低脂氧化酶1大麦
本申请是于2001年1月22日,指定所有的国家,以一个丹麦公司——嘉士伯研究实验室(Carlsberg Rearch Laboratory)的名义作为PCT国际专利申请提出申请的。
发明领域
本申请是在植物生物技术领域。更具体地说,本申请涉及编码具有剧烈降低的9-过氧羟基十八烷酸形成活性之酶的突变型大麦脂氧化酶1基因(lox-1)。本发明还涉及在酿酒过程中lox-1纯合的大麦品种用于减少酿制成的产品例如啤酒在贮藏过程中异味(off-flavor)的形成的用途。
发明背景
脂氧化酶是催化游离的和酯化的多不饱和脂肪酸的双氧化作用的一个酶家族(EC1.13.11.12),该多不饱和脂肪酸含有1(Z)、4(Z)-戊二烯构型。长期以来,脂氧化酶催化反应的产物被怀疑为在植物谷粒/种子和从谷粒/种子获得的食品中腐败味出现的主要原因(Robinson等,1995,Food Chem.,54:33-43)。脂氧化酶涉及在大豆加工过程中产生的挥发性己醛的生产,它具有不受欢迎的香气,限制了在食品中大豆蛋白的使用。人们认为三种在大豆种子中表达的脂氧化酶同工酶有助于脂类氧化和己醛形成。为了减少己醛形成和改善它们的香味稳定性,已生产了缺乏一种或多种这些同工酶的大豆突变体。这一方法的成果已由Hildebrand等,1990,J.Agric.Food Chem.38:1934-1936所评估。缺乏大豆脂氧化酶3的突变体产生较高的己醛水平,表明该同工酶转化由脂类氧化产生的13-过氧羟基十八烷酸为非挥发性的产物。缺乏所有三种种子脂氧化酶的三-零大豆品系(triplenullsoybean line)的野外使用特性显示这些酶对于正常的农学和种子特性不是必需的(Narvel等,1998,Crop Sci.38:926-928)。
脂氧化酶还涉及水稻异味的产生,这可在谷物贮藏过程中发生。在贮藏的谷物中可检测到游离脂肪酸的释放,这是甘油三酯储备代谢的表现。发现水稻品种Daw Dam累积低水平的戊醛和己醛,该戊醛和己醛在贮存中给予较好的香味稳定性(Susu ki等,1999,J.Agric.Food Chem.47:1119-1124)。这一希望的表型归因于水稻酯氧化酶-3的缺乏,该酯氧化酶-3氧化不饱和脂酰基链以形成9-羟基过氧化十八烷酸定位异构体。
人们认为脂氧化酶途径与许多分支途径结合,并且它在植物生长和生理等许多方面的作用尚未完全了解。建议使用在种子作物中改变9-氢过氧化作用活性的修饰的脂氧化酶途径,调节它们对由曲霉属真菌(Aspergillus spp.)引起的真菌毒素的污染的敏感性(WO9726364),这与植物病原体抗性中包含的这种途径一致,但与这里描述的本发明的目的无关。
在许多促成啤酒香味的植物香味挥发物中,具有6-12个碳链的高度不饱和醛具有特别低的器官感觉香味阈(Meilgaard 1975,MBAATech.Quart.12:151-168)。反式-2-壬烯醛,是该类群的一个成员,具有极低的0.01ppb的香味阈值并给予啤酒令人不快的稻草样、“硬纸板”味道。由反式-2-壬烯醛引起的特征性异味是贮存1-3个月或更长的啤酒的普遍特征,并对于窖藏啤酒的香味特别有害,该窖藏啤酒是用浅色麦芽酿制并具有柔和的香味。
人们早已知道亚硫酸盐改善啤酒的香味稳定性,其不仅通过结合氧并充当抗氧化剂,并且还通过形成存在于啤酒中的具有醛和酮的挥发性亚硫酸氢盐加成化合物。啤酒中亚硫酸盐的两个主要的来源是通过发酵过程中酵母经硫的同化途径产生亚硫酸盐,和在包装前将亚硫酸盐加入啤酒。增强酵母亚硫酸盐生产和分泌的发酵条件将允许从存在于麦芽汁中的羰基形成亚硫酸盐-羰基加合物,并防止它们通过酵母的进一步代谢(Dufour1991,Proc.Eur.Brew.Conv.Congr.,Lisbon,209-216页)以这种形式羰基(例如乙醛和双乙酰)可转移到啤酒中。在啤酒熟化过程中,亚硫酸盐防止羰基化合物反式-2-壬烯醛出现的能力已通过使用一个酵母菌株酿造啤酒得以证实,其中所述酵母的硫同化途径被阻断(Johannesen等,1999,Proc.Eur.Brew.Conv.Congr.,Nice,655-662)。装瓶后,通过在37℃贮存7天强制熟化啤酒,这以后发现反式-2-壬烯醛的水平大大超过味觉阈限。如果在装瓶前向低亚硫酸盐的啤酒中加入10PPm的亚硫酸盐,在强制熟化过程中反式-2-壬烯醛的出现明显减少。亚硫酸盐和羰基化合物之间的反应是可逆的,并且在热力学和动力学的控制下。亚硫酸盐化合物的表观平衡常数从羰基化合物的10-6M(例如乙醛、己醛、和癸醛),到双乙酰和丙酮酸盐的10-3(Dufour 1991,出处同前)。在啤酒贮藏过程中,由包装而产生的气体交换将允许氧气进入啤酒造成亚硫酸盐损失,这样较弱的亚硫酸氢盐加合物将分解,使游离羰基出现在啤酒中。而亚硫酸盐毫无疑问地增强啤酒香味稳定性,特别是在短期内,在包装的啤酒中其保持力极大程度上取决于由包装而产生的气体交换和温度。在酿制的啤酒中,发酵过程中生产的亚硫酸盐的自然水平是可变的,并且装瓶前亚硫酸盐的加入不是一个普遍接受的方法。由于这些原因,亚硫酸盐独自不能提供在全世界所使用的不同的啤酒贮藏条件下增强啤酒长期的香味稳定性的可靠方法。
通常公认在啤酒中发现的反式-2-壬烯醛产生自从大麦谷粒脂类衍生的多不饱和脂肪酸的氧化,其中18-碳链脂肪酸,亚油酸[分类为18;2,n-6多不饱和脂肪酸(Broun,Gettner和Sommerville 1999,Annu.Rev.Nutr.19:197-216)]是最丰富的。然而,由此形成反式-2-壬烯醛的机制在文献中很少一致。已提出导致反式-2-壬烯醛从多-不饱和脂肪酸形成的酶促途径的存在,但是从没有在大麦谷粒中或麦粒发芽的过程中实验证明各个酶促步骤(Gardner 1988,Adv.Cereal Sci.Technol.9:161-215)。已提出将使用反义或共抑制基因技术减少大麦谷粒中的脂氧化酶-1的水平,并因此控制9-羟基过氧化和在制成的大麦谷粒中减少乙醛和乙醇的水平的概念作为控制异味形成的方法,但是没有报道这种方法的结果(McElroy和Jacobsen,1995,Bio/Technology13:245-249)。
已开发了一种强制试验作为评估啤酒反式-2-壬烯醛效能的方法,其中在麦芽汁或啤酒中反式-2-壬烯醛的形成是通过在减低的pH的条件下提高样品温度诱导的(100℃,pH4.0持续2小时)。把麦芽汁和酿制的啤酒中的反式-2-壬烯醛的效能与干麦芽中的脂氧化酶活性的总水平联系起来的努力显示,脂氧化酶可促成反式-2-壬烯醛在熟化啤酒中的出现(Drost等,1990,J.Am.Soc.Brew.Chem.48:124-131)。然而,大麦麦芽中的脂氧化酶活性在麦粒发芽过程结束时由窑干过程中酶失活的程度所调节的事实严格限制了从这一研究得出的结论。因此,仅检测了残余的麦芽脂氧化酶活性对得到的麦芽汁和酿制的啤酒中的反式-2-壬烯醛效能的影响。该研究没能评估在大麦谷粒的发育和麦粒发芽过程中催化脂氧化酶酶促途径中第一步的脂氧化酶,以及它们作为在啤酒中发现的反式-2-壬烯醛水平的决定因素的作用。实际上,缺失一种或多种脂氧化酶同工酶的大麦栽培变种的缺乏造成不可能提供关于在大麦麦芽中控制反式-2-壬烯醛的形成之脂氧化酶途径的作用的令人信服的证据。当与自动氧化/化学途径比较时,需要这样的实验以评估酶促途径对啤酒中反式-2-壬烯醛形成的贡献。酿酒过程包括麦芽汁沸腾的高温步骤,其中提出会发生这些非酶促反应(Noel等,1999,J.Agric.Food Chem.47:4323-4326)。
发明综述
本发明提供了一种具有大大降低的脂氧化酶-1活性的大麦栽培变种。在一个实施方案中,本发明的大麦植物含有一个表达水平大大降低了的异构酶脂氧化酶-1的突变型lox-1基因。在另一个实施方案中,大麦植物含有表达野生型lox-1之反义序列的异源核酸序列,因此降低了酶的活性。
如本文所显示的,从本发明的降低了脂氧化酶的大麦生产的麦芽和麦芽汁(例如,从纯合突变型lox-1基因的大麦栽培变种生产的),可用于生产具有明显增强的香味稳定性和反式-2-壬烯醛水平降低的啤酒,特别是在已知促进T2N出现的条件下。本发明证实大麦麦芽脂氧化酶-1生产9-羟过氧化-十八双酸(9-HPOD)的能力,与啤酒中反式-2-壬烯醛的存在之间的关系。本发明进一步证实在酿酒过程中纯合突变型lox-1基因的大麦,在贮存过程中和暴露于提高的贮存温度的条件下,改善了啤酒的香味稳定性的用途。这些特性提高了啤酒的质量,并用于延长货架期和减少在转运和贮存过程中制冷啤酒的需要。
本发明提供了具有降低的脂氧化酶-1活性的大麦植物及其部分,包括表达本文所描述的突变型LOX-1蛋白质的大麦植物,以及用于生产这些大麦植物、植物部分、植物产品的方法,和具体的从本发明的大麦植物生产的麦芽和啤酒产品。
附图简述
图1是显示抑制剂去甲二氢愈创木酸(NDGA)对从发芽3天的大麦谷粒的胚纯化的脂氧化酶1和2的活性之免疫亲和性的影响的曲线图。
图2是显示从开花后5天到完全成熟(FM)的过程中,品系G和栽培变种Vintage发育中的谷粒之鲜重的曲线图。每个测定是取自6个穗状花序的平均单个谷粒重。
图3是显示从开花后5天到完全成熟(FM)的过程中,品系G和栽培变种Vintage的发育中的谷粒的干重的曲线图。每个测定是取自5个谷粒中的三个样品的平均单个谷粒重。
图4是显示从开花后5天到完全成熟(FM)的过程中,品系G和栽培变种Vintage的发育中谷粒中的总脂氧化酶活性的曲线图。
图5是显示在品系G之发育中的谷粒中由脂氧化酶活性氧化亚油酸的9-HPOD和13-HPOD产物的曲线图。
图6是显示品系G和栽培变种Vintage的发芽谷粒的胚中的总脂氧化酶活性,表达为μmol/min/10个胚(U/10个胚)的曲线图。
图7是显示在品系G和栽培变种Vintage的发芽谷粒的胚中由脂氧化酶活性氧化亚油酸的9-HPOD和13-HPOD产物,显示9-HPOD和13-HPOD水平的曲线图。
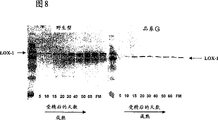
图8是显示脂氧化酶1的免疫检测的蛋白质印迹,该脂氧化酶存在于从开花后5天到完全成熟(FM)的品系G和栽培变种Vintage[wt]的发育谷粒的胚中。
图9是显示在发芽0-6天的品系G和栽培变种Vintage[野生型]的谷粒胚中脂氧化酶-1的免疫检测的蛋白质印迹。
图10是用lox-1 cDNA的3’非转录区探测的Northern印迹,显示在从开花后5天到完全成熟(FM)的品系G和栽培变种Vintage[wt]的发育谷粒中检测的脂氧化酶1转录物。
图11是用lox-1cDNA的3,非转录区探测的Northern印迹,显示在发芽0-6天的品系G和栽培变种Vintage[w-t]的谷粒胚中检测的脂氧化酶1转录物。
图12A-12G是lox-1野生型栽培变种Vintage等位基因(WT)和品系G等位基因(LG)的启动子和转录区的核苷酸序列比对。在基因序列中的转录起始位点(+1)、ATG起始密码子(+69)和翻译终止密码子(+4231)下面划线。在品系G等位基因中鉴别的核苷酸突变以粗斜体显示并用星号指出。
图13是栽培变种Vintage(野生型)的lox-1基因和品系G的突变型lot-1基因的示意图。从+1到+4375的转录物由7个外显子(画点的框)和6个内含子(白色框)组成。显示了lox-1基因中的两个突变
图14是用于野生型lox-1cDNA和lox-1基因以及品系G的突变型lox-1基因的瞬时表达的基因盒的示意图。在具有内含子1(Ubi-1)的组成型玉米泛素启动子和nos终止子之间克隆脂氧化酶编码序列。
图15是显示用包含野生型lox-1cDNA、来自品系G的突变型lox-1基因、WTlox-1基因和一个对照GUS报道基因的基因盒转染的大麦糊粉原生质体中的脂氧化酶活性的条线图。通过KI的氧化和分光光度计确定数量在微量滴定板中分析转染的原生质体提取物中的脂氧化酶活性。提取物中脂氧化酶1的活性表示为单位/μg蛋白质,并显示2份重复试验的3次测定的平均值。
图16是一个序列比对,该序列比对显示野生型和突变型lox-1基因之间的RFLP是由于在核苷酸2347位点上的点突变,产生一个额外的AatII限制位点。
图17是在聚合酶链反应-切割扩增的多态位点(PCR-CAPS)分析中,扩增和切割的lox-1PCR片段的示意图。PCR引物的位置由箭头指示,AatII位点显示于基因之上(序列位置)。PCR产物中的外显子和内含子区域分别通过点和白色框区分,并给出AatII消化片段的大小。
图18是一个电泳的琼脂糖凝胶,该凝胶显示在PCR-CAPS分析的第一步中从品系G和栽培变种Vintage基因组DNA扩增的lox-1PCR片段(652bp)。
图19是一个电泳的琼脂糖凝胶,该凝胶显示在野生型和突变型lox-1基因中由PCR-CAPS检测的RFLP。突变型基因的AatII消化片段包括唯一的313bp的限制性片段,用星号指示。
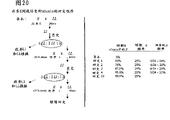
图20是一个表格,该表格显示品系G对栽培变种Alexis的单隐性基因对ll(低脂氧化酶特性)的回交程序。LL基因型是表达野生型脂氧化酶活性(显性等位基因)的植物,ll基因型是表达低脂氧化酶(隐性等位基因)的植物。L1是含有野生型和低脂氧化酶等位基因的杂合植物。既然低脂氧化酶特性是一种隐性特性,L1植物显示野生型脂氧化酶活性。在每一轮回交后(包括自授粉—),预期ll子代占25%的子代。显示了观察到的低脂氧化酶活性的频率。具有杂合的低脂氧化酶等位基因的栽培变种Alexis遗传背景的计算百分比以%Alexis表示。
图21是一个电泳的琼脂糖凝胶,该凝胶显示在品系G-Alexis回交程序的ll子代中的突变型lox-1基因的PCR-CAPS检测。对品系G(泳道2)、栽培变种Vintage(泳道3)、第三(泳道4)和第四次回交(泳道5-9)的ll子代的基因组DNA进行PCR-CAPS分析。对照,回交的高lox品系(泳道10)。
图22A-22B是大豆脂氧化酶LOX-1(Gm1)、LOX-2(Gm2)、LOX-3(Gm3)和大麦脂氧化酶LOX-1(Hv1)和LOX-2(Hv2)的氨基酸序列的比较性比对。
发明的详细描述
根据本发明,提供了用于生产例如啤酒的一种饮料的植物原料、植物产物和方法,该饮料具有降低含量的异味化合物反式-2-壬烯醛,这样,在贮藏过程中和暴露于提高温度的条件下的饮料,例如啤酒,相对于对照饮料,香味稳定性得到改善。更具体地说,本发明提供的大麦品种,其发育和发芽的谷粒产生酶的活性水平大大降低的脂氧化酶-1,称之为LOX-1。这种大麦品种,例如,当在啤酒酿造过程中使用时,与对照的大麦品种相比较,产生具有降低的反式-2-壬烯醛水平的啤酒。
下文描述了用于生产、表征、和证实具有大大降低的LOX-1活性的大麦品种的方法,以及这种类型的大麦用于生产香味稳定的啤酒的用途。
1.定义
如本文所使用的,下列术语具有指示的定义:
“植物部分”是指植物或植物的特定部分,例如茎、叶、根、花、种子、谷粒、果实或芽。
“LOX-1”是指脂氧化酶-1蛋白质;“lox-1”是指编码LOX-1的基因。
“突变型大麦lox-1”是指编码突变型脂氧化酶1多肽的诱变处理的大麦基因。
“未突变对照”是指植物、核酸、基因、多肽、植物部分、或包含野生型基因或蛋白质的植物产品。
“异源的”是指非天然序列,例如衍生自另一种类的序列,或基因工程重组的序列或不同于天然序列的合成序列。
“植物产物”是指从植物或植物部分加工得来的产物,包括例如麦芽和麦芽汁。
“酸性氨基酸”是指天冬氨酸或谷氨酸。
“碱性氨基酸”是指组氨酸、赖氨酸或精氨酸。
“极性氨基酸”是指苏氨酸、丝氨酸、酪氨酸、色氨酸、天冬酰胺、或谷氨酰胺。
“器官感觉的特性”是指吸引嗅觉和味觉的特性,例如训练有素的品尝小组分析它。
“酿造的产品”是指通过捣碎、煮沸和发酵制成的产品,例如啤酒。
“减少的反式-2-壬烯醛”是指与野生型(对照)的情况相比少于约50%。
2.脂氧化酶活性
脂氧化酶催化多不饱和脂肪酸的氧化。在大麦中,已知异构酶LOX-1和LOX-2。LOX-1主要催化多不饱和十八烷脂肪酸的9-氢过氧化作用,而LOX-2主要催化多不饱和十八烷脂肪酸的13-氢过氧化作用。下面的实施例中显示的数据表明大麦LOX-19-氢过氧化活性和啤酒中反式-2-壬烯醛存在之间的关系。因此,具有降低了LOX-1活性的大麦可用于生产与对照相比具有减少的反式-2-壬烯醛水平和/或效力的啤酒。
3.低脂氧化酶大麦的生产
可使用已知的各种遗传学方法生产本发明的植物,即,以稳定的、可遗传的形式降低在大麦植物中表达的脂氧化酶1的酶活水平。这些方法包括,但不限于反义技术和突变,例如化学和辐射诱导的突变,以及定点突变。
大麦转化。可用设计用于操纵lox-1基因表达或改变lox-1基因结构的多种核酸分子转化大麦。各种方法例如,根癌农杆菌介导的转移(Tingay等,1997,Plant J.11:1369-1376),粒子轰击(Wan和Lemaux,1994,Plant Physiol.,104:37-48,或聚乙二醇(PEG)-介导的DNA摄入(Funatsuki和Kihare,1995,Theor.Appl.Genet.,91:707-712),可用于成功引导核酸进入大麦细胞,例如进入原生质体、愈伤组织或胚。
可使用各种启动子驱动感兴趣的基因的表达。对于包括反义序列之包含lox-1的载体表达,可使用天然lox-1启动子区域。Lox-1的启动子序列包含于核苷酸2602-3511,它包括编号为U83904的EMBL的5’UTR。换句话说,可使用组成型驱动感兴趣的基因表达的启动子,例如Ubi.1玉米遍在蛋白启动子(Wan和Lemaux,出处同前;Kj aerulff等,P.Mathis,编,1995,Photosynthesis:from Light to Biosphere,II卷,151-154)。表达载体还可含有一个转录终止区,例如胭脂碱合酶基因的3’终止子(3’-nos)(Bevan,等,1983,Nuci.Acids Res.,11:369-358)与在转基因大麦中表达的基因融合(Wan和Lemaux,出处同前;Funatsuki和Kihara,出处同前)。
表达载体还可包含一个基因,该基因允许当载体成功整合进入细胞时,经转化细胞的筛选。这些基因可编码抗生素或除草剂抗性基因,例如新霉素磷酸转移酶(npt)或膦丝菌素乙酰基转移酶(bar)基因。当表达时,这些抗性基因允许转化的细胞在分别含有新霉素或bialaphos的培养基中生长(见例如,Wan和Lemaux,出处同前;Funatsu ki和Kihara,出处同前;Kj aerulff等,在P.Mathis中,出处同前)。
转化后,细胞可在选择性培养基中生长一段时间,然后培养以使苗形成,然后是根系统,而后形成小植株。Funatsuki和Kihara开发了成功的大麦转化方法(出处同前),其中用含有新霉素磷酸转移酶的表达载体经PEG转化大麦原生质体,随后在新霉素中选择,产生含有转基因的可育植物。显示转基因整合进入基因组,并且大多数转基因植物表达由转基因编码的蛋白质。这些转基因植物杂交后还能传递和表达转基因。
应当理解各种转化方法、表达载体、启动子、选择性标记等是已知的并对大麦的转化是有用的。
大麦诱变。使用嵌合的RNA/DNA寡核苷酸,lox-1基因可被定向用于位点专一诱变。这些嵌合的RNA/DNA寡核苷酸已显示在希望的位点成功引导突变进入植物细胞(Zhu等,1999,Proc.Natl.Acad.S ci.96:8768-8773;和Beetham等,1999,Proc.Natl.Acad.Sci.96:8774-8778)和哺乳动物细胞(Yoon等,1999,Proc.Natl.Acad.Sci.93:2071-2076)。可以各种方法将嵌合的RNA/DNA寡核苷酸转化进入大麦原生质体或感兴趣的细胞,例如使用上文描述的PEG介导的或粒子轰击介导的转化方法。然后可通过组织培养使各个原生质体或细胞再生至完整的可育植物,并可例如使用下文实施例中详细描述的以PCR为基础的方法证实和跟踪突变的发生。
这种定点诱变的方法可用于lox-1基因中的突变特异性残基。Lox-1基因可在启动子区域的一个或多个核苷酸位置突变,以下调或消除lox-1的转录。特异性诱变还可用于向lox-1编码区导入改变,例如,降低酶的活性。这种突变包括,但不限于,导致移码的插入、缺失和取代,LOX-1蛋白的截短、和/或酶底物腔的中性或疏水性特征的改变。
反义表达。Lox-1表达的减少还可通过在大麦细胞中的lox-1反义构建物的表达完成。已报道了在大麦中反义构建物的表达以减少靶蛋白质表达的方法,例如,Gilpin,M.J.等,1998,在:光合作用:机制和效应(Photosynthesis:Mechanisms and Effect),G.Garab,编,IV卷,2983-2986中;Kjaerulff等,1995在:光合作用:从光到生物圈(Photosynthesis:from Light to Biosphere),P.Mathis,编、II卷,151-154中。
大麦细胞可以用含有反义核酸序列的表达构建物转化。该表达构建物产生一种反义RNA分子,该反义RNA分子能够通过互补碱基配对,特异性地与从野生型lox-1基因产生的至少一部分的mRNA结合,并能够打断前一mRNA的剪接或这种mRNA的翻译。一个组成型或组织/时序特异性启动子,例如,上文描述的大麦lox-1启动子能够驱动反义核酸序列的表达。
化学诱变。化学诱变剂叠氮化钠(NaN3)通常用于大麦的诱变,并已知在大麦基因组的DNA(脱氧核糖核酸)序列中诱导稳定的突变(Olsen等,1993,Proc.Natl.Acad.Sci.USA,90:8043-8047)。其它化学诱变剂,例如,甲磺酸乙酯(EMS)、叠氮化甘油(AG,3-叠氮-1,2-丙二醇)、甲硝脲(MNU)、和马来酰肼(MH)也可用于诱导DNA突变(Rank,J.等,1997,Mutat.Res.390:121-7),也可使用紫外线照射。
如下文实施例中所显示的,大麦栽培变种(cv)Vintage和栽培变种Caruso的谷粒用叠氮化钠处理并通过自体受精繁殖到第三代(M3)。
4.低脂氧化酶大麦的鉴定和选择。
例如可通过脂氧化酶活性的分析实现在谷粒中具有降低的脂氧化酶异构酶活性的大麦植物的鉴定和选择。酶促分析可用于确定已知存在于成熟或发芽谷粒中的两种主要的脂氧化酶LOX-1和LOX2的活性。这种分析可区别LOX-1和LOX-2的活性。
LOX-1和LOX2的一种选择性分析是基于通过脂氧化酶的多不饱和脂肪酸的氧化作用,和这种氧化作用的氢过氧化产物的分光光度计检测。相对于LOX-2,LOX-1的这种分析之特异性是利用了LOX-1对于抑制剂的比较不敏感性,例如对NDGA。
选择性分析还可通过使用免疫沉淀选择性地从试验中除去LOX-1或LOX-2完成。特异性抗-LOX-1和抗-LOX-2抗体,例如,单克隆抗体,可如Holtman等,1996,出处同前,中描述的从纯化的LOX-1或LOX-2中制备。
可使这些分析方法适合于微滴定板试验步骤,或其它已知允许许多样品的快速筛选的可重复的、高通量的试验形式。这些试验可有效地以非破坏性形式筛选发芽谷粒的叶尖,使得在筛选中选择的籽苗可进一步繁殖。
在推定的突变中LOX-1活性的损失可通过酶活分析证实。例如,谷粒提取物可与亚油酸一起培养,并且例如使用反向HPLC分析亚油酸氧化产物。从亚油酸形成的9-HPOD和13-HPOD的相对含量提供了LOX-1活性的测定方法,其主要产物是9-HPOD。
如下文实施例所显示的,通过抑制剂存在的情况下的氧化分析以及通过免疫沉淀分析,筛选诱变处理的栽培变种Vintage和栽培变种Caruso的M3代的约20,000颗谷粒的LOX-1和LOX-2的活性。使用这些筛选方法,发现在栽培变种Vintage中具有一个LOX-1活性较大降低的突变体,并称之为品系G(Line G)。该突变体的表型可在M4和M5代遗传。
国立工业和海洋微生物保藏有限公司(National Collections ofIndustrial,Food and Marine Bacteria)(NCIMB),23St.MacharDrive,Avberdeen,AB243RY,Scotland,UK,在布达佩斯条约的条款下,于2001年1月4日保藏了从品系G大麦生产的种子,保藏号为:NCIMB41078。
5.遗传序列
用于说明在本发明大麦植物中低脂氧化酶表型的基因型改变的精确描述,在育种计划中对于鉴定具有这种遗传改变的植物和对于使这种遗传特性杂交进入其它大麦栽培变种是有用的。可使用各种已知的分子和生物化学方法确定低脂氧化酶表型的遗传基础。
通常认为顺式作用和反式作用遗传序列均可决定给定的基因在基因组中的表达以及该基因产物的活性。基因表达中的控制点包括时程调节、组织特异性和基因转录速度、转录物的稳定性和转录物翻译的速度。基因表达和稳定性的水平以及编码的酶的比活性水平将决定在组织中检测的酶活水平。
植物基因序列的改变可通过已知的基因组相关部分的DNA测序确定,而Northern印迹分析提供了一个在给定的植物组织中监测稳定的转录物水平的方法。可通过从组织中提取酶并测定酶活评估在植物组织中表达的酶。
如下文实施例所显示的,决定在栽培变种Vintage中诱导的品系G突变体之低脂氧化酶表型的遗传改变的同一性可以下面的方式确定。存在于亲本栽培变种Vintage和品系G中的编码LOX-1蛋白的结构基因通过聚合酶链反应(PCR)扩增,并且测定调节基因表达的上游启动子序列以及含有内含子和外显子的整个编码序列。
来自品系G和来自野生型栽培变种Vintage的lox-1基因的核苷酸序列的比较揭示在2个外显子中的2个核苷酸取代,其中之一(在位置+2347)导致在表达的蛋白质中非保守性氨基酸替换(甘氨酸368变为天冬氨酸)。
图22显示大豆(Gm:Glycine maxL)脂氧化酶LOX-1(编号P08170)、LOX-2(编号P08170)、LOX-3(编号AAB41272)和大麦(hv:Hordeum vulgare)脂氧化酶LOX-1(编号P29114)和LOX-2(编号AAB70865.1)的序列对比。保守的氨基酸残基和带电残基的保守性替换以粗体显示。在序列比对之上显示了大豆Glycine max的LOX-3的二级结构排布,其中H=α螺旋,E=β链,并且显示了与酶功能相关的残基(通过箭头或实心圆识别),如Skrzypczak-Jankun等,1997,Proteins29:15-31所描述的。
在大豆LOX-3中参与非血红素铁结合或催化作用所必须的氨基酸残基(*)包括:H518、H523、H709[3N原子];N713,I857。大麦LOX1中的等同残基是H517、H522、H708、N712和I862。在催化中具有预测作用的大豆LOX-3残基是(●):H266、H513、H776、F264、F272、F714、W519、R552、R726、D766、D779、K278。大麦LOX1中的等同残基是H261、H512、H775、F259、F267、F713、W518、R551、R725、D778和K273。
位于蛋白质二级结构的环和螺旋-加帽位置的脯氨酸(P86、109、167、171、223、234、291、311、324、343、345、371、381、382、486、541、548、600、616、627、685、726、734788、829、833839、857)和甘氨酸(G49、67、68、70、91、107、137、187、192、210、217、218、260、306、307、336、392、409、458、474、490、569、607、674、676、720、736、783、828、850、855)残基(+),可促进肽主链的急剧扭转和折叠。
相关植物的脂氧化酶的序列比对显示大麦LOX-1中的甘氨酸-368是强烈保守的。而且,对应于大豆LOX-1中的甘氨酸-353的这一残基是全部58个残基中的35个高度保守的残基之一,从其晶体结构可见,所述58个残基沿酶的底物腔II排列。在图22所显示的植物脂氧化酶序列比对中用框突出显示了这些保守残基(Minor等,1996,Biochemistry35:10687-10701),包括如下大麦LOX-1残基:Y224、L268、W355、E364、G368、V369、N370、I374、L424、L499、K501、A502、V504、D508、S509、H512、Q513、L514、H517、W518、H522、I556、L559、A560、L564、I565、I570、T574、S585、Q715、Y718、N724、R725、P726、T727、L772和I862。35个保守性残基中除7个外全部是中性或疏水性残基。大麦中甘氨酸-368位置上的带电残基或在另一个沿底物腔II排列的保守性中性或疏水性残基的置换,可能扰乱酶的结构和功能特性。大麦品系G LOX1(◆)中G变为D368的突变位于α-螺旋H6和β-链E12之间。
如图22所示,脂氧化酶家族的酶具有高度的序列保守性,这反映在对于包括大豆LOX1和LOX3的植物脂氧化酶家族的一些成员测定的保守的二级结构中,(Skrzypczak-Jankun等,1997,出处同前)。大麦LOX1与大豆LOX3具有56.9%的序列同一性,和67.8%的序列相似性。在大豆LOX3异构酶中的一些氨基酸残基被鉴定为非血红素铁的配体,或表明对它的活性是必须的(表示为*●)。由于大麦LOX1和大豆LOX3之间高度的序列保守性,有理由预测存在于与鉴定的对于LOX3的功能是重要的序列同源之大麦LOX1序列中的残基,对酶的活性也是关键的。因此,在任何这些位置上的非保守的氨基酸置换,包括对应于沿底物腔排列的大豆LOX3的35个高度保守残基的大麦LOX1中的那些残基的置换,以及在对酶活必需的其它位置上的置换,很可能降低脂氧化酶的活性。
已知当它们位于二级结构元件之间时,氨基酸残基脯氨酸和甘氨酸促进肽主链的扭转,这使蛋白质呈现折叠的三级结构。在螺旋加帽基序中脯氨酸和甘氨酸残基也是常见的(Parker和Hefford,1997,Protein Eng.,10:487-496,http://www.expasy.ch)。在品系G LOX1中的单一非保守置换,其中位于两个预测的结构元件之间的甘氨酸为天冬氨酸取代,则导致酶活性的明显损失。因此预测:位于结构元件外部的区域中,引起大麦LOX1中在一个或多个脯氨酸或甘氨酸残基上的非保守性氨基酸置换的LOX-1基因中的突变,可同样地防止天然蛋白质的折叠并因此降低编码的酶的活性。
因此,在一个实施方案中,具有降低是脂氧化酶1活性的本发明的有用的突变型大麦植物包含一个突变的核酸序列,该序列通过插入一个或多个酸性、碱性或极性氨基酸改变酶之底物腔的中性或疏水特性。例如,一个有用的核酸序列[SEQ ID NO:11编码大麦LOX-1蛋白质[SEQ ID NO:12]包含在氨基酸368从甘氨酸到Xaa的置换,其中Xaa是一个酸性、碱性、或极性氨基酸。本发明的大麦突变型LOX-1的一个特异性氨基酸序列是其中Xaa为天冬氨酸,例如品系G。
如下文实施例中所显示的,品系G中基因型的改变对lox-1基因的表达没有可察觉的影响,但是在品系G成熟和发芽谷粒中检测到的LOX-1活性约为亲本品系栽培变种Vintage的谷粒中检测到的9%。为了提供在品系G的LOX-1中的氨基酸突变是造成低-lox-1表型之原因的直接证据,在取自大麦糊粉的原生质体中,品系G lox-1和栽培变种Vintage lox-1的编码序列被瞬时表达,并且与野生型LOX-1酶相比显示出突变型LOX-1酶的活性大大降低了。
6.繁殖品系之间的转移
本发明基因型的大麦植物的遗传特征改变之检测用于鉴定在大麦品系中特定遗传特征的存在,并用于促进育种计划中繁殖品系之间该特征的转移。可利用各种分子方法检测染色体组序列中的改变。这些方法包括,但不限于,限制性片段长度多态性的检测(Gebhardt和Salamini1992,Int.Rev.Cytology.,135:201-237)和基于定量PCR的检测方法,例如使用荧光引物进行扩增,例如,TaqMan引物探测系统(Ibraham等,1998,Anal.Chem70,2013-2017)。检测方法的选择将取决于特定遗传特征,但较佳地应是快速的并提供清晰的可解释的数据。
如下文中实施例所显示的,提供了一个PCR-切割扩增的多态位点试验(PCR-Cleavage Amplified Polymorphic Site assay)(PCR-CAPS)用于品系G的突变型脂氧化酶-1基因的检测。在品系G的lox-1基因中位置+2347上核苷酸的置换导入由AatII限制性内切酶识别的额外的位点,这可通过PCR-CAPS试验检测到。适当的Lox-1检测方法不限于本试验,但可同样地基于TaqMan技术,以及其它已知的检测方法。
如下文的实施例所显示的,PCR-CAPS分析应用于来自回交程序的4代繁殖材料,其中品系G中低脂氧化酶表型系统地回交进入栽培变种Alexis。显示低脂氧化酶的表型随lox-1基因的遗传而遗传,并且该表型被鉴定为隐性的并只在对lox-1基因纯合的品系中见到。
因此,本发明的植物子代包括例如在回交程序中获得的、含有突变型lox-1和表达一个低脂氧化酶表型的繁殖品系。
7.酿造
本文证实本发明的大麦植物,包括具有低的脂氧化酶1活性的植物部分、植物子代、谷粒和例如麦芽和麦芽汁的植物产物,与由野生型对照大麦种类产生的饮料相比,可用于制备对于在一段有限的时间期限内,或在提高了贮藏温度的条件下,具有降低水平的游离反式-2-壬烯醛的饮料的生产。为了这些比较的目的,啤酒的亚硫酸盐含量控制在5ppm或以下,因为认为装瓶时较高的亚硫酸盐水平将暂时延缓游离反式-2-壬烯醛的出现。例如,从本发明描述的突变的大麦品系G得来的麦芽酿造的啤酒,与从对照的、非突变的大麦获得的麦芽酿造的啤酒相比,在一段有限的时间里具有稳定的器官感觉特性。
酿造试验和瓶装啤酒的评估提供了评估不同成分对成品啤酒的质量和稳定性的影响的最好的方法。为了检测不同大麦麦芽的影响,需要足够的大麦谷粒进行麦粒发芽和在中试规模和半生产规模上的酿造试验。在大麦增殖过程中,可评估大麦品系的野外使用特性。大麦品系的发芽特性可在中试或工业生产规模的发芽过程中评估,并较佳地应基于国家麦芽制造的质量推荐,例如欧洲酿酒协定(EuropeanBrewing Convention)对于麦芽制造质量的推荐(Analytica-EBC/European Brewing Convention,1998,Publ.Hans Carl Getranke-Fachverlag,Nurnberg,Germany)。中试或半工业生产规模酿造后,啤酒装入棕色瓶子中并冷却至5℃以最佳保存。在这一阶段,可由经过培训的、能够检测包括异味化合物反式-2-壬烯醛的特定啤酒风味的品尝小组分析鲜啤酒。另外,对于包括反式-2-壬烯醛的主要风味成分采用化学方法分析啤酒。然后在已知显示啤酒的长期贮藏稳定性的各种贮藏条件下,对啤酒重复这些啤酒质量分析的方法,例如强制熟化处理。
如下文实施例所显示的,为了在一个工业化麦芽制造坊获得10吨该品系的麦芽,经几个季节在野外增殖品系G的大麦。均具有野生型LOX-1表型的对照大麦品种栽培变种Vintage和栽培变种Nevada在相似的条件下进行发芽。从品系G和对照的大麦栽培变种获得的烘干的麦芽属于半工业酿酒试验所需的规格范围。
以30-h1的规模进行酿造试验,新装瓶啤酒的评估显示从品系G和对照栽培变种的麦芽酿造的啤酒含有的反式-2-壬烯醛的含量低于味觉阈,并且品尝小组认为是满意的。两种强制熟化处理(在37℃贮藏7天或在30℃贮藏6-12周)用于评估啤酒的香味稳定性。对于品尝小组的评价以及游离反式-2-壬烯醛的水平而言,发现从品系G的麦芽酿造的啤酒的香味稳定性比对照麦芽生产的啤酒的香味稳定性好,并发现该改进具有统计学显著性。
实施例
在下文的实施例中进一步详细说明本发明。应理解当所示优选的实施方案时,这些实施例仅以举例说明的方式给出。
实施例1 从诱变处理的大麦筛选和选择脂氧化酶异构酶突变体
1.大麦诱变
根据公开的方法用叠氮化钠诱变处理大麦Hordeum vulgare栽培变种Vintage和栽培变种Caruso的谷粒(Kleinhofs等,1978突变研究(Mutation Research)51:29-35)。突变处理将点突变导入基因组DNA,例如,可在编码的蛋白质中产生氨基酸的变化。突变的M1谷粒在温室中经过两代而增殖,收集M3谷粒用于筛选。根据Kleinhofs等,1978,出处同前,在M2代中观察到的单个基因形状突变的频率为每10,000个M2代的谷粒1.0-2.7个突变。既然大多数基因突变是隐性的并且仅在纯合状态下可被检测到,在M3代中筛选突变诱导的类群,其中预期的纯合突变体谷粒的比例将比较高。在M3突变诱导的材料中预期每10,000谷粒中0.9-2.3的突变频率。
2.在M3突变诱导的谷粒中脂氧化酶1(LOX-1)和脂氧化酶2(LOX-2)的活性的非破坏性分析
开发了一种用于检测具有降低的LOX-1活性的突变型大麦谷粒的快速筛选方法,并具有如下标准:筛选步骤应不阻止谷粒/籽苗的增殖;选择的谷粒/籽苗组织应表达可以计量的脂氧化酶活性水平;该试验应区别LOX-1活性和LOX-2的活性;试验步骤应包含多重样本。
发芽谷粒的不同组织中,即枝条、根、和胚的盾片组织以及胚乳,总脂氧化酶活性水平如下进行分析:通过在含有2mM NaN3,和0.5mM苯甲基磺酰基氟(PMSF)的冰冷的20mM Tris-HCl,pH7.5中匀浆化制备大麦籽苗组织的提取物,随后通过在1000g离心10分钟除去不溶解的物质。在25℃,通过加入2.9ml的20mM亚油酸底物分析100μl提取物中脂氧化酶的活性,该亚油酸(游离酸,L-1376,Sigma,USA)底物通过在5ml含有1%的Tween20的水中扩散35μl的亚油酸制备。随后进行分光光度反应,其中由于在氢过氧化物的产物中共轭二烯的形成,在234nm处吸光度(A234nm)增加的速度与存在的酶活性成比例。一个单位的脂氧化酶活性定义为ΔA234=在3-ml的反应中每分钟0.001,等价于0.12微摩尔亚油酸的氧化。
在黑暗中发芽4天的谷粒的叶组织具有可检测的脂氧化酶活性的最高水平(Holtman等,1996,植物生理Plant Physiology111:569-576)。这样选择4-天籽苗的叶尖用于非破坏性脂氧化酶筛选分析。在pH4.5和pH9.0之间测定总大麦脂氧化酶活性的最适pH,发现为pH6.5。因此,选择含有0.2M硼酸的25mM HEPES缓冲液(pH6.5)用于筛选分析。
既然在4日龄籽苗的枝条中免疫检测到LOX-1和LOX-2酶(Holtman等,1996,出处同前),使用LOX-1和LOX-2特异性分析试验。发现由Es kin等,1977,Crit.Rev.Food,科学与营养Science andNutrition9:1-40所鉴定的脂氧化酶抑制剂去甲二氢愈创木酸(NDGA)是大麦脂氧化酶的一个选择性的抑制剂。NDGA在1×10-5M时,强烈地抑制纯化的大麦LOX-2,而LOX-1保留了47%的活性(图1)。通过测定分别由LOX-1和LOX-2的亚油酸氧化产生的9-过氧羟基十八烷酸(9-HPOD)与13-过氧羟基十八烷酸(13-HPOD)的比,在叶尖分析中检验这种抑制剂的选择。在栽培变种Vintage叶尖的脂氧化酶分析中,加入1×10-5M NDGA,形成的13-HPOD的比例从24.5%下降至9.5%。
叶尖提取物中的LOX-2活性的选择性分析基于LOX-1-特异性单克隆抗体(5D2)的使用(Holtman等,1996,出处同前)以免疫沉淀存在于提取物中的LOX-1。LOX-1沉淀后在提取物中检测到的残余的脂氧化酶活性提供了LOX-2活性的测定方法。使用LOX-1和LOX-2的特异性单克隆抗体(分别表示为5D2和5.8),通过用ELISA分析定量提取物上清液中残余的LOX-1和OOX-2来评估该免疫沉淀的效率(Holtman等,1996,出处同前)。栽培变种Vintage叶尖提取物的LOX-1免疫沉淀除去了85%的LOX-1蛋白质和15%的LOX-2蛋白质。
在V底的96孔平板中,通过向20μl的各个叶尖提取物加入5μl5D2-包被的Dynabeads(Dynal)和75μl缓冲液[20mM Tris-HCl pH7.5,1%v/v,胎牛血清(HyClone)]进行免疫沉淀。该平板在滴定板摇床上(MTS4,IKA,Labor Technik)4℃下培养1小时。通过在Sigma302-K离心机上以2000rpm、4℃下离心10分钟得到免疫沉淀的沉淀。如下文所述,但要加入100μl分析缓冲液(25mM HEPES,0.2M硼酸,pH6.5),在平底96孔平板中分析从各个样本获得的上清液(70μl)的脂氧化酶活性。
使LOX-1和LOX-2分析适合于高流通量筛选方法。从8个4天龄发芽谷粒得到的叶尖(1cm)在150μl冰冷缓冲液(20mM Tris-HCl,pH7.5)中,在多孔匀浆器中分别匀浆化2×30秒(Berg等,1992,Electrophoresis13:76-81)。3000rpm离心15分钟后,40μl的各个提取物的上清液转移到平底96孔平板上。向各个孔中加入170μl缓冲液(25mM HEPES,0.2M、pH6.5的硼酸,1×10-5M NDGA)和10μl的底物(20mM亚油酸),然后在25℃培养20分钟。通过加入20μl饱和的碘化钾溶液(KI)使该反应终止并在25℃进一步培养8分钟。过氧羟二烯和KI之间的氧化还原反应产生I2,它通过在一个微量培养板读数器的350nm处的消光最大值得以监控(Multiskan MCC/340)。
3.诱变处理大麦的M3和M4谷粒中潜在的脂氧化酶1突变型的鉴定
栽培变种Vintage和栽培变种Caruso的M3代的谷粒在45℃贮存6.5天以打破休眠,保证95%的发芽频率。使栽培变种Vintage(9318)和栽培变种Caruso(9633)的M3谷粒发芽并筛选品系,该品系的LOX-1活性为15%或少于野生型谷粒。增殖推定的突变型品系(50栽培变种Vintage和42栽培变种Caruso品系)到M4代,收获之,并重新筛选发芽的谷粒。经过在从各个品系中获得的5个叶尖的提取物中测定脂氧化酶活性后,在一个栽培变种Vintage品系和六个栽培变种Caruso品系中证实了突变体LOX-1表型。当检验这7个推定的突变型的发芽胚中的LOX-1和LOX-2的活性时,只有栽培变种Vintage突变型(表示为品系G)显示LOX-1的大量减少。在成熟休眠的谷粒中,存在于胚中的脂氧化酶活性几乎全部为LOX-1的活性,归因于两种异构酶的不同表达形式(Schmitt和van Mechelen,1997,Plant Sci.128:141-150)。取自品系G成熟干谷粒(M5代)的胚提取物中总脂氧化酶活性与在栽培变种Vintage胚提取物中0.74±0.44U/mg蛋白质相比为0.06±0.04U/mg蛋白质,如实施例1的部分2中描述的使用分光光度法的脂氧化酶分析进行测定。发现品系G的M4和M5代的成熟胚中残余的脂氧化酶活性约为亲本品系的9%。
实施例2 品系G是具有低脂氧化酶表型的栽培变种Vintage突变体
在M5代的材料中分析品系G的农学特性和突变体表型。进行最初的分析以证实所分析的M5的材料为突变体表型纯合。在M3代检测到的品系G中低LOX-1表型可产生显性或隐性突变。如果在M3代选择的品系G对显性突变是杂合的,那么在其随后的几代中将显示表型的分离。测定取自M5代的休眠谷粒之26个品系G胚的脂氧化酶活性,并与栽培变种Vintage野生型胚比较。在所有的品系G胚中的脂氧化酶活性非常低,平均为每毫克蛋白质0.06±0.04U脂氧化酶,与之相比较在野生型栽培变种Vintage胚中平均为每毫克蛋白质0.74±0.44U脂氧化酶。这些数据证实M5代中的品系G对于低脂氧化酶的性状是纯合的。
1.品系G具有野生型植物生长生理学和谷粒发育
使品系G和栽培变种Vintage谷粒发芽并在人工气候室中,相对湿度为80%时,在有光照的条件下15℃培育16小时,及在黑暗的条件下12℃培育8小时。就植物的高度、每棵植物上分蘖枝的数量、开花的起始和每棵穗状花序上的谷粒数量而言,品系G和栽培变种Vintage植物的生长特征相似。从开花后(DAF)5天直到完全成熟(约90DAF)的发育过程中,品系G和野生型栽培变种Vintage的谷粒的鲜重(图2)和干重(图3)非常相近。
2.在整个发育过程中品系G的谷粒具有低脂氧化酶1表型
测定了品系G(M5代)和野生型栽培变种Vintage的发育大麦谷粒提取物中的脂氧化酶活性。谷粒在含有0.1%(v/v)Nonidet P-40(一种增强脂氧化酶提取的非离子去污剂)的冰冷的、pH7.5的20mMTris-HCl缓冲液中匀浆,并在15,000g离心20分钟除去不溶解的物质。提取物中脂氧化酶活性通过极谱分析方法,在含有1.2mM亚油酸的200μl氧饱和缓冲液(0.2M硼酸、25mM HEPES、pH6.5)中,25℃时,使用Clark型电极测定氧消耗而测定。在品系G和野生型谷粒的谷粒发育的最初20天脂氧化酶活性增加,但是在谷粒突变的过程中只有品系G的脂氧化酶的活性水平下降(图4)。
在亚油酸氧化的过程中形成的9-HPOD和13-HPOD的相对含量提供了在谷物提取物中测定LOX-1和LOX-2活性水平的方法。在这种情况下,从谷物提取缓冲液中略去Nonidet P-40,以避免氢过氧化物-消耗酶的共提取。100μl的该提取物与含有200μM亚油酸的10ml50mM pH6.5的磷酸缓冲液混合,培养20分钟。通过调整pH到3.5终止反应,加入一个内标物。试验中形成的氢过氧化物结合到一个十八烷基固相柱上(Bakerbond,Baker)并用甲醇洗提。然后,以由Aarle等,1991,FEBS Letters280:159-162描述的方法,0.5ml/min的流速,通过在C-18柱上的反相HPLC分离9-HPOD和13-HPOD,该C-18柱采用恒溶剂洗脱溶液(四氢呋喃∶甲醇∶H2O∶乙酸;25:30:44.9:0.1(v/v)用浓氨水调节到pH5.5)洗脱。在234nm检测到氢过氧化物,依照内标物前列腺素B2校正HPOD峰值。
图5显示在最初20DAF过程中13-HPOD是存在于谷粒中的脂氧化酶活性的主要产物,而在谷粒成熟过程中由脂氧化酶活性形成9-HPOD。而品系G和野生型谷粒的提取物都具有相似的13-HPOD合成活性的特征,品系G在9-HPOD合成活性上没有显示野生型的增加。这些数据与品系G大麦谷粒成熟过程中LOX-1活性的损失是一致的。
3.品系G谷粒在发芽时具有低脂氧化酶1表型
如实施例1所描述的在15℃时分析发芽谷粒的胚提取物中总脂氧化酶的活性。存在于休眠的野生型谷粒中的脂氧化酶活性在发芽的最初4天下降,然后提高(图6)。在品系G中,休眠谷粒中的脂氧化酶活性非常低,但4天后提高。
在发芽中的胚中由脂氧化酶活性形成的HPOD的分析显示,9-HPOD是存在于休眠野生型谷粒中的脂氧化酶的主要产物(图7)。随着提取物中脂氧化酶活性的减少9-HPOD形成的水平下降。4天后脂氧化酶活性的提高伴随9-HPOD和13-HPOD的形成。品系G休眠谷粒中低脂氧化酶活性与HPOD形成的缺乏有关,而4天后活性的提高主要产生13-HPOD。这些数据提供了如下证据:导致9-HPOD形成的LOX-1活性在品系G的发育、休眠和发芽的大麦谷粒的胚中大大减少,而导致13-HPOD形成的LOX-2活性在品系G中没有变化。
实施例3 品系G具有引起低脂氧化酶表型的突变型脂氧化酶1基因 (lox-1)
为了提供突变体的完整的描述,调查研究了品系G的低-LOX-1表型的分子基础。进行下面的分析以提供完整的表型特征:
1.在正在发育和正在发芽的品系G的谷粒中合成脂氧化酶-1
如实施例2所描述的,取自发育中和发芽中的大麦谷粒的胚提取物的蛋白质印迹分析与脂氧化酶活性的测定平行进行。根据Laemmli,1970,Nature227:680-685,使用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)分离粗提物。根据Towbin等,(1979)Proc.Natl.Acad.Sci.USA,76:4350-4354,使用半干印迹将分离的蛋白质转移到硝化纤维。如Holtman等,1996,Plant Physiology111:569-576所描述的,以500倍稀释的LOX-1特异性单克隆抗体5D2探测印迹,随后用与碱性磷酸酶偶联的山羊抗-小鼠抗体培养,并如Holtman等,1996,Plant Physiology111:569-576所描述的,用碱性磷酸酶底物硝基蓝四唑和5-溴-4-氯-3吲哚-磷酸盐检测。蛋白质印迹分析显示在栽培变种Vintage胚中从10DAF开始,在发育的谷粒只能感检测到LOX-1蛋白质,并且其水平在谷物成熟的过程中增长(图8)。该蛋白质也存在于栽培变种Vintage休眠的谷粒的胚中,但在发芽过程中缓慢下降(图9)。尽管在品系G胚的提取物中识别出LOX-1,并且作为与栽培变种Vintage LOX-1相似大小的蛋白质在SDS-PAGE中迁移,品系G中蛋白质的免疫可检测水平比栽培变种Vintage中的略低。
2.在发育和发芽的品系-G的谷粒中表达lox-1基因
根据Hensgens和van Os-Ruygrok,1989,Rice Genet.Newslett,6:163-168,的方法从发育中和发芽中的大麦谷粒的胚分离总RNA,同时如实施例2中描述的平行测定脂氧化酶的活性。在变性琼脂糖凝胶上分离RNA样品(7.5μg),并如Sambrook等,1989,在分子克隆(Molecular Cloning),实验室手册(Laboratory Manual),ColdSpring Harbour Laboratory Press,NY,所描述的进行Northern印迹分析。如Holtman等,1996 Plant Physiol.111:569-576所描述的,使用Amersham Random Prime Kit,使印迹与从大麦3’未翻译区,lox1cDNA(EMBL编号:L35931)的核苷酸2659-2801[SEQ ID NO:1]产生的32p-标记的探针杂交。
从30DAF开始,在发育中的和成熟栽培变种Vintage和品系G谷粒的胚中检测编码LOX-1的lox-1转录物(图10)。在栽培变种Vintage和品系G的胚中,发芽过程中lox-1转录物的水平增加,表明lox-1基因的从头表达(图11)。因为lox-1转录物可检测的水平在品系G和栽培变种Vintage胚中是相似的,lox-1转录的减少或转录物稳定性的降低都不能解释品系G的低脂氧化酶表型。
3.品系G的lox-1基因编码一个脂氧化酶-1的突变型
分析并比较品系G和栽培变种Vintage的lox-1基因的核苷酸序列以测定品系G的低-LOX-1表型的分子基础,其特征是lox-1基因正常转录,但减少谷粒中表达的脂氧化酶的累积和活性。
根据Pich和Schubert1993,Nucleic Acids Res.21:3328描述的方法,从籽苗的叶组织中分离品系G和野生型栽培变种Vintage基因组DNA。使用基于大麦lox-1基因的序列的引物,通过聚合酶链反应(PCR)扩增基因组DNA制剂中的lox-1基因(van Mechelen等,1995,BBA1254:221-225;Rouster等,1997,Plant J.11:513-523)。在图12中表示的用于扩增lox-1启动子和编码区的寡核苷酸引物的位置和序列如下:
正向引物5’-GAA AAG CTT GGA GGT AGA CGC TGC-3’[SEQID NO:2]和反向引物5’-TAT AGG ATC CTT GTT CTT GGC CTCCTC TCC TCG-3’[SEQ ID NO:3]用于PCR扩增品系G和栽培变种Vintage lox-1的lox-1启动子结构域(-361到+68)。
正向引物5’-AGT GAA AAA CAG TGT GCT GGT G-3,[SEQ IDNO:4]和反向引物5’-GGC TTA AAG AGC AACTGC TGA-3’[SEQID NO:5]用于PCR扩增品系Glox-1编码区。
正向引物5’-CAA G AT GCA TAT GCT GCT GGG AG-3’[SEQ IDNO:6]和反向引物5’-CGA TGG TTT AAA TTA GAT GGA GAT GCTGT-3’[SEQ ID NO:7]用于PCR扩增栽培变种Vintagelox-1编码区。
根据酶提供者的说明,PCR反应由含有50pmol引物的50μl体积中的250ng基因组DNA和2U Pfu DNA聚合酶(Promega)组成。在Stratagene Robocycler中进行PCR扩增:94℃1分钟,1轮;94℃1分钟、62℃2分钟和72℃5分钟,30轮;72℃10分钟,1轮。在1.2%的琼脂糖凝胶上分离PCR产物。在长度上对应于扩增区域的DNA片段使用Qiax II凝胶提取试剂盒(Qiagen)纯化,并克隆进入质粒pcDNA2.1(Invitrogen)。用特异性寡核苷酸引物,使用双脱氧核苷酸链终止反应测定克隆的lox-1启动子和编码区的链的核苷酸序列,并在ABI PRISM310Genetic Analyzer(PE Biosystems)上分析。使用DNA STAR序列分析软件包(DNA STAR Inc.,USA)进行序列比较。
大麦lox-1编码区的启动子区和内含子-外显子结构显示于图13中,并从野生型lox-1基因组的核苷酸序列与cDNA序列的比较推断得到(图12)。从-363到+68的lox-1启动子区的测定序列的区域(相对于测定的转录起始位点编号;van Mechelen等,1995,BBA1254:221-225)足以指导天然基因的胚特异性和时序调节的基因表达特征(Rouster等,1998,Plant J.15:435-440)。野生型lox-1基因的启动子和转录的区域[SEQ ID NO:8]长度为4663nt,并含有长度在82nt和634nt之间的6个内含子,它们不存在于各自的cDNA中[SEQ IDNO:10],因此在RNA转录剪接过程中必须除去。
品系G的lox-1的核苷酸序列与野生型的核苷酸序列比较(图12)显示品系G lox-1等位基因具有两个点突变。一个是在外显子1中的位置221上沉默型C到T的置换,第二个是在外显子3中的位置2347上G到A的置换(图13)。野生型大麦lox-1基因编码具有862个氨基酸残基的蛋白质[SEQ ID NO:9],而在品系G的lox-1等位基因的位置2347上的突变引起在编码的蛋白质中残基368上甘氨酸到天冬氨酸的氨基酸置换。
相关植物的脂氧化酶的序列比对显示,大麦LOX-1中的甘氨酸-368是强烈保守的。而且,对应于大豆LOX-1中的甘氨酸353的该残基是51个中性或疏水性残基之一,从其晶体结构上看,该中性或疏水性残基沿酶的底物腔排列(Minor等,1996,Biochemistry 35:10687-10701)并在图22中显示。因此,在该位置上带电氨基酸残基的插入可能扰乱该酶的结构和功能特性。
4.由品系G的lox-1等位基因编码的突变的LOX-1蛋白质具有低酶活,并是品系G的低脂氧化酶表型的原因。
在品系G中诱导突变的lox-1等位基因的栽培变种Vintage谷粒的叠氮化钠的诱变处理,可能在品系G的基因组中诱导了额外的突变。使用了两个实验方法以证实品系G中突变型lox-1等位基因,而不是基因组中的其它突变,是其低脂氧化酶表型的原因。为了证实在突变的酶中甘氨酸到天冬氨酸的置换引起稳定性和活性的降低,测定了由突变体和野生型lox-1等位基因编码的LOX-1的酶活性。在从吸涨的成熟谷粒中分离的糊粉原生质体中瞬时表达两个lox-1基因,因为预期在这些细胞中内源脂氧化酶的表达的水平低于检出限。在糊粉组织中没有检测到任何在发芽大麦中表达的经鉴定的大麦脂氧化酶基因(vanMechelen等,1999,出处同前)。为了指导在糊粉原生质体中lox-1基因的瞬时表达,其编码区域与已知在这些原生质体中有活性的组成型启动子翻译性融合。
在质粒pcDNA2.1中克隆的突变体编码区(序列位置+1到+4350)和野生型lox-1基因的编码区(序列位置+69到+4230)以及野生型lox-1cDNA的编码区(序列位置+69到+2654)(见部分3)在载体的多位点接头中通过KpnI和EcoRV的酶切消化切除。在pUBARN质粒中(Jensen等,1998,Hereditas129:215-225),组成型活化的玉米泛素Ubi启动子(如美国专利005510474A中所描述的)和Nos终止子之间克隆编码区,取代编码膦丝菌素乙酰基转移酶的bar基因。
根据Skriver等,1991,Proc.Natl.Acad.Sci.USA,88:7266-7270的方法从吸涨的大麦喜玛拉雅栽培变种(Hordeum vulgare cvHimalaya)的糊粉组织中分离原生质体。在0℃时,用100μg质粒DNA(等摩尔量的各个质粒)通过聚乙二醇(PEG)介导的DNA吸收转染等分试样的2×105个原生质体(Lee等,1997,Plant Mol.Biol.13:21-29),然后如前面所描述的(Skriver等,1991,出处同前)在25℃下,培养于糊粉原生质体培养基中。经过48小时的培养后,仔细地除去培养基,重悬浮原生质体,并在300μl的脂氧化酶分析缓冲液中(0.2mM硼酸、25mM HEPES、pH6.5)匀化。匀浆液以15,000g离心5分钟以沉淀不溶解的物质,随后使用实施例1中第1节描述的快速筛选方法(但除去NDGA抑制剂),分析上清液(10μl)的总脂氧化酶活性。原生质体提取物中的蛋白质含量通过由Bio-RadLaboratories,Hercules,California,USA提供的Bradford染料-结合实验测定(Bradford1976,Anal.Biochem.,72:248),并且脂氧化酶活性表达为提取物中每毫克蛋白质的活性。
用对照的质粒,pUBI-GUS转染的原生质体(其中玉米泛素-1启动子指导β-葡糖醛酸酶报道基因的表达)没有给出任何可检测的脂氧化酶活性。野生型lox-1基因和原生质体中的cDNA的瞬时表达都在原生质体提取物中给出高水平的脂氧化酶活性(图15)。与基因组序列相比,野生型lox-1 cDNA较高的表达可能是由于对于较小的lox-1cDNA表达质粒较高的转染频率(4929bp对6505bp的lox-1基因构建物)。突变型lox-1基因的瞬时表达给出低水平的脂氧化酶活性,是野生型脂氧化酶活性的~10%。这些数据清楚表明品系G中的突变型lox-1基因编码具有大大降低活性的脂氧化酶,这是低脂氧化酶表型的原因。
实施例4 扩增的PCR-切割多态性位点(PCR-CAPS)分析:用于突变型lox-1基因鉴定的一种方法
基于PCR-CAPS分析,开发了在任何遗传背景下鉴别品系G突变型lox-1基因的分析方法。该分析方法包括基因组DNA片段的PCR扩增,随后通过用特异性的限制性内切核酸酶消化扩增的序列,显示限制性片段的长度多态性(RFLP)。
突变型lox-1基因的编码序列包含两个点突变(见实施例3),其中在位置2347的突变(图12)引入一个附加的Aat II限制性内切核酸酶切割位点,这在野生型lox-1基因中没有发现(图16)。基于由在lox-1基因中的该限制性位点的存在产生的多态性,显示随后的PCR-CAPS分析,以区别野生型lox-1基因和突变的lox-1基因。
根据Pich和Schubert(1993,出处同前)的方法从大麦(Hordeumvulgare,L.)栽培变种Vintage和品系G的M6籽苗的嫩叶分离基因组DNA。使用对lox-1基因特异性的引物通过PCR扩增包含位置2347的DNA序列(品系G的lox-1基因突变位点)[SEQ ID NO:8]。通过选择的正向引物5’-CGCTACGACGTCTACAACGA-3’[SEQ IDNO:13]和反向引物5’-CAGACTACTTTTTGGCGGGA-3’扩增的DNA片段显示于图17中。根据提供者的说明,使用存在于含有50pmol的各个引物和1单位Taq DNA聚合酶(Promega)的50μl体积中的250ng基因组DNA进行PCR反应。在Stratagene Robocycler上进行PCR扩增:94℃1分钟,1轮;94℃1分钟,60℃1.5分钟,72℃2分钟,30轮;72℃10分钟,1轮。突变体和野生型lox-1基因的扩增的片段为~650bp(图18),对应于预期的大小(图17)。在自旋柱(Qiagen)上纯化的PCR产物在37℃用25单位的AatII限制性内切酶消化24小时,并在1.2%的琼脂糖凝胶上分析。
野生型lox-1PCR产物的消化产生10,179和462bp的DNA片段,从突变型lox-1PCR产物得到的片段为10,149,179和313bp,其中额外的DNA片段是由于lox-1PCR产物的部分消化(图19)。该片段型式与从这一突变产生的预期的RFLP一致,其中313bp的片段对于突变型lox-1是独特的。该PCR-CAPS分析提供了用于在大麦中鉴定lox-1等位基因的可重复的和特异性的手段,并因此能够用于以引导该基因进入新的大麦品种为目的的大麦育种计划。
实施例5 回交品系G的低脂氧化酶表型到栽培变种Alexis显示对突变型lox-1基因的遗传连锁
重复回交用于从品系G转移低脂氧化酶表型进入回归亲本(在这种情况下是cv.Alexis)。图20中显示了该回交程序,与低脂氧化酶表型的选择结合,使用回归亲本基因组渐进置换品系G的基因组。而且,将除去叠氮化钠诱变处理过程中引入品系G基因组的其它突变。在纯合的低脂氧化酶品系G(表示为基因型ll)与栽培变种Alexis(表示为基因型LL)的第一次回交中,子代品系将是杂合的(表示为基因型Ll)。归因于隐性突变的低脂氧化酶表型在杂合品系中将检测不到突变。该后代自花传粉并将给出正常的孟德尔分离种群,即1LL:2Ll:1ll。从第一次回交产生的低lox杂合ll基因型将具有50%的栽培变种Alexis遗传背景。经过10轮的回交,回归亲本的背景将达到约99.9%。
大麦(Hordeum vulgare,L.)栽培变种Alexis和品系G在温室中通过回交程序增殖。回交的子代谷粒在培养皿中滤纸上发芽,用4ml的水浸泡,在22℃黑暗中3天。如实施例1中描述的,通过测定取自发芽籽苗的胚鞘(顶端7mm)的提取物中总脂氧化酶活性筛选低脂氧化酶品系。还使用实施例4中描述的PCR-CAPS试验分析第三次和第四次回交的子代的突变型lox-1基因的遗传。
四次回交后代的分离子代中低脂氧化酶表型的预期的频率对于隐性突变是25%。观察到的四次回交后代的子代(24个谷粒)中低脂氧化酶活性的频率与预期的频率一致(图20)。当使用PCR-CAPS试验分析具有低lox纯合ll基因型的第三次和第四次回交子代时,发现它们都具有诊断性313bp的片段,而具有野生型脂氧化酶活性的子代缺乏这一片段(图21)。
该回交程序证实突变型lox-1等位基因可转移到新的遗传背景,并在孟德尔分离后以隐性单因子形式遗传。既然在第四次回交子代中回归亲本的背景是93.8%,突变型lox-1基因和低脂氧化酶表型的共遗传提供了它们遗传连锁的证据。
实施例6 从品系G的大麦麦芽酿造的啤酒在贮藏过程中累积较少的反式-2-壬烯醛,给予改善的风味稳定性
为了提供充足的谷粒用于工业化发麦芽,大麦栽培变种Vintage和品系G野外增殖几个季节。随后进行工业规模的麦芽制造和酿造试验以及制成啤酒的分析有以证实品系G低脂氧化酶大麦对于改善香味稳定性的评价。
1.品系G和栽培变种Vintage的工业化发麦芽和焙烤
在一个工业的麦芽坊中以如下两个试验进行10吨范围的麦芽制造:
试验1:品系G大麦谷粒(1996年收获)
浸泡条件:8小时湿润;14小时干燥;8小时湿润;10小时干燥;在16℃的浸泡水中4小时湿润。发芽条件:18℃12小时;16℃24小时;14℃24小时;12℃60小时。焙烤条件:60℃12小时;68℃3小时;74℃4小时;80℃3小时。
试验2:栽培变种Vintage和品系G(1996年/1997年收获)
浸泡条件:8小时湿润;10小时干燥;6小时湿润;15小时干燥;在15℃的浸泡水中4小时湿润。发芽条件:在15℃使用入口空气5天并喷雾保持湿气水平。焙烤条件:50℃10小时;60℃2小时;80℃2.5小时。
从试验1获得的品系G麦芽的2个样本与对照麦芽栽培变种Nevada(表1)比较和从试验2获得的品系G麦芽与栽培变种Vintage比较的发芽分析证实品系G麦芽适合于酿造试验。
表1
| 发麦芽试验1 |
|
|
|
|
| 大麦品种 |
|
栽培变种Nevada |
品系G |
品系G |
| 收割年份 |
|
1996 |
1996 |
1996 |
|
|
|
|
|
|
| 麦芽分析 |
|
|
|
|
| 水分含量 |
% |
4.7 |
4.3 |
4.4 |
| 提取物fine as is |
%ai. |
76.9 |
76.1 |
75.3 |
| 提取物fine d.m. |
%dm. |
81.4 |
79.5 |
78.7 |
| 糖化作用时间 |
分钟 |
<10 |
<10 |
<10 |
| 糖化力 |
%WK |
252 |
373 |
365 |
| 颜色 |
EBC |
2.8 |
4.4 |
3.8 |
| pH |
|
6.16 |
5.97 |
5.99 |
| 混浊度 |
EBC |
9.0 |
2.5 |
2.4 |
| 总蛋白d.m. |
% |
10.35 |
10.74 |
12.03 |
| 可溶性氮 |
mg/l |
647 |
787 |
765 |
| 可溶性蛋白质%麦芽d.m. |
%dm. |
3.7 |
4.4 |
4.3 |
| Kolbach |
|
35.3 |
40.8 |
35.4 |
| 游离氨基氮 |
mg/l |
97 |
125 |
118 |
| 易碎性 |
% |
89.5 |
85.6 |
89.5 |
| 完全未修饰谷粒 |
% |
1.1 |
0.6 |
0.5 |
| 部分未修饰谷粒 |
% |
2.3 |
1.0 |
0.6 |
| 麦芽汁中的β-葡聚糖 |
mg/l |
114 |
66 |
36 |
| 麦芽中的β-葡聚糖 |
%w/w |
0.24 |
0.11 |
0.05 |
| S-甲基甲硫氨酸/DMS eq |
μg/g |
2.4 |
6.4 |
8.4 |
| 游离DMS |
μg/g |
1.0 |
6.6 |
4.7 |
| HDNA |
μg/kg |
n.d. |
0.3/0.6 |
0.3/0.3 |
表2
| 发麦芽试验2 |
|
|
|
|
| 大麦品种 |
|
Vintage |
品系G |
品系G |
| 收割年份 |
|
1996 |
1996 |
1997 |
|
|
|
|
|
|
| 水分含量 |
% |
4.1 |
4.1 |
4.3 |
| 提取物fine as is |
%ai. |
77.0 |
75.6 |
77.5 |
| 提取物fine d.m. |
%dm. |
80.3 |
78.8 |
80.9 |
| 精细/粗糙差别 |
%dm. |
0.7 |
1.6 |
1.7 |
| 糖化作用时间 |
分钟 |
- |
- |
<10 |
| 糖化力 |
%WK |
343 |
342 |
268 |
| 颜色 |
EBC |
2.5 |
2.8 |
3.4 |
| pH |
|
6.05 |
6.01 |
6.12 |
| 混浊度 |
EBC |
1.5 |
1.3 |
2.5 |
| 总蛋白d.m. |
% |
10.98 |
12.22 |
9.82 |
| 可溶性氮 |
mg/l |
696 |
741 |
610 |
| 可溶性蛋白质%麦芽d.m. |
%dm. |
3.9 |
4.1 |
3.4 |
| Kolbach |
|
35.2 |
33.7 |
34.6 |
| 游离氨基氮 |
mg/l |
110 |
117 |
100 |
| 易碎性 |
% |
91.3 |
81.8 |
89.5 |
| 完全未修饰谷粒 |
% |
0.7 |
1.1 |
1.3 |
| 部分未修饰谷粒 |
% |
1.0 |
2.7 |
2.7 |
| 麦芽汁中的β-葡聚糖 |
mg/l |
97 |
172 |
117 |
| 麦芽中的β-葡聚糖 |
%w/w |
- |
- |
0.3 |
| S-甲基甲硫氨酸/DMS eq |
μg/g |
- |
- |
- |
| 游离DMS |
μg/g |
- |
|
- |
2.用品系G、栽培变种Vintage麦芽和对照麦芽栽培变种Nevada进行工业酿造
使用从试验1中的品系G和对照麦芽栽培变种Nevada麦芽以及试验2中的品系G和对照麦芽栽培变种Vintage制备的麦芽汁进行两批酿造试验。
根据如下方案使用475kg麦芽以30-h1的工业规模酿造啤酒:在50℃捣碎;50℃30分钟;从50℃加热到70℃,30分钟;70℃15分钟。一部分麦芽汁从70℃加热到100℃20分钟,并在100℃5分钟,而主要部分的麦芽汁再在70℃保持25分钟,然后两部分麦芽汁混合并在76℃保持10分钟。麦芽汁的沸腾、余下谷粒的涡流分离、冷却、发酵、窖藏和包装入棕色玻璃瓶中等酿酒步骤是根据标准酿造方法进行的。
3.品系G、栽培变种Vintage麦芽和对照麦芽栽培变种Nevada酿制的啤酒的风味稳定性和T2N的含量
新装瓶的啤酒在5℃贮存,并在生产2个月内分析。遵循两个不同类型的啤酒贮藏条件,在两个独立的实验室中评估新鲜啤酒和贮藏啤酒的香味稳定性。在实验室A中使啤酒经过强制熟化过程,即该啤酒在37℃贮藏7天的时间,而在试验室B中该啤酒在30℃贮藏6和12周。基本上如Gronqvist等,1993,Proceedings of the 24th EBCCongress,Oslo,421-428所描述的,在用O-(2,3,4,5,6-五氟苯甲基)-羟胺的羰基衍生作用后,使用气相色谱分析和质谱检测测定啤酒中反式-2-壬烯醛水平。一个经过培训的啤酒品尝小组评估了啤酒全面的味道得分,包括显示啤酒中游离反式-2-壬烯醛存在的纸板味的检测。
实验室A:强制熟化耐受性
在第一次酿酒试验中,从品系G酿制的啤酒与对照麦芽,栽培变种Nevada酿制的啤酒的比较(表3)证明从品系G酿制的啤酒具有更强的香味稳定性,并且强制熟化后当与对照比较时,具有较低的反式-2-壬烯醛含量。第二个试验,比较从品系G麦芽酿制的啤酒和从栽培变种Vintage麦芽酿制的啤酒,该亲本栽培变种证实了原始数据(表4)。
表3
| 酿造试验1 |
|
|
| 大麦麦芽 |
栽培变种Nevada |
品系G |
| SO<sub>2</sub>含量(mg/ml) |
1 |
1 |
| T2N<sup>**</sup>(ppb)-新鲜啤酒 |
0.009 |
0.005 |
| T2N<sup>**</sup>(ppb)-熟化啤酒(37℃/7天) |
0.117 |
0.025 |
| 香味<sup>*</sup>-新鲜啤酒 |
5.9 |
5.3 |
| 香味<sup>*</sup>-熟化啤酒(37℃/7天) |
1.3 |
5.1 |
*增加质量的香味评估范围1-10;**反式-2-壬烯醛。
表4
| 酿造试验2 |
|
|
| 大麦麦芽 |
栽培变种Vintage |
品系G |
| SO<sub>2</sub>含量(mg/ml) |
2 |
2.5 |
| T2N<sup>**</sup>(ppb)-新鲜啤酒 |
0.023 |
0.019 |
| T2N<sup>**</sup>(ppb)-熟化啤酒(37℃/7天) |
0.078 |
0.035 |
| 香味<sup>*</sup>-新鲜啤酒 |
5.9 |
6.1 |
| 香味<sup>*</sup>-熟化啤酒(37℃/7天) |
2.9 |
5.9 |
*增加质量的香味评估范围1-10;**反式-2-壬烯醛。
实验室B:30℃贮藏耐受性
在贮藏温度提高到30℃的条件下放置6和12周后,当与从任何两个参考麦芽酿制的啤酒比较时,从品系G麦芽酿制的啤酒具有较低的反式-2-壬烯醛水平(表5和6),并且由品尝小组判定具有较好的香味稳定性。在这些分析的啤酒中,反式-2-壬烯醛的味觉阈接近0.08ppb。
表5
| 酿造实验1 |
|
|
| 大麦麦芽 |
栽培变种Nevada |
品系G |
| 反式-2-壬烯醛(ppb)—新鲜啤酒 |
0.050 |
0.044 |
| 反式-2-壬烯醛(ppb)-30℃/6周 |
0.072 |
0.037 |
| 反式-2-壬烯醛(ppb)-30℃/12周 |
0.095 |
0.046 |
| 反式-2-壬烯醛味道<sup>*</sup>—新鲜啤酒 |
0.6 |
0.3 |
| 反式-2-壬烯醛味道*—30℃/6周 |
3.7 |
1.4 |
| 反式-2-壬烯醛味道<sup>*</sup>—30℃/12周 |
2.5 |
0.6 |
*反式-2-壬烯醛味道检测范围在1-10的范围内。
表6
| 酿造实验2 |
|
|
| 大麦麦芽 |
栽培变种Vintage |
品系G |
| 反式-2-壬烯醛(ppb)—新鲜啤酒 |
0.070 |
0.062 |
| 反式-2-壬烯醛(ppb)-30℃/6周 |
0.093 |
0.070 |
| 反式-2-壬烯醛(ppb)-30℃/12周 |
0.133 |
0.080 |
| 反式-2-壬烯醛味道*—新鲜啤酒 |
0.3 |
0.9 |
| 反式-2-壬烯醛味道<sup>*</sup>—30℃/6周 |
2.5 |
1.7 |
| 反式-2-壬烯醛味道<sup>*</sup>—30℃/12周 |
2.2 |
1.3 |
*反式-2-壬烯醛味道检测范围在1-10的范围内。
从组合的酿造实验数据可见,如通过在30℃贮存后检测啤酒中反式-2-壬烯醛的水平测定的从品系G麦芽酿制的啤酒的改进的香味稳定性,显示了统计学的显著性(表7)。
表7
贮藏啤酒中的反式-2-壬烯醛
| 新鲜啤酒 |
平均值 |
标准偏差 |
差异 |
2-tailed p |
显著性(p<0.05) |
| 参考 |
0.060 |
0.012 |
0.007 |
0.34 |
无 |
| 品系G |
0.053 |
0.011 |
|
|
|
| 6周,30℃ |
平均值 |
标准偏差 |
差异 |
2-tailed p |
显著性(p<0.05) |
| 参考参考 |
0.083 |
0.013 |
0.029 |
0.031 |
有有 |
| 品系G |
0.054 |
0.021 |
|
|
|
| 12周,30℃ |
平均值 |
标准偏差 |
差异 |
2-tailed p |
显著性(p<0.05) |
| 参考 |
0.114 |
0.023 |
0.051 |
0.003 |
有 |
| 品系G |
0.063 |
0.020 |
|
|
|
因为在两个酿造试验中天然亚硫酸盐的水平很低,熟化啤酒中游离反式-2-壬烯醛水平将密切反映不同啤酒的反式-2-壬烯醛的潜能,即存在于新鲜啤酒中的反式-2-壬烯醛加合物的水平。通过与游离反式-2-壬烯醛混合,添加亚硫酸盐可暂时延缓熟化过程,直到通过包装的气体交换造成的氧化作用使亚硫酸盐的水平下降。
该描述的用低-LOX-1大麦麦芽进行的酿酒试验提供了第一次明确的证据,即在发芽和酿造过程中,大麦中LOX-1的活性是熟化啤酒中异味化合物反式-2-壬烯醛出现的关键性决定因素。
上述说明包括引用大量的发表的文献。如充分阐明的,为了本发明的全部目的,本文引用每一篇发表的文献作为参考。
序列表
<110>Douma, Anneke
Doderer,Albert
Cameron-Mills,Verena
Skadhauge,Birgitte
Bech,Lene
<120>低脂氧化酶1大麦
<130>11225.12W001
<160>18
<170>PatentIn version3.O
<210>1
<211>143
<212>DNA
D<213>大麦(Hordeum Vulgare)
<400>1
<210>2
<210>24
<212>DNA
<213>大麦
<400>2
<210>3
<211>33
<212>DNA
<213>大麦
<400>3
<210>4
<211>22
<212>DNA
<213>大麦
<400>4
<210>5
<211>21
<212>DNA
<213>大麦
<400>5
<210>6
<211>23
<212>DNA
<213>大麦
<400>6
<210>7
<211>29
<212>DNA
<213>大麦
<400>7
<210>8
<211>4663
<212>DNA
<213>大麦
<400>8
<210>9
(211>862
(212>PRT
<213>大麦
<400>9
<210>10
<211>2818
<212>DNA
<213>大麦
<400>10
<210>11
<211>4663
<212>DNA
<213>大麦
<220>
<221>variation
<222> (2346)..(2348)
<223>“n”is a,c,t,or g encoding an acidic,basic,or polar aminoa(
1
<400>11
<210>12
<211>862
<212>PRT
<213>大麦
<220>
<221>variation
<222> (368)..(368)
<223>“xaa”is an acidic,basic,or polar amino acid
<400>12
<210>13
<211>20
<212>DNA
<213>大麦
<400>13
<210>14
<211>20
<212>DNA
<213>大麦
<400>14
<210>15
<211>839
<212>PRT
<213>Glycine max
<400>15
<210>16
<211>865
<212>PRT
<213>Glycine max
<400>16
Ile
865
<210>17
<211>857
<212>PRT
<213>Glycine max
<400>17
<210>18
<211>864
<212>PRT
<213>大麦
<400>18