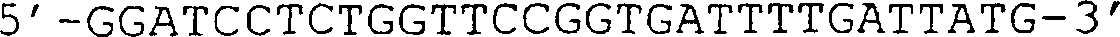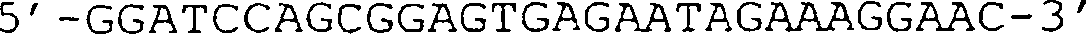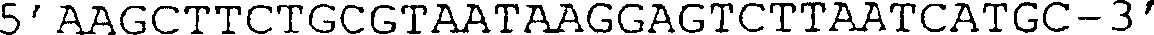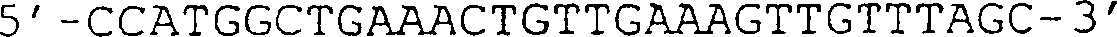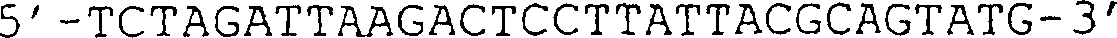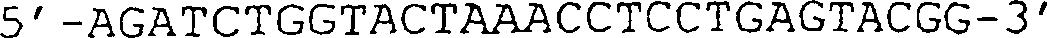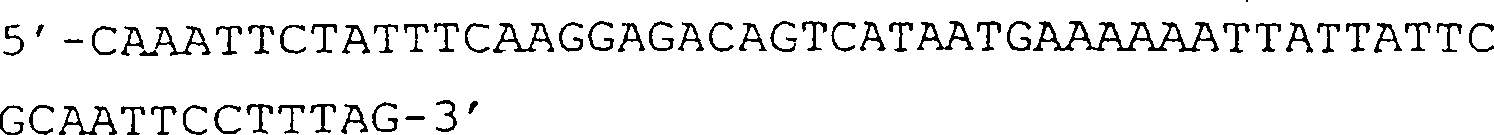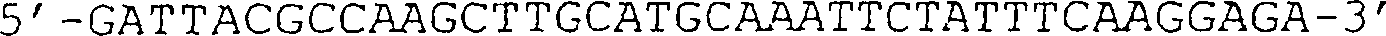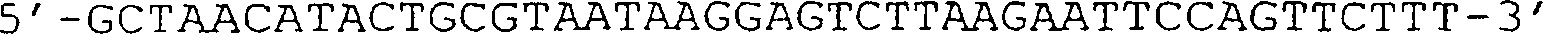-
Gebiet der
Erfindung
-
Die
vorliegende Erfindung bezieht sich auf die Gebiete der Molekularbiologie
und Immunologie. Die vorliegende Erfindung bezieht sich insbesondere
auf das Gebiet der Erzeugung von Helferphagen und von Phagen-Display-Bibliotheken
zur Identifizierung von Bindungsmolekülen.
-
Hintergrund
der Erfindung
-
Ein
Individuum muss ein dynamisches Immunsystem haben, das sich schnell
anpassen kann, um adäquat
auf potentiell schädliche
Mikroorganismen reagieren zu können
und auf die Einflüsse
einer mannigfaltigen und ständig
wechselnden Umgebung reagieren zu können. Höhere Organismen haben spezialisierte
molekulare Mechanismen entwickelt, um den Einsatz von klonal verteilten,
mannigfaltigen Repertoires von Antigen-Rezeptor-Molekülen, die
von Zellen des Immunsystems exprimiert werden, zu gewährleisten:
Immunglobulin(Ig)-Moleküle
auf B-Lymphocyten und T-Zell-Rezeptoren auf T-Lymphocyten. Ein primäres Repertoire
von (im Allgemeinen niederaffinen) Ig-Rezeptoren wird während der
B-Zell-Differenzierung im Knochenmark als Ergebnis der Neuanordnung
von keimliniencodierten Gensegmenten etabliert. Die weitere Verfeinerung
der Ig-Rezeptor-Spezifität und -Affinität erfolgt
in peripheren lymphatischen Organen, wo antigenstimulierte B-Lymphocyten
eine somatische Hypermutationsmaschinerie aktivieren, die spezifisch
auf die variablen (V) Bereiche der Immunglobuline abzielt. Während dieses
Vorgangs werden B-Zell-Klone mit mutanten Ig-Rezeptoren mit höherer Affinität zu dem
auslösenden
Antigen zur klonalen Vermehrung und Reifung zu antikörpersezernierenden
Plasmazellen stimuliert (Übersicht
in Berek und Milstein, 1987).
-
DNA-Rekombinationstechnik
wird verwendet, um viele Aspekte der Vorgänge nachzubilden, die die Erzeugung
und Selektion von natürlichen
humanen Antikörperrepertoires
beherrschen (Übersicht
in Winter und Milstein, 1991; Vaughan et al., 1998). Der Aufbau
großer
Repertoires von Antikörperfragmenten
(wie Fab-Fragmente oder Einzelketten-Fv-Fragmente, scFv), die auf
der Oberfläche
von filamentösen
Phagenpartikeln exprimiert werden, und die Selektion solcher Phagen
durch "Panning" mit Antigenen wurde
als vielseitiges und schnelles Verfahren entwickelt, um Antikörper mit
gewünschten
Spezifitäten
zu erhalten (Übersicht
in Burton und Barbas, 1994). Eine anschließende Optimierung der Affinität von einzelnen
Phagenantikörpern wurde
erreicht, indem man Repertoires mutanter Antikörper der selektierten Phagen
erzeugte und durch Selektion in Bezug auf die Bindung an das Antigen
unter stringenteren Bedingungen auf Nachkommen mit höherer Affinität testete
(Übersicht
in Hoogenboom, 1994).
-
M13
und von M13 abgeleitete Phagen (zuweilen auch Viren genannt) sind
filamentöse
Phagen, die F-Pili tragende (F-positive) Escherichia-coli-(E. coli)-Zellen
selektiv infizieren können.
Im Phagengenom sind 11 Proteine codiert, während die Phagenhülle selbst
aus 5 dieser Proteine besteht: Gen-3-, -6-, -7-, -8- und -9-(g3,
g6, g7, g8 und g9)-Proteine, die an die (zirkuläre) einzelsträngige DNA
(ssDNA) des viralen Genoms gebunden sind und diese schützen. Der
Lebenszyklus des Virus kann in unterschiedliche Phasen unterteilt werden.
Das g3-Protein (g3p) von M13-Phagen und M13-Derivaten umfasst drei
funktionelle Domänen:
D1, D2 und D3, die über
zwei glycinreiche Linker miteinander verknüpft sind. Eine alternative
Nomenklatur für g3p-Domänen wurde
ebenfalls allgemein akzeptiert; D1, D2 und D3 werden dabei N1, N2
bzw. CT genannt. Die N-terminalen
D1/D2-Bereiche treten in Wechselwirkung mit dem C-terminalen D3-Bereich, wie von
Chatellier et al. (1999) unter Verwendung von mehreren Deletionsmutanten
von g3p herausgefunden wurde. Da die Funktionalität einer
D3-Domäne
des Proteins für
den Zusammenbau stabiler Phagen erforderlich ist, betrifft die Mutation
einer weniger oder nicht infektiösen
Mutante des Phagenhüllproteins
vorzugsweise einen D3-Bereich des g3p oder einen funktionellen Teil,
ein Derivat und/oder Analogon dieses D3-Bereichs. Die D3-Domäne bindet
vermutlich an DNA innerhalb des Viruspartikels. Der Verlust der
D3-Domäne führt funktionell
zu seltenen phagenartigen Partikeln, die sehr lang und sehr zerbrechlich
sind (Pratt et al., 1969; Crissman und Smith, 1984; Rakonjac und
Model, 1998). Die D1- und die D2-Domäne treten vermutlich miteinander
in Wechselwirkung, bis der Phage an die Bakterien bindet, während D1/D2
in bestimmten Stadien auch mit D3 in Wechselwirkung treten (Chatellier
et al., 1999). Die Linker, die in g3p zwischen D1, D2 und D3 vorhanden
sind, spielen anscheinend auch eine Rolle bei der Infektiosität des Phagenpartikels
(Nilsson et al., 2000). Studien, in denen eine Protease-Spaltungsstelle
zwischen D1 und D2 eingeführt
wurde, zeigten, dass der Phagenpartikel nach der Spaltung nicht
mehr infektiös
war (Kristensen und Winter, 1998). Eine funktionelle Analyse von g3p
zeigte, dass von den N-terminalen Bereichen von g3p die D1-Domäne für die Infektion
essentiell ist. Ein Verlust dieser Domäne führt zu Phagen, die keine Bakterien
infizieren können
(Lubkowski et al., 1998; Nelson et al., 1981; Deng et al., 1999;
Riechmann und Holliger, 1997; Holliger und Riechmann, 1997). Es
hat sich gezeigt, dass die D2-Domäne auf dem Phagen mit der D1-Domäne von g3p
in Wechselwirkung tritt (1). Aufgrund der Konkurrenz
von Proteinen, die sich auf dem F-Pilus (auf F-positiven Bakterien)
befinden und eine höhere
Affinität
zu D2 als zu D1 haben, dissoziieren die D1- und die D2-Domäne von g3p
voneinander ab. Die Bindung von D2 an den F-Pilus führt zu einem
Vorgang, der zum Rückzug
des F-Pilus zur Zellmembran von E. coli führt. Aufgrund dieses Vorgangs
kommt der Phagenpartikel in engen Kontakt mit der Bakterienmembran.
Jetzt kann die dissoziierte D1-Domäne mit bakteriellen Proteinen,
wie dem TolA-Rezeptor, in Wechselwirkung treten, was zur Einführung der
Phagen-DNA in die E.-coli-Zelle führt (Lubkowski et al., 1999).
Die Tatsache, dass die Entfernung der D2-Domäne die Infektion nicht verhindert,
aber Phagen in die Lage versetzt, E. coli ohne F-Pili zu infizieren
(Riechmann und Holliger, 1997; Deng et al., 1999), zeigt, dass die
Anwesenheit der D2-Domäne
die Spezifität
erhöht
und dass D2 eine wichtige Rolle bei der Verhinderung von Infektionen, die
von F-Pili unabhängig
sind, spielt. Die Bindung von D1 an die spezifischen Rezeptoren
auf der Oberfläche der
E.-coli-Zelle (ein Merkmal, das nicht F+-spezifisch
ist) ist in 2 dargestellt. Dieser Vorgang
löst die
Injektion des viralen Genoms in das Bakterium aus (wie in 3 gezeigt
ist). Obwohl der Verlust der D2-Domäne zur Bildung von Phagenpartikeln
führt,
die E. coli in etwas reduzierter spezifi scher Weise infizieren können, ist das
Ausmaß der
Infektionen durch eine solche Population von Phagen anscheinend
erheblich reduziert. Nach der Infektion von E. coli durch einen
Phagenpartikel wird die ssDNA des Virus aufgrund der Einwirkung
von mehreren bakteriellen Enzymen doppelsträngig. Das doppelsträngige Phagengenom
dient jetzt als Matrize für die
Transcription und Translation aller 11 Gene, die sich auf dem Phagengenom
befinden. Neben diesen proteincodierenden Bereichen enthält das Phagengenom
auch einen intergenischen Bereich: den F1-Replikationsstartpunkt
(F1-ORI). Die DNA-Sequenz dieses F1-ORI kann in 2 getrennte Teilbereiche
unterteilt werden. Ein Teilbereich ist für die Einleitung und Beendigung
der Synthese von ssDNA über
den sogenannten "Mechanismus
des rollenden Kreises" verantwortlich,
und der andere Teilbereich ist für
die Einleitung der Verpackung der gebildeten zirkulären ssDNA
verantwortlich, die zur Bildung und Freisetzung neuer Viruspartikel
führt.
-
Es
hat sich gezeigt, dass Polypeptide, wie Stücke von Aminosäuren, Proteinteile
oder auch ganze Proteine, mit Hilfe der Molekulargenetik an die
terminalen Enden von mehreren Partikelhüllproteinen addiert werden
können,
ohne die Funktionalität
dieser Proteine im Lebenszyklus des Phagen zu stören (Smith, 1985; Cwirla et
al., 1990; Devlin et al., 1990; Bass et al., 1990; Felici et al.,
1993; Luzzago et al., 1993).
-
Dieses
Merkmal ermöglicht
es Forschern, Peptide oder Proteine auf Phagen zu präsentieren,
was zur Erzeugung von Peptid- oder Proteinexpressions-Phagen-Display-Bibliotheken
führt.
Eines der Proteine, die in der Technik zum Fusionieren mit Polypeptiden
für Phagen-Display-Zwecke
verwendet werden, ist das g3-Protein
(g3p), bei dem es sich um ein Hüllprotein
handelt, das für
eine effiziente und effektive Infektion und anschließenden Eintritt
des viralen Genoms in die E.-coli-Zelle
erforderlich ist.
-
Für die Herstellung
von Phagen, die mit dem g3p-Hüllprotein
fusionierte Polypeptide präsentieren, führten auf
diesem Gebiet bewanderte Forscher ein Plasmid zusammen mit dem Phagengenom
in E.-coli-Zellen ein. Dieses Plasmid enthält einen aktiven Promotor stromaufwärts einer
rastererhaltenden Fusion zwischen dem g3-codierenden Gen und einem
interessierenden Gen (X), das zum Beispiel Polypeptide, wie Proteine,
wie Antikörper
oder Fragmente, wie Fab-Fragmente
oder scFvs, codiert. Die Einführung
dieses Plasmids zusammen mit dem Genom des Helferphagen in eine
E.-coli-Zelle führt
zur Erzeugung von Phagen, die auf ihrer Hülle entweder das Wildtyp-g3p
aus dem viralen Genom, das Fusionsprodukt g3p-X aus dem Plasmid oder
ein Gemisch der beiden enthalten, da ein Phagenpartikel fünf g3ps
auf seiner Oberfläche
trägt.
Der Vorgang des Einbaus von g3p oder g3p-X erfolgt im Allgemeinen
zufällig.
Die Anwesenheit einer F1-ORI-Sequenz im g3p-X-Expressionsvektor
(Plasmid) führt
die Phagensynthesemaschinerie so in die Irre, dass zwei Arten von
zirkulärer
ssDNA gebildet werden: eine ist vom Genom des Phagen abgeleitet,
und die andere ist vom Expressionsvektor abgeleitet. Während der
Synthese von neuen Phagen ist die Maschinerie nicht in der Lage, den
Unterschied zwischen diesen beiden Formen von ssDNA zu erkennen,
was zur Synthese einer gemischten Population von Phagen führt, wobei
ein Teil das Phagengenom enthält
und ein Teil die Vektor-DNA beherbergt. Aufgrund dieser Vorgänge enthält das Gemisch
wenigstens einige Phagen, bei denen die phänotypische Information auf
der Außenseite
(des g3p-X-Fusionsproteins) innerhalb der genotypischen Information
im Innern des Partikels (dem g3p-X-Expressionsvektor) konserviert
wird. Ein infektiöser
Wildtypphage und ein Phage, der ein an g3P gebundenes Fusionsprotein
trägt,
sind in 4 abgebildet. Gemäß der einschlägigen Lehre gibt
es mehrere Probleme, die die Verwendung dieser Grundanordnungen
betreffen.
-
Der
hohe Anteil der genotypischen Wildtypphagen in Phagenpopulationen,
die in Bakterien gezüchtet wurden,
die sowohl das Phagengenom als auch den Expressionsvektor enthalten,
brachten Forscher dazu, mutante F1-ORI-Sequenzen in M13-Genomen
zu entwerfen. Solche mutanten M13-Stämme sind in Bezug auf den Einbau
ihres Genoms in Phagenpartikel während
des Zusammenbaus der Phagen weniger effektiv, was zu einem erhöhten Prozentsatz
an Phagen führt,
die Vektorsequenzen enthalten, wenn sie coexprimiert werden. Diese
mutanten Phagen, wie die kommerziell erhältlichen Stämme R408, VCSM13 und M13KO7,
werden "Helferphagen" genannt. Das Genom
dieser Helferphagen kann Gene enthalten, die erforderlich sind,
um neue (Helfer-)Phagen in E. coli zusammen zubauen und anschließend neues
F-Pili exprimierendes E. coli zu infizieren. Sowohl VCSM13 als auch
M13K07 wurden mit einem Replikationsstartpunkt (ORI) des P15A-Typs
versehen, der zur Vervielfältigung
des viralen Genoms in E. coli führt.
Außerdem
gewährleistet
die ORI-Einführung, dass
das alte und das neu gebildete E. coli nach der Zellteilung wenigstens
eine Kopie des viralen Genoms enthält.
-
Es
wurde vorgeschlagen und schließlich
von mehreren Forschern bewiesen, dass die Einführung von Plasmiden, die ein
g3p-scFv-Fusionsprodukt zusammen mit dem Genom der Helferphagen
enthalten, in E.-coli-Zellen zu ungefähr 99% neu gebildeter Phagen
führt,
die das g3p-scFv-Fusionsprotein-Expressionsplasmid beherbergen,
denen aber die g3p-scFv-Fusion auf der Oberfläche fehlt (Beekwilder et al.,
1999). Das Fehlen von g3p-X ist ein erheblicher Nachteil bei der
Verwendung von Display-Bibliotheken für die Identifizierung von spezifischen
Proteinen oder Peptiden, wie scFv, die an ein interessierendes Ziel
(wie Tumorantigene) binden. Dies impliziert, dass im Falle von Phagen-Display-Bibliotheken
in einem Experiment wenigstens ein 100facher Überschuss an produzierten Phagen
verwendet werden muss, um eine Selektion mit allen möglichen
anwesenden Fusionsproteinen durchzuführen. Gemäß der einschlägigen Lehre
führt diese Überlastung
mit relativ nutzlosen Phagen in einem Experiment zu (zu) vielen
falsch positiven Ergebnissen. Zum Beispiel sollten bei einem Panning-Experiment wenigstens
1012 Phagen hinzugefügt werden, damit 1 Kopie von
jeder möglichen Fusion
in dem Experiment vorhanden ist, da eine solche Bibliothek ungefähr 1010 verschiedene g3p-scFv-Fusionen (1%) enthält. Die
Phagen in diesen ungefähr
1% exprimieren auf ihrer Hülle
im Allgemeinen nur eine einzige g3p-scFv-Fusion zusammen mit vier
normalen g3ps (keine Fusionen), während der Rest der Helferphagen
(ungefähr
99%) fünf
g3ps und keine g3p-scFv-Fusionen exprimiert. Um in einem Panning-Experiment theoretisch
die Anwesenheit von 100 Kopien jedes einzelnen Fusionsproteins zu
gewährleisten,
muss man in einem solchen Experiment ungefähr 1014 Phagen
verwenden. Der Fachmann versucht im Allgemeinen, einen wenigstens
100fachen Überschuss
jedes einzelnen einzigartigen Fusionsproteins zu verwenden, um die
Anwesenheit einer ausreichenden Anzahl jeder einzelnen Fusion zu
gewährleisten
und keine relevanten Bindungsmoleküle zu schnell in den ersten
Panning-Durchläufen
zu verlieren. Die Zahl der Phagen (1014)
entspricht mehr oder weniger der Höchstzahl an Phagenpartikeln,
die in einem Milliliter (ml) enthalten sein können. Die Viskosität einer
solchen Lösung
ist äußerst hoch,
und daher ist sie relativ nutzlos. Insbesondere wenn ELISA-Panning-Strategien
verwendet werden (bei denen das Volumen eines Napfes nur 200 μl beträgt), können solche
Bibliotheken nicht verwendet werden.
-
Zusätzlich zu
diesen Problemen wird angenommen, dass je nach dem Antigen und der
Stringenz der Waschverfahren im Allgemeinen durchschnittlich einer
von jeweils 107 Phagen aufgrund von unspezifischer Bindung
an das Antigen bindet. Für
die Anwendung von 1012 scFv-exprimierenden
Ausgangsphagen (1%) auf ein Panning-Verfahren muss man also ungefähr 1014 Phagen hinzufügen (von denen 99% kein scFv-Fragment exprimieren).
Es wird allgemein angenommen, dass von diesen 1012 Phagen
ungefähr
104 Partikel mutmaßlich interessierende Phagen
sein könnten.
Je nach den Waschbedingungen betrug die Zahl der berechneten Hintergrundphagen,
die man bei Verwendung von in der Technik vorhandenen Bibliotheken
normalerweise nach einem Panning-Durchlauf findet, ungefähr 106–107, während
nur wenige dieser Phagen relevante Bindungseigenschaften zu haben
scheinen. Dies ist eines der wichtigsten Probleme, die man in der
Technik erkannt hat: Nach dem ersten Panning-Durchlauf treten zu
viele Hintergrundphagen als anfängliche
Bindungsmoleküle
im Phagengemisch auf, während
nur wenige signifikante und interessierende Bindungsmoleküle in diesem
Gemisch vorhanden sind. Daher ist die absolute Zahl von isolierten
Phagen nach einem Panning-Durchlauf eindeutig zu hoch (106–107). Außerdem
bleiben auch in anschließenden
Panning-Durchläufen
unspezifische Hintergrundphagen vorhanden. In Bibliotheken, die
in der Technik verwendet werden, vermehren sich die meisten dieser
unspezifischen Bindungsmoleküle
bei der Vermehrung auf Bakterien und treten so weiterhin in einem zweiten
Panning-Durchlauf
auf. Daher sind gemäß der einschlägigen Lehre
das Hintergrundniveau von unspezifisch bindenden Phagen und die
Gesamtzahl der Phagen pro ml in diesen Typen von Bibliotheken unannehmbar
hoch und bleibt auch in anschließenden Panning-Durchläufen hoch.
-
Eine
Möglichkeit,
die von einschlägigen
Forschern als Lösung
des Problems, dass man zu viele Hintergrundphagen erhält, denen
eine g3p-X-Fusion fehlt, vorgeschlagen wurde, bestand darin, das
g3p-codierende Gen ganz aus dem Helferphagengenom zu entfernen.
Im Prinzip gewährleistet
dieses System, dass während
der Phagensynthese in einer E.-coli-Zelle (die das g3-lose Phagengenom
und einen g3p-X-Fusionsprotein-Expressionsvektor erhielt) nur g3p-X-Proteine in die neu
gebildete Phagenhülle
eingebaut werden. Dadurch exprimiert jeder synthetisierte Phage
fünf Kopien
des g3p-X-Fusionsprodukts, und es werden kaum Phagen synthetisiert,
die g3p allein exprimieren oder die weniger als fünf g3p-X-Fusionen
exprimieren. R408-d3 und M13MDΔD3
sind zwei Beispiele für
g3-negative Helferphagen (Dueñas
und Borrebaeck, 1995; Rakonjac et al., 1997). Da das Genom dieser
Phagen keine g3p-Synthese unterstützen kann, können kaum
Phagenpartikel gebildet werden, die weniger als fünf g3p-X-Fusionsproteine tragen,
oder wenn sie gebildet werden, erweisen sie sich aufgrund von Instabilität als nichtinfektiös, da gemäß der einschlägigen Lehre
fünf g3ps
notwendig sind, um einen stabilen Phagenpartikel zu gewährleisten.
-
In
der Technik wurde anerkannt, dass zur Herstellung von Helferphagen,
die das g3-Gen nicht enthalten, aber dennoch infektiös sind und
zur Erzeugung von Bibliotheken von Phagen, die fünf g3p-X-Fusionsproteine tragen,
verwendet werden können
und die keine Phagen mit weniger als fünf g3p-X-Fusionen umfassen, eine
externe Quelle für
g3p erforderlich ist. Eine solche Quelle kann ein Vektor ohne F1-ORI
sein, der aber dennoch einen aktiven Promotor stromaufwärts des
vollen offenen Leserasters (ORF) von g3 enthält. Ein Hauptproblem, das vom
Fachmann anerkannt wird, besteht darin, dass die Ausbeute nach dem
Erzeugungsschritt zur Produktion neu gebildeter Helferphagen, denen
ein g3-Gen fehlt,
drastisch gering ist. Tatsächlich liegt
die Ausbeute aller beschriebenen Systeme unter 1010 Phagen
pro Liter, was bedeutet, dass für
eine Bibliothek von 1010 einzelnen Klonen
wenigstens 100 Liter Helferphagenkultur notwendig sind (NB: die
Helferphagen müssen
gereinigt werden), um die Bibliothek einmal zu kultivieren. Gemäß der einschlägigen Lehre
sind also Phagenbibliotheken, die mit so geringen Titern von Helferphagen
erzeugt werden, für
Phagen-Display-Zwecke
nicht geeignet, und daher können
diese Bibliotheken nicht für
Panning- Experimente
verwendet werden. Ein Verfahren zur Komplementierung von g3p-Deletions-Phagen
wurde vor kurzem vorgestellt; dabei wurde Wildtyp-g3p durch eine
Nucleinsäure,
die das Wildtyp-g3p codiert, bereitgestellt, wobei die Nucleinsäure stabil
in das Wirtszellgenom eingebaut war (Rondot et al., 2001).
-
Phagen,
die deletierte g3ps exprimieren, die mit heterologen Proteinen fusioniert
sind, wurden erzeugt. Für
den Aufbau der meisten herkömmlichen
Fab-Bibliotheken
und einiger scFv-Phagen-Display-Bibliotheken wurden die D1-Domäne und Teile
der D2-Domäne
entfernt, um ein kürzeres
Fusionsprotein zu gewährleisten,
das in der Technik als ein Produkt galt, das leichter translatiert
werden konnte als ein g3p in voller Länge, das mit einem Fab-Fragment
in voller Länge
verknüpft
ist. Der kürzere
g3p-Teil würde
die Erzeugung eines lebensfähigen
und geeigneten Helferphagen nicht verhindern. Selbstverständlich sind
solche Phagen noch von g3ps mit voller Länge abhängig, die für eine funktionelle Infektion
von E.-coli-Zellen auf ihrer Oberfläche neben der deletierten g3p-Fusion mit dem Fab-Fragment
vorhanden sind. Außerdem
wurden Phagen erzeugt, die deletierte g3ps, die mit ligandenbindenden
Proteinen fusioniert sind, exprimieren; ihre Infektionsfähigkeit hängt von
Antigenen ab, die mit den Teilen von g3p fusioniert wurden, die
im nichtinfektiösen
Phagenpartikel fehlen (Krebber et al., 1997; Spada et al., 1997).
Die Infektiosität
dieser Partikel hängt
von einer Wechselwirkung zwischen dem ligandbindenden Protein, wie
einem Antikörper
oder einem Fragment davon, und ihrem jeweiligen Liganden (oder Antigen)
ab. Diese Wechselwirkungsabhängigkeit
reduziert jedoch die Effizienz der Infektion aufgrund des Wegfalls
einer direkten Verknüpfung
zwischen den g3p-Domänen
und einer allgemeinen hemmenden Wirkung des löslichen N-terminalen Teils von g3p, der mit dem
Antigen gekoppelt ist.
-
Den
oben genannten g3-negativen Helferphagen R408-d3 und M13MDΔD3 fehlt
in ihrem Genom ein bakterieller ORI und ein Selektionsmarker. Das
Fehlen eines Selektionsmarkers in den g3-negativen Genomen hat eine
erhebliche Wirkung auf den Produktionsmaßstab von Helferphagen, da
es zu einer Überwucherung mit
Bakterien führt,
die das Helferphagengenom nicht enthalten. Es ist bekannt, dass
Bakterien langsamer wachsen, wenn sie mit dem Helferphagen oder
Virus infiziert sind. Daher überwuchern
Bakterien, denen das Phagengenom fehlt, schnell die anderen Bakterien,
die das Genom enthalten. Eine weitere Wirkung des Fehlens eines
ORI oder eines Selektionsmarkers besteht darin, dass in sich teilenden
Bakterien während
der Produktion und Expansion von Phagen-Display-Bibliotheken keine g3-negativen Phagengenome
gehalten werden können.
Dies ist ein sehr wichtiges negatives Merkmal, da eine Überwucherung
mit Bakterien, die das Phagengenom verloren haben oder die nie eines
erhielten, anscheinend einen Wachstumsvorteil gegenüber Bakterien
hat, die das Phagengenom enthalten. Außerdem sind solche "leeren" Bakterien selbstverständlich auch
nicht in der Lage, einen Phagen zu erzeugen, und daher sind die
Phagen-Display-Vektoren,
die Fusionsproteinfragmente enthalten, in solchen Helferphagen,
denen Bakterien fehlen, permanent verloren.
-
Wie
bereits erwähnt,
sind die g3ps vermutlich essentiell für den Zusammenbau von stabilen
M13-artigen Phagen, und wegen ihrer entscheidenden Rolle bei der
Infektion sollten g3ps in sonstiger Weise bereitgestellt werden,
wenn g3-negative Helferphagen erzeugt werden sollen. In der Technik
gibt es ein Vorurteil gegen die Herstellung von Phagen-Display-Bibliotheken,
denen g3ps fehlen, da Phagen, denen g3ps fehlen, nicht stabil sind.
Rakonjac et al. (1997) konstruierten einen g3-negativen Helferphagen
VCSM13 parallel zu einem g3-negativen Helferphagen R408 und verendeten
Helferplasmide mit entweder dem psp- oder dem lac-Promotor stromaufwärts einer
g3-Sequenz mit voller Länge,
so dass g3 während
der Helferphagensynthese substituiert wurde (Model et al., 1997).
Gemäß der einschlägigen Lehre
hat der lac-Promotor jedoch den Nachteil, dass er nicht vollständig abgeschaltet
werden kann, auch nicht in Gegenwart von hohen Konzentrationen an Glucose
(3–5%)
im Medium (Rakonjac und Model, 1998). Ein zusätzliches Problem, das in der
Technik wohlbekannt ist, besteht darin, dass auch sehr geringe Konzentrationen
von g3p in E. coli die Infektion von M13-artigen Phagen blockieren
können.
Außerdem
hat sich gezeigt, dass eine gemeinsame Verkapselung von Plasmiden
zusammen mit dem Phagengenom auftreten kann (Russel und Model, 1989;
Krebber et al., 1995; Rakonjac et al., 1997). Wenn eine gemeinsame
Verkapselung mit dem lac-gesteuerten Helferplasmid erfolgt, konkurriert
sie mit den lac-gesteuerten Vektoren, die im Phagen-Display verwendet
werden, was zur effizienten Produktion von infektiösen Phagenpartikeln
führt,
die das g3p-X-Fusionsprodukt nicht enthalten. Alles in allem ist
der lac-Promotor gemäß der einschlägigen Lehre
also nicht der beste in Frage kommende Promotor im Helferplasmidsystem.
Der psp-Promotor hat den Vorteil, in E. coli bis zur Infektion relativ
inaktiv zu sein (Rakonjac et al., 1997). Nach Infektion mit einem
Phagen der M13-Klasse wird der psp-Promotor aktiviert, und jetzt
erzeugt das Helferplasmid g3-Proteine. Der Nachteil dieses Promotors
besteht jedoch darin, dass das Ausmaß der RNA-Produktion nicht
durch externe Faktoren reguliert werden kann, sondern entweder durch
Mutierenlassen (und Ändern
der Aktivität)
des Promotors oder durch Ändern
der ribosomalen Bindungsstelle (RBS) oder anderer Elemente, die
die Promotoraktivität
beeinflussen, reguliert werden muss. Das ideale Niveau der Promotoraktivität in einem
speziellen E.-coli-Stamm herauszufinden, kann zeitraubend sein,
und es muss für jeden
E.-coli-Stamm getrennt optimiert werden. Gemäß der einschlägigen Lehre
ist das psp-Promotorsystem auch aufgrund der Inflexibilität von E.-coli-Stämmen, der
zeitraubenden Optimierung und des signifikant niedrigen Niveaus
der Helferphagenproduktion für
eine Produktion von Helferphagen in großem Maßstab nicht sehr attraktiv.
-
Ein
erhebliches problematisches Merkmal aller beschriebenen Helferphagensysteme
ist das Auftreten unerwünschter
Rekombinationsereignisse zwischen dem Helfergenom und den (Helfer-)Plasmiden.
Das Problem, dem sich einschlägige
Forscher gegenübersehen,
besteht darin, dass die g3-DNA-Sequenzen in den Helferphagen zu
den g3-Sequenzen im Phagen-Display-Vektor und/oder im Helferphagenplasmid
homolog sind. Dies führt
in vielen Fällen
zu einer Rekombination zwischen den beiden DNA-Stämmen und
daher zu einem Verlust der Funktionalität der Bibliothek als Ganzes.
-
Es
ist ein Ziel der vorliegenden Erfindung, einige der oben beschriebenen,
aus der Technik bekannten Probleme und Nachteile betreffend die
Erzeugung von Phagenpartikeln und Helferphagen, die Verwendung von
Helferphagen bei der Produktion von Phagen-Display-Bibliotheken
und die Probleme und Nachteile, die für die Identifizierung von relevanten
Bindungsmolekülen
unter Verwendung solcher Bibliotheken bekannt sind, anzugehen.
-
Kurzbeschreibung
der Figuren
-
1.
Schematische Darstellung des g3-Proteins (g3p), das in der Hülle von
M13-Phagen vorhanden ist. Die D3-Domäne von g3p ist über das
g8-Protein (g8p) mit der einzelsträngigen DNA innerhalb des Partikels verknüpft, während die
D1- und D2-Domänen
außerhalb
des Partikels miteinander in Wechselwirkung treten und für die Fusion
zum Beispiel mit scFv verwendet werden können.
-
2.
Schematische Darstellung der Wechselwirkung zwischen der D2-Domäne von g3p
und dem F-Pilus auf der Oberfläche
von E. coli mit anschließender
Wechselwirkung der D1-Domäne
mit anderen Komponenten der bakteriellen Oberfläche.
-
3.
Schematische Darstellung der Wechselwirkung der D2-Domäne von g3p
mit dem F-Pilus (links) und der D1-Domäne von g3p mit dem TolA-Rezeptor
(siehe 2) und des anschließenden Eintritts (rechts) des
Phagengenoms in das Cytoplasma des Bakteriums.
-
4.
Schematische Darstellung eines Wildtypphagen, der an seinem infektiösen Ende
fünf g3ps
exprimiert (links), und eines rekombinanten Phagen, der an seinem
infektiösen
Ende vier Wildtyp-g3ps und ein g3p-X-Fusionsprotein exprimiert (rechts).
Der rekombinante Phage beherbergt auch die genetische Information
des auf der Oberfläche
vorhandenen Fusionsproteins.
-
5.
Schematische Darstellung des pBAD/gIII-g3-Helfervektors, der das
g3-Gen in voller
Länge unter
der Kontrolle des AraC/BAD-Promotors beherbergt und weiterhin ein
Ampicillin-Resistenz-Gen und einen ColE1-Replikationsstartpunkt
(ORI) beherbergt.
-
6. (A) Schematische Darstellung des VCSM13-Helferphagengenoms.
Die 11 Gene sowie das Kanamycin-Resistenz-Gen (KanR), das Verpackungssignal (PS)
und das große
und das kleine Fragment des ursprünglichen intergenischen Bereichs
(IG) sind angezeigt. (B) Sequenz des in (A) gezeigten VCSM13-Genoms. Die Translation
des g3-Gens ist im Ein-Buchstaben-Code angegeben. (C) Schematische
Darstellung des von VCSM13 abgeleiteten g3-negativen Helferphagengenoms,
aus dem das offene Leseraster (ORF) des g3-Gens deletiert wurde.
(D) Sequenz des Teils des Helferphagengenoms, der die Position der
in (C) abgebildeten g3-Deletion umgibt. Die HindIII-Stelle auf Position
3431 ist unterstrichen, und danach folgen die 6 halbfett gedruckten
Codons stromaufwärts
des TAA-Stopcodons.
-
7. (A) Schematische Darstellung des von
VCSM13 abgeleiteten D3-Helferphagengenoms, aus dem die D1- und die
D2-Domäne
des g3-Gens deletiert sind. Der D3-Teil des g3-Gens codiert den
carboxyterminalen Teil des g3-Proteins,
der die Erzeugung von stabilen, aber im Wesentlichen nichtinfektiösen Helferphagen
ermöglicht.
Das D3-Helferphagengenom ist mit dem CT-Helferphagengenom identisch.
(B) Sequenz der D3-Domäne
von g3, die in der in (A) gezeigten Nucleinsäure vorhanden ist. Die BamHI-Stelle
auf Position 3488 sowie das GTG-Start- und das TAA-Stopcodon sind
unterstrichen.
-
8.
Anpassung der Codonverwendung im g3-Gen und der D3-Domäne, um eine
homologe Rekombination während
der Helferphagenproduktion und während
der Amplifikation der Phagen-Display-Bibliothek zu verhindern. Die
linke Spalte zeigt die erhaltenen Aminosäuren im Ein-Buchstaben-Code,
und die rechte Spalte zeigt die optimalen Codons.
-
9.
Schematische Darstellung des pUC-g3-Helferplasmids, der das g3-Gen
in voller Länge
unter der Kontrolle des lac-Promotors beherbergt und weiterhin ein
Ampicillin-Resistenz-Gen (AmpR) und einen ColE1-Replikationsstartpunkt
(ORI) beherbergt.
-
10.
Schematische Darstellung des von VCSM13 abgeleiteten N2CT-Helferphagengenoms,
aus dem die N1(D1)-Domäne
des g3-Gens, die dem Phagen Infektiosität verleiht, deletiert wurde.
-
11.
Schematische Darstellung des von VCSM13 abgeleiteten p3-negativen
Helferphagengenoms, das die g3-Leadersequenz exprimiert, auf die
nur sieben Aminosäuren
folgen. Auf dem DNA-Niveau sind ein großer Teil der N2(D2)-Domäne und die
gesamte CT(D3)-Domäne
noch vorhanden. Der frühere
C-terminale Bereich
von Gen III ist vorhanden, codiert aber wegen Rasterverschiebung
kein funktionelles Protein und beinhaltet daher viele ins Raster
passende Stopcodons.
-
Kurzbeschreibung
der Erfindung
-
Die
vorliegende Erfindung stellt Verfahren und Mittel bereit, die im
Wesentlichen nicht die oben skizzierten Nachteile aufweisen und
die dadurch gekennzeichnet sind, dass sie neue Phagenpartikel, wie
chimärische
Phagen, neue Helferphagen, Bibliotheken, die die chimärischen
Phagen umfassen, sowie Verfahren und Mittel zur Herstellung der
chimärischen
Phagen und der Helferphagen liefern.
-
Die
Erfindung stellt einen chimärischen
Phagen bereit, der eine Hülle
aufweist, die ein Gemisch von Proteinen umfasst, wobei das Gemisch
ein Fusionsprotein umfasst, bei dem ein proteinartiges Molekül mit einer
funktionellen Form eines Phagenhüllproteins
fusioniert ist, wobei das Gemisch weiterhin eine mutante Form des
Phagenhüllproteins
umfasst, wobei die Bindung der mutanten Form an einen Wirtszellrezeptor
beeinträchtigt
ist. Die Erfindung stellt auch einen chimärischen Phagen bereit, der
eine Hülle
aufweist, die ein Gemisch von Proteinen umfasst, wobei das Gemisch
ein Fusionsprotein umfasst, bei dem ein proteinartiges Molekül mit einem
Phagenhüllprotein
oder einem Fragment oder Derivat davon fusioniert ist, und wobei
das Fusionsprotein funktionsfähig
in Bezug darauf ist, dass es den chimärischen Phagen infektiös macht,
wobei das Gemisch weiterhin eine mutante Form des Phagenhüllproteins
umfasst, wobei die Bindung der mutanten Form an einen Wirtszellrezeptor
beeinträchtigt
ist.
-
In
einer Ausführungsform
stellt die Erfindung einen Helferphagen bereit, der eine Nucleinsäure umfasst,
die Phagenproteine oder funktionelle Äquivalente davon codiert, die
für den
Zusammenbau des Helferphagen wesentlich sind, wobei die Nucleinsäure weiterhin
eine mutante Form eines Phagenhüllproteins
codiert, wobei die Bindung der mutanten Form an einen Wirtszellrezeptor
beeinträchtigt
ist und wobei der Helferphage keine Nucleinsäure umfasst, die eine funktionelle
Form des Phagenhüllproteins
codiert.
-
Die
Erfindung stellt weiterhin Verfahren und Mittel bereit, um Phagenpartikel,
chimärische
Phagen, infektiöse
Phagen und Helferphagen gemäß der Erfindung
herzustellen. Die Erfindung stellt auch Phagensammlungen bereit,
wie Phagen-Display-Bibliotheken,
die chimärische
Phagen und/oder infektiöse
Phagen gemäß der Erfindung
umfassen.
-
Ausführliche
Beschreibung der Erfindung
-
Die
vorliegende Erfindung stellt einen chimärischen Phagen bereit, der
eine Hülle
aufweist, die ein Gemisch von Proteinen umfasst, wobei das Gemisch
ein Fusionsprotein umfasst, bei dem ein proteinartiges Molekül mit einer
funktionellen Form eines Phagenhüllproteins
fusioniert ist, wobei das Gemisch weiterhin eine mutante Form des
Phagenhüllproteins
umfasst, wobei die mutante Form dadurch gekennzeichnet ist, dass
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form und keine Kopien der funktionellen Form umfasst, weniger infektiös ist als
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form sowie wenigstens eine Kopie der funktionellen Form umfasst.
In einer Ausführungsform
der Erfindung ist die mutante Form dadurch gekennzeichnet, dass
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der die mutante Form und keine
Kopien des Fusionsproteins trägt,
weniger infektiös
ist als ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der zusätzlich zu der mutanten Form
wenigstens eine Kopie des Fusionsproteins trägt. Vorzugsweise ist die mutante
Form dadurch gekennzeichnet, dass ein Phage, der kein Wildtyp-Phagenhüllprotein
umfasst, von dem die mutante Form abgeleitet ist, und der die mutante Form
und keine Kopien des Fusionsproteins oder der funktionellen Form
trägt,
nichtinfektiös
ist. Besonders bevorzugt ist die mutante Form weiter dadurch gekennzeichnet,
dass ein Phage, der eine Hülle
aufweist, die die mutante Form in Anwesenheit oder Abwesenheit von
Kopien der funktionellen Form umfasst, stabil ist. "Stabil", wie der Ausdruck
hier verwendet wird, bedeutet, dass der Teil von g3p, der in der
mutanten Form noch vorhanden ist (und der auch in der funktionellen
Form vorhanden ist), Merkmale wie DNA-Bindung und Steifigkeit des Phagen gewährleistet,
aber nicht zur Infektiosität
des Phagen beiträgt
wie die Domänen
in der funktionellen Form, die in der mutanten Form nicht vorhanden
sind. Die Erfindung stellt auch einen infektiösen Phagen bereit, der wenigstens
eine Kopie einer mutanten Form eines Phagenhüllproteins enthält, wobei
die mutante Form die Fähigkeit,
die Infektion eines natürlichen
Wirts durch den infektiösen
Phagen zu vermitteln, verloren hat.
-
In
einer bevorzugten Ausführungsform
ist das Phagenhüllprotein
das g3-Protein (g3p), das in der Hülle von Phagen wie M13 und
R408 vorhanden ist. Besonders bevorzugt umfasst die mutante Form
eine Mutation im D1- und/oder D2-Bereich von g3p. In einem anderen
bevorzugten Aspekt der Erfindung sind der chimärische Phage und/oder der infektiöse Phage
Bestandteil einer Phagensammlung, wie einer Phagen-Display-Bibliothek.
In einem besonders bevorzugten Aspekt der Erfindung besteht eine
solche Phagensammlung im Wesentlichen aus chimärischen Phage oder infektiösen Phagen,
die durch die Erfindung bereitgestellt werden. Ebenso bevorzugt
sind chimärische
Phagen oder infektiöse
Phagen gemäß der Erfindung,
die bindende Struktureinheiten, wie Antikörper oder Fragmente davon,
als Bestandteil des Fusionsproteins umfassen.
-
In
einer anderen Ausführungsform
stellt die Erfindung ein Verfahren zur Herstellung eines Phagenpartikels
bereit, das die folgenden Schritte umfasst: das Versehen einer Wirtszelle
mit einer ersten Nucleinsäure, die
ein Fusionsprotein codiert, wobei das Fusionsprotein ein proteinartiges
Molekül
umfasst, das mit einer funktionellen Form eines Phagenhüllproteins
fusioniert ist, das Versehen der Wirtszelle mit einer zweiten Nucleinsäure, die
eine mutante Form des Phagenhüllproteins
codiert, wobei die mutante Form dadurch gekennzeichnet ist, dass
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form und keine Kopien der funktionellen Form umfasst, weniger infektiös ist als
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form sowie wenigstens eine Kopie der funktionellen Form umfasst,
und wobei die Wirtszelle eine zusätzliche Nucleinsäuresequenz
umfasst, die wenigstens alle anderen Proteine oder funktionellen Äquivalente
davon codiert, die für
den Zusammenbau des Phagenpartikels in der Wirtszelle wesentlich
sind, und das Kultivieren der Wirtszelle, so dass der Phagenpartikel
zusammengebaut werden kann. Die Erfindung stellt ein Verfahren zur
Herstellung eines Phagenpartikels bereit, wobei das Verfahren die
folgenden Schritte umfasst: das Versehen einer Wirtszelle mit einer
ersten Nucleinsäure,
die ein Fusionsprotein codiert, wobei das Fusionsprotein ein proteinartiges
Molekül
umfasst, das mit einer funktionellen Form eines Phagenhüllproteins
fusioniert ist, das Versehen der Wirtszelle mit einer zweiten Nucleinsäure, die eine
mutante Form des Phagenhüllproteins
codiert, wobei die Bindung der mutanten Form an einen Wirtszellrezeptor
beeinträchtigt
ist, und das Kultivieren der Wirtszelle, so dass der Phagenpartikel
zusammengebaut werden kann. Vorzugsweise werden diese Verfahren
gemäß der Erfindung
zur Herstellung eines Phagenpartikels angewendet, um den chimärischen
Phagen und/oder den infektiösen
Phagen herzustellen. Besonders bevorzugt wird das Verfahren angewendet,
um einen Phagenpartikel, wie einen chimärischen Phagen oder einen infektiösen Phagen
herzustellen, der durch die Erfindung bereitgestellt wird und der
Nucleinsäure
umfasst, die die mutante Form unter der Kontrolle eines regulierbaren
Promotors, wie des AraC/BAD-Promotors, des lac-Promotors oder des
psp-Promotors, codiert.
-
In
einer anderen Ausführungsform
stellt die Erfindung einen Helferphagen bereit, der eine Nucleinsäure umfasst,
die Phagenproteine oder funktionelle Äquivalente davon codiert, die
für den
Zusammenbau des Helferphagen wesentlich sind, wobei die Nucleinsäure weiterhin
eine mutante Form eines Phagenhüllproteins codiert,
wobei die mutante Form dadurch gekennzeichnet ist, dass ein Phage,
der kein Wildtyp-Phagenhüllprotein
umfasst, von dem die mutante Form abgeleitet ist, und der eine Hülle aufweist,
die die mutante Form und keine Kopien einer funktionellen Form des
Phagenhüllproteins
umfasst, weniger infektiös
ist als ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die wenigstens
eine Kopie der funktionellen Form umfasst, wobei die funktionelle
Form dadurch gekennzeichnet ist, dass sie einen Phagenpartikel,
der die funktionelle Form in seiner Hülle trägt, infektiös macht, und wobei der Helferphage
keine exprimierbare Nucleinsäure
umfasst, die die funktionelle Form codiert. Die Erfindung stellt
einen Helferphagen bereit, der eine Nucleinsäure umfasst, die Phagenproteine
oder funktionelle Äquivalente
davon codiert, die für
den Zusammenbau des Helferphagen wesentlich sind, wobei die Nucleinsäure weiterhin
eine mutante Form eines Phagenhüllproteins
codiert, wobei die Bindung der mutanten Form an einen Wirtszellrezeptor
beeinträchtigt
ist, und wobei der Helferphage keine Nucleinsäure umfasst, die eine funktionelle
Form des Phagenhüllproteins
codiert.
-
In
noch einer anderen Ausführungsform
stellt die Erfindung ein Verfahren zur Herstellung eines Helferphagen
bereit, das die folgenden Schritte umfasst: das Versehen einer Wirtszelle
mit einer ersten Nucleinsäure,
die eine funktionelle Form eines Phagenhüllproteins codiert, das Versehen
der Wirtszelle mit einer zweiten Nucleinsäure, die eine mutante Form
des Phagenhüllproteins
codiert, wobei die mutante Form dadurch gekennzeichnet ist, dass
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form umfasst, weniger infektiös
ist als ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die wenigstens
eine Kopie der funktionellen Form umfasst, wobei die Wirtszelle
eine zusätzliche
Nucleinsäuresequenz
umfasst, die wenigstens alle anderen Proteine oder funktionellen Äquivalente
davon codiert, die für den
Zusammenbau des Helferphagen in der Wirtszelle wesentlich sind,
und das Kultivieren der Wirtszelle, so dass der Helferphage zusammengebaut
werden kann. Die Erfindung stellt auch ein Verfahren zur Herstellung eines
Helferphagen bereit, das die folgenden Schritte umfasst: das Versehen
einer Wirtszelle mit einer ersten Nucleinsäure, die eine funktionelle
Form eines Phagenhüllproteins
codiert, das Versehen der Wirtszelle mit einer zweiten Nucleinsäure, die
eine mutante Form des Phagenhüllproteins codiert,
wobei die Bindung der mutanten Form an einen Wirtszellrezeptor beeinträchtigt ist,
wobei die Wirtszelle eine zusätzliche
Nucleinsäuresequenz
umfasst, die wenigstens alle anderen Proteine oder funktionellen Äquivalente
davon codiert, die für den
Zusammenbau des Helferphagen in der Wirtszelle wesentlich sind,
und das Kultivieren der Wirtszelle, so dass der Helferphage zusammengebaut
werden kann.
-
In
einem anderen Aspekt stellt die Erfindung Verfahren und Mittel bereit,
um einen Phagenpartikel, wie einen chimärischen Phagen, einen infektiösen Phagen
oder einen Helferphagen gemäß der Erfindung
herzustellen, wobei getrennte Nucleinsäuren, die entweder (1) eine
funktionelle Form des Phagenhüllproteins
allein oder mit einem proteinartigen Molekül fusioniert oder (2) eine
mutante Form des Phagenhüllproteins
codieren, jeweils Codons in den überlappenden
Bereichen zwischen den proteincodierenden Teilen umfassen, die im Wesentlichen
kein Ereignis der homologen Rekombination zwischen den getrennten
Nucleinsäuren
verursachen. In einem bevorzugten Aspekt umfassen die getrennten
Nucleinsäuren
jeweils einander nicht störende Replikationsstartpunkte
und einzigartige Selektionsmarker.
-
In
einer anderen Ausführungsform
stellt die Erfindung ein Verfahren zur Anreicherung eines ersten Partners
eines Bindungspaars in einem Repertoire aus ersten Partnern von
Bindungspaaren, die aus der Gruppe ausgewählt sind, die aus einem Antikörper, einem
Antikörperfragment,
einem Einzelketten-Fv-Fragment, einem Fab-Fragment, einem variablen
Bereich, einem CDR-Bereich, einem Immunglobulin oder einem funktionellen
Teil davon besteht, wobei der erste Partner des Bindungspaars spezifisch
für einen
zweiten Partner des Bindungspaars ist, bereit, wobei das Verfahren
die folgenden Schritte umfasst: das In-Kontakt-Bringen einer Phagensammlung,
die chimärische
oder infektiöse
Phagen gemäß der Erfindung
umfasst, mit Material, das den zweiten Partner des Bindungspaars
umfasst, unter Bedingungen, die eine spezifische Bindung ermöglichen,
das Entfernen von unspezifischen Bindungsmolekülen, und das Gewinnen von spezifischen
Bindungsmolekülen,
wobei die spezifischen Bindungsmoleküle den ersten Partner des Bindungspaars
umfassen. Das Material kann zweite Partner von Bindungspaaren, wie
gereinigte Proteine, rekombinante Proteine und/oder in oder auf
Zellen vorhandene Proteine, umfassen. In einer bevorzugten Ausführungsform
stellt die Erfindung ein Verfahren zur Anreicherung eines ersten
Partners eines Bindungspaars bereit, wobei das Verfahren weiterhin
die folgenden Schritte umfasst: Gewinnen einer DNA-Sequenz, die
den ersten Partner des Bindungspaars codiert, aus einem Phagen,
das Subklonieren der DNA-Sequenz in einem geeigneten Expressionsvektor,
das Exprimieren der DNA-Sequenz in einem geeigneten Wirt und das
Kultivieren des geeigneten Wirts unter Bedingungen, bei denen der
erste Partner des spezifischen Bindungspaars produziert wird. Ein
geeigneter Expressionsvektor kann ein Plasmidvektor sein, der einen
aktiven Promotor umfasst, welcher die Expression des ersten Partners
des spezifischen Bindungspaars in geeigneten Wirten, wie eukaryontischen
Zellen, wie Hefezellen oder Säugerzellen,
reguliert.
-
In
einem anderen Aspekt stellt die Erfindung ein Nucleinsäuremolekül bereit,
das eine Sequenz umfasst, die eine mutante Form eines Phagenhüllproteins
codiert, wobei die mutante Form dadurch gekennzeichnet ist, dass
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form und keine funktionelle Form des Phagenhüllproteins umfasst, weniger
infektiös
ist als ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form sowie wenigstens eine Kopie der funktionellen Form des Phagenhüllproteins
umfasst, wobei die funktionelle Form dadurch gekennzeichnet ist,
dass sie einen Phagen, der die funktionelle Form in seiner Hülle trägt, infektiös macht.
Das Nucleinsäuremolekül kann weiterhin
alle relevanten Nucleinsäuren
umfassen, die Proteine codieren, welche für den Zusammenbau eines Phagen
in einer Wirtszelle erforderlich sind.
-
Die
Erfindung stellt einen chimärischen
Phagen bereit, der eine Hülle
aufweist, die ein Gemisch von Proteinen umfasst, wobei das Gemisch
ein Fusionsprotein umfasst, bei dem ein proteinartiges Molekül mit einer
funktionellen Form eines Phagenhüllproteins
fusioniert ist, wobei das Gemisch weiterhin eine mutante Form des
Phagenhüllproteins
umfasst, wobei die mutante Form dadurch gekenn zeichnet ist, dass
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die die mutante
Form des Phagenhüllproteins
und keine Kopie oder Kopien der funktionellen Form des Phagenhüllproteins
umfasst, weniger infektiös
ist als ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die wenigstens
eine funktionelle Form des Phagenhüllproteins umfasst. Der hier
verwendete Ausdruck "funktionelle
Form" bezieht sich
auf ein Phagenhüllprotein,
das erheblich zur Infektiosität
des Partikels beiträgt,
an den es gebunden ist. Das Phagenhüllprotein selbst ist nicht
infektiös,
aber die funktionelle Form macht den Phagenpartikel, an den sie
gebunden ist, infektiös.
Neben dem Beitrag zur Infektiosität des Phagenpartikels unterstützt das
Phagenhüllprotein
auch andere Funktionen, wie die Stabilisierung des Phagenpartikels.
Eine mutante Form eines Phagenhüllproteins gemäß der Erfindung
kann den Phagenpartikel weniger infektiös oder nichtinfektiös machen,
sollte aber immer noch andere Funktionen des Phagenhüllproteins,
wie die Stabilisierung des Phagen, unterstützen. Ein Phage, der kein Wildtyp-Phagenhüllprotein
umfasst, von dem die mutante Form abgeleitet ist, und der keine
funktionellen Formen des Phagenhüllproteins
umfasst, sondern nur mutante Formen des Phagenhüllproteins umfasst, ist weniger
infektiös
als ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine oder mehrere funktionelle
Formen des Phagenhüllproteins
neben den mutanten Formen des Phagenhüllproteins in seiner Hülle umfasst.
Der hier verwendete Ausdruck "weniger
infektiös" kann auch nichtinfektiös bedeuten.
Obwohl ein chimärischer
Phage der Erfindung wenigstens eine Kopie der mutanten Form des
Phagenhüllproteins
in seiner Hülle
umfasst, besitzt der chimärische
Phage Infektionsfähigkeit,
da er auch wenigstens eine funktionelle Form des Phagenhüllproteins
in seiner Hülle
umfasst. Der hier verwendete Ausdruck "funktionelle Form eines Phagenhüllproteins" bedeutet auch einen
Teil, ein Derivat und/oder ein Analogon davon, das noch insofern
Funktionalität
beinhaltet, als es den Phagen, an den es gebunden ist, infektiös macht.
Der hier verwendete Ausdruck "mutante
Form eines Phagenhüllproteins" bedeutet auch einen
Teil, ein Derivat und/oder ein Analogon der mutanten Form, wobei
die mutante Form dadurch gekennzeichnet ist, dass ein Phage, der
kein Wildtyp- Phagenhüllprotein
umfasst, von dem die mutante Form abgeleitet ist, und der eine Hülle aufweist,
die nur mutante Formen des Phagenhüllproteins oder Teile, Derivate und/oder
Analoga davon umfasst, weniger infektiös oder nichtinfektiös ist im
Vergleich zu einem Phagen, der kein Wildtyp-Phagenhüllprotein
umfasst, von dem die mutante Form abgeleitet ist, und der eine Hülle aufweist, die
wenigstens eine funktionelle Form des Phagenhüllproteins umfasst.
-
In
einer bevorzugten Ausführungsform
ist das Phagenhüllprotein
das g3-Protein (g3p), das in einer Wildtyp- oder funktionellen Form
einen Phagen, an den es gebunden ist, infektiös macht. Wie in 2 und 3 skizziert
ist, sind bestimmte Teile des g3-Proteins an der Erkennung von Wirtszellrezeptoren
beteiligt. Die Rezeptorbindung einer mutanten Form ist "beeinträchtigt", wenn eine Veränderung
im g3-Protein dafür sorgt,
dass der Rezeptor vom g3-Protein oder Teilen davon in geringerem
Maße erkannt
und gebunden wird, als wenn keine Veränderung vorhanden ist. Daher
bedeutet der hier verwendete Ausdruck "beeinträchtigt", dass das Hüllprotein, wie g3p, den Wirtszellrezeptor
weniger effizient bindet oder dass g3p seine Fähigkeit, den Wirtszellrezeptor
zu binden und/oder zu erkennen, vollständig verloren hat. In einer
bevorzugten Ausführungsform
umfasst die mutante Form des Phagenhüllproteins eine Veränderung
im g3-Protein, die
aus einer Mutation im D1-Bereich, im D2-Bereich oder in beiden besteht.
Der hier verwendete Ausdruck "Veränderung" oder "Mutation" bedeutet eine oder
mehrere Punktmutationen, Abschnitte von Mutationen, Deletionen,
Substitutionen, Ersetzungen und/oder Austausch von Teilen. In einer
besonders bevorzugten Ausführungsform
ist die Veränderung
im g3-Protein eine Deletion im Wesentlichen des gesamten D1- und/oder
D2-Bereichs. Diese Veränderung
kann auch eine Substitution des deletierten g3-Proteinteils durch
ein Protein oder Peptid bedeuten, das nicht zur Infektiosität des Helferphagen,
des chimärischen
Phagen, des infektiösen
Phagen oder des Phagenpartikels beiträgt.
-
In
einer besonders bevorzugten Ausführungsform
der Erfindung umfasst ein chimärischer
Phage oder ein infektiöser
Phage gemäß der Erfindung
eine Nucleinsäure,
die ein Fusionsprotein codiert, bei dem ein proteinartiges Molekül mit einer
funktionellen Form des Phagenhüllproteins
fusioniert ist, wobei der chimärische Phage
der Erfindung einen M13-, M13K07, VCSM13- oder R408-Stamm oder eine Mutante,
ein Derivat oder einen analogen Stamm, die von einem dieser Stämme abgeleitet
sind, umfasst. Ein proteinartiges Molekül gemäß der Erfindung ist mit der
funktionellen Form des Phagenhüllproteins
fusioniert und umfasst ein Protein wie eine ligandenbindende Struktureinheit
oder ein Immunglobulin (wie einen Antikörper). Ein proteinartiges Molekül kann auch
ein Peptid wie eine zufällige
Sequenz von Aminosäuren
oder eine nichtzufällige
Sequenz von Aminosäuren
bedeuten, wie ein Antikörperfragment
oder Derivate davon (Fab-Fragment, Einzelketten-Fv-Fragment (scFv),
variabler Bereich oder CDR-Bereich). Ein proteinartiges Molekül kann auch
einen ersten Partner eines spezifischen Bindungspaars bedeuten oder
kann Fusionen zwischen verschiedenen Arten von (Fragmenten von)
Proteinen und/oder Fusionen zwischen (Fragmenten von) Proteinen
und (statistischen oder nichtstatistischen) Peptiden, wie von Antikörpern erkannten
Markern, bedeuten.
-
Ein
chimärischer
Phage der Erfindung hängt
in Bezug auf die Infektion erheblich von der funktionellen Form
des Phagenhüllproteins
g3p und von der Anwesenheit eines Teils oder von Teilen des Phagenhüllproteins,
die zur Infektiosität
des Phagen beitragen, ab. Die mutante Form des Phagenhüllproteins
ist in denjenigen Teilen des Phagenhüllproteins mutiert, die den
Phagen infektiös
machen. Beispiele für
die Mutation sind unter anderem Deletionen, Substitutionen von Resten
oder Fragmenten, Austausche und/oder Ersetzungen durch andere Proteinfragmente,
die ihn weniger infektiös
machen. Die Proteinfragmente können
mit Phagenhüllproteinen
oder Fragmenten davon verwandt sein oder auch nicht und sind im
Wesentlichen nicht in der Lage, eine Infektion einer Wirtszelle
durch den Phagenpartikel zu induzieren.
-
In
einer anderen Ausführungsform
stellt die Erfindung eine Phagensammlung bereit, die einen chimärischen
Phagen oder einen infektiösen
Phagen gemäß der Erfindung
umfasst. Phagen der vorliegenden Erfindung sind insbesondere für die Erzeugung
von Phagen-Display-Bibliotheken geeignet. Daher ist die Phagensammlung
in einer besonders bevorzugten Ausführungsform eine Phagen-Display-Bibliothek.
In einer ganz besonders bevorzugten Ausführungsform besteht die Phagensammlung
im Wesentlichen aus chimärischen Phagen
oder infektiösen
Phagen der Erfindung. Ein proteinartiges Molekül, wie (statistische oder nichtstatistische)
Sequenzen von Aminosäuren,
Peptide, Proteinteile oder auch ganze Proteine, können mit
den Phagenhüllproteinen
fusioniert werden und einen ersten Partner eines spezifischen Bindungspaars
bilden. Diese Fusion erfolgt typischerweise an den terminalen Enden
des Hüllproteins.
Die Addition beeinflusst die Funktion des Phagenhüllproteins
typischerweise nicht. Außerdem
stört sie
häufig
auch nicht die Funktion der addierten Struktureinheit. So ist es
möglich,
Bibliotheken zu erzeugen, die zum Beispiel verwendet werden können, um spezifische
Bindungsmoleküle
zu lokalisieren und zu klonieren. Solche Bibliotheken können Peptide
oder größere Moleküle umfassen.
Vorzugsweise umfassen die größeren Moleküle ein Protein,
wie einen Antikörper oder
einen funktionellen Teil, ein Derivat und/oder Analogon davon, wie
schwere und/oder leichte Ketten eines Immunglobulinmoleküls in voller
Länge oder
Fragmente von Immunglobulinen, wie Fab-Fragmente, Einzelketten-Fv(scFv)-Fragmente,
CDR-Bereiche, einzelne
variable Bereiche und/oder Kombinationen der obigen.
-
In
einem anderen Aspekt stellt die Erfindung ein Verfahren zur Herstellung
eines Phagenpartikels bereit, das die folgenden Schritte umfasst:
das Versehen einer Wirtszelle mit einer ersten Nucleinsäure, die
ein Fusionsprotein codiert, wobei das Fusionsprotein ein proteinartiges
Molekül
umfasst, das mit einer funktionellen Form eines Phagenhüllproteins
oder einem funktionellen Teil, Derivat und/oder Analogon des Phagenhüllproteins
fusioniert ist, das Versehen der Wirtszelle mit einer zweiten Nucleinsäure, die
eine mutante Form des Phagenhüllproteins
codiert, wobei die mutante Form durch das oben beschriebene gekennzeichnet
ist und wobei "weniger
infektiös" auch nichtinfektiös bedeuten
kann, und das Kultivieren der Zelle, so dass der Phage zusammengebaut
werden kann, wobei die Wirtszelle ansonsten oder zusätzlich Nucleinsäuren umfasst,
die wenigstens alle wesentlichen Proteine oder funktionelle Äquivalente
der wesentlichen Proteine für
den Zusammenbau des Phagenpartikels codieren. In einer bevorzugten
Ausführungsform
stellt die Erfindung ein Verfahren bereit, bei dem die Nucleinsäure, die
wenigstens alle anderen Proteine oder funktionellen Äquivalente
davon, die für
den Zusammenbau des Phagenpartikels in der Wirtszelle wesentlich
sind, codiert, aus einem Helferphagen besteht und der Helferphage
verwendet wird, um die Nucleinsäure
an die Wirtszelle abzugeben. In einer besonders bevorzugten Ausführungsform
umfasst die Nucleinsäure,
die durch den Helferphagen abgegeben wird, auch die zweite Nucleinsäure, die
die mutante Form des Phagenhüllproteins
codiert. In einer ganz besonders bevorzugten Ausführungsform
sind die erste und die zweite Nucleinsäure getrennte Nucleinsäuren, die
jeweils getrennte einzigartige Selektionsmarker umfassen können, um
zu gewährleisten,
dass die Wirtszelle wenigstens eine Kopie von jeder getrennten Nucleinsäure umfasst
und dass jede getrennte einzigartige Replikationsstartpunkte umfasst,
um zu gewährleisten,
dass es während
der Replikation keine Störungen
gibt. In einem anderen Aspekt der Erfindung ist die Zahl der möglichen
homologen Rekombinationsereignisse zwischen überlappenden Stücken von
Nucleinsäuresequenzen
zwischen den getrennten Nucleinsäuren
aufgrund der Verwendung unterschiedlicher Codons innerhalb jeder
Nucleinsäure
reduziert.
-
Unter
Verwendung eines Verfahrens der Erfindung ist es möglich, Phagenpartikel
zu erzeugen, die wenigstens zwei Varianten umfassen, die von demselben
Hüllprotein
abgeleitet sind. Solche Phagenpartikel wurden in der Technik bisher
nicht verwendet. Gemäß der einschlägigen Lehre
werden Phagenpartikel verwendet, die eine Hülle haben, die entweder deletierte
g3-Proteine, mit heterologen Proteinen verknüpfte g3-Proteine, mit heterologen
Proteinen verknüpfte
deletierte g3-Proteine oder eine begrenzte Zahl von g3-Proteinen,
aber keine Gemische davon enthält.
Die vorliegende Erfindung stellt neue Phagenpartikel (chimärische Phagen,
infektiöse
Phagen und Helferphagen) bereit, bei denen die relative Zahl der
in einer Hülle
vorhandenen g3p-Varianten (mit einer heterologen Struktureinheit
fusioniert oder auch nicht) variieren kann. Typischerweise möchte man
die relative Menge der verschiedenen Varianten in der Phagenhülle beeinflussen.
Zu diesem Zweck ist die Expression des Fusionsproteins und/oder
die Expression der mutanten Form des Phagenhüllproteins vorzugsweise regulierbar
(steuerbar). Vorzugsweise wird dies erreicht, indem man die Expression
des Gens, das das Phagenhüllprotein
codiert, auf dem Transcriptionsniveau reguliert. So steht die Expression
des Fusionsproteins und/oder der mutanten Form des Phagenhüllproteins
vorzugsweise unter der Kontrolle eines Promotors, der gut steuerbar
ist. Beispiele für
solche Promotoren sind der lac-Promotor, der psp-Promotor und der AraC/BAD-Promotor,
wobei letzterer durch die Konzentration von Glucose oder Arabinose
im Medium beeinflusst wird. Besonders vorteilhaft ist der AraC/BAD-Promotor.
Der AraC/BAD-Promotor wird bevorzugt, weil es sich um einen Promotor
handelt, der sich sehr genau steuern lässt. Dieser Promotor ist in
Anwesenheit von Glucose für
alle praktischen Zwecke inaktiv und in Abwesenheit von Glucose nur
geringfügig
aktiv. Dies bedeutet, dass eine geringe Konzentration oder die Abwesenheit
von Glucose den Promotor aktiviert, was zur Hochregulation des interessierenden
Gens führt,
das sich unter der Kontrolle des Promotors befindet. Wenn zum Beispiel
die Deletionsmutante von g3 (der die infektiösen Bereiche D1 und D2 fehlen)
unter der Kontrolle eines solchen Promotors steht, wäre die relative
Zahl von Deletionsmutanten im Vergleich zu g3-Fusionsproteinen der
vollen Länge
(oder wenigstens funktionellen g3-Fusionsproteinen) gering, wenn
die Glucosekonzentration relativ hoch ist. Im Prinzip führt dieses
System zu einer genauen Regulation der Anzahl der Deletionsmutanten
und funktionellen Hüllproteine,
wie g3, auf der Hülle
eines Phagenpartikels, und daher kann der Prozentsatz solcher Hüllproteine
und Deletionsmutanten reguliert werden. Außerdem kann die Aktivität des AraC/BAD-Promotors durch die
Zugabe von Arabinose zum Medium sehr genau reguliert werden. Die
verwendete Arabinosekonzentration bestimmt das Expressionsniveau
des Proteins in E.-coli-Zellen. Daher wird eine optimale Regulation
des Gehalts an Phagenhüllprotein
erreicht, indem man diesen AraC/BAD-Promotor verwendet und die Kulturbedingungen
der Wirtszelle verändert.
Die Verwendung eines Promotors, der von Arabinose abhängt, wie
der AraC/BAD-Promotor, anstatt von IPTG, wie das lac-Operon, verhindert
mögliche
Probleme, die aufgrund der gemeinsamen Verkapselung mit dem Helferplasmid
in Viruspartikeln während
der Helferphagensynthese auftreten. Wie oben beschrieben wurde,
hat sich gezeigt, dass eine gemeinsame Verkapselung von Plasmiden
mit dem Phagengenom erfolgt (Russel und Model, 1989; Krebber et
al., 1995; Rakonjac et al., 1997). Wenn eine gemeinsame Verkapselung
mit einem lac-gesteuerten Helferplasmid erfolgt, kann dieses mit
den lac-gesteuerten Vektoren konkurrieren, die im Allgemeinen für das Phagen-Display
verwendet werden, was zur Produktion von infektiösen Phagenpartikeln führt, die
das g3p-X-Fusionsprodukt nicht enthalten. Dieses Problem tritt am
wahrscheinlichsten dann nicht auf, wenn der AraC/BAD-Promotor verwendet
wird. In den Studien, die durch die vorliegende Erfindung vorgestellt
werden, wurden solche Konkurrenzprobleme mit dem lac-Promotor jedoch
nicht beobachtet. Daher könne
auch andere regulierbare Promotoren, wie der psp- und der lac-Promotor,
verwendet werden, um Phagen gemäß der Erfindung
zu erzeugen. Es ist daher auch Bestandteil der Erfindung, solche
Promotoren als Alternative zum AraC/BAD-Promotor zu verwenden.
-
Ein
Verfahren der Erfindung zur Herstellung eines Phagenpartikels wird
vorzugsweise verwendet, um einen chimärischen Phagen gemäß der Erfindung
herzustellen. Der Wirt kann mit jeder geeigneten Methode mit Nucleinsäure versehen
werden, die ein Phagenprotein codiert. Vorzugsweise jedoch wird
der Wirt mit einem Helferphagen gemäß der Erfindung versehen. Ein
Helferphage gemäß der Erfindung
umfasst Nucleinsäure,
die andere Phagenproteine oder funktionelle Äquivalente davon, die für den Zusammenbau
des Helferphagen wesentlich sind, codiert, wobei die Nucleinsäure weiterhin
eine mutante Form eines Phagenhüllproteins codiert,
wobei die mutante Form dadurch gekennzeichnet ist, dass ein Phage,
der kein Wildtyp-Phagenhüllprotein
umfasst, von dem die mutante Form abgeleitet ist, und der eine Hülle aufweist,
die mutante Formen des Phagenhüllproteins
und keine funktionellen Formen des Phagenhüllproteins umfasst, weniger
infektiös
ist als ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die wenigstens
eine funktionelle Form des Phagenhüllproteins umfasst, und wobei
der Helferphage keine Nucleinsäure
umfasst, die eine funktionelle Form des Phagenhüllproteins codiert. "Andere Phagenproteine" bedeutet andere
als die funktionelle Form des Phagenhüllproteins und als die mutante
Form des Phagenhüllproteins.
Mit Nucleinsäure,
die letztere codiert, kann die Wirtszelle in einer alternativen
Weise versehen werden. Der Helferphage kann jedoch weiterhin Nucleinsäure umfassen,
die die funktionelle Form des Phagenhüllproteins oder die mutante
Form oder beide codiert. Vorzugsweise umfasst der Helferphage keine Nucleinsäure, die
das Fusionsprotein codiert. Auf diese Weise ist der Helferphage
gleichmäßig und
kann verwendet werden, um Phagen zu produzieren, die präferentiell
Nucleinsäure,
die das Fusionsprotein codiert, in Abwesenheit von Nucleinsäure, die
ein anderes erforderliches Helferphagenprotein codiert, umfassen.
Somit werden das Fusionsprotein und die mutante Form des Phagenhüllproteins
vorzugsweise von getrennten Nucleinsäuren codiert. Vorzugsweise
umfasst jede der getrennten Nucleinsäuren einen einzigartigen Selektionsmarker.
Vorzugsweise umfassen die getrennten Nucleinsäuren einander nicht störende Replikationsstartpunkte,
wobei die Replikationsstartpunkte nicht miteinander konkurrieren,
was zu Bakterienzellen führt,
die die getrennten Nucleinsäuren
für die
Erzeugung von neuen Phagenpartikeln tolerieren.
-
Gemäß der einschlägigen Lehre
ist es sehr schwierig, Chargen von Helferphagen zu erzeugen, bei denen
die Helferphagen Nucleinsäure
beherbergen, die alle wesentlichen Proteine codiert, die für den Zusammenbau
eines Phagenpartikels in einer bakteriellen Wirtszelle erforderlich
sind, und wobei der Nucleinsäure ein
Gen, das g3p codiert, fehlt. Anschließend ist es gemäß der einschlägigen Lehre
schwierig, unter Verwendung solcher Helferphagen Phagenbibliotheken
zu produzieren. In der Technik sind mehrere Schwierigkeiten bekannt,
die eine geeignete Erzeugung solcher Chargen von Helferphagen behindern.
Die vorliegende Erfindung stellt Verfahren und Mittel sowie eine
gute Kombination von Merkmalen, wie die Verwendung von spezifischen
Replikationsstartpunkten, Selektionsmarkern und Codons in überlappenden
DNA-Sequenzen bereit, die die Produktion von Phagenchargen ermöglichen,
welche hohe Anteile von geeigneten Helferphagen enthalten und die
anschließend
für die
Erzeugung von chimärischen
oder infektiösen
Phagen gemäß der Erfindung
angewendet werden können.
Daher stellt die Erfindung in einer Ausführungsform Verfahren und Mittel zur
Herstellung von Helferphagen bereit, die funktionelle Formen und/oder
Wildtypformen eines Phagenhüllproteins
in ihrer Hülle
tragen, denen aber dennoch Nucleinsäure fehlt, die für die funktionelle
Form und/oder die Wildtypform des Phagenhüllproteins codiert. Die Erfindung
stellt ein Verfahren zur Herstellung eines Helferphagen bereit,
das die folgenden Schritte umfasst: das Versehen einer Wirtszelle
mit einer ersten Nucleinsäure,
die eine funktionelle Form eines Phagenhüllproteins codiert, das Versehen
der Wirtszelle mit einer zweiten Nucleinsäure, die eine mutante Form
des Phagenhüllproteins
codiert, wobei die mutante Form dadurch gekennzeichnet ist, dass
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die mutante Formen
des Phagenhüllproteins
und keine Kopien der funktionellen Form umfasst, weniger infektiös ist als
ein Phage, der kein Wildtyp-Phagenhüllprotein umfasst, von dem
die mutante Form abgeleitet ist, und der eine Hülle aufweist, die wenigstens
eine funktionelle Form des Phagenhüllproteins umfasst, und das
Kultivieren der Wirtszelle, so dass der Helferphage zusammengebaut
werden kann, wobei die Wirtszelle zusätzlich Nucleinsäure umfasst,
die wenigstens alle anderen Proteine oder funktionellen Äquivalente
davon codiert, die für
den Zusammenbau des Helferphagen in der Wirtszelle wesentlich sind.
Vorzugsweise ist das Phagenhüllprotein
das g3-Protein,
das in der Hülle
der meisten, wenn nicht aller Bakteriophagen vorhanden ist. Vorzugsweise
werden die anderen Proteine von der zweiten Nucleinsäure codiert,
wobei die Expression der mutanten Form des Phagenhüllproteins
und/oder die Expression der funktionellen Form reguliert wird, indem
man die Kulturbedingungen der Wirtszelle verändert, und sie sich vorzugsweise
unter der Kontrolle eines regulierbaren Promotors, wie des AraC/BAD-,
psp- und lac-Promotors,
befindet, wie es oben beschrieben ist.
-
Man
sagt, ein Phagenhüllprotein
trage erheblich zur Infektiosität
eines Phagen bei, wenn es nach Einbau in einen Phagen den Phagen
in die Lage versetzt, in einer Weise, die mit einer Wildtypversion
des Phagenhüllproteins
vergleichbar ist, einen bakteriellen Wirt zu erkennen, zu binden
und/oder zu infizieren. Die hier verwendeten Ausdrücke "weniger infektiös" oder "nichtinfektiös" beziehen sich auf
einen Phagen, der keine funktionelle oder Wildtypform eines Phagenhüllproteins,
vorzugsweise g3p, trägt
und der im Vergleich zum Wildtypphagen (Partikel) eine erheblich
beeinträchtigte,
reduzierte Infektionsfähigkeit
aufweist, was zum Beispiel in Plaque-Assays, die dem Fachmann wohlbekannt
sind, bestimmt werden kann. Die hier verwendeten Ausdrücke "weniger infektiös" und "nichtinfektiös" können sich
auch auf eine Abnahme der Wirtszellspezifität beziehen. Im Allgemeinen
ist ein nichtinfektiöser
Phage nicht in der Lage, eine Bakterienzelle zu erkennen, zu binden
und/oder in sie einzutreten, wie es für Wildtypphagen in 2 und 3 skizziert
ist, während
ein weniger infektiöser
Phage dies mit geringerer Effizienz kann. Vorzugsweise infiziert
ein Phage, der mutante Formen aufweist, die Wirtszelle mit einer
Effizienz, die kleiner als 50% ist im Vergleich zu einem Phagen,
der wenigstens eine funktionelle Form trägt, besonders bevorzugt kleiner
als 10%, ganz besonders bevorzugt kleiner als 1%, unter ansonsten
vergleichbaren Bedingungen.
-
In
Ausführungsformen,
bei denen das Phagenhüllprotein
ein g3p umfasst, umfasst die mutante Form vorzugsweise wenigstens
einen strukturellen Teil, ein Derivat und/oder Analogon des D3-Bereichs.
Der strukturelle Teil, das Derivat und/oder Analogon umfasst wenigstens
die Funktionalität
von g3p, den Zusammenbau von stabilen Phagenpartikeln zu ermöglichen.
-
Ein
Derivat eines Proteins, wie der Ausdruck hier verwendet wird, umfasst
dieselbe Aktivität
wie das Protein, von dem das Derivat abgeleitet ist. Wenn das Protein
eine funktionelle Form eines Phagenhüllproteins ist, umfasst das
Derivat dieselbe Funktionalität.
Wenn das Protein eine mutante Form eines Phagenhüllproteins ist, ist das Derivat
ebenfalls eine mutante Form des Phagenhüllproteins. Wenn das Protein
eine mutante Form eines Phagenhüllproteins
ist, die einen Phagen, der nur mutante Formen des Phagenhüllproteins
trägt, weniger
infektiös
macht, so ist das Derivat ebenfalls ein Phagenhüllprotein, das einen Phagen,
der nur Derivate und/oder mutante Formen des Phagenhüllproteins
trägt,
weniger infektiös
macht. Typische Derivate sind Proteine, die eine oder mehrere konservative
Aminosäuresubstitutionen
umfassen. Derivate können
jedoch auch Insertionen und/oder Deletionen umfassen. Weiterhin
können
Derivate auch Austausche von Aminosäuren oder von Sequenzen von
Aminosäuren
innerhalb desselben Proteins oder zwischen zwei oder mehr miteinander
verwandten und/oder nicht miteinander verwandten Proteinen umfassen.
Ein Analogon eines Proteins, wie der Ausdruck hier verwendet wird,
umfasst im Wesentlichen dieselbe Aktivität wie das Protein, zu dem das Analogon
analog ist. Wenn das Protein eine funktionelle Form eines Phagenhüllproteins
ist, umfasst das Analogon dieselbe Funktionalität.
-
Vor
der Erfindung war es schwierig, eine ausreichende Zahl von Helferphagen
zu erzeugen, denen ein funktionelles g3-Gen oder ein Teil, Derivat
und/oder Analogon davon in ihrem Genom fehlt, die aber dennoch in
einem Infektionsdurchlauf bei E.-coli-Zellen infektiös sind.
Der vorliegenden Erfindung gelingt es, solche Helferphagen effizient
zu erzeugen. Solche Helferphagen tragen funktionelle Formen und/oder
Wildtyp-g3-Proteine (g3ps) auf ihrer Oberfläche, aber dennoch fehlt ihnen
eine Nucleinsäure,
die die funktionelle Form und/oder das Wildtyp-g3-Protein codiert.
Die Erfindung stellt weiterhin Verfahren und Mittel bereit, um mit
Hilfe solcher Helferphagen Bibliotheken zu erzeugen, die chimärische Phagen
umfassen. Vorzugsweise werden Phagen-Display-Bibliotheken erzeugt.
Besonders bevorzugt präsentieren
Bibliotheken eine Vielzahl von Einzelketten-Fv(scFv)-Fragmenten.
Unter Verwendung der Mittel und Verfahren der Erfindung können Phagenbibliotheken
erzeugt werden, die eine Zahl von infektiösen Phagen enthalten, die im
Vergleich zu der Zahl der nicht oder weniger infektiösen Phagen
erheblich größer ist,
als im Stand der Technik beschrieben und vorhanden. Gleichzeitig
werden die Helferphagen, die zur Herstellung solcher Bibliotheken
verwendet werden, (über
einen Infektionsdurchlauf in E.-coli-Zellen) im Wesentlichen nichtinfektiös, da das
g3-Gen in den Phagen nicht in einer zur Infektiosität beitragenden
Form vorhanden ist. Nach der Infektion eines bakteriellen Wirts
können
sich diese Phagen also nicht auf andere Bakterienzellen ausbreiten,
außer
natürlich
durch Teilung des bereits infizierten Wirtes zu Tochterzellen. Solche
Bibliotheken werden daher ebenfalls durch die Erfindung bereitgestellt. Die
erzeugten Bibliotheken sind besonders nützlich für Panning-Experimente, da die
Titer von Phagen pro Milliliter erheblich höher sind, als sie bis zur vorliegenden
Erfindung in der Technik verwendet wurden. Außerdem präsentieren Bibliotheken der
Erfindung weniger unspezifische Klebrigkeit. Die Bibliotheken präsentieren
also weniger falsch positive Ergebnisse als Bibliotheken im Stand
der Technik. Außerdem
können
nach einem Durchlauf des Panning nur Phagen, die eine funktionelle
Form auf ihrer Oberfläche
präsentieren
(vorzugsweise mit einem proteinartigen Molekül fusioniert), in E.-coli-Zellen
vermehrt werden, während
Phagen, die keine funktionellen Formen des Phagenhüllproteins
tragen, sondern nur mutante Formen des Phagenhüllproteins tragen, im Wesentlichen
nichtinfektiös
sind und in E.-coli-Zellen nicht vermehrt werden können. Daher
wird die Zahl der verbleibenden Phagen, die für einen zweiten Panning-Durchlauf
verwendet werden können,
erheblich reduziert. Infolge der Verwendung der chimärischen
Phagen der Erfindung, die in den durch die vorliegende Erfindung
bereitgestellten Bibliotheken vorhanden sind, nimmt die Zahl der
Panning-Durchläufe
ab, und die Zahl der relevanten Bindungsmoleküle wird viel effizienter erhalten,
als es vor der vorliegenden Erfindung möglich war.
-
In
einem anderen Aspekt stellt die Erfindung Nucleinsäuren und
Helferphagen, die die Nucleinsäuren umfassen,
welche genomische DNA-Sequenzen umfassen, bereit, wobei wenigstens
die Domänen
von g3p, die für
die Infektion verantwortlich sind und zu dieser beitragen, funktionell
entfernt sind. Die Erfindung stellt auch genomische Helferphagen-DNA
bereit, bei der die Leadersequenz und wenigstens die D3-Domäne unberührt und
miteinander fusioniert sind. Diese Nucleinsäuren beruhen vorzugsweise auf
genomischen Sequenzen von VCSM13 und M13K07. Aufgrund des Fehlens
einer funktionellen D1-Domäne
sind Phagenpartikel, die durch diese Nucleinsäuren erzeugt wurden, im Wesentlichen
nichtinfektiös. "Im Wesentlichen" bedeutet in diesem
Zusammenhang, dass keine Ausbreitung oder wenigstens erheblich weniger
Ausbreitung der Phagen auf andere Bakterienzellen als das Produktionsbakterium
durch infektiöse
Merkmale, die von g3p geliefert werden, erfolgt. Dieses Fehlen einer
Ausbreitung auf andere Bakterienzellen ist auf das Fehlen einer funktionellen
Form von g3p zurückzuführen. Wenn
während
der Produktion eine Quelle für
die Wildtypform oder funktionelle Form von g3p bereitgestellt wird,
können
die erzeugten Phagen ein Bakterium infizieren. Wenn das Bakterium
jedoch infolge einer Infektion Phagen produziert, ist der resultierende
Phagenpartikel nicht in der Lage, ein anderes Bakterium zu infizieren,
es sei denn, dass wiederum während
der Produktion eine Quelle für
infektiöses
g3p bereitgestellt wird. Ein chimärischer Phage oder ein infektiöser Phage
der Erfindung umfasst vorzugsweise einen Teil von g3p, der nach
einem Infektionsdurchlauf in E.-coli-Zellen die Erzeugung eines
stabilen Phagenpartikels gewährleistet.
Zu diesem Zweck umfasst der Helferphage vorzugsweise eine Nucleinsäure, die
eine mutante Form des g3p codiert. Vorzugsweise umfasst die mutante
Form D3 oder einen funktionellen Teil, ein Derivat und/oder ein
Analogon davon.
-
Eine
Phagen-Display-Bibliothek kann zum Beispiel erzeugt werden, indem
man eine Sammlung von Bakterien mit einer Bibliothek von Nucleinsäuren versieht,
die g3p codieren, das mit einer Reihe von verschiedenen proteinhaltigen
Molekülen
fusioniert ist, und die Bakterien mit Helferphagen der Erfindung
infiziert. In einer besonders bevorzugten Ausführungsform enthält eine
Bibliothek von Phagen-Display-Partikeln, die mit diesen Helferphagen
erzeugt wird, Phagen, die keinen infektiösen Teile von g3p auf ihrer
Oberfläche
tragen, und Phagen, die eine oder zwei g3p-X-Fusionen in voller
Länge tragen,
neben nichtinfektiösen
oder weniger infektiösen
Teilen von g3p-Deletionsproteinen. Phagen in diesen Bibliotheksgemischen,
die keine g3p-X-Fusionsproteine exprimieren, können keine Bakterien mehr infizieren,
da sie in Abwesenheit von infektiösen g3p-Teilen, die nicht mit
X fusioniert sind, erzeugt wurden (X umfasst ein interessierendes
proteinartiges Molekül
oder einen interessierenden Fusionspartner, wie Immunglobuline oder
Fragmente von Immunglobulinen, wie Fab-Fragmente oder scFv-Fragmente).
-
In
einem Aspekt stellt die Erfindung Helferphagen bereit, die die Anwesenheit
eines Selektionsmarkers mit der Anwesenheit eines bakteriellen Replikationsstartpunkts
(ORI) kombinieren, um die beschriebenen Probleme bei der Produktion
von g3-negativen Helferphagen und anschließend bei der Erzeugung von
Phagen-Display-Bibliotheken zu überwinden.
Die Anwesenheit einer solchen Kombination gewährleistet die Produktion von
großen
Mengen an Helferphagen und/oder Helfergenomen. g3-negative Helferphagen
mit einem ORI und einem Resistenzmarker können aus den Helferphagen VCSM13
und M13K07 hergestellt werden. Diese Helferphagen enthalten im Unterschied
zu M13 oder R408 ein Kanamycin-Resistenz-Gen aus dem Tn903-Transposon
und einen P15A-ORI, die beide in den intergenischen Bereich des
Phagengenoms eingesetzt werden. Ein weiterer Aspekt der Erfindung
ist die Tatsache, dass diese Helferphagen wegen dieses Resistenz-Gens
und der Anwesenheit dieses besonderen ORI leicht in großen Mengen
wachsen können,
während
leere oder kein Plasmid oder kein Genom enthaltende Bakterien unter
dem Selektionsdruck entfernt werden, und dass keine Störungen zwischen
ORIs aus dem Phagengenom und dem Helferplasmid oder zwischen ORIs
aus dem Phagengenom und den Plasmiden der Display-Bibliothek auftreten,
wenn beide Nucleinsäuren in
derselben bakteriellen Wirtszelle vorhanden sind. VCSM13 und M13K07
enthalten den P15A-ORI. Um das Verschwinden des Helfergenoms oder
des Helfervektors zu verhindern, sollten die ORIs keine Störungen verursachen,
und daher werden von P15A abgeleitete ORIs im Vektor nicht verwendet.
Für den
auf die Produktion von Helferphagen angewendeten Vektor wurde der
ColE1-ORI gewählt.
Neben den oben genannten Merkmalen und Effekten ist es auch wichtig,
dass der Helfervektor für
die Erzeugung von Helferphagen keinen F1-ORI trägt, um den Einbau des Helfervektors
anstatt des viralen Genoms in den Phagenpartikel zu verhindern.
-
Die
Erfindung stellt auch Vektoren bereit, die eine regulierte Expression
der mutanten Form und/oder der funktionellen Form von g3p durch
die Verwendung eines regulierbaren Promotors ermöglichen, die weiterhin ein
Resistenz-Gen enthalten, das von dem im Helfergenom vorhandenen
Kanamycin-Resistenz-Gen verschieden ist. Diese komplementäre Resistenz
wird hier durch das beta-Lactamase(Ampicillin)-Gen
bereitgestellt, da dessen Produkt relativ stabil ist und ein vollständiges Abtöten von
Bakterien gewährleistet,
die das Genprodukt nicht exprimieren.
-
Der
pBAD/gIII-Vektor (Invitrogen) kann als Gerüstvektor für die Produktion von Helfervektoren
der Erfindung verwendet werden. Alternativ dazu können Vektoren
wie pUC19 verwendet werden. Vorzugsweise bestehen weitere Merkmale
dieser Grundhelfervektoren darin, dass Bereiche mit Sequenzhomologie
minimiert sind, was die Möglichkeit
der homologen Rekombination erheblich reduziert.
-
Die
Erfindung gibt weiterhin die Verwendung der bakteriellen Wirtszellen
TOP10, LMG und/oder XL1blue für
die Herstellung von Helferphagen an, die ein g3-negatives Genom
enthalten, aber aufgrund des g3p, das auf den Phagen vorhanden ist,
da es vom Helfervektor geliefert wurde, dennoch infektiös sind.
Der Genotyp der TOP10- und LMG-Bakterien gewährleistet, dass sie Arabinose
in die Zelle transportieren können, aber
dass sie diese nicht metabolisieren können (Genotyp: araABCD– und araEFGH+).
Außerdem
sind die TOP10-Bakterien recA- und endA-defizitär, was die Wahrscheinlichkeit
einer Rekombination und Mutation verringert. Weiterhin sind die
TOP10-Bakterien F-negativ, wodurch sie resistent gegen Phagen sind,
die interessierende Phagenchargen kontaminieren könnten.
-
Die
vorliegende Erfindung gibt auch die Sequenz des VCSM13-Phagengenoms
und deren Verwendung zum Zwecke des Aufbaus einer Phagen-Display-Bibliothek
an und stellt Klonierungsphagen und Plasmidmutanten bereit, die
bei allgemeinen molekularbiologischen Verfahren und Mitteln nützlich sind.
-
Die
vorliegende Erfindung stellt weiterhin ein partiell deletiertes
g3-Gen bereit, das im Helferphagengenom noch vorhanden ist, um stabile,
aber im Wesentlichen nichtinfektiöse Helferphagen bereitzustellen,
die infektiöse
g3ps auf ihrer Hülle
beherbergen. Die Erfindung beschreibt dieses partiell deletierte
g3-Gen, das synthetisch hergestellt wird, indem man synthetische
Primer in einer solchen Weise verwendet, dass das funktionell deletierte
g3-Gen auf dem Aminosäureniveau
dasselbe Protein codiert wie der andere Teil des g3-Gens, der in
demselben Bakterium vorhanden ist, aber die verwendeten Codons nicht
zu homologen Rekombinationsereignissen führen. Da die Leadersequenzen
in den verschiedenen Ausführungen
sehr unterschiedlich sind, brauchen diese Bereiche im Helfergenom,
Phagen-Display-Vektor oder Helfervektor (mit den scFv-codierenden
Genen) nicht verändert
zu werden. Im Prinzip ist es nicht wichtig, ob das g3-Gen im Helfergenom,
im Phagen-Display-Vektor oder im Helfervektor verändert wurde,
solange die beiden überlappenden
(im Aminosäuregehalt)
und vorher homologen g3-Teile, die in eine E.-coli-Zelle eingeführt werden,
nicht zueinander passen.
-
Die
vorliegende Erfindung beschreibt die Verwendung von Codonveränderungen
im g3-Gen für
die Herstellung von Helferphagen, die aufgrund von g3ps, die in
den Helfervektoren codiert sind, infektiös sind, denen aber ein Wildtyp-
oder wenigstens ein infektiöses
g3-Gen in ihrem Genom fehlt, und für die Herstellung von chimärischen
Phagen gemäß der Erfindung.
Die Verwendung von Codonveränderungen
gewährleistet eine
reduzierte Wahrscheinlichkeit für
homologe Rekombinationseffekte, die während des Vorgangs der Erzeugung
von Helferphagen auftreten könnten.
Die Erfindung gibt vorzugsweise die Verwendung von Codonveränderungen
im g3-Gen oder Teilen davon für
die Herstellung von Phagen-Display-Bibliotheken an, bei denen das
Helferphagengenom, das zusammen mit Nucleinsäuren, die für g3p-X-Fusionsproteine codieren,
in eine E.-coli-Zelle gebracht wird, nicht zu dem g3-Gen homolog
ist, das in der DNA vorhanden ist, die für das g3p-X-Fusionsprotein
codiert. Diese Codonveränderungen
gewährleisten,
dass die Wahrscheinlichkeit für
homologe Rekombinationsereignisse bei der Herstellung von Phagen-Display-Bibliotheken
erheblich abnimmt, wodurch die Qualität dieser Bibliotheken und ihre
Verwendungen erheblich verbessert werden.
-
Experimentelle Verfahren
-
Primer
-
Die
folgenden Primer (von Genset oder Invitrogen erhalten) wurden bei
der Erzeugung der verschiedenen Vektoren und genomischen Helferphagenkonstrukte
verwendet. Die meisten Restriktionsenzyme bauen DNA kaum oder gar
nicht ab, wenn sich ihr entsprechendes Palindrom in der Nähe des Endes
der DNA befindet. Daher wurde eine Sequenz von 8 Nucleotiden zum
5'-Ende des D3-,
g3-minus- und g3-ORF-Primers addiert, wobei diese Sequenz ein A/T-reiches
nichthybridisierendes Octamer ist.
-
D3-Primer D3
BamHI Forward
-
-
g3-minus-Primer g3
minus HindIII Forward
-
g3
minus HindIII Backward
-
g3-ORF-Primer g3
ORF NcoI Forward
-
-
CT-
und N2CT-Primer SnaBIclon
-
-
-
-
-
-
-
-
PCR-Reaktionen
und Produktisolierung
-
PCR-Reaktionen
mit D3-, g3-minus- und g3-ORF-Primern wurden standardmäßig (abgesehen
von der Verlängerungszeit
des DNA-Synthesezyklusschritts) unter Verwendung des folgenden 50-μl-Heißstart-PCR-Schemas
und des AmpliTaq-PCR-Kits
von Perkin Elmer durchgeführt:
1 μl 10
mM dNTP (Roche Diagnostics), 4 μl
25 mM MgCl2, 5 μl 10 × PCR-Puffer, der mit dem Kit
mitgeliefert wird, 5 μl
2,5 μM Forward-Primer,
5 μl 2,5 μM Backward-Primer,
0,3 μl 5
Einheiten/μl
AmpliTaq, 10–50
ng Matrize, steriles bidestilliertes Wasser. Alle Komponenten wurden
auf Eis gehalten, bis sie in den vorgeheizten PCR-Block gegeben
wurden. Das Standardprogramm war wie folgt. 12 Zyklen von 25 s bei
94°C, 52°C Assoziation
während
25 s, 72°C
Polymerisation, am Ende ein Zyklus von 72°C während 7 min und dann Lagerung
bei 4°C.
Die Zeit der Polymerisation für
die Synthese von neuen Helfergenomen betrug 12 min, und für die g3-Amplifikation
und für
die Amplifikation des AraC-Gens und des AraC/BAD-Promotors wurde
sie auf 90 s eingestellt. PCR-Reaktionen mit pUC-g3-Primern wurden
wie oben durchgeführt,
aber mit Pwo-Polymerase (Boehringer Mannheim) anstatt AmpliTaq und
mit dem folgenden Programm: 30 Zyklen von 45 s bei 94°C, 50°C Assoziation
während
30 s und 72°C
Polymerisation während
1 min, am Ende ein Zyklus von 68°C
während
8 min und dann Lagerung bei 4°C. PCR-Reaktionen
mit CT- und N2CT-Primern wurden wie oben durchgeführt, aber
mit Taq-Polymerase
(Gibco) anstelle von AmpliTaq und mit dem folgenden Programm: 25
Zyklen von 30 s bei 96°C,
53°C Assoziation
während
30 s und 72°C
Polymerisation während
2 min, am Ende ein Zyklus von 72°C
während
10 min und dann Lagerung bei 4°C.
Eine Ausnahme war die PCR-Reaktion mit den N2CT-Primern BglN2 und
PacIclon, die mit Hilfe von rekombinanter Taq-DNA-Polymerase (Invitrogen)
mit dem folgenden Programm durchgeführt wurde: 30 Zyklen von 45
s bei 94°C,
55°C Assoziation
während
30 s und 72°C
Polymerisation während
2,5 min, am Ende ein Zyklus von 72°C während 10 min und dann Lagerung
bei 4°C.
-
Alle
PCR-Produkte wurden auf 0,5%–1%
TBE-Agarose-Gelen, die 100 ng/ml Ethidiumbromid enthielten, aufgetrennt.
Nach der Bildaufzeichnung wurden die gewünschten Fragmente unter Verwendung
von sterilen chirurgischen Einwegmessern herausgeschnitten und mit
dem Gel-Reinigungskit von Qiagen gemäß den beiliegenden Anweisungen
isoliert.
-
Ligierungsreaktionen
-
Ligierungsreaktionen
wurden standardmäßig in den
folgenden Reaktionsgemischen durchgeführt:
50 ng Vektor- oder
Helfergenom
25 ng Insert
4 μl 5 × Ligierungspuffer (Gibco BRL)
1 μl T4-Lipase
(Gibco BRL, 200 Einheiten/μl)
steriles
bidestilliertes Wasser auf 20 μl
-
Für den Aufbau
des pUC-g3-Helferplasmids wurde jedoch T4-DNA-Lipase (Roche, 1 Einheit/μl) verwendet,
während
beim Aufbau der Helferphagengenome CT, N2CT und p3-minus T4-DNA-Ligase
(NEB, 400 Einheiten/μl)
verwendet wurde. Die Gemische wurden über Nacht bei 6–16°C inkubiert.
Dann wurden 30 μl
steriles Wasser, 5 μl
3 M Kaliumacetat, pH 4,8 mit Essigsäure eingestellt (KAc), 1 μl 10 mg/ml
Glycogen und 50 μl
Isopropanol oder 96% Ethanol hinzugefügt und gründlich gemischt. Nach 15 min
Fällung
wurden die Röhrchen
10 min lang bei 4°C
mit maximaler Geschwindigkeit zentrifugiert. Das Sediment wurde
einmal mit 1 ml 70%igem Ethanol gewaschen und nach dem Trocknen
in 10 μl
sterilem Wasser gelöst.
Die Hälfte
dieses Volumens wurde zusammen mit 50 μl kompetenten Zellen für die Elektroporation
verwendet.
-
Sequenzierung
-
Die
Sequenzierung der Klone erfolgte gemäß den Anweisungen, die mit
dem ABI PRISM BigDye Terminator Cycle Sequencing Kit (Applied Biosystems)
mitgeschickt wurden, bei einer Assoziationstemperatur von 50°C. Alle Klone
wurden sequenziert, um die Richtigkeit der Produkte zu überprüfen. Die
Sequenz des VCSM13-Helferphagengenoms wurde nach dem Primer-Walking-Verfahren
auf beiden Strängen
bestimmt, einem Verfahren, das dem Fachmann allgemein bekannt ist.
-
Elektroporation
-
Alle
Bakterienstämme
außer
denjenigen, die für
die Produktion von Helferphagen verwendet wurden, wurden von Herstellern
als elektroporationskompetente Zellen mit der höchsten verfügbaren Kompetenz erworben und
nach den Anweisungen des Herstellers unter Verwendung von 0,1-cm-Küvetten (BioRad)
transformiert. Die Produktion von Helferphagen hängt jedoch von TOP10- oder
LMG-Zellen (Stratagene) ab, die das Helferplasmid (pBAD/gIII-g3)
enthalten. Diese Zellen wurden wie folgt kompetent gemacht und bis
zur Verwendung bei –80°C gelagert:
Eine Kolonie der Bakterien wurde verwendet, um 10 ml 2 × TY mit
Ampicillin (100 μg/ml)
und für
LMG auch mit Tetracyclin (10 μg/ml)
zu beimpfen, und durch kräftiges
Schütteln
bei 30°C über Nacht
kultiviert. Dann wurden die Kulturen 5 min lang mit 3000 U/min sedimentiert.
Das Sediment wurde in 500 ml frischem 2 × YT einschließlich der
Antibiotika resuspendiert und in einem 2-l-Erlenmeyerkolben auf
einem Schüttelbrett
bei 37°C
bis OD 0,5 kultiviert. Diese Zellen wurden 45 min lang auf Eiswasser
abkühlen
gelassen und 25 min lang in einer Sorvall-Zentrifuge unter Verwendung
eines GLA-3000-Rotors
in vorgekühlten
Bechern und Rotor mit 3000 U/min bei 4°C zentrifugiert. Der Überstand
wurde verworfen, und die Zellen wurden langsam und vorsichtig in
100 ml eiskaltem 10%igem Glycerin resuspendiert. Die Zentrifugations-
und Glycerinschritte wurden zweimal wiederholt. Das endgültige Sediment
wurde sehr vorsichtig in 5 ml 10%igem eiskaltem Glycerin aufgenommen
und in vorgekühlten
Eppendorf-Röhrchen
aliquotiert. Dann wurden diese Röhrchen
5 min lang in ein Gemisch von Ethanol und Trockeneis eingetaucht,
um ein sehr schnelles Gefrieren der Zellen zu gewährleisten.
Die Röhrchen,
die die elektrokompetenten Zellen enthielten, wurden bis zur Verwendung
bei –80°C gelagert.
Ein vergleichbares Verfahren wurde verwendet, um kompetente Zellen
von XL-1 (Stratagene) herzustellen, die das Helferplasmid pUC-g3
enthielten.
-
Phagenproduktion
-
Der
gewünschte
F-positive E.-coli-Stamm wird in 2 × YT-Medium, das die erforderlichen
Antibiotika enthielt, eingeimpft und bei 37°C mit 220 U/min bis OD 0,2 kultiviert.
Der (Helfer-)Phage wird zu der Kultur gegeben und 30–45 min
lang bei 37°C
in einem nichtschüttelnden
Wasserbad inkubiert. Dann wird Kanamycin (50 μg/ml) zu den Zellen gegeben,
und die Zellen werden 30–45
min lang bei 37°C
mit 220 U/min weiterinkubiert. Anschließend wird diese Lösung 15
min lang bei Raumtemperatur mit 3500 U/min zentrifugiert. Der Überstand
wird vorsichtig entfernt, und das Sediment wird auf das gewünschte Volumen
von 2 × YT-Medium,
das alle erforderlichen Antibiotika enthält, gebracht. Die Zellen werden
für eine
Phagen-Display-Bibliothek über Nacht
bei 30°C
und für
reguläre
(Helfer-)Phagen bei 37°C
auf einem Schüttelbrett
kultiviert.
-
Titerbestimmung
-
Eine
Kolonie von Xl1blue (Stratagene) wird in 5 ml 2 × YT, das 10 μg Tetracyclin
pro ml enthält
(YT-T), in einem 50-ml-Röhrchen
(Falcon) eingeimpft und über
Nacht bei 37°C
mit 220 U/min kultiviert. 200 μl
dieser Kultur werden zu 5 ml YT-T gegeben und bis OD 0,2 kultiviert.
Dann wird die Phagen-Stammlösung
in YT verdünnt,
und nach Bedarf wird eine Verdünnungsreihe
hergestellt, um die Zahl der plaquebildenden Einheiten zu bestimmen.
Für jeden
Verdünnungsschritt
werden 100 μl
der XL1Blue-Kultur mit OD 0,2 entnommen und zu 100 μl der Phagen
gegeben. Dieses Gemisch wird 25 min lang bei 37°C in einem Wasserbad (nichtschüttelnd) inkubiert.
Die 200 μl
Bakterienzellen werden auf eine Platte mit 2 × YT Brühe, die die erforderlichen
Antibiotika enthält,
pipettiert. Die Suspension wird unter Verwendung eines sterilen
Glasstabs ausgebreitet. Nach dem Trocknen der Platten werden sie
umgedreht und in einen Inkubator von 37°C übergeführt. Nach Kultur über Nacht
werden die Kolonien gezählt.
Jede Kolonie zeigt die Gegenwart von 1 infektiösen Phagenpartikel in der ursprünglichen
Phagenlösung
an. Die Zahl der infektiösen
Partikel pro ml der analysierten Stammlösung wird bestimmt. Die Phagenpartikel
werden gemäß den mitgeliefer ten
Anweisungen einem ELISA-Test mit Anti-M13 und Anti-M13-HRP-Konjugat
(Pharmacia) unterzogen.
-
Isolierung
von DNA aus Phagen
-
Eine
Phagenkultur wird über
Nacht kultiviert, wie es oben beschrieben ist. Wenn eine Isolierung
von DNA im großen
Maßstab
erforderlich war, wurde der BioRad Plasmid Maxi Prep Kit gemäß den Anweisungen des
Herstellers verwendet, abgesehen vom Elutionsschritt, der 10 min
lang mit 10 mM Tris pH 8,5 bei 65°C durchgeführt wird.
Isolierungen im kleinen oder mittleren Maßstab wurden unter Verwendung
des Mini-Prep-Kits von Qiagen gemäß den mit dem Kit mitgelieferten
Anweisungen durchgeführt,
abgesehen vom Elutionsschritt. Der Elutionsschritt wurde hier 10
min lang bei 65°C
durchgeführt.
-
PEG-Fällung
-
Das
Medium, das Bakterien und Phagen enthält, wird in 450-ml-Bechern
aufgefangen. Das Gemisch wird 20 min lang in einer vorgekühlten Sorvall-Zentrifuge
unter Verwendung eines GSA-3000-Rotors mit 8000 U/min zentrifugiert.
Dann werden 90 ml 20% PEG/2,5 M NaCl in saubere 450-ml-Becher pipettiert.
360 ml des Überstands
des zentrifugierten Mediums, der die Phagen enthält, werden in die PEG-haltigen
Becher gegeben und gut gemischt. Das Gemisch wird 2 h lang auf Eiswasser
oder über
Nacht in den Kühlschrank
gestellt. Der Niederschlag wird durch 20 min Zentrifugation in einer
vorgekühlten
Sorvall-Zentrifuge mit 8000 U/min sedimentiert. Der Überstand
wird dekantiert, und die Becher werden 5 min lang austropfen gelassen,
um so viel Fällungspuffer
wie möglich
zu entfernen. Anschließend
werden 32 ml PBS/1% Rinderserumalbumin (BSA) zu den 450-ml-Bechern,
die die sedimentierten Phagen enthalten, gegeben, und die Becher
werden 15 min lang auf einem Flaschenroller rotieren gelassen. Die
Lösung
wird in ein SS-34-kompatibles Zentrifugenröhrchen übergeführt und 25 min lang in einer
vorgekühlten
Sorvall-Zentrifuge, die einen SS-34-Rotor (oder eine äquivalente
Ausrüstung)
enthält,
mit 13 000 U/min zentrifugiert. In diesem Schritt werden Trümmer und
kleine Bakterien aller Art entfernt. Inzwi schen wird der Kolben
aus einer 50-ml-Spritze entfernt und an einem 0,45-μm-Filter (Whatman)
befestigt. Der zentrifugierte Überstand
wird in die Spritze übergeführt und
durch den Filter gedrückt.
In diesem Schritt werden alle kleinen Bakterien und anderen Zellen
entfernt. 8 ml 20% PEG/2,5 M NaCl werden hinzugefügt, und
es wird gut gemischt. Die Röhrchen
werden 1 h lang auf Eis gestellt. Der Hochgeschwindigkeitszentrifugationsschritt
wird so wiederholt, wie es oben beschrieben ist. Der Überstand
wird dekantiert, und das Röhrchen
wird 5 min lang auf einem Papiertuch austropfen gelassen. Das Phagensediment wird
in 5 ml PBS/1% BSA gelöst.
Dann werden 5 ml 100%iges Glycerin zu der Phagenlösung gegeben,
und es wird gut gemischt. Die Phagen werden bei –20°C gelagert. Typischerweise enthält die Lösung ungefähr 2 bis
5 × 1013 infektiöse Phagenpartikel pro ml.
-
Beispiele
-
Zur
Erläuterung
der Erfindung werden die folgenden Beispiele angegeben, die den
Umfang der Erfindung nicht einschränken sollen.
-
Beispiel 1
-
Klonierung des pBAD/9III-g3-Helfervektors
-
Das
volle offene Leseraster (ORF) des g3-Gens wurde erzeugt, indem man
M13K07-DNA (Gibco-BRL) als Matrize in einer Standard-PCR-Reaktion
zusammen mit den Primern g3 ORF NcoI Forward und g3 ORF XbaI Backward
verwendete. Das gereinigte PCR-Produkt und der pBAD/gIII-Vektor
wurden beide gleichzeitig 4 h lang bei 37°C in Puffer H (Roche Diagnostics)
mit NcoI (NEB) und XbaI (Roche Diagnostics) abgebaut. Nach der Ligierung,
Isolierung und Elektroporation in TOP10-(Stratagene) und LMG-Zellen
(Stratagene) wurden zwei korrekte Klone durch Sequenzieren ausgewählt und
im großen
Maßstab
kultiviert, und dann wurde die DNA isoliert. Die DNA wurde mit 70%
Ethanol in Gegenwart von KAc erneut gefällt, und der Niederschlag wurde
zweimal mit 70% Ethanol gewaschen. Nach dem Trocknen wurde die DNA
in sterilem bidestilliertem Wasser gelöst und bis zur Verwendung bei –20°C aufbewahrt.
Das resultie rende Plasmid pBAD/gIII-g3 ist in 5 gezeigt.
Dieser Helfervektor enthält
als wichtigste Merkmale das g3-Gen in voller Größe unter der Kontrolle des
AraC/BAD-Promotors, ein Ampicillin-Resistenz-Gen und einen ColE1-ORI.
-
Beispiel 2
-
Klonierung des g3-negativen
Helferphagengenoms
-
VCSM13
(Stratagene) ist ein verbreitet verwendeter Helferphage, doch seine
Sequenz ist nicht öffentlich
zugänglich.
Die volle DNA-Sequenz des VCSM13-Genoms
wurde bestimmt und ist schematisch in 6A und
als Nucleotidsequenz in 6B gezeigt.
Die Verwendung der Primer g3 minus HindIII Forward und g3 minus
HindIII Backward sowie M13K07 und VCSM13 als Matrizen in einer Standard-PCR-Reaktion
führte
zur Bildung eines PCR-Produkts, das HindIII-Stellen an beiden Enden der DNA enthielt.
Nach der Trennung, Gelisolierung und Reinigung, Abbau mit HindIII
(Roche Diagnostics) und erneuter Reinigung der DNA wurde das Produkt
unter Standard-Ligierungsbedingungen selbstligiert, und das resultierende
Helferphagengenom, das g3-minus genannt wurde, wurde durch Elektroporation
in XL1Blue-Zellen (Stratagene) eingeführt. Die transformierten Zellen
wurden in 5 ml 2TY-Medium resuspendiert und 1 h lang einer Schüttelkultur
bei 37°C
unterzogen. Kanamycin wurde bis zu einer Endkonzentration von 50 μg/ml hinzugefügt, und
die Zellen wurden weitere 5 h lang unter denselben Bedingungen wachsen
gelassen. Die Kultur wurde 15 min lang mit 3000 U/min zentrifugiert,
und der Überstand
wurde durch ein 0,22-μm-Filter
passiert, um Bakterien zu entfernen. Gleichzeitig wurde eine Kultur
von exponentiell wachsenden XL-1-Blue-Bakterien hergestellt. Fraktionen
des Filtrats (50–1000 μl), die Phagenpartikel
enthielten, wurden zu 5 ml XL-1-Blue-Bakterien gegeben und 30 min
lang bei 37°C
ohne Schütteln
inkubiert. Die Kultur wurde erneut zentrifugiert, der Überstand
wurde verworfen, und die Zellen wurden auf 2 × YT-K-T-Platten ausgestrichen
und zum Wachstum über
Nacht in einen Inkubator von 37°C übergeführt. Acht
korrekte Klone, denen der g3-ORF fehlt (über die BamHI-Stelle überprüft) und
die die eingeführte
HindIII-Stelle enthalten, wurden isoliert und für die Produktion von g3-losen
Helferphagen in Gegenwart des pBAD/gIII-g3-Helferplasmids verwendet.
Nur zwei Klone, die in Gegenwart des Helferplasmids Phagen bilden
konnten, wurden behalten. Aus diesen Klonen wurde eine große Menge
DNA isoliert und für weitere
Experimente aufbewahrt. Das erhaltene, von VCSM13 abgeleitete g3-negative
Helferphagengenom, in dem das gesamte g3-Gen außer den letzten sechs Codons
durch eine HindIII-Stelle ersetzt wurde, ist schematisch in 6C gezeigt,
während
die g3-negative Sequenz in 6D gezeigt
ist.
-
Beispiel 3
-
Klonierung des D3-Helferphagengenoms
mit einem partiell deletierten g3-Gen
-
Die
Konstruktion eines Helferphagengenoms, das nur den D3-Teil des g3-Gens
exprimiert, war mit den oben beschriebenen g3-negativen Helferphagen
vergleichbar, außer
dass die verwendeten Primer D3 BamHI Forward und D3 BamHI Backward
waren, um das neue Genom zu erzeugen. Alle anderen Verfahren waren dieselben
wie bei dem g3-Negativ-Verfahren, abgesehen von der Verwendung von
BamHI anstelle von HindIII. Am Ende wurde die DNA von zwei korrekten
Klonen behalten und bei –20°C aufbewahrt.
Das endgültige,
von VCSM13 abgeleitete Konstrukt des Helferphagengenoms (genannt
D3: es exprimiert nur diesen Teil des g3-Gens, der nicht zur Infektiosität des Phagenpartikels
beiträgt)
ist in 7A gezeigt, während die
D3-Sequenz in 7B gezeigt ist.
-
Beispiel 4
-
Produktion von infektiösen Helferphagen,
die kein Gen tragen, das das Wildtyp-g3p codiert
-
Die
Verfahren zur Erzeugung von g3-losen und partiell deletiertes g3
(oder D3) exprimierenden Helferphagen sind identisch. Gefrorene
kompetente TOP10- oder LMG-Zellen, die das pBAD/gIII-g3-Helferplasmid
enthielten, wurden einer Elektroporation mit Hilfe von 100 ng Helferphagen-DNA
unterzogen. Nach der Gewinnung wurden die Zellen in 4 × 250 ml
2 × YT-K-A-Medium übergeführt, das
mit 0,05% Arabinose (Sigma) angereichert war. Phagen wurden während der
Kultur über
Nacht bei 37°C
und unter kräftigem
Schütteln
produziert. Am nächsten
Tag wurden die Phagen gemäß den oben
beschriebenen Standardverfahren für die Fällung und Lagerung gereinigt
und gelagert. Die Zahl der infektiösen Partikel und die Zahl der
Phagen wurden durch Titrations- und ELISA-Verfahren, die ebenfalls
oben beschrieben wurden, bestimmt. Für g3-lose Helferphagen wurden
ungefähr
5 × 1011 infektiöse Partikel synthetisiert,
während
für D3
unter Verwendung dieser Verfahren ungefähr 5 × 1013 infektiöse Phagen
gebildet wurden.
-
Beispiel 5
-
Vermehrung und Ernte von
Phagen-Display-Bibliotheken, die infektiöse Phagen, welche g3p-scFv-Fusionen tragen,
und nichtinfektiöse
Helferphagen enthalten, mit Hilfe von g3-negativen und partiell
g3-deletierten Helferphagen
-
Eine
gefrorene Bibliothek wurde wie folgt eingeimpft. Im Allgemeinen
wurden ungefähr
5–10 μl konzentrierte
Stammlösungen
in 25 ml 2 × YT,
das die erforderlichen Antibiotika und 5% Glucose enthielt, eingeimpft
und bei 37°C
unter kräftigem
Schütteln
gezüchtet.
Bei OD 0,3–0,4
(nach etwa 2–3
h) wurde ein 500- bis 1000facher Überschuss
an Helferphage hinzugefügt.
Das Medium, das die Helferphagen und Bakterien enthielt, wurde 25
min lang in einem Wasserbad von 37°C ohne Schütteln inkubiert. Die Entfernung
von toten Zellen und des Überschusses
von Phagenpartikeln erfolgte nach einem Zentrifugationsschritt mit
3000 U/min während
15 min. Das Sediment wurde in 250 ml 2 × YT mit Antibiotika, aber
ohne Glucose, resuspendiert und über
Nacht bei 30°C
mit guter Durchlüftung
kultiviert. Am nächsten
Tag wurden die gebildeten Phagen unter Verwendung des Standard-PEG/NaCl-Verfahrens
isoliert und gelagert.
-
Beispiel 6
-
Codonverwendung in g3
und dem partiell deletierten g3-Gen (D3) zur Verhinderung einer
homologen Rekombination während
der Helferphagenproduktion und Vermehrung der Bibliothek
-
Um
mögliche
Rekombinationen zwischen einer genomischen Nucleinsäure, die
den Helferphagen-g3-Proteinbereich (oder einen Teil davon, wie die
D3-Domäne)
codiert, und anderen Nucleinsäuren
(wie dem Phagen-Display-Vektor und dem AraC/BAD-Helfervektor) zu
verhindern, wird eine Reihe von Helferphagen entworfen, die innerhalb
des g3p-Bereichs veränderte
Codons enthalten. Neu translatierte g3ps sind mit dem Wildtyp-g3-Protein
oder -Proteinteil (D3) identisch. Aufgrund dieser Veränderungen
können
g3-ORF codierende DNA-Domänen
nicht oder kaum mit den Phagen-Display-Vektoren oder dem AraC/BAD-g3-Helfervektor
rekombinieren. Die Codons, die verwendet werden, um nichthomologe
g3-Gene zu erzeugen, sind in 8 abgebildet
und sind für
die Transcriptionsmaschinerie von E. coli optimal. Die PCR-Erzeugung
von Helferphagengenomen (VCSM13, M13K07, D3, g3-minus oder AraC/BAD)
mit g3-Kopf-Rückwärts- und
g3-Ende-Vorwärts-Primern
mit NotI-Restriktionsstellen gewährleisten
die Erzeugung von PCR-Produkten, die alle Helferphagenkomponenten
und -gene außer
dem g3-ORF enthalten. Neue g3-Bereiche werden mit überlappenden
Primern konstruiert und in Helferphagen eingesetzt. Das durch PCR
erzeugte g3p oder Teile davon werden mit NotI abgebaut und in das
mit NotI angebaute, durch PCR erzeugte Helferphagengenom ligiert. Nach
der Transformation und Selektion von korrekten Helfergenomen (mit
einem neuen g3-Gen) werden Helferphagen so gezüchtet, wie es beschrieben wurde.
-
Beispiel 7
-
Selektion von mit Thyroglobulin
wechselwirkenden Phagen unter Verwendung einer Bibliothek, die mit
Helferphagen vermehrt wurde, die nur den D3-Teil von g3p in ihrem
Genom umfassen
-
Um
die D3-Helferphagen in Standard-Phagenselektionen zu bewerten, wurde
eine Selektion durchgeführt,
bei der eine Antikörper-Phagen-Display-Bibliothek verwendet
wurde, die mit Hilfe der D3-Helferphagen vermehrt wurde, wie es
oben beschrieben ist. Die verwendeten Verfahren entsprachen im Wesentlichen
den von De Kruif et al. (1995a) beschriebenen. Kurz gesagt, ein
Kunststoffröhrchen
wurde mit Thyroglobulin beschichtet. Das Röhrchen wurde in PBS, das 2%
Milch enthielt (MPBS), blockiert, und danach wurde die ebenfalls
in MPBS blockierte Antikörper-Phagen-Display-Bibliothek
in das Röhrchen
gegeben. Die Phagen wurden 2 h lang binden gelassen, und danach
wurden nichtbindende Phagen entfernt, indem man das Röhrchen in PBS,
das 0,1% Tween-20 enthielt, wusch, wie es von De Kruif et al. (1995a)
beschrieben wurde. Die bindenden Phagen wurden in 50 mM Glycin/HCl,
pH 2,2, eluiert (10 min bei RT) und verwendet, um frisch gezüchtete XL-1-Blue-Bakterien
zu infizieren. Die Bakterien wurden auf 2TY-Agarplatten, die die
geeigneten Antibiotika und Glucose enthielten, ausgestrichen, über Nacht
bei 37°C
inkubiert und verwendet, um eine angereicherte Phagen-Display-Bibliothek
herzustellen; der Phage D3 wurde wiederum als Helferphage verwendet.
Das Verfahren wurde einmal wiederholt, und danach wurden einzelne
E.-coli-Kolonien verwendet, um monoklonale Phagenantikörper herzustellen.
Diese monoklonalen Phagenantikörper
wurden in ELISA auf ihre Fähigkeit
getestet, spezifisch an das Thyroglobulin-Antigen zu binden. Die
Ergebnisse zeigen, dass nach zwei Selektionsdurchläufen 25/46
Kolonien eine positive Bindung an Thyroglobulin zeigen. Vorher fanden
wir heraus, dass bei Verwendung eines allgemeinen VCS-M13 als Helferphage
in diesem Selektionsformat wenigstens 3 Selektionsdurchläufe erforderlich
waren, um spezifische Bindungsmoleküle zu erhalten.
-
Beispiel 8
-
Selektion von mit Myelomzellen
wechselwirkenden Phagen unter Verwendung einer Bibliothek, die mit
Helferphagen vermehrt wurde, die nur den D3-Teil von g3p in ihrem
Genom umfassen
-
Um
die D3-Helferphagen in Selektionen an intakten Zellen zu bewerten,
wurde eine Selektion durchgeführt,
bei der eine Antikörper-Phagen-Display-Bibliothek
verwendet wurde, die mit Hilfe des D3-Helferphagen vermehrt wurde.
Die verwendeten Verfahren entsprachen im Wesentlichen den von De
Kruif et al. (1995a und 1995b) beschriebenen. Kurz gesagt, Myelomzellen
(AML, CD33+, CD34+) wurden aus dem Blut eines Patienten erhalten,
der an der Utrechter Universitätsklinik
(Niederlande) behandelt wurde. 0,5 ml Phagenbibliothek wurden in
3 ml RPMI-Medium, das 10% FCS enthielt (RPMIS), gegeben und 15 min
auf Eis inkubiert. Die Myelomzellen wurden hinzugefügt, und
die Zellsuspension wurde 2 h lang bei 4°C rotieren gelassen. Die Zellen
wurden fünfmal
in 50 ml eisgekühltem
RPMIS gewaschen, und danach wurden die bindenden Phagen eluiert
(in 50 mM Glycin/HCl, pH 2,2, 10 min bei RT) und verwendet, um frisch
gezüchtete
XL-1-Blue-Bakterien zu infizieren. Die Bakterien wurden auf 2TY-Agarplatten,
die die geeigneten Antibiotika und Glucose enthielten, ausgestrichen, über Nacht
bei 37°C
inkubiert und verwendet, um eine angereicherte Phagen-Display-Bibliothek
herzustellen. Wiederum wurden die D3 exprimierenden Helferphagen
als Helferphagen verwendet. Das Verfahren wurde einmal wiederholt,
und danach wurden einzelne E.-coli-Kolonien verwendet, um monoklonale Phagenantikörper herzustellen.
Diese monoklonalen Phagenantikörper
wurden in FACS-Verfahren auf ihre Fähigkeit getestet, Myelomzellen
zu binden. Die Ergebnisse zeigen, dass 23 von 41 getesteten Klonen
spezifisch an Epitope banden, die auf den Myelomzellen exprimiert
werden. Im Allgemeinen sind drei oder mehr Selektionsdurchläufe erforderlich,
um eine ähnliche
Zahl von bindenden Phagen zu erhalten, wobei identische Verfahren
verwendet werden, mit der Ausnahme, dass VCS-M13-Helferphagen anstelle
der in der Erfindung beschriebenen Helferphagen verwendet wurden.
-
Beispiel 9
-
Klonierung des pUC-g3-Helferplasmids
-
Als
Alternative zu dem oben beschriebenen pBAD/gIII-g3-Helferplasmid
wurde ein auf pUC19 basierendes Helferplasmid für die Expression von Wildtyp-g3p
unter der Kontrolle eines lac-Promotors konstruiert. Die Verwendung
der Primer H3leadA und p3endEco sowie VCSM13 als Matrize in einer
oben ausführlich
beschriebenen PCR-Reaktion führte
zur Bildung eines PCR-Produkts, das die g3-Gensequenz mit 28 zusätzlichen
Nucleotiden stromaufwärts
des Gens und 15 zusätzlichen
Nucleotiden einschließlich
einer eingeführten EcoRI-Stelle
stromab wärts
enthielt. Die Verwendung der Primer H3leadB und p3endEco sowie dieses PCR-Produkts
als Matrize in einer oben ausführlich
beschriebenen PCR-Reaktion führte
zur Bildung eines verlängerten
PCR-Produkts, das die g3-Gensequenz mit eingeführten HindIII- und EcoRI-Stellen
stromaufwärts bzw.
stromabwärts
des Gens enthielt. Das verlängerte
PCR-Produkt wurde in das Plasmid pCR4-BluntII-TOPO einkloniert, wobei man den Zero
Blunt TOPO PCR Cloning Kit (Invitrogen) verwendete, und die Sequenz des
Inserts wurde überprüft. Das
Plasmid pUC19 (New England Biolabs) und das pCR4-BluntII-TOPO-Derivat, das
das g3-Gen enthielt, wurden beide mit HindIII und EcoRI abgebaut.
Das Fragment, das das g3-Gen aus dem pCR4-TOPO-Derivat enthielt,
und das Vektorfragment von pUC19 wurden aus Gel gereinigt und ligiert. Das
resultierende Plasmid, das pUC-g3
genannt wurde, wurde durch Elektroporation in XL-1-Zellen (Stratagene)
eingeführt,
und ein korrekter Klon wurde ausgewählt, indem man DNA sequenzierte,
die aus den transformierten Zellen isoliert wurde. Der Helfervektor
pUC-g3, der das
g3-Gen unter der Kontrolle des lac-Promotors enthält, ist
schematisch in 9 gezeigt.
-
Beispiel 10
-
Alternative Klonierung
von Helferphagengenomen mit einem partiell deletierten g3-Gen
-
Bei
den in Beispiel 2 und 3 beschriebenen Klonierungsverfahren wird
ein sehr großer
Bereich des Phagengenoms durch PCR amplifiziert, mit der Gefahr,
aufgrund der eingeschränkten
Kopiertreue der DNA-Polymerase künstliche
Mutationen einzuführen.
Eine alternative Strategie wurde angewendet, um Helfergenome mit
einem partiell deletierten g3-Gen zu erzeugen. Bei dieser Strategie
wurden ein Teil der g3-Sequenz, der den g3-Leader codiert, und ein
Teil der g3-Sequenz, der einen C-terminalen Teil des g3-Proteins
codiert, getrennt amplifiziert. Die beiden Fragmente wurden in einem
Shuttle-Vektor miteinander kombiniert und anschließend in
das Phagengenom kloniert. Um die resultierenden Konstrukte von dem
in Beispiel 3 beschriebenen D3-Helferphagen zu unterscheiden, wurde
die allgemein akzeptierte alternative Nomenklatur für die g3-Domänen verwendet,
wobei D1, D2 und D3 als N1, N2 bzw. CT bezeich net werden. Die Verwendung
der Primer SnaBIclon und Bamlead sowie VCSM13 als Matrize in einer
oben ausführlich
beschriebenen PCR-Reaktion führte zur
Bildung eines PCR-Produkts, das die g3-Leadersequenz mit einer nativen
SnaBI-Stelle stromaufwärts und einer
eingeführten
BamHI-Stelle stromabwärts
enthielt. Die Verwendung der Primer BamCT und PacIclon sowie VCSM13
als Matrize in einer oben ausführlich
beschriebenen PCR-Reaktion führte
zur Bildung eines PCR-Produkts, das die g3-CT-Domäne mit einer
eingeführten
BamHI-Stelle stromaufwärts
und einer nativen PacI-Stelle stromabwärts enthielt. Die Verwendung
der Primer BglN2 und PacIclon sowie VCSM13 als Matrize in einer
oben ausführlich
beschriebenen PCR-Reaktion führte
zur Bildung eines PCR-Produkts, das die g3-N2- und -CT-Domänen mit
einer eingeführten
BglII-Stelle stromaufwärts
und einer nativen PacI-Stelle stromabwärts enthielt. Alle PCR-Produkte
wurden in das Plasmid pCR4-TOPO kloniert, wobei man den TOPO TA
Cloning Kit for Sequencing (Invitrogen) verwendete, und die Sequenzen
der Inserts wurden überprüft. Das pCR4-TOPO-Derivat,
das die Leadersequenz enthielt, wurde mit NotI und BamHI abgebaut,
und das Insert wurde aus Agarose-Gel isoliert. Das pCR4-TOPO-Derivat,
das die CT-Domäne
enthielt, wurde ebenfalls mit NotI und BamHI abgebaut, und das Vektorfragment
wurde isoliert. Das Insert wurde mit dem Vektorfragment ligiert,
was zu einem pCR4-TOPO-Derivat führte,
das das g3-Gen enthält,
bei dem die N1- und die N2-Domäne deletiert
sind. Dieses Plasmid und VCSM13 wurden beide mit SnaBI und PacI
abgebaut. Nach der Isolierung wurde das Plasmid-Insert mit dem VCSM13-Vektorfragment
ligiert. Die Sequenz des Inserts in dem resultierenden Phagemid,
das CT genannt wird und mit dem in 7A gezeigten
D3 identisch ist, wurde überprüft. Ein
vergleichbares Verfahren wurde verwendet, um ein Phagemid zu konstruieren,
dem nur die N1-Domäne fehlt.
Für dieses
Konstrukt wurde das pCR4-TOPO-Derivat,
das die N2- und CT-Domänen
enthielt, mit NotI und BglII abgebaut, und das Vektorfragment wurde
isoliert. Das Insert, das die Leadersequenz enthielt, wurde mit
dem Vektorfragment ligiert (die klebrigen Enden von BglII und BamHI
sind kompatibel), was zu einem pCR4-TOPO-Derivat führte, das
das g3-Gen enthielt, bei dem die N1-Domäne deletiert ist. Dieses Gen
wurde in VCSM13 subkloniert, wie es oben beschrieben wurde, und
die Sequenz des Inserts in dem resultierenden Phagemid, das N2CT
genannt wird, wurde überprüft. Das
von VCSM13 abgeleitete Helferphagengenom N2CT ist schematisch in 10 gezeigt.
-
Beispiel 11
-
Klonierung des p3-negativen
Helferphagengenoms
-
Bei
der Klonierung des in Beispiel 2 beschriebenen g3-negativen Helferphagengenoms
wurde fast die gesamte g3-Sequenz deletiert. Das g3-Gen überlappt
mit der Ribosomenbindungsstelle des g6-Gens und kann noch andere
wichtige Merkmale enthalten. Eine alternative Klonierungsstrategie
wurde angewendet, bei der ein Teil der N2-Domäne und die gesamte CT-Domäne auf dem
DNA-Niveau beibehalten
werden, diese Domänen
aber auf dem Proteinniveau durch eine Rasterverschiebung deletiert
werden. Das pCR4-TOPO-Derivat, das den g3-Leader enthält, der mit der g3-CT-Domäne (siehe
oben) fusioniert ist, und VCSM13 wurden beide mit BamHI und SnaBI
geschnitten. Nach der Isolierung aus Gel wurde das Insertfragment
des pCR4-TOPO-Derivats, das den g3-Leader enthält, mit dem Vektorfragment
von VCSM13 ligiert, das einen C-terminalen Teil der N2-Domäne und die
gesamte CT-Domäne
von g3 enthält.
Die Sequenz des Inserts im resultierenden Phagemid, das p3-minus
genannt wird, wurde überprüft. In diesem
Konstrukt tritt eine Rasterverschiebung an der BamHI-Stelle auf, die sich
zwischen dem g3-Leader und dem C-terminalen Teil der N2-Domäne befindet, was
zur Einführung
von über
30 Stopcodons stromabwärts
der BamHI-Stelle führt.
Das von VCSM13 abgeleitete Helferphagengenom p3-minus ist in 11 schematisch
gezeigt.
-
Literatur
-
- Balint RF and Larrick JW (1993) Antibody engineering by
parsimonious mutagenesis. Gene 137: 109
- Barbas CF, Hu D, Dunlop N, Sawyer L, Cababa D, Hendry RM, Nara
PL, Burton DR (1994) In vitro evolution of a neutralizing human
antibody to human immunodeficiency virus type 1 to enhance affinity
and broaden strain cross-reactivity. Proc Natl Acad Sci USA. 91:
3809
- Bass SH, Greene R and Wells JA (1990) Hormone phage: An enrichment
method for variant proteins with altered binding properties. Proteins
8: 309–314
- Beekwilder J, Rakonjac J, Jongsma M, Bosch D (1999) A phagemid
vector using the E. coli phage shock promoter facilitates phage
display of toxic proteins. Gene 288: 23–31
- Berek C, and Milstein C (1987) Mutation drift and repertoire
shift in the maturation of the immune response. Immunol Rev 96:
23
- Burton DR and Barbas CF (1994) Human antibodies from combinatorial
libraries. Adv Immunol 57: 191
- Chatellier J, Hartley O, Griffiths AD, Fersht AR, Winter G,
Riechmann L (1999) Interdomain interactions within the gene 3 protein
of filamentous phage. FEBS Letters 463: 371–374
- Crissman JW and Smith GP (1984) Gene-III protein of filamentous
phage: evidence for a carboxyl-terminal domain with a role in morphogenesis.
Virology 132: 445–455
- Cwirla SE, Peters EA, Barrett RE and Dower WJ (1990) Peptides
of phage: A vast library of peptides for identifying ligands. Proc
Natl Acad Sci USA 87: 6378–638
- De Kruif J, Boel E and Logtenberg T (1995a) Selection and application
of human single chain Fv antibody fragments from a semi-synthetic
phage antibody display library with designed CDR3 regions. J Mol
Biol 248: 97–105
- De Kruif J, Terstappen L, Boel E and Logtenberg T (1995b) Rapid
selection of cell subpopulation-specific human monoclonal antibodies
from a synthetic phage antibody library. Proc Natl Acad Sci USA
92: 3938–3942
- Deng LW, Malik P, Perham RN (1999) Interaction of the globular
domains of pIII protein of filamentous bacteriophage fd with the
F-pilus of Escherichia coli. Virology 253: 271–277
- Devlin JJ, Panganiban LC and Devlin PE (1990) Random peptide
libraries: A source of specific protein binding molecules. Science
249: 404–406
- Dueñas
M, Borrebaeck CA (1995) Novel helper phage design: intergenic region
affects the assembly of bacteriophage and the size of antibody libraries.
FEMS microbiology letters 125: 317–322
- Felici F, Luzzago A and Cortese R (1993) Mimicking of discontinuous
epitopes by phage displayed proteins. II. Selection of clones recognized
by a protective monoclonal antibody against the Bordetella pertussis
toxin from phage peptide libraries. Gene 128: 21–27
- Hawkins RE, Russel SJ, Winter G (1992) Selection of phage antibodies
by binding affinity: mimicking affinity maturation. J Mol Biol 226:
889
- Holliger P, Riechmann L (1997) A conserved infection pathway
for filamentous bacteriophage is suggested by the structure of the
membrane penetration domain of the minor coat protein g3p from phage
fd. Structure 5: 265–275
- Hoogenboom HR (1994) Designing and optimizing library selection
strategies for generating high-affinity antibodies. Trends in Biotechnol.
15: 62
- Krebber C, Spada S, Desplancq D, Pluckthun A (1995) Coselection
of cognate antibody-antigen pairs by selectively-infective phage.
FEBS Lett. 377: 227–31
- Krebber C, Spada S, Desplancq D, Krebber A, Ge L, Plückthun (1997)
Selectively-infective Phage (SIP): A mechanistic dissection of a
novel in vivo selection for protein-ligand interactions. J Mol Biol
268: 607–618
- Kristensen P, Winter G (1998) Proteolytic selection for protein
folding using filamentous bacteriophage. Folding and Design 3: 321–328
- Low NM, Holliger PH, Winter G (1996) Mimicking somatic hypermutation:
affinity maturation of antibodies displayed on bacteriophage using
a bacterial mutator strain. J Mol Biol 260: 359
- Lubkowski J, Hennecke F, Pluckthun A, Wlodawer A (1998) The
structural basis of phage display elucidated by the crystal structure
of the N-terminal domains of g3p. Nat Struct Biol 5: 140–147
- Lubkowski J, Hennecke F, Pluckthun A, Wlodawer A (1999) Filamentous
phage infection: crystal structure of g3p in complex with its coreceptor,
the C-terminal domain of TolA. Structure 7: 711–722
- Luzzago A, Felici F, Tramontano A, Pessi A and Cortese R (1993)
Mimicking of discontinuous epitopes by phage displayed proteins.
I. Epitope mapping of human H ferritin using a phage display library
of constrained peptides. Gene 128: 51–57
- Lopez J and Webster RE (1983) Morphogenesis of filamentous bacteriophage
f1: orientation of extrusion and production of polyphage. Virology
127: 177–193
- Model P, Jovanovic G, Dworkin J. (1997) The Escherichia coli
phage shock protein (psp) operon. Mol Microbiol 24: 255–261
- Nelson FK, Friedman SM and Smith GP (1981) Filamentous phage
DNA cloning vectors: A noninfective mutant with a nonpolar deletion
in gene III. Virology 108: 338–350
- Nilsson N, Malmborg A-C, Borrebaeck CA (2000) The phage infection
process: a functional role for the distal linker region of bacteriophage
protein 3. J Virol 74: 4229–4235
- Pratt D, Tzagoloff H and Beaudoin J (1969) Conditional lethal
mutants of the small filamentous coliphage. II. Two genes for coat
proteins. Virology 39: 42–53
- Rakonjac J, Jovanovic G, Model P. (1997) Filamentous phage infection-mediated
gene expression: construction and propagation of the gIII deletion
mutant helper phage R408D3. Gene 198: 99–103
- Rakonjac J, Model P. (1998) Roles of pIII in filamentous phage
assembly. J Mol Biol 282: 25–41
- Riechmann L, Holliger P (1997) The C-terminal domain of TolA
is the coreceptor for filamentous phage infection of E. coli. Cell
90: 351–360
- Rondot S, Koch J, Breitling F, Dübel S (2001) A helper phage
to improve single-chain antibody presentation in phage display.
Nature Biotech 19: 75–78
- Russel M, Model P (1989) Genetic analysis of the filamentous
bacteriophage packaging signal and of the proteins that interact
with it. J Virol 63: 3284–3295
- Smith GP (1985) Filamentous fusion phage: Novel expression vectors
that display cloned antigens on the surface of the virion. Science
228: 1315–1317
- Spada S, Krebber C and Pluckthun A (1997) Selectively infective
phages (SIP). Biol Chem 378: 445–456
- Vaughan TJ, Osbourn JK, Tempest PR (1998) Human antibodies by
design. Nat Biotechnol 16: 535
- Winter G, Milstein C (1991) Man-made antibodies. Nature 349:
293
- Yang W-P, Green K, Pinz-Sweeney S, Briones AT, Burton DR, Barbas
CF (1995) CDR walking mutagenesis for the affinity maturation of
a potent human ant-HIV-1 antibody into the picomolar range. J Mol
Biol 254: 392