JP7246726B2 - 重量が増大した植物の製造方法 - Google Patents
重量が増大した植物の製造方法 Download PDFInfo
- Publication number
- JP7246726B2 JP7246726B2 JP2019219693A JP2019219693A JP7246726B2 JP 7246726 B2 JP7246726 B2 JP 7246726B2 JP 2019219693 A JP2019219693 A JP 2019219693A JP 2019219693 A JP2019219693 A JP 2019219693A JP 7246726 B2 JP7246726 B2 JP 7246726B2
- Authority
- JP
- Japan
- Prior art keywords
- plant
- gene
- phytochrome
- plants
- rice
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images

Description
<1> 重量が増大した植物の製造方法であって、フィトクロムA遺伝子の機能が抑制された植物を、通常の栽培条件における栄養素の濃度と比して少なくとも1.5倍高い濃度にて栽培する、方法。
<2> 前記栄養素は、窒素、リン酸及びカリウムである、<1>に記載の方法。
本発明において重量増大の対象となる「植物」は、フィトクロムA遺伝子の機能が抑制されれば、特に制限はなく、例えば、単子葉植物(例えば、イネ、トウモロコシ、ソルガム、コムギ等のイネ科植物)及び双子葉植物(例えば、トマト、ジャガイモ等のナス科植物)を含む被子植物、裸子植物、コケ植物、シダ植物、草本植物、並びに木本植物が挙げられる。さらに、これら植物の遺伝子組み換え体やゲノム編集体(例えば、除草剤耐性作物、害虫耐性作物、病害耐性作物、食味向上作物、保存性向上作物、収量向上作物)であっても良い。
本発明において「フィトクロムA遺伝子」とは、光受容タンパク質の1種であるフィトクロムA(phyA)をコードする遺伝子であり、例えば、下記表に示すアミノ酸配列を含むタンパク質をコードする遺伝子(下記表に示すヌクレオチド配列を含むタンパク質をコードする遺伝子が挙げられる。なお、表1において「遺伝子数」は、各植物種が有するフィトクロムA遺伝子の遺伝子数(パラロガス遺伝子の数)を示す。

本発明において、「フィトクロムA遺伝子の機能の抑制」には、該機能の完全な抑制(阻害)及び部分的な抑制の双方が含まれる。また、複数のフィトクロムA遺伝子(phyA1~phyA5等のパラロガス遺伝子)を有する植物種も存在する(例えば、表1に示すコムギにおいて、AEA40430、AEA40439及びAEA40449にて示されるアミノ酸配列をコードする遺伝子は、各々phyA1遺伝子、phyA2遺伝子及びphyA3遺伝子である)。かかる植物種の場合、少なくとも1のフィトクロムA遺伝子(例えば、コムギにおいてphyA1遺伝子のみ)の機能が抑制されていればよいが、全てのフィトクロムA遺伝子(例えば、コムギにおいてはphyA1遺伝子、phyA2遺伝子及びphyA3遺伝子)の機能が抑制されていることが好ましい。また、フィトクロムA遺伝子の機能の抑制には、フィトクロムA遺伝子の発現抑制の他、フィトクロムA遺伝子がコードするタンパク質の活性の抑制が含まれる。そして、かかる抑制は、例えば、フィトクロムA遺伝子のコード領域、非コード領域、転写制御領域(プロモーター領域)等に変異を導入することによって行なうことができる。
後述の実施例に示すとおり、本発明の製造方法において、上述のフィトクロムA遺伝子の機能が抑制された植物を、通常の栽培条件における栄養素の濃度と比して少なくとも1.5倍高い濃度にて栽培することによって、重量が増大した植物を得ることができる。

phyAを欠損したイネ(phyA欠損イネ)及びその親品種である日本晴を、以下の実験に供した。phyA欠損イネは、旧農業生物資源研究所で作製されたレトロトランスポゾンTos17による突然変異体集団(ミュータントパネル)より選抜されたイネである(非特許文献1及び2 参照)。後述の実施例1及び4においては、phyA欠損イネ 系統A♯1を用い、実施例2においては、系統A♯1とは独立した系統であるphyA欠損イネ 系統A#2を用いた。実施例3においては前記A♯1とA♯2の2系統を用いた。
330mLの下記高栄養又は低栄養の水耕液あたり、6本のイネを用い、水耕栽培を実施した。水耕液は2日ごとに新しい液に交換し、イネは5葉期になるまで(おおよそ3週間)育てた。水耕液は、以下のとおりに、各種ストック液を混合して調製した。
I液(500mLストック)
KNO3(硝酸カリウム、MW=101.1) 101.1g(2M)
NH4H2PO4(りん酸二水素アンモニウム、MW=115.03) 57.5g(1M)
II液(200mLストック)
MgSO4-7H2O(硫酸マグネシウム七水和物、MW=246.48) 49.3g(1M)
III液(100mLストック)
CaCl2-2H2O(塩化カルシウム二水和物、MW=147.01) 73.5g(5M)
IV液(200mLストック)
H3BO3(ホウ酸、MW=61.83) 0.572g(46.26mM)
MnSO4-5H2O(硫酸マンガン(II)五水和物、MW=241.08) 0.307g(6.37mM)
CuSO4-5H2O(硫酸銅(II)五水和物、MW=249.7) 0.012g(0.24mM)
ZnSO4-7H2O(硫酸亜鉛(II)七水和物、MW=287.56) 0.156g(2.71mM)
K2MoO4(モリブデン酸カリウム、MW=238.13) 0.023g(0.48mM)
V液(100mLストック)
EDTA-Na-Fe(II)-H2O(Fe-EDTA、MW=385.06) 0.193g(50mM)。
後述の実施例2においては、2005年5月中旬にイネ幼植物(4から5葉期のイネ)を水田に移植した。肥料(基肥、穂肥)は通常の施肥量で実施した。具体的には、表2に示す日本晴の普通作物栽培基準に基づく施肥量にて栽培を行なった。イネ幼植物は30センチ(疎植)又は15センチ(密植)の間隔で水田に移植した。
新しい水耕液交換後4、24、48時間後にサンプリングした水耕液を13AI 0.2μm クロマトディスク(ジーエルサイエンス社製)でろ過処理し、各栄養素含量をイオンクロマトグラフ(ダイオネックス社製)を用いて測定した。
上述のとおり、水耕液を用い、低栄養条件又は高栄養条件(低栄養条件と比べて栄養素が2.5倍濃い)にて、phyA欠損イネ及び親品種を栽培した。そして、得られた各幼苗を、地上部と根に分けて乾燥重量を測定した。得られた結果を表3に示す。

上述のとおり、水田において、疎植(株間:30cm、高栄養条件)又は密植(株間:15cm、通常の栄養条件)にてphyA欠損イネ及び親品種を籾が成熟するまで栽培した。そして、得られた全植物体及び籾の乾燥重量を測定した。得られた結果を表4に示す。
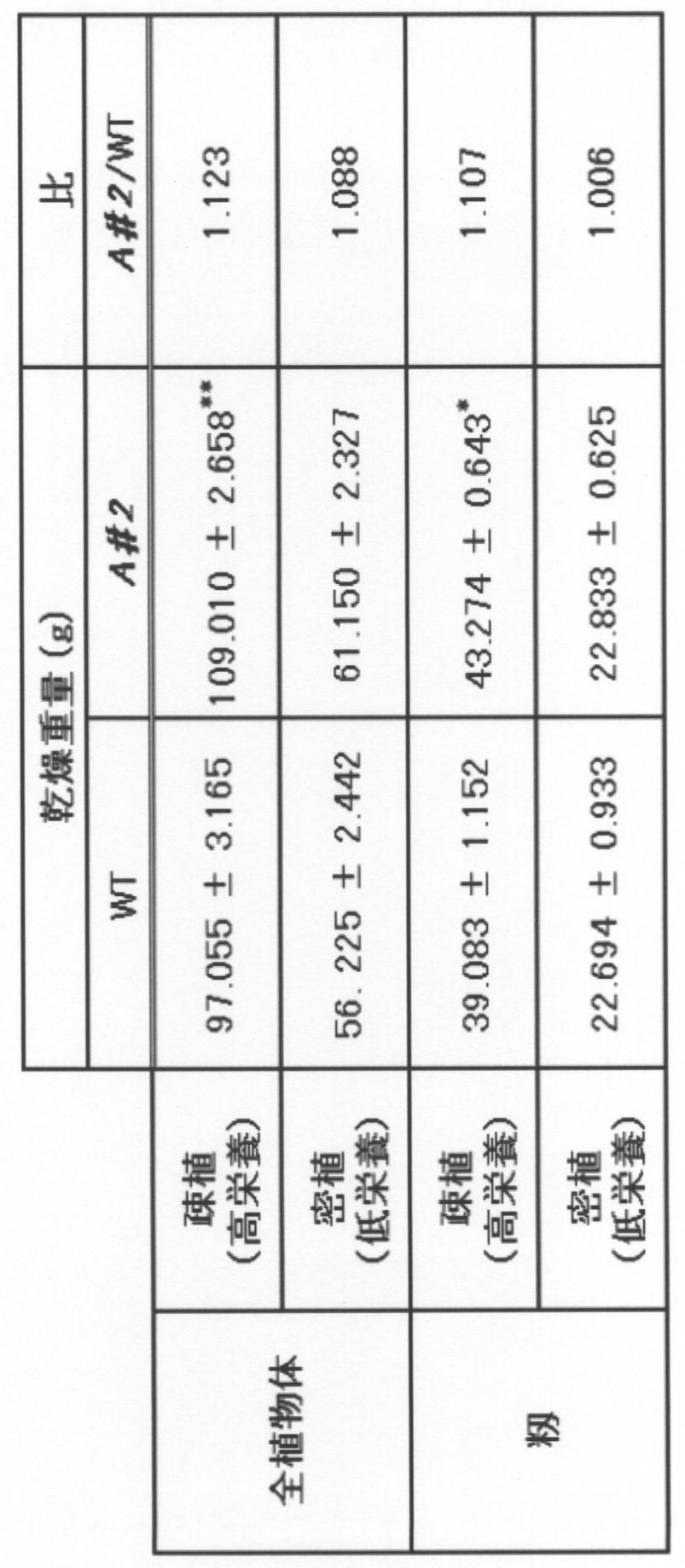
上述のとおり、水田において、通常の栄養条件下又は高栄養条件(施肥量2倍)にてphyA欠損イネ(2系統)及び親品種を籾が成熟するまで栽培した。そして、得られたイネの穂及び地上部(穂以外の部分)の乾燥重量を測定した。得られた結果を表5に示す。

実施例1~3に示すとおり、高栄養条件にてphyA欠損イネを栽培した結果、その重量は親品種と比して増大した。このことから、phyA欠損イネの栄養素の吸収能力が、高栄養条件下において促進することが予想される。
Claims (2)
- 重量が増大した植物の製造方法であって、
フィトクロムA遺伝子の機能が抑制された植物を、通常の栽培条件における栄養素の濃度と比して少なくとも1.5倍高い濃度にて栽培する、方法。 - 前記栄養素は、窒素、リン酸及びカリウムである、請求項1に記載の方法。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2019027742 | 2019-02-19 | ||
| JP2019027742 | 2019-02-19 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2020130174A JP2020130174A (ja) | 2020-08-31 |
| JP7246726B2 true JP7246726B2 (ja) | 2023-03-28 |
Family
ID=72276840
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2019219693A Active JP7246726B2 (ja) | 2019-02-19 | 2019-12-04 | 重量が増大した植物の製造方法 |
Country Status (1)
| Country | Link |
|---|---|
| JP (1) | JP7246726B2 (ja) |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2003020935A1 (fr) | 2001-09-03 | 2003-03-13 | National Institute Of Agrobiological Sciences | Regulation du temps de floraison de vegetaux a travers l'expression du phytochrome c |
| JP2004520822A (ja) | 2000-12-06 | 2004-07-15 | シンジェンタ リミテッド | トマト(tomato)由来のHP−1突然変異体の活性を模倣する、組換え植物 |
| JP2007145614A (ja) | 2005-11-24 | 2007-06-14 | Sumika Takeda Engei Kk | 高濃度液体肥料組成物 |
| JP2010233509A (ja) | 2009-03-31 | 2010-10-21 | National Institute Of Agrobiological Sciences | 青色ledを利用した、イネを短期間で低コストに育成収穫する方法、及びこの方法に適した系統の選抜 |
-
2019
- 2019-12-04 JP JP2019219693A patent/JP7246726B2/ja active Active
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2004520822A (ja) | 2000-12-06 | 2004-07-15 | シンジェンタ リミテッド | トマト(tomato)由来のHP−1突然変異体の活性を模倣する、組換え植物 |
| WO2003020935A1 (fr) | 2001-09-03 | 2003-03-13 | National Institute Of Agrobiological Sciences | Regulation du temps de floraison de vegetaux a travers l'expression du phytochrome c |
| JP2007145614A (ja) | 2005-11-24 | 2007-06-14 | Sumika Takeda Engei Kk | 高濃度液体肥料組成物 |
| JP2010233509A (ja) | 2009-03-31 | 2010-10-21 | National Institute Of Agrobiological Sciences | 青色ledを利用した、イネを短期間で低コストに育成収穫する方法、及びこの方法に適した系統の選抜 |
Also Published As
| Publication number | Publication date |
|---|---|
| JP2020130174A (ja) | 2020-08-31 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Kieu et al. | Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes | |
| JP6375398B2 (ja) | Alsインヒビター除草剤耐性ベータ・ブルガリス突然変異体 | |
| Wang et al. | Mutagenesis reveals that the OsPPa6 gene is required for enhancing the alkaline tolerance in rice | |
| Zhao et al. | Over-expression of a tobacco nitrate reductase gene in wheat (Triticum aestivum L.) increases seed protein content and weight without augmenting nitrogen supplying | |
| WO2019024534A1 (zh) | 使植物具有除草剂抗性的水稻als突变型蛋白及其应用 | |
| CN108368515A (zh) | 耐旱玉米 | |
| Wang et al. | Enhanced soluble sugar content in tomato fruit using CRISPR/Cas9-mediated SlINVINH1 and SlVPE5 gene editing | |
| BR112020016016A2 (pt) | Composições e métodos para aprimorar rendimentos de cultura através do empilhamento de traços | |
| Park et al. | Applications of CRISPR/Cas9 as new strategies for short breeding to drought gene in rice | |
| Abe et al. | Gene overexpression resources in cereals for functional genomics and discovery of useful genes | |
| Lata et al. | Engineering cereal crops for enhanced abiotic stress tolerance | |
| CN110881367A (zh) | 一种玉米事件t抗-4及其使用方法 | |
| JP2015508663A (ja) | 種子活力のモジュレーション | |
| JP7246726B2 (ja) | 重量が増大した植物の製造方法 | |
| CN105802931A (zh) | Crk4蛋白及其编码基因在调控植物茎叶生长中的应用 | |
| JP6874956B2 (ja) | セシウム吸収を制御する遺伝子およびセシウム低吸収性植物 | |
| Ali et al. | An assessment on CRISPR Cas as a novel asset in mitigating drought stress | |
| US20230279419A1 (en) | Enhancement of productivity in c3 plants | |
| CN111269924B (zh) | 水稻半胱氨酸合酶编码基因OsASTOL1突变体及其应用 | |
| US20200347399A1 (en) | Method for promoting an increase in plant biomass, productivity, and drought resistance | |
| Yang et al. | Phylogenetic and expression analysis of pear yellow stripe-like transporters and functional verification of PbrYSL4 in pear pollen | |
| CN116121298B (zh) | 抑制hsrp1基因的表达在提高植物耐热性中的应用 | |
| CN104450739B (zh) | 一种水稻源抗虫相关基因OsHR1及其编码产物与应用 | |
| Shahwar et al. | Mutagenesis-based Plant Breeding Approaches and Genome Engineering: A Review Focused on Tomato | |
| CN115851821B (zh) | Bbx16基因在提高植物盐耐受性中的应用 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20220628 |
|
| A977 | Report on retrieval |
Free format text: JAPANESE INTERMEDIATE CODE: A971007 Effective date: 20230224 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20230302 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20230308 |
|
| R150 | Certificate of patent or registration of utility model |
Ref document number: 7246726 Country of ref document: JP Free format text: JAPANESE INTERMEDIATE CODE: R150 |