CN112839958A - 抗体变体组合及其用途 - Google Patents
抗体变体组合及其用途 Download PDFInfo
- Publication number
- CN112839958A CN112839958A CN201980037949.XA CN201980037949A CN112839958A CN 112839958 A CN112839958 A CN 112839958A CN 201980037949 A CN201980037949 A CN 201980037949A CN 112839958 A CN112839958 A CN 112839958A
- Authority
- CN
- China
- Prior art keywords
- antibody
- substitution
- region
- binding
- igg1
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 238000006467 substitution reaction Methods 0.000 claims abstract description 738
- 239000000203 mixture Substances 0.000 claims abstract description 250
- 125000003275 alpha amino acid group Chemical group 0.000 claims abstract description 106
- 230000027455 binding Effects 0.000 claims description 620
- 210000004027 cell Anatomy 0.000 claims description 418
- 239000000427 antigen Substances 0.000 claims description 273
- 108091007433 antigens Proteins 0.000 claims description 272
- 102000036639 antigens Human genes 0.000 claims description 272
- 239000003814 drug Substances 0.000 claims description 65
- 238000000034 method Methods 0.000 claims description 56
- 102100035360 Cerebellar degeneration-related antigen 1 Human genes 0.000 claims description 51
- 239000004337 magnesium citrate Substances 0.000 claims description 50
- 101100112922 Candida albicans CDR3 gene Proteins 0.000 claims description 49
- 102100035361 Cerebellar degeneration-related protein 2 Human genes 0.000 claims description 49
- 101000737793 Homo sapiens Cerebellar degeneration-related antigen 1 Proteins 0.000 claims description 49
- 101000737796 Homo sapiens Cerebellar degeneration-related protein 2 Proteins 0.000 claims description 49
- 239000001825 Polyoxyethene (8) stearate Substances 0.000 claims description 47
- 210000003719 b-lymphocyte Anatomy 0.000 claims description 35
- 210000004369 blood Anatomy 0.000 claims description 30
- 239000008280 blood Substances 0.000 claims description 30
- 210000001744 T-lymphocyte Anatomy 0.000 claims description 25
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 claims description 24
- 201000010099 disease Diseases 0.000 claims description 22
- 102100024217 CAMPATH-1 antigen Human genes 0.000 claims description 17
- 108010065524 CD52 Antigen Proteins 0.000 claims description 17
- 102100022005 B-lymphocyte antigen CD20 Human genes 0.000 claims description 15
- 101000897405 Homo sapiens B-lymphocyte antigen CD20 Proteins 0.000 claims description 15
- 206010028980 Neoplasm Diseases 0.000 claims description 15
- 210000004881 tumor cell Anatomy 0.000 claims description 14
- 201000011510 cancer Diseases 0.000 claims description 11
- 210000004698 lymphocyte Anatomy 0.000 claims description 11
- 102000006354 HLA-DR Antigens Human genes 0.000 claims description 10
- 108010058597 HLA-DR Antigens Proteins 0.000 claims description 10
- 210000000265 leukocyte Anatomy 0.000 claims description 10
- 101000777628 Homo sapiens Leukocyte antigen CD37 Proteins 0.000 claims description 6
- 101000610605 Homo sapiens Tumor necrosis factor receptor superfamily member 10A Proteins 0.000 claims description 6
- 101000610604 Homo sapiens Tumor necrosis factor receptor superfamily member 10B Proteins 0.000 claims description 6
- 102100031586 Leukocyte antigen CD37 Human genes 0.000 claims description 6
- 102100040113 Tumor necrosis factor receptor superfamily member 10A Human genes 0.000 claims description 6
- 102100040112 Tumor necrosis factor receptor superfamily member 10B Human genes 0.000 claims description 6
- 210000000822 natural killer cell Anatomy 0.000 claims description 6
- 229940124597 therapeutic agent Drugs 0.000 claims description 6
- 208000023275 Autoimmune disease Diseases 0.000 claims description 5
- 108050005493 CD3 protein, epsilon/gamma/delta subunit Proteins 0.000 claims description 5
- 102000017420 CD3 protein, epsilon/gamma/delta subunit Human genes 0.000 claims description 5
- 208000035473 Communicable disease Diseases 0.000 claims description 5
- 230000000779 depleting effect Effects 0.000 claims description 5
- 230000001939 inductive effect Effects 0.000 claims description 5
- 210000003289 regulatory T cell Anatomy 0.000 claims description 5
- 101000934341 Homo sapiens T-cell surface glycoprotein CD5 Proteins 0.000 claims description 4
- 102100025244 T-cell surface glycoprotein CD5 Human genes 0.000 claims description 4
- 208000037979 autoimmune inflammatory disease Diseases 0.000 claims description 4
- 208000019691 hematopoietic and lymphoid cell neoplasm Diseases 0.000 claims description 4
- 239000008194 pharmaceutical composition Substances 0.000 claims description 3
- 230000035755 proliferation Effects 0.000 claims description 3
- 210000004981 tumor-associated macrophage Anatomy 0.000 claims description 2
- 210000004985 myeloid-derived suppressor cell Anatomy 0.000 claims 2
- 101000851370 Homo sapiens Tumor necrosis factor receptor superfamily member 9 Proteins 0.000 claims 1
- 102100040678 Programmed cell death protein 1 Human genes 0.000 claims 1
- 101710089372 Programmed cell death protein 1 Proteins 0.000 claims 1
- 102100036856 Tumor necrosis factor receptor superfamily member 9 Human genes 0.000 claims 1
- 239000003937 drug carrier Substances 0.000 claims 1
- 239000000546 pharmaceutical excipient Substances 0.000 claims 1
- 239000012636 effector Substances 0.000 abstract description 78
- 230000006870 function Effects 0.000 abstract description 68
- 230000010056 antibody-dependent cellular cytotoxicity Effects 0.000 abstract description 60
- 230000001419 dependent effect Effects 0.000 abstract description 46
- 230000004913 activation Effects 0.000 abstract description 14
- 230000001270 agonistic effect Effects 0.000 abstract description 12
- 230000035772 mutation Effects 0.000 description 298
- 230000004540 complement-dependent cytotoxicity Effects 0.000 description 235
- 238000006384 oligomerization reaction Methods 0.000 description 115
- 230000002401 inhibitory effect Effects 0.000 description 111
- 230000000694 effects Effects 0.000 description 109
- 235000001014 amino acid Nutrition 0.000 description 106
- 230000002708 enhancing effect Effects 0.000 description 106
- 230000003993 interaction Effects 0.000 description 87
- XJMOSONTPMZWPB-UHFFFAOYSA-M propidium iodide Chemical compound [I-].[I-].C12=CC(N)=CC=C2C2=CC=C(N)C=C2[N+](CCC[N+](C)(CC)CC)=C1C1=CC=CC=C1 XJMOSONTPMZWPB-UHFFFAOYSA-M 0.000 description 63
- 229920001184 polypeptide Polymers 0.000 description 55
- 108090000765 processed proteins & peptides Proteins 0.000 description 55
- 102000004196 processed proteins & peptides Human genes 0.000 description 55
- 102000005962 receptors Human genes 0.000 description 54
- 108020003175 receptors Proteins 0.000 description 54
- 230000009089 cytolysis Effects 0.000 description 45
- 108060003951 Immunoglobulin Proteins 0.000 description 44
- 102000018358 immunoglobulin Human genes 0.000 description 44
- 108090000623 proteins and genes Proteins 0.000 description 44
- 238000010606 normalization Methods 0.000 description 39
- 102000004169 proteins and genes Human genes 0.000 description 39
- 235000018102 proteins Nutrition 0.000 description 38
- 238000011534 incubation Methods 0.000 description 29
- 238000004020 luminiscence type Methods 0.000 description 27
- 150000001413 amino acids Chemical class 0.000 description 26
- 101710099301 Low affinity immunoglobulin gamma Fc region receptor III-A Proteins 0.000 description 20
- 102100029193 Low affinity immunoglobulin gamma Fc region receptor III-A Human genes 0.000 description 20
- 238000000684 flow cytometry Methods 0.000 description 20
- 239000000523 sample Substances 0.000 description 20
- 230000000295 complement effect Effects 0.000 description 19
- 108010021468 Fc gamma receptor IIA Proteins 0.000 description 18
- 102100026120 IgG receptor FcRn large subunit p51 Human genes 0.000 description 18
- 102100029204 Low affinity immunoglobulin gamma Fc region receptor II-a Human genes 0.000 description 18
- 230000005764 inhibitory process Effects 0.000 description 18
- 239000013598 vector Substances 0.000 description 18
- 101710177940 IgG receptor FcRn large subunit p51 Proteins 0.000 description 17
- 238000003556 assay Methods 0.000 description 17
- 239000003795 chemical substances by application Substances 0.000 description 17
- 230000000875 corresponding effect Effects 0.000 description 17
- 230000001404 mediated effect Effects 0.000 description 17
- 206010025323 Lymphomas Diseases 0.000 description 16
- 238000010790 dilution Methods 0.000 description 16
- 239000012895 dilution Substances 0.000 description 16
- 230000014509 gene expression Effects 0.000 description 16
- 238000000338 in vitro Methods 0.000 description 16
- 108010068617 neonatal Fc receptor Proteins 0.000 description 16
- 230000024203 complement activation Effects 0.000 description 15
- 210000002845 virion Anatomy 0.000 description 15
- 238000001994 activation Methods 0.000 description 14
- 239000013642 negative control Substances 0.000 description 14
- 238000002965 ELISA Methods 0.000 description 13
- 230000001965 increasing effect Effects 0.000 description 13
- ZTOJFFHGPLIVKC-YAFCTCPESA-N (2e)-3-ethyl-2-[(z)-(3-ethyl-6-sulfo-1,3-benzothiazol-2-ylidene)hydrazinylidene]-1,3-benzothiazole-6-sulfonic acid Chemical compound S\1C2=CC(S(O)(=O)=O)=CC=C2N(CC)C/1=N/N=C1/SC2=CC(S(O)(=O)=O)=CC=C2N1CC ZTOJFFHGPLIVKC-YAFCTCPESA-N 0.000 description 12
- 108010073807 IgG Receptors Proteins 0.000 description 12
- 102000009490 IgG Receptors Human genes 0.000 description 12
- 241000699670 Mus sp. Species 0.000 description 12
- 238000001514 detection method Methods 0.000 description 12
- 238000003782 apoptosis assay Methods 0.000 description 11
- 230000005522 programmed cell death Effects 0.000 description 11
- 230000001105 regulatory effect Effects 0.000 description 11
- 210000001519 tissue Anatomy 0.000 description 11
- 238000002474 experimental method Methods 0.000 description 10
- 239000012634 fragment Substances 0.000 description 10
- 230000004044 response Effects 0.000 description 10
- 210000001266 CD8-positive T-lymphocyte Anatomy 0.000 description 9
- 238000002835 absorbance Methods 0.000 description 9
- 230000015572 biosynthetic process Effects 0.000 description 9
- 230000006037 cell lysis Effects 0.000 description 9
- 210000002966 serum Anatomy 0.000 description 9
- 102000009109 Fc receptors Human genes 0.000 description 8
- 108010087819 Fc receptors Proteins 0.000 description 8
- 125000000539 amino acid group Chemical group 0.000 description 8
- 238000004166 bioassay Methods 0.000 description 8
- 102000006834 complement receptors Human genes 0.000 description 8
- 108010047295 complement receptors Proteins 0.000 description 8
- HKSZLNNOFSGOKW-UHFFFAOYSA-N ent-staurosporine Natural products C12=C3N4C5=CC=CC=C5C3=C3CNC(=O)C3=C2C2=CC=CC=C2N1C1CC(NC)C(OC)C4(C)O1 HKSZLNNOFSGOKW-UHFFFAOYSA-N 0.000 description 8
- 102000039446 nucleic acids Human genes 0.000 description 8
- 108020004707 nucleic acids Proteins 0.000 description 8
- 150000007523 nucleic acids Chemical class 0.000 description 8
- HKSZLNNOFSGOKW-FYTWVXJKSA-N staurosporine Chemical compound C12=C3N4C5=CC=CC=C5C3=C3CNC(=O)C3=C2C2=CC=CC=C2N1[C@H]1C[C@@H](NC)[C@@H](OC)[C@]4(C)O1 HKSZLNNOFSGOKW-FYTWVXJKSA-N 0.000 description 8
- CGPUWJWCVCFERF-UHFFFAOYSA-N staurosporine Natural products C12=C3N4C5=CC=CC=C5C3=C3CNC(=O)C3=C2C2=CC=CC=C2N1C1CC(NC)C(OC)C4(OC)O1 CGPUWJWCVCFERF-UHFFFAOYSA-N 0.000 description 8
- 208000003950 B-cell lymphoma Diseases 0.000 description 7
- 208000011691 Burkitt lymphomas Diseases 0.000 description 7
- 241000283707 Capra Species 0.000 description 7
- 108010047041 Complementarity Determining Regions Proteins 0.000 description 7
- 238000012286 ELISA Assay Methods 0.000 description 7
- 108060008683 Tumor Necrosis Factor Receptor Proteins 0.000 description 7
- 239000000556 agonist Substances 0.000 description 7
- 229940049595 antibody-drug conjugate Drugs 0.000 description 7
- 210000000170 cell membrane Anatomy 0.000 description 7
- 238000003776 cleavage reaction Methods 0.000 description 7
- 230000009827 complement-dependent cellular cytotoxicity Effects 0.000 description 7
- 239000000758 substrate Substances 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- 102000003298 tumor necrosis factor receptor Human genes 0.000 description 7
- 208000010839 B-cell chronic lymphocytic leukemia Diseases 0.000 description 6
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 6
- 108010021472 Fc gamma receptor IIB Proteins 0.000 description 6
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 6
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 6
- 102100029205 Low affinity immunoglobulin gamma Fc region receptor II-b Human genes 0.000 description 6
- 208000031422 Lymphocytic Chronic B-Cell Leukemia Diseases 0.000 description 6
- 206010061902 Pancreatic neoplasm Diseases 0.000 description 6
- 238000011579 SCID mouse model Methods 0.000 description 6
- 239000000611 antibody drug conjugate Substances 0.000 description 6
- 208000032852 chronic lymphocytic leukemia Diseases 0.000 description 6
- 231100000673 dose–response relationship Toxicity 0.000 description 6
- 239000013604 expression vector Substances 0.000 description 6
- 238000006206 glycosylation reaction Methods 0.000 description 6
- 208000015486 malignant pancreatic neoplasm Diseases 0.000 description 6
- 230000004048 modification Effects 0.000 description 6
- 238000012986 modification Methods 0.000 description 6
- 201000002528 pancreatic cancer Diseases 0.000 description 6
- 208000008443 pancreatic carcinoma Diseases 0.000 description 6
- 230000008569 process Effects 0.000 description 6
- 230000002829 reductive effect Effects 0.000 description 6
- 230000007017 scission Effects 0.000 description 6
- 230000001225 therapeutic effect Effects 0.000 description 6
- 230000035899 viability Effects 0.000 description 6
- 241000894006 Bacteria Species 0.000 description 5
- 108090000331 Firefly luciferases Proteins 0.000 description 5
- 102000002702 GPI-Linked Proteins Human genes 0.000 description 5
- 108010043685 GPI-Linked Proteins Proteins 0.000 description 5
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 5
- 108060001084 Luciferase Proteins 0.000 description 5
- 239000005089 Luciferase Substances 0.000 description 5
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 5
- 108091027981 Response element Proteins 0.000 description 5
- 238000006243 chemical reaction Methods 0.000 description 5
- 230000004927 fusion Effects 0.000 description 5
- 230000013595 glycosylation Effects 0.000 description 5
- 239000003446 ligand Substances 0.000 description 5
- 230000014207 opsonization Effects 0.000 description 5
- 239000013612 plasmid Substances 0.000 description 5
- 102000000844 Cell Surface Receptors Human genes 0.000 description 4
- 108010001857 Cell Surface Receptors Proteins 0.000 description 4
- 229910052693 Europium Inorganic materials 0.000 description 4
- 102000003688 G-Protein-Coupled Receptors Human genes 0.000 description 4
- 108090000045 G-Protein-Coupled Receptors Proteins 0.000 description 4
- 108010021625 Immunoglobulin Fragments Proteins 0.000 description 4
- 102000008394 Immunoglobulin Fragments Human genes 0.000 description 4
- 102000006496 Immunoglobulin Heavy Chains Human genes 0.000 description 4
- 108010019476 Immunoglobulin Heavy Chains Proteins 0.000 description 4
- 108090000467 Interferon-beta Proteins 0.000 description 4
- 102000003996 Interferon-beta Human genes 0.000 description 4
- 208000015914 Non-Hodgkin lymphomas Diseases 0.000 description 4
- 206010057249 Phagocytosis Diseases 0.000 description 4
- 206010035226 Plasma cell myeloma Diseases 0.000 description 4
- 102000010780 Platelet-Derived Growth Factor Human genes 0.000 description 4
- 108010038512 Platelet-Derived Growth Factor Proteins 0.000 description 4
- 208000006664 Precursor Cell Lymphoblastic Leukemia-Lymphoma Diseases 0.000 description 4
- 206010054094 Tumour necrosis Diseases 0.000 description 4
- 230000002378 acidificating effect Effects 0.000 description 4
- 230000001154 acute effect Effects 0.000 description 4
- 230000003833 cell viability Effects 0.000 description 4
- 239000013522 chelant Substances 0.000 description 4
- 102000003675 cytokine receptors Human genes 0.000 description 4
- 108010057085 cytokine receptors Proteins 0.000 description 4
- 230000003013 cytotoxicity Effects 0.000 description 4
- 231100000135 cytotoxicity Toxicity 0.000 description 4
- 229940079593 drug Drugs 0.000 description 4
- OGPBJKLSAFTDLK-UHFFFAOYSA-N europium atom Chemical compound [Eu] OGPBJKLSAFTDLK-UHFFFAOYSA-N 0.000 description 4
- 210000004602 germ cell Anatomy 0.000 description 4
- 229940072221 immunoglobulins Drugs 0.000 description 4
- 229960001388 interferon-beta Drugs 0.000 description 4
- 230000002147 killing effect Effects 0.000 description 4
- 201000011649 lymphoblastic lymphoma Diseases 0.000 description 4
- 238000004519 manufacturing process Methods 0.000 description 4
- 230000007246 mechanism Effects 0.000 description 4
- 201000000050 myeloid neoplasm Diseases 0.000 description 4
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 4
- 230000008782 phagocytosis Effects 0.000 description 4
- 239000012071 phase Substances 0.000 description 4
- 239000000047 product Substances 0.000 description 4
- 241000894007 species Species 0.000 description 4
- 231100000747 viability assay Toxicity 0.000 description 4
- 238000003026 viability measurement method Methods 0.000 description 4
- TZCPCKNHXULUIY-RGULYWFUSA-N 1,2-distearoyl-sn-glycero-3-phosphoserine Chemical compound CCCCCCCCCCCCCCCCCC(=O)OC[C@H](COP(O)(=O)OC[C@H](N)C(O)=O)OC(=O)CCCCCCCCCCCCCCCCC TZCPCKNHXULUIY-RGULYWFUSA-N 0.000 description 3
- NFGXHKASABOEEW-UHFFFAOYSA-N 1-methylethyl 11-methoxy-3,7,11-trimethyl-2,4-dodecadienoate Chemical compound COC(C)(C)CCCC(C)CC=CC(C)=CC(=O)OC(C)C NFGXHKASABOEEW-UHFFFAOYSA-N 0.000 description 3
- 239000004475 Arginine Substances 0.000 description 3
- 206010009944 Colon cancer Diseases 0.000 description 3
- 108020004414 DNA Proteins 0.000 description 3
- ZWZWYGMENQVNFU-UHFFFAOYSA-N Glycerophosphorylserin Natural products OC(=O)C(N)COP(O)(=O)OCC(O)CO ZWZWYGMENQVNFU-UHFFFAOYSA-N 0.000 description 3
- 102000003886 Glycoproteins Human genes 0.000 description 3
- 108090000288 Glycoproteins Proteins 0.000 description 3
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 description 3
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 description 3
- 102000008070 Interferon-gamma Human genes 0.000 description 3
- 108010074328 Interferon-gamma Proteins 0.000 description 3
- KDXKERNSBIXSRK-YFKPBYRVSA-N L-lysine Chemical compound NCCCC[C@H](N)C(O)=O KDXKERNSBIXSRK-YFKPBYRVSA-N 0.000 description 3
- 239000004472 Lysine Substances 0.000 description 3
- 241000124008 Mammalia Species 0.000 description 3
- 241001436793 Meru Species 0.000 description 3
- MUBZPKHOEPUJKR-UHFFFAOYSA-N Oxalic acid Chemical compound OC(=O)C(O)=O MUBZPKHOEPUJKR-UHFFFAOYSA-N 0.000 description 3
- 101710120037 Toxin CcdB Proteins 0.000 description 3
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 3
- 239000002246 antineoplastic agent Substances 0.000 description 3
- 230000006907 apoptotic process Effects 0.000 description 3
- ODKSFYDXXFIFQN-UHFFFAOYSA-N arginine Natural products OC(=O)C(N)CCCNC(N)=N ODKSFYDXXFIFQN-UHFFFAOYSA-N 0.000 description 3
- 230000001580 bacterial effect Effects 0.000 description 3
- 108010051210 beta-Fructofuranosidase Proteins 0.000 description 3
- 210000000601 blood cell Anatomy 0.000 description 3
- 239000000872 buffer Substances 0.000 description 3
- 210000004899 c-terminal region Anatomy 0.000 description 3
- 238000012054 celltiter-glo Methods 0.000 description 3
- 208000029742 colonic neoplasm Diseases 0.000 description 3
- 229940127089 cytotoxic agent Drugs 0.000 description 3
- 230000007423 decrease Effects 0.000 description 3
- 238000005516 engineering process Methods 0.000 description 3
- 125000001495 ethyl group Chemical group [H]C([H])([H])C([H])([H])* 0.000 description 3
- 108020001507 fusion proteins Proteins 0.000 description 3
- 102000037865 fusion proteins Human genes 0.000 description 3
- 238000011102 hetero oligomerization reaction Methods 0.000 description 3
- 230000001900 immune effect Effects 0.000 description 3
- 210000000987 immune system Anatomy 0.000 description 3
- 229960003130 interferon gamma Drugs 0.000 description 3
- 235000011073 invertase Nutrition 0.000 description 3
- 239000001573 invertase Substances 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 210000002540 macrophage Anatomy 0.000 description 3
- 230000003211 malignant effect Effects 0.000 description 3
- 210000004379 membrane Anatomy 0.000 description 3
- 239000012528 membrane Substances 0.000 description 3
- 239000000178 monomer Substances 0.000 description 3
- 239000002773 nucleotide Substances 0.000 description 3
- 125000003729 nucleotide group Chemical group 0.000 description 3
- 239000013641 positive control Substances 0.000 description 3
- 230000004481 post-translational protein modification Effects 0.000 description 3
- 238000002360 preparation method Methods 0.000 description 3
- 238000003259 recombinant expression Methods 0.000 description 3
- 230000010076 replication Effects 0.000 description 3
- 238000013207 serial dilution Methods 0.000 description 3
- 230000011664 signaling Effects 0.000 description 3
- 230000001629 suppression Effects 0.000 description 3
- 238000003786 synthesis reaction Methods 0.000 description 3
- 230000008685 targeting Effects 0.000 description 3
- 230000001988 toxicity Effects 0.000 description 3
- 231100000419 toxicity Toxicity 0.000 description 3
- 102000003390 tumor necrosis factor Human genes 0.000 description 3
- 108090000672 Annexin A5 Proteins 0.000 description 2
- 102000004121 Annexin A5 Human genes 0.000 description 2
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 2
- 108010005939 Ciliary Neurotrophic Factor Proteins 0.000 description 2
- 102100031614 Ciliary neurotrophic factor Human genes 0.000 description 2
- 108010034753 Complement Membrane Attack Complex Proteins 0.000 description 2
- 102000004127 Cytokines Human genes 0.000 description 2
- 108090000695 Cytokines Proteins 0.000 description 2
- AOJJSUZBOXZQNB-TZSSRYMLSA-N Doxorubicin Chemical compound O([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)[C@H](O)[C@H](C)O1 AOJJSUZBOXZQNB-TZSSRYMLSA-N 0.000 description 2
- 241000196324 Embryophyta Species 0.000 description 2
- 102000003951 Erythropoietin Human genes 0.000 description 2
- 108090000394 Erythropoietin Proteins 0.000 description 2
- 241000588724 Escherichia coli Species 0.000 description 2
- -1 FR3 Proteins 0.000 description 2
- 108050007372 Fibroblast Growth Factor Proteins 0.000 description 2
- 102000018233 Fibroblast Growth Factor Human genes 0.000 description 2
- 241000233866 Fungi Species 0.000 description 2
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 2
- 102100039620 Granulocyte-macrophage colony-stimulating factor Human genes 0.000 description 2
- 238000011993 High Performance Size Exclusion Chromatography Methods 0.000 description 2
- 241000282412 Homo Species 0.000 description 2
- 102000003746 Insulin Receptor Human genes 0.000 description 2
- 108010001127 Insulin Receptor Proteins 0.000 description 2
- 108010086140 Interferon alpha-beta Receptor Proteins 0.000 description 2
- 102000007438 Interferon alpha-beta Receptor Human genes 0.000 description 2
- 102000006992 Interferon-alpha Human genes 0.000 description 2
- 108010047761 Interferon-alpha Proteins 0.000 description 2
- 102000000589 Interleukin-1 Human genes 0.000 description 2
- 108010002352 Interleukin-1 Proteins 0.000 description 2
- 102000003814 Interleukin-10 Human genes 0.000 description 2
- 108090000174 Interleukin-10 Proteins 0.000 description 2
- 102000000588 Interleukin-2 Human genes 0.000 description 2
- 108010002350 Interleukin-2 Proteins 0.000 description 2
- 102000000646 Interleukin-3 Human genes 0.000 description 2
- 108010002386 Interleukin-3 Proteins 0.000 description 2
- 102000004388 Interleukin-4 Human genes 0.000 description 2
- 108090000978 Interleukin-4 Proteins 0.000 description 2
- 102100039897 Interleukin-5 Human genes 0.000 description 2
- 108010002616 Interleukin-5 Proteins 0.000 description 2
- 102000004889 Interleukin-6 Human genes 0.000 description 2
- 108090001005 Interleukin-6 Proteins 0.000 description 2
- 102100021592 Interleukin-7 Human genes 0.000 description 2
- 108010002586 Interleukin-7 Proteins 0.000 description 2
- 102000000585 Interleukin-9 Human genes 0.000 description 2
- 108010002335 Interleukin-9 Proteins 0.000 description 2
- 241000235058 Komagataella pastoris Species 0.000 description 2
- ODKSFYDXXFIFQN-BYPYZUCNSA-P L-argininium(2+) Chemical compound NC(=[NH2+])NCCC[C@H]([NH3+])C(O)=O ODKSFYDXXFIFQN-BYPYZUCNSA-P 0.000 description 2
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 2
- AYFVYJQAPQTCCC-GBXIJSLDSA-N L-threonine Chemical compound C[C@@H](O)[C@H](N)C(O)=O AYFVYJQAPQTCCC-GBXIJSLDSA-N 0.000 description 2
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 2
- 102000007651 Macrophage Colony-Stimulating Factor Human genes 0.000 description 2
- 108010046938 Macrophage Colony-Stimulating Factor Proteins 0.000 description 2
- 102000018697 Membrane Proteins Human genes 0.000 description 2
- 108010052285 Membrane Proteins Proteins 0.000 description 2
- 241000699666 Mus <mouse, genus> Species 0.000 description 2
- 102000007339 Nerve Growth Factor Receptors Human genes 0.000 description 2
- 108010032605 Nerve Growth Factor Receptors Proteins 0.000 description 2
- 239000004793 Polystyrene Substances 0.000 description 2
- 241000283984 Rodentia Species 0.000 description 2
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 2
- 241000191967 Staphylococcus aureus Species 0.000 description 2
- 108700031126 Tetraspanins Proteins 0.000 description 2
- 102000043977 Tetraspanins Human genes 0.000 description 2
- 102000036693 Thrombopoietin Human genes 0.000 description 2
- 108010041111 Thrombopoietin Proteins 0.000 description 2
- 102000004887 Transforming Growth Factor beta Human genes 0.000 description 2
- 108090001012 Transforming Growth Factor beta Proteins 0.000 description 2
- 229940098773 bovine serum albumin Drugs 0.000 description 2
- 230000030833 cell death Effects 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 210000004978 chinese hamster ovary cell Anatomy 0.000 description 2
- 230000004154 complement system Effects 0.000 description 2
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 2
- 238000000151 deposition Methods 0.000 description 2
- 238000002405 diagnostic procedure Methods 0.000 description 2
- 208000035475 disorder Diseases 0.000 description 2
- 238000010494 dissociation reaction Methods 0.000 description 2
- 230000005593 dissociations Effects 0.000 description 2
- 238000009826 distribution Methods 0.000 description 2
- 230000009977 dual effect Effects 0.000 description 2
- 229940105423 erythropoietin Drugs 0.000 description 2
- 229940126864 fibroblast growth factor Drugs 0.000 description 2
- 238000013467 fragmentation Methods 0.000 description 2
- 238000006062 fragmentation reaction Methods 0.000 description 2
- 230000005714 functional activity Effects 0.000 description 2
- 235000013922 glutamic acid Nutrition 0.000 description 2
- 239000004220 glutamic acid Substances 0.000 description 2
- 230000003394 haemopoietic effect Effects 0.000 description 2
- 108091008039 hormone receptors Proteins 0.000 description 2
- 210000005260 human cell Anatomy 0.000 description 2
- 238000003384 imaging method Methods 0.000 description 2
- 210000002865 immune cell Anatomy 0.000 description 2
- 230000028993 immune response Effects 0.000 description 2
- 238000001727 in vivo Methods 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- 108010085650 interferon gamma receptor Proteins 0.000 description 2
- 229940076144 interleukin-10 Drugs 0.000 description 2
- 229940076264 interleukin-3 Drugs 0.000 description 2
- 229940028885 interleukin-4 Drugs 0.000 description 2
- 229940100602 interleukin-5 Drugs 0.000 description 2
- 229940100601 interleukin-6 Drugs 0.000 description 2
- 229940100994 interleukin-7 Drugs 0.000 description 2
- 229940118526 interleukin-9 Drugs 0.000 description 2
- 230000003834 intracellular effect Effects 0.000 description 2
- 230000004068 intracellular signaling Effects 0.000 description 2
- 125000003588 lysine group Chemical group [H]N([H])C([H])([H])C([H])([H])C([H])([H])C([H])([H])C([H])(N([H])[H])C(*)=O 0.000 description 2
- 210000001616 monocyte Anatomy 0.000 description 2
- 230000007935 neutral effect Effects 0.000 description 2
- 210000000440 neutrophil Anatomy 0.000 description 2
- 239000002245 particle Substances 0.000 description 2
- 230000037361 pathway Effects 0.000 description 2
- 210000000680 phagosome Anatomy 0.000 description 2
- 229920002223 polystyrene Polymers 0.000 description 2
- 239000011148 porous material Substances 0.000 description 2
- OXCMYAYHXIHQOA-UHFFFAOYSA-N potassium;[2-butyl-5-chloro-3-[[4-[2-(1,2,4-triaza-3-azanidacyclopenta-1,4-dien-5-yl)phenyl]phenyl]methyl]imidazol-4-yl]methanol Chemical compound [K+].CCCCC1=NC(Cl)=C(CO)N1CC1=CC=C(C=2C(=CC=CC=2)C2=N[N-]N=N2)C=C1 OXCMYAYHXIHQOA-UHFFFAOYSA-N 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 230000009467 reduction Effects 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 229910052708 sodium Inorganic materials 0.000 description 2
- 239000011734 sodium Substances 0.000 description 2
- HEMHJVSKTPXQMS-UHFFFAOYSA-M sodium hydroxide Inorganic materials [OH-].[Na+] HEMHJVSKTPXQMS-UHFFFAOYSA-M 0.000 description 2
- 230000000638 stimulation Effects 0.000 description 2
- ZRKFYGHZFMAOKI-QMGMOQQFSA-N tgfbeta Chemical compound C([C@H](NC(=O)[C@H](C(C)C)NC(=O)CNC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC(C)C)NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](C)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N1[C@@H](CCC1)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O)C1=CC=C(O)C=C1 ZRKFYGHZFMAOKI-QMGMOQQFSA-N 0.000 description 2
- 238000002560 therapeutic procedure Methods 0.000 description 2
- 231100000331 toxic Toxicity 0.000 description 2
- 230000002588 toxic effect Effects 0.000 description 2
- 230000009261 transgenic effect Effects 0.000 description 2
- 230000032258 transport Effects 0.000 description 2
- 229910052721 tungsten Inorganic materials 0.000 description 2
- VBEQCZHXXJYVRD-GACYYNSASA-N uroanthelone Chemical compound C([C@@H](C(=O)N[C@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O)C(C)C)[C@@H](C)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)NC(=O)[C@@H](NC(=O)CNC(=O)CNC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@H](CO)NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CS)NC(=O)CNC(=O)[C@H]1N(CCC1)C(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O)C(C)C)[C@@H](C)CC)C1=CC=C(O)C=C1 VBEQCZHXXJYVRD-GACYYNSASA-N 0.000 description 2
- 239000013603 viral vector Substances 0.000 description 2
- 230000003612 virological effect Effects 0.000 description 2
- HBZBAMXERPYTFS-SECBINFHSA-N (4S)-2-(6,7-dihydro-5H-pyrrolo[3,2-f][1,3]benzothiazol-2-yl)-4,5-dihydro-1,3-thiazole-4-carboxylic acid Chemical compound OC(=O)[C@H]1CSC(=N1)c1nc2cc3CCNc3cc2s1 HBZBAMXERPYTFS-SECBINFHSA-N 0.000 description 1
- ZTOJFFHGPLIVKC-UHFFFAOYSA-N 3-ethyl-2-[(3-ethyl-6-sulfo-1,3-benzothiazol-2-ylidene)hydrazinylidene]-1,3-benzothiazole-6-sulfonic acid Chemical compound S1C2=CC(S(O)(=O)=O)=CC=C2N(CC)C1=NN=C1SC2=CC(S(O)(=O)=O)=CC=C2N1CC ZTOJFFHGPLIVKC-UHFFFAOYSA-N 0.000 description 1
- 206010069754 Acquired gene mutation Diseases 0.000 description 1
- 102000000412 Annexin Human genes 0.000 description 1
- 108050008874 Annexin Proteins 0.000 description 1
- 108010083359 Antigen Receptors Proteins 0.000 description 1
- 102000006306 Antigen Receptors Human genes 0.000 description 1
- MLDQJTXFUGDVEO-UHFFFAOYSA-N BAY-43-9006 Chemical compound C1=NC(C(=O)NC)=CC(OC=2C=CC(NC(=O)NC=3C=C(C(Cl)=CC=3)C(F)(F)F)=CC=2)=C1 MLDQJTXFUGDVEO-UHFFFAOYSA-N 0.000 description 1
- 208000035143 Bacterial infection Diseases 0.000 description 1
- 102100023995 Beta-nerve growth factor Human genes 0.000 description 1
- 102000003930 C-Type Lectins Human genes 0.000 description 1
- 108090000342 C-Type Lectins Proteins 0.000 description 1
- 101150013553 CD40 gene Proteins 0.000 description 1
- 101100009017 Caenorhabditis elegans dcr-1 gene Proteins 0.000 description 1
- 241000282832 Camelidae Species 0.000 description 1
- 102000014914 Carrier Proteins Human genes 0.000 description 1
- 108010067225 Cell Adhesion Molecules Proteins 0.000 description 1
- 102000016289 Cell Adhesion Molecules Human genes 0.000 description 1
- 238000003734 CellTiter-Glo Luminescent Cell Viability Assay Methods 0.000 description 1
- 102000009410 Chemokine receptor Human genes 0.000 description 1
- 108050000299 Chemokine receptor Proteins 0.000 description 1
- 108010028780 Complement C3 Proteins 0.000 description 1
- 102000016918 Complement C3 Human genes 0.000 description 1
- 102000016574 Complement C3-C5 Convertases Human genes 0.000 description 1
- 108010067641 Complement C3-C5 Convertases Proteins 0.000 description 1
- 241000557626 Corvus corax Species 0.000 description 1
- 102000053602 DNA Human genes 0.000 description 1
- 102000010170 Death domains Human genes 0.000 description 1
- 108050001718 Death domains Proteins 0.000 description 1
- 241000702421 Dependoparvovirus Species 0.000 description 1
- SNRUBQQJIBEYMU-UHFFFAOYSA-N Dodecane Natural products CCCCCCCCCCCC SNRUBQQJIBEYMU-UHFFFAOYSA-N 0.000 description 1
- 101100009019 Drosophila melanogaster Dcr-1 gene Proteins 0.000 description 1
- 101100012887 Drosophila melanogaster btl gene Proteins 0.000 description 1
- 101100012878 Drosophila melanogaster htl gene Proteins 0.000 description 1
- 102000001301 EGF receptor Human genes 0.000 description 1
- 108060006698 EGF receptor Proteins 0.000 description 1
- 102000004190 Enzymes Human genes 0.000 description 1
- 108090000790 Enzymes Proteins 0.000 description 1
- 102400001368 Epidermal growth factor Human genes 0.000 description 1
- 101800003838 Epidermal growth factor Proteins 0.000 description 1
- HKVAMNSJSFKALM-GKUWKFKPSA-N Everolimus Chemical compound C1C[C@@H](OCCO)[C@H](OC)C[C@@H]1C[C@@H](C)[C@H]1OC(=O)[C@@H]2CCCCN2C(=O)C(=O)[C@](O)(O2)[C@H](C)CC[C@H]2C[C@H](OC)/C(C)=C/C=C/C=C/[C@@H](C)C[C@@H](C)C(=O)[C@H](OC)[C@H](O)/C(C)=C/[C@@H](C)C(=O)C1 HKVAMNSJSFKALM-GKUWKFKPSA-N 0.000 description 1
- GHASVSINZRGABV-UHFFFAOYSA-N Fluorouracil Chemical compound FC1=CNC(=O)NC1=O GHASVSINZRGABV-UHFFFAOYSA-N 0.000 description 1
- 206010017533 Fungal infection Diseases 0.000 description 1
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Natural products NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 229930186217 Glycolipid Natural products 0.000 description 1
- 102000009465 Growth Factor Receptors Human genes 0.000 description 1
- 108010009202 Growth Factor Receptors Proteins 0.000 description 1
- 108010051696 Growth Hormone Proteins 0.000 description 1
- 101001057612 Homo sapiens Dual specificity protein phosphatase 5 Proteins 0.000 description 1
- 101001059454 Homo sapiens Serine/threonine-protein kinase MARK2 Proteins 0.000 description 1
- 101000830565 Homo sapiens Tumor necrosis factor ligand superfamily member 10 Proteins 0.000 description 1
- 101000920026 Homo sapiens Tumor necrosis factor receptor superfamily member EDAR Proteins 0.000 description 1
- 102000004157 Hydrolases Human genes 0.000 description 1
- 108090000604 Hydrolases Proteins 0.000 description 1
- DGAQECJNVWCQMB-PUAWFVPOSA-M Ilexoside XXIX Chemical compound C[C@@H]1CC[C@@]2(CC[C@@]3(C(=CC[C@H]4[C@]3(CC[C@@H]5[C@@]4(CC[C@@H](C5(C)C)OS(=O)(=O)[O-])C)C)[C@@H]2[C@]1(C)O)C)C(=O)O[C@H]6[C@@H]([C@H]([C@@H]([C@H](O6)CO)O)O)O.[Na+] DGAQECJNVWCQMB-PUAWFVPOSA-M 0.000 description 1
- 102000003815 Interleukin-11 Human genes 0.000 description 1
- 108090000177 Interleukin-11 Proteins 0.000 description 1
- 102000004553 Interleukin-11 Receptors Human genes 0.000 description 1
- 108010017521 Interleukin-11 Receptors Proteins 0.000 description 1
- 102000013462 Interleukin-12 Human genes 0.000 description 1
- 108010065805 Interleukin-12 Proteins 0.000 description 1
- 102000004560 Interleukin-12 Receptors Human genes 0.000 description 1
- 108010017515 Interleukin-12 Receptors Proteins 0.000 description 1
- 102000003816 Interleukin-13 Human genes 0.000 description 1
- 108090000176 Interleukin-13 Proteins 0.000 description 1
- 102000004559 Interleukin-13 Receptors Human genes 0.000 description 1
- 108010017511 Interleukin-13 Receptors Proteins 0.000 description 1
- 102000003812 Interleukin-15 Human genes 0.000 description 1
- 108090000172 Interleukin-15 Proteins 0.000 description 1
- 102000004556 Interleukin-15 Receptors Human genes 0.000 description 1
- 108010017535 Interleukin-15 Receptors Proteins 0.000 description 1
- 102000010787 Interleukin-4 Receptors Human genes 0.000 description 1
- 108010038486 Interleukin-4 Receptors Proteins 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- 239000002147 L01XE04 - Sunitinib Substances 0.000 description 1
- 239000005511 L01XE05 - Sorafenib Substances 0.000 description 1
- VKVDRTGWLVZJOM-DCAQKATOSA-N Leu-Val-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O VKVDRTGWLVZJOM-DCAQKATOSA-N 0.000 description 1
- 108090001030 Lipoproteins Proteins 0.000 description 1
- 102000004895 Lipoproteins Human genes 0.000 description 1
- 239000012515 MabSelect SuRe Substances 0.000 description 1
- 108010061593 Member 14 Tumor Necrosis Factor Receptors Proteins 0.000 description 1
- 241000699660 Mus musculus Species 0.000 description 1
- 101100153533 Mus musculus Ltbr gene Proteins 0.000 description 1
- 208000031888 Mycoses Diseases 0.000 description 1
- 108010025020 Nerve Growth Factor Proteins 0.000 description 1
- 108091028043 Nucleic acid sequence Proteins 0.000 description 1
- 102000003987 Oncostatin M Receptors Human genes 0.000 description 1
- 108010082522 Oncostatin M Receptors Proteins 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 229930012538 Paclitaxel Natural products 0.000 description 1
- 108090000526 Papain Proteins 0.000 description 1
- 102000004160 Phosphoric Monoester Hydrolases Human genes 0.000 description 1
- 108090000608 Phosphoric Monoester Hydrolases Proteins 0.000 description 1
- 239000004365 Protease Substances 0.000 description 1
- 229940079156 Proteasome inhibitor Drugs 0.000 description 1
- 102000002727 Protein Tyrosine Phosphatase Human genes 0.000 description 1
- 102000004022 Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000412 Protein-Tyrosine Kinases Proteins 0.000 description 1
- 229940078123 Ras inhibitor Drugs 0.000 description 1
- 102000004278 Receptor Protein-Tyrosine Kinases Human genes 0.000 description 1
- 108090000873 Receptor Protein-Tyrosine Kinases Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 108700008625 Reporter Genes Proteins 0.000 description 1
- 239000006146 Roswell Park Memorial Institute medium Substances 0.000 description 1
- 241000239226 Scorpiones Species 0.000 description 1
- 102000012479 Serine Proteases Human genes 0.000 description 1
- 108010022999 Serine Proteases Proteins 0.000 description 1
- 102100028904 Serine/threonine-protein kinase MARK2 Human genes 0.000 description 1
- 102100038803 Somatotropin Human genes 0.000 description 1
- 108010068542 Somatotropin Receptors Proteins 0.000 description 1
- BPEGJWRSRHCHSN-UHFFFAOYSA-N Temozolomide Chemical compound O=C1N(C)N=NC2=C(C(N)=O)N=CN21 BPEGJWRSRHCHSN-UHFFFAOYSA-N 0.000 description 1
- 102100024554 Tetranectin Human genes 0.000 description 1
- AYFVYJQAPQTCCC-UHFFFAOYSA-N Threonine Natural products CC(O)C(N)C(O)=O AYFVYJQAPQTCCC-UHFFFAOYSA-N 0.000 description 1
- 239000004473 Threonine Substances 0.000 description 1
- 102000002689 Toll-like receptor Human genes 0.000 description 1
- 108020000411 Toll-like receptor Proteins 0.000 description 1
- 108700019146 Transgenes Proteins 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- 102100028785 Tumor necrosis factor receptor superfamily member 14 Human genes 0.000 description 1
- 101710187743 Tumor necrosis factor receptor superfamily member 1A Proteins 0.000 description 1
- 102100033732 Tumor necrosis factor receptor superfamily member 1A Human genes 0.000 description 1
- 102100040245 Tumor necrosis factor receptor superfamily member 5 Human genes 0.000 description 1
- 102100030810 Tumor necrosis factor receptor superfamily member EDAR Human genes 0.000 description 1
- 208000036142 Viral infection Diseases 0.000 description 1
- 230000009471 action Effects 0.000 description 1
- 230000021917 activation of membrane attack complex Effects 0.000 description 1
- 238000001042 affinity chromatography Methods 0.000 description 1
- 230000008484 agonism Effects 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 125000001931 aliphatic group Chemical group 0.000 description 1
- 150000003862 amino acid derivatives Chemical class 0.000 description 1
- 239000004037 angiogenesis inhibitor Substances 0.000 description 1
- 238000011091 antibody purification Methods 0.000 description 1
- 238000009175 antibody therapy Methods 0.000 description 1
- 230000005888 antibody-dependent cellular phagocytosis Effects 0.000 description 1
- 210000000612 antigen-presenting cell Anatomy 0.000 description 1
- 229940076005 apoptosis modulator Drugs 0.000 description 1
- 150000001483 arginine derivatives Chemical class 0.000 description 1
- 125000003118 aryl group Chemical group 0.000 description 1
- 208000022362 bacterial infectious disease Diseases 0.000 description 1
- 210000003651 basophil Anatomy 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- 108091008324 binding proteins Proteins 0.000 description 1
- 230000033228 biological regulation Effects 0.000 description 1
- 230000008512 biological response Effects 0.000 description 1
- 229960001467 bortezomib Drugs 0.000 description 1
- GXJABQQUPOEUTA-RDJZCZTQSA-N bortezomib Chemical compound C([C@@H](C(=O)N[C@@H](CC(C)C)B(O)O)NC(=O)C=1N=CC=NC=1)C1=CC=CC=C1 GXJABQQUPOEUTA-RDJZCZTQSA-N 0.000 description 1
- 238000005251 capillar electrophoresis Methods 0.000 description 1
- 229960004562 carboplatin Drugs 0.000 description 1
- 190000008236 carboplatin Chemical compound 0.000 description 1
- 238000004113 cell culture Methods 0.000 description 1
- 230000022534 cell killing Effects 0.000 description 1
- 230000004663 cell proliferation Effects 0.000 description 1
- 210000002421 cell wall Anatomy 0.000 description 1
- 230000033077 cellular process Effects 0.000 description 1
- 238000005119 centrifugation Methods 0.000 description 1
- 239000003153 chemical reaction reagent Substances 0.000 description 1
- 230000010428 chromatin condensation Effects 0.000 description 1
- 239000013611 chromosomal DNA Substances 0.000 description 1
- DQLATGHUWYMOKM-UHFFFAOYSA-L cisplatin Chemical compound N[Pt](N)(Cl)Cl DQLATGHUWYMOKM-UHFFFAOYSA-L 0.000 description 1
- 229960004316 cisplatin Drugs 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 238000002648 combination therapy Methods 0.000 description 1
- 150000001875 compounds Chemical class 0.000 description 1
- 230000008878 coupling Effects 0.000 description 1
- 238000010168 coupling process Methods 0.000 description 1
- 238000005859 coupling reaction Methods 0.000 description 1
- 239000012228 culture supernatant Substances 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 1
- 239000002254 cytotoxic agent Substances 0.000 description 1
- 231100000599 cytotoxic agent Toxicity 0.000 description 1
- 230000001472 cytotoxic effect Effects 0.000 description 1
- 238000002784 cytotoxicity assay Methods 0.000 description 1
- 231100000263 cytotoxicity test Toxicity 0.000 description 1
- 230000034994 death Effects 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 230000002950 deficient Effects 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 210000004443 dendritic cell Anatomy 0.000 description 1
- 230000008021 deposition Effects 0.000 description 1
- 238000011033 desalting Methods 0.000 description 1
- 230000001627 detrimental effect Effects 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 239000000539 dimer Substances 0.000 description 1
- 208000037765 diseases and disorders Diseases 0.000 description 1
- 239000002552 dosage form Substances 0.000 description 1
- 229960004679 doxorubicin Drugs 0.000 description 1
- 230000008030 elimination Effects 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 230000002121 endocytic effect Effects 0.000 description 1
- 230000012202 endocytosis Effects 0.000 description 1
- 210000001163 endosome Anatomy 0.000 description 1
- 230000003511 endothelial effect Effects 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 230000002255 enzymatic effect Effects 0.000 description 1
- 229940088598 enzyme Drugs 0.000 description 1
- 210000003979 eosinophil Anatomy 0.000 description 1
- 229940116977 epidermal growth factor Drugs 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 229960005167 everolimus Drugs 0.000 description 1
- 210000003754 fetus Anatomy 0.000 description 1
- 229960002949 fluorouracil Drugs 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- SDUQYLNIPVEERB-QPPQHZFASA-N gemcitabine Chemical compound O=C1N=C(N)C=CN1[C@H]1C(F)(F)[C@H](O)[C@@H](CO)O1 SDUQYLNIPVEERB-QPPQHZFASA-N 0.000 description 1
- 229960005277 gemcitabine Drugs 0.000 description 1
- 125000000291 glutamic acid group Chemical group N[C@@H](CCC(O)=O)C(=O)* 0.000 description 1
- ZDXPYRJPNDTMRX-UHFFFAOYSA-N glutamine Natural products OC(=O)C(N)CCC(N)=O ZDXPYRJPNDTMRX-UHFFFAOYSA-N 0.000 description 1
- 125000003630 glycyl group Chemical group [H]N([H])C([H])([H])C(*)=O 0.000 description 1
- 210000003714 granulocyte Anatomy 0.000 description 1
- 239000000122 growth hormone Substances 0.000 description 1
- 210000003630 histaminocyte Anatomy 0.000 description 1
- 229940121372 histone deacetylase inhibitor Drugs 0.000 description 1
- 239000003276 histone deacetylase inhibitor Substances 0.000 description 1
- 102000043381 human DUSP5 Human genes 0.000 description 1
- 102000044949 human TNFSF10 Human genes 0.000 description 1
- 235000020256 human milk Nutrition 0.000 description 1
- 210000004251 human milk Anatomy 0.000 description 1
- 210000004408 hybridoma Anatomy 0.000 description 1
- 230000005931 immune cell recruitment Effects 0.000 description 1
- 230000036737 immune function Effects 0.000 description 1
- 102000028557 immunoglobulin binding proteins Human genes 0.000 description 1
- 108091009323 immunoglobulin binding proteins Proteins 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 238000002513 implantation Methods 0.000 description 1
- 238000000099 in vitro assay Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 230000006882 induction of apoptosis Effects 0.000 description 1
- 208000015181 infectious disease Diseases 0.000 description 1
- 208000027866 inflammatory disease Diseases 0.000 description 1
- 102000006495 integrins Human genes 0.000 description 1
- 108010044426 integrins Proteins 0.000 description 1
- 230000002452 interceptive effect Effects 0.000 description 1
- 102000002467 interleukin receptors Human genes 0.000 description 1
- 108010093036 interleukin receptors Proteins 0.000 description 1
- 229940074383 interleukin-11 Drugs 0.000 description 1
- 229940117681 interleukin-12 Drugs 0.000 description 1
- 230000000968 intestinal effect Effects 0.000 description 1
- 210000004347 intestinal mucosa Anatomy 0.000 description 1
- 210000000936 intestine Anatomy 0.000 description 1
- 230000006662 intracellular pathway Effects 0.000 description 1
- 230000031146 intracellular signal transduction Effects 0.000 description 1
- 229960004768 irinotecan Drugs 0.000 description 1
- UWKQSNNFCGGAFS-XIFFEERXSA-N irinotecan Chemical compound C1=C2C(CC)=C3CN(C(C4=C([C@@](C(=O)OC4)(O)CC)C=4)=O)C=4C3=NC2=CC=C1OC(=O)N(CC1)CCC1N1CCCCC1 UWKQSNNFCGGAFS-XIFFEERXSA-N 0.000 description 1
- 229940043355 kinase inhibitor Drugs 0.000 description 1
- 210000001865 kupffer cell Anatomy 0.000 description 1
- 230000006674 lysosomal degradation Effects 0.000 description 1
- 210000003712 lysosome Anatomy 0.000 description 1
- 230000001868 lysosomic effect Effects 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 230000008774 maternal effect Effects 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- 230000005012 migration Effects 0.000 description 1
- 238000013508 migration Methods 0.000 description 1
- 238000012544 monitoring process Methods 0.000 description 1
- 210000000066 myeloid cell Anatomy 0.000 description 1
- 229940053128 nerve growth factor Drugs 0.000 description 1
- 238000010899 nucleation Methods 0.000 description 1
- 239000002417 nutraceutical Substances 0.000 description 1
- 235000021436 nutraceutical agent Nutrition 0.000 description 1
- 230000001662 opsonic effect Effects 0.000 description 1
- 235000006408 oxalic acid Nutrition 0.000 description 1
- DWAFYCQODLXJNR-BNTLRKBRSA-L oxaliplatin Chemical compound O1C(=O)C(=O)O[Pt]11N[C@@H]2CCCC[C@H]2N1 DWAFYCQODLXJNR-BNTLRKBRSA-L 0.000 description 1
- 229960001756 oxaliplatin Drugs 0.000 description 1
- 229960001592 paclitaxel Drugs 0.000 description 1
- 229940055729 papain Drugs 0.000 description 1
- 235000019834 papain Nutrition 0.000 description 1
- 230000036961 partial effect Effects 0.000 description 1
- 229960005079 pemetrexed Drugs 0.000 description 1
- QOFFJEBXNKRSPX-ZDUSSCGKSA-N pemetrexed Chemical compound C1=N[C]2NC(N)=NC(=O)C2=C1CCC1=CC=C(C(=O)N[C@@H](CCC(O)=O)C(O)=O)C=C1 QOFFJEBXNKRSPX-ZDUSSCGKSA-N 0.000 description 1
- 238000010647 peptide synthesis reaction Methods 0.000 description 1
- 210000001539 phagocyte Anatomy 0.000 description 1
- 239000003757 phosphotransferase inhibitor Substances 0.000 description 1
- 210000002826 placenta Anatomy 0.000 description 1
- 230000036470 plasma concentration Effects 0.000 description 1
- 229920002401 polyacrylamide Polymers 0.000 description 1
- 229920000642 polymer Polymers 0.000 description 1
- 238000006116 polymerization reaction Methods 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 239000003207 proteasome inhibitor Substances 0.000 description 1
- 108020000494 protein-tyrosine phosphatase Proteins 0.000 description 1
- 238000004445 quantitative analysis Methods 0.000 description 1
- 238000001959 radiotherapy Methods 0.000 description 1
- 238000002708 random mutagenesis Methods 0.000 description 1
- 230000019491 signal transduction Effects 0.000 description 1
- 230000009131 signaling function Effects 0.000 description 1
- 238000002741 site-directed mutagenesis Methods 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- 229910000162 sodium phosphate Inorganic materials 0.000 description 1
- 239000000243 solution Substances 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 230000037439 somatic mutation Effects 0.000 description 1
- 229960003787 sorafenib Drugs 0.000 description 1
- 101150065015 spa gene Proteins 0.000 description 1
- 230000000087 stabilizing effect Effects 0.000 description 1
- 238000010186 staining Methods 0.000 description 1
- 210000000130 stem cell Anatomy 0.000 description 1
- 229960001796 sunitinib Drugs 0.000 description 1
- WINHZLLDWRZWRT-ATVHPVEESA-N sunitinib Chemical compound CCN(CC)CCNC(=O)C1=C(C)NC(\C=C/2C3=CC(F)=CC=C3NC\2=O)=C1C WINHZLLDWRZWRT-ATVHPVEESA-N 0.000 description 1
- 238000001356 surgical procedure Methods 0.000 description 1
- 208000024891 symptom Diseases 0.000 description 1
- RCINICONZNJXQF-MZXODVADSA-N taxol Chemical compound O([C@@H]1[C@@]2(C[C@@H](C(C)=C(C2(C)C)[C@H](C([C@]2(C)[C@@H](O)C[C@H]3OC[C@]3([C@H]21)OC(C)=O)=O)OC(=O)C)OC(=O)[C@H](O)[C@@H](NC(=O)C=1C=CC=CC=1)C=1C=CC=CC=1)O)C(=O)C1=CC=CC=C1 RCINICONZNJXQF-MZXODVADSA-N 0.000 description 1
- 229960004964 temozolomide Drugs 0.000 description 1
- 108010013645 tetranectin Proteins 0.000 description 1
- 238000011287 therapeutic dose Methods 0.000 description 1
- 231100000816 toxic dose Toxicity 0.000 description 1
- 231100000440 toxicity profile Toxicity 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 238000011830 transgenic mouse model Methods 0.000 description 1
- 230000010474 transient expression Effects 0.000 description 1
- 239000013638 trimer Substances 0.000 description 1
- 230000007306 turnover Effects 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 241000701161 unidentified adenovirus Species 0.000 description 1
- 241001430294 unidentified retrovirus Species 0.000 description 1
- 230000002792 vascular Effects 0.000 description 1
- 238000010865 video microscopy Methods 0.000 description 1
- 239000000277 virosome Substances 0.000 description 1
- WAEXFXRVDQXREF-UHFFFAOYSA-N vorinostat Chemical compound ONC(=O)CCCCCCC(=O)NC1=CC=CC=C1 WAEXFXRVDQXREF-UHFFFAOYSA-N 0.000 description 1
- 229960000237 vorinostat Drugs 0.000 description 1
- 210000005253 yeast cell Anatomy 0.000 description 1
Images




















































































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2878—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the NGF-receptor/TNF-receptor superfamily, e.g. CD27, CD30, CD40, CD95
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K45/00—Medicinal preparations containing active ingredients not provided for in groups A61K31/00 - A61K41/00
- A61K45/06—Mixtures of active ingredients without chemical characterisation, e.g. antiphlogistics and cardiaca
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2809—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against the T-cell receptor (TcR)-CD3 complex
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2818—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against CD28 or CD152
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2803—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily
- C07K16/2833—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the immunoglobulin superfamily against MHC-molecules, e.g. HLA-molecules
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2887—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against CD20
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2893—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against CD52
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2896—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against molecules with a "CD"-designation, not provided for elsewhere
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/46—Hybrid immunoglobulins
- C07K16/468—Immunoglobulins having two or more different antigen binding sites, e.g. multifunctional antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
- A61K2039/507—Comprising a combination of two or more separate antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/31—Immunoglobulins specific features characterized by aspects of specificity or valency multispecific
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/71—Decreased effector function due to an Fc-modification
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/72—Increased effector function due to an Fc-modification
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
- C07K2317/732—Antibody-dependent cellular cytotoxicity [ADCC]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/73—Inducing cell death, e.g. apoptosis, necrosis or inhibition of cell proliferation
- C07K2317/734—Complement-dependent cytotoxicity [CDC]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/94—Stability, e.g. half-life, pH, temperature or enzyme-resistance
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Genetics & Genomics (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Pharmacology & Pharmacy (AREA)
- Epidemiology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Peptides Or Proteins (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Acyclic And Carbocyclic Compounds In Medicinal Compositions (AREA)
- Nitrogen Condensed Heterocyclic Rings (AREA)
- Pyridine Compounds (AREA)
- Saccharide Compounds (AREA)
- Nitrogen And Oxygen Or Sulfur-Condensed Heterocyclic Ring Systems (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Epoxy Compounds (AREA)
- Plural Heterocyclic Compounds (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
Abstract
本文提供了具有由Fc区中的氨基酸取代产生的经修饰的Fc效应器功能的第一抗体和第二抗体的组合,该氨基酸取代允许共依赖性激活效应器功能如CDC和/或ADCC。还提供了具有由Fc区中的氨基酸取代产生的激动活性或增强的激动活性的第一抗体和第二抗体的组合物,其中激动活性是第一抗体和第二抗体两者共依赖的。
Description
发明领域
本发明涉及具有由Fc区中氨基酸取代产生的经修饰的Fc效应器功能的抗体,即如增加的Fc效应器功能或降低的Fc效应器功能,以及此类抗体的组合以及包含此类抗体的组合物的用途。
发明背景
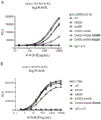
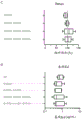
单克隆抗体的Fc介导的效应器功能,如补体依赖性细胞毒性(CDC)、抗体依赖性细胞介导的细胞毒性(ADCC)和抗体依赖性细胞介导的吞噬作用(ADCP)有助于由功效和毒性定义的治疗窗。CDC由C1q与抗体Fc区的结合来启动。C1q是一种多聚体蛋白质,其由6个附着到茎的球状结合头组成。单个球状结合头对IgG的亲和力低;C1q必须通过结合细胞表面上的多个IgG1分子来触发经典补体途径来获得亲合力。ADCC和ADCP通过将IgG Fc区与效应细胞上的Fcγ受体(FcγR)结合而启动。
已经显示靶标结合在细胞表面上后的IgG六聚化支持亲合的C1q结合。六聚化通过分子间非共价Fc-Fc相互作用介导,并且可以通过CH3域中的点突变,包括E345R和E430G来增强Fc-Fc相互作用。
WO2013/004842公开了包含具有一个或多个氨基酸修饰的变体Fc区的抗体或多肽,所述氨基酸修饰导致经修饰的效应器功能,如补体依赖性细胞毒性(CDC)。
WO2014/108198公开了包含具有一个或多个氨基酸修饰的变体Fc区的多肽如抗体,所述修饰导致增加的补体依赖性细胞毒性(CDC)。
WO2012/130831涉及具有由于多肽Fc区中一个或多个氨基酸取代而改变的效应器功能的包含Fc区的多肽。与包含野生型IgG Fc区的多肽相比,这些多肽对人FcγRIIIa和/或FcγRIIa和/或FcγRI的亲和力降低,并且显示由所述多肽诱导的ADCC降低至包含野生型IgG Fc区的多肽诱导的ADCC的至少20%。WO2012/130831未公开具有增强的Fc-Fc相互作用和/或增强的形成六聚体的能力的抗体。
如上所述,先前在增强抗体之间的Fc-Fc相互作用上的努力具有增强效应器功能的作用,例如增强的CDC和/或ADCC,这可能导致抗体结合的靶细胞的细胞死亡。
然而,如果靶抗原在体内健康细胞和引起疾病的细胞上都普遍表达,那么抗体可能会通过杀死健康细胞而变得有毒性。因此,需要使具有增强的效应器功能的抗体的效应器功能依赖于另一种抗体,该抗体的组合对引起疾病的细胞具有选择性,从而防止杀伤健康细胞。
发明目的
具有增强的效应器功能的单独起作用的抗体仅依赖于抗体的抗原结合区来实现对其靶细胞的特异性,这可能将合适的靶标限制为在患病细胞上高度选择性表达的那些靶标。具有效应器功能降低突变的单独起作用的抗体可以不伤害表达那些抗体的抗原靶标的健康细胞,但是其效能可以受到效应器功能降低突变的限制。因此,显然需要制备抗体的组合,其中每种单独的抗体都可以结合引起疾病的细胞和健康细胞,但是只有两种抗体都结合同一引起疾病的细胞时,增强的效应器功能才被激活或优先被激活。如果效应器功能的激活对同时被抗体组合结合的细胞具有选择性,则效应器功能激活与单独靶标结合的这种解偶联能够产生结合靶标的抗体,该靶标由于对健康细胞的不想要的毒性或由于对疾病细胞的功效不足而至今不能以最佳方式用于抗体治疗。
因此,本发明的目的是提供第一抗体和第二抗体,所述第一抗体和第二抗体经工程改造以对被两种抗体同时结合的靶细胞提供最大活性,其中与对被两种抗体同时结合的细胞的活性相比,第一抗体对仅被第一抗体结合的靶细胞不提供活性或提供最小活性,且其中第二抗体对仅被第二抗体结合的靶细胞提供最小或降低的活性。
本发明的另一目的是提供第一抗体和第二抗体,所述第一抗体和第二抗体都包含人IgG的Fc区和抗原结合区,两者均具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,并且第一抗体还具有用于降低效应器功能如CDC和/或ADCC的取代,其中与没有所述用于降低效应器功能的取代的第一亲本抗体相比,第一抗体具有降低的效应器功能如CDC和/或ADCC,其中单独与第一抗体和第二抗体相比,第一抗体和第二抗体的组合具有增强的效应器功能。
本发明的又一目的是提供第一抗体和第二抗体,所述第一抗体包含人IgG的Fc区和抗原结合区,与第一亲本抗体相比,第一抗体具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,以及用于降低效应器功能如CDC和/或ADCC的取代;所述第二抗体包含人IgG的Fc区和抗原结合区,与第二亲本抗体相比,所述第二抗体具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,以及用于增强效应器功能如CDC和/或ADCC的取代;其中第一抗体和第二抗体的活性通过互补的自身寡聚化抑制取代而变得共依赖。
本发明的另一目的是提供第一抗体和第二抗体,所述第一抗体包含人IgG的Fc区和抗原结合区,与第一亲本抗体相比,第一抗体具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,以及用于降低效应器功能如CDC和/或ADCC的取代;所述第二抗体包含人IgG的Fc区和抗原结合区,与第二亲本抗体相比,第二抗体具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,以及诱导激动活性的取代,例如在结合时增强靶受体的激活;其中第一抗体和第二抗体的活性通过互补的自身寡聚化抑制取代而变得共依赖。
本发明的又一目的是提供第一抗体和第二抗体,所述第一抗体包含人IgG的Fc区和抗原结合区,与第一亲本抗体相比,所述第一抗体具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,以及用于降低激动活性的取代,例如在结合时降低靶受体的激活;所述第二抗体包含人IgG的Fc区和抗原结合区,与第二亲本抗体相比,所述第二抗体具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,以及用于降低激动活性的取代,如在结合时降低靶受体的激活;其中第一抗体和第二种抗体的组合具有通过互补的自身寡聚化抑制取代而变得共依赖性增强的激动活性。
本发明的另一目的是提供第一抗体和第二抗体,所述第一抗体包含人IgG的Fc区和抗原结合区,与第一亲本抗体相比,所述第一抗体具有增强Fc-Fc相互作用的取代,抑制自身寡聚化的取代,以及降低效应器功能如CDC和/或ADCC的取代;所述第二抗体包含人IgG的Fc区和抗原结合区,与第二亲本抗体相比,所述第二抗体具有抑制自身寡聚化的取代,增强Fc-Fc相互作用的取代以及抗体的抗原结合区与相应的抗原结合时激活信号传导,任选诱导增强的信号传导的取代;其中第一抗体和第二抗体的活性通过互补的自身寡聚化抑制取代而变得共依赖。
发明概述
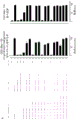
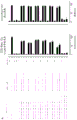
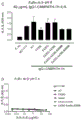
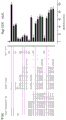
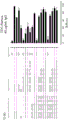
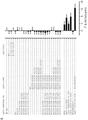
如本文所述,本发明涉及第一抗体和第二抗体的组合,所述第一抗体具有Fc区和抗原结合区,其中该Fc区具有一个Fc-Fc增强性取代和降低效应器功能如CDC和/或ADCC的一个或多个取代;所述第二抗体具有Fc区和抗原结合区,其中该Fc区具有一个Fc-Fc增强性取代,并且任选具有增强Fc效应器功能如CDC和/或ADCC的一个或多个取代。第一抗体和第二抗体还具有互补的自身寡聚化抑制取代,其使第一抗体和第二抗体异寡聚化变得共依赖。
不受理论的限制,认为本发明的第一抗体和第二抗体的组合能够降低组合的毒性并增加组合的治疗窗口,该组合具有互补的取代,该取代使第一抗体和第二抗体的效应器功能或信号传导功能,例如激动活性变得共依赖。此外,可将本发明的第一抗体和第二抗体的组合用于特异性消耗(deplete)表达被第一抗体和第二抗体识别的抗原的细胞群。因此,可将本发明的第一抗体和第二抗体的组合用于特异性消耗表达被第一抗体和第二抗体识别的第一抗原和第二抗原的肿瘤细胞群,而不消耗表达仅被第一抗体和第二抗体识别的第一抗原或第二抗原的健康细胞群或组织。
一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.G237位置处氨基酸的一个取代,或者选自G236R、G236K、K322A、E269K、K322E和P329R的一个或多个取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
f.如果第一Fc区包含K322E或P329R取代,则选自下组的一个或多个取代:K326A、K326W、E333A和E333S;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统(Edelmanet al.,Proc Natl Acad Sci U S A.1969 May;63(1):78-85;Kabat et al.,Sequencesof Proteins of Immunological Interest,Fifth Edition.1991 NIH Publication NO:.91-3242)的人IgG1。
一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.选自L234、L235、G237、G236的一个或多个氨基酸取代,或者选自K322A和E269K的一个或多个取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
在一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.G237位置处氨基酸的一个取代,或选自G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E的一个或多个取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.P329位置处氨基酸的一个取代,或K322E取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
f.选自下组的一个或多个取代:K326A、K326W、E333A和E333S;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
另一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.K248E和T437R取代;和
b.K439E或S440K取代;和
c.G237或P329位置处氨基酸的一个取代,或选自下组的一个或多个取代:G236R、G236K、K322A、K332E、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.K248E和T437R取代,和
e.K439E或S440K;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
根据本发明,对应于E430、E345或S440Y或S440W取代的位置处的取代被认为是Fc-Fc增强性取代。
根据本发明,K439E或S440K取代被认为是互补的自身寡聚化抑制取代。也就是说,具有如K439E的第一抗体与具有K439E取代的另一抗体不可以形成寡聚体,然而具有K439E取代的抗体与具有S440K取代的另一抗体可以形成寡聚体。具有S440K取代的抗体与具有S440K取代的另一抗体不可以形成寡聚体。
根据本发明,G237位置处氨基酸的取代,或选自G236R、G236K、K322A、E269K、K322E、P329R、L234A、L234F、L235A、L235Q和L235E的一个或多个取代被认为是C1q结合位点调节取代,并且降低效应器功能如CDC和/或ADCC。
根据本发明,选自K326A、K326W、E333A和E333S的一个或多个取代被认为是C1q结合位点调节取代,并且增强效应器功能如CDC和/或ADCC。
在一实施方案中,第二Fc区包含G237A取代。
根据本发明,G237A取代被认为是Fc-gamma受体调节取代,并且减少Fc-gamma受体结合。
也就是说,本发明的发明人在本发明的第一方面发现,通过组合第一抗体和第二抗体,其中第一抗体具有Fc-Fc增强性取代,自身寡聚化抑制取代以及降低效应器功能如CDC和/或ADCC的一个或多个取代,第二抗体具有Fc-Fc增强性取代,自身寡聚化抑制取代以及任选增强效应器功能如CDC和/或ADCC的一个或多个取代,并且第一抗体和第二抗体具有互补的自身寡聚化抑制取代,从而使第一抗体和第二抗体异寡聚化变得共依赖。
与具有Fc-Fc增强和自身寡聚化抑制取代但无C1q调节取代的抗体组合相比时,抗体组合可能可以改善对患病细胞的效应器功能活性相比对健康细胞的效应器功能活性之间的安全界限。
另一方面,本发明提供了抗体,其包含人IgG的Fc区和能够结合抗原的抗原结合区,其中所述Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,取代不为S440K,和
c.G237位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E。
另一方面,本发明提供了抗体,其包含人IgG的Fc区和能够结合抗原的抗原结合区,其中所述Fc区包含
a.K248E和T437R取代,和
b.K248E或S440K取代,和
c.G237或P329位置处氨基酸的一个取代,或者选自G236R、G236K、K322的一个或多个取代,
d.K332E、E269K、L234A、L234F、L235A、L235Q、L235E、K326A、K326W、E333A和E333S。
一方面,本发明提供了包含第一抗体和第二抗体的组合物,其中第一抗体包含第一抗原结合区和根据本文所述任何实施方案或方面的第一Fc区,并且第二抗体包含第二抗原结合区和根据本文所述任何实施方案或方面的第二Fc区。
一方面,本发明提供了包含第一抗体和第二抗体的组合物,其中所述第一抗体包含能够结合第一抗原的第一抗原结合区和人IgG的第一Fc区,第二抗体包含能够结合第二抗原的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.G237位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、E269K、K322E和P329R;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,和
f.如果第一Fc区包含K322E或P329R取代,则选自K326A、K326W、E333A和E333S中的一个或多个取代;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
一方面,本发明提供了包含第一抗体和第二抗体的组合物,其中第一抗体包含能够结合第一抗原的第一抗原结合区和人IgG的第一Fc区,第二抗体包含能够结合第二抗原的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.G237位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
一方面,本发明提供了包含第一抗体和第二抗体的组合物,其中第一抗体包含能够结合第一抗原的第一抗原结合区和人IgG的第一Fc区,第二抗体包含能够结合第二抗原的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.P329位置处氨基酸的一个取代,或者K322E取代;
以及所述第二Fc包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
f.选自K326A、K326W、E333A和E333S中的一个或多个取代;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
另一方面,本发明提供了包含第一抗体和第二抗体的组合物,其中第一抗体包含能够结合第一抗原的第一抗原结合区和人IgG的第一Fc区,并且第二抗体包含能够结合第二抗原的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
a.K248E和T437R取代,和
b.K439E或S440K取代,和
c.G237或P329位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、K332E、E269K、L234A、L234F、L235A、L235Q和L235E;
以及第二Fc区包含
d.K248E和T437R取代,和
e.K439E或S440K的一个取代;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
另一方面,本发明涉及消耗表达第一抗原和第二抗原的细胞群的方法,该方法包括使所述细胞群与第一抗体和第二抗体或根据任何第一抗体和第二抗体的组合物或本文定义的组合物接触。
另一方面,本发明涉及治疗患有疾病的个体的方法,该方法包括向所述个体施用有效量的如本文所述的权利要求的第一抗体和第二抗体或有效量的如本文所述的组合物。
另一方面,本发明涉及试剂盒,其包括第一容器和第二容器,所述第一容器包含如本文定义的第一抗体,所述第二容器包含如本文定义的第二抗体。
另一方面,本发明涉及用于治疗癌症、自身免疫性疾病、炎性疾病或传染病的本文所述的第一抗体和第二抗体或组合物。
在另一方面,本发明涉及治疗患有疾病的个体的方法,该方法包括向所述个体施用有效量的本文所述的第一抗体和第二抗体或组合物。
以下进一步详细描述本发明的这些和其他方面,特别是第一抗体和第二抗体的各种用途和治疗应用。
附图简述
图1显示通过引入P329R突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%合并的正常人血清(NHS)存在下,将Wien 133细胞与浓度抗体浓度系列一起温育。CDC效力表示为(A)由碘化丙啶(PI)阳性细胞百分比确定的裂解百分比,和(B)剂量反应-反应曲线下面积(AUC),相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图2显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为(A)由PI阳性细胞百分比确定的裂解百分比,和(B)AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图3显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20-11B8-E430G-S440K混合抗体变体对不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特(Burkitt)氏淋巴瘤细胞系Wien 133(A),Daudi(B),Raji(C)和Ramos(D),急性成淋巴细胞性淋巴瘤(ALL)细胞系REH(E),骨髓瘤细胞系U266B1(F)和B细胞淋巴瘤细胞系U-698-M(G),用30μg/mL抗体进行体外CDC测定。CDC活性表示为通过每种细胞系相对于非结合对照抗体IgG1-b12(0%),以及针对REH,U266B1和Wien 133为IgG1-CAMPATH-1H-E430G(100%),或针对Daudi,Raji,Ramos和U-698-M为IgG1-11B8-E430G(100%)标准化的PI阳性细胞百分比确定的裂解百分比。
图4显示具有C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)的抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图5显示具有CD1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图6显示具有C1q结合抑制突变G236R(A)、K322A(B)、E269K(C)、K322E(D)或P329R(E)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有C1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图7显示具有C1q结合抑制突变G236R或K322A(A,B)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(C,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人淋巴瘤细胞系Wien 133(A,C)和Raji(B,D)的结合。通过流式细胞术测试抗体结合。结合表示为荧光强度的几何平均值(MFI)。使用没有一抗或非结合抗gp120抗体IgG1-b12的样品作为结合的阴性对照。
图8显示具有C1q结合抑制突变G236R或K322A(A,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(B,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体的ADCC能力。(A,B)进行了ADCC报告物生物测定(ADCC reporterBioassay),其中Raji靶细胞与抗体稀释系列与稳定表达高亲和力FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的NFAT反应元件的Jurkat人T细胞一起温育。萤光素酶产生通过发光读数定量。(C,D)进行体外铕TDA(EuTDA)ADCC测定,其中将Wien 133靶细胞与抗体稀释系列和人PBMC(E:T 100:1)一起温育。细胞裂解通过测量上清液中EuTDA荧光螯合物的信号来确定。
图9显示具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-K439E+具有或不具有C1q结合增强突变E333S的Fc-Fc相互作用增强的抗CD20 IgG1-11B8-S440K变体混合抗体变体的CDC活性的选择性。所测试的Fc-Fc相互作用增强突变是E430G、E345K、E345R和E345R-E430G。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图10显示了G236位置处(G236R或G236K)或G237位置处(G237A、G237T、G237Q或G237R)具有C1q结合调节突变或双突变G237A-K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合调节突变E333S、E333A、K326A、K326W-E333S、G237A或G237A-E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图11显示具有或不具有C1q结合抑制突变G236R的抗CD52 CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 11B8-E430G-S440K混合抗体同种型变体(IgG1,IgG2,IgG3和铰链稳定的IgG4)的CDC活性的选择性。在20%NHS存在下,将Wien133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图12显示具有或不具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD37 IgG1-CD37-37.3-E430G-S440K混合抗体变体对(A)Daudi和(B)Wien 133细胞的CDC活性的选择性。在20%NHS存在下,将靶细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-CD37-37.3-E430G混合物(100%)进行标准化。
图13显示BxPC-3人胰腺癌细胞上(A)单一抗体变体或(B)具有或不具有C1q结合抑制突变G236R的抗DR5 IgG1-DR5-01-G56T-E430G-K439E+具有或不具有C1q结合增强突变E333S的IgG1-DR5-05-E430G-S440K的抗体变体的激动剂混合物的DR5介导的细胞毒性。进行三天存活力测定,并使用CellTiter-Glo试剂盒确定细胞存活力。
图14显示具有C1q结合调节突变G237A、G236R、A327K、K322E或P329R的抗CD52IgG1-CAMPATH-1H-E430G的抗体变体的CDC活性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12-S440K(0%;未显示)和IgG1-CAMPATH-1H-E430G(100%)进行标准化。
图15显示具有C1q结合抑制突变G236R或K322A(A,B,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(D,E,F)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人FcRn的结合。用5μg/mL包被的重组人FcRn细胞外域(FcRnhsECDHis-B2M-BIO)和抗体稀释液系列进行FcRn ELISA。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图16显示SCID小鼠中500μg静脉内施用的抗体的清除率。(A-C)通过ELISA测定血浆样品的总人IgG,并针对(A)IgG1-CAMPATH-1H变体,(B)IgG1-11B8变体和(C)IgG1-CAMPATH-1H变体+IgG1-11B8变体的组合物,绘制浓度对时间的曲线。每个数据点代表一式三份样品的平均值+/-标准差。(D)抗体施用后直至第21天的清除率,用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1,000/AUC确定。
图17显示如ELISA测定所检测,具有C1q结合抑制突变G236R或G237T的固定的IgG1-CAMPATH-1H-E430G-K439E变体和具有C1q结合增强突变K326A或E333S的IgG1-11B8-E430G-S440K变体与FcγRIIA同种异型131H(A),FcγRIIA同种异型131R(B),FcγRIIB(C),FcγRIIIA同种异型158V(D)和FcγRIIIA同种异型158F(E)的加His标签的二聚生物素化ECD的结合。相对于无抗体对照(背景)和与IgG1-11B8-E430G-S440K的结合(100%),对20μg/mL抗体样品呈现结合。使用链霉亲合素-polyHRP和ABTS进行检测。
图18显示具有或不具有C1q结合抑制突变(G236R或G237T)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的具有FcγR结合抑制突变G237A的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mL IgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图19显示通过血细胞的流式细胞术分析确定的全血中IgG1-CAMPATH-1H和IgG1-11B8的变体组合的选择性活性。Y轴:在效应细胞存在下进行过夜(o/n)温育后,总淋巴细胞群(CD66b阴性)的B细胞(CD19阳性/CD3阴性)分数或T细胞(CD19阴性/CD3阳性)分数。X轴:不同的治疗组。符号代表来自五个不同健康供体的细胞,每个供体在两次单独的温育中进行了测试。(A)IgG1-CAMPATH-1H-G236R-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(B)包含额外G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(C)包含额外G236R或G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合包含额外C1q结合增强E333S突变的IgG1-11B8-G237A-E430G-S440K的选择性活性。(D)与具有额外突变G236R、G237Q或G237R的IgG1-CAMPATH-1H-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的共依赖性抗体组合相比,不同B细胞靶向抗体的B细胞消耗的深度。Y轴:如上确定的B细胞分数的对数标度表示。
图20显示具有抗CD52组分中C1q结合抑制突变以及抗CD20组分中C1q结合增强突变的IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体的组合对具有CD20和CD52的不同表达水平的不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特氏淋巴瘤细胞系Daudi(A),Raji(B)和Ramos(C),ALL细胞系REH(D)和B细胞淋巴瘤细胞系U-698-M(E),用0.01-40μg/mL抗体进行体外CDC测定。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图21显示具有或不具有G236R C1q结合抑制突变的抗-CD37 IgG1-CD37-37.3-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将(A)Daudi细胞和(B)WIL2-S细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mLIgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CD37-37.3-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图22显示(A)具有Fc:Fc相互作用增强突变E430G或E345R,自身寡聚化抑制突变K439E组合任何FcγR结合抑制和C1q结合调节突变G236R、G237A或G237T的抗CD52 IgG1-CAMPATH-1H抗体变体与人淋巴瘤细胞系Wien 133的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-Campath-1H(100%)的Bmax值标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(B)具有突变E430G和K439E组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-Campath-1H的结合标准化显示。(C)具有E430G和S440K突变组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞结合的表观Kd值。
图23显示具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K组合任何C1q结合调节突变K326A或E333A(A)或E333S、G237A或G237A-E333S(B)的抗CD20 IgG1-11B8抗体变体与人淋巴瘤细胞系Raji的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-11B8的Bmax值(100%)标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(C,D)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(C)或E333S、G237A或G237A-E333S(D)的IgG1-11B8抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-11B8的结合标准化显示。(E,F)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(E)或E333S、G237A或G237A-E333S(F)的IgG1-11B8抗体变体与Raji细胞结合的表观Kd值。
图24显示抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的FcRn结合。(A,C)显示使用40μg/ml抗体浓度于(A)pH 6.0或(C)pH 7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E和C1q结合调节突变G237A或G237T的抗CD52抗体IgG1-CAMPATH-1H的变体与人FcRn的结合。(B,D)显示使用40μg/ml抗体浓度于(B)pH 6.0或(D)pH7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和C1q结合调节突变K326A、E333A、G237A或G237A-E333S的抗CD20抗体IgG1-11B8的变体与人FcRn的结合。FcRnELISA用2μg/mL包被的重组人FcRn胞外域(FcRnECDHis-B2M-BIO)和抗体稀释系列进行。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图25显示从用抗CD52 IgG1-CAMPATH-1H或抗-IgG1-11B8抗体变体或其混合物注射的小鼠收集的血液样品中测量的总人IgG(hIgG)浓度。(A)从用野生型IgG1-CAMPATH-1H,IgG1-CAMPATH-1H-E430G-K439E-G237Q或IgG1-CAMPATH-1H-E430G-K439E-G236R注射的小鼠收集的血液样品的总hIgG浓度。(B)从用野生型IgG1-11B8,IgG1-11B8-E430G-S440K-G237A或IgG1-11B8-E430G-S440K-E333S注射的小鼠收集的血液样品的总hIgG浓度。(C)从用野生型IgG1-CAMPATH-1H+IgG1-11B8混合物或带有(A)和(B)中突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物注射的小鼠收集的血液样品的总hIgG浓度。在所有图中,虚线表示SCID小鼠中野生型IgG1抗体随时间预测的IgG1浓度。(D)抗体施用后直至第21天的清除率用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1000/AUC确定。
图26显示减去在不含抗体的阴性对照样品中检测的平均C4d浓度后,用带有突变E430G、K439E或S440K和G236R、G237A、G237Q或G237R的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12的抗体变体温育的样品中检测的C4d浓度。阳性对照样品包括带有E345R、E430G和S440Y Fc-Fc相互作用增强突变(RGY)的抗体变体。
图27显示用带有E430G、K439E或S440K和G236R、G237T、K326A或E333S突变的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12抗体变体调理后,通过流式细胞术检测C1q与Wien 133细胞的结合,所述Wien 133细胞于冰上用正常人血清作为补体来源温育。将平均荧光强度值相对于无抗体的对照反应(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物的最高水平(100%)标准化,通过拟合对数激动剂反应模型进行估算。(A)对照反应的C1q结合。(B-D)(B)IgG1-CAMPATH-E430G-K439E,(C)IgG1-CAMPATH-E430G-K439E-G236R和(D)IgG1-CAMPATH-E430G-K439E-G237R混合非结合对照IgG1-b12或不同的IgG1-11B8变体的C1q结合。
图28显示带有自身寡聚化抑制突变K439E或S440K和C1q结合调节突变的IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G抗体变体的FcγR结合。(A-E)如ELISA测定所检测,固定化抗体变体与(A)高亲和力同种异型FcγRIIA 131H,(B)低亲和力同种异型FcγRIIA131R,(C)FcγRIIB,(D)高亲和力同种异型FcγRIIIA 158V,或(E)低亲和力同种异型FcγRIIIA158F的加His标签的二聚生物素化ECD的结合。(F)ELISA检测的固定化FcγRIa与抗体变体的结合。表示了20μg/mL抗体样品的结合,并且每个实验在减去不与一抗温育的孔中的信号后相对于野生型IgG1-CAMPATH-1H所观测的平均信号(100%)标准化。使用链霉亲合素-polyHRP和ABTS进行检测。
图29显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的抗CD52IgG1-CAMPATH-1H,抗CD20 IgG1-11B8和抗CD52 IgG1-h2E8混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。在三个独立实验中测量了CDC效力,并表示为(A)平均AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化,和(B)在40μg/ml的抗体浓度时,由碘化丙啶阳性确定的平均裂解百分比。
图30(A,B)显示单一试剂和组合的带有所示自身寡聚化抑制突变和C1q结合调节突变的抗CD52 IgG1-CAMPATH-1H-E430G,抗CD20 IgG1-11B8-E430G和非抗原结合IgG1-b12-E430G抗体变体的CDC效力。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图31显示与Raji淋巴瘤细胞和不同浓度的IgG1-CAMPATH-1H和IgG1-11B8抗体变体共培养时,通过发光水平(RLU)测定的稳定表达(A)FcγRIIa或(B)FcγRIIIa的Jurkat报告物细胞系的激活。每个实验的发光值相对于IgG1-b12(0%)和野生型IgG1-Campath-1H+野生型IgG1-11B8(100%)观测到的值标准化,然后求三个(FcγRIIa)或两个(FcγRIIIa)实验重复的平均值。
图32显示由非等摩尔比的带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物诱导的对Wien 133细胞的共依赖性CDC。(A)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。(B)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。
图33显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H抗体变体与抗HLA-DR IgG1-HLA-DR-huL243变体(A)或抗HLA-DRIgG1-HLA-DR-1D09C3变体(B)混合物的CDC活性的选择性。在20%NHS存在下,将Oci-Ly17细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和(A)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-huL243-E430G(100%)或(B)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-1D09C3-E430G混合物(100%)标准化。
图34显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。显示具有突变L234A、L234A-L235A、L234F、L234F-L235E、L235A、L235Q、G236R或G237Q的IgG1-CAMPATH-1H-E430G-K439E变体以及这些变体与非结合对照抗体IgG1-b12或IgG1-11B8-E430G-S440K混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图35显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。(A)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345K、E345Q、E345R或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(B)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(C)带有匹配的Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8的抗体变体作为单一试剂或其混合物的CDC效力。(D)带有Fc-Fc相互作用增强突变E430G或K248E-T437R,自身寡聚化抑制突变K439E或S440K,和C1q结合调节突变G236R、G237A或E333S的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图36显示由带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CD37-37-3和IgG1-11B8抗体变体混合物诱导的对Raji淋巴瘤细胞的共依赖性CDC。(A)由稀释的所示抗体变体或其混合物诱导的细胞裂解的曲线下相对面积(AUC),相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CD37-37-3-E430G+IgG1-11B8-E430G混合物为100%)标准化。(B)由所示抗体变体及其混合物诱导的最大裂解百分比。
图37显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。在20%NHS存在下,将患者CLL样品与抗体浓度系列一起温育。CDC效力表示为与抗体变体温育后活的B细胞的百分比。示出使用来自患者1(A),2(B)和3(C)的CLL样品的结果。
图38显示全血样品与带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM抗体变体混合物温育后,通过流式细胞术检测的B细胞,CD4+ T细胞和CD8+ T细胞的分数。在与所示IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(A)B细胞,(B)CD4+ T细胞和(C)CD8+ T细胞的百分比。在与所示IgG1-CAMPATH-1H,IgG1-CD5-INSERM和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(D)B细胞,(E)CD4+ T细胞和(F)CD8+ T细胞的百分比。分数计算为[100%x(样品的细胞计数/“无Ab样品”的细胞计数)x(“无Ab样品”的粒细胞计数/样品的粒细胞计数)]。
图39显示了带有增强Fc-Fc相互作用,抑制自身寡聚化和抑制(G237T)或增强(K326W-E333S)C1q结合的突变的抗DR4和抗DR5抗体变体在癌细胞中协同激活程序性细胞死亡。(A)与所示抗体变体温育72小时后,BxPC-3人胰腺癌细胞的存活力。(B)与所示抗体变体温育72小时后,COLO 205人结肠癌细胞的存活力。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
发明详述
在描述本发明的实施方案时,为了清楚起见将采用特定术语。然而,本发明不意图限于如此选择的特定术语,并且应当理解,每个特定术语包括以类似方式操作以实现类似目的所有技术等同方案。
定义
术语“亲本抗体”应当理解为下述抗体,其与根据本发明的多肽或抗体相同,但是其中亲本抗体不具有根据本发明的C1q结合调节取代。因此,亲本抗体可以具有增加Fc-Fc的取代和抑制自身寡聚化的取代。术语“C1q结合调节取代”应理解为可抑制C1q结合的取代,例如G237位氨基酸的一个取代,或选自G236R,G236K,K322A,E269K,K322E和P329R的一个或多个取代,或可以增强C1q结合的取代,例如选自K326A,K326W,E333A和E333S的一个或多个取代。
术语“包含免疫球蛋白的Fc区和结合区的多肽”在本发明的背景中指包含免疫球蛋白的Fc区和能够结合例如存在于细胞、细菌或病毒体上的任何分子如多肽的结合区的多肽。免疫球蛋白的Fc区定义为通常在用木瓜蛋白酶(其为本领域技术人员已知)消化抗体后产生的抗体片段,其包括免疫球蛋白的两个CH2-CH3区和连接区,例如铰链区。抗体重链的恒定域定义抗体同种型,例如IgG1、IgG2、IgG3、IgG4、IgA1、IgA2、IgM、IgD、或IgE。Fc区凭借称为Fc受体的细胞表面受体和补体系统的蛋白质介导抗体的效应器功能。结合区可以是能够结合细胞、细菌或病毒体的多肽序列,如蛋白质、蛋白质配体、受体、抗原结合区、或配体结合区。若结合区是例如受体,则“包含免疫球蛋白的Fc区和结合区的多肽”可以制备为免疫球蛋白的Fc区和所述结合区的融合蛋白。若结合区是抗原结合区,则“包含免疫球蛋白的Fc区和结合区的多肽”可以是抗体,如嵌合抗体、人源化抗体或人抗体或仅重链抗体或ScFv-Fc融合物。包含免疫球蛋白的Fc区和结合区的多肽通常可以包含连接区,例如铰链区和免疫球蛋白重链的两个CH2-CH3区,因此“包含免疫球蛋白的Fc区和结合区的多肽”可以是“至少包含免疫球蛋白的Fc区和结合区的多肽”。术语“免疫球蛋白的Fc区”在本发明的背景中意指根据抗体亚型的连接区,例如铰链区,并且存在免疫球蛋白的CH2和CH3区,例如,人IgG1、IgG2、IgG3、IgG4、IgD、IgA1、IgGA2、IgM或IgE。多肽不限于人起源,但是可以是任何起源的,例如小鼠或猕猴起源。术语“野生型Fc区”在本发明的背景中指具有天然存在的氨基酸序列的免疫球蛋白Fc区。
如本文所用,术语“铰链区”意指免疫球蛋白重链的铰链区。因此,例如,根据EU编号,人IgG1抗体的铰链区对应于氨基酸216-230。
如本文所用,术语“CH2区”或“CH2域”意指免疫球蛋白重链的CH2区。因此,例如,根据EU编号,人IgG1抗体的CH2区对应于氨基酸231-340。然而,CH2区也可以是如本文所述的任何其他亚型。
如本文所用,术语“CH3区”或“CH3域”意指免疫球蛋白重链的CH3区。因此,例如,根据EU编号,人IgG1抗体的CH3区对应于氨基酸341-447。然而,CH3区也可以是如本文所述的任何其他亚型。
术语“免疫球蛋白”是指由两对多肽链,即一对轻(L)低分子量链和一对重(H)链组成的一类结构相关的糖蛋白,所有四条链潜在通过二硫键互相连接。免疫球蛋白的结构是已经完善建立的。参见例如Fundamental Immunology第7章(Paul,W.编,第2章RavenPress,N.Y.(1989))。简而言之,每条重链通常包含重链可变区(本文中简写为VH)和重链恒定区。重链恒定区通常包含三个域:CH1、CH2和CH3。重链经由二硫键在所谓的“铰链区”中互相连接。每条轻链通常包含轻链可变区(本文中简写为VL)和轻链恒定区。轻链恒定区通常包含一个域CL。VH和VL区可以进一步再分成高可变区(或超变区,其在序列上和/或结构上限定的环的形式中高度可变),也称作互补决定区(CDR),散布有称作框架区(FR)的更保守的区域。每个VH和VL通常由三个CDR和四个FR构成,以如下的次序从氨基端向羧基端排列:FR1,CDR1,FR2,CDR2,FR3,CDR3,FR4(亦见Chothia和Lesk J.Mol.Biol.196,901 917(1987))。除非另外叙述或与背景相冲突,使用DomainGapAlign(Lefranc MP.,NucleicAcids Research 1999;27:209-212和Ehrenmann F.,Kaas Q.和Lefranc M.-P.NucleicAcids Res.,38,D301-307(2010);还可见因特网http地址www.imgt.org/)根据IMGT规则鉴定本文中的CDR序列。除非另外叙述或与背景相冲突,根据EU-numbering(Edelman et al.,Proc Natl Acad Sci U S A.1969 May;63(1):78-85;Kabat et al.,Sequences ofproteins of immunological interest.5th Edition-1991 NIH Publication NO:.91-3242)提及本发明中的Fc区/Fc域中的氨基酸位置。
术语“抗体(Ab)”在本发明的背景中指具有特异性结合抗原的能力的免疫球蛋白分子、免疫球蛋白分子的片段、或其任一者的衍生物。本发明的抗体包含免疫球蛋白的Fc域和抗原结合区。抗体通常含有两个CH2-CH3区和连接区,例如铰链区,例如至少Fc域。因此,本发明的抗体可以包含Fc区和抗原结合区。免疫球蛋白分子的重链和轻链的可变区含有与抗原相互作用的结合域。抗体的恒定区或“Fc”区可介导免疫球蛋白对宿主组织或因子的结合,包括免疫系统的各种细胞(如效应细胞)和补体系统的组分如C1q(补体激活的经典途径中的第一组分)。抗体还可以是多特异性抗体,如双特异性抗体或类似的分子。术语“双特异性抗体”指对至少两种不同的、通常不重叠的表位具有特异性的抗体。这类表位可以在相同或不同的靶标上。若表位在不同的靶标上,这类靶标可以在相同细胞或不同细胞或细胞类型中。如上文指示的,除非另外说明或与背景清楚地冲突,术语抗体在本文中包括抗体的片段,其至少包含Fc区的一部分且保留特异性结合抗原的能力。这类片段可以通过任何已知的技术提供,如酶促切割、肽合成和重组表达技术。已经显示抗体的抗原结合功能可通过全长抗体的片段实施。涵盖在术语“Ab”或“抗体”内的结合片段的实例包括但不限于单价抗体(由Genmab在WO2007059782中描述);重链抗体,其仅由两条重链组成并且在例如骆驼中天然存在(例如,Hamers-Casterman(1993)Nature 363:446);ThioMabs(Roche,WO2011069104),链交换工程化域(SEED或Seed-body),其是不对称的且双特异性抗体样的分子(Merck,WO2007110205);Triomab(Pharma/Fresenius Biotech,Lindhofer etal.1995 J Immunol 155:219;WO2002020039);FcΔAdp(Regeneron,WO2010151792),Azymetric Scaffold(Zymeworks/Merck,WO2012/058768),mAb-Fv(Xencor,WO2011/028952),Xmab(Xencor),双重可变域免疫球蛋白(Abbott,DVD-Ig,美国专利NO:.7,612,181);双重域双头抗体(Unilever;Sanofi Aventis,WO20100226923),双二抗体(Di-diabody)(ImClone/Eli Lilly),突出-入-空穴(Knobs-into-holes)抗体形式(Genentech,WO9850431);DuoBody(Genmab,WO 2011/131746);双特异性IgG1和IgG2(Pfizer/Rinat,WO11143545),DuetMab(MedImmune,US2014/0348839),静电转向(Electrostaticsteering)抗体形式(Amgen,EP1870459和WO 2009089004;Chugai,US201000155133;Oncomed,WO2010129304A2);双特异性IgG1和IgG2(Rinat neurosciences Corporation,WO11143545),CrossMAbs(Roche,WO2011117329),LUZ-Y(Genentech),Biclonic(Merus,WO2013157953),双重靶向域抗体(GSK/Domantis),识别两种靶标的二合一抗体或双重作用Fab(Genentech,NovImmune,Adimab),交联的Mab(Karmanos Cancer Center),共价融合的单抗(AIMM),CovX-body(CovX/Pfizer),FynomAb(Covagen/Janssen ilag),DutaMab(Dutalys/Roche),iMab(MedImmune),IgG样双特异性(ImClone/Eli Lilly,Shen,J.,etal.J Immunol Methods,2007.318(1-2):p.65-74),TIG-body,DIG-body和PIG-body(Pharmabcine),双重亲和力再靶向分子(Fc-DART或Ig-DART,Macrogenics,WO/2008/157379,WO/2010/080538),BEAT(Glenmark),Zybodies(Zyngenia),用共同轻链的方法(Crucell/Merus,US7262028)或用共同重链的方法(κλBodies,NovImmune,WO2012023053),以及包含与含有Fc区的抗体片段融合的多肽序列的融合蛋白,如scFv融合物,如ZymoGenetics/BMS的BsAb,Biogen Idec的HERCULES(US007951918),EmergentBioSolutions/Trubion和Zymogenetics/BMS的SCORPIONS,Ts2Ab(MedImmune/AZ(Dimasi,N.,et al.J Mol Biol,2009.393(3):p.672-92),Genetech/Roche的scFv融合物,Novartis的scFv融合物,Immunomedics的scFv融合物,Changzhou Adam Biotech Inc的scFv融合物(CN 102250246),Roche的TvAb(WO 2012025525,WO 2012025530),f-Star的mAb2(WO2008/003116)和双重scFv融合物。还应当理解,除非另有规定,术语抗体也包括多克隆抗体、单克隆抗体(如人单克隆抗体)、抗体混合物(重组多克隆)例如通过Symphogen和Merus(Oligoclonics)开发的技术生成的,如WO2015/158867中描述的多聚体Fc蛋白,如WO2014/031646中描述的融合蛋白和抗体样多肽,如嵌合抗体和人源化抗体。所生成的抗体可潜在地拥有任何同种型。
在本文中使用时,术语“全长抗体”指含有与在该同种型的野生型抗体中通常找到的那些重链和轻链恒定域和可变域对应的全部重链和轻链恒定域和可变域的抗体(例如亲本抗体)。
如本文中所用,术语“人抗体”意图包括具有源自人种系免疫球蛋白序列的可变和恒定区的抗体。本发明的人抗体可以包含不由人种系免疫球蛋白序列编码的氨基酸残基(例如通过体外随机或定点诱变或者通过体内体细胞突变引入的突变、插入或缺失)。然而,如本文中所用,术语“人抗体”不意图包括如下的抗体,其中源自另一哺乳动物物种(例如小鼠)的种系的CDR序列被嫁接到人框架序列上。
如本文所用,术语“嵌合抗体”指其中的两种链类型(即重链和轻链)由于抗体工程而是嵌合的抗体。嵌合链是含有与人起源恒定区连接的外来可变域(源自非人物种,或合成的或从包括人的任何物种工程化改造而来)的链。
如本文所用,术语“人源化抗体”指其中的两种链类型由于抗体工程而人源化的抗体。人源化链通常是其中的可变域的互补决定区(CDR)为外来的(源自除人以外的物种或合成的),而链的剩余部分是人起源的链。人源化评估基于所得的氨基酸序列,而不是基于方法本身,这允许使用除植入之外的方案。
如本文所用,术语“单克隆抗体”、“单克隆Ab”、单克隆抗体组合物”、“单抗”等指具有单个分子组成的抗体分子制备物。单克隆抗体组合物显示对特定表位的单一结合特异性和亲和性。因此,术语“人单克隆抗体”指显示单个结合特异性的抗体,其具有源自人种系免疫球蛋白序列的可变区和恒定区。人单抗可以通过杂交瘤产生,所述杂交瘤包括从转基因或转染色体非人动物(例如转基因小鼠)获得的B细胞,该B细胞具有包含人重链转基因全集和轻链转基因全集的基因组(经重排以产生功能性人抗体),并与永生化细胞融合。
如本文中所用,“同种型”指由重链恒定区基因编码的免疫球蛋白类(例如IgG1、IgG2、IgG3、IgG4、IgD、IgA1、IgGA2、IgE或IgM或其任何同种异型如IgG1m(za)和IgG1m(f))。此外,每个重链同种型可与kappa(κ)或lambda(λ)轻链组合。本文使用的术语“混合同种型”指通过将一种同种型的结构特征与来自另一同种型的类似区域组合,从而产生杂合同种型而产生的免疫球蛋白的Fc区。混合同种型可以包含Fc区,所述Fc区具有包含两种或更多种同种型的序列,所述同种型选自以下IgG1、IgG2、IgG3、IgG4、IgD、IgA1、IgGA2、IgE、或IgM,从而产生组合,诸如例如IgG1/IgG3、IgG1/IgG4、IgG2/IgG3、IgG2/IgG4或IgG1/IgA。
如本文所用,术语“抗原结合区”、“抗原结合区”、“结合区”或抗原结合域指能够结合抗原的抗体区域。此结合区通常由抗体的VH和VL域定义,它们可以进一步细分为高变性区(或高变区,其可以是序列和/或结构上定义的环形式上高变的),也称为互补决定区(CDR),散布着更保守的区域,称为框架区(FR)。抗原可以是任何分子,如多肽,例如存在于细胞、细菌或病毒体上。
如本文所用,术语“靶标/靶”指与抗体的抗原结合区结合的分子。靶标包括制备的抗体所针对的任何抗原。就抗体而言,术语“抗原”和“靶标”可以互换使用,并且就本发明的任何方面或实施方案而言构成相同的含义和目的。
术语“表位”指能够特异性结合抗体可变域的蛋白质决定簇。表位通常由分子,如氨基酸、糖侧链或其组合的表面分组组成,并且通常具有特定的三维结构特征以及特定的电荷特征。构象和非构象表位的区别在于,在变性溶剂的存在下,与前者的结合而非后者的结合丧失。表位可以包含直接参与结合的氨基酸残基(也称为表位的免疫显性组分)和其他不直接参与结合的氨基酸残基。
本发明的“抗体”或“抗体变体”或“亲本抗体的变体”是与“亲本抗体”相比包含一个或多个突变的抗体分子。不同的术语可以互换使用,并就本发明的任何方面或实施方案而言构成相同的含义和目的。示例性亲本抗体形式包括但不限于野生型抗体、全长抗体或含Fc的抗体片段、双特异性抗体、人抗体、人源化抗体、嵌合抗体或其任何组合。不同的术语可以互换使用,并且就本发明的任何方面或实施方案而言构成相同的含义和目的。氨基酸取代可以将天然氨基酸交换为另一种天然存在的氨基酸,或者非天然存在的氨基酸衍生物。氨基酸取代可以是保守的或非保守的。在本发明的背景中,保守取代可以通过以下三个表中的一个或多个中反映的氨基酸类别内的取代来定义:
用于保守取代的氨基酸残基类别
| 酸性残基 | Asp(D)和Glu(E) |
| 碱性残基 | Lys(K),Arg(R)和His(H) |
| 亲水性不带电荷的残基 | Ser(S),Thr(T),Asn(N)和Gln(Q) |
| 脂肪族不带电荷的残基 | Gly(G),Ala(A),Val(V),Leu(L)和Ile(I) |
| 非极性不带电荷的残基 | Cys(C),Met(M)和Pro(P) |
| 芳香族残基 | Phe(F),Tyr(Y)和Trp(W) |
备选的保守氨基酸残基取代类别
| 1 | A | S | T |
| 2 | D | E | |
| 3 | N | Q | |
| 4 | R | K | |
| 5 | I | L | M |
| 6 | F | Y | W |
氨基酸残基的备选物理和功能分类

在本发明的背景中,变体中的取代表示为:
初始氨基酸-位置-取代的氨基酸
使用三字母码或单字母码,包括代码Xaa和X来表示氨基酸残基。因此,标记“E345R”或“Glu345Arg”意为在对应于亲本抗体中的345位氨基酸的变体氨基酸位置中包含以精氨酸取代谷氨酸的取代的变体。
当位置本身不存在于抗体中,但变体包含氨基酸插入时,例如:
位置-取代的氨基酸;使用标记,例如“448E”。
此类符号就一系列同源多肽或抗体中的修饰而言特别相关。
类似地,当取代氨基酸残基的身份不重要时:
初始氨基酸-位置;或“E345”。
对于初始氨基酸和/或取代的氨基酸可以包含超过1个但非全部氨基酸的修饰,345位处谷氨酸取代为精氨酸、赖氨酸或色氨酸:
可以在本发明的背景中互换使用“Glu345Arg、Lys、Trp”或“E345R、K、W”或“E345R/K/W”或“E345至R、K或W”。
此外,术语“取代”涵盖取代为其他19种天然氨基酸的任一种,或取代为其他氨基酸,如非天然氨基酸。例如,345位氨基酸E的取代包括下列取代的每种:345A、345C、345D、345G、345H、345F、345I、345K、345L、345M、345N、345P、345Q、345R、345S、345T、345V、345W和345Y。这相当于名称345X,其中X指任意氨基酸。这些取代也可以被称为E345A、E345C等,或E345A、C等,或E345A/C/等。这类似适用于本发明中提及的每一个位置,以在本文中明确包括此类取代的任一个。
如本文所用,术语“效应细胞”指与免疫应答的识别和激活阶段相反参与免疫应答的效应器阶段的免疫细胞。示例性免疫细胞包括髓样或淋巴样起源的细胞,例如淋巴细胞(如B细胞和T细胞,包括溶细胞T细胞(CTL))、杀伤细胞、天然杀伤细胞、巨噬细胞、单核细胞、嗜曙红细胞、多形核细胞,例如嗜中性粒细胞、粒细胞、肥大细胞和嗜碱性粒细胞。一些效应细胞表达Fc受体(FcR)或补体受体并发挥特定的免疫功能。在一些实施方案中,效应细胞诸如例如天然杀伤细胞能够诱导ADCC。例如,表达FcR的单核细胞、巨噬细胞、嗜中性粒细胞、树突细胞和Kupffer细胞参与靶细胞的特异性杀伤并将抗原呈递给免疫系统的其他组分,或与呈递抗原的细胞结合。在一些实施方案中,ADCC可以通过抗体驱动的经典补体激活进一步增强,导致激活的C3片段沉积在靶细胞上。C3切割产物是在髓样细胞上表达的补体受体(CR)(例如CR3)的配体。效应细胞上CR对补体片段的识别可以促进增强的Fc受体介导的ADCC。在一些实施方案中,抗体驱动的经典补体激活导致靶细胞上的C3片段。这些C3切割产物可以促进直接补体依赖性细胞性细胞毒性(CDCC)。在一些实施方案中,效应细胞可以吞噬靶抗原、靶颗粒或靶细胞。效应细胞上特定FcR或补体受体的表达可以通过体液因子如细胞因子调节。例如,已经发现FcγRI的表达被干扰素γ(IFNγ)和/或G-CSF上调。此种增强的表达增加携带FcγRI的细胞对靶标的细胞毒性活性。效应细胞可以吞噬靶抗原或吞噬或裂解靶细胞。在一些实施方案中,抗体驱动的经典补体激活导致靶细胞上的C3片段。这些C3切割产物可以通过效应细胞促进直接吞噬作用或通过增强抗体介导的吞噬作用间接促进。
如本文所用,术语“Fc效应器功能”或“Fc介导的效应器功能”意指作为使多肽或抗体与其在细胞膜上靶标(如抗原)结合的结果的功能,其中Fc效应器功能可归因于多肽或抗体的Fc区。Fc效应器功能的实例包括(i)C1q结合,(ii)补体激活,(iii)补体依赖性细胞毒性(CDC),(iv)抗体依赖性细胞介导的细胞毒性(ADCC),(v)Fc-γ受体结合,(vi)抗体依赖性细胞吞噬作用(ADCP),(vii)补体依赖性细胞性细胞毒性(CDCC),(viii)补体增强的细胞毒性,(ix)由抗体介导的调理抗体对补体受体的结合,(x)调理作用,和(xi)(i)至(x)中任一项的组合。
如本文所用,术语“聚簇依赖性功能”意指在多肽或抗体与其抗原结合,任选在细胞上,细胞膜上,在病毒体上寡聚化后形成抗原复合物的结果的功能。,或在另一个粒子上。聚类依赖性效应器功能的实例包括(i)抗体寡聚体形成,(ii)抗体寡聚体稳定性,(iii)抗原寡聚体形成,(iv)抗原寡聚体稳定性,(v)细胞凋亡诱导,(vi)增殖调节,例如作为增殖减少,抑制或刺激,和(vii)(i)至(vi)中任一项的组合。
如本文所用,术语“激动性”理解为细胞膜上受体的刺激或激活,导致生物应答,如胞内信号传导。此类激动效力可以导致凋亡(程序性细胞死亡)的诱导或免疫细胞激活,或细胞内途径的激活。
如实施例16中所述,使用以下步骤,可以在针对表达胞内死亡域的靶标的抗体的存活力测定法中测定激动活性或增加的激动活性:
i)在聚苯乙烯96孔平底板中接种表达响应抗体的靶标(例如DR5)的细胞系,37℃过夜,
ii)添加一定范围(0.0003至20,000ng/mL)内的抗体(例如抗DR5抗体)的连续稀释,并且37℃温育3天,
iii)通过量化ATP的存在来测定细胞存活力,例如通过使用CellTiter-Glo发光细胞存活力测定法进行,
iv)使用以下公式计算活细胞:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
如实施例29、30、31和32中所述,使用以下步骤,可以在针对激活胞内信号传导途径的靶标的抗体的报告物测定法中测定激动活性或增加的激动活性:
i)接种用靶标和NFAT响应元件表达下游的萤光素酶报告物基因稳定转染的Jurkat细胞,将细胞在96孔平底板中于37℃温育,
ii)添加范围为例如19.5至5,000ng/mL的抗体连续稀释物,并温育5小时,
iii)向细胞添加萤火虫萤光素酶底物(5’-氟萤光素)并温育5-10分钟,
iv)使用Envision MultiLable读板仪测定发光。
如本文中所用,术语“载体”意指能够诱导连接到载体中的核酸区段的转录的核酸分子。一种类型的载体是“质粒”,其是一种环形双链DNA环的形式。另一种类型的载体是病毒载体,其中核酸区段可以连接到病毒基因组中。某些载体能够它们被导入的宿主细胞内自主复制(例如具有细菌复制起点的细菌载体和附加体型哺乳动物载体)。其它载体(如非附加体型哺乳动物载体)可以在导入宿主细胞内后整合到宿主细胞的基因组中,并且借此与宿主基因组一起复制。此外,某些载体能够指导与之可操作连接的基因的表达。这类载体在本文中称作“重组表达载体”(或简称为“表达载体”)。一般地,在重组DNA技术中有用的表达载体经常是质粒的形式。在本说明书中,“质粒”和“载体”可以互换使用,因为质粒是最常用的载体形式。然而,本发明意图包含表达载体的这类其它形式,例如病毒载体(例如复制缺陷型逆转录病毒、腺病毒和腺相关病毒),其发挥同等的功能。
如本文中所用,术语“重组宿主细胞”(或简称作“宿主细胞”)意指其中引入了表达载体的细胞。应当理解,这类术语意图不仅指特定的受试者细胞,还指这类细胞的后代。因为在连续传代过程中由于突变或环境影响可能发生某些修饰,所以这类后代可能实际上与亲本细胞不相同,但是仍然包含如本文所用的术语“宿主细胞”的范围内。重组宿主细胞包括例如转染瘤,例如CHO细胞、HEK-293细胞、PER.C6、NS0细胞和淋巴细胞,以及原核细胞如大肠杆菌(E.coli)和其他真核宿主如植物细胞和真菌。
如本文中所用,术语“转染瘤”包括表达抗体或靶抗原的重组真核宿主细胞,例如CHO细胞、PER.C6、NS0细胞、HEK-293细胞、植物细胞或真菌,包括酵母细胞。
术语“制剂”指抗体变体的制备物和不同抗体变体混合物,其在与同细胞(例如,在细胞表面上表达的抗原)、细胞膜、病毒体或其他结构相关的抗原相互作用时具有增加的形成寡聚体的能力,这可导致增强的由抗原进行的信号传导和/或激活。
如本文中所用,术语“亲和力”是一个分子(例如抗体)与另一个(例如靶标或抗原)在单个位点处的结合强度,如抗体的个别抗原结合位点与抗原的单价结合。
如本文中所用,术语“亲合力”是两个结构之间的多个结合位点的组合强度,如同时与靶标相互作用的抗体的多个抗原结合位点之间,或例如抗体与C1q之间。当存在超过一个结合相互作用时,仅当所有结合位点解离时两个结构才会解离,因此,解离速率会比个别结合位点的解离速率慢,并由此与个别结合位点结合的强度(亲和力)相比提供更大的有效总结合强度(亲合力)。
如本文所用,术语“寡聚体”指与至少原则上由无限数目的单体组成的聚合物相反,由超过一个但是有限数目的单体单元(例如抗体)组成的分子。示例性的寡聚体是二聚体、三聚体、四聚体、五聚体和六聚体。希腊前缀通常用于表示寡聚体中单体单元的数目,例如四聚体由四个单元组成,六聚体由六个单元组成。
如本文所用,术语“寡聚化”意指将单体转化为有限的聚合度的过程。在本文中,观测到包含根据本发明的靶结合区的抗体可以在靶标结合后例如在细胞表面通过Fc区的非共价缔合形成寡聚体,例如六聚体。在本申请的背景中,“自身寡聚化”或“自动寡聚化”旨在指具有相同蛋白质序列(不考虑翻译后修饰)的抗体分子之间的寡聚化过程。如本文所用,术语“异源寡聚化”意指具有不同蛋白质序列(不考虑翻译后修饰)的抗体分子之间的寡聚过程。参与异源寡聚化的不同抗体可以例如结合不同的抗原,例如不同的靶蛋白、糖蛋白、聚糖或糖脂。
术语“抑制自身寡聚化的取代”或“自身寡聚化抑制性取代”意指包含免疫球蛋白的Fc区和抗原结合区的抗体中抑制具有相同蛋白质序列(不考虑翻译后修饰)的抗体分子之间的寡聚化过程的取代。根据实施例2和15中所述的方法所测量,抑制自身寡聚化可以例如导致多肽的CDC活性的EC50升高或最大CDC裂解活性降低。抑制自身寡聚化的取代的实例为K439E和S440K。
如本文所用,术语“聚簇”意指通过非共价相互作用使抗体、多肽、抗原或其他蛋白质寡聚化。
如本文所用,术语“Fc-Fc增强”意指增加两个含Fc区的抗体或多肽的Fc区之间的结合强度或稳定所述Fc区之间的相互作用,使得多肽在靶标结合后形成寡聚体。
如本文所用,Fc-Fc增强性取代指根据EU编号的对应于人IgG1的以下位置处的取代:E430、E345或S440,条件是S440位的取代为S440Y或S440W。因此,如本文所用的Fc-Fc增强性取代指以下氨基酸取代E430G、E345K、E430S、E430F、E430T、E345Q、E345R、E345Y、S440W和S440Y。在优选实施方案中,Fc-Fc增强性取代为E430G、E345K或E345R。
当在本文背景中使用两种抗原时,术语“共定位”或其语法变体一方面旨在指两种抗原在同一细胞上共表达的情况。抗原在细胞上可能已经彼此相邻,或者抗原可以通过本发明的结合多肽,例如抗体的寡聚化而聚集在一起。此外,术语“共定位”还旨在指两种抗原在不同细胞上表达,但是此类细胞在位置上是彼此紧邻的情况。
如本文所用,术语“共依赖性”意指依赖于两种或更多种具有自身寡聚化抑制性取代的不同的含Fc域的多肽同时结合到同一靶标、细胞或病毒体的功能作用。在本发明的背景中,可以为共依赖性的功能作用包括成簇依赖性功能,Fc介导的效应器功能,以及效应分子例如FcγR或C1的结合,但不一定是含Fc域的多肽单独结合至它们的靶抗原。如本文所用,具有抑制自身寡聚化的取代的含Fc域的不同多肽可以各自单独结合不同的靶标、细胞或病毒体,但是共依赖性功能的结果取决于两种或更多种不同组分同时结合到同一靶标、细胞或病毒体。如本文中所使用的,借助于共依赖性的含Fc域的多肽混合物中不同组分之间的非共价Fc-Fc相互作用的恢复,通过具有自身寡聚化抑制性取代的两个或多个不同的含Fc域的多肽特异性地恢复共依赖性功能作用。
如本文所用,术语“安全界限(safety margin)”意指药物的有效性和安全性指标,并且定义为药物的最小治疗剂量(对患病组织的功效)和最小毒性剂量(对健康组织的功效)之间的范围。
如本文所用,术语“C1q结合”意指C1q与抗体之间的直接相互作用。直接C1q结合可以例如通过在人工表面上使用固定化抗体来评估。当与细胞或病毒体表面上的预先确定的抗原结合时,可以评估导致C1q与抗体寡聚体的高亲合力结合的多价相互作用。
在ELISA测定法中可使用以下步骤测定C1q与多肽或抗体的结合:i)用100μl PBS中的1μg/mL多肽或抗体将96-孔Microlon ELISA板于4℃包被过夜,ii)将该板与100μL/孔的C1q连续稀释系列于37℃一起温育1小时,以3倍稀释度,最终C1q浓度范围30-0.01μg/mL,iii)将该板与100μl/孔的兔抗人C1q于室温一起温育1小时,iv)将板与100μl/孔猪抗兔IgG-HRP于室温一起温育1小时,v)于室温将具有100μL/孔底物的板与1mg/mL 2,2'-连氮基-二(3-乙基苯并噻唑啉-6-磺酸)温育15分钟,vi)通过添加100μL 2%草酸/孔终止反应。用BioTek EL808酶标仪在405nm处测量吸光度。
如本文中所用,术语C1q结合取代意指包含免疫球蛋白Fc区和抗原结合区的多肽中增强与C1q直接相互作用的取代。增强C1q结合可以例如导致C1q与包含免疫球蛋白的Fc区和抗原结合区的多肽之间的相互作用的EC50降低,其根据上述测定C1q结合的方法测量。
如本文中所用,术语“补体激活”是指经典补体途径的激活,其由称为C1的大型大分子复合物与表面上的抗体-抗原复合物结合而起始。C1是复合物,其由6个识别蛋白C1q和丝氨酸蛋白酶的异四聚体C1r2C1s2组成。C1是涉及一系列切割反应的经典补体级联早期事件中的第一个蛋白质复合物,所述一系列切割反应开始于C4切割成C4a和C4b以及C2切割成C2a和C2b。C4b被沉积并且与C2a一起形成称为C3转化酶的酶促活性转化酶,该转化酶将补体成分C3切割成C3b和C3a,其形成C5转化酶。此种C5转化酶将C5两分为C5a和C5b,并且最后一种成分被沉积在膜上,并且继而触发补体激活的晚期事件,其中末端补体成分C5b、C6、C7、C8和C9组装成膜攻击复合物(MAC)。补体级联导致在细胞膜中创建孔,这引起细胞裂解,也称为补体依赖性细胞毒性(CDC)。可以通过使用C1q功效、CDC动力学CDC测定法(如WO2013/004842,WO2014/108198中所述),或通过Beurskens等(Journal of Immunology,2012 vol.188NO:.7,April 1,3532-3541)描述的C3b和C4b的细胞沉积方法来评估补体激活。
如本文中所用,术语“补体依赖性细胞毒性”(“CDC”)意指抗体介导的补体激活过程,当抗体与其在细胞或病毒体上的靶标结合时由于由MAC装配体创建的膜中的孔而导致细胞或病毒体的裂解。
如本文中所用,术语“抗体依赖性细胞介导的细胞毒性”(“ADCC”)意指通过表达识别结合抗体的恒定区的Fc受体的细胞杀死抗体包被的靶细胞或病毒体的机制。如本文中所用,术语“抗体依赖性细胞吞噬”(“ADCP”)意指通过吞噬细胞内化消除抗体包被的靶细胞或病毒体的机制。内化的抗体包被的靶细胞或病毒体包含在称为吞噬体的囊泡中,然后该囊泡与一个或多个溶酶体融合以形成吞噬溶酶体。可以通过使用用巨噬细胞作为效应细胞的体外细胞毒性测定法和视频显微术来评估ADCP,如van Bij等(Journal of HepatologyVolume 53,Issue 4,October 2010,Pages 677–685)描述。
如本文中所用,术语“补体依赖性细胞性细胞毒性”(“CDCC”)意指由于抗体介导的补体激活由表达识别与靶细胞或病毒体共价结合的补体3(C3)切割产物的补体受体的细胞杀死靶细胞或病毒体的机制。可以以与针对ADCC描述的类似方式评估CDCC。
如本文中所用,术语“血浆半衰期”指在消除期间(分布阶段后)将血浆中多肽浓度降低至其初始浓度的一半所花费的时间。对于抗体,分布阶段通常为1-3天,在此期间,由于血浆和组织之间的重新分布,血浆浓度降低约50%。血浆半衰期可通过本领域公知的方法测量。
如本文中所用,术语“血浆清除率”是在对活生物体施用时多肽从血液中除去的速率的定量测量。血浆清除率可以计算为剂量/AUC(mL/天/kg),其中AUC值(曲线下面积)从浓度-时间曲线测定。
如本文中所用,术语“抗体-药物缀合物”指对至少一种类型的恶性细胞具有特异性的抗体或含Fc的多肽、药物和将药物与例如抗体偶联的接头。接头在存在恶性细胞的情况下是可切割的或不可切割的;其中抗体-药物缀合物杀死恶性细胞。
如本文中所用,术语“抗体-药物缀合物摄取”指抗体-药物缀合物与细胞上的靶标结合,然后被细胞膜摄取/吞噬,从而被吸入细胞中的过程。如WO 2011/157741中所述,抗体-药物缀合物摄取可以作为“在体外杀伤测定法中通过抗TF ADC进行的抗体介导的内化和细胞杀伤”评估。
如本文中所用,术语“凋亡”指可在细胞中发生的程序性细胞死亡(PCD)的过程。生化事件导致特征性细胞变化(形态学)和死亡。这些变化包括起泡、细胞皱缩、核碎裂、染色质浓缩和染色体DNA片段化。抗体对某种受体的结合可以诱导凋亡。
如本文中所用,术语“程序性细胞死亡”或“PCD”指由胞内程序介导的任何形式的细胞死亡。存在不同形式的PCD,各种类型的PCD的共同之处在于它们通过活性细胞过程来执行,所述活性细胞过程可以通过干扰胞内信号传导而被截获。在一个具体实施方案中,可以通过用缀合的膜联蛋白V染色细胞或组织,与磷脂酰丝氨酸暴露相关来测定细胞或组织中任何形式的PCD的发生。
如本文所用,术语“膜联蛋白V”指膜联蛋白组中结合细胞表面上的磷脂酰丝氨酸(PS)的蛋白质。
如本文中所用,术语“FcRn”意指为Fc受体的新生儿Fc受体。它在啮齿类中以独特的受体第一次被发现,所述独特的受体能够将IgG从母乳中通过新生啮齿类肠道上皮细胞转运到新生儿血流中。进一步的研究揭示了人中的类似受体。然而,在人中,在胎盘中发现它帮助促进母体IgG向生长的胎儿的转运,并且其也已经显示在监测IgG周转中发挥作用。FcRn在6.0-6.5的酸性pH,但不在中性或更高的pH结合IgG。因此,FcRn可以从微酸性pH的肠腔(肠的内部)中结合IgG,并确保向pH中性到碱性(pH7.0-7.5)的基底侧有效单向转运。此受体在IgG成人救助中也通过其在内皮细胞内吞途径中的发生起作用。酸性内体中的FcRn受体结合通过胞饮作用内化的IgG,将其再循环到细胞表面,于血液的碱性pH将其释放,由此防止其经历溶酶体降解。此机制可以提供了与其他同种型相比更大的血液中IgG半衰期的解释。
如本文中所用,术语“蛋白A”意指最初在细菌金黄色葡萄球菌(Staphylococcusaureus)的细胞壁中发现的56kDa MSCRAMM表面蛋白。它由spa基因编码,并且其调节受DNA拓扑结构、细胞渗透压和称为ArlS-ArlR的双组分系统控制。由于其结合免疫球蛋白的能力,它也在生物化学研究中得到应用。它由折叠到三螺旋束中的5个同源Ig结合域组成。每个域能结合来自许多哺乳动物物种的蛋白,最显著的是IgG。它结合大多数免疫球蛋白的重链Fc区(与FcRn受体的保守结合位点重叠),并也与人VH3家族的Fab区域相互作用。通过血清中的这些相互作用,IgG分子通过其Fc区而不是单纯通过其Fab区域结合细菌,由此细菌破坏调理、补体激活和吞噬作用。
如本文中所用,术语“蛋白G”意指在组C和G链球菌细菌中表达的免疫球蛋白结合蛋白,其很类似蛋白A但有不同的特异性。它是65kDa(G148蛋白G)和58kDa(C40蛋白G)细胞表面蛋白,其已经在通过其对Fc区的结合纯化抗体中得到应用。
本发明的具体实施方案
如本文所述,第一方面,本发明涉及与第二抗体组合用作药物的第一抗体,其中第一抗体和第二抗体的活性是共依赖的,并且第一抗体具有Fc区,该Fc区具有一个Fc-Fc增强,自身寡聚化抑制的取代和减少效应器功能如CDC和/或ADCC的一个或多个取代,第二抗体具有Fc区,该Fc区具有一个Fc-Fc增强,自身寡聚化抑制的取代和任选增强Fc效应器功能如CDC和/或ADCC的一个或多个取代。因此,第一抗体和第二抗体的此类组合可以增加使用第一抗体和第二抗体的此类组合的安全界限,其中第一抗体的作用取决于第二抗体的存在,并且第二抗体的作用取决于第一抗体的存在。
如本发明发明人所显示,第一抗体具有Fc区,该Fc区具有一个Fc-Fc增强性取代和降低效应器功能如CDC和/或ADCC的一个或多个取代,以及互补的自身寡聚化抑制取代,该抗体用作单个抗体时显示没有或只有非常有限的CDC活性。类似地,根据本发明的第二抗体,其具有一个Fc-Fc增强性取代和增强Fc效应器功能如CDC和/或ADCC的一个或多个取代,以及互补的自身寡聚化抑制取代,该抗体用作单个抗体时显示只有有限的CDC活性。然而,令人惊讶地,根据本发明的第一抗体和第二抗体的组合能够恢复诱导的CDC水平,而单一试剂显示没有或有限的诱导CDC的能力。
一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.选自L234、L235、G237、G236的一个或多个取代,或选自K322A和E269K的一个或多个取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
另一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.选自G237位置处氨基酸的一个取代,或选自下组的一个或多个取代:G236R、G236K、K322A、E269K、K322E、P329R、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,和
f.如果第一Fc区包含K322E或P329R取代,则选自K326A、K326W、E333A和E333S的一个或多个取代;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统(Edelmanet al.,Proc Natl Acad Sci U S A.1969May;63(1):78-85;Kabat et al.,Sequences ofProteins of Immunological Interest,Fifth Edition.1991 NIH Publication NO:.91-3242)的人IgG1。
一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.选自G237位置处氨基酸的一个取代,或选自下组的一个或多个取代:G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.选自E430,E345和S440位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.P329位置处氨基酸的一个取代,或K322E取代;
以及所述第二Fc区包含
d.选自E430,E345和S440位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
f.选自K326A、K326W、E333A和E333S的一个或多个取代;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
根据本发明,对应于E430、E345或S440Y或S440W取代位置处的取代被认为是Fc-Fc增强性取代,这样的取代在多肽或抗体中引入了增强Fc-Fc相互作用和寡聚化的作用。当抗体的抗原结合区与相应的靶抗原结合时,发生增强的寡聚化。增强的寡聚化产生寡聚体,例如六聚体。诸如六聚体的寡聚结构的产生具有通过增加多肽的C1q结合亲和力来增强Fc效应器功能如CDC的作用。
在一个实施方案中,第一抗体包含对应于E430、E345或S440Y或S440W取代的位置处至多一个取代。在一个实施方案中,第二抗体包含对应于E430、E345或S440Y或S440W取代的位置处至多一个取代。因此,在一个实施方案中,Fc区包含对应于E430、E345或S440Y或S440W取代的位置处至多一个取代。本发明的一个实施方案中,第一Fc区和第二Fc区包含选自下组的取代:E430G、E345K、E430S、E430F、E430T、E345Q、E345R、E345Y、S440W和S440Y。因此,在一个实施方案中,第一Fc区可以具有E430G取代且第二Fc区可以具有E345K取代。在另一实施方案中,第一Fc区可以具有E345K取代且第二抗体可以具有E430G取代。可以独立地选择第一Fc区和第二Fc区中的取代形成Fc-Fc增强性取代的组。
本发明的一个实施方案中,第一Fc区和第二Fc区包含选自E430G、E345K和E345R的取代。
本发明的一个实施方案中,第一Fc区和第二Fc区包含E430G取代。本发明的一个实施方案中,第一Fc区和第二Fc区包含E345K取代。本发明的一个实施方案中,第一Fc区和第二Fc区包含E345R取代。本发明的一个实施方案中,第一Fc区和第二Fc区包含S440Y取代。
根据本发明,第一Fc区和第二Fc区还包含K439E或S440K取代,其被认为是互补的寡聚化抑制取代。也就是说,具有如K439E的第一抗体不可以与具有K439E取代的另一抗体形成寡聚体,但是具有K439E取代的抗体可以与具有S440K取代的另一抗体形成寡聚体。具有S440K取代的抗体不可以与具有S440K取代的另一抗体形成寡聚体,但是可以与具有K439E取代的抗体形成寡聚体。因此,本发明的一个实施方案中,第一Fc区包含K439E取代且第二Fc区包含S440K取代。本发明的一个实施方案中,第一Fc区包含S440K取代且第二Fc区包含K439E取代。当Fc区包含S440K寡聚化抑制取代时,则该Fc区可以不包含Fc-Fc增强性取代S440Y或S440W。因此,具有S440K寡聚化抑制取代的Fc区可以在对应于E430或E345的氨基酸位置处具有Fc-Fc增强性取代。
另一方面,本发明提供了与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.K248E和T437R取代;和
b.K439E或S440K取代;和
c.G237或P329位置处氨基酸的一个取代,或选自下组的一个或多个取代:G236R、G236K、K322A、K332E、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.K248E和T437R取代,和
e.K439E或S440K取代;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
根据本文的任何方面或实施方案,对应于E430、E345或S440Y或S440W取代的位置处的Fc-Fc增强性取代可以取代以下两个取代:T248E和T437R。
本发明的一个实施方案中,第一Fc区包含降低效应器功能如CDC和/或ADCC的取代。因此,本发明的一个实施方案中,第一Fc区包含降低CDC的取代。本发明的一个实施方案中,第一Fc区包含降低ADCC的取代。本发明的一个实施方案中,第一Fc区包含降低CDC和ADCC的取代。
本发明的一个实施方案中,第一Fc区包含G237位置处的一个氨基酸取代。G237位置处氨基酸的取代可以具有降低抗体诱导CDC的能力的作用。本文提供了实施方案,其中第一Fc区包含这样的取代,其除了降低CDC活性外,还可以降低Fc-gamma受体结合,从而降低Fc-gamma受体介导的效应器功能。
本发明的一个实施方案中,第一Fc区包含选自下组的一个取代:G237A、G237T、G237Q、G237R、G237S、G237N、G237D、G237E、G237K、G237V、G237M、G237I、G237L、G237H、G237F、G237Y、G237W和G237P。
发明人发现,通过用代表天然存在氨基酸的各种类别的氨基酸,如丙氨酸、苏氨酸、谷氨酰胺或精氨酸取代对应于人IgG1中237位置处的甘氨酸,降低了抗体诱导CDC的能力。因此,认为另一种天然氨基酸任意取代G237都会降低抗体诱导CDC的能力。G237位置处氨基酸的取代可以降低具有Fc-Fc增强性取代和自身寡聚化抑制取代的抗体自行诱导CDC的能力。
本发明的一个实施方案中,第一Fc区包含选自G237T、G237A、G237Q和G237R的一个取代。在一个实施方案中,第一Fc区包含G237T取代。本发明的一个实施方案中,第一Fc区包含G237A取代。本发明的一个实施方案中,第一Fc区包含G237S取代。本发明的一个实施方案中,Fc区包含G237Q取代。本发明的一个实施方案中,第一Fc区包含G237R取代。
本发明的一个实施方案中,第一Fc区包含选自G236R、G236K、E269K和P329R的一个或多个取代。本发明的一个实施方案中,第一Fc区包含L234F和L235E取代。在此提供了实施方案,其中第一Fc区包含这样的取代,其除了降低CDC活性外,还可以降低Fc-gamma受体的结合,从而降低Fc-gamma受体介导的效应器功能。
本发明的一个实施方案中,第一Fc区包含选自G236R、G236K和E269K的一个或多个取代。本发明的一个实施方案中,第一Fc区包含选自G236R和E269K的一个或多个取代。本发明的一个实施方案中,第一Fc区包含G263R或G236K取代。本发明的一个实施方案中,第一Fc区包含G236R取代。本发明的一个实施方案中,第一Fc区包含G236K取代。本发明的实施方案中,第一Fc区包含G269K取代。
本发明的一个实施方案中,第一Fc区包含选自K322A和K322E的一个取代。本文提供了实施方案,其中第一Fc区包含这样的取代,其可以降低抗体诱导CDC活性的能力,同时保留抗体结合Fc-gamma受体的能力。
本发明的一个实施方案中,第一Fc区包含K322A取代。
本发明的一个实施方案中,第一Fc区包含K322E取代。
本发明的一个实施方案中,第一Fc区包含P329位置处的氨基酸取代。
本发明的一个实施方案中,第一Fc区包含选自P329R、P329K、P329E、P329D和P329A的一个取代。本发明的一个实施方案中,第一Fc区包含P329R取代。本发明的一个实施方案中,第一Fc区包含P329R取代。本发明的一个实施方案中,第一Fc区包含P329K取代。本发明的一个实施方案中,第一Fc区包含P329E取代。本发明的一个实施方案中,第一Fc区包含一个P329D取代。本发明的一个实施方案中,该第一Fc区包含一个P329A取代。
本文提供了实施方案,其中第一抗体包含第一Fc区,该Fc区具有Fc-Fc增强性取代,其引入增强抗体的Fc-Fc相互作用和寡聚化的作用。当抗体的抗原结合区与相应的靶抗原结合时,发生增强的寡聚化。增强的寡聚化产生寡聚体如六聚体。诸如六聚体的寡聚结构的产生通过增加抗体的C1q结合亲合力而具有增强Fc效应器功能如CDC的作用。然而,通过引入自身寡聚化抑制取代和降低效应器功能如CDC和/或ADCC的取代,产生的抗体具有降低的效应器功能如CDC和/或ADCC,这可以改善与第二抗体组合的抗体的毒性特征(profile)的控制。第一抗体具有降低的效应器功能如CDC和/或ADCC,应当在将第一抗体与亲本抗体相比时理解,该亲本抗体具有相同的Fc-Fc增强性取代和自身寡聚化抑制取代,但不具有降低效应器功能的取代。
本发明的一个实施方案中,第二Fc区包含增强效应器功能如CDC和/或ADCC的取代。因此,在一个实施方案中,第二Fc区包含增强CDC的取代。在一个实施方案中,第二Fc区包含增强ADCC的取代。在一个实施方案中,第二Fc区包含增强CDC和ADCC的取代。第二抗体具有增强的效应器功能如CDC和/或ADCC,应当在将第二抗体与亲本抗体相比时理解,该亲本抗体具有相同的Fc-Fc增强性取代和自身寡聚化抑制取代,但不具有增强效应器功能的取代。
本发明的一个实施方案中,第二Fc区包含选自下组的一个或多个取代:G237A、K326A、K326W、E333A和E333S。
本发明的一个实施方案中,第二Fc区包含G237A取代。本发明的一个实施方案中,第二Fc区包含降低Fc-gamma受体结合的取代,例如G237A。
本发明的一个实施方案中,第二Fc区包含选自下组的一个或多个取代:K326A、K326W、E333A和E333S。
本发明的一个实施方案中,第二Fc区包含选自下组的一个取代:K326A、K326W、E333A和E333S。
本发明的一个实施方案中,第二Fc区包含选自下组的一个取代:K326A、K326W、E333A和E333S。本发明的一个实施方案中,第二Fc区包含K326A取代。本发明的一个实施方案中,第二Fc区包含E333A取代。本发明的第二Fc区包含E333S取代。
本发明的一个实施方案中,第二Fc区包含选自下组的两个取代:K326A、K326W、E333A和E333S。
本发明的一个实施方案中,第二Fc区包含K326W和E333S取代。本发明的一个实施方案中,第二Fc区包含K326A和E333A取代。
本发明的一个实施方案中,第二Fc区包含G237A和E333S取代。
本发明的一个实施方案中,第二Fc区包含K326A取代。
本发明的一个实施方案中,第二Fc区包含E333S取代。
本文提供了实施方案,其中第二抗体包含第二Fc区,该Fc区具有Fc-Fc增强性取代,其引入增强抗体的Fc-Fc相互作用和寡聚化的作用。当抗体的抗原结合区与相应的靶抗原结合时,发生增加的寡聚化。增加的寡聚化产生寡聚体,例如六聚体。诸如六聚体的寡聚结构的产生通过增加抗体的C1q结合亲合力而具有增强Fc效应器功能如CDC的作用。但是,通过引入增强效应器功能如CDC和/或ADCC的取代,生成的抗体具有增加的寡聚化和增强的效应器功能如CDC和/或ADCC,这可以改善与第一抗体组合的抗体的功效。第二抗体具有增强的效应器功能如CDC和/或ADCC,应当在将第二抗体与亲本抗体相比时理解,该亲本抗体具有相同的Fc-Fc增强性取代和自身寡聚化抑制取代,但不具有增强效应器功能的取代。
下表提供了实施方案的非限制性列表,描述了具有特定取代的第一多肽和第二多肽的组合。因此,例如,下表的实施方案1为包含第一抗体和第二抗体的组合,其中第一抗体包含分别对应于人IgG中E430G、K439E和G236R位置处的取代,第二抗体分别包含人IgG中E430G和S440K取代。如本文所述,所有实施方案1至177的第一抗体和第二抗体可任选包含其他取代。








下表提供了实施方案的优选列表,描述了具有特定取代的第一抗体和第二抗体的组合,因此,例如,下表的实施方案1为第一抗体和第二抗体的组合,其中第一抗体包含分别对应于人IgG中E430G,K439E和G236R位置处的取代,第二抗体分别包含人IgG中E430G和S440K取代。如本文所述,所有以下实施方案1至36的第一抗体和第二抗体可任选包含其他取代。



在一个实施方案中,第一Fc区包含E430G、K439E和G236R取代,并且第二Fc区包含E430G和S440K取代。在一个实施方案中,第一Fc区包含E430G、K439E和G237Q取代,并且第二Fc该区包含E430G和S440K取代。
在一个实施方案中,第一Fc区包含E430G和K439E取代,并且第二Fc区包含E430G、S440K和G236R取代。在一个实施方案中,第一Fc区包含E430G和K439E取代,并且第二Fc区包含E430G、S440K和G237Q取代。在一个实施方案中,第一Fc区包含E430G、K439E和G236R取代,并且第二Fc区包含E430G、S440K和G237A取代。在一个实施方案中,第一Fc区包含E430G、K439E和G236R取代,并且第二Fc区包含E430G、S440K和E333S取代。在一个实施方案中,第一Fc区包含E430G、K439E和G237A取代,并且第二Fc区包含E430G、S440K和G236R取代。在一个实施方案中,第一Fc区包含E430G、K439E和E333S取代,并且第二Fc区包含E430G、S440K和G236R取代。在一个实施方案中,第一Fc区包含E430G、K439E和G237Q取代,并且第二Fc区包含E430G、S440K和G237A取代。在一个实施方案中,第一Fc区包含E430G、K439E和G237Q取代,并且第二Fc区包含E430G、S440K和E333S取代。在一个实施方案中,第一Fc区包含E430G、K439E和G237A取代,并且第二Fc区包含E430G、S440K和G237Q取代。在一个实施方案中,第一Fc区包含E430G、K439E和E333S取代,并且第二Fc区包含E430G、S440K和G237Q取代。在一个实施方案中,第一Fc区包含E345K、K439E和G236R取代,并且第二Fc区包含E345K、S440K和E333S取代。在一个实施方案中,第一Fc区包含E345R、K439E和G236R取代,并且第二Fc区包含E345K、S440K和E333S取代。在一个实施方案中,第一Fc区包含E345R、K439E和G237Q取代,并且第二Fc区包含E345K和S440K取代。
本发明的一个实施方案中,第一抗体和/或第二抗体是人的,人源化的或嵌合的。本发明的一个实施方案中,第一抗体和第二抗体是人的,人源化的或嵌合的。本发明的一个实施方案中,第一抗体和第二抗体是人的。本发明的一个实施方案中,第一抗体和第二抗体是人源化的。本发明的一个实施方案中,第一抗体是人的,而第二抗体是人源化的。本发明的一个实施方案中,第一抗体是人源化的,而第二抗体是人抗体。
本发明的一个实施方案中,第一抗体和/或第二抗体是单克隆抗体。本发明的一个实施方案中,第一抗体和第二抗体是单克隆抗体。本发明的一个实施方案中,第一抗体和第二抗体是双特异性抗体。在一个实施方案中,第一抗体是单克隆抗体,而第二抗体是双特异性抗体。在一个实施方案中,第一抗体是双特异性抗体,而第二抗体是单克隆抗体。
应当理解,本文所述的实施方案提及抗体是指包含免疫球蛋白的Fc区和抗原结合区的抗体,该抗体也可以是多特异性抗体,例如双特异性抗体,其具有免疫球蛋白的第一Fc区和第一抗原结合区,以及具有免疫球蛋白的第二Fc区和第二抗原结合区的第二多肽或抗体。
本发明的一个实施方案中,第一抗体和/或第二抗体是IgG1,IgG2,IgG3或IgG4同种型。本发明的一个实施方案中,第一抗体和第二抗体是IgG1,IgG2,IgG3或IgG4同种型。本发明的一个实施方案中,第一抗体和第二抗体是人IgG1,IgG2,IgG3或IgG4同种型。本发明的一个实施方案中,第一抗体和第二抗体是IgG1,IgG2或IgG4同种型。本发明的一个实施方案中,第一抗体和第二抗体是人IgG1,IgG2或IgG4同种型。本发明的一个实施方案中,第一抗体和/或第二抗体是IgG1同种型。本发明的一个实施方案中,第一抗体和/或第二抗体是人IgG1同种型。本发明的一个实施方案中,第一抗体和第二抗体是IgG1同种型。在一个实施方案中,第一抗体和第二抗体是IgG2同种型。本发明的一个实施方案中,第一抗体和第二抗体是IgG4同种型。本发明的一个实施方案中,第一抗体是IgG1同种型,而第二抗体是IgG2同种型。本发明的一个实施方案中,第一抗体是IgG2同种型,而第二抗体是IgG1同种型。
本发明的一个实施方案中,第一抗体包含IgG1同种型的重链。本发明的一个实施方案中,第二抗体包含IgG1同种型的重链。本发明的一个实施方案中,第一抗体包含IgG2同种型的重链。本发明的一个实施方案中,第二抗体包含IgG2同种型的重链。本发明的一个实施方案中,第一抗体包含IgG3同种型的重链。本发明的一个实施方案中,第二抗体包含IgG3同种型的重链。本发明的一个实施方案中,第一抗体包含IgG4同种型的重链。本发明的一个实施方案中,第二抗体包含IgG4同种型的重链。
本发明的一个实施方案中,第一抗体包含IgG1同种型的重链,而第二抗体包含IgG1同种型的重链。
在优选的实施方案中,所述第一抗体包含人IgG1同种型的第一Fc区和/或所述第二抗体包含人IgG1同种型的第二Fc区。
本发明的一个实施方案中,抗体或第一抗体和/或第二抗体包含第一恒定区和/或第二恒定区,其包含选自表1的序列。
本发明的一个实施方案中,抗体或第一抗体和/或第二抗体包含第一恒定区和/或第二恒定区,其包含选自SEQ ID NO:63-122、135-138、140-145的序列。
本发明的一个实施方案中,第一抗体和/或第二抗体包含第一重链恒定区和/或第二重链恒定区,其包含选自SEQ ID NO:63-122、135-138、140-145的序列,其中第一重链序列和第二重链序列独立地选自该组。
本发明的一个实施方案中,抗体或第一抗体和/或第二抗体包含第一恒定区和/或第二恒定区,其包含选自SEQ ID NO:63-122、135-138和140-145的序列,其中引入至多5个另外的取代,例如至多4个另外的取代,例如至多3个另外的取代,例如至多2个另外的取代,例如至多1个另外的取代。
本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:84所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:87所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体包含恒定区,其包含SEQ ID NO:101所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:107所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:105所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:103所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:85所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:104所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:82所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:66所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:68所示的序列。本发明的一个实施方案中,抗体,例如第一抗体或第二抗体,包含恒定区,其包含SEQ ID NO:73所示的序列。
本发明的一个实施方案中,第一抗原和第二抗原都是细胞表面暴露的分子。在一个实施方案中,第一抗原和第二抗原在同一细胞上表达。在一个实施方案中,第一抗原和第二抗原在同一组织中表达。
本发明的一个实施方案中,第一抗原和第二抗原共定位于细胞或组织中,所述细胞或组织是用于待治疗的疾病或病症的靶细胞或靶组织。本发明的一个实施方案中,第一抗原和第二抗原是不同的。
本发明的一个实施方案中,第一抗体和第二抗体消耗表达第一抗原和第二抗原的细胞群。
本发明的一个实施方案中,细胞群是肿瘤细胞。
本发明的一个实施方案中,细胞群是血液肿瘤细胞或实体瘤细胞。
本发明的一个实施方案中,细胞群是白细胞,例如白细胞细胞群。
本发明的一个实施方案中,细胞群是淋巴细胞,例如淋巴细胞细胞群。
本发明的一个实施方案中,细胞群是B细胞,例如B细胞群。本发明的一个实施方案中,细胞群是B细胞群的亚群。
本发明的一个实施方案中,细胞群是T细胞,例如T细胞群。本发明的一个实施方案中,细胞群是T细胞群的亚群。本发明的一个实施方案中,细胞群是调节性T细胞。
本发明的一个实施方案中,细胞群是NK细胞。本发明的一个实施方案中,细胞群是髓样来源的抑制细胞。本发明的一个实施方案中,细胞群是肿瘤相关的巨噬细胞。
本文描述了实施方案,其中根据本发明的第一抗体和第二抗体用作药物以消耗表达被第一抗体和第二抗体识别的第一抗原和第二抗原的特定细胞群。因此,根据本发明的第一抗体和第二抗体可以用于消耗表达被第一抗体和第二抗体识别的第一抗原和第二抗原的肿瘤细胞,而第一抗体和第二抗体可以不消耗仅表达第一抗原或第二抗原的健康组织。根据本发明的第一抗体和第二抗体还可在消耗免疫系统的特定细胞群中特别有用,例如淋巴细胞的特定亚群,例如B细胞或T细胞,甚至B细胞的亚群或T细胞的亚群。
本发明的一个实施方案中,抗体是单特异性抗体,双特异性抗体或多特异性抗体。本发明的一个实施方案是天然抗体。
具有天然的如人Fc域的抗体还可以是具有除本发明突变以外的其他突变的抗体,例如影响糖基化或使抗体成为双特异性抗体的突变。术语“天然抗体”是指不包含任何遗传引入的突变的任何抗体。因此在本发明的意义上,包含天然存在的修饰的抗体,例如不同的同种异型被理解为“天然抗体”,从而可以被理解为亲本抗体。天然抗体可以用作根据本发明的第一抗体中的至少三个取代或第二抗体中的至少两个取代的模板,从而提供本发明的第一抗体和第二抗体。包含除本发明之外的其他取代的亲本抗体的实例是如WO2011/131746(Genmab)中所述的双特异性抗体,其利用还原条件促进包含IgG4-样CH3区的两种抗体的半分子交换,从而形成双特异性抗体而不会伴随形成聚集体。亲本抗体的其他实例包括但不限于双特异性抗体,例如异二聚双特异性:Triomabs(Fresenius);双特异性IgG1和IgG2(Rinat neurosciences公司);FcΔAdp(Regeneron);Knobs-into-holes(Genentech);静电转向(Amgen,Chugai,Oncomed);SEEDbody(Merck);酶学支架(Zymeworks);mAb-Fv(Xencor);以及LUZ-Y(Genentech)。其他示例性亲本抗体形式包括但不限于野生型抗体,全长抗体或含Fc的抗体片段,人抗体,人源化抗体,嵌合抗体或其任何组合。
在本发明的一方面,抗体包含人IgG的Fc区和能够结合抗原的抗原结合区,其中所述Fc区包含
选自E430,E345和S440位置处氨基酸的一个取代,条件是S440中的取代为S440Y或S440W,和
K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
G237位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E。
另一方面,抗体包含人IgG的Fc区和能够结合抗原的抗原结合区,其中所述Fc区包含
K248E和T437R取代,和
K439E或S440K取代,和
G237或P329位置处氨基酸的一个取代,或选自下组的一个或多个取代:G236R、G236K、K322A、K332E、E269K、L234A、L234F、L235A、L235Q、L235E、K326A、K326W、E333A和E333S。
本发明的一个实施方案中,Fc区包含选自下组的取代:E430G、E345K、E430S、E430F、E430T、E345Q、E345R、E345Y、S440W和S440Y。
本发明的一个实施方案中,Fc区包含选自E430G、E345K和E345R的取代。
本发明的一个实施方案中,Fc区包含E430G取代。
本发明的一个实施方案中,Fc区包含选自E430、E345和S440位置处至多一个取代,条件是S440中的取代为S440Y或S440W。
本发明的一个实施方案中,Fc区包含选自下组的一个取代:G237A、G237T、G237Q、G237R、G237S、G237N、G237D、G237E、G237K、G237V、G237M、G237I、G237L、G237H、G237F、G237Y、G237W、G237P。
本发明的一个实施方案中,Fc区包含选自下组的一个取代:G237A、G237T、G237S、G237Q、G237R。本发明的一个实施方案中,Fc区包含G237Q取代。
本发明的一个实施方案中,Fc区包含选自G236R和E269K的一个或多个取代。
本发明的一个实施方案中,Fc区包含G236R取代。
本发明的一个实施方案中,其中Fc区包含E269K取代。
本发明的一个实施方案中,Fc区包含K322A取代。
靶标和应用方法
根据本发明的第一和/或第二抗体可以结合在同一细胞上表达的靶标。在一个实施方案中,靶标是激活,抑制,调控和/或调节信号转导途径的靶标。
可以特别适合用作根据本发明的靶标的实例是细胞表面受体和配体。
以下蛋白质类别也可以特别适合用作根据本发明的第一抗体和/或第二抗体的抗原结合靶标:肿瘤坏死受体超家族,GPI锚定蛋白,脂质蛋白,水解酶(EC 3.)和调节剂超家族,B7家族相关蛋白,免疫球蛋白超家族,白介素受体家族,整联蛋白,Ig样细胞粘附分子家族,受体型蛋白酪氨酸磷酸酶,C型凝集素,四跨膜蛋白,跨膜域4(Membrane spanning 4-domains),激活白细胞免疫球蛋白样受体,C-C基序趋化因子受体,G蛋白偶联受体,Toll样受体,酪氨酸激酶。本发明的一个实施方案中,第一抗原结合区和第二抗原结合区能够结合相同蛋白质类别的靶抗原。本发明的一个实施方案中,第一抗原结合区和第二抗原结合区能够结合来自不同的蛋白质类别的靶抗原。
本发明的一个实施方案中,第一抗原结合区能够结合来自GPI锚定蛋白的蛋白质类别的靶抗原,而第二抗原结合区能够结合来自四跨膜蛋白的蛋白质类别的靶抗原。本发明的一个实施方案中,第一抗原结合区能够结合来自四联蛋白的蛋白质类别的靶抗原,而第二抗原结合区能够结合来自GPI锚定蛋白的蛋白质类别的靶抗原。
本发明的一个实施方案中,第一抗原结合区能够结合来自GPI锚定蛋白的蛋白质类别的靶抗原,而第二抗原结合区能够结合来自跨膜域4的蛋白质类别的靶抗原。本发明的一个实施方案中,第一抗原结合区能够结合来自跨膜域4的蛋白质类别的靶抗原,而第二抗原结合区能够结合来自GPI锚定蛋白的蛋白质类别的靶抗原。
本发明的一个实施方案中,第一抗原结合区能够结合来自肿瘤坏死受体超家族的蛋白质类别的靶抗原,而第二抗原结合区能够结合来自肿瘤坏死受体超家族的蛋白质类别的靶抗原。
本发明的一个实施方案中,第一抗原结合区能够结合来自肿瘤坏死受体超家族的蛋白质类别的靶抗原,而第二抗原结合区能够结合来自免疫球蛋白超家族的蛋白类别的靶抗原。
例如,细胞表面受体包括属于受体家族的受体,例如造血因子受体家族、细胞因子受体家族、酪氨酸激酶受体家族、丝氨酸/苏氨酸激酶受体家族、TNF受体家族、G蛋白偶联受体家族、GPI锚定受体家族、酪氨酸磷酸酶受体家族、粘附因子家族和激素受体家族。涉及属于这些受体家族的受体及其特征的各种参考文献是可获得的,包括例如Cooke B A.,KingR J B.,van der Molen H J.ed.New Comprehensive Biochemistry Vol.18B"Hormones和their Actions PartΠ"pp.1-46(1988)Elsevier Science Publishers BV.,New York,USA;Patthy L.(1990)Cell,61:13-14;Ullrich A.,et al.(1990)Cell,61:203-212;Massagul J.(1992)Cell,69:1067-1070;Miyajima A.,et al.(1992)Annu.Rev.Immunol.,10:295-331;Taga T.和Kishimoto T.(1992)FASEB J.,7:3387-3396;Fantl W I.,et al.(1993)Annu.Rev.Biochem.,62:453-481;Smith C A.,et al.(1994)Cell,76:959-962;Flower D R.(1999)Biochim.Biophys.Acta,1422:207-234;以及M.Miyasaka编,CellTechnology,增补卷,手册系列,"Handbook for Adhesion Factors"(1994)(Shujunsha,Tokyo,Japan)。
本发明的一个实施方案中,抗体包含抗原结合区,其中抗原结合区结合肿瘤坏死因子受体超家族(TNFR-SF)或G蛋白偶联受体(GPCR)超家族的成员。
本发明的一个实施方案中,所述第一抗体和/或第二抗体结合至细胞表面受体,例如激素受体和细胞因子受体。示例性细胞因子受体包括例如造血因子受体、淋巴因子受体、生长因子受体、分化控制因子受体等。细胞因子受体的实例是促红细胞生成素(EPO)受体、血小板生成素(TPO)受体、粒细胞集落刺激因子(G-CSF)受体、巨噬细胞集落刺激因子(M-CSF)受体、粒状巨噬细胞集落刺激因子(GM-CSF)受体、肿瘤坏死因子(TNF)受体、白细胞介素-1(IL-1)受体、白细胞介素-2(IL-2)受体、白细胞介素-3(IL-3)受体、白细胞介素-4(IL-4)受体、白细胞介素-5(IL-5)受体、白细胞介素-6(IL-6)受体、白细胞介素-7(IL-7)受体、白细胞介素-9(IL-9)受体、白细胞介素-10(IL-10)受体、白细胞介素-11(IL-11)受体、白细胞介素-12(IL-12)受体、白细胞介素-13(IL-13)受体、白细胞介素-15(IL-15)受体、干扰素-α(IFN-α)受体、干扰素-β(IFN-β)受体、干扰素-γ(IFN-γ)受体、生长激素(GH)受体、胰岛素受体、血液干细胞增殖因子(SCF)受体、血管表皮生长因子(VEGF)受体、表皮细胞生长因子(EGF)受体、神经生长因子(NGF)受体、成纤维细胞生长因子(FGF)受体、血小板衍生生长因子(PDGF)受体、转化生长因子-β(TGF-β)受体、白细胞迁移抑制因子(LIF)受体、睫状神经营养因子(CNTF)受体、制瘤蛋白M(OSM)受体和Notch家族受体。
肿瘤坏死因子受体超家族(TNFRSF)是一组受体,其特征在于能够通过细胞外富含半胱氨酸的域结合肿瘤坏死因子超家族(TNFSF)的配体。TNF受体在质膜中形成三聚体复合物。TNFRSF包括以下29种蛋白质;TNFR1(Uniprot P19438),FAS(Uniprot P25445),DR3(Uniprot Q93038),DR4(Uniprot O00220),DR5(Uniprot O14763),DR6(Uniprot O75509),NGFR(Uniprot P08138),EDAR(Uniprot Q9UNE0),DcR1 Uniprot O14798),DcR2(UniprotQ9UBN6),DcR3(Uniprot O95407),OPG(Uniprot O00300),TROY(Uniprot Q9NS68),XEDAR(Uniprot Q9HAV5),LTbR(Uniprot P36941),HVEM(Uniprot Q92956)),CD120b(UniprotP20333),OX40(Uniprot P43489),CD40(Uniprot P25942),CD27(Uniprot P26842),CD30(Uniprot P28908),4-1BB(Uniprot Q07011),RANK(Uniprot Q9Y6Q6),TACI(UniprotO14836)),BLySR(Uniprot Q96RJ3),BCMA(Uniprot Q02223),GITR(Uniprot Q9Y5U5),RELT(Uniprot Q969Z4)。
本发明的一个实施方案中,抗体、第一抗体和/或第二抗体包含能够结合选自下组的抗原的抗原结合区:DR4、DR5、CD20、CD37、CD52、HLA-DR、CD3和CD5。
在一个实施方案中,抗原结合区能够结合DR4。在一个实施方案中,抗原结合区能够结合DR5。在一实施方案中,抗原结合区能够结合CD20。在一实施方案中,抗原结合区能够结合CD37。在一实施方案中,抗原结合区能够结合CD52。在一实施方案中,抗原结合区能够结合HLA-DR。在一实施方案中,抗原结合区能够结合CD3。在一个实施方案中,抗原结合区能够结合CD5。
本发明的一个实施方案中,抗体、或第一抗体和/或第二抗体包含含有以下的抗原结合区:
VH区,其包含SEQ ID NO:196所示的CDR1序列,SEQ ID NO:196所示的CDR2序列,和SEQ ID NO:198所示的CDR3序列,以及VL区,其包含SEQ ID NO:200所示的CDR1序列,AAT所示的CDR2序列,和SEQ ID NO:201所示的CDR3序列[DR4];
VH区,其包含SEQ ID NO:50所示的CDR1序列,SEQ ID NO:51所示的CDR2序列,和SEQ ID NO:52所示的CDR3序列,以及VL区,其包含SEQ ID NO:54所示的CDR1序列,FAS所示的CDR2序列,和SEQ ID NO:55所示的CDR3序列[DR5-01-G56T];
VH区,其包含SEQ ID NO:57所示的CDR1序列,SEQ ID NO:58所示的CDR2序列,和SEQ ID NO:59所示的CDR3序列,以及VL区,其包含SEQ ID NO:61所示的CDR1序列,RTS所示的CDR2序列,和SEQ ID NO:62所示的CDR3序列[DR5-05];
VH区,其包含SEQ ID NO:36所示的CDR1序列,SEQ ID NO:37所示的CDR2序列,和SEQ ID NO:38所示的CDR3序列,以及VL区,其包含SEQ ID NO:40所示的CDR1序列,DAS所示的CDR2序列,和SEQ ID NO:41所示的CDR3序列[CD20,7D8];
VH区,其包含SEQ ID NO:9所示的CDR1序列,SEQ ID NO:10所示的CDR2序列,和SEQID NO:11所示的CDR3序列,以及VL区,其包含SEQ ID NO:13所示的CDR1序列,DAS所示的CDR2序列,和SEQ ID NO:14所示的CDR3序列[CD20,11B8];
VH区,其包含SEQ ID NO:43所示的CDR1序列,SEQ ID NO:44所示的CDR2序列,和SEQ ID NO:45所示的CDR3序列,以及VL区,其包含SEQ ID NO:47所示的CDR1序列,VAT所示的CDR2序列,和SEQ ID NO:48所示的CDR3序列[CD37];
VH区,其包含SEQ ID NO:2所示的CDR1序列,SEQ ID NO:3所示的CDR2序列,和SEQID NO:4所示的CDR3序列,以及VL区,其包含SEQ ID NO:6所示的CDR1序列,NTN所示的CDR2序列,和SEQ ID NO:7所示的CDR3序列[CD52,CAMPATH-1H];
VH区,其包含SEQ ID NO:161所示的CDR1序列,SEQ ID NO:162所示的CDR2序列,和SEQ ID NO:163所示的CDR3序列,以及VL区,其包含SEQ ID NO:165所示的CDR1序列,LVS所示的CDR2序列,和SEQ ID NO:166所示的CDR3序列[CD52,h2E8];
VH区,其包含SEQ ID NO:168所示的CDR1序列,SEQ ID NO:169所示的CDR2序列,和SEQ ID NO:170所示的CDR3序列,以及VL区,其包含SEQ ID NO:172所示的CDR1序列,SEQ IDNO:AAS所示的CDR2序列,和SEQ ID NO:173所示的CDR3序列[HLA-DR,hul243];
VH区,其包含SEQ ID NO:175所示的CDR1序列,SEQ ID NO:176所示的CDR2序列,和SEQ ID NO:177所示的CDR3序列,以及VL区,其包含SEQ ID NO:179所示的CDR1序列,DNN所示的CDR2序列,和SEQ ID NO:180所示的CDR3序列[HLA-DR,1D09C3];
VH区,其包含SEQ ID NO:182所示的CDR1序列,SEQ ID NO:183所示的CDR2序列,和SEQ ID NO:184所示的CDR3序列,以及VL区,其包含SEQ ID NO:186所示的CDR1序列,SEQ IDNO:DTS所示的CDR2序列,和SEQ ID NO:187所示的CDR3序列[CD3,huCLB T3/4];
VH区,其包含SEQ ID NO:189所示的CDR1序列,SEQ ID NO:190所示的CDR2序列,和SEQ ID NO:191所示的CDR3序列,以及VL区,其包含SEQ ID NO:193所示的CDR1序列,ATS所示的CDR2序列,和SEQ ID NO:194所示的CDR3序列[CD5]。
一方面,本发明涉及包含第一抗体和第二抗体的组合物,其中第一抗体包含根据本文公开的任何实施方案的第一抗原结合区和第一Fc区,第二抗体包含根据本文公开的任何方面或实施方案的第二抗原结合区和第二Fc区。
另一方面,本发明涉及包含第一抗体和第二抗体的组合物,其中第一抗体包含能够与第一抗原结合的第一抗原结合区和人IgG的第一Fc区,第二抗体包含能够与第二抗原结合的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
G237位置处氨基酸的一个取代,或选自下组的一个或多个取代:G236R、G236K、K322A、E269K、K322E、P329R、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,和
如果第一Fc区包含K322E或P329R取代,则选自下组的一个或多个取代:K326A,K326W,E333A和E333S;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
一方面,本发明提供了包含第一抗体和第二抗体的组合物,其中第一抗体包含能够与第一抗原结合的第一抗原结合区和人IgG的第一Fc区,第二抗体包含能够与第二抗原结合的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
G237位置处氨基酸的一个取代,或选自下组的一个或多个取代:G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,和
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
一方面,本发明提供了包含第一抗体和第二抗体的组合物,其中第一抗体包含能够与第一抗原结合的第一抗原结合区和人IgG的第一Fc区,第二抗体包含能够与第二抗原结合的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
P329位置处氨基酸的一个取代,或者K322E取代;
以及所述第二Fc区包含
选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,和
选自K326A、K326W、E333A和E333S的一个或多个取代;
其中第一Fc区具有K439E取代且第二Fc区具有S440K取代,或者第一Fc区具有S440K取代且第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
本发明的一个实施方案中,包含第一Fc区和第二Fc区的组合物包含选自下组的取代:E430G、E345K、E430S、E430F、E430T、E345Q、E345R、E345Y、S440W和S440Y。
本发明的一个实施方案中,包含第一Fc区和第二Fc区的组合物包含选自下组的取代:E430G、E345K和E345R。
本发明的一个实施方案中,包含第一Fc区和第二Fc区的组合物包含E430G取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自下组的一个取代:G237A、G237T、G237Q、G237R、G237S、G237N、G237D、G237E、G237K、G237V、G237M、G237I、G237L、G237H、G237F、G237Y、G237W和G237P。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自下组的一个取代:G237A、G237T、G237Q、G237R和G237S。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自下组的一个取代:G237A、G237T、G237Q和G237R。
根据权利要求43至50中任一项所述的组合物,其中所述第一Fc区包含G237Q取代。
根据权利要求43至50中任一项所述的组合物,其中所述第一Fc区包含G237T取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自下组的一个或多个取代:G236R和G236K。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自G236R、G236K和E269K的一个或多个取代。本发明的一个实施方案中,包含第一Fc区的组合物包含选自G236R和E269K的一个或多个取代。本发明的一个实施方案中,包含第一Fc区的组合物包含G263R或G236K取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自K322E和K322A的一个取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含P329R取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含G236R取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含P329位置处氨基酸的一个取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自下组的一个取代:P329R、P329A、P329T、P329Q、P329R、P329S、P329N、P329D、P329E、P329K、P329V、P329M、P329I、P329L、P329H、P329F、P329Y、P329W和P329P。
本发明的一个实施方案中,包含第一Fc区的组合物包含选自下组的一个取代:P329R、P329K、P329E、P329D和P329A。
本发明的一个实施方案中,包含第一Fc区的组合物包含P329R取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含K322A取代。
本发明的一个实施方案中,包含第一Fc区的组合物包含E269K取代。
本发明的一个实施方案中,包含第二Fc区的组合物包含选自下组的一个或多个取代:G237A、K326A、K326W、E333A和E333S。
本发明的一个实施方案中,包含第二Fc区的组合物包含选自下组的一个或多个取代:K326A、K326W、E333A和E333S。
本发明的一个实施方案中,包含第二Fc区的组合物包含选自由以下的一个取代:K326A、K326W、E333A和E333S。
本发明的一个实施方案中,包含第二Fc区的组合物包含G237A取代。
本发明的一个实施方案中,包含第二Fc区的组合物包含K326A取代。
本发明的一个实施方案中,包含第二Fc区的组合物包含E333S取代。
本发明的一个实施方案中,包含第二Fc区的组合物包含选自下组的两个取代:K326A、K326W、E333A和E333S。
本发明的一个实施方案中,包含第二Fc区的组合物包含K326W和E333S取代。
本发明的一个实施方案中,包含第二Fc区的组合物包含K326A和E333A取代。
本发明的一个实施方案中,包含第二Fc区的组合物包含G237A和E333S取代。
本发明的一个实施方案中,组合物包含的第一抗体和第二抗体以1:50至50:1摩尔比,例如1:1摩尔比、1:2摩尔比、1:3摩尔比、1:4摩尔比、1:5摩尔比、1:6摩尔比、1:7摩尔比、1:8摩尔比、1:9摩尔比、1:10摩尔比、1:15摩尔比、1:20摩尔比、1:25摩尔比、1:30摩尔比、1:35摩尔比、1:40摩尔比、1:45摩尔比、1:50摩尔比、50:1摩尔比、45:1摩尔比、40:1摩尔比、35:1摩尔比、30:1摩尔比、25:1摩尔比、20:1摩尔比、15:1摩尔比、10:1摩尔比、9:1摩尔比、8:1摩尔比、7:1摩尔比、6:1摩尔比、5:1摩尔比、4:1摩尔比、3:1摩尔比、2:1摩尔比存在于组合物中。
本发明的一个实施方案中,组合物包含的第一抗体和第二抗体以约1:50至50:1摩尔比,如约1:40至40:1摩尔比,如约1:30至30:1摩尔比,约1:20至20:1摩尔比,如约1:10至10:1摩尔比,如约1:9至9:1摩尔比,如约1:5至5:1摩尔比存在于组合物中。
本发明的一个实施方案中,组合物包含的第一抗体和第二抗体和/或任何其他多肽以等摩尔比存在于组合物中。
本发明的一个实施方案中,根据任何方面或实施方案的组合物为药物组合物。
一方面,本发明涉及消耗表达第一抗原和第二抗原的细胞群的方法,该方法包括使所述细胞群与根据本文所公开的任何方面或实施方案的第一抗体和第二抗体或组合物接触。
本发明的一个实施方案中,细胞群是肿瘤细胞群,例如血液肿瘤细胞群或实体瘤细胞群。
本发明的一个实施方案中,细胞群存在于血液中。
本发明的一个实施方案中,细胞群是白细胞,例如白细胞细胞群。
本发明的一个实施方案中,细胞群是白细胞群的亚群。
本发明的一个实施方案中,细胞群是淋巴细胞细胞群。
本发明的一个实施方案中,细胞群是B细胞群。本发明的一个实施方案中,细胞群是B细胞群的亚群。
本发明的一个实施方案中,细胞群是T细胞群。本发明的一个实施方案中,细胞群是T细胞群的亚群。本发明的一个实施方案中,细胞群是调节性T细胞,例如调节性T细胞群。
本发明的一个实施方案中,细胞群是NK细胞群。
本发明的一个实施方案中,细胞群是髓样来源的抑制细胞。
治疗应用
根据本发明的任何方面或实施方案的第一抗体和第二抗体、双特异性抗体或组合物可以用作药物,即用于治疗应用。
一方面,本发明提供了根据本文公开的任何方面或实施方案的第一抗体和第二抗体或组合物,其用作药物。
另一方面,本发明提供了根据本文公开的任何方面或实施方案的抗体或组合物,其用于治疗癌症、自身免疫性疾病、炎性疾病或传染病。
又一方面,本发明涉及治疗患有疾病的个体的方法,包括向个体施用有效量的根据本文公开的任何方面或实施方案的第一抗体和第二抗体或组合物。
再一方面,本发明涉及治疗患有疾病的个体的方法,该方法包括向个体施用有效量的根据本文所述的任何方面或实施方案的第一抗体和第二抗体,或有效量的根据本文所述的任何方面或实施方案的组合物。
本发明的实施方案中,疾病选自:癌症、自身免疫性疾病、炎性疾病和传染病。
本发明的实施方案中,所述方法包括施用另外的治疗剂。
本发明的实施方案中,根据本文所述的任何方面或实施方案的方法包括进一步施用另外的治疗剂。
本发明的实施方案中,另外的治疗剂是一种或多种选自下组的抗癌剂:化学治疗剂(包括但不限于紫杉醇、替莫唑胺、顺铂、卡铂、奥沙利铂、伊立替康、多柔比星、吉西他滨、5-氟尿嘧啶、培美曲塞)、激酶抑制剂(包括但不限于索拉非尼、舒尼替尼或依维莫司)、凋亡调节剂(包括但不限于重组人TRAIL或birinapant)、RAS抑制剂、蛋白酶体抑制剂(包括但不限于bortezomib)、组蛋白去乙酰化酶抑制剂(包括但不限于伏立诺他)、营养保健品、细胞因子(包括但不限于IFN-γ)、抗体或抗体模拟物(包括但不限于抗EGFR、抗IGF-1R、抗-VEGF、抗CD20、抗CD38、抗HER2、抗PD-1、抗PD-L1、抗-CTLA4、抗-CD40、抗-CD137、抗-GITR抗体和抗体模拟物)、抗体-药物缀合物。
试剂盒
应当理解,下文描述的实施方案提及第一抗体和第二抗体是指包含免疫球蛋白的Fc区和抗原结合区的抗体。
本发明还涉及用于在治疗中同时、分开或依次使用的试剂盒,其包含本文所述的第一抗体和第二抗体。此外,可以根据本文描述的任何方法获得此类第一抗体和第二抗体。
一方面,本发明涉及包含根据本文所述任何方面或实施方案的抗体或组合物的试剂盒,其中所述第一抗体和第二抗体或组合物在一个或多个容器如小瓶中。
本发明的一个实施方案中,试剂盒包含根据本文所述任何方面或实施方案的第一抗体和第二抗体或组合物,其用于在治疗中同时、分开或依次使用。
另一方面,本发明涉及根据本文所述任何实施方案的第一抗体和第二抗体、组合物或试剂盒用于诊断方法的用途。
另一方面,本发明涉及诊断方法,其包括将根据本文所述任何实施方案的第一抗体和第二抗体、组合物或试剂盒施用于人或其他哺乳动物的身体的至少一部分。
另一方面,本发明涉及根据本文所述任何实施方案的第一抗体和第二抗体、组合物或试剂盒在对人或其他哺乳动物身体的至少一部分进行成像中的用途。
另一方面,本发明涉及用于对人或其他哺乳动物身体的至少一部分进行成像的方法,其包括施用根据本文所述任何实施方案的第一抗体和第二抗体、组合物或试剂盒。
进一步应用
应当理解,下文描述的实施方案提及第一抗体和第二抗体是指各自包含免疫球蛋白的Fc区和抗原结合区的第一抗体和第二抗体。
另一方面,本发明涉及如上所述的本发明的第一抗体和第二抗体,其用作药物,特别是用作治疗疾病或病症的药物。此类疾病和病症的实例包括但不限于癌症、自身免疫性疾病、炎性疾病、传染病、细菌性、病毒性或真菌性感染。
另一方面,本发明涉及本文所述的第一抗体和第二抗体、双特异性抗体、组合物和试剂盒,其用于治疗疾病如癌症。
另一方面,本发明涉及治疗人疾病的方法,包括施用本文所述的第一抗体和第二抗体、组合物或试剂盒。
另一方面,本发明涉及治疗人癌症的方法,包括施用第一抗体和第二抗体、组合物或试剂盒。
“治疗”是指以缓解、改善、阻止或根除(治愈)症状或疾病状态为目的施用有效量的本发明的治疗活性化合物。
“有效量”或“治疗有效量”是指在必要的剂量和时间段内有效实现期望治疗结果的量。抗体的治疗有效量可以根据诸如疾病状态、个体的年龄、性别和体重以及抗体在个体中引发期望响应的能力等因素而变化。治疗有效量也是其中治疗有益效果超过抗体或抗体部分的任何毒性或有害效果的量。
剂量
应当理解,下文描述的实施方案提及多肽或抗体是指包含免疫球蛋白的Fc区和抗原结合区的抗体,抗体也可以是多特异性抗体如双特异性抗体,其具有免疫球蛋白的第一Fc区和第一抗原结合区,以及具有免疫球蛋白的第二Fc区和第二抗原结合区的第二抗体。
抗体的有效剂量和剂量方案取决于待治疗的疾病或病症,并且可由本领域技术人员确定。本发明抗体的治疗有效量的示例性非限制性范围为约0.1至100mg/kg,例如约0.1至50mg/kg,例如约0.1至20mg/kg,例如约0.1至10mg/kg,例如约0.5,约0.3,约1,约3,约5或约8mg/kg。
本发明的抗体也可以在联合疗法中施用,即与对于要治疗的疾病或病况相关的其他治疗剂组合施用。因此,在一个实施方案中,含有抗体的药物用于与一种或多种其他治疗剂组合,例如细胞毒剂、化学治疗剂或抗血管生成剂。这样的组合施用可以是同时的、分开的或依次的。
在另一实施方案中,本发明提供治疗或预防疾病如癌症的方法,该方法包括向有此需要的受试者与放射治疗和/或手术组合施用治疗有效量的本发明的变体或药物组合物。
制备方法
应当理解,下文描述的实施方案提及多肽或抗体是指包含免疫球蛋白的Fc区和抗原结合区的抗体,抗体也可以是多特异性多肽或抗体,其具有免疫球蛋白的第一Fc区和第一抗原结合区,以及具有免疫球蛋白的第二Fc区和第二抗原结合区的第二多肽或抗体。
本发明还提供了编码根据上述任一方面的抗体的分离的核酸和载体,以及编码所述变体的载体和表达系统。用于抗体及其变体的合适的核酸构建体、载体和表达系统是本领域已知的,并且在实施例中描述。在变体抗体不仅包含重链(或其含有Fc的片段)而且还包含轻链的实施方案中,编码重链和轻链部分的核苷酸序列可以存在于相同或不同的核酸或载体上。
本发明还提供了在宿主细胞中产生根据上述任一方面的抗体的方法,其中所述抗体至少包含重链的Fc区,所述方法包括以下步骤:
a)提供编码所述变体的所述Fc区的核苷酸构建体,
b)在宿主细胞中表达所述核苷酸构建体,和
c)从所述宿主细胞的细胞培养物中回收所述抗体变体。
在一些实施方案中,抗体是重链抗体。然而,在大多数实施方案中,抗体还含有轻链,因此所述宿主细胞在相同或不同的载体上进一步表达轻链编码构建体。
适用于抗体重组表达的宿主细胞是本领域公知的,包括CHO、HEK-293、Expi293、PER-C6、NS/0和Sp2/0细胞。在一个实施方案中,所述宿主细胞是能够进行蛋白质的Asn连接糖基化的细胞,例如真核细胞,如哺乳动物细胞,例如人细胞。在另一个实施方案中,所述宿主细胞是非人细胞,其经遗传工程改造以产生具有人样或人糖基化的糖蛋白。此类细胞的实例是遗传修饰的巴斯德毕赤酵母(Pichia pastoris)((Hamilton et al.,Science 301(2003)1244-1246;Potgieter et al.,J.Biotechnology 139(2009)318-325))和遗传修饰的浮萍(Lemna minor)(Cox et al.,Nature Biotechnology 12(2006)1591-1597)。
在一个实施方案中,所述宿主细胞是不能从抗体重链有效除去C-末端赖氨酸K447残基的宿主细胞。例如,Liu et al.(2008)J Pharm Sci 97:2426(通过引用并入本文)的表2列出了许多此类抗体生产系统,例如,Sp2/0,NS/0或转基因乳腺(山羊),其中仅获得C-末端赖氨酸部分除去。在一个实施方案中,宿主细胞是具有改变的糖基化机制的宿主细胞。此类细胞已在本领域中描述,并且可用作宿主细胞,在其中表达本发明的变体,从而产生具有改变的糖基化的抗体。参见例如Shields,R.L.et al.(2002)J.Biol.Chem.277:26733-26740;Umana et al.(1999)Nat.Biotech.17:176-1以及EP1176195;WO03/035835;和WO99/54342。用于产生工程化糖型的其他方法是本领域已知的,包括但不限于Davies et al.,2001,Biotechnol Bioeng 74:288-294;Shields et al,2002,J Biol Chem 277:26733-26740;Shinkawa et al.,2003,J Biol Chem 278:3466-3473),US6602684,WO00/61739A1;WO01/292246A1;WO02/311140A1;WO 02/30954A1;PotelligentTM技术(Biowa,Inc.Princeton,N.J.);GlycoMAbTM糖基化工程化技术(GLYCART biotechnology AG,Zurich,Switzerland);US 20030115614;Okazaki et al.,2004,JMB,336:1239-49中描述的那些。
本发明还涉及通过上述本发明的方法获得或可获得的抗体。
另一方面,本发明涉及能够产生本发明抗体的宿主细胞。在一个实施方案中,宿主细胞已经用本发明的核苷酸构建体转化或转染。
通过以下实例进一步说明本发明,这些实施例不应解释为进一步的限制。
表1序列表
























定义实施例5-23中测试的取代的表


定义自身寡聚化抑制性取代的表
| 包含第一Fc区的多肽 | 包含第二Fc区的多肽 |
| K439E | S440K |
| S440K | K439E |
定义FcGammaR结合抑制性和C1q结合抑制性取代的表


实施例
实施例1
抗体表达构建体
为了表达本文使用的人和人源化抗体,通过基因合成(GeneArt Gene Synthesis;ThermoFisher Scientific)制备了可变重链(VH)和可变轻链(VL),并克隆到pcDNA3.3表达载体(ThermoFisher Scientific)中,该表达载体包含人IgG重链(HC)恒定区(恒定区人IgG1m(f)HC:SEQ ID NO:22;恒定区人IgG2HC:SEQ ID NO:31;恒定区人IgG3 HC:SEQ IDNO:32;或恒定区人IgG4HC:SEQ ID NO:33)和或人kappa轻链(LC)恒定区:SEQ ID NO:34。通过基因合成引入所需的突变。本申请中的CD20抗体变体具有源自先前描述的CD20抗体IgG1-CD20-11B8的VH和VL序列(WO2004/035607;VH:SEQ ID NO:8;VL:SEQ ID NO:12)。本申请中的CD52抗体变体具有源自先前描述的CD52抗体CAMPATH-1H的VH和VL序列(Crowe等,1992Clin Exp Immunol.87(1):105-110;VH:SEQ ID NO:1;VL:SEQ ID No.5)和来自先前描述的CD52抗体h2E8的VH和VL序列(US2014/0127236;VH:SEQ ID NO:160;VL:SEQ ID NO:164)。本申请中的CD37抗体变体具有源自先前描述的CD37抗体IgG1-CD37-37.3的VH和VL序列(WO2011/112978;VH:SEQ ID NO:42;VL:SEQ ID NO:46)。本申请中的DR4抗体变体具有源自先前描述的DR4抗体DR4-chCTB007的VH和VL序列(US2009/0136503A1;VH:SEQ ID NO:200;VL:SEQ ID NO:204)。本申请中的DR5抗体变体具有源自先前描述的DR5抗体DR5-01-G56T(WO 2017/093447;VH:SEQ ID NO:49;VL:SEQ ID NO:53)和DR5-05(WO2014/009358;VH:SEQ ID NO:56;VL:SEQ ID NO:60)的VH和VL序列。本申请中的HLA-DR抗体变体具有源自先前描述的HLA-DR抗体HLA-DR-hul243(US8722047B2;VH:SEQ ID NO:168;VL:SEQ ID NO:172)和源自先前描述的HLA-DR抗体HLA-DR-1D09C3(US7521047B2;VH:SEQ ID NO:176;VL:SEQ ID NO:180)的VH和VL序列。本申请中的CD3抗体变体具有源自先前描述的CD3抗体huCLB-T3/4的VH和VL序列(Parren等,Res.Immunol 1991Nov-Dec;142(9):749-63;VH:SEQID NO:184;VL:SEQ ID NO:188)。本申请中的CD5抗体变体具有源自先前描述的CD5抗体CD5-INSERM的VH和VL序列(WO2010145895;VH:SEQ ID NO:192;VL:SEQ ID NO:196)。在一些实验中,人IgG1抗体b12(HIV gp120特异性抗体)被用作阴性对照(Barbas et al.,J MolBiol.1993 Apr 5;230(3):812-23;VH:SEQ ID NO:15;VL:SEQ ID NO:19)。
瞬时表达抗体构建体
基本如Vink等(Vink et al.,2014Methods 65(1):5-10)所述,使用293fectin(Life Technologies)在Expi293F细胞(Gibco,目录号A14635)中瞬时转染编码抗体的重链和轻链的质粒DNA混合物。上清液中的抗体浓度通过在280nm处的吸光度测量。含抗体的上清液可直接用于体外测定,或按以下所述纯化抗体。
抗体纯化和定量分析
通过蛋白A亲和层析纯化抗体。将培养物上清液用0.20μM死端滤器过滤,并加载到5mL MabSelect SuRe柱(GE Healthcare)上,用0.02M柠檬酸钠-NaOH(pH3)清洗并洗脱。在纯化后立即将洗脱液加载到HiPrep脱盐柱(GE Healthcare)上,并且将抗体缓冲液交换到12.6mM NaH2PO4,140mM NaCl,pH 7.4缓冲液(B.Braun或Thermo Fisher)中。缓冲液交换后,将样品用0.20μM死端滤器无菌过滤。通过许多生物分析测定分析纯化的蛋白质,包括用十二烷基硫酸钠-聚丙烯酰胺凝胶(CE-SDS)的毛细管电泳和高效大小排阻层析(HP-SEC)。通过280nm处的吸光度测量浓度。将纯化的抗体于2-8℃保存。
实施例2:通过在具有Fc-Fc相互作用增强突变E430G的抗CD52 IgG1-CAMPATH-1H-K439E+抗CD20 IgG1-11B8-S440K中引入P329R突变的混合抗体变体的CDC活性的选择性
使用抗CD20抗体IgG1-11B8和抗CD52抗体IgG1-CAMPATH-1H的变体混合物测试了P329R突变对体外CDC效力的影响。在20%NHS下对Wien 133细胞进行体外CDC测定,测试了不同浓度的纯化抗体(最终浓度范围为0.001-60.0μg/mL)。在抗体IgG1-11B8和IgG1-CAMPATH-1H中引入了不同的突变:诱导增强Fc-Fc相互作用的E430G;抑制C1q直接与抗体结合的P329R;以及K439E或S440K突变,其均通过抑制分子间Fc-Fc相互作用抑制同源六聚体抗体复合物的形成,并通过交叉互补的Fc-Fc相互作用促进异源六聚体抗体复合物的形成。作为对照,将单个抗体与非结合同种型对照抗体IgG1-b12或IgG1-b12-E430G同样以1:1混合,以能够直接比较各个组分及由其组成混合物的浓度。对于CDC测定,将具有0.2%牛血清白蛋白(BSA;Roche,目录号10735086001)的RPMI(Lonza,目录号BE12-115F)中的0.1x 106个Wien 133细胞(由英国牛津BioAnaLab Limited的Geoff Hale博士友情提供)与纯化抗体系列浓度以80μL总体积在聚苯乙烯圆底96孔板(Greiner bio-one目录号650101)中用摇动器于室温预温育15分钟。接着,添加20μL正常人血清(NHS;Sanquin,参考号M0008)作为补体来源,并于37℃培养箱温育45分钟(最终NHS浓度为20%;以3倍稀释度,最终抗体浓度为0.001-10.0μg/mL)。通过将板置于冰上来终止反应,然后通过离心沉淀细胞并置换上清液,用20μL的2μg/mL碘化丙啶溶液(PI;Sigma Aldrich,目录号P4170)置换。由IntellicytiQue筛选器(Westburg)通过流式细胞术确定PI阳性细胞数目,并以(PI阳性细胞数目/细胞总数目)x100%计算裂解百分比。使用GraphPad PRISM中经对数转换的浓度,使用非线性剂量反应拟合的最佳拟合值分析数据,并计算三个实验重复的剂量反应曲线下面积。相对曲线下面积(AUC)值表示相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物为100%)标准化。
抗CD52 IgG1-CAMPATH-1H-E430G+抗CD20 IgG1-11B8-E430G(均包含SEQ ID NO:26)的1:1混合物诱导Wiens 133细胞的高效细胞裂解(图1显示通过引入P329R突变,抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%合并的正常人血清(NHS)存在下,将Wien 133细胞与浓度抗体浓度系列一起温育。CDC效力表示为(A)由碘化丙啶(PI)阳性细胞百分比确定的裂解百分比,和(B)剂量反应-反应曲线下面积(AUC),相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图2显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为(A)由PI阳性细胞百分比确定的裂解百分比,和(B)AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图3显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20-11B8-E430G-S440K混合抗体变体对不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特(Burkitt)氏淋巴瘤细胞系Wien 133(A),Daudi(B),Raji(C)和Ramos(D),急性成淋巴细胞性淋巴瘤(ALL)细胞系REH(E),骨髓瘤细胞系U266B1(F)和B细胞淋巴瘤细胞系U-698-M(G),用30μg/mL抗体进行体外CDC测定。CDC活性表示为通过每种细胞系相对于非结合对照抗体IgG1-b12(0%),以及针对REH,U266B1和Wien 133为IgG1-CAMPATH-1H-E430G(100%),或针对Daudi,Raji,Ramos和U-698-M为IgG1-11B8-E430G(100%)标准化的PI阳性细胞百分比确定的裂解百分比。
图4显示具有C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)的抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图5显示具有CD1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图6显示具有C1q结合抑制突变G236R(A)、K322A(B)、E269K(C)、K322E(D)或P329R(E)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有C1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图7显示具有C1q结合抑制突变G236R或K322A(A,B)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(C,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人淋巴瘤细胞系Wien 133(A,C)和Raji(B,D)的结合。通过流式细胞术测试抗体结合。结合表示为荧光强度的几何平均值(MFI)。使用没有一抗或非结合抗gp120抗体IgG1-b12的样品作为结合的阴性对照。
图8显示具有C1q结合抑制突变G236R或K322A(A,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(B,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体的ADCC能力。(A,B)进行了ADCC报告物生物测定(ADCC reporterBioassay),其中Raji靶细胞与抗体稀释系列与稳定表达高亲和力FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的NFAT反应元件的Jurkat人T细胞一起温育。萤光素酶产生通过发光读数定量。(C,D)进行体外铕TDA(EuTDA)ADCC测定,其中将Wien 133靶细胞与抗体稀释系列和人PBMC(E:T 100:1)一起温育。细胞裂解通过测量上清液中EuTDA荧光螯合物的信号来确定。
图9显示具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-K439E+具有或不具有C1q结合增强突变E333S的Fc-Fc相互作用增强的抗CD20 IgG1-11B8-S440K变体混合抗体变体的CDC活性的选择性。所测试的Fc-Fc相互作用增强突变是E430G、E345K、E345R和E345R-E430G。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图10显示了G236位置处(G236R或G236K)或G237位置处(G237A、G237T、G237Q或G237R)具有C1q结合调节突变或双突变G237A-K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合调节突变E333S、E333A、K326A、K326W-E333S、G237A或G237A-E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图11显示具有或不具有C1q结合抑制突变G236R的抗CD52 CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 11B8-E430G-S440K混合抗体同种型变体(IgG1,IgG2,IgG3和铰链稳定的IgG4)的CDC活性的选择性。在20%NHS存在下,将Wien133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图12显示具有或不具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD37 IgG1-CD37-37.3-E430G-S440K混合抗体变体对(A)Daudi和(B)Wien 133细胞的CDC活性的选择性。在20%NHS存在下,将靶细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-CD37-37.3-E430G混合物(100%)进行标准化。
图13显示BxPC-3人胰腺癌细胞上(A)单一抗体变体或(B)具有或不具有C1q结合抑制突变G236R的抗DR5 IgG1-DR5-01-G56T-E430G-K439E+具有或不具有C1q结合增强突变E333S的IgG1-DR5-05-E430G-S440K的抗体变体的激动剂混合物的DR5介导的细胞毒性。进行三天存活力测定,并使用CellTiter-Glo试剂盒确定细胞存活力。
图14显示具有C1q结合调节突变G237A、G236R、A327K、K322E或P329R的抗CD52IgG1-CAMPATH-1H-E430G的抗体变体的CDC活性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12-S440K(0%;未显示)和IgG1-CAMPATH-1H-E430G(100%)进行标准化。
图15显示具有C1q结合抑制突变G236R或K322A(A,B,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(D,E,F)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人FcRn的结合。用5μg/mL包被的重组人FcRn细胞外域(FcRnhsECDHis-B2M-BIO)和抗体稀释液系列进行FcRn ELISA。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图16显示SCID小鼠中500μg静脉内施用的抗体的清除率。(A-C)通过ELISA测定血浆样品的总人IgG,并针对(A)IgG1-CAMPATH-1H变体,(B)IgG1-11B8变体和(C)IgG1-CAMPATH-1H变体+IgG1-11B8变体的组合物,绘制浓度对时间的曲线。每个数据点代表一式三份样品的平均值+/-标准差。(D)抗体施用后直至第21天的清除率,用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1,000/AUC确定。
图17显示如ELISA测定所检测,具有C1q结合抑制突变G236R或G237T的固定的IgG1-CAMPATH-1H-E430G-K439E变体和具有C1q结合增强突变K326A或E333S的IgG1-11B8-E430G-S440K变体与FcγRIIA同种异型131H(A),FcγRIIA同种异型131R(B),FcγRIIB(C),FcγRIIIA同种异型158V(D)和FcγRIIIA同种异型158F(E)的加His标签的二聚生物素化ECD的结合。相对于无抗体对照(背景)和与IgG1-11B8-E430G-S440K的结合(100%),对20μg/mL抗体样品呈现结合。使用链霉亲合素-polyHRP和ABTS进行检测。
图18显示具有或不具有C1q结合抑制突变(G236R或G237T)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的具有FcγR结合抑制突变G237A的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mL IgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图19显示通过血细胞的流式细胞术分析确定的全血中IgG1-CAMPATH-1H和IgG1-11B8的变体组合的选择性活性。Y轴:在效应细胞存在下进行过夜(o/n)温育后,总淋巴细胞群(CD66b阴性)的B细胞(CD19阳性/CD3阴性)分数或T细胞(CD19阴性/CD3阳性)分数。X轴:不同的治疗组。符号代表来自五个不同健康供体的细胞,每个供体在两次单独的温育中进行了测试。(A)IgG1-CAMPATH-1H-G236R-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(B)包含额外G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(C)包含额外G236R或G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合包含额外C1q结合增强E333S突变的IgG1-11B8-G237A-E430G-S440K的选择性活性。(D)与具有额外突变G236R、G237Q或G237R的IgG1-CAMPATH-1H-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的共依赖性抗体组合相比,不同B细胞靶向抗体的B细胞消耗的深度。Y轴:如上确定的B细胞分数的对数标度表示。
图20显示具有抗CD52组分中C1q结合抑制突变以及抗CD20组分中C1q结合增强突变的IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体的组合对具有CD20和CD52的不同表达水平的不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特氏淋巴瘤细胞系Daudi(A),Raji(B)和Ramos(C),ALL细胞系REH(D)和B细胞淋巴瘤细胞系U-698-M(E),用0.01-40μg/mL抗体进行体外CDC测定。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图21显示具有或不具有G236R C1q结合抑制突变的抗-CD37 IgG1-CD37-37.3-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将(A)Daudi细胞和(B)WIL2-S细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mLIgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CD37-37.3-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图22显示(A)具有Fc:Fc相互作用增强突变E430G或E345R,自身寡聚化抑制突变K439E组合任何FcγR结合抑制和C1q结合调节突变G236R、G237A或G237T的抗CD52 IgG1-CAMPATH-1H抗体变体与人淋巴瘤细胞系Wien 133的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-Campath-1H(100%)的Bmax值标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(B)具有突变E430G和K439E组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-Campath-1H的结合标准化显示。(C)具有E430G和S440K突变组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞结合的表观Kd值。
图23显示具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K组合任何C1q结合调节突变K326A或E333A(A)或E333S、G237A或G237A-E333S(B)的抗CD20 IgG1-11B8抗体变体与人淋巴瘤细胞系Raji的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-11B8的Bmax值(100%)标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(C,D)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(C)或E333S、G237A或G237A-E333S(D)的IgG1-11B8抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-11B8的结合标准化显示。(E,F)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(E)或E333S、G237A或G237A-E333S(F)的IgG1-11B8抗体变体与Raji细胞结合的表观Kd值。
图24显示抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的FcRn结合。(A,C)显示使用40μg/ml抗体浓度于(A)pH 6.0或(C)pH 7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E和C1q结合调节突变G237A或G237T的抗CD52抗体IgG1-CAMPATH-1H的变体与人FcRn的结合。(B,D)显示使用40μg/ml抗体浓度于(B)pH 6.0或(D)pH7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和C1q结合调节突变K326A、E333A、G237A或G237A-E333S的抗CD20抗体IgG1-11B8的变体与人FcRn的结合。FcRnELISA用2μg/mL包被的重组人FcRn胞外域(FcRnECDHis-B2M-BIO)和抗体稀释系列进行。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图25显示从用抗CD52 IgG1-CAMPATH-1H或抗-IgG1-11B8抗体变体或其混合物注射的小鼠收集的血液样品中测量的总人IgG(hIgG)浓度。(A)从用野生型IgG1-CAMPATH-1H,IgG1-CAMPATH-1H-E430G-K439E-G237Q或IgG1-CAMPATH-1H-E430G-K439E-G236R注射的小鼠收集的血液样品的总hIgG浓度。(B)从用野生型IgG1-11B8,IgG1-11B8-E430G-S440K-G237A或IgG1-11B8-E430G-S440K-E333S注射的小鼠收集的血液样品的总hIgG浓度。(C)从用野生型IgG1-CAMPATH-1H+IgG1-11B8混合物或带有(A)和(B)中突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物注射的小鼠收集的血液样品的总hIgG浓度。在所有图中,虚线表示SCID小鼠中野生型IgG1抗体随时间预测的IgG1浓度。(D)抗体施用后直至第21天的清除率用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1000/AUC确定。
图26显示减去在不含抗体的阴性对照样品中检测的平均C4d浓度后,用带有突变E430G、K439E或S440K和G236R、G237A、G237Q或G237R的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12的抗体变体温育的样品中检测的C4d浓度。阳性对照样品包括带有E345R、E430G和S440Y Fc-Fc相互作用增强突变(RGY)的抗体变体。
图27显示用带有E430G、K439E或S440K和G236R、G237T、K326A或E333S突变的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12抗体变体调理后,通过流式细胞术检测C1q与Wien 133细胞的结合,所述Wien 133细胞于冰上用正常人血清作为补体来源温育。将平均荧光强度值相对于无抗体的对照反应(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物的最高水平(100%)标准化,通过拟合对数激动剂反应模型进行估算。(A)对照反应的C1q结合。(B-D)(B)IgG1-CAMPATH-E430G-K439E,(C)IgG1-CAMPATH-E430G-K439E-G236R和(D)IgG1-CAMPATH-E430G-K439E-G237R混合非结合对照IgG1-b12或不同的IgG1-11B8变体的C1q结合。
图28显示带有自身寡聚化抑制突变K439E或S440K和C1q结合调节突变的IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G抗体变体的FcγR结合。(A-E)如ELISA测定所检测,固定化抗体变体与(A)高亲和力同种异型FcγRIIA 131H,(B)低亲和力同种异型FcγRIIA131R,(C)FcγRIIB,(D)高亲和力同种异型FcγRIIIA 158V,或(E)低亲和力同种异型FcγRIIIA158F的加His标签的二聚生物素化ECD的结合。(F)ELISA检测的固定化FcγRIa与抗体变体的结合。表示了20μg/mL抗体样品的结合,并且每个实验在减去不与一抗温育的孔中的信号后相对于野生型IgG1-CAMPATH-1H所观测的平均信号(100%)标准化。使用链霉亲合素-polyHRP和ABTS进行检测。
图29显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的抗CD52IgG1-CAMPATH-1H,抗CD20 IgG1-11B8和抗CD52 IgG1-h2E8混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。在三个独立实验中测量了CDC效力,并表示为(A)平均AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化,和(B)在40μg/ml的抗体浓度时,由碘化丙啶阳性确定的平均裂解百分比。
图30(A,B)显示单一试剂和组合的带有所示自身寡聚化抑制突变和C1q结合调节突变的抗CD52 IgG1-CAMPATH-1H-E430G,抗CD20 IgG1-11B8-E430G和非抗原结合IgG1-b12-E430G抗体变体的CDC效力。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图31显示与Raji淋巴瘤细胞和不同浓度的IgG1-CAMPATH-1H和IgG1-11B8抗体变体共培养时,通过发光水平(RLU)测定的稳定表达(A)FcγRIIa或(B)FcγRIIIa的Jurkat报告物细胞系的激活。每个实验的发光值相对于IgG1-b12(0%)和野生型IgG1-Campath-1H+野生型IgG1-11B8(100%)观测到的值标准化,然后求三个(FcγRIIa)或两个(FcγRIIIa)实验重复的平均值。
图32显示由非等摩尔比的带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物诱导的对Wien 133细胞的共依赖性CDC。(A)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。(B)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。
图33显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H抗体变体与抗HLA-DR IgG1-HLA-DR-huL243变体(A)或抗HLA-DRIgG1-HLA-DR-1D09C3变体(B)混合物的CDC活性的选择性。在20%NHS存在下,将Oci-Ly17细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和(A)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-huL243-E430G(100%)或(B)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-1D09C3-E430G混合物(100%)标准化。
图34显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。显示具有突变L234A、L234A-L235A、L234F、L234F-L235E、L235A、L235Q、G236R或G237Q的IgG1-CAMPATH-1H-E430G-K439E变体以及这些变体与非结合对照抗体IgG1-b12或IgG1-11B8-E430G-S440K混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图35显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。(A)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345K、E345Q、E345R或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(B)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(C)带有匹配的Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8的抗体变体作为单一试剂或其混合物的CDC效力。(D)带有Fc-Fc相互作用增强突变E430G或K248E-T437R,自身寡聚化抑制突变K439E或S440K,和C1q结合调节突变G236R、G237A或E333S的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图36显示由带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CD37-37-3和IgG1-11B8抗体变体混合物诱导的对Raji淋巴瘤细胞的共依赖性CDC。(A)由稀释的所示抗体变体或其混合物诱导的细胞裂解的曲线下相对面积(AUC),相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CD37-37-3-E430G+IgG1-11B8-E430G混合物为100%)标准化。(B)由所示抗体变体及其混合物诱导的最大裂解百分比。
图37显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。在20%NHS存在下,将患者CLL样品与抗体浓度系列一起温育。CDC效力表示为与抗体变体温育后活的B细胞的百分比。示出使用来自患者1(A),2(B)和3(C)的CLL样品的结果。
图38显示全血样品与带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM抗体变体混合物温育后,通过流式细胞术检测的B细胞,CD4+ T细胞和CD8+ T细胞的分数。在与所示IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(A)B细胞,(B)CD4+ T细胞和(C)CD8+ T细胞的百分比。在与所示IgG1-CAMPATH-1H,IgG1-CD5-INSERM和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(D)B细胞,(E)CD4+ T细胞和(F)CD8+ T细胞的百分比。分数计算为[100%x(样品的细胞计数/“无Ab样品”的细胞计数)x(“无Ab样品”的粒细胞计数/样品的粒细胞计数)]。
图39显示了带有增强Fc-Fc相互作用,抑制自身寡聚化和抑制(G237T)或增强(K326W-E333S)C1q结合的突变的抗DR4和抗DR5抗体变体在癌细胞中协同激活程序性细胞死亡。(A)与所示抗体变体温育72小时后,BxPC-3人胰腺癌细胞的存活力。(B)与所示抗体变体温育72小时后,COLO 205人结肠癌细胞的存活力。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
A;设为100%,图1显示通过引入P329R突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%合并的正常人血清(NHS)存在下,将Wien 133细胞与浓度抗体浓度系列一起温育。CDC效力表示为(A)由碘化丙啶(PI)阳性细胞百分比确定的裂解百分比,和(B)剂量反应-反应曲线下面积(AUC),相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图2显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为(A)由PI阳性细胞百分比确定的裂解百分比,和(B)AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图3显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20-11B8-E430G-S440K混合抗体变体对不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特(Burkitt)氏淋巴瘤细胞系Wien 133(A),Daudi(B),Raji(C)和Ramos(D),急性成淋巴细胞性淋巴瘤(ALL)细胞系REH(E),骨髓瘤细胞系U266B1(F)和B细胞淋巴瘤细胞系U-698-M(G),用30μg/mL抗体进行体外CDC测定。CDC活性表示为通过每种细胞系相对于非结合对照抗体IgG1-b12(0%),以及针对REH,U266B1和Wien 133为IgG1-CAMPATH-1H-E430G(100%),或针对Daudi,Raji,Ramos和U-698-M为IgG1-11B8-E430G(100%)标准化的PI阳性细胞百分比确定的裂解百分比。
图4显示具有C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)的抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图5显示具有CD1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图6显示具有C1q结合抑制突变G236R(A)、K322A(B)、E269K(C)、K322E(D)或P329R(E)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有C1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图7显示具有C1q结合抑制突变G236R或K322A(A,B)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(C,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人淋巴瘤细胞系Wien 133(A,C)和Raji(B,D)的结合。通过流式细胞术测试抗体结合。结合表示为荧光强度的几何平均值(MFI)。使用没有一抗或非结合抗gp120抗体IgG1-b12的样品作为结合的阴性对照。
图8显示具有C1q结合抑制突变G236R或K322A(A,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(B,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体的ADCC能力。(A,B)进行了ADCC报告物生物测定(ADCC reporterBioassay),其中Raji靶细胞与抗体稀释系列与稳定表达高亲和力FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的NFAT反应元件的Jurkat人T细胞一起温育。萤光素酶产生通过发光读数定量。(C,D)进行体外铕TDA(EuTDA)ADCC测定,其中将Wien 133靶细胞与抗体稀释系列和人PBMC(E:T 100:1)一起温育。细胞裂解通过测量上清液中EuTDA荧光螯合物的信号来确定。
图9显示具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-K439E+具有或不具有C1q结合增强突变E333S的Fc-Fc相互作用增强的抗CD20 IgG1-11B8-S440K变体混合抗体变体的CDC活性的选择性。所测试的Fc-Fc相互作用增强突变是E430G、E345K、E345R和E345R-E430G。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图10显示了G236位置处(G236R或G236K)或G237位置处(G237A、G237T、G237Q或G237R)具有C1q结合调节突变或双突变G237A-K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合调节突变E333S、E333A、K326A、K326W-E333S、G237A或G237A-E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图11显示具有或不具有C1q结合抑制突变G236R的抗CD52 CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 11B8-E430G-S440K混合抗体同种型变体(IgG1,IgG2,IgG3和铰链稳定的IgG4)的CDC活性的选择性。在20%NHS存在下,将Wien133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图12显示具有或不具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD37 IgG1-CD37-37.3-E430G-S440K混合抗体变体对(A)Daudi和(B)Wien 133细胞的CDC活性的选择性。在20%NHS存在下,将靶细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-CD37-37.3-E430G混合物(100%)进行标准化。
图13显示BxPC-3人胰腺癌细胞上(A)单一抗体变体或(B)具有或不具有C1q结合抑制突变G236R的抗DR5 IgG1-DR5-01-G56T-E430G-K439E+具有或不具有C1q结合增强突变E333S的IgG1-DR5-05-E430G-S440K的抗体变体的激动剂混合物的DR5介导的细胞毒性。进行三天存活力测定,并使用CellTiter-Glo试剂盒确定细胞存活力。
图14显示具有C1q结合调节突变G237A、G236R、A327K、K322E或P329R的抗CD52IgG1-CAMPATH-1H-E430G的抗体变体的CDC活性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12-S440K(0%;未显示)和IgG1-CAMPATH-1H-E430G(100%)进行标准化。
图15显示具有C1q结合抑制突变G236R或K322A(A,B,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(D,E,F)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人FcRn的结合。用5μg/mL包被的重组人FcRn细胞外域(FcRnhsECDHis-B2M-BIO)和抗体稀释液系列进行FcRn ELISA。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图16显示SCID小鼠中500μg静脉内施用的抗体的清除率。(A-C)通过ELISA测定血浆样品的总人IgG,并针对(A)IgG1-CAMPATH-1H变体,(B)IgG1-11B8变体和(C)IgG1-CAMPATH-1H变体+IgG1-11B8变体的组合物,绘制浓度对时间的曲线。每个数据点代表一式三份样品的平均值+/-标准差。(D)抗体施用后直至第21天的清除率,用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1,000/AUC确定。
图17显示如ELISA测定所检测,具有C1q结合抑制突变G236R或G237T的固定的IgG1-CAMPATH-1H-E430G-K439E变体和具有C1q结合增强突变K326A或E333S的IgG1-11B8-E430G-S440K变体与FcγRIIA同种异型131H(A),FcγRIIA同种异型131R(B),FcγRIIB(C),FcγRIIIA同种异型158V(D)和FcγRIIIA同种异型158F(E)的加His标签的二聚生物素化ECD的结合。相对于无抗体对照(背景)和与IgG1-11B8-E430G-S440K的结合(100%),对20μg/mL抗体样品呈现结合。使用链霉亲合素-polyHRP和ABTS进行检测。
图18显示具有或不具有C1q结合抑制突变(G236R或G237T)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的具有FcγR结合抑制突变G237A的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mL IgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图19显示通过血细胞的流式细胞术分析确定的全血中IgG1-CAMPATH-1H和IgG1-11B8的变体组合的选择性活性。Y轴:在效应细胞存在下进行过夜(o/n)温育后,总淋巴细胞群(CD66b阴性)的B细胞(CD19阳性/CD3阴性)分数或T细胞(CD19阴性/CD3阳性)分数。X轴:不同的治疗组。符号代表来自五个不同健康供体的细胞,每个供体在两次单独的温育中进行了测试。(A)IgG1-CAMPATH-1H-G236R-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(B)包含额外G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(C)包含额外G236R或G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合包含额外C1q结合增强E333S突变的IgG1-11B8-G237A-E430G-S440K的选择性活性。(D)与具有额外突变G236R、G237Q或G237R的IgG1-CAMPATH-1H-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的共依赖性抗体组合相比,不同B细胞靶向抗体的B细胞消除的深度。Y轴:如上确定的B细胞分数的对数标度表示。
图20显示具有抗CD52组分中C1q结合抑制突变以及抗CD20组分中C1q结合增强突变的IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体的组合对具有CD20和CD52的不同表达水平的不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特氏淋巴瘤细胞系Daudi(A),Raji(B)和Ramos(C),ALL细胞系REH(D)和B细胞淋巴瘤细胞系U-698-M(E),用0.01-40μg/mL抗体进行体外CDC测定。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图21显示具有或不具有G236R C1q结合抑制突变的抗-CD37 IgG1-CD37-37.3-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将(A)Daudi细胞和(B)WIL2-S细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mLIgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CD37-37.3-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图22显示(A)具有Fc:Fc相互作用增强突变E430G或E345R,自身寡聚化抑制突变K439E组合任何FcγR结合抑制和C1q结合调节突变G236R、G237A或G237T的抗CD52 IgG1-CAMPATH-1H抗体变体与人淋巴瘤细胞系Wien 133的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-Campath-1H(100%)的Bmax值标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(B)具有突变E430G和K439E组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-Campath-1H的结合标准化显示。(C)具有E430G和S440K突变组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞结合的表观Kd值。
图23显示具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K组合任何C1q结合调节突变K326A或E333A(A)或E333S、G237A或G237A-E333S(B)的抗CD20 IgG1-11B8抗体变体与人淋巴瘤细胞系Raji的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-11B8的Bmax值(100%)标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(C,D)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(C)或E333S、G237A或G237A-E333S(D)的IgG1-11B8抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-11B8的结合标准化显示。(E,F)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(E)或E333S、G237A或G237A-E333S(F)的IgG1-11B8抗体变体与Raji细胞结合的表观Kd值。
图24显示抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的FcRn结合。(A,C)显示使用40μg/ml抗体浓度于(A)pH 6.0或(C)pH 7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E和C1q结合调节突变G237A或G237T的抗CD52抗体IgG1-CAMPATH-1H的变体与人FcRn的结合。(B,D)显示使用40μg/ml抗体浓度于(B)pH 6.0或(D)pH7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和C1q结合调节突变K326A、E333A、G237A或G237A-E333S的抗CD20抗体IgG1-11B8的变体与人FcRn的结合。FcRnELISA用2μg/mL包被的重组人FcRn胞外域(FcRnECDHis-B2M-BIO)和抗体稀释系列进行。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图25显示从用抗CD52 IgG1-CAMPATH-1H或抗-IgG1-11B8抗体变体或其混合物注射的小鼠收集的血液样品中测量的总人IgG(hIgG)浓度。(A)从用野生型IgG1-CAMPATH-1H,IgG1-CAMPATH-1H-E430G-K439E-G237Q或IgG1-CAMPATH-1H-E430G-K439E-G236R注射的小鼠收集的血液样品的总hIgG浓度。(B)从用野生型IgG1-11B8,IgG1-11B8-E430G-S440K-G237A或IgG1-11B8-E430G-S440K-E333S注射的小鼠收集的血液样品的总hIgG浓度。(C)从用野生型IgG1-CAMPATH-1H+IgG1-11B8混合物或带有(A)和(B)中突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物注射的小鼠收集的血液样品的总hIgG浓度。在所有图中,虚线表示SCID小鼠中野生型IgG1抗体随时间预测的IgG1浓度。(D)抗体施用后直至第21天的清除率用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1000/AUC确定。
图26显示减去在不含抗体的阴性对照样品中检测的平均C4d浓度后,用带有突变E430G、K439E或S440K和G236R、G237A、G237Q或G237R的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12的抗体变体温育的样品中检测的C4d浓度。阳性对照样品包括带有E345R、E430G和S440Y Fc-Fc相互作用增强突变(RGY)的抗体变体。
图27显示用带有E430G、K439E或S440K和G236R、G237T、K326A或E333S突变的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12抗体变体调理后,通过流式细胞术检测C1q与Wien 133细胞的结合,所述Wien 133细胞于冰上用正常人血清作为补体来源温育。将平均荧光强度值相对于无抗体的对照反应(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物的最高水平(100%)标准化,通过拟合对数激动剂反应模型进行估算。(A)对照反应的C1q结合。(B-D)(B)IgG1-CAMPATH-E430G-K439E,(C)IgG1-CAMPATH-E430G-K439E-G236R和(D)IgG1-CAMPATH-E430G-K439E-G237R混合非结合对照IgG1-b12或不同的IgG1-11B8变体的C1q结合。
图28显示带有自身寡聚化抑制突变K439E或S440K和C1q结合调节突变的IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G抗体变体的FcγR结合。(A-E)如ELISA测定所检测,固定化抗体变体与(A)高亲和力同种异型FcγRIIA 131H,(B)低亲和力同种异型FcγRIIA131R,(C)FcγRIIB,(D)高亲和力同种异型FcγRIIIA 158V,或(E)低亲和力同种异型FcγRIIIA 158F的加His标签的二聚生物素化ECD的结合。(F)ELISA检测的固定化FcγRIa与抗体变体的结合。表示了20μg/mL抗体样品的结合,并且每个实验在减去不与一抗温育的孔中的信号后相对于野生型IgG1-CAMPATH-1H所观测的平均信号(100%)标准化。使用链霉亲合素-polyHRP和ABTS进行检测。
图29显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的抗CD52IgG1-CAMPATH-1H,抗CD20 IgG1-11B8和抗CD52 IgG1-h2E8混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。在三个独立实验中测量了CDC效力,并表示为(A)平均AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化,和(B)在40μg/ml的抗体浓度时,由碘化丙啶阳性确定的平均裂解百分比。
图30(A,B)显示单一试剂和组合的带有所示自身寡聚化抑制突变和C1q结合调节突变的抗CD52 IgG1-CAMPATH-1H-E430G,抗CD20 IgG1-11B8-E430G和非抗原结合IgG1-b12-E430G抗体变体的CDC效力。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图31显示与Raji淋巴瘤细胞和不同浓度的IgG1-CAMPATH-1H和IgG1-11B8抗体变体共培养时,通过发光水平(RLU)测定的稳定表达(A)FcγRIIa或(B)FcγRIIIa的Jurkat报告物细胞系的激活。每个实验的发光值相对于IgG1-b12(0%)和野生型IgG1-Campath-1H+野生型IgG1-11B8(100%)观测到的值标准化,然后求三个(FcγRIIa)或两个(FcγRIIIa)实验重复的平均值。
图32显示由非等摩尔比的带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物诱导的对Wien 133细胞的共依赖性CDC。(A)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。(B)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。
图33显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H抗体变体与抗HLA-DR IgG1-HLA-DR-huL243变体(A)或抗HLA-DRIgG1-HLA-DR-1D09C3变体(B)混合物的CDC活性的选择性。在20%NHS存在下,将Oci-Ly17细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和(A)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-huL243-E430G(100%)或(B)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-1D09C3-E430G混合物(100%)标准化。
图34显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。显示具有突变L234A、L234A-L235A、L234F、L234F-L235E、L235A、L235Q、G236R或G237Q的IgG1-CAMPATH-1H-E430G-K439E变体以及这些变体与非结合对照抗体IgG1-b12或IgG1-11B8-E430G-S440K混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图35显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。(A)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345K、E345Q、E345R或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(B)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(C)带有匹配的Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8的抗体变体作为单一试剂或其混合物的CDC效力。(D)带有Fc-Fc相互作用增强突变E430G或K248E-T437R,自身寡聚化抑制突变K439E或S440K,和C1q结合调节突变G236R、G237A或E333S的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图36显示由带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CD37-37-3和IgG1-11B8抗体变体混合物诱导的对Raji淋巴瘤细胞的共依赖性CDC。(A)由稀释的所示抗体变体或其混合物诱导的细胞裂解的曲线下相对面积(AUC),相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CD37-37-3-E430G+IgG1-11B8-E430G混合物为100%)标准化。(B)由所示抗体变体及其混合物诱导的最大裂解百分比。
图37显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。在20%NHS存在下,将患者CLL样品与抗体浓度系列一起温育。CDC效力表示为与抗体变体温育后活的B细胞的百分比。示出使用来自患者1(A),2(B)和3(C)的CLL样品的结果。
图38显示全血样品与带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM抗体变体混合物温育后,通过流式细胞术检测的B细胞,CD4+ T细胞和CD8+T细胞的分数。在与所示IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(A)B细胞,(B)CD4+ T细胞和(C)CD8+ T细胞的百分比。在与所示IgG1-CAMPATH-1H,IgG1-CD5-INSERM和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(D)B细胞,(E)CD4+ T细胞和(F)CD8+ T细胞的百分比。分数计算为[100%x(样品的细胞计数/“无Ab样品”的细胞计数)x(“无Ab样品”的粒细胞计数/样品的粒细胞计数)]。
图39显示了带有增强Fc-Fc相互作用,抑制自身寡聚化和抑制(G237T)或增强(K326W-E333S)C1q结合的突变的抗DR4和抗DR5抗体变体在癌细胞中协同激活程序性细胞死亡。(A)与所示抗体变体温育72小时后,BxPC-3人胰腺癌细胞的存活力。(B)与所示抗体变体温育72小时后,COLO 205人结肠癌细胞的存活力。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
B),其被发现比单独的单个抗体更有效。(图1显示通过引入P329R突变,抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%合并的正常人血清(NHS)存在下,将Wien 133细胞与浓度抗体浓度系列一起温育。CDC效力表示为(A)由碘化丙啶(PI)阳性细胞百分比确定的裂解百分比,和(B)剂量反应-反应曲线下面积(AUC),相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图2显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为(A)由PI阳性细胞百分比确定的裂解百分比,和(B)AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图3显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20-11B8-E430G-S440K混合抗体变体对不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特(Burkitt)氏淋巴瘤细胞系Wien 133(A),Daudi(B),Raji(C)和Ramos(D),急性成淋巴细胞性淋巴瘤(ALL)细胞系REH(E),骨髓瘤细胞系U266B1(F)和B细胞淋巴瘤细胞系U-698-M(G),用30μg/mL抗体进行体外CDC测定。CDC活性表示为通过每种细胞系相对于非结合对照抗体IgG1-b12(0%),以及针对REH,U266B1和Wien 133为IgG1-CAMPATH-1H-E430G(100%),或针对Daudi,Raji,Ramos和U-698-M为IgG1-11B8-E430G(100%)标准化的PI阳性细胞百分比确定的裂解百分比。
图4显示具有C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)的抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图5显示具有CD1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图6显示具有C1q结合抑制突变G236R(A)、K322A(B)、E269K(C)、K322E(D)或P329R(E)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有C1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图7显示具有C1q结合抑制突变G236R或K322A(A,B)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(C,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人淋巴瘤细胞系Wien 133(A,C)和Raji(B,D)的结合。通过流式细胞术测试抗体结合。结合表示为荧光强度的几何平均值(MFI)。使用没有一抗或非结合抗gp120抗体IgG1-b12的样品作为结合的阴性对照。
图8显示具有C1q结合抑制突变G236R或K322A(A,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(B,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体的ADCC能力。(A,B)进行了ADCC报告物生物测定(ADCC reporterBioassay),其中Raji靶细胞与抗体稀释系列与稳定表达高亲和力FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的NFAT反应元件的Jurkat人T细胞一起温育。萤光素酶产生通过发光读数定量。(C,D)进行体外铕TDA(EuTDA)ADCC测定,其中将Wien 133靶细胞与抗体稀释系列和人PBMC(E:T 100:1)一起温育。细胞裂解通过测量上清液中EuTDA荧光螯合物的信号来确定。
图9显示具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-K439E+具有或不具有C1q结合增强突变E333S的Fc-Fc相互作用增强的抗CD20 IgG1-11B8-S440K变体混合抗体变体的CDC活性的选择性。所测试的Fc-Fc相互作用增强突变是E430G、E345K、E345R和E345R-E430G。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图10显示了G236位置处(G236R或G236K)或G237位置处(G237A、G237T、G237Q或G237R)具有C1q结合调节突变或双突变G237A-K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合调节突变E333S、E333A、K326A、K326W-E333S、G237A或G237A-E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图11显示具有或不具有C1q结合抑制突变G236R的抗CD52 CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 11B8-E430G-S440K混合抗体同种型变体(IgG1,IgG2,IgG3和铰链稳定的IgG4)的CDC活性的选择性。在20%NHS存在下,将Wien133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图12显示具有或不具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD37 IgG1-CD37-37.3-E430G-S440K混合抗体变体对(A)Daudi和(B)Wien 133细胞的CDC活性的选择性。在20%NHS存在下,将靶细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-CD37-37.3-E430G混合物(100%)进行标准化。
图13显示BxPC-3人胰腺癌细胞上(A)单一抗体变体或(B)具有或不具有C1q结合抑制突变G236R的抗DR5 IgG1-DR5-01-G56T-E430G-K439E+具有或不具有C1q结合增强突变E333S的IgG1-DR5-05-E430G-S440K的抗体变体的激动剂混合物的DR5介导的细胞毒性。进行三天存活力测定,并使用CellTiter-Glo试剂盒确定细胞存活力。
图14显示具有C1q结合调节突变G237A、G236R、A327K、K322E或P329R的抗CD52IgG1-CAMPATH-1H-E430G的抗体变体的CDC活性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12-S440K(0%;未显示)和IgG1-CAMPATH-1H-E430G(100%)进行标准化。
图15显示具有C1q结合抑制突变G236R或K322A(A,B,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(D,E,F)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人FcRn的结合。用5μg/mL包被的重组人FcRn细胞外域(FcRnhsECDHis-B2M-BIO)和抗体稀释液系列进行FcRn ELISA。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图16显示SCID小鼠中500μg静脉内施用的抗体的清除率。(A-C)通过ELISA测定血浆样品的总人IgG,并针对(A)IgG1-CAMPATH-1H变体,(B)IgG1-11B8变体和(C)IgG1-CAMPATH-1H变体+IgG1-11B8变体的组合物,绘制浓度对时间的曲线。每个数据点代表一式三份样品的平均值+/-标准差。(D)抗体施用后直至第21天的清除率,用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1,000/AUC确定。
图17显示如ELISA测定所检测,具有C1q结合抑制突变G236R或G237T的固定的IgG1-CAMPATH-1H-E430G-K439E变体和具有C1q结合增强突变K326A或E333S的IgG1-11B8-E430G-S440K变体与FcγRIIA同种异型131H(A),FcγRIIA同种异型131R(B),FcγRIIB(C),FcγRIIIA同种异型158V(D)和FcγRIIIA同种异型158F(E)的加His标签的二聚生物素化ECD的结合。相对于无抗体对照(背景)和与IgG1-11B8-E430G-S440K的结合(100%),对20μg/mL抗体样品呈现结合。使用链霉亲合素-polyHRP和ABTS进行检测。
图18显示具有或不具有C1q结合抑制突变(G236R或G237T)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的具有FcγR结合抑制突变G237A的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mL IgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图19显示通过血细胞的流式细胞术分析确定的全血中IgG1-CAMPATH-1H和IgG1-11B8的变体组合的选择性活性。Y轴:在效应细胞存在下进行过夜(o/n)温育后,总淋巴细胞群(CD66b阴性)的B细胞(CD19阳性/CD3阴性)分数或T细胞(CD19阴性/CD3阳性)分数。X轴:不同的治疗组。符号代表来自五个不同健康供体的细胞,每个供体在两次单独的温育中进行了测试。(A)IgG1-CAMPATH-1H-G236R-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(B)包含额外G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(C)包含额外G236R或G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合包含额外C1q结合增强E333S突变的IgG1-11B8-G237A-E430G-S440K的选择性活性。(D)与具有额外突变G236R、G237Q或G237R的IgG1-CAMPATH-1H-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的共依赖性抗体组合相比,不同B细胞靶向抗体的B细胞消耗的深度。Y轴:如上确定的B细胞分数的对数标度表示。
图20显示具有抗CD52组分中C1q结合抑制突变以及抗CD20组分中C1q结合增强突变的IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体的组合对具有CD20和CD52的不同表达水平的不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特氏淋巴瘤细胞系Daudi(A),Raji(B)和Ramos(C),ALL细胞系REH(D)和B细胞淋巴瘤细胞系U-698-M(E),用0.01-40μg/mL抗体进行体外CDC测定。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图21显示具有或不具有G236R C1q结合抑制突变的抗-CD37 IgG1-CD37-37.3-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将(A)Daudi细胞和(B)WIL2-S细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mLIgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CD37-37.3-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图22显示(A)具有Fc:Fc相互作用增强突变E430G或E345R,自身寡聚化抑制突变K439E组合任何FcγR结合抑制和C1q结合调节突变G236R、G237A或G237T的抗CD52 IgG1-CAMPATH-1H抗体变体与人淋巴瘤细胞系Wien 133的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-Campath-1H(100%)的Bmax值标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(B)具有突变E430G和K439E组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-Campath-1H的结合标准化显示。(C)具有E430G和S440K突变组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞结合的表观Kd值。
图23显示具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K组合任何C1q结合调节突变K326A或E333A(A)或E333S、G237A或G237A-E333S(B)的抗CD20 IgG1-11B8抗体变体与人淋巴瘤细胞系Raji的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-11B8的Bmax值(100%)标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(C,D)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(C)或E333S、G237A或G237A-E333S(D)的IgG1-11B8抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-11B8的结合标准化显示。(E,F)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(E)或E333S、G237A或G237A-E333S(F)的IgG1-11B8抗体变体与Raji细胞结合的表观Kd值。
图24显示抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的FcRn结合。(A,C)显示使用40μg/ml抗体浓度于(A)pH 6.0或(C)pH 7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E和C1q结合调节突变G237A或G237T的抗CD52抗体IgG1-CAMPATH-1H的变体与人FcRn的结合。(B,D)显示使用40μg/ml抗体浓度于(B)pH 6.0或(D)pH7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和C1q结合调节突变K326A、E333A、G237A或G237A-E333S的抗CD20抗体IgG1-11B8的变体与人FcRn的结合。FcRnELISA用2μg/mL包被的重组人FcRn胞外域(FcRnECDHis-B2M-BIO)和抗体稀释系列进行。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图25显示从用抗CD52 IgG1-CAMPATH-1H或抗-IgG1-11B8抗体变体或其混合物注射的小鼠收集的血液样品中测量的总人IgG(hIgG)浓度。(A)从用野生型IgG1-CAMPATH-1H,IgG1-CAMPATH-1H-E430G-K439E-G237Q或IgG1-CAMPATH-1H-E430G-K439E-G236R注射的小鼠收集的血液样品的总hIgG浓度。(B)从用野生型IgG1-11B8,IgG1-11B8-E430G-S440K-G237A或IgG1-11B8-E430G-S440K-E333S注射的小鼠收集的血液样品的总hIgG浓度。(C)从用野生型IgG1-CAMPATH-1H+IgG1-11B8混合物或带有(A)和(B)中突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物注射的小鼠收集的血液样品的总hIgG浓度。在所有图中,虚线表示SCID小鼠中野生型IgG1抗体随时间预测的IgG1浓度。(D)抗体施用后直至第21天的清除率用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1000/AUC确定。
图26显示减去在不含抗体的阴性对照样品中检测的平均C4d浓度后,用带有突变E430G、K439E或S440K和G236R、G237A、G237Q或G237R的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12的抗体变体温育的样品中检测的C4d浓度。阳性对照样品包括带有E345R、E430G和S440Y Fc-Fc相互作用增强突变(RGY)的抗体变体。
图27显示用带有E430G、K439E或S440K和G236R、G237T、K326A或E333S突变的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12抗体变体调理后,通过流式细胞术检测C1q与Wien 133细胞的结合,所述Wien 133细胞于冰上用正常人血清作为补体来源温育。将平均荧光强度值相对于无抗体的对照反应(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物的最高水平(100%)标准化,通过拟合对数激动剂反应模型进行估算。(A)对照反应的C1q结合。(B-D)(B)IgG1-CAMPATH-E430G-K439E,(C)IgG1-CAMPATH-E430G-K439E-G236R和(D)IgG1-CAMPATH-E430G-K439E-G237R混合非结合对照IgG1-b12或不同的IgG1-11B8变体的C1q结合。
图28显示带有自身寡聚化抑制突变K439E或S440K和C1q结合调节突变的IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G抗体变体的FcγR结合。(A-E)如ELISA测定所检测,固定化抗体变体与(A)高亲和力同种异型FcγRIIA 131H,(B)低亲和力同种异型FcγRIIA131R,(C)FcγRIIB,(D)高亲和力同种异型FcγRIIIA 158V,或(E)低亲和力同种异型FcγRIIIA158F的加His标签的二聚生物素化ECD的结合。(F)ELISA检测的固定化FcγRIa与抗体变体的结合。表示了20μg/mL抗体样品的结合,并且每个实验在减去不与一抗温育的孔中的信号后相对于野生型IgG1-CAMPATH-1H所观测的平均信号(100%)标准化。使用链霉亲合素-polyHRP和ABTS进行检测。
图29显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的抗CD52IgG1-CAMPATH-1H,抗CD20 IgG1-11B8和抗CD52 IgG1-h2E8混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。在三个独立实验中测量了CDC效力,并表示为(A)平均AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化,和(B)在40μg/ml的抗体浓度时,由碘化丙啶阳性确定的平均裂解百分比。
图30(A,B)显示单一试剂和组合的带有所示自身寡聚化抑制突变和C1q结合调节突变的抗CD52 IgG1-CAMPATH-1H-E430G,抗CD20 IgG1-11B8-E430G和非抗原结合IgG1-b12-E430G抗体变体的CDC效力。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图31显示与Raji淋巴瘤细胞和不同浓度的IgG1-CAMPATH-1H和IgG1-11B8抗体变体共培养时,通过发光水平(RLU)测定的稳定表达(A)FcγRIIa或(B)FcγRIIIa的Jurkat报告物细胞系的激活。每个实验的发光值相对于IgG1-b12(0%)和野生型IgG1-Campath-1H+野生型IgG1-11B8(100%)观测到的值标准化,然后求三个(FcγRIIa)或两个(FcγRIIIa)实验重复的平均值。
图32显示由非等摩尔比的带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物诱导的对Wien 133细胞的共依赖性CDC。(A)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。(B)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。
图33显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H抗体变体与抗HLA-DR IgG1-HLA-DR-huL243变体(A)或抗HLA-DRIgG1-HLA-DR-1D09C3变体(B)混合物的CDC活性的选择性。在20%NHS存在下,将Oci-Ly17细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和(A)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-huL243-E430G(100%)或(B)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-1D09C3-E430G混合物(100%)标准化。
图34显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。显示具有突变L234A、L234A-L235A、L234F、L234F-L235E、L235A、L235Q、G236R或G237Q的IgG1-CAMPATH-1H-E430G-K439E变体以及这些变体与非结合对照抗体IgG1-b12或IgG1-11B8-E430G-S440K混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图35显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。(A)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345K、E345Q、E345R或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(B)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(C)带有匹配的Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8的抗体变体作为单一试剂或其混合物的CDC效力。(D)带有Fc-Fc相互作用增强突变E430G或K248E-T437R,自身寡聚化抑制突变K439E或S440K,和C1q结合调节突变G236R、G237A或E333S的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图36显示由带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CD37-37-3和IgG1-11B8抗体变体混合物诱导的对Raji淋巴瘤细胞的共依赖性CDC。(A)由稀释的所示抗体变体或其混合物诱导的细胞裂解的曲线下相对面积(AUC),相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CD37-37-3-E430G+IgG1-11B8-E430G混合物为100%)标准化。(B)由所示抗体变体及其混合物诱导的最大裂解百分比。
图37显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。在20%NHS存在下,将患者CLL样品与抗体浓度系列一起温育。CDC效力表示为与抗体变体温育后活的B细胞的百分比。示出使用来自患者1(A),2(B)和3(C)的CLL样品的结果。
图38显示全血样品与带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM抗体变体混合物温育后,通过流式细胞术检测的B细胞,CD4+ T细胞和CD8+T细胞的分数。在与所示IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(A)B细胞,(B)CD4+ T细胞和(C)CD8+ T细胞的百分比。在与所示IgG1-CAMPATH-1H,IgG1-CD5-INSERM和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(D)B细胞,(E)CD4+ T细胞和(F)CD8+ T细胞的百分比。分数计算为[100%x(样品的细胞计数/“无Ab样品”的细胞计数)x(“无Ab样品”的粒细胞计数/样品的粒细胞计数)]。
图39显示了带有增强Fc-Fc相互作用,抑制自身寡聚化和抑制(G237T)或增强(K326W-E333S)C1q结合的突变的抗DR4和抗DR5抗体变体在癌细胞中协同激活程序性细胞死亡。(A)与所示抗体变体温育72小时后,BxPC-3人胰腺癌细胞的存活力。(B)与所示抗体变体温育72小时后,COLO 205人结肠癌细胞的存活力。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
B).将IgG1-CAMPATH-1H-E430G作为单一试剂进行测试时,引入K439E突变(SEQ IDNO:80)导致CDC效力降低,而K439E-P329R(SEQ ID NO:100)导致CDC活性完全丧失。(图1显示通过引入P329R突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%合并的正常人血清(NHS)存在下,将Wien 133细胞与浓度抗体浓度系列一起温育。CDC效力表示为(A)由碘化丙啶(PI)阳性细胞百分比确定的裂解百分比,和(B)剂量反应-反应曲线下面积(AUC),相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图2显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为(A)由PI阳性细胞百分比确定的裂解百分比,和(B)AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图3显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20-11B8-E430G-S440K混合抗体变体对不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特(Burkitt)氏淋巴瘤细胞系Wien 133(A),Daudi(B),Raji(C)和Ramos(D),急性成淋巴细胞性淋巴瘤(ALL)细胞系REH(E),骨髓瘤细胞系U266B1(F)和B细胞淋巴瘤细胞系U-698-M(G),用30μg/mL抗体进行体外CDC测定。CDC活性表示为通过每种细胞系相对于非结合对照抗体IgG1-b12(0%),以及针对REH,U266B1和Wien 133为IgG1-CAMPATH-1H-E430G(100%),或针对Daudi,Raji,Ramos和U-698-M为IgG1-11B8-E430G(100%)标准化的PI阳性细胞百分比确定的裂解百分比。
图4显示具有C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)的抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图5显示具有CD1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图6显示具有C1q结合抑制突变G236R(A)、K322A(B)、E269K(C)、K322E(D)或P329R(E)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有C1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图7显示具有C1q结合抑制突变G236R或K322A(A,B)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(C,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人淋巴瘤细胞系Wien 133(A,C)和Raji(B,D)的结合。通过流式细胞术测试抗体结合。结合表示为荧光强度的几何平均值(MFI)。使用没有一抗或非结合抗gp120抗体IgG1-b12的样品作为结合的阴性对照。
图8显示具有C1q结合抑制突变G236R或K322A(A,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(B,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体的ADCC能力。(A,B)进行了ADCC报告物生物测定(ADCC reporterBioassay),其中Raji靶细胞与抗体稀释系列与稳定表达高亲和力FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的NFAT反应元件的Jurkat人T细胞一起温育。萤光素酶产生通过发光读数定量。(C,D)进行体外铕TDA(EuTDA)ADCC测定,其中将Wien 133靶细胞与抗体稀释系列和人PBMC(E:T 100:1)一起温育。细胞裂解通过测量上清液中EuTDA荧光螯合物的信号来确定。
图9显示具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-K439E+具有或不具有C1q结合增强突变E333S的Fc-Fc相互作用增强的抗CD20 IgG1-11B8-S440K变体混合抗体变体的CDC活性的选择性。所测试的Fc-Fc相互作用增强突变是E430G、E345K、E345R和E345R-E430G。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图10显示了G236位置处(G236R或G236K)或G237位置处(G237A、G237T、G237Q或G237R)具有C1q结合调节突变或双突变G237A-K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合调节突变E333S、E333A、K326A、K326W-E333S、G237A或G237A-E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图11显示具有或不具有C1q结合抑制突变G236R的抗CD52 CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 11B8-E430G-S440K混合抗体同种型变体(IgG1,IgG2,IgG3和铰链稳定的IgG4)的CDC活性的选择性。在20%NHS存在下,将Wien133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图12显示具有或不具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD37 IgG1-CD37-37.3-E430G-S440K混合抗体变体对(A)Daudi和(B)Wien 133细胞的CDC活性的选择性。在20%NHS存在下,将靶细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-CD37-37.3-E430G混合物(100%)进行标准化。
图13显示BxPC-3人胰腺癌细胞上(A)单一抗体变体或(B)具有或不具有C1q结合抑制突变G236R的抗DR5 IgG1-DR5-01-G56T-E430G-K439E+具有或不具有C1q结合增强突变E333S的IgG1-DR5-05-E430G-S440K的抗体变体的激动剂混合物的DR5介导的细胞毒性。进行三天存活力测定,并使用CellTiter-Glo试剂盒确定细胞存活力。
图14显示具有C1q结合调节突变G237A、G236R、A327K、K322E或P329R的抗CD52IgG1-CAMPATH-1H-E430G的抗体变体的CDC活性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12-S440K(0%;未显示)和IgG1-CAMPATH-1H-E430G(100%)进行标准化。
图15显示具有C1q结合抑制突变G236R或K322A(A,B,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(D,E,F)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人FcRn的结合。用5μg/mL包被的重组人FcRn细胞外域(FcRnhsECDHis-B2M-BIO)和抗体稀释液系列进行FcRn ELISA。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图16显示SCID小鼠中500μg静脉内施用的抗体的清除率。(A-C)通过ELISA测定血浆样品的总人IgG,并针对(A)IgG1-CAMPATH-1H变体,(B)IgG1-11B8变体和(C)IgG1-CAMPATH-1H变体+IgG1-11B8变体的组合物,绘制浓度对时间的曲线。每个数据点代表一式三份样品的平均值+/-标准差。(D)抗体施用后直至第21天的清除率,用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1,000/AUC确定。
图17显示如ELISA测定所检测,具有C1q结合抑制突变G236R或G237T的固定的IgG1-CAMPATH-1H-E430G-K439E变体和具有C1q结合增强突变K326A或E333S的IgG1-11B8-E430G-S440K变体与FcγRIIA同种异型131H(A),FcγRIIA同种异型131R(B),FcγRIIB(C),FcγRIIIA同种异型158V(D)和FcγRIIIA同种异型158F(E)的加His标签的二聚生物素化ECD的结合。相对于无抗体对照(背景)和与IgG1-11B8-E430G-S440K的结合(100%),对20μg/mL抗体样品呈现结合。使用链霉亲合素-polyHRP和ABTS进行检测。
图18显示具有或不具有C1q结合抑制突变(G236R或G237T)的抗CD52IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的具有FcγR结合抑制突变G237A的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mL IgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图19显示通过血细胞的流式细胞术分析确定的全血中IgG1-CAMPATH-1H和IgG1-11B8的变体组合的选择性活性。Y轴:在效应细胞存在下进行过夜(o/n)温育后,总淋巴细胞群(CD66b阴性)的B细胞(CD19阳性/CD3阴性)分数或T细胞(CD19阴性/CD3阳性)分数。X轴:不同的治疗组。符号代表来自五个不同健康供体的细胞,每个供体在两次单独的温育中进行了测试。(A)IgG1-CAMPATH-1H-G236R-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(B)包含额外G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(C)包含额外G236R或G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合包含额外C1q结合增强E333S突变的IgG1-11B8-G237A-E430G-S440K的选择性活性。(D)与具有额外突变G236R、G237Q或G237R的IgG1-CAMPATH-1H-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的共依赖性抗体组合相比,不同B细胞靶向抗体的B细胞消耗的深度。Y轴:如上确定的B细胞分数的对数标度表示。
图20显示具有抗CD52组分中C1q结合抑制突变以及抗CD20组分中C1q结合增强突变的IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体的组合对具有CD20和CD52的不同表达水平的不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特氏淋巴瘤细胞系Daudi(A),Raji(B)和Ramos(C),ALL细胞系REH(D)和B细胞淋巴瘤细胞系U-698-M(E),用0.01-40μg/mL抗体进行体外CDC测定。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图21显示具有或不具有G236R C1q结合抑制突变的抗-CD37 IgG1-CD37-37.3-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将(A)Daudi细胞和(B)WIL2-S细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mLIgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CD37-37.3-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图22显示(A)具有Fc:Fc相互作用增强突变E430G或E345R,自身寡聚化抑制突变K439E组合任何FcγR结合抑制和C1q结合调节突变G236R、G237A或G237T的抗CD52 IgG1-CAMPATH-1H抗体变体与人淋巴瘤细胞系Wien 133的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-Campath-1H(100%)的Bmax值标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(B)具有突变E430G和K439E组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-Campath-1H的结合标准化显示。(C)具有E430G和S440K突变组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞结合的表观Kd值。
图23显示具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K组合任何C1q结合调节突变K326A或E333A(A)或E333S、G237A或G237A-E333S(B)的抗CD20 IgG1-11B8抗体变体与人淋巴瘤细胞系Raji的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-11B8的Bmax值(100%)标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(C,D)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(C)或E333S、G237A或G237A-E333S(D)的IgG1-11B8抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-11B8的结合标准化显示。(E,F)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(E)或E333S、G237A或G237A-E333S(F)的IgG1-11B8抗体变体与Raji细胞结合的表观Kd值。
图24显示抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的FcRn结合。(A,C)显示使用40μg/ml抗体浓度于(A)pH 6.0或(C)pH 7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E和C1q结合调节突变G237A或G237T的抗CD52抗体IgG1-CAMPATH-1H的变体与人FcRn的结合。(B,D)显示使用40μg/ml抗体浓度于(B)pH 6.0或(D)pH7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和C1q结合调节突变K326A、E333A、G237A或G237A-E333S的抗CD20抗体IgG1-11B8的变体与人FcRn的结合。FcRnELISA用2μg/mL包被的重组人FcRn胞外域(FcRnECDHis-B2M-BIO)和抗体稀释系列进行。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图25显示从用抗CD52 IgG1-CAMPATH-1H或抗-IgG1-11B8抗体变体或其混合物注射的小鼠收集的血液样品中测量的总人IgG(hIgG)浓度。(A)从用野生型IgG1-CAMPATH-1H,IgG1-CAMPATH-1H-E430G-K439E-G237Q或IgG1-CAMPATH-1H-E430G-K439E-G236R注射的小鼠收集的血液样品的总hIgG浓度。(B)从用野生型IgG1-11B8,IgG1-11B8-E430G-S440K-G237A或IgG1-11B8-E430G-S440K-E333S注射的小鼠收集的血液样品的总hIgG浓度。(C)从用野生型IgG1-CAMPATH-1H+IgG1-11B8混合物或带有(A)和(B)中突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物注射的小鼠收集的血液样品的总hIgG浓度。在所有图中,虚线表示SCID小鼠中野生型IgG1抗体随时间预测的IgG1浓度。(D)抗体施用后直至第21天的清除率用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1000/AUC确定。
图26显示减去在不含抗体的阴性对照样品中检测的平均C4d浓度后,用带有突变E430G、K439E或S440K和G236R、G237A、G237Q或G237R的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12的抗体变体温育的样品中检测的C4d浓度。阳性对照样品包括带有E345R、E430G和S440Y Fc-Fc相互作用增强突变(RGY)的抗体变体。
图27显示用带有E430G、K439E或S440K和G236R、G237T、K326A或E333S突变的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12抗体变体调理后,通过流式细胞术检测C1q与Wien 133细胞的结合,所述Wien 133细胞于冰上用正常人血清作为补体来源温育。将平均荧光强度值相对于无抗体的对照反应(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物的最高水平(100%)标准化,通过拟合对数激动剂反应模型进行估算。(A)对照反应的C1q结合。(B-D)(B)IgG1-CAMPATH-E430G-K439E,(C)IgG1-CAMPATH-E430G-K439E-G236R和(D)IgG1-CAMPATH-E430G-K439E-G237R混合非结合对照IgG1-b12或不同的IgG1-11B8变体的C1q结合。
图28显示带有自身寡聚化抑制突变K439E或S440K和C1q结合调节突变的IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G抗体变体的FcγR结合。(A-E)如ELISA测定所检测,固定化抗体变体与(A)高亲和力同种异型FcγRIIA 131H,(B)低亲和力同种异型FcγRIIA131R,(C)FcγRIIB,(D)高亲和力同种异型FcγRIIIA158V,或(E)低亲和力同种异型FcγRIIIA158F的加His标签的二聚生物素化ECD的结合。(F)ELISA检测的固定化FcγRIa与抗体变体的结合。表示了20μg/mL抗体样品的结合,并且每个实验在减去不与一抗温育的孔中的信号后相对于野生型IgG1-CAMPATH-1H所观测的平均信号(100%)标准化。使用链霉亲合素-polyHRP和ABTS进行检测。
图29显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的抗CD52IgG1-CAMPATH-1H,抗CD20 IgG1-11B8和抗CD52 IgG1-h2E8混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。在三个独立实验中测量了CDC效力,并表示为(A)平均AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化,和(B)在40μg/ml的抗体浓度时,由碘化丙啶阳性确定的平均裂解百分比。
图30(A,B)显示单一试剂和组合的带有所示自身寡聚化抑制突变和C1q结合调节突变的抗CD52 IgG1-CAMPATH-1H-E430G,抗CD20 IgG1-11B8-E430G和非抗原结合IgG1-b12-E430G抗体变体的CDC效力。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图31显示与Raji淋巴瘤细胞和不同浓度的IgG1-CAMPATH-1H和IgG1-11B8抗体变体共培养时,通过发光水平(RLU)测定的稳定表达(A)FcγRIIa或(B)FcγRIIIa的Jurkat报告物细胞系的激活。每个实验的发光值相对于IgG1-b12(0%)和野生型IgG1-Campath-1H+野生型IgG1-11B8(100%)观测到的值标准化,然后求三个(FcγRIIa)或两个(FcγRIIIa)实验重复的平均值。
图32显示由非等摩尔比的带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物诱导的对Wien 133细胞的共依赖性CDC。(A)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。(B)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。
图33显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H抗体变体与抗HLA-DR IgG1-HLA-DR-huL243变体(A)或抗HLA-DRIgG1-HLA-DR-1D09C3变体(B)混合物的CDC活性的选择性。在20%NHS存在下,将Oci-Ly17细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和(A)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-huL243-E430G(100%)或(B)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-1D09C3-E430G混合物(100%)标准化。
图34显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。显示具有突变L234A、L234A-L235A、L234F、L234F-L235E、L235A、L235Q、G236R或G237Q的IgG1-CAMPATH-1H-E430G-K439E变体以及这些变体与非结合对照抗体IgG1-b12或IgG1-11B8-E430G-S440K混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图35显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。(A)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345K、E345Q、E345R或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(B)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(C)带有匹配的Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8的抗体变体作为单一试剂或其混合物的CDC效力。(D)带有Fc-Fc相互作用增强突变E430G或K248E-T437R,自身寡聚化抑制突变K439E或S440K,和C1q结合调节突变G236R、G237A或E333S的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图36显示由带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CD37-37-3和IgG1-11B8抗体变体混合物诱导的对Raji淋巴瘤细胞的共依赖性CDC。(A)由稀释的所示抗体变体或其混合物诱导的细胞裂解的曲线下相对面积(AUC),相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CD37-37-3-E430G+IgG1-11B8-E430G混合物为100%)标准化。(B)由所示抗体变体及其混合物诱导的最大裂解百分比。
图37显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。在20%NHS存在下,将患者CLL样品与抗体浓度系列一起温育。CDC效力表示为与抗体变体温育后活的B细胞的百分比。示出使用来自患者1(A),2(B)和3(C)的CLL样品的结果。
图38显示全血样品与带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM抗体变体混合物温育后,通过流式细胞术检测的B细胞,CD4+ T细胞和CD8+T细胞的分数。在与所示IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(A)B细胞,(B)CD4+ T细胞和(C)CD8+ T细胞的百分比。在与所示IgG1-CAMPATH-1H,IgG1-CD5-INSERM和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(D)B细胞,(E)CD4+ T细胞和(F)CD8+ T细胞的百分比。分数计算为[100%x(样品的细胞计数/“无Ab样品”的细胞计数)x(“无Ab样品”的粒细胞计数/样品的粒细胞计数)]。
图39显示了带有增强Fc-Fc相互作用,抑制自身寡聚化和抑制(G237T)或增强(K326W-E333S)C1q结合的突变的抗DR4和抗DR5抗体变体在癌细胞中协同激活程序性细胞死亡。(A)与所示抗体变体温育72小时后,BxPC-3人胰腺癌细胞的存活力。(B)与所示抗体变体温育72小时后,COLO 205人结肠癌细胞的存活力。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
A-B).对于IgG1-11B8-E430G,引入S440K突变(SEQ ID NO:101)或S440K-P329R(SEQ ID NO:114)导致CDC效力的丧失。(图1显示通过引入P329R突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%合并的正常人血清(NHS)存在下,将Wien 133细胞与浓度抗体浓度系列一起温育。CDC效力表示为(A)由碘化丙啶(PI)阳性细胞百分比确定的裂解百分比,和(B)剂量反应-反应曲线下面积(AUC),相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图2显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为(A)由PI阳性细胞百分比确定的裂解百分比,和(B)AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图3显示通过引入K322E突变,抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20-11B8-E430G-S440K混合抗体变体对不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特(Burkitt)氏淋巴瘤细胞系Wien 133(A),Daudi(B),Raji(C)和Ramos(D),急性成淋巴细胞性淋巴瘤(ALL)细胞系REH(E),骨髓瘤细胞系U266B1(F)和B细胞淋巴瘤细胞系U-698-M(G),用30μg/mL抗体进行体外CDC测定。CDC活性表示为通过每种细胞系相对于非结合对照抗体IgG1-b12(0%),以及针对REH,U266B1和Wien 133为IgG1-CAMPATH-1H-E430G(100%),或针对Daudi,Raji,Ramos和U-698-M为IgG1-11B8-E430G(100%)标准化的PI阳性细胞百分比确定的裂解百分比。
图4显示具有C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)的抗CD52IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图5显示具有CD1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图6显示具有C1q结合抑制突变G236R(A)、K322A(B)、E269K(C)、K322E(D)或P329R(E)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有C1q结合增强突变(E333S、K326W或K326A/E333A)的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体混合物IgG1-b12-K439E+IgG1-b12-S440K(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图7显示具有C1q结合抑制突变G236R或K322A(A,B)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(C,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人淋巴瘤细胞系Wien 133(A,C)和Raji(B,D)的结合。通过流式细胞术测试抗体结合。结合表示为荧光强度的几何平均值(MFI)。使用没有一抗或非结合抗gp120抗体IgG1-b12的样品作为结合的阴性对照。
图8显示具有C1q结合抑制突变G236R或K322A(A,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(B,D)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体的ADCC能力。(A,B)进行了ADCC报告物生物测定(ADCC reporterBioassay),其中Raji靶细胞与抗体稀释系列与稳定表达高亲和力FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的NFAT反应元件的Jurkat人T细胞一起温育。萤光素酶产生通过发光读数定量。(C,D)进行体外铕TDA(EuTDA)ADCC测定,其中将Wien 133靶细胞与抗体稀释系列和人PBMC(E:T 100:1)一起温育。细胞裂解通过测量上清液中EuTDA荧光螯合物的信号来确定。
图9显示具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-K439E+具有或不具有C1q结合增强突变E333S的Fc-Fc相互作用增强的抗CD20 IgG1-11B8-S440K变体混合抗体变体的CDC活性的选择性。所测试的Fc-Fc相互作用增强突变是E430G、E345K、E345R和E345R-E430G。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图10显示了G236位置处(G236R或G236K)或G237位置处(G237A、G237T、G237Q或G237R)具有C1q结合调节突变或双突变G237A-K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合调节突变E333S、E333A、K326A、K326W-E333S、G237A或G237A-E333S的抗CD20IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图11显示具有或不具有C1q结合抑制突变G236R的抗CD52 CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 11B8-E430G-S440K混合抗体同种型变体(IgG1,IgG2,IgG3和铰链稳定的IgG4)的CDC活性的选择性。在20%NHS存在下,将Wien133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图12显示具有或不具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD37 IgG1-CD37-37.3-E430G-S440K混合抗体变体对(A)Daudi和(B)Wien 133细胞的CDC活性的选择性。在20%NHS存在下,将靶细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-CD37-37.3-E430G混合物(100%)进行标准化。
图13显示BxPC-3人胰腺癌细胞上(A)单一抗体变体或(B)具有或不具有C1q结合抑制突变G236R的抗DR5 IgG1-DR5-01-G56T-E430G-K439E+具有或不具有C1q结合增强突变E333S的IgG1-DR5-05-E430G-S440K的抗体变体的激动剂混合物的DR5介导的细胞毒性。进行三天存活力测定,并使用CellTiter-Glo试剂盒确定细胞存活力。
图14显示具有C1q结合调节突变G237A、G236R、A327K、K322E或P329R的抗CD52IgG1-CAMPATH-1H-E430G的抗体变体的CDC活性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC。相对于非结合对照抗体IgG1-b12-S440K(0%;未显示)和IgG1-CAMPATH-1H-E430G(100%)进行标准化。
图15显示具有C1q结合抑制突变G236R或K322A(A,B,C)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E的抗体变体和具有C1q结合增强突变E333S(D,E,F)的抗CD20 IgG1-11B8-E430G-S440K的抗体变体与人FcRn的结合。用5μg/mL包被的重组人FcRn细胞外域(FcRnhsECDHis-B2M-BIO)和抗体稀释液系列进行FcRn ELISA。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图16显示SCID小鼠中500μg静脉内施用的抗体的清除率。(A-C)通过ELISA测定血浆样品的总人IgG,并针对(A)IgG1-CAMPATH-1H变体,(B)IgG1-11B8变体和(C)IgG1-CAMPATH-1H变体+IgG1-11B8变体的组合物,绘制浓度对时间的曲线。每个数据点代表一式三份样品的平均值+/-标准差。(D)抗体施用后直至第21天的清除率,用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1,000/AUC确定。
图17显示如ELISA测定所检测,具有C1q结合抑制突变G236R或G237T的固定的IgG1-CAMPATH-1H-E430G-K439E变体和具有C1q结合增强突变K326A或E333S的IgG1-11B8-E430G-S440K变体与FcγRIIA同种异型131H(A),FcγRIIA同种异型131R(B),FcγRIIB(C),FcγRIIIA同种异型158V(D)和FcγRIIIA同种异型158F(E)的加His标签的二聚生物素化ECD的结合。相对于无抗体对照(背景)和与IgG1-11B8-E430G-S440K的结合(100%),对20μg/mL抗体样品呈现结合。使用链霉亲合素-polyHRP和ABTS进行检测。
图18显示具有或不具有C1q结合抑制突变(G236R或G237T)的抗CD52 IgG1-CAMPATH-1H-E430G-K439E+具有或不具有C1q结合增强突变E333S的具有FcγR结合抑制突变G237A的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mL IgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图19显示通过血细胞的流式细胞术分析确定的全血中IgG1-CAMPATH-1H和IgG1-11B8的变体组合的选择性活性。Y轴:在效应细胞存在下进行过夜(o/n)温育后,总淋巴细胞群(CD66b阴性)的B细胞(CD19阳性/CD3阴性)分数或T细胞(CD19阴性/CD3阳性)分数。X轴:不同的治疗组。符号代表来自五个不同健康供体的细胞,每个供体在两次单独的温育中进行了测试。(A)IgG1-CAMPATH-1H-G236R-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(B)包含额外G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合IgG1-11B8-G237A-E430G-S440K的选择性活性。(C)包含额外G236R或G237突变的IgG1-CAMPATH-1H-E430G-K439E变体混合包含额外C1q结合增强E333S突变的IgG1-11B8-G237A-E430G-S440K的选择性活性。(D)与具有额外突变G236R、G237Q或G237R的IgG1-CAMPATH-1H-E430G-K439E混合IgG1-11B8-G237A-E430G-S440K的共依赖性抗体组合相比,不同B细胞靶向抗体的B细胞消耗的深度。Y轴:如上确定的B细胞分数的对数标度表示。
图20显示具有抗CD52组分中C1q结合抑制突变以及抗CD20组分中C1q结合增强突变的IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体的组合对具有CD20和CD52的不同表达水平的不同细胞系的CDC活性的选择性。在20%NHS存在下,使用伯基特氏淋巴瘤细胞系Daudi(A),Raji(B)和Ramos(C),ALL细胞系REH(D)和B细胞淋巴瘤细胞系U-698-M(E),用0.01-40μg/mL抗体进行体外CDC测定。CDC效力表示为PI阳性细胞百分比的标准化AUC和最大裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图21显示具有或不具有G236R C1q结合抑制突变的抗-CD37 IgG1-CD37-37.3-E430G-K439E+具有或不具有C1q结合增强突变E333S的抗CD20 IgG1-11B8-E430G-S440K混合抗体变体的CDC活性的选择性。在20%NHS存在下,将(A)Daudi细胞和(B)WIL2-S细胞与抗体浓度系列一起温育。CDC效力表示为PI阳性细胞百分比的标准化AUC,并表示为40μg/mLIgG时的裂解。相对于非结合对照抗体IgG1-b12(0%)和IgG1-CD37-37.3-E430G+IgG1-11B8-E430G混合物(100%)进行标准化。
图22显示(A)具有Fc:Fc相互作用增强突变E430G或E345R,自身寡聚化抑制突变K439E组合任何FcγR结合抑制和C1q结合调节突变G236R、G237A或G237T的抗CD52 IgG1-CAMPATH-1H抗体变体与人淋巴瘤细胞系Wien 133的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-Campath-1H(100%)的Bmax值标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(B)具有突变E430G和K439E组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-Campath-1H的结合标准化显示。(C)具有E430G和S440K突变组合任何C1q结合调节突变G236R、G237A或G237T的IgG1-Campath-1H抗体变体与Raji细胞结合的表观Kd值。
图23显示具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K组合任何C1q结合调节突变K326A或E333A(A)或E333S、G237A或G237A-E333S(B)的抗CD20 IgG1-11B8抗体变体与人淋巴瘤细胞系Raji的结合。抗体结合通过流式细胞术测试,并相对于野生型IgG1-11B8的Bmax值(100%)标准化表示。使用非结合抗gp120抗体IgG1-b12作为结合的阴性对照。(C,D)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(C)或E333S、G237A或G237A-E333S(D)的IgG1-11B8抗体变体与Raji细胞的最大结合(Bmax)相对于野生型IgG1-11B8的结合标准化显示。(E,F)具有突变E430G和S440K组合任何C1q结合调节突变K326A或E333A(E)或E333S、G237A或G237A-E333S(F)的IgG1-11B8抗体变体与Raji细胞结合的表观Kd值。
图24显示抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的FcRn结合。(A,C)显示使用40μg/ml抗体浓度于(A)pH 6.0或(C)pH 7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E和C1q结合调节突变G237A或G237T的抗CD52抗体IgG1-CAMPATH-1H的变体与人FcRn的结合。(B,D)显示使用40μg/ml抗体浓度于(B)pH 6.0或(D)pH7.4时具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和C1q结合调节突变K326A、E333A、G237A或G237A-E333S的抗CD20抗体IgG1-11B8的变体与人FcRn的结合。FcRnELISA用2μg/mL包被的重组人FcRn胞外域(FcRnECDHis-B2M-BIO)和抗体稀释系列进行。结合抗体的量用缀合HRP的山羊抗人IgG1抗体和化学发光底物ABTS确定。在405nm处测量吸光度。
图25显示从用抗CD52 IgG1-CAMPATH-1H或抗-IgG1-11B8抗体变体或其混合物注射的小鼠收集的血液样品中测量的总人IgG(hIgG)浓度。(A)从用野生型IgG1-CAMPATH-1H,IgG1-CAMPATH-1H-E430G-K439E-G237Q或IgG1-CAMPATH-1H-E430G-K439E-G236R注射的小鼠收集的血液样品的总hIgG浓度。(B)从用野生型IgG1-11B8,IgG1-11B8-E430G-S440K-G237A或IgG1-11B8-E430G-S440K-E333S注射的小鼠收集的血液样品的总hIgG浓度。(C)从用野生型IgG1-CAMPATH-1H+IgG1-11B8混合物或带有(A)和(B)中突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物注射的小鼠收集的血液样品的总hIgG浓度。在所有图中,虚线表示SCID小鼠中野生型IgG1抗体随时间预测的IgG1浓度。(D)抗体施用后直至第21天的清除率用D,注射剂量和AUC(浓度-时间曲线的曲线下面积),按照式D*1000/AUC确定。
图26显示减去在不含抗体的阴性对照样品中检测的平均C4d浓度后,用带有突变E430G、K439E或S440K和G236R、G237A、G237Q或G237R的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12的抗体变体温育的样品中检测的C4d浓度。阳性对照样品包括带有E345R、E430G和S440Y Fc-Fc相互作用增强突变(RGY)的抗体变体。
图27显示用带有E430G、K439E或S440K和G236R、G237T、K326A或E333S突变的IgG1-CAMPATH-1H,IgG1-11B8和IgG1-b12抗体变体调理后,通过流式细胞术检测C1q与Wien 133细胞的结合,所述Wien 133细胞于冰上用正常人血清作为补体来源温育。将平均荧光强度值相对于无抗体的对照反应(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物的最高水平(100%)标准化,通过拟合对数激动剂反应模型进行估算。(A)对照反应的C1q结合。(B-D)(B)IgG1-CAMPATH-E430G-K439E,(C)IgG1-CAMPATH-E430G-K439E-G236R和(D)IgG1-CAMPATH-E430G-K439E-G237R混合非结合对照IgG1-b12或不同的IgG1-11B8变体的C1q结合。
图28显示带有自身寡聚化抑制突变K439E或S440K和C1q结合调节突变的IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G抗体变体的FcγR结合。(A-E)如ELISA测定所检测,固定化抗体变体与(A)高亲和力同种异型FcγRIIA 131H,(B)低亲和力同种异型FcγRIIA131R,(C)FcγRIIB,(D)高亲和力同种异型FcγRIIIA 158V,或(E)低亲和力同种异型FcγRIIIA158F的加His标签的二聚生物素化ECD的结合。(F)ELISA检测的固定化FcγRIa与抗体变体的结合。表示了20μg/mL抗体样品的结合,并且每个实验在减去不与一抗温育的孔中的信号后相对于野生型IgG1-CAMPATH-1H所观测的平均信号(100%)标准化。使用链霉亲合素-polyHRP和ABTS进行检测。
图29显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的抗CD52IgG1-CAMPATH-1H,抗CD20 IgG1-11B8和抗CD52 IgG1-h2E8混合抗体变体的CDC活性的选择性。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。在三个独立实验中测量了CDC效力,并表示为(A)平均AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化,和(B)在40μg/ml的抗体浓度时,由碘化丙啶阳性确定的平均裂解百分比。
图30(A,B)显示单一试剂和组合的带有所示自身寡聚化抑制突变和C1q结合调节突变的抗CD52 IgG1-CAMPATH-1H-E430G,抗CD20 IgG1-11B8-E430G和非抗原结合IgG1-b12-E430G抗体变体的CDC效力。在20%NHS存在下,将Wien 133细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图31显示与Raji淋巴瘤细胞和不同浓度的IgG1-CAMPATH-1H和IgG1-11B8抗体变体共培养时,通过发光水平(RLU)测定的稳定表达(A)FcγRIIa或(B)FcγRIIIa的Jurkat报告物细胞系的激活。每个实验的发光值相对于IgG1-b12(0%)和野生型IgG1-Campath-1H+野生型IgG1-11B8(100%)观测到的值标准化,然后求三个(FcγRIIa)或两个(FcγRIIIa)实验重复的平均值。
图32显示由非等摩尔比的带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的IgG1-CAMPATH-1H和IgG1-11B8抗体变体混合物诱导的对Wien 133细胞的共依赖性CDC。(A)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。(B)由包含等摩尔和非等摩尔浓度比的IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A的混合物诱导的共依赖性CDC。
图33显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H抗体变体与抗HLA-DR IgG1-HLA-DR-huL243变体(A)或抗HLA-DRIgG1-HLA-DR-1D09C3变体(B)混合物的CDC活性的选择性。在20%NHS存在下,将Oci-Ly17细胞与抗体浓度系列一起温育。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和(A)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-huL243-E430G(100%)或(B)IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-1D09C3-E430G混合物(100%)标准化。
图34显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。显示具有突变L234A、L234A-L235A、L234F、L234F-L235E、L235A、L235Q、G236R或G237Q的IgG1-CAMPATH-1H-E430G-K439E变体以及这些变体与非结合对照抗体IgG1-b12或IgG1-11B8-E430G-S440K混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图35显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和抑制C1q结合的突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。(A)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345K、E345Q、E345R或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(B)带有Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。(C)带有匹配的Fc-Fc相互作用增强突变E430G、E430N、E430T、E430V、E430Y、E345A、E345Q、E345V或E345Y,自身寡聚化抑制突变K439E或S440K,和C1q结合抑制突变G236R或G237A的IgG1-CAMPATH-1H和IgG1-11B8的抗体变体作为单一试剂或其混合物的CDC效力。(D)带有Fc-Fc相互作用增强突变E430G或K248E-T437R,自身寡聚化抑制突变K439E或S440K,和C1q结合调节突变G236R、G237A或E333S的IgG1-CAMPATH-1H和IgG1-11B8抗体变体作为单一试剂或其混合物的CDC效力。CDC效力表示为AUC,相对于非结合对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物(100%)标准化。
图36显示由带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CD37-37-3和IgG1-11B8抗体变体混合物诱导的对Raji淋巴瘤细胞的共依赖性CDC。(A)由稀释的所示抗体变体或其混合物诱导的细胞裂解的曲线下相对面积(AUC),相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CD37-37-3-E430G+IgG1-11B8-E430G混合物为100%)标准化。(B)由所示抗体变体及其混合物诱导的最大裂解百分比。
图37显示通过引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变,抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8混合抗体变体的选择性共依赖性CDC活性。在20%NHS存在下,将患者CLL样品与抗体浓度系列一起温育。CDC效力表示为与抗体变体温育后活的B细胞的百分比。示出使用来自患者1(A),2(B)和3(C)的CLL样品的结果。
图38显示全血样品与带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合突变的IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM抗体变体混合物温育后,通过流式细胞术检测的B细胞,CD4+ T细胞和CD8+T细胞的分数。在与所示IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(A)B细胞,(B)CD4+ T细胞和(C)CD8+ T细胞的百分比。在与所示IgG1-CAMPATH-1H,IgG1-CD5-INSERM和IgG1-b12抗体变体温育后,在4个供体的全血样品中检测的(D)B细胞,(E)CD4+ T细胞和(F)CD8+ T细胞的百分比。分数计算为[100%x(样品的细胞计数/“无Ab样品”的细胞计数)x(“无Ab样品”的粒细胞计数/样品的粒细胞计数)]。
图39显示了带有增强Fc-Fc相互作用,抑制自身寡聚化和抑制(G237T)或增强(K326W-E333S)C1q结合的突变的抗DR4和抗DR5抗体变体在癌细胞中协同激活程序性细胞死亡。(A)与所示抗体变体温育72小时后,BxPC-3人胰腺癌细胞的存活力。(B)与所示抗体变体温育72小时后,COLO 205人结肠癌细胞的存活力。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
A-B).
抗CD20 IgG1-11B8-E430G-S440K(无单一试剂CDC活性)+抗CD52 IgG1-CAMPATH-1H-E430G-K439E(部分单一试剂CDC活性)混合物完全恢复了对Wien 133的最大CDC活性,类似于靶向CD20和CD52的混合物IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G的水平(图1A中为相似的剂量反应;图1B中为100%)。将抗CD20 IgG1-11B8-E430G-S440K(无单一试剂CDC活性)添加至抗CD52 IgG1-CAMPATH-1H-P329R-E430G-K439E(无单一试剂CDC活性),在Wien133上部分恢复细胞裂解(图1A中饱和靶标结合时为56%;图1B中为28%)。
将抗CD20 IgG1-11B8-P329R-E430G-S440K(无单一试剂CDC活性)添加到抗CD52IgG1-CAMPATH-1H-E430G-K439E(部分单一试剂CDC活性),在Wien 133上部分恢复CDC活性至低于IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物的水平(图1A中较高的EC50和较低的最大杀伤力;图1B中为47%),但适度高于抗CD52 IgG1-CAMPATH-1H-E430G-K439E+对照抗体(IgG1-b12)混合物,其具有相似的最大杀伤力,但较高的EC50(图1A)和较低的相对AUC(图1B中为36%)。相反,将抗CD20 IgG1-11B8-P329R-E430G-S440K(无单一试剂CDC活性)添加至抗CD52 IgG1-CAMPATH-1H-P329R-E430G-K439E(无单一试剂CDC活性)不恢复任何CDC活性(图1A-B)。
这些数据说明,引入抑制直接C1q结合的突变P329R可以进一步抑制具有增强Fc-Fc相互作用(E430G)的K439E+S440K抗体混合物中单个组分的CDC活性。出乎意料的是,观测到对于作为单一试剂均未显示可检测CDC活性的两种针对两种不同靶标的抗体,对于两种抗体中仅一种包含C1q结合抑制突变K322E的混合物可部分恢复CDC活性。这些数据表明,通过引入C1q结合抑制突变P329R以降低或抑制单一试剂活性,可将抗CD20 IgG-E430G-S440K+抗CD52 IgG-E430G-K439E抗体混合物应用于产生对同时表达两种不同靶标的细胞的CDC活性的选择性。不受理论的限制,异源六聚体IgG组件中的三个抗体中C1q结合位点的C1q亲合力可能高得足以恢复部分CDC活性。相反,所有六个抗体中失去所有六个C1q结合位点,例如均包含P329R突变的两种抗体混合物中,导致CDC活性丧失。
实施例3:通过在具有Fc-Fc相互作用增强突变E430G的抗CD52 IgG1-CAMPATH-1H-K439E+抗CD20 IgG1-11B8-S440K中引入K322E突变的混合抗体变体的CDC活性的选择性
使用抗CD20抗体IgG1-11B8和抗CD52抗体IgG1-CAMPATH-1H的变体混合物测试了突变K322E对体外CDC效力的影响。基本如实施例2所述,在含20%NHS的情况下对Wien 133细胞进行的体外CDC测定中测试了不同浓度的纯化抗体(最终浓度范围为0.001-30.0μg/mL)。在抗体IgG1-11B8和IgG1-CAMPATH-1H:E430G中引入了不同的突变:诱导增强Fc-Fc相互作用的E430G;抑制C1q直接与抗体结合的K322E;以及突变K439E或S440K,其通过抑制分子间Fc-Fc相互作用抑制同源六聚体抗体复合物形成和通过交叉互补的Fc-Fc相互作用促进异源六聚体抗体复合物形成。由PI阳性细胞数目计算裂解百分比。使用对数转化的浓度,使用非线性剂量反应拟合的最佳拟合值分析数据,并计算三个实验重复的剂量反应曲线下面积。相对曲线下面积(AUC)值表示相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物为100%)标准化。
抗CD52 IgG1-CAMPATH-1H-E430G+抗CD20 IgG1-11B8-E430G的1:1混合物诱导Wien 133细胞的高效细胞裂解(错误!未找到参考源。A;设为100%错误!找不到参考源。B)。将IgG1-CAMPATH-1H-E430G作为单一试剂测试时,引入K439E突变导致CDC效力降低,而K439E-K322E(SEQ ID NO:91)导致CDC活性完全丧失(错误!未找到参考源。A-B)。对于IgG1-11B8-E430G,引入S440K突变或S440K-K322E(SEQ ID NO:109)导致CDC效力丧失(错误!未找到参考源。A-B)。
抗CD20 IgG1-11B8-E430G-S440K(无单一试剂CDC活性)+抗CD52 IgG1-CAMPATH-1H-E430G-K439E(部分单一试剂CDC活性)混合物恢复对Wien 133细胞的高CDC活性,与CD20和CD52靶向性混合物IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G的水平相当(图2A中相当的剂量反应;图2B中为87%)。将抗CD20 IgG1-11B8-E430G-S440K(无单一试剂CDC活性)添加至抗CD52 IgG1-CAMPATH-1H-K322E-E430G-K439E(无单一试剂CDC活性)部分恢复对Wien133的细胞裂解(图2A中比IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物更高的EC50;图2B中为60%)。
将抗CD20 IgG1-11B8-K322E-E430G-S440K(无单一试剂CDC活性)添加到抗CD52IgG1-CAMPATH-1H-E430G-K439E(部分单一试剂CDC活性)部分恢复对Wien 133的CDC活性(图2A中比IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物更高的EC50;图2B中为66%)。相反,将抗CD20 IgG1-11B8-K322E-E430G-S440K(无单一试剂CDC活性)添加到抗CD52IgG1-CAMPATH-1H-K322E-E430G-K439E(无单一试剂CDC活性)只能恢复最小的CDC活性(在图2A中高EC50且最大杀伤约31%;在图2B为10%)。
这些数据表明,引入抑制直接C1q结合的突变K322E,可以进一步抑制具有增强Fc-Fc相互作用(E430G)的K439E+S440K抗体混合物中单个组分的CDC活性。出乎意料的是,观察到对于作为单一试剂均未显示可检测CDC活性的两种针对两种不同靶标的抗体,两种抗体中仅一种包含C1q结合抑制突变K322E的混合物,可恢复CDC活性至接近最大细胞裂解。这些数据表明,通过引入C1q结合抑制突变K322E以降低或抑制单一试剂活性,可将抗CD20IgG-E430G-S440K+抗CD52 IgG-E430G-K439E抗体混合物应用于产生对同时表达两种不同靶标的细胞的CDC活性的选择性。
实施例4:通过在具有Fc-Fc相互作用增强突变E430G的抗CD20+抗CD52抗体中引入K322E突变的混合抗体变体在不同细胞系上的CDC活性的选择性
实施例2和实施例3描述了Fc-Fc相互作用增强的抗CD20和抗CD52抗体的特定组合物在两种组分同时存在时显示对Wien 133细胞的最有效的CDC活性,条件是每种抗体都含有K439E或S440K突变,其通过Fc-Fc相互作用阻止同源寡聚化。混合物的选择性活性与其单独组分相比通过在抗CD52抗体IgG1-CAPMATH-1H-E430G-S440K中引入C1q结合抑制突变P329R(实施例2)或K322E(实施例3)以抑制其作为单一组分的残留CDC活性得到改善。在此,在7种不同的人细胞系上测试了具有K322E突变的抗CD20 IgG1-11B8-E430G-S440K+抗CD52IgG1-CAMPATH-1H-E430G-K439E抗体变体混合物对细胞表面表达足够水平的CD20和CD52的细胞的CDC活性的选择性:伯基特氏淋巴瘤细胞系Wien 133,Daudi(ATCC,目录号CCL-213),Raji(ATCC,目录号CCL-86)和Ramos(ATCC,目录号CRL-1596),急性成淋巴细胞性淋巴瘤(ALL)细胞系REH(DSMZ,目录号ACC-22),骨髓瘤细胞系U266B1(ATCC,目录号TIB-196),以及细胞淋巴瘤细胞系U-698-M(DSMZ,目录号ACC-4)。体外CDC测定采用20%NHS和30.0μg/mL的最终抗体浓度进行,基本如实施例2所述。细胞裂解由PI阳性细胞数目计算得到,取三个实验重复的平均值,每种细胞系相对于阴性对照抗体IgG1-b12(0%)和IgG1-CAMPATH-1H-E430G(100%,针对REH,U266B1和Wien 133)或IgG1-11B8-E430G(100%,针对Daudi,Raji,Ramos和U-698-M)所测量的细胞裂解标准化,取决于哪种抗体诱导最高的裂解。
使用人IgG校准试剂盒(Biocytex,目录号CP010),通过间接免疫荧光测定CD52和CD20的细胞表面表达水平。将100,000个细胞/孔接种到聚苯乙烯96孔圆底板(GreinerBio-One,目录号650101)。接下来的步骤于4℃进行。将细胞以300xg离心3分钟进行沉淀,并重悬于包含饱和浓度10μg/mL IgG1-CAMPATH-1H(抗CD52)或IgG1-11B8(抗-CD20)的50μLPBS(B Braun,目录号3623140)中。于4℃温育30分钟后,将细胞以300xg离心3分钟进行沉淀,并重悬于150μL FACS缓冲液(PBS+0.1%(w/v)牛血清白蛋白(BSA)+0.02%(w/v)叠氮化钠)。根据制造商说明,将设置和校准珠添加到板。将平行的细胞和珠用150μL FACS缓冲液再清洗两次,重悬于50μL缀合有FITC的小鼠IgG吸附的山羊抗人IgG(BioCytex)中,并于4℃温育30分钟。将细胞和珠清洗两次,并重悬于150μL FACS缓冲液中。将细胞重悬于固定液(BioCytex)中,并在避光条件下于4℃温育5至60分钟。通过记录活细胞群内10,000个事件,用FACS Canto ll(BD Biosciences)通过流式细胞术测量免疫荧光。使用GraphPad Prism软件,将校准珠的荧光强度几何平均值用于计算被迫经过零强度和零浓度的校准曲线。对于每种细胞系,使用GraphPad软件,基于校正曲线方程(标准曲线中未知数的插值),代表CD52和CD20细胞表面表达的IgG1-CAMPATH-1H和IgG1-11B8的抗体结合能力(ABC)分别使用几何平均荧光强度,接着减去没有一抗温育的孔中测定的背景值来计算。从两次独立实验对ABC值得出平均值(总结于表2中)。
图3显示靶标饱和时IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G的CDC活性随靶标表达水平而变化:U266B1(CD20 ABC<20,000;CD52 ABC>1x106)和REH(CD20 ABC<20,000;CD52 ABC>100,000)细胞对抗CD20抗体IgG1-11B8-E430G的溶解具有抗性(resilient),但对抗CD52抗体IgG1-CAMPATH-1H-E430G敏感。反之亦然,Daudi细胞(CD52 ABC<75,000;CD20ABC>100,000)对IgG1-CAMPATH-1H-E430G有抗性,但对IgG1-11B8-E430G敏感。其他四种测试的细胞系对IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G均敏感:Wien 133(CD20ABC>100,000;CD52 ABC>300,000),Raji(CD20 ABC>100,000;CD52 ABC>85,000),U-698-M(CD20ABC>70,000;CD52:ABC>90,000)和Ramos(CD20 ABC>80,000;CD52 ABC>175,000)。
对于IgG1-CAMPATH-1H-E430G,引入K439E突变导致对U-698-M和Raji细胞的单一试剂CDC活性降低(CD52 ABC<100,000),但对细胞系U266B1(CD52 ABC>1x106)细胞,Wien133(CD52 ABC>300,000),Ramos(CD52 ABC>175,000)和REH细胞(CD52 ABC>135,000)没有明显影响。通过在IgG1-CAMPATH-1H-E430G-K439E中引入K322E的C1q结合抑制进一步消除对响应于IgG1-CAMPATH-1H-E430G的所有测试细胞系的单一试剂CDC活性。对于IgG1-11B8-E430G,仅引入S440K突变或引入S440K和K322E突变两者导致完全抑制对IgG1-11B8-E430G敏感的所有测试细胞系的单一试剂CDC活性:Daudi(CD20 ABC<115,000),Wien 133(CD20ABC<110,000),Raji(CD20 ABC<120,000),U-698-M(CD20 ABC<75,000)和Ramos(CD20 ABC<85,000)。
IgG1-CAMPATH-1H-E430G-K439E+IgG1-11B8-E430G-S440K混合物对所有测试细胞系均显示CDC活性,而与CD20和CD52表面表达水平无关(图3)。与之形成鲜明对比的是,IgG1-CAMPATH-1H-E430G-K439E-K322E+IgG1-11B8-E430G-S440K混合物仅显示选择性裂解展示CD20和CD52两者的充足表面表达水平的那些细胞系:Wien 133(CD20 ABC>100,000;CD52 ABC>300,000),Ramos(CD20 ABC>80,000;CD52 ABC>175,000),U-698-M(CD20 ABC>70,000;CD52 ABC>90,000)和Raji(CD20 ABC>100,000;CD52 ABC>85,000)。
当CD20或CD52仅以极低的水平表达时,IgG1-CAMPATH-1H-E430G-K439E-K322E+IgG1-11B8-E430G-S440K混合物未观测到CDC活性:Daudi(CD52 ABC<75,000),U266B1U266B1(CD20 ABC<20,000)和REH(CD20 ABC<20,000)。当在两种抗体(IgG1-CAMPATH-1H-E430G-K439E-K322E+IgG1-11B8-E430G-S440K-K322E)中均引入C1q结合抑制突变K322E时,未观测到CDC活性。总之,这些数据表明,IgG1-CAMPATH-1H-E430G-K439E-K322E的C1q募集和CDC效力取决于其与IgG1-11B8-E430G-S440K的异源寡聚化。
总之,使用IgG1-CAMPATH-1H-E430G-K439E-K322E+IgG1-11B8-E430G-S440K混合物可以选择性杀伤表达足够水平的CD20和CD52两者的细胞。相反,这种混合物在表达如此低的CD20或CD52表达水平以至于它们不能分别被IgG1-11B8-E430G-S440K和IgG1-CAMPATH-1H-E430G-K439E-K322E的单一试剂CDC活性杀死的细胞系中显示背景裂解水平。
表2:CD52和CD20在血液癌症细胞系的细胞表面表达

1细胞表面表达表示为每种细胞特异性抗体结合能力(ABC)的数目,使用人IgG校准试剂盒(Biocytex)确定。
实施例5:抗CD52抗体IgG1-CAMPATH-1H-E430G-K439E中不同C1q结合抑制突变对具有抗CD20抗体IgG1-11B8-E430G-S440K的抗体混合物的选择性CDC活性的影响的分析
上面已经描述P329R(错误!未找到参考源)或K322E(错误!未找到参考源)的引入。C1q结合抑制突变导致抑制IgG1-CAMPATH-1H-E430G-K439E在Wien 133细胞上的残留CDC活性,而与IgG1-11B8-E430G-S440K混合时,CDC活性完全恢复、部分恢复或完全没有恢复,这取决于所测试的细胞系(错误!未找到参考源)。在此,用与IgG1-11B8-E430G-S440K混合物中的IgG1-CAMPATH-1H-E430G-K439E变体(包含G236R(SEQ ID NO:84),K322A(SEQ ID NO:90),E269K(SEQ ID NO:81),C1q结合抑制突变K322E或P329R(在此按预期降低的C1q结合亲和力列出)),在使用Wien 133细胞的体外CDC测定中比较不同C1q结合抑制突变的影响。使用Wien 33细胞的体外CDC测定利用20%NHS和系列稀释抗体浓度(以4倍稀释度,最终浓度范围为0.002-40.0μg/mL)进行,基本上如所述错误!找不到参考源。细胞裂解由IntellicytiQueTM筛选器通过流式细胞术测量的PI阳性细胞数目计算,取三个实验重复的平均值。相对曲线下面积(AUC)值代表相对于最小裂解(对于阴性对照IgG1-b12-K439E+IgG1-b12-S440K为0%)和最大裂解(对于IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G为100%)标准化。错误!找不到参考源。显示对Wien 133细胞的剂量反应CDC活性曲线的AUC值的条形图。引入任一C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)导致完全抑制IgG1-CAMPATH-1H-E430G-K439E Wien 133的残留单一试剂CDC活性。在具有IgG1-11B8-E430G-S440K的混合物中,使用含有单个C1q结合抑制突变P329R、K322E、E269K、K322A或G236R的IgG1-CAMPATH-1H-E430G-K439E变体,观测到CDC活性恢复的水平提高。这些数据表明,IgG1-CAMPATH-1H-E430G-K439E变体(P329R、K322E、E269K、K322A或G236R)的C1q结合亲和力与具有IgG1-11B8-E430G-S440K的混合物的CDC活性恢复之间存在直接相关性。
实施例6:抗CD20抗体IgG1-11B8-E430G-S440K中不同C1q结合增强突变对具有抗CD52抗体IgG1-CAMPATH-1H-E430G-K439E的抗体混合物的选择性CDC活性的影响的分析
用具有IgG1-CAMPATH-1H-E430G-K439E的混合物中的IgG1-11B8-E430G-S440K变体(包含C1q结合增强突变E333S(SEQ ID NO:103),K326W(SEQ ID NO:112)或K326A/E333A(SEQ ID NO:111;此处以预期增加的C1q结合亲和力列出)),在使用Wien 133细胞的体外CDC测定中比较不同C1q结合增强突变的效果。使用Wien 133细胞的体外CDC测定,利用20%NHS和系列稀释的抗体浓度(以4倍稀释度,最终浓度范围为0.002-40.0μg/mL)进行,基本如所述错误!找不到参考源。细胞裂解由Intellicyt iQueTM筛选器通过流式细胞术测量的PI阳性细胞数目计算,取三个实验重复的平均值。相对曲线下面积(AUC)值代表相对于最小裂解(对于阴性对照IgG1-b12-K439E+IgG1-b12-S440K为0%)和最大裂解(对于IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物为100%)标准化。错误!找不到参考源。显示对Wien 133细胞的剂量反应CDC活性曲线的AUC值的条形图。引入C1q结合增强突变E333S、K326W或K326A/E333A导致增加IgG1-11B8-E430G-S440K对Wien 133的单一试剂CDC活性(相比无C1q结合增强突变的情况下1%,E333S为7%;K326W为18%;K326A/E333A为25%)。在具有IgG1-CAMPATH-1H-E430G-K439E的混合物中,利用包含C1q结合增强突变K326A/E333A、K326W或E333S的IgG1-11B8-E430G-S440K变体观测到CDC活性水平增加。这些数据表明,IgG1-11B8-E430G-S440K变体(K326A/E333A、K326W或E333S)的C1q结合亲和力与具有IgG1-CAMPATH-1H-E430G-K439E的混合物的CDC活性之间存在直接相关性。
实施例7:抗CD52抗体IgG1-CAMPATH-1H-E430G-K439E中不同C1q结合抑制突变和抗CD20抗体IgG1-11B8-E430G-S440K中C1q结合增强突变对抗CD52和抗CD20抗体变体的抗体混合物的选择性CDC活性的影响的分析
用具有包含C1q结合增强突变E333S、K326W或K326A/E333A的IgG1-11B8-E430G-S440K变体(实施例6所述)的混合物中的包含C1q结合抑制突变G236R、K322A、E269K、K322E或P329R的IgG1-CAMPATH-1H-E430G-K439E变体(实施例5所述),在使用Wien 133细胞的体外CDC测定中比较影响C1q结合亲和力的不同突变的作用。使用Wien 133细胞的体外CDC测定,利用20%NHS和系列稀释的抗体浓度(以4倍稀释度,最终浓度范围为0.002-40.0μg/mL)进行,基本如实施例2所述。细胞裂解由Intellicyt iQueTM筛选器通过流式细胞术所测量的PI阳性细胞数目计算,取三个实验重复的平均值。相对曲线下面积(AUC)值代表相对于最小裂解(对于阴性对照IgG1-b12-K439E+IgG1-b12-S440K为0%)和最大裂解(对于IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G为100%)标准化。图6显示对Wien 133细胞的剂量反应CDC活性曲线的AUC值的条形图。与具有无C1q结合增强突变的IgG1-11B8-E430G-S440K的混合物相比,具有C1q结合增强突变的抗CD20 IgG1-11B8-E430G-S440K混合具有C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H-E430G-K439E抗体变体的混合物显示对Wien 133细胞的CDC活性的改善的恢复。与包含C1q结合抑制突变的IgG1-CAMPATH-1H-E430G-K439E变体混合时,IgG1-11B8-E430G-S440K变体中的C1q结合亲和力的预期强度(K326A/E333A>K326W>E333S)与CDC活性恢复水平之间存在相关性,其中对于G236R,最大裂解恢复略有提高(图6A;表3),对于K322A(图6B;表3),E269K(图6C;表3),K322E(图6D;表3),最大裂解恢复与增加的EC50值,或对于P329R,最大裂解在EC50值增加的情况下保持降低(图6E;表3)。
表3


实施例8:抗CD52 IgG1-CAMPATH-1H抗体变体和抗CD20 IgG1-11B8抗体变体在细胞上的靶标结合
通过流式细胞术分析具有E430G、K439E和C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H和具有E430G、S440K和C1q结合增强突变E333S的抗CD20 IgG1-11B8与Wien 133和Raji淋巴瘤细胞的结合。将细胞悬浮液用FACS缓冲液清洗并以2.5x106个细胞/mL的浓度重悬于FACS缓冲液[PBS+0.1%(w/v)牛血清白蛋白(BSA)+0.02%(w/v)叠氮化钠]中。将40μL细胞悬浮液样品(100,000个细胞/孔)接种到聚苯乙烯96孔圆底培养板(Greiner Bio-One;Cat nr 650101),并与40μL抗体样品(以3倍稀释度,终浓度为0.001-30μg/mL)于4℃温育30分钟。将细胞于4℃以300xg离心3分钟进行沉淀,并用150μL FACS缓冲液清洗两次。将细胞与50μL二抗R-藻红蛋白(R-PE)缀合的山羊抗人IgG F(ab')2(JacksonImmunoResearch,目录号109-116-098,1:100)于4℃避光温育30分钟。将细胞用150μL FACS缓冲液清洗两次,重悬于150μL FACS缓冲液中,然后用Intellicyt iQue筛选器通过流式细胞术分析抗体结合。使用GraphPad Prism软件,结合曲线利用非线性回归分析(具有可变斜率的S形剂量反应)进行分析。
图7显示对于IgG1-CAMPATH-1H,所有测试的抗体变体与Wien 133(图7A)和Raji细胞(图7B)均显示相似的剂量依赖性结合。这些数据表明,引入单突变E430G和K439E以及双突变E430G-K439E对细胞表面上的靶标结合没有影响。同样,引入另外的突变G236R或K322A对细胞表面上的CD52靶标结合没有影响。
图7显示对于IgG1-11B8,所有测试的抗体变体与Wien 133(图7C)和Raji细胞(图7D)均显示相似的的剂量依赖性结合。这些数据表明,引入单突变E430G和S440K以及双突变E430G-S440K对细胞表面上的靶标结合没有影响。同样,引入另外的突变E333S对细胞表面上的CD20靶标结合没有影响。
实施例9:使用抗CD52和抗CD20抗体变体混合物分析抗CD52抗体IgG1-CAMPATH-1H-E430G-K439E中C1q结合抑制突变和抗CD20抗体IgG1-11B8-E430G-S440K中C1q结合增强突变对FcγR介导的效应器功能的影响
在对Raji细胞的ADCC报告物生物测定和对Wien 133细胞利用人外周血单个核细胞(PBMC)的体外铕TDA(EuTDA)ADCC测定中,测试IgG1-CAMPATH-1H-E430G-K439E变体中引入C1q结合抑制突变G236R或K322A和IgG1-11B-E430G-S440K中引入C1q结合增强突变E333S对抗体依赖性细胞性细胞毒性(ADCC)的影响。
对于ADCC报告物生物测定,在Raji细胞上使用Bio-Glo萤光素酶测定系统(Promega,目录号G7940)测试IgG1-CAMPATH-1H(WT、E430G、K439E(SEQ ID NO:29),E430G-K439E,E430G-K439E-G236R和E430G-K439E-K322A)和IgG1-11B8(WT、E430G、S440K(SEQ IDNO:30),E430G-S440K、E430G-S440K-E333S)变体。该试剂盒包含Jurkat人T细胞作为效应细胞,其经工程改造以稳定表达高亲和力FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的激活T细胞核因子(NFAT)反应元件。简而言之,将Raji细胞(5.000个细胞/孔)接种于384孔白色OptiPlates(Perkin Elmer目录号6007290)中,在补充有4%低IgG血清(Promega,目录号G711A)的ADCC测定缓冲液[RPMI-1640培养基(Lonza,目录号BE12-115F)]中,并在包含抗体浓度系列(以3.5倍稀释度,最终浓度为0.4-10,000ng/mL)和解冻的ADCC生物测定效应细胞(Promega,目录号G701A)的30μL总体积于37℃/5%CO2温育6小时。将板于室温(RT)温育15分钟后,添加30μL Bio Glo萤光素酶试剂[Bio-Glo萤光素酶测定缓冲液(Promega,目录号G719A)中的Bio-Glo萤光素酶测定底物(Promega目录号G720A)],并于室温温育5分钟。用EnVision Multilabel Reader(Perkin Elmer)通过发光读数来定量萤光素酶的产生。
在ADCC报告物生物测定中,IgG1-CAMPATH-E430G-K439E中引入C1q结合抑制突变G236在Raji细胞上的单个抗CD52抗体结合后导致效应细胞中的FcγRIIIa激活的完全抑制,而C1q结合抑制突变K322A导致部分抑制FcγRIIIa激活(图8A)。在Raji细胞上结合单个抗CD20抗体后,所有测试的IgG1-11B8抗体变体均显示效应细胞中相当大的FcγRIIIa激活,而包含S440K和/或E430G和/或E333S突变的IgG1-11B8变体之间无显著差异(图8B)。
在Wien 133细胞上进行了体外EuTDA ADCC测定(DELFIA EuTDA细胞毒性测定,Perkin Elmer,目录号AD0116),利用来自三个不同健康供体的新鲜分离的PBMC作为效应细胞,以测试IgG1-CAMPATH-1H抗体变体(WT、K439E、E430G-K439E、G236R-E430G-K439E和K322A-E430G-K439E)和IgG1-11B8抗体变体(WT、S440K、E430G-S440K和E333S-E430G-S440K)作为单一试剂或组合(IgG1-CAMPATH-1H-G236R-E430G-K439E或-K322A-E430G-K439E和IgG1-11B8-E333S-E430G-S440K)的ADCC活性。根据制造商说明书,使用淋巴细胞分离培养基(Lonza,目录号17-829E)从血沉棕黄层(Sanquin,Amsterdam,The Netherlands)中分离PBMC,用于标准Ficoll密度离心。将细胞重悬于补充有10%含铁的供体牛血清(DBSI,ThermoFischer,目录号10371029)和青霉素/链霉素(Pen/Strep,Lonza,目录号DE17-603E)的RPMI-1640培养基(Lonza,目录号BE12-115F)中后,细胞通过台盼蓝(SigmaAldrich,目录号T8154)排除进行计数,并浓缩至2x107个细胞/mL。
将Wien 133细胞收获,清洗(PBS中两次,1200rpm,5分钟),收集在补充有10%DBSI和Pen/Strep的RPMI-1640培养基中,浓度为1x106个细胞/mL,向其中加入15μLDELFIABATDA试剂(Perkin Elmer,目录号C136-100,5μL/3x106个细胞)。将混合物在水浴中于37℃温育20分钟。清洗细胞后(在50mL PBS中五次,1200rpm,5分钟),将细胞重悬于补充有10%DBSI和Pen/Strep的RPMI-1640培养基中,通过台盼蓝排除法计数,并稀释至2x105个细胞/mL的浓度。
对于ADCC实验,将50μL BATDA标记的Wien 133细胞(10,000个细胞/孔)与IgG1-CAMPATH-1H和/或IgG1-11B8抗体变体的浓度系列(以10倍稀释度,最终浓度为0.01-10,000ng/mL)在96孔V底微量滴定板(Greiner Bio-One;目录号651101)的总体积150μL补充有10%DBSI和Pen/Strep的RPMI-1640培养基中一起预温育。室温下15分钟后,添加50μL PBMC(1x106个细胞),得到100:1的效应物与靶标比率(E:T),并于37℃/5%CO2温育2小时。为了确定最大的细胞裂解量,根据制造商建议,将50μL BATDA标记的Wien 133细胞(10,000个细胞/孔)与补充有10%DBSI,Pen/Strep和DELFIA裂解缓冲液(0.03%Digitonin和19%DMSO,Perkin Elmer,目录号AD0116-A)的150μL RPMI-1640培养基一起温育。为了确定自发裂解的量,将50μL BATDA标记的Wien 133细胞(10,000个细胞/孔)在不含任何抗体或效应细胞的150μL培养基中温育。
为了测量释放的BATDA试剂的量,将板离心(500xg,10分钟),将20μL上清液转移至DELFIA微量滴定板,然后添加200μL DELFIA铕溶液(Perkin Elmer,目录号AD0116-B)。随后,将板于室温摇动温育15分钟。用时间分辨荧光计(Perkin Elmer EnVision 2104MultiDetection Microplate Reader)测量铕-BATDA(EuTDA),并用于如下所示计算抗体介导的裂解百分比:(样品释放-自发释放)/(最大释放-自发释放)x100%。
作为单一试剂测试时,在WT IgG1-CAMPATH-1H中引入E430G和/或K439E突变对剂量反应性ADCC介导的Wien 133细胞杀伤没有影响(图8C)。在IgG1-CAMPATH-E430G-K439E中引入C1q结合抑制突变G236R导致完全抑制单一试剂的ADCC活性,而C1q结合抑制突变K322A对单一试剂IgG1-CAMPATH-E430G-K439E对Wien 13细胞的ADCC活性没有影响(图8C)。在Wien 133细胞上,所有测试的IgG1-11B8抗体变体作为单一试剂测试时均显示相当大的ADCC效力,而在包含S440K和/或E430G和/或E333S突变的IgG1-11B8变体之间没有显著差异(图8D)。在Wien 133细胞上,IgG1-11B8-E333S-E430G-S440K与IgG1-CAMPATH-1H-G236R-E430G-K439E或IgG1-CAMPATH-1H-K322A-E430G-K439E的混合物均显示相当大的ADCC效力(图8D)。
总之,在发光ADCC报告物生物测定和体外EuTDA ADCC测定两者中,突变G236R均强烈抑制IgG-CAMPATH-E430G-K439E的ADCC活性。具有K322A突变的IgG-CAMPATH-E430G-K439E在发光ADCC报告物生物测定中显示出降低的活性,但在体外EuTDA ADCC测定中显示出保留实质的ADCC活性。
实施例10:针对具有C1q结合抑制突变的IgG1-CAMPATH-K439E变体和具有或不具有C1q结合增强突变的IgG1-11B8-S440K变体的混合物对Wien 133细胞的选择性CDC活性分析不同Fc-Fc相互作用增强突变
实施例5描述了IgG1-CAMPATH-1H-E430G-K439E中引入C1q结合抑制突变(G236R、K322A、E269K、K322E或P329R)导致完全抑制单一试剂对Wien133的CDC活性,与IgG1-11B8-E430G-S440K混合后可恢复CDC活性。实施例6描述了含有C1q结合增强突变(如E333S)的IgG1-11B8-E430G-S440K,当使用Wien 133细胞在体外CDC测定中作为单一试剂测试时,显示出非常有限的CDC活性,但是与IgG1-CAMPATH-1H-E430G-K439E混合后显示CDC活性恢复。实施例7描述了含有C1q结合抑制突变G236R或K322A的抗CD52抗体IgG1-CAMPATH-1H-E430G-K439E与含有增强C1q结合突变E333S的抗CD20抗体IgG1-11B8-E430G-S440K的组合在Wien 133细胞上显示选择性CDC活性,而单一试剂未观测到CDC活性。在此,在Wien 133细胞的体外CDC测定中,分析了其他Fc-Fc增强突变如E345K、E345R和E345R-E430G的这个原理,即具有Fc-Fc增强突变(如E430G),K439E突变和C1q结合增强突变(如G236R或K322A)的IgG1-CAMPATH-1H变体与具有Fc-Fc增强突变(如E430G),S440K突变和任选C1q结合增强突变(如E333S)的IgG1-11B8抗体变体的混合物选择性诱导CDC活性。使用Wien 133细胞的体外CDC测定,利用20%NHS和抗体浓度系列(以3倍稀释度,最终浓度范围为0.003-10.0μg/mL),基本如实施例2所述。如实施例2所述,细胞裂解和相对曲线下面积(AUC)值由两次实验重复的PI阳性细胞数目计算。将AUC相对于阴性对照抗体IgG1-b12(0%)和阳性对照IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G(100%)的值标准化,同时提供的最大裂解数据反映10μg/mL IgG时非标准化的细胞裂解。
错误!找不到参考源。显示对Wien 133细胞的剂量反应CDC活性曲线AUC值和最大裂解的条形图。在具有Fc-Fc相互作用增强突变E430G、E345K(分别为SEQ ID NO:66或67)或E345R(分别为SEQ ID NO:74或73)之一的IgG1-CAMPATH-1H-K439E中引入C1q结合抑制突变G236R(错误!未找到参考源。A)或K322A(错误!未找到参考来源.B),在Wien133细胞上作为单一靶向药剂测试时导致CDC活性丧失。相反,具有两个Fc-Fc相互作用增强突变E345R-E430G和C1q结合抑制突变G236R(SEQ ID NO:71)的IgG1-CAMPATH-1H-K439E变体在Wien133细胞上显示出残留的单一试剂CDC活性(错误!找不到参考源。A)。包含Fc-Fc相互作用增强突变E430G、E345K或E345R的IgG1-CAMPATH-1H-K439E-G236R(错误!未找到参考来源.A)或IgG1-CAMPATH-1H-K439E-K322A(错误!未找到参考来源.B)与IgG1-11B8-E430G-S440K或IgG1-11B8-E430G-S440K-E333S的混合物在Wien 133细胞上显示CDC活性的恢复,而所有这些变体作为单一试剂均未诱导任何CDC。在使用Wien 133细胞的体外CDC测定中,包含Fc-Fc相互作用增强突变E430G或E345K(SEQ ID NO:68)的IgG1-11B8-S440K-E333S抗体变体作为单一试剂进行测试时,不诱导显著的CDC活性,而使用包含两个Fc-Fc相互作用增强突变E345R-E430G(错误!未找到参考来源。C)的IgG1-11B8-E345R-E430G-S440K-E333S(SEQ IDNO:72)观测到单一试剂活性。含有Fc-Fc相互作用增强突变E345K或E430G的IgG1-11B8-S440K-E333S和IgG1-CAMPATH-1H-E430G-K439E-G236R的混合物在Wien 133细胞上显示CDC活性恢复,而所有这些变体作为单一试剂均未诱导任何CDC(错误!未找到参考来源。C)。同样,均包含Fc-Fc相互作用增强突变E345K的IgG1-11B8-E345K-S440K-E333S与IgG1-CAMPATH-1H-E345K-K439E-G236R或IgG1-CAMPATH-1H-E345K-K439E-K322A的混合物,在Wien 133细胞上显示CDC活性恢复,而单一试剂则没有诱导CDC(错误!未找到参考来源。C)。
总之,这些数据表明,可以通过包含Fc-Fc相互作用增强突变之一如E430G、E345K或E345R和C1q结合抑制突变G236R或K322A的IgG1-CAMPATH-1H-K439E变体混合包含Fc-Fc相互作用增强突变之一且具有或不具有C1q结合增强突变如E333S的IgG1-11B8-S440K变体的组合物来实现对CD52阳性/CD20阳性Wien 133细胞的选择性CDC活性。总之,具有不同Fc-Fc相互作用增强突变的单个抗体的CDC活性可通过引入自身寡聚化抑制突变并调节C1q结合强度来控制。补体活性在将具有互补的自身寡聚化抑制突变的两种此类抗体混合后恢复,这允许在同时被两种抗体结合的细胞上进行选择性异源寡聚化。
实施例11:针对抗CD52 IgG1-CAMPATH-E430G-K439E和抗CD20 IgG1-11B8-E430G-S440K抗体变体的混合物对Wien 133细胞的选择性CDC活性分析不同C1q结合调节突变
实施例7描述了包含C1q结合抑制突变如G236R或K322A的抗CD52抗体IgG1-CAMPATH-1H-E430G-K439E与包含C1q结合增强突变如E333S、K326W或K326A-E333A的抗CD20抗体IgG1-11B8-E430G-S440K的组合在Wien 133细胞上显示选择性CDC活性,而单一试剂几乎没有观测到CDC活性。在此,在IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体的不同组合中测试了替代的C1q结合调节突变,目的是抑制单一试剂的CDC活性,并通过抗CD52和抗CD20抗体变体的混合物使选择性CDC活性能够最大恢复。使用Wien 133细胞的体外CDC测定,利用20%NHS和抗体浓度系列(以3倍稀释度,最终浓度范围为0.003-10.0μg/mL)进行,基本如实施例2所述。如实施例2所述,细胞裂解和相对AUC值由两次实验重复的PI阳性细胞数目计算。将AUC相对于阴性对照抗体IgG1-b12(0%)和阳性对照IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G(100%)的值标准化,同时呈现的最大裂解数据反映10μg/mL IgG时非标准化的细胞裂解。
首先,包含氨基酸G236位置处(G236R或G236K(SEQ ID NO:83))或G237位置处(G237A(SEQ ID NO:85),G237T(SEQ ID NO:89),G237Q(SEQ ID NO:87)或G237R(SEQ IDNO:88))取代或双突变G237A-K322A(SEQ ID NO:86)的IgG1-CAMPATH-1H-E430G-K439E抗体变体与具有或不具有C1q结合增强突变E333S的IgG1-11B8-E430G-S440K组合。如错误!未找到参考来源A所示,IgG1-CAMPATH-1H-E430G-K439E中的G236或G237位置处所有测试的C1q结合调节突变均导致与具有或不具有C1q结合增强突变E333S的IgG1-11B8-E430G-S440K的混合物中的选择性CDC活性,而作为单一试剂在Wien 133细胞上几乎没有CDC活性。相反,包含双突变G237A-K322A的IgG1-CAMPATH-1H-E430G-K439E变体与具有或不具有C1q结合增强突变E333S的IgG1-11B8-E430G-S440K组合时,未显示CDC活性的恢复。
其次,包含C1q结合增强突变K326A(SEQ ID NO:110)、E333A(SEQ ID NO:102)、E333S或K236W-E333S(SEQ ID NO:157)的IgG1-11B8-E430G-S440K抗体变体与包含C1q结合抑制突变G236R或K322A的IgG1-CAMPATH-1H-E430G-K439E组合。如错误!未找到参考来源B所示,在Wien 133细胞上,包含单个C1q结合增强突变K326A、E333A或E333S的IgG1-11B8-E430G-S440K抗体变体作为单一试剂保持没有CDC活性,而包含双突变K236W-E333S的IgG1-11B8-E430G-S440K变体作为单一试剂显示出明显更高的CDC活性。在Wien 133细胞上,包含单个C1q结合增强突变K326A、E333A或E333S的IgG1-11B8-E430G-S440K抗体变体与包含C1q结合抑制突变G236R或K322A的IgG1-CAMPATH-1H-E430G-K439E变体的所有测试的组合均显示CDC活性的恢复,而作为单一试剂显示几乎没有CDC活性。
还将包含C1q结合增强突变K326A或E333A的IgG1-11B8-E430G-S440K抗体变体与包含在G237位置处C1q结合调节突变(G237A、G237T、G237Q或G237R)的IgG1-CAMPATH-1H-E430G-K439E组合。如图10B所示,在Wien 133细胞上,IgG1-11B8-E430G-S440K-K326A或IgG1-11B8-E430G-S440K-E333A与G237位置处包含所测试突变之一的IgG1-CAMPATH-1H-E430G-K439E抗体变体的所有测试的组合导致选择性CDC活性,而所有单一试剂显示几乎没有CDC活性。
将突变G237A引入IgG1-11B8-E430G-S440K抗体变体中,以抑制S440K组分(SEQ IDNO:105)中FcγR介导的效应器功能。为了补偿可能减少的C1q结合,在与C1q结合增强突变E333S(SEQ ID NO:106)的组合中测试G237A。将IgG1-11B8变体与IgG1-CAMPATH-1H-E430G-K439E-G236R组合。如错误!未找到参考来源B所示,在Wien 133细胞上,这些组合也显示选择性CDC活性,而单一试剂显示几乎没有CDC活性。
总之,具有Fc-Fc相互作用增强突变E430G的单个抗体的CDC活性可通过引入自身寡聚化抑制突变组合抑制C1q结合的不同突变来控制。通过与包含不同C1q结合增强突变和互补的自身寡聚化抑制突变的抗体混合(这允许在同时被两种抗体结合的细胞上进行选择性异源寡聚化),使用此类抗体可实现补体活性的最大恢复。
实施例12:分析不同人IgG同种型骨架中抗CD52抗体和抗CD20抗体变体的混合物对Wen 133细胞的选择性CDC活性
将抗CD52 CAMPATH-1H的VH序列克隆至包含E430G-K439E-G236R突变的人IgG1、IgG2、IgG3和铰链稳定的IgG4(S228P)Fc骨架中,并将抗CD2011B8的VH序列克隆至包含E430G-S440K-E333S突变的人IgG1、IgG2、IgG3和铰链稳定的IgG4(S228P)Fc骨架中。测试这些抗CD52和抗CD20同种型变体的不同组合的选择性CDC活性。使用Wien 133细胞的体外CDC测定,利用20%NHS和抗体浓度系列(以3倍稀释度,最终浓度范围为0.003-10.0μg/mL)进行,基本如实施例2所述。如实施例2所述,细胞裂解和相对AUC值由两个实验重复的PI阳性细胞数目计算。将AUC相对于阴性对照抗体IgG1-b12(0%)和阳性对照IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G(100%)的值标准化,同时呈现的最大裂解数据反映10μg/mL IgG时非标准化的细胞裂解。
IgG2-CAMPATH-1H-E430G-K439E-G236R和IgG2-11B8-E430G-S440K或IgG2-11B8-E430G-S440K-E333S混合物显示恢复CDC活性,但与相应的IgG1抗体变体混合物相比,CDC功效(AUC)降低(图11)。IgG2-CAMPATH-1H-E430G-K439E-G236R和IgG2-11B8-E430G-S440K未显示单一试剂CDC活性,而IgG2-11B8-E430G-S440K-E333S中引入C1q结合增强突变E333S导致单一试剂在Wien 133细胞上诱导一些CDC活性(图11)。
对于任何测试的IgG3同种型变体的单一试剂或混合物均未观测到CDC活性(图11)。对于测试的IgG4同种型变体的单一试剂或混合物也均未观测到CDC活性(图11)。
令人惊讶地,IgG2-CAMPATH-1H-E430G-K439E-G236R与IgG1-11B8-E430G-S440K-E333S的混合物以及铰链稳定的IgG4-CAMPATH-1H-E430G-K439E-G236R(SEQ ID NO:146)与IgG1-11B8-E430G-S440K-E333S的混合物显示CDC功效的部分恢复,而各个单一试剂在Wien133细胞上均未诱导CDC(图11)。
总之,包含Fc-Fc相互作用增强突变E430G的具有不同IgG骨架的单个抗体的CDC活性可通过引入自身寡聚化抑制突变组合调节C1q结合的突变来控制。通过混合包含互补的自身寡聚化抑制突变的这类抗体,该类抗体的CDC活性可通过同时被两种抗体结合的细胞上的选择性异源寡聚化来恢复。
实施例13:具有不同靶标表达水平的细胞系上抗CD52 IgG1-CAMPATH-1H抗体变体和抗CD37 IgG1-CD37-37.3抗体变体的混合物的CDC活性的选择性
如实施例4所述,IgG1-CAMPATH-1H-E430G-K439E-K322E+IgG1-11B8-E430G-S440K混合物的选择性CDC活性仅可以在表达足够水平的两个靶标(即CD20和CD52)的细胞上实现。此外,实施例7中描述了使用包含C1q结合抑制突变如G236R或K322A的IgG1-CAMPATH-1H-E430G-K439E和包含C1q结合增强突变如E333S的IgG1-11B8-E430G-S440K,实现了在抗体组合具有保留的选择性情况下的Wien 133细胞最大杀伤。在此,在使用Daudi和Wien 133细胞的体外CDC测定中,测试了抗CD52抗体变体和抗CD37抗体变体的组合的选择性CDC活性。使用Daudi和Wien 133细胞的体外CDC测定,利用20%NHS和抗体浓度系列(以3倍稀释度,最终浓度范围为0.003-10.0μg/mL),基本如实施例2所述。如实施例2所述,细胞裂解和相对AUC值由两个实验重复(Daudi)和一个实验(Wien 133)的PI阳性细胞数目计算。AUC相对于阴性对照抗体IgG1-b12(0%)和阳性对照IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G(100%)的值标准化。
发现表达低CD52水平的Daudi细胞(表2)对由IgG1-CAMPATH-1H变体(WT、E430G、E430G-K439E、E430G-K439E-G236R和E430G-K439E-K322A)作为单一试剂测试时诱导的CDC具有抗性(图12A)。IgG1-CD37-37.3-E430G在Daudi细胞(表达高CD37水平,数据未显示)上的CDC活性通过引入S440K突变抑制,并且通过另外引入C1q结合增强突变E333S而部分恢复。WT IgG1-CD37-37.3在Daudi细胞上不诱导CDC活性。对于IgG1-CD37-37.3-E430G-S440K与包含G236R或K322A的IgG1-CAMPATH-1H-E430G-K439E变体的混合物,观测到的CDC活性类似于单一试剂IgG1-CD37-37.3-E430G-S440K的CDC活性,表明低CD52表达时与IgG1-CAMPATH-1H变体缺乏协同性。对于IgG1-CD37-37.3-E430G-S440K-E333S与包含C1q抑制突变G236R或K322A的IgG1-CAMPATH-1H-E430G-K439E变体的混合物,观测到低协同性。这些数据表明,IgG1-CAMPATH-1H-E430G-K439E中的C1q结合抑制突变G236R抑制了表达低水平CD52的细胞的虚假共激活。
发现表达适度CD37水平(数据未显示)的Wien 133细胞对由所有IgG1-CD37-37.3变体(WT、E430G、E430G-S440K、E430G-S440K-E333S)作为单一试剂测试时诱导的CDC具有抗性(图12B)。IgG1-CAMPATH-1H-E430G在表达高CD52水平的Wien 133细胞(ABC>300,000;表2)上的CDC活性通过引入K439E突变降低至WT IgG1-CAMPATH-1H的水平。进一步引入C1q结合抑制突变(IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-CAMPATH-1H-E430G-K439E-K322A)导致完全抑制单一试剂在Wien133细胞上的CDC活性。对于IgG1-CAMPATH-1H-E430G-K439E-G236R或IgG1-CAMPATH-1H-E430G-K439E-K322A与IgG1-CD37-37.3-E430G-S440K或IgG1-CD37-37.3-E430G-S440K-E333S的混合物,观测到CDC活性的恢复水平优于WT IgG1-CAMPATH-1H的水平,而所有这些变体作为单一试剂在Wien 133细胞上均未显示CDC活性。
总之,可以在表达明显水平的CD37和CD52两者的Wien 133细胞上建立IgG1-CAMPATH-1H-E430G和IgG1-CD37-37.3-E430G抗体变体混合物的CDC效力的选择性恢复,而表达低水平CD52的Daudi细胞则可以得到保护而免于包含自身寡聚化抑制突变和C1q结合抑制突变G236R或K322A两者的IgG1-CAMPATH-1H-E430G变体存在的影响。
实施例14:BxPC-3细胞上两种非交叉阻断抗DR5抗体的混合物的选择性DR5激动剂活性
两种非交叉阻断抗死亡受体5(DR5)抗体IgG1-DR5-01-G56T-E430G+IgG1-DR5-05-E430G混合物充当DR5激动剂来诱导杀伤DR5阳性癌细胞(WO17093447)。在此进行了存活力测定,以研究IgG1-DR5-01-G56T-E430G抗体变体(K439E、K439E-G236R)与IgG1-DR5-05-E430G抗体变体(S440K、S440K-E333S)的组合诱导杀伤表达低水平DR5(数据未显示)的人BxPC-3胰腺癌细胞(ATCC,目录号CRL-1687)的能力。BxPC-3细胞通过胰蛋白酶消化收获并通过细胞过滤器。细胞以1200rpm离心5分钟进行沉淀,并以0.5x105个细胞/mL的浓度重悬于培养基(RPMI1640,含25mM Hepes和L-谷氨酰胺(Lonza)+10%DBSI(Life Technologies,目录号10371-029)+Pen/Strep)中。将100μL单细胞悬浮液(5,000个细胞/孔)接种到聚苯乙烯96孔平底板(Greiner Bio-One,目录号655182)中,并允许于37℃过夜贴壁。第二天,添加50μL抗体稀释系列的样品(以3倍稀释度,最终浓度范围为0.003-20μg/mL)和10μL纯化的人C1q储备液(Quidel,目录号A400,终浓度为2.5μg/mL),并于37℃温育3天。将细胞与5μM星形孢菌素(Sigma Aldrich,目录号S6942)一起温育作为阳性对照。用CellTiter-Glo发光细胞存活力测定法(Promega,目录号G7571)确定细胞培养物的存活力,该测定法量化存在的ATP,这是代谢活性细胞的指标。自试剂盒,每孔添加15μL萤光素溶液试剂,并以500rpm摇动板2分钟进行混合。接着,将板于37℃温育1.5小时。将100μL上清液转移至白色OptiPlate-96(Perkin Elmer,目录号6005299),并用EnVision Multilabel Reader(PerkinElmer)测量发光。使用GraphPad Prism软件,数据利用非线性回归(具有可变斜率的S形剂量反应)进行分析和绘图。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
测试的单一试剂均未诱导杀伤BxPC-3细胞(图13A),而IgG1-DR5-01-G56T-E430G+IgG1-DR5-05-E430G的组合诱导剂量依赖性杀伤BxPC-3细胞(图13B)。这种选择性杀伤保留在IgG1-DR5-01-G56T-E430G-K439E+IgG1-DR5-05-E430G-S440K的组合中,表明两种抗体均形成异源六聚体。通过调节C1q结合位点,观测到DR5激动剂活性的BxPc-3细胞杀伤的诱导利用IgG1-DR5-01-G56T-E430G-K439E(无C1q结合抑制突变)+IgG1-DR5-05-E430G-S440K-E333S(具有C1q结合增强突变E333S)的组合最为有效,利用IgG1-DR5-01-G56T-E430G-K439E-G236R(具有C1q结合抑制突变G236R)+IgG1-DR5-05-E430G+S440K(无C1q结合增强突变)稍微降低。IgG1-DR5-01-G56T-E430G-K439E-E333S(具有C1q结合抑制突变)+IgG1-DR5-05-E430G-S440K-E333S(具有E333S C1q结合增强突变)的其他测试组合显示与IgG1-DR5-01-G56T-E430G+IgG1-DR5-05-E430G类似的效力。
总之,引入C1q调节突变不损伤抗DR5抗体的异源低聚化混合物对BxPC-3细胞的选择性杀伤。而是,混合物的功效与DR5结合后在BxPC-3细胞形成的异源寡聚体抗体结构的预期组合的C1q结合亲合力的功效成比例。
实施例15:抗CD52抗体IgG1-CAMPATH-1H-E430G中不同C1q结合抑制突变对CDC活性的影响的分析
在使用Wien 133细胞的体外CDC测定中,比较了在IgG1-CAMPATH-1H-E430G中引入不同C1q结合抑制突变(G237A、G236R、A327K、K322E或P329R)的影响。使用Wien 133细胞的体外CDC测定,利用20%NHS和序列稀释抗体浓度(以4倍稀释度,最终浓度范围为0.002-40.0μg/mL)进行,基本如错误!找不到参考源。细胞裂解由Intellicyt iQueTM筛选器通过流式细胞术测量的PI阳性细胞数目计算,取三个实验重复的平均值。相对曲线下面积(AUC)代表相对于最小裂解(使用阴性对照IgG1-b12-K439E+IgG1-b12-S440K(未显示)为0%)和最大裂解(使用IgG1-CAMPATH-1H-E430G为100%)标准化。错误!找不到参考源。显示G237A突变(SEQ ID NO:124)对IgG1-CAMPATH-1H-E430G对Wien 133细胞的CDC功效没有影响。引入G236R(SEQ ID NO:123)或A327K(SEQ ID NO:63)突变,导致CDC活性部分降低,而K322E(SEQ ID NO:132)突变导致CDC活性降低至低于WT IgG1-CAMPATH-1H的水平的残留水平。仅P329R突变(SEQ ID NO:133)导致完全抑制IgG1-CAMPATH-1H-E430G-P329R对Wien133细胞的CDC活性。
实施例16:抗CD52 IgG1-CAMPATH-1H抗体变体和抗CD20 IgG1-11B8抗体变体的FcRn结合
新生儿Fc受体(FcRn)通过保护IgG免受降解来负责延长IgG的血浆半衰期。抗体内化后,FcRn结合核内体中的抗体Fc区,其中相互作用在弱酸性环境(pH 6.0)中稳定。循环至环境为中性(pH 7.4)的质膜后相互作用消失,并且抗体释放回到循环中。这影响IgG的血浆半衰期。
进行FcRn结合酶联免疫吸附测定(ELISA),以评估人FcRn与具有E430G、K439E和C1q结合抑制突变G236R或K322A的抗CD52 IgG1-CAMPATH-1H和具有E430G、S440K和C1q结合增强突变E333S的抗CD20 IgG1-11B8的结合。于室温(RT)在摇动的情况下将Streptawell96孔板(Roche,目录号1734776001)用在加有0.2%BSA的补充有0.05%Tween 20的PBS(PBST)中稀释的5μg/mL(100μL/孔)重组产生的人FcRn生物素化细胞外域[FcRnhsECDHis-B2M-BIO,即具有C端His标签的人FcRn细胞外域(FcRnhsECDHis;SEQ ID NO:155),作为与β2微球蛋白(B2M;SEQ ID NO:156)形成的二聚体]包被2小时。将板用PBST清洗3次。加入系列稀释抗体样品(pH6.0或pH7.4的PBST/0.2%BSA中以5倍稀释度,最终浓度范围为0.0005-40μg/mL),并于室温摇动温育1小时。用pH 6.0或pH7.4的PBST/0.2%BSA清洗板。加入在pH6.0或pH 7.4的PBST/0.2%BSA中稀释的辣根过氧化物酶(HRP)缀合的多克隆山羊抗人类kappa轻链(1:5,000;Sigma,目录号A-7164),并将板于室温摇动温育1小时。用pH 6.0或pH7.4的PBST/0.2%BSA清洗后,添加100μL 2,2'-连氮基-双(3-乙基苯并噻唑啉-6-磺酸(ABTS;1mg/mL;Roche目录号11112422001和11112597001))作为底物,并将板于室温避光温育10分钟,然后使用100μL 2%草酸(Riedel de Haen,目录号33506)终止反应,于室温温育10分钟,并使用ELISA读数器在405nm处测量吸光度。使用GraphPad Prism软件,对数转换的数据通过拟合具有可变斜率的S形剂量反应曲线进行分析。
所有测试的IgG1-CAMPATH-1H抗体变体显示pH 7.4时均不与人FcRn结合(图15A),pH 6.0时则高效的FcRn结合(图15B)。考虑到IgG1-CAMPATH-1H变体K439E(表观下限端)和变体E430G-K439E(表观上限端)的结合曲线之间的展开(spread),以及具有带FcRn结合位点的野生型Fc域的IgG1-b12和IgG1-CAMPATH-1H的最大FcRn结合之间的展开(图15B,C),测试的IgG1-CAMPATH-1H变体之间结合的表观差异被解读是不显著的。所有测试的IgG1-11B8抗体变体显示pH 7.4时均不与人FcRn结合(图15D),pH 6.0时则高效的FcRn结合(图15E)。IgG1-11B8-E430G-S440K中引入C1q结合增强突变E333S对与人FcRn的结合没有影响(图15E,F)。这些数据共同表明,具有E430G、K439E和C1q结合抑制突变G236R或K322A的抗CD52IgG1-CAMPATH-1H和具有E430G、S440K和C1q结合增强突变E333S的抗CD20 IgG1-11B8显示与人FcRn的正常结合。
实施例17:包含E430G、K439E、S440K和/或K322E突变及其组合的IgG1-CAMPATH-1H和IgG1-11B8抗体变体的药代动力学(PK)分析
在SCID小鼠的PK实验中,研究了K439E、S440K、E430G和K322E突变对IgG1-CAMPATH-1H和/或IgG1-11B8清除率的影响。将IgG1-CAMPATH-1H-E430G、-K439E、-E430G-K439E和-K322-E430G-K439E的清除率与WT IgG1-CAMPATH-1H的清除率进行比较,并将IgG1-11B8-E430G、-S440K、-E430G-S440K和-K322-E430G-S440K的清除率与WT IgG1-11B8的清除率进行比较。此外,还确定了如图16C和图16D所示的IgG1-CAMPATH-1H抗体变体与IgG1-11B8抗体变体的组合的清除率,并与WT IgG1-CAMPATH-1H和WT IgG1-11B8的组合的清除率进行比较。
本研究中,将小鼠安置于中央实验室动物设施(Utrecht,The Netherlands),并按照FELASA定义的良好动物规范在AAALAC和ISO 9001:2000认可的动物设施(GDL)中进行处理。所有实验均遵照由自指令(2010/63/EU)转换的荷兰动物保护法(WoD)进行,并经荷兰动物伦理委员会(CCD)和当地动物福利机构批准。向11-12周大的雌性SCID(C.B-17/ Hsd-Prkdcscid,Envigo)小鼠(每组3只小鼠)静脉内注射500μg抗体(单一试剂500μg;抗体混合物250μg+250μg)(25mg/kg),注射量为200μL。抗体施用后10分钟,4小时,1天,2天,7天,14天和21天,从隐静脉和颊静脉(cheek vein)交替穿刺收集50μL血液样品。将血液收集到含肝素的小瓶中,并以14,000xg离心10分钟。用380μL PBS稀释20μL血浆样品,并在测定抗体浓度之前于-20℃保存。使用夹心ELISA测定人总IgG浓度。小鼠抗人IgG-kappa mAb克隆MH16(CLB Sanquin,目录号M1268)被用作捕获抗体,并于4℃在100μL中以PBS中2μg/mL的浓度过夜包被到96孔ELISA微孔板(Greiner,目录号655092)。通过在板振荡器上与补充有0.2%BSA的PBS于室温温育1小时来封闭板。清洗后,加入100μL稀释的血浆样品,并在板振荡器上于室温温育1小时。将板用300μL PBST清洗3次,然后在板振荡器上与100μL过氧化物酶标记的山羊抗人IgG免疫球蛋白(Jackson,目录号109-035-098;在补充有0.2%BSA的PBST中1:10.000)于室温温育1小时。将板再次用300μL PBST清洗3次,然后于室温与100μL底物ABTS一起避光温育15分钟。加入100μL 2%草酸终止反应,并于室温温育10分钟。用酶标仪(Biotek,Winooski,VT)在405nm处测量吸光度。通过使用注射材料作为参考曲线来计算浓度。包括含有IgG1 kappa(结合位点,目录号BP078)的人骨髓瘤蛋白作为板对照。绘制人IgG浓度(以μg/mL为单位)(图16A为IgG1-CAMPATH-1H变体,图16B为IgG1-11B8变体,图16C为组合),使用Graphpad Prism软件计算曲线下面积(AUC)。到采血最后一天(第21天)为止的清除率通过公式确定:D*1,000/AUC,其中D是注射剂量(25mg/kg)(图16D)。
Hsd-Prkdcscid,Envigo)小鼠(每组3只小鼠)静脉内注射500μg抗体(单一试剂500μg;抗体混合物250μg+250μg)(25mg/kg),注射量为200μL。抗体施用后10分钟,4小时,1天,2天,7天,14天和21天,从隐静脉和颊静脉(cheek vein)交替穿刺收集50μL血液样品。将血液收集到含肝素的小瓶中,并以14,000xg离心10分钟。用380μL PBS稀释20μL血浆样品,并在测定抗体浓度之前于-20℃保存。使用夹心ELISA测定人总IgG浓度。小鼠抗人IgG-kappa mAb克隆MH16(CLB Sanquin,目录号M1268)被用作捕获抗体,并于4℃在100μL中以PBS中2μg/mL的浓度过夜包被到96孔ELISA微孔板(Greiner,目录号655092)。通过在板振荡器上与补充有0.2%BSA的PBS于室温温育1小时来封闭板。清洗后,加入100μL稀释的血浆样品,并在板振荡器上于室温温育1小时。将板用300μL PBST清洗3次,然后在板振荡器上与100μL过氧化物酶标记的山羊抗人IgG免疫球蛋白(Jackson,目录号109-035-098;在补充有0.2%BSA的PBST中1:10.000)于室温温育1小时。将板再次用300μL PBST清洗3次,然后于室温与100μL底物ABTS一起避光温育15分钟。加入100μL 2%草酸终止反应,并于室温温育10分钟。用酶标仪(Biotek,Winooski,VT)在405nm处测量吸光度。通过使用注射材料作为参考曲线来计算浓度。包括含有IgG1 kappa(结合位点,目录号BP078)的人骨髓瘤蛋白作为板对照。绘制人IgG浓度(以μg/mL为单位)(图16A为IgG1-CAMPATH-1H变体,图16B为IgG1-11B8变体,图16C为组合),使用Graphpad Prism软件计算曲线下面积(AUC)。到采血最后一天(第21天)为止的清除率通过公式确定:D*1,000/AUC,其中D是注射剂量(25mg/kg)(图16D)。
所有IgG1-CAMPATH-1变体(包括WT)的清除率比IgG1-11B8变体和预测的IgG1曲线(基于2室模型)的清除率更快一些。
在IgG1-CAMPATH-1H和IgG1-11B8中引入E430G突变导致这些抗体的清除率的少量增加。在IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G中引入K439E或S440K和/或K322E突变导致清除率分别相当于WT IgG1-CAMPATH-1H和WT IgG1-11B8的清除率(图16D)。所有测试的IgG1-CAMPATH-1H变体与IgG1-11B8变体的组合显示清除率与WT IgG1-CAMPATH-1H+IgG1-11B8抗体的组合的清除率相当。
这些数据一起表明,具有E430G、K439E、E430G-K439E或E430G-K439E-K322E突变的抗CD52 IgG1-CAMPATH-1H和具有E430G、S440K、E430G-S440K或E430G-S440K-K322E突变的抗CD20 IgG1-11B8及其混合物显示的清除率类似于WT IgG1抗体的清除率。
实施例18:G236R、G237T、K326A或E333S突变对具有六聚化增强突变和K439E或S440K的抗CD52或抗CD20抗体的体外FcγR结合的影响
在ELISA测定中使用纯化的抗体,测试IgG1-CAMPATH-1H或IgG1-11B8抗体变体与FcγRIIA同种异型131H、FcγRIIA同种异型131R、FcγRIIB、FcγRIIIA同种异型158F和FcγRIIIA同种异型158V的二聚ECD的结合。为了检测与二聚体FcγR变体的结合,将96孔Microlon ELISA板(Greiner,Germany)用PBS中的山羊F(ab’)2-抗人IgG-F(ab’)2(JacksonLaboratory,109-006-097,1μg/mL)于4℃包被过夜,清洗,并用200μL/孔PBS/0.2%BSA于室温(RT)封闭1小时。两次温育之间进行清洗,将板依次与以下温育:PBST/0.2%BSA中的IgG1-CAMPATH-1H或IgG1-11B8抗体变体稀释系列(以五倍的步骤,0.0013-20μg/mL)的100μL/孔于室温1小时,PBST/0.2%BSA中的加His标签的C端生物素化的二聚FcγRECD变体(1μg/mL)的100μL/孔于室温1小时,以及PBST/0.2%BSA中的100μL/孔链霉亲合素-polyHRP(CLB,M2032,1:10.000)作为检测抗体于室温30分钟。用1mg/mL ABTS(Roche,Mannheim,Germany)进行显影约20分钟(IIA-131H)或30分钟(IIA-131R、IIB、IIIA-158V、IIIA-158F)。为了终止反应,加入100μL 2%草酸。用酶标仪(BioTek,Winooski,VT)在405nm处测量吸光度。绘制了20μg/mL抗体浓度下的FcγR结合。数据基于三个独立的重复,每个实验相对于ELISA中的背景信号(无抗体对照,0%)和内标IgG1-11B8-E430G-S440K(设置为100%)进行标准化。
由于FcγR介导的效应器功能对IgG六聚体调节的敏感性可能不如CDC,因此由六聚体调节的共依赖性混合物的细胞毒性的完全选择性可能需要抑制FcγR与混合物中每个单个抗体的结合,特别是存在表达FcγR受体的效应细胞时。将抑制C1q结合的突变G236R或G237T引入抗体IgG1-CAMPATH-1H-E430G-K439E时,如通过ELISA所检测的,与FcγR变体IIA、IIB和IIIA的结合被强烈抑制(图17)。将增强C1q结合的突变K326A或E333S引入IgG1-11B8-E430G-S440K时,突变K326A导致与所有测试的FcγR变体的结合增加,而突变E333S分别将与FcγR变体IIA/B和IIIA的结合降低约50%和约20%(图17)。FcγR与IgG1-11B8-E430G-S440K、IgG1-11B8-K326A-E430G-S440K或IgG1-11B8-E333S-E430G-S440K的结合的进一步抑制可能需要另外的突变,例如实施例19所测试的突变G237A。
总之,尽管具有K326A或E333S突变的IgG1-11B8-E430G-S440K变体保留了FcγR结合,但是包含G236R或G237T突变的IgG1-CAMPATH-1H-E430G-K439E变体未显示可检测的与FcγR变体IIA、IIB和IIIA的结合。
实施例19:针对抗CD52 IgG1-CAMPATH-1H-E430G-K439E和抗CD20 IgG1-11B8-E430G-S440K-G237A抗体变体的混合物对Wien 133细胞的选择性CDC活性分析不同C1q结合调节突变
实施例11中显示,在具有或不具有C1q结合增强突变E333S的IgG1-11B8-E430G-S440K变体中引入FcγR结合抑制突变G237A与包含C1q结合抑制突变G236R的IgG1-CAMPATH-1H-E430G-K439E变体组合时在10μg/mL IgG时不损害CDC活性。在此,将另一个C1q结合抑制突变(G237T)引入IgG1-CAMPATH-1H-E430G-K439E中,并与IgG1-11B8-E430G-S440K-G237A和IgG1-11B8-E430G-S440K-G237A-E333S组合进行测试。使用Wien 133细胞的体外CDC测定,利用20%NHS和抗体浓度系列(以3.3倍稀释度,最终浓度范围为0.01-40.0μg/mL),基本如实施例2所述。如实施例2所述,细胞裂解和相对AUC值由三个实验重复的PI阳性细胞数目来计算。将AUC相对于阴性对照抗体IgG1-b12(0%)和阳性对照IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G(100%)的值标准化,同时呈现的最大裂解数据代表40μg/mLIgG时非标准化的细胞裂解。
由于FcγR介导的效应器功能对IgG六聚体调节的敏感性可能不如CDC,因此对六聚体调节的共依赖性混合物的细胞毒性的完全选择性可能需要抑制FcγR与混合物中每个单个抗体的结合,特别是存在表达FcγR受体的效应细胞时。将突变G236R和G237T引入IgG1-Campath-E430G-K439E强烈抑制FcγR结合(实施例18),但是IgG1-11B8-E430G-S440K显示常规的FcγR结合。为了抑制测试混合物中11B8组分的FcγR结合,将突变G237A引入具有或不具有另外的E333S突变的抗体IgG1-11B8-E430G-S440K中。如图18所示,在Wien 133细胞上,引入C1q结合抑制突变G237R或G237T有效降低IgG1-CAMPATH-1H-E430G-K439E组分的单一试剂活性,而与IgG1-11B8-E430G-S440K-G237A或IgG1-11B8-E430G-S440K-G237A-E333S组合保持完整的CDC活性。
总之,通过在抗CD52 K439E组分中引入C1q结合抑制突变G236R或G237T,增加了分别具有Fc-Fc相互作用增强突变E430G和自身寡聚化抑制突变K439E或S440K的抗CD52IgG1-CAMPATH-1H+抗CD20 IgG1-11B8-G237A抗体变体的CDC活性的选择性。在11B8变体中引入C1q结合增强突变E333S不增加组合中CDC活性的恢复,因为使用IgG1-11B8-E430G-S440K-G237A(具有G237A用于FcγR结合抑制)已经实现了补体活性的最大恢复。
实施例20:通过共依赖性抗体组合消耗血液细胞亚群
实施例19中,显示设计为以共依赖性方式起作用的抗CD52和抗CD20抗体变体的混合物对Wien 133细胞的CDC活性。在全血细胞毒性测定中,测试了共依赖性抗体混合物的特异性。CD52在T细胞和B细胞上均表达,而CD20在B细胞上表达但基本不在T细胞上表达。因此,预期独立起作用的CD52和CD20靶向抗体的混合物将消耗T细胞和B细胞两者,而预期严格共依赖性Ab混合物仅消耗B细胞,因为它们表达CD52和CD20两者。因此,该实验的目的是测试哪种抗CD52和抗CD20抗体变体混合物可以消除B细胞而不影响T细胞群。
在乌得勒支大学医学中心(University Medical Center Utrecht)获得用水蛭素处理的匿名血液样本。第一天期间,在补充有0.2%BSA的RPMI 1640培养基中制备CD52,CD20和gp120定向抗体的混合物。将抗体转移至包含RPMI/0.2%BSA中的30μL水蛭素处理的血液的板中,终体积为100μL,最终抗体浓度为10μg/mL,并于37℃和5%CO2下温育18小时。第二天,将等体积的红细胞(RBC)裂解缓冲液添加到混合物中,然后通过离心(300rcf下3分钟)收集细胞。重复该步骤,直到所有红细胞都被溶解为止,这通过混合物的透明度来评估。
随后,添加针对用荧光染料标记的不同血液亚群的检测抗体的混合物,并将该混合物于4℃温育30分钟。用FACS缓冲液进行三个清洗步骤后,将细胞重悬于FACS缓冲液中,并使用LSR Fortessa X20 FACS(BD)通过流式细胞术确定剩余细胞群的相对丰度。平行地,使用单个免疫检测试剂或可固定存活力染料(FVS)对补偿对照珠(UltraComp eBeads,Thermo Scientific目录号01-2222)进行染色的程序,并使用流式细胞术检测。所有步骤均在冰上进行。所用的检测试剂是用efluor 450标记的抗CD3(e-biosciences目录号48-0037),用BV711标记的CD19(Biolegend目录号302245),用PE-Cy7标记的CD66b(Biolegend目录号305115)和FVS存活力染色剂BV510(BD目录号564406)。简而言之,通过排除FVS阳性细胞和双联体(doublet)来对活细胞进行门控。在活细胞群中,髓样细胞由CD66b高表达进行门控,而淋巴样细胞由CD3高/CD19低/CD66低表达分出T细胞,由CD3低/CD19高/CD66低表达分出B细胞;其余的CD3低/CD19低/CD66低淋巴样细胞可以包含NK细胞,但在这些实验中未作进一步表征。在三个独立实验中测试了五个不同健康供体的血液样本。
与对肿瘤细胞系的CDC测定相反,全血测定是在可介导ADCC和ADCP的表达FcγR的效应细胞存在下进行的,因此除CDC以外,血细胞的消耗还可通过不同的效应机制介导。由于FcγR介导的效应器功能对IgG六聚体的敏感性可能不如CDC,因此可能需要对FcγR介导的效应器功能进行额外抑制,以使细胞毒性共依赖于非常接近的形成异源六聚体的两种抗体试剂的结合。在不同的供体中,淋巴细胞群中B细胞和T细胞的相对丰度范围分别为约1.5-6%和20-40%(图19A)。实际上,CD52定向的IgG1-Campath-1H-E430G和CD20定向的IgG1-11B8-E430G混合物消耗了所有T细胞(表达CD52)和B细胞(表达CD52和CD20)。基于WO2013004842中公开的突变,IgG1-Campath-1H-E430G-K439E+IgG1-11B8-E430G-S440K混合物在该测定设置中不是共依赖的(图19A)。IgG1-Campath-1H-E430G-K439E在B细胞和T细胞上均显示大量单一试剂活性,这可通过在肿瘤细胞系上高IgG浓度下观测到的FcγR介导的活性和残留CDC活性两者来解释(实施例4)。IgG1-11B8-E430G-S440K在B细胞上显示单一试剂活性,这可能由FcγR介导的活性解释。确实,IgG1-Campath-1H-E430G-K439E+IgG1-11B8-E430G-S440K混合物导致B细胞和T细胞两者的大量消耗。与之形成鲜明对比的是,IgG1-Campath-1H-G236R-E430G-K439E+IgG1-11B8-G237A-E430G-S440K混合物显示以严格共依赖性方式仅选择性消耗B细胞而不是T细胞(图19A)。实际上,IgG1-Campath-1H-G236R-E430G-K439E在B细胞或T细胞上均未显示明显的单一试剂活性,这表明除了如在前的实施例5、7和19中所观测到的抑制单一试剂CDC活性外,引入突变G236R消除FcγR介导的活性,如实施例18所预期。存在于IgG1-11B8-G237A-E430G-S440K中突变G237A的引入消除了B细胞上残留的单一试剂活性,这最可能是通过抑制残留的FcγR介导的活性。
包含G237位置处额外的突变,即G237A、G237T、G237Q或G237R的IgG1-CD52-Campath-E430G-K439E变体显示改变水平的单一试剂活性(图19B)。变体IgG1-CD52-Campath-G237A-E430G-K439E和IgG1-CD52-Campath-G237T-E430G-K439E作为单一试剂诱导T细胞数目减少。相反地,变体IgG1-CD52-Campath-G237Q-E430G-K439E和IgG1-CD52-Campath-G237R-E430G-K439E未显示可察觉的单一试剂活性,既不对B细胞也不对T细胞。没有单一试剂活性的IgG1-11B8-G237A-E430G-S440K与IgG1-CD52-Campath-G237Q-E430G-K439E或与IgG1-CD52-Campath-G237R-E430G-K439E混合时,观测到B细胞而非T细胞的有力且选择性消耗(图19B)。
包含增强C1q亲和力的突变E333S的IgG1-11B8-G237A-E333S-E430G-S440K存在下获得类似结果:该组分在B细胞或T细胞上都未显示可察觉的单一试剂活性,而与IgG1-CD52-Campath-G236R-E430G-K439E,IgG1-CD52-Campath-G237Q-E430G-K439E或IgG1-CD52-Campath-G237R-E430G-K439E混合时导致B细胞,而非T细胞的有力且选择性消耗(图19C)。相反地,将IgG1-11B8-G237A-E333S-E430G-S440K与IgG1-CD52-Campath-G237A-E430G-K439E或IgG1-CD52-Campath-G237T-E430G-K439E混合时,观测到可检测的T细胞消耗(图19C),与图19B中的结果一致。
还比较了靶向表达CD52和CD20两者的B细胞的共依赖性抗体混合物的功效与独立B细胞靶向性抗体利妥昔单抗(rituximab)、奥滨尤妥珠单抗(obinutuzumab)、IgG1-11B8和IgG1-11B8-E430G的功效。令人感兴趣的是,靶向CD52和CD20两者的严格共依赖性抗体混合物的B细胞消耗深度超过了利妥昔单抗、奥滨尤妥珠单抗、IgG1-11B8和IgG1-11B8-E430G的深度(图19D)。这对于IgG1-11B8-G237A-E430G-S440K与IgG1-CD52-Campath-G236R-E430G-K439E,IgG1-CD52-Campath-G237Q-E430G-K439E或IgG1-CD52-Campath-G237R-E430G-K439E的混合物也是如此,所有这些都显示了B细胞而非T细胞的选择性消耗(图19A/B)。
总之,这些数据表明,靶向两种不同细胞表面靶标的两种抗体的组合可以被迫以严格共依赖性方式发挥作用。结果,从CD52表达细胞的总群体中实现了选择性消耗血液亚群,此处是表达CD20的B细胞。这些严格共依赖性抗体组合由除了G236R、G237Q或G237T突变外具有Fc域突变E430G和K439E的一种抗体和具有Fc域突变G237A、E430G和S440K和可选的额外突变E333S的一种抗体组成。
实施例21:与抗CD20 IgG1-11B8-E430G-S440K变体组合时,通过在抗CD52 IgG1-CAMPATH-1H-E430G K439E中引入C1q结合抑制突变G236R或G237T混合的抗体变体对多种细胞系的CDC活性的增强的选择性
前面实施例中,通过与具有或不具有C1q结合增强突变E333S和/或FcγR结合抑制突变G237A的IgG1-11B8-E430G-S440K抗体变体组合将C1q结合抑制突变引入IgG1-CAMPATH-1H-E430G-K439E(实施例3中为K322E,实施例19中为G236R或G237T)显示CDC活性的增强的选择性。在此,在具有CD52和CD20不同表达水平的人肿瘤细胞系上(参见下面的表4),测试了将C1q结合抑制突变G236R或G237T引入IgG1-CAMPATH-1H-E430G-K439E后对CD20/CD52双阳性细胞的CDC活性的选择性。基本如实施例2所述,使用Daudi,Raji,Ramos,REH和U-698-M细胞系的体外CDC测定,利用20%NHS和抗体浓度系列(以3.3倍稀释度,最终浓度范围为0.01-40.0μg/mL)进行。如实施例2所述,细胞裂解和相对AUC值由三个实验重复的PI阳性细胞数目计算。将AUC相对于阴性对照抗体IgG1-b12(0%)和阳性对照IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G(100%)的值标准化,而呈现的最大裂解数据反映40μg/mL IgG时非标准化的细胞裂解浓度。
表4:不同细胞系上的CD20和CD52表达水平

*通过QIFIKIT分析确定的ABC抗体结合能力(数据未显示)
具有C1q结合抑制突变G237R或G237T的IgG1-CAMPATH-1H-E430G-K439E变体在具有不同CD52表达水平的所有测试细胞系上均未显示单一试剂活性(与背景水平相当):Daudi(图20A),Raji(图20B),Ramos(图20C),REH(图20D)和U-698-M(图20E)。对于细胞系Daudi,Ramos,Raji和U-698-M,与具有测试的C1q结合增强突变E333S(具有或不具有FcγR抑制突变G237A)或K326A的IgG1-11B8-E430G-S440K抗体变体组合时,观测到增加的CDC活性恢复。当CD20表达水平较低(REH:19,000ABC CD20)时,在CD20靶向性11B8中C1q增强突变K326A或E333S的影响不太明显。与靶标表达水平一致,低CD20表达性细胞系REH的CDC高度依赖于CD52靶向药剂的存在,而低CD52表达性细胞系Daudi的CDC高度依赖于CD20靶向药剂的存在。实际上,在细胞系REH和Daudi上,IgG1-G236R-E430G-K439E+IgG1-11B8-E430G-S440K诱导的CDC AUC和最大裂解大大低于IgG1-Campath-1H-E430G+IgG1-11B8-E430G组合诱导的CDC AUC和最大裂解,说明该混合物对CD20/CD52双阳性细胞系Ramos,Raji和U-698-M的相对选择性。通过在CD20靶向药剂中引入的C1q增强突变K326A、E333S或G237A-E333S的存在调节低CD52表达性Daudi细胞的CDC敏感性。
总之,通过在抗-CD52 E430G-K439E组分中引入C1q结合抑制突变如G236R或G237T,可以建立IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K组合的CDC活性的选择性,而与靶标在不同细胞系中的表达水平无关;通过在抗CD20 E430G-S440K组分中引入C1q结合增强突变如E333S或K326A来调节混合物的功效。
实施例22:实施例15:抗CD37 IgG1-CD37-37.3抗体变体和抗CD20 IgG1-11B8抗体的混合物在具有不同靶标表达水平的细胞系上的CDC活性的选择性
前面实施例中,显示了抗CD52与抗CD20抗体变体(错误!未找到参考源。错误!未找到参考源。错误!未找到参考源。)以及抗-CD52与抗CD37抗体变体(错误!找不到参考源。)的组合的选择性CDC活性。在此,在使用Daudi和WIL2-S细胞的体外CDC测定中,测试了抗CD37抗体变体与抗CD20抗体变体的组合的选择性CDC活性。使用Daudi和B淋巴母细胞WIL2-S细胞(ATCC,CRL-8885)的体外CDC测定,利用20%NHS和抗体浓度系列(以3倍稀释度,最终浓度范围为0.005-10.0μg/mL)进行,基本如所述错误!找不到参考源。WIL2-S细胞的培养基由以下组成:具有25mM Hepes和L-谷氨酰胺的RPMI 1640(Lonza,目录号BE12-115F),补充有+10%热灭活的DBSI,1mM丙酮酸钠(Lonza,目录号BE13-115E)和50U/mL的Pen/Strep。细胞裂解由三个实验重复平均的PI阳性细胞数目计算,并相对于阴性对照抗体IgG1-b12(0%)和10μg/mL IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G(100%)所测量的细胞裂解标准化。
作为单一试剂测试时,Daudi细胞对WT抗CD37抗体IgG1-CD37-37.3和WT II型抗CD20抗体IgG1-11B8的CDC诱导具有抗性,而两种WT抗体混合物观测到一些CDC活性(图21A)。引入Fc-Fc相互作用增强突变作为单一试剂测试时导致两种抗体的CDC活性诱导,作为混合物测试时则导致诱导强的CDC活性。IgG1-CD37-37.3-E430G的单一试剂CDC活性通过引入K439E突变强烈抑制,并且通过额外引入C1q结合抑制突变G236R则完全抑制。IgG1-11B8-E430G的单一试剂CDC活性通过引入S440K突变抑制,也在存在另外的C1q结合增强突变E333S的情况下。对于所有测试的IgG1-CD37-37.3抗体变体(包含E430G、E430G-K439E或E430G-K439E-G326R)与IgG1-11B8抗体变体(包含E430G、E430G-S440K或E430G-S440K-E333S)的混合物,在Daudi细胞上观测到强的CDC活性。
在WIL2-S细胞上,所有IgG1-CD37-37.3抗体变体(WT、E430G、E430G-K439E和E430G-K439E-E333S)作为单一试剂测试时的CDC活性都非常低(图21B)。在II型抗CD20抗体IgG1-11B8中引入E430G突变导致对WIL2-S细胞的CDC活性诱导。引入S440K突变强烈抑制IgG1-11B8-E430G的单一试剂CDC活性,而引入另外的C1q结合增强突变E333S则导致IgG1-11B8-E430G-S440K-E333S的低CDC活性(图21B)。对于所有测试的IgG1-CD37-37.3抗体变体(包含E430G、E430G-K439E或E430G-K439E-G326R)与IgG1-11B8抗体变体(包含E430G、E430G-S440K或E430G-S440K-E333S)的混合物,在WIL2-S细胞上观测到强的CDC活性。
总之,这些数据表明测试的IgG1-CD37-37.3-E430G抗体变体(K439E和K439E-G236R)与IgG1-11B8-E430G抗体变体(S440K和S440K-E333S)的混合物对Daudi和WIL2-S细胞的选择性CDC活性,而当作为单一试剂测试时,单个抗体几乎观测不到CDC活性。
实施例23:抗CD37 IgG1-CD37-37.3抗体变体和抗CD20 IgG1-11B8抗体变体以不同比例混合的组合物对Daudi细胞的CDC活性
实施例22中,在使用Daudi细胞的体外CDC测定中,显示与抗CD20抗体变体以1:1比例混合的抗CD37抗体变体的选择性CDC活性。在此,检测了改变混合物中存在的两种共依赖性抗体组分之间的比例的影响。在全因子设计中生成包含浓度范围为0.0005-10μg/mL的IgG1-CD37-37.3-G236R-E430G-K439E和浓度范围为0.013-10μg/mL的IgG1-11B8-E430G-S440K的组合物,并在体外CDC测定中进行测试。基本如实施例2所述,使用Daudi细胞的体外CDC测定,利用20%NHS进行。细胞裂解由PI阳性细胞数目计算。
Daudi细胞通过IgG1-CD37-37.3-G236R-E430G-K439E和IgG1-11B8-E430G-S440K的混合物以共依赖性方式裂解。单一试剂显示有限的活性:与非结合对照抗体IgG1-b12混合的高达10μg/mL的IgG1-CD37-37.3-G236R-E430G-K439E诱导最多8%的裂解,而与IgG1-b12混合的高达10μg/mL的IgG1-11B8-E430G-S440K诱导最多7%的裂解。相反地,对于代表大范围的不同抗体比例的组合物,IgG1-CD37-37.3-G236R-E430G-K439E和IgG1-11B8-E430G-S440K的混合物诱导Daudi细胞的有效裂解(表5)。
值得注意的是,通过提供10μg/mL的IgG1-11B8-E430G-S440K使CD20结合饱和时,添加≥0.12μg/mL IgG1-CD37-37.3-G236R-E430G-K439E导致≥70%的Daudi细胞裂解。对于包含10μg/mL的IgG1-11B8-E430G-S440K和≥0.013μg/mL的IgG1-CD37-37.3-G236R-E430G-K439E的组合物,观测到可检测的裂解。类似地,使用10μg/mL的IgG1-CD37-37.3-G236R-E430G-K439E使CD37结合饱和时,≥1.1μg/mL的IgG1-11B8-E430G-S440K诱导≥69%的Daudi细胞裂解。对于包含10μg/mL的IgG1-CD37-37.3-G236R-E430G-K439E和≥0.12μg/mL的IgG1-11B8-E430G-S440K的组合物,观测到可检测的裂解。
这意味着包含比例范围为约1:1000-1:1,或1:1-1:100的IgG1-CD37-37.3-G236R-E430G-K439E和IgG1-11B8-E430G-S440K的抗体组合物产生可检测的Daudi细胞裂解,只要两个靶标CD37和CD20中至少一个的结合是饱和的。包含比例范围为约1:100-1:1,或1:1-1:10的IgG1-CD37-37.3-G236R-E430G-K439E和IgG1-11B8-E430G-S440K的抗体组合物产生>69%的Daudi细胞裂解,只要两个靶标中至少一个的结合是饱和的。
总之,这些数据表明,比例相差很大的抗体组合物产生可检测至高效的CDC对Daudi细胞的裂解,尤其是在两个靶标中至少一个的结合饱和的情况下,而第二种组分不存在或以低丰度存在时则未观测到可检测的裂解。
表5显示以全因子设计混合的浓度范围为0.0005-10μg/mL的IgG1-CD37-37.3-G236R-E430G-K439E和浓度范围为0.013-10μg/mL的IgG1-11B8-E430G-S440K的组合物的CDC活性。在20%NHS存在下,将Daudi细胞与抗体混合物一起温育。Daudi细胞的裂解由PI阳性细胞分数计算。

实施例24:抗-CD52 IgG1-CAMPATH-1H抗体变体在Wien 133细胞上的靶标结合
通过流式细胞术分析具有Fc-Fc相互作用增强突变E430G或E345R,自身寡聚化抑制突变K439E以及FcγR结合抑制和C1q结合调节突变G236R、G237A或G237T之任一个的抗CD52 IgG1-CAMPATH-1H抗体变体与Wien133淋巴瘤细胞的结合。细胞悬浮液用PBS清洗并以2x106个细胞/mL的浓度重悬于FACS缓冲液[PBS+0.1%(w/v)牛血清白蛋白(BSA)+0.02%(w/v)叠氮化钠]中。将50μL细胞悬浮液样品(100,000个细胞/孔)接种到聚苯乙烯96孔圆底板(Greiner Bio-One;目录号650261)中,并与50μL抗体样品(以3倍稀释度,最终浓度为0.0002-10μg/mL)于4℃温育30分钟。细胞通过于4℃以300x g离心3分钟进行沉淀,并用150μL FACS缓冲液清洗3次。将细胞与50μL二抗R-藻红蛋白(R-PE)缀合的山羊抗人IgG F(ab')2(Jackson ImmunoResearch,目录号109-116-098,1:200)于4℃避光温育30分钟。将细胞用150μL FACS缓冲液清洗两次,重悬于30μL FACS缓冲液中,并用Intellicyt iQue筛选器通过流式细胞术分析抗体结合。减去仅与二抗温育的细胞观测到的背景信号后,使用GraphPad Prism软件版本8中提供的Hill斜率模型,通过非线性回归分析具有特异性结合的结合曲线确定最大结合水平(Bmax)和表观Kd。将每个实验的结合数据相对于针对野生型IgG1-11B8计算的最大结合水平(Bmax)进行标准化。
图22A显示在代表性实验中对IgG1-CAMPATH-1H变体观测到的结合,其中所有测试的抗体变体均显示与Wien 133细胞的相似剂量依赖性结合。这些数据表明,引入突变E430G和K439E对细胞表面上的靶标结合没有影响。当引入突变E345R而不是突变E430G时,观测到相当的靶标结合。同样,在IgG1-CAMPATH-1H-E430G-K439E中引入另外的突变G236R、G237A或G237T之任一个对细胞表面上的CD52靶标结合均无影响。比较从三个独立实验收集的数据时,抗体变体与野生型抗体相比,观测到平均最大结合(Bmax;图22B)无差异,并观测到平均表观Kd无差异(图22C)。
实施例25:包含E430G-S440K突变的抗CD20 IgG1-11B8抗体变体在Raji细胞上的靶标结合
通过流式细胞术分析具有Fc:Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和C1q结合调节突变的抗CD20 IgG1-11B8抗体变体与Raji淋巴瘤细胞的结合。基本如实施例24所述,使用Raji细胞的体外结合测定,利用连续稀释抗体浓度(以3倍稀释度,最终浓度范围为0.0007-40μg/ml)进行。用Intellicyt iQue筛选器通过流式细胞术分析抗体结合。减去仅与二抗温育的细胞观测到的背景信号后,使用GraphPad Prism软件版本8中提供的Hill斜率模型,通过非线性回归分析具有特异性结合的结合曲线来确定最大结合水平(Bmax)和表观Kd。每个实验的结合数据相对于对野生型IgG1-11B8计算的最大结合水平(Bmax)进行标准化。
图23A和B显示代表性结合实验,表明所有测试的IgG1-11B8抗体变体显示相似的与Raji细胞的剂量依赖性结合。取三个独立实验的平均值时,带有上述任何突变的IgG1-11B8抗体变体的最大结合(Bmax)值和平均表观Kd值与野生型IgG1-11B8的那些相似(图23C-F)。这些数据表明,引入单个突变或组合突变E430G和S440K对细胞表面上的靶标结合没有影响。在IgG1-11B8-E430G-S440K中引入另外的C1q结合增强突变K326A、E333A或E333S之任一种,C1q结合抑制突变G237A或C1q结合调节突变G237A-E333S的组合对细胞表面上的CD20靶标结合均没有影响。
实施例26:包含E430G和K439E或S440K突变的抗CD52 IgG1-CAMPATH-1H和抗CD20IgG1-11B8抗体变体的FcRn结合
新生儿Fc受体(FcRn)通过保护IgG免受降解来负责延长IgG的血浆半衰期。抗体内化后,FcRn结合核内体中的抗体Fc区,其中相互作用在弱酸性环境(pH 6.0)中稳定。循环至环境为中性(pH 7.4)的质膜后相互作用消失,并且抗体释放回到循环中。这影响IgG的血浆半衰期。
进行FcRn结合酶联免疫吸附测定(ELISA),以评估人FcRn与具有抗Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E(在IgG1-CAMPATH-1H的情况下)或S440K(在IgG1-11B8的情况下),以及C1q结合调节突变G237A、G237T、K326A、E333A或G237A-E333S之任一的抗CD52 IgG1-CAMPATH-1H或抗CD20 IgG1-11B8的结合。于室温(RT)在摇动情况下将Streptawell 96孔板(Roche,目录号1734776001)用在PBS中稀释的2μg/mL(100μL/孔)重组产生的人FcRn生物素化细胞外域[FcRnECDHis-B2M-BIO,即具有C端His标签的人FcRn细胞外域(FcRnECDHis;SEQ ID NO:155),作为与β2微球蛋白(B2M;SEQ ID NO:156)形成的二聚体]包被2小时。将板用PBST清洗3次。加入pH6.0或pH7.4的PBST/0.2%BSA中最终浓度为40μg/mL的抗体样品,并于室温摇动温育1小时。将板用pH6.0或pH7.4的PBST/0.2%BSA清洗3次。加入用pH 6.0或pH 7.4的PBST/0.2%BSA稀释的辣根过氧化物酶(HRP)缀合的多克隆山羊抗人类kappa轻链(1:5,000;Sigma,目录号A-7164),并于室温摇动温育1小时。用pH 6.0或pH 7.4的PBST/0.2%BSA清洗3次后,加入100μL 2,2'-连氮基双(3-乙基苯并噻唑啉-6-磺酸(ABTS;1mg/mL;Roche目录号11112422001)作为底物,并将板于室温避光温育10分钟。使用100μL 2%草酸(Riedel de Haen,目录号33506)终止反应,于室温温育10分钟,并使用ELISA读数器在405nm处测量吸光度。
pH 6.0时,由野生型IgG1-CAMPATH-1H和带有突变E430G或E430G-K439E的IgG1-CAMPATH-1H抗体变体,以及带有C1q结合调节突变G237A或G237T的IgG1-CAMPATH-1H-E430G-K439E抗体变体观测到高度相似的FcRn结合(图24A)。同样,pH 6.0时,野生型IgG1-11B8,带有突变E430G或E430G-S440K的IgG1-11B8抗体变体,和带有C1q结合调节突变K326A、E333A、G237A或G237A-E333S的IgG1-11B8-E430G-S440K抗体变体显示相似的FcRn结合(图24B)。pH 7.4时,与测试的其他抗体相比,IgG1-b12显示与FcRn的低残留结合,所有所述其他抗体均显示与在没有抗体的情况下温育的孔所记录的背景信号基本相似的结合(图24C,D)。总之,这些结果显示,具有六聚化增强突变E430G,自身寡聚化抑制突变K439E或S440K,以及C1q结合调节突变K326A、G237A、G237T、E333A或G237A-E333S的抗CD52IgG1-CAMPATH-1H和抗CD20 IgG1-11B8显示与人FcRn的正常结合。
实施例27:带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变的抗体变体的药代动力学(PK)分析
在小鼠研究中,分析了具有Fc-Fc相互作用增强突变,自身寡聚化抑制突变和C1q结合调节突变的抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的药代动力学特性。抗体变体既作为单个抗体又作为混合物进行测试。
本研究中,将小鼠安置于中央实验室动物设施(Utrecht,The Netherlands),并保持在单独通风的笼子中,随意提供水和食物。所有实验均遵照由自指令(2010/63/EU)转化的荷兰动物保护法(WoD)进行,并经荷兰中央动物实验委员会(CCD)和地方伦理委员会批准。使用每组3只小鼠,向SCID小鼠(C.B-17/ Hsd-Prkdc<scid,Envigo)静脉内注射500μg抗体(野生型IgG1-CAMPATH-1H,其含有E430G和K439E突变和G237Q或G236R突变的变体,野生型IgG1-11B8,其含有E430G和S440K突变和G237A或E333S突变的变体,或IgG1-CAMPATH-1H和IgG1-11B8抗体变体的组合)。抗体施用后10分钟,4小时,1天,2天,7天,14天和21天,从面静脉收集50μL血液样品。将血液收集到含肝素的小瓶中,并以10,000g离心5分钟。将血浆保持于–20℃,直至测定抗体浓度。
Hsd-Prkdc<scid,Envigo)静脉内注射500μg抗体(野生型IgG1-CAMPATH-1H,其含有E430G和K439E突变和G237Q或G236R突变的变体,野生型IgG1-11B8,其含有E430G和S440K突变和G237A或E333S突变的变体,或IgG1-CAMPATH-1H和IgG1-11B8抗体变体的组合)。抗体施用后10分钟,4小时,1天,2天,7天,14天和21天,从面静脉收集50μL血液样品。将血液收集到含肝素的小瓶中,并以10,000g离心5分钟。将血浆保持于–20℃,直至测定抗体浓度。
使用总hIgG ELISA确定特异性人IgG浓度。将以2μg/mL浓度包被于96孔MicrolonELISA板(Greiner,Germany)的小鼠抗人IgG IgG2amm-1015-6A05-Fab(内部生产的抗体,批次3233-025-EP,Genmab,The Netherlands)用作捕获抗体。用补充有0.2%牛血清白蛋白的PBS封闭板后,添加样品,在ELISA缓冲液(补充有0.05%Tween 20和0.2%牛血清白蛋白的PBS)中系列稀释,并在板摇动器上于室温(RT)温育1小时。随后将板与山羊抗人IgG免疫球蛋白(#109-035-098,Jackson,West Grace,PA)一起温育,并用2,2'-连氮基双(3-乙基苯并噻唑啉-6-磺酸)(ABTS;Roche,Mannheim,Germany)显影。用于注射的相应材料用作参考曲线。用酶标仪(Biotek,Winooski,VT)在405nm处测量吸光度。
尽管小鼠中注射的所有IgG1-CAMPATH-1H抗体变体的观测到的IgG浓度比SCID小鼠中野生型IgG1抗体的基于2室模型的预期浓度曲线更低,但野生型抗体IgG1-CAMPATH-1H与抗体变体IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-CAMPATH-1H-E430G-K439E-G237Q的清除率相当(图25A)。野生型抗体IgG1-11B8及其变体IgG1-11B8-E430G-S440K-G237A和IgG1-11B8-E430G-S440K-E333S的清除率彼此相当(图25B)。在IgG1-11B8及其变体的情况下,总IgG1浓度曲线类似于SCID小鼠中野生型IgG1的预测浓度曲线。与单个抗体的观测结果一致,引入增强Fc-Fc相互作用,抑制自身寡聚化或调节C1q结合的突变不影响IgG1-CAMPATH-1H和IgG1-11B8抗体混合物的药代动力学(图25C)。混合物的总IgG浓度曲线与SCID小鼠中野生型IgG1的预测浓度曲线相似,但略低。
具有突变E430G-S440K-G237A的IgG1-11B8变体显示与野生型IgG1-11B8相似的清除率(图25D)。具有突变E430G-K439E-G236R或E430G-K439E-G237Q的IgG1-CAMPATH-1H变体显示的清除率与野生型IgG1-CAMPATH-1H的清除率相似(图25D),而所有测试的IgG1-CAMPATH-1H变体的清除率比测试的IgG1-11B8变体的清除率更高。所有测试的IgG1-CAMPATH-1H变体与IgG1-11B8变体的组合显示的清除率与WT IgG1-CAMPATH-1H+IgG1-11B8抗体组合物的清除率相当。总之,引入调节Fc-Fc相互作用,自身寡聚化和C1q结合的突变不影响IgG1-CAMPATH-1H和IgG1-11B8抗体或其混合物在小鼠中的药代动力学特征。
实施例28:带有Fc-Fc相互作用增强突变,自身寡聚化抑制突变和C1q结合调节突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体的液相补体激活
抗体寡聚化后,补体因子C1可与抗体多聚体结合,引发补体级联反应的进一步激活。在经典补体途径激活后,激活的C1s将C4切割成片段C4a和C4b,其被因子I进一步加工成C4d。因此,C4d的血清浓度可用作经典补体途径激活的度量。在此,在具有完整补体(未加热灭活)的人血清中温育抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体后,测量C4d血清浓度,以确定引入增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变是否诱导经典补体途径激活,而不依赖于靶标结合。
将单个抗体与90%正常人AB血清(NHS)混合,最终重组IgG总终浓度为100μg/ml。然后将抗体于37℃温育60分钟,随后置于冰上。接着,使用样品稀释液(来自Quidel,MicroVue Complement C4d Fragment EIA试剂盒)将样品稀释100倍。根据制造商说明书,将样品和试剂盒提供的标准液应用到96孔微量滴定板,该板已经使用250-300μl清洗缓冲液进行3次板清洗步骤来预处理,且在每个步骤后印干(blot drying)。将板在摇动器上于室温温育30分钟。接下来,将微量滴定板倒空并用清洗缓冲液清洗5次,并在每个步骤后印干。每孔加入50μl试剂盒提供的C4d缀合物,并且于室温温育30分钟。再次用清洗缓冲液清洗所有孔5次,并且每个步骤后印干。然后,每孔加入100μl试剂盒提供的底物,并于室温温育30分钟。为了终止酶促反应,将50μl试剂盒提供的终止溶液添加到所有孔,并使用ELISA读数器(Biotek,Winooski,VT)在405nm波长处测定吸光度。在用于这些测定的血清中检测到约11至13μg/ml的背景C4d水平。因此,将不包含抗体的两个样品用作阴性对照,并从与抗体变体温育的样品中测量的C4d浓度中减去这些样品中测量的C4d平均浓度。根据三个独立实验中记录的所有值计算样品平均值和标准差。
对于包含E345R、E430G,S440Y(RGY)突变的抗体变体,检测如通过血清中C4d浓度确定的补体激活(图26)。在重组Her2ECDHis(SEQ ID NO:159)与Her2特异性抗体变体IgG1-1014-005-E345R-E430G-S440Y(IgG1-1014-005-RGY;SEQ ID NO:158)温育后,检测到最强的补体激活,而重组Her2ECDHis不诱导补体激活。对于IgG1-CAMPATH-1H、IgG1-11B8或IgG1-b12抗体变体的任一种或其混合物,均未检测到与背景C4d水平显著不同的C4d浓度。
总之,在IgG1-CAMPATH-1H、IgG1-11B8或IgG1-b12抗体变体中引入Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K和C1q结合调节突变G236R、G237A、G237Q或G237R不导致溶液中的补体激活。
实施例29:包含E430G和K439E或S440K突变的抗体变体在细胞上的C1q结合
进行流式细胞分析术以分析与带有增强Fc-Fc相互作用,抑制自身寡聚化或调节C1q结合的突变的抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体温育的Wien 133细胞的C1q结合。对于每个样品,将0.1x 106个Wien 133细胞与以2.5倍稀释度步骤的浓度范围为20至0.033μg/ml的抗体变体的稀释液于37℃温育15分钟。接着,将板在冰上冷却至0℃,放置30分钟。加入冷却的正常人AB血清(NHS)至20%的终浓度,并将板于0℃在冰上温育10-20分钟。将细胞用冰冷的FACS缓冲液(PBS+0.1%BSA+0.02%NaN3)清洗两次,然后与20μg/ml兔抗人C1q-FITC(Dako;目录号F0254)于4℃温育30分钟。在FACS缓冲液中清洗细胞两次后,将细胞悬浮于30μl FACS缓冲液中,并用iQue流式细胞仪(Intellicyt)进行分析。使用GraphPad Prism版本8,在浓度轴的对数转换后,将作为平均荧光强度值测量的C1q结合用对数激动剂反应模型拟合,并且相对于无抗体温育的孔的平均MFI(0%)和阳性对照IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G的剂量-反应曲线的拟合上限值(100%)进行标准化。
将Wien 133细胞与IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G混合物温育后,观测到高效的C1q结合(图27A)。IgG1-CAMPATH-1H-E430G的单一试剂C1q结合活性接近阳性对照混合物诱导的水平,而IgG1-11B8-E430G显示低的单一试剂C1q结合活性。
在IgG1-CAMPATH-1H-E430G中引入自身寡聚化抑制突变K439E导致C1q结合效率的部分消除,而在IgG1-11B8-E430G中引入自身寡聚化抑制突变S440K导致完全丧失C1q结合(图27B)。与阳性对照混合物相比,通过混合IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K,与C1q结合的能力恢复至中等水平。通过在IgG1-11B8-E430G-S440K中引入C1q结合增强突变K326A或E333S实现单一试剂C1q结合的温和恢复。将后两种抗体变体之任一种与IgG1-CAMPATH-1H-E430G-K439E混合,导致C1q结合的恢复类似于IgG1-CAMPATH-1H-E430G-K439E+IgG1-11B8-E430G-S440K混合物的水平。
通过在IgG1-CAMPATH-1H-E430G-K439E中引入抑制C1q结合的突变G236R(图27C)或G237T(图27D)的任一种完全消除了C1q结合。将IgG1-CAMPATH-1H-E430G-K439E-G236R与带有S440K、S440K-K326A或S440K-E333S突变的任一种的IgG1-11B8-E430G抗体变体混合后,观测到C1q结合的部分恢复(图27C)。同样,通过将IgG1-CAMPATH-1H-E430G-K439E-G237T与带有S440K、S440K-K326A或S440K-E333S突变之任一种的IgG1-11B8-E430G抗体变体混合,观测到C1q结合的部分恢复(图27D)。
总之,虽然引入自身寡聚化抑制突变K439E或S440K部分消除带有Fc-Fc增强突变E430G的抗体的C1q结合能力,但是通过引入另外的C1q结合抑制突变可以实现C1q结合的完全消除。通过混合具有互补突变K439E和S440K的抗体使C1q结合恢复至不同水平,取决于单个抗体中引入的C1q调节突变。
实施例30:C1q结合调节突变对具有六聚化增强突变和自身寡聚化抑制突变的抗CD52和CD20抗体的体外FcγR结合的影响
使用纯化的抗体,在ELISA测定中测试IgG1-CAMPATH-1H和IgG1-11B8抗体变体与FcγRIA(FCGR1AECDHis;SEQ ID NO:149)的单体胞外域(ECD)和与FcγRIIA同种异型131H(diFCGR2AH-HisBAP;SEQ ID NO:150),FcγRIIA同种异型131R(diFCGR2AR-HisBAP;SEQ IDNO:151),FcγRIIB(diFCGR2B-HisBAP;SEQ ID NO:152),FcγRIIIA同种异型158F(diFCGR3AF-HisBAP;SEQ ID NO:153)及FcγRIIIA同种异型158V(diFCGR3AV-HisBAP;SEQID NO:154)的二聚体胞外域的结合。为了检测与二聚体FcγR变体的结合,将96孔MicrolonELISA板(Greiner,Germany)用PBS中的山羊F(ab’)2-抗人-IgG-F(ab’)2(JacksonLaboratory,109-006-097,1μg/mL)于4℃包被过夜,清洗,并且用200μL/孔的PBS/0.2%BSA于室温(RT)封闭1小时。在温育间进行清洗,将板依次与100μL/孔PBST/0.2%BSA中的20μg/ml的IgG1-CAMPATH-1H抗体变体于室温在摇动的情况下温育1小时,于室温与100μL/孔PBST/0.2%BSA中的加His标签的C端生物素化的二聚体FcγRECD变体(1μg/mL)摇动情况下温育1小时,以及与100μL/孔PBST/0.2%BSA中的链霉亲合素-polyHRP(CLB,M2032,1:10.000)作为检测抗体于室温摇动温育30分钟。用1mg/mL ABTS(Roche,Mannheim,Germany)进行显影约10分钟(IIAH,IIAR,IIIAV),20分钟(IIIAF)或30分钟(IIB)。为了终止反应,以100μL/孔添加2%草酸。用酶标仪(BioTek,Winooski,VT)在405nm处测量吸光度。为了检测与FcγRIa的结合,将96孔Microlon ELISA板(Greiner,Germany)用PBS中的FCGR1AECD-His(1μg/mL)于4℃包被过夜,清洗并且用PBS/0.2%BSA以200μL/孔于室温(RT)封闭1小时。在温育之间进行清洗,将板依次与PBST/0.2%BSA中的20μg/ml IgG1-CAMPATH-1H抗体变体以100μL/孔于室温摇动温育1小时,与PBST/0.2%BSA中的山羊抗人-kappaLC-HRP(1:5000)以100μL/孔于室温摇动温育30分钟。用1mg/mL ABTS(Roche,Mannheim,Germany)进行显影约10分钟。为了终止反应,以100μL/孔添加2%草酸。用酶标仪(BioTek,Winooski,VT)在405nm处测量吸光度。
通过相对于ELISA中的背景信号(无抗体对照,0%)和内标IgG1-CAMPATH-1H(设为100%)标准化每块板,将来自三个(FcγRI,FcγRIIB,FcγRIIIA)或两个(FcγRIIA)独立重复的数据合并在一起,然后取所有实验的平均值。还预期对抗体变体引入以调节C1q结合的突变影响FcγR结合。通过ELISA评估具有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K和C1q结合调节突变G236R、G237A、K322A、G237T、G237Q、G237R或G237A-E333S之任一种的IgG1-CAMPATH-1H和IgG1-11B8抗体变体与FcγRIIa、FcγRIIb、FcγRIIIa和FcγRIa的结合揭示在IgG1-CAMPATH-1H-E430G-K439E中引入突变G236R、G237A、G237T、G237Q或G237R强烈抑制与所有测试的FcγR变体的结合(图28A-F)。对于IgG1-CAMPATH-1H-E430G-G237A,观测到与FcγRIIb和低亲和力FcγRIIa和FcγRIIIa受体的低残留结合。还观测到具有突变G236R、G237T、G237Q和G237R的IgG1-CAMPATH-1H-E430G-K439E变体与FcγRIa的低残留结合。野生型IgG1-11B8和IgG1-11B8-E430G-S440K显示相似的与所有测试的FcγR的强烈结合,其通过在IgG1-11B8-E430G-S440K中引入G237A或G237A-E333S抑制。后两种抗体变体显示与FcγRIa、FcγRIIb和低亲和力和FcγRIIa受体的低残留结合。引入突变K322A轻度抑制IgG1-CAMPATH-1H-E430G-K439E与所有测试的FcγR的结合。
总之,通过在IgG1-CAMPATH-1H-E430G-K439E中引入C1q结合调节突变G236R、G237A、G237T、G237Q和G237R,强烈抑制FcγR结合。类似地,在IgG1-11B8-E430G-S440K中引入突变G237A或G237A-E333S强烈抑制FcγR结合。
实施例31:通过在具有Fc-Fc相互作用增强突变E430G的抗CD52 IgG1-h2E8,IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体中引入C1q结合调节突变得到的混合抗体变体的CDC活性的选择性
与IgG1-CAMPATH-1H类似,人源化抗体IgG1-h2E8靶向人CD52。在此,使用Wien 133淋巴瘤细胞研究了在带有Fc-Fc相互作用增强突变和自身寡聚化抑制突变的IgG1-CAMPATH-1H、IgG1-h2E8、IgG1-11B8抗体变体中引入C1q结合调节突变对CDC效力的影响。
在抗体IgG1-CAMPATH-1H、IgG1-h2E8、IgG1-11B8和IgG1-b12中引入不同的突变:诱导增强的Fc-Fc相互作用的E430G;突变K439E或S440K之任一种,其通过抑制分子间Fc-Fc相互作用来抑制同源六聚体抗体复合物的形成,并通过交叉互补的Fc-Fc相互作用来促进异源六聚体抗体复合物的形成;抑制C1q与异源六聚体抗体复合物结合并抑制与Fcγ受体结合的G236R、G237A、G237R、G237T或G237Q。作为对照,还将单个抗体与非结合同种型对照抗体IgG1-b12或与IgG1-b12-E430G-S440K以1:1混合,以能够直接比较单个组分及由其组成的混合物的浓度。基本如实施例2中所述,在含20%NHS的情况下对Wien 133细胞的体外CDC测定中,测试了一系列浓度的纯化抗体(以3.3倍稀释度,最终浓度范围为0.009-40.0μg/mL)。细胞裂解百分比计算为(PI阳性细胞数目/细胞总数目)×100%。利用GraphPadPRISM中的对数变换浓度,使用非线性激动剂反应模型的最佳拟合值来分析数据,并计算三个实验重复的剂量反应曲线下面积。相对曲线下面积(AUC)值表示相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物为100%)标准化。
由IgG1-11B8-E430G诱导的中等单一试剂CDC效力通过引入突变S440K或双突变S440K-G237A而消除(图29A)。与IgG1-11B8-E430G相比,IgG1-CAMPATH-1H-E430G诱导更强的单一试剂CDC效力,这通过引入突变K439E-G236R而完全消除。可以通过将IgG1-CAMPATH-1H-E430G-K439E-G236R与IgG1-11B8-E430G-S440K混合使CDC效力部分恢复至阳性对照混合物IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G功效(相对AUC)的约80%。
与野生型IgG1-CAMPATH-1H类似,在Wien 133细胞中野生型IgG1-h2E8诱导的CDC具有中等效力,这可以通过引入Fc-Fc相互作用增强突变E430G来增强至IgG1-CAMPATH-1H-E430G诱导的相同水平。将IgG1-h2E8-E430G与IgG1-11B8-E430G混合与IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G的混合物以相当的效力诱导CDC。IgG1-h2E8-E430G的单一试剂CDC活性可以通过引入自身寡聚化抑制突变K439E来部分抑制。由40μg/ml单一试剂IgG1-h2E8-E430G-K439E诱导的最大裂解接近IgG1-h2E8-E430G+IgG1-11B8-E430G混合物的水平(图29B)。通过将IgG1-h2E8-E430G-K439E与IgG1-11B8-E430G-S440K混合来恢复CDC效力(恢复阳性对照混合物IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G功效的约90%),但是与非抗原结合IgG1-b12-E430G-S440K混合则不恢复CDC效力。在后一种混合物中的IgG1-11B8-E430G-S440K中引入突变G237A不影响混合物的CDC功效,无论是以AUC还是以40μg/ml时的最大裂解测量(图29B)。
在IgG1-h2E8-E430G-K439E中引入突变G236R完全消除残留的单一试剂CDC活性,而可以通过与IgG1-11B8-E430G-S440K混合恢复与对照混合物相比CDC效力的约79%。在IgG1-11B8-E430G-S440K中引入突变G237A轻度抑制在与IgG1-h2E8-E430G-K439E-G236R混合后恢复的CDC效力。在IgG1-h2E8-E430G-K439E中引入突变G237T抑制但不完全消除单一试剂CDC效力。与IgG1-h2E8-E430G-K439E-G236R和IgG1-11B8-E430G-S440K抗体变体的混合物类似,IgG1-h2E8-E430G-K439E-G237T的CDC效力可以通过与IgG1-11B8-E430G-S440K混合恢复至对照混合物功效的约88%,或与IgG1-11B8-E430G-S440K-G237A混合恢复(至对照混合物功效的约79%)。与引入突变G236R类似,在IgG1-h2E8-E430G-K439E中引入G237Q完全消除残留单一试剂CDC活性。通过与IgG1-11B8-E430G-S440K混合,可使CDC功效恢复至对照混合物功效的约84%。在后一种混合物中的IgG1-11B8-E430G-S440K中引入突变G237A诱导CDC功效的轻度抑制至对照混合物功效的约74%。40μg/ml时由具有任一C1q结合调节突变G236R、G237T或G237Q的IgG1-h2E8-E430G-K439E抗体变体与IgG1-11B8-E430G-S440K或IgG1-11B8-E430G-S440K-G237A的所有抗体混合物诱导的最大裂解与由IgG1-h2E8-E430G-K439E和IgG1-11B8-E430G-S440K的混合物诱导的最大裂解相当(图29B)。
总之,带有E430G突变,自身寡聚化抑制突变K439E和C1q结合调节突变的抗CD52抗体IgG1-h2E8变体当与抗CD20 IgG1-11B8抗体变体混合时显示与带有相同突变的IgG1-CAMPATH-1H抗体变体相当的在Wien 133细胞中的选择性共依赖性CDC效力,显示所观测到的由突变诱导的效果并非是IgG1-CAMPATH-1H特异的,而是也适用于其他CD52靶向性抗体。
实施例32:包含增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变的抗体变体的不依赖于靶标结合的募集
前面实施例中,证明了带有Fc-Fc相互作用增强突变和自身寡聚化抑制突变的抗原结合抗体变体的单一试剂CDC活性可以通过引入C1q结合抑制突变而进一步降低。将具有降低的单一试剂活性的抗体变体与带有C1q结合调节突变的互补抗原结合抗体变体混合后,观测到CDC效力的恢复。在此,我们测试了通过将抗原结合抗体变体与带有所述突变的非结合抗体变体混合是否也可以诱导共依赖性六聚体和CDC。
在抗CD52 IgG1-CAMPATH-1H抗体,抗CD20 IgG1-11B8抗体和非结合对照抗体IgG1-b12中引入了不同的突变:诱导增强Fc-Fc相互作用的E430G,突变K439E或S440K之任一种,它们通过抑制分子间Fc-Fc相互作用来抑制同源六聚体抗体复合物的形成,并通过交叉互补的Fc-Fc相互作用来促进异源六聚体抗体复合物的形成;抑制C1q与异源六聚体抗体复合物结合的G236R或G237A。基本如实施例2所述,在20%NHS的情况下对Wien 133细胞进行的体外CDC测定中,测试了一系列浓度的纯化抗体(以3.3倍稀释度,最终浓度范围为0.009-40.0μg/mL)。细胞裂解百分比计算为(PI阳性细胞数目/细胞总数目)×100%。利用GraphPad PRISM中的对数转换浓度,使用非线性激动剂剂量反应模型的最佳拟合值来分析数据,并计算三个实验重复的剂量反应曲线下面积。相对曲线下面积(AUC)值代表相对于同种型对照抗体IgG1-b12观测到的AUC值(0%)和IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G的混合物的AUC值(100%)标准化的值。
图30A,B显示,对于带有E430G突变与突变K439E或S440K之任一种的组合的非结合性对照抗体IgG1-b12的抗体变体混合物,未观测到单一试剂活性。另外,如前面实施例所述,未观测到抗体IgG1-CAMPATH-1H-E430G-K439E-G236R,IgG1-CAMPATH-1H-E430G-K439E-G237Q,IgG1-CAMPATH-1H-E430G-K439E-G237R或IgG1-11B8-E430G-S440K的单一试剂活性。将IgG1-CAMPATH-1H-E430G-K439E-G237T与非抗原结合的IgG1-b12-E430G-S440K混合时,检测到CDC具有阳性对照混合物(IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G)的CDC功效的9%。将IgG1-11B8-E430G-S440K与非结合性IgG1-b12-E430G-K439E温育时,可以检测到适度的CDC。相反,与CD20抗原结合的IgG1-11B8-E430G-S440K混合后,所测试的所有四个CAMPATH变体都可以有效地恢复CDC。
通过比较与CD52结合抗体IgG1-CAMPATH-1H或非结合对照抗体IgG1-b12的变体(均包含E430G,K439E和G236R突变)混合后的CDC效力,在单独的实验中更详细地分析了CD20定向性IgG1-11B8变体对非细胞结合抗体的潜在募集(图30C,D)。对于带有E430G突变与K439E或S440K突变之任一种和G236R或G237A突变之任一种的组合的非结合对照抗体IgG1-b12的抗体变体,未观测到单一试剂活性。混合都带有E430G突变,K439E或S440K突变之任一种和G236R或G237A突变之任一种的两个非抗原结合IgG1-b12抗体变体时,未观测到CDC效力。将非抗原结合抗体IgG1-b12-E430G-K439E-G236R与IgG1-11B8-E430G-S440K或IgG1-11B8-E430G-S440K-G237A之任一种混合后分别观测到CDC效力部分恢复至阳性对照混合物CDC功效的约15%和19%。通过将IgG1-CAMPATH-1H-E430G-K439E-G236R与IgG1-11B8-E430G-S440K或IgG1-11B8-E430G-S440K-G237A之任一种混合,CDC活性可以分别恢复至阳性对照混合物诱导的水平的约65%和71%。将IgG1-CAMPATH-1H-E430G-K439E-G236R与非靶标结合抗体IgG1-b12-E430G-S440K或IgG1-b12-E430G-S440K-G237A之任一种混合时,不诱导这种恢复。
这些数据表明,带有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变S440K和任选C1q结合抑制突变G237A的抗原结合抗体变体可以募集带有互补性突变E430G、K439E和G236R的非抗原结合抗体变体,最终导致诱导CDC。如果抗原结合抗体除E430G-K439E外还包含C1q结合抑制突变G236R、G237Q或G237R之任一种,则强烈抑制互补抗体E430G-S440K的不依赖于抗原结合的募集,而G237T无法阻断互补抗体的不依赖于抗原结合的募集。
实施例33:通过带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变的抗体变体激活FcγRIIa和FcγRIIIa的能力
实施例31中,研究了带有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K之任一种和C1q结合调节突变G236R、G237A、G237T、G237Q,G237R或K322A之任一种的抗体变体与FcγRIa、FcγRIIa、FcγRIIb和FcγRIIIa的结合。在G236或G237位置处引入C1q结合调节突变导致抑制FcγR结合。在此,通过使用表达人FcγRIIa或人FcγRIIIa的高亲和力同种异型变体的表达靶标的Raji细胞和Jurkat报告物细胞系在Promega报告物测定中确定cγRIIa和FcγRIIIa介导的激活,我们研究了在抗CD52IgG1-CAMPATH-1H-E430G-K439E和抗CD20 IgG1-11B8-E430G-S440K中引入C1q结合调节突变G236R、G237A、G237A-E333S或G237Q是否影响ADCC的诱导。
对Raji细胞使用ADCC报告物生物测定(Promega,FcγRIIa:目录号G9995;FcγRIIIa:目录号G7018)根据生产商说明书,定量上述IgG1-CAMPATH-1H和IgG1-11B8抗体变体对FcγR介导的信号传导的激活。作为效应细胞,该试剂盒包含Jurkat人T细胞,该细胞经工程改造以稳定表达高亲和力的FcγRIIa(FcγRII-H)或FcγRIIIa(V158)和驱动萤火虫萤光素酶表达的激活T细胞核因子(NFAT)响应元件。简而言之,将Raji细胞(5.000个细胞/孔)接种到384孔白色OptiPlates(Perkin Elmer目录号6007290)中在补充有12%低IgG血清(Promega;目录号G711A)的ADCC测定缓冲液(Promega,目录号G719A)中,并以含抗体浓度系列(以4倍稀释度,0.0002-40μg/mL终浓度)的30μL的总体积于37℃/5%CO2温育6小时,然后解冻ADCC生物测定效应细胞。于室温(RT)温育板15分钟后,添加30μLBio Glo Assay萤光素酶试剂,并于RT温育5分钟。通过在EnVision Multilabel Reader(Perkin Elmer)上的发光读数来定量萤光素酶的产生。通过减去自仅有培养基的样品(无Raji细胞,无抗体,无效应细胞)测定的背景发光信号,将发光信号标准化。利用GraphPad PRISM中的对数转换浓度,使用非线性激动剂剂量反应模型的最佳拟合值来分析数据,并计算三个(FcγRIIa)或两个(FcγRIIIa)实验重复的剂量反应曲线下面积。每个实验相对于与非结合对照IgG1-b12温育的细胞观测到的报告物活性(0%)以及野生型IgG1-Campath-1H+野生型IgG1-11B8混合物的活性(100%),将AUC值标准化,然后对实验重复取平均值。
作为单一试剂,抗体变体IgG1-11B8-E430G-S440K诱导FcγRIIa介导的激活(图31A),这与实施例31中显示该变体的FcγRIIa结合的结果一致。相反,在IgG1-CAMPATH-1H-E430G-K439E中引入C1q结合调节突变G236R或G237Q,或在IgG1-11B8-E430G-S440K中引入突变G237A或G237A-E333S时,未观测到单一试剂抗体变体的FcγRIIa-介导的激活。带有突变G236R或G237Q的IgG1-CAMPATH-1H-E430G-K439E变体和带有突变G237A或G237A-E333S的IgG1-11B8-E430G-S440K变体的混合物也不诱导FcγRIIa-介导的激活。但是,IgG1-11B8-E430G-S440K和IgG1-CAMPATH-1H-E430G-K439E-G236R或IgG1-CAMPATH-1H-E430G-K439E-G237Q之任一种的混合物确实诱导FcγRIIa-介导的激活。测试了相同的抗体变体诱导FcγRIIIa-介导的激活的能力(图31B)。在此,IgG1-11B8-E430G-S440K单独或与带有G236R或G237Q突变的IgG1-CAMPATH-1H-E430G-K439E变体混合时,观测到FcγRIIIa-介导的激活。具有或不具有突变G236R的IgG1-CAMPATH-1H-E430G-K439E分别诱导低或中等的FcγRIIIa-介导的激活。当IgG1-CAMPATH-1H-E430G-K439E-G236R或IgG1-CAMPATH-1H-E430G-K439E-G237Q与IgG1-11B8-E430G-S440K变体G237A或G237A-E333S混合时,无法检测到FcγRIIIa-介导的激活。
总之,带有Fc-Fc相互作用增强和自身寡聚化抑制突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体诱导FcγRIIa-或FcγRIIIa-介导的激活的能力(用作ADCC的替代量度),通过引入C1q结合调节突变G236R、G237A,G237A-E333S或G237Q有效消除。
实施例34:滴定具有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变的抗CD52 IgG1-CAMPATH-1H和抗CD20 IgG1-11B8抗体变体的混合物组分后对Wien 133细胞的CDC活性的选择性
前面实施例中,将带有增强Fc-Fc相互作用的突变以及抑制自身寡聚化的突变和调节C1q结合的突变的抗体变体以1:1的比例混合。在此,我们测试了通过以非等摩尔比混合两种抗体变体是否也获得选择性共依赖性CDC活性。
基本如实施例2所述,使用Wien 133细胞的体外CDC测定,利用20%NHS进行。将单个抗体以3.3倍稀释度(最终浓度范围为0.005-20.0μg/mL)滴定。应用于抗体混合物时,滴定一种组分(以5倍稀释度,最终浓度范围为0.0003-20.0μg/mL),而另一种组分以20μg/mL的固定浓度使用。如实施例2所述,细胞裂解由PI阳性细胞数目计算。
通过IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A的滴定混合物(1:1比例)诱导对Wien 133细胞的有效CDC活性(图32A)。将Wien 133细胞与抗体变体混合物一起温育后,甚至更有效地诱导CDC,该混合物中滴定一种抗体变体,而另一种抗体变体以20μg/mL的固定浓度应用。将滴定的IgG1-CAMPATH-1H-E430G-K439E-G236R与20μg/mL非靶标结合抗体IgG1-b12或其带有E430G-S440K-G237A突变的变体混合后,未检测到CDC活性,表明即使在使CD20结合饱和的IgG浓度下,异源六聚体化保持靶标结合依赖性。在没有血清的情况下混合IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-11B8-E430G-S440K-G237A时也未观测到CDC,表明C1q是稳定六聚体复合物所需的。对于IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A混合物,获得了高度相似的结果(图32B)。CDC仅由IgG1-CAMPATH-1H-E430G-K439E-G237Q和IgG1-11B8-E430G-S440K-G237A的混合物诱导,在这两种抗体变体都被滴定的混合物中,或在一种抗体变体被滴定而另一种抗体变体以20μg/mL的固定浓度应用的混合物中。IgG1-CAMPATH-1H-E430G-K439E-G237Q和非靶标结合抗体IgG1-b12或其带有E430G-S440K-G237A突变的变体的混合物不诱导CDC。
总之,通过带有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K之任一种和C1q结合调节突变G236R、G237A或G237Q之任一种的IgG1-CAMPATH-1H和IgG1-11B8抗体变体在大范围的非等摩尔比内可以共依赖地诱导对Wien 133细胞的高效的CDC:CAMPATH:11B8比例范围从100:3降至1:100时观测到>50%裂解,条件是使与两种靶标中的至少一种的结合饱和。
实施例35:带有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K之任一种和C1q结合调节突变P329R或G236R的抗CD52和抗HLA-DR抗体变体的选择性
在此,在Oci-Ly17淋巴瘤细胞上研究了带有Fc-Fc相互作用增强突变,自身寡聚化抑制突变和C1q结合调节突变的抗CD52和抗HLA-DR抗体变体的选择性共依赖性CDC效力。
在抗CD52抗体IgG1-CAMPATH-1H和抗HLA-DR抗体IgG1-HLA-DR-huL243和IgG1-HLA-DR-1D09C3中引入不同的突变:诱导增强的Fc-Fc相互作用的E430G,自身寡聚化抑制突变K439E或S440K之任一种,以及抑制C1q结合的突变G236或P329R之任一种。作为对照,还将单个抗体与非结合同种型对照抗体IgG1-b12或IgG1-b12-E430G-S440K以1:1混合,以能够直接比较单个组分及由其组成的混合物的浓度。基本如实施例2进一步所述,在20%NHS的情况下每种条件对30,000个Oci-Ly17淋巴瘤细胞的体外CDC测定中测试了一系列浓度的纯化抗体(以3.3倍稀释度,最终浓度范围为0.009-40.0μg/mL)。细胞裂解百分比计算为(PI阳性细胞数目/细胞总数目)×100%。利用GraphPad PRISM中的对数转换浓度,使用非线性激动剂剂量反应模型的最佳拟合值来分析数据,并计算三个实验重复的剂量反应曲线下面积。相对曲线下面积(AUC)值代表相对于最小裂解(对于IgG1-b12为0%)和最大裂解(对于IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-huL243-E430G混合物为100%,图33A;或对于IgG1-CAMPATH-1H-E430G+IgG1-HLA-DR-1D09C3-E430G混合物为100%,图33B)标准化。
IgG1-CAMPATH-1H-E430G和IgG1-HLA-DR-huL243-E430G的混合物诱导Oci-Ly17细胞的有效CDC(阳性对照混合物;设为100%,图33A)。后一种混合物的单一试剂诱导CDC至阳性对照混合物功效的约83%(IgG1-CAMPATH-1H-E430G)和80%(IgG1-HLA-DR-huL243-E430G)。在IgG1-CAMPATH-1H-E430G中引入突变S440K将诱导CDC的能力降低至对照混合物功效的约24%,而在IgG1-HLA-DR-huL243-E430G中引入双突变K439E-P329R降低诱导CDC的能力至对照混合物功效的约8.5%。通过混合IgG1-HLA-DR-huL243-E430G-K439E-P329R和IgG1-CAMPATH-1H-E430G-S440K,使诱导CDC的能力恢复至对照混合物功效的约38%。通过引入双突变K439E-G236R,消除了IgG1-HLA-DR-huL243-E430G的单一试剂CDC功效。通过将IgG1-HLA-DR-huL243-E430G-K439E-G236R与IgG1-CAMPATH-1H-E430G-S440K混合,可以获得CDC效力恢复至对照混合物功效的约75%,但是与非靶标结合抗体变体IgG1-b12-E430G-S440K混合则不会。
测试抗HLA-DR抗体IgG1-HLA-DR-1D09C3而非IgG1-HLA-DR-huL243变体时,观测到相当的结果(图33B)。IgG1-CAMPATH-1H-E430G和IgG1-HLA-DR-1D09C3-E430G的混合物诱导Oci-Ly17细胞的高效CDC(阳性对照混合物;设为100%,图33B)。后一种混合物的单一试剂诱导CDC至对照混合物功效的约70%(IgG1-CAMPATH-1H-E430G)和95%(IgG1-HLA-DR-1D09C3-E430G)。在IgG1-CAMPATH-1H-E430G中引入突变S440K将诱导CDC的能力降低至对照混合物功效的20%,而在IgG1-HLA-DR-1D09C3-E430G中引入双突变K439E-P329R则降低诱导CDC的能力至对照混合物功效的约7%。通过混合IgG1-HLA-DR-huL243-E430G-K439E-P329R和IgG1-CAMPATH-1H-E430G-S440K,使诱导CDC的能力恢复至对照混合物功效诱导水平的约35%。通过引入双突变K439E-G236R,消除了IgG1-HLA-DR-1D09C3-E430G的单一试剂CDC效力。可以通过将IgG1-HLA-DR-1D09C3-E430G-K439E-G236R与IgG1-CAMPATH-1H-E430G-S440K混合,获得CDC效力恢复至对照混合物功效的约50%,而与非靶标结合抗体变体IgG1-b12-E430G-S440K混合则不会。
总之,这些数据显示,通过混合通过引入Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K和C1q结合抑制突变P329R或G236R得到的抗CD52 IgG1-CAMPATH-1H的抗体变体与抗-HLA-DR IgG1-HLA-DR-huL243或IgG1-HLA-DR-1D09C3的抗体变体,可以诱导Oci-Ly17淋巴瘤细胞的选择性共依赖性CDC。
实施例36:包含L234和L235位置处抑制FcγR结合和C1q结合的突变的抗体变体的选择性
实施例5和12中,描述了在抗体变体中引入调节C1q结合的突变后对CDC效力的影响。在IgG1-CAMPATH-1H-E430G-K439E中引入突变G236R和G237Q导致对Wien 133细胞的单一试剂CDC活性的消除。在此,研究了L234和L235位置处引入C1q结合调节突变后,IgG1-CAMPATH-1H-E430G-K439E和IgG1-11B8-E430G-S440K抗体变体对Wien 133细胞上的CDC效力的影响。
在抗体IgG1-CAMPATH-1H和IgG1-11B8中引入了不同的突变:诱导Fc-Fc相互作用增强的E430G;自身寡聚化抑制突变K439E或S440K之任一种;和抑制C1q与异源六聚体抗体复合物结合的L234A、L234A-L235A、L234F、L234F-L235E、L235A、L235Q、G236R或G237Q。作为对照,还将单个抗体与非结合同种型对照抗体IgG1-b12以1:1混合,以能够直接比较单个组分及由其组成的混合物的浓度。在此,将来自瞬时转染的EXPI293上清液的未纯化的上清液用作包含L234、L235或其组合处的取代的IgG1-CAMPATH-1H变体的IgG来源。基本如实施例2所述,在20%NHS的情况下对Wien 133细胞的体外CDC测定中,测试一系列浓度的抗体(以3.3倍稀释度,最终浓度范围为0.009-40.0μg/mL)。将细胞裂解百分比计算为(PI阳性细胞数目/细胞总数目)×100%。利用GraphPad PRISM中的对数转换浓度,使用非线性激动剂剂量反应模型的最佳拟合值来分析数据,并计算三个实验重复的剂量反应曲线下面积。相对曲线下面积(AUC)值代表相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物为100%)标准化。
模拟转染的EXPI293上清液未诱导Wien 133细胞裂解,排除所用未纯化上清液中物质的贡献。引入突变L234A、L234A-L235A、L234F-L235E、L235A、L235Q、G236R或G237Q中的任何一个后,完全消除了IgG1-CAMPATH-1H-E430G-K439E对Wien 133细胞的单一试剂CDC效力,而观测到IgG1-CAMPATH-1H-E430G-K439E-L234F的低残留CDC活性(图34)。对于所有这些抗体变体,与IgG1-11B8-E430G-S440K混合后,可以使CDC效力部分恢复。CDC恢复的效率范围为阳性对照混合物IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G的AUC定义的功效的约70%(IgG1-CAMPATH-1H-E430G-K439E-G236R+IgG1-11B8-E430G-S440K)至约84%(IgG1-CAMPATH-1H-E430G-K439E-L234F+IgG1-11B8-E430G-S440K)。
总之,通过引入C1q结合抑制突变L234A、L234A-L235A、L234F、L234F-L235E、L235A和L235Q可以消除IgG1-CAMPATH-1H-E430G-K439E对Wien 133细胞的单一试剂活性,与引入突变G236R或G237Q的效果相当。通过将这些抗体变体与IgG1-11B8-E430G-S440K混合,可以实现CDC效力的恢复。因此,观测到前面实施例和本实施例中测试的任何C1q结合抑制取代可以改善本发明共依赖地起作用的抗体的选择性。在这一大类中,G236K/R、G237、L234和L235取代是优选的取代,因为它们相对温和地调节C1q结合,同时抑制FcγR-介导的效应器功能。不受理论的限制,如果在被两种抗体结合的细胞上本发明的两种抗体之间形成的异源六聚体复合物具有足够的C1q结合亲合力,则这可以使得有可能恢复有效的寡聚化依赖性活性,例如CDC,从而在被两种抗体结合的细胞上选择性地显示最大活性。
实施例37:除抑制自身寡聚化和调节C1q结合的突变外,E345和E430位置处具有备选Fc-Fc相互作用增强突变的抗体变体的选择性
前面实施例中,将Fc-Fc相互作用增强突变E345K、E345R和E430G引入共依赖性抗体变体中。在此,我们研究了在带有抑制自身寡聚化和调节C1q结合的突变的IgG1-CAMPATH-1H和IgG1-11B8抗体变体中的E430和E345位置处引入备选取代是否导致对Wien133细胞的CDC的共依赖性诱导。
在抗体IgG1-CAMPATH-1H和IgG1-11B8中引入不同的突变。选择E430取代E430G、E430N、E430T、E430V和E430Y来代表广泛不同的取代,例如小的,亲水的,疏水的和大的/芳香族的突变。同样,选择引入E345中的突变E345A、E345K、E345Q、E345R、E345V,E345Y代表较小的,碱性/带电荷的,亲水的,疏水的和大的/芳香族的突变。先前建议突变组合K248E-T437R通过Fc-Fc相互作用来调节IgG寡聚化(WO2018031258)。此外,抗体包含自身寡聚化抑制突变K439E或S440K,和/或C1q调节突变G236R、G237A或E333S。作为对照,还将单个抗体与非结合同种型对照抗体IgG1-b12以1:1混合,以能够直接比较各个组分及由其组成的混合物的浓度。在此,将瞬时转染的EXPI293上清液中未纯化的上清液用作不含E430G、E345K或E345R取代的抗体变体的IgG来源。基本如实施例2所述,在20%NHS的情况下对Wien 133细胞的体外CDC测定中,测试了一系列浓度的抗体(最终浓度范围0.009-40.0μg/mL;以3.3倍稀释度)。细胞裂解百分比计算为(PI阳性细胞数目/细胞总数目)x100%。利用GraphPadPRISM中的对数转换浓度,使用非线性激动剂剂量反应模型的最佳拟合值来分析数据,并计算两次实验重复的剂量反应曲线下面积。以相对曲线下面积(AUC)值呈现的CDC功效/效力代表每个板相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物为100%)标准化的数值,它们然后在两个独立的实验重复中取平均值。
模拟转染的EXPI293上清液未诱导Wien 133细胞裂解,排除所用未纯化上清液中物质的贡献。如前面实施例所述,在IgG1-11B8中引入突变E430G-S440K或E430G-S440K-G237A消除了单一试剂CDC效力(图35A),而通过引入突变E430G-K439E-G236R可消除IgG1-CAMPATH-1H的单一试剂CDC活性。通过将IgG1-CAMPATH-1H-E430G-K439E-G236R与具有E430G-S440K或E430G-S440K-G237A突变之任一种的IgG1-11B8抗体变体混合,使CDC效力分别恢复至阳性对照混合物IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G所测量功效的约82%和91%。
在IgG1-CAMPATH-1H-K439E-G236R中引入突变E430N(得到SEQ ID NO:115),E430T(得到SEQ ID NO:117),E430V(得到SEQ ID NO:119),E345A(得到SEQ ID NO:64),E345K(得到SEQ ID NO:66),E345Q(得到SEQ ID NO:69),E345R(得到SEQ ID NO:73)或E345Y(得到SEQ ID NO:77)代替E430G,获得与对E430G观测的那些结果相当的结果:带有所提及任何一种E430取代的IgG1-CAMPATH-1H抗体变体的单一试剂CDC活性处于背景水平,而通过混合IgG1-CAMPATH-1H变体与IgG1-11B8-E430G-S440K或IgG1-11B8-E430G-S440K-G237A获得CDC活性的恢复。由混合物得到的CDC效力恢复范围为对照混合物功效的约78%(IgG1-CAMPATH-1H-E345Q-K439E-G236R+IgG1-11B8-E430G-S440K)至约92%(IgG1-CAMPATH-1H-E345R-K439E-G236R+IgG1-11B8-E430G-S440K-G237A)。抗体变体IgG1-CAMPATH-1H-E430Y-K439E-G236R(SEQ ID NO:121)诱导对照混合物功效的约44%的残留单一试剂CDC活性。CDC效力恢复至通过混合IgG1-CAMPATH-1H-E430Y-K439E-G236R和IgG1-11B8-S440K-G237A的阳性对照混合物诱导水平的约94%。
在IgG1-11B8-E430G-S440K-G237A中用突变E430N(得到SEQ ID NO:116),E430T(得到SEQ ID NO:118),E430V(得到SEQ ID NO:120),E345A(得到SEQ ID NO:65),E345Q(得到SEQ ID NO:70),E345V(得到SEQ ID NO:76)或E345Y(得到SEQ ID NO:78)中的任何一个代替E430G突变时,获得与对E430G观测的那些结果相当的结果:带有所提及任何一个E430取代的IgG1-11B8抗体变体的单一试剂CDC活性处于背景水平,而通过混合IgG1-11B8变体与IgG1-CAMPATH-1H-E430G-K439E-G236R获得CDC活性的恢复(图35B)。由混合物得到的CDC效力恢复范围为对照混合物功效的约80%(IgG1-CAMPATH-1H-E430G-K439E-G236R+IgG1-11B8-E430V-S440K-G237A至约86%(IgG1-CAMPATH-1H-E430G-K439E-G236R+IgG1-11B8-E345Q-S440K-G237A)。对于抗体变体IgG1-11B8-E430Y-S440K-G237A(SEQ ID NO:122),观测到阳性对照混合物诱导的水平的约21%残留单一试剂CDC活性,而CDC效力可恢复至由通过混合IgG1-11B8-E430Y-S440K-G237A与IgG1-CAMPATH-1H-E430G-K439E-G236R得到的阳性对照混合物诱导的水平的约91%。
混合IgG1-CAMPATH-1H-K439E-G236R和IgG1-11B8-S440K-G237A抗体变体后,还观测到CDC效力的有效恢复,其中将相同的Fc-Fc相互作用调节突变引入两种抗体,选自以下突变:E430N、E430T、E430V、E345A、E345Q、E345V、E345Y(图35C)。如图35A和B所示,这些抗体变体的单一试剂活性处于背景水平。CDC效力的恢复范围为由阳性对照混合物诱导的水平的约70%(IgG1-CAMPATH-1H-E430V-K439E-G236R+IgG1-11B8-E430V-S440K-G237A)至约89%(IgG1-CAMPATH-1H-E430T-K439E-G236R+IgG1-11B8-E430T-S440-G237A)。CDC效力的最高恢复是通过引入E430Y突变的IgG1-CAMPATH-1H-K439E-G236R和IgG1-11B8-S440K-G237A抗体变体的混合物获得的(由阳性对照混合物诱导的水平的约97%),而两种抗体变体均诱导残留单一试剂CDC活性,如上所述。
已经描述了在抗体变体中引入双突变K248E-T437R来促进Fc-Fc相互作用。如图35D所示,在IgG1-11B8-K248E-T437R中引入突变S440K-G237A(得到SEQ ID NO:131)消除了对Wien 133细胞的单一试剂CDC效力。在IgG1-11B8-K248E-T437R中引入突变S440K-E333S(得到SEQ ID NO:130)后,观测到单一试剂CDC活性的部分抑制。将IgG1-11B8-K248E-T437R-S440K-G237A与IgG1-CAMPATH-1H-E430G-K439E-G236R混合时,观测到CDC效力有效恢复至对照混合物功效的约87%。此外,在还带有K439E-G236R突变(得到SEQ ID NO:128)的IgG1-CAMPATH-1H抗体变体中引入突变K248E-T437R得到没有任何单一试剂CDC活性的抗体变体。通过将IgG1-CAMPATH-1H-K248E-T437R-K439E-G236R与带有E430G-S440K-G237A、K248E-T437R-S440K-G237A或K248E-T437R-S440K-E333S突变的IgG1-11B8抗体变体之任一种混合,CDC效力可分别有效恢复至对照混合物功效的约87%、84%和97%。同样,在还具有K439E-G237Q突变(得到SEQ ID NO:129)的IgG1-CAMPATH-1H抗体变体中引入突变K248E-T437R得到没有任何单一试剂CDC活性的抗体变体。通过将IgG1-CAMPATH-1H-K248E-T437R-K439E-G237Q与带有K248E-T437R-S440K-G237A或K248E-T437R-S440K-E333S突变的IgG1-11B8抗体变体混合,可以获得CDC效力分别恢复至对照混合物功效的约86%和98%。
总之,通过混合在抑制自身寡聚化和调节C1q结合的突变外带有不同Fc-Fc相互作用调节突变,包括所有测试的E430取代(E430G、E430N、E430T、E430V、E430Y),所有测试的E345取代(E345A、E345K、E345Q、E345R、E345V、E345Y)和K248E-T437R的IgG1-CAMPATH-1H和IgG1-11B8抗体变体可以获得对Wien 133细胞的选择性共依赖性CDC效力。此外,当混合的抗体包含不同的Fc-Fc相互作用增强突变时,观测到共依赖的活性。
因此,可以预期的是,任何E430或任何E345取代都可应用于本发明的共依赖地起作用的抗体,条件是它们还包含自身寡聚化抑制突变K439E或S440K,并且条件是一种抗体还包含C1q结合调节突变,例如G236R、G237Q或G237A。Fc-Fc相互作用增强取代E430Y和E430F由于其残留单一试剂活性,对于本发明的抗体而言可能是不太优选的取代。
实施例38:由带有E430G-S440K突变和C1q结合抑制突变的抗体变体和带有E430G-K439E突变和C1q结合增强突变的抗体变体组成的混合物在Raji淋巴瘤细胞上的CDC活性的选择性
前面实施例中,测量了包含两种抗体变体的共依赖性混合物诱导CDC的能力,所述抗体变体之一是带有Fc-Fc相互作用增强突变和自身寡聚化抑制突变K439E与C1q结合抑制突变的组合的抗体变体,另一种是带有Fc-Fc相互作用增强突变和自身寡聚化抑制突变S440K任选与C1q结合增强突变的组合的抗体变体。在此,我们测试了Raji细胞上的选择性共依赖性CDC诱导是否还可以被反向诱导,即通过将带有轻度C1q结合抑制或增强突变的IgG1-CD37-37-3-E430G-K439E抗体变体与带有C1q结合抑制突变的IgG1-11B8-E430G-S440K抗体变体混合。
在抗体IgG1-CD37-37-3和IgG1-11B8中引入不同的突变:诱导增强Fc-Fc相互作用的E430G;自身寡聚化抑制突变K439E或S440K之任一种;抑制C1q与异源六聚体抗体复合物结合的G236R、G237A、G237Q;增强C1q与异源六聚体抗体复合物结合的E333S。作为对照,还将单个抗体与非结合同种型对照抗体IgG1-b12以1:1混合,以能够直接比较各个组分及由其组成的混合物的浓度。在此,还将来自瞬时转染的EXPI293上清液的未纯化上清液用作IgG来源。基本如实施例2所述,在20%NHS情况下对Raji细胞的体外CDC测定中,测试了一系列浓度的纯化抗体(最终浓度范围为0.009-40.0μg/mL;3.3倍稀释度)。细胞裂解百分比计算为(PI阳性细胞数目/细胞总数目)×100%。利用GraphPad PRISM中的对数转换浓度,使用非线性剂量-反应拟合的最佳拟合值分析数据,并计算三个实验重复的剂量-反应曲线下面积。以相对曲线下面积(AUC)值呈现的CDC效力代表每个板相对于最小裂解(IgG1-b12为0%)和最大裂解(IgG1-CD37-37-3-E430G+IgG1-11B8-E430G混合物为100%)的标准化的数值,它们随后在两个独立的实验重复中取平均值。IgG1-CD37-37-3-E430G和IgG1-11B8-E430G的阳性对照混合物在Raji细胞上诱导有效的CDC(图36A)。引入突变K439E可以强烈抑制IgG1-11B8-E430G抗体变体的单一试剂活性,其不受额外引入C1q结合抑制(G237A)或C1q结合增强(E333S)突变的进一步影响。CDC效力还可以通过在具有或不具有C1q结合抑制突变G237A的情况下引入突变S440K来强烈抑制。尽管在IgG1-CD37-37-3-E430G中引入突变K439E-G236R也消除单一试剂CDC效力,但可通过将IgG1-CD37-37-3-E430G-K439E-G236R与IgG1-11B8-E430G-S440K或IgG1-11B8-E430G-S440K-G237A之任一种混合使CDC效力分别恢复至阳性对照混合物诱导的水平的约72%和62%。同样,在IgG1-CD37-37-3-E430G中引入突变K439E-G237A、K439E-E333S(SEQ ID NO:82)、S440K-G236R(SEQ ID NO:104)、S440K-G237A或S440K-G237Q(SEQ ID NO:107)没有产生单一试剂活性,而这些抗体与带有互补性自身寡聚化抑制突变和C1q结合抑制(G237A)或增强(E333S)突变之任一种的IgG1-11B8抗体变体的混合物使CDC效力部分恢复(范围为对照混合物功效的约43%至94%)。由IgG1-CD37-37-3和IgG1-11B8抗体变体的共依赖性混合物诱导的40μg/mL浓度时的最大裂解是强烈的,范围为对照混合物功效的76%至93%(图36B)。
总之,尽管带有E430G突变的IgG1-CD37-37-3和IgG1-11B8抗体变体的单一试剂CDC活性可通过引入K439E或S440K突变之任一种和C1q结合调节突变G236R、G237A、G237Q、E333S之任一种来消除,此类抗体变体混合物使CDC效力恢复,而与在两种抗体之任一种中引入哪种自身寡聚化抑制突变和C1q结合抑制或增强突变的组合无关。
实施例39:带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变的抗CD52和抗CD20抗体变体的共依赖性组合对患者CLL细胞的选择性补体依赖性细胞毒性
前面实施例中,使用体外培养的肿瘤细胞系来测试抗体变体组合的CDC效力。在此,使用来自慢性淋巴白血病(CLL)患者的肿瘤细胞,通过混合带有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K之任一种和C1q结合性调节突变G236R或E333S之任一种的抗CD20 IgG1-11B8和抗CD52 IgG1-CAMPATH-1H抗体变体,研究了CDC的选择性共依赖性诱导。作为对照,还将单个抗体与非结合同种型对照抗体IgG1-b12以1:1混合,以能够直接比较各个组分及由其组成的混合物的浓度。在20%NHS的情况下对源自三名CLL患者的肿瘤细胞的体外CDC测定中,测试了一系列浓度的纯化抗体(最终浓度范围为0.003-10.0μg/mL;3.3倍稀释度)。将来自三名CLL患者的外周血单个核细胞(PBMC;ConversantBio)于37℃解冻,在补充有10%供体牛血清和铁(DBSI;Life Technologies,目录号20371)的RPMI 1640(Life technologies)中清洗,并使其在RPMI+0.2%BSA中的浓度达到3.33x 106个PBMC/mL。向96孔板的每个孔中添加30μL(相当于100,000个细胞/孔)以及50μL 2倍浓度的抗体稀释系列。在摇动器上于室温温育15分钟后,向每个孔中添加20μL非人血清,并于37℃温育45分钟。然后将细胞离心并用FACS缓冲液(PBS+0.1%BSA+0.02%叠氮化钠)清洗。向每个孔中加入50μL染色混合物,并于4℃温育30分钟。使用FACS缓冲液清洗板两次之后,用Fortessa流式细胞仪(BD)测量活细胞百分比。
使用QIFI试剂盒(DAKO;目录号K0078),通过定量流式细胞术程序确定CD20和CD52的靶标表达水平。将三名患者的CLL患者PBMC以100,000个细胞/孔接种在96孔圆底板中,并离心进行细胞沉淀。弃去上清液后,将一抗小鼠抗人CD20(IgG1-mm-IgG1-7D8-K409R)和抗人CD52(Tebu Bio;目录号MAB0944)抗体或同种型对照抗体加入每个孔中,并于4℃温育30-45分钟。在单独的孔中,加入50μL试剂盒提供的设置和校准珠。离心细胞并弃去上清液后,将细胞在FACS缓冲液中清洗两次。然后,将细胞重悬于50μL/孔的二抗山羊抗小鼠IgG-FITC缀合物(DAKO;目录号F0479)中,并于4℃黑暗温育30-45分钟。将细胞清洗两次,最后重悬于FACS缓冲液中,用于在Fortessa流式细胞仪上进行分析。根据制造商指南,通过基于校准曲线计算抗体结合能力来确定抗原量。
在所有三个CLL患者样品中,最强CDC效力是由IgG1-CAMPATH-1H-E430G和IgG1-11B8-E430G混合物诱导的(图37A-C)。IgG1-CAMPATH-1H-E430G的单一试剂CDC活性接近由IgG1-CAMPATH-1H-E430G+IgG1-11B8-E430G混合物诱导的水平(患者1,2)或与其相当(患者3)。IgG1-11B8-E430G作为单一试剂诱导CDC的能力因患者而异,从无CDC活性(患者3)和中等效力(患者1)到接近由单一试剂IgG1-CAMPATH-1H-E430G诱导的水平的CDC效力(患者2)。IgG1-11B8抗体变体的单一试剂活性与不同样品的CD20表达水平相关(患者1:48E+03;患者2:172E+03;患者3:39E+03)。在IgG1-CAMPATH-1H-E430G中引入突变K439E-G236R和在IgG1-11B8-E430G中引入突变S440K或S440K-E333S后,单一试剂活性被完全消除。但是,通过将IgG1-CAMPATH-1H-E430G-K439E-G236R与IgG1-11B8-E430G-S440K或IgG1-11B8-E430G-S440K-E333S混合,可以恢复CDC效力。CDC效力恢复的程度因患者样品而异。
总之,带有突变E430G、K439E或S440K和G236R或E333S的抗CD20和抗CD52抗体变体混合物可以以共依赖的方式诱导患者CLL样品的CDC。
实施例40:外周血单个核细胞样品中T-细胞群的选择性消耗
共依赖性抗体混合物的产生允许从全血样品中选择性消耗特定血液亚群,如针对B细胞的实施例20中所示,而使T细胞保持不变。在此,我们测试了带有Fc-Fc相互作用增强突变,自身寡聚化抑制突变和C1q结合调节突变的共依赖性抗体变体的混合物是否可以选择性地消耗来自全血样品的T细胞,而保持B细胞分数不变。
将源自四名健康人供体(UMC Utrecht)的30μL全血样品(用水蛭素保存以防止凝固)等分取样于96孔圆底板(Greiner Bio-One;目录号650101)中,并补充50μL RPMI+0/2%BSA(Lonza;目录号BE12-115F/U1)。向每个孔中加入20μL具有以下突变的IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM抗体的两种抗体变体的混合物(最终浓度为10μg/mL):诱导增强Fc-Fc相互作用的E430G;自身寡聚化抑制突变K439E或S440K之任一种;和C1q调节突变G236R、G237A或K326A。作为对照,将单个抗体与带有E430G、K439E、S440K、G236R或G237A突变的非结合同种型对照抗体IgG1-b12的变体以1:1混合,从而能够直接比较各个组分及由其组成的混合物的浓度。于37℃和5%CO2温育过夜后,将板离心,并将细胞用150μLPBS(B.Braun;目录号220/12257874/1110)清洗一次。接着,将细胞重悬于80μL以1:1000稀释于PBS的胺反应存活力染料(Invitrogen;目录号A10346A)中,并于4℃黑暗温育30分钟。然后将细胞离心,并且将沉淀重悬于可固定存活力染料510(FVS510,BD;目录号564406)外的80μL针对CD3(eBioscience;目录号48-0037),CD4(eBioscience;目录号47-0048),CD8(BioLegend;目录号301028),CD19(BioLegend,目录号302245),CD45(BioLegend;目录号368505),CD56(BD;目录号564849)和CD66b(BioLegend;目录号305115)的荧光标记的抗体的混合物中。将细胞于4℃黑暗温育30分钟,然后离心,接着使用冷RBC裂解缓冲液裂解红细胞。清洗细胞并在FACS缓冲液(PBS+0.1%BSA+0.02%NaN3)中吸收,然后用流式细胞仪(BDFortessa)以75μL固定体积样品进行分析。使用以下公式计算B细胞,CD4+T细胞和CD8+T细胞分数:恢复分数(靶标群体,%)=100%*(靶标细胞计数(样品)/靶标细胞计数(无Ab对照))*(粒细胞计数(无Ab对照)/粒细胞计数(样品)。在此,通过在无裂解诱导抗体情况下温育的对照样品的细胞计数进行比较,我们测量了靶定细胞的相对恢复,使用粒细胞群作为内部对照(因为它不被裂解诱导抗体靶向)针对方法诱导的样品间变化校正。
将抗CD52抗体IgG1-CAMPATH-1H和抗CD3抗体IgG1-huCLB-T3/4的变体用于从全血样品中选择性消耗T细胞而不影响B细胞。当以单一试剂形式应用时,IgG1-CAMPATH-1H-E430G强烈消耗B细胞,CD4+T细胞和CD8+T细胞,而单一试剂IgG1-huCLB-T3/4-E430G部分消耗CD4+T细胞(图38A,B,C)。未观测到非靶标结合抗体变体IgG1-b12-E430G-K439E-G236R和IgG1-b12-E430G-S440K-G237A的混合物对淋巴细胞亚群的消耗。当将后者抗体变体之任一种与互补性IgG1-CAMPATH-1H-E430G-K439E-G236R或IgG1-huCLB-T3/4-E430G-S440K-K326A混合时,也未观测到消耗。相反,IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-huCLB-T3/4-E430G-S440K-K326A的混合物以共依赖的方式强烈消耗CD4+T细胞和在较小程度上消耗CD8+T细胞,而不影响这些样品中计数的B细胞。
此外,将抗CD52抗体IgG1-CAMPATH-1H和抗CD5抗体IgG1-CD5-INSERM的变体用于从全血样本中选择性消耗T细胞,而不影响B细胞。再次,单一试剂IgG1-CAMPATH-1H-E430G诱导B细胞,CD4+T细胞和CD8+T细胞的强烈消耗,而单一试剂IgG1-CD5-INSERM-E430G未消耗任何分析的淋巴细胞群(图38D,E,F)。未观测到非靶标结合抗体变体IgG1-b12-E430G-K439E-G236R和IgG1-b12-E430G-S440K-G237A混合物对淋巴细胞亚群的消耗。当将后者抗体变体之任一种与互补性IgG1-CAMPATH-1H-E430G-K439E-G236R或IgG1-CD5-INSERM-E430G-S440K-K326A混合时,也未观测到消耗。相反,IgG1-CAMPATH-1H-E430G-K439E-G236R和IgG1-CD5-INSERM-E430G-S440K-K326A混合物以共依赖的方式强烈且选择性地消耗CD4+T细胞,但不消耗B细胞。IgG1-CAMPATH-1H-E430G-K439E-K326A和IgG1-CD5-INSERM-E430G-S440K-G237A的混合物对CD8+T细胞的影响因供体而异。
总之,带有Fc-Fc相互作用增强突变E430G,自身寡聚化抑制突变K439E或S440K和C1q结合调节突变G236R或G237A的抗CD52IgG1-CAMPATH-1H,IgG1-huCLB-T3/4和IgG1-CD5-INSERM的抗体变体混合物可用于从健康供体全血样品中选择性且共依赖性地消耗T细胞群,而使B细胞群不受损害。
与实施例20的结果相结合,这表明使用靶标组合,可将本发明公开的包含带有突变的Fc域的抗体组合用于选择性地消耗B细胞、CD8+T细胞或CD4+T-细胞之外的不同血液亚群,所述靶标组合的共表达是靶定的血液亚群的选择性标志。
实施例41:带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变的抗DR4和抗DR5抗体变体的混合物共依赖性诱导程序性细胞死亡
进行存活力分析以研究带有增强Fc-Fc相互作用,抑制自身寡聚化和调节C1q结合的突变的抗DR4和抗DR5抗体变体混合物是否可以共依赖性诱导BxPC-3人胰腺癌细胞和COLO 205结肠癌细胞的程序性细胞死亡(PCD)。
在抗DR4抗体IgG1-DR4-chCTB007和抗DR5抗体IgG1-DR5-01中引入不同的突变:诱导增强Fc-Fc相互作用的E430G;自身寡聚化抑制突变K439E或S440K之任一种;C1q结合调节突变G237T、K326W或E333S,其抑制(G237T)或增强(K326W、E333S)C1q的结合。作为对照,还将单个抗体与具有E430G-K439E或E430G-S440K突变的非结合同种型对照抗体IgG1-b12的变体以1:1混合,从而直接比较各个组分及由其组成的混合物的浓度。基本如实施例14所述进行存活力测定。简而言之,将90μL单细胞悬浮液(5,000个细胞/孔)接种于聚苯乙烯96孔平底板(Greiner Bio-One,目录号655182),并且于37℃温育过夜。添加连续稀释系列的20μL抗体混合物(以5倍稀释度,最终浓度范围为0.0003至20μg/mL)和10μL C1q(Quidel;最终浓度为2.5μg/mL),并于37℃温育3天。作为诱导细胞死亡的阴性和阳性对照,将细胞分别在无抗体的情况下温育或与5μM星形孢菌素一起温育。如实施例14所述,通过CellTiter-Glo发光细胞存活力测定法确定所培养细胞的存活力。使用GraphPad Prism软件,利用非线性回归(具有可变斜率的S形剂量反应)进行数据分析和绘图。使用以下公式计算活细胞百分比:活细胞%=[(抗体样品发光值-星形孢菌素样品发光值)/(无抗体样品发光值-星形孢菌素样品发光值)]*100。
IgG1-DR5-01-E430G-K439E-G237T在作为单一试剂应用时,在BxPC-3或COLO 205细胞中未观测到程序性细胞死亡(图39A,B)。IgG1-DR4-chCTB007-E430G-S440K-K326W-E333S在作为单一试剂应用时,确实诱导程序性细胞死亡,如在使用的最高浓度下在BxPC-3和COLO 205测定中剩下的约67%和60%的活细胞所反映。IgG1-DR4-chCTB007-E430G-S440K-K326W-E333S诱导程序性细胞死亡的能力可通过将其与IgG1-DR5-01-E430G-K439E-G237T混合而增强,使用这两种细胞系观测到的效应。
总之,BxPC-3和COLO 205癌细胞中程序性细胞死亡的共依赖性激活可通过带有增强Fc-Fc相互作用,抑制自身寡聚化和抑制(G237T)或增强(K326W-E333S)C1q结合的突变的抗DR4和抗DR5抗体变体的混合物来实现。
序列表
<110> 根马布私人有限公司
<120> 抗体变体组合及其用途
<130> P/0121-WO-PCT[2]
<150> DK PA 2018 00195
<151> 2018-05-03
<150> DK PA 2018 00644
<151> 2018-09-26
<160> 201
<170> PatentIn version 3.5
<210> 1
<211> 121
<212> PRT
<213> 人工序列
<220>
<223> 人源化, VH区
<400> 1
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Asn Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Phe Ile Arg Asp Lys Ala Lys Gly Tyr Thr Thr Glu Tyr Asn Pro
50 55 60
Ser Val Lys Gly Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn Gln
65 70 75 80
Phe Ser Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Gly His Thr Ala Ala Pro Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Ser Leu Val Thr Val Ser Ser
115 120
<210> 2
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 2
Gly Phe Thr Phe Thr Asp Phe Tyr
1 5
<210> 3
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 3
Ile Arg Asp Lys Ala Lys Gly Tyr Thr Thr
1 5 10
<210> 4
<211> 12
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 4
Ala Arg Glu Gly His Thr Ala Ala Pro Phe Asp Tyr
1 5 10
<210> 5
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区
<400> 5
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Ile Asp Lys Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Thr Asn Asn Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln His Ile Ser Arg Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 6
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 6
Gln Asn Ile Asp Lys Tyr
1 5
<210> 7
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR3
<400> 7
Leu Gln His Ile Ser Arg Pro Arg Thr
1 5
<210> 8
<211> 125
<212> PRT
<213> 人工序列
<220>
<223> VH区
<400> 8
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val His Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Gly Ser Gly Phe Thr Phe Ser Tyr His
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Gly Thr Gly Gly Val Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Met Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Tyr Gly Ala Gly Ser Phe Tyr Asp Gly Leu Tyr Gly Met
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 9
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 9
Gly Phe Thr Phe Ser Tyr His Ala
1 5
<210> 10
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 10
Ile Gly Thr Gly Gly Val Thr
1 5
<210> 11
<211> 19
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR3
<400> 11
Ala Arg Asp Tyr Tyr Gly Ala Gly Ser Phe Tyr Asp Gly Leu Tyr Gly
1 5 10 15
Met Asp Val
<210> 12
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> VL区
<400> 12
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asp Trp Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 13
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 13
Gln Ser Val Ser Ser Tyr
1 5
<210> 14
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR3
<400> 14
Gln Gln Arg Ser Asp Trp Pro Leu Thr
1 5
<210> 15
<211> 127
<212> PRT
<213> 人工序列
<220>
<223> VH区
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Gln Ala Ser Gly Tyr Arg Phe Ser Asn Phe
20 25 30
Val Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Phe Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Tyr Asn Gly Asn Lys Glu Phe Ser Ala Lys Phe
50 55 60
Gln Asp Arg Val Thr Phe Thr Ala Asp Thr Ser Ala Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Gly Pro Tyr Ser Trp Asp Asp Ser Pro Gln Asp Asn Tyr
100 105 110
Tyr Met Asp Val Trp Gly Lys Gly Thr Thr Val Ile Val Ser Ser
115 120 125
<210> 16
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 16
Gly Tyr Arg Phe Ser Asn Phe Val
1 5
<210> 17
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 17
Ile Asn Pro Tyr Asn Gly Asn Lys
1 5
<210> 18
<211> 20
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR3
<400> 18
Ala Arg Val Gly Pro Tyr Ser Trp Asp Asp Ser Pro Gln Asp Asn Tyr
1 5 10 15
Tyr Met Asp Val
20
<210> 19
<211> 108
<212> PRT
<213> 人工序列
<220>
<223> VL区
<400> 19
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Phe Ser Cys Arg Ser Ser His Ser Ile Arg Ser Arg
20 25 30
Arg Val Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro Arg Leu Val
35 40 45
Ile His Gly Val Ser Asn Arg Ala Ser Gly Ile Ser Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Thr Arg Val Glu
65 70 75 80
Pro Glu Asp Phe Ala Leu Tyr Tyr Cys Gln Val Tyr Gly Ala Ser Ser
85 90 95
Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Arg Lys
100 105
<210> 20
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 20
His Ser Ile Arg Ser Arg Arg
1 5
<210> 21
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 21
Gln Val Tyr Gly Ala Ser Ser Tyr Thr
1 5
<210> 22
<211> 330
<212> PRT
<213> 人
<400> 22
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 23
<211> 330
<212> PRT
<213> 人
<400> 23
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 24
<211> 330
<212> PRT
<213> 人
<400> 24
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Pro Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 25
<211> 330
<212> PRT
<213> 人
<400> 25
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Pro Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Gly Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 26
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 26
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 27
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 27
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Lys Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 28
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 28
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Arg Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 29
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 29
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 30
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 30
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 31
<211> 326
<212> PRT
<213> 人
<400> 31
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 32
<211> 377
<212> PRT
<213> 人
<400> 32
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 33
<211> 327
<212> PRT
<213> 人
<400> 33
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 34
<211> 107
<212> PRT
<213> 人
<400> 34
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 35
<211> 122
<212> PRT
<213> 人工序列
<220>
<223> VH区
<400> 35
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Asp Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe His Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Ser Trp Asn Ser Gly Thr Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys Asp Ile Gln Tyr Gly Asn Tyr Tyr Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 36
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 36
Gly Phe Thr Phe His Asp Tyr Ala
1 5
<210> 37
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 37
Ile Ser Trp Asn Ser Gly Thr Ile
1 5
<210> 38
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR3
<400> 38
Ala Lys Asp Ile Gln Tyr Gly Asn Tyr Tyr Tyr Gly Met Asp Val
1 5 10 15
<210> 39
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> VL区
<400> 39
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 40
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 40
Gln Ser Val Ser Ser Tyr
1 5
<210> 41
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR3
<400> 41
Gln Gln Arg Ser Asn Trp Pro Ile Thr
1 5
<210> 42
<211> 115
<212> PRT
<213> 人工序列
<220>
<223> VH区
<400> 42
Gln Val Gln Val Lys Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Ser
20 25 30
Gly Val Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Gly Asp Gly Ser Thr Asn Tyr His Ser Ala Leu Lys
50 55 60
Ser Arg Leu Ser Ile Lys Lys Asp His Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Leu Asn Ser Leu Gln Thr Asp Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Lys Gly Gly Tyr Ser Leu Ala His Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ala
115
<210> 43
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 43
Gly Phe Ser Leu Thr Thr Ser Gly
1 5
<210> 44
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 44
Ile Trp Gly Asp Gly Ser Thr
1 5
<210> 45
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR3
<400> 45
Ala Lys Gly Gly Tyr Ser Leu Ala His
1 5
<210> 46
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> VL区
<400> 46
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Val Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Arg Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Val
35 40 45
Asn Val Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Leu Lys Ile Asn Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Gly Thr Tyr Tyr Cys Gln His Tyr Trp Gly Thr Thr Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 47
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 47
Glu Asn Ile Arg Ser Asn
1 5
<210> 48
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 48
Gln His Tyr Trp Gly Thr Thr Trp Thr
1 5
<210> 49
<211> 118
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区
<400> 49
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Phe Ile His Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Thr Asn Thr Lys Tyr Asp Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Thr Thr Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Gly Leu Tyr Thr Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 50
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR1
<400> 50
Gly Phe Asn Ile Lys Asp Thr Phe
1 5
<210> 51
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR2
<400> 51
Ile Asp Pro Ala Asn Thr Asn Thr
1 5
<210> 52
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR3
<400> 52
Val Arg Gly Leu Tyr Thr Tyr Tyr Phe Asp Tyr
1 5 10
<210> 53
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区
<400> 53
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asn Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Phe Ala Ser Gln Ser Ile Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Asn Ser Trp Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 54
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR1
<400> 54
Gln Ser Ile Ser Asn Asn
1 5
<210> 55
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR3
<400> 55
Gln Gln Gly Asn Ser Trp Pro Tyr Thr
1 5
<210> 56
<211> 118
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区
<400> 56
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
His Met His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Thr Glu Tyr Asp Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Val Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Thr Asn Val Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 57
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR1
<400> 57
Gly Phe Asn Ile Lys Asp Thr His
1 5
<210> 58
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化, VH区 CDR2
<400> 58
Ile Asp Pro Ala Asn Gly Asn Thr
1 5
<210> 59
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 人源化, VH区 CDR3
<400> 59
Ala Arg Trp Gly Thr Asn Val Tyr Phe Ala Tyr
1 5 10
<210> 60
<211> 106
<212> PRT
<213> 人工序列
<220>
<223> 人源化, VL区
<400> 60
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Trp Ile Tyr
35 40 45
Arg Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr His Ser Tyr Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 61
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 人源化, VL区 CDR1
<400> 61
Ser Ser Val Ser Tyr
1 5
<210> 62
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> 人源化, VL区 CDR3
<400> 62
Gln Gln Tyr His Ser Tyr Pro Pro Thr
1 5
<210> 63
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 63
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Lys Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 64
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 64
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Ala Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 65
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 65
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Ala Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 66
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 66
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Lys Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 67
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 67
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Ala Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Lys Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 68
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 68
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Lys Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 69
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 69
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Gln Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 70
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 70
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Gln Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 71
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 71
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Arg Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 72
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 72
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Arg Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 73
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 73
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Arg Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 74
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 74
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Ala Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Arg Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 75
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 75
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Val Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 76
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 76
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Val Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 77
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 77
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Tyr Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 78
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 78
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Tyr Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 79
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 79
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 80
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 80
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 81
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 81
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Lys Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 82
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 82
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 83
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 83
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Lys Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 84
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 84
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 85
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 85
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 86
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 86
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Ala Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 87
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 87
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gln Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 88
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 88
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Arg Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 89
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 89
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Thr Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 90
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 90
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Ala Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 91
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 91
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Glu Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 92
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 92
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 93
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 93
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 94
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 94
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 95
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 95
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 96
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 96
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 97
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 97
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 98
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 98
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Gln Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 99
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 99
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Gln Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 100
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 100
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Arg Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 101
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 101
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 102
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 102
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Ala Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 103
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 103
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 104
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 104
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 105
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 105
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 106
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 106
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 107
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 107
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gln Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 108
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 108
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Arg Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 109
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 109
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Glu Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 110
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Ala Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 111
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 111
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Ala Ala Leu Pro Ala Pro Ile Ala Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 112
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 112
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Trp Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 113
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 113
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 114
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 114
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Arg Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 115
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 115
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Asn Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 116
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 116
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Asn Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 117
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 117
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Thr Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 118
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 118
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Thr Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 119
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 119
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Val Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 120
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 120
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Val Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 121
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 121
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Tyr Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 122
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 122
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Tyr Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 123
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 123
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 124
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 124
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 125
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 125
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gln Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 126
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 126
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Arg Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 127
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 127
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Thr Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 128
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 128
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Glu Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Arg
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 129
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 129
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gln Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Glu Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Arg
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 130
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 130
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Glu Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Arg
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 131
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 131
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Ala Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Glu Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Arg
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 132
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 132
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Glu Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 133
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 133
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Arg Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 134
<211> 326
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 134
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 135
<211> 326
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 135
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 136
<211> 326
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 136
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Arg Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Glu Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 137
<211> 326
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 137
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Lys Lys Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 138
<211> 326
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 138
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Ser Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Lys Lys Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 139
<211> 377
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 139
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 140
<211> 377
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 140
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Glu Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 141
<211> 377
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 141
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Glu Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 142
<211> 377
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 142
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Lys Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 143
<211> 377
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 143
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Lys Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 144
<211> 327
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 144
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 145
<211> 327
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 145
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Glu Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 146
<211> 327
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 146
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Arg Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Glu Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 147
<211> 327
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 147
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Lys Lys
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 148
<211> 327
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 148
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr Gln Lys Lys
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 149
<211> 298
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc gamma受体
<400> 149
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
Val Trp Phe His His His His His His His
290 295
<210> 150
<211> 418
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc gamma受体
<400> 150
Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu Trp Leu
1 5 10 15
Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp Ser Gln
20 25 30
Ala Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp Ile
35 40 45
Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala Arg
50 55 60
Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile
65 70 75 80
Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp
85 90 95
Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro
100 105 110
Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His
115 120 125
Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser Trp
130 135 140
Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser
145 150 155 160
Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala Asn
165 170 175
His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr
180 185 190
Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser Met
195 200 205
Gly Ser Ser Ser Pro Val Ala Pro Pro Lys Ala Val Leu Lys Leu Glu
210 215 220
Pro Pro Trp Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys
225 230 235 240
Gln Gly Ala Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn
245 250 255
Gly Asn Leu Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala
260 265 270
Asn Asn Asn Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser
275 280 285
Leu Ser Asp Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu
290 295 300
Gln Thr Pro His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg
305 310 315 320
Cys His Ser Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln
325 330 335
Asn Gly Lys Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile
340 345 350
Pro Gln Ala Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn
355 360 365
Ile Gly Tyr Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln
370 375 380
Val Pro Ser Met Gly Pro Gly Ser Ser Ser His His His His His His
385 390 395 400
Pro Gly Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp
405 410 415
His Glu
<210> 151
<211> 405
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc gamma受体
<400> 151
Met Val Leu Ser Leu Leu Tyr Leu Leu Thr Ala Leu Pro Gly Ile Leu
1 5 10 15
Ser Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp Ile
20 25 30
Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala Arg
35 40 45
Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile
50 55 60
Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp
65 70 75 80
Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro
85 90 95
Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His
100 105 110
Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser Trp
115 120 125
Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser
130 135 140
Gln Lys Phe Ser Arg Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala Asn
145 150 155 160
His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr
165 170 175
Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser Met
180 185 190
Gly Ser Ser Ser Pro Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu
195 200 205
Pro Pro Trp Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys
210 215 220
Gln Gly Ala Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn
225 230 235 240
Gly Asn Leu Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala
245 250 255
Asn Asn Asn Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser
260 265 270
Leu Ser Asp Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu
275 280 285
Gln Thr Pro His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg
290 295 300
Cys His Ser Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln
305 310 315 320
Asn Gly Lys Ser Gln Lys Phe Ser Arg Leu Asp Pro Thr Phe Ser Ile
325 330 335
Pro Gln Ala Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn
340 345 350
Ile Gly Tyr Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln
355 360 365
Val Pro Ser Met Gly Ser Ser Ser Pro Gly Ser Ser Ser His His His
370 375 380
His His His Pro Gly Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
385 390 395 400
Ile Glu Trp His Glu
405
<210> 152
<211> 405
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc gamma受体
<400> 152
Met Val Leu Ser Leu Leu Tyr Leu Leu Thr Ala Leu Pro Gly Ile Leu
1 5 10 15
Ser Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile
20 25 30
Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His
35 40 45
Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile
50 55 60
Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp
65 70 75 80
Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro
85 90 95
Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His
100 105 110
Leu Glu Phe Gln Glu Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp
115 120 125
Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser
130 135 140
Lys Lys Phe Ser Arg Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn
145 150 155 160
His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr
165 170 175
Leu Tyr Ser Ser Lys Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser
180 185 190
Ser Pro Met Gly Pro Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu
195 200 205
Pro Gln Trp Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys
210 215 220
Arg Gly Thr His Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn
225 230 235 240
Gly Asn Leu Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala
245 250 255
Asn Asn Asn Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser
260 265 270
Leu Ser Asp Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu
275 280 285
Gln Thr Pro His Leu Glu Phe Gln Glu Gly Glu Thr Ile Val Leu Arg
290 295 300
Cys His Ser Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln
305 310 315 320
Asn Gly Lys Ser Lys Lys Phe Ser Arg Ser Asp Pro Asn Phe Ser Ile
325 330 335
Pro Gln Ala Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn
340 345 350
Ile Gly Tyr Thr Leu Tyr Ser Ser Lys Pro Val Thr Ile Thr Val Gln
355 360 365
Ala Pro Ser Ser Ser Pro Met Gly Pro Gly Ser Ser Ser His His His
370 375 380
His His His Pro Gly Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
385 390 395 400
Ile Glu Trp His Glu
405
<210> 153
<211> 410
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc gamma受体
<400> 153
Met Val Leu Ser Leu Leu Tyr Leu Leu Thr Ala Leu Pro Gly Ile Ser
1 5 10 15
Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro Gln Trp Tyr
20 25 30
Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln Gly Ala Tyr
35 40 45
Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu Ser Leu Ile
50 55 60
Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr Val Asp Asp
65 70 75 80
Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu Ser Asp Pro
85 90 95
Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln Ala Pro Arg
100 105 110
Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys His Ser Trp
115 120 125
Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn Gly Lys Gly
130 135 140
Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro Lys Ala Thr
145 150 155 160
Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Phe Gly Ser Lys
165 170 175
Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln Gly Pro Ser
180 185 190
Met Gly Ser Ser Ser Pro Ser Glu Asp Leu Pro Lys Ala Val Val Phe
195 200 205
Leu Glu Pro Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu
210 215 220
Lys Cys Gln Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe
225 230 235 240
His Asn Glu Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp
245 250 255
Ala Ala Thr Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu
260 265 270
Ser Thr Leu Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu
275 280 285
Leu Leu Gln Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His
290 295 300
Leu Arg Cys His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr
305 310 315 320
Leu Gln Asn Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe
325 330 335
Tyr Ile Pro Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg
340 345 350
Gly Leu Phe Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr
355 360 365
Ile Thr Gln Gly Pro Ser Met Gly Ser Ser Ser Pro Gly Pro Gly Ser
370 375 380
Ser Ser His His His His His His Pro Gly Gly Gly Leu Asn Asp Ile
385 390 395 400
Phe Glu Ala Gln Lys Ile Glu Trp His Glu
405 410
<210> 154
<211> 410
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc gamma受体
<400> 154
Met Val Leu Ser Leu Leu Tyr Leu Leu Thr Ala Leu Pro Gly Ile Ser
1 5 10 15
Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro Gln Trp Tyr
20 25 30
Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln Gly Ala Tyr
35 40 45
Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu Ser Leu Ile
50 55 60
Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr Val Asp Asp
65 70 75 80
Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu Ser Asp Pro
85 90 95
Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln Ala Pro Arg
100 105 110
Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys His Ser Trp
115 120 125
Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn Gly Lys Gly
130 135 140
Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro Lys Ala Thr
145 150 155 160
Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val Gly Ser Lys
165 170 175
Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln Gly Pro Ser
180 185 190
Met Gly Ser Ser Ser Pro Ser Glu Asp Leu Pro Lys Ala Val Val Phe
195 200 205
Leu Glu Pro Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu
210 215 220
Lys Cys Gln Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe
225 230 235 240
His Asn Glu Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp
245 250 255
Ala Ala Thr Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu
260 265 270
Ser Thr Leu Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu
275 280 285
Leu Leu Gln Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His
290 295 300
Leu Arg Cys His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr
305 310 315 320
Leu Gln Asn Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe
325 330 335
Tyr Ile Pro Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg
340 345 350
Gly Leu Val Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr
355 360 365
Ile Thr Gln Gly Pro Ser Met Gly Ser Ser Ser Pro Gly Pro Gly Ser
370 375 380
Ser Ser His His His His His His Pro Gly Gly Gly Leu Asn Asp Ile
385 390 395 400
Phe Glu Ala Gln Lys Ile Glu Trp His Glu
405 410
<210> 155
<211> 280
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 155
Ala Glu Ser His Leu Ser Leu Leu Tyr His Leu Thr Ala Val Ser Ser
1 5 10 15
Pro Ala Pro Gly Thr Pro Ala Phe Trp Val Ser Gly Trp Leu Gly Pro
20 25 30
Gln Gln Tyr Leu Ser Tyr Asn Ser Leu Arg Gly Glu Ala Glu Pro Cys
35 40 45
Gly Ala Trp Val Trp Glu Asn Gln Val Ser Trp Tyr Trp Glu Lys Glu
50 55 60
Thr Thr Asp Leu Arg Ile Lys Glu Lys Leu Phe Leu Glu Ala Phe Lys
65 70 75 80
Ala Leu Gly Gly Lys Gly Pro Tyr Thr Leu Gln Gly Leu Leu Gly Cys
85 90 95
Glu Leu Gly Pro Asp Asn Thr Ser Val Pro Thr Ala Lys Phe Ala Leu
100 105 110
Asn Gly Glu Glu Phe Met Asn Phe Asp Leu Lys Gln Gly Thr Trp Gly
115 120 125
Gly Asp Trp Pro Glu Ala Leu Ala Ile Ser Gln Arg Trp Gln Gln Gln
130 135 140
Asp Lys Ala Ala Asn Lys Glu Leu Thr Phe Leu Leu Phe Ser Cys Pro
145 150 155 160
His Arg Leu Arg Glu His Leu Glu Arg Gly Arg Gly Asn Leu Glu Trp
165 170 175
Lys Glu Pro Pro Ser Met Arg Leu Lys Ala Arg Pro Ser Ser Pro Gly
180 185 190
Phe Ser Val Leu Thr Cys Ser Ala Phe Ser Phe Tyr Pro Pro Glu Leu
195 200 205
Gln Leu Arg Phe Leu Arg Asn Gly Leu Ala Ala Gly Thr Gly Gln Gly
210 215 220
Asp Phe Gly Pro Asn Ser Asp Gly Ser Phe His Ala Ser Ser Ser Leu
225 230 235 240
Thr Val Lys Ser Gly Asp Glu His His Tyr Cys Cys Ile Val Gln His
245 250 255
Ala Gly Leu Ala Gln Pro Leu Arg Val Glu Leu Glu Ser Pro Ala Lys
260 265 270
Ser Ser His His His His His His
275 280
<210> 156
<211> 99
<212> PRT
<213> 人
<400> 156
Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg His Pro Ala Glu
1 5 10 15
Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His Pro
20 25 30
Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu Lys
35 40 45
Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr Leu
50 55 60
Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala Cys
65 70 75 80
Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp Asp
85 90 95
Arg Asp Met
<210> 157
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 157
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Trp Ala Leu Pro Ala Pro Ile Ser Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Lys Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 158
<211> 330
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Fc区
<400> 158
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Arg Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Gly Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Tyr Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 159
<211> 653
<212> PRT
<213> 人工序列
<220>
<223> 修饰的Her2
<400> 159
Met Glu Leu Ala Ala Leu Cys Arg Trp Gly Leu Leu Leu Ala Leu Leu
1 5 10 15
Pro Pro Gly Ala Ala Ser Thr Gln Val Cys Thr Gly Thr Asp Met Lys
20 25 30
Leu Arg Leu Pro Ala Ser Pro Glu Thr His Leu Asp Met Leu Arg His
35 40 45
Leu Tyr Gln Gly Cys Gln Val Val Gln Gly Asn Leu Glu Leu Thr Tyr
50 55 60
Leu Pro Thr Asn Ala Ser Leu Ser Phe Leu Gln Asp Ile Gln Glu Val
65 70 75 80
Gln Gly Tyr Val Leu Ile Ala His Asn Gln Val Arg Gln Val Pro Leu
85 90 95
Gln Arg Leu Arg Ile Val Arg Gly Thr Gln Leu Phe Glu Asp Asn Tyr
100 105 110
Ala Leu Ala Val Leu Asp Asn Gly Asp Pro Leu Asn Asn Thr Thr Pro
115 120 125
Val Thr Gly Ala Ser Pro Gly Gly Leu Arg Glu Leu Gln Leu Arg Ser
130 135 140
Leu Thr Glu Ile Leu Lys Gly Gly Val Leu Ile Gln Arg Asn Pro Gln
145 150 155 160
Leu Cys Tyr Gln Asp Thr Ile Leu Trp Lys Asp Ile Phe His Lys Asn
165 170 175
Asn Gln Leu Ala Leu Thr Leu Ile Asp Thr Asn Arg Ser Arg Ala Cys
180 185 190
His Pro Cys Ser Pro Met Cys Lys Gly Ser Arg Cys Trp Gly Glu Ser
195 200 205
Ser Glu Asp Cys Gln Ser Leu Thr Arg Thr Val Cys Ala Gly Gly Cys
210 215 220
Ala Arg Cys Lys Gly Pro Leu Pro Thr Asp Cys Cys His Glu Gln Cys
225 230 235 240
Ala Ala Gly Cys Thr Gly Pro Lys His Ser Asp Cys Leu Ala Cys Leu
245 250 255
His Phe Asn His Ser Gly Ile Cys Glu Leu His Cys Pro Ala Leu Val
260 265 270
Thr Tyr Asn Thr Asp Thr Phe Glu Ser Met Pro Asn Pro Glu Gly Arg
275 280 285
Tyr Thr Phe Gly Ala Ser Cys Val Thr Ala Cys Pro Tyr Asn Tyr Leu
290 295 300
Ser Thr Asp Val Gly Ser Cys Thr Leu Val Cys Pro Leu His Asn Gln
305 310 315 320
Glu Val Thr Ala Glu Asp Gly Thr Gln Arg Cys Glu Lys Cys Ser Lys
325 330 335
Pro Cys Ala Arg Val Cys Tyr Gly Leu Gly Met Glu His Leu Arg Glu
340 345 350
Val Arg Ala Val Thr Ser Ala Asn Ile Gln Glu Phe Ala Gly Cys Lys
355 360 365
Lys Ile Phe Gly Ser Leu Ala Phe Leu Pro Glu Ser Phe Asp Gly Asp
370 375 380
Pro Ala Ser Asn Thr Ala Pro Leu Gln Pro Glu Gln Leu Gln Val Phe
385 390 395 400
Glu Thr Leu Glu Glu Ile Thr Gly Tyr Leu Tyr Ile Ser Ala Trp Pro
405 410 415
Asp Ser Leu Pro Asp Leu Ser Val Phe Gln Asn Leu Gln Val Ile Arg
420 425 430
Gly Arg Ile Leu His Asn Gly Ala Tyr Ser Leu Thr Leu Gln Gly Leu
435 440 445
Gly Ile Ser Trp Leu Gly Leu Arg Ser Leu Arg Glu Leu Gly Ser Gly
450 455 460
Leu Ala Leu Ile His His Asn Thr His Leu Cys Phe Val His Thr Val
465 470 475 480
Pro Trp Asp Gln Leu Phe Arg Asn Pro His Gln Ala Leu Leu His Thr
485 490 495
Ala Asn Arg Pro Glu Asp Glu Cys Val Gly Glu Gly Leu Ala Cys His
500 505 510
Gln Leu Cys Ala Arg Gly His Cys Trp Gly Pro Gly Pro Thr Gln Cys
515 520 525
Val Asn Cys Ser Gln Phe Leu Arg Gly Gln Glu Cys Val Glu Glu Cys
530 535 540
Arg Val Leu Gln Gly Leu Pro Arg Glu Tyr Val Asn Ala Arg His Cys
545 550 555 560
Leu Pro Cys His Pro Glu Cys Gln Pro Gln Asn Gly Ser Val Thr Cys
565 570 575
Phe Gly Pro Glu Ala Asp Gln Cys Val Ala Cys Ala His Tyr Lys Asp
580 585 590
Pro Pro Phe Cys Val Ala Arg Cys Pro Ser Gly Val Lys Pro Asp Leu
595 600 605
Ser Tyr Met Pro Ile Trp Lys Phe Pro Asp Glu Glu Gly Ala Cys Gln
610 615 620
Pro Cys Pro Ile Asn Cys Thr His Ser Cys Val Asp Leu Asp Asp Lys
625 630 635 640
Gly Cys Pro Ala Glu Gln Arg His His His His His His
645 650
<210> 160
<211> 113
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区
<400> 160
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Val
35 40 45
Ala Met Met Lys Thr Lys Gly Gly Arg Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Ser Asp Gly Tyr Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
100 105 110
Ser
<210> 161
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR1
<400> 161
Gly Phe Thr Phe Ser Arg Tyr Gly
1 5
<210> 162
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR2
<400> 162
Met Lys Thr Lys Gly Gly Arg Thr
1 5
<210> 163
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR3
<400> 163
Ala Ser Asp Gly Tyr Tyr
1 5
<210> 164
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区
<400> 164
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Leu Val Ser Lys Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Leu Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 165
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR1
<400> 165
Gln Ser Leu Leu His Ser Asp Gly Lys Thr Tyr
1 5 10
<210> 166
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR3
<400> 166
Trp Gln Gly Thr His Leu Trp Thr
1 5
<210> 167
<211> 121
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区
<400> 167
Gln Val Gln Leu Gln Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Arg Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Asp Ile Thr Ala Val Val Pro Thr Gly Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Ser Leu Val Thr Val Ser Ser
115 120
<210> 168
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 168
Gly Phe Thr Phe Thr Asn Tyr Gly
1 5
<210> 169
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR2
<400> 169
Ile Asn Thr Tyr Thr Arg Glu Pro
1 5
<210> 170
<211> 14
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR3
<400> 170
Ala Arg Asp Ile Thr Ala Val Val Pro Thr Gly Phe Asp Tyr
1 5 10
<210> 171
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区
<400> 171
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Ser Asn
20 25 30
Leu Ala Trp Tyr Arg Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Val
35 40 45
Phe Ala Ala Ser Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln His Phe Trp Thr Thr Pro Trp
85 90 95
Ala Phe Gly Gly Gly Thr Lys Leu Gln Ile Lys
100 105
<210> 172
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR1
<400> 172
Glu Asn Ile Tyr Ser Asn
1 5
<210> 173
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR3
<400> 173
Gln His Phe Trp Thr Thr Pro Trp Ala
1 5
<210> 174
<211> 120
<212> PRT
<213> 人工序列
<220>
<223> VH区
<400> 174
Gln Val Gln Leu Lys Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Val Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Leu Ile Asp Trp Asp Asp Asp Lys Tyr Tyr Ser Thr Ser
50 55 60
Leu Lys Thr Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Ser Pro Arg Tyr Arg Gly Ala Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 175
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 175
Gly Phe Ser Leu Ser Thr Ser Gly Val Gly
1 5 10
<210> 176
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 176
Ile Asp Trp Asp Asp Asp Lys
1 5
<210> 177
<211> 12
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR3
<400> 177
Ala Arg Ser Pro Arg Tyr Arg Gly Ala Phe Asp Tyr
1 5 10
<210> 178
<211> 108
<212> PRT
<213> 人工序列
<220>
<223> VL区
<400> 178
Asp Ile Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Glu Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Met Asn Val
85 90 95
His Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 179
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 179
Glu Ser Asn Ile Gly Asn Asn Tyr
1 5
<210> 180
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR3
<400> 180
Gln Ser Tyr Asp Met Asn Val His Val
1 5
<210> 181
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区
<400> 181
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Tyr Ser Arg Tyr Ile Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Pro Leu Tyr Gly Ser Ser Pro Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 182
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR1
<400> 182
Gly Phe Thr Phe Ser Ser Tyr Gly
1 5
<210> 183
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR2
<400> 183
Ile Ser Arg Tyr Ser Arg Tyr Ile
1 5
<210> 184
<211> 12
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR3
<400> 184
Ala Arg Arg Pro Leu Tyr Gly Ser Ser Pro Asp Tyr
1 5 10
<210> 185
<211> 106
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区
<400> 185
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Thr Tyr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Phe Gln Gly Ser Gly Tyr Pro Leu Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Met Arg
100 105
<210> 186
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR1
<400> 186
Ser Ser Val Thr Tyr
1 5
<210> 187
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR3
<400> 187
Phe Gln Gly Ser Gly Tyr Pro Leu Thr
1 5
<210> 188
<211> 118
<212> PRT
<213> 人工序列
<220>
<223> VH区
<400> 188
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Thr Gly Tyr Ser Ile Thr Ser Gly
20 25 30
Tyr Tyr Trp His Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Gly Tyr Ile Ser Tyr Ser Gly Phe Thr Asn Tyr Lys Thr Ser Leu
50 55 60
Ile Asn Arg Ile Ser Ile Thr His Asp Thr Ser Glu Asn Gln Phe Phe
65 70 75 80
Leu Asn Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Asp Arg Thr Gly Ser Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 189
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR1
<400> 189
Gly Tyr Ser Ile Thr Ser Gly Tyr Tyr
1 5
<210> 190
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR2
<400> 190
Ile Ser Tyr Ser Gly Phe Thr
1 5
<210> 191
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR3
<400> 191
Ala Gly Asp Arg Thr Gly Ser Trp Phe Ala Tyr
1 5 10
<210> 192
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> VL区
<400> 192
Asp Ile Gln Val Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Ile Ser Leu Thr Cys Arg Thr Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Phe Gln Gln Lys Pro Asp Gly Thr Phe Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Ala Asp Tyr Tyr Cys Leu Gln Tyr Ala Ser Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 193
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR1
<400> 193
Gln Asp Ile Ser Asn Tyr
1 5
<210> 194
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> VL区 CDR3
<400> 194
Leu Gln Tyr Ala Ser Tyr Pro Phe Thr
1 5
<210> 195
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区
<400> 195
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Thr Lys Tyr Asp Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Tyr Tyr Tyr Val Ser Asn Ala Trp Phe Thr Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala
115
<210> 196
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR1
<400> 196
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 197
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VH区 CDR2
<400> 197
Ile Asp Pro Ala Asn Gly Asn Thr
1 5
<210> 198
<211> 12
<212> PRT
<213> 人工序列
<220>
<223> VH区 CDR3
<400> 198
Ala Tyr Tyr Tyr Val Ser Asn Ala Trp Phe Thr Tyr
1 5 10
<210> 199
<211> 106
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区
<400> 199
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Val Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Ser Asn
20 25 30
Leu Glu Trp Tyr Gln Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Val
35 40 45
Tyr Ala Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Leu Lys Ile Asn Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Gly Ser Tyr Tyr Cys Gln His Phe Trp Gly Thr Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 200
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR1
<400> 200
Glu Asn Ile Tyr Ser Asn
1 5
<210> 201
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 人源化 VL区 CDR3
<400> 201
Gln His Phe Trp Gly Thr Trp Thr
1 5
Claims (97)
1.与第二抗体组合用作药物的第一抗体,所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则所述取代不为S440K,和
c.G237位置处氨基酸的一个取代,或者选自G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E的一个或多个取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则取代不为S440K,
其中所述第一Fc区具有K439E取代且所述第二Fc区具有S440K取代,或者所述第一Fc区具有S440K取代且所述第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
2.与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则所述取代不为S440K,和
c.P329位置处氨基酸的一个取代,或者K322E取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则所述取代不为S440K,
f.选自下组的一个或多个取代:K326A、K326W、E333A和E333S;
其中所述第一Fc区具有K439E取代且所述第二Fc区具有S440K取代,或者所述第一Fc区具有S440K取代且所述第二Fc区具有K439E取代;
其中氨基酸位置对应于根据Eu编号系统的人IgG1。
3.与第二抗体组合用作药物的第一抗体,其中所述第一抗体包含人IgG的第一Fc区和能够与第一抗原结合的第一抗原结合区,所述第二抗体包含人IgG的第二Fc区和能够与第二抗原结合的第二抗原结合区,并且所述第一Fc区包含
a.K248E和T437R取代;和
b.K439E或S440K取代;和
c.G237或P329位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、K332E、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.K248E和T437R取代,和
e.K439E或S440K取代;
其中所述第一Fc区具有K439E取代且所述第二Fc区具有S440K取代,或者所述第一Fc区具有S440K取代且所述第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
4.根据权利要求1和2中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区和第二Fc区包含选自下组的取代:E430G、E345K、E430S、E430F、E430T、E345Q、E345R、E345Y、S440W和S440Y。
5.根据权利要求1至2和4中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区和第二Fc区包含选自下组的取代:E430G、E345K和E345R。
6.根据权利要求1至2和4至5中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区和第二Fc区包含E430G取代。
7.根据权利要求1和3至6中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含选自下组的一个取代:G237A、G237T、G237Q、G237R、G237S、G237N、G237D、G237E、G237K、G237V、G237M、G237I、G237L、G237H、G237F、G237Y、G237W、G237P。
8.根据权利要求1和3至7中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含选自下组的一个取代:G237A、G237T、G237S、G237Q、G237R。
9.根据权利要求1和3至8中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含G237Q取代。
10.根据权利要求1和3至6中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含选自下组的一个或多个取代:G236R和E269K。
11.根据权利要求1,3至6和10中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含G236R取代。
12.根据权利要求1,3至6和10中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含E269K取代。
13.根据权利要求1和3至6中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含K322A取代。
14.根据权利要求2至6中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含P329R取代。
15.根据权利要求2至6中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一Fc区包含K322E取代。
16.根据权利要求1和3至13中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含选自下组的一个或多个取代:G237A、K326A、K326W、E333A和E333S。
17.根据权利要求1,3至13和16中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含G237A取代。
18.根据权利要求1至15中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含选自下组的一个或多个取代:K326A、K326W、E333A和E333S。
19.根据权利要求1至15和18中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含选自下组的一个取代:K326A、K326W、E333A和E333S。
20.根据权利要求1,3至13和16至17中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含G237A和E333S取代。
21.根据权利要求1至19中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含K326A取代。
22.根据权利要求1至19中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含E333S取代。
23.根据权利要求1至16和18中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含选自下组的两个取代:K326A、K326W、E333A和E333S。
24.根据权利要求1至16,18和23中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含K326W和E333S取代。
25.根据权利要求1至16,18和23中任一项的与第二抗体组合用作药物的第一抗体,其中所述第二Fc区包含K326A和E333A取代。
26.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一抗体和/或第二抗体为人的,人源化的或嵌合的。
27.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一抗体和/或所述第二抗体为单克隆抗体。
28.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一抗体和/或所述第二抗体为人IgG1、IgG2、IgG3或IgG4同种型。
29.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一抗体和/或所述第二抗体为人IgG1同种型。
30.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一抗原和第二抗原都是细胞表面暴露的分子。
31.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一抗原和第二抗原共定位于细胞或组织中,所述细胞或组织是待治疗的疾病或病症的靶细胞或靶组织。
32.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述第一抗原和第二抗原是不同的。
33.根据前述权利要求中任一项的与第二抗体组合用作药物的第一抗体,其中所述药物消耗表达所述第一抗原和第二抗原的细胞群。
34.根据前述权利要求33的与第二抗体组合用作药物的第一抗体,其中细胞群是肿瘤细胞。
35.根据权利要求33至34中任一项的与第二抗体组合用作药物的第一抗体,其中细胞群是血液肿瘤细胞或实体瘤细胞。
36.根据权利要求33至34中任一项的与第二抗体组合用作药物的第一抗体,其中细胞群是白细胞、淋巴细胞、B细胞、T细胞、调节性T细胞、NK细胞、髓样来源的抑制细胞、肿瘤相关的巨噬细胞群。
37.抗体,其包含人IgG的Fc区和能够结合抗原的抗原结合区,其中所述Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则取代不为S440K,和
c.G237位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E。
38.抗体,其包含人IgG的Fc区和能够结合抗原的抗原结合区,其中所述Fc区包含
a.K248E和T437R取代,和
b.K439E或S440K取代,和
c.G237或P329位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A,
d.K332E、E269K、L234A、L234F、L235A、L235Q、L235E、K326A、K326W、E333A和E333S。
39.根据权利要求37的抗体,其中所述Fc区包含选自下组的取代:E430G、E345K、E430S、E430F、E430T、E345Q、E345R、E345Y、S440W和S440Y。
40.根据权利要求37和39中任一项的抗体,其中所述Fc区包含选自下组的取代:E430G、E345K和E345R。
41.根据权利要求37和39至41中任一项的抗体,其中所述Fc区包含E430G取代。
42.根据权利要求37至41中任一项的抗体,其中所述Fc区包含选自下组的一个取代:G237A、G237T、G237Q、G237R、G237S、G237N、G237D、G237E、G237K、G237V、G237M、G237I、G237L、G237H、G237F、G237Y、G237W、G237P。
43.根据权利要求37至42中任一项的抗体,其中所述Fc区包含选自下组的一个取代:G237A、G237T、G237S、G237Q、G237R。
44.根据权利要求37至43中任一项的抗体,其中所述Fc区包含G237Q或G237A取代。
45.根据权利要求37至41中任一项的抗体,其中所述Fc区包含选自下组的一个或多个取代:G236R和E269K。
46.根据权利要求37至41和45中任一项的抗体,其中所述Fc区包含G236R取代。
47.根据权利要求37至41和45中任一项的抗体,其中所述Fc区包含E269K取代。
48.根据权利要求37至41中任一项的抗体,其中所述Fc区包含K322A取代。
49.根据权利要求37至48中任一项的抗体,其中所述抗体为人IgG1、IgG2、IgG3或IgG4同种型。
50.根据权利要求37至49中任一项的抗体,其中所述抗体为人IgG1同种型。
51.组合物,其包含第一抗体和第二抗体,其中所述第一抗体包含第一抗原结合区和根据权利要求1至15和26至36中任一项的第一Fc区,并且所述第二抗体包含第二抗原结合区和根据权利要求1至6和16至36中任一项的第二Fc区。
52.组合物,其包含第一抗体和第二抗体,其中所述第一抗体包含能够结合第一抗原的第一抗原结合区和人IgG的第一Fc区,并且所述第二抗体包含能够结合第二抗原的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则所述取代不为S440K,和
c.G237位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则所述取代不为S440K,
其中所述第一Fc区具有K439E取代且所述第二Fc区具有S440K取代,或者所述第一Fc区具有S440K取代且所述第二Fc区具有K439E取代,
其中氨基酸位置对应于根据Eu编号系统的人IgG1。
53.组合物,其包含第一抗体和第二抗体,其中所述第一抗体包含能够结合第一抗原的第一抗原结合区和人IgG的第一Fc区,并且所述第二抗体包含能够结合第二抗原的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
a.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
b.K439E或S440K取代,条件是如果(a)中的取代为S440Y或S440W,则所述取代不为S440K,和
c.P329位置处氨基酸的一个取代,或者K322E取代;
以及所述第二Fc区包含
d.选自E430、E345和S440的位置处氨基酸的一个取代,条件是S440处的取代为S440Y或S440W,和
e.K439E或S440K取代,条件是如果(d)中的取代为S440Y或S440W,则所述取代不为S440K,
f.选自K326A、K326W、E333A和E333S中的一个或多个取代;
其中所述第一Fc区具有K439E取代且所述第二Fc区具有S440K取代,或者所述第一Fc区具有S440K取代且所述第二Fc区具有K439E取代,其中氨基酸位置对应于根据Eu编号系统的人IgG1。
54.组合物,其包含第一抗体和第二抗体,其中所述第一抗体包含能够结合第一抗原的第一抗原结合区和人IgG的第一Fc区,并且所述第二抗体包含能够结合第二抗原的第二抗原结合区和人IgG的第二Fc区,其中所述第一Fc区包含
a.K248E和T437R取代,和
b.K439E或S440K取代,和
c.G237或P329位置处氨基酸的一个取代,或者选自下组的一个或多个取代:G236R、G236K、K322A、K332E、E269K、L234A、L234F、L235A、L235Q和L235E;
以及所述第二Fc区包含
d.K248E和T437R取代,和
e.一个K439E或S440K取代;
其中所述第一Fc区具有K439E取代且所述第二Fc区具有S440K取代,或者所述第一Fc区具有S440K取代且所述第二Fc区具有K439E取代;其中氨基酸位置对应于根据Eu编号系统的人IgG1。
55.根据权利要求52至53中任一项的组合物,其中所述第一Fc区和第二Fc区包含选自下组的取代:E430G、E345K、E430S、E430F、E430T、E345Q、E345R、E345Y、S440W和S440Y。
56.根据权利要求52至53和55中任一项的组合物,其中所述第一Fc区和第二Fc区包含选自下组的取代:E430G、E345K和E345R。
57.根据权利要求52至53和55至56中任一项的组合物,其中所述第一Fc区和第二Fc区包含E430G取代。
58.根据权利要求52至57中任一项的组合物,其中所述第一Fc区包含选自下组的一个取代:G237A、G237T、G237Q、G237R、G237S、G237N、G237D、G237E、G237K、G237V、G237M、G237I、G237L、G237H、G237F、G237Y、G237W、G237P。
59.根据权利要求52至58中任一项的组合物,其中所述第一Fc区包含选自下组的一个取代:G237A、G237T、G237Q、G237R和G237S。
60.根据权利要求52至59中任一项的组合物,其中所述第一Fc区包含G237Q取代。
61.根据权利要求52至57中任一项的组合物,其中所述第一Fc区包含选自下组的一个或多个取代:G236R和E269K。
62.根据权利要求52至57和61中任一项的组合物,其中所述第一Fc区包含G236R取代。
63.根据权利要求52至57和61中任一项的组合物,其中所述第一Fc区包含E269K取代。
64.根据权利要求52,54至57中任一项的组合物,其中所述第一Fc区包含K322A取代。
65.根据权利要求53至57中任一项的组合物,其中所述第一Fc区包含P329R取代。
66.根据权利要求53至57中任一项的组合物,其中所述第一Fc区包含K322E取代。
67.根据权利要求52,54至64中任一项的组合物,其中所述第二Fc区包含选自下组的一个或多个取代:G237A、K326A、K326W、E333A和E333S。
68.根据权利要求52,54至64和67中任一项的组合物,其中所述第二Fc区包含G237A取代。
69.根据权利要求52至67中任一项的组合物,其中所述第二Fc区包含选自下组的一个取代:K326A、K326W、E333A和E333S。
70.根据权利要求52至67和69中任一项的组合物,其中所述第二Fc区包含选自下组的两个取代:K326A、K326W、E333A和E333S。
71.根据权利要求52至67和69至70中任一项的组合物,其中所述第二Fc区包含K326W和E333S取代。
72.根据权利要求52至67和69至70中任一项的组合物,其中所述第二Fc区包含K326A和E333A取代。
73.根据权利要求52至67和69中任一项的组合物,其中所述第二Fc区包含E333S取代。
74.根据权利要求52,54至67中任一项的组合物,其中所述第二Fc区包含G237A和E333S取代。
75.根据权利要求51至74中任一项的组合物,其中所述第一抗原和第二抗原都是细胞表面暴露的分子。
76.根据权利要求51至75中任一项的组合物,其中所述第一抗原和第二抗原是不同的。
77.根据权利要求51至76中任一项的组合物,其中所述第一抗体和所述第二抗体以约1:50至50:1摩尔比,如约1:1摩尔比、约1:2摩尔比、约1:3摩尔比、约1:4摩尔比、约1:5摩尔比、约1:6摩尔比、约1:7摩尔比、约1:8摩尔比、约1:9摩尔比、约1:10摩尔比、约1:15摩尔比、约1:20摩尔比、约1:25摩尔比、约1:30摩尔比、约1:35摩尔比、约1:40摩尔比、约1:45摩尔比、约1:50摩尔比、约50:1摩尔比、约45:1摩尔比、约40:1摩尔比、约35:1摩尔比、约30:1摩尔比、25:1摩尔比、20:1摩尔比、15:1摩尔比、10:1摩尔比、9:1摩尔比、8:1摩尔比、7:1摩尔比、6:1摩尔比、5:1摩尔比、4:1摩尔比、3:1摩尔比、2:1摩尔比存在于所述组合物中。
78.根据权利要求51至77中任一项的组合物,其中所述第一抗体和所述第二抗体以约1:50至50:1摩尔比,如约1:40至40:1摩尔比,如约1:30至30:1摩尔比,如约1:20至20:1摩尔比,如约1:10至10:1摩尔比,如约1:9至9:1摩尔比,如约1:5至5:1摩尔比存在于所述组合物中。
79.根据权利要求51至78中任一项的组合物,其中所述第一抗体和所述第二抗体以1:1摩尔比存在于所述组合物中。
80.根据权利要求51至79中任一项的组合物,其中所述组合物还包含药物载体或赋形剂。
81.根据权利要求51至80中任一项的组合物,其中所述组合物为药物组合物。
82.根据前述权利要求中任一项的组合物,其用作药物。
83.根据权利要求1至36中任一项的第一抗体或第二抗体,根据权利要求37至50中任一项的抗体,或根据权利要求51至82中任一项的组合物,其中所述抗原结合区能够结合选自下组的抗原:DR4、DR5、CD20、CD37、CD52、HLA-DR、CD3、CD5、4-1BB、PD1。
84.根据权利要求1至36和83中任一项的第一抗体或第二抗体,根据权利要求37至50和83中任一项的抗体,或根据权利要求51至83中任一项的组合物,其中所述抗原结合区包含:
a.VH区,其包含SEQ ID NO:196所示的CDR1序列,SEQ ID NO:196所示的CDR2序列,和SEQ ID NO:198所示的CDR3序列,以及VL区,其包含SEQ ID NO:200所示的CDR1序列,AAT所示的CDR2序列,和SEQ ID NO:201所示的CDR3序列[DR4];
b.VH区,其包含SEQ ID NO:50所示的CDR1序列,SEQ ID NO:51所示的CDR2序列,和SEQID NO:52所示的CDR3序列,以及VL区,其包含SEQ ID NO:54所示的CDR1序列,FAS所示的CDR2序列,和SEQ ID NO:55所示的CDR3序列[DR5-01-G56T];
c.VH区,其包含SEQ ID NO:57所示的CDR1序列,SEQ ID NO:58所示的CDR2序列,和SEQID NO:59所示的CDR3序列,以及VL区,其包含SEQ ID NO:61所示的CDR1序列,RTS所示的CDR2序列,和SEQ ID NO:62所示的CDR3序列[DR5-05];
d.VH区,其包含SEQ ID NO:36所示的CDR1序列,SEQ ID NO:37所示的CDR2序列,和SEQID NO:38所示的CDR3序列,以及VL区,其包含SEQ ID NO:40所示的CDR1序列,DAS所示的CDR2序列,和SEQ ID NO:41所示的CDR3序列[CD20,7D8];
e.VH区,其包含SEQ ID NO:9所示的CDR1序列,SEQ ID NO:10所示的CDR2序列,和SEQID NO:11所示的CDR3序列,以及VL区,其包含SEQ ID NO:13所示的CDR1序列,DAS所示的CDR2序列,和SEQ ID NO:14所示的CDR3序列[CD20,11B8];
f.VH区,其包含SEQ ID NO:43所示的CDR1序列,SEQ ID NO:44所示的CDR2序列,和SEQID NO:45所示的CDR3序列,以及VL区,其包含SEQ ID NO:47所示的CDR1序列,VAT所示的CDR2序列,和SEQ ID NO:48所示的CDR3序列[CD37];
g.VH区,其包含SEQ ID NO:2所示的CDR1序列,SEQ ID NO:3所示的CDR2序列,和SEQ IDNO:4所示的CDR3序列,以及VL区,其包含SEQ ID NO:6所示的CDR1序列,NTN所示的CDR2序列,和SEQ ID NO:7所示的CDR3序列[CD52,CAMPATH-1H];
h.VH区,其包含SEQ ID NO:161所示的CDR1序列,SEQ ID NO:162所示的CDR2序列,和SEQ ID NO:163所示的CDR3序列,以及VL区,其包含SEQ ID NO:165所示的CDR1序列,LVS所示的CDR2序列,和SEQ ID NO:166所示的CDR3序列[CD52,h2E8];
i.VH区,其包含SEQ ID NO:168所示的CDR1序列,SEQ ID NO:169所示的CDR2序列,和SEQ ID NO:170所示的CDR3序列,以及VL区,其包含SEQ ID NO:172所示的CDR1序列,SEQ IDNO:AAS所示的CDR2序列,和SEQ ID NO:173所示的CDR3序列[HLA-DR,hul243];
j.VH区,其包含SEQ ID NO:175所示的CDR1序列,SEQ ID NO:176所示的CDR2序列,和SEQ ID NO:177所示的CDR3序列,以及VL区,其包含SEQ ID NO:179所示的CDR1序列,DNN所示的CDR2序列,和SEQ ID NO:180所示的CDR3序列[HLA-DR,1D09C3];
k.VH区,其包含SEQ ID NO:182所示的CDR1序列,SEQ ID NO:183所示的CDR2序列,和SEQ ID NO:184所示的CDR3序列,以及VL区,其包含SEQ ID NO:186所示的CDR1序列,SEQ IDNO:DTS所示的CDR2序列,和SEQ ID NO:187所示的CDR3序列[CD3,huCLB T3/4];或者
l.VH区,其包含SEQ ID NO:189所示的CDR1序列,SEQ ID NO:190所示的CDR2序列,和SEQ ID NO:191所示的CDR3序列,以及VL区,其包含SEQ ID NO:193所示的CDR1序列,ATS所示的CDR2序列,和SEQ ID NO:194所示的CDR3序列[CD5]。
85.治疗患有疾病的个体的方法,其包括向所述个体施用有效量的根据权利要求1至37和83至84中任一项的第一抗体和第二抗体,根据权利要求37至50和83至84中任一项的抗体,或者根据权利要求51至84中任一项的组合物。
86.根据权利要求85的方法,其中所述疾病选自:癌症、自身免疫性疾病、炎症性疾病和传染性疾病。
87.根据权利要求85至86中任一项的方法,其中所述方法包括施用另外的治疗剂。
88.消耗表达第一抗原和第二抗原的细胞群的方法,该方法包括使所述细胞群与根据权利要求1至84的第一抗体和第二抗体,抗体或组合物接触。
89.根据权利要求88的方法,其中所述细胞群是肿瘤细胞群,例如血液肿瘤细胞群或实体瘤细胞群。
90.在表达第一抗原和第二抗原的细胞群中诱导增殖的方法,该方法包括使所述细胞群与根据权利要求1至84的第一抗体和第二抗体,抗体或组合物接触。
91.根据权利要求88至90中任一项的方法,其中所述细胞群存在于血液中。
92.根据权利要求88至91中任一项的方法,其中所述细胞群是白细胞。
93.根据权利要求88至92中任一项的方法,其中所述细胞群是白细胞细胞群的亚群。
94.根据权利要求88至93中任一项的方法,其中所述细胞群是淋巴细胞细胞群。
95.根据权利要求94的方法,其中所述细胞群是B细胞群、T细胞群、NK细胞群、调节性T细胞群、髓样来源的抑制细胞群。
96.根据权利要求85至95中任一项的方法,其中所述第一抗原和/或第二抗原为TNFR-SF的成员。
97.试剂盒,其包含第一容器和第二容器,所述第一容器包含权利要求1至15,26至36和83至84中任一项所定义的第一抗体,所述第二容器包含权利要求1至6,16至36和83至84中任一项所定义的第二抗体。
Applications Claiming Priority (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| DKPA201800195 | 2018-05-03 | ||
| DKPA201800195 | 2018-05-03 | ||
| DKPA201800644 | 2018-09-26 | ||
| DKPA201800644 | 2018-09-26 | ||
| PCT/EP2019/061455 WO2019211472A1 (en) | 2018-05-03 | 2019-05-03 | Antibody variant combinations and uses thereof |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN112839958A true CN112839958A (zh) | 2021-05-25 |
Family
ID=66476613
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201980037949.XA Pending CN112839958A (zh) | 2018-05-03 | 2019-05-03 | 抗体变体组合及其用途 |
Country Status (13)
| Country | Link |
|---|---|
| US (1) | US20210238296A1 (zh) |
| EP (1) | EP3788074A1 (zh) |
| JP (1) | JP2021522303A (zh) |
| KR (1) | KR20210008380A (zh) |
| CN (1) | CN112839958A (zh) |
| AU (1) | AU2019264217A1 (zh) |
| BR (1) | BR112020022179A2 (zh) |
| CA (1) | CA3098486A1 (zh) |
| MA (1) | MA52485A (zh) |
| MX (1) | MX2020011552A (zh) |
| PH (1) | PH12020551808A1 (zh) |
| SG (1) | SG11202010601VA (zh) |
| WO (1) | WO2019211472A1 (zh) |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB2595299B (en) * | 2020-05-21 | 2022-08-03 | Mabsolve Ltd | Modified immunoglobulin FC regions |
| KR20230156921A (ko) * | 2021-03-12 | 2023-11-15 | 젠맵 에이/에스 | 비-활성화 항체 변이체 |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2014006217A1 (en) * | 2012-07-06 | 2014-01-09 | Genmab B.V. | Dimeric protein with triple mutations |
| CN105229026A (zh) * | 2013-01-10 | 2016-01-06 | 根马布私人有限公司 | 人类IgG1 FC区变体及其用途 |
| CN107709364A (zh) * | 2015-04-07 | 2018-02-16 | 豪夫迈·罗氏有限公司 | 具有激动剂活性的抗原结合复合体及使用方法 |
Family Cites Families (51)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO1998050431A2 (en) | 1997-05-02 | 1998-11-12 | Genentech, Inc. | A method for making multispecific antibodies having heteromultimeric and common components |
| DK2180007T4 (da) | 1998-04-20 | 2017-11-27 | Roche Glycart Ag | Glycosyleringsteknik for antistoffer til forbedring af antistofafhængig cellecytotoxicitet |
| EP2275540B1 (en) | 1999-04-09 | 2016-03-23 | Kyowa Hakko Kirin Co., Ltd. | Method for controlling the activity of immunologically functional molecule |
| JP4668498B2 (ja) | 1999-10-19 | 2011-04-13 | 協和発酵キリン株式会社 | ポリペプチドの製造方法 |
| US7521047B2 (en) | 2000-05-12 | 2009-04-21 | Gpc Biotech Ag | Human polypeptides causing or leading to the killing of cells including lymphoid tumor cells |
| DE10043437A1 (de) | 2000-09-04 | 2002-03-28 | Horst Lindhofer | Verwendung von trifunktionellen bispezifischen und trispezifischen Antikörpern zur Behandlung von malignem Aszites |
| US6946292B2 (en) | 2000-10-06 | 2005-09-20 | Kyowa Hakko Kogyo Co., Ltd. | Cells producing antibody compositions with increased antibody dependent cytotoxic activity |
| CA2424977C (en) | 2000-10-06 | 2008-03-18 | Kyowa Hakko Kogyo Co., Ltd. | Process for purifying antibody |
| MXPA03002974A (es) | 2000-10-06 | 2004-05-05 | Kyowa Hakko Kogyo Kk | Celulas que producen composiciones de anticuerpo. |
| BR0213761A (pt) | 2001-10-25 | 2005-04-12 | Genentech Inc | Composições, preparação farmacêutica, artigo industrializado, método de tratamento de mamìferos, célula hospedeira, método para a produção de uma glicoproteìna e uso da composição |
| CN101537180B (zh) | 2002-07-18 | 2016-02-10 | 莫鲁斯有限公司 | 抗体混合物的重组生产 |
| PL216630B1 (pl) | 2002-10-17 | 2014-04-30 | Genmab As | Izolowane ludzkie przeciwciało monoklonalne wiążące ludzki CD20, związane z tym przeciwciałem transfektoma, komórka gospodarza, transgeniczne zwierzę lub roślina, kompozycja, immunokoniugat, cząsteczka bispecyficzna, wektor ekspresyjny, kompozycja farmaceutyczna, zastosowanie medyczne, zestaw oraz przeciwciało antyidiotypowe i jego zastosowanie |
| US7741568B2 (en) | 2005-01-13 | 2010-06-22 | The Wiremold Company | Downward facing receptacle assembly for cable raceway |
| AU2006218454B2 (en) | 2005-03-03 | 2011-11-17 | Immunomedics, Inc. | Humanized L243 antibodies |
| WO2006106905A1 (ja) | 2005-03-31 | 2006-10-12 | Chugai Seiyaku Kabushiki Kaisha | 会合制御によるポリペプチド製造方法 |
| US7612181B2 (en) | 2005-08-19 | 2009-11-03 | Abbott Laboratories | Dual variable domain immunoglobulin and uses thereof |
| KR101866623B1 (ko) | 2005-11-28 | 2018-07-04 | 젠맵 에이/에스 | 재조합 1가 항체 및 그의 제조 방법 |
| BRPI0709598A8 (pt) | 2006-03-17 | 2019-01-08 | Biogen Idec Inc | composições de polipeptídeos estabilizados |
| ES2395969T3 (es) | 2006-03-24 | 2013-02-18 | Merck Patent Gmbh | Dominios de proteínas heterodiméricas genéticamente modificados |
| CN101074261A (zh) | 2006-04-30 | 2007-11-21 | 北京同为时代生物技术有限公司 | Trail受体1和/或trail受体2特异性抗体及其应用 |
| AT503902B1 (de) | 2006-07-05 | 2008-06-15 | F Star Biotech Forsch & Entw | Verfahren zur manipulation von immunglobulinen |
| EP2158221B1 (en) | 2007-06-21 | 2018-08-29 | MacroGenics, Inc. | Covalent diabodies and uses thereof |
| EP2050764A1 (en) | 2007-10-15 | 2009-04-22 | sanofi-aventis | Novel polyvalent bispecific antibody format and uses thereof |
| PT2235064E (pt) | 2008-01-07 | 2016-03-01 | Amgen Inc | Método de preparação de moléculas heterodiméricas de fc de anticorpos utilizando efeitos de indução eletrostática |
| HUE043084T2 (hu) | 2008-12-19 | 2019-08-28 | Macrogenics Inc | Kovalens diatestek és alkalmazásaik |
| WO2010129304A2 (en) | 2009-04-27 | 2010-11-11 | Oncomed Pharmaceuticals, Inc. | Method for making heteromultimeric molecules |
| DK2430051T3 (en) | 2009-05-14 | 2016-07-04 | Inst Nat De La Santé Et De La Rech Medicale | Compositions containing antibodies for the treatment of CD5 + HLA-DR + B- or T-cell-related diseases |
| EP2975051B1 (en) | 2009-06-26 | 2021-04-14 | Regeneron Pharmaceuticals, Inc. | Readily isolated bispecific antibodies with native immunoglobulin format |
| WO2011028952A1 (en) | 2009-09-02 | 2011-03-10 | Xencor, Inc. | Compositions and methods for simultaneous bivalent and monovalent co-engagement of antigens |
| MX2012006406A (es) | 2009-12-04 | 2012-07-25 | Genentech Inc | Anticuerpos multiespecificos, analogos de anticuerpo, composiciones y metodos. |
| RU2610662C9 (ru) | 2010-03-12 | 2017-07-24 | Иммьюноджен, Инк. | Cd37-связывающие молекулы и их иммуноконъюгаты |
| TWI426920B (zh) | 2010-03-26 | 2014-02-21 | Hoffmann La Roche | 雙專一性、雙價抗-vegf/抗-ang-2抗體 |
| US9150663B2 (en) | 2010-04-20 | 2015-10-06 | Genmab A/S | Heterodimeric antibody Fc-containing proteins and methods for production thereof |
| JP6022444B2 (ja) | 2010-05-14 | 2016-11-09 | ライナット ニューロサイエンス コーポレイション | ヘテロ二量体タンパク質ならびにそれを生産および精製するための方法 |
| AU2011267106B2 (en) | 2010-06-15 | 2015-05-14 | Genmab A/S | Human antibody drug conjugates against tissue factor |
| PL2606064T3 (pl) | 2010-08-16 | 2015-07-31 | Novimmune Sa | Sposoby wytwarzania wieloswoistych i wielowartościowych przeciwciał |
| CA2807269A1 (en) | 2010-08-24 | 2012-03-01 | Roche Glycart Ag | Activatable bispecific antibodies |
| JP5758004B2 (ja) | 2010-08-24 | 2015-08-05 | エフ.ホフマン−ラ ロシュ アーゲーF. Hoffmann−La Roche Aktiengesellschaft | ジスルフィドによって安定化されたFv断片を含む二重特異性抗体 |
| JP6167040B2 (ja) | 2010-11-05 | 2017-07-19 | ザイムワークス,インコーポレイテッド | Fcドメイン中に突然変異を有する、安定したヘテロ二量体抗体の設計 |
| SG193554A1 (en) | 2011-03-29 | 2013-11-29 | Roche Glycart Ag | Antibody fc variants |
| GB201109238D0 (en) | 2011-06-01 | 2011-07-13 | Antitope Ltd | Antibodies |
| CN102250246A (zh) | 2011-06-10 | 2011-11-23 | 常州亚当生物技术有限公司 | 抗VEGF/PDGFRβ双特异性抗体及其应用 |
| UA117901C2 (uk) * | 2011-07-06 | 2018-10-25 | Ґенмаб Б.В. | Спосіб посилення ефекторної функції вихідного поліпептиду, його варіанти та їх застосування |
| WO2013096291A2 (en) | 2011-12-20 | 2013-06-27 | Medimmune, Llc | Modified polypeptides for bispecific antibody scaffolds |
| PT2838918T (pt) | 2012-04-20 | 2019-08-23 | Merus Nv | Métodos e meios para a produção de moléculas heterrodiméricas do tipo ig |
| EP2684896A1 (en) | 2012-07-09 | 2014-01-15 | International-Drug-Development-Biotech | Anti-DR5 family antibodies, bispecific or multivalent anti-DR5 family antibodies and methods of use thereof |
| AU2013305885B2 (en) | 2012-08-20 | 2017-12-21 | Gliknik Inc. | Molecules with antigen binding and polyvalent Fc gamma receptor binding activity |
| CN106255704A (zh) | 2014-04-16 | 2016-12-21 | Ucb生物制药私人有限公司 | 多聚体Fc蛋白 |
| MX2018006333A (es) | 2015-12-01 | 2018-08-01 | Genmab Bv | Anticuerpos anti-receptor de muerte 5 (dr5) y metodos de uso de los mismos. |
| WO2017117179A1 (en) * | 2015-12-28 | 2017-07-06 | Massachusetts Institute Of Technology | Bispecific antibodies having constant region mutations and uses therefor |
| US10669344B2 (en) | 2016-08-12 | 2020-06-02 | Janssen Biotech, Inc. | Engineered antibodies and other Fc-domain containing molecules with enhanced agonism and effector functions |
-
2019
- 2019-05-03 AU AU2019264217A patent/AU2019264217A1/en active Pending
- 2019-05-03 BR BR112020022179-1A patent/BR112020022179A2/pt unknown
- 2019-05-03 SG SG11202010601VA patent/SG11202010601VA/en unknown
- 2019-05-03 MX MX2020011552A patent/MX2020011552A/es unknown
- 2019-05-03 WO PCT/EP2019/061455 patent/WO2019211472A1/en active Application Filing
- 2019-05-03 KR KR1020207034871A patent/KR20210008380A/ko not_active Application Discontinuation
- 2019-05-03 EP EP19723054.3A patent/EP3788074A1/en active Pending
- 2019-05-03 CN CN201980037949.XA patent/CN112839958A/zh active Pending
- 2019-05-03 CA CA3098486A patent/CA3098486A1/en active Pending
- 2019-05-03 MA MA052485A patent/MA52485A/fr unknown
- 2019-05-03 US US17/051,205 patent/US20210238296A1/en active Pending
- 2019-05-03 JP JP2020561788A patent/JP2021522303A/ja active Pending
-
2020
- 2020-10-29 PH PH12020551808A patent/PH12020551808A1/en unknown
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2014006217A1 (en) * | 2012-07-06 | 2014-01-09 | Genmab B.V. | Dimeric protein with triple mutations |
| CN104736174A (zh) * | 2012-07-06 | 2015-06-24 | 根马布私人有限公司 | 具有三重突变的二聚体蛋白质 |
| CN105229026A (zh) * | 2013-01-10 | 2016-01-06 | 根马布私人有限公司 | 人类IgG1 FC区变体及其用途 |
| CN107709364A (zh) * | 2015-04-07 | 2018-02-16 | 豪夫迈·罗氏有限公司 | 具有激动剂活性的抗原结合复合体及使用方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| MA52485A (fr) | 2021-03-10 |
| SG11202010601VA (en) | 2020-11-27 |
| KR20210008380A (ko) | 2021-01-21 |
| PH12020551808A1 (en) | 2021-07-26 |
| WO2019211472A1 (en) | 2019-11-07 |
| BR112020022179A2 (pt) | 2021-02-02 |
| CA3098486A1 (en) | 2019-11-07 |
| US20210238296A1 (en) | 2021-08-05 |
| EP3788074A1 (en) | 2021-03-10 |
| AU2019264217A1 (en) | 2020-12-17 |
| MX2020011552A (es) | 2020-11-24 |
| JP2021522303A (ja) | 2021-08-30 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR102346336B1 (ko) | Pd1 및 lag3에 특이적으로 결합하는 이중특이적 항체 | |
| KR102632202B1 (ko) | Cd40 및 cd137에 대한 다중특이적 항체 | |
| US20230107363A1 (en) | Polypeptide variants and uses thereof | |
| AU2024200261A1 (en) | Anti-DR5 antibodies and methods of use thereof | |
| US20200181277A1 (en) | Polypeptide variants and uses thereof | |
| JP2021524276A (ja) | Cd38抗体のバリアントおよびその使用 | |
| CN112839958A (zh) | 抗体变体组合及其用途 | |
| JP2021526845A (ja) | Cd38抗体を使用したトロゴサイトーシスを介した治療 | |
| US20220411529A1 (en) | Antibody variant combinations and uses thereof | |
| US20230365714A1 (en) | Antibodies capable of binding to ror2 and bispecific antibodies binding to ror2 and cd3 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |