WO2022102595A1 - 形質転換体並びにそれを用いるカロテノイド組成物の製造方法 - Google Patents
形質転換体並びにそれを用いるカロテノイド組成物の製造方法 Download PDFInfo
- Publication number
- WO2022102595A1 WO2022102595A1 PCT/JP2021/041098 JP2021041098W WO2022102595A1 WO 2022102595 A1 WO2022102595 A1 WO 2022102595A1 JP 2021041098 W JP2021041098 W JP 2021041098W WO 2022102595 A1 WO2022102595 A1 WO 2022102595A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- gene
- promoter
- capsanthin
- tag
- carotenoid
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Images











Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/52—Genes encoding for enzymes or proenzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N5/00—Undifferentiated human, animal or plant cells, e.g. cell lines; Tissues; Cultivation or maintenance thereof; Culture media therefor
- C12N5/10—Cells modified by introduction of foreign genetic material
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/001—Oxidoreductases (1.) acting on the CH-CH group of donors (1.3)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0071—Oxidoreductases (1.) acting on paired donors with incorporation of molecular oxygen (1.14)
- C12N9/0073—Oxidoreductases (1.) acting on paired donors with incorporation of molecular oxygen (1.14) with NADH or NADPH as one donor, and incorporation of one atom of oxygen 1.14.13
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0071—Oxidoreductases (1.) acting on paired donors with incorporation of molecular oxygen (1.14)
- C12N9/0077—Oxidoreductases (1.) acting on paired donors with incorporation of molecular oxygen (1.14) with a reduced iron-sulfur protein as one donor (1.14.15)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1085—Transferases (2.) transferring alkyl or aryl groups other than methyl groups (2.5)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/90—Isomerases (5.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P23/00—Preparation of compounds containing a cyclohexene ring having an unsaturated side chain containing at least ten carbon atoms bound by conjugated double bonds, e.g. carotenes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y103/00—Oxidoreductases acting on the CH-CH group of donors (1.3)
- C12Y103/99—Oxidoreductases acting on the CH-CH group of donors (1.3) with other acceptors (1.3.99)
- C12Y103/99031—Phytoene desaturase (lycopene-forming) (1.3.99.31)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y114/00—Oxidoreductases acting on paired donors, with incorporation or reduction of molecular oxygen (1.14)
- C12Y114/13—Oxidoreductases acting on paired donors, with incorporation or reduction of molecular oxygen (1.14) with NADH or NADPH as one donor, and incorporation of one atom of oxygen (1.14.13)
- C12Y114/1309—Zeaxanthin epoxidase (1.14.13.90)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y114/00—Oxidoreductases acting on paired donors, with incorporation or reduction of molecular oxygen (1.14)
- C12Y114/13—Oxidoreductases acting on paired donors, with incorporation or reduction of molecular oxygen (1.14) with NADH or NADPH as one donor, and incorporation of one atom of oxygen (1.14.13)
- C12Y114/13129—Beta-carotene 3-hydroxylase (1.14.13.129)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y205/00—Transferases transferring alkyl or aryl groups, other than methyl groups (2.5)
- C12Y205/01—Transferases transferring alkyl or aryl groups, other than methyl groups (2.5) transferring alkyl or aryl groups, other than methyl groups (2.5.1)
- C12Y205/01029—Geranylgeranyl diphosphate synthase (2.5.1.29)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y205/00—Transferases transferring alkyl or aryl groups, other than methyl groups (2.5)
- C12Y205/01—Transferases transferring alkyl or aryl groups, other than methyl groups (2.5) transferring alkyl or aryl groups, other than methyl groups (2.5.1)
- C12Y205/01032—15-Cis-phytoene synthase (2.5.1.32)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y503/00—Intramolecular oxidoreductases (5.3)
- C12Y503/99—Other intramolecular oxidoreductases (5.3.99)
- C12Y503/99008—Capsanthin/capsorubin synthase (5.3.99.8)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y505/00—Intramolecular lyases (5.5)
- C12Y505/01—Intramolecular lyases (5.5.1)
- C12Y505/01019—Lycopene beta-cyclase (5.5.1.19)
Definitions
- the present invention relates to a transformant and a method for producing a carotenoid composition using the transformant.
- Carotenoids are natural pigments produced by plants, algae, bacteria, etc., and are an important group of compounds that control various physiological functions in living organisms. Many carotenoids have been isolated so far, and the health function of carotenoids, which are present only in trace amounts in nature, has begun to attract attention, but the trace amount makes commercial production costly and difficult.
- Escherichia coli The most studied microorganism as a host for carotenoid production is Escherichia coli.
- Escherichia coli synthesizes isopentenyl diphosphate (IPP) and its isomer dimethylallyl diphosphate (DMAPP) by the non-mevalonate pathway (MEP pathway) as the basic metabolic pathway of isoprenoids including carotenoids (DMAPP).
- MEP pathway non-mevalonate pathway
- DMAPP non-mevalonate pathway
- Non-Patent Document 1 The synthesized IPP and DMAPP are metabolized to geranyl diphosphate (GPP) and farnesyl diphosphate (FPP) by a continuous condensation reaction mediated by prenyltransferase.
- GPP geranyl diphosphate
- FPP farnesyl diphosphate
- Escherichia coli is a non-carotenoid-producing bacterium and does not possess the enzyme genes required for carotenoid synthesis after FPP, but synthesizes geranylgeranylpyrophosphate (GGPP) synthase gene (crtE), phytoen synthase gene (crtB), and lycopene.
- GGPP geranylgeranylpyrophosphate
- crtE geranylgeranylpyrophosphate
- crtB phytoen synthase gene
- lycopene such as the phytoen unsaturated enzyme gene (crtI), the lycopene cyclase gene (crtY) that synthesizes ⁇ -carotene, and the ⁇ -carotenoid hydroxylase gene (crtZ) that synthesizes ⁇ cryptoxanthin and zeaxanthin.
- the pathway engineering approach is roughly divided into a method of modifying an endogenous metabolic pathway including a MEP pathway and a method of introducing a mevalonic acid (MVA) pathway gene cluster derived from a heterologous organism.
- MEP pathway a mevalonic acid pathway gene cluster derived from a heterologous organism.
- Non-Patent Document 2 a method for significantly increasing carotenoid production in Escherichia coli in which a type 1 IPP isomerase gene (idi) derived from S. cerevisiae or green alga (Haematococcus pluvialis) is highly expressed.
- idi type 1 IPP isomerase gene
- CrtB type 1 IPP isomerase gene derived from S. cerevisiae or green alga
- Non-Patent Document 4 Escherichia coli in which the MVA pathway gene group (including type 2 IPP isomerase gene) derived from the actinomycete Streptomyces sp. CL190 strain is functionally expressed is used, and dehydrogenated D-MVA lactone is used as a substrate.
- dehydrogenated zeaxanthin is synthesized (Non-Patent Document 4). This approach is considered to be a powerful and effective tool in pathway engineering research for carotenoid production in E. coli as a host (Non-Patent Document 5).
- Non-Patent Document 6 zeaxanthin epoxidizing enzyme (ZEP) gene synthesizes antheraxanthin from zeaxanthin, and the capsanthin-capsanthin synthase (CCS) gene synthesizes capsanthin from antheraxanthin.
- ZEP zeaxanthin epoxidizing enzyme
- CCS capsanthin-capsanthin synthase
- capsanthin could not be synthesized by previously known methods using recombinant microorganisms.
- the present inventor introduced the CrtE gene, CrtB gene, CrtI gene, CrtY gene, CrtZ gene, ZEP gene, and CCS gene into enterobacteria together with the IDI gene, no capsanthin was produced.
- An object of the present invention is to provide a technique for producing a composition containing a carotenoid, such as capsanthin, which could not be synthesized so far, by a gene recombination technique.
- the present inventor came up with a novel idea of controlling the expression level of only some of the capsanthin biosynthetic genes so that the expression level is relatively high. Therefore, in order to obtain a relatively large amount of capsanthin substrate, a recombinant vector was constructed so that the expression level of the upstream gene in the capsanthin biosynthetic gene cluster was relatively high, and the recombinant vector was transformed. I tried to synthesize capsanthin. However, this method did not produce capsanthin at all when idi was incorporated.
- the transformant produced capsanthin. I found that it can be generated. Further, when the ZEP gene and the CCS gene are incorporated into a recombinant vector so as to be expressed so that ZEP and CCS indirectly bind to each other via a multimer of a predetermined affinity protein, capsorbin and kukurubita are used. It was also unexpectedly found that various other carotenoids such as xanthine A can also be produced.
- the present invention is an invention completed by further studies based on these findings.
- Item 1 An upstream gene of a carotenoid biosynthetic gene, including crtY and crtZ, operably linked to the first promoter and said first promoter. A second promoter having a higher promoter strength than the first promoter, and the ZEP gene and CCS gene operably linked to the second promoter. Is a transformant introduced into a host cell. Item 2. Item 2. The transformant according to Item 1, wherein the difference in promoter intensity between the first promoter and the second promoter is equal to or greater than the difference in promoter intensity between the promoter peculiar to the carotenoid biosynthesis gene and PBAD. Item 3.
- Item 4. Item 6. The transformant according to any one of Items 1 to 3, wherein the first promoter is a promoter specific to a biosynthetic gene of Plac or a carotenoid. Item 5.
- Item 6. The transformant according to any one of Items 1 to 4, wherein the second promoter is PBAD or Ptac. Item 6. Further, the S protein gene, the S tag gene and the linker gene have been introduced into the host cell. The S tag gene is ligated to each of the ZEP gene and the CCS gene. Item 2.
- the carotenoid composition comprises capsanthin, capsanthin 3'-acetate, capsanthin 3-acetate, kukurubitaxanthin A, capsanthin diacetate, capsanthin 3,6-epoxide, and / or capsanthin 3,6-epoxide 3'-acetate.
- Item 2 The production method according to Item 10 or 11.
- the carotenoid biosynthetic pathway and the enzymes that mediate it are shown. It is a schematic diagram of the recombinant vector prepared in Comparative Example 1. It is a schematic diagram of the recombinant vector prepared in Comparative Example 2 (Comparative Example 2-1 and Comparative Example 2-2). It is an HPLC chromatogram of the carotenoid composition obtained in Comparative Example 2 (Comparative Example 2-1 and Comparative Example 2-2). It is a schematic diagram of the recombinant vector prepared in Example 1. 6 is an HPLC chromatogram of the carotenoid composition obtained in Example 1.
- the UPLC chromatogram (a) of the carotenoid composition obtained in Example 1, the absorption spectrum of capsanthin (b), and the mass spectrum of capsanthin (c) are shown. It is a schematic diagram of the recombinant vector prepared in Example 2 (Example 2-1 and Example 2-2). The HPLC chromatogram of the carotenoid composition obtained in Example 2 (Example 2-1 and Example 2-2) is shown. The amount of capsanthin produced in the carotenoid composition obtained in Example 2 (Example 2-1 and Example 2-2) is shown. It is a schematic diagram of the recombinant vector prepared in Example 3 (Example 3-1 to Example 3-3).
- Example 3-3 The cooperative system of CCS and ZEP using S protein constructed by the expression system according to Example 3 (Examples 3-1 to 3-3) is schematically shown.
- the analysis result of the carotenoid composition obtained in Example 3 (Example 3-1 to Example 3-3) is shown.
- the analysis result of the carotenoid composition obtained in Example 3 (Example 3-2) is shown.
- the transformants of the present invention are a first promoter and an upstream gene of a carotenoid biosynthesis gene, including crtY and crtZ, operably linked to the first promoter; said first promoter. It is characterized in that a second promoter having a higher promoter strength and a ZEP gene and a CCS gene operably linked to the second promoter are introduced into a host cell. This allows the carotenoid biosynthetic enzyme to be expressed in the host cell, allowing the production of carotenoid compositions.
- FIG. 1 shows a carotenoid biosynthetic pathway and a group of enzymes that mediate it.
- FIG. 1 gives an example of biosynthesis of carotenoids using recombinant Escherichia coli.
- CrtE is a GGPP synthase that synthesizes geranylgeranylpyrophosphate (GGPP) from farnesyl diphosphate (FPP), and the gene encoding CrtE will be referred to as "crtE" below.
- CrtB is a phytoene synthase that synthesizes phytoene from GGPP, and in the following, the gene encoding CrtB will be referred to as "crtB".
- CrtI is a phytoene desaturase that synthesizes lycopene from phytoene, and in the following, the gene encoding CrtI will be referred to as "crtI".
- CrtY is a lycopene cyclase that synthesizes ⁇ -carotene from lycopene, and in the following, the gene encoding CrtY will be referred to as "crtY".
- CrtZ is a ⁇ -carotene hydroxylase that synthesizes ⁇ -cryptoxanthin and zeaxanthin from ⁇ -carotene, and the gene encoding CrtZ is hereinafter referred to as “crtZ”.
- ZEP is a zeaxanthin epoxidizing enzyme that synthesizes antheraxanthin and violaxanthin from zeaxanthin.
- CCS is a capsanthin / capsanthin synthase known to synthesize capsanthin and capsanthin from antheraxanthin and violaxanthin, respectively. Further, in the findings found by the present inventor, CCS synthesizes capsanthin 3'-acetate and kukurubitaxanthin A from antheraxanthin, and from violaxanthin, capsanthin 3-acetate, capsanthin diacetate, capsanthin 3,6. -Epoxide, capsanthin 3,6-epoxide 3'-It is also a synthetic enzyme that synthesizes acetate.
- the above-mentioned gene encoding carotenoid biosynthetic enzyme is a known gene, and its specific sequence can be appropriately obtained from an official database or the like.
- the organism from which these carotenoid biosynthesis genes are derived is not particularly limited, and is selected without particular limitation from organisms that produce carotenoids corresponding to these genes.
- Specific origin organisms include plants and microorganisms, and specifically, the genus Capsicum (for example, Capsicum annuum), the genus Lilium (for example, Lilyum lancifolia), the genus Arabidopsis (for example, Arabidopsis thaliana), and the genus Zea (for example, Zea).
- the genus Prunus eg Prunus armeniaca
- the genus Ipomoea eg Ipomoea nil
- the genus Gentiana eg Gentiana lutea
- the genus Marchantia eg the genus Marchantia , Heamatococcus, Monorapidium, Phaffia (Xanthophyllomyces), Paracoccus, Brevundimonas, Gordonia (Rhodococcus), Nostoccus , Pantoea genus.
- the organism from which each gene is derived is not limited, but the same organism is preferable.
- the carotenoid biosynthetic gene that can be used in the present invention includes not only a base sequence obtained from an official database and a codon-optimized sequence thereof, but also an enzyme having carotenoid biosynthetic enzyme activity.
- an enzyme having carotenoid biosynthetic enzyme activity for example, (I-2), (II-2), (III-2) with respect to the base sequences of (I-1), (II-1), and (III-1) below that are similar to the sequence.
- (I-3), (II-3), and (III-3) are also included.
- Preferred examples of the base sequence of the above-mentioned carotenoid biosynthesis gene include the following.
- (III-1) Nucleotide sequence of CCS gene shown by DDBJ accession number X76165 (I-2)
- an enzyme having phytoen synthase activity for synthesizing phytoen from GGPP and lycopene from phytoen.
- Enzymes with phytoen unsaturated enzyme activity, enzymes with lycopen cyclase activity that synthesizes ⁇ -carotene from lycopene, and enzymes with ⁇ -carotene hydroxylase activity that synthesize ⁇ cryptoxanthin and zeaxanthin from ⁇ -carotene are encoded respectively.
- (II-2) A nucleic acid encoding an enzyme having a zeaxanthin epoxidizing enzyme activity that synthesizes anteraxanthin and violaxanthin from zeaxanthin, and having a base sequence complementary to the base sequence shown in (II-1) above.
- Nucleic acid sequence that hybridizes under stringent conditions, (III-2) Capsanthin and capsanthin are synthesized from antheraxanthin and violaxanthin, respectively, and in addition, capsanthin, capsanthin 3'-acetate, capsanthin 3-acetate, kukurubitaxanthin A, capsanthin diacetate, capsanthin 3,6-epoxide.
- Capsanthin 3,6-epoxide 3'-A gene encoding an enzymatic activity that synthesizes acetate, and under stringent conditions with a nucleic acid consisting of a base sequence complementary to the base sequence shown in (III-1) above. Nucleic acid sequence to hybridize, (I-3)
- Enzymes with phytoen unsaturated enzyme activity, enzymes with lycopen cyclase activity that synthesizes ⁇ -carotene from lycopene, and enzymes with ⁇ -carotene hydroxylase activity that synthesize ⁇ cryptoxanthin and zeaxanthin from ⁇ -carotene are encoded respectively.
- II-3 A gene encoding an enzyme having zeaxanthin epoxidizing enzyme activity that synthesizes antheraxanthin and violaxanthin from zeaxanthin, and has 90% or more homology with the base sequence shown in (II-1) above.
- Capsanthin and capsanthin are synthesized from antheraxanthin and violaxanthin, respectively, and in addition, capsanthin, capsanthin 3'-acetate, capsanthin 3-acetate, kukurubitaxanthin A, capsanthin diacetate, capsanthin 3,6-epoxide. , Capsanthin 3,6-epoxide 3'-A gene encoding an enzymatic activity for synthesizing acetate, which has 90% or more homology with the base sequence shown in (III-1) above.
- stringent conditions means 0.5% SDS, 5 ⁇ Denhartz's, 0.1% bovine serum. 6 ⁇ SSC (1 ⁇ SSC is 0.15M NaCl, 0.015M sodium citrate, pH 7) containing albumin (BSA), 0.1% polyvinylpyrrolidone, 0.1% ficol 400] and 100 ⁇ g / ml salmon sperm DNA. In 0.0), it refers to the condition of keeping warm at 50 ° C to 65 ° C for 4 hours to overnight. Hybridization under stringent conditions is specifically performed by the following method.
- a nylon membrane on which a DNA library or cDNA library was immobilized was prepared, and in a prehybridization solution containing 6 ⁇ SSC, 0.5% SDS, 5 ⁇ Denhardz, 100 ⁇ g / ml salmon sperm DNA, at 65 ° C. Block the nylon membrane. Then add each probe labeled at 32 P and insulate at 65 ° C. overnight. This nylon film was placed in 6 ⁇ SSC at room temperature for 10 minutes, in 2 ⁇ SSC containing 0.1% SDS, at room temperature for 10 minutes, in 0.2 ⁇ SSC containing 0.1% SDS, at 45 ° C. for 30 minutes. After washing, autoradiography can be taken to detect nucleic acids that specifically hybridize to the probe.
- the homology may be 90% or more, preferably 95% or more, more preferably 98% or more, still more preferably 98. 5.5% or more, more preferably 99% or more, even more preferably 99.5% or more, particularly preferably 99.9% or more.
- "Homology" is calculated using publicly available or commercially available software with an algorithm that compares the reference sequence as a query sequence. Specifically, BLAST, FASTA, GENETYX (manufactured by Genetics Co., Ltd.) and the like can be used, and these may be used by setting them to default parameters.
- the above-mentioned carotenoid biosynthesis gene may be integrated into the recombinant vector with the full-length sequence integrated into the recombinant vector, or may be integrated into the recombinant vector with the sequence unnecessary for the expression of the target enzyme in the host removed. ..
- the organism from which the carotenoid biosynthesis gene is derived is a plant and the host is a non-plant cell, it is incorporated into the recombinant vector with the signal sequence removed from the viewpoint of further improving the production of capsanthin. Is preferable.
- the signal sequence of each plant-derived gene can also be easily obtained by using an official database or various gene analysis tools.
- crtE, crtB, crtI, crtY and crtZ are classified as “upstream genes", and the ZEP gene and CCS gene are classified as “downstream genes”.
- the upstream gene of the carotenoid biosynthesis gene is operably linked to the first promoter, and the downstream gene of the carotenoid biosynthesis gene is operably linked to the second promoter. That is, the first promoter is a region that controls transcription of upstream genes, and the second promoter is a region that controls transcription of downstream genes.
- operably linked means that the regulatory factor and the above-mentioned gene are linked in a state in which they can act in a host cell.
- the second promoter is designed so that the promoter strength is higher than that of the first promoter.
- the promoter strength of the second promoter is three times or more the promoter strength of the first promoter.
- the promoter intensity refers to the frequency of transcription initiation, and is also referred to as, for example, Pbla unit (Pbla unit; Deuschle et al., EMBO J., 5: 2987-2994 (1986)) and mirror unit (Miller units; ⁇ galactosidase unit).
- the expression level of the upstream gene of the carotenoid biosynthesis gene is controlled to be relatively low and the expression level of the downstream gene is controlled to be relatively high by the first promoter and the second promoter. , Allows the synthesis of carotenoids.
- the difference between the promoter strength between the first promoter and the second promoter is inherently provided in the promoter peculiar to the carotenoid biosynthesis gene (preferably Pantoea ananatis). It is preferable that the promoter strength is equal to or greater than the difference in promoter strength between the promoter, more preferably the promoter contained in DDBJ accession number D90087) and the PBAD promoter, and the promoter inherent in the carotenoid biosynthetic gene (preferably Pantoea ananatis). , More preferably, it is more than the difference in promoter strength between the promoter of DDBJ accession number D90087) and the promoter.
- the first promoter and the second promoter are not particularly limited as long as the second promoter is designed so that the promoter strength is relatively higher, and the recombination incorporating these promoters is not particularly limited. It is appropriately selected according to the type of host into which the vector is introduced.
- the first promoter and the second promoter may be a natural promoter or an artificial promoter.
- the promoter used as the first promoter and the second promoter in prokaryotic organisms such as Escherichia coli, the lac promoter (Plac), the promoter peculiar to the carotenoid biosynthesis gene, the BAD promoter (PBAD), and the tac promoter ( Ptac), trc promoter, T7 promoter, T7lac promoter, T5 promoter, UV5lac promoter, L8-UV5lac promoter, cspA promoter, CAG promoter, CMV promoter, RSV promoter, etc.
- the lac promoter the promoter peculiar to the carotenoid biosynthesis gene
- PBAD BAD promoter
- Ptac tac promoter
- GAL1 promoter In yeast, GAL1 promoter, NMT1 promoter, TEF1 promoter, ADH1 Examples thereof include a promoter, TPI1 promoter, HXT7 promoter, TDH3 promoter, PGK1 promoter, PYK1 promoter, CAG promoter, CMV promoter, RSV promoter and the like.
- the promoter unique to the carotenoid biosynthesis gene is a promoter inherent in carotenoid biosynthesis.
- the promoter peculiar to the carotenoid biosynthesis gene it can be used without particular limitation, and in the organism from which the above-mentioned carotenoid biosynthesis gene is derived, the promoter naturally provided as the promoter of the carotenoid biosynthesis gene should be used. Can be done.
- a gene placed under the control of a promoter specific to the biosynthetic gene of carotenoid is derived from Pantoea ananatis, it is also inherently provided in Pantoea ananatis as a promoter specific to the biosynthetic gene of the carotenoid.
- a promoter more preferably a promoter included in DDBJ accession number D90087, can be used.
- two types having different promoter strengths can be selected, the one with the lower promoter strength is selected as the first promoter, and the one with the lower promoter strength is selected as the second promoter.
- a combination of two types of promoters having a promoter strength difference in the promoter strength is selected from various promoters including the above examples, and the promoter is selected.
- the one with the lower strength can be selected as the first promoter, and the one with the lower strength can be selected as the second promoter.
- the first promoter includes a promoter specific to the biosynthetic gene of Plac and carotenoid
- the second promoter includes PBAD and Ptac, and more preferably Ptac.
- the control of the expression level by using the first promoter and the second promoter having different promoter intensities as described above suppresses the reaction of synthesizing ⁇ -carotene from lycopene by CCS. Therefore, the upstream gene expressed by the first promoter may contain at least crtY and crtZ.
- upstream genes may be arranged under the control of the first promoter as in crtY and crtZ, or may be arranged under the control of other promoters.
- the other upstream genes are placed under the control of the first promoter.
- Specific examples of the case where another upstream gene is placed under the control of the first promoter include (i) crtI; (ii) crtI and crtB; and (iii). ) CrtI, crtB and crtE may be included.
- the IPP isomerase gene (idi) may be contained together with other upstream genes, that is, (iv) crtI, crtB, crtE and idi may be further arranged under the control of the first promoter.
- the first promoter and the gene arranged under its control and the second promoter and the gene arranged under its control are both one recombinant vector by being integrated into one expression vector. Or may be incorporated into a separate expression vector to form a first recombinant vector and a second recombinant vector, respectively.
- the gene CCS for S-protein-mediated expression of the coordinated system of CCS and ZEP has the lycopene ⁇ -cyclase activity of CrtY in addition to the capsanthin / capsanthin synthesis activity. Even if the CCS gene is efficiently expressed in E. coli due to such dual activity of CCS, it acts in collaboration with the phytoene desaturase CrtI instead of ZEP to receive lycopene from CrtI as a substrate and receive ⁇ -carotene.
- the reaction to be synthesized may be prioritized. Therefore, in order to further increase the amount of reaction product produced by CCS by selectively coordinating CCS with ZEP, the CCS gene and ZEP gene are coordinated with CCS and ZEP via S protein as follows. It is preferably introduced into the host cell in a manner that expresses the system. A diagram schematically illustrating the cooperative system is shown in FIG.
- the S protein gene, S tag gene and linker gene are further introduced into the host cell, and the ZEP gene and the CCS gene are respectively introduced.
- the S-tag gene is ligated to, and the linker gene is ligated between the ZEP gene and the S-tag gene and / or between the CCS gene and the S-tag gene.
- the S tag and the S protein have an affinity tag and an affinity protein relationship specific to each other, and are polypeptides constituting one RNase A molecule.
- the N-terminal 20 amino acid fragment obtained by cleaving RNase A with satylicin is an S tag, and the remaining fragment consisting of about 100 amino acids is an S protein.
- the S tag and S protein can be non-covalently bound to reproduce the original higher-order structure, resulting in RNase activity (Lopez-Alonso, J. P., Bruix, M., Font, J). ., Ribo, M., Vilanova, M., Rico, M., Gotte, G., Libonati, M., Gonzalez, C. and Laurents, D. V. (2006) J. Biol. Chem. 281, 9400 -9406).
- the ZEP gene and the CCS gene to which the S tag gene is linked are expressed as S tag fusion ZEP and S tag fusion CCS in the host cell, respectively. Furthermore, since the linker gene is linked between the ZEP gene and the S tag gene and / or between the CCS gene and the S tag gene, the S tag fusion ZEP and S tag expressed in the host cell are linked.
- the fusion CCS takes the form of a fusion protein via a linker between the S tag and the enzyme.
- both S-tag fusion ZEP and S-tag fusion CCS are expressed in the form of a fusion protein via a linker between the S tag and the enzyme (“S tag + linker +” in the figure. "CCS" and "S tag + linker + ZEP”) are shown with examples.
- S-tag fusion ZEP and S-tag fusion CCS are co-expressed with S-protein, as shown in FIG. 12, a complex in which fusion ZEP and fusion CCS are indirectly bound to the S-protein multimer via S-tag. Form the body.
- ZEP and CCS physically approach each other in the host cell, so that CCS can selectively collaborate with ZEP. That is, the CCS can smoothly receive the metabolites from ZEP. This makes it possible to obtain more carotenoids produced by CCS metabolism.
- capsanthin 3,6-epoxide 3'-acetate produced improves the amount of capsanthin produced and / or capsanthin, capsanthin 3'-acetate, capsanthin 3-acetate, kukurubitaxanthin A, capsanthin diacetate, capsanthin 3,6-epoxide, and / Alternatively, the amount of capsanthin 3,6-epoxide 3'-acetate produced can be improved.
- S protein is known as a cleavage product consisting of about 100 amino acids by satilysin of RNase A as described above, its amino acid sequence is known (specifically, SEQ ID NO: 2 is mentioned).
- the base sequence of the S protein gene is also known (specifically, SEQ ID NO: 1 is mentioned). Examples of the base sequence of the S protein gene that can be used in the present invention include the following.
- (IV-1) The base sequence shown by SEQ ID NO: 1, (IV-2) Nucleic acid and stringent, which is a gene encoding a protein that forms a multimer and specifically binds to an S tag, and has a base sequence complementary to the base sequence shown in (IV-1) above. Nucleic acid sequence that hybridizes under various conditions, and (IV-3) a gene that encodes a protein that forms a multimer and specifically binds to an S tag, and is the base shown in (IV-1) above. A base sequence having 90% or more homology with the sequence.
- the S tag is known as an N-terminal 20-amino acid cleavage product of RNase A by satylysin as described above, its amino acid sequence is known (specifically, the 12th sequence of SEQ ID NO: 4 is phenylalanine. Alternatively, an amino acid sequence that is histidine can be mentioned), and the base sequence of the S protein gene is also known (specifically, the 34th to 36th sequences of SEQ ID NO: 3 encode histidine instead of the sequence encoding phenylalanine. A base sequence which is a sequence can be mentioned.).
- S tag examples include any of the following.
- the sequence identity other than the 12th sequence to the amino acid sequence shown in SEQ ID NO: 4 is 90% or more, preferably 95% or more.
- a polypeptide that specifically binds to S protein is 90% or more, preferably 95% or more.
- amino acid sequence shown in SEQ ID NO: 4 is the 12th wild-type S tag for the purpose of suppressing the effect on RNase activity caused by the binding of the S tag to the S protein in the host cell. It has been replaced by phenylalanine from histidine.
- sequence identity is defined as BLASTPACKAGE [sgi32 bit edition, Version 2.0.12; available from National Center for Biotechnology Information (NCBI)] bl2sevo The value of the identity of the amino acid sequence obtained by Lett., Vol. 174, p247-250, 1999) is shown.
- the parameters may be set to Gap insertion Cost value: 11 and Gap extension Cost value: 1.
- the base sequence of the S tag gene that can be used in the present invention is not particularly limited as long as it is the base sequence encoding the above S tag, but the following are preferable.
- V-1 The base sequence shown in SEQ ID NO: 3 (that is, the base sequence encoding the amino acid sequence shown in SEQ ID NO: 4), (V-2) A gene encoding a peptide that specifically binds to S protein, which is the base sequence shown in (V-1) above (however, the base sequence other than the 34th to 36th sequences of SEQ ID NO: 3) and 90.
- the 34th to 36th sequences of SEQ ID NO: 3 in the above (V-1) to (V-2) of the S tag gene are RNase activities due to the binding of the expressed S tag to the S protein in the host cell.
- the histidine-encoding sequence present in the wild-type S tag has been replaced with a phenylalanine-encoding sequence for the purpose of suppressing the effect on phenylalanine.
- the binding position of the S tag gene is not particularly limited, but the S tag binds to the N-terminal of the enzyme (ZEP and / or CCS) from the viewpoint of reducing the influence on the activity of the enzyme fused with the S tag. Can be designed to.
- the linker gene is particularly limited as long as the linker (peptide) has an appropriate distance between the S tag and the enzyme in the fusion protein so that the degree of freedom can be guaranteed between CCS and ZEP in the complex. Not done.
- the preferred sequence of the linker (peptide) in the fusion protein is a sequence in which non-polar amino acids and polar amino acids are alternately linked from the viewpoint of eliminating the bias of the polarity of the linker (peptide) itself and suppressing undesired adsorption.
- the combination of the non-polar amino acid and the polar amino acid is preferably a combination of glycine and serine.
- Examples of the length of the linker (peptide) include 15 to 25 amino acids, preferably 18 to 22 amino acids.
- the intervening position of the linker gene may be at least one between the ZEP gene and the S tag gene and between the CCS gene and the S tag gene, but preferably at least the ZEP gene and the S tag gene. And particularly preferably only between the ZEP gene and the S-tag gene (no interposition between the CCS gene and the S-tag gene).
- other recombinant vectors may contain regulatory sequences other than the first promoter and the second promoter.
- regulatory sequences are located upstream (5'side), within the sequence, or downstream (3'side) of the upstream gene sequence of the carotenoid biosynthetic gene and / or the downstream gene sequence of the carotenoid biosynthetic gene. Positioned and affects transcription, RNA processing, stability, etc.
- Regulatory sequences include translation leader sequences, introns, polyadenylation recognition sequences, RNA processing sites, effector binding sites, stem-loop structures and the like.
- Expression vector a vector constructed for gene recombination from a plasmid, virus, phage, transposon, IS element, fasmid, cosmid, etc. that can grow autonomously in the host can be used.
- expression vectors for example, pET, pLG, pACYC, pBR, pUC, pKC, pRep, pHS, pKK, pDHE, pPLc, pMBL, pUR, pIN, ⁇ gt, pBdCI, pUB, pC, pBD; pSA, Examples thereof include pAJ, YEp, YRp, YIp, pYAC, pCDM, pMT2PC, adenovirus vector, adeno-associated virus vector, retrovirus vector, herpesvirus vector and the like.
- the underlying expression vector may be the same or different between the first recombinant vector and the second recombinant vector. Further, the expression vector may be used by selecting an appropriate combination with the host cell. For example, as a preferable example when Escherichia coli is used as the host cell, a combination of pACYC vector and pUC vector and JM101 (DE3) Escherichia coli strain and the like can be used. Can be mentioned.
- the host is not particularly limited as long as the recombinant vector is stable, can autonomously proliferate, and can express the trait of a foreign gene, and may be a carotenoid-producing cell or a carotenoid-non-producing cell.
- Preferred examples of the host include bacteria belonging to the genus Escherichia such as Escherichia coli, the genus Bacillus such as Bacillus subtilis, and the genus Yeast belonging to the genus Pseudomonas putida, for example. , Saccharomyces cerevisiae, fission yeast (S.
- pombe Trula yeast (Candida utilis), methanol-utilizing yeast (Pichia pastoris), red yeast (Xanthophyllomyces dendrhos, etc.), etc. It may be a cell, an insect cell, a plant or the like. Among these, Escherichia coli is a particularly preferable host.
- the transformant of the present invention can be obtained by introducing a recombinant vector into a host, and the conditions for introducing the recombinant vector into the host can be appropriately set according to the type of host and the like. good.
- a bacterium for example, a heat shock method, a method using competent cells treated with calcium ions, an electroporation method, and the like can be mentioned.
- yeast for example, an electroporation method, a spheroplast method, a lithium acetate method and the like can be mentioned.
- the host is an animal cell, for example, an electroporation method, a calcium phosphate method, a lipofection method and the like can be mentioned.
- the host is an insect cell, for example, a calcium phosphate method, a lipofection method, an electroporation method and the like can be mentioned.
- a plant cell for example, an electroporation method, an Agrobacterium method, a particle gun method, a PEG method and the like can be mentioned.
- Whether or not the recombinant vector has been incorporated into the host can be confirmed by the PCR method, the Southern hybridization method, the Northern hybridization method, or the like.
- the recombinant vector may be separated and purified from the transformant.
- Separation and purification of the recombinant vector can be performed, for example, when the host is a bacterium, based on the lysed product obtained by lysing the bacterium.
- a method of lysis treatment is performed with a lytic enzyme such as lysozyme, and if necessary, a protease and other enzymes and a surfactant such as sodium lauryl sulfate (SDS) are used in combination.
- a lytic enzyme such as lysozyme
- a protease and other enzymes and a surfactant such as sodium lauryl sulfate (SDS) are used in combination.
- SDS sodium lauryl sulfate
- Separation and purification of DNA from the lysate can be performed, for example, by appropriately combining a phenol treatment, a protein removal treatment by a protease treatment, a ribonuclease treatment, an alcohol precipitation treatment, and a commercially available kit.
- DNA can be cleaved according to a conventional method, for example, using restriction enzyme treatment.
- restriction enzyme for example, a type II restriction enzyme that acts on a specific nucleotide sequence can be used.
- the binding between the DNA and the expression vector can be performed using, for example, DNA ligase.
- the separated / purified DNA was cleaved with a specific restriction enzyme, subjected to agarose gel electrophoresis, polyacrylamide gel electrophoresis, capillary electrophoresis, etc., and stained with ethidium bromide, SYBR Green solution, etc., and the detected band. From the long pattern, it can be confirmed that the target has been transformed.
- transformation has been performed by designing a primer and performing PCR using the separated / purified DNA as a template.
- the amplified product obtained by PCR is subjected to agarose gel electrophoresis, polyacrylamide gel electrophoresis, capillary electrophoresis, etc., stained with ethidium bromide, SYBR Green solution, etc., and the amplified product is detected as a band. You can confirm that it has been converted.
- amplification product it is also possible to detect the amplification product by performing PCR using a primer labeled in advance with a fluorescent dye or the like. Further, a method of binding the amplification product to a solid phase such as a microplate and confirming the amplification product by fluorescence, an enzymatic reaction or the like may also be adopted.
- the method for producing a carotenoid composition of the present invention can be obtained by a production method including a step of culturing a transformant of the present invention.
- the culture conditions of the transformant may be appropriately set in consideration of the nutritional and physiological properties of the host, but liquid culture is preferable. Further, in the case of industrial production, aeration stirring culture is preferable.
- the carbon source may be any carbon compound that can be assimilated, and examples thereof include molasses, glucose, fructose, maltose, sucrose, lactose, sucrose, starch, lactose, glycerol, and pyruvate.
- the nitrogen source may be any assimilating nitrogen compound, for example, natural components such as corn steep liquor, peptone (animal peptone, casein peptone, soybean peptone, etc.), extract (meat extract, yeast extract, etc.); ammonium acetate. , Ammonium salts such as ammonium chloride and ammonium sulfate; amino acids such as glutamate, asvaraginic acid and glycine can be mentioned.
- natural components such as corn steep liquor, peptone (animal peptone, casein peptone, soybean peptone, etc.), extract (meat extract, yeast extract, etc.); ammonium acetate.
- Ammonium salts such as ammonium chloride and ammonium sulfate
- amino acids such as glutamate, asvaraginic acid and glycine can be mentioned.
- salts such as phosphates, carbonates, sulfates, magnesium, calcium, potassium, iron, manganese and zinc, and specific vitamins may be used as needed. good.
- the culture temperature can be appropriately set as long as the transformant of the present invention can grow and the transformant of the present invention produces the polypeptide of the present invention, but is preferably about 20 to 40 ° C., preferably about 20 to 40 ° C.
- the temperature is about 30 to 37 ° C.
- the culturing may be completed at an appropriate time in anticipation of the time when the polypeptide of the present invention reaches the maximum yield, and examples of the culturing time include about 24 to 200 hours, preferably about 60 to 90 hours.
- Examples of the method for recovering the bacterial cells after the completion of the culture include filtration, decantation, and centrifugation of the culture solution.
- the recovered cells can be treated with water, sodium chloride solution, dimethylformamide or the like, if necessary.
- the carotenoid composition obtained in the production method of the present invention typically contains capsanthin, preferably further capsanthin, capsanthin 3'-acetate, capsanthin 3-acetate, cucurbitaxanthin A, capsanthbin diacetate. , Capsanthin 3,6-epoxide, and / or capsanthin 3,6-epoxide 3'-acetate.
- An appropriate organic solvent can be used for the extraction of the carotenoid composition from the cells.
- the organic solvent include methanol, ethanol, isopropyl alcohol, acetone, hexane, diethyl ether, tetrahydrofuran, methyl ethyl ketone, methyl isobutyl ketone, dichloromethane, chloroform, dimethylformamide, dimethyl sulfoxide, methyl acetate, ethyl acetate and the like.
- One or more kinds can be selected from the solvent of.
- acetone, methanol and diethyl ether are preferable.
- the carotenoid composition fraction obtained as described above may be concentrated and dried as it is as a composition, or each carotenoid can be purified, concentrated and dried.
- the purification method can be performed by, for example, appropriately combining methods such as gel filtration, adsorption chromatography, ion exchange chromatography, and affinity chromatography.
- drying method include freeze-drying, vacuum drying, spray drying and the like.
- the promoters used in the following examples and comparative examples are promoters inherent in Plac and carotenoid biosynthetic genes (promoters inherent in Pantoea ananatis, and are specifically included in DDBJ accession number D90087. Promoter), PBAD, or Ptac, and the relationship of promoter intensity is Plac ⁇ promoter peculiar to the biosynthetic gene of the carotenoid ⁇ PBAD ⁇ Ptac, and promoter intensity of PBAD is the promoter peculiar to the biosynthetic gene of the carotenoid. It is more than 3 times the strength of the promoter.
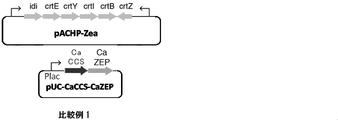
- pACHP-Zea shown in FIG. 2 was constructed as the first recombination vector to be incorporated, and pUC-Plac-CaCCS-CaZEP shown in FIG. 2 was constructed as the second recombination vector.
- These recombinant vectors were designed so that the expression level of the upstream gene was relatively high due to the promoter peculiar to the carotenoid biosynthesis gene (derived from Pantoea ananatis), and the expression level of the downstream gene was relatively low due to Plac.
- Table 1 shows an outline of the first recombinant vector and the second recombinant vector of this comparative example.
- PACHP-Zea as the first recombinant vector is a vector for zeaxanthin synthesis used in the report of Non-Patent Document 3.
- pACHP-Zea is a gene group required for zeaxanthin biosynthesis (Pantoea ananatis-derived crtE, crtY, crtI, crtJc gene, crtY, crtD The number D90087)) is inserted and configured.
- the pUC-Plac-CaCCS-CaZEP as the second recombination vector is a Capsicum annuum-derived ZEP gene (hereinafter referred to as “CaZEP”; DDBJ accession number XM_016705616) and a Capsicum annuum-derived CCS gene (hereinafter referred to as “CaCCS”).
- DDBJ accession number X76165 was prepared by placing it downstream of the Plac promoter of the pUC vector.
- Both the first recombinant vector and the second recombinant vector were introduced into Escherichia coli JM109 strain by the heat shock method.
- the single colony on the LB medium was transplanted to 5 mL of the LB liquid medium and cultured with shaking at 200 rpm and 37 ° C. for 16 hours. 100 ⁇ L of the culture solution was transplanted into 10 mL of TB liquid medium, and after shaking culture at 200 rpm and 30 ° C. for 24 hours, 10 ⁇ L of isopropyl- ⁇ -D-galactopyranoside (IPTG) was added to induce gene expression, and further culture was performed. Continued for 48 hours. In addition, as antibiotics, chloramphenicol (30 mg / L) and kanamycin (40 mg / L) were added to the medium.
- IPTG isopropyl- ⁇ -D-galactopyranoside
- 1 mL of the obtained culture solution was centrifuged at 8,000 g for 1 minute, and the cell fraction was collected by removing the supernatant.
- the cell fraction was suspended in 1% sodium chloride solution, centrifuged at 8,000 g for 1 minute, the supernatant was removed, and the cells were collected.
- 0.5 mL of acetone and 1 mL of diethyl ether / hexane (1: 1 (volume basis)) were added, and the cells were vigorously stirred with vortex. After adding 1 mL of water and stirring, the mixture was allowed to stand for 5 minutes, the upper layer was collected in a 1.5 mL centrifuge tube, and dried in a vacuum centrifugal concentrator.
- the obtained dry matter was subjected to separation of carotenoids using Aquity H-class UPLC (Waters).
- Aquity UPLC BEH C18 volume (2.1x100 mm, 1.7 ⁇ m, Waters) as a column, acetonitrile: ultrapure water (85:15 (volume basis)) as mobile phase A, acetonitrile: methanol (65:35 (65:35) as mobile phase B). Volume-based)) was used, and the liquid was sent by a gradient for 8 minutes in which mobile phase A: mobile phase B changed from 100: 0 to 0: 100. The carotenoids were then separated by holding at 0: 100 for 5 minutes.
- the prepared first recombinant vector pAC-HiEBIY-Z-CaZEP was introduced into Escherichia coli JM101 (DE3) and cultured in the same manner as in Comparative Example 1 to obtain a supernatant.
- the obtained supernatant was subjected to separation of carotenoids using Hitachi HPLC Chromaster system.
- Inertsil ODS-3 (4.6x150 mm, 5 ⁇ m, GL Science) was used as a column, acetonitrile was used as mobile phase A, and isopropanol was used as mobile phase B, and mobile phase A: mobile phase B was sent at 100: 0 for 5 minutes.
- Mobile phase A The mobile phase B was delivered by a gradient for 5 minutes from 100: 0 to 50:50 (volume basis), and 50:50 was held for another 5 minutes to separate the carotenoid.
- FIG. 4 shows the obtained HPLC chromatogram. As shown in FIG. 4, it was confirmed that zeaxanthin and ⁇ -carotene and many other carotenoid intermediates were produced in addition to violaxanthin and antheraxanthin.
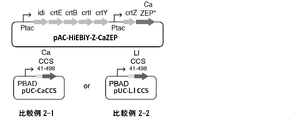
- Comparative Example 2-1 of Comparative Example 2-1 and Comparative Example 2-2 it was investigated whether capsanthin and capsanthin could be produced by co-expressing CaCCS in this carotenoid-producing Escherichia coli strain.
- the N-terminal chloroplast signal sequence of CaCCS was predicted, and CaCCS 41-498 with the N-terminal amino acid removed was designed.
- a plasmid pUC-CaCCS in which CaCCS 41-498 was placed downstream of the PBAD promoter of the pUC vector was prepared as a second recombinant vector and introduced into the above-mentioned carotenoid-producing Escherichia coli strain.
- Comparative Example 2-2 of Comparative Example 2-1 and Comparative Example 2-2 CCS derived from Lilium recombination (hereinafter referred to as “LlCCS”; DDBJ accession number GU443955) was introduced instead of CaCCS.
- LlCCS CCS derived from Lilium recombination
- PUC-LlCCS was prepared as a second recombinant vector in the same manner as in Comparative Example 2-1 and introduced into the above-mentioned carotenoid-producing Escherichia coli strain.
- Comparative Example 2-1 and Comparative Example 2-2 after introducing each of the above second recombinant vectors, the single colonies on the LB medium were transplanted into 3 mL of LB liquid medium, and cultured with shaking at 200 rpm and 37 ° C. for 16 hours. 100 ⁇ L of the obtained culture medium was transplanted to 10 mL of TB liquid medium, and after shaking culture at 200 rpm and 30 ° C. for 24 hours, 10 ⁇ L of isopropyl- ⁇ -D-galactopyranoside (IPTG) was added to induce gene expression, and further. Culture was continued for 48 hours. In addition, as antibiotics, chloramphenicol (30 mg / L) and kanamycin (40 mg / L) were added to the medium.
- IPTG isopropyl- ⁇ -D-galactopyranoside
- the obtained supernatant was subjected to separation of carotenoids using Hitachi HPLC Chromaster system.
- Inertsil ODS-3 (4.6x150 mm, 5 ⁇ m, GL Science) was used as a column, acetonitrile was used as mobile phase A, and isopropanol was used as mobile phase B, and mobile phase A: mobile phase B was sent at 100: 0 for 5 minutes.
- Mobile phase A The mobile phase B was sent by a gradient for 5 minutes from 100: 0 to 50:50 (volume basis), and 50:50 was held for another 5 minutes.
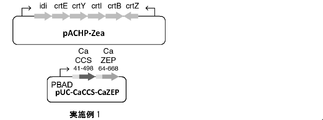
- Example 1 In this example, pACHP-Zea shown in FIG. 5 was constructed as the first recombination vector to be incorporated, and pUC-CaCCS-CaZEP shown in FIG. 5 was constructed as the second recombination vector.
- These recombinant vectors were designed so that the expression level of the upstream gene was relatively low due to the promoter peculiar to the carotenoid biosynthesis gene (derived from Pantoea ananatis), and the expression level of the downstream gene was relatively high due to PBAD.
- Table 3 shows an outline of the first recombinant vector and the second recombinant vector of this example.
- PACHP-Zea as the first recombinant vector was prepared in the same manner as in Comparative Example 1. According to the report of Non-Patent Document 3, this pACHP-Zea is known that zeaxanthin occupies the total carotenoid without ⁇ -carotene being detected in the carotenoid composition by the transformant.
- the pUC-CaCCS-CaZEP as the second recombinant vector was prepared by arranging CaZEP 64-668 and CaCCS 41-498 downstream of the PBAD promoter of the pUC vector.
- Both the first recombinant vector and the second recombinant vector were introduced into Escherichia coli JM101 (DE3) in the same manner as in Comparative Example 1.
- the single colony on the LB medium was transplanted to 3 mL of the LB liquid medium and cultured with shaking at 200 rpm and 37 ° C. for 16 hours.
- 100 ⁇ L of the culture solution was transplanted into 10 mL of TB liquid medium, and after shaking culture at 200 rpm and 30 ° C. for 24 hours, 10 ⁇ L of isopropyl- ⁇ -D-galactopyranoside (IPTG) was added to induce gene expression, and further culture was performed.
- IPTG isopropyl- ⁇ -D-galactopyranoside
- the obtained supernatant was subjected to separation of carotenoids using Hitachi HPLC Chromaster system.
- Inertsil ODS-3 (4.6x150 mm, 5 ⁇ m, GL Science) was used as a column, acetonitrile was used as mobile phase A, and isopropanol was used as mobile phase B, and mobile phase A: mobile phase B was sent at 100: 0 for 5 minutes.
- Mobile phase A The mobile phase B was delivered by a gradient for 5 minutes from 100: 0 to 50:50 (volume basis), and 50:50 was held for another 5 minutes to separate the carotenoid.
- the obtained supernatant (carotenoid) was also subjected to Aquity H-class UPLC (Waters).
- Aquity UPLC BEH C18 volume (2.1x100 mm, 1.7 ⁇ m, Waters) as a column, acetonitrile: ultrapure water (85:15 (volume ratio)) as mobile phase A, acetonitrile: methanol (65:35 (65:35) as mobile phase B) Volume ratio))
- A: B from 100: 0 to 0: 100
- the carotenoid was separated by a program in which 0: 100 was held for 5 minutes.
- FIG. 6 the HPLC chromatogram when the first expression vector is expressed alone (top) and the HPLC chromatograph when the first expression vector and the second expression vector are co-expressed (Example 1).
- the results with grams (bottom) and the absorption spectra of peaks 1, 2, 3 and 5 detected in the latter (Example 1) are shown.
- a new peak 5 was confirmed by co-expression of the first expression vector and the second expression vector.
- FIG. 7a shows the results of analyzing the same supernatant (sample) with UPLC and comparing it with the standard.
- a new peak 5 was confirmed by co-expression of the first expression vector and the second expression vector (Example 1).
- the absorption spectrum of peak 5 is shown in FIG. 7b.
- the peak 5 confirmed by the UPLC chromatogram of FIG. 7a showed a retention time and an absorption spectrum similar to those of capsanthin.
- the amount of capsanthin produced per culture solution was 292 ⁇ g / L.
- LC / MS analysis of peak 5 was performed using a Waters XEvo G2S Q TOF mass spectrometer (Waters) equipped with an Accuracy UPLC system.
- the analysis conditions by the time-of-flight mass spectrometer are a capillary voltage of 3.2 kV, a cone voltage of 20 eV, 120 ° C., and a nitrogen gas of 30 L / h, and scan m / z 100 to 1,500. bottom.
- the MS / MS spectrum was measured with a quadrupole TOF MS / MS device using argon with a collision energy of 20 V.
- UV-VIS absorption spectra were recorded from 200 nm to 600 nm using a photodiode array.
- HPLC separation uses an Accuracy 1.7 ⁇ m BEH UPLC C18 column (Waters), and a gradient program (flow rate 0) from acetonitrile: ultrapure water (85:15 (volume basis)) to acetonitrile: methanol (65:35 (volume basis)). .4 mL / min, 15 min) was used.
- the results are shown in FIG. 7c.
- the molecular ion peak shown in FIG. 7c was in agreement with the calculated molecular weight of capsanthin.
- Example 2 (Example 2-1 and Example 2-2)
- pACHP-Zea shown in FIG. 8 was constructed as the first recombination vector to be incorporated
- pUC-CaCCS-CaZEP (two types) shown in FIG. 8 was constructed as the second recombination vector.
- These recombinant vectors were designed so that the expression level of the upstream gene was relatively low due to the promoter peculiar to the carotenoid biosynthesis gene (derived from Pantoea ananatis), and the expression level of the downstream gene was relatively high due to Ptac.
- Table 4 shows an outline of the first recombinant vector and the second recombinant vector of this example.
- PACHP-Zea as the first recombinant vector was prepared in the same manner as in Comparative Example 1. According to the report of Non-Patent Document 3, this pACHP-Zea is known that zeaxanthin occupies the total carotenoid without ⁇ -carotene being detected in the carotenoid composition by the transformant.
- the pUC-CaCCS-CaZEP as the second recombinant vector is a pUC vector containing CaZEP (without signal sequence removed) and CaCCS (without signal sequence removed) or CaCCS 41-498 . It was manufactured by arranging it downstream of the Ptac promoter.
- Both the first recombinant vector and the second recombinant vector were introduced into Escherichia coli JM101 (DE3) in the same manner as in Comparative Example 1.
- the single colony on the LB medium was transplanted to 0.5 mL of the LB liquid medium and cultured with shaking at 1000 rpm and 37 ° C. for 16 hours.
- 20 ⁇ L of the culture solution was transplanted into 2 mL of TB liquid medium, and after shaking culture at 1000 rpm and 30 ° C. for 24 hours, 2 ⁇ L of 1M isopropyl- ⁇ -D-galactopyranoside (IPTG) was added to induce gene expression.
- IPTG isopropyl- ⁇ -D-galactopyranoside
- the cells were further cultured for 48 hours.
- antibiotics chloramphenicol (30 mg / L) and kanamycin (40 mg / L) were added to the medium.
- the obtained supernatant was subjected to separation of carotenoids using Hitachi HPLC Chromaster system.
- Inertsil ODS-3 (4.6x150 mm, 5 ⁇ m, GL Science) was used as a column, acetonitrile was used as mobile phase A, and isopropanol was used as mobile phase B, and mobile phase A: mobile phase B was sent at 100: 0 for 5 minutes.
- Mobile phase A The mobile phase B was delivered by a gradient for 5 minutes from 100: 0 to 50:50 (volume basis), and 50:50 was held for another 5 minutes to separate the carotenoid.
- FIG. 9 The analysis result is shown in FIG. In FIG. 9, the HPLC chromatogram (top) in which the first expression vector and the second expression vector for comparison are co-expressed, and the first expression vector and the second expression vector (full-length CaCCS are incorporated). What: The HPLC chromatogram (middle) in which Example 2-2) was co-expressed, and the first expression vector and the second expression vector (CaCCS 41-498 from which the signal sequence was removed) were incorporated.
- FIG. 10 shows a graph comparing the intensities of the peak 5 of capsanthin in FIG.
- a second expression vector incorporating CCS 41-498 from which the signal sequence has been removed is used as compared with the case where the second expression vector incorporating the full-length CCS is used (Example 2-2). (Example 2-1) produced more capsanthin.
- the second expression vector incorporating the full-length CCS was used (Example 2-2) per culture medium.
- the capsanthin production was 326 ⁇ g / L, and the capsanthin production per culture medium was 452 ⁇ g / L when the second expression vector incorporating CCS 41-498 from which the signal sequence was removed was used (Example 2-1). ..
- Example 3 (Example 3-1 and Example 3-2, Example 3-3)
- the same pACHP-Zea as in Comparative Example 1 was used as the first recombinant vector.
- this pACHP-Zea is known that zeaxanthin occupies the total carotenoid without ⁇ -carotene being detected in the carotenoid composition by the transformant.
- this second recombinant vector is a protein in which CCS and ZEP are fused with S-Tag, respectively (in FIG. 12, “S tag + linker + CCS” and “CCS, respectively”. After being expressed as "S tag + linker + ZEP"), it binds indirectly via a multimerized S Protein (indicated as “multimer” in FIG. 12) and is a complex (Fig. 12). In 12, it was designed to form a "complex"). The aspect of the second recombinant vector thus designed is also referred to as "incorporated with S-Tag".
- the three second recombinant vectors pUC-S / slC / sZ, pUC-S / sC / slZ, and pUC-S / slC / slZ shown in FIG. 11 are between S-Tag and CCS. And / or the combination of presence or absence of a linker between S-Tag and ZEP was designed to have different configurations. By thus differentiating the position of introduction of the linker in the second recombinant vector into which S-Tag was incorporated, three conditions with different degrees of freedom between CCS and ZEP after expression were constructed.
- the linker consists of 20 amino acid residues in which a unit (GS) consisting of the non-polar amino acid glycine (G) and the polar amino acid serine (S) is continuous by 10 units. ( ⁇ GS ⁇ 10 linker) was used.
- the fusion position of S-Tag was set to the N-terminal of each of CaCCS and CaZEP.
- pUClac-ptac-CaCCSM40-CaZEP (pMF541) (Furubayashi, M., Kubo, A., Takemura, M., Otani, Y., Maoka, T., Terada, Y., Yaoi, K., Ohdan, K., Misawa, N. and Mitani, Y. (2021) J. Agric. Food Chem., 69, 5076-5085) were used.
- Example 2 After introducing the above-mentioned first recombinant vector and second recombinant vector in the same manner as in Example 1, a single colony on LB medium was transplanted into 3 mL of 2YT liquid medium, and Shaking manufactured by Yamato Kagaku Co., Ltd. Bath BW201 was used for culturing with shaking at 105 rpm and 28 ° C. for 16 hours. 20 ⁇ L of the culture solution was transplanted into 2.1 mL of TB liquid medium, and 0.1 mM isopropyl- ⁇ -D-galactopira after shaking culture at 180 rpm for 4 hours at 30 ° C. using Kuhner's Lab-Therm LT-X.
- the obtained supernatant was subjected to separation of carotenoids using Hitachi HPLC Chromaster system.
- Inertsil ODS-3 (4.6x150 mm, 5 ⁇ m, GL Science) was used as a column, acetonitrile was used as mobile phase A, and isopropanol was used as mobile phase B, and mobile phase A: mobile phase B was sent at 100: 0 for 5 minutes.
- Mobile phase A The mobile phase B was delivered by a gradient for 5 minutes from 100: 0 to 50:50 (volume basis), and 50:50 was held for another 5 minutes to separate carotenoids.
- FIG. x A graph comparing the peak intensities of each carotenoid in the HPLC analysis is shown in FIG. x
- the second recombinant vector pUC-CaCCS 41-498 -CaZEP (indicated as “C / Z” in the figure) used in Example 2-1 in which S-Tag is not incorporated is shown. The results when used are also shown.
- the case where the second recombinant vector in which S-Tag was incorporated was used rather than the case where S-Tag was not incorporated (Example 2-1) (Examples 3-1 to 3-1). In Example 3-3), more capsanthin was accumulated.
- Table 7 shows the results calculated from the peak intensity based on the molar extinction coefficient.
- the second recombinant vector in which S-tag was incorporated was used.
- the amount of capsanthin produced per culture solution was improved.
- the amount of capsanthin produced per culture medium is high. Not only was it significantly improved, but the ratio of capsanthin to the total amount of carotenoids shown in Table 7 was also significantly improved.
- FIG. 14 shows the results of UPLC analysis of the carotenoid obtained when pUC-S / sC / slZ was used as the second vector (Example 3-2) by the same method as in Example 1.
- each peak of capsanthin 3'-acetate, capsanthin 3,6-epoxide, capsanthin 3,6-epoxide 3'-acetate, capsanthin 3-acetate, capsorbindiacetate, and kukurubitaxanthin A was detected.
- the molecular ion peak was in agreement with the calculated value of the molecular weight of each carotenoid.
- Table 8 shows the results of calculating the ratio of each carotenoid from the peak intensity of UPLC analysis.
- SEQ ID NO: 3 the 34th to 36th sequences of the base sequence encoding S-Tag are replaced with the sequence encoding phenylalanine.
- SEQ ID NO: 4 is the amino acid sequence of S-Tag in which the 12th position is replaced with phenylalanine.
- SEQ ID NOs: 5 and 6 are the nucleotide sequence and amino acid sequence of CaCCS into which S-Tag has been introduced, respectively.
- SEQ ID NOs: 7 and 8 are the nucleotide sequence and amino acid sequence of CaCCS into which S-Tag is introduced via a linker, respectively.
- SEQ ID NOs: 9 and 10 are the nucleotide sequence and amino acid sequence of CaZEP into which S-Tag has been introduced, respectively.
- SEQ ID NOs: 11 and 12 are the nucleotide sequence and amino acid sequence of CaZEP into which S-Tag has been introduced via a linker, respectively.
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Cell Biology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Priority Applications (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US18/036,611 US20230416759A1 (en) | 2020-11-12 | 2021-11-09 | Transformant and method for producing carotenoid composition using same |
| EP21891841.5A EP4245843A4 (en) | 2020-11-12 | 2021-11-09 | TRANSFORMANT AND METHOD FOR PRODUCING A CAROTINOID COMPOSITION THEREOF |
| CN202180075145.6A CN116457454A (zh) | 2020-11-12 | 2021-11-09 | 转化体以及使用其的类胡萝卜素组合物的制造方法 |
| JP2022561924A JP7813719B2 (ja) | 2020-11-12 | 2021-11-09 | 形質転換体並びにそれを用いるカロテノイド組成物の製造方法 |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2020188462 | 2020-11-12 | ||
| JP2020-188462 | 2020-11-12 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2022102595A1 true WO2022102595A1 (ja) | 2022-05-19 |
Family
ID=81601269
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP2021/041098 Ceased WO2022102595A1 (ja) | 2020-11-12 | 2021-11-09 | 形質転換体並びにそれを用いるカロテノイド組成物の製造方法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US20230416759A1 (https=) |
| EP (1) | EP4245843A4 (https=) |
| JP (1) | JP7813719B2 (https=) |
| CN (1) | CN116457454A (https=) |
| WO (1) | WO2022102595A1 (https=) |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH09510608A (ja) * | 1994-03-01 | 1997-10-28 | サントル・ナショナル・ドゥ・ラ・ルシェルシェ・シャンティフィク | Dna構築物、それから誘導した細胞及び植物 |
| JP2008532492A (ja) * | 2005-02-11 | 2008-08-21 | カラマズー・ホールディングス・インコーポレイテッド | ゼアキサンチンの過剰蓄積を示すトウガラシ属品種およびそこから得られる産物 |
| US20140380516A1 (en) * | 2013-06-21 | 2014-12-25 | Seminis Vegetable Seeds, Inc. | Selection of mature fruit color in pepper plants |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR101424123B1 (ko) * | 2012-11-05 | 2014-08-20 | 대한민국 | 캡산틴을 생성하는 형질전환 컬러쌀 및 이의 제조방법 |
-
2021
- 2021-11-09 CN CN202180075145.6A patent/CN116457454A/zh active Pending
- 2021-11-09 US US18/036,611 patent/US20230416759A1/en active Pending
- 2021-11-09 EP EP21891841.5A patent/EP4245843A4/en active Pending
- 2021-11-09 WO PCT/JP2021/041098 patent/WO2022102595A1/ja not_active Ceased
- 2021-11-09 JP JP2022561924A patent/JP7813719B2/ja active Active
Patent Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH09510608A (ja) * | 1994-03-01 | 1997-10-28 | サントル・ナショナル・ドゥ・ラ・ルシェルシェ・シャンティフィク | Dna構築物、それから誘導した細胞及び植物 |
| JP2008532492A (ja) * | 2005-02-11 | 2008-08-21 | カラマズー・ホールディングス・インコーポレイテッド | ゼアキサンチンの過剰蓄積を示すトウガラシ属品種およびそこから得られる産物 |
| US20140380516A1 (en) * | 2013-06-21 | 2014-12-25 | Seminis Vegetable Seeds, Inc. | Selection of mature fruit color in pepper plants |
Non-Patent Citations (17)
| Title |
|---|
| BOUVIER, F. ET AL., THE PLANT JOURNAL, vol. 6, no. 1, 1994, pages 45 |
| EDITH H. M. LEMPENS, INGRID VAN BAAL, JOOST L. J. VAN DONGEN, TILMAN M. HACKENG, : "Noncovalent Synthesis of Protein Dendrimers", CHEMISTRY - A EUROPEAN JOURNAL, JOHN WILEY & SONS, INC, DE, vol. 15, no. 35, 7 September 2009 (2009-09-07), DE, pages 8760 - 8767, XP055349052, ISSN: 0947-6539, DOI: 10.1002/chem.200900748 * |
| FURUBAYASHI MAIKO, KUBO AKIKO, TAKEMURA MIHO, OTANI YUKO, MAOKA TAKASHI, TERADA YOSHINOBU, YAOI KATSURO, OHDAN KOHJI, MISAWA NORIH: "Capsanthin Production in Escherichia coli by Overexpression of Capsanthin/Capsorubin Synthase from Capsicum annuum", JOURNAL OF AGRICULTURAL AND FOOD CHEMISTRY, AMERICAN CHEMICAL SOCIETY, US, vol. 69, no. 17, 5 May 2021 (2021-05-05), US , pages 5076 - 5085, XP055929965, ISSN: 0021-8561, DOI: 10.1021/acs.jafc.1c00083 * |
| FURUBAYASHI, M.KUBO, A.TAKEMURA, M.OTANI, Y.MAOKA, T.TERADA, Y.YAOI, K.OHDAN, K.MISAWA, N.MITANI, Y., J.AGRIC.FOOD CHEM., vol. 69, 2021, pages 5076 - 5085 |
| JEKNIĆ ZORAN, MORRÉ JEFFREY T., JEKNIĆ STEVAN, JEVREMOVIĆ SLAĐANA, SUBOTIĆ ANGELINA, CHEN TONY H.H.: "Cloning and Functional Characterization of a Gene for Capsanthin-Capsorubin Synthase from Tiger Lily (Lilium lancifolium Thunb. ‘Splendens’)", PLANT AND CELL PHSIOLOGY, OXFORD UNIVERSITY PRESS, UK, vol. 53, no. 11, 1 November 2012 (2012-11-01), UK , pages 1899 - 1912, XP055929969, ISSN: 0032-0781, DOI: 10.1093/pcp/pcs128 * |
| JOURNAL OF THE SOCIETY FOR BIOTECHNOLOGY, vol. 93, no. 7, 2015 |
| KAJIWARA, S. ET AL., BIOCHEM. J., vol. 324, 1997, pages 421 |
| KAKINUMA, K. ET AL., J. AM. CHEM. SOC., vol. 123, 2001, pages 1238 |
| KUMAGAI MONTO H., YVES KELLER, FLORENCE BOUVIER, DAVID CLARY, BILAL CAMARA: "Functional integration of non-native carotenoids into chloroplasts by viral-derived expression of capsanthin-capsorubin synthase in Nicotiana benthamiana", THE PLANT JOURNAL, vol. 14, no. 3, 31 May 1998 (1998-05-31), pages 305 - 315, XP055929968, DOI: 10.1046/j.1365-313X.1998.00128.x * |
| LANZER ET AL., PNAS USA, vol. 85, 1988, pages 8973 - 8977 |
| LOPEZ-ALONSO, J.P.BRUIX, M.FONT, J.RIBO, M.VILANOVA, M.RICO, M.GOTTE, G.LIBONATI, M.GONZALEZ, C.LAURENTS, D.V., J.BIOL.CHEM., vol. 281, 2006, pages 9400 - 9406 |
| MEYER ET AL., NATURE CHEMICAL BIOLOGY, vol. 15, 2019, pages 196 - 204 |
| PBLA UNITDEUSCHLE ET AL., EMBO J., vol. 5, 1986, pages 2987 - 2994 |
| ROHMER, M. ET AL., BIOCHEM. J., vol. 295, 1993, pages 517 |
| See also references of EP4245843A4 |
| TAKEMURA, M. ET AL., TETRAHEDRON LETTERS, vol. 56, 2015, pages 6063 |
| TATIANA A. TATSUSOVATHOMAS L. MADDEN, FEMS MICROBIOL. LETT., vol. 174, 1999, pages 247 - 250 |
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2022102595A1 (https=) | 2022-05-19 |
| CN116457454A (zh) | 2023-07-18 |
| EP4245843A1 (en) | 2023-09-20 |
| EP4245843A4 (en) | 2025-01-29 |
| JP7813719B2 (ja) | 2026-02-13 |
| US20230416759A1 (en) | 2023-12-28 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Heider et al. | Carotenoid biosynthesis and overproduction in Corynebacterium glutamicum | |
| CN114502734A (zh) | 微生物产生植物大麻素和植物大麻素前体的方法和细胞 | |
| US20220325313A1 (en) | Biosynthesis of alpha-ionone and beta-ionone | |
| CA2996711C (en) | Method of fermentative alpha-ionone production | |
| CN117143951A (zh) | 类胡萝卜素和脱辅基类胡萝卜素的生产 | |
| Xinrui et al. | Metabolic engineering of Escherichia coli for high-level production of violaxanthin | |
| CN111527203B (zh) | 细胞色素p450单加氧酶催化的倍半萜的氧化 | |
| CN118813433A (zh) | 一种生产β-胡萝卜素的基因工程菌及其制备方法和应用 | |
| WO2006091924A2 (en) | Producing carotenoids | |
| US20240228986A1 (en) | Engineered cells, enzymes, and methods for producing cannabinoids | |
| WO2021050371A1 (en) | Biotin synthases for efficient production of biotin | |
| KR20210033100A (ko) | 효소반응을 이용한 크로신-4의 생산방법 | |
| JP7813719B2 (ja) | 形質転換体並びにそれを用いるカロテノイド組成物の製造方法 | |
| CN108949788B (zh) | 番茄红素合成相关基因及其应用 | |
| CN103052709B (zh) | 原伊鲁烯合酶 | |
| JP6635535B1 (ja) | Efpタンパク質を発現する大腸菌およびそれを用いたフラボノイド化合物製造方法 | |
| US7241611B2 (en) | Methods for synthesis of holo-photoactive yellow protein | |
| CN113528471A (zh) | 一种从头合成黄烷酮的三功能酶及其合成方法和应用 | |
| KR101400274B1 (ko) | P450 효소의 촉매활성을 증대시키는 cpr 유전자를 포함하는 재조합 벡터, 이에 의하여 형질전환된 세균 및 이를 이용한 p450 촉매반응 화합물의 제조방법 | |
| KR20210033101A (ko) | 효소반응을 이용한 크로신-2의 생산방법 | |
| WO2004101746A2 (en) | Production of carotenoids in microorganisms | |
| KR20250075325A (ko) | 크로신 및 전구체 생산용 재조합 효모 | |
| KR20230012321A (ko) | 레티노익산의 미생물 생합성 | |
| WO2023222879A2 (en) | Methods for producing monoterpene indole alkaloids | |
| CN117730147A (zh) | 苯丙素类化合物的生物合成 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application |
Ref document number: 21891841 Country of ref document: EP Kind code of ref document: A1 |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 202180075145.6 Country of ref document: CN |
|
| ENP | Entry into the national phase |
Ref document number: 2022561924 Country of ref document: JP Kind code of ref document: A |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 18036611 Country of ref document: US |
|
| NENP | Non-entry into the national phase |
Ref country code: DE |
|
| ENP | Entry into the national phase |
Ref document number: 2021891841 Country of ref document: EP Effective date: 20230612 |