CN106170554B - 过表达n-糖基化途径调节基因以调节重组蛋白的糖基化 - Google Patents
过表达n-糖基化途径调节基因以调节重组蛋白的糖基化 Download PDFInfo
- Publication number
- CN106170554B CN106170554B CN201480074585.XA CN201480074585A CN106170554B CN 106170554 B CN106170554 B CN 106170554B CN 201480074585 A CN201480074585 A CN 201480074585A CN 106170554 B CN106170554 B CN 106170554B
- Authority
- CN
- China
- Prior art keywords
- cell
- day
- culture
- protein
- cells
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images





Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1048—Glycosyltransferases (2.4)
- C12N9/1051—Hexosyltransferases (2.4.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/005—Glycopeptides, glycoproteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y204/00—Glycosyltransferases (2.4)
- C12Y204/01—Hexosyltransferases (2.4.1)
- C12Y204/01101—Alpha-1,3-mannosyl-glycoprotein 2-beta-N-acetylglucosaminyltransferase (2.4.1.101)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/40—Immunoglobulins specific features characterized by post-translational modification
- C07K2317/41—Glycosylation, sialylation, or fucosylation
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y204/00—Glycosyltransferases (2.4)
- C12Y204/01—Hexosyltransferases (2.4.1)
- C12Y204/01094—Protein N-acetylglucosaminyltransferase (2.4.1.94)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y204/00—Glycosyltransferases (2.4)
- C12Y204/01—Hexosyltransferases (2.4.1)
- C12Y204/01143—Alpha-1,6-mannosyl-glycoprotein 2-beta-N-acetylglucosaminyltransferase (2.4.1.143)
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Medicinal Chemistry (AREA)
- Biophysics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Immunology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Plant Pathology (AREA)
- Physics & Mathematics (AREA)
- Cell Biology (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Enzymes And Modification Thereof (AREA)
- Saccharide Compounds (AREA)
Abstract
提供了调节表达目标蛋白的细胞培养物的特性的方法。在不同实施方案中,所述方法与涉及N‑糖基化途径的蛋白质的过表达有关。
Description
发明领域
本发明总体上涉及用于调节由细胞培养物(包括哺乳动物细胞细胞培养物,诸如CHO细胞培养物)产生的重组蛋白的一个或多个特性的方法。
相关申请的交叉引用
本申请要求2014年1月29日提交的美国临时申请号61/933,192的权益,所述申请以引用的方式并入本文。
发明背景
糖基化是哺乳动物细胞中常见的翻译后修饰;正常人免疫球蛋白和中国仓鼠卵巢(CHO)细胞中产生的治疗性单克隆抗体(mAb)都是糖蛋白。治疗性mAb的药代动力学特性和效应功能都可受糖基化影响。末端糖诸如岩藻糖和半乳糖可影响抗体依赖性细胞毒性(ADCC)和补体依赖性细胞毒性(CDC;Wright,A.和S.L.Morrison,Trends Biotechnol(1997)15:26-32)。高甘露聚糖可增加某些mAb的血清清除率,因此潜在地影响了功效(Goetze等,(2011)Glycobiology 21:949-59)。另选地,高甘露糖糖型可增加Fc γIII受体的抗体亲和性,因此增加了某些抗体的ADCC活性(Yu等(2012)MAb 4:475-87)。因此,对于每个重组mAb,需维持某些最好地支持mAb治疗潜力的糖基化特性。
用于在细胞培养中操控蛋白质的高甘露糖糖型含量的方法包括改变培养基组成、重量摩尔渗透压浓度、pH、温度等(Yu等,同上,Pacis等,同上,Chee Furng Wong等(2005)Biotechnol Bioeng 89:164-177;Ahn等(2008)Biotechnol Bioeng 101:1234-44)。这些方法的有效性对于细胞系、分子类型和培养基环境是特异的,并且通常通过试错法来获得。另选地,这些方法可能也会改变抗体生产率、细胞培养物行为和其他抗体质量属性。
仍存在鉴定可在没有降低CHO生产培养物性能和抗体产率的情况下调控mAb上的高甘露糖糖型(特别是甘露糖5)的机制的需要。这种方法将有益于治疗性蛋白质的工艺过程开发。本发明提供一种通过操控涉及N-糖基化途径的蛋白质的表达水平来调控高甘露糖糖型含量的方法。
发明概述
本发明提供一种用于在哺乳动物细胞培养过程期间调控重组蛋白的高甘露糖糖型含量的方法,其包括转化宿主细胞以过表达涉及N-糖基化途径的蛋白质。本发明还提供一种用于在哺乳动物细胞培养过程期间减少重组蛋白的高甘露糖糖型含量的方法。在一个实施方案中,蛋白质是N-乙酰基-葡萄糖胺转移酶-1(由Mgat1编码);在本发明的另一个实施方案中,蛋白质是N-乙酰基-葡萄糖胺转移酶-2(由Mgat2编码)。在本发明的再一个实施方案中,蛋白质是UDP-半乳糖转运蛋白(由Slc35a2编码)。
本发明的附加实施方案包括转化宿主细胞以过表达两种或更多种涉及N-糖基化途径的蛋白质,包括上述蛋白质的组合。在一个实施方案中,用Mgat1和Mgat2转化宿主细胞;在其他实施方案中,用Mgat1和Slc35a2、用Mgat2和Slc35a2或者用Mgat1、Mgat2和Slc35a2转化宿主细胞。
本发明还提供先前已经过转染以表达重组蛋白的宿主细胞系的转染。在一个实施方案中,重组蛋白是包含抗体Fc区的蛋白质。再一个实施方案包括表达选自由Fc融合蛋白、抗体、免疫球蛋白和肽体组成的组的重组蛋白的宿主细胞。
在再一个实施方案中,首先转染宿主细胞以过表达Mgat1、Mgat2和Slc35a2中的一种或多种,然后进行转染以表达重组蛋白。在一个实施方案中,重组蛋白是包含抗体Fc区的蛋白质。再一个实施方案包括选自由Fc融合蛋白、抗体、免疫球蛋白和肽体组成的组的重组蛋白的表达。
任选地,本发明还包括收获由细胞培养物产生的重组蛋白的步骤。在再一个实施方案中,将宿主细胞产生的重组蛋白纯化并配制成药学上可接受的制剂。
在再一个实施方案中,重组蛋白的高甘露糖糖型含量与由细胞未通过转染以过表达涉及N-连接糖基化的蛋白质来进行操控的培养物产生的重组蛋白相比是减少的。在一个实施方案中,高甘露聚糖物质为甘露糖5(Man5)。在另一个实施方案中,高甘露聚糖物质为甘露糖6(Man6)、甘露糖7(Man7)、甘露糖8(包括甘露糖8a和8b;Man8a和8b或甘露糖9(Man9)。在再一个实施方案中,高甘露聚糖物质包括Man5、Man6、Man7、Man8a、Man8b和/或Man9的混合物。
本发明提供重组蛋白高甘露糖糖型含量减少的再一个实施方案。在再一个实施方案中,重组蛋白的高甘露糖糖型含量小于或等于5%。在另一个实施方案中,重组蛋白的高甘露糖糖型含量小于或等于10%。在再一个实施方案中,由本发明的细胞培养物产生的重组蛋白的高甘露糖糖型含量小于6%、7%、8%、9%或10%。在又一个实施方案中,由本发明的细胞培养物产生的重组蛋白的高甘露糖糖型含量小于0.5%、1%、2%、3%、4%或5%。再一个实施方案包括小于12%、小于15%、小于20%或小于30%、40%或50%的高甘露糖糖型含量。
附加的实施方案包括使用分批或补料分批培养物和使用灌流培养物。在一个实施方案中,使用交替式切向流(ATF)灌流所述培养物。
与本文描述的本发明的任何实施方案相结合,根据需要还可向培养容器中添加消泡剂。可选地或此外,使用1M碳酸钠或另一种适合的碱来使pH维持在所需设定值。
如本文所述,在本发明的一个方面,可通过灌流维持细胞培养物。在一个实施方案中,灌流开始于细胞培养的第1天或大约第1天至第9天或大约第9天。在一个相关实施方案中,灌流开始于细胞培养的第3天或大约第3天至第7天或大约第7天。在一个实施方案中,灌流在细胞已达到生产期时开始。在本发明的再一个实施方案中,灌流通过交替式切向流来完成。在一个相关实施方案中,通过交替式切向流使用超滤器或微滤器来完成灌流。
在本发明的再一个实施方案中提供了连续灌流;在又一个实施方案中,灌流速率是恒定的。在本发明的一个实施方案中提供了以每天小于或等于1.0个工作容积的速率进行的灌流。在一个相关实施方案中,在细胞培养期间以在生产期期间从每天0.25个工作容积增加至每天1.0个工作容积的速率进行灌流。在另一个相关实施方案中,在细胞培养的第9天至第11天以达到每天1.0个工作容积的速率进行灌流。在另一个相关实施方案中,在细胞培养的第10天以达到每天1.0个工作容积的速率进行灌流。
在一个实施方案中,细胞培养物在培养的第3天-第7天之前接受灌注细胞培养基补料。
在本发明的又一方面,通过分批补料来维持细胞培养物。在补料分批培养的一个实施方案中,所述培养物在生产期间补料三次。在再一个实施方案中,所述培养物在第二天与第四天之间的一天、在第5天与第7天之间的一天以及在第8天与第10天之间的一天补料。另一个实施方案提供了所述培养物在生产期间补料四次的补料分批法。在又一个实施方案中,所述培养物在第二天与第四天之间的一天、在第5天与第6天之间的一天、在第7天与第8天之间的一天以及在第8天与第10天或更迟之间的一天补料。
根据本发明的一个实施方案,哺乳动物细胞培养物通过用无血清培养基中的至少0.5x 106个细胞/mL至3.0x 106个细胞/mL接种生物反应器来建立。在一个可选或再一个实施方案中,哺乳动物细胞培养物通过用无血清培养基中的至少0.5x 106个细胞/mL至1.5x106个细胞/mL接种生物反应器来建立。
本发明还可包括培养期间的温度变化。在一个实施方案中,温度变化为36℃至31℃。在一个实施方案中,本发明还包括36℃至33℃的温度变化。在一个相关实施方案中,温度变化发生在生长期与生产期之间的过渡处。在一个相关实施方案中,温度变化在生产期期间发生。
在另一个实施方案中,本发明还包括通过L-天冬酰胺饥饿随后用具有5mM或更少的L-天冬酰胺浓度的无血清灌流培养基灌流来诱导细胞生长停滞。在另一个实施方案中,本发明还包括通过用具有5mM或更少的L-天冬酰胺浓度的无血清灌流培养基灌流来诱导细胞生长停滞。在一个相关实施方案中,无血清灌流培养基中L-天冬酰胺的浓度小于或等于5mM。在一个相关实施方案中,无血清灌流培养基中L-天冬酰胺的浓度小于或等于4.0mM。在一个相关实施方案中,无血清灌流培养基中L-天冬酰胺的浓度小于或等于3.0mM。在一个相关实施方案中,无血清灌流培养基中L-天冬酰胺的浓度小于或等于2.0mM。在一个相关实施方案中,无血清灌流培养基中L-天冬酰胺的浓度小于或等于1.0mM。在一个相关实施方案中,无血清灌流培养基中L-天冬酰胺的浓度为0mM。在一个相关实施方案中,在L-天冬酰胺饥饿之前或期间监控细胞培养基的L-天冬酰胺浓度。
在又一个实施方案中,本发明包括在生产期期间细胞压积小于或等于35%。在一个相关实施方案中,所述细胞压积小于或等于35%。在一个相关实施方案中,所述细胞压积小于或等于30%。
在一个相关实施方案中,在小于或等于35%的细胞压积时的哺乳动物细胞培养物的活细胞密度为10x106个活细胞/ml至80x106个活细胞/ml。在另一个实施方案中,所述哺乳动物细胞培养物的活细胞密度为20x106个活细胞/ml至30x106个活细胞/ml。
在又一个实施方案中,生物反应器的容量为至少500L。在又一个实施方案中,生物反应器的容量为至少500L至2000L。在又一个实施方案中,生物反应器的容量为至少1000L至2000L。
在又一个实施方案中,所述哺乳动物细胞是中国仓鼠卵巢(CHO)细胞。在又一个实施方案中,重组蛋白选自由人抗体、人源化抗体、嵌合抗体、重组融合蛋白或细胞因子组成的组。
附图简述
图1展示在用于实施例2中第一组补料分批培养物的细胞系的传代期间倍增时间的克隆可变性。在这个实施例中,使来自表达MAb B的细胞系的细胞转化以过表达Mgat1、Mgat2和/或Slc35A2。M1M2表示过表达Mgat1和Mgat2(由实心圆代表的个别克隆)的细胞系;M1M2S表示过表达Mgat1、Mgat2和Slc35a2(由空心三角形代表的个别克隆)的细胞系;并且S表示过表达Slc35A2(由实心正方形代表的个别克隆)的细胞系。用空载体转化对照细胞(由空心正方形代表的个别克隆)。中央方框涵盖第一四分位数(Q1)至第三四分位数(Q3),并且方框的高度为四分位数间距(IQR),方框内的条带是中值,如果没有值大于Q3+1.5IQR,则上线(top whisker)从Q3延伸至低于Q3+1.5IQR的最大值或极大值。如果没有值小于Q1-1.5IQR,则下线(bottom whisker)从Q1延伸至高于Q1-1.5IQR的最小值或极小值。
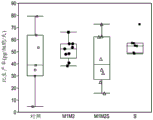
图2展示在用于实施例2中第二组补料分批细胞的细胞系的传代期间倍增时间的克隆可变性。在这个实施例中,使来自表达MAb B的细胞系的细胞转化以过表达Mgat1、Mgat2和/或Slc35A2。M1M2表示过表达Mgat1和Mgat2的细胞系;M1M2S表示过表达Mgat1、Mgat2和Slc35a2的细胞系;并且S表示过表达Slc35A2的细胞系。用空载体转化对照细胞。如对图1所描述的,表示个别克隆;各方框的参数如对图1所描述的一样。
图3呈现与实施例2描述的第二补料分批实验第10天的对照相比,就活细胞密度而言显示的生长对于所有过表达的细胞系(M1M2、M1M2S和S)是相似的。各方框中的个别值再次说明了观察到的克隆可变性。如对图1所描述的,表示个别克隆;各方框的参数如对图1所描述的一样。
图4呈现在实施例2描述的第二补料分批实验期间产生的抗体的效价的克隆可变性比较。如对图1所描述的,表示个别克隆;各方框的参数如对图1所描述的一样。
图5呈现在实施例2描述的第二补料分批实验期间产生的抗体的比产率的克隆可变性。如对图1所描述的,表示个别克隆;各方框的参数如对图1所描述的一样。
图6提供在实施例2描述的第二补料分批实验期间产生的抗体的HM百分比的克隆可变性指示。如对图1所描述的,表示个别克隆;各方框的参数如对图1所描述的一样。
发明详述
虽然在本申请中使用的术语在本领域中是标准的,但是本文提供了某些术语的定义以确保权利要求意思的清楚和明确。单位、前缀和符号可以他们的SI(国际单位制)接受的形式来表示。本文列举的数值范围包括界定了范围的数字,并且包括并支持所界定的范围内的每个整数。除非另外指示,否则本文描述的方法和技术一般根据本领域中所熟知且如在整篇本说明书中所引用并论述的各种一般参考文献和更特定参考文献中所述的常规方法来进行。参见,例如,Sambrook等Molecular Cloning:A Laboratory Manual,第3版,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(2001)和Ausubel等,Current Protocols in Molecular Biology,Greene Publishing Associates(1992)以及Harlow和Lane Antibodies:A Laboratory Manual Cold Spring Harbor LaboratoryPress,Cold Spring Harbor,N.Y.(1990)。
公开的方法适用于在搅拌釜反应器(包括传统分批细胞培养物和补料分批细胞培养物,其可能但不必包括自旋滤器)、灌流系统(包括交替式切向流(“ATF”)培养物、声学(acoustic)灌流系统、深度滤器灌流系统及其他系统)、中空纤维生物反应器(HFB,其在一些情况下可用于灌流工艺)以及各种其他细胞培养方法中生长的贴壁培养物或悬浮培养物(参见,例如,以引用的方式整体并入本文的Tao等,(2003)Biotechnol.Bioeng.82:751-65;Kuystermans和Al-Rubeai,(2011)“Bioreactor Systems for Producing Antibody fromMammalian Cells”in Antibody Expression and Production,Cell Engineering 7:25-52,Al-Rubeai(ed)Springer;Catapano等,(2009)“Bioreactor Design and Scale-Up”inCell and Tissue Reaction Engineering:Principles and Practice,Eibl等(编)Springer-Verlag)。
本申请中引用的所有文件或文件的部分,包括但不限于专利、专利申请、文章、书籍和论文,均特此明确地以引用的方式并入。本发明的一个实施方案中描述的可与本发明的其他实施方案组合。
定义
如本文所使用,术语“一个(a)”和“一种(an)”意指一个(种)或多个(种),除非另外确切地指出。此外,除非另外为上下文所需,否则单数术语将包括复数,并且复数术语将包括单数。通常,与本文所述的细胞和组织培养、分子生物学、免疫学、微生物学、遗传学和蛋白质及核酸化学以及杂交关联使用的命名以及细胞和组织培养、分子生物学、免疫学、微生物学、遗传学和蛋白质及核酸化学以及杂交的技术是本领域中熟知和常用的那些命名和技术。
本公开提供调节表达“目标蛋白”的细胞培养物的特性的方法;“目标蛋白”包括天然存在的蛋白质、重组蛋白和改造的蛋白质(例如,不存在于自然中且已由人类设计和/或创造的蛋白质)。目标蛋白可为但不必为已知或疑似是治疗学上相关的蛋白质。目标蛋白的具体实例包括抗原结合蛋白(如本文所描述或定义的)、肽体(即,包含直接或间接与其他分子诸如抗体的Fc结构域融合的肽,其中肽部分特异性地与所需靶标结合;例如,如以引用的方式整体并入本文的美国专利申请公布号US2006/0140934所述的,所述肽可与Fc区融合或者嵌入Fc-环或修饰的Fc分子中)、融合蛋白(例如,Fc融合蛋白,其中Fc片段与蛋白质或肽(包括肽体)融合)、细胞因子、生长因子、激素和其他天然存在的分泌蛋白以及天然存在的蛋白质的突变形式。
术语“抗原结合蛋白”被以其广义使用并且意指包含与抗原或靶标结合的一部分并且任选地包含允许抗原结合部分采用促进抗原结合蛋白与抗原结合的构型的构架或框架部分的蛋白质。抗原结合蛋白的实例包括人抗体、人源化抗体;嵌合抗体;重组抗体;单链抗体;双功能抗体;三功能抗体;四功能抗体;Fab片段;F(ab’)2片段;IgD抗体;IgE抗体;IgM抗体;IgG1抗体;IgG2抗体;IgG3抗体;或IgG4抗体以及其片段。抗原结合蛋白可以包括,例如具有移植的CDR或CDR衍生物的替代蛋白构架或人工构架。此类构架包括,但不限于:抗体来源的构架,其包含被引入以便例如使抗原结合蛋白的三维结构稳定的突变;以及完全合成的构架,其包含例如生物相容性聚合物。参见,例如,Korndorfer等,2003,Proteins:Structure,Function,and Bioinformatics,53(1):121-129(2003);Roque等,Biotechnol.Prog.20:639-654(2004)。此外,可使用肽抗体模拟物(“PAM”),也可使用基于抗体模拟物的构架,所述抗体模拟物利用纤连蛋白组分作为构架。
抗原结合蛋白可以具有例如天然存在的免疫球蛋白的结构。“免疫球蛋白”是四聚体分子。在天然存在的免疫球蛋白中,每个四聚体主要由两个相同的多肽链对组成,每对具有一个“轻”链(约25kDa)和一个“重”链(约50kDa-70kDa)。每条链的氨基端部分包括主要负责抗原识别的具有约100至110个或更多个氨基酸的可变区。每条链的羧基端部分限定了主要负责效应功能的恒定区。人轻链被分类为κ轻链和λ轻链。重链被分类为μ、δ、γ、α或ε,并且将抗体的同种型分别定义为IgM、IgD、IgG、IgA以及IgE。
天然存在的免疫球蛋白链展现出由三个高变区连接的相对保守的框架区(FR)的相同通用结构,所述高变区又称为互补决定区或CDR。从N端至C端,轻链和重链均包含结构域FR1、CDR1、FR2、CDR2、FR3、CDR3以及FR4。可根据Kabat等in Sequences of Proteins ofImmunological Interest,第5版,US Dept.of Health and Human Services,PHS,NIH,NIH出版号91-3242,(1991)的定义来进行每个结构域的氨基酸分配。按照要求,也可根据另选命名方案诸如Chothia的命名方案(参见,Chothia和Lesk,(1987)J.Mol.Biol.196:901-917;Chothia等,(1989)Nature 342:878-883或者Honegger和Pluckthun,(2001)J.Mol.Biol.309:657-670)重新定义CDR。
在本公开的上下文中,当离解数(KD)≤10-8M时,称抗原结合蛋白“特异性地结合”或“选择性地结合”其靶标抗原。当KD≤5x 10-9M时,抗体以“高亲和性”特异性地结合抗原,并且当KD≤5x 10-10M时,抗体以“极高亲和性”特异性地结合抗原。
除非另外说明,否则术语“抗体”包括对任何同种型或亚类的糖基化的和非糖基化的免疫球蛋白或与完整抗体竞争以特异性结合的其抗原结合区的提及。此外,除非另外说明,否则术语“抗体”是指完整免疫球蛋白或与完整抗体竞争以特异性结合的其抗原结合部分。抗原结合部分可以通过重组DNA技术或通过完整抗体的酶促裂解或化学裂解来产生,并且可形成目标蛋白的元件。抗原结合部分尤其包括Fab、Fab’、F(ab’)2、Fv、结构域抗体(dAb)、包括互补决定区(CDR)的片段、单链抗体(scFv)、嵌合抗体、双功能抗体、三功能抗体、四功能抗体以及含有足以赋予与多肽的特异性抗原结合的至少一部分免疫球蛋白的多肽。
Fab片段是具有VL、VH、CL和CH1结构域的单价片段;F(ab’)2是具有两个由铰链区二硫键连接的Fab片段的二价片段;Fd片段具有VH和CH1结构域;Fv片段具有抗体的单臂的VL和VH结构域;并且dAb片段具有VH结构域、VL结构域或者VH或VL结构域的抗原结合片段(美国专利号6,846,634、6,696,245、美国申请公布号05/0202512、04/0202995、04/0038291、04/0009507、03/0039958,Ward等,(1989)Nature 341:544-546)。
单链抗体(scFv)是VL区和VH区通过接头(例如,具有氨基酸残基的合成序列)连接以形成连续的蛋白质链的抗体,其中接头足够长以允许蛋白质链在其自身上向后折叠并形成单价抗原结合位点(参见,例如,Bird等,Science 242:423-26(1988)以及Huston等,(1988)Proc.Natl.Acad.Sci.USA 85:5879-83)。双功能抗体是包含两个多肽链的二价抗体,其中每个多肽链包含通过接头连接的VH结构域和VL结构域,所述接头太短以致不能允许同一链上的两个结构域之间成对,由此允许每个结构域与另一个多肽链上的互补结构域成对(参见,例如,Holliger等,(1993)Proc.Natl.Acad.Sci.USA 90:6444-48;以及Poljak等,(1994)Structure 2:1121-23)。如果双功能抗体的两个多肽链是相同的,那么由它们成对而产生的双功能抗体将具有两个相同的抗原结合位点。具有不同序列的多肽链可以周来制造具有两个不同抗原结合位点的双功能抗体。类似地,三功能抗体和四功能抗体是分别包含三个和四个多肽链并且分别形成三个和四个抗原结合位点的抗体,所述抗原结合位点可以是相同的或不同的。
可将一个或多个CDR共价地或非共价地并入到分子中以使所述分子成为抗原结合蛋白。抗原结合蛋白可以并入一个或多个CDR作为更大的多肽链的部分,可以使一个或多个CDR与另一个多肽链共价地连接,或可以非共价地并入一个或多个CDR。CDR容许抗原结合蛋白特异性地与特定的目标抗原结合。
抗原结合蛋白可以具有一个或多个结合位点。如果存在多于一个结合位点,则结合位点可以彼此相同或可以不同。例如,天然存在的人免疫球蛋白通常具有两个相同的结合位点,而“双特异性”或“双功能性”抗体具有两个不同的结合位点。
为了清楚的目的,并且如本文所述,应当指出抗原结合蛋白可但不必为人源的(例如,人抗体),并且在一些情况下将包括非人蛋白例如大鼠蛋白或小鼠蛋白,并且在其他情况下,抗原结合蛋白可包括人蛋白和非人蛋白的杂交种(例如,人源化抗体)。
目标蛋白可包括人抗体。术语“人抗体”包括具有一个或多个源自人免疫球蛋白序列的可变区和恒定区的所有抗体。在一个实施方案中,所有的可变结构域和恒定结构域均源自人免疫球蛋白序列(完全人抗体)。此类抗体可以各种方式(包括通过与小鼠的目标抗原的免疫作用)来制备,所述小鼠,诸如源自 UltiMabTM或
UltiMabTM或 系统的小鼠,被遗传修饰以表达源自人重链编码基因和/或轻链编码基因的抗体。还可采用基于噬菌体的方法。
系统的小鼠,被遗传修饰以表达源自人重链编码基因和/或轻链编码基因的抗体。还可采用基于噬菌体的方法。
另选地,目标蛋白可包括人源化抗体。“人源化抗体”具有因一个或多个氨基酸置换、缺失和/或添加而与源自非人物种的抗体的序列不同的序列,这样使得与非人物种抗体相比,当向人类受试者施用时人源化抗体不太可能诱导免疫应答和/或诱导不太严重的免疫应答。在一个实施方案中,非人物种抗体的重链和/或轻链的框架结构域和恒定结构域中的某些氨基酸被突变以产生人源化抗体。在另一个实施方案中,使来自人抗体的一个或多个恒定结构域与非人物种的一个或多个可变结构域融合。如何制得人源化抗体的实例可见于美国专利号6,054,297、5,886,152和5,877,293中。
如本文所用,“Fc”区包含两个重链片段,所述重链片段包含抗体的CH2和CH3结构域。两个重链片段通过两个或更多个二硫键且通过CH3结构域的疏水性相互作用结合在一起。包含Fc区的目标蛋白(包括抗原结合蛋白和Fc融合蛋白)形成了本公开的另一个方面。
“半抗体(hemibody)”是包含完整重链、完整轻链以及与完整重链的Fc区成对的第二重链Fc区的免疫功能性免疫球蛋白构建体。可但不必采用接头来连接重链Fc区和第二重链Fc区。在具体实施方案中,半抗体为本文所公开的抗原结合蛋白的单价形式。在其他实施方案中,可采用带电的残基对来使一个Fc区与第二Fc区缔合。在本公开的上下文中,半抗体可为目标蛋白。
术语“宿主细胞”意指已用或能够用核酸序列转化并且因此表达目标基因的细胞。所述术语包括亲本细胞的子代,无论子代在形态或基因组成方面是否与原始亲本细胞相同,只要存在目标基因即可。细胞培养物可包含一个或多个宿主细胞。
术语“杂交瘤”意指由永生化细胞与抗体产生细胞的融合而产生的细胞或细胞的子代。产生的杂交瘤是产生抗体的永生化细胞。用来创建杂交瘤的单个细胞可来自任何哺乳动物源,包括但不限于仓鼠、大鼠、猪、兔、绵羊、山羊和人。所述术语还包括当异杂交骨髓瘤融合时产生的三源杂交瘤细胞系,三源杂交瘤细胞系是人细胞和小鼠骨髓瘤细胞系之间的融合的产物,随后与浆细胞融合。所述术语旨在包括产生抗体的任何永生化杂交细胞系,例如像四源杂交瘤(参见,例如,Milstein等,(1983)Nature,537:3053)。
术语“培养物”和“细胞培养物”可互换使用,并且是指在培养基中在适于细胞群的存活和/或生长的条件下维持的细胞群。本领域技术人员将清楚,这些术语是指包含细胞群和所述群所悬浮于的培养基的组合。
术语“多肽”和“蛋白质”(例如,如在目标蛋白或目标多肽的上下文中所用)在本文可互换使用以指氨基酸残基的聚合物。所述术语还适用于一个或多个氨基酸残基是相应天然存在的氨基酸的类似物或模拟物的氨基酸聚合物以及适用于天然存在的氨基酸聚合物。所述术语还可包括已被修饰(例如,通过添加碳水化合物残基以形成糖蛋白)或被磷酸化的氨基酸聚合物。多肽和蛋白质可由天然存在的和非重组的细胞产生,或者多肽和蛋白质可由基因工程改造的或重组的细胞产生。多肽和蛋白质可包含具有天然蛋白质的氨基酸序列的分子,或者具有缺失、添加和/或置换的一个或多个氨基酸的天然序列的分子。
术语“多肽”和“蛋白质”包括只包含天然存在的氨基酸的分子,以及包含非天然存在的氨基酸的分子。非天然存在的氨基酸的实例(按照要求,其可置换存在于本文公开的任何序列中的任何天然存在的氨基酸)包括:4-羟脯氨酸、γ-羧基谷氨酸、ε-N,N,N-三甲基赖氨酸、ε-N-乙酰赖氨酸、O-磷酸丝氨酸、N-乙酰丝氨酸、N-甲酰甲硫氨酸、3-甲基组氨酸、5-羟赖氨酸、σ-N-甲基精氨酸及其他类似的氨基酸和亚氨基酸(例如,4-羟脯氨酸)。根据标准用法和惯例,在本文所用的多肽符号中,左手方向是氨基末端方向且右手方向是羧基末端方向。
可被插入蛋白质或多肽序列或者置换蛋白质或多肽序列中的野生型残基的非天然存在的氨基酸的实例的非限制性列表包括β-氨基酸、高氨基酸、环状氨基酸以及具有衍生侧链的氨基酸。实例包括(呈L-型或D-型;如括号中缩写的):瓜氨酸(Cit)、高瓜氨酸(hCit)、Nα-甲基瓜氨酸(NMeCit)、Nα-甲基高瓜氨酸(Nα-MeHoCit)、鸟氨酸(Orn)、Nα-甲基鸟氨酸(Nα-MeOrn或NMeOrn)、肌氨酸(Sar)、高赖氨酸(hLys或hK)、高精氨酸(hArg或hR)、高谷氨酰胺(hQ)、Nα-甲基精氨酸(NMeR)、Nα-甲基亮氨酸(Na-MeL或NMeL)、N-甲基高赖氨酸(NMeHoK)、Nα-甲基谷氨酰胺(NMeQ)、正亮氨酸(Nle)、正缬氨酸(Nva)、1,2,3,4-四氢异喹啉(Tic)、八氢吲哚-2-羧酸(Oic)、3-(1-萘基)丙氨酸(1-Nal)、3-(2-萘基)丙氨酸(2-Nal)、1,2,3,4-四氢异喹啉(Tic)、2-茚满基甘氨酸(IgI)、对-吲哚苯丙氨酸(pI-Phe)、对-氨基苯丙氨酸(4AmP或4-氨基-Phe)、4-胍基苯丙氨酸(Guf)、甘氨酰赖氨酸(缩写“K(Nε-甘氨酰)”或“K(甘氨酰)”或“K(gly)”)、硝基苯丙氨酸(硝基phe)、氨基苯丙氨酸(氨基phe或氨基-Phe)、苄基苯丙氨酸(苄基phe)、γ-羧基谷氨酸(γ-羧基glu)、羧脯氨酸(羟基pro)、对羧基-苯丙氨酸(Cpa)、α-氨基己二酸(Aad)、Nα-甲基缬氨酸(NMeVal)、N-α-甲基亮氨酸(NMeLeu)、Nα-甲基正亮氨酸(NMeNle)、环戊基甘氨酸(Cpg)、环己基甘氨酸(Chg)、乙酰精氨酸(乙酰arg)、α,β-二氨基丙酸(Dpr)、α,γ-二氨基丁酸(Dab)、二氨基丙酸(Dap)、环己基丙氨酸(Cha)、4-甲基-苯丙氨酸(MePhe)、β,β-二苯基-丙氨酸(BiPhA)、氨基丁酸(Abu)、4-苯基-苯丙氨酸(或二苯基丙氨酸;4Bip)、α-氨基-异丁酸(Aib)、β-丙氨酸、β-氨基丙酸、哌啶酸、氨基己酸、氨基庚酸、氨基庚二酸、锁链素、二氨基庚二酸、N-乙基甘氨酸、N-乙基天冬酰胺、羟赖氨酸、别-羟赖氨酸、异锁链素、别-异亮氨酸、N-甲基甘氨酸、N-甲基异亮氨酸、N-甲基缬氨酸、4-羟脯氨酸(Hyp)、γ-羧基谷氨酸、ε-N,N,N-三甲基赖氨酸、ε-N-乙酰赖氨酸、O-磷酸丝氨酸、N-乙酰丝氨酸、N-甲酰甲硫氨酸、3-甲基组氨酸、5-羟赖氨酸、ω-甲基精氨酸、4-氨基-O-邻苯二甲酸(4APA)以及其他类似氨基酸和明确列出的那些氨基酸中的任一种的衍生形式。
“细胞培养”或“培养”指的是多细胞生物或组织外的细胞的生长和增殖。用于哺乳动物细胞的适合的培养条件是本领域已知的。参见,例如,Animal cell culture:APractical Approach,D.Rickwood,编,Oxford University Press,New York(1992)。哺乳动物细胞可在悬浮液中或当附着到固体基质上时培养。可使用具有或没有微载体的流化床生物反应器、中空纤维生物反应器、转瓶、摇瓶或搅拌釜生物反应器。在一个实施方案中,使用500L至2000L生物反应器。在一个实施方案中,使用1000L至2000L生物反应器。
术语“细胞培养基”(也称为“培养基”、“细胞培养基(cell culture media)”、“组织培养基”)是指用于使细胞例如动物或哺乳动物细胞生长的任何营养液,并且其通常提供来自以下的至少一种或多种组分:能量来源(通常呈碳水化合物的形式,诸如葡萄糖);所有必需氨基酸中的一种或多种,并且通常为20种基本氨基酸,外加半胱氨酸;通常以低浓度需要的维生素和/或其他有机化合物;脂质或游离脂肪酸;以及微量元素,例如通常以极低的浓度(常在微摩尔浓度范围内)需要的无机化合物或天然存在的元素。
取决于要培养的细胞的需要和/或所需的细胞培养参数,所述营养液可任选地增补有附加的任选组分以使细胞生长优化,诸如激素和其他生长因子例如胰岛素、转铁蛋白、表皮生长因子、血清等;盐类,例如钙、镁和磷酸盐以及缓冲液,例如HEPES;核苷和碱基,例如腺苷、胸苷、次黄嘌呤;以及蛋白质和组织水解产物,例如水解的动物蛋白或植物蛋白(蛋白胨或蛋白胨混合物,其可获得自动物副产品、精制明胶或植物物质);抗生素,例如庆大霉素;细胞保护剂或表面活性剂,诸如 F68(还被称为
F68(还被称为 F68和
F68和 P188;非离子三嵌段共聚物,其由侧接聚氧化乙烯(聚环氧乙烷)的两个亲水链的聚氧化丙烯(聚环氧丙烷)的中央疏水链构成);聚胺,例如腐胺、亚精胺和精胺(参见,例如,WIPO公布号WO 2008/154014)以及丙酮酸盐(参见,例如,美国专利号8053238)。
P188;非离子三嵌段共聚物,其由侧接聚氧化乙烯(聚环氧乙烷)的两个亲水链的聚氧化丙烯(聚环氧丙烷)的中央疏水链构成);聚胺,例如腐胺、亚精胺和精胺(参见,例如,WIPO公布号WO 2008/154014)以及丙酮酸盐(参见,例如,美国专利号8053238)。
细胞培养基包括通常在任何细胞培养方法中采用的和/或已知用于任何细胞培养方法的那些,诸如但不限于细胞的分批培养、延长分批培养、补料分批培养和/或灌流培养或连续培养。
“基础”(或分批)细胞培养基是指通常用来启动细胞培养,并且足够完全支持细胞培养的细胞培养基。
“生长”细胞培养基是指通常用于指数生长时期(生长期)期间的细胞培养中并且足够完全支持此阶段期间的细胞培养的细胞培养基。生长细胞培养基还可含有赋予对并入宿主细胞系内的选择标记的抗性或存活性的选择剂。此类选择剂包括但不限于遗传霉素(G4118)、新霉素、潮霉素B、嘌呤霉素、博莱霉素、蛋氨酸亚砜亚胺、氨甲蝶呤、无谷氨酰胺细胞培养基、缺乏甘氨酸、次黄嘌呤和胸苷或单独缺乏胸苷的细胞培养基。
“生产”细胞培养基是指通常在指数生长将要结束且蛋白生产接管“过渡”期和/或“产物”期时用于过渡期间的细胞培养中并且足够完全维持此阶段期间所需的细胞密度、细胞活力和/或产物效价的细胞培养基。
“灌流”细胞培养基是指通常用于通过灌流培养或连续培养方法维持的细胞培养中,并且足够完全支持此过程期间的细胞培养的细胞培养基。灌流细胞培养基制剂可更为富集或比基础细胞培养基制剂更为浓缩以适应用来移除废培养基的方法。灌流细胞培养基可在生长期和生产期期间使用。
浓缩细胞培养基可含有一些或所有维持细胞培养所需的营养物;特别是可含有鉴定为或已知是在细胞培养的生产期过程期间消耗的营养物的浓缩培养基。浓缩培养基可基于几乎任何细胞培养基制剂。此类浓缩补料培养基可含有例如约2X、3X、4X、5X、6X、7X、8X、9X、10X、12X、14X、16X、20X、30X、50X、100x、200X、400X、600X、800X乃至1000X它们的正常量的细胞培养基的一些或所有组分。
用来制备细胞培养基的组分可完全地碾磨成粉末培养基制剂;部分地与液体增补物一起碾磨,按需添加到细胞培养基中;或以完全液体形式向细胞培养物中添加。
细胞培养物还可增补有可能难以制备或在细胞培养中快速耗乏的特定营养物的独立浓缩补料。此类营养物可为氨基酸,诸如酪氨酸、半胱氨酸和/或胱氨酸(参见,例如,WIPO公布号2012/145682)。在一个实施方案中,独立地向在含有酪氨酸的细胞培养基中生长的细胞培养物中补料浓缩的酪氨酸溶液,以使酪氨酸在细胞培养物中的浓度不超过8mM。在另一个实施方案中,独立地向在缺乏酪氨酸、胱氨酸或半胱氨酸的细胞培养基中生长的细胞培养物中补料酪氨酸和胱氨酸的浓缩溶液。所述独立补料可在生产期之前或生产期开始时开始。所述独立补料可在与浓缩补料培养基相同或不同的日子通过细胞培养基的分批补料来实现。所述独立补料还可在与灌流培养基相同或不同的日子进行灌流。
“无血清”应用于不含动物血清诸如胎牛血清的细胞培养基中。在其他情况中,各种组织培养基(包括确定的培养基)是可商购获得的,例如,可使用下列细胞培养基中的任何一种或组合:RPMI-1640培养基、RPMI-1641培养基、Dulbecco氏改良的Eagle氏培养基(DMEM)、最低必需Eagle培养基、F-12K培养基、Ham氏F12培养基、Iscove氏改良的Dulbecco氏培养基、McCoy氏5A培养基、Leibovitz氏L-15培养基以及无血清培养基,诸如EX-CELLTM300系列(JRHBiosciences,Lenexa,Kansas)。此类培养基的无血清版也是可获得的。取决于要培养的细胞的需要和/或所需的细胞培养参数,细胞培养基可增补有附加的或增加浓度的组分,诸如氨基酸、盐类、糖类、维生素、激素、生长因子、缓冲液、抗生素、脂类、微量元素等。
术语“生物反应器”意指用于细胞培养物的生长的任何容器。本公开的细胞培养物可在生物反应器中生长,可基于由生长于所述生物反应器中的细胞产生的目标蛋白的应用而选择所述生物反应器。生物反应器可为任何尺寸,只要它对细胞培养有用;通常,生物反应器的尺寸适于在其之内生长的细胞培养物的容积。通常,生物反应器会是至少1升,并且可为2、5、10、50、100、200、250、500、1,000、1500、2000、2,500、5,000、8,000、10,000、12,000升或更多或在中间的任何容积。可在培养期期间控制生物反应器的内部条件,包括但不限于pH和温度。本领域技术人员将知道并且将能够基于相关考虑而选择用于实践本发明的适合的生物反应器。
“细胞密度”是指给定容积的培养基中的细胞数量。“活细胞密度”是指如通过标准活力测定(诸如台盼蓝染料排斥方法)所确定的给定容积的培养基中的活细胞数量。
术语“细胞活力”意指培养中的细胞在给定组的培养条件或实验变量下生存的能力。所述术语还指在当时的培养中在特定时间相对于总数的细胞(活的和死的)活着的细胞的部分。
“细胞压积”(PCV),也称为“细胞压积百分比”(%PCV),是细胞所占容积与细胞培养物的总容积的比率,表示为百分比(参见,Stettler等,(2006)Biotechnol Bioeng.Dec20:95(6):1228-33)。细胞压积是细胞密度和细胞直径的函数;细胞压积的增加可由细胞密度或细胞直径或两者的增加而引起。细胞压积是细胞培养物中固体含量的量度。在收获和下游纯化期间去除固体。更多的固体意味着在收获和下游纯化步骤期间从所需产物中分离所述固体物料的更多的努力。另外,在收获过程期间所需产物可困于所述固体中并损失,导致产物收率减少。因为宿主细胞大小不同并且细胞培养物还可含有死的和垂死的细胞以及其他细胞碎片,所以细胞压积比起细胞密度或活细胞密度是描述细胞培养物内固体含量的更加精确的方式。例如,取决于细胞大小,具有50x106个细胞/ml的细胞密度的2000L培养物将具有迥然不同的细胞压积。此外,一些细胞当处于生长停滞状态时将在大小方面增加,因此由于细胞大小增加而导致生物量增加,使得在生长停滞和后生长停滞之前的细胞压积很可能会不同。
“生长停滞”,其还被称为“细胞生长停滞”,是细胞在数量上停止增加或当细胞周期不再进行时的时刻。可通过测定细胞培养物的活细胞密度来监控生长停滞。处于生长停滞状态的一些细胞可在大小而非数量方面增加,因此生长停滞的培养物的细胞压积可能增加。如果细胞不处于健康下降中,那么可通过反转导致生长停滞的条件来在某种程度上反转生长停滞。
术语“效价”意指由给定量的培养基体积中的细胞培养物产生的目标多肽或目标蛋白(其可为天然存在的或重组的目标蛋白)的总量。效价可以每毫升(或其他体积的量度)培养基毫克或微克多肽或蛋白质的单位表示。“累积效价”是由细胞在培养过程期间产生的效价,并且可例如通过测量每日效价并利用这些值来计算累积效价而确定。
术语“补料分批培养”是指悬浮培养的形式,并且意指继培养过程开始之后一次或多次向培养物中提供附加组分的培养细胞的方法。提供的组分通常包含已在培养过程期间耗乏的用于细胞的营养补充剂。此外或可选地,附加组分可包括补充性组分(例如,细胞周期抑制性化合物)。通常在某点处停止补料分批培养,并且将培养基中的细胞和/或组分收获并任选地纯化。
术语“综合活细胞密度”或“IVCD”可互换使用,并且意指培养过程中的平均活细胞密度乘以培养已运行的时间量。
通过两个时间点之间的平均活细胞密度(VCD)乘以这两个时间点之间的持续时间来计算“累积活细胞密度”(CVCD)。CVCD是由VCD对时间绘图形成的曲线下的面积。
细胞培养过程的描述
在重组蛋白产生期间,期望具有使细胞生长至所需密度的可控系统,然后使细胞的生理状态转换至细胞利用能量和底物来产生目标重组蛋白而不是形成更多细胞的生长停滞的高生产率状态。用于到达这个目标的各种方法存在并且包括温度变化和氨基酸饥饿以及使用细胞周期抑制剂或可停滞细胞生长而不引起细胞死亡的其他分子。
重组蛋白的生产开始于在培养皿、培养瓶、培养试管、生物反应器或其他适合的容器中建立表达蛋白质的哺乳动物细胞生产型细胞培养物。通常使用较小的生产型生物反应器,在一个实施方案中,所述生物反应器为500L至2000L。在另一个实施方案中,使用1000L-2000L生物反应器。用来接种生物反应器的种子细胞密度可对产生的重组蛋白水平具有积极影响。在一个实施方案中,用无血清培养基中的至少0.5x106个活细胞/mL至和超过3.0x106个活细胞/mL接种生物反应器。在另一个实施方案中,接种为1.0x106个活细胞/mL。
哺乳动物细胞然后经受指数生长期。可在没有补充性补料的情况下维持细胞培养物直至达到所需细胞密度。在一个实施方案中,在有或没有补充性补料的情况下维持细胞培养物持续多达三天。在另一个实施方案中,可按照所需细胞密度接种培养物以开始生产期,而没有简短的生长期。在本文的任何实施方案中,还可通过任何前述的方法来启动生长期至生产期的转换。
在生长期与生产期之间的过渡处以及在生产期期间,细胞压积百分比(%PCV)等于或小于35%。生产期期间维持的所需细胞压积等于或小于35%。在一个实施方案中,所述细胞压积等于或小于30%。在另一个实施方案中,所述细胞压积等于或小于20%。在又一个实施方案中,所述细胞压积等于或小于15%。在再一个实施方案中,所述细胞压积等于或小于10%。
在生长期与生产期之间的过渡处的和在生产期期间维持的所需活细胞密度可根据项目而不同。它可基于来自历史数据的当量细胞压积来确定。在一个实施方案中,所述活细胞密度为至少约10x106个活细胞/mL至80x106个活细胞/mL。在一个实施方案中,所述活细胞密度为至少约10x106个活细胞/mL至70x106个活细胞/mL。在一个实施方案中,所述活细胞密度为至少约10x106个活细胞/mL至60x106个活细胞/mL。在一个实施方案中,所述活细胞密度为至少约10x106个活细胞/mL至50x106个活细胞/mL。在一个实施方案中,所述活细胞密度为至少约10x106个活细胞/mL至40x106个活细胞/mL。在另一个实施方案中,所述活细胞密度为至少约10x106个活细胞/mL至30x106个活细胞/mL。在另一个实施方案中,所述活细胞密度为至少约10x106个活细胞/mL至20x106个活细胞/mL。在另一个实施方案中,所述活细胞密度为至少约20x106个活细胞/mL至30x106个活细胞/mL。在另一个实施方案中,所述活细胞密度为至少约20x106个活细胞/mL至至少约25x106个活细胞/mL,或者至少约20x106个活细胞/mL。
生产期期间较低的细胞压积有助于缓解可阻碍更高细胞密度灌流培养的溶解氧鼓泡问题。较低的细胞压积还允许较小的培养基体积,所述较小的培养基体积允许使用较小的培养基储存容器并且可与较小流速组合。与较高的细胞生物量培养物相比,较小的细胞压积还对收获和下游处理具有较小影响。所有这些都降低了与生产重组蛋白治疗剂相关的成本。
通常在用于通过哺乳动物细胞培养来生产重组蛋白的商业过程中使用三种方法:分批培养、补料分批培养和灌流培养。分批培养是使细胞在固定体积的培养基中生长持续较短时间段随后全部收获的不连续的方法。利用分批法生长的培养物经历细胞密度的增加直至达到最大细胞密度,随后随着培养基组分消耗和代谢副产物(诸如乳酸盐和氨)的水平积累,活细胞密度下降。通常收获发生在当达到最大细胞密度(取决于培养基制剂、细胞系等,通常为5x106个细胞/mL-10x106个细胞/mL)时的时刻。分批法是最简单的培养方法,然而活细胞密度受养分有效性的限制,并且一旦细胞处于最大密度,培养物就衰退且生产就减少。不存在延长生产期的能力,因为废物积累和营养耗乏快速地导致了培养物衰退,(通常大约3天至7天)。
补料分批培养通过提供灌注或连续培养基补料以补充已消耗的那些培养基组分来改进分批法。因为补料分批培养物在整个操作过程中始终接受附加的营养物,所以当与分批法相比时它们具有达到较高的细胞密度(取决于培养基制剂、细胞系等,>10x106个细胞/ml至30x106个细胞/ml))和增加的产物效价的潜力。不同于分批法,双相培养可通过操控补料策略和培养基制剂来建立和维持,以区分达成所需细胞密度(生长期)的细胞增殖时期与暂停的或缓慢的细胞生长时期(生产期)。如此,与分批培养相比,补料分批培养具有达到较高产物效价的潜力。通常,在生长期使用分批法并且在生产期使用补料分批法,但是可在整个过程中使用补料分批补料策略。然而,不同于分批法,生物反应器容积是限制补料量的限制因素。另外,如同分批法一样,代谢副产物积累会导致培养物衰退,这限制了生产期的持续时间,约1.5周至3周。补料分批培养是不连续的,并且收获通常在代谢副产物水平或培养物活力达到预定水平时发生。当与没有补料发生的分批培养相比时,补料分批培养可产生更大量的重组蛋白。参见,例如,美国专利号5,672,502。
灌流法通过添加新鲜培养基并同时移除废培养基而提供了优于分批法和补料分批法的潜在改进。典型的大规模商业细胞培养策略努力达到几乎三分之一至超过二分之一的反应器容积为生物量的高细胞密度,60x106个细胞/mL-90(+)x106个细胞/mL。用灌流培养已达到了>1x108个细胞/mL的极限细胞密度,并且甚至预测了更高的密度。典型的灌流培养开始于持续一天或两天的分批培养启动,随后在培养的整个生长期和生产期中向培养物中连续、逐步和/或间歇添加新鲜补料培养基并同时移除废培养基并保留细胞和另外的高分子量化合物诸如蛋白质(基于滤器分子量截断值)。诸如沉降、离心或过滤的各种方法可用来移除废培养基,同时维持细胞密度。已报告了每天一部分工作容积至每天许多多个工作容积的灌流流速。
灌流法的优势为比起分批培养方法或补料分批培养方法,生产培养可维持较长期间。然而,需要增加的培养基制备、使用、存储和处理以支持长期灌流培养,特别是具有高细胞密度的那些灌流培养,其还需要甚至更多的营养物,并且与分批法或补料分批法相比,这一切驱使生产成本甚至更高。此外,更高的细胞密度可引起生产期间的问题,诸如维持溶解氧水平和具有增加的放气的问题,其包括提供更多氧气并去除更多二氧化碳,这将导致更多发泡和对消泡剂策略的改变的需要;以及收获和下游处理期间的问题,其中去除过多的细胞材料所需的努力可导致产物损失,由于增加的细胞团块而否定了增加的效价的效益。
还提供组合了生长期期间的补料分批补料随后是生产期期间的连续灌流的大规模细胞培养策略。所述方法把以小于或等于35%细胞压积维持细胞培养的生产期作为目标。
在一个实施方案中,在生长期期间使用具有灌注补料的补料分批培养来维持细胞培养。然后可在生产期期间使用灌流补料。在一个实施方案中,灌流在细胞已达到生产期时开始。在另一个实施方案中,灌流开始于细胞培养的第3天或大约第3天至第9天或大约第9天。在另一个实施方案中,灌流开始于细胞培养的第5天或大约第5天至第7天或大约第7天。
生长期期间使用灌注补料允许细胞过渡至生产期中,引起了作为启动并控制生产期的手段的对温度变化更少的依赖性,然而36℃至31℃的温度变化可在生长期与生产期之间发生。在一个实施方案中,变化为36℃至33℃。在另一个实施方案中,可通过使补料分批培养物暴露于细胞周期抑制剂中来启动细胞生长停滞在补料分批培养中的启动。在另一个实施方案中,可通过用包含细胞周期抑制剂的无血清灌流培养基灌流来实现细胞生长停滞在补料分批培养中的启动。
如本文所述,可用至少0.5x106个活细胞/mL至和超过3.0x106个活细胞/mL在无血清培养基中接种生物反应器,例如1.0x106个活细胞/mL。
灌流培养是细胞培养物接受新鲜灌流补料培养基并同时移除废培养基的一种培养。灌流可为连续的、逐步的、间歇的或者这些中的任何的任一种或全部的组合。灌流速率可为每天小于一个工作容积至每天许多个工作容积。细胞在培养物中保留并且移除的废培养基基本不含细胞或比所述培养物具有显著较少的细胞。由细胞培养物表达的重组蛋白也可在培养物中保留。灌流可通过包括离心、沉降或过滤的若干手段来实现,参见,例如,Voisard等,(2003),Biotechnology and Bioengineering 82:751-65。过滤法的一个实例为交替式切向流过滤。交替式切向流通过将培养基泵送通过中空纤维滤器组件来维持。参见,例如,美国专利号6,544,424;Furey(2002)Gen.Eng.News.22(7),62-63。
“灌流流速”是在给定时间内从生物反应器穿过(添加和移除)的培养基的量,通常表现为一部分或多倍的工作容积。“工作容积”是指用于细胞培养的生物反应器容积的量。在一个实施方案中,所述灌流流速为每天一个工作容积或更少。可配制灌流补料培养基以使灌流营养浓度最大化,使灌流速率最小化。
可用含有在细胞培养的生产期过程期间消耗的组分诸如营养物和氨基酸的浓缩补料培养基增补细胞培养物。浓缩补料培养基可基于几乎任何细胞培养基制剂。这种浓缩补料培养基可含有例如约5X、6X、7X、8X、9X、10X、12X、14X、16X、20X、30X、50X、100x、200X、400X、600X、800X乃至1000X它们的正常量的细胞培养基的大部分组分。浓缩补料培养基经常用于补料分批培养过程中。
根据本发明的方法可用来在多阶段培养过程中改进重组蛋白的生产。在多阶段过程中,以两个或更多个不同阶段培养细胞。例如,细胞可首先以一个或多个生长期在使细胞增殖和活力最大化的环境条件下培养,然后在使蛋白质生产最大化的条件下转移至生产期。在用于通过哺乳动物细胞生产蛋白质的商业过程中,在最终生产培养之前,通常有存在于不同培养容器中的多个(例如,至少约2、3、4、5、6、7、8、9或10个)生长期。
一个或多个过渡期可先于或隔开生长期和生产期。在多阶段过程中,可至少在商业细胞培养最终生产期的生长期和生产期期间采用根据本发明的方法,然而它还可在前生长期中采用。可以大规模进行生产期。可以至少约100、500、1000、2000、3000、5000、7000、8000、10,000、15,000、20,000升的容积进行大规模过程。在一个实施方案中,生产在500L、1000L和/或2000L生物反应器中进行。
生长期可在比生产期更高的温度下存在。例如,生长期可在约35℃至约38℃的第一温度下存在,并且生产期可在约29℃至37℃,任选地约30℃至约36℃或约30℃至约34℃的第二温度下存在。此外,可在温度变化的同时、之前和/或之后添加蛋白生产的化学诱导物,例如像咖啡因、丁酸盐和六亚甲基二乙酰胺(HMBA)。如果在温度变化后添加诱导物,它们可在温度变化后一小时至五天,任选地在温度变化后一至两天添加。当细胞产生所需蛋白质时,细胞培养物可维持数天乃至数周。
可使用本领域已知的任何分析技术监控并评估来自细胞培养物的样品。可在培养期间监控包括重组蛋白和培养基质量和特征的多种参数。可以合意的频率间歇地获取并监控样品,包括实时或近实时的连续监控。
通常,最终生产培养物(N-x to N-1)之前的细胞培养物用来产生将用来接种生产型生物反应器(N-1培养物)的种子细胞。种子细胞密度可对产生的重组蛋白水平具有积极影响。产物水平倾向于随着增加种子密度而升高。效价的改善不但受制于更高的种子密度,而且很可能受制于生产中的细胞的代谢状态和细胞周期状态的影响。
可通过任何培养方法来生产种子细胞。一个这样的方法是使用交替式切向流过滤的灌流培养。可使用交替式切向流过滤操作N-1生物反应器以提供高密度的细胞来接种生产型生物反应器。N-1阶段可用来使细胞生长至>90x 106个细胞/mL的密度。N-1生物反应器可用来产生灌注种子培养物或可用作滚种储备培养物(rolling seed stock culture),可维持所述滚种储备培养物来以高种子细胞密度接种多生产型生物反应器。生产的生长阶段持续时间的范围可为7至14天,并且可被设计以便在接种生产型生物反应器之前使细胞维持指数生长。优化灌流速率、培养基制剂和定时以使细胞生长并将他们以最有益于优化他们的生产的状态递送至生产型生物反应器。对于接种生产型生物反应器,可达到>15x106个细胞/mL的种子细胞密度。接种时更高的种子细胞密度可减少乃至消除达到所需生产密度所需的时间。
本发明发现了在调控重组蛋白糖基化的存在和/或量方面的特定效用。本发明所用的细胞系(也称为“宿主细胞”)经过基因工程改造以表达具有商业或科学价值的多肽。细胞系通常从起源于原代培养物的谱系得到,所述原代培养物可在培养中维持无限时间。基因工程改造的细胞系涉及用重组多核苷酸分子转染、转化或转导所述细胞和/或另外的改变(例如,通过同源重组和基因活化或重组细胞与非重组细胞的融合),以便引起宿主细胞表达所需重组多肽。用于基因工程改造细胞和/或细胞系以表达目标多肽的方法和载体为本领域技术人员所熟知;例如,Current Protocols in Molecular Biology,Ausubel等,编.(Wiley和Sons,New York,1988,和季刊更新);Sambrook等,Molecular Cloning:ALaboratory Manual(Cold Spring Laboratory Press,1989);Kaufman,R.J.,Large ScaleMammalian Cell Culture,1990,pp.15-69阐明了各种技术。
动物细胞系来源于祖细胞是来源于多细胞动物的细胞。一种动物细胞系为哺乳动物细胞系。适于在培养中生长的多种多样的哺乳动物细胞系购自美国种质保存中心(Manassas,Va.)和商业供应商。工业中常用的细胞系的实例包括VERO、BHK、HeLa、CV1(包括Cos)、MDCK、293、3T3、骨髓瘤细胞系(例如,NSO、NS1)、PC12、WI38细胞和中国仓鼠卵巢(CHO)细胞。CHO细胞广泛用于生产复合重组蛋白,例如,细胞因子、凝血因子和抗体(Brasel等(1996),Blood88:2004-2012;Kaufman等(1988),J.Biol Chem 263:6352-6362;McKinnon等(1991),J Mol Endocrinol 6:231-239;Wood等(1990),J.Immunol.145:3011-3016)。二氢叶酸还原酶(DHFR)缺陷型突变细胞系(Urlaub等(1980),Proc Natl Acad Sci USA 77:4216-4220)、DXB11和DG-44是合意的CHO宿主细胞系,因为有效的DHFR可选择和可扩增基因表达系统允许这些细胞中的高水平重组蛋白表达(Kaufman R.J.(1990),Meth Enzymol185:537-566)。此外,这些细胞易于操控作为贴壁培养物或悬浮培养物并且显示出相对良好的遗传稳定性。CHO细胞和在它们中重组表达的蛋白已被广泛表征并且已被监管机构批准用于临床商业生产中。
在另一方面,本发明提供已引入重组表达载体的宿主细胞。宿主细胞可为任何原核细胞(例如大肠杆菌(E.coli))或真核细胞(例如酵母、昆虫或哺乳动物细胞(例加,CHO细胞))。可通过常规转化或转柒技术将载体DNA引入原核或真核细胞中。很多转染方法是本领域已知的,并且包括了使用脂质(例如, )、磷酸钙、阳离子聚合物、DEAE-葡聚糖、活化的树状聚合物和磁珠。另外的转染方法利用基于仪器的技术。实例包括电穿孔、生物射弹技术、显微注射和激光转染/光注射,所述激光转染/光注射使用光(例如,激光)以将核酸引入宿主细胞中。
)、磷酸钙、阳离子聚合物、DEAE-葡聚糖、活化的树状聚合物和磁珠。另外的转染方法利用基于仪器的技术。实例包括电穿孔、生物射弹技术、显微注射和激光转染/光注射,所述激光转染/光注射使用光(例如,激光)以将核酸引入宿主细胞中。
对于哺乳动物细胞的稳定转染,众所周知,根据使用的表达载体和转染技术,仅一小部分细胞可将外源DNA整合至它们的基因组中。为了鉴定和选择这些整合体,通常将编码选择标记(例加,对于对抗生素的抗性)的基因连同目标基因一起引入宿主细胞中。优选的选择标记包括赋予对药物诸如G418、潮霉素和氨甲蝶呤的抗性的那些选择标记。在其他方法中,可通过药物选择鉴定稳定转染了引入的核酸的细胞(例加,已并入选择标记基因的细胞将存活,而其它细胞死亡)。
目标蛋白
本发明的方法可用来培养表达目标重组蛋白的细胞。表达的重组蛋白可分泌至培养基中,可从所述培养基中回收和/或收集它们。此外,可利用已知的方法和从商业供应商获得的产品来纯化或部分地纯化来自此类培养物或组分(例如,来自培养基)的蛋白质。然后可将纯化的蛋白质“配制”(意指缓冲液交换)、灭菌、成批包装和/或包装以用于最终用户。用于药物组合物的适合制剂包括在Remington′s Pharmaceutical Sciences,第18版.1995,Mack Publishing Company,Easton,PA中描述的那些制剂。
可用本发明方法来生产的多肽的实例包括了包含与以下蛋白质之一的全部或部分相同或基本相似的氨基酸序列的蛋白质:肿瘤坏死因子(TNF)、flt3配体(WO 94/28391)、促红细胞生成素(erythropoeitin)、促血小板生成素(thrombopoeitin)、降钙素、IL-2、血管生成素-2(Maisonpierre等(1997),Science 277(5322):55-60)、NF-κ B的受体激活物的配体(RANKL,WO 01/36637)、肿瘤坏死因子(TNF)相关的凋亡诱导配体(TRAIL,WO 97/01633)、胸腺基质淋巴细胞生成素、粒细胞集落刺激因子、粒细胞-巨噬细胞集落刺激因子(GM-CSF,澳大利亚专利号588819)、肥大细胞生长因子、干细胞生长因子(美国专利号6,204,363)、表皮生长因子、角质化细胞生长因子、巨核细胞(megakaryote)生长和发育因子、RANTES、人纤维蛋白原样2蛋白(FGL2;NCBI登录号NM_00682;Rüegg和Pytela(1995),Gene160:257-62)、生长激素、胰岛素、促胰岛素、胰岛素样生长因子、甲状旁腺素、包括α-干扰素、γ-干扰素和共有序列干扰素(consensus interferon)的干扰素(美国专利号4,695,623和4,897471)、神经生长因子、脑源性神经营养因子、突触结合蛋白样蛋白(SLP 1-5)、神经营养因子-3、胰高血糖素、白细胞介素、集落刺激因子、淋巴毒素-B、白血病抑制因子和制瘤素-M。对可根据本发明方法生产的蛋白质的描述可见于例如Human Cytokines:Handbook for Basic and Clinical Research,全册(Aggarwal和Gutterman,编BlackwellSciences,Cambridge,MA,1998);Growth Factors:A Practical Approach(McKay和Leigh,编,Oxford University Press Inc.,New York,1993);以及The Cytokine Handbook,第1 册和第2册(Thompson和Lotze编,Academic Press,San Diego,CA,2003)中。
此外,本发明的方法将用来生产包含任何上述蛋白质的受体的全部或部分氨基酸序列的蛋白质、此类受体或任何上述蛋白质的拮抗剂和/或基本类似于此类受体或拮抗剂的蛋白质。这些受体和拮抗剂包括:两种形式的肿瘤坏死因子受体(TNFR,也称为p55和p75,美国专利号5,395,760和美国专利号5,610,279)、白细胞介素-1(IL-1)受体(I型和II型;欧洲专利号0460846、美国专利号4,968,607和美国专利号5,767,064,)、IL-1受体拮抗剂(美国专利号6,337,072)、IL-1拮抗剂或抑制剂(美国专利号5,981,713、6,096,728和5,075,222)、IL-2受体、IL-4受体(欧洲专利号0 367 566和美国专利号5,856,296)、IL-15受体、IL-17受体、IL-18受体、Fc受体、粒细胞-巨噬细胞集落刺激因子受体、粒细胞集落刺激因子受体、制瘤素-M的受体和白血病抑制因子的受体、NF-κ B的受体激活物(RANK,WO 01/36637和美国专利号6,271,349)、骨保护素(美国专利号6,015,938)、TRAIL的受体(包括TRAIL受体1、2、3和4)以及包含死亡结构域的受体,诸如Fas或凋亡诱导受体(AIR)。
可利用本发明来生产的其他蛋白质包括包含分化抗原(也称为CD蛋白)或它们的配体或基本类似于这两者之一的蛋白质的全部或部分氨基酸序列的蛋白质。在Leukocyte Typing VI(Proceedings of the VIth International Workshop and Conference,Kishimoto,Kikutani等人,编,Kobe,Japan,1996)中公开了此类抗原。在后续的研讨会中公开了类似的CD蛋白。此类抗原的实例包括CD22、CD27、CD30、CD39、CD40和另外配体(CD27配体、CD30配体等)。若干CD抗原是TNF受体家族的成员,所述TNF家族还包括41BB和OX40。配体通常是TNF家族的成员,41BB配体和OX40配体也是。
也可利用本发明生产酶促活性蛋白或它们的配体。实例包括包含以下蛋白质或它们的配体或基本类似于这两者之一的蛋白质中的一个的全部或部分的蛋白质:包括TNF-α转化酶的解联蛋白和金属蛋白酶结构域家族成员、各种激酶、葡糖脑苷脂酶、超氧化物歧化酶、组织纤溶酶原激活物、因子VIII、因子IX、载脂蛋白E、载脂蛋白A-I、珠蛋白、IL-2拮抗剂、α-1抗胰蛋白酶、任何上述酶类的配体和为数众多的其他酶类和它们的配体。
可生产的抗体的实例包括但不限于识别包括但不限于上述蛋白质和/或以下抗原的蛋白质中的任何一种或组合的那些抗体:CD2、CD3、CD4、CD8、CD11a、CD14、CD18、CD20、CD22、CD23、CD25、CD33、CD40、CD44、CD52、CD80(B7.1)、CD86(B7.2)、CD147、IL-1α、IL-1β、IL-2、IL-3、IL-7、IL-4、IL-5、IL-8、IL-10、IL-1受体、IL-2受体、IL-4受体、IL-6受体、IL-13受体、IL-18受体亚基、FGL2、PDGF-β及其类似物(参见,美国专利号5,272,064和5,149,792)、VEGF、TGF、TGF-β2、TGF-β1、EGF受体(参见,美国专利号6,235,883)、VEGF受体、肝细胞生长因子、骨保护素配体、干扰素γ、B淋巴细胞刺激物(BlyS,也称为BAFF、THANK、TALL-1和zTNF4;参见,Do和Chen-Kiang(2002),Cytokine Growth Factor Rev.13(1):19-25)、C5补体、IgE、肿瘤抗原CA125、肿瘤抗原MUC1、PEM抗原、LCG(其是表达的与肺癌相关的基因产物)、HER-2、HER-3、肿瘤相关糖蛋白TAG-72、SK-1抗原、在结肠癌和/或胰腺癌患者血清中以升高的水平存在的肿瘤相关表位、在乳腺、结肠、鳞状细胞、前列腺、胰腺、肺和/或肾癌细胞和/或黑色素瘤、胶质瘤或成神经细胞瘤细胞上表达的癌症相关表位或蛋白、肿瘤坏死中心、整联蛋白α4β7、整联蛋白VLA-4、整联蛋白(包括包含了α4β7的整联蛋白)、TRAIL受体1、2、3和4、RANK、RANK配体、TNF-α、黏附分子VAP-1、上皮细胞黏附分子(EpCAM)、细胞间黏附分子-3(ICAM-3)、白细胞整联蛋白粘附素、血小板糖蛋白gp IIb/IIIa、心肌肌球蛋白重链、甲状旁腺素、rNAPc2(其是因子VIIa组织因子的抑制剂)、MHC I、癌胚抗原(CEA)、甲胎蛋白(AFP)、肿瘤坏死因子(TNF)、CTLA-4(其是细胞毒性T淋巴细胞相关抗原)、Fc-γ-1受体、HLA-DR 10β、HLA-DR抗原、硬化蛋白、L-选择素、呼吸融合病毒、人类免疫缺陷病毒(HIV)、乙型肝炎病毒(HBV)、变异链球菌(Streptococcus mutans)和金黄色葡萄球菌(Staphlycoccus aureus)。
可利用本发明方法来生产的已知抗体的具体实例包括但不限于阿达木单抗、贝伐单抗、英夫利昔单抗、阿昔单抗、阿仑单抗、巴匹珠单抗、巴利昔单抗、贝利木单抗、布雷奴单抗、布罗达单抗(brodalumab)、康纳单抗、培舍珠单抗、西妥昔单抗、可那木单抗(conatumumab)、地诺单抗(denosumab)、依库珠单抗、etrolizumab、evolocumab、吉妥珠单抗奥唑米星、戈利木单抗(golimumab)、替伊莫单抗、拉贝珠单抗、马帕木单抗、马妥珠单抗、美泊利单抗、莫他珠单抗、莫罗单抗-CD3、那他珠单抗、帕利珠单抗、奥法木单抗、奥马珠单抗、奥戈伏单抗(oregovomab)、帕利珠单抗、帕尼单抗、pemtumomab、培妥珠单抗、兰尼单抗、利妥昔单抗、罗维珠单抗、塔西单抗、托西莫单抗、曲妥珠单抗、优特克单抗、维多珠单抗(vedolizomab)、扎芦木单抗和扎木单抗。
本发明还可用来生产包含例如任何上述蛋白质的重组融合蛋白。例如,可利用本发明的方法来生产包含上述蛋白质之一加上多聚化结构域诸如亮氨酸拉链、卷曲螺旋、免疫球蛋白或基本类似的蛋白质的Fc部分的重组融合蛋白。参见,例如,WO94/10308;Lovejoy等(1993),Science 259:1288-1293;Harbury等(1993),Science 262:1401-05;Harbury等(1994),Nature 371:80-83; 等(1999),Structure7:255-64。明确地包括在此类重组融合蛋白中的是受体的部分融合到抗体诸如依那西普(p75 TNFR:Fc)、阿巴西普和贝拉西普(CTLA4:Fc)的Fc部分的蛋白质。
等(1999),Structure7:255-64。明确地包括在此类重组融合蛋白中的是受体的部分融合到抗体诸如依那西普(p75 TNFR:Fc)、阿巴西普和贝拉西普(CTLA4:Fc)的Fc部分的蛋白质。
本发明的范围不受本文所述的具体实施方案限制,所述具体实施方案意欲作为本发明的个别方面的说明,并且功能上等同的方法和组分构成本发明的方面。确实,除了本文示出和描述的那些之外,根据以上描述和所附附图,本发明的各种修改将对于本领域技术人员而言变得显而易见。此类修改旨在处于所附权利要求书的范围内。
实施例
实施例1
将表达以往显示出低水平高甘露糖(HM)表达的重组人抗体(MAb A)的产生单克隆抗体的中国仓鼠卵巢(CHO)细胞系用于siRNA实验。使用基于DHFR的选择克隆获得所述细胞系;对于常规培养,将细胞悬浮培养于含有氨甲蝶呤(MTX)的选择培养基中。在36℃、5%CO2和85%相对湿度下,将培养物维持在放空的125mL或250mL锥形摇瓶(Corning LifeSciences,Lowell,MA)或50mL放空的自旋试管(TPP,Trasadingen,Switzerland)中。将锥形瓶以120rpm用25mm轨道直径在大容量自动CO2培养箱(Thermo Fisher Scientific,Waltham,MA)中振荡,并且将自旋试管以225rpm、50mm轨道直径在大容量ISF4-X培养箱(Kuhner AG,Basel,Switzerland)中振荡。
检测八种不同的19聚体siRNA的Mgat1、Mgat2和Slc35a2。根据制造商的方案使用 RNAiMAX(Invitrogen;Life Technologies;一种基于脂质的转染试剂,其与核酸复合并促进细胞与核酸的转染)将siRNA瞬时转染至MAb A细胞系中。简言之,在转染日用500微升培养基将细胞以每孔2×105个接种在Corning公司的六孔板上。对于转染,使10pmol siRNA在100微升Opti-Mem I培养基中与1.5微升Lipofectamine RNAiMAX复合,并在室温下孵育10分钟。将RNAi-Lipofectamine RNAiMax试剂复合物添加至各孔中。将板在36℃下于CO2培养箱中孵育3天。使用1X裂解缓冲液(Affymetrix Inc.,Sanata Clara,CA)在50℃下将细胞裂解1小时。通过
RNAiMAX(Invitrogen;Life Technologies;一种基于脂质的转染试剂,其与核酸复合并促进细胞与核酸的转染)将siRNA瞬时转染至MAb A细胞系中。简言之,在转染日用500微升培养基将细胞以每孔2×105个接种在Corning公司的六孔板上。对于转染,使10pmol siRNA在100微升Opti-Mem I培养基中与1.5微升Lipofectamine RNAiMAX复合,并在室温下孵育10分钟。将RNAi-Lipofectamine RNAiMax试剂复合物添加至各孔中。将板在36℃下于CO2培养箱中孵育3天。使用1X裂解缓冲液(Affymetrix Inc.,Sanata Clara,CA)在50℃下将细胞裂解1小时。通过 多路复用测定使用FLEXMAP
多路复用测定使用FLEXMAP 系统(Luminex,Austin TX;蛋白质和基因组多路复用珠阵列测定)来将裂解物用于mRNA表达分析。
系统(Luminex,Austin TX;蛋白质和基因组多路复用珠阵列测定)来将裂解物用于mRNA表达分析。
对于mRNA表达分析,利用QuantiGene Plex 2.0试剂系统(Affymetrix Inc.,Santa Clara,CA)。简言之,使用增补有蛋白激酶K(储备浓度50mg/mL)的1X裂解缓冲液(QS0100)(Affymetrix Inc.,Santa Clara,CA)将来自5x 105个活细胞的细胞团裂解并在50℃下孵育1小时。将细胞裂解物在-80℃下储存直至准备使用。使用靶向Mgat1、Mgat2和Slc35a3以及归一化基因的定制的基因特异性探针组(Affymetrix,Inc.Santa Clara,CA)。将冷冻的裂解物解冻并使用Affymetrix公司的标准方案处理而用于mRNA表达水平分析。
将来源于测量来自每基因每孔100珠的中值报告基因荧光的数据测定并表示为中值荧光强度(MFI)。将本底信号在缺乏来自空白样品的靶mRNA的情况下测定并从在靶mRNA存在下获得的信号中减去。针对两种持家基因:GusB和TBP将目标基因的荧光强度归一化。

将各靶RNA的测定强度通过测定检测极限(LOD)来评估,定义为信号是三个高于本底的标准差的靶浓度。变异系数(CV)测量试验精密度,并且是标准差与平均数的比率。
将所有样品重复操作三次,并且将裂解缓冲液用作空白。使用Luminex公司的Bio-Plex 3D酶标仪分析样品,并且使用Bio-Plex数据管理器5.0(Bio-Rad Laboratories,Hercules,CA)获得数据。
对于各基因,选择达到>85%敲除而没有任何显著脱靶效应的两种siRNA。然后将所选的siRNA转染至MAb A细胞系中,并且进行10天的补料分批抗体生产操作。在补料分批生产研究中,将细胞以3.5x105个细胞/mL接种至生产培养基中。将3mL工作容积用于24深孔板(Axygen Scientific,Union City,CA)中,或者将25mL工作容积用于125mL放空的摇瓶中。在第3天、第6天和第8天将培养物补料7%的最初培养容积的快速灌注补料。在第3天、第6天和第8天将葡萄糖补料至10g/L的目标。在生产操作的第10天收获离心的条件培养基。还在第3天、第6天、第8天和第10天获取样品的生长、活力和代谢数据,并且在第6天、第8天和第10天获取样品的效价和HM分析。
在第3天、第6天、第8天和第10天将细胞团样品从培养物中取出以用于mRNA表达分析;下表1示出了结果。
表1:通过siRNA处理的Mgat1、Mgat2和Slc35a2的水平的减少

mRNA表达分析显示在10天培养操作中Mgat1和Mgat2转录物水平减少>50%。Slc35a2的表达水平从第3天至第6天减少>50%,而到第8天和第10天只看到7%-14%的减少。
在补料分批的第十天测定抗体效价和%HM。由亲和蛋白A POROS PA ID传感器盒通过使用Waters UPLC来测量效价。使用Caliper GX II HM试验(Caliper Life SciencesInc.,PerkinElmer公司)或使用UPLC HILIC(亲水相互作用色谱)(装备有与Acquity UPLCBEH Glycan Column一起使用的UPLC Fluorescence(FLR)检测器的Waters Acquity UPLC)来测量高甘露糖含量。下表2示出了结果(结果表示为平均值加/减标准差)。
表2:用siRNA处理的细胞系的效价和%HM
| 样品 | 效价(g/L) | %HM |
| 无siRNA | 0.75±0.09 | 0.81±0.07 |
| siRNA MGAT1 | 0.73±0.05 | 72.42±3.28 |
| siRNA MGAT2 | 0.87±0.27 | 1.98±0.31 |
| siRNA Slc35A2 | 0.57±0.36 | 1.56±0.29 |
这些结果的分析表明在用Mgatl siRNA处理的细胞中,HM的水平降低70%,然而Mgat2或Slc35a2的敲除不显著影响HM。然而到第10天,Slc35a2的水平已恢复至对照值的90%,因此对于此实验,不可能排除这种基因在调节HM中的作用。用siRNA处理未观察到产生的抗体的效价的显著变化。此外,siRNA处理似乎不影响生产率或细胞活力,表明了在减少的Mgat1 mRNA表达的情况下观察到的升高的HM水平很可能直接与减少的Mgat1活性有关。
实施例2
将表达以往显示出高水平(即,>10%)高甘露糖型聚糖的重组人抗体(MAb B)的产生单克隆抗体的中国仓鼠卵巢(CHO)细胞系用于转柒实验。使用基于DHFR的选择克隆获得所述细胞系;对于常规培养,将细胞悬浮培养于含有MTX的选择培养基中。基本如先前所述,将培养物维持在放空的125mL或250mL锥形摇瓶(Corning Life Sciences,Lowell,MA)或50mL放空的自旋试管(TPP,Trasadingen,Switzerland)中。
将MAb B细胞用任一以下载体来转染:零表达载体对照、含有用弗林蛋白酶pep2A连接的Mgat1和Mgat2的双顺反子表达载体(M1M2)、含有Slc35a2的载体(S)或共转染的Mgat1、Mgat2和Slc35a2载体(M1M2S)。这些细胞在选择培养基中恢复至大于80%活力后,使用流式细胞术将它们单细胞克隆。对于来源于单细胞的那些细胞系,分析目标基因的表达水平。对于四种不同载体中的每个,分析大于40个克隆的构型的目标基因的表达并且基于此分析而选择二十种过表达细胞系和十种对照细胞系以用于基本如先前所述进行的两个独立10天补料分批生产操作中的进一步表征。
在第一补料分批生产开始时,平均来说,过表达细胞系的倍增时间较高(>25小时),并且与具有较高PDL的对照(<25小时)相比,PDL较低(图1)。因此,为全部显示出相似倍增时间的较小克隆亚组进行第二10天补料分批实验(图2)。
来自第二补料分批实验的所选克隆中的mRNA表达的统计分析确认了与对照相比过表达的倍数变化较为显著。与Mgat1基因相比,Mgat2基因具有较大的表达增加倍数。下表3示出了结果。
表3:三组过表达克隆的Mgat1、Mgat2和Slc35a2转录物水平的平均增加倍数。
| 样品 | Mgat1 | Mgat2 | Slc35A2 |
| M1M2 | 6.24 | 26.29 | 0.99 |
| M1M2S | 1.45 | 7.76 | 37.28 |
| S | 0.83 | 0.99 | 27.1 |
为了进一步研究诱导的过表达水平,使用液相色谱串联质谱法(LC-MS/MS)量化Mgat1和Mgat2的蛋白质水平。分析用空载体(EV)转染的对照细胞系以及过表达Mgat1和Mgat2的两种系(B1和B2)。下表4示出了结果。以百万分率(ppm)测量相对蛋白质表达。
表4:第10天对照细胞系和过表达Mgat1和Mgat2的两种细胞系中的归一化mRNA和蛋白质水平

如表4所示,EV展现出Mgat1和Mgat2的基础表达水平,然而在B1和B2细胞系中这些蛋白质的表达水平显著升高。此外,在B1和B2细胞系中,与Mgat1相比,Mgat2显示了升高的表达水平。这些数据与对于第10天Mgat1和Mgat2的mRNA表达水平所观察到的那些数据密切相关。
相比之下,在第8天和第10天(分析蛋白质表达的两天)对于这三种细胞系,持家对照蛋白的表达水平(GAPDH;数据未示出)保持恒定。
图3、4和5示出了生长和比生产率在所述组之间相似;然而,与另外两组相比,对于过表达Mgat1和2的克隆,效价显著增加。与对照细胞(分别为70%和29%)相比,过表达Mgat1和Mgat2的细胞系(M1M2)以及过表达全部三种基因的那些克隆(M1M2S)显示了高甘露糖水平的减少(图6)。然而,当与对照相比时过表达Slc35a2的细胞系(S)没有显示出统计学上显著的变化(图6)。因为UDP-半乳糖转运蛋白(由Slc35A2编码的蛋白质)的作用是将核苷酸糖底物(包括UDP-GlcNAc)转运至高尔基体腔中,所以如果UDP-GlcNAc水平是限制性的,则升高的Slc35a2水平将不会影响随后的聚糖加工。
为每种过表达的细胞系(M1M2、M1M2S和S)评估分泌的重组MAb B的糖型特性。与对照细胞系相比,M1M2细胞系显示了所有HM类诸如M5、M6、M7和M8b的显著减少。结果展示于下表5中。
表5:在补料分批的第10天过表达Mgat1、Mgat2和/或Slc35a2的细胞系中产生的抗体的糖基化特性

在对照空载体细胞系中,A2G0F是评估的八种复合聚糖种类之中的优势种类(49.17%),其次是A2G1F(16.46%)和其他复合糖型。对于M1M2和M1M2S,看到相对于不同种类百分比的相似趋势。然而,在M1M2细胞系的情况下,与对照相比,A2G0F糖型的量显著增加27%(显著性值p=0.0076)。这暗示随着Mgat2的表达存在着杂合聚糖(A1G0M5)至A2G0的有效转化,并且因此与对照细胞系相比,更多底物可用于岩藻糖基转移酶8(Fut8)以制造更多A2G0F产物。虽然在此实验中Slc35a2的过表达似乎不引起复合聚糖水平的显著升高,但是这些结果表明Mgat1和Mgat2的过表达可增加HM聚糖至复合糖型的转化,因此降低HM水平。
实施例3
将CHO宿主细胞系单独用Mgat1或Mgat2转柒,或用Mgat1和Mgat2表达载体共转染。这些细胞在选择培养基中恢复至大于80%活力后,使用流式细胞术将它们单细胞克隆。分析总计291个克隆的Mgat1和Mgat2基因的表达。其中,基于良好生长和活力而选择表达了高于重组CHO细胞系中检测的水平的Mgat1和Mgat2水平的48个克隆,所述重组CHO细胞系表达具有以往低水平(即,<5%)高甘露糖类型聚糖的人单克隆抗体(MAb A)。
使全部48个克隆生长至少60个PDL(群体倍增水平),并且在此时间过程期间在三个不同时间点处分析Mgat 1和Mgat2的mRNA表达水平。基于各自mRNA的稳定表达水平而选择十六个克隆。基本如Remy和Michnick(1999),Proc.Natl.Acad.Sci.,96:5394-5399所述,为了进一步评估这些克隆的转柒能力,将全部48个克隆用含有绿色荧光蛋白(GFP)的载体在蛋白质片断互补测定中进行瞬时转染。
选择展现出最高转染率的七个克隆以用于进一步分析。表6示出前七个克隆相对于对照CHO的Mgat1和Mgat2 mRNA倍数变化。
表6:Mgat1和Mgat2的转录物水平的平均增加倍数

将以往展示出高水平(即,>15%)高甘露糖类型聚糖的单克隆抗体(MAb C)用于过表达Mgat1和/或Mgat2的改造的宿主细胞或者对照非改造的CHO宿主细胞中的转柒。如先前所述,创建稳定细胞群并选择以用于10天补料分批生产操作中的进一步表征。为每种过表达的细胞系评估分泌的重组MAb C的糖型特性和效价。与对照宿主相比,对于MAb C,在宿主38C2中检测到显著较低水平的高甘露聚糖,而没有影响生产率(即,效价)。表7示出了结果;数值反映出从补料分批生产试验获得的第10天效价和聚糖水平。
表7:表达MAb C的非扩增克隆的效价和糖基化特性

类似地,使150nM和300nM扩增群形成并在10天补料分批生产试验中分析。在150nM群的情况下,与对照宿主细胞相比,所有过表达的宿主细胞展示出显著降低的高甘露糖%,而没有影响效价。表8和9示出了结果。
表8:表达MAb C的150nM扩增克隆的效价和糖基化特性

表8续

*表示P值显著
如表9所示,随着300nM扩增,对于五种群(61A9、45F2、63C5、38C2和2B8)检测到显著减小水平的高甘露聚糖水平。
表9:300nM扩增MAb C的效价和糖基化特性

表9续

*表示P值显著
这些结果表明了经过转化以过表达Mgat1和/或Mgat2的宿主细胞可用来制备HM聚糖至复合糖型的转化增加的重组蛋白,并且因此降低HM水平。
Claims (30)
1.一种在哺乳动物细胞培养过程期间调控重组蛋白的高甘露糖糖型含量的方法,其包括转染哺乳动物宿主细胞以过表达涉及N-糖基化途径的蛋白质,其中所述涉及N-糖基化途径的蛋白质选自Mgat1和/或Mgat2;以及Mgat1、Mgat2和Slc35a2。
2.根据权利要求1所述的方法,其中所述宿主细胞先前已经过转染以表达重组蛋白,并且随后进行转染以表达涉及N-糖基化途径的蛋白质。
3.根据权利要求1所述的方法,其中所述宿主细胞首先用涉及N-糖基化途径的蛋白质转染,随后进行转染以表达重组蛋白。
4.根据权利要求1或2所述的方法,其中所述重组蛋白是包含抗体Fc区的蛋白质。
5.根据权利要求1或2所述的方法,其中所述重组蛋白为Fc融合蛋白。
6.根据权利要求1或2所述的方法,其中所述重组蛋白为抗体。
7.根据权利要求1或2所述的方法,其中所述重组蛋白为免疫球蛋白。
8.根据权利要求1或2所述的方法,其中所述重组蛋白为肽体。
9.根据权利要求1或2所述的方法,其中在经过转染以过表达涉及N-糖基化途径的蛋白质的所述细胞中表达的重组蛋白的高甘露糖糖型含量与由细胞未通过转染以过表达涉及N-糖基化途径的蛋白质进行操控的培养物产生的所述重组蛋白相比是减少的。
10.根据权利要求1或2所述的方法,其中所述高甘露糖糖型选自甘露糖5 (Man5);甘露糖6 (Man6);甘露糖7 (Man7);甘露糖8 (Man8);或甘露糖9 (Man9);及其组合。
11.根据权利要求9所述的方法,其中重组蛋白的高甘露糖糖型含量调控为小于或等于10%。
12.根据权利要求9所述的方法,其中重组蛋白的高甘露糖糖型含量调控为小于或等于5%。
13.根据权利要求1或2所述的方法,其中所述哺乳动物细胞培养过程利用补料分批培养方法、灌流培养方法及其组合。
14.根据权利要求13所述的方法,其中使用交替式切向流(ATF)灌流培养物。
15.根据权利要求14所述的方法,其中灌流开始于细胞培养的第1天至第9天。
16.根据权利要求14所述的方法,其中灌流开始于细胞培养的第3天至第7天。
17.根据权利要求14-16中任一项所述的方法,其中灌流在细胞已到达生产期时开始。
18.根据权利要求17所述的方法,其中通过交替式切向流使用超滤器或微滤器来完成灌流。
19.根据权利要求13所述的方法,其中通过分批补料来维持细胞培养物。
20.根据权利要求19所述的方法,其中所述培养物在生产期间补料三次。
21.根据权利要求20所述的方法,其中所述培养物在第二天与第四天之间的一天、在第5天与第7天之间的一天以及在第8天与第10天之间的一天补料。
22.根据权利要求19所述的方法,其中所述培养物在生产期间补料四次。
23.根据权利要求22所述的方法,其中所述培养物在第二天与第四天之间的一天、在第5天与第6天之间的一天、在第7天与第8天之间的一天以及在第8天与第10天或更迟之间的一天补料。
24.根据权利要求13所述的方法,其中调控的高甘露糖糖型含量与未通过转染以过表达涉及N-糖基化途径的蛋白质来进行操控的哺乳动物宿主细胞相比是减少的。
25.根据权利要求1或2所述的方法,其中哺乳动物细胞培养物通过用无血清培养基中的至少0.5 x 106个细胞/mL至3.0 x 106个细胞/mL接种生物反应器来建立。
26.根据权利要求1或2所述的方法,其中所述哺乳动物细胞是中国仓鼠卵巢(CHO)细胞。
27.根据权利要求25所述的方法,其中所述方法减少了所述重组蛋白的高甘露糖糖型含量。
28.根据权利要求1或2所述的方法,其中所述方法还包括用于所述重组蛋白的收获步骤。
29.根据权利要求28所述的方法,其中将所述收获的重组蛋白纯化并配制成药学上可接受的制剂。
30.通过权利要求29所述的方法获得的经过纯化、配制的重组蛋白。
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN202110902965.XA CN113774084B (zh) | 2014-01-29 | 2014-12-11 | 过表达n-糖基化途径调节基因以调节重组蛋白的糖基化 |
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201461933137P | 2014-01-29 | 2014-01-29 | |
| US61/933,137 | 2014-01-29 | ||
| PCT/US2014/069744 WO2015116315A1 (en) | 2014-01-29 | 2014-12-11 | Overexpression of n-glycosylation pathway regulators to modulate glycosylation of recombinant proteins |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN202110902965.XA Division CN113774084B (zh) | 2014-01-29 | 2014-12-11 | 过表达n-糖基化途径调节基因以调节重组蛋白的糖基化 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN106170554A CN106170554A (zh) | 2016-11-30 |
| CN106170554B true CN106170554B (zh) | 2021-08-17 |
Family
ID=53757626
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201480074585.XA Active CN106170554B (zh) | 2014-01-29 | 2014-12-11 | 过表达n-糖基化途径调节基因以调节重组蛋白的糖基化 |
| CN202110902965.XA Active CN113774084B (zh) | 2014-01-29 | 2014-12-11 | 过表达n-糖基化途径调节基因以调节重组蛋白的糖基化 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN202110902965.XA Active CN113774084B (zh) | 2014-01-29 | 2014-12-11 | 过表达n-糖基化途径调节基因以调节重组蛋白的糖基化 |
Country Status (14)
| Country | Link |
|---|---|
| US (3) | US10227627B2 (zh) |
| EP (1) | EP3099806A1 (zh) |
| JP (4) | JP6474420B2 (zh) |
| KR (3) | KR102519540B1 (zh) |
| CN (2) | CN106170554B (zh) |
| AU (3) | AU2014380174B2 (zh) |
| BR (1) | BR112016017606A2 (zh) |
| CA (1) | CA2938079C (zh) |
| CL (1) | CL2016001900A1 (zh) |
| EA (2) | EA037565B1 (zh) |
| IL (3) | IL282517B (zh) |
| MX (2) | MX2016009911A (zh) |
| SG (1) | SG10201912081RA (zh) |
| WO (1) | WO2015116315A1 (zh) |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR101936049B1 (ko) | 2015-10-15 | 2019-01-08 | (주)알테오젠 | IgG Fc 도메인을 가지는 융합 단백질의 생산방법 |
| MA47775A (fr) * | 2017-03-14 | 2020-01-22 | Amgen Inc | Contrôle des glycoformes afucosylées totales d'anticorps produits en culture cellulaire |
| KR20210005689A (ko) | 2018-05-01 | 2021-01-14 | 암젠 인크 | 조절된 글리칸 프로파일을 갖는 항체 |
| CN109576212B (zh) * | 2018-12-14 | 2021-06-22 | 杭州奕安济世生物药业有限公司 | 高密度接种培养中种子细胞的培养方法及其应用 |
| EP4326766A1 (en) | 2021-04-23 | 2024-02-28 | Amgen Inc. | Anti-tslp antibody compositions and uses thereof |
| CN113684322B (zh) * | 2021-09-09 | 2024-01-23 | 上海药明生物技术有限公司 | 一种降低抗体类蛋白高聚甘露糖型水平的方法 |
| CN114525252B (zh) * | 2022-03-10 | 2023-12-01 | 广州源井生物科技有限公司 | 一种提高mda-mb-231单细胞克隆形成率的单克隆增强培养基与培养方法及其应用 |
| TW202426485A (zh) | 2022-10-26 | 2024-07-01 | 美商安進公司 | 抗tslp抗體組成物及其用途 |
Family Cites Families (43)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6936694B1 (en) | 1982-05-06 | 2005-08-30 | Intermune, Inc. | Manufacture and expression of large structural genes |
| AU588819B2 (en) | 1984-10-29 | 1989-09-28 | Immunex Corporation | Cloning of human granulocyte-macrophage colony stimulating factor gene |
| US5672502A (en) | 1985-06-28 | 1997-09-30 | Celltech Therapeutics Limited | Animal cell culture |
| US4968607A (en) | 1987-11-25 | 1990-11-06 | Immunex Corporation | Interleukin-1 receptors |
| US5075222A (en) | 1988-05-27 | 1991-12-24 | Synergen, Inc. | Interleukin-1 inhibitors |
| AU643427B2 (en) | 1988-10-31 | 1993-11-18 | Immunex Corporation | Interleukin-4 receptors |
| US5395760A (en) | 1989-09-05 | 1995-03-07 | Immunex Corporation | DNA encoding tumor necrosis factor-α and -β receptors |
| EP1132471A3 (de) | 1989-09-12 | 2001-11-28 | F. Hoffmann-La Roche Ag | TNF-bindende Proteine |
| US6204363B1 (en) | 1989-10-16 | 2001-03-20 | Amgen Inc. | Stem cell factor |
| US5149792A (en) | 1989-12-19 | 1992-09-22 | Amgen Inc. | Platelet-derived growth factor B chain analogs |
| US5272064A (en) | 1989-12-19 | 1993-12-21 | Amgen Inc. | DNA molecules encoding platelet-derived growth factor B chain analogs and method for expression thereof |
| WO1991018982A1 (en) | 1990-06-05 | 1991-12-12 | Immunex Corporation | Type ii interleukin-1 receptors |
| US5350683A (en) | 1990-06-05 | 1994-09-27 | Immunex Corporation | DNA encoding type II interleukin-1 receptors |
| GB9014932D0 (en) | 1990-07-05 | 1990-08-22 | Celltech Ltd | Recombinant dna product and method |
| WO1994004679A1 (en) | 1991-06-14 | 1994-03-03 | Genentech, Inc. | Method for making humanized antibodies |
| JPH05244982A (ja) | 1991-12-06 | 1993-09-24 | Sumitomo Chem Co Ltd | 擬人化b−b10 |
| PT672141E (pt) | 1992-10-23 | 2003-09-30 | Immunex Corp | Metodos de preparacao de proteinas oligomericas soluveis |
| US5981713A (en) | 1994-10-13 | 1999-11-09 | Applied Research Systems Ars Holding N.V. | Antibodies to intereleukin-1 antagonists |
| EP0835305B1 (en) | 1995-06-29 | 2005-11-23 | Immunex Corporation | Cytokine that induces apoptosis |
| US6613544B1 (en) | 1995-12-22 | 2003-09-02 | Amgen Inc. | Osteoprotegerin |
| US6096728A (en) | 1996-02-09 | 2000-08-01 | Amgen Inc. | Composition and method for treating inflammatory diseases |
| US6271349B1 (en) | 1996-12-23 | 2001-08-07 | Immunex Corporation | Receptor activator of NF-κB |
| US6235883B1 (en) | 1997-05-05 | 2001-05-22 | Abgenix, Inc. | Human monoclonal antibodies to epidermal growth factor receptor |
| GB9722131D0 (en) | 1997-10-20 | 1997-12-17 | Medical Res Council | Method |
| US6337072B1 (en) | 1998-04-03 | 2002-01-08 | Hyseq, Inc. | Interleukin-1 receptor antagonist and recombinant production thereof |
| AU1920301A (en) | 1999-11-17 | 2001-05-30 | Immunex Corporation | Receptor activator of nf-kappa b |
| US6544424B1 (en) | 1999-12-03 | 2003-04-08 | Refined Technology Company | Fluid filtration system |
| GB9928787D0 (en) | 1999-12-03 | 2000-02-02 | Medical Res Council | Direct screening method |
| GB0025144D0 (en) | 2000-10-13 | 2000-11-29 | Medical Res Council | Concatenated nucleic acid sequences |
| US20040202995A1 (en) | 2003-04-09 | 2004-10-14 | Domantis | Nucleic acids, proteins, and screening methods |
| US20050186577A1 (en) * | 2004-02-20 | 2005-08-25 | Yixin Wang | Breast cancer prognostics |
| EP2308958A3 (en) | 2004-03-05 | 2011-08-10 | DSM IP Assets B.V. | Process for cell culturing by continuous perfusion and alternating tangential flow |
| JP5017116B2 (ja) | 2004-09-24 | 2012-09-05 | アムジエン・インコーポレーテツド | 修飾Fc分子 |
| US8053238B2 (en) | 2005-10-31 | 2011-11-08 | Unhwa Corporation | Isolated population of plant single cells and method of preparing the same |
| US20070190057A1 (en) * | 2006-01-23 | 2007-08-16 | Jian Wu | Methods for modulating mannose content of recombinant proteins |
| US20100145032A1 (en) * | 2007-01-18 | 2010-06-10 | Suomen Punainen Risti, Veripalelu | Novel carbohydrate profile compositions from human cells and methods for analysis and modification thereof |
| WO2008154014A2 (en) * | 2007-06-11 | 2008-12-18 | Amgen Inc. | A method for culturing mammalian cells to improve recombinant protein production |
| TW201028431A (en) * | 2008-10-31 | 2010-08-01 | Lonza Ag | Novel tools for the production of glycosylated proteins in host cells |
| EP2421892A1 (en) * | 2009-04-20 | 2012-02-29 | Pfizer Inc. | Control of protein glycosylation and compositions and methods relating thereto |
| WO2011109600A1 (en) * | 2010-03-05 | 2011-09-09 | Alnylam Pharmaceuticals, Inc. | Compositions and methods for modifying the glycosylation pattern of a polypeptide |
| ES2602108T3 (es) * | 2010-04-07 | 2017-02-17 | Momenta Pharmaceuticals, Inc. | Método para cuantificar glicoformas que contienen alta manosa |
| TW201209160A (en) * | 2010-04-27 | 2012-03-01 | Lonza Ag | Improved glycosylation of proteins in host cells |
| WO2012145682A1 (en) | 2011-04-21 | 2012-10-26 | Amgen Inc. | A method for culturing mammalian cells to improve recombinant protein production |
-
2014
- 2014-12-11 KR KR1020227019897A patent/KR102519540B1/ko active IP Right Grant
- 2014-12-11 EP EP14821424.0A patent/EP3099806A1/en active Pending
- 2014-12-11 SG SG10201912081RA patent/SG10201912081RA/en unknown
- 2014-12-11 CN CN201480074585.XA patent/CN106170554B/zh active Active
- 2014-12-11 IL IL282517A patent/IL282517B/en unknown
- 2014-12-11 KR KR1020167021676A patent/KR102301034B1/ko active IP Right Grant
- 2014-12-11 EA EA201691532A patent/EA037565B1/ru unknown
- 2014-12-11 BR BR112016017606A patent/BR112016017606A2/pt not_active Application Discontinuation
- 2014-12-11 MX MX2016009911A patent/MX2016009911A/es active IP Right Grant
- 2014-12-11 WO PCT/US2014/069744 patent/WO2015116315A1/en active Application Filing
- 2014-12-11 EA EA202190244A patent/EA202190244A3/ru unknown
- 2014-12-11 JP JP2016549077A patent/JP6474420B2/ja active Active
- 2014-12-11 KR KR1020217028516A patent/KR102410393B1/ko active IP Right Grant
- 2014-12-11 CA CA2938079A patent/CA2938079C/en active Active
- 2014-12-11 CN CN202110902965.XA patent/CN113774084B/zh active Active
- 2014-12-11 AU AU2014380174A patent/AU2014380174B2/en active Active
-
2016
- 2016-07-24 IL IL246914A patent/IL246914B/en active IP Right Grant
- 2016-07-26 CL CL2016001900A patent/CL2016001900A1/es unknown
- 2016-07-29 MX MX2020012356A patent/MX2020012356A/es unknown
-
2018
- 2018-09-13 US US16/130,879 patent/US10227627B2/en active Active
- 2018-11-15 AU AU2018264100A patent/AU2018264100B2/en active Active
-
2019
- 2019-01-28 JP JP2019011871A patent/JP6797950B2/ja active Active
- 2019-01-29 US US16/261,311 patent/US10655156B2/en active Active
- 2019-11-24 IL IL270861A patent/IL270861B/en active IP Right Grant
-
2020
- 2020-05-15 US US16/875,832 patent/US10907186B2/en active Active
- 2020-09-23 AU AU2020239681A patent/AU2020239681B2/en active Active
- 2020-11-17 JP JP2020190707A patent/JP7034236B2/ja active Active
-
2022
- 2022-02-28 JP JP2022029077A patent/JP2022068365A/ja active Pending
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| AU2020239681B2 (en) | Overexpression Of N-Glycosylation Pathway Regulators To Modulate Glycosylation Of Recombinant Proteins | |
| US11299760B2 (en) | Use of monensin to regulate glycosylation of recombinant proteins | |
| US10106829B2 (en) | Overexpression of N-glycosylation pathway regulators to modulate glycosylation of recombinant proteins | |
| EA047078B1 (ru) | Сверхэкспрессия регуляторов пути n-гликозилирования для модуляции гликозилирования рекомбинантных белков |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| REG | Reference to a national code |
Ref country code: HK Ref legal event code: DE Ref document number: 1231928 Country of ref document: HK |
|
| GR01 | Patent grant | ||
| GR01 | Patent grant |