KR0132425B1 - 신규 폴리펩타이드 및 그 제조방법 - Google Patents
신규 폴리펩타이드 및 그 제조방법Info
- Publication number
- KR0132425B1 KR0132425B1 KR1019900003202A KR900003202A KR0132425B1 KR 0132425 B1 KR0132425 B1 KR 0132425B1 KR 1019900003202 A KR1019900003202 A KR 1019900003202A KR 900003202 A KR900003202 A KR 900003202A KR 0132425 B1 KR0132425 B1 KR 0132425B1
- Authority
- KR
- South Korea
- Prior art keywords
- polypeptide
- dna
- cells
- amino acid
- acid sequence
- Prior art date
Links
Images













Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/575—Hormones
- C07K14/655—Somatostatins
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/22—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against growth factors ; against growth regulators
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P17/00—Drugs for dermatological disorders
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P43/00—Drugs for specific purposes, not provided for in groups A61P1/00-A61P41/00
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/475—Growth factors; Growth regulators
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/475—Growth factors; Growth regulators
- C07K14/48—Nerve growth factor [NGF]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Zoology (AREA)
- Engineering & Computer Science (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Pharmacology & Pharmacy (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Public Health (AREA)
- Animal Behavior & Ethology (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Wood Science & Technology (AREA)
- Immunology (AREA)
- General Engineering & Computer Science (AREA)
- Veterinary Medicine (AREA)
- Biotechnology (AREA)
- Physics & Mathematics (AREA)
- Endocrinology (AREA)
- Plant Pathology (AREA)
- Microbiology (AREA)
- Dermatology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
Abstract
내용 없음.
Description
제1도은 실시예1에서 얻어진 플라스미드 pUNK5 내의 폴리펩타이드(I) cDNA를 포함한 DNA 의 제한 효소지도이며,
제2도는 실시예1에서 얻어진 플라스미드 pUNK5 내의 폴리펩타이드(I) cDNA를 포함하는 DNA 의 뉴틀레오티드 서열 및 그로부터 해독된 아미노산서열을 나타내며,
제3도는 실시예1에서 얻어진 본 발명의 폴리펩타이드(I) 의 아미노산서열 (윗줄)과 사람 βNGF의 아미노산서열 (아랫줄) 과의 비교를 나타내며,
제4도는 실시예2에서 얻어진 프라스미드 pHNT2 내의 폴리펩타이드(I) cDNA를 포함하는 DNA 의 제한효소지도이며,
제5도는 실시예 2 에서 얻어진 플라스미드 pHNT2 내의 폴리펩타이드(I) cDNA를 포함하는 DNA 의 뉴클레오티드서열 및 그로부터 해독된 아미노산서열을 나타내며,
제6도는 실시예3에서 얻어진 에스케리키아 콜라이 ( Escherichia coli) 에 대한 폴리펩타이드(I) 발현벡터의 구축을 나타내는 도식적 표현이며,
제7도는 실시예 5에서 얻어진 동물세포에 대한 폴리펩타이드(I) 발현벡터 pNTK 26 및 pNTL 145 의 구축을 나타내는 도식적 표현이며,
제8도는 실시예6에서 얻어진 동물세포에 대한 폴리펩타이드(I) 발현벡터 pNTS 101 의 구축을 나타내는 도식적 표현이며,
제9도는 실시예8에서 얻어진 효모에 대한 폴리펩타이드(I) 발현벡터 pANT 341T 의 구축을 나타내는 도식적 표현이며,
제10도는 실시예12에서 얻어진 닭의 태아지각 신경세포의 생존에 대한 각 시료의 영향을 나타내는 그래프이며,
제11도는 실시예13에서 얻어진 생쥐 폴리펩타이드(I) 유전자의 뉴클레오티드서열 및, 그로부터 해독된 아미노산서열을 나타내고,
제12도는 실시예15에서 얻어진 폴리펩타이드(I) 발현벡터 pTB 1139 의 구축을 나타내는 도식적 표현이다.
본 발명은 신규폴리펩타이드, 그에 대한 DNA 서열코오딩 및 그 용도에 관한 것이다.
표피 생장인자 ( 이후부터 EGF 로 표기) 및 신경생장인자 ( 이후부터 NGF 로 표기)의 발견 이후로 많은 세포생장인자들이 분리되고 그 구조가 밝혀져 왔다.
세포생장인자들은 세포분화 메카니즘 및 세포증식 메카니즘의 해명에 유용하고 이들중 일부는 ( 사람 EGF를 포함) 약제로서 유용한 것으로 개대된다. 따라서, 이들에 대한 연구는 근년간 널리 증가되어 왔다.
사람 NGF 유전자가 분리되었지만, DNA재조합 기술을 이용하여 사람 NGF를 대량제조하는 것에 관해서는 보고된 바가 없다.
동물세포의 생장을 촉진하는 신규폴리펩타이드가 얻어진다면, 그것에 의 해 새로운 탐구를 할 수 있다. 공지의 생장인자와 유사한 활성을 갖는 그러한 신규폴리펩타이드는 또한 약제로서 활용할 수도 있을 것이다.
이러한 신규폴리펩타이드를 발견하기 위하여, 본 발명자들은 DNA서열코오딩 NGF를 프로우브로서 사용하고 사람신경교종의 cDNA 라이브러리로부터 그와 함께 혼성화할 수 있는 DNA서열을 클로닝하였다.
그결과, 본 발명자들은 신규폴리펩타이드에 대한 DNA (cDNA)서열코오딩을 얻는데 성공하였다.
본 발명의 cDNA는 신규폴리펩타이드를 제조하는 숙주세포로 표현될 수 있다. 이 폴리펩타이드는 또한 동물세포의 분화, 생장 및 생존에 관한 연구용 시약으로서 사용될 수 있다. 이 폴리펩타이드는 또한 약제로서 사용 가능하다. 본 발명자들은 상기 기재한 정보에 바탕하여 더 이상의 연구를 수행하여 마침내 본 발명을 완성하였다.
본 발명의 목적은 따라서 연구조사용 시약 또는 약제로서 유용한 신규 폴리펩타이드를 제공하는 것이다. 기타 목적은 하기의 기술내용 및 첨부된 도면으로부터 자명해질 것이다.
본 발명은 :
(1) 그 분자내에 하기의 아미노산서열 (II)을 포함하는 폴리펩타이드 :
TyrAlaGluHisLysSerHisArgGlyGluTyrSerValCys
AspSerGluSerLeuTrpValThrAspLysSerSerAlaIle
AspIleArgGlyHisGlnValThrValLeuGlyGluIleLys
ThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArg
CysLysGluAlaArgProValLysAsnGlyCysArgGlyIle
AspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThr
TyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGly
TrpArgTrpIleArgIleAspThrSerCysValCysAlaLeu
SerArgLysIleGlyArg (II)
(이 아미노산서열은 이후부터 식(II)로 간단히 표기함),
(2) (1)에 기재된 폴리펩타이드에 대한 DNA 코오딩,
(3) (2)에 기재된 DNA서열을 포함하는 벡터,
(4) (3)에 기재된 중개물에 의해 형질전화된 형질전환체 및,
(5) 배지내에서 (4)에 기재된 형질전환체을 배양하여 (1)에 기재된 폴리펩타이드를 배지내에서 제조 및 축적하는 것으로 이루어지는 폴리펩타이드(I)의 제조방법을 제공한다.
본 발명의 폴리펩타이드(I)은 식(II)의 아미노산서열을 갖는 폴리펩타이드와 또한 식(II)의 아미노산서열의 C- 말단에 첨가된 트레오닌잔기를 갖는 폴리펩타이드를 포함한다. 상기 폴리펩타이드에 덧붙여, 본 발명의 폴리펩타이드와 동일한 활성을 갖는 상기 폴리펩타이드의 부분, 및 상기아미노산서열의 부분이 하나 또는 그 이상의 다른 아미노산 또는 아미노산서열로 치환된, 또는 상기 아미노산서열 속에 하나 또는 그 이상의 다른 아미노산 또는 아미노산서열의 첨가 또는 삽입된 폴리펩타이드 및 상기 폴리펩타이드와 동일한 활성을 갖는 폴리펩타이드를 포함한다.
아미노산서열(II) 의 C- 말단에 Thr 이 첨가된 하기의 아미노산서열(II')를 갖는 폴리펩타이드(I)이 하기에 언급된 실시예에서 E. 콜라이 (E. coli) 형질전환체에 의해 제조되었다.
TyrAlaGluHisLysSerHisArgGlyGluTyrSerValCys
AspSerGluSerLeuTrpValThrAspLysSerSerAlaIle
AspIleArgGlyHisGlnValThrValLeuGlyGluIleLys
ThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArg
CysLysGluAlaArgProValLysAsnGlyCysArgGlyIle
AspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThr
TyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGly
TrpArgTrpIleArgIleAspThrSerCysValCysAlaLeu
SerArgLysIleGlyArgThr (II')
아미노산서열(II) 또는 (II')를 갖는 폴리펩타이드(I)은 또한 동물세포 형질전환체를 사용하여 발현될 수 있다.
DNA 재조합기술을 사용하여 폴리펩타이드(I)을 제조할 때, 폴리펩타이드(I)에 대한 유전자 코오딩으로부터
개시코오돈 ATG상류에 해당되는 메티오닌잔기를 폴리펩타이드(I)의 N- 말단에 첨가할 수 있다.
본 발명의 폴리펩타이드(I)에 대한 DNA 코오딩은 예를 들어, 하기 방법으로 얻을 수 있다 :
(1) 메신저 RNA (mRNA)를 폴리펩타이드(I) -생성세포로부터 분리한다.
(2) mRNA 로부터 1가닥 상보 DNA(cDNA)를 합성한다음, 이중가닥 DNA를 합성한다.
(3) 상보 DNA를 플라스미드 또는 파아지 속에 도입한다.
(4) 숙주세포를 그렇게해서 수득한 재조합 파아지 또는 플라스미드로써 형질전환시킨다.
(5) 그렇게해서 수득한 형질전환체의 배양 후, 원하는 DNA를 함유하는 플라스미드 또는 파아지를 플라그(plague) 혼성화 또는 군락혼성화 같은 적절한 방법에 의해 형질전환체로부터 분리시킨다.
(6) 원하는 클로닝된 DNA 서열을 플라스미드 또는 파아지로부터 절단한다.
(7) 클로닝된 DNA를 적절한 플라스미드 속에 서브클로닝 한다.
폴리펩타이드(I)에 대한 mRNA 코오딩은 폴리펩타이드(I)-생성세포, 예컨데 사람 및 쥐 등의 동물의 세포, 조직 및 기관, 특히 사람신경교종, 사람신경교세포, 사람태반, 쥐신경교종, 신장, 간, 심장, 뇌, 비장, 흉신, 허파 및 하악선에 의해 얻어질 수 있다.
폴리펩타이드(I)-생성세포로부터 mRNA를 제조하는 적당한 방법에는 구아니딘 티오시아네이트법(J.M. Chirgwin et al., Biochemistry, 1s, 5294 (1979))등이 포함된다.
그렇게해서 수득한 mRNA를 형(template)으로서 사용하여, 예컨데 H. 오까야마 일행의 방법(Molecular and Cellular Biology, 2, 161 (1979) ; ibid. 3, 280 (1989)) 또는 U. 구블러 및 B.J. 호프만의 방법 (Gene 25,263, (1983))에 따라서 역전사효소를 사용하여 cDNA를 합성한다. 그렇게해서 수득한 cDNA를 플라스미드 속에 도입하여 사람 cDNA 라이브러리를 제조한다.
cDNA 가 도입될 수 있는 플라스미드에는 예를 들어, pBR 322(Gene 2, 95 (1977)), pBR 325(Gene 4, 121 (1978)), pUCI 2 (Gene 19, 259 (1982)), pUCI 3(Gene 19, 259, (1982)), pUCI 8(Gene 33, 103 (1985)), pUCI 9(Gene 33, 103 (1985)), pUCI 18 (Method in Enzymology 153, 3 (1987)), pUCI 19(Method in Enzymology 153, (1987))이 포함된다. 그러나, 숙주세포내에서 모사될 수 있고, 지지될 수 있는 한 다른 어떠한 플라스미드도 사용가능하다. cDNA가 도입될 수 있는 파아지 벡터로는 예컨데 λgtill (R. Young and R. Davis, Proc. Natl. Acad. Sci. U.S.A. 80, 1194 (1983))이 포함된다. 그러나, 숙주세포내에서 성장할 수 있다면, 어떠한 다른 파아지벡터로 적당하다.
플라스미드 속에 cDNA를 삽입하는 방법에는 예컨데, T. 마니아티스 일행의 (Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory , p.239 (1982))에 기재된 방법이 포함된다. 파아지벡터 속에 cDNA를 삽입시키는 방법에는 예컨데, T.V. 휸 일행의 방법 (DNA Cloning, a Practical Approach 1, 49 (1985))이 포함된다. 그렇게해서 얻어진 플라스미드 또는 파아지벡터를 E. 콜라이 (E. coli) 와 같은 적절한 세포내에 도입한다.
상기 E. 콜라이 (E. coli)의 예로는 E. 콜라이(E. coli) K12DH1 (Proc. Natl. Acad. Sci. U.S.A. 60, 160 (1968)), JM 103 (Nucl, Acids Res. 9, 309 (1981)), JA 221 ( journal of Molecular Biology 120, 517 (1978)), HB 101 (Journal of Molecular Biology 41, 459,(1969)) 및 C 600 (Genetics 39, 440 (1954)) 이 포함된다.
플라스미드에 의해 숙주세포를 형질전환하는 적당한 방법에는 예를들어, T. 마니아티스 일행의 (Molecular Cloning, Cold Spring Harbor Laboratory, p.249 (1982))에 기재된 염화칼슘법 또는 염화칼슘/염화로듐법이 포함된다. 또한, 예를들어 파아지벡터는 실험실내(in vitro)패키지법을 사용하여 증식된 E. 콜라이(E. coli)속에 인공적항원 변환될 수 있다.
폴리펩타이드(I) cDNA를 함유하는 cDNA 라이브러리는 상기 언급한 방법들에 의해 수득할 수 있다. 그러나, 이들을 상업적제품으로도 구입할 수 있다. 예를들어, 사람신경교종-유도 cDNA 라이브러리와 사람태반- 유도 cDNA 라이브러리는 미합중국의 콜로테그라보라토리스,Inc. 로부터 구입가능하다. cDNA 라이브러리로부터 폴리펩타이드(I) cDNA를 클로닝하는 적당한 방법에는 표지된 프로우브를 사용하는 플라그 혼성화법 또는 군락 혼성화법 (T. Maniatis 일행, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory (1982)) 이다. 폴리펩타이드(I)를 위한 DNA코오딩과 혼성가능하다면 어떠한 DNA도 상기 혼성화에 프로우브로서 사용되는 DNA 로 사용가능하다.
이러한 DNA로는 예를 들어, NGF의 전체 또는 부분에 대한 cDNA 코오딩, 게놈 DNA, 화학적으로 합성된 DNA 및, NGF의 아미노산서열에 바탕하여 화학적으로 합성된 올리고뉴클레오티드가 포함된다.
상기 언급한 NGF 의 예로는 생쥐NGF (Proc. Natl. Acad. U.S.A. 68, 2417 (1971), Nature 302, 538 (1983)), 사람NGF(Nature 303, 821 (1983)) 및 기타동물의 NGF가 포함된다.
그렇게해서 클로닝된 폴리펩타이드(I) cDNA 는 필요하다면, 폴리펩타이드(I) cDNA를 발현시키기 위하여 예컨데, pBR 322, pUCI 2, pUCI 3, pUCI 8, pUCI 9, pUCI 18, pUCI 19 속에 서브클로닝될 수 있다.
그렇게해서 얻어진 DNA의 뉴클레오티드 서열을 예컨데, 막삼-길버트법 (A. M. Maxam and W Gilbert, Proc. Natl. Acad. Sci. U.S.A. 74, 560 (1977)) 또는 디데옥시법 ( J. Messing 일행, Nucleic Acids Reseach 2, 309, (1981))으로 결정하여 폴리펩타이드(I) cDNA의 존재를 확인할 수 있다. 그 결과, 폴리펩타이드(I)에 대한 전체지역 코오딩이 파악되지 않은 겅우, cDNA는 DNA 단편을 프로우브로서 사용하는 플라그 혼성화 또는 군락 혼성화에 의해 다시 클로닝되어 파악되지 않은 다른 지역을 얻을 수 있다.
상기 기대된 바와 같이, 폴리펩타이드(I)에 대한 DNA코오딩을 얻을 수 있다.
상기 방법들에 덧붙여, 본발명의 폴리펩타이드(I)에 대한 DNA 분절코오딩을 포함하는 DNA는 또한 사람, 쥐, 생쥐 등의 게놈 DNA 라이브러리부터 클로닝함으로써 수득할 수 있다.
또한, 폴리펩타이드(I)에 대한 DNA 코오딩은, 그렇게 해서 콜로닝된 DNA의 뉴클레오티드 서열로부터 해명된 폴리펩타이드(I)의 아미노산서열에 바탕한 화학적 분석에 의해 얻을 수 있다.
본 발명의 폴리펩타이드(I)에 대한 DNA코오딩으로서 사용되는 어떠한 DNA도 폴리펩타이드(I)에 대해 코오딩되는 한, 사용될 수 있다. 실례로는 하기식 (III)의 뉴클레오티드 서열에 의해 표시되는 DNA와 하기식 (III)의 뉴클레오티드 서열의 3' - 말단에 ACA가 더 첨가된 DNA가 포함된다. :
TACGCGGAGC ATAAGAGTCA CCGAGGGGAG TACTCGGTAT
GAGACAGTGA GAGTCTGTGG GTGACCGACA AGTCATCGGC
CATCGACATT CGGGGACACC AGGTCACGGT GCTGGGGGAG
ATCAAAACGG GCAACTCTCC CGTCAAACAA TATTTTTATG
AAACGCGATG TAAGGAAGCC AGGCCGGTCA AAAACGGTTG
CAGGGGTATT GATGATAAAC ACTGGAACTC TCAGTGCAAA
ACATCCCAAA CCTACGTCCG AGCACTGACT TCAGAGAACA
ATAAACTCGT GGGCTGGCGG TGGATACGGA TAGACACGTC
CTGTGTGTGT GCCTTGTCGA GAAAAATCGG AAGA (III)
( 이 뉴클레오티드 서열은 이후부터 또한 간략히 식 (III) 으로 표기함.)
몇몇 경우에, 이 DNA를 구성하는 뉴클레오티드 서열의 부분이 제거되거나 치환될 수 있다. 또한, 하나 또는 그 이상의 부가적 염기를 이 DNA에 첨가 또는 삽입할 수 있다. 염기의 제거, 치환 또는 첨가를 상응하는 아미노산 또는 아미노산들의 발현에 해당되는 코오돈 단위에 의해 수행하는 것이 바람직하다.
그렇게해서 얻어진 폴리펩타이드(I) 에 대한 DNA 코오딩은 그대로 사용되거나, 또는 바람직하다면, 원하는 용도에 따라서 제한효소에 의해 절단될 수 있다.
본 발명의 폴리펩타이드(I)을 수득하는 적당한 방법에는 (1) 사람을 포함한 동물의 조직으로부터 폴리펩타이드(I)의 분리, (2) 펩타이드 합성에 의한 폴리펩타이드(I)의제조 및 (3) 유전자 재조합을 이용한 폴리펩타이드(I)의 합성이 포함된다. 세 번째 방법이 공업적으로 바람직하다.
DNA재조합기술을 이용한 폴리펩타이드(I)의 제조를 위한 발현시스템(숙주-벡터시스템)의 예로는 박테리아, 방선균류, 효모, 곰팡이, 곤충세포 및 동물세포의 발현시스템이 포함된다.
적당한 발현 방법으로는 (a) 세포내에서 유전자 생성물을 제조 및 축적하는 것, (b) 유전자 생성물을 세포 밖으로 분비시키고 이들을 배지내에 추적하는 것, 및 (c) 유전자 생성물을 페리플라즘 속으로 분비하는 것이 포함된다.
상기 (b) 및 (c) 방법에 있어서 폴리펩타이드(I)을 분비하기 위하여, 시그날 펩타이드에 대한 DNA코오딩 또는 시그날 펩타이드 및 프로펩타이드(prepro)에 대한 DNA코오딩을 폴리펩타이드에 대한 DNA코오딩의 5' - 말단에 연결할 수 있다. 폴리펩타이드(I)의 분비를 유도할 수 있는 한 어떠한 펩타이드도 상기 언급한 시그날 펩타이드로서 사용가능하다. 이러한 시그날 펩타이드의 예로는 E. 콜라이 (E. coli) 장독소 및 그 돌연변이체의 시그날 펩타이드, 바실루스 아밀로리퀴파시엔스( Bacillus amyloliquefaciens)d 중성 프로베아제 및 - 아밀라제의 시그날 펩타이드, 바실루스 브레비스 (Bacillus brevis) 중간벽 단백질의 시그날 펩타이드, 사카 로마이세스 세레비사애 (Saccharomyces cerevisiae) 인베르타아제, 포스파타아제, α-인자 및 킬러 인자의 시그날 펩타이드, 아스페르키루스 아와모리( Aspergillus awamori ) 글로코 아밀라아제의 시그날 펩타이드, 폴리펩타이드(I) 의 시그날 펩타이드, 달걀 흰자위 리소집 및 그 돌연변이체의 시그날 펩타이드, 사람 인터로 이킨-2 의 시그날 펩타이드, 및 사람, 생쥐, 쥐, 닭 및 소의 NGF 의 시그날 펩타이드가 포함된다. 적당한 프로펩타이드의 예로는 S. 세레비시애 (S. cerevisiae) α- 인자 및 킬러 인자의 프로펩타이드, A. 아와모리 (A. awamori) 글루코아밀라아제의 프로펩타이드, 폴리펩타이드(I)의 프로펩타이드 및 사람 엔도셀린 (endothelin), 사람, 생쥐, 쥐, 닭, 및 소의 NGF의 프로펩타이드가 포함된다.
상기 방법들에 덧붙여, 폴리펩타이드(I)은 또한, 폴리펩타이드(I)과 또다른 단백질의 융합단백질을 제조한 후에 적절한 프로테아제로 이것을 절단하는 것에 의해서도 수득할 수 있다.
개시 코오돈은 폴리펩타이드(I)에 대한 DNA 코오딩, 시그날 펩타이드 및 폴리펩타이드(I)에 대한 DNA 코오딩과 같은 폴리펩타이드(I)에 대한 DNA 분질 코오딩을 함유하는 상기 DNA 의 5'- 말단에 첨가될 수 있고, 종말 코오돈은 그 하류에 삽입할 수 있다. 얻어진 DNA를 벡터내의 프로모터로부터 하류에 삽입할 수 있으며, 이로써 폴리펩타이드(I) 발현벡터가 구축된다.
폴리펩타이드(I)의 발현에 사용되는 벡터로서는, 선택된 숙주세포내에서 작용하는 한 어떠한 벡터도 사용가능하다. E. 콜라이 (E. coli) 발현벡터의 예로는 pBR 322, pBR 325, pUCI 2, pUCI 3, pUCI 8, pUCI 9, pUCI 18, pUCI 19 및 그 유도체들이 포함된다. 바실루스 서브틸리스(Bacillus subtilis) 발현벡터의 예로는 pUB1 10, pC1 94, pE1 94, pTB 5 및 그 유도체들이 포함되고, B. 브레비스 (B. brevis) 발현벡터의 예로는 pUB1 10, pHY 481, pC1 94, pHY 500, pNU 200 및 그 유도체들이 포함된다. S. 세레비시애 (S. cerevisiae) 발현벡터의 예로는 pSH 19, pSH 15 및 그 유도체들이 포함되고, 스키조사카로 마이세스 폼베(Schizosaccharomyces pombe) 발현벡터의 예로는 pDB 248, pPA-4 및 그 유도체들이 포함된다. 동물세포발현벡터의 예로는 테트로비루스 벡터, 바시니아 비루스벡터, 보빈파필로마 비루스 벡터 및 SV 40- 계열벡터 (예컨데 pKSV-10, pSV 2-dhfr 및 pTB 389)가 포함된다.
선택된 숙주세포내에서 작용하는 한 어떠한 프로모터도 적합하다.
예를 들어, E. 콜라이 (E. coli) 벡터를 사용하는 경우, 적당한 프로모터는 trp 프로모터, lac 프로모터, tac프로모터, λPL프로모터, rec A프로모터 및 T7 프로모터가 포함된다.
B. 서브틸리스(B. subtilis) 벡터를 사용하는 경우, 적당한 프로모터의 예로는 SP01 프로모터, P1 프로모터 및 중성 프로테아제 유전자 프로모터가 포함된다. B. 브레비스( B. brevis)벡터를 사용하는 경우, 적당한 프로모터의 예로는 세포의 주단백질 유전자 프로모터 및 SP01 프로모터가 포함된다. S. 세레비시애(S.cerevisiae) 벡터를 사용하는 경우, 적당한 프로모터의 예로는 GLD프로모터, PH05프로모터, GAL 10프로모터, GAL 7프로모터, PGK프로모터 및 α- 인자프로모터가 포함된다. S. 폼베(S. pombe) 벡터를 사용하는 경우, 적당한 프로모터의 예로는 GLD프로모터 및 SV 40프로모터가 포함된다.
동물세포벡터를 사용하는 경우, 적당한 프로모터의 예로는 SV40프로모터, LTR 프로모터 및 메탈로리오네인 프로모터가 포함된다.
발현효율을 증가시키기 위해서는 , 효모에 있어서는 폴리펩타이드(I) 에 대한 DNA 코오딩으로부터 하류에 터미네이터( 예컨데 PGK 터미네이터)를 사용하는 것이 바람직하고, 동물세포에 있어서는 인핸서(enhancer), RNA 스플리싱 시그날, 폴리 A 첨가 시그날 또는 선택된 마커(marker)를 사용하는 거싱 바람직하다.
본 발명의 발현벡터를 구축하는 방법은 그 자체가 공지되어 있고, 예를 들면, 문헌 (Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory (1982))에 기재되어 있다.
그렇게해서 제조된 폴리펩타이드 발현벡터를 사용하여, 숙주세포를 형질전환시킬 수 있다.
적당한 숙주세포로는 E.콜라이 (E. coli) B.서브틸리스(B. subtilis) 및 B.브레비스(B. brevis)와 같은 박테리아, 스트렙토마이세스 리비단스 (Streptomyces lividans)와 같은 방선균류, S. 세레비시애 (S. cerevisiae), 스키조사카로마이세스 폼베(Schizosaccaromyces pombe) 및 피치아 파스토리스(pichia pastoris)와 같은 효모, 아스페르길루스 오리자애(aspergillus orizae), 아스페르길루스 니듈란스(aspergillus nidulans) 및 아스페르길루스 니거(Aspergillus niger) 와 같은 곰팡이 및 원숭이 세포 COS-7 세포, 베로세포, 중국산 햄스터 난소 세포(CHO) 및 생쥐 L 세포와 같은 동물세포가 포함된다.
더욱 특정하게는, 적당한 E.콜라이(E. coli) 균주로는 DH1, JM 103, JA 221, HB 101, C600, MV1184 및 그 돌연변이체들이 포함된다. 적당한 B. 서브틸리스 (B. subtilis)균주로는 MI 114, 1A 274 및 그 돌연변이체들이 포함된다. 적당한 B. 브레비스 (B. brevis) 균주로는 47, 47-5, HPD 31 및 그 돌연변이체들이 포함된다. 적당한 S. 세레비시애 (S. cerevisiae)균주로는 AH 22R-, NA 47-3Aρ-, TB39ρ- 및 그 돌연변이체들이 포함된다. 적당한 S. 폼베(S. pombe)균주로는 ATCC 38399, TH168 및 그 돌연변이체들이 포함된다.
폴리펩타이드(I) 발현 플라스미드와 같은 본 발명의 DNA서열을 사용하는 숙주세포의 형질전환방법은 이 분야에 공지되어 있다. E. 콜라이 (E. coli)는 예컨데 코헨일행의 방법(Proc. Natl. Acad. Sci. U.S.A. 69, 2110 (1972))에 의해 형질전환시킬 수 있다.
B. 서브틸리스 (B. subtilis)는 예컨데 프로토플라스트법(Molecular General Genetics 168, 111 (1979)) 또는 콤페턴트법(J. Mo1. Bio1. 56, 209 (1971))에 의해 형질전환시킬 수 있다. B. 브레비스 (B. brevis)는 예컨데 다까하시일행의 방법(J. Bacterio1. 156, 1130, (1983))에 의해 형질전환시킬 수 있다. S. 세레비시애 (S. cerevisiae) 및 S.폼베(S. pombe) 는 예컨데 히넨의 방법(Proc. NAtl. Acad. Sci. U.S.A. 75, 1929 (1978)) 또는 리튬법(J.Bacterio1 153, 163(1983))에 의해 형질전환시킬 수 있다. 동물세포는 예컨데 그램의 방법(Virology 52, 456 (1973))에 의해 형질전환시킬 수 있다.
이상 기재된 바와 같이, 폴리펩타이드(I)에 대한 DNA분절코오딩을 함유하는 DNA로써 형질전환된 형질전환체를 본 발명에 따라서 수득할 수 있다.
숙주세포가 박테리아, 방선균류, 효모 또는 곰팡이인 형질전환체를 배양하는 경우, 배양에 사용되는 배지로는 액체배지가 적당하다. 탄소 공급원, 질소 공급원, 무기화합물 및 기타 형질전환체의 생장에 필요한 영양물이 그 속에 함유된다. 적당한 탄소 공급원으로는 예컨데 포도당, 덱스트린, 가용성녹말 및 슈크로오스가 포함된다. 적당한 질소원천에는 암모늄염, 질산염, 아미노산, 옥수수스팁(steep)액, 펩톤, 카제인, 육류추출액, 콩죽 및 감자추출용액과 같은 무기 또는 유기물질이 포함된다. 적당한 유기화합물로는 예컨데 염화칼슘, 인산이수소나트륨 및 염화마그네슘이 포함된다.
배지의 pH는 약 5 ∼ 8 바람직하다.
숙주가 E.콜라이(E. coli)인 경우, 배양에 사용되는 배지는 예컨데, 포도당과 카사미노산을 함유하는 M9배지 (Miller, Journal of Experiments in Molecular Genetics, 431∼ 433, Cold Spring Harbor Laboratory, New York (1972))가 바람직하다. 배양은 보통 14 ∼ 43 ℃에서 약 3 ∼ 24 시간동안, 필요하다면 통기 또는 흔들면서 수행한다.
숙주가 바실루스(Bacillus)인 경우, 배양은 보통 약 30 ∼ 40도씨에서 약 16 ∼ 96시간 동안, 필요하다면, 통기 또는 교반하면서 수행한다.
효모 형질전환체를 배양하는 경우, 적당한 배지의 예로는 부르크홀더 최소배지 (K. L. Bostian 일행, Proc. NAtl. Acad. Sci. U.S.A. 77, 4505 (1980))가 포함된다.
배지의 pH를 약 5 ∼ 8로 조절하는 것이 바람직하다. 배양은 보통 약 20 ∼ 35℃에서 약 24 ∼ 144시간동안, 필요하다면 통기 또는 흔들면서 수행한다.
동물세포 형질전환체를 배양하는 경우, 적당한 배지의 예로는 약 5 ∼ 20 % 태아송아지 혈청을 함유하는 MEM배지 (Science 122, 501 (1952)), DMEM배지 (Virology 8, 396 (1959)), RPMI 1640 배지(J. Am. Med. Assoc. 199, 519 (1967)) 및 199 배지(Proc. Soc. Exp. Bio1. Med. 73, 1 (1950))가 포함된다.
pH는 약 6 ∼ 8이 바람직하다. 배양은 보통 약 30 ∼ 40℃에서 약 15 ∼ 60시간동안, 필요하다면 통기 또는 흔들면서 수행한다.
본 발명의 폴리펩타이드(I)은 세포의 내부 또는 외부에서 생성 및 축적될 수 있다.
배양된 세포로부터 세포간 폴리펩타이드(I)을 추출하는 경우, 이 분야 공지방법에 의해 배양후에 세포를 수집한다. 다음에, 수집한 세포를 우레아 또는 구아니딘 염산염과 같은 단백질 변성제 또는 트리톤 X-100과 같은 계면활성제를 함유하는 적절한 완충용액내에 현탁시킨 후, 원심분리함으로써 폴리펩타이드(I)을 함유하는 상충액을 수득한다. 또 다르게는 수집된 세포를 초음파처리, 리소짐과 같은 효소에 의한 처리 또는 냉동-용해에 이어서 원심분리함에 의해 분열시켜 폴리펩타이드(I)을 함유하는 상충액을 수득할 수 있다.
배양상충액내에 함유된 또는 세포내에 생성 및 축적된 폴리펩타이드(I)의 정제는공지 정제 방법을 적절히 조합하여 수행할 수 있다. 이들 공지 정제법으로는 염석출 및 용매석출과 같은 용해도차를 이용하는 방법; 투석, 한의여과, 겔투과크로마토그래피 및 SDS-폴리아크릴아미드겔전기영동법과 같은, 분자량의 차이를 주로 이용하는방법; 이온교환컬럼크로마토그래피와 같이 전하의 차이를 이용하는 방법; 친화도크로마토그래피와 같이 특수친화도를 이용하는 방법; 역상고속액체크로마토그래피와 같이 소수성의 차이를 이용하는 방법; 및 등전집점 전기영동법과 같이 등전접의 차이를 이용하는 방법들이 포함된다.
그렇게 해서 수득한 펩타이드(I)이 활성을 갖는다면, 그대로 사용할 수 있다. 만약 활성을 나타내지 않는다면, 효소적 또는 비효소적 방법에 의한 활성화 후에 사용할 수 있다.
본 발명의 폴리펩타이드(I)의 활성은 효소면역분석, 방사선면역분석 등에 의해 측정할 수 있다.
폴리펩타이드(I)은 동물세포의 분화 및 생장의 촉진, 동물세포의 생존촉진, 유전자발현의 촉진 및, 단백질 및 효소의 생성유도의 기능을 갖는다. 따라서, 이들 기능을 지표로 하여 폴리펩타이드(I)의 활성을 분석할 수 있다. NGF와의 동족관계로 인하여, 폴리펩타이드(I)은 NGF와 유사한 활성 및 기능을 가질 수 있다. 이러한 활성 및 기능의 실례로는 PC 12세포내에서의 축색외부생장의 촉진 (L. A. Greene, Brain Research 133, 350 (1977); R. Heumann 일행, Experimental Cell Research 145, 179 (1983))과 닭 태아 지각신경절 (배근 신경절 )의 생존촉진 기능 (A. M. Davies R. M. Lindsay, Developmental Biology 111, 62(1985))이 포함된다.
본 발명의 폴리펩타이드(I)은 동물세포의 분화, 생장 및 생존에 관한 연구용 시약으로서 유용하다. 폴리펩타이드(I)을 이들 연구에 사용하는 경우, 바람직하게는 동물세포용 배지에 폴리펩타이드(I)을 첨가하여 최종농도가 약 0.1 ∼ 1,000 mg/ml, 더욱 바람직하게는 약 1 ∼ 100 mg/ml가 되게 한다. 동물세포를 폴리펩타이드(I)을 함유하는 배지내에서 배양하는 것에 의해 동물세포의 분화, 생장 및 생존을 측정할 수 있다.
폴리펩타이드(I)은 또한 손상된 조직 및 기관의 복구기능을 할 수 있으며, 따라서 폴리펩타이드(I)은 약제로서 유용하다.
또한, 폴리펩타이드(I)에 대한 DNA 코오딩은 폴리펩타이드(I)의 검출 및 측적 및, NGF 유전자의 클로닝을 위한 프로우브로서 이용할 수 있다.
폴리펩타이드(I)을 코오딩하는 DNA를 프로우브를 사용할 때, 예를 들어, 폴리펩타이드(I)에 대해 코오딩하는 DNA 0.5㎍ (약 300bp)을 아머샴(Amersham)에서 공급하는 미크 해독키트 (약 107 cpm)를 사용하여 (α- 32p) dCTP ( 400 Ci/mmo1)로서 표지한다. 플라스 혼성화에 의한 클로닝에 있어서, 혼성화는 필터당 0.005㎍ (105 cpm) 상기 표지된 프로우브를 사용하여 수행한다.
본 명세서 및 도면에서 염기, 아미노산 등을 약어로써 표시하는 경우, 생화학명명법에 관한 IUPAC-IUB위원회에서 채택되거난 이 분야에서 통상적으로 사용되는 약어들을 사용한다. 아미노산에 대하여 광학이성질체가 존재할 수 있는 경우, 달리 명시하지 않는 한 L-형을 나타낸다.
DNA : 데옥시리보핵산
A : 아데닌
C : 시토신
G : 구아닌
T : 리민
Ala : 알라닌
Arg : 아르기닌
Asn : 아스파라긴
Asp : 아스파라긴산
Cys : 시스테인
Gln : 글루타민
Glu : 글루탐산
Gly : 글리신
His : 히스티딘
Ile : 이소로이신
Leu : 로이신
Lys : 리신
Met : 메티오닌
Phe : 페틸알라닌
Pro : 프롤린
Ser : 세린
Thr : 트레오닌
Trp : 트립토판
Tyr : 티로신
Val : 발린
Boc : t - 부틸옥시카르보닐
MeBzl : p - 메틸벤질
Bzl : 벤질
-P : 펩타이드의 고체합성용 폴리스티렌수지
PAM : p - 옥시메틸페닐아세트아미도메틸수지
AcOH : 아세트산
OBzl : 벤질에스테르
Tos : 토실
Br-z : 2 - 브로모벤질옥시카르보닐
Cl-z : 2 - 클로로벤질옥시카르보닐
하기 참고예1에서 수득한 미생물과 하기 실시예들에서 수득한 형질전환체는 부다페스트 조약에 따라서 일본국 오오사까 발효연구소(IFO) 와 또한 일본국 국재통상 및 산업성 산업과학 기술청 발효학술연구소 (FRI)에 기탁하였다. 이들의 수탁번호 및 기탁일은 표 1에 주어진다.

본 발명을 이하에서 하기 참고예와 실시예로써 더욱 상세히 기술할 것이다. 이들 참고예 및 실시예는 본 발명의 영역을 제한하려는 목적이 없음은 물론이다.
참고예 1 (S. 세레비시애 (S. cerevisiae) TB39ρ- 의 제조) S. 세레비시애 (S. cerevisiae) NA 74-3A(a, pho9, his4, leu2) (IFO 10430, FERM BP-1947) (EP-317, 209A에 상응하는 일분국 특허출원(공개) 제63-283716/1989 참고)를 S. 세레비시애 (S. cerevisiae) DK-13D (α, leu2, trp1, his3 ) (Molecular and Cellular Biology 4, 771 (1984))와 교배시킨다. 수득된 균주를 에티듐 브로마이드로 처리하여 무호흡 균주 S. 세러비시애 (S. cerevisiae) TB 39ρ-(α, MAta, leu2, his3, pho9, P-) (IFO 10467, FERM BP-2399)를 수득한다.
참고예 2 (항-N-말단펩타이드 항체의 제조)
(1) H-Tyr-Ala-Glu-His-Lys-Ser-His-Arg-Gly-Glu-Tyr-Ser-Val-Cys-OH의 합성
이 펩타이드의 합성은 자동펩타이드 합성기 모델430A(Applied Biosystems)를 사용하는 고체합성법에 의한다. 프로그램으로, 스탠다드 1을 사용한다. 합성은 기본적으로 문헌 (R. B. Merrifield, Adv. Enzymo1. 32, 221 296 (1969))에 기재된 방법에 따라서 수행한다. Boc-Cys(MeBzl)PAM-P(0.5mmol/g)를 수지로 사용하고, 카르복실말단에서부터 연속적으로 합성을 수행한다. Boc-아미노산으로서는, Boc-Val, Box-Ser(Bzl), Boc-Tyr(Br-Z), Boc-Glu(OBzl), Boc-Gly, Boc-Arg(Tos), Boc-His(Tos), Boc-Lys(Cl-Z) 및 Boc-Ala를 사용한다. 펩타이드 수지를 아미노말단 Try까지 합성한 다음, 합성기로부터 꺼내고 건조시킨다.
1g의 펩타이드 수지에 1.5ml의 P-크레졸과 0.5ml의 1,2-에탄디티올을 첨가하고, 약 8ml의 액체불화수소를 여기에 또 첨가한 후, 0℃에서 2시간동안 반응시킨다. 반응완결 후, 건조기내에서 감압하에 불화수소를 제거하고 0.1% 머캅토 에탄올의 디에틸에테르용액으로 씻은 후, 디에틸에테르로 씻음으로써 대부분의 포함된 시약을 제거한다. 3% 아세트산 10ml로 펩타이드를 추출하고, 추출용액내에 포함된 수지를 여과로써 제거한다. 여액을 세파덱스(Sephadex) G-25 칼럼을 사용하여 겔투과크로마토그래피하여 정제한다. 겔투과크로마토그래피의 조건은 다음과 같다.
칼럼칫수 : 2.8 x 60cm ; 검출 파장 : 280 nm ;
용매 : 3% 아세트산 ; 유속 : 40 ml/hr
펩타이드를 함유하는 분획들을 수집하고 동결건조시켜 분말시료를 수득한다. 얻어진 분말시료를 하기 조건하에 역상고속액체크로마토그래피로 더 정제한다.
칼럼 : YMC pack, A-324 ODS 10 x 250nm ;
칼럼온도 : 25℃
용리제 A : 0.1% 트리플루오로아세트산 - 99.9% 증류수 ;
용리제 B : 0.1% 트리플루오로아세트산 - 99.9% 아세토니트릴 ;
용출프로그램 : 0분 (90%A + 10%B),
30분 (60%A + 40%B);
용출속도 : 2ml/분 ;
검출파장 : 230 nm
상기 조건하에서 23.0분의 체류시간에서 용출된 주 피크분획들을 수집하고, 바이오래드(Bio RAD AG1 x 8 칼럼 (AcOH 형, 1.8 x 5 cm)에 통과시킨다. 세척을 또한 수행한다. 다음에, 증류에 이어 동결건조시켜 아세토니트릴을 제거한다. 그리하여 56mg의 백색분말이 수득된다. 얻어진 생성물은 상기언급한 고속액체크로마토그래피에서와 같은 조건하에 23.0분에서 예리한 단일피크를 나타낸다.
문헌 (G. L. Elman, Arch. Biochem. Biophys. 82, 70 ∼ 77 (1959))에 기재된 방법에 의한 유리 SH기의 측정 : 114%
아미노산 분석치 : Ser 1.65(2) ; Glu 2.13(2) ; Gly 1.00(1) ; Ala 1.04(1) ; 1/2 Cys 0.82(1) ; Val 1.03(1) ; Tyr 1.97(2) ; Lys 0.95(1) ; His 1.72(2) ; Arg 1.00(1)
회수율 : 74%
1/2 Cys는 퍼포트믹 산화법으로 측정한다.괄호안의 수치는 어른치를 나타낸다.
(2) N-말단펩타이드와 헤모시아닌의 접합물의 제조
4ml의 0.2M 인산염 완충액(pH 7.3)에 상기 (1)에서 수득한 -말단펩타이드 5mg과 헤모시아닌 10mg을 용해시키고, 2.5% 글루타르알데히드의 냉각수 용액 400μl를 여기에 교반하면서 적가한다. 3시간동안 얼음냉각하면서 교반한 후, 증류수에 대한 투석을 수행하여 N-말단펩타이드와 헤모시아닌의 접합물을 제조한다.
(3) N-말단펩타이드와 소 혈청알부민의 접합물의 제조
3ml의 0.1M 인산염 완충액(pH7.5)에 132mg의 소 혈청알부민(BSA) (A 용액)을 가한다. 200μl의 디메틸포름아미드에 11.2mg의 N-(γ-말레이미드 부틸옥시) 숙신이미드(GMBS)(B 용액)를 가한다. B 용액을 교반기로 교반하면서 A 용액에 적가하고, 혼합용액을 30 ℃에서 30분간 반응시킨다. 그 다음, 용리제로서 0.1M 인산염 완충액(pH 6.5) - 0.1M NaCl을 사용하여 세파덱스(Sephadex) G-25컬럼에 의해 정제하여 말레이미드기가 도입된 소혈청알부민을 수득한다.
0.1M 인산염 완충액 -5mM EDTA에 상기 (1)에서 수득한 펩타이드 5mg과 말레이미드기 - 도입된 소혈청알부민 20mg을 첨가한 후 (총부피는 5ml 이하임), 30도씨에서 60분간 반응시킨다. 그 다음, 여기에 총부피가 12ml가 될 때까지 PBS(인산염-완충염수)를 첨가하므로써, N-말단펩타이드와 소혈청알부민의 접합물을 수득한다.
(4) 항 - 폴리펩타이드(I) N-말단 펩타이드의 제조
상기 (2)에서 수득한 N-말단 펩타이드와 헤모시아닌의 접합물을 프로인드보조액과 철저히 혼합하고, 얻어진 혼합물을 토끼에게 피하주사한다. 그 다음, 2주간격으로 N-말단 펩타이드와 상기 (3)에서 수득한 소혈청알부민의 접합물을 프로인드 보조액과 혼합하고, 얻어진 혼합물을 동일한 토끼에게 주사한다.
상기와 같이 면역된 토끼로부터 수집한 혈액을 원심분리하여 항-폴리펩타이드(I) N-말단 펩타이드 항체를 수득한다.
실시예1 (폴리펩타이드(I) cDNA의 클로닝)
에스케리키아 콜라이(Escherichia coli) Y 1090을 사람신경교종 - 유도λgtill cDNA 라이브러리 (Clontech Laboratories, Inc.)로 감염시킨 다음, 문헌 (Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory(1982))에 기재된 NZCYM 배지를 함유하는 아가플레이트 위에 6 x 105 파아지를 펼쳐놓은 후, 37℃에서 5시간동안 배양한다. 그 다음, 나일론 막을 플레이트위에 놓고, 1분동안 둔 후에 제거한다. 이 나일론 막을 0.5M NaOH - 1.5M NaCl, 다음에 1.5M NaCl-0.5M 트리스 -HCl(pH8.0) 및 또한 2 x SSC (Molecular Cloning, A Laboratory Manual, cold Spring Harbor Laboratory (1982) 참고) 에 적신다. 공기건조후, 막을 80℃에 2시간동안 둔다.
사람 βNGF (Nature 303, 821 (1983) 참고)에 대한 DNA (약 0.38kb) 코오딩을 화학적으로 합성하고 니크 해독에 의해 (α-32p) dCTP 로써 표지함으로써, 프로우브를 제조한다.
상기 수득한 나일론 막과 프로우브를 사용하여, 문헌 (Molecular Cloning, A Laboratory Manual, Cold Spring Harbor LAboratory (1982))에 기재된 방법에 따라서 혼성화를 실시한다. 즉, 프로우브를 함유하는 혼성화용액에 나일론 막을 담그고 65℃에 16시간동안 유지한다.
나일론 막을 실온의 2 x SSC - 0.1% SDS 로 씻은 다음 60℃의 1 x SSC-0.1% SDS로 씻는다. 그 다음, 방사선자기법에 의해 포지티브클론을 검출한다.
그렇게해서 수득한 클론 βGN 1321로부터 EcoRI 로써 cDNA 부분을 절단해내고 플라스미드 pUCI 18 (Takara Shuzo)의 EcoRI 부위에 삽입하여 플라스미드 pUNK5를 수득한다. 그렇게해서 수득한 플라스미드 pUNK5를 사용하여, 코헨 일행의 방법 (상기 기재됨)으로 E.콜라이 (E. coli) MV 1184 (Takara Shuzo)를 형질전환시켜 형질전환체 E. 콜라이 (E. coli) MV 1184/pUNK5 (IFO 14832, FERM BP-2304)를 수득한다.
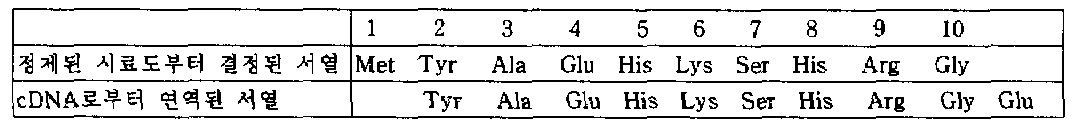
제 1도는 플라스미드 pUNK5내에 함유된 폴리펩타이드(I) cDNA를 포함하고 전체길이가 약 0.78kb인 cDNA의 제한 효소지도를 나타낸다. 제 1도에서, □는 비해독 지역을, ▨는 프로펩타이드 코오드 지역을, ▥는 식 (II)의 아미노산서열의 C-말단에서 트레오닌잔기를 더 갖는 폴리펩타이드에 대해 코오딩하는 지역을 나타낸다.
상기에서 수득한 cDNA부분의 뉴클레오티드 서열을 디데옥시법 (Messing 일행, Nucl. Acid. Res 9, 309 (1981))으로 결정한다. 제 2도에서, 위치 1에서부터 아미노산 서열의 N-말단으로까지 걸친 지역은 포르펩타이드의 일부분이고, 위치 1 ∼ 118 또는 위치 1 ∼ 119의 지역은 식(II)의 아미노산 서열을 갖는 폴리펩타이드와 식(II)의 아미노산 서열의 C-말단에서 트레오닌잔기를 더 갖는 폴리펩타이드를 나타낸다.
제 3도는 문헌 (Ullrich 일행, Nature 303, 821 (1983))에 기재된 사람 βNGF의 아미노산서열과 비교하여, 상기방법에 의해 결정된 폴리펩타이드(I)의 아미노산 서열을 나타낸다. 제 3도에서, 윗줄은 폴리펩타이드(I)의 119 아미노산의 서열을 가리키고, 아랫줄은 사람 βNGF의 아미노산 서열을 가리킨다. 동일한 아미노산 잔기 부분은 네모칸속에 넣어진다. 이 도면에서, __는 단지 화학적 결합을 나타낸다.
이 비교로부터 분명한 바와 같이, 본 발명의 폴리펩타이드(I)의 119 아미노산의 서열은 상기 사람 βNGF의 아미노산 서열과 약 60%동족성(homology)을 갖는다.
또한, 상기한 바대로 결정된 폴리펩타이드(I)의 119 아미노산의 서열을 문헌 (Angeletti 일행, Proceedings of National Academy of Sciences, U.S.A. 68, 2417 (1971)) 및 (Scott 일행,Nature 302, 538 (1983))에 나와있는 생쥐 βNGF의 아미노산서열과 비교할 때, 약 60%의 동족성을 갖는다.
상기 비교로부터, 본 발명의 폴리펩타이드(I)은 신규 폴리펩타이드인 것으로 판명된다.
실시예 2 (폴리펩타이드(I) cDNA의 리클로닝(recloning))
실시예1에서 수득한 pUNK5에 함유된 폴리펩타이드(I) cDNA부분의 5'- 말단부위를 함유하는 EcoRI-AhaIII 단편을 프로우브로서 사용하여, 사람 신경교종-유도 cDNA 라이브러리 (Clontech Laboratories, Inc.) 중 하나를 실시예1에서와 유사한 방식으로 클로닝한다.
그렇게해서 수득한 많은 포지티브클론 λHNT 31중 하나로부터, EcoRI로써 cDNA 부분을 절단해내고 플라스미드 pUCI 19 (Takara Shuzo)의 EcoRI부위속에 삽입하여 플라스미드 pHNH 2를 수득한다. 제 4도는 플라스미드 pHNT 2 속에 삽입된 폴리펩타이드(I) cDNA (약 1.1kb) 의 제한효소지도를 나타낸다.
제 4도에서, ▥는 단일펩타이드 코오드 지역을 나타내고, ▨는 프로펩타이드 코오드 지역을 나타내고, ▤는 식(II)으 아미노산서열의 C-말단에서 트레오닌잔기를 더 갖는 폴리펩타이드에 대해 코오딩하는 지역을 나타낸다.
상기에서 수득한 cDNA 부분의 뉴클레오티드 서열을 디에옥시법(상기 기술됨)에 의해 결정한다. 제 5도는 결정된 뉴클레오티드 서열과 그에 의해 해독된 아미노산 서열을 나타낸다. 제 5도에서 시그날은 시그날 펩타이드를 가리키고, 프로는 프로펩타이드를 가리키고 숙성은 폴리펩타이드(I) (숙성 단백질)을 가리킨다.
실시예 3 (에스케리키아 콜라이 (Escherichia coli)에 대한 폴리펩타이드(I) 발현벡터의 구축)실시예 1에서 수득한 플라스미드 pUNK 5 속에 삽입된 폴리펩타이드(I) cDNA는 폴리 펩타이드(I)의 N-말단으로부터 11번째 티토신 잔기에 대해 코오딩하는 지역부근에 ScaI 부위를 갖고, 50염기에 의한 폴리펩타이드(I)의 종말 코오돈으로부터 하류에 NsiI 부위를 갖는다. (제2도, 4 및 5도 참고). 0.3kb ScaI - NsiI 분절을 플라스미드 pUNK 5 로부터 분리하고, 여기에 어댑터 NGFTE - 1 (35 mer), NGFTE - 2 ( 33 mer), NGFTE - 3 (7mer) 및 NGFTE - 4 (15 mer)를 T4 DNA 리가아제로써 결찰시킨 후, 제한 효소 NdeI 및 BamHI로써 처리한다. 그리하여, 0.3 - kb NdeI - BamHI 단편을 수득한다. (제 6도 참고).
이들 어댑터는 다음과 같다 :
NGFTE-1 : 5' TATGTACGCGGAGCATAAGAGTCACCGAGGGGAGT 3' 35 mer
NGFTE-2 : 5' ACTCCCCTCGGTGACTCTTATGCTCCGCGTACA 3' 33 mer
NGFTE-3 : 5' TGCCAGG 3' 7 mer
NGFTE-4 : 5' GATCCCTGGATGCA 3' 15 mer
T7 프로모터를 갖는 발현벡터 pET - 3C ( Rosenberg 일행, Gene 56, 125 (1987))를 유사하게 NdeI 및 BamHI로써 절단하여 4.4 - kb NdeI - BamHI 단편을 분리한다.
상기 수득한 4.4 - kb NdeI-BamHI 단편을 T4 DNA 리가아제로써 0.3 - kb NdeI - BamHI 단편에 결찰시킨 다음, 결찰된 단편을 E. 콜라이 (E. coli)속에 삽입하여 형질전환체를 제조한다 얻어진 암피실린 - 내성 형질전환체 E.콜라이(E. coli) DH 1/pENGFT 102 로부터 분리된 플라스미드는 pENGFT 102라 명한다. (제 6도).
실시예4(형질전환체의 분리 및 발현)
실시예 3에서 수득된 폴리펩타이드(I) 발현벡터pENGFT 102를 사용하여 E. 콜라이(E. coli) BL21 (DE3) (GEne 56, 125(1987)) 을 형질전환시켜 형질전환체 E. 콜라이(E. coil) BL21 (DE3) / pENGFT 102 (IFO 14874, FERM BP-2420) 를 수득한다.
형질 전환체 E. 콜라이(E. coil) BL21 (DE3) / pENGFT 102 를 시험관 내에서 50μg/ml 암피실린과 0.2% 포도당을 함유하는 LB배지 5ml상에서 37。C에서 16시간 동안 배양한다. 얻어진 배양용액 1ml를 갖는 배지 20ml를 함유하는 200ml 플라스크 속에 옮기고 37。C에서 배양한다. 클레트(Klett) 값이 170∼200에 도달할 때, IPTG를 첨가하여 최종농도가 0.4mM이 되게 하고, 배양을 3시간 동안 더 계속한다. 얻어진 배양용액 30μl 로부터 수집한 세포를 시료 완충액 [50mM 트리스-HCL(pH 6.8) , 2mM EDTA, 1%SDS, 1% 머캅토에탄올, 8% 글리세롤, 0.025% 브로모페놀블루]에 현탁시키고, 5분간 가열한 후, 0.1%SDS를 함유하는 16% 폴리아크릴아미드 겔 상에 전기영동한다. 전기 영홍 후, 코오 마씨 브릴리안트 블루(Coomassie Brilliant Blue)로써 겔을 염색한다. 그 결과, 상기 벡터 pET-3C를 사용하여 E. 콜라이(E. coil) BL21 (DE3)를 형질전환시켜 얻은 E. 콜라이(E. coil) BL21 (DE3)/pET-3C내에서 검출되지 않은 15-킬로달톤(kDa) 단백질이 E. 콜라이(E. coil) BL21 (DE3) /pENGFT102내에서 검출된다. 생성된 15 kDa단백질의 양은 전체 단백질의 약 10%이다. 이 단백질은 또한 토끼 항 -생쥐 NGF항체를 사용하는 웨스턴 블로팅 법(Western blotting method)(Collaborative Research, Inc. U.S.A) 에 의해서도 검출된다.
실시예5 (동물세포에 대한 폴리펩타이드 (Ⅰ) 발현 벡터의 구축)
폴리펩타이드 ( I ) cDNA 를 함유하는 1.1kb EcoRI 단편을 실시예2에서 수득한 플라스미드 pHNT2로부터 분리시킨다. 발현벡터 pTB389 (EP-251, 244A에 상응하는 일본국 특허 공개 공보 제 64-2572/1989에 기재되어 있음)를 EcoRI로써 유사하게 절단한다. 얻어진 단편을 폴리펩타이드 (Ⅰ) cDNA를 함유하는 상기 1.1 kbEcoRI 단편에 T4DNA리가아제로써 결찰시킨 다음, 결찰 혼합물을 E. 콜라이(E. coil) DH1의 형질 전환에 사용한다. (Molecular Cloning A Laboratory Manual1, Cold Spring Harbor Laboratory, p.505, 1982). 얻어진 암피실린-내성 형질 전환체 E. coil DH 1 / pNTK 26 로부터 플라스미드를 분리하고 이 플라스미드는 pNTK 26이라 명한다.
아벨슨 생쥐 백혈병 비루스 (A-MuLV) LTR) 지역을 함유하는 1.1kb ClaI-HindШ 단편을 플라스미드 pTB 505로부터 분리한다 (EP-225, 701A 에 상응하는 일본국 특허 공개 공보 제 62-175182/1987에 기재됨). 플라스미드 pNTK26을 제한 효소ClaI 및 HindШ 으로써 유사하게 절단하고, 작은 단편을 제거한다. 그 다음 얻어진 단편을 A-MuLV LTR지역을 함유하는 상기 1.1kb ClaI-HindШ 단편에 T4 DNA 리가아제로써 결찰시키고, 결찰 혼합물을 E. 콜라이(E. coil) DH1의 형질 전환에 사용하여 암피실린-내성형질전환체 E.콜라이(E. coli) DH 1/pNTL 145(IFO 14873,FERM BP -2421) 를 수득한다. 그리하여 수득된 형질전환체로부터 플라스미드 pNTL145를 분리한다. (제 7도)
실시예6 (동물세포에 대한 폴리펩타이드 (Ⅰ) 발현벡터의 구축)
폴리펩타이드 (Ⅰ) cDNA 내에서 시그날 펩타이드, 프로펩타이드 및 폴리펩타이드 (Ⅰ) 에 대해 코오딩하는 지역을 함유하는 0.83-kb EcoRI-AhaⅢ 단편을 실시예2 에서 수득한 플라스미드 pHNT2로부터 분리시킨다.
(AhaⅢ부위의 위치에 대해서는 제 4도 및 5도를 참고).
얻어진 단편의 5`-말단 (EcoRI)을 클레노우(Klenow) 단편으로 염기쌍 말단으로 만든 다음, XhoI 링커 pCCTCGAGG를 T4 리가아제로써 그 각각의 말단에 결찰시킨 후, XhoI 로 처리한다. 그리하여, 0.86-kb XohI 로 처리한다.
동물세포에 대한 발현벡터 pKSV-10 (Pharmacia) 를 제한효소 BglⅡ로 절단한 다음에 얻어진 단편의 양 말단 (XhoI)을 클레노우단편으로써 염기쌍 말단으로 만든다. XhoI 링커(상기 기재됨)을 여기에 첨가하고, 이 단편을 상기 0.86kb XhoI 단편에 T4 DNA리가아제로써 결찰시킨다.
결찰된 단편을 사용하여 E. 콜라이 ( E. coli ) DH1을 형질전환시킨다. 얻어진 암피실린-내성 형질 전환체 E. 콜라이 ( E. coli ) DH1/pNTS101로부터 플라스미드 pNTS101을 분리한다.(제 8도)
실시예7 (동물세포 내에서 폴리펩타이드 (Ⅰ) cDNA의 발현)
10%태아 송아지 혈청을 함유하는 둘벡코 벤형 이글스 배지 (DMEM 배지) (Flow Laboratories) 내에서 단일층으로 배양한 후, 동일 배지에 대해 배지를 교환한다. 교환한지 4시간 후, 각각 발현벡터 pTB 389, 10μg의 폴리펩타이드 (Ⅰ) 발현벡터 pNTK 26 및 폴리펩타이드 (Ⅰ) 발현 벡터 pNTL 145를 함유하는 인산칼슘겔을 공지방법[Graham 일행, Virology 52, 456 (1973) ]에 따라서 제조하고, 세포에 첨가하여 각각 형질전환체 COS-7/pTB389, COs-7/pNTK26 및 COS-7/pNTL145를 수득한다. 이들 세포를 이산화탄소 항온 기내에서 4시간 동안 배양한 다음 글리세롤로 처리한 후 [ Gorman 일행, Science 221, 551 (1983) ]. 3일 동안 배양한다. 배양후 배양물을 원심분리하여 배양 상충액을 수득한다. PC12세포를 문헌[Brain Esearch 133, 350(1977)] 및 [Experimental Cell Research 145,179 (1983)] 에 기재된 방법에 따라서 각상충액의 존재하에 배양하고, 축색이 세포직경의 2배 이상인 세포의 비율을 계산한다. 결과가 표 2에 주어진다.

상기 기재된 바와 유사한 방법으로 수득한 배양상충액을 사용하여, 공(co-) 배양된 중격 및 기저 전뇌 뉴우런 [M.Kakihama and Suno, Nerve Chemistry 27, 166(1998) ]의 아세틸콜린 (ACh) 함량에 대한 영향을 조사하였다.
17일 된 태아 뇌로부터 중격 및 긱저 전뇌를 절개하고 , 그로부터 하따나까 일행의 방법[ Develope. Brain Res. 30, 47(1986)]에 따라서 신경세포를 분리한다. 세포를 , 약 1×106세포/cm²/웰의 밀도로 100μg/ml의 폴리-L-오르니틴으로 선처리한 48-웰 플레이트 상에 심고, 500㎕의 무혈청 DME/F12/N2 배지내에서 24시간 동안 배양한다. 흡입에 의해 제거한 후, 500㎕의 DME/F12/10% FCS와 표본상의 상충액을 더한다. 2일 후, 배양용액을 동일한 배양용액 750ml로 교환하고, 상충액을 다시 더한 후, 2일간 배양한다. 상충액은 2종류 방식으로 첨가한다. 즉, 50μl의 상충액을 처음 2일간 더하고 75μl를 나중 2일간 더하여 최종 농도가 10%가 되게 한다. 와꼬준 야꾸사로 부터 구입한 생쥐 NGF (7S-NGF) 를 사용하는 경우, 그것을 0.1% 오브알부민/PBS로 희석하고 10μl를 더한다.
상충액 첨가로부터 4일 후, 흡입에 의해 상충액을 제거하고 ACh측정을 위해 20 ∼ 60pmol/20μl 의 ENC (에틸호모콜린)와 냉각된 0.3N PCA 500μl를 여기에 첨가한다. 부드럽게 교반한 후, 500μl 의 용액을 에펜돌프 마이크로튜브에 옮긴다. 상기 보고된 방법에 따라서 후속 조작을 수행하고, HPLC/ECD(고색액체크로마토그래피/전기화학 검출계)를 사용하여 ACh양을 측정한다. ACh추출 후, 세포를 500μl의 1N NaOH에 용해시키고, 단백질의 양을 결정한다( Bio-RAD protein assay). 두 네트 t-테스트를 통계처리에 사용한다.
결과가 표 3에 주어진다.

실시예 8(효모에 대한 폴리펩타이드(Ⅰ)발현 벡터의 구축)
사람 리소좀 발현 벡터 PGEL125( 유럽특허 공보 제 255, 233호에 기재된 방법에 따라 제조)를 HindⅢ으로 절단하고, 얻어진 단편을 클레노우 단편으로 염기쌍 말단으로 만든 후, T4 DNA 리가아제로 결찰시켜 HindⅢ부위를 전혀 갖지 않는 플라스미드 pGEL 125H를 수득한다. 그 다음, 플라스미드 pGEL125H를 XhoI로써 절단하고, 얻어진 단편을 T4 DNA 리가아제로써 XhoI- HindⅢ 어댑터 결찰시켜8.3-kb HindⅢ-BamHⅠ 단편을 수득한다. 1.6-kb EcoRI 단편-함유-α-인자 유전자를 프라스미드 p69A [Cell 30 , 993(1982)]로부터 분리하고 클레노우 단편으로 염기쌍 말단으로 만든다. 그 다음, 얻어진 단편을 T4 DNA리가아제로써 BamHⅠ링커 5`CCGGATCCGG3` 에 결찰시킨 후, BamHⅠ 및 HindⅢ으로 처리한다. 그렇게 해서 수득한 0.9kb BamHⅠ-HindⅢ 단편( α-인자 유전자의 프로모터와 프레프로 지역에 대한 DNA 코오딩을 함유)을 상기 8.3kb HindⅢ-BamH 단편에 T4 DNA 리가아제로써 결찰시키고 이 반응 혼합물을 사용하여 E, 콜라이( E, coil) DH1 을 형질 전환시킨다. 얻어진 암피실린- 내성 형질 전환체로부터 분리된 플라스미드 pALFA103(9.2kb)이라 명한다.
결찰시켜8.3-kb HindⅢ-BamHⅠ 단편을 수득한다. 1.6-kb EcoRI 단편-함유-α-인자 유전자를 프라스미드 p69A [Cell 30 , 993(1982)]로부터 분리하고 클레노우 단편으로 염기쌍 말단으로 만든다. 그 다음, 얻어진 단편을 T4 DNA리가아제로써 BamHⅠ링커 5`CCGGATCCGG3` 에 결찰시킨 후, BamHⅠ 및 HindⅢ으로 처리한다. 그렇게 해서 수득한 0.9kb BamHⅠ-HindⅢ 단편( α-인자 유전자의 프로모터와 프레프로 지역에 대한 DNA 코오딩을 함유)을 상기 8.3kb HindⅢ-BamH 단편에 T4 DNA 리가아제로써 결찰시키고 이 반응 혼합물을 사용하여 E, 콜라이( E, coil) DH1 을 형질 전환시킨다. 얻어진 암피실린- 내성 형질 전환체로부터 분리된 플라스미드 pALFA103(9.2kb)이라 명한다.
α- 인자 유전자의 프로모터와 플레프로지역에 대해 코오딩하는 DNA서열을 함유하는 0.9-kb 단편을 분리한 다음에 파아지벡터 M13mp18 속에 삽입한다. 24염기로써 α-인자의 프로지역에 대해 코오딩하는 DNA의 3`- 말단(HindⅢ부위)으로부터 상류의 신규HindⅢ부위를 제조하기 위하여 프로지역의 81번째 위치의 세린의 콩코오돈 TCT를 AGC로 전환시켰다. 즉, 프라이머 5`TTTATCCAAGCTTACCCCTTC 3` 와, 0.9-kb BamHⅠ-HindⅢ 단편을 함유하는 상기 파아지벡터 M13mp18을 사용하여 , 아미샴키트에 의해 부위-지향 돌연변이 유발을 수행하여 원하는 클론을 수득한다. 돌연변이 유발 이전보다 24bp짧은 0.9-kb BamHⅠ-HindⅢ을 수득된 클론으로부터 분리하고, pGEL125H로부터 파생된 8.3-kb BamHⅠ-HindⅢ 단편(상기 기재됨)에 결찰시켜 플라스미드 pALFA 310을 수득한다.
PGK 터미네이터를 함유하는 0.29-kb AhaⅢ-SaⅡ 단편을 플라스미드 pGLDp31-RcT (유럽특허공보 제 0235430 호) 로 부터 분리한다. XhoI 링커 pCCTCGAGG 를 T4 DNA리가아제로써 이 단편에 결찰시킨 후에, XhoI 및 SaⅢ로써 처리하여 PGK터미네이터를 함유하는 0.29-kb XhoI-SaⅡ단편을 수득한다. 이 0.29-kb XhoI-SaⅡ단편을 플라스미드 pALFA310 (상기 기재됨) 내의 α-인자의 프로지역에 대해 코오딩하는 DNA로부터 하류에 위치한 XhoI부위 속에 삽입한다. 그리하여 플라스미드 pALFA 310T가 수득된다.
폴리펩타이드(Ⅰ) cDNA를 함유하는 1.1-kb EcoRI 단편을 실시예2에서 수득한 플라스미드 pHNT2로부터 분리한다. 이 1.1-kb EcoRI 단편을 AhaⅢ으로 절단한 후, XhoI 링커를 첨가한다. 얻어진 단편을 ScaI로 절단하여 0.36-kb ScaI-XhoI 단편을 수득한다. 이 0.36-kb-ScaI-XhoI 단편에 하기 합성 DNA와XhoI-HindⅢ 어댑터 (상기 기재됨)을 결절시킨 후, HIⅡ으로 처리한다.
5·GCTTGGATAAAAGATACGCGGAGCATAAGAGTCACCGAGGGGAGT3'
3'ACCTATTTTCTATGCGCCTCGTATTCTCAGTGGTCCCCTCA5'
HindⅢ ScaⅠ
그리하여, 폴리펩타이드 ( I ) 에 대해 코오팅하는 0.4-kb HindIII 단편을 수득한다.
폴리펩타이드(I)에 대해 코오딩하는 0.4-kb HinrIII 단편을 플라스미드 pALFA310T의 - 인자의 프레프로지역에 대해 코오딩하는 DNA의 3' - 말단에 위치한 HindIII 부위속에 삽입시켜,폴리펩타이드(I) 발현 벡터 pANT341T를 수득한다(제 9도)
실시예 9 : (형질전환체의 분리와 폴리펩타이드(I) cDNA의 발현)
실시예8에서 수득한 폴리펩타이드(I) 발현 벡터 pANT314T를 사용하여, 참고예1에서 수득한 S. 세레비시애(S. cerevisiae) TB 39ρ-(IFO 10467, FERM BP -2399)를 리튬법 [J.Bacterial. 153, 163 (1983)]에 따라서 형질전환시켜, 형질전환체 S. 세레비시애(S. cerevisiae) TB 39ρ-/pANT314(IFO 10475, FERM BP -2530)를 수득한다.
형질전환체 S. 세레비시에 (S. cerevisiae) TB 39ρ-/pANT331T를 시험관 내의 5ml의 변형 부르크홀더 배지(리터당 89g 의 슈크로오스, 11g의 글르코오스, 3.6g의 아스파라긴 및 0.44g의 KH2PO4를 함유함)[Amer. J. Bot. 30, 206(1943)] 속에 접종하고 30℃에서 3일간 흔들면서 배양한다. 1ml 의 얻어진 혼합물을 상기 배지 4ml를 함유하는 시험관 내로 옮기고, 30℃에서 3일간 흔들면서 배양한다. 이 배양물 2ml를 상기 배지 18ml를 담은 200ml 삼각 플라스크 속에 또한 옮기고 30℃에서 3일간 흔들면서 배양한다.
그렇게 해서 수득한 배양물을 원심분리하고, 그 상층액 750μl에 트리콜로로아세트산을 가하여 단백질을 침전시킨다. 침전물을 시료완충액 [Laemmli, Nature 227, 680(1970)]내에 용해시키고, 100℃에서 5분간 가열한 후, 0.5% SDS를 함유하는 15% 폴리아크릴아미드겔 상에서 전기 영동한다. 겔상의 단백질을 베네트법에 따라서 니트로셀룰로오스 막에 옮긴다. [Analytical Biochemistry, 195(1981)].
토끼의 항 - 생쥐항체(Collaborative Research Inc. U.S.A) 및 친화도 - 정제된 HRP - 연결된 산양의 항 - 토끼 IgG(Bio RAD, U.S.A)를 사용하여 웨스턴 블로팅을 실시한다. 그 결과, 폴리펩타이드(I)의 분자량 약 15킬로달튼(kDa)에 해당하는 띠가 검출된다. 반면, S. 세레비시애(S. cerevisiae) TB 39ρ-/pALFA 310T의 상층액에 대해서는, 이 띠가 검출되지 않는다.
실시예10: (E. 콜라이 (E. coli)에 의한 폴리펩타이드(I)의 제조)
실시예3에서 수득한 폴리펩타이드(I) 발현켁터 pENGFT102 와 T7 리소짐 발현 벡터 pLysS를 사용하여 에스케리키아콜라이 BL21(DE3)을 형질전환시켜 형질전환체 E. 콜라이 (E. coli) BL21/(DE3)/pLysS, pENGFT102(IFO 14903, FERM BP-2529)를 수득한다.
형질전환체 E. 콜라이(E. coli) BL21(DE3)/ pLysS. pENGFT 102를, 50μg/ml의 암피실린, 10ug/ml의 콜로로암페니콜 및 0.2% 글루코스롤 함유하는 LB 배지 [1% 트립톤 (Difco), 0.5% 효모추출액)] 내에서 37℃에서 16시간동안 흔들어주면서 배양한다. 배양물(12.5 ml)을 동일한 배지 250ml를 함유하는 1리터 삼각 플라스크속에 옮기고, 30℃에서 3시간동안 흔들어 주면서 배양한다. 그 즉시, 배양 용액의 콜레트(Klett)값이 170 에 도달한다. 이 배양물에 이소프로필 - β - D (-) - 티오갈락토피로노시드를 최종 농도가 0.1mM이 되게 첨가하고, 30℃에서 3시간동안 흔들어 주면서 배양한다. 그렇게해서 수득한 30ul의 배양물로부터 수집한 세포를 30μl의 시료 완충액[Laemmli, Nature 227, 680(1970)] 내에 현탁시키고, 100℃에서 5분간 가열한 후, 0.1% SDS를 함유하는 16% 폴리아크릴아미드 겔에 대해 전기 영동 시킨다. 겔 상의 단백질을 버네트법 [Analytical Biochemistry 112, 195(1981)]에 따라서 니트로 셀룰로오스 막에 옮긴 다음, 토끼의 항- 생쥐 NGF 항체 (Collaborative Research Inc, U.S.A)와 친화도 -정제된 HRP - 연결된 산양의 항- 토끼 IgG (Bio RAD, U.S.A)를 사용하여 웨스턴 블로팅을 수행한다. 그 결과, 15킬로달톤 (kDa)의 분자량을 갖는 폴리펩타이드(I)이 검출된다.
상기 기재된 바와 유사한 방식으로 수득되고 전기 영동한 겔을 코오마씨블릴리안트 불루로써 염색하면, 폴리펩타이드(I)에 해당하는 15-kDa 단백질이 검출되고, 그 생성량은 단백질의 총량을 기준으로 약 10%인 것으로 추정된다.
실시예11: 실시예 10에서 수득한 형질전환체 E. 콜라이(E, Coli) BL21 (DE3)/pLysS, pENGFT 102의 배양물(3.75리터)을 원심분리하여 17g(wet)의 세포를 수득한다. 세포를 375ml의 50mM 트리 - HCL(pH 8.0)에 현탁시키고 냉동 - 용해시킨후, 음파 진동기 (Kaijo Denki, 2A, 2분 )로 3회 처리한다. 부숴진 세포현탁액을 원심분리하고, 얻어진 침전물을 60ml의 5M 구아니딘 염산염 - 5mM EDTA - 1mM PMSF - 0.1mM APMSF -20mM 디티오트레이톨(DTT) - 50mM 인산나트륨 완충액(pH 6.0)내에 용해시킨다. 그렇게해서 수득한 용액을 2M 구아니딘 염산염 - 5mM EDTA - 0.1mM APMSF - 5mM DTT - 25mM 인산나트륨 완충액 (pH 6.0)으로 평형화된 세파크릴 S-200 칼럼에 적용하고, 위스턴 볼로팅법( 상기 기재됨) 으로 폴리펩타이드(I)이 검출되는 분획들을 수집한다. (부피 = 300ml). 이 용액을 YM5 막을 설치한 한외여과기 (Amicon)를 하용하여 농축하고, 얻어진 농축용액 50ml를 상기 기재된 바와같은 세파크릴 S-200컬럼에 적용한다. 그리하여, 328mg의 정제된 폴리펩타이드(I)을 함유하는 용액 164ml를 수득한다. 순도는 SDS - 폴리아크릴아미드겔전기영동법으로 조사한다. 그 결과, 얻어진 정제된 폴리펩타이드(I)는 실질적으로 균질하다는 것이 확인된다.
상기 정제된 폴리펩타이드(I)을 함유하는 용액을 울트라포어 RPSC 칼럼(0.46 x 7.5 cm , Altex)상에 부하하고, 트리플루오로아세트산 - 아세토니트릴용리제 용매계를 사용하여 고속액체크로마토그래피(HPLC) 하여 균질 폴리펩타이드(I)을 수득한다. 얻어진 폴리펩타이드(I)의 N-말단 아미노산 서열을 기체상 단백질 서열화장치 (Model 470A, Applied Biosystems)로써 결정한다. 결과적으로, 정제된 폴리펩타이드(I)의 N-말단 아미노산 서열은 메티오닌잔기가 N-말단에 첨가된 것을 제외하고 cDNA의 뉴클레오티드 서열로부터 연역된 폴리펩타이드 (I)의 N-말단 아미노산 서열과 일치한다. (표 4 ).
상기 수득된 정제된 시료의 아미노산 조성을 닌히드린법에 의해 결정된다. 그 결과, 관찰된 값은 N-말단에 메티오닌잔기가 첨가된 폴리펩타이드(I)로부터 계산된 이론치와 거의 일치한다. ( 표 5 ).

1) Glu를 11로 취하여 계산
2) 폴리펩타이드 (I)의 N-말단에 메티오닌잔기가 첨가된 경우의 계산
상기 정제된 폴리펩타이드(I)을 합유하는 용액 (단백질 농도 : 2mg/ml )을 2mM 구아니딘 염산염 - 5mM EDTA - 0.1mM APMSF - 5mM DTT - 25mM 인산나트륨완충액 (pH 6.0) 으로 회석하여 단백질 농도가 10μg/ml 가 되게한다. 희석된 용액을 50배량의 1mM EDTA - 50mM NaHCO3- Na2CO3(pH 10.0) 에 대하여 4℃에서 16시간동안 투석하고 동일 완충액에 대하여 4시간동안 더 투석한다. 얻어진 투석된 액체의 PC12 세포에 대한 영향을 문헌 [Brain Research 133. 350 (1979) ] 및 [Experimental Cell Research 145, 179(1983)]에 기재된 방법에 따라서 시험한다. 그 결과, PC12 세포의 6%가 내부투석액의 첨가에 의해 축색을 가지고, 그의 2%는 길이가 세포체 직경의 적어도 2배인 것이 관찰된다. 반면, 대조용으로서 1mM EDTA - 50mM NaHCO3- NaCO3(pH 10.0) 에 대해서는 0.5% 이하의 세포가 축색을 가진다. 상기 기재된 바와 유사한 방법에 의해 수득된 정재된 폴리펩타이드(I)은 닭래아 지각 뉴우턴( 배근 신경질)의 생존촉진 활성을 갖는다.(상기 기재됨). 실시예12(동물세포내에서 폴리펩타이드(I) cDNA 의 발현) 이산회탄소 항온기내에서 10% 태아 송아지 혈청을 함유하는 둘벡코 변형 이글스 배지(DMEM 배지) 내에서 원숭이 COS-7 세포를 단일층으로 배양한 후, 동일배지로 배지를 교환한다. 교환한지 4시간후, 10μg의 pTB 389 (EP-251, 244A 에 해당하는 일본국 특허 공개 제64-2572/1989호에 기재됨) 또는 10μg의 pNTL 145 (실시예5를 참고)를 공지방법 [Graham 일행, Virology 52, 456 (1973)]에 따라서 제조하고 세포에 첨가한다. 이들 세포를 4시간동안 배양한 다음, 글리세롤[Gorman 일행, Science 221, 551(1983)]로 처리한 후에 10% 태아 송아지 혈청을 함유하는 DMEM 내에서 16시간동안 배양한다. 배지를 0.5% 태아 송아지 혈청을 함유하는 DMEM으로 교환한후, 세포를 2 일간 더 배양하고, 얻어진 배양물을 원심분리한다. 그렇게 해서 수득한 pTE 389로 형질 감염된 COS-7 세포의 배양물 상층액(시료1)과 pNTL145 로 형질감염된 COS-7세포의 배양물 상층액(시료2)을 하기 실험에 사용한다.
트리클로로아세트산 0.5ml의 각 시료에 첨가하여 단백질을 침전시킨다. 얻어진 침전물을 시료완충액 [Laemmli, Nature 227, 680(1970)]내에 용해시키고 100℃에서 2분간 가열한 후 에, 0.5% SDS를 함유하는 17% 폴리아크릴아미드겔상에서 전기영동 시킨다. 겔 상의 단백질을 버네트법[Analytical Biochemistry 112, 195(1981)]에 따라서 니트로셀룰로오스막에 옮긴다. 참고 예2에서 수득한 항 - 펩타이드(I) N - 말단 펩타이드 항체와 친화도 - 정제된 HRP - 연결된 산양의 항 - 토끼 IgG(Bio RAD, U.S.A)를 사용하여 웨스턴 볼로팅을 실시한다. 그 결과, pNTL 145로 형질감염된 COS-7 세포의 배양물상층액 (시료2)에 대하여, 폴리펩타이드(I)에 해당하는 분자량 약 15킬로달튼(kDa)의 띠가 검출된다. 그러나, pTB 389로 형질전환된 COS-7 세포의 배양물 상층액 (시료1)에 대하여, 폴리펩타이드(I)에 해당하는 띠가 검출되지 않는다. 항 - 생쥐 NGF 항체 (Collaborative Research, U.S.A)를 상기 항 - 폴리핍타이드(I) N - 말단 펩타이드 항체 대신에 사용하는 경우, 펩타이드(I)에 해당하는 띠가 검출된다.
지각 뉴우런 (배근 신경질)을 8일된 닭의 배로부터 분리하고 37℃에서 20분간 CMF (무 칼슘 - 마그네슘) - PBS 내의 1% 트립신( 돼지 췌장 결정화된 트립신, Wako Junyaku)으로 처리하여 세포를 분산시킨다. 플라스틱 디쉬상에서 10% 태아 송아지 혈청(FCS)를 함유하는 DMEM 내에서 세포의 선배양을 실시하여 비신경 세포를 점착 시킨다. 그 다음, 점착되지 않은 세포를 원심분리로써 수집하고 104∼105세포/웰의 밀도로 폴리 - L - 오르니틴으로 코우팅된 24-웰 폴레이트상에 심는다. DMEM에 대하여 투석된 각 시료를 배양을 위해, 배양 배지로서 혼합 배지(10% FCS, 1μM Ara-C 및 50μg/ml 의 카나마이신을 함유하는 DMEM : 햄스 (Ham's) F12= 1:1) 여기에 첨가한다. 4일간의 배양 후, 세포가 평활한 표면과 세포체 직경의 적어도 2개 길이의 축색을 갖는 것을 척도로 하여, 웰당 10시야에 대해 생존 신경세포의 수를 결정한다.
배양을 개시하여 3일후에, 신경세포의 외관을 서로 비교한다. 각 세포내에 105세포를 놓고 각 시료를 배양용약을 기준으로 10부피 %의 양으로 첨가하는 경우, 시료2에 있어서, 수많은 생존 신경 세포가 관찰되고 축색은 빽빽히 분포된다. 대조적으로, 시료1에 있어서는 수많은 죽은 세포(윤곽이 고르지 않고 축색이 없는 부유하는 세포)가 관찰되고 생존세포의 수는 실시에 2에서 보다 적다.
104 세포/웰의 밀도로 심어진 세포를 배양하는 경우, 모든 시료가 5% 및 10%의 시료 농도에서 축색의 높은 발달을 나타낸다. 그러나, 2%의 시료농도에서, 고 발달된 축색을 갖는 신경세포가 실시예2에서 관찰된 한편, 실시예1에서는 축색의 발달이 나쁘고 세포체는 작다.
104 세포/웰의 밀도로 심어진 세포를 배양하는 경우, 배양을 개시한지 4일후에 생존 신경세포의 수를 센다. (제10도). 제 10도에 대해 설명하면, 빈 원 (○)은 pTB 389로 형질감염된 COS-7 세포의 배양물 상층액 (시료1)을 나타내고, 검은 원 (●)은 pNT145로 형질감염된 COS-7 세포의 배양물 상층액( 시료1 )을 나타낸다. 시료2는 지각신경세포의 생존을 증가시키며 농도에 따른 그 효과를 시료1과 비교한다. 실시예13(쥐 폴리펩타이드(I) 유전자의 콜로닝):
폴리펩타이드(I)cDNA를 함유하는 1·1-kb EcoRI DNA 단편을 실시예2에서 수득한 플라스미드 pHNT2 로부터 분리하고 올리고 표지화 반응(Nippon Gene)에 의해 표지하여 프로우브를 수득한다.
5주된 쥐의 각 기관으로부터 구아니딘-CsCl 법에 의해 전체 RNA를 제조하고, 올리고 - dt 셀룰로오스를 사용하여 폴리(A) RNA를 수득한다. 상기 프로우브를 사용하여, 각 조직으로부터 수득한 폴리(A) RNA의 노오든 블로팅(Northern blotting)을 실시한다.
결과적으로, 신장, 간, 심장, 뇌, 췌장, 흉신, 허파 및 하악선에서 폴리펩타이드(I)의 1, 4 - kb 메신저 RNA(mRNA) 가 검출된다. 상기 결과는 폴리펩타이드(I) 유전자가 쥐에게도 존재하고 많은 조직중에서 발현됨을 제시한다.
사람 폴리펩타이드(I)에 대해 코오딩하는 0.45 - kb EcoRI - Aha Ⅲ 단편을 실시예 1에서 수득한 폴라스미드 pUNK5 로부터 분리하고, 이 단편을 프로우브로 사용하여 쥐 게놈 DNA의 사우든 (Southern) 혼성화를 실시한다. 이 프로우브는 약 7.4-kb EcoRI 단편, 약 3.8 - kb BglⅡ 단편 및 약 3.3 kb HindⅢ 단편에 대해 혼성화하며, 이는 폴리펩타이드(I) 유전자가 쥐에도 또한 존재함을 제시한다.
그 다음, 실시예2에서 수득한 폴라스미드 pHNT2로부터 폴리펩타이드(I) cDNA를 함유하는 1·1 - kb EcoRI 분리하고, 쥐의 폴리펩타이드(I) 유전자를 이 단편을 프로우브로서 사용하여 클로닝한다. 암컷 쥐(Sprague-Dawley)의 간으로부터 유도된 DNA를 부분적으로 섭취시키고 얻어진 단편을 파론(Charon) 4A 파아지 속에 도입시켜 구축한, 클로닝에 사용되는 쥐 게놈 DNA 라이브러리는 콜론테크(Clontech)사로부터 구입한 것이다. E. 콜라이(E. coli) LE 362를 이 파아지 라이크러리로 감염시켜 폴레이트 당 약 5 x 105 플라그를 형성한다. 10개의 개별적 플레이트로부터 파아지 DNA를 공지 방법 T. maniatis일행, Molecular Clining, A Laboratory Manual에 따라서 니트로셀룰로오스막에 옮기고, 상기 프로우브와 혼성화 시킨다. 그 결과, 7 포지티브 클론이 수득된다. 한 포지티브 클론 (λ rNGF 2- 8)은 약 12 - kb 삽입된 DNA 단편을 합유한다. 사우든 혼성화의 결과로부터, DNA 단편 중에서 0.95 - kb BglⅡ - HindⅢ 단편내에 폴리펩타이드(I)에 대해 코오딩하는 지역이 존재한다는 것이 추론된다. 그 다음, 0.95 - kb BglⅡ - HindⅢ 단편을 플라스미드 pUCl18 (Takara Shuzo) 내에서 서브콜로닝하여 플라스미드 pRNT18을 수득한다. 플라스미드 pRNT18을 사용하여, E. 콜라이(E. coli) DHl을 형질전환시켜 형질전환체 E. 콜라이 (E. coli) DHl/pRNT18(IFO 14943, FERM BP - 2618)을 수득한다.
상기 0.95 - kb BglⅡ - HindⅢ 단편을 다양한 제한 효소로써 절단하고, 얻어진 단편을 각각 pUCl18, M13mp18 등내에서 서브콜로닝한다. 그 다음, 시쿼나아제(Toyobo)를 사용하여 이들의 뉴콜레오티드 서열을 결정한다. (제 11도). 결과적으로, 0.95 - kb BglⅡ - HindⅢ 단편은 시그날펩타이드에 대해 코오딩하는 지역, 프로지역, 및 쥐폴리펩타이드(I)의 숙성 프로태인을 함유함과 인트론은 존재하지 않음이 드러난다.
뉴콜레오티드 서열로부터 유도된 쥐의 폴리펩타이드(I)의 아미노산 서열을 사람의 폴리펩타이드(I)의 것과 비교하면, 시그날 펩타이드 및 프로지역의 11째 잔기에서 차이가 관찰되나, 숙성 단백질 ( 폴리펩다이트(I))에 대해서는 차이가 없다. 따라서 쥐 폴리펩타이드(I)의 아미노산서열은 사람 폴리펩타이드의 경우와 완전히 일치함이 입증된다. 실시예14(폴리펩타이드(I) cDNA의 콜로닝) :
시그날 서열, 프로지역 및 폴리펩타이드(I)에대해 코오딩하는 0.83 - kb DNA 단편을 폴리펩타이드(I) cDNA로부터 프로우브를 제조한다. 얻어진 프로우브를 사용하여, 실시에 1 및 2 에서와 유사한 방식으로 사람 태반 라이브러리(Clontech Laboratories, Inc)로부터 0.73 - kb 및 1·1 - kb 폴리펩타이드(I) cDNA를 클로닝한다. 그렇게 해서 수득한 폴리펩타이드(I) cDNA의 뉴콜레오티드 서열은 실시예 1 및 2에서 콜로닝된 폴리펩타이드(I) cDNA의 뉴콜레오티드 서열과 일치한다. 실시예 15(폴리펩타이드 (I) 발현 벡터의 도입에 의한 폴리펩타이드(I) - 생성 동물세포균주의 설립)
(1) 발현벡터의 구축
폴리펩타이드(I) cDNA의 시그날 펩타이드, 프로펩타이드 및 폴리펩타이드(I)에 대해 코오딩하는 지역을 함유하는 0.86 - kb Ecori - Aha Ⅲ 단편을 실시에2에서 수득한 폴라스미드 pHNT2 로부터 분리한다. 다른 한편, 인터로이킨 (IL)-cDNA의 발현을 위한 플라스미드 pTB399 [Cell Struct.Funct.12,205(1987)]를 BglⅡ로써 절단한 다음에 DNA폴리미라아제 콜레노우 단편으로 처리한후, EcoRI로써 더 절단하여 단편 (약 3.8kb)을 수득하고 그로부터 IL-cDNA 부분을 제거한다. 이 단편에 T4 DNA 리가아제 반응에 의해 상기 0.86 - kb EcoRI - AhaⅢ 단편을 결찰시켜 플라스미드 pTB1091을 수득한다.
그 다음, 플라스미드 pLG 89 [Gene 25, 179(1983)]으로부터 히드로마이신 B - 내성 유전자를 함유하는 1.0 - kb BamHI 단편을 분리하고, pTB 86 의 네오마이신 - 내성 유전자 [Cell Struct. Funct. 12, 205(1987)]를 함유하는 지역 (1.0 -kbBglⅡ - Sma I)으로 대치한다. 그리하여, HSV TK 유전자 프로모터를 갖는 히그로마이신 -내성 유전자 발현벡터 pTB 681을 구축한다. 플라스미드 pTB 681을 PvuⅡ로써 절단하여 수득한 1.8 - kb 단편에 Hind Ⅲ 링커를 첨가한 다음, 얻어진 단편을 상기 수득한 폴리펩다이드(I) 발현 벡터 pTB 1091의 HindⅢ 부위속에 삽입하여 히그로마이신 - 내성 유전자를 갖는 폴리펩다이트 (I) 발현 벡터 pTB 1139를 구축한다. (제12도). (2) 폴리펩다이드(I) - 생성 동물세포균주의 설립 생쥐 L 세로 (TK - 결핍균주)를 직경 6cm 의 팔콘 스케일상에 (7 x 105셀/ 스케일)심고, 10% FCS를 함유하는 이글스 MEM내에서 배양한다. 다음날, 그램 일행의 방법 [Virology 52, 456 (1973)]에 따라서 10μg의 발현 벡터 pTB 1139로 세포를 감염시킨후, 상기 배지내에서 2일간 배양한다. 트립신으로 처리한후 , 얻어진 세포를 새 스케일상에 다시 싣고, 500μg/ml 의 히그로마이신 B(시그마)를 함유하는 10% FCS-MEM내에서 배양을 계속한다. 2 내지 3 주후, 군락형태로 증식된 히그로마이신-내성세포를 수득한다. 그리하여 수득한 히그로마이신 - 내성 L 세포를 제한 희석법과 같은 공지방법에 따라서 클로닝하여 콜론 L-HI-1, L-H6-1, L-Hl1-1, L-H13-1, L-H14-1 (IFO 50223, FERM BP-2754), L-H18-1, L-H19-1, L-H35-1, L-H36-1 및 L-H43-1을 수득한다. 각 클론의 세포를 24-웰 플레이트상에 심고 배양한다. 셀이 합류하는 경우, 배지를 0.1% FCS를 함유하는 0.5ml/웰의 MEM 배지로 교환한다. 2일간의 배양 후, 상층액을 SDS-폴리아크릴아미드 겔 전기영동을 시키고, 참고예2에서 제조딘 폴리펩타이드(I) N-말단 펩타이드 항체를 사용하는 웨스턴 블로팅에 의해 폴리펩타이드(I)을 검출한다.
그 결과, 상기 각 콜론의 배지내에서 약 1mg의 폴리펩타이드(I)가 생성되었음이 밝혀진다.
Claims (10)
- 분자내에 하기 아미노산 서열 (Ⅱ)를 포함하는 폴리펩타이드(I)TyrAlaGluHisLysSerHisArgGlyGluTyrSerValCysAspSerGluSerLeuTrpValThrAspLysSerSerAlaIleAspIleArgGlyHisGlnValThrValLeuGlyGluIleLysThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArgCysLysGluAlaArgProValLysAsnGlyCysArgGlyIleAspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThrTyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGlyTrpArgTrpIleArgIleAspThrSerCysValCysAlaLeuSerArgLysIleGlyArg (Ⅱ)
- 제1항에 있어서, 하기 아미노산 서열(Ⅱ')을 갖는 폴리펩타이드TyrAlaGluHisLysSerHisArgGlyGluTyrSerValCysAspSerGluSerLeuTrpValThrAspLysSerSerAlaIleAspIleArgGlyHisGlnValThrValLeuGlyGluIleLysThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArgCysLysGluAlaArgProValLysAsnGlyCysArgGlyIleAspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThrTyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGlyTrpArgTrpIleArgIleAspThrSerCysValCysAlaLeuSerArgLysIleGlyArgThr (Ⅱ')
- 분자내에 하기 아미노산 서열(Ⅱ)을 포함하는 폴리펩타이드(I)에 대해 코오딩하는DNATyrAlaGluHisLysSerHisArgGlyGluTyrSerValCysAspSerGluSerLeuTrpValThrAspLysSerSerAlaIleAspIleArgGlyHisGlnValThrValLeuGlyGluIleLysThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArgCysLysGluAlaArgProValLysAsnGlyCysArgGlyIleAspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThrTyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGlyTrpArgTrpIleArgIleAspThrSerCysValCysAlaLeuSerArgLysIleGlyArg (Ⅱ)
- 분자내에하기 아미노산 서열(Ⅱ)을 포함하는 폴리펩타이드(Ⅰ)에 대해 코오딩하는 DNA를 함유하는 벡터TyrAlaGluHisLysSerHisArgGlyGluTyrSerValCysAspSerGluSerLeuTrpValThrAspLysSerSerAlaIleAspIleArgGlyHisGlnValThrValLeuGlyGluIleLysThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArgCysLysGluAlaArgProValLysAsnGlyCysArgGlyIleAspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThrTyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGlyTrpArgTrpIleArgIleAspThrSerCysValCysAlaLeuSerArgLysIleGlyArg (Ⅱ)
- 분자내에 하기 아미노산 서열 (Ⅱ)을 포함하는 폴리펩타이드(I)에 대해 포오딩하는 DNA를 함유하는 벡터에 의해 형질전환된 형질 전환체TyrAlaGluHisLysSerHisArgGlyGluTyrSerValCysAspSerGluSerLeuTrpValThrAspLysSerSerAlaIleAspIleArgGlyHisGlnValThrValLeuGlyGluIleLysThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArgCysLysGluAlaArgProValLysAsnGlyCysArgGlyIleAspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThrTyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGlyTrpArgTrpIleArgIleAspThrSerCysValCysAlaLeuSerArgLysIleGlyArg (Ⅱ)
- 제5항에 따른 에스케리키아 콜라이 BL21(DE3)/pENGFT102 (FERM BP-2420).
- 제5항에 따른 사카로마이세스 세레비시애 TB39ρ-/pANT 341T (FERM BP-2530)
- 제5항에 따른 에스케리키아 콜라이 BL21(DE3)/pLysS, pENGFT102 (FERM BP-2529).
- 제5항에 따른 L-H14-l (FERM BP -2754)
- 폴리펩타이드(I)에 대해 코오딩하는 DNA를 함유하는 벡터에 의해 형질전환된 형질 전환체를 배지 내에서 배양하고폴리펩타이드르 축적하고폴리펩타이드를 수집하는 것으로 구성되는 ,분자내에 하기 아미노산 서열 (Ⅱ)을 포함하는 폴리펩타이드(I)의 제조 방법TyrAlaGluHisLysSerHisArgGlyGluTyrSerValCysAspSerGluSerLeuTrpValThrAspLysSerSerAlaIleAspIleArgGlyHisGlnValThrValLeuGlyGluIleLysThrGlyAsnSerProValLysGlnTyrPheTyrGluThrArgCysLysGluAlaArgProValLysAsnGlyCysArgGlyIleAspAspLysHisTrpAsnSerGlnCysLysThrSerGlnThrTyrValArgAlaLeuThrSerGluAsnAsnLysLeuValGlyTrpArgTrpIleArgIleAspThrSerCysValCysAlaLeuSerArgLysIleGlyArg (Ⅱ)
Applications Claiming Priority (8)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP5898389 | 1989-03-10 | ||
| JP1-58983 | 1989-03-10 | ||
| JP127710/1989 | 1989-05-19 | ||
| JP12771089 | 1989-05-19 | ||
| JP193654/1989 | 1989-07-26 | ||
| JP19365489 | 1989-07-26 | ||
| JP263613/1989 | 1989-10-09 | ||
| JP26361389 | 1989-10-09 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| KR900014428A KR900014428A (ko) | 1990-10-23 |
| KR0132425B1 true KR0132425B1 (ko) | 1998-04-14 |
Family
ID=27463714
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1019900003202A KR0132425B1 (ko) | 1989-03-10 | 1990-03-10 | 신규 폴리펩타이드 및 그 제조방법 |
Country Status (11)
| Country | Link |
|---|---|
| US (1) | US6709837B1 (ko) |
| EP (1) | EP0386752B1 (ko) |
| JP (1) | JPH0780904B2 (ko) |
| KR (1) | KR0132425B1 (ko) |
| AT (1) | ATE111521T1 (ko) |
| CA (1) | CA2011833C (ko) |
| DE (1) | DE69012377T2 (ko) |
| DK (1) | DK0386752T3 (ko) |
| ES (1) | ES2059854T3 (ko) |
| HK (1) | HK34896A (ko) |
| IE (1) | IE65361B1 (ko) |
Families Citing this family (16)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5656435A (en) * | 1989-08-28 | 1997-08-12 | Takeda Chemical Industries, Ltd. | Antibodies to peptides having NGF-like activity, said antibodies having no substantial cross-reactivity with NGF, and use thereof |
| US5180820A (en) * | 1989-08-30 | 1993-01-19 | Barde Yves Alain | Brain-derived neurotrophic factor |
| US6933276B1 (en) | 1989-08-30 | 2005-08-23 | Regeneron Pharmaceuticals, Inc. | Methods of treating peripheral neuropathies using neurotrophin-3 |
| GR1000980B (el) * | 1989-08-30 | 1993-03-31 | Max Planck Gesellschaft | Νευροτροφινη-3,ενας νεος neυροτροφικος παραγοντας που σχετιζεται με τον παραγοντα αναπτυξης νευρων και τον νευροτροφικο παραγοντα που προερχεται απο τον εγκεφαλο. |
| US5235043A (en) * | 1990-04-06 | 1993-08-10 | Synergen, Inc. | Production of biologically active, recombinant members of the ngf/bdnf family of neurotrophic proteins |
| US6511823B1 (en) * | 1990-08-20 | 2003-01-28 | American Cyanamid Corporation | Heparin binding neurotrophic factor gene sequence |
| DK0499993T3 (da) * | 1991-02-18 | 1997-12-01 | Takeda Chemical Industries Ltd | Fremgangsmåde til fremstilling af human nervevækstfaktor-2 |
| US5389529A (en) * | 1991-06-12 | 1995-02-14 | Regeneron Pharmaceuticals, Inc. | Modified lamβ signal sequence and processes for producing recombinant neurotrophins |
| EP0573904B1 (en) * | 1992-06-08 | 2001-09-26 | Takeda Chemical Industries, Ltd. | Therapeutic agent for neutropenia |
| AU5684594A (en) * | 1992-12-02 | 1994-06-22 | Novo Nordisk A/S | Novel human amyloid protein precursor homologue and kunitz-type inhibitors |
| JP2732005B2 (ja) * | 1992-12-14 | 1998-03-25 | 眞一 富永 | ヒトst2をコードするdna、該dnaの発現産物、該dnaを発現させることによる発現産物の製造方法 |
| JPH07246040A (ja) * | 1993-07-06 | 1995-09-26 | Takeda Chem Ind Ltd | ニューロトロフィン−3遺伝子が不活性化された胚幹細胞および該遺伝子発現不全動物 |
| US5877016A (en) * | 1994-03-18 | 1999-03-02 | Genentech, Inc. | Human trk receptors and neurotrophic factor inhibitors |
| US5681931A (en) * | 1995-03-15 | 1997-10-28 | Becton, Dickinson And Company | Human restrictin |
| US5864018A (en) * | 1996-04-16 | 1999-01-26 | Schering Aktiengesellschaft | Antibodies to advanced glycosylation end-product receptor polypeptides and uses therefor |
| AU7301698A (en) * | 1996-11-22 | 1998-06-10 | Human Genome Sciences, Inc. | Thermostable polymerases having altered fidelity |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DK161152C (da) | 1983-03-03 | 1991-11-11 | Genentech Inc | Polypeptid med egenskaber som human beta-nervevaekstfaktor og fremgangsmaade til fremstilling deraf, dna-isolat omfattende en sekvens som koder for polypeptidet, replicerbar udtrykkelsesvektor for dna-sekvensen, rekombinant vaertscelle transformeret med vektoren, farmaceutisk praeparat indeholdende polypeptidet og fremg. der omfatter anvendelsen af polypeptidet til fremst. af et farmaceutisk praeparat |
| US5180820A (en) * | 1989-08-30 | 1993-01-19 | Barde Yves Alain | Brain-derived neurotrophic factor |
-
1990
- 1990-03-05 US US07/488,696 patent/US6709837B1/en not_active Expired - Fee Related
- 1990-03-06 IE IE78590A patent/IE65361B1/en not_active IP Right Cessation
- 1990-03-08 DK DK90104419.8T patent/DK0386752T3/da active
- 1990-03-08 DE DE69012377T patent/DE69012377T2/de not_active Expired - Lifetime
- 1990-03-08 JP JP2057751A patent/JPH0780904B2/ja not_active Expired - Lifetime
- 1990-03-08 AT AT90104419T patent/ATE111521T1/de not_active IP Right Cessation
- 1990-03-08 EP EP90104419A patent/EP0386752B1/en not_active Expired - Lifetime
- 1990-03-08 ES ES90104419T patent/ES2059854T3/es not_active Expired - Lifetime
- 1990-03-09 CA CA002011833A patent/CA2011833C/en not_active Expired - Lifetime
- 1990-03-10 KR KR1019900003202A patent/KR0132425B1/ko not_active IP Right Cessation
-
1996
- 1996-02-29 HK HK34896A patent/HK34896A/xx not_active IP Right Cessation
Also Published As
| Publication number | Publication date |
|---|---|
| US6709837B1 (en) | 2004-03-23 |
| DK0386752T3 (da) | 1994-12-05 |
| JPH0780904B2 (ja) | 1995-08-30 |
| DE69012377T2 (de) | 1995-05-04 |
| ATE111521T1 (de) | 1994-09-15 |
| EP0386752B1 (en) | 1994-09-14 |
| DE69012377D1 (de) | 1994-10-20 |
| JPH03204897A (ja) | 1991-09-06 |
| HK34896A (en) | 1996-03-08 |
| CA2011833C (en) | 2002-11-05 |
| CA2011833A1 (en) | 1990-09-10 |
| IE900785L (en) | 1990-09-10 |
| EP0386752A1 (en) | 1990-09-12 |
| IE65361B1 (en) | 1995-10-18 |
| KR900014428A (ko) | 1990-10-23 |
| ES2059854T3 (es) | 1994-11-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR0132425B1 (ko) | 신규 폴리펩타이드 및 그 제조방법 | |
| US5141856A (en) | Expression of purified ciliary neurotrophic factor | |
| Takahashi et al. | Cloning and sequencing of cDNA of Sarcophaga peregrina humoral lectin induced on injury of the body wall. | |
| US5221620A (en) | Cloning and expression of transforming growth factor β2 | |
| US4994559A (en) | Human basic fibroblast growth factor | |
| GB2214185A (en) | Proteins and methods for their production | |
| WO1985002198A1 (en) | Microbial expression of type i transforming growth factor, polypeptide analogs thereof and hybrid egf/tgf polypeptides | |
| EP0226181B1 (en) | Human placenta angiogenic factor capable of stimulating capillary endothelial cell protease synthesis, DNA synthesis and migration | |
| EP0414151B1 (en) | Production of human nerve growth factor proteins | |
| US4732972A (en) | Polypeptides having growth hormone releasing activity | |
| US6312688B1 (en) | Tyrosine-phosphatase-related protein | |
| JPH05103675A (ja) | ヒト神経成長因子2の製造法 | |
| JPH06317587A (ja) | 抗体およびその用途 | |
| JP3232415B2 (ja) | モノクローナル抗体,その製造法および用途 | |
| HU211309A9 (hu) | Új polipeptid és előállítása | |
| EP0421059B1 (en) | Purified ciliary neurotrophic factor | |
| AU642858C (en) | Purified ciliary neurotrophic factor | |
| JPH04128300A (ja) | ヒト神経成長因子蛋白質およびその製造法 | |
| JPH03139285A (ja) | 酵母によるヒト神経成長因子の製造法 | |
| CA2113076A1 (en) | Recombinant human interleukin 6 with homogeneous n-terminus and production thereof | |
| JPS62130697A (ja) | ペプチドの製造法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| E701 | Decision to grant or registration of patent right | ||
| GRNT | Written decision to grant | ||
| FPAY | Annual fee payment |
Payment date: 20101124 Year of fee payment: 14 |
|
| LAPS | Lapse due to unpaid annual fee |