KR20190078574A - 조립된 당단백질 - Google Patents
조립된 당단백질 Download PDFInfo
- Publication number
- KR20190078574A KR20190078574A KR1020197011934A KR20197011934A KR20190078574A KR 20190078574 A KR20190078574 A KR 20190078574A KR 1020197011934 A KR1020197011934 A KR 1020197011934A KR 20197011934 A KR20197011934 A KR 20197011934A KR 20190078574 A KR20190078574 A KR 20190078574A
- Authority
- KR
- South Korea
- Prior art keywords
- gly
- leu
- thr
- ala
- ser
- Prior art date
Links
- 102000003886 Glycoproteins Human genes 0.000 title description 42
- 108090000288 Glycoproteins Proteins 0.000 title description 42
- 239000000427 antigen Substances 0.000 claims abstract description 305
- 108091007433 antigens Proteins 0.000 claims abstract description 305
- 102000036639 antigens Human genes 0.000 claims abstract description 305
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 claims abstract description 160
- 238000000034 method Methods 0.000 claims abstract description 128
- 230000027455 binding Effects 0.000 claims abstract description 102
- 230000003472 neutralizing effect Effects 0.000 claims abstract description 77
- 230000002829 reductive effect Effects 0.000 claims abstract description 69
- 235000018417 cysteine Nutrition 0.000 claims abstract description 40
- 239000003638 chemical reducing agent Substances 0.000 claims abstract description 38
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 claims abstract description 29
- 239000002671 adjuvant Substances 0.000 claims abstract description 28
- 210000003719 b-lymphocyte Anatomy 0.000 claims abstract description 27
- 210000002865 immune cell Anatomy 0.000 claims abstract description 27
- 230000028993 immune response Effects 0.000 claims abstract description 23
- 239000007800 oxidant agent Substances 0.000 claims abstract description 15
- 239000003085 diluting agent Substances 0.000 claims abstract description 12
- 238000007865 diluting Methods 0.000 claims abstract description 11
- 241000711549 Hepacivirus C Species 0.000 claims description 227
- 239000000203 mixture Substances 0.000 claims description 115
- 210000004027 cell Anatomy 0.000 claims description 95
- 238000004519 manufacturing process Methods 0.000 claims description 49
- 208000015181 infectious disease Diseases 0.000 claims description 39
- 229960005486 vaccine Drugs 0.000 claims description 38
- 241000700605 Viruses Species 0.000 claims description 35
- 238000011282 treatment Methods 0.000 claims description 21
- 206010028980 Neoplasm Diseases 0.000 claims description 18
- 108010003533 Viral Envelope Proteins Proteins 0.000 claims description 18
- 208000024891 symptom Diseases 0.000 claims description 16
- 210000004779 membrane envelope Anatomy 0.000 claims description 15
- 238000000746 purification Methods 0.000 claims description 15
- 238000004458 analytical method Methods 0.000 claims description 14
- 108020003175 receptors Proteins 0.000 claims description 14
- 102000005962 receptors Human genes 0.000 claims description 14
- 125000000539 amino acid group Chemical group 0.000 claims description 13
- 102000008482 12E7 Antigen Human genes 0.000 claims description 12
- 108010020567 12E7 Antigen Proteins 0.000 claims description 12
- 201000011510 cancer Diseases 0.000 claims description 12
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 claims description 11
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 claims description 10
- 239000007787 solid Substances 0.000 claims description 10
- 230000008569 process Effects 0.000 claims description 9
- 230000003053 immunization Effects 0.000 claims description 8
- 230000035772 mutation Effects 0.000 claims description 8
- 230000002265 prevention Effects 0.000 claims description 8
- 238000001514 detection method Methods 0.000 claims description 7
- 238000012544 monitoring process Methods 0.000 claims description 6
- 150000001875 compounds Chemical class 0.000 claims description 5
- 239000003814 drug Substances 0.000 claims description 5
- 238000002955 isolation Methods 0.000 claims description 5
- 230000001225 therapeutic effect Effects 0.000 claims description 5
- 208000006454 hepatitis Diseases 0.000 claims description 4
- 238000002372 labelling Methods 0.000 claims description 4
- 238000006467 substitution reaction Methods 0.000 claims description 4
- 239000000758 substrate Substances 0.000 claims description 4
- 238000003745 diagnosis Methods 0.000 claims description 3
- 239000000032 diagnostic agent Substances 0.000 claims description 3
- 229940039227 diagnostic agent Drugs 0.000 claims description 3
- 231100000283 hepatitis Toxicity 0.000 claims description 3
- 230000003452 intergenotypic effect Effects 0.000 claims 1
- 230000003834 intracellular effect Effects 0.000 abstract 1
- 239000000178 monomer Substances 0.000 description 116
- 102000004169 proteins and genes Human genes 0.000 description 102
- 108090000623 proteins and genes Proteins 0.000 description 102
- 235000018102 proteins Nutrition 0.000 description 94
- VHJLVAABSRFDPM-QWWZWVQMSA-N dithiothreitol Chemical compound SC[C@@H](O)[C@H](O)CS VHJLVAABSRFDPM-QWWZWVQMSA-N 0.000 description 70
- 206010057190 Respiratory tract infections Diseases 0.000 description 62
- 230000009467 reduction Effects 0.000 description 60
- 241001465754 Metazoa Species 0.000 description 43
- 239000000539 dimer Substances 0.000 description 43
- RWSXRVCMGQZWBV-WDSKDSINSA-N glutathione Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-WDSKDSINSA-N 0.000 description 41
- 239000002953 phosphate buffered saline Substances 0.000 description 35
- 102100027221 CD81 antigen Human genes 0.000 description 30
- 101000914479 Homo sapiens CD81 antigen Proteins 0.000 description 30
- PBVAJRFEEOIAGW-UHFFFAOYSA-N 3-[bis(2-carboxyethyl)phosphanyl]propanoic acid;hydrochloride Chemical compound Cl.OC(=O)CCP(CCC(O)=O)CCC(O)=O PBVAJRFEEOIAGW-UHFFFAOYSA-N 0.000 description 28
- 241000725303 Human immunodeficiency virus Species 0.000 description 28
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 28
- 108010016616 cysteinylglycine Proteins 0.000 description 27
- 238000010790 dilution Methods 0.000 description 26
- 239000012895 dilution Substances 0.000 description 26
- 238000001641 gel filtration chromatography Methods 0.000 description 26
- 241000894007 species Species 0.000 description 26
- 108090000765 processed proteins & peptides Proteins 0.000 description 24
- 108010061238 threonyl-glycine Proteins 0.000 description 24
- 239000000243 solution Substances 0.000 description 22
- 150000001413 amino acids Chemical class 0.000 description 20
- 230000015572 biosynthetic process Effects 0.000 description 20
- 239000000872 buffer Substances 0.000 description 20
- 108010050848 glycylleucine Proteins 0.000 description 20
- 210000002966 serum Anatomy 0.000 description 20
- 238000004113 cell culture Methods 0.000 description 18
- 239000003795 chemical substances by application Substances 0.000 description 18
- 229960003180 glutathione Drugs 0.000 description 18
- VPZXBVLAVMBEQI-UHFFFAOYSA-N glycyl-DL-alpha-alanine Natural products OC(=O)C(C)NC(=O)CN VPZXBVLAVMBEQI-UHFFFAOYSA-N 0.000 description 18
- 102000004196 processed proteins & peptides Human genes 0.000 description 18
- 108010029384 tryptophyl-histidine Proteins 0.000 description 18
- RVRLFABOQXZUJX-UHFFFAOYSA-N 1-[1-(2,5-dioxopyrrol-1-yl)ethyl]pyrrole-2,5-dione Chemical compound O=C1C=CC(=O)N1C(C)N1C(=O)C=CC1=O RVRLFABOQXZUJX-UHFFFAOYSA-N 0.000 description 17
- 238000003556 assay Methods 0.000 description 16
- 108020004414 DNA Proteins 0.000 description 15
- 102100038132 Endogenous retrovirus group K member 6 Pro protein Human genes 0.000 description 15
- 239000011545 carbonate/bicarbonate buffer Substances 0.000 description 15
- 108010078326 glycyl-glycyl-valine Proteins 0.000 description 15
- 238000013207 serial dilution Methods 0.000 description 15
- 239000013598 vector Substances 0.000 description 15
- 108010001336 Horseradish Peroxidase Proteins 0.000 description 14
- 108010060199 cysteinylproline Proteins 0.000 description 14
- 238000002474 experimental method Methods 0.000 description 14
- 239000000499 gel Substances 0.000 description 14
- 238000002523 gelfiltration Methods 0.000 description 14
- 108010009111 arginyl-glycyl-glutamic acid Proteins 0.000 description 13
- 238000006243 chemical reaction Methods 0.000 description 13
- 239000012228 culture supernatant Substances 0.000 description 13
- 239000013612 plasmid Substances 0.000 description 13
- 239000000523 sample Substances 0.000 description 13
- 125000003396 thiol group Chemical group [H]S* 0.000 description 13
- 230000003612 virological effect Effects 0.000 description 13
- QTBSBXVTEAMEQO-UHFFFAOYSA-N Acetic acid Chemical compound CC(O)=O QTBSBXVTEAMEQO-UHFFFAOYSA-N 0.000 description 12
- 108010024636 Glutathione Proteins 0.000 description 12
- HNDVDQJCIGZPNO-UHFFFAOYSA-N Histidine Chemical compound OC(=O)C(N)CC1=CN=CN1 HNDVDQJCIGZPNO-UHFFFAOYSA-N 0.000 description 12
- KZNQNBZMBZJQJO-UHFFFAOYSA-N N-glycyl-L-proline Natural products NCC(=O)N1CCCC1C(O)=O KZNQNBZMBZJQJO-UHFFFAOYSA-N 0.000 description 12
- AJHCSUXXECOXOY-UHFFFAOYSA-N N-glycyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)CN)C(O)=O)=CNC2=C1 AJHCSUXXECOXOY-UHFFFAOYSA-N 0.000 description 12
- 239000000463 material Substances 0.000 description 12
- 230000009257 reactivity Effects 0.000 description 12
- HLTLEIXYIJDFOY-ZLUOBGJFSA-N Asn-Cys-Asn Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(O)=O HLTLEIXYIJDFOY-ZLUOBGJFSA-N 0.000 description 11
- 238000002965 ELISA Methods 0.000 description 11
- 102000004190 Enzymes Human genes 0.000 description 11
- 108090000790 Enzymes Proteins 0.000 description 11
- BQVUABVGYYSDCJ-UHFFFAOYSA-N Nalpha-L-Leucyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)CC(C)C)C(O)=O)=CNC2=C1 BQVUABVGYYSDCJ-UHFFFAOYSA-N 0.000 description 11
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 11
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 11
- BSNZTJXVDOINSR-JXUBOQSCSA-N Thr-Ala-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O BSNZTJXVDOINSR-JXUBOQSCSA-N 0.000 description 11
- 238000001042 affinity chromatography Methods 0.000 description 11
- 108010047495 alanylglycine Proteins 0.000 description 11
- 108010087924 alanylproline Proteins 0.000 description 11
- 108010062796 arginyllysine Proteins 0.000 description 11
- 150000001945 cysteines Chemical class 0.000 description 11
- 238000001943 fluorescence-activated cell sorting Methods 0.000 description 11
- 238000009472 formulation Methods 0.000 description 11
- 108010089804 glycyl-threonine Proteins 0.000 description 11
- 238000011534 incubation Methods 0.000 description 11
- 230000036961 partial effect Effects 0.000 description 11
- 229920001184 polypeptide Polymers 0.000 description 11
- 238000007619 statistical method Methods 0.000 description 11
- 238000001890 transfection Methods 0.000 description 11
- 238000005406 washing Methods 0.000 description 11
- VKCOHFFSTKCXEQ-OLHMAJIHSA-N Asn-Asn-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VKCOHFFSTKCXEQ-OLHMAJIHSA-N 0.000 description 10
- 101710121417 Envelope glycoprotein Proteins 0.000 description 10
- ISQOVWDWRUONJH-YESZJQIVSA-N His-Tyr-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=C(C=C2)O)NC(=O)[C@H](CC3=CN=CN3)N)C(=O)O ISQOVWDWRUONJH-YESZJQIVSA-N 0.000 description 10
- MVVSHHJKJRZVNY-ACRUOGEOSA-N Leu-Phe-Tyr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O MVVSHHJKJRZVNY-ACRUOGEOSA-N 0.000 description 10
- 239000012505 Superdex™ Substances 0.000 description 10
- 230000005847 immunogenicity Effects 0.000 description 10
- 230000001965 increasing effect Effects 0.000 description 10
- 108010057821 leucylproline Proteins 0.000 description 10
- 108010038320 lysylphenylalanine Proteins 0.000 description 10
- 230000003287 optical effect Effects 0.000 description 10
- 238000002360 preparation method Methods 0.000 description 10
- HNXWVVHIGTZTBO-LKXGYXEUSA-N Asn-Ser-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC(N)=O HNXWVVHIGTZTBO-LKXGYXEUSA-N 0.000 description 9
- 241000700199 Cavia porcellus Species 0.000 description 9
- PSERKXGRRADTKA-MNXVOIDGSA-N Gln-Leu-Ile Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O PSERKXGRRADTKA-MNXVOIDGSA-N 0.000 description 9
- WHUUTDBJXJRKMK-UHFFFAOYSA-N Glutamic acid Natural products OC(=O)C(N)CCC(O)=O WHUUTDBJXJRKMK-UHFFFAOYSA-N 0.000 description 9
- IMRNSEPSPFQNHF-STQMWFEESA-N Gly-Ser-Trp Chemical compound NCC(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CNC2=CC=CC=C12)C(=O)O IMRNSEPSPFQNHF-STQMWFEESA-N 0.000 description 9
- JBCLFWXMTIKCCB-UHFFFAOYSA-N H-Gly-Phe-OH Natural products NCC(=O)NC(C(O)=O)CC1=CC=CC=C1 JBCLFWXMTIKCCB-UHFFFAOYSA-N 0.000 description 9
- 241000880493 Leptailurus serval Species 0.000 description 9
- WNGVUZWBXZKQES-YUMQZZPRSA-N Leu-Ala-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O WNGVUZWBXZKQES-YUMQZZPRSA-N 0.000 description 9
- KLSUAWUZBMAZCL-RHYQMDGZSA-N Leu-Thr-Pro Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(O)=O KLSUAWUZBMAZCL-RHYQMDGZSA-N 0.000 description 9
- OKKJLVBELUTLKV-UHFFFAOYSA-N Methanol Chemical compound OC OKKJLVBELUTLKV-UHFFFAOYSA-N 0.000 description 9
- CDNPIRSCAFMMBE-SRVKXCTJSA-N Phe-Asn-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O CDNPIRSCAFMMBE-SRVKXCTJSA-N 0.000 description 9
- QUBVFEANYYWBTM-VEVYYDQMSA-N Pro-Thr-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(O)=O QUBVFEANYYWBTM-VEVYYDQMSA-N 0.000 description 9
- 229940024606 amino acid Drugs 0.000 description 9
- 235000001014 amino acid Nutrition 0.000 description 9
- 230000000890 antigenic effect Effects 0.000 description 9
- 108010029539 arginyl-prolyl-proline Proteins 0.000 description 9
- 230000000694 effects Effects 0.000 description 9
- 244000052769 pathogen Species 0.000 description 9
- 108010004914 prolylarginine Proteins 0.000 description 9
- 108010053725 prolylvaline Proteins 0.000 description 9
- 239000006228 supernatant Substances 0.000 description 9
- 108010020532 tyrosyl-proline Proteins 0.000 description 9
- AUFACLFHBAGZEN-ZLUOBGJFSA-N Ala-Ser-Cys Chemical compound N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O AUFACLFHBAGZEN-ZLUOBGJFSA-N 0.000 description 8
- 101100269553 Arabidopsis thaliana ALA7 gene Proteins 0.000 description 8
- BWGNESOTFCXPMA-UHFFFAOYSA-N Dihydrogen disulfide Chemical compound SS BWGNESOTFCXPMA-UHFFFAOYSA-N 0.000 description 8
- WMOMPXKOKASNBK-PEFMBERDSA-N Gln-Asn-Ile Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WMOMPXKOKASNBK-PEFMBERDSA-N 0.000 description 8
- 108010053070 Glutathione Disulfide Proteins 0.000 description 8
- GNLJXWBNLAIPEP-MELADBBJSA-N Lys-His-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CN=CN2)NC(=O)[C@H](CCCCN)N)C(=O)O GNLJXWBNLAIPEP-MELADBBJSA-N 0.000 description 8
- 108091028043 Nucleic acid sequence Proteins 0.000 description 8
- 108010093581 aspartyl-proline Proteins 0.000 description 8
- 239000012634 fragment Substances 0.000 description 8
- YPZRWBKMTBYPTK-BJDJZHNGSA-N glutathione disulfide Chemical compound OC(=O)[C@@H](N)CCC(=O)N[C@H](C(=O)NCC(O)=O)CSSC[C@@H](C(=O)NCC(O)=O)NC(=O)CC[C@H](N)C(O)=O YPZRWBKMTBYPTK-BJDJZHNGSA-N 0.000 description 8
- 238000003018 immunoassay Methods 0.000 description 8
- 108010034529 leucyl-lysine Proteins 0.000 description 8
- 239000002609 medium Substances 0.000 description 8
- 239000013642 negative control Substances 0.000 description 8
- 230000001717 pathogenic effect Effects 0.000 description 8
- 238000003118 sandwich ELISA Methods 0.000 description 8
- 238000012360 testing method Methods 0.000 description 8
- 238000003146 transient transfection Methods 0.000 description 8
- TZDNWXDLYFIFPT-BJDJZHNGSA-N Ala-Ile-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O TZDNWXDLYFIFPT-BJDJZHNGSA-N 0.000 description 7
- MDNAVFBZPROEHO-UHFFFAOYSA-N Ala-Lys-Val Natural products CC(C)C(C(O)=O)NC(=O)C(NC(=O)C(C)N)CCCCN MDNAVFBZPROEHO-UHFFFAOYSA-N 0.000 description 7
- FOQFHANLUJDQEE-GUBZILKMSA-N Arg-Pro-Cys Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CCCN=C(N)N)N)C(=O)N[C@@H](CS)C(=O)O FOQFHANLUJDQEE-GUBZILKMSA-N 0.000 description 7
- SNYCNNPOFYBCEK-ZLUOBGJFSA-N Asn-Ser-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O SNYCNNPOFYBCEK-ZLUOBGJFSA-N 0.000 description 7
- QYRMBFWDSFGSFC-OLHMAJIHSA-N Asn-Thr-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N)O QYRMBFWDSFGSFC-OLHMAJIHSA-N 0.000 description 7
- UDDITVWSXPEAIQ-IHRRRGAJSA-N Cys-Phe-Arg Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O UDDITVWSXPEAIQ-IHRRRGAJSA-N 0.000 description 7
- QQOWCDCBFFBRQH-IXOXFDKPSA-N Cys-Phe-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)[C@H](CS)N)O QQOWCDCBFFBRQH-IXOXFDKPSA-N 0.000 description 7
- DHDOADIPGZTAHT-YUMQZZPRSA-N Gly-Glu-Arg Chemical compound NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N DHDOADIPGZTAHT-YUMQZZPRSA-N 0.000 description 7
- RXGLHDWAZQECBI-SRVKXCTJSA-N Leu-Leu-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O RXGLHDWAZQECBI-SRVKXCTJSA-N 0.000 description 7
- FHJQROWZEJFZPO-SRVKXCTJSA-N Pro-Val-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H]1CCCN1 FHJQROWZEJFZPO-SRVKXCTJSA-N 0.000 description 7
- 108010076504 Protein Sorting Signals Proteins 0.000 description 7
- SVWQEIRZHHNBIO-WHFBIAKZSA-N Ser-Gly-Cys Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CS)C(O)=O SVWQEIRZHHNBIO-WHFBIAKZSA-N 0.000 description 7
- PIFJAFRUVWZRKR-QMMMGPOBSA-N Val-Gly-Gly Chemical compound CC(C)[C@H]([NH3+])C(=O)NCC(=O)NCC([O-])=O PIFJAFRUVWZRKR-QMMMGPOBSA-N 0.000 description 7
- 210000004369 blood Anatomy 0.000 description 7
- 239000008280 blood Substances 0.000 description 7
- 210000004899 c-terminal region Anatomy 0.000 description 7
- 238000001914 filtration Methods 0.000 description 7
- 108010082286 glycyl-seryl-alanine Proteins 0.000 description 7
- 108010025306 histidylleucine Proteins 0.000 description 7
- 230000001976 improved effect Effects 0.000 description 7
- 108010017391 lysylvaline Proteins 0.000 description 7
- 238000006386 neutralization reaction Methods 0.000 description 7
- 108010025488 pinealon Proteins 0.000 description 7
- 239000013641 positive control Substances 0.000 description 7
- 108010026333 seryl-proline Proteins 0.000 description 7
- 239000003826 tablet Substances 0.000 description 7
- HFBFSOAKPUZCCO-ZLUOBGJFSA-N Ala-Cys-Asn Chemical compound C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)N)C(=O)O)N HFBFSOAKPUZCCO-ZLUOBGJFSA-N 0.000 description 6
- OGSQONVYSTZIJB-WDSOQIARSA-N Arg-Leu-Trp Chemical compound CC(C)C[C@H](NC(=O)[C@@H](N)CCCN=C(N)N)C(=O)N[C@@H](Cc1c[nH]c2ccccc12)C(O)=O OGSQONVYSTZIJB-WDSOQIARSA-N 0.000 description 6
- ISJWBVIYRBAXEB-CIUDSAMLSA-N Arg-Ser-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O ISJWBVIYRBAXEB-CIUDSAMLSA-N 0.000 description 6
- JAHCWGSVNZXHRR-SVSWQMSJSA-N Cys-Thr-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CS)N JAHCWGSVNZXHRR-SVSWQMSJSA-N 0.000 description 6
- NNXIQPMZGZUFJJ-AVGNSLFASA-N Gln-His-Lys Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)N)N NNXIQPMZGZUFJJ-AVGNSLFASA-N 0.000 description 6
- QPRZKNOOOBWXSU-CIUDSAMLSA-N Glu-Asp-Arg Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N QPRZKNOOOBWXSU-CIUDSAMLSA-N 0.000 description 6
- BMWFDYIYBAFROD-WPRPVWTQSA-N Gly-Pro-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)CN BMWFDYIYBAFROD-WPRPVWTQSA-N 0.000 description 6
- WCORRBXVISTKQL-WHFBIAKZSA-N Gly-Ser-Ser Chemical compound NCC(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O WCORRBXVISTKQL-WHFBIAKZSA-N 0.000 description 6
- DNVDEMWIYLVIQU-RCOVLWMOSA-N Gly-Val-Asp Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O DNVDEMWIYLVIQU-RCOVLWMOSA-N 0.000 description 6
- VJJSDSNFXCWCEJ-DJFWLOJKSA-N His-Ile-Asn Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O VJJSDSNFXCWCEJ-DJFWLOJKSA-N 0.000 description 6
- 241000713772 Human immunodeficiency virus 1 Species 0.000 description 6
- HDODQNPMSHDXJT-GHCJXIJMSA-N Ile-Asn-Ser Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O HDODQNPMSHDXJT-GHCJXIJMSA-N 0.000 description 6
- SBANPBVRHYIMRR-UHFFFAOYSA-N Leu-Ser-Pro Natural products CC(C)CC(N)C(=O)NC(CO)C(=O)N1CCCC1C(O)=O SBANPBVRHYIMRR-UHFFFAOYSA-N 0.000 description 6
- 101100342977 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) leu-1 gene Proteins 0.000 description 6
- GPSMLZQVIIYLDK-ULQDDVLXSA-N Phe-Lys-Val Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O GPSMLZQVIIYLDK-ULQDDVLXSA-N 0.000 description 6
- UQFYNFTYDHUIMI-WHFBIAKZSA-N Ser-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)[C@@H](N)CO UQFYNFTYDHUIMI-WHFBIAKZSA-N 0.000 description 6
- TYVAWPFQYFPSBR-BFHQHQDPSA-N Thr-Ala-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(=O)NCC(O)=O TYVAWPFQYFPSBR-BFHQHQDPSA-N 0.000 description 6
- FRQRWAMUESPWMT-HSHDSVGOSA-N Thr-Trp-Met Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CCSC)C(=O)O)N)O FRQRWAMUESPWMT-HSHDSVGOSA-N 0.000 description 6
- OAZLRFLMQASGNW-PMVMPFDFSA-N Trp-His-Tyr Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CN=CN3)C(=O)N[C@@H](CC4=CC=C(C=C4)O)C(=O)O)N OAZLRFLMQASGNW-PMVMPFDFSA-N 0.000 description 6
- QHWMVGCEQAPQDK-UMPQAUOISA-N Trp-Thr-Arg Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N)O QHWMVGCEQAPQDK-UMPQAUOISA-N 0.000 description 6
- VPEFOFYNHBWFNQ-UFYCRDLUSA-N Tyr-Pro-Tyr Chemical compound C([C@H](N)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=C(O)C=C1 VPEFOFYNHBWFNQ-UFYCRDLUSA-N 0.000 description 6
- FPCIBLUVDNXPJO-XPUUQOCRSA-N Val-Cys-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CS)C(=O)NCC(O)=O FPCIBLUVDNXPJO-XPUUQOCRSA-N 0.000 description 6
- YDPFWRVQHFWBKI-GVXVVHGQSA-N Val-Glu-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N YDPFWRVQHFWBKI-GVXVVHGQSA-N 0.000 description 6
- XBJKAZATRJBDCU-GUBZILKMSA-N Val-Pro-Ala Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O XBJKAZATRJBDCU-GUBZILKMSA-N 0.000 description 6
- 230000002776 aggregation Effects 0.000 description 6
- 238000004220 aggregation Methods 0.000 description 6
- 108010044940 alanylglutamine Proteins 0.000 description 6
- 239000011324 bead Substances 0.000 description 6
- 238000010367 cloning Methods 0.000 description 6
- 239000013604 expression vector Substances 0.000 description 6
- 239000011544 gradient gel Substances 0.000 description 6
- 230000036541 health Effects 0.000 description 6
- 238000002826 magnetic-activated cell sorting Methods 0.000 description 6
- 229920002401 polyacrylamide Polymers 0.000 description 6
- 229920000642 polymer Polymers 0.000 description 6
- 108010029020 prolylglycine Proteins 0.000 description 6
- 230000000069 prophylactic effect Effects 0.000 description 6
- 238000011002 quantification Methods 0.000 description 6
- 238000001542 size-exclusion chromatography Methods 0.000 description 6
- 108010080629 tryptophan-leucine Proteins 0.000 description 6
- 108010038745 tryptophylglycine Proteins 0.000 description 6
- 208000030507 AIDS Diseases 0.000 description 5
- XYTNPQNAZREREP-XQXXSGGOSA-N Ala-Glu-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XYTNPQNAZREREP-XQXXSGGOSA-N 0.000 description 5
- SOBIAADAMRHGKH-CIUDSAMLSA-N Ala-Leu-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O SOBIAADAMRHGKH-CIUDSAMLSA-N 0.000 description 5
- YLVGUOGAFAJMKP-JYJNAYRXSA-N Arg-Met-Tyr Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N YLVGUOGAFAJMKP-JYJNAYRXSA-N 0.000 description 5
- FTCGGKNCJZOPNB-WHFBIAKZSA-N Asn-Gly-Ser Chemical compound NC(=O)C[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O FTCGGKNCJZOPNB-WHFBIAKZSA-N 0.000 description 5
- IBLAOXSULLECQZ-IUKAMOBKSA-N Asn-Ile-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CC(N)=O IBLAOXSULLECQZ-IUKAMOBKSA-N 0.000 description 5
- FMNBYVSGRCXWEK-FOHZUACHSA-N Asn-Thr-Gly Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O FMNBYVSGRCXWEK-FOHZUACHSA-N 0.000 description 5
- IPPFAOCLQSGHJV-WFBYXXMGSA-N Asn-Trp-Ala Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](C)C(O)=O IPPFAOCLQSGHJV-WFBYXXMGSA-N 0.000 description 5
- XABFFGOGKOORCG-CIUDSAMLSA-N Cys-Asp-Leu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O XABFFGOGKOORCG-CIUDSAMLSA-N 0.000 description 5
- TXGDWPBLUFQODU-XGEHTFHBSA-N Cys-Pro-Thr Chemical compound [H]N[C@@H](CS)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O TXGDWPBLUFQODU-XGEHTFHBSA-N 0.000 description 5
- NXPXQIZKDOXIHH-JSGCOSHPSA-N Gln-Gly-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CCC(=O)N)N NXPXQIZKDOXIHH-JSGCOSHPSA-N 0.000 description 5
- FYBSCGZLICNOBA-XQXXSGGOSA-N Glu-Ala-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O FYBSCGZLICNOBA-XQXXSGGOSA-N 0.000 description 5
- KKCUFHUTMKQQCF-SRVKXCTJSA-N Glu-Arg-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O KKCUFHUTMKQQCF-SRVKXCTJSA-N 0.000 description 5
- DDXZHOHEABQXSE-NKIYYHGXSA-N Glu-Thr-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)O DDXZHOHEABQXSE-NKIYYHGXSA-N 0.000 description 5
- JBRBACJPBZNFMF-YUMQZZPRSA-N Gly-Ala-Lys Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN JBRBACJPBZNFMF-YUMQZZPRSA-N 0.000 description 5
- QXPRJQPCFXMCIY-NKWVEPMBSA-N Gly-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)CN QXPRJQPCFXMCIY-NKWVEPMBSA-N 0.000 description 5
- DTPOVRRYXPJJAZ-FJXKBIBVSA-N Gly-Arg-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCN=C(N)N DTPOVRRYXPJJAZ-FJXKBIBVSA-N 0.000 description 5
- OLPPXYMMIARYAL-QMMMGPOBSA-N Gly-Gly-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)CNC(=O)CN OLPPXYMMIARYAL-QMMMGPOBSA-N 0.000 description 5
- COVXELOAORHTND-LSJOCFKGSA-N Gly-Ile-Val Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O COVXELOAORHTND-LSJOCFKGSA-N 0.000 description 5
- TVTZEOHWHUVYCG-KYNKHSRBSA-N Gly-Thr-Thr Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O TVTZEOHWHUVYCG-KYNKHSRBSA-N 0.000 description 5
- VYMGAXSNYUFVCK-GUBZILKMSA-N His-Gln-Asn Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC(=O)N)C(=O)O)N VYMGAXSNYUFVCK-GUBZILKMSA-N 0.000 description 5
- HGUUMQWGYCVPKG-DCAQKATOSA-N Leu-Pro-Cys Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CS)C(=O)O)N HGUUMQWGYCVPKG-DCAQKATOSA-N 0.000 description 5
- AAKRWBIIGKPOKQ-ONGXEEELSA-N Leu-Val-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O AAKRWBIIGKPOKQ-ONGXEEELSA-N 0.000 description 5
- 241000699666 Mus <mouse, genus> Species 0.000 description 5
- YBAFDPFAUTYYRW-UHFFFAOYSA-N N-L-alpha-glutamyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=O YBAFDPFAUTYYRW-UHFFFAOYSA-N 0.000 description 5
- AUEJLPRZGVVDNU-UHFFFAOYSA-N N-L-tyrosyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CC1=CC=C(O)C=C1 AUEJLPRZGVVDNU-UHFFFAOYSA-N 0.000 description 5
- XYSXOCIWCPFOCG-IHRRRGAJSA-N Pro-Leu-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O XYSXOCIWCPFOCG-IHRRRGAJSA-N 0.000 description 5
- CGSOWZUPLOKYOR-AVGNSLFASA-N Pro-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 CGSOWZUPLOKYOR-AVGNSLFASA-N 0.000 description 5
- SNGZLPOXVRTNMB-LPEHRKFASA-N Pro-Ser-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CO)C(=O)N2CCC[C@@H]2C(=O)O SNGZLPOXVRTNMB-LPEHRKFASA-N 0.000 description 5
- BVTYXOFTHDXSNI-IHRRRGAJSA-N Pro-Tyr-Cys Chemical compound C([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H]1NCCC1)C1=CC=C(O)C=C1 BVTYXOFTHDXSNI-IHRRRGAJSA-N 0.000 description 5
- PGSWNLRYYONGPE-JYJNAYRXSA-N Pro-Val-Tyr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O PGSWNLRYYONGPE-JYJNAYRXSA-N 0.000 description 5
- CKDXFSPMIDSMGV-GUBZILKMSA-N Ser-Pro-Val Chemical compound [H]N[C@@H](CO)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=O CKDXFSPMIDSMGV-GUBZILKMSA-N 0.000 description 5
- PURRNJBBXDDWLX-ZDLURKLDSA-N Ser-Thr-Gly Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CO)N)O PURRNJBBXDDWLX-ZDLURKLDSA-N 0.000 description 5
- PIQRHJQWEPWFJG-UWJYBYFXSA-N Ser-Tyr-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C)C(O)=O PIQRHJQWEPWFJG-UWJYBYFXSA-N 0.000 description 5
- 238000000692 Student's t-test Methods 0.000 description 5
- 210000001744 T-lymphocyte Anatomy 0.000 description 5
- ZTPXSEUVYNNZRB-CDMKHQONSA-N Thr-Gly-Phe Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O ZTPXSEUVYNNZRB-CDMKHQONSA-N 0.000 description 5
- MUAFDCVOHYAFNG-RCWTZXSCSA-N Thr-Pro-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(O)=O MUAFDCVOHYAFNG-RCWTZXSCSA-N 0.000 description 5
- ADBDQGBDNUTRDB-ULQDDVLXSA-N Tyr-Arg-Leu Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O ADBDQGBDNUTRDB-ULQDDVLXSA-N 0.000 description 5
- URIRWLJVWHYLET-ONGXEEELSA-N Val-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)C(C)C URIRWLJVWHYLET-ONGXEEELSA-N 0.000 description 5
- 108010005233 alanylglutamic acid Proteins 0.000 description 5
- 230000005875 antibody response Effects 0.000 description 5
- 108010060035 arginylproline Proteins 0.000 description 5
- 239000003153 chemical reaction reagent Substances 0.000 description 5
- 230000002950 deficient Effects 0.000 description 5
- 238000001739 density measurement Methods 0.000 description 5
- 108010049041 glutamylalanine Proteins 0.000 description 5
- 108010077515 glycylproline Proteins 0.000 description 5
- 108010084389 glycyltryptophan Proteins 0.000 description 5
- 230000002163 immunogen Effects 0.000 description 5
- 230000003993 interaction Effects 0.000 description 5
- 108010003700 lysyl aspartic acid Proteins 0.000 description 5
- 108020004707 nucleic acids Proteins 0.000 description 5
- 102000039446 nucleic acids Human genes 0.000 description 5
- 150000007523 nucleic acids Chemical class 0.000 description 5
- 239000000137 peptide hydrolase inhibitor Substances 0.000 description 5
- 230000036515 potency Effects 0.000 description 5
- 239000000047 product Substances 0.000 description 5
- 150000003839 salts Chemical class 0.000 description 5
- 238000010186 staining Methods 0.000 description 5
- 238000012353 t test Methods 0.000 description 5
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 5
- UWQJHXKARZWDIJ-ZLUOBGJFSA-N Ala-Ala-Cys Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CS)C(O)=O UWQJHXKARZWDIJ-ZLUOBGJFSA-N 0.000 description 4
- RLMISHABBKUNFO-WHFBIAKZSA-N Ala-Ala-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O RLMISHABBKUNFO-WHFBIAKZSA-N 0.000 description 4
- OLVCTPPSXNRGKV-GUBZILKMSA-N Ala-Pro-Pro Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 OLVCTPPSXNRGKV-GUBZILKMSA-N 0.000 description 4
- FFZJHQODAYHGPO-KZVJFYERSA-N Ala-Pro-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](C)N FFZJHQODAYHGPO-KZVJFYERSA-N 0.000 description 4
- IASNWHAGGYTEKX-IUCAKERBSA-N Arg-Arg-Gly Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)NCC(O)=O IASNWHAGGYTEKX-IUCAKERBSA-N 0.000 description 4
- YUGFLWBWAJFGKY-BQBZGAKWSA-N Arg-Cys-Gly Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CS)C(=O)NCC(O)=O YUGFLWBWAJFGKY-BQBZGAKWSA-N 0.000 description 4
- AUFHLLPVPSMEOG-YUMQZZPRSA-N Arg-Gly-Glu Chemical compound NC(N)=NCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O AUFHLLPVPSMEOG-YUMQZZPRSA-N 0.000 description 4
- JEOCWTUOMKEEMF-RHYQMDGZSA-N Arg-Leu-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JEOCWTUOMKEEMF-RHYQMDGZSA-N 0.000 description 4
- DIIGDGJKTMLQQW-IHRRRGAJSA-N Arg-Lys-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCCN=C(N)N)N DIIGDGJKTMLQQW-IHRRRGAJSA-N 0.000 description 4
- YCYXHLZRUSJITQ-SRVKXCTJSA-N Arg-Pro-Pro Chemical compound NC(=N)NCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 YCYXHLZRUSJITQ-SRVKXCTJSA-N 0.000 description 4
- OWSMKCJUBAPHED-JYJNAYRXSA-N Arg-Pro-Tyr Chemical compound NC(N)=NCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 OWSMKCJUBAPHED-JYJNAYRXSA-N 0.000 description 4
- GFFRWIJAFFMQGM-NUMRIWBASA-N Asn-Glu-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O GFFRWIJAFFMQGM-NUMRIWBASA-N 0.000 description 4
- WEDGJJRCJNHYSF-SRVKXCTJSA-N Asp-Cys-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC(=O)O)N WEDGJJRCJNHYSF-SRVKXCTJSA-N 0.000 description 4
- GYWQGGUCMDCUJE-DLOVCJGASA-N Asp-Phe-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](C)C(O)=O GYWQGGUCMDCUJE-DLOVCJGASA-N 0.000 description 4
- QSFHZPQUAAQHAQ-CIUDSAMLSA-N Asp-Ser-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O QSFHZPQUAAQHAQ-CIUDSAMLSA-N 0.000 description 4
- 102000012406 Carcinoembryonic Antigen Human genes 0.000 description 4
- 108010022366 Carcinoembryonic Antigen Proteins 0.000 description 4
- 208000034628 Celiac artery compression syndrome Diseases 0.000 description 4
- CVLIHKBUPSFRQP-WHFBIAKZSA-N Cys-Gly-Ala Chemical compound [H]N[C@@H](CS)C(=O)NCC(=O)N[C@@H](C)C(O)=O CVLIHKBUPSFRQP-WHFBIAKZSA-N 0.000 description 4
- XTHUKRLJRUVVBF-WHFBIAKZSA-N Cys-Gly-Ser Chemical compound SC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O XTHUKRLJRUVVBF-WHFBIAKZSA-N 0.000 description 4
- RWVBNRYBHAGYSG-GUBZILKMSA-N Cys-Met-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CS)N RWVBNRYBHAGYSG-GUBZILKMSA-N 0.000 description 4
- NGOIQDYZMIKCOK-NAKRPEOUSA-N Cys-Val-Ile Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O NGOIQDYZMIKCOK-NAKRPEOUSA-N 0.000 description 4
- 101710091045 Envelope protein Proteins 0.000 description 4
- JKDBRTNMYXYLHO-JYJNAYRXSA-N Gln-Tyr-Leu Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=C(O)C=C1 JKDBRTNMYXYLHO-JYJNAYRXSA-N 0.000 description 4
- IDEODOAVGCMUQV-GUBZILKMSA-N Glu-Ser-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O IDEODOAVGCMUQV-GUBZILKMSA-N 0.000 description 4
- UEGIPZAXNBYCCP-NKWVEPMBSA-N Gly-Cys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CS)NC(=O)CN)C(=O)O UEGIPZAXNBYCCP-NKWVEPMBSA-N 0.000 description 4
- LRQXRHGQEVWGPV-NHCYSSNCSA-N Gly-Leu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)CN LRQXRHGQEVWGPV-NHCYSSNCSA-N 0.000 description 4
- WNZOCXUOGVYYBJ-CDMKHQONSA-N Gly-Phe-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)CN)O WNZOCXUOGVYYBJ-CDMKHQONSA-N 0.000 description 4
- SSFWXSNOKDZNHY-QXEWZRGKSA-N Gly-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)CN SSFWXSNOKDZNHY-QXEWZRGKSA-N 0.000 description 4
- ISSDODCYBOWWIP-GJZGRUSLSA-N Gly-Pro-Trp Chemical compound [H]NCC(=O)N1CCC[C@H]1C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O ISSDODCYBOWWIP-GJZGRUSLSA-N 0.000 description 4
- SOEGEPHNZOISMT-BYPYZUCNSA-N Gly-Ser-Gly Chemical compound NCC(=O)N[C@@H](CO)C(=O)NCC(O)=O SOEGEPHNZOISMT-BYPYZUCNSA-N 0.000 description 4
- GWCJMBNBFYBQCV-XPUUQOCRSA-N Gly-Val-Ala Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O GWCJMBNBFYBQCV-XPUUQOCRSA-N 0.000 description 4
- BCISUQVFDGYZBO-QSFUFRPTSA-N Ile-Val-Asp Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(O)=O BCISUQVFDGYZBO-QSFUFRPTSA-N 0.000 description 4
- DZQMXBALGUHGJT-GUBZILKMSA-N Leu-Glu-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O DZQMXBALGUHGJT-GUBZILKMSA-N 0.000 description 4
- OXRLYTYUXAQTHP-YUMQZZPRSA-N Leu-Gly-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](C)C(O)=O OXRLYTYUXAQTHP-YUMQZZPRSA-N 0.000 description 4
- USLNHQZCDQJBOV-ZPFDUUQYSA-N Leu-Ile-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O USLNHQZCDQJBOV-ZPFDUUQYSA-N 0.000 description 4
- JFSGIJSCJFQGSZ-MXAVVETBSA-N Leu-Ile-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CC(C)C)N JFSGIJSCJFQGSZ-MXAVVETBSA-N 0.000 description 4
- IFMPDNRWZZEZSL-SRVKXCTJSA-N Leu-Leu-Cys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CS)C(O)=O IFMPDNRWZZEZSL-SRVKXCTJSA-N 0.000 description 4
- ZDBMWELMUCLUPL-QEJZJMRPSA-N Leu-Phe-Ala Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](C)C(O)=O)CC1=CC=CC=C1 ZDBMWELMUCLUPL-QEJZJMRPSA-N 0.000 description 4
- SBANPBVRHYIMRR-GARJFASQSA-N Leu-Ser-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CO)C(=O)N1CCC[C@@H]1C(=O)O)N SBANPBVRHYIMRR-GARJFASQSA-N 0.000 description 4
- SITLTJHOQZFJGG-UHFFFAOYSA-N N-L-alpha-glutamyl-L-valine Natural products CC(C)C(C(O)=O)NC(=O)C(N)CCC(O)=O SITLTJHOQZFJGG-UHFFFAOYSA-N 0.000 description 4
- PESQCPHRXOFIPX-UHFFFAOYSA-N N-L-methionyl-L-tyrosine Natural products CSCCC(N)C(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 PESQCPHRXOFIPX-UHFFFAOYSA-N 0.000 description 4
- 108010079364 N-glycylalanine Proteins 0.000 description 4
- 241000283973 Oryctolagus cuniculus Species 0.000 description 4
- JTKGCYOOJLUETJ-ULQDDVLXSA-N Phe-Val-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC1=CC=CC=C1 JTKGCYOOJLUETJ-ULQDDVLXSA-N 0.000 description 4
- IFMDQWDAJUMMJC-DCAQKATOSA-N Pro-Ala-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O IFMDQWDAJUMMJC-DCAQKATOSA-N 0.000 description 4
- OLHDPZMYUSBGDE-GUBZILKMSA-N Pro-Arg-Cys Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CS)C(=O)O OLHDPZMYUSBGDE-GUBZILKMSA-N 0.000 description 4
- XJROSHJRQTXWAE-XGEHTFHBSA-N Pro-Cys-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CS)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XJROSHJRQTXWAE-XGEHTFHBSA-N 0.000 description 4
- VVAWNPIOYXAMAL-KJEVXHAQSA-N Pro-Thr-Tyr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O VVAWNPIOYXAMAL-KJEVXHAQSA-N 0.000 description 4
- VVEQUISRWJDGMX-VKOGCVSHSA-N Pro-Trp-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@@H]3CCCN3 VVEQUISRWJDGMX-VKOGCVSHSA-N 0.000 description 4
- 229940124158 Protease/peptidase inhibitor Drugs 0.000 description 4
- 101710188315 Protein X Proteins 0.000 description 4
- KDGARKCAKHBEDB-NKWVEPMBSA-N Ser-Gly-Pro Chemical compound C1C[C@@H](N(C1)C(=O)CNC(=O)[C@H](CO)N)C(=O)O KDGARKCAKHBEDB-NKWVEPMBSA-N 0.000 description 4
- BCAVNDNYOGTQMQ-AAEUAGOBSA-N Ser-Trp-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)NCC(O)=O BCAVNDNYOGTQMQ-AAEUAGOBSA-N 0.000 description 4
- XTWXRUWACCXBMU-XIRDDKMYSA-N Ser-Trp-His Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CN=CN3)C(=O)O)NC(=O)[C@H](CO)N XTWXRUWACCXBMU-XIRDDKMYSA-N 0.000 description 4
- SGZVZUCRAVSPKQ-FXQIFTODSA-N Ser-Val-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CO)N SGZVZUCRAVSPKQ-FXQIFTODSA-N 0.000 description 4
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 4
- GZYNMZQXFRWDFH-YTWAJWBKSA-N Thr-Arg-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N1CCC[C@@H]1C(=O)O)N)O GZYNMZQXFRWDFH-YTWAJWBKSA-N 0.000 description 4
- TZKPNGDGUVREEB-FOHZUACHSA-N Thr-Asn-Gly Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O TZKPNGDGUVREEB-FOHZUACHSA-N 0.000 description 4
- VXMHQKHDKCATDV-VEVYYDQMSA-N Thr-Asp-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O VXMHQKHDKCATDV-VEVYYDQMSA-N 0.000 description 4
- RKDFEMGVMMYYNG-WDCWCFNPSA-N Thr-Gln-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O RKDFEMGVMMYYNG-WDCWCFNPSA-N 0.000 description 4
- QQWNRERCGGZOKG-WEDXCCLWSA-N Thr-Gly-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O QQWNRERCGGZOKG-WEDXCCLWSA-N 0.000 description 4
- NCXVJIQMWSGRHY-KXNHARMFSA-N Thr-Leu-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N)O NCXVJIQMWSGRHY-KXNHARMFSA-N 0.000 description 4
- KERCOYANYUPLHJ-XGEHTFHBSA-N Thr-Pro-Ser Chemical compound C[C@@H](O)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O KERCOYANYUPLHJ-XGEHTFHBSA-N 0.000 description 4
- YRJOLUDFVAUXLI-GSSVUCPTSA-N Thr-Thr-Asp Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC(O)=O YRJOLUDFVAUXLI-GSSVUCPTSA-N 0.000 description 4
- CYCGARJWIQWPQM-YJRXYDGGSA-N Thr-Tyr-Ser Chemical compound C[C@@H](O)[C@H]([NH3+])C(=O)N[C@H](C(=O)N[C@@H](CO)C([O-])=O)CC1=CC=C(O)C=C1 CYCGARJWIQWPQM-YJRXYDGGSA-N 0.000 description 4
- 102000003978 Tissue Plasminogen Activator Human genes 0.000 description 4
- 108090000373 Tissue Plasminogen Activator Proteins 0.000 description 4
- YDTKYBHPRULROG-LTHWPDAASA-N Trp-Ile-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N YDTKYBHPRULROG-LTHWPDAASA-N 0.000 description 4
- ACGIVBXINJFALS-HKUYNNGSSA-N Trp-Phe-Gly Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CC2=CNC3=CC=CC=C32)N ACGIVBXINJFALS-HKUYNNGSSA-N 0.000 description 4
- BVDHHLMIZFCAAU-BZSNNMDCSA-N Tyr-Cys-Phe Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O BVDHHLMIZFCAAU-BZSNNMDCSA-N 0.000 description 4
- VXFXIBCCVLJCJT-JYJNAYRXSA-N Tyr-Pro-Pro Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N1CCC[C@H]1C(=O)N1CCC[C@H]1C(O)=O VXFXIBCCVLJCJT-JYJNAYRXSA-N 0.000 description 4
- SCBITHMBEJNRHC-LSJOCFKGSA-N Val-Asp-Val Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](C(C)C)C(=O)O)N SCBITHMBEJNRHC-LSJOCFKGSA-N 0.000 description 4
- KZKMBGXCNLPYKD-YEPSODPASA-N Val-Gly-Thr Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(O)=O KZKMBGXCNLPYKD-YEPSODPASA-N 0.000 description 4
- AEFJNECXZCODJM-UWVGGRQHSA-N Val-Val-Gly Chemical compound CC(C)[C@H]([NH3+])C(=O)N[C@@H](C(C)C)C(=O)NCC([O-])=O AEFJNECXZCODJM-UWVGGRQHSA-N 0.000 description 4
- 108010024078 alanyl-glycyl-serine Proteins 0.000 description 4
- 108010070944 alanylhistidine Proteins 0.000 description 4
- 108010068265 aspartyltyrosine Proteins 0.000 description 4
- 230000009286 beneficial effect Effects 0.000 description 4
- 230000008901 benefit Effects 0.000 description 4
- 239000002775 capsule Substances 0.000 description 4
- 230000001413 cellular effect Effects 0.000 description 4
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 4
- 239000003431 cross linking reagent Substances 0.000 description 4
- 230000007423 decrease Effects 0.000 description 4
- 238000004925 denaturation Methods 0.000 description 4
- 230000036425 denaturation Effects 0.000 description 4
- 238000010828 elution Methods 0.000 description 4
- 108010063718 gamma-glutamylaspartic acid Proteins 0.000 description 4
- XKUKSGPZAADMRA-UHFFFAOYSA-N glycyl-glycyl-glycine Chemical compound NCC(=O)NCC(=O)NCC(O)=O XKUKSGPZAADMRA-UHFFFAOYSA-N 0.000 description 4
- 108010015792 glycyllysine Proteins 0.000 description 4
- 108010085325 histidylproline Proteins 0.000 description 4
- 230000036039 immunity Effects 0.000 description 4
- 230000006872 improvement Effects 0.000 description 4
- 230000000977 initiatory effect Effects 0.000 description 4
- 238000004255 ion exchange chromatography Methods 0.000 description 4
- 108010078274 isoleucylvaline Proteins 0.000 description 4
- 108010053037 kyotorphin Proteins 0.000 description 4
- 239000007788 liquid Substances 0.000 description 4
- 238000000569 multi-angle light scattering Methods 0.000 description 4
- 230000003647 oxidation Effects 0.000 description 4
- 238000007254 oxidation reaction Methods 0.000 description 4
- 239000008188 pellet Substances 0.000 description 4
- 108010051242 phenylalanylserine Proteins 0.000 description 4
- 108091033319 polynucleotide Proteins 0.000 description 4
- 102000040430 polynucleotide Human genes 0.000 description 4
- 239000002157 polynucleotide Substances 0.000 description 4
- 108010079317 prolyl-tyrosine Proteins 0.000 description 4
- 108010090894 prolylleucine Proteins 0.000 description 4
- 230000030788 protein refolding Effects 0.000 description 4
- 239000000126 substance Substances 0.000 description 4
- 239000000725 suspension Substances 0.000 description 4
- 229960000187 tissue plasminogen activator Drugs 0.000 description 4
- 239000013638 trimer Substances 0.000 description 4
- 108010078580 tyrosylleucine Proteins 0.000 description 4
- 108010073969 valyllysine Proteins 0.000 description 4
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N (+)-Biotin Chemical compound N1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21 YBJHBAHKTGYVGT-ZKWXMUAHSA-N 0.000 description 3
- XVZCXCTYGHPNEM-IHRRRGAJSA-N (2s)-1-[(2s)-2-[[(2s)-2-amino-4-methylpentanoyl]amino]-4-methylpentanoyl]pyrrolidine-2-carboxylic acid Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(O)=O XVZCXCTYGHPNEM-IHRRRGAJSA-N 0.000 description 3
- PKOHVHWNGUHYRE-ZFWWWQNUSA-N (2s)-1-[2-[[(2s)-2-amino-3-(1h-indol-3-yl)propanoyl]amino]acetyl]pyrrolidine-2-carboxylic acid Chemical compound O=C([C@H](CC=1C2=CC=CC=C2NC=1)N)NCC(=O)N1CCC[C@H]1C(O)=O PKOHVHWNGUHYRE-ZFWWWQNUSA-N 0.000 description 3
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 3
- -1 ADHl Proteins 0.000 description 3
- HRPVXLWXLXDGHG-UHFFFAOYSA-N Acrylamide Chemical compound NC(=O)C=C HRPVXLWXLXDGHG-UHFFFAOYSA-N 0.000 description 3
- GFBLJMHGHAXGNY-ZLUOBGJFSA-N Ala-Asn-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O GFBLJMHGHAXGNY-ZLUOBGJFSA-N 0.000 description 3
- NINQYGGNRIBFSC-CIUDSAMLSA-N Ala-Lys-Ser Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CO)C(O)=O NINQYGGNRIBFSC-CIUDSAMLSA-N 0.000 description 3
- KTXKIYXZQFWJKB-VZFHVOOUSA-N Ala-Thr-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O KTXKIYXZQFWJKB-VZFHVOOUSA-N 0.000 description 3
- YCTIYBUTCKNOTI-UWJYBYFXSA-N Ala-Tyr-Asp Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N YCTIYBUTCKNOTI-UWJYBYFXSA-N 0.000 description 3
- YJHKTAMKPGFJCT-NRPADANISA-N Ala-Val-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O YJHKTAMKPGFJCT-NRPADANISA-N 0.000 description 3
- XPSGESXVBSQZPL-SRVKXCTJSA-N Arg-Arg-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O XPSGESXVBSQZPL-SRVKXCTJSA-N 0.000 description 3
- IGULQRCJLQQPSM-DCAQKATOSA-N Arg-Cys-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(C)C)C(O)=O IGULQRCJLQQPSM-DCAQKATOSA-N 0.000 description 3
- DNLQVHBBMPZUGJ-BQBZGAKWSA-N Arg-Ser-Gly Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O DNLQVHBBMPZUGJ-BQBZGAKWSA-N 0.000 description 3
- FTMRPIVPSDVGCC-GUBZILKMSA-N Arg-Val-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N FTMRPIVPSDVGCC-GUBZILKMSA-N 0.000 description 3
- XWGJDUSDTRPQRK-ZLUOBGJFSA-N Asn-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC(N)=O XWGJDUSDTRPQRK-ZLUOBGJFSA-N 0.000 description 3
- XQQVCUIBGYFKDC-OLHMAJIHSA-N Asn-Asp-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XQQVCUIBGYFKDC-OLHMAJIHSA-N 0.000 description 3
- RAQMSGVCGSJKCL-FOHZUACHSA-N Asn-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(N)=O RAQMSGVCGSJKCL-FOHZUACHSA-N 0.000 description 3
- AYOAHKWVQLNPDM-HJGDQZAQSA-N Asn-Lys-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O AYOAHKWVQLNPDM-HJGDQZAQSA-N 0.000 description 3
- BKFXFUPYETWGGA-XVSYOHENSA-N Asn-Phe-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O BKFXFUPYETWGGA-XVSYOHENSA-N 0.000 description 3
- JBDLMLZNDRLDIX-HJGDQZAQSA-N Asn-Thr-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O JBDLMLZNDRLDIX-HJGDQZAQSA-N 0.000 description 3
- ULZOQOKFYMXHPZ-AQZXSJQPSA-N Asn-Trp-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ULZOQOKFYMXHPZ-AQZXSJQPSA-N 0.000 description 3
- XEGZSHSPQNDNRH-JRQIVUDYSA-N Asn-Tyr-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XEGZSHSPQNDNRH-JRQIVUDYSA-N 0.000 description 3
- ZLGKHJHFYSRUBH-FXQIFTODSA-N Asp-Arg-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O ZLGKHJHFYSRUBH-FXQIFTODSA-N 0.000 description 3
- VWWAFGHMPWBKEP-GMOBBJLQSA-N Asp-Met-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CC(=O)O)N VWWAFGHMPWBKEP-GMOBBJLQSA-N 0.000 description 3
- IWLZBRTUIVXZJD-OLHMAJIHSA-N Asp-Thr-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(O)=O IWLZBRTUIVXZJD-OLHMAJIHSA-N 0.000 description 3
- 241000228212 Aspergillus Species 0.000 description 3
- 241000193830 Bacillus <bacterium> Species 0.000 description 3
- 241000283707 Capra Species 0.000 description 3
- 241000700198 Cavia Species 0.000 description 3
- 108020004705 Codon Proteins 0.000 description 3
- 241000711573 Coronaviridae Species 0.000 description 3
- ZWNFOZNJYNDNGM-UBHSHLNASA-N Cys-Asn-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CS)N ZWNFOZNJYNDNGM-UBHSHLNASA-N 0.000 description 3
- UPURLDIGQGTUPJ-ZKWXMUAHSA-N Cys-Gly-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CS)N UPURLDIGQGTUPJ-ZKWXMUAHSA-N 0.000 description 3
- HJXSYJVCMUOUNY-SRVKXCTJSA-N Cys-Ser-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CS)N HJXSYJVCMUOUNY-SRVKXCTJSA-N 0.000 description 3
- FANFRJOFTYCNRG-JYBASQMISA-N Cys-Thr-Trp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CS)N)O FANFRJOFTYCNRG-JYBASQMISA-N 0.000 description 3
- PNEAWXSKCKCHDK-XIRDDKMYSA-N Cys-Trp-His Chemical compound C([C@H](NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CS)N)C(O)=O)C1=CN=CN1 PNEAWXSKCKCHDK-XIRDDKMYSA-N 0.000 description 3
- 241000196324 Embryophyta Species 0.000 description 3
- 241000588914 Enterobacter Species 0.000 description 3
- 241000710831 Flavivirus Species 0.000 description 3
- INKFLNZBTSNFON-CIUDSAMLSA-N Gln-Ala-Arg Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O INKFLNZBTSNFON-CIUDSAMLSA-N 0.000 description 3
- BRKUZSLQMPNVFN-SRVKXCTJSA-N Glu-His-Arg Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O BRKUZSLQMPNVFN-SRVKXCTJSA-N 0.000 description 3
- FBEJIDRSQCGFJI-GUBZILKMSA-N Glu-Leu-Ser Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O FBEJIDRSQCGFJI-GUBZILKMSA-N 0.000 description 3
- SOYWRINXUSUWEQ-DLOVCJGASA-N Glu-Val-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCC(O)=O SOYWRINXUSUWEQ-DLOVCJGASA-N 0.000 description 3
- PYTZFYUXZZHOAD-WHFBIAKZSA-N Gly-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)CN PYTZFYUXZZHOAD-WHFBIAKZSA-N 0.000 description 3
- BRFJMRSRMOMIMU-WHFBIAKZSA-N Gly-Ala-Asn Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(O)=O BRFJMRSRMOMIMU-WHFBIAKZSA-N 0.000 description 3
- UGVQELHRNUDMAA-BYPYZUCNSA-N Gly-Ala-Gly Chemical compound [NH3+]CC(=O)N[C@@H](C)C(=O)NCC([O-])=O UGVQELHRNUDMAA-BYPYZUCNSA-N 0.000 description 3
- GNBMOZPQUXTCRW-STQMWFEESA-N Gly-Asn-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CC(N)=O)NC(=O)CN)C(O)=O)=CNC2=C1 GNBMOZPQUXTCRW-STQMWFEESA-N 0.000 description 3
- JSNNHGHYGYMVCK-XVKPBYJWSA-N Gly-Glu-Val Chemical compound [H]NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O JSNNHGHYGYMVCK-XVKPBYJWSA-N 0.000 description 3
- UFPXDFOYHVEIPI-BYPYZUCNSA-N Gly-Gly-Asp Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CC(O)=O UFPXDFOYHVEIPI-BYPYZUCNSA-N 0.000 description 3
- YWAQATDNEKZFFK-BYPYZUCNSA-N Gly-Gly-Ser Chemical compound NCC(=O)NCC(=O)N[C@@H](CO)C(O)=O YWAQATDNEKZFFK-BYPYZUCNSA-N 0.000 description 3
- HAOUOFNNJJLVNS-BQBZGAKWSA-N Gly-Pro-Ser Chemical compound NCC(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O HAOUOFNNJJLVNS-BQBZGAKWSA-N 0.000 description 3
- IRJWAYCXIYUHQE-WHFBIAKZSA-N Gly-Ser-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)CN IRJWAYCXIYUHQE-WHFBIAKZSA-N 0.000 description 3
- WNGHUXFWEWTKAO-YUMQZZPRSA-N Gly-Ser-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)CN WNGHUXFWEWTKAO-YUMQZZPRSA-N 0.000 description 3
- 241000238631 Hexapoda Species 0.000 description 3
- DLTCGJZBNFOWFL-LKTVYLICSA-N His-Tyr-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)NC(=O)[C@H](CC2=CN=CN2)N DLTCGJZBNFOWFL-LKTVYLICSA-N 0.000 description 3
- 241000282412 Homo Species 0.000 description 3
- DMHGKBGOUAJRHU-UHFFFAOYSA-N Ile-Arg-Pro Natural products CCC(C)C(N)C(=O)NC(CCCN=C(N)N)C(=O)N1CCCC1C(O)=O DMHGKBGOUAJRHU-UHFFFAOYSA-N 0.000 description 3
- RQZFWBLDTBDEOF-RNJOBUHISA-N Ile-Val-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N RQZFWBLDTBDEOF-RNJOBUHISA-N 0.000 description 3
- 108060003951 Immunoglobulin Proteins 0.000 description 3
- 108010065920 Insulin Lispro Proteins 0.000 description 3
- 102000013462 Interleukin-12 Human genes 0.000 description 3
- 108010065805 Interleukin-12 Proteins 0.000 description 3
- 102000004889 Interleukin-6 Human genes 0.000 description 3
- 108090001005 Interleukin-6 Proteins 0.000 description 3
- 108020004684 Internal Ribosome Entry Sites Proteins 0.000 description 3
- PWWVAXIEGOYWEE-UHFFFAOYSA-N Isophenergan Chemical compound C1=CC=C2N(CC(C)N(C)C)C3=CC=CC=C3SC2=C1 PWWVAXIEGOYWEE-UHFFFAOYSA-N 0.000 description 3
- FADYJNXDPBKVCA-UHFFFAOYSA-N L-Phenylalanyl-L-lysin Natural products NCCCCC(C(O)=O)NC(=O)C(N)CC1=CC=CC=C1 FADYJNXDPBKVCA-UHFFFAOYSA-N 0.000 description 3
- BQSLGJHIAGOZCD-CIUDSAMLSA-N Leu-Ala-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O BQSLGJHIAGOZCD-CIUDSAMLSA-N 0.000 description 3
- ILJREDZFPHTUIE-GUBZILKMSA-N Leu-Asp-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O ILJREDZFPHTUIE-GUBZILKMSA-N 0.000 description 3
- FMEICTQWUKNAGC-YUMQZZPRSA-N Leu-Gly-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O FMEICTQWUKNAGC-YUMQZZPRSA-N 0.000 description 3
- BKTXKJMNTSMJDQ-AVGNSLFASA-N Leu-His-Gln Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N BKTXKJMNTSMJDQ-AVGNSLFASA-N 0.000 description 3
- IAJFFZORSWOZPQ-SRVKXCTJSA-N Leu-Leu-Asn Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O IAJFFZORSWOZPQ-SRVKXCTJSA-N 0.000 description 3
- HVHRPWQEQHIQJF-AVGNSLFASA-N Leu-Lys-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O HVHRPWQEQHIQJF-AVGNSLFASA-N 0.000 description 3
- PPGBXYKMUMHFBF-KATARQTJSA-N Leu-Ser-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O PPGBXYKMUMHFBF-KATARQTJSA-N 0.000 description 3
- WFCKERTZVCQXKH-KBPBESRZSA-N Leu-Tyr-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(O)=O WFCKERTZVCQXKH-KBPBESRZSA-N 0.000 description 3
- 241000219745 Lupinus Species 0.000 description 3
- QFSYGUMEANRNJE-DCAQKATOSA-N Lys-Val-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCCN)N QFSYGUMEANRNJE-DCAQKATOSA-N 0.000 description 3
- CAODKDAPYGUMLK-FXQIFTODSA-N Met-Asn-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O CAODKDAPYGUMLK-FXQIFTODSA-N 0.000 description 3
- OVTOTTGZBWXLFU-QXEWZRGKSA-N Met-Val-Asp Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(O)=O OVTOTTGZBWXLFU-QXEWZRGKSA-N 0.000 description 3
- 241000699670 Mus sp. Species 0.000 description 3
- XZFYRXDAULDNFX-UHFFFAOYSA-N N-L-cysteinyl-L-phenylalanine Natural products SCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XZFYRXDAULDNFX-UHFFFAOYSA-N 0.000 description 3
- XMBSYZWANAQXEV-UHFFFAOYSA-N N-alpha-L-glutamyl-L-phenylalanine Natural products OC(=O)CCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XMBSYZWANAQXEV-UHFFFAOYSA-N 0.000 description 3
- 241000244206 Nematoda Species 0.000 description 3
- 108010065395 Neuropep-1 Proteins 0.000 description 3
- 101800001020 Non-structural protein 4A Proteins 0.000 description 3
- 241000606860 Pasteurella Species 0.000 description 3
- 239000001888 Peptone Substances 0.000 description 3
- 108010080698 Peptones Proteins 0.000 description 3
- MMYUOSCXBJFUNV-QWRGUYRKSA-N Phe-Gly-Cys Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)NCC(=O)N[C@@H](CS)C(=O)O)N MMYUOSCXBJFUNV-QWRGUYRKSA-N 0.000 description 3
- VZFPYFRVHMSSNA-JURCDPSOSA-N Phe-Ile-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CC1=CC=CC=C1 VZFPYFRVHMSSNA-JURCDPSOSA-N 0.000 description 3
- PTDAGKJHZBGDKD-OEAJRASXSA-N Phe-Thr-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N)O PTDAGKJHZBGDKD-OEAJRASXSA-N 0.000 description 3
- FYQSMXKJYTZYRP-DCAQKATOSA-N Pro-Ala-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 FYQSMXKJYTZYRP-DCAQKATOSA-N 0.000 description 3
- ICTZKEXYDDZZFP-SRVKXCTJSA-N Pro-Arg-Pro Chemical compound N([C@@H](CCCN=C(N)N)C(=O)N1[C@@H](CCC1)C(O)=O)C(=O)[C@@H]1CCCN1 ICTZKEXYDDZZFP-SRVKXCTJSA-N 0.000 description 3
- HQVPQXMCQKXARZ-FXQIFTODSA-N Pro-Cys-Ser Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CS)C(=O)N[C@@H](CO)C(=O)O HQVPQXMCQKXARZ-FXQIFTODSA-N 0.000 description 3
- OZAPWFHRPINHND-GUBZILKMSA-N Pro-Cys-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(O)=O OZAPWFHRPINHND-GUBZILKMSA-N 0.000 description 3
- LSIWVWRUTKPXDS-DCAQKATOSA-N Pro-Gln-Arg Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O LSIWVWRUTKPXDS-DCAQKATOSA-N 0.000 description 3
- PULPZRAHVFBVTO-DCAQKATOSA-N Pro-Glu-Arg Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O PULPZRAHVFBVTO-DCAQKATOSA-N 0.000 description 3
- 241000125945 Protoparvovirus Species 0.000 description 3
- 208000035415 Reinfection Diseases 0.000 description 3
- KCNSGAMPBPYUAI-CIUDSAMLSA-N Ser-Leu-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O KCNSGAMPBPYUAI-CIUDSAMLSA-N 0.000 description 3
- FBLNYDYPCLFTSP-IXOXFDKPSA-N Ser-Phe-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O FBLNYDYPCLFTSP-IXOXFDKPSA-N 0.000 description 3
- DBMJMQXJHONAFJ-UHFFFAOYSA-M Sodium laurylsulphate Chemical compound [Na+].CCCCCCCCCCCCOS([O-])(=O)=O DBMJMQXJHONAFJ-UHFFFAOYSA-M 0.000 description 3
- 241000282898 Sus scrofa Species 0.000 description 3
- 230000024932 T cell mediated immunity Effects 0.000 description 3
- LMMDEZPNUTZJAY-GCJQMDKQSA-N Thr-Asp-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O LMMDEZPNUTZJAY-GCJQMDKQSA-N 0.000 description 3
- ZUUDNCOCILSYAM-KKHAAJSZSA-N Thr-Asp-Val Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O ZUUDNCOCILSYAM-KKHAAJSZSA-N 0.000 description 3
- YSXYEJWDHBCTDJ-DVJZZOLTSA-N Thr-Gly-Trp Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N)O YSXYEJWDHBCTDJ-DVJZZOLTSA-N 0.000 description 3
- UYTYTDMCDBPDSC-URLPEUOOSA-N Thr-Ile-Phe Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N UYTYTDMCDBPDSC-URLPEUOOSA-N 0.000 description 3
- NQQMWWVVGIXUOX-SVSWQMSJSA-N Thr-Ser-Ile Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O NQQMWWVVGIXUOX-SVSWQMSJSA-N 0.000 description 3
- 241000223996 Toxoplasma Species 0.000 description 3
- FEZASNVQLJQBHW-CABZTGNLSA-N Trp-Gly-Ala Chemical compound C1=CC=C2C(C[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O)=CNC2=C1 FEZASNVQLJQBHW-CABZTGNLSA-N 0.000 description 3
- DZIKVMCFXIIETR-JSGCOSHPSA-N Trp-Gly-Glu Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O DZIKVMCFXIIETR-JSGCOSHPSA-N 0.000 description 3
- RCMHSGRBJCMFLR-BPUTZDHNSA-N Trp-Met-Asn Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(N)=O)C(O)=O)=CNC2=C1 RCMHSGRBJCMFLR-BPUTZDHNSA-N 0.000 description 3
- OOEUVMFKKZYSRX-LEWSCRJBSA-N Tyr-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CC=C(C=C2)O)N OOEUVMFKKZYSRX-LEWSCRJBSA-N 0.000 description 3
- CTDPLKMBVALCGN-JSGCOSHPSA-N Tyr-Gly-Val Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O CTDPLKMBVALCGN-JSGCOSHPSA-N 0.000 description 3
- NVZVJIUDICCMHZ-BZSNNMDCSA-N Tyr-Phe-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O NVZVJIUDICCMHZ-BZSNNMDCSA-N 0.000 description 3
- PLVVHGFEMSDRET-IHPCNDPISA-N Tyr-Ser-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC3=CC=C(C=C3)O)N PLVVHGFEMSDRET-IHPCNDPISA-N 0.000 description 3
- NZBSVMQZQMEUHI-WZLNRYEVSA-N Tyr-Thr-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N NZBSVMQZQMEUHI-WZLNRYEVSA-N 0.000 description 3
- HJSLDXZAZGFPDK-ULQDDVLXSA-N Val-Phe-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)[C@H](C(C)C)N HJSLDXZAZGFPDK-ULQDDVLXSA-N 0.000 description 3
- YQYFYUSYEDNLSD-YEPSODPASA-N Val-Thr-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O YQYFYUSYEDNLSD-YEPSODPASA-N 0.000 description 3
- DVLWZWNAQUBZBC-ZNSHCXBVSA-N Val-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](C(C)C)N)O DVLWZWNAQUBZBC-ZNSHCXBVSA-N 0.000 description 3
- OFTXTCGQJXTNQS-XGEHTFHBSA-N Val-Thr-Ser Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](C(C)C)N)O OFTXTCGQJXTNQS-XGEHTFHBSA-N 0.000 description 3
- QTXGUIMEHKCPBH-FHWLQOOXSA-N Val-Trp-Lys Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](N)C(C)C)C(=O)N[C@@H](CCCCN)C(O)=O)=CNC2=C1 QTXGUIMEHKCPBH-FHWLQOOXSA-N 0.000 description 3
- LLJLBRRXKZTTRD-GUBZILKMSA-N Val-Val-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(=O)O)N LLJLBRRXKZTTRD-GUBZILKMSA-N 0.000 description 3
- 208000036142 Viral infection Diseases 0.000 description 3
- 238000000246 agarose gel electrophoresis Methods 0.000 description 3
- 108010045350 alanyl-tyrosyl-alanine Proteins 0.000 description 3
- 239000011230 binding agent Substances 0.000 description 3
- 238000009835 boiling Methods 0.000 description 3
- 235000014633 carbohydrates Nutrition 0.000 description 3
- 150000001720 carbohydrates Chemical class 0.000 description 3
- 238000005119 centrifugation Methods 0.000 description 3
- 238000004587 chromatography analysis Methods 0.000 description 3
- 229910017052 cobalt Inorganic materials 0.000 description 3
- 239000010941 cobalt Substances 0.000 description 3
- GUTLYIVDDKVIGB-UHFFFAOYSA-N cobalt atom Chemical compound [Co] GUTLYIVDDKVIGB-UHFFFAOYSA-N 0.000 description 3
- 230000000052 comparative effect Effects 0.000 description 3
- 230000021615 conjugation Effects 0.000 description 3
- 238000004132 cross linking Methods 0.000 description 3
- 230000003111 delayed effect Effects 0.000 description 3
- 238000012217 deletion Methods 0.000 description 3
- 230000037430 deletion Effects 0.000 description 3
- 230000001419 dependent effect Effects 0.000 description 3
- 239000003937 drug carrier Substances 0.000 description 3
- 230000001605 fetal effect Effects 0.000 description 3
- 230000006870 function Effects 0.000 description 3
- 244000053095 fungal pathogen Species 0.000 description 3
- 108010055341 glutamyl-glutamic acid Proteins 0.000 description 3
- JYPCXBJRLBHWME-UHFFFAOYSA-N glycyl-L-prolyl-L-arginine Natural products NCC(=O)N1CCCC1C(=O)NC(CCCN=C(N)N)C(O)=O JYPCXBJRLBHWME-UHFFFAOYSA-N 0.000 description 3
- XBGGUPMXALFZOT-UHFFFAOYSA-N glycyl-L-tyrosine hemihydrate Natural products NCC(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 XBGGUPMXALFZOT-UHFFFAOYSA-N 0.000 description 3
- 108010010147 glycylglutamine Proteins 0.000 description 3
- 108010037850 glycylvaline Proteins 0.000 description 3
- 208000010710 hepatitis C virus infection Diseases 0.000 description 3
- 108010018006 histidylserine Proteins 0.000 description 3
- RAXXELZNTBOGNW-UHFFFAOYSA-N imidazole Natural products C1=CNC=N1 RAXXELZNTBOGNW-UHFFFAOYSA-N 0.000 description 3
- 238000002649 immunization Methods 0.000 description 3
- 102000018358 immunoglobulin Human genes 0.000 description 3
- 108010090333 leucyl-lysyl-proline Proteins 0.000 description 3
- 230000000670 limiting effect Effects 0.000 description 3
- 239000002502 liposome Substances 0.000 description 3
- 239000011159 matrix material Substances 0.000 description 3
- 230000001404 mediated effect Effects 0.000 description 3
- 201000001441 melanoma Diseases 0.000 description 3
- 230000004048 modification Effects 0.000 description 3
- 238000012986 modification Methods 0.000 description 3
- 229940035032 monophosphoryl lipid a Drugs 0.000 description 3
- 235000019319 peptone Nutrition 0.000 description 3
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 3
- 239000008194 pharmaceutical composition Substances 0.000 description 3
- 108010073101 phenylalanylleucine Proteins 0.000 description 3
- 229920005862 polyol Polymers 0.000 description 3
- 150000003077 polyols Chemical class 0.000 description 3
- 239000000843 powder Substances 0.000 description 3
- 108010031719 prolyl-serine Proteins 0.000 description 3
- 229940021993 prophylactic vaccine Drugs 0.000 description 3
- 239000001397 quillaja saponaria molina bark Substances 0.000 description 3
- 230000004044 response Effects 0.000 description 3
- 230000000717 retained effect Effects 0.000 description 3
- 229930182490 saponin Natural products 0.000 description 3
- 150000007949 saponins Chemical class 0.000 description 3
- 230000028327 secretion Effects 0.000 description 3
- 238000000926 separation method Methods 0.000 description 3
- 230000002269 spontaneous effect Effects 0.000 description 3
- 238000003153 stable transfection Methods 0.000 description 3
- 230000008685 targeting Effects 0.000 description 3
- 210000001519 tissue Anatomy 0.000 description 3
- 108010051110 tyrosyl-lysine Proteins 0.000 description 3
- 241001430294 unidentified retrovirus Species 0.000 description 3
- 238000002255 vaccination Methods 0.000 description 3
- 230000029812 viral genome replication Effects 0.000 description 3
- 230000009385 viral infection Effects 0.000 description 3
- YYGNTYWPHWGJRM-UHFFFAOYSA-N (6E,10E,14E,18E)-2,6,10,15,19,23-hexamethyltetracosa-2,6,10,14,18,22-hexaene Chemical compound CC(C)=CCCC(C)=CCCC(C)=CCCC=C(C)CCC=C(C)CCC=C(C)C YYGNTYWPHWGJRM-UHFFFAOYSA-N 0.000 description 2
- UHPMCKVQTMMPCG-UHFFFAOYSA-N 5,8-dihydroxy-2-methoxy-6-methyl-7-(2-oxopropyl)naphthalene-1,4-dione Chemical compound CC1=C(CC(C)=O)C(O)=C2C(=O)C(OC)=CC(=O)C2=C1O UHPMCKVQTMMPCG-UHFFFAOYSA-N 0.000 description 2
- 108010042708 Acetylmuramyl-Alanyl-Isoglutamine Proteins 0.000 description 2
- 241000589291 Acinetobacter Species 0.000 description 2
- YYSWCHMLFJLLBJ-ZLUOBGJFSA-N Ala-Ala-Ser Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O YYSWCHMLFJLLBJ-ZLUOBGJFSA-N 0.000 description 2
- JBVSSSZFNTXJDX-YTLHQDLWSA-N Ala-Ala-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](C)N JBVSSSZFNTXJDX-YTLHQDLWSA-N 0.000 description 2
- JBGSZRYCXBPWGX-BQBZGAKWSA-N Ala-Arg-Gly Chemical compound OC(=O)CNC(=O)[C@@H](NC(=O)[C@@H](N)C)CCCN=C(N)N JBGSZRYCXBPWGX-BQBZGAKWSA-N 0.000 description 2
- KIUYPHAMDKDICO-WHFBIAKZSA-N Ala-Asp-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O KIUYPHAMDKDICO-WHFBIAKZSA-N 0.000 description 2
- ZIWWTZWAKYBUOB-CIUDSAMLSA-N Ala-Asp-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O ZIWWTZWAKYBUOB-CIUDSAMLSA-N 0.000 description 2
- MIPWEZAIMPYQST-FXQIFTODSA-N Ala-Cys-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(O)=O MIPWEZAIMPYQST-FXQIFTODSA-N 0.000 description 2
- SFNFGFDRYJKZKN-XQXXSGGOSA-N Ala-Gln-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](C)N)O SFNFGFDRYJKZKN-XQXXSGGOSA-N 0.000 description 2
- WKOBSJOZRJJVRZ-FXQIFTODSA-N Ala-Glu-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O WKOBSJOZRJJVRZ-FXQIFTODSA-N 0.000 description 2
- ZVFVBBGVOILKPO-WHFBIAKZSA-N Ala-Gly-Ala Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O ZVFVBBGVOILKPO-WHFBIAKZSA-N 0.000 description 2
- PCIFXPRIFWKWLK-YUMQZZPRSA-N Ala-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@H](C)N PCIFXPRIFWKWLK-YUMQZZPRSA-N 0.000 description 2
- HHRAXZAYZFFRAM-CIUDSAMLSA-N Ala-Leu-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O HHRAXZAYZFFRAM-CIUDSAMLSA-N 0.000 description 2
- MNZHHDPWDWQJCQ-YUMQZZPRSA-N Ala-Leu-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O MNZHHDPWDWQJCQ-YUMQZZPRSA-N 0.000 description 2
- OYJCVIGKMXUVKB-GARJFASQSA-N Ala-Leu-Pro Chemical compound C[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N OYJCVIGKMXUVKB-GARJFASQSA-N 0.000 description 2
- AJBVYEYZVYPFCF-CIUDSAMLSA-N Ala-Lys-Asn Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(O)=O AJBVYEYZVYPFCF-CIUDSAMLSA-N 0.000 description 2
- CYBJZLQSUJEMAS-LFSVMHDDSA-N Ala-Phe-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)[C@H](C)N)O CYBJZLQSUJEMAS-LFSVMHDDSA-N 0.000 description 2
- VHAQSYHSDKERBS-XPUUQOCRSA-N Ala-Val-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O VHAQSYHSDKERBS-XPUUQOCRSA-N 0.000 description 2
- NLYYHIKRBRMAJV-AEJSXWLSSA-N Ala-Val-Pro Chemical compound C[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N NLYYHIKRBRMAJV-AEJSXWLSSA-N 0.000 description 2
- REWSWYIDQIELBE-FXQIFTODSA-N Ala-Val-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O REWSWYIDQIELBE-FXQIFTODSA-N 0.000 description 2
- OMSKGWFGWCQFBD-KZVJFYERSA-N Ala-Val-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OMSKGWFGWCQFBD-KZVJFYERSA-N 0.000 description 2
- OVVUNXXROOFSIM-SDDRHHMPSA-N Arg-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCCN=C(N)N)N)C(=O)O OVVUNXXROOFSIM-SDDRHHMPSA-N 0.000 description 2
- XVLLUZMFSAYKJV-GUBZILKMSA-N Arg-Asp-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O XVLLUZMFSAYKJV-GUBZILKMSA-N 0.000 description 2
- DXQIQUIQYAGRCC-CIUDSAMLSA-N Arg-Asp-Gln Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N)CN=C(N)N DXQIQUIQYAGRCC-CIUDSAMLSA-N 0.000 description 2
- JTWOBPNAVBESFW-FXQIFTODSA-N Arg-Cys-Asp Chemical compound C(C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)O)C(=O)O)N)CN=C(N)N JTWOBPNAVBESFW-FXQIFTODSA-N 0.000 description 2
- FEZJJKXNPSEYEV-CIUDSAMLSA-N Arg-Gln-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O FEZJJKXNPSEYEV-CIUDSAMLSA-N 0.000 description 2
- LKDHUGLXOHYINY-XUXIUFHCSA-N Arg-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N LKDHUGLXOHYINY-XUXIUFHCSA-N 0.000 description 2
- YKZJPIPFKGYHKY-DCAQKATOSA-N Arg-Leu-Asp Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O YKZJPIPFKGYHKY-DCAQKATOSA-N 0.000 description 2
- OTZMRMHZCMZOJZ-SRVKXCTJSA-N Arg-Leu-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O OTZMRMHZCMZOJZ-SRVKXCTJSA-N 0.000 description 2
- DNUKXVMPARLPFN-XUXIUFHCSA-N Arg-Leu-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O DNUKXVMPARLPFN-XUXIUFHCSA-N 0.000 description 2
- FSNVAJOPUDVQAR-AVGNSLFASA-N Arg-Lys-Arg Chemical compound NC(=N)NCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O FSNVAJOPUDVQAR-AVGNSLFASA-N 0.000 description 2
- NPAVRDPEFVKELR-DCAQKATOSA-N Arg-Lys-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(O)=O NPAVRDPEFVKELR-DCAQKATOSA-N 0.000 description 2
- ATABBWFGOHKROJ-GUBZILKMSA-N Arg-Pro-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(O)=O ATABBWFGOHKROJ-GUBZILKMSA-N 0.000 description 2
- CGWVCWFQGXOUSJ-ULQDDVLXSA-N Arg-Tyr-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(O)=O CGWVCWFQGXOUSJ-ULQDDVLXSA-N 0.000 description 2
- ULBHWNVWSCJLCO-NHCYSSNCSA-N Arg-Val-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCN=C(N)N ULBHWNVWSCJLCO-NHCYSSNCSA-N 0.000 description 2
- VYZBPPBKFCHCIS-WPRPVWTQSA-N Arg-Val-Gly Chemical compound OC(=O)CNC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCN=C(N)N VYZBPPBKFCHCIS-WPRPVWTQSA-N 0.000 description 2
- 241001533466 Asfivirus Species 0.000 description 2
- DAPLJWATMAXPPZ-CIUDSAMLSA-N Asn-Asn-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CC(N)=O DAPLJWATMAXPPZ-CIUDSAMLSA-N 0.000 description 2
- SPIPSJXLZVTXJL-ZLUOBGJFSA-N Asn-Cys-Ser Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CO)C(O)=O SPIPSJXLZVTXJL-ZLUOBGJFSA-N 0.000 description 2
- QRHYAUYXBVVDSB-LKXGYXEUSA-N Asn-Cys-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CS)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QRHYAUYXBVVDSB-LKXGYXEUSA-N 0.000 description 2
- PHJPKNUWWHRAOC-PEFMBERDSA-N Asn-Ile-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N PHJPKNUWWHRAOC-PEFMBERDSA-N 0.000 description 2
- NCFJQJRLQJEECD-NHCYSSNCSA-N Asn-Leu-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O NCFJQJRLQJEECD-NHCYSSNCSA-N 0.000 description 2
- RTFWCVDISAMGEQ-SRVKXCTJSA-N Asn-Phe-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N RTFWCVDISAMGEQ-SRVKXCTJSA-N 0.000 description 2
- CPYHLXSGDBDULY-IHPCNDPISA-N Asn-Trp-Phe Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O CPYHLXSGDBDULY-IHPCNDPISA-N 0.000 description 2
- KRXIWXCXOARFNT-ZLUOBGJFSA-N Asp-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC(O)=O KRXIWXCXOARFNT-ZLUOBGJFSA-N 0.000 description 2
- VPPXTHJNTYDNFJ-CIUDSAMLSA-N Asp-Ala-Lys Chemical compound C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)O)N VPPXTHJNTYDNFJ-CIUDSAMLSA-N 0.000 description 2
- FAEIQWHBRBWUBN-FXQIFTODSA-N Asp-Arg-Ser Chemical compound C(C[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC(=O)O)N)CN=C(N)N FAEIQWHBRBWUBN-FXQIFTODSA-N 0.000 description 2
- XYBJLTKSGFBLCS-QXEWZRGKSA-N Asp-Arg-Val Chemical compound NC(N)=NCCC[C@@H](C(=O)N[C@@H](C(C)C)C(O)=O)NC(=O)[C@@H](N)CC(O)=O XYBJLTKSGFBLCS-QXEWZRGKSA-N 0.000 description 2
- RYEWQKQXRJCHIO-SRVKXCTJSA-N Asp-Asn-Tyr Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 RYEWQKQXRJCHIO-SRVKXCTJSA-N 0.000 description 2
- JGDBHIVECJGXJA-FXQIFTODSA-N Asp-Asp-Arg Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O JGDBHIVECJGXJA-FXQIFTODSA-N 0.000 description 2
- VHQOCWWKXIOAQI-WDSKDSINSA-N Asp-Gln-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O VHQOCWWKXIOAQI-WDSKDSINSA-N 0.000 description 2
- NHSDEZURHWEZPN-SXTJYALSSA-N Asp-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)NC(=O)[C@H](CC(=O)O)N NHSDEZURHWEZPN-SXTJYALSSA-N 0.000 description 2
- UJGRZQYSNYTCAX-SRVKXCTJSA-N Asp-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC(O)=O UJGRZQYSNYTCAX-SRVKXCTJSA-N 0.000 description 2
- GWWSUMLEWKQHLR-NUMRIWBASA-N Asp-Thr-Glu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CC(=O)O)N)O GWWSUMLEWKQHLR-NUMRIWBASA-N 0.000 description 2
- VHUKCUHLFMRHOD-MELADBBJSA-N Asp-Tyr-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=C(C=C2)O)NC(=O)[C@H](CC(=O)O)N)C(=O)O VHUKCUHLFMRHOD-MELADBBJSA-N 0.000 description 2
- XWKBWZXGNXTDKY-ZKWXMUAHSA-N Asp-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC(O)=O XWKBWZXGNXTDKY-ZKWXMUAHSA-N 0.000 description 2
- SFJUYBCDQBAYAJ-YDHLFZDLSA-N Asp-Val-Phe Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 SFJUYBCDQBAYAJ-YDHLFZDLSA-N 0.000 description 2
- 241000894006 Bacteria Species 0.000 description 2
- 241000588807 Bordetella Species 0.000 description 2
- 241001453380 Burkholderia Species 0.000 description 2
- 101100022187 Caenorhabditis elegans mab-10 gene Proteins 0.000 description 2
- 241000589876 Campylobacter Species 0.000 description 2
- 241000253343 Chromadorea Species 0.000 description 2
- 241000193403 Clostridium Species 0.000 description 2
- 108091026890 Coding region Proteins 0.000 description 2
- 206010009944 Colon cancer Diseases 0.000 description 2
- 241000186216 Corynebacterium Species 0.000 description 2
- 239000004971 Cross linker Substances 0.000 description 2
- 241000223935 Cryptosporidium Species 0.000 description 2
- OJQJUQUBJGTCRY-WFBYXXMGSA-N Cys-Ala-Trp Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CS)N OJQJUQUBJGTCRY-WFBYXXMGSA-N 0.000 description 2
- DCXGXDGGXVZVMY-GHCJXIJMSA-N Cys-Asn-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CS DCXGXDGGXVZVMY-GHCJXIJMSA-N 0.000 description 2
- CPTUXCUWQIBZIF-ZLUOBGJFSA-N Cys-Asn-Ser Chemical compound SC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O CPTUXCUWQIBZIF-ZLUOBGJFSA-N 0.000 description 2
- NQSUTVRXXBGVDQ-LKXGYXEUSA-N Cys-Asn-Thr Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NQSUTVRXXBGVDQ-LKXGYXEUSA-N 0.000 description 2
- KOHBWQDSVCARMI-BWBBJGPYSA-N Cys-Cys-Thr Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CS)C(=O)N[C@@H]([C@@H](C)O)C(O)=O KOHBWQDSVCARMI-BWBBJGPYSA-N 0.000 description 2
- DZLQXIFVQFTFJY-BYPYZUCNSA-N Cys-Gly-Gly Chemical compound SC[C@H](N)C(=O)NCC(=O)NCC(O)=O DZLQXIFVQFTFJY-BYPYZUCNSA-N 0.000 description 2
- SKSJPIBFNFPTJB-NKWVEPMBSA-N Cys-Gly-Pro Chemical compound C1C[C@@H](N(C1)C(=O)CNC(=O)[C@H](CS)N)C(=O)O SKSJPIBFNFPTJB-NKWVEPMBSA-N 0.000 description 2
- WAJDEKCJRKGRPG-CIUDSAMLSA-N Cys-His-Ser Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CS)N WAJDEKCJRKGRPG-CIUDSAMLSA-N 0.000 description 2
- HMWBPUDETPKSSS-DCAQKATOSA-N Cys-Pro-Lys Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CS)N)C(=O)N[C@@H](CCCCN)C(=O)O HMWBPUDETPKSSS-DCAQKATOSA-N 0.000 description 2
- NXQCSPVUPLUTJH-WHFBIAKZSA-N Cys-Ser-Gly Chemical compound SC[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O NXQCSPVUPLUTJH-WHFBIAKZSA-N 0.000 description 2
- NRVQLLDIJJEIIZ-VZFHVOOUSA-N Cys-Thr-Ala Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](C)C(=O)O)NC(=O)[C@H](CS)N)O NRVQLLDIJJEIIZ-VZFHVOOUSA-N 0.000 description 2
- 102000004127 Cytokines Human genes 0.000 description 2
- 108090000695 Cytokines Proteins 0.000 description 2
- 241001533413 Deltavirus Species 0.000 description 2
- 241000725619 Dengue virus Species 0.000 description 2
- IAZDPXIOMUYVGZ-UHFFFAOYSA-N Dimethylsulphoxide Chemical compound CS(C)=O IAZDPXIOMUYVGZ-UHFFFAOYSA-N 0.000 description 2
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 2
- 201000011001 Ebola Hemorrhagic Fever Diseases 0.000 description 2
- 201000009051 Embryonal Carcinoma Diseases 0.000 description 2
- 241000588722 Escherichia Species 0.000 description 2
- 241000711950 Filoviridae Species 0.000 description 2
- 241000223218 Fusarium Species 0.000 description 2
- RZSLYUUFFVHFRQ-FXQIFTODSA-N Gln-Ala-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O RZSLYUUFFVHFRQ-FXQIFTODSA-N 0.000 description 2
- PRBLYKYHAJEABA-SRVKXCTJSA-N Gln-Arg-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O PRBLYKYHAJEABA-SRVKXCTJSA-N 0.000 description 2
- AAOBFSKXAVIORT-GUBZILKMSA-N Gln-Asn-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O AAOBFSKXAVIORT-GUBZILKMSA-N 0.000 description 2
- PKVWNYGXMNWJSI-CIUDSAMLSA-N Gln-Gln-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O PKVWNYGXMNWJSI-CIUDSAMLSA-N 0.000 description 2
- XFKUFUJECJUQTQ-CIUDSAMLSA-N Gln-Gln-Glu Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O XFKUFUJECJUQTQ-CIUDSAMLSA-N 0.000 description 2
- IWUFOVSLWADEJC-AVGNSLFASA-N Gln-His-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(C)C)C(O)=O IWUFOVSLWADEJC-AVGNSLFASA-N 0.000 description 2
- FFVXLVGUJBCKRX-UKJIMTQDSA-N Gln-Ile-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](CCC(=O)N)N FFVXLVGUJBCKRX-UKJIMTQDSA-N 0.000 description 2
- LGIKBBLQVSWUGK-DCAQKATOSA-N Gln-Leu-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O LGIKBBLQVSWUGK-DCAQKATOSA-N 0.000 description 2
- XFAUJGNLHIGXET-AVGNSLFASA-N Gln-Leu-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O XFAUJGNLHIGXET-AVGNSLFASA-N 0.000 description 2
- YPMDZWPZFOZYFG-GUBZILKMSA-N Gln-Leu-Ser Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O YPMDZWPZFOZYFG-GUBZILKMSA-N 0.000 description 2
- IOFDDSNZJDIGPB-GVXVVHGQSA-N Gln-Leu-Val Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O IOFDDSNZJDIGPB-GVXVVHGQSA-N 0.000 description 2
- ININBLZFFVOQIO-JHEQGTHGSA-N Gln-Thr-Gly Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](CCC(=O)N)N)O ININBLZFFVOQIO-JHEQGTHGSA-N 0.000 description 2
- VEYGCDYMOXHJLS-GVXVVHGQSA-N Gln-Val-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O VEYGCDYMOXHJLS-GVXVVHGQSA-N 0.000 description 2
- RUFHOVYUYSNDNY-ACZMJKKPSA-N Glu-Ala-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(O)=O RUFHOVYUYSNDNY-ACZMJKKPSA-N 0.000 description 2
- CGOHAEBMDSEKFB-FXQIFTODSA-N Glu-Glu-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O CGOHAEBMDSEKFB-FXQIFTODSA-N 0.000 description 2
- QJCKNLPMTPXXEM-AUTRQRHGSA-N Glu-Glu-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=O QJCKNLPMTPXXEM-AUTRQRHGSA-N 0.000 description 2
- HVYWQYLBVXMXSV-GUBZILKMSA-N Glu-Leu-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O HVYWQYLBVXMXSV-GUBZILKMSA-N 0.000 description 2
- IOUQWHIEQYQVFD-JYJNAYRXSA-N Glu-Leu-Tyr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O IOUQWHIEQYQVFD-JYJNAYRXSA-N 0.000 description 2
- JYXKPJVDCAWMDG-ZPFDUUQYSA-N Glu-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CCC(=O)O)N JYXKPJVDCAWMDG-ZPFDUUQYSA-N 0.000 description 2
- SYWCGQOIIARSIX-SRVKXCTJSA-N Glu-Pro-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O SYWCGQOIIARSIX-SRVKXCTJSA-N 0.000 description 2
- MRWYPDWDZSLWJM-ACZMJKKPSA-N Glu-Ser-Asp Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O MRWYPDWDZSLWJM-ACZMJKKPSA-N 0.000 description 2
- CQGBSALYGOXQPE-HTUGSXCWSA-N Glu-Thr-Phe Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)O CQGBSALYGOXQPE-HTUGSXCWSA-N 0.000 description 2
- HBMRTXJZQDVRFT-DZKIICNBSA-N Glu-Tyr-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(O)=O HBMRTXJZQDVRFT-DZKIICNBSA-N 0.000 description 2
- HQTDNEZTGZUWSY-XVKPBYJWSA-N Glu-Val-Gly Chemical compound CC(C)[C@H](NC(=O)[C@@H](N)CCC(O)=O)C(=O)NCC(O)=O HQTDNEZTGZUWSY-XVKPBYJWSA-N 0.000 description 2
- WGYHAAXZWPEBDQ-IFFSRLJSSA-N Glu-Val-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O WGYHAAXZWPEBDQ-IFFSRLJSSA-N 0.000 description 2
- VSVZIEVNUYDAFR-YUMQZZPRSA-N Gly-Ala-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)CN VSVZIEVNUYDAFR-YUMQZZPRSA-N 0.000 description 2
- BGVYNAQWHSTTSP-BYULHYEWSA-N Gly-Asn-Ile Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O BGVYNAQWHSTTSP-BYULHYEWSA-N 0.000 description 2
- XRTDOIOIBMAXCT-NKWVEPMBSA-N Gly-Asn-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)N)NC(=O)CN)C(=O)O XRTDOIOIBMAXCT-NKWVEPMBSA-N 0.000 description 2
- XBWMTPAIUQIWKA-BYULHYEWSA-N Gly-Asp-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)CN XBWMTPAIUQIWKA-BYULHYEWSA-N 0.000 description 2
- QCTLGOYODITHPQ-WHFBIAKZSA-N Gly-Cys-Ser Chemical compound [H]NCC(=O)N[C@@H](CS)C(=O)N[C@@H](CO)C(O)=O QCTLGOYODITHPQ-WHFBIAKZSA-N 0.000 description 2
- VNBNZUAPOYGRDB-ZDLURKLDSA-N Gly-Cys-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)CN)O VNBNZUAPOYGRDB-ZDLURKLDSA-N 0.000 description 2
- PEZZSFLFXXFUQD-XPUUQOCRSA-N Gly-Cys-Val Chemical compound [H]NCC(=O)N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(O)=O PEZZSFLFXXFUQD-XPUUQOCRSA-N 0.000 description 2
- BPQYBFAXRGMGGY-LAEOZQHASA-N Gly-Gln-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)CN BPQYBFAXRGMGGY-LAEOZQHASA-N 0.000 description 2
- HDNXXTBKOJKWNN-WDSKDSINSA-N Gly-Glu-Asn Chemical compound NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O HDNXXTBKOJKWNN-WDSKDSINSA-N 0.000 description 2
- XTQFHTHIAKKCTM-YFKPBYRVSA-N Gly-Glu-Gly Chemical compound NCC(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O XTQFHTHIAKKCTM-YFKPBYRVSA-N 0.000 description 2
- HQRHFUYMGCHHJS-LURJTMIESA-N Gly-Gly-Arg Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N HQRHFUYMGCHHJS-LURJTMIESA-N 0.000 description 2
- KMSGYZQRXPUKGI-BYPYZUCNSA-N Gly-Gly-Asn Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CC(N)=O KMSGYZQRXPUKGI-BYPYZUCNSA-N 0.000 description 2
- ITZOBNKQDZEOCE-NHCYSSNCSA-N Gly-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)CN ITZOBNKQDZEOCE-NHCYSSNCSA-N 0.000 description 2
- UHPAZODVFFYEEL-QWRGUYRKSA-N Gly-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)CN UHPAZODVFFYEEL-QWRGUYRKSA-N 0.000 description 2
- QSQXZZCGPXQBPP-BQBZGAKWSA-N Gly-Pro-Cys Chemical compound C1C[C@H](N(C1)C(=O)CN)C(=O)N[C@@H](CS)C(=O)O QSQXZZCGPXQBPP-BQBZGAKWSA-N 0.000 description 2
- LCRDMSSAKLTKBU-ZDLURKLDSA-N Gly-Ser-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)CN LCRDMSSAKLTKBU-ZDLURKLDSA-N 0.000 description 2
- DBUNZBWUWCIELX-JHEQGTHGSA-N Gly-Thr-Glu Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O DBUNZBWUWCIELX-JHEQGTHGSA-N 0.000 description 2
- GWNIGUKSRJBIHX-STQMWFEESA-N Gly-Tyr-Arg Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)CN)O GWNIGUKSRJBIHX-STQMWFEESA-N 0.000 description 2
- GJHWILMUOANXTG-WPRPVWTQSA-N Gly-Val-Arg Chemical compound [H]NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O GJHWILMUOANXTG-WPRPVWTQSA-N 0.000 description 2
- SYOJVRNQCXYEOV-XVKPBYJWSA-N Gly-Val-Glu Chemical compound [H]NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O SYOJVRNQCXYEOV-XVKPBYJWSA-N 0.000 description 2
- RYAOJUMWLWUGNW-QMMMGPOBSA-N Gly-Val-Gly Chemical compound NCC(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O RYAOJUMWLWUGNW-QMMMGPOBSA-N 0.000 description 2
- YGHSQRJSHKYUJY-SCZZXKLOSA-N Gly-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)CN YGHSQRJSHKYUJY-SCZZXKLOSA-N 0.000 description 2
- KSOBNUBCYHGUKH-UWVGGRQHSA-N Gly-Val-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)CN KSOBNUBCYHGUKH-UWVGGRQHSA-N 0.000 description 2
- DHMQDGOQFOQNFH-UHFFFAOYSA-N Glycine Chemical compound NCC(O)=O DHMQDGOQFOQNFH-UHFFFAOYSA-N 0.000 description 2
- 208000031886 HIV Infections Diseases 0.000 description 2
- 241000606790 Haemophilus Species 0.000 description 2
- 208000005176 Hepatitis C Diseases 0.000 description 2
- AWHJQEYGWRKPHE-LSJOCFKGSA-N His-Ala-Arg Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O AWHJQEYGWRKPHE-LSJOCFKGSA-N 0.000 description 2
- SYMSVYVUSPSAAO-IHRRRGAJSA-N His-Arg-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O SYMSVYVUSPSAAO-IHRRRGAJSA-N 0.000 description 2
- SDTPKSOWFXBACN-GUBZILKMSA-N His-Glu-Asp Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O SDTPKSOWFXBACN-GUBZILKMSA-N 0.000 description 2
- BPOHQCZZSFBSON-KKUMJFAQSA-N His-Leu-His Chemical compound CC(C)C[C@H](NC(=O)[C@@H](N)Cc1cnc[nH]1)C(=O)N[C@@H](Cc1cnc[nH]1)C(O)=O BPOHQCZZSFBSON-KKUMJFAQSA-N 0.000 description 2
- IGBBXBFSLKRHJB-BZSNNMDCSA-N His-Lys-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CN=CN1 IGBBXBFSLKRHJB-BZSNNMDCSA-N 0.000 description 2
- ILUVWFTXAUYOBW-CUJWVEQBSA-N His-Ser-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CC1=CN=CN1)N)O ILUVWFTXAUYOBW-CUJWVEQBSA-N 0.000 description 2
- BCSGDNGNHKBRRJ-ULQDDVLXSA-N His-Tyr-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)NC(=O)[C@H](CC2=CN=CN2)N BCSGDNGNHKBRRJ-ULQDDVLXSA-N 0.000 description 2
- GBMSSORHVHAYLU-QTKMDUPCSA-N His-Val-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CC1=CN=CN1)N)O GBMSSORHVHAYLU-QTKMDUPCSA-N 0.000 description 2
- 241000713340 Human immunodeficiency virus 2 Species 0.000 description 2
- VEXZGXHMUGYJMC-UHFFFAOYSA-N Hydrochloric acid Chemical compound Cl VEXZGXHMUGYJMC-UHFFFAOYSA-N 0.000 description 2
- HDOYNXLPTRQLAD-JBDRJPRFSA-N Ile-Ala-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(=O)O)N HDOYNXLPTRQLAD-JBDRJPRFSA-N 0.000 description 2
- QTUSJASXLGLJSR-OSUNSFLBSA-N Ile-Arg-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N QTUSJASXLGLJSR-OSUNSFLBSA-N 0.000 description 2
- QADCTXFNLZBZAB-GHCJXIJMSA-N Ile-Asn-Ala Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](C)C(=O)O)N QADCTXFNLZBZAB-GHCJXIJMSA-N 0.000 description 2
- YKRIXHPEIZUDDY-GMOBBJLQSA-N Ile-Asn-Arg Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N YKRIXHPEIZUDDY-GMOBBJLQSA-N 0.000 description 2
- KOPIAUWNLKKELG-SIGLWIIPSA-N Ile-His-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)N KOPIAUWNLKKELG-SIGLWIIPSA-N 0.000 description 2
- CMNMPCTVCWWYHY-MXAVVETBSA-N Ile-His-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CC(C)C)C(=O)O)N CMNMPCTVCWWYHY-MXAVVETBSA-N 0.000 description 2
- SVBAHOMTJRFSIC-SXTJYALSSA-N Ile-Ile-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(=O)N)C(=O)O)N SVBAHOMTJRFSIC-SXTJYALSSA-N 0.000 description 2
- PFPUFNLHBXKPHY-HTFCKZLJSA-N Ile-Ile-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(=O)O)N PFPUFNLHBXKPHY-HTFCKZLJSA-N 0.000 description 2
- 102000006992 Interferon-alpha Human genes 0.000 description 2
- 108010047761 Interferon-alpha Proteins 0.000 description 2
- 102000000588 Interleukin-2 Human genes 0.000 description 2
- 108010002350 Interleukin-2 Proteins 0.000 description 2
- 241000588748 Klebsiella Species 0.000 description 2
- 241000235058 Komagataella pastoris Species 0.000 description 2
- PMGDADKJMCOXHX-UHFFFAOYSA-N L-Arginyl-L-glutamin-acetat Natural products NC(=N)NCCCC(N)C(=O)NC(CCC(N)=O)C(O)=O PMGDADKJMCOXHX-UHFFFAOYSA-N 0.000 description 2
- RWSXRVCMGQZWBV-PHDIDXHHSA-N L-Glutathione Natural products OC(=O)[C@H](N)CCC(=O)N[C@H](CS)C(=O)NCC(O)=O RWSXRVCMGQZWBV-PHDIDXHHSA-N 0.000 description 2
- IBMVEYRWAWIOTN-UHFFFAOYSA-N L-Leucyl-L-Arginyl-L-Proline Natural products CC(C)CC(N)C(=O)NC(CCCN=C(N)N)C(=O)N1CCCC1C(O)=O IBMVEYRWAWIOTN-UHFFFAOYSA-N 0.000 description 2
- WHUUTDBJXJRKMK-VKHMYHEASA-N L-glutamic acid Chemical compound OC(=O)[C@@H](N)CCC(O)=O WHUUTDBJXJRKMK-VKHMYHEASA-N 0.000 description 2
- SENJXOPIZNYLHU-UHFFFAOYSA-N L-leucyl-L-arginine Natural products CC(C)CC(N)C(=O)NC(C(O)=O)CCCN=C(N)N SENJXOPIZNYLHU-UHFFFAOYSA-N 0.000 description 2
- 239000004166 Lanolin Substances 0.000 description 2
- 241000713666 Lentivirus Species 0.000 description 2
- 241000589902 Leptospira Species 0.000 description 2
- LJHGALIOHLRRQN-DCAQKATOSA-N Leu-Ala-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N LJHGALIOHLRRQN-DCAQKATOSA-N 0.000 description 2
- ZRLUISBDKUWAIZ-CIUDSAMLSA-N Leu-Ala-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC(O)=O ZRLUISBDKUWAIZ-CIUDSAMLSA-N 0.000 description 2
- XBBKIIGCUMBKCO-JXUBOQSCSA-N Leu-Ala-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O XBBKIIGCUMBKCO-JXUBOQSCSA-N 0.000 description 2
- HBJZFCIVFIBNSV-DCAQKATOSA-N Leu-Arg-Asn Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CC(N)=O)C(O)=O HBJZFCIVFIBNSV-DCAQKATOSA-N 0.000 description 2
- YOZCKMXHBYKOMQ-IHRRRGAJSA-N Leu-Arg-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCCN)C(=O)O)N YOZCKMXHBYKOMQ-IHRRRGAJSA-N 0.000 description 2
- WGNOPSQMIQERPK-UHFFFAOYSA-N Leu-Asn-Pro Natural products CC(C)CC(N)C(=O)NC(CC(=O)N)C(=O)N1CCCC1C(=O)O WGNOPSQMIQERPK-UHFFFAOYSA-N 0.000 description 2
- OGCQGUIWMSBHRZ-CIUDSAMLSA-N Leu-Asn-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O OGCQGUIWMSBHRZ-CIUDSAMLSA-N 0.000 description 2
- QCSFMCFHVGTLFF-NHCYSSNCSA-N Leu-Asp-Val Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O QCSFMCFHVGTLFF-NHCYSSNCSA-N 0.000 description 2
- LOLUPZNNADDTAA-AVGNSLFASA-N Leu-Gln-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O LOLUPZNNADDTAA-AVGNSLFASA-N 0.000 description 2
- APFJUBGRZGMQFF-QWRGUYRKSA-N Leu-Gly-Lys Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCCN APFJUBGRZGMQFF-QWRGUYRKSA-N 0.000 description 2
- YFBBUHJJUXXZOF-UWVGGRQHSA-N Leu-Gly-Pro Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N1CCC[C@H]1C(O)=O YFBBUHJJUXXZOF-UWVGGRQHSA-N 0.000 description 2
- HRTRLSRYZZKPCO-BJDJZHNGSA-N Leu-Ile-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O HRTRLSRYZZKPCO-BJDJZHNGSA-N 0.000 description 2
- LIINDKYIGYTDLG-PPCPHDFISA-N Leu-Ile-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LIINDKYIGYTDLG-PPCPHDFISA-N 0.000 description 2
- DSFYPIUSAMSERP-IHRRRGAJSA-N Leu-Leu-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N DSFYPIUSAMSERP-IHRRRGAJSA-N 0.000 description 2
- KYIIALJHAOIAHF-KKUMJFAQSA-N Leu-Leu-His Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CN=CN1 KYIIALJHAOIAHF-KKUMJFAQSA-N 0.000 description 2
- UBZGNBKMIJHOHL-BZSNNMDCSA-N Leu-Leu-Phe Chemical compound CC(C)C[C@H]([NH3+])C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C([O-])=O)CC1=CC=CC=C1 UBZGNBKMIJHOHL-BZSNNMDCSA-N 0.000 description 2
- IEWBEPKLKUXQBU-VOAKCMCISA-N Leu-Leu-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O IEWBEPKLKUXQBU-VOAKCMCISA-N 0.000 description 2
- JVTYXRRFZCEPPK-RHYQMDGZSA-N Leu-Met-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CC(C)C)N)O JVTYXRRFZCEPPK-RHYQMDGZSA-N 0.000 description 2
- PTRKPHUGYULXPU-KKUMJFAQSA-N Leu-Phe-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O PTRKPHUGYULXPU-KKUMJFAQSA-N 0.000 description 2
- ILDSIMPXNFWKLH-KATARQTJSA-N Leu-Thr-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O ILDSIMPXNFWKLH-KATARQTJSA-N 0.000 description 2
- XOEDPXDZJHBQIX-ULQDDVLXSA-N Leu-Val-Phe Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 XOEDPXDZJHBQIX-ULQDDVLXSA-N 0.000 description 2
- WQWZXKWOEVSGQM-DCAQKATOSA-N Lys-Ala-Met Chemical compound CSCC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN WQWZXKWOEVSGQM-DCAQKATOSA-N 0.000 description 2
- QYOXSYXPHUHOJR-GUBZILKMSA-N Lys-Asn-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O QYOXSYXPHUHOJR-GUBZILKMSA-N 0.000 description 2
- LZWNAOIMTLNMDW-NHCYSSNCSA-N Lys-Asn-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CCCCN)N LZWNAOIMTLNMDW-NHCYSSNCSA-N 0.000 description 2
- MWVUEPNEPWMFBD-SRVKXCTJSA-N Lys-Cys-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@H](C(O)=O)CCCCN MWVUEPNEPWMFBD-SRVKXCTJSA-N 0.000 description 2
- OIQSIMFSVLLWBX-VOAKCMCISA-N Lys-Leu-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OIQSIMFSVLLWBX-VOAKCMCISA-N 0.000 description 2
- IOQWIOPSKJOEKI-SRVKXCTJSA-N Lys-Ser-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O IOQWIOPSKJOEKI-SRVKXCTJSA-N 0.000 description 2
- ZUGVARDEGWMMLK-SRVKXCTJSA-N Lys-Ser-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCCN ZUGVARDEGWMMLK-SRVKXCTJSA-N 0.000 description 2
- BVRNWWHJYNPJDG-XIRDDKMYSA-N Lys-Trp-Asn Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCCCN)N BVRNWWHJYNPJDG-XIRDDKMYSA-N 0.000 description 2
- OHXUUQDOBQKSNB-AVGNSLFASA-N Lys-Val-Arg Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O OHXUUQDOBQKSNB-AVGNSLFASA-N 0.000 description 2
- RPWQJSBMXJSCPD-XUXIUFHCSA-N Lys-Val-Ile Chemical compound CC[C@H](C)[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CCCCN)C(C)C)C(O)=O RPWQJSBMXJSCPD-XUXIUFHCSA-N 0.000 description 2
- DRRXXZBXDMLGFC-IHRRRGAJSA-N Lys-Val-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCCN DRRXXZBXDMLGFC-IHRRRGAJSA-N 0.000 description 2
- 108700018351 Major Histocompatibility Complex Proteins 0.000 description 2
- 101710175625 Maltose/maltodextrin-binding periplasmic protein Proteins 0.000 description 2
- GAELMDJMQDUDLJ-BQBZGAKWSA-N Met-Ala-Gly Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O GAELMDJMQDUDLJ-BQBZGAKWSA-N 0.000 description 2
- UOENBSHXYCHSAU-YUMQZZPRSA-N Met-Gln-Gly Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O UOENBSHXYCHSAU-YUMQZZPRSA-N 0.000 description 2
- IUYCGMNKIZDRQI-BQBZGAKWSA-N Met-Gly-Ala Chemical compound CSCC[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O IUYCGMNKIZDRQI-BQBZGAKWSA-N 0.000 description 2
- RKRFGIBULDYDPF-XIRDDKMYSA-N Met-Trp-Gln Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N RKRFGIBULDYDPF-XIRDDKMYSA-N 0.000 description 2
- 108010008707 Mucin-1 Proteins 0.000 description 2
- 102000007298 Mucin-1 Human genes 0.000 description 2
- 241000186359 Mycobacterium Species 0.000 description 2
- WUGMRIBZSVSJNP-UHFFFAOYSA-N N-L-alanyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)C)C(O)=O)=CNC2=C1 WUGMRIBZSVSJNP-UHFFFAOYSA-N 0.000 description 2
- WYBVBIHNJWOLCJ-UHFFFAOYSA-N N-L-arginyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCCN=C(N)N WYBVBIHNJWOLCJ-UHFFFAOYSA-N 0.000 description 2
- 101800001014 Non-structural protein 5A Proteins 0.000 description 2
- 241001631646 Papillomaviridae Species 0.000 description 2
- 241000711504 Paramyxoviridae Species 0.000 description 2
- HHOOEUSPFGPZFP-QWRGUYRKSA-N Phe-Asn-Gly Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O HHOOEUSPFGPZFP-QWRGUYRKSA-N 0.000 description 2
- WPTYDQPGBMDUBI-QWRGUYRKSA-N Phe-Gly-Asn Chemical compound N[C@@H](Cc1ccccc1)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O WPTYDQPGBMDUBI-QWRGUYRKSA-N 0.000 description 2
- KXUZHWXENMYOHC-QEJZJMRPSA-N Phe-Leu-Ala Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O KXUZHWXENMYOHC-QEJZJMRPSA-N 0.000 description 2
- INHMISZWLJZQGH-ULQDDVLXSA-N Phe-Leu-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC1=CC=CC=C1 INHMISZWLJZQGH-ULQDDVLXSA-N 0.000 description 2
- SHUFSZDAIPLZLF-BEAPCOKYSA-N Phe-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CC=CC=C2)N)O SHUFSZDAIPLZLF-BEAPCOKYSA-N 0.000 description 2
- CDHURCQGUDNBMA-UBHSHLNASA-N Phe-Val-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC1=CC=CC=C1 CDHURCQGUDNBMA-UBHSHLNASA-N 0.000 description 2
- 241000709664 Picornaviridae Species 0.000 description 2
- RVGRUAULSDPKGF-UHFFFAOYSA-N Poloxamer Chemical compound C1CO1.CC1CO1 RVGRUAULSDPKGF-UHFFFAOYSA-N 0.000 description 2
- 239000002202 Polyethylene glycol Substances 0.000 description 2
- KIZQGKLMXKGDIV-BQBZGAKWSA-N Pro-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 KIZQGKLMXKGDIV-BQBZGAKWSA-N 0.000 description 2
- BNBBNGZZKQUWCD-IUCAKERBSA-N Pro-Arg-Gly Chemical compound NC(N)=NCCC[C@@H](C(=O)NCC(O)=O)NC(=O)[C@@H]1CCCN1 BNBBNGZZKQUWCD-IUCAKERBSA-N 0.000 description 2
- XWYXZPHPYKRYPA-GMOBBJLQSA-N Pro-Asn-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O XWYXZPHPYKRYPA-GMOBBJLQSA-N 0.000 description 2
- LUGOKRWYNMDGTD-FXQIFTODSA-N Pro-Cys-Asn Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)N)C(=O)O LUGOKRWYNMDGTD-FXQIFTODSA-N 0.000 description 2
- LCWXSALTPTZKNM-CIUDSAMLSA-N Pro-Cys-Glu Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(=O)O)C(=O)O LCWXSALTPTZKNM-CIUDSAMLSA-N 0.000 description 2
- SZZBUDVXWZZPDH-BQBZGAKWSA-N Pro-Cys-Gly Chemical compound OC(=O)CNC(=O)[C@H](CS)NC(=O)[C@@H]1CCCN1 SZZBUDVXWZZPDH-BQBZGAKWSA-N 0.000 description 2
- ZPPVJIJMIKTERM-YUMQZZPRSA-N Pro-Gln-Gly Chemical compound OC(=O)CNC(=O)[C@H](CCC(=O)N)NC(=O)[C@@H]1CCCN1 ZPPVJIJMIKTERM-YUMQZZPRSA-N 0.000 description 2
- LHALYDBUDCWMDY-CIUDSAMLSA-N Pro-Glu-Ala Chemical compound C[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H]1CCCN1)C(O)=O LHALYDBUDCWMDY-CIUDSAMLSA-N 0.000 description 2
- WVOXLKUUVCCCSU-ZPFDUUQYSA-N Pro-Glu-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WVOXLKUUVCCCSU-ZPFDUUQYSA-N 0.000 description 2
- CLNJSLSHKJECME-BQBZGAKWSA-N Pro-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)[C@@H]1CCCN1 CLNJSLSHKJECME-BQBZGAKWSA-N 0.000 description 2
- ULIWFCCJIOEHMU-BQBZGAKWSA-N Pro-Gly-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H]1CCCN1 ULIWFCCJIOEHMU-BQBZGAKWSA-N 0.000 description 2
- VYWNORHENYEQDW-YUMQZZPRSA-N Pro-Gly-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H]1CCCN1 VYWNORHENYEQDW-YUMQZZPRSA-N 0.000 description 2
- NFLNBHLMLYALOO-DCAQKATOSA-N Pro-Leu-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@@H]1CCCN1 NFLNBHLMLYALOO-DCAQKATOSA-N 0.000 description 2
- JLMZKEQFMVORMA-SRVKXCTJSA-N Pro-Pro-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 JLMZKEQFMVORMA-SRVKXCTJSA-N 0.000 description 2
- HWLKHNDRXWTFTN-GUBZILKMSA-N Pro-Pro-Cys Chemical compound C1C[C@H](NC1)C(=O)N2CCC[C@H]2C(=O)N[C@@H](CS)C(=O)O HWLKHNDRXWTFTN-GUBZILKMSA-N 0.000 description 2
- SBVPYBFMIGDIDX-SRVKXCTJSA-N Pro-Pro-Pro Chemical compound OC(=O)[C@@H]1CCCN1C(=O)[C@H]1N(C(=O)[C@H]2NCCC2)CCC1 SBVPYBFMIGDIDX-SRVKXCTJSA-N 0.000 description 2
- DCHQYSOGURGJST-FJXKBIBVSA-N Pro-Thr-Gly Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O DCHQYSOGURGJST-FJXKBIBVSA-N 0.000 description 2
- AJJDPGVVNPUZCR-RHYQMDGZSA-N Pro-Thr-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@@H]1CCCN1)O AJJDPGVVNPUZCR-RHYQMDGZSA-N 0.000 description 2
- ZYJMLBCDFPIGNL-JYJNAYRXSA-N Pro-Tyr-Arg Chemical compound NC(=N)NCCC[C@H](NC(=O)[C@H](Cc1ccc(O)cc1)NC(=O)[C@@H]1CCCN1)C(O)=O ZYJMLBCDFPIGNL-JYJNAYRXSA-N 0.000 description 2
- 241000588769 Proteus <enterobacteria> Species 0.000 description 2
- 241000588768 Providencia Species 0.000 description 2
- 241000589516 Pseudomonas Species 0.000 description 2
- 241000725643 Respiratory syncytial virus Species 0.000 description 2
- 241000607142 Salmonella Species 0.000 description 2
- BKOKTRCZXRIQPX-ZLUOBGJFSA-N Ser-Ala-Cys Chemical compound C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CO)N BKOKTRCZXRIQPX-ZLUOBGJFSA-N 0.000 description 2
- YQHZVYJAGWMHES-ZLUOBGJFSA-N Ser-Ala-Ser Chemical compound OC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O YQHZVYJAGWMHES-ZLUOBGJFSA-N 0.000 description 2
- JJKSSJVYOVRJMZ-FXQIFTODSA-N Ser-Arg-Cys Chemical compound C(C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CO)N)CN=C(N)N JJKSSJVYOVRJMZ-FXQIFTODSA-N 0.000 description 2
- HQTKVSCNCDLXSX-BQBZGAKWSA-N Ser-Arg-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O HQTKVSCNCDLXSX-BQBZGAKWSA-N 0.000 description 2
- DSSOYPJWSWFOLK-CIUDSAMLSA-N Ser-Cys-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(C)C)C(O)=O DSSOYPJWSWFOLK-CIUDSAMLSA-N 0.000 description 2
- YRBGKVIWMNEVCZ-WDSKDSINSA-N Ser-Glu-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O YRBGKVIWMNEVCZ-WDSKDSINSA-N 0.000 description 2
- YMTLKLXDFCSCNX-BYPYZUCNSA-N Ser-Gly-Gly Chemical compound OC[C@H](N)C(=O)NCC(=O)NCC(O)=O YMTLKLXDFCSCNX-BYPYZUCNSA-N 0.000 description 2
- UBRMZSHOOIVJPW-SRVKXCTJSA-N Ser-Leu-Lys Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(O)=O UBRMZSHOOIVJPW-SRVKXCTJSA-N 0.000 description 2
- FKYWFUYPVKLJLP-DCAQKATOSA-N Ser-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CO FKYWFUYPVKLJLP-DCAQKATOSA-N 0.000 description 2
- FLONGDPORFIVQW-XGEHTFHBSA-N Ser-Pro-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CO FLONGDPORFIVQW-XGEHTFHBSA-N 0.000 description 2
- SRSPTFBENMJHMR-WHFBIAKZSA-N Ser-Ser-Gly Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O SRSPTFBENMJHMR-WHFBIAKZSA-N 0.000 description 2
- XQJCEKXQUJQNNK-ZLUOBGJFSA-N Ser-Ser-Ser Chemical compound OC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O XQJCEKXQUJQNNK-ZLUOBGJFSA-N 0.000 description 2
- VGQVAVQWKJLIRM-FXQIFTODSA-N Ser-Ser-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O VGQVAVQWKJLIRM-FXQIFTODSA-N 0.000 description 2
- XJDMUQCLVSCRSJ-VZFHVOOUSA-N Ser-Thr-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O XJDMUQCLVSCRSJ-VZFHVOOUSA-N 0.000 description 2
- SOACHCFYJMCMHC-BWBBJGPYSA-N Ser-Thr-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CO)N)O SOACHCFYJMCMHC-BWBBJGPYSA-N 0.000 description 2
- SZRNDHWMVSFPSP-XKBZYTNZSA-N Ser-Thr-Gln Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CO)N)O SZRNDHWMVSFPSP-XKBZYTNZSA-N 0.000 description 2
- BDMWLJLPPUCLNV-XGEHTFHBSA-N Ser-Thr-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O BDMWLJLPPUCLNV-XGEHTFHBSA-N 0.000 description 2
- FVFUOQIYDPAIJR-XIRDDKMYSA-N Ser-Trp-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CO)N FVFUOQIYDPAIJR-XIRDDKMYSA-N 0.000 description 2
- ZWSZBWAFDZRBNM-UBHSHLNASA-N Ser-Trp-Ser Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CO)C(O)=O ZWSZBWAFDZRBNM-UBHSHLNASA-N 0.000 description 2
- BEBVVQPDSHHWQL-NRPADANISA-N Ser-Val-Glu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O BEBVVQPDSHHWQL-NRPADANISA-N 0.000 description 2
- MFQMZDPAZRZAPV-NAKRPEOUSA-N Ser-Val-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CO)N MFQMZDPAZRZAPV-NAKRPEOUSA-N 0.000 description 2
- JGUWRQWULDWNCM-FXQIFTODSA-N Ser-Val-Ser Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O JGUWRQWULDWNCM-FXQIFTODSA-N 0.000 description 2
- 241000607768 Shigella Species 0.000 description 2
- 241000191940 Staphylococcus Species 0.000 description 2
- 241000194017 Streptococcus Species 0.000 description 2
- 230000005867 T cell response Effects 0.000 description 2
- BHEOSNUKNHRBNM-UHFFFAOYSA-N Tetramethylsqualene Natural products CC(=C)C(C)CCC(=C)C(C)CCC(C)=CCCC=C(C)CCC(C)C(=C)CCC(C)C(C)=C BHEOSNUKNHRBNM-UHFFFAOYSA-N 0.000 description 2
- TWLMXDWFVNEFFK-FJXKBIBVSA-N Thr-Arg-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O TWLMXDWFVNEFFK-FJXKBIBVSA-N 0.000 description 2
- MFEBUIFJVPNZLO-OLHMAJIHSA-N Thr-Asp-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O MFEBUIFJVPNZLO-OLHMAJIHSA-N 0.000 description 2
- OYTNZCBFDXGQGE-XQXXSGGOSA-N Thr-Gln-Ala Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](C)C(=O)O)N)O OYTNZCBFDXGQGE-XQXXSGGOSA-N 0.000 description 2
- LIXBDERDAGNVAV-XKBZYTNZSA-N Thr-Gln-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(O)=O LIXBDERDAGNVAV-XKBZYTNZSA-N 0.000 description 2
- KGKWKSSSQGGYAU-SUSMZKCASA-N Thr-Gln-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N)O KGKWKSSSQGGYAU-SUSMZKCASA-N 0.000 description 2
- DXNUZQGVOMCGNS-SWRJLBSHSA-N Thr-Gln-Trp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N)O DXNUZQGVOMCGNS-SWRJLBSHSA-N 0.000 description 2
- KCRQEJSKXAIULJ-FJXKBIBVSA-N Thr-Gly-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(O)=O KCRQEJSKXAIULJ-FJXKBIBVSA-N 0.000 description 2
- WYKJENSCCRJLRC-ZDLURKLDSA-N Thr-Gly-Cys Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)N[C@@H](CS)C(=O)O)N)O WYKJENSCCRJLRC-ZDLURKLDSA-N 0.000 description 2
- XPNSAQMEAVSQRD-FBCQKBJTSA-N Thr-Gly-Gly Chemical compound C[C@@H](O)[C@H](N)C(=O)NCC(=O)NCC(O)=O XPNSAQMEAVSQRD-FBCQKBJTSA-N 0.000 description 2
- VRUFCJZQDACGLH-UVOCVTCTSA-N Thr-Leu-Thr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VRUFCJZQDACGLH-UVOCVTCTSA-N 0.000 description 2
- JWQNAFHCXKVZKZ-UVOCVTCTSA-N Thr-Lys-Thr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JWQNAFHCXKVZKZ-UVOCVTCTSA-N 0.000 description 2
- WTMPKZWHRCMMMT-KZVJFYERSA-N Thr-Pro-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O WTMPKZWHRCMMMT-KZVJFYERSA-N 0.000 description 2
- DEGCBBCMYWNJNA-RHYQMDGZSA-N Thr-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)[C@@H](C)O DEGCBBCMYWNJNA-RHYQMDGZSA-N 0.000 description 2
- AHERARIZBPOMNU-KATARQTJSA-N Thr-Ser-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O AHERARIZBPOMNU-KATARQTJSA-N 0.000 description 2
- MFMGPEKYBXFIRF-SUSMZKCASA-N Thr-Thr-Gln Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(N)=O)C(O)=O MFMGPEKYBXFIRF-SUSMZKCASA-N 0.000 description 2
- UQCNIMDPYICBTR-KYNKHSRBSA-N Thr-Thr-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O UQCNIMDPYICBTR-KYNKHSRBSA-N 0.000 description 2
- BBPCSGKKPJUYRB-UVOCVTCTSA-N Thr-Thr-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O BBPCSGKKPJUYRB-UVOCVTCTSA-N 0.000 description 2
- ZESGVALRVJIVLZ-VFCFLDTKSA-N Thr-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@@H]1C(=O)O)N)O ZESGVALRVJIVLZ-VFCFLDTKSA-N 0.000 description 2
- LVRFMARKDGGZMX-IZPVPAKOSA-N Thr-Tyr-Thr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)O)C(O)=O)CC1=CC=C(O)C=C1 LVRFMARKDGGZMX-IZPVPAKOSA-N 0.000 description 2
- BTAJAOWZCWOHBU-HSHDSVGOSA-N Thr-Val-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)[C@@H](C)O)C(C)C)C(O)=O)=CNC2=C1 BTAJAOWZCWOHBU-HSHDSVGOSA-N 0.000 description 2
- TWJDQTTXXZDJKV-BPUTZDHNSA-N Trp-Arg-Ser Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(O)=O TWJDQTTXXZDJKV-BPUTZDHNSA-N 0.000 description 2
- YEGMNOHLZNGOCG-UBHSHLNASA-N Trp-Asn-Asn Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O YEGMNOHLZNGOCG-UBHSHLNASA-N 0.000 description 2
- ADBFWLXCCKIXBQ-XIRDDKMYSA-N Trp-Asn-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N ADBFWLXCCKIXBQ-XIRDDKMYSA-N 0.000 description 2
- VKMOGXREKGVZAF-QEJZJMRPSA-N Trp-Asp-Gln Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N VKMOGXREKGVZAF-QEJZJMRPSA-N 0.000 description 2
- YWXMGBUGMLJMIP-IHPCNDPISA-N Tyr-Cys-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC3=CC=C(C=C3)O)N YWXMGBUGMLJMIP-IHPCNDPISA-N 0.000 description 2
- WVGKPKDWYQXWLU-BZSNNMDCSA-N Tyr-His-Lys Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)N[C@@H](CCCCN)C(=O)O)N)O WVGKPKDWYQXWLU-BZSNNMDCSA-N 0.000 description 2
- PMHLLBKTDHQMCY-ULQDDVLXSA-N Tyr-Lys-Val Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O PMHLLBKTDHQMCY-ULQDDVLXSA-N 0.000 description 2
- OKDNSNWJEXAMSU-IRXDYDNUSA-N Tyr-Phe-Gly Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)NCC(O)=O)C1=CC=C(O)C=C1 OKDNSNWJEXAMSU-IRXDYDNUSA-N 0.000 description 2
- KLQPIEVIKOQRAW-IZPVPAKOSA-N Tyr-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N)O KLQPIEVIKOQRAW-IZPVPAKOSA-N 0.000 description 2
- AKKYBQGHUAWPJR-MNSWYVGCSA-N Tyr-Thr-Trp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CC3=CC=C(C=C3)O)N)O AKKYBQGHUAWPJR-MNSWYVGCSA-N 0.000 description 2
- NWEGIYMHTZXVBP-JSGCOSHPSA-N Tyr-Val-Gly Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O NWEGIYMHTZXVBP-JSGCOSHPSA-N 0.000 description 2
- ZLFHAAGHGQBQQN-GUBZILKMSA-N Val-Ala-Pro Natural products CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O ZLFHAAGHGQBQQN-GUBZILKMSA-N 0.000 description 2
- XPYNXORPPVTVQK-SRVKXCTJSA-N Val-Arg-Met Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCSC)C(=O)O)N XPYNXORPPVTVQK-SRVKXCTJSA-N 0.000 description 2
- DNOOLPROHJWCSQ-RCWTZXSCSA-N Val-Arg-Thr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O DNOOLPROHJWCSQ-RCWTZXSCSA-N 0.000 description 2
- QGFPYRPIUXBYGR-YDHLFZDLSA-N Val-Asn-Phe Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N QGFPYRPIUXBYGR-YDHLFZDLSA-N 0.000 description 2
- ISERLACIZUGCDX-ZKWXMUAHSA-N Val-Asp-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](C(C)C)N ISERLACIZUGCDX-ZKWXMUAHSA-N 0.000 description 2
- COSLEEOIYRPTHD-YDHLFZDLSA-N Val-Asp-Tyr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 COSLEEOIYRPTHD-YDHLFZDLSA-N 0.000 description 2
- YTPLVNUZZOBFFC-SCZZXKLOSA-N Val-Gly-Pro Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N1CCC[C@@H]1C(O)=O YTPLVNUZZOBFFC-SCZZXKLOSA-N 0.000 description 2
- LAYSXAOGWHKNED-XPUUQOCRSA-N Val-Gly-Ser Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O LAYSXAOGWHKNED-XPUUQOCRSA-N 0.000 description 2
- JPPXDMBGXJBTIB-ULQDDVLXSA-N Val-His-Tyr Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)O)N JPPXDMBGXJBTIB-ULQDDVLXSA-N 0.000 description 2
- FEXILLGKGGTLRI-NHCYSSNCSA-N Val-Leu-Asn Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](C(C)C)N FEXILLGKGGTLRI-NHCYSSNCSA-N 0.000 description 2
- LYERIXUFCYVFFX-GVXVVHGQSA-N Val-Leu-Glu Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](C(C)C)N LYERIXUFCYVFFX-GVXVVHGQSA-N 0.000 description 2
- BTWMICVCQLKKNR-DCAQKATOSA-N Val-Leu-Ser Chemical compound CC(C)[C@H]([NH3+])C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C([O-])=O BTWMICVCQLKKNR-DCAQKATOSA-N 0.000 description 2
- SYSWVVCYSXBVJG-RHYQMDGZSA-N Val-Leu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](C(C)C)N)O SYSWVVCYSXBVJG-RHYQMDGZSA-N 0.000 description 2
- SBJCTAZFSZXWSR-AVGNSLFASA-N Val-Met-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N SBJCTAZFSZXWSR-AVGNSLFASA-N 0.000 description 2
- NZGOVKLVQNOEKP-YDHLFZDLSA-N Val-Phe-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(=O)N)C(=O)O)N NZGOVKLVQNOEKP-YDHLFZDLSA-N 0.000 description 2
- MJOUSKQHAIARKI-JYJNAYRXSA-N Val-Phe-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](C(C)C)C(O)=O)CC1=CC=CC=C1 MJOUSKQHAIARKI-JYJNAYRXSA-N 0.000 description 2
- DOFAQXCYFQKSHT-SRVKXCTJSA-N Val-Pro-Pro Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 DOFAQXCYFQKSHT-SRVKXCTJSA-N 0.000 description 2
- DEGUERSKQBRZMZ-FXQIFTODSA-N Val-Ser-Ala Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O DEGUERSKQBRZMZ-FXQIFTODSA-N 0.000 description 2
- QZKVWWIUSQGWMY-IHRRRGAJSA-N Val-Ser-Phe Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 QZKVWWIUSQGWMY-IHRRRGAJSA-N 0.000 description 2
- UVHFONIHVHLDDQ-IFFSRLJSSA-N Val-Thr-Glu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](C(C)C)N)O UVHFONIHVHLDDQ-IFFSRLJSSA-N 0.000 description 2
- CFIBZQOLUDURST-IHRRRGAJSA-N Val-Tyr-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CS)C(=O)O)N CFIBZQOLUDURST-IHRRRGAJSA-N 0.000 description 2
- VVIZITNVZUAEMI-DLOVCJGASA-N Val-Val-Gln Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCC(N)=O VVIZITNVZUAEMI-DLOVCJGASA-N 0.000 description 2
- 241000607598 Vibrio Species 0.000 description 2
- 241000607734 Yersinia <bacteria> Species 0.000 description 2
- 238000002835 absorbance Methods 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 239000004480 active ingredient Substances 0.000 description 2
- 208000009956 adenocarcinoma Diseases 0.000 description 2
- 239000003463 adsorbent Substances 0.000 description 2
- 239000000443 aerosol Substances 0.000 description 2
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 2
- 229940037003 alum Drugs 0.000 description 2
- AVKUERGKIZMTKX-NJBDSQKTSA-N ampicillin Chemical compound C1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1 AVKUERGKIZMTKX-NJBDSQKTSA-N 0.000 description 2
- 229960000723 ampicillin Drugs 0.000 description 2
- 230000000840 anti-viral effect Effects 0.000 description 2
- 238000013459 approach Methods 0.000 description 2
- 108010069926 arginyl-glycyl-serine Proteins 0.000 description 2
- 108010068380 arginylarginine Proteins 0.000 description 2
- 108010038633 aspartylglutamate Proteins 0.000 description 2
- 108010047857 aspartylglycine Proteins 0.000 description 2
- 244000309743 astrovirus Species 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000004071 biological effect Effects 0.000 description 2
- BBWBEZAMXFGUGK-UHFFFAOYSA-N bis(dodecylsulfanyl)-methylarsane Chemical compound CCCCCCCCCCCCS[As](C)SCCCCCCCCCCCC BBWBEZAMXFGUGK-UHFFFAOYSA-N 0.000 description 2
- 210000002798 bone marrow cell Anatomy 0.000 description 2
- DQXBYHZEEUGOBF-UHFFFAOYSA-N but-3-enoic acid;ethene Chemical compound C=C.OC(=O)CC=C DQXBYHZEEUGOBF-UHFFFAOYSA-N 0.000 description 2
- 230000022534 cell killing Effects 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 235000012000 cholesterol Nutrition 0.000 description 2
- 208000029742 colonic neoplasm Diseases 0.000 description 2
- 239000003086 colorant Substances 0.000 description 2
- 239000000356 contaminant Substances 0.000 description 2
- 108010069495 cysteinyltyrosine Proteins 0.000 description 2
- 210000005220 cytoplasmic tail Anatomy 0.000 description 2
- 239000003405 delayed action preparation Substances 0.000 description 2
- 238000011161 development Methods 0.000 description 2
- 230000018109 developmental process Effects 0.000 description 2
- 238000003113 dilution method Methods 0.000 description 2
- FSXRLASFHBWESK-UHFFFAOYSA-N dipeptide phenylalanyl-tyrosine Natural products C=1C=C(O)C=CC=1CC(C(O)=O)NC(=O)C(N)CC1=CC=CC=C1 FSXRLASFHBWESK-UHFFFAOYSA-N 0.000 description 2
- 201000010099 disease Diseases 0.000 description 2
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 2
- 150000002019 disulfides Chemical class 0.000 description 2
- PRAKJMSDJKAYCZ-UHFFFAOYSA-N dodecahydrosqualene Natural products CC(C)CCCC(C)CCCC(C)CCCCC(C)CCCC(C)CCCC(C)C PRAKJMSDJKAYCZ-UHFFFAOYSA-N 0.000 description 2
- 239000000975 dye Substances 0.000 description 2
- 238000001962 electrophoresis Methods 0.000 description 2
- 230000008030 elimination Effects 0.000 description 2
- 239000000839 emulsion Substances 0.000 description 2
- 239000002158 endotoxin Substances 0.000 description 2
- 210000003743 erythrocyte Anatomy 0.000 description 2
- 239000005038 ethylene vinyl acetate Substances 0.000 description 2
- 238000011156 evaluation Methods 0.000 description 2
- 238000000684 flow cytometry Methods 0.000 description 2
- 150000002270 gangliosides Chemical class 0.000 description 2
- 201000006585 gastric adenocarcinoma Diseases 0.000 description 2
- 230000002496 gastric effect Effects 0.000 description 2
- 238000001502 gel electrophoresis Methods 0.000 description 2
- 108010085059 glutamyl-arginyl-proline Proteins 0.000 description 2
- 108010008237 glutamyl-valyl-glycine Proteins 0.000 description 2
- 108010000434 glycyl-alanyl-leucine Proteins 0.000 description 2
- 108010027668 glycyl-alanyl-valine Proteins 0.000 description 2
- 108010062266 glycyl-glycyl-argininal Proteins 0.000 description 2
- 108010051307 glycyl-glycyl-proline Proteins 0.000 description 2
- 108010025801 glycyl-prolyl-arginine Proteins 0.000 description 2
- 108010081551 glycylphenylalanine Proteins 0.000 description 2
- 239000008241 heterogeneous mixture Substances 0.000 description 2
- 230000028996 humoral immune response Effects 0.000 description 2
- 239000000017 hydrogel Substances 0.000 description 2
- 230000002209 hydrophobic effect Effects 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 230000002458 infectious effect Effects 0.000 description 2
- 230000002401 inhibitory effect Effects 0.000 description 2
- 108010027338 isoleucylcysteine Proteins 0.000 description 2
- 229940039717 lanolin Drugs 0.000 description 2
- 235000019388 lanolin Nutrition 0.000 description 2
- 108010083708 leucyl-aspartyl-valine Proteins 0.000 description 2
- 108010051673 leucyl-glycyl-phenylalanine Proteins 0.000 description 2
- 108010073472 leucyl-prolyl-proline Proteins 0.000 description 2
- 108010000761 leucylarginine Proteins 0.000 description 2
- 208000032839 leukemia Diseases 0.000 description 2
- 238000012417 linear regression Methods 0.000 description 2
- 150000002632 lipids Chemical class 0.000 description 2
- 229920006008 lipopolysaccharide Polymers 0.000 description 2
- 238000011068 loading method Methods 0.000 description 2
- 239000000314 lubricant Substances 0.000 description 2
- 210000004698 lymphocyte Anatomy 0.000 description 2
- 108010009298 lysylglutamic acid Proteins 0.000 description 2
- 108010064235 lysylglycine Proteins 0.000 description 2
- 108010054155 lysyllysine Proteins 0.000 description 2
- HQKMJHAJHXVSDF-UHFFFAOYSA-L magnesium stearate Chemical compound [Mg+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O HQKMJHAJHXVSDF-UHFFFAOYSA-L 0.000 description 2
- 230000014759 maintenance of location Effects 0.000 description 2
- 210000004962 mammalian cell Anatomy 0.000 description 2
- 239000012528 membrane Substances 0.000 description 2
- 238000005374 membrane filtration Methods 0.000 description 2
- 108010056582 methionylglutamic acid Proteins 0.000 description 2
- 108010034507 methionyltryptophan Proteins 0.000 description 2
- 230000005012 migration Effects 0.000 description 2
- 238000013508 migration Methods 0.000 description 2
- 238000010369 molecular cloning Methods 0.000 description 2
- 208000005871 monkeypox Diseases 0.000 description 2
- BSOQXXWZTUDTEL-ZUYCGGNHSA-N muramyl dipeptide Chemical compound OC(=O)CC[C@H](C(N)=O)NC(=O)[C@H](C)NC(=O)[C@@H](C)O[C@H]1[C@H](O)[C@@H](CO)O[C@@H](O)[C@@H]1NC(C)=O BSOQXXWZTUDTEL-ZUYCGGNHSA-N 0.000 description 2
- 239000003921 oil Substances 0.000 description 2
- 239000000546 pharmaceutical excipient Substances 0.000 description 2
- 108010064486 phenylalanyl-leucyl-valine Proteins 0.000 description 2
- 108010012581 phenylalanylglutamate Proteins 0.000 description 2
- 108010073025 phenylalanylphenylalanine Proteins 0.000 description 2
- 229920001993 poloxamer 188 Polymers 0.000 description 2
- 229920001200 poly(ethylene-vinyl acetate) Polymers 0.000 description 2
- 238000002264 polyacrylamide gel electrophoresis Methods 0.000 description 2
- 229920001223 polyethylene glycol Polymers 0.000 description 2
- 239000003755 preservative agent Substances 0.000 description 2
- 108010020755 prolyl-glycyl-glycine Proteins 0.000 description 2
- 108700042769 prolyl-leucyl-glycine Proteins 0.000 description 2
- 230000004845 protein aggregation Effects 0.000 description 2
- 238000001742 protein purification Methods 0.000 description 2
- 238000011160 research Methods 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 239000012723 sample buffer Substances 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 108010048818 seryl-histidine Proteins 0.000 description 2
- 108010071207 serylmethionine Proteins 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 239000001488 sodium phosphate Substances 0.000 description 2
- 229910000162 sodium phosphate Inorganic materials 0.000 description 2
- 229940031439 squalene Drugs 0.000 description 2
- TUHBEKDERLKLEC-UHFFFAOYSA-N squalene Natural products CC(=CCCC(=CCCC(=CCCC=C(/C)CCC=C(/C)CC=C(C)C)C)C)C TUHBEKDERLKLEC-UHFFFAOYSA-N 0.000 description 2
- 230000006641 stabilisation Effects 0.000 description 2
- 238000011105 stabilization Methods 0.000 description 2
- 235000000346 sugar Nutrition 0.000 description 2
- 230000020382 suppression by virus of host antigen processing and presentation of peptide antigen via MHC class I Effects 0.000 description 2
- 230000002459 sustained effect Effects 0.000 description 2
- 239000012730 sustained-release form Substances 0.000 description 2
- 238000003786 synthesis reaction Methods 0.000 description 2
- 229940124597 therapeutic agent Drugs 0.000 description 2
- CWERGRDVMFNCDR-UHFFFAOYSA-N thioglycolic acid Chemical compound OC(=O)CS CWERGRDVMFNCDR-UHFFFAOYSA-N 0.000 description 2
- 230000014616 translation Effects 0.000 description 2
- 238000013519 translation Methods 0.000 description 2
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 2
- 210000004881 tumor cell Anatomy 0.000 description 2
- 108010003137 tyrosyltyrosine Proteins 0.000 description 2
- 238000000108 ultra-filtration Methods 0.000 description 2
- 241000701161 unidentified adenovirus Species 0.000 description 2
- 241001529453 unidentified herpesvirus Species 0.000 description 2
- 229940125575 vaccine candidate Drugs 0.000 description 2
- 108010015385 valyl-prolyl-proline Proteins 0.000 description 2
- 239000003981 vehicle Substances 0.000 description 2
- 108010027345 wheylin-1 peptide Proteins 0.000 description 2
- DMWMUMWKGKGSNW-OPMCLZTFSA-N (2S)-6-amino-2-[[(2S)-2-[[(2S)-6-amino-2-[[(2S)-2-[[(2S)-2-[[(2S,3S)-2-[[(2S)-4-amino-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[(2S)-4-amino-2-[[(2S)-4-amino-2-[[2-[[(2R)-2-amino-3-[(2R)-2,3-di(hexadecanoyloxy)propyl]sulfanylpropanoyl]amino]acetyl]amino]-4-oxobutanoyl]amino]-4-oxobutanoyl]amino]-3-carboxypropanoyl]amino]-4-carboxybutanoyl]amino]-3-hydroxypropanoyl]amino]-4-oxobutanoyl]amino]-3-methylpentanoyl]amino]-3-hydroxypropanoyl]amino]-3-phenylpropanoyl]amino]hexanoyl]amino]-4-carboxybutanoyl]amino]hexanoic acid Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](CSC[C@H](N)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(=O)N[C@@H](Cc1ccccc1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(O)=O)OC(=O)CCCCCCCCCCCCCCC DMWMUMWKGKGSNW-OPMCLZTFSA-N 0.000 description 1
- OMDQUFIYNPYJFM-XKDAHURESA-N (2r,3r,4s,5r,6s)-2-(hydroxymethyl)-6-[[(2r,3s,4r,5s,6r)-4,5,6-trihydroxy-3-[(2s,3s,4s,5s,6r)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyoxan-2-yl]methoxy]oxane-3,4,5-triol Chemical compound O[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@@H]1OC[C@@H]1[C@@H](O[C@H]2[C@H]([C@@H](O)[C@H](O)[C@@H](CO)O2)O)[C@H](O)[C@H](O)[C@H](O)O1 OMDQUFIYNPYJFM-XKDAHURESA-N 0.000 description 1
- BRPMXFSTKXXNHF-IUCAKERBSA-N (2s)-1-[2-[[(2s)-pyrrolidine-2-carbonyl]amino]acetyl]pyrrolidine-2-carboxylic acid Chemical compound OC(=O)[C@@H]1CCCN1C(=O)CNC(=O)[C@H]1NCCC1 BRPMXFSTKXXNHF-IUCAKERBSA-N 0.000 description 1
- ZCPBEAHAVUJKAE-UHTWSYAYSA-N (2s)-2-[[(2s)-2-[[(2r)-2-[(2-aminoacetyl)amino]-3-phenylpropanoyl]amino]propanoyl]amino]butanedioic acid Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](NC(=O)CN)CC1=CC=CC=C1 ZCPBEAHAVUJKAE-UHTWSYAYSA-N 0.000 description 1
- LUEWUZLMQUOBSB-FSKGGBMCSA-N (2s,3s,4s,5s,6r)-2-[(2r,3s,4r,5r,6s)-6-[(2r,3s,4r,5s,6s)-4,5-dihydroxy-2-(hydroxymethyl)-6-[(2r,4r,5s,6r)-4,5,6-trihydroxy-2-(hydroxymethyl)oxan-3-yl]oxyoxan-3-yl]oxy-4,5-dihydroxy-2-(hydroxymethyl)oxan-3-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol Chemical compound O[C@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@H]1O[C@@H]1[C@@H](CO)O[C@@H](O[C@@H]2[C@H](O[C@@H](OC3[C@H](O[C@@H](O)[C@@H](O)[C@H]3O)CO)[C@@H](O)[C@H]2O)CO)[C@H](O)[C@H]1O LUEWUZLMQUOBSB-FSKGGBMCSA-N 0.000 description 1
- FQVLRGLGWNWPSS-BXBUPLCLSA-N (4r,7s,10s,13s,16r)-16-acetamido-13-(1h-imidazol-5-ylmethyl)-10-methyl-6,9,12,15-tetraoxo-7-propan-2-yl-1,2-dithia-5,8,11,14-tetrazacycloheptadecane-4-carboxamide Chemical compound N1C(=O)[C@@H](NC(C)=O)CSSC[C@@H](C(N)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C)NC(=O)[C@@H]1CC1=CN=CN1 FQVLRGLGWNWPSS-BXBUPLCLSA-N 0.000 description 1
- XBBVURRQGJPTHH-UHFFFAOYSA-N 2-hydroxyacetic acid;2-hydroxypropanoic acid Chemical compound OCC(O)=O.CC(O)C(O)=O XBBVURRQGJPTHH-UHFFFAOYSA-N 0.000 description 1
- UAIUNKRWKOVEES-UHFFFAOYSA-N 3,3',5,5'-tetramethylbenzidine Chemical compound CC1=C(N)C(C)=CC(C=2C=C(C)C(N)=C(C)C=2)=C1 UAIUNKRWKOVEES-UHFFFAOYSA-N 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- 102100030310 5,6-dihydroxyindole-2-carboxylic acid oxidase Human genes 0.000 description 1
- 241000235389 Absidia Species 0.000 description 1
- 241000224422 Acanthamoeba Species 0.000 description 1
- 241000606750 Actinobacillus Species 0.000 description 1
- 241000186361 Actinobacteria <class> Species 0.000 description 1
- 241000186046 Actinomyces Species 0.000 description 1
- 101800001401 Activation peptide Proteins 0.000 description 1
- 102400000069 Activation peptide Human genes 0.000 description 1
- 241000701242 Adenoviridae Species 0.000 description 1
- 241000607534 Aeromonas Species 0.000 description 1
- 229920001817 Agar Polymers 0.000 description 1
- BUANFPRKJKJSRR-ACZMJKKPSA-N Ala-Ala-Gln Chemical compound C[C@H]([NH3+])C(=O)N[C@@H](C)C(=O)N[C@H](C([O-])=O)CCC(N)=O BUANFPRKJKJSRR-ACZMJKKPSA-N 0.000 description 1
- PIPTUBPKYFRLCP-NHCYSSNCSA-N Ala-Ala-Phe Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 PIPTUBPKYFRLCP-NHCYSSNCSA-N 0.000 description 1
- KQFRUSHJPKXBMB-BHDSKKPTSA-N Ala-Ala-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](C)NC(=O)[C@@H](N)C)C(O)=O)=CNC2=C1 KQFRUSHJPKXBMB-BHDSKKPTSA-N 0.000 description 1
- ODWSTKXGQGYHSH-FXQIFTODSA-N Ala-Arg-Ala Chemical compound C[C@H](N)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(O)=O ODWSTKXGQGYHSH-FXQIFTODSA-N 0.000 description 1
- TTXMOJWKNRJWQJ-FXQIFTODSA-N Ala-Arg-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CCCN=C(N)N TTXMOJWKNRJWQJ-FXQIFTODSA-N 0.000 description 1
- NXSFUECZFORGOG-CIUDSAMLSA-N Ala-Asn-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O NXSFUECZFORGOG-CIUDSAMLSA-N 0.000 description 1
- GORKKVHIBWAQHM-GCJQMDKQSA-N Ala-Asn-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O GORKKVHIBWAQHM-GCJQMDKQSA-N 0.000 description 1
- MKZCBYZBCINNJN-DLOVCJGASA-N Ala-Asp-Phe Chemical compound C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 MKZCBYZBCINNJN-DLOVCJGASA-N 0.000 description 1
- RCQRKPUXJAGEEC-ZLUOBGJFSA-N Ala-Cys-Cys Chemical compound C[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CS)C(O)=O RCQRKPUXJAGEEC-ZLUOBGJFSA-N 0.000 description 1
- XAGIMRPOEJSYER-CIUDSAMLSA-N Ala-Cys-Lys Chemical compound C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CCCCN)C(=O)O)N XAGIMRPOEJSYER-CIUDSAMLSA-N 0.000 description 1
- HJCMDXDYPOUFDY-WHFBIAKZSA-N Ala-Gln Chemical compound C[C@H](N)C(=O)N[C@H](C(O)=O)CCC(N)=O HJCMDXDYPOUFDY-WHFBIAKZSA-N 0.000 description 1
- LGFCAXJBAZESCF-ACZMJKKPSA-N Ala-Gln-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O LGFCAXJBAZESCF-ACZMJKKPSA-N 0.000 description 1
- IFTVANMRTIHKML-WDSKDSINSA-N Ala-Gln-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O IFTVANMRTIHKML-WDSKDSINSA-N 0.000 description 1
- BLGHHPHXVJWCNK-GUBZILKMSA-N Ala-Gln-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O BLGHHPHXVJWCNK-GUBZILKMSA-N 0.000 description 1
- ZDYNWWQXFRUOEO-XDTLVQLUSA-N Ala-Gln-Tyr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O ZDYNWWQXFRUOEO-XDTLVQLUSA-N 0.000 description 1
- GGNHBHYDMUDXQB-KBIXCLLPSA-N Ala-Glu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)N GGNHBHYDMUDXQB-KBIXCLLPSA-N 0.000 description 1
- HXNNRBHASOSVPG-GUBZILKMSA-N Ala-Glu-Leu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O HXNNRBHASOSVPG-GUBZILKMSA-N 0.000 description 1
- VGPWRRFOPXVGOH-BYPYZUCNSA-N Ala-Gly-Gly Chemical compound C[C@H](N)C(=O)NCC(=O)NCC(O)=O VGPWRRFOPXVGOH-BYPYZUCNSA-N 0.000 description 1
- BLIMFWGRQKRCGT-YUMQZZPRSA-N Ala-Gly-Lys Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCCN BLIMFWGRQKRCGT-YUMQZZPRSA-N 0.000 description 1
- QHASENCZLDHBGX-ONGXEEELSA-N Ala-Gly-Phe Chemical compound C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 QHASENCZLDHBGX-ONGXEEELSA-N 0.000 description 1
- FAJIYNONGXEXAI-CQDKDKBSSA-N Ala-His-Phe Chemical compound C([C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CNC=N1 FAJIYNONGXEXAI-CQDKDKBSSA-N 0.000 description 1
- CBCCCLMNOBLBSC-XVYDVKMFSA-N Ala-His-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CO)C(O)=O CBCCCLMNOBLBSC-XVYDVKMFSA-N 0.000 description 1
- NJWJSLCQEDMGNC-MBLNEYKQSA-N Ala-His-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](C)N)O NJWJSLCQEDMGNC-MBLNEYKQSA-N 0.000 description 1
- PNALXAODQKTNLV-JBDRJPRFSA-N Ala-Ile-Ala Chemical compound C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O PNALXAODQKTNLV-JBDRJPRFSA-N 0.000 description 1
- FOHXUHGZZKETFI-JBDRJPRFSA-N Ala-Ile-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](C)N FOHXUHGZZKETFI-JBDRJPRFSA-N 0.000 description 1
- DVJSJDDYCYSMFR-ZKWXMUAHSA-N Ala-Ile-Gly Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(O)=O DVJSJDDYCYSMFR-ZKWXMUAHSA-N 0.000 description 1
- RZZMZYZXNJRPOJ-BJDJZHNGSA-N Ala-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](C)N RZZMZYZXNJRPOJ-BJDJZHNGSA-N 0.000 description 1
- XCZXVTHYGSMQGH-NAKRPEOUSA-N Ala-Ile-Met Chemical compound C[C@H]([NH3+])C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCSC)C([O-])=O XCZXVTHYGSMQGH-NAKRPEOUSA-N 0.000 description 1
- OKIKVSXTXVVFDV-MMWGEVLESA-N Ala-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](C)N OKIKVSXTXVVFDV-MMWGEVLESA-N 0.000 description 1
- CCDFBRZVTDDJNM-GUBZILKMSA-N Ala-Leu-Glu Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O CCDFBRZVTDDJNM-GUBZILKMSA-N 0.000 description 1
- ZKEHTYWGPMMGBC-XUXIUFHCSA-N Ala-Leu-Leu-Ser Chemical compound C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O ZKEHTYWGPMMGBC-XUXIUFHCSA-N 0.000 description 1
- VHVVPYOJIIQCKS-QEJZJMRPSA-N Ala-Leu-Phe Chemical compound C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 VHVVPYOJIIQCKS-QEJZJMRPSA-N 0.000 description 1
- RGQCNKIDEQJEBT-CQDKDKBSSA-N Ala-Leu-Tyr Chemical compound C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 RGQCNKIDEQJEBT-CQDKDKBSSA-N 0.000 description 1
- IAUSCRHURCZUJP-CIUDSAMLSA-N Ala-Lys-Cys Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](CS)C(O)=O IAUSCRHURCZUJP-CIUDSAMLSA-N 0.000 description 1
- PIXQDIGKDNNOOV-GUBZILKMSA-N Ala-Lys-Gln Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(O)=O PIXQDIGKDNNOOV-GUBZILKMSA-N 0.000 description 1
- OQWQTGBOFPJOIF-DLOVCJGASA-N Ala-Lys-His Chemical compound C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N OQWQTGBOFPJOIF-DLOVCJGASA-N 0.000 description 1
- PMQXMXAASGFUDX-SRVKXCTJSA-N Ala-Lys-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](C)N)CCCCN PMQXMXAASGFUDX-SRVKXCTJSA-N 0.000 description 1
- OINVDEKBKBCPLX-JXUBOQSCSA-N Ala-Lys-Thr Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OINVDEKBKBCPLX-JXUBOQSCSA-N 0.000 description 1
- MDNAVFBZPROEHO-DCAQKATOSA-N Ala-Lys-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O MDNAVFBZPROEHO-DCAQKATOSA-N 0.000 description 1
- DGLQWAFPIXDKRL-UBHSHLNASA-N Ala-Met-Phe Chemical compound C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N DGLQWAFPIXDKRL-UBHSHLNASA-N 0.000 description 1
- 108010011667 Ala-Phe-Ala Proteins 0.000 description 1
- XRUJOVRWNMBAAA-NHCYSSNCSA-N Ala-Phe-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=CC=C1 XRUJOVRWNMBAAA-NHCYSSNCSA-N 0.000 description 1
- PEIBBAXIKUAYGN-UBHSHLNASA-N Ala-Phe-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=CC=C1 PEIBBAXIKUAYGN-UBHSHLNASA-N 0.000 description 1
- WEZNQZHACPSMEF-QEJZJMRPSA-N Ala-Phe-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=CC=C1 WEZNQZHACPSMEF-QEJZJMRPSA-N 0.000 description 1
- IHMCQESUJVZTKW-UBHSHLNASA-N Ala-Phe-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](C)N)CC1=CC=CC=C1 IHMCQESUJVZTKW-UBHSHLNASA-N 0.000 description 1
- ADSGHMXEAZJJNF-DCAQKATOSA-N Ala-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](C)N ADSGHMXEAZJJNF-DCAQKATOSA-N 0.000 description 1
- RTZCUEHYUQZIDE-WHFBIAKZSA-N Ala-Ser-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O RTZCUEHYUQZIDE-WHFBIAKZSA-N 0.000 description 1
- DYXOFPBJBAHWFY-JBDRJPRFSA-N Ala-Ser-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@H](C)N DYXOFPBJBAHWFY-JBDRJPRFSA-N 0.000 description 1
- HOVPGJUNRLMIOZ-CIUDSAMLSA-N Ala-Ser-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@H](C)N HOVPGJUNRLMIOZ-CIUDSAMLSA-N 0.000 description 1
- UCDOXFBTMLKASE-HERUPUMHSA-N Ala-Ser-Trp Chemical compound C[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N UCDOXFBTMLKASE-HERUPUMHSA-N 0.000 description 1
- VRTOMXFZHGWHIJ-KZVJFYERSA-N Ala-Thr-Arg Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O VRTOMXFZHGWHIJ-KZVJFYERSA-N 0.000 description 1
- AAWLEICNDUHIJM-MBLNEYKQSA-N Ala-Thr-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](C)N)O AAWLEICNDUHIJM-MBLNEYKQSA-N 0.000 description 1
- IOFVWPYSRSCWHI-JXUBOQSCSA-N Ala-Thr-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@H](C)N IOFVWPYSRSCWHI-JXUBOQSCSA-N 0.000 description 1
- SAHQGRZIQVEJPF-JXUBOQSCSA-N Ala-Thr-Lys Chemical compound C[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@H](C(O)=O)CCCCN SAHQGRZIQVEJPF-JXUBOQSCSA-N 0.000 description 1
- QOIGKCBMXUCDQU-KDXUFGMBSA-N Ala-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](C)N)O QOIGKCBMXUCDQU-KDXUFGMBSA-N 0.000 description 1
- KUFVXLQLDHJVOG-SHGPDSBTSA-N Ala-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](C)N)O KUFVXLQLDHJVOG-SHGPDSBTSA-N 0.000 description 1
- IETUUAHKCHOQHP-KZVJFYERSA-N Ala-Thr-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@H](C)N)[C@@H](C)O)C(O)=O IETUUAHKCHOQHP-KZVJFYERSA-N 0.000 description 1
- ZVWXMTTZJKBJCI-BHDSKKPTSA-N Ala-Trp-Ala Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](N)C)C(=O)N[C@@H](C)C(O)=O)=CNC2=C1 ZVWXMTTZJKBJCI-BHDSKKPTSA-N 0.000 description 1
- VQBULXOHAZSTQY-GKCIPKSASA-N Ala-Trp-Phe Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O VQBULXOHAZSTQY-GKCIPKSASA-N 0.000 description 1
- QDGMZAOSMNGBLP-MRFFXTKBSA-N Ala-Trp-Tyr Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)O)N QDGMZAOSMNGBLP-MRFFXTKBSA-N 0.000 description 1
- KLKARCOHVHLAJP-UWJYBYFXSA-N Ala-Tyr-Cys Chemical compound C[C@H](N)C(=O)N[C@@H](Cc1ccc(O)cc1)C(=O)N[C@@H](CS)C(O)=O KLKARCOHVHLAJP-UWJYBYFXSA-N 0.000 description 1
- BHFOJPDOQPWJRN-XDTLVQLUSA-N Ala-Tyr-Gln Chemical compound C[C@H](N)C(=O)N[C@@H](Cc1ccc(O)cc1)C(=O)N[C@@H](CCC(N)=O)C(O)=O BHFOJPDOQPWJRN-XDTLVQLUSA-N 0.000 description 1
- XKXAZPSREVUCRT-BPNCWPANSA-N Ala-Tyr-Met Chemical compound CSCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](C)N)CC1=CC=C(O)C=C1 XKXAZPSREVUCRT-BPNCWPANSA-N 0.000 description 1
- BVLPIIBTWIYOML-ZKWXMUAHSA-N Ala-Val-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O BVLPIIBTWIYOML-ZKWXMUAHSA-N 0.000 description 1
- BOKLLPVAQDSLHC-FXQIFTODSA-N Ala-Val-Cys Chemical compound C[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CS)C(=O)O)N BOKLLPVAQDSLHC-FXQIFTODSA-N 0.000 description 1
- ZCUFMRIQCPNOHZ-NRPADANISA-N Ala-Val-Gln Chemical compound C[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N ZCUFMRIQCPNOHZ-NRPADANISA-N 0.000 description 1
- LYILPUNCKACNGF-NAKRPEOUSA-N Ala-Val-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C)N LYILPUNCKACNGF-NAKRPEOUSA-N 0.000 description 1
- DHONNEYAZPNGSG-UBHSHLNASA-N Ala-Val-Phe Chemical compound C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 DHONNEYAZPNGSG-UBHSHLNASA-N 0.000 description 1
- 102100034035 Alcohol dehydrogenase 1A Human genes 0.000 description 1
- 241000710929 Alphavirus Species 0.000 description 1
- 241001147657 Ancylostoma Species 0.000 description 1
- 108020005544 Antisense RNA Proteins 0.000 description 1
- 108010039627 Aprotinin Proteins 0.000 description 1
- 241000712892 Arenaviridae Species 0.000 description 1
- 241000712891 Arenavirus Species 0.000 description 1
- DBKNLHKEVPZVQC-LPEHRKFASA-N Arg-Ala-Pro Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@@H]1C(O)=O DBKNLHKEVPZVQC-LPEHRKFASA-N 0.000 description 1
- OLDOLPWZEMHNIA-PJODQICGSA-N Arg-Ala-Trp Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O OLDOLPWZEMHNIA-PJODQICGSA-N 0.000 description 1
- MUXONAMCEUBVGA-DCAQKATOSA-N Arg-Arg-Gln Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCC(N)=O)C(O)=O MUXONAMCEUBVGA-DCAQKATOSA-N 0.000 description 1
- JGDGLDNAQJJGJI-AVGNSLFASA-N Arg-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCCN=C(N)N)N JGDGLDNAQJJGJI-AVGNSLFASA-N 0.000 description 1
- DPXDVGDLWJYZBH-GUBZILKMSA-N Arg-Asn-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O DPXDVGDLWJYZBH-GUBZILKMSA-N 0.000 description 1
- DPNHSNLIULPOBH-GUBZILKMSA-N Arg-Asn-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CCCN=C(N)N)N DPNHSNLIULPOBH-GUBZILKMSA-N 0.000 description 1
- ZTKHZAXGTFXUDD-VEVYYDQMSA-N Arg-Asn-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ZTKHZAXGTFXUDD-VEVYYDQMSA-N 0.000 description 1
- PQWTZSNVWSOFFK-FXQIFTODSA-N Arg-Asp-Asn Chemical compound C(C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)CN=C(N)N PQWTZSNVWSOFFK-FXQIFTODSA-N 0.000 description 1
- JSHVMZANPXCDTL-GMOBBJLQSA-N Arg-Asp-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JSHVMZANPXCDTL-GMOBBJLQSA-N 0.000 description 1
- SQKPKIJVWHAWNF-DCAQKATOSA-N Arg-Asp-Lys Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(O)=O SQKPKIJVWHAWNF-DCAQKATOSA-N 0.000 description 1
- MFAMTAVAFBPXDC-LPEHRKFASA-N Arg-Asp-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCCN=C(N)N)N)C(=O)O MFAMTAVAFBPXDC-LPEHRKFASA-N 0.000 description 1
- ASQYTJJWAMDISW-BPUTZDHNSA-N Arg-Asp-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCCN=C(N)N)N ASQYTJJWAMDISW-BPUTZDHNSA-N 0.000 description 1
- HJAICMSAKODKRF-GUBZILKMSA-N Arg-Cys-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O HJAICMSAKODKRF-GUBZILKMSA-N 0.000 description 1
- NAARDJBSSPUGCF-FXQIFTODSA-N Arg-Cys-Asn Chemical compound C(C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)CN=C(N)N NAARDJBSSPUGCF-FXQIFTODSA-N 0.000 description 1
- GDVDRMUYICMNFJ-CIUDSAMLSA-N Arg-Cys-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(O)=O)C(O)=O GDVDRMUYICMNFJ-CIUDSAMLSA-N 0.000 description 1
- VSPLYCLMFAUZRF-GUBZILKMSA-N Arg-Cys-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CCCN=C(N)N)N VSPLYCLMFAUZRF-GUBZILKMSA-N 0.000 description 1
- BBYTXXRNSFUOOX-IHRRRGAJSA-N Arg-Cys-Phe Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O BBYTXXRNSFUOOX-IHRRRGAJSA-N 0.000 description 1
- JVMKBJNSRZWDBO-FXQIFTODSA-N Arg-Cys-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CO)C(O)=O JVMKBJNSRZWDBO-FXQIFTODSA-N 0.000 description 1
- YHSNASXGBPAHRL-BPUTZDHNSA-N Arg-Cys-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CCCN=C(N)N)N YHSNASXGBPAHRL-BPUTZDHNSA-N 0.000 description 1
- JUWQNWXEGDYCIE-YUMQZZPRSA-N Arg-Gln-Gly Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O JUWQNWXEGDYCIE-YUMQZZPRSA-N 0.000 description 1
- PBSOQGZLPFVXPU-YUMQZZPRSA-N Arg-Glu-Gly Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O PBSOQGZLPFVXPU-YUMQZZPRSA-N 0.000 description 1
- OHYQKYUTLIPFOX-ZPFDUUQYSA-N Arg-Glu-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O OHYQKYUTLIPFOX-ZPFDUUQYSA-N 0.000 description 1
- GOWZVQXTHUCNSQ-NHCYSSNCSA-N Arg-Glu-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O GOWZVQXTHUCNSQ-NHCYSSNCSA-N 0.000 description 1
- IYMAXBFPHPZYIK-BQBZGAKWSA-N Arg-Gly-Asp Chemical compound NC(N)=NCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(O)=O IYMAXBFPHPZYIK-BQBZGAKWSA-N 0.000 description 1
- 108010010777 Arg-Gly-Asp-Gly Proteins 0.000 description 1
- CYXCAHZVPFREJD-LURJTMIESA-N Arg-Gly-Gly Chemical compound NC(=N)NCCC[C@H](N)C(=O)NCC(=O)NCC(O)=O CYXCAHZVPFREJD-LURJTMIESA-N 0.000 description 1
- NKNILFJYKKHBKE-WPRPVWTQSA-N Arg-Gly-Val Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O NKNILFJYKKHBKE-WPRPVWTQSA-N 0.000 description 1
- SLNCSSWAIDUUGF-LSJOCFKGSA-N Arg-His-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C)C(O)=O SLNCSSWAIDUUGF-LSJOCFKGSA-N 0.000 description 1
- YBIAYFFIVAZXPK-AVGNSLFASA-N Arg-His-Arg Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O YBIAYFFIVAZXPK-AVGNSLFASA-N 0.000 description 1
- MMGCRPZQZWTZTA-IHRRRGAJSA-N Arg-His-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N MMGCRPZQZWTZTA-IHRRRGAJSA-N 0.000 description 1
- ZJEDSBGPBXVBMP-PYJNHQTQSA-N Arg-His-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O ZJEDSBGPBXVBMP-PYJNHQTQSA-N 0.000 description 1
- CVKOQHYVDVYJSI-QTKMDUPCSA-N Arg-His-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CCCN=C(N)N)N)O CVKOQHYVDVYJSI-QTKMDUPCSA-N 0.000 description 1
- HCIUUZGFTDTEGM-NAKRPEOUSA-N Arg-Ile-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N HCIUUZGFTDTEGM-NAKRPEOUSA-N 0.000 description 1
- LVMUGODRNHFGRA-AVGNSLFASA-N Arg-Leu-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O LVMUGODRNHFGRA-AVGNSLFASA-N 0.000 description 1
- GMFAGHNRXPSSJS-SRVKXCTJSA-N Arg-Leu-Gln Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O GMFAGHNRXPSSJS-SRVKXCTJSA-N 0.000 description 1
- WMEVEPXNCMKNGH-IHRRRGAJSA-N Arg-Leu-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N WMEVEPXNCMKNGH-IHRRRGAJSA-N 0.000 description 1
- PZBSKYJGKNNYNK-ULQDDVLXSA-N Arg-Leu-Tyr Chemical compound CC(C)C[C@H](NC(=O)[C@@H](N)CCCN=C(N)N)C(=O)N[C@@H](Cc1ccc(O)cc1)C(O)=O PZBSKYJGKNNYNK-ULQDDVLXSA-N 0.000 description 1
- JOADBFCFJGNIKF-GUBZILKMSA-N Arg-Met-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C)C(O)=O JOADBFCFJGNIKF-GUBZILKMSA-N 0.000 description 1
- JBIRFLWXWDSDTR-CYDGBPFRSA-N Arg-Met-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCSC)NC(=O)[C@H](CCCN=C(N)N)N JBIRFLWXWDSDTR-CYDGBPFRSA-N 0.000 description 1
- DTBPLQNKYCYUOM-JYJNAYRXSA-N Arg-Met-Phe Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 DTBPLQNKYCYUOM-JYJNAYRXSA-N 0.000 description 1
- PRLPSDIHSRITSF-UNQGMJICSA-N Arg-Phe-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O PRLPSDIHSRITSF-UNQGMJICSA-N 0.000 description 1
- HNJNAMGZQZPSRE-GUBZILKMSA-N Arg-Pro-Asn Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O HNJNAMGZQZPSRE-GUBZILKMSA-N 0.000 description 1
- OVQJAKFLFTZDNC-GUBZILKMSA-N Arg-Pro-Asp Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O OVQJAKFLFTZDNC-GUBZILKMSA-N 0.000 description 1
- HGKHPCFTRQDHCU-IUCAKERBSA-N Arg-Pro-Gly Chemical compound NC(N)=NCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O HGKHPCFTRQDHCU-IUCAKERBSA-N 0.000 description 1
- UULLJGQFCDXVTQ-CYDGBPFRSA-N Arg-Pro-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(O)=O UULLJGQFCDXVTQ-CYDGBPFRSA-N 0.000 description 1
- NGYHSXDNNOFHNE-AVGNSLFASA-N Arg-Pro-Leu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O NGYHSXDNNOFHNE-AVGNSLFASA-N 0.000 description 1
- VUGWHBXPMAHEGZ-SRVKXCTJSA-N Arg-Pro-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCN=C(N)N VUGWHBXPMAHEGZ-SRVKXCTJSA-N 0.000 description 1
- VRTWYUYCJGNFES-CIUDSAMLSA-N Arg-Ser-Gln Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(O)=O VRTWYUYCJGNFES-CIUDSAMLSA-N 0.000 description 1
- URAUIUGLHBRPMF-NAKRPEOUSA-N Arg-Ser-Ile Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O URAUIUGLHBRPMF-NAKRPEOUSA-N 0.000 description 1
- ICRHGPYYXMWHIE-LPEHRKFASA-N Arg-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)[C@H](CCCN=C(N)N)N)C(=O)O ICRHGPYYXMWHIE-LPEHRKFASA-N 0.000 description 1
- FRBAHXABMQXSJQ-FXQIFTODSA-N Arg-Ser-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O FRBAHXABMQXSJQ-FXQIFTODSA-N 0.000 description 1
- AIFHRTPABBBHKU-RCWTZXSCSA-N Arg-Thr-Arg Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O AIFHRTPABBBHKU-RCWTZXSCSA-N 0.000 description 1
- HRCIIMCTUIAKQB-XGEHTFHBSA-N Arg-Thr-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N)O HRCIIMCTUIAKQB-XGEHTFHBSA-N 0.000 description 1
- LYJXHXGPWDTLKW-HJGDQZAQSA-N Arg-Thr-Gln Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N)O LYJXHXGPWDTLKW-HJGDQZAQSA-N 0.000 description 1
- UZSQXCMNUPKLCC-FJXKBIBVSA-N Arg-Thr-Gly Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O UZSQXCMNUPKLCC-FJXKBIBVSA-N 0.000 description 1
- ZPWMEWYQBWSGAO-ZJDVBMNYSA-N Arg-Thr-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ZPWMEWYQBWSGAO-ZJDVBMNYSA-N 0.000 description 1
- XRNXPIGJPQHCPC-RCWTZXSCSA-N Arg-Thr-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CCCNC(N)=N)[C@@H](C)O)C(O)=O XRNXPIGJPQHCPC-RCWTZXSCSA-N 0.000 description 1
- LOVIQNMIPQVIGT-BVSLBCMMSA-N Arg-Trp-Phe Chemical compound C([C@H](NC(=O)[C@H](CC=1C2=CC=CC=C2NC=1)NC(=O)[C@H](CCCN=C(N)N)N)C(O)=O)C1=CC=CC=C1 LOVIQNMIPQVIGT-BVSLBCMMSA-N 0.000 description 1
- ZCSHHTFOZULVLN-SZMVWBNQSA-N Arg-Trp-Val Chemical compound C1=CC=C2C(C[C@@H](C(=O)N[C@@H](C(C)C)C(O)=O)NC(=O)[C@@H](N)CCCN=C(N)N)=CNC2=C1 ZCSHHTFOZULVLN-SZMVWBNQSA-N 0.000 description 1
- LLQIAIUAKGNOSE-NHCYSSNCSA-N Arg-Val-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CCCN=C(N)N LLQIAIUAKGNOSE-NHCYSSNCSA-N 0.000 description 1
- QLSRIZIDQXDQHK-RCWTZXSCSA-N Arg-Val-Thr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QLSRIZIDQXDQHK-RCWTZXSCSA-N 0.000 description 1
- WHLDJYNHXOMGMU-JYJNAYRXSA-N Arg-Val-Tyr Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 WHLDJYNHXOMGMU-JYJNAYRXSA-N 0.000 description 1
- ANAHQDPQQBDOBM-UHFFFAOYSA-N Arg-Val-Tyr Natural products CC(C)C(NC(=O)C(N)CCNC(=N)N)C(=O)NC(Cc1ccc(O)cc1)C(=O)O ANAHQDPQQBDOBM-UHFFFAOYSA-N 0.000 description 1
- 241000244186 Ascaris Species 0.000 description 1
- ACRYGQFHAQHDSF-ZLUOBGJFSA-N Asn-Asn-Asn Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ACRYGQFHAQHDSF-ZLUOBGJFSA-N 0.000 description 1
- PCKRJVZAQZWNKM-WHFBIAKZSA-N Asn-Asn-Gly Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O PCKRJVZAQZWNKM-WHFBIAKZSA-N 0.000 description 1
- AYZAWXAPBAYCHO-CIUDSAMLSA-N Asn-Asn-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC(=O)N)N AYZAWXAPBAYCHO-CIUDSAMLSA-N 0.000 description 1
- DXZNJWFECGJCQR-FXQIFTODSA-N Asn-Asn-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC(=O)N)N DXZNJWFECGJCQR-FXQIFTODSA-N 0.000 description 1
- PIWWUBYJNONVTJ-ZLUOBGJFSA-N Asn-Asp-Asn Chemical compound C([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)C(=O)N PIWWUBYJNONVTJ-ZLUOBGJFSA-N 0.000 description 1
- GMCOADLDNLGOFE-ZLUOBGJFSA-N Asn-Asp-Cys Chemical compound C([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CS)C(=O)O)N)C(=O)N GMCOADLDNLGOFE-ZLUOBGJFSA-N 0.000 description 1
- UPALZCBCKAMGIY-PEFMBERDSA-N Asn-Gln-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O UPALZCBCKAMGIY-PEFMBERDSA-N 0.000 description 1
- KUYKVGODHGHFDI-ACZMJKKPSA-N Asn-Gln-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(O)=O KUYKVGODHGHFDI-ACZMJKKPSA-N 0.000 description 1
- JREOBWLIZLXRIS-GUBZILKMSA-N Asn-Glu-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O JREOBWLIZLXRIS-GUBZILKMSA-N 0.000 description 1
- OOWSBIOUKIUWLO-RCOVLWMOSA-N Asn-Gly-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O OOWSBIOUKIUWLO-RCOVLWMOSA-N 0.000 description 1
- ODBSSLHUFPJRED-CIUDSAMLSA-N Asn-His-Asn Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N ODBSSLHUFPJRED-CIUDSAMLSA-N 0.000 description 1
- XLHLPYFMXGOASD-CIUDSAMLSA-N Asn-His-Asp Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CC(=O)N)N XLHLPYFMXGOASD-CIUDSAMLSA-N 0.000 description 1
- IKLAUGBIDCDFOY-SRVKXCTJSA-N Asn-His-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(C)C)C(O)=O IKLAUGBIDCDFOY-SRVKXCTJSA-N 0.000 description 1
- UYXXMIZGHYKYAT-NHCYSSNCSA-N Asn-His-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CC(=O)N)N UYXXMIZGHYKYAT-NHCYSSNCSA-N 0.000 description 1
- SEKBHZJLARBNPB-GHCJXIJMSA-N Asn-Ile-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O SEKBHZJLARBNPB-GHCJXIJMSA-N 0.000 description 1
- SPCONPVIDFMDJI-QSFUFRPTSA-N Asn-Ile-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O SPCONPVIDFMDJI-QSFUFRPTSA-N 0.000 description 1
- GLWFAWNYGWBMOC-SRVKXCTJSA-N Asn-Leu-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O GLWFAWNYGWBMOC-SRVKXCTJSA-N 0.000 description 1
- NLDNNZKUSLAYFW-NHCYSSNCSA-N Asn-Lys-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O NLDNNZKUSLAYFW-NHCYSSNCSA-N 0.000 description 1
- PBFXCUOEGVJTMV-QXEWZRGKSA-N Asn-Met-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C(C)C)C(O)=O PBFXCUOEGVJTMV-QXEWZRGKSA-N 0.000 description 1
- ZJIFRAPZHAGLGR-MELADBBJSA-N Asn-Phe-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=CC=C2)NC(=O)[C@H](CC(=O)N)N)C(=O)O ZJIFRAPZHAGLGR-MELADBBJSA-N 0.000 description 1
- YXVAESUIQFDBHN-SRVKXCTJSA-N Asn-Phe-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O YXVAESUIQFDBHN-SRVKXCTJSA-N 0.000 description 1
- XMHFCUKJRCQXGI-CIUDSAMLSA-N Asn-Pro-Gln Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CC(=O)N)N)C(=O)N[C@@H](CCC(=O)N)C(=O)O XMHFCUKJRCQXGI-CIUDSAMLSA-N 0.000 description 1
- AWXDRZJQCVHCIT-DCAQKATOSA-N Asn-Pro-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CC(N)=O AWXDRZJQCVHCIT-DCAQKATOSA-N 0.000 description 1
- IDUUACUJKUXKKD-VEVYYDQMSA-N Asn-Pro-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O IDUUACUJKUXKKD-VEVYYDQMSA-N 0.000 description 1
- REQUGIWGOGSOEZ-ZLUOBGJFSA-N Asn-Ser-Asn Chemical compound C([C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)C(=O)N REQUGIWGOGSOEZ-ZLUOBGJFSA-N 0.000 description 1
- MKJBPDLENBUHQU-CIUDSAMLSA-N Asn-Ser-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O MKJBPDLENBUHQU-CIUDSAMLSA-N 0.000 description 1
- JXMREEPBRANWBY-VEVYYDQMSA-N Asn-Thr-Arg Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O JXMREEPBRANWBY-VEVYYDQMSA-N 0.000 description 1
- WUQXMTITJLFXAU-JIOCBJNQSA-N Asn-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(=O)N)N)O WUQXMTITJLFXAU-JIOCBJNQSA-N 0.000 description 1
- AMGQTNHANMRPOE-LKXGYXEUSA-N Asn-Thr-Ser Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O AMGQTNHANMRPOE-LKXGYXEUSA-N 0.000 description 1
- TZQWZQSMHDVLQL-QEJZJMRPSA-N Asn-Trp-Gln Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N TZQWZQSMHDVLQL-QEJZJMRPSA-N 0.000 description 1
- RDLYUKRPEJERMM-XIRDDKMYSA-N Asn-Trp-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC(C)C)C(O)=O RDLYUKRPEJERMM-XIRDDKMYSA-N 0.000 description 1
- RTFXPCYMDYBZNQ-SRVKXCTJSA-N Asn-Tyr-Asn Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(N)=O)C(O)=O RTFXPCYMDYBZNQ-SRVKXCTJSA-N 0.000 description 1
- QNNBHTFDFFFHGC-KKUMJFAQSA-N Asn-Tyr-Lys Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)N)N)O QNNBHTFDFFFHGC-KKUMJFAQSA-N 0.000 description 1
- XLDMSQYOYXINSZ-QXEWZRGKSA-N Asn-Val-Arg Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](CC(=O)N)N XLDMSQYOYXINSZ-QXEWZRGKSA-N 0.000 description 1
- QXNGSPZMGFEZNO-QRTARXTBSA-N Asn-Val-Trp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O QXNGSPZMGFEZNO-QRTARXTBSA-N 0.000 description 1
- HPNDBHLITCHRSO-WHFBIAKZSA-N Asp-Ala-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)NCC(O)=O HPNDBHLITCHRSO-WHFBIAKZSA-N 0.000 description 1
- XBQSLMACWDXWLJ-GHCJXIJMSA-N Asp-Ala-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O XBQSLMACWDXWLJ-GHCJXIJMSA-N 0.000 description 1
- OERMIMJQPQUIPK-FXQIFTODSA-N Asp-Arg-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(O)=O OERMIMJQPQUIPK-FXQIFTODSA-N 0.000 description 1
- SDHFVYLZFBDSQT-DCAQKATOSA-N Asp-Arg-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(=O)O)N SDHFVYLZFBDSQT-DCAQKATOSA-N 0.000 description 1
- DBWYWXNMZZYIRY-LPEHRKFASA-N Asp-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(=O)O)N)C(=O)O DBWYWXNMZZYIRY-LPEHRKFASA-N 0.000 description 1
- YNQIDCRRTWGHJD-ZLUOBGJFSA-N Asp-Asn-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CC(O)=O YNQIDCRRTWGHJD-ZLUOBGJFSA-N 0.000 description 1
- QRULNKJGYQQZMW-ZLUOBGJFSA-N Asp-Asn-Asp Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O QRULNKJGYQQZMW-ZLUOBGJFSA-N 0.000 description 1
- RDRMWJBLOSRRAW-BYULHYEWSA-N Asp-Asn-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O RDRMWJBLOSRRAW-BYULHYEWSA-N 0.000 description 1
- CELPEWWLSXMVPH-CIUDSAMLSA-N Asp-Asp-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CC(O)=O CELPEWWLSXMVPH-CIUDSAMLSA-N 0.000 description 1
- ZRAOLTNMSCSCLN-ZLUOBGJFSA-N Asp-Cys-Asn Chemical compound C([C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)C(=O)O ZRAOLTNMSCSCLN-ZLUOBGJFSA-N 0.000 description 1
- RSMIHCFQDCVVBR-CIUDSAMLSA-N Asp-Gln-Arg Chemical compound OC(=O)C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(O)=O)CCCNC(N)=N RSMIHCFQDCVVBR-CIUDSAMLSA-N 0.000 description 1
- BKXPJCBEHWFSTF-ACZMJKKPSA-N Asp-Gln-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O BKXPJCBEHWFSTF-ACZMJKKPSA-N 0.000 description 1
- HRGGPWBIMIQANI-GUBZILKMSA-N Asp-Gln-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O HRGGPWBIMIQANI-GUBZILKMSA-N 0.000 description 1
- IJHUZMGJRGNXIW-CIUDSAMLSA-N Asp-Glu-Arg Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O IJHUZMGJRGNXIW-CIUDSAMLSA-N 0.000 description 1
- OMMIEVATLAGRCK-BYPYZUCNSA-N Asp-Gly-Gly Chemical compound OC(=O)C[C@H](N)C(=O)NCC(=O)NCC(O)=O OMMIEVATLAGRCK-BYPYZUCNSA-N 0.000 description 1
- PZXPWHFYZXTFBI-YUMQZZPRSA-N Asp-Gly-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(O)=O PZXPWHFYZXTFBI-YUMQZZPRSA-N 0.000 description 1
- SNDBKTFJWVEVPO-WHFBIAKZSA-N Asp-Gly-Ser Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](CO)C(O)=O SNDBKTFJWVEVPO-WHFBIAKZSA-N 0.000 description 1
- YRBGRUOSJROZEI-NHCYSSNCSA-N Asp-His-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C(C)C)C(O)=O YRBGRUOSJROZEI-NHCYSSNCSA-N 0.000 description 1
- YFSLJHLQOALGSY-ZPFDUUQYSA-N Asp-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)O)N YFSLJHLQOALGSY-ZPFDUUQYSA-N 0.000 description 1
- SPWXXPFDTMYTRI-IUKAMOBKSA-N Asp-Ile-Thr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SPWXXPFDTMYTRI-IUKAMOBKSA-N 0.000 description 1
- AITKTFCQOBRJTG-CIUDSAMLSA-N Asp-Leu-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CC(=O)O)N AITKTFCQOBRJTG-CIUDSAMLSA-N 0.000 description 1
- DWOGMPWRQQWPPF-GUBZILKMSA-N Asp-Leu-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O DWOGMPWRQQWPPF-GUBZILKMSA-N 0.000 description 1
- XSXVLWBWIPKUSN-UHFFFAOYSA-N Asp-Leu-Glu-Asp Chemical compound OC(=O)CC(N)C(=O)NC(CC(C)C)C(=O)NC(CCC(O)=O)C(=O)NC(CC(O)=O)C(O)=O XSXVLWBWIPKUSN-UHFFFAOYSA-N 0.000 description 1
- AYFVRYXNDHBECD-YUMQZZPRSA-N Asp-Leu-Gly Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O AYFVRYXNDHBECD-YUMQZZPRSA-N 0.000 description 1
- IVPNEDNYYYFAGI-GARJFASQSA-N Asp-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(=O)O)N IVPNEDNYYYFAGI-GARJFASQSA-N 0.000 description 1
- UMHUHHJMEXNSIV-CIUDSAMLSA-N Asp-Leu-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC(O)=O UMHUHHJMEXNSIV-CIUDSAMLSA-N 0.000 description 1
- MYOHQBFRJQFIDZ-KKUMJFAQSA-N Asp-Leu-Tyr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O MYOHQBFRJQFIDZ-KKUMJFAQSA-N 0.000 description 1
- XWSIYTYNLKCLJB-CIUDSAMLSA-N Asp-Lys-Asn Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(O)=O XWSIYTYNLKCLJB-CIUDSAMLSA-N 0.000 description 1
- NVFSJIXJZCDICF-SRVKXCTJSA-N Asp-Lys-Lys Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(=O)O)N NVFSJIXJZCDICF-SRVKXCTJSA-N 0.000 description 1
- MYLZFUMPZCPJCJ-NHCYSSNCSA-N Asp-Lys-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O MYLZFUMPZCPJCJ-NHCYSSNCSA-N 0.000 description 1
- WDMNFNXKGSLIOB-GUBZILKMSA-N Asp-Met-Met Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CC(=O)O)N WDMNFNXKGSLIOB-GUBZILKMSA-N 0.000 description 1
- IOXWDLNHXZOXQP-FXQIFTODSA-N Asp-Met-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC(=O)O)N IOXWDLNHXZOXQP-FXQIFTODSA-N 0.000 description 1
- WOPJVEMFXYHZEE-SRVKXCTJSA-N Asp-Phe-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O WOPJVEMFXYHZEE-SRVKXCTJSA-N 0.000 description 1
- KRQFMDNIUOVRIF-KKUMJFAQSA-N Asp-Phe-His Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)NC(=O)[C@H](CC(=O)O)N KRQFMDNIUOVRIF-KKUMJFAQSA-N 0.000 description 1
- JUWISGAGWSDGDH-KKUMJFAQSA-N Asp-Phe-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)CC(O)=O)CC1=CC=CC=C1 JUWISGAGWSDGDH-KKUMJFAQSA-N 0.000 description 1
- AHWRSSLYSGLBGD-CIUDSAMLSA-N Asp-Pro-Glu Chemical compound OC(=O)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O AHWRSSLYSGLBGD-CIUDSAMLSA-N 0.000 description 1
- YFGUZQQCSDZRBN-DCAQKATOSA-N Asp-Pro-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O YFGUZQQCSDZRBN-DCAQKATOSA-N 0.000 description 1
- ZBYLEBZCVKLPCY-FXQIFTODSA-N Asp-Ser-Arg Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O ZBYLEBZCVKLPCY-FXQIFTODSA-N 0.000 description 1
- ZQFRDAZBTSFGGW-SRVKXCTJSA-N Asp-Ser-Phe Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O ZQFRDAZBTSFGGW-SRVKXCTJSA-N 0.000 description 1
- MGSVBZIBCCKGCY-ZLUOBGJFSA-N Asp-Ser-Ser Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O MGSVBZIBCCKGCY-ZLUOBGJFSA-N 0.000 description 1
- XYPJXLLXNSAWHZ-SRVKXCTJSA-N Asp-Ser-Tyr Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O XYPJXLLXNSAWHZ-SRVKXCTJSA-N 0.000 description 1
- MNQMTYSEKZHIDF-GCJQMDKQSA-N Asp-Thr-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O MNQMTYSEKZHIDF-GCJQMDKQSA-N 0.000 description 1
- KBJVTFWQWXCYCQ-IUKAMOBKSA-N Asp-Thr-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O KBJVTFWQWXCYCQ-IUKAMOBKSA-N 0.000 description 1
- PLNJUJGNLDSFOP-UWJYBYFXSA-N Asp-Tyr-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C)C(O)=O PLNJUJGNLDSFOP-UWJYBYFXSA-N 0.000 description 1
- KNDCWFXCFKSEBM-AVGNSLFASA-N Asp-Tyr-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O KNDCWFXCFKSEBM-AVGNSLFASA-N 0.000 description 1
- MFDPBZAFCRKYEY-LAEOZQHASA-N Asp-Val-Gln Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O MFDPBZAFCRKYEY-LAEOZQHASA-N 0.000 description 1
- UXRVDHVARNBOIO-QSFUFRPTSA-N Asp-Val-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CC(=O)O)N UXRVDHVARNBOIO-QSFUFRPTSA-N 0.000 description 1
- GIKOVDMXBAFXDF-NHCYSSNCSA-N Asp-Val-Leu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O GIKOVDMXBAFXDF-NHCYSSNCSA-N 0.000 description 1
- ZUNMTUPRQMWMHX-LSJOCFKGSA-N Asp-Val-Val Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C(C)C)C(O)=O ZUNMTUPRQMWMHX-LSJOCFKGSA-N 0.000 description 1
- 241000416162 Astragalus gummifer Species 0.000 description 1
- 241001533362 Astroviridae Species 0.000 description 1
- 102100024222 B-lymphocyte antigen CD19 Human genes 0.000 description 1
- 241000606125 Bacteroides Species 0.000 description 1
- 241000701412 Baculoviridae Species 0.000 description 1
- 241001235574 Balantidium Species 0.000 description 1
- 241000702628 Birnaviridae Species 0.000 description 1
- 241000335423 Blastomyces Species 0.000 description 1
- 241000228405 Blastomyces dermatitidis Species 0.000 description 1
- 241000589968 Borrelia Species 0.000 description 1
- 108091003079 Bovine Serum Albumin Proteins 0.000 description 1
- 206010006187 Breast cancer Diseases 0.000 description 1
- 208000026310 Breast neoplasm Diseases 0.000 description 1
- 241000589562 Brucella Species 0.000 description 1
- 206010006500 Brucellosis Diseases 0.000 description 1
- 241000244036 Brugia Species 0.000 description 1
- 241000714198 Caliciviridae Species 0.000 description 1
- 241000222120 Candida <Saccharomycetales> Species 0.000 description 1
- 241000222122 Candida albicans Species 0.000 description 1
- 206010007134 Candida infections Diseases 0.000 description 1
- BVKZGUZCCUSVTD-UHFFFAOYSA-L Carbonate Chemical compound [O-]C([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-L 0.000 description 1
- 201000009030 Carcinoma Diseases 0.000 description 1
- 108010001857 Cell Surface Receptors Proteins 0.000 description 1
- 102000000844 Cell Surface Receptors Human genes 0.000 description 1
- 241000282994 Cervidae Species 0.000 description 1
- 206010008631 Cholera Diseases 0.000 description 1
- 241001533399 Circoviridae Species 0.000 description 1
- 241000588923 Citrobacter Species 0.000 description 1
- 241001327942 Clonorchis Species 0.000 description 1
- 241000223205 Coccidioides immitis Species 0.000 description 1
- 208000001333 Colorectal Neoplasms Diseases 0.000 description 1
- 208000035473 Communicable disease Diseases 0.000 description 1
- 229920002261 Corn starch Polymers 0.000 description 1
- 201000007336 Cryptococcosis Diseases 0.000 description 1
- 241001527609 Cryptococcus Species 0.000 description 1
- 241000221204 Cryptococcus neoformans Species 0.000 description 1
- 241000195493 Cryptophyta Species 0.000 description 1
- 241000235555 Cunninghamella Species 0.000 description 1
- 241000244152 Cyclophyllidea Species 0.000 description 1
- HAYVTMHUNMMXCV-IMJSIDKUSA-N Cys-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H](N)CS HAYVTMHUNMMXCV-IMJSIDKUSA-N 0.000 description 1
- TVYMKYUSZSVOAG-ZLUOBGJFSA-N Cys-Ala-Ala Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O TVYMKYUSZSVOAG-ZLUOBGJFSA-N 0.000 description 1
- AMRLSQGGERHDHJ-FXQIFTODSA-N Cys-Ala-Arg Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O AMRLSQGGERHDHJ-FXQIFTODSA-N 0.000 description 1
- PRXCTTWKGJAPMT-ZLUOBGJFSA-N Cys-Ala-Ser Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O PRXCTTWKGJAPMT-ZLUOBGJFSA-N 0.000 description 1
- UKVGHFORADMBEN-GUBZILKMSA-N Cys-Arg-Arg Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O UKVGHFORADMBEN-GUBZILKMSA-N 0.000 description 1
- DEVDFMRWZASYOF-ZLUOBGJFSA-N Cys-Asn-Asp Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O DEVDFMRWZASYOF-ZLUOBGJFSA-N 0.000 description 1
- KLLFLHBKSJAUMZ-ACZMJKKPSA-N Cys-Asn-Glu Chemical compound C(CC(=O)O)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CS)N KLLFLHBKSJAUMZ-ACZMJKKPSA-N 0.000 description 1
- GSNRZJNHMVMOFV-ACZMJKKPSA-N Cys-Asp-Glu Chemical compound C(CC(=O)O)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CS)N GSNRZJNHMVMOFV-ACZMJKKPSA-N 0.000 description 1
- GGIHYKLJUIZYGH-ZLUOBGJFSA-N Cys-Cys-Asp Chemical compound C([C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CS)N)C(=O)O GGIHYKLJUIZYGH-ZLUOBGJFSA-N 0.000 description 1
- ATPDEYTYWVMINF-ZLUOBGJFSA-N Cys-Cys-Ser Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CS)C(=O)N[C@@H](CO)C(O)=O ATPDEYTYWVMINF-ZLUOBGJFSA-N 0.000 description 1
- ZVNFONSZVUBRAV-CIUDSAMLSA-N Cys-Gln-Arg Chemical compound C(C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CS)N)CN=C(N)N ZVNFONSZVUBRAV-CIUDSAMLSA-N 0.000 description 1
- RFHGRMMADHHQSA-KBIXCLLPSA-N Cys-Gln-Ile Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O RFHGRMMADHHQSA-KBIXCLLPSA-N 0.000 description 1
- GUKYYUFHWYRMEU-WHFBIAKZSA-N Cys-Gly-Asp Chemical compound [H]N[C@@H](CS)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(O)=O GUKYYUFHWYRMEU-WHFBIAKZSA-N 0.000 description 1
- LBOLGUYQEPZSKM-YUMQZZPRSA-N Cys-Gly-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CS)N LBOLGUYQEPZSKM-YUMQZZPRSA-N 0.000 description 1
- XELISBQUZZAPQK-CIUDSAMLSA-N Cys-His-Cys Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CS)N XELISBQUZZAPQK-CIUDSAMLSA-N 0.000 description 1
- CUXIOFHFFXNUGG-HTFCKZLJSA-N Cys-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)NC(=O)[C@H](CS)N CUXIOFHFFXNUGG-HTFCKZLJSA-N 0.000 description 1
- VPQZSNQICFCCSO-BJDJZHNGSA-N Cys-Leu-Ile Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O VPQZSNQICFCCSO-BJDJZHNGSA-N 0.000 description 1
- WVLZTXGTNGHPBO-SRVKXCTJSA-N Cys-Leu-Leu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O WVLZTXGTNGHPBO-SRVKXCTJSA-N 0.000 description 1
- XZKJEOMFLDVXJG-KATARQTJSA-N Cys-Leu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CS)N)O XZKJEOMFLDVXJG-KATARQTJSA-N 0.000 description 1
- MKMKILWCRQLDFJ-DCAQKATOSA-N Cys-Lys-Arg Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O MKMKILWCRQLDFJ-DCAQKATOSA-N 0.000 description 1
- XMVZMBGFIOQONW-GARJFASQSA-N Cys-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CS)N)C(=O)O XMVZMBGFIOQONW-GARJFASQSA-N 0.000 description 1
- HJGUQJJJXQGXGJ-FXQIFTODSA-N Cys-Met-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CS)N HJGUQJJJXQGXGJ-FXQIFTODSA-N 0.000 description 1
- SDDJEOCJUFKAPV-BPUTZDHNSA-N Cys-Met-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CS)CCSC)C(O)=O)=CNC2=C1 SDDJEOCJUFKAPV-BPUTZDHNSA-N 0.000 description 1
- JEKIARHEWURQRJ-BZSNNMDCSA-N Cys-Phe-Tyr Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)O)NC(=O)[C@H](CS)N JEKIARHEWURQRJ-BZSNNMDCSA-N 0.000 description 1
- SMEYEQDCCBHTEF-FXQIFTODSA-N Cys-Pro-Ala Chemical compound [H]N[C@@H](CS)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O SMEYEQDCCBHTEF-FXQIFTODSA-N 0.000 description 1
- MBRWOKXNHTUJMB-CIUDSAMLSA-N Cys-Pro-Glu Chemical compound [H]N[C@@H](CS)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O MBRWOKXNHTUJMB-CIUDSAMLSA-N 0.000 description 1
- DQUWSUWXPWGTQT-DCAQKATOSA-N Cys-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CS DQUWSUWXPWGTQT-DCAQKATOSA-N 0.000 description 1
- ZGERHCJBLPQPGV-ACZMJKKPSA-N Cys-Ser-Gln Chemical compound C(CC(=O)N)[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CS)N ZGERHCJBLPQPGV-ACZMJKKPSA-N 0.000 description 1
- GGRDJANMZPGMNS-CIUDSAMLSA-N Cys-Ser-Leu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O GGRDJANMZPGMNS-CIUDSAMLSA-N 0.000 description 1
- YNJBLTDKTMKEET-ZLUOBGJFSA-N Cys-Ser-Ser Chemical compound SC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O YNJBLTDKTMKEET-ZLUOBGJFSA-N 0.000 description 1
- XWTGTTNUCCEFJI-UBHSHLNASA-N Cys-Ser-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CS)N XWTGTTNUCCEFJI-UBHSHLNASA-N 0.000 description 1
- IXPSSIBVVKSOIE-SRVKXCTJSA-N Cys-Ser-Tyr Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](CS)N)O IXPSSIBVVKSOIE-SRVKXCTJSA-N 0.000 description 1
- NDNZRWUDUMTITL-FXQIFTODSA-N Cys-Ser-Val Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O NDNZRWUDUMTITL-FXQIFTODSA-N 0.000 description 1
- ZLFRUAFDAIFNHN-LKXGYXEUSA-N Cys-Thr-Asp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CS)N)O ZLFRUAFDAIFNHN-LKXGYXEUSA-N 0.000 description 1
- FTTZLFIEUQHLHH-BWBBJGPYSA-N Cys-Thr-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CS)N)O FTTZLFIEUQHLHH-BWBBJGPYSA-N 0.000 description 1
- UKHNKRGNFKSHCG-CUJWVEQBSA-N Cys-Thr-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CS)N)O UKHNKRGNFKSHCG-CUJWVEQBSA-N 0.000 description 1
- GFAPBMCRSMSGDZ-XGEHTFHBSA-N Cys-Thr-Met Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CS)N)O GFAPBMCRSMSGDZ-XGEHTFHBSA-N 0.000 description 1
- IQXSTXKVEMRMMB-XAVMHZPKSA-N Cys-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CS)N)O IQXSTXKVEMRMMB-XAVMHZPKSA-N 0.000 description 1
- KFYPRIGJTICABD-XGEHTFHBSA-N Cys-Thr-Val Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](CS)N)O KFYPRIGJTICABD-XGEHTFHBSA-N 0.000 description 1
- KARBMKZDLYMMOW-JYBASQMISA-N Cys-Trp-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CS)N)O KARBMKZDLYMMOW-JYBASQMISA-N 0.000 description 1
- IOLWXFWVYYCVTJ-NRPADANISA-N Cys-Val-Gln Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CS)N IOLWXFWVYYCVTJ-NRPADANISA-N 0.000 description 1
- KZZYVYWSXMFYEC-DCAQKATOSA-N Cys-Val-Leu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O KZZYVYWSXMFYEC-DCAQKATOSA-N 0.000 description 1
- ATFSDBMHRCDLBV-BPUTZDHNSA-N Cys-Val-Trp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CS)N ATFSDBMHRCDLBV-BPUTZDHNSA-N 0.000 description 1
- 102000012410 DNA Ligases Human genes 0.000 description 1
- 108010061982 DNA Ligases Proteins 0.000 description 1
- 101710088194 Dehydrogenase Proteins 0.000 description 1
- 229920002307 Dextran Polymers 0.000 description 1
- 108010053187 Diphtheria Toxin Proteins 0.000 description 1
- 102000016607 Diphtheria Toxin Human genes 0.000 description 1
- 241001137876 Diphyllobothrium Species 0.000 description 1
- 235000003550 Dracunculus Nutrition 0.000 description 1
- 241000316827 Dracunculus <angiosperm> Species 0.000 description 1
- 102000001301 EGF receptor Human genes 0.000 description 1
- 108060006698 EGF receptor Proteins 0.000 description 1
- 241001126301 Echinostoma Species 0.000 description 1
- 241000224431 Entamoeba Species 0.000 description 1
- 241000194033 Enterococcus Species 0.000 description 1
- 101710204837 Envelope small membrane protein Proteins 0.000 description 1
- 241000186811 Erysipelothrix Species 0.000 description 1
- 241000588724 Escherichia coli Species 0.000 description 1
- 241001126309 Fasciolopsis Species 0.000 description 1
- 201000008808 Fibrosarcoma Diseases 0.000 description 1
- 241000710781 Flaviviridae Species 0.000 description 1
- 241000589601 Francisella Species 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- 101150038242 GAL10 gene Proteins 0.000 description 1
- 229920000926 Galactomannan Polymers 0.000 description 1
- 102100024637 Galectin-10 Human genes 0.000 description 1
- 102100039555 Galectin-7 Human genes 0.000 description 1
- 108010010803 Gelatin Proteins 0.000 description 1
- 206010060891 General symptom Diseases 0.000 description 1
- 101000892220 Geobacillus thermodenitrificans (strain NG80-2) Long-chain-alcohol dehydrogenase 1 Proteins 0.000 description 1
- 241000232864 Geospiza Species 0.000 description 1
- 241000224466 Giardia Species 0.000 description 1
- YJIUYQKQBBQYHZ-ACZMJKKPSA-N Gln-Ala-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O YJIUYQKQBBQYHZ-ACZMJKKPSA-N 0.000 description 1
- UWZLBXOBVKRUFE-HGNGGELXSA-N Gln-Ala-His Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCC(=O)N)N UWZLBXOBVKRUFE-HGNGGELXSA-N 0.000 description 1
- SHERTACNJPYHAR-ACZMJKKPSA-N Gln-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCC(N)=O SHERTACNJPYHAR-ACZMJKKPSA-N 0.000 description 1
- OVQXQLWWJSNYFV-XEGUGMAKSA-N Gln-Ala-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)CCC(N)=O)C)C(O)=O)=CNC2=C1 OVQXQLWWJSNYFV-XEGUGMAKSA-N 0.000 description 1
- RGXXLQWXBFNXTG-CIUDSAMLSA-N Gln-Arg-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(O)=O RGXXLQWXBFNXTG-CIUDSAMLSA-N 0.000 description 1
- WOACHWLUOFZLGJ-GUBZILKMSA-N Gln-Arg-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(O)=O WOACHWLUOFZLGJ-GUBZILKMSA-N 0.000 description 1
- SXIJQMBEVYWAQT-GUBZILKMSA-N Gln-Asp-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCC(=O)N)N SXIJQMBEVYWAQT-GUBZILKMSA-N 0.000 description 1
- IXFVOPOHSRKJNG-LAEOZQHASA-N Gln-Asp-Val Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O IXFVOPOHSRKJNG-LAEOZQHASA-N 0.000 description 1
- ALUBSZXSNSPDQV-WDSKDSINSA-N Gln-Cys-Gly Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@@H](CS)C(=O)NCC(O)=O ALUBSZXSNSPDQV-WDSKDSINSA-N 0.000 description 1
- COYGBRTZEVWZBW-XKBZYTNZSA-N Gln-Cys-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CS)NC(=O)[C@@H](N)CCC(N)=O COYGBRTZEVWZBW-XKBZYTNZSA-N 0.000 description 1
- PNENQZWRFMUZOM-DCAQKATOSA-N Gln-Glu-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O PNENQZWRFMUZOM-DCAQKATOSA-N 0.000 description 1
- XJKAKYXMFHUIHT-AUTRQRHGSA-N Gln-Glu-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CCC(=O)N)N XJKAKYXMFHUIHT-AUTRQRHGSA-N 0.000 description 1
- VSXBYIJUAXPAAL-WDSKDSINSA-N Gln-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)[C@@H](N)CCC(N)=O VSXBYIJUAXPAAL-WDSKDSINSA-N 0.000 description 1
- QQAPDATZKKTBIY-YUMQZZPRSA-N Gln-Gly-Met Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)NCC(=O)N[C@@H](CCSC)C(O)=O QQAPDATZKKTBIY-YUMQZZPRSA-N 0.000 description 1
- SBHVGKBYOQKAEA-SDDRHHMPSA-N Gln-His-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CN=CN2)NC(=O)[C@H](CCC(=O)N)N)C(=O)O SBHVGKBYOQKAEA-SDDRHHMPSA-N 0.000 description 1
- HDUDGCZEOZEFOA-KBIXCLLPSA-N Gln-Ile-Ala Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)O)NC(=O)[C@H](CCC(=O)N)N HDUDGCZEOZEFOA-KBIXCLLPSA-N 0.000 description 1
- TWTWUBHEWQPMQW-ZPFDUUQYSA-N Gln-Ile-Arg Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O TWTWUBHEWQPMQW-ZPFDUUQYSA-N 0.000 description 1
- KHGGWBRVRPHFMH-PEFMBERDSA-N Gln-Ile-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CCC(=O)N)N KHGGWBRVRPHFMH-PEFMBERDSA-N 0.000 description 1
- HYPVLWGNBIYTNA-GUBZILKMSA-N Gln-Leu-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O HYPVLWGNBIYTNA-GUBZILKMSA-N 0.000 description 1
- HWEINOMSWQSJDC-SRVKXCTJSA-N Gln-Leu-Arg Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O HWEINOMSWQSJDC-SRVKXCTJSA-N 0.000 description 1
- SHAUZYVSXAMYAZ-JYJNAYRXSA-N Gln-Leu-Phe Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)NC(=O)[C@H](CCC(=O)N)N SHAUZYVSXAMYAZ-JYJNAYRXSA-N 0.000 description 1
- ZBKUIQNCRIYVGH-SDDRHHMPSA-N Gln-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)N)N ZBKUIQNCRIYVGH-SDDRHHMPSA-N 0.000 description 1
- LURQDGKYBFWWJA-MNXVOIDGSA-N Gln-Lys-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(=O)N)N LURQDGKYBFWWJA-MNXVOIDGSA-N 0.000 description 1
- FKXCBKCOSVIGCT-AVGNSLFASA-N Gln-Lys-Leu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O FKXCBKCOSVIGCT-AVGNSLFASA-N 0.000 description 1
- KLKYKPXITJBSNI-CIUDSAMLSA-N Gln-Met-Ala Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C)C(O)=O KLKYKPXITJBSNI-CIUDSAMLSA-N 0.000 description 1
- CULXMOZETKLBDI-XIRDDKMYSA-N Gln-Met-Trp Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CCC(=O)N)N CULXMOZETKLBDI-XIRDDKMYSA-N 0.000 description 1
- GQTNWYFWSUFFRA-KKUMJFAQSA-N Gln-Met-Tyr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O GQTNWYFWSUFFRA-KKUMJFAQSA-N 0.000 description 1
- FQCILXROGNOZON-YUMQZZPRSA-N Gln-Pro-Gly Chemical compound NC(=O)CC[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O FQCILXROGNOZON-YUMQZZPRSA-N 0.000 description 1
- OKARHJKJTKFQBM-ACZMJKKPSA-N Gln-Ser-Asn Chemical compound C(CC(=O)N)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(=O)N)C(=O)O)N OKARHJKJTKFQBM-ACZMJKKPSA-N 0.000 description 1
- JILRMFFFCHUUTJ-ACZMJKKPSA-N Gln-Ser-Ser Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O JILRMFFFCHUUTJ-ACZMJKKPSA-N 0.000 description 1
- DYVMTEWCGAVKSE-HJGDQZAQSA-N Gln-Thr-Arg Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](CCC(=O)N)N)O DYVMTEWCGAVKSE-HJGDQZAQSA-N 0.000 description 1
- STHSGOZLFLFGSS-SUSMZKCASA-N Gln-Thr-Thr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O STHSGOZLFLFGSS-SUSMZKCASA-N 0.000 description 1
- WIMVKDYAKRAUCG-IHRRRGAJSA-N Gln-Tyr-Glu Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CCC(=O)N)N)O WIMVKDYAKRAUCG-IHRRRGAJSA-N 0.000 description 1
- UBRQJXFDVZNYJP-AVGNSLFASA-N Gln-Tyr-Ser Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CCC(=O)N)N)O UBRQJXFDVZNYJP-AVGNSLFASA-N 0.000 description 1
- OACPJRQRAHMQEQ-NHCYSSNCSA-N Gln-Val-Arg Chemical compound NC(=O)CC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O OACPJRQRAHMQEQ-NHCYSSNCSA-N 0.000 description 1
- BBFCMGBMYIAGRS-AUTRQRHGSA-N Gln-Val-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O BBFCMGBMYIAGRS-AUTRQRHGSA-N 0.000 description 1
- OGMQXTXGLDNBSS-FXQIFTODSA-N Glu-Ala-Gln Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(N)=O)C(O)=O OGMQXTXGLDNBSS-FXQIFTODSA-N 0.000 description 1
- HUWSBFYAGXCXKC-CIUDSAMLSA-N Glu-Ala-Met Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCSC)C(O)=O HUWSBFYAGXCXKC-CIUDSAMLSA-N 0.000 description 1
- ATRHMOJQJWPVBQ-DRZSPHRISA-N Glu-Ala-Phe Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O ATRHMOJQJWPVBQ-DRZSPHRISA-N 0.000 description 1
- IYAUFWMUCGBFMQ-CIUDSAMLSA-N Glu-Arg-Cys Chemical compound C(C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)CN=C(N)N IYAUFWMUCGBFMQ-CIUDSAMLSA-N 0.000 description 1
- CGYDXNKRIMJMLV-GUBZILKMSA-N Glu-Arg-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=O CGYDXNKRIMJMLV-GUBZILKMSA-N 0.000 description 1
- KEBACWCLVOXFNC-DCAQKATOSA-N Glu-Arg-Met Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCSC)C(O)=O KEBACWCLVOXFNC-DCAQKATOSA-N 0.000 description 1
- VPKBCVUDBNINAH-GARJFASQSA-N Glu-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCC(=O)O)N)C(=O)O VPKBCVUDBNINAH-GARJFASQSA-N 0.000 description 1
- NKSGKPWXSWBRRX-ACZMJKKPSA-N Glu-Asn-Cys Chemical compound C(CC(=O)O)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CS)C(=O)O)N NKSGKPWXSWBRRX-ACZMJKKPSA-N 0.000 description 1
- YYOBUPFZLKQUAX-FXQIFTODSA-N Glu-Asn-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O YYOBUPFZLKQUAX-FXQIFTODSA-N 0.000 description 1
- ZOXBSICWUDAOHX-GUBZILKMSA-N Glu-Asn-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@@H](N)CCC(O)=O ZOXBSICWUDAOHX-GUBZILKMSA-N 0.000 description 1
- XXCDTYBVGMPIOA-FXQIFTODSA-N Glu-Asp-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O XXCDTYBVGMPIOA-FXQIFTODSA-N 0.000 description 1
- GZWOBWMOMPFPCD-CIUDSAMLSA-N Glu-Asp-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCC(=O)O)N GZWOBWMOMPFPCD-CIUDSAMLSA-N 0.000 description 1
- JRCUFCXYZLPSDZ-ACZMJKKPSA-N Glu-Asp-Ser Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O JRCUFCXYZLPSDZ-ACZMJKKPSA-N 0.000 description 1
- KIMXNQXJJWWVIN-AVGNSLFASA-N Glu-Cys-Tyr Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CCC(=O)O)N)O KIMXNQXJJWWVIN-AVGNSLFASA-N 0.000 description 1
- OXEMJGCAJFFREE-FXQIFTODSA-N Glu-Gln-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O OXEMJGCAJFFREE-FXQIFTODSA-N 0.000 description 1
- GYCPQVFKCPPRQB-GUBZILKMSA-N Glu-Gln-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCC(=O)O)N GYCPQVFKCPPRQB-GUBZILKMSA-N 0.000 description 1
- RFDHKPSHTXZKLL-IHRRRGAJSA-N Glu-Gln-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CCC(=O)O)N RFDHKPSHTXZKLL-IHRRRGAJSA-N 0.000 description 1
- HNVFSTLPVJWIDV-CIUDSAMLSA-N Glu-Glu-Gln Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O HNVFSTLPVJWIDV-CIUDSAMLSA-N 0.000 description 1
- PXXGVUVQWQGGIG-YUMQZZPRSA-N Glu-Gly-Arg Chemical compound OC(=O)CC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N PXXGVUVQWQGGIG-YUMQZZPRSA-N 0.000 description 1
- UHVIQGKBMXEVGN-WDSKDSINSA-N Glu-Gly-Asn Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O UHVIQGKBMXEVGN-WDSKDSINSA-N 0.000 description 1
- OGNJZUXUTPQVBR-BQBZGAKWSA-N Glu-Gly-Glu Chemical compound OC(=O)CC[C@H](N)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O OGNJZUXUTPQVBR-BQBZGAKWSA-N 0.000 description 1
- QIQABBIDHGQXGA-ZPFDUUQYSA-N Glu-Ile-Arg Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O QIQABBIDHGQXGA-ZPFDUUQYSA-N 0.000 description 1
- QXDXIXFSFHUYAX-MNXVOIDGSA-N Glu-Ile-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCC(O)=O QXDXIXFSFHUYAX-MNXVOIDGSA-N 0.000 description 1
- XTZDZAXYPDISRR-MNXVOIDGSA-N Glu-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N XTZDZAXYPDISRR-MNXVOIDGSA-N 0.000 description 1
- GXMXPCXXKVWOSM-KQXIARHKSA-N Glu-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)O)N GXMXPCXXKVWOSM-KQXIARHKSA-N 0.000 description 1
- KRRFFAHEAOCBCQ-SIUGBPQLSA-N Glu-Ile-Tyr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O KRRFFAHEAOCBCQ-SIUGBPQLSA-N 0.000 description 1
- PJBVXVBTTFZPHJ-GUBZILKMSA-N Glu-Leu-Asp Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CCC(=O)O)N PJBVXVBTTFZPHJ-GUBZILKMSA-N 0.000 description 1
- MWMJCGBSIORNCD-AVGNSLFASA-N Glu-Leu-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O MWMJCGBSIORNCD-AVGNSLFASA-N 0.000 description 1
- ZGEJRLJEAMPEDV-SRVKXCTJSA-N Glu-Lys-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(=O)O)N ZGEJRLJEAMPEDV-SRVKXCTJSA-N 0.000 description 1
- QMOSCLNJVKSHHU-YUMQZZPRSA-N Glu-Met-Gly Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCSC)C(=O)NCC(O)=O QMOSCLNJVKSHHU-YUMQZZPRSA-N 0.000 description 1
- HQOGXFLBAKJUMH-CIUDSAMLSA-N Glu-Met-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CCC(=O)O)N HQOGXFLBAKJUMH-CIUDSAMLSA-N 0.000 description 1
- KJBGAZSLZAQDPV-KKUMJFAQSA-N Glu-Phe-Arg Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](CCC(=O)O)N KJBGAZSLZAQDPV-KKUMJFAQSA-N 0.000 description 1
- YTRBQAQSUDSIQE-FHWLQOOXSA-N Glu-Phe-Phe Chemical compound C([C@H](NC(=O)[C@H](CCC(O)=O)N)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=CC=C1 YTRBQAQSUDSIQE-FHWLQOOXSA-N 0.000 description 1
- BPLNJYHNAJVLRT-ACZMJKKPSA-N Glu-Ser-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O BPLNJYHNAJVLRT-ACZMJKKPSA-N 0.000 description 1
- DAHLWSFUXOHMIA-FXQIFTODSA-N Glu-Ser-Gln Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(O)=O DAHLWSFUXOHMIA-FXQIFTODSA-N 0.000 description 1
- GMVCSRBOSIUTFC-FXQIFTODSA-N Glu-Ser-Glu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O GMVCSRBOSIUTFC-FXQIFTODSA-N 0.000 description 1
- SYAYROHMAIHWFB-KBIXCLLPSA-N Glu-Ser-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O SYAYROHMAIHWFB-KBIXCLLPSA-N 0.000 description 1
- WXONSNSSBYQGNN-AVGNSLFASA-N Glu-Ser-Tyr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O WXONSNSSBYQGNN-AVGNSLFASA-N 0.000 description 1
- HZISRJBYZAODRV-XQXXSGGOSA-N Glu-Thr-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O HZISRJBYZAODRV-XQXXSGGOSA-N 0.000 description 1
- BDISFWMLMNBTGP-NUMRIWBASA-N Glu-Thr-Asp Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(O)=O BDISFWMLMNBTGP-NUMRIWBASA-N 0.000 description 1
- DTLLNDVORUEOTM-WDCWCFNPSA-N Glu-Thr-Lys Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(O)=O DTLLNDVORUEOTM-WDCWCFNPSA-N 0.000 description 1
- CAQXJMUDOLSBPF-SUSMZKCASA-N Glu-Thr-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O CAQXJMUDOLSBPF-SUSMZKCASA-N 0.000 description 1
- UQULNJAARAXSPO-ZCWPNWOLSA-N Glu-Thr-Thr-Tyr Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 UQULNJAARAXSPO-ZCWPNWOLSA-N 0.000 description 1
- QVXWAFZDWRLXTI-NWLDYVSISA-N Glu-Thr-Trp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CCC(=O)O)N)O QVXWAFZDWRLXTI-NWLDYVSISA-N 0.000 description 1
- BPCLDCNZBUYGOD-BPUTZDHNSA-N Glu-Trp-Glu Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCC(O)=O)N)C(=O)N[C@@H](CCC(O)=O)C(O)=O)=CNC2=C1 BPCLDCNZBUYGOD-BPUTZDHNSA-N 0.000 description 1
- KIEICAOUSNYOLM-NRPADANISA-N Glu-Val-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O KIEICAOUSNYOLM-NRPADANISA-N 0.000 description 1
- ZALGPUWUVHOGAE-GVXVVHGQSA-N Glu-Val-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCC(=O)O)N ZALGPUWUVHOGAE-GVXVVHGQSA-N 0.000 description 1
- FGGKGJHCVMYGCD-UKJIMTQDSA-N Glu-Val-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O FGGKGJHCVMYGCD-UKJIMTQDSA-N 0.000 description 1
- VIPDPMHGICREIS-GVXVVHGQSA-N Glu-Val-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O VIPDPMHGICREIS-GVXVVHGQSA-N 0.000 description 1
- FVGOGEGGQLNZGH-DZKIICNBSA-N Glu-Val-Phe Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 FVGOGEGGQLNZGH-DZKIICNBSA-N 0.000 description 1
- 229920001503 Glucan Polymers 0.000 description 1
- 229920002581 Glucomannan Polymers 0.000 description 1
- 102100041003 Glutamate carboxypeptidase 2 Human genes 0.000 description 1
- PUUYVMYCMIWHFE-BQBZGAKWSA-N Gly-Ala-Arg Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N PUUYVMYCMIWHFE-BQBZGAKWSA-N 0.000 description 1
- GQGAFTPXAPKSCF-WHFBIAKZSA-N Gly-Ala-Cys Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CS)C(=O)O GQGAFTPXAPKSCF-WHFBIAKZSA-N 0.000 description 1
- MFVQGXGQRIXBPK-WDSKDSINSA-N Gly-Ala-Glu Chemical compound NCC(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O MFVQGXGQRIXBPK-WDSKDSINSA-N 0.000 description 1
- LERGJIVJIIODPZ-ZANVPECISA-N Gly-Ala-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](NC(=O)CN)C)C(O)=O)=CNC2=C1 LERGJIVJIIODPZ-ZANVPECISA-N 0.000 description 1
- JRDYDYXZKFNNRQ-XPUUQOCRSA-N Gly-Ala-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)CN JRDYDYXZKFNNRQ-XPUUQOCRSA-N 0.000 description 1
- RJIVPOXLQFJRTG-LURJTMIESA-N Gly-Arg-Gly Chemical compound OC(=O)CNC(=O)[C@@H](NC(=O)CN)CCCN=C(N)N RJIVPOXLQFJRTG-LURJTMIESA-N 0.000 description 1
- JPXNYFOHTHSREU-UWVGGRQHSA-N Gly-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)CN JPXNYFOHTHSREU-UWVGGRQHSA-N 0.000 description 1
- OCQUNKSFDYDXBG-QXEWZRGKSA-N Gly-Arg-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCN=C(N)N OCQUNKSFDYDXBG-QXEWZRGKSA-N 0.000 description 1
- OVSKVOOUFAKODB-UWVGGRQHSA-N Gly-Arg-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CCCN=C(N)N OVSKVOOUFAKODB-UWVGGRQHSA-N 0.000 description 1
- VXKCPBPQEKKERH-IUCAKERBSA-N Gly-Arg-Pro Chemical compound NC(N)=NCCC[C@H](NC(=O)CN)C(=O)N1CCC[C@H]1C(O)=O VXKCPBPQEKKERH-IUCAKERBSA-N 0.000 description 1
- KKBWDNZXYLGJEY-UHFFFAOYSA-N Gly-Arg-Pro Natural products NCC(=O)NC(CCNC(=N)N)C(=O)N1CCCC1C(=O)O KKBWDNZXYLGJEY-UHFFFAOYSA-N 0.000 description 1
- GWCRIHNSVMOBEQ-BQBZGAKWSA-N Gly-Arg-Ser Chemical compound [H]NCC(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(O)=O GWCRIHNSVMOBEQ-BQBZGAKWSA-N 0.000 description 1
- UXJHNZODTMHWRD-WHFBIAKZSA-N Gly-Asn-Ala Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(O)=O UXJHNZODTMHWRD-WHFBIAKZSA-N 0.000 description 1
- CIMULJZTTOBOPN-WHFBIAKZSA-N Gly-Asn-Asn Chemical compound NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O CIMULJZTTOBOPN-WHFBIAKZSA-N 0.000 description 1
- AIJAPFVDBFYNKN-WHFBIAKZSA-N Gly-Asn-Asp Chemical compound C([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)CN)C(=O)N AIJAPFVDBFYNKN-WHFBIAKZSA-N 0.000 description 1
- NZAFOTBEULLEQB-WDSKDSINSA-N Gly-Asn-Glu Chemical compound C(CC(=O)O)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)CN NZAFOTBEULLEQB-WDSKDSINSA-N 0.000 description 1
- JVWPPCWUDRJGAE-YUMQZZPRSA-N Gly-Asn-Leu Chemical compound [H]NCC(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(C)C)C(O)=O JVWPPCWUDRJGAE-YUMQZZPRSA-N 0.000 description 1
- FMNHBTKMRFVGRO-FOHZUACHSA-N Gly-Asn-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](CC(N)=O)NC(=O)CN FMNHBTKMRFVGRO-FOHZUACHSA-N 0.000 description 1
- XQHSBNVACKQWAV-WHFBIAKZSA-N Gly-Asp-Asn Chemical compound [H]NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O XQHSBNVACKQWAV-WHFBIAKZSA-N 0.000 description 1
- FUTAPPOITCCWTH-WHFBIAKZSA-N Gly-Asp-Asp Chemical compound [H]NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O FUTAPPOITCCWTH-WHFBIAKZSA-N 0.000 description 1
- ZRZILYKEJBMFHY-BQBZGAKWSA-N Gly-Asp-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)CN ZRZILYKEJBMFHY-BQBZGAKWSA-N 0.000 description 1
- CEXINUGNTZFNRY-BYPYZUCNSA-N Gly-Cys-Gly Chemical compound [NH3+]CC(=O)N[C@@H](CS)C(=O)NCC([O-])=O CEXINUGNTZFNRY-BYPYZUCNSA-N 0.000 description 1
- GHHAMXVMWXMGSV-STQMWFEESA-N Gly-Cys-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CS)NC(=O)CN)C(O)=O)=CNC2=C1 GHHAMXVMWXMGSV-STQMWFEESA-N 0.000 description 1
- BYYNJRSNDARRBX-YFKPBYRVSA-N Gly-Gln-Gly Chemical compound NCC(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O BYYNJRSNDARRBX-YFKPBYRVSA-N 0.000 description 1
- NPSWCZIRBAYNSB-JHEQGTHGSA-N Gly-Gln-Thr Chemical compound [H]NCC(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NPSWCZIRBAYNSB-JHEQGTHGSA-N 0.000 description 1
- CCQOOWAONKGYKQ-BYPYZUCNSA-N Gly-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)CN CCQOOWAONKGYKQ-BYPYZUCNSA-N 0.000 description 1
- GDOZQTNZPCUARW-YFKPBYRVSA-N Gly-Gly-Glu Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CCC(O)=O GDOZQTNZPCUARW-YFKPBYRVSA-N 0.000 description 1
- BUEFQXUHTUZXHR-LURJTMIESA-N Gly-Gly-Pro zwitterion Chemical compound NCC(=O)NCC(=O)N1CCC[C@H]1C(O)=O BUEFQXUHTUZXHR-LURJTMIESA-N 0.000 description 1
- FQKKPCWTZZEDIC-XPUUQOCRSA-N Gly-His-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H](NC(=O)CN)CC1=CN=CN1 FQKKPCWTZZEDIC-XPUUQOCRSA-N 0.000 description 1
- HHSOPSCKAZKQHQ-PEXQALLHSA-N Gly-His-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)CN HHSOPSCKAZKQHQ-PEXQALLHSA-N 0.000 description 1
- SWQALSGKVLYKDT-UHFFFAOYSA-N Gly-Ile-Ala Natural products NCC(=O)NC(C(C)CC)C(=O)NC(C)C(O)=O SWQALSGKVLYKDT-UHFFFAOYSA-N 0.000 description 1
- SXJHOPPTOJACOA-QXEWZRGKSA-N Gly-Ile-Arg Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CCCN=C(N)N SXJHOPPTOJACOA-QXEWZRGKSA-N 0.000 description 1
- VIIBEIQMLJEUJG-LAEOZQHASA-N Gly-Ile-Gln Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(N)=O)C(O)=O VIIBEIQMLJEUJG-LAEOZQHASA-N 0.000 description 1
- HKSNHPVETYYJBK-LAEOZQHASA-N Gly-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)CN HKSNHPVETYYJBK-LAEOZQHASA-N 0.000 description 1
- ZOTGXWMKUFSKEU-QXEWZRGKSA-N Gly-Ile-Met Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCSC)C(O)=O ZOTGXWMKUFSKEU-QXEWZRGKSA-N 0.000 description 1
- BHPQOIPBLYJNAW-NGZCFLSTSA-N Gly-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)CN BHPQOIPBLYJNAW-NGZCFLSTSA-N 0.000 description 1
- SCWYHUQOOFRVHP-MBLNEYKQSA-N Gly-Ile-Thr Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SCWYHUQOOFRVHP-MBLNEYKQSA-N 0.000 description 1
- UYPPAMNTTMJHJW-KCTSRDHCSA-N Gly-Ile-Trp Chemical compound [H]NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O UYPPAMNTTMJHJW-KCTSRDHCSA-N 0.000 description 1
- XVYKMNXXJXQKME-XEGUGMAKSA-N Gly-Ile-Tyr Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 XVYKMNXXJXQKME-XEGUGMAKSA-N 0.000 description 1
- YTSVAIMKVLZUDU-YUMQZZPRSA-N Gly-Leu-Asp Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O YTSVAIMKVLZUDU-YUMQZZPRSA-N 0.000 description 1
- YIFUFYZELCMPJP-YUMQZZPRSA-N Gly-Leu-Cys Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CS)C(O)=O YIFUFYZELCMPJP-YUMQZZPRSA-N 0.000 description 1
- ULZCYBYDTUMHNF-IUCAKERBSA-N Gly-Leu-Glu Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O ULZCYBYDTUMHNF-IUCAKERBSA-N 0.000 description 1
- UUYBFNKHOCJCHT-VHSXEESVSA-N Gly-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)CN UUYBFNKHOCJCHT-VHSXEESVSA-N 0.000 description 1
- NNCSJUBVFBDDLC-YUMQZZPRSA-N Gly-Leu-Ser Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O NNCSJUBVFBDDLC-YUMQZZPRSA-N 0.000 description 1
- LHYJCVCQPWRMKZ-WEDXCCLWSA-N Gly-Leu-Thr Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LHYJCVCQPWRMKZ-WEDXCCLWSA-N 0.000 description 1
- AFWYPMDMDYCKMD-KBPBESRZSA-N Gly-Leu-Tyr Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 AFWYPMDMDYCKMD-KBPBESRZSA-N 0.000 description 1
- MHXKHKWHPNETGG-QWRGUYRKSA-N Gly-Lys-Leu Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O MHXKHKWHPNETGG-QWRGUYRKSA-N 0.000 description 1
- MHZXESQPPXOING-KBPBESRZSA-N Gly-Lys-Phe Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O MHZXESQPPXOING-KBPBESRZSA-N 0.000 description 1
- WDEHMRNSGHVNOH-VHSXEESVSA-N Gly-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)CN)C(=O)O WDEHMRNSGHVNOH-VHSXEESVSA-N 0.000 description 1
- QGDOOCIPHSSADO-STQMWFEESA-N Gly-Met-Phe Chemical compound [H]NCC(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O QGDOOCIPHSSADO-STQMWFEESA-N 0.000 description 1
- UWQDKRIZSROAKS-FJXKBIBVSA-N Gly-Met-Thr Chemical compound [H]NCC(=O)N[C@@H](CCSC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O UWQDKRIZSROAKS-FJXKBIBVSA-N 0.000 description 1
- NSVOVKWEKGEOQB-LURJTMIESA-N Gly-Pro-Gly Chemical compound NCC(=O)N1CCC[C@H]1C(=O)NCC(O)=O NSVOVKWEKGEOQB-LURJTMIESA-N 0.000 description 1
- HFPVRZWORNJRRC-UWVGGRQHSA-N Gly-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)CN HFPVRZWORNJRRC-UWVGGRQHSA-N 0.000 description 1
- ABPRMMYHROQBLY-NKWVEPMBSA-N Gly-Ser-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CO)NC(=O)CN)C(=O)O ABPRMMYHROQBLY-NKWVEPMBSA-N 0.000 description 1
- ZLCLYFGMKFCDCN-XPUUQOCRSA-N Gly-Ser-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CO)NC(=O)CN)C(O)=O ZLCLYFGMKFCDCN-XPUUQOCRSA-N 0.000 description 1
- CUVBTVWFVIIDOC-YEPSODPASA-N Gly-Thr-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)O)NC(=O)CN CUVBTVWFVIIDOC-YEPSODPASA-N 0.000 description 1
- PYFIQROSWQERAS-LBPRGKRZSA-N Gly-Trp-Gly Chemical compound C1=CC=C2C(C[C@H](NC(=O)CN)C(=O)NCC(O)=O)=CNC2=C1 PYFIQROSWQERAS-LBPRGKRZSA-N 0.000 description 1
- SFOXOSKVTLDEDM-HOTGVXAUSA-N Gly-Trp-Leu Chemical compound C1=CC=C2C(C[C@@H](C(=O)N[C@@H](CC(C)C)C(O)=O)NC(=O)CN)=CNC2=C1 SFOXOSKVTLDEDM-HOTGVXAUSA-N 0.000 description 1
- UMRIXLHPZZIOML-OALUTQOASA-N Gly-Trp-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CC2=CNC3=CC=CC=C32)NC(=O)CN UMRIXLHPZZIOML-OALUTQOASA-N 0.000 description 1
- MREVELMMFOLESM-HOCLYGCPSA-N Gly-Trp-Val Chemical compound [H]NCC(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](C(C)C)C(O)=O MREVELMMFOLESM-HOCLYGCPSA-N 0.000 description 1
- DNAZKGFYFRGZIH-QWRGUYRKSA-N Gly-Tyr-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)CN)CC1=CC=C(O)C=C1 DNAZKGFYFRGZIH-QWRGUYRKSA-N 0.000 description 1
- YDIDLLVFCYSXNY-RCOVLWMOSA-N Gly-Val-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)CN YDIDLLVFCYSXNY-RCOVLWMOSA-N 0.000 description 1
- BAYQNCWLXIDLHX-ONGXEEELSA-N Gly-Val-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)CN BAYQNCWLXIDLHX-ONGXEEELSA-N 0.000 description 1
- COZMNNJEGNPDED-HOCLYGCPSA-N Gly-Val-Trp Chemical compound [H]NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O COZMNNJEGNPDED-HOCLYGCPSA-N 0.000 description 1
- IZVICCORZOSGPT-JSGCOSHPSA-N Gly-Val-Tyr Chemical compound [H]NCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O IZVICCORZOSGPT-JSGCOSHPSA-N 0.000 description 1
- 239000004471 Glycine Substances 0.000 description 1
- 229930186217 Glycolipid Natural products 0.000 description 1
- 102000009465 Growth Factor Receptors Human genes 0.000 description 1
- 108010009202 Growth Factor Receptors Proteins 0.000 description 1
- RVKIPWVMZANZLI-UHFFFAOYSA-N H-Lys-Trp-OH Natural products C1=CC=C2C(CC(NC(=O)C(N)CCCCN)C(O)=O)=CNC2=C1 RVKIPWVMZANZLI-UHFFFAOYSA-N 0.000 description 1
- 208000037357 HIV infectious disease Diseases 0.000 description 1
- 241000589989 Helicobacter Species 0.000 description 1
- 241000590002 Helicobacter pylori Species 0.000 description 1
- 241000711557 Hepacivirus Species 0.000 description 1
- 241000700739 Hepadnaviridae Species 0.000 description 1
- 241000700721 Hepatitis B virus Species 0.000 description 1
- 108010073141 Hepatitis C virus glycoprotein E2 Proteins 0.000 description 1
- 208000037319 Hepatitis infectious Diseases 0.000 description 1
- 241000700586 Herpesviridae Species 0.000 description 1
- UCDWNBFOZCZSNV-AVGNSLFASA-N His-Arg-Met Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCSC)C(O)=O UCDWNBFOZCZSNV-AVGNSLFASA-N 0.000 description 1
- CJGDTAHEMXLRMB-ULQDDVLXSA-N His-Arg-Phe Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O CJGDTAHEMXLRMB-ULQDDVLXSA-N 0.000 description 1
- NOQPTNXSGNPJNS-YUMQZZPRSA-N His-Asn-Gly Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O NOQPTNXSGNPJNS-YUMQZZPRSA-N 0.000 description 1
- MDBYBTWRMOAJAY-NHCYSSNCSA-N His-Asn-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC1=CN=CN1)N MDBYBTWRMOAJAY-NHCYSSNCSA-N 0.000 description 1
- LSQHWKPPOFDHHZ-YUMQZZPRSA-N His-Asp-Gly Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)NCC(=O)O)N LSQHWKPPOFDHHZ-YUMQZZPRSA-N 0.000 description 1
- RBOOOLVEKJHUNA-CIUDSAMLSA-N His-Cys-Asn Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(O)=O RBOOOLVEKJHUNA-CIUDSAMLSA-N 0.000 description 1
- WGHJXSONOOTTCZ-JYJNAYRXSA-N His-Glu-Tyr Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O WGHJXSONOOTTCZ-JYJNAYRXSA-N 0.000 description 1
- PGTISAJTWZPFGN-PEXQALLHSA-N His-Gly-Ile Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)NCC(=O)N[C@@H]([C@@H](C)CC)C(O)=O PGTISAJTWZPFGN-PEXQALLHSA-N 0.000 description 1
- RGPWUJOMKFYFSR-QWRGUYRKSA-N His-Gly-Leu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)NCC(=O)N[C@@H](CC(C)C)C(O)=O RGPWUJOMKFYFSR-QWRGUYRKSA-N 0.000 description 1
- NTXIJPDAHXSHNL-ONGXEEELSA-N His-Gly-Val Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O NTXIJPDAHXSHNL-ONGXEEELSA-N 0.000 description 1
- 108010093488 His-His-His-His-His-His Proteins 0.000 description 1
- VTZYMXGGXOFBMX-DJFWLOJKSA-N His-Ile-Asp Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O VTZYMXGGXOFBMX-DJFWLOJKSA-N 0.000 description 1
- SKOKHBGDXGTDDP-MELADBBJSA-N His-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CN=CN2)N SKOKHBGDXGTDDP-MELADBBJSA-N 0.000 description 1
- FHGVHXCQMJWQPK-SRVKXCTJSA-N His-Lys-Asn Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(O)=O FHGVHXCQMJWQPK-SRVKXCTJSA-N 0.000 description 1
- SGLXGEDPYJPGIQ-ACRUOGEOSA-N His-Phe-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)O)NC(=O)[C@H](CC3=CN=CN3)N SGLXGEDPYJPGIQ-ACRUOGEOSA-N 0.000 description 1
- ULRFSEJGSHYLQI-YESZJQIVSA-N His-Phe-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=CC=C2)NC(=O)[C@H](CC3=CN=CN3)N)C(=O)O ULRFSEJGSHYLQI-YESZJQIVSA-N 0.000 description 1
- PYNPBMCLAKTHJL-SRVKXCTJSA-N His-Pro-Glu Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O PYNPBMCLAKTHJL-SRVKXCTJSA-N 0.000 description 1
- WCHONUZTYDQMBY-PYJNHQTQSA-N His-Pro-Ile Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WCHONUZTYDQMBY-PYJNHQTQSA-N 0.000 description 1
- OWYIDJCNRWRSJY-QTKMDUPCSA-N His-Pro-Thr Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O OWYIDJCNRWRSJY-QTKMDUPCSA-N 0.000 description 1
- KAXZXLSXFWSNNZ-XVYDVKMFSA-N His-Ser-Ala Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O KAXZXLSXFWSNNZ-XVYDVKMFSA-N 0.000 description 1
- FCPSGEVYIVXPPO-QTKMDUPCSA-N His-Thr-Arg Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O FCPSGEVYIVXPPO-QTKMDUPCSA-N 0.000 description 1
- MDOBWSFNSNPENN-PMVVWTBXSA-N His-Thr-Gly Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O MDOBWSFNSNPENN-PMVVWTBXSA-N 0.000 description 1
- NBWATNYAUVSAEQ-ZEILLAHLSA-N His-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CC1=CN=CN1)N)O NBWATNYAUVSAEQ-ZEILLAHLSA-N 0.000 description 1
- VXZZUXWAOMWWJH-QTKMDUPCSA-N His-Thr-Val Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O VXZZUXWAOMWWJH-QTKMDUPCSA-N 0.000 description 1
- YERBCFWVWITTEJ-NAZCDGGXSA-N His-Trp-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CC3=CN=CN3)N)O YERBCFWVWITTEJ-NAZCDGGXSA-N 0.000 description 1
- WSWAUVHXQREQQG-JYJNAYRXSA-N His-Tyr-Gln Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O WSWAUVHXQREQQG-JYJNAYRXSA-N 0.000 description 1
- XGBVLRJLHUVCNK-DCAQKATOSA-N His-Val-Ser Chemical compound [H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O XGBVLRJLHUVCNK-DCAQKATOSA-N 0.000 description 1
- CMPHFUWXKBPNRS-WDSOQIARSA-N His-Val-Trp Chemical compound C([C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)C1=CNC=N1 CMPHFUWXKBPNRS-WDSOQIARSA-N 0.000 description 1
- 108091006054 His-tagged proteins Proteins 0.000 description 1
- 241000228402 Histoplasma Species 0.000 description 1
- 241000228404 Histoplasma capsulatum Species 0.000 description 1
- 101000773083 Homo sapiens 5,6-dihydroxyindole-2-carboxylic acid oxidase Proteins 0.000 description 1
- 101000780443 Homo sapiens Alcohol dehydrogenase 1A Proteins 0.000 description 1
- 101000980825 Homo sapiens B-lymphocyte antigen CD19 Proteins 0.000 description 1
- 101000608772 Homo sapiens Galectin-7 Proteins 0.000 description 1
- 101000892862 Homo sapiens Glutamate carboxypeptidase 2 Proteins 0.000 description 1
- 101000798109 Homo sapiens Melanotransferrin Proteins 0.000 description 1
- 101000934338 Homo sapiens Myeloid cell surface antigen CD33 Proteins 0.000 description 1
- 101001012157 Homo sapiens Receptor tyrosine-protein kinase erbB-2 Proteins 0.000 description 1
- 241000701024 Human betaherpesvirus 5 Species 0.000 description 1
- 241000711920 Human orthopneumovirus Species 0.000 description 1
- 241000701806 Human papillomavirus Species 0.000 description 1
- 241000702617 Human parvovirus B19 Species 0.000 description 1
- 206010020751 Hypersensitivity Diseases 0.000 description 1
- NKVZTQVGUNLLQW-JBDRJPRFSA-N Ile-Ala-Ala Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(=O)O)N NKVZTQVGUNLLQW-JBDRJPRFSA-N 0.000 description 1
- JXUGDUWBMKIJDC-NAKRPEOUSA-N Ile-Ala-Arg Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O JXUGDUWBMKIJDC-NAKRPEOUSA-N 0.000 description 1
- HERITAGIPLEJMT-GVARAGBVSA-N Ile-Ala-Tyr Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 HERITAGIPLEJMT-GVARAGBVSA-N 0.000 description 1
- QLRMMMQNCWBNPQ-QXEWZRGKSA-N Ile-Arg-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)NCC(=O)O)N QLRMMMQNCWBNPQ-QXEWZRGKSA-N 0.000 description 1
- WECYRWOMWSCWNX-XUXIUFHCSA-N Ile-Arg-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CC(C)C)C(O)=O WECYRWOMWSCWNX-XUXIUFHCSA-N 0.000 description 1
- UNDGQKWQNSTPPW-CYDGBPFRSA-N Ile-Arg-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCSC)C(=O)O)N UNDGQKWQNSTPPW-CYDGBPFRSA-N 0.000 description 1
- DMHGKBGOUAJRHU-RVMXOQNASA-N Ile-Arg-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N1CCC[C@@H]1C(=O)O)N DMHGKBGOUAJRHU-RVMXOQNASA-N 0.000 description 1
- NULSANWBUWLTKN-NAKRPEOUSA-N Ile-Arg-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CO)C(=O)O)N NULSANWBUWLTKN-NAKRPEOUSA-N 0.000 description 1
- GYAFMRQGWHXMII-IUKAMOBKSA-N Ile-Asp-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N GYAFMRQGWHXMII-IUKAMOBKSA-N 0.000 description 1
- AQTWDZDISVGCAC-CFMVVWHZSA-N Ile-Asp-Tyr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N AQTWDZDISVGCAC-CFMVVWHZSA-N 0.000 description 1
- LDRALPZEVHVXEK-KBIXCLLPSA-N Ile-Cys-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N LDRALPZEVHVXEK-KBIXCLLPSA-N 0.000 description 1
- WZDCVAWMBUNDDY-KBIXCLLPSA-N Ile-Glu-Ala Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](C)C(=O)O)N WZDCVAWMBUNDDY-KBIXCLLPSA-N 0.000 description 1
- PHIXPNQDGGILMP-YVNDNENWSA-N Ile-Glu-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N PHIXPNQDGGILMP-YVNDNENWSA-N 0.000 description 1
- XLCZWMJPVGRWHJ-KQXIARHKSA-N Ile-Glu-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N XLCZWMJPVGRWHJ-KQXIARHKSA-N 0.000 description 1
- NZOCIWKZUVUNDW-ZKWXMUAHSA-N Ile-Gly-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O NZOCIWKZUVUNDW-ZKWXMUAHSA-N 0.000 description 1
- CDGLBYSAZFIIJO-RCOVLWMOSA-N Ile-Gly-Gly Chemical compound CC[C@H](C)[C@H]([NH3+])C(=O)NCC(=O)NCC([O-])=O CDGLBYSAZFIIJO-RCOVLWMOSA-N 0.000 description 1
- VNDQNDYEPSXHLU-JUKXBJQTSA-N Ile-His-Tyr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)O)N VNDQNDYEPSXHLU-JUKXBJQTSA-N 0.000 description 1
- BBQABUDWDUKJMB-LZXPERKUSA-N Ile-Ile-Ile Chemical compound CC[C@H](C)[C@H]([NH3+])C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C([O-])=O BBQABUDWDUKJMB-LZXPERKUSA-N 0.000 description 1
- CSQNHSGHAPRGPQ-YTFOTSKYSA-N Ile-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCCN)C(=O)O)N CSQNHSGHAPRGPQ-YTFOTSKYSA-N 0.000 description 1
- QZZIBQZLWBOOJH-PEDHHIEDSA-N Ile-Ile-Val Chemical compound N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(=O)O QZZIBQZLWBOOJH-PEDHHIEDSA-N 0.000 description 1
- TWYOYAKMLHWMOJ-ZPFDUUQYSA-N Ile-Leu-Asn Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O TWYOYAKMLHWMOJ-ZPFDUUQYSA-N 0.000 description 1
- FZWVCYCYWCLQDH-NHCYSSNCSA-N Ile-Leu-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)NCC(=O)O)N FZWVCYCYWCLQDH-NHCYSSNCSA-N 0.000 description 1
- HPCFRQWLTRDGHT-AJNGGQMLSA-N Ile-Leu-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O HPCFRQWLTRDGHT-AJNGGQMLSA-N 0.000 description 1
- OVDKXUDMKXAZIV-ZPFDUUQYSA-N Ile-Lys-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(=O)N)C(=O)O)N OVDKXUDMKXAZIV-ZPFDUUQYSA-N 0.000 description 1
- PARSHQDZROHERM-NHCYSSNCSA-N Ile-Lys-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)NCC(=O)O)N PARSHQDZROHERM-NHCYSSNCSA-N 0.000 description 1
- CEPIAEUVRKGPGP-DSYPUSFNSA-N Ile-Lys-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)[C@@H](C)CC)C(O)=O)=CNC2=C1 CEPIAEUVRKGPGP-DSYPUSFNSA-N 0.000 description 1
- DNKDIDZHXZAGRY-HJWJTTGWSA-N Ile-Met-Phe Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N DNKDIDZHXZAGRY-HJWJTTGWSA-N 0.000 description 1
- NPAYJTAXWXJKLO-NAKRPEOUSA-N Ile-Met-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CO)C(=O)O)N NPAYJTAXWXJKLO-NAKRPEOUSA-N 0.000 description 1
- BKPPWVSPSIUXHZ-OSUNSFLBSA-N Ile-Met-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N BKPPWVSPSIUXHZ-OSUNSFLBSA-N 0.000 description 1
- XLXPYSDGMXTTNQ-DKIMLUQUSA-N Ile-Phe-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](Cc1ccccc1)C(=O)N[C@@H](CC(C)C)C(O)=O XLXPYSDGMXTTNQ-DKIMLUQUSA-N 0.000 description 1
- XLXPYSDGMXTTNQ-UHFFFAOYSA-N Ile-Phe-Leu Natural products CCC(C)C(N)C(=O)NC(C(=O)NC(CC(C)C)C(O)=O)CC1=CC=CC=C1 XLXPYSDGMXTTNQ-UHFFFAOYSA-N 0.000 description 1
- LRAUKBMYHHNADU-DKIMLUQUSA-N Ile-Phe-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)[C@@H](C)CC)CC1=CC=CC=C1 LRAUKBMYHHNADU-DKIMLUQUSA-N 0.000 description 1
- BATWGBRIZANGPN-ZPFDUUQYSA-N Ile-Pro-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(=O)N)C(=O)O)N BATWGBRIZANGPN-ZPFDUUQYSA-N 0.000 description 1
- NLZVTPYXYXMCIP-XUXIUFHCSA-N Ile-Pro-Lys Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(O)=O NLZVTPYXYXMCIP-XUXIUFHCSA-N 0.000 description 1
- CAHCWMVNBZJVAW-NAKRPEOUSA-N Ile-Pro-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)O)N CAHCWMVNBZJVAW-NAKRPEOUSA-N 0.000 description 1
- JHNJNTMTZHEDLJ-NAKRPEOUSA-N Ile-Ser-Arg Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O JHNJNTMTZHEDLJ-NAKRPEOUSA-N 0.000 description 1
- PELCGFMHLZXWBQ-BJDJZHNGSA-N Ile-Ser-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)O)N PELCGFMHLZXWBQ-BJDJZHNGSA-N 0.000 description 1
- ZDNNDIJTUHQCAM-MXAVVETBSA-N Ile-Ser-Phe Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N ZDNNDIJTUHQCAM-MXAVVETBSA-N 0.000 description 1
- JDCQDJVYUXNCGF-SPOWBLRKSA-N Ile-Ser-Trp Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N JDCQDJVYUXNCGF-SPOWBLRKSA-N 0.000 description 1
- CNMOKANDJMLAIF-CIQUZCHMSA-N Ile-Thr-Ala Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O CNMOKANDJMLAIF-CIQUZCHMSA-N 0.000 description 1
- YBKKLDBBPFIXBQ-MBLNEYKQSA-N Ile-Thr-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)O)N YBKKLDBBPFIXBQ-MBLNEYKQSA-N 0.000 description 1
- NURNJECQNNCRBK-FLBSBUHZSA-N Ile-Thr-Thr Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NURNJECQNNCRBK-FLBSBUHZSA-N 0.000 description 1
- DGTOKVBDZXJHNZ-WZLNRYEVSA-N Ile-Thr-Tyr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N DGTOKVBDZXJHNZ-WZLNRYEVSA-N 0.000 description 1
- WKSHBPRUIRGWRZ-KCTSRDHCSA-N Ile-Trp-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)NCC(=O)O)N WKSHBPRUIRGWRZ-KCTSRDHCSA-N 0.000 description 1
- HQLSBZFLOUHQJK-STECZYCISA-N Ile-Tyr-Arg Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N HQLSBZFLOUHQJK-STECZYCISA-N 0.000 description 1
- GNXGAVNTVNOCLL-SIUGBPQLSA-N Ile-Tyr-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N GNXGAVNTVNOCLL-SIUGBPQLSA-N 0.000 description 1
- PRTZQMBYUZFSFA-XEGUGMAKSA-N Ile-Tyr-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)NCC(=O)O)N PRTZQMBYUZFSFA-XEGUGMAKSA-N 0.000 description 1
- JSLIXOUMAOUGBN-JUKXBJQTSA-N Ile-Tyr-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N JSLIXOUMAOUGBN-JUKXBJQTSA-N 0.000 description 1
- NSPNUMNLZNOPAQ-SJWGOKEGSA-N Ile-Tyr-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N2CCC[C@@H]2C(=O)O)N NSPNUMNLZNOPAQ-SJWGOKEGSA-N 0.000 description 1
- NGKPIPCGMLWHBX-WZLNRYEVSA-N Ile-Tyr-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N NGKPIPCGMLWHBX-WZLNRYEVSA-N 0.000 description 1
- AUIYHFRUOOKTGX-UKJIMTQDSA-N Ile-Val-Gln Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N AUIYHFRUOOKTGX-UKJIMTQDSA-N 0.000 description 1
- ZSESFIFAYQEKRD-CYDGBPFRSA-N Ile-Val-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCSC)C(=O)O)N ZSESFIFAYQEKRD-CYDGBPFRSA-N 0.000 description 1
- WIYDLTIBHZSPKY-HJWJTTGWSA-N Ile-Val-Phe Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 WIYDLTIBHZSPKY-HJWJTTGWSA-N 0.000 description 1
- QSXSHZIRKTUXNG-STECZYCISA-N Ile-Val-Tyr Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 QSXSHZIRKTUXNG-STECZYCISA-N 0.000 description 1
- 206010061598 Immunodeficiency Diseases 0.000 description 1
- 206010062016 Immunosuppression Diseases 0.000 description 1
- 101710125507 Integrase/recombinase Proteins 0.000 description 1
- 102000003996 Interferon-beta Human genes 0.000 description 1
- 108090000467 Interferon-beta Proteins 0.000 description 1
- 102000014150 Interferons Human genes 0.000 description 1
- 108010050904 Interferons Proteins 0.000 description 1
- 102000000589 Interleukin-1 Human genes 0.000 description 1
- 108010002352 Interleukin-1 Proteins 0.000 description 1
- 102000004388 Interleukin-4 Human genes 0.000 description 1
- 108090000978 Interleukin-4 Proteins 0.000 description 1
- 241000567229 Isospora Species 0.000 description 1
- 244000285963 Kluyveromyces fragilis Species 0.000 description 1
- 235000014663 Kluyveromyces fragilis Nutrition 0.000 description 1
- 241001138401 Kluyveromyces lactis Species 0.000 description 1
- HGCNKOLVKRAVHD-UHFFFAOYSA-N L-Met-L-Phe Natural products CSCCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 HGCNKOLVKRAVHD-UHFFFAOYSA-N 0.000 description 1
- SITWEMZOJNKJCH-UHFFFAOYSA-N L-alanine-L-arginine Natural products CC(N)C(=O)NC(C(O)=O)CCCNC(N)=N SITWEMZOJNKJCH-UHFFFAOYSA-N 0.000 description 1
- UGTHTQWIQKEDEH-BQBZGAKWSA-N L-alanyl-L-prolylglycine zwitterion Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O UGTHTQWIQKEDEH-BQBZGAKWSA-N 0.000 description 1
- LHSGPCFBGJHPCY-UHFFFAOYSA-N L-leucine-L-tyrosine Natural products CC(C)CC(N)C(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 LHSGPCFBGJHPCY-UHFFFAOYSA-N 0.000 description 1
- KFKWRHQBZQICHA-STQMWFEESA-N L-leucyl-L-phenylalanine Natural products CC(C)C[C@H](N)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 KFKWRHQBZQICHA-STQMWFEESA-N 0.000 description 1
- QIVBCDIJIAJPQS-VIFPVBQESA-N L-tryptophane Chemical compound C1=CC=C2C(C[C@H](N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-VIFPVBQESA-N 0.000 description 1
- OUYCCCASQSFEME-QMMMGPOBSA-N L-tyrosine Chemical compound OC(=O)[C@@H](N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-QMMMGPOBSA-N 0.000 description 1
- LZDNBBYBDGBADK-UHFFFAOYSA-N L-valyl-L-tryptophan Natural products C1=CC=C2C(CC(NC(=O)C(N)C(C)C)C(O)=O)=CNC2=C1 LZDNBBYBDGBADK-UHFFFAOYSA-N 0.000 description 1
- 206010023927 Lassa fever Diseases 0.000 description 1
- 241000589248 Legionella Species 0.000 description 1
- 208000007764 Legionnaires' Disease Diseases 0.000 description 1
- 241000222722 Leishmania <genus> Species 0.000 description 1
- 101710180643 Leishmanolysin Proteins 0.000 description 1
- 229920001491 Lentinan Polymers 0.000 description 1
- CZCSUZMIRKFFFA-CIUDSAMLSA-N Leu-Ala-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(O)=O CZCSUZMIRKFFFA-CIUDSAMLSA-N 0.000 description 1
- CQQGCWPXDHTTNF-GUBZILKMSA-N Leu-Ala-Glu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(O)=O CQQGCWPXDHTTNF-GUBZILKMSA-N 0.000 description 1
- PBCHMHROGNUXMK-DLOVCJGASA-N Leu-Ala-His Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CN=CN1 PBCHMHROGNUXMK-DLOVCJGASA-N 0.000 description 1
- QPRQGENIBFLVEB-BJDJZHNGSA-N Leu-Ala-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O QPRQGENIBFLVEB-BJDJZHNGSA-N 0.000 description 1
- WSGXUIQTEZDVHJ-GARJFASQSA-N Leu-Ala-Pro Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@@H]1C(O)=O WSGXUIQTEZDVHJ-GARJFASQSA-N 0.000 description 1
- JUWJEAPUNARGCF-DCAQKATOSA-N Leu-Arg-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(O)=O JUWJEAPUNARGCF-DCAQKATOSA-N 0.000 description 1
- NTRAGDHVSGKUSF-AVGNSLFASA-N Leu-Arg-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O NTRAGDHVSGKUSF-AVGNSLFASA-N 0.000 description 1
- REPPKAMYTOJTFC-DCAQKATOSA-N Leu-Arg-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O REPPKAMYTOJTFC-DCAQKATOSA-N 0.000 description 1
- IBMVEYRWAWIOTN-RWMBFGLXSA-N Leu-Arg-Pro Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N1CCC[C@@H]1C(O)=O IBMVEYRWAWIOTN-RWMBFGLXSA-N 0.000 description 1
- DBVWMYGBVFCRBE-CIUDSAMLSA-N Leu-Asn-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O DBVWMYGBVFCRBE-CIUDSAMLSA-N 0.000 description 1
- BAJIJEGGUYXZGC-CIUDSAMLSA-N Leu-Asn-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CS)C(=O)O)N BAJIJEGGUYXZGC-CIUDSAMLSA-N 0.000 description 1
- WGNOPSQMIQERPK-GARJFASQSA-N Leu-Asn-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N1CCC[C@@H]1C(=O)O)N WGNOPSQMIQERPK-GARJFASQSA-N 0.000 description 1
- FIJMQLGQLBLBOL-HJGDQZAQSA-N Leu-Asn-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O FIJMQLGQLBLBOL-HJGDQZAQSA-N 0.000 description 1
- WXHFZJFZWNCDNB-KKUMJFAQSA-N Leu-Asn-Tyr Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 WXHFZJFZWNCDNB-KKUMJFAQSA-N 0.000 description 1
- PJYSOYLLTJKZHC-GUBZILKMSA-N Leu-Asp-Gln Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CCC(N)=O PJYSOYLLTJKZHC-GUBZILKMSA-N 0.000 description 1
- ULXYQAJWJGLCNR-YUMQZZPRSA-N Leu-Asp-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O ULXYQAJWJGLCNR-YUMQZZPRSA-N 0.000 description 1
- MMEDVBWCMGRKKC-GARJFASQSA-N Leu-Asp-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N MMEDVBWCMGRKKC-GARJFASQSA-N 0.000 description 1
- PVMPDMIKUVNOBD-CIUDSAMLSA-N Leu-Asp-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O PVMPDMIKUVNOBD-CIUDSAMLSA-N 0.000 description 1
- HUEBCHPSXSQUGN-GARJFASQSA-N Leu-Cys-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CS)C(=O)N1CCC[C@@H]1C(=O)O)N HUEBCHPSXSQUGN-GARJFASQSA-N 0.000 description 1
- ZYLJULGXQDNXDK-GUBZILKMSA-N Leu-Gln-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O ZYLJULGXQDNXDK-GUBZILKMSA-N 0.000 description 1
- YVKSMSDXKMSIRX-GUBZILKMSA-N Leu-Glu-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O YVKSMSDXKMSIRX-GUBZILKMSA-N 0.000 description 1
- HFBCHNRFRYLZNV-GUBZILKMSA-N Leu-Glu-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O HFBCHNRFRYLZNV-GUBZILKMSA-N 0.000 description 1
- QVFGXCVIXXBFHO-AVGNSLFASA-N Leu-Glu-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O QVFGXCVIXXBFHO-AVGNSLFASA-N 0.000 description 1
- LAGPXKYZCCTSGQ-JYJNAYRXSA-N Leu-Glu-Phe Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O LAGPXKYZCCTSGQ-JYJNAYRXSA-N 0.000 description 1
- WQWSMEOYXJTFRU-GUBZILKMSA-N Leu-Glu-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O WQWSMEOYXJTFRU-GUBZILKMSA-N 0.000 description 1
- ZFNLIDNJUWNIJL-WDCWCFNPSA-N Leu-Glu-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ZFNLIDNJUWNIJL-WDCWCFNPSA-N 0.000 description 1
- CCQLQKZTXZBXTN-NHCYSSNCSA-N Leu-Gly-Ile Chemical compound [H]N[C@@H](CC(C)C)C(=O)NCC(=O)N[C@@H]([C@@H](C)CC)C(O)=O CCQLQKZTXZBXTN-NHCYSSNCSA-N 0.000 description 1
- KEVYYIMVELOXCT-KBPBESRZSA-N Leu-Gly-Phe Chemical compound CC(C)C[C@H]([NH3+])C(=O)NCC(=O)N[C@H](C([O-])=O)CC1=CC=CC=C1 KEVYYIMVELOXCT-KBPBESRZSA-N 0.000 description 1
- PBGDOSARRIJMEV-DLOVCJGASA-N Leu-His-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](C)C(O)=O PBGDOSARRIJMEV-DLOVCJGASA-N 0.000 description 1
- YWYQSLOTVIRCFE-SRVKXCTJSA-N Leu-His-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(O)=O)C(O)=O YWYQSLOTVIRCFE-SRVKXCTJSA-N 0.000 description 1
- CFZZDVMBRYFFNU-QWRGUYRKSA-N Leu-His-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)NCC(O)=O CFZZDVMBRYFFNU-QWRGUYRKSA-N 0.000 description 1
- OHZIZVWQXJPBJS-IXOXFDKPSA-N Leu-His-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OHZIZVWQXJPBJS-IXOXFDKPSA-N 0.000 description 1
- HGFGEMSVBMCFKK-MNXVOIDGSA-N Leu-Ile-Glu Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCC(O)=O)C(O)=O HGFGEMSVBMCFKK-MNXVOIDGSA-N 0.000 description 1
- ZALAVHVPPOHAOL-XUXIUFHCSA-N Leu-Ile-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CC(C)C)N ZALAVHVPPOHAOL-XUXIUFHCSA-N 0.000 description 1
- QLDHBYRUNQZIJQ-DKIMLUQUSA-N Leu-Ile-Phe Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O QLDHBYRUNQZIJQ-DKIMLUQUSA-N 0.000 description 1
- JNDYEOUZBLOVOF-AVGNSLFASA-N Leu-Leu-Gln Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O JNDYEOUZBLOVOF-AVGNSLFASA-N 0.000 description 1
- YOKVEHGYYQEQOP-QWRGUYRKSA-N Leu-Leu-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O YOKVEHGYYQEQOP-QWRGUYRKSA-N 0.000 description 1
- FAELBUXXFQLUAX-AJNGGQMLSA-N Leu-Leu-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC(C)C FAELBUXXFQLUAX-AJNGGQMLSA-N 0.000 description 1
- LXKNSJLSGPNHSK-KKUMJFAQSA-N Leu-Leu-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)O)N LXKNSJLSGPNHSK-KKUMJFAQSA-N 0.000 description 1
- XVZCXCTYGHPNEM-UHFFFAOYSA-N Leu-Leu-Pro Natural products CC(C)CC(N)C(=O)NC(CC(C)C)C(=O)N1CCCC1C(O)=O XVZCXCTYGHPNEM-UHFFFAOYSA-N 0.000 description 1
- UCNNZELZXFXXJQ-BZSNNMDCSA-N Leu-Leu-Tyr Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 UCNNZELZXFXXJQ-BZSNNMDCSA-N 0.000 description 1
- DCGXHWINSHEPIR-SRVKXCTJSA-N Leu-Lys-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CS)C(=O)O)N DCGXHWINSHEPIR-SRVKXCTJSA-N 0.000 description 1
- LVTJJOJKDCVZGP-QWRGUYRKSA-N Leu-Lys-Gly Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)NCC(O)=O LVTJJOJKDCVZGP-QWRGUYRKSA-N 0.000 description 1
- OVZLLFONXILPDZ-VOAKCMCISA-N Leu-Lys-Thr Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OVZLLFONXILPDZ-VOAKCMCISA-N 0.000 description 1
- CPONGMJGVIAWEH-DCAQKATOSA-N Leu-Met-Ala Chemical compound CSCC[C@H](NC(=O)[C@@H](N)CC(C)C)C(=O)N[C@@H](C)C(O)=O CPONGMJGVIAWEH-DCAQKATOSA-N 0.000 description 1
- NHRINZSPIUXYQZ-DCAQKATOSA-N Leu-Met-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CS)C(=O)O)N NHRINZSPIUXYQZ-DCAQKATOSA-N 0.000 description 1
- FLNPJLDPGMLWAU-UWVGGRQHSA-N Leu-Met-Gly Chemical compound OC(=O)CNC(=O)[C@H](CCSC)NC(=O)[C@@H](N)CC(C)C FLNPJLDPGMLWAU-UWVGGRQHSA-N 0.000 description 1
- GNRPTBRHRRZCMA-RWMBFGLXSA-N Leu-Met-Pro Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCSC)C(=O)N1CCC[C@@H]1C(=O)O)N GNRPTBRHRRZCMA-RWMBFGLXSA-N 0.000 description 1
- HDHQQEDVWQGBEE-DCAQKATOSA-N Leu-Met-Ser Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CO)C(O)=O HDHQQEDVWQGBEE-DCAQKATOSA-N 0.000 description 1
- BIZNDKMFQHDOIE-KKUMJFAQSA-N Leu-Phe-Asn Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(N)=O)C(O)=O)CC1=CC=CC=C1 BIZNDKMFQHDOIE-KKUMJFAQSA-N 0.000 description 1
- ZAVCJRJOQKIOJW-KKUMJFAQSA-N Leu-Phe-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(O)=O)C(O)=O)CC1=CC=CC=C1 ZAVCJRJOQKIOJW-KKUMJFAQSA-N 0.000 description 1
- YESNGRDJQWDYLH-KKUMJFAQSA-N Leu-Phe-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CS)C(=O)O)N YESNGRDJQWDYLH-KKUMJFAQSA-N 0.000 description 1
- YWKNKRAKOCLOLH-OEAJRASXSA-N Leu-Phe-Thr Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)O)C(O)=O)CC1=CC=CC=C1 YWKNKRAKOCLOLH-OEAJRASXSA-N 0.000 description 1
- WMIOEVKKYIMVKI-DCAQKATOSA-N Leu-Pro-Ala Chemical compound [H]N[C@@H](CC(C)C)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O WMIOEVKKYIMVKI-DCAQKATOSA-N 0.000 description 1
- KWLWZYMNUZJKMZ-IHRRRGAJSA-N Leu-Pro-Leu Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O KWLWZYMNUZJKMZ-IHRRRGAJSA-N 0.000 description 1
- PWPBLZXWFXJFHE-RHYQMDGZSA-N Leu-Pro-Thr Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O PWPBLZXWFXJFHE-RHYQMDGZSA-N 0.000 description 1
- KZZCOWMDDXDKSS-CIUDSAMLSA-N Leu-Ser-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O KZZCOWMDDXDKSS-CIUDSAMLSA-N 0.000 description 1
- AKVBOOKXVAMKSS-GUBZILKMSA-N Leu-Ser-Gln Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(O)=O AKVBOOKXVAMKSS-GUBZILKMSA-N 0.000 description 1
- BRTVHXHCUSXYRI-CIUDSAMLSA-N Leu-Ser-Ser Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O BRTVHXHCUSXYRI-CIUDSAMLSA-N 0.000 description 1
- SVBJIZVVYJYGLA-DCAQKATOSA-N Leu-Ser-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O SVBJIZVVYJYGLA-DCAQKATOSA-N 0.000 description 1
- ZDJQVSIPFLMNOX-RHYQMDGZSA-N Leu-Thr-Arg Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CCCN=C(N)N ZDJQVSIPFLMNOX-RHYQMDGZSA-N 0.000 description 1
- AEDWWMMHUGYIFD-HJGDQZAQSA-N Leu-Thr-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(N)=O)C(O)=O AEDWWMMHUGYIFD-HJGDQZAQSA-N 0.000 description 1
- ICYRCNICGBJLGM-HJGDQZAQSA-N Leu-Thr-Asp Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC(O)=O ICYRCNICGBJLGM-HJGDQZAQSA-N 0.000 description 1
- LINKCQUOMUDLKN-KATARQTJSA-N Leu-Thr-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CC(C)C)N)O LINKCQUOMUDLKN-KATARQTJSA-N 0.000 description 1
- LFSQWRSVPNKJGP-WDCWCFNPSA-N Leu-Thr-Glu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CCC(O)=O LFSQWRSVPNKJGP-WDCWCFNPSA-N 0.000 description 1
- QWWPYKKLXWOITQ-VOAKCMCISA-N Leu-Thr-Leu Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC(C)C QWWPYKKLXWOITQ-VOAKCMCISA-N 0.000 description 1
- DAYQSYGBCUKVKT-VOAKCMCISA-N Leu-Thr-Lys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(O)=O DAYQSYGBCUKVKT-VOAKCMCISA-N 0.000 description 1
- ODRREERHVHMIPT-OEAJRASXSA-N Leu-Thr-Phe Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 ODRREERHVHMIPT-OEAJRASXSA-N 0.000 description 1
- AIQWYVFNBNNOLU-RHYQMDGZSA-N Leu-Thr-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O AIQWYVFNBNNOLU-RHYQMDGZSA-N 0.000 description 1
- CNWDWAMPKVYJJB-NUTKFTJISA-N Leu-Trp-Ala Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](N)CC(C)C)C(=O)N[C@@H](C)C(O)=O)=CNC2=C1 CNWDWAMPKVYJJB-NUTKFTJISA-N 0.000 description 1
- URJUVJDTPXCQFL-IHPCNDPISA-N Leu-Trp-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC3=CN=CN3)C(=O)O)N URJUVJDTPXCQFL-IHPCNDPISA-N 0.000 description 1
- HQBOMRTVKVKFMN-WDSOQIARSA-N Leu-Trp-Val Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](C(C)C)C(O)=O HQBOMRTVKVKFMN-WDSOQIARSA-N 0.000 description 1
- UCRJTSIIAYHOHE-ULQDDVLXSA-N Leu-Tyr-Arg Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N UCRJTSIIAYHOHE-ULQDDVLXSA-N 0.000 description 1
- VQHUBNVKFFLWRP-ULQDDVLXSA-N Leu-Tyr-Val Chemical compound CC(C)C[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](C(C)C)C(O)=O)CC1=CC=C(O)C=C1 VQHUBNVKFFLWRP-ULQDDVLXSA-N 0.000 description 1
- XZNJZXJZBMBGGS-NHCYSSNCSA-N Leu-Val-Asn Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O XZNJZXJZBMBGGS-NHCYSSNCSA-N 0.000 description 1
- TUIOUEWKFFVNLH-DCAQKATOSA-N Leu-Val-Cys Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CS)C(O)=O TUIOUEWKFFVNLH-DCAQKATOSA-N 0.000 description 1
- QESXLSQLQHHTIX-RHYQMDGZSA-N Leu-Val-Thr Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O QESXLSQLQHHTIX-RHYQMDGZSA-N 0.000 description 1
- GDBQQVLCIARPGH-UHFFFAOYSA-N Leupeptin Natural products CC(C)CC(NC(C)=O)C(=O)NC(CC(C)C)C(=O)NC(C=O)CCCN=C(N)N GDBQQVLCIARPGH-UHFFFAOYSA-N 0.000 description 1
- 241000186781 Listeria Species 0.000 description 1
- 241000186779 Listeria monocytogenes Species 0.000 description 1
- 206010058467 Lung neoplasm malignant Diseases 0.000 description 1
- 241001625930 Luria Species 0.000 description 1
- 239000006137 Luria-Bertani broth Substances 0.000 description 1
- 208000016604 Lyme disease Diseases 0.000 description 1
- BTSXLXFPMZXVPR-DLOVCJGASA-N Lys-Ala-His Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCCCN)N BTSXLXFPMZXVPR-DLOVCJGASA-N 0.000 description 1
- NFLFJGGKOHYZJF-BJDJZHNGSA-N Lys-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN NFLFJGGKOHYZJF-BJDJZHNGSA-N 0.000 description 1
- WSXTWLJHTLRFLW-SRVKXCTJSA-N Lys-Ala-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O WSXTWLJHTLRFLW-SRVKXCTJSA-N 0.000 description 1
- KNKHAVVBVXKOGX-JXUBOQSCSA-N Lys-Ala-Thr Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O KNKHAVVBVXKOGX-JXUBOQSCSA-N 0.000 description 1
- IRNSXVOWSXSULE-DCAQKATOSA-N Lys-Ala-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN IRNSXVOWSXSULE-DCAQKATOSA-N 0.000 description 1
- CLBGMWIYPYAZPR-AVGNSLFASA-N Lys-Arg-Arg Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O CLBGMWIYPYAZPR-AVGNSLFASA-N 0.000 description 1
- GAOJCVKPIGHTGO-UWVGGRQHSA-N Lys-Arg-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O GAOJCVKPIGHTGO-UWVGGRQHSA-N 0.000 description 1
- GGAPIOORBXHMNY-ULQDDVLXSA-N Lys-Arg-Tyr Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCCCN)N)O GGAPIOORBXHMNY-ULQDDVLXSA-N 0.000 description 1
- HQVDJTYKCMIWJP-YUMQZZPRSA-N Lys-Asn-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(O)=O HQVDJTYKCMIWJP-YUMQZZPRSA-N 0.000 description 1
- DEFGUIIUYAUEDU-ZPFDUUQYSA-N Lys-Asn-Ile Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O DEFGUIIUYAUEDU-ZPFDUUQYSA-N 0.000 description 1
- GKFNXYMAMKJSKD-NHCYSSNCSA-N Lys-Asp-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O GKFNXYMAMKJSKD-NHCYSSNCSA-N 0.000 description 1
- RDIILCRAWOSDOQ-CIUDSAMLSA-N Lys-Cys-Asp Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)O)C(=O)O)N RDIILCRAWOSDOQ-CIUDSAMLSA-N 0.000 description 1
- RLZDUFRBMQNYIJ-YUMQZZPRSA-N Lys-Cys-Gly Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CS)C(=O)NCC(=O)O)N RLZDUFRBMQNYIJ-YUMQZZPRSA-N 0.000 description 1
- SSYOBDBNBQBSQE-SRVKXCTJSA-N Lys-Cys-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(C)C)C(O)=O SSYOBDBNBQBSQE-SRVKXCTJSA-N 0.000 description 1
- KSFQPRLZAUXXPT-GARJFASQSA-N Lys-Cys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CS)NC(=O)[C@H](CCCCN)N)C(=O)O KSFQPRLZAUXXPT-GARJFASQSA-N 0.000 description 1
- QQUJSUFWEDZQQY-AVGNSLFASA-N Lys-Gln-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(O)=O)CCCCN QQUJSUFWEDZQQY-AVGNSLFASA-N 0.000 description 1
- GRADYHMSAUIKPS-DCAQKATOSA-N Lys-Glu-Gln Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(N)=O)C(O)=O GRADYHMSAUIKPS-DCAQKATOSA-N 0.000 description 1
- ULUQBUKAPDUKOC-GVXVVHGQSA-N Lys-Glu-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O ULUQBUKAPDUKOC-GVXVVHGQSA-N 0.000 description 1
- FHIAJWBDZVHLAH-YUMQZZPRSA-N Lys-Gly-Ser Chemical compound NCCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O FHIAJWBDZVHLAH-YUMQZZPRSA-N 0.000 description 1
- ZASPELYMPSACER-HOCLYGCPSA-N Lys-Gly-Trp Chemical compound [H]N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O ZASPELYMPSACER-HOCLYGCPSA-N 0.000 description 1
- SKRGVGLIRUGANF-AVGNSLFASA-N Lys-Leu-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(O)=O SKRGVGLIRUGANF-AVGNSLFASA-N 0.000 description 1
- WVJNGSFKBKOKRV-AJNGGQMLSA-N Lys-Leu-Ile Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O WVJNGSFKBKOKRV-AJNGGQMLSA-N 0.000 description 1
- AIRZWUMAHCDDHR-KKUMJFAQSA-N Lys-Leu-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O AIRZWUMAHCDDHR-KKUMJFAQSA-N 0.000 description 1
- RBEATVHTWHTHTJ-KKUMJFAQSA-N Lys-Leu-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(O)=O RBEATVHTWHTHTJ-KKUMJFAQSA-N 0.000 description 1
- YPLVCBKEPJPBDQ-MELADBBJSA-N Lys-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCCN)N YPLVCBKEPJPBDQ-MELADBBJSA-N 0.000 description 1
- ZJWIXBZTAAJERF-IHRRRGAJSA-N Lys-Lys-Arg Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@H](C(O)=O)CCCN=C(N)N ZJWIXBZTAAJERF-IHRRRGAJSA-N 0.000 description 1
- ALGGDNMLQNFVIZ-SRVKXCTJSA-N Lys-Lys-Asp Chemical compound C(CCN)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(=O)O)C(=O)O)N ALGGDNMLQNFVIZ-SRVKXCTJSA-N 0.000 description 1
- ATNKHRAIZCMCCN-BZSNNMDCSA-N Lys-Lys-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CCCCN)N ATNKHRAIZCMCCN-BZSNNMDCSA-N 0.000 description 1
- YXPJCVNIDDKGOE-MELADBBJSA-N Lys-Lys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CCCCN)N)C(=O)O YXPJCVNIDDKGOE-MELADBBJSA-N 0.000 description 1
- QQPSCXKFDSORFT-IHRRRGAJSA-N Lys-Lys-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)CCCCN QQPSCXKFDSORFT-IHRRRGAJSA-N 0.000 description 1
- AEIIJFBQVGYVEV-YESZJQIVSA-N Lys-Phe-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=CC=C2)NC(=O)[C@H](CCCCN)N)C(=O)O AEIIJFBQVGYVEV-YESZJQIVSA-N 0.000 description 1
- BOJYMMBYBNOOGG-DCAQKATOSA-N Lys-Pro-Ala Chemical compound [H]N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O BOJYMMBYBNOOGG-DCAQKATOSA-N 0.000 description 1
- PDIDTSZKKFEDMB-UWVGGRQHSA-N Lys-Pro-Gly Chemical compound [H]N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O PDIDTSZKKFEDMB-UWVGGRQHSA-N 0.000 description 1
- QBHGXFQJFPWJIH-XUXIUFHCSA-N Lys-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCCN QBHGXFQJFPWJIH-XUXIUFHCSA-N 0.000 description 1
- HYSVGEAWTGPMOA-IHRRRGAJSA-N Lys-Pro-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O HYSVGEAWTGPMOA-IHRRRGAJSA-N 0.000 description 1
- LOGFVTREOLYCPF-RHYQMDGZSA-N Lys-Pro-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)CCCCN LOGFVTREOLYCPF-RHYQMDGZSA-N 0.000 description 1
- SQXZLVXQXWILKW-KKUMJFAQSA-N Lys-Ser-Phe Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O SQXZLVXQXWILKW-KKUMJFAQSA-N 0.000 description 1
- MEQLGHAMAUPOSJ-DCAQKATOSA-N Lys-Ser-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O MEQLGHAMAUPOSJ-DCAQKATOSA-N 0.000 description 1
- TVOOGUNBIWAURO-KATARQTJSA-N Lys-Thr-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCCCN)N)O TVOOGUNBIWAURO-KATARQTJSA-N 0.000 description 1
- RPWTZTBIFGENIA-VOAKCMCISA-N Lys-Thr-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O RPWTZTBIFGENIA-VOAKCMCISA-N 0.000 description 1
- YKBSXQFZWFXFIB-VOAKCMCISA-N Lys-Thr-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CCCCN)C(O)=O YKBSXQFZWFXFIB-VOAKCMCISA-N 0.000 description 1
- BDFHWFUAQLIMJO-KXNHARMFSA-N Lys-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCCCN)N)O BDFHWFUAQLIMJO-KXNHARMFSA-N 0.000 description 1
- WINFHLHJTRGLCV-BZSNNMDCSA-N Lys-Tyr-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CCCCN)C(O)=O)CC1=CC=C(O)C=C1 WINFHLHJTRGLCV-BZSNNMDCSA-N 0.000 description 1
- VWPJQIHBBOJWDN-DCAQKATOSA-N Lys-Val-Ala Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O VWPJQIHBBOJWDN-DCAQKATOSA-N 0.000 description 1
- NYTDJEZBAAFLLG-IHRRRGAJSA-N Lys-Val-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(O)=O NYTDJEZBAAFLLG-IHRRRGAJSA-N 0.000 description 1
- IKXQOBUBZSOWDY-AVGNSLFASA-N Lys-Val-Val Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](CCCCN)N IKXQOBUBZSOWDY-AVGNSLFASA-N 0.000 description 1
- 101710145006 Lysis protein Proteins 0.000 description 1
- 241001444195 Madurella Species 0.000 description 1
- 241000555676 Malassezia Species 0.000 description 1
- PEEHTFAAVSWFBL-UHFFFAOYSA-N Maleimide Chemical compound O=C1NC(=O)C=C1 PEEHTFAAVSWFBL-UHFFFAOYSA-N 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- 206010026749 Mania Diseases 0.000 description 1
- 241000712079 Measles morbillivirus Species 0.000 description 1
- 102100032239 Melanotransferrin Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- DTICLBJHRYSJLH-GUBZILKMSA-N Met-Ala-Val Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](C(C)C)C(O)=O DTICLBJHRYSJLH-GUBZILKMSA-N 0.000 description 1
- CWFYZYQMUDWGTI-GUBZILKMSA-N Met-Arg-Asp Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O CWFYZYQMUDWGTI-GUBZILKMSA-N 0.000 description 1
- DSWOTZCVCBEPOU-IUCAKERBSA-N Met-Arg-Gly Chemical compound CSCC[C@H](N)C(=O)N[C@H](C(=O)NCC(O)=O)CCCNC(N)=N DSWOTZCVCBEPOU-IUCAKERBSA-N 0.000 description 1
- ZEDVFJPQNNBMST-CYDGBPFRSA-N Met-Arg-Ile Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O ZEDVFJPQNNBMST-CYDGBPFRSA-N 0.000 description 1
- IHITVQKJXQQGLJ-LPEHRKFASA-N Met-Asn-Pro Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N1CCC[C@@H]1C(=O)O)N IHITVQKJXQQGLJ-LPEHRKFASA-N 0.000 description 1
- HDNOQCZWJGGHSS-VEVYYDQMSA-N Met-Asn-Thr Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O HDNOQCZWJGGHSS-VEVYYDQMSA-N 0.000 description 1
- QWTGQXGNNMIUCW-BPUTZDHNSA-N Met-Asn-Trp Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O QWTGQXGNNMIUCW-BPUTZDHNSA-N 0.000 description 1
- UZVWDRPUTHXQAM-FXQIFTODSA-N Met-Asp-Ala Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(O)=O UZVWDRPUTHXQAM-FXQIFTODSA-N 0.000 description 1
- YLLWCSDBVGZLOW-CIUDSAMLSA-N Met-Gln-Ala Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(O)=O YLLWCSDBVGZLOW-CIUDSAMLSA-N 0.000 description 1
- GPAHWYRSHCKICP-GUBZILKMSA-N Met-Glu-Glu Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O GPAHWYRSHCKICP-GUBZILKMSA-N 0.000 description 1
- YAWKHFKCNSXYDS-XIRDDKMYSA-N Met-Glu-Trp Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N YAWKHFKCNSXYDS-XIRDDKMYSA-N 0.000 description 1
- LQMHZERGCQJKAH-STQMWFEESA-N Met-Gly-Phe Chemical compound CSCC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 LQMHZERGCQJKAH-STQMWFEESA-N 0.000 description 1
- MVBZBRKNZVJEKK-DTWKUNHWSA-N Met-Gly-Pro Chemical compound CSCC[C@@H](C(=O)NCC(=O)N1CCC[C@@H]1C(=O)O)N MVBZBRKNZVJEKK-DTWKUNHWSA-N 0.000 description 1
- FZUNSVYYPYJYAP-NAKRPEOUSA-N Met-Ile-Ala Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O FZUNSVYYPYJYAP-NAKRPEOUSA-N 0.000 description 1
- AFFKUNVPPLQUGA-DCAQKATOSA-N Met-Leu-Ala Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O AFFKUNVPPLQUGA-DCAQKATOSA-N 0.000 description 1
- BEZJTLKUMFMITF-AVGNSLFASA-N Met-Lys-Arg Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@H](C(O)=O)CCCNC(N)=N BEZJTLKUMFMITF-AVGNSLFASA-N 0.000 description 1
- WUYLWZRHRLLEGB-AVGNSLFASA-N Met-Met-Leu Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(O)=O WUYLWZRHRLLEGB-AVGNSLFASA-N 0.000 description 1
- OIFHHODAXVWKJN-ULQDDVLXSA-N Met-Phe-Leu Chemical compound CSCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CC(C)C)C(O)=O)CC1=CC=CC=C1 OIFHHODAXVWKJN-ULQDDVLXSA-N 0.000 description 1
- JQHYVIKEFYETEW-IHRRRGAJSA-N Met-Phe-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H](CO)C(O)=O)CC1=CC=CC=C1 JQHYVIKEFYETEW-IHRRRGAJSA-N 0.000 description 1
- DBMLDOWSVHMQQN-XGEHTFHBSA-N Met-Ser-Thr Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O DBMLDOWSVHMQQN-XGEHTFHBSA-N 0.000 description 1
- SOAYQFDWEIWPPR-IHRRRGAJSA-N Met-Ser-Tyr Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O SOAYQFDWEIWPPR-IHRRRGAJSA-N 0.000 description 1
- XLTSAUGGDYRFLS-UMPQAUOISA-N Met-Thr-Trp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)NC(=O)[C@H](CCSC)N)O XLTSAUGGDYRFLS-UMPQAUOISA-N 0.000 description 1
- WYNIRYZIFZGWQD-BPUTZDHNSA-N Met-Trp-Asn Chemical compound CSCC[C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC(=O)N)C(=O)O)N WYNIRYZIFZGWQD-BPUTZDHNSA-N 0.000 description 1
- YDKYJRZWRJTILC-WDSOQIARSA-N Met-Trp-Lys Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](N)CCSC)C(=O)N[C@@H](CCCCN)C(O)=O)=CNC2=C1 YDKYJRZWRJTILC-WDSOQIARSA-N 0.000 description 1
- CULGJGUDIJATIP-STQMWFEESA-N Met-Tyr-Gly Chemical compound CSCC[C@H](N)C(=O)N[C@H](C(=O)NCC(O)=O)CC1=CC=C(O)C=C1 CULGJGUDIJATIP-STQMWFEESA-N 0.000 description 1
- MUDYEFAKNSTFAI-JYJNAYRXSA-N Met-Tyr-Val Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(O)=O MUDYEFAKNSTFAI-JYJNAYRXSA-N 0.000 description 1
- CQRGINSEMFBACV-WPRPVWTQSA-N Met-Val-Gly Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O CQRGINSEMFBACV-WPRPVWTQSA-N 0.000 description 1
- LBSWWNKMVPAXOI-GUBZILKMSA-N Met-Val-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O LBSWWNKMVPAXOI-GUBZILKMSA-N 0.000 description 1
- 108010006035 Metalloproteases Proteins 0.000 description 1
- 102000005741 Metalloproteases Human genes 0.000 description 1
- 206010027476 Metastases Diseases 0.000 description 1
- 108060004795 Methyltransferase Proteins 0.000 description 1
- 241000192041 Micrococcus Species 0.000 description 1
- 241001480037 Microsporum Species 0.000 description 1
- 239000012901 Milli-Q water Substances 0.000 description 1
- 241000588621 Moraxella Species 0.000 description 1
- 241000588771 Morganella <proteobacterium> Species 0.000 description 1
- 102000015728 Mucins Human genes 0.000 description 1
- 108010063954 Mucins Proteins 0.000 description 1
- 241000235395 Mucor Species 0.000 description 1
- 241000711386 Mumps virus Species 0.000 description 1
- 241001529936 Murinae Species 0.000 description 1
- 241000204031 Mycoplasma Species 0.000 description 1
- 102100025243 Myeloid cell surface antigen CD33 Human genes 0.000 description 1
- 108010002311 N-glycylglutamic acid Proteins 0.000 description 1
- 108010066427 N-valyltryptophan Proteins 0.000 description 1
- 108010087066 N2-tryptophyllysine Proteins 0.000 description 1
- 241000224436 Naegleria Species 0.000 description 1
- 241000588653 Neisseria Species 0.000 description 1
- 206010061309 Neoplasm progression Diseases 0.000 description 1
- 241000187654 Nocardia Species 0.000 description 1
- 241001263478 Norovirus Species 0.000 description 1
- XDMCWZFLLGVIID-SXPRBRBTSA-N O-(3-O-D-galactosyl-N-acetyl-beta-D-galactosaminyl)-L-serine Chemical compound CC(=O)N[C@H]1[C@H](OC[C@H]([NH3+])C([O-])=O)O[C@H](CO)[C@H](O)[C@@H]1OC1[C@H](O)[C@@H](O)[C@@H](O)[C@@H](CO)O1 XDMCWZFLLGVIID-SXPRBRBTSA-N 0.000 description 1
- 241000320412 Ogataea angusta Species 0.000 description 1
- 108091034117 Oligonucleotide Proteins 0.000 description 1
- 241000243981 Onchocerca Species 0.000 description 1
- 241000233654 Oomycetes Species 0.000 description 1
- 241000242716 Opisthorchis Species 0.000 description 1
- 239000012124 Opti-MEM Substances 0.000 description 1
- 241000700635 Orf virus Species 0.000 description 1
- 241000712464 Orthomyxoviridae Species 0.000 description 1
- 108700006640 OspA Proteins 0.000 description 1
- 101710160107 Outer membrane protein A Proteins 0.000 description 1
- 206010033128 Ovarian cancer Diseases 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 241001236817 Paecilomyces <Clavicipitaceae> Species 0.000 description 1
- 241000282579 Pan Species 0.000 description 1
- 241000526686 Paracoccidioides brasiliensis Species 0.000 description 1
- 241001480233 Paragonimus Species 0.000 description 1
- 208000002606 Paramyxoviridae Infections Diseases 0.000 description 1
- 241000701945 Parvoviridae Species 0.000 description 1
- 241000228143 Penicillium Species 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 241000150350 Peribunyaviridae Species 0.000 description 1
- 108010081690 Pertussis Toxin Proteins 0.000 description 1
- AJOKKVTWEMXZHC-DRZSPHRISA-N Phe-Ala-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=CC=C1 AJOKKVTWEMXZHC-DRZSPHRISA-N 0.000 description 1
- FPTXMUIBLMGTQH-ONGXEEELSA-N Phe-Ala-Gly Chemical compound OC(=O)CNC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=CC=C1 FPTXMUIBLMGTQH-ONGXEEELSA-N 0.000 description 1
- BBDSZDHUCPSYAC-QEJZJMRPSA-N Phe-Ala-Leu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O BBDSZDHUCPSYAC-QEJZJMRPSA-N 0.000 description 1
- DPUOLKQSMYLRDR-UBHSHLNASA-N Phe-Arg-Ala Chemical compound NC(N)=NCCC[C@@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1 DPUOLKQSMYLRDR-UBHSHLNASA-N 0.000 description 1
- YYRCPTVAPLQRNC-ULQDDVLXSA-N Phe-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H](N)CC1=CC=CC=C1 YYRCPTVAPLQRNC-ULQDDVLXSA-N 0.000 description 1
- UUWCIPUVJJIEEP-SRVKXCTJSA-N Phe-Asn-Cys Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CS)C(=O)O)N UUWCIPUVJJIEEP-SRVKXCTJSA-N 0.000 description 1
- DDYIRGBOZVKRFR-AVGNSLFASA-N Phe-Asp-Glu Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N DDYIRGBOZVKRFR-AVGNSLFASA-N 0.000 description 1
- OJUMUUXGSXUZJZ-SRVKXCTJSA-N Phe-Asp-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CO)C(O)=O OJUMUUXGSXUZJZ-SRVKXCTJSA-N 0.000 description 1
- UEHNWRNADDPYNK-DLOVCJGASA-N Phe-Cys-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC1=CC=CC=C1)N UEHNWRNADDPYNK-DLOVCJGASA-N 0.000 description 1
- HPECNYCQLSVCHH-BZSNNMDCSA-N Phe-Cys-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC2=CC=CC=C2)C(=O)O)N HPECNYCQLSVCHH-BZSNNMDCSA-N 0.000 description 1
- AKJAKCBHLJGRBU-JYJNAYRXSA-N Phe-Glu-His Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N AKJAKCBHLJGRBU-JYJNAYRXSA-N 0.000 description 1
- JJHVFCUWLSKADD-ONGXEEELSA-N Phe-Gly-Ala Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)NCC(=O)N[C@@H](C)C(O)=O JJHVFCUWLSKADD-ONGXEEELSA-N 0.000 description 1
- WFHRXJOZEXUKLV-IRXDYDNUSA-N Phe-Gly-Tyr Chemical compound C([C@H](N)C(=O)NCC(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=CC=C1 WFHRXJOZEXUKLV-IRXDYDNUSA-N 0.000 description 1
- ONORAGIFHNAADN-LLLHUVSDSA-N Phe-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CC=CC=C2)N ONORAGIFHNAADN-LLLHUVSDSA-N 0.000 description 1
- CWFGECHCRMGPPT-MXAVVETBSA-N Phe-Ile-Ser Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O CWFGECHCRMGPPT-MXAVVETBSA-N 0.000 description 1
- YTILBRIUASDGBL-BZSNNMDCSA-N Phe-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC1=CC=CC=C1 YTILBRIUASDGBL-BZSNNMDCSA-N 0.000 description 1
- CMHTUJQZQXFNTQ-OEAJRASXSA-N Phe-Leu-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC1=CC=CC=C1)N)O CMHTUJQZQXFNTQ-OEAJRASXSA-N 0.000 description 1
- ACJULKNZOCRWEI-ULQDDVLXSA-N Phe-Met-Leu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(O)=O ACJULKNZOCRWEI-ULQDDVLXSA-N 0.000 description 1
- DSXPMZMSJHOKKK-HJOGWXRNSA-N Phe-Phe-Tyr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O DSXPMZMSJHOKKK-HJOGWXRNSA-N 0.000 description 1
- GKRCCTYAGQPMMP-IHRRRGAJSA-N Phe-Ser-Met Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCSC)C(O)=O GKRCCTYAGQPMMP-IHRRRGAJSA-N 0.000 description 1
- IAOZOFPONWDXNT-IXOXFDKPSA-N Phe-Ser-Thr Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O IAOZOFPONWDXNT-IXOXFDKPSA-N 0.000 description 1
- RAGOJJCBGXARPO-XVSYOHENSA-N Phe-Thr-Asp Chemical compound OC(=O)C[C@@H](C(O)=O)NC(=O)[C@H]([C@H](O)C)NC(=O)[C@@H](N)CC1=CC=CC=C1 RAGOJJCBGXARPO-XVSYOHENSA-N 0.000 description 1
- XNQMZHLAYFWSGJ-HTUGSXCWSA-N Phe-Thr-Glu Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O XNQMZHLAYFWSGJ-HTUGSXCWSA-N 0.000 description 1
- BSKMOCNNLNDIMU-CDMKHQONSA-N Phe-Thr-Gly Chemical compound [H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(O)=O BSKMOCNNLNDIMU-CDMKHQONSA-N 0.000 description 1
- YFXXRYFWJFQAFW-JHYOHUSXSA-N Phe-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N)O YFXXRYFWJFQAFW-JHYOHUSXSA-N 0.000 description 1
- NWVMQNAELALJFW-RNXOBYDBSA-N Phe-Trp-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=CC=C1 NWVMQNAELALJFW-RNXOBYDBSA-N 0.000 description 1
- VFDRDMOMHBJGKD-UFYCRDLUSA-N Phe-Tyr-Arg Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N VFDRDMOMHBJGKD-UFYCRDLUSA-N 0.000 description 1
- QUUCAHIYARMNBL-FHWLQOOXSA-N Phe-Tyr-Gln Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N QUUCAHIYARMNBL-FHWLQOOXSA-N 0.000 description 1
- GCFNFKNPCMBHNT-IRXDYDNUSA-N Phe-Tyr-Gly Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)NCC(=O)O)N GCFNFKNPCMBHNT-IRXDYDNUSA-N 0.000 description 1
- MHNBYYFXWDUGBW-RPTUDFQQSA-N Phe-Tyr-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)NC(=O)[C@H](CC2=CC=CC=C2)N)O MHNBYYFXWDUGBW-RPTUDFQQSA-N 0.000 description 1
- XALFIVXGQUEGKV-JSGCOSHPSA-N Phe-Val-Gly Chemical compound OC(=O)CNC(=O)[C@H](C(C)C)NC(=O)[C@@H](N)CC1=CC=CC=C1 XALFIVXGQUEGKV-JSGCOSHPSA-N 0.000 description 1
- IEIFEYBAYFSRBQ-IHRRRGAJSA-N Phe-Val-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)N IEIFEYBAYFSRBQ-IHRRRGAJSA-N 0.000 description 1
- 206010035226 Plasma cell myeloma Diseases 0.000 description 1
- 102000013566 Plasminogen Human genes 0.000 description 1
- 108010051456 Plasminogen Proteins 0.000 description 1
- 241000224016 Plasmodium Species 0.000 description 1
- 241000607000 Plesiomonas Species 0.000 description 1
- 241000142787 Pneumocystis jirovecii Species 0.000 description 1
- 229920001213 Polysorbate 20 Polymers 0.000 description 1
- 239000004793 Polystyrene Substances 0.000 description 1
- 241000700625 Poxviridae Species 0.000 description 1
- 108091000054 Prion Proteins 0.000 description 1
- 102000029797 Prion Human genes 0.000 description 1
- VXCHGLYSIOOZIS-GUBZILKMSA-N Pro-Ala-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 VXCHGLYSIOOZIS-GUBZILKMSA-N 0.000 description 1
- CGBYDGAJHSOGFQ-LPEHRKFASA-N Pro-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@@H]2CCCN2 CGBYDGAJHSOGFQ-LPEHRKFASA-N 0.000 description 1
- NHDVNAKDACFHPX-GUBZILKMSA-N Pro-Arg-Ala Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(O)=O NHDVNAKDACFHPX-GUBZILKMSA-N 0.000 description 1
- LNLNHXIQPGKRJQ-SRVKXCTJSA-N Pro-Arg-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@@H]1CCCN1 LNLNHXIQPGKRJQ-SRVKXCTJSA-N 0.000 description 1
- VCYJKOLZYPYGJV-AVGNSLFASA-N Pro-Arg-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O VCYJKOLZYPYGJV-AVGNSLFASA-N 0.000 description 1
- NUZHSNLQJDYSRW-BZSNNMDCSA-N Pro-Arg-Trp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O NUZHSNLQJDYSRW-BZSNNMDCSA-N 0.000 description 1
- TXPUNZXZDVJUJQ-LPEHRKFASA-N Pro-Asn-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CC(=O)N)C(=O)N2CCC[C@@H]2C(=O)O TXPUNZXZDVJUJQ-LPEHRKFASA-N 0.000 description 1
- UTAUEDINXUMHLG-FXQIFTODSA-N Pro-Asp-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@@H]1CCCN1 UTAUEDINXUMHLG-FXQIFTODSA-N 0.000 description 1
- KPDRZQUWJKTMBP-DCAQKATOSA-N Pro-Asp-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@@H]1CCCN1 KPDRZQUWJKTMBP-DCAQKATOSA-N 0.000 description 1
- SFECXGVELZFBFJ-VEVYYDQMSA-N Pro-Asp-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SFECXGVELZFBFJ-VEVYYDQMSA-N 0.000 description 1
- DIZLUAZLNDFDPR-CIUDSAMLSA-N Pro-Cys-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CS)NC(=O)[C@@H]1CCCN1 DIZLUAZLNDFDPR-CIUDSAMLSA-N 0.000 description 1
- OLTFZQIYCNOBLI-DCAQKATOSA-N Pro-Cys-Lys Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCCCN)C(=O)O OLTFZQIYCNOBLI-DCAQKATOSA-N 0.000 description 1
- PZSCUPVOJGKHEP-CIUDSAMLSA-N Pro-Gln-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O PZSCUPVOJGKHEP-CIUDSAMLSA-N 0.000 description 1
- VOZIBWWZSBIXQN-SRVKXCTJSA-N Pro-Glu-Lys Chemical compound NCCCC[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H]1CCCN1)C(O)=O VOZIBWWZSBIXQN-SRVKXCTJSA-N 0.000 description 1
- WFHYFCWBLSKEMS-KKUMJFAQSA-N Pro-Glu-Phe Chemical compound N([C@@H](CCC(=O)O)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C(=O)[C@@H]1CCCN1 WFHYFCWBLSKEMS-KKUMJFAQSA-N 0.000 description 1
- LGSANCBHSMDFDY-GARJFASQSA-N Pro-Glu-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCC(=O)O)C(=O)N2CCC[C@@H]2C(=O)O LGSANCBHSMDFDY-GARJFASQSA-N 0.000 description 1
- LXVLKXPFIDDHJG-CIUDSAMLSA-N Pro-Glu-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O LXVLKXPFIDDHJG-CIUDSAMLSA-N 0.000 description 1
- QGOZJLYCGRYYRW-KKUMJFAQSA-N Pro-Glu-Tyr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O QGOZJLYCGRYYRW-KKUMJFAQSA-N 0.000 description 1
- DMKWYMWNEKIPFC-IUCAKERBSA-N Pro-Gly-Arg Chemical compound [H]N1CCC[C@H]1C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(O)=O DMKWYMWNEKIPFC-IUCAKERBSA-N 0.000 description 1
- QNZLIVROMORQFH-BQBZGAKWSA-N Pro-Gly-Cys Chemical compound C1C[C@H](NC1)C(=O)NCC(=O)N[C@@H](CS)C(=O)O QNZLIVROMORQFH-BQBZGAKWSA-N 0.000 description 1
- JMVQDLDPDBXAAX-YUMQZZPRSA-N Pro-Gly-Gln Chemical compound NC(=O)CC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H]1CCCN1 JMVQDLDPDBXAAX-YUMQZZPRSA-N 0.000 description 1
- HAAQQNHQZBOWFO-LURJTMIESA-N Pro-Gly-Gly Chemical compound OC(=O)CNC(=O)CNC(=O)[C@@H]1CCCN1 HAAQQNHQZBOWFO-LURJTMIESA-N 0.000 description 1
- VWXGFAIZUQBBBG-UWVGGRQHSA-N Pro-His-Gly Chemical compound C([C@@H](C(=O)NCC(=O)[O-])NC(=O)[C@H]1[NH2+]CCC1)C1=CN=CN1 VWXGFAIZUQBBBG-UWVGGRQHSA-N 0.000 description 1
- SOACYAXADBWDDT-CYDGBPFRSA-N Pro-Ile-Arg Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O SOACYAXADBWDDT-CYDGBPFRSA-N 0.000 description 1
- AQGUSRZKDZYGGV-GMOBBJLQSA-N Pro-Ile-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O AQGUSRZKDZYGGV-GMOBBJLQSA-N 0.000 description 1
- FJLODLCIOJUDRG-PYJNHQTQSA-N Pro-Ile-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@@H]2CCCN2 FJLODLCIOJUDRG-PYJNHQTQSA-N 0.000 description 1
- LNOWDSPAYBWJOR-PEDHHIEDSA-N Pro-Ile-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O LNOWDSPAYBWJOR-PEDHHIEDSA-N 0.000 description 1
- UREQLMJCKFLLHM-NAKRPEOUSA-N Pro-Ile-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CO)C(O)=O UREQLMJCKFLLHM-NAKRPEOUSA-N 0.000 description 1
- FMLRRBDLBJLJIK-DCAQKATOSA-N Pro-Leu-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 FMLRRBDLBJLJIK-DCAQKATOSA-N 0.000 description 1
- HFNPOYOKIPGAEI-SRVKXCTJSA-N Pro-Leu-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 HFNPOYOKIPGAEI-SRVKXCTJSA-N 0.000 description 1
- FXGIMYRVJJEIIM-UWVGGRQHSA-N Pro-Leu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 FXGIMYRVJJEIIM-UWVGGRQHSA-N 0.000 description 1
- CPRLKHJUFAXVTD-ULQDDVLXSA-N Pro-Leu-Tyr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O CPRLKHJUFAXVTD-ULQDDVLXSA-N 0.000 description 1
- SUENWIFTSTWUKD-AVGNSLFASA-N Pro-Leu-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C(C)C)C(O)=O SUENWIFTSTWUKD-AVGNSLFASA-N 0.000 description 1
- ABSSTGUCBCDKMU-UWVGGRQHSA-N Pro-Lys-Gly Chemical compound NCCCC[C@@H](C(=O)NCC(O)=O)NC(=O)[C@@H]1CCCN1 ABSSTGUCBCDKMU-UWVGGRQHSA-N 0.000 description 1
- HBBBLSVBQGZKOZ-GUBZILKMSA-N Pro-Met-Ala Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CCSC)C(=O)N[C@@H](C)C(O)=O HBBBLSVBQGZKOZ-GUBZILKMSA-N 0.000 description 1
- KDBHVPXBQADZKY-GUBZILKMSA-N Pro-Pro-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 KDBHVPXBQADZKY-GUBZILKMSA-N 0.000 description 1
- FHZJRBVMLGOHBX-GUBZILKMSA-N Pro-Pro-Asp Chemical compound OC(=O)C[C@H](NC(=O)[C@@H]1CCCN1C(=O)[C@@H]1CCCN1)C(O)=O FHZJRBVMLGOHBX-GUBZILKMSA-N 0.000 description 1
- FYKUEXMZYFIZKA-DCAQKATOSA-N Pro-Pro-Gln Chemical compound [H]N1CCC[C@H]1C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O FYKUEXMZYFIZKA-DCAQKATOSA-N 0.000 description 1
- KBUAPZAZPWNYSW-SRVKXCTJSA-N Pro-Pro-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 KBUAPZAZPWNYSW-SRVKXCTJSA-N 0.000 description 1
- GMJDSFYVTAMIBF-FXQIFTODSA-N Pro-Ser-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O GMJDSFYVTAMIBF-FXQIFTODSA-N 0.000 description 1
- BGWKULMLUIUPKY-BQBZGAKWSA-N Pro-Ser-Gly Chemical compound OC(=O)CNC(=O)[C@H](CO)NC(=O)[C@@H]1CCCN1 BGWKULMLUIUPKY-BQBZGAKWSA-N 0.000 description 1
- LNICFEXCAHIJOR-DCAQKATOSA-N Pro-Ser-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O LNICFEXCAHIJOR-DCAQKATOSA-N 0.000 description 1
- KWMZPPWYBVZIER-XGEHTFHBSA-N Pro-Ser-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O KWMZPPWYBVZIER-XGEHTFHBSA-N 0.000 description 1
- PRKWBYCXBBSLSK-GUBZILKMSA-N Pro-Ser-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(O)=O PRKWBYCXBBSLSK-GUBZILKMSA-N 0.000 description 1
- JDJMFMVVJHLWDP-UNQGMJICSA-N Pro-Thr-Phe Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O JDJMFMVVJHLWDP-UNQGMJICSA-N 0.000 description 1
- AIOWVDNPESPXRB-YTWAJWBKSA-N Pro-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@@H]2CCCN2)O AIOWVDNPESPXRB-YTWAJWBKSA-N 0.000 description 1
- CXGLFEOYCJFKPR-RCWTZXSCSA-N Pro-Thr-Val Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O CXGLFEOYCJFKPR-RCWTZXSCSA-N 0.000 description 1
- YIPFBJGBRCJJJD-FHWLQOOXSA-N Pro-Trp-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@@H]3CCCN3 YIPFBJGBRCJJJD-FHWLQOOXSA-N 0.000 description 1
- QDDJNKWPTJHROJ-UFYCRDLUSA-N Pro-Tyr-Tyr Chemical compound C([C@@H](C(=O)O)NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)[C@H]1NCCC1)C1=CC=C(O)C=C1 QDDJNKWPTJHROJ-UFYCRDLUSA-N 0.000 description 1
- WWXNZNWZNZPDIF-SRVKXCTJSA-N Pro-Val-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H]1CCCN1 WWXNZNWZNZPDIF-SRVKXCTJSA-N 0.000 description 1
- ZAUHSLVPDLNTRZ-QXEWZRGKSA-N Pro-Val-Asn Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O ZAUHSLVPDLNTRZ-QXEWZRGKSA-N 0.000 description 1
- STGVYUTZKGPRCI-GUBZILKMSA-N Pro-Val-Cys Chemical compound SC[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H]1CCCN1 STGVYUTZKGPRCI-GUBZILKMSA-N 0.000 description 1
- FUOGXAQMNJMBFG-WPRPVWTQSA-N Pro-Val-Gly Chemical compound OC(=O)CNC(=O)[C@H](C(C)C)NC(=O)[C@@H]1CCCN1 FUOGXAQMNJMBFG-WPRPVWTQSA-N 0.000 description 1
- OQSGBXGNAFQGGS-CYDGBPFRSA-N Pro-Val-Ile Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O OQSGBXGNAFQGGS-CYDGBPFRSA-N 0.000 description 1
- ZMLRZBWCXPQADC-TUAOUCFPSA-N Pro-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@@H]2CCCN2 ZMLRZBWCXPQADC-TUAOUCFPSA-N 0.000 description 1
- 241000282330 Procyon lotor Species 0.000 description 1
- 241000186429 Propionibacterium Species 0.000 description 1
- 108010072866 Prostate-Specific Antigen Proteins 0.000 description 1
- 102100038358 Prostate-specific antigen Human genes 0.000 description 1
- 239000004365 Protease Substances 0.000 description 1
- 241000196250 Prototheca Species 0.000 description 1
- 241000223596 Pseudallescheria Species 0.000 description 1
- 241001137874 Pseudophyllidea Species 0.000 description 1
- 101800001554 RNA-directed RNA polymerase Proteins 0.000 description 1
- 241000711798 Rabies lyssavirus Species 0.000 description 1
- 102100030086 Receptor tyrosine-protein kinase erbB-2 Human genes 0.000 description 1
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 1
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 1
- 208000015634 Rectal Neoplasms Diseases 0.000 description 1
- 241000702247 Reoviridae Species 0.000 description 1
- 241000712907 Retroviridae Species 0.000 description 1
- 241000244200 Rhabditida Species 0.000 description 1
- 241000711931 Rhabdoviridae Species 0.000 description 1
- 241000235527 Rhizopus Species 0.000 description 1
- 241000220010 Rhode Species 0.000 description 1
- 241000316848 Rhodococcus <scale insect> Species 0.000 description 1
- 241000223252 Rhodotorula Species 0.000 description 1
- IWUCXVSUMQZMFG-AFCXAGJDSA-N Ribavirin Chemical compound N1=C(C(=O)N)N=CN1[C@H]1[C@H](O)[C@H](O)[C@@H](CO)O1 IWUCXVSUMQZMFG-AFCXAGJDSA-N 0.000 description 1
- 241000702670 Rotavirus Species 0.000 description 1
- 241000132889 Scedosporium Species 0.000 description 1
- 241000242678 Schistosoma Species 0.000 description 1
- 241000235347 Schizosaccharomyces pombe Species 0.000 description 1
- 229920002684 Sepharose Polymers 0.000 description 1
- MWMKFWJYRRGXOR-ZLUOBGJFSA-N Ser-Ala-Asn Chemical compound N[C@H](C(=O)N[C@H](C(=O)N[C@H](C(=O)O)CC(N)=O)C)CO MWMKFWJYRRGXOR-ZLUOBGJFSA-N 0.000 description 1
- SRTCFKGBYBZRHA-ACZMJKKPSA-N Ser-Ala-Glu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(O)=O SRTCFKGBYBZRHA-ACZMJKKPSA-N 0.000 description 1
- BTKUIVBNGBFTTP-WHFBIAKZSA-N Ser-Ala-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)NCC(O)=O BTKUIVBNGBFTTP-WHFBIAKZSA-N 0.000 description 1
- HRNQLKCLPVKZNE-CIUDSAMLSA-N Ser-Ala-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H](CC(C)C)C(O)=O HRNQLKCLPVKZNE-CIUDSAMLSA-N 0.000 description 1
- IDQFQFVEWMWRQQ-DLOVCJGASA-N Ser-Ala-Phe Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O IDQFQFVEWMWRQQ-DLOVCJGASA-N 0.000 description 1
- BRKHVZNDAOMAHX-BIIVOSGPSA-N Ser-Ala-Pro Chemical compound C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N BRKHVZNDAOMAHX-BIIVOSGPSA-N 0.000 description 1
- GXXTUIUYTWGPMV-FXQIFTODSA-N Ser-Arg-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(O)=O GXXTUIUYTWGPMV-FXQIFTODSA-N 0.000 description 1
- NLQUOHDCLSFABG-GUBZILKMSA-N Ser-Arg-Arg Chemical compound NC(N)=NCCC[C@H](NC(=O)[C@H](CO)N)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O NLQUOHDCLSFABG-GUBZILKMSA-N 0.000 description 1
- RZUOXAKGNHXZTB-GUBZILKMSA-N Ser-Arg-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCSC)C(O)=O RZUOXAKGNHXZTB-GUBZILKMSA-N 0.000 description 1
- NRCJWSGXMAPYQX-LPEHRKFASA-N Ser-Arg-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CO)N)C(=O)O NRCJWSGXMAPYQX-LPEHRKFASA-N 0.000 description 1
- OYEDZGNMSBZCIM-XGEHTFHBSA-N Ser-Arg-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(O)=O OYEDZGNMSBZCIM-XGEHTFHBSA-N 0.000 description 1
- KAAPNMOKUUPKOE-SRVKXCTJSA-N Ser-Asn-Phe Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 KAAPNMOKUUPKOE-SRVKXCTJSA-N 0.000 description 1
- TYYBJUYSTWJHGO-ZKWXMUAHSA-N Ser-Asn-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O TYYBJUYSTWJHGO-ZKWXMUAHSA-N 0.000 description 1
- BNFVPSRLHHPQKS-WHFBIAKZSA-N Ser-Asp-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O BNFVPSRLHHPQKS-WHFBIAKZSA-N 0.000 description 1
- BGOWRLSWJCVYAQ-CIUDSAMLSA-N Ser-Asp-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O BGOWRLSWJCVYAQ-CIUDSAMLSA-N 0.000 description 1
- DBIDZNUXSLXVRG-FXQIFTODSA-N Ser-Asp-Met Chemical compound CSCC[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@H](CO)N DBIDZNUXSLXVRG-FXQIFTODSA-N 0.000 description 1
- OLIJLNWFEQEFDM-SRVKXCTJSA-N Ser-Asp-Phe Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 OLIJLNWFEQEFDM-SRVKXCTJSA-N 0.000 description 1
- SWSRFJZZMNLMLY-ZKWXMUAHSA-N Ser-Asp-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O SWSRFJZZMNLMLY-ZKWXMUAHSA-N 0.000 description 1
- RNFKSBPHLTZHLU-WHFBIAKZSA-N Ser-Cys-Gly Chemical compound C([C@@H](C(=O)N[C@@H](CS)C(=O)NCC(=O)O)N)O RNFKSBPHLTZHLU-WHFBIAKZSA-N 0.000 description 1
- MOVJSUIKUNCVMG-ZLUOBGJFSA-N Ser-Cys-Ser Chemical compound C([C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CO)C(=O)O)N)O MOVJSUIKUNCVMG-ZLUOBGJFSA-N 0.000 description 1
- CDVFZMOFNJPUDD-ACZMJKKPSA-N Ser-Gln-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O CDVFZMOFNJPUDD-ACZMJKKPSA-N 0.000 description 1
- OJPHFSOMBZKQKQ-GUBZILKMSA-N Ser-Gln-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](N)CO OJPHFSOMBZKQKQ-GUBZILKMSA-N 0.000 description 1
- PVDTYLHUWAEYGY-CIUDSAMLSA-N Ser-Glu-Arg Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O PVDTYLHUWAEYGY-CIUDSAMLSA-N 0.000 description 1
- SQBLRDDJTUJDMV-ACZMJKKPSA-N Ser-Glu-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O SQBLRDDJTUJDMV-ACZMJKKPSA-N 0.000 description 1
- GYXVUTAOICLGKJ-ACZMJKKPSA-N Ser-Glu-Cys Chemical compound C(CC(=O)O)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CO)N GYXVUTAOICLGKJ-ACZMJKKPSA-N 0.000 description 1
- LALNXSXEYFUUDD-GUBZILKMSA-N Ser-Glu-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O LALNXSXEYFUUDD-GUBZILKMSA-N 0.000 description 1
- UFKPDBLKLOBMRH-XHNCKOQMSA-N Ser-Glu-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CO)N)C(=O)O UFKPDBLKLOBMRH-XHNCKOQMSA-N 0.000 description 1
- MUARUIBTKQJKFY-WHFBIAKZSA-N Ser-Gly-Asp Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(O)=O MUARUIBTKQJKFY-WHFBIAKZSA-N 0.000 description 1
- MIJWOJAXARLEHA-WDSKDSINSA-N Ser-Gly-Glu Chemical compound OC[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CCC(O)=O MIJWOJAXARLEHA-WDSKDSINSA-N 0.000 description 1
- WSTIOCFMWXNOCX-YUMQZZPRSA-N Ser-Gly-Lys Chemical compound C(CCN)C[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CO)N WSTIOCFMWXNOCX-YUMQZZPRSA-N 0.000 description 1
- JFWDJFULOLKQFY-QWRGUYRKSA-N Ser-Gly-Phe Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O JFWDJFULOLKQFY-QWRGUYRKSA-N 0.000 description 1
- UIGMAMGZOJVTDN-WHFBIAKZSA-N Ser-Gly-Ser Chemical compound OC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O UIGMAMGZOJVTDN-WHFBIAKZSA-N 0.000 description 1
- UGHCUDLCCVVIJR-VGDYDELISA-N Ser-His-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CO)N UGHCUDLCCVVIJR-VGDYDELISA-N 0.000 description 1
- SFTZTYBXIXLRGQ-JBDRJPRFSA-N Ser-Ile-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O SFTZTYBXIXLRGQ-JBDRJPRFSA-N 0.000 description 1
- RIAKPZVSNBBNRE-BJDJZHNGSA-N Ser-Ile-Leu Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O RIAKPZVSNBBNRE-BJDJZHNGSA-N 0.000 description 1
- ZOPISOXXPQNOCO-SVSWQMSJSA-N Ser-Ile-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](CO)N ZOPISOXXPQNOCO-SVSWQMSJSA-N 0.000 description 1
- QYSFWUIXDFJUDW-DCAQKATOSA-N Ser-Leu-Arg Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O QYSFWUIXDFJUDW-DCAQKATOSA-N 0.000 description 1
- NLOAIFSWUUFQFR-CIUDSAMLSA-N Ser-Leu-Asp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O NLOAIFSWUUFQFR-CIUDSAMLSA-N 0.000 description 1
- GJFYFGOEWLDQGW-GUBZILKMSA-N Ser-Leu-Gln Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CO)N GJFYFGOEWLDQGW-GUBZILKMSA-N 0.000 description 1
- IUXGJEIKJBYKOO-SRVKXCTJSA-N Ser-Leu-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CO)N IUXGJEIKJBYKOO-SRVKXCTJSA-N 0.000 description 1
- VMLONWHIORGALA-SRVKXCTJSA-N Ser-Leu-Leu Chemical compound CC(C)C[C@@H](C([O-])=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]([NH3+])CO VMLONWHIORGALA-SRVKXCTJSA-N 0.000 description 1
- XXNYYSXNXCJYKX-DCAQKATOSA-N Ser-Leu-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCSC)C(O)=O XXNYYSXNXCJYKX-DCAQKATOSA-N 0.000 description 1
- XNXRTQZTFVMJIJ-DCAQKATOSA-N Ser-Met-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CC(C)C)C(O)=O XNXRTQZTFVMJIJ-DCAQKATOSA-N 0.000 description 1
- JAWGSPUJAXYXJA-IHRRRGAJSA-N Ser-Phe-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@H](CO)N)CC1=CC=CC=C1 JAWGSPUJAXYXJA-IHRRRGAJSA-N 0.000 description 1
- GDUZTEQRAOXYJS-SRVKXCTJSA-N Ser-Phe-Asn Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CO)N GDUZTEQRAOXYJS-SRVKXCTJSA-N 0.000 description 1
- FZEUTKVQGMVGHW-AVGNSLFASA-N Ser-Phe-Gln Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O FZEUTKVQGMVGHW-AVGNSLFASA-N 0.000 description 1
- MQUZANJDFOQOBX-SRVKXCTJSA-N Ser-Phe-Ser Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O MQUZANJDFOQOBX-SRVKXCTJSA-N 0.000 description 1
- PJIQEIFXZPCWOJ-FXQIFTODSA-N Ser-Pro-Asp Chemical compound [H]N[C@@H](CO)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(O)=O PJIQEIFXZPCWOJ-FXQIFTODSA-N 0.000 description 1
- DINQYZRMXGWWTG-GUBZILKMSA-N Ser-Pro-Pro Chemical compound OC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 DINQYZRMXGWWTG-GUBZILKMSA-N 0.000 description 1
- HHJFMHQYEAAOBM-ZLUOBGJFSA-N Ser-Ser-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O HHJFMHQYEAAOBM-ZLUOBGJFSA-N 0.000 description 1
- WLJPJRGQRNCIQS-ZLUOBGJFSA-N Ser-Ser-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O WLJPJRGQRNCIQS-ZLUOBGJFSA-N 0.000 description 1
- VFWQQZMRKFOGLE-ZLUOBGJFSA-N Ser-Ser-Cys Chemical compound C([C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O)N)O VFWQQZMRKFOGLE-ZLUOBGJFSA-N 0.000 description 1
- JCLAFVNDBJMLBC-JBDRJPRFSA-N Ser-Ser-Ile Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JCLAFVNDBJMLBC-JBDRJPRFSA-N 0.000 description 1
- BMKNXTJLHFIAAH-CIUDSAMLSA-N Ser-Ser-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O BMKNXTJLHFIAAH-CIUDSAMLSA-N 0.000 description 1
- AABIBDJHSKIMJK-FXQIFTODSA-N Ser-Ser-Met Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCSC)C(O)=O AABIBDJHSKIMJK-FXQIFTODSA-N 0.000 description 1
- OLKICIBQRVSQMA-SRVKXCTJSA-N Ser-Ser-Tyr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O OLKICIBQRVSQMA-SRVKXCTJSA-N 0.000 description 1
- RXUOAOOZIWABBW-XGEHTFHBSA-N Ser-Thr-Arg Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N RXUOAOOZIWABBW-XGEHTFHBSA-N 0.000 description 1
- KKKVOZNCLALMPV-XKBZYTNZSA-N Ser-Thr-Glu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=O KKKVOZNCLALMPV-XKBZYTNZSA-N 0.000 description 1
- NADLKBTYNKUJEP-KATARQTJSA-N Ser-Thr-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O NADLKBTYNKUJEP-KATARQTJSA-N 0.000 description 1
- PCJLFYBAQZQOFE-KATARQTJSA-N Ser-Thr-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CO)N)O PCJLFYBAQZQOFE-KATARQTJSA-N 0.000 description 1
- UYLKOSODXYSWMQ-XGEHTFHBSA-N Ser-Thr-Met Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CO)N)O UYLKOSODXYSWMQ-XGEHTFHBSA-N 0.000 description 1
- SNXUIBACCONSOH-BWBBJGPYSA-N Ser-Thr-Ser Chemical compound OC[C@H](N)C(=O)N[C@@H]([C@H](O)C)C(=O)N[C@@H](CO)C(O)=O SNXUIBACCONSOH-BWBBJGPYSA-N 0.000 description 1
- VLMIUSLQONKLDV-HEIBUPTGSA-N Ser-Thr-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VLMIUSLQONKLDV-HEIBUPTGSA-N 0.000 description 1
- OJFFAQFRCVPHNN-JYBASQMISA-N Ser-Thr-Trp Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(O)=O OJFFAQFRCVPHNN-JYBASQMISA-N 0.000 description 1
- ZKOKTQPHFMRSJP-YJRXYDGGSA-N Ser-Thr-Tyr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O ZKOKTQPHFMRSJP-YJRXYDGGSA-N 0.000 description 1
- STIAINRLUUKYKM-WFBYXXMGSA-N Ser-Trp-Ala Chemical compound C1=CC=C2C(C[C@@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@@H](N)CO)=CNC2=C1 STIAINRLUUKYKM-WFBYXXMGSA-N 0.000 description 1
- UQGAAZXSCGWMFU-UBHSHLNASA-N Ser-Trp-Asp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CO)N UQGAAZXSCGWMFU-UBHSHLNASA-N 0.000 description 1
- VAIWUNAAPZZGRI-IHPCNDPISA-N Ser-Trp-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CC2=CNC3=CC=CC=C32)NC(=O)[C@H](CO)N VAIWUNAAPZZGRI-IHPCNDPISA-N 0.000 description 1
- NERYDXBVARJIQS-JYBASQMISA-N Ser-Trp-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)NC(=O)[C@H](CO)N)O NERYDXBVARJIQS-JYBASQMISA-N 0.000 description 1
- GSCVDSBEYVGMJQ-SRVKXCTJSA-N Ser-Tyr-Asp Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](CO)N)O GSCVDSBEYVGMJQ-SRVKXCTJSA-N 0.000 description 1
- PLQWGQUNUPMNOD-KKUMJFAQSA-N Ser-Tyr-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(O)=O PLQWGQUNUPMNOD-KKUMJFAQSA-N 0.000 description 1
- VVKVHAOOUGNDPJ-SRVKXCTJSA-N Ser-Tyr-Ser Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CO)C(O)=O VVKVHAOOUGNDPJ-SRVKXCTJSA-N 0.000 description 1
- PMTWIUBUQRGCSB-FXQIFTODSA-N Ser-Val-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C)C(O)=O PMTWIUBUQRGCSB-FXQIFTODSA-N 0.000 description 1
- JZRYFUGREMECBH-XPUUQOCRSA-N Ser-Val-Gly Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)NCC(O)=O JZRYFUGREMECBH-XPUUQOCRSA-N 0.000 description 1
- YEDSOSIKVUMIJE-DCAQKATOSA-N Ser-Val-Leu Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O YEDSOSIKVUMIJE-DCAQKATOSA-N 0.000 description 1
- ANOQEBQWIAYIMV-AEJSXWLSSA-N Ser-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CO)N ANOQEBQWIAYIMV-AEJSXWLSSA-N 0.000 description 1
- SIEBDTCABMZCLF-XGEHTFHBSA-N Ser-Val-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SIEBDTCABMZCLF-XGEHTFHBSA-N 0.000 description 1
- 101800001838 Serine protease/helicase NS3 Proteins 0.000 description 1
- 241000607720 Serratia Species 0.000 description 1
- 241000607715 Serratia marcescens Species 0.000 description 1
- 241000700584 Simplexvirus Species 0.000 description 1
- 108010003723 Single-Domain Antibodies Proteins 0.000 description 1
- 244000061458 Solanum melongena Species 0.000 description 1
- 235000002597 Solanum melongena Nutrition 0.000 description 1
- 241000244042 Spirurida Species 0.000 description 1
- 241001149962 Sporothrix Species 0.000 description 1
- 229920002472 Starch Polymers 0.000 description 1
- 241000122971 Stenotrophomonas Species 0.000 description 1
- 229930182558 Sterol Natural products 0.000 description 1
- 208000005718 Stomach Neoplasms Diseases 0.000 description 1
- 241000243788 Strongylida Species 0.000 description 1
- 241000244174 Strongyloides Species 0.000 description 1
- 108700005078 Synthetic Genes Proteins 0.000 description 1
- 230000029662 T-helper 1 type immune response Effects 0.000 description 1
- PZBFGYYEXUXCOF-UHFFFAOYSA-N TCEP Chemical compound OC(=O)CCP(CCC(O)=O)CCC(O)=O PZBFGYYEXUXCOF-UHFFFAOYSA-N 0.000 description 1
- 102000003141 Tachykinin Human genes 0.000 description 1
- 241000244155 Taenia Species 0.000 description 1
- 241001523006 Talaromyces marneffei Species 0.000 description 1
- 102000007000 Tenascin Human genes 0.000 description 1
- 108010008125 Tenascin Proteins 0.000 description 1
- 108700031126 Tetraspanins Proteins 0.000 description 1
- 102000043977 Tetraspanins Human genes 0.000 description 1
- IGROJMCBGRFRGI-YTLHQDLWSA-N Thr-Ala-Ala Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](C)C(O)=O IGROJMCBGRFRGI-YTLHQDLWSA-N 0.000 description 1
- DWYAUVCQDTZIJI-VZFHVOOUSA-N Thr-Ala-Ser Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O DWYAUVCQDTZIJI-VZFHVOOUSA-N 0.000 description 1
- GLQFKOVWXPPFTP-VEVYYDQMSA-N Thr-Arg-Asp Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O GLQFKOVWXPPFTP-VEVYYDQMSA-N 0.000 description 1
- JMQUAZXYFAEOIH-XGEHTFHBSA-N Thr-Arg-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CS)C(=O)O)N)O JMQUAZXYFAEOIH-XGEHTFHBSA-N 0.000 description 1
- NAXBBCLCEOTAIG-RHYQMDGZSA-N Thr-Arg-Lys Chemical compound NC(N)=NCCC[C@H](NC(=O)[C@@H](N)[C@H](O)C)C(=O)N[C@@H](CCCCN)C(O)=O NAXBBCLCEOTAIG-RHYQMDGZSA-N 0.000 description 1
- WFUAUEQXPVNAEF-ZJDVBMNYSA-N Thr-Arg-Thr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)O)C(O)=O)CCCN=C(N)N WFUAUEQXPVNAEF-ZJDVBMNYSA-N 0.000 description 1
- UNURFMVMXLENAZ-KJEVXHAQSA-N Thr-Arg-Tyr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O UNURFMVMXLENAZ-KJEVXHAQSA-N 0.000 description 1
- JNQZPAWOPBZGIX-RCWTZXSCSA-N Thr-Arg-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)[C@@H](C)O)CCCN=C(N)N JNQZPAWOPBZGIX-RCWTZXSCSA-N 0.000 description 1
- LXWZOMSOUAMOIA-JIOCBJNQSA-N Thr-Asn-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N1CCC[C@@H]1C(=O)O)N)O LXWZOMSOUAMOIA-JIOCBJNQSA-N 0.000 description 1
- PZVGOVRNGKEFCB-KKHAAJSZSA-N Thr-Asn-Val Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](C(C)C)C(=O)O)N)O PZVGOVRNGKEFCB-KKHAAJSZSA-N 0.000 description 1
- NLJKZUGAIIRWJN-LKXGYXEUSA-N Thr-Asp-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CS)C(=O)O)N)O NLJKZUGAIIRWJN-LKXGYXEUSA-N 0.000 description 1
- NLSNVZAREYQMGR-HJGDQZAQSA-N Thr-Asp-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O NLSNVZAREYQMGR-HJGDQZAQSA-N 0.000 description 1
- JXKMXEBNZCKSDY-JIOCBJNQSA-N Thr-Asp-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N1CCC[C@@H]1C(=O)O)N)O JXKMXEBNZCKSDY-JIOCBJNQSA-N 0.000 description 1
- LYGKYFKSZTUXGZ-ZDLURKLDSA-N Thr-Cys-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CS)C(=O)NCC(O)=O LYGKYFKSZTUXGZ-ZDLURKLDSA-N 0.000 description 1
- ZLNWJMRLHLGKFX-SVSWQMSJSA-N Thr-Cys-Ile Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CS)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O ZLNWJMRLHLGKFX-SVSWQMSJSA-N 0.000 description 1
- DSLHSTIUAPKERR-XGEHTFHBSA-N Thr-Cys-Val Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CS)C(=O)N[C@@H](C(C)C)C(O)=O DSLHSTIUAPKERR-XGEHTFHBSA-N 0.000 description 1
- WLDUCKSCDRIVLJ-NUMRIWBASA-N Thr-Gln-Asp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC(=O)O)C(=O)O)N)O WLDUCKSCDRIVLJ-NUMRIWBASA-N 0.000 description 1
- RCEHMXVEMNXRIW-IRIUXVKKSA-N Thr-Gln-Tyr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N)O RCEHMXVEMNXRIW-IRIUXVKKSA-N 0.000 description 1
- FHDLKMFZKRUQCE-HJGDQZAQSA-N Thr-Glu-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O FHDLKMFZKRUQCE-HJGDQZAQSA-N 0.000 description 1
- LHEZGZQRLDBSRR-WDCWCFNPSA-N Thr-Glu-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O LHEZGZQRLDBSRR-WDCWCFNPSA-N 0.000 description 1
- VULNJDORNLBPNG-SWRJLBSHSA-N Thr-Glu-Trp Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N)O VULNJDORNLBPNG-SWRJLBSHSA-N 0.000 description 1
- BNGDYRRHRGOPHX-IFFSRLJSSA-N Thr-Glu-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)[C@@H](C)O)C(O)=O BNGDYRRHRGOPHX-IFFSRLJSSA-N 0.000 description 1
- SLUWOCTZVGMURC-BFHQHQDPSA-N Thr-Gly-Ala Chemical compound C[C@@H](O)[C@H](N)C(=O)NCC(=O)N[C@@H](C)C(O)=O SLUWOCTZVGMURC-BFHQHQDPSA-N 0.000 description 1
- XFTYVCHLARBHBQ-FOHZUACHSA-N Thr-Gly-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O XFTYVCHLARBHBQ-FOHZUACHSA-N 0.000 description 1
- YZUWGFXVVZQJEI-PMVVWTBXSA-N Thr-Gly-His Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N)O YZUWGFXVVZQJEI-PMVVWTBXSA-N 0.000 description 1
- MSIYNSBKKVMGFO-BHNWBGBOSA-N Thr-Gly-Pro Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)N1CCC[C@@H]1C(=O)O)N)O MSIYNSBKKVMGFO-BHNWBGBOSA-N 0.000 description 1
- KBBRNEDOYWMIJP-KYNKHSRBSA-N Thr-Gly-Thr Chemical compound C[C@H]([C@@H](C(=O)NCC(=O)N[C@@H]([C@@H](C)O)C(=O)O)N)O KBBRNEDOYWMIJP-KYNKHSRBSA-N 0.000 description 1
- NQVDGKYAUHTCME-QTKMDUPCSA-N Thr-His-Arg Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N)O NQVDGKYAUHTCME-QTKMDUPCSA-N 0.000 description 1
- FKIGTIXHSRNKJU-IXOXFDKPSA-N Thr-His-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)[C@H](O)C)CC1=CN=CN1 FKIGTIXHSRNKJU-IXOXFDKPSA-N 0.000 description 1
- KRGDDWVBBDLPSJ-CUJWVEQBSA-N Thr-His-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CO)C(O)=O KRGDDWVBBDLPSJ-CUJWVEQBSA-N 0.000 description 1
- YUPVPKZBKCLFLT-QTKMDUPCSA-N Thr-His-Val Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)N[C@@H](C(C)C)C(=O)O)N)O YUPVPKZBKCLFLT-QTKMDUPCSA-N 0.000 description 1
- PAXANSWUSVPFNK-IUKAMOBKSA-N Thr-Ile-Asn Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N PAXANSWUSVPFNK-IUKAMOBKSA-N 0.000 description 1
- XTCNBOBTROGWMW-RWRJDSDZSA-N Thr-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N XTCNBOBTROGWMW-RWRJDSDZSA-N 0.000 description 1
- URPSJRMWHQTARR-MBLNEYKQSA-N Thr-Ile-Gly Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(O)=O URPSJRMWHQTARR-MBLNEYKQSA-N 0.000 description 1
- JRAUIKJSEAKTGD-TUBUOCAGSA-N Thr-Ile-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N JRAUIKJSEAKTGD-TUBUOCAGSA-N 0.000 description 1
- LCCSEJSPBWKBNT-OSUNSFLBSA-N Thr-Ile-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H]([C@@H](C)O)N LCCSEJSPBWKBNT-OSUNSFLBSA-N 0.000 description 1
- GXUWHVZYDAHFSV-FLBSBUHZSA-N Thr-Ile-Thr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)O)C(O)=O GXUWHVZYDAHFSV-FLBSBUHZSA-N 0.000 description 1
- IMDMLDSVUSMAEJ-HJGDQZAQSA-N Thr-Leu-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=O IMDMLDSVUSMAEJ-HJGDQZAQSA-N 0.000 description 1
- XIULAFZYEKSGAJ-IXOXFDKPSA-N Thr-Leu-His Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CC1=CNC=N1 XIULAFZYEKSGAJ-IXOXFDKPSA-N 0.000 description 1
- MEJHFIOYJHTWMK-VOAKCMCISA-N Thr-Leu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)[C@@H](C)O MEJHFIOYJHTWMK-VOAKCMCISA-N 0.000 description 1
- PRNGXSILMXSWQQ-OEAJRASXSA-N Thr-Leu-Phe Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O PRNGXSILMXSWQQ-OEAJRASXSA-N 0.000 description 1
- CJXURNZYNHCYFD-WDCWCFNPSA-N Thr-Lys-Gln Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N)O CJXURNZYNHCYFD-WDCWCFNPSA-N 0.000 description 1
- SPVHQURZJCUDQC-VOAKCMCISA-N Thr-Lys-Leu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O SPVHQURZJCUDQC-VOAKCMCISA-N 0.000 description 1
- KKPOGALELPLJTL-MEYUZBJRSA-N Thr-Lys-Tyr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 KKPOGALELPLJTL-MEYUZBJRSA-N 0.000 description 1
- DXPURPNJDFCKKO-RHYQMDGZSA-N Thr-Lys-Val Chemical compound CC(C)[C@H](NC(=O)[C@H](CCCCN)NC(=O)[C@@H](N)[C@@H](C)O)C(O)=O DXPURPNJDFCKKO-RHYQMDGZSA-N 0.000 description 1
- OHDXOXIZXSFCDN-RCWTZXSCSA-N Thr-Met-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O OHDXOXIZXSFCDN-RCWTZXSCSA-N 0.000 description 1
- WYLAVUAWOUVUCA-XVSYOHENSA-N Thr-Phe-Asp Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(O)=O)C(O)=O WYLAVUAWOUVUCA-XVSYOHENSA-N 0.000 description 1
- JMBRNXUOLJFURW-BEAPCOKYSA-N Thr-Phe-Pro Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N2CCC[C@@H]2C(=O)O)N)O JMBRNXUOLJFURW-BEAPCOKYSA-N 0.000 description 1
- ABWNZPOIUJMNKT-IXOXFDKPSA-N Thr-Phe-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O ABWNZPOIUJMNKT-IXOXFDKPSA-N 0.000 description 1
- MXDOAJQRJBMGMO-FJXKBIBVSA-N Thr-Pro-Gly Chemical compound C[C@@H](O)[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O MXDOAJQRJBMGMO-FJXKBIBVSA-N 0.000 description 1
- MROIJTGJGIDEEJ-RCWTZXSCSA-N Thr-Pro-Pro Chemical compound C[C@@H](O)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 MROIJTGJGIDEEJ-RCWTZXSCSA-N 0.000 description 1
- GVMXJJAJLIEASL-ZJDVBMNYSA-N Thr-Pro-Thr Chemical compound C[C@@H](O)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=O GVMXJJAJLIEASL-ZJDVBMNYSA-N 0.000 description 1
- PRTHQBSMXILLPC-XGEHTFHBSA-N Thr-Ser-Arg Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O PRTHQBSMXILLPC-XGEHTFHBSA-N 0.000 description 1
- NBIIPOKZPUGATB-BWBBJGPYSA-N Thr-Ser-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CS)C(=O)O)N)O NBIIPOKZPUGATB-BWBBJGPYSA-N 0.000 description 1
- XHWCDRUPDNSDAZ-XKBZYTNZSA-N Thr-Ser-Glu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N)O XHWCDRUPDNSDAZ-XKBZYTNZSA-N 0.000 description 1
- HUPLKEHTTQBXSC-YJRXYDGGSA-N Thr-Ser-Tyr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 HUPLKEHTTQBXSC-YJRXYDGGSA-N 0.000 description 1
- NDZYTIMDOZMECO-SHGPDSBTSA-N Thr-Thr-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C)C(O)=O NDZYTIMDOZMECO-SHGPDSBTSA-N 0.000 description 1
- ZMYCLHFLHRVOEA-HEIBUPTGSA-N Thr-Thr-Ser Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(O)=O ZMYCLHFLHRVOEA-HEIBUPTGSA-N 0.000 description 1
- FBQHKSPOIAFUEI-OWLDWWDNSA-N Thr-Trp-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](C)C(O)=O FBQHKSPOIAFUEI-OWLDWWDNSA-N 0.000 description 1
- BJJRNAVDQGREGC-HOUAVDHOSA-N Thr-Trp-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC(=O)N)C(=O)O)N)O BJJRNAVDQGREGC-HOUAVDHOSA-N 0.000 description 1
- XEVHXNLPUBVQEX-DVJZZOLTSA-N Thr-Trp-Gly Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)NCC(=O)O)N)O XEVHXNLPUBVQEX-DVJZZOLTSA-N 0.000 description 1
- VEENWOSZGWWKHW-SZZJOZGLSA-N Thr-Trp-His Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC3=CN=CN3)C(=O)O)N)O VEENWOSZGWWKHW-SZZJOZGLSA-N 0.000 description 1
- NLWDSYKZUPRMBJ-IEGACIPQSA-N Thr-Trp-Leu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)N[C@@H](CC(C)C)C(=O)O)N)O NLWDSYKZUPRMBJ-IEGACIPQSA-N 0.000 description 1
- NJGMALCNYAMYCB-JRQIVUDYSA-N Thr-Tyr-Asn Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(N)=O)C(O)=O NJGMALCNYAMYCB-JRQIVUDYSA-N 0.000 description 1
- KAJRRNHOVMZYBL-IRIUXVKKSA-N Thr-Tyr-Gln Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(N)=O)C(O)=O KAJRRNHOVMZYBL-IRIUXVKKSA-N 0.000 description 1
- JAWUQFCGNVEDRN-MEYUZBJRSA-N Thr-Tyr-Leu Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC(C)C)C(=O)O)N)O JAWUQFCGNVEDRN-MEYUZBJRSA-N 0.000 description 1
- KZTLZZQTJMCGIP-ZJDVBMNYSA-N Thr-Val-Thr Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O KZTLZZQTJMCGIP-ZJDVBMNYSA-N 0.000 description 1
- BPGDJSUFQKWUBK-KJEVXHAQSA-N Thr-Val-Tyr Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 BPGDJSUFQKWUBK-KJEVXHAQSA-N 0.000 description 1
- VYVBSMCZNHOZGD-RCWTZXSCSA-N Thr-Val-Val Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](C(C)C)C(O)=O VYVBSMCZNHOZGD-RCWTZXSCSA-N 0.000 description 1
- 241000244031 Toxocara Species 0.000 description 1
- 229920001615 Tragacanth Polymers 0.000 description 1
- 241000242541 Trematoda Species 0.000 description 1
- 241000589886 Treponema Species 0.000 description 1
- 241000243774 Trichinella Species 0.000 description 1
- 241000224526 Trichomonas Species 0.000 description 1
- 208000005448 Trichomonas Infections Diseases 0.000 description 1
- 206010044620 Trichomoniasis Diseases 0.000 description 1
- 241000223238 Trichophyton Species 0.000 description 1
- 241000223230 Trichosporon Species 0.000 description 1
- 241000223231 Trichosporon beigelii Species 0.000 description 1
- 241001489151 Trichuris Species 0.000 description 1
- 239000007983 Tris buffer Substances 0.000 description 1
- OHGNSVACHBZKSS-KWQFWETISA-N Trp-Ala Chemical compound C1=CC=C2C(C[C@H]([NH3+])C(=O)N[C@@H](C)C([O-])=O)=CNC2=C1 OHGNSVACHBZKSS-KWQFWETISA-N 0.000 description 1
- CXUFDWZBHKUGKK-CABZTGNLSA-N Trp-Ala-Gly Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O)=CNC2=C1 CXUFDWZBHKUGKK-CABZTGNLSA-N 0.000 description 1
- HYVLNORXQGKONN-NUTKFTJISA-N Trp-Ala-Lys Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O)=CNC2=C1 HYVLNORXQGKONN-NUTKFTJISA-N 0.000 description 1
- PEYSVKMXSLPQRU-FJHTZYQYSA-N Trp-Ala-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N)O PEYSVKMXSLPQRU-FJHTZYQYSA-N 0.000 description 1
- RSUXQZNWAOTBQF-XIRDDKMYSA-N Trp-Arg-Gln Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N RSUXQZNWAOTBQF-XIRDDKMYSA-N 0.000 description 1
- ICNFHVUVCNWUAB-SZMVWBNQSA-N Trp-Arg-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N ICNFHVUVCNWUAB-SZMVWBNQSA-N 0.000 description 1
- RNFZZCMCRDFNAE-WFBYXXMGSA-N Trp-Asn-Ala Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(O)=O RNFZZCMCRDFNAE-WFBYXXMGSA-N 0.000 description 1
- UTQBQJNSNXJNIH-IHPCNDPISA-N Trp-Asn-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC2=CNC3=CC=CC=C32)N UTQBQJNSNXJNIH-IHPCNDPISA-N 0.000 description 1
- DEZKIRSBKKXUEV-NYVOZVTQSA-N Trp-Asp-Trp Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC3=CNC4=CC=CC=C43)C(=O)O)N DEZKIRSBKKXUEV-NYVOZVTQSA-N 0.000 description 1
- LGEPIBQBGZTBHL-SXNHZJKMSA-N Trp-Gln-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N LGEPIBQBGZTBHL-SXNHZJKMSA-N 0.000 description 1
- JLTQXEOXIJMCLZ-ZVZYQTTQSA-N Trp-Gln-Val Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C(C)C)C(O)=O)=CNC2=C1 JLTQXEOXIJMCLZ-ZVZYQTTQSA-N 0.000 description 1
- VMBBTANKMSRJSS-JSGCOSHPSA-N Trp-Glu-Gly Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O VMBBTANKMSRJSS-JSGCOSHPSA-N 0.000 description 1
- HXNVJPQADLRHGR-JBACZVJFSA-N Trp-Glu-Tyr Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)O)N HXNVJPQADLRHGR-JBACZVJFSA-N 0.000 description 1
- OGXQLUCMJZSJPW-LYSGOOTNSA-N Trp-Gly-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CC1=CNC2=CC=CC=C21)N)O OGXQLUCMJZSJPW-LYSGOOTNSA-N 0.000 description 1
- WSGPBCAGEGHKQJ-BBRMVZONSA-N Trp-Gly-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)CNC(=O)[C@H](CC1=CNC2=CC=CC=C21)N WSGPBCAGEGHKQJ-BBRMVZONSA-N 0.000 description 1
- HNIWONZFMIPCCT-SIXJUCDHSA-N Trp-His-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CC2=CNC3=CC=CC=C32)N HNIWONZFMIPCCT-SIXJUCDHSA-N 0.000 description 1
- KULBQAVOXHQLIY-HSCHXYMDSA-N Trp-Ile-Leu Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O)=CNC2=C1 KULBQAVOXHQLIY-HSCHXYMDSA-N 0.000 description 1
- SAKLWFSRZTZQAJ-GQGQLFGLSA-N Trp-Ile-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N SAKLWFSRZTZQAJ-GQGQLFGLSA-N 0.000 description 1
- CXPJPTFWKXNDKV-NUTKFTJISA-N Trp-Leu-Ala Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=O)=CNC2=C1 CXPJPTFWKXNDKV-NUTKFTJISA-N 0.000 description 1
- YVXIAOOYAKBAAI-SZMVWBNQSA-N Trp-Leu-Gln Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O)=CNC2=C1 YVXIAOOYAKBAAI-SZMVWBNQSA-N 0.000 description 1
- YPBYQWFZAAQMGW-XIRDDKMYSA-N Trp-Lys-Asn Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(=O)N)C(=O)O)N YPBYQWFZAAQMGW-XIRDDKMYSA-N 0.000 description 1
- ABEVJDLMFPTGPS-SZMVWBNQSA-N Trp-Met-Met Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCSC)C(O)=O ABEVJDLMFPTGPS-SZMVWBNQSA-N 0.000 description 1
- XGFOXYJQBRTJPO-PJODQICGSA-N Trp-Pro-Ala Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O XGFOXYJQBRTJPO-PJODQICGSA-N 0.000 description 1
- XOLLWQIBBLBAHQ-WDSOQIARSA-N Trp-Pro-Leu Chemical compound [H]N[C@@H](CC1=CNC2=C1C=CC=C2)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(C)C)C(O)=O XOLLWQIBBLBAHQ-WDSOQIARSA-N 0.000 description 1
- KBKTUNYBNJWFRL-UBHSHLNASA-N Trp-Ser-Asn Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(N)=O)C(O)=O)=CNC2=C1 KBKTUNYBNJWFRL-UBHSHLNASA-N 0.000 description 1
- UJGDFQRPYGJBEH-AAEUAGOBSA-N Trp-Ser-Gly Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CO)C(=O)NCC(=O)O)N UJGDFQRPYGJBEH-AAEUAGOBSA-N 0.000 description 1
- SGQSAIFDESQBRA-IHPCNDPISA-N Trp-Tyr-Asn Chemical compound C1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC3=CC=C(C=C3)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N SGQSAIFDESQBRA-IHPCNDPISA-N 0.000 description 1
- RWTFCAMQLFNPTK-UMPQAUOISA-N Trp-Val-Thr Chemical compound C1=CC=C2C(C[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O)=CNC2=C1 RWTFCAMQLFNPTK-UMPQAUOISA-N 0.000 description 1
- 241000223104 Trypanosoma Species 0.000 description 1
- 102000018690 Trypsinogen Human genes 0.000 description 1
- 108010027252 Trypsinogen Proteins 0.000 description 1
- QIVBCDIJIAJPQS-UHFFFAOYSA-N Tryptophan Natural products C1=CC=C2C(CC(N)C(O)=O)=CNC2=C1 QIVBCDIJIAJPQS-UHFFFAOYSA-N 0.000 description 1
- JONPRIHUYSPIMA-UWJYBYFXSA-N Tyr-Ala-Asn Chemical compound NC(=O)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 JONPRIHUYSPIMA-UWJYBYFXSA-N 0.000 description 1
- XLMDWQNAOKLKCP-XDTLVQLUSA-N Tyr-Ala-Gln Chemical compound C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N XLMDWQNAOKLKCP-XDTLVQLUSA-N 0.000 description 1
- TVOGEPLDNYTAHD-CQDKDKBSSA-N Tyr-Ala-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 TVOGEPLDNYTAHD-CQDKDKBSSA-N 0.000 description 1
- XGEUYEOEZYFHRL-KKXDTOCCSA-N Tyr-Ala-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=C(O)C=C1 XGEUYEOEZYFHRL-KKXDTOCCSA-N 0.000 description 1
- NOXKHHXSHQFSGJ-FQPOAREZSA-N Tyr-Ala-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 NOXKHHXSHQFSGJ-FQPOAREZSA-N 0.000 description 1
- AKXBNSZMYAOGLS-STQMWFEESA-N Tyr-Arg-Gly Chemical compound NC(N)=NCCC[C@@H](C(=O)NCC(O)=O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 AKXBNSZMYAOGLS-STQMWFEESA-N 0.000 description 1
- VTFWAGGJDRSQFG-MELADBBJSA-N Tyr-Asn-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)N)NC(=O)[C@H](CC2=CC=C(C=C2)O)N)C(=O)O VTFWAGGJDRSQFG-MELADBBJSA-N 0.000 description 1
- UMXSDHPSMROQRB-YJRXYDGGSA-N Tyr-Cys-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](CC1=CC=C(C=C1)O)N)O UMXSDHPSMROQRB-YJRXYDGGSA-N 0.000 description 1
- IYHNBRUWVBIVJR-IHRRRGAJSA-N Tyr-Gln-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 IYHNBRUWVBIVJR-IHRRRGAJSA-N 0.000 description 1
- NZFCWALTLNFHHC-JYJNAYRXSA-N Tyr-Glu-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 NZFCWALTLNFHHC-JYJNAYRXSA-N 0.000 description 1
- CNLKDWSAORJEMW-KWQFWETISA-N Tyr-Gly-Ala Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(=O)N[C@@H](C)C(O)=O CNLKDWSAORJEMW-KWQFWETISA-N 0.000 description 1
- OLWFDNLLBWQWCP-STQMWFEESA-N Tyr-Gly-Met Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(=O)N[C@@H](CCSC)C(O)=O OLWFDNLLBWQWCP-STQMWFEESA-N 0.000 description 1
- ILTXFANLDMJWPR-SIUGBPQLSA-N Tyr-Ile-Glu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N ILTXFANLDMJWPR-SIUGBPQLSA-N 0.000 description 1
- GGXUDPQWAWRINY-XEGUGMAKSA-N Tyr-Ile-Gly Chemical compound OC(=O)CNC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 GGXUDPQWAWRINY-XEGUGMAKSA-N 0.000 description 1
- WSFXJLFSJSXGMQ-MGHWNKPDSA-N Tyr-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N WSFXJLFSJSXGMQ-MGHWNKPDSA-N 0.000 description 1
- OHOVFPKXPZODHS-SJWGOKEGSA-N Tyr-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC2=CC=C(C=C2)O)N OHOVFPKXPZODHS-SJWGOKEGSA-N 0.000 description 1
- GULIUBBXCYPDJU-CQDKDKBSSA-N Tyr-Leu-Ala Chemical compound [O-]C(=O)[C@H](C)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]([NH3+])CC1=CC=C(O)C=C1 GULIUBBXCYPDJU-CQDKDKBSSA-N 0.000 description 1
- WDGDKHLSDIOXQC-ACRUOGEOSA-N Tyr-Leu-Phe Chemical compound C([C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(O)=O)C1=CC=C(O)C=C1 WDGDKHLSDIOXQC-ACRUOGEOSA-N 0.000 description 1
- DMWNPLOERDAHSY-MEYUZBJRSA-N Tyr-Leu-Thr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O DMWNPLOERDAHSY-MEYUZBJRSA-N 0.000 description 1
- OLYXUGBVBGSZDN-ACRUOGEOSA-N Tyr-Leu-Tyr Chemical compound C([C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(O)=O)C1=CC=C(O)C=C1 OLYXUGBVBGSZDN-ACRUOGEOSA-N 0.000 description 1
- CDKZJGMPZHPAJC-ULQDDVLXSA-N Tyr-Leu-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 CDKZJGMPZHPAJC-ULQDDVLXSA-N 0.000 description 1
- OGPKMBOPMDTEDM-IHRRRGAJSA-N Tyr-Met-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N OGPKMBOPMDTEDM-IHRRRGAJSA-N 0.000 description 1
- VYTUETMEZZLJFU-IHRRRGAJSA-N Tyr-Pro-Cys Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CC2=CC=C(C=C2)O)N)C(=O)N[C@@H](CS)C(=O)O VYTUETMEZZLJFU-IHRRRGAJSA-N 0.000 description 1
- FRMFMFNMGQGMNB-BVSLBCMMSA-N Tyr-Pro-Trp Chemical compound C([C@H](N)C(=O)N1[C@@H](CCC1)C(=O)N[C@@H](CC=1C2=CC=CC=C2NC=1)C(O)=O)C1=CC=C(O)C=C1 FRMFMFNMGQGMNB-BVSLBCMMSA-N 0.000 description 1
- RWOKVQUCENPXGE-IHRRRGAJSA-N Tyr-Ser-Arg Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O RWOKVQUCENPXGE-IHRRRGAJSA-N 0.000 description 1
- QPOUERMDWKKZEG-HJPIBITLSA-N Tyr-Ser-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CO)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 QPOUERMDWKKZEG-HJPIBITLSA-N 0.000 description 1
- LUMQYLVYUIRHHU-YJRXYDGGSA-N Tyr-Ser-Thr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O LUMQYLVYUIRHHU-YJRXYDGGSA-N 0.000 description 1
- PWKMJDQXKCENMF-MEYUZBJRSA-N Tyr-Thr-Leu Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O PWKMJDQXKCENMF-MEYUZBJRSA-N 0.000 description 1
- CLEGSEJVGBYZBJ-MEYUZBJRSA-N Tyr-Thr-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H]([C@H](O)C)NC(=O)[C@@H](N)CC1=CC=C(O)C=C1 CLEGSEJVGBYZBJ-MEYUZBJRSA-N 0.000 description 1
- MWUYSCVVPVITMW-IGNZVWTISA-N Tyr-Tyr-Ala Chemical compound C([C@@H](C(=O)N[C@@H](C)C(O)=O)NC(=O)[C@@H](N)CC=1C=CC(O)=CC=1)C1=CC=C(O)C=C1 MWUYSCVVPVITMW-IGNZVWTISA-N 0.000 description 1
- AGDDLOQMXUQPDY-BZSNNMDCSA-N Tyr-Tyr-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CO)C(O)=O AGDDLOQMXUQPDY-BZSNNMDCSA-N 0.000 description 1
- RVGVIWNHABGIFH-IHRRRGAJSA-N Tyr-Val-Ser Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(O)=O RVGVIWNHABGIFH-IHRRRGAJSA-N 0.000 description 1
- ABSXSJZNRAQDDI-KJEVXHAQSA-N Tyr-Val-Thr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O ABSXSJZNRAQDDI-KJEVXHAQSA-N 0.000 description 1
- YKBUNNNRNZZUID-UFYCRDLUSA-N Tyr-Val-Tyr Chemical compound [H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O YKBUNNNRNZZUID-UFYCRDLUSA-N 0.000 description 1
- 208000007097 Urinary Bladder Neoplasms Diseases 0.000 description 1
- 108010064997 VPY tripeptide Proteins 0.000 description 1
- IZFVRRYRMQFVGX-NRPADANISA-N Val-Ala-Gln Chemical compound C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](C(C)C)N IZFVRRYRMQFVGX-NRPADANISA-N 0.000 description 1
- YFOCMOVJBQDBCE-NRPADANISA-N Val-Ala-Glu Chemical compound C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](C(C)C)N YFOCMOVJBQDBCE-NRPADANISA-N 0.000 description 1
- REJBPZVUHYNMEN-LSJOCFKGSA-N Val-Ala-His Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](C(C)C)N REJBPZVUHYNMEN-LSJOCFKGSA-N 0.000 description 1
- RUCNAYOMFXRIKJ-DCAQKATOSA-N Val-Ala-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCCCN RUCNAYOMFXRIKJ-DCAQKATOSA-N 0.000 description 1
- AZSHAZJLOZQYAY-FXQIFTODSA-N Val-Ala-Ser Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O AZSHAZJLOZQYAY-FXQIFTODSA-N 0.000 description 1
- LABUITCFCAABSV-BPNCWPANSA-N Val-Ala-Tyr Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 LABUITCFCAABSV-BPNCWPANSA-N 0.000 description 1
- LABUITCFCAABSV-UHFFFAOYSA-N Val-Ala-Tyr Natural products CC(C)C(N)C(=O)NC(C)C(=O)NC(C(O)=O)CC1=CC=C(O)C=C1 LABUITCFCAABSV-UHFFFAOYSA-N 0.000 description 1
- JOQSQZFKFYJKKJ-GUBZILKMSA-N Val-Arg-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CS)C(=O)O)N JOQSQZFKFYJKKJ-GUBZILKMSA-N 0.000 description 1
- COYSIHFOCOMGCF-UHFFFAOYSA-N Val-Arg-Gly Natural products CC(C)C(N)C(=O)NC(C(=O)NCC(O)=O)CCCN=C(N)N COYSIHFOCOMGCF-UHFFFAOYSA-N 0.000 description 1
- PVPAOIGJYHVWBT-KKHAAJSZSA-N Val-Asn-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](C(C)C)N)O PVPAOIGJYHVWBT-KKHAAJSZSA-N 0.000 description 1
- IQQYYFPCWKWUHW-YDHLFZDLSA-N Val-Asn-Tyr Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N IQQYYFPCWKWUHW-YDHLFZDLSA-N 0.000 description 1
- XLDYBRXERHITNH-QSFUFRPTSA-N Val-Asp-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)C(C)C XLDYBRXERHITNH-QSFUFRPTSA-N 0.000 description 1
- HHSILIQTHXABKM-YDHLFZDLSA-N Val-Asp-Phe Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](Cc1ccccc1)C(O)=O HHSILIQTHXABKM-YDHLFZDLSA-N 0.000 description 1
- CWSIBTLMMQLPPZ-FXQIFTODSA-N Val-Cys-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CS)NC(=O)[C@H](C(C)C)N CWSIBTLMMQLPPZ-FXQIFTODSA-N 0.000 description 1
- OUUBKKIJQIAPRI-LAEOZQHASA-N Val-Gln-Asn Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O OUUBKKIJQIAPRI-LAEOZQHASA-N 0.000 description 1
- QHFQQRKNGCXTHL-AUTRQRHGSA-N Val-Gln-Glu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=O QHFQQRKNGCXTHL-AUTRQRHGSA-N 0.000 description 1
- JXGWQYWDUOWQHA-DZKIICNBSA-N Val-Gln-Phe Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)O)N JXGWQYWDUOWQHA-DZKIICNBSA-N 0.000 description 1
- NYTKXWLZSNRILS-IFFSRLJSSA-N Val-Gln-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)N)NC(=O)[C@H](C(C)C)N)O NYTKXWLZSNRILS-IFFSRLJSSA-N 0.000 description 1
- AAOPYWQQBXHINJ-DZKIICNBSA-N Val-Gln-Tyr Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)O)N AAOPYWQQBXHINJ-DZKIICNBSA-N 0.000 description 1
- CVIXTAITYJQMPE-LAEOZQHASA-N Val-Glu-Asn Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O CVIXTAITYJQMPE-LAEOZQHASA-N 0.000 description 1
- VLDMQVZZWDOKQF-AUTRQRHGSA-N Val-Glu-Gln Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N VLDMQVZZWDOKQF-AUTRQRHGSA-N 0.000 description 1
- NXRAUQGGHPCJIB-RCOVLWMOSA-N Val-Gly-Asn Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O NXRAUQGGHPCJIB-RCOVLWMOSA-N 0.000 description 1
- JVYIGCARISMLMV-HOCLYGCPSA-N Val-Gly-Trp Chemical compound CC(C)[C@@H](C(=O)NCC(=O)N[C@@H](CC1=CNC2=CC=CC=C21)C(=O)O)N JVYIGCARISMLMV-HOCLYGCPSA-N 0.000 description 1
- XXROXFHCMVXETG-UWVGGRQHSA-N Val-Gly-Val Chemical compound CC(C)[C@H](N)C(=O)NCC(=O)N[C@@H](C(C)C)C(O)=O XXROXFHCMVXETG-UWVGGRQHSA-N 0.000 description 1
- PTFPUAXGIKTVNN-ONGXEEELSA-N Val-His-Gly Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)NCC(=O)O)N PTFPUAXGIKTVNN-ONGXEEELSA-N 0.000 description 1
- LKUDRJSNRWVGMS-QSFUFRPTSA-N Val-Ile-Asp Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)O)NC(=O)[C@H](C(C)C)N LKUDRJSNRWVGMS-QSFUFRPTSA-N 0.000 description 1
- PYPZMFDMCCWNST-NAKRPEOUSA-N Val-Ile-Cys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](C(C)C)N PYPZMFDMCCWNST-NAKRPEOUSA-N 0.000 description 1
- UKEVLVBHRKWECS-LSJOCFKGSA-N Val-Ile-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](C(C)C)N UKEVLVBHRKWECS-LSJOCFKGSA-N 0.000 description 1
- FTKXYXACXYOHND-XUXIUFHCSA-N Val-Ile-Leu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(C)C)C(O)=O FTKXYXACXYOHND-XUXIUFHCSA-N 0.000 description 1
- DJQIUOKSNRBTSV-CYDGBPFRSA-N Val-Ile-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](C(C)C)N DJQIUOKSNRBTSV-CYDGBPFRSA-N 0.000 description 1
- BMOFUVHDBROBSE-DCAQKATOSA-N Val-Leu-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](C(C)C)N BMOFUVHDBROBSE-DCAQKATOSA-N 0.000 description 1
- QRVPEKJBBRYISE-XUXIUFHCSA-N Val-Lys-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C(C)C)N QRVPEKJBBRYISE-XUXIUFHCSA-N 0.000 description 1
- YMTOEGGOCHVGEH-IHRRRGAJSA-N Val-Lys-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(O)=O YMTOEGGOCHVGEH-IHRRRGAJSA-N 0.000 description 1
- XPKCFQZDQGVJCX-RHYQMDGZSA-N Val-Lys-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C(C)C)N)O XPKCFQZDQGVJCX-RHYQMDGZSA-N 0.000 description 1
- JVGHIFMSFBZDHH-WPRPVWTQSA-N Val-Met-Gly Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)NCC(=O)O)N JVGHIFMSFBZDHH-WPRPVWTQSA-N 0.000 description 1
- VCIYTVOBLZHFSC-XHSDSOJGSA-N Val-Phe-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N2CCC[C@@H]2C(=O)O)N VCIYTVOBLZHFSC-XHSDSOJGSA-N 0.000 description 1
- YKNOJPJWNVHORX-UNQGMJICSA-N Val-Phe-Thr Chemical compound CC(C)[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@@H](C)O)C(O)=O)CC1=CC=CC=C1 YKNOJPJWNVHORX-UNQGMJICSA-N 0.000 description 1
- JMCOXFSCTGKLLB-FKBYEOEOSA-N Val-Phe-Trp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC2=CNC3=CC=CC=C32)C(=O)O)N JMCOXFSCTGKLLB-FKBYEOEOSA-N 0.000 description 1
- GQMNEJMFMCJJTD-NHCYSSNCSA-N Val-Pro-Gln Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(N)=O)C(O)=O GQMNEJMFMCJJTD-NHCYSSNCSA-N 0.000 description 1
- SJRUJQFQVLMZFW-WPRPVWTQSA-N Val-Pro-Gly Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)NCC(O)=O SJRUJQFQVLMZFW-WPRPVWTQSA-N 0.000 description 1
- USLVEJAHTBLSIL-CYDGBPFRSA-N Val-Pro-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)C(C)C USLVEJAHTBLSIL-CYDGBPFRSA-N 0.000 description 1
- NHXZRXLFOBFMDM-AVGNSLFASA-N Val-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@@H](N)C(C)C NHXZRXLFOBFMDM-AVGNSLFASA-N 0.000 description 1
- MIKHIIQMRFYVOR-RCWTZXSCSA-N Val-Pro-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](C(C)C)N)O MIKHIIQMRFYVOR-RCWTZXSCSA-N 0.000 description 1
- QWCZXKIFPWPQHR-JYJNAYRXSA-N Val-Pro-Tyr Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 QWCZXKIFPWPQHR-JYJNAYRXSA-N 0.000 description 1
- UGFMVXRXULGLNO-XPUUQOCRSA-N Val-Ser-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O UGFMVXRXULGLNO-XPUUQOCRSA-N 0.000 description 1
- KRAHMIJVUPUOTQ-DCAQKATOSA-N Val-Ser-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N KRAHMIJVUPUOTQ-DCAQKATOSA-N 0.000 description 1
- VHIZXDZMTDVFGX-DCAQKATOSA-N Val-Ser-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](C(C)C)N VHIZXDZMTDVFGX-DCAQKATOSA-N 0.000 description 1
- QTPQHINADBYBNA-DCAQKATOSA-N Val-Ser-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCCN QTPQHINADBYBNA-DCAQKATOSA-N 0.000 description 1
- GBIUHAYJGWVNLN-UHFFFAOYSA-N Val-Ser-Pro Natural products CC(C)C(N)C(=O)NC(CO)C(=O)N1CCCC1C(O)=O GBIUHAYJGWVNLN-UHFFFAOYSA-N 0.000 description 1
- UJMCYJKPDFQLHX-XGEHTFHBSA-N Val-Ser-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](CO)NC(=O)[C@H](C(C)C)N)O UJMCYJKPDFQLHX-XGEHTFHBSA-N 0.000 description 1
- LCHZBEUVGAVMKS-RHYQMDGZSA-N Val-Thr-Leu Chemical compound CC(C)C[C@H](NC(=O)[C@@H](NC(=O)[C@@H](N)C(C)C)[C@@H](C)O)C(O)=O LCHZBEUVGAVMKS-RHYQMDGZSA-N 0.000 description 1
- IRAUYEAFPFPVND-UVBJJODRSA-N Val-Trp-Ala Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@@H](N)C(C)C)C(=O)N[C@@H](C)C(O)=O)=CNC2=C1 IRAUYEAFPFPVND-UVBJJODRSA-N 0.000 description 1
- JPBGMZDTPVGGMQ-ULQDDVLXSA-N Val-Tyr-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC2=CN=CN2)C(=O)O)N JPBGMZDTPVGGMQ-ULQDDVLXSA-N 0.000 description 1
- OWFGFHQMSBTKLX-UFYCRDLUSA-N Val-Tyr-Tyr Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC1=CC=C(C=C1)O)C(=O)N[C@@H](CC2=CC=C(C=C2)O)C(=O)O)N OWFGFHQMSBTKLX-UFYCRDLUSA-N 0.000 description 1
- AOILQMZPNLUXCM-AVGNSLFASA-N Val-Val-Lys Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCCN AOILQMZPNLUXCM-AVGNSLFASA-N 0.000 description 1
- IOUPEELXVYPCPG-UHFFFAOYSA-N Valylglycine Chemical compound CC(C)C(N)C(=O)NCC(O)=O IOUPEELXVYPCPG-UHFFFAOYSA-N 0.000 description 1
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 1
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 1
- 208000028227 Viral hemorrhagic fever Diseases 0.000 description 1
- 108700039655 Viroporin Proteins Proteins 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- YVNQAIFQFWTPLQ-UHFFFAOYSA-O [4-[[4-(4-ethoxyanilino)phenyl]-[4-[ethyl-[(3-sulfophenyl)methyl]amino]-2-methylphenyl]methylidene]-3-methylcyclohexa-2,5-dien-1-ylidene]-ethyl-[(3-sulfophenyl)methyl]azanium Chemical compound C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C(=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S(O)(=O)=O)C)C=2C(=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S(O)(=O)=O)C)C=C1 YVNQAIFQFWTPLQ-UHFFFAOYSA-O 0.000 description 1
- 238000010521 absorption reaction Methods 0.000 description 1
- 108010081404 acein-2 Proteins 0.000 description 1
- 108010036951 achatin I Proteins 0.000 description 1
- 239000003929 acidic solution Substances 0.000 description 1
- 239000008186 active pharmaceutical agent Substances 0.000 description 1
- 239000013543 active substance Substances 0.000 description 1
- 239000000654 additive Substances 0.000 description 1
- 108700010877 adenoviridae proteins Proteins 0.000 description 1
- 239000008272 agar Substances 0.000 description 1
- 108010076324 alanyl-glycyl-glycine Proteins 0.000 description 1
- 108010069020 alanyl-prolyl-glycine Proteins 0.000 description 1
- 108010086434 alanyl-seryl-glycine Proteins 0.000 description 1
- 108010078114 alanyl-tryptophyl-alanine Proteins 0.000 description 1
- 108010041407 alanylaspartic acid Proteins 0.000 description 1
- 150000001298 alcohols Chemical class 0.000 description 1
- 239000000783 alginic acid Substances 0.000 description 1
- 235000010443 alginic acid Nutrition 0.000 description 1
- 229920000615 alginic acid Polymers 0.000 description 1
- 229960001126 alginic acid Drugs 0.000 description 1
- 150000004781 alginic acids Chemical class 0.000 description 1
- ANBQYFIVLNNZCU-CQCLMDPOSA-N alpha-L-Fucp-(1->2)-[alpha-D-GalpNAc-(1->3)]-beta-D-Galp-(1->3)-[alpha-L-Fucp-(1->4)]-beta-D-GlcpNAc-(1->3)-beta-D-Galp Chemical compound O[C@H]1[C@H](O)[C@H](O)[C@H](C)O[C@H]1O[C@H]1[C@H](O[C@H]2[C@@H]([C@@H](O[C@@H]3[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O3)NC(C)=O)[C@@H](O)[C@@H](CO)O2)O[C@H]2[C@H]([C@H](O)[C@H](O)[C@H](C)O2)O)[C@@H](NC(C)=O)[C@H](O[C@H]2[C@H]([C@@H](CO)O[C@@H](O)[C@@H]2O)O)O[C@@H]1CO ANBQYFIVLNNZCU-CQCLMDPOSA-N 0.000 description 1
- SRHNADOZAAWYLV-XLMUYGLTSA-N alpha-L-Fucp-(1->2)-beta-D-Galp-(1->4)-[alpha-L-Fucp-(1->3)]-beta-D-GlcpNAc Chemical compound O[C@H]1[C@H](O)[C@H](O)[C@H](C)O[C@H]1O[C@H]1[C@H](O[C@H]2[C@@H]([C@@H](NC(C)=O)[C@H](O)O[C@@H]2CO)O[C@H]2[C@H]([C@H](O)[C@H](O)[C@H](C)O2)O)O[C@H](CO)[C@H](O)[C@@H]1O SRHNADOZAAWYLV-XLMUYGLTSA-N 0.000 description 1
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 1
- ILRRQNADMUWWFW-UHFFFAOYSA-K aluminium phosphate Chemical compound O1[Al]2OP1(=O)O2 ILRRQNADMUWWFW-UHFFFAOYSA-K 0.000 description 1
- 229940103272 aluminum potassium sulfate Drugs 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 239000003242 anti bacterial agent Substances 0.000 description 1
- 230000000844 anti-bacterial effect Effects 0.000 description 1
- 230000001745 anti-biotin effect Effects 0.000 description 1
- 238000011091 antibody purification Methods 0.000 description 1
- 239000003429 antifungal agent Substances 0.000 description 1
- 229940121375 antifungal agent Drugs 0.000 description 1
- 229960004405 aprotinin Drugs 0.000 description 1
- 108010008355 arginyl-glutamine Proteins 0.000 description 1
- 108010072041 arginyl-glycyl-aspartic acid Proteins 0.000 description 1
- 108010089442 arginyl-leucyl-alanyl-arginine Proteins 0.000 description 1
- 108010043240 arginyl-leucyl-glycine Proteins 0.000 description 1
- 108010018691 arginyl-threonyl-arginine Proteins 0.000 description 1
- 108010059459 arginyl-threonyl-phenylalanine Proteins 0.000 description 1
- 238000000149 argon plasma sintering Methods 0.000 description 1
- 108010010430 asparagine-proline-alanine Proteins 0.000 description 1
- 108010040443 aspartyl-aspartic acid Proteins 0.000 description 1
- 108010069205 aspartyl-phenylalanine Proteins 0.000 description 1
- 230000004888 barrier function Effects 0.000 description 1
- 210000003323 beak Anatomy 0.000 description 1
- CXQCLLQQYTUUKJ-ALWAHNIESA-N beta-D-GalpNAc-(1->4)-[alpha-Neup5Ac-(2->8)-alpha-Neup5Ac-(2->3)]-beta-D-Galp-(1->4)-beta-D-Glcp-(1<->1')-Cer(d18:1/18:0) Chemical compound O[C@@H]1[C@@H](O)[C@H](OC[C@H](NC(=O)CCCCCCCCCCCCCCCCC)[C@H](O)\C=C\CCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@@H](CO)O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)C(O)=O)[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](CO)O1 CXQCLLQQYTUUKJ-ALWAHNIESA-N 0.000 description 1
- 239000012148 binding buffer Substances 0.000 description 1
- 230000008033 biological extinction Effects 0.000 description 1
- 229960002685 biotin Drugs 0.000 description 1
- 235000020958 biotin Nutrition 0.000 description 1
- 239000011616 biotin Substances 0.000 description 1
- 210000002459 blastocyst Anatomy 0.000 description 1
- 230000000903 blocking effect Effects 0.000 description 1
- 210000001124 body fluid Anatomy 0.000 description 1
- 239000010839 body fluid Substances 0.000 description 1
- 238000006664 bond formation reaction Methods 0.000 description 1
- 229940098773 bovine serum albumin Drugs 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 201000003984 candidiasis Diseases 0.000 description 1
- 210000000845 cartilage Anatomy 0.000 description 1
- 239000006143 cell culture medium Substances 0.000 description 1
- 230000003915 cell function Effects 0.000 description 1
- 239000013592 cell lysate Substances 0.000 description 1
- 230000003833 cell viability Effects 0.000 description 1
- 230000007969 cellular immunity Effects 0.000 description 1
- 210000003850 cellular structure Anatomy 0.000 description 1
- CETPSERCERDGAM-UHFFFAOYSA-N ceric oxide Chemical compound O=[Ce]=O CETPSERCERDGAM-UHFFFAOYSA-N 0.000 description 1
- 229910000422 cerium(IV) oxide Inorganic materials 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 239000002738 chelating agent Substances 0.000 description 1
- 238000002512 chemotherapy Methods 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 239000011248 coating agent Substances 0.000 description 1
- 238000000576 coating method Methods 0.000 description 1
- 210000001072 colon Anatomy 0.000 description 1
- 201000010897 colon adenocarcinoma Diseases 0.000 description 1
- 230000002860 competitive effect Effects 0.000 description 1
- 239000003184 complementary RNA Substances 0.000 description 1
- 238000011109 contamination Methods 0.000 description 1
- NKLPQNGYXWVELD-UHFFFAOYSA-M coomassie brilliant blue Chemical compound [Na+].C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=2C=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=C1 NKLPQNGYXWVELD-UHFFFAOYSA-M 0.000 description 1
- 150000004696 coordination complex Chemical class 0.000 description 1
- 229920001577 copolymer Polymers 0.000 description 1
- 239000008120 corn starch Substances 0.000 description 1
- 239000007822 coupling agent Substances 0.000 description 1
- 230000009260 cross reactivity Effects 0.000 description 1
- 239000012531 culture fluid Substances 0.000 description 1
- 108010004073 cysteinylcysteine Proteins 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 229940068840 d-biotin Drugs 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000034994 death Effects 0.000 description 1
- 229960002086 dextran Drugs 0.000 description 1
- 238000000502 dialysis Methods 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 1
- 108010054813 diprotin B Proteins 0.000 description 1
- 229940042399 direct acting antivirals protease inhibitors Drugs 0.000 description 1
- 239000007884 disintegrant Substances 0.000 description 1
- 239000002612 dispersion medium Substances 0.000 description 1
- 230000006334 disulfide bridging Effects 0.000 description 1
- 125000002228 disulfide group Chemical group 0.000 description 1
- 239000002552 dosage form Substances 0.000 description 1
- 241001493065 dsRNA viruses Species 0.000 description 1
- 238000004043 dyeing Methods 0.000 description 1
- 229940056913 eftilagimod alfa Drugs 0.000 description 1
- 238000003379 elimination reaction Methods 0.000 description 1
- 239000012149 elution buffer Substances 0.000 description 1
- 210000002257 embryonic structure Anatomy 0.000 description 1
- 239000003995 emulsifying agent Substances 0.000 description 1
- 210000001900 endoderm Anatomy 0.000 description 1
- 230000008472 epithelial growth Effects 0.000 description 1
- 230000017188 evasion or tolerance of host immune response Effects 0.000 description 1
- 230000001747 exhibiting effect Effects 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 238000005189 flocculation Methods 0.000 description 1
- 230000016615 flocculation Effects 0.000 description 1
- 239000012530 fluid Substances 0.000 description 1
- 239000003205 fragrance Substances 0.000 description 1
- 238000007710 freezing Methods 0.000 description 1
- 230000008014 freezing Effects 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- UIWYJDYFSGRHKR-UHFFFAOYSA-N gadolinium atom Chemical compound [Gd] UIWYJDYFSGRHKR-UHFFFAOYSA-N 0.000 description 1
- GIVLTTJNORAZON-HDBOBKCLSA-N ganglioside GM2 (18:0) Chemical compound O[C@@H]1[C@@H](O)[C@H](OC[C@H](NC(=O)CCCCCCCCCCCCCCCCC)[C@H](O)\C=C\CCCCCCCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)[C@@H](O[C@H]2[C@@H]([C@@H](O)[C@@H](O)[C@@H](CO)O2)NC(C)=O)[C@@H](CO)O1 GIVLTTJNORAZON-HDBOBKCLSA-N 0.000 description 1
- PFJKOHUKELZMLE-VEUXDRLPSA-N ganglioside GM3 Chemical compound O[C@@H]1[C@@H](O)[C@H](OC[C@@H]([C@H](O)/C=C/CCCCCCCCCCCCC)NC(=O)CCCCCCCCCCCCC\C=C/CCCCCCCC)O[C@H](CO)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@]2(O[C@H]([C@H](NC(C)=O)[C@@H](O)C2)[C@H](O)[C@H](O)CO)C(O)=O)[C@@H](O)[C@@H](CO)O1 PFJKOHUKELZMLE-VEUXDRLPSA-N 0.000 description 1
- 206010017758 gastric cancer Diseases 0.000 description 1
- 210000001035 gastrointestinal tract Anatomy 0.000 description 1
- 239000008273 gelatin Substances 0.000 description 1
- 229920000159 gelatin Polymers 0.000 description 1
- 235000019322 gelatine Nutrition 0.000 description 1
- 235000011852 gelatine desserts Nutrition 0.000 description 1
- 238000007429 general method Methods 0.000 description 1
- 229940046240 glucomannan Drugs 0.000 description 1
- 229960002989 glutamic acid Drugs 0.000 description 1
- 108010078144 glutaminyl-glycine Proteins 0.000 description 1
- 108010080575 glutamyl-aspartyl-alanine Proteins 0.000 description 1
- 108010073628 glutamyl-valyl-phenylalanine Proteins 0.000 description 1
- 150000004676 glycans Chemical group 0.000 description 1
- 125000003976 glyceryl group Chemical group [H]C([*])([H])C(O[H])([H])C(O[H])([H])[H] 0.000 description 1
- 150000002334 glycols Chemical class 0.000 description 1
- 108010067216 glycyl-glycyl-glycine Proteins 0.000 description 1
- 108010026364 glycyl-glycyl-leucine Proteins 0.000 description 1
- 108010066198 glycyl-leucyl-phenylalanine Proteins 0.000 description 1
- 108010050475 glycyl-leucyl-tyrosine Proteins 0.000 description 1
- 108010087823 glycyltyrosine Proteins 0.000 description 1
- 239000003979 granulating agent Substances 0.000 description 1
- 230000012010 growth Effects 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 238000003306 harvesting Methods 0.000 description 1
- IXCSERBJSXMMFS-UHFFFAOYSA-N hcl hcl Chemical compound Cl.Cl IXCSERBJSXMMFS-UHFFFAOYSA-N 0.000 description 1
- SHFKGANKURXVMY-LCWPZEQJSA-N hcv e2 protein Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H](CCC(N)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H]([C@@H](C)O)NC(=O)[C@@H](NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CO)NC(=O)[C@H](C)NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](NC(=O)[C@@H](NC(=O)[C@H](CC=1NC=NC=1)NC(=O)[C@H](C)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](C)NC(=O)CNC(=O)CNC(=O)[C@@H](NC(=O)[C@@H](NC(=O)[C@@H](NC(=O)[C@@H](NC(=O)[C@H](CC=1C=CC(O)=CC=1)NC(=O)CNC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](NC(=O)CNC(=O)[C@H](C)NC(=O)[C@H](CC=1C=CC=CC=1)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H](N)CCSC)C(C)C)[C@@H](C)O)[C@@H](C)O)C(C)C)[C@@H](C)O)[C@@H](C)O)[C@@H](C)O)CC1=CC=CC=C1 SHFKGANKURXVMY-LCWPZEQJSA-N 0.000 description 1
- 230000005802 health problem Effects 0.000 description 1
- 229940037467 helicobacter pylori Drugs 0.000 description 1
- 210000002443 helper t lymphocyte Anatomy 0.000 description 1
- 238000011134 hematopoietic stem cell transplantation Methods 0.000 description 1
- 208000005252 hepatitis A Diseases 0.000 description 1
- 239000000833 heterodimer Substances 0.000 description 1
- 108010040030 histidinoalanine Proteins 0.000 description 1
- 125000000487 histidyl group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C([H])=N1 0.000 description 1
- 108010036413 histidylglycine Proteins 0.000 description 1
- 208000033519 human immunodeficiency virus infectious disease Diseases 0.000 description 1
- 235000020256 human milk Nutrition 0.000 description 1
- 210000004251 human milk Anatomy 0.000 description 1
- 230000004727 humoral immunity Effects 0.000 description 1
- 229920001600 hydrophobic polymer Polymers 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- DOUYETYNHWVLEO-UHFFFAOYSA-N imiquimod Chemical compound C1=CC=CC2=C3N(CC(C)C)C=NC3=C(N)N=C21 DOUYETYNHWVLEO-UHFFFAOYSA-N 0.000 description 1
- 229960002751 imiquimod Drugs 0.000 description 1
- 230000002519 immonomodulatory effect Effects 0.000 description 1
- 230000036737 immune function Effects 0.000 description 1
- 210000004201 immune sera Anatomy 0.000 description 1
- 229940042743 immune sera Drugs 0.000 description 1
- 230000000984 immunochemical effect Effects 0.000 description 1
- 230000016784 immunoglobulin production Effects 0.000 description 1
- 230000001506 immunosuppresive effect Effects 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 239000007943 implant Substances 0.000 description 1
- 238000002513 implantation Methods 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 238000001727 in vivo Methods 0.000 description 1
- 238000005462 in vivo assay Methods 0.000 description 1
- 210000003000 inclusion body Anatomy 0.000 description 1
- 238000010348 incorporation Methods 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 230000001939 inductive effect Effects 0.000 description 1
- 206010022000 influenza Diseases 0.000 description 1
- 238000003331 infrared imaging Methods 0.000 description 1
- 239000003112 inhibitor Substances 0.000 description 1
- 230000005764 inhibitory process Effects 0.000 description 1
- ZPNFWUPYTFPOJU-LPYSRVMUSA-N iniprol Chemical compound C([C@H]1C(=O)NCC(=O)NCC(=O)N[C@H]2CSSC[C@H]3C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@H](C(N[C@H](C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC=4C=CC=CC=4)C(=O)N[C@@H](CC=4C=CC(O)=CC=4)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](C)NC(=O)[C@H](CO)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC=4C=CC=CC=4)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CC(N)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C)NC(=O)[C@H](CCCNC(N)=N)NC2=O)C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CSSC[C@H](NC(=O)[C@H](CC=2C=CC=CC=2)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H]2N(CCC2)C(=O)[C@@H](N)CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(O)=O)C(=O)N2[C@@H](CCC2)C(=O)N2[C@@H](CCC2)C(=O)N[C@@H](CC=2C=CC(O)=CC=2)C(=O)N[C@@H]([C@@H](C)O)C(=O)NCC(=O)N2[C@@H](CCC2)C(=O)N3)C(=O)NCC(=O)NCC(=O)N[C@@H](C)C(O)=O)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@H](C(=O)N[C@@H](CC=2C=CC=CC=2)C(=O)N[C@H](C(=O)N1)C(C)C)[C@@H](C)O)[C@@H](C)CC)=O)[C@@H](C)CC)C1=CC=C(O)C=C1 ZPNFWUPYTFPOJU-LPYSRVMUSA-N 0.000 description 1
- 238000011221 initial treatment Methods 0.000 description 1
- 230000002452 interceptive effect Effects 0.000 description 1
- 229940079322 interferon Drugs 0.000 description 1
- 229960001388 interferon-beta Drugs 0.000 description 1
- 229940117681 interleukin-12 Drugs 0.000 description 1
- 229940100601 interleukin-6 Drugs 0.000 description 1
- 230000009878 intermolecular interaction Effects 0.000 description 1
- 238000007918 intramuscular administration Methods 0.000 description 1
- 238000007912 intraperitoneal administration Methods 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 230000009545 invasion Effects 0.000 description 1
- 108010060857 isoleucyl-valyl-tyrosine Proteins 0.000 description 1
- 210000003734 kidney Anatomy 0.000 description 1
- 238000002386 leaching Methods 0.000 description 1
- 229940115286 lentinan Drugs 0.000 description 1
- 108010044311 leucyl-glycyl-glycine Proteins 0.000 description 1
- 108010044056 leucyl-phenylalanine Proteins 0.000 description 1
- 108010012058 leucyltyrosine Proteins 0.000 description 1
- 210000000265 leukocyte Anatomy 0.000 description 1
- GDBQQVLCIARPGH-ULQDDVLXSA-N leupeptin Chemical compound CC(C)C[C@H](NC(C)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C=O)CCCN=C(N)N GDBQQVLCIARPGH-ULQDDVLXSA-N 0.000 description 1
- 108010052968 leupeptin Proteins 0.000 description 1
- 239000003446 ligand Substances 0.000 description 1
- 238000004811 liquid chromatography Methods 0.000 description 1
- 210000005229 liver cell Anatomy 0.000 description 1
- 208000019423 liver disease Diseases 0.000 description 1
- 239000007937 lozenge Substances 0.000 description 1
- 201000005296 lung carcinoma Diseases 0.000 description 1
- 239000006166 lysate Substances 0.000 description 1
- 235000019359 magnesium stearate Nutrition 0.000 description 1
- 230000010874 maintenance of protein location Effects 0.000 description 1
- 230000003211 malignant effect Effects 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 230000035800 maturation Effects 0.000 description 1
- 238000005259 measurement Methods 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 229910052751 metal Inorganic materials 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- 230000009401 metastasis Effects 0.000 description 1
- 108010085203 methionylmethionine Proteins 0.000 description 1
- 108010068488 methionylphenylalanine Proteins 0.000 description 1
- 239000000693 micelle Substances 0.000 description 1
- 238000001471 micro-filtration Methods 0.000 description 1
- 244000005700 microbiome Species 0.000 description 1
- 239000003094 microcapsule Substances 0.000 description 1
- 239000004005 microsphere Substances 0.000 description 1
- 108010071421 milk fat globule Proteins 0.000 description 1
- 239000002480 mineral oil Substances 0.000 description 1
- 235000010446 mineral oil Nutrition 0.000 description 1
- 239000007758 minimum essential medium Substances 0.000 description 1
- 239000003595 mist Substances 0.000 description 1
- 150000004712 monophosphates Chemical class 0.000 description 1
- UXOUKMQIEVGVLY-UHFFFAOYSA-N morin Natural products OC1=CC(O)=CC(C2=C(C(=O)C3=C(O)C=C(O)C=C3O2)O)=C1 UXOUKMQIEVGVLY-UHFFFAOYSA-N 0.000 description 1
- 238000002703 mutagenesis Methods 0.000 description 1
- 231100000350 mutagenesis Toxicity 0.000 description 1
- 201000000050 myeloid neoplasm Diseases 0.000 description 1
- 239000006199 nebulizer Substances 0.000 description 1
- 208000004235 neutropenia Diseases 0.000 description 1
- 231100000252 nontoxic Toxicity 0.000 description 1
- 230000003000 nontoxic effect Effects 0.000 description 1
- 239000002773 nucleotide Substances 0.000 description 1
- 125000003729 nucleotide group Chemical group 0.000 description 1
- 238000011369 optimal treatment Methods 0.000 description 1
- 239000006186 oral dosage form Substances 0.000 description 1
- 230000008520 organization Effects 0.000 description 1
- WRUUGTRCQOWXEG-UHFFFAOYSA-N pamidronate Chemical compound NCCC(O)(P(O)(O)=O)P(O)(O)=O WRUUGTRCQOWXEG-UHFFFAOYSA-N 0.000 description 1
- 208000003154 papilloma Diseases 0.000 description 1
- 239000012188 paraffin wax Substances 0.000 description 1
- 244000045947 parasite Species 0.000 description 1
- 238000007911 parenteral administration Methods 0.000 description 1
- 239000002245 particle Substances 0.000 description 1
- 239000001814 pectin Substances 0.000 description 1
- 235000010987 pectin Nutrition 0.000 description 1
- 229920001277 pectin Polymers 0.000 description 1
- 230000010412 perfusion Effects 0.000 description 1
- 238000002823 phage display Methods 0.000 description 1
- 238000009520 phase I clinical trial Methods 0.000 description 1
- 108010084572 phenylalanyl-valine Proteins 0.000 description 1
- 108010018625 phenylalanylarginine Proteins 0.000 description 1
- 108010083476 phenylalanyltryptophan Proteins 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 230000035479 physiological effects, processes and functions Effects 0.000 description 1
- 239000006187 pill Substances 0.000 description 1
- 210000004180 plasmocyte Anatomy 0.000 description 1
- 229920000747 poly(lactic acid) Polymers 0.000 description 1
- 229920000728 polyester Polymers 0.000 description 1
- 229920002338 polyhydroxyethylmethacrylate Polymers 0.000 description 1
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 1
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 1
- 229920002223 polystyrene Polymers 0.000 description 1
- 229920002451 polyvinyl alcohol Polymers 0.000 description 1
- GRLPQNLYRHEGIJ-UHFFFAOYSA-J potassium aluminium sulfate Chemical compound [Al+3].[K+].[O-]S([O-])(=O)=O.[O-]S([O-])(=O)=O GRLPQNLYRHEGIJ-UHFFFAOYSA-J 0.000 description 1
- 230000003389 potentiating effect Effects 0.000 description 1
- 238000001556 precipitation Methods 0.000 description 1
- 238000004237 preparative chromatography Methods 0.000 description 1
- 230000003449 preventive effect Effects 0.000 description 1
- 230000037452 priming Effects 0.000 description 1
- 239000013587 production medium Substances 0.000 description 1
- 238000004393 prognosis Methods 0.000 description 1
- 108010014614 prolyl-glycyl-proline Proteins 0.000 description 1
- 108010025826 prolyl-leucyl-arginine Proteins 0.000 description 1
- 108010077112 prolyl-proline Proteins 0.000 description 1
- 108010087846 prolyl-prolyl-glycine Proteins 0.000 description 1
- 108010015796 prolylisoleucine Proteins 0.000 description 1
- 239000003380 propellant Substances 0.000 description 1
- 125000002568 propynyl group Chemical group [*]C#CC([H])([H])[H] 0.000 description 1
- 210000002307 prostate Anatomy 0.000 description 1
- 229940121649 protein inhibitor Drugs 0.000 description 1
- 239000012268 protein inhibitor Substances 0.000 description 1
- 230000005180 public health Effects 0.000 description 1
- 230000002685 pulmonary effect Effects 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 208000013718 rectal benign neoplasm Diseases 0.000 description 1
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 1
- 238000010405 reoxidation reaction Methods 0.000 description 1
- 230000010076 replication Effects 0.000 description 1
- 239000011347 resin Substances 0.000 description 1
- 229920005989 resin Polymers 0.000 description 1
- 230000002441 reversible effect Effects 0.000 description 1
- 229960000329 ribavirin Drugs 0.000 description 1
- HZCAHMRRMINHDJ-DBRKOABJSA-N ribavirin Natural products O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1N=CN=C1 HZCAHMRRMINHDJ-DBRKOABJSA-N 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 230000011218 segmentation Effects 0.000 description 1
- 108010048397 seryl-lysyl-leucine Proteins 0.000 description 1
- 230000035939 shock Effects 0.000 description 1
- 239000002356 single layer Substances 0.000 description 1
- 239000010802 sludge Substances 0.000 description 1
- 239000002904 solvent Substances 0.000 description 1
- 238000002798 spectrophotometry method Methods 0.000 description 1
- 150000003408 sphingolipids Chemical class 0.000 description 1
- 230000007480 spreading Effects 0.000 description 1
- 238000003892 spreading Methods 0.000 description 1
- 239000003381 stabilizer Substances 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 235000019698 starch Nutrition 0.000 description 1
- 239000007858 starting material Substances 0.000 description 1
- 230000000707 stereoselective effect Effects 0.000 description 1
- 150000003432 sterols Chemical class 0.000 description 1
- 235000003702 sterols Nutrition 0.000 description 1
- 239000011550 stock solution Substances 0.000 description 1
- 201000011549 stomach cancer Diseases 0.000 description 1
- 238000005728 strengthening Methods 0.000 description 1
- 238000007920 subcutaneous administration Methods 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 239000000829 suppository Substances 0.000 description 1
- 230000004083 survival effect Effects 0.000 description 1
- 239000000375 suspending agent Substances 0.000 description 1
- 238000013268 sustained release Methods 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 108060008037 tachykinin Proteins 0.000 description 1
- 101150047061 tag-72 gene Proteins 0.000 description 1
- 238000002560 therapeutic procedure Methods 0.000 description 1
- 229940021747 therapeutic vaccine Drugs 0.000 description 1
- 150000003568 thioethers Chemical class 0.000 description 1
- 230000000699 topical effect Effects 0.000 description 1
- 239000003053 toxin Substances 0.000 description 1
- 231100000765 toxin Toxicity 0.000 description 1
- 239000000196 tragacanth Substances 0.000 description 1
- 235000010487 tragacanth Nutrition 0.000 description 1
- 229940116362 tragacanth Drugs 0.000 description 1
- 230000002103 transcriptional effect Effects 0.000 description 1
- 239000012096 transfection reagent Substances 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 230000009261 transgenic effect Effects 0.000 description 1
- 230000007704 transition Effects 0.000 description 1
- 108091005703 transmembrane proteins Proteins 0.000 description 1
- 102000035160 transmembrane proteins Human genes 0.000 description 1
- 230000014599 transmission of virus Effects 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- 238000011269 treatment regimen Methods 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 108010036387 trimethionine Proteins 0.000 description 1
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 1
- 238000003211 trypan blue cell staining Methods 0.000 description 1
- 108010084932 tryptophyl-proline Proteins 0.000 description 1
- 108010044292 tryptophyltyrosine Proteins 0.000 description 1
- 201000008827 tuberculosis Diseases 0.000 description 1
- 230000005751 tumor progression Effects 0.000 description 1
- 230000029069 type 2 immune response Effects 0.000 description 1
- OUYCCCASQSFEME-UHFFFAOYSA-N tyrosine Natural products OC(=O)C(N)CC1=CC=C(O)C=C1 OUYCCCASQSFEME-UHFFFAOYSA-N 0.000 description 1
- 108010035534 tyrosyl-leucyl-alanine Proteins 0.000 description 1
- 238000005199 ultracentrifugation Methods 0.000 description 1
- 241000701447 unidentified baculovirus Species 0.000 description 1
- 241000712461 unidentified influenza virus Species 0.000 description 1
- 238000011870 unpaired t-test Methods 0.000 description 1
- 108010043941 valyl-glutamyl-isoleucyl-asparaginyl-cysteinyl-threonyl-arginine Proteins 0.000 description 1
- 230000002792 vascular Effects 0.000 description 1
- 230000006656 viral protein synthesis Effects 0.000 description 1
- 239000013603 viral vector Substances 0.000 description 1
- 210000002845 virion Anatomy 0.000 description 1
- 239000011534 wash buffer Substances 0.000 description 1
- 239000000080 wetting agent Substances 0.000 description 1
Images



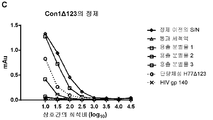
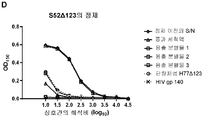

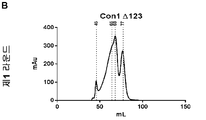
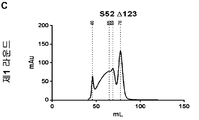



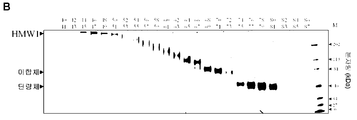









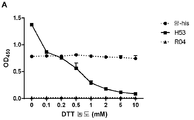

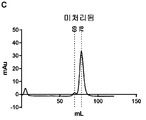
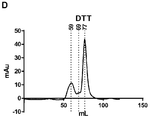
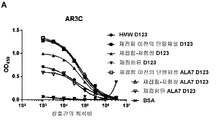
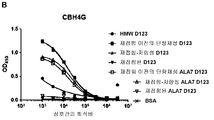



















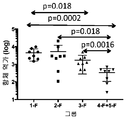
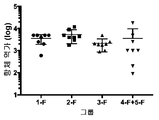
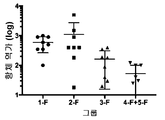
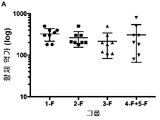
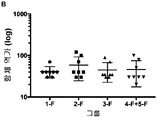
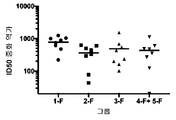
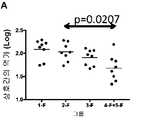

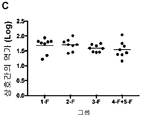






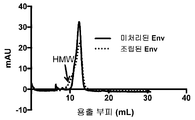
Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/12—Antivirals
- A61P31/14—Antivirals for RNA viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K1/00—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length
- C07K1/107—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length by chemical modification of precursor peptides
- C07K1/113—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length by chemical modification of precursor peptides without change of the primary structure
- C07K1/1133—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length by chemical modification of precursor peptides without change of the primary structure by redox-reactions involving cystein/cystin side chains
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/29—Hepatitis virus
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K1/00—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length
- C07K1/13—Labelling of peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K1/00—General methods for the preparation of peptides, i.e. processes for the organic chemical preparation of peptides or proteins of any length
- C07K1/14—Extraction; Separation; Purification
- C07K1/16—Extraction; Separation; Purification by chromatography
- C07K1/22—Affinity chromatography or related techniques based upon selective absorption processes
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/08—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses
- C07K16/10—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from viruses from RNA viruses
- C07K16/1081—Togaviridae, e.g. flavivirus, rubella virus, hog cholera virus
- C07K16/109—Hepatitis C virus; Hepatitis G virus
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5044—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics involving specific cell types
- G01N33/5047—Cells of the immune system
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/5005—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells
- G01N33/5008—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics
- G01N33/5044—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing involving human or animal cells for testing or evaluating the effect of chemical or biological compounds, e.g. drugs, cosmetics involving specific cell types
- G01N33/5047—Cells of the immune system
- G01N33/5052—Cells of the immune system involving B-cells
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/569—Immunoassay; Biospecific binding assay; Materials therefor for microorganisms, e.g. protozoa, bacteria, viruses
- G01N33/56966—Animal cells
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N33/00—Investigating or analysing materials by specific methods not covered by groups G01N1/00 - G01N31/00
- G01N33/48—Biological material, e.g. blood, urine; Haemocytometers
- G01N33/50—Chemical analysis of biological material, e.g. blood, urine; Testing involving biospecific ligand binding methods; Immunological testing
- G01N33/53—Immunoassay; Biospecific binding assay; Materials therefor
- G01N33/576—Immunoassay; Biospecific binding assay; Materials therefor for hepatitis
- G01N33/5767—Immunoassay; Biospecific binding assay; Materials therefor for hepatitis non-A, non-B hepatitis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/64—Medicinal preparations containing antigens or antibodies characterised by the architecture of the carrier-antigen complex, e.g. repetition of carrier-antigen units
- A61K2039/645—Dendrimers; Multiple antigen peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/33—Crossreactivity, e.g. for species or epitope, or lack of said crossreactivity
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/24011—Flaviviridae
- C12N2770/24211—Hepacivirus, e.g. hepatitis C virus, hepatitis G virus
- C12N2770/24222—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2770/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssRNA viruses positive-sense
- C12N2770/00011—Details
- C12N2770/24011—Flaviviridae
- C12N2770/24211—Hepacivirus, e.g. hepatitis C virus, hepatitis G virus
- C12N2770/24234—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Immunology (AREA)
- Molecular Biology (AREA)
- Engineering & Computer Science (AREA)
- Organic Chemistry (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Biochemistry (AREA)
- Hematology (AREA)
- Urology & Nephrology (AREA)
- Analytical Chemistry (AREA)
- Virology (AREA)
- Cell Biology (AREA)
- Genetics & Genomics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biophysics (AREA)
- Microbiology (AREA)
- Physics & Mathematics (AREA)
- Biotechnology (AREA)
- Pathology (AREA)
- General Physics & Mathematics (AREA)
- Food Science & Technology (AREA)
- Communicable Diseases (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Tropical Medicine & Parasitology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- General Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Toxicology (AREA)
- Mycology (AREA)
- Epidemiology (AREA)
- Gastroenterology & Hepatology (AREA)
- Oncology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
Abstract
(i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 항원을 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 낮은 급수의 항원을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 항원을 높은 급수의 항원으로 조립시키는 단계를 포함하고, 여기서 단계 (ii)에서 낮은 급수의 항원의 적어도 10%가 높은 급수의 항원으로 전환되고, 이로써 조립된 높은 급수의 항원이 낮은 급수의 항원과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타내고 적어도 하나의 중화 항체에 대한 결합을 보유하는, 고유의 낮은 급수의 항원으로부터 조립된 높은 급수의 항원을 세포밖에서 제조하는 방법. 상기 방법의 단계들을 수행하는 단계, 및 조립된 높은 급수의 항원을 약학적으로 또는 생리학적으로 허용가능한 희석제, 담체 또는 보조제와 혼합하는 단계를 포함하는 백신 조성물의 제조 방법. 조립된 항원이 고유의 대조적인 높은 급수의 항원과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타내는, 높은 급수의 세포밖에서 조립된 항원을 포함하는 조성물. 면역 반응을 자극하거나, 또는 항원에 특이적인 면역 세포, 예컨대 B-세포를 검출하고/하거나 단리하기 위한 조립된 높은 급수의 항원의 용도.
Description
본 명세서는 일반적으로 백신 및 진단용 조성물에 관한 것이다. 특별히, 명세서는 관심있는 항원, 예컨대 HCV 엔벨롭(envelope) 2(E2) 당단백질 및 HIV 엔벨롭 단백질의 높은 급수(higher order) 형태의 생산을 용이하게 한다. 높은 급수의 항원은 백신 생산 및 생체외 결합 적용을 위해 적합하다.
본 명세서에서 참고문헌에 대한 서지 세부사항은 또한 명세서 말미에 열거된다.
본 명세서에서 임의의 선행 기술에 대한 언급은, 이러한 선행 기술이 임의의 국가에서 일반적 상식의 일부를 형성함을 인정하거나 시사하는 것이 아니고 그렇게 취급되어서도 안된다.
간염 C 바이러스는 세계 인구의 3%를 만성적으로 감염시키고 만성 간 질환의 주요 원인인 고도로 다양화된 병원체이다. HCV는 7가지의 고도로 분기된 유전자형(G1-G7) 및 67개 이상의 아형(a, b, c 등)으로서 퍼지고, 이에 대하여 이용가능한 예방적 백신은 존재하지 않는다. 최근에, 직접 작용하는 항바이러스제(DAA: direct acting antiviral)는 치료된 개별체의 > 95%에서 바이러스 제거를 가능하게 하는 임상 실험에 도달하였다. 그러나, DAA는 재감염을 예방할 수 없고, 이들의 높은 비용은 의료 제도에 막대한 경제적 부담을 준다. 더욱이, 대략 5000만명이 감염으로 진단되지 않아서, 지속적으로 바이러스를 확산시키는 수단을 제공한다. 예방적 백신은 새로운 감염, 및 가능하게는 재감염을 방지하고, DAA의 사용이 포함된 제거 프로그램을 상당히 증대시킬 것이다. 그러므로, 세계 보건 기구는 개발해야 할 우선 사항으로서 예방적 백신을 인식하고 있다.
주 표면 당단백질, E2는 비리온(virion)을 숙주 세포 수용체 CD81에 부착시키고, 유전자형 특이적이고 광범위한 (교차-유전자형 보호용) 중화 항체(bNAb: broadly neutralizing antibody)를 생성한다. 자연 감염시, 개개인중 30%는 그들의 감염을 자발적으로 물리치고, 이는 bNAb의 신속한 유도 및 광범위한 반응성 T 세포 반응과 상관된다. 추가적으로, 광범위한 중화 단일클론성 항체(NMAb: neutralizing monoclonal antibody)의 칵테일(cocktail)은 HCV의 작은 동물 모델에서 확립된 HCV 감염을 예방하고 물리칠 수 있다. 그러나, 재조합 HCV 당단백질의 상 I 임상 실험 및 E2의 전장 또는 절단 형태를 함유하는 백신을 사용하여 동물에서 수행된 이전의 백신접종 연구는 높은 역가의 NAb를 이끌어내지 못했고, 매우 제한된 교차-유전형 중화가 관찰되었다. HCV E2는 고도로 글리코실화되고, 신속한 서열 변화를 겪으며, 다중의 가변성 영역을 소유하는데, 이는 모두 면역 회피(immune evasion)에 관여된다. 비-중화 항체는 중화 항체의 결합을 방해하는 것으로 제시되어 왔다.
개선된 HCV 백신을 개발하기 위한 선행 기술 접근법은 CD81 결합을 보유하면서도 가변성 영역을 제거하도록 E2를 변형시키는데 초점을 두었다. 최근에, 발명자들은 원하는 광범위 중화 항체 반응을 생성하는데 있어서 올리고머, 특별히 고분자량 올리고머(HMW 형태)가 단량체에 비해 더 우수함을 알아내었다. 이러한 접근법은 유망하지만, 야생형 HCV E2 및 과가변성 영역이 없는 변형된 E2(예를 들어 델타123 HCV E2)의 고분자량(HMW) 형태는 일반적으로 낮은 수율로 발현되고 구조적으로 이종성인데, 이는 백신 생산에 있어서 난제를 만들어 낸다. 현재, 올리고머는 E2를 발현하는 플라스미드를 숙주 세포내로 형질감염시키고, 세포에 의해 E2가 고유의 올리고머로 접힐 수 있도록 함으로써 생산된다. 이어서 올리고머는 필요에 따라서 다른 성분으로부터 분리된다.
이러한 배경으로 인해, 본 발명자들은 상업용 백신 제조 목적을 위한 몇몇 잠재적 이점, 예컨대 몇몇 실시태양에서, 비용 및 정제의 복잡성이 감소된 E2의 올리고머 형태를 생산하기 위한 전략을 개발하였다. 이러한 방법은 관심있는 임의의 항원, 및 항원의 조립된 높은 급수 형태의 생산을 위해 광범위하게 적용될 수 있다.
하나의 양태에서, 본 개시내용은 (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 항원을 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 낮은 급수의 항원을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 항원을 높은 급수의 항원으로 조립시키는 단계를 포함하는, 고유의 낮은 급수의 항원으로부터 세포밖에서 조립된 높은 급수의 항원을 제조하는 방법을 가능하게 한다. 방법의 하나의 실시태양에서, 단계 (ii)에서 낮은 급수의 항원의 적어도 10%는 높은 급수의 항원으로 전환된다. 방법의 하나의 실시태양에서, 조립된 높은 급수의 항원은 낮은 급수의 항원과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타낸다. 방법의 하나의 실시태양에서, 조립된 높은 급수의 항원은 적어도 하나의 중화 항체에 대한 결합을 보유한다.
하나의 실시태양에서, 항원은 바이러스성 엔벨롭 항원이다. 하나의 실시태양에서, 바이러스성 엔벨롭 항원은 HCV 또는 HIV이다.
또 다른 실시태양에서, 항원은 암 항원이다.
하나의 실시태양에서, 높은 급수의 암 항원은 4합체(tetramer), 5합체(pentamer), 6합체(hexamer), 10합체(decamer) 등, 23합체까지이다.
하나의 실시태양에서, (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 항원을 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 낮은 급수의 항원을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 항원을 높은 급수의 항원으로 조립시키는 단계를 포함하고, 여기서 단계 (ii)에서 낮은 급수의 항원의 적어도 10%가 높은 급수의 항원으로 전환되고, 이로써 조립된 높은 급수의 항원이 낮은 급수의 항원과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타내고 적어도 하나의 중화 항체에 대한 결합을 보유하는, 고유의 낮은 급수의 항원으로부터 세포밖에서 조립된 높은 급수의 항원을 제조하는 방법이 제공된다.
하나의 실시태양에서, 높은 급수의 항원은 예방적 또는 치료학적 백신으로서 피험체에게 전달되기 이전에 정제되고 약학적으로 허용가능한 희석제, 담체 또는 보조제와 혼합된다.
하나의 실시태양에서, 본 개시내용은 (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 간염 C 바이러스(HCV) 엔벨롭 당단백질 2(E2)를 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 E2를 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 E2를 높은 급수의 HCV E2로 조립시키는 단계를 포함하는, 고유의 낮은 급수의 HCV E2로부터 세포밖에서 조립된 높은 급수의 HCV E2 항원을 제조하는 방법을 가능하게 한다. 방법의 하나의 실시태양에서, 단계 (ii)에서 낮은 급수의 E2 항원의 적어도 20%는 높은 급수의 항원으로 조립된다. 하나의 실시태양에서, 낮은 급수의 HCV E2와 비교하여 조립된 높은 급수의 HCV E2는 적어도 감소된 비-중화 항체에 대한 결합을 나타내고, 적어도 하나의 중화 항체에 대한 결합을 보유한다.
따라서, 하나의 실시태양에서, 본 개시내용은 (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 간염 C 바이러스(HCV) 엔벨롭 당단백질 2(E2)를 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 E2를 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 E2를 높은 급수의 HCV E2으로 조립시키는 단계를 포함하고, 여기서 단계 (ii)에서 낮은 급수의 항원의 적어도 20%가 높은 급수의 항원으로 전환되고, 이로써 조립된 높은 급수의 HCV E2가 낮은 급수의 항원과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타내고 적어도 하나의 중화 항체에 대한 결합을 보유하는, 고유의 낮은 급수의 HCV E2로부터 세포밖에서 조립된 높은 급수의 HCV E2 항원을 제조하는 방법을 가능하게 한다.
또 다른 하나의 실시태양에서, 개시내용은 (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 HIV env를 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 Env을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 Env을 높은 급수의 HCV Env으로 조립시키는 단계를 포함하고, 여기서 단계 (ii)에서 낮은 급수의 항원의 적어도 10%가 높은 급수의 항원으로 전환되는, 고유의 낮은 급수의 HIV env로부터 세포밖에서 조립된 높은 급수의 HIV 엔벨롭 당단백질 항원을 제조하는 방법을 가능하게 한다.
하나의 실시태양에서, 낮은 급수의 항원을 높은 급수의 항원으로 조립하는 방법의 효율을 개선시키기 위해, 단계 (ii)로부터 남아있는 낮은 급수의 항원을 포함하는 용액에 의해 처음 단계 (i) 및 (ii)를 반복한다.
놀랍게도, 심지어 HCV E2 단량체의 시스테인 변형된 버전(version), 예컨대 Ala7 작성물이 또한 본원에 기재된 방법을 사용하여 높은 급수의 형태로 조립된다. 시스테인 변형된 형태, 예컨대 Ala7은 우세하게 단량체 형태로 재조합적으로 발현되므로, 예를 들면, HCV E2 RBD 형태 또는 델타123 형태에 의해 대체적으로 생산되는 경우 상이한 형태의 혼합물로부터 단량체 형태를 정제하기 보다는 상기 물질로부터 높은 급수의 형태를 생산하는 것이 유용할 것이다. 이와 같이, 시스테인 변형된 형태가 또한 본 방법에서 조립된다는 발견은 낮은 급수의 또는 단량체의 유용한 공급원을 제공함으로써 백신 제작을 용이하게 한다. 더욱이, 본원에서 결정된 바와 같이 높은 급수의 형태로 초기에 조립되지 않는 낮은 급수의 항원은 방법이 반복될 경우 높은 급수의 형태를 형성할 수 있어서, 높은 급수의 형태의 더욱 더 높은 수율을 가능하게 한다.
방법의 하나의 실시태양에서, 단계 (i)에서 또는 단계 (i) 이전에, 낮은 급수의 항원을 포함하는 용액에는 고유의 올리고머 또는 높은 급수의 항원이 실질적으로 고갈되어 있다.
하나의 실시태양에서, 낮은 급수의 항원의 적어도 25%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60% 또는 적어도 70%, 또는 적어도 80%, 또는 적어도 90% 또는 적어도 95% 또는 그 이상이 높은 급수의 항원으로 전환된다.
또 다른 실시태양에서, 발명자들은 높은 급수의 항원의 수율에 있어서 유전자형 특이적 차이를 발견하였다. 따라서, 하나의 실시태양에서, 방법은 조립된 올리고머성 항원의 가장 큰 수율을 생성하는 바이러스 또는 항원 유전자형을 선택하는 단계를 추가로 포함한다.
하나의 실시태양에서, 조립된 높은 급수의 항원은 하나 이상의 중화 항체에 결합하거나 이를 이끌어내는 고유의 대조적인 높은 급수의 항원의 능력을 보유하거나 이를 초과한다.
하나의 실시태양에서, 조립된 높은 급수의 항원은 HCV E2의 수용체-결합 도메인(RBD)이다.
하나의 실시태양에서, 조립된 높은 급수의 HCV E2 항원은 과가변성 영역의 전부 또는 일부, 예컨대 과가변성 영역 1(HVR1) 또는 이의 일부, 과가변성 영역 2(HVR2) 또는 이의 일부 및/또는 상호유전자형 가변성 영역(igVR/VR3) 또는 이의 일부중 하나 이상이 결핍된다.
하나의 실시태양에서, 조립된 올리고머성 항원은 C581, C585, C652, C677, C494, C486, C459, C452, C564, C597, 및 C569로 구성된 군에서 선택된 하나 이상의 아미노산 잔기에서 비-시스테인 치환 또는 돌연변이를 포함한다. 본원에서 논의된 바와 같이, 시스테인 변형된 형태의 사용은 놀랍게도 높은 급수의 형태로 조립될 수 있는 낮은 급수의 항원의 공급원을 제공한다.
또 다른 양태에서, 본 개시내용은 (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 항원을 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 낮은 급수의 항원을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 항원을 높은 급수의 항원으로 조립시키고, 이후 조립된 높은 급수의 항원을 약학적으로 또는 생리학적으로 허용가능한 희석제, 담체 또는 보조제와 혼합하는 단계를 포함하는, 본원에 개시된 조립 방법을 포함하는 백신 조성물의 제조 방법을 제공한다.
몇몇 실시태양에서, 항원은 바이러스성 엔벨롭 항원 또는 암 항원이다.
몇몇 실시태양에서, 바이러스성 엔벨롭 항원은 간염 바이러스 항원 또는 HIV 엔벨롭 항원이다.
또 다른 양태에서, 본 개시내용은 (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 항원을 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 낮은 급수의 항원을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 항원을 높은 급수의 항원으로 조립시키는 단계를 포함하는 본원에 개시된 방법에 의해 낮은 급수의 항원으로부터 생산되는, 조립된 높은 급수의 항원, 또는 이를 포함하는 조성물을 제공한다.
하나의 실시태양에서, 높은 급수의 세포밖에서 조립된 항원을 포함하는 조성물이 가능하다. 하나의 실시태양에서, 높은 급수의 조립된 항원은 숙주 세포에서 생산된 대응하는 대조 항원과 비교하여 더욱 유리한 면역원성 프로파일(profile)을 나타낸다. 하나의 실시태양에서, 유리한 면역원성 프로파일은, 바이러스성 엔벨롭 항원의 하나 이상의 비-중화 에피토프의 더 큰 폐쇄 및/또는 하나 이상의 중화 에피토프의 더 큰 노출을 포함한다. 하나의 실시태양에서, 유리한 면역원성 프로파일은, 숙주 세포의 환경에서 높은 급수의 항원으로 조립되는 대응하는 대조 항원과 비교하여 더욱 효과적인 면역 반응을 결정하는 조립된 항원을 포함한다. 하나의 실시태양에서, 유리한 면역원성 프로파일은 피험체에서 종양 진행을 감소시키거나 예방할 수 있는 더욱 강력한 면역 반응을 포함한다. 하나의 실시태양에서, 조립된 항원은 고유의 대조적인 높은 급수의 (올리고머성) 항원과 비교하여 감소된 비-중화 항체에 대한 결합을 나타냄으로써 고유의 대조적인 높은 급수의 항원과 구별가능하다. 감소된 결합은 검출불가능한 결합을 포함한다(에피토프는 조립된 항원에서 폐쇄되어 비-중화 항체와 결합되지 않는 반면, 에피토프는 비-중화 항체에 결합하는 고유의 대조 항원에서 폐쇄되지 않는다). 특별히, 항체 CBH4G에 의해 인식되는 에피토프는 본원에 기재된 바와 같은 조립된 E2에서 폐쇄된다.
또 다른 실시태양에서, 본 개시내용은 높은 급수의 세포밖에서 조립된 HCV E2 당단백질 항원을 포함하고, 여기서 조립된 E2가 비-중화 항체에 대해 감소된 결합을 나타내는 조성물을 가능하게 한다. 특별히, 비-중화 항체 CBH4G는 고유의 대조적인 올리고머성 HCV E2와 비교하여 감소된 조립된 E2에 대한 결합을 나타내었다. 이러한 항체는 문헌, 예를 들면, 케크(Keck) 등의 문헌[PLos Pathogens: 8(4) e1002653, April 2012]에 기재되어 있다. 또한, 항체 2A12는 고유의 대조적인 올리고머성 HCV E2와 비교하여 감소된 조립된 E2에 대한 결합을 나타내었다. 항체 패널 및 이들을 생성하는 방법은 본원에 참고로 인용된 비에티어(Vietheer) 등의 문헌[Hepatology: 65(4), 1117-1131, 2017], 및 본원에 언급된 참고자료, 예컨대 참고자료 5, 33-36, 17 및 37에 기재되어 있고, 보충 자료는 출판사로부터 이용가능하다.
이와 같이, 하나의 실시태양에서 조립된 항원은 항체 2A12 또는 항체 CBH4G에 의해 결합되는 에피토프 및/또는 에피토프 잔기가 덜 노출되거나, 당분야에 공지된 비-중화 항체, 예컨대 항체 2A12 또는 항체 CBH4G에 의해 인식되지 않는다.
또 다른 실시태양에서, 본 개시내용은 높은 급수의 세포밖에서 조립된 높은 급수의 바이러스성 엔벨롭 항원을 포함하고, 여기서 조립된 바이러스성 env 항원이 고유의 대조적인 높은 급수의 바이러스성 엔벨롭 항원과 비교하여 감소된 비-중화 항체에 대한 결합을 나타내는 조성물을 가능하게 한다.
한가지 실시태양에서, 재접힘된(refolded) 올리고머성 항원은 하기로 구성된 군에서 선택된 적어도 하나의 특징을 나타낸다:
(i) 고유의 대조 항원 형태 또는 단량체성 형태와 비교하여 감소된 비-중화 항체에 대한 결합;
(ii) 고유의 대조 항원 형태와 비교하여 적어도 실질적으로 동일한 중화 항체에 대한 결합;
(iii) 고유의 대조 항원 형태 또는 단량체성 항원과 비교하여 더 낮은 비-중화 항체의 역가의 생산을 이끌어냄;
(iv) 중화 항체의 생산을 이끌어냄;
(v) 광범위 중화 항체의 생산을 이끌어냄;
(vi) 임의적으로 중화 항체의 더 높은 역가의 생산을 이끌어냄; 및
(vii) 임의적으로 광범위 중화 항체의 더 높은 역가의 생산을 이끌어냄.
또 다른 실시태양에서, 본 발명의 개시내용은 높은 급수의 세포밖에서 조립된 높은 급수의 HIV 엔벨롭 항원을 포함하고, 여기서 조립된 HIV env 항원이 고유의 대조적인 높은 급수의 HIV 엔벨롭 항원과 비교하여 감소된 비-중화 항체에 대한 결합을 나타내는 조성물을 가능하게 한다.
적합한 대조군은 본 개시내용의 내용에서 숙련가에 의해 잘 이해될 것이다. 하나의 실시태양에서, 대조 항원은 숙주 세포, 예컨대 세균 또는 포유동물, 효모, 식물 또는 곤충 세포에서 재조합적으로 생산되는, 관심있는 검사 항원과 동일한 유전자형이다.
하나의 실시태양에서, 조립된 높은 급수의 항원은 HCV E2의 수용체-결합 도메인(RBD)이다.
하나의 실시태양에서, HCV E2 항원은 과가변성 영역의 전부 또는 일부, 예컨대 과가변성 영역 1(HVR1) 또는 이의 일부, 과가변성 영역 2(HVR2) 또는 이의 일부 및/또는 상호유전자형 가변성 영역(igVR/VR3) 또는 이의 일부중 하나 이상이 결핍된다. HVR1, HVR2 및 igVR/VR3 서열의 예는 도 12에 제시된다.
몇몇 실시태양에서, E2 항원은 C581, C585, C652, C677, C494, C486, C459, C452, C564, C597, 및 C569로 구성된 군에서 선택된 하나 이상의 아미노산 잔기에서 비-시스테인 치환 또는 돌연변이를 포함한다. 하나의 실시태양에서, 이와 관련하여 하나 이상의 아미노산에 대한 언급은 2, 3, 4, 5, 6, 또는 7개의 아미노산이 비-시스테인 치환되거나, 또는 결실되거나, 달리는 돌연변이됨을 의미한다.
하나의 실시태양에서, 본 개시내용은 조립된 높은 급수의 HCV E2 항원(단량체성 E2로부터 조립됨) 및 약학적으로 또는 생리학적으로 허용가능한 담체 및/또는 희석제를 포함하는 조성물을 제공한다.
하나의 실시태양에서, 조성물은 약학적으로 또는 생리학적으로 허용가능한 희석제, 담체 또는 보조제를 포함한다. 하나의 실시태양에서, 항원은 백신 조성물에 적합한 임의의 형태의 담체내에 존재할 수 있다.
하나의 실시태양에서, 개시내용은 HCV 또는 HIV 감염과 같은 바이러스 감염, 또는 암, 또는 바이러스성 엔벨롭 또는 암 항원과 연관된 증상의 치료 또는 예방을 위한 약제의 제조에 있어서의 본원에 기재된 바와 같은 조성물의 용도를 제공한다.
하나의 실시태양에서, 개시내용은 항원, 예컨대 E2, HCV 감염과 연관된 증상을 진단 또는 모니터링하거나, 또는 항-HCV 치료 프로토콜(protocol)을 모니터링하기 위한 진단용 제제의 제조에 있어서의 조립된 높은 급수의 항원, 또는 조성물의 용도를 제공한다.
하나의 실시태양에서, 개시내용은 면역 반응을 이끌어내기에 충분한 조건하에 일정 시간 동안 효과량의 본원에 기재된 조립된 높은 급수의 항원, 또는 본원에 기재된 바와 같은 조성물을 피험체 또는 환자에게 투여하는 단계를 포함하는, 피험체 또는 환자에서 면역 반응을 이끌어내는 방법을 제공한다.
하나의 실시태양에서, 개시내용은 본원에 기재된 바와 같은 조립된 올리고머성 항원, 또는 조성물을 피험체에게 투여하는 단계를 포함하는, HCV로부터의 감염에 대해 피험체를 면역화시키는 방법을 제공한다.
하나의 실시태양에서, 개시내용은 HCV 감염을 치료하거나 예방하기에 충분한 조건하에 일정 시간 동안 기재된 바와 같은 조립된 올리고머성 항원, 또는 조성물을 피험체에게 투여하는 단계를 포함하는, HCV 감염을 치료하거나 예방하는 방법을 제공한다.
하나의 실시태양에서, 조성물은 추가로 상이한 병원체로부터의 높은 급수의 2차 항원을 포함한다.
하나의 실시태양에서, 조립된 높은 급수의 항원은 검출가능한 또는 정제 태그(tag)를 포함한다.
하나의 실시태양에서, 개시내용은 효과량의 항원을 피험체에게 투여하는 단계 및 생산된 항체를 정제하는 단계를 포함하는, 본원에 기재된 바와 같은 높은 급수의 조립된 항원에 대한 정제된 항체를 생산하는 방법을 제공한다.
하나의 실시태양에서, 본원에 기재된 바와 같은 조립된 높은 급수의 항원/E2 항원을 특이적으로 인식하는 항체가 제공된다. 다르게는 또는 추가적으로, 고유적이지만 조립되지 않은 항원, 또는 조립되지만 고유적이지 않은 항원 상에 노출된 에피토프를 인식하는 항체가 식별될 수 있다.
하나의 실시태양에서, 본원에 기재된 바와 같은 조립된 높은 급수의 항원, 또는 본원에 기재된 바와 같은 조성물을 포함하는 키트, 또는 고체 또는 반-고체 기질이 제공된다.
하나의 실시태양에서, 개시내용은 (i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수의 항원을 접촉시키는 단계; 및 (ii) 환원제를 제거 또는 희석하거나 환원된 낮은 급수의 항원을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 항원을 높은 급수의 항원으로 조립시키는 단계를 포함하는 본원에 개시된 방법에 의해 낮은 급수의 항원으로부터 생산된 조립된 높은 급수의 항원, 또는 높은 급수의 세포밖에서 조립된 높은 급수의 암 항원 또는 바이러스성 엔벨롭 항원을 포함하는 조성물(여기서 조립된 바이러스성 env 항원이 고유의 대조적인 높은 급수의 바이러스성 엔벨롭 항원과 비교하여 감소된 비-중화 항체에 대한 결합을 나타냄)의, 항원 특이적 면역 세포에 결합하는/이를 검출하는 용도를 제공한다.
하나의 실시태양에서, 본 발명의 높은 급수의 조립된 항원은 항원에 특이적인 면역 세포, 예컨대 B-세포의 검출 및/또는 단리를 위해, 또는 이를 위해 사용될 경우 적합하다. 한가지 실시태양에서, 면역 세포는 항원에 특이적인 T-세포이다.
또 다른 실시태양에서, 개시내용은 i) 항원에 특이적인 면역 세포/B-세포를 본원에 기재된 바와 같은 조립된 높은 급수의 항원으로 표지화하는 단계; 및 ii) 표지화된 면역 세포/B-세포를 세포분석에 의해 검출하고/하거나 단리하는 단계를 포함하는, 항원에 특이적인 면역 세포/B-세포를 검출하고/하거나 단리하는 방법을 가능하게 한다.
또 다른 실시태양에서, 개시내용은 i) 항원, 예컨대 HCV 또는 HIV에 특이적인 면역 세포/B-세포를 조립된 높은 급수의 항원으로 표지화하는 단계; 및 ii) 표지화된 면역 세포/B-세포를 세포분석에 의해 검출하고/하거나 단리하는 단계를 포함하는, 항원, 예컨대 HCV 또는 HIV에 특이적인 면역 세포/B-세포를 검출하고/하거나 단리하는 방법을 가능하게 한다.
이러한 요약은 명세서에 기재된 모든 실시태양을 빠뜨림없이 열거한 것이 아니다.
몇몇 도면은 색 표시 또는 도면 요소를 함유한다. 도면의 색채 버전은 요청시 특허권자로부터 또는 적절한 특허청으로부터 입수가능하다. 만일 특허청으로부터 수득될 경우 요금이 부과될 수 있다.
도 1은 Δ123-함유 플라스미드의 아가로스 겔 전기영동을 보여준다. 밴드의 상단 행은 벡터 주쇄를 나타내고, 밴드의 하단 행은 Δ123 삽입물을 나타내는데, 이는 제한 효소에 의한 소화에 의해 벡터로부터 단리되었다. pcDNA-H77cΔ123-HIS 및 pcDNA-S52Δ123-HIS는 둘 다 NheI 및 XbaI에 의해 소화된 반면, pcDNA-Con1Δ123-HIS는 NheI 및 XhoI에 의해 소화되었다. H77c, Con1 및 S52 Δ123-함유 플라스미드로부터 방출된 삽입물의 예상 길이는 각각 762 bp, 759 및 710 bp이다. 진룰러(GeneRuler) 1 kb DNA 래더(DNA ladder)가 크기 표지(M)로서 사용되었다.
도 2는 Δ123의 형질감염 및 정제를 보여준다. (A-B) FS293F 세포 내로의 pcDNA-Con1Δ123-HIS(A) 및 pcDNA-S52Δ123-HIS(B)의 일시적 형질감염으로부터의 단백질 발현은 샌드위치(sandwich) ELISA를 통해 분석되었다. 형질감염 이후 3, 5, 7 및 9 일째 수거된 세포 배양 상청액이 2배 희석된 후 이들의 반-로그식(half-logarithmic) 연속 희석물이 이합체성 MBP-CD81-LEL113-201-코팅된 효소 면역검정 플레이트에 적용되었다. 단량체성 H77c Δ123(1 ㎍/㎖)의 반-로그식 연속 희석물 및 HIV gp140이 함유된 세포 배양 상청액(이는 6xHIS 태그가 결핍됨)이 또한 각각 양성 및 음성 대조군으로서 포함되었다. 이후 1/1000로 희석된 토끼 항-HIS 항체가 첨가되고, 1/1000로 희석된 HRP-접합된(conjugated) 염소 항-토끼 면역글로불린의 단일 희석물에 의해 검출되었다. (C-D) 친화도 크로마토그래피 이후 Con1 Δ123(C) 및 S52 Δ123(D)은 동일한 방식으로 수행되는 샌드위치 ELISA를 통해 분석되었다. 세포 배양 상청액에 함유된 단백질은 코발트-충전된 탈론(TALON) 비이드에 적용되어 6x HIS 태그 부착 Δ123의 결합을 허용하고, 탈론 상호작용 이후 상청액은 통과액으로서 수집되었다. 이어서 단백질은 세척되고 3가지 별도의 분별물로 용출되었다. 모든 샘플은 ELISA를 위해 본래의 제조물에 대하여 이들의 양을 동등히 하기 위해 적절할 경우 희석되었다.
도 3은 Δ123의 겔 여과 프로파일을 보여준다. (A) 친화도-정제된 H77c Δ123 당단백질은 FS293F 세포에서 안정한 형질감염으로부터 생산되었고, 수퍼덱스(Superdex) 200을 사용하여 겔 여과 크로마토그래피에 의해 분석되었다. 점선 및 피크 높이는 단량체(78), 이합체(70), HMW2(59) 및 HMW1(46) Δ123에 상응하는 분별물을 지시한다. (B-C)는 FS293F 세포로의 일시적 형질감염의 1 라운드(round)로부터 생산된 친화도-정제된 Con1 Δ123(B) 및 S52 Δ123(C)의 수퍼덱스 200에 의한 겔 여과 크로마토그래피를 보여준다. 점선 및 피크 높이는 각각 Con1 및 S52에 대하여 숫자 77, 68, 66, 46, 및 78, 69, 65, 46에 의해 각각 표지화된 단량체, 이합체, HMW2 및 HMW1에 상응하는 분별물을 지시한다. (D-E)는 FS293F 세포로의 Con1 Δ123(D) 및 S52 Δ123(E)의 일시적 형질감염이 반복되었음을 보여주고(즉 라운드 2), 생산된 Δ123 당단백질은 친화도-정제된 후, 라운드 1과 동일한 방식으로 겔 여과 크로마토그래피에 의해 분석되었다.
도 4는 SDS-PAGE를 통해 분석된 분별화된 Δ123의 겔 여과를 보여준다. 겔 여과 분별화된 (A) Con1 Δ123 및 (B) S52 Δ123의 비-환원 SDS-PAGE 및 쿠마시(coomassie) 염색: 각각의 2가지 연속 분별물로부터 20 uL을 모아서 5-12% 폴리아크릴아미드 구배 겔 상으로 적재하였다. 겔 여과 분별화된 (C) Con1 Δ123 및 (D) S52 Δ123의 환원 SDS-PAGE 및 쿠마시 염색: 비-환원 SDS-PAGE와 동일한 부피가 12% 아크릴아미드 겔 상으로 적재되었다. 바이오-래드(Bio-Rad) 광범위 SDS-PAGE 표준물이 표지(M)로서 적재되었다.
도 5(A)는 단량체성 및 HMW1 Δ123에 결합하는 MAb의 능력의 일예를 보여준다. 단량체성 Δ123(적색)(2 ㎍/㎖) 및 HMW1 Δ123(청색)(5 ㎍/㎖) 및 음성 대조군으로서의 BSA(녹색)는 효소 면역검정 플레이트 상으로 코팅되었다. 이어서 1차 항체의 반-로그식 연속 희석물이 첨가되었고, 결합은 적절한 HRP-접합된 2차 항체에 의해 검출되었다. 이러한 기법을 사용하여, HMW1 Δ123에 대한 패널 MAb의 상대적 결합은 이후 각각의 결합 곡선의 중간점을 비교함으로써 계산되었고, 이러한 중간점은 가장 높은 광학 밀도 값의 절반으로서 취해졌다. 결합에서 배수 차이는 표 7의 작성을 위해 단량체 및 H77c HMW1 Δ123에 상대적으로 계산되었다. (B)는 MAb의 패널에 대한 Δ123의 상이한 균주의 반응성을 보여준다. 단일 희석점 평가를 수행하였고, 여기서 H77c, Con1 및 S52 Δ123 단량체(5 ㎍/㎖) 뿐만 아니라 음성 대조군으로서의 BSA(5 ㎍/㎖)는 효소 면역검정 플레이트 상에 코팅되었다. 1차 항체의 단일 희석물이 첨가되었고, HRP-접합된 2차 항체에 특이적인 적절한 종에 의해 결합이 검출되었다.
도 6(A)는 소규모의 트리스(2-카복시에틸)포스핀 하이드로클로라이드(TCEP) 환원을 보여준다. 효소 면역검정 플레이트 상에 코팅된 H77 Δ123 단량체는 PBS pH 9.6에서 제조된 상이한 농도의 TCEP(0 내지 500mM)에 의해 30 분 동안 37℃하에 환원되었다. 입체배좌(conformation)-의존적 마우스 H53(1 ㎍/㎖) 뿐만 아니라 각각 양성 및 음성 대조군으로서의 토끼 항-HIS(1/1000) 및 인간 항-CMV R04(1/1000)를 포함하여, 1차 항체의 단일 희석물이 첨가되었다. 결합은 적절한 HRP-접합된 2차 항체에 의해 검출되었다. 각각의 1차 항체에 대한 반응성은 3중으로 검사되었고, 평균 광학 밀도 값은 표준 편차를 나타내는 오차 막대를 갖는 선 도표로서 도표화되었다. (B)는 TCEP 및 BMOE에 의해 처리된 단량체성 H77c Δ123을 보여준다. 1 내지 7로 표지화된 상이한 조건하에 TCEP 환원되고 BMOE 가교결합제에 의해 재접힘된 후의 단량체성 H77c Δ123(10 ㎍)의 비-환원 SDS-PAGE 및 쿠마시 염색: 이는 표 8에 기재되어 있다. Δ123 샘플은 대조군으로서의 단량체성 및 HMW1 H77c Δ123(각각 5 ㎍) 뿐만 아니라 광범위 SDS-PAGE 표지(M)와 함께 5-12% 폴리아크릴아미드 구배 겔 상에 적재되었다.
도 7(A)는 글루타티온(glutathione)으로 처리된 단량체성 H77c Δ123을 보여준다. 1 내지 5로 표지화된 상이한 조건하에 산화환원-셔플링(redox-shuffling) 시스템을 사용하여 GSH 및 GSSG에 의해 재접힘된 후의 단량체성 H77c Δ123(10 ㎍)의 비-환원 SDS-PAGE 및 쿠마시 염색: 이는 표 9에 기재되어 있다. H77c Δ123 샘플은 대조군으로서의 단량체성 및 HMW1 H77c Δ123(각각 5 ㎍) 뿐만 아니라 광범위 SDS-PAGE 표지(M)와 함께 5-12% 폴리아크릴아미드 구배 겔 상에 적재되었다. (B)는 글루타티온에 의해 처리된 단량체성 H77c Δ123의 겔 여과 프로파일 및 미처리된 단량체성 H77c Δ123의 겔 여과 크로마토그래피를 보여준다(수퍼덱스 200 사용). (C)는 10 ㎍/㎕의 당단백질이 2mM GSH 및 0.4mM GSSG에 의해 처리된 이후의 H77c Δ123의 겔 여과 크로마토그래피를 보여준다(수퍼덱스 200 사용). 점선 및 피크 높이는 단량체(78) 및 이합체(69)에 상응하는 분별물을 지시한다.
도 8(A)는 소규모의 DTT 환원을 보여준다. 효소 면역검정 플레이트 상에 코팅된 H77c Δ123 단량체는 탄산염-중탄산염 완충액 pH 9.6에서 제조된 상이한 농도의 DTT(0 내지 1OmM)에 의해 30분 동안 37℃에서 환원되었다. 입체배좌-의존적 마우스 H53(1 ㎍/㎖) 뿐만 아니라, 각각 양성 및 음성 대조군을 나타내는 토끼 항-HIS(1/1000) 및 인간 항-CMV R04를 포함하여, 1차 항체의 단일 희석물을 첨가하였다. 결합은 적절한 HRP-접합된 2차 항체에 의해 검출되었다. 각각의 1차 항체에 대한 반응성은 3중으로 검사되었고, 평균 광학 밀도 값은 표준 편차를 나타내는 오차 막대를 갖는 선 도표로서 도표화되었다. (B)는 DTT에 의해 처리된 단량체성 H77c Δ123을 보여준다. 상이한 조건하에서의 DTT 환원(1 내지 12로 표지화되고 표 3.6에 기재됨) 및 느린 희석 방법에 의한 재접힘 이후 단량체성 H77c Δ123(10 ㎍)의 비-환원 SDS-PAGE 및 쿠마시 염색. H77c Δ123 샘플은, 대조군으로서의 단량체성 및 HMW1 H77c Δ123(각각 5 ㎍) 뿐만 아니라 광범위 SDS-PAGE 표지(M)와 함께, 5-12% 폴리아크릴아미드 구배 겔 상에 적재되었다. (C)는 미처리된 단량체성 H77c Δ123의 겔 여과 크로마토그래피(수퍼덱스 200 사용)를 보여준다. (D)는 1 ㎍/㎕의 당단백질이 1 mM DTT에 의해 처리된 이후의 H77c Δ123의 겔 여과 크로마토그래피(수퍼덱스 200 사용)를 보여준다. 점선 및 피크 높이는 단량체(77-78), 이합체(69) 및 HMW Δ123(59)에 상응하는 분별물을 지시한다.
도 9는 DTT-처리된 Δ123 및 ALA7 Δ123의 항원 특징을 보여준다. DTT-처리된 Δ123 및 ALA7 Δ123은 실시예 15에 기재된 바와 같이 항체 (A) AR3C (B) CBH4G (C) HC84.27 및 (D) HCV1과의 결합을 평가함으로써 특징지워졌다.
도 10은 겔 여과 크로마토그래피를 사용하여 DTT-처리된 Δ123이 재접힘됨으로써 생성된 생성물의 분석을 보여준다. 미처리된 단량체의 프로파일이 제시되고, 이후 실시예 14에 기재된 바와 같이 DTT에 의한 처리의 횟수(1차, 2차, 또는 3차 처리)를 증가시킨다.
도 11은 실시예 14에 기재된 바와 같이 겔 여과 크로마토그래피를 사용하여 재접힘되지 않은 Δ123 단량체의 재접힘 및 프로테아제 저해제의 존재하의 Δ123의 재접힘으로부터 생성된 생성물의 분석을 보여준다.
도 12는 사용된 HCV 단리물의 상응하는 E2661 영역의 클러스탈(Clustal)W 정렬을 제공한다. H77c(AF009606, Gla, 서열 번호 11), J6(AF177036, G2a, 서열 번호 16), s52(GU814263, G3a, 서열 번호 13), ED43(GU814265, G4a, 서열 번호 10), SA13(AF064490, G5a, 서열 번호 12), EUHK2(Y12083, G6a, 서열 번호 14), QC69 (EF108306, G7a, 서열 번호 15). HVR1, HVR2 및 igVR/VR3은 적색/주황색으로 제시된다. ED43의 아미노산 잔기 GFLASLFY, YTWGENETD 및 YRLWHF에 상응하는 잔기는 CD81 결합 영역이다.
도 13은 코돈 최적화된 Δ123Ala7에 대한 뉴클레오티드 작성물(밑줄친 영역은 제한 효소 자리에 상응함, GGTACC = KpnI, GGATCC = BamHI, CTCGAG = XhoI) 및 Δ123Ala7에 대한 단백질 서열을 보여준다.
도 14 내지 도 14cb는 하기 단백질 서열의 클러스탈오메가(ClustalOmega) 아미노산 정렬을 표시한다: AF009606 코딩 서열(서열 번호 6), AF009606 전장 E2(서열 번호 7), AF009606 E2661(서열 번호 8), WT_E2661(서열 번호 4) 및 Δ123(서열 번호 3). 밑줄친 영역은 잔기 630-635에 상응한다.
도 15(A)는 상이한 농도의 β-머캅토에탄올에 의해 환원되고 조립된 샘플의 비-환원 SDS-PAGE를 보여준다. 정밀 크기 표지는 kDa으로 지시되는 분자량에 의해 좌측에 제시된다. 단량체, 이합체 및 고분자량 형태를 나타내는 크기는 우측에 제시된다. (B)는 100 mM BME에 의해 처리된 후(파선) 또는 미처리된(실선) 단량체성 Δ123의 수퍼덱스 200에 의한 겔 여과 크로마토그래피를 보여준다.
도 16은 조립된 HCV 단백질에 의해 백신접종된 동물에서의 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 단량체성 Δ123 단백질에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 2-F 및 4-F+5-F 사이의 항체 역가에서의 차이는 만-휘트니(Mann-Whitney) 비대응(unpaired) t-테스트를 사용할 경우 통계적으로 유의적이었다(p=0.0281). 프리즘(Prism) v7.0.
도 17은 조립된 HCV 단백질에 의해 백신접종된 동물에서 에피토프 I에 대한 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 유전자형 1a H77c 서열의 잔기 409-428을 포괄하는 합성 펩타이드에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 18은 조립된 HCV 단백질에 의해 백신접종된 동물에서 에피토프 III에 대한 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 유전자형 1a H77c 서열의 잔기 523-549를 포괄하는 합성 펩타이드에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 19는 조립된 HCV 단백질에 의해 백신접종된 동물에서 유전자형 2a 에피토프 I에 대한 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 유전자형 2a J6 서열의 잔기 409-428을 포괄하는 합성 펩타이드에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 20은 HCV E2 및 그의 세포 수용체 CD81 사이의 상호작용을 저해하는 면역 혈청의 능력을 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 (A) H77c G1a E2 및 CD81, 및 (B) J6 G2a E2 및 CD81 사이의 결합을 저해하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 21은 유전자형 1a 바이러스에 의한 간 세포의 감염을 예방하는 면역 혈청의 능력을 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 G1a HCVpp에 의한 감염을 예방하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이다.
도 22는 HMW1 및 단량체 면역 혈청의 특이성을 보여준다. 기니아 피그(guinea pig) 혈청의 연속 희석물이 일정한 양의 HCV1(A), HC84-27(B), AR3C(C) 및 2A12(D)에 첨가되었다. 항체가 단량체성 Δ123에 첨가되었고, 결합된 MAb는 항-인간 Fab2에 의해 검출되었다. 그룹은 만-휘트니 t-테스트를 사용하여 비교되었다(프리즘 v7).
도 23은 조립된 Δ123을 사용하여 분류된 B 세포 개체군의 FACS 도표를 보여준다. CD 19 양성 및 항-E2 양성 B 세포는 항-CD19 Cy7 항체 및 조립된 Δ123에 의해 검출되었다.
도 24는 조립된 단백질의 크기를 결정하는 크기-배제 크로마토그래피-다각도 광산란을 보여준다. 조립된 Δ123A7(청색) 및 조립된 Δ123(적색) 샘플의 UV(A280 nm) 신호의 중첩 및 몰 질량.
도 25는 DTT에 의해 처리된 후, HMW 형태로 조립되지 않은 Δ123A7 단량체(이는 조립이 수반되는 DTT에 의한 변성의 제2 라운드를 거치지 않을 것임)의 크기-배제 크로마토그래피를 보여준다. 단량체 및 HMW 종은 각각 표에 제시된 화살표 및 %에 의해 지시된다.
도 26은 조립이 수반되는 DTT에 의한 변성을 거친 RBD 단량체의 크기-배제 크로마토그래피를 보여준다. 단량체 및 HMW 종은 각각 표에 제시된 화살표 및 %에 의해 지시된다.
도 27은 조립이 수반되는 DTT에 의한 변성을 거친 RBDA7 단량체의 크기-배제 크로마토그래피를 보여준다. 단량체 및 HMW 종은 각각 표에 제시된 화살표 및 %에 의해 지시된다.
도 28은 조립이 수반되는 DTT에 의한 변성을 거친 env 단량체의 크기-배제 크로마토그래피를 보여준다. HMW 종은 화살표에 의해 지시된다.
도 1은 Δ123-함유 플라스미드의 아가로스 겔 전기영동을 보여준다. 밴드의 상단 행은 벡터 주쇄를 나타내고, 밴드의 하단 행은 Δ123 삽입물을 나타내는데, 이는 제한 효소에 의한 소화에 의해 벡터로부터 단리되었다. pcDNA-H77cΔ123-HIS 및 pcDNA-S52Δ123-HIS는 둘 다 NheI 및 XbaI에 의해 소화된 반면, pcDNA-Con1Δ123-HIS는 NheI 및 XhoI에 의해 소화되었다. H77c, Con1 및 S52 Δ123-함유 플라스미드로부터 방출된 삽입물의 예상 길이는 각각 762 bp, 759 및 710 bp이다. 진룰러(GeneRuler) 1 kb DNA 래더(DNA ladder)가 크기 표지(M)로서 사용되었다.
도 2는 Δ123의 형질감염 및 정제를 보여준다. (A-B) FS293F 세포 내로의 pcDNA-Con1Δ123-HIS(A) 및 pcDNA-S52Δ123-HIS(B)의 일시적 형질감염으로부터의 단백질 발현은 샌드위치(sandwich) ELISA를 통해 분석되었다. 형질감염 이후 3, 5, 7 및 9 일째 수거된 세포 배양 상청액이 2배 희석된 후 이들의 반-로그식(half-logarithmic) 연속 희석물이 이합체성 MBP-CD81-LEL113-201-코팅된 효소 면역검정 플레이트에 적용되었다. 단량체성 H77c Δ123(1 ㎍/㎖)의 반-로그식 연속 희석물 및 HIV gp140이 함유된 세포 배양 상청액(이는 6xHIS 태그가 결핍됨)이 또한 각각 양성 및 음성 대조군으로서 포함되었다. 이후 1/1000로 희석된 토끼 항-HIS 항체가 첨가되고, 1/1000로 희석된 HRP-접합된(conjugated) 염소 항-토끼 면역글로불린의 단일 희석물에 의해 검출되었다. (C-D) 친화도 크로마토그래피 이후 Con1 Δ123(C) 및 S52 Δ123(D)은 동일한 방식으로 수행되는 샌드위치 ELISA를 통해 분석되었다. 세포 배양 상청액에 함유된 단백질은 코발트-충전된 탈론(TALON) 비이드에 적용되어 6x HIS 태그 부착 Δ123의 결합을 허용하고, 탈론 상호작용 이후 상청액은 통과액으로서 수집되었다. 이어서 단백질은 세척되고 3가지 별도의 분별물로 용출되었다. 모든 샘플은 ELISA를 위해 본래의 제조물에 대하여 이들의 양을 동등히 하기 위해 적절할 경우 희석되었다.
도 3은 Δ123의 겔 여과 프로파일을 보여준다. (A) 친화도-정제된 H77c Δ123 당단백질은 FS293F 세포에서 안정한 형질감염으로부터 생산되었고, 수퍼덱스(Superdex) 200을 사용하여 겔 여과 크로마토그래피에 의해 분석되었다. 점선 및 피크 높이는 단량체(78), 이합체(70), HMW2(59) 및 HMW1(46) Δ123에 상응하는 분별물을 지시한다. (B-C)는 FS293F 세포로의 일시적 형질감염의 1 라운드(round)로부터 생산된 친화도-정제된 Con1 Δ123(B) 및 S52 Δ123(C)의 수퍼덱스 200에 의한 겔 여과 크로마토그래피를 보여준다. 점선 및 피크 높이는 각각 Con1 및 S52에 대하여 숫자 77, 68, 66, 46, 및 78, 69, 65, 46에 의해 각각 표지화된 단량체, 이합체, HMW2 및 HMW1에 상응하는 분별물을 지시한다. (D-E)는 FS293F 세포로의 Con1 Δ123(D) 및 S52 Δ123(E)의 일시적 형질감염이 반복되었음을 보여주고(즉 라운드 2), 생산된 Δ123 당단백질은 친화도-정제된 후, 라운드 1과 동일한 방식으로 겔 여과 크로마토그래피에 의해 분석되었다.
도 4는 SDS-PAGE를 통해 분석된 분별화된 Δ123의 겔 여과를 보여준다. 겔 여과 분별화된 (A) Con1 Δ123 및 (B) S52 Δ123의 비-환원 SDS-PAGE 및 쿠마시(coomassie) 염색: 각각의 2가지 연속 분별물로부터 20 uL을 모아서 5-12% 폴리아크릴아미드 구배 겔 상으로 적재하였다. 겔 여과 분별화된 (C) Con1 Δ123 및 (D) S52 Δ123의 환원 SDS-PAGE 및 쿠마시 염색: 비-환원 SDS-PAGE와 동일한 부피가 12% 아크릴아미드 겔 상으로 적재되었다. 바이오-래드(Bio-Rad) 광범위 SDS-PAGE 표준물이 표지(M)로서 적재되었다.
도 5(A)는 단량체성 및 HMW1 Δ123에 결합하는 MAb의 능력의 일예를 보여준다. 단량체성 Δ123(적색)(2 ㎍/㎖) 및 HMW1 Δ123(청색)(5 ㎍/㎖) 및 음성 대조군으로서의 BSA(녹색)는 효소 면역검정 플레이트 상으로 코팅되었다. 이어서 1차 항체의 반-로그식 연속 희석물이 첨가되었고, 결합은 적절한 HRP-접합된 2차 항체에 의해 검출되었다. 이러한 기법을 사용하여, HMW1 Δ123에 대한 패널 MAb의 상대적 결합은 이후 각각의 결합 곡선의 중간점을 비교함으로써 계산되었고, 이러한 중간점은 가장 높은 광학 밀도 값의 절반으로서 취해졌다. 결합에서 배수 차이는 표 7의 작성을 위해 단량체 및 H77c HMW1 Δ123에 상대적으로 계산되었다. (B)는 MAb의 패널에 대한 Δ123의 상이한 균주의 반응성을 보여준다. 단일 희석점 평가를 수행하였고, 여기서 H77c, Con1 및 S52 Δ123 단량체(5 ㎍/㎖) 뿐만 아니라 음성 대조군으로서의 BSA(5 ㎍/㎖)는 효소 면역검정 플레이트 상에 코팅되었다. 1차 항체의 단일 희석물이 첨가되었고, HRP-접합된 2차 항체에 특이적인 적절한 종에 의해 결합이 검출되었다.
도 6(A)는 소규모의 트리스(2-카복시에틸)포스핀 하이드로클로라이드(TCEP) 환원을 보여준다. 효소 면역검정 플레이트 상에 코팅된 H77 Δ123 단량체는 PBS pH 9.6에서 제조된 상이한 농도의 TCEP(0 내지 500mM)에 의해 30 분 동안 37℃하에 환원되었다. 입체배좌(conformation)-의존적 마우스 H53(1 ㎍/㎖) 뿐만 아니라 각각 양성 및 음성 대조군으로서의 토끼 항-HIS(1/1000) 및 인간 항-CMV R04(1/1000)를 포함하여, 1차 항체의 단일 희석물이 첨가되었다. 결합은 적절한 HRP-접합된 2차 항체에 의해 검출되었다. 각각의 1차 항체에 대한 반응성은 3중으로 검사되었고, 평균 광학 밀도 값은 표준 편차를 나타내는 오차 막대를 갖는 선 도표로서 도표화되었다. (B)는 TCEP 및 BMOE에 의해 처리된 단량체성 H77c Δ123을 보여준다. 1 내지 7로 표지화된 상이한 조건하에 TCEP 환원되고 BMOE 가교결합제에 의해 재접힘된 후의 단량체성 H77c Δ123(10 ㎍)의 비-환원 SDS-PAGE 및 쿠마시 염색: 이는 표 8에 기재되어 있다. Δ123 샘플은 대조군으로서의 단량체성 및 HMW1 H77c Δ123(각각 5 ㎍) 뿐만 아니라 광범위 SDS-PAGE 표지(M)와 함께 5-12% 폴리아크릴아미드 구배 겔 상에 적재되었다.
도 7(A)는 글루타티온(glutathione)으로 처리된 단량체성 H77c Δ123을 보여준다. 1 내지 5로 표지화된 상이한 조건하에 산화환원-셔플링(redox-shuffling) 시스템을 사용하여 GSH 및 GSSG에 의해 재접힘된 후의 단량체성 H77c Δ123(10 ㎍)의 비-환원 SDS-PAGE 및 쿠마시 염색: 이는 표 9에 기재되어 있다. H77c Δ123 샘플은 대조군으로서의 단량체성 및 HMW1 H77c Δ123(각각 5 ㎍) 뿐만 아니라 광범위 SDS-PAGE 표지(M)와 함께 5-12% 폴리아크릴아미드 구배 겔 상에 적재되었다. (B)는 글루타티온에 의해 처리된 단량체성 H77c Δ123의 겔 여과 프로파일 및 미처리된 단량체성 H77c Δ123의 겔 여과 크로마토그래피를 보여준다(수퍼덱스 200 사용). (C)는 10 ㎍/㎕의 당단백질이 2mM GSH 및 0.4mM GSSG에 의해 처리된 이후의 H77c Δ123의 겔 여과 크로마토그래피를 보여준다(수퍼덱스 200 사용). 점선 및 피크 높이는 단량체(78) 및 이합체(69)에 상응하는 분별물을 지시한다.
도 8(A)는 소규모의 DTT 환원을 보여준다. 효소 면역검정 플레이트 상에 코팅된 H77c Δ123 단량체는 탄산염-중탄산염 완충액 pH 9.6에서 제조된 상이한 농도의 DTT(0 내지 1OmM)에 의해 30분 동안 37℃에서 환원되었다. 입체배좌-의존적 마우스 H53(1 ㎍/㎖) 뿐만 아니라, 각각 양성 및 음성 대조군을 나타내는 토끼 항-HIS(1/1000) 및 인간 항-CMV R04를 포함하여, 1차 항체의 단일 희석물을 첨가하였다. 결합은 적절한 HRP-접합된 2차 항체에 의해 검출되었다. 각각의 1차 항체에 대한 반응성은 3중으로 검사되었고, 평균 광학 밀도 값은 표준 편차를 나타내는 오차 막대를 갖는 선 도표로서 도표화되었다. (B)는 DTT에 의해 처리된 단량체성 H77c Δ123을 보여준다. 상이한 조건하에서의 DTT 환원(1 내지 12로 표지화되고 표 3.6에 기재됨) 및 느린 희석 방법에 의한 재접힘 이후 단량체성 H77c Δ123(10 ㎍)의 비-환원 SDS-PAGE 및 쿠마시 염색. H77c Δ123 샘플은, 대조군으로서의 단량체성 및 HMW1 H77c Δ123(각각 5 ㎍) 뿐만 아니라 광범위 SDS-PAGE 표지(M)와 함께, 5-12% 폴리아크릴아미드 구배 겔 상에 적재되었다. (C)는 미처리된 단량체성 H77c Δ123의 겔 여과 크로마토그래피(수퍼덱스 200 사용)를 보여준다. (D)는 1 ㎍/㎕의 당단백질이 1 mM DTT에 의해 처리된 이후의 H77c Δ123의 겔 여과 크로마토그래피(수퍼덱스 200 사용)를 보여준다. 점선 및 피크 높이는 단량체(77-78), 이합체(69) 및 HMW Δ123(59)에 상응하는 분별물을 지시한다.
도 9는 DTT-처리된 Δ123 및 ALA7 Δ123의 항원 특징을 보여준다. DTT-처리된 Δ123 및 ALA7 Δ123은 실시예 15에 기재된 바와 같이 항체 (A) AR3C (B) CBH4G (C) HC84.27 및 (D) HCV1과의 결합을 평가함으로써 특징지워졌다.
도 10은 겔 여과 크로마토그래피를 사용하여 DTT-처리된 Δ123이 재접힘됨으로써 생성된 생성물의 분석을 보여준다. 미처리된 단량체의 프로파일이 제시되고, 이후 실시예 14에 기재된 바와 같이 DTT에 의한 처리의 횟수(1차, 2차, 또는 3차 처리)를 증가시킨다.
도 11은 실시예 14에 기재된 바와 같이 겔 여과 크로마토그래피를 사용하여 재접힘되지 않은 Δ123 단량체의 재접힘 및 프로테아제 저해제의 존재하의 Δ123의 재접힘으로부터 생성된 생성물의 분석을 보여준다.
도 12는 사용된 HCV 단리물의 상응하는 E2661 영역의 클러스탈(Clustal)W 정렬을 제공한다. H77c(AF009606, Gla, 서열 번호 11), J6(AF177036, G2a, 서열 번호 16), s52(GU814263, G3a, 서열 번호 13), ED43(GU814265, G4a, 서열 번호 10), SA13(AF064490, G5a, 서열 번호 12), EUHK2(Y12083, G6a, 서열 번호 14), QC69 (EF108306, G7a, 서열 번호 15). HVR1, HVR2 및 igVR/VR3은 적색/주황색으로 제시된다. ED43의 아미노산 잔기 GFLASLFY, YTWGENETD 및 YRLWHF에 상응하는 잔기는 CD81 결합 영역이다.
도 13은 코돈 최적화된 Δ123Ala7에 대한 뉴클레오티드 작성물(밑줄친 영역은 제한 효소 자리에 상응함, GGTACC = KpnI, GGATCC = BamHI, CTCGAG = XhoI) 및 Δ123Ala7에 대한 단백질 서열을 보여준다.
도 14 내지 도 14cb는 하기 단백질 서열의 클러스탈오메가(ClustalOmega) 아미노산 정렬을 표시한다: AF009606 코딩 서열(서열 번호 6), AF009606 전장 E2(서열 번호 7), AF009606 E2661(서열 번호 8), WT_E2661(서열 번호 4) 및 Δ123(서열 번호 3). 밑줄친 영역은 잔기 630-635에 상응한다.
도 15(A)는 상이한 농도의 β-머캅토에탄올에 의해 환원되고 조립된 샘플의 비-환원 SDS-PAGE를 보여준다. 정밀 크기 표지는 kDa으로 지시되는 분자량에 의해 좌측에 제시된다. 단량체, 이합체 및 고분자량 형태를 나타내는 크기는 우측에 제시된다. (B)는 100 mM BME에 의해 처리된 후(파선) 또는 미처리된(실선) 단량체성 Δ123의 수퍼덱스 200에 의한 겔 여과 크로마토그래피를 보여준다.
도 16은 조립된 HCV 단백질에 의해 백신접종된 동물에서의 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 단량체성 Δ123 단백질에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 2-F 및 4-F+5-F 사이의 항체 역가에서의 차이는 만-휘트니(Mann-Whitney) 비대응(unpaired) t-테스트를 사용할 경우 통계적으로 유의적이었다(p=0.0281). 프리즘(Prism) v7.0.
도 17은 조립된 HCV 단백질에 의해 백신접종된 동물에서 에피토프 I에 대한 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 유전자형 1a H77c 서열의 잔기 409-428을 포괄하는 합성 펩타이드에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 18은 조립된 HCV 단백질에 의해 백신접종된 동물에서 에피토프 III에 대한 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 유전자형 1a H77c 서열의 잔기 523-549를 포괄하는 합성 펩타이드에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 19는 조립된 HCV 단백질에 의해 백신접종된 동물에서 유전자형 2a 에피토프 I에 대한 항체 역가를 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 유전자형 2a J6 서열의 잔기 409-428을 포괄하는 합성 펩타이드에 결합하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 20은 HCV E2 및 그의 세포 수용체 CD81 사이의 상호작용을 저해하는 면역 혈청의 능력을 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 (A) H77c G1a E2 및 CD81, 및 (B) J6 G2a E2 및 CD81 사이의 결합을 저해하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이고, 상위 및 하위 막대는 표준 편차를 나타낸다. 그룹간의 항체 역가에서의 차이는 만-휘트니 비대응 t-테스트를 사용하여 결정되었다. 프리즘 v7.0.
도 21은 유전자형 1a 바이러스에 의한 간 세포의 감염을 예방하는 면역 혈청의 능력을 보여준다. 표 15에 열거된 항원에 의해 백신접종된 동물로부터의 최종 채혈은 G1a HCVpp에 의한 감염을 예방하는 이들의 능력에 대해 분석되었다. 각각의 그룹에 대한 상호간의 항체 역가가 제시된다. 수평선은 평균이다.
도 22는 HMW1 및 단량체 면역 혈청의 특이성을 보여준다. 기니아 피그(guinea pig) 혈청의 연속 희석물이 일정한 양의 HCV1(A), HC84-27(B), AR3C(C) 및 2A12(D)에 첨가되었다. 항체가 단량체성 Δ123에 첨가되었고, 결합된 MAb는 항-인간 Fab2에 의해 검출되었다. 그룹은 만-휘트니 t-테스트를 사용하여 비교되었다(프리즘 v7).
도 23은 조립된 Δ123을 사용하여 분류된 B 세포 개체군의 FACS 도표를 보여준다. CD 19 양성 및 항-E2 양성 B 세포는 항-CD19 Cy7 항체 및 조립된 Δ123에 의해 검출되었다.
도 24는 조립된 단백질의 크기를 결정하는 크기-배제 크로마토그래피-다각도 광산란을 보여준다. 조립된 Δ123A7(청색) 및 조립된 Δ123(적색) 샘플의 UV(A280 nm) 신호의 중첩 및 몰 질량.
도 25는 DTT에 의해 처리된 후, HMW 형태로 조립되지 않은 Δ123A7 단량체(이는 조립이 수반되는 DTT에 의한 변성의 제2 라운드를 거치지 않을 것임)의 크기-배제 크로마토그래피를 보여준다. 단량체 및 HMW 종은 각각 표에 제시된 화살표 및 %에 의해 지시된다.
도 26은 조립이 수반되는 DTT에 의한 변성을 거친 RBD 단량체의 크기-배제 크로마토그래피를 보여준다. 단량체 및 HMW 종은 각각 표에 제시된 화살표 및 %에 의해 지시된다.
도 27은 조립이 수반되는 DTT에 의한 변성을 거친 RBDA7 단량체의 크기-배제 크로마토그래피를 보여준다. 단량체 및 HMW 종은 각각 표에 제시된 화살표 및 %에 의해 지시된다.
도 28은 조립이 수반되는 DTT에 의한 변성을 거친 env 단량체의 크기-배제 크로마토그래피를 보여준다. HMW 종은 화살표에 의해 지시된다.
표의 간단한 설명
표 1은 일시적 형질감염 증상을 보여준다.
표 2는 MAb의 목록을 보여준다.
표 3은 DTT 환원을 위한 조건을 보여준다.
표 4는 글루타티온에 의한 단백질 재점힘을 위한 조건을 보여준다.
표 5는 TCEP 환원을 위한 조건을 보여준다.
표 6은 Δ123의 상이한 올리고머성 형태의 발현을 보여준다. 단량체, 이합체, HMW2 및 HMW1의 백분율은 겔 여과 곡선(도 3B-E) 상에서 이들의 상응하는 피크하의 면적을 곡선하의 총 면적에 의해 나눠줌으로써 계산되었다. 곡선하의 면적은 지이 헬쓰케어 라이프 사이언시즈(GE Healthcare Life Sciences)에 의한 유니콘(UNICORN) 제어 소프트웨어를 사용하여 정량화되었다.
표 7은 HMW1 Δ123의 항원 특징을 보여준다.
표 8은 Δ123의 TCEP 환원 및 BMOE-중재된 재접힘을 위한 조건을 보여준다.
표 9는 글루타티온에 의한 H77c Δ123 재접힘을 위한 조건을 보여준다. 단량체 대 이합체의 비는, 글루타티온 처리된 H77c Δ123의 비-환원 SDS-PAGE 분석으로부터 이합체의 밀도측정값에 의해 단량체의 밀도측정값을 나눠줌으로써 계산되었다(도 7A). 밀도측정값은 LI-COR 오디세이(Odyssey) 시스템을 사용하여 정량화되었다.
표 10은 글루타티온-처리된 단량체성 H77c Δ123으로부터의 다합체 형성의 정량화를 보여준다. 겔 여과 곡선(도 7B) 상의 단량체 및 이합체 피크하의 면적은 곡선하의 총 면적의 백분율로서 계산되었고, 단량체 대 다합체의 비는 단량체 피크하의 면적을 이합체 피크하의 면적에 의해 나눠줌으로써 계산되었다.
표 11은 DTT-처리된 단량체성 H77c Δ123으로부터의 다합체 형성의 정량화를 보여준다. 겔 여과 곡선(도 7C-D) 상의 단량체 및 HMW 피크하의 면적은 곡선하의 총 면적의 백분율로서 계산되었고, 단량체(76 분) 대 다합체(58 분)의 비는 단량체 피크하의 면적을 다합체 피크하의 면적에 의해 나눠줌으로써 계산되었다.
표 12는 DTT-처리된 단량체성 H77c Δ123으로부터의 다합체 형성의 정량화를 보여준다. 겔 여과 곡선(도 7B) 상의 단량체 및 다합체 피크하의 면적은 곡선하의 총 면적의 백분율로서 계산되었고, 단량체 대 다합체의 비는 단량체 피크하의 면적을 다합체 피크하의 면적에 의해 나눠줌으로써 계산되었다.
표 13은 항원 Δ123을 사용하여 재접힘 단백질을 생성하는 방법을 보여준다.
표 14는 상이한 재접힘 방법에 의해 생성된 재접힘 단백질%를 보여준다.
표 15는 면역화 그룹을 보여준다.
표 16은 단량체성 Δ123에 대한 면역 혈청 반응성의 통계학적 분석을 보여준다.
표 17은 H77c 에피토프 I에 결합하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 18은 H77c 에피토프 III에 결합하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 19는 J6 에피토프 I에 결합하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 20은 CD81로의 H77c G1a E2 결합을 차단하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 21은 CD81로의 JFH-1 G2a E2 결합을 차단하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 22는 H77c HCV 바이러스가 간 세포를 감염시키는 것을 예방하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 23은 HCV1의 결합을 예방하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 24는 HC84-27의 결합을 예방하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 25는 AR3C의 결합을 예방하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 26은 2A12의 결합을 예방하는 면역 혈청 능력의 통계학적 분석을 보여준다.
표 27은 재조립된 단백질의 SEC-MALS를 보여준다.
서열 목록에 대한 핵심
서열 번호 1: 코돈 최적화된 Δ123Ala7을 위한 DNA 작성물.
서열 번호 2: 코돈 최적화된 Δ123Ala7을 인코딩하는 DNA 서열.
서열 번호 3: 아미노산 서열 Δ123Ala7.
서열 번호 4: WT E2661(RBD)을 인코딩하는 아미노산 서열.
서열 번호 5: Δ123 E2661.
서열 번호 6: AF009606 코딩 서열에 상응하는 아미노산 잔기.
서열 번호 7: AF009606 전장 E2 단백질 서열에 상응하는 아미노산 잔기.
서열 번호 8: AF009606 E2661에 상응하는 아미노산 잔기.
서열 번호 9: N-말단 신호 서열.
서열 번호 10: 아미노산 서열 ED43.
서열 번호 11: 아미노산 서열 H77c.
서열 번호 12: 아미노산 서열 SA13.
서열 번호 13: 아미노산 서열 s52.
서열 번호 14: 아미노산 서열 EUHK2.
서열 번호 15: 아미노산 서열 QC69.
서열 번호 16: 아미노산 서열 J6.
서열 번호 17: C-말단 막관통 도메인 및 세포질 테일(cytoplasmic tail)이 결핍된 HIV env에 상응하는 아미노산 잔기.
서열 번호 18: N-말단 리더(leader) 서열을 갖는 HIV env에 상응하는 아미노산 잔기.
서열 번호 19: 코돈 H77c Δ123을 인코딩하는 DNA 서열.
서열 번호 20: Con1 Δ123을 인코딩하는 DNA 서열.
서열 번호 21: S52 Δ123을 인코딩하는 DNA 서열.
서열 번호 22: 인간 트립시노겐(trypsinogen) 서열 펩타이드.
서열 번호 23: 인간 조직 플라스미노겐(plasminogen) 활성화제 신호 펩타이드(tPA).
서열 번호 24: 6개의 His 태그.
발명의 상세한 설명
본 개시내용은 제제를 위한 특별한 선별 절차, 제제의 특정 제형, 및 다양한 의학적 방법론에 제한되지 않고, 이처럼 다양할 수 있다.
본 명세서에서 각각의 실시태양은, 달리 명백히 지시되지 않는 한, 필요한 부분만 약간 수정하여 모든 다른 실시태양에 적용된다.
달리 정의되지 않는 한, 본원에 사용된 모든 기술적 및 과학적 용어는 본 발명이 속하는 분야의 숙련가에 의해 통상적으로 이해되는 바와 동일한 의미를 갖는다. 본원에 기재된 재료 및 방법과 유사하거나 동등한 임의의 재료 및 방법이 본 발명을 실행하거나 검사하기 위해 사용될 수 있다. 실행자는 당분야의 숙련가에게 공지된 기술 및 기타 방법의 정의 및 용어를 위해 특별히 상기 샘브룩(Sambrook) 등의 문헌(1989), 콜리간(Coligan) 등의 문헌[Current Protocols In Protein Science, John Wiley & Sons, Inc., 1995-1997](특별히 1장, 5장 및 6장), 및 어수벨(Ausubel) 등의 문헌[Current Protocols in Molecular Biology, Supplement 47, John Wiley & Sons, New York, 1999]; 콜로위크(Colowick) 및 카플란(Kaplan)(편집)의 문헌[Methods In Enzymology, Academic Press, Inc.]; 웨이어(Weir) 및 블랙웰(Blackwell)의 문헌[Handbook of Experimental Immunology, Vols. I-IV, Blackwell Scientific Publications, 1986]; 조클릭(Joklik)(편집)의 문헌[Virology, 3rd Edition, 1988]; 필즈(Fields) 및 니페(Knipe)(편집)의 문헌[Fundamental Virology, 2nd Edition, 1991]; 필즈(Fields) 등의 문헌[Virology, 3rd Edition, Lippincott-Raven, Philadelphia, Pa., 1996]을 참조한다. 또한, 스태비(Staby), 라토레(Rathore) 및 아후가(Ahuga)(편집)의 문헌[Preparative Chromatography for Separation of Proteins; Whiley, 2017](특별히 3장 및 7장)을 참고한다. 또한 웬(Wen), 엘리스(Ellis), 푸자르(Pujar)(편집)의 문헌[Vaccine Development and Manufacturing, Wiley, 2014](특별히 4장, 6장, 8장, 11장)을 참고한다. 또한 방법 및 재료를 위해 국제 특허출원 공개공보 제WO 2008022401호, 제WO 2012016290호 및 제WO 2012068637호를 참고할 수 있다.
정의
본 명세서 전반에 걸쳐, 문맥상 달리 정의되어야 하지 않는 한, "포함하다", 또는 변형어, 예컨대 "포함하고" 또는 "포함하는"이라는 단어는, 언급된 요소 또는 정수, 또는 요소들 또는 정수들의 그룹의 포함을 내포하지만, 임의의 다른 요소 또는 정수, 또는 요소들 또는 정수들의 그룹을 배제하지 않는 것으로 이해될 것이다. "구성되는"은 "구성되는"이라는 어구 뒤에 수반되는 어떤것이라도 포함하고 이에 제한됨을 의미한다. 이와 같이, "구성되는"이라는 어구는 열거된 요소들이 필수적이거나 의무적이고 다른 요소가 존재하지 않을 수 있음을 지시한다. "본질적으로 구성되는"은 이러한 어구 뒤에 열거되는 임의의 요소를 포함하고, 열거된 요소에 대한 개시내용에서 특정화된 활성 또는 작용에 간섭하지 않거나 이에 기여하는 다른 요소로 제한됨을 의미한다.
본원에서 사용되는 경우, 단수 형태는 문맥상 명백히 달리 지시되지 않는 한 복수의 양태를 포함한다. 이와 같이, 예를 들면, "조성물"에 대한 언급은 단일 조성물, 뿐만 아니라 둘 이상의 조성물을 포함하고; "제제"에 대한 언급은 하나의 제제 뿐만 아니라, 둘 이상의 제제를 포함하며; "개시내용"에 대한 언급은 개시내용의 단일 양태 및 다중 양태를 포함하는 등등이다.
실시예는 단량체성 HCV E2로부터의 올리고머성 HCV E2의 조립 및 단량체성 HIV env로부터의 올리고머 HIV env의 조립을 예시하지만, 본 개시내용은 이들 특별한 실시예에 제한되지 않고, 백신 생산을 위해 또는 생체외 결합 적용에 있어서 바이러스성 엔벨롭 단백질 및 암 항원의 조립으로 확장된다. 이러한 관점에서, 방법은 낮은 급수의 항원으로부터 높은 급수의 항원의 생산을 제공한다. "높은 급수의" 항원에 대한 언급은 3합체 또는 더 큰 다합체를 의미하는 반면, "낮은 급수의 형태"는 단량체 또는 이합체를 의미한다. 용어 "항원" "종" 및 "형태"는 상호교환적으로 사용된다.
"고유의"에 대한 언급은 항원이 세포내에서 세포 기작을 사용하여 조립됨을 의미한다. 용어 "조립된" 및 "접힌" 또는 "재접힌"은 상호교환적으로 사용되지만, "조립된"은 조립된 항원의 부분적인 드 노보(de novo) 성질을 전달하기 위해 사용된다. "조립된"에 대한 언급은 "세포-부재" 조립을 의미한다. 하나의 실시태양에서, 조립된 항원은 이들의 고유의 대응물과 동일하게 효과적이거나 이에 비해 더욱 효과적이다.
본원에서 사용되는 경우 "비-중화 항체"는 바이러스성 항원에 결합하지만 바이러스의 진입을 감소시키거나 방해하지 않는 항체를 지칭한다. 하나의 실시태양에서, 이는 E2에 결합하지만 바이러스의 진입을 감소시키거나 방해하지 않는 항체를 지칭한다. 암 항원과 관련하여, 비-중화 항체에 대한 언급은 종양 세포 살해를 중재하지 않는 항체를 의미한다.
본원에서 사용되는 경우 "중화 항체"는 바이러스성 항원에 결합하고 결합될 경우 바이러스의 진입을 저해하는 항체를 지칭한다. 하나의 실시태양에서, 이는 E2에 결합하고, E2에 결합할 경우 바이러스의 진입을 저해하는 항체를 지칭한다. 암 항원과 관련하여, 중화 항체에 대한 언급은 종양 세포 살해를 직접적으로 또는 간접적으로 중재하는 항체를 의미한다.
본원에서 사용되는 경우 "광범위 중화 항체"는 항원/HCV 항원의 다중의 유전자형 또는 하위유형에 대해 교차 보호를 제공하는 항체를 지칭한다.
높은 급수의 또는 올리고머성 형태는 항원의 구조적 형태이고, 3합체 및 더 큰 접힌 형태를 포함한다. 높은 급수의 항원 또는 형태 또는 종은 낮은 급수의 형태, 항원 또는 종으로부터 조립된다. 낮은 급수의 형태로는 항원의 단량체성 또는 이합체성 형태가 포함된다.
"대조"에 대한 언급은 당분야의 숙련가에게 이해될 것이고, 본 발명과 관련하여 의미있는 결과를 생성하거나 생성하기 쉬운 비교자 또는 비교자들을 사용함을 의미한다. 전형적으로, 대조군은 숙주 세포 또는 세포 부재 발현 시스템에서 생산되는 대응물이다.
임의의 바이러스성 엔벨롭 항원은 본 명세서에 기재된 방법을 사용하여 조작될 수 있다. 바이러스 과의 비-제한적 예로는 아데노바이러스과(Adenoviridae), 아프리카 돼지 열병-유사 바이러스, 아레나바이러스과(Arenaviridae), 아테리바이러스(Arterivirus), 아스트로바이러스과(Astroviridae), 바큘로바이러스과(Baculoviridae), 비르나바이러스과(Birnaviridae), 부니아바이러스과(Bunyaviridae), 칼리시바이러스과(Caliciviridae), 시르코바이러스과(Circoviridae), 코로나바이러스과(Coronaviridae), 델타바이러스(Deltavirus), 필로바이러스과(Filoviridae), 플라비바이러스과(Flaviviridae), 헤파드나바이러스과(Hepadnaviridae), 헤페바이러스과(Hepeviridae), 헤르페스바이러스과(Herpesviridae), 오르토믹소바이러스과(Orthomyxoviridae), 파라믹소바이러스과(Paramyxoviridae), 피코르나바이러스과(Picornaviridae), 폭스바이러스과(Poxviridae), 레오바이러스과(Reoviridae), 레트로바이러스과(Retroviridae) 및 랍도바이러스과(Rhabdoviridae)가 포함된다. 특별한 바이러스 엔벨롭 항원은 파라믹소바이러스과, 레트로바이러스과 및 필로바이러스과로부터이다.
바이러스성 엔벨롭 항원의 비-제한적 예는 병원체성 바이러스, 예컨대 인플루엔자 헤마글루티닌(HA: haemagglutinin); 렌티바이러스(lentivirus), 예컨대 R2 하위유형 또는 HIV-2 gp125를 비롯한 HIV-1 당단백질(gp) 120; 코로나바이러스, 예컨대 SARS Si 당단백질; 파라믹소바이러스, 예컨대 호흡기 세포융합 바이러스(RSV: respiratory syncytial virus) F2; 또는 플라비바이러스, 예컨대 뎅기열(Dengue) 바이러스 E 단백질로부터 유래된다.
항원의 한가지 중요한 그룹은 병원체, 예컨대 인간의 원발성 전신 진균류 병원체로부터 유래되고, 예컨대 콕치디오이데스 임미티스(Coccidioides immitis), 히스토플라스마 캅술라툼(Histoplasma capsulatum), 블라스토미세스 데르마티티디스(Blastomyces dermatitidis), 및 파라콕치디오이데스 브라실리엔시스(Paracoccidioides brasiliensis)이다. 면역력이 약화된 숙주에 의존하는 경향이 있는 중요한 기회감염성 진균류 병원체로는 크립토코쿠스 네오포르만스(Cryptococcus neoformans), 뉴모시스티스 위로베시(Pneumocystis jiroveci), 칸디다 아종(Candida spp.), 아스페르길루스 아종(Aspergillus spp.), 페니실리움 마르네페이(Penicillium marneffei), 및 지고미세테스(Zygomycetes), 트리코스포론 베이겔리이(Trichosporon beigelii), 및 푸사리움 아종(Fusarium spp.)이 포함된다. 여러 가지 병원체성 진균류는 면역력이 약화된 피험체, 예컨대, 무엇보다도, AIDS에 걸리거나, 화학요법 유도된 호중구 감소증을 앓는 피험체, 또는 조혈 모세포 이식을 겪은 환자와 연관된다.
몇몇 실시태양에서, 항원은 세균, 진균류, 바이러스, 조류(algae), 기생충(외부-기생충 및 내부-기생충 포함), 프리온(prion), 난균류(oomycete), 슬라임(slime), 곰팡이(mould), 선충, 미코플라스마(mycoplasma) 등을 비롯한 미생물로부터 유래된다. 비-제한적 예에 의해, 미생물은 다음의 목, 속 또는 종들중 하나 이상으로부터 선택된다: 아시네토박터(Acinetobacter), 악티노바실러스(Actinobacillus), 악티노미세테스(Actinomycetes), 악티노미세스(Actinomyces), 에로모나스(Aeromonas), 바실루스(Bacillus), 박테로이데스(Bacteroides), 보르데텔라(Bordetella), 보렐리아(Borrelia), 브루셀라(Brucella), 부르크홀데리아(Burkholderia), 캄필로박터(Campylobacter), 시트로박터(Citrobacter), 클로스트리디움(Clostridium), 코리네박테리움(Corynebacterium), 엔테로박터(Enterobacter), 엔테로코쿠스(Enterococcus), 에리시펠로트릭스(Erysipelothrix), 에스케리키아(Escherichia), 프란시셀라(Francisella), 헤모필루스(Haemophilus), 헬리코박터(Helicobacter), 클레브시엘라(Klebsiella), 레기오넬라(Legionella), 렙토스피라(Leptospira), 리스테리아(Listeria), 미크로코쿠스(Micrococcus), 모락셀라(Moraxella), 모르가넬라(Morganella), 미코박테리움(Mycobacterium)(결핵), 노카르디아(Nocardia), 네이세리아(Neisseria), 파스테우렐라(Pasteurella), 플레시오모나스(Plesiomonas), 프로피오니박테리움(Propionibacterium), 프로테우스(Proteus), 프로비덴시아(Providencia), 슈도모나스(Pseudomonas), 로도코쿠스(Rhodococcus), 살모넬라(Salmonella), 세라티아(Serratia), 시겔라(Shigella), 스타필로코쿠스(Staphylococcus), 스테노트로포모나스(Stenotrophomonas), 스트렙토코쿠스(Streptococcus), 트레포네마(Treponema), 비브리오(Vibrio)(콜레라) 및 예르시니아(Yersinia)(흑사병), 아데노바이러스과, 아프리카 돼지 열병-유사 바이러스, 아레나바이러스과[예컨대 바이러스성 출혈열, 라사열(Lassa fever)], 아스트로바이러스과(아스트로바이러스), 부니아바이러스과[라 크로세(La Crosse)], 칼리시바이러스과(노로바이러스), 코로나바이러스과(코로나 바이러스), 필로바이러스과[예컨대 에볼라(Ebola) 바이러스, 마르부르그(Marburg) 바이러스], 파르보바이러스과(Parvoviridae)(B19 바이러스), 플라비바이러스과(예컨대 간염 C 바이러스, 뎅기열 바이러스), 헤파드나바이러스과(예컨대 간염 B 바이러스, 델타바이러스), 헤르페스바이러스과(단순 포진 바이러스, 수두 대상포진 바이러스), 오르토믹소바이러스과(인플루엔자 바이러스), 파포바바이러스과(Papovaviridae)(유두종 바이러스), 파라믹소바이러스과(예컨대 인간 파라인플루엔자 바이러스, 유행성 이하선염 바이러스, 홍역 바이러스, 인간 호흡기 세포융합 바이러스), 피코르나바이러스과(감기 바이러스), 폭스바이러스과[천연두 바이러스, 오르프(orf) 바이러스, 원숭이 두창(monkey pox) 바이러스], 레오바이러스과(로타바이러스), 레트로바이러스과(인간 면역결핍 바이러스), 파르보바이러스과(파르보바이러스), 파필로마바이러스과(Papillomaviridae)(유두종바이러스), 알파바이러스, 랍도바이러스과(광견병 바이러스), 트리파노소마(Trypanosoma), 레이쉬마니아(Leishmania), 기아르디아(Giardia), 트리코모나스(Trichomonas), 엔타모에바(Entamoeba), 네글레리아(Naegleria), 아칸타모에바(Acanthamoeba), 플라스모디움(Plasmodium), 톡소플라스마(Toxoplasma), 크립토스포리디움(Cryptosporidium), 이소스포라(Isospora), 발란티디움(Balantidium), 스키스토소마(Schistosoma), 에키노스토마(Echinostoma), 파스키올롭시스(Fasciolopsis), 클로노르키스(Clonorchis), 파스키올라(Fasciola), 오피스토르키스(Opisthorchis), 파라고니무스(Paragonimus), 의엽류(Pseudophyllidea)[예를 들어, 디필로보트리움(Diphyllobothrium)], 원엽목(Cyclophyllidea)[예를 들어, 테니아(Taenia)]. 병원체성 선충류로는 하기 목으로부터의 종이 포함된다; 간선충목(Rhabditida)[예를 들어, 스트롱길로이데스(Strongyloides)], 원선충목(Strongylida)[예를 들어, 안실로스토마(Ancylostoma)], 회충류(Ascarida)[예를 들어, 아스카리스(Ascaris), 톡소카라(Toxocara)], 시미선충류(Spirurida)[예를 들어, 드라쿤쿨루스(Dracunculus), 브루기아(Brugia), 옹코세르카(Onchocerca), 우케리아(Wucheria)], 및 쌍기충강(Adenophorea)[예를 들어, 트리쿠리스(Trichuris) 및 트리키넬라(Trichinella)], 프로토테카(Prototheca), 티에스테리아(Ptiesteria), 압시디아(Absidia), 아스페르길루스, 블라스토미세스(Blastomyces), 칸디다(효모), 클라도피알로페라(Cladophialophera), 콕치디오이데스, 크립토코쿠스(Cryptococcus), 쿤닝하멜라(Cunninghamella), 푸사리움, 히스토플라스마(Histoplasma), 마두렐라(Madurella), 말라세지아(Malassezia), 미크로스포룸(Microsporum), 무코(Mucor), 페실로미세스(Paecilomyces), 파라콕치디오이데스, 페니실리움(Penicillium), 뉴모시스티스, 슈달레스케리아(Pseudallescheria), 리조푸스(Rhizopus), 로도토룰라(Rhodotorula), 스케도스포리움(Scedosporium), 스포로트릭스(Sporothrix), 트리코피톤(Trichophyton) 및 트리코스포론(Trichosporon). 혼동을 피하기 위해, 병원체는 신생 병원체를 포함할 수 있다.
예시적인 암 항원으로는 CD 항원, 당단백질, 당지질[강글리오시드(ganglioside)], 탄수화물[루이스(Lewis)-Y] 혈관 표적(VEGF/R), 성장 인자, 및 기조직(stromal) 또는 세포외 바탕질 항원[FAP, 테나신(Tenascin)] 등이 포함된다. 예를 들면, 다음과 같이 열거된다: KS ¼ pan-암종 항원, 난소 암종 항원(CA125), 전립선 산 인산염, 전립선 특이적 항원, 흑색종-연관된 항원 p97, 흑색종 항원 gp75, 고분자량 흑색종 항원(HMW-MAA), 전립선 특이적 막 항원, 암배아 항원(CEA: carcinoembryonic antigen), 다형태성 상피 뮤신 항원, 인간 유지방구(milk fat globule) 항원, 결장직장 종양-연관된 항원, CEA, TAG-72, LEA, 버킷(Burkitt)의 림프종 항원-38.13, CD19, 인간 B-림프종 항원-CD20, CD33, 흑색종 특이적 항원, 강글리오시드 GD2, 강글리오시드 GD3, 강글리오시드 GM2, 강글리오시드 GM3, 세포-표면 항원의 종양-특이적 이식 유형(TSTA), 바이러스에 의해 유도된 종양 항원, T-항원 DNA 종양 바이러스, RNA 종양 바이러스의 엔벨롭 항원, 종양태아 항원-알파-태아단백질, 결장의 CEA, 방광 종양 종양태아 항원, 분화 항원, 인간 폐 암종 항원 L6, L20, 섬유육종의 항원, 인간 백혈병 T 세포 항원-Gp37, 신생당단백질, 스핑고지질(sphingolipid), 유방암 항원, EGFR(상피 성장 인자 수용체), HER2 항원, 다형태성 상피 뮤신, 악성 인간 림프구 항원-APO-1, 분화 항원(태아 적혈구에서 발견되는 I 항원 포함), 초기 내배엽, 성인 적혈구에서 발견되는 I 항원, 착상전 배아, 위 선암에서 발견되는 I(Ma), M18, 유방 상피에서 발견되는 M39, 골수 세포에서 발견되는 SSEA-1, VEP8, VEP9, Myl, VIM-D5, 결장직장암에서 발견되는 Du56-22, TRA-1-85(혈액군 H), 결장 선암에서 발견되는 C14, 폐 선암에서 발견되는 F3, 위암에서 발견되는 AH6, Y 합텐(hapten), 배아 암종 세포에서 발견되는 LeY, TL5(혈액군 A), A431 세포에서 발견되는 EGF 수용체, 췌장암에서 발견되는 E1 시리즈(혈액군 B), 배아 암종 세포에서 발견되는 FC10.2, 위 선암 항원, 선암에서 발견되는 CO-514(혈액군 Lea), 선암에서 발견되는 NS-10, CO-43(혈액군 Leb), A431 세포의 EGF 수용체에서 발견되는 G49, 결장 선암에서 발견되는 MH2(혈액군 ALeb/Ley), 결장암에서 발견되는 19.9, 위암 뮤신, 골수 세포에서 발견되는 TsA7, 흑색종에서 발견되는 R24, 배아 암종 세포에서 발견되는 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2, 및 M 1:22:25:8, 및 4 내지 8-세포 단계 배아에서 발견되는 SSEA-3 및 SSEA-4.
본원에서 사용되는 경우, 용어 "인간 면역결핍 바이러스" 또는 "HIV"는 렌티바이러스 속 및 레트로바이러스과의 일부의, 엔벨롭을 갖는 양성 단일-가닥의 RNA 일원을 지칭한다. 시간 초과시 HIV는 후천적 면역결핍 증후군(AIDS)을 일으킨다. 본원에서 사용되는 경우, 이 용어는 임의의 HIV 유전자형을 지칭하고, 예를 들면, 제한되지 않지만 HIV1 또는 HIV2 또는 이의 임의의 그룹 또는 하위유형이다. 한가지 실시태양에서, HIV-1은 그룹 M, N, O 또는 P로부터이다. 한가지 실시태양에서, HIV-1은 A, B, C, D, E, F, G, H, J, K 또는 이의 순환성 재조합 형태(CRF: circulating recombinant form)로부터 선택된 하위유형이다. HIV는 엔벨롭 단백질 당단백질(gp) 120 및 env를 인코딩한다.
본원에서 사용되는 경우, 용어 "간염 C 바이러스" 또는 "HCV"는 플라비바이러스과의 헤파시바이러스(Hepacivirus) 속에 속하는, 엔벨롭을 갖는 양성 센스(sense), 단일-가닥의 RNA 바이러스를 지칭한다. 본원에서 사용되는 경우, 이 용어는 임의의 유전자형의 HCV를 지칭하고, 예를 들면, 제한되지 않지만 HCV 유전자형 1(G1), HCV 유전자형 2(G2), HCV 유전자형 3(G3), HCV 유전자형 4(G4), HCV 유전자형 5(G5), HCV 유전자형 6(G6), HCV 유전자형 7(G7)의 균주이고, 이의 임의의 하위유형, 예를 들어 하위유형 a, b, c, d, e 등을 포함할 수 있다. HCV는 숙주 세포내로의 바이러스의 진입에 필요한 2개의 당단백질 E1 및 E2를 인코딩한다.
본원에서 사용되는 경우, 또한 "E2"로서 지칭되는 "HCV E2"는 HCV의 임의의 유전자형/하위유형으로부터의 E2 폴리펩타이드를 포함한다. 한가지 실시태양에서, E2는 HCV 유전자형 G1, G2, G3, G4, G5, G6, G7, 또는 이의 키메라 버전으로부터 유래된다. "유래되는"은 이들 유전자형중 하나 이상에 직접적으로 또는 간접적으로 기초함을 의미한다. 유전자형은 자연적으로 다양하고, 인간에 의해 추가로 변형될 수 있으며, 전형적으로 보존적 돌연변이를 포함하는 이러한 작용성 변형체가 포괄된다. 하나 이상의 아미노산 돌연변이를 포함하는 작용성 변형체는 숙련가에게 공지되고, 재조합 E2 엑토도메인(ectodomain)을 포함하는 작용성 변형체를 포함할 수 있다. 이 용어는, 예를 들면, 입체배좌 또는 다른 에피토프를 인식하고/하거나 E1E2 이합체 형성을 중재하는 하나 이상의 항체에 의한 항체 결합, 수용체 결합을 중재하는 전장 E2 폴리펩타이드의 일부를 포함하는 변형체를 추가로 포함한다. 이러한 용어는 E2의 변형된 형태, 예컨대 면역원성(델타 123 형태) 또는 단량체 생산(예를 들어 "Ala7")을 증가시키기 위한 변형을 포함한다.
하나의 예시적인 모 HCV E2 폴리펩타이드는 유전자형 H77 1a(E2 661 또는 E2e)의 아미노산 384-661 또는 또 다른 HCV 유전자형으로부터의 상응하는 부분을 포함하는 E2 폴리펩타이드의 수용체 결합 부분이다. 따라서, 활성화된 E2 폴리펩타이드는 막관통 도메인 부재의 CD81-결합을 위해 요구되는 엑토도메인의 전부 또는 일부를 포함한다. 추가의 변형체는 분할 또는 분비를 위해 필수적인 서열의 부가 또는 결실/파괴를 포함할 수 있다. 예를 들면, E384TH는 서열 펩타이드 분할 및 당단백질 분비를 변형시키기 위해 포함되거나, 결실되거나, 변형될 수 있다[맥카프레이(McCaffrey) 등의 문헌(2007)]. 한가지 실시태양에서, E2 폴리펩타이드는 하나 이상의 과가변성 영역 또는 이의 일부가 결핍된다. 한가지 실시태양에서, E2는 과가변성 영역, 예컨대 과가변성 영역 1(HVR1) 또는 이의 일부, 과가변성 영역 2(HVR2) 또는 이의 일부, 및 상호유전자형 가변성 영역(igVR/VR3) 또는 이의 일부중 하나 이상이 결핍된다.
한가지 실시태양에서, E2는 HVR1, HVR2 및 igVR/VR3이 결핍된다. 한가지 실시태양에서, E2는 Δ123이다. 한가지 실시태양에서, E2는 서열 번호 3, 4, 5, 6, 7, 또는 8에 제시된 서열; 또는 CD81 결합 활성을 보유하는 이의 단편; 또는 이에 대해 적어도 50%, 또는 적어도 55%, 또는 적어도 60%, 또는 적어도 65%, 또는 적어도 70%, 또는 적어도 75%, 또는 적어도 80%, 또는 적어도 85%, 또는 적어도 90%, 또는 적어도 95%, 또는 적어도 97%, 또는 적어도 98%, 또는 적어도 99% 동일한 서열을 포함한다.
한가지 실시태양에서, E2는 돌연변이되거나 파괴되는 시스테인을 0개 또는 하나 이상 포함한다. 이와 같이, 하나의 실시태양에서, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 또는 15개의 시스테인이 결실되거나 파괴된다. 하나의 실시태양에서, 이들은 C581, C585, C652, C677, C494, C486, C459, C452, C564, C597, C569 및/또는 C620으로부터 선택된다. 한가지 실시태양에서, 돌연변이되거나 파괴된 시스테인은 C486, C581 및 C652이다. 한가지 실시태양에서, 돌연변이되거나 파괴된 시스테인은 C581, C585 및 C652이다. 한가지 실시태양에서, 돌연변이되거나 파괴된 시스테인은 C452, C486, C581 및 C652이다. 한가지 실시태양에서, 돌연변이되거나 파괴된 시스테인은 C569, C581, C585 및 C652이다. 한가지 실시태양에서, 돌연변이되거나 파괴된 시스테인은 C486, C581 및 C652이다. 한가지 실시태양에서, 돌연변이되거나 파괴된 시스테인은 C452, C486, C569, C581, C585 및 C652이다. 한가지 실시태양에서, 돌연변이되거나 파괴된 시스테인은 C452, C486, C569, C581, C585, C597 및 C652이다. 특허청구범위를 비롯하여 본 명세서 전반에 걸쳐, HCV 당단백질 E2의 폴리펩타이드 잔기에 대한 모든 넘버링(numbering)은 도 14에 제시된 원형(prototype) HCV-H77 폴리펩타이드 서열, 유전자은행(Genbank) 등록 번호 AF009606(서열 번호 6)에 기초한다. E2의 성숙한 형태는 서열 번호 6의 아미노산 잔기 384 내지 746에 의해 포괄되고, 별도로 서열 번호 7에 제시된다. 본원에서 지칭된 변형은 서열 번호 6에 제시되고 및 도 14에 제시된 바와 같은 아미노산 넘버링을 참고하여 이루어진다.
E2661의 하나의 예시적인 시스테인 돌연변이된 버전은 하기 시스테인의 돌연변이 또는 파괴를 포함한다: C581, C585, C652, C486, C452, C597, 및 C569. 이러한 돌연변이체는 "Ala7"로서 지칭된다. 추가의 E2의 시스테인 변형된 버전은 본원에 그의 전체가 인용된 국제 특허출원 공개공보 제WO 2012/016290호에 기재되어 있다.
본원에서 사용되는 경우 "CD81"은 분화 클러스터(cluster of differentiation) 81을 지칭하고, 이는 테트라스파닌(tetraspanin) 상과의 막관통 단백질이고, HCV 숙주 수용체이다.
HCV E2의 수용체 결합 도메인(RBD)은 CD81-결합 모티프(motif) 및 접힘부를 포함하고 각각 상이한 디설파이드 및 글리칸(glycan) 배열을 함유하는 다양한 상이한 종으로 올리고머화된다. 본 발명자(들)는 단량체성 및 이합체성 WT E2661 및 Δ123 E2661 상의 디설파이드 결합 배열을 식별하기 위해 상당한 작업을 시행하였고, 이는 심지어 단량체성 단백질로서의 이들 단백질 둘 다가 실제로 이종성이고, 하나 건너 하나씩의 다중의 분자내 디설파이드 결합된 형태로 존재함을 시사한다.
HCV E2가 재조합적으로 생산되는 경우, 일반적으로 20 내지 30%가 올리고머성이고 대략 70%가 단량체성인데, 이는 (본원에서 결정되는 경우) 이용되는 유전자형에 의존한다. 더 많은 단량체가 시스테인 변형된 형태, 예컨대 Ala7에 의해 생산된다.
일예로서, 델타123의 유전자형 1a H77c 서열의 FS293F 세포로의 안정한 형질감염으로부터 생성된 "델타123" 또는 "Δ123"로서 지칭되는, 과가변성 영역 1(HVR1), 과가변성 영역 2(HVR2) 및 상호유전자형 가변성 영역(igVR 또는 VR3)이 제거된 E2 수용체 결합 도메인(잔기 384-661)의 형태의 대략 20%만이 HMW 형태이고, 단량체 경우의 64.9%와 비교된다(도 3A). 본원에 제시된 바와 같이 이러한 백분율은 증가된 양의 올리고머성 형태를 생산하는 유전자형을 선택함으로써 증가될 수 있다.
관례적으로, 원치않는 응집체로부터 낮은 급수의 종을 재생하기 위해 환원 및 재접힘 방법이 이용된다. 본 발명에 따라서, 환원 및 재접힘은 단량체로부터 올리고머를, 또는 단량체 및 올리고머로부터 올리고머를 생성하기 위해 이용된다.
하나의 실시태양에서, 본 발명은 고유의 HCV E2로부터 재접힘된 재조합 올리고머성 간염 C 바이러스(HCV) 엔벨롭 당단백질 2(E2)를 제조하는 방법을 제공하고, 이러한 방법은: (i) 하나 이상의 고유의 시스테인(또는 디설파이드 결합)을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 고유의 E2를 접촉시키는 단계; 및 (ii) 환원제를 제거하거나 환원된 고유의 E2를 산화제와 접촉시켜서 환원된 단량체성 E2의 올리고머성 HCV E2로의 재접힘을 이끌어내는 단계를 포함하고,
여기서 단계 (ii)에서 단량체의 적어도 20%는 올리고머로 전환되고, 재접힘된 올리고머성 HCV E2는 단량체성 E2와 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타낸다.
한가지 실시태양에서, 단계 (i)은 단계 (ii) 이전에 2회 수행된다. 한가지 실시태양에서, 단계 (ii)는 단계 (ii) 이전에 3회 이상 수행된다. 몇몇 실시태양에서, 단계 (i) 및 (ii)는 2회 이상 반복된다.
한가지 실시태양에서, 재접힘된 올리고머성 HCV E2는 하기로 구성된 군에서 선택된 적어도 하나의 특징을 나타낸다:
(i) 고유의 대조 HCV E2 형태 또는 단량체성 E2와 비교하여 감소된 비-중화 항체에 대한 결합;
(ii) 고유의 대조 HCV E2 형태와 비교하여 적어도 실질적으로 동일한 중화 항체에 대한 결합;
(iii) 고유의 대조 HCV E2 형태 또는 단량체성 E2와 비교하여 더 낮은 비-중화 항체의 역가의 생산을 이끌어냄;
(iv) 중화 항체의 생산을 이끌어냄;
(v) 광범위 중화 항체의 생산을 이끌어냄;
(vi) 임의적으로 중화 항체의 더 높은 역가의 생산을 이끌어냄; 및
(vii) 임의적으로 광범위 중화 항체의 더 높은 역가의 생산을 이끌어냄.
고유의 HCV E2 단량체는 상이한 HCV E2 종의 혼합물로부터 효율적으로 생산되고 효과적으로 정제될 수 있다. 고유의 단량체성 HCV E2는 본원에 기재된 바와 같이 또는 당분야에 공지된 바와 같이 생산된다. 고유의 단량체성 및 올리고머성 HCV E2는 본원에 기재된 바와 같이 또는 당분야에 공지된 바와 같이 생산된다. 전형적으로, 단백질은 HCV E2를 인코딩하는 적합한 발현 벡터에 의해 형질전환된 숙주 세포에서 재조합적으로 생산된다.
적합한 포유동물 세포주로는, 제한되는 것은 아니지만, BHK, VERO, HT1080, 293, 293T, FS293F, Expi293, RD, COS-7, CHO, 주르캣(Jurkat), HUT, SUPT, C8166, MOLT4/클론8, MT-2, MT-4, H9, PML, CEM, 골수종 세포(예를 들어, SB20 세포) 및 CEMX174(예를 들면, ATCC로부터 입수가능함)가 포함된다. 기타 숙주 세포로는 제한없이 효모, 예를 들어 피키아 파스토리스(Pichia pastoris), 또는 곤충 세포, 예컨대 Sf9 세포가 포함된다.
세포는 500 ㎖, 1 ℓ, 1.5 ℓ, 2 ℓ, 2.5 ℓ 또는 3 ℓ 부피에서 배양될 수 있다. 하나의 예에서, 세포는 회분식 세포 배양 공정을 사용하여 배양된다. 하나의 예에서, 세포는 관류(perfusion) 세포 배양 공정을 사용하여 배양된다. 하나의 예에서, 세포는 종자 배지 및 생산 배지에서 배양된다. 하나의 예에서, 세포는 교반되는-탱크 반응기에서 배양된다. 하나의 예에서, 반응기의 부피는 약 1 ℓ 내지 약 2500 ℓ이다. 하나의 예에서, 반응기는 1 ℓ 반응기, 1.5 ℓ 반응기, 2 ℓ 반응기, 2.5 ℓ 반응기 또는 3 ℓ 반응기이다. 하나의 예에서, 세포는 파장 생물반응기(wave reactor)에서 배양된다. 하나의 예에서, 세포는 세포 공장 시스템, 예를 들어 눈크(Nunc) 세포 공장 시스템에서 배양된다.
합성 DNA는 적합한 프로모터(promoter) 및 다른 적절한 전사 조절 요소를 함유하는 발현 벡터내로의 분자 클로닝에 의해 재조합적으로 발현되고, 원핵 또는 진핵 숙주 세포내로 형질감염되어 재조합 단백질을 생산한다. 이러한 조작을 위한 기법은 샘브룩(Sambrook) 등의 문헌[Molecular Cloning: A Laboratory Manual (2nd ed.). Cold Spring Harbour Laboratory, Cold Spring Harbour, N.Y., 1989]; 어수벨(Ausubel) 등의 문헌[Current Protocols in Molecular Biology, Green Pub. Associates and Wiley-Interscience, New York, 1988]에 기재되어 있다.
예를 들면, 효모에서의 발현을 위한 작성물은 바람직하게는 합성 유전자를, 이에 작동적으로 연결된 관련된 전사 및 번역 제어 서열, 예컨대 프로모터(예컨대 GAL10, GAL7, ADH1, TDH3 또는 PGK), 및 종결 서열[예컨대, 사카로미세스 세레비지에(Saccharomyces cerevisiae) ADH1 종결자]와 함께 함유한다. 효모는 사카로미세스 세레비지에, 한세눌라 폴리오르파(Hansenula polymorpha), 피키아 파스토리스, 클루이베로미세스 프라길리스(Kluyveromyces fragilis), 클루이베로미세스 락티스(Kluyveromyces lactis), 및 스키조사카로미세스 폼베(Schizosaccharomyces pombe)로 구성된 군에서 선택될 수 있다. 또한 로즈(Rose) 등의 문헌[Yeast Genetics: A Laboratory Course Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1990]을 참조한다. 핵산 분자는 당분야에 공지된 바와 같이 효모에서 발현하기 위해 코돈 최적화될 수 있다[샤프(Sharp) 및 코웨(Cowe)의 문헌 "Yeast, 7: 657-678, 1991" 참조]. 임의의 소정의 세포 유형을 위해 적절한 벡터 및 제어 요소는, 발현 벡터에 대하여 당분야에 공지된 정보 및 본 명세서의 교시내용을 고려하여 당분야의 숙련가에 의해 선택될 수 있다.
숙주 세포주에서의 클로닝 및 발현을 위해 이용될 수 있는 벡터는 당분야에 잘 공지되어 있고, 제한되는 것은 아니지만, 포유동물 세포주 또는 효모(진균류) 세포에서의 클로닝 및 발현을 위한 벡터, 세균 세포주에서의 클로닝 및 발현을 위한 벡터, 파아지에서의 클로닝 및 발현을 위한 벡터, 및 곤충 세포주에서의 클로닝 및 발현을 위한 벡터가 포함된다. 발현된 단백질은 표준 단백질 정제 방법을 사용하여 회수될 수 있다. 번역 제어 요소는 코자크(M. Kozak)에 의해 검토되었다[예를 들어, 코자크의 문헌 "Mamm Genome, 7(8): 563-74, 1996"; 코자크의 문헌 "Biochimie., 76(9): 815-21, 1994"; 코자크의 문헌 "J Cell Biol, 108(2): 229-241, 1989"; 코자크 및 샤트킨(Shatkin)의 문헌 "Methods Enzymol, 60: 360-375, 1979"].
HCV E2를 인코딩하는 예시적인 폴리뉴클레오티드는 서열 목록에 제공되고, 서열 번호 1 또는 2에 제시된 폴리뉴클레오티드 서열을 포함한다.
고유의 HCV E2는 단량체성, 이합체성, 3합체성, 4합체성, 5합체성 이상 등23-합체성까지일 수 있고, 1OO kDa 이상 또는 200 kDa 이상의 분자량을 갖는 형태(예컨대 HMW1, 또는 HMW2 형태)를 포함한다. 단량체성 및 올리고머성 형태는 크기, 겔 여과 특징, 항체 반응성 등에 기초하여 선택될 수 있다. 발현된 단백질은 친화도 크로마토그래피에 의해, 예컨대 항체 친화도 크로마토그래피에 의해 세포 성분들로부터 정제될 수 있다.
하나의 실시태양에서, 방법은 본원에 기재된 바와 같은 방법에 의해 처리되기 이전의 고유의 HCV E2와 비교하여 재접힘된 올리고머성 HCV E2의 순도를 증가시킨다. 하나의 실시태양에서, 방법은 세포 배양액으로부터 단리된 고유의 올리고머성 HCV E2와 비교하여 재접힘된 올리고머성 HCV E2의 순도를 증가시킨다. 본원에서 사용되는 경우, 용어 "정제된" 또는 "순도"는 오염원, 즉 세포 또는 바이러스성 오염원, 예컨대 제한되지 않지만, 단백질, 지질, 핵산 및 탄수화물로부터의 올리고머성 HCV E2의 분리를 지칭한다.
더욱이, 하나의 실시태양에서 방법은 샘플중의 올리고머성 HCV E2의 농도를 증가시키고, 즉 단량체성 HCV E2를 함유하는 샘플로부터 본원에 기재된 바와 같은 방법에 의한 샘플의 처리는 올리고머성 HCV E2의 농도를 적어도 10%, 또는 적어도 20%, 또는 적어도 30%, 또는 적어도 40%, 또는 적어도 50%, 또는 적어도 60%, 또는 적어도 70%, 또는 적어도 80%, 또는 적어도 90%, 또는 적어도 95% 증가시킨다. 한가지 실시태양에서, 방법은 이합체의 농도를 적어도 10%, 또는 적어도 20%, 또는 적어도 30%, 또는 적어도 40%, 또는 적어도 50%, 또는 적어도 60%, 또는 적어도 70%, 또는 적어도 80%, 또는 적어도 90%, 또는 적어도 95% 증가시킨다. 한가지 실시태양에서, 방법은 3합체 및/또는 더 높은 급수의 형태의 농도를 적어도 10%, 또는 적어도 20%, 또는 적어도 30%, 또는 적어도 40%, 또는 적어도 50%, 또는 적어도 60%, 또는 적어도 70%, 또는 적어도 80%, 또는 적어도 90%, 또는 적어도 95% 증가시킨다.
하나의 실시태양에서, 재접힘된 올리고머성 HCV E2는 고유의 대조적인 올리고머성 HCV E2와 비교하여 변형된 안정성, 즉 변형된 열역학적 안정성 및/또는 운동학적 안정성을 나타낸다. 하나의 실시태양에서, 재접힘된 올리고머성 HCV E2는 고유의 대조적인 올리고머성 HCV E2와 비교하여 증가된 안정성을 나타낸다.
하나의 실시태양에서, 재접힘된 올리고머성 HCV E2는 비-중화 항체에 대하여 고유의 올리고머성 HCV E2와 동등한 결합을 나타낸다. 한가지 실시태양에서, 올리고머는 이합체, 또는 3합체, 또는 4합체, 또는 5합체 내지 23합체이다.
HCV E2 단량체의 부분적 환원이 바람직하다. 부분적 환원은 입체배좌적 에피토프를 인식하는 항체에 의한 일부 항체 반응성이 보유되는 경우 달성된다. 예를 들면, 단백질 입체배좌의 10%, 20% 또는 30%가 본원에 사용된 부분적 환원 프로토콜에서 보유될 수 있다.
하나의 실시태양에서, 환원 조건은 사용되는 환원제에 의존하여 약 7 이상의 pH를 포함한다.
용어 "올리고머" 또는 "다합체"는 3합체, 4합체 등, 또는 고분자량(HMW) 형태로서 접히는 항원 또는 E2의 버전을 포함한다. 상이한 형태는, 예를 들면, 비-환원 겔 전기영동에서의 이들의 이동 패턴에 의해, 또는 겔 여과 크로마토그래피에 의해 또는 이들의 항체 반응성에 의해 식별된다. 하나의 실시태양에서, 단량체, 이합체, HMW2 및 HMW 델타123 형태는 각각 약 46, 97, 239 및 2400의 분자량을 갖는다.
하나의 구체적인 실시태양에서, 낮은 급수의 형태는 이합체성 형태를 포함하거나, 또는 낮은 급수의 형태는 단량체성 형태를 포함한다.
하나의 실시태양에서, 올리고머성 형태는 3합체성 형태이다.
하나의 실시태양에서, 올리고머성 형태는 3합체성 및/또는 더 높은 급수의 형태이다.
하나의 실시태양에서, 올리고머성 형태는 높은 급수의 형태, 예컨대 본원에 기재된 바와 같은 HMW2 또는 HMW, 또는 본원에 기재된 바와 같은 이의 변형된 형태이다.
하나의 실시태양에서, 고유의 E2는 수용체 결합 도메인을 포함하고, 스템(stem) 영역 및 막관통 도메인이 결핍된 E2의 변형된 형태이다.
당분야에 공지된 바와 같이, 막관통 도메인은 잔기 715-746이고 스템 영역은 잔기 662 내지 714이다. 하나의 실시태양에서, 이러한 형태는 WT E2661이다.
본원에 기재된 바와 같이, HCV E2의 모든 고유의 형태는 상이한 분자내 및 분자간(이합체 및 그 이상의 경우) 디설파이드 결합된 형태를 포함한다. 본 개시내용의 하나의 실시태양에 따라서, 단량체는 환원되고 올리고머로 재접힘되거나, 올리고머는 환원되고 올리고머로 재접힘되거나, 또는 단량체 및 올리고머는 환원되고 이합체 또는 올리고머로 재접힘된다.
방법의 하나의 실시태양에서, 단량체의 적어도 25%, 적어도 30%, 적어도 40%, 적어도 50%, 적어도 60% 또는 적어도 70%가 올리고머로 전환된다.
또 다른 실시태양에서, 단량체의 적어도 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 또는 적어도 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 97%, 98%, 99%가 올리고머로 전환된다.
하나의 실시태양에서, 올리고머는 1OO kDa 이상의 겉보기 분자량을 갖는다. 하나의 실시태양에서, 올리고머는 200 kDa 이상의 겉보기 분자량을 갖는다. 하나의 실시태양에서, 올리고머는 2000 kDa 이상의 겉보기 분자량을 갖는다.
하나의 실시태양에서, 방법은 재접힘된 올리고머성 HCV E2의 가장 큰 수율을 생성하는 HCV 유전자형을 선택하는 단계를 추가로 포함한다. 한가지 실시태양에서, HCV 유전자형은 Gl, G2, G3, G4, G5, G6, G7 및/또는 기타 HCV 유전자형으로부터 선택된다. 예시적인 비-제한적 실시태양에서, HCV 유전자형은 Con1 또는 S52이다.
하나의 실시태양에서, 재접힘된 올리고머성 HCV E2는 단량체 재접힘에 의해 제조되지 않는 대조적 올리고머성 HCV E2와 비교하여 감소된 항체 CBH4G에 대한 결합을 나타낸다. 고유의 올리고머는 세포외 단량체 재접힘에 의해 제조되지 않는 대조적 올리고머성 HCV E2의 일예이다.
방법의 하나의 실시태양에서, 재접힘된 올리고머성 HCV E2는 중화 항체에 결합하는 대조적 올리고머성 HCV E2의 능력을 실질적으로 보유한다. 방법의 하나의 실시태양에서, 올리고머성 HCV E2는 광범위 중화 항체에 결합하는 대조적 올리고머성 HCV E2의 능력을 실질적으로 보유한다.
하나의 실시태양에서, 환원제는 가역성 환원제이고, 예를 들면 DTT 또는 2-머캅토에탄올, 트리스(2-카복시에틸)포스핀, 환원된 글루타티온, 티오글리콜산 또는 기타 티올 함유 제제로부터 선택된다.
방법의 하나의 실시태양에서, 환원제는 고유의 E2의 부분적 환원을 일으키기 위해 약 0.1 내지 1.O mM에서 사용되는 디티오트레이톨이다.
방법의 하나의 실시태양에서, 용액은 pH > 7 내지 9의 완충액이다.
하나의 실시태양에서, 항원 농도는 탄산염 완충액 pH 9.6에서 1 mg/㎖이다. 0.6mM DTT(최종 농도)가 사용되고 항온처리는 1 시간 동안 37℃에서이다. 용액은 PBS pH 6.8에 의해 느리게 희석되고, 항원 농도 및 완충액은 PBS pH 6.8에 의해 교환된다.
하나의 실시태양에서, 고유의 E2(및 결과적으로 재접힘된 올리고머)는 과가변성 영역의 전부 또는 일부, 예컨대 과가변성 영역 1(HVR1) 또는 이의 일부, 과가변성 영역 2(HVR2) 또는 이의 일부 및/또는 상호유전자형 가변성 영역(igVR/VR3) 또는 이의 일부중 하나 이상이 결핍된다. 수용체 결합 도메인은, 바이러스 침습을 위해 필수적이지 않은 영역 상에서의 면역 반응에 집중하고 광범위 중화 항체의 에피토프를 차폐함으로써 면역 회피에서 일정 역할을 담당하는 가변성 영역 HVR1, HVR2 및 igVR을 포함한다. HVR2 및/또는 igVR 및/또는 HVR1의 적어도 일부의 결실은 광범위 중화 항체를 이끌어내는 보존적 에피토프 상에서의 면역 반응에 집중하도록 영향을 받는다.
RBD를 포함하고 모든 3개의 가변성 영역이 결핍된 작성물은 국제 특허출원 공개공보 제WO 2008/022401호에 기재된 바와 같이 E2661 델타123 또는 Δ123으로 지칭된다. 결실된 영역은 국제 특허출원 공개공보 제WO 08/022401호(본원에 그의 전체가 인용됨)에 기재된 가요성 연결기 서열, 예컨대 Gly, Ser, Ala 및 Arg으로 구성된 군에서 선택되는 잔기를 포함하는 20개 이하의 아미노산의 펩타이드 서열(예를 들면 GSSG 또는 ETHGSSG)에 의해 임의적으로 대체된다.
한가지 실시태양에서, 고유의 E2는 정제를 돕기 위한 신호 서열 또는 단백질 태그를 포함한다. 한가지 실시태양에서, 신호 서열은 세포로부터 단백질을 분비시킨다. 한가지 실시태양에서, 신호 서열은 서열 번호 9에 제공된 아미노산 서열 "MNPLLILTFVAAALA"을 포함하는 N-말단 신호 서열이다. 한가지 실시태양에서, 단백질 태그는 C-말단 His-태그이다. 한가지 실시태양에서, C-말단 His-태그는 아미노산 서열 "HHHHHH"를 포함한다.
본 발명자들은 단량체성 E2 조차도 이종성이고 상이한 분자내 디설파이드 배열을 나타냄을 발견하였다. 이들은 단량체의 환원이 더 많은 동종성 올리고머의 재접힘을 위한 우수한 출발점을 제공할 것으로 판단하였다.
하나의 실시태양에서, 고유의 E2는 C581, C585, C652, C677, C494, C486, C459, C452, C564, C597, C569 및/또는 C620으로부터 선택된 하나 이상의 아미노산 잔기에서 비-시스테인 치환 또는 돌연변이를 포함한다. 놀랍게도, 비-시스테인 잔기에 대해 치환된 7개의 시스테인 잔기, 즉 C452, C486, C569, C597, C581, C585, C652 비-시스테인 치환을 갖는 HCV E2 단량체("Ala7"로서 지칭됨) 조차도 더 높은 수준의 올리고머로 재접힘된다.
E2는 당단백질의 3차 구조를 스캐폴딩(scaffolding)하는, ER에서 9개의 분자내 디설파이드 결합을 형성하는 18개의 고도로 보존된 시스테인 잔기를 포함한다. 국제 특허출원 공개공보 제WO 12/016290호(본원에 참고로 그의 전체가 인용됨)에 기재된 바와 같이, 이들 디설파이드중 일부는 CD81 결합 및 MAb H53 결합을 위해 불필요하다. 또한, 7개의 시스테인 잔기의 제거를 포함하여, 시스테인 변형된 형태는 유의적으로 단량체 생산을 증가시키고 올리고머의 생산을 감소시킨다. 정상적으로, E2가 재조합적으로 숙주 세포에서 생산될 경우, 이는 상이한 단량체 및 올리고머성 형태의 혼합물을 생산한다. 국제 특허출원 공개공보 제WO 12/016290호는 다양한 시스테인 변형된 HCV E2의 생산을 기재하였다. 국제 특허출원 공개공보 제WO 12/016290호에 개시된 다른 작성물이 본원에서 고려되고, 예컨대 C452, C486, C569, 및 C597로부터 선택된 2, 3, 또는 4개의 시스테인이 돌연변이되거나 파괴된 Ala 치환된 형태이다. E2661은 C677에서 시스테인이 결핍되고 17개의 시스테인을 갖는다.
몇몇 실시태양에서, 고유의 HCV E2는 HCV E2의 시스테인 변형된 형태, 예컨대 Ala4, Ala5, Ala6 또는 Ala7 변형된 형태이고, 이는 재조합적으로 발현될 경우 적어도 40%의 단량체 및 70% 미만의 올리고머, 또는 적어도 50%의 단량체 및 50% 미만의 올리고머를 고유의 형태로 생산한다.
본원에서 결정된 바와 같이, C620-Ala의 돌연변이생성은 고분자량 올리고머 형성을 차단한다. 따라서 높은 급수의 형태의 생산을 위한 고유의 HCV E2 형태는 일반적으로 C620이다.
또 다른 광범위한 양태에서, 본 기재내용은: (i) 하나 이상의 디설파이드 결합을 부분적으로 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 고유의 E2를 접촉시키는 단계; 및 (ii) 환원제를 제거하거나 (i)로부터의 E2를 산화제와 접촉시켜서 단량체성 E2의 올리고머성 엔벨롭 당단백질로의 재접힘을 이끌어내는 단계를 포함하고, 여기서 단계 (ii)에서 단량체의 적어도 20%가 올리고머로 전환되고, 올리고머가 단량체성 당단백질과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타내는, 재조합 올리고머성 바이러스성 엔벨롭 당단백질을 단량체성 엔벨롭 당단백질로부터 제조하는 방법을 제공한다.
한가지 실시태양에서, 단계 (i)은 단계 (ii) 이전에 2회 수행된다. 한가지 실시태양에서, 단계 (ii)는 단계 (ii) 이전에 3회 수행된다.
또 다른 실시태양에서, 본 명세서는 앞서 본원에 기재된 환원 및 재접힘 방법에 의해 생산된 재접힘된 재조합 올리고머성 간염 C 바이러스(HCV) E2 당단백질을 포함하는 조성물을 가능하게 한다.
하나의 실시태양에서, 재접힘된 올리고머성 단백질은 서열 번호 3, 4, 5, 6, 7 및 8중 하나에 제시된 아미노산, 또는 이의 절단되거나 변형된 버전, 또는 작용성 변형체를 포함한다. 작용성 변형체 및 변형된 형태는 변형전 형태 또는 임의의 다른 적합한 대조군과 비교하여 향상된 면역원성을 나타낼 수 있다.
하나의 실시태양에서, 조성물은 본원에 기재된 환원 및 재접힘 방법에 의해 생산된 재접힘된 재조합 올리고머성 간염 C 바이러스(HCV) E2 당단백질을 포함하고, 여기서 올리고머성 HCV E2는 단량체 환원 및 재접힘에 의해 제조되지 않은 대조군 HMW E2와 비교하여 감소된 비-중화 항체에 대한 결합을 나타낸다.
하나의 실시태양에서, 조성물은 본원에 기재된 환원 및 재접힘 방법에 의해 생산된 재접힘된 재조합 올리고머성 간염 C 바이러스(HCV) E2 당단백질을 포함하고, 여기서 재접힘된 올리고머성 HCV E2는 단량체 환원 및 재접힘에 의해 제조되지 않은 대조군 HMW E2와 비교하여 감소된 항체 CBH4G 또는 AR3C에 대한 결합을 나타낸다.
하나의 실시태양에서, 조성물은 재접힘된 올리고머성 HCV E2를 포함하고, 여기서 재접힘된 올리고머성 HCV E2는 단량체 환원 및 재접힘에 의해 제조되지 않은 대조군 HMW E2와 비교하여 감소된 항체 CBH4G에 대한 결합을 나타낸다. 하나의 실시태양에서, 조성물은 재접힘된 올리고머성 HCV E2를 포함하고, 여기서 재접힘된 올리고머성 HCV E2는 단량체 환원 및 재접힘에 의해 제조되지 않은 대조군 HMW E2와 비교하여 감소된 항체 AR3C에 대한 결합을 나타낸다.
하나의 실시태양에서, 조성물은 단량체 재접힘된 올리고머성 HCV E2 및 약학적으로 또는 생리학적으로 허용가능한 담체 및/또는 희석제를 포함한다.
하나의 실시태양에서, 조성물은 추가로 보조제를 포함한다.
당분야의 숙련가라면 보조제는 면역 반응을 유도하는 조성물의 능력을 향상시키는 임의의 제제일 수 있음을 인식할 것이다. 한가지 실시태양에서, 보조제는 항원에 대한 면역 반응을 증가시킴으로써 작용할 수 있다. 한가지 실시태양에서, 보조제는 Th1 및/또는 Th2 면역 반응을 증가시킬 수 있다. 한가지 실시태양에서, 보조제는 명반 염(alum salt) 또는 기타 무기질 보조제; 텐소액티브 제제(tensoactive agent); 세균성 유도체; 비히클(vehicle) 또는 지효성 물질 또는 사이토킨(cytokine), 예컨대 페트로브스키(Petrovsky) 등의 문헌(2004) 및 윌슨-웰더(Wilson-Welder) 등의 문헌(2009)에 기재된 물질일 수 있다. 하나의 실시태양에서, 보조제는 인산 알루미늄, 수산화 알루미늄, 황산 알루미늄 칼륨(명반), 인산 칼슘, 프런드(Freund)의 완전 보조제, 프런드의 불완전 보조제, MF-59, 사포닌, QS-21, 지질다당류(LPS: lipopolysaccharide), 모노포스포릴 리피드 A(MPLA: monophosphoryl lipid A), Th1 활성화 펩타이드(예를 들어 IMP321), TLR-2 리간드[예를 들어 OspA, 뮤라밀 디펩타이드(MDP: muramyl dipeptide), 대식세포 활성화 지질펩타이드-2(MALP-2: macrophage activating lipopeptide-2)], CpG 보조제, 백일해 독소, 열 불안정성 독소(LTK63 및 LT-R192G), 디프테리아 독소, 이미퀴모드(Imiquimod), 아다백스(Addavax), 이스코마트릭스(ISCOMATRIX), 과립구 대식세포-콜로니 자극 인자(GM-CSF: granulocyte macrophage-colony stimulating factor), IL-12, IL-6, IL-4, IL-2, IL-1, IFN-g, AS04(MPLA 및 QS-21을 함유하는 리포솜 제형), 글리세릴, 및 오일 유화액, 예컨대 파라핀, 무기질유, 라놀린(lanolin), 스쿠알렌(squalene), ISA-70, 및 몬타나이드(montanide)로부터 선택될 수 있다. 하나의 예시적인 실시태양에서, 보조제는 사포닌 기제 보조제이다. 관련된 양태에서, 보조제는 추가로 콜레스테롤 및 스테롤을 포함하는 사포닌 기제 보조제이고, 이의 예시적인 일예는 이스코마트릭스 보조제이다. 하나의 예시적인 실시태양에서, 보조제는 MF59이다. 하나의 예시적인 실시태양에서, 보조제는 아다백스이다. 하나의 실시태양에서, 보조제는 탄수화물 보조제, 예를 들면, 글루칸, 덱스트란, 레티난(lentinan), 글루코만난(glucomannan) 또는 갈락토만난(galactomannan)이다.
하나의 실시태양에서, 본 명세서는 HCV 감염의 치료 또는 예방에 있어서의, 또는 이를 위한 약제의 제조에 있어서의, 재접힘된 올리고머성 HCV E2를 포함하는 조성물의 용도를 제공한다.
하나의 실시태양에서, 본 명세서는 HCV 감염의 진단 또는 모니터링, 또는 항-HCV 치료 프로토콜의 모니터링에 있어서의, 또는 이를 위한 진단용 제제의 제조에 있어서의, 재접힘된 올리고머성 HCV E2를 포함하는 조성물의 용도를 제공한다.
하나의 실시태양에서, 조성물은 3합체성, 또는 3합체성 이상의 높은 급수의 형태의 HCV E2를 70 중량% 이상 또는 80 중량% 이상으로 포함한다.
하나의 실시태양에서, 조성물은 이합체성 HCV E2를 70 중량% 이상 또는 80 중량% 이상으로 포함한다.
하나의 실시태양에서, 조성물은 더 높은 급수의 형태의 HCV E2 당단백질을 70 중량% 이상 또는 80 중량% 이상으로 포함한다.
하나의 실시태양에서, 본 명세서는 면역 반응을 이끌어내기에 충분한 조건하에 일정한 시간 동안 본원에 기재된 바와 같은 재접힘된 올리고머성 HCV E2를 포함하는 효과량의 조성물을 피험체 또는 환자에게 투여하는 단계를 포함하는, 피험체 또는 환자에서 면역 반응을 이끌어내는 방법을 제공한다.
이들 실시태양에 따라서, 조성물은 일반적으로 광범위 중화 항체의 생성을 포함하는 면역 반응을 이끌어내기에 충분한 조건하에 일정한 시간 동안 투여된다. 본 발명의 조성물은 단일 투약 또는 적용으로서 투여될 수 있다. 다르게는, 조성물은 반복 투약 또는 적용을 포함할 수 있고, 예를 들면 조성물은 2, 3, 4, 5, 6, 7, 8, 9, 또는 10회 이상 투여될 수 있다.
본 발명은 추가로 본원에 기재된 바와 같은 재접힘된 올리고머성 HCV E2를 포함하는 효과량의 조성물을 피험체 또는 환자에게 투여하는 단계를 포함하는, 피험체 또는 환자에서 면역 반응을 이끌어내는 방법을 추가로 제공한다. 본원에 기재된 바와 같은 재접힘된 올리고머성 HCV E2를 포함하는, 간염 C 바이러스로부터의 감염에 대해 피험체를 면역화하기 위한 조성물, 특별히 백신 조성물이 또한 본 발명에 의해 고려된다.
용어 "백신"은, 본원에서 사용되는 경우, 피험체에서 면역학적 반응을 유도하는 적어도 하나의 면역학적으로 활성인 성분, 및 가능하지만 필수적이지 않은, 이러한 활성 성분의 면역학적 활성을 향상시키는 하나 이상의 추가적인 성분(예를 들면 보조제)을 포함하는 약학 조성물을 지칭한다. 백신은 약학 조성물에 전형적인 추가의 성분을 추가적으로 포함할 수 있다. 백신의 면역학적 활성 성분은 높은 급수의 조립된 항원, 예컨대 재접힘된 올리고머성 HCV E2 조성물을 포함한다. 용어 "백신" 및 "백신 조성물"은 본 발명에서 상호교환적으로 사용된다.
본 발명에서 고려되는 "피험체"는 인간 또는 동물이고, 실험실 또는 기술분야에서 용인된 검사 또는 비히클 동물이 포함된다. "환자"는 치료 또는 예방이 필요한 인간 피험체를 포함한다.
하나의 실시태양에서, 본 명세서는 본원에 기재된 바와 같은 조립된 또는 재접힘된 올리고머성 항원을 포함하는 조성물을 피험체에게 투여하는 단계를 포함하는, 항원과 연관된 증상에 대해 피험체를 면역화하는 방법을 또한 가능하게 한다.
하나의 실시태양에서, 본 명세서는 본원에 기재된 바와 같은 세포밖에서 재접힘된 올리고머성 HCV E2를 포함하는 조성물을 피험체에게 투여하는 단계를 포함하는, HCV로부터의 감염에 대해 피험체를 면역화하는 방법을 또한 가능하게 한다.
하나의 실시태양에서, 본 명세서는 HCV 감염을 치료하거나 예방하기에 충분한 조건하에 일정 시간 동안 본원에 기재된 바와 같은 세포밖에서 재접힘된 올리고머성 HCV E2를 포함하는 조성물을 피험체에게 투여하는 단계를 포함하는, HCV 감염을 치료하거나 예방하는 방법을 또한 가능하게 한다.
"치료학적 효과량" 및 "예방학적 효과량"을 비롯한 용어 "효과량"은, 본원에서 사용되는 경우, 원하는 치료학적, 예방적, 또는 생리학적 효과를 일부 피험체에서 제공하는 일련의 방출 또는 지효성 시스템의 일부로서의 또는 단일 용량으로서의 본 발명의 조성물의 충분한 양을 의미한다. 원치않는 효과, 예를 들어 부작용은 때때로 원하는 치료학적 효과와 함께 나타날 수 있고; 이에 따라, 의사는 적절한 "효과량"을 결정하는데 있어서 잠재적인 위험에 대한 점재적인 이점의 균형을 맞춘다. 필요한 조성물의 정확한 양은, 피험체의 종, 연령 및 일반적 증상, 투여 방식 등에 의존하여 피험체마다 다를 것이다. 이와 같이, 정확한 "효과량"을 특정화하는 것은 불가능할 수 있다. 그러나, 임의의 개별적 경우에 "적절한 효과량"은 일상적 기술 또는 실험을 사용하여 당분야의 숙련가에 의해 결정될 수 있다. 당분야의 숙련가라면 조성물 또는 다른 제제의 이전 투여, 피험체의 크기, 피험체 증후의 위중성, 또는 감염된 개체군에서의 증후의 위중성, 바이러스 양, 및 특별한 조성물 또는 선택된 투여 경로와 같은 인자에 기초하여 필요한 양을 결정할 수 있을 것이다.
용어 "치료"는 항원, 예컨대 E2 항원 및 HCV 감염과 연관된 증상의 하나 이상의 증후에 있어서, 또는 HCV의 진전된 증후의 발전 위험 또는 HCV를 전염시킬 위험에 있어서의 적어도 일부 피험체에서의 임의의 측정가능하거나 통계적으로 유의적인 개선을 지칭한다.
용어 "방지" 및 "예방" 등은 상호교환적으로 사용되고, 후속적인 감염을 예방 또는 약화시키거나, 감염될 위험을 감소시키거나, 또는 HCV 감염과 연관된 증상 또는 증상의 징후의 위중성 또는 개시를 감소시킬 목적으로 HCV 에 의해 감염된 것으로 알려지지 않은 피험체에게 본 발명의 조성물을 투여함을 포함한다.
백신 조성물의 투여는 일반적으로 예방학적 목적을 위한 것이다. 조성물의 예방학적 투여는 임의의 후속적인 감염을 예방하거나 약화시키는 작용을 한다. 한가지 실시태양에서, 백신 조성물은 병원체에 의한 피험체의 재감염을 예방하기 위한 것이다. "약학적으로 허용가능한" 조성물은 수여 환자에 의해 용인되는 것이다. 효과량의 백신이 투여되는 것이 고려된다. "효과량"은 원하는 생물학적 효과를 달성하기 위해, 예컨대 충분한 체액 면역 또는 세포 면역을 유도하기 위해 충분한 양이다. 이는 백신의 유형, 수여자의 연령, 성별, 건강 및 체중에 따라 달라질 수 있다. 원하는 생물학적 효과의 예로는, 제한되는 것은 아니지만, 증후의 미발현, 증후의 감소, 조직 및 코 분비물에서 바이러스 역가의 감소, 간염 C 바이러스에 의한 감염에 대한 완전한 보호, 및 간염 C 바이러스에 의한 감염에 대한 부분적 보호가 포함된다.
몇몇 실시태양에서, 본 발명의 백신 또는 조성물은, 이의 존재가 수여 환자의 생리학에 있어서 검출가능한 변화를 초래하여 감염성 간염 C 바이러스의 적어도 하나의 균주에 대하여 적어도 하나의 1차 또는 2차 체액 면역 또는 세포 면역 반응을 향상시키거나 이러한 반응에 있어서 향상을 나타낸다면 생리학적으로 유의적이다. 백신 조성물은 바이러스 감염에 대하여 보호하기 위해 투여된다. "보호"는 절대적일 필요는 없고, 즉 간염 C 감염은 대조 모집단 또는 환자의 집단과 비교하여 통계적으로 유의적인 개선이 존재한다면 완전히 예방되거나 근절될 필요는 없다. 보호는 간염 C 바이러스 감염의 증후의 위중성 또는 이의 개시의 신속성을 감소시키는 것으로 제한될 수 있다.
하나의 실시태양에서, 본 발명의 백신 조성물은 감염의 개시 이전(예상되는 감염을 예방하거나 약화시키기 위함) 또는 감염의 개시 이후에 피험체에게 제공되고, 이에 따라 바이러스 감염에 대해 보호한다. 몇몇 실시태양에서, 본 발명의 백신 조성물은 피험체 사이의 바이러스 전염을 감소시키기 위해 감염의 개시 전 또는 후에 피험체에게 제공된다.
추가로, 본 발명의 조성물은 유일한 활성 약학 제제로서 투여되거나, 또는 바이러스성 또는 암 증상, 예컨대 간염 C 감염 또는 HCV 감염과 연관된 증후를 치료하거나 예방하기 위한 하나 이상의 제제와 함께 사용될 수 있음을 인식할 것이다. 본 발명의 조성물 또는 조성물의 조합물과 함께 투여되는 다른 제제는 HCV 감염에 의해 야기되는 질병을 위한 치료제, 또는 직접적 또는 간접적 기작에 의해 HCV 바이러스 복제를 억제하는 치료제를 포함한다. 이들 제제로는, 제한되는 것은 아니지만, 숙주 면역 조절인자[예를 들면, 인터페론(interferon)-알파, 페길화된(pegylated) 인터페론-알파, 컨센서스(consensus) 인터페론, 인터페론-베타, 인터페론-감마, CpG 올리고뉴클레오티드 등]; 숙주 세포 기능을 저해하는 항바이러스 화합물, 예컨대 일인산이노신 탈수소효소[예를 들면, 리바비린(ribavirin) 등]; 면역 기능을 조절하는 사이토킨[예를 들면, 인터류킨(interleukin) 2, 인터류킨 6, 및 인터류킨 12]; 유형 1 헬퍼(helper) T 세포 반응의 진행을 향상시키는 화합물; 간섭 RNA; 안티센스(anti-sense) RNA; HCV에 대해 유도되는 HCV 항원 또는 항원 보조제 조합물을 포함하는 백신; HCV 바이러스 복제의 내부 리보솜 진입 자리(IRES: internal ribosome entry site) 개시된 번역 단계를 저해함으로써 바이러스 단백질 합성을 차단하거나, 또는 바이러스의 입자 성숙 및 방출을 차단하기 위해 숙주 세포 성분과 상호작용하는 제제(비로포린(viroporin) 계열의 막 단백질, 예컨대, 예를 들면, HCV P7 등에 대해 표적화되는 제제); 및 바이러스 복제에 연루된 바이러스 게놈의 다른 단백질을 표적화함으로써 HCV의 복제를 저해하고/하거나 다른 바이러스 표적의 기능을 간섭하는 임의의 제제 또는 제제의 조합물, 예컨대 NS3/NS4A 프로테아제, NS3 헬리카아제(helicase), NS5B 폴리머라아제, NS4A 단백질 및 NS5A 단백질의 저해제가 포함된다.
역시 또 다른 실시태양에 따라서, 본 발명의 약학 조성물은 HCV 수명 주기에서, 제한되지 않지만, 헬리카아제, 폴리머라아제, 메탈로프로테아제(metalloprotease), NS4A 단백질, NS5A 단백질, 및 내부 리보솜 진입 자리(IRES)를 비롯한 표적에 대한 다른 저해제(들)를 추가로 포함할 수 있다.
투여는 일반적으로 E2-특이적 항체의 생산을 포함하는 면역 반응 또는 세포 면역 반응을 이끌어내기에 충분한 조건하에 일정한 시간 동안이다. 면역원성 조성물은, 편리한 방식으로, 예컨대 폐, 경구, 정맥내(여기서 수용성), 복강내, 근육내, 피하, 피내, 척추강내 또는 좌약 경로 또는 이식(예를 들어 지효성 제형 사용)에 의해 투여될 수 있다. 투여는 전신적이거나 국부적일 수 있지만, 전신적 투여가 더욱 편리하다. 고려되는 다른 투여 경로는 패치(patch), 세포 전달, 임플란트, 설하, 안내, 국소적, 경구적, 직장내, 질내, 비강내 또는 경피적이다.
본원에서 사용되는 경우, "면역 반응"은 본 발명의 조성물의 존재에 대한 전체적인 신체의 반응을 지칭하고, 이는 항체를 만들고 조성물에 대해 면역력을 발달시킴을 포함한다. 그러므로, 항원에 대한 면역 반응은 또한 관심있는 항원에 대한 체액 및/또는 세포 면역 반응의 피험체에서의 발달을 포함한다. "체액 면역 반응"은 혈장 세포에 의해 생산된 항체에 의해 중재된다. "세포 면역 반응"은 T 림프구 및/또는 기타 백혈구에 의해 중재되는 반응이다. 본원에서 사용되는 경우, "항체 역가"는 각각의 피험체에 대한 면역전 샘플에 비해 더 큰 값을 초래하는 면역후 혈청에서 가장 높은 희석비로서 정의될 수 있다.
본 발명의 실시태양은 재접힘된 올리고머성 항원, 예컨대 HCV E2에 대한 면역 반응을 평가하기 위한 검정을 또한 제공한다. 이러한 검정은 생체내 검정, 예컨대 항체 반응 및 지연된 유형의 과민성 반응을 측정하기 위한 검정을 포함할 수 있다. 한가지 실시태양에서, 항체 반응을 주로 측정하는 검정은 B-세포 기능 뿐만 아니라 B-세포/T-세포 상호작용을 측정할 수 있다. 항체 반응 검정의 경우, 혈액중 항체 역가는 항원 시험감염(challenge) 이후 비교될 수 있다.
본 명세서는 효과량의 재접힘된 올리고머성 항원, 예컨대 HCV E2를 피험체에게 투여하는 단계; 및 생산된 항체를 정제하는 단계를 포함하는, 본원에 기재된 바와 같은 재접힘된 올리고머성 항원, 예컨대 HCV E2에 대한 정제된 항체를 생산하는 방법을 제공한다.
또 다른 실시태양에서, 본 발명은 재접힘된 올리고머성 항원, 예컨대 HCV E2에 대해 야기된 항체를 제공한다. 특이적 항체는 조립된 항원을 인식하는 반면, 고유의 항원을 인식하지 못하고/못하거나, 또는 그 반대이다. 항체는 다중클론성이거나 단일클론성일 수 있다. 추가로, 항체는 진단학적, 예후적, 치료학적, 예방학적, 및 선별 목적을 위해, 전형적으로 관련 분야의 숙련가에게 공지된 준거를 사용하여 선택될 수 있다.
용어 "항체" 및 "항체들"은 다중클론성 및 단일클론성 항체 및 단일클론성 항체로부터 유래된 모든 다양한 형태를 포함하고, 제한되지 않지만 전장 항체(예를 들어 온전한 Fc 영역을 갖는 항체), 항원-결합 단편, 예를 들면, Fv, Fab, Fab' 및 F(ab').sub.2 단편; 및 재조합 방법을 사용하여 생산된 항체-유래된 폴리펩타이드, 예컨대 단일 쇄 항체가 포함된다. 용어 "항체" 및 "항체들"은, 본원에서 사용되는 경우, 예를 들면 유전자이식 동물에서 또는 파이지 디스플레이(phage display)를 통해 생산된 인간 항체, 뿐만 아니라 항체, 인간 또는 인간화된 항체, 영장류화된 항체 또는 탈면역화된(deimmunized) 항체를 또한 지칭한다. 이는 또한 치료학적으로 허용가능한 항체의 다른 형태, 및 이의 항원-결합 단편, 예를 들면 연골 해양 동물 또는 낙타과 동물로부터 유래된 단일 도메인 항체, 또는 이러한 항체에 기초한 라이브러리로부터의 항체를 포함한다. 항체의 단편화되거나 변형된 형태의 선택은 또한 단편 또는 변형된 형태가 항체 또는 단편의 반감기에 미치는 임의의 영향을 고려함을 포함한다.
몇몇 실시태양에서, 항체는 약학적으로 또는 약리학적으로 허용가능한 담체, 희석제 또는 부형제와 함께 제공된다. 다른 실시태양에서, 항체는 진단 또는 예후를 위해 선택된다. 몇몇 실시태양에서, HCV E2 당단백질 항체의 재접힘된 올리고머성 형태를 포함하는 키트가 제공된다.
"약학적으로 허용가능한 담체 및/또는 희석제"는, 별도로 바람직하지 않지는 않은, 즉 그 자체로 또는 활성 조성물과 함께 실질적인 부작용을 초래할 것 같지 않은 물질로 구성된 약학 비히클이다. 담체는 모든 용매, 분산 매질, 코팅물, 항균 및 항진균제, 긴장성(tonicity)을 조정하기 위한 제제, 흡수 및 소거 속도를 증가 또는 감소시키기 위한 제제, pH를 유지하기 위한 완충액, 킬레이트화제, 막 또는 장벽 통과제를 포함할 수 있다. 약학적으로 허용가능한 염은 별도로 바람직하지 않지는 않은 염이다. 제제 또는 제제를 포함하는 조성물은 약학적으로 허용가능한 무독성 염, 예컨대 산 부가 염 또는 금속 착체의 형태로 투여될 수 있다.
경구 투여를 위해, 조성물은 고체 또는 액체 제제, 예컨대 캡슐, 환제, 정제, 로젠지(lozenge), 분말, 현탁액 또는 유화액으로 제형화될 수 있다. 경구 투여형으로 조성물을 제조함에 있어서, 임의의 통상적인 약학 매질이 이용될 수 있고, 예컨대, 예를 들면, 경구 액체 제제(예컨대, 예를 들면, 현탁액, 엑릭시르(elixir) 및 용액)의 경우 물, 글리콜, 오일, 알코올, 방향제, 보존제, 착색제, 현탁제 등이거나; 또는 경구 고체 제제(예컨대, 예를 들면, 분말, 캡슐 및 정제)의 경우 담체, 예컨대 전분, 당분, 희석제, 과립화제, 윤활제, 결합제, 붕해제 등이다. 정제 및 캡슐은 투여시 이들의 용이성으로 인해 가장 유리한 경구 단위 투여형을 대표하고, 이 경우 고체 약학 담체가 명백히 이용된다. 정제는 결합제, 예컨대 트라가칸트(tragacanth), 옥수수 전분 또는 젤라틴; 붕해제, 예컨대 알긴산; 및 윤활제, 예컨대 스테아르산 마그네슘을 함유할 수 있다. 원한다면, 정제는 표준 기법에 의해 당분-코팅되거나 장성(enteric)-코팅될 수 있다. 활성 조성물은 조성물이 위장관을 통해 안정하게 통과하도록 캡슐화될 수 있다. 예를 들면, 국제 특허출원 공개공보 제WO 96/11698호를 참조한다.
비경구 투여를 위해, 조성물은 담체에 용해되어 용액 또는 현탁액으로서 투여될 수 있다. 경점막 또는 경피(패치 포함) 전달을 위해, 당분야에 공지된 적절한 침투제가 조성물을 전달하기 위해 사용된다. 흡입을 위해, 임의의 편리한 시스템, 예컨대 건조 분말 에어로졸(aerosol), 액체 전달 시스템, 에어 제트 분무기(air jet nebulizer), 추진제 시스템이 전달에 사용된다. 예를 들면, 제형은 에어로졸 또는 미스트(mist) 형태로 투여될 수 있다. 조성물은 또한 지속된 전달 형식 또는 서방성 형식으로 전달될 수 있다. 예를 들면, 지속된 전달을 가능하게 하는 생분해성 미소구체(microsphere) 또는 캡슐 또는 기타 중합체 입체구조물이 제형에 포함될 수 있다. 제형은 약물동력학 및 생체내분포(biodistribution)를 바꾸기 위해 변형될 수 있다. 약물동력학의 일반적 논의를 위해, 예를 들어, 문헌[Remington's Pharmaceutical Sciences, 18th 'Ed., Mack Publishing Company, Easton, PA, U.S.A.1990 (supra)]을 참고한다. 몇몇 실시태양에서, 제형은 액체 단층 또는 이중층, 예컨대 리포솜 또는 미셀(micelle)에 혼입될 수 있다. 당분야에 공지된 표적화 요법은 제제를 특정 유형의 세포 또는 조직에 더욱 특이적으로 전달하기 위해 사용될 수 있다.
투여되는 활성화제의 실제량 및 투여 속도 및 시간-경과는 질병의 성질 및 위중성에 따라 달라질 것이다. 치료의 처방, 예를 들어 투여량, 시기 선택 등에 대한 결정은 의사 또는 전문가의 책임하에 있고, 전형적으로 개별 환자의 증상, 전달 부위, 투여 방법 및 의사에게 공지된 다른 요인을 고려한다. 기법 및 프로토콜의 예는 문헌[Remington's Pharmaceutical Sciences, 1990 (supra)]에서 찾아볼 수 있다.
제조될 수 있는 서방성 제제는 면역 반응을 유도하기 위해 특별히 편리하다. 서방성 제제의 예는 폴리펩타이드가 함유된 고체 소수성 중합체의 반투과성 매트릭스를 포함하고, 이러한 매트릭스는 성형 제품, 예를 들어, 필름, 또는 마이크로캡슐의 형태로 존재한다. 서방성 매트릭스의 예로는 폴리에스테르, 하이드로겔[예를 들면, 폴리(2-하이드록시에틸-메타크릴레이트), 또는 폴리(비닐알코올)], 폴리락티드, L-글루탐산 및 에틸-L-글루타메이트의 공중합체, 비-분해성 에틸렌-비닐 아세테이트, 분해성 락트산-글리콜산 공중합체, 및 폴리-D-(-)-3-하이드록시부티르산이 포함된다. 중합체, 예컨대 에틸렌-비닐 아세테이트 및 락트산-글리콜산은 100 일에 걸쳐 분자를 방출시킬 수 있는 반면, 어떤 하이드로겔은 더 짧은 기간 동안 단백질을 방출시킨다. 지질 함량이 약 30%를 초과하는 콜레스테롤인 작은(약 200 내지 800 옹스트롬) 단일층판상(unilamellar) 유형의 리포솜이 사용될 수 있고, 선택되는 비율은 최적 치료법을 위해 조정된다.
단백질의 안정화는 설프하이드릴 잔기를 변형시키고, 산성 용액으로부터 동결건조시키며, 수분 함량을 제어하고, 적절한 첨가제를 사용하며, 특정 중합체 매트릭스 조성물을 개발함으로써 달성될 수 있다. 단백질의 생체내 반감기는 당분야에 공지된 기법을 사용하여 연장될 수 있고, 예를 들면, 다른 요소, 예컨대 폴리에틸렌글리콜(PEG) 기의 결합에 의한 기법이 포함된다.
당분야에 개시된 바와 같은 프라임-부스트(prime-boost) 면역화 전략이 고려된다. 예를 들면 국제 특허출원 공개공보 제WO/2003/047617호를 참조한다. 이와 같이, 조성물은 백신, 프라이밍(priming) 또는 부스팅(boosting) 제제의 형태일 수 있다.
하나의 실시태양에서, 본 명세서는 본원에 기재된 바와 같은 환원되고 재접힘된 올리고머성 HCV E2 형태를 포함하는 조성물을 포함하는 키트, 또는 고체 또는 반고체 기질을 제공한다.
용어 "단리된"은 정상적으로는 고유의 상태에서 동반되는 성분들이 실질적으로 또는 본질적으로 존재하지 않는 물질을 의미한다. 예를 들면, "단리된 핵산 분자"는 자연-발생 상태에서 옆에 배치된 서열로부터 단리된 핵산 또는 폴리뉴클레오티드, 예를 들어, 정상적으로 DNA 단편에 인접한 서열로부터 제거되어진 그러한 DNA 단편을 지칭한다. 제한없이, 단리된 핵산, 폴리뉴클레오티드, 펩타이드, 또는 폴리펩타이드는 정제에 의해 단리되는 고유의 서열, 또는 재조합 또는 합성 수단에 의해 생산되는 서열을 지칭할 수 있다. 몇몇 실시태양에서, 정제된 재접힘된 올리고머성 HCV E2는 적어도 95 내지 99% 순수하다.
당분야의 숙련가라면 본원에 기재된 바와 같은 방법에 의해 생산된 재접힘된 올리고머성 HCV E2는, 예를 들면 하나 이상의 하기 단계를 포함하는 당분야의 숙련가에게 공지된 임의의 방법에 의해 정제될 수 있음을 인식할 것이다: 원심분리, 미소여과, 항체 정제, 심층 여과, 한외여과, 투석여과, 침전, 비이드 크로마토그래피(예를 들면 크기 배제 크로마토그래피, 이온 교환 크로마토그래피 또는 친화도 크로마토그래피), 막 흡착제(예를 들면 이온 교환 크로마토그래피 또는 친화도 크로마토그래피). 하나의 예에서, 본원에 기재된 바와 같은 방법에 의해 생산된 재접힘된 올리고머성 HCV E2는 단백질 태그, 예컨대 재접힘된 올리고머성 HCV E2의 정제를 돕는 HIS-태그를 포함한다.
하나의 실시태양에서, 본 명세서는 재접힘된 재조합 올리고머성 간염 C 바이러스(HCV) 엔벨롭 당단백질 2(E2)를 재조합 HCV E2로부터 제조하는 방법을 가능하게 하고, 여기서 재조합 E2는 단량체성 E2이거나, 또는 단량체성 및 올리고머성 재조합 E2의 혼합물을 포함하고, 상기 방법은 하기 단계를 포함한다: (i) 하나 이상의 단량체 디설파이드 결합을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 재조합 E2 접촉시키는 단계; 및 (ii) 환원제를 제거하거나 단량체를 산화제와 접촉시켜서 재조합 E2의 올리고머성 HCV E2로의 재접힘을 이끌어내는 단계; 여기서 재접힘된 재조합 올리고머성 E2는 재접힘(ii) 이전의 재조합 E2에 비해 적어도 20% 이상의 올리고머를 포함하고, 재접힘된 올리고머성 HCV E2는 재조합 단량체성 E2와 비교하여 감소된 비-중화 항체에 대한 결합을 나타낸다.
몇몇 실시태양에서, 재접힘된 올리고머성 E2는 E2의 이합체, 3합체, 및/또는 더 높은 급수의 형태를 포함한다.
또 다른 양태에서, 본 명세서는 재접힘된 재조합 올리고머성 바이러스성 엔벨롭 당단백질을 고유의 엔벨롭 당단백질로부터 제조하는 더욱 일반적인 방법을 제공하고, 상기 공정은 하기 단계를 포함한다: (i) 하나 이상의 단량체 디설파이드 결합 또는 고유의 시스테인을 부분적으로 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 고유의 E2를 접촉시키는 단계; 및 (ii) 환원제를 제거하거나 환원된 고유의 HCV E2를 산화제와 접촉시켜서 환원된 단량체성 엔벨롭 당단백질의 올리고머성 엔벨롭 당단백질로의 재접힘을 이끌어내는 단계; 여기서 단계 (ii)에서 단량체의 적어도 20%는 접힘된 올리고머로 전환되고, 올리고머는 단량체성 당단백질과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타낸다.
당분야의 숙련가라면 기재된 바와 같이 일단 생산된 높은 급수의 항원은 당분야의 숙련가에게 공지된 임의의 방법에 의해 정제될 수 있음을 인식할 것이다. 예를 들면 "정제하는" 또는 "정제"는 하기 단계들중 하나 이상을 포함할 수 있다: 원심분리(예를 들어 초원심분리), 크로마토그래피(예를 들어 크기 배제 크로마토그래피, 이온 교환 크로마토그래피 또는 친화도 크로마토그래피), 및/또는 막 흡착제(예를 들면 이온 교환 크로마토그래피 또는 친화도 크로마토그래피), 여과(예를 들어 막 여과, 한외여과 또는 투석여과).
당분야의 숙련가라면 정제된 높은 급수의 항원이 약학 제형, 예컨대 백신 조성물로 제형화될 수 있음을 인식할 것이다. 이러한 조성물은: 보조제, 부형제, 결합제, 보존제, 담체 커플링제, 완충제, 안정화제, 유화제, 습윤제, 비-바이러스성 벡터 및 형질감염 촉진 화합물중 하나 이상을 포함할 수 있다. 숙련가라면 이러한 백신 조성물이 동결건조될 수 있음을 추가로 이해할 것이다. 하나의 실시태양에서, 생산된 백신 조성물은 인간 용도에 적합하다. 또 다른 실시태양에서, 백신은 수의학적 용도에 적합하다.
본원에 기재된 바와 같은 높은 급수의 항원 또는 조성물은 항원 특이적 면역 세포에 결합하기 위해/이를 검출하기 위해 사용될 수 있다. 한가지 실시태양에서, 면역 세포는 B-세포이다. 한가지 실시태양에서, 면역 세포는 T-세포이다. 항원성 면역 세포의 검출은, 피험체가 항원과 연관된 병원체에 의해 감염되고/되거나, 피험체가 항원에 의해 백신접종됨을 지시한다. 한가지 실시태양에서, 본원에 기재된 바와 같은 높은 급수의 항원, 또는 본원에 기재된 바와 같은 조성물은 엔벨롭을 갖는 바이러스에 특이적인 B-세포의 검출 및/또는 단리를 위해 사용될 수 있다. 한가지 실시태양에서, 엔벨롭을 갖는 바이러스는 HCV이다. 한가지 실시태양에서, 엔벨롭을 갖는 바이러스는 HIV이다.
당분야의 숙련가라면 본 발명의 높은 급수의 항원이 면역 세포의 진단용 검정 및/또는 단리에서 사용하기에 적합함을 인식할 것이다. 한가지 실시태양에서, 높은 급수의 항원은 유세포분석에서 면역 세포를 위한 착색제 또는 표지로서 사용될 수 있다. 한가지 실시태양에서, 높은 급수의 항원은 표지, 염료 또는 형광단과 착화되어 유세포분석을 사용하여 검출을 허용한다. 한가지 실시태양에서, 표지는 d-비오틴이다.
면역 세포/들의 단리는, 예를 들면 알트만(Altman) 등의 문헌(1996), 볼러스(Vollers) 등의 문헌(2008) 및 돌턴(Dolton) 등의 문헌(2015)에 기재된 바와 같은 "4합체 검정" 또는 "4합체 착색"을 포함하고, 여기서 본원에 기재된 바와 같은 높은 급수의 항원은 검정에서 주요 조직적합성 복합체(MHC: major histocompatibility complex) 4합체의 역할을 대체한다.
한가지 실시태양에서, 본 발명은 i) HCV에 특이적인 면역 세포/B-세포를 본원에 기재된 바와 같은 높은 급수의 항원에 의해 표지화하는 단계; 및 ii) 표지화된 면역 세포/B-세포를 세포분석에 의해 검출하고/하거나 단리하는 단계를 포함하는, HCV에 대해 특이적인 면역 세포/B-세포를 검출하고/하거나 단리하는 방법을 제공한다.
변형체에 대한 언급은 부분, 유도체 및 화학적 유사체를 포함한다. 고려되는 화학적 유사체는 측쇄의 변형, 합성 동안의 비천연 아미노산 및/또는 이들의 유도체의 혼입 및 연결기 또는 다교결합기, 또는 무엇보다도 입체배좌의 구속을 부과하는 기타 방법의 사용을 포함한다.
본 개시내용은 조립된 높은 급수의 항원의 특징 및 조립 방법에 대한 하기의 비제한적 실시예에 의해 완성된다. 상기 주지된 바와 같이, 비록 개시내용이 간염 및 HIV 엔벨롭 항원을 사용하여 설명되지만, 본 발명은 항원의 항원성 및 면역원성을 향상시키기 위해 낮은 급수의 항원으로부터 높은 급수의 올리고머를 형성하는 본 발명의 방법을 사용하여 생산된 항원으로 확장된다. 본원에 기재된 바와 같이, 높은 급수의 항원은 당분야에 공지된 여러 가지 기법을 사용하여 추가로 특성규명될 수 있다. 그러나 데이터에 대한 분석은 환원에 의해 생산된 조립된 올리고머를 보여주고, 본원에 기재된 조립 방법은 세포내에서 생산된 대조 항원에 의해 나타나지 않는 항체 결합 프로파일을 나타내며, 따라서 본 발명의 조성물은 이를 바탕으로 선행 기술의 항원과 구별될 수 있다.
실시예
실시예 1: 발현 벡터 및 플라스미드
클로닝. C-말단 6x 히스티딘(HIS) 태그와 융합된 HCV E2 Δ123의 DNA 서열 (서열 번호 24)을 pcDNA3.1 포유동물 발현 벡터[인비트로겐(Invitrogen)]내로 클로닝하였다. Con1 유전자형 1b(G1b) Δ123(서열 번호 20)을 CSL에 의해 클로닝하였다. H77c 유전자형 1a(G1a) Δ123(서열 번호 19)을 또한 CSL을 사용하지만 특허등록된 CSL-변형된 pcDNA3.1 벡터내로 클로닝하였다. H77c Δ123 및 Con1 Δ123을 포함한 생성된 플라스미드를 각각 pcDNA-H77cΔ123-HIS 및 pcDNA-Con1Δ123-HIS로서 명명하였다. S52 G3a Δ123(서열 번호 21)의 DNA 서열을 pcDNA3.1의 NheI 및 XbaI 자리로 T4 DNA 리가아제(ligase)에 의해 결찰시켜 pcDNA-S52Δ123-HIS를 생산하였다.
DNA 발현 및 정제. pcDNA-Con1Δ123-HIS 및 pcDNA-S52Δ123-HIS를 DH5α 에스케리치아 콜라이(Escherichia coli)내로 열 충격 방법에 의해 형질전환시키고[프로거(Froger) 및 홀(Hall)의 문헌(2007)], 이어서 루리아-베르타니(LB: Luria-Bertani) 암피실린(ampicillin)(100 ㎍/㎖) 한천 플레이트 상에서 성장시켰다. 단일 콜로니를 암피실린-함유(100 ㎍/㎖) LB 브로스(broth)에서 배양하고, DNA 플라스미드를 퀴아겐(QIAGEN) 플라스미드 맥시 프렙 키트(maxi prep kit)를 사용하여 제조업체의 권고에 따라 추출하였다. 클로닝된 플라스미드의 성공적인 결찰 및 규모 확장을 확인하기 위해, 맥시 프렙으로부터의 pcDNA-Con1Δ123-HIS 및 pcDNA-S52Δ123-HIS를 각각 NheI 및 XhoI; 및 NheI 및 XbaI 제한 효소에 의해 소화시켰다. 이어서 생성된 소화물을 아가로스 겔 전기영동하고, 겔 독(Gel Doc) XR+ 시스템[바이오-래드 래보레이토리즈(Bio-Rad Laboratories)] 및 퀀터티 원(Quantity One) 1-D 분석 소프트웨어(바이오-래드 래보레이토리즈)를 사용하여 겔을 관찰함으로써 예상된 제한 패턴을 확인하였다. 또한 빅다이 터미네이터(BigDye Terminator) v3.1 케미스트리(chemistry)를 사용하여 플라스미드를 서열화하고[마이크로몬(Micromon)에 의함], CLC 서열 뷰어(Viewer)(퀴아겐) 및 핀치티비(FinchTV)[게오스피자(Geospiza)]를 사용하여 서열을 분석하였다.
실시예 2: 단백질 발현 및 정제
일시적 형질감염. 인간 배아 신장 293 세포주로부터 유래된 프리스타일(FreeStyle) 293-F(FS293F) 세포(인비트로겐)를 1 ℓ 들이 엘렌메이어(Erlenmeyer) 세포 배양 플라스크[코닝(Corning)]에서 150 ㎖중 1 x 106 생존가능 세포/㎖로 접종하고, 프리스타일 293(FS293) 발현 배지(인비트로겐)에서 유지시켰다. 세포를 37℃에서 8% C02를 갖는 습윤 대기하에, 4 x 상대 원심력(RCF: relative centrifugal force)으로 회전하는 회전 진탕기 상에서 스테리-사이클(Steri-Cycle) C02 항온처리기[써모 일렉트론 코포레이션(Thermo Electron Corporation)]에 의해 항온처리하였다. 293펙틴(fectin) 형질감염 시약(인비트로겐)을 제조업체의 지시에 따라 사용하고, 표 1에 제시된 부피를 이용하여 pcDNA-Con1Δ123-HIS 및 pcDNA-S52Δ123-HIS를 FS293F 세포내로 일시적으로 형질감염시켰다. 형질감염 후 1 일째, 100 ㎖의 FS293 발현 배지를 첨가하여, 총 형질감염 부피를 250 ㎖로 상승시켰다. 또한 세포 배양액에 0.5% 루핀 펩톤(lupin peptone)[솔라비아 바이오테크놀로지(Solabia Biotechnology)] 및 0.02% 플루오닉(Pluronic) F-68[깁코(Gibco)]를 보충하였다. 형질감염 후 3, 5 및 7 일째 배지 절반을 교체하였고, 이로써 세포 배양 상청액의 절반(125 ㎖)을 300 x RCF에서 5 분 동안 원심분리에 의해 수거하였다. 펠릿화된 세포를 0.5% 루핀 펩톤 및 0.02% 플루오닉 F-68이 또한 보충된 125 ㎖의 신선한 FS293 발현 배지에 재현탁시키고, 이어서 세포 배양 플라스크로 되돌려보냈다. 최종적으로, 세포 배양 상청액(250 ㎖)의 전체 수거를 형질감염 후 9 일째 수행하였다. 모든 수거된 상청액을 15,344 x RCF에서 30 분 동안 추가로 원심분리하고, 잔여 세포 및 세포 찌꺼기를 제거하기 위해 펠릿을 폐기하였다. 각각의 수거일에 트립판 블루(trypan blue) 염료 배제 방법 및 혈구계(hemocytometer)를 사용하여 세포 계수를 수행함으로써 세포 밀도 및 생존도를 결정하였다.
친화도 크로마토그래피. 분비된 HIS-태그 부착 Δ123 당단백질을 함유하는 세포 배양 상청액을 로커(rocker) 상에서 10 ㎖(즉 1 칼럼 부피)의 코발트-충전된 탈론(TALON) 금속 친화도 수지[클론테크(ClonTech)]에 2 시간 동안 실온(RT)에서 적용하여, HIS-태그 부착 단백질의 결합을 허용하였다. 비이드를 20 칼럼 부피의 세척 완충액(50mM 인산 나트륨, 300mM 염화 나트륨, pH 7.0)에 의해 5 ㎖/분의 유속으로 2회 세척하고,5 칼럼 부피의 용출 완충액(50mM 인산 나트륨, 300mM 염화 나트륨, 200mM 이미다졸, pH 7.0)에 의해 1 ㎖/분의 유속으로 용출시켰다. 30 kDa의 분자량 컷-오프(MWCO: molecular weight cut-off)를 갖는 아미콘 초원심분리 필터 유닛(Amicon Ultra centrifugal filter unit)[메르크 밀리포어(Merck Millipore))]을 사용하여 용출액에 함유된 단백질을 농축시키고, 염산(HC1)을 사용하여 pH 6.8으로 조정된 인산염-완충된 염수(PBS, 깁코)(PBS pH 6.8)에서 세척하고, 사용할 때까지 4℃에서 저장하였다.
실시예 3: 겔 여과 크로마토그래피
수퍼덱스 200 제조용 등급(prep grade) 16/600 칼럼[지이 헬쓰케어(GE Healthcare)] 상에서 AKTA 고속 단백질 액체 크로마토그래피(FPLC: fast protein liquid chromatography) 시스템(지이 헬쓰케어)을 사용하여 단백질을 겔 여과 크로마토그래피하였다. 이동 완충액으로서 여과되고 탈기된 PBS(pH 6.8)를 사용하여 0.5 ㎖/분의 유속으로 운행시켰다. 원하는 올리고머성 종을 함유하는 분별물을 모은 다음, 10 kDa의 MWCO를 갖는 아미콘 초원심분리 필터 유닛을 사용하여 농축시키고, 사용할 때까지 PBS pH 6.8에서 저장하였다. 1 cm의 통로 길이를 갖는 크벳(cuvette)을 사용하여 280 nm의 광학 밀도(OD280)에서 분광측광에 의해 단백질 농도를 결정하고, 하기 식을 사용하여 mg/㎖로 계산하였다:
흡광 계수 × OD280에서의 흡광도
Δ123의 아미노산 서열을 사용하여 흡광 계수를 결정하였고, 하기 식을 사용하여 이를 계산하였다:
분자량(Da)/((5690 × #트립토판) + (1280 × #티로신))
실시예 4: 폴리아크릴아미드 겔 전기영동(PAGE)
적절하다면 환원 또는 비-환원 조건하에 황산 도데실 나트륨(SDS)-PAGE를 통해 단백질을 분석하였다. 환원 SDS-PAGE의 경우, 단백질 샘플을 2%(부피/부피) β-머캅토에탄올이 함유된 1x 샘플 완충액중에서 100℃에서 5 분 동안 가열하고, 이어서 광범위한 SDS-PAGE 표준물(바이오-래드 래보레이토리즈)과 함께 12% 아크릴아미드 겔 상으로 적재하였다. 수직 전기영동 장치(CLP)를 사용하여 1x 이동 완충액(25mM 트리스 염기, 192mM 글리신, 0.1% SDS, pH 8.3)에서 180V하에 ∼1.5 시간 동안 전기영동을 수행하였다. 4-12% 폴리아크릴아미드 구배 겔을 사용하고 샘플 완충액중 β-머캅토에탄올이 없는 것을 제외하고 동일한 방식으로 비-환원 SDS-PAGE를 수행하였다. 겔을 0.25%(부피/부피) 쿠마시 브릴리언트 블루(Coomassie brilliant blue) R-250(바이오-래드 래보레이토리즈), 10%(부피/부피) 아세트산 및 50%(부피/부피) 메탄올로 염색한 다음, 10%(부피/부피) 아세트산 및 50%(부피/부피) 메탄올로 탈색시키고, 오디세이 적외선 영상촬영 시스템(LI-COR)을 사용하여 스캐닝하였다. 밴드 강도를 또한 오디세이 시스템에 의해 정량화하였다.
실시예 5: 항체
단일클론성 항체(MAb) AR3A, AR3B, AR3C 및 AR3D는 더 스크립스 리서치 인스티튜트(The Scripps Research Institute)의 만순 로우 박사(Dr Mansun Law)의 기증물이었다[로우(Law) 등의 문헌(2008)]. CBH-4B, CBH-4D, HC-11 및 HCV-1은 스탠포드 대학의 스티븐 펑 박사(Dr. Steven Foung)의 기증물이었다. H52 및 H53은 진 두뷔슨 박사(Dr. Jean Dubuisson) 및 해리 그린버그 박사(Dr. Harry Greenberg)의 기증물이었다. MAb 1, 7, 10, 12, 16, 20, 24 및 60을 CSL 리미티드(CSL Ltd)와 협력하여 본 발명자들의 실험실에서 생산하였다(표 2). 적합한 항체 패널은 문헌, 예를 들면, 케크(Keck) 등의 문헌[PLos Pathogens: 8(4) e1002653, April 2012]에 기재되어 있다. 또한, 항체 2A12는 고유의 대조적인 올리고머성 HCV E2와 비교하여 감소된 조립된 E2에 대한 결합을 나타내었다. 항체 패널 및 이들의 생성 방법은 본원에 참고로 인용된 비에티어(Vietheer P.) 등의 문헌[Hepatology: 65(4), 1117-1131, 2017], 및 이에 언급된 참고문헌, 예컨대 참고문헌 5, 33-36, 17 및 37, 및 출판사로부터 입수가능한 보충 자료에 기재되어 있다.
실시예 6: 효소 연결된 면역흡착 검정(ELISA)
샌드위치 ELISA. 성공적인 단백질 발현 및 정제를 확인하기 위해, 맥시소브(Maxisorb) 평저 96 웰 플레이트(눈크)를 사용하여 샌드위치 ELISA를 수행하였다. 웰을 50mM 탄산염-중탄산염 완충액 pH 9.6중 5 ㎍/㎖의 이합체성 말토스 결합 단백질(MBP)-CD81-LEL113-201로 코팅시키고, 이어서 하룻밤 4℃에서 항온처리하였다. 여기서 및 후속 세척 단계에서 플레이트를 0.05% 트윈 20(PBST)이 함유된 PBS에서 4회 세척하고, 이어서 PBS중 10 mg/㎖의 소 혈청 알부민[BSA, 시그마-알드리치(Sigma-Aldrich)](BSA10PBS)에 의해 1 시간 동안 RT에서 차단하였다. 세척 후, 조직 배양 상청액의 반-로그식 연속 희석을 5 mg/㎖ BSA가 함유된 PBST(BSA5PBST)에서 수행하고, 이어서 1 시간 동안 RT에서 항온처리하였다. 세척 후, BSA5PBST중 토끼 항-6x HIS 에피토프 태그 항체[락랜드 이뮤노케미칼스(Rockland Immunochemicals)]의 단일 희석물을 첨가하고, 1 시간 동안 RT에서 항온처리하였다. 세척한 다음, 호스 래디쉬 퍼옥시다아제(HRP: horse radish peroxidase)에 접합된 염소 항-토끼 면역글로불린[다코(Dako)]의 단일 희석물을 첨가하였다. 세척후 3,3',5,5'-테트라메틸벤지딘(TMB) 기질(시그마-알드리치)을 사용하여 제조업체의 지시에 따라 ELISA를 진행하고, 반응을 1M HCl에 의해 중단시켰다. 최종적으로, 멀티스칸 어센트(Multiskan Ascent) 마이크로플레이트 판독기(써모 일렉트론 코포레이션)를 사용하여 광학 밀도를 450 nm에서 측정하였다.
단일 희석점 평가 및 직접적 ELISA. 당분야에 공지된 항체의 패널, 예컨대 실시예 5/표 2에 열거된 항체의 패널에 대한 Δ123의 반응성을 평가하기 위해, 단일 희석점 평가를 초기에 수행하였고, 여기서 맥시소브 평저 96 웰 플레이트를 5 ㎍/㎖의 단량체성 Δ123으로 직접 코팅한 다음 하룻밤 4℃에서 항온처리하였다. 세척하고 BSA10PBS에 의해 차단한 후, 1차 항체의 단일 희석물을 첨가하고, 이후 2차 항체(다코)를 첨가하였고, 이때 각각의 첨가 사이에 세척하고 1 시간 동안 RT에서 항온처리하였다. 플레이트를 전개시키고 샌드위치 ELISA에 대해 상기 기재된 바와 같이 측정하였다. 직접적 ELISA를 동일한 방법으로 수행하였으나, 단 웰을 2 ㎍/㎖의 단량체성 Δ123 또는 5㎍/㎖의 HMW1 Δ123중 적절한 것으로 코팅하였고, 1차 항체를 BSA5PBST에서 반-로그식 연속 희석하였다.
실시예 7: 단백질 환원 및 재접힘
소규모 환원. 단백질 환원에서 사용하기 위한 최적의 DTT[피어스(Pierce)] 농도를 결정하기 위해, 맥시소브 평저 96 웰 플레이트를 50mM 탄산염-중탄산염 완충액 pH 9.6중 5 ㎍/㎖의 단량체성 H77c Δ123으로 코팅하고, 실온에서 최소한 2 시간 동안 항온처리하였다. PBST에 의해 4회 세척한 후, 탄산염-중탄산염 완충액에서 제조된 상이한 농도의 DTT(0 내지 1OmM)를 첨가하고 37℃에서 30 분 동안 항온처리하였다. 이어서 플레이트를 즉시 6 회 PBST에서 세척하여 DTT를 제거하고, 이후 BSA10PBS에 의해 차단하였다. 실시예 6에 기재된 바와 같이, 항-HIS, H53 및 R04를 포함하는 1차 항체를 첨가하고, 이어서 적절한 2차 항체를 첨가하였다. 최종적으로, 결합된 항체를 검출하고 실시예 6에 기재된 바와 같이 측정하였다. 환원제로서 TCEP를 사용하여 동일한 실험을 반복하였다.
용액중 DTT에 의한 환원 및 재접힘. 탄산염-중탄산염 완충액에서 제조된 H77c Δ123 단량체를 상이한 조건(표 3)하에 DTT 환원시켜서 최적의 HMW Δ123 형성에 필요한 요건을 결정하였다. 단백질 재접힘을 느린 희석에 의해 수행하였고, 이는 초기 샘플 부피의 절반인 일정한 부피로 PBS pH 6.8을 3회 단계식으로 첨가함으로써 달성되었다. 예를 들면, 4x 200 ㎕의 PBS pH 6.8를 400 ㎕의 샘플에 첨가한다. 각각의 첨가 후 15 분 동안 RT에서 항온처리하였다. 10 kDa의 MWCO를 갖는 아미콘 초원심분리 필터 유닛을 사용하여 DTT를 제거하고, 이어서 PBS pH 6.8에서 2회 세정하였다. 이어서, 재접힘된 단백질을 실시예 4에 기재된 바와 같이 비-환원 SDS-PAGE에 의해 분석하였다. 다합체가 성공적으로 형성된 경우 더 큰 규모의 재접힘을 수행하였고, 실시예 3에 기재된 바와 같이 겔 여과 크로마토그래피에 의해 분석하였다.
산화환원-셔플링 시스템을 사용하는 환원 및 재접힘. 1:5 비의 산화된 L-글루타티온(GSSG, 피어스) 대 환원된 L-글루타티온(GSH, 피어스)을 조사된 상이한 조건으로(표 4) 탄산염-중탄산염 완충액에서 제조된 H77c Δ123 단량체에 첨가하였다. L-글루타티온을 제거함으로써 반응을 중단시키고, 이어서 재접힘된 단백질을 실시예 7에 기재된 바와 같이 분석하였다. 다합체가 성공적으로 형성된 경우 더 큰 규모의 재접힘을 수행하였고, 실시예 3에 기재된 바와 같이 겔 여과 크로마토그래피에 의해 분석하였다.
트리스(2-카복시에틸)포스핀 하이드로클로라이드(TCEP)-환원된 단백질과 비스말레이미도에탄(BMOE)의 가교결합. 상이한 농도의 TCEP를 상이한 농도(표 5)로 PBS pH 6.8에서 제조된 H77c Δ123 단량체에 첨가하고, 30 분 동안 37℃에서 항온처리하였다. 이어서, 사용 직전에 디메틸 설폭사이드(시그마)에 용해된 BMOE(시그마)를 0.2mM의 최종 농도로 첨가하고, 제조업체에 의해 지시된 바와 같이 1 시간 동안 RT에서 항온처리하였다. BMOE 및 TCEP를 제거함으로써 반응을 중단시키고, 이어서 재접힘된 단백질을 실시예 7에 기재된 바와 같이 분석하였다.
실시예 8: Δ123의 생산
균주 H77c, Con1 및 S52로부터의 Δ123의 DNA 서열을 pcDNA3.1 기반의 벡터내로 혼입하여, 하기 플라스미드를 생성하였다: pcDNA-H77cΔ123-HIS, pcDNA-Con1Δ123-HIS 및 pcDNA-S52Δ123-HIS. 이들 플라스미드는 아가로스 겔 전기영동 상에서 예상된 제한 패턴을 나타내었고, H77c, Con1 및 S52 Δ123 삽입물을 나타내는 밴드 모두는 각각 762, 759 및 710 bp의 그들의 예상된 길이에 상응하였다(도 1). H77c Δ123을 특허등록된 CSL-변형된 pcDNA3.1 벡터내로 클로닝하였고, 이는 Con1 및 S52가 함유된 표준 인비트로겐 벡터와 비교하여 관찰된 더 큰 벡터 크기(이는 각각 동일한 수평 위치로 이동됨)를 설명한다(도 1).
Con1 및 S52 Δ123 플라스미드를 포유동물 FS293F 세포내로 일시적으로 형질감염시켜서 Δ123 당단백질을 생산하였다. 동유체력에 대해 세포를 보호하기 위한 플루오닉 F-68, 및 성장 효율을 증진시키기 위한 루핀 펩톤이 보충된 FS293 발현 배지에서 세포를 현탁액으로 유지시켰다. 1 x 106 생존가능 세포/㎖로 세포를 접종하였고, 세포 생존도는 형질감염 후 0 내지 9일 동안 대략 90%로 유지되었다. Δ123 당단백질의 성공적인 발현을 확인하기 위해, 샌드위치 ELISA를 통해 세포 배양 상청액을 분석하였다(도 2A-B). 형질감염 후 3, 5, 7 및 9일째 수거된 상청액의 반-로그식 연속 희석물을 이합체성 MBP-CD81-LEL113-201-코팅된 효소 면역검정 플레이트에 적용하고, 포획된 Δ123 당단백질을 토끼 항-his 항체 및 HRP-접합된 염소 항-토끼 면역글로불린에 의해 검출하였다. 제 1 웰에서 1 ㎍/㎖ 농도의 단량체성 H77c Δ123은 양성 대조군으로서 작용하는 반면, 제1 웰에서 1/2로 희석된 HIV 엔벨롭 단백질 gp140-함유 상청액은 음성 대조군으로서 작용한다. 모든 세포 배양 상청액은 CD81에 대해 양성 결합을 나타내었는데, 이는 상청액중 Δ123 당단백질의 존재, 및 이에 따라 성공적인 단백질 발현을 지시한다. 더욱이, 세포 배양 상청액을 나타내는 곡선이 양성 대조군 위에 나타났고, 이는 Δ123 당단백질의 1 ㎍/㎖ 초과의 수율(Con1 및 S52의 경우 각각 대략 4 ㎍/㎖ 및 20 ㎍/㎖)을 지시한다. 광학 밀도(OD) 값은 5 및 7 일째 수행된 수거의 경우 가장 높았고, 이는 단백질 발현이 이들 중간 시점에서 어느 정도 더욱 효율적이었음을 시사한다. 또한 S52 Δ123 형질감염으로부터의 상청액이 Con1 Δ123에 비해 더 큰 수준의 CD81 결합을 나타내었음을 주지해야 하는데, 이는 S52 균주에 대한 단백질 발현이 더욱 효율적이거나, S52 Δ123이 CD81에 대해 더욱 강력한 친화도를 가짐을 시사한다.
세포 배양 상청액으로부터 Δ123 당단백질을 단리하기 위해, 친화도 크로마토그래피를 탈론 비이드를 사용하여 수행하였고, 세파로스 매트릭스는 코발트로 충전되었고, 이는 종래의 니켈-친화도 지지체에 비해 더욱 높은 특이성으로 his-태그 부착 단백질에 효율적으로 결합한다. 또한 단백질 정제의 유효성을 세포 배양 상청액에 대해 수행된 것과 동일한 방식으로 샌드위치 ELISA에 의해 분석하였다(도 2C-D). 용출액을 3개의 별도의 분별물로 순차적으로 수집하였고, 이때 제1 분별물에서 용출된 단백질은 정제 이전의 상청액과 유사한 수준의 CD81 결합을 보여주는데, 이는 분별물 1이 대부분의 Δ123 당단백질을 포함하였음을 지시한다. 분별물 2에서 용출된 단백질은 감소된 CD81 결합을 나타내었는데, 이는 더 적은 양으로 Δ123이 존재하였음을 지시하고, 한편 분별물 3은 무시할만한 CD81 결합을 나타내었고 HIV gp140 음성 대조군과 정렬된 곡선을 생산하였다. S52 Δ123 정제로부터의 세척액 및 정제 이후의 상청액(즉, 통과액)은 양성 대조군과 거의 유사한 CD81 결합을 나타낸 반면, Con1 Δ123의 경우에 낮은 수준이 관찰되었는데, 이는 Con1 Δ123 정제가 더욱 효과적으로 수행되었음을 지시한다.
실시예 9: Δ123 발현 프로파일
겔 여과 크로마토그래피에 의한 친화도-정제된 H77c Δ123 발현의 균주로부터의 Δ123의 DNA 서열에 대한 이전 분석은, H77c Δ123이 크기가 상이한 다양한 종들로서 나타남을 보여주었다(도 3A). 이들은 단량체, 이합체, HMW2 및 HMW1 Δ123으로 구성되고, 이는 각각 46, 97, 239 및 2402 kDa의 분자량을 갖고 이들의 항원성 뿐만 아니라 면역원성이 다양하다[드루머(H. Drummer), 미공개 데이터]. HMW 종은 bNAb를 포함할 수 있는 반면, 단량체는 유형-특이적 NAb를 유도한다(드루머, 미공개 데이터). 그러나, H77c Δ123의 FS293F 세포로의 안정한 형질감염[이러한 프로젝트의 착수 이전에 닥터 로브 제이 센터(Dr. Rob J Center)에 의해 수행됨]으로부터 생성된 Δ123의 단지 23.13%만이 HMW 형태였고, 이는 단량체의 경우의 64.90%와 비교된다(도 3A 및 표 6).
임의의 유전자형으로부터의 Δ123은, 이것이 E2의 가장 보존적인 영역을 포함하므로 동등하게 bNAb를 이끌어낼 수 있어야 한다. 그러나, WT E2 및 Δ123의 발현 프로파일은 상이한 유전자형 사이에서 다양할 것이다. Con1 Δ123 및/또는 S52 Δ123이 HMW 종을 더 높은 수율로 발현하는지의 여부를 결정하기 위해, FS293F 세포로의 일시적 형질감염로부터의 친화도-정제된 Δ123 당단백질을, AKTA FPLC 시스템을 사용하는 수퍼덱스 200 제조용 등급 16/600 칼럼 상에서 겔 여과 크로마토그래피에 의해 분석하였다. 유니콘 소프트웨어를 사용하여 피크하의 면적을 정량화한 결과, Con1(도 3B) 및 S52(도 3C) 균주 둘 다가 실제로 더 높은 수율로 HMW Δ123을 생성함이 밝혀졌는데, HMW 형태(HMW1 및 HMW2 조합)의 생성된 모든 Δ123 당단백질의 46.02%(Conl) 및 44.88%(S52)이었고, 이는 H77c의 경우에 23.13%인 것과 비교된다(표 6).
이들 결과의 재현성을 평가하기 위해, Δ123의 일시적 형질감염 및 정제를 반복하였고, Δ123 발현을 분석하기 위해 다시 겔 여과 크로마토그래피를 수행하였다. y-축 상의 척도로 알 수 있듯이, 제2 라운드에서 Con1 Δ123의 발현 수준은 라운드 1의 수준의 대략 2배였던 반면, S52 Δ123의 수준은 상대적으로 안정하게 유지되었다(도 3D-E). 발현 수준에서의 편차에도 불구하고, Con1 및 S52 Δ123의 발현 프로파일이 라운드 1 및 라운드 2 둘 다의 경우 유사하게 유지되었음을 주지하는 것은 중요하였다. H77c와 비교하여, HMW Δ123의 수율에서의 개선이 다시 관찰되었고, HMW 종의 유사한 비율이 라운드 2에서 생성되었다 - Con1 및 S52의 경우 각각 46.98% 및 41.56%(표 6).
겔 여과 분별화된 Δ123 당단백질을 SDS-PAGE에 의해 분석하였다. 5-12% 폴리아크릴아미드 구배 겔 상의 비-환원 SDS-PAGE는 겔 여과 크로마토그래피로부터의 결과를 확증하였고, 다시, Δ123이 다양한 디설파이드 연결된 올리고머성 형태로 존재함을 입증하였고 밴드 강도 및 분자량 사이에 반비례 관계를 나타내었다(도 4A-B). 또한, 디설파이드 결합을 방해하는(이는 차례로 또한 소수성 결합을 방해하는) 환원제인 β-머캅토에탄올을 사용하여 12% 아크릴아미드 겔 상에서 환원성 SDS-PAGE 분석을 수행하였다. 결과로부터, 디설파이드 결합이 높은 급수의 구조물의 형성 및 안정화를 위해 필수불가결함을 알 수 있는데, 이는 일단 이들 디설파이드가 β-머캅토에탄올에 의해 분할되면 이들 모두 단량체로서 이동되기 때문이다(도 4C-D). 소수성 상호작용은 높은 급수 종의 형성에 기여할 수 있다.
이들 데이터는 Con1 및 S52 균주가 H77c와 비교하여 실제로 더 높은 수율로 HMW Δ123을 발현함을 보여주고, HMW Δ123 생산을 개선시키는 하나의 가능한 방법을 입증한다.
실시예 10: HMW Δ123의 항원적 특징
Con1 및 S52 균주에 의해 생성된 HMW Δ123이 H77c의 HMW Δ123의 항원 패턴을 재현하는지의 여부를 평가하기 위해, 모든 3가지 균주로부터의 겔 여과 분별화된 단량체성 및 HMW1 Δ123을 입체배좌 감수성 및 비감수성 MAb의 패널에 대하여 ELISA 형식으로 검사하였다(도 5A). 단일 희석점 평가(도 5B)를 초기에 신속한 검사로서 수행하여, 어떠한 MAb가 H77c, 및 Con1 및 S52중 하나 또는 둘 다의 Δ123에 대해 양성 반응성을 나타내는지를 결정하였고, 이로써 균주 사이의 항원성을 비교할 수 있었다. 이러한 기준을 충족시키는 MAb를 직접적 ELISA에서 추가로 검사하였고, 여기서 MAb의 반-로그식 연속 희석물을 단량체성 및 HMW1 Δ123에 의해 별도로 코팅된 효소 면역검정 플레이트에 적용하고, 이후 적절한 HRP-접합된 2차 항체에 의해 검출하였다. 이어서 각각의 결합 곡선의 중심점을 비교함으로써 HMW1 Δ123에 대한 각각의 MAb의 상대적 결합을 계산하였고(표 7), 이러한 중심점은 가장 높은 광학 밀도 값의 절반으로서 취해졌다.
단일 희석점 평가는 입체배좌 감수성 마우스 H53 및 MAb 1, 7, 12, 16 및 20 모두가 H77c Δ123에 대해 양성 반응성을 나타내었음을 보여준 반면, Con1 및 S52 Δ123에 대한 무시할만한 반응성이 입증되었는데, 이는 MAb의 표적 에피토프를 인식불가능하게 만들고 MAb가 유형-특이적이 되는 방식으로 이들 2가지 균주의 에피토프 완전성이 변경되었음을 지시한다. 입체배좌 감수성 인간 AR3A에 의해서는 상반되었고, 여기서 Con1 및 S52 Δ123에 대한 양성 반응이 있었지만 H77c에 대해서는 양성 반응이 없었다(도 5B).
직접적 ELISA에서 추가로 검사된 11가지의 MAb중에서, 8가지(AR3B, AR3C, AR3D, CBH-4B, CBH-4D, HC-11, MAb 10 및 60)가 단량체성 형태와 비교될 경우 Δ123의 HMW 형태에 대한 감소된 결합을 나타내었는데(표 7), 이는 이들의 표적 에피토프가 더욱 폐쇄되었음을 지시한다. 단지 마우스 H52만이 모든 3가지 균주에 대한 HMW1 Δ123에 결합하는 능력이 현저히 향상되었는데(표 7), 이는 잔기 C652로의 개선된 접근성을 시사한다.
Con1 및 S52의 HMW 형태를 H77c의 HMW 형태에 결합시키는 각각의 MAb의 능력을 비교할 경우, 검사된 11가지의 MAb중에서 5가지(HCV1, AR3B, AR3C, CBH-4D, HC-11)가 증가되거나 동등한 수준의 결합을 나타내었다. 증가된 결합은 또한 AR3D 및 H52에 의해 입증되었지만 단지 Con1 Δ123에 대한 것이었고, 반면 MAb 10 및 24는 균주 둘 다에 대해 감소된 결합 수준을 나타내었다(표 7).
실시예 11: TCEP 및 BMOE를 사용하는 Δ123의 환원 및 재접힘
단량체성 Δ123을 부분적으로 환원시킴으로써 Δ123 단량체를 단량체로 다시 접어 유리 설프하이드릴을 생성하고, 이후 이들 설프하이드릴을 분자간 디설파이드 결합으로 재-접합시킴으로써 다합체 형성을 중재하는 능력을 평가하였다.
TCEP, 즉 설프하이드릴-부재 환원제를 사용하는 Δ123의 소규모 환원을 신속한 검사로서 시험하여, Δ123의 부분적 환원을 위해 필요한 TCEP의 최적의 농도를 결정하였다(도 6). 이는, 효소 면역검정 플레이트 상으로 코팅된 단량체성 H77c Δ123을 상이한 농도의 TCEP(0-500mM)에 30 분 동안 37℃에서 노출시켜서 환원이 발생하기 위한 시간을 허용함으로써 실행되었다. TCEP 환원으로부터 야기된 구조적 변화의 정도를 결정하기 위해, 입체배좌-의존적 마우스 H53 항체의 단일 희석물을 플레이트에 적용하고, HRP-접합된 항-마우스 항체에 의해 결합을 검출하였다. 10 내지 500mM TCEP로 처리된 H77c Δ123은 H53에 대한 결합의 점진적 감소를 나타내었고, 이때 모든 광학 밀도 값은 R04 음성 대조군(인간 사이토메갈로바이러스에 대한 인간 MAb)의 값에 비해 더 컸다. 이들 결과로부터 Δ123의 입체배좌는 H53 인식이 어느 정도 보유되었으므로 완전히 파괴되지 않았음을 알 수 있는데, 이는 부분적 환원이 달성되었고, 환원이 TCEP 농도의 증가에 따라 점진적으로 증가되었음을 지시한다. 더욱이, 양성 항-HIS 결합은 상이한 농도의 TCEP의 존재하에 일관되게 발생하여, 결합된 Δ123의 손실이 감소된 H53 결합을 설명할 수 있는 가능성을 제거하였다. Δ123 입체배좌의 보유가 Δ123의 항원성 및 면역원성 특성을 보존하고 HMW Δ123으로의 후속적인 재형성(reformation)을 허용하기 위해 필수적인 것으로 고려되므로, 10 내지 200mM 사이의 TCEP 농도가 추가의 단백질 환원 실험에 적합한 것으로 결정되었다.
효소 면역검정 플레이트-결합된 단백질 상에서의 환원은 다수의 시약 농도의 신속한 검사를 허용한다. 그러나, 용액에서의 환원이 잠재적 백신 생산과 관련하여 더욱 적절하다. 따라서, TCEP가 6 내지 8의 pH 범위에서 가장 효과적이므로 H77c Δ123 단량체를 PBS pH 6.8에서 제조하였다. 다양한 농도의 Δ123을 또한 제조하여 초기에 존재하는 상이한 양의 단백질이 단량체의 HMW 형태로의 전환 효율에 영향을 미치는지의 여부를 조사하였다. 이어서 TCEP를 이전의 소규모 환원 실험에 의해 안내된 농도로 첨가하였다. 부분적으로 환원된 Δ123 단량체를 HMW Δ123으로 후속적으로 전환시키기 위해, 환원 이후 단백질 재접힘, 즉 디설파이드 재형성 및 재셔플링(reshuffling)이 관여하는 공정을 수행하였다. 이는, 6.5 내지 7.5의 pH 범위에서 유리 설프하이드릴과 함께 티오에테르 결합을 형성하는 짧은 동종이작용성의 말레이미드 가교결합제인 BMOE를 첨가함으로써 촉진되었다. 결과로서 설프하이드릴 기가 접합되고, 이에 따라 디설파이드 재형성을 모방한다. 더욱이, TCEP는 설프하이드릴-부재이고, 따라서 설프하이드릴-반응성 BMOE 가교결합제와 함께 사용하기에 적절하여, 본 실험에서 환원제로서의 TCEP의 사용을 정당화시킨다. 재접힘된 Δ123 당단백질을 비-환원 SDS-PAGE에 의해 분석하였다(도 6B). 모든 샘플은 단량체 대조군 및 모의(mock) 대조군과 유사한 이동 패턴을 나타내었고, 이는 TCEP 및 BMOE의 부재하에 환원되고 재접힘되었다(도 6B 및 표 8 상의 래인 7). HMW Δ123 대조군의 밴드와 정렬되는 밴드는 없었는데, 이는 TCEP 환원 및 BMOE를 사용하는 재접힘이 단량체성 Δ123를 HMW Δ123으로 전환시키는데 성공하지 않았음을 보여준다.
실시예 12: 산화환원-셔플링 시스템을 사용하는 환원 및 재접힘
산화환원-셔플링 시스템을 이용하여 환원된 설프하이드릴 사이의 산화를 촉진시켰고, 이로써 환원 및 재접힘을 환원제(GSH) 및 산화제(GSSG)의 존재하에 동시에 수행하였다. 탄산염-중탄산염 완충액 pH 9.6에서 H77c Δ123 단량체를 제조함으로써 이를 실행하였는데, 높은 pH 환경이 GSH 및 GSSG의 환원 및 산화 능력을 각각 촉진시키기 때문이다. 응집 효율에 미치는 단백질 농도의 영향을 조사하기 위해, H77c Δ123 단량체를 또한 다양한 농도에서 제조하였다(표 9). 이어서, 5 GSH: 1 GSSG의 비를 첨가하여 2mM GSH 및 0.4mM GSSG의 최종 농도가 되고, 이는 전형적으로 상업용 단백질 재접힘 키트에서 사용되는 농도로서, 90% 초과의 단백질이 재접힘되는 것으로 보고되었다[써모 피셔 사이언티픽(Thermo Fisher Scientific)에 의해 제공된 정보]. 이어서 이들 Δ123 샘플을 37℃에서 다양한 시간 동안 항온처리하였고(표 9), 이후 즉시 PBS pH 6.8에 의해 50배 희석하고, 반응을 종결시키기 위해 원심분리 필터 유닛을 사용하여 글루타티온을 제거하였다.
Δ123 샘플을 비-환원 SDS-PAGE에 의해 분석하였고(도 7A), 이는, 이합체 밴드의 등장 및 이합체에 비해 크기가 더 큰 높은 급수의 구조물의 존재에 의해 제시되는 바와 같이, 단량체성 Δ123의 이합체성 Δ123 및 더 높은 급수의 종으로의 성공적인 재접힘을 보여주었다. 단량체 및 모의 대조군 각각은 예상된 바와 같이 단량체성 Δ123의 크기에 상응하는 밴드를 생산하였다. 그러나, 희미한 이합체 밴드가 또한 모의 대조군의 경우 관찰되었다(도 7A 상의 래인 1). 이는 단량체성 Δ123에 대한 겔 여과 프로파일과 일치하고(도 7B), 이는 또한 소량의 오염 이합체성 Δ123을 나타내었다. 이합체의 검출은 또한 공기 또는 더 높은 온도로의 노출의 결과로서 자발적인 단백질 응집을 시사할 수 있는데, 이는 미처리된 Δ123 단량체가 이전에 4℃에서 뚜껑덮힌 관에서 저장되었기 때문이다. 추가적으로, 겔에 적재하기 이전에 샘플을 비등시키는 것은 응집을 촉진시킬 수 있다. LI-COR 오디세이 시스템을 사용하여 단량체 및 이합체 밴드 밀도측정값을 정량화한 결과, 모의 대조군은 이합체에 비해 4.65배 더 많은 단량체를 함유하는 것으로 나타났다. 대부분의 단량체는 1 ㎍/㎕의 단량체성 Δ123을 글루타티온으로 처리할 경우 높은 급수의 종으로 재접힘되지 못하였고, 이때 이합체에 비해 6.89배 더 많은 단량체를 가졌으며 이는 모의 대조군에 대해 기록된 것 보다 더 높았다. 대조적으로, 더 높은 농도(5 및 10 ㎍/㎕)의 사용은 더 많은 이합체의 형성을 초래하였는데, 이는 다량의 단백질의 존재하에서의 재접힘이 Δ123 응집 효율을 향상시켰음을 지시한다. 또한 2 시간에서 24 시간으로의 항온처리 시간의 증가는 더 많은 이합체의 형성을 유도하였는데, 이는 도 7A 상의 병치 조건 3 및 5에 의해 입증된다. 모든 검사된 조건중에서, 단량체의 이합체로의 전환은 10 ㎍/㎕의 단량체성 Δ123을 24 시간 동안 글루타티온으로 처리할 경우 가장 효율적이었다. 그러므로, 이들 조건을 이용하는 실험을 규모 확장시켜 반복하였고, 겔 여과 크로마토그래피에 의해 분석하였다.
반복 실험으로부터의 Δ123의 겔 여과 프로파일은 단량체 및 이합체 피크를 나타내었고(도 7C), 이는 비-환원 SDS-PAGE의 결과와 일치한다(도 7A). 이합체 피크 상에 숄더(shoulder)가 또한 관찰되었는데, 이는 높은 급수의 종의 일부 형성을 지시한다. 유니콘 소프트웨어를 사용하여 곡선하의 면적을 정량화한 결과, Δ123 단량체의 거의 절반이 다합체로 재접힘된 것으로 밝혀졌고, 이때 이합체는 대부분의 단백질을 포함하였다(표 10). 전체적으로, 이들 결과는 산화환원-셔플링 시스템의 사용이 실제로 글루타티온-환원된 Δ123 단량체 사이에 분자간 디설파이드 결합 형성을 유도하였음을 입증하였다. 우세하게, 이들 실험은 이합체의 형성을 초래하였다.
실시예 13: 디티오트레이톨(DTT)을 사용한 환원 및 느린 희석을 통한 재접힘
Δ123 응집에 대한 본 발명자들의 연구를 확장시키기 위해, 상이한 환원제 및 재접힘 기법을 이용하였다: 각각 DTT 및 느린 희석. TCEP와 마찬가지로, ELISA 형식으로 Δ123의 소규모의 DTT 환원을 초기에 시험하여 Δ123의 부분적 환원을 달성하기 위해 필요한 최적의 DTT 농도를 결정하였다(도 8A). 이는 효소 면역검정 플레이트 상으로 코팅된 H77c Δ123 단량체를 상이한 농도의 DTT(0-1OmM)에 30 분 동안 37℃에서 노출시킴으로써 실행되었다. 이어서 플레이트를 세척하여 DTT를 제거함으로써 추가의 환원을 중단시킨 후, 차단하고, 1차 항체(항-HIS, H53 및 R04)를 첨가하고, 적절한 HRP-접합된 2차 항체에 의해 검출하였다. Δ123의 사실상 완전한 환원이 > 2mM DTT에 의해 달성되었다. 0.1 내지 1.OmM DTT로 처리된 Δ123은 점진적으로 감소된 H53 결합을 나타내었고, 따라서 이들 농도를 추가로 후속적인 DTT 환원 실험에서 조사하였다. 일정한 항-HIS 결합이 모든 DTT 농도에서 관찰되었고, 감소된 H53 결합에 대한 요인으로서 결합된 Δ123의 손실을 제거하였다.
이어서, 용액중의 DTT 환원을 조사하였고, 여기서 다양한 농도의 H77c Δ123 단량체(표 11)를 탄산염-중탄산염 완충액 pH 9.6에서 제조하고, 7을 초과하는 pH에서 환원 능력을 보유하는 DTT에 노출시켰다. 환원된 설프하이드릴은 결국 환원제의 부재하에 자발적 산화를 겪게 되어, 디설파이드 결합의 재형성을 유도할 것이다. 이러한 과정이 발생될 수 있도록 하기 위해, 환원된 Δ123 당단백질을 후속적으로 느린 희석에 의해 재접힘되도록 하고, 이러한 희석은 PBS pH 6.8의 Δ123 샘플로의 단계식 첨가를 포함한다. 더욱이, 이러한 절차는 DTT를 희석하고, pH 환경을 점진적으로 중화시켜 그의 활성을 약화시킴으로써 설프하이드릴의 산화를 촉진시킨다.
재접힘된 Δ123 당단백질을 비-환원 SDS-PAGE에 의해 분석하였다(도 8B). 모의 대조군은 예상대로의 단량체 밴드, 및 이합체성 Δ123에 상응하는 대략 100 kDa에서 희미한 밴드를 생산하였는데, 이는 이합체 밴드가 단량체 대조군에서는 관찰되지 않았으므로 이합체 또는 아마도 자발적 단백질 응집에 의한 낮은 수준의 오염을 지시한다. 단량체 및 이합체 밴드 밀도측정값을 정량화한 결과, 모의 대조군이 이합체성 단백질에 비해 3.41배 더 많은 단량체성 단백질을 함유하는 것으로 나타났다(도 8B 및 표 11 상의 래인 1). 모든 검사 조건하에서 환원되고 재접힘된 Δ123 샘플의 경우 올리고머의 증가된 양이 기록되었는데(도 8B), 이는 일정 비율의 Δ123 단량체가 성공적인 분자간 디설파이드 결합 형성의 결과로서 실제로 다합체로 전환되었음을 보여준다. 단량체 대 이합체 비의 유의적인 감소가 DTT 첨가 후 30 분 동안 항온처리된 샘플에 대해 기록되었고, 이는 이합체성 단백질에 비해 2 배 미만으로 더 많은 단량체성 단백질을 생성하였다. 2 시간 이상의 항온처리가 30 분간의 항온처리에 비해 더 많은 다합체 형성을 일으키지 않았으므로 항온처리 기간의 연장은 Δ123 응집을 향상시키지 못하였다. 1 ㎍/㎕의 단량체성 Δ123이 DTT에 의해 처리될 경우 단량체 대 이합체의 낮은 비가 기록되었고, 이는 글루타티온 처리로부터의 결과와 상반되었다. 1 ㎍/㎕의 단량체성 Δ123을 O.1mM DTT에 의해 30 분 동안 처리할 경우 1.33의 최저치가 기록되었는데(도 8B 및 표 11 상의 래인 11), 이는 이합체로의 전환이 이들 조건을 사용함으로써 가장 효율적이었음을 지시한다. 높은 급수의 종의 형성은 1 ㎍/㎕의 단량체성 Δ123을 1mM DTT에 의해 30 분 동안 처리한 경우 비-환원 SDS-PAGE 상에서 매우 분명하였다(도 8B 상의 래인 9).
다합체 형성을 위한 최적의 조건(1mM DTT, 30 분 동안)을 사용하여 규모 확장 반복 실험을 수행하였다. 이어서 재접힘된 Δ123 당단백질을 겔 여과 크로마토그래피에 의해 분석하여, 비-환원 SDS-PAGE에서의 관측 결과를 확증하였고 생산된 각각의 올리고머성 종에 대한 더욱 정확한 측정값을 수득하였다(도 8C-D). DTT 처리 이후 단량체 피크의 보유에 의해 제시되는 바와 같이 단량체의 비율은 영향받지 않았다. 겔 여과 프로파일은 또한 이합체성 Δ123의 일부 형성을 지시하는데, 이는 소량의 단백질이 이합체성 Δ123에 상응하는 면적내에서 용출되었기 때문이고, 이는 처리 이전의 겔 여과 프로파일에 비해 더욱 현저하다. 매우 중요하게는, 200kDa 이상의 크기에 상응하는 넓은 피크가 HMW2 Δ123에 대해 관찰되는 용출 시간과 유사한 대략 59 분에서 용출되었고, 이는 Δ123의 더 큰 다합체로의 성공적인 응집을 확인해 준다. 피크하 면적의 정량화로부터, 단량체의 대략 1/5이 HMW Δ123으로 응집되었음이 밝혀졌다. 느린 희석이 수반되는 DTT 환원은 단량체성 Δ123을 HMW Δ123으로 전환시키기 위해 사용된 3가지 방법들중 가장 효과적이었다.
실시예 14: DTT-처리된 Δ123의 재접힘으로부터 생성된 생성물의 크기 분석
하기 프로토콜을 사용하여 DTT-처리된 Δ123의 재접힘으로부터 생성된 생성물의 크기를 분석하였다.
재료:

단량체성 Δ123을 환원시키기 위해, 1 mg의 단량체성 Δ123을 탄산염-중탄산염 완충액중 총 1 ㎖의 부피로 5 ㎖의 황색의 뚜껑있는 관에서 제조하였다. 프로테아제 저해제의 존재하에 재접힘되는 경우, 5 ㎍/㎖의 아프로티닌(aprotinin), 8 ㎍/㎖의 류펩틴(leupeptin) 및 5μM 키포이넨신(kifeunensine)을 또한 첨가하였다. DTT를 밀리(Milli)-Q 워터에 용해시키고, 0.3mM의 최종 농도(즉 3 ㎕의 10OmM DTT)로 관에 첨가하였다. 그런 다음, 혼합물을 37℃에서 30 분 동안 항온처리하였다. DTT에 의해 2회 처리하기 위해, 항온처리 후, 3 ㎕의 1OOmM DTT를 관에 첨가하고 혼합물을 37℃에서 30 분 동안 항온처리하였다. DTT에 의해 3회 처리하기 위해, 제2의 항온처리 후, 3 ㎕의 1OOmM DTT를 관에 첨가하고 혼합물을 37℃에서 30 분 동안 항온처리하였다.
재접힘을 위해, 0.5 ㎖의 PBS를 첨가하고 단백질을 실온에서 15 분 동안 항온처리하였다. 이러한 단계를 3회 반복하였다. 아미콘 초원심분리 필터 유닛(10k MWCO)을 사용하여 DTT를 제거하고, 단백질을 PBS에서 2회 세정하였다.
크기 분석을 위해, 단백질을 1.5 ㎖의 에펜도르프 관으로 옮기고 13,000 rpm하에 4℃에서 10 분 동안 원심분리하였다. 이어서 16/600 수퍼덱스 200 제조용 등급 칼럼을 사용하여 겔 여과 크로마토그래피를 수행하였다.
DTT 처리로부터, 단량체성 Δ123이 높은 급수의 종으로 재접힘되어, 59 및 54 분의 용출 시간에 피크를 형성함이 밝혀졌다(도 10). DTT 처리의 수를 증가시킨 결과 HMW-유사 피크는 좌측으로, 즉 1차 처리에서 관찰된 59 분의 용출 시간으로부터 2차 또는 3차 처리 이후의 54 분(도 10)으로 전이되었다. 한편, 우측으로의 약간의 전이가 단량체 피크에 대해 다음과 같이 관찰되었다; 1차, 2차 및 3차 처리의 경우 각각 77에서 76에서 75의 용출 시간으로 전이됨. 프로테아제 저해제를 첨가한 결과 잔여 단량체 피크는 75 분에 용출되었고(도 11, 하부 예) 재접힘된 다합체는 54 분에 용출되었는데, 이는 프로테아제 저해제가 이러한 공정에서 유익할 수 있음을 시사한다.
재접힘되지 않은 잔여 단량체성 단백질이 재접힘될 수 있는지의 여부를 검사하기 위해, DTT로 2회 처리된 상기 기재된 실시예로부터의 잔여 단량체성 종(겔 여과 분별물 72 내지 81을 모으고 아미콘 초원심분리 필터 유닛(10k MWCO)을 사용하여 농축시킴)을 수집하였다. 샘플을 상기 기재된 바와 같이 DTT에 의해 2회 다시 처리하여, 재접힘을 수행하고 단백질을 크기 분석하였다.
재접힘되지 않은 단량체성 Δ123의 재접힘 결과(도 11 , 상부 예), 37%의 이합체가 형성되고 HMW-유사체 형성은 거의 없었던 반면, 63%가 단량체로서 남아있었는데, 이는 Δ123 단량체의 하위-모집단이 재접힘에 저항함을 지시한다. 또한, 단량체 피크의 우측에 비교적 짧은 피크가 형성되었다. 그러나, 프로테아제 저해제의 존재하에서의 재접힘은 이러한 피크의 크기 감소를 초래하였다.
결과는 적어도 60 내지 70% 수율의 올리고머(> 100 MW 또는 200 MW)가 본 방법을 사용하여 생산될 수 있음을 입증한다.
실시예 15: DTT-처리된 Δ123 및 ALA7 Δ123의 항원성 특징
하기 프로토콜에 따라 하기 기재된 항체의 패널을 사용하여 DTT-처리된 Δ123 및 ALA7 Δ123 사이의 항원성을 비교하였다.
재료:
시약:

단백질:

항체:

눈크 맥시소브 평저 96 웰 플레이트를 50mM 탄산염-중탄산염 완충액 pH9.6에서 제조된 5 ㎍/㎖의 상기 열거된 단백질에 의해 코팅하고, 하룻밤(o/n) 4℃에서 항온처리하였다. 다음날, 플레이트를 PBST에 의해 4회 세척하고 100 ㎕/웰의 BSA1OPBS에 의해 1 시간 동안 RT에서 차단하였다. 그런 다음, 플레이트를 PBST에 의해 4회 세척하고, '재료'에서 열거된 1차 항체의 반-로그식 연속 희석을 수행하고, 1 시간 동안 RT에서 항온처리하였다. 그런 다음, 플레이트를 PBST에 의해 4회 세척하고, '재료'에서 열거된 50 ㎕/웰의 2차 항체를 첨가하고, 1 시간 동안 RT에서 항온처리하였다. 그런 다음, 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 TMB에 의해 전개시킨 후, 50 ㎕/웰의 HCl을 첨가하였다. 결과는 도 9에 제시된 바와 같다. 재접힘된 Δ123은 단량체성 Δ123 대조군 및 재접힘 저항성 Δ123 대조군과 비교하여 감소된 AR3C 및 CBH4G에 대한 결합을 나타내었는데, 이는 표적 에피토프가 재접힘된 Δ123에서 더욱 폐쇄되었음을 지시하고, HMW Δ123의 전형적인 프로파일이다. 재접힘된 ALA7 Δ123은 단량체성 ALA7 Δ123 대조군 및 재접힘 저항성 ALA7 Δ123 대조군과 비교하여 감소된 AR3C 및 CBH4G에 대한 결합을 나타내었고, 이는 표적 에피토프가 재접힘된 ALA7 Δ123에서 더욱 폐쇄되었음을 지시하고, 단백질의 고분자량 형태의 전형적인 프로파일이다. 재접힘된 Δ123 및 재접힘된 ALA7Δ123에 대한 CBH4G의 감소된 반응성은 비-중화 에피토프가 고유의 HMW 물질에서에 비해 재접힘된 물질에서 더욱 폐쇄됨을 지시할 수 있다. 이는, 비-중화 항체 반응의 생성이 억제되기 쉽기 때문에, 재접힘된 물질이 면역원으로서 사용되는 경우 이점을 제공할 수 있다.
실시예 16: 재접힘 실험
상기 수행된 추가의 재접힘 실험의 결과는 표 13 및 14에 요약되어 있다.
요약하면, 친화도-정제된 Δ123의 겔 여과 크로마토그래피는 우세한 단량체성 형태에 더하여 디설파이드 연결된 이합체 및 높은 급수의 HMW 다합체의 이종성 혼합물의 존재를 보여준다. HMW Δ123은 매력적인 백신 후보이다. 그러나, Δ123의 HMW 형태의 매력은 상대적으로 낮은 발현의 수율 및 오염 단백질의 존재로 인해 다소 경감된다. 백신 제조를 위해 유익한 HMW Δ123 생산의 수율을 개선시키는 몇몇 방법을 조사하였다.
인간-유래된 FS293F 세포에서 일시적 형질감염으로부터의 Δ123의 발현을 평가하고, Δ123을 포획하기 위해 CD81-LEL113-201을 이용하는 샌드위치 ELISA를 통해 성공적인 Δ123 생산 및 정제를 입증하였다.
겔 여과 크로마토그래피로부터 수득된 결과에 의해 Con1 및 S52 균주 둘 다가 더 높은 비율의 HMW Δ123을 일관되게 발현하는 것으로 밝혀졌고, 이때 Con1 및 S52의 경우 각각 대략 23% 및 20% 증가하였고, H77c 균주에 의해 생성된 HMW Δ123의 비율의 사실상 두배였다. 더욱이, 이들 결과로부터 상이한 HCV 균주가 다양한 양의 HMW Δ123을 생산함이 밝혀졌다. 결과는 형질감염 및 정제의 독립적인 제2 라운드로부터 생성된 Con1 및 S52 Δ123 둘 다에 대해 재현가능하였다.
비록 코발트-충전된 탈론 비이드를 사용하는 친화도 크로마토그래피가 분비된 Δ123 당단백질을 세포 배양 상청액으로부터 효율적으로 정제하지만, 이들 비이드로부터의 코발트의 침출은 백신 생산을 위해 사용될 경우 건강 문제를 야기할 수 있다. Δ123을 정제할 경우 더욱 안전한 대안은 항체 친화도 크로마토그래피의 사용일 수 있다.
HMW Δ123의 구조가 한정되지 않은채로 남아있지만, 본원에 기재된 결과는 이전에 WT E2 및 단량체성 Δ123에서 접근가능한 비-특이적 및 유형-특이적 중화 영역을 폐쇄하는 특유의 접힘 배열을 분자간 디설파이드 결합이 중재함을 시사한다. H77c Δ123에 대한 면역원성 연구는, 단량체가 bNAb를 생성하지 못하는 반면, HMW 형태가 모든 7가지 HCV 유전자형을 중화시킬 수 있는 항체를 이끌어낼 수 있음을 증명함으로써 상기 주장을 지지하는데, 이는 이들 항체 반응이 이들 영역의 폐쇄로 인해 비-중화 에피토프로부터 멀리 인도되고, 이에 따라 그 대신 광범위 중화 에피토프를 표적으로 함을 지시한다[드루머, 미공개 데이터].
단량체가 상이한 Δ123 종의 이종성 혼합물로부터 효율적으로 생산되고 효과적으로 정제될 수 있으므로, 단량체성 Δ123의 HMW 형태로의 재접힘을 위한 전략이 평가되었다. 유리 설프하이드릴을 생성하기 위한 단량체의 부분적 환원, 및 이후의 이들 설프하이드릴의 접합 또는 재산화가 H77c HMW Δ123에서 디설파이드 배열을 재현하는 방식으로 디설파이드 결합의 재셔플링을 유도할 것이라는 가설이 세워졌다. 관례적으로, 원치않는 단백질 응집체로부터, 특별히 세균성 봉입체(inclusion body)로부터 낮은 급수의 종(단량체/이합체/3합체)을 재생하기 위해 환원 및 재접힘 방법을 사용한다[싱(Singh) 및 판다(Panda)의 문헌(2005)].
동종이작용성 BMOE 가교결합제는 유리 설프하이드릴에 반응하여 이들의 접합을 초래한다. 더욱이, 접합이 개별적인 Δ123 서브유닛 사이에서 일어난다면, HMW Δ123으로의 응집을 중재하는 분자간 상호작용이 달성될 것이다. 그러나, 재접힘 과정에서의 BMOE 가교결합제의 활용은 분자간 디설파이드 결합 형성을 촉진시키지 못하였다. 이는 BMOE가 아마도 입체 장해 또는 실험적 오차에 기인하여 Δ123 단량체를 시험된 조건에서 가교결합시키지 못하였음을 시사한다. 더욱이, BMOE-처리된 양성 대조군의 부재는 이들 BMOE 재접힘 실험의 유효성을 평가하는 것을 극히 어렵게 만들었다. 이는 다른 가교결합제가 가교결합된 다합체를 생성하지 않을 것이고 이들의 생산을 위한 유용한 방법이 아닐 것이라고 말하는 것은 아니다. 역으로, 느린 희석 방법이 수반되는 DTT 환원, 뿐만 아니라 글루타티온-기반의 산화환원-셔플링 시스템의 사용은 이합체 및 높은 급수의 종을 형성하였는데, 이는 유리 설프하이드릴이 분자간 디설파이드 결합으로 산화되었음을 지시한다. 구체적으로, 시스테인 잔기 452, 486, 569 및 597에서의 설프하이드릴은 개별 Δ123 서브유닛을 함께 가교화하는데 직접적으로 관여될 수 있는데, 이들 잔기가 E2 RBD의 구조적 완전성에 필수적이지 않기 때문이다.
DTT를 사용하는 환원 및 느린 희석을 통한 재접힘은 시험된 3가지 방법들중 단량체성 Δ123을 HMW 종으로 전환시키는데 있어서 가장 효과적인 방법이었다. 이 방법은 Δ123 단량체의 대략 1/5을 HMW 종으로 전환시킬 수 있었다. 산화환원-셔플링 시스템의 사용은, 비록 이합체가 될지라도, Δ123 단량체의 대략 50%로 재접힘을 달성하였다.
실시예 17: 베타-머캅토에탄올(βME)을 사용하는 환원 및 조립
βME를 사용한 부분적 환원에 의한 Δ123 단량체의 환원 및 고분자량 올리고머로의 조립을 평가하였다. 단량체성 Δ123(20 ㎍)을 각각의 관에서 PBS중 20 ㎕의 총 부피로 제조하였다. 0 내지 1OOmM의 최종 농도가 되도록 βME을 첨가하고 15 분 동안 RT에서 항온처리하였다. 단백질을 재조립하기 위해, 10 ㎕의 PBS를 첨가하고 단백질을 RT에서 7 분 동안 항온처리하고 2회 반복하였다. 아미콘 초원심분리 필터 유닛(30k MWCO)을 사용하여 βME를 제거하고, 단백질을 PBS에서 2회 세정하였다. 단백질을 비-환원 SDS-PAGE에서 분석하였다. βME-처리된 Δ123Δ1233 및 HMW1 Δ123의 샘플 및 2 ㎕의 단백질 표준물을 별도로 웰에 적재하였고, 이어서 겔 전기영동을 120V에서 수행하였다. 일단 분리 겔의 상부에 염료 전방부가 이동되면, 전압을 160V로 상승시켰다. 겔을 쿠마시(0.25% 쿠마시 브릴리언트 블루 G-250, 50% 메탄올 및 10% 아세트산)로 1 시간 동안 착색시키고, 이어서 겔의 배경이 투명해질 때까지 탈색시켰다(50% 메탄올 및 10% 아세트산). 최종적으로, Li-코르오디세이(CorODyssey) 영상촬영 시스템을 사용하여 겔을 관찰하였다.
처리된 단백질 및 미처리된 단백질을 비-환원 조건하에 SDS-PAGE에서 분석하였다. 결과(도 15A)로부터 미처리된 단량체성 Δ123이 대부분 단량체로서 이동됨을 알 수 있고, 이때 아마도 샘플을 SDS-PAGE에 첨가하기 이전에 비등시킨 결과로서 소량의 이합체가 존재한다. βME의 고농도에서, 더 큰 HMW 올리고머가 도 15A에 지시된 바와 같이 관찰될 수 있다. 이들의 존재를 확인하기 위해, Δ123 단량체를 1OOmM βME에 의해 환원시키고 조립한 후 겔 여과 크로마토그래피 상에서 분석하였다. 결과(도 15B)로부터, 음영처리된 영역에 의해 지시되는 바와 같이 1OOmM βME로 처리된 Δ123 단량체 상에서 작은 숄더가 존재하였음을 알 수 있다. 이는 비효율적이긴 하지만 Δ123을 높은 급수의 올리고머로 재조립시킬 수 있음을 시사한다.
실시예 18: 조립된 당단백질의 면역원성
조립된 단백질이 단량체성 항원과 비교하여 동물에서 더욱 면역원성인지의 여부를 결정하기 위해, 이계교배된(outbred) 기니아 피그를 100 ㎍의 단백질에 의해 4회 백신접종하고, 3주 뒤 보조제로서 아다백스(Addavax)를 사용하였다(표 15). 기니아 피그에게 최종 백신접종을 한지 2주 후 이를 채혈하고 면역 혈청을 분석하여 Δ123 단량체성 단백질에 대한 항체 역가를 결정하였다.
눈크 맥시소프 평저 96 웰 플레이트를 50 ㎕/웰의 50mM 탄산염-중탄산염 완충액 pH 9.6에서 제조된 5 ㎍/㎖의 단량체성 H77c E2 Δ123에 의해 코팅하고 하룻밤 4℃에서 항온처리하였다. 다음 날, 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 BSA10PBS에 의해 1 시간 동안 실온에서 차단하였다. PBST에 의해 4회 세척한 후, 1/100에서 시작된 기니아 피그 혈청의 반-로그식 연속 희석물을 플레이트에 2 시간 동안 실온에서 첨가하였다. 플레이트를 PBST에 의해 4회 세척하고, 항-HIS 항체를 1/1000로 첨가하고 1 시간 동안 실온에서 항온처리하였다. 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 TMB에 의해 전개시킨 후, 50 ㎕/웰의 HCl을 첨가하였다. 450 nm 내지 650 nm에서의 광학 밀도를 연속 희석비에 대해 도표화하고(y-축), 0.5 유닛의 광학 밀도를 제공하기 위해 필요한 상호간의 희석비를 사용하여 항체 역가를 결정하였다. 4-F 및 5-F 그룹이 각각 4 마리의 동물을 가지므로, 둘 다 HCV 단백질의 단량체성 버전을 수여받았고, 이들 그룹으로부터의 결과를 다른 그룹과의 비교를 위해 조합하였다.
결과로부터 재접힘된 Δ123 및 Δ123A7 단백질이 백신접종된 동물은 이들 단백질의 단량체성 형태가 백신접종된 동물에 비해 더 높은 항체 역가를 생성하였음을 알 수 있다(Δ123=41826, Δ123A7=28686 대 조합된 Δ123 및 Δ123A7 단량체 그룹=21756)(도 16). 이들 그룹의 평균 역가는 또한 더 높았다. 조립된 Δ123 그룹의 경우, 항체 역가의 기하 평균은 고유의 Δ123 HMW 백신접종된 그룹에 비해 더 높았다(표 16). 조립된 Δ123을 수여받은 동물은 단량체성 항원을 수여받은 동물에 비해 통계적으로 유의적인 더 높은 항체 역가를 나타내었다(p=0.0286)(도 16).
실시예 19: 항체 반응의 특이성
조립된 항원이 항체의 상이한 특이성을 생성하였는지의 여부를 결정하기 위해, HCV에 의한 감염을 예방하는 광범위 중화 항체의 표적인 E2 당단백질의 주요 항원 영역을 나타내는 합성 펩타이드에 대하여 혈청을 검사하였다. 에피토프 I은잔기 412-423에 걸쳐 있고, 광범위 중화 항체의 에피토프, 예컨대 마우스 AP33[오우시안카(Owsianka) 등의 문헌(2001); 오우시안카 등의 문헌(2005); 타르(Tarr) 등의 문헌(2006)]; 래트 3/11[타르 등의 문헌(2006); 플린트(Flint) 등의 문헌(2009)]; 인간 HCV1 및 95-2[브로에링(Broering) 등의 문헌(2009)]; 인간 HC33.1 및 관련 항체[케크(Keck) 등의 문헌(2013)]; H77.39[사보(Sabo) 등의 문헌(2011)]; MRCT10.v362 및 hu5B3.v3[판투아(Pantua) 등의 문헌(2013)]; 및 MAb24[알함마드(Alhammad) 등의 문헌(2015)]를 포괄한다. 합성 펩타이드를 사용하여 이러한 특이성 및 잔기 408-428을 검출하였고, 이는 이전에 기니아 피그 혈청에서 다중클론성 항체 반응성을 특징짓기 위해 사용되었다[비에티어(Vietheer) 등의 문헌(2017)].
눈크 맥시소프 평저 96 웰 플레이트를 50 ㎕/웰의 50mM 탄산염-중탄산염 완충액 pH 9.6에서 제조된 5 ㎍/㎖의 H77c 에피토프 I 펩타이드(408-428)에 의해 코팅하고 하룻밤 4℃에서 항온처리하였다. 다음 날, 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 BSA10PBS에 의해 1 시간 동안 RT에서 차단하였다. PBST에 의해 4회 세척한 후, 1/5에서 시작된 기니아 피그 혈청의 반-로그식 연속 희석물을 첨가하고 1 시간 동안 결합하도록 허용하였다. PBST에 의해 4회 세척한 후, 50 ㎕/웰의 항-기니아 피그 HRP 항체를 첨가하고 1 시간 동안 RT에서 항온처리하였다. 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 TMB에 의해 전개시킨 후, 50 ㎕/웰의 HCl을 첨가하였다.
결과로부터 조립된 Δ123 및 Δ123A7에 의해 백신접종된 동물은 이들 항원의 단량체성 형태에 의해 백신접종된 동물에 비해 유의적으로 더 높은 항체 역가를 가졌음을 알 수 있다(도 17). 조립된 Δ123의 경우, 평균 역가는 단량체성 그룹에 비해 15배 더 높았고 고유의 Δ123 백신접종된 그룹에 비해 1.15배 더 높았다. 조립된 Δ123A7의 경우 평균 역가는 단량체성 그룹에 비해 5배 높았다. 이러한 데이터는, 본 실시예에서 HCV E2의 단량체인 항원을 고유적으로 접힌 상태로부터 높은 급수의 올리고머로 항원을 조립하는 것이 광범위 중화 항체 에피토프로 인도되는 항체의 역가를 증가시켜서 항원의 면역원성을 유의적으로 개선시켰음을 시사한다.
실시예 20: 항체 반응-에피토프 III의 특이성
조립된 항원이 항체의 상이한 특이성을 생성하였는지의 여부를 결정하기 위해, 잔기 523-549에 걸친 CD81 결합 루프(loop)를 나타내는 합성 펩타이드에 대해 혈청을 검사하였다. 이러한 항원 영역은 항원 영역 3의 일부를 포함하고, 중화 항체에 대한 표적이다.
눈크 맥시소프 평저 96 웰 플레이트를 50 ㎕/웰의 50mM 탄산염-중탄산염 완충액 pH 9.6에서 제조된 5 ㎍/㎖의 H77c 에피토프 III 펩타이드(523-549)에 의해 코팅하고 하룻밤 4℃에서 항온처리하였다. 다음 날, 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 BSA10PBS에 의해 1 시간 동안 RT에서 차단하였다. PBST에 의해 4회 세척한 후, 1/5에서 시작된 기니아 피그 혈청의 반-로그식 연속 희석물을 첨가하고 1 시간 동안 결합하도록 허용하였다. PBST에 의해 4회 세척한 후, 50 ㎕/웰의 항-기니아 피그 HRP 항체를 첨가하고 1 시간 동안 RT에서 항온처리하였다. 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 TMB에 의해 전개시킨 후, 50 ㎕/웰의 HCl을 첨가하였다.
결과(도 18)로부터, 조립된 Δ123 및 Δ123A7을 수여받은 동물은 H77c 에피토프 III에 대해 항체의 높은 역가를 생성하였고, 이때 Δ123에 의해 백신접종된 동물의 평균 및 기하 평균은 고유적으로 접힌 단량체성 물질을 수여받은 동물에 비해 더 높음을 알 수 있다(표 18). 추가적으로, 조립된 Δ123 및 Δ123A7 단백질을 수여받은 동물에서의 항체 역가는, 단량체성 물질을 수여받은 동물에서 관찰되는 광범위한 항체 역가와는 대조적으로 일관되게 높았다(Δ123 및 Δ123A7의 경우 역가에서 5 내지 6배 범위인 반면 단량체의 경우 200배임). 이는, 조립된 단백질이 백신접종된 동물에서 일정한 면역 반응을 유도할 수 있는 반면 단량체성 출발 물질이 이러한 영역에 특이적인 항체를 비연속적으로 생산하였음을 시사한다.
실시예 21: 에피토프 I에 대한 항체 반응의 교차 반응성
H77c 에피토프 I에 대해 생성된 항체가 상이한 유전자형의 동등한 서열과 교차-반응성인지를 결정하기 위해, 잔기 408-428에 걸친 유전자형 2a J6 펩타이드를 사용하여 ELISA를 수행하였다.
눈크 맥시소프 평저 96 웰 플레이트를 50 ㎕/웰의 50mM 탄산염-중탄산염 완충액 pH 9.6에서 제조된 5 ㎍/㎖의 J6 에피토프 I 펩타이드에 의해 코팅하고 하룻밤 4℃에서 항온처리하였다. 다음 날, 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 BSA10PBS에 의해 1 시간 동안 RT에서 차단하였다. PBST에 의해 4회 세척한 후, 1/5에서 시작된 기니아 피그 혈청의 반-로그식 연속 희석물을 첨가하고 1 시간 동안 결합하도록 허용하였다. PBST에 의해 4회 세척한 후, 50 ㎕/웰의 항-기니아 피그 HRP 항체를 첨가하고 1 시간 동안 RT에서 항온처리하였다. 플레이트를 PBST에 의해 4회 세척하고, 100 ㎕/웰의 TMB에 의해 전개시킨 후, 50 ㎕/웰의 HCl을 첨가하였다.
결과(도 19)로부터, Δ123 및 Δ123A7에 의해 백신접종된 동물에서 생성된 항체는 에피토프 I의 유전자형 2a 균주 J6 서열과 교차-반응성이었고, 이들 그룹에서 항체의 평균 및 기하 평균 역가는 이들 항원의 고유의 단량체성 형태에 의한 백신접종에 의해 달성된 역가에 비해 더 높았음을 알 수 있다(표 19). Δ123에 의해 백신접종된 동물의 평균 항체 역가는, Δ123 단백질을 발현하는 플라스미드에 의해 형질감염된 293 FS 세포으로부터 직접 수거된 Δ123의 고유의 고분자량 형태에 의한 백신접종을 통해 달성된 역가에 비해 거의 2배 더 높았다(표 19).
실시예 22: HCV E2 CD81 저해의 특성규명
HCV E2는 세포 수용체 CD81을 사용하여 세포에 부착된다. 본 발명자들의 실험실에서, 본 발명자들은 가용성 E2의 재조합 CD81로의 결합을 방지하는 면역 혈청에 존재하는 항체의 능력을 결정하기 위한 검정을 이전에 이용하였다.
눈크 맥시소프 평저 플레이트를 50mM 탄산염-중탄산염 완충액 pH 9.6에서 제조된 5 ㎍/㎖의 MBP-CD81LEL에 의해 코팅하고 하룻밤 4℃에서 저장하였다. 다음 날, 플레이트를 100 ㎕/웰의 BSA10PBS로 1 시간 동안 차단하고 PBST에 의해 4회 세척하였다. 1/5에서 시작된 기니아 피그 혈청의 반-로그식 연속 희석물 50㎕를 50 ㎕의 분별화되지 않은 E2 RBD(별도의 Gla H77c 및 G2a JFH1)에 1 ㎍/㎖로 모든 웰에 첨가한 다음, 1 시간 동안 RT에서 항온처리하였다. 80 ㎕의 E2 RBD-혈청 혼합물을 CD81-코팅된 플레이트에 첨가하고, 4℃에서 하룻밤 항온처리하였다. 플레이트를 PBST에 의해 4회 세척한 후, 50 ㎕/웰의 마우스 항-HIS를 1/500으로 첨가하고 1 시간 동안 RT에서 항온처리하였다. 플레이트를 PBST에 의해 4회 세척하고 100 ㎕/웰의 TMB에 의해 전개시킨 후, 50 ㎕/웰의 1M HCl을 첨가하여 반응을 중단시켰다. 결과로부터, 조립된 Δ123 및 Δ123A7에 의해 백신접종된 동물은 H77c G1a E2 및 CD81 사이의 상호작용을 방지할 수 있는 항체의 높은 역가를 생성하였고, 고유의 Δ123 HMW 단백질을 수여받은 마우스에서 존재하는 것과 유사한 수준으로 및 이들 항원의 단량체성 형태에 의해 백신접종된 마우스에서 달성된 평균 수준과 유사한 수준으로 존재하였음을 알 수 있다(도 20A 및 표 20).
CD81에 결합하는 G2a E2의 결합을 저해하는 면역 혈청의 능력을 평가하였다(도 20B). 조립된 Δ123 및 Δ123A7에 의해 백신접종된 동물에서의 항체 역가는 이들 항원의 단량체성 형태에 의해 백신접종된 동물 및 고유의 Δ123 HMW에 의해 백신접종된 동물의 역가와 유사하였다. 이들 데이터는, HCV의 이종성 유전자형이 세포 표면 수용체 CD81에 결합하는 것을 방지하는 교차 반응성 항체를 생성하는데 있어서 Δ123 및 Δ123A7의 조립된 형태가 더욱 유리함을 시사한다.
실시예 23: 동종성 유전자형 1a 바이러스의 중화
생성된 항체가 HCV에 의한 간 세포주의 감염을 방지하는 능력을 일으키는지의 여부를 결정하기 위해, 동종성 유전자형 1a 슈도타입(pseudotyped) 바이러스를 사용하여 중화 검정을 수행하였다.
G1a로부터의 E1E2 이종이합체를 혼입한 HCVpp를 생산하기 위한 발현 벡터는 pE1E2H77c였다[드루머(Drummer) 등의 문헌(2003)]. pNL4-3.Luc.R-.E-(pNL43)을, 나타니엘 란다우 박사(Dr. Nathaniel Landau)에 의해 미국 국립보건원(NIH) 산하 국립 알러지 및 감염병 연구소(NIAID)의 AIDS 분과의 NIH AIDS 시약 프로그램으로부터 수득하였다[콘노어(Connor) 등의 문헌(1995); 헤(He) 등의 문헌(1995)]. 1.5 ㎖의 에펜도르프 관에서 6 ㎍의 E1E2H77c DNA를 6 ㎍의 pNL43 DNA와 혼합하였다. 빈 벡터 대조군의 경우, 1 ㎍의 pcDNA4c를 1 ㎍의 pNL43 DNA와 혼합하였다. 312 ㎕의 FuGENE 6을 5200 ㎕의 옵티-멤(Opti-MEM)[+글루타맥스(glutamax)]에 첨가하고, 이어서 RT에서 5 분 동안 항온처리하고, 600 ㎕의 이러한 혼합물을 DNA 혼합물에 첨가하였다. 100 ㎕를 빈 대조군에 첨가하였다. 이들을 RT에서 20 분 동안 항온처리하였다. PC3 실험실에서, 8개 관의 내용물을 단계 1로부터의 별도의 293T 세포-함유 접시에 적가하였다. 빈 대조군을 단계 1로부터의 293T 세포-함유 플레이트의 1개의 웰에 첨가하였다. 후겐(Fugene) 6/옵티멤/DNA 혼합물을 37℃ 및 5% C02에서 20 분 동안 항온처리한 후, 293T 세포에 적가하였다. 세포를 37℃에서 5% C02하에 3 일 동안 항온처리하였다. 3 일 후, 감염성 E1E2 슈도타입 바이러스(HCVPP) 또는 빈 벡터 슈도타입 바이러스(Emptypp)가 함유된 상청액 유체를 제거하고, 중화 검정에서 사용하기 위해 준비된 0.45uM 필터를 통해 여과시켰다.
열에 의해 불활성화된 기니아 피그 혈청의 반-로그식 연속 희석을 1/20에서 시작하면서 수행하였다. 250 ㎕의 혈청을 250 ㎕의 HCVpp에 첨가하고 1 시간 동안 항온처리하였다. 150 ㎕/웰의 혈청-바이러스 혼합물을 Huh7.5 세포에 3중으로 첨가하였다. 150 ㎕/웰의 빈 대조군을 항원 그룹이 없는 플레이트의 제일 아래열에 3중으로 첨가하였다. 이어서 플레이트를 37℃ 및 5% C02에서 항온처리하였다.
42 시간 후 배지를 Huh7.5 세포로부터 제거하고, 이어서 이를 1x PBS에서 1회 세척하였다. 50 ㎕/웰의 1x 세포 배양 용해 시약을 첨가하고 세포 용해물을 u-형 바닥의 96 웰 플레이트로 옮겼다. 5 ㎕의 용해물을 384 웰 백색 플레이트로 옮기고 클라리오스타(CLARIOstar) 플레이트 판독기에서 판독하였다. 3중 값의 평균을 혈청 희석 비에 대해 도표화하고, 비-선형 회귀 분석에 의해 곡선을 정합시키고, 세포를 감염시키는 바이러스에 대한 50% 저해 용량을 달성하는데 필요한 혈청의 상호간의 희석비를 계산하였다(프리즘 v7).
결과(도 21)로부터, 조립된 Δ123 및 Δ123A7에 의해 백신접종된 8/8 마리의 동물이 G1a HCVpp에 의한 감염을 방지할 수 있는 항체를 생성하였음을 알 수 있다. 대조적으로, 단량체성 항원을 수여받은 그룹에서는 단지 7/8 마리의 동물이 중화 항체를 생성하였다. Δ123A7을 수여받은 그룹에서 평균 및 기하 평균 항체 역가는 단량체성 항원을 사용하여 달성된 항체 역가에 비해 더 높았고, 이는 조립 방법을 통한 항원의 크기 증가가 그의 면역원성을 개선시킴을 시사한다(표 22). 조립을 통해 달성된 항체 중화 역가는 항원의 고유의 HMW를 사용하여 달성되는 역가와 유사한 범위에 속하였고(표 22), 이는 상기 방법이 단량체성 항원과 비교하여 HCV의 동종성 균주에 대해 효과적인 중화 항체를 생성하는 적어도 동등한 능력을 갖고 이에 비해 개선된 항원을 생산함을 시사한다.
실시예 24: E2에 대한 인간 단일클론성 항체의 결합을 저해하는 면역 혈청의 능력
기니아 피그에서 생성된 항체가 공지된 인간의 광범위 중화 항체의 에피토프와 유사한 에피토프를 인식하는지의 여부에 관하여 항체 반응의 특이성을 추가로 조사하기 위해, 경쟁적 효소 면역검정을 수행하였다.
인간 항체 HCV-1[브로에링(Broering) 등의 문헌(2009)], HC84-27[크레이( Krey) 등의 문헌(2013)], AR3C[로우(Law) 등의 문헌(2008)] 및 2A12[칸(Khan) 등의 문헌(2014)]를 VH 및 VL 영역[진아트(Geneart)]의 합성에 의해 생산하였고, 조직 플라스미노겐 활성화제(tPA) 리더의 인도하에 IgG1을 발현하기 위해 각각 pcDNA3-tPA-LC 및 pcDNA3-tPA-HC내로 클로닝하였다. 중쇄 및 경쇄 인코딩 플라스미드를 IgG 생산을 위해 FS293F 세포내로 형질감염하였다.
일정한 양의 MAb 및 각각의 기니아 피그 혈청의 반-로그식 연속 희석물을 차단된 웰에 첨가하고, 2 시간 동안 실온에서 항온처리한 후, 단량체성 Δ123이 결합된 플레이트에 첨가하였다. 잔여 MAb 결합을 항-인간 Fab2에 의해 검출하였다. 곡선을 비-선형 회귀에 의해 정합하고, 각각의 혈청 샘플에 대한 ID50을 결정하기 위해 사용하였다. 혈청 샘플이 검사된 가장 높은 농도에서 ID50을 달성하지 못하였을때(1:10 희석, log1O1), log1O0.5의 값을 그 혈청에 할당하였다.
결과로부터, 조립된 Δ123 및 Δ123A7에 의해 백신접종된 동물이 412-423 영역을 인식하는 항체 HCV1의 에피토프(에피토프 I)와 중첩되는 항체의 더 높은 평균 및 기하 평균 역가를 나타내었음을 알 수 있다(표 23 및 도 22A). HCV1 항체는 침팬지에서 HCV를 예방하고 치료하는 것으로 보여졌다[모린(Morin) 등의 문헌(2012)]. 추가적으로, 역가는 고유의 HMW Δ123을 사용하여 달성되는 역가와 유사하였고, 이는 상기 방법이 등가의 항원을 생산함을 보여준다. Δ123을 수여받은 동물에서의 항체 역가는 단량체 백신접종된 그룹에서의 역가에 비해 통계학적으로 더 높았다(도 22A).
결과로부터, 단량체성 백신 또는 고유의 HMW 백신과 비교하여, 조립된 Δ123에 의해 백신접종된 동물은 E2의 430-446 영역을 인식하고 세포 수용체 CD81과 접촉된 영역의 일부를 포함하는 항체 HC 84-27의 에피토프와 중첩되는 항체의 더 높은 평균 및 기하 평균 역가를 나타내었음을 알 수 있다(표 24 및 도 22B). 이러한 항체 특이성은 광범위하게 중화하고, HCV 도피 돌연변이체(escape mutant)의 생성을 방지한다[크레이(Krey) 등의 문헌(2013); 케크(Keck) 등의 문헌(2012)]. 이러한 데이터로부터, 조립 방법이 HC84.27에 의해 인식되는 에피토프와 중첩되는 항체의 생성에 있어서 개선된 항원을 생산함을 알 수 있다. Δ123을 수여받은 동물에서의 항체 역가는 단량체 백신접종된 그룹에서의 역가에 비해 통계학적으로 더 높았다(도 22B).
결과로부터, 단량체성 백신과 비교하여, 조립된 Δ123 및 Δ123A7에 의해 백신접종된 동물은 항체 AR3C의 에피토프와 중첩되는 항체의 더 높은 평균 및 기하 평균 역가를 나타내었음을 알 수 있다(표 25 및 도 22C). Δ123의 경우, 평균 및 기하 평균 역가는 동등한 고유의 HMW 항원에 의해 생성된 역가에 비해 더 높았다(표 25).
결과로부터, 단량체성 백신과 비교하여, 조립된 Δ123 및 Δ123A7에 의해 백신접종된 동물은 비-중화 항체 2A12의 결합을 방지할 수 있는 항체의 더 낮은 평균 및 기하 평균 역가를 나타내었음을 알 수 있다(표 26 및 도 22D). 데이터는 단량체성 Δ123 및 Δ123A7의 조립이 HCV의 중화[장(Zhang) 등의 문헌(2009)] 및 중화 항체의 생성[비에티어(Vietheer) 등의 문헌(2017)]을 방해하는 것으로 보이는 비-중화 에피토프를 폐쇄함을 시사한다.
실시예 25: E2 특이적 B 세포를 검출하기 위한 조립된 단백질의 용도
항원에 특이적인 항체에 의해 면역 세포를 식별하기 위해 조립된 HCV 단백질이 사용될 수 있는지의 여부를 결정하기 위해, 만성적으로 감염된 인간으로부터 단리된 PBMC를 사용하여 다음을 수행하였다.
동결 PBMC로부터의 B 세포의 강화
시약

HCV 감염된 개별체로부터의 PBMC를 실온에서 해동시켰다. 사세포 제거 키트(#130-090-101 밀테니 바이오텍)를 사용하여 사세포를 제거하였다. 107의 전체 세포당, 0.25 ㎖의 20x 결합 완충액 스톡 용액(Stock Solution)을 4.75 ㎖의 멸균수로 희석시켰다. 107의 전체 세포당 1OO ul의 사세포 제거 마이크로비이드를 첨가하고, 혼합하고, 15 분 동안 실온에서 항온처리하였다. 사세포를 MACS 분리기 상에서 제거하였다. 세포를 MACS 완충액(PBS, 0.5% BSA, 2mM EDTA)에 재현탁시켰다: 50 ㎖의 차가운 PBS + 0.25 g의 BSA 및 0.2 ㎖의 0.5M EDTA. 107의 전체 세포당 40 ul의 MACS 완충액 및 107의 전체 세포당 10 ul의 비오틴-항체 칵테일을 첨가하고, 10 분 동안 냉장고에서 항온처리하였다. 107의 전체 세포당 30 ul의 MACS 완충액을 첨가하였다. 20 ul의 항-비오틴 마이크로비이드를 20분 동안 냉장고에서 첨가하였다. 500 ul의 MACS 완충액에 재현탁시키고 MACS 분리기의 MS 칼럼 상에서 비-B 세포 개체군을 제거하였다. FACS 아리아 분류(Aria Sort)를 위해 세포를 다음과 같이 착색시켰다:

FACS 완충액에서 1OO ul의 CD19 PE-Cy7 항체(1:50)에 의해 세포를 표지화하였다. 웰을 혼합하고 30 분 동안 냉장고에서 항온처리하였다. FACS 완충액에서 2회 세척한 후, 현탁된 세포 펠릿을 FACS 완충액중 1OO ul의 정제된 CD81 항체(1:50)에 의해 30 분 동안 4℃에서 착색시켰다. 2회 세척한 후, 재현탁된 세포 펠릿을 20 ug의 델타3 단백질과 함께 1OO ul의 FACS 완충액에서 30 분 동안 4℃에서 항온처리하였다. 세포를 500 ul의 FACS 완충액에 의해 300 g에서 5 분 동안 2회 세척하였다. 재현탁된 세포 펠릿을 1OO ul의 CD81 APC 항체(1:50) 및 펜타-HIS 488(1:100)에 의해 FACS 완충액에서 30 분 동안 4℃에서 착색시켰다. 세포를 500 ul의 FACS 완충액에 의해 300 g에서 5 분 동안 2회 세척한 후, 세포를 200 ul의 FACS 완충액에 재현탁시키고, 즉시 FACS 분류를 진행하였다.

결과(도 23)로부터, C-말단 his 태그의 다중의 복사물이 함유된 조립된 Δ123이 E2-반응성 B 세포를 효율적으로 검출할 수 있음을 알 수 있다. B 세포 개체군의 대략 0.5%가 E2 특이적이었다. 이는 면역 표지 검출을 위한 다가 단백질로서 작용하는 조립된 단백질의 용도를 입증한다.
실시예 26: 조립된 단백질의 크기
SEC-MALS 분석을 사용하여 조립된 Δ123A7 및 조립된 Δ123 단백질의 몰 질량을 결정하였다.
샘플을 적재하기 이전에, 와이어트(Wyatt) WTC-030-N5 4.6/300 칼럼을 MT-PBS에 의해 평형화시켰다. 유속은 0.2 ㎖/분이었다. 던 헬레오스(DAWN Heleos) MALS 검출기를 아길렌트(Agilent) 1200 시리즈 UV 다이오드 어레이(diode array) 검출기 및 옵티랩(Optilab) T-rEx RI 검출기와 직렬로 사용하였다. MALS 검출기를 BSA에 의해 정규화시켰다.
262 내지 675 kDa 범위에 걸친 MW 종이 함유된 조립된 Δ123A7 샘플은 다중분산된다. 중량 평균 몰 질량은 409 kDa이다. 210 내지 744 kDa 범위에 걸친 MW 종이 함유된 조립된 Δ123 샘플은 다중분산된다. 중량 평균 몰 질량은 408.7 kDa이다. 조립된 Δ123A7 샘플에서 E2 프로모터의 범위는 5 내지 15이고, Δ123에서는 4 내지 16이며, 이때 각 경우 평균은 9이다.
실시예 27: 단백질의 조립
환원제 DTT를 사용하여 달성될 수 있는 조립의 범위를 검사하기 위해, 다중의 독립적 조립 실험을 수행하였다. 결과로부터, 단량체성 Δ123A7이 80%까지의 효율로 재접힘될 수 있는 반면, 71%까지의 Δ123이 HMW 형태로 조립될 수 있음을 알 수 있다.
잔여 단량체성 종이 DTT에 의해 처리되어 HMW 형태로 조립될 수 있는지의 여부를 평가하였다. 이전 실험으로부터 HMW 형태로 조립되지 않은 단량체성 Δ123A7의 1 mg/㎖ 용액을 0.6mM의 최종 농도의 DTT에 의해 37℃에서 30 분 동안 처리하였다. 추가의 3 ul의 10OmM DTT를 단백질 제조물에 첨가하고, 혼합하고, 또 다른 30 분 동안 37℃에서 항온처리하였다. 이어서 250 ul의 1x PBS pH 6.8을 첨가하고, 실온에서 15 분 동안 항온처리하고, 2회 이상 반복하였다. 이어서 단백질을 완충액 교환하고, 500 ul의 부피가 달성될 때까지 4 ㎖의 아미콘 초원심분리 장치(30K MWCO)를 사용하여 농축시키고 1x PBS pH 6.8에 의해 2회 세척한 후 에펜도르프 관으로 옮겼다. 처리된 단백질을 겔 여과 크로마토그래피에 의해 분석하였다. 결과로부터, 잔여 단량체성 Δ123A7의 30%가 HMW 형태로 조립될 수 있었음을 알 수 있다(도 25). 이는, 본 방법을 사용하여 단량체성 HCV E2 단백질을 HMW 형태로 조립하는 능력은 제한되지 않고, 실제로 단량체성 형태의 100%까지 조립된 HMW 형태로 전환될 수 있음을 시사한다.
실시예 28: 다른 단백질의 조립
동일한 방법을 사용하여 HCV E2의 다른 형태가 HMW 종으로 조립될 수 있는지의 여부를 검사하였다. 잔기 384-661을 포함하는 단량체성 H77c E2를 실시예 25에 기재된 바와 같이 DTT에 의해 처리하고 HMW 형태로 조립시켰다. 겔 여과 크로마토그래피 결과, 대략 40%가 HMW 형태로 조립된 것으로 밝혀졌다(도 26). RBDA7 단백질을 사용하여 동일한 과정을 수행하였고, 여기서 C581, C585, C652, C677, C494, C486, C459, C452, C564, C597, 및 C569(A7)에서 Cys-Ala 돌연변이를 갖는 잔기 384-661을 포함한 단량체성 H77c E2를 동일한 방식으로 처리하였다. 본 실시예에서, 대략 10%가 HMW 형태로 조립되었다(도 27).
본 방법이 다른 단백질로부터 높은 급수의 종을 조립하는데 광범위하게 적용될 수 있는지의 여부를 조사하기 위해, 상기 방법을 사용하여 AD8 서열로부터 C-말단 막관통 도메인 및 세포질 테일을 제거하도록 절단된 주로 단량체성인 HIV 엔벨롭 단백질(Env)을 사용하였다. DTT로 처리되고 조립된 후, 겔 여과 프로파일에서 env의 HMW 종의 형성에 상응하는 작은 전이가 관찰되었다(도 28). 높은 급수의 올리고머, 특별히 3합체의 형성은 HIV에 대해 바람직한데, 이들이 광범위 중화 항체의 결합을 보유하는 것으로 보이고 광범위 중화 항체의 생산을 위한 선호되는 백신 후보이기 때문이다[드 태예(de Taeye) 등의 문헌(2015)]. 이러한 데이터는, 단량체 또는 낮은 급수의 종으로부터 높은 급수의 올리고머가 형성되는 것이 항원 생산을 위해 바람직한 경우, 본 방법이 다른 단백질에 적용될 수 있음을 시사한다.



























참고문헌


SEQUENCE LISTING
<110> Macfarlane Burnet Institute for Medical Research and Public
Health Limited
<120> Assembled glycoproteins
<130> 526133
<140> PCT/AU2017/051037
<141> 2017-09-22
<150> AU 2016903961
<151> 2016-09-29
<160> 24
<170> PatentIn version 3.5
<210> 1
<211> 774
<212> DNA
<213> Artificial Sequence
<220>
<223> Modified viral sequence
<400> 1
ggtaccgcta gcgccaccat gaaccccctg ctgatcctga cctttgtggc cgctgccctg 60
gccgagacac accagaacat ccagctgatc aacaccaacg gcagctggca catcaacagc 120
accgccctga actgcaacga gagcctgaac acaggctggc tggccggcct gttctaccag 180
cacaagttca acagcagcgg agcccccgag agactggcct cttgtggatc ttctggcgcc 240
tggcactacc cccctagacc ttgtggaatc gtgcccgcca agagcgtgtg cggccctgtg 300
tactgcttca cccctagccc tgtggtcgtg ggcaccaccg atagatctgg cgcccctacc 360
tattcctggg gcgccaacga caccgacgtg ttcgtgctga acaacacccg gccacccctg 420
ggcaattggt tcggctgcac ctggatgaac tccaccggct tcaccaaagt gtgcggcgct 480
cctcctgccg gatccagcgg agcacctacc gacgccttca gaaagcaccc cgaggccacc 540
tactctagag ccggatctgg cccctggatc acccccagat gcatggtgga ctacccctac 600
cggctgtggc actatccctg caccatcaac tacaccatct tcaaagtgcg gatgtacgtg 660
ggcggcgtgg aacacagact ggaagccgcc tgcaactgga ccagaggcga gagagccgac 720
ctggaagatc gggacagaag cgagcaccac caccatcacc actgatgact cgag 774
<210> 2
<211> 747
<212> DNA
<213> Artificial Sequence
<220>
<223> Modified viral sequence
<400> 2
atgaaccccc tgctgatcct gacctttgtg gccgctgccc tggccgagac acaccagaac 60
atccagctga tcaacaccaa cggcagctgg cacatcaaca gcaccgccct gaactgcaac 120
gagagcctga acacaggctg gctggccggc ctgttctacc agcacaagtt caacagcagc 180
ggagcccccg agagactggc ctcttgtgga tcttctggcg cctggcacta cccccctaga 240
ccttgtggaa tcgtgcccgc caagagcgtg tgcggccctg tgtactgctt cacccctagc 300
cctgtggtcg tgggcaccac cgatagatct ggcgccccta cctattcctg gggcgccaac 360
gacaccgacg tgttcgtgct gaacaacacc cggccacccc tgggcaattg gttcggctgc 420
acctggatga actccaccgg cttcaccaaa gtgtgcggcg ctcctcctgc cggatccagc 480
ggagcaccta ccgacgcctt cagaaagcac cccgaggcca cctactctag agccggatct 540
ggcccctgga tcacccccag atgcatggtg gactacccct accggctgtg gcactatccc 600
tgcaccatca actacaccat cttcaaagtg cggatgtacg tgggcggcgt ggaacacaga 660
ctggaagccg cctgcaactg gaccagaggc gagagagccg acctggaaga tcgggacaga 720
agcgagcacc accaccatca ccactga 747
<210> 3
<211> 227
<212> PRT
<213> Artificial Sequence
<220>
<223> Modified viral sequence
<400> 3
Glu Thr His Gln Asn Ile Gln Leu Ile Asn Thr Asn Gly Ser Trp His
1 5 10 15
Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu Ser Leu Asn Thr Gly Trp
20 25 30
Leu Ala Gly Leu Phe Tyr Gln His Lys Phe Asn Ser Ser Gly Ala Pro
35 40 45
Glu Arg Leu Ala Ser Cys Gly Ser Ser Gly Ala Trp His Tyr Pro Pro
50 55 60
Arg Pro Cys Gly Ile Val Pro Ala Lys Ser Val Cys Gly Pro Val Tyr
65 70 75 80
Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr Asp Arg Ser Gly
85 90 95
Ala Pro Thr Tyr Ser Trp Gly Ala Asn Asp Thr Asp Val Phe Val Leu
100 105 110
Asn Asn Thr Arg Pro Pro Leu Gly Asn Trp Phe Gly Cys Thr Trp Met
115 120 125
Asn Ser Thr Gly Phe Thr Lys Val Cys Gly Ala Pro Pro Ala Gly Ser
130 135 140
Ser Gly Ala Pro Thr Asp Ala Phe Arg Lys His Pro Glu Ala Thr Tyr
145 150 155 160
Ser Arg Ala Gly Ser Gly Pro Trp Ile Thr Pro Arg Cys Met Val Asp
165 170 175
Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Ile Asn Tyr Thr Ile
180 185 190
Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg Leu Glu Ala
195 200 205
Ala Cys Asn Trp Thr Arg Gly Glu Arg Ala Asp Leu Glu Asp Arg Asp
210 215 220
Arg Ser Glu
225
<210> 4
<211> 278
<212> PRT
<213> Hepatitis C virus
<400> 4
Glu Thr His Val Thr Gly Gly Asn Ala Gly Arg Thr Thr Ala Gly Leu
1 5 10 15
Val Gly Leu Leu Thr Pro Gly Ala Lys Gln Asn Ile Gln Leu Ile Asn
20 25 30
Thr Asn Gly Ser Trp His Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu
35 40 45
Ser Leu Asn Thr Gly Trp Leu Ala Gly Leu Phe Tyr Gln His Lys Phe
50 55 60
Asn Ser Ser Gly Cys Pro Glu Arg Leu Ala Ser Cys Arg Arg Leu Thr
65 70 75 80
Asp Phe Ala Gln Gly Trp Gly Pro Ile Ser Tyr Ala Asn Gly Ser Gly
85 90 95
Leu Asp Glu Arg Pro Tyr Cys Trp His Tyr Pro Pro Arg Pro Cys Gly
100 105 110
Ile Val Pro Ala Lys Ser Val Cys Gly Pro Val Tyr Cys Phe Thr Pro
115 120 125
Ser Pro Val Val Val Gly Thr Thr Asp Arg Ser Gly Ala Pro Thr Tyr
130 135 140
Ser Trp Gly Ala Asn Asp Thr Asp Val Phe Val Leu Asn Asn Thr Arg
145 150 155 160
Pro Pro Leu Gly Asn Trp Phe Gly Cys Thr Trp Met Asn Ser Thr Gly
165 170 175
Phe Thr Lys Val Cys Gly Ala Pro Pro Cys Val Ile Gly Gly Val Gly
180 185 190
Asn Asn Thr Leu Leu Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu
195 200 205
Ala Thr Tyr Ser Arg Cys Gly Ser Gly Pro Trp Ile Thr Pro Arg Cys
210 215 220
Met Val Asp Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Ile Asn
225 230 235 240
Tyr Thr Ile Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg
245 250 255
Leu Glu Ala Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asp Leu Glu
260 265 270
Asp Arg Asp Arg Ser Glu
275
<210> 5
<211> 227
<212> PRT
<213> Artificial Sequence
<220>
<223> modified E2
<400> 5
Glu Thr His Gln Asn Ile Gln Leu Ile Asn Thr Asn Gly Ser Trp His
1 5 10 15
Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu Ser Leu Asn Thr Gly Trp
20 25 30
Leu Ala Gly Leu Phe Tyr Gln His Lys Phe Asn Ser Ser Gly Cys Pro
35 40 45
Glu Arg Leu Ala Ser Cys Gly Ser Ser Gly Cys Trp His Tyr Pro Pro
50 55 60
Arg Pro Cys Gly Ile Val Pro Ala Lys Ser Val Cys Gly Pro Val Tyr
65 70 75 80
Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr Asp Arg Ser Gly
85 90 95
Ala Pro Thr Tyr Ser Trp Gly Ala Asn Asp Thr Asp Val Phe Val Leu
100 105 110
Asn Asn Thr Arg Pro Pro Leu Gly Asn Trp Phe Gly Cys Thr Trp Met
115 120 125
Asn Ser Thr Gly Phe Thr Lys Val Cys Gly Ala Pro Pro Cys Gly Ser
130 135 140
Ser Gly Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu Ala Thr Tyr
145 150 155 160
Ser Arg Cys Gly Ser Gly Pro Trp Ile Thr Pro Arg Cys Met Val Asp
165 170 175
Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Ile Asn Tyr Thr Ile
180 185 190
Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg Leu Glu Ala
195 200 205
Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asp Leu Glu Asp Arg Asp
210 215 220
Arg Ser Glu
225
<210> 6
<211> 3011
<212> PRT
<213> Hepatitis C virus
<400> 6
Met Ser Thr Asn Pro Lys Pro Gln Arg Lys Thr Lys Arg Asn Thr Asn
1 5 10 15
Arg Arg Pro Gln Asp Val Lys Phe Pro Gly Gly Gly Gln Ile Val Gly
20 25 30
Gly Val Tyr Leu Leu Pro Arg Arg Gly Pro Arg Leu Gly Val Arg Ala
35 40 45
Thr Arg Lys Thr Ser Glu Arg Ser Gln Pro Arg Gly Arg Arg Gln Pro
50 55 60
Ile Pro Lys Ala Arg Arg Pro Glu Gly Arg Thr Trp Ala Gln Pro Gly
65 70 75 80
Tyr Pro Trp Pro Leu Tyr Gly Asn Glu Gly Cys Gly Trp Ala Gly Trp
85 90 95
Leu Leu Ser Pro Arg Gly Ser Arg Pro Ser Trp Gly Pro Thr Asp Pro
100 105 110
Arg Arg Arg Ser Arg Asn Leu Gly Lys Val Ile Asp Thr Leu Thr Cys
115 120 125
Gly Phe Ala Asp Leu Met Gly Tyr Ile Pro Leu Val Gly Ala Pro Leu
130 135 140
Gly Gly Ala Ala Arg Ala Leu Ala His Gly Val Arg Val Leu Glu Asp
145 150 155 160
Gly Val Asn Tyr Ala Thr Gly Asn Leu Pro Gly Cys Ser Phe Ser Ile
165 170 175
Phe Leu Leu Ala Leu Leu Ser Cys Leu Thr Val Pro Ala Ser Ala Tyr
180 185 190
Gln Val Arg Asn Ser Ser Gly Leu Tyr His Val Thr Asn Asp Cys Pro
195 200 205
Asn Ser Ser Ile Val Tyr Glu Ala Ala Asp Ala Ile Leu His Thr Pro
210 215 220
Gly Cys Val Pro Cys Val Arg Glu Gly Asn Ala Ser Arg Cys Trp Val
225 230 235 240
Ala Val Thr Pro Thr Val Ala Thr Arg Asp Gly Lys Leu Pro Thr Thr
245 250 255
Gln Leu Arg Arg His Ile Asp Leu Leu Val Gly Ser Ala Thr Leu Cys
260 265 270
Ser Ala Leu Tyr Val Gly Asp Leu Cys Gly Ser Val Phe Leu Val Gly
275 280 285
Gln Leu Phe Thr Phe Ser Pro Arg Arg His Trp Thr Thr Gln Asp Cys
290 295 300
Asn Cys Ser Ile Tyr Pro Gly His Ile Thr Gly His Arg Met Ala Trp
305 310 315 320
Asp Met Met Met Asn Trp Ser Pro Thr Ala Ala Leu Val Val Ala Gln
325 330 335
Leu Leu Arg Ile Pro Gln Ala Ile Met Asp Met Ile Ala Gly Ala His
340 345 350
Trp Gly Val Leu Ala Gly Ile Ala Tyr Phe Ser Met Val Gly Asn Trp
355 360 365
Ala Lys Val Leu Val Val Leu Leu Leu Phe Ala Gly Val Asp Ala Glu
370 375 380
Thr His Val Thr Gly Gly Ser Ala Gly Arg Thr Thr Ala Gly Leu Val
385 390 395 400
Gly Leu Leu Thr Pro Gly Ala Lys Gln Asn Ile Gln Leu Ile Asn Thr
405 410 415
Asn Gly Ser Trp His Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu Ser
420 425 430
Leu Asn Thr Gly Trp Leu Ala Gly Leu Phe Tyr Gln His Lys Phe Asn
435 440 445
Ser Ser Gly Cys Pro Glu Arg Leu Ala Ser Cys Arg Arg Leu Thr Asp
450 455 460
Phe Ala Gln Gly Trp Gly Pro Ile Ser Tyr Ala Asn Gly Ser Gly Leu
465 470 475 480
Asp Glu Arg Pro Tyr Cys Trp His Tyr Pro Pro Arg Pro Cys Gly Ile
485 490 495
Val Pro Ala Lys Ser Val Cys Gly Pro Val Tyr Cys Phe Thr Pro Ser
500 505 510
Pro Val Val Val Gly Thr Thr Asp Arg Ser Gly Ala Pro Thr Tyr Ser
515 520 525
Trp Gly Ala Asn Asp Thr Asp Val Phe Val Leu Asn Asn Thr Arg Pro
530 535 540
Pro Leu Gly Asn Trp Phe Gly Cys Thr Trp Met Asn Ser Thr Gly Phe
545 550 555 560
Thr Lys Val Cys Gly Ala Pro Pro Cys Val Ile Gly Gly Val Gly Asn
565 570 575
Asn Thr Leu Leu Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu Ala
580 585 590
Thr Tyr Ser Arg Cys Gly Ser Gly Pro Trp Ile Thr Pro Arg Cys Met
595 600 605
Val Asp Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Ile Asn Tyr
610 615 620
Thr Ile Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg Leu
625 630 635 640
Glu Ala Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asp Leu Glu Asp
645 650 655
Arg Asp Arg Ser Glu Leu Ser Pro Leu Leu Leu Ser Thr Thr Gln Trp
660 665 670
Gln Val Leu Pro Cys Ser Phe Thr Thr Leu Pro Ala Leu Ser Thr Gly
675 680 685
Leu Ile His Leu His Gln Asn Ile Val Asp Val Gln Tyr Leu Tyr Gly
690 695 700
Val Gly Ser Ser Ile Ala Ser Trp Ala Ile Lys Trp Glu Tyr Val Val
705 710 715 720
Leu Leu Phe Leu Leu Leu Ala Asp Ala Arg Val Cys Ser Cys Leu Trp
725 730 735
Met Met Leu Leu Ile Ser Gln Ala Glu Ala Ala Leu Glu Asn Leu Val
740 745 750
Ile Leu Asn Ala Ala Ser Leu Ala Gly Thr His Gly Leu Val Ser Phe
755 760 765
Leu Val Phe Phe Cys Phe Ala Trp Tyr Leu Lys Gly Arg Trp Val Pro
770 775 780
Gly Ala Val Tyr Ala Phe Tyr Gly Met Trp Pro Leu Leu Leu Leu Leu
785 790 795 800
Leu Ala Leu Pro Gln Arg Ala Tyr Ala Leu Asp Thr Glu Val Ala Ala
805 810 815
Ser Cys Gly Gly Val Val Leu Val Gly Leu Met Ala Leu Thr Leu Ser
820 825 830
Pro Tyr Tyr Lys Arg Tyr Ile Ser Trp Cys Met Trp Trp Leu Gln Tyr
835 840 845
Phe Leu Thr Arg Val Glu Ala Gln Leu His Val Trp Val Pro Pro Leu
850 855 860
Asn Val Arg Gly Gly Arg Asp Ala Val Ile Leu Leu Met Cys Val Val
865 870 875 880
His Pro Thr Leu Val Phe Asp Ile Thr Lys Leu Leu Leu Ala Ile Phe
885 890 895
Gly Pro Leu Trp Ile Leu Gln Ala Ser Leu Leu Lys Val Pro Tyr Phe
900 905 910
Val Arg Val Gln Gly Leu Leu Arg Ile Cys Ala Leu Ala Arg Lys Ile
915 920 925
Ala Gly Gly His Tyr Val Gln Met Ala Ile Ile Lys Leu Gly Ala Leu
930 935 940
Thr Gly Thr Tyr Val Tyr Asn His Leu Thr Pro Leu Arg Asp Trp Ala
945 950 955 960
His Asn Gly Leu Arg Asp Leu Ala Val Ala Val Glu Pro Val Val Phe
965 970 975
Ser Arg Met Glu Thr Lys Leu Ile Thr Trp Gly Ala Asp Thr Ala Ala
980 985 990
Cys Gly Asp Ile Ile Asn Gly Leu Pro Val Ser Ala Arg Arg Gly Gln
995 1000 1005
Glu Ile Leu Leu Gly Pro Ala Asp Gly Met Val Ser Lys Gly Trp
1010 1015 1020
Arg Leu Leu Ala Pro Ile Thr Ala Tyr Ala Gln Gln Thr Arg Gly
1025 1030 1035
Leu Leu Gly Cys Ile Ile Thr Ser Leu Thr Gly Arg Asp Lys Asn
1040 1045 1050
Gln Val Glu Gly Glu Val Gln Ile Val Ser Thr Ala Thr Gln Thr
1055 1060 1065
Phe Leu Ala Thr Cys Ile Asn Gly Val Cys Trp Thr Val Tyr His
1070 1075 1080
Gly Ala Gly Thr Arg Thr Ile Ala Ser Pro Lys Gly Pro Val Ile
1085 1090 1095
Gln Met Tyr Thr Asn Val Asp Gln Asp Leu Val Gly Trp Pro Ala
1100 1105 1110
Pro Gln Gly Ser Arg Ser Leu Thr Pro Cys Thr Cys Gly Ser Ser
1115 1120 1125
Asp Leu Tyr Leu Val Thr Arg His Ala Asp Val Ile Pro Val Arg
1130 1135 1140
Arg Arg Gly Asp Ser Arg Gly Ser Leu Leu Ser Pro Arg Pro Ile
1145 1150 1155
Ser Tyr Leu Lys Gly Ser Ser Gly Gly Pro Leu Leu Cys Pro Ala
1160 1165 1170
Gly His Ala Val Gly Leu Phe Arg Ala Ala Val Cys Thr Arg Gly
1175 1180 1185
Val Ala Lys Ala Val Asp Phe Ile Pro Val Glu Asn Leu Glu Thr
1190 1195 1200
Thr Met Arg Ser Pro Val Phe Thr Asp Asn Ser Ser Pro Pro Ala
1205 1210 1215
Val Pro Gln Ser Phe Gln Val Ala His Leu His Ala Pro Thr Gly
1220 1225 1230
Ser Gly Lys Ser Thr Lys Val Pro Ala Ala Tyr Ala Ala Gln Gly
1235 1240 1245
Tyr Lys Val Leu Val Leu Asn Pro Ser Val Ala Ala Thr Leu Gly
1250 1255 1260
Phe Gly Ala Tyr Met Ser Lys Ala His Gly Val Asp Pro Asn Ile
1265 1270 1275
Arg Thr Gly Val Arg Thr Ile Thr Thr Gly Ser Pro Ile Thr Tyr
1280 1285 1290
Ser Thr Tyr Gly Lys Phe Leu Ala Asp Gly Gly Cys Ser Gly Gly
1295 1300 1305
Ala Tyr Asp Ile Ile Ile Cys Asp Glu Cys His Ser Thr Asp Ala
1310 1315 1320
Thr Ser Ile Leu Gly Ile Gly Thr Val Leu Asp Gln Ala Glu Thr
1325 1330 1335
Ala Gly Ala Arg Leu Val Val Leu Ala Thr Ala Thr Pro Pro Gly
1340 1345 1350
Ser Val Thr Val Ser His Pro Asn Ile Glu Glu Val Ala Leu Ser
1355 1360 1365
Thr Thr Gly Glu Ile Pro Phe Tyr Gly Lys Ala Ile Pro Leu Glu
1370 1375 1380
Val Ile Lys Gly Gly Arg His Leu Ile Phe Cys His Ser Lys Lys
1385 1390 1395
Lys Cys Asp Glu Leu Ala Ala Lys Leu Val Ala Leu Gly Ile Asn
1400 1405 1410
Ala Val Ala Tyr Tyr Arg Gly Leu Asp Val Ser Val Ile Pro Thr
1415 1420 1425
Ser Gly Asp Val Val Val Val Ser Thr Asp Ala Leu Met Thr Gly
1430 1435 1440
Phe Thr Gly Asp Phe Asp Ser Val Ile Asp Cys Asn Thr Cys Val
1445 1450 1455
Thr Gln Thr Val Asp Phe Ser Leu Asp Pro Thr Phe Thr Ile Glu
1460 1465 1470
Thr Thr Thr Leu Pro Gln Asp Ala Val Ser Arg Thr Gln Arg Arg
1475 1480 1485
Gly Arg Thr Gly Arg Gly Lys Pro Gly Ile Tyr Arg Phe Val Ala
1490 1495 1500
Pro Gly Glu Arg Pro Ser Gly Met Phe Asp Ser Ser Val Leu Cys
1505 1510 1515
Glu Cys Tyr Asp Ala Gly Cys Ala Trp Tyr Glu Leu Thr Pro Ala
1520 1525 1530
Glu Thr Thr Val Arg Leu Arg Ala Tyr Met Asn Thr Pro Gly Leu
1535 1540 1545
Pro Val Cys Gln Asp His Leu Glu Phe Trp Glu Gly Val Phe Thr
1550 1555 1560
Gly Leu Thr His Ile Asp Ala His Phe Leu Ser Gln Thr Lys Gln
1565 1570 1575
Ser Gly Glu Asn Phe Pro Tyr Leu Val Ala Tyr Gln Ala Thr Val
1580 1585 1590
Cys Ala Arg Ala Gln Ala Pro Pro Pro Ser Trp Asp Gln Met Trp
1595 1600 1605
Lys Cys Leu Ile Arg Leu Lys Pro Thr Leu His Gly Pro Thr Pro
1610 1615 1620
Leu Leu Tyr Arg Leu Gly Ala Val Gln Asn Glu Val Thr Leu Thr
1625 1630 1635
His Pro Ile Thr Lys Tyr Ile Met Thr Cys Met Ser Ala Asp Leu
1640 1645 1650
Glu Val Val Thr Ser Thr Trp Val Leu Val Gly Gly Val Leu Ala
1655 1660 1665
Ala Leu Ala Ala Tyr Cys Leu Ser Thr Gly Cys Val Val Ile Val
1670 1675 1680
Gly Arg Ile Val Leu Ser Gly Lys Pro Ala Ile Ile Pro Asp Arg
1685 1690 1695
Glu Val Leu Tyr Gln Glu Phe Asp Glu Met Glu Glu Cys Ser Gln
1700 1705 1710
His Leu Pro Tyr Ile Glu Gln Gly Met Met Leu Ala Glu Gln Phe
1715 1720 1725
Lys Gln Lys Ala Leu Gly Leu Leu Gln Thr Ala Ser Arg Gln Ala
1730 1735 1740
Glu Val Ile Thr Pro Ala Val Gln Thr Asn Trp Gln Lys Leu Glu
1745 1750 1755
Val Phe Trp Ala Lys His Met Trp Asn Phe Ile Ser Gly Ile Gln
1760 1765 1770
Tyr Leu Ala Gly Leu Ser Thr Leu Pro Gly Asn Pro Ala Ile Ala
1775 1780 1785
Ser Leu Met Ala Phe Thr Ala Ala Val Thr Ser Pro Leu Thr Thr
1790 1795 1800
Gly Gln Thr Leu Leu Phe Asn Ile Leu Gly Gly Trp Val Ala Ala
1805 1810 1815
Gln Leu Ala Ala Pro Gly Ala Ala Thr Ala Phe Val Gly Ala Gly
1820 1825 1830
Leu Ala Gly Ala Ala Ile Gly Ser Val Gly Leu Gly Lys Val Leu
1835 1840 1845
Val Asp Ile Leu Ala Gly Tyr Gly Ala Gly Val Ala Gly Ala Leu
1850 1855 1860
Val Ala Phe Lys Ile Met Ser Gly Glu Val Pro Ser Thr Glu Asp
1865 1870 1875
Leu Val Asn Leu Leu Pro Ala Ile Leu Ser Pro Gly Ala Leu Val
1880 1885 1890
Val Gly Val Val Cys Ala Ala Ile Leu Arg Arg His Val Gly Pro
1895 1900 1905
Gly Glu Gly Ala Val Gln Trp Met Asn Arg Leu Ile Ala Phe Ala
1910 1915 1920
Ser Arg Gly Asn His Val Ser Pro Thr His Tyr Val Pro Glu Ser
1925 1930 1935
Asp Ala Ala Ala Arg Val Thr Ala Ile Leu Ser Ser Leu Thr Val
1940 1945 1950
Thr Gln Leu Leu Arg Arg Leu His Gln Trp Ile Ser Ser Glu Cys
1955 1960 1965
Thr Thr Pro Cys Ser Gly Ser Trp Leu Arg Asp Ile Trp Asp Trp
1970 1975 1980
Ile Cys Glu Val Leu Ser Asp Phe Lys Thr Trp Leu Lys Ala Lys
1985 1990 1995
Leu Met Pro Gln Leu Pro Gly Ile Pro Phe Val Ser Cys Gln Arg
2000 2005 2010
Gly Tyr Arg Gly Val Trp Arg Gly Asp Gly Ile Met His Thr Arg
2015 2020 2025
Cys His Cys Gly Ala Glu Ile Thr Gly His Val Lys Asn Gly Thr
2030 2035 2040
Met Arg Ile Val Gly Pro Arg Thr Cys Arg Asn Met Trp Ser Gly
2045 2050 2055
Thr Phe Pro Ile Asn Ala Tyr Thr Thr Gly Pro Cys Thr Pro Leu
2060 2065 2070
Pro Ala Pro Asn Tyr Lys Phe Ala Leu Trp Arg Val Ser Ala Glu
2075 2080 2085
Glu Tyr Val Glu Ile Arg Arg Val Gly Asp Phe His Tyr Val Ser
2090 2095 2100
Gly Met Thr Thr Asp Asn Leu Lys Cys Pro Cys Gln Ile Pro Ser
2105 2110 2115
Pro Glu Phe Phe Thr Glu Leu Asp Gly Val Arg Leu His Arg Phe
2120 2125 2130
Ala Pro Pro Cys Lys Pro Leu Leu Arg Glu Glu Val Ser Phe Arg
2135 2140 2145
Val Gly Leu His Glu Tyr Pro Val Gly Ser Gln Leu Pro Cys Glu
2150 2155 2160
Pro Glu Pro Asp Val Ala Val Leu Thr Ser Met Leu Thr Asp Pro
2165 2170 2175
Ser His Ile Thr Ala Glu Ala Ala Gly Arg Arg Leu Ala Arg Gly
2180 2185 2190
Ser Pro Pro Ser Met Ala Ser Ser Ser Ala Ser Gln Leu Ser Ala
2195 2200 2205
Pro Ser Leu Lys Ala Thr Cys Thr Ala Asn His Asp Ser Pro Asp
2210 2215 2220
Ala Glu Leu Ile Glu Ala Asn Leu Leu Trp Arg Gln Glu Met Gly
2225 2230 2235
Gly Asn Ile Thr Arg Val Glu Ser Glu Asn Lys Val Val Ile Leu
2240 2245 2250
Asp Ser Phe Asp Pro Leu Val Ala Glu Glu Asp Glu Arg Glu Val
2255 2260 2265
Ser Val Pro Ala Glu Ile Leu Arg Lys Ser Arg Arg Phe Ala Arg
2270 2275 2280
Ala Leu Pro Val Trp Ala Arg Pro Asp Tyr Asn Pro Pro Leu Val
2285 2290 2295
Glu Thr Trp Lys Lys Pro Asp Tyr Glu Pro Pro Val Val His Gly
2300 2305 2310
Cys Pro Leu Pro Pro Pro Arg Ser Pro Pro Val Pro Pro Pro Arg
2315 2320 2325
Lys Lys Arg Thr Val Val Leu Thr Glu Ser Thr Leu Ser Thr Ala
2330 2335 2340
Leu Ala Glu Leu Ala Thr Lys Ser Phe Gly Ser Ser Ser Thr Ser
2345 2350 2355
Gly Ile Thr Gly Asp Asn Thr Thr Thr Ser Ser Glu Pro Ala Pro
2360 2365 2370
Ser Gly Cys Pro Pro Asp Ser Asp Val Glu Ser Tyr Ser Ser Met
2375 2380 2385
Pro Pro Leu Glu Gly Glu Pro Gly Asp Pro Asp Leu Ser Asp Gly
2390 2395 2400
Ser Trp Ser Thr Val Ser Ser Gly Ala Asp Thr Glu Asp Val Val
2405 2410 2415
Cys Cys Ser Met Ser Tyr Ser Trp Thr Gly Ala Leu Val Thr Pro
2420 2425 2430
Cys Ala Ala Glu Glu Gln Lys Leu Pro Ile Asn Ala Leu Ser Asn
2435 2440 2445
Ser Leu Leu Arg His His Asn Leu Val Tyr Ser Thr Thr Ser Arg
2450 2455 2460
Ser Ala Cys Gln Arg Gln Lys Lys Val Thr Phe Asp Arg Leu Gln
2465 2470 2475
Val Leu Asp Ser His Tyr Gln Asp Val Leu Lys Glu Val Lys Ala
2480 2485 2490
Ala Ala Ser Lys Val Lys Ala Asn Leu Leu Ser Val Glu Glu Ala
2495 2500 2505
Cys Ser Leu Thr Pro Pro His Ser Ala Lys Ser Lys Phe Gly Tyr
2510 2515 2520
Gly Ala Lys Asp Val Arg Cys His Ala Arg Lys Ala Val Ala His
2525 2530 2535
Ile Asn Ser Val Trp Lys Asp Leu Leu Glu Asp Ser Val Thr Pro
2540 2545 2550
Ile Asp Thr Thr Ile Met Ala Lys Asn Glu Val Phe Cys Val Gln
2555 2560 2565
Pro Glu Lys Gly Gly Arg Lys Pro Ala Arg Leu Ile Val Phe Pro
2570 2575 2580
Asp Leu Gly Val Arg Val Cys Glu Lys Met Ala Leu Tyr Asp Val
2585 2590 2595
Val Ser Lys Leu Pro Leu Ala Val Met Gly Ser Ser Tyr Gly Phe
2600 2605 2610
Gln Tyr Ser Pro Gly Gln Arg Val Glu Phe Leu Val Gln Ala Trp
2615 2620 2625
Lys Ser Lys Lys Thr Pro Met Gly Phe Ser Tyr Asp Thr Arg Cys
2630 2635 2640
Phe Asp Ser Thr Val Thr Glu Ser Asp Ile Arg Thr Glu Glu Ala
2645 2650 2655
Ile Tyr Gln Cys Cys Asp Leu Asp Pro Gln Ala Arg Val Ala Ile
2660 2665 2670
Lys Ser Leu Thr Glu Arg Leu Tyr Val Gly Gly Pro Leu Thr Asn
2675 2680 2685
Ser Arg Gly Glu Asn Cys Gly Tyr Arg Arg Cys Arg Ala Ser Gly
2690 2695 2700
Val Leu Thr Thr Ser Cys Gly Asn Thr Leu Thr Cys Tyr Ile Lys
2705 2710 2715
Ala Arg Ala Ala Cys Arg Ala Ala Gly Leu Gln Asp Cys Thr Met
2720 2725 2730
Leu Val Cys Gly Asp Asp Leu Val Val Ile Cys Glu Ser Ala Gly
2735 2740 2745
Val Gln Glu Asp Ala Ala Ser Leu Arg Ala Phe Thr Glu Ala Met
2750 2755 2760
Thr Arg Tyr Ser Ala Pro Pro Gly Asp Pro Pro Gln Pro Glu Tyr
2765 2770 2775
Asp Leu Glu Leu Ile Thr Ser Cys Ser Ser Asn Val Ser Val Ala
2780 2785 2790
His Asp Gly Ala Gly Lys Arg Val Tyr Tyr Leu Thr Arg Asp Pro
2795 2800 2805
Thr Thr Pro Leu Ala Arg Ala Ala Trp Glu Thr Ala Arg His Thr
2810 2815 2820
Pro Val Asn Ser Trp Leu Gly Asn Ile Ile Met Phe Ala Pro Thr
2825 2830 2835
Leu Trp Ala Arg Met Ile Leu Met Thr His Phe Phe Ser Val Leu
2840 2845 2850
Ile Ala Arg Asp Gln Leu Glu Gln Ala Leu Asn Cys Glu Ile Tyr
2855 2860 2865
Gly Ala Cys Tyr Ser Ile Glu Pro Leu Asp Leu Pro Pro Ile Ile
2870 2875 2880
Gln Arg Leu His Gly Leu Ser Ala Phe Ser Leu His Ser Tyr Ser
2885 2890 2895
Pro Gly Glu Ile Asn Arg Val Ala Ala Cys Leu Arg Lys Leu Gly
2900 2905 2910
Val Pro Pro Leu Arg Ala Trp Arg His Arg Ala Arg Ser Val Arg
2915 2920 2925
Ala Arg Leu Leu Ser Arg Gly Gly Arg Ala Ala Ile Cys Gly Lys
2930 2935 2940
Tyr Leu Phe Asn Trp Ala Val Arg Thr Lys Leu Lys Leu Thr Pro
2945 2950 2955
Ile Ala Ala Ala Gly Arg Leu Asp Leu Ser Gly Trp Phe Thr Ala
2960 2965 2970
Gly Tyr Ser Gly Gly Asp Ile Tyr His Ser Val Ser His Ala Arg
2975 2980 2985
Pro Arg Trp Phe Trp Phe Cys Leu Leu Leu Leu Ala Ala Gly Val
2990 2995 3000
Gly Ile Tyr Leu Leu Pro Asn Arg
3005 3010
<210> 7
<211> 363
<212> PRT
<213> Hepatitis C virus
<400> 7
Glu Thr His Val Thr Gly Gly Ser Ala Gly Arg Thr Thr Ala Gly Leu
1 5 10 15
Val Gly Leu Leu Thr Pro Gly Ala Lys Gln Asn Ile Gln Leu Ile Asn
20 25 30
Thr Asn Gly Ser Trp His Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu
35 40 45
Ser Leu Asn Thr Gly Trp Leu Ala Gly Leu Phe Tyr Gln His Lys Phe
50 55 60
Asn Ser Ser Gly Cys Pro Glu Arg Leu Ala Ser Cys Arg Arg Leu Thr
65 70 75 80
Asp Phe Ala Gln Gly Trp Gly Pro Ile Ser Tyr Ala Asn Gly Ser Gly
85 90 95
Leu Asp Glu Arg Pro Tyr Cys Trp His Tyr Pro Pro Arg Pro Cys Gly
100 105 110
Ile Val Pro Ala Lys Ser Val Cys Gly Pro Val Tyr Cys Phe Thr Pro
115 120 125
Ser Pro Val Val Val Gly Thr Thr Asp Arg Ser Gly Ala Pro Thr Tyr
130 135 140
Ser Trp Gly Ala Asn Asp Thr Asp Val Phe Val Leu Asn Asn Thr Arg
145 150 155 160
Pro Pro Leu Gly Asn Trp Phe Gly Cys Thr Trp Met Asn Ser Thr Gly
165 170 175
Phe Thr Lys Val Cys Gly Ala Pro Pro Cys Val Ile Gly Gly Val Gly
180 185 190
Asn Asn Thr Leu Leu Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu
195 200 205
Ala Thr Tyr Ser Arg Cys Gly Ser Gly Pro Trp Ile Thr Pro Arg Cys
210 215 220
Met Val Asp Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Ile Asn
225 230 235 240
Tyr Thr Ile Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg
245 250 255
Leu Glu Ala Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asp Leu Glu
260 265 270
Asp Arg Asp Arg Ser Glu Leu Ser Pro Leu Leu Leu Ser Thr Thr Gln
275 280 285
Trp Gln Val Leu Pro Cys Ser Phe Thr Thr Leu Pro Ala Leu Ser Thr
290 295 300
Gly Leu Ile His Leu His Gln Asn Ile Val Asp Val Gln Tyr Leu Tyr
305 310 315 320
Gly Val Gly Ser Ser Ile Ala Ser Trp Ala Ile Lys Trp Glu Tyr Val
325 330 335
Val Leu Leu Phe Leu Leu Leu Ala Asp Ala Arg Val Cys Ser Cys Leu
340 345 350
Trp Met Met Leu Leu Ile Ser Gln Ala Glu Ala
355 360
<210> 8
<211> 278
<212> PRT
<213> Hepatitis C virus
<400> 8
Glu Thr His Val Thr Gly Gly Ser Ala Gly Arg Thr Thr Ala Gly Leu
1 5 10 15
Val Gly Leu Leu Thr Pro Gly Ala Lys Gln Asn Ile Gln Leu Ile Asn
20 25 30
Thr Asn Gly Ser Trp His Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu
35 40 45
Ser Leu Asn Thr Gly Trp Leu Ala Gly Leu Phe Tyr Gln His Lys Phe
50 55 60
Asn Ser Ser Gly Cys Pro Glu Arg Leu Ala Ser Cys Arg Arg Leu Thr
65 70 75 80
Asp Phe Ala Gln Gly Trp Gly Pro Ile Ser Tyr Ala Asn Gly Ser Gly
85 90 95
Leu Asp Glu Arg Pro Tyr Cys Trp His Tyr Pro Pro Arg Pro Cys Gly
100 105 110
Ile Val Pro Ala Lys Ser Val Cys Gly Pro Val Tyr Cys Phe Thr Pro
115 120 125
Ser Pro Val Val Val Gly Thr Thr Asp Arg Ser Gly Ala Pro Thr Tyr
130 135 140
Ser Trp Gly Ala Asn Asp Thr Asp Val Phe Val Leu Asn Asn Thr Arg
145 150 155 160
Pro Pro Leu Gly Asn Trp Phe Gly Cys Thr Trp Met Asn Ser Thr Gly
165 170 175
Phe Thr Lys Val Cys Gly Ala Pro Pro Cys Val Ile Gly Gly Val Gly
180 185 190
Asn Asn Thr Leu Leu Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu
195 200 205
Ala Thr Tyr Ser Arg Cys Gly Ser Gly Pro Trp Ile Thr Pro Arg Cys
210 215 220
Met Val Asp Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Ile Asn
225 230 235 240
Tyr Thr Ile Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg
245 250 255
Leu Glu Ala Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asp Leu Glu
260 265 270
Asp Arg Asp Arg Ser Glu
275
<210> 9
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> N-Terminal signal sequence
<400> 9
Met Asn Pro Leu Leu Ile Leu Thr Phe Val Ala Ala Ala Leu Ala
1 5 10 15
<210> 10
<211> 353
<212> PRT
<213> Hepatitis C virus
<400> 10
Tyr Phe Ser Met Gln Ala Asn Trp Ala Lys Val Ile Leu Val Leu Phe
1 5 10 15
Leu Phe Ala Gly Val Asp Ala Glu Thr His Val Ser Gly Ala Ala Val
20 25 30
Gly Arg Ser Thr Ala Gly Leu Ala Asn Leu Phe Ser Ser Gly Ser Lys
35 40 45
Gln Asn Leu Gln Leu Ile Asn Ser Asn Gly Ser Trp His Ile Asn Arg
50 55 60
Thr Ala Leu Asn Cys Asn Asp Ser Leu Asn Thr Gly Phe Leu Ala Ser
65 70 75 80
Leu Phe Tyr Thr His Lys Phe Asn Ser Ser Gly Cys Ser Glu Arg Leu
85 90 95
Ala Cys Cys Lys Ser Leu Asp Ser Tyr Gly Gln Gly Trp Gly Pro Leu
100 105 110
Gly Val Ala Asn Ile Ser Gly Ser Ser Asp Asp Arg Pro Tyr Cys Trp
115 120 125
His Tyr Ala Pro Arg Pro Cys Gly Ile Val Pro Ala Ser Ser Val Cys
130 135 140
Gly Pro Val Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr
145 150 155 160
Asp His Val Gly Val Pro Thr Tyr Thr Trp Gly Glu Asn Glu Thr Asp
165 170 175
Val Phe Leu Leu Asn Ser Thr Arg Pro Pro His Gly Ala Trp Phe Gly
180 185 190
Cys Val Trp Met Asn Ser Thr Gly Phe Thr Lys Thr Cys Gly Ala Pro
195 200 205
Pro Cys Glu Val Asn Thr Asn Asn Gly Thr Trp His Cys Pro Thr Asp
210 215 220
Cys Phe Arg Lys His Pro Glu Thr Thr Tyr Ala Lys Cys Gly Ser Gly
225 230 235 240
Pro Trp Ile Thr Pro Arg Cys Leu Ile Asp Tyr Pro Tyr Arg Leu Trp
245 250 255
His Phe Pro Cys Thr Ala Asn Phe Ser Val Phe Asn Ile Arg Thr Phe
260 265 270
Val Gly Gly Ile Glu His Arg Met Gln Ala Ala Cys Asn Trp Thr Arg
275 280 285
Gly Glu Val Cys Gly Leu Glu His Arg Asp Arg Val Glu Leu Ser Pro
290 295 300
Leu Leu Leu Thr Thr Thr Ala Trp Gln Ile Leu Pro Cys Ser Phe Thr
305 310 315 320
Thr Leu Pro Ala Leu Ser Thr Gly Leu Ile His Leu His Gln Asn Ile
325 330 335
Val Asp Val Gln Tyr Leu Tyr Gly Val Gly Ser Ala Val Val Ser Trp
340 345 350
Ala
<210> 11
<211> 353
<212> PRT
<213> Hepatitis C virus
<400> 11
Tyr Phe Ser Met Val Gly Asn Trp Ala Lys Val Leu Val Val Leu Leu
1 5 10 15
Leu Phe Ala Gly Val Asp Ala Glu Thr His Val Thr Gly Gly Asn Ala
20 25 30
Gly Arg Thr Thr Ala Gly Leu Val Gly Leu Leu Thr Pro Gly Ala Lys
35 40 45
Gln Asn Ile Gln Leu Ile Asn Thr Asn Gly Ser Trp His Ile Asn Ser
50 55 60
Thr Ala Leu Asn Cys Asn Glu Ser Leu Asn Thr Gly Trp Leu Ala Gly
65 70 75 80
Leu Phe Tyr Gln His Lys Phe Asn Ser Ser Gly Cys Pro Glu Arg Leu
85 90 95
Ala Ser Cys Arg Arg Leu Thr Asp Phe Ala Gln Gly Trp Gly Pro Ile
100 105 110
Ser Tyr Ala Asn Gly Ser Gly Leu Asp Glu Arg Pro Tyr Cys Trp His
115 120 125
Tyr Pro Pro Arg Pro Cys Gly Ile Val Pro Ala Lys Ser Val Cys Gly
130 135 140
Pro Val Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr Asp
145 150 155 160
Arg Ser Gly Ala Pro Thr Tyr Ser Trp Gly Ala Asn Asp Thr Asp Val
165 170 175
Phe Val Leu Asn Asn Thr Arg Pro Pro Leu Gly Asn Trp Phe Gly Cys
180 185 190
Thr Trp Met Asn Ser Thr Gly Phe Thr Lys Val Cys Gly Ala Pro Pro
195 200 205
Cys Val Ile Gly Gly Val Gly Asn Asn Thr Leu Leu Cys Pro Thr Asp
210 215 220
Cys Phe Arg Lys His Pro Glu Ala Thr Tyr Ser Arg Cys Gly Ser Gly
225 230 235 240
Pro Trp Ile Thr Pro Arg Cys Met Val Asp Tyr Pro Tyr Arg Leu Trp
245 250 255
His Tyr Pro Cys Thr Ile Asn Tyr Thr Ile Phe Lys Val Arg Met Tyr
260 265 270
Val Gly Gly Val Glu His Arg Leu Glu Ala Ala Cys Asn Trp Thr Arg
275 280 285
Gly Glu Arg Cys Asp Leu Glu Asp Arg Asp Arg Ser Glu Leu Ser Pro
290 295 300
Leu Leu Leu Ser Thr Thr Gln Trp Gln Val Leu Pro Cys Ser Phe Thr
305 310 315 320
Thr Leu Pro Ala Leu Ser Thr Gly Leu Ile His Leu His Gln Asn Ile
325 330 335
Val Asp Val Gln Tyr Leu Tyr Gly Val Gly Ser Ser Ile Ala Ser Trp
340 345 350
Ala
<210> 12
<211> 354
<212> PRT
<213> Hepatitis C virus
<400> 12
Tyr Tyr Ala Ser Ala Ala Asn Trp Ala Lys Val Val Leu Val Leu Phe
1 5 10 15
Leu Phe Ala Gly Val Asp Ala Asn Thr Arg Thr Val Gly Gly Ser Ala
20 25 30
Ala Gln Gly Ala Arg Gly Leu Ala Ser Leu Phe Thr Pro Gly Pro Gln
35 40 45
Gln Asn Leu Gln Leu Ile Asn Thr Asn Gly Ser Trp His Ile Asn Arg
50 55 60
Thr Ala Leu Asn Cys Asn Asp Ser Leu Gln Thr Gly Phe Val Ala Gly
65 70 75 80
Leu Leu Tyr Tyr His Lys Phe Asn Ser Thr Gly Cys Pro Gln Arg Met
85 90 95
Ala Ser Cys Arg Pro Leu Ala Ala Phe Asp Gln Gly Trp Gly Thr Ile
100 105 110
Ser Tyr Ala Ala Val Ser Gly Pro Ser Asp Asp Lys Pro Tyr Cys Trp
115 120 125
His Tyr Pro Pro Arg Pro Cys Gly Ile Val Pro Ala Arg Gly Val Cys
130 135 140
Gly Pro Val Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr
145 150 155 160
Asp Arg Lys Gly Asn Pro Thr Tyr Ser Trp Gly Glu Asn Glu Thr Asp
165 170 175
Ile Phe Leu Leu Asn Asn Thr Arg Pro Pro Thr Gly Asn Trp Phe Gly
180 185 190
Cys Thr Trp Met Asn Ser Thr Gly Phe Val Lys Thr Cys Gly Ala Pro
195 200 205
Pro Cys Asn Leu Gly Pro Thr Gly Asn Asn Ser Leu Lys Cys Pro Thr
210 215 220
Asp Cys Phe Arg Lys His Pro Asp Ala Thr Tyr Thr Lys Cys Gly Ser
225 230 235 240
Gly Pro Trp Leu Thr Pro Arg Cys Leu Val His Tyr Pro Tyr Arg Leu
245 250 255
Trp His Tyr Pro Cys Thr Leu Asn Tyr Thr Ile Phe Lys Val Arg Met
260 265 270
Tyr Ile Gly Gly Leu Glu His Arg Leu Glu Val Ala Cys Asn Trp Thr
275 280 285
Arg Gly Glu Arg Cys Asp Leu Glu Asp Arg Asp Arg Ala Glu Leu Ser
290 295 300
Pro Leu Leu His Thr Thr Thr Gln Trp Ala Ile Leu Pro Cys Ser Phe
305 310 315 320
Thr Pro Thr Pro Ala Leu Ser Thr Gly Leu Ile His Leu His Gln Asn
325 330 335
Ile Val Asp Thr Gln Tyr Leu Tyr Gly Leu Ser Ser Ser Ile Val Ser
340 345 350
Trp Ala
<210> 13
<211> 359
<212> PRT
<213> Hepatitis C virus
<400> 13
Tyr Tyr Ser Met Gln Gly Asn Trp Ala Lys Val Ala Ile Val Met Ile
1 5 10 15
Met Phe Ser Gly Val Asp Ala Glu Thr Tyr Val Thr Gly Gly Ser Val
20 25 30
Ala His Ser Ala Arg Gly Leu Thr Ser Leu Phe Ser Met Gly Ala Lys
35 40 45
Gln Lys Leu Gln Leu Val Asn Thr Asn Gly Ser Trp His Ile Asn Ser
50 55 60
Thr Ala Leu Asn Cys Asn Glu Ser Ile Asn Thr Gly Phe Ile Ala Gly
65 70 75 80
Leu Phe Tyr Tyr His Lys Phe Asn Ser Thr Gly Cys Pro Gln Arg Leu
85 90 95
Ser Ser Cys Lys Pro Ile Ile Ser Phe Arg Gln Gly Trp Gly Pro Leu
100 105 110
Thr Asp Ala Asn Ile Thr Gly Pro Ser Asp Asp Arg Pro Tyr Cys Trp
115 120 125
His Tyr Ala Pro Arg Pro Cys Ser Val Val Pro Ala Ser Ser Val Cys
130 135 140
Gly Pro Val Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr
145 150 155 160
Asp Ile Lys Gly Arg Pro Thr Tyr Asn Trp Gly Glu Asn Glu Thr Asp
165 170 175
Val Phe Leu Leu Glu Ser Leu Arg Pro Pro Ser Gly Arg Trp Phe Gly
180 185 190
Cys Ala Trp Met Asn Ser Thr Gly Phe Leu Lys Thr Cys Gly Ala Pro
195 200 205
Pro Cys Asn Ile Tyr Gly Gly Glu Gly Asp Pro Glu Asn Glu Thr Asp
210 215 220
Leu Phe Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu Ala Thr Tyr
225 230 235 240
Ser Arg Cys Gly Ala Gly Pro Trp Leu Thr Pro Arg Cys Met Val Asp
245 250 255
Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Val Asn Phe Thr Leu
260 265 270
Phe Lys Val Arg Met Phe Val Gly Gly Phe Glu His Arg Phe Thr Ala
275 280 285
Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asn Ile Glu Asp Arg Asp
290 295 300
Arg Ser Glu Gln His Pro Leu Leu His Ser Thr Thr Glu Leu Ala Ile
305 310 315 320
Leu Pro Cys Ser Phe Thr Pro Met Pro Ala Leu Ser Thr Gly Leu Ile
325 330 335
His Leu His Gln Asn Ile Val Asp Val Gln Tyr Leu Tyr Gly Val Gly
340 345 350
Ser Asp Met Val Gly Trp Ala
355
<210> 14
<211> 357
<212> PRT
<213> Hepatitis C virus
<400> 14
Tyr Phe Gly Met Ala Gly Asn Trp Leu Lys Val Leu Ala Val Leu Phe
1 5 10 15
Leu Phe Ala Gly Val Glu Ala Gln Thr Met Ile Ala His Gly Val Ser
20 25 30
Gln Thr Thr Ser Gly Phe Ala Ser Leu Leu Thr Pro Gly Ala Lys Gln
35 40 45
Asn Ile Gln Leu Ile Asn Thr Asn Gly Ser Trp His Ile Asn Arg Thr
50 55 60
Ala Leu Asn Cys Asn Asp Ser Leu Gln Thr Gly Phe Leu Ala Ser Leu
65 70 75 80
Phe Tyr Thr His Lys Phe Asn Ser Ser Gly Cys Pro Glu Arg Met Ala
85 90 95
Ala Cys Lys Pro Leu Ala Glu Phe Arg Gln Gly Trp Gly Gln Ile Thr
100 105 110
His Lys Asn Val Ser Gly Pro Ser Asp Asp Arg Pro Tyr Cys Trp His
115 120 125
Tyr Ala Pro Arg Pro Cys Glu Val Val Pro Ala Arg Ser Val Cys Gly
130 135 140
Pro Val Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr Asp
145 150 155 160
Lys Arg Gly Asn Pro Thr Tyr Thr Trp Gly Glu Asn Glu Thr Asp Val
165 170 175
Phe Met Leu Glu Ser Leu Arg Pro Pro Thr Gly Gly Trp Phe Gly Cys
180 185 190
Thr Trp Met Asn Ser Thr Gly Phe Thr Lys Thr Cys Gly Ala Pro Pro
195 200 205
Cys Gln Ile Val Pro Gly Asn Tyr Asn Ser Ser Ala Asn Glu Leu Leu
210 215 220
Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu Ala Thr Tyr Gln Arg
225 230 235 240
Cys Gly Ser Gly Pro Trp Val Thr Pro Arg Cys Leu Val Asp Tyr Ala
245 250 255
Tyr Arg Leu Trp His Tyr Pro Cys Thr Val Asn Phe Thr Leu His Lys
260 265 270
Val Arg Met Phe Val Gly Gly Thr Glu His Arg Phe Asp Val Ala Cys
275 280 285
Asn Trp Thr Arg Gly Glu Arg Cys Glu Leu His Asp Arg Asn Arg Ile
290 295 300
Glu Met Ser Pro Leu Leu Phe Ser Thr Thr Gln Leu Ser Ile Leu Pro
305 310 315 320
Cys Ser Phe Ser Thr Met Pro Ala Leu Ser Thr Gly Leu Ile His Leu
325 330 335
His Gln Asn Ile Val Asp Val Gln Tyr Leu Tyr Gly Val Ser Thr Asn
340 345 350
Val Thr Ser Trp Val
355
<210> 15
<211> 357
<212> PRT
<213> Hepatitis C virus
<400> 15
Tyr Phe Gly Met Ala Gly Asn Trp Ala Lys Val Ile Leu Ile Met Leu
1 5 10 15
Leu Met Ser Gly Val Asp Ala Glu Thr Met Ala Val Gly Ala Arg Ala
20 25 30
Ala His Thr Thr Gly Ala Leu Val Ser Leu Leu Asn Pro Gly Pro Ser
35 40 45
Gln Arg Leu Gln Leu Ile Asn Thr Asn Gly Ser Trp His Ile Asn Arg
50 55 60
Thr Ala Leu Asn Cys Asn Asp Ser Leu Gln Thr Gly Phe Ile Ala Ala
65 70 75 80
Leu Phe Tyr Thr His Arg Phe Asn Ser Ser Gly Cys Pro Glu Arg Met
85 90 95
Ala Ser Cys Lys Pro Leu Ser Asp Phe Asp Gln Gly Trp Gly Pro Leu
100 105 110
Trp Tyr Asn Ser Thr Glu Arg Pro Ser Asp Gln Arg Pro Tyr Cys Trp
115 120 125
His Tyr Ala Pro Ser Pro Cys Gly Ile Val Pro Ala Lys Asp Val Cys
130 135 140
Gly Pro Val Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr
145 150 155 160
Asp Arg Arg Gly Val Pro Thr Tyr Thr Trp Gly Glu Asn Glu Ser Asp
165 170 175
Val Phe Leu Leu Asn Ser Thr Arg Pro Pro Gln Gly Ser Trp Phe Gly
180 185 190
Cys Ser Trp Met Asn Thr Thr Gly Phe Thr Lys Thr Cys Gly Gly Pro
195 200 205
Pro Cys Lys Ile Arg Pro Gln Gly Ala Gln Ser Asn Thr Ser Leu Thr
210 215 220
Cys Pro Thr Asp Cys Phe Arg Lys His Pro Arg Ala Thr Tyr Ser Ala
225 230 235 240
Cys Gly Ser Gly Pro Trp Leu Thr Pro Arg Cys Met Val His Tyr Pro
245 250 255
Tyr Arg Leu Trp His Tyr Pro Cys Thr Val Asn Phe Thr Ile His Lys
260 265 270
Val Arg Leu Tyr Ile Gly Gly Val Glu His Arg Leu Asp Ala Ala Cys
275 280 285
Asn Trp Thr Arg Gly Glu Arg Cys Asp Leu Glu Asp Arg Asp Arg Val
290 295 300
Asp Met Ser Pro Leu Leu His Ser Thr Thr Glu Leu Ala Ile Leu Pro
305 310 315 320
Cys Ser Phe Val Pro Leu Pro Ala Leu Ser Thr Gly Leu Ile His Leu
325 330 335
His Gln Asn Ile Val Asp Ala Gln Tyr Leu Tyr Gly Leu Ser Pro Ala
340 345 350
Ile Ile Ser Trp Ala
355
<210> 16
<211> 357
<212> PRT
<213> Hepatitis C virus
<400> 16
Tyr Phe Ser Met Gln Gly Ala Trp Ala Lys Val Val Val Ile Leu Leu
1 5 10 15
Leu Ala Ala Gly Val Asp Ala Arg Thr His Thr Val Gly Gly Ser Ala
20 25 30
Ala Gln Thr Thr Gly Arg Leu Thr Ser Leu Phe Asp Met Gly Pro Arg
35 40 45
Gln Lys Ile Gln Leu Val Asn Thr Asn Gly Ser Trp His Ile Asn Arg
50 55 60
Thr Ala Leu Asn Cys Asn Asp Ser Leu His Thr Gly Phe Ile Ala Ser
65 70 75 80
Leu Phe Tyr Thr His Ser Phe Asn Ser Ser Gly Cys Pro Glu Arg Met
85 90 95
Ser Ala Cys Arg Ser Ile Glu Ala Phe Arg Val Gly Trp Gly Ala Leu
100 105 110
Gln Tyr Glu Asp Asn Val Thr Asn Pro Glu Asp Met Arg Pro Tyr Cys
115 120 125
Trp His Tyr Pro Pro Arg Gln Cys Gly Val Val Ser Ala Lys Thr Val
130 135 140
Cys Gly Pro Val Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr
145 150 155 160
Thr Asp Arg Leu Gly Ala Pro Thr Tyr Thr Trp Gly Glu Asn Glu Thr
165 170 175
Asp Val Phe Leu Leu Asn Ser Thr Arg Pro Pro Leu Gly Ser Trp Phe
180 185 190
Gly Cys Thr Trp Met Asn Ser Ser Gly Tyr Thr Lys Thr Cys Gly Ala
195 200 205
Pro Pro Cys Arg Thr Arg Ala Asp Phe Asn Ala Ser Thr Asp Leu Leu
210 215 220
Cys Pro Thr Asp Cys Phe Arg Lys His Pro Asp Thr Thr Tyr Leu Lys
225 230 235 240
Cys Gly Ser Gly Pro Trp Leu Thr Pro Arg Cys Leu Ile Asp Tyr Pro
245 250 255
Tyr Arg Leu Trp His Tyr Pro Cys Thr Val Asn Tyr Thr Ile Phe Lys
260 265 270
Ile Arg Met Tyr Val Gly Gly Val Glu His Arg Leu Thr Ala Ala Cys
275 280 285
Asn Phe Thr Arg Gly Asp Arg Cys Asn Leu Glu Asp Arg Asp Arg Ser
290 295 300
Gln Leu Ser Pro Leu Leu His Ser Thr Thr Glu Trp Ala Ile Leu Pro
305 310 315 320
Cys Ser Tyr Ser Asp Leu Pro Ala Leu Ser Thr Gly Leu Leu His Leu
325 330 335
His Gln Asn Ile Val Asp Val Gln Phe Met Tyr Gly Leu Ser Pro Ala
340 345 350
Leu Thr Lys Tyr Ile
355
<210> 17
<211> 632
<212> PRT
<213> Human immunodeficiency virus type 1
<400> 17
Glu Asn Leu Trp Val Thr Val Tyr Tyr Gly Val Pro Val Trp Lys Glu
1 5 10 15
Ala Thr Thr Thr Leu Phe Cys Ala Ser Asp Ala Lys Ala Tyr Asp Thr
20 25 30
Glu Val His Asn Val Trp Ala Thr His Ala Cys Val Pro Thr Asp Pro
35 40 45
Asn Pro Gln Glu Val Val Leu Glu Asn Val Thr Glu Asn Phe Asn Met
50 55 60
Trp Lys Asn Asn Met Val Glu Gln Met His Glu Asp Ile Ile Ser Leu
65 70 75 80
Trp Asp Gln Ser Leu Lys Pro Cys Val Lys Leu Thr Pro Leu Cys Val
85 90 95
Thr Leu Asn Cys Thr Asp Leu Arg Asn Val Thr Ser Ser Glu Gly Met
100 105 110
Arg Gly Glu Ile Lys Asn Cys Ser Phe Asn Ile Thr Thr Ser Ile Arg
115 120 125
Asp Lys Val Lys Lys Asp Tyr Ala Leu Phe Tyr Arg Leu Asp Val Val
130 135 140
Pro Ile Asp Asn Asp Asn Thr Ser Tyr Arg Leu Ile Asn Cys Asn Thr
145 150 155 160
Ser Thr Cys Thr Gln Ala Cys Pro Lys Val Ser Phe Glu Pro Ile Pro
165 170 175
Ile His Tyr Cys Thr Pro Ala Gly Phe Ala Ile Leu Lys Cys Lys Asp
180 185 190
Lys Lys Phe Asn Gly Thr Gly Pro Cys Lys Asn Val Ser Thr Val Gln
195 200 205
Cys Thr His Gly Ile Arg Pro Val Val Ser Thr Gln Leu Leu Leu Asn
210 215 220
Gly Ser Leu Ala Glu Glu Glu Val Val Ile Arg Ser Ser Asn Phe Thr
225 230 235 240
Asp Asn Ala Lys Asn Ile Ile Val Gln Leu Lys Glu Ser Val Glu Ile
245 250 255
Asn Cys Thr Arg Pro Asn Asn Asn Thr Arg Lys Ser Ile His Ile Gly
260 265 270
Pro Gly Arg Cys Phe Tyr Thr Thr Gly Asp Ile Ile Gly Asp Ile Arg
275 280 285
Gln Ala His Cys Asn Ile Ser Arg Thr Lys Trp Asn Asn Thr Leu Asn
290 295 300
Gln Ile Ala Thr Lys Leu Lys Glu Gln Phe Gly Asn Asn Lys Thr Ile
305 310 315 320
Val Phe Asn Gln Ser Ser Gly Gly Asp Pro Glu Ile Val Met His Ser
325 330 335
Phe Asn Cys Gly Gly Glu Phe Phe Tyr Cys Asn Ser Thr Gln Leu Phe
340 345 350
Asn Ser Thr Trp Asn Phe Asn Gly Thr Trp Asn Leu Thr Gln Ser Asn
355 360 365
Gly Thr Glu Gly Asn Asp Thr Ile Thr Leu Pro Cys Arg Ile Lys Gln
370 375 380
Ile Ile Asn Met Trp Gln Glu Val Gly Lys Ala Met Tyr Ala Pro Pro
385 390 395 400
Ile Arg Gly Gln Ile Arg Cys Ser Ser Asn Ile Thr Gly Leu Ile Leu
405 410 415
Thr Arg Asp Gly Gly Asn Asn His Asn Asn Asp Thr Glu Thr Phe Arg
420 425 430
Pro Gly Gly Gly Asp Met Arg Asp Asn Trp Arg Ser Glu Leu Tyr Lys
435 440 445
Tyr Lys Val Val Lys Ile Glu Pro Leu Gly Val Ala Pro Thr Lys Cys
450 455 460
Lys Arg Arg Val Val Gln Arg Arg Arg Arg Lys Arg Ala Val Gly Thr
465 470 475 480
Ile Gly Ala Met Phe Leu Gly Phe Leu Gly Ala Ala Gly Ser Thr Met
485 490 495
Gly Ala Ala Ser Ile Thr Leu Thr Val Gln Ala Arg Leu Leu Leu Ser
500 505 510
Gly Ile Val Gln Gln Gln Asn Asn Leu Leu Arg Ala Pro Glu Ala Gln
515 520 525
Gln His Leu Leu Gln Leu Thr Val Trp Gly Ile Lys Gln Leu Gln Ala
530 535 540
Arg Val Leu Ala Val Glu Arg Tyr Leu Arg Asp Gln Gln Leu Leu Gly
545 550 555 560
Ile Trp Gly Cys Ser Gly Lys Leu Ile Cys Cys Thr Ala Val Pro Trp
565 570 575
Asn Ala Ser Trp Ser Asn Lys Thr Leu Asp Met Ile Trp Asn Asn Met
580 585 590
Thr Trp Met Glu Trp Glu Arg Glu Ile Asp Asn Tyr Thr Gly Leu Ile
595 600 605
Tyr Thr Leu Ile Glu Glu Ser Gln Asn Gln Gln Glu Lys Asn Glu Gln
610 615 620
Glu Leu Leu Glu Leu Asp Gly Ser
625 630
<210> 18
<211> 661
<212> PRT
<213> Artificial Sequence
<220>
<223> Amino acid residues corresponding to HIV1 env with N-terminal
leader sequence and C-terminal HIS tag
<400> 18
Met Asp Ala Met Lys Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Ala Ser Glu Asn Leu Trp Val Thr Val Tyr Tyr
20 25 30
Gly Val Pro Val Trp Lys Glu Ala Thr Thr Thr Leu Phe Cys Ala Ser
35 40 45
Asp Ala Lys Ala Tyr Asp Thr Glu Val His Asn Val Trp Ala Thr His
50 55 60
Ala Cys Val Pro Thr Asp Pro Asn Pro Gln Glu Val Val Leu Glu Asn
65 70 75 80
Val Thr Glu Asn Phe Asn Met Trp Lys Asn Asn Met Val Glu Gln Met
85 90 95
His Glu Asp Ile Ile Ser Leu Trp Asp Gln Ser Leu Lys Pro Cys Val
100 105 110
Lys Leu Thr Pro Leu Cys Val Thr Leu Asn Cys Thr Asp Leu Arg Asn
115 120 125
Val Thr Ser Ser Glu Gly Met Arg Gly Glu Ile Lys Asn Cys Ser Phe
130 135 140
Asn Ile Thr Thr Ser Ile Arg Asp Lys Val Lys Lys Asp Tyr Ala Leu
145 150 155 160
Phe Tyr Arg Leu Asp Val Val Pro Ile Asp Asn Asp Asn Thr Ser Tyr
165 170 175
Arg Leu Ile Asn Cys Asn Thr Ser Thr Cys Thr Gln Ala Cys Pro Lys
180 185 190
Val Ser Phe Glu Pro Ile Pro Ile His Tyr Cys Thr Pro Ala Gly Phe
195 200 205
Ala Ile Leu Lys Cys Lys Asp Lys Lys Phe Asn Gly Thr Gly Pro Cys
210 215 220
Lys Asn Val Ser Thr Val Gln Cys Thr His Gly Ile Arg Pro Val Val
225 230 235 240
Ser Thr Gln Leu Leu Leu Asn Gly Ser Leu Ala Glu Glu Glu Val Val
245 250 255
Ile Arg Ser Ser Asn Phe Thr Asp Asn Ala Lys Asn Ile Ile Val Gln
260 265 270
Leu Lys Glu Ser Val Glu Ile Asn Cys Thr Arg Pro Asn Asn Asn Thr
275 280 285
Arg Lys Ser Ile His Ile Gly Pro Gly Arg Cys Phe Tyr Thr Thr Gly
290 295 300
Asp Ile Ile Gly Asp Ile Arg Gln Ala His Cys Asn Ile Ser Arg Thr
305 310 315 320
Lys Trp Asn Asn Thr Leu Asn Gln Ile Ala Thr Lys Leu Lys Glu Gln
325 330 335
Phe Gly Asn Asn Lys Thr Ile Val Phe Asn Gln Ser Ser Gly Gly Asp
340 345 350
Pro Glu Ile Val Met His Ser Phe Asn Cys Gly Gly Glu Phe Phe Tyr
355 360 365
Cys Asn Ser Thr Gln Leu Phe Asn Ser Thr Trp Asn Phe Asn Gly Thr
370 375 380
Trp Asn Leu Thr Gln Ser Asn Gly Thr Glu Gly Asn Asp Thr Ile Thr
385 390 395 400
Leu Pro Cys Arg Ile Lys Gln Ile Ile Asn Met Trp Gln Glu Val Gly
405 410 415
Lys Ala Met Tyr Ala Pro Pro Ile Arg Gly Gln Ile Arg Cys Ser Ser
420 425 430
Asn Ile Thr Gly Leu Ile Leu Thr Arg Asp Gly Gly Asn Asn His Asn
435 440 445
Asn Asp Thr Glu Thr Phe Arg Pro Gly Gly Gly Asp Met Arg Asp Asn
450 455 460
Trp Arg Ser Glu Leu Tyr Lys Tyr Lys Val Val Lys Ile Glu Pro Leu
465 470 475 480
Gly Val Ala Pro Thr Lys Cys Lys Arg Arg Val Val Gln Arg Arg Arg
485 490 495
Arg Lys Arg Ala Val Gly Thr Ile Gly Ala Met Phe Leu Gly Phe Leu
500 505 510
Gly Ala Ala Gly Ser Thr Met Gly Ala Ala Ser Ile Thr Leu Thr Val
515 520 525
Gln Ala Arg Leu Leu Leu Ser Gly Ile Val Gln Gln Gln Asn Asn Leu
530 535 540
Leu Arg Ala Pro Glu Ala Gln Gln His Leu Leu Gln Leu Thr Val Trp
545 550 555 560
Gly Ile Lys Gln Leu Gln Ala Arg Val Leu Ala Val Glu Arg Tyr Leu
565 570 575
Arg Asp Gln Gln Leu Leu Gly Ile Trp Gly Cys Ser Gly Lys Leu Ile
580 585 590
Cys Cys Thr Ala Val Pro Trp Asn Ala Ser Trp Ser Asn Lys Thr Leu
595 600 605
Asp Met Ile Trp Asn Asn Met Thr Trp Met Glu Trp Glu Arg Glu Ile
610 615 620
Asp Asn Tyr Thr Gly Leu Ile Tyr Thr Leu Ile Glu Glu Ser Gln Asn
625 630 635 640
Gln Gln Glu Lys Asn Glu Gln Glu Leu Leu Glu Leu Asp Gly Ser His
645 650 655
His His His His His
660
<210> 19
<211> 233
<212> PRT
<213> Artificial Sequence
<220>
<223> H77cdelta123
<400> 19
Glu Thr His Gln Asn Ile Gln Leu Ile Asn Thr Asn Gly Ser Trp His
1 5 10 15
Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu Ser Leu Asn Thr Gly Trp
20 25 30
Leu Ala Gly Leu Phe Tyr Gln His Lys Phe Asn Ser Ser Gly Cys Pro
35 40 45
Glu Arg Leu Ala Ser Cys Gly Ser Ser Gly Cys Trp His Tyr Pro Pro
50 55 60
Arg Pro Cys Gly Ile Val Pro Ala Lys Ser Val Cys Gly Pro Val Tyr
65 70 75 80
Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr Asp Arg Ser Gly
85 90 95
Ala Pro Thr Tyr Ser Trp Gly Ala Asn Asp Thr Asp Val Phe Val Leu
100 105 110
Asn Asn Thr Arg Pro Pro Leu Gly Asn Trp Phe Gly Cys Thr Trp Met
115 120 125
Asn Ser Thr Gly Phe Thr Lys Val Cys Gly Ala Pro Pro Cys Gly Ser
130 135 140
Ser Gly Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu Ala Thr Tyr
145 150 155 160
Ser Arg Cys Gly Ser Gly Pro Trp Ile Thr Pro Arg Cys Met Val Asp
165 170 175
Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Ile Asn Tyr Thr Ile
180 185 190
Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg Leu Glu Ala
195 200 205
Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asp Leu Glu Asp Arg Asp
210 215 220
Arg Ser Glu His His His His His His
225 230
<210> 20
<211> 228
<212> PRT
<213> Artificial Sequence
<220>
<223> Con1delta123
<400> 20
Gly Thr Tyr Gly Ser Ser Gly Gln Leu Val Asn Thr Asn Gly Ser Trp
1 5 10 15
His Ile Asn Arg Thr Ala Leu Asn Cys Asn Asp Ser Leu Asn Thr Gly
20 25 30
Phe Leu Ala Ala Leu Phe Tyr Val His Lys Phe Asn Ser Ser Gly Cys
35 40 45
Pro Glu Arg Met Ala Ser Cys Gly Ser Ser Gly Cys Trp His Tyr Ala
50 55 60
Pro Arg Pro Cys Gly Ile Val Pro Ala Ala Gln Val Cys Gly Pro Val
65 70 75 80
Tyr Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr Asp Arg Phe
85 90 95
Gly Val Pro Thr Tyr Ser Trp Gly Glu Asn Glu Thr Asp Val Leu Leu
100 105 110
Leu Asn Asn Thr Arg Pro Pro Gln Gly Asn Trp Phe Gly Cys Thr Trp
115 120 125
Met Asn Ser Thr Gly Phe Thr Lys Thr Cys Gly Gly Pro Pro Cys Gly
130 135 140
Ser Ser Gly Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu Ala Thr
145 150 155 160
Tyr Thr Lys Cys Gly Ser Gly Pro Trp Leu Thr Pro Arg Cys Leu Val
165 170 175
His Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Val Asn Phe Thr
180 185 190
Ile Phe Lys Val Arg Met Tyr Val Gly Gly Val Glu His Arg Leu Glu
195 200 205
Ala Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asn Leu Glu Asp Arg
210 215 220
Asp Arg Ser Glu
225
<210> 21
<211> 227
<212> PRT
<213> Artificial Sequence
<220>
<223> S52delta123
<400> 21
Glu Thr His Gln Lys Leu Gln Leu Val Asn Thr Asn Gly Ser Trp His
1 5 10 15
Ile Asn Ser Thr Ala Leu Asn Cys Asn Glu Ser Ile Asn Thr Gly Phe
20 25 30
Ile Ala Gly Leu Phe Tyr Tyr His Lys Phe Asn Ser Thr Gly Cys Pro
35 40 45
Gln Arg Leu Ser Ser Cys Gly Ser Ser Gly Cys Trp His Tyr Ala Pro
50 55 60
Arg Pro Cys Ser Val Val Pro Ala Ser Ser Val Cys Gly Pro Val Tyr
65 70 75 80
Cys Phe Thr Pro Ser Pro Val Val Val Gly Thr Thr Asp Ile Lys Gly
85 90 95
Lys Pro Thr Tyr Asn Trp Gly Glu Asn Glu Thr Asp Val Phe Leu Leu
100 105 110
Glu Ser Leu Arg Pro Pro Ser Gly Arg Trp Phe Gly Cys Ala Trp Met
115 120 125
Asn Ser Thr Gly Phe Leu Lys Thr Cys Gly Ala Pro Pro Cys Gly Ser
130 135 140
Ser Gly Cys Pro Thr Asp Cys Phe Arg Lys His Pro Glu Ala Thr Tyr
145 150 155 160
Ser Arg Cys Gly Ala Gly Pro Trp Leu Thr Pro Arg Cys Met Val Asp
165 170 175
Tyr Pro Tyr Arg Leu Trp His Tyr Pro Cys Thr Val Asn Phe Thr Leu
180 185 190
Phe Lys Val Arg Met Phe Val Gly Gly Phe Glu His Arg Phe Thr Ala
195 200 205
Ala Cys Asn Trp Thr Arg Gly Glu Arg Cys Asn Ile Glu Asp Arg Asp
210 215 220
Arg Ser Glu
225
<210> 22
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> human trypsinogen signal peptide
<400> 22
Met Asn Pro Leu Leu Ile Leu Thr Phe Val Ala Ala Ala Leu Ala
1 5 10 15
<210> 23
<211> 23
<212> PRT
<213> Artificial Sequence
<220>
<223> human tissue plasminogen activator signal peptide (tPA)
<400> 23
Met Asp Ala Met Lys Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Ala Ser
20
<210> 24
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> 6 HIS tag
<400> 24
His His His His His His
1 5
Claims (29)
- 고유의 낮은 급수의 항원으로부터 세포밖에서 조립된 높은 급수의 항원을 제조하는 방법으로서,
(i) 하나 이상의 고유의 시스테인을 환원시키기에 충분한 조건하에 일정 시간 동안 환원제를 포함하는 용액과 낮은 급수(lower order)의 항원을 접촉시키는 단계; 및
(ii) 환원제를 제거 또는 희석하거나 환원된 낮은 급수의 항원을 산화제와 접촉시켜서 (i)로부터의 낮은 급수의 항원을 높은 급수(higher order)의 항원으로 조립시키는 단계
를 포함하되, 단계 (ii)에서 낮은 급수의 항원의 10% 이상이 높은 급수의 항원으로 전환되고, 이로써 조립된 높은 급수의 항원이 낮은 급수의 항원과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타내고 하나 이상의 중화 항체에 대한 결합을 보유하는, 방법. - 제1항에 있어서,
낮은 급수의 항원을 높은 급수의 항원으로 조립하는 방법의 효율을 개선시키기 위해, 단계 (ii)로부터 남아있는 낮은 급수의 항원을 포함하는 용액에 의해 처음 단계 (i) 및 (ii)를 반복하는 방법. - 제1항 또는 제2항에 있어서,
단계 (i)에서 또는 단계 (i)의 이전에, 낮은 급수의 항원을 포함하는 용액에서 고유의 올리고머 또는 높은 급수의 항원이 실질적으로 고갈된 방법. - 제1항 내지 제3항중 어느 한 항에 있어서,
낮은 급수의 항원의 25% 이상, 30% 이상, 40% 이상, 50% 이상, 60% 이상, 70% 이상, 80% 이상, 90% 이상 또는 95% 이상이 높은 급수의 항원으로 전환되는 방법. - 제1항 내지 제4항중 어느 한 항에 있어서,
조립된 높은 급수의 항원이, 하나 이상의 중화 항체에 결합하거나 이를 이끌어내는 고유의 대조적인 높은 급수의 항원의 능력을 보유하거나 이를 초과하는 방법. - 제1항 내지 제5항중 어느 한 항에 있어서,
조립된 높은 급수의 항원이 간염 C 바이러스(HCV) 엔벨롭(envelope) 2(E2)의 수용체-결합 도메인(RBD: receptor-binding domain)인 방법. - 제6항에 있어서,
조립된 높은 급수의 항원에서 과가변성 영역의 전부 또는 일부, 예컨대 과가변성 영역 1(HVR1) 또는 이의 일부, 과가변성 영역 2(HVR2) 또는 이의 일부 및/또는 상호유전자형(intergenotypic) 가변성 영역(igVR/VR3) 또는 이의 일부중 하나 이상이 결핍된 방법. - 제6항 또는 제7항에 있어서,
조립된 올리고머성 항원이 C581, C585, C652, C677, C494, C486, C459, C452, C564, C597, 및 C569로 구성된 군에서 선택된 하나 이상의 아미노산 잔기에 비-시스테인 치환 또는 돌연변이를 포함하는 방법. - 제1항 내지 제8항중 어느 한 항에 따른 방법을 포함하고, 조립된 높은 급수의 항원을 약학적으로 또는 생리학적으로 허용가능한 희석제, 담체 또는 보조제와 혼합하는, 백신 조성물의 제조 방법.
- 제1항에 있어서,
항원이 바이러스성 엔벨롭 항원 또는 암 항원인 방법. - 제10항에 있어서,
바이러스성 엔벨롭 항원이 간염 바이러스 항원 또는 HIV 엔벨롭 항원인 방법. - 제1항 내지 제11항중 어느 한 항에 따른 방법에 의해 낮은 급수의 항원으로부터 생산된, 조립된 높은 급수의 항원, 또는 이를 포함하는 조성물.
- 높은 급수의 세포밖에서 조립된 항원을 포함하고, 여기서 조립된 항원이 고유의 대조적인 높은 급수의 항원과 비교하여 적어도 감소된 비-중화 항체에 대한 결합을 나타내는, 조성물.
- 제13항에 있어서,
조립된 높은 급수의 항원이 HCV E2이거나 HCV E2의 수용체-결합 도메인(RBD)인 조성물. - 제14항에 있어서,
E2 항원에서 과가변성 영역의 전부 또는 일부, 예컨대 과가변성 영역 1(HVR1) 또는 이의 일부, 과가변성 영역 2(HVR2) 또는 이의 일부 및/또는 상호유전자형 가변성 영역(igVR/VR3) 또는 이의 일부중 하나 이상이 결핍된 조성물. - 제14항 또는 제15항에 있어서,
E2 항원이 C581, C585, C652, C677, C494, C486, C459, C452, C564, C597, 및 C569로 구성된 군에서 선택된 하나 이상의 아미노산 잔기에 비-시스테인 치환 또는 돌연변이를 포함하는 조성물. - 제12항 내지 제16항중 어느 한 항에 있어서,
약학적으로 또는 생리학적으로 허용가능한 희석제, 담체 또는 보조제를 포함하는 조성물. - 항원 또는 HCV 감염과 연관된 증상의 치료 또는 예방에 있어서의, 또는 이를 위한 약제의 제조에 있어서의 제12항 내지 제17항중 어느 한 항의 조성물의 용도.
- HCV 감염의 진단 또는 모니터링, 또는 항-HCV 치료 프로토콜(protocol)의 모니터링을 위한 진단용 제제의 제조에 있어서의 제12항의 조립된 높은 급수의 항원, 또는 제13항 내지 제17항중 어느 한 항의 조성물의 용도.
- 면역 반응을 이끌어내기에 충분한 조건하에 일정 시간 동안 효과량의 제12항의 조립된 높은 급수의 항원, 또는 제13항 내지 제17항중 어느 한 항의 조성물을 피험체 또는 환자에게 투여하는 단계를 포함하는, 피험체 또는 환자에서 면역 반응을 이끌어내는 방법.
- 제12항의 조립된 올리고머성 E2 항원, 또는 제13항 내지 제17항중 어느 한 항의 조성물을 피험체에게 투여하는 단계를 포함하는, HCV로부터의 감염에 대해 피험체를 면역화시키는 방법.
- 항원, 예컨대 HCV E2 및 HCV 감염과 연관된 증상을 치료하거나 예방하기에 충분한 조건하에 일정 시간 동안 제12항의 조립된 올리고머성 항원, 또는 제13항 내지 제17항중 어느 한 항의 조성물을 피험체에게 투여하는 단계를 포함하는, 피험체에서 HCV 감염을 치료하거나 예방하는 방법.
- 제13항 내지 제16항중 어느 한 항에 있어서,
조립된 높은 급수의 항원이 검출가능한 또는 정제 태그(tag)를 포함하는 조성물. - 제13항 내지 제16항중 어느 한 항의 높은 급수의 조립된 항원에 대한 정제된 항체를 생산하는 방법으로서,
효과량의 상기 항원을 피험체에게 투여하는 단계, 및
생산된 항체를 정제하는 단계
를 포함하는, 방법. - 제12항 내지 제16항중 어느 한 항의 조립된 항원을 특이적으로 인식하는 항체.
- 제12항의 조립된 높은 급수의 항원, 또는 제13항 내지 제16항중 어느 한 항의 조성물을 포함하는 키트, 또는 고체 또는 반-고체 기질.
- 항원 특이적 면역 세포에 결합하는/이를 검출하는, 제12항의 조립된 높은 급수의 항원, 또는 제13항 내지 제16항중 어느 한 항의 조성물의 용도.
- 항원에 특이적인 면역 세포, 예컨대 B-세포의 검출 및/또는 단리를 위한, 제12항의 조립된 높은 급수의 항원, 또는 제13항 내지 제16항중 어느 한 항의 조성물의 용도.
- i) HCV에 특이적인 면역 세포/B-세포를 제13항 내지 제16항중 어느 한 항의 조립된 높은 급수의 E2 항원으로 표지화하는 단계; 및
ii) 표지화된 면역 세포/B-세포를 세포분석에 의해 검출하고/하거나 단리하는 단계
를 포함하는, HCV에 특이적인 면역 세포/B-세포를 검출하고/하거나 단리하는 방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| AU2016903961 | 2016-09-29 | ||
| AU2016903961A AU2016903961A0 (en) | 2016-09-29 | Refolded glycoproteins for vaccine production | |
| PCT/AU2017/051037 WO2018058177A1 (en) | 2016-09-29 | 2017-09-22 | Assembled glycoproteins |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20190078574A true KR20190078574A (ko) | 2019-07-04 |
Family
ID=61762303
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020197011934A KR20190078574A (ko) | 2016-09-29 | 2017-09-22 | 조립된 당단백질 |
Country Status (8)
| Country | Link |
|---|---|
| US (1) | US20190284230A1 (ko) |
| EP (1) | EP3519424A4 (ko) |
| JP (1) | JP7068284B2 (ko) |
| KR (1) | KR20190078574A (ko) |
| CN (1) | CN110049993A (ko) |
| AU (1) | AU2017334263A1 (ko) |
| IL (1) | IL265524B2 (ko) |
| WO (1) | WO2018058177A1 (ko) |
Families Citing this family (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN112285361B (zh) * | 2020-09-27 | 2023-12-05 | 中国人民解放军空军军医大学 | 排除抗-cd38单克隆抗体药物对抗人球蛋白检测干扰的试剂 |
| CN115389681B (zh) * | 2022-11-01 | 2023-01-20 | 常州百瑞吉生物医药有限公司 | 一种巯基化透明质酸衍生物中二硫苏糖醇残留的检测方法 |
Family Cites Families (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ES2174957T5 (es) * | 1994-07-29 | 2006-12-16 | Innogenetics N.V. | Proteinas purificadas de envoltura de virus de la hepatitis c para uso diagnostico y terapeutico. |
| EP1463521A4 (en) * | 2001-12-17 | 2009-06-24 | Us Gov Health & Human Serv | GP41 INHIBITORS |
| RU2319505C2 (ru) * | 2001-12-18 | 2008-03-20 | Иннодженетикс Н.В. | Очищенные оболочечные белки вируса гепатита с для диагностического и терапевтического применения |
| EP2061805B1 (en) | 2006-08-25 | 2020-02-12 | Macfarlane Burnet Institute for Medical Research and Public Health Limited | Recombinant hcv e2 glycoprotein |
| EP2600894B1 (en) | 2010-08-04 | 2017-02-01 | The Macfarlane Burnet Institute For Medical Research And Public Health Ltd | Modified hepatitis c virus proteins |
| JP5897024B2 (ja) | 2010-11-26 | 2016-03-30 | ザ マクファーレーン バーネット インスティテュート フォー メディカル リサーチ アンド パブリック ヘルス リミテッドThe Macfarlane Burnet Institute for Medical Research and Public Health Ltd | 組成物および方法 |
-
2017
- 2017-09-22 US US16/337,900 patent/US20190284230A1/en active Pending
- 2017-09-22 AU AU2017334263A patent/AU2017334263A1/en active Pending
- 2017-09-22 JP JP2019517034A patent/JP7068284B2/ja active Active
- 2017-09-22 EP EP17854239.5A patent/EP3519424A4/en active Pending
- 2017-09-22 KR KR1020197011934A patent/KR20190078574A/ko not_active IP Right Cessation
- 2017-09-22 WO PCT/AU2017/051037 patent/WO2018058177A1/en unknown
- 2017-09-22 CN CN201780063166.XA patent/CN110049993A/zh active Pending
-
2019
- 2019-03-20 IL IL265524A patent/IL265524B2/en unknown
Also Published As
| Publication number | Publication date |
|---|---|
| AU2017334263A1 (en) | 2019-04-04 |
| IL265524B (en) | 2022-10-01 |
| US20190284230A1 (en) | 2019-09-19 |
| WO2018058177A1 (en) | 2018-04-05 |
| EP3519424A1 (en) | 2019-08-07 |
| IL265524A (en) | 2019-05-30 |
| JP7068284B2 (ja) | 2022-05-16 |
| EP3519424A4 (en) | 2020-06-03 |
| JP2020500837A (ja) | 2020-01-16 |
| CN110049993A (zh) | 2019-07-23 |
| IL265524B2 (en) | 2023-02-01 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| RU2247729C2 (ru) | Олигомерная частица, индуцирующая иммунитет против вируса гепатита с, способ получения олигомерной частицы, композиция, специфическое антитело, набор (варианты), иммунологический анализ и вакцина против вируса гепатита с | |
| JP2023513693A (ja) | SARS-CoV-2ワクチン | |
| JP4173741B2 (ja) | コアグリコシル化されたhcvエンベロープタンパク質 | |
| US11578103B2 (en) | Nucleic acids encoding Zika virus-like particles and their use in Zika virus vaccines and diagnostic assays | |
| US11179460B2 (en) | Virus-like particles comprising zika antigen | |
| CN108290960B (zh) | 丙型肝炎病毒e1/e2异二聚体及其生产方法 | |
| KR20190078574A (ko) | 조립된 당단백질 | |
| EP3188755B1 (en) | Vaccine | |
| US20030021805A1 (en) | Generation of HCV-like particles and chimeric HCV virus | |
| CN107648601B (zh) | 一种丙型肝炎病毒三价亚单位疫苗的制备及应用 | |
| CZ20004802A3 (cs) | Částice obsahující obalové proteiny HCV a jejich použití pro očkování |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| AMND | Amendment | ||
| E902 | Notification of reason for refusal | ||
| AMND | Amendment | ||
| E601 | Decision to refuse application | ||
| AMND | Amendment | ||
| X601 | Decision of rejection after re-examination |