, Sojabohne,
Erdnuss, Kokosnuss, Ölpalme und
Mais.
-
Die
Erfindung umfasst ein Triglyceridöl aus einer Pflanze, die ausgewählt ist
aus der Gruppe bestehend aus Raps, Crambe, Brassica juncea, Canola,
Flachs, Sonnenblume, Distel, Baumwolle, Cuphea, Sojabohne, Erdnuss,
Kokosnuss, Ölpalme
oder Mais, wobei die Fettsäurezusammensetzung
des Öls
dahingehend modifiziert wurde, dass sie hydroxylierte Fettsäuren enthält, durch
ein Verfahren, bei dem man eine Pflanzenzelle, die ein in ihr Genom
integriertes DNA-Konstrukt aufweist, welches eine für eine Hydroxylase
kodierende Sequenz gemäß der vorliegenden
Erfindung umfasst, unter Bedingungen anzüchtet, die die Tran skription
und Translation der besagten Pflanzen-Hydroxylase in den Pflanzenzellen
erlauben.
-
Die
Erfindung umfasst ein Verfahren zur Isolierung von Nukleinsäurefragmenten,
die für
eine Fettsäure-Hydroxylase
kodieren, bei dem man:
- (a) SEQ ID NO: 4 und
andere Fettsäure-Hydroxylase-Sequenzen
und Fettsäure-Desaturasen
vergleicht;
- (b) konservierte Sequenzen von 4 oder mehr Aminosäuren, die
in Schritt (a) erhalten werden, identifiziert;
- (c) auf Basis der in Schritt (b) identifizierten, konservierten
Sequenzen degenerierte Oligomere erstellt;
- (d) die degenerierten Oligomere von Schritt (c) verwendet, um
durch Sequenz-abhängige
Methoden Sequenzen zu isolieren, die für Fettsäure-Hydroxylasen kodieren;
- (e) ausgehend von der Nukleotidsequenz des Gens die abgeleitete
Aminosäuresequenz
des kodierten Genprodukts erhält;
und
- (f) durch Analyse von Aminosäuresequenz-Unterschieden
zwischen Fettsäure-Desaturasen
und Fettsäure-Hydroxylasen
Hydroxylase-Gene von einem Desaturase-Gen unterscheidet.
-
KURZE BESCHREIBUNG
DER ZEICHNUNGEN
-
Die 1A–D zeigen
die Massenspektren von Hydroxyfettsäure-Standards (1A,
O-TMS-Methyl-Rizinoleat; 1B, O-TMS-Methyl-Densipoleat; 1C,
O-TMS-Methyl-Lesqueroleat; und 1D, O-TMS-Methyl-Auricoleat.
-
2 zeigt
das Fragmentierungsmuster von Trimethylsilylierten Methylestern
von Hydroxyfettsäuren.
-
3A zeigt das Gaschromatogramm von Fettsäuren, die
aus Samen von Wildtyp-Arabidopsis-Pflanzen extrahiert wurden. 3B zeigt das Gaschromatogramm von Fettsäuren, die
aus Samen von transgenen Arabidopsis-Pflanzen extrahiert wurden,
die das fah12-Hydroxylase-Gen enthielten. Die Zahlen verweisen auf die
folgenden Fettsäuren:
[1] 16:0; [2] 18:0; [3] 18:1cisΔ9;
[4] 18:2cisΔ9,12;
[5] 20:0; [6] 20:1cisΔ11; [7] 18:3cisΔ9,12,15;
[8] 22:1cisΔ13;
m [9] 24:1cisΔ13;
[10] Rizinolsäure
[11] Densipolsäu
re, [12] Lesquerolsäure;
[13] Auricolsäure.
-
Die 4A–D zeigen
die Massenspektren von neuen Fettsäuren, die in Samen von transgenen
Pflanzen gefunden wurden. 4A zeigt
das Massenspektrum des Peaks 10 aus 3B. 4B zeigt
das Massenspektrum des Peaks 11 aus 3B. 4C zeigt
das Massenspektrum von Peak 12 aus 3B. 4D zeigt
das Massenspektrum von Peak 14 aus 3B.
-
5 zeigt
die Nukleotidsequenz von pLesg2 (SEQ ID NO: 1).
-
6 zeigt
die Nukleotidsequenz von pLesg3 (SEQ ID NO: 2).
-
7 zeigt
einen Northernblot von Gesamt-RNA aus Samen von L. fendleri, die
mit pLesq2 oder pLesq3 als Sonde untersucht wurden. S verweist darauf,
dass die RNA aus Samen stammt; L verweist darauf, dass die RNA aus
Blättern
stammt.
-
Die 8A–B zeigen
die Nukleotidsequenz eines genomischen Klons, der für pLesq-HYD
(SEQ ID NO: 3) kodiert sowie die davon abgeleitete Aminosäuresequenz
des Hydroxylase-Enzyms, das durch dieses Gen kodiert wird (SEQ ID
NO: 4).
-
Die 9A–B zeigen
verschiedene Sequenz-Alignments von abgeleiteten Aminosäuresequenzen
für kappa-Hydroxylasen
und mikrosomale Δ12-Desaturasen.
Die Abkürzungen
sind die Folgenden: Rcfah12, fah12, Hydroxylase-Gen aus R. communis
(van de Loo et al., 1995); lffah12, kappa-Hydroxylase-Gen aus L. fendleri;
Atfad2, fad2, Desaturase aus Arabidopsis thaliana (Okuley et al.,
1994); Gmfad2-1, fad2 Desaturase aus Glycine max (GenBank Zugriffsnummer
L43920); Gmfad2-2, fad2 Desaturase aus Glycine max (GenBank Zugriffsnummer
L43921); Zmfad2, fad2 Desaturase aus Zea mays (WO 94/11516); Rcfad2,
Fragment der fad2 Desaturase aus R. communis (WO 94/11516); Bnfad2,
fad2 Desaturase aus Brassica napus (WO 94/11516); LFFAH12.AMI, SEQ
ID NO: 4; FAH12.AMI, SEQ ID NO: 5; ATFAC2.AMI, SEQ ID NO: 6; BNFAD2.AMI,
SEQ ID NO: 7; GMFAD2-1.AMI, SEQ ID NO: 8; GMFAD-2.AMI, SEQ ID NO:
9; ZMFAD2.AMI, SEQ ID NO: 10; und RCFAD2.AMI, SEQ ID NO: 11.
-
10 zeigt
einen Southernblot von genomischer DNA aus L. fendleri, die mit
pLesq-HYD als Sonde untersucht wurde. E=EcoRI, H=HindIII, X=XbaI.
-
11 zeigt
eine Kartierung des binären
Ti-Plasmids pSLJ44024.
-
Wie
oben dargestellt ist, werden gemäß einer
ersten Ausführungsform
bestimmte rekombinante DNA-Konstrukte bereitgestellt, die für die Transkription
oder für
die Transkription und Translation (Expression) der kappa-Hydroxylasesequenz
aus Pflanzen sorgen können.
Es sind insbesondere Konstrukte bevorzugt, die zur Transkription
oder zur Transkription und Translation in Pflanzen-Wirtszellen befähigt sind.
Solche Konstrukte können
eine Vielzahl von regulatorischen Regionen umfassen, einschließlich transkriptionaler
Initiationsregionen, die von Genen erhalten werden, die vorzugsweise
im Samengewebe von Pflanzen exprimiert werden. Gemäß eines
zweiten Aspekts werden solche Konstrukte in Pflanzenzellen bereitgestellt,
insbesondere in Pflanzenzellen, die eine exprimierte Pflanzenkappa-Hydroxylase
umfassen.
-
Eine
solche Pflanzen-kappa-Hydroxylase kann in einer Wirtszelle oder
in Nachkommen derselben mittels der Expression eines Konstrukts
in der Zelle produziert werden. Zellen, die eine Pflanzen-kappa-Hydroxylase
als Ergebnis der Produktion der für die Pflanzen-kappa-Hydroxylase
kodierenden Sequenz umfassen, werden vorliegend ebenfalls vorgeschlagen.
-
Gemäß einer
weiteren Ausführungsform
werden Verfahren zur Verwendung einer für eine Pflanzen-kappa-Hydroxylase
kodierenden DNA-Sequenz zur Modifikation eines Teils der hydroxylierten
Fettsäuren, die
in einer Zelle gebildet werden, insbesondere einer Pflanzenzelle,
bereitgestellt. Pflanzenzellen, die eine solche modifizierte Zusammensetzung
bezüglich
der hydroxylierten Fettsäuren
aufweisen, werden vorliegend ebenfalls vorgeschlagen.
-
Pflanzen-kappa-Hydroxylase-Proteine
und damit verwandte Sequenzen, einschließlich Aminosäure- und
Nukleinsäuresequenzen
werden vorgeschlagen. Die vorliegend beispielhaft angeführte Pflanzen-kappa-Hydroxylase
umfasst eine Fettsäure-Hydroxylase
aus Lesquerella fendleri. Diese beispielhaft angeführte Fettsäure-Hydroxylase
kann verwendet werden, um andere erfindungsgemäße Pflanzen-Fettsäure-Hydroxylasen
zu erhalten.
-
Gemäß eines
weiteren Aspektes dieser Erfindung wird eine Nukleinsäuresequenz
beschrieben, die die Samen-spezifische Expression einer für ein assoziiertes
Polypeptid kodierenden Sequenz steuert. Die Verwendung dieser Nukleinsäuresequenz
oder von Fragmenten, die sich davon ableiten, um eine Samenspezifische
Expression einer beliebigen kodierenden Sequenz in höheren Pflanzen
zu erhalten, wird vorliegend vorgeschlagen.
-
DETAILLIERTE
BESCHREIBUNG DER ERFINDUNG
-
Eine
erfindungsgemäße genetisch
transformierte Pflanze, die hydroxylierte Fettsäuren akkumuliert, kann durch
Expression der doppelsträngigen
DNA-Moleküle,
die in dieser Anmeldung beschrieben werden, erhalten werden.
-
Eine
erfindungsgemäße Pflanzen-Fettsäure-Hydroxylase
umfasst eine beliebige Aminosäuresequenz (wie
beispielsweise ein Protein, Peptid oder Peptidfragment oder Nukleinsäuresequenzen,
die für
solche Polypeptide kodieren), die aus einer Pflanzen-Bezugsquelle erhältlich ist
und die Fähigkeit
zeigt, die Herstellung von Rizinolsäure, Lesquerolsäure, Hydroxyerucasäure (16-Hydroxy-cis-13-docosensäure) oder
Hydroxypalmitoleinsäure
(12-Hydroxy-cis-9-hexadecensäure)
aus CoA, ACP oder Lipidgebundenen Monoen-Fettsäuresubstraten unter für ein Pflanzen-Enzym reaktiven Bedingungen
zu katalysieren. Mit "für ein Enzym
reaktive Bedingungen" ist
gemeint, dass sämtliche
erforderlichen Bedingungen in einer Umgebung verfügbar sind
(d.h. solche Faktoren wie Temperatur, pH, Fehlen von inhibierenden
Substanzen), die dem Enzym seine Funktionsfähigkeit erlauben.
-
Die
bevorzugte Aktivität
einer Pflanzen-Fettsäure-Hydroxylase
gegenüber
einem bestimmten Fettsäuresubstrat
wird durch Vergleich der Produktmengen an hydroxylierten Fettsäure, die
für verschiedene
Fettsäuresubstrate
erhalten werden, bestimmt. Beispielsweise ist mit "Oleat bevorzugend" gemeint, dass die
Hydroxylase-Aktivität
der Enzympräparation
eine Präferenz
für Oleat-enthaltende
Substrate im Vergleich mit anderen Substraten zeigt. Obwohl das
genaue Substrat für
die Fettsäure-Hydroxylase aus Rizinus
nicht bekannt ist, wird angenommen, dass es sich um einen einfach
ungesättigten
Fettsäurerest
handelt, der mit einem Phospholipid wie Phosphatidylcholin verestert
ist. Es ist jedoch auch möglich,
dass die einfach unge sättigten
Fettsäuren
mit Phosphatidylethanolamin verestert sind; Phosphatidsäure oder
ein neutrales Lipid wie Diacylglycerol oder ein Coenzym A-Thioester
könnten
ebenfalls Substrate sein. Wie oben angemerkt, wurde eine signifikante
Aktivität
in radioaktiven Markierungsuntersuchungen beobachtet, in denen andere
Fettsäuresubstrate als
Oleate verwendet wurden (Howling et al., 1972), was darauf verweist,
dass die Substratspezifität
für eine Familie
von verwandten Fettsäure-Verbindungen
existiert. Da die Hydroxylase aus Rizinus die Hydroxy-Gruppen 3
Kohlenstoffatome von einer Doppelbindung entfernt einbringt, proximal
zum Methyl-Kohlenstoff der Fettsäure,
haben wir dieses Enzym der Einfachheit halber als kappa-Hydroxylase
bezeichnet. Wir glauben interessanterweise, dass die Rizinus-kappa-Hydroxylase
zur Herstellung von 12-Hydroxy-9-octadecensäure (Rizinoleat), 12-Hydroxy-9-hexadecensäure, 14-Hydroxy-11-eicosensäure, 16-Hydroxy-13-docosansäure, 9-Hydroxy-6-octadecensäure verwendet
werden könnte,
indem man sie in Pflanzen-Spezies exprimiert, die die nicht-hydroxylierten Vorstufen
produzieren. Wir streben ferner die Herstellung von zusätzlich modifizierten Fettsäuren, wie
12-Hydroxy-9,15-octadecadiensäure
an, die aus der Desaturierung von hydroxylierten Fettsäuren (z.B.
12-Hydroxy-9-octadecensäure in diesem
Beispiel) resultieren.
-
Wir
nehmen ferner an, dass zukünftige
Fortschritte in der Pflanzengenetik zur Herstellung von Substratfettsäuren wie
Icosensäureester
und Palmitoleinsäureester
in Pflanzen führen
werden, die normalerweise solche Fettsäuren nicht akkumulieren. Wir
nehmen ferner an, dass die vorliegend beschriebene Erfindung im Zusammenhang
mit solchen künftigen
Verbesserungen verwendet werden kann, um die erfindungsgemäßen hydroxylierten
Fettsäuren
in beliebigen Pflanzen-Spezies herzustellen, die geeignet sind,
die genetische Modifikation zu steuern. Daher ist die Anwendbarkeit
dieser Erfindung nach unserer Vorstel lung nicht nur auf diejenigen
Spezies beschränkt,
die gegenwärtig
geeignete Substrate akkumulieren.
-
Wie
oben erwähnt
wird eine erfindungsgemäße Pflanzen-kappa-Hydroxylase
eine Aktivität
gegenüber verschiedenen
Fettsäuresubstraten
zeigen. Während
der Biosynthese von Fetten in einer Pflanzenzelle sind die Fettsäuren üblicherweise
kovalent an Acyl-Trägerproteine
(ACP), Coenzym A (CoA) oder verschiedene zelluläre Lipide gebunden. Pflanzen-kappa-Hydroxylasen,
die eine bevorzugte Aktivität
gegenüber
Lipid-gebundenen Acylsubstraten aufweisen, sind besonders bevorzugt,
weil sie wahrscheinlich eng mit dem normalen Weg der Synthese von
Speicherlipiden in unreifen Embryos assoziiert sind. Eine Aktivität gegenüber Acyl-CoA-Substraten
und anderen synthetischen Substraten wird jedoch beispielsweise
vorliegend ebenfalls vorgeschlagen.
-
Andere
Pflanzen-kappa-Hydroxylasen sind ausgehend von den spezifischen,
beispielhaft angeführten Sequenzen,
die vorliegend bereitgestellt werden, erhältlich. Des Weiteren wird es
ersichtlich sein, dass man natürliche
und synthetische Pflanzen-kappa-Hydroxylasen,
einschließlich
modifizierter Aminosäuresequenzen und
Startmaterialien, für
das synthetische Protein-Modeling
ausgehend von den beispielhaft angeführten Pflanzen-kappa-Hydroxylasen
und den Pflanzen-kappa-Hydroxylasen, die durch Verwendung solcher
beispielhaft angeführten
Sequenzen erhalten werden, erhalten kann. Modifizierte Aminosäuresequenzen
umfassen Sequenzen die mutiert, trunkiert, vergrößert usw. wurden, wobei solche
Sequenzen partiell oder vollständig
synthetisiert sein können.
Sequenzen, die tatsächlich
aus Pflanzen-Präparationen
gereinigt werden, oder die mit diesen identisch sind oder für identische
Proteine kodieren, werden unabhängig
von dem verwendeten Verfahren zum Erhalt des Proteins oder der Sequenz
gleichermaßen
als natürlich
erhalten angesehen.
-
Der
Fachmann wird somit problemlos erkennen, dass Antikörperpräparation,
Nukleinsäure-Sonden (DNA
oder RNA) u.ä.
hergestellt und verwendet werden können, um "homologe" oder "verwandte" kappa-Hydroxylasen aus einer Vielzahl
pflanzlicher Bezugsquellen durchzumustern (Screening) und zu gewinnen. Üblicherweise
werden Nukleinsäure-Sonden
markiert, um die Detektion zu erlauben, vorzugsweise mittels Radioaktivität, obwohl
Enzyme oder andere Verfahren ebenfalls verwendet werden können. Für immunologische Screening-Verfahren
werden entweder monoklonale oder polyklonale Antikörper-Präparationen
verwendet. Polyklonale Antikörper
sind üblicherweise
bei der Isolierung von Genen nützlicher,
obwohl sie weniger spezifisch sind. Bei der Detektion wird der Antikörper unter
Verwendung von Radioaktivität
oder mit einem beliebigen aus einer Vielzahl von Systemen aus sekundärem Antikörper/Enzymkonjugat
markiert, die kommerziell verfügbar
sind.
-
Homologe
Sequenzen werden gefunden, wenn Sequenzidentität vorliegt, und sie können durch
Vergleich der Sequenzinformation (Nukleinsäure oder Aminosäure) oder
durch Hybridisierungsreaktionen zwischen einer bekannten kappa-Hydroxylase
und einer Kandidaten-Bezugsquelle bestimmt werden. Konservative
Austausche, wie Glu/Asp, Val/Ile, Ser/Thr, Arg/Lys und Gln/Asn können ebenfalls
bei Bestimmung der Sequenzhomologie berücksichtigt werden. Üblicherweise
kann eine längere
Nukleinsäuresequenz
eine geringe Sequenzidentität
von nur 50 bis 60 %, und stärker
bevorzugt eine Sequenzidentität
von mindestens etwa 70 %, zwischen der Zielsequenz und der gegebenen
Pflanzen-kappa-Hydroxylase
von Interesse aufweisen (wobei jegliche Deletionen, die vorhanden
sein können,
nicht eingeschlossen sind) und dennoch als verwandt gelten. Aminosäuresequenzen
werden bei einer Sequenzidentität
von nur 25 % zwischen zwei vollständig reifen Proteinen als homolog
angesehen (für
einen allgemeinen Überblick
siehe Doolittle, R.F., OF URFS and ORFS, University Science Books,
CA, 1986).
-
Eine
genomische oder eine andere geeignete Bibliothek, die ausgehend
von einer Kandidatenpflanzen-Bezugsquelle von Interesse hergestellt
worden ist, kann mit konservierten Sequenzen der Pflanzen-kappa-Hydroxylase
als Sonde untersucht werden, um homolog verwandte Sequenzen zu identifizieren.
Die Verwendung einer gesamten cDNA oder einer anderen Sequenz ist
möglich,
wenn kürzere
Sonden-Sequenzen nicht identifiziert wurden. Positive Klone werden
anschließend
durch Restriktionsenzymverdau und/oder Sequenzierung analysiert.
Wenn eine genomische Bibliothek verwendet wird, können eine
oder mehrere Sequenzen identifiziert werden, wobei sowohl die kodierende
Region als auch die transkriptionalen regulatorischen Elemente des
kappa-Hydroxylase-Gens
aus einer solchen Pflanzen-Bezugsquelle bereitgestellt werden. Sonden
können
auch beträchtlich
kürzer
sein als die gesamte Sequenz. Beispielsweise können Oligonukleotide verwendet
werden, wobei diese mindestens etwa 10, vorzugsweise mindestens
etwa 15, stärker
bevorzugt mindestens 20 Nukleotide lang sein sollten. Wenn Regionen
kürzerer
Länge für den Vergleich
verwendet werden, ist ein höherer
Grad von Identität
erforderlich als bei längeren
Sequenzen. Kürzere
Sonden sind oftmals insbesondere nützlich bei einer Polymerase-Kettenreaktion
(PCR), insbesondere wenn hochkonservierte Sequenzen identifiziert
werden können
(siehe Gould et al., 1989 für
Beispiele der Verwendung einer PCR zur Isolierung von homologen
Genen aus taxonomisch verschiedenen Spezies).
-
Wenn
längere
Nukleinsäurefragmente
als Sonden eingesetzt werden (> 100
bp), insbesondere bei Verwendung von vollständigen oder großen cDNA-Sequenzen,
würde man
ein Screening bei geringer Stringenz durchführen (beispielsweise bei 40
bis 50°C
unter der Schmelztemperatur der Sonde), um ein Signal von der Zielprobe
zu erhalten, welche eine Abweichung von 20 bis 50 % aufweist, d.h.
homologe Sequenzen (Beltz et al., 1983).
-
Gemäß einer
bevorzugten Ausführungsform
wird eine Pflanzen-kappa-Hydroxylase
gemäß der vorliegenden
Erfindung insgesamt mindestens 60 % Aminosäuresequenz-Ähnlichkeit mit der beispielhaft
angeführten
Pflanzen-kappa-Hydroxylase aufweisen. Insbesondere werden kappa-Hydroxylasen
bevorzugt, die von einer Aminosäure-
oder Nukleinsäuresequenz
aus Rizinus oder Lesquerella erhältlich
sind. Die Pflanzen-kappa-Hydroxylasen können eine bevorzugte Aktivität gegenüber längerkettigen
oder kürzerkettigen
Fettsäuresubstraten
aufweisen. Pflanzen-Fettsäure-Hydroxylasen,
die eine Oleat-12-Hydroxylase-Aktivität und eine Eicosenoat-14-Hydroxylase-Aktivität aufweisen,
werden beide aufgrund eines in vitro-Belegs (Howling et al., 1972)
und aufgrund des vorliegend erbrachten Belegs, dass die Rizinus-kappa-Hydroxylase
auf beide Substrate wirkt, als homolog verwandte Proteine angesehen.
Die hydroxylierten Fettsäuren
können
weiteren enzymatischen Modifikationen durch andere Enzyme unterworfen
werden, die normalerweise vorhanden sind oder durch biotechnologische
Verfahren eingebracht werden. So wird beispielsweise angenommen,
dass 14-Hydroxy-11,17-eicosadiensäure, die
in einigen Lesquerella-Spezies vorhanden ist (Smith, 1985), durch
Desaturierung von 14-Hydroxy-11-eicosensäure hergestellt
wird.
-
Es
können
nicht nur Gen-Klone und Materialien, die sich von diesen ableiten,
zur Identifizierung von homologen Pflanzen-Fettsäure-Hydroxylasen verwendet
werden, sondern die resultierenden Sequenzen, die von diesen erhalten
werden, können
auch ein weiteres Verfahren zum Erhalt von Pflanzen-Fettsäure-Hydroxylasen aus
anderen pflanzlichen Bezugsquellen bereitstellen. Die PCR könnte eine
besonders nützliche
Methode sein, um ausgehend von den vorliegend bereitgestellten Sequenzdaten
verwandte Pflanzen-Fettsäure-Hydroxylasen
zu erhalten. Der Fachmann wird in der Lage sein, Oligonukleotid-Sonden
zu erstellen, die auf Sequenzvergleichen oder Regionen basieren,
die eine eine hochkonservierte Sequenz aufweisen. Von besonderem
Interesse sind Polymerase-Kettenreaktions-Primer, die auf den konservierten
Aminosäuresequenz-Regionen
zwischen der Rizinus-kappa-Hydroxylase und der L. fendleri-Hydroxylase
(SEQ ID NO: 4) basieren. Details zum Erstellen von Sonden sowie
zu Verfahren für
eine PCR-Reaktion unter Verwendung dieser Sonden werden in den Beispielen
näher beschrieben.
-
Es
sollte ferner angemerkt werden, dass die Fettsäure-Hydroxylasen von einer Vielzahl von
Bezugsquellen dazu verwendet werden können, Fettsäure-Hydroxylierungs-Ereignisse
in einer großen
Vielzahl von Pflanzen und in vivo-Applikationen zu untersuchen.
Da alle Pflanzen Fettsäuren über einen
gemeinsamen metabolischen Weg synthetisieren, kann die Untersuchung
und/oder Anwendung einer Pflanzen-Fettsäure-Hydroxylase mit einem heterologen
Pflanzen-Wirt problemlos in einer Vielzahl von Spezies erreicht
werden.
-
Sobald
die Nukleinsäuresequenz
erhalten wurde, ist die Transkription oder die Transkription und Translation
(Expression) der Pflanzen-Fettsäure-Hydroxylasen
in einer Wirtszelle angestrebt, um eine einfache Bezugsquelle für das Enzym
herzustellen und/oder die Zusammensetzung der Fettsäuren zu
modifizieren, die in dieser in Form von freien Fettsäuren, Estern
(insbesondere mit Glycerol-Lipiden verestert oder als Bestandteile
von Wachs-Estern), Estoliden oder Ethern vorgefunden werden. Andere
nützliche
Anwendungen können
gefunden werden, wenn die Wirtszelle eine Pflanzen-Wirtszelle ist,
in vitro und in vivo.
-
Beispielsweise
kann durch Erhöhen
der Menge an kappa-Hydroxylase,
die der Pflanze zugänglich
ist, ein erhöhter
Prozentsatz von Rizinoleat oder Lesqueroleat (14-Hydroxy-11-eicosensäure) bereitgestellt
werden.
-
Kappa-Hydroxylase
-
Die
vorliegende Erfindung zeigt einen Mechanismus der Biosynthese von
Rizinolsäure
in Pflanzen; es wird nämlich
gezeigt, dass eine spezifische Pflanzen-kappa-Hydroxylase mit bevorzugter
Aktivität
gegenüber Fettsäuresubstraten
zumindest in einigen Pflanzen-Spezies an der Akkumulierung von hydroxylierten
Fettsäuren
beteiligt ist. Die Verwendung der Begriffe Rizinoleat oder Rizinolsäure (oder
Lesqueroleat oder Lesquerolsäure,
Densipoleat usw.) soll die freien Säuren, die ACP- und CoA-Ester,
die Salze dieser Säuren,
die Glycerol-Lipidester (insbesondere die Triacylglycerolester),
die Wachs-Ester, die Estoliden und die Etherderivate dieser Säuren umfassen.
-
Die
Feststellung, dass Pflanzen-Fettsäure-Hydroxylasen an der in
vivo-Produktion von hydroxylierten Fettsäuren aktiv beteiligt sind,
verweist auf mehrere Möglichkeiten
hinsichtlich der Bezugsquellen für
das Pflanzenenzym. Tatsächlich
werden hydroxylierte Fettsäuren
in einigen natürlichen
Pflanzen-Spezies im Überschuss
vorgefunden. Beispielsweise kommen drei Hydroxy-Fettsäuren, die mit Rizinoleat verwandt
sind, in großen
Mengen in Samenölen
verschiedener Lesquerella-Spezies vor. Besonders interessant ist
die Lesquerolsäure,
ein Homolog zu Rizinoleat von 20 Kohlenstoffatomen mit zwei zusätzlichen
Kohlenstoffatomen am Carboxyl-Ende der Kette (Smith, 1985). Andere
natürliche
Pflanzen-Bezugsquellen für
hydroxylierte Fettsäuren
umfassen (sind jedoch nicht beschränkt auf) Samen der Gattung
Linum, Samen von Wrightia-Spezies, Lycopodium-Spezies, Strophanthus-Spezies,
Convolvulaces-Spezies, Calendula-Spezies und zahlreichen anderen
(van der Loo et al., 1993).
-
Pflanzen
mit signifikantem Vorhandensein von Rizinoleat oder Lesqueroleat
oder desaturierten anderen oder modifizierten Derivaten dieser Fettsäuren sind
bevorzugte Kandidaten, um na türlich
abgeleitete kappa-Hydroxylasen zu erhalten. Beispielsweise enthält Lesquerella
densipila eine zweifach ungesättigte
Fettsäure
mit 18 Kohlenstoffatomen mit einer Hydroxyl-Gruppe (van de Loo et
al., 1993), von der gemäß der Theorie, auf
der diese Erfindung basiert, angenommen wird, dass sie von einem
Enzym produziert wird, das mit der Rizinus-kappa-Hydroxylase eng
verwandt ist. Darüber
hinaus kann ein Vergleich zwischen kappa-Hydroxylasen und zwischen
Pflanzen-Fettsäure-Hydroxylasen,
die Hydroxyl-Gruppen an anderen Positionen als dem 12-Kohlenstoffatom des
Oleats oder dem 14-Kohlenstoffatom des Lesqueroleats einbringen,
oder Hydroxyl-Gruppen bei anderen Substraten als Ölsäure und
Icosensäure
einbringen, Einblicke für
das Protein-Modeling und für
andere Modifikationen zur Erzeugung synthetischer Hydroxylasen geben,
wie oben diskutiert wird. Beispielsweise streben wir an, auf Basis
der Informationen, welche aus den strukturellen Vergleichen der Δ12-Desaturasen
und der kappa-Hydroxylase erhalten werden, genetische Modifikationen
in den Struktur-Genen der Δ12-Desaturasen
vorzunehmen, die diese Desaturasen zu kappa-Hydroxylasen umwandeln.
Wir streben ferner an, Veränderungen
in Δ15-Hydroxylasen
vorzunehmen, die diese zu Hydroxylasen umwandeln, welche eine Substrat-Spezifität aufweisen,
die mit der der Desaturasen vergleichbar ist (z. B. Konversion von 18:2Δ9,12 zu
15OH-18:2Δ9,12.
Da der Unterschied zwischen einer Hydroxylase und einer Desaturase
die Anordnung eines Protons betrifft, denken wir, dass diese Veränderung
durch systematische Veränderung
der geladenen Gruppen im Bereich nahe des aktiven Zentrums des Enzyms
bewirkt werden kann.
-
Von
besonderem Interesse sind Fettsäure-Hydroxylasen,
die eine Aktivität
gegenüber
anderen Fettsäure-Substraten
als Oleat zeigen oder die Hydroxyl-Gruppe an einer anderen Stelle
als dem C12-Kohlenstoffatom einbringen. Wie oben beschrieben ist,
können
auch andere Pflanzen-Bezugsquellen Bezugsquellen für diese
Enzyme bereitstellen, z.B. durch die Verwendung von Pro teinreinigung,
Nukleinsäure-Sonden,
Antikörper-Präparationen,
Protein-Modeling oder Sequenzvergleichen. Von besonderem Interesse
sind die jeweiligen Aminosäure-
und Nukleinsäuresequenzen,
die solchen Pflanzen-Fettsäure-Hydroxylasen
entsprechen. Wie ebenfalls zuvor beschrieben wurde, können – sobald
eine Nukleinsäuresequenz
für eine
gegebene Pflanzen-Hydroxylase
erhalten wird – weitere
pflanzliche Sequenzen verglichen und/oder mit Sonden untersucht werden,
um zu dieser Nukleinsäuresequenz
homolog verwandte DNA-Sequenzen zu erhalten usw.
-
Verwendungen
in Gentechnik-Anwendungen
-
Wie
im Stand der Technik hinreichend bekannt ist, kann ein für eine Pflanzen-kappa-Hydroxylase
kodierender cDNA-Klon, sobald er erhalten wurde, dazu verwendet
werden, die entsprechenden genomischen Nukleinsäuresequenzen zu diesem zu erhalten.
-
Die
Nukleinsäuresequenzen,
die für
die Pflanzen-kappa-Hydroxylasen kodieren, können in verschiedenen Konstrukten
verwendet werden, z.B. als Sonden, um weitere Sequenzen von derselben
oder von anderen Spezies zu erhalten. Alternativ dazu können die
Sequenzen in Verbindung mit geeigneten regulatorischen Sequenzen
verwendet werden, um die Mengen der jeweiligen Hydroxylase von Interesse
in einer Wirtszelle für
die Produktion von hydroxylierten Fettsäuren zu erhöhen, oder zur Untersuchung
des Enzyms in vitro oder in vivo, oder um die Mengen der jeweiligen
Hydroxylase von Interesse bei einigen Anwendungen zu verringern oder
zu erhöhen,
in denen die Wirtszelle eine Pflanzen-Einheit ist, einschließlich Pflanzenzellen,
Pflanzenteilen (einschließlich
jedoch nicht beschränkt
auf Samen, Schnitte oder Gewebe) und Pflanzen.
-
Eine
Nukleinsäuresequenz,
die für
eine Pflanzen-kappa-Hydroxylase
gemäß der vorliegenden
Erfindung kodiert, kann eine genomische, eine cDNA- oder eine mRNA-Sequenz
umfassen. Mit "kodierend" ist gemeint, dass
die Sequenz einer bestimmten Aminosäuresequenz entspricht, entweder
in einer Sense-Orientierung
oder in einer Antisense-Orientierung. Mit "rekombinant" ist gemeint, dass die Sequenz eine
Modifikation enthält,
die gentechnisch durch Manipulation mittels Mutagenese, Restriktionsenzyme
und Ähnliches
erzeugt wurde. Eine cDNA-Sequenz
kann für
Prä-Prozessierungssequenzen,
wie beispielsweise Transit- oder Signalpeptid-Sequenzen, kodieren
oder auch nicht. Transit- oder Signalpeptid-Sequenzen ermöglichen
die Bereitstellung des Proteins an ein gegebenes Organell und werden
oftmals bei Eintritt in das Organell von dem Polypeptid abgespalten,
wobei die "reife" Sequenz freigesetzt
wird. Die Verwendung der Vorläufer-DNA-Sequenz ist
bei pflanzlichen Zell-Expressions-Kassetten bevorzugt.
-
Ferner
kann wie oben erwähnt
die vollständige
genomische Sequenz der Pflanzen-kappa-Hydroxylase durch Screening
einer genomischen Bibliothek mit einer Sonde, wie beispielsweise
einer cDNA-Sonde, und Isolierung derjenigen Sequenzen, die die Expression
in Samen-Gewebe regulieren, erhalten werden. Auf diese Weise können Transkriptions-
und Translations-Initiationsregionen, Introns, und/oder Transkript-Terminationsregionen
der Pflanzen-kappa-Hydroxylase für
die Verwendung in einer Vielzahl von DNA-Konstrukten erhalten werden,
mit oder ohne kappa-Hydroxylase-Struktur-Gen. Somit können Nukleinsäuresequenzen,
die der erfindungsgemäßen Pflanzen-kappa-Hydroxylase
entsprechen, auch Signalsequenzen bereitstellen, die für den direkten
Transport in ein Organell nützlich
sind, 5'-stromaufwärts gelegene
nicht-kodierende regulatorische Regionen (Promotoren) mit nützlichen
Gewebe- und Timing-Eigenschaften, 3'-stromabwärts gelegene nicht-kodierende
regulatorische Regionen, welche als transkriptionale und translationa le
regulatorische Regionen nützlich
sind, und sie können
Einblick in andere Eigenschaften des Gens vermitteln.
-
Sobald
die gewünschte
Nukleinsäuresequenz
der Pflanzen-kappa-Hydroxylase
erhalten wird, kann sie auf vielfältige Weise manipuliert werden.
Wenn die Sequenz nicht-kodierende flankierende Regionen umfaßt, können die
flankierenden Regionen einer Resektion, einer Mutagenese usw. unterworfen
werden. Somit können
Transitionen, Transversionen, Deletionen und Insertionen bei der
natürlich
vorkommenden Sequenz durchgeführt
werden. Darüber
hinaus kann die gesamte Sequenz oder ein Teil der Sequenz synthetisiert
werden. Ein oder mehrere Kodons im Struktur-Gen können modifiziert
werden, um eine modifizierte Aminosäuresequenz bereitzustellen,
oder es können
ein oder mehrere Kodon-Mutationen eingebracht werden, um für geeignete
Restriktionsschnittstellen zu sorgen, oder aus anderen Gründen, die
bei der Konstruktion oder Expression eine Rolle spielen. Das Struktur-Gen
kann ferner durch Verwendung synthetischer Adapter oder durch Verwendung
von Verbindungsmolekülen
zur Einbringung einer oder mehrerer geeigneter Restriktionsschnittstellen
o.ä. modifiziert
werden.
-
Die
Nukleinsäure-
oder Aminosäuresequenz,
die für
eine Pflanzen-kappa-Hydroxylase gemäß der vorliegenden Erfindung
kodiert, kann auf vielfältige
Weise mit anderen nicht-nativen (oder "heterologen") Sequenzen kombiniert werden. Mit "heterologen" Sequenzen ist jede
beliebige Sequenz gemeint, die natürlicherweise nicht in Verbindung
mit der Pflanzen-kappa-Hydroxylase
vorgefunden wird, einschließlich
z.B. eine Kombination von Nukleinsäuresequenzen aus derselben
Pflanze, die natürlicherweise
nicht miteinander verbunden vorgefunden werden.
-
Die
DNA-Sequenz, die für
eine Pflanzen-kappa-Hydroxylase gemäß der vorliegenden Erfindung
kodiert, kann in Verbindung mit al len oder einem Teil der Gen-Sequenzen
verwendet werden, die normalerweise mit der kappa-Hydroxylase assoziiert
sind. Eine DNA-Sequenz, die für
eine kappa-Hydroxylase kodiert, wird in ihren Einzelteilen in ein
DNA-Konstrukt kombiniert, das in 5'-3'-Richtung
der Transkription eine Transkriptions-Initiations-Kontrollregion umfasst,
die in der Lage ist, die Transkription und Translation in einer
Wirtszelle zu fördern,
die DNA-Sequenz, die für
die Pflanzen-kappa-Hydroxylase kodiert, sowie eine die Transkription oder
Translation terminierende Region.
-
Potentielle
Wirtszellen umfassen prokaryotische und eukaryotische Zellen. Eine
Wirtszelle kann – abhängig von
der beabsichtigten Verwendung – einzellig
sein oder in einem vielzellig differenzierten oder undifferenzierten
Organismus vorkommen. Zellen gemäß dieser
Erfindung können
dadurch unterschieden werden, dass sie eine Pflanzen-kappa-Hydroxylase
aufweisen, die in Bezug auf die Wildtypzelle, in der diese vorliegt, fremd
ist, beispielsweise, indem sie ein rekombinantes Nukleinsäurekonstrukt
aufweist, das für
eine Pflanzen-kappa-Hydroxylase
kodiert.
-
Abhängig vom
Wirt werden die regulatorischen Regionen variieren, wobei Regionen
aus viralen Genen, aus Plasmid-Genen oder aus chromosomalen Genen
oder Ähnlichen
eingeschlossen sind. Für
die Expression in prokaryotischen oder eukaryotischen Mikroorganismen,
insbesondere in einzelligen Wirten, kann eine große Vielzahl
von konstitutiven oder regulierbaren Promotoren eingesetzt werden.
Die Expression in einem Mikroorganismus kann eine einfache Bezugsquelle
für das
Pflanzen-Enzym bereitstellen. Unter den beschriebenen Transkriptions-Initiations-Regionen sind Regionen
aus bakteriellen Wirten und aus Hefe-Wirten, wie beispielsweise E. coli,
B. subtilis, Saccharomyces cerevisiae, wobei Gene wie β-Galaktosidase,
T7-Polymerase, Tryptophan E und Ähnliche
eingeschlossen sind.
-
Die
Konstrukte werden meist regulatorische Regionen umfassen, die in
Pflanzen funktionstüchtig
sind, was für
die modifizierte Produktion von Pflanzen-kappa-Hydroxylase sorgt
und zu einer Modifikation der Fettsäurezusammensetzung führt. Der
offene Leserahmen, der für
die Pflanzen-kappa-Hydroxylase oder ein funktionstüchtiges
Fragment derselben kodiert, wird an seinem 5'-Ende mit einer regulatorischen Transkriptions-Initiations-Region, wie z.B.
der Wildtypsequenz, die natürlicherweise
5'-stromaufwärts vom
kappa-Hydroxylase-Struktur-Gen vorkommt, zusammengefügt. Zahlreiche
andere Transkriptions-Initiations-Regionen sind verfügbar, die für eine große Vielzahl von konstitutiven
oder regulierbaren (z.B. induzierbaren) Funktionen der Transkription
des Struktur-Gens sorgen. Unter den Transkriptions-Initiations-Regionen,
die für
Pflanzen verwendet werden, sind solche Regionen, die beispielsweise
mit den Struktur-Genen der Nopalin-Synthase und der Mannopin-Synthase
assoziiert sind, oder mit Napin, Sojabohnen-β-conglycinin, Oleosin, dem 12S-Speicherprotein,
den 35S-Promotoren des Blumenkohl-Mosaikvirus und Ähnlichen.
Die Transkription/Translation-Initiations-Regionen, die solchen
Struktur-Genen entsprechen, werden unmittelbar 5'-stromaufwärts vom jeweiligen Startkodon
gefunden. In Ausführungsformen,
in denen die Expression des kappa-Hydroxylase-Proteins in einem
Pflanzenwirt erwünscht
ist, wird die Verwendung des gesamten oder eines Teils des vollständigen Pflanzen-kappa-Hydroxylase-Gens
angestrebt; es können
dementsprechend alle oder ein Teil der 5'-stromaufwärts gelegenen nicht-kodierenden
Regionen (Promotor) gemeinsam mit der Sequenz des Struktur-Gens
und den 3'-stromabwärts gelegenen
nicht-kodierenden Regionen verwendet werden. Wenn ein anderer Promotor
erwünscht
ist (wie beispielsweise ein Promotor der in Bezug auf den Pflanzen-Wirt
von Interesse nativ ist, oder ein modifizierter Promotor, d.h. mit
Transkriptions-Initiations-Regionen, die von einer Gen-Bezugsquelle stammen,
und Translations-Initiations-Regionen, die von einer anderen Gen-Bezugsquelle stammen,
einschließlich
der Sequenz, die für
die Pflanzen-kappa-Hydroxylase von Interesse kodiert, oder verstärkende Promotoren,
wie die Double-35S-CaMV-Promotoren,
können
die Sequenzen unter Verwendung von Standardverfahren zusammengefügt werden.
-
Bei
Anwendungen, in denen die 5'-stromaufwärts gelegenen
nicht-kodierenden Regionen von anderen während der Samenreifung regulierten
Genen erhalten werden, sind vorzugsweise solche erwünscht, die in
Embryo-Gewebe von Pflanzen exprimiert werden, wie die Transkriptions-Initiations-Kontrollregionen
des Napin-Gens aus B. napus oder des 12S-Speicherproteins von Arabidopsis,
oder des β-conglycinins
aus Sojabohnen (Bray et al., 1987), oder der kappa-Hydroxylase-Promotor
aus L. fendleri, der vorliegend beschrieben wird. Transkriptions-Initiations-Regionen,
die vorzugsweise in Samen-Gewebe exprimiert werden, d.h. die in anderen
Pflanzen-Teilen nicht nachgewiesen werden können, werden für Fettsäuremodifikationen
als wünschenswert
erachtet, um alle störenden
oder nachteiligen Auswirkungen auf das Gen-Produkt zu minimieren.
-
Regulatorische
Transkript-Terminations-Regionen können ebenfalls in den erfindungsgemäßen DNA-Konstrukten
vorgesehen werden. Transkript-Terminations-Regionen können durch
die DNA-Sequenz
bereitgestellt werden, die für
die Pflanzen-kappa-Hydroxylase
kodiert, oder es kann eine vorteilhafte Transkriptions-Terminations-Region
von einer anderen Gen-Bezugsquelle abgeleitet werden, beispielsweise
von der Transkript-Terminations-Region,
die natürlicherweise
mit der Transkript-Initiations-Region
assoziiert ist. Wenn die Transkript-Terminations-Region von einer anderen
Gen-Bezugsquelle stammt, wird sie mindestens etwa 0,5 kb, vorzugsweise
etwa 1 bis 3 kb, der Sequenz 3' vom
Struktur-Gen enthalten, von dem die Terminations-Region abgeleitet
ist.
-
Pflanzen-Expressions-
oder Transkriptions-Konstrukte mit einer Pflanzen-kappa-Hydroxylase
als DNA-Sequenz von Interesse für
die gesteigerte oder verringerte Expression derselben können mit
einer großen
Vielzahl von pflanzlichen Lebensformen verwendet werden, insbesondere
mit pflanzlichen Lebensformen, die bei der Herstellung von pflanzlichen Ölen für Nahrungszwecke
und industrielle Anwendungen beteiligt sind. Am stärksten bevorzugt
sind gemäßigte Ölsamen-Nutzpflanzen.
Pflanzen von Interesse umfassen (sind jedoch nicht beschränkt auf)
Raps (Canola und Sorten mit hohem Erucasäure-Gehalt), Crambe, Brassica juncea,
Brassica nigra, Sumpfblume (meadowfom), Flachs, Sonnenblume, Distel,
Baumwolle, Cuphea, Sojabohne, Erdnuss, Kokosnuss und Ölpalmen
und Mais. Ein wichtiges Kriterium bei der Auswahl geeigneter Pflanzen
für die
Einbringung einer kappa-Hydroxylase
ist das Vorhandensein eines geeigneten Substrats für die Hydroxylase
in der Wirtspflanze. Daher wird beispielsweise die Produktion von
Rizinolsäure
am besten in Pflanzen erreicht, die normalerweise hohe Mengen Ölsäure in den
Samen-Lipiden aufweisen.
Die Produktion von Lesquerolsäure
wird am besten in Pflanzen erreicht, die hohe Mengen Icosensäure in den
Samen-Lipiden aufweisen.
-
Abhängig von
den Verfahren zur Einbringung der rekombinanten Konstrukte in die
Wirtszelle können weitere
DNA-Sequenzen erforderlich sein. Es ist wichtig, dass die Erfindung
bei dikotylen und monokotylen Spezies gleichermaßen angewendet werden kann
und problemlos auf neue und/oder verbesserte Transformations- und
Regulationsverfahren anwendbar ist. Das Verfahren der Transformation
ist für
die vorliegende Erfindung nicht entscheidend; verschiedene Verfahren
der Transformation von Pflanzen sind gegenwärtig verfügbar. Sofern neuere Verfahren
zur Transformation von Nutzpflanzen verfügbar sind, können diese
direkt im vorliegenden Rahmen verwendet werden. Beispielsweise können zahlreiche
Pflanzen-Spezies, die von Natur aus für eine Agrobacterium-Infektion
empfänglich
sind, erfolg reich durch Verfahren der Agrobacterium-vermittelten
Transformation unter Verwendung von binären oder dreiteiligen Vektoren
transformiert werden. Zusätzliche
Verfahren von Mikroinjektion, DNA-Partikelbombardment und Elektroporation
sind entwikkelt worden, die die Transformation verschiedener monokotylen
und dikotylen Pflanzen-Spezies erlauben.
-
Bei
der Entwicklung der DNA-Konstrukte werden die verschiedenen Bestandteile
des Konstruktes oder Fragmente desselben in einen geeigneten Klonierungsvektor
insertiert, der in der Lage ist, sich in einem bakteriellen Wirt
zu replizieren, z.B. in E. coli. Es existieren zahlreiche Vektoren,
die in der Literatur beschrieben wurden. Nach jeder Klonierung kann
das Plasmid isoliert und weiteren Manipulationen unterzogen werden,
wie beispielsweise Restriktion, Insertion von neuen Fragmenten,
Ligation, Deletion, Insertion, Resektion etc., sodass die Bestandteile
der gewünschten
Sequenz zurechtgeschnitten werden. Sobald das Konstrukt vollständig ist,
kann es gemäß der Art
der Wirtszellen-Transformation für
die weitere Manipulation in einen geeigneten Vektor eingebracht
werden.
-
Normalerweise
wird das DNA-Konstrukt ein Struktur-Gen mit den notwendigen regulatorischen
Regionen für
die Expression in einem Wirt und die Selektion von transformierten
Zellen umfassen. Das Gen kann für
eine Resistenz gegen ein cytotoxisches Mittel, z.B. ein Antibiotikum,
ein Schwermetall, ein Toxin usw. sorgen, für eine Komplementierung, die
für eine
Prototrophie in einem auxotrophen Wirt sorgt, für virale Immunität und Ähnliches.
Abhängig
von der Anzahl der verschiedenen Wirts-Spezies, in die das Expressionskonstrukt oder
Bestandteile desselben eingebracht wird, können ein oder mehrere Marker
verwendet werden, wobei verschiedene Selektionbedingungen bei den
verschiedenen Wirten angewendet werden.
-
Es
wird angemerkt, dass die Degeneration des DNA-Codes dafür sorgt,
dass einige Kodon-Substitutionen bei DNA-Sequenzen zulässig sind,
ohne irgendeine entsprechende Modifikation bei der Aminosäuresequenz
zu verursachen.
-
Wie
oben erwähnt,
ist die Art, auf die das DNA-Konstrukt in die Pflanzenzelle eingebracht
wird, für
die vorliegende Erfindung nicht entscheidend. Jedes beliebige Verfahren,
das für
eine effiziente Transformation sorgt, kann verwendet werden. Verschiedene
Verfahren der Transformation von Pflanzenzellen umfassen die Verwendung
von Ti- oder Ri-Plasmiden, Mikroinjektion, Elektroporation, Infiltration,
Imbibition, DNA-Partikelbombardement,
Liposomenfusion, DNA-Bombardement oder Ähnliches. In vielen Fällen wird
es erwünscht sein,
dass das Konstrukt auf einer oder auf beiden Seiten der T-DNA eingegrenzt
ist; es wird besonders erwünscht
sein, dass die linke und die rechte Grenze vorhanden sind, insbesondere
die rechte Grenze. Dies ist besonders nützlich, wenn das Konstrukt
A. tumefaciens oder A. rhizogenes als Art der Transformation verwendet,
obgleich die T-DNA-Grenzen auch bei anderen Arten der Transformation
Anwendung finden können.
-
Wenn
Agrobacterium für
die Transformtion der Pflanzenzellen verwendet wird, kann ein Vektor
verwendet werden, der zum Zwecke der homologen Kombination mit der
T-DNA oder dem Ti- oder Ri-Plasmid, die in dem Agrobacterium-Wirt
vorhanden sind, in den Agrobacterium-Wirt eingebracht wird. Das
Ti- oder Ri-Plasmid, welches die T-DNA für die Rekombination umfasst,
kann „bewaffnet" (armed) sein (fähig, die
Gallen-Bildung hervorzurufen) oder „unbewaffnet" (disarmed) sein
(unfähig,
Gallen hervorzurufen), wobei letzteres erlaubt ist, solange die
vir-Gene in dem transformierten Agrobacterium-Wirt vorhanden sind.
Das „bewaffnete" Plasmid kann zu
einer Mischung von normalen Pflanzenzellen und Gallen führen.
-
In
einigen Fällen,
in denen Agrobacterium als Vehikel zur Transformation von Pflanzenzellen
verwendet wird, wird das von der (den) T-DNA-Grenze(n) eingefasste
Expressionskonstrukt in einen Vektor mit breitem Wirtsspektrum insertiert,
wobei Vektoren mit breitem Wirtsspektrum in der Literatur beschrieben
sind. Oftmals wird pRK2 oder Derivate desselben verwendet. Siehe
beispielsweise Ditta et al. (1980). Umfasst vom Expressionskonstrukt
und der T-DNA werden ein oder mehrere Marker sein, die die Selektion
von transformiertem Agrobacterium und transformierten Pflanzenzellen
erlauben. Mehrere Marker sind für
die Verwendung in Pflanzenzellen entwickelt worden, wie z.B. die
Resistenz gegen Kanamycin, das Aminoglykosid G418, Hygromycin und Ähnliche.
Abhängig
vom jeweiligen Wirt und der Art der Konstruktion werden verschiedene
Marker bevorzugt werden.
-
Bei
der Transformation von Pflanzenzellen unter Verwendung von Agrobacterium
können
Explantate kombiniert und mit dem transformierten Agrobacterium
für eine
ausreichende Transformationsdauer inkubiert werden. Die Bakterien
werden getötet
und die Pflanzenzellen werden in einem geeigneten selektiven Medium kultiviert.
Sobald sich der Callus gebildet hat, kann die Sprossbildung durch
Verwendung geeigneter Pflanzenhormone nach bekannten Verfahren unterstützt werden,
und die Sprösslinge
können
zur Regeneration der Pflanzen auf Medium zur Wurzelbildung übertragen
werden. Die Pflanzen können
anschließend
angezüchtet werden,
um Samen zu bilden, und die Samen können verwendet werden, um repetitive
Generationen zu etablieren, und um pflanzliche Öle zu isolieren.
-
Die
nunmehr allgemein beschriebene Erfindung wird leichter durch Verweis
auf die nachfolgenden Beispiele verständlich sein, wobei diese lediglich
zum Zwecke der Illustration enthalten sind und die vorliegende Erfindung
nicht beschränken
sollen.
-
BEISPIELE
-
In
der nachfolgenden experimentellen Offenbarung werden alle Temperaturen
in Grad Celsius (°)
angegeben, Gewichte werden in Gramm (g), Milligramm (mg) oder Mikrogramm
(μg) angegeben,
Konzentrationen werden in molar (M), millimolar (mM) oder mikromolar
(μM) angegeben,
und alle Volumen werden in Litern (l), Mikrolitern (μl) oder Millilitern
(ml) angegeben, sofern nicht auf anderes verwiesen wird.
-
BEISPIEL 1 – HERSTELLUNG
VON NEUEN HYDROXYLIERTEN FETTSÄUREN
IN ARABIDOPSIS THALIANA
-
Übersicht
-
Die
kappa-Hydroxylase, die durch das zuvor beschriebene fah12-Gen von Rizinus kodiert
wird, wurde verwendet, um Rizinolsäure, Lesquerolsäure, Densipolsäure und
Auricolsäure
in transgenen Arabidopsis-Pflanzen herzustellen. Dieses Beispiel
setzt das in der vorhergehenden Anmeldung gelehrte Beispiel 2 in die
Praxis um.
-
Herstellung
von transgenen Pflanzen
-
Eine
Vielzahl von Verfahren ist entwickelt worden, um eine DNA-Sequenz
von Interesse in das Genom eines Pflanzen-Wirts zu insertieren,
um die Transkription und Translation der Sequenz zur Verursachung
einer phänotypischen
Veränderung
zu erhalten. Die nachfolgenden Verfahren stellen lediglich eines
von mehreren äquivalenten
Mitteln zur Herstellung transgener Pflanzen und zum Bewirken der
Expression des Hydroxylase-Gens dar.
-
Arabidopsis-Pflanzen
wurden durch Agrobacterium-vermittelte Transformation transformiert,
wobei die kappa-Hydroxylase durch das fah12-Gen von Rizinus auf
dem binären
Ti-Plasmid pB6 kodiert wurde. Dieses Plasmid wurde zuvor verwendet,
um Nicotiana tabacum zum Zwecke der Herstellung von Rizinolsäure zu transformieren.
-
Material
zum Animpfen des Agrobacterium tumefaciens-Stamms GV3101, der das
binäre
Ti-Plasmid pB6 enthielt, wurde auf L-Nährmedium-Platten ausplattiert,
die 50 μg/ml
Kanamycin enthielten, und für
2 Tage bei 30°C
inkubiert. Einzelne Kolonien wurden verwendet, um große Flüssigkeitskulturen
(L-Nährmedium
mit 50 mg/l Rifampicin, 110 mg/l Gentamycin und 200 mg/l Kanamycin)
anzuimpfen, die für
die Transformation von Arabidopsis-Pflanzen verwendet wurden.
-
Die
Arabidopsis-Pflanzen wurden mittels der in planta-Transformationsmethode
transformiert, wie sie im Wesentlichen von Bechtold et al., 1993)
beschrieben wurde. Zellen von A. tumefaciens GV3101 (pB6) wurden
durch Zentrifugation aus Flüssigkulturen
geerntet, anschließend
bei einer OD600 = 0,8 in Infiltrationsmedium
resuspendiert (Infiltrationsmedium war Murashige und Skoog Makro-
und Mikronährstoffmedium
(Sigma Chemical Co., St. Louis, MO), das 10 mg/l 6-Benzylaminopurin
und 5 % Glucose enthielt). Gruppen von 12–15 Pflanzen wurden für 3 bis
4 Wochen bei natürlichem
Licht bei einer durchschnittlichen täglichen Temperatur von ungefähr 25°C in Töpfen von
8,89 cm (3,5 Inch), die Erde enthielten, angezüchtet. Die intakten Pflanzen wurden
in die bakterielle Suspension eingetaucht und anschließend in
einer Vakuumkammer eingebracht und unter ein Vakuum von 600 mm gesetzt,
das von einer Laborvakuumpumpe produziert wurde, bis die Gewebe einheitlich
mit Wasser durchtränkt
erschienen (ungefähr
10 Minuten). Die Pflanzen wurden bei 25°C für 4 Wochen bei kontinuierlichem
Licht angezüchtet
(100 μmol
m–2 s–1 Bestrahlung
im Bereich von 400 bis 700 nm). Die Samen, die von allen Pflanzen
in einem Topf erhalten wurden, wurden als eigene Gruppe geerntet.
Die Samen wurden durch sequentielle Behandlung für 2 Minuten mit Ethanol, gefolgt
von 10 Minuten mit einer Mischung von Haushaltsbleiche (Chlorox),
Wasser und Tween-80 (50 %, 50 %, 0,05 %) sterilisiert und anschließend gründlich mit
sterilem Wasser gespült.
Die Samen wurden in 100 ml Petrischalen in einer hohen Dichte (2000
bis 4000 pro Platte) auf Agar-verfestigtem Medium ausplattiert,
das 1/2 × Murashige
und Skoog-Salzmedium enthielt, mit B5-Vitaminen (Sigma Chemical
Co., St. Louis, MO) angereichert war und 50 mg/l Kanamycin enthielt.
Nach Inkubation für
48 Stunden bei 4°C
zur Stimulation der Keimung wurden die Keimlinge für einen
Zeitraum von 7 Tagen angezüchtet,
bis die Transformanten eindeutig als gesunde grüne Keimlinge gegen einen Hintergrund
von chlorotischen, Kanamycin-sensitiven Keimlingen identifiziert
werden konnten. Die Transformanten wurden für 2 Wochen auf Erde übertragen,
bevor Blattgewebe für
die DNA- und Lipid-Analyse benutzt werden konnte. Mehr als 20 Transformanten
wurden erhalten.
-
Die
DNA wurde aus jungen Blättern
von Transformanten extrahiert, um das Vorhandensein eines intakten
fah12-Gens zu verifizieren. Das Vorhandensein des Transgens in mehreren
vermeintlichen transgenen Linien wurde unter Verwendung der Polymerase-Kettenreaktion
zur Amplifizierung des Inserts von pB6 verifiziert. Bei den verwendeten
Primer handelte es sich um HF2 = GCTCTTTTGTGCGCTCATTC (SEQ ID NO:
12) und HR1 = CGGTACCAGAAAACGCCTTG (SEQ ID NO: 13), die erstellt
wurden, um die Amplifikation eines 700 bp-Fragments zu erlauben.
Ungefähr
100 ng genomische DNA wurden zu einer Lösung gegeben, die 25 pmol von
jedem Primer enthielt, 1,5 U Taq-Polymerase (Boehringer Mannheim),
200 μM dNTPs,
50 mM KCl, 10 mM Tris-HCl (pH 9), 0,1 % (v/v) Triton X-00, 1,5 mM
MgCl2, 3 % (v/v) Formamid, das Endvolumen
betrug 50 μl.
Die Amplifikationsbedingungen waren: 4 min Denaturierungsschritt
bei 94°C,
gefolgt von 30 Zyklen bei 92°C
für 1 min,
55°C für 1 min,
72°C für 2 min.
Ein finaler Extensionsschritt bei 72°C für 5 min schloss das Programm
ab. Die Transformanten konnten nach Sichtbarmachen eines charakteristischen
amplifizierten Fragments von 1 kb auf einem Ethidiumbromid-gefärbten Agarosegel
in positiver Hinsicht identifiziert werden. Alle transgenen Linien,
die getestet wurden, ergaben ein PCR-Produkt einer Größe, die
mit dem erwarteten Genotyp übereinstimmte,
was bestätigte,
dass die Linien in der Tat transgen waren. Alle weiteren Experimente wurden
mit drei repräsentativen
transgenen Linien des Wildtyps durchgeführt, die als 1-3, 4D, 7-4 bezeichnet wurden,
sowie mit einer Linie der fad2-Mutantenlinie JB12. Die transgene
JB12-Linie wurde einbezogen, um zu testen, ob die erhöhte Akkumulierung
von Ölsäure in dieser
Mutante eine Auswirkung auf die Menge der Rizinolsäure hat,
die in transgenen Pflanzen akkumuliert wird.
-
Analyse von
transgenen Pflanzen
-
Blätter und
Samen von fah12-transgenen Arabidopsis-Pflanzen wurden unter Verwendung
der Gaschromatographie auf das Vorhandensein von hydroxylierten
Fettsäuren
analysiert. Die Lipide wurden aus 100–200 mg Blattgewebe oder aus
50 Samen extrahiert. Es wurden Fettsäuremethylester (FAMES) hergestellt,
indem Gewebe in einem 13 × 100
mm Glasbehälter
mit Schraubverschluss, der mit einer mit Teflon ausgekleideten Kappe
versehen war, in 1,5 ml 1,0 M methanolischem HCl (Supelco Co.) eingebracht
und für
2 Stunden auf 80°C
erhitzt wurde. Beim Abkühlen
wurde 1 ml Petroleumether zugegeben, und die FAMES wurden entfernt,
indem die Etherphase abgedampft wurde, welche anschließend in
einem Glasbehälter
in einem Stickstoff-Strom getrocknet wurde. 100 μl N,O-Bis(trimethylsilyl)-trifluoracetamid
(BSTFA; Pierce Chemical Co.) und 200 μl Acetonitril wurden zugegeben,
um die Hydroxyl-Gruppen zu derivatisieren. Die Reaktion wurde bei
70°C für 15 Minuten
ausgeführt.
Die Produkte wurden in Stickstoff getrocknet, in 100 μl Chloroform
gelöst und
in ein Gaschromatograph-Behältnis
gegeben. 2 μl
je der Probe wurden auf einer SP2340 Fused-Silica-Kapillarsäule (30
m, 0,75 mm ID, 0,20 mm Film, Supelco) analysiert, wobei ein Hewlett-Packard-Gaschromatograph
der Serie 5890 II verwendet wurde. Die Proben wurden nicht aufgeteilt,
das Temperaturprogramm betrug 195°C
für 18
min, wurde mit 25°C/min
auf 230°C
erhöht,
bei 230°C
für 5 min
gehalten und anschließend
mit 25°C/min
auf 195°C
abgesenkt, und es wurden Flammenionisations-Detektoren verwendet.
-
Die
chromatographische Elutionszeit von Methylestern und O-TMS-Derivaten
von Rizinolsäure,
Lesquerolsäure
und Auricolsäure
wurde durch GC-MS der Lipidproben von Samen von L. fendleri und
Vergleich mit veröffentlichten
Chromatogrammen von Fettsäuren
dieser Spezies etabliert (Carlson et al., 1990). Es wurde ein O-TMS-Methyl-Rizinoleat-Standard
aus Rizinolsäure
hergestellt, die von Sigma Chemicals Co (St. Louis, MO) erhalten
wurde. O-TMS-Methyl-Lesqueroleat-Standards und O-TMS-Methyl-Auricolat-Standards
wurden ausgehend von Triacylglycerolen, die aus Samen von L. fendleri
gereinigt worden waren, hergestellt. Das Massenspektrum von O-TMS-Methyl-Rizinoleat,
O-TMS-Methyl-Densipoleat, O-TMS-Methyl-Lesqueroleat und O-TMS-Methyl-Auricoleat
sind jeweils in den 1A–D gezeigt. Die Struktur der
charakteristischen Ionen, die während
der Massenspektrometrie dieser Derivate erzeugt werden, sind in 2 gezeigt.
-
Die
aus dem transgenen Gewebe extrahierten Lipide wurden durch Gaschromatographie
und Massenspektrometrie auf das Vorhandensein von hydroxylierten
Fettsäuren
untersucht. Zum Zwecke der Referenz sei darauf verwiesen, dass die
durchschnittliche Fettsäurezusammensetzung
von Blättern
in einer Arabidopsis-Wildtyp-Linie
und einer fad2-Mutanten-Linien von Miguel und Browse (1992) beschrieben
wurden. Gachromatogramme von methylierten und silylierten Fettsäuren von
Samen vom Wildtyp und von einer fah12-transgenen Wildtypflanze sind
in den 3A bzw. 3B gezeigt.
Die Profile sind sehr ähnlich,
mit Ausnahme des Vorhandenseins von drei kleinen aber getrennten
Peaks bei 14,3, 15,9 und 18,9 Minuten. Ein sehr kleiner Peak bei
20,15 Minuten war ebenfalls zu erkennen. Die Elutionszeit der Peaks
bei 14,3 und 18,9 Minuten entsprach exakt denen der zum Vergleich
hergestellten Rizinolsäure-
bzw. Lesquerolsäure-Standards. In Lipidextrakten von
Blättern
oder Wurzeln des Wildtyps und der fah12-transgenen Wildtyplinien
wurden keine signifikanten Unterschiede beobachtet (Tabelle 1).
Obwohl das fah12-Gen in der ganzen Pflanze exprimiert wird, beobachteten wir
Auswirkungen auf die Fettsäurezusammensetzung
lediglich im Samengewebe. Eine ähnliche
Beobachtung wurde bei transgenem fah12-Tabak gemacht.
-
Tabelle
1: Fettsäurezusammensetzung
von Lipiden aus transgenem Arabidopsis und Wildtyp-Arabidopsis.
Die Werte sind Mittelwerte, die aus der Analyse von Proben aus drei
unabhängigen
transgenen Linien oder drei unabhängigen Proben von Wildtyp- und fad2-Linien
erhalten wurden.
-
Um
zu bestätigen,
dass die beobachteten neuen Peaks in den transgenen Linien den Derivaten
von Rizinolsäure,
Lesquerolsäure,
Densipolsäure
und Auricolsäure
entsprachen, wurde eine Massenspektrometrie verwendet. Die Fettsäure-Derivate
wurden wie oben beschrieben mittels Gaschromatographie aufgetrennt,
mit der Ausnahme, dass ein massenselektiver Hewlett-Packard Detektor
der 5971-Serie anstelle des Flammenionisationsdetektor aus dem vorherigen
Experiment verwendet wurde. Die Spektren der vier neuen Peaks in 3B (Peaknummern 10, 11, 12 und 13)
sind jeweils in den 4A–D gezeigt. Der Vergleich des
Spektrums, das für
die Standards erhalten wurde, mit dem Spektrum, das für die vier
Peaks aus den transgenen Linien erhalten wurde, bestätigte die
Identität
der vier neuen Peaks. Auf Basis der drei charakteristischen Peaks
bei M/Z 187, 270 und 299 wird Peak 10 unzweifelhaft als
O-TMS-Methyl-Rizinoleat
identifiziert. Auf Basis der drei charakteristischen Peaks bei M/Z
185, 270 und 299 wird Peak 11 unzweifelhaft als O-TMS-Methyl-Densipoleat identifiziert.
Auf Basis der drei charakteristischen Peaks bei M/Z 187, 298 und
327 wird Peak 12 unzweifelhaft als O-TMS-Methyl-Lesqueroleat
identifiziert. Auf Basis der drei charakteristischen Peaks bei M/Z
185, 298 und 327 wird Peak 13 unzweifelhaft als O-TMS-Methyl-Auricoleat
identifiziert. Diese Ergebnisse zeigen eindeutig, dass die fah12-cDNA
für eine
Hydroxylase kodiert, welche sowohl Ölsäure hydroxyliert, wobei Rizinolsäure entsteht,
als auch Icosensäure
hydroxyliert, wobei Lesquerolsäure
entsteht. Diese Ergebnisse liefern einen weiteren Beleg dafür, dass
die Hydroxylase in einer heterologen Pflanzen-Spezies auf so exprimiert
werden kann, dass das Enzym katalytisch funkti onstüchtig ist.
Diese Ergebnisse zeigen ferner, dass die Expression dieses Hydroxylase-Gens
zur Akkumulierung von Rizinolsäure,
Lesquerolsäure,
Densipolsäure
und Auricolsäure
in einer Pflanzen-Spezies führt,
die normalerweise keine hydroxylierten Fettsäuren in extrahierbaren Lipiden
akkumuliert.
-
Das
Vorhandensein von Lesquerolsäure
in den transgenen Pflanzen wurde in der vorhergehenden Patentanmeldung
(Nr. 08/320,982) auf Basis des biochemischen Hinweises, der eine
breite Substratspezifität
der kappa-Hydroxylase verriet, angenommen. Im Gegensatz dazu war
die Akkumulierung von Densipol- und
Auricolsäure
schlechter vorhersagbar. Da Arabidopsis normalerweise keine signifikanten
Mengen des nicht-hydroxylierten Vorläufers dieser Fettsäuren umfasst,
die als Substrate für
die Hydroxylase dienen können, scheint
es, dass ein oder mehrere der drei in Arabidopsis bekannten n-3-Fettsäure-Desaturasen (z.B.
fad3, fad7, fad8; Überblick
in Gibson et al., 1995) in der Lage sind, die hydroxylierten Verbindungen
an der n-3-Position zu desaturieren. Dies bedeutet, dass Densipolsäure durch
die Wirkung einer n-3-Desaturase auf Rizinolsäure produziert wird. Auricolsäure wird
durch die Wirkung einer n-3-Desaturase auf Lesquerolsäure produziert.
Die fad3-Desaturase
ist mit annähernder
Sicherheit dafür
verantwortlich, da sie im endoplasmatischen Retikulum lokalisiert
ist. Dies kann zukünftig
getestet werden, indem fah12-enthaltene transgene Pflanzen aus der
fad3-defizienten Mutante von Arabidopsis hergestellt werden (ähnliche
Experimente können
mit fad7 und fad8 durchgeführt
werden). Es ist ferner formell möglich,
dass die Enzyme, die normalerweise 18:1cisΔ9 zu 20:1cisΔ11 elongieren
12OH-18:1cisΔ9 zu
14OH-20:1cisΔ11 elongieren
und 12OH-18:2cisΔ9,15 zu
14OH-20:2cisΔ11,17.
-
Die
Menge der verschiedenen Fettsäuren
in Lipiden von Samen, Blatt und Wurzel der Kontrolle und der transgenen
Pflanzen ist in Tabelle 1 dargestellt. Obwohl die Menge an hydroxylierten Fettsäuren, die
in diesem Beispiel produziert wurden, geringer ist, als es für die kommerzielle
Produktion von Rizinoleat und anderen hydroxylierten Fettsäuren aus
Pflanzen erwünscht
ist, streben wir zahlreiche Verbesserungen dieser Erfindung an,
die das Niveau der Akkumulierung von hydroxylierten Fettsäuren in
Pflanzen, welche das fah12-Hydroxylase-Gen oder verwandte Hydroxylase-Gene
exprimieren, erhöhen
werden. Verbesserungen beim Ausmaß und bei der Gewebespezifität der Expression
des Hydroxylase-Gens sind angestrebt. Verfahren, die dies durch
Verwendung eines starken, samenspezifischen Promotors, wie dem Napin-Promotor
aus B. napus, oder durch Verwendung der nativen Promotoren des fah12-Gens
aus Rizinus oder des entsprechenden Hydroxylase-Gens von L. fendleri,
erreichen werden, werden für
den Fachmann offensichtlich sein. Zusätzliche angestrebte Verbesserungen
betreffen die Modifikation der Enzyme, welche die hydroxylierten
Fettsäuren von
Phosphatidylcholin abspalten, die Verringerung der Aktivität von Enzymen,
die hydroxylierte Fettsäuren abbauen,
sowie das Ersetzen von Acyltransferasen, die hydroxylierte Fettsäuren zu
den sn-1-, sn-2- und sn-3-Positionen
der Glycerol-Lipide transferieren. Obwohl Gene für diese Enzyme in der wissenschaftlichen Literatur
nicht beschrieben worden sind, kann ihre Nützlichkeit bei der Verbesserung
des Ausmaßes
der Produktion von hydroxylierten Fettsäuren ohne weiteres basierend
auf den Ergebnissen der biochemischen Untersuchungen der Rizinoleat-Synthese
abgesehen werden.
-
Obgleich
Arabidopsis keine ökonomisch
wichtige Pflanzen-Spezies
ist, ist sie in weitem Umfang bei Pflanzenbiologen als Modell für höhere Pflanzen
anerkannt. Daher soll die Aufnahme dieses Beispiels die allgemeine
Nützlichkeit
der vorliegend beschriebenen Erfindung zeigen, um die Öl-Zusammensetzung
in höheren
Pflanzen zu modifizieren. Ein Vorteil der Untersuchung der Expression
des neuen Gens in Arabidopsis besteht darin, dass beträchtliches
Wissen über
den Lipid-Metabolismus in diesem System besteht, und eine Sammlung
von Mutanten verfügbar ist,
die dazu verwendet werden kann, nützliche Informationen in Bezug
auf die Biochemie der Fettsäure-Hydroxylierung
in Pflanzen-Spezies zu erlangen. Ein weiterer Vorteile besteht darin,
dass alle Informationen, die über
den Rizinoleat-Metabolismus
in Arabidopsis erhalten wurden, leicht auf eng verwandte Spezies,
wie beispielsweise die Nutzpflanzen Brassica napus, Brassica juncea
oder Crambe abyssinica, übertragen
werden können,
um Rizinoleat, Lesqueroleat und andere hydroxylierte Fettsäuren für die industrielle
Verwendung in großen
Mengen zu produzieren. Die kappa-Hydroxylase ist für die Produktion von
Rizinoleat oder Lesqueroleat in jeder beliebigen Pflanzen-Spezies
nützlich,
welche signifikante Mengen der Vorläufer, Ölsäure und Icosensäure, akkumuliert.
Von besonderem Interesse sind genetisch modifizierte Varianten,
die große
Mengen Ölsäure akkumulieren.
Solche Varianten sind gegenwärtig
für Sonnenblume
und Canola verfügbar.
Die Produktion von Lesquerolsäure
und verwandten Hydroxy-Fettsäuren
kann in Spezies erreicht werden, die große Mengen an Icosensäure und
anderen langkettigen Monoensäuren
akkumulieren. Solche Pflanzen könnten
in der Zukunft mittels Gentechnik aus Pflanzen hergestellt werden,
die solche Vorläufer
normalerweise nicht produzieren. Wir nehmen daher an, dass die Verwendung
von kappa-Hydroxylase von allgemeiner Nützlichkeit ist.
-
BEISPIEL 2: ISOLIERUNG
EINES GENOMISCHEN KLONS FÜR
DIE LESQUERELLA-KAPPA-HYDROXYLASE
-
Überblick
-
Nukleotidsequenzregionen,
die sowohl in der Rizinus-kappa-Hydroxylase
als auch in der Arabidopsis-fad2-Δ12-Fettsäure-Desaturase konserviert
sind, wurden verwendet, um Oligonukleotid-Primer zu erstellen. Diese
wurden mit genomischer DNA von Lesquerella fendleri verwendet, um
Fragmente mehrerer homolo ger Gene zu amplifizieren. Diese amplifizierten
Fragmente wurden anschließend
als Hybridisierungssonden verwendet, um genomische Klone vollständiger Länge aus
einer genomischen Bibliothek von L. fendleri zu identifizieren.
Hydroxylierte Fettsäuren
sind für
das Samengewebe von Lesquerella sp. spezifisch und werden in vegetativen
Geweben nicht in einem nennenswerten Umfang gefunden. Eines der
zwei Gene, die durch dieses Verfahren identifiziert wurden, wurde
sowohl in Blättern
als auch in sich entwickelnden Samen exprimiert, und es wird daher
angenommen, dass es der Δ12-Fettsäure-Desaturase
entspricht. Das andere Gen wurde in sich entwickelnden Samen im
hohen Ausmaß exprimiert,
es wurde jedoch nicht oder nur in sehr geringem Ausmaß in Blättern exprimiert;
es handelt sich um die kappa-Hydroxylase
dieser Spezies. Die Identität
dieses Gens wird durch Einbringen des Gens in transgene Arabidopsis-Pflanzen
und Nachweis, dass dies die Akkumulierung von Rizinolsäure, Lesquerolsäure, Densipolsäure und
Auricolsäure
in den Lipiden des Samens bewirkt, nachgewiesen. Der Promotor dieses
Gens ist ebenfalls von Nutzen, da er in der Lage ist, die Expression eines
Gens in sich entwickelnden Samen zu einer Zeit zu steuern, zu der
Speicherlipide akkumuliert werden. Dieser Promotor ist somit von
großem
Nutzen für
zahlreiche Anwendungen bei der Genmanipulation von Samen, insbesondere
bei Mitgliedern der Brassicaceen.
-
Die
verschiedenen Schritte, die an diesem Vorgang beteiligt sind, werden
nachfolgend im Detail beschrieben. Sofern nicht anders angegeben,
werden zur Manipulation von Nukleinsäuren, Bakterien und Phagen
Routineverfahren angewendet wie sie in Sambrook et al. (1989) beschrieben
werden.
-
Isolierung
eines Fragments des kappa-Hydroxylase-Gens aus Lesquerella
-
Oligonukleotid-Primer
für die
Amplifikation der kappa-Hydroxylase
aus L. fendleri wurden erstellt, indem Regionen mit hoher Homologie
der abgeleiteten Aminosäuresequenz
zwischen der Rizinus-kappa-Hydroxylase und der Arabidopsis-Δ12-Desaturase (fad2)
ausgewählt
wurden. Da die meisten Aminosäuren
durch mehrere unterschiedliche Kodons kodiert werden, wurden diese
Oligonukleotide so erstellt, dass sie alle möglichen Kodons umfassten, die
für die
entsprechenden Aminosäuren
kodieren können.
Die Sequenz dieser gemischten Oligonukleotide war:
Oligo 1:
TAYWSNCAYMGNMGNCAYCA (SEQ ID NO: 14)
Oligo 2: RTGRTGNGCNACRTGNGTRTC
(SEQ ID NO: 15)
(wobei: Y = C + T; W = A + T; S = G + C; N
= A + G + C + T; M = A + C; R = A + G ist).
-
Diese
Oligonukleotide wurde dazu verwendet, ein DNA-Fragment der genomischen
DNA von L. fendleri durch Polymerase-Kettenreaktion (PCR) unter Verwendung
der folgenden Bedingungen zu amplifizieren: Ungefähr 100 ng
genomischer DNA wurden zu einer Lösung gegeben, die 25 pmol jedes
Primers, 1,5 U Taq-Polymerase
(Boehringer Mannheim), 200 μM
dNTPs, 50 mM KCl, 10 mM Tris-HCl (pH 9), 0,1 % (v/v) Triton X-100,
1,5 mM MgCl2, 3 % (v/v) Formamid enthielt,
wobei ein Endvolumen von 50 μl
erhalten wurde. Die Amplifikationsbedingungen waren: 4 min Denaturierungsschritt
bei 94°C,
gefolgt von 30 Zyklen bei 92°C
für 1 min,
55°C für 1 min,
72°C für 2 min.
Ein finaler Extensionsschritt bei 72°C für 5 min schloss das Programm
ab.
-
Nach
elektrophoretischer Trennung der Produkte der PCR-Reaktion in Agarosegelen
wurden PCR-Produkte von ungefähr 540
bp beobachtet. Zwei dieser Fragmente wurden in pBluescript (Stratagene) kloniert,
wodurch die Plasmide pLesq2 und pLesq3 entstanden. Die Sequenz der
Inserts in diesen zwei Plasmiden wurde durch Kettenabbruchverfahren
bestimmt. Die Sequenz des Inserts von pLesq2 ist als 5 dargestellt
(SEQ ID NO: 1) und die Sequenz des Inserts von pLesq3 ist als 6 dargestellt
(SEQ ID NO: 2). Der hohe Grad an Sequenzidentität zwischen den zwei Klonen
verweist darauf, dass diese potentielle Kandidaten waren, entweder
eine Δ12-Desaturase
oder eine gamma-Hydroxylase zu sein.
-
Northern-Analyse
-
In
L. fendleri werden hydroxylierte Fettsäuren in großen Mengen in Samenölen gefunden,
jedoch nicht in nennenswerten Mengen in den Blättern. Daher besteht ein wichtiges
Kriterium bei der Unterscheidung zwischen einer Fettsäure-Desaturase
und einer kappa-Hydroxylase darin, dass man erwartet, dass das kappa-Hydroxylase-Gen
in Geweben mit einem hohen Gehalt an hydroxylierten Fettsäuren stärker exprimiert
wird als in anderen Geweben, wohingegen alle Pflanzengewebe mRNA
für eine ω6-Fettsäure-Desaturase
enthalten sollten, da zweifach ungesättigte Fettsäuren in
den Lipiden aller Gewebe in den meisten oder in allen Pflanzen gefunden
werden. Daher war es von großem
Interesse zu bestimmen, ob das Gen, welches pLesq2 entspricht, ebenfalls
nur in Samen exprimiert wird, oder ob es auch in anderen Geweben
exprimiert wird. Dieser Frage wurde nachgegangen, indem man die
Hybridisierung von pLesq2 an RNA, die aus sich entwickelnden Samen
oder von Blättern
erhalten wurde, untersuchte.
-
Gesamt-RNA
wurde aus sich entwickelnden Samen und jungen Blättern von L. fendleri unter
Verwendung eines Rneasy-RNA-Extraktions-Kits
(Qiagen) gemäß den Anweisungen
des Herstellers gereinigt. Die RNA-Konzentrationen wurden durch
UV- Spektrophotometrie
bei λ =
260 und 280 nm quantifiziert. Um sicherzustellen, dass das für den Northernblot
verwendete Gel gleichmäßig beladen
wurde, wurden die RNA-Konzentrationen weiter angepasst, nachdem
die Fluoreszenz der mit Ethidiumbromid gefärbten und auf einem denaturierenden
Testgel getrennten RNA-Proben unter UV-Licht aufgenommen worden
war.
-
Die
wie oben beschrieben präparierte
Gesamt-DNA aus Blättern
und sich entwickelnden Samen wurde elektrophoretisch auf einem Agarosegel
getrennt, das Formaldehyd enthielt (Iba et al., 1993). Eine gleiche Menge
(10 μg)
RNA wurde in beide Spuren geladen, und RNA-Standards (0,16 bis 1,77
kb-Leiter, Gibco-BRL) wurden
in die dritte Spur geladen. Nach der Elektrophorese wurde die RNA
vom Gel auf eine Nylonmembran (Hybond N+, Amersham) übertragen
und durch Einwirken von UV-Licht auf dem Filter fixiert. Eine 32P-markierte Sonde wurde durch Random-Priming aus der Insert-DNA
des Klons pLesq2 hergestellt und über Nacht bei 52°C mit der
Membran hybridisiert, nachdem diese für 2 Stunden vorhybridisiert
worden war. Die Vorhybridisierungslösung enthielt 5X SSC, 10X Denhardt-Lösung, 0,1
% SDS, 0,1 M KPO4 pH 6,8, 100 μg/ml Lachssperma-DNA.
Die Hybridisierungslösung
hatte dieselbe Basiszusammensetzung, enthielt jedoch kein SDS und enthielt
10 % Dextransulfat und 30 % Formamid. Der Blot wurde einmal in 2X
SSC, 0,5 % SDS bei 65°C
und anschließend
in 1X SSC bei derselben Temperatur gewaschen.
-
Ein
kurzes Einwirken (30 min) des Blots auf einen Röntgenfilm zeigte, dass die
Sonde pLesq2 mit einer einzelnen Bande hybridisiert, die lediglich
in der Spur mit der Samen-RNA vorhanden war (7). Der
Blot wurde erneut mit dem Insert des pLesq3-Gens als Sonde untersucht,
was zu Banden ähnlicher
Intensität
in den Spuren der Samen und der Blätter führte (7).
-
Diese
Ergebnisse zeigen, dass das Gen, welches dem Klon pLesq2 entspricht,
in hohem Maße
und spezifisch in den Samen von L. fendleri exprimiert wird. In
Verbindung mit der Kenntnis der Nukleotidsequenz und der abgeleiteten
Aminosäuresequenz
ist eine Samen-spezifische Expression des Gens, das dem Insert in pLesq2
entspricht, ein überzeugender
Indikator einer Rolle des Enzyms bei der Synthese von hydroxylierten Fettsäuren in
Samenöl.
-
Charakterisierung
eines genomischen Klons für
die gamma-Hydroxylase
-
Genomische
DNA wurde wie von Murray und Thompson (1980) beschrieben aus jungen
Blättern
von L. fendleri isoliert. Eine genomische Bibliothek aus einem partiellem
Sau3AI-Verdau, die in dem Vektor λDashII (Stratagene,
11011 North Toorey Pines Road, La Jolla, CA 92037) konstruiert wurde,
wurde durch partielles Verdauen von 500 μg DANN, Größenselektion der DNA in einem
Sucrosegradient (Sambrook et al., 1989) und Ligieren der DNA (durchschnittliche
Größe 12 kb)
mit dem BamHI-verdauten Arm von λDashII
hergestellt. Die gesamte Ligation wurde gemäß den Bedingungen des Herstellers
gesammelt und auf den E. coli Stamm XL1-Blue MRA-P2 (Stratagene)
ausplattiert. Dies ergab 5 × 105 primäre
rekombinante Klone. Die Bibliothek wurde anschließend gemäß den Bedingungen
des Herstellers amplifiziert. Eine Fraktion der genomischen Bibliothek
wurde auf E. coli XL1-Blue ausplattiert, und die resultierenden
Plaques (150000) wurden gemäß den Bedingungen
des Herstellers auf geladene Nylonmembranen (Hybond N+, Amersham) überführt. Die
DNA wurde mittels UV in einem Stratalinker (Stratagene) mit den
Filtern quervernetzt.
-
Mehrere
Klone, die genomische DNA-Sequenzen trugen, welche der Hydroxylase
von L. fendleri entsprachen, wurden isoliert, indem man die Membranen
mit dem Insert von pLesq2 als Sonde un tersuchte, das mittels PCR
mit internen Primer amplifiziert und mit 32P
durch Random-Priming markiert worden war. Die Filter wurden für 2 Stunden
bei 65°C
in 7 % SDS, 1 mM EDTA, 0,25 M Na2HPO4 (pH 7,2), 1 % BSA prähybridisiert und für 16 Stunden
in derselben Lösung
mit der Sonde hybridisiert. Die Filter wurden nacheinander bei 65°C in Lösungen gewaschen,
die 2X SSC, 1X SSC, 0,5 X SSC und zusätzlich 0,1 % SDS enthielten.
Ein 2,6 kb-XbaI-Fragment, das die vollständige kodierende Sequenz für die gamma-Hydroxylase
und ungefähr
1 kb der 5'-stromaufwärts gelegenen
Region enthielt, wurde in die entsprechende Stelle des pBluescript
KS subkloniert, wobei das Plasmid pLesq-Hyd entstand, und die Sequenz
wurde vollständig
durch das Dideoxy-Kettenbabbruchs-Verfahren unter Verwendung eines
automatischen Sequenziergerätes
bestimmt. Die Sequenzdaten wurden unter Verwendung des Programms
DNASIS (Hitachi Company) untersucht.
-
Die
Sequenz des Inserts von Klon pLesq-Hyd ist in den 8A–B gezeigt.
Die Sequenz enthält
1855 bp kontinuierliche DNA-Sequenz (SEQ ID NO: 3). Der Klon kodiert
eine 5'-nicht-translatierte Region
von 401 bp (d.h. Nukleotide, die dem ersten ATG-Kodon voranstehen),
einen offenen Leserahmen von 1152 bp sowie eine 3'-untranslatierte
Region von 302 bp. Der offene Leserahmen kodiert ein Protein von
384 Aminosäuren mit
einem vorhergesagten Molekulargewicht von 44370 (SEQ ID NO: 4).
Der Amino-Terminus weist keine Merkmale eines typischen Signalpeptids
auf (von Heijne, 1985).
-
Das
genaue Translations-Initiations-Methionin ist experimentell nicht
bestimmt worden; man nimmt jedoch auf Basis der abgeleiteten Aminosäuresequenz-Homologie
zu der Rizinus-kappa-Hydroxylase
(unten angemerkt) an, dass es sich um das Methionin handelt, welches
von dem ersten ATG-Kodon bei Nukleotid 402 kodiert wird.
-
Der
Vergleich der abgeleiteten Aminosäuresequenz von pLesq-Hyd mit
Sequenzen Membran-gebundener Desaturasen und der Rizinus-Hydroxylase (9A–B) zeigt,
dass pLesq-Hyd homolog zu diesen Genen ist. Die Figur zeigt ein
Alignment der Hydroxylase von L. fendleri (SEQ ID NO: 4) mit der
Hydroxylase von Rizinus (van de Loo et al., 1995), der Arabidopsis
fad2-cDNA, die eine im endoplasmatischen Retikulum lokalisierte Δ12-Desaturase (fad2
genannt) kodiert (Okuley et al., 1994), zwei fad2-Desaturase-Klonen
aus der Sojabohne, einem Brassica napus-fad2-Klon, einem Zea mays-fad2-Klon
und der partiellen Sequenz eines fad2-Klons aus R. communis. Der
hohe Grad an Sequenzhomologie verweist darauf, dass die Genprodukte ähnliche
Funktionen aufweisen. So beträgt
beispielsweise die Gesamthomologie zwischen der Lesquerella-Hydroxylase
und der Arabidopsis-fad2-Desaturase 92,2 % Ähnlichkeit und 84,8 % Identität, und die
zwei Sequenzen unterscheiden sich hinsichtlich ihrer Länge lediglich
in einer Aminosäure.
-
Southern-Hybridisierung
-
Die
Southernanalyse wurde verwendet, um die Kopienzahl der Gene im Genom
von L. fendleri zu bestimmen, die dem pLesq-Hyd-Klon entsprachen. Genomische DNA (5 μg) wurde
mit EcoRI, HindIII und XbaI verdaut und auf einem 0,9%igen Agarosegel
getrennt. Die DNA wurde gemäß dem Protokoll
des Herstellers mittels Alkali-Blot auf eine geladene Nylonmembran
(Hybond N+. Amersham) übertragen.
Der Blot wurde für 2
Stunden bei 65°C
in 7 % SDS, 1 mM EDTA, 0, 25 M Na2HPO4 (pH 7,2), 1 % BSA vorhybridisiert und für 16 Stunden
in derselben Lösung
mit der Sonde hybridisiert, wobei das pLesq-Hyd-Insert mit internen
Primern PCR-amplifiziert und durch Random-Priming mit 32P
markiert worden war. Die Filter wurden nacheinander bei 65°C in Lösungen gewaschen,
die 2X SSC, 1X SSC, 0,5X SSC und zusätzlich 0,1 % SDS enthielten
und anschließend
einem Röntgenfilm
ausgesetzt.
-
Die
Sonde hybridisierte mit einer einzelnen Bande in jedem Verdau der
DNA von L. fendleri (10), was darauf verweist, dass
das Gen, von dem pLesq-Hyd transkribiert wird, in einer einzelnen
Kopie im Genom von L. fendleri vorliegt.
-
Expression
von pLesq-Hyd in transgenen Pflanzen
-
Es
gibt eine große
Vielzahl von Pflanzen-Promotorsequenzen, die verwendet werden können, um
eine Gewebe-spezifische Expression von klonierten Genen in transgenen
Pflanzen zu bewirken. Der Napin-Promotor und die Promotoren des
Acylcarrierproteins sind beispielsweise bereits zuvor bei der Modifikation
der Zusammensetzung von Samenöl
durch Expression einer Antisense-Form einer Desaturase verwendet
worden (Knutsen et al., 1992). Von dem Promotor der β-Untereinheit
des Sojabohnen-β-Conglycinin
ist gezeigt worden, dass er in hohem Maße aktiv ist und zu einer Gewebe-spezifischen
Expression in transgenen Pflanzen anderer Spezies als der Sojabohne
führt (Bray
et al., 1987). Obwohl wir die Verwendung des kappa-Hydroxylase-Promotors aus L.
fendleri in den vorleiegenden Beispielen beschrieben haben, können auch
andere Promotoren für
die Herstellung einer modifizierter Samenöl-Zusammensetzungen verwendet
werden, die zu einer Samen-spezifischer Expression führen. Solche
Modifikationen der vorliegend beschriebenen Erfindung werden für den Fachmann
offensichtlich sein.
-
Konstrukte
für die
Expression der kappa-Hydroxylase von L. fendleri in Pflanzenzellen
werden wie folgt hergestellt: Ein 13 kb-SalI-Fragment, welches das
pLesq-Hyg-Gen enthielt, wurde in die XhoI-Schnittstelle des binären Ti-Plasmid-Vektors
pSLJ44026 (Jones et al., 1992) (11) kloniert,
wobei das Plasmid pTi-Hyd entstand, und durch Elektroporation in
den Agrobacterium tumefaciens-Stamm GV3101 transformiert. Der Stamm
GV3101 (Koncz und Schell, 1986) enthält ein „unbewaffnetes" (disarmed) Ti-Plasmid.
Zellen für die
Elektroporation wurden wie folgt hergestellt. GV3101 wurde in LB-Medium
mit verringertem NaCl (5 g/l) angezüchtet. Eine 250 ml-Kultur wurde
bis zu einer OD600 = 0,6 angezüchtet und
anschließend
bei 4000 UpM für 15
Minuten zentrifugiert (Sorvall GS-A-Rotor). Der Überstand wurde unverzüglich von
dem lockeren Pellet abgezogen, und das Pellet wurde vorsichtig in
500 ml eiskaltem Wasser resuspendiert. Die Zellen wurden wie zuvor
zentrifugiert, in 30 ml eiskaltem Wasser resuspendiert, und in ein
30 ml-Gefäß transferiert
und bei 5000 UpM für
5 min zentrifugiert (Sorvall SS-34-Rotor). Dies wurde dreimal wiederholt,
wobei die Zellen nacheinander in 30 ml eiskaltem Wasser, 30 ml eiskaltem
10%igem Glycerol und schließlich
in 0,75 ml eiskaltem 10%igen Glycerol resuspendiert wurden. Die
Zellen wurden aliquotiert, in flüssigem
Stickstoff gefroren und bei –80 °C gelagert.
-
Bei
den Elektroporationen wurde ein Biorad-Gene-Pulsar-Instrument unter
Verwendung kalter 2 ml-gap-Küvetten,
die 40 μl
Zellen und 1 μl
DNA in Wasser enthielten, bei einer Spannung von 2,5 KV und 200 Ohm
Widerstand eingesetzt. Die elektroporierten Zellen wurden mit 1
ml SOC-Medium (Sambrook et al., 1898, Seite A2) verdünnt und
bei 28°C
für 2–4 Stunden
inkubiert, bevor sie auf Kanamycin-enthaltendem Medium (50 mg/l)
ausplattiert wurden.
-
Arabidopsis
thaliana kann mit den Agrobacterium-Zellen transformiert werden,
die das pTi-Hyd entahlten, wie oben in Beispiel 1 beschrieben ist.
Das Vorhandensein von hydroxylierten Fettsäuren in transgenen Arabidopsis-Pflanzen
kann ebenfalls durch die oben in Beispiel 1 beschriebenen Verfahren
nachgewiesen werden.
-
Konstitutive Expression
der Hydroxylase aus L. fendleri in transgenen Pflanzen
-
Ein
1,5 kb-EcoRI-Fragment von pLesq-Hyg, das die gesamte kodierende
Region der Hydroxylase umfasste, wurde aus dem Gel gereinigt, und
anschließend
in die entsprechende Schnittstelle von pBluescript KS (Stratagene)
kloniert. Die Plasmid-DNA von mehreren rekombinanten Klonen wurde
anschließend
mit PstI geschnitten, das jeweils nur einmal in dem Insert und einmal
in der Polylinkersequenz des Vektors schneiden sollte. Die Freisetzung
eines 920 bp-Fragments mit PstI zeigte die richtige Orientierung
des Inserts für
weitere Manipulationen. DNA aus einem solchen Klon wurde weiter
mit SalI geschnitten, die 5'-Überhänge wurden
mit dem Klenow-Fragment der DNA-Polymerase I aufgefüllt und
anschließend
mit SacI geschnitten. Das Insert-Fragment wurde aus dem Gel gereinigt
und zwischen die SmaI- und SacI-Schnittstellen von pBI121 (Clontech)
hinter den Blumenkohl-Mosaikvirus 35S-Promotor kloniert. Nachdem
geprüft
worden war, dass die Sequenz der Verbindung zwischen Insert- und Vektor-DNA passend
war, wurde Plasmid-DNA eines rekombinanten Klons verwendet, um A.
tumefaciens (GV3101) zu transformieren. Kanamycin-resistente Kolonien
wurden anschließend
wie zuvor beschrieben für
eine in planta-Transformation von A. thaliana verwendet.
-
DNA
aus Kanamycin-resistenten Keimlingen wurde extrahiert und dazu verwendet,
ausgewählte
Fragmente der Hydroxylase unter Verwendung von nested-Primern mittels
PCR zu amplifizieren. Wenn Fragmente der erwarteten Größe amplifiziert
werden konnten, wurden die entsprechenden Pflanzen in einem Gewächshaus
oder auf Agarplatten angezüchtet,
und die Fettsäuren
wurden von vollständig
ausgebildeten Blättern, Wurzeln
und getrockneten Samen extrahiert. Es wurde anschließend eine
GC-MS-Analyse wie zuvor beschrieben durchgeführt, um die unterschiedlichen Fettsäure-Arten
zu charakterisieren und die Akkumulierung von Hydroxyfettsäuren in
transgenen Geweben zu detektieren.
-
BEISPIEL 3 – ERHALT
VON ANDEREN PFLANZEN-FETTSÄURE-HYDROXYLASEN
-
In
einer vorhergehenden Patentanmeldung haben wir Wege beschrieben,
auf denen die Rizinus-fah12-Sequenz verwendet werden kann, um andere
kappa-Hydroxylasen durch Verfahren wie PCR und heterologe Hybridisierung
zu identifizieren. Aufgrund des hohen Grads an Sequenzähnlichkeit
zwischen Δ12-Desaturasen
und kappa-Hydroxylasen stellt der Stand der Technik jedoch keine
Lehre bereit, wie zwischen den beiden Arten von Enzymen ohne funktionalen
Test, wie beispielsweise Nachweis der Aktivität in transgenen Pflanzen oder
in einem anderen geeigneten Wirt (z.B. transgene mikrobielle oder
tierische Zellen), unterschieden werden kann. Die Identifizierung
der Hydroxylase aus L. fendleri sorgt für die Entwicklung von Kriterien,
mittels derer eine Hydroxylase und eine Desaturase allein auf Basis
der abgeleiteten Aminosäuresequenz-Information
voneinander unterschieden werden können.
-
Die 9A–B zeigen
ein Sequenz-Alignment der Hydroxylase-Sequenzen aus Rizinus und L. fendleri mit
der Hydroxylase-Sequenz
aus Rizinus und allen öffentlich
verfügbaren
Sequenzen aller mikrosomaler Δ12-Fettsäure-Desaturasen
aus Pflanzen. Von den 384 Aminosäureresten
in der Rizinus-Hydroxylase-Sequenz sind mehr als 95 % identisch
mit dem entsprechenden Rest in mindestens einer der Desaturase-Sequenzen.
Daher ist keiner dieser Reste für
die katalytischen Unterschiede zwischen Hydroxylase und Desaturase
verantwortlich. Von den verbleibenden 16 Resten in der Rizinus-Hydroxylase
und den 14 Resten in der Lesquerella-Hydroxylase stellen alle bis
auf sechs Fälle
dar, in denen die Hydroxylase-Sequenz einen konservativen Austausch
im Vergleich mit einer oder mehreren der Desaturase-Sequenzen aufweist,
oder es gibt eine große
Variabilität
hinsichtlich der Aminosäure
in dieser Position in den verschiedenen Desaturasen. Mit konservativ
meinen wir, dass die folgenden Aminosäuren funktionell äquivalent
sind: Ser/Thr, Ile/Leu/Val/Met, Asp/Glu. Diese strukturellen Unterschiede
können
somit nicht zu den katalytischen Unterschieden zwischen Desaturasen
und Hydroxylasen beitragen. Dies lässt lediglich sechs Aminosäurereste übrig, in
denen sowohl die Rizinus-Hydroxylase
als auch die Lesquerella-Hydroxylase von allen bekannten Desaturasen
abweichen, und in denen alle bekannten mikrosomalen Δ12-Desaturasen
einen identischen Aminosäurerest
aufweisen. Diese Reste treten an den Positionen 69, 111, 155, 226,
304 und 331 des Alignments in 9 auf.
Daher unterscheiden diese sechs Reste Hydroxylasen von Desaturasen.
Basierend auf dieser Analyse behaupten wir, dass ein beliebiges
Enzym mit einer Sequenzidentität
von mehr als 60 % zu einem der Enzyme, die in 9 aufgeführt sind,
als Hydroxylase klassifiziert werden kann, wenn es sich von der
Sequenz der Desaturasen an diesen sechs Positionen unterscheidet.
Aufgrund leichter Unterschiede bezüglich der Anzahl der Reste
in einem bestimmten Protein kann die Nummerierung von Protein zu
Protein abweichen; der Sinn des Nummerierungssystems wird jedoch
ersichtlich sein, wenn das betreffende Protein mit der Rizinus-Hydroxylase
unter Verwendung des vorliegend gezeigten Nummerierungssystems in
einem Alignment gegenübergestellt
wird. Somit zeigen die vorliegend offenbarten strukturellen Kriterien
in Zusammenhang mit den Verfahren zur Verwendung des Lesquerella-Hydroxylase-Gens
für die
Isolierung homologer Gene, wie Pflanzen-kappa-Hydroxylase-Gene zum
Zwecke der genetischen Modifizierung einer Fettsäurezusammensetzung isoliert
und identifiziert werden können.
-
Wenn
man überlegt,
welche der sechs Substitutionen allein oder primär für die Unterschiede bezüglich der
katalytischen Aktivität
der erfindungsgemäßen Hydroxylasen
und der Desaturasen verantwortlich ist, sehen wir es als wahrscheinlich
an, dass das Ersetzen eines Phe durch ein Tyr in Position 226 wegen
der bekannten Teilnahme von Tyrosin-Radikalen bei der enzymatischen
Katalyse ausschließlich
für diesen
Unterschied bezüglich
der katalytischen Aktivität
verantwortlich sein könnte.
Andere Substitutionen wie Ala durch Ser in Position 331 können Auswirkungen
beim Modulieren der Gesamtgeschwindigkeit der Reaktion haben. Auf
dieser Basis streben wir die Erzeugung neuer kappa-Hydroxylasen
durch ortsspezifische Mutagenese von Δ12-Desaturasen an. Wir streben ferner die
Umwandlung von Δ15-Desaturasen und Δ9-Desaturasen
zu Hydroxylasen durch eine ähnliche
Anwendung der ortsspezifischen Mutagenese an.
-
ABSCHLIESSENDE
BEMERKUNGEN
-
Durch
die obigen Beispiele wird der Nachweis entscheidener Faktoren bei
der Herstellung von neuen hydroxylierten Fettsäuren durch Expression eines
kappa-Hydroxylase-Gens aus Rizinus in transgenen Pflanzen beschrieben.
Darüber
hinaus wird eine vollständige
cDNA-Sequenz der kappa-Hydroxylase von Lesquerella fendleri bereitgestellt.
Eine vollständige
Sequenz der Rizinus-Hydroxylase wird ebenfalls in verschiedenen
Konstrukten zur Verwendung in Wirtszellen bereitgestellt. Durch
diese Erfindung kann man für
eine Vielzahl von Anwendungen die Aminosäure- und Nukleinsäuresequenzen
erhalten, die für
Pflanzen-Fettsäure-Hydroxylasen
vielfältigen
Ursprungs kodieren.
-
Literaturhinweise
-
- Beltz, G.A., Jacobs, K.A., Eickbuch, T.H.,
Cherbas, P.T., Kafatos, F.C. (1983) Isolation of multigene families
and determination of homologies by filter hybridization methods.
Methods in Enzymology 100, 266–285.
- Bray, E.A., Naito, S., Pan, N.S., Anderson, E., Dube, P., Beachy,
R. N. (1987) Expressian of the β-subunit
of β-conglycinin in seeds
of transgenic plants. Planta 172:364–370.
- Carlson, K.D., Chaudhry, A., Bagby, M.O (1990) Analysis of oil
and meal from lesquerella fendleri seed. J. Am. Oil Chem. Soc. 67,
438–442.
- Ditta, G., Stanfield, S., Corbin, D., Helinski, D.R. (1980)
Broad host range DNA cloning system for gram-negative bacteria:
Construction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad.
Sci. USA 77,7347–7351.
- Gould, S.J., Subramani, S., Scheffler, I.E. (1989) Use of the
DNA polymerase chain reaction for homology probing. Proc. Natl.
Acad. Sci. USA 86, 1934–1938.
- Hirsinger, F. (1989) New oil crops, in Oil Crops of the World,
Robbelen, G., Downey, K.R., and Ashri, A., Eds., McGraw-Hill, New
York, pp. 518–533.
- Howling, D., Morris, L.J., Gurr, M.I., James, A.T. (1972) The
specificity of fatty acid desaturases and hydroxylases. The dehydrogenation
and hydroxylation of monoenoic acids, Biochim. Biophys. Acta 260,
10.
- Huyuh, T.V., Young, R.A., Davis, R.W. (1985) Constructing and
screening cDNA libraries in λgt10
and λgt11.
In DNA Cloning, Vol. 1: A Practical Approach, (ed) D.M. Glover.
IRL Press, Washington DC pp 49–77.
- Iba, K., Gibson, S., Nishiuchi, T., Fuse, T., Nishimura, M.,
Arondel, V., Hugly, S., and Somerville, C. (1993) A gene encoding
a chloroplast omega-3 fatty acid desaturase complements alterations
in fatty acid desaturation and chloroplast copy number of the fad7
mutant of Arabidopsis thaliana. J. Biol. Chem. 268, 24099–24105.
- Jones, J.D.G., Shlumukov, L., Carland, F., English, J., Scofield,
S., Bishop, G.J., Harrison, K. (1992) Effective vectors for transformation,
expression of heterologous genes, and assaying transposon excision
in transgenic plants. Transgenic Res. 1, 285–297.
- Knutson, D.S., Thompson, G.A., Radke, S.E., Johnson, W.B., Knauf,
V.C., Kridl, J.C. (1992) Proc. Natl. Acad. Sci. USA 89, 2624–2628.
- Koncz, C., Schell, J. (1986) The promoter of Tl-DNA
gene 5 controls the tissue-specific expression of chimeric genes
carried by a novel type of Agrobacterium binary vector. Mol. Gen.
Genet. 204, 383–396.
- Miquel, M. Browse, J. (1992) Arabidopsis mutants deficient in
polyunsaturated fatty acid synthesis. J. Biol. Chem. 267, 1502–1509.
- Murray, M.G., Thompson, W. F. (1980) Rapid isolation of high
molecular weight plant DNA. Nucl. Acid Res. 8, 4321–4325.
- Okukey, J., Lightner, J., Feldman, K., Yadav, N., Lark, E.,
Browse, J. (1994) Arabidopsis FAD2 gene encodes the enzyme that
is essential for polyunsaturated lipid
- Sambrook, J., Fritsch, E.F., and Maniatis, T., Molecular Cloning:
a Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory Press,
1989.
- Smith C.R., Jr. (1985) Unusual seed oils and their fatty acids,
in Fatty Acids, Pryde E.H., Ed., American Oil Chemists' Society, Champaign,
Second edition, pp 29–47.
- van de Loo, F.J., Fox, B.G., Somerville, C. (1993) Unusual fatty
acids, in Lipid Metabolism in Plants, T.S. Moore Jr., Ed., CRC Press,
Boca Raton, pp91–126.
- van de Loo, F.N., Turner, S., Somerville, C.R. (1995) An oleate
12-hydraxylase from castor (Ricinus communis L.) is a fatty acyl
desaturase homolog. Proc. Natl. Acad. Sci. USA 92, 6743–6747.
- von Heijne, G. (1985) Signal sequences. J. Mol. Biol. 184, 99–105
- Atsmon, D. (1989) Castor, in Oil Crops of the World, Robbelen,
G., Downey, K.R., and Ashri, A., Eds., McGraw-Hill, New York, pp.
438–447.
- Bechtold, N., Ellis, J. and Pelletier, G. (1993) In Planta Agrobacterium
mediated gene transfer by infiltration of adult Arabidopsis thaliana
plants. C.R. Acad. Sci. Paris 316, 1194–1199.
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-