KR20160121702A - 짠맛 펩타이드 - Google Patents
짠맛 펩타이드 Download PDFInfo
- Publication number
- KR20160121702A KR20160121702A KR1020150050771A KR20150050771A KR20160121702A KR 20160121702 A KR20160121702 A KR 20160121702A KR 1020150050771 A KR1020150050771 A KR 1020150050771A KR 20150050771 A KR20150050771 A KR 20150050771A KR 20160121702 A KR20160121702 A KR 20160121702A
- Authority
- KR
- South Korea
- Prior art keywords
- taste
- peptide
- salty
- binding
- peptides
- Prior art date
Links
Images




















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K5/00—Peptides containing up to four amino acids in a fully defined sequence; Derivatives thereof
- C07K5/04—Peptides containing up to four amino acids in a fully defined sequence; Derivatives thereof containing only normal peptide links
- C07K5/10—Tetrapeptides
- C07K5/1021—Tetrapeptides with the first amino acid being acidic
-
- A—HUMAN NECESSITIES
- A23—FOODS OR FOODSTUFFS; TREATMENT THEREOF, NOT COVERED BY OTHER CLASSES
- A23L—FOODS, FOODSTUFFS, OR NON-ALCOHOLIC BEVERAGES, NOT COVERED BY SUBCLASSES A21D OR A23B-A23J; THEIR PREPARATION OR TREATMENT, e.g. COOKING, MODIFICATION OF NUTRITIVE QUALITIES, PHYSICAL TREATMENT; PRESERVATION OF FOODS OR FOODSTUFFS, IN GENERAL
- A23L27/00—Spices; Flavouring agents or condiments; Artificial sweetening agents; Table salts; Dietetic salt substitutes; Preparation or treatment thereof
- A23L27/30—Artificial sweetening agents
- A23L27/31—Artificial sweetening agents containing amino acids, nucleotides, peptides or derivatives
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K7/00—Peptides having 5 to 20 amino acids in a fully defined sequence; Derivatives thereof
- C07K7/04—Linear peptides containing only normal peptide links
- C07K7/06—Linear peptides containing only normal peptide links having 5 to 11 amino acids
Abstract
본 발명은 짠맛 펩타이드에 관한 것으로, 더욱 상세하게는 4~5개의 아미노산으로 이루어진 짧은 펩타이드로 짠맛을 나타내는 것을 확인하였으며, 소금 대체 물질로서 짠맛 식품 첨가물 개발이 가능할 것으로 기대된다.
Description
본 발명은 짠맛 펩타이드에 관한 것이다.
염(염화나트륨)은 음식물의 조미 및 가공에 있어서, 음식물에의 맛 부여, 음식물의 보존성 향상, 음식물의 물성 개선 등의 중요한 역할을 하고 있다. 식염은 특히 맛있다고 느끼게 하는 맛(식염미)을 음식물에 부여하며, 그 구성성분인 나트륨과 염소는 인체의 필수영양소이다.
그러나, 식염의 구성성분인 나트륨의 과잉섭취는 많은 건강문제, 예로서 고혈압 등의 심장병이나 혈관계 질환의 위험인자가 되는 것으로 여겨지고 있다. 일본은 물론 여러 선진국에서는 이들 질환에 걸리기 쉬운 고령자층의 증가에 따라 식염, 특히 나트륨의 섭취량의 감소가 강하게 요구되고 있다.
식염섭취량의 감소를 위해서는 음식물의 조미 및 가공에 있어서 식염의 사용량을 감소시키는 것이 가장 단순한 방법이다. 그러나, 가정 조리식품이든 가공음식물이든 음식물에 함유되는 식염량을 10% 이상 감소시키면 그 음식물의 미감은 일반적으로 손상받게 된다.
식염미를 손상시키지 않고 식염, 특히 나트륨의 섭취량을 감소시키는 방법, 즉 일반적으로 감염 방법이라고 하는 것으로는, 그 자신이 식염미를 띠는 물질(이하, 식염대체물질이라고 함)을 사용하는 방법과, 그 자신이 식염미를 띠지는 않지만 식염과 공존시 그 식염미를 증강하는 물질(이하, 식염미 증강물질이라고 함)을 사용하는 방법 등이 알려져 있다.
식염대체물질로는 예로서 칼륨염, 암모늄염, 염기성 아미노산, 염기성 아미노산으로 이루어지는 펩타이드 및 글루콘산의 알칼리금속염 등이 알려져 있다. 식염미 증강물질은 식염을 대체할 수는 없으나 식염의 식염미를 증강시킴으로써 식염의 사용량을 감소시켜, 감염(減鹽)을 가능하게 하는 물질이다.
식염미 증강물질로는 예로서 분자량이 50,000 달톤 이하의 콜라겐을 가수분해하여 얻어지는 펩타이드 (특허문헌 1), 감미 단백질인 토마틴(thaumatin) (특허문헌 2), 시트르산 생산능을 갖는 흑국균(Aspergillus niger)으로 제국된 흑국, 및 황국균(Aspergillus oryzae)으로 제국된 황국의 혼합물을 소화시켜 수득한 분해 용액 (특허문헌 3) 등이 알려져 있다.
또한, 국내에서는 재래 간장으로부터 짠맛 단백질을 발견한 바 있으나, 이 짠맛 단백질은 분자량이 500 Da에서 10,000 Da에 이르며, 만노스(mannose)와 N-아세틸-글루코사민(N-acetyl-glucosamin)의 당이 수식된 고분자로서, 구체적인 서열을 밝혀지지 않았으며 숙성과정을 거친 후에 추출해야 얻을 수 있어 제조과정이 번거롭다[특허문헌 4, 5].
본 발명자들은 안정되고 높은 단맛을 가지는 새로운 감미 펩타이드를 개발하고자, 단맛을 나타내는 단백질의 구조적, 단백질 공학적 연구를 통해 밝혀진 중요 부위의 서열로부터 디자인된 펩아티드 중 일부에서 짠맛을 나타내는 펩타이드를 확인함으로써 본 발명을 완성하게 되었다.
따라서, 본 발명은 새롭게 디자인된 짠맛 펩타이드를 제공하는데 그 목적이 있다.
상기 과제를 해결하기 위한 수단으로서, 본 발명은 신규 짠맛 펩타이드를 제공한다.
또한, 상기 과제를 해결하기 위한 다른 수단으로서, 본 발명은 상기 짠맛 펩타이드를 포함하는 식염 대체제 및/또는 식염 식품 첨가제를 제공한다.
본 발명에 따른 짠맛 펩타이드는 4~5개 아미노산으로 이루어진 짧은 펩타이드로, 당 수식이 되지 않은 상태에서도 짠맛을 나타내는 것을 확인하였다. 본 발명의 짠맛 펩타이드를 응용하여 소금을 대체할 수 있는 새로운 식염 대체제로 개발 가능하므로 현대 질병에서 가장 큰 문제가 되는 성인병을 예방할 수 있는 우수한 대체제로 사용되리라 기대된다.
도 1은 탄수화물 수크로스의 메커니즘(좌)과 합성 감미료 사카린의 메커니즘(우)이 다른 것을 나타낸 것이다.
도 2는 TRPV1(transient receptor potential cation channel subfamily V member 1) 구조를 나타낸 것이다.
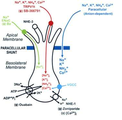
도 3은 용상 유도 맛 수용체 세포 내 나트륨 이온 수송 및 전방의 혀 내 짠맛 전달 모델을 나타낸 것이다.
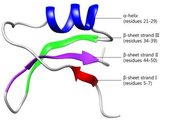
도 4는 브라제인의 3차원 구조를 나타낸 것이다.
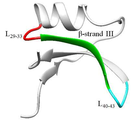
도 5는 브라제인의 중요 부위(29~43번째 잔기)를 나타낸 것이다.
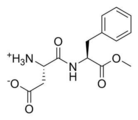
도 6은 아스파탐의 화학구조식을 나타낸 것이다.
도 7은 겔 크로마토그래피를 이용한 펩타이드 정제를 나타낸 것이다.
도 8은 브라제인과 T1R2의 결합 포켓을 예측한 것이다.
도 9는 단맛 수용체 T1R2와 브라제인의 도킹 모델을 예측한 것이다.
도 10은 단맛 수용체 T1R2와 BZ1의 도킹 모델을 예측한 것이다.
도 11은 단맛 수용체 T1R2와 BZ2의 도킹 모델을 예측한 것이다.
도 12는 단맛 수용체 T1R2와 BZ3의 도킹 모델을 예측한 것이다.
도 13은 단맛 수용체 T1R2와 BZ4의 도킹 모델을 예측한 것이다.
도 14는 단맛 수용체 T1R2와 BZ5의 도킹 모델을 예측한 것이다.
도 15는 단맛 수용체 T1R2와 BZ6의 도킹 모델을 예측한 것이다.
도 16은 단맛 수용체 T1R2와 BZ7의 도킹 모델을 예측한 것이다.
도 17은 단맛 수용체 T1R2와 BZ8의 도킹 모델을 예측한 것이다.
도 18은 단맛 수용체 T1R2와 BZ9의 도킹 모델을 예측한 것이다.
도 19는 단맛 수용체 T1R2와 BZ10의 도킹 모델을 예측한 것이다.
도 20은 TRPV1 구조와 리간드 도킹 모델을 나타낸 것이다
도 21은 T2R1와 BZ10의 도킹 모델을 예측한 것이다.
도 22는 펩타이드의 CD 스펙트라를 나타낸 것이다[BZ1: solid line, BZ2: dotted line, BZ5: dashed line, BZ6: dot-dashed line].
도 23은 브라제인과 T1R2 사이의 수소결합을 나타낸 것이다.
도 24는 짠맛 펩타이드(BZ3~5) 및 TRPV1 간의 상호작용을 나타낸 것이다[a: BZ3(DKHAR); b: BZ4(KKRAR); c: BZ5(DEKR)].
도 2는 TRPV1(transient receptor potential cation channel subfamily V member 1) 구조를 나타낸 것이다.
도 3은 용상 유도 맛 수용체 세포 내 나트륨 이온 수송 및 전방의 혀 내 짠맛 전달 모델을 나타낸 것이다.
도 4는 브라제인의 3차원 구조를 나타낸 것이다.
도 5는 브라제인의 중요 부위(29~43번째 잔기)를 나타낸 것이다.
도 6은 아스파탐의 화학구조식을 나타낸 것이다.
도 7은 겔 크로마토그래피를 이용한 펩타이드 정제를 나타낸 것이다.
도 8은 브라제인과 T1R2의 결합 포켓을 예측한 것이다.
도 9는 단맛 수용체 T1R2와 브라제인의 도킹 모델을 예측한 것이다.
도 10은 단맛 수용체 T1R2와 BZ1의 도킹 모델을 예측한 것이다.
도 11은 단맛 수용체 T1R2와 BZ2의 도킹 모델을 예측한 것이다.
도 12는 단맛 수용체 T1R2와 BZ3의 도킹 모델을 예측한 것이다.
도 13은 단맛 수용체 T1R2와 BZ4의 도킹 모델을 예측한 것이다.
도 14는 단맛 수용체 T1R2와 BZ5의 도킹 모델을 예측한 것이다.
도 15는 단맛 수용체 T1R2와 BZ6의 도킹 모델을 예측한 것이다.
도 16은 단맛 수용체 T1R2와 BZ7의 도킹 모델을 예측한 것이다.
도 17은 단맛 수용체 T1R2와 BZ8의 도킹 모델을 예측한 것이다.
도 18은 단맛 수용체 T1R2와 BZ9의 도킹 모델을 예측한 것이다.
도 19는 단맛 수용체 T1R2와 BZ10의 도킹 모델을 예측한 것이다.
도 20은 TRPV1 구조와 리간드 도킹 모델을 나타낸 것이다
도 21은 T2R1와 BZ10의 도킹 모델을 예측한 것이다.
도 22는 펩타이드의 CD 스펙트라를 나타낸 것이다[BZ1: solid line, BZ2: dotted line, BZ5: dashed line, BZ6: dot-dashed line].
도 23은 브라제인과 T1R2 사이의 수소결합을 나타낸 것이다.
도 24는 짠맛 펩타이드(BZ3~5) 및 TRPV1 간의 상호작용을 나타낸 것이다[a: BZ3(DKHAR); b: BZ4(KKRAR); c: BZ5(DEKR)].
본 발명은 짠맛 펩타이드에 관한 것이다.
상기 펩타이드는 서열번호 3, 4 및/또는 5로 표시되는 서열로 이루어진다.
서열번호 3: DKHAR (브라제인 유래)
서열번호 4: KKRAR (인공서열)
서열번호 5: DEKR (브라제인 유래)
본 발명의 펩타이드는 기존의 당이 수식된 고분자 펩타이드(분자량이 500~10000Da)와는 달리 4~5개의 아미노산으로 이루어진 짧은 펩타이드로, 제조가 용이할 뿐만 아니라 염을 직접 섭취하지 않기 때문에 나트륨의 섭취를 줄일 수 있고, 칼로리가 거의 없기 때문에 저칼로리 음식으로 응용할 수 있는 장점을 가진다. 또한, 본 발명에서 당 수식이 되지 않은 상태에서도 짠맛을 나타내는 것을 확인하였다.
본 발명은 또한, 상기 짠맛 펩타이드를 포함하는 식염 대체제 및/또는 식품 첨가제를 제공한다.
본 발명의 식염미 대체제 또는 식품 첨가제의 대상이 되는 식품으로는 식염을 함유하지 않는 음식물이든 식염을 함유하는 음식물이든, 먹을 때 식염이 함유되는 식품이라면 특별한 제한은 없다. 식품으로 예를 들면, 국, 찌개, 탕 등의 국물, 고추장, 청국장, 된장, 간장, 소스, 드레싱, 마요네즈, 토마토 케찹 등의 조미료, 일본식 맑은 국과 같은 맑은 국, 콘소메 스프, 계란 스프, 미역 스프, 상어 지느러미 스프, 포타쥬(potage), 된장국 등의 스프류, 면류 (메밀 국수, 우동, 라면, 파스타 등)용 국물 및 소스류, 죽, 야채 및 쌀의 포리지(porridge), 및 찻물에 말은 밥 등의 쌀 조리 식품, 햄, 소시지, 치즈 등의 축산 가공품, 어묵, 건어물, 젓갈, 진미(chinmi) 등의 수산 가공품, 피클 등의 야채 가공품, 포테이토 칩, 전병, 쿠키 등의 스낵류, 가열 식품, 튀긴 식품, 구운 식품, 카레 등의 조리된 식품 등을 들 수 있다.
상기 식염미 증강제 및/또는 식품 첨가제는 염기성 물질 및/또는 숙신산을 함유하고, 더 필요하다면 무기염, 산, 아미노산류, 핵산, 당류, 부형제 등의 음식물에 사용할 수 있는 각종 첨가물을 함유할 수 있다.
상기 무기염으로는 식염, 염화칼륨, 염화암모늄 등을 들 수 있다. 상기 산으로는 아스코르브산, 푸마르산, 말산, 주석산, 시트르산, 지방산 등과 같은 카르복실산 및 이들의 염 등을 들 수 있다. 상기 염으로는 나트륨 및 칼륨염을 들 수 있다. 상기 아미노산으로는 글루타민산 나트륨, 글리신 등을 들 수 있다. 상기 핵산으로는 이노신산 나트륨, 구아닐산 나트륨 등을 들 수 있다. 상기 당류로는 자당, 포도당, 유당 등을 들 수 있다. 상기 부형제로는 전분 가수분해물인 덱스트린, 각종 전분 등을 들 수 있다. 이들의 사용량은 사용목적에 따라 적절히 설정할 수 있는데, 예로서 펩타이드 100 중량부 당 0.1 내지 500 중량부 사용할 수 있다.
이하, 본 발명에 따르는 실시예를 통하여 본 발명을 보다 상세히 설명하나, 본 발명의 범위가 하기 제시된 실시예에 의해 제한되는 것은 아니다.
[
실시예
]
I. 참조
1. 맛 인지 메커니즘
맛은 혀의 맛 유두(taste papillae)에서 인지된다(Mombaerts, 2004). 맛 유두는 혀 뒤쪽 편에 존재하는 성곽유두(circumvallate papillae), 뒤쪽 측면 가장자리에 있는 잎새 유두(foliate papillae) 그리고 혀끝과 혀의 측면에 존재하는 버섯유두(fungiform papilla)의 세 부분으로 구분할 수 있다. 세 종류의 맛 유두는 맛 봉오리(taste bud)로 구성되어 있으며, 다시 각 맛 봉오리는 50-150개의 특별한 맛 수용세포(taste receptor cells, TRCs)로 구성되어 있다.
TRCs는 Type I에서 IV까지 4종류로 분류할 수 있다. Type I 맛 수용세포는 ecto-ATPase를 발현하여 그룹 외의 세포까지 ATP가 확산되는 것을 막아준다. Type II 맛 수용세포는, 맛 수용체인 T1Rs, T2Rs를 포함한 GPCR(G protein coupled receptors)를 발현시키는 감각 수용세포로 알려져 있으며, 4종류 맛 수용세포에서 유일하게 리간드에 의해 자극이 유발된다. Type III 맛 수용세포는 시냅스를 형성하며, Type IV 맛 수용세포는 맛 봉오리 아래에 위치하여 다른 TRCs로 분화될 수 있는 기저세포의 역할을 한다. 하나의 맛 수용세포는 하나의 맛만 인지할 수 있도록 프로그램 되어 있다.
TRCs에서 발현되는 GPCRs는 7개의 알파-나선구조의 막 관통 도메인 (seven trans-membrane domain, 7TM domain)으로 구성되며 N-말단은 세포 밖에, C-말단은 세포 내에 위치한다. 막 관통 부위와 N-말단은 리간드와 결합하며, 3번째 세포 내 loop와 C- 말단 부위에는 G 단백질이 결합된다. 단맛, 우마미맛, 쓴맛은 GPCR 수용체에 의해 인지된다 (Kitagawa et al., 2001). 반면 짠맛과 신맛은 맛 봉오리에 위치한 이온 채널(ion channel) 또는 이온 교환자(ion exchanger)를 통해 나트륨/칼륨 이온, 수소 이온을 통해 감지된다(Mombaerts, 2004). 단맛과 우마미맛은 class C GPCR 수용체에 해당되는 T1Rs에 의해 인지되며 (Shi and Zhang, 2006), 쓴맛은 class A GPCR 수용체에 속하는 T2Rs에 의해 인지된다 (Huang et al., 2007).
2. 단맛 수용체
T1R2
/
T1R3
인간의 단맛 수용체는 class C GPCR에 속하는 두 개의 소단위체(subunit)이 이합체를 형성하여 작용한다. 단맛을 인지하는 수용체는 T1R2와 T1R3로써 각각 Tasr2, Tasr3 유전자에서 발현된다. T1R2/T1R3는 ATD(amino-terminal ectodomain; VFTM(venus-flytrap binding domain)), CRD(cystein rich domain), TMD(trans-membrane domain)의 3개의 도메인으로 구성되어 있. T1R2와 T1R3의 ATD(T1R2, 1-484; T1R3, 1-462)의 VFTM은 리간드와 결합하며, CRD(T1R2, 491-544; T1R3, 495-547)는 ATD와 TMD를 연결하여 구조를 유지하고, TMD(T1R2, 575-815; T1R3, 590-817)은 막을 관통하여 VFTM이 리간드와 결합함으로써 생기는 구조적 변화를 G 단백질에 전달하는 역할을 한다.
T1R2/T1R3의 구조는 아직 밝혀지지 않았으며, 가장 높은 상동성을 갖는 mGluR(glutamate receptor)로부터 기능을 예측할 수 있다. mGluR은 T1R1/T1R3의 이종이합체로 작용하며, N- 말단 부위에 글루탐산과 결합하여 복합체를 형성할 경우 상당한 구조적 변화를 유발한다 (Lopez Cascales et al., 2010). mGluR의 리간드 결합 부위인 m1-LBR은 리간드의 부재시 휴지형인 자유형 I의 형태를 갖고, 리간드가 결합한 경우 활성형인 자유형 II의 복합체를 형성한다고 예상되어 진다 (Temussi et al ., 2002). 이와 마찬가지로 단맛 수용체인 T1R2/T1R3 역시 리간드와 결합하여 활성형의 복합체를 형성할 것으로 예측된다.
리간드가 당일 경우 T1R2/T1R3와 복합체를 형성함으로써 구조적 변화가 일어나고, 분리된 Gαgust를 통해 AC(adenylate cyclase)를 활성화시킴으로써 ATP를 cAMP로 전환시킨다. 전환된 cAMP는 직접 cyclic nucleotide-dependent ion channels을 통해 양이온을 유입시킴으로써 탈분극을 일으키거나, PKA(protein kinase A)를 활성화시키고 기저막의 칼륨 채널을 인산화하여 닫음으로써 칼슘 이온의 유입을 유도하여 탈분극을 일으킨다.
반면 당이 아닌 감미료로 인한 단맛은 Gqα, Gqβγ로 구성된 Gq 단백질이 PLC β2(phospholipase C β2)를 자극하여 PIP2 (phosphatidylinositol 4,5-bisphosphate)를 DAG (diacylglycerol)와 IP3로 가수분해하고(Margolskee, 2002), 높아진 IP3 농도로 인해 맛 인지세포로부터 칼슘 이온을 방출하여 탈분극을 유발한다 (Ohkuri et al., 2009) (도 1).
3.
TRPV1
에 의한 짠맛 인지
많은 연구 결과를 통해, 짠맛은 나트륨 이온이 맛 수용세포의 나트륨 이온 채널(ENaC)에 유입되면서 인지되는 것이 밝혀졌다. 설치류를 이용한 전기 생리학적 연구 결과에 따르면, ENaC를 블록킹한다고 알려진 아밀로라이드(amiloride)를 구강 투여한 쥐의 경우 CT (chorda tympani)의 NaCl에 대한 신경반응이 현저하게 감소하는 것이 밝혀졌다. 하지만 아밀로라이드가 NaCl에 대한 모든 CT 반응을 제거하지 못하였고, 이에 따라 또 다른 맛 전달 과정이 있음이 예상되었다 (Smith et al, 2012).
TRPV1은 비선택성 양이온 채널로서 캡사이신(capsaicin), 열, 산에 의해 활성화 되는 통각 수용체이다. 맛 봉오리에 다양한 종류의 TRPV1t가 존재함이 밝혀졌다 (Liu et al., 2001). 나트륨, 칼륨, 암모늄 이온에 대한 CT 반응이 TRPV1의 작용제에 의해 증가함이 밝혀졌으며, ENaC와 TRPV1을 모두 blocking한 설치류는 NaCl에 대한 CT 반응이 말소됨이 확인되었다. 이와 같은 전기 생리학적 연구를 통해 TRPV1 채널은 중요한 짠맛 인식 경로임이 밝혀졌다 (도 3). 메일라드 펩타이드(maillard reacted peptide, MRPs)는 열과 당에 의해 가수분해와 당 수식이 된 형태의 단백질이다. Katsumata 등의 연구 결과에 따르면 MRPs에 의해 TRPV1이 제거된 쥐의 NaCl에 대한 CT 신경 반응이 감지되며, MRPs 자체만으로도 짠 맛이 유도되는 것이 밝혀졌다 (Katsumata et al ., 2008). 이와 관련하여 2012년에는 한국식품연구원의 류미라 박사팀이 재래간장으로부터 추출한 MRPs인 KFRI-LHe가 TRPV1과의 결합을 통해 짠맛을 나타내는 것을 발견하였다.
TRPV1의 고차 구조는 Liao 등에 의해 밝혀졌으며 Cryo-EM을 이용하여 3.4Å해상도로 고차구조 및 활성화된 구조를 밝혀냈으며 (Liao et al., 2013), 본 발명을 통해 밝혀진 TRPV1 구조를 사용하여 docking을 실시하였다. TRPV1 은 동일한 네 개의 소단위체로 이루어져 있으며 이온통로를 중심으로 네 개의 단위체가 배치되어 있다. 하나의 소단위체에는 Transmembrane segment (S5-S6)과 측면에 있는 S1-S4 전위수용 부위 사이에 pore loop이 존재하며 이들은 S4-S5 linker를 통해 연결되어 있다. 특히, S4-S5 linker 부위는 TRP 부위와 상호작용하여 입체성 변조 (allosteric modulation)에 관여하는 것으로 알려져 있다. TRPV1의 세포내 조립에 관련하여, 소단위체들의 배치는 세포질 내 부위인 N-말단 ankryin repeats의 상호작용으로부터 촉진되며 이 영역들을 통해 TRP 채널의 기능적인 측면을 더 많이 파악할 수 있을 것으로 예상된다(도 2).
4. 단백질
브라제인
브라제인(brazzein)은 서아프리카의 P. brazzeana의 열매에서 추출된 감미단백질로, 수크로오스와 비교하였을 때 약 500배에서 2000배 이상 단맛을 나타낸다. 브라제인은 major 타입과 minor 타입의 형태가 있으며, 식물에서 추출한 브라제인은 대부분 major 타입으로 존재한다. Major 타입은 N-말단에 pyroglutamic acid 잔기를 포함하여 54개의 아미노산을 가지는 반면 minor 타입은 N-말단의 pyroglutamic acid가 결손된 53개의 아미노산을 가지며 major 타입보다 약 2배의 단맛을 나타낸다. 브라제인은 감미단백질 중 가장 작은 크기로써 약 6.5 kDa의 분자량을 갖고 있는 단량체(monomer)이다. NMR을 통해 밝혀진 브라제인의 3차 구조는 1개의 α-helix (residues 21-29)와 3개의 β-sheet (strand I, residues 5-7; strand II, residues 44-50, strand III, residues 34-39)로 구성되어있으며(도 4), 8개의 시스테인 잔기(cysteine)가 4개의 이황화 결합(disulfide bond)을 형성하여 열과 pH에 대한 안정성이 매우 높다 (Gao et al., 1999).
본 연구에서는 연구의 목적의 달성을 위해, 감미 단백질인 브라제인의 루프부분을 기반으로 10개의 펩타이드를 디자인하고, 관능검사를 하였으며 컴퓨터 모델링을 통해 그 결과를 분석하였다.
II
. 기기 및 시약
1. 기기 및 시약
펩타이드의 맛 시험을 위해 BZ1, BZ2, BZ6은 AbClon사(Seoul, Korea)에 의뢰하여, BZ3-BZ5, BZ7-BZ10은 Peptron사(Daejeon, Korea)에 의뢰하여 합성하였다. 합성된 펩타이드는 GE Healthcare Life Sciences사(Pittsburgh, PA, USA)의 Sephadex G-10 컬럼을 사용하여 정제하였으며, 정제된 펩타이드의 농도 측정을 위해 UV/VIS 분광광도계인 HITACH사(Japan)의 U-2000 제품을 사용하였다.
2. 구조 예측 및 컴퓨터 도킹 프로그램
디자인한 펩타이드의 가상 구조 시각화를 위해 Diderot Universite(Paris, France)에서 운영하는 인터넷 웹 서버 PEP-FOLD 프로그램을 이용하였다. 또한, 리간드와 리셉터의 결합 구조 예측을 위하여 California University의 UCSF Chimera v1.7 프로그램과 미국 The Scripps Research Institute의 Autodock Vina 프로그램을 연계하여 사용하였다.
Ⅲ. 실험방법
1.
펩타이드의
디자인
1.1
펩타이드
BZ1
,
BZ2
의 디자인
변이체 연구를 통해 브라제인의 29-43번 아미노산 서열의 β-sheet 구조(도 5)가 브라제인의 단맛에 중요한 부분임을 밝혀졌다(Jin et al., 2003). 따라서 이 서열을 기준으로 BZ1을 디자인하였으며, 합성한 펩타이드의 서열은‘DKHARSGECFYDEKR’로서 총 15개의 아미노산으로 구성된다.
다양한 브라제인 변이체 연구는 단맛을 증가시키기도, 감소시키기도 하였다. 브라제인의 29번 Arg 잔기를 Lys 잔기로(Jin et al., 2003), 31번 His 잔기를 Arg 잔기로 변이시켜 각각 단맛이 증가하였으며 (Lee J.W et al ., 2013), 36번 Glu 잔기를 Asp 잔기로 변이시키고 (Do H.D et al ., 2011), 41번 Glu 잔기를 Ala 잔기로 변이시킨 경우에도 각각 야생형 대비 단맛이 증가하였다 (Lee J.W et al ., 2013). 이러한 변이체 연구 결과를 토대로 BZ1의 29, 31, 36, 41번 잔기를 다음서열‘KKRARSGDCFYDAKR’과 같이 변형하여 디자인하였다 (표 1).
| Name | Sub name | Sequence |
| BZ1 | BZ 29-43 wild | DKHARSGECFYDEKR (서열번호 1) |
| BZ2 | BZ 29-43 mutant | KKRARSGDCFYDAKR (서열번호 2) |
1.2
펩타이드
BZ3
-
BZ6
의 디자인
아스파탐(aspartame)은 Asp 잔기와 Phe 잔기로 구성된 dipeptide의 methyl ester 형태로(도 6), 음료와 식품의 인공 감미료로 사용되고 있다 (Ager et al ., 1998). 단 2개의 아미노산으로 구성된 아스파탐이 단맛을 나타내는 것처럼, 브라제인의 짧은 단편이 단맛을 나타낼 것으로 기대하여 BZ1과 BZ2를 세분화하여 BZ3-BZ6을 디자인하였다. 브라제인에는 3개의 loop가 존재한다. Loop I은 13-15번 부위이며, loop II는 30-33번 서열에 해당된다. 마지막으로 Loop III은 40-43번의 서열로써 BZ1의 서열로 선정된 29-43번 서열에는 Loop II와 Loop III가 존재한다 (도 5). 이러한 단맛 단백질의 Loop는 단맛 단백질의 단맛을 나타낸다고 예상되는 ‘sweet finger’모델이 될 가능성이 있다 (Temussi et al ., 2002). 따라서 BZ1의 Loop II에 해당되는 29-33번 부위‘DKHAR’를 BZ3로 선정하였으며. BZ4는 BZ3의 첫 번째 잔기인 Asp를 BZ2와 같이 Lys으로 바꿈으로써 펩타이드가 양성을 띄도록 디자인 하였다. 선행연구에서 보고되었듯이 Loop III의 Glu41이 T1R2의 Glu252와 근접할 뿐 아니라 Arg43가 중요 잔기이므로 (Yoon et al ., 2011), Loop III에 해당되는 40-43번 부위인 ‘DEKR’를 BZ5로 선정하였으며, BZ6 역시 BZ2와 같이 Glu 잔기를 Ala으로 바꿈으로써 음성 잔기가 중성이 되도록 하였다 (표 2).
| Name | Sub name | Sequence |
| BZ3 | BZ 29-33 wild | DKHAR (서열번호 3) |
| BZ4 | BZ 29-33 mutant | KKRAR (서열번호 4) |
| BZ5 | BZ 40-43 wild | DEKR (서열번호 5) |
| BZ6 | BZ 40-43 mutant | DAKR (서열번호 6) |
1.3
펩타이드
BZ7
-
BZ10
의 디자인
Loop II를 기반으로 디자인한 BZ3에서 음성 잔기에 해당되는 Asp를 제외한 4개의 서열인‘KHAR’을 BZ7로 선정함으로써, Loop II의 단맛에 미치는 영향과 전기적 성질에 따른 단맛 변화, 더 짧은 펩타이드에서도 단맛이 나타나는지를 연구하고자 디자인하였다.
반면 Loop III에 해당되는 BZ5는 양 옆으로 2개씩 잔기를 추가하여 BZ5의 맛과 비교하여 주변 잔기인‘FY’,‘VL’의 역할을 확인하고자 하였으며, 이에 따라‘FYDEKRVL’이 BZ8의 서열로 선정되었다.
BZ8의 loop 구조를 유지함으로써 단맛 수용체와 올바르게 결합할 수 있도록, BZ9, BZ10를 디자인하였다. BZ9는 BZ8 서열 가운데 Pro 잔기를 삽입함으로써 loop 구조가 유지되도록 디자인되었으며, 그 서열은‘FYDPEKRVL’와 같다. BZ10은 BZ9의 서열 양 끝에 Cys 잔기를 추가함으로써 이황화 결합을 통해 loop 구조가 형성되도록 하였으며, 그 서열은‘CFYDEKRVLC’와 같다 (표 3).
| Name | Sub name | Sequence |
| BZ7 | BZ 30-33 wild | KHAR [서열번호 7] |
| BZ8 | BZ 38-45 wild | FYDEKRVL [서열번호 8] |
| BZ9 | BZ 38-45 mutant | FYDPEKRVL [서열번호 9] |
| BZ10 | BZ 38-45 mutant2 | CFYDEKRVLC [서열번호 10] |
2.
펩타이드의
합성 및 정제
BZ1, BZ2, BZ6은 AbClon사에 의뢰하여 80% 이상의 순도로 HPLC(high performance liquid chromatography)를 통해 정제하여 각 5 mg씩 합성하였다. BZ3~BZ5, BZ7~BZ10은 Peptron사에 의뢰하여 95% 이상의 순도로 HPLC 정제하여 각 10 mg씩 합성하였다.
합성과정과 HPLC 정제 과정에서 첨가되어 신맛을 나게 할 수 있는 TFA, 불순물을 제거하기 위해 Sephadex G-10 (GE Healthcare)를 이용해 펩타이드를 정제하였다. 정제 전 시료는 3 mg/ml의 농도로 3차 증류수를 이용하여 용해되었다. Sephadex G-10 컬럼은 bed volume의 10배 이상의 3차 증류수를 흘려주어 파장 205 nm에서의 흡광도의 변화가 나타나지 않을 때까지 세척하였다. 흡광도의 변화가 없는 것을 확인한 후에, 앞서 3차 증류수에 용해시킨 1 ml의 시료를 컬럼에 주입하였다. Bed의 표면이 보일 때쯤, 자유낙하를 통해 0.5 ml/min의 유속으로 3차 증류수로 목적 펩타이드를 용출시킨다. 최종적으로 정제된 펩타이드 용액은 freeze dryer를 사용하여 동결건조하였다.
3.
펩타이드의
맛 시험
총 14명의 훈련된 패널을 대상으로 맛 시험을 실시하였다. 패널들은 건강한 남녀로서 흡연을 하지 않는 대상으로 선정하였다. 정확한 맛의 판단을 위해 식전에 실시하였으며, 시험 전 3차 증류수로 입을 세척하였다. 합성된 펩타이드의 안전성이 알려져 있지 않으므로 맛을 본 후에는 바로 뱉고 증류수로 입을 세척하였다. 맛에 대한 결과는 블라인드 테스트로 하되, 심리적 요인을 제거하기 위하여 자유롭게 표현하도록 하였다.
4.
펩타이드의
구조 예측
화학적 합성을 통해 합성된 펩타이드의 구조는, 해당 서열의 브라제인과 타우마틴에서의 구조와 많은 차이가 있을 것이다. 따라서 합성된 펩타이드가 가장 안정한 에너지를 갖는 구조를 예측하고, 실제 브라제인과 타우마틴에서의 구조와 비교하였다.
프랑스 Paris Diderot 대학에서 제공하는 PEP-FOLD ver. 1.5.3 (http://bioserv.rpbs.univ-paris-diderot.fr/PEP-FOLD/)를 통해 디자인한 펩타이드의 구조를 예측하였다 (Thevenet et al., 2012). 프로그램을 통해 작성된 pdb 파일은 미국 California 대학에서 제공하는 UCSF Chimera ver. 1.9 프로그램을 통해 구현되었으며 (Pettersen et al., 2004), 웹페이지(https://www.cgl.ucsf.edu/chimera/download.htmlPEP-FOLD)를 통해 다운로드하였다. PEP-FOLD에서 제공하는 예측 프로그램은 9개에서 36개의 아미노산 서열에 한하여 예측이 가능하다. 따라서 아미노산이 9개 이하로 구성된 BZ3-BZ8의 경우 C-말단에 Ala 잔기를 추가하여 9개가 되도록 한 후 PEP-FOLD, USCF Chimera 프로그램을 통해 추가된 Ala 잔기를 제거하였다. 또한, 이황화 결합이 포함된 펩타이드인 BZ10의 경우 이황화결합 옵션을 선택함으로써 이황화결합이 도입되도록 하였다.
5.
펩타이드와
맛 인지 수용체의 결합 예측
펩타이드와 맛 인지 수용체와의 결합부위와 에너지를 예측하기 위해 Autodock vina 프로그램을 사용하였다 (Trott et al., 2010). 본 과정은 Intel(R) Core(TM) i3-3220 3.30 GHz CPU, memory 4 GB, Windows 7 사양의 컴퓨터에서 운용되었다. 구조가 아직 밝혀지지 않은 수용체인 T1R2, T2R1 구조는 스위스 Basel 대학의 분자생물학센터에서 제공하는 SWISS-MODEL 웹 서버를 통해 예측되었으며 (Biasini et al., 2014), 웹페이지 (http://swissmodel.expasy.org/)에서 운용이 가능하다. T1R2의 구조는 GPCR familly에 해당하는 human glutamate receptor (PDB ID: 2E4U)를, T2R1은 human delta opioid 7tm receptor (PDB ID: 4N6H)를 기반으로 예측하였다. 모든 pdb 단백질 구조 파일은 AutoDockTools (ADT) 프로그램을 활용하여 Autodock vina 도킹을 위한 pdbqt 파일로 추출하였다 (Morris et al., 2009). ADT 프로그램을 활용한 준비과정에는 물 분자 제거, 수소 원자 추가, 수용체의 결합 예측 공간 선정을 위한 Grid box 설정, 리간드의 torsion 선택, AMBER FF99와 Gasteiger-Marsili 계산법을 통한 charge 부여 과정이 포함된다. 수용체와 펩타이드의 도킹을 위해 선정된 범위(Grid box)를 표 4에 명시하였다. ADT와 Autodock vina 프로그램은 웹사이트 (http://autodock.scripps.edu/downloads)에서 다운로드하였다.
| T1R2 | T1R3 | T2R1 | TRPV1 | |
| center_x | 24.73 | 26.03 | -6.02 | 0.075 |
| center_y | -5.771 | 12.94 | -73.57 | 0.000 |
| center_z | 53.92 | 51.58 | 85.02 | 7.475 |
| size_x | 59.77 | 63.12 | 34.50 | 120.0 |
| size_y | 64.78 | 68.50 | 47.78 | 126.0 |
| size_z | 74.49 | 65.61 | 67.12 | 100.0 |
Ⅳ. 결과
1.
펩타이드의
합성 및 정제
브라제인으로부터 디자인된 10개의 합성된 펩타이드는 HPLC 정제 순도는 대체로 90% 이상이지만 짠맛 펩타이드의 경우 95% 이상으로 높은 수준의 순도를 보였다 (표 5).
더불어 Sephadex-G10 컬럼을 이용하여 불순물 및 합성과 HPLC 정제에 사용되어 맛 시험에 방해가 되는 TFA를 제거하였다. 정제된 펩타이드는 파장 205 nm에서 흡광도를 측정하여 정량하였으며 (도 7), 펩타이드가 용출된 E1~E5 분획을 모아 동결건조하였다.
| Peptide | Purity (%) | Peptide | Purity (%) |
| BZ1 | 80.04 | BZ2 | 96.84 |
| BZ3 | 97.32 | BZ4 | 99.87 |
| BZ5 | 96.79 | BZ6 | 86.11 |
| BZ7 | 97.39 | BZ8 | 99.89 |
| BZ9 | 98.59 | BZ10 | 96.86 |
2.
펩타이드의
맛 시험
BZ1부터 BZ10까지 합성된 펩타이드에 대하여 14명의 패널이 그 맛을 시험하였다. 모든 패널은 주관식 응답지에 느끼는 맛을 자유롭게 표현하도록 하였다. 그 결과 짠맛을 나타내었던 BZ3, BZ4, BZ5와 쓴맛이 느껴진 BZ10을 제외하고는 아무런 맛을 나타내지 못하였다. 특히 BZ4는 짠맛이 강하며 후미가 길게 남아있는 특징을 가졌으며, BZ10에서는 역겨운 쓴맛이 느껴졌다 (표 6).
| N ame | Sub Name | Taste | Intensity |
| BZ1 | BZ 29-43 wild | none | - |
| BZ2 | BZ 29-43 mutant | none | - |
| BZ3 | BZ 29-33 wild | salty | ++ |
| BZ4 | BZ 29-33 mutant | salty | +++++ |
| BZ5 | BZ 40-43 wild | salty | ++ |
| BZ6 | BZ 40-43 mutant | none | - |
| BZ7 | BZ 30-33 wild | none | - |
| BZ8 | BZ 38-45 wild | none | - |
| BZ9 | BZ 38-45 mutant | none | - |
| BZ10 | BZ 38-45 mutant2 | bitter | +++++ |
3.
펩타이드의
구조 예측
합성된 10개의 펩타이드와 실제 단백질 상에서의 구조와의 비교를 통해 구조적 차이를 확인하고, 예측된 펩타이드 구조 파일을 활용하여 맛 인지 수용체와의 결합 모델을 확인하기 위해 PEP-FOLD Web sever를 통해 디자인한 10개의 펩타이드의 구조를 예측하였다.
펩타이드의 구조 예측 결과와 브라제인 상의 구조를 비교한 결과, BZ1은 원래의 구조를 잘 유지한 반면, BZ2는 다소 구부러진 형태로 존재하는 것을 알 수 있다. BZ3와 BZ4는 브라제인의 29-33번 서열의 β-turn 구조를 매우 잘 유지하고 있으며, BZ5와 BZ6의 경우 브라제인의 40-43번 서열의 구부러진 모양에 대하여 오히려 BZ6이 더 잘 유지함을 알 수 있다 (표 7).
BZ7, BZ8 그리고 BZ9의 경우 각각의 브라제인 서열에 해당되는 30-33번 서열, 38-45번 서열에 대하여 그 구조가 매우 잘 유지된다 (표 8).


4.
펩타이드와
맛 인지 수용체의 결합 예측
4.1
펩타이드와
단맛 인지 수용체
T1R2
와의 결합
단맛 수용체의 부위 특이적 변이법 연구와 키메라 형성 연구를 통해 T1R2의 VFT 모듈이 단맛 물질의 수용에 중요함이 밝혀진 바 있다 (Assadi-Porte et al., 2010). 또한, mGlu 수용체와의 상동성 연구를 통해 T1R3보다는 T1R2에 브라제인이 결합할 것으로 예측되었다 (Walters et al., 2006). 따라서 본 연구에서는 10개의 디자인된 펩타이드와 T1R2를 컴퓨터 모델링을 통해 결합함으로써 결합 위치와 결합에너지를 예측하였다. 또한 컴퓨터 모델링 이전에, CASTp Sever를 통해 T1R2에 존재할 수 있는 모든 결합포켓(binding pocket)를 예측하였다 (Dundas et al ., 2006). 브라제인과 T1R2의 상호작용에 집중하기 위하여, 96개의 예상 결합포켓에서, 브라제인의 직경인 35 Å 보다 짧은 결합부위를 제외한 21개의 결합포켓을 선정하였다 (표 9).
단맛을 나타내는 브라제인의 결합부위와 펩타이드의 결합 부위를 비교하기 위해 T1R2와 브라제인의 결합부위를 예측하였다. 그 결과 브라제인은 POC ID (pocket identification) 90번(검정)과 POC ID 92번(회색)에 해당되는 (도 8), VFT 모듈에 위치한 cleft 부분의 중앙에 정확하게 결합함을 보였다 (도 9의 A). 결합 모델의 단면을 통해서도 브라제인의 N-말단과 L40 -43 부위가 T1R2에 결합함을 알 수 있다 (도 9의 B). 브라제인과 T1R2의 결합 에너지는 -14.0 kcal/mol이다. BZ1과 BZ2의 T1R2와의 결합 예측부위는 각각 도 10, 도 11과 같다. 그 결과 BZ1, BZ2 모두 브라제인의 결합부위인 POC ID 90, 92 중 브라제인의 L40 -43이 결합한 부위인 POC ID 90에 결합한 것을 알 수 있다 (도 10, 도 11). 이때의 결합력은 각각 -12.4 kcal/mol, -11.4 kcal/mol이다.
| POC IDa | Length(Å)b | The functionally relevant residues |
| 73 | 36 | M194, V195, M198, I206, V208, L223, V227, I232, C233, I234, V273, I299 |
| 74 | 40 | M194, L197, M198, I299, A300, L323, I325, I450 |
| 77 | 35 | L350, T353, S354, C359, N360, Q361, D364, N365, C366, L367 |
| 78 | 46 | Y389, S390, Y393, A394, H397, K426, N428, I436, F437, F438 |
| 79 | 43 | V208, V210, R217, G220, Q221, L223, G224, I234, Q237, E238, T239 |
| 80 | 54 | D26, F27, Y28, Q55, E97, I98, V99, P110, Y113, F114 |
| 81 | 43 | L54, Q55, M58, D100, V101, C102, I104, N106, N107, Q109, P110 |
| 82 | 58 | V136, A137, P160, V395, A398, L399, Q419, L402, V415, Y416, P417, Q419, L420, E422, E423 |
| 83 | 54 | N292, F293, T294, E315, L316, R317, H318, G320, T321, W453, W455, W483, H484, T485, I486 |
| 84 | 55 | R202, N204, W205, D231, M494, C495, S496, Y506, V508, V512, C513, F515, Y533 |
| 85 | 64 | D26, F27, L41, H42, F53, Q55, V56, I98, D100, P347, P348, S351 |
| 86 | 98 | L158, P160, Q161, I162, K174, P178, A179, L180, L181, P417, W418, L420, L421, I424, W425, F438, D439, P440, Q441, G442 |
| 87 | 106 | T183, T184, P185, I327, Q328, S329, Y386, V330, Y386, S387, S390, L432, D433, H434, Q435, I436, F437, F438, D439, D443, V444, A445, H447, Y469 |
| 88 | 108 | D32, Y33, L34, L89, R134, V135, V136, L399, L402, L403, D406, K407, K412, R413, V414, V415, Y416, Q419 |
| 89 | 130 | S40, E63, V66, I67, Y103, D142, N143, S144, S165, A166, I167, T184, Y215, P277, D278, L279, E302, S303, A305, I306, D307, T326, I327, E 382, R383, V384 |
| 90 | 109 | K65, I306, D307, P308, H311, N312, E315, I376, L377, R378, L379, S380, G381, E382, R383, W453, R457, S458, Q459, N460, F462 |
| 91 | 178 | V64, K65, Y69, N70, Q73, S336, F338, R339, E340, W341, G342, P343, P349, R352, T353, S356, Y357, C359, N365, A369, T370, L371, S372, F373, N374, T375, I376, L377, R378, L379, S380 |
| 92 | 195 | A43, N44, M45, K46, G47, K60, E61, Y62, E63, V64, K65, V66, Y103, I104, S105, N106, E145, S211, S212, D213, L240, P241, T242, L243, Q244, P245, N246, Q255, L257, P277, D278, L279, T280, Y282, H283, Q355, S356, T358 |
a. POC ID: pocket identification; b. All the selected ones among total 92 potential binding pockets are longer than 35Å which is the length of brazzein.
브라제인의 29-33번 부위를 기반으로 디자인한 BZ3와 BZ4의 T1R2와의 결합 예측부위는 각각 도 12, 도 13과 같다. 합성 및 정제된 BZ3는 약한 짠맛을, BZ4는 강한 짠맛을 나타냈다. BZ3와 BZ4 모두 cleft의 왼쪽 부위에 결합하였으며, POC ID 92에 해당된다. 이때의 결합에너지는 각각 -7.6 kcal/mol, -5.4 kcal/mol이다.
BZ5와 BZ6은 브라제인의 40-43번 서열을 기반으로 디자인하였으며, BZ5는 약한 짠맛을, BZ6은 아무런 맛도 나지 않았다. T1R2와 BZ5, BZ6의 결합 예측부위는 각각 도 14, 도 15에 나타내었으며, BZ5는 cleft의 왼쪽 윗부분인 POC ID 85 및 92에, BZ6는 cleft의 오른쪽인 POC ID 90에 결합한 것을 알 수 있다. 이때의 결합 에너지는 각각 -8.3 kcal/mol, -7.6 kcal/mol이다.
브라제인의 30-33번 서열을 기반으로 디자인한 BZ7의 T1R2와의 결합 예측부위는 도 16과 같다. 결합 부위 예측 결과, cleft의 왼쪽 부분인 POC ID 92에 -6.9 kcal/mol의 결합 에너지로 결합하였다.
BZ8, BZ9, BZ10은 브라제인의 38-45번 서열을 기반으로 디자인하였으며, 모두 단맛을 나타내지 못하였다. BZ8과 BZ9의 경우 자극적인 맛이며 세기가 굉장히 약했으나, BZ10의 경우 매우 강한 쓴맛을 나타내었다. T1R2와의 결합 예측부위는 각각 도 17, 도 18, 도 19에 나타내었다. 결합 예측 결과 BZ8과 BZ10은 cleft의 우측에 해당되는 POC ID 92에 결합하였으며, BZ9는 cleft에서 벗어난 좌측면 (POC ID 81)에 결합하였다. 이때의 결합력은 BZ8부터 BZ10까지 순서대로 -7..2, -6.9, -7.1 kcal/mol이다.
4.2
펩타이드와
통각 수용채널
TRPV1
과의 결합
짠맛의 인지를 위한 pathway에는 나트륨 이온 채널인 ENaC와 통각 수용체인 TRPV1의 자극을 통한 2가지가 존재한다. 펩타이드는 ENaC를 통해서 탈분극을 일으키지 못하므로, 통각 수용채널인 TRPV1과 결합하여 짠맛을 전달 및 증가시킬 가능성이 있다. 본 결합 예측프로그램에 사용된 TRPV1은 electron cryo-microscopy 방법으로 그 구조가 밝혀진 Rattus norvegicus의 TRPV1으로서 (Liao et al., 2013) PDI ID는 3J5R이다 (도 20의 A). Homo-tetramer로 구성된 TRPV1의 모든 chain을 대상으로 동시에 도킹을 실시하였다.
합성 및 정제된 10개의 펩타이드 중에서 짠맛을 나타내는 BZ3, BZ4, BZ5와 TRPV1의 결합을 예측하였다 (도 20). 그 결과 Chain B의 막 관통부위에서 S1-S4 helix 구조 사이의 pore 부분에 결합되었으며, 결합 에너지는 각각 -7.2 kcal/mol, -8.0 kcal/mol, -6.5 kcal/mol이다.
본 결과는 TRPV1(transient receptor potential cation channel subfamily V member 1)에 결합하여 통각을 전달하는 물질인 캡사이신이 S3-S6 helix 구조의 pore에 결합한 결과를 나타낸 lee 등의 (Lee et al., 2011) 결과와 차이를 보이지만, 사실상 S3, S4 helix에 결합한 것에 있어서는 비슷한 결과라고 할 수 있다. 합성된 펩타이드의 도킹 결과를 볼 때, 캡사이신과 유사하게 S1-S4 helix 구조 사이에 결합하였다. 하지만, 캡사이신의 결과와 다르게 펩타이드들은 모두 S5 helix 와 추가적인 상호작용을 하는 것을 볼 수 있다. 이는 펩타이드와 캡사이신의 구조적인 차이로 인해 생긴 결과로 예상되며 안정한 결합을 유지하기 위해 S5 helix 와 추가적인 상호작용하는 것으로 여겨진다. 펩타이드들은 S1-S4 helix 구조, S4-S5 linker 및 S5 helix 와 상호작용하는 것으로 나타나 캡사이신과 달리 넓은 결합부위를 가지고 있는 것으로 보이고 이 차이로 인해 맛의 차이가 나타나는 것으로 추정된다(도 20).
V. 토의
결합부위를 분석함으로써 단맛이 나타나지 않는 이유와 짠맛과 쓴맛을 나타내는 이유를 확인할 수 있었다.
| Ligand | Receptor |
Binding
energy
?Gb ( kcal / mol ) |
Taste |
Binding
Site
( POC ID ) |
| Brazzein | T1R2 | -14.0 | sweet | center of cleft (90, 92) |
| Aspartame | T1R2 | -5.6 | sweet | center of cleft (90, 92) |
| BZ1 | T1R2 | -12.4 | none | right side of cleft (90) |
| BZ2 | T1R2 | -11.4 | none | right side of cleft (90) |
| BZ3 | T1R2 | -7.6 | salty | left side of cleft (92) |
| BZ4 | T1R2 | -5.4 | salty | left side of cleft (92) |
| BZ5 | T1R2 | -5.6 | salty | left side of cleft (85, 92) |
| BZ6 | T1R2 | -8.3 | none | right side of cleft (90) |
| BZ7 | T1R2 | -7.6 | none | left side of cleft (92) |
| BZ8 | T1R2 | -7.2 | none | right side of cleft (90) |
| BZ9 | T1R2 | -6.9 | none | left side of VFT module (81) |
| BZ10 | T1R2 | -7.1 | bitter | right side of cleft (90) |
| BZ3 | TRPV1 | -8.4 | salty | TM1-4 pore |
| BZ4 | TRPV1 | -8.3 | salty | TM1-4 pore |
| BZ5 | TRPV1 | -8.1 | salty | TM1-4 pore |
| BZ10 | T2R1 | -7.6 | bitter | N- terminal |
합성한 펩타이드로부터 단맛이 나지 않는 첫 번째 이유로 예상할 수 있는 것은, 브라제인에서의 구조가 합성한 펩타이드에서 완벽하게 유지되지 않았다는 것이다. PEP-FOLD 구조 예측을 통해 예측된 펩타이드는 비교적 브라제인에서의 구조와 비슷한 것으로 보이며, 실제 합성된 펩타이드의 CD(circular dichroism) spectra 결과 역시 예측된 구조가 실제에서의 구조의 유효성을 잘 설명해 줄 수 있었다. 예측된 구조의 유효성 확인을 위한 CD spectra 측정 과정을 위해 인하대학교 공동기기원에서 보유한 Jasco사의 J-815 CD spectrascometer를 활용하였으며 10개의 펩타이드 중 BZ1, BZ2, BZ6, BZ7의 CD spectra를 측정하였다 (도 22).
BZ1, 2, 6, 7의 CD 결과로부터 2차 구조를 예측하기 위해 CDSSTR 알고리즘을 활용하여 분석하였다 (Johnson, 1999). 56개의 단백질을 reference로 갖고 있는 SMP56 reference set을 기반으로 CD 결과를 분석한 결과, BZ1과 BZ2는 β-sheet 구조의 비율이, BZ3와 BZ4는 turn의 비율이 높은 것으로 나타났다 (Sreerama et al., 2004). 또한, BZ2 보다 BZ1의 β-sheet 구조의 비율이 더 높은 것과, BZ4가 BZ3 보다 turn 구조가 높게 나온 것을 통해, 예측된 결과가 실제 펩타이드의 구조를 잘 예측하고 있다는 것을 알 수 있다 (표 11). 하지만 비록 CD spectra 분석 결과와 PEP-FOLD 구조 예측 결과가 상관성 있게 나타났다 할지라도, PEP-FOLD 구조 예측 결과 얻은 펩타이드의 구조가 브라제인에서의 구조와 완벽하게 동일하지 않을뿐더러, 수용액 상태에서 자유자재로 결합의 회전이 일어나는 펩타이드 구조가 가질 수 있는 경우의 수는 무한대와 같다. 따라서 브라제인의 단맛을 나타내는 펩타이드를 합성하였다 할지라도 브라제인에서의 본래 구조를 가질 확률이 매우 낮아 단맛을 나타내지 못하였을 수 있다.

디자인한 펩타이드에서 단맛이 나타나지 않은 결과에 대해 예상되는 두 번째 이유는, 브라제인이 결합해야 하는 다중 결합 부위(multi-binding site)가 충족되지 않았다는 것이다. 단맛을 인식하는 GPCR C 계열 수용체인 T1R2/T1R3는 당을 비롯한 인공 감미료와 결합하여 G 단백질을 통해 뇌로 단맛의 신호를 전달한다. 이는 수용체와 리간드의 결합 부위 및 에너지가 단맛 전달의 중요한 요소가 된다는 것을 말한다. T1R2와 T1R3 중에서 감미단백질과 결합하는 것으로 알려진 T1R2과 브라제인의 결합을 컴퓨터 모델링을 통해 결합한 결과 도 9와 같이 T1R2의 cleft 전체 부분에 결합한 것을 확인할 수 있었으며 그때의 POC ID는 90과 92였다. 또한, -14 kcal/mol의 높은 결합 에너지로 결합하는 것을 확인할 수 있었다. 또한, 브라제인과 T1R2의 수소결합을 분석함으로써 단맛을 나타내기 위해 결합에 관여하는 잔기와 (도 23) 수소결합 하는 잔기를 확인하였다 (표 12).

상기 표 12에서 볼 수 있듯이, 브라제인은 11개의 수소결합을 갖고 강하게 결합하며, N- 말단과 36-54번 잔기에 해당하는 loop가 T1R2와의 결합에 관여하는 것을 확인할 수 있었다. 또한, 46, 60, 255, 312-314, 458번 잔기에 해당되는 T1R2의 POC ID 90, 92 부분이 브라제인과 강한 수소결합을 통해 결합하는 것을 알 수 있다. 이와 비교하여 BZ1와 BZ2는 브라제인의 loop가 결합하는 POC ID 90에는 결합하지만 N-말단이 결합하는 POC ID 92에는 결합하지 못하였다. 반면 짠맛을 나타내었던 BZ3, BZ4, BZ5는 브라제인의 POC ID 92이 결합하는 부위에는 결합하였으나, loop가 결합하는 부위에는 결합하지 못하였다. 마찬가지로 BZ6, BZ8, BZ10은 loop가 결합하는 부위에는 결합하였으나 N-말단 결합 부위에는 결합하지 못하였다. 그 외에도 BZ7, BZ9는 N-말단 결합부위에 결합하는 결과를 보임으로써, 디자인한 10개의 펩타이드 중에서 N-말단 결합부위와 loop 결합 부위를 둘 다 충족시키는 것은 없었다. 본 결과를 통해 내릴 수 있는 결론은 브라제인은 T1R2에 다중 결합 부위로 결합하며, 그 중에 한 부분이라도 충족되지 않는다면 단맛을 나타낼 수 없다는 것이다.
펩타이드와 상호작용에 관여하는 TRPV1 chain B의 공통적인 아미노산이 존재하는 것을 도킹 결과를 통해 예측할 수 있었다(도 24). Glu513, Val567, Gln700, Ile703 의 경우, 3개의 펩타이드 모두에서 상호작용에 관련하는 아미노산으로 나타났다. 특히, Val567 은 S5 helix에 포함된 아미노산으로 S5 helix 와 상호작용이 공통적으로 일어난 것으로 보아 S5 helix 의 Val567과 그 주변 아미노산과 결합이 이루어졌을 때 짠맛을 나타낼 수 있는 가능성을 시사한다. 합성된 펩타이드가 위치한 S1-S4 helix pore 구조의 내부는 극성 아미노산으로 이루어져 있다. 이는 극성 아미노산을 많이 포함하고 있는 펩타이드의 결합 안정성에 큰 기여를 할 것으로 예상되고 도킹 결과 통해 S1-S4 helix pore 에 가장 낮은 에너지로 결합함을 볼 수 있었다. 이와 관련하여 BZ4 가 가장 짠맛을 나타낸 것도 가장 낮은 에너지로 가장 안정한 결합을 하고 있기 때문일 것이라고 추정할 수 있다.
아스파탐의 발견 이후 수많은 짧은 펩타이드의 연구 및 개발이 진행되었다. Tamura 그룹에서 개발된 펩타이드는 디펩타이드(dipeptide), 트리펩타이드(tripeptide)로서 우마미 맛을 지녀 육수의 우마미 맛의 주원인으로 생각되어지고 있다(Tamura et al ., 1989). 여기서 주목할 점은, 개발된 디펩타이드 및 트리펩타이드는 우마미맛 뿐만 아니라 짠맛까지 함께 동반한 것들이 존재하였는데, 이들은 주로 음전하를 띠는 아스파르트산(aspartic acid) 및 글루탐산(glutamic acid)으로 이루어졌다는 것이다. 본 연구에서 개발된 가장 짠맛을 지닌 KKRAR은 주로 양전하를 띄는 리신(lysine)과 아르기닌(arginine) 포함하고 있어 기존에 개발되었던 음전하를 띈 펩타이드와 반대 전하를 띈 점에서 짠맛을 가지는 조건은 전하와 크게 무관하다는 것을 유추할 수 있다.
BZ3, BZ4, BZ5가 짠맛을 나타낸 것과 통각 수용체 TRPV1와의 도킹 결과는 우리에게 중요한 2가지 사항을 시사한다. 짧은 펩타이드로 짠맛을 나타낼 수 있다는 것이 그 첫 번째 시사점이다. 2012년 한국식품연구원의 류미라 박사팀이 재래 간장으로부터 발견한 짠맛 단백질의 분자량은 500 Da에서 10,000 Da에 이를 뿐 아니라 만노스와 N-아세틸-글루코사민의 당 수식이 된 고분자이며, 숙성과정을 거친 후에 추출해야 얻을 수 있다 (Rhyu et al., 2012). 반면 BZ3~5의 3가지 펩타이드는 4~5개 아미노산으로 이루어진 짧은 펩타이드이며, 당 수식이 되지 않은 상태에서도 짠맛을 나타내는 것을 본 실험을 통하여 확인하였다. 즉, 소금을 대체할 수 있는 펩타이드성 짠맛 식품 첨가물의 개발이 가능하다는 것을 말한다. 두 번째 시사점은 리간드가 통각 수용체 TRPV1에 결합하는 부위에 따라 전달되는 신호의 종류가 다양해 질 수 있다는 것이다. 매운 맛의 통증을 전달하는 캡사이신은 TRPV1의 S3, S4에 결합하면서 S1-S4 helix pore 가운데 위치하였다. BZ3~5 역시 S3, S4에 결합하였으나 S1-S4 helix pore 기저부에 결합하였다는 것과 S5 helix와 추가적인 상호작용을 한 것이 캡사이신의 결합양상과의 차이점이다. 즉, Voltage sensor domain에 속하는 S3, S4의 측면에 결합한 경우 매운맛을, 기저부에 결합하고 S5 helix와 상호작용 할 경우 짠맛을 전달함을 도킹 결과를 통해 예상할 수 있다.
참고문헌
Assadi-Porter, F. M., Maillet, E. L., Radek, J. T., Quijada, J., Markely, J. L., Max, M. (2010) Key amino acid residues involved in multi-point binding interactions between brazzein, a sweet protein, and the T1R2-T1R3 human sweet receptor. J. Mol . Biol ., 398, 584-599.
Biasini, M., Bienert, S., Waterhouse, A., Arnold, K., Studer, G., Schmidt, T., Kiefer, F., Cassarino, T. G., Bertoni, M., Bordoli, L., Schwede, T. (2014) SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res., 42, 1-7.
Do, H. D., Jo, H. J., Jo, D. H., Kong, K. H. (2011) Mutagenesis of critical amino acid residues in α-helix and β-sheet structures of brazzein. Bull. Kor . Chem . Soc ., 32, 4106-4108.
Dundas, J., Ouyang, Z., Tseng, J., Binkowski, A., Turpaz, Y., Liang J. (2006) CASTp: computed atlas of surface topography of proteins with structural and topographical mapping of functionally annotated residues. Nuc . Acid Res ., 34, 116-118.
Huang, Y. j., Maruyama, Y., Dvoryanchikov, G., Pereira, E., Chaudhari, N., Roper, S. D. (2007) The role of pannexin 1 hemichannels in ATP release and cell-cell communication in mouse taste buds. PNAS, vol. 104, no. 15, 6436-6441.
Jin, Z., Danilova, V., Assadi-Porter, F. M., Aceti D. J., Markley, J. L., Hellekant, G. (2003) Critical regions for the sweetness of brazzein. FEBS Lett ., 544, 33-37.
Johnson, W. C. (1999) Analyzing protein circular dichroism spectra for accurate secondary structures. Proteins, 35, 307-312.
Katsumata, T., Nakakuki, H., Tokunaga, C., Fujii, N., Egi, M., Phan, T. H., Mummalaneni, S., DeSimone, J. A., Lyall, V. (2008) Effect of maillard reacted peptides on human salt taste and the amiloride-insensitive salt taste receptor (TRPV1t). Chem . Senses , 33, 665-680.
Kitagawa, M., Kusakabe, Y., Miura, H., Ninomiya, Y., Hino, A. (2001) Molecular genetic identification of a candidate receptor gene for sweet taste. Biochem . Biophys . Res . Commun ., 283, 236-242.
Lee, J. H., Lee, Y., Ryu, H., Kang, D. W., Lee, J., Lazar, J., Pearce, L. V., Pavlyukovets, V. A., Blumberg, P. M., Choi, S. (2011) Structural insights into transient receptor potential vanilloid type 1 (TRPV1) from homology modeling, flexible docking, and mutational studies. J. Comput . Aided Mol. Des ., 25, 317-327.
Lee, J. W., Cha, J. E., Jo, H. J., Kong, K. H. (2013) Multiple mutations of the critical amino acid residues for the sweetness of the sweet-tasting protein, brazzein. Food Chem ., 138, 1370-1373.
Liu L., Simon S. A. (2001) Acidic stimuli activates two distinct pathways in taste receptor cells from rat fungiform papillae. Brain Res ., 923, 58-70.
Lopez Cascales, J. J., Oliveira Costa, S. D., Bert L. de Groot, Walters, D. E. (2010) Binding of glutamate to the umami receptor. Biophys . Chem., 152, 139-144.
Margolskee, R. F. (2002) Molecular mechanisms of bitter and sweet taste transduction. J. Biol . Chem ., 277, 1-4.
Mombaerts, P. (2004) Genes and ligands for odorant, vomeronasal and taste receptors. Nat . Rev . Neurosci ., 5, 263-278.
Morris, G. M., Huey, R., Lindstrom, W., Sanner, M. F., Belew, R. K., Goodsell, D. S. and Olson, A. J. (2009) Autodock4 and AutoDockTools4: automated docking with selective receptor flexiblity. J. Comput . Chem ., 16, 2785-2791.
Pettersen, E. F., Goddard, T. D., Huang, C. C., Couch, G. S, Greenblatt, D. M., Meng, E. C., Ferrin, T. E. (2004) UCSF Chimera-visualization system for exploratory research and analysis. J. Comput . Chem ., 25, 1605-1612.
Ohkuri, T., Yasumatsu, K., Horio, N., Jyotaki, M., Margolskee, R. F., Ninomiya, Y. (2009) Multiple sweet receptors and transduction pathways revealed in knockout mice by temperature dependence and gurmarin sensitivity. Am. J. Physiol . Regul . Integr . Comp . Physiol ., 296, 960-971.
Shi, P., Zhang, J. (2006) Contrasting modes of evolution between vertebrate sweet/umami receptor genes and bitter receptor genes. Mol . Biol . Evol., 23, 292-300.
Smith, K. R., Treesukosol, Y., Brennan, A. B., Contreras, R. J., Spector, A. C. (2012) Contribution of the TRPV1 channel to salt taste quality in mice as assessed by conditioned taste aversion generalization and chorda tympani nerve responses. Am . J. Physiol . Regul . Integr . Comp . Physiol ., 303, 1195-1205.
Sreerama, N., Woody, R. W. (2004) On the analysis of membrane protein circular dichroism spectra. Protein Sci., 13, 100.
Temussi, P. A. (2002) Why are sweet proteins sweet? Interaction of brazzein, monellin and thaumatin with the T1R2-T1R3 receptor. FEBS Lett ., 526, 1-4.
Thevenet, P., Shen, Y., Maupetit, J., Guyon, F., Derreumaux, P., Tuffery, P. (2012) PEP-FOLD: an updated de novo structure prediction server for both linear and disulfide bonded cyclic peptides. Nucleic Acids Res., 40, 288-293.
Trott, O., Olson, A. J. (2010) AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput . Chem ., 31, 455-461.
Walters, D. E., Hellekant G. J. (2006) Interactions of the sweet protein brazzein with the sweet taste receptor. Agric . Food Chem ., 54, 10129-10133.
Yoon, S. Y., Kong, J. N., Jo, D. H., Kong. K. H. (2011) Residue mutations in the sweetness loops for the sweet-tasting protein brazzein. Food Chem., 129, 1327-1330.
<110> Chung-Ang University Industry-Academy Cooperation Foundation
<120> Salty peptide
<130> P15U21C0062
<160> 10
<170> KopatentIn 2.0
<210> 1
<211> 15
<212> PRT
<213> pentadiplandra brazzeana
<400> 1
Asp Lys His Ala Arg Ser Gly Glu Cys Phe Tyr Asp Glu Lys Arg
1 5 10 15
<210> 2
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> BZ 29-43 mutant
<400> 2
Lys Lys Arg Ala Arg Ser Gly Asp Cys Phe Tyr Asp Ala Lys Arg
1 5 10 15
<210> 3
<211> 5
<212> PRT
<213> pentadiplandra brazzeana
<400> 3
Asp Lys His Ala Arg
1 5
<210> 4
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> BZ 29-33 mutant
<400> 4
Lys Lys Arg Ala Arg
1 5
<210> 5
<211> 4
<212> PRT
<213> pentadiplandra brazzeana
<400> 5
Asp Glu Lys Arg
1
<210> 6
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<223> BZ 40-43 mutant
<400> 6
Asp Ala Lys Arg
1
<210> 7
<211> 4
<212> PRT
<213> pentadiplandra brazzeana
<400> 7
Lys His Ala Arg
1
<210> 8
<211> 8
<212> PRT
<213> pentadiplandra brazzeana
<400> 8
Phe Tyr Asp Glu Lys Arg Val Leu
1 5
<210> 9
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> BZ 38-45 mutant
<400> 9
Phe Tyr Asp Pro Glu Lys Arg Val Leu
1 5
<210> 10
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> BZ 38-45 mutant2
<400> 10
Cys Phe Tyr Asp Glu Lys Arg Val Leu Cys
1 5 10
Claims (5)
- 서열번호 3으로 표시되는 짠맛 펩타이드.
- 서열번호 4로 표시되는 짠맛 펩타이드.
- 서열번호 5로 표시되는 짠맛 펩타이드.
- 제 1 항 내지 제 3 항 중에서 선택된 하나의 항의 펩타이드를 포함하는 식염 대체제.
- 제 1 항 내지 제 3 항 중에서 선택된 하나의 항의 펩타이드를 포함하는 식품 첨가제.
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020150050771A KR101682803B1 (ko) | 2015-04-10 | 2015-04-10 | 짠맛 펩타이드 |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| KR1020150050771A KR101682803B1 (ko) | 2015-04-10 | 2015-04-10 | 짠맛 펩타이드 |
Related Child Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020160135017A Division KR101682865B1 (ko) | 2016-10-18 | 2016-10-18 | 짠맛 펩타이드 |
| KR1020160135015A Division KR101682864B1 (ko) | 2016-10-18 | 2016-10-18 | 짠맛 펩타이드 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| KR20160121702A true KR20160121702A (ko) | 2016-10-20 |
| KR101682803B1 KR101682803B1 (ko) | 2016-12-06 |
Family
ID=57251172
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020150050771A KR101682803B1 (ko) | 2015-04-10 | 2015-04-10 | 짠맛 펩타이드 |
Country Status (1)
| Country | Link |
|---|---|
| KR (1) | KR101682803B1 (ko) |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR102315036B1 (ko) * | 2020-07-24 | 2021-10-21 | 중앙대학교 산학협력단 | 염미성 펩타이드 발현용 재조합 발현벡터 및 이를 이용한 생산 방법 |
Citations (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS633766A (ja) | 1986-06-14 | 1988-01-08 | ドイチエ ジエラテイネ−フアブリケン シユテス ウント コンパニ− ゲゼルシヤフト ミツト ベシユレンクテル ハフツング | 減塩食品の製造方法 |
| JPS63137658A (ja) | 1986-11-28 | 1988-06-09 | San Ei Chem Ind Ltd | 食塩の塩辛度の増加法 |
| JPH0253456A (ja) | 1988-08-19 | 1990-02-22 | Nakano Vinegar Co Ltd | 塩辛味増強剤 |
| KR20010101729A (ko) * | 1999-11-29 | 2001-11-14 | 히라타 다다시 | 식염미 증강방법, 식염미 증강제, 식염미 조미료 및식염미 증강식품 |
| JP2009113563A (ja) * | 2007-11-02 | 2009-05-28 | Tokai Rubber Ind Ltd | 衝撃吸収アセンブリおよびその取付方法 |
| KR101126163B1 (ko) | 2009-11-06 | 2012-03-22 | 한국식품연구원 | 장기 숙성 재래 간장에서 분리한 메일라드 펩타이드를 유효성분으로 함유하는 trpv1 활성 관련 질환 또는 염증 관련 질환의 예방 또는 치료용 약학적 조성물 |
| WO2013000922A1 (en) * | 2011-06-27 | 2013-01-03 | Universite Pierre Et Marie Curie (Paris 6) | Ccr2 antagonist peptides |
| KR20130057403A (ko) | 2011-11-23 | 2013-05-31 | 한국식품연구원 | 감미 증강제 |
| KR20130076052A (ko) * | 2011-12-28 | 2013-07-08 | 대상 주식회사 | L-글루탐산 및 l-라이신을 포함하는 아미노산 조미료 조성물 |
-
2015
- 2015-04-10 KR KR1020150050771A patent/KR101682803B1/ko active IP Right Grant
Patent Citations (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS633766A (ja) | 1986-06-14 | 1988-01-08 | ドイチエ ジエラテイネ−フアブリケン シユテス ウント コンパニ− ゲゼルシヤフト ミツト ベシユレンクテル ハフツング | 減塩食品の製造方法 |
| JPS63137658A (ja) | 1986-11-28 | 1988-06-09 | San Ei Chem Ind Ltd | 食塩の塩辛度の増加法 |
| JPH0253456A (ja) | 1988-08-19 | 1990-02-22 | Nakano Vinegar Co Ltd | 塩辛味増強剤 |
| KR20010101729A (ko) * | 1999-11-29 | 2001-11-14 | 히라타 다다시 | 식염미 증강방법, 식염미 증강제, 식염미 조미료 및식염미 증강식품 |
| JP2009113563A (ja) * | 2007-11-02 | 2009-05-28 | Tokai Rubber Ind Ltd | 衝撃吸収アセンブリおよびその取付方法 |
| KR101126163B1 (ko) | 2009-11-06 | 2012-03-22 | 한국식품연구원 | 장기 숙성 재래 간장에서 분리한 메일라드 펩타이드를 유효성분으로 함유하는 trpv1 활성 관련 질환 또는 염증 관련 질환의 예방 또는 치료용 약학적 조성물 |
| WO2013000922A1 (en) * | 2011-06-27 | 2013-01-03 | Universite Pierre Et Marie Curie (Paris 6) | Ccr2 antagonist peptides |
| KR20130057403A (ko) | 2011-11-23 | 2013-05-31 | 한국식품연구원 | 감미 증강제 |
| KR20130076052A (ko) * | 2011-12-28 | 2013-07-08 | 대상 주식회사 | L-글루탐산 및 l-라이신을 포함하는 아미노산 조미료 조성물 |
Cited By (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| KR102315036B1 (ko) * | 2020-07-24 | 2021-10-21 | 중앙대학교 산학협력단 | 염미성 펩타이드 발현용 재조합 발현벡터 및 이를 이용한 생산 방법 |
Also Published As
| Publication number | Publication date |
|---|---|
| KR101682803B1 (ko) | 2016-12-06 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Shiyan et al. | Novel umami peptides from tilapia lower jaw and molecular docking to the taste receptor T1R1/T1R3 | |
| Kong et al. | Comparison of non-volatile umami components in chicken soup and chicken enzymatic hydrolysate | |
| Zhang et al. | Identification and virtual screening of novel umami peptides from chicken soup by molecular docking | |
| Hellfritsch et al. | Human psychometric and taste receptor responses to steviol glycosides | |
| Schindler et al. | Discovery of salt taste enhancing arginyl dipeptides in protein digests and fermented fish sauces by means of a sensomics approach | |
| CN102481006B (zh) | 肽用于赋予浓味的用途 | |
| Chen et al. | Characterization of novel umami-active peptides from Stropharia rugoso-annulata mushroom and in silico study on action mechanism | |
| Zhang et al. | Umami peptides screened based on peptidomics and virtual screening from Ruditapes philippinarum and Mactra veneriformis clams | |
| Rega et al. | Design of sweet protein based sweeteners: Hints from structure–function relationships | |
| Yu et al. | Identification of Oncorhynchus mykiss nebulin-derived peptides as bitter taste receptor TAS2R14 blockers by in silico screening and molecular docking | |
| Shen et al. | Identification of novel saltiness-enhancing peptides from yeast extract and their mechanism of action for transmembrane channel-like 4 (TMC4) protein through experimental and integrated computational modeling | |
| Mollica et al. | Exploring the biological consequences of conformational changes in aspartame models containing constrained analogues of phenylalanine | |
| Shen et al. | Isolation, taste characterization and molecular docking study of novel umami peptides from Lactarius volemus (Fr.) | |
| Diepeveen et al. | Molecular insights into human taste perception and umami tastants: A review | |
| Wang et al. | Identification of umami peptides based on virtual screening and molecular docking from Atlantic cod (Gadus morhua) | |
| Zhao et al. | Revealing the secret of umami taste of peptides derived from fermented broad bean paste | |
| Newcomb et al. | Heritable differences in chemosensory ability among humans | |
| An et al. | Identification, taste characterization, and molecular docking study of a novel microbiota-derived umami peptide | |
| Zhao et al. | In silico identification of novel small molecule umami peptide from ovotransferrin | |
| Paladino et al. | Functional hypothesis on miraculin’sweetness by a molecular dynamics approach | |
| Zhao et al. | Taste characteristics and umami mechanism of novel umami peptides from hen egg proteins | |
| KR101682803B1 (ko) | 짠맛 펩타이드 | |
| Zhao et al. | Combining molecular docking and molecular dynamics simulation to discover four novel umami peptides from tuna skeletal myosin with sensory evaluation validation | |
| Gasymov et al. | Evaluation of anti-cancer and anti-covid-19 properties of cationic pentapeptide Glu-Gln-Arg-Pro-Arg, from rice bran protein and its d-isomer analogs through molecular docking simulations | |
| Wang et al. | Identification of novel umami peptides from yeast extract and the mechanism against T1R1/T1R3 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A107 | Divisional application of patent | ||
| E701 | Decision to grant or registration of patent right | ||
| FPAY | Annual fee payment |
Payment date: 20191001 Year of fee payment: 4 |